Перекрестная ссылка на родственные заявки
В этой заявке заявлен приоритет к предварительной заявке на патент США № 61/653,522, поданной 5 апреля 2018, озаглавленной “СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР, И РОДСТВЕННЫЕ КОМПОЗИЦИИ”, содержание которой включено сюда в качестве ссылки полностью.
Включение в качестве ссылки списка последовательностей
Данная заявка подана вместе со списком последовательностей в электронном формате. Список последовательностей представлен в виде файла, озаглавленного 735042012740SeqList.txt, созданного 3 апреля 2019, размером 179 килобайтов. Информация в электронном формате списка последовательностей включена сюда в качестве ссылки полностью.
Область техники
Данное изобретение относится к способам инжиниринга иммунных клеток, клеточных композиций, содержащих сконструированные иммунные клетки, наборам и готовым изделиям для целенаправленного воздействия на последовательность нуклеиновых кислот, кодирующую рекомбинантный рецептор до конкретного геномного локуса и/или для модулирования экспрессии гена в геномном локусе, и их применению в связи с противораковой иммунотерапией, включающей адоптивный перенос сконструированных T клеток.
Уровень техники
Адоптивные клеточные терапии, в которых применяют рекомбинантно экспрессированные Т-клеточные рецепторы (TCR) или другие антигенные рецепторы (например, химерные антигенные рецепторы (CAR)) для распознавания опухолевых антигенов, представляют собой привлекательные терапевтические методы для лечения раков и других заболеваний. Экспрессия и функция рекомбинантных TCR или других антигенных рецепторов могут быть ограниченными и/или гетерогенными в популяции клеток. Улучшенные стратегии необходимы для достижения высоких и/или гомогенных уровней экспрессии и функционирования рекомбинантных рецепторов. Эти стратегии могут способствовать созданию клеток, демонстрирующих желаемые уровни экспрессии и/или свойства для применения в адоптивной иммунотерапии, например, при лечении рака, инфекционных заболеваний и аутоиммунных заболеваний. Представлены способы, клетки, композиции и наборы для применения в способах, которые отвечают таким нуждам.
Сущность изобретения
Представлены генетически сконструированные клетки, которые содержат генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), и трансген, кодирующий рекомбинантный рецептор, такой как Т-клеточный рецептор (TCR) или химерный антигенный рецептор (CAR), который интегрирован, через репарацию, направляемую гомологией (HDR), в или рядом с одним или более целевыми сайтами, и композиция, содержащая сконструированные клетки, способы получения сконструированных клеток и родственные способы и применения. В некоторых аспектах, посредством генетического разрушения и направленной интеграции трансгенных последовательностей, одну или более из эндогенных TCR цепей уменьшают или инактивируют в экспрессии. В некоторых из любых таких вариантов, рекомбинантный рецептор может связываться с антигеном, который ассоциирован с клеткой или тканью заболевания, расстройства или состояния. В некоторых из любых таких вариантов, рекомбинантный рецептор может связываться с антигеном, который является специфическим к клетке или ткани заболевания, расстройства или состояния. В некоторых из любых таких вариантов, рекомбинантный рецептор может связываться с антигеном, который экспрессирован на клетке или ткани, ассоциированной с заболеванием, расстройством или состоянием.
Также представлена композиция, содержащая сконструированную клетку или множество сконструированных клеток, описанных здесь. В конкретных вариантах, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции содержат генетическое разрушение, по меньшей мере, одного целевого сайта в гене, кодирующем домен или область гена альфа константы Т-клеточного рецептора (TRAC) и/или гена бета константы Т-клеточного рецептора (TRBC). В определенных вариантах, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции экспрессируют рекомбинантный рецептор или его антигенсвязывающий фрагмент и/или демонстрируют антигенное связывание. В некоторых из любых таких вариантов, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции экспрессируют рекомбинантный рецептор или его антигенсвязывающий фрагмент и/или демонстрируют связывание с антигеном.
Представлены композиции, содержащие множество сконструированных T клеток. В некоторых из любых таких вариантов, композиция, содержащая множество сконструированных T клеток, содержащих рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, кодированный трансгеном, и генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), где рекомбинантный рецептор способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния, и где, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции содержат генетическое разрушение, по меньшей мере, одного целевого сайта в гене TRAC и/или гене TRBC; и/или, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции экспрессируют рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, и/или демонстрируют связывание с антигеном; и трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, интегрирован в или рядом с одним из, по меньшей мере, одним целевым сайтом через репарацию, направляемую гомологией (HDR).
В некоторых вариантах, коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора или его антигенсвязывающего фрагмента или цепи среди множества клеток ниже, чем 0,70, 0,65, 0,60, 0,55, 0,50, 0,45, 0,40, 0,35 или 0,30 или менее. В конкретных вариантах, коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора или его антигенсвязывающего фрагмента или цепи среди множества клеток на, по меньшей мере, 100%, 95%, 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20% или 10% ниже, чем коэффициент вариации экспрессии и/или антигенного связывания того же рекомбинантного рецептора, который интегрирован в геном случайной интеграцией. В некоторых из любых таких вариантов, рекомбинантный рецептор способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния.
В определенных вариантах, экспрессию и/или антигенное связывание рекомбинантного рецептора или его антигенсвязывающего фрагмента оценивают через контакт клеток в композиции со связывающим реагентом, специфическим к TCRα цепи или TCRβ цепи и оценку связывания реагента с клетками. В некоторых вариантах, связывающим реагентом является анти-TCR Vβ антитело или анти-TCR Vα антитело, которое специфически распознает определенное семейство Vβ или Vα цепей.
В конкретных вариантах, связывающим агентом является пептидный антиген-MHC комплекс, который необязательно является тетрамером. В определенных вариантах, композиция, описанная здесь, дополнительно содержит фармацевтически приемлемый носитель. Представлена композиция, содержащая множество сконструированных Т клеток, содержащих рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, кодированные трансгеном, и генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), где, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции содержат генетическое разрушение, по меньшей мере, одного целевого сайта в гене TRAC и/или гене TRBC; и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелены для интеграции в или рядом с, по меньшей мере, одним целевым сайтом через репарацию, направляемую гомологией (HDR).
Также здесь представлена композиция, содержащая множество сконструированных Т клеток, содержащих рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, кодированные трансгеном, и генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), где, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции экспрессируют рекомбинантный рецептор или его антигенсвязывающий фрагмент и/или демонстрируют антигенное связывание; и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелены для интеграции в или рядом с, по меньшей мере, одним целевым сайтом через репарацию, направляемую гомологией (HDR).
Также представлена композиция, содержащая множество сконструированных Т клеток, содержащих рекомбинантный рецептор или его антигенсвязывающий фрагмент, кодированные трансгеном, и генетическое разрушение, по меньшей мере, одного целевого сайта в Гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), где коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора среди множества клеток ниже, чем 0,70, 0,65, 0,60, 0,55, 0,50, 0,45, 0,40, 0,35 или 0,30 или менее.
Также здесь представлена композиция, содержащая множество сконструированных Т клеток, содержащих рекомбинантный рецептор или его антигенсвязывающий фрагмент, кодированные трансгеном, и генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), где коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора среди множества клеток на, по меньшей мере, 100%, 95%, 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20% или 10% ниже, чем коэффициент вариации экспрессии и/или антигенного связывания того же рекомбинантного рецептора, который интегрирован в геном случайной интеграцией.
В некоторых вариантах, композицию получают: (a) введением во множество T клеток одного или более агентов, где каждый из одного или более агентов независимо способны вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), тем самым вызывая генетическое разрушение, по меньшей мере, одного целевого сайта; и (b) введением во множество T клеток матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или цепь, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию на или рядом с, по меньшей мере, одним целевым сайтом через репарацию, направляемую гомологией (HDR). В конкретных вариантах, экспрессию и/или антигенное связывание рекомбинантного рецептора или его антигенсвязывающего фрагмента оценивают через контакт клеток в композиции со связывающим реагентом, специфическим к TCRα цепи или TCRβ цепи и оценку связывания реагента с клетками.
В некоторых из любых таких вариантов, сконструированные Т клетки содержат, по меньшей мере, одно генетическое разрушение в гене TRAC. В некоторых из любых таких вариантов, сконструированные Т клетки содержат, по меньшей мере, одно генетическое разрушение в гене TRBC. В некоторых из любых таких вариантов, сконструированные Т клетки содержат, по меньшей мере, одно генетическое разрушение целевого сайта в гене TRAC и, по меньшей мере, одно генетическое разрушение целевого сайта в гене TRBC.
В определенных вариантах, связывающим реагентом является анти-TCR Vβ антитело или является анти-TCR Vα антитело, которое специфически распознает конкретное семейство Vβ или Vα цепей. В некоторых вариантах, связывающим агентом является пептидный антиген-МНС комплекс, который необязательно является тетрамером. В конкретных вариантах, по меньшей мере, один из одного или более агентов способен вызывать генетическое разрушение целевого сайта в гене TRAC. В определенных вариантах, по меньшей мере, один из одного или более агентов способен вызывать генетическое разрушение целевого сайта в гене TRBC. В некоторых вариантах, один или более агентов содержат, по меньшей мере, один агент, который способен вызывать генетическое разрушение целевого сайта в гене TRAC и, по меньшей мере, один агент, который способен вызывать генетическое разрушение целевого сайта в гене TRBC. В конкретных вариантах, геном TRBC является один или оба из генов бета константы Т-клеточного рецептора 1 (TRBC1) или бета константы Т-клеточного рецептора 2 (TRBC2). В определенных вариантах, один или более агентов, способных вызывать генетическое разрушение, содержат ДНК-связывающий белок или ДНК-связывающую нуклеиновую кислоту, которые специфически связываются с или гибридизируются в целевой сайт. В некоторых вариантах, один или более агент, способный вызывать генетическое разрушение, содержит (a) слитый белок, содержащий ДНК-направленный белок и нуклеазу или (b) РНК-направляемую нуклеазу.
В конкретных вариантах, ДНК-направленный белок или РНК-направляемая нуклеаза содержит белок «цинковый палец» (ZFP), TAL белок или связанную с кластеризованной регулярной промежуточной короткой палиндромной нуклеиновой кислотой (CRISPR) нуклеазу (Cas), специфическую к целевому сайту. В определенных вариантах, один или более агент содержит нуклеазу «цинковый палец» нуклеазу «цинковый палец» (ZFN), TAL-эффекторную нуклеазу (TALEN) и/или CRISPR-Cas9 сочетание, которые специфически связываются с, распознают или гибридизируются в целевой сайт. В некоторых вариантах, каждый из одного или более агентов содержит направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту. В конкретных вариантах, один или более агентов вводят в виде рибонуклеопротеинового (RNP) комплекса, содержащего нРНК и Cas9 белок. В определенных вариантах, RNP вводят через электропорацию, генную пушку, трансфекцию фосфата кальция, сгущение или сжатие клеток. В некоторых вариантах, RNP вводят электропорацией. В конкретных вариантах, один или более агентов вводят в виде одного или более полинуклеотидов, кодирующих нРНК и/или Cas9 белок. В определенных вариантах, по меньшей мере, один целевой сайт находится внутри экзона TRAC, TRBC1 и/или TRBC2 гена.
В некоторых из любых таких вариантов, генетическое разрушение производится нуклеазой «цинковый палец» (ZFN), TAL-эффекторной нуклеазой (TALEN) или сочетанием CRISPR-Cas9, которые специфически связываются с, распознают или гибридизируются в целевой сайт. В некоторых из любых таких вариантов, генетическое разрушение CRISPR-Cas9 сочетанием включает направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту. В некоторых из любых таких вариантов, CRISPR-Cas9 сочетанием является рибонуклеопротеиновый (RNP) комплекс, содержащий нРНК и Cas9 белок. В некоторых из любых таких вариантов, RNP вводят через электропорацию. В некоторых из любых таких вариантов, по меньшей мере, один целевой сайт находится внутри экзона TRAC, TRBC1 и/или TRBC2 гена.
В некоторых вариантах, нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в TRAC гене и содержит последовательность, выбранную из группы, состоящей из UCUCUCAGCUGGUACACGGC (SEQ ID NO:28), UGGAUUUAGAGUCUCUCAGC (SEQ ID NO:29), ACACGGCAGGGUCAGGGUUC (SEQ ID NO:30), GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31), GCUGGUACACGGCAGGGUCA (SEQ ID NO:32), CUCAGCUGGUACACGGC (SEQ ID NO:33), UGGUACACGGCAGGGUC (SEQ ID NO:34), GCUAGACAUGAGGUCUA (SEQ ID NO:35), GUCAGAUUUGUUGCUCC (SEQ ID NO:36), UCAGCUGGUACACGGCA (SEQ ID NO:37), GCAGACAGACUUGUCAC (SEQ ID NO:38), GGUACACGGCAGGGUCA (SEQ ID NO:39), CUUCAAGAGCAACAGUGCUG (SEQ ID NO:40), AGAGCAACAGUGCUGUGGCC (SEQ ID NO:41), AAAGUCAGAUUUGUUGCUCC (SEQ ID NO:42), ACAAAACUGUGCUAGACAUG (SEQ ID NO:43), AAACUGUGCUAGACAUG (SEQ ID NO:44), UGUGCUAGACAUGAGGUCUA (SEQ ID NO:45), GGCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:46), GCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:47), GGGGAAGAAGGUGUCUUC (SEQ ID NO:48), GUUUUGUCUGUGAUAUACACAU (SEQ ID NO:49), GGCAGACAGACUUGUCACUGGAUU (SEQ ID NO:50), GCAGACAGACUUGUCACUGGAUU (SEQ ID NO:51), GACAGACUUGUCACUGGAUU (SEQ ID NO:52), GUGAAUAGGCAGACAGACUUGUCA (SEQ ID NO:53), GAAUAGGCAGACAGACUUGUCA (SEQ ID NO:54), GAGUCUCUCAGCUGGUACACGG (SEQ ID NO:55), GUCUCUCAGCUGGUACACGG (SEQ ID NO:56), GGUACACGGCAGGGUCAGGGUU (SEQ ID NO:57) и GUACACGGCAGGGUCAGGGUU (SEQ ID NO:58). В конкретных вариантах, нРНК имеет домен целенаправленного воздействия, содержащий последовательность GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31).
В определенных вариантах, нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в одном или обоих TRBC1 и TRBC2 генах и содержит последовательность, выбранную из группы, состоящей из CACCCAGAUCGUCAGCGCCG (SEQ ID NO:59), CAAACACAGCGACCUCGGGU (SEQ ID NO:60), UGACGAGUGGACCCAGGAUA (SEQ ID NO:61), GGCUCUCGGAGAAUGACGAG (SEQ ID NO:62), GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63), GAAAAACGUGUUCCCACCCG (SEQ ID NO:64), AUGACGAGUGGACCCAGGAU (SEQ ID NO:65), AGUCCAGUUCUACGGGCUCU (SEQ ID NO:66), CGCUGUCAAGUCCAGUUCUA (SEQ ID NO:67), AUCGUCAGCGCCGAGGCCUG (SEQ ID NO:68), UCAAACACAGCGACCUCGGG (SEQ ID NO:69), CGUAGAACUGGACUUGACAG (SEQ ID NO:70), AGGCCUCGGCGCUGACGAUC (SEQ ID NO:71), UGACAGCGGAAGUGGUUGCG (SEQ ID NO:72), UUGACAGCGGAAGUGGUUGC (SEQ ID NO:73), UCUCCGAGAGCCCGUAGAAC (SEQ ID NO:74), CGGGUGGGAACACGUUUUUC (SEQ ID NO:75), GACAGGUUUGGCCCUAUCCU (SEQ ID NO:76), GAUCGUCAGCGCCGAGGCCU (SEQ ID NO:77), GGCUCAAACACAGCGACCUC (SEQ ID NO:78), UGAGGGUCUCGGCCACCUUC (SEQ ID NO:79), AGGCUUCUACCCCGACCACG (SEQ ID NO:80), CCGACCACGUGGAGCUGAGC (SEQ ID NO:81), UGACAGGUUUGGCCCUAUCC (SEQ ID NO:82), CUUGACAGCGGAAGUGGUUG (SEQ ID NO:83), AGAUCGUCAGCGCCGAGGCC (SEQ ID NO:84), GCGCUGACGAUCUGGGUGAC (SEQ ID NO:85), UGAGGGCGGGCUGCUCCUUG (SEQ ID NO:86), GUUGCGGGGGUUCUGCCAGA (SEQ ID NO:87), AGCUCAGCUCCACGUGGUCG (SEQ ID NO:88), GCGGCUGCUCAGGCAGUAUC (SEQ ID NO:89), GCGGGGGUUCUGCCAGAAGG (SEQ ID NO:90), UGGCUCAAACACAGCGACCU (SEQ ID NO:91), ACUGGACUUGACAGCGGAAG (SEQ ID NO:92), GACAGCGGAAGUGGUUGCGG (SEQ ID NO:93), GCUGUCAAGUCCAGUUCUAC (SEQ ID NO:94), GUAUCUGGAGUCAUUGAGGG (SEQ ID NO:95), CUCGGCGCUGACGAUCU (SEQ ID NO:96), CCUCGGCGCUGACGAUC (SEQ ID NO:97), CCGAGAGCCCGUAGAAC (SEQ ID NO:98), CCAGAUCGUCAGCGCCG (SEQ ID NO:99), GAAUGACGAGUGGACCC (SEQ ID NO:100), GGGUGACAGGUUUGGCCCUAUC (SEQ ID NO:101), GGUGACAGGUUUGGCCCUAUC (SEQ ID NO:102), GUGACAGGUUUGGCCCUAUC (SEQ ID NO:103), GACAGGUUUGGCCCUAUC (SEQ ID NO:104), GAUACUGCCUGAGCAGCCGCCU (SEQ ID NO:105), GACCACGUGGAGCUGAGCUGGUGG (SEQ ID NO:106), GUGGAGCUGAGCUGGUGG (SEQ ID NO:107), GGGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:108), GGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:109), GCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:110), GGGCUGCUCCUUGAGGGGCU (SEQ ID NO:111), GGCUGCUCCUUGAGGGGCU (SEQ ID NO:112), GCUGCUCCUUGAGGGGCU (SEQ ID NO:113), GGUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:114), GUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:115) и GAAUGGGAAGGAGGUGCACAG (SEQ ID NO:116). В некоторых вариантах, нРНК имеет домен целенаправленного воздействия, содержащий последовательность GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63).
В некоторых из любых таких вариантов, трансген интегрирован матричным полинуклеотидом, введенным в каждую из множества T клеток. В конкретных вариантах, матричный полинуклеотид содержит структуру [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии]. В определенных вариантах, 5’ плечо гомологии и 3’ плечо гомологии содержит последовательности нуклеиновых кислот, гомологичные к последовательностям нуклеиновых кислот, окружающих, по меньшей мере, один целевой сайт. В некоторых вариантах, 5’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот 5’ целевого сайта. В конкретных вариантах, 3’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот 3’ целевого сайта. В определенных вариантах, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет, по меньшей мере, или, по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов. В некоторых вариантах, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 нуклеотидов. В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от или около 50 и от или около 100 нуклеотидов в длину, от или около 100 и от или около 250 нуклеотидов в длину, от или около 250 и от или около 500 нуклеотидов в длину, от или около 500 и от или около 750 нуклеотидов в длину, от или около 750 и от или около 1000 нуклеотидов в длину или от или около 1000 и от или около 2000 нуклеотидов в длину.
В конкретных вариантах, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от или от около 100 до 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов. В конкретных вариантах, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от или около 100 до от или около 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов в длину.
В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от или около 200, 300, 400, 500, 600, 700 или 800 нуклеотидов в длину или любое значение между любыми из вышеописанных. В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет более чем от или около 300 нуклеотидов в длину. В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от или около 400, 500 или 600 нуклеотидов в длину или любое значение между любыми из вышеописанных. В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от или около 500 и от или около 600 нуклеотидов в длину. В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет более чем от или около 300 нуклеотидов в длину.
В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, интегрирован в или рядом с целевым сайтом в TRAC гене. В некоторых вариантах, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, интегрирован в или рядом с целевым сайтом в одном или обоих TRBC1 и TRBC2 генах.
В некоторых из любых таких вариантов, рекомбинантным рецептором является химерный антигенный рецептор (CAR). В некоторых из любых таких вариантов, CAR содержит внеклеточный домен, содержащий антигенсвязывающий домен, специфический к антигену. В некоторых из любых таких вариантов, антигенсвязывающим доменом является scFv; трансмембранный домен; цитоплазматический сигнальный домен, полученный из костимулирующей молекулы и цитоплазматический сигнальный домен, полученный из первичной сигнальной ITAM-содержащей молекулы. В некоторых из любых таких вариантов, CAR дополнительно содержит спейсер между трансмембранным доменом и антигенсвязывающим доменом. В некоторых из любых таких вариантов, костимулирующей молекулой является или она содержит 4-1BB, необязательно, человеческий 4-1BB. В некоторых из любых таких вариантов, ITAM-содержащей молекулой является или она содержит CD3дзэта сигнальный домен. В некоторых из любых таких вариантов, ITAM-содержащей молекулой является человеческий CD3дзэта сигнальный домен.
В некоторых из любых таких вариантов, рекомбинантным рецептором является рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь. В некоторых из любых таких вариантов, рекомбинантным рецептором является рекомбинантный TCR, содержащий альфа (TCRα) цепь и бета (TCRβ) цепь, и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, содержит последовательность нуклеиновых кислот, кодирующую TCRα цепь и последовательность нуклеиновых кислот, кодирующую TCRβ цепь. В некоторых из любых таких вариантов, трансген дополнительно содержит один или более мультицистронных элементов, и мультицистронные элементы расположены между последовательностью нуклеиновых кислот, кодирующей TCRα, или его часть, и последовательностью нуклеиновых кислот, кодирующей TCRβ или его часть. В некоторых из любых таких вариантов, мультицистронные элементы содержат последовательность, кодирующую элемент проскока рибосомы, выбранный из T2A, P2A, E2A или F2A или внутреннего участка посадки рибосомы (IRES).
В некоторых из любых таких вариантов, сконструированные клетки дополнительно содержат один или более вторых трансгенов, где второй трансген интегрируют в или рядом с, по меньшей мере, одним целевым сайтом через репарацию, направляемую гомологией (HDR). В некоторых из любых таких вариантов, рекомбинантным рецептором является рекомбинантный TCR, и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, содержит последовательность нуклеиновых кислот, кодирующую одну цепь рекомбинантного TCR, и второй трансген содержит последовательность нуклеиновых кислот, кодирующую другую цепь рекомбинантного TCR. В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, содержит последовательность нуклеиновых кислот, кодирующую TCRα цепь, и второй трансген содержит последовательность нуклеиновых кислот, кодирующую TCRβ цепь или ее часть. В некоторых из любых таких вариантов, интеграцию второго трансгена проводят вторым матричным полинуклеотидом, введенным в каждую из множества T клеток, где указанный второй матричный полинуклеотид содержит структуру [второе 5’ плечо гомологии]-[один или более второй трансген]-[второй 3’ плечо гомологии].
В определенных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене. В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в одном или обоих TRBC1 и TRBC2 генах. В конкретных вариантах, композицию получают дополнительным введением в иммунную клетку одного или более вторых матричных полинуклеотидов, содержащих один или более вторых трансгенов, где второй трансген нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
В определенных вариантах, второй матричный полинуклеотид содержит структуру [второе 5’ плечо гомологии]-[один или более второй трансген]-[второе 3’ плечо гомологии]. В некоторых вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии содержат последовательности нуклеиновых кислот, гомологичные последовательностям нуклеиновых кислот, окружающим, по меньшей мере, один целевой сайт. В конкретных вариантах, второе 5’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот второго 5’ целевого сайта. В определенных вариантах, второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот второго 3’ целевого сайта.
В некоторых вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет, по меньшей мере, или, по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов. В конкретных вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет от около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 нуклеотидов. В определенных вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет от или от около 100 до 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов.
В некоторых вариантах, один или более вторых трансгенов нацелены на интеграцию в или рядом с целевым сайтом в TRAC гене. В конкретных вариантах, одни или более вторых трансгенов нацелены на интеграцию в или рядом с целевым сайтом в TRBC1 или TRBC2 гене. В определенных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, TRBC1 гене или TRBC2 гене, и один или более вторых трансгенов нацелены на интеграцию в или рядом с одним или более целевыми сайтами, на которые не нацелен трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь,.
В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, и одни или более вторых трансгенов нацелены на интеграцию в или рядом с одним или более целевыми сайтами в TRBC1 гене и/или TRBC2 гене. В конкретных вариантах, один или более вторых трансгенов кодируют молекулу, выбранную из костимулирующего лиганда, цитокина, растворимого однонитевого переменного фрагмента (scFv), иммуномодулирующего слитого белка, химерного рецептора переключения (CSR) или корецептора. В определенных вариантах, кодированной молекулой является костимулирующий лиганд, необязательно выбранный из лиганда фактора некроза опухоли (TNF), выбранного из 4-1BBL, OX40L, CD70, LIGHT и CD30L, или лиганд суперсемейства иммуноглобулина (Ig), выбранный из CD80 и CD86.
В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, интегрирован в или рядом с целевым сайтом в TRAC гене, и один или более вторых трансгенов интегрированы в или рядом с одним или более другими целевыми сайтами среди TRAC гена, TRBC1 гена или TRBC2 гена, и который не интегрирован трансгеном, кодирующим рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь. В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, интегрированы в или рядом с целевым сайтом в TRAC гене, и один или более вторых трансгенов интегрированы в или рядом с одним или более целевыми сайтами в TRBC1 гене и/или TRBC2 гене. В некоторых из любых таких вариантов, один или более вторых трансгенов кодируют молекулы, выбранные из костимулирующего лиганда, цитокина, растворимого однонитевого переменного фрагмента (scFv), иммуномодулирующего слитого белка, химерного рецептора переключения (CSR) или корецептора.
В некоторых вариантах, кодированной молекулой является цитокин, необязательно выбранный из IL-2, IL-3, IL-6, IL-11, IL-12, IL-7, IL-15, IL-21, гранулоцитарно-моноцитарного колониестимулирующего фактора (GM-CSF), интерферона альфа (IFN-α), интерферона бета (IFN-β) или интерферона гамма (IFN-γ) и эритропоэтина. В конкретных вариантах, кодированной молекулой является растворимый однонитевой переменный фрагмент (scFv), который необязательно связывает полипептид, который имеет иммунодепрессивное действие или иммуностимулирующее действие, выбранный из CD47, PD-1, CTLA-4 и их лигандов или CD28, OX-40, 4-1BB и их лигандов.
В определенных вариантах, кодированной молекулой является иммуномодулирующий слитый белок, необязательно содержащий: (a) внеклеточный связывающий домен, который специфически связывает антиген, полученный из CD200R, SIRPα, CD279 (PD-1), CD2, CD95 (Fas), CD152 (CTLA4), CD223 (LAG3), CD272 (BTLA), A2aR, KIR, TIM3, CD300 или LPA5; (b) внутриклеточный связывающий домен, полученный из CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD47, CD79A, CD79B, CD134 (OX40), CD137 (4-1BB), CD150 (SLAMF1), CD278 (ICOS), CD357 (GITR), ХАРD11, DAP10, DAP12, FcRα, FcRβ, FcRγ, Fyn, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, ROR2, Ryk, Slp76, pTα, TCRα, TCRβ, TRFM, Zap70, PTCH2 или любого их сочетания; и (c) гидрофобный трансмембранный домен, полученный из CD2, CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD79A, CD79B, CD80, CD86, CD95 (Fas), CD134 (OX40), CD137 (4-1BB), CD150 (SLAMF1), CD152 (CTLA4), CD200R, CD223 (LAG3), CD270 (HVEM), CD272 (BTLA), CD273 (PD-L2), CD274 (PD-L1), CD278 (ICOS), CD279 (PD-1), CD300, CD357 (GITR), A2aR, DAP10, FcRα, FcRβ, FcRγ, Fyn, GAL9, KIR, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, PTCH2, ROR2, Ryk, Slp76, SIRPα, pTα, TCRα, TCRβ, TIM3, TRIM, LPA5 или Zap70. В некоторых вариантах, кодированной молекулой является химерный рецептор переключения (CSR), который необязательно содержат усеченный внеклеточный домен PD1 и трансмембранный и цитоплазматический сигнальные домены CD28. В конкретных вариантах, кодированной молекулой является корецептор, необязательно выбранный из CD4 или CD8.
В определенных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует одну цепь рекомбинантного TCR, и второй трансген кодирует другую цепь рекомбинантного TCR. В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует альфа (TCRα) цепь рекомбинантного TCR, и второй трансген кодирует бета (TCRβ) цепь рекомбинантного TCR. В конкретных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо дополнительно содержат регулирующий или контрольный элемент.
В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, дополнительно содержит гетерологичный регулирующий или контрольный элемент. В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо дополнительно содержит гетерологичный регулирующий или контрольный элемент. В некоторых из любых таких вариантов, гетерологичный регулирующий или контрольный элемент содержит гетерологичный промотор. В некоторых из любых таких вариантов, гетерологичный промотор является или содержит промотор человеческого фактора элонгации 1 альфа (EF1α) или MND промотор или его вариант. В некоторых из любых таких вариантов, гетерологичным промотором является индуцируемый промотор или репрессируемый промотор.
В определенных вариантах, регулирующий или контрольный элемент содержит промотор, энхансер, интрон, сигнал полиаденилирования, консенсусную последовательность Козака, последовательность акцептора сплайсинга или последовательность донора сплайсинга. В некоторых вариантах, регулирующий или контрольный элемент содержит промотор. В конкретных вариантах, промотор выбирают из конститутивного промотора, индуцируемого промотора, репрессируемого промотора и/или ткань-специфического промотора. В определенных вариантах, промотор выбирают из РНК pol I, pol II или pol III промотора. В некоторых вариантах, промотор выбирают из: pol III промотора, который является U6 или H1 промотором; или pol II промотора, который является промотором CMV, SV40 ранней области или аденовирусным основным поздним промотором. В конкретных вариантах, промотором является или он содержит промотор человеческого фактора элонгации 1 альфа (EF1α) или MND промотор или его вариант. В определенных вариантах, промотором является индуцируемый промотор или репрессируемый промотор. В некоторых вариантах, промотор содержит последовательность оператора Lac, последовательность оператора тетрациклина, последовательность оператора галактозы или последовательность оператора доксициклина или их аналог, или он способен связываться или распознаваться репрессором Lac или тетрациклиновым репрессором или их аналогом. В конкретных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо содержит один или более мультицистронных элементов.
В определенных вариантах, один или более мультицистронных элементов находятся против хода транскрипции трансгена, кодирующего рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или одного или более вторых трансгенов. В некоторых вариантах, мультицистронные элементы расположены между трансгеном, кодирующим рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и одним или более вторыми трансгенами. В конкретных вариантах, мультицистронные элементы расположены между последовательностью нуклеиновых кислот, кодирующей TCRα или его часть и последовательностью нуклеиновых кислот, кодирующей TCRβ или его часть. В определенных вариантах, мультицистронные элементы содержат последовательность, кодирующую рибопартикулярный элемент проскока, выбранный из T2A, P2A, E2A или F2A или внутренний участок посадки рибосомы (IRES).
В некоторых из любых таких вариантов, TCRα цепь содержит постоянную область (Cα), содержащую введение одного или более цистеиновых остатков, и/или TCRβ цепь содержит Cβ область, содержащую введение одного или более цистеиновых остатков, где один или более введенных цистеиновых остатков способны образовывать один или более ненативных дисульфидных мостиков между альфа цепью и бета цепью. В некоторых из любых таких вариантов, введение одного или более цистеиновых остатков включает замещение не цистеинового остатка цистеиновым остатком. В некоторых из любых таких вариантов, Cα область содержит цистеин в положении, соответствующем положению 48 с нумерацией, как указано в любой из SEQ ID NO: 24; и/или Cβ область содержит цистеин в положении, соответствующем положению 57 с нумерацией, как указано в SEQ ID NO: 20.
В определенных вариантах, последовательность, кодирующая рибопартикулярный элемент проскока, нацелена на то, чтобы быть внутри рамки с геном в целевом сайте. В некоторых вариантах, при HDR, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо функционально связаны с эндогенным промотором гена в целевом сайте. В определенных вариантах, рекомбинантный TCR способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессируется на клетке или ткани заболевания, расстройства или состояния. В конкретных вариантах, заболеванием, расстройством или состоянием является инфекционное заболевание или расстройство, аутоиммунное заболевание, воспалительное заболевание или опухоль или рак. В конкретных вариантах, антигеном является опухолевый антиген или патогенный антиген. В определенных вариантах, патогенным антигеном является бактериальный антиген или вирусный антиген.
В некоторых вариантах, антигеном является вирусный антиген, и вирусный антиген выбирают из вируса гепатита A, гепатита B, гепатита C (HCV), папилломавируса человека (HPV), вирусной инфекции гепатита, вируса Эпштейн-Барр (EBV), герпесвируса человека 8 (HHV-8), вируса-1 T-клеточного лейкоза человека (HTLV-1), вируса-2 T-клеточного лейкоза человека (HTLV-2) или цитомегаловируса (CMV). В конкретных вариантах, антигеном является антиген из HPV, выбранный из HPV-16, HPV-18, HPV-31, HPV-33 и HPV-35. В определенных вариантах, антигеном является HPV-16 антиген, который является HPV-16 E6 или HPV-16 E7 антигеном. В некоторых вариантах, вирусным антигеном является EBV антиген, выбранный из ядерного антигена Эпштейн-Барр (EBNA)-1, EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, EBNA-лидерного белка (EBNA-LP), латентных мембранных белков LMP-1, LMP-2A и LMP-2B, EBV-EA, EBV-MA и EBV-VCA. В конкретных вариантах, вирусным антигеном является HTLV-антиген, который является TAX. В определенных вариантах, вирусным антигеном является HBV антиген, который является ядерным антигеном гепатита B или оболочечным антигеном гепатита B. В некоторых вариантах, антигеном является опухолевый антиген.
В конкретных вариантах, антиген выбирают из глиома-ассоциированного антигена, β-человеческого хорионного гонадотропина, альфафетопротеина (AFP), лектин-реактивного AFP, тироглобулина, RAGE-1, MN-CA IX, обратной транскриптазы теломеразы человека, RU1, RU2 (AS), кишечной карбоксилэстеразы, mut hsp70-2, M-CSF, Меланин-A/MART-1, WT-1, S-100, MBP, CD63, MUC1 (например, MUC1-8), p53, Ras, циклина B1, HER-2/neu, карциноэмбрионального антигена (CEA), gp100, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11, MAGE-A11, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-C1, BAGE, GAGE-1, GAGE-2, pl5, тирозиназы, тирозиназа-родственного белка 1 (TRP-1), тирозиназа-родственного белка 2 (TRP-2), β-катенина, NY-ESO-1, LAGE-1a, PP1, MDM2, MDM4, EGVFvIII, Tax, SSX2, теломеразы, TARP, pp65, CDK4, виментина, S100, eIF-4A1, IFN-индуцируемого p78, меланотрансферрина (p97), Уроплакина II, простата-специфического антигена (PSA), человеческого калликреина (huK2), простата-специфического мембранного антигена (PSM) и простатической кислой фосфатазы (PAP), эластазы нейтрофилов, эфрина B2, BA-46, Bcr-abl, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, Каспазы 8, FRa, CD24, CD44, CD133, CD 166, epCAM, CA-125, HE4, Oval, рецептора эстрогена, рецептора прогестерона, uPA, PAI-1, CD19, CD20, CD22, ROR1, CD33/IL3Ra, c-Met, PSMA, Гликолипида F77, GD-2, инсулинового фактора роста (IGF)-I, IGF-II, рецептора IGF-I и мезотелина.
В определенных вариантах, T клеткой является CD8+ T клетка или ее подтипы. В некоторых вариантах, T клеткой является CD4+ T клетка или ее подтипы. В конкретных вариантах, Т клетка аутологична субъекту. В определенных вариантах, Т клетка аллогенна субъекту. В некоторых вариантах, первый матричный полинуклеотид, один или более вторых матричных полинуклеотидов и/или один или более полинуклеотидов, кодирующих нРНК и/или Cas9 белок содержатся в одном или более векторах, которые необязательно являются вирусными векторами. В конкретных вариантах, вектором является AAV вектор. В определенных вариантах, AAV вектор выбирают из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 или AAV8 вектора. В некоторых вариантах, AAV вектором является AAV2 или AAV6 вектор. В конкретных вариантах, вирусным вектором является ретровирусный вектор. В определенных вариантах, вирусным вектором является лентивирусный вектор.
В некоторых из любых таких вариантов, T клетки содержат CD8+ T клетку и/или CD4+ T клетки или их подтипы. В некоторых из любых таких вариантов, T клетки являются аутологичными субъекту. В некоторых из любых таких вариантов, T клетки являются аллогенными субъекту. В некоторых из любых таких вариантов, описанная здесь композиция дополнительно содержит фармацевтически приемлемый носитель.
В некоторых вариантах, введение одного или более агентов, способных вызывать генетическое разрушение, и введение матричного полинуклеотида проводят одновременно или последовательно, в любом порядке. В конкретных вариантах, введение матричного полинуклеотида проводят после введения одного или более агентов, способных вызывать генетическое разрушение. В определенных вариантах, матричный полинуклеотид вводят сразу же после или в течение около 30 секунд, 1 минуты, 2 минут, 3 минут, 4 минут, 5 минут, 6 минут, 6 минут, 8 минут, 9 минут, 10 минут, 15 минут, 20 минут, 30 минут, 40 минут, 50 минут, 60 минут, 90 минут, 2 часов, 3 часов или 4 часов после введения одного или более агентов, способных вызывать генетическое разрушение.
В некоторых вариантах, введение матричного полинуклеотида и введение одного или более вторых матричных полинуклеотидов проводят одновременно или последовательно, в любом порядке. В конкретных вариантах, введение одного или более агента, способного вызывать генетическое разрушение, и введение матричного полинуклеотида проводят в одной экспериментальной реакции. В определенных вариантах, введение одного или более агента, способного вызывать генетическое разрушение, и введение матричного полинуклеотида и вторых матричных полинуклеотидов проводят в одной экспериментальной реакции. В некоторых вариантах, описанная здесь композиция дополнительно содержит фармацевтически приемлемый носитель.
В некоторых вариантах, представлены способы получения генетически сконструированной иммунной клетки, которые включают (a) введение в иммунную клетку одного или более агентов, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), тем самым вызывая генетическое разрушение, по меньшей мере, одного целевого сайта; и (b) введение в иммунную клетку матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
В некоторых из любых таких вариантов, представлены способы получения генетически сконструированной иммунной клетки, которые включают (a) введение в иммунную клетку одного или более агентов, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), тем самым вызывая генетическое разрушение, по меньшей мере, одного целевого сайта; и (b) введение в иммунную клетку матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или его цепь, где указанный рекомбинантный рецептор способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния, где трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR), где введение матричного полинуклеотида проводят после введения одного или более агента, способного вызывать генетическое разрушение.
В некоторых вариантах, также представлены способы получения генетически сконструированной иммунной клетки, которые включают введение в иммунную клетку, имеющую генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где генетическое разрушение вызвано одним или более агентами, где каждый из одного или более агентов независимо способны вызывать генетическое разрушение, и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого положения через репарацию, направляемую гомологией (HDR).
В некоторых из любых таких вариантов, также представлены способы получения генетически сконструированной иммунной клетки, которые включают введение в иммунную клетку, имеющую генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или его цепь, где указанный рекомбинантный рецептор способе связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния, где генетическое разрушение вызвано одним или более агентами, где каждый из одного или более агентов независимо способны вызывать генетическое разрушение, и трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
В некоторых из любых таких вариантов, матричный полинуклеотид вводят сразу же после или в течение от или около 30 секунд, 1 минуты, 2 минут, 3 минут, 4 минут, 5 минут, 6 минут, 6 минут, 8 минут, 9 минут, 10 минут, 15 минут, 20 минут, 30 минут, 40 минут, 50 минут, 60 минут, 90 минут, 2 часов, 3 часов или 4 часов после введения одного или более агентов, способных вызывать генетическое разрушение. В некоторых из любых таких вариантов, матричный полинуклеотид вводят в или около 2 часов после введения одного или более агентов.
В некоторых из любых таких вариантов, одна или более иммунных клеток содержат T клетки. В некоторых из любых таких вариантов, T клетки содержат CD4+ T клетки, CD8+ T клетки или CD4+ и CD8+ T клетки. В некоторых из любых таких вариантов, T клетки содержат CD4+ и CD8+ T клетки и соотношение CD4+ к CD8+ T клеткам составляет от около 1:3 до около 3:1. В некоторых из любых таких вариантов, необязательно от около 1:2 до около 2:1, соотношение CD4+ к CD8+ T клеткам составляет около 1:1.
В некоторых из любых таких вариантов, один или более агентов содержат CRISPR-Cas9 сочетание, и CRISPR-Cas9 сочетание содержит направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту. В некоторых из любых таких вариантов, CRISPR-Cas9 сочетанием является рибонуклеопротеиновый (RNP) комплекс, содержащий нРНК и Cas9 белок. В некоторых из любых таких вариантов, концентрация RNP составляет от около 1 мкМ до около 5 мкМ. В некоторых из любых таких вариантов, концентрация RNP составляет около 2 мкМ.
В некоторых из любых таких вариантов, один или более агентов вводят электропорацией. В некоторых из любых таких вариантов, матричный полинуклеотид содержится в вирусных векторах, и введение матричного полинуклеотида проводят трансдукцией. В некоторых из любых таких вариантов, вектором является AAV вектор.
В некоторых из любых таких вариантов, способ включает инкубирование клеток in vitro со стимулирующими агентами в условиях для стимулирования или активации одной или более иммунных клеток и до введения одного или более агентов. В некоторых из любых таких вариантов, стимулирующие агенты содержат и анти-CD3 и/или анти-CD28 антитела, необязательно, анти-CD3/анти-CD28 сферы. В некоторых из любых таких вариантов, соотношение сфер к клеткам составляет около 1:1. В некоторых из любых таких вариантов, стимулирующие агенты удаляют из иммунной клетки до введения одного или более агентов.
В некоторых из любых таких вариантов, способ дополнительно включает инкубирование клеток до, во время или после введения одного или более агентов и/или введения матричного полинуклеотида с одним или более рекомбинантными цитокинами. В некоторых из любых таких вариантов, один или более рекомбинантных цитокинов выбирают из группы, состоящей из IL-2, IL-7 и IL-15. В некоторых из любых таких вариантов, один или более рекомбинантных цитокинов добавляют в концентрации, выбранной из концентрации IL-2 от около 10 Ед/мл до около 200 Ед/мл. В некоторых из любых таких вариантов, концентрация составляет от около 50 МЕд/мл до около 100 Ед/мл; IL-7 в концентрации от 0,5 нг/мл до 50 нг/мл. В некоторых из любых таких вариантов, концентрация составляет от около 5 нг/мл до около 10 нг/мл и/или IL-15 в концентрации от 0,1 нг/мл до 20 нг/мл, необязательно, от около 0,5 нг/мл до около 5 нг/мл. В некоторых из любых таких вариантов, инкубирование проводят после введения одного или более агентов и введения матричного полинуклеотида в течение вплоть до или приблизительно 24 часов, 36 часов, 48 часов, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или 21 дней, необязательно вплоть до или около 7 дней.
В некоторых из любых таких вариантов, рекомбинантным рецептором является химерный антигенный рецептор (CAR). В некоторых из любых таких вариантов, CAR содержит внеклеточный домен, содержащий антигенсвязывающий домен, специфический к антигену; трансмембранный домен; цитоплазматический сигнальный домен, полученный из костимулирующей молекулы; и цитоплазматический сигнальный домен, полученный из первичной сигнальной ITAM-содержащей молекулы. В некоторых из любых таких вариантов, CAR дополнительно содержит спейсер между трансмембранным доменом и антигенсвязывающим доменом. В некоторых из любых таких вариантов, антигенсвязывающим доменом является scFv. В некоторых из любых таких вариантов, костимулирующей молекулой является или она содержит 4-1BB. В некоторых из любых таких вариантов, костимулирующей молекулой является человеческий 4-1BB. В некоторых из любых таких вариантов, ITAM-содержащей молекулой является или она содержит CD3дзэта сигнальный домен. В некоторых из любых таких вариантов, ITAM-содержащей молекулой является человеческий CD3дзэта сигнальный домен.
В некоторых из любых таких вариантов, рекомбинантным рецептором является рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь. В некоторых из любых таких вариантов, рекомбинантным рецептором является рекомбинантный TCR, содержащий альфа (TCRα) цепь и бета (TCRβ) цепь, и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, содержит последовательность нуклеиновых кислот, кодирующую TCRα цепь и последовательность нуклеиновых кислот, кодирующую TCRβ цепь.
В некоторых из любых таких вариантов, представлены способы получения генетически сконструированной иммунной клетки, включающие (a) введение в иммунную клетку одного или более агентов, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), тем самым вызывая генетическое разрушение, по меньшей мере, одного целевого сайта; и (b) введение в иммунную клетку матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный рецептор, который является рекомбинантным Т-клеточным рецептором (TCR) или его антигенсвязывающий фрагмент или его цепь, где указанный трансген содержит гетерологичный промотор, и где трансген нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
В некоторых из любых таких вариантов, представлены способы получения генетически сконструированной иммунной клетки, включающие введение в иммунную клетку, имеющую генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный рецептор, которым является рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где указанный трансген содержит гетерологичный промотор, где генетическое разрушение вызвано одним или более агентами, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение, и трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
В некоторых вариантах, по меньшей мере, один из одного или более агентов способен вызывать генетическое разрушение целевого сайта в гене TRAC. В конкретных вариантах, по меньшей мере, один из одного или более агентов способен вызывать генетическое разрушение целевого сайта в гене TRBC. В определенных вариантах, один или более агентов содержат, по меньшей мере, один агент, который способен вызывать генетическое разрушение целевого сайта в гене TRAC и, по меньшей мере, один агент, который способен вызывать генетическое разрушение целевого сайта в гене TRBC. В некоторых вариантах, геном TRBC является один или оба из генов бета константы Т-клеточного рецептора 1 (TRBC1) или бета константы Т-клеточного рецептора 2 (TRBC2).
Представлен способы получения генетически сконструированной иммунной клетки, включающий: (a) введение в иммунную клетку одного или более агентов, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), тем самым вызывая генетическое разрушение, по меньшей мере, одного целевого сайта; и (b) введение в иммунную клетку матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом через репарацию, направляемую гомологией (HDR).
Также представлены способы получения генетически сконструированной иммунной клетки, включающие: введение в иммунную клетку, имеющую генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и гене бета константы Т-клеточного рецептора (TRBC), матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где генетическое разрушение вызвано одним или более агентами, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение, и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR). В конкретных вариантах, TRBC геном является один или оба из гена бета константы Т-клеточного рецептора 1 (TRBC1) или бета константы Т-клеточного рецептора 2 (TRBC2).
В некоторых из любых таких вариантов, также представлены способы получения генетически сконструированной иммунной клетки, включающие (a) введение в иммунную клетку, по меньшей мере, одного агента, который способен вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и, по меньшей мере, одного агента, который способен вызывать генетическое разрушение целевого сайта в гене бета константы Т-клеточного рецептора (TRBC), тем самым вызывая генетическое разрушение целевых сайтов; и (b) введение в иммунную клетку матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный рецептор, которым является рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
В некоторых из любых таких вариантов, также представлены способы получения генетически сконструированной иммунной клетки, включающие введение в иммунную клетку, имеющую генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC), и генетическое разрушение, по меньшей мере, одного целевого сайта в гене бета константы Т-клеточного рецептора (TRBC), матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный рецептор, которым является рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где генетические разрушения вызваны, по меньшей мере, одним агентом, который способен вызывать генетическое разрушение целевого сайта в TRAC гене, и, по меньшей мере, одним агентом, который способен вызывать генетическое разрушение в TRBC гене, и трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR). В некоторых из любых таких вариантов, TRBC геном является один или оба из генов бета константы Т-клеточного рецептора 1 (TRBC1) или бета константы Т-клеточного рецептора 2 (TRBC2).
В определенных вариантах, один или более агент, способный вызывать генетическое разрушение содержит ДНК-связывающий белок или ДНК-связывающую нуклеиновую кислоту, которые специфически связываются с или гибридизируются в целевой сайт. В некоторых вариантах, один или более агент, способный вызывать генетическое разрушение содержит (a) слитый белок, содержащий ДНК-направленный белок и нуклеазу, или (b) РНК-направляемую нуклеазу. В конкретных вариантах, ДНК-направленный белок или РНК-направляемая нуклеаза содержит белок «цинковый палец» (ZFP), TAL белок или связанную с кластеризованной регулярной промежуточной короткой падлиндромной нуклеиновой кислотой (CRISPR) нуклеазу (Cas), специфическую для целевого сайта.
В определенных вариантах, один или более агент содержит нуклеазу «цинковый палец» (ZFN), TAL-эффекторную нуклеазу (TALEN) и/или CRISPR-Cas9 сочетание, которые специфически связываются с, распознают или гибридизируются с целевым сайтом. В некоторых вариантах, каждый из одного или более агентов содержит направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту. В некоторых из любых таких вариантов, каждый из одного или более агентов содержит CRISPR-Cas9 сочетание, и CRISPR-Cas9 сочетание содержит направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, один целевой сайт. В конкретных вариантах, один или более агентов вводят в виде рибонуклеопротеинового (RNP) комплекса, содержащего нРНК и Cas9 белок. В некоторых из любых таких вариантов, CRISPR-Cas9 сочетанием является рибонуклеопротеиновый (RNP) комплекс, содержащий нРНК и Cas9 белок. В некоторых из любых таких вариантов, концентрация RNP составляет от около 1 мкМ до около 5 мкМ. В некоторых из любых таких вариантов, концентрация RNP составляет около 2 мкМ.
В определенных вариантах, RNP вводят через электропорацию, генную пушку, трансфекцию фосфата кальция, сгущение или сжатие клеток. В некоторых вариантах, RNP вводят через электропорацию. В конкретных вариантах, один или более агентов вводят в виде одного или более полинуклеотидов, кодирующих нРНК и/или Cas9 белок. В определенных вариантах, по меньшей мере, один целевой сайт находится внутри экзона TRAC, TRBC1 и/или TRBC2 гена. В некоторых из любых таких вариантов, по меньшей мере, один целевой сайт находится внутри экзона TRAC и экзона с TRBC1 или TRBC2 гена.
В некоторых вариантах, нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в TRAC гене и содержит последовательность, выбранную из группы, состоящей из UCUCUCAGCUGGUACACGGC (SEQ ID NO:28), UGGAUUUAGAGUCUCUCAGC (SEQ ID NO:29), ACACGGCAGGGUCAGGGUUC (SEQ ID NO:30), GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31), GCUGGUACACGGCAGGGUCA (SEQ ID NO:32), CUCAGCUGGUACACGGC (SEQ ID NO:33), UGGUACACGGCAGGGUC (SEQ ID NO:34), GCUAGACAUGAGGUCUA (SEQ ID NO:35), GUCAGAUUUGUUGCUCC (SEQ ID NO:36), UCAGCUGGUACACGGCA (SEQ ID NO:37), GCAGACAGACUUGUCAC (SEQ ID NO:38), GGUACACGGCAGGGUCA (SEQ ID NO:39), CUUCAAGAGCAACAGUGCUG (SEQ ID NO:40), AGAGCAACAGUGCUGUGGCC (SEQ ID NO:41), AAAGUCAGAUUUGUUGCUCC (SEQ ID NO:42), ACAAAACUGUGCUAGACAUG (SEQ ID NO:43), AAACUGUGCUAGACAUG (SEQ ID NO:44), UGUGCUAGACAUGAGGUCUA (SEQ ID NO:45), GGCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:46), GCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:47), GGGGAAGAAGGUGUCUUC (SEQ ID NO:48), GUUUUGUCUGUGAUAUACACAU (SEQ ID NO:49), GGCAGACAGACUUGUCACUGGAUU (SEQ ID NO:50), GCAGACAGACUUGUCACUGGAUU (SEQ ID NO:51), GACAGACUUGUCACUGGAUU (SEQ ID NO:52), GUGAAUAGGCAGACAGACUUGUCA (SEQ ID NO:53), GAAUAGGCAGACAGACUUGUCA (SEQ ID NO:54), GAGUCUCUCAGCUGGUACACGG (SEQ ID NO:55), GUCUCUCAGCUGGUACACGG (SEQ ID NO:56), GGUACACGGCAGGGUCAGGGUU (SEQ ID NO:57) и GUACACGGCAGGGUCAGGGUU (SEQ ID NO:58). В конкретных вариантах, нРНК имеет домен целенаправленного воздействия, содержащий последовательность GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31).
В определенных вариантах, нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в одном или обоих TRBC1 и TRBC2 генах и содержит последовательность, выбранную из группы, состоящей из CACCCAGAUCGUCAGCGCCG (SEQ ID NO:59), CAAACACAGCGACCUCGGGU (SEQ ID NO:60), UGACGAGUGGACCCAGGAUA (SEQ ID NO:61), GGCUCUCGGAGAAUGACGAG (SEQ ID NO:62), GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63), GAAAAACGUGUUCCCACCCG (SEQ ID NO:64), AUGACGAGUGGACCCAGGAU (SEQ ID NO:65), AGUCCAGUUCUACGGGCUCU (SEQ ID NO:66), CGCUGUCAAGUCCAGUUCUA (SEQ ID NO:67), AUCGUCAGCGCCGAGGCCUG (SEQ ID NO:68), UCAAACACAGCGACCUCGGG (SEQ ID NO:69), CGUAGAACUGGACUUGACAG (SEQ ID NO:70), AGGCCUCGGCGCUGACGAUC (SEQ ID NO:71), UGACAGCGGAAGUGGUUGCG (SEQ ID NO:72), UUGACAGCGGAAGUGGUUGC (SEQ ID NO:73), UCUCCGAGAGCCCGUAGAAC (SEQ ID NO:74), CGGGUGGGAACACGUUUUUC (SEQ ID NO:75), GACAGGUUUGGCCCUAUCCU (SEQ ID NO:76), GAUCGUCAGCGCCGAGGCCU (SEQ ID NO:77), GGCUCAAACACAGCGACCUC (SEQ ID NO:78), UGAGGGUCUCGGCCACCUUC (SEQ ID NO:79), AGGCUUCUACCCCGACCACG (SEQ ID NO:80), CCGACCACGUGGAGCUGAGC (SEQ ID NO:81), UGACAGGUUUGGCCCUAUCC (SEQ ID NO:82), CUUGACAGCGGAAGUGGUUG (SEQ ID NO:83), AGAUCGUCAGCGCCGAGGCC (SEQ ID NO:84), GCGCUGACGAUCUGGGUGAC (SEQ ID NO:85), UGAGGGCGGGCUGCUCCUUG (SEQ ID NO:86), GUUGCGGGGGUUCUGCCAGA (SEQ ID NO:87), AGCUCAGCUCCACGUGGUCG (SEQ ID NO:88), GCGGCUGCUCAGGCAGUAUC (SEQ ID NO:89), GCGGGGGUUCUGCCAGAAGG (SEQ ID NO:90), UGGCUCAAACACAGCGACCU (SEQ ID NO:91), ACUGGACUUGACAGCGGAAG (SEQ ID NO:92), GACAGCGGAAGUGGUUGCGG (SEQ ID NO:93), GCUGUCAAGUCCAGUUCUAC (SEQ ID NO:94), GUAUCUGGAGUCAUUGAGGG (SEQ ID NO:95), CUCGGCGCUGACGAUCU (SEQ ID NO:96), CCUCGGCGCUGACGAUC (SEQ ID NO:97), CCGAGAGCCCGUAGAAC (SEQ ID NO:98), CCAGAUCGUCAGCGCCG (SEQ ID NO:99), GAAUGACGAGUGGACCC (SEQ ID NO:100), GGGUGACAGGUUUGGCCCUAUC (SEQ ID NO:101), GGUGACAGGUUUGGCCCUAUC (SEQ ID NO:102), GUGACAGGUUUGGCCCUAUC (SEQ ID NO:103), GACAGGUUUGGCCCUAUC (SEQ ID NO:104), GAUACUGCCUGAGCAGCCGCCU (SEQ ID NO:105), GACCACGUGGAGCUGAGCUGGUGG (SEQ ID NO:106), GUGGAGCUGAGCUGGUGG (SEQ ID NO:107), GGGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:108), GGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:109), GCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:110), GGGCUGCUCCUUGAGGGGCU (SEQ ID NO:111), GGCUGCUCCUUGAGGGGCU (SEQ ID NO:112), GCUGCUCCUUGAGGGGCU (SEQ ID NO:113), GGUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:114), GUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:115) и GAAUGGGAAGGAGGUGCACAG (SEQ ID NO:116). В некоторых вариантах, нРНК имеет домен целенаправленного воздействия, содержащий последовательность GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63).
В конкретных вариантах, матричный полинуклеотид содержит структуру [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии]. В определенных вариантах, 5’ плечо гомологии и 3’ плечо гомологии содержит последовательности нуклеиновых кислот гомологичные последовательностям нуклеиновых кислот, окружающим, по меньшей мере, один целевой сайт. В некоторых вариантах, 5’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот 5’ целевого сайта. В конкретных вариантах, 3’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот 3’ целевого сайта.
В определенных вариантах, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет, по меньшей мере, или, по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов. В некоторых вариантах, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 нуклеотидов. В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от около 50 и от около 100 нуклеотидов в длину, от около 100 и от около 250 нуклеотидов в длину, от около 250 и от около 500 нуклеотидов в длину, от около 500 и от около 750 нуклеотидов в длину, от около 750 и от около 1000 нуклеотидов в длину или от около 1000 и от около 2000 нуклеотидов в длину.
В конкретных вариантах, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от или от около 100 до 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов. В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от около 100 до около 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов в длину.
В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от около 200, 300, 400, 500, 600, 700 или 800 нуклеотидов в длину или любое значение между любыми из вышеописанных. В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет более чем от около 300 нуклеотидов в длину, необязательно где 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от около 400, 500 или 600 нуклеотидов в длину или любое значение между любыми из вышеописанных. В некоторых из любых таких вариантов, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет более чем от около 300 нуклеотидов в длину.
В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене. В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в одном или обоих TRBC1 и TRBC2 генах. В некоторых из любых таких вариантов, рекомбинантным рецептором является рекомбинантный TCR, содержащий альфа (TCRα) цепь и бета (TCRβ) цепь, и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, содержит последовательность нуклеиновых кислот, кодирующую TCRα цепь и последовательность нуклеиновых кислот, кодирующую TCRβ цепь. В некоторых из любых таких вариантов, трансген дополнительно содержит один или более мультицистронных элементов, и мультицистронные элементы расположены между последовательностью нуклеиновых кислот, кодирующей TCRα или его часть и последовательностью нуклеиновых кислот, кодирующей TCRβ или его часть. В некоторых из любых таких вариантов, мультицистронные элементы содержат последовательность, кодирующую элемент проскока рибосомы, выбранный из T2A, P2A, E2A или F2A или внутренний участок посадки рибосомы (IRES).
В некоторых из любых таких вариантов, рекомбинантным рецептором является рекомбинантный TCR и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, содержит последовательность нуклеиновых кислот, кодирующую одну цепь рекомбинантного TCR, и второй трансген содержит последовательность нуклеиновых кислот, кодирующую другую цепь рекомбинантного TCR. В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, содержит последовательность нуклеиновых кислот, кодирующую TCRα цепь, и второй трансген содержит последовательность нуклеиновых кислот, кодирующую TCRβ цепь или ее часть.
В определенных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене. В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в одном или обоих TRBC1 и TRBC2 генах. В конкретных вариантах, включающих введение в иммунную клетку одного или более вторых матричных полинуклеотидов, содержащих один или более вторые трансгены, где второй трансген нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
В определенных вариантах, второй матричный полинуклеотид содержит структуру [второе 5’ плечо гомологии]-[один или более второй трансген]-[второе 3’ плечо гомологии]. В некоторых вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот, гомологичные последовательностям нуклеиновых кислот, окружающим, по меньшей мере, один целевой сайт. В конкретных вариантах, второе 5’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот второго 5’ целевого сайта.
В определенных вариантах, второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот второго 3’ целевого сайта. В некоторых вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет, по меньшей мере, или, по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов. В конкретных вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет от около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 нуклеотидов. В определенных вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет от или от около 100 до 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов.
В некоторых вариантах, один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене. В конкретных вариантах, один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRBC1 или TRBC2 гене. В определенных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, TRBC1 гене или TRBC2 гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами, на которые не нацелен трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь,. В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, TRBC1 гене или TRBC2 гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более другими целевыми сайтами среди TRAC гена, TRBC1 гена или TRBC2 гена, и на которые не нацелен трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь,. В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами в TRBC1 гене и/или TRBC2 гене. В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами в TRBC1 гене и/или TRBC2 гене.
В конкретных вариантах, один или более вторых трансгенов кодирует молекулу, выбранную из костимулирующего лиганда, цитокина, растворимого однонитевого переменного фрагмента (scFv), иммуномодулирующего слитого белка, химерного рецептора переключения (CSR) или корецептора. В определенных вариантах, кодированной молекулой является костимулирующий лиганд, необязательно выбранный из лиганда фактора некроза опухоли (TNF), выбранного из 4-1BBL, OX40L, CD70, LIGHT и CD30L или лиганда суперсемейства иммуноглобулина (Ig), выбранного из CD80 и CD86. В некоторых вариантах, кодированной молекулой является цитокин, необязательно выбранный из IL-2, IL-3, IL-6, IL-11, IL-12, IL-7, IL-15, IL-21, гранулоцитарно-моноцитарного колониестимулирующего фактора (GM-CSF), интерферона альфа (IFN-α), интерферона бета (IFN-β) или интерферона гамма (IFN-γ) и эритропоэтина. В конкретных вариантах, кодированной молекулой является растворимый однонитевой переменный фрагмент (scFv), который необязательно связывает полипептид, который обладает иммунодепрессивным действием или иммуностимулирующим действием, выбранный из CD47, PD-1, CTLA-4 и их лигандов или CD28, OX-40, 4-1BB и их лигандов.
В определенных вариантах, кодированной молекулой является иммуномодулирующий слитый белок, необязательно содержащий: (a) внеклеточный связывающий домен, который специфически связывает антиген, полученный из CD200R, SIRPα, CD279 (PD-1), CD2, CD95 (Fas), CD152 (CTLA4), CD223 (LAG3), CD272 (BTLA), A2aR, KIR, TIM3, CD300 или LPA5; (b) внутриклеточный связывающий домен, полученный из CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD47, CD79A, CD79B, CD134 (OX40), CD137 (4-1BB), CD150 (SLAMF1), CD278 (ICOS), CD357 (GITR), ХАРD11, DAP10, DAP12, FcRα, FcRβ, FcRγ, Fyn, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, ROR2, Ryk, Slp76, pTα, TCRα, TCRβ, TRFM, Zap70, PTCH2 или любого их сочетания; и (c) гидрофобный трансмембранный домен, полученный из CD2, CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD79A, CD79B, CD80, CD86, CD95 (Fas), CD134 (OX40), CD137 (4-1BB), CD150 (SLAMF1), CD152 (CTLA4), CD200R, CD223 (LAG3), CD270 (HVEM), CD272 (BTLA), CD273 (PD-L2), CD274 (PD-L1), CD278 (ICOS), CD279 (PD-1), CD300, CD357 (GITR), A2aR, DAP10, FcRα, FcRβ, FcRγ, Fyn, GAL9, KIR, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, PTCH2, ROR2, Ryk, Slp76, SIRPα, pTα, TCRα, TCRβ, TIM3, TRIM, LPA5 или Zap70. В некоторых вариантах, кодированной молекулой является химерный рецептор переключения (CSR), который необязательно содержит усеченный внеклеточный домен PD1 и трансмембранный и цитоплазматический сигнальные домены CD28. В конкретных вариантах, кодированной молекулой является корецептор, необязательно выбранный из CD4 или CD8.
В определенных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует одну цепь рекомбинантного TCR, и второй трансген кодирует другую цепь рекомбинантного TCR. В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует альфа (TCRα) цепь рекомбинантного TCR и второй трансген кодирует бета (TCRβ) цепь рекомбинантного TCR. В конкретных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо дополнительно содержат регулирующий или контрольный элемент. В определенных вариантах, регулирующий или контрольный элемент содержит промотор, энхансер, интрон, сигнал полиаденилирования, консенсусную последовательность Козака, последовательность акцептора сплайсинга или последовательность донора сплайсинга. В некоторых вариантах, регулирующий или контрольный элемент содержит промотор. В конкретных вариантах, промотор выбирают из конститутивного промотора, индуцируемого промотора, репрессируемого промотора и/или ткань-специфического промотора. В некоторых вариантах, промотор выбирают из РНК pol I, pol II или pol III промотора.
В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, дополнительно содержит регулирующий или контрольный элемент. В некоторых из любых таких вариантов, трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо дополнительно содержат гетерологичный регулирующий или контрольный элемент. В некоторых из любых таких вариантов, гетерологичный регулирующий или контрольный элемент содержит гетерологичный промотор. В некоторых из любых таких вариантов, гетерологичным промотором является или он содержит промотор человеческого фактора элонгации 1 альфа (EF1α) или MND промотор или их вариант. В некоторых из любых таких вариантов, гетерологичным промотором является индуцируемый промотор или репрессируемый промотор.
В конкретных вариантах, промотор выбирают из: pol III промотора, который является U6 или H1 промотором; или pol II промотора, который является промотором CMV, SV40 ранней области или основной поздней области аденовируса. В определенных вариантах, промотором является или он содержит промотор человеческого фактора элонгации 1 альфа (EF1α) или MND промотор или их вариант. В некоторых вариантах, промотором является индуцируемый промотор или репрессируемый промотор. В конкретных вариантах, промотор содержит Lac последовательность оператора, тетрациклиновую последовательность оператора, галактозную последовательность оператора или доксициклиновую последовательность оператора или их аналог, или способен связываться или распознаваться Lac репрессором или тетрациклиновым репрессором или их аналогом.
В определенных вариантах, трансген? кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо содержат один или более мультицистронных элементов. В некоторых вариантах, один или более мультицистронных элементов расположены против хода транскрипции трансгена, кодирующего рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов. В конкретных вариантах, мультицистронные элементы расположены между трансгеном, кодирующим рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и одним или более вторыми трансгенами. В определенных вариантах, мультицистронные элементы расположены между последовательностью нуклеиновых кислот, кодирующей TCRα или его часть, и последовательностью нуклеиновых кислот, кодирующей TCRβ или его часть. В некоторых вариантах, мультицистронные элементы содержат последовательность, кодирующую рибопартикулярный элемент проскока, выбранный из T2A, P2A, E2A или F2A или участок внутренней посадки рибосомы (IRES). В некоторых вариантах, последовательность, кодирующая рибопартикулярный элемент проскока, нацелена на то, чтобы быть внутри рамки с геном в целевом сайте.
В определенных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо функционально связан с эндогенным промотором гена на целевом сайте.
В некоторых из любых таких вариантов, TCRα цепь содержит постоянную область (Cα), содержащую введение одного или более цистеиновых остатков и/или TCRβ цепь содержит область Cβ, содержащую введение одного или более цистеиновых остатков, где один или более введенных цистеиновых остатков способны образовывать один или более не нативных дисульфидных мостиков между альфа цепью и бета цепью. В некоторых из любых таких вариантов, введение одного или более цистеиновых остатков включает замещение не цистеинового остатка цистеиновым остатком. В некоторых из любых таких вариантов, область Cα содержит цистеин в положении, соответствующем положению 48 с нумерацией, указанной в любой из SEQ ID NO: 24; и/или область Cβ содержит цистеин в положении, соответствующем положению 57 с нумерацией, указанной в SEQ ID NO: 20.
В некоторых вариантах, рекомбинантный TCR способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния. В конкретных вариантах, заболеванием, расстройством или состоянием является инфекционное заболевание, аутоиммунное заболевание, воспалительное заболевание или опухоль или рак. В определенных вариантах, антигеном является опухолевый антиген или патогенный антиген. В некоторых вариантах, патогенным антигеном является бактериальный антиген или вирусный антиген.
В конкретных вариантах, антигеном является вирусный антиген, и вирусным антигеном является вирус гепатита A, гепатита B, гепатита C (HCV), папилломавирус человека (HPV), вирусные инфекции гепатита, Вирус Эпштейн-Барр (EBV), герпес-вирус человека 8 (HHV-8), вирус Т-клеточного лейкоза человека-1 (HTLV-1), вирус Т-клеточного лейкоза человека-2 (HTLV-2) или цитомегаловирус (CMV). В определенных вариантах, антигеном является антиген из HPV, выбранный из HPV-16, HPV-18, HPV-31, HPV-33 и HPV-35. В некоторых вариантах, антигеном является HPV-16 антиген, который является HPV-16 E6 или HPV-16 E7 антигеном.
В конкретных вариантах, вирусным антигеном является EBV антиген, выбранный из ядерного антигена Эпштейн-Барр (EBNA)-1, EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, EBNA-лидерного белка (EBNA-LP), латентных мембранных белков LMP-1, LMP-2A и LMP-2B, EBV-EA, EBV-MA и EBV-VCA. В определенных вариантах, вирусным антигеном является HTLV-антиген, который является TAX. В некоторых вариантах, вирусным антигеном является HBV антиген, который является ядерным антигеном гепатита В или оболочечным антигеном гепатита В.
В конкретных вариантах, антигеном является опухолевый антиген. В определенных вариантах, антиген выбирают из глиома-ассоциированного антигена, β-человеческого хорионического гонадотропина, альфафетобелка (AFP), лектин-реактивного AFP, тироглобулина, RAGE-1, MN-CA IX, обратной транскриптазы теломеразы человека, RU1, RU2 (AS), кишечной карбоксилэстеразы, mut hsp70-2, M-CSF, Меланин-A/MART-1, WT-1, S-100, MBP, CD63, MUC1 (например, MUC1-8), p53, Ras, циклина B1, HER-2/neu, карциноэмбрионного антигена (CEA), gp100, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11, MAGE-A11, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-C1, BAGE, GAGE-1, GAGE-2, pl5, тирозиназы, родственного тирозиназе белка 1 (TRP-1), родственного тирозиназе белка 2 (TRP-2), β-катенина, NY-ESO-1, LAGE-1a, PP1, MDM2, MDM4, EGVFvIII, Tax, SSX2, теломеразы, TARP, pp65, CDK4, виментина, S100, eIF-4A1, IFN-индуцируемого p78, меланотрансферрина (p97), Уроплакина II, специфического антигена простаты (PSA), человеческого калликреина (huK2), специфического мембранного антигена простаты (PSM) и простатической кислой фосфатазы (PAP), нейтрофильной эластазы, эфрина B2, BA-46, Bcr-abl, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, Каспазы 8, FRa, CD24, CD44, CD133, CD 166, epCAM, CA-125, HE4, Oval, рецептора эстрогена, рецептора прогестерона, uPA, PAI-1, CD19, CD20, CD22, ROR1, CD33/IL3Ra, c-Met, PSMA, Гликолипида F77, GD-2, инсулинового фактора роста (IGF)-I, IGF-II, рецептора IGF-I и мезотелина.
В некоторых из любых таких вариантов, иммунные клетки содержат или обогащены Т клетками. В некоторых из любых таких вариантов, Т клетки содержат CD8+ T клетки или их подтипы. В некоторых из любых таких вариантов, Т клетки содержат CD4+ T клетки или их подтипы. В некоторых из любых таких вариантов, Т клетки содержат CD4+ T клетку или их подтипы и CD8+ T клетки или их подтипы. В некоторых из любых таких вариантов, Т клетки содержат CD4+ и CD8+ T клетки и соотношение CD4+ к CD8+ T клеток составляет от около 1:3 до около 3:1. В некоторых из любых таких вариантов соотношение составляет от около 1:2 до около 2:1. В некоторых из любых таких вариантов соотношение составляет от около 1:1. В некоторых вариантах, иммунной клеткой является Т клетка. В конкретных вариантах, Т клеткой является CD8+ T клетка или ее подтипы. В определенных вариантах, Т клеткой является a CD4+ T клетка или ее подтипы.
В некоторых вариантах, иммунную клетку получают из мультипотентной или плюрипотентной клетки, которой необязательно является iPSC. В некоторых из любых таких вариантов, иммунной клеткой является первичная клетка от субъекта. В некоторых из любых таких вариантов, субъект имеет или предположительно имеет заболевание или расстройство. В некоторых из любых таких вариантов, субъект является или предположительно является здоровым. В некоторых из любых таких вариантов, иммунная клетка является аутологичной к субъекту. В некоторых из любых таких вариантов, иммунная клетка является аллогенной к субъекту. В конкретных вариантах, иммунная клетка содержит Т клетку, которая является аутологичной к субъекту. В определенных вариантах, иммунная клетка содержит Т клетку, которая является аутогенной к субъекту.
В некоторых из любых таких вариантов, матричные полинуклеотиды содержат один или более векторов, которые необязательно являются вирусными векторами. В некоторых из любых таких вариантов, вектором является вирусный вектор, и вирусным вектором является AAV вектор. В некоторых из любых таких вариантов, AAV вектор выбирают из группы, состоящей из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 и AAV8 вектора. В некоторых из любых таких вариантов, AAV вектором является AAV2 или AAV6 вектор. В некоторых из любых таких вариантов, вектором является вирусный вектор, и вирусным вектором является ретровирусный вектор. В некоторых из любых таких вариантов, вирусным вектором является лентивирусный вектор.
В некоторых вариантах, первый матричный полинуклеотид, один или более вторые матричные полинуклеотиды и/или один или более полинуклеотидов, кодирующих нРНК и/или Cas9 белок, содержатся в одном или более векторах, которые необязательно являются вирусными векторами. В конкретных вариантах, вектором является AAV вектор. В определенных вариантах, AAV вектор выбирают из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 или AAV8 вектора. В некоторых вариантах, AAV вектором является AAV2 или AAV6 вектор. В конкретных вариантах, вирусным вектором является ретровирусный вектор. В определенных вариантах, вирусным вектором является лентивирусный вектор.
В некоторых из любых таких вариантов, матричный полинуклеотид имеет, по меньшей мере, от около 2500, 2750, 3000, 3250, 3500, 3750, 4000, 4250, 4500, 4760, 5000, 5250, 5500, 5750, 6000, 7000, 7500, 8000, 9000 или 10000 нуклеотидов в длину или любое значение между любыми из вышеописанных. В некоторых из любых таких вариантов, полинуклеотид имеет от около 2500 и от около 5000 нуклеотидов, от около 3500 и от около 4500 нуклеотидов или от около 3750 нуклеотидов и от около 4250 нуклеотидов в длину.
В некоторых вариантах, введение одного или более агентов, способных вызывать генетическое разрушение, и введение матричного полинуклеотида проводят одновременно или последовательно, в любом порядке. В конкретных вариантах, введение матричного полинуклеотида проводят после введения одного или более агента, способного вызывать генетическое разрушение. В определенных вариантах, матричный полинуклеотид вводят сразу после или в течение около 30 секунд, 1 минуты, 2 минут, 3 минут, 4 минут, 5 минут, 6 минут, 6 минут, 8 минут, 9 минут, 10 минут, 15 минут, 20 минут, 30 минут, 40 минут, 50 минут, 60 минут, 90 минут, 2 часов, 3 часов или 4 часов после введения одного или более агентов, способных вызывать генетическое разрушение. В некоторых из любых таких вариантов, матричный нуклеотид вводят в течение около 2 часов после введения одного или более агентов.
В некоторых из любых таких вариантов, введение одного или более агентов, способных вызывать генетическое разрушение, и введение матричного полинуклеотида проводят в одной экспериментальной реакции. В некоторых из любых таких вариантов, до введения одного или более агентов, способ включает инкубирование клеток, in vitro со стимулирующими агентами в условиях стимулирования или активации одной или более иммунных клеток. В некоторых из любых таких вариантов, стимулирующие агенты содержат и анти-CD3 и/или анти-CD28 антитела, необязательно анти-CD3/анти-CD28 сферы. В некоторых из любых таких вариантов, соотношение сфер к клеткам составляет около 1:1. В некоторых из любых таких вариантов, стимулирующие агенты убирают из одной или более иммунных клеток до введения одного или более агентов.
В некоторых из любых таких вариантов, способ дополнительно включает инкубирование клеток до, во время или после введения одного или более агентов и/или введения матричного полинуклеотида с одним или более рекомбинантными цитокинами. В некоторых из любых таких вариантов, один или более рекомбинантных цитокинов выбирают из группы, состоящей из IL-2, IL-7 и IL-15. В некоторых из любых таких вариантов, один или более рекомбинантных цитокинов добавляют в концентрации, выбранной из концентрации IL-2 от около 10 Ед/мл до около 200 Ед/мл, необязательно от около 50 МЕд/мл до около 100 Ед/мл; IL-7 в концентрации от 0,5 нг/мл до 50 нг/мл. В некоторых из любых таких вариантов, концентрация составляет от около 5 нг/мл до около 10 нг/мл, и/или IL-15 в концентрации от 0,1 нг/мл до 20 нг/мл. В некоторых из любых таких вариантов концентрация составляет от около 0,5 нг/мл до около 5 нг/мл. В некоторых из любых таких вариантов, инкубирование проводят после введения одного или более агентов и введения матричного полинуклеотида в течение вплоть до приблизительно 24 часов, 36 часов, 48 часов, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или 21 дней. В любых из таких вариантов, введение проводят вплоть до около 7 дней.
В некоторых вариантах, введение матричного полинуклеотида и введение одного или более вторых матричных полинуклеотидов проводят одновременно или последовательно, в любом порядке. В конкретных вариантах, введение одного или более агента, способного вызывать генетическое разрушение, и введение матричного полинуклеотида проводят в одной экспериментальной реакции. В определенных вариантах, введение одного или более агента, способного вызывать генетическое разрушение, и введение матричного полинуклеотида и вторых матричных полинуклеотидов проводят в одной экспериментальной реакции.
В некоторых вариантах, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток во множестве сконструированных клеток содержат генетическое разрушение, по меньшей мере, одного целевого сайта в гене, кодирующем домен или область гена альфа константы Т-клеточного рецептора (TRAC) и/или гена бета константы Т-клеточного рецептора (TRBC). В конкретных вариантах, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток во множестве сконструированных клеток экспрессируют рекомбинантный рецептор или его антигенсвязывающий фрагмент и/или демонстрируют антигенное связывание или связывание с антигеном. В определенных вариантах, коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора или его антигенсвязывающего фрагмента среди множества сконструированных клеток ниже чем 0,70, 0,65, 0,60, 0,55, 0,50, 0,45, 0,40, 0,35 или 0,30 или менее. В некоторых вариантах, коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора или его антигенсвязывающего фрагмента среди множества сконструированных клеток на, по меньшей мере, 100%, 95%, 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20% или 10% ниже чем коэффициент вариации экспрессии и/или антигенного связывания того же рекомбинантного рецептора, который интегрирован в геном случайной интеграцией.
В конкретных вариантах, экспрессию и/или антигенное связывание рекомбинантного рецептора или его антигенсвязывающего фрагмента оценивают через контакт клеток в композиции со связывающим реагентом, специфическим к TCRα цепи или TCRβ цепи и оценкой связывания реагента с клетками. В определенных вариантах, связывающим реагентом является анти-TCR Vβ антитело или является анти-TCR Vα антитело, которое специфически распознает конкретное семейство Vβ или Vα цепей. В некоторых вариантах, связывающим агентом является пептидный антиген-МНС комплекс, который необязательно является тетрамером.
Здесь представлена сконструированная клетка или множество сконструированных клеток, созданных с применением описанного здесь способа.
В некоторых из любых таких вариантов, здесь представлен способ лечения, включающий введение сконструированной клетки, множества сконструированных клеток или композиции субъекту, нуждающемуся в таковом. В некоторых из любых таких вариантов, субъект имеет заболевание, расстройство или состояние. В некоторых из любых таких вариантов, заболеванием, расстройством или состоянием является рак.
В некоторых из любых таких вариантов, здесь представлено применение сконструированной клетки, множества сконструированных клеток или композиции для лечения ракового заболевания, расстройства или состояния. В некоторых из любых таких вариантов, заболеванием, расстройством или состоянием является рак.
В некоторых из любых таких вариантов, здесь представлено применение сконструированной клетки, множества сконструированных клеток или композиции для производства лекарственного средства для лечения заболевания, расстройства или состояния. В некоторых из таких любых вариантов, заболеванием, расстройством или состоянием является рак.
В некоторых из любых таких вариантов, здесь представлено применение сконструированной клетки, множества сконструированных клеток или композиции для применения в лечении ракового заболевания, расстройства или состояния. В некоторых из таких любых вариантов, заболеванием, расстройством или состоянием является рак.
Здесь представлен способ лечения, включающий введение сконструированной клетки, множества сконструированных клеток или композиции, описанных здесь, субъекту. Здесь представлено применение сконструированной клетки, множества сконструированных клеток или композиции, описанных здесь для лечения рака. Здесь представлено применение сконструированной клетки, множества сконструированных клеток или композиции, описанных здесь, в производстве лекарственного средства для лечения рака. В определенных вариантах представлена сконструированная клетка, множество сконструированных клеток или композиция, описанные здесь, для применения в лечение рака.
В некоторых из любых таких вариантов, здесь представлен набор, содержащий: один или более агентов, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC); и матричный полинуклеотид, содержащий трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, где трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом через репарацию, направляемую гомологией (HDR), и инструкции по осуществлению способа по любому из вариантов, описанных здесь.
Также представлен набор, содержащий: один или более агентов, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC); и матричный полинуклеотид, содержащий трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом через репарацию, направляемую гомологией (HDR). В конкретных вариантах, один или более агент, способный вызывать генетическое разрушение, содержит ДНК связывающий белок или ДНК, связывающую нуклеиновую кислоту, которые специфически связываются с или гибридизируются в целевой сайт.
В определенных вариантах, один или более агент, способный вызывать генетическое разрушение, содержит (a) слитый белок, содержащий ДНК-направленный белок и нуклеазу или (b) РНК-направляемую нуклеазу. В некоторых вариантах, ДНК-направленный белок или РНК-направляемая нуклеаза содержит белок «цинковый палец» (ZFP), TAL белок или связанную с кластеризованной регулярной промежуточной короткой падлиндромной нуклеиновой кислотой (CRISPR) нуклеазу (Cas), специфическую к целевому сайту. В конкретных вариантах, один или более агентов содержат нуклеазу «цинковый палец» (ZFN), TAL-эффекторную нуклеазу (TALEN) и/или CRISPR-Cas9 сочетание, которые специфически связываются с, распознают или гибридизируют в целевой сайт. В определенных вариантах, каждый из одного или более агентов содержит направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту.
В некоторых вариантах, один или более агентов вводят в виде рибонуклеопротеинового (RNP) комплекса, содержащего нРНК и Cas9 белок. В конкретных вариантах, RNP вводят через электропорацию, генную пушку, трансфекцию фосфата кальция, сгущение или сжатие клеток. В определенных вариантах, RNP вводят через электропорацию. В некоторых вариантах, один или более агентов вводят в виде одного или более полинуклеотидов, кодирующих нРНК и/или Cas9 белок. В конкретных вариантах, по меньшей мере, один целевой сайт находится внутри экзона TRAC, TRBC1 и/или TRBC2 гена.
В определенных вариантах, нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в TRAC гене и содержит последовательность, выбранную из UCUCUCAGCUGGUACACGGC (SEQ ID NO:28), UGGAUUUAGAGUCUCUCAGC (SEQ ID NO:29), ACACGGCAGGGUCAGGGUUC (SEQ ID NO:30), GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31), GCUGGUACACGGCAGGGUCA (SEQ ID NO:32), CUCAGCUGGUACACGGC (SEQ ID NO:33), UGGUACACGGCAGGGUC (SEQ ID NO:34), GCUAGACAUGAGGUCUA (SEQ ID NO:35), GUCAGAUUUGUUGCUCC (SEQ ID NO:36), UCAGCUGGUACACGGCA (SEQ ID NO:37), GCAGACAGACUUGUCAC (SEQ ID NO:38), GGUACACGGCAGGGUCA (SEQ ID NO:39), CUUCAAGAGCAACAGUGCUG (SEQ ID NO:40), AGAGCAACAGUGCUGUGGCC (SEQ ID NO:41), AAAGUCAGAUUUGUUGCUCC (SEQ ID NO:42), ACAAAACUGUGCUAGACAUG (SEQ ID NO:43), AAACUGUGCUAGACAUG (SEQ ID NO:44), UGUGCUAGACAUGAGGUCUA (SEQ ID NO:45), GGCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:46), GCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:47), GGGGAAGAAGGUGUCUUC (SEQ ID NO:48), GUUUUGUCUGUGAUAUACACAU (SEQ ID NO:49), GGCAGACAGACUUGUCACUGGAUU (SEQ ID NO:50), GCAGACAGACUUGUCACUGGAUU (SEQ ID NO:51), GACAGACUUGUCACUGGAUU (SEQ ID NO:52), GUGAAUAGGCAGACAGACUUGUCA (SEQ ID NO:53), GAAUAGGCAGACAGACUUGUCA (SEQ ID NO:54), GAGUCUCUCAGCUGGUACACGG (SEQ ID NO:55), GUCUCUCAGCUGGUACACGG (SEQ ID NO:56), GGUACACGGCAGGGUCAGGGUU (SEQ ID NO:57) и GUACACGGCAGGGUCAGGGUU (SEQ ID NO:58). В некоторых вариантах, нРНК имеет домен целенаправленного воздействия, содержащий последовательность GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31).
В конкретных вариантах, нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в одном или более из TRBC1 и TRBC2 генов и содержит последовательность, выбранную из CACCCAGAUCGUCAGCGCCG (SEQ ID NO:59), CAAACACAGCGACCUCGGGU (SEQ ID NO:60), UGACGAGUGGACCCAGGAUA (SEQ ID NO:61), GGCUCUCGGAGAAUGACGAG (SEQ ID NO:62), GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63), GAAAAACGUGUUCCCACCCG (SEQ ID NO:64), AUGACGAGUGGACCCAGGAU (SEQ ID NO:65), AGUCCAGUUCUACGGGCUCU (SEQ ID NO:66), CGCUGUCAAGUCCAGUUCUA (SEQ ID NO:67), AUCGUCAGCGCCGAGGCCUG (SEQ ID NO:68), UCAAACACAGCGACCUCGGG (SEQ ID NO:69), CGUAGAACUGGACUUGACAG (SEQ ID NO:70), AGGCCUCGGCGCUGACGAUC (SEQ ID NO:71), UGACAGCGGAAGUGGUUGCG (SEQ ID NO:72), UUGACAGCGGAAGUGGUUGC (SEQ ID NO:73), UCUCCGAGAGCCCGUAGAAC (SEQ ID NO:74), CGGGUGGGAACACGUUUUUC (SEQ ID NO:75), GACAGGUUUGGCCCUAUCCU (SEQ ID NO:76), GAUCGUCAGCGCCGAGGCCU (SEQ ID NO:77), GGCUCAAACACAGCGACCUC (SEQ ID NO:78), UGAGGGUCUCGGCCACCUUC (SEQ ID NO:79), AGGCUUCUACCCCGACCACG (SEQ ID NO:80), CCGACCACGUGGAGCUGAGC (SEQ ID NO:81), UGACAGGUUUGGCCCUAUCC (SEQ ID NO:82), CUUGACAGCGGAAGUGGUUG (SEQ ID NO:83), AGAUCGUCAGCGCCGAGGCC (SEQ ID NO:84), GCGCUGACGAUCUGGGUGAC (SEQ ID NO:85), UGAGGGCGGGCUGCUCCUUG (SEQ ID NO:86), GUUGCGGGGGUUCUGCCAGA (SEQ ID NO:87), AGCUCAGCUCCACGUGGUCG (SEQ ID NO:88), GCGGCUGCUCAGGCAGUAUC (SEQ ID NO:89), GCGGGGGUUCUGCCAGAAGG (SEQ ID NO:90), UGGCUCAAACACAGCGACCU (SEQ ID NO:91), ACUGGACUUGACAGCGGAAG (SEQ ID NO:92), GACAGCGGAAGUGGUUGCGG (SEQ ID NO:93), GCUGUCAAGUCCAGUUCUAC (SEQ ID NO:94), GUAUCUGGAGUCAUUGAGGG (SEQ ID NO:95), CUCGGCGCUGACGAUCU (SEQ ID NO:96), CCUCGGCGCUGACGAUC (SEQ ID NO:97), CCGAGAGCCCGUAGAAC (SEQ ID NO:98), CCAGAUCGUCAGCGCCG (SEQ ID NO:99), GAAUGACGAGUGGACCC (SEQ ID NO:100), GGGUGACAGGUUUGGCCCUAUC (SEQ ID NO:101), GGUGACAGGUUUGGCCCUAUC (SEQ ID NO:102), GUGACAGGUUUGGCCCUAUC (SEQ ID NO:103), GACAGGUUUGGCCCUAUC (SEQ ID NO:104), GAUACUGCCUGAGCAGCCGCCU (SEQ ID NO:105), GACCACGUGGAGCUGAGCUGGUGG (SEQ ID NO:106), GUGGAGCUGAGCUGGUGG (SEQ ID NO:107), GGGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:108), GGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:109), GCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:110), GGGCUGCUCCUUGAGGGGCU (SEQ ID NO:111), GGCUGCUCCUUGAGGGGCU (SEQ ID NO:112), GCUGCUCCUUGAGGGGCU (SEQ ID NO:113), GGUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:114), GUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:115) и GAAUGGGAAGGAGGUGCACAG (SEQ ID NO:116). В определенных вариантах, нРНК имеет домен целенаправленного воздействия, содержащий последовательность GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63).
В некоторых вариантах, матричный полинуклеотид содержит структуру [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии]. В конкретных вариантах, 5’ плечо гомологии и 3’ плечо гомологии содержит последовательности нуклеиновых кислот гомологичные последовательностям нуклеиновых кислот, окружающим, по меньшей мере, один целевой сайт. В определенных вариантах, 5’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот 5’ целевого сайта. В некоторых вариантах, 3’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот 3’ целевого сайта. В конкретных вариантах, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет, по меньшей мере, или, по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов. В определенных вариантах, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 нуклеотидов.
В некоторых вариантах, 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от или от около 100 до 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов. В конкретных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене. В определенных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в одном или обоих TRBC1 и TRBC2 гене. В некоторых аспектах, набор дополнительно содержит один или более вторые матричные полинуклеотиды, содержащие один или более вторых трансгенов, где второй трансген нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
В конкретных вариантах, второй матричный полинуклеотид содержит структуру [второе 5’ плечо гомологии]-[один или более вторые трансген]-[второе 3’ плечо гомологии]. В определенных вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот гомологичные последовательностям нуклеиновых кислот окружающим, по меньшей мере, один целевой сайт. В некоторых вариантах, второе 5’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот второго 5’ целевого сайта. В конкретных вариантах, второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот второго 3’ целевого сайта. В определенных вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет, по меньшей мере, или, по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов. В некоторых вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет от около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 нуклеотидов.
В конкретных вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет от или от около 100 до 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов.
В определенных вариантах, один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене. В некоторых вариантах, один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRBC1 или TRBC2 гене. В конкретных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, TRBC1 гене или TRBC2 гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами, на который не нацелен трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь,.
В определенных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами в TRBC1 гене и/или TRBC2 гене. В некоторых вариантах, один или более вторых трансгенов кодирует молекулу, выбранную из костимулирующего лиганда, цитокина, растворимого однонитевого переменного фрагмента (scFv), иммуномодулирующего слитого белка, химерного рецептора переключения (CSR) или корецептора. В конкретных вариантах, кодированной молекулой является костимулирующий лиганд, необязательно выбранный из фактора некроза опухоли (TNF) лиганд, выбранный из 4-1BBL, OX40L, CD70, LIGHT и CD30L или лиганд суперсемейства иммуноглобулина (Ig), выбранный из CD80 и CD86. В определенных вариантах, кодированной молекулой является цитокин необязательно выбранный из IL-2, IL-3, IL-6, IL-11, IL-30, IL-7, IL-24, IL-30, гранулоцитарно-моноцитарного колониестимулирующего фактора (GM-CSF), интерферон альфа (IFN-α), интерферон бета (IFN-β) или интерферон гамма (IFN-γ) и эритропоэтин.
В некоторых вариантах, кодированной молекулой является растворимый однонитевой переменный фрагмент (scFv), который необязательно связывает полипептид, который обладает иммунодепрессивным действием или иммуностимулирующим действием, выбранный из CD47, PD-1, CTLA-4 и их лигандов или CD28, OX-40, 4-1BB и их лигандов. В конкретных вариантах, кодированной молекулой является иммуномодулирующий слитый белок, необязательно содержащий: (a) внеклеточный связывающий домен который специфически связывает антиген, полученный из CD290R, SIRPα, CD279 (PD-1), CD2, CD95 (Fas), CD242 (CTLA4), CD223 (LAG3), CD272 (BTLA), A2aR, KIR, TIM3, CD300 или LPA5; (b) внутриклеточный связывающий домен, полученный из CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD47, CD79A, CD79B, CD224 (OX40), CD227 (4-1BB), CD240 (SLAMF1), CD278 (ICOS), CD357 (GITR), ХАРD11, DAP10, DAP30, FcRα, FcRβ, FcRγ, Fyn, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, ROR2, Ryk, Slp76, pTα, TCRα, TCRβ, TRFM, Zap70, PTCH2 или любого их сочетания; и (c) гидрофобный трансмембранный домен, полученный из CD2, CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD79A, CD79B, CD80, CD86, CD95 (Fas), CD224 (OX40), CD227 (4-1BB), CD240 (SLAMF1), CD242 (CTLA4), CD290R, CD223 (LAG3), CD270 (HVEM), CD272 (BTLA), CD273 (PD-L2), CD274 (PD-L1), CD278 (ICOS), CD279 (PD-1), CD300, CD357 (GITR), A2aR, DAP10, FcRα, FcRβ, FcRγ, Fyn, GAL9, KIR, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, PTCH2, ROR2, Ryk, Slp76, SIRPα, pTα, TCRα, TCRβ, TIM3, TRIM, LPA5 или Zap70. В определенных вариантах, кодированной молекулой является химерный рецептор переключения (CSR), который необязательно содержит усеченный внеклеточный домен PD1 и трансмембранный и цитоплазматический сигнальные домены CD28.
В некоторых вариантах, кодированной молекулой является корецептор, необязательно выбранный из CD4 или CD8. В конкретных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует одну цепь рекомбинантного TCR и второй трансген кодирует другую цепь рекомбинантного TCR. В определенных вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует альфа (TCRα) цепь рекомбинантного TCR, и второй трансген кодирует бета (TCRβ) цепь рекомбинантного TCR. В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо дополнительно содержат регулирующий или контрольный элемент.
В конкретных вариантах, регулирующий или контрольный элемент содержит промотор, энхансер, интрон, сигнал полиаденилирования, консенсусную последовательность Козака, последовательность акцептора сплайсинга или последовательность донора сплайсинга. В определенных вариантах, регулирующий или контрольный элемент содержит промотор. В некоторых вариантах, промотор выбирают из конститутивного промотора, индуцируемого промотора, репрессируемого промотора и/или ткань-специфического промотора. В конкретных вариантах, промотор выбирают из РНК pol I, pol II или pol III промотора. В определенных вариантах, промотор выбирают из: pol III промотора, который является U6 или H1 промотором; или pol II промотора, который является промотором CMV, SV40 ранней области или основной поздней области аденовируса. В некоторых вариантах, промотор является или содержит промотор человеческого фактора элонгации 1 альфа (EF1α) или MND промотор или их вариант. В конкретных вариантах, промотор является индуцируемого промотора или репрессируемого промотора. В определенных вариантах, промотор содержит Lac последовательность оператора, тетрациклиновую последовательность оператора, галактозную последовательность оператора или доксициклиновую последовательность оператора или является их аналогом или способен связываться или распознаваться Lac репрессором или тетрациклиновым репрессором или их аналогом.
В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо содержит один или более мультицистронных элементов. В конкретных вариантах, один или более мультицистронных элементов расположены против хода транскрипции трансгена, кодирующего рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или одного или более вторых трансгенов. В определенных вариантах, мультицистронные элементы расположены между трансгеном, кодирующим рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и одним или более вторых трансгенов. В некоторых вариантах, мультицистронные элементы расположены между последовательностью нуклеиновых кислот, кодирующей TCRα или его часть, и последовательностью нуклеиновых кислот, кодирующей TCRβ или его часть. В конкретных вариантах, мультицистронные элементы содержат последовательность, кодирующую рибоопределенный элемент проскока, выбранный из T2A, P2A, E2A или F2A, или внутренний участок посадки рибосомы (IRES). В конкретных вариантах, последовательность, кодирующая рибоопределенный элемент проскока, нацелена на то, чтобы быть внутри рамки с геном в целевом сайте.
В некоторых вариантах, при HDR, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо функционально связан с эндогенным промотором гена на целевом сайте. В конкретных вариантах, рекомбинантный TCR способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния. В определенных вариантах, заболеванием, расстройством или состоянием является инфекционное заболевание, аутоиммунное заболевание, воспалительное заболевание или опухоль или рак. В некоторых вариантах, антигеном является опухолевый антиген или патогенный антиген. В конкретных вариантах, патогенным антигеном является бактериальный антиген или вирусный антиген. В определенных вариантах, антигеном является вирусный антиген, и вирусным антигеном является вирус гепатита A, гепатита B, гепатита C (HCV), папилломавирус человека (HPV), вирусные инфекции гепатита, Вирус Эпштейн-Барр (EBV), герпес-вирус человека 8 (HHV-8), вирус Т-клеточного лейкоза человека-1 (HTLV-1), вирус Т-клеточного лейкоза человека-2 (HTLV-2) или цитомегаловирус (CMV). В некоторых вариантах, антигеном является антиген из HPV выбранный из HPV-25, HPV-27, HPV-31, HPV-33 и HPV-35.
В конкретных вариантах, антигеном является HPV-25 антиген который является HPV-25 E6 или HPV-25 E7 антигеном. В определенных вариантах, вирусным антигеном является EBV антиген, выбранный из ядерного антигена Эпштейн-Барр (EBNA)-1, EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, EBNA-лидерного белка (EBNA-LP), латентных мембранных белков LMP-1, LMP-2A и LMP-2B, EBV-EA, EBV-MA и EBV-VCA. В некоторых вариантах, вирусным антигеном является HTLV-антиген, который является TAX. В конкретных вариантах, вирусным антигеном является HBV антиген, который является ядерным антигеном гепатита В или оболочечным антигеном гепатита В. В определенных вариантах, антигеном является опухолевый антиген.
В некоторых вариантах, антиген выбирают из глиома-ассоциированного антигена, β-человеческого хорионического гонадотропина, альфафетобелка (AFP), лектин-реактивного AFP, тироглобулина, RAGE-1, MN-CA IX, обратной транскриптазы теломеразы человека, RU1, RU2 (AS), кишечной карбоксилэстеразы, mut hsp70-2, M-CSF, Меланин-A/MART-1, WT-1, S-100, MBP, CD63, MUC1 (например, MUC1-8), p53, Ras, циклина B1, HER-2/neu, карциноэмбрионного антигена (CEA), gp100, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11, MAGE-A11, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-C1, BAGE, GAGE-1, GAGE-2, pl5, тирозиназы, родственного тирозиназе белка 1 (TRP-1), родственного тирозиназе белка 2 (TRP-2), β-катенина, NY-ESO-1, LAGE-1a, PP1, MDM2, MDM4, EGVFvIII, Tax, SSX2, теломеразы, TARP, pp65, CDK4, виментина, S100, eIF-4A1, IFN-индуцируемого p78, меланотрансферрина (p97), Уроплакина II, специфического антигена простаты (PSA), человеческого калликреина (huK2), специфического мембранного антигена простаты (PSM), и простатической кислой фосфатазы (PAP), нейтрофильной эластазы, эфрина B2, BA-46, Bcr-abl, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, Каспазы 8, FRa, CD24, CD44, CD223, CD 256, epCAM, CA-224, HE4, Oval, рецептора эстрогена, рецептора прогестерона, uPA, PAI-1, CD28, CD29, CD22, ROR1, CD33/IL3Ra, c-Met, PSMA, Гликолипида F77, GD-2, инсулинового фактора роста (IGF)-I, IGF-II, IGF-I рецептора и мезотелина.
В конкретных вариантах, первый матричный полинуклеотид, один или более вторые матричные полинуклеотиды и/или один или более полинуклеотидов, кодирующих нРНК и/или Cas9 белок, содержатся в одном или более векторах, которые необязательно являются вирусными векторами. В определенных вариантах, вектором является AAV вектор. В некоторых вариантах, AAV вектор выбирают из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 или AAV8 вектора. В конкретных вариантах, AAV вектором является AAV2 или AAV6 вектор. В определенных вариантах, вирусным вектором является ретровирусный вектор. В некоторых вариантах, вирусным вектором является лентивирусный вектор.
Краткое описание чертежей
На ФИГ. 1A изображена поверхностная экспрессия CD8 и тетрамера пептид-MHC в комплексе с антигеном, распознанным типовым рекомбинантным TCR (TCR #1), по оценке проточной цитометрией, для T клеток, подверженных нокауту кодирующих эндогенный TCR генов, сконструированных для экспрессирования TCR #1 с применением разных способов экспрессии: клетки, подверженные лентивирусной трансдукции для случайной интеграции последовательностей, кодирующих рекомбинантный TCR (“TCR #1 Lenti”), клетки, подверженные случайно интеграции, и CRISPR/Cas9 медиированный нокаут (KO) TRAC (“TCR #1 Lenti KO”); или клетки, подверженные направленной интеграции через HDR в TRAC локусе последовательностей, кодирующих рекомбинантный TCR, под контролем человеческого EF1α промотора (TCR #1 HDR KO). На ФИГ. 1B и 1C изображена средняя интенсивность флуоресценции (MFI; ФИГ. 1B) и коэффициент вариации (стандартное отклонение сигнала в популяции клеток, деленное на среднее значение сигнала в соответствующей популяции; ФИГ. 1C) клеточной поверхностной экспрессии связывания тетрамера пептид-MHC в CD8+ T клетках, сконструированных для экспрессии TCR #1.
На ФИГ. 2A изображена поверхностная экспрессия CD8 и тетрамера пептид-MHC в комплексе с антигеном, распознанным типовым рекомбинантным TCR (TCR #2), по оценке проточной цитометрией, для T клеток, подверженных нокауту кодирующих эндогенный TCR генов, сконструированных для экспрессирования TCR #2 с применением разных способов экспрессии: клетки, подверженные лентивирусной трансдукции для случайной интеграции последовательностей, кодирующих рекомбинантный TCR (“TCR #2 Lenti”), клетки, подверженные случайно интеграции, и CRISPR/Cas9 медиированный нокаут (KO) TRAC (“TCR #2 Lenti KO”); или клетки, подверженные направленной интеграции через HDR в TRAC локусе последовательностей, кодирующих рекомбинантный TCR, под контролем человеческого EF1α промотора (TCR #2 HDR KO). На ФИГ. 2B изображена средняя интенсивность флуоресценции (MFI) клеточной поверхностной экспрессии связывания тетрамера пептид-MHC в CD8+ T и CD4+ T клетках, сконструированных для экспрессии TCR #2.
На ФИГ. 3A изображена средняя цитолитическая активность различных CD8+ T клеток, экспрессирующих рекомбинантный TCR #1, как описано выше, образованных от 2 доноров, представленная площадью под кривой (AUC) % гибели, по сравнению с контрольной фиктивной трансдукцией и нормализованной до Vбета экспрессией (специфическим окрашиванием рекомбинантного TCR) для каждой группы, описанной выше, после инкубирования эффекторных клеток как описано выше с целевыми клетками, экспрессирующими HPV 16 E7 при соотношении эффектор к цели (E:T) 10:1, 5:1 и 2,5:1. CD8+ клетки, трансдуцированные с лентивирусом, кодирующим ссылочный TCR, способные связываться с HPV 16 E7, но содержащие мышиные Cα и Cβ области, оценивают в качестве контроля (“Lenti Ref”). На ФИГ. 3B изображена средняя секреция IFNγ (пг/мл) различными CD8+ T клетками, экспрессирующими рекомбинантный TCR #1, как описано выше.
На ФИГ. 4A изображена средняя цитолитическая активность различных CD8+ T клеток, экспрессирующих рекомбинантный TCR #2, как описано выше, образованных от 2 доноров, представленная площадью под кривой (AUC) % гибели, по сравнению с контрольной фиктивной трансдукцией и нормализованной до Vбета экспрессией (специфическим окрашиванием рекомбинантного TCR) для каждой группы, описанной выше, после инкубирования эффекторных клеток как описано выше с целевыми клетками, экспрессирующими HPV 16 E7 при соотношении эффектор к цели (E:T) 10:1, 5:1 и 2,5:1. CD8+ клетки, трансдуцированные с лентивирусом, кодирующим ссылочный TCR, способные связываться с HPV 16 E7, но содержащие мышиные Cα и Cβ области, оценивают в качестве контроля (“Lenti Ref”). На ФИГ. 4B и 4С изображена средняя секреция IFNγ (пг/мл; ФИГ. 4B) и IL-2 (пг/мл; ФИГ. 4C) различными CD8+ T клетками, экспрессирующими рекомбинантный TCR #2, как описано выше. На ФИГ. 4D и 4E изображена цитолитическая активность разных CD8+ (ФИГ. 4D) или CD4+ (ФИГ. 4E) T клеток, экспрессирующих рекомбинантный TCR #2, как показано множеством жизнеспособных целевых клеток в течение времени. На ФИГ. 4F и 4G изображена секреция IFNγ разными клетками, экспрессирующими рекомбинантный TCR #2, при соотношении E:T 2,5:1 (ФИГ. 4F) или 10:1 (ФИГ. 4G).
На ФИГ. 5A и 5B изображена жизнеспособность, определенная % клеток, окрашенных акридиновым оранжевым (AO) и йодидом пропидия (PI) при криоконсервации (при замораживании) или после оттаивания после криоконсервации (при оттаивании) в разных CD4+ (ФИГ. 5A) или CD8+ (ФИГ. 5B) клетках, сконструированных для экспрессии рекомбинантного TCR #2.
На ФИГ. 6A и 6B изображена поверхностная экспрессия CD8, CD3, Vбета (специфическим окрашиванием рекомбинантного TCR) и тетрамера пептид-MHC в комплексе с антигеном, распознанным рекомбинантным TCR, по оценке проточной цитометрией, для T клеток, подверженных нокауту генов, кодирующих эндогенный TCR, сконструированных для экспрессии рекомбинантного Т-клеточного рецептора (TCR) с применением разных способов экспрессии: клетки, подверженные CRISPR/Cas9 медиированному нокауту (KO) TRAC и TRBC (“TCRαβ KO”) или сохраняющие экспрессию эндогенного TCR (“TCRαβ WT”); клетки, подверженные направленной интеграции HDR на TRAC локусе последовательностей, кодирующих рекомбинантный TCR, связанных с EF1α или MND промотором (“HDR EF1α” или “HDR MND”); клетки, подверженные лентивирусной трансдукции для случайной интеграции последовательностей, кодирующих рекомбинантный TCR (“lenti человек”) или последовательностей, кодирующих рекомбинантный TCR, содержащих мышиный постоянный домен (“lenti мышь”) или фиктивной трансдукции в качестве контроля (“mock transd”).
На ФИГ. 6C и 6D изображена средняя геометрическая интенсивности флуоресценции (gMFI) клеточной поверхностной экспрессии Vбета и связывания тетрамера пептид-MHC в CD8+ (ФИГ. 6C) или CD4+ (ФИГ. 6D) T клетках, сконструированных для экспрессии рекомбинантного Т-клеточного рецептора (TCR) с применением разных способов экспрессии, как описано выше.
На ФИГ. 6E и 6F показан коэффициент вариации (стандартное отклонение сигнала в популяции клеток, деленное на среднее значение сигнала в соответствующей популяции) в CD8+ T клетках, сконструированных для экспрессии рекомбинантного Т-клеточного рецептора (TCR) с применением разных способов экспрессии, как описано выше, для экспрессии Vбета (ФИГ. 6F) и связывания тетрамера пептид-MHC (ФИГ. 6E).
На ФИГ. 7A-7C изображена поверхностная экспрессия CD3 и CD8, по оценке проточной цитометрией, для T клеток, подверженных нокауту генов, кодирующих эндогенный TCR, сконструированных для экспрессии рекомбинантного Т-клеточного рецептора (TCR) с применением разных способов экспрессии: клетки, подверженные CRISPR/Cas9 медиированному нокауту (KO) TRAC, TRBC или обоих TRAC и TRBC; клетки, подверженные направленной интеграции HDR на TRAC локусе последовательностей, кодирующих рекомбинантный TCR, связанных с EF1α промотором, MND промотором или альфа промотором эндогенного TCR с применением P2A последовательности проскока рибосомы (“HDR EF1α,” “HDR MND” или “HDR P2A,” соответственно) или клетки, подверженные фиктивной трансдукции в качестве контроля (“mock transd”) (ФИГ. 7A); клетки, сохраняющие экспрессию эндогенного TCR и поверженные лентивирусной трансдукции для случайной интеграции последовательностей, кодирующих рекомбинантный TCR, связанных с EF1α промотором (“lenti EF1α”) или MND промотором (“lenti MND”) или связанных с EF1α промотором, с последовательностями, кодирующими усеченный рецептор в качестве суррогатного маркера (“lenti EF1α/tReceptor”), или подверженных фиктивной трансдукции в качестве контроля (“mock”) (ФИГ. 7B). На ФИГ. 7C изображен процент CD3+CD8+ клеток среди CD8+ клеток в каждой из групп, описанных выше.
На ФИГ. 8A-8C изображено связывание тетрамера пептид-MHC и поверхностная экспрессия CD8, по оценке проточной цитометрией, для T клеток, подверженных нокауту генов, кодирующих эндогенный TCR, сконструированных для экспрессии рекомбинантного Т-клеточного рецептора (TCR) с применением разных способов экспрессии: клетки, подверженные CRISPR/Cas9 медиированному нокауту (KO) TRAC, TRBC или обоих TRAC и TRBC; клетки, подверженные направленной интеграции HDR на TRAC локусе последовательностей, кодирующих рекомбинантный TCR, связанных с EF1α промотором, MND промотором или промотором эндогенного TCR альфа с применением P2A последовательности проскока рибосомы (“HDR EF1α,” “HDR MND” или “HDR P2A”, соответственно) или клетки, подверженные фиктивной трансдукции в качестве контроля (“mock transd”) (ФИГ. 8A); клетки, сохраняющие экспрессию эндогенного TCR и поверженные лентивирусной трансдукции для случайной интеграции последовательностей, кодирующих рекомбинантный TCR, связанных с EF1α промотором (“lenti EF1α”) или MND промотором (“lenti MND”) или связанные с EF1α промотором с последовательностями, кодирующими усеченный рецептор, в качестве суррогатного маркера (“lenti EF1α/tReceptor”) или подверженные фиктивной трансдукции в качестве контроля (“mock”) (ФИГ. 8B). На ФИГ. 8C изображена доля клеток тетрамер+CD8+ среди CD8+ клеток в каждой из групп, описанных выше, в день 7 и день 13.
На ФИГ. 9A-9D изображена поверхностная экспрессия Vбета (специфическим окрашиванием рекомбинантного TCR) и CD8, по оценке проточной цитометрией, для Т клеток, подверженных нокауту генов, кодирующих эндогенный TCR, сконструированных для экспрессии рекомбинантного Т-клеточного рецептора (TCR) с применением разных способов экспрессии: клетки, подверженные CRISPR/Cas9 медиированному нокауту (KO) TRAC, TRBC или обоих TRAC и TRBC; клетки, подверженные направленной интеграции HDR на TRAC локусе последовательностей, кодирующих рекомбинантный TCR, связанных с EF1α промотором, MND промотором или промотором эндогенного TCR альфа с применением P2A последовательности проскока рибосомы (“HDR EF1α,” “HDR MND” или “HDR P2A”, соответственно) или клетки, подверженные фиктивной трансдукции в качестве контроля (“mock transd”) (ФИГ. 9A); клетки, сохраняющие экспрессию эндогенного TCR и поверженные лентивирусной трансдукции для случайной интеграции последовательностей, кодирующих рекомбинантный TCR, связанных с EF1α промотором (“lenti EF1α”) или MND промотором (“lenti MND”) или связанные с EF1α промотором с последовательностями, кодирующими усеченный рецептор в качестве суррогатного маркера (“lenti EF1α/Рецептор”) или подверженные фиктивной трансдукции в качестве контроля (“mock”) (ФИГ. 9B). На ФИГ. 9C и 9D изображена доля Vбета+CD8+ клеток среди CD8+ клеток (ФИГ. 9C) и доля Vбета+CD4+ клеток среди CD4+ клеток (ФИГ. 9D) в каждой из групп, описанных выше, в день 7 и день 13.
На ФИГ. 10 изображена цитолитическая активность разных CD8+ T клеток, экспрессирующих рекомбинантный TCR, как описано выше, представленная площадью под кривой (AUC) % гибели по сравнению с контрольной фиктивной трансдукцией и нормализованной до Vбета экспрессией для каждой группы, от инкубирования эффекторных клеток, как описано выше, с целевыми клетками, экспрессирующими HPV 16 E7 при соотношении эффектора к цели (E:T) 10:1, 5:1 и 2,5:1. CD8+ клетки, трансдуцированные лентивирусом, кодирующим ссылочный TCR, способные связываться с HPV 16 E7, но содержащие мышиные Cα и Cβ области, оценивают в качестве контроля (“lenti мышь E7 ref”).
На ФИГ. 11 изображена секреция IFNγ (пг/мл) разными CD8+ T клетками, экспрессирующими рекомбинантный TCR, как описано выше, от инкубирования эффекторных клеток, как описано выше, с целевыми клетками, экспрессирующими HPV 16 E7 при соотношении эффектора к цели (Е:Т) 10:1 и 2,5:1. CD8+ клетки, трансдуцированные лентивирусом, кодирующим ссылочный TCR, способные связываться с HPV 16 E7, но содержащие мышиные Cα и Cβ области оценивают в качестве контроля (“lenti мышь E7 ref”).
На ФИГ. 12 изображена теплокарта, показывающая относительную активность разных T клеток, экспрессирующих рекомбинантный TCR, как описано выше, в разных функциональных анализах: AUC % гибели при соотношениях E:T 10:1, 5:1 и 2,5:1 (“AUC”), связывание тетрамера в CD8+ клетках в дни 7 и 13 (“тетрамер CD8”), анализ пролиферации (“CTV импульс”) с применением SCC152 клеток или T2 целевых клеток, и в которые вводили антигенный пептид, и секреция IFNγ из CD8+ клеток (“CD8, секретирующие IFNγ”).
На ФИГ. 13A-13B изображены результаты изменений объема опухоли в течение времени в мышиной модели UPCI:SCC152 плоскоклеточной карциномы, которой вводили CD4+ и CD8+ клетки, сконструированные для экспрессии типового рекомбинантного TCR #2, вызванных разными способами: TCR#2, контролируемый промотором человеческого фактора элонгации 1 альфа (EF1α), нацеленные для интеграции на TRAC локусе через HDR (TCR #2 HDR KO EF1α); TCR#2, контролируемый промотором эндогенного TRAC (расположенным против хода транскрипции вставленным в рамку P2A элементом проскока рибосомы), нацеленные для интеграции на TRAC локусе через HDR (TCR #2 HDR KO P2A); TCR#2, случайно интегрированный с применением лентивирусного конструкта (TCR #2 Lenti); TCR#2, случайно интегрированный с применением лентивирусного конструкта в клетки, содержащие нокаут эндогенного TRAC (TCR #2 Lenti KO); и ссылочный TCR, способный связываться с HPV 16 E7, но содержащий мышиные Cα и Cβ области, случайно интегрированные с применением лентивирусного конструкта (Lenti ref), по сравнению с мышами, которые не получали сконструированные клетки (только опухоль) или которым вводили клетки, процессированные в тех же условиях, которые применяют для электропорации, но без добавления RNP (mock KO), в дозе 6×106 (ФИГ. 13A) или 3×106 (ФИГ. 13B) клеток, экспрессирующих TCR.
На ФИГ. 14A-14B изображена кривая выживания мышей из каждой группы, описанной выше, для мышей, получавших дозу 6×106 (ФИГ. 14A) или 3×106 (ФИГ. 14B) клеток, экспрессирующих рекомбинантный TCR.
На ФИГ. 15A-15B изображен % изменения массы тела в течение времени у мышей в каждой группе, описанной выше, для мышей, получающих дозу 6×106 (ФИГ. 15A) или 3×106 (ФИГ. 15B) клеток, экспрессирующих рекомбинантный TCR.
На ФИГ. 16A-16B изображены результаты интеграции в различные моменты времени для разных тестированных длин плеча гомологии, по оценке изменений в GFP шаблонов через 24,48 и 72 часа (ФИГ. 16A), и через 96 часов или 7 дней (ФИГ. 16B) после трансдукции с препаратами AAV, содержащими HDR матричные полинуклеотиды.
На ФИГ. 17A-17B изображены изменения соотношения интеграции для HDR с применением разных длин плеча гомологии, через 24, 48, 72 и 96 часов или 7 дней для четырех разных доноров, доноров 1 и 2 (ФИГ. 17A) и доноров 3 и 4 (ФИГ. 17B).
На ФИГ. 18A-18B изображены результаты оценки экспрессии и активности типовых анти-CD19 CAR, в клетках, сконструированных интеграцией последовательности нуклеиновых кислот в локус эндогенного TRAC. На ФИГ. 18A изображена поверхностная экспрессия CD3 и анти-CD19 CAR (определенная окрашиванием анти-идиотипическим (анти-ID) антителом, которое специфически распознает CAR) по оценке проточной цитометрией, для Т клеток, подверженных нокауту генов, кодирующих эндогенный TCR, сконструированных для экспрессии анти-CD19 CAR с применением разных способов экспрессии: клетки, подверженные ретровирусной трансдукции для случайной интеграции последовательностей, кодирующих рекомбинантный TCR (“Только ретровирус”), клетки, подверженные целевой интеграции через HDR в TRAC локус последовательностей, кодирующих рекомбинантный TCR, под контролем человеческого EF1α промотора (EF1α) или эндогенного TRAC промотора, с применением P2A последовательности проскока рибосомы (P2A). На ФИГ. 18B изображена экспрессия, по оценке проточной цитометрией, типовых Т клеток, экспрессирующих анти-CD19 CAR, для разных способов экспрессии, описанных выше, подверженных электропорации с рибонуклеопротеиновыми (RNP) комплексами, содержащими нРНК TRAC-целенаправленного воздействия или TRBC-целенаправленного воздействия.
На ФИГ. 19A-19C изображена экспрессия и антиген-специфическая функция клеток, экспрессирующих типовой анти-CD19 CAR, сконструированных с применением разных способов экспрессии после повторных раундов антигенной стимуляции с целевыми клетками. На ФИГ. 19A изображена доля клеток, экспрессирующих CAR, наблюдаемая после 3 раундов стимулирования целевыми клетками. На ФИГ. 19B изображена средняя интенсивность флуоресценции (MFI) и на ФИГ. 19C изображен коэффициент вариации (стандартное отклонение сигнала в популяции клеток, деленное на среднее значение сигнала в соответствующей популяции) для Т клеток, сконструированных для экспрессии анти-CD19 CAR, после 3 раундов стимулирования.
На ФИГ. 20A-20B изображена секреция IFNγ (ФИГ. 20A; пг/мл) и цитолитическая активность (ФИГ. 20B) клеток, экспрессирующих типовой анти-CD19 CAR, с применением разных способов конструирования, инкубированных с K562 целевыми клетками, сконструированными для экспрессии CD19 (K562-CD19) или не сконструированными K562 (исходными) при соотношении эффектора к цели (Е:Т) 2:1.
На ФИГ. 21A-21B изображены результаты оценки экспрессии и активности типового анти-BCMA CAR в клетках, сконструированных интеграцией последовательностей нуклеиновых кислот в локус эндогенного TRAC. На ФИГ. 21A изображена поверхностная экспрессия CD3 и анти-BCMA CAR (распознанных BCMA-Fc слитым белком), по оценке проточной цитометрией, для Т клеток, сконструированных для экспрессии анти-BCMA CAR, с применением разных способов экспрессии: клетки, подверженные ретровирусной трансдукции для случайной интеграции последовательностей, кодирующих рекомбинантный TCR (“Только лентивирус”), клетки, подверженные целевой интеграции через HDR в TRAC локус последовательностей, кодирующих рекомбинантный TCR, под контролем человеческого EF1α промотора (EF1α) или эндогенного TCR альфа промотора, с применением P2A последовательности проскока рибосомы (P2A). На ФИГ. 21B изображена экспрессия, по оценке проточной цитометрией, типовых Т клеток, экспрессирующих анти-BCMA CAR, для разных способов экспрессии, описанных выше, подверженных электропорации с рибонуклеопротеиновыми (RNP) комплексами, содержащими нРНК TRAC-целенаправленного воздействия или TRBC-целенаправленного воздействия.
На ФИГ. 22A-22B изображена экспрессия и антиген-специфическая функция клеток, экспрессирующих типовой анти-BCMA CAR, сконструированных с применением разных способов экспрессии, после повторных раундов антигенной стимуляции целевыми клетками. На ФИГ. 22A изображена доля клеток, экспрессирующих CAR, наблюдаемая после 3 раундов стимулирования целевыми клетками. На ФИГ. 22B показан уровень секреции IFNγ (верхняя панель, пг/мл) и интерлейкина-2 (IL-2; нижняя панель).
Подробное описание
Здесь представлены способы получения генетически сконструированных иммунных клеток, экспрессирующих рекомбинантный рецептор, такой как рекомбинантный Т-клеточный рецептор (TCR). Также представлены генетически сконструированные иммунные клетки, экспрессирующие рекомбинантный рецептор, такой как рекомбинантный Т-клеточный рецептор (TCR) и композиции, содержащие такие клетки. Представленные варианты включают специфически нацеленные последовательности нуклеиновых кислот, кодирующие рекомбинантный рецептор, в конкретный локус, например, на одном или более локусах гена эндогенного TCR. В некоторых контекстах, представленные варианты включают вызов направленного генетического разрушения, например, создание разрыва ДНК, с применением методов генной инженерии и репарации, направляемой гомологией (HDR) для целевых нуклеиновых кислот, кодирующих рекомбинантный рецептор в локусах гена эндогенного TCR, тем самым снижая или исключая экспрессию генов эндогенного TCR и способствуя однородной или гомогенной экспрессии рекомбинантного рецептора в клеточной популяции. Также представлены родственные клеточные композиции, нуклеиновые кислоты и наборы для применения в способах, представленных здесь.
Терапии на основе T клеток, такие как адоптивные Т клеточные терапии (включая те, которые вовлекают использование сконструированных клеток, экспрессирующих рекомбинантные рецепторы, специфические для рассматриваемого заболевания или расстройства, такие как TCR, CAR и/или другие рекомбинантные антигенные рецепторы) могут быть эффективны в лечении рака и других заболеваний и расстройств. В определенных контекстах, доступные подходы к созданию сконструированных клеток для адоптивной клеточной терапии не всегда могут быть полностью удовлетворительными. В некоторых контекстах, оптимальная эффективность может зависеть от способности введенных клеток экспрессировать рекомбинантный рецептор, и для рекомбинантного рецептора, способности распознавать и связываться с целью, например, целевым антигеном, в субъекте, опухолях и их окружающих средах, и способности к однородной, гомогенной и/или стабильной экспрессии рецепторов между клетками, такими как популяция иммунных клеток и/или клетки в терапевтической клеточной композиции.
В некоторых случаях, доступные в настоящее время способы, например, случайная интеграция последовательностей, кодирующих рекомбинантный рецептор, не являются полностью удовлетворительными по одному или более этим аспектам. В некоторых аспектах, вариабельная интеграция последовательностей, кодирующих рекомбинантный рецептор, может дать нестабильную экспрессию, вариабельное количество копий нуклеиновых кислот, возможный инсерционный мутагенез и/или вариабельность экспрессии рецептора и/или генетическое разрушение в клеточной композиции, такой как терапевтическая клеточная композиция. В некоторых аспектах, применение конкретных векторов случайной интеграции, таких как определенные лентивирусные векторы, требует проведения анализа репликационно-компетентного лентивируса (RCL).
В некоторых случаях, стабильность и/или эффективность экспрессии рекомбинантного рецептора ограничена в определенных клетках или определенных клеточных популяциях, сконструированных с применением доступных на сегодня способов. В некоторых вариантах, рекомбинантный рецептор экспрессируется только в определенных клетках, и уровень экспрессии или антигенного связывания рекомбинантным рецептором широко варьируется среди клеток в популяции. В конкретных аспектах, уровень экспрессии рекомбинантного рецептора может быть сложно предсказать, контролировать и/или регулировать. В некоторых случаях, полуслучайная или случайная интеграция трансгена, кодирующего рецептор, в геном клетки может, в некоторых случаях, вызвать побочные и/или нежелательные эффекты из-за интеграции последовательности нуклеиновых кислот в нежелательную локацию в геноме, например, в незаменимый ген или ген, критический для регулирования активности клетки. В некоторых случаях, случайная интеграция последовательности нуклеиновых кислот, кодирующей рецептор, может вызвать нестабильную, нерегулируемую, неконтролируемую и/или субоптимальную экспрессию или антигенное связывание, онкогенную трансформацию и транскрипционное выключение последовательности нуклеиновых кислот, в зависимости от места интеграции и/или числа копий последовательности нуклеиновых кислот. В других случаях, в частности, для рекомбинантных TCR, субоптимальная экспрессия сконструированного или рекомбинантного TCR, которая может возникать из-за экспрессии одной или более цепей эндогенного TCR в сконструированных клетках, может вызвать нарушение комплементарности между цепью рекомбинантного TCRα или β и цепью эндогенного TCRα или β. В некоторых аспектах, неправильно спаренные TCR могут привести к нежелательному нацеливанию клеток и потенциальным побочным эффектам. В некоторых аспектах, неправильно спаренные TCR могут конкурировать за инвариантные CD3 сигнальные молекулы, которые вовлечены в позволение экспрессии рекомбинантного TCR комплекса на поверхности клетки, тем самым снижая экспрессию рекомбинантного TCR на поверхности клетки и/или способность распознавать и связываться с целью, например, целевым антигеном.
В некоторых вариантах, направленное генетическое разрушение одного или более локусов гена эндогенного TCR может привести к снижению риска или возможности нарушения комплементарности между цепями сконструированного или рекомбинантного TCR и эндогенного TCR. Неправильно спаренные TCR могут, в некоторых аспектах, создавать новые TCR, которые потенциально могут вызвать больший риск нежелательного или ненамеренного распознавания антигена и/или побочные эффекты, и/или могут снизить уровни экспрессии желательных сконструированных или рекомбинантных TCR. В некоторых аспектах, снижение или предотвращение экспрессии эндогенного TCR может повысить экспрессию сконструированного или рекомбинантного TCR в T клетки или T клеточные композиции, по сравнению с клетками, в которых экспрессия TCR не снижена или не предотвращена. В некоторых вариантах, экспрессия рекомбинантного TCR может быть повышена в 1,5 раза, 2 раза, 3 раза, 4 раза, 5 раз или более. Например, в некоторых случаях, субоптимальная экспрессия сконструированного или рекомбинантного TCR может возникать из-за конкуренции с эндогенным TCR и/или с TCR, имеющими неправильно спаренные цепи, для сигнальных молекул и/или доменов, таких как инвариантные CD3 сигнальные молекулы (например, доступность соэкспрессированной соэкспрессии CD3 δ, ε, γ и ζ цепей), которые вовлечены в разрешение экспрессии комплекса на поверхности клетки. В некоторых аспектах, доступные CD3ζ молекулы могут ограничивать экспрессию и функцию TCR в клетках. В некоторых аспектах, доступные в настоящее время способов доставки трансгенов, например, кодирование рекомбинантных рецепторов, таких как рекомбинантные TCR, могут показать недостаточную интеграцию и/или пониженную экспрессию рекомбинантных рецепторов. В некоторых аспектах, эффективность интеграции и/или экспрессии рекомбинантного рецептора в популяции может быть низкой и/или варьироваться.
В некоторых аспектах, развитие гуманизированного и/или полностью человеческого рекомбинантного TCR представляет технические проблемы. Например, в некоторых аспектах, рецептор гуманизированного и/или полностью человеческого рекомбинантного TCR конкурирует с комплексами эндогенного TCR и может сформировать нарушение комплементарности с цепями эндогенного TCRα и/или TCRβ, которое может, в определенных аспектах, снижать подачу сигнала, активность и/или экспрессию рекомбинантного TCR, и, в конце концов, вызвать снижение активности сконструированных клеток. Одним из способов решения этих проблем является создание рекомбинантных TCR с мышиными постоянными доменами для предотвращения нарушения комплементарности с цепями эндогенного человеческого TCRα или TCRβ. Однако применение рекомбинантных TCR с мышиными последовательностями может, в некоторых аспектах, представлять риск иммунного ответа. Представленные полинуклеотиды, реагенты, готовые изделия, наборы и способы решают эти проблемы вставкой последовательностей, кодирующих весь или часть рекомбинантного TCR в эндогенном гене, кодирующем одну или более цепей TCR. В конкретных аспектах, такая вставка служит для нарушения экспрессии гена эндогенного TCR, при этом позволяя экспрессию полностью гуманизированного и/или человеческого рекомбинантного TCR, снижая вероятность конкуренции от или нарушения комплементарности с цепями эндогенного TCR или применения мышиных последовательностей, которые потенциально могут быть иммуногенными.
В некоторых контекстах, доступные подходы к конструированию множества и/или популяции клеток дают гетерогенную, не однородную и/или неравномерную экспрессию рекомбинантного рецептора из-за разницы в эффективности введения нуклеиновой кислоты, разницы в геномной локации интеграции и/или копийности, нарушения комплементарности и/или конкуренции с цепями эндогенного TCR и/или другими факторами. В некоторых контекстах, доступные подходы к конструированию дают популяцию клеток, которая является гетерогенной в смысле экспрессии рекомбинантного рецептора и/или нокаута конкретных локусов. В некоторых аспектах, гетерогенная и не однородная экспрессия в популяции клеток может привести к снижению общего уровня экспрессии, стабильности экспрессии и/или антигенного связывания рекомбинантным рецептором, снижению функционирования сконструированных клеток и/или неоднородному лекарственному продукту, тем самым снижая эффективность сконструированных клеток.
В некоторых вариантах, здесь представлены способы образования или получения генетически сконструированных клеток, которые содержат TRAC и/или TRBC локус, включающий последовательности нуклеиновых кислот, кодирующие рекомбинантный TCR или его фрагмент. В некоторых аспектах, TRAC и/или TRBC локус в генетически сконструированных клеток содержит трансгенную последовательность (также называемую здесь экзогенной или гетерологичной последовательностью нуклеиновых кислот), кодирующую весь или часть рекомбинантного TCR, интегрированного в эндогенный TRAC и/или TRBC локус, который обычно кодирует TCRα или TCRβ постоянные домены. В некоторых вариантах, способы включают инициацию направленного генетического разрушения и репарации, направляемой гомологией (HDR), с применением одного или более матричных полинуклеотидов, содержащих трансген, кодирующий весь или часть рекомбинантного TCR, тем самым нацеленной интеграции трансгена на TRAC и/или TRBC локус. Также представлены клетки с клеточные композиции, созданные способами. В некоторых аспектах, исключение экспрессии цепей эндогенного TCRα и/или TCRβ может снизить нарушение комплементарности между эндогенными и сконструированными или рекомбинантными цепями.
В некоторых вариантах, представленные полинуклеотиды, трансгены и/или векторы при доставке в иммунные клетки вызывают экспрессию рекомбинантных рецепторов, например, TCR, которые могут модулировать активность T клетки и, в некоторых случаях, могут модулировать дифференциацию T клетки или гомеостаз. Полученные генетически сконструированные клетки или клеточные композиции могут применяться в способах адоптивной клеточной терапии.
В некоторых аспектах, по сравнению с обычными способами получения генетически сконструированных иммунных клеток, экспрессирующих рекомбинантный рецептор, такой как рекомбинантный Т-клеточный рецептор (TCR) или химерный антигенный рецептор (CAR), представленные способы позволяют более высокую, более стабильную и/или намного более однородную или гомогенную экспрессию рекомбинантного рецептора. В некоторых аспектах, представленные варианты предлагают преимущества в получении сконструированных Т клеток с улучшенной, однородной, гомогенной, устойчивой и/или стабильной экспрессией рекомбинантного рецептора, при минимизации возможного нарушения комплементарности, нецеленаправленного воздействия, полуслучайной или случайной интеграции трансгена и/или конкуренции от эндогенных TCR. В некоторых аспектах, представленные варианты позволяют предсказуемую и стабильную интеграцию на одном целевом генном локусе или множестве целевых генных локусов, дают стабильную копийность (обычно 1 или 2) нуклеиновых кислот, имеют пониженную, низкую или отсутствующую возможность инсерционного мутагенеза, обеспечивает стабильность в экспрессии рекомбинантного рецептора и экспрессии генов эндогенного рецептора в клеточной популяции, и убирают необходимость в анализе RCL. В некоторых аспектах, представленные варианты основаны на наблюдениях, что целевой нокаут нуклеиновых кислот, кодирующих рекомбинантный рецептор на одном или более локусах гена эндогенного TCR, что снижает или убирает экспрессию генов эндогенного TCR, давая более высокий общий уровень экспрессии, более однородную и стабильную экспрессию и/или антигенное связывание, и улучшенную функцию сконструированных клеток, включая улучшенные противоопухолевые эффекты.
Представленные варианты также предлагают преимущества в получении сконструированных Т клеток, где во всех клетках, которые экспрессируют рекомбинантный рецептор, также нокаутирована, снижена и/или устранена экспрессия одного или более локусов генов эндогенного TCR (таких как эндогенные гены, кодирующие TCRα и/или TCRβ цепи) через редактирование генома и HDR. По сравнению с подходами, которые могут дать гетерогенную смесь, где некоторые клетки, которые экспрессируют рекомбинантный рецептор, могут быть нокаутированы для локусов гена эндогенного TCR, в то время как другие клетки, которые экспрессируют рекомбинантный рецептор, могут сохранять локус гена эндогенного TCR, представленные варианты могут применяться для создания по существу более гомогенной и однородной популяции клеток, например, где все клетки, которые экспрессируют рекомбинантный рецептор, содержат нокаут одного или более локусов гена эндогенного TCR.
В некоторых аспектах, представленные варианты основаны на наблюдениях улучшенной эффективности интеграции и экспрессии и антигенного связывания TCR с применением подхода направленного нокина. Направленный нокаут одного или более локусов гена эндогенного TCR (таких как эндогенные гены, кодирующие TCRα и/или TCRβ цепи) через редактирование генома, объединенный с целевым нокином нуклеиновых кислот кодирующего рекомбинантного рецептора (такого как рекомбинантный TCR или CAR) посредством репарации, направляемой гомологией (HDR), может способствовать образованию сконструированных Т клеток, в которых улучшена экспрессия, функция и однородность экспрессии и/или другие желаемые характеристики или свойства, и крайне высока эффективность.
Также представлены способы конструирования, приготовления и получения сконструированных клеток и наборы и устройства для создания или получения сконструированных клеток. Представлены полинуклеотиды, например, вирусные векторы, которые содержат последовательность нуклеиновых кислот, кодирующую рекомбинантный рецептор или его часть, и способы введения таких полинуклеотидов в клетки, такие как трансдукция, или физическая доставка, такая как электропорация. Также представлены композиции, содержащие сконструированные клетки, способы, наборы и устройства для введения клеток и композиций субъектам, например, для адоптивной клеточной терапии.
Все публикации, включая патентные документы, научные статьи и базы данных, упомянутые в этой заявке, включены в качестве ссылки полностью для всех целей в той же степени, как если бы каждая отдельная публикация была индивидуально включена в качестве ссылки. Если определение, данное здесь, противоречит или иным образом не согласуется с определением, данным в патентах, заявках, опубликованных заявках и других публикациях, которые включены сюда в качестве ссылки, определение, изложенное здесь, преобладает над определением, которое включено сюда в качестве ссылки.
Заголовки разделов, используемые здесь, предназначены только для организационных целей и не должны рассматриваться как ограничение описываемого предмета.
I. СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР, РЕПАТАЦИЕЙ, НАПРАВЛЕННОЙ ГОМОЛОГИЕЙ (HDR)
Здесь представлены способы получения генетически сконструированной иммунной клетки, например, генетически сконструированной Т клетки, для адоптивной клеточной терапии, родственные композиции, способы, применения и наборы и готовые изделия для проведения способов. Иммунные клетки обычно конструируют для экспрессии рекомбинантной молекулы, такой как рекомбинантный рецептор, например, рекомбинантный Т-клеточный рецептор (TCR) или химерный антигенный рецептор (CAR). В некоторых вариантах, также представлены композиции, содержащие популяцию клеток, которые сконструированы для экспрессии рекомбинантного рецептора, например, TCR или CAR, такую как популяция клеток, которая демонстрирует более улучшенную, однородную, гомогенную и/или стабильную экспрессию и/или антигенное связывание рекомбинантным рецептором, включая генетически сконструированные иммунные клетки, полученные любым из представленных способов. В некоторых вариантах, представленные композиции демонстрируют пониженный коэффициент вариации экспрессии и/или антигенного связывания по сравнению с этим показателем для клеточной популяции и/или композиций, созданных обычными способами. В некоторых вариантах, также представлены способы и применения композиции и/или клеток для терапии, включая те, в которые включено введение композиции и/или клеток.
В некоторых вариантах, представлены способы получения генетически сконструированной иммунной клетки, например, генетически сконструированной Т клетки для адоптивной клеточной терапии. В некоторых вариантах, представленные способы включают введение в иммунную клетку одного или более агентов, способных вызывать генетическое разрушение одного или боле целевых сайтов (также известных как “целевое положение”, “целевая последовательность ДНК” или “целевая локация”) в гене, кодирующем домен или область цепи Т-клеточного рецептора альфа (TCRα) и/или одном или более генах, кодирующих домен или область цепи Т-клеточного рецептора бета (TCRβ) (также называемых в описании “один или более агентов” или “агенты” со ссылкой на аспекты представленных способов); и введение в иммунную клетку полинуклеотида, например, матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный рецептор или его цепь, где трансген, кодирующий рекомбинантный рецептор или его цепь нацелен на или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
В некоторых вариантах, представлены способы создания или получения генетически сконструированных клеток, которые содержат локус TRAC и/или TRBC, включающий последовательности нуклеиновых кислот, кодирующие рекомбинантный TCR или его фрагмент. В некоторых аспектах, локус TRAC и/или TRBC в генетически сконструированных клетках содержит трансгенную последовательность (также обозначенную здесь как экзогенная или гетерологичная последовательности нуклеиновых кислот), кодирующую весь или часть рекомбинантного TCR, интегрированную в эндогенный локус TRAC и/или TRBC, который обычно кодирует TCRα или TCRβ постоянные домены. В некоторых вариантах, способы включают инициацию направленного генетического разрушения и репарацию, направляемую гомологией (HDR) с применением одного или более матричных полинуклеотидов, содержащих трансген, кодирующий весь или часть рекомбинантного TCR, следовательно, направленную интеграцию трансгена на локус TRAC и/или TRBC. Также представлены клетки и клеточные композиции, созданные способами.
В конкретных вариантах, трансгенная последовательность, кодирующая весь или часть рекомбинантного TCR, содержит последовательность нуклеотидов, кодирующую TCRα цепь и/или TCRβ цепь. В некоторых вариантах, может применяться один или более полинуклеотидов, например, матричных полинуклеотидов. В некоторых вариантах, каждый полинуклеотид, например, матричный полинуклеотид, может содержать последовательность нуклеотидов, кодирующую либо TCRα цепь, либо TCRβ цепь. В некоторых вариантах, полинуклеотид, например, матричный полинуклеотид, содержит последовательность нуклеиновых кислот, кодирующую весь или часть рекомбинантного рецептора или цепи, например, рекомбинантного TCR или его цепи. В определенных вариантах, последовательность нуклеиновых кислот нацелена в целевой сайт, который находится в локусе гена, который кодирует эндогенный рецептор, например, одного или более генов, кодирующих цепь эндогенного TCR или его части. В определенных вариантах, последовательность нуклеиновых кислот нацелена на интеграцию в локус эндогенного гена. В определенных вариантах, интеграция генетически прерывает экспрессию эндогенного рецептора, кодированного геном в целевом сайте. В конкретных вариантах, трансген, кодирующий часть рекомбинантного рецептора, нацелен на локус гена через HDR.
В некоторых вариантах, представленные способы включают введение в иммунную клетку одного или более агентов, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), тем самым вызывая генетическое разрушение, по меньшей мере, одного целевого сайта; и введение в иммунную клетку матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR). В конкретных вариантах, интеграция в или рядом с целевым сайтом проводится в часть кодирующей последовательности TRAC и/или TRBC гена, например, часть кодирующей последовательности ниже, или 3’ целевого сайта.
В некоторых вариантах, один из, по меньшей мере, одного из целевых сайтов находится в гене альфа константы Т-клеточного рецептора (TRAC). В некоторых вариантах, один из, по меньшей мере, одного из целевых сайтов находится в гене бета константы Т-клеточного рецептора 1 (TRBC1) или бета константы Т-клеточного рецептора 2 (TRBC2). В некоторых вариантах, один или более целевых сайтов находится в TRAC гене и одном или обоих TRBC1 и TRBC2 генах.
В некоторых вариантах, представленные способы включают введение в иммунную клетку, имеющую генетическое разрушение одного или более целевых сайтов в гене, кодирующем домен или область цепи Т-клеточного рецептора альфа (TCRα) и/или одном или более генов, кодирующих домен или область цепи Т-клеточного рецептора бета (TCRβ), матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный рецептор, где трансген, кодирующий рекомбинантный рецептор или его цепь нацелен на или рядом с, по меньшей мере, одним целевым сайтом через HDR.
В представленных вариантах термин “введение” охватывает множество способов введения ДНК в клетку, либо in vitro, либо in vivo, где такие способы включают превращение, трансдукцию, трансфекцию (например, электропорацию) и инфекцию. Векторы применяют для введения ДНК, кодирующей молекулы, в клетки. Возможные векторы включают плазмидные векторы и вирусные векторы. Вирусные векторы включают ретровирусные векторы, лентивирусные векторы и другие векторы, такие как аденовирусные векторы или аденоассоциированные векторы. Способы, такие как электропорация, также могут применяться для введения или доставки белка или рибонуклеопротеина (RNP), например, содержащего Cas9 белок в комплексе с целевой нРНК, в рассматриваемые клетки.
В некоторых случаях, представленные здесь варианты включают одно или более направленное генетическое разрушение, например, ДНК разрыв, в одном или более локусах гена эндогенного TCR (такого как гены, кодирующие TCRα и/или TCRβ цепи) методами редактирования генома, объединенными с целевым нокином нуклеиновых кислот, кодирующих рекомбинантный рецептор (такой как рекомбинантный TCR или CAR), репарацией, направляемой гомологией (HDR). В некоторых вариантах, стадия HDR требует разрыва, например, двухнитевого разрыва в ДНК в целевой геномной локации. В некоторых вариантах, разрыв ДНК происходит в результате стадии редактирования генома, например, ДНК разрывы, созданные целевыми нуклеазами, применяемыми при редактировании генома.
В некоторых вариантах, варианты включают создание направленного разрыва ДНК с применением методов редактирования генома и/или целевых нуклеаз, с последующей HDR на основе одного или более матричных полинуклеотидов, например, матричных полинуклеотидов, которые содержат гомологические последовательности и один или более трансгенов, например, нуклеиновых кислот, кодирующих рекомбинантный рецептор или его цепь и/или других экзогенных или рекомбинантных нуклеиновых кислот, для специфического нацеливания и интеграции последовательностей нуклеиновых кислот, кодирующих рекомбинантный рецептор или его цепь и/или другие экзогенные или рекомбинантные нуклеиновые кислоты на или рядом с разрывом ДНК.
В некоторых вариантах, направленное генетическое разрушение и направленная интеграция нуклеиновых кислот, кодирующих рекомбинантный рецептор, посредством HDR происходит на одном или более целевых сайтах (также называемых “целевым положением”, “целевой последовательностью ДНК” или “целевой локацией”) эндогенных генов, которые кодируют один или более доменов, областей и/или цепей эндогенного Т-клеточного рецептора (TCR). В некоторых вариантах, направленное генетическое разрушение вызывается в TCRα гене. В некоторых вариантах, направленное генетическое разрушение вызывается в TCRβ гене. В некоторых вариантах, направленное генетическое разрушение вызывается в эндогенном TCRα гене и эндогенном TCRβ гене. Эндогенные TCR гены могут включать один или более генов, кодирующих TCRα постоянный домен (кодированный TRAC у человека) и/или TCRβ постоянный домен (кодированный TRBC1 или TRBC2 у человека).
В некоторых вариантах, направленное генетическое разрушение одного или более локусов гена эндогенного TCR может привести к снижению риска или вероятности нарушения комплементарности между цепями сконструированного или рекомбинантного TCR и эндогенного TCR. Неправильно спаренные TCR могут создать новый TCR, который потенциально может вызвать повышенный риск нежелательного или ненамеренного распознавания антигена и/или побочных эффектов, и/или может снизить уровни экспрессии желаемых сконструированных или рекомбинантных TCR. В некоторых аспектах, снижение или предотвращение экспрессии эндогенного TCR может повысить экспрессию сконструированного или рекомбинантного TCR в T клетках или T клеточных композициях по сравнению с клетками, в которых экспрессия TCR не понижена или предотвращена. В некоторых вариантах, экспрессия рекомбинантного TCR может быть повышена в 1,5 раза, 2 раза, 3 раза, 4 раза, 5 раз или более. Например, в некоторых случаях, субоптимальная экспрессия сконструированных или рекомбинантных TCR может происходить из-за конкуренции с эндогенным TCR и/или с TCR, имеющими неправильно спаренные цепи, для сигнальных доменов, таких как инвариантные CD3 сигнальные молекулы, которые вовлечены в разрешение экспрессии комплекса на поверхности клеток.
В некоторых вариантах, матричный полинуклеотид вводят в сконструированные клетки до, одновременно с или после введения агентов, способных вызывать одно или более направленное генетическое разрушение. В присутствии одного или более направленных генетических разрушений, например, разрыва ДНК, матричный полинуклеотид может применяться в качестве матрицы репарации ДНК, для эффективного копирования и интеграции трансгенов, например, последовательности нуклеиновых кислот, кодирующей рекомбинантный рецептор, рядом с сайтом направленного генетического разрушения посредством HDR, основанной на гомологии между последовательностью эндогенного гена, окружающей целевой сайт, и 5’ и/или 3’ плечами гомологии, включенными в матричный полинуклеотид.
В некоторых вариантах, стадии редактирования генома и HDR проводят одновременно и/или в одной экспериментальной реакции. В некоторых вариантах, стадии редактирования генома и HDR проводят непрерывно или последовательно, в одной или последовательных экспериментальных реакциях. В некоторых вариантах, стадии редактирования генома и HDR проводят в отдельных экспериментальных реакциях, одновременно или в разное время.
Иммунные клетки могут включать популяцию клеток, содержащих T клетки. Такими клетками могут быть клетки, которые получают от субъекта, такие, которые получены из образца мононуклеаров периферической крови (PBMC), образца нефракционированных Т клеток, образца лимфоцитов, образца белых кровяных клеток, продукта афереза или продукта лейкафереза. В некоторых вариантах, T клетки могут быть разделены или выбраны для обогащения T клеток в популяции с применением способов положительной селекции или отрицательной селекции и обогащения. В некоторых вариантах, популяция содержит CD4+, CD8+ или CD4+ и CD8+ T клетки. В некоторых вариантах, стадия введения полинуклеотидной матрицы и стадия введения агента (например, Cas9/нРНК RNP) могут проводиться одновременно или последовательно в любом порядке. В конкретных вариантах, полинуклеотидную матрицу вводят в иммунные клетки после вызова генетического разрушения стадией введения агентов (например, Cas9/нРНК RNP). В некоторых вариантах, до, во время или после введения полинуклеотидной матрицы и одного или более агентов (например, Cas9/нРНК RNP), клетки культивируют или инкубируют в условиях стимулирования размножения и/или пролиферации клеток.
В конкретных вариантах представленных способов, введение матричного полинуклеотида проводят после введения одного или более агента, способного вызывать генетическое разрушение. Может применяться любой описанный способ введения одного или более агентов, в зависимости от конкретного агента, применяемого для вызова генетического разрушения. В некоторых аспектах, разрушение проводят редактированием генома, например, с применением РНК-направляемой нуклеазы, такой как система кластерных коротких палиндромных нуклеиновых кислот, разделенных регулярными промежутками (CRISPR)-Cas, такой как система CRISPR-Cas9, специфическая к разрушаемому TRAC или TRBC локусу. В некоторых вариантах, агент, содержащий Cas9 и направляющую РНК (нРНК), содержащую домен целенаправленного воздействия, который нацелен на область TRAC или TRBC локуса, вводят в клетку. В некоторых вариантах, агент является или содержит рибонуклеопротеиновый (RNP) комплекс Cas9 и нРНК, содержащей нацеленный на TRAC/TRBC домен целенаправленного воздействия (Cas9/нРНК RNP). В некоторых вариантах, введение включает контакт агента или его части с клетками in vitro, который может включать культивирование или инкубирование клетки и агента в течение вплоть до 24, 36 или 48 часов или 3, 4, 5, 6, 7 или 8 дней. В некоторых вариантах, введение дополнительно может включать проведение доставки агента в клетки. В разных вариантах, способы, композиции и клетки в соответствии с данным изобретением применяют прямую доставку рибонуклеопротеиновых (RNP) комплексов Cas9 и нРНК в клетки, например, электропорацией. В некоторых вариантах, RNP комплексы включают нРНК, которая модифицирована так, чтобы включать 3’ поли-A хвост и 5’ Анти-Реверсный аналог кэпа (ARCA) кэп. В некоторых случаях, электропорация модифицируемых клеток включает шоковую обработку клеток холодом, клетки, например, при 32°C, с последующей электропорацией клеток и до высевания в планшеты.
В таких аспектах представленных способов, матричный полинуклеотид вводят в клетки после введения одного или более агентов, таких как Cas9/нРНК RNP, например, которые были введены электропорацией. В некоторых вариантах, матричный полинуклеотид вводят сразу же после введения одного или более агентов, способных вызывать генетическое разрушение. В некоторых вариантах, матричный полинуклеотид вводят в клетки в пределах или около 30 секунд, в пределах или около 1 минуты, в пределах или около 2 минут, в пределах или около 3 минут, в пределах или около 4 минут, в пределах или около 5 минут, в пределах или около 6 минут, в пределах или около 6 минут, в пределах или около 8 минут, в пределах или около 9 минут, в пределах или около 10 минут, в пределах или около 15 минут, в пределах или около 20 минут, в пределах или около 30 минут, в пределах или около 40 минут, в пределах или около 50 минут, в пределах или около 60 минут, в пределах или около 90 минут, в пределах или около 2 часов, в пределах или около 3 часов или в пределах или около 4 часов после введения одного или более агентов, способных вызывать генетическое разрушение. В некоторых вариантах, матричный полинуклеотид вводят в клетки в период времени от около 15 минут до около 4 часов после введения одного или более агентов, например, в течение от около 15 минут до около 3 часов, от около 15 минут до около 2 часов, от около 15 минут до около 1 часа, от около 15 минут до около 30 минут, от около 30 минут до около 4 часов, от около 30 минут до около 3 часов, от около 30 минут до около 2 часов, от около 30 минут до около 1 часа, от около 1 часа до около 4 часов, от около 1 часа до около 3 часов, от около 1 часа до около 2 часов, от около 2 часов до около 4 часов, от около 2 часов до около 3 часов или от около 3 часов до около 4 часов. В некоторых вариантах, матричный полинуклеотид вводят клетки в течение от около 2 часов после введения одного или более агентов, таких как Cas9/нРНК RNP, например, которые были введены электропорацией.
Может применяться любой способ введения матричного полинуклеотида, в зависимости от конкретных способов, применяемых для доставки матричного полинуклеотида в клетки. Типовые способы включают способы для переноса нуклеиновых кислот, кодирующих рецепторы, включая вирусную, например, ретровирусную или лентивирусную, трансдукцию, транспозоны и электропорацию. В конкретных вариантах, применяют способы вирусной трансдукции. В некоторых вариантах, матричные полинуклеотиды могут быть перенесены или введены в клетки в частицах рекомбинантного инфекционного вируса, таких как, например, векторы, полученные из полученный из вируса обезьян 40 (SV40), аденовирусов, адено-ассоциированного вируса (AAV). В некоторых вариантах, рекомбинантные нуклеиновые кислоты переносят в T клетки с применением рекомбинантных лентивирусных векторов или ретровирусных векторов, таких как гамма-ретровирусные векторы (см., например, Koste et al. (2014) Gene Therapy 2014 Apr 3. doi: 10,1038/gt,2014,25; Carlens et al. (2000) Exp Hematol 28(10): 1137-46; Alonso-Camino et al. (2013) Mol Ther Nucl Acids 2, e93; Park et al., Trends Biotechnol. 2011 November 29(11): 550-557. В конкретных вариантах, вирусным вектором является AAV, такой как AAV2 или AAV6.
В некоторых вариантах, до, во время или после контакта агента с клетками и/или до, во время или после проведения доставки (например, электропорацией), представленные способы включают инкубирование клеток в присутствии цитокина, стимулирующего агента и/или агента, который способен вызывать пролиферацию, стимулирование или активацию иммунных клеток (например, T клеток). В некоторых вариантах, по меньшей мере, часть инкубирования проводят в присутствии стимулирующего агента, который является или содержит антитело, специфическое к CD3, антитело, специфическое к CD28 и/или цитокин, такой как анти-CD3/анти-CD28 сферы. В некоторых вариантах, по меньшей мере, часть инкубирования проводят в присутствии цитокина, такого как один или более из рекомбинантного IL-2, рекомбинантного IL-7 и/или рекомбинантного IL-15. В некоторых вариантах, инкубирование проводят в течение вплоть до 8 дней часов до или после введения одного или более агентов, таких как Cas9/нРНК RNP, например, через электропорацию, и матричного полинуклеотида, например, вплоть до 24 часов, 36 часов или 48 часов или 3, 4, 5, 6, 7 или 8 дней.
В некоторых вариантах, способ включает активацию или стимулирование клеток стимулирующим агентом (например, анти-CD3/анти-CD28 антителами) до введения агента, например, Cas9/нРНК RNP, и полинуклеотидной матрицы. В некоторых вариантах, инкубирование в присутствии стимулирующего агента (например, анти-CD3/анти-CD28) проводят в течение от 6 часов до 96 часов, например, за 24-48 часов или 24-36 часов до введения одного или более агентов, таких как Cas9/нРНК RNP, например, через электропорацию. В некоторых вариантах, инкубирование со стимулирующими агентами может дополнительно включать присутствие цитокина, такого как одни или более из рекомбинантного IL-2, рекомбинантного IL-7 и/или рекомбинантного IL-15. В некоторых вариантах, инкубирование проводят в присутствии рекомбинантного цитокина, такого как IL-2 (например, от 1 Ед/мл до 500 Ед/мл, например, от 10 Ед/мл до 200 Ед/мл, например, по меньшей мере, или около 50 Ед/мл или 100 Ед/мл), IL-7 (например, от 0,5 нг/мл до 50 нг/мл, например, от 1 нг/мл до 20 нг/мл, например, по меньшей мере, или около 5 нг/мл или 10 нг/мл) или IL-15 (например, от 0,1 нг/мл до 50 нг/мл, например, от 0,5 нг/мл до 25 нг/мл, например, по меньшей мере, или около 1 нг/мл или 5 нг/мл). В некоторых вариантах стимулирующий агент (например, анти-CD3/анти-CD28 антитела) промывают или удаляют из клеток до введения или доставки в клетки агентов, способных вызывать генетическое разрушение Cas9/нРНК RNP и/или полинуклеотидной матрицы. В некоторых вариантах, до введения агентов клеткам дают отдыхать, например, удаляя любой стимулирующий или активирующий агент. В некоторых вариантах, до введения агентов, стимулирующий или активирующий агент и/или цитокины не удаляют.
В некоторых вариантах, после введения агентов, например, Cas9/нРНК, и/или полинуклеотидной матрицы клетки инкубируют, культивируют или выращивают в питательной среде в присутствии рекомбинантного цитокина, такого как один или более из рекомбинантного IL-2, рекомбинантного IL-7 и/или рекомбинантного IL-15. В некоторых вариантах, инкубирование проводят в присутствии рекомбинантного цитокина, такого как IL-2 (например, от 1 Ед/мл до 500 Ед/мл, например, от 10 Ед/мл до 200 Ед/мл, например, по меньшей мере, или около 50 Ед/мл или 100 Ед/мл), IL-7 (например, от 0,5 нг/мл до 50 нг/мл, например, от 1 нг/мл до 20 нг/мл, например, по меньшей мере, или около 5 нг/мл или 10 нг/мл) или IL-15 (например, от 0,1 нг/мл до 50 нг/мл, например, от 0,5 нг/мл до 25 нг/мл, например, по меньшей мере, или около 1 нг/мл или 5 нг/мл). Клетки могут быть инкубированы или культивированы в условиях, вызывающих пролиферацию или размножение клеток. В некоторых вариантах, клетки могут быть инкубированы или культивированы до достижения порогового количества клеток для сбора, например, терапевтически эффективной дозы.
В некоторых вариантах, инкубирование во время любой стадии способа или всего способа может проходить при температуре от 30°C±2°C до 39°C±2°C, например, по меньшей мере, или около, по меньшей мере, 30°C±2°C, 32°C±2°C, 34°C±2°C или 37°C±2°C. В некоторых вариантах, по меньшей мере, часть инкубирования проводят при 30°C±2°C и, по меньшей мере, часть инкубирования проводят при 37°C±2°C.
А. Генетическое разрушение
В некоторых вариантах, одно или более направленных генетических разрушений вызывают на эндогенном TCRα гене и/или эндогенном TCRβ гене. В некоторых вариантах, направленное генетическое разрушение вызывают на одном или более генах, кодирующих постоянный домен TCRα (также известном как постоянная область TCRα; кодированная TRAC у человека) и/или постоянный домен TCRβ (также известный как постоянная область TCRβ; кодированная TRBC1 или TRBC2 у человека). В некоторых вариантах, направленное генетическое разрушение вызывают на локусах TRAC, TRBC1 и TRBC2.
В некоторых вариантах, направленное генетическое разрушение дает разрыв или ник ДНК. В некоторых вариантах, в месте разрыва ДНК действие клеточных механизмов репарации ДНК может дать нокаут, вставку, миссенс или знаковую мутацию, такую как биаллельная знаковая мутация, делецию всего или части гена. В некоторых вариантах, генетическое разрушение может быть направлено на одни или более экзонов гена или его части, такие как первый или второй экзон. В некоторых вариантах, связывающий ДНК белок или связывающая ДНК нуклеиновая кислота, которые специфически связываются с или гибридизируются с последовательностями в области рядом с одним из, по меньшей мере, одного целевого сайта, применяют для направленного разрушения. В некоторых аспектах, при отсутствии экзогенных матричных полинуклеотидов для HDR, разрушение, направленное генетическое разрушение вызывает делецию, мутацию или вставку в экзоне гена. В некоторых вариантах, матричные полинуклеотиды, например, матричные полинуклеотиды, которые включают последовательности нуклеиновых кислот, кодирующие рекомбинантный рецептор, и гомологические последовательности, могут быть введены для направленной интеграции последовательностей, кодирующих рекомбинантный рецептор на или рядом с сайтом генетического разрушения через HDR (см. раздел I.B. здесь).
В некоторых вариантах, генетическое разрушение проводят введением одного или более агентов, способных вызывать генетическое разрушение. В некоторых вариантах, такие агенты содержат связывающий ДНК белок или связывающую ДНК нуклеиновую кислоту, которые специфически связываются с или гибридизируются с геном. В некоторых вариантах, агент содержит разные компоненты, такие как слитый белок, содержащий ДНК-направленный белок и нуклеазу или РНК-направляемую нуклеазу. В некоторых вариантах, агенты могут быть направлены на одну или более целевых локаций, например, на TRAC гене и одном или обоих TRBC1 и TRBC2 генах.
В некоторых вариантах, генетическое разрушение происходит на целевом сайте (также называемом и/или известном как “целевое положение”, “целевая последовательность ДНК” или “целевая локация”). В некоторых вариантах, целевым сайтом является или он включает сайт на целевой ДНК (например, геномной ДНК), который модифицируют одним или более агентами, способными вызывать генетическое разрушение, например, Cas9 молекулой в комплексе с нРНК, которая определяет целевой сайт. Например, в некоторых вариантах, целевой сайт может включать локации в ДНК, например, локус эндогенного TRAC, TRBC1 и/или TRBC2, где происходит отщепление или разрывы ДНК. В некоторых аспектах, интеграция последовательности нуклеиновых кислот через HDR может происходить на или рядом с целевым сайтом или целевой последовательностью. В некоторых вариантах, целевым сайтом может быть сайт между двумя нуклеотидами, например, соседними нуклеотидами, на ДНК, в которую добавляют один или более нуклеотидов. Целевой сайт может содержать один или более нуклеотидов, которые изменяются матричным полинуклеотидом. В некоторых вариантах, целевой сайт находится в целевой последовательности (например, последовательности, с которой связывается нРНК). В некоторых вариантах, целевой сайт находится выше или ниже целевой последовательности.
1. Целевые сайты на генах, кодирующих эндогенный Т-клеточный рецептор (TCR)
В некоторых вариантах, направленное генетическое разрушение происходит на эндогенных генах, которые кодируют один или более доменов, областей и/или цепей эндогенного Т-клеточного рецептора (TCR). В некоторых вариантах, генетическое разрушение направлено на локус эндогенного гена, который кодирует TCRα и/или TCRβ. В некоторых вариантах, генетическое разрушение направлено на ген, кодирующий TCRα постоянный домен (TRAC у человека) и/или TCRβ постоянный домен (TRBC1 или TRBC2 у человека).
В некоторых вариантах, “Т-клеточный рецептор” или “TCR”, включая эндогенные TCR, является молекулой, которая содержит переменные α и β цепи (также известной как TCRα и TCRβ, соответственно) или переменные γ и δ цепи (также известной как TCRγ и TCRδ, соответственно) или их антигенсвязывающие части, и которая способна специфически связываться с пептидной связью на MHC молекуле. В некоторых вариантах, TCR является αβ формой. Обычно, TCR, которые существуют в αβ и γδ формах, генетически структурно похожи, но T клетки, экспрессирующие их, могут иметь разные анатомические локации или функции. Обычно одна T клетка экспрессирует один тип TCR. TCR может быть найден на поверхности клетки или а растворимой форме. Обычно TCR находят на поверхности T клетки (или T лимфоцитов), где он обычно отвечает за распознавание антигенов, связанных с молекулами главного комплекса гистосовместимости (MHC).
В некоторых вариантах, TCR может содержать переменный домен и постоянный домен (также известный как постоянная область), трансмембранный домен и/или короткий цитоплазматический концевой сегмент (см., например, Janeway et al., Immunobiology: The Immune System in Health и Disease, 3rd Ed., Current Biology Publications, p. 4:33, 1997). В некоторых вариантах, TCR цепь содержит один или более постоянных доменов. Например, внеклеточная часть данной TCR цепи (например, TCRα цепь или TCRβ цепь) может содержать два иммуноголобулиноподобных домена, таких как переменные домены (например, Vα или Vβ; обычно аминокислоты 1-116 на основе номенклатуры Кэбота, Kabat et al., “Sequences of Proteins of Immunological Interest, US Dept. Health и Human Services, Public Health Service National Institutes of Health, 1991, 5th ed.) и постоянный домен (например, постоянный домен α цепи или TCR Cα, обычно положения 117-259 цепи на основе номенклатуры Кэбота, или постоянный домен β цепи или TCR Cβ, обычно положения 117-295 цепи на основе номенклатуры Кэбота), расположенный рядом с клеточной мембраной. Например, в некоторых случаях, внеклеточная часть TCR, образованная двумя цепями, содержит два мембрана-проксимальных постоянных домена и два мембрана-дистальных переменных домена.
В некоторых вариантах, эндогенным TCR Cα кодирован TRAC геном (номенклатура IMGT). Типовая последовательность человеческого локуса гена постоянного домена альфа цепи Т-клеточного рецептора (TRAC) представлена в SEQ ID NO:1 (Ссылочная последовательность NCBI: NG_001332,3, TRAC). В некоторых вариантах, кодированный эндогенный Cα содержит последовательность аминокислот, представленную в SEQ ID NO: 19 или 24 (UniProtKB Образец № P01848 или Genbank Образец № CAA26636,1). В определенных вариантах, генетическое разрушение направлено на, рядом или в TRAC локус. В конкретных вариантах, генетическое разрушение направлено на, рядом с или внутри открытой рамки считывания TRAC локуса. В определенных вариантах, генетическое разрушение направлено на, рядом или в открытую рамку считывания, которая кодирует TCRα постоянный домен. В некоторых вариантах, генетическое разрушение направлено на, рядом или в локус, имеющий последовательность нуклеиновых кислот, представленную в SEQ ID NO: 1, или последовательность, имеющую от, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,9% идентичности последовательности со всеми или частью, например, от, по меньшей мере, 500, 1000, 1500, 2000, 2500, 3000, 3500 или 4000 соседних нуклеотидов последовательности нуклеиновых кислот, представленной в SEQ ID NO: 1.
У человека, типовой геномный локус TRAC содержит открытую рамку считывания, которая содержит 4 экзона и 3 интрона. Типовой мРНК транскрипт TRAC может охватывать последовательность, соответствующую координатам Хромосомы 14: 22,547,506-22,552,154, на прямой нити, со ссылкой на версию человеческого генома GRCh38 (UCSC Genome Browser on Human Dec. 2013 (GRCh38/hg38) Assembly). В таблице 1 ниже представлены координаты экзонов и интронов открытых рамок считывания и нетранслированных областей транскрипта типового человеческого TRAC локуса.
Таблица 1. Координаты экзонов и интронов типового человеческого TRAC локуса (GRCh38, хромосома 14, прямая нить).
В некоторых вариантах, эндогенным TCR Cβ являются кодированные TRBC1 или TRBC2 гены (номенклатура IMGT). Типовая последовательность человеческого локуса гена постоянного домена 1 цепи Т-клеточного рецептора бета (TRBC1) представлена в SEQ ID NO:2 (ссылочная последовательность NCBI: NG_001333,2, TRBC1); и типовой последовательностью человеческого локуса гена постоянного домена 2 цепи Т-клеточного рецептора бета (TRBC2) представлена в SEQ ID NO:3 (Ссылочная последовательность NCBI: NG_001333,2, TRBC2). В некоторых вариантах, кодированный Cβ имеет или содержит последовательность аминокислот, представленную в SEQ ID NO:20, 21 или 25 (Uniprot Образец № P01850, A0A5B9 или A0A0G2JNG9). В некоторых вариантах, генетическое разрушение направлено на, рядом или в локус TRBC1 гена. В конкретных вариантах, генетическое разрушение направлено на, рядом или в открытую рамку считывания TRBC1 локуса. В определенных вариантах, генетическое разрушение направлено на, рядом или в открытую рамку считывания, которая кодирует TCRβ постоянный домен. В некоторых вариантах, генетическое разрушение направлено на, рядом или в локус, имеющий последовательность нуклеиновых кислот, представленную в SEQ ID NO: 2 или последовательность, имеющую от, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,9% идентичность последовательности со всеми или частью, например, от, по меньшей мере, 500, 1000, 1500, 2000, 2500, 3000, 3500 или 4000 соседних нуклеотидов последовательности нуклеиновых кислот, представленной в SEQ ID NO: 2.
У человека, типовой геномный локус TRBC1 содержит открытую рамку считывания, которая содержит 4 экзона и 3 интрона. Типовой мРНК транскрипт TRBC1 может охватывать последовательность, соответствующую координатам Хромосомы 7: 142,791,694-142,793,368, на прямой нити, со ссылкой на версию человеческого генома GRCh38 (UCSC Genome Browser on Human Dec. 2013 (GRCh38/hg38) Assembly). В таблице 2 ниже представлены координаты экзонов и интронов открытых рамок считывания и нетранслированных областей транскрипта типового человеческого TRBC1 локуса.
Таблица 2. Координаты экзонов и интронов типового человеческого TRBC1 локуса (GRCh38, хромосома 7, прямая нить).
В конкретных вариантах, генетическое разрушение направлено на, рядом или в TRBC2 локус. В конкретных вариантах, генетическое разрушение направлено на, рядом или в открытую рамку считывания TRBC2 локуса. В определенных вариантах, генетическое разрушение направлено на, рядом или в открытую рамку считывания, которая кодирует TCRβ постоянный домен. В некоторых вариантах, генетическое разрушение направлено на, рядом или в локус, имеющий последовательность нуклеиновых кислот, представленную в SEQ ID NO: 3 или последовательность, имеющую от, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,9% идентичность последовательности со всеми или частью, например, от, по меньшей мере, 500, 1000, 1500, 2000, 2500, 3000, 3500 или 4000 соседних нуклеотидов последовательности нуклеиновых кислот, представленной в SEQ ID NO: 3.
У человека, типовой геномный локус TRBC2 содержит открытую рамку считывания, которая содержит 4 экзона и 3 интрона. Типовой мРНК транскрипт TRBC2 может охватывать последовательность, соответствующую координатам Хромосомы 7: 142,801,041-142,802,748, на прямой нити, со ссылкой на версию человеческого генома GRCh38 (UCSC Genome Browser on Human Dec. 2013 (GRCh38/hg38) Assembly). В таблице 3 ниже представлены координаты экзонов и интронов открытых рамок считывания и нетранслированных областей транскрипта типового человеческого TRBC2 локуса.
Таблица 3. Координаты экзонов и интронов типового человеческого TRBC2 локуса (GRCh38, хромосома 7, прямая нить).
В некоторых аспектах, трансген (например, экзогенные последовательности нуклеиновых кислот) в матричном полинуклеотиде могут применяться для определения локации целевых сайтов и/или плечей гомологии. В некоторых аспектах, целевой сайт генетического разрушения может применяться в качестве гида для создания матричных полинуклеотидов и/или плечей гомологии, применяемых для HDR. В некоторых вариантах, генетическое разрушение может быть направлено рядом с желаемым сайтом направленной интеграции трансгенных последовательностей (например, кодирующим рекомбинантный TCR или его части). В некоторых аспектах, целевой сайт находится внутри экзона открытой рамки считывания TRAC, TRBC1 и/или TRBC2 локуса. В некоторых аспектах, целевой сайт находится внутри интрона открытой рамки считывания TRAC, TRBC1 и/или TRBC2 локуса.
В некоторых вариантах, генетическое разрушение, например, разрыв ДНК, нацелено в непосредственной близости к началу кодирующей области (например, ранней кодирующей области, например, в пределах 500 п.о. от инициирующего кодона или оставшейся кодирующей последовательности, например, ниже первых 500 п.о. от инициирующего кодона). В некоторых вариантах, генетическое разрушение, например, Разрыв ДНК, направлено на раннюю кодирующую область рассматриваемого гена, например, TRAC, TRBC1 и/или TRBC2, включая последовательность, расположенную сразу после сайта начала транскрипции, в пределах первого экзона кодирующей последовательности или в пределах 500 п.о. от сайта начала транскрипции (например, менее 500, 450, 400, 350, 300, 250, 200, 150, 100 или 50 п.о.) или в пределах 500 п.о. от инициирующего кодона (например, менее 500, 450, 400, 350, 300, 250, 200, 150, 100 или 50 п.о.).
В некоторых вариантах, целевой сайт находится внутри экзона эндогенного TRAC, TRBC1, и/или TRBC2 локуса. В определенных вариантах, целевой сайт находится в пределах интрона эндогенного TRAC, TRBC1, и/или TRBC2 локуса. В некоторых аспектах, целевой сайт находится в пределах регулирующего или контролирующего элемента, например, промотора, 5’ нетранслированной области (UTR) или 3’ UTR TRAC, TRBC1, и/или TRBC2 локуса. В определенных вариантах, целевой сайт находится в пределах открытой рамки считывания эндогенного TRAC, TRBC1, и/или TRBC2 локуса. В конкретных вариантах, целевой сайт находится в пределах экзона в открытой рамке считывания TRAC, TRBC1 и/или TRBC2 локуса.
В конкретных вариантах, генетическое разрушение, например, разрыв ДНК, направлен на или в пределы открытой рамки считывания рассматриваемого гена или локуса, например, TRAC, TRBC1 и/или TRBC2. В некоторых вариантах, генетическое разрушение направлено на или в пределах интрона в открытой рамке считывания рассматриваемого гена или локуса. В некоторых вариантах, генетическое разрушение направлено в пределах экзона в открытой рамке считывания рассматриваемого гена или локуса.
В конкретных вариантах, генетическое разрушение, например, разрыв ДНК, направлено на или в пределах интрона. В определенных вариантах, генетическое разрушение, например, разрыв ДНК, направлено на или в пределах экзона. В некоторых вариантах, генетическое разрушение, например, разрыв ДНК, направлено на или в пределах экзона рассматриваемого гена, например, TRAC, TRBC1 и/или TRBC2.
В некоторых вариантах, генетическое разрушение, например, разрыв ДНК, направлено в пределах экзона TRAC гена, открытой рамки считывания или локуса. В определенных вариантах, генетическое разрушение находится в первом экзоне, втором экзоне, третьем экзоне или четвертом экзоне TRAC гена, открытой рамки считывания или локуса. В конкретных вариантах, генетическое разрушение находится в первом экзоне TRAC гена, открытой рамки считывания или локуса. В некоторых вариантах, генетическое разрушение находится в пределах 500 пар оснований (п.о.) ниже 5’ окончания первого экзона в TRAC гене, открытой рамке считывания или локусе. В конкретных вариантах, генетическое разрушение находится между наибольшим 5’ нуклеотидом экзона 1 и выше наибольшего 3’ нуклеотида экзона 1. В определенных вариантах, генетическое разрушение находится в пределах 400 п.о., 350 п.о., 300 п.о., 250 п.о., 200 п.о., 150 п.о., 100 п.о. или 50 п.о. ниже 5’ окончания первого экзона TRAC гена, открытой рамки считывания или локуса. В конкретных вариантах, генетическое разрушение находится между 1 п.о. и 400 п.о., между 50 и 300 п.о., между 100 п.о. и 200 п.о. или между 100 п.о. и 150 п.о. ниже 5’ окончания первого экзона TRAC гена, открытой рамки считывания или локуса, все включительно. В определенных вариантах, генетическое разрушение находится между 100 п.о. и 150 п.о. ниже 5’ окончания первого экзона TRAC гена, открытой рамки считывания или локуса, включительно.
В конкретных вариантах, генетическое разрушение, например, разрыв ДНК, направлено в пределах экзона TRBC гена, открытой рамки считывания или локуса, например, TRBC1 и/или TRBC2. В определенных вариантах, генетическое разрушение находится в первом экзоне, втором экзоне, третьем экзоне или четвертом экзоне TRBC1 и/или TRBC2 гена, открытой рамки считывания или локуса. В некоторых вариантах, генетическое разрушение находится в первом экзоне TRBC1 и/или TRBC2 гена, открытой рамки считывания или локуса. В определенных вариантах, генетическое разрушение находится в первом экзоне, втором экзоне, третьем экзоне или четвертом экзоне TRBC1 и/или TRBC2 гена, открытой рамки считывания или локуса. В некоторых вариантах, генетическое разрушение находится между наибольшим 5’ нуклеотидом экзона 1 и выше наибольшего 3’ нуклеотида экзона 1. В конкретных вариантах, генетическое разрушение находится в первом экзоне TRBC гена, открытой рамки считывания или локуса. В некоторых вариантах, генетическое разрушение находится в пределах 400 п.о., 350 п.о., 300 п.о., 250 п.о., 200 п.о., 150 п.о., 100 п.о. или 50 п.о. ниже 5’ окончания первого экзона TRBC1 и/или TRBC2 гена, открытой рамки считывания или локуса. В конкретных вариантах, генетическое разрушение находится между 1 п.о. и 400 п.о., между 50 и 300 п.о., между 100 п.о. и 200 п.о. или между 100 п.о. и 150 п.о. ниже 5’ окончания первого экзона TRBC1 и/или TRBC2 гена, открытой рамки считывания или локуса, все включительно. В определенных вариантах, генетическое разрушение находится между 100 п.о. и 150 п.о. ниже 5’ окончания первого экзона TRBC1 и/или TRBC2 гена, открытой рамки считывания или локуса, включительно.
2. Способы генетического разрушения
Способы создания генетического разрушения, включая описанные здесь, могут включать применение одного или более агентов, способных вызывать генетическое разрушение, таких как сконструированные системы для вызова генетического разрушения, расщепления и/или двухнитевого разрыва (DSB) или ника в целевом сайте или целевом положении в эндогенной ДНК, так, что репарация разрыва ошибочным процессом, таким как негомологичное соединение концов (NHEJ) или репарация с применением матрицы репарации HDR, может дать нокаут гена и/или вставку рассматриваемой последовательности (например, экзогенных последовательностей нуклеиновых кислот или трансгена, кодирующего часть химерного рецептора) рядом с целевым сайтом или положением. Также представлены один или более агентов, способных вызывать генетическое разрушение, для применения в представленных здесь способах. В некоторых аспектах, один или более агентов могут применяться в сочетании с матричными нуклеотидами, представленными здесь, для медиированной репарацией, направляемой гомологией (HDR), направленной интеграции трансгенных последовательностей (например, описанных здесь в разделе I.B).
В некоторых вариантах, один или более агентов, способных вызывать генетическое разрушение, содержит ДНК связывающий белок или ДНК связывающую нуклеиновую кислоту, которые специфически связываются с или гибридизируются с конкретным сайтом или положением в геноме, например, целевом сайте или целевом положении. В некоторых аспектах, направленное генетическое разрушение, например, разрыв ДНК или отщепление, эндогенных генов, кодирующих TCR, достигается с применением белка или нуклеиновой кислоты, сочетаемых или образующих комплексы с нуклеазой редактирования генома, такого как химерный или слитый белок. В некоторых вариантах, один или более агентов, способных вызывать генетическое разрушение, содержит РНК-направляемую нуклеазу или слитый белок, содержащий ДНК-направленный белок и нуклеазу.
В некоторых вариантах, агент содержит разные компоненты, такие как РНК-направляемая нуклеаза или слитый белок, содержащий ДНК-направленный белок и нуклеазу. В некоторых вариантах, направленное генетическое разрушение проводят с применением молекулы целенаправленного воздействия на ДНК, которая включает связывающийся с ДНК белок, такой как один или более белок «цинковый палец» (ZFP) или эффекторы, подобные активаторам транскрипции (TALE), слитый с нуклеазой, такой как эндонуклеаза. В некоторых вариантах, направленное генетическое разрушение проводят с применением РНК-направляемых нуклеаз, таких как связанная с кластеризованной регулярной промежуточной короткой падлиндромной нуклеиновой кислотой (CRISPR) (Cas) система (включая Cas и/или Cfp1). В некоторых вариантах, направленное генетическое разрушение проводят с применением агентов, способных вызывать генетическое разрушение, таких как специфические к последовательность или направленные нуклеазы, включая связывающиеся с ДНК направленные нуклеазы и нуклеазы редактирования генома, такие как нуклеазы «цинковый палец» (ZFN) и эффекторные нуклеазы, подобные активаторам транскрипции (TALEN) и РНК-направляемые нуклеазы, такие как CRISPR-ассоциированная нуклеазная (Cas) система, специфически созданная для направления на, по меньшей мере, один целевой сайт, последовательность гена или его части. Типовые ZFN, TALE и TALEN описаны в, например, Lloyd et al., Frontiers in Immunology, 4(221): 1-7 (2013).
Белки «цинковый палец» (ZFP), эффекторы, подобные активаторам транскрипции (TALE) и домены связывания CRISPR системы могут быть “сконструированы” для связывания с определенной нуклеотидной последовательностью, например, через конструирование (изменение одной или более аминокислот) области распознающей спирали существующего в природе ZFP или TALE белка. Сконструированные связывающие ДНК белки (ZFP или TALE) являются белками, которые не существуют в природе. Разумные критерии для разработки включают применение правил замещения и компьютеризированных алгоритмов для обработки информации в базе данных, хранящей информацию о дизайнах и данных связывания существующих ZFP и/или TALE. См., например, патенты США №№ 6,140,081; 6,453,242; и 6,534,261; см. также WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496 и публикацию США № 20110301073.
В некоторых вариантах, один или более агентов специфически направлен на, по меньшей мере, один целевой сайт, например, рядом с рассматриваемым геном, например, TRAC, TRBC1 и/или TRBC2. В некоторых вариантах, агент содержит ZFN, TALEN или CRISPR/Cas9 сочетание, которые специфически связываются с, распознают или гибридизируют в целевые сайты. В некоторых вариантах, CRISPR/Cas9 система включает сконструированную crРНК/АКТРr РНК (“одиночную направляющую РНК”) для проведения специфического отщепления. В некоторых вариантах, агент содержит нуклеазы, основанные на системе Argonaute (например, из T. thermophilus, известные как ‘TtAgo’, (Swarts et al. (2014) Nature 507(7491): 258-261). Направленное расщепление с применением любых нуклеазных систем, описанных здесь, может применяться для вставки последовательностей трансгена, например, последовательностей нуклеиновых кислот, кодирующих рекомбинантный рецептор, в определенную целевую локацию, например, на эндогенных TCR генах, с применением либо HDR, либо NHEJ-медиированных способов.
В некоторых вариантах, “связывающий ЛНК белок цинковый палец” (или связывающий домен) является белком или доменом в большем белке, который связывает ДНК сиквенс-специфичным методом через один или более цинковых пальцев, которые являются областями последовательности аминокислот в связывающем домене, структура которого стабилизирована через координацию иона цинка. Термин связывающий ДНК белок цинковый палец часто сокращают до белок «цинковый палец» или ZFP. Среди ZFP имеются искусственные ZFP домены целенаправленного воздействия определенных ДНК последовательностей, обычно длиной 9-18 нуклеотидов, созданные сборкой отдельных пальцев. ZFP включают такие, в которых отдельный пальцевый домен имеет приблизительно 30 аминокислот в длину и содержит альфа спираль, содержащую два инвариантных гистидиновых остатка, координированных через цинк с двумя цистеинами одинарного бета изгиба, и имеющих два, три, четыре или пять пальцев. В общем, сиквенс-специфичность ZFP может быть изменена проведением аминокислотных замещений в четырех положениях спирали (−1, 2, 3, и 6) на распознающей спирали цинкового пальца. Таким образом, например, ZFP или ZFP-содержащая молекула не существует в природе, например, конструируется для связывания с выбранным целевым сайтом.
В некоторых случаях, ДНК-направленная молекула является или содержит ДНК связывающий домен цинкового пальца, слитый с ДНК расщепляющим доменом с получением нуклеазы цинкового пальца (ZFN). Например, слитые белки содержат расщепляющий домен (или расщепляющий полудомен) из, по меньшей мере, одной рестрикционной нуклеазы типа IIS и одного или более связывающих доменов цинкового пальца, которые могут быть или могут не быть сконструированы. В некоторых случаях, расщепляющий домен берут из рестрикционной эндонуклеазы типа IIS FokI, которая обычно катализирует двухнитевое расщепление ДНК, на 9 нуклеотидах от ее сайта распознавания на одной цепи и 13 нуклеотидах от ее сайта распознавания на другой. См., например, Патенты США №№ 5,356,802; 5,436,150 и 5,487,994; Li et al. (1992) Proc. Natl. Acad. Sci. USA 89:4275-4279; Li et al. (1993) Proc. Natl. Acad. Sci. USA 90:2764-2768; Kim et al. (1994a) Proc. Natl. Acad. Sci. USA 91:883-887; Kim et al. (1994b) J. Biol. Chem. 269:978-982. Некоторые ген-специфические сконструированные цинковые пальцы доступны коммерчески. Например, доступна платформа, называемая CompoZr, для конструирования цинковых пальцев, которая дает специфически направленные цинковые пальцы для тысяч целей. См., например, Gaj et al., Trends in Biotechnology, 2013, 31(7), 397-405. В некоторых случаях, применяют коммерчески доступные цинковые пальцы или создают под заказ.
В некоторых вариантах, один или более целевых сайтов, например, в пределах TRAC, TRBC1 и/или TRBC2 генов, могут быть направлены на генетическое разрушение сконструированными ZFN. Типовые ZFN, который направлены на гены эндогенного Т-клеточного рецептора (TCR), включают описанные в, например, US 2015/0164954, US 2011/0158957, US 2015/0056705, US 8956828 и Torikawa et al. (2012) Blood 119:5697-5705, описания которых включены с качестве ссылки полностью, или те, которые представлены в любой из SEQ ID NOS:213-224 (TRAC) или SEQ ID NOS: 225 и 226 (TRBC).
Эффектор, подобный активаторам транскрипции (TALE) является белком из бактериальных видов Xanthomonas, содержащим множество повторяющихся последовательностей, где каждый повтор содержит ди-остатки в положении 12 и 13 (RVD), которые являются специфическими к каждому нуклеотидному основанию направленной последовательности нуклеиновых кислот. Связывающие домены с подобными модульными свойствами связывания нуклеиновой кислоты основание-к-основанию (MBBBD), также могут быть получены из разных бактериальных видов. Новые модульные белки обладают преимуществом демонстрации большей вариабельности последовательности, чем повторы TAL. В некоторых вариантах, RVD, связанные с распознаванием разных нуклеотидов, являются HD для распознавания C, NG для распознавания T, NI для распознавания A, NN для распознавания G или A, NS для распознавания A, C, G или T, HG для распознавания T, IG для распознавания T, NK для распознавания G, HA для распознавания C, ND для распознавания C, HI для распознавания C, HN для распознавания G, NA для распознавания G, SN для распознавания G или A и YG для распознавания T, TL для распознавания A, VT для распознавания A или G и SW для распознавания A. В некоторых вариантах, критические аминокислоты 12 и 13 могут быть мутированы до других аминокислотных остатков для модулирования их специфичности к нуклеотидам A, T, C и G и, в частности, для улучшения такой специфичности.
В некоторых вариантах, “TALE ДНК связывающий домен” или “TALE” является полипептидом, содержащим один или более TALE повторных доменов/единиц. Повторные домены, каждый из которых содержит пару вариабельных аминокислотных остатков (RVD), вовлечены в связывание TALE с его родственной целевой ДНК последовательностью. Отдельная “повторная единица” (также называемая “повтор”) обычно имеет 33-35 аминокислот в длину и демонстрирует, по меньшей мере, некоторую гомологию последовательности с другими TALE повторными последовательностями в существующем в природе белке TALE. TALE белки могут быть сконструированы для связывания с целевым сайтом с применением канонических или не канонических RVD в повторных единицах. См., например, патенты США №№ 8,586,526 и 9,458,205.
В некоторых вариантах, “TALE-нуклеазой” (TALEN) является слитый белок, содержащий связывающий домен нуклеиновой кислоты, обычно полученный из эффектора, подобного активаторам транскрипции (TALE) и каталитического домена нуклеазы, который расщепляет целевую последовательность нуклеиновой кислоты. Каталитический домен содержит домен нуклеазы или домен, имеющий активность эндонуклеазы, такой как, например, I-TevI, ColE7, NucA и Fok-I. В конкретном варианте, TALE домен может быть слит с мегануклеазой, такой как, например, I-CreI и I-OnuI, или ее функциональным вариантом. В некоторых вариантах, TALEN является мономерной TALEN. Мономерной TALEN является TALEN, которая не требует димеризации для специфического распознавания и расщепления, такого как сливание сконструированных TAL повторов с каталитическим доменом I-TevI, описанное в WO2012138927. TALEN была описана и применена для направленного воздействия на гены и модификации генов (см., например, Boch et al. (2009) Science 326(5959): 1509-12.; Moscou и Bogdanove (2009) Science 326(5959): 1501; Christian et al. (2010) Genetics 186(2): 757-61; Li et al. (2011) Nucleic Acids Res 39(1): 359-72).
В некоторых вариантах, TRAC, TRBC1 и/или TRBC2 гены могут быть направлены на генетическое разрушение сконструированной TALEN. Типовые TALEN, который направлены на гены эндогенного Т-клеточного рецептора (TCR), включают описанные в, например, WO 2017/070429, WO 2015/136001, US20170016025 и US20150203817, описания которых включено сюда в качестве ссылки полностью.
В некоторых вариантах, “TtAgo” является прокариотическим белком Argonaute, который считается вовлеченным в выключение генов. TtAgo получают из бактерии Thermus thermophilus. См, например, Swarts et al, (2014) Nature 507(7491): 258-261, Sheng et al., (2013) Proc. Natl. Acad. Sci. U.S.A. 111, 652). “TtAgo система” включает все компоненты, требующие включения, например, направляющих ДНК для расщепления ферментом TtAgo.
В некоторых вариантах, сконструированный белок «цинковый палец», TALE белок или CRISPR/Cas система не существуют в природе, и их получение в основном является результатом эмпирического процесса, такого как фаговый дисплей, ловушка взаимодействия или гибридная селекция. См., например, Патент США № 5,789,538; Патент США № 5,925,523; Патент США № 6,007,988; Патент США № 6,013,453; Патент США № 6,200,759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970; WO 01/88197 и WO 02/099084.
Цинковый палец и TALE ДНК-связывающие домены могут быть сконструированы для связывания определенной нуклеотидной последовательности, например через конструирование (изменение одной или более аминокислот) области распознающей спирали природного белка «цинковый палец» или конструированием аминокислот, вовлеченных в связывание ДНК (пары вариабельных аминокислотных остатков или RVD области). Поэтому сконструированные белки «цинковый палец» или TALE белки являются белками, которые не существуют в природе. Не ограничивающие примеры способов конструирования белков «цинковый палец» и TALE включают дизайн и селекцию. Сконструированным белком является белок, не существующий в природе, дизайн/композиция которого является результатом в основном разумных критериев. Разумные критерии для дизайна включают применение правил замещения и компьютеризированных алгоритмов для обработки информации в базе данных, хранящей информацию о дизайнах и данных связывания существующих ZFP или TALE дизайнов (канонических и не канонических RVD) и данных связывания. См., например, Патенты США №№ 9,458,205; 8,586,526; 6,140,081; 6,453,242; и 6,534,261; см. также WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
Различные способы и композиции для направленного расщепления могут применяться, например, для вызова направленного мутагенеза, вызова направленных делений последовательностей клеточной ДНК, и поддержки направленной рекомбинации в определенном хромосомном локусе. См., например, Патенты США №№ 9,255,250; 9,200,266; 9,045,763; 9,005,973; 9,150,847; 8,956,828; 8,945,868; 8,703,489; 8,586,526; 6,534,261; 6,599,692; 6,503,717; 6,689,558; 7,067,317; 7,262,054; 7,888,121; 7,972,854; 7,914,796; 7,951,925; 8,110,379; 8,409,861; Публикации патентов США 20030232410; 20050208489; 20050026157; 20050064474; 20060063231; 20080159996; 201000218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983; 20130196373; 20140120622; 20150056705; 20150335708; 20160030477 и 20160024474, описания которых включены сюда в качестве ссылок полностью. Также представлены один или более агентов, способных вызывать генетическое разрушение. Также представлены полинуклеотиды (например, молекулы нуклеиновой кислоты), кодирующие один или боле компонентов одного или более агентов, способных вызывать генетическое разрушение.
а. Crispr/Cas9
В некоторых вариантах, направленное генетическое разрушение, например, разрыв ДНК, эндогенных генов, кодирующих TCR, такие как TRAC и TRBC1 или TRBC2 у человека, проводят с применением кластеризованных регулярных промежуточных коротких палиндромных повторов (CRISPR) и CRISPR-ассоциированных (Cas) белков. См. Sander и Joung, (2014) Nature Biotechnology, 32(4): 347-355.
В общем, “CRISPR система” относится совокупно к транскриптам и другим элементам, вовлеченным в экспрессию или направление активности CRISPR-ассоциированных (“Cas”) генов, включая последовательности, кодирующие Cas ген, tracr (транс-активирующую CRISPR) последовательность (например, tracrРНК или активную частичную tracrРНК), tracr-парную последовательность (охватывающую “прямой повтор” и tracrРНК-процессированный частичный прямой повтор в контексте эндогенной CRISPR системы), направляющую последовательность (также называемую “спейсер” в контексте эндогенной CRISPR системы) и/или другие последовательности и транскрипты из CRISPR локуса.
В некоторых аспектах, CRISPR/Cas нуклеаза или CRISPR/Cas нуклеазная система включает не кодирующую направляющую РНК (нРНК), которая секвенс-спцифичски связывается с ДНК, и Cas белок (например, Cas9), с функциональностью нуклеазы.
1) Направляющая РНК (нРНК)
В некоторых вариантах, один или более агентов содержит, по меньшей мере, одну из: направляющей РНК (нРНК), имеющей домен целенаправленного воздействия, который является комплементарным с целевым сайтом TRAC гена; нРНК, имеющую домен целенаправленного воздействия, который является комплементарным с целевым сайтом одного или обоих TRBC1 и TRBC2 генов; или, по меньшей мере, одной нуклеиновой кислоты, кодирующей нРНК.
В некоторых аспектах, “нРНК молекула” относится к нуклеиновой кислоте, которая способствует специфическому целенаправленному воздействию или хомингу нРНК молекулы/Cas9 молекулярного комплекса в целевую нуклеиновую кислоту, такую как локус на геномной ДНК клетки. нРНК молекулы могут быть мономолекулярными (имеющими одну молекулу РНК), иногда называемыми здесь “химерными” нРНК, или модульными (содержащими более одной, и обычно две отдельные молекулы РНК). В общем, направляющей последовательностью, например, направляющей РНК, является любая полинуклеотидная последовательность, содержащая, по меньшей мере, часть последовательности, которая имеет достаточную комплементарность с целевой полинуклеотидной последовательностью, такой как TRAC, TRBC1 и/или TRBC2 гены у человека, для гибридизации с целевой последовательностью в целевом сайте и прямого сиквенс-специфического связывания CRISPR комплекса с целевой последовательностью. В некоторых вариантах, в контексте образования CRISPR комплекса, “целевая последовательность” в общем относится к последовательности, с которой имеет комплементарность сконструированная направляющая последовательность, где гибридизация между целевой последовательностью и доменом, например, доменом целенаправленного воздействия, направляющей РНК способствует образованию CRISPR комплекса. Полная комплементарность не обязательно требуется, при условии, что существует достаточная комплементарность для гибридизации с помощь образованию CRISPR комплекса. В общем, направляющую последовательность выбирают так, чтобы снизить степень вторичной структуры в направляющей последовательности. Вторичная структура может быть определена любым подходящим алгоритмом предсказания полинуклеотида.
В некоторых вариантах, направляющую РНК (нРНК), специфическую к рассматриваемому локусу (например, на TRAC, TRBC1 и/или TRBC2 локусах у человека) применяют для РНК-направляемых нуклеаз, например, Cas, для вызова разрыва ДНК на целевом сайте или в целевом положении. Способы конструирования нРНК и типовых доменов целенаправленного воздействия могут включать описанные, например, в WO2015/161276, WO2017/193107, WO2017/093969, US2016/272999 и US2015/056705.
Несколько типовых нРНК структур, с указанными здесь доменами, описаны в WO2015/161276, например, на ФИГ. 1A-1G здесь. Не желая быть связанными теорией, в отношении трехмерной формы или внутри- и межнитевых взаимодействий активной формы нРНК, области высокой комплементарности иногда показаны как дуплексы в WO2015/161276, например, на ФИГ. 1A-1G здесь и других изображениях, представленных здесь.
В некоторых случаях, нРНК является мономолекулярной или химерной нРНК, содержащей от 5’ до 3’: домен целенаправленного воздействия, который направлен на целевой сайт или положение, например, в последовательности из TRAC локуса (типовая нуклеотидная последовательность локуса человеческого TRAC гена, представленная в SEQ ID NO:1; NCBI Ссылочная последовательность: NG_001332,3, TRAC; типовая геномная последовательность, описанная в таблице 1 здесь); первый домен комплементарности; связывающий домен; второй домен комплементарности (который является комплементарным с первым доменом комплементарности); проксимальный домен; и, необязательно, хвостовой домен. В некоторых случаях, нРНК является мономолекулярной или химерной нРНК, содержащей, от 5’ до 3’: домен целенаправленного воздействия, который нацелен на целевой сайт или положение, например, в последовательности от TRBC1 или TRBC2 локуса (типовая нуклеотидная последовательность локуса человеческого TRBC1 гена, представленная в SEQ ID NO:2; NCBI Ссылочная последовательность: NG_001333,2, TRBC1; типовая геномная последовательность, описанная в таблице 2 здесь; типовая нуклеотидная последовательность локуса человеческого TRBC2 гена, представленная в SEQ ID NO:3; NCBI Ссылочная последовательность: NG_001333,2, TRBC2; типовая геномная последовательность, описанная в таблице 3 здесь); первый домен комплементарности; связывающий домен; второй домен комплементарности (который является комплементарным с первым доменом комплементарности); проксимальный домен; и, необязательно, хвостовой домен.
В других случаях, нРНК является модульной нРНК, содержащей первую и вторую нити. В этих случаях, первая нить предпочтительно включает от 5’ до 3’: домен целенаправленного воздействия (который нацелен на целевой сайт или положение, например, в последовательности от TRAC локуса (типовая нуклеотидная последовательность локуса человеческого TRAC гена, представленная в SEQ ID NO:1; NCBI Ссылочная последовательность: NG_001332,3, TRAC; типовая геномная последовательность, описанная в таблице 1 здесь) или TRBC1 или TRBC2 локуса (типовая нуклеотидная последовательность локуса человеческого TRBC1 гена, представленная в SEQ ID NO:2; NCBI Ссылочная последовательность: NG_001333,2, БКТР11; типовая геномная последовательность, описанная в таблице 2 здесь; типовая нуклеотидная последовательность локуса человеческого TRBC2 гена, представленная в SEQ ID NO:3; NCBI Ссылочная последовательность: NG_001333,2, TRBC2); и первый домен комплементарности. Вторая нить обычно включает, от 5’ до 3’: необязательно, 5’ удлиняющий домен; второй домен комплементарности; проксимальный домен; и необязательно, хвостовой домен.
А) Домен целенаправленного воздействия
Примеры размещения доменов целенаправленного воздействия включают такие, которые описаны в WO2015/161276, например, на ФИГ. 1A-1G здесь. Домен целенаправленного воздействия содержит нуклеотидную последовательность, которая комплементарна, например, по меньшей мере, 80, 85, 90, 95, 98 или 99% комплементарности, например, полностью комплементарна, целевой последовательности на целевой нуклеиновой кислоте. Нить целевой нуклеиновой кислоты, содержащая целевую последовательность, называют здесь “комплементарной нитью” целевой нуклеиновой кислоты. Руководство по выбору доменов целенаправленного воздействия может быть найдено, например, в Fu Y et al., Nat Biotechnol 2014 (doi: 10,1038/nbt,2808) и Sternberg SH et al., Nature 2014 (doi: 10,1038/nature13011).
Домен целенаправленного воздействия является частью РНК молекулы и поэтому содержит основной урацил (U), в любая ДНК, кодирующая молекулу нРНК, будет содержать основной тимин (T). Не желая быть связанными теорией, в некоторых вариантах, полагают, что комплементарность домена целенаправленного воздействия с целевой последовательностью играет роль в специфичности взаимодействия комплекса нРНК молекулы/Cas9 молекулы с целевой нуклеиновой кислотой. Понятно, что в паре домен целенаправленного воздействия и целевая последовательность, урациловые основания в домене целенаправленного воздействия будут образовывать пары с адениновыми основаниями в целевой последовательности. В некоторых вариантах, сам целевой домен содержит в 5’-3’ направлении необязательный второй домен и коровый домен. В некоторых вариантах, коровый домен полностью комплементарен с целевой последовательностью. В некоторых вариантах, домен целенаправленного воздействия имеет 5-50 нуклеотидов в длину. Нить целевой нуклеиновой кислоты, с которой комплементарен домен целенаправленного воздействия, называют здесь комплементарной нитью. Некоторые или все нуклеотиды домена могут иметь модификацию, например, для того, чтобы сделать его менее подверженным разложению, улучшения биосовместимости и т.д. В качестве неограничивающего примера, остов целевого домена может быть модифицирован фосфоротиоатом или другими модификациями. В некоторых случаях, нуклеотид домена целенаправленного воздействия может содержать 2’ модификацию, например, 2-ацетилирование, например, 2’ метилирование или другие модификации.
В разных вариантах, домен целенаправленного воздействия имеет 16-26 нуклеотидов в длину (т.е. он имеет 16 нуклеотидов в длину или 17 нуклеотидов в длину или 18, 19, 20, 21, 22, 23, 24, 25 или 26 нуклеотидов в длину).
В) Типовые домены целенаправленного воздействия
Типовые домены целенаправленного воздействия, содержащиеся в нРНК для направленного генетического разрушения человеческого TRAC, TRBC1 или TRBC2, включают те, которые описаны в, например, WO2015/161276, WO2017/193107, WO2017/093969, US2016/272999 и US2015/056705, или домен целенаправленного воздействия, который может связываться с целевыми последовательностями, описанными далее. Типовые домены целенаправленного воздействия, содержащиеся в нРНК для направленного генетического разрушения человеческого TRAC локуса с применением S. pyogenes или S. aureus Cas9, могут включать любые из представленных в
Таблица 4. Типовые последовательности домена целенаправленного воздействия TRAC нРНК
Типовые домены целенаправленного воздействия, содержащиеся в нРНК для направленного генетического разрушения человеческого TRBC1 или TRBC2 локуса с применением S. pyogenes или S. aureus Cas9 могут включать любые из представленных в таблице 5.
Таблица 5. Типовые последовательности домена целенаправленного воздействия TRBC1 или TRBC2 нРНК
В некоторых вариантах, нРНК для целенаправленного воздействия на TRAC, TRBC1 и/или TRBC2 может быть любая, которая описана здесь или где-либо еще, например, в WO2015/161276, WO2017/193107, WO2017/093969, US2016/272999 и US2015/056705, или домен целенаправленного воздействия, который связывается с направленными последовательностями, описанными далее. В некоторых вариантах, последовательность, на которую направлена CRISPR/Cas9 нРНК в локусе TRAC гена представлена в SEQ ID NOS: 117, 163 и 165-211, например, GAGAATCAAAATCGGTGAAT (SEQ ID NO:163) или ATTCACCGATTTTGATTCTC (SEQ ID NO:117). В некоторых вариантах, последовательность, на которую направлена CRISPR/Cas9 нРНК в локусах TRBC1 и/или TRBC2 гена, представлена в SEQ ID NOS: 118, 164 и 212, например, GGCCTCGGCGCTGACGATCT (SEQ ID NO:164) или AGATCGTCAGCGCCGAGGCC (SEQ ID NO:118). В некоторых вариантах, нРНК последовательностью домена целенаправленного воздействия для целенаправленного воздействия на целевой сайт в локусе TRAC гена является GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31). В некоторых вариантах, нРНК последовательность домена целенаправленного воздействия для целенаправленного воздействия на целевой сайт в локусах TRBC1 и/или TRBC2 гена является GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63).
В некоторых вариантах, нРНК для целенаправленного воздействия на локус TRAC гена может быть получен in vitro транскрипцией последовательности AGCGCTCTCGTACAGAGTTGGCATTATAATACGACTCACTATAGGGGAGAATCAAAATCGGTGAATGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTTTTT (представленной в SEQ ID NO:26; подчеркнутая жирной линией часть комплементарна с целевым сайтом в TRAC локусе) или химическим синтезом, где нРНК имеет последовательность 5’- GAG AAU CAA AAU CGG UGA AUG UUU UAG AGC UAG AAA UAG CAA GUU AAA AUA AGG CUA GUC CGU UAU CAA CUU GAA AAA GUG GCA CCG AGU CGG UGC UUU U -3’ (представленную в SEQ ID NO:27; см. Osborn et al., Mol Ther. 24(3):570-581 (2016)). Другие типовые нРНК последовательности для получения генетического разрушения эндогенных генов, кодирующих TCR домены или области, например, TRAC, TRBC1 и/или TRBC2, описаны, например, в публикации международной заявки на патент № WO2015/161276. Типовые способы редактирования генома локусов эндогенного TCR включают те, которые описаны в, например, публикациях США №№ US2011/0158957, US2014/0301990, US2015/0098954,US2016/0208243; US2016/272999 и US2015/056705; международных публикациях РСТ № WO2014/191128, WO2015/136001, WO2015/161276, WO2016/069283, WO2016/016341, WO2017/193107, и WO2017/093969; и Osborn et al. (2016) Mol. Ther. 24(3):570-581. Любые из известных способов, которые могут применяться для получения генетического разрушения эндогенных генов, кодирующих TCR домены или области, могут применяться в представленных здесь вариантах.
В некоторых вариантах, домены целенаправленного воздействия включают домены для введения генетического разрушения на локусах TRAC, TRBC1 и/или TRBC2 с применением S. pyogenes Cas9 или с применением N. meningitidis Cas9. В некоторых вариантах, домены целенаправленного воздействия включают домены для введения генетического разрушения в локусах TRAC, TRBC1 и/или TRBC2 с применением S. pyogenes Cas9. Любые из доменов целенаправленного воздействия могут применяться с S. pyogenes Cas9 молекулой, которая создает двухнитевой разрыв (Cas9 нуклеаза) или однонитевой разрыв (Cas9 никаза).
В некоторых вариантах, двойное целенаправленное воздействие применяют для создания двух ников на противоположных нитях ДНК с применением S. pyogenes Cas9 никаз с двумя доменами целенаправленного воздействия, которые комплементарны с противоположными нитями ДНК, например, нРНК, содержащая любой домен целенаправленного воздействия минус нити, может быть спарена с любой нРНК, содержащей домен целенаправленного воздействия плюс нити. В некоторых вариантах, две нРНК ориентированы на ДНК, такую как PAM, повернутую наружу, и расстояние между 5’ концами нРНК составляет 0-50 п.о. В некоторых вариантах, две нРНК используют для нацеливания на две Cas9 нуклеазы или две Cas9 никазы, например, с применением пары комплекса Cas9 молекулы/нРНК молекулы, направляемого двумя разными молекулами нРНК для расщепления целевого домена с двумя однонитевыми разрывами на противоположных нитях целевого домена. В некоторых вариантах, две Cas9 никазы могут включать молекулу, имеющую HNH активность, например, Cas9 молекулу, имеющую RuvC активность, инактивированную, например, Cas9 молекулой, имеющей мутацию на D10, например, D10A мутацию, молекулу, имеющую RuvC активность, например, Cas9 молекулу, имеющую HNH активность, инактивированную, например, Cas9 молекулой, имеющей мутацию на H840, например, H840A, или молекулу, имеющую RuvC активность, например, Cas9 молекулу, имеющую HNH активность, инактивированную, например, Cas9 молекулой, имеющей мутацию на N863, например, N863A. В некоторых вариантах, каждая из двух нРНК образует комплекс с D10A Cas9 никазой.
В некоторых вариантах, целевая последовательность (целевой домен) расположена рядом с TRAC, TRBC1 и/или TRBC2 локусом, например, любой частью TRAC, TRBC1 и/или TRBC2 кодирующей последовательности, представленной в SEQ ID NO: 1-3 или описанной в таблицах 1-3 здесь. В некоторых вариантах, целевая нуклеиновая кислота, комплементарная с доменом целенаправленного воздействия, расположена на ранней кодирующей области рассматриваемого гена, такого как TRAC, TRBC1 и/или TRBC2. Целенаправленное воздействие ранней кодирующей области может применяться для генетического разрушения (т.е. устранения экспрессии) рассматриваемого гена. В некоторых вариантах, ранняя кодирующая область рассматриваемого гена включает последовательность, расположенную сразу же после инициирующего кодона (например, ATG) или в пределах 500 п.о. от инициирующего кодона (например, менее 500, 450, 400, 350, 300, 250, 200, 150, 100, 50 п.о., 40 п.о., 30 п.о., 20 п.о. или 10 п.о.). В конкретных примерах, целевая нуклеиновая кислота находится в пределах 200 п.о., 150 п.о., 100 п.о., 50 п.о., 40 п.о., 30 п.о., 20 п.о. или 10 п.о. инициирующего кодона. В некоторых примерах, домен целенаправленного воздействия нРНК комплементарен, например, по меньшей мере, на 80, 85, 90, 95, 98 или 99% комплементарности, например, полностью комплементарен целевой последовательности на целевой нуклеиновой кислоте, такой как целевая нуклеиновая кислота в TRAC, TRBC1 и/или TRBC2 локусе.
В некоторых аспектах, нРНК может быть направлена на сайт в пределах экзона открытой рамки считывания эндогенного TRAC, TRBC1 и/или TRBC2 локуса. В некоторых аспектах, нРНК может быть нацелена на сайт в пределах интрона открытой рамки считывания TRAC, TRBC1 и/или TRBC2 локуса. В некоторых аспектах, нРНК может быть нацелена на сайт в пределах регулирующего или контрольного элемента, например, промотора, TRAC, TRBC1 и/или TRBC2 локуса. В некоторых аспектах, целевым сайтом на TRAC, TRBC1 и/или TRBC2 локусе, на который направлена нРНК, может быть любой целевой сайт, описанный здесь, например, в разделе I.A.1. В некоторых вариантах, нРНК может быть нацелена на сайт в пределах или близко к экзонам, соответствующим ранней кодирующей области, например, экзону 1, 2 или 3 открытой рамки считывания эндогенного TRAC, TRBC1 и/или TRBC2 локуса, или включающим последовательность сразу же за сайтом начала транскрипции, в пределах экзона 1, 2 или 3 или в пределах менее 500, 450, 400, 350, 300, 250, 200, 150, 100 или 50 п.о. от экзона 1, 2 или 3. В некоторых вариантах, нРНК может быть нацелена на сайт рядом с экзоном 2 эндогенного TRAC, TRBC1 и/или TRBC2 локуса или в пределах менее 500, 450, 400, 350, 300, 250, 200, 150, 100 или 50 п.о. от экзона 2.
С) Первый домен комплементарности
Примеры первого домена комплементарности включают такие, которые описаны в WO2015/161276, например, на ФИГ. 1A-1G здесь. Первый домен комплементарности комплементарен со вторым доменом комплементарности, описанным здесь, и обычно имеет достаточную комплементарность со вторым доменом комплементарности для образования дуплексной области в, по меньшей мере, некоторых физиологических условиях. Первый домен комплементарности обычно имеет 5-30 нуклеотидов в длину, и может иметь 5-25 нуклеотидов в длину, 7-25 нуклеотидов в длину, 7-22 нуклеотидов в длину, 7-18 нуклеотидов в длину или 7-15 нуклеотидов в длину. В разных вариантах, первый домен комплементарности имеет 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеотидов в длину.
Обычно первый домен комплементарности не имеет точной комплементарности с целью второго домена комплементарности. В некоторых вариантах, первый домен комплементарности может иметь 1, 2, 3, 4 или 5 нуклеотидов, которые не комплементарны с соответствующим нуклеотидом второго домена комплементарности. Например, сегмент из 1, 2, 3, 4, 5 или 6, (например, 3) нуклеотидов первого домена комплементарности может не спариваться в дуплекс, и может образовывать не дуплексную или внепетлевую область. В некоторых случаях, непарная или внепетлевая область, например, 3 нуклеотида вне петли, присутствует на втором домене комплементарности. Эта непарная область необязательно начинается через 1, 2, 3, 4, 5 или 6, например, 4, нуклеотида от 5’ конца второго домена комплементарности.
Первый домен комплементарности может включать 3 субдомена, которые в направлении 5’-3’ являются: 5’ субдоменом, центральным субдоменом и 3’ субдоменом. В некоторых вариантах, 5’ субдомен имеет 4-9, например, 4, 5, 6, 7, 8 или 9 нуклеотидов в длину. В некоторых вариантах, центральный субдомен имеет 1, 2 или 3, например, 1, нуклеотид в длину. В некоторых вариантах, 3’ субдомен имеет 3-25, например, 4-22, 4-18 или 4-10 или 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеотидов в длину.
В некоторых вариантах, первый и второй домены комплементарности, когда образуют дуплекс, содержат 11 парных нуклеотидов, например, в нРНК последовательности (одна парная нить подчеркнута, одна выделена жирным шрифтом): NNNNNNNNNNNNNNNNNNNNGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (SEQ ID NO:142).
В некоторых вариантах, первый и второй домены комплементарности, когда образуют дуплекс, содержат 15 парных нуклеотидов, например, в нРНК последовательности (одна парная нить подчеркнута, одна выделена жирным шрифтом): NNNNNNNNNNNNNNNNNNNNGUUUUAGAGCUAUGCUGAAAAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (SEQ ID NO:143).
В некоторых вариантах первый и второй домены комплементарности, когда образуют дуплекс, содержат 16 парных нуклеотидов, например, в нРНК последовательности (одна парная нить подчеркнута, одна выделена жирным шрифтом): NNNNNNNNNNNNNNNNNNNNGUUUUAGAGCUAUGCUGGAAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (SEQ ID NO:144).
В некоторых вариантах первый и второй домены комплементарности, когда образуют дуплекс, содержат 21 парных нуклеотидов, например, в нРНК последовательности (одна парная нить подчеркнута, одна выделена жирным шрифтом): NNNNNNNNNNNNNNNNNNNNGUUUUAGAGCUAUGCUGUUUUGGAAACAAAACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (SEQ ID NO:145).
В некоторых вариантах, нуклеотиды заменяют для удаления поли-U АКТРt, например в нРНК последовательностях (замененные нуклеотиды подчеркнуты): NNNNNNNNNNNNNNNNNNNNGUAUUAGAGCUAGAAAUAGCAAGUUAAUAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (SEQ ID NO:146); NNNNNNNNNNNNNNNNNNNNGUUUAAGAGCUAGAAAUAGCAAGUUUAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (SEQ ID NO:147); и NNNNNNNNNNNNNNNNNNNNGUAUUAGAGCUAUGCUGUAUUGGAAACAAUACAGCAUAGCAAGUUAAUAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC (SEQ ID NO:148).
Первый домен комплементарности может разделять гомологию с, или быть полученным из существующего в природе первого домена комплементарности. В некоторых вариантах, он имеет, по меньшей мере, 50% гомологию с первым доменом комплементарности, описанным здесь, например, S. pyogenes, S. aureus, N. meningtidis или S. thermophilus, первый домен комплементарности.
Необходимо отметить, что один или более, или даже все нуклеотиды первого домена комплементарности могут иметь модификацию вместе с линиями, описанными здесь для домена целенаправленного воздействия.
D) Связывающий домен
Примеры связывающих доменов включают те, которые описаны в WO2015/161276, например, на ФИГ. 1A-1G здесь. В мономолекулярной или химерной нРНК, связывающий домен служит для связи первого домена комплементарности со вторым доменом комплементарности мономолекулярной нРНК. Связывающий домен может связывать первый и второй домены комплементарности ковалентно или не ковалентно. В некоторых вариантах, связь является ковалентной. В некоторых вариантах, связывающий домен ковалентно связывает первый и второй домены комплементарности, см., например, WO2015/161276, например, на ФИГ. 1B-1E здесь. В некоторых вариантах, связывающий домен является или содержит ковалентную связь, расположенную между первым доменом комплементарности и вторым доменом комплементарности. Обычно связывающий домен содержит один или более, например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 нуклеотидов, но в разных вариантах линкер может иметь 20, 30, 40, 50 или даже 100 нуклеотидов в длину.
В модульных нРНК молекулах, две молекулы, связанные посредством гибридизации доменов комплементарности и связывающего домена, могут не присутствовать. См., например, WO2015/161276, например, на ФИГ. 1A здесь.
Множество связывающих доменов подходит для применения в мономолекулярных нРНК молекулах. Связывающие домены могут состоять из ковалентной связи или быть короткими, то есть иметь один или более нуклеотидов, например, 1, 2, 3, 4 или 5 нуклеотидов в длину. В некоторых вариантах, связывающий домен имеет 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20 или 25 или более нуклеотидов в длину. В некоторых вариантах, связывающий домен имеет 2-50, 2-40, 2-30, 2-20, 2-10 или 2-5 нуклеотидов в длину. В некоторых вариантах, связывающий домен разделяет гомологию с, или получен из природной последовательности, например, последовательности tracrРНК, которая составляет 5’ ко второму домену комплементарности. В некоторых вариантах, связывающий домен имеет, по меньшей мере, 50% гомологию со связывающим доменом, описанным здесь.
Как описано здесь для первого домена комплементарности, некоторые или все нуклеотиды связывающего домена могут включать модификацию.
Е) 5’ Расширяющий домен
В некоторых случаях, модулярная нРНК может содержать дополнительную последовательность, 5’ ко второму домену комплементарности, названную здесь 5’ расширяющий домен, WO2015/161276, например, на ФИГ. 1A здесь. В некоторых вариантах, 5’ расширяющий домен имеет 2-10, 2-9, 2-8, 2-7, 2-6, 2-5 или 2-4 нуклеотидов в длину. В некоторых вариантах, 5’ расширяющий домен имеет 2, 3, 4, 5, 6, 7, 8, 9 или 10 или более нуклеотидов в длину.
F) Второй домен комплементарности
Примеры второго домена комплементарности включает такие, которые описаны в WO2015/161276, например, на ФИГ. 1A-1G здесь. Второй домен комплементарности комплементарен с первым доменом комплементарности, и обычно имеет достаточную комплементарность со вторым доменом комплементарности с получением дуплексной области в, по меньшей мере, некоторых физиологических условиях. В некоторых случаях, например, как показано в WO2015/161276, например, на ФИГ. 1A-1B здесь, второй домен комплементарности может включать последовательность, которая не имеет комплементарности с первым доменом комплементарности, например, последовательности, которая выпадает из дуплексной области.
Второй домен комплементарности может иметь 5-27 нуклеотидов в длину, и в некоторых случаях может быть длиннее первой области комплементарности. Например, второй домен комплементарности может иметь 7-27 нуклеотидов в длину, 7-25 нуклеотидов в длину, 7-20 нуклеотидов в длину или 7-17 нуклеотидов в длину. Более конкретно, домен комплементарности может иметь 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 нуклеотидов в длину.
В некоторых вариантах, второй домен комплементарности содержит 3 субдомена, которые, в направлении 5’-3’, включают: 5’ субдомен, центральный субдомен, и 3’ субдомен. В некоторых вариантах, 5’ субдомен имеет 3-25, например, 4-22, 4-18 или 4-10 или 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 нуклеотидов в длину. В некоторых вариантах, центральный субдомен имеет 1, 2, 3, 4 или 5, например, 3, нуклеотидов в длину. В некоторых вариантах, 3’ субдомен имеет 4-9, например, 4, 5, 6, 7, 8 или 9 нуклеотидов в длину.
В некоторых вариантах, 5’ субдомен и 3’ субдомен первого домена комплементарности, соответственно, являются комплементарными, например, полностью комплементарными с 3’ субдоменом и 5’ субдоменом второго домена комплементарности.
Второй домен комплементарности может разделять гомологию с или быть полученным из природного второго домена комплементарности. В некоторых вариантах, он имеет, по меньшей мере, 50% гомологию со вторым доменом комплементарности, описанным здесь, например, S. pyogenes, S. aureus, N. meningtidis или S. thermophilus, первым доменом комплементарности.
Некоторые или все нуклеотиды второго домена комплементарности могут иметь модификацию, например, описанную здесь модификацию.
G) Проксимальный домен
Примеры проксимальных доменов включают такие, которые описаны в WO2015/161276, например, на ФИГ. 1A-1G здесь. В некоторых вариантах, проксимальный домен имеет 5-20 нуклеотидов в длину. В некоторых вариантах, проксимальный домен может разделять гомологию с или быть полученным из природного проксимального домена. В некоторых вариантах, он имеет, по меньшей мере, 50% гомологию с проксимальным доменом, описанным здесь, например, S. pyogenes, S. aureus, N. meningtidis или S. thermophilus, проксимальным доменом.
Некоторые или все нуклеотиды проксимального домена могут иметь модификацию вместе с линиями, описанными здесь.
Н) Хвостовой домен
Примеры хвостовых доменов включают те, которые описаны в WO2015/161276, например, на ФИГ. 1A-1G здесь. Как можно увидеть при проверке хвостовых доменов в WO2015/161276, например, на ФИГ. 1A и ФИГ. 1B-1F здесь, широкий спектр хвостовых доменов подходит для применения в молекулах нРНК. В разных вариантах, хвостовой домен имеет 0 (отсутствует), 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 нуклеотидов в длину. В определенных вариантах, нуклеотиды хвостового домена получают из или разделяют гомологию с последовательностью из 5’ конца природного хвостового домена, см., например, WO2015/161276, например, на ФИГ. 1D или 1E здесь. Хвостовой домен также необязательно включает последовательности, которые комплементарны друг к другу, и которые, по меньшей мере, в некоторых физиологических условиях, образуют дуплексную область.
Хвостовые домены могут разделять гомологию с или быть полученными из природных проксимальных хвостовых доменов. В качестве не ограничивающего примера, данный хвостовой домен согласно разным вариантам данного изобретения может разделять, по меньшей мере, 50% гомологию с природным хвостовым доменом, раскрытым здесь, например, S. pyogenes, S. aureus, N. meningtidis или S. thermophilus хвостовым доменом.
В определенных случаях, хвостовой домен включает нуклеотиды на 3’ конце, относящиеся к способу in vitro или in vivo транскрипции. Если T7 промотор применяют для in vitro транскрипции нРНК, этими нуклеотидами могут быть любые нуклеотиды, присутствующие до 3’ конца ДНК матрицы. Если U6 промотор применяют для in vivo транскрипции, этими нуклеотидами может быть последовательность UUUUUU. Если применяют альтернативные pol-III промоторы, этими нуклеотидами могут быть разные числа или урациловые основаниям, или они могут включать альтернативные основания.
В качестве не ограничивающего примера, в разных вариантах, проксимальный и хвостовой домены, взятые вместе, включают следующие последовательности: AAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCU (SEQ ID NO:149), AAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC (SEQ ID NO:150), AAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCGGAUC (SEQ ID NO:151), AAGGCUAGUCCGUUAUCAACUUGAAAAAGUG (SEQ ID NO:152), AAGGCUAGUCCGUUAUCA (SEQ ID NO:153) или AAGGCUAGUCCG (SEQ ID NO:154).
В некоторых вариантах, хвостовой домен содержит 3’ последовательность UUUUUU, например, если U6 промотор применяют для транскрипции. В некоторых вариантах, хвостовой домен содержит 3’ последовательность UUUU, например, если H1 промотор применяют для транскрипции. В некоторых вариантах, хвостовой домен содержит переменное число 3’U, в зависимости от, например, стоп-кодона применяемого pol-III промотора. В некоторых вариантах, хвостовой домен содержит переменную 3’ последовательность, полученную из матрицы ДНК, если применяют T7 промотор. В некоторых вариантах, хвостовой домен содержит переменную 3’ последовательность, полученную из матрицы ДНК, например, если in vitro транскрипцию применяют для создания РНК молекулы. В некоторых вариантах, хвостовой домен содержит переменную 3’ последовательность, полученную из матрицы ДНК, например, если pol-II промотор применяют для проведения транскрипции.
В некоторых вариантах нРНК имеет следующую структуру: 5’ [домен целенаправленного воздействия]-[первый домен комплементарности]-[связывающий домен]-[второй домен комплементарности]-[проксимальный домен]-[хвостовой домен]-3’, где домен целенаправленного воздействия содержит коровый домен и, необязательно, вторичный домен, и имеет 10-50 нуклеотидов в длину; первый домен комплементарности имеет 5-25 нуклеотидов в длину и, в некоторых вариантах, имеет, по меньшей мере, 50, 60, 70, 80, 85, 90, 95, 98 или 99% гомологию со ссылочным первым доменом комплементарности, описанным здесь; связывающий домен имеет 1-5 нуклеотидов в длину; проксимальный домен имеет 5-20 нуклеотидов в длину и, в некоторых вариантах, имеет, по меньшей мере, 50, 60, 70, 80, 85, 90, 95, 98 или 99% гомологию со ссылочным проксимальным доменом, описанным здесь; и хвостовой домен отсутствует или имеет нуклеотидную последовательность 1-50 нуклеотидов в длину и, в некоторых вариантах, имеет, по меньшей мере, 50, 60, 70, 80, 85, 90, 95, 98 или 99% гомологию со ссылочным хвостовым доменом, описанным здесь.
I) Типовые химерные нРНК
В некоторых вариантах, мономолекулярная или химерная, нРНК содержит, предпочтительно, от 5’ до 3’: домен целенаправленного воздействия, например, содержащий 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 нуклеотидов (которые комплементарны к целевой нуклеиновой кислоте); первый домен комплементарности; связывающий домен; второй домен комплементарности (который комплементарен первому домену комплементарности); проксимальный домен; и хвостовой домен, где (a) проксимальный и хвостовой домены, взятые вместе, содержат, по меньшей мере, 15, 18, 20, 25, 30, 31, 35, 40, 45, 49, 50 или 53 нуклеотидов; (b) имеется, по меньшей мере, 15, 18, 20, 25, 30, 31, 35, 40, 45, 49, 50 или 53 нуклеотидов от 3’ до последнего нуклеотида второго домена комплементарности; или (c) имеется, по меньшей мере, 16, 19, 21, 26, 31, 32, 36, 41, 46, 50, 51 или 54 нуклеотидов от 3’ до последнего нуклеотида второго домена комплементарности, которые комплементарны их соответствующим нуклеотидам первого домена комплементарности.
В некоторых вариантах, последовательности из (a), (b) или (c), имеют, по меньшей мере, 60, 75, 80, 85, 90, 95 или 99% гомологию с соответствующими последовательностями природной нРНК или с нРНК, описанной здесь. В некоторых вариантах, проксимальный и хвостовой домены, взятые вместе, содержат по меньшей мере, 15, 18, 20, 25, 30, 31, 35, 40, 45, 49, 50 или 53 нуклеотидов. В некоторых вариантах, имеется, по меньшей мере, 15, 18, 20, 25, 30, 31, 35, 40, 45, 49, 50 или 53 нуклеотидов от 3’ до последнего нуклеотида второго домена комплементарности. В некоторых вариантах, имеется, по меньшей мере, 16, 19, 21, 26, 31, 32, 36, 41, 46, 50, 51 или 54 нуклеотидов от 3’ до последнего нуклеотида второго домена комплементарности, которые комплементарны их соответствующим нуклеотидам первого домена комплементарности. В некоторых вариантах, домен целенаправленного воздействия содержит, имеет или состоит из 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 нуклеотидов (например, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 последовательных нуклеотидов), имеющих комплементарность с целевым доменом, например, домен целенаправленного воздействия имеет 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 нуклеотидов в длину.
В некоторых вариантах, мономолекулярная или химерная, нРНК молекула (содержащая домен целенаправленного воздействия, первый домен комплементарности, связывающий домен, второй домен комплементарности, проксимальный домен и, необязательно, хвостовой домен) содержит следующую последовательность, в которой домен целенаправленного воздействия изображен как 20 N, но может быть любой последовательностью и имеет в длину от 16 до 26 нуклеотидов, и в которой после нРНК последовательности следуют 6 U, которые служат в качестве стоп-кодона для U6 промотора, но которые могут либо отсутствовать, либо иметь меньшее количество: NNNNNNNNNNNNNNNNNNNNGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUUUU (SEQ ID NO:155). В некоторых вариантах, мономолекулярная или химерная, нРНК молекула является S. pyogenes нРНК молекулой.
В некоторых вариантах, мономолекулярная или химерная, нРНК молекула (содержащая домен целенаправленного воздействия, первый домен комплементарности, связывающий домен, второй домен комплементарности, проксимальный домен и, необязательно, хвостовой домен) содержит следующую последовательность, в которой домен целенаправленного воздействия изображен как 20 N, но может быть любой последовательностью и имеет в длину от 16 до 26 нуклеотидов, и в которой после нРНК последовательности следуют 6 U, которые служат в качестве стоп-кодона для U6 промотора, но которые могут либо отсутствовать, либо иметь меньшее количество: NNNNNNNNNNNNNNNNNNNNGUUUUAGUACUCUGGAAACAGAAUCUACUAAAACAAGGCAAAAUGCCGUGUUUAUCUCGUCAACUUGUUGGCGAGAUUUUUU (SEQ ID NO:156). В некоторых вариантах, мономолекулярная или химерная, нРНК молекула является S. aureus нРНК молекулой. Последовательности и структуры типовых химерных нРНК также показаны в WO2015/161276, например, на ФИГ. 10A-10B здесь.
J) Типовые модульные нРНК
В некоторых вариантах, модульная нРНК содержит первую и вторую нити. Первая нить содержит, предпочтительно, от 5’ до 3’; домен целенаправленного воздействия, например, содержащий 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 нуклеотидов; первый домен комплементарности. Вторая нить содержит, предпочтительно, от 5’ до 3’: необязательно 5’ расширяющий домен; второй домен комплементарности; проксимальный домен; и хвостовой домен, где: (a) проксимальный и хвостовой домены, взятые вместе, содержат, по меньшей мере, 15, 18, 20, 25, 30, 31, 35, 40, 45, 49, 50 или 53 нуклеотидов; (b) имеется, по меньшей мере, 15, 18, 20, 25, 30, 31, 35, 40, 45, 49, 50 или 53 нуклеотидов от 3’ до последнего нуклеотида второго домена комплементарности; или (c) имеется, по меньшей мере, 16, 19, 21, 26, 31, 32, 36, 41, 46, 50, 51 или 54 нуклеотидов от 3’ до последнего нуклеотида второго домена комплементарности, которые комплементарны их соответствующим нуклеотидам первого домена комплементарности.
В некоторых вариантах, последовательности из (a), (b) или (c), имеют, по меньшей мере, 60, 75, 80, 85, 90, 95 или 99% гомологию с соответствующими последовательностями природной нРНК или с нРНК, описанной здесь. В некоторых вариантах, проксимальный и хвостовой домены, взятые вместе, содержат по меньшей мере, 15, 18, 20, 25, 30, 31, 35, 40, 45, 49, 50 или 53 нуклеотидов. В некоторых вариантах, имеется, по меньшей мере, 15, 18, 20, 25, 30, 31, 35, 40, 45, 49, 50 или 53 нуклеотидов от 3’ до последнего нуклеотида второго домена комплементарности.
В некоторых вариантах, имеется, по меньшей мере, 16, 19, 21, 26, 31, 32, 36, 41, 46, 50, 51 или 54 нуклеотидов от 3’ до последнего нуклеотида второго домена комплементарности, которые комплементарны их соответствующим нуклеотидам первого домена комплементарности.
В некоторых вариантах, домен целенаправленного воздействия содержит, имеет или состоит из 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 нуклеотидов (например, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 последовательных нуклеотидов), имеющих комплементарность с целевым доменом, например, домен целенаправленного воздействия имеет 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 нуклеотидов в длину.
К) Способы конструирования нРНК
Способы конструирования нРНК описаны здесь, включая способы выбора, конструирования и обоснования доменов целенаправленного воздействия. Типовые домены целенаправленного воздействия также представлены здесь. Домены целенаправленного воздействия, обсуждаемые здесь, могут быть введены в нРНК, описанные здесь.
В некоторых вариантах, направляющую РНК (нРНК), специфическую к целевому гену (например, TRAC, TRBC1 и/или TRBC2 у человека), применяют для РНК-направляемых нуклеаз, например, Cas, для вызова разрыва ДНК на целевом сайте или целевом положении. Способы конструирования нРНК и типовых доменов целенаправленного воздействия могут включать те, которые описаны в, например, международной публикации PCT № WO2015/161276. Домены целенаправленного воздействия могут быть введены в нРНК, которую применяют для целенаправленного воздействия Cas9 нуклеазы на целевой сайт или целевое положение.
Способы выбора и обоснования целевых последовательностей, а также нецелевые анализы описаны, например, в Mali et al., 2013 Science 339(6121): 823-826; Hsu et al. Nat Biotechnol, 31(9): 827-32; Fu et al., 2014 Nat Biotechnol, doi: 10,1038/nbt,2808. PubMed PMID: 24463574; Heigwer et al., 2014 Nat Methods 11(2):122-3. doi: 10,1038/nmeth,2812. PubMed PMID: 24481216; Bae et al., 2014 Bioinformatics PubMed PMID: 24463181; Xiao A et al., 2014 Bioinformatics PubMed PMID: 24389662.
В некоторых вариантах, программные средства могут применяться для оптимизации выбора нРНК в пределах пользовательских целевых последовательностей, например, для минимизации общей нецелевой активности по геному. Нецелевая активность может отличаться от расщепления. Например, для каждого возможного выбора нРНК с применением S. pyogenes Cas9, программные средства могут идентифицировать все потенциальные нецелевые последовательности (предшествующие либо NAG, либо NGG PAM) по геному, который содержит вплоть до определенного количества (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) ошибочно спаренных пар оснований. Эффективность расщепления на каждой нецелевой последовательности может быть спрогнозирована, например, с применением полученной в эксперименте схемы взвешивания. Каждая возможная нРНК затем может быть ранжирована в соответствии с ее общим спрогнозированным нецелевым расщеплением; ведущие нРНК представляют такие, которые вероятно имеют наибольшее целевое и наименьшее нецелевое расщепление. Другие функции, например, автоматизированное конструирование реагента для конструирования вектора нРНК, конструирование праймера для целевого анализа Surveyor, и конструирование праймера для обнаружения с высокой пропускной способностью и количественной оценки нецелевого расщепления через секвенирование следующего поколения также могут быть включены в средство. Потенциальные нРНК молекулы могут быть оценены известными в данной области техники способами или как описано здесь.
В некоторых вариантах, нРНК для применения с S. pyogenes, S. aureus и N. meningitidis Cas9s идентифицируют с применением алгоритма поиска ДНК последовательности, например, с применением заказного программного обеспечения для конструирования нРНК на основе общедоступного средства Cas-OFFinder (Bae et al. Bioinformatics. 2014; 30(10): 1473-1475). Заказное программное обеспечение для конструирования нРНК оценивает гидов после расчета их полногеномной нецелевой предрасположенности. Обычно пары, ранжированные от превосходного спаривания до 7 ошибочных спариваний, рассматривают для гидов длиной от 17 до 24. В некоторых аспектах, как только компьютер определит нецелевые сайты, для каждого гида рассчитывают совокупную оценку и выводят в виде таблицы с помощью веб-интерфейса. Кроме идентификации потенциальных нРНК сайтов, соседних к PAM последовательностям, программа также может идентифицировать все PAM соседние последовательности, которые отличаются 1, 2, 3 или более нуклеотидами из выбранных сайтов нРНК. В некоторых вариантах, геномные ДНК последовательности для каждого гена получают из UCSC Геномного браузера, и последовательности могут быть проверены на присутствие повторяющихся элементов с применением общедоступной программы RepeatMasker. RepeatMasker исследует входящие ДНК последовательности на присутствие повторных элементов и областей низкой сложности. На выходе получают детальную аннотацию повторов, присутствующих в данной искомой последовательности.
После идентификации, нРНК могут быть ранжированы в ряды на основе одного или более из их расстояний до целевого сайта, их ортогональности и присутствия 5’ G (на основе идентификации близких совпадений в человеческом геноме, содержащем соответствующую PAM, например, в случае S. pyogenes, NGG PAM, в случае S. aureus, NNGRR (например, NNGRRT или NNGRRV) PAM, и в случае N. meningtidis, NNNNGATT или NNNNGCTT PAM). Ортогональность относится к количеству последовательностей в человеческом геноме, которые содержат минимальное количество ошибочных спариваний с целевой последовательностью. “Высокий уровень ортогональности” или “хорошая ортогональность” может, например, относится к 20-mer доменам целенаправленного воздействия, которые не имеют идентичных последовательностей в человеческом геноме, кроме намеченной цели, и не имеют последовательностей, которые содержат одно или два ошибочных спаривания в целевой последовательности. Домены целенаправленного воздействия с хорошей ортогональностью выбирают для минимизации нецелевого расщепления ДНК. Должно быть понятно, что это неограничивающий пример, и что множество стратегий может применяться для идентификации нРНК для применения с S. pyogenes, S. aureus и N. meningitidis или другими Cas9 ферментами.
В некоторых вариантах, нРНК для применения с S. pyogenes Cas9 могут быть идентифицированы с применением общедоступного веб-сервера ZiFiT (Fu et al., Improving CRISPR-Cas nuclease specificity using truncated guide RNAs. Nat Biotechnol. 2014 Jan 26. doi: 10,1038/nbt,2808. PubMed PMID: 24463574, оригинальные ссылки представлены в Sander et al., 2007, NAR 35:W599-605; Sander et al., 2010, NAR 38: W462-8). Кроме идентификации потенциальных нРНК сайтов, соседних с PAM последовательностями, программа также идентифицирует все PAM соседние последовательности, которые отличаются 1, 2, 3 или более нуклеотидами из выбранных нРНК сайтов. В некоторых аспектах, геномные ДНК последовательности для каждого гена могут быть получены из UCSC Геномного браузера, и последовательности могут быть проверены на присутствие повторяющихся элементов с применением общедоступной программы RepeatMasker. RepeatMasker исследует входящие ДНК последовательности на присутствие повторных элементов и областей низкой сложности. На выходе получают детальную аннотацию повторов, присутствующих в данной искомой последовательности.
После идентификации, нРНК для применения с S. pyogenes Cas9, могут быть ранжированы на ряды, например, на 5 рядов. В некоторых вариантах, домены целенаправленного воздействия для первого ряда нРНК молекул выбирают на основе их расстояния до целевого сайта, их ортогональности и присутствия 5’ G (на основе ZiFiT идентификации близких совпадений в человеческом геноме, содержащем NGG PAM). В некоторых вариантах, обе, 17-mer и 20-mer нРНК, сконструированы для целей. В некоторых аспектах, нРНК также выбирают для стратегии разрезания одинарной-нРНК нуклеазы и для двойной нРНК никазы. Критерии для выбора нРНК и определения того, какие нРНК могут применяться для какой стратегии, могут быть основаны на нескольких соображениях. В некоторых вариантах, идентифицируют нРНК для расщепления одинарной-нРНК нуклеазы и для стратегии двойной-нРНК спаренной “никазы”. В некоторых вариантах, для выбора нРНК, включая определение того, какие нРНК могут применяться для стратегии двойной-нРНК спаренной “никазы”, пары нРНК должны быть ориентированы на ДНК, что PAM, направленные наружу и усеченные с D10A Cas9 никазой, дадут 5’ липких концов. В некоторых аспектах, можно предположить, что расщепление двойными парами никазы приведет к делеции всей промежуточной последовательности с разумной частотой. Однако расщепление двойными парами никаз также может часто приводить к инсерционно-делеционным мутациям только в одной из нРНК. Потенциальные члены пары могут быть тестированы на предмет того, насколько эффективно они удаляют всю последовательность по сравнению с просто вызовом инсерционно-делеционных мутаций на сайте одной нРНК.
В некоторых вариантах, домены целенаправленного воздействия первого ряда нРНК молекул могут быть выбраны на основе (1) разумного расстояния до целевого положения, например, в пределах первых 500 п.о. кодирующей последовательности ниже инициирующего кодона, (2) высокого уровня ортогональности, и (3) присутствия 5’ G. В некоторых вариантах, для выбора второго ряда нРНК, требование 5’G можно исключить, но ограничения по расстоянию требуется, и высокий уровень ортогональности требуется. В некоторых вариантах, при выборе третьего ряда применяют те же ограничения по расстоянию и требование 5’G, но исключают требование хорошей ортогональности. В некоторых вариантах, при выборе четвертого ряда применяют те же ограничения по расстоянию, но исключают требование хорошей ортогональности и начала с 5’G. В некоторых вариантах, при выборе пятого ряда исключают требование хорошей ортогональности и 5’G, и сканируют более длинные последовательности (например, остальные кодирующие последовательности, например, дополнительные 500 п.о. выше или ниже транскрипции целевого сайта). В определенных случаях, никакие нРНК не идентифицируют по критериям конкретного ряда.
В некоторых вариантах, нРНК идентифицируют для расщепления одинарной-нРНК нуклеазы, а также для стратегии двойной-нРНК спаренной “никазы”.
В некоторых аспектах, нРНК для применения с N. meningitidis и S. aureus Cas9s могут быть идентифицированы вручную сканированием геномной ДНК последовательности на присутствие PAM последовательностей. Эти нРНК могут быть разделены на два ряда. В некоторых вариантах, для нРНК первого ряда, домены целенаправленного воздействия выбирают в пределах первых 500 п.о. кодирующей последовательности ниже инициирующего кодона. В некоторых вариантах, для нРНК второго ряда, домены целенаправленного воздействия выбирают в пределах оставшейся кодирующей последовательности (ниже первых 500 п.о.). В определенных вариантах, никакие нРНК не идентифицируют на основе критерия конкретного ряда.
В некоторых вариантах, в другой стратегии идентификации направляющих РНК (нРНК) для применения с S. pyogenes, S. aureus и N. meningtidis Cas9s может применяться алгоритм поиска ДНК последовательности. В некоторых аспектах, конструирование направляющей РНК проводят с применением заказной программы для конструирования направляющей РНК на основе общедоступного средства Cas-OFFinder (Bae et al. Bioinformatics. 2014; 30(10): 1473-1475). Заказное программное обеспечение для конструирования нРНК оценивает гидов после расчета их полногеномной нецелевой предрасположенности. Обычно пары, ранжированные от превосходного спаривания до 7 ошибочных спариваний, рассматривают для гидов длиной от 17 до 24. Как только компьютер определит нецелевые сайты, для каждого гида рассчитывают совокупную оценку и выводят в виде таблицы с помощью веб-интерфейса. Кроме идентификации потенциальных нРНК сайтов, соседних к PAM последовательностям, программа также может идентифицировать все PAM соседние последовательности, которые отличаются 1, 2, 3 или более нуклеотидами из выбранных сайтов нРНК. В некоторых вариантах, геномные ДНК последовательности для каждого гена получают из UCSC Геномного браузера, и последовательности могут быть проверены на присутствие повторяющихся элементов с применением общедоступной программы RepeatMasker. RepeatMasker исследует входящие ДНК последовательности на присутствие повторных элементов и областей низкой сложности. На выходе получают детальную аннотацию повторов, присутствующих в данной искомой последовательности.
В некоторых вариантах, после идентификации нРНК ранжируют на ряды на основе их расстояния до целевого сайта или их ортогональности (на основе идентификации близких совпадений в человеческом геноме, содержащем соответствующую PAM, например, в случае S. pyogenes, NGG PAM, в случае S. aureus, NNGRR (например, NNGRRT или NNGRRV) PAM, и в случае N. meningtidis, NNNNGATT или NNNNGCTT PAM. В некоторых аспектах, домен целенаправленного воздействия с хорошей ортогональностью выбирают для минимизации нецелевого расщепления ДНК.
В качестве примера, для S. pyogenes и N. meningtidis целей, 17-mer или 20-mer нРНК могут быть сконструированы. В качестве другого примера, для S. aureus целей, 18-mer, 19-mer, 20-mer, 21-mer, 22-mer, 23-mer и 24-mer нРНК могут быть сконструированы.
В некоторых вариантах, нРНК идентифицируют и для расщепления одинарной-нРНК нуклеазы и для стратегии двоной-нРНК спаренной “никазы”. В некоторых вариантах для выбора нРНК, включая определение того, какая нРНК может применяться для стратегии двойной-нРНК спаренной “никазы”, нРНК пары должны быть ориентированы на ДНК так, что PAM, направленные наружу и усеченные с D10A Cas9 никазой, дадут 5’ липких концов. В некоторых аспектах, можно предположить, что расщепление двойными парами никазы приведет к делеции всей промежуточной последовательности с разумной частотой. Однако расщепление двойными парами никаз также может часто приводить к инсерционно-делеционным мутациям только в одной из нРНК. Потенциальные члены пары могут быть тестированы на предмет того, насколько эффективно они удаляют всю последовательность по сравнению с просто вызовом инсерционно-делеционных мутаций на сайте одной нРНК.
Для конструирования стратегий для генетического разрушения, в некоторых вариантах, домены целенаправленного воздействия для ряда 1 нРНК молекул для S. pyogenes выбирают на основе их расстояния до целевого сайта и их ортогональности (PAM является NGG). В некоторых случаях, домены целенаправленного воздействия для ряда 1 нРНК молекул выбирают на основе (1) разумного расстояния до целевого положения, например, в пределах первых 500 п.о. кодирующей последовательности ниже инициирующего кодона и (2) высокого уровня ортогональности. В некоторых аспектах, для выбора ряда 2 нРНК, высокий уровень ортогональности не требуется. В некоторых случаях, ряд 3 нРНК снимает требование хорошей ортогональности и может быть сканирована более длинная последовательность (например, оставшаяся кодирующая последовательность). В определенных случаях, никакие нРНК не идентифицируют на основе критериев конкретного ряда.
Для конструирования стратегий для генетического разрушения, в некоторых вариантах, домен целенаправленного воздействия для ряда 1 нРНК молекул для N. meningtidis выбирают в пределах первых 500 п.о. кодирующей последовательности, и он имеет высокий уровень ортогональности. Домен целенаправленного воздействия для ряда 2 нРНК молекул для N. meningtidis выбирают в пределах первых 500 п.о. кодирующей последовательности, и он не требуют высокой ортогональности. Домен целенаправленного воздействия для ряда 3 нРНК молекул для N. meningtidis выбирают в пределах остатка кодирующей последовательности ниже 500 п.о. Необходимо отметить, что ряды являются не включительными (каждая нРНК перечислена только один раз). В определенных случаях, никакие нРНК не идентифицируют на основе критериев конкретного ряда.
Для конструирования стратегий для генетического разрушения, в некоторых вариантах, домен целенаправленного воздействия для ряда 1 нРНК молекул для S. aureus выбирают в пределах первых 500 п.о. кодирующей последовательности, он имеет высокий уровень ортогональности и содержит NNGRRT PAM. В некоторых вариантах, домен целенаправленного воздействия для ряда 2 нРНК молекул для S. aureus выбирают в пределах первых 500 п.о. кодирующей последовательности, уровень ортогональности не требуется, и он содержит NNGRRT PAM. В некоторых вариантах, домен целенаправленного воздействия для ряда 3 нРНК молекул для S. aureus выбирают в пределах остатка кодирующей последовательности ниже, и он содержит NNGRRT PAM. В некоторых вариантах, домен целенаправленного воздействия для ряда 4 нРНК молекул для S. aureus выбирают в пределах первых 500 п.о. кодирующей последовательности, и он содержит NNGRRV PAM. В некоторых вариантах, домен целенаправленного воздействия для ряда 5 нРНК молекул для S. aureus выбирают в пределах остатка кодирующей последовательности ниже, и он содержит NNGRRV PAM. В определенных случаях, никакие нРНК не идентифицируют на основе критериев конкретного ряда.
2) Cas9
Cas9 молекулы множества видов могут применяться в способах и композициях, описанных здесь. Хотя S. pyogenes, S. aureus, N. meningitidis и S. thermophilus Cas9 молекулы являются предметом большей части раскрытия здесь, также могут применяться Cas9 молекулы, полученные из или основанные на Cas9 белках других видов, перечисленных здесь. Другими словами, хотя в большей части описания здесь применяют S. pyogenes, S. aureus, N. meningitidis и S. thermophilus Cas9 молекулы, Cas9 молекулы из других видов могут их заменить. Такие виды включают: Acidovorax avenae, Actinobacillus pleuropneumoniae, Actinobacillus succinogenes, Actinobacillus suis, Actinomyces sp., Cycliphilusdenitrificans, Aminomonas paucivorans, Bacillus cereus, Bacillus smithii, Bacillus thuringiensis, Bacteroides sp., Blastopirellula marina, Bradyrhizobium sp., Brevibacillus laterosporus, Campylobacter coli, Campylobacter jejuni, Campylobacter lari, Candidatus puniceispirillum, Clostridium cellulolyticum, Clostridium perfringens, Corynebacterium accolens, Corynebacterium diphtheria, Corynebacterium matruchotii, Dinoroseobacter shibae, Eubacterium dolichum, Gammaproteobacterium, Gluconacetobacter diazotrophicus, Haemophilus parainfluenzae, Haemophilus sputorum, Helicobacter canadensis, Helicobacter cinaedi, Helicobacter mustelae, Ilyobacter polytropus, Kingella kingae, Lactobacillus crispatus, Listeria ivanovii, Listeria monocytogenes, Listeriaceae bacterium, Methylocystis sp., Methylosinus trichosporium, Mobiluncus mulieris, Neisseria bacilliformis, Neisseria cinerea, Neisseria flavescens, Neisseria lactamica, Neisseria meningitidis, Neisseria sp., Neisseria wadsworthii, Nitrosomonas sp., Parvibaculum lavamentivorans, Pasteurella multocida, Phascolarctobacterium succinatutens, Ralstonia syzygii, Rhodopseudomonas palustris, Rhodovulum sp., Simonsiella muelleri, Sphingomonas sp., Sporolactobacillus vineae, Staphylococcus aureus, Staphylococcus lugdunensis, Streptococcus sp., Subdoligranulum sp., Tistrella mobilis, Treponema sp. или Verminephrobacter eiseniae. Примеры Cas9 молекул могут включать те, которые описаны в, например, WO2015/161276, WO2017/193107, WO2017/093969, US2016/272999 и US2015/056705.
Cas9 молекула или Cas9 полипептид, как этот термин применяется здесь, относится к молекуле или полипептиду, который может взаимодействовать с нРНК молекулой и, совместно с нРНК молекулой, находится или локализуется на сайте, который содержит целевой домен и PAM последовательность. Cas9 молекула и Cas9 полипептид, как эти термины применяются здесь, относится к природным Cas9 молекулам и к сконструированным, измененным или модифицированным Cas9 молекулам или Cas9 полипептидам, которые отличаются, например, по меньшей мере, одним аминокислотным остатком, от ссылочной последовательности, например, наиболее похожей природной Cas9 молекулы.
Кристаллические структуры определены для двух разных природных бактериальных Cas9 молекул (Jinek et al., Science, 343(6176):1247997, 2014) и для S. pyogenes Cas9 с направляющей РНК (например, синтетического слияния crРНК и tracrРНК) (Nishimasu et al., Cell, 156:935-949, 2014; и Anders et al., Nature, 2014, doi: 10,1038/nature13579).
Природная Cas9 молекула содержит две доли: распознающую (REC) долю и нуклеазную (NUC) долю; каждая из которых дополнительно содержит домены, описанные здесь. Типовая схема организации важных Cas9 доменов в первичной структуре описана в WO2015/161276, например, на ФИГ. 8A-8B здесь. Номенклатура домена и нумерация аминокислотных остатков, охватываемых каждым доменом, применяемая в этом раскрытии, такая, как описана в Nishimasu et al. Нумерация аминокислотных остатков дана со ссылкой на Cas9 из S. pyogenes.
REC доля содержит насыщенную аргинином мостиковую спираль (BH), REC1 домен и REC2 домен. REC доля не имеет структурной схожести с другими известными белками, что указывает на то, что она является Cas9-специфическим функциональным доменом. BH домен является длинной α-спиралью и насыщенной аргинином областью, и содержит аминокислоты 60-93 последовательности S. pyogenes Cas9. REC1 домен является важным для распознавания дуплекса повтор:антиповтор, например, нРНК или tracrРНК, и поэтому является критичным для активности Cas9 через распознавание целевой последовательности. REC1 домен содержит два REC1 мотива на аминокислотах 94-179 и 308-717 последовательности S. pyogenes Cas9. Эти два REC1 домена, хотя и разделены REC2 доменом в линейной первичной структуре, собираются в третичную структуру для образования REC1 домена. REC2 домен или его части также могут играть роль в распознавании дуплекса повтор:антиповтор. REC2 домен содержит аминокислоты 180-307 последовательности S. pyogenes Cas9.
NUC доля содержит RuvC домен (также называемый здесь RuvC-подобный домен), HNH домен (также называемый здесь HNH-подобный домен) и PAM-взаимодействующий (PI) домен. RuvC домен имеет структурное сходство с членами суперсемейства ретровирусной интегразы и расщепляет одну нить, например, не-комплементарную нить целевой молекулы нуклеиновой кислоты. RuvC домен собран из трех разделенных RuvC мотивов (RuvC I, RuvCII и RuvCIII, который часто в общем обозначены как RuvCI домен или N-концевой RuvC домен, RuvCII домен и RuvCIII домен) на аминокислотах 1-59, 718-769 и 909-1098, соответственно, последовательности S. pyogenes Cas9. Так же как и REC1 домен, три RuvC мотива линейно разделены другими доменами в первичной структуре, однако в третичной структуре три RuvC мотива собираются и образуют RuvC домен. HNH домен имеет структурное сходство с HNH эндонуклеазами и расщепляет одну нить, например, комплементарную нить целевой молекулы нуклеиновой кислоты. HNH домен лежит между RuvC II-III мотивами и содержит аминокислоты 775-908 последовательности S. pyogenes Cas9. PI домен взаимодействует с PAM целевой молекулы нуклеиновой кислоты и содержит аминокислоты 1099-1368 последовательности S. pyogenes Cas9.
A) RuvC-подобный домен и HNH-подобный домен
В некоторых вариантах, Cas9 молекула или Cas9 полипептид содержит HNH-подобный домен и RuvC-подобный домен. В некоторых вариантах, активность расщепления зависит от RuvC-подобного домена и HNH-подобного домена. Cas9 молекула или Cas9 полипептид, например, eaCas9 молекула или eaCas9 полипептид, могут содержать один или более из следующих доменов: RuvC-подобный домен и HNH-подобный домен. В некоторых вариантах, Cas9 молекулой или Cas9 полипептидом является eaCas9 молекула или eaCas9 полипептид, и eaCas9 молекула или eaCas9 полипептид содержит RuvC-подобный домен, например, RuvC-подобный домен, описанный здесь, и/или HNH-подобный домен, например, HNH-подобный домен, описанный здесь.
В) RuvC-подобные домены
В некоторых вариантах, RuvC-подобный домен расщепляет одну нить, например, не-комплементарную нить целевой молекулы нуклеиновой кислоты. Cas9 молекула или Cas9 полипептид могут включать более одного RuvC-подобного домена (например, один, два, три или более RuvC-подобных доменов). В некоторых вариантах, RuvC-подобный домен имеет, по меньшей мере, 5, 6, 7, 8 аминокислот в длину, но не более 20, 19, 18, 17, 16 или 15 аминокислот в длину. В некоторых вариантах, Cas9 молекула или Cas9 полипептид содержит N-концевой RuvC-подобный домен, имеющий около 10-20 аминокислот, например, около 15 аминокислот в длину.
С) N-концевые RuvC-подобные домены
Некоторые природные Cas9 молекулы содержат более одного RuvC-подобного домена, где расщепление зависит от N-концевого RuvC-подобного домена. Следовательно, Cas9 молекула или Cas9 полипептид может содержать N-концевой RuvC-подобный домен.
В варианте, N-концевой RuvC-подобный домен является расщепление-компенентным.
В варианте, N-концевой RuvC-подобный домен является расщепление-некомпететным.
В некоторых вариантах, N-концевой RuvC-подобный домен отличается от последовательности N-концевого RuvC-подобного домена, описанного здесь, например, в WO2015/161276, например, на ФИГ. 3A-3B или ФИГ. 7A-7B здесь, не менее 1, но не более чем 2, 3, 4 или 5 остатками. В некоторых вариантах, присутствуют 1, 2 или все 3 из высококонсервативных остатков, идентифицированных в WO2015/161276, например, на ФИГ. 3A-3B или ФИГ. 7A-7B здесь.
В некоторых вариантах, N-концевой RuvC-подобный домен отличается от последовательности N-концевого RuvC-подобного домена, описанного здесь, например, в WO2015/161276, например, на ФИГ. 3A-3B или ФИГ. 7A-7B здесь, не менее 1, но не более чем 2, 3, 4 или 5 остатками. В некоторых вариантах, присутствуют 1, 2, 3 или все 4 из высококонсервативных остатков, идентифицированных в WO2015/161276, например, на ФИГ. 4A-4B или ФИГ. 7A-7B здесь.
D) Дополнительные RuvC-подобные домены
В дополнение к N-концевому RuvC-подобному домену, Cas9 молекула или Cas9 полипептид, например, eaCas9 молекула или eaCas9 полипептид, могут содержать один или более дополнительных RuvC-подобных доменов. В некоторых вариантах, Cas9 молекула или Cas9 полипептид может содержать два дополнительных RuvC-подобных домена. Предпочтительно, дополнительные RuvC-подобный домен имеет, по меньшей мере, 5 аминокислот в длину и, например, менее 15 аминокислот в длину, например, 5-10 аминокислот в длину, например, 8 аминокислот в длину.
Е) HNH-подобные домены
В некоторых вариантах, HNH-подобный домен расщепляет однонитевой комплементаный домен, например, комплементарную нить двухнитевой молекулы нуклеиновой кислоты. В некоторых вариантах, HNH-подобный домен составляет, по меньшей мере, 15, 20, 25 аминокислот в длину, но не более 40, 35 или 30 аминокислот в длину, например, 20-35 аминокислот в длину, например, 25-30 аминокислот в длину. Типовые HNH-подобные домены описаны здесь.
В некоторых вариантах, HNH-подобный домен является расщепление-компенентным.
В некоторых вариантах, HNH-подобный домен является расщепление-некомпенентным.
В некоторых вариантах, HNH-подобный домен отличается от последовательности HNH-подобного домена, описанного здесь, например, в WO2015/161276, например, на ФИГ. 5A-5C или ФИГ. 7A-7B здесь, не менее 1, но не более 2, 3, 4 или 5 остатками. В некоторых вариантах, присутствует 1 или оба высококонсервативных остатка, идентифицированных в WO2015/161276, например, на ФИГ. 5A-5C или ФИГ. 7A-7B здесь.
В некоторых вариантах, HNH-подобный домен отличается от последовательности HNH-подобного домена, описанного здесь, например, в WO2015/161276, например, на ФИГ. 5A-5C или ФИГ. 7A-7B здесь, не менее 1, но не более 2, 3, 4 или 5 остатками. В некоторых вариантах, присутствует 1, 2, все 3 высококонсервативных остатка, идентифицированных в WO2015/161276, например, в ФИГ. 6A-6B или ФИГ. 7A-7B здесь.
F) Активность нуклеазы и хеликазы
В некоторых вариантах, Cas9 молекула или Cas9 полипептид способны расщеплять целевую молекулу нуклеиновой кислоты. Обычно дикие Cas9 молекулы расщепляют обе нити целевой молекулы нуклеиновой кислоты. Cas9 молекулы и Cas9 полипептиды могут быть сконструированы для изменения расщепления нуклеазы (или других свойств), например, для получения Cas9 молекулы или Cas9 полипептида, которые являются никазой или с отсутствием способности расщеплять целевую нуклеиновую кислоту. Cas9 молекула или Cas9 полипептид, которые способны расщеплять целевую молекулу нуклеиновой кислоты, называют здесь eaCas9 молекулой или eaCas9 полипептидом.
В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид имеет одну или более из следующих активностей: активность никазы, например, способность расщеплять одну нить, например, не-комплементарную нитью или комплементарную нить, молекулы нуклеиновой кислоты; активность двухнитевой нуклеазы, т.е. способность расщеплять обе нити двухнитевой нуклеиновой кислоты и создавать двухнитевой разрыв, что, в некоторых вариантах, является присутствием активности двух никаз; активность эндонуклеазы; активность экзонуклеазы; и активность геликазы, т.е. способность раскручивать спиральную структуру двухнитевой нуклеиновой кислоты.
В некоторых вариантах, ферментативно активные, или eaCas9 молекула или eaCas9 полипептид расщепляют обе нити и вызывают двухнитевой разрыв. В некоторых вариантах, eaCas9 молекула расщепляет только одну нить, например, нить, к которой гибридизируется нРНК, или нить, комплементарную к нити нРНК, с которой гибридизируется. В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид имеет активность расщепления, связанную с HNH-подобным доменом. В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид имеет активность расщепления, связанную с N-концевым RuvC-подобным доменом. В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид имеет активность расщепления, связанную с HNH-подобным доменом, и активность расщепления, связанную с N-концевым RuvC-подобным доменом. В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид содержит активный или расщепление-компететный HNH-подобный домен, и неактивный или расщепление-некомпетентный N-концевой RuvC-подобный домен. В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид содержит неактивный или расщепление-некомпетентный HNH-подобный домен и активный или расщепление-компететный N-концевой RuvC-подобный домен.
Некоторые Cas9 молекулы или Cas9 полипептиды обладают способностью взаимодействовать с нРНК молекулой, и в сочетании с нРНК молекулой, локализоваться на коровом целевом домене, но не способны расщеплять целевую нуклеиновую кислоту или не способна расщеплять при эффективных степенях. Cas9 молекулы, не имеющие или по существу не имеющие активность расщепления называют здесь eiCas9 молекула или eiCas9 полипептид. Например, eiCas9 молекула или eiCas9 полипептид может не иметь активность расщепления или иметь по существу меньшую, например, менее 20, 10, 5, 1 или 0,1% активности расщепления по сравнению со ссылочной Cas9 молекулой или eiCas9 полипептидом, измеренную описанным здесь анализом.
G) Целенаправленное воздействие и PAM
Cas9 молекулой или Cas9 полипептидом является полипептид, который может взаимодействовать с направляющей РНК (нРНК) молекулой и, в сочетании с нРНК молекулой, локализоваться на сайте, который содержит целевой домен и PAM последовательность.
В некоторых вариантах, способность eaCas9 молекулы или eaCas9 полипептида взаимодействовать с и расщеплять целевую нуклеиновую кислоту зависит от PAM последовательности. PAM последовательностью является последовательность в целевой нуклеиновой кислоте. В некоторых вариантах, расщепление целевой нуклеиновой кислоты происходит выше PAM последовательности. EaCas9 молекулы из разных бактериальных видов могут распознавать разные мотивы последовательности (например, PAM последовательности). В некоторых вариантах, eaCas9 молекула S. pyogenes распознает мотив последовательности NGG, NAG, NGA и направляет расщепление целевой последовательности нуклеиновых кислот 1-10, например, 3-5, пар оснований выше этой последовательности. См., например, Mali et al., Science 2013; 339(6121): 823-826. В некоторых вариантах, eaCas9 молекула S. thermophilus распознает мотив последовательности NGGNG и/или NNAGAAW (W=A или T) и направляет расщепление целевой последовательности нуклеиновых кислот 1-10, например, 3-5, пар оснований выше этих последовательностей. См., например, Horvath et al., Science 2010; 327(5962):167-170, и Deveau et al., J Bacteriol 2008; 190(4): 1390-1400. В некоторых вариантах, eaCas9 молекула S. mutans распознает мотив последовательности NGG и/или NAAR (R=A или G)) и направляет расщепление коровой целевой последовательности нуклеиновых кислот 1-10, например, 3-5 пар оснований, выше этой последовательности. См., например, Deveau et al., J Bacteriol 2008; 190(4): 1390-1400. В некоторых вариантах, eaCas9 молекула S. aureus распознает мотив последовательности NNGRR (R=A или G) и направляет расщепление целевой последовательности нуклеиновых кислот 1-10, например, 3-5, пар оснований выше этой последовательности. В некоторых вариантах, eaCas9 молекула S. aureus распознает мотив последовательности NNGRRT (R=A или G) и направляет расщепление целевой последовательности нуклеиновых кислот 1-10, например, 3-5, пар оснований выше этой последовательности. В некоторых вариантах, eaCas9 молекула S. aureus распознает мотив последовательности NNGRRV (R=A или G) и направляет расщепление целевой последовательности нуклеиновых кислот 1-10, например, 3-5, пар оснований выше этой последовательности. В некоторых вариантах, eaCas9 молекула N. meningitidis распознает мотив последовательности NNNNGATT или NNNGCTT (R=A или G, V=A, G или C и направляет расщепление целевой последовательности нуклеиновых кислот 1-10, например, 3-5, пар оснований выше этой последовательности. См., например, Hou et al., PNAS Early Edition 2013, 1-6. Способность Cas9 молекулы распознавать PAM последовательность может быть определена, например, с применением анализа превращения, описанного в Jinek et al., Science 2012 337:816. В указанных выше вариантах, N может быть любым нуклеотидным остатком, например, любым из A, G, C или T.
Как обсуждается здесь, Cas9 молекулы могут быть сконструированы для изменения PAM специфичности Cas9 молекулы.
Типовые природные Cas9 молекулы описаны в Chylinski et al., РНК Biology 2013 10:5, 727-737. Такие Cas9 молекулы включают Cas9 молекулы кластера 1-78 бактериального семейства.
Типовые природные Cas9 молекулы включают Cas9 молекулу кластера 1 бактериального семейства. Примеры включают Cas9 молекулу: S. pyogenes (например, штамма SF370, MGAS10270, MGAS10750, MGAS2096, MGAS315, MGAS5005, MGAS6180, MGAS9429, NZ131 и SSI-1), S. thermophilus (например, штамма LMD-9), S. pseudoporcinus (например, штамма SPIN 20026), S. mutans (например, штамма UA159, NN2025), S. macacae (например, штамма NCTC11558), S. gallolyticus (например, штамма UCN34, ATCC BAA-2069), S. equines (например, штамма ATCC 9812, MGCS 124), S. dysdalactiae (например, штамма GGS 124), S. bovis (например, штамма ATCC 700338), S. anginosus (например, штамма F0211), S. agalactiae (например, штамма NEM316, A909), Listeria monocytogenes (например, штамма F6854), Listeria innocua (L. innocua, например, штамма Clip11262), Enterococcus italicus (например, штамма DSM 15952) или Enterococcus faecium (например, штамма 1,231,408). Другой типовой Cas9 молекулой является Cas9 молекула Neisseria meningitidis (Hou et al., PNAS Early Edition 2013, 1-6).
В некоторых вариантах, Cas9 молекула или Cas9 полипептид, например, eaCas9 молекула или eaCas9 полипептид, содержит последовательность аминокислот: имеющую 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% гомологию с; отличающуюся не более чем 2, 5, 10, 15, 20, 30 или 40% аминокислотных остатков по сравнению с; отличающуюся, по меньшей мере, 1, 2, 5, 10 или 20 аминокислотами, но не больше чем 100, 80, 70, 60, 50, 40 или 30 аминокислотами от; или идентичную последовательности любой Cas9 молекулы, описанной здесь или последовательности природной Cas9 молекулы, например, Cas9 молекулы от видов, перечисленных здесь (например, SEQ ID NOS:157-162) или описанных в Chylinski et al., RNA Biology 2013 10:5, 727-737; Hou et al., PNAS Early Edition 2013, 1-6. В некоторых вариантах, Cas9 молекула или Cas9 полипептид имеет одну или более из следующих активностей: активность никазы; активность двухнитевого расщепления (например, активность эндонуклеаза и/или экзонуклеазы); активность геликазы; или способность, вместе с нРНК молекулой, располагаться на целевой нуклеиновой кислоте.
В некоторых вариантах, Cas9 молекула или Cas9 полипептид содержит аминокислотную последовательность консенсусной последовательности из WO2015/161276, например, на ФИГ. 2A-2G здесь, где “*” означает любую аминокислоту, найденную в соответствующем положении аминокислотной последовательности Cas9 молекулы S. pyogenes, S. thermophilus, S. mutans и L. innocua, и “-” означает любую аминокислоту. В некоторых вариантах, Cas9 молекула или Cas9 полипептид отличается от последовательности консенсусной последовательности SEQ ID NOS: 157-162 или консенсусной последовательности, описанной в WO2015/161276, например, на ФИГ. 2A-2G здесь, на, по меньшей мере, 1, но не более чем 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных остатков. В некоторых вариантах, Cas9 молекула или Cas9 полипептид содержит аминокислотную последовательность SEQ ID NO:162 или как описана в WO2015/161276, например, на ФИГ. 7A-7B здесь, где “*” означает любую аминокислоту, найденную в соответствующем положении аминокислотной последовательности Cas9 молекулы S. pyogenes или N. meningitidis, “-” означает любую аминокислоту и “-” означает любую аминокислоту или ее отсутствие. В некоторых вариантах, Cas9 молекула или Cas9 полипептид отличается от последовательности SEQ ID NO:161 или 162 или как описано в WO2015/161276, например, на ФИГ. 7A-7B здесь, на, по меньшей мере, 1, но не более чем 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных остатков.
Сравнение последовательностей множества Cas9 молекул показывает, что определенные области являются консервативными. Они идентифицированы как: область 1 (остатки 1-180 или в случае области 1’, остатки 120-180); область 2 (остатки 360-480); область 3 (остатки 660-720); область 4 (остатки 817-900); и область 5 (остатки 900-960).
В некоторых вариантах, Cas9 молекула или Cas9 полипептид содержит области 1-5, вместе с достаточными дополнительными последовательностями Cas9 молекулы для получения биологически активной молекулы, например, Cas9 молекулы, имеющей, по меньшей мере, одну описанную здесь активность. В некоторых вариантах, каждая из областей 1-6, независимо, имеет 50%, 60%, 70% или 80% гомологию с соответствующими остатками Cas9 молекулы или Cas9 полипептида, описанных здесь, например, представленных в SEQ ID NOS:157-162, или a последовательностью, описанными в WO2015/161276, например, из ФИГ. 2A-2G или из ФИГ. 7A-7B здесь.
Н) Сконструированные или измененные Cas9 молекулы и Cas9 полипептиды
Cas9 молекулы и Cas9 полипептиды, описанные здесь, например, природные Cas9 молекулы, могут обладать любым количеством свойств, включая: активность никазы, активность нуклеазы (например, активность эндонуклеазы и/или экзонуклеазы); активность геликазы; способность функционально связываться с нРНК молекулой; и способность нацеливаться (или локализоваться на) сайте на нуклеиновой кислоте (например, PAM распознавание и специфичность). В некоторых вариантах, Cas9 молекула или Cas9 полипептид может включать все или подмножество этих свойств. В типовых вариантах, Cas9 молекула или Cas9 полипептид обладает способностью взаимодействовать с нРНК молекулой и, в сочетании с нРНК молекулой, локализоваться на сайте в нуклеиновой кислоте. Другие активности, например, PAM специфичность, расщепляющая активность или активность геликазы, могут более широко варьироваться в Cas9 молекулах и Cas9 полипептидах.
Cas9 молекулы включают сконструированные Cas9 молекулы и сконструированные Cas9 полипептиды (“сконструированные” в данном контексте означает, что Cas9 молекула или Cas9 полипептид отличается от ссылочных последовательностей, и не предполагает ограничение процесса или происхождения). Сконструированная Cas9 молекула или Cas9 полипептид может иметь измененные ферментные свойства, например, измененную активность нуклеазы (по сравнению с природной или другой ссылочной Cas9 молекулой) или измененную активность геликазы. Как обсуждается здесь, сконструированная Cas9 молекула или Cas9 полипептид могут иметь активность никазы (противоположную активности двухнитевой нуклеазы). В некоторых вариантах, сконструированная Cas9 молекула или Cas9 полипептид могут иметь изменения, которые меняют ее размер, например, делецию последовательности аминокислоты, которая снижает ее размер, например, без значительного влияния на одну или более или любую Cas9 активность. В некоторых вариантах, сконструированная Cas9 молекула или Cas9 полипептид может содержать изменение, которое влияет на PAM распознавание. Например, сконструированная Cas9 молекула может быть изменена для распознавания PAM последовательности, отличной от той, которая распознается эндогенным диким PI доменом. В некоторых вариантах Cas9 молекула или Cas9 полипептид может отличаться последовательностью от природной Cas9 молекулы, но не иметь значительных изменений в одной или более активностях Cas9.
Cas9 молекулы или Cas9 полипептиды с желаемыми свойствами могут быть получены множеством путей, например, изменением исходных, например, природных Cas9 молекул или Cas9 полипептидов, с получением измененной Cas9 молекулы или Cas9 полипептида, имеющих желаемое свойство. Например, может быть введена одна или более мутаций или изменений относительно исходной Cas9 молекулы, например, природной или сконструированной Cas9 молекулы. Такие мутации и изменения содержат: замещения (например, консервативные замещения ли замещения не незаменимых аминокислот); вставки; или делеции. В некоторых вариантах, Cas9 молекула или Cas9 полипептид может содержать одну или более мутаций или отличий, например, по меньшей мере, 1, 2, 3, 4, 5, 10, 15, 20, 30, 40 или 50 мутаций, но менее 200, 100 или 80 мутаций относительно ссылочной, например, исходной Cas9 молекулы.
В некоторых вариантах, мутация или мутации не оказывают существенного действия на активность Cas9, например, описанную здесь активность Cas9. В некоторых вариантах, мутация или мутации оказывают значительное действие на активность Cas9, например, описанную здесь активность Cas9.
I) Cas9 молекулы и Cas9 полипептиды, не расщепляющие и с модифицированным расщеплением
В некоторых вариантах, Cas9 молекула или Cas9 полипептид обладает свойством расщепления, которое отличается от природных Cas9 молекул, например, которое отличается от природной Cas9 молекулы, имеющей ближайшую гомологию. Например, Cas9 молекула или Cas9 полипептид может отличаться от природных Cas9 молекул, например, Cas9 молекулы S. pyogenes, следующим: способностью модулировать, например, снижать или повышать, расщепление двухнтевой нуклеиновой кислоты (активностью эндонуклеазы и/или экзонуклеазы), например, по сравнению с природной Cas9 молекулой (например, Cas9 молекулой S. pyogenes); способностью модулировать, например, снижать или повышать, расщепление одной нити нуклеиновой кислоты, например, не комплементарной нити молекулы нуклеиновой кислоты или комплементарной нити молекулы нуклеиновой кислоты (активностью никазы), например, по сравнению с природной Cas9 молекулой (например, Cas9 молекулой S. pyogenes); или способностью расщеплять молекулу нуклеиновой кислоты, например, двухнитевую или однонитевую молекулу нуклеиновой кислоты, которые могут быть убраны.
J) Модифицированное расщепление eaCas9 молекулами и eaCas9 полипептидами
В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид имеет одну или более из следующих активностей: расщепляющая активность, связанная с N-концевым RuvC-подобным доменом; расщепляющая активность, связанная с HNH-подобным доменом; расщепляющая активность, связанная с HNH-подобным доменом и расщепляющая активность, связанная с N-концевым RuvC-подобным доменом.
В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид содержит активный или расщепление-компететный, HNH-подобный домен и неактивный или расщепление-некомпететный, N-концевой RuvC-подобный домен. Типовой неактивный или расщепление-некомпететный N-концевой RuvC-подобный домен может иметь мутацию аспарагиновой кислоты в N-концевом RuvC-подобном домене, например, аспарагиновой кислоты в положении 9 консенсусной последовательности SEQ ID NOS: 157-162 или консенсусной последовательности, раскрытой в WO2015/161276, например, на ФИГ. 2A-2G здесь, или аспарагиновой кислоты в положении 10 SEQ ID NO:162, например, которая может быть заменена аланином. В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид отличается от дикого типа в N-концевом RuvC-подобном домене и не расщепляет целевую нуклеиновую кислоту или расщепляет со значительно меньшей эффективностью, например, менее 20, 10, 5, 1 или 1% от расщепляющей активности ссылочной Cas9 молекулы, например, измеренной в анализе, описанном здесь. Ссылочной Cas9 молекулой может быть природная немодифицированная Cas9 молекула, например, природная Cas9 молекула, такая как Cas9 молекула S. pyogenes или S. thermophilus. В некоторых вариантах, ссылочной Cas9 молекулой может быть природная Cas9 молекула, имеющая ближайшую идентичность или гомологию последовательности.
В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид содержит неактивный или расщепление-некомпететный HNH домен и активный или расщепление-компететный N-концевой RuvC-подобный домен. Типовые неактивные или расщепление-некомпететные HNH-подобные домены могут иметь мутацию в одном или более: гистидине в HNH-подобном домене, например, гистидине, показанном в положении 856 консенсусной последовательности SEQ ID NOS:157-162 или консенсусной последовательности, раскрытой в WO2015/161276, например, на ФИГ. 2A-2G здесь, например, он может быть замещен аланином; и одном или более аспарагинах в HNH-подобном домене, например, аспарагине, показанном в положении 870 консенсусной последовательности SEQ ID NOS:157-162 или консенсусной последовательности, раскрытой в WO2015/161276, например, на ФИГ. 2A-2G здесь и/или в положении 879 консенсусной последовательности SEQ ID NOS:157-162 или консенсусной последовательности, раскрытой в WO2015/161276, например, на ФИГ. 2A-2G здесь, например, которые могут быть замещены аланином. В некоторых вариантах, eaCas9 отличается от дикого типа в HNH-подобном домене и не расщепляет целевую нуклеиновую кислоту или расщепляет со значительно меньшей эффективностью, например, менее 20, 10, 5, 1 или 1% от расщепляющей активности ссылочной Cas9 молекулы, например, измеренной в анализе, описанном здесь. Ссылочной Cas9 молекулой может быть природная немодифицированная Cas9 молекула, например, природная Cas9 молекула, такая как Cas9 молекула S. pyogenes или S. thermophilus. В некоторых вариантах, ссылочной Cas9 молекулой может быть природная Cas9 молекула, имеющая ближайшую идентичность или гомологию последовательности.
В некоторых вариантах, eaCas9 молекула или eaCas9 полипептид содержит неактивный или расщепление-некомпететный HNH домен и активный или расщепление-компететный N-концевой RuvC-подобный домен. Типовые неактивные или расщепление-некомпететные HNH-подобные домены могут иметь мутацию в одном или более: гистидине в HNH-подобном домене, например, гистидине, показанном в положении 856 консенсусной последовательности SEQ ID NOS:157-162 или консенсусной последовательности, раскрытой в WO2015/161276, например, на ФИГ. 2A-2G здесь, например, он может быть замещен аланином; и одном или более аспарагинах в HNH-подобном домене, например, аспарагине, показанном в положении 870 консенсусной последовательности SEQ ID NOS:157-162 или консенсусной последовательности, раскрытой в WO2015/161276, например, на ФИГ. 2A-2G здесь и/или в положении 879 консенсусной последовательности SEQ ID NOS:157-162 или консенсусной последовательности, раскрытой в WO2015/161276, например, на ФИГ. 2A-2G здесь, например, которые могут быть замещены аланином. В некоторых вариантах, eaCas9 отличается от дикого типа в HNH-подобном домене и не расщепляет целевую нуклеиновую кислоту или расщепляет со значительно меньшей эффективностью, например, менее 20, 10, 5, 1 или 1% от расщепляющей активности ссылочной Cas9 молекулы, например, измеренной в анализе, описанном здесь. Ссылочной Cas9 молекулой может быть природная немодифицированная Cas9 молекула, например, природная Cas9 молекула, такая как Cas9 молекула S. pyogenes или S. thermophilus. В некоторых вариантах, ссылочной Cas9 молекулой может быть природная Cas9 молекула, имеющая ближайшую идентичность или гомологию последовательности.
К) Изменения способность расщеплять одну или обе нити целевой нуклеиновой кислоты
В некоторых вариантах, типовая активность Cas9 включает одну или более из PAM специфичности, расщепляющей активности и активности геликазы. Мутации могут присутствовать, например, в: одном или более RuvC-подобных доменах, например, N-концевом RuvC-подобном домене; HNH-подобном домене; области вне RuvC-подобных доменов и HNH-подобного домена. В некоторых вариантах, мутации присутствуют в RuvC-подобном домене, например, N-концевом RuvC-подобном. В некоторых вариантах, мутации присутствуют в HNH-подобном домене. В некоторых вариантах, мутации присутствуют в обоих RuvC-подобном домене, например, N-концевом RuvC-подобном домене, и HNH-подобном домене.
Типовые мутации, которые могут быть проведены в RuvC домене или HNH домене со ссылкой на S. pyogenes последовательность, включают: D10A, E762A, H840A, N854A, N863A и/или D986A.
В некоторых вариантах, Cas9 молекулой или Cas9 полипептидом является eiCas9 молекула или eiCas9 полипептид, содержащий одно или более отличий в RuvC домене и/или в HNH домене по сравнению со ссылочной Cas9 молекулой, и eiCas9 молекула или eiCas9 полипептид не расщепляют нуклеиновую кислоту или расщепляют со значительно меньшей эффективностью, чем дикий тип, например, по сравнению с диким типом в анализе расщепления, например, как описано здесь, расщепляет менее 50, 25, 10 или 1% по сравнению со ссылочной Cas9 молекулой, как измерено в описанном здесь анализе.
Может ли или нет конкретная последовательность, например, замещение, влиять на одну или более активностей, таких как целенаправленное воздействие, активность расщепления и т.д., может быть оценено или предсказано, например, путем оценки консервативности мутации. В некоторых вариантах, “не незаменимым” аминокислотным остатком, как применяется в контексте Cas9 молекулы, является остаток, который может быть изменен в дикой последовательности Cas9 молекулы, например, природной Cas9 молекулы, например, eaCas9 молекулы, без прекращения, или более предпочтительно, без существенного изменения Cas9 активности (например, расщепляющей активности), в то время как изменение “незаменимого” аминокислотного остатка может вызвать значительную потерю активности (например, расщепляющей активности).
В некоторых вариантах, Cas9 молекула или Cas9 полипептид имеет расщепляющее свойство, которое отличается от природных Cas9 молекул, например, которое отличается от природной Cas9 молекулы, имеющей ближайшую гомологию. Например, Cas9 молекула или Cas9 полипептид может отличаться от природных Cas9 молекул, например, Cas9 молекулы S aureus, S. pyogenes или C. jejuni, следующим: способностью модулировать, например, снижать или повышать, расщепление двухнитевого разрыва (активность эндонуклеазы и/или экзонуклеазы), например, по сравнению с природной Cas9 молекулой (например, Cas9 молекулой S aureus, S. pyogenes или C. jejuni); способностью модулировать, например, снижать или повышать, расщепление одной нити нуклеиновой кислоты, например, не комплементарной нити молекулы нуклеиновой кислоты или комплементарной нити молекулы нуклеиновой кислоты (активность никазы), например, по сравнению с природной Cas9 молекулой (например, Cas9 молекулой S aureus, S. pyogenes или C. jejuni); или способность расщеплять молекулу нуклеиновой кислоты, например, двухнитевую или однонитевую молекулу нуклеиновой кислоты, которая может быть исключена.
В некоторых вариантах, измененной Cas9 молекулой или Cas9 полипептидом является eaCas9 молекула или eaCas9 полипептид, имеющие одной или более из следующих активностей: расщепляющая активность, связанная с RuvC доменом; расщепляющая активность, связанная с HNH доменом; расщепляющая активность, связанная с HNH доменом и расщепляющая активность, связанная с RuvC доменом.
В некоторых вариантах, измененной Cas9 молекулой или Cas9 полипептидом является eiCas9 молекула или eaCas9 полипептид, который не расщепляет молекулу нуклеиновой кислоты (либо двухнитевую, либо однонитевую молекулу нуклеиновой кислоты) или расщепляет молекулу нуклеиновой кислоты со значительно меньшей эффективностью, например, менее 20, 10, 5, 1 или 0,1% от расщепляющей активности ссылочной Cas9 молекулы, например, как измерено описанным здесь анализом. Ссылочной Cas9 молекулой может быть природная немодифицированная Cas9 молекула, например, природная Cas9 молекула, такая как Cas9 молекула S. pyogenes, S. thermophilus, S. aureus, C. jejuni или N. meningitidis. В некоторых вариантах, ссылочной Cas9 молекулой является природная Cas9 молекула, имеющая ближайшую идентичность или гомологию последовательности. В некоторых вариантах, eiCas9 молекула или eiCas9 полипептид не имеет значительной расщепляющей активности, связанной с RuvC доменом, и расщепляющей активности, связанной с HNH доменом.
В некоторых вариантах, измененной Cas9 молекулой или Cas9 полипептидом является eaCas9 молекула или eaCas9 полипептид, содержащие фиксированные аминокислотные остатки S. pyogenes, показанные в консенсусной последовательности, раскрытой в WO2015/161276, например, в ФИГ. 2A-2G здесь, и имеющие одну или более аминокислот, которые отличаются от аминокислотной последовательности S. pyogenes (например, имеют замещение) на одном или более остатках (например, 2, 3, 5, 10, 15, 20, 30, 50, 70, 80, 90, 100, 200 аминокислотных остатках) в SEQ ID NO:162 или остатке, представленном “-” в консенсусной последовательности, описанной в WO2015/161276, например, в ФИГ. 2A-2G здесь.
В некоторых вариантах, измененной Cas9 молекулой или Cas9 полипептидом, например, eaCas9 молекулой, может быть слитая, например, из двух или более разных Cas9 молекул или Cas9 полипептидов, например, двух или более природных Cas9 молекул разных видов. Например, фрагмент природной Cas9 молекулы одного вида может быть слит с фрагментом Cas9 молекулы второго вида. В качестве примера, фрагмент Cas9 молекулы S. pyogenes, содержащий N-концевой RuvC-подобный домен, может быть слит с фрагментом Cas9 молекулы видов, отличных от S. pyogenes (например, S. thermophilus), содержащих HNH-подобный домен.
L) Cas9 Молекулы с измененным распознаванием PAM или без распознавания PAM
Существующие в природе Cas9 молекулы могут распознавать PAM последовательности, например распознавание PAM последовательностей описано здесь, например, для S. pyogenes, S. thermophilus, S. mutans, S. aureus и N. meningitidis.
В некоторых вариантах, Cas9 молекула или Cas9 полипептид имеют такую же специфичность к PAM, как существующая в природе Cas9 молекула. В других вариантах, Cas9 молекула или Cas9 полипептид имеет специфичность к PAM не связанную с существующей в природе Cas9 молекулой, или специфичность к PAM, не связанную с существующей в природе Cas9 молекулой, с которой она имеет наибольшую гомологию последовательности. Например, существующая в природе Cas9 молекула может быть изменена, например, для изменения распознавания PAM, например, для изменения PAM последовательности, которую распознает Cas9 молекула или Cas9 полипептид, для снижения целевых сайтов и/или улучшения специфичности; или устранения требования в распознавании PAM. В некоторых вариантах, Cas9 молекула может быть изменена, например, для увеличения длины последовательности распознавания PAM и/или улучшения специфичности Cas9 до высокого уровня идентичности, например, для снижения целевых сайтов и повышения специфичности. В некоторых вариантах, длина последовательности распознавания PAM составляет, по меньшей мере, 4, 5, 6, 7, 8, 9, 10 или 15 аминокислот в длину.
Cas9 молекулы или Cas9 полипептиды, которые распознают разные PAM последовательности и/или снижают нецелевую активность могут быть созданы с применением направленного развития. Типовые способы и системы, которые могут применяться для направленного развития Cas9 молекул, описаны, например, в Svelte et al. Nature 2011, 472(7344): 499-503. Кандидатные Cas9 молекулы могут быть оценены, например, описанными здесь способами.
Изменения PI домена, который медиирует распознавание PAM, обсуждается здесь.
М) Синтетические Cas9 молекулы и Cas9 полипептиды с измененными PI доменами
Современные способы редактирования генома ограничены разнообразием целевых последовательностей, на которые могут быть направлены PAM последовательности, распознаваемые применяемой Cas9 молекулой. Синтетическая Cas9 молекулы (или Syn-Cas9 молекула) или синтетическая Cas9 полипептид (или Syn-Cas9 полипептид), как этот термин используется здесь, относится к Cas9 молекуле или Cas9 полипептиду, который содержит Cas9 который домен от одних бактериальных видов и функциональный измененный PI домен, т.е., PI домен, отличный от того, который связан с природным Cas9 коровым доменом, например, от других бактериальных видов.
В некоторых вариантах, измененный PI домен распознает PAM последовательность, которая отличается от PAM последовательности, распознаваемой существующей в природе Cas9, из которой получен Cas9 коровый домен. В некоторых вариантах, измененный PI домен распознает ту же PAM последовательность, которая распознается существующей в природе Cas9, из которое получен Cas9 коровый домен, но с другой аффинностью или специфичностью. Syn-Cas9 молекула или Syn-Cas9 полипептид могут быть, соответственно, Syn-eaCas9 молекулой или Syn-eaCas9 полипептидом или Syn-eiCas9 молекулой, Syn-eiCas9 полипептидом.
Типовая Syn-Cas9 молекула или Syn-Cas9 полипептид содержит: a) Cas9 коровый домен, например, Cas9 коровый домен, например, S. aureus, S. pyogenes или C. jejuni Cas9 коровый домен; и b) измененный PI домен из Cas9 последовательности видов X.
В некоторых вариантах, RKR мотив (PAM связывающий мотив) такого измененного PI домена содержит: различия в 1, 2 или 3 аминокислотных остатках; различие в аминокислотной последовательности в первом, втором или третьем положении; различия в аминокислотной последовательности в первом и втором положениях, первом и третьем положениях или втором и третьем положениях; по сравнению с последовательностью RKR мотива природного или эндогенного PI домена, связанного с Cas9 коровым доменом.
В некоторых вариантах, Syn-Cas9 молекула или Syn-Cas9 полипептид также могут быть оптимизированы по размеру, например, Syn-Cas9 молекула или Syn-Cas9 полипептид содержит одну или более делеций, и необязательно, один или более линкеров, расположенных между аминокислотными остатками, прилегающими к делециям. В некоторых вариантах, Syn-Cas9 молекула или Syn-Cas9 полипептид содержит REC делецию.
N) Оптимизированные по размеру Cas9 Молекулы и Cas9 Полипептиды
Сконструированные Cas9 молекулы и сконструированные Cas9 полипептиды, описанные здесь, включают Cas9 молекулу или Cas9 полипептид, содержащие делецию, которая снижает размер молекулы при сохранении желаемых свойств Cas9, например, по существу, природную конформацию, Cas9 активность нуклеазы и/или целевое распознавание молекул нуклеиновой кислоты. Здесь представлены Cas9 молекулы или Cas9 полипептиды, содержащие одну или более делеций и необязательно один или более линкеров, где линкер расположен между аминокислотными остатками, которые расположены рядом с делецией. Способы идентификации подходящих делеций в ссылочной Cas9 молекуле, способы создания Cas9 молекул с делецией и линкером, и способы применения таких Cas9 молекул будут очевидны при обзоре этого документа.
Cas9 молекула, например, S. aureus, S. pyogenes или C. jejuni Cas9 молекула, имеющая делецию, меньше, например, имеет пониженное число аминокислот, чем соответствующая существующая в природе Cas9 молекула. Меньший размер Cas9 молекул позволяет повысить гибкость для способов доставки и, тем самым, увеличивает применимость для редактирования генома. Cas9 молекула или Cas9 полипептид могут содержать одну или более делеций, которые существенно не влияют или не снижают активность полученных Cas9 молекул или Cas9 полипептидов, описанных здесь. Активности, сохраняющиеся в Cas9 молекулах или Cas9 полипептидах, содержащих делецию как описано здесь, включают одну или более из следующих: активность никазы, т.е., способность расщеплять одну нить, например, не-комплементарную нить или комплементарную нить, молекулы нуклеиновой кислоты; двухнитевая активность нуклеазы, т.е., способность расщеплять обе нити двухнитевой нуклеиновой кислоты и создавать двухнитевой разрыв, которая, в некоторых вариантах является присутствием активностей двух никаз; эндоактивность нуклеазы; активность экзонуклеазы; активность геликазы, т.е., способность разматывать спиральную структуру двухнитевой нуклеиновой кислоты; и активность распознавания молекулы нуклеиновой кислоты, например, целевой нуклеинов кислот или нРНК.
Активность Cas9 молекул или Cas9 полипептидов, описанная здесь, может быть оценена с применением анализов активности, описанных здесь или известных.
О) Идентифицирующие области, подходящие для делеции
Подходящие области Cas9 молекул для делеции могут быть идентифицированы множеством способов. Существующие в природе ортологические Cas9 молекулы из разных бактериальных видов могут быть смоделированы в кристаллическую структуру S. pyogenes Cas9 (Nishimasu et al., Cell, 156:935-949, 2014) для проверки уровня превращения выбранных Cas9 ортологов по отношению к трехмерной конформации белка. Менее консервированные или неконсервированные области, которые пространственно удалены от областей, вовлеченных в активность Cas9, например, интерфейс с целевой молекулой нуклеиновой кислоты и/или нРНК, представляют области или домены, которые являются кандидатами для делеции без значительного влияния или снижения активности Cas9.
Р) REC-оптимизированные Cas9 молекулы и Cas9 полипептиды
REC-оптимизированная Cas9 молекула или REC-оптимизированный Cas9 полипептид, как этот термин используется здесь, относится к Cas9 молекуле или Cas9 полипептиду, который содержит делецию в одном или обоих REC2 доменах и RE1CT домене (вместе REC делецию), где делеция содержит, по меньшей мере, 10% аминокислотных остатков в когнатном домене. REC-оптимизированной Cas9 молекулой или Cas9 полипептидом может быть eaCas9 молекула или eaCas9 полипептид или eiCas9 молекула или eiCas9 полипептид. Типовая REC-оптимизированная Cas9 молекула или REC-оптимизированный Cas9 полипептид содержит: a) делецию, выбранную из: i) REC2 делеции; ii) REC1CT делеции; или iii) REC1SUB делеции.
Необязательно, линкер расположен между аминокислотными остатками, которые окружают делецию. В некоторых вариантах, Cas9 молекула или Cas9 полипептид включают только одну делецию или только две делеции. Cas9 молекула или Cas9 полипептид может содержать REC2 делецию и REC1CT делецию. Cas9 молекула или Cas9 полипептид может содержать REC2 делецию и REC1SUB делецию.
В общем, делеция будет содержать, по меньшей мере, 10% аминокислот в когнатном домене, например, REC2 делеция будет включать, по меньшей мере, 10% аминокислот REC2 домене. Делеция может содержать: по меньшей мере, 10, 20, 30, 40, 50, 60, 70, 80 или 90% аминокислотных остатков ее когнатного домена; все аминокислотные остатки ее когнатного домена; аминокислотный остаток вне ее когнатного домена; множество аминокислотных остатков вне ее когнатного домена; аминокислотный остаток непосредственно N-концевой к ее когнатному домену; аминокислотный остаток непосредственно C-концевой ее когнатному домену; аминокислотный остаток, непосредственно N-концевой ее когнатному, и аминокислотный остаток, непосредственно C-концевой ее когнатному домену; множество, например, вплоть до 5, 10, 15 или 20 аминокислотных остатков N-концевых к ее когнатному домену; множество, например, вплоть до 5, 10, 15 или 20 аминокислотных остатков C-концевых к ее когнатному домену; множество, например, вплоть до 5, 10, 15 или 20 аминокислотных остатков N концевых к ее когнатному домену и множество, например, вплоть до 5, 10, 15 или 20 аминокислотных остатков C концевых к ее когнатному домену.
В некоторых вариантах, делеция не выходит за рамки: ее когнатного домена; N-концевого аминокислотного остатка ее когнатного домена; C-концевого аминокислотного остатка ее когнатного домена.
REC-оптимизированная Cas9 молекула или REC-оптимизированный Cas9 полипептид может включать линкер, расположенный между аминокислотными остатками, которые окружают делецию. Подходящие линкеры для применения между аминокислотными остатками, которые окружают REC делецию в REC-оптимизированной Cas9 молекуле, описаны здесь.
В некоторых вариантах, REC-оптимизированная Cas9 молекула или REC-оптимизированный Cas9 полипептид содержит аминокислотную последовательность, кроме любых REC делеций и связанного линкера, имеющую, по меньшей мере, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 99 или 100% гомологию с аминокислотной последовательностью существующей в природе Cas9, например, S. aureus Cas9 молекулы, S. pyogenes Cas9 молекулы или C. jejuni Cas9 молекулы.
В некоторых вариантах, REC-оптимизированная Cas9 молекула или REC-оптимизированный Cas9 полипептид содержит аминокислотную последовательность, которая, кроме любых REC делеций и связанного линкера, отличается не более чем 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20 или 25 аминокислотными остатками из аминокислотной последовательности существующей в природе Cas9, например, S. aureus Cas9 молекулы, S. pyogenes Cas9 молекулы или C. jejuni Cas9 молекулы.
В некоторых вариантах, REC-оптимизированная Cas9 молекула или REC-оптимизированный Cas9 полипептид содержит аминокислотную последовательность, которая, кроме любой REC делеции и связанного линкера, отличается не более чем на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20 или 25% аминокислотных остатков от аминокислотной последовательности существующей в природе Cas9, например, S. aureus Cas9 молекулы, S. pyogenes Cas9 молекулы или C. jejuni Cas9 молекулы.
Для сравнения последовательностей обычно одна последовательность является ссылочной последовательностью, с которой сравнивают тестируемые последовательности. При применении алгоритма сравнения последовательностей, тестируемую и ссылочную последовательность вводят в компьютер, обозначают координаты подпоследовательности, при необходимости, и обозначают параметры программы алгоритма последовательности. Могут применяться параметры программы «по умолчанию» или могут быть обозначены альтернативные параметры. Алгоритм сравнения последовательности затем рассчитывает процент идентичности последовательности для тестируемых последовательностей относительно ссылочной последовательности, на основе параметров программы. Способы выравнивания последовательностей для сравнения хорошо известны. Оптимальное выравнивание последовательностей для сравнения может быть проведено, например, алгоритмом локальной гомологии из Smith and Waterman, (1970) Adv. Appl. Math. 2:482c, алгоритмом выравнивания гомологии Needleman and Wunsch, (1970) J. Mol. Biol. 48:443, поиском для метода подобия из Pearson and Lipman, (1988) Proc. Nat’l. Acad. Sci. USA 85:2444, компьютеризированными вариантами этих алгоритмов (GAP, BESTFIT, FASTA, и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI) или ручным выравниванием и визуальной проверкой (см., например, Brent et al., (2003) Current Protocols in Molecular Biology).
Два примера алгоритмов, которые подходят для определения процента идентичности последовательностей и сходства последовательностей, включают алгоритмы BLAST и BLAST 2,0, которые описаны в Altschul et al., (1977) Nuc. Acids Res. 25:3389-3402; и Altschul et al., (1990) J. Mol. Biol. 215:403-410, соответственно. Программа для проведения анализов BLAST общедоступна через National Center for Biotechnology Information.
Процент идентичности между двумя аминокислотными последовательностями также может быть определен с применением алгоритма из E. Meyers and W. Miller, (1988) Comput. Appl. Biosci. 4:11-17), который включен в программу ALIGN (версия 2,0), с применением таблицы весов замен остатков PAM120, штрафа за удлинение гэпа 12 и штрафа за гэп 4. Кроме того, процент идентичности между двумя аминокислотными последовательностями может быть определен с применением алгоритма Needleman and Wunsch (1970) J. Mol. Biol. 48:444-453), который включен в программу GAP в программном пакете GCG (доступном на www.gcg.com), с применением либо матрицы Blossom 62, либо матрицы PAM250, и штрафа за открытие гэпа 16, 14, 12, 10, 8, 6 или 4 и штрафа за продолжение гэпа 1, 2, 3, 4, 5 или 6.
Информация о последовательностях для типовых REC делеций представлена для 83 существующих в природе Cas9 ортологов, описанных в, например, международных публикациях PCT №№ WO2015/161276, WO2017/193107 и WO2017/093969.
Q) Нуклеиновые кислоты, кодирующие Cas9 молекулы
Нуклеиновые кислоты, кодирующие Cas9 молекулы или Cas9 полипептиды, например, eaCas9 молекулу или eaCas9 полипептид, могут применяться в связи с любым из вариантов, представленных здесь.
Типовые нуклеиновые кислоты, кодирующие Cas9 молекулы или Cas9 полипептиды, описаны в Cong et al., Science 2013, 399(6121):819-823; Wang et al., Cell 2013, 153(4):910-918; Mali et al., Science 2013, 399(6121):823-826; Jinek et al., Science 2012, 337(6096):816-821, и WO2015/161276, например, на ФИГ. 8 здесь.
В некоторых вариантах, нуклеиновой кислотой, кодирующей Cas9 молекулу или Cas9 полипептид, может быть синтетическая последовательность нуклеиновых кислот. Например, синтетическая молекула нуклеиновой кислоты может быть химически модифицирована. В некоторых вариантах, Cas9 мРНК имеет одно или более (например, все) из следующих свойств: она кэпирована, полиаденилирована, замещена 5-метилцитидином и/или псевдоуридином.
В дополнение или альтернативно, синтетическая последовательность нуклеиновых кислот может быть оптимизирована кодоном, например, по меньшей мере, один не обычный кодон или менее обычный кодон может быть замещен обычным кодоном. Например, синтетическая нуклеиновая кислота может направлять синтез оптимизированного мессенджера мРНК, например, оптимизированного для экспрессии в системе экспрессии млекопитающего, например, описанной здесь.
В дополнение или альтернативно, нуклеиновая кислота, кодирующая Cas9 молекулу или Cas9 полипептид, может содержать последовательность ядерной локализации (ПЯЛ). Последовательность ядерной локализации известна.
Если любая из Cas9 последовательностей слита с пептидом или полипептидом на C-окончании, понятно, что стоп-кодон будет удален.
R) Другие Cas молекулы и Cas полипептиды
Разные типы Cas молекул или Cas полипептидов могут применяться для практики изобретений, раскрытых здесь. В некоторых вариантах, могут применяться Cas молекулы II типа Cas систем. В других вариантах, применяют Cas молекулы других Cas систем. Например, могут применяться Cas молекулы I типа или III типа. Типовые Cas молекулы (и Cas системы) описаны, например, в Haft et al., PLoS Computational Biology 2005, 1(6): e60 и Makarova et al., Nature Review Microbiology 2011, 9:467-477, содержание обеих ссылок включено сюда в качестве ссылки полностью. Типовые Cas молекулы (и Cas системы) также показаны в таблице 6.
Таблица 6. Cas системы
• Подтип III-B
3) Cpf1
В некоторых вариантах, направляющая РНК или нРНК способствует специфической ассоциации целенаправленного воздействия РНК-направляемой нуклеазы, такой как Cas9 или Cpf1, на целевую последовательность, такую как геномная или эписомная последовательность в клетке. В общем, нРНК могут быть мономолекулярными (содержащими одну РНК молекулу, и обозначаемыми альтернативно химерными) или модулярными (содержащими более одной, обычно две, отдельные РНК молекулы, такие как crРНК и tracrРНК, которые обычно связаны друг с другом, например, дуплексированием). нРНК и их части описаны в литературе, например, в Briner et al. (Molecular Cell 56(2), 333-339, October 23, 2014 (Briner), которая включена сюда в качестве ссылки), и в Cotta-Ramusino.
Направляющие РНК, мономолекулярные или модулярные, обычно включают домен целенаправленного воздействия, который полностью или частично комплементарен целевому, и обычно имеет 10-30 нуклеотидов в длину, и в определенных вариантах, имеет 16-24 нуклеотидов в длину (например, 16, 17, 18, 19, 20, 21, 22, 23 или 24 нуклеотидов в длину). В некоторых аспектах, домены целенаправленного воздействия находятся на или рядом с 5’ окончанием нРНК в случае Cas9 нРНК, и на или рядом с 3’ окончанием в случае Cpf1 нРНК. Хотя предшествующее описание сфокусировано на нРНК для применения с Cas9, должно быть понятно, что другие РНК-направляемые нуклеазы были (или могут быть) обнаружены или изобретены, которые применяют нРНК, которые отличаются некоторым образом от тех, которые описаны до настоящего момента. Например, Cpf1 (“CRISPR от Prevotella and Franciscella 1”) является недавно обнаруженной РНК-направляемой нуклеазой, которая не требует tracrРНК для функционирования. (Zetsche et al., 2015, Cell 163, 759-771 October 22, 2015 (Zetsche I), включенная сюда в качестве ссылки). нРНК для применения в системе редактирования Cpf1 генома обычно включает домен целенаправленного воздействия и домен комплементарности (альтернативно названный “ручка”). Также необходимо отметить, что в нРНК для применения с Cpf1, домен целенаправленного воздействия обычно присутствует на или рядом с 3’ окончанием, а не с 5’ окончанием, как описано выше для Cas9 нРНК (ручка находится рядом с 5’ окончанием Cpf1 нРНК).
Хотя структурные различия могут существовать между нРНК из разных прокариотных видов или между Cpf1 и Cas9 нРНК, принципы работы нРНК обычно согласуются. Из-за согласованности их работы, нРНК могут быть определены, в широком смысле, их последовательностями домена целенаправленного воздействия, и специалисты в данной области техники поймут, что данная последовательность домена целенаправленного воздействия может быть введена в любую подходящую нРНК, включая мономолекулярную или химерную нРНК или нРНК, которая включает одну или более химических модификаций и/или модификаций последовательности (замещения, дополнительные нуклеотиды, усечения и т.д.). Таким образом, в некоторых аспектах данного раскрытия, нРНК могут быть описаны исключительно с точки зрения их последовательностей домена целенаправленного воздействия.
Более обще, некоторые аспекты данного раскрытия относятся к системам, способам и композициям, которые могут быть использованы с применением множества РНК-направляемых нуклеаз. Если не указано иначе, термин нРНК должен пониматься как охватывающий любые подходящие нРНК, которые могут применяться с любой РНК-направляемой нуклеазой, в не только такие нРНК, которые совместимы с конкретными видами Cas9 или Cpf1. В качестве иллюстрации, термин нРНК может, в определенных вариантах, включать нРНК для применения с любой РНК-направляемой нуклеазой, существующей в классе 2 CRISPR системы, такой как CRISPR система II типа или V типа, или РНК-направляемой нуклеазой, полученной или адаптированной из нее.
Определенные типовые модификации, обсуждаемые в этом разделе, могут быть включены в любое положение в нРНК последовательности включая, без ограничений, от около 5’ окончания (например, в пределах 1-10, 1-5 или 1-2 нуклеотидов от 5’ окончания) и/или от около 3’ окончания (например, в пределах 1-10, 1-5 или 1-2 нуклеотидов от 3’ окончания). В некоторых случаях, модификации расположены в функциональных мотивах, таких как дуплекс повтор-анти-повтор Cas9 нРНК, структура «петля-на-стебле» Cas9 или Cpf1 нРНК и/или a домен целенаправленного воздействия нРНК.
РНК-направляемые нуклеазы включают, но не ограничены ими, существующие в природе нуклеазы класса 2 CRISPR, такие как Cas9, и Cpf1, а также другие нуклеазы, производные или полученные из них. В функциональном плане, РНК-направляемые нуклеазы определены как такие нуклеазы, которые: (a) взаимодействуют с (например, образуют комплекс с) нРНК; и (b) вместе с нРНК, связанной с ними, и необязательно расщепляют или модифицируют целевую область ДНК, которая включает (i) последовательность, комплементарную домену целенаправленного воздействия нРНК и, необязательно, (ii) дополнительную последовательность, названную “мотив, прилегающий к протоспейсеру” или “PAM”, который более подробно описан ниже. Как будет показано в следующих примерах, РНК-направляемые нуклеазы могут быть определены, в широком смысле, их PAM специфичностью и расщепляющей активностью, даже если вариации могут существовать между отдельными РНК-направляемыми нуклеазами, которые имеют общую PAM специфичность или расщепляющую активность. Специалисты в данной области техники поймут, что некоторые аспекты данного раскрытия относятся к системам, способам и композициям, которые могут применяться с применением любых подходящих РНК-направляемых нуклеаз, имеющих определенную PAM специфичность и/или расщепляющую активность. По этой причине, если не указано иначе, термин РНК-направляемая нуклеаза должен пониматься как общий термин, и не ограничиваться каким-либо конкретным типом (например, Cas9 к Cpf1), видами (например, S. pyogenes к S. aureus) или вариацией (например, полноразмерная к усеченной или расщепленной; существующая в природе специфичность к PAM к сконструированной специфичности к PAM, и т.д.) РНК-направляемой нуклеазы.
В дополнение к распознающим специфическим последовательностным ориентациям PAM и протоспейсеров, РНК-направляемые нуклеазы в некоторых вариантах также могут распознавать специфические PAM последовательности. S. aureus Cas9, например, обычно распознает PAM последовательность NNGRRT или NNGRRV, где N остатки находятся сразу же после 3’ области, распознанной нРНК доменом целенаправленного воздействия. S. pyogenes Cas9 обычно распознает NGG PAM последовательности. И F. novicida Cpf1 обычно распознает TTN PAM последовательность.
Кристаллическая структура Acidaminococcus sp. Cpf1 в комплексе с crРНК и двухнитевой (дн) ДНК целью, включая TTTN PAM последовательность, была решена Yamano et al. (Cell. 2016 May 5; 165(4): 949-962 (Yamano), включенной сюда в качестве ссылки). Cpf1, как и Cas9, имеет две доли: REC (распознающую) долю и NUC (нуклеазную) долю. REC доля включает REC1 и REC2 домены, которые не похожи ни на какие известные белковые структуры. NUC доля, тем временем, включает три RuvC домена (RuvC-I, -II и -III) и BH домен. Однако, в отличие от Cas9, Cpf1 REC доля не имеет HNH домена и включает другие домены, которые также не имеют сходства с известными белковыми структурами: структурно уникальный PI домен, три Wedge (WED) домена (WED-I, -II и -III) и нуклеазный (Nuc) домен.
Хотя Cas9 и Cpf1 имеют сходство в структуре и функционировании, должно быть понятно, что определенные активности Cpf1 медиированы структурными доменами, которые не аналогичны каким-либо Cas9 доменам. Например, расщепление комплементарной нити целевой ДНК вероятно медиируется Nuc доменом, который отличается последовательностно и пространственно от HNH домена Cas9. Дополнительно, часть не целенаправленного воздействия Cpf1 нРНК (ручка) принимает структуру псевдоузла, а не структуру «петля на стебле», образованную дуплексом повтор:антиповтор в Cas9 нРНК.
Нуклеиновые кислоты, кодирующие РНК-направляемые нуклеазы, например, Cas9, Cpf1 или их функциональные фрагменты, представлены здесь. Типовые нуклеиновые кислоты, кодирующие РНК-направляемые нуклеазы, описаны ранее (см., например, Cong 2013; Wang 2013; Mali 2013; Jinek 2012).
3. Доставка агентов для генетического разрушения
В некоторых вариантах, направленное генетическое разрушение, например, разрыв ДНК, эндогенных генов, кодирующих TCR, такую, как TRAC и TRBC1 или TRBC2 у человека, проводят доставкой или введением одного или более агентов, способных вызывать генетическое разрушение, например, Cas9 и/или нРНК компонентов, в клетку с применением любого количество известных способов доставки или носителей для введения или переноса в клетки, например, с применением лентивирусных векторов доставки или любых известных способов или носителей для доставки Cas9 молекул и нРНК. Типовые способы описаны в, например, Wang et al. (2012) J. Immunother. 35(9): 689-701; Cooper et al. (2003) Blood. 101:1637-1644; Verhoeyen et al. (2009) Methods Mol Biol. 506: 97-114; и Cavalieri et al. (2003) Blood. 102(2): 497-505. В некоторых вариантах, последовательности нуклеиновых кислот, кодирующие один или более компонентов одного или более агентов, способных вызывать генетическое разрушение, например, разрыв ДНК, вводят в клетки, например, любыми способами для введения нуклеиновых кислот в клетку, описанными здесь или известными. В некоторых вариантах, вектор, кодирующий компоненты одного или более агентов, способных вызывать генетическое разрушение, такой как CRISPR направляющая РНК и/или Cas9 фермент, может быть доставлен в клетку.
В некоторых вариантах, один или более агентов, способных вызывать генетическое разрушение, например, один или более агенты, которые являются Cas9/нРНК, вводят в клетку в виде рибонуклеопротеинового (RNP) комплекса. RNP комплексы включают последовательность рибонуклеотидов, таких как РНК или нРНК молекула, и белок, такой как Cas9 белок или его вариант. Например, Cas9 белок доставляют в виде RNP комплекса, который содержит Cas9 белок и нРНК молекулу целенаправленного воздействия на целевую последовательность, например, с применением электропорации или другого физического способа. В некоторых вариантах, RNP доставляют в клетку через электропорацию или другие физические средства, например, генную пушку, трансфекцию фосфата кальция, сгущение или сжатие клеток. В некоторых вариантах, RNP может проходить через плазматическую мембрану клетки без необходимости в дополнительных агентах доставки (например, низкомолекулярных агентов, жиров и т.д.). В некоторых вариантах, доставка одного или более агентов, способных вызывать генетическое разрушение, например, CRISPR/Cas9, в виде RNP имеет преимущество в том, что целевое разрушение происходит временно, например, в клетках, в которые введен RNP, без размножения агента до клеточных потомков. Например, доставка RNP минимизирует передачу агента его потомкам, тем самым снижая шанс нецелевого генетического разрушения в потомках. В таких случаях, генетическое разрушение и интеграцию трансгена (обсуждаемые далее в разделе I.B) могут наследоваться клетками потомками, но без самого агента, что может дальше вызвать нецелевые генетические разрушения, переданные клеткам наследникам.
Агенты и компоненты, способные вызывать генетическое разрушение, например, Cas9 молекула и нРНК молекула, могут быть введены в клетки во множестве форм с применением множества способов доставки и составов, как представлено в таблицах 7 и 8 или способах, описанных в, например, WO 2015/161276; US 2015/0056705, US 2016/0272999, US 2017/0211075; или US 2017/0016027. Как описано далее, способы доставки и составы могут применяться для доставки матричных полинуклеотидов и/или других агентов в клетки на предыдущих или последующих стадиях описанных здесь способов.
Таблица 7. Типовые способы доставки
Таблица 8. Сравнение типовых способов доставки
В некоторых вариантах, ДНК кодирующие Cas9 молекулы и/или нРНК молекулы или RNP комплексы, содержащие Cas9 молекулу и/или нРНК молекулы, могут быть доставлены в клетки известными способами или как описано здесь. Например, Cas9-кодирующие и/или нРНК-кодирующие ДНК могут быть доставлены, например, векторами (например, вирусными или не вирусными векторами), не векторными способами (например, с применением голых ДНК или ДНК комплексов) или их сочетаний. В некоторых вариантах, полинуклеотид, содержащий агенты и/или их компоненты, доставляют вектором (например, вирусным вектором/вирусом или плазмидой). Вектор может быть любым, описанным здесь.
В некоторых аспектах, CRISPR фермент (например, Cas9 нуклеаза) в сочетании с (и, необязательно, в комплексе с) направляющей последовательностью доставляют в клетку. Например, один или более элементов CRISPR системы получают из типа I, типа II или типа III CRISPR системы. Например, один или более элементов CRISPR системы получают из конкретного организма, содержащего эндогенную CRISPR систему, такого как Streptococcus pyogenes, Staphylococcus aureus или Neisseria meningitides.
В некоторых вариантах, Cas9 нуклеазу (например, которая кодирована мРНК из Staphylococcus aureus или из Streptococcus pyogenes, например, pCW-Cas9, Addgene #50661, Wang et al. (2014) Science, 3:343-80-4; или нуклеазные или никазные лентивирусные векторы, доступные от Applied Biological Materials (ABM; Canada) под кат. №№ K002, K003, K005 или K006) и направляющую РНК, специфическую к целевому гену (например, TRAC, TRBC1 и/или TRBC2 у человека) вводят в клетки. В некоторых вариантах, нРНК последовательности которые являются или содержат домен целенаправленного воздействия последовательности целенаправленного воздействия на целевой сайт в конкретном гене, таком как TRAC, TRBC1 и/или TRBC2 гены, обозначают или идентифицируют. Полногеномная база данных нРНК для редактирования CRISPR генома общедоступна, она содержит типовые последовательности одинарных направляющих РНК (онРНК) для целенаправленного воздействия на конститутивные экзоны генов в человеческом геноме или мышином геноме (см., например, genescript.com/gRNA-database.html; см. также Sanjana et al. (2014) Nat. Methods, 11:783-4). В некоторых аспектах, нРНК последовательность является или содержит последовательность с минимальным нецелевым связыванием с нецелевым сайтом или положением.
В некоторых вариантах, полинуклеотид, содержащий агенты и/или их компоненты, или RNP комплекс доставляют не векторным способом (например, с применением голых ДНК или ДНК комплексов). Например, ДНК или РНК или белки или их сочетание, например, рибонуклеопротеиновые (RNP) комплексы, могут быть доставлены, например, органически модифицированным диоксидом кремния или силикатом (Ormosil), электропорацией, временным сгущением или сжатием клеток (например, как описано у Lee, et al. (2012) Nano Lett 12: 6322-27, Kollmannsperger et al. (2016) Nat Comm 7, 10372 doi:10,1038/ncomms10372).), генной пушки, сонопорацией, магнитофекцией, медиированной жиром трансфекцией, дендримерами, неорганическими наночастицами, фосфатами кальция или их сочетанием.
В некоторых вариантах, доставка электропорацией включает смешивание клеток с Cas9- и/или нРНК-кодирующей ДНК или RNP комплексом в картридже, камере или кювете, и применение одного или более электрических импульсов определенной длительности и амплитуды. В некоторых вариантах, доставку электропорацией проводят с применением системы, в которой клетки смешивают с Cas9- и/или нРНК-кодирующей ДНК в сосуде, соединенном с устройством (например, помпой), которая закачивает смесь в картридж, камеру или кювету, где применяют один или более электрических импульсов определенной длительности и амплитуды, после чего клетки доставляют во второй сосуд.
В некоторых вариантах, средством доставки является не вирусный вектор. В некоторых вариантах, не вирусным вектором является неорганическая наночастица. Типовые неорганические наночастицы включают, например, магнитные наночастицы (например, Fe3MnO2) и диоксид кремния. Внешняя поверхность наночастицы может быть конъюгирована с положительно заряженным полимером (например, полиэтиленимином, полилизином, полисерином), который позволяет присоединение (например, конъюгирование или улавливание) нагрузки. В некоторых вариантах, не вирусным вектором является органическая наночастица. типовые органические наночастицы включают, например, SNALP липосомы, которые содержат катионные жиры вместе с нейтральными хелперными жирами, которые покрыты полиэтиленгликолем (ПЭГ), и протамин-гуклеиновыми кислотными комплексами, покрытыми жиром. Типовые жиры и/или полимеры известны и могут применяться в представленных вариантах.
В некоторых вариантах, носитель имеет целевые модификации для повышения целевого клеточного обновления наночастиц и липосомов, например, клеточноспецифических антигенов, моноклональных антител, одноцепочечных антител, аптамеров, полимеров, сахаров и проникающих пептидов (например, описанных в US 2016/0272999). В некоторых вариантах, в носителе применяют фузогенные и эндосом-дестабилизирующие пептиды/полимеры. В некоторых вариантах, носитель переносит стимулированные кислотой конформационные изменения (например, для усиления эндосомального высвобождения карго). В некоторых вариантах, применяют расщепляемый стимулом полимер, например, для выделения в клеточный компартмент. Например, могут применяться катионные полимеры на основе дисульфида, которые расщепляют в восстанавливающей клеточной среде.
В некоторых вариантах, средством доставки является биологическое не вирусное средство доставки. В некоторых вариантах, средством является ослабленная бактерия (например, природная или искусственно сконструированная для того, чтобы быть инвазивной, но ослабленной для предотвращения патогенеза и экспрессирования трансгена (например, Listeria monocytogenes, определенные штаммы Salmonella, Bifidobacterium longum, и модифицированная Escherichia coli), бактерия, имеющая питательный и ткань-специфический тропизм к целевым специфическим клеткам, бактерия, имеющая модифицированные поверхностные белки для изменения специфичности целевой клетки). В некоторых вариантах, носителем является генетически модифицированный бактериофаг (например, сконструированные фаги, имеющие большую упаковочную способность, меньшую иммуногенность, содержащие поддерживающие последовательности плазмиды млекопитающих и в которые введены лиганды целенаправленного воздействия). В некоторых вариантах, носителем является вирусоподобная частица млекопитающего. Например, модифицированные вирусные частицы могут быть созданы (например, очисткой “пустых” частиц после ex vivo сборки вируса с желаемыми карго). Носитель также может быть сконструирован для введения лигандов целенаправленного воздействия для изменения целевой специфичности к ткани. В некоторых вариантах, носителем является биологический липосом. Например, биологическим липосомом является частица на основе фосфолипида, полученная из человеческой клетки, например, тени эритроцита, которая является красной кровяной клеткой, разбитой на сферические структуры, полученные у субъекта (например, целенаправленное воздействие на ткань может быть достигнуто присоединением разных лигандов, специфических к ткани или клетке) или секреторные экзосомы - полученные у субъекта связанные с мембраной нановезикулы (30-100 нм) эндоцитарного происхождения (например, которые могут быть получены из различных типов клеток и поэтому могут быть приняты клетками без необходимости лигандов целенаправленного воздействия).
В некоторых вариантах, РНК, кодирующие Cas9 молекулы и/или нРНК молекулы, могут быть доставлены в клетки, например, целевые клетки, описанные здесь, известными способами или как описано здесь. Например, Cas9-кодирующая и/или нРНК-кодирующая РНК может быть доставлена, например, микроинъекцией, электропорацией, временным сгущением или сжатием клеток (например, как описано у Lee, et al.(2012) Nano Lett 12: 6322-27), медиированной жиром трансфекцией, медиированной пептидом доставкой или их сочетанием.
В некоторых вариантах, доставка электропорацией включает смешивание клеток с РНК, кодирующей Cas9 молекулы и/или нРНК молекулы в картридж, камеру или кювету и применение одного или более электрических импульсов определенной длительности и амплитуды. В некоторых вариантах, доставку электропорацией проводят с применением системы, в которой клетки смешивают с РНК, кодирующей Cas9 молекулы и/или нРНК молекулы в сосуде, соединенном с устройством (например, помпой), которая закачивает смесь в картридж, камеру или кювету, где применяют один или более электрических импульсов определенной длительности и амплитуды, после чего клетки доставляют во второй сосуд.
В некоторых вариантах, Cas9 молекулы могут быть доставлены в клетки известными методами или как описано здесь. Например, молекулы Cas9 белка могут быть доставлены, например, микроинъекцией, электропорацией, временным сгущением или сжатием клеток (например, как описано у Lee, et al. (2012) Nano Lett 12: 6322-27), медиированной жиром трансфекцией, медиированной пептидом доставкой или их сочетанием. Доставка может сопровождаться ДНК, кодирующей нРНК или нРНК.
В некоторых вариантах, доставка электропорацией включает смешивание клетки с Cas9 молекулами с или без нРНК молекул в картридже, камере или кювете, с применением одного или более электрических импульсов определенной длительности и амплитуды. В некоторых вариантах, доставку электропорацией проводят с применением системы, в которой клетки смешивают с Cas9 молекулами с или без нРНК молекул в сосуде, соединенном с устройством (например, помпой), которая закачивает смесь в картридж, камеру или кювету, где применяют один или более электрических импульсов определенной длительности и амплитуды, после чего клетки доставляют во второй сосуд.
В некоторых вариантах, полинуклеотид, содержащий агенты и/или их компоненты доставляют сочетанием векторных и не-векторных способов. Например, виросом содержит липосом, объединенный с инактивированным вирусом (например, вирусом ВИЧ или гриппа), который может обеспечить более эффективный перенос гена, чем либо вирусный, либо липосомный способ по отдельности.
В некоторых вариантах, более одного агента или их компонентов доставляют в клетку. Например, в некоторых вариантах, агенты, способные вызывать генетическое разрушение двух или более локаций в геноме, TRAC, TRBC1 и/или TRBC2 локусов, доставляют в клетку. В некоторых вариантах, агенты и их компоненты доставляют с применением одного способа. Например, в некоторых вариантах, агенты, вызывающие генетическое разрушение TRAC, TRBC1 и/или TRBC2 локусов, доставляют в виде полинуклеотидов, кодирующих компоненты для генетического разрушения. В некоторых вариантах, один полинуклеотид может кодировать агенты, которые направлены на TRAC, TRBC1 и/или TRBC2 локусы. В некоторых вариантах, два или более разных нуклеотида могут кодировать агенты, которые направлены на TRAC, TRBC1 и/или TRBC2 локусы. В некоторых вариантах, агенты, способные вызывать генетическое разрушение, могут быть доставлены в виде рибонуклеопротеиновых (RNP) комплексов, и два или более разных RNP комплексов могут быть доставлены вместе в виде смеси или отдельно.
В некоторых вариантах, доставляют одну или более молекул нуклеиновой кислоты, отличных от одного или более агентов, способных вызывать генетическое разрушение, и/или их компонентов, например, компонента Cas9 молекулы и/или компонента нРНК молекулы, таких как матричный полинуклеотид для HDR-направленной интеграции (например, описанной в разделе I.B. здесь). В некоторых вариантах, молекулу нуклеиновой кислоты, например, матричный полинуклеотид, доставляют в то же время, что и один или более компонентов Cas системы. В некоторых вариантах, молекулу нуклеиновой кислоты доставляют до или после (например, менее около 1 минуты, 5 минут, 10 минут, 15 минут, 30 минут, 1 часа, 2 часов, 3 часов, 6 часов, 9 часов, 12 часов, 1 дня, 2 дней, 3 дней, 1 недели, 2 недель или 4 недель) доставки одного или более компонентов Cas системы. В некоторых вариантах, молекулу нуклеиновой кислоты, например, матричный полинуклеотид, доставляют средствами, отличающимися от одного или более компонентов Cas системы, например, компонента Cas9 молекулы и/или компонента нРНК молекулы. Молекула нуклеиновой кислоты, например, матричный полинуклеотид, может быть доставлена любым из способов доставки, описанных здесь. Например, молекула нуклеиновой кислоты, например, матричный полинуклеотид, может быть доставлена вирусным вектором, например, ретровирусом или лентивирусом, и компонент Cas9 молекулы и/или компонент нРНК молекулы может быть доставлен электропорацией. В некоторых вариантах, молекула нуклеиновой кислоты, например, матричный полинуклеотид, включает один или более трансгенов, например, трансгенов, которые кодируют рекомбинантный TCR, рекомбинантный CAR и/или другие генные продукты.
В. Направленная интеграция через репарацию, направляемую гомологией (HDR)
В некоторых из вариантов, представленных здесь, репарация, направляемая гомологией (HDR), может применяться для направленной интеграции определенной части матричного полинуклеотида, содержащего трансген, например, последовательность нуклеиновых кислот, кодирующую рекомбинантный рецептор, в определенную локацию в геноме, например, TRAC, TRBC1 и/или TRBC2 локусе. В некоторых вариантах, присутствие генетического разрушения (например, разрыва ДНК, такого как описан в разделе I.A) и матричного полинуклеотида, содержащего одно или более плеч гомологии (например, содержащего последовательности нуклеиновых кислот, гомологичные последовательностям, окружающим генетическое разрушение) может вызвать или направить HDR, с гомологичными последовательностями, действующими как матрица для репарации ДНК. На основе гомологии между последовательностью эндогенного гена, окружающей генетическое разрушение, и 5’ и/или 3’ плечами гомологии, включенными в матричный полинуклеотид, механизм клеточной репарации ДНК может применять матричный полинуклеотид для репарации разрыва ДНК и пересинтезирования генетической информации в сайте генетического разрушения, тем самым эффективно вставляя или интегрируя трансгенные последовательности в матричном полинуклеотиде на или рядом с сайтом генетического разрушения. В некоторых вариантах, генетическое разрушение, например, TRAC, TRBC1 и/или TRBC2 локус, может быть создано любыми способами для получения направленного генетического разрушения, описанного здесь.
Также представлены полинуклеотиды, например, матричные полинуклеотиды, описанные здесь. В некоторых вариантах, представленные полинуклеотиды могут применяться в способах, описанных здесь, например, включающих HDR, для нацеливания на трансгенные последовательности, кодирующие часть рекомбинантного рецептора, например, рекомбинантного TCR, в локусе эндогенного TRAC, TRBC1 и/или TRBC2.
В некоторых вариантах, матричный полинуклеотид имеет, или содержит полинуклеотид, содержащий трансген (экзогенные или гетерологические последовательности нуклеиновых кислот), кодирующий рекомбинантный рецептор или его часть (например, одну или более цепей, областей или доменов рекомбинантного рецептора), и гомологические последовательности (например, плечи гомологии), которые гомологичны последовательностям на или рядом с эндогенным геномным сайтом, например, на локусе эндогенного TRAC, TRBC1 и/или TRBC2. В некоторых аспектах, матричный полинуклеотид вводят в виде линейного ДНК фрагмента или он содержится в векторе. В некоторых аспектах, стадию вызова генетического разрушения и стадию направленной интеграции (например, введением матричного полинуклеотида) проводят одновременно или последовательно.
1. Репарация, направляемая гомологией (HDR)
В некоторых вариантах, репарация, направляемая гомологией (HDR), может применяться для направленной интеграции или вставки одной или более последовательностей нуклеиновых кислот, например, трансгенных последовательностей, в одном или более целевых сайтах в геноме, например, TRAC, TRBC1 и/или TRBC2 локусе. В некоторых вариантах, вызванная нуклеазой HDR может применяться для изменения целевой последовательности, интеграции трансгена в конкретную целевую локацию и/или для редактирования или репарации мутации в конкретном целевом гене.
Изменение последовательности нуклеиновых кислот на целевом сайт может проводиться HDR с экзогенно полученными матричными полинуклеотидами (также обозначенными как донорный полинуклеотид или матричная последовательность). Например, матричный полинуклеотид обеспечивает изменение целевой последовательности, такое как вставка трансгена, содержащегося в матричном полинуклеотиде. В некоторых вариантах, плазмида или вектор может применяться в качестве матрицы для гомологичной рекомбинации. В некоторых вариантах, линейный ДНК фрагмент может применяться в качестве матрицы для гомологичной рекомбинации. В некоторых вариантах, однонитевой матричный полинуклеотид может применяться в качестве матрицы для изменения целевой последовательности альтернативными способами репарации, направляемой гомологией (например, однонитевым отжигом) между целевой последовательностью и матричным полинуклеотидом. Вызванное матричным полинуклеотидом изменение целевой последовательности зависит от расщепления нуклеазой, например, целевой нуклеазой, такой как CRISPR/Cas9. Расщепление нуклеазой может включать двухнитевой разрыв или два однонитевых разрыва.
В некоторых вариантах, “рекомбинация” относится к процессу обмена генетической информацией между двумя полинуклеотидами. В некоторых вариантах, “гомологичная рекомбинация (ГР)” относится к специализированной форме такого обмена, который происходит, например, во время репарации двухнитевых разрывов в клетках через механизмы репарации, направляемой гомологией. Эти способы требуют гомологии нуклеотидной последовательности, применения матричного полинуклеотида для матричной репарации целевой ДНК (т.е., такой, в которой применяют двухнитевой разрыв, например, целевого сайта в эндогенном гене), и по разному известной как “не кроссоверная конверсия генов” или “короткая трактовая конверсия гена” так как она приводит к переносу генетической информации из матричного полинуклеотида в цель. В некоторых вариантах, такой перенос может включать коррекцию ошибки спаривания гетеродуплексной ДНК, которая образуется между разорванной целью и матричным полинуклеотидом, и/или “синтез-зависимый отжиг нити”, при котором матричный полинуклеотид применяют для пересинтеза генетической информации, которая становится частью цели, и/или родственные способы. Такие специализированные ГР часто дают изменение последовательности целевой молекулы так, что часть или вся последовательность матричного полинуклеотида введена в целевой полинуклеотид.
В некоторых вариантах, матричный полинуклеотид, например, полинуклеотид-содержащий трансген, интегрируют в геном клетки через независимые от гомологии механизмы. Способы включают создание двухнитевого разрыва (DSB) в геноме клетки и расщепление молекулы матричного полинуклеотида с применением нуклеазы так, что матричный полинуклеотид интегрируют в сайт DSB. В некоторых вариантах, матричный полинуклеотид интегрируют не зависимыми от гомологии способами (например, NHEJ). При in vivo расщеплении матричные полинуклеотиды могут быть интегрированы целевой методикой в геном клетки в локации DSB. Матричный полинуклеотид может включать один или более одинаковых целевых сайтов для одной или более нуклеаз, применяемых для создания DSB. Таким образом, матричный полинуклеотид может быть расщеплен одной или более одинаковыми нуклеазами, применяемыми для расщепления эндогенного гена, в который желательна интеграция. В некоторых вариантах, матричный полинуклеотид включает разные целевые сайты нуклеазы из нуклеазы, применяемой для вызова DSB. Как описано здесь, генетическое разрушение целевого сайта или целевого положения может быть создано любыми механизмами, такими как ZFN, TALEN, CRISPR/Cas9 система или TtAgo нуклеазы.
В некоторых вариантах, механизмы репарации ДНК могут быть вызваны нуклеазой после (1) одного двухнитевого разрыва, (2) двух однонитевых разрывов, (3) двух двухнитевых разрывов с разрывом, возникающим на каждой стороне целевого сайта, (4) одного двухнитевого разрыва и двух однонитевых разрывов, где двухнитевой разрыв и два однонитевых разрыва возникают на каждой стороне целевого сайта, (5) четырех однонитевых разрывов с парой однонитевых разрывов, возникающих на каждой стороне целевого сайта или (6) одного однонитевого разрыва. В некоторых вариантах, применяют однонитевой матричный полинуклеотид, и целевой сайт может быть изменен альтернативной HDR.
Вызванное матричным полинуклеотидом изменение целевого сайта зависит от расщепления молекулой нуклеазы. Расщепление нуклеазой может содержать ник, духнитевой разрыв или два однонитевых разрыва, например, на каждой нити ДНК в целевом сайте. После введения разрывов на целевом сайте, резекция возникает на разорванных концах в однонитевых выступающих ДНК областях.
В канонической HDR, вводят двухнитевой матричный полинуклеотид, содержащий гомологичную последовательность, в целевой сайт, который либо непосредственно вводят в целевой сайт, либо применяют в качестве матрицы для вставки трансгена или коррекции последовательности целевого сайта. После резекции на разрыве, репарация может развиваться двумя разными путями, например, моделью двойной структуры Холлидея (или путем репарации двухнитевого разрыва, РДНР) или путем синтез-зависимого отжига нити (SDSA).
В модели двойной структуры Холлидея происходит встраивание в нить двух однонитевых липких концов целевого сайта в гомологичную последовательность в матричном полинуклеотиде, вызывая образование промежуточного результата с двумя структурами Холлидея. Структуры мигрируют по мере того, как новая ДНК синтезируется из концов встраиваемой нити для заполнения гэпа, получаемого при резекции. Конец свежей синтезированной ДНК лигируют до резецированного конца, и структуры распадаются с получением вставки в целевой сайт, например, вставки трансгена в матричный полинуклеотид. Кроссовер с матричным полинуклеотидом может возникать при распаде структур.
При пути SDSA, только однонитевой липкий конец внедряется в матричный полинуклеотид, и новую ДНК синтезируют от конца внедряемой нити для заполнения гэпа, полученного при резекции. Свежесинтезированную ДНК затем отжигают до оставшегося однонитевого липкого конца, новую ДНК синтезируют для заполнения гэпа, и нити лигируют с получением модифицированного ДНК дуплекса.
В альтернативной HDR, вводят однонитевой матричный полинуклеотид, например, матричный полинуклеотид. Ник, однонитевой разрыв или двухнитевой разрыв на целевом сайте, для изменения желаемого целевого сайта, медиируют молекулой нуклеазы, и резекция на разрыве происходит для раскрытия однонитевых липких концов. Введение последовательности матричного полинуклеотида для коррекции или изменения целевого сайта ДНК обычно происходит путем SDSA, как описано здесь.
“Альтернативная HDR” или альтернативная репарация, направляемая гомологией, в некоторых вариантах, относится к способу репарации ДНК повреждения с применением гомологичной нуклеиновой кислоты (например, эндогенной гомологичной последовательности, например, сестринской хроматиды или экзогенных нуклеиновых кислот, например, матричного полинуклеотида). Альтернативная HDR отличается от канонической HDR тем, что в способе применяют пути, отличающиеся от канонической HDR, и может быть ингибирована медиаторами канонической HDR, RAD51 и BRCA2. также, в альтернативной HDR применяется однонитевая или содержащая разрыв гомологичная нуклеиновая кислота для репарации разрыва. “Каноническая HDR” или каноническая репарация, направляемая гомологией, в некоторых вариантах, относится к способу репарации повреждения ДНК с применением гомологичной нуклеиновой кислоты (например, эндогенной гомологичной последовательности, например, сестринской хроматиды, или экзогенной нуклеиновой кислоты, например, матричной нуклеиновой кислоты). Каноническая HDR обычно действует, если имеется значительная резекция на двухнитевом разрыве, образуются, по меньшей мере, одну однонитевую часть ДНК. В нормальной клетке HDR обычно включает ряд стадий, таких как распознавание разрыва, стабилизация разрыва, резекция, стабилизация однонитевой ДНК, образование промежуточного ДНК кроссовера, разделение промежуточного кроссовера и лигирование. Способ требует RAD51 и BRCA2, и гомологичная нуклеиновая кислота обычно является двухнитевой. Если не указано иначе, термин “HDR” в некоторых вариантах охватывает каноническую HDR и альтернативную HDR.
В некоторых вариантах, двухнитевой расщепление проводят нуклеазой, например, Cas9 молекулой, обладающей расщепляющей активностью, связанной с HNH-подобным доменом, и расщепляющей активностью, связанной с RuvC-подобным доменом, например, N-концевым RuvC-подобным доменом, например, диким типом Cas9. Такие варианты требуют только одной нРНК.
В некоторых вариантах, однонитевой разрыв или ник производится молекулой нуклеазы, имеющей активность никазы, например, Cas9 никазы. ДНК, содержащая разрыв, может быть субстратом для альтернативной HDR.
В некоторых вариантах, двухнитевые разрывы или ники проводятся нуклеазой, например, Cas9 молекулой, имеющей активность никазы, например, расщепляющую активность, связанную с HNH-подобным доменом, или расщепляющую активность, связанную с N-концевым RuvC-подобным доменом. Такие варианты обычно требуют двух нРНК, одну для размещения каждого однонитевого разрыва. В некоторых вариантах, Cas9 молекула, имеющая активность никазы, расщепляет нить, с которой гибридизируется нРНК, но не нить, которая комплементарна с нитью, с которой гибридизируется нРНК. В некоторых вариантах, Cas9 молекула, имеющая активность никазы, не расщепляет нить, с которой гибридизируется нРНК, но скорее расщепляет нить, которая является комплементарной к нити, с которой гибридизируется нРНК. В некоторых вариантах, никаза имеет HNH активность, например, Cas9 молекулу, имеющую инактивированную RuvC активность, например, Cas9 молекулу, имеющую мутацию на D10, например, D10A мутацию. D10A инкативирует RuvC; поэтому Cas9 никаза имеет (только) HNH активность и разрывает нить, с которой гибридизируется нРНК (например, комплементарную нить, которая не имеет NGG PAM на ней). В некоторых вариантах, Cas9 молекула, имеющая H840, например, H840A, мутацию, может применяться в качестве никазы. H840A инкативирует HNH; поэтому Cas9 никаза имеет (только) RuvC активность и разрывает не комплементарную нить (например, нить, которая имеет NGG PAM, и последовательность которой идентична нРНК). В некоторых вариантах, Cas9 молекулой является никаза N-концевого RuvC-подобного домена, например, Cas9 молекула содержит мутацию на N863, например, N863A.
В некоторых вариантах, в которых никазу и две нРНК применяют для расположения двух однонитевых разрывов, один разрыв находится на+нити, и один разрыв находится на - нити целевой ДНК. PAM развернута вовне. нРНК могут быть выбраны так, что нРНК разделяют от около 0-50, 0-100 или 0-200 нуклеотидов. В некоторых вариантах, не существует перекрытия между целевыми последовательностями, которые комплементарны доменам целенаправленного воздействия двух нРНК. В некоторых вариантах, нРНК не перекрываются и разделены не менее 50, 100 или 200 нуклеотидов. В некоторых вариантах, применение двух нРНК может повысить специфичность, например, через снижение нецелевого связывания (Ran et al., Cell 2013).
В некоторых вариантах, одинарный разрыв может применяться для того, чтобы вызвать HDR, например, альтернативной HDR. Здесь рассматривается, что одинарный разрыв может применяться для повышения соотношения HR к NHEJ в данном сайте расщепления, например, целевом сайте. В некоторых вариантах, однонитевой разрыв формируют на нити ДНК на целевом сайте, к которому комплементарен домен целенаправленного воздействия указанной нРНК. В другом варианте, однонитевой разрыв формируют на нити ДНК на целевом сайте, отличной от нити, к которой комплементарен домен целенаправленного воздействия указанной нРНК.
В некоторых вариантах, другие пути репарации ДНК, такие как однонитевой отжиг (SSA), репарация однонитевого разрыва (SSBR), репарация неправильного спаривания (MMR), эксцизионная репарация оснований (BER), эксцизионная репарация нуклеотида (NER), внутринитевое поперечная сшивка (ICL), синтез «через/сквозь» поврежденные участки (TLS), безошибочная пострепликативная репарация (PRR) может применяться клеткой для репарации двухнитевого или однонитевого разрыва, созданного нуклеазой.
2. Размещение генетического разрушения (например, разрыва нити ДНК)
Направленная интеграция дает трансген, интегрируемый в определенный ген или локус в геноме. Трансген может быть интегрирован в любом месте рядом с одним из, по меньшей мере, одного из целевых сайтов или сайта в геноме. В некоторых вариантах, трансген интегрирован в или рядом с одним из, по меньшей мере, одного из целевых сайтов, например, в пределах 300, 250, 200, 150, 100, 50, 10, 5, 4, 3, 2, 1 или меньше пар оснований до или после сайта расщепления, например, в пределах 100, 50, 10, 5, 4, 3, 2, 1 пар оснований с любой стороны целевого сайта, например, в пределах 50, 10, 5, 4, 3, 2, 1 пар оснований с любой стороны целевого сайта. В некоторых вариантах, интегрированная последовательность, содержащая трансген, не включает любые векторы последовательностей (например, вирусные векторы последовательностей). В некоторых вариантах, интегрированная последовательность включает часть векторов последовательностей (например, вирусных векторов последовательностей).
Двухнитевой разрыв или однонитевой разрыв на одной из нитей должен быть достаточно близким к сайту для направленной интеграцией, так, что изменение происходит в желаемой области, например, происходит вставка трансгена или коррекция мутации. В некоторых вариантах, расстояние составляет не более 10, 25, 50, 100, 200, 300, 350, 400 или 500 нуклеотидов. В некоторых вариантах, полагают, что разрыв должен быть достаточно близким к сайту для направленной интеграции так, что разрыв происходит в области, которая подвержена медиированному экзонуклеазой удаления во время конечной резекции. В некоторых вариантах, домен целенаправленного воздействия сконфигурирован так, что агент расщепления, например, двухнитевой или однонитевой разрыв, расположен в пределах 1, 2, 3, 4, 5, 10, 15, 20, 25, 30, 35, 40, 45,50, 60, 70, 80, 90, 100, 150, 200, 300, 350, 400 или 500 нуклеотидов от области, которую желательно изменить, например, сайта для направленной вставки. Разрыв, например, двухнитевой или однонитевой разрыв, может быть расположен до или после области, которую желательно изменить, например, сайта для направленной вставки. В некоторых вариантах, разрыв расположен в области, которую желательно изменить, например, в пределах области, определенной, по меньшей мере, двумя мутантными нуклеотидами. В некоторых вариантах, разрыв расположен непосредственно рядом с областью, которую желательно изменить, например, непосредственно до или после сайта для направленной интеграции.
В некоторых вариантах, однонитевой разрыв сопровождается дополнительным однонитевым разрывом, расположенным на второй нРНК молекуле. Например, домены целенаправленного воздействия сконфигурированы так, что событие расщепления, например, два однонитевых разрыва, расположены в пределах 1, 2, 3, 4, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 150, 200, 300, 350, 400 или 500 нуклеотидов от сайта направленной интеграции. В некоторых вариантах, первая и вторая нРНК молекулы сконфигурированы так, что при направлении Cas9 никазы однонитевой разрыв будет сопровождаться дополнительным однонитевым разрывом, расположенным второй нРНК, достаточно близкой друг к другу, что дает изменение желаемой области. В некоторых вариантах, первая и вторая нРНК молекулы сконфигурированы так, что однонитевой разрыв, расположенный указанной второй нРНК, расположен в пределах 10, 20, 30, 40 или 50 нуклеотидов от разрыва, расположенного указанной первой нРНК молекулой, например, если Cas9 является никаза. В некоторых вариантах, две нРНК молекулы сконфигурированы так, чтобы расположить разрывы в том же положении или в пределах нескольких нуклеотидов друг от друга, на разных нитях, например, существенно имитируя двухнитевой разрыв.
В некоторых вариантах, в которых нРНК (мономолекулярная (или химерная) или модулярная нРНК) и Cas9 нуклеаза вызывает двухнитевой разрыв для целей вызова HDR-медиированной вставки трансгена или коррекции, сайт расщепления находится между 0-200 п.о. (например, 0-175, 0-150, 0-125, 0-100, 0-75, 0-50, 0-25, 25-200, 25-175, 25-150, 25-125, 25-100, 25-75, 25-50, 50-200, 50-175, 50-150, 50-125, 50-100, 50-75, 75-200, 75-175, 75-150, 75-125, 75-100 п.о.) от сайта для направленной интеграции. В некоторых вариантах, сайт расщепления находится между 0-100 п.о. (например, 0-75, 0-50, 0-25, 25-100, 25-75, 25-50, 50-100, 50-75 или 75-100 п.о.) от сайта для направленной интеграции.
В некоторых вариантах, можно способствовать HDR через применение никаз для создания разрыва с липкими концами. В некоторых вариантах, однонитевая природа липких концов может усилить вероятность клетки к репарации разрыва через HDR, в отличие от, например, NHEJ.
Более конкретно, в некоторых вариантах, HDR способствует выбор первой нРНК, которая направляет первую никазу на первый целевой сайт, и второй нРНК, которая направляет вторую никазу на второй целевой сайт, который находится на противоположной нити ДНК от первого целевого сайта и в стороне от первого ника. В некоторых вариантах, домен целенаправленного воздействия нРНК молекулы сконфигурирован так, чтобы располагать событие расщепления достаточно далеко от предварительно выбранного нуклеотида, например, нуклеотида кодирующей области, так, чтобы нуклеотид не изменялся. В некоторых вариантах, домен целенаправленного воздействия нРНК молекулы сконфигурирован так, чтобы располагать событие интронного расщепления достаточно далеко от границы интрон/экзон или существующего в природе сигнала сплайсинга для того, чтобы избежать изменения экзонной последовательности или нежелательных событий сплайсинга. В некоторых вариантах, домен целенаправленного воздействия нРНК молекулы сконфигурирован так, чтобы располагать ранний экзон так, чтобы позволить делецию или нокаут эндогенного гена, и/или позволить интеграцию внутри рамки трансгена на или рядом с, по меньшей мере, одним из целевых сайтов.
В некоторых вариантах, двухнитевой разрыв может сопровождаться дополнительным двухнитевым разрывом, расположенным на второй нРНК молекулой. В некоторых вариантах, двухнитевой разрыв может сопровождаться двумя дополнительными однонитевыми разрывами, расположенными на второй нРНК молекуле и третьей нРНК молекуле.
В некоторых вариантах, две нРНК, например, независимо, мономолекулярная (или химерная) или модулярная нРНК, сконфигурированы так, чтобы расположить двухнитевой разрыв на обеих сторонах сайта для направленной интеграции.
3. Матричные полинуклеотиды
Матричный полинуклеотид, имеющий гомологию с последовательностями рядом с одним или более целевыми сайтами в эндогенной ДНК, может применяться для изменения структуры целевой ДНК, например, направленной вставки трансгена. В некоторых вариантах, матричный полинуклеотид содержит гомологические последовательности (например, плечи гомологии) расположенные вокруг трансгена, например, последовательности нуклеиновых кислот, кодирующей рекомбинантный рецептор, для направленной вставки. В некоторых вариантах, гомологические последовательности направлены на трансген в одном или более локусах TRAC, TRBC1 и/или TRBC2. В некоторых вариантах, матричный полинуклеотид включает дополнительные последовательности (кодирующие или не кодирующие последовательности) между плечами гомологии, такие как регулирующие последовательности, такие как промоторы и/или энхансеры, донорные и/или акцепторные сайты сплайсинга, внутренний участок посадки рибосомы (IRES), последовательности, кодирующие элемент проскока рибосомы (например, 2A пептиды), маркеры и/или SA сайты, и/или один или более дополнительных трансгенов.
Рассматриваемые последовательности в матричном полинуклеотиде могут содержать одну или более последовательностей, кодирующих функциональный полипептид (например, кДНК), с или без промотора.
В некоторых вариантах, трансген, содержащийся в матричном полинуклеотиде, содержит последовательность, кодирующую рецептор поверхности клетки (например, рекомбинантный рецептор) или его цепь, антитело, антиген, фермент, фактор роста, ядерный рецептор, гормон, лимфокин, цитокин, репортер, функциональные фрагменты или функциональные варианты любого из представленных здесь, и сочетание представленных здесь. Трансген может кодировать один или более белков, применяемых в противораковой терапии, например один или более химерных антигенных рецепторов (CAR) и/или рекомбинантный Т-клеточный рецептор (TCR). В некоторых вариантах, трансген может кодировать любые рекомбинантные рецепторы, как описано в разделе IV здесь, или любые их цепи, области и/или домены. В некоторых вариантах, трансген кодирует рекомбинантный Т-клеточный рецептор (TCR) или любые его цепи, области и/или домены.
В определенных вариантах, полинуклеотид, например, матричный полинуклеотид содержит и/или включает трансген, кодирующий весь или часть рекомбинантного рецептора, например, рекомбинантного TCR или его цепи. В конкретных вариантах, трансген направлен на целевые сайты, которые находятся внутри гена, локуса или открытой рамки считывания, которые кодируют эндогенный рецептор, например, эндогенный ген, кодирующий одну или более областей, цепей или частей TCR.
В определенных вариантах, матричный полинуклеотид включает или содержит трансген, часть трансгена и/или нуклеиновую кислоту, кодирующие рекомбинантный рецептор, который является рекомбинантным TCR или его цепью, которая содержит один или более переменных доменов и один или более постоянных доменов. В определенных вариантах, рекомбинантный TCR или его цепь содержит один или более постоянных доменов, которые имеет полную, например, от около 100% идентичность со всем или частью и/или фрагментом постоянного домена эндогенного TCR. В некоторых вариантах, трансген кодирует сечь или часть постоянного домена, например, часть или фрагмент постоянного домена, который полностью или частично идентичен постоянному домену эндогенного TCR. В некоторых вариантах, трансген содержит нуклеотиды с последовательностью, имеющей, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99%, 99,5% или 99,9% идентичность последовательности со всей или частью последовательности нуклеиновых кислот, представленной в SEQ ID NOS: 1, 2 или 3.
В некоторых вариантах, трансген содержит последовательность, кодирующую TCRα и/или TCRβ цепь или ее часть, которая кодон-оптимизирована. В некоторых вариантах, трансген кодирует часть TCRα и/или TCRβ цепи при менее 100% идентичности аминокислотной последовательности с соответствующей частью природной или эндогенной TCRα и/или TCRβ цепи. В некоторых вариантах, кодированная TCRα и/или TCRβ цепь содержит аминокислотную последовательность с, с около или с, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более чем 99% идентичностью но менее 100% идентичностью с соответствующей природной или эндогенной TCRα и/или TCRβ цепью. В конкретных вариантах, трансген кодирует TCRα и/или TCRβ постоянный домен или его часть с менее 100% идентичностью аминокислотной последовательности с соответствующим природным или эндогенным TCRα и/или TCRβ постоянным доменом. В некоторых вариантах, TCRα и/или TCRβ постоянный домен содержит аминокислотную последовательность с, с около или с, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или более чем 99% идентичностью, но менее 100% идентичностью с соответствующей природной или эндогенной TCRα и/или TCRβ цепью.
В определенных вариантах, трансген содержит одну или более модификаций для введения одного или более цистеиновых остатков, которые способны образовывать один или более не природных дисульфидных мостиков между TCRα цепью и TCRβ цепью. В некоторых вариантах, трансген кодирует TCRα цепь или ее часть, содержащую TCRα постоянный домен, содержащий цистеин в положении, соответствующем положению 48 с нумерацией, указанной в SEQ ID NO: 24. В некоторых вариантах, TCRα постоянный домен имеет аминокислотную последовательность, представленную в любой из SEQ ID NOS: 19 или 24, или последовательность аминокислот, которая имеет, имеет около или имеет, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99% идентичность последовательности к ней, содержащую один или более цистеиновых остатков, способных образовывать не природную дисульфидную связь с TCRβ цепью. В некоторых вариантах, трансген кодирует TCRβ цепь или ее часть, содержащую TCRβ постоянный домен, содержащий цистеин в положении, соответствующем положению 57 с нумерацией, указанной в SEQ ID NO: 20. В некоторых вариантах, TCRβ постоянный домен имеет аминокислотную последовательность, представленную в любом из SEQ ID NOS: 20, 21 или 25, или последовательность аминокислот, которая имеет, имеет около или имеет, по меньшей мере, 70%, 75%, 80%, 85% 90%, 95%, 97%, 98%, 99% идентичность последовательности к ней, содержащую один или более цистеиновых остатков, способных образовывать не природную дисульфидную связь с TCRα цепью.
В конкретных вариантах, трансген кодирует TCRα и/или TCRβ цепь и/или постоянные домены TCRα и/или TCRβ цепи, содержащие одну или более модификаций для введения одной или более дисульфидных связей. В некоторых вариантах, трансген кодирует TCRα и/или TCRβ цепь и/или TCRα и/или TCRβ с одной или более модификаций для удаления или предотвращения природной дисульфидной связи, например, между TCRα кодированным трансгеном, и эндогенной TCRβ цепью, или между TCRβ, кодированным трансгеном, и эндогенной TCRα цепью. В некоторых вариантах, один или более природных цистеинов, которые образуют и/или способны образовывать природную межцепочечную дисульфидную связь, замещены другим остатком, например, серином или аланином. В некоторых вариантах, TCRα и/или TCRβ цепь и/или постоянные домены TCRα и/или TCRβ цепи модифицированы для замещения одного или более не цистеиновых остатков на цистеин. В некоторых вариантах, один или более не природных цистеиновых остатков способны образовывать не природные дисульфидные связи, например, между рекомбинантной TCRα и TCRβ цепями, кодированными трансгеном. В некоторых вариантах, цистеин вводят на одном или более остатков Thr48, Thr45, Tyr10, Thr45 и Ser15 со ссылкой на нумерацию TCRα постоянного домена, представленную в SEQ ID NO: 24. В определенных вариантах, цистеины могут быть введены на остатках Ser57, Ser77, Ser17, Asp59 и Glu15 TCRβ цепи со ссылкой на нумерацию TCRβ цепи, представленную в SEQ ID NO: 20. Типовые не природные дисульфидные связи TCR описаны в опубликованной международной заявке РСТ № WO2006/000830, WO2006/037960 и Kuball et al. (2007) Blood, 109:2331-2338. В некоторых вариантах, трансген кодирует часть TCRα цепи и/или TCRα постоянного домена, содержащую одну или более модификаций для введения одной или более дисульфидных связей.
В некоторых вариантах, трансген кодирует всю или часть TCRα цепи и/или TCRα постоянного домена с одной или более модификациями для удаления или предотвращения природной дисульфидной связи, например, между TCRα цепью, кодированной трансгеном, и эндогенной TCRβ цепью. В некоторых вариантах, один или более природных цистеинов, которые образуют и/или способны образовывать природную межцепочечную дисульфидную связь, замещены на другой остаток, например, серин или аланин. В некоторых вариантах, часть TCRα цепи и/или TCRα постоянного домена модифицируют для замены одного или более не цистеиновых остатков на цистеин. В некоторых вариантах, один или более не природных цистеиновых остатков способны образовывать не природные дисульфидные связи, например, с TCRβ цепью, кодированной трансгеном. В некоторых вариантах, трансген кодирует всю или часть TCRβ цепи и/или TCRβ постоянного домена с одной или более модификациями для удаления или предотвращения природной дисульфидной связи, например, между TCRβ цепью, кодированной трансгеном, и эндогенной TCRα цепью. В некоторых вариантах, один или более природных цистеинов, которые образуют и/или способны образовывать природную межцепочечную дисульфидную связь, замещены на другой остаток, например, серин или аланин. В некоторых вариантах, часть TCRβ цепи и/или TCRβ постоянного домена модифицируют для замены одного или более не цистеиновых остатков на цистеин.
В некоторых вариантах, один или более не природных цистеиновых остатков и способны образовывать не природные дисульфидные связи, например, с TCRα цепи, кодированной трансгеном. В некоторых вариантах, один или более разных матричных полинуклеотидов применяют для направленной интеграции трансгена в один или более разных целевых сайтов. Для направленной интеграции в разные целевые сайты, одно или более генетических разрушений (например, разрывы ДНК), создают на одном или более целевых сайтах; и одну или более разных гомологических последовательностей применяют для направленной интеграции трансгена в соответствующий целевой сайт. В некоторых вариантах, трансген, вставляемый в каждый сайт, является одинаковым или по существу одинаковым. В некоторых вариантах, трансген, вставляемый в каждый сайт, является разным. В некоторых вариантах, два или более разных трансгена, кодирующих две или более разных доменов или цепей белка, вставляют на одном или более целевых сайтах. В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует одну цепь рекомбинантного TCR, и второй трансген кодирует другую цепь рекомбинантного TCR. В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует альфа (TCRα) цепь рекомбинантного TCR, и второй трансген кодирует бета (TCRβ) цепь рекомбинантного TCR. В некоторых вариантах, два или более трансгена, кодирующих разные домены рекомбинантных рецепторов, направлены для интеграции в двух или более целевых сайтах. Например, в некоторых вариантах, трансген, кодирующий альфа цепь рекомбинантного TCR, направлен для интеграции в TRAC локус, и трансген, кодирующий бета цепь рекомбинантного TCR, направлен для интеграции в TRBC1 и/или TRBC2 локусы.
В некоторых вариантах, два или более разных матричных полинуклеотидов применяют для нацеливания двух или более трансгенов для интеграции в два или более разных локуса эндогенного гена. В некоторых вариантах, первый матричный полинуклеотид включает трансген, кодирующий рекомбинантный рецептор. В некоторых вариантах, второй матричный полинуклеотид включает один или более вторых трансгенов, например, один или более вторых трансгенов, кодирующих одну или более разных молекулу, полипептидов и/или факторов. Любое описание или характеризация матричного полинуклеотида, представленные здесь, также могут применяться к одному или более вторыми матричными полинуклеотидами.
В некоторых вариантах, один или более вторых трансгенов нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта в TRAC гене. В некоторых вариантах, один или более вторых трансгенов нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевой сайта в TRBC1 или TRBC2 гене. В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевой сайта в TRAC гене, TRBC1 гене или TRBC2 гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами, на который не нацелен трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь.
В некоторых вариантах, молекулой, полипептидом или фактором, кодированными одним или более вторыми трансгенами, является молекула, полипептид, фактор или агент, которые могут обеспечивать костимулирующий сигнал в иммунную клетку, например, T клетку. В некоторых вариантах, молекулой, полипептидом, фактором или агентом, кодированными вторым трансгеном, является дополнительный рецептор, например, дополнительный рекомбинантный рецептор. В некоторых вариантах, дополнительный рецептор может обеспечивать костимулирующий сигнал и/или импульсы или обратно изменять ингибирующий сигнал.
В некоторых вариантах, один или более вторых трансгенов кодирует молекулу, выбранную из костимулирующего лиганда, цитокина, растворимого однонитевого переменного фрагмента (scFv), иммуномодулирующего слитого белка, химерного рецептора переключения (CSR) или корецептора.
В некоторых вариантах, молекулой, полипептидом или фактором, кодированным одним или более вторыми трансгенами является костимулирующий лиганд. Типовые костимулирующие лиганды включают лиганд фактора некроза опухоли (TNF) или лиганд суперсемейства иммуноглобулина (Ig). В некоторых вариантах, типовые TNF лиганды включают 4-1BBL, OX40L, CD70, LIGHT, и CD30L. В некоторых вариантах, типовые лиганды суперсемейства Ig включают CD80 и CD86. В некоторых вариантах, костимулирующий лиганд включает CD3, CD27, CD28, CD83, CD127, 4-1BB, PD-1 или PDIL. В некоторых вариантах, молекулой, полипептидом или фактором, кодированным одним или более вторыми трансгенами является цитокин, такой как IL-2, IL-3, IL-6, IL-11, IL-12, IL7, IL-15, IL-21, гранулоцитарно-моноцитарный колониестимулирующий фактор (GM-CSF), интерферон альфа (IFN-α), интерферон бета (IFN-β) или интерферон гамма (IFN-γ) и эритропоэтин. Типовые костимулирующие лиганды и цитокины, которые могут быть кодированы одним или более вторыми трансгенами, включают такие, которые описаны в, например, WO 2008121420.
В некоторых вариантах, молекулой, полипептидом или фактором, кодированным одним или более вторыми трансгенами, является растворимый однонитевой переменный фрагмент (scFv), такой как scFv, который связывает полипептид, который обладает иммунодепрессивным действием или иммуностимулирующим действием, такой как CD47, PD-1, CTLA-4 и их лиганды или CD28, OX-40, 4-1BB и их лиганды. Типовые scFv, которые могут быть кодированы одним или более вторыми трансгенами, включают те, которые описаны в, например, WO 2014134165.
В некоторых вариантах, молекулой, полипептидом или фактором, кодированным одним или более вторыми трансгенами, является иммуномодулирующий слитый белок или химерный рецептор переключения (CSR). В некоторых вариантах, кодированный иммуномодулирующий слитый белок содержит (a) внеклеточный компонент, состоящий из связывающего домена, который специфически связывает цель, (b) внутриклеточный компонент, состоящий из внутриклеточного связывающего домена, и (c) гидрофобный компонент, соединяющий внеклеточный и внутриклеточный компоненты. В некоторых вариантах, кодированный иммуномодулирующий слитый белок содержит (a) внеклеточный связывающий домен, который специфически связывает антиген, полученный из CD200R, SIRPα, CD279 (PD-1), CD2, CD95 (Fas), CD152 (CTLA4), CD223 (LAG3), CD272 (BTLA), A2aR, KIR, TIM3, CD300 или LPA5; (b) внутриклеточный связывающий домен, полученный из CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD47, CD79A, CD79B, CD134 (OX40), CD137 (4-1BB), CD150 (SLAMF1), CD278 (ICOS), CD357 (GITR), ХАРD11, DAP10, DAP12, FcRα, FcRβ, FcRγ, Fyn, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, ROR2, Ryk, Slp76, pTα, TCRα, TCRβ, TRFM, Zap70, PTCH2 или любого их сочетания; и (c) гидрофобный трансмембранный домен, полученный из CD2, CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD79A, CD79B, CD80, CD86, CD95 (Fas), CD134 (OX40), CD137 (4-1BB), CD150 (SLAMF1), CD152 (CTLA4), CD200R, CD223 (LAG3), CD270 (HVEM), CD272 (BTLA), CD273 (PD-L2), CD274 (PD-L1), CD278 (ICOS), CD279 (PD-1), CD300, CD357 (GITR), A2aR, DAP10, FcRα, FcRβ, FcRγ, Fyn, GAL9, KIR, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, PTCH2, ROR2, Ryk, Slp76, SIRPα, pTα, TCRα, TCRβ, TIM3, TRIM, LPA5 или Zap70. В некоторых вариантах, молекулой, полипептидом или фактором, кодированным одним или более вторыми трансгенами, является химерный рецептор переключения (CSR), такой как CSR, содержащий усеченный внеклеточный домен PD1 и трансмембранный и цитоплазматический сигнальные домены CD28. Типовые иммуномодулирующие слитые белки или CSR, которые могут быть кодированы один или более вторыми трансгенами, включают такие, которые описаны в, например, WO 2014134165, US 2014/0219975, WO 2013/019615 и Liu et al., Cancer Res. (2016) 76(6):1578-90.
В некоторых вариантах, молекулой, полипептидом или фактором, кодированным одним или более вторыми трансгенами, является корецептор. В некоторых вариантах, типовые корецепторы включают CD4 или CD8.
В некоторых вариантах, один или более целевых сайтов расположены на или рядом с одним или более из TRAC, TRBC1 и/или TRBC2 локусов. В некоторых вариантах, первый целевой сайт находится на или рядом с кодирующей последовательностью локуса TRAC гена, и второй целевой сайт находится на или рядом с кодирующей последовательностью локуса TRBC1 гена. В некоторых вариантах, первый целевой сайт находится на или рядом с кодирующей последовательностью локуса TRAC гена, и второй целевой сайт находится на или рядом с кодирующей последовательностью локуса TRBC2 гена. В некоторых вариантах, первый целевой сайт находится на или рядом с кодирующей последовательностью локуса TRAC гена, и второй целевой сайт обоих локусов TRBC1 и TRBC2, например, на последовательности, которая законсервирована между TRBC1 и TRBC2.
В некоторых вариантах, один или более разных ДНК сайтов, например, TRAC, TRBC1 и/или TRBC2 локусы, являются целями, и один или более трансгенов вставлены в каждый сайт. В некоторых вариантах, трансген, вставленный в каждый сайт, является одинаковым или по существу одинаковым. В некоторых вариантах, трансген, вставленный в каждый сайт, является разным. В некоторых вариантах, трансген вставлен только в один целевой сайт (например, TRAC локус), и другой целевой сайт направлен для редактирования генома (например, нокаут).
В некоторых вариантах, любые из длин и положений плеч гомологии и родственных положений к целевым сайтам, такие как любые, описанные здесь, также могут применяться к одному или более вторым матричным полинуклеотидам.
В некоторых вариантах, вызванная нуклеазой HDR дает вставку трансгена (также называемого “экзогенная последовательность” или “трансгенная последовательность”) для экспрессии трансгена для целевой вставки. Матричная полинуклеотидная последовательность обычно не идентична геномной последовательности, где она размещена. Матричная полинуклеотидная последовательность может содержать не гомологичную последовательность, окруженную двумя областями гомологии, для того, чтобы позволить эффективную в рассматриваемой локации. Дополнительно, матричная полинуклеотидная последовательность может содержать векторную молекулу, содержащую последовательности, которые не гомологичны рассматриваемой области в клеточном хроматине. Матричная полинуклеотидная последовательность может содержать несколько периодических областей гомологии к клеточному хроматину. Например, для целевой вставки последовательностей, обычно не присутствующих в рассматриваемой области, указанные последовательности могут присутствовать в трансгене и быть окруженными областями гомологии к последовательностям в рассматриваемой области.
В некоторых аспектах, рассматриваемые последовательности нуклеиновых кислот, включая кодирующие и/или не кодирующие последовательности и/или частичные кодирующие последовательности, которые вставлены или интегрированы в целевую локацию в геноме, также могут называться “трансгеном”, “трансгенными последовательностями”, “экзогенными последовательностями нуклеиновых кислот”, “гетерологическими последовательностями” или “донорными последовательностями”. В некоторых аспектах, трансгеном является последовательность нуклеиновых кислот, которая является экзогенной или гетерологической к эндогенным геномным последовательностям, таким как эндогенные геномные последовательности в специфическом целевом локусе или целевой локации в геноме, T клетки, например, человеческой T клетки. В некоторых аспектах, трансгеном является последовательность, которая модифицирована или изменена по сравнению с эндогенной геномной последовательностью в целевом локусе или целевой локации T клетки, например, человеческой T клетки. В некоторых аспектах, трансгеном является последовательность нуклеиновых кислот, которая происходит из или модифицирована по сравнению с последовательностью нуклеиновых кислот от разных генов, видов и/или происхождения. В некоторых аспектах, трансгеном является последовательность, которая получена из последовательности из другого локуса, например, другой геномной области или другого гена, тех же видов.
Полинуклеотиды для вставки также могут называться “трансгены” или “экзогенные последовательности” или “донорные” полинуклеотиды или молекулы. Матричным полинуклеотидом может быть ДНК, однонитевая и/или двухтнитевая, и она может быть введена в клетку в линейной или кольцевой форме. Матричным полинуклеотидом может быть РНК, однонитевая и/или двухнитевая, и она может быть введена в виде РНК молекулы (например, части РНК вируса). См. также Публикации патентов США №№ 20100047805 и 20110207221. Матричный полинуклеотид также может быть введен в ДНК форме, которая может быть введена в клетку в кольцевой или линейной форме. Если она введена в линейной форме, концы матричного полинуклеотида могут быть защищены (например, от экзонуклеолитического разрушения) известными способами. Например, один или более дидеоксинуклеотидных остатков добавляют к 3’ окончанию линейной молекулы и/или самокомплементарные олигонуклеотиды лигируют на один или оба конца. См., например, Chang et al. (1987) Proc. Natl. Acad. Sci. USA 84:4959-4963; Nehls et al. (1996) Science 272:886-889. Дополнительные способы защиты экзогенных полинуклеотидов от разрушения включают, но не ограничены ими, добавление концевых аминогрупп и применением модифицированных межнуклеотидных связей, таких как, например, фосфоротиоаты, фосфорамидаты и O-метилрибозные или деоксирибозные остатки. Если он введен в двухнитевой форме, матричный полинуклеотид может включать один или более целевых сайтов нуклеазы, например, целевые сайты нуклеазы, окружающие трансген, интегрируемый в геном клетки. См., например, Публикацию патента США №. 20130326645.
В некоторых вариантах, двухнитевой матричный полинуклеотид включает последовательности (также называемые трансген) более чем 1 т.н. в длину, например между 2 и 200 т.н., между 2 и 10 т.н. (или любое промежуточное значение). Двухнитевой матричный полинуклеотид также включает, по меньшей мере, один целевой сайт нуклеазы, например. В некоторых вариантах, матричный полинуклеотид включает, по меньшей мере, 2 целевых сайта, например, для пары ZFN или TALEN. Обычно целевые сайты нуклеазы находятся вне трансгенных последовательностей, например, 5’ и/или 3’ до трансгенных последовательностей, для расщепления трансгена. Сайты расщепления нуклеазы могут быть для любых нуклеаз. В некоторых вариантах, целевые сайты нуклеаза, содержащиеся в двухнитевом матричном полинуклеотиде, предназначены для тех же нуклеаз, которые применяют для расщепления эндогенной цели, в которую интегрирован расщепленный матричный полинуклеотид способами, не зависимыми от гомологии.
В некоторых вариантах, матричная система нуклеиновой кислоты является двухнитевой. В некоторых вариантах, матричная система нуклеиновой кислоты является однонитевой. В некоторых вариантах, матричная система нуклеиновой кислоты содержит однонитевую часть и двухнитевую часть.
В некоторых вариантах, матричный полинуклеотид содержит трансген, например, кодирующие рекомбинантный рецептор последовательности нуклеиновых кислот, окруженные гомологическими последовательностями (также называемыми “плечи гомологии”) на 5’ и 3’ концах, что позволяет запустить механизм репарации ДНК, например, механизм гомологичной рекомбинации, для применения матричного полинуклеотида в виде матрицы для репарации, эффективной вставки трансгена в целевой сайт интеграции в геноме. Плечо гомологии должно распространяться, по меньшей мере, так далеко, как область, в которой может происходить конечная резекция, например, для того, чтобы позволить иссеченному однонитевому липкому концу найти комплементарную область в матричном полинуклеотиде. Общая длина должна быть ограничена параметрами, такими как размер плазмиды или пределы упаковки вируса. В некоторых вариантах, плечо гомологии не продлевается в повторяющиеся элементы, например, ALU повторы или LINE повторы.
Типовая длина плеча гомологии включает, по меньшей мере, или, по меньшей мере, около, или составляет, или составляет около 50, 100, 200, 250, 300, 400, 500, 600, 700, 750, 800, 900, 1000, 2000, 3000, 4000 или 5000 нуклеотидов. В некоторых вариантах, длина плеча гомологии составляет 50-100, 100-250, 250-500, 500-750, 750-1000, 1000-2000, 2000-3000, 3000-4000 или 4000-5000 нуклеотидов. типовые длины плеча гомологии включают менее или менее чем около, или составляют или составляют около 50, 100, 200, 250, 300, 400, 500, 600, 700, 750, 800, 900, 1000, 2000, 3000, 4000 или 5000 нуклеотидов. В некоторых вариантах, длина плеча гомологии составляет 50-100, 100-250, 250-500, 500-750, 750-1000, 1000-2000, 2000-3000, 3000-4000 или 4000-5000 нуклеотидов.
Целевой сайт (также известный как “целевое положение”, “целевая последовательность ДНК” или “целевая локация”) в некоторых вариантах относится к сайту на целевой ДНК (например, хромосоме), который модифицирован одним или более агентами, способными вызывать генетическое разрушение, например, Cas9 молекулой. Например, целевой сайт может быть модифицирован расщеплением ДНК Cas9 молекулой на целевом сайте и модификацией, направленной матричным полинуклеотидом, например, целевой вставкой трансгена, на целевом сайте. В некоторых вариантах, целевым сайтом может быть сайт между двумя нуклеотидами, например, соседними нуклеотидами, на ДНК, в которую добавляют один или более нуклеотидов. Целевой сайт может содержать один или более нуклеотидов, которые изменены матричным полинуклеотидом. В некоторых вариантах, целевой сайт находится в пределах целевой последовательности (например, последовательности, с которой связана нРНК). В некоторых вариантах, целевой сайт находится выше или ниже целевой последовательности (например, последовательности, с которой связана нРНК). В некоторых аспектах, может быть создана пара однонитевых разрывов (например, ников) на каждой стороне целевого сайта.
В некоторых вариантах, матричный полинуклеотид содержит около 500 до 1000, например, 600 до 900 или 700 до 800, пар оснований гомологии на каждой стороне целевого сайта на эндогенном гене. В некоторых вариантах, матричный полинуклеотид содержит, по меньшей мере, или менее или около 200, 300, 400, 500, 600, 700, 800, 900 или 1000 пар оснований гомологии 5’ целевого сайта, 3’ целевого сайта или обоих 5’ и 3’ целевого сайта, например, в TRAC, TRBC1, и/или TRBC2 гене, локусе или открытой рамке считывания (например, описанных в таблицах 1-3 здесь).
В некоторых вариантах, матричный полинуклеотид содержит около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 3000, 4000 или 5000 пар оснований гомологии 3’ целевого сайта. В некоторых вариантах, матричный полинуклеотид содержит около 100 до 500, 200 до 400 или 250 до 350, пар оснований гомологии 3’ трансгена и/или целевого сайта. В некоторых вариантах, матричный полинуклеотид содержит менее около 100, 90, 80, 70, 60, 50, 40, 30, 20, 15 или 10 пар оснований гомологии 5’ целевого сайта, например, в TRAC, TRBC1, и/или TRBC2 гене, локусе или открытой рамке считывания (например, описанных в таблицах 1-3 здесь).
В некоторых вариантах, матричный полинуклеотид содержит около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 3000, 4000 или 5000 пар оснований гомологии 5’ целевого сайта. В некоторых вариантах, матричный полинуклеотид содержит около 100 до 500, 200 до 400 или 250 до 350, пар оснований гомологии 5’ трансгена и/или целевого сайта. В некоторых вариантах, матричный полинуклеотид содержит менее около 100, 90, 80, 70, 60, 50, 40, 30, 20, 15 или 10 пар оснований гомологии 3’ целевого сайта, например, в TRAC, TRBC1, и/или TRBC2 гене, локусе или открытой рамке считывания (например, описанных в таблицах 1-3 здесь).
В некоторых вариантах, матричным полинуклеотидом является последовательность нуклеиновых кислот, которая может применяться в сочетании с одним или более агентами, способными вызывать генетическое разрушение для изменения структуры целевого сайта. В некоторых вариантах, целевой сайт модифицируют так, чтобы он имеет некоторые или все последовательности матричного полинуклеотида, обычно рядом с сайтами расщепления. В некоторых вариантах, матричный полинуклеотид является однонитевым. В некоторых вариантах, матричный полинуклеотид является двухнитевым. В некоторых вариантах, матричный полинуклеотид является ДНК, например, двухнитевой ДНК. В некоторых вариантах, матричный полинуклеотид является однонитевой ДНК. В некоторых вариантах, матричный полинуклеотид кодирован на том же векторном скелете, например, AAV геноме, плазмидной ДНК, такой как Cas9 и нРНК. В некоторых вариантах, матричный полинуклеотид иссекают из векторного скелета in vivo, например, он окружен последовательностями распознавания нРНК. В некоторых вариантах, матричный полинуклеотид находится на отдельной полинуклеотидной молекуле, такой как Cas9 и нРНК. В некоторых вариантах, Cas9 и нРНК вводят в форме рибонуклеопротеинового (RNP) комплекса, и матричный полинуклеотид вводят в виде полинуклеотидной молекулы, например, в векторе.
В некоторых вариантах, матричный полинуклеотид изменяет структуру целевого сайта, например, вставкой трансгена, участием в репарации, направляемой гомологией. В некоторых вариантах, матричный полинуклеотид изменяет последовательность целевого сайта.
В некоторых вариантах, матричный полинуклеотид включает последовательность, которая соответствует сайту на целевой последовательности, которая расщепляется одним или более агентами, способными вызывать генетическое разрушение. В некоторых вариантах, матричный полинуклеотид включает последовательность, которая соответствует обоим, первому сайту на целевой последовательности, который расщепляется первым агентом, способным вызвать генетическое разрушение, и второму сайту на целевой последовательности, который расщепляется вторым агентом, способным вызывать генетическое разрушение.
В некоторых вариантах, матричный полинуклеотид содержит следующие компоненты: [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии]. Плечи гомологии представлены для рекомбинации в хромосому, тем самым вставляя трансген в ДНК на или рядом с сайтом расщепления, например, целевыми сайтами. В некоторых вариантах, плечи гомологии окружают наиболее отдаленные целевые сайты.
В некоторых вариантах, 3’ окончание 5’ плеча гомологии находится в положении, следующем за 5’ окончанием трансгена. В некоторых вариантах, 5’ плечо гомологии может распространяться на, по меньшей мере, 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 3000, 4000 или 5000 нуклеотидов 5’ от 5’ окончания трансгена.
В некоторых вариантах, 5’ окончание 3’ плеча гомологии находится в положении, следующем за 3’ окончанием трансгена. В некоторых вариантах, 3’ плечо гомологии может распространяться на, по меньшей мере, 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 3000, 4000 или 5000 нуклеотидов 3’ от 3’ окончания трансгена.
В некоторых вариантах, для направленной вставки, плечи гомологии, например, 5’ и 3’ плечи гомологии, могут каждое содержать около 1000 пар оснований (п.о.) последовательности, окружающей наиболее отдаленные нРНК (например, 1000 п.о. от последовательности на любой стороне мутации).
В некоторых вариантах, один или более вторых матричных полинуклеотидов, содержащих один или более вторых трансгенов, могут быть введены. В некоторых вариантах, один или более вторых трансгенов нацелены на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
В некоторых вариантах, один или более вторых матричных полинуклеотидов содержит структуру [второе 5’ плечо гомологии]-[один или более вторые трансген]-[второе 3’ плечо гомологии]. Плечи гомологии обеспечивают рекомбинацию в хромосомы, следовательно, вставку трансгена в ДНК на или рядом с сайтом расщепления, например, целевым сайтом. В некоторых вариантах, плечи гомологии окружают наиболее отдаленные места расщепления. В некоторых вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот, гомологичные последовательностям нуклеиновых кислот окружающим, по меньшей мере, один целевой сайт. В некоторых вариантах, второе 5’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот второго 5’ целевого сайта. В некоторых вариантах, второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот второго 3’ целевого сайта. В некоторых вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет, по меньшей мере, или, по меньшей мере, около, или составляет или составляет около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований, или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований. В некоторых вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет между около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 пар оснований. В некоторых вариантах, второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований.
В некоторых вариантах, один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене (например, описанном в таблице 1 здесь). В некоторых вариантах, один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRBC1 или TRBC2 гене (например, описанных в таблицах 2-3 здесь).
Здесь предполагается, что одно или оба плеча гомологии могут быть сокращены для избежания включения определенных повторных элементов последовательности, например, Alu повторов или LINE элементов. Например, 5’ плечо гомологии может быть сокращено для избежания повторных элементов последовательности. В некоторых вариантах, 3’ плечо гомологии может быть сокращено для избежания повторных элементов последовательности. В некоторых вариантах, оба 5’ и 3’ плеча гомологии могут быть сокращены для избежания включения определенных повторных элементов последовательности. Здесь предполагается, что матричные полинуклеотиды для целевой вставки могут быть созданы для применения в виде однонитевого олигонуклеотида, например, однонитевого олигодеоксинуклеотида (ssODN). При применении ssODN, 5’ и 3’ плечи гомологии могут варьироваться вплоть до около 200 пар оснований (п.о.) в длину, например, по меньшей мере, 25, 50, 75, 100, 125, 150, 175 или 200 п.о. в длину. Более длинные плечи гомологии также рассматриваются для ssODN в качестве улучшений в продолжающемся синтезе олигонуклеотида. В некоторых вариантах, долее длинное плечо гомологии получают способом, отличным от химического синтеза, например, денатурацией длинной двухнитевой нуклеиновой кислоты и очисткой одной из нитей, например, аффинностью для цепь-специфической последовательности, прикрепленной к твердому субстрату.
В некоторых вариантах, альтернативная HDR проходит более эффективно, когда матричный полинуклеотид продлевает гомологию 5’ до целевого сайта (т.е., в 5’ направлении нити целевого сайта). Следовательно, в некоторых вариантах, матричный полинуклеотид имеет более длинное плечо гомологии и более короткое плечо гомологии, где длинное плечо гомологии может ренатурировать 5’ к целевому сайту. В некоторых вариантах, плечо, которое может ренатурировать 5’ к целевому сайту составляет, по меньшей мере, 25, 50, 75, 100, 125, 150, 175 или 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 3000, 4000 или 5000 нуклеотидов от целевого сайта или 5’ или 3’ окончаний трансгена. В некоторых вариантах, плечо, которое может ренатурировать 5’ к целевому сайту на, по меньшей мере, 10%, 20%, 30%, 40% или 50% длиннее, чем плечо, которое может ренатурировать 3’ к целевому сайту. В некоторых вариантах, плечо, которое может ренатурировать 5’ к целевому сайту на, по меньшей мере, 2x, 3x, 4x или 5x длиннее, чем плечо, которое может ренатурировать 3’ к целевому сайту. В зависимости от того, где матрица онДНК может ренатурировать к нетронутой нити или нити целевого сайта, плечо гомологии, которое ренатурирует 5’ к целевому сайту, может быть на 5’ окончании матрицы онДНК или 3’ окончании матрицы онДНК, соответственно.
Также, в некоторых вариантах, матричный полинуклеотид имеет 5’ плечо гомологии, трансген и 3’ плечо гомологии, так, что матричный полинуклеотид содержит расширенную гомологию к 5’ целевого сайта. Например, 5’ плечо гомологии и 3’ плечо гомологии могут иметь по существу одинаковую длину, но трансген может продлевать скорее 5’ целевого сайта, чем 3’ целевого сайта. В некоторых вариантах, плечо гомологии продлевается, по меньшей мере, 10%, 20%, 30%, 40%, 50%, 2x, 3x, 4x или 5x скорее до 5’ окончания целевого сайта, чем до 3’ окончания целевого сайта.
В некоторых вариантах, альтернативная HDR проходит более эффективно, если матричный полинуклеотид расположен по центру целевого сайта. Следовательно, в некоторых вариантах, матричный полинуклеотид имеет два плеча гомологии, которые по существу имеют одинаковый размер.
Например, первое плечо гомологии матричного полинуклеотида имеет длину, которая составляет пределах 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% от второго плеча гомологии матричного полинуклеотида.
Также, в некоторых вариантах, матричный полинуклеотид имеет 5’ плечо гомологии, трансген и 3’ плечо гомологии, так, что матричный полинуклеотид расширяется по существу на одинаковое расстояние на любой стороне целевого сайта. Например, плечи гомологии могут иметь разные длины, но трансген может быть выбран так, чтобы компенсировать это. Например, трансген может быть дальше 5’ от целевого сайта, чем 3’ целевого сайта, но плечо гомологии 5’ целевого сайта короче, чем плечо гомологии 3’ целевого сайта, для компенсации. Возможно и обратное, то, что трансген может быть дальше 3’ от целевого сайта, чем 5’ от целевого сайта, но плечо гомологии 3’ целевого сайта короче, чем плечо гомологии 5’ целевого сайта, для компенсации.
В некоторых вариантах, матричный полинуклеотид является однонитевой нуклеиновой кислотой. В другом варианте, матричный полинуклеотид является двухнитевой нуклеиновой кислотой. В некоторых вариантах, матричный полинуклеотид содержит нуклеотидную последовательность, например, из одного или более нуклеотидов, которые будут добавлены к или создадут матрицу изменения в целевой ДНК. В некоторых вариантах, матричный полинуклеотид содержит нуклеотидную последовательность, которая может применяться для модификации целевого сайта. В некоторых вариантах, матричный полинуклеотид содержит нуклеотидную последовательность, например, из одного или более нуклеотидов, которая соответствует дикому типу последовательности целевой ДНК, например, целевого сайта.
Матричный полинуклеотид может содержать трансген. В некоторых вариантах, матричный полинуклеотид содержит 5’ плечо гомологии. В некоторых вариантах, матрица нуклеиновой кислоты содержит 3’ плечо гомологии.
В некоторых вариантах, матричный полинуклеотид является двухнитевой ДНК. Длина может быть, например, около 200-5000 пар оснований, например, около 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600, 1800, 2000, 2500, 3000, 4000 или 5000 пар оснований. Длина может быть, например, по меньшей мере, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600, 1800, 2000, 2500, 3000, 4000 или 5000 пар оснований. В некоторых вариантах, длина составляет не более чем 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600, 1800, 2000, 2500, 3000, 4000 или 5000 пар оснований. В некоторых вариантах, двухнитевой матричный полинуклеотид имеет длину около 160 пар оснований, например, около 200-4000, 300-3500, 400-3000, 500-2500, 600-2000, 700-1900, 800-1800, 900-1700, 1000-1600, 1100-1500 или 1200-1400 пар оснований.
Матричный полинуклеотид может быть линейной однонитевой ДНК. В некоторых вариантах, матричный полинуклеотид имеет, (i) линейную однонитевую ДНК, которая может ренатурировать с никованной нитью целевой ДНК, (ii) линейную однонитевую ДНК, которая может ренатурировать с нетронутой нитью целевой ДНК, (iii) линейную однонитевую ДНК, которая может ренатурировать со считанной нитью целевой ДНК, (iv) линейную однонитевую ДНК, которая может ренатурировать с не считанной нитью целевой ДНК или более одной из вышеописанных.
Длина может составлять, например, около 200-5000 пар оснований, например, около 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600, 1800, 2000, 2500, 3000, 4000 или 5000 нуклеотидов. Длина может составлять, например, по меньшей мере, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600, 1800, 2000, 2500, 3000, 4000 или 5000 нуклеотидов. В некоторых вариантах, длина составляет не более чем 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600, 1800, 2000, 2500, 3000, 4000 или 5000 нуклеотидов. В некоторых вариантах, однонитевой матричный полинуклеотид имеет длину около 160 нуклеотидов, например, около 200-4000, 300-3500, 400-3000, 500-2500, 600-2000, 700-1900, 800-1800, 900-1700, 1000-1600, 1100-1500 или 1200-1400 нуклеотидов.
В некоторых вариантах, матричный полинуклеотид является круговой двухнитевой ДНК, например, плазмидой. В некоторых вариантах, матричный полинуклеотид содержит около 500 до 1000 пар оснований гомологии на любой стороне трансгена и/или целевого сайта. В некоторых вариантах, матричный полинуклеотид содержит около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований гомологии 5’ целевого сайта или трансгена, 3’ целевого сайта или трансгена или оба 5’ и 3’ целевого сайта или трансгена. В некоторых вариантах, матричный полинуклеотид содержит, по меньшей мере, 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований гомологии 5’ целевого сайта или трансгена, 3’ целевого сайта или трансгена или оба 5’ и 3’ целевого сайта или трансгена. В некоторых вариантах, матричный полинуклеотид содержит не более 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований гомологии 5’ целевого сайта или трансгена, 3’ целевого сайта или трансгена или оба 5’ и 3’ целевого сайта или трансгена.
В некоторых вариантах, длина любого из полинуклеотидов, например, матричных полинуклеотидов, может быть, например, от около 200-10000 нуклеотидов, например, от около 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600, 1800, 2000, 2500, 3000, 4000, 5000, 6000, 7000, 8000, 9000 или 10000 нуклеотидов или значение между любыми из вышеописанных. В некоторых вариантах, длина может быть, например, по меньшей мере, от около 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600, 1800, 2000, 2500, 3000, 4000, 5000, 6000, 7000, 8000, 9000 или 10000 нуклеотидов или значение между любыми из вышеописанных. В некоторых вариантах, длина составляет не более чем от около 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600, 1800, 2000, 2500, 3000, 4000, 5000, 6000, 7000, 8000, 9000 или 10000 нуклеотидов. В некоторых вариантах, длина составляет от около 200-4000, 300-3500, 400-3000, 500-2500, 600-2000, 700-1900, 800-1800, 900-1700, 1000-1600, 1100-1500 или 1200-1400 нуклеотидов. В некоторых вариантах, полинуклеотид составляет, по меньшей мере, от около 2500, 2750, 3000, 3250, 3500, 3750, 4000, 4250, 4500, 4760, 5000, 5250, 5500, 5750, 6000, 7000, 7500, 8000, 9000 или 10000 нуклеотидов в длину или любое значение между любыми из вышеописанных. В некоторых вариантах, полинуклеотид имеет от около 2500 и от около 5000 нуклеотидов, от около 3500 и от около 4500 нуклеотидов или от около 3750 нуклеотидов и от около 4250 нуклеотидов в длину. В некоторых вариантах, полинуклеотид составляет от около 2500, 2750, 3000, 3250, 3500, 3750, 4000, 4250, 4500, 4760, 5000, 5250, 5500, 5750, 6000, 7000, 7500, 8000, 9000 или 10000 нуклеотидов в длину.
В некоторых вариантах, матричный полинуклеотид содержит плечи гомологии для целенаправленного воздействия на эндогенный TRAC локус (типовая нуклеотидная последовательность локус человеческого TRAC гена представлена в SEQ ID NO:1; NCBI Ссылочная последовательность: NG_001332,3, TRAC или описана в таблице 1 здесь). В некоторых вариантах, генетическое разрушение TRAC локуса вводят в раннюю кодирующую область гена, включающую последовательность, расположенную непосредственно после сайта начала транскрипции, в пределах первого экзона кодирующей последовательности или в пределах 500 п.о. от сайта начала транскрипции (например, менее 500, 450, 400, 350, 300, 250, 200, 150, 100 или 50 п.о.) или в пределах 500 п.о. от инициирующего кодона (например, менее 500, 450, 400, 350, 300, 250, 200, 150, 100 или 50 п.о.). В некоторых вариантах, генетическое разрушение вводят с применением любой из направленных нуклеаз и/или нРНК, описанных в разделе I.A здесь. В некоторых вариантах, матричный полинуклеотид содержит около 500 до 1000, например, 600 до 900 или 700 до 800, пар оснований гомологии на любой стороне генетического разрушения, введенного направленной нуклеазой и/или нРНК. В некоторых вариантах, матричный полинуклеотид содержит около 500, 600, 700, 800, 900 или 1000 пар оснований 5’ последовательностей плеча гомологии, которые гомологичны 500, 600, 700, 800, 900 или 1000 парам оснований последовательностей 5’ генетического разрушения (например, на TRAC локусе), трансгена, и около 500, 600, 700, 800, 900 или 1000 пар оснований 3’ последовательностей плеча гомологии, которые гомологичны 500, 600, 700, 800, 900 или 1000 парам оснований последовательностей 3’ генетического разрушения (например, на TRAC локусе). В некоторых вариантах, типовые 5’ и 3’ плечи гомологии для направленной интеграции на TRAC локусе представлены в SEQ ID NO: 124 и 125, соответственно. В некоторых вариантах, типовые 5’ и 3’ плечи гомологии для направленной интеграции на TRAC локусе представлены в SEQ ID NOS: 227-233 и 234-240, соответственно.
В некоторых вариантах, матричный полинуклеотид содержит плечи гомологии для целенаправленного воздействия эндогенного TRBC1 или TRBC2 локуса (типовая нуклеотидная последовательность локуса человеческого TRBC1 гена представлена в SEQ ID NO:2; NCBI Ссылочная последовательность: NG_001333,2, TRBC1, описана в таблице 2 здесь; типовая нуклеотидная последовательность локуса человеческого TRBC2 гена представлена в SEQ ID NO:3; NCBI Ссылочная последовательность: NG_001333,2, TRBC2, описана в таблице 3 здесь). В некоторых вариантах, генетическое разрушение TRBC1 или TRBC2 локуса вводят на ранней кодирующей области гена, включая последовательность, расположенную непосредственно после сайта начала транскрипции, в пределах первого экзона кодирующей последовательности или в пределах 500 п.о. сайта начала транскрипции (например, менее 500, 450, 400, 350, 300, 250, 200, 150, 100 или 50 п.о.) или в пределах 500 п.о. от инициирующего кодона (например, менее 500, 450, 400, 350, 300, 250, 200, 150, 100 или 50 п.о.). В некоторых вариантах, генетическое разрушение вводят с применением любой из направленных нуклеазы и/или нРНК, описанных в разделе I.A здесь. В некоторых вариантах, матричный полинуклеотид содержит около 500 до 1000, например, 600 до 900 или 700 до 800, пар оснований гомологии на любой стороне генетического разрушения, введенного направленными нуклеазой и/или нРНК. В некоторых вариантах, матричный полинуклеотид содержит около 500, 600, 700, 800, 900 или 1000 пар оснований 5’ последовательностей плеча гомологии, которые гомологичны 500, 600, 700, 800, 900 или 1000 парам оснований последовательностей 5’ генетического разрушения (например, на TRBC1 или TRBC2 локусе), трансген и около 500, 600, 700, 800, 900 или 1000 пар оснований 3’ последовательностей плеча гомологии, которые гомологичны 500, 600, 700, 800, 900 или 1000 пар оснований последовательностей 3’ генетического разрушения (например, на TRBC1 или TRBC2 локусе).
В некоторых вариантах, любые длины и положения плеч гомологии и родственного положения к целевым сайтам, такие как описаны здесь, также могут применяться для одного или более вторых матричных полинуклеотидов.
В некоторых случаях, матричный полинуклеотид содержит промотор, например, промотор, который является экзогенным и/или не присутствует на или рядом с целевым локусом. В некоторых вариантах, промотор управляет экспрессией только в клетке определенного типа (например, специфический к T клетке или B клетке или NK клетке промотор). В некоторых вариантах, в которых функциональные полипептидные кодирующие последовательности не содержат промотор, экспрессия интегрированного трансгена обеспечивается транскрипцией, управляемой эндогенным промотором или другим контрольным элементом в рассматриваемой области.
Трансген, включая трансген, кодирующий рекомбинантный рецептор или антигенное связывание, его часть или его цепь и/или один или более вторых трансгенов, может быть вставлен так, что экспрессия управляется эндогенным промотором в месте интеграции, а именно, промотором, который управляет экспрессией эндогенного гена, в который вставлен трансген (например, TRAC, TRBC1 и/или TRBC2). Например, кодирующие последовательности в трансгене могут быть вставлены без промотора, но внутри рамки с кодирующей последовательностью эндогенного целевого гена, так, что экспрессия интегрированного трансгена контролируется транскрипцией эндогенного промотора в сайте интеграции. В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо функционально связан с эндогенным промотором гена на целевом сайте. В некоторых вариантах, элемент проскока рибосомы/элемент саморасщепления, такой как 2A элемент, помещают выше трансгенной кодирующей последовательности так, что элемент проскока рибосомы/элемент саморасщепления помещают внутри рамки с эндогенным геном так, что экспрессия трансгена, кодирующего рекомбинантный или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов функционально связан с промотором эндогенного TCRα.
В некоторых вариантах, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо содержит один или более мультицистронных элементов. В некоторых вариантах, один или более мультицистронных элементов расположены против хода транскрипции трансгена, кодирующего рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов. В некоторых вариантах, мультицистронные элементы расположены между трансгеном, кодирующим рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и одним или более вторыми трансгенами. В некоторых вариантах, мультицистронные элементы расположены между последовательностью нуклеиновых кислот, кодирующей TCRα или его часть, и последовательностью нуклеиновых кислот, кодирующей TCRβ или его часть. В некоторых вариантах, элемент проскока рибосомы содержит последовательность, кодирующую элемент проскока рибосомы, выбранный из T2A, P2A, E2A или F2A или внутренний участок посадки рибосомы (IRES).
В некоторых вариантах, кодированная TCRα цепь и TCRβ цепь разделены линкерную или спейсерную область. В некоторых вариантах, включена линкерная последовательность, которая связывает TCRα и TCRβ цепи с образованием одной полипептидной нити. В некоторых вариантах, линкер имеет достаточную длину для того, чтобы охватывать расстояние между C окончанием α цепи и N окончанием β цепи или наоборот, также обеспечивая то, что длина линкера не настолько большая, чтобы блокировать или снижать связывание с целевым комплексом пептид-MHC. В некоторых вариантах, линкером может быть любой линкер, способный образовывать одну полипептидную нить, сохраняя специфичность к TCR связыванию. В некоторых вариантах, линкер может содержать от или от около 10 до 45 аминокислот, например, от 10 до 30 аминокислот или от 26 до 41 аминокислотных остатков, например 29, 30, 31 или 32 аминокислот. В некоторых вариантах, линкер имеет формулу -PGGG-(SGGGG)n-P-, где n равно 5 или 6 и P является пролином, G является глицином и S является серином (SEQ ID NO: 22). В некоторых вариантах, линкер имеет последовательность GSADDAKKDAAKKDGKS (SEQ ID NO: 23). В некоторых вариантах, линкер или спейсер между TCRα цепью или ее частью, и TCRβ цепью или ее частью, которые распознаются и/или могут быть расщеплены протеазой. В определенных вариантах, линкер или спейсер между TCRα цепью или ее частью и TCRβ цепью или ее частью содержит элемент проскока рибосомы или элемент саморасщепления.
В некоторых вариантах, трансгеном является или включает последовательность нуклеотидов, которая является или включает структуру [TCRβ цепь]-[линкер]-[TCRα цепь]. В конкретных вариантах, трансгеном является или включает последовательность нуклеотидов, которая является или включает структуру [TCRβ цепь]-[элемент саморасщепления]-[TCRα цепь]. В определенных вариантах, трансген является или включает последовательность нуклеотидов, которая является или включает структуру [TCRβ цепь]-[последовательность проскока рибосомы]-[TCRα цепь]. В некоторых вариантах, трансген является или включает последовательность нуклеотидов, которая является или включает структуру [TCRα цепь]-[линкер]-[TCRβ цепь]. В конкретных вариантах, трансген является или включает последовательность нуклеотидов, которая является или включает структуру [TCRα цепь]-[элемент саморасщепления]-[TCRβ цепь]. В определенных вариантах, трансген является или включает последовательность нуклеотидов, которая является или включает структуру [TCRα цепь]-[последовательность проскока рибосомы]-[TCRβ цепь]. В некоторых вариантах, структурами является кодированная полинуклеотидная нить или одно- или двухнитевой полинуклеотид, в 5’-3’ направлении.
В некоторых случаях, элемент проскока рибосомы/элемент саморасщепления, такой как T2A, может привести к тому, что рибосома проскочит (проскок рибосомы) синтез пептидной связи на C-окончании 2A элемента, что приведет к разделению между концом 2A последовательности и следующего пептида ниже (см., например, de Felipe, Genetic Vaccines и Ther. 2:13 (2004) и de Felipe et al. Traffic 5:616-626 (2004)). Это позволяет контроль вставленного трансгена транскрипцией эндогенного промотора на сайте интеграции, например, TRAC, TRBC1 и/или TRBC2 промотора. Типовой элемент проскока рибосомы/элемент саморасщепления включает 2A последовательности из вируса ящера (F2A, например, SEQ ID NO: 11), A вируса лошадиного ринита (E2A, например, SEQ ID NO: 10), вируса Thosea asigna (T2A, например, SEQ ID NO: 6 или 7) и свиного тешовируса-1 (P2A, например, SEQ ID NO: 8 или 9) как описано в Публикации патента США №. 20070116690. В некоторых вариантах, матричный полинуклеотид включает P2A элемент проскока рибосомы (последовательность, представленную в SEQ ID NO: 8 или 9) выше трансгена, например, рекомбинантного рецептора, кодирующего нуклеиновые кислоты.
В некоторых вариантах, трансген может содержать промотор и/или энхансер, например конститутивный промотор или индуцируемый или ткань-специфический промотор. В некоторых вариантах, промотор является или содержит конститутивный промотор. Типовые конститутивные промоторы включают, например, ранний промотор вируса обезьян 40 (SV40), предранний промотор цитомегаловируса (CMV), промотор человеческого убиквитина C (UBC), промотор человеческого фактора элонгации 1α (EF1α), промотор мышиной фосфоглицераткиназы 1 (PGK) и промотор куриного β-активна, сопряженный с ранним энхансером CMV (CAGG). В некоторых вариантах, конститутивным промотором является синтетический или модифицированный промотор. В некоторых вариантах, промотор является или содержит MND промотор, синтетический промотор, который содержит U3 область модифицированного MoMuLV LTR с энхансером вируса миелопролиферативной саркомы (последовательность, представленная в SEQ ID NO:18 или 126; см. Challita et al. (1995) J. Virol. 69(2):748-755). В некоторых вариантах, промотором является ткань-специфический промотор. В другом варианте, промотором является вирусный промотор. В другом варианте, промотором является не вирусный промотор. В некоторых случаях, промотор выбирают из промотора человеческого фактора элонгации 1 альфа (EF1α) (последовательность, представленная в SEQ ID NO:4 или 5) или его модифицированная форма (EF1α промотор с HTLV1 энхансером; последовательность, представленная в SEQ ID NO: 127) или MND промотор (последовательность, представленная в SEQ ID NO:18 или 126). В некоторых вариантах, трансген не включает регулирующий элемент, например, промотор.
В некоторых вариантах, “тандемная” кассета интегрирована в выбранный сайт. В некоторых вариантах, одна или более “тандемных” кассет кодирует один или более полипептид или факторы, каждый из которых независимо контролируется регулирующим элементом или все которые контролируются как мультицистронная система экспрессии. В некоторых вариантах, например, в которых полинуклеотид содержит первую и вторую последовательность нуклеиновых кислот, кодирующие последовательности, кодирующие каждую из разных полипептидных цепей, могут быть функционально связаны промотором, который может быть одинаковым или разным. В некоторых вариантах, молекула нуклеиновой кислоты может содержать промотор, который управляет экспрессией двух или более разных полипептидных цепей. В некоторых вариантах, такие молекулы нуклеиновой кислоты могут быть мультицистронными (бицистронными или трицистронными, см., например, патент США № 6,060,273). В некоторых вариантах, транскрипционные единицы могут быть сконструированы как бицистронная единица, содержащая IRES (внутренний участок посадки рибосомы), которая позволяет coэкспрессию генных продуктов через мРНК от одного промотора. Альтернативно, в некоторых случаях, один промотор может направлять экспрессию РНК, которая содержит, в одной открытой рамке считывания (ORF), два или три полипептида, разделенных друг с другом последовательностями, кодирующими пептид саморасщепления (например, 2A последовательности) или сайт распознавания протеазы (например, фурин), как описано здесь. Таким образом, ORF кодирует один полипептид, который, либо во время (в случае 2A), либо после трансляции перерабатывается на отдельные белки. В некоторых вариантах, “тандемная кассета” включает первый компонент кассеты, содержащий последовательность без промотора, затем последовательность превращения транскрипции и вторую последовательность, кодирующую кассету автономной экспрессии или последовательность мультицистронной экспрессии. В некоторых вариантах, тандемная кассета кодирует два или более разных полипептида или фактора, например, две или более цепей или доменов рекомбинантного рецептора. В некоторых вариантах, последовательности нуклеиновых кислот, кодирующие две или более цепей или доменов рекомбинантного рецептора, введены в виде кассет тандемной экспрессии или би- или мультицистронных кассет, в целевой сайт интеграции ДНК.
Трансген может быть вставлен в эндогенный ген, так, что экспрессируются все, некоторые или никакие эндогенные гены. В некоторых вариантах, трансген (например, с или без кодирующих пептид последовательностей) интегрируют в любой эндогенный локус. В некоторых вариантах, трансген интегрируют в локусы TRAC, TRBC1 и/или TRBC2 гена.
В некоторых вариантах, экзогенные последовательности могут также включать транскрипционные или трансляционные регулирующие последовательности, например, промоторы, энхансеры, инсуляторы, внутренние участки посадки рибосомы, последовательности, кодирующие 2A пептиды и/или сигналы полиаденилирования. Далее, контрольные элементы рассматриваемых генов могут быть функционально связаны с генами репортерами для создания химерных генов (например, кассет экспрессии репортера). Дополнительно, могут быть включены последовательности акцептора сплайсинга. Типовые известные последовательности сайтов акцептора сплайсинга включают, например, CTGACCTCTTCTCTTCCTCCCACAG, (SEQ ID NO:119) (из человеческого HBB гена) и TTTCTCTCCACAG (SEQ ID NO:120) (из человеческого иммуноглобулин-гамма гена).
В типовом варианте, матричный полинуклеотид включает плечи гомологии для целенаправленного воздействия на TRAC локус, регулирующие последовательности, например, промотор, и последовательности нуклеиновых кислот, кодирующие рекомбинантный рецептор, например, TCR. В типовом варианте, применяют дополнительный матричный полинуклеотид, который включает плечи гомологии для целенаправленного воздействия на TRBC1 и/или TRBC2 локусы, регулирующие последовательности, например, промотор, и последовательности нуклеиновых кислот, кодирующие другой фактор.
В некоторых вариантах, типовые матричные полинуклеотиды содержат трансген, кодирующий рекомбинантный Т-клеточный рецептор под функциональным контролем промотора человеческого фактора элонгации 1 альфа (EF1α) с HTLV1 энхансером (последовательность, представленная в SEQ ID NO:127) или MND промотора (последовательность, представленная в SEQ ID NO:126) или связанный с последовательностями нуклеиновых кислот, кодирующими P2A элемент проскока рибосомы (последовательность, представленная в SEQ ID NO:8) для управления экспрессией рекомбинантного TCR из эндогенного целевого локуса гена (например, TRAC), 5’ плеча гомологии последовательности приблизительно 600 п.о. (например, представленной в SEQ ID NO:124), 3’ плеча гомологии последовательности приблизительно 600 п.о. (например, представленной в SEQ ID NO:125), которые гомологичны последовательностям окружающим целевой сайт интеграции в экзоне 1 человеческой TCR α постоянной области (TRAC) гена. В некоторых вариантах, матричный полинуклеотид дополнительно содержит другие последовательности нуклеиновых кислот, например, последовательности нуклеиновых кислот, кодирующие маркер, например, поверхностный маркер или маркер отбора. В некоторых вариантах, матричный полинуклеотид дополнительно содержит вирусный вектор последовательностей, например, вектор последовательностей адено-ассоциированного вируса (AAV).
Трансген, содержащийся на матричном полинуклеотиде, описанном здесь, может быть выделен из плазмид, клеток или других источников с применением известных стандартных методов, таких как ПЦР. Матричный полинуклеотид для применения может включать разные типы топологии, включая круговые сверхспиральные, круговые расслабленные, линейные и подобные. Альтернативно, они могут быть химически синтезированы с применением стандартных методов синтеза олигонуклеотида. Кроме того, матричные полинуклеотиды могут быть метилированы или не иметь метилирования. Матричные полинуклеотиды могут быть в форме бактериальных или дрожжевых искусственных хромосом (BAC или YAC).
Полинуклеотид может быть введен в клетку как часть векторной молекулы, имеющей дополнительные последовательности, такие как, например, точки начала репликации, промоторы и гены, кодирующие резистентность к антибиотикам. Более того, матричные полинуклеотиды могут быть введены в виде депротеинизированных нуклеиновых кислот, в виде нуклеиновых кислот в комплексе с материалами, такими как липосомы, наночастицы или полоксамер, или могут быть доставлены вирусами (например, аденовирусом, AAV, герпесвирусом, ретровирусом, лентивирусом и интеграза-дефективным лентивирусом (IDLV)).
В других аспектах, матричный полинуклеотид доставляет вирусными и/или не вирусными методами переноса гена. В некоторых вариантах, матричный полинуклеотид доставляют в клетку через адено-ассоциированный вирус (AAV). Может применяться любой AAV, включая, но не ограничиваясь ими, AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 и их сочетания. В некоторых случаях, AAV содержит LTR, которые являются гетерологическим серотипом по сравнению с капсидным серотипом (например, AAV2 ITRs с AAV5, AAV6 или AAV8 капсидами). Матричный полинуклеотид может быть доставлен с применением той же системы переноса гена, которую применяют для доставки нуклеазы (включая на том же векторе) или может быть доставлена с применением разных систем доставки с той, которой доставляют нуклеазу. В некоторых вариантах, матричный полинуклеотид доставляют с применением вирусного вектора (например, AAV) и нуклеазы доставляют в форме мРНК. Клетка также может быть процессирована одной или более молекулами, которые ингибируют связывание вирусного вектора с рецептором поверхности клетки, как описано здесь, до, одновременно с и/или после доставки вирусного вектора (например, несущего нуклеазы и/или матричный полинуклеотид).
В некоторых вариантах, матричный полинуклеотид содержится в вирусном векторе и составляет, по меньшей мере, от около 2500, 2750, 3000, 3250, 3500, 3750, 4000, 4250, 4500, 4760, 5000, 5250, 5500, 5750, 6000, 7000, 7500, 8000, 9000 или 10000 нуклеотидов в длину или любое значение между любыми из вышеописанных. В некоторых вариантах, полинуклеотид содержится в вирусном векторе и имеет от около 2500 и от около 5000 нуклеотидов, от около 3500 и от около 4500 нуклеотидов или от около 3750 нуклеотидов и от около 4250 нуклеотидов в длину. В некоторых вариантах, полинуклеотид содержится в вирусном векторе и составляет от около 2500, 2750, 3000, 3250, 3500, 3750, 4000, 4250, 4500, 4760, 5000, 5250, 5500, 5750, 6000, 7000, 7500, 8000, 9000 или 10000 нуклеотидов в длину.
В некоторых вариантах, матричный полинуклеотид является аденовирусным вектором, например, AAV вектором, например, онДНК молекулой с длиной и последовательностью, которые позволяют упаковать ее в AAV капсид. Вектор может быть, например, менее 5 т.н. и может содержать ITR последовательность, которая способствует упаковке в капсид. Вектор может иметь дефицит интеграции. В некоторых вариантах, матричный полинуклеотид содержит около 150 до 1000 нуклеотидов гомологии на каждой стороне трансгенного и/или целевого сайта. В некоторых вариантах, матричный полинуклеотид содержит около 100, 150, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов 5’ целевого сайта или трансгена, 3’ целевого сайта или трансгена или обоих 5’ и 3’ целевого сайта или трансгена. В некоторых вариантах, матричный полинуклеотид содержит, по меньшей мере, 100, 150, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов 5’ целевого сайта или трансгена, 3’ целевого сайта или трансгена или обоих 5’ и 3’ целевого сайта или трансгена. В некоторых вариантах, матричный полинуклеотид содержит не более 100, 150, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов 5’ целевого сайта или трансгена, 3’ целевого сайта или трансгена или обоих 5’ и 3’ целевого сайта или трансгена.
В некоторых вариантах, матричный полинуклеотид является лентивирусным вектором, например, IDLV (интеграция дефицитным лентивирусом). В некоторых вариантах, матричный полинуклеотид содержит около 500 до 1000 пар оснований гомологии на каждой стороне трансгена и/или целевого сайта. В некоторых вариантах, матричный полинуклеотид содержит около 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований гомологии 5’ целевого сайта или трансгена, 3’ целевого сайта или трансгена или обоих 5’ и 3’ целевого сайта или трансгена. В некоторых вариантах, матричный полинуклеотид содержит, по меньшей мере, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований гомологии 5’ целевого сайта или трансгена, 3’ целевого сайта или трансгена или обоих 5’ и 3’ целевого сайта или трансгена. В некоторых вариантах, матричный полинуклеотид содержит не более 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований гомологии 5’ целевого сайта или трансгена, 3’ целевого сайта или трансгена или обоих 5’ и 3’ целевого сайта или трансгена. В некоторых вариантах, матричный полинуклеотид содержит одну или более мутаций, например, молчащих мутаций, которые предотвращают распознавание и расщепление Cas9 матричным полинуклеотидом. Матричный полинуклеотид может содержать, например, по меньшей мере, 1, 2, 3, 4, 5, 10, 20 или 30 молчащих мутаций относительно соответствующей последовательности в геноме изменяемой клетки. В некоторых вариантах, матричный полинуклеотид содержит не более 2, 3, 4, 5, 10, 20, 30 или 50 молчащих мутаций относительно соответствующей последовательности в геноме изменяемой клетки. В некоторых вариантах, кДНК содержит одну или более мутаций, например, молчащих мутаций, которые предотвращают распознавание и расщепление Cas9 матричным полинуклеотидом. Матричный полинуклеотид может содержать, например, по меньшей мере, 1, 2, 3, 4, 5, 10, 20 или 30 молчащих мутаций относительно соответствующей последовательности в геноме изменяемой клетки. В некоторых вариантах, матричный полинуклеотид содержит не более 2, 3, 4, 5, 10, 20, 30 или 50 молчащих мутаций относительно соответствующей последовательности в геноме изменяемой клетки.
Двухнитевые матричные полинуклеотиды, описанные здесь, могут включать одно или более не природных оснований и/или скелетов. В частности, вставка матричного полинуклеотида с метилированными цитозинами может проводиться с применением способов, описанных здесь для достижения состояния транскрипционной неподвижности в рассматриваемой области.
Матричный полинуклеотид может содержать любой рассматриваемый трансген (экзогенную последовательность). Типовые экзогенные последовательности включают, но не ограничены ими, любой полипептид, кодирующий последовательность (например, кДНК или ее фрагменты), промотор последовательностей, энхансер последовательностей, эпитопные метки, маркерные гены, сайты распознавания расщепления фермента и разные типы конструктов экспрессии. Маркерные гены включают, но не ограничены ими, последовательности, кодирующие белок, который медиирует резистентность к антибиотику (например, резистентность к ампициллину, резистентность к неомицину, резистентность к G418, резистентность к пиромицину), последовательности, кодирующие окрашенные или флуоресцентные или люминесцентные белки (например, зеленый флуоресцентный белок, усиленный зеленый флуоресцентный белок, красный флуоресцентный белок, люциферазу) и белки, которые медиируют усиленный рост клетки и/или амплификацию гена (например, дигидрофолатредуктазы). Эпитопные метки включают, например, одну или более копий FLAG, His, myc, Tap, HA или любой определяемой аминокислотной последовательности.
В некоторых вариантах, трансген содержит полинуклеотид, кодирующий любой полипептид, экспрессия которого в клетку желательна, включая, но не ограничиваясь ими, антитела, антигены, ферменты, рецепторы (клеточной поверхности или ядерные), гормоны, лимфокины, цитокины, репортерные полипептиды, факторы роста и функциональные фрагменты любых из вышеперечисленных. В некоторых вариантах, экзогенная последовательность (трансген) содержит полинуклеотид, кодирующий один или более рекомбинантных рецепторов, например, функциональных не-TCR антигенных рецепторов, химерных антигенных рецепторов (CAR) и Т-клеточных рецепторов (TCR), таких как трансгенные TCR, сконструированные TCR или рекомбинантные TCR, и компоненты любых из вышеперечисленных.
В некоторых вариантах, кодирующими последовательностями могут быть, например, кДНК. Экзогенные последовательности также могут быть фрагментом трансгена для связывания с рассматриваемой эндогенной последовательности гена. Например, фрагмент трансгена, содержащий последовательность на 3’ окончании рассматриваемого гена, может применяться для коррекции, через вставку или замещение, последовательности, кодирующей мутацию в 3’ окончании эндогенной последовательности гена. Также, фрагмент может содержать последовательности, похожие с 5’ окончанием эндогенного гена для вставки/замещения эндогенных последовательностей для коррекции или модификации такой эндогенной последовательности. Дополнительно, фрагмент может кодировать рассматриваемый функциональный домен (каталитический, секреторный или подобный) для связывания in situ с последовательностью эндогенного гена с получением слитого белка.
В некоторых вариантах, трансген дополнительно кодирует один или более маркеров. В некоторых вариантах, одним или более маркерами являются маркер трансдукции, суррогатный маркер и/или маркер отбора.
В некоторых вариантах, маркером является маркер трансдукции или суррогатный маркер. Маркер трансдукции или суррогатный маркер могут применяться для определения клеток, которые введены с полинуклеотидом, например, полинуклеотидом, кодирующим рекомбинантный рецептор. В некоторых вариантах, маркер трансдукции может показывать или подтверждать модификацию клетки. В некоторых вариантах, суррогатным маркером является белок, который получают так, чтобы он соэкспрессировался на поверхности клетки с рекомбинантным рецептором, например, TCR или CAR. В конкретных вариантах, таким суррогатным маркером является белок поверхности, который модифицирован так, чтобы иметь незначительную или не иметь активности. В определенных вариантах, суррогатный маркер кодирован на том же полинуклеотиде, который кодирует рекомбинантный рецептор. В некоторых вариантах, последовательность нуклеиновых кислот, кодирующая рекомбинантный рецептор, функционально связана с последовательностью нуклеиновых кислот, кодирующей маркер, необязательно разделенный внутренним участком посадки рибосомы (IRES) или нуклеиновой кислотой, кодирующей пептид саморасщепления или пептид, который вызывает проскок рибосомы, такой как 2A последовательность, такая как T2A, P2A, E2A или F2A. Гены внешних маркеров, в некоторых случаях, могут применяться в сочетании со сконструированными клетками чтобы разрешить определение или отбор клеток, и, в некоторых случаях, также способствуют суициду клетки.
Типовые суррогатные маркеры могут включать усеченные формы полипептидов клеточной поверхности, такие как усеченные формы, которые не функциональны и не трансдуцируют или не способны к трансдукции сигнала или сигнала, обычно трансдуцируемого полной формой полипептида клеточной поверхности, и/или не делают или не способны к интернализации. Типовые усеченные полипептиды клеточной поверхности включают усеченные формы факторов роста или других рецепторов, таких как усеченный рецептор 2 человеческого эпидермального фактора роста (tHER2), усеченный рецептор эпидермального фактора (tEGFR, типовая последовательность tEGFR представлена в SEQ ID NO:12 или 13) или простатический специфический мембранный антиген (PSMA) или их модифицированные формы. tEGFR может содержать эпитоп, распознанный антителом цетуксимабом (Erbitux®) или другим терапевтическим анти-EGFR антителом или связывающей молекулой, которая может применяться для идентификации или выбора клеток, которые были сконструированы с tEGFR конструктом и кодированным экзогенным белком, и/или исключения или отделения клеток, экспрессирующих кодированный экзогенный белок. См. патент США № 8,802,374 и Liu et al., Nature Biotech. 2016 April; 34(4): 430-434). В некоторых аспектах, маркер, например, суррогатный маркер, включает все или часть (например, усеченную форму) CD34, NGFR, CD19 или усеченного CD19, например, усеченного рецептора не человеческого CD19 или эпидермального фактора роста (например, tEGFR). В некоторых вариантах, маркер содержит флуоресцентный белок, такой как зеленый флуоресцентный белок (GFP), усиленный зеленый флуоресцентный белок (EGFP), такой как супер-свернутый GFP (sfGFP), красный флуоресцентный белок (RFP), такой как tdTomato, mCherry, mStrawberry, AsRed2, DsRed или DsRed2, голубой флуоресцентный белок (CFP), сине-зеленый флуоресцентный белок (BFP), усиленный синий флуоресцентный белок (EBFP) и желтый флуоресцентный белок (YFP), и их варианты, включая варианты видов, мономерные варианты и кодон-оптимизированные и/или усиленные варианты флуоресцентных белков. В некоторых вариантах, маркер является или содержит фермент, такой как люцифераза, lacZ ген из E. coli, щелочная фосфатаза, секретированная эмбрионная щелочная фосфатаза (SEAP), хлорамфениколацетилтрансфераза (CAT). Типовые испускающие свет гены-репортеры включают люциферазу (luc), β-галактозидазу, хлорамфениколацетилтрансферазу (CAT), β-глюкуронидазу (GUS) или из варианты.
В некоторых вариантах, маркером является маркер отбора. В некоторых вариантах, маркер отбора является или содержит полипептид, который предоставляет резистентность к экзогенным агентами или лекарственным средствам. В некоторых вариантах, маркером отбора является ген резистентности к антибиотику. В некоторых вариантах, маркером отбора является ген резистентности к антибиотику, предоставляющий резистентность к антибиотику клеткам млекопитающих. В некоторых вариантах, маркер отбора является или содержит ген резистентности к Пиромицину, ген резистентности к Гигромицину, ген резистентности к Бластицидину, ген резистентности к Неомицину, ген резистентности к Генетицину или ген резистентности к Зеоцину или их модифицрованные формы.
В некоторых вариантах, нуклеиновая кислота, кодирующая маркер, функционально связана с полинуклеотидом, кодирующим для линкерной последовательности, такой как расщепляемая линкерная последовательность, например, T2A. Например, маркер и, необязательно, линкерная последовательность могут быть такими, как описаны в публикации РСТ № WO2014031687. Например, маркером может быть усеченный EGFR (tEGFR) который, необязательно, связан с линкерной последовательностью, такой как T2A расщепляемая линкерная последовательность. Типовой полипептид для усеченного EGFR (например, tEGFR) содержит последовательность аминокислот, представленную в SEQ ID NO: 12 или 13 или последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности с SEQ ID NO: 12 или 13.
В некоторых вариантах, маркером является молекула, например, белок поверхности клетки, не найденный в природе на T клетке или не найденный в природе на поверхности T клетки или ее части.
В некоторых вариантах, молекулой является не своя молекула, например, не свой белок, т.е. тот, который не распознается как “свой” иммунной системой хозяина, в который адоптивно переносят клетки.
В некоторых вариантах, маркер не имеет терапевтической функции и/или не оказывает действие, отличное от применения в качестве маркера для генной инженерии, например, для выбора клеток, успешно сконструированных. В других вариантах, маркером может быть терапевтическая молекула или молекула, другим образом оказывающая некий желаемый эффект, такая как лиганд для клетки, обнаруженной in vivo, такой как костимулирующая молекула или иммунная контрольная точка для улучшения и/или ослабления ответа клеток при адоптивном переносе и столкновении с лигандом.
В некоторых вариантах, такой трансген дополнительно включает T2A элемент проскока рибосомы и/или последовательность, кодирующую маркер, такую как tEGFR последовательность, например, ниже TCR или CAR, такую, как представлена в SEQ ID NO: 12 или 13, соответственно, или последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности с SEQ ID NO: 12 или 13.
В некоторых вариантах, матричный полинуклеотид кодирует рекомбинантный рецептор, который служит для направления функции T клетки. Химерными антигенными рецепторами (CAR) являются молекулы, сконструированные для направления иммунных клеток на определенные молекулярные цели, экспрессированных на поверхностях клеток. В наиболее основной форме, они являются рецепторами, введенными в клетку, которые сочетают домен специфичности, экспрессированный на внешней стороне клетки, с сигнальными путями внутри клетки так, что когда домен специфичности взаимодействует с его целью, клетки активируются. Другие CAR получают из вариантов рецептором T-клетки (TCR), где домен специфичности, такой как scFv или некоторый тип рецептора, сливают с сигнальным доменом TCR. Эти конструкты затем вводят в T клетку, позволяя T клетке активироваться в присутствии клетки, экспрессирующей целевой антиген, что вызывает атаку целевой клетки активированной T клеткой независимым от MHC образом (см. Chicaybam et at (2011) Int Rev Immunol 30:294-311). Альтернативно, кассеты экспрессии CAR могут быть введены в иммунную клетку для дальнейшего энграфтмента так, что CAR кассета находится под контролем специфического промотора T клетки (например, FOXP3 промотора, см. Mantel et. al (2006) J. Immunol 176: 3593-3602).
В качестве типового примера, матричный полинуклеотид включен в виде конструкта вектора адено-ассоциированного вируса (AAV), содержащего последовательность нуклеиновых кислот, кодирующую рекомбинантные TCR α и TCR β цепь и под контролем конститутивного промотора, окруженного плечами гомологии около 600 пар оснований каждый от 5’ и 3’ стороны последовательности нуклеиновых кислот, кодирующей рекомбинантный TCR, для целенаправленного воздействия на экзон 1 эндогенного TRAC гена. Типовое 5’ плечо гомологии для целенаправленного воздействия на TRAC включает последовательность, представленную в SEQ ID NO:124. Типовое 3’ плечо гомологии для целенаправленного воздействия на TRAC включают последовательность, представленную в SEQ ID NO:125.
При конструировании таких кассет экспрессии, следуя идеям данной спецификации, применяют методики, хорошо известные в области молекулярной биологии (см, например, Ausubel или Maniatis). До применения кассеты экспрессии для создания трансгенного животного, отвечаемость кассеты экспрессии на вызов стресса, связанного с выбранными контрольными элементами, может быть тестирована введением кассеты экспрессии в подходящую колонию клеток (например, первичные клетки, трансформированные клетки или бессмертные колонии клеток).
Направленная вставка не кодирующей последовательности нуклеиновых кислот также может быть проведена. Последовательности кодирующие антисмысловые РНК, РНКi, shРНК и микро РНК (miРНК) также могут применяться для направленных вставок. В дополнительных вариантах, матричный полинуклеотид может содержать не кодирующие последовательности, которые являются специфическими целевыми сайтами для дополнительных дизайнов нуклеазы. Затем, дополнительные нуклеазы могут быть экспрессированы в клетки так, что исходный матричный полинуклеотид расщепляется и модифицируется вставкой другого рассматриваемого матричного полинуклеотида. Таким образом, могут быть получены реитеративные интеграции матричных полинуклеотидов, позволяющие прамидирование признаков в конкретном рассматриваемом локусе, например, локусах TRAC, TRBC1 и/или TRBC2 гена.
В некоторых вариантах, полинуклеотид содержит структуру: [5′ плечо гомологии]-[трансгенная последовательность]-[3′ плечо гомологии]. В некоторых вариантах, полинуклеотид содержит структуру: [5′ плечо гомологии]-[мультицистронный элемент]-[трансгенная последовательность]-[3′ плечо гомологии]. В некоторых вариантах, полинуклеотид содержит структуру: [5′ плечо гомологии]-[промотор]-[трансгенная последовательность]-[3′ плечо гомологии].
4. Доставка матричных полинуклеотидов
В некоторых вариантах, полинуклеотид, например, полинуклеотид, такой как матричный полинуклеотид, кодирующий химерный рецептор, вводят в клетки в форме нуклеотида, например, в виде полинуклеотида или вектора. В конкретных вариантах, полинуклеотид содержит трансген, который кодирует химерный рецептор или его часть.
В некоторых вариантах, матричный полинуклеотид вводят в клетку для конструирования, в дополнение к агентам, способным вызывать направленное генетическое разрушение, например, нуклеазе и/или нРНК. В некоторых вариантах, матричные полинуклеотиды могут быть доставлены до, одновременно с или после агентов, способных вызывать направленное генетическое разрушение, вводимых в клетку. В некоторых вариантах, матричные полинуклеотиды доставляют одновременно в агентами. В некоторых вариантах, матричные полинуклеотиды доставляют до агентов, например, от секунд до часов до дней до агентов, включая, но не ограничиваясь ими, за 1-60 минут (или любое время между) до агентов, за 1-24 часа (или любое время между) до агентов или более чем за 24 часа до агентов. В некоторых вариантах, матричные полинуклеотиды доставляют после агентов, от секунд до часов до дней после агентов, включая сразу же после доставки агента, например, между или между около 30 секунд до 4 часов, например, около 30 секунд, 1 минуты, 2 минут, 3 минут, 4 минут, 5 минут, 6 минут, 6 минут, 8 минут, 9 минут, 10 минут, 15 минут, 20 минут, 30 минут, 40 минут, 50 минут, 60 минут, 90 минут, 2 часов, 3 часов или 4 часов после доставки агентов и/или, предпочтительно, в пределах 4 часов после доставки агентов. В некоторых вариантах, матричный полинуклеотид доставляют более чем через 4 часа после доставки агентов. В некоторых вариантах, матричные полинуклеотиды доставляют после агентов, например, включая, но не ограничиваясь ими, в пределах от 1 секунды до 60 минут (или любое время между) после агентов, от 1 до 4 часов (или любое время между) после агентов или более чем через 4 часа после агентов.
В некоторых вариантах, матричные полинуклеотиды могут быть доставлены с применением тех же систем доставки, что и агенты, способные вызывать направленное генетическое разрушение, например, нуклеаза и/или нРНК. В некоторых вариантах, матричные полинуклеотиды могут быть доставлены с применением систем, отличных от систем доставки агентов, способных вызывать направленное генетическое разрушение, например, нуклеазы и/или нРНК. В некоторых вариантах, матричный полинуклеотид доставляют одновременно с агентами. В других вариантах, матричный полинуклеотид доставляют в другое время, до или после доставки агентов. Любой из способов доставки, описанных здесь в разделе I.A.3 (например, в таблицах 7 и 8) для доставки нуклеиновых кислот в агенты, способные вызывать направленное генетическое разрушение, например, нуклеазу и/или нРНК, могут применяться для доставки матричного полинуклеотида.
В некоторых вариантах, один или более агентов и матричный полинуклеотид доставляют в одном и той же формате или способе. Например, в некоторых вариантах, и один или более агентов и матричный полинуклеотид содержатся в векторе, например, вирусном векторе. В некоторых вариантах, матричный полинуклеотид кодирован на том же остове вектора, например, AAV геноме, плазмиде ДНК, такой как Cas9 и нРНК. В некоторых аспектах, один или более агентов и матричный полинуклеотид находятся в разных форматах, например, комплекса рибонуклеиновая кислота-белок (RNP) для Cas9-нРНК агента и линейной ДНК для матричного полинуклеотида, но их доставляют с применением одинакового способа. В некоторых аспектах, один или более агентов и матричный полинуклеотид находятся в разных форматах, например, комплекса рибонуклеиновая кислота-белок (RNP) для Cas9-нРНК агента, и матричный полинуклеотид содержится в AAV векторе, и RNP доставляют с применением физического способа доставки (например, электропорации) и матричный полинуклеотид доставляют через трансдукцию AAV вирусных препаратов. В некоторых аспектах, матричный полинуклеотид доставляют сразу же после, например, в пределах около 1, 2, 3, 4, 5, 10, 20, 30, 40, 50 или 60 минут после доставки одного или более агентов.
В некоторых вариантах, матричным полинуклеотидом является линейная или кольцевая молекула нуклеиновой кислоты, такая как линейная или кольцевая ДНК или линейная РНК, и он может доставляться с применением любых способов, описанных в разделе I.A.3 здесь (например, таблицах 7 и 8) для доставки молекул нуклеиновой кислоты в клетку.
В конкретных вариантах, полинуклеотид, например, матричный полинуклеотид, вводят в клетки нуклеотидной форме, например, в виде или в пределах не вирусного вектора. В некоторых вариантах, не вирусный вектор является или включает полинуклеотид, например, ДНК или РНК полинуклеотид, который подходит для трансдукции и/или трансфекции любым подходящим и/или известным не вирусным способом доставки гена, таким как, но не ограниченным ими, микроинъекция, электропорация, временное сгущение или сжатие клеток (например, как описано у Lee, et al. (2012) Nano Lett 12: 6322-27), медиированная жирами трансфекция, медиированная пептидом доставка, например, проникающие в клетки пептиды, или их сочетание. В некоторых вариантах, не вирусный полинуклеотид доставляют в клетку не вирусным способом, описанным здесь, таким как не вирусные способы, перечисленные в таблице 8 здесь.
В некоторых вариантах, матричная полинуклеотидная последовательность может содержаться в векторе молекулы, содержащей последовательности, которые не гомологичны рассматриваемой области в геномной ДНК. В некоторых вариантах, вирусом является ДНК вирус (например, днДНК или онДНК вирус). В некоторых вариантах, вирусом является РНК вирус (например, онРНК или днРНК вирус). Типовые вирусные векторы/вирусы включают, например, ретровирусы, лентивирусы, аденовирус, адено-ассоциированный вирус (AAV), вирусы осповакцины, поксвирусы и вирусы простого герпеса, или любые из вирусов, описанных здесь.
В некоторых вариантах, матричный полинуклеотид может быть перенесен в клетки с применением рекомбинантных инфекционных вирусных частиц, таких как, например, векторы, полученные из вируса обезьян 40 (SV40), аденовирусов, адено-ассоциированных вирусов (AAV). В некоторых вариантах, матричный полинуклеотид переносят в T клетки с применением рекомбинантных лентивирусных векторов или ретровирусных векторов, таких как гамма-ретровирусные векторы (см., например, Koste et al. (2014) Gene Therapy 2014 Apr 3. doi: 10,1038/gt,2014,25; Carlens et al. (2000) Exp Hematol 28(10): 1137-46; Alonso-Camino et al. (2013) Mol Ther Nucl Acids 2, e93; Park et al., Trends Biotechnol. 2011 November 29(11): 550-557) или полученные из ВИЧ-1 лентивирусные векторы.
В некоторых вариантах, ретровирусный вектор имеет длинный концевой повтор (LTR), например, ретровирусный вектор, полученный из вируса мышиного лейкоза Молони (MoMLV), вируса миелопролиферативной саркомы (MPSV), вируса мышиных эмбриональных стволовых клеток (MESV), вируса мышиных стволовых клеток (MSCV) или вируса некроза селезенки (SFFV). Большинство ретровирусных векторов получают из мышиных ретровирусов. В некоторых вариантах, ретровирусы включают те, которые получены из любого источника клеток птиц или млекопитающих. Ретровирусы обычно являются амфотропными, что означает, что они способны заражать клетки хозяина нескольких видов, включая человека. В одном варианте, экспрессируемый ген заменяет ретровирусные gag, pol и/или env последовательности. Множество иллюстративных ретровирусных систем описано (например, Патенты США №№ 5,219,740; 6,207,453; 5,219,740; Miller and Rosman (1989) BioTechniques 7:980-990; Miller, A. D. (1990) Human Gene Therapy 1:5-14; Scarpa et al. (1991) Virology 180:849-852; Burns et al. (1993) Proc. Natl. Acad. Sci. USA 90:8033-8037; и Boris-Lawrie and Temin (1993) Cur. Opin. Genet. Develop. 3:102-109).
В некоторых вариантах, матричные полинуклеотиды и нуклеазы могут быть на одном и том же векторе, например, AAV векторе (например, AAV6). В некоторых вариантах, матричные полинуклеотиды доставляют с применением AAV вектора, и агенты, способные вызывать направленное генетическое разрушение, например, нуклеазу и/или нРНК, доставляют в другой форме, например, в виде мРНК кодирующей нуклеазы и/или нРНК. В некоторых вариантах, матричные полинуклеотиды и нуклеазы доставляют с применением одного и того же типа способа, например, вирусным вектором, но на отдельных векторах. В некоторых вариантах, матричные полинуклеотиды доставляют в разных системах доставки как агенты, способные вызывать генетическое разрушение, например, нуклеазы и/или нРНК. В некоторых вариантах, матричный полинуклеотид иссекают из остова вектора in vivo, например, он окружен последовательностями распознавания нРНК. В некоторых вариантах, матричный полинуклеотид находится на отдельной полинуклеотидной молекуле, такой как Cas9 и нРНК. В некоторых вариантах, Cas9 и нРНК вводят в форме рибонуклеопротеинового (RNP) комплекса, и матричный полинуклеотид вводят в виде полинуклеотидной молекулы, например, в векторе или линейной молекуле нуклеиновой кислоты, например, линейной ДНК. Типы или нуклеиновые кислоты и векторы для доставки включают любые из тех, которые описаны в разделе III здесь.
С. Оценка сконструированных Т клеток и композиций
В некоторых вариантах, способы включают оценку T клеток или T клеточных композиций, сконструированных для экспрессии рекомбинантных TCR для конкретных свойств. Например, способы включают оценку T клеток или T клеточных композиций для экспрессии рекомбинантного TCR на поверхности клеток и/или для распознавания пептида в контексте MHC молекулы. Например, в любом из представленных здесь вариантов, функциональные анализы могут быть проведены на T клетках или T клеточных композициях, экспрессирующих экзогенный рекомбинантный TCR, сконструированный или полученный с применением любых способов, описанных здесь. В некоторых вариантах, также могут быть проведены анализы для определения функциональности TCR и активности подачи сигнала TCR.
В некоторых вариантах, T клетки или T клеточные композиции оценивают на экспрессию рекомбинантного TCR на поверхности клетки, например, на возможность или способность экспрессировать функциональный TCR, такой как TCRαβ, на поверхности клетки. В некоторых вариантах, T клетки или T клеточные композиции оценивают на возможность или способность экспрессированных TCR распознавать пептид в контексте MHC молекулы, например, связывающих антигенов или эпитопов в контексте MHC молекулы. В некоторых вариантах, способы включают оценку T клеток или T клеточных композиций на T клеточную активность и/или функциональность. В некоторых вариантах, T клетки или T клеточные композиции оценивают на экспрессию маркера для трансдукции или введения трансгена.
В некоторых вариантах, T клетки или T клеточные композиции оценивают на экспрессию рекомбинантного TCR на поверхности клеток, например, на возможность или способность экспрессировать функциональный TCR, такой как TCRαβ, на поверхности клетки. В некоторых вариантах, оценка поверхностной экспрессии TCR включает контакт клеток каждой T клеточной композиции со связывающим реагентом, специфическим для TCRα цепи или TCRβ цепи, и оценку связывания реагента с клетками. В некоторых вариантах, связывающим реагентом является антитело. В некоторых вариантах, связывающий реагент является выявляемо меченным, необязательно, флуоресцентно меченным, прямо или косвенно. В некоторых вариантах, связывающий реагент является флуоресцентно меченным антителом, таким как антитело, меченное прямо или косвенно. В некоторых вариантах, связывающим реагентом является анти-pan-TCR Vβ антитело или является анти-pan-TCR Vα антитело. В некоторых вариантах, связывающий реагент распознает специфическое семейство цепей. В некоторых вариантах, связывающим реагентом является анти-TCR Vβ или анти-TCR Vα антитело, которое распознает или связывается со специфическим семейством, таким как анти-TCR Vβ22 антитело или анти-TCR Vβ2 антитело. В некоторых вариантах, экспрессия определяется с применением антител против одной или более общих частей, например, внеклеточных частей, TCR. Например, экспрессия TCR на поверхности клетки может быть определена с применением pan-реактивного анти-TCR антитела, такого как pan-реактивное TCR Vβ антитело или pan-реактивное TCR Vα антитело. Pan-реактивные антитела могут определять области TCR независимо от их специфичности связывания антигена или эпитопа. В некоторых вариантах, клетки окрашивают с применением связывающего реагента, например, меченного антитела, которое распознает экспрессию TCR на поверхности клетки, такого как меченное pan-реактивное TCR Vα антитело или его антигенсвязывающий фрагмент, и определяют с применением флуоресцентной микроскопии, проточной цитометрии или сортировки флуоресцентно-активированных клеток (FACS). В некоторых вариантах, идентифицируют и/или выбирают T клетки или T клеточные композиции, которые экспрессируют TCR на поверхности клетки, например, положительные к окрашиванию с применением pan-реактивных анти-TCR антител, таких как pan-реактивное TCR Vβ антитело или pan-реактивное TCR Vα антитело.
В некоторых вариантах, T клетки или T клеточные композиции оценивают на возможность или способность экпрессированных TCR распознавать пептид в контексте MHC молекулы, например, связывания антигенов или эпитопов в контексте MHC молекулы. Например, в некоторых вариантах, оценка T клеток или T клеточных композиций на распознавания пептида в контексте MHC молекулы включает: (1) контакт клеток или клеток T клеточной композиции с целевым антигеном, содержащим пептид-MHC комплекс, и (2) определение присутствия или отсутствия связывания пептид-MHC комплекса с клетками и/или определение присутствия или отсутствия активации T клетки TCR-экспрессирующих клеток при взаимодействии с пептид-MHC комплексом.
В некоторых вариантах, T клетки или T клеточные композиции, в которые вводят последовательности нуклеиновых кислот, кодирующие рекомбинантные TCR, тестируют подтверждением того, что рекомбинантные TCR связываются с желаемыми или известными антигенами, такими как TCR лиганд (MHC-пептидный комплекс). В некоторых вариантах, связывание клеток с антигеном или эпитопом может быть определено множеством способов. В некоторых способах, конкретный антиген, например, MHC-пептидный комплекс, может быть определяемо меченным так, чтобы можно было визуализировать связывание с рецептором, например, TCR. В некоторых вариантах, антиген может быть растворимым или экспрессироваться в растворимой форме. В некоторых вариантах, TCR лигандом может быть пептид-MHC тетрамер, и в некоторых случаях, пептид-MHC тетрамер может быть определяемо помечен, например, помечен флуоресцентной меткой. Пептид-MHC тетрамер может быть помечен прямо или косвенно. В некоторых вариантах, флуоресцентная метка может быть определена с применением проточной цитометрии или сортировки флуоресцентно-активированных клеток (FACS) или флуоресцентной микроскопией. В некоторых вариантах, способы включают идентификацию одной или более T клеток или T клеточных композиций, которые распознают пептид в контексте MHC молекулы, т.е. пептид-MHC комплекс.
В некоторых случаях, связывание TCR, такого как рекомбинантный TCR, с пептидным эпитопом, например, в комплексе с MHC, дает или запускает функциональное свойство взаимодействия. Например, T клетка, экспрессирующая TCR, такой как рекомбинантный TCR, при специфическом связывании с MHC-пептидный комплексом, может индуцировать путь трансдукции сигналов в клетке, вызывая клеточную экспрессию или секрецию эффекторной молекулы (например, цитокина), репортера или другого определяемого считывания взаимодействия, или вызывает T клеточную активацию или T клеточный ответ, такой как T клеточная пролиферация, образование цитокина, цитотоксический ответ T клетки или другие ответы. В некоторых вариантах, TCR, такой как рекомбинантный TCR, может специфически связываться с, и иммунологически распознавать пептидный эпитоп, так, что связывание с пептидным эпитопом вызывает иммунный ответ.
Способы тестирования TCR на способность распознавать пептидный эпитоп целевого полипептида и антигенную специфичность известны. В некоторых вариантах, T клетки или T клеточные композиции, полученные представленными способами, контактируют с пептид-MHC комплексом, либо в растворимой форме, либо через сокультивирование с активированными пептидами антигенпрезентирующими клетками (например, T2 клетками или другими известными антигенпрезентирующими клетками, которые совпадают с MHC аллелью рекомбинантного TCR). Типовые антигены и MHC аллели рекомбинантного TCR описаны в разделе III. В некоторых вариантах, способы включают оценку свойств, таких как функциональные свойства, экзогенного рекомбинантного TCR. В некоторых вариантах, способ включает оценку активации T клетки через экзогенный рекомбинантный TCR, например, определение присутствия или отсутствия активации T клетки TCR-экспрессирующими клетками при взаимодействии с пептид-MHC комплексом. В некоторых вариантах, считывание активации T клетки такими способами включает выделение цитокинов (например, интерферона-γ, гранулоцитарного/моноцитарного колониестимулирующего фактора (GM-CSF), фактора некроза опухоли (TNF-α) или интерлейкина 2 (IL-2)). Кроме того, функция TCR может быть оценена измерение клеточной цитотоксичности, как описано в Zhao et al., J. Immunol., 174:4415-4423 (2005).
В некоторых вариантах, оценка активации T клетки включает оценку активности или экспрессии молекул нуклеиновой кислоты, кодирующих репортер, например, репортер активации T клетки, оценивая выделение цитокинов, и/или оценивая функциональную активность T клетки.
В некоторых вариантах, один или более анализов включают один или более измерительных приборов, тип результата или анализа и/или считывание данных. В некоторых вариантах, один или более анализов проводят с применением флуоресцентно меченных реагентов, таких как антитела, прямо или косвенно меченных флуорофорами, и определяют с применением инструмента проточной цитометрии или сортировки флуоресцентно-активированных клеток (FACS). Например, для проточной цитометрии или FACS, может быть определено множество разных флуорофоров, которые имеют разные пики длин волн возбуждения и испускания. Таким образом, множество флуорофорных меток может применяться для оценки множества свойств, например, экспрессии TCR, распознавания пептида в контексте MHC молекулы и/или экспрессии репортера активации T клетки, в одной экспериментальной реакции. В некоторых вариантах, один или более анализов проводят с высокой пропускной способностью, мультиплексированным и/или крупномасштабным методом.
В некоторых вариантах, способы дополнительно включают аспекты оценки активации T клетки, такие как оценка выделения цитокинов и/или оценка функциональной активности T клетки, например, цитолитической активности и/или хелперной T клеточной активности. В некоторых вариантах, оценки могут быть проведены для T клеток или T клеточных композиций, полученных с применением описанных здесь вариантов.
В некоторых вариантах, функциональные анализы проводят на первичных T клетках, таких, которые выделены непосредственно у субъекта и/или выделены у субъекта и заморожены, таких как первичные CD4+ и/или CD8+ T клетки, которые были сконструированы с применением представленных здесь вариантов.
В некоторых вариантах, способы включают проведение функциональных анализов или определение функции TCR или T клетки. Например, функциональные анализы для определения активности TCR или активности T клетки включают определение секреции цитокина, цитолитической активности и/или хелперной T клеточной активности. Например, оценка T клеточной активации включает оценку выделения цитокинов и/или оценку функциональной активности T клетки. В некоторых вариантах, при связывании TCR с антигеном или эпитопом, цитоплазматический домен или внутриклеточный связывающий домен TCR активирует, по меньшей мере, одну из нормальных эффекторных функций или ответов иммунной клетки, например, T клетки, сконструированной для экспрессии TCR. Например, в некоторых контекстах, TCR вызывает функционирование T клетки, такое как цитолитическая активность и/или хелперная Т клеточная активность, такая как секреция цитокинов или других факторов. В некоторых вариантах, внутриклеточный связывающий домен или домены включают цитоплазматические последовательности Т-клеточного рецептора (TCR), и в некоторых аспектах, также со-рецепторов, которые в природном контексте действуют согласованно с таким рецептором для инициации трансдукции сигнала после взаимодействия антигенного рецептора, и/или любого производного или варианта таких молекул, и/или любой синтетической последовательности, которая имеет такую же функциональную способность.
В некоторых вариантах, T клетки или T клеточные композиции, содержащие экзогенные рекомбинантные TCR, оценивают для иммунологического считывания, например, с применением анализа T клетки. В некоторых вариантах, TCR-экспрессирующие клетки могут активировать ответ CD8+ T клетки. В некоторых вариантах, ответы CD8+ T клетки могут быть оценены отслеживанием реакционной способности CTL с применением анализов, которые включают, но не ограничены ими, целевой лизис клеток через выделение 51Cr, анализы целевого лизиса клеток с применением реагентов визуализации в реальном масштабе времени, анализы целевого лизиса клеток с применением реагента определения апопотоза (например, реагента Каспазы 3/7) или определение выделения интерферона гамма, например, твердофазным ИФА (ELISA), внутриклеточным цитокиновым окрашиванием или ELISPOT. В некоторых вариантах, TCR-экспрессирующие клетки могут активировать ответ CD4+ T клетки. В некоторых аспектах, ответы CD4+ T клетки могут быть оценены анализами, которые измеряют пролиферацию, например, введением [3H]-тимидина в клеточную ДНК и/или образование цитокинов, например, ELISA, внутриклеточным окрашиванием цитокина или ELISPOT. В некоторых случаях, цитокин может включать, например, интерлейкин-2 (IL-2), интерферон-гамма (IFN-гамма), интерлейкин-4 (IL-4), TNF-α, интерлейкин-6 (IL-6), интерлейкин-10 (IL-10), интерлейкин-12 (IL-12) или TGF β. В некоторых вариантах, распознавание или связывание пептидного эпитопа, такого как MHC класса I или класса II эпитопа, через TCR, может вызвать или активировать ответ CD8+ T клетки и/или ответ CD4+ T клетки.
II. КЛЕТКИ ДЛЯ ГЕННОЙ ИНЖЕНЕРИИ
В некоторых из представленных вариантов, клетками для конструирования являются иммунные клетки, такие как T клетки. Представлены генетически сконструированные клетки или популяции клеток, где одна или более из клеток содержит нокаут одного или более эндогенных TCR генов и кодирующие рекомбинантный рецептор нуклеиновые кислоты и/или другой трансген, которые интегрированы в одни или более генов эндогенного TCR. Также представлены популяции или композиции таких клеток, композиции, содержащие такие клетки и/или обогащенные клетками, которые сконструированы с применением представленных способов.
В некоторых вариантах, клетки для конструирования включают один или более подмножеств T клеток или других типов клеток, таких как популяции полных T клеток, CD4+ клетки, CD8+ клетки и их субпопуляции, такие, которые определены функцией, состоянием активации, зрелостью, потенциалом для дифференциации, экспансией, рециркуляцией, локализацией и/или потенциалом жизнестойкости, антиген-специфичностью, типом антигенного рецептора, присутствием конкретном органе или отделе, профилем выбора маркера или цитокина и/или степенью дифференциации. Со ссылкой на подлежащего лечению субъекта, клетки могут быть аллогенными и/или аутологичными. В число способов входят доступные способы. В некоторых аспектах, таких как доступные методики, клетки являются плюрипотентными и/или мультипотентными, такими как стволовые клетки, такие индуцированные плюрипотентные стволовые клетки (иПСК). В некоторых вариантах, способы включают выделение клеток у субъекта, их приготовление, обработку, культивирование и/или конструирование, как описано здесь, и повторное введение тому же пациенту, до или после криоконсервации.
Подтипы и субпопуляции T клеток и/или CD4+ и/или CD8+ T клеток включают наивные T (TN) клетки, эффекторные T клетки (TEFF), T клетки памяти и их подтипы, такие как стволовые T клетки памяти (TSCM), центральные T клетки памяти (TCM), эффекторные T клетки памяти (TEM) или терминально-дифференцированные эффекторные T клетки памяти, инфильтрующие опухоли лимфоциты (TIL), незрелые T клетки, зрелые T клетки, хелперные T клетки, цитотоксические T клетки, мукозальные инвариантные T (MAIT) клетки, существующие в природе и адаптивные регулирующие T (Treg) клетки, хелперные T клетки, такие как TH1 клетки, TH2 клетки, TH3 клетки, TH17 клетки, TH9 клетки, TH22 клетки, фолликулярные хелперные T клетки, альфа/бета T клетки, и дельта/гамма T клетки. В некоторых вариантах, клеткой является регулирующая T клетка (Treg). В некоторых вариантах, клетка дополнительно содержит рекомбинантные FOXP3 или их вариант.
В некоторых вариантах, клетки включают одну или более нуклеиновых кислот, введенных генной инженерией, и поэтому экспрессирующих рекомбинантные или генетически сконструированные продукты таких нуклеиновых кислот. В некоторых вариантах, нуклеиновые кислоты являются гетерологичными, т.е. обычно не присутствующими в клетке или образце, полученном из клетки, такими, которые получены из другого организма или клетки, которые, например, обычно не находят в сконструированной клетке, и/или организме, из которого такая клетка получена. В некоторых вариантах, нуклеиновые кислоты являются не существующими в природе, например, нуклеиновой кислотой, не найденной в природе, включая такую, которая содержит химерные сочетания нуклеиновых кислот, кодирующих разные домены из множества разных типов клеток.
В некоторых вариантах, получение сконструированных клеток включает одну или более стадий культивирования и/или приготовления. Клетки для конструирования могут быть выделены из образца, такого как биологический образец, например, полученный из или производный из субъекта. В некоторых вариантах, субъектом, из которого выделяют клетки, является такой, который имеет заболевание или состояние или нуждается в клеточной терапии, или которому вводят клеточную терапию. Субъектом в некоторых вариантах является человек, нуждающийся в конкретном терапевтическом вмешательстве, таком как адоптивная клеточная терапия, для которой выделены, процессированы и/или сконструированы клетки.
Следовательно, клетками в некоторых вариантах являются первичные клетки, например, первичные человеческие клетки. Образцы включают ткань, жидкость и другие образцы, взятые непосредственно у субъекта, а также образцы, полученные на одной или более стадиях обработки, таких как разделение, центрифугирование, генная инженерия (например, трансдукция с вирусным вектором), промывание и/или инкубирование. Биологическим образцом может быть образец, полученный непосредственно из биологического источника, или образец, который процессирован. Биологические образцы включают, но не ограничены ими, жидкости тела, такие как кровь, плазма, сыворотка, спинномозговая жидкость, синовиальная жидкость, моча и пот, образцы ткани и органов, включая процессированные образцы, полученные из них.
В некоторых аспектах, образцом, из которого получают или выделяют клетки, является кровь или полученный из крови образец, или является или является полученным из афереза лейкафереза продукт. Типовые образцы включают цельную кровь, мононуклеары периферической крови (PBMC), лейкоциты, костный мозг, вилочковую железу, биопсию ткани, опухоль, лейкоз, лимфому, лимфатический узел, кишечную лимфоидную ткань, лимфоидную ткань слизистых оболочек, печень, легкое, желудок, кишечник, толстую кишку, почки, поджелудочную железу, молочную железу, кость, простату, шейку матки, яички, яичники, гланды или другой орган, и/или полученные из них клетки. Образцы включают, в контексте клеточной терапии, например, адоптивной клеточной терапии, образцы от аутологичных и аллогенных источников.
В некоторых вариантах, клетки получают из колоний клеток, например, колоний T клеток. Клетки в некоторых вариантах получают из ксеногенного источника, например, от мыши, крысы, не человекообразного примата или свиньи.
В некоторых вариантах, выделение клеток включает одну или более из стадий приготовления и/или неаффинного клеточного разделения. В некоторых примерах, клетки промывают, центрифугируют и/или инкубируют в присутствии одного или более реагентов, например, для удаления нежелательных компонентов, обогащения желательными компонентами, лизирования или удаления клеток, чувствительных к конкретным реагентам. В некоторых примерах, клетки разделяют на основе одного или более свойств, таких как плотность, свойства адгерентности, размер, чувствительность и/или резистентность к конкретным компонентам.
В некоторых примерах, клетки из кровотока субъекта получают, например, аферезом или лейкаферезом. Образцы, в некоторых аспектах, содержит лимфоциты, включающие T клетки, моноциты, гранулоциты, B клетки, другие ядросодержащие белые кровяные клетки, красные кровяные клетки и/или тромбоциты, и в некоторых аспектах, содержат клетки, отличные от красных кровяных клеток и тромбоцитов.
В некоторых вариантах, кровяные клетки, собранные у субъекта, промывают, например, для удаления фракции плазмы и для помещения клеток в подходящий буфер или среду для дальнейшей обработки. В некоторых вариантах, клетки промывают физиологическим раствором с фосфатным буфером (PBS). В некоторых вариантах, промывочные растворы не содержат кальций и/или магний и/или многие или все двухвалентные катионы. В некоторых аспектах, стадию промывания проводят в полуавтоматической “проточной” центрифуге (например, Cobe 2991 cell processor, Baxter) согласно инструкциям производителя. В некоторых аспектах, стадию промывания осуществляют тангенциальной поточной фильтрацией (TFF) согласно инструкциям производителя. В некоторых вариантах, клетки ресуспендируют во множестве биосовместимых буферов после промывки, таких как, например, не содержащий Ca++/Mg++ PBS. В определенных вариантах, компоненты образца клеток крови удаляют, и клетки ресуспендируют непосредственно в культуральной среде.
В некоторых вариантах, способы включают способы разделения клеток на основе плотности, такие как получение белых кровяных клеток из периферической крови лизированием красных кровяных клеток и центрифугированием через градиент Перколла или Фиколла.
В некоторых вариантах, способы выделения включают разделение разных типов клеток на основе экспрессии или присутствия в клетке одной или более специфических молекул, таких как поверхностные маркеры, например, поверхностные белки, внутриклеточные маркеры или нуклеиновые кислоты. В некоторых вариантах, может применяться любой способ разделения на основе таких маркеров. В некоторых вариантах, разделением является разделение на основе аффинности или иммуноаффинности. Например, выделение в некоторых аспектах включает разделение клеток и клеточных популяций на основе экспрессии клеток или уровня экспрессии одного или более маркеров, обычно клеточных поверхностных маркеров, например, инкубированием с антителом или связывающим партнером, который специфически связывается с такими маркерами, обычно с последующими стадиями промывания и разделения клеток, связанных с антителом или связывающим партнером, от клеток, не имеющих связи с антителом или связывающим партнером.
Такие стадии разделения могут быть основаны на положительной селекции, при которой клетки, связывающие реагенты, остаются для дальнейшего использования, и/или отрицательной селекции, при которой остаются клетки, не имеющие связи с антителом или связывающим партнером. В некоторых примерах, обе фракции остаются для последующего применения. В некоторых аспектах, отрицательная селекция может быть особенно полезна, если недоступно антитело, которое специфически идентифицирует тип клетки в гетерогенной популяции, так как такое разделение наилучшим образом проводят на основе маркеров, экспрессированных клетками, отличными от желаемой популяции.
Разделение не обязательно дает 100% обогащение или удаление конкретной популяции клеток или клеток, экспрессирующих конкретный маркер. Например, положительная селекция или обогащение клетками конкретного типа, такими, которые экспрессируют маркер, относится к увеличению количества или доли таких клеток, но не обязательно дает полное отсутствие клеток, не экспрессирующих маркер. Также, отрицательная селекция, удаление или истощение клеток конкретного типа, таких, которые экспрессируют маркер, относится к снижению количества или доли таких клеток, но не обязательно дает полное удаление всех таких клеток.
В некоторых примерах проводится несколько раундов стадий разделения, где положительно или отрицательно выбранная фракция с одной стадии подвергается другой стадии разделения, например последующей положительной или отрицательной селекции. В некоторых примерах, одна стадия разделения может истощить клетки, экспрессирующие множество маркеров одновременно, например, путем инкубации клеток с множеством антител или связывающих партнеров, каждый из которых специфичен для маркера, нацеленный на отрицательную селекцию. Аналогичным образом, множество типов клеток могут быть одновременно положительно выбраны путем инкубации клеток с множеством антител или связывающих партнеров, экспрессированных на различных типах клеток.
Например, в некоторых аспектах, конкретные субпопуляции T клеток, таких как клетки положительные или экспрессирующие высокие уровни одного или более поверхностных маркеров, например, CD28+, CD62L+, CCR7+, CD27+, CD127+, CD4+, CD8+, CD45RA+, и/или CD45RO+ T клетки, выделяют методами положительной или отрицательной селекции.
Например, CD3+, CD28+ T клетки могут быть положительно выбраны с применением анти-CD3/анти-CD28 конъюгированных магнитных сфер (например, DYNABEADS® M-450 CD3/CD28 T Cell Expander).
В некоторых вариантах, выделение проводят обогащением конкретной популяции клеток положительной селекцией или обеднением конкретной популяции клеток отрицательной селекцией. В некоторых вариантах, положительную или отрицательную селекцию. проводят инкубированием клеток с одним или более антителами или другим связывающим агентом, который специфически связывается с одним или более поверхностными маркерами, экспрессированными или экспрессированными (маркер+) при относительно высоком уровне (маркервысокий) положительно или отрицательно выбранных клеток, соответственно.
В некоторых вариантах, T клетки отделяют от образца PBMC отрицательным выбором маркеров, экспрессированных на не T клетках, таких как B клетки, моноциты или другие белые кровяные клетки, такие как CD14. В некоторых аспектах, стадию выбора CD4+ или CD8+ применяют для отделения CD4+ хелпера и CD8+ цитотоксических T клеток. Такие CD4+ и CD8+ популяции могут быть далее отсортированы на субпопуляции положительной или отрицательной селекцией для маркеров, экспрессированных или экспрессированных до относительно высокой степени на одной или более субпопуляций наивных, клеток памяти и/или эффекторных T клеток.
В некоторых вариантах, CD8+ клетки дополнительно обогащают или обедняют наивными, центральными клетками памяти, эффекторными клетками памяти и/или центральными стволовыми клетками памяти, например, положительной или отрицательной селекцией на основе поверхностных антигенов, ассоциированных с соответствующей субпопуляцией. В некоторых вариантах, обогащение центральными T клетками памяти (TCM) проводят для повышения эффективности, например, для улучшения долговременной выживаемости, размножения и/или энграфтмента после введения, что в некоторых аспектах особенно устойчиво в таких субпопуляциях. См. Terakura et al. (2012) Blood,1:72-82; Wang et al. (2012) J Immunother. 35(9):689-701. В некоторых вариантах, объединение TCM-обогащенных CD8+ T клеток и CD4+ T клеток дополнительно увеличивает эффективность.
В вариантах, T клетки памяти присутствуют в обоих CD62L+ и CD62L- подмножествах CD8+ лимфоцитов периферической крови. PBMC могут быть обогащены или обеднены CD62L-CD8+ и/или CD62L+CD8+ фракциями, например, с применением анти-CD8 и анти-CD62L антител.
В некоторых вариантах, обогащение центральными T клетками памяти (TCM) основано на положительной или высокой поверхностной экспрессии CD45RO, CD62L, CCR7, CD28, CD3, и/или CD 127; в некоторых аспектах, оно основано на отрицательной селекции для клеток, экспрессирующих или высоко экспрессирующих CD45RA и/или гранзим B. В некоторых аспектах, выделение CD8+ популяции, обогащенной TCM клетками, проводят обеднением клеток, экспрессирующих CD4, CD14, CD45RA, и положительной селекцией или обогащением клеток, экспрессирующих CD62L. В одном аспекте, обогащение центральными T клетками памяти (TCM) клетки проводят, начиная с отрицательной фракции клеток, выбранных на основе CD4 экспрессии, которую подвергают отрицательной селекции на основе экспрессии CD14 и CD45RA, и положительно селекции на основе CD62L. Такие изменения в некоторых аспектах проводят одновременно, и в других аспектах, поводят последовательно, в любом порядке. В некоторых аспектах, одинаковую стадию селекции на основе экспрессии CD4, применяемую для получения популяции или субпопуляции CD8+ клеток, также применяют для создания популяции или субпопуляции CD4+ клеток, так, что обе, положительная и отрицательная фракции после сепарации на основе CD4, остаются и применяются на последующих стадиях способов, необязательно после одной или более дополнительных стадий положительной или отрицательной селекции.
В конкретном примере, образец PBMC или образец других белых кровяных клеток подвергают селекции CD4+ клеток, где остаются и отрицательные и положительные фракции. Отрицательную фракцию затем подвергают отрицательной селекции на основе экспрессии CD14 и CD45RA или ROR1, и положительной селекции на основе маркерных характеристик центральных T клеток памяти, таких как CD62L или CCR7, где положительные и отрицательные селекции проводят в любом порядке.
CD4+ T хелперные клетки сортируют на наивные, центральные клетки памяти и эффекторные клетки идентификацией клеточных популяций, которые имеют антигены на поверхности клетки. CD4+ лимфоциты могут быть получены стандартными методами. В некоторых вариантах, наивными CD4+ T лимфоцитами являются CD45RO-, CD45RA+, CD62L+, CD4+ T клетки. В некоторых вариантах, центральными клетками памяти CD4+ являются CD62L+ и CD45RO+. В некоторых вариантах, эффекторными CD4+ клетками являются CD62L- и CD45RO-.
В одном примере, для обогащения CD4+ клеток отрицательной селекцией, коктейль моноклональных антител обычно включает антитела к CD14, CD20, CD11b, CD16, HLA-DR и CD8. В некоторых вариантах, антитело или связывающий партнер связывается с твердой подложкой или матрицей, такой как магнитные сферы или парамагнитные сферы, чтобы позволить разделение клеток для положительной и/или отрицательной селекции. Например, в некоторых вариантах, клетки и популяции клеток разделяют или изолируют с применением методик иммуномагнитного (или аффинномагнитного) разделения (обзор представлен в Methods in Molecular Medicine, vol. 58: Metastasis Research Protocols, Vol. 2: Cell Behavior In vitro и In vivo, p 17-25 Edited by: S. A. Brooks и U. Schumacher © Humana Press Inc., Totowa, NJ).
В некоторых аспектах, образец или композицию отделяемых клеток инкубируют с небольшим намагничиваемым или магнитно восприимчивым материалом, таким как магнитно восприимчивые частицы или микрочастицы, такие как парамагнитные (например, такие как Dynalbeads или MACS сферы). Магнитно восприимчивый материал, например, частица, в основном прямо или косвенно присоединен к связывающему партнеру, например, антителу, которое специфически связывается с молекулой, например, поверхностным маркером, присутствующем на клетке, клетках или популяции клеток, которые желательно отделить, например, которые желательно отрицательно или положительно выбрать.
В некоторых вариантах, магнитная частица или сфера содержит магнитно восприимчивый материал, связанный со специфическим связывающим членом, таким как антитело или другой связывающий партнер. Существует множество хорошо известных магнитно восприимчивых материалов, применяемых в способах магнитного разделения. Подходящие магнитные частицы включают такие, которые описаны у Molday, Патент США № 4,452,773, и в спецификации европейского патента EP 452342 B, которые включены сюда в качестве ссылки. Коллоидные отсортированные частицы, такие как описаны у Owen, Патент США № 4,795,698, и Liberti et al., Патент США № 5,200,084 являются другими примерами.
Инкубирование обычно проводят в условиях, где антитела или связывающие партнеры или молекулы, такие как вторичные антитела или другие реагенты, которые специфически связываются с такими антителами или связывающими партнерами, которые присоединены в магнитной частице или сфере, специфически связываются с молекулами клеточной поверхности, если присутствует на клетках в образце.
В некоторых аспектах, образец помещают в магнитное поле, и клетки, имеющие магнитно восприимчивые или намагничиваемые частицы, присоединенные к нему, будут привлекаться к магниту и отделяться от не меченных клеток. Для положительной селекции, клетки, которые привлечены к магниту, остаются; для отрицательной селекции клетки, которые не привлечены (не меченные клетки) остаются. В некоторых аспектах, сочетание положительной и отрицательной селекции проводят во время одной и той же стадии селекции, где положительные и отрицательные фракции сохраняются и далее обрабатываются или подвергаются дальнейшему разделению.
В определенных вариантах, магнитно восприимчивые частицы покрыты первичными антителами или другими связывающими партнерами, вторичными антителами, лектинами ферментами или стрептавидином. В определенных вариантах, магнитные частицы присоединены к клеткам через покрытие первичными антителами, специфическими для одного или более маркеров. В определенных вариантах, клетки, скорее чем сферы, мечены первичным антителом или связывающим партнером, и затем добавляют покрытые специфическими к клеточному типу вторичными антителами или другим связывающим партнером (например, стрептавидином) магнитные частицы. В определенных вариантах, покрытые стрептавидином магнитные частицы применяют в сочетании с биотинилированными первичными или вторичными антителами.
В некоторых вариантах, магнитно восприимчивые частицы остаются присоединенными к клеткам, которые затем инкубируют, культивируют и/или конструируют; в некоторых аспектах, частицы остаются присоединенными к клеткам для введения пациенту. В некоторых вариантах, намагничиваемые или магнитно восприимчивые частицы удаляют с клеток. Способы удаления намагничиваемых частиц с клеток известны и включают, например, применение конкурирующих не меченных антител, намагничиваемых частиц или антител, конъюгированных с расщепляемыми линкерами, и т.д. В некоторых вариантах, намагничиваемые частицы являются биоразлагаемыми.
В некоторых вариантах, селекцию на основе аффинности проводят через активируемую магнитным полем сортировку клеток (MACS) (Miltenyi Biotec, Auburn, CA). Системы активируемой магнитным полем сортировки клеток (MACS) способны с высокой чистотой выбирать клетки, имеющие намагничиваемые частицы, присоединенные к ним. В определенных вариантах, MACS работает в режиме, где не целевые и целевые виды далее элюируют после применения внешнего магнитного поля. То есть клетки, присоединенные к намагничиваемым частицам, хранят на месте, пока не присоединенные виды элюируют. Затем, после завершения этой первой стадии элюирования, виды, которые захвачены магнитным полем и сохранены от элюирования, высвобождают любым способом так, чтобы элюировать и восстановить их. В определенных аспектах, не целевые клетки мечены и обеднены от гетерогенной популяции клеток.
В определенных вариантах, выделение или разделение проводят с применением системы, устройства или аппарата, которые проводят одну или более из стадий выделения, клеточного препарата, разделения, обработки, культивирования и/или составления способов. В некоторых аспектах, систему применяют для проведения каждой из этих стадий в закрытой или стерильной окружающей среде, например, для минимизации ошибки, ручных манипуляций и/или загрязнения. В одном примере, системой является система, описанная в международной публикации PCT № WO2009/072003 или US 20110003380 A1.
В некоторых вариантах, система или аппарат проводит одну или более, например, все стадии выделения, обработки, конструирования и составления в интегрированной или замкнутой системе и/или в автоматизированном или программируемом режиме. В некоторых аспектах, система или аппарат включает компьютер и/или компьютерную программу, связанную с системой или аппаратом, что позволяет пользователю программировать, контролировать, оценивать результат и/или корректировать разные аспекты стадий обработки, выделения, конструирования или составления.
В некоторых аспектах, разделение и/или другие стадии проводят с применением системы CliniMACS (Miltenyi Biotec), например, для автоматизированного разделения клеток в клиническом масштабе в закрытой и стерильной системе. Компоненты могут включать встроенный микрокомпьютер, блок магнитного разделения, перистальтический насос и разные запорные клапаны. Встроенный компьютер в некоторых аспектах контролирует все компоненты инструмента и направляет систему на проведение повторяющихся процедур в стандартизированной последовательности. Блок магнитного разделения в некоторых аспектах включает подвижный постоянный магнит и держатель для разделяющей колонки. Перистальтический насос контролирует скорость потока чрез комплект трубок и, вместе с запорными клапанами, обеспечивает контролируемый поток буфера через систему и непрерывную суспензию клеток.
Система CliniMACS в некоторых аспектах применяет сопряженные с антителом намагничиваемые частицы, которые подают в стерильном непирогенном растворе. В некоторых вариантах, после мечения клеток магнитными частицами, клетки промывают для удаления избыточных частиц. Мешок для клеточного препарата затем присоединяют к комплекту трубок, который затем присоединяют к мешку, содержащему буфер и мешку для сбора клеток. Комплект трубок состоит из предварительно собранных стерильных трубок, включающих предколонку и разделительную колонку, и предназначен только для однократного применения. После начала программы разделения, система автоматически наносит образец клеток на делительную колонку. Меченные клетки остаются внутри колонки, а не меченные клетки удаляют несколькими стадиями промывки. В некоторых вариантах, популяции клеток для применения в способах, описанных здесь, не мечены и не остаются в колонке. В некоторых вариантах, популяции клеток для применения в способах, описанных здесь, мечены и остаются в колонке. В некоторых вариантах, популяции клеток для применения в способах, описанных здесь, элюируют из колонки после удаления магнитного поля и собирают в мешок для сбора клеток.
В определенных вариантах, разделение и/или другие стадии проводят с применением системы CliniMACS Prodigy (Miltenyi Biotec). Система CliniMACS Prodigy в некоторых аспектах оборудована блоком обработки клеток, который позволяет автоматизированное промывание и фракционирование клеток центрифугированием. Система CliniMACS Prodigy также может включать встроенную камеру и программу для распознавания изображений, которая определяет оптимальную конечную точку фракционирования клеток через распознавание макроскопических слоев источника клеточного продукта. Например, периферическая кровь может быть автоматически разделена на слои эритроцитов, белых кровяных клеток и плазмы. Система CliniMACS Prodigy может также включать встроенную камеру для культивирования клеток, которая выполняет протоколы клеточных культур, такие как, например, дифференциация и разрастание клеток, загрузка антигена и долгосрочное культивирование клеток. Входные порты могут позволить стерильное удаление и пополнение среды, и клетки можно отслеживать с применением встроенного микроскопа. См., например, Klebanoff et al. (2012) J Immunother. 35(9): 651-660, Terakura et al. (2012) Blood, 1:72-82, и Wang et al. (2012) J Immunother.35(9):689-701.
В некоторых вариантах, описанную здесь клеточную популяцию собирают и обогащают (или обедняют) проточной цитометрией, в которой клетки, окрашенные для множества маркеров поверхности клетки, вводят в жидкий поток. В некоторых вариантах, описанную здесь клеточную популяцию собирают и обогащают (или обедняют) препаративной (FACS) сортировкой. В определенных вариантах, описанную здесь клеточную популяцию собирают и обогащают (или обедняют) применением чипов микроэлектромеханических систем (MEMS) в сочетании с системой определения на основе FACS (см., например, WO 2010/033140, Cho et al. (2010) Lab Chip 10:1567-1573; и Godin et al. (2008) J Biophoton. 1(5):355-376). В обоих случаях, клетки могут быть мечены множеством маркеров, позволяющих выделение хорошо определенных подмножеств T клеток с высокой чистотой.
В некоторых вариантах, антитела или связывающие партнеры мечены одним или более определяемыми маркерами для того, чтобы способствовать разделению для положительной и/или отрицательной селекции. Например, разделение может быть основано на связывании с флуоресцентно меченными антителами. В некоторых примерах, разделение клеток на основе связывания антител или других связывающих партнеров, специфических для одного или более поверхностных маркеров проводят в жидком потоке, таком как сортировка флуоресцентно активированных клеток (FACS), включая препаративную (FACS) и/или чипы микроэлектромеханических систем (MEMS), например, в сочетании с проточно-цитометрической системой определения. Такие способы позволяют положительную и отрицательную селекцию на основе множества маркеров одновременно.
В некоторых вариантах, способы получения включают стадии замораживания, например, криоконсервации клеток, до или после выделения, инкубирования и/или конструирования. В некоторых вариантах, стадия замораживания и последующего оттаивания удаляет гранулоциты и, до некоторой степени, моноциты в клеточной популяции. В некоторых вариантах, клетки суспендируют в замораживающем растворе, например, после стадии промывания для удаления плазмы и тромбоцитов. В некоторых аспектах могут применяться любые из известных замораживающих растворов и параметров. Один их примеров включает применение PBS, содержащего 20% ДМСО и 8% альбумин человеческой сыворотки (HSA) или другую подходящую среду для замораживания клеток. Его затем разбавляют 1:1 средой так, что конечная концентрация ДМСО и HSA составляет 10% и 4%, соответственно. Затем клетки замораживают до −80°C со скоростью 1° в минуту и хранят в паровой фазе в резервуаре для хранения в жидком азоте.
В некоторых вариантах, представленные способы включают стадии выращивания, инкубирования, культивирования и/или генной инженерии. Например, в некоторых вариантах, представлены способы инкубирования и/или конструирования обедненных клеточных популяций и инициирующих культивирование составов.
Таким образом, в некоторых вариантах, клеточные популяции инкубируют в инициирующей культивацию композиции. Инкубирование и/или конструирование может проводиться в культуральном сосуде, таком как блок, камера, ячейка, колонка, пробирка, комплект трубок, клапан, флакон, чашка для культивирования, мешок или другой контейнер для культивирования или выращивания клеток.
В некоторых вариантах, клетки инкубируют и/или культивируют до или в сочетании с генной инженерией. Стадии инкубирования могут включать культивирование, выращивание, стимулирование, активацию и/или размножение. В некоторых вариантах, композиции или клетки инкубируют в присутствии стимулирующих условий или стимулирующего агента. Такие условия включают условия, разработанные для вызова пролиферации, размножения, активации и/или выживания клеток в популяции, для имитации воздействия антигеном и/или для примирования клеток для генной инженерии, такой как введение нуклеиновых кислот, кодирующих рекомбинантный рецептор, например, рекомбинантный TCR.
Условия могут включать одно или более из конкретной среды, температуры, содержания кислорода, содержания диоксида углерода, времени, агентов, например, питательных веществ, аминокислот, антибиотиков, ионов и/или стимулирующих факторов, таких как цитокины, хемокины, антигены, связывающие партнеры, слитые белки, рекомбинантные растворимые рецепторы и другие агенты, созданные для активации клеток.
В некоторых вариантах, стимулирующие условия или агенты включают один или более агент, например, лиганд, который способен активировать внутриклеточную сигнальную область TCR комплекса. В некоторых аспектах, агент включает ли инициирует TCR/CD3 внутриклеточный сигнальный каскад в T клетке. Такие агенты включают антитела, такие как антитела, специфические к TCR, например, анти-CD3. В некоторых вариантах, стимулирующие условия включают один или более агент, например, лиганд, который способен стимулировать костимулирующий рецептор, например, анти-CD28. В некоторых вариантах, такие агенты и/или лиганды могут быть связаны с твердой подложкой, такой как сферы, и/или одним или более цитокинами. Необязательно, способ размножения может дополнительно включать стадию добавления анти-CD3 и/или анти-CD28 антитела в культуральную среду (например, в концентрации, по меньшей мере, около 0,5 нг/мл). В некоторых вариантах, стимулирующие агенты включают IL-2, IL-15 и/или IL-7. В некоторых аспектах, концентрация IL-2 составляет, по меньшей мере, около 10 единиц/мл.
В некоторых аспектах, инкубирование проводят в соответствии с описанным в патенте США № 6,040,177 Riddell et al., Klebanoff et al. (2012) J Immunother. 35(9): 651-660, Terakura et al. (2012) Blood,1:72-82, и/или Wang et al. (2012) J Immunother. 35(9):689-701.
В некоторых вариантах, T клетки размножают добавлением инициирующей культивацию композиции поддерживающих клеток, таких как не делящиеся мононуклеары периферической крови (PBMC), (например, так, чтобы полученная популяция клеток содержала, по меньшей мере, около 5, 10, 20 или 40 или более PBMC поддерживающих клеток для каждого T лимфоцита в исходной популяции, которую размножают); и инкубированием культуры (например, в течение времени, достаточного для размножения количества T клеток). В некоторых аспектах, не делящиеся поддерживающие клетки могут содержать гамма-облученные PBMC поддерживающие клетки. В некоторых вариантах, PBMC облучают гамма лучами в интервале от около 3000 до 3600 рад для предотвращения деления клеток. В некоторых аспектах, поддерживающие клетки добавляют в культуральную вреду до добавления популяций T клеток.
В некоторых вариантах, стимулирующие условия включают температуру, подходящую для роста человеческих T лимфоцитов, например, по меньшей мере, около 25 градусов Цельсия, в общем, по меньшей мере, около 30 градусов, и в общем, от около 37 градусов Цельсия. Необязательно, инкубирование может дополнительно включать добавление не делящихся EBV-трансформированных лимфобластоидных клеток (LCL) в качестве поддерживающих клеток. LCL могут быть облучены гамма лучами в интервале от около 6000 до 10000 рад. LCL поддерживающие клетки в некоторых аспектах представлены в любом подходящем количестве, таком как соотношение LCL поддерживающих клеток к исходным T лимфоцитам, по меньшей мере, около 10:1.
В некоторых вариантах, антиген-специфические T клетки, такие как антиген-специфические CD4+ и/или CD8+ T клетки, получают стимулированием наивных или антиген-спцифических Т лимфоцитов с антигеном. Например, колонии или клоны антиген-специфических T клеток могут быть созданы для цитомегаловирусных антигенов выделением T клеток у зараженных субъектов и стимулированием клеток in vitro с тем же антигеном.
Разные способы введения генетически сконструированных компонентов, например, агентов для вызова генетического разрушения и/или нуклеиновых кислот, кодирующих рекомбинантные рецепторы, например, CAR или TCR, известны и могут применяться с представленными способами и композициями. Типовые способы включают способы для переноса нуклеиновых кислот, кодирующих полипептиды или рецепторы, включая перенос через вирусные векторы, например, ретровирусные или лентивирусные, не вирусные векторы или транспозоны, например, систему транспозон Sleeping Beauty. Способы переноса генов могут включать трансдукцию, электропорацию или другой способ, который дает перенос гена в клетку, или любые способы доставки, описанные в разделе I.A здесь. Другие подходы и векторы для переноса нуклеиновых кислот, кодирующих рекомбинантные продукты, описаны, например, в WO2014055668 и патенте США № 7,446,190.
В некоторых вариантах, рекомбинантные нуклеиновые кислоты переносят в T клетки через электропорацию (см., например, Chicaybam et al, (2013) PLoS ONE8(3):e60298 и Van Tedeloo et al. (2000) Gene Therapy 7(16): 1431-1437). В некоторых вариантах, рекомбинантные нуклеиновые кислоты переносят в T клетки через транспозицию (см., например, Manuri et al. (2010) Hum Gene Ther 21(4): 427-437; Sharma et al. (2013) Molec Ther Nucl Acids 2, e74; и Huang et al. (2009) Methods Mol Biol 506: 115-126). Другие способы введения и экспрессии генетического материала в иммунные клетки включают трансфекцию фосфата кальция (например, как описано в Current Protocols in Molecular Biology, John Wiley & Sons, New York. N.Y.), слияние протопласта, катионную трансфекцию, медиированную липосомой; баллистическую трансфекцию микрочастиц, усиленную частицами вольфрама (Johnston, Nature, 346: 776-777 (1990)); и совместное осаждение фосфата стронция и ДНК (Brash et al., Mol. Cell Biol., 7: 2031-2034 (1987)).
В некоторых вариантах, перенос гена осуществляют сначала стимулированием клетки, таким как объединение ее со стимулом, который вызывает ответ, такой как пролиферация, выживание и/или активация, например, измеренная экспрессией цитокина или маркера активации, после трансдукции активированных клеток, и размножением в культуре до количеств, достаточных для клинического применения.
В некоторых контекстах, может потребоваться защита от возможности того, что сверхэкспрессия стимулирующего фактора (например, лимфокина или цитокина) потенциально может дать нежелательный результат или уменьшить эффективность у субъекта, такого, как фактор, ассоциированный с токсичностью у субъекта. Таким образом, в некоторых контекстах, сконструированные клетки включают генные сегменты, которые делают клетки подверженными отрицательной селекции in vivo, например, при введении при адоптивной иммунотерапии. Например в некоторых аспектах, клетки сконструированы так, что они могут быть исключены в результате изменения in vivo состояния пациента, которому их вводят. Отрицательно выбираемый фенотип может получиться при вставке гена, который придает чувствительность вводимому агенту, например, соединению. Отрицательно выбираемые гены включают ген тимидинкиназы вируса простого герпеса I типа (HSV-I TK) (Wigler et al., Cell 11:223, 1977), который придает чувствительность к ганцикловиру; ген клеточной гипоксантинфосфорибозилтрансферазы (HPRT), ген клеточной аденинфосфорибозилтрансферазы (APRT), бактериальную цитозиндеаминазу (Mullen et al., Proc. Natl. Acad. Sci. USA. 89:33 (1992)).
В некоторых вариантах, клетки, например, T клетки, могут быть сконструированы либо во время, либо после размножения. Такое конструирование для введения гена желаемого полипептида или рецептора может проводиться с любым подходящим ретровирусным вектором, например. Генетически модифицированная клеточная популяция затем может быть освобождена из исходного стимула (CD3/CD28 стимула, например) и затем быть стимулирована вторым типом стимула (например, через de novo введенный рецептор). Этот второй тип стимула может включать антигенный стимул в форме пептида/MHC молекулы, когнатного (поперечно-сшивающего) лиганда генетически введенного рецептора (например, природного лиганда CAR) или любого лиганда (такого как антитело), который непосредственно связывается с рамкой нового рецептора (например, через распознавание постоянных областей рецептором). См., например, Cheadle et al, “Chimeric antigen receptors for T-cell based therapy” Methods Mol Biol. 2012; 907:645-66 или Barrett et al., Chimeric Antigen Receptor Therapy for Cancer Annual Review of Medicine Vol. 65: 333-347 (2014).
Среди дополнительных нуклеиновых кислот, например, генов для введения, находятся такие, которые улучшают эффективность терапии, способствуя жизнеспособности и/или функционированию перенесенных клеток; гены, обеспечивающие генный маркер для выбора и/или оценки клеток, например, для оценки in vivo выживания или локализации; гены для улучшения безопасности, например, делающие клетки восприимчивыми к отрицательной селекции in vivo как описано у Lupton S. D. et al., Mol. and Cell Biol., 11:6 (1991); и Riddell et al., Human Gene Therapy 3:319-338 (1992); см. также публикации PCT/US91/08442 и PCT/US94/05601 Lupton et al., описывающие применение бифункциональных отбираемых слитых генов, полученных слиянием доминантного положительного отбираемого маркера с отрицательными отбираемым маркером. См., например, Riddell et al., патент США № 6,040,177, в столбцах 14-17.
Как описано здесь, в некоторых вариантах, клетки инкубируют и/или культивируют до или в сочетании с генной инженерией. Стадии инкубирования могут включать культивирование, выращивание, стимулирование, активацию, размножение и/или замораживание для консервации, например, криоконсервации.
III. НУКЛЕИНОВЫЕ КИСЛОТЫ, ВЕКТОРЫ И ДОСТАВКА
В некоторых вариантах, один или более агентов для генетического разрушения и/или матричных полинуклеотидов, например, матричных полинуклеотидов, содержащих трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, или один или более вторых матричных полинуклеотидов, вводят в клетки в форме нуклеиновой кислоты, например, в виде полинуклеотидов и/или векторов. Как описано в разделе I здесь, компоненты для конструирования могут быть доставлены в разных формах с применением разных способов доставки, включая полинуклеотидные кодирующие компоненты. Также представлены один или более полинуклеотидов (например, молекулы нуклеиновой кислоты), кодирующих один или более компонентов одного или более агентов, способных вызывать генетическое разрушение, и/или одного или более матричных полинуклеотидов, содержащих трансген, и векторов для генетически сконструированных клеток для направленной интеграции трансгена.
В некоторых вариантах, представлены матричные полинуклеотиды, например, матричные полинуклеотиды для целенаправленного воздействия трансгена на определенную геномную целевую локацию, например, на TRAC, TRBC1 и/или TRBC2 локус. В некоторых вариантах, представлены любые матричные полинуклеотиды, описанные в разделе I.B здесь. В некоторых вариантах, матричный полинуклеотид содержит трансген, который включает последовательности нуклеиновых кислот, которые кодируют рекомбинантный рецептор или другие полипептиды и/или факторы, и плечи гомологии для направленной интеграции. В некоторых вариантах, матричный полинуклеотид может содержаться в векторе.
В некоторых вариантах, агенты, способные вызывать генетическое разрушение, могут быть кодированы в одном или более полинуклеотидах. В некоторых вариантах, компонент агентов, например, Cas9 молекулы и/или нРНК молекулы, может быть кодирован в одном или более полинуклеотидах, и введены в клетки. В некоторых вариантах, полинуклеотид, кодирующий один или более компонентов агентов, может быть включен в вектор.
В некоторых вариантах, вектор может содержать последовательность, которая кодирует Cas9 молекулу и/или нРНК молекулу и/или матричные полинуклеотиды. Вектор также может содержать последовательность, кодирующую сигнальный пептид (например, для ядерной локализации, ядрышковой локализации, митохондриальной локализации), слитую, например, с последовательностью Cas9 молекулы. Например, вектор может содержать последовательность ядерной локализации (например, от SV40), слитую с последовательностью, кодирующей Cas9 молекулу.
Один или более регулирующих/контрольных элементов, например, промотор, энхансер, интрон, сигнал полиаденилирования, консенсусная последовательность Козака, внутренний участок посадки рибосомы (IRES), 2A последовательность и акцептор или донов сплайсинга могут быть включены в векторы. В некоторых вариантах, промотор выбирают из РНК pol I, pol II или pol III промотора. В некоторых вариантах, промотор распознается РНК полимеразой II (например, CMV, промотором SV40 ранней области или основной поздней области аденовируса). В другом варианте, промотор распознается РНК полимеразой III (например, U6 или H1 промотор).
В другом варианте, промотор является регулированным промотором (например, индуцируемым промотором). В некоторых вариантах, промотор является индуцируемым промотором или репрессируемым промотором. В некоторых вариантах, промотор содержит Lac последовательность оператора, тетрациклиновую последовательность оператора, галактозную последовательность оператора или доксициклиновую последовательность оператора или является их аналогом или способен связываться или распознаваться Lac репрессором или тетрациклиновым репрессором или их аналогом.
В некоторых вариантах, промотор является или содержит конститутивный промотор. Типовые конститутивные промоторы включают, например, ранний промотор вируса обезьян 40 (SV40), средне-ранний промотор цитомегаловируса (CMV), промотор человеческого Убиквитина C (UBC), промотор человеческого фактора элонгации 1α (EF1α), промотор мышиной фосфоглицераткиназы 1 (PGK) и промотор куриного β-Актина, сопряженный с ранним энхансером CMV (CAGG). В некоторых вариантах, конститутивным промотором является синтетический или модифицированный промотор. В некоторых вариантах, промотором является или содержит MND промотор, синтетический промотор, который сдержит U3 область модифицированного MoMuLV LTR с энхансером вируса миелопролиферативной саркомы (последовательность, представленную в SEQ ID NO:18 или 126; см. Challita et al. (1995) J. Virol. 69(2):748-755). В некоторых вариантах, промотор является ткань-специфическим промотором. В другом варианте, промотор является вирусным промотором. В другом варианте, промотор является не вирусным промотором. В некоторых вариантах, типовые промоторы могут включать, но не ограничены ими, промотор человеческого фактора элонгации 1 альфа (EF1α) (последовательность, представленную в SEQ ID NO:4 или 5) или его модифицированную форму (EF1α промотор с HTLV1 энхансером; последовательность, представленную в SEQ ID NO: 127) или MND промотор (последовательность, представленную в SEQ ID NO:18 или 126). В некоторых вариантах, полинуклеотид и/или вектор не включает регулирующий элемент, например, промотор.
В некоторых вариантах, вектором или средством доставки является вирусный вектор (например, для создания рекомбинантных вирусов). В некоторых вариантах, вирусом является ДНК вирус (например, днДНК или онДНК вирус). В некоторых вариантах, вирусом является РНК вирус (например, онРНК вирус). Типовые вирусные векторы/вирусы включают, например, ретровирусы, лентивирусы, аденовирусы, адено-ассоциированный вирус (AAV), вирусы осповакцины, поксвирусы и вирусы простого герпеса или любые из вирусов, описанных здесь повсюду.
В некоторых вариантах, вирус заражает делящиеся клетки. В другом варианте, вирус заражает не делящиеся клетки. В другом варианте, вирус заражает и делящиеся и не делящиеся клетки. В другом варианте, вирус может интегрироваться в геном хозяина. В другом варианте, вирус сконструирован так, чтобы иметь пониженную иммунность, например, у человека. В другом варианте, вирус способен реплицироваться. В другом варианте, репликация вируса нарушена, например, он имеет одну или более кодирующих областей для генов, необходимых для дополнительных раундов репликации вириона и/или упаковки, замещенных другими генами или удаленными. В другом варианте, вирус вызывает временную экспрессию Cas9 молекулы и/или нРНК молекулы для целей временного вызова генетического разрушения. В другом варианте, вирус вызывает длительную, например, по меньшей мере, 1 неделю, 2 недели, 1 месяц, 2 месяца, 3 месяца, 6 месяцев, 9 месяцев, 1 год, 2 года или постоянную экспрессию Cas9 молекулы и/или нРНК молекулы. Упаковочная способность вирусов может варьироваться, например, от, по меньшей мере, около 4 т.н. до, по меньшей мере, около 30 т.н., например, по меньшей мере, около 5 т.н., 10 т.н., 15 т.н., 20 т.н., 25 т.н., 30 т.н., 35 т.н., 40 т.н., 45 т.н. или 50 т.н.
В некоторых вариантах, полинуклеотид, содержащий агенты и/или матричный полинуклеотид доставляют рекомбинантным ретровирусом. В другом варианте, ретровирус (например, вирус мышиного лейкоза Молони) содержит обратную транскриптазу, например, которая позволяет интеграцию в геном хозяина. В некоторых вариантах, ретровирус способен к репликации. В другом варианте, ретровирус не способен к репликации, например, имеет одну или более кодирующих областей для генов, необходимых для дополнительных раундов репликации вириона и упаковки, замещенных другими генами или удаленными.
В некоторых вариантах, полинуклеотид, содержащий агенты и/или матричный полинуклеотид, доставляют рекомбинантным лентивирусом. Например, лентивирус не способен к репликации, например, не содержит один или более генов, требуемых для вирусной репликации. В некоторых вариантах, лентивирусом является лентивирус, полученный из ВИЧ.
В некоторых вариантах, полинуклеотид содержащий агенты и/или матричный полинуклеотид, доставляют рекомбинантным аденовирусом. В другом варианте, аденовирус сконструирован так, чтобы иметь пониженную иммунность у человека.
В некоторых вариантах, полинуклеотид, содержащий агенты и/или матричный полинуклеотид, доставляют рекомбинантным AAV. В некоторых вариантах, AAV может вводить свой геном в геном клетки хозяина, например, целевой клетки, как описано здесь. В другом варианте, AAV является самокомплементарным адено-ассоциированным вирусом (scAAV), например, scAAV, который упаковывает обе нити, которые ренатурируют вместе с образованием двухнитевой ДНК. AAV серотипы, которые могут применяться в раскрытых способах, включают AAV1, AAV2, модифицированный AAV2 (например, модификации в Y444F, Y500F, Y730F и/или S662V), AAV3, модифицированный AAV3 (например, модификации в Y705F, Y731F и/или T492V), AAV4, AAV5, AAV6, модифицированный AAV6 (например, модификации в S663V и/или T492V), AAV7, AAV8, AAV 8.2, AAV9, AAV.rh10, модифицированный AAV.rh10, AAV.rh32/33, модифицированный AAV.rh32/33, AAV.rh43, модифицированный AAV.rh43, AAV.rh64R1, модифицированный AAV.rh64R1, и псевдотипированный AAV, такой как AAV2/8, AAV2/5 и AAV2/6, которые также могут применяться в раскрытых способах.
В некоторых вариантах, полинуклеотид, содержащий агенты и/или матричный полинуклеотид, доставляют гибридным вирусом, например, гибридом одного или более вирусов, описанных здесь.
Упаковывающую клетку применяют для образования частиц вируса, которые способны заражать целевую клетку. Такая клетка включает 293 клетку, которая может упаковывать аденовирус, и ψ2 клетку или PA317 клетку, которая может упаковывать ретровирус. Вирусный вектор, применяемый в генной терапии, обычно создается клеточной линией-продуцентом, которая упаковывает вектор нуклеиновых кислот в вирусную частицу. Вектор обычно содержит минимальные вирусные последовательности, требуемые для упаковки и последующей интеграции в хозяина или целевую клетку (если применяется), с другими вирусными последовательностями, замененными кассетой экспрессии, кодирующими экспрессируемый белок, например, Cas9. Например, AAV вектор, применяемый в генной терапии, обычно обладает только последовательностью инвертированного концевого повтора (ITR) от AAV генома, который требуется для упаковки и генной экспрессии в клетке хозяина или целевой клетке. Недостающие вирусные функции передаются in trans клеточной линией упаковки. С этого момента, вирусную ДНК упаковывают в колонию клеток, которая содержит хелперную плазмиду, кодирующую другие AAV гены, а именно, rep и cap, но не имеет ITR последовательности. Колонию клеток также заражают аденовирусом в качестве хелпера. Хелперный вирус способствует репликации AAV вектора и экспрессии AAV генов из хелперной плазмиды. Хелперная плазмида не упакована в значительных количествах из-за отсутствия ITR последовательностей. Загрязнение аденовирусом может быть снижено, например, тепловой обработкой, к которой аденовирус более чувствителен, чем AAV.
В некоторых вариантах, вирусный вектор обладает способностью распознавания типа клеток. Например, вирусный вектор может быть псевдотипирован с разными/альтернативными гликопротеинами вирусной оболочки; сконструирован с клеточным тип-специфическим рецептором (например, генетической модификацией гликопротеинов вирусной оболочки для введения лигандов целенаправленного воздействия, таких как пептидный лиганд, однонитевое антитело, фактор роста); и/или сконструирован так, чтобы иметь молекулярный мостик с двойной специфичностью, с одним концом, распознающим вирусный гликопротеин, и другим концом, распознающим группу поверхности целевой клетки (например, лиганд-рецептор, моноклональное антитело, авидин-биотин и химическое конъюгирование).
В некоторых вариантах, вирусный вектор достигает специфической к типу клетки экспрессии. Например, ткань-специфический промотор может быть сконструирован так, чтобы ограничивать экспрессию трансгена (Cas9 и нРНК) только в специфической целевой клетке. Специфичность вектора также может быть медиирована микроРНК-зависимым контролем трансгенной экспрессии. В некоторых вариантах, вирусный вектор повышает эффективность слияния вирусного вектора и мембраны целевой клетки. Например, слитый белок, такой как способный к слиянию геммаглутинин (HA), может быть введен для повышения вирусного поглощения в клетки. В некоторых вариантах, вирусный вектор обладает способностью ядерной локализации. Например, вирус, который требует расщепления ядерной мембраны (во время деления клетки) и поэтому не будет заражать не делящуюся клетку, может быть изменен для введения пептида ядерной локализации в матричный белок вируса, тем самым позволяя трансдукцию не пролиферирующих клеток.
IV. РЕКОМБИНАНТНЫЕ РЕЦЕПТОРЫ
В некоторых вариантах, трансген для направленной интеграции кодирует рекомбинантный рецептор или его антигенсвязывающий фрагмент или его цепь. В некоторых вариантах, рекомбинантным рецептором является рекомбинантный антигенный рецептор или рекомбинантный рецептор, который связывает антиген. В некоторых вариантах, рекомбинантным рецептором является рекомбинантный или сконструированный Т-клеточный рецептор (TCR), который отличается от эндогенного TCR, кодированного T клеткой. В некоторых вариантах, рекомбинантным рецептором является химерный антигенный рецептор (CAR) или TCR-подобный CAR. В некоторых вариантах, трансген может кодировать домен, область или цепь рекомбинантного рецептора, и один или более вторых трансгенов могут кодировать другие домены, области или цепи рекомбинантного рецептора. В некоторых вариантах, представленные полинуклеотиды, векторы, композиции, способы, готовые изделия и/или наборы применяют для конструирования клеток, которые экспрессируют рекомбинантный TCR или его антигенсвязывающий фрагмент.
В некоторых вариантах, представленные рекомбинантные рецепторы, например, TCR или CAR, способны связываться с или распознавать, например, специфически связываться или распознавать, антиген, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния, такого как рак или опухоль. В некоторых аспектах, антиген имеет форму пептида, например, является пептидным антигеном или пептидным эпитопом. В некоторых вариантах, представленные TCR связываются с, например, специфически связываются с антигеном, который является пептидом, в контексте основной гистосовместимости (MHC) молекулы.
Наблюдение, что рекомбинантный рецептор связывается с антигеном, например, пептидным антигеном или специфически связывается с антигеном, например, пептидным антигеном, не обязательно означает, что он связывается с антигеном любого вида. Например, в некоторых вариантах, характеристики связывания с антигеном, например, пептидным антигеном в контексте MHC, такие как способность специфически связываться с ним и/или конкурировать для связывания с ним со ссылочной связывающей молекулой или рецептором, и/или связываться с конкретной аффинностью или конкурировать до определенной степени, в некоторых вариантах, относятся к способности по отношению к человеческому антигену, и рекомбинантный рецептор может не иметь этой характеристики по отношению к антигену от другого вида, такого как мышь. В некоторых аспектах, степень связывания рекомбинантного рецептора или его антигенсвязывающего фрагмента с неродственным антигеном или белком, таким как неродственный пептидный антиген, составляет менее около 10% связывания рекомбинантного рецептора или его антигенсвязывающего фрагмента с антигеном, например, когнатным антигеном, как измерено, например, радиоиммуноанализом (РИА), анализом пептидного титрования или анализом рецептора.
А. Т-клеточные рецепторы (TCR)
В некоторых вариантах, рекомбинантным рецептором, который вводят в клетку, является Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент.
В некоторых вариантах, “Т-клеточным рецептором” или “TCR” является молекула, которая содержит α и β цепи (также известные как TCRα и TCRβ, соответственно) или γ и δ цепи (также известные как TCRγ и TCRδ, соответственно) или их антигенсвязывающие части, и которая способна специфически связываться с антигеном, например, пептидным антигеном или пептидным эпитопом, связанным с MHC молекулой. В некоторых вариантах, TCR имеет αβ форму. Обычно, TCR, который существует в αβ и γδ формах, в общем структурно схожи, но T клетки, экспрессирующие их, могут иметь разные анатомические локации или функции. TCR может быть найден на поверхности клетки или в растворимой форме. В общем, TCR найден на поверхности T клетки (или T лимфоцитов), где он обычно отвечает за распознавание антигенов, связанных с молекулами основного комплекса гистосовместимости (MHC).
Обычно специфическое связывание рекомбинантного рецептора, например, TCR, с пептидным эпитопом, например, в комплексе с MHC, управляется присутствием антигенсвязывающего сайта, содержащего одну или более областей определения комплементарности (CDR). В общем, понятно, что специфическое связывание не означает, что конкретный пептидный эпитоп, например, в комплексе с MHC, является единственным, с которым может связываться Молекула MHC-пептида, так как не специфическое связывание с другими молекулами также может происходить. В некоторых вариантах, связывание рекомбинантного рецептора с пептидом в контексте MHC молекулы происходит с более высоким сродством, чем связывание с такими другими молекулами, например, другим пептидом в контексте MHC молекулы или нецелевым (контрольным) пептидом в контексте MHC молекулы, например, по меньшей мере, около 2-кратным, по меньшей мере, около 10-кратным, по меньшей мере, около 20-кратным, по меньшей мере, около 50-кратным или по меньшей мере, около 100-кратным по сравнению со сродством связывания с такими другими молекулами.
В некоторых вариантах, рекомбинантный рецептор, например, TCR, может быть оценен на безопасность или активность нецелевого связывания с применением любого из множества известных скриннинговых исследований. В некоторых вариантах, создание иммунного ответа к конкретному рекомбинантному рецептору, например, TCR, может быть измерено в присутствии клетки, которая известна как экспрессирующая целевой пептидный эпитоп, такой как клетка, полученная из нормальных тканей, аллогенных колоний клеток, которые экспрессируют один или более разных типов MHC, или других источников ткани или клетки. В некоторых вариантах, клетки или ткани включают нормальные клетки или ткани. В некоторых вариантах, связывание с клетками может быть тестировано в 2-мерных культурах. В некоторых вариантах, связывание с клеткой может быть тестировано в 3-мерных культурах. В некоторых вариантах, в качестве контроля, тканями или клетками могут быть такие, которые известны как экспрессирующие целевой эпитоп. Иммунный ответ может быть оценен прямо или косвенно, например, через оценку активации иммунных клеток, таких как T клетки (например, цитотоксической активности), образования цитокина (например, интерферона гамма) или активации сигнального каскада, такого как анализ репортерного гена.
Если не указано иначе, термин “TCR” должен пониматься как охватывающий полные TCR, а также их антигенсвязывающие части или антигенсвязывающие фрагменты. В некоторых вариантах, TCR является нативным или полноразмерным TCR, таким как TCR, содержащий альфа (TCRα) цепь и бета (TCRβ) цепь. В некоторых вариантах, TCR является антигенсвязывающей частью, которая меньше полноразмерного TCR, но которая связывается со специфической пептидной связью в MHC молекуле, такой, которая связывается с MHC-пептидным комплексом. В некоторых случаях, антигенсвязывающая часть или фрагмент TCR может содержать только часть структурных доменов полноразмерного или нативного TCR, но также способна связывать пептидный эпитоп, такой как MHC-пептидный комплекс, с которым связывается полноразмерный TCR. В некоторых случаях, антигенсвязывающая часть содержит переменные домены TCR, такие как переменная α (Vα) цепь и переменная β (Vβ) цепь TCR или его антигенсвязывающих фрагментов, достаточных для образования сайта связывания со специфическим MHC-пептидным комплексом.
В некоторых вариантах, переменные домены TCR содержат области, определяющие комплементарность (CDR), которые обычно являются первичными участниками антигенного распознавания и способностей к связыванию и специфичности пептида, MHC и/или MHC-пептидного комплекса. В некоторых вариантах, CDR TCR или их сочетание образует весь или по существу весь антигенсвязывающий сайт данной TCR молекулы. Разные CDR в пределах переменной области TCR цепи обычно разделены каркасными областями (FR), которые обычно демонстрируют меньшую вариабельность среди TCR молекул по сравнению с CDR (см., например, Jores et al., Proc. Nat'l Acad. Sci. U.S.A. 87:9138, 1990; Chothia et al., EMBO J. 7:3745, 1988; см. также Lefranc et al., Dev. Comp. Immunol. 27:55, 2003). В некоторых вариантах, CDR3 является основной CDR, отвечающей за антигенное связывание или специфичность, или является наиболее важной среди CDR на данной переменной области TCR для распознавания антигена, и/или для взаимодействия с процессированной пептидной частью пептид-MHC комплекса. В некоторых контекстах, CDR1 α цепи может взаимодействовать с N-концевой частью определенных антигенных пептидов. В некоторых контекстах, CDR1 β цепи может взаимодействовать с C-концевой частью пептида. В некоторых контекстах, CDR2 вносит наибольший вклад в или является первичной CDR, отвечающей за взаимодействие м или распознавание MHC части MHC-пептидный комплекса. В некоторых вариантах, переменная область β-цепи может содержать дополнительную гиперпеременную область (CDR4 или HVR4), которая обычно вовлечена в суперантигенное связывание, но не антигенное распознавание (Kotb (1995) Clinical Microbiology Reviews, 8:411-426).
В некоторых вариантах, α-цепь и/или β-цепь TCR также может содержать постоянный домен, трансмембранный домен и/или короткий цитоплазматический хвост (см., например, Janeway et al., Immunobiology: The Immune System in Health и Disease, 3rd Ed., Current Biology Publications, p. 4:33, 1997). В некоторых аспектах, каждая цепь (например, альфа или бета) TCR может обладать одним N-концевым иммуноглобулиновым переменным доменом, одним иммуноглобулиновым постоянным доменом, трансмембранной областью и коротким цитоплазматических хвостом на C-концевой части. В некоторых вариантах, TCR, например через цитоплазматический хвост, ассоциирован с инвариантными белками CD3 комплекса, вовлеченными в медиирование трансдукции сигнала. В некоторых случаях, структура позволяет TCR ассоциироваться с другими молекулами, такими как CD3 и их подъединицы. Например, TCR, содержащие постоянные домены с трансмембранной областью, может прикреплять белок в клеточной мембране и ассоциировать с инвариантными подъединицами CD3 сигнального аппарата или комплекса. Внутриклеточные хвосты CD3 сигнальных подъединиц (например, CD3γ, CD3δ, CD3ε и CD3ζ цепи) содержат один или более основанных на иммунорецепторе тирозина мотивов активации или ITAM и обычно вовлечены в сигнальную способность TCR комплекса.
В некоторых вариантах, могут быть определены разные домены или области TCR. В некоторых случаях, точный локус домена или области может варьироваться в зависимости от конкретного структурного или гомологичного моделирования или других характеристик, применяемых для описания конкретного домена. Понятно, что ссылка на аминокислоты, включая специфические последовательности, указанные как SEQ ID NO, применяемые для описания организации домена рекомбинантного рецептора, например, TCR, даны для иллюстративных целей и не предназначены для ограничения объема представленных вариантов. В некоторых случаях, специфический домен (например, переменный или постоянный) может быть на несколько аминокислот (например, одну, две, три или четыре) длиннее или короче. В некоторых аспектах, остатки TCR известны или могут быть идентифицированы согласно системе нумерации International Immunogenetics Information System (IMGT) (см., например, www.imgt.org; см. также Lefranc et al. (2003) Developmental и Comparative Immunology, 2&;55-77; и The T Cell Factsbook 2nd Edition, Lefranc и LeFranc Academic Press 2001). С применением этой системы, CDR1 последовательности в пределах TCR Vα цепей и/или Vβ цепи соответствуют аминокислотам, присутствующим между остатками №№ 27-38, включительно, CDR2 последовательности в пределах TCR Vα цепи и/или Vβ цепи соответствуют аминокислотам, присутствующим между остатками №№ 56-65, включительно, и CDR3 последовательности в пределах TCR Vα цепи и/или Vβ цепи соответствуют аминокислотам, присутствующим между остатками №№ 105-117, включительно.
В некоторых вариантах, каждая из α цепи и β цепи TCR содержит постоянный домен. В некоторых вариантах, постоянный домен α цепи (Cα) и постоянный домен β цепи (Cβ) по отдельности являются постоянным доменом млекопитающих, таких как человек или мышь. В некоторых вариантах, постоянный домен находится рядом с клеточной мембраной. Например, в некоторых случаях, внеклеточная часть TCR, образованная двумя цепями, содержит два мембрана-проксимальных постоянных домена, и два мембрана-дистальных переменных домена, где каждый переменный домен содержит CDR.
В некоторых вариантах, каждый из Cα и Cβ доменов является человеческим. В некоторых вариантах, Cα кодирован TRAC геном (IMGT номенклатура) или является его вариантом. В некоторых вариантах, Cα имеет или содержит последовательность аминокислот, представленную в SEQ ID NO: 19, или последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности с SEQ ID NO: 19. В некоторых вариантах, Cα имеет или содержит последовательность аминокислот, представленную в любом из SEQ ID NO:19. В некоторых вариантах, Cα имеет или содержит последовательность аминокислот, например, зрелый полипептид, кодированную последовательностью нуклеиновых кислот, представленной в SEQ ID NO:1, или последовательностью аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность к последовательности аминокислот, например, зрелому полипептиду, кодированному последовательностью нуклеиновых кислот, представленной в SEQ ID NO:1. В некоторых вариантах, Cβ кодирован TRBC1 или TRBC2 генами (IMGT номенклатура) или является его вариантом. В некоторых вариантах, Cβ имеет или содержит последовательность аминокислот, представленную в SEQ ID NO:20 или 21, или последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность к последовательности SEQ ID NO: 20 или 21. В некоторых вариантах, Cβ имеет или содержит последовательность аминокислот, представленную в SEQ ID NO: 20 или 21. В некоторых вариантах, Cβ имеет или содержит последовательность аминокислот, например, зрелый полипептид, кодированный последовательностью нуклеиновых кислот, представленной в SEQ ID NO:2 или 3, или последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность к последовательности аминокислот, например, зрелому полипептиду, кодированному последовательностью нуклеиновых кислот, представленной в SEQ ID NO:2 или 3.
В некоторых вариантах, любой из представленных TCR или их антигенсвязывающих фрагментов может быть человеческим/мышиным химерным TCR. В некоторых случаях, TCR или его антигенсвязывающий фрагмент имеет α цепь и/или β цепь, содержащую мышиную постоянную область. В некоторых аспектах, Cα и/или Cβ области являются мышиными постоянными областями. В некоторых вариантах, Cα является мышиной постоянной областью, которая является или содержит последовательность аминокислот, представленную в SEQ ID NO: 14, 15, 121 или 122, или последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности к SEQ ID NO: 14, 15, 121 или 122. В некоторых вариантах, Cα является или содержит последовательность аминокислот, представленную в SEQ ID NO: 14, 15, 121 или 122. В некоторых вариантах, Cβ является мышиной постоянной областью, которая является или содержит последовательность аминокислот, представленную в SEQ ID NO: 16, 17 или 123, или последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности к SEQ ID NO: 16, 17 или 123. В некоторых вариантах, Cβ является или содержит последовательность аминокислот, представленную в SEQ ID NO: 16, 17 или 123.
В некоторых из любых таких вариантов, TCR или его антигенсвязывающий фрагмент содержат одну или более модификаций в α цепи и/или β цепи, такие, при которых при экспрессировании TCR или его антигенсвязывающего фрагмента в клетке, частота нарушения комплементарности между TCR α цепью и β цепью и эндогенной TCR α цепью и β цепью снижается, экспрессия TCR α цепи и β цепи повышается и/или стабильность TCR α цепи и β цепи повышается. В некоторых вариантах, одна или более модификаций включает замещение, делецию или вставку одной или более аминокислот в Cα области и/или Cβ области. В некоторых аспектах, одна или более модификаций содержит замещения для введения одного или более цистеиновых остатков, которые способны образовывать один или более ненативных дисульфидных мостиков между α цепью и β цепью.
В некоторых из любых таких вариантов, TCR или его антигенсвязывающий фрагмент содержит Cα область, содержащую цистеин в положении, соответствующем положению 48 с нумерацией, указанной в SEQ ID NO: 24, и/или Cβ область, содержащую цистеин в положении, соответствующем положению 57 с нумерацией, указанной в SEQ ID NO: 20. В некоторых вариантах, указанная Cα область содержит аминокислотную последовательность, представленную в любой из SEQ ID NOS: 19 или 24, или последовательность аминокислот, которая имеет, по меньшей мере, 90% идентичность последовательности к ней, содержащую один или более цистеиновых остатков, способных образовывать ненативную дисульфидную связь с β цепью; и/или указанная Cβ область содержит аминокислотную последовательность, представленную в любой из SEQ ID NOS: 20, 21 или 25, или последовательность аминокислот, которая имеет, по меньшей мере, 90% идентичность последовательности к ней, содержащую один или более цистеиновых остатков, способных образовывать ненативную дисульфидную связь с α цепью.
В некоторых из любых таких вариантов, TCR или его антигенсвязывающий фрагмент кодирован нуклеотидной последовательностью, которая кодон-оптимизирована.
В некоторых из любых таких вариантов, связывающая молекула или TCR или его антигенсвязывающий фрагмент выделен или очищен или является рекомбинантным. В некоторых из любых таких вариантов, связывающая молекула или TCR или его антигенсвязывающий фрагмент является человеческим.
В некоторых вариантах, TCR может быть гетеродимером из двух цепей, α и β, которые связаны, например, дисульфидной связью или дисульфидными связями. В некоторых вариантах, постоянный домен TCR может содержать короткие связывающие последовательности, в которых цистеиновые остатки образуют дисульфидную связь, тем самым связывая две цепи TCR. В некоторых вариантах, TCR может иметь дополнительный цистеиновый остаток на каждой из α и β цепях, так, что TCR содержит две дисульфидные связи в постоянных доменах. В некоторых вариантах, каждый из постоянного и переменного доменов содержит дисульфидные связи, образованные цистеиновыми остатками.
В некоторых вариантах, TCR может содержать введенную дисульфидную связь или связи. В некоторых вариантах, нативные дисульфидные связи не присутствуют. В некоторых вариантах, один или более нативных цистеинов (например, в постоянном домене α цепи и β цепи), которые образуют нативные межцепочечные дисульфидные связи, замещены другим остатком, таким как серин или аланин. В некоторых вариантах, введенные дисульфидные связи могут быть образованы мутацией не-цистеиновых остатков на альфа и β цепях, таких как в постоянном домене α цепи и β цепи, до цистеина. В некоторых вариантах, присутствие ненативных цистеиновых остатков (например, дающих одну или более ненативных дисульфидных связей) в рекомбинантном TCR может благоприятствовать образованию желаемого рекомбинантного TCR в клетке, в которую его вводят, над экспрессией ошибочно спаренных TCR пар, содержащих нативную TCR цепь.
Типовые ненативные дисульфидные связи TCR описаны в опубликованной международной заявке РСТ № WO2006/000830, WO2006037960 и Kuball et al. (2007) Blood, 109:2331-2338. В некоторых вариантах, цистеины могут быть введены на остатке Thr48 Cα цепи и Ser57 Cβ цепи, на остатке Thr45 Cα цепи и Ser77 Cβ цепи, на остатке Tyr10 Cα цепи и Ser17 Cβ цепи, на остатке Thr45 Cα цепи и Asp59 Cβ цепи и/или на остатке Ser15 Cα цепи и Glu15 Cβ цепи со ссылкой на нумерацию Cα, представленную в SEQ ID NO: 24, или Cβ, представленную в SEQ ID NO:20. В некоторых вариантах, любая из представленных цистеиновых мутаций может быть сделана в соответствующем положении в другой последовательности, например, в мышиных Cα и Cβ последовательностях, описанных здесь. Термин “соответствующее” со ссылкой на положения белка, например, при описании того, что положения аминокислот “соответствуют” положениям аминокислот в раскрытой последовательности, такой как указана в Списке последовательностей, относится к положениям аминокислот, идентифицированных при выравнивании с раскрытой последовательностью на основе выравнивания структурной последовательности или применении стандартного алгоритма выравнивания, такого как GAP алгоритм. Например, соответствующие остатки могут быть определены выравниванием ссылочной последовательности с Cα последовательностью, представленной в любой из SEQ ID NO: 24, или Cβ последовательностью представленной в SEQ ID NO: 20, способами структурного выравнивания, описанными здесь. Выравниванием последовательностей, соответствующие остатки могут быть идентифицированы, например, с применением консервативных и идентичных аминокислотных остатков в качестве направляющих.
Типовые последовательности (например, CDR, Vα и/или Vβ и постоянная область последовательностей) представленных TCR описаны здесь.
В некоторых вариантах, рекомбинантный TCR или его антигенсвязывающая часть (или другая связывающая Молекула MHC-пептида, такая как TCR-подобное антитело) известен как, или вероятно может, или может распознавать пептидный эпитоп или T клеточный эпитоп целевого полипептида, когда представлен клетками в контексте MHC молекулы, т.е. MHC-пептидный комплекса целевого полипептида. В некоторых вариантах, рекомбинантный TCR (или другая связывающая Молекула MHC-пептида, такая как TCR-подобное антитело) известен как, или вероятно демонстрирует специфическое связывание для эпитопа T клетки целевого полипептида, например, при воспроизведении как MHC-пептидный комплекс. Способы оценки связывания или взаимодействия связывающей MHC-пептид молекулы (например, TCR или TCR-подобного антитела) известны, включая любой из типовых способов, описанных здесь.
В некоторых вариантах, MHC молекулой является MHC класса I или MHC класса II молекула. В некоторых вариантах, MHC содержит сайт связывания полиморфного пептида или полость связывания, которая может, в некоторых случаях, образовывать комплекс с пептидными эпитопами полипептидов, включая пептидные эпитопы, процессированные клеточным механизмом. В некоторых случаях, MHC молекулы могут воспроизводиться или экспрессироваться на клеточной поверхности, включая комплекс с пептидом, т.е. MHC-пептидный комплекс, для представления антигена в конформации, распознаваемой TCR на T клетках или других связывающих MHC-пептид молекулах. В общем, MHC молекулы класса I являются гетеродимерами, имеющими трансмембранную α цепь, в некоторых случаях с тремя α доменами, и не ковалентно связанный β2 микроглобулин. В общем, MHC молекулы класса II состоят из двух трансмембранных гликопротеинов, α и β, оба которых обычно охватывают мембрану. MHC молекула может включать эффективную часть MHC, которая содержит антигенсвязывающий сайт или сайты для связывания пептида и последовательностей, необходимых для распознавания подходящей связывающей молекулой, такой как TCR. В некоторых вариантах, MHC молекулы класса I доставляют пептиды, возникающие в цитозоле, на поверхность клетки, где пептид:MHC комплекс распознается T клетками, такими как, обычно CD8+ T клетки, но в некоторых случаях, CD4+ T клетки. В некоторых вариантах, MHC молекулы класса II доставляют пептиды, возникающие в везикулярной системе, на поверхности клетки, где они обычно распознаются CD4+ T клетками. Обычно MHC молекулы кодированы группой связанных локусов, которые вместе называют H-2 в мышином и человеческом лейкоцитарном антигене (HLA) у человека. В некоторых аспектах, человеческий MHC также может называться человеческий лейкоцитарный антиген (HLA).
В некоторых вариантах, пептидным эпитопом или T клеточным эпитопом является пептид, который может быть получен из, или основан на фрагменте большей биологической молекулы, такой как полипептид или белок, и который способен ассоциироваться с или образовывать комплекс с MHC молекулой. В некоторых вариантах, пептид имеет от около 8 до около 24 аминокислот в длину. В некоторых вариантах, пептид имеет длину от или от около 9-22 аминокислот для распознавания в MHC класса II комплексе. В некоторых вариантах, пептид имеет длину от или от около 8-13 аминокислот для распознавания в MHC класса I комплексе. В некоторых вариантах, MHC молекула и пептидный эпитоп или T клеточный эпитоп образуют комплекс или ассоциируются через нековалентные взаимодействия пептида в полости связывания или щели MHC молекулы.
В некоторых вариантах, MHC-пептидный комплекс присутствует или отображается на поверхности клетки. В некоторых вариантах, MHC-пептидный комплекс может специфически распознаваться TCR или его антигенсвязывающей частью или другой связывающей MHC-пептид молекулой. В некоторых вариантах, T клеточный эпитоп или пептидный эпитоп способны вызывать иммунный ответ у животного через его характеристики связывания с MHC молекулами. В некоторых вариантах, при распознавании T клеточного эпитопа, такого как MHC-пептидный комплекс, TCR (или другая связывающая MHC-пептид молекула) дает или запускает сигнал активации в T клетку, который вызывает ответ T клетки, такой как пролиферация T клетки, образование цитокина, цитотоксичный ответ T клетки или другой ответ.
В некоторых вариантах, TCR или другая связывающая MHC-пептид молекула, распознает или потенциально распознает T клеточный эпитоп в контексте MHC молекулы класса I. MHC белки класса I экспрессируются во всех ядросодержащих клетках высших позвоночных. MHC молекулой класса I является гетеродимер, состоящий из 46-кДа тяжелой цепи, которая не ковалентно связана с 12-кДа легкой цепью β-2 микроглобулина. У человека имеется несколько MHC аллелей, таких как, например, HLA-A2, HLA-A1, HLA-A3, HLA-A24, HLA-A28, HLA-A31, HLA-A33, HLA-A34, HLA-B7, HLA-B45 и HLA-Cw8. Последовательности MHC аллелей известны и могут быть найдены, например, в базе данных IMGT/HLA, доступной на www.ebi.ac.uk/ipd/imgt/hla. В некоторых вариантах, MHC аллелью класса I является HLA-A2 аллель, которая в некоторых популяциях экспрессируется приблизительно 50% популяции. В некоторых вариантах, HLA-A2 аллелью может быть HLA-A*0201, *0202, *0203, *0206 или *0207 генный продукт. В некоторых случаях, могут иметься различия в частоте подтипов среди разных популяций. Например, в некоторых вариантах, более 95% HLA-A2 положительной кавказской популяции является HLA-A*0201, а в китайской популяции частота описана как приблизительно 23% HLA-A*0201, 45% HLA-A*0207, 8% HLA-A*0206 и 23% HLA-A*0203.
В некоторых вариантах, MHC-класса I рестриктированные пептиды имеют 8-15 аминокислот в длину, например, 8-10 аминокислот в длину. В некоторых вариантах, MHC молекулы класса I связывают пептиды, полученные из эндогенных антигенов, таких как белки опухоли, вирусные или бактериальные белки, полученные в болезненных или зараженных клетках, которые процессируют в цитоплазме клетки через цитозольный путь. В некоторых вариантах, MHC класса I-пептид комплексы, отображенные на поверхности клетки, обычно распознаются TCR, экспрессированных на CD8+ T клетках, таких как цитотоксические T клетки. В некоторых вариантах, MHC класса I-пептид комплексы могут быть распознаны TCR, экспрессированными на CD4+ T клетках, таких как TCR, демонстрирующие CD8- или частично CD8- независимое связывание.
В некоторых вариантах, TCR или другие связывающие MHC-пептид молекулы распознают или потенциально распознают T клеточный эпитоп в контексте MHC молекулы класса II. MHC белки класса II экспрессированы в подмножестве ядросодержащих клеток позвоночных, обычно называемых антиген-презентующие клетки (АПК). У человека имеется несколько MHC класса II аллелей, таких как, например, DR1, DR3, DR4, DR7, DR52, DQ1, DQ2, DQ4, DQ8 и DP1. В некоторых вариантах, MHC класса II аллель является HLA-DRB1*0101, HLA-DRB*0301, HLA-DRB*0701, HLA-DRB*0401, HLA-DQB1*0201. Последовательности MHC аллелей известны и могут быть найдены, например, в базе данных IMGT/HLA, доступной на www.ebi.ac.uk/ipd/imgt/hla.
В некоторых вариантах, MHC-класса II рестрикртированные пептиды обычно имеют около 9-25 остатков в длину, например, 15-25 остатков или 13-18 остатков в длину, и, в некоторых случаях, содержат связывающую коровую область из около 9 аминокислот или около 12 аминокислот. В некоторых вариантах, MHC молекулы класса II связывают пептиды, полученные из экзогенных антигенов, которые интернализированы фагоцитозом или эндоцитозом и процессированы в эндосомальном/лизосомальном пути. В некоторых вариантах, MHC класса II-пептид комплексы, воспроизведенные на поверхности клеток, обычно распознаются CD4+ клетками, такими как хелперные T клетки. В некоторых вариантах, воспроизведенные MHC класса II-пептид комплексы могут распознаваться TCR, экспрессированными на CD8+ T клетках.
Обычно пептидным эпитопом или T клеточным эпитопом является пептидная часть антигена. В некоторых вариантах, антиген известен, и в некоторых случаях пептидный эпитоп, распознаваемый TCR или его антигенсвязывающей частью (или другими связывающими MHC-пептид молекулами), также может быть известен, например, известен до проведения представленного способа.
В некоторых вариантах, антигеном является ассоциированный с опухолью антиген, антиген, экспрессированный в конкретном типе клетки, ассоциированной с аутоиммунным или воспалительным заболеванием, или антиген, полученный из вирусного патогена или бактериального патогена. В некоторых вариантах, антигеном является антиген, вовлеченный в заболевание. В некоторых вариантах, заболевание может быть вызвано злокачественностью или превращением клеток, таким как рак. В некоторых вариантах, антигеном может быть антиген внутриклеточного белка из опухолевой или раковой клетки, такой как ассоциированный с опухолью антиген. В некоторых случаях, из-за того, что основная часть раковых антигенов получена из внутриклеточных белков, которые могут быть поражены только на поверхности клетки в контексте MHC молекулы, TCR являются идеальными кандидатами для терапии, так как они вовлечены в распознавание этого класса антигенов. В некоторых вариантах, заболевание может быть вызвано заражением, например, бактериальной или вирусной инфекцией. В некоторых вариантах, антигеном является ассоциированный с вирусом раковый антиген. В некоторых случаях, рекомбинантный TCR или его антигенсвязывающая часть (и другие Связывающие MHC-пептид молекулы) распознают или потенциально распознают пептиды, полученные из вирусных белков, которые в природе процессируются в зараженных клетках и воспроизводятся MHC молекулой на поверхности клетки. В некоторых вариантах, заболеванием может быть аутоиммунное заболевание. Другие цели включают перечисленные HLA Factsbook (Marsh et al. (2000)) и другие известные.
В некоторых вариантах, антигеном является такой, который ассоциирован с опухолью или раком. В некоторых вариантах, опухолевым или раковым антигеном является такой, который может быть найден на злокачественной клетке, найден внутри злокачественной клетки или является медиатором роста опухолевой клетки. В некоторых вариантах, опухолевым или раковым антигеном является такой, который преимущественно экспрессируется или сверхэкспрессируется опухолевой клеткой или раковой клеткой. Множество опухолевых антигенов идентифицировано и известно, включая MHC-рестриктированные, определенные T клеткой опухолевые антигены (см., например, cancerimmunity.org/peptide/; Boon и Old (1997) Curr Opin Immunol, 9:681 -3; Cheever et al. (2009) Clin Cancer Res, 15:5323-37). В некоторых вариантах, опухолевые антигены включают, но не ограничены ими, мутированные пептиды, антигены дифференциации и сверхэкспрессированные антигены, все которые служат в качестве целей для терапии.
В некоторых вариантах, опухолевым или раковым антигеном является антиген лимфомы, (например, неходжкинской лимфомы или лимфомы Ходжкина), раковый антиген B-клеточной лимфомы, антиген лейкоза, антиген миеломы (т.е., множественной миеломы или плазмоклеточной миеломы), антиген острого лимфобластного лейкоза, антиген хронического миелоидного лейкоза или антиген острого миелогенного лейкоза. В некоторых вариантах, раковым антигеном является антиген, который сверхэкспрессируется или ассоциирован с раком, которым являются аденокарциномы, такие как рак поджелудочной железы, толстой кишки, молочной железы, яичников, легкого, простаты, головы и шеи, включая множественные миеломы и B клеточные лимфомы. В некоторых вариантах, антиген ассоциирован с раком, таким как рак простаты, рак легких, рак груди, рак яичников, рак поджелудочной железы, рак кожи, рак печени (например, печеночноклеточная аденокарцинома), рак кишечника или рак мочевого пузыря.
В некоторых вариантах, антигеном является опухолевый антиген, который может быть глиома-ассоциированным антигеном, β-человеческим хорионическим гонадотропином, альфафетобелком (AFP), антиген созревания B-клетки (BCMA, BCM), рецептором фактора активации B-клетки (BAFFR, BR3) и/или трансмембранным активатором и партнером CAML (TACI), Fc Рецептор-подобным 5 (FCRL5, FcRH5), лектин-реактивным AFP, тироглобулином, RAGE-1, MN-CA IX, обратной транскриптазой теломеразы человека, RU1, RU2 (AS), кишечной карбоксилэстеразой, mut hsp70-2, M-CSF, Меланин-A/MART-1, WT-1, S-100, MBP, CD63, MUC1 (например, MUC1-8), p53, Ras, циклином B1, HER-2/neu, карциноэмбрионным антигеном (CEA), gp100, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11, MAGE-A11, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-C1, BAGE, GAGE-1, GAGE-2, pl5, тирозиназой, родственным тирозиназе белком 1 (TRP-1), родственным тирозиназе белком 2 (TRP-2), β-катенином, NY-ESO-1, LAGE-1a, PP1, MDM2, MDM4, EGVFvIII, Tax, SSX2, теломеразой, TARP, pp65, CDK4, виментином, S100, eIF-4A1, IFN-индуцируемым p78 и меланотрансферрином (p97), Уроплакином II, специфическим антигеном простаты (PSA), человеческим калликреином (huK2), специфическим мембранным антигеном простаты (PSM) и простатической кислой фосфатазой (PAP), нейтрофильной эластазой, эфрином B2, BA-46, бета-катенином, Bcr-abl, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, Каспазой 8 или B-Raf антигеном. Другие опухолевые антигены могут включать любые, полученные из FRa, CD24, CD44, CD133, CD 166, epCAM, CA-125, HE4, Oval, рецептора эстрогена, рецептора прогестерона, uPA, PAI-1, CD19, CD20, CD22, ROR1, мезотелина, CD33/IL3Ra, c-Met, PSMA, Гликолипида F77, GD-2, инсулинового фактора роста (IGF)-I, IGF-II и IGF-I рецептора. Специфические ассоциированные с опухолью антигены или T клеточные эпитопы известны (см., например, van der Bruggen et al. (2013) Cancer Immun, доступную на www.cancerimmunity.org/peptide/; Cheever et al. (2009) Clin Cancer Res, 15, 5323-37).
В некоторых вариантах, антигеном является вирусный антиген. Множество вирусных антигенных целей идентифицированы и известны, включая пептиды, полученные из вирусных геномов в ВИЧ, HTLV и других вирусах (см., например, Addo et al. (2007) PLoS ONE, 2, e321; Tsomides et al. (1994) J Exp Med, 180, 1283-93; Utz et al. (1996) J Virol, 70, 843-51). типовые вирусные антигены включают, но не ограничены ими, антиген из гепатита A, гепатита B (например, HBV которые и поверхностные антигены (HBVc, HBVs)), гепатита C (HCV), Вируса Эпштейн-Барр (например, EBVA), человеческого папилломавируса (HPV; например, E6 и E7), вируса иммунодефицита человека типа-1 (ВИЧ1), герпесвируса саркомы Капоши (KSHV), папилломавируса человека (HPV), вируса гриппа, вируса Ласса, HTLN-1, HIN-1, HIN-II, CMN, EBN или HPN. В некоторых вариантах, целевым белком является бактериальный антиген или другой патогенный антиген, такой как антигены Mycobacterium tuberculosis (MT), антигены трипаносомы, например, Tiypansoma cruzi (T. cruzi), такие как поверхностный антиген (TSA) или малярийные антигены. специфические вирусные антигены или эпитопы или другие патогенные антигены или T клеточные эпитопы известны (см., например, Addo et al. (2007) PLoS ONE, 2:e321; Anikeeva et al. (2009) Clin Immunol, 130:98-109).
В некоторых вариантах, антигеном является антиген, полученный из вируса, ассоциированного с раком, такого как онкогенный вирус. Например, онкогенным вирусом является такой, для которого заражение определенными вирусами приводит к развитию разных типов рака, например, антигены вируса гепатита A, гепатита B (например, HBV коровые и поверхностные антигены (HBVc, HBVs)), гепатита C (HCV), папилломавируса человека (HPV), вирусных инфекций гепатита, вируса Эпштейн-Барр (EBV), герпес-вируса человека 8 (HHV-8), вируса Т-клеточного лейкоза человека-1 (HTLV-1), вируса Т-клеточного лейкоза человека-2 (HTLV-2) или цитомегаловируса (CMV).
В некоторых вариантах, вирусным антигеном является HPV антиген, который, в некоторых случаях, может привести к повышению риска развития рака шейки матки. В некоторых вариантах, антигеном может быть HPV-16 антиген, HPV-18 антиген, HPV-31 антиген, HPV-33 антиген или HPV-35 антиген. В некоторых вариантах, вирусным антигеном является HPV-16 антиген (например, серореактивные области E1, E2, E6 и/или E7 белков HPV-16, см., например, Патент США № 6,531,127) или HPV-18 антиген (например, серореактивные области L1 и/или L2 белков HPV-18, такие как описаны в Патенте США № 5,840,306). В некоторых вариантах, вирусным антигеном является HPV-16 антиген из E6 и/или E7 белков HPV-16. В некоторых вариантах, TCR является TCR, направленный против HPV-16 E6 или HPV-16 E7. В некоторых вариантах, TCR является TCR, описанный в, например, WO 2015/184228, WO 2015/009604 и WO 2015/009606.
В некоторых вариантах, вирусным антигеном является HBV или HCV антиген, который, в некоторых случаях, может привести к большему риску развития рака печени, чем у HBV или HCV отрицательных субъектов. Например, в некоторых вариантах, гетерологичным антигеном является HBV антиген, такой как ядерный антиген гепатита В или оболочечный антиген гепатита В (US2012/0308580).
В некоторых вариантах, вирусным антигеном является EBV антиген, который, в некоторых случаях, может привести к большему риску развития лимфомы Буркитта, носоглоточной карциномы и болезни Ходжкина, чем EBV отрицательных субъектов. Например, EBV является герпес-вирусом человека, который, в некоторых случаях, связан с многочисленными опухолями человека различного тканевого происхождения. Хотя в первую очередь он обнаруживается в виде бессимптомной инфекции, EBV-положительные опухоли могут быть охарактеризованы активной экспрессией вирусных генных продуктов, таких как EBNA-1, LMP-1 и LMP-2A. В некоторых вариантах, гетерологичным антигеном является EBV антиген, который может включать ядерный антиген Эпштейн-Барр (EBNA)-1, EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, EBNA-лидерный белок (EBNA-LP), латентные мембранные белки LMP-1, LMP-2A и LMP-2B, EBV-EA, EBV-MA или EBV-VCA.
В некоторых вариантах, вирусным антигеном является HTLV-1 или HTLV-2 антиген, который, в некоторых случаях, может привести к большему риску развития T-клеточного лейкоза, чем у HTLV-1 или HTLV-2 отрицательных субъектов. Например, в некоторых вариантах, гетерологичным антигеном является HTLV-антиген, такой как TAX.
В некоторых вариантах, вирусным антиген является HHV-8 антиген, который, в некоторых случаях, может привести к большему риску развития саркомы Капоши, чем у HHV-8 отрицательных субъектов. В некоторых вариантах, гетерологичным антигеном является CMV антиген, такой как pp65 или pp64 (см. патент США № 8,361,473).
В некоторых вариантах, антигеном является аутоантиген, такой как антиген полипептида, ассоциированного с аутоиммунным заболеванием или расстройством. В некоторых вариантах, аутоиммунным заболеванием или расстройством может быть рассеянный склероз (MS), ревматоидный артрит (RA), синдром Шегрена, склеродермия, полиомиозит, дерматомиозит, системная красная волчанка, юношеский ревматоидный артрит, анкилозирующий спондилит, миастения гравис (MG), буллезный пемфигоид (антитела к базальной мембране на дермо-эпидермальном соединении), пузырчатка (антитела к белковому комплексу мукополисахарида или внутриклеточному цементирующему веществу), гломерулонефрит (антитела к клубочковой базальной мембране), синдром Гудпасчера, аутоиммунная гемолитическая анемия (антитела к эритроцитам), болезнь Хашимото (антитела к щитовидной железе), пернициозная анемия (антитела к внутреннему фактору), идиопатическая тромбоцитопеническая пурпура (антитела к тромбоцитам), болезнь Грейвса или болезнь Аддисона (антитела к тироглобулину). В некоторых вариантах, аутоантигеном, таким как аутоантиген, ассоциированный с одним или более из вышепредставленных аутоиммунных заболеваний, может быть коллаген, такой как коллаген II типа, микобактериальный белок теплового шока, тироглобулин, рецептор ацетилхолина (AcHR), основной миелиновый белок (MBP) или протеолипидный белок (PLP). Специфические аутоиммунные ассоциированные эпитопы или антигены известны (см., например, Bulek et al. (2012) Nat Immunol, 13:283-9; Harkiolaki et al. (2009) Immunity, 30:348-57; Skowera et al. (2008) J Clin Invest, 1(18): 3390-402).
В некоторых вариантах, идентичность пептидного эпитопа целевого антигена известна, что, в некоторых случаях, может применяться для получения или создания рассматриваемого TCR или при оценке функциональной активности или свойства, в том числе в связи с представленными способами. В некоторых вариантах, пептидные эпитопы могут быть определены или идентифицированы на основе HLA-рестриктированного мотива в целевом рассматриваемом антигене. В некоторых вариантах, пептиды идентифицируют с применением известных компьютерных моделей прогноза. В некоторых вариантах, для прогноза сайтов связывания MHC класса I, такие модели включают, но не ограничены ими, ProPred1 (Singh and Raghava (2001) Bioinformatics 17(12):1236-1237, и SYFPEITHI (см. Schuler et al. (2007) Immunoinformatics Methods in Molecular Biology, 409(1): 75-93 2007). В некоторых вариантах, MHC-рестриктированным эпитопом является HLA-A0201, который экспрессируется у приблизительно 39-46% всех кавказцев и, поэтому, представляет подходящий выбор MHC антигена для применения при получении TCR или другой связывающей MHC-пептид молекулы. В некоторых аспектах, связывающие HLA-A*0201 мотивы и сайты расщепления для протеасом и иммунных протеасом с применением компьютерных моделей прогнозов известны. Для прогнозирования сайтов связывания MHC класса I, такие модели включают, но не ограничены ими, ProPred1 (более подробно описанную в Singh and Raghava, ProPred: prediction of HLA-DR binding sites. BIOINFORMATICS 17(12):1236-1237 2001), и SYFPEITHI (см. Schuler et al. SYFPEITHI, Database for Searching и T-Cell Epitope Prediction. in Immunoinformatics Methods in Molecular Biology, vol 409(1): 75-93 2007). Представлены способы скриннинга и клетки, применяемые в способах скриннинга, такие как T клетки, который распознают антиген или эпитоп, в контексте молекулы основного комплекса гистосовместимости (MHC).
В некоторых вариантах, MHC содержит сайт связывания или полость связывания полиморфного пептида, который может, в некоторых случаях, образовывать комплекс с пептидными эпитопами полипептидов, включая пептидные эпитопы, процессированные клеточным механизмом. В некоторых случаях, MHC молекулы могут быть воспроизведены или экспрессированы на поверхности клетки, в том числе в виде комплекса с пептидом, т.е. MHC-пептидный комплекса, для представления антигена в структуре, распознаваемой TCR на T клетках или других связывающих Молекула MHC-пептидах. В общем, MHC молекулы класса I являются гетеродимерами, имеющими трансмембранную α цепь, в некоторых случаях с тремя α доменами, и не ковалентно ассоциированный β2 микроглобулин. В основном, MHC молекулы класса II состоят из двух трансмембранных гликопротеинов, α и β, оба которых обычно продлевают мембрану. MHC молекула может включать эффективную часть MHC, которая содержит сайт связывания эпитопа или сайты связывания пептида, и последовательность, необходимую для распознавания подходящей связывающей молекулы, такой как TCR. В некоторых вариантах, MHC молекулы класса I доставляют пептиды, возникающие в цитозоле, на поверхность клетки, где пептид:MHC комплекс распознается T клетками, такими как в основном CD8+ T клетки, но в некоторых случаях, CD4+ T клетки. В некоторых вариантах, MHC молекулы класса II доставляют пептиды, возникающие в везикулярной системе, на поверхность клетки, где они обычно распознаются CD4+ T клетками. В основном, MHC молекулы кодированы группой связанных локусов, которые вместе называют H-2 в мышином и человеческом лейкоцитарном антигене (HLA) у человека. В некоторых аспектах, человеческий MHC также может быть назван человеческим лейкоцитарным антигеном (HLA).
В некоторых вариантах, пептидным эпитопом или T клеточным эпитопом является пептид, который может быть получен из или на основе фрагмента большей биологической молекулы, такой как полипептид или белок, и который способен ассоциироваться с или образовывать комплекс с MHC молекулой. В некоторых вариантах, пептид имеет от около 8 до около 24 аминокислот в длину. В некоторых вариантах, пептид имеет длину от или от около 9-22 аминокислот для распознавания в MHC класса II комплексе. В некоторых вариантах, пептид имеет длину от или от около 8-13 аминокислот для распознавания в MHC класса I комплексе. В некоторых вариантах, MHC молекула и пептидный эпитоп или T клеточный эпитоп образуют комплекс или ассоциируются через не ковалентные взаимодействия пептида в полости связывания или щели MHC молекулы.
В некоторых вариантах, MHC-пептидный комплекс присутствует или воспроизводится на поверхности клетки. В некоторых вариантах, MHC-пептидный комплекс может специфически распознаваться TCR или его антигенсвязывающей частью или другой связывающей MHC-пептид молекулой. В некоторых вариантах, T клеточный эпитоп или пептидный эпитоп способны вызывать иммунный ответ у животного через его характеристики связывания с MHC молекулами. В некоторых вариантах, при распознавании T клеточного эпитопа, такого как MHC-пептидный комплекс, TCR (или другая Связывающая MHC-пептид молекула) вызывает или запускает сигнал активации к T клетке, который вызывает ответ T клетки, такой как пролиферация T клетки, образование цитокина, цитотоксический ответ T клетки или другой ответ.
В некоторых вариантах, связывающей MHC-пептид молекулой является TCR или его связывающий эпитоп фрагмент. В некоторых вариантах, MHC-пептид связывающей молекулой является TCR-подобный CAR, который содержит антитело или его связывающий эпитоп фрагмент, такое как TCR-подобное антитело, такое, как сконструировано для связывания с MHC-пептидный комплексами. В некоторых вариантах, такая связывающая молекула связывается со связывающей последовательностью, такой как Т клеточный эпитоп, содержащий аминокислотную последовательность или антиген целевого полипептида. В некоторых вариантах, связывающая последовательность целевого пептида или целевого полипептида известна. В некоторых вариантах, MHC-пептид связывающей молекулой может быть получена из природных источников, или может быть частично или полностью синтетической или рекомбинантной.
В некоторых вариантах, MHC-пептид связывающей молекулой является молекула или ее часть, которая обладает способностью связываться, например, специфически связываться с пептидным эпитопом, который присутствует или воспроизведен в контексте MHC молекулы, т.е. MHC-пептидный комплексом, например, на поверхности клетки. В некоторых вариантах, связывающая молекула может включать любую существующую в природе, синтетическую, полусинтетическую или рекомбинантно полученную молекулу, которая может связываться, например, специфически связываться с MHC-пептидный комплексом. Типовые Связывающие MHC-пептид молекулы включают Т-клеточные рецепторы или антитела или их антигенсвязывающие части, включая отдельные цепи переменных областей иммуноглобулина (например, scTCR, scFv), которые демонстрируют специфическую способность связываться с MHC-пептидный комплексом.
В некоторых вариантах, TCR является полноразмерный TCR. В некоторых вариантах, TCR является антигенсвязывающей частью. В некоторых вариантах, TCR является димерным TCR (dTCR). В некоторых вариантах, TCR является однонитевым TCR (sc-TCR). TCR может быть связан с клеткой или в растворимой форме. В некоторых вариантах, TCR имеет связанную с клеткой форму, экпрессированную на поверхности клетки.
В некоторых вариантах dTCR содержит первый полипептид, где последовательность, соответствующая представленной переменной области последовательности TCR α цепи слита с N окончанием последовательности, соответствующей постоянной области внеклеточной последовательности TCR α цепи, и второй полипептид, где последовательность, соответствующая представленной последовательности переменной области TCR β цепи слита с N окончанием последовательности, соответствующей постоянной области внеклеточной последовательности TCR β, где первый и второй полипептиды связаны дисульфидной связью. В некоторых вариантах, связь может соответствовать нативной межнитевой дисульфидной связи, присутствующей в нативных димерных αβ TCR. В некоторых вариантах, межнитевые дисульфидные связи не присутствуют в нативном TCR. Например, в некоторых вариантах, один или более цистеинов могут быть введены в постоянную область внеклеточных последовательностей dTCR полипептидной пары. В некоторых случаях, обе, нативная и не нативная дисульфидные связи могут быть желательны. В некоторых вариантах, TCR содержит трансмембранную последовательность для прикрепления к мембране.
В некоторых вариантах, dTCR содержит представленную TCR α цепь, содержащую переменный α домен, постоянный α домен и первый мотив димеризации, присоединенный к C-окончанию постоянного α домена, и представленную β цепь, содержащую переменный β домен, постоянный β домен и первый мотив димеризации, присоединенный к C-окончанию постоянного β домена, где первый и второй мотивы димеризации могут взаимодействовать с образованием ковалентной связи между аминокислотами в первом мотиве димеризации и аминокислотами во втором мотиве димеризации, связывающей TCR α цепь и TCR β цепь вместе.
В некоторых вариантах, TCR является scTCR, который является одинарной аминокислотной нитью, содержащей α цепь и β цепь, которые способны связываться с MHC-пептидный комплексами. Обычно scTCR может быть получен с применением известных способов, см., например, международные опубликованные PCT №№ WO 96/13593, WO 96/18105, WO99/18129, WO 04/033685, WO2006/037960, WO2011/044186; патент США № 7,569,664; и Schlueter, C. J. et al. J. Mol. Biol. 256, 859 (1996).
В некоторых вариантах, scTCR содержит первый сегмент, образованный аминокислотной последовательностью, соответствующей последовательности представленной переменной области TCR α цепи, второй сегмент, образованный аминокислотной последовательностью, соответствующей последовательности представленной переменной области TCR β цепи, слитый с N окончанием аминокислотной последовательности, соответствующей постоянному домену внеклеточной последовательности TCR β цепи, и линкер последовательности, связывающей C окончание первого сегмента с N окончанием второго сегмента.
В некоторых вариантах, scTCR содержит первый сегмент, образованный аминокислотной последовательностью, соответствующей последовательности представленной переменной области TCR β цепи, второй сегмент, образованный аминокислотной последовательностью, соответствующей последовательности представленной переменной области TCR α цепи, слитый с N окончанием аминокислотной последовательности, соответствующей постоянному домену внеклеточной последовательности TCR α цепи и линкер последовательности, связывающей C окончание первого сегмента с N окончанием второго сегмента.
В некоторых вариантах, scTCR содержит первый сегмент, образованный представленной переменной областью последовательности α цепи, слитый с N концом постоянного домена внеклеточной последовательности α цепи, и второй сегмент, образованный переменной областью последовательности β цепи, слитый с N концом последовательности внеклеточной постоянной и трансмембранной последовательности β цепи, и, необязательно, линкерную последовательность, связывающую C окончание первого сегмента с N окончанием второго сегмента.
В некоторых вариантах, scTCR содержит первый сегмент, образованный представленной переменной областью последовательности TCR β цепи, слитый с N концом постоянного домена внеклеточной последовательности β цепи, и второй сегмент, образованный переменной областью последовательности α цепи, слитый с N концом последовательности внеклеточной постоянной и трансмембранной последовательности α цепи, и, необязательно, линкерную последовательность, связывающую C окончание первого сегмента с N окончанием второго сегмента.
В некоторых вариантах, для scTCR для связывания MHC-пептидный комплекса, α и β цепи должны быть спарены так, чтобы их переменные области последовательностей были ориентированы для такого связывания. Разные способы облегчения спаривания α и β в scTCR известны. В некоторых вариантах, включают линкерную последовательность, которая связывает α и β цепи с получением одной полипептидной нити. В некоторых вариантах, линкер должен иметь достаточную длину для охватывания расстояния между C окончанием α цепи и N окончанием β цепи или наоборот, а также для гарантии того, что длина линкера не настолько велика, чтобы он блокировал или снижал связывание scTCR с целевым пептид-MHC комплексом.
В некоторых вариантах, линкером scTCR, который связывает первые и второй сегменты TCR, может быть любой линкер, способный образовывать одинарную полипептидную нить, при этом сохраняя специфичность TCR связывания. В некоторых вариантах, линкерная последовательность может, например, иметь формулу -P-AA-P-, где P является пролином и AA представляет аминокислотные последовательности, где аминокислотами являются глицин и серин. В некоторых вариантах, первый и второй сегменты спарены так, чтобы переменная область их последовательностей были ориентированы для такого связывания. Следовательно, в некоторых случаях, линкер имеет достаточную длину для охватывания расстояния между C окончанием первого сегмента и N окончанием второго сегмента или наоборот, но не настолько большую, чтобы он блокировал или снижал связывание scTCR с целевым лигандом. В некоторых вариантах, линкер может содержать от или от около 10-45 аминокислот, например, 10-30 аминокислот или 26-41 аминокислотных остатков, например 29, 30, 31 или 32 аминокислот. В некоторых вариантах, линкер имеет формулу -PGGG-(SGGGG)n-P-, где n равно 5 или 6 и P является пролином, G является глицином и S является серином (SEQ ID NO: 22). В некоторых вариантах, линкер имеет последовательность GSADDAKКДАAKKDGKS (SEQ ID NO: 23).
В некоторых вариантах, scTCR содержит дисульфидную связь между остатками одинарной аминокислотной нити, в некоторых случаях, может способствовать стабильности спаривания между α и β областями однонитевой молекулы (см., например, патент США № 7,569,664). В некоторых вариантах, scTCR содержит ковалентную дисульфидную связь, связывающую остаток иммуноглобулиновой области постоянного домена α цепи с остатком иммуноглобулиновой области постоянного домена β цепи однонитевой молекулы. В некоторых вариантах, дисульфидная связь соответствует нативной дисульфидной связи, присутствующей в нативном dTCR. В некоторых вариантах, дисульфидная связь в нативном TCR не присутствует. В некоторых вариантах, дисульфидной связью является введенная не нативная дисульфидная связь, например, путем введения одного или более цистеинов в постоянную область внеклеточной последовательности первой и второй цепи областей scTCR полипептида. Типовые цистеиновые мутации включают любые, описанные здесь. В некоторых случаях, и нативная и не нативная дисульфидная связь может присутствовать.
В некоторых вариантах, scTCR является не связанным дисульфидом усеченным TCR, в котором гетерологичные лейциновые «молнии», слитые с их C-окончаниями, способствуют ассоциации цепи (см., например, международную опубликованную PCT № WO99/60120). В некоторых вариантах, scTCR содержит TCRα переменный домен, ковалентно связанный с TCRβ переменным доменом через пептидный линкер (см., например, международную опубликованную PCT № WO99/18129).
В некоторых вариантах, любой из представленных TCR, включая dTCR или scTCR, может быть связан с сигнальными доменами, что дает активный TCR на поверхности T клетки. В некоторых вариантах, TCR экспрессируется на поверхности клетки. В некоторых вариантах, TCR не содержит последовательность, соответствующую трансмембранной последовательности. В некоторых вариантах, трансмембранный домен положительно заряжен. В некоторых вариантах, трансмембранный домен может быть Cα или Cβ трансмембранным доменом. В некоторых вариантах, трансмембранный домен может происходить не из TCR, например, трансмембранная область из CD3z, CD28 или B7.1. В некоторых вариантах, TCR не содержит последовательность, соответствующую цитоплазматическим последовательностям. В некоторых вариантах, TCR содержит CD3z сигнальный домен. В некоторых вариантах, TCR способен образовывать TCR комплекс с CD3.
В некоторых вариантах, TCR является растворимый TCR. В некоторых вариантах, растворимый TCR имеет структуру, описанную в WO99/60120 или WO 03/020763. В некоторых вариантах, TCR не содержит последовательность, соответствующую трансмембранной последовательности, например, чтобы позволить мембране закрепиться в клетке, в которой он экспрессируется. В некоторых вариантах, TCR не содержит последовательность, соответствующую цитоплазматическим последовательностям.
В некоторых вариантах, рекомбинантный рецептор, например, TCR или его антигенсвязывающий фрагмент, является или был модифицирован по сравнению с известным рекомбинантным рецептором. В определенных вариантах, рекомбинантные рецепторы, например, TCR или его антигенсвязывающие фрагменты, включают одну или более аминокислотных вариаций, например, замещений, делеций, вставок и/или мутаций по сравнению с последовательностью рекомбинантного рецептора, например, TCR, описанного здесь или известного. Типовые варианты включают такие, которые созданы для улучшения сродства связывания и/или других биологических свойств связывающей молекулы. Варианты аминокислотной последовательности связывающей молекулы могут быть получены введением подходящих модификаций в нуклеотидную последовательность, кодирующую связывающую молекулу, или пептидным синтезом. Такие модификации включают, например, делеции из и/или вставки в и/или замещения остатков в аминокислотных последовательностях связывающей молекулы. Любые сочетания делеции, вставки и замещения могут быть проведены для получения конечного конструкта, при условии что конечный конструкт обладает желаемыми характеристиками, например, антигенным связыванием.
В некоторых вариантах, способы направленного развития применяют для создания TCR с измененными свойствами, такими как более высокая аффинность к определенному пептиду в контексте MHC молекулы. В некоторых вариантах, направленное развитие достигается способами дисплея, включающими, но не ограниченными ими, дрожжевой дисплей (Holler et al. (2003) Nat Immunol, 4, 55-62; Holler et al. (2000) Proc Natl Acad Sci U S A, 97, 5387-92), фаговый дисплей (Li et al. (2005) Nat Biotechnol, 23, 349-54) или T клеточный дисплей (Chervin et al. (2008) J Immunol Methods, 339, 175-84). В некоторых вариантах, подходы дисплея включают конструирование или модификацию известных, исходных или ссылочных TCR. Например, в некоторых случаях, ссылочный TCR, такой как любой из представленных здесь, может применяться в качестве матрицы для получения мутагенизированных TCR, в которых один или более остатков CDR мутированы, и выбирают мутанты с желаемым измененным свойством, таким как более высокая аффинность для пептидного эпитопа в контексте MHC молекулы.
В определенных вариантах, рекомбинантные рецепторы, например, TCR или их антигенсвязывающие фрагменты, включают одно или более аминокислотных замещений, например, по сравнению с рекомбинантным рецептором, например, TCR, последовательности по сравнению с последовательностью природного спектра, например, человеческого спектра. Исследуемые сайты для заместительного мутагенеза включают CDR, FR и/или постоянные области. Аминокислотные замещения могут быть введены в рассматриваемую связывающую молекулу, и продукты подвергают скринингу желаемой активности, например, сохраненной/улучшенной антигенной аффинности или авидности, пониженной иммуногенности, улучшенного периода полужизни, CD8-независимого связывания или активности, поверхностной экспрессии, содействия спариванию TCR цепи и/или других улучшенных свойства или функций.
В некоторых вариантах, один или более остатков в пределах CDR рекомбинантного рецептора, например, TCR, замещены. В некоторых вариантах, замещение проводят для свертывания последовательности или положения в последовательности до генеративной линии последовательности, такой как последовательность связывающей молекулы, найденная в генеративной линии (например, генеративной линии человека), например, для снижения вероятности иммуногенности, например, при введении человеку.
В определенных вариантах, замещения, вставки или делеции могут проводиться в пределах одной или более CDR до той степени, пока такие изменения по существу не снизят способность рекомбинантного рецептора, например, TCR или его антигенсвязывающего фрагмента, связывать антиген. Например, консервативные изменения (например, консервативные замещения, представленные здесь), которые по существу не снижают сродство связывания, могут быть проведены в CDR. Такие изменения, например, могут быть вне контактирующих с антигеном остатков в CDR. В определенных вариантах переменных последовательностей, представленных здесь, каждая CDR либо не изменена, либо содержит не более одного, двух или трех аминокислотных замещений.
Вставки аминокислотных последовательностей включают амино- и/или карбоксил-концевые слияния, варьирующиеся в длину от одного остатка до полипептидов, содержащих сотни или более остатков, а также вставки внутри последовательностей одного или множества аминокислотных остатков.
В некоторых аспектах, TCR или его антигенсвязывающий фрагмент может содержать одну или более модификаций в α цепи и/или β цепи так, что если TCR или его антигенсвязывающий фрагмент экспрессируется в клетке, частота ошибочного спаривания между α цепью и β цепью TCR и α цепью и β цепью эндогенного TCR снижается, экспрессия α цепи и β цепи TCR повышается и/или стабильность α цепи и β цепи TCR повышается.
В некоторых вариантах, TCR содержит один или более не нативных остатков для введения ковалентной дисульфидной связи, связывающей остаток иммуноглобулиновой области постоянного домена α цепи с остатком иммуноглобулиновой области постоянного домена β цепи. В некоторых вариантах, один или более цистеинов могут быть введены в постоянную область внеклеточных последовательностей первого и второго сегментов TCR полипептида. типовые не ограничивающие модификации в TCR для введения не нативных цистеиновых остатков описаны здесь (см. также международную PCT № WO2006/000830 и WO2006037960). В некоторых случаях, и нативные и не нативные дисульфидные связи могут быть желательны. В некоторых вариантах, TCR или его антигенсвязывающий фрагмент модифицированы так, что межнитевые дисульфидные связи в нативном TCR не присутствуют.
В некоторых вариантах, трансмембранный домен постоянной области TCR может быть модифицирован так, чтобы содержать большее число гидрофобных остатков (см., например, Haga-Friedman et al. (2012) Journal of Immunology, 188:5538-5546). В некоторых вариантах, трансмембранная область TCR α цепи содержит одну или более мутаций, соответствующих S116L, G119V или F120L, со ссылкой на нумерацию Cα, представленную в SEQ ID NO:24.
В некоторых вариантах, TCR или его антигенсвязывающий фрагмент кодирован нуклеотидной последовательностью, которая является или была кодон-оптимизированной. Типовые кодон-оптимизированные варианты описаны здесь везде.
В. Химерные антигенные рецепторы (CAR)
В некоторых вариантах, рекомбинантным рецептором, который вводят в клетку, является химерный антигенный рецептор (CAR) или его антигенсвязывающий фрагмент. В некоторых вариантах представлены сконструированные клетки, такие как T клетки, которые экспрессируют CAR со специфичностью к конкретному антигену (или маркеру или лиганду), такому как антиген, экспрессированный на поверхности конкретного типа клетки. В некоторых вариантах, антигеном является полипептид. В некоторых вариантах, им является углевод или другая молекула. В некоторых вариантах, антиген селективно экспрессирован или сверхэкспрессирован на клетке заболевания или состояния, например, опухоли или патогенных клеток, по сравнению с нормальными или не таргетными клетками или тканями. В других вариантах, антиген экспрессирован на нормальных клетках и/или экспрессирован на сконструированных клетках.
В конкретных вариантах, рекомбинантный рецептор, такой как химерный рецептор, содержит внутриклеточную сигнальную область, которая включает цитоплазматический сигнальный домен (также взаимозаменяемо называемый внутриклеточный связывающий домен), такой как цитоплазматическая (внутриклеточная) область, способная вызывать первичный сигнал активации в Т клетке, например, цитоплазматический сигнальный домен компонента Т-клеточного рецептора (TCR) (например, цитоплазматический сигнальный домен дзэта цепи CD3-дзэта (CD3ζ) цепи или его функциональный вариант или сигнальная часть) и/или который содержит иммунорецепторный тирозиновый активирующий мотив (ITAM).
В некоторых вариантах, химерный рецептор дополнительно содержит внеклеточный лиганд-связывающий домен, который специфически связывается с лигандом (например, антигеном) антигена. В некоторых вариантах, химерным рецептором является CAR, который содержит внеклеточный антиген-распознающий домен, который специфически связывается с антигеном. В некоторых вариантах, лигандом, таким как антиген, является белок, экспрессированный на поверхности клеток. В некоторых вариантах, CAR является TCR-подобный CAR, и антигеном является процессированный пептидный антиген, такой как пептидный антиген внутриклеточного белка, который, как и TCR, распознается на поверхности клетки в контексте молекулы главного комплекса гистосовместимости (MHC).
Типовые антигенные рецепторы, включающие CAR, и способы конструирования и введения таких рецепторов в клетки, включают такие, которые описаны, например, в публикациях заявок на международный патент №№ WO200014257, WO2013126726, WO2012/129514, WO2014031687, WO2013/166321, WO2013/071154, WO2013/123061, публикациях заявок на патент США №№ US2002131960, US2013287748, US20130149337, патентах США №№: 6,451,995, 7,446,190, 8,252,592, 8,339,645, 8,398,282, 7,446,179, 6,410,319, 7,070,995, 7,265,209, 7,354,762, 7,446,191, 8,324,353, и 8,479,118, и заявке на европейский патент № EP2537416, и/или такие, который описаны у Sadelain et al., Cancer Discov. 2013 April; 3(4): 388-398; Davila et al. (2013) PLoS ONE 8(4): e61338; Turtle et al., Curr. Opin. Immunol., 2012 October; 24(5): 633-39; Wu et al., Cancer, 2012 March 18(2): 160-75. В некоторых аспектах, антигенные рецепторы включают CAR, описанные в патенте США №: 7,446,190, и такие, которые описаны в Публикации заявки на международный патент №: WO/2014055668 A1. Примеры CAR включают CAR, описанные в любой из вышеуказанных публикаций, таких как WO2014031687, US 8,339,645, US 7,446,179, US 2013/0149337, Патент США №: 7,446,190, Патент США №: 8,389,282, Kochenderfer et al., 2013, Nature Reviews Clinical Oncology, 10, 267-276 (2013); Wang et al. (2012) J. Immunother. 35(9): 689-701; и Brentjens et al., Sci Transl Med. 2013 5(177). См. также WO2014031687, US 8,339,645, US 7,446,179, US 2013/0149337, Патент США №: 7,446,190, и Патент США №: 8,389,282.
В некоторых вариантах, CAR сконструирован со специфичностью к конкретному антигену (или маркеру или лиганду), такому как антиген, экспрессированный в конкретном типе клетки, являющийся целью адоптивной терапии, например, раковый маркер, и/или антиген, предназначенный для вызова ослабляющего ответа, такой как антиген, экспрессированный на нормальном или не болезненном типе клетки. Таким образом, CAR обычно включает в его внеклеточной части одну или более антигенсвязывающих молекул, таких как один или более антигенсвязывающи фрагмент, домен или часть, или один или более переменных доменов антитела, и/или молекул антитела. В некоторых вариантах, CAR включает антигенсвязывающую часть или части молекулы антитела, такую как однонитевой фрагмент антитела (scFv), полученный из переменной тяжелой (VH) и переменной легкой (VL) цепей моноклонального антитела (mAb).
В некоторых вариантах, антитело или его антигенсвязывающая часть экспрессируется на клетках как часть рекомбинантного рецептора, такого как антигенный рецептор. Антигенные рецепторы включают функциональные не-TCR антигенные рецепторы, такие как химерные антигенные рецепторы (CAR). В общем, CAR, содержащий антитело или антигенсвязывающий фрагмент, который демонстрирует TCR-подобную специфичность, направленное против пептид-MHC комплексов, также может называться TCR-подобный CAR. В некоторых вариантах, внеклеточный антигенсвязывающий домен, специфический для MHC-пептидного комплекса TCR-подобного CAR, связан с одним или более внутриклеточными сигнальными компонентами, в некоторых аспектах, через линкеры и/или трансмембранные домены. В некоторых вариантах, такие молекулы обычно могут имитировать или приблизительно соответствовать сигналу через природный антигенный рецептор, такой как TCR, и, необязательно, сигнал через такой рецептор в сочетании с костимулирующим рецептором.
В некоторых вариантах, рекомбинантный рецептор, такой как химерный рецептор (например, CAR), включает лигандсвязывающий домен, который связывается, например, специфически связывается с антигеном (или лигандом). Антигены, на которые направлены химерные рецепторы, включают такие, которые экспрессированы в контексте заболевания, состояния или клеточного типа, на которые направлена адоптивная клеточная терапия. Заболевания и состояния включают пролиферативные, неопластические и злокачественные заболевания и расстройства, включая рак и опухоли, включая гемобластоз, раки иммунной системы, такие как лимфомы, лейкозы и/или миеломы, такие как B, T и миелоидные лейкозы, лимфомы и миеломы.
В некоторых вариантах, антигеном (или лигандом) является полипептид. В некоторых вариантах, он является углеводом или другой молекулой. В некоторых вариантах, антиген (или лиганд) селективно экспрессируется или сверхэкспрессируется на клетках заболевания или состояния, например, опухолевых или патогенных клетках, по сравнению с нормальными или не таргетными клетками или тканями. В других вариантах, антиген экспрессируется на нормальных клетках и/или экспрессируется на сконструированных клетках.
В некоторых вариантах, CAR содержит антитело или антигенсвязывающий фрагмент (например, scFv), который специфически распознает антиген, такой как нативный антиген, экспрессированный на поверхности клетки.
В некоторых вариантах, антигеном (или лигандом) является опухолевый антиген или раковый маркер. В некоторых вариантах, антигеном (или лигандом) является или включает αvβ6 интегрин (avb6 интегрин), антиген созревания B клетки (BCMA), B7-H3, B7-H6, карбонангидразу 9 (CA9, также известную как CAIX или G250), раково-тестикулярный антиген, раково-тестикулярный антиген 1B (CTAG, также известный как NY-ESO-1 и LAGE-2), карциноэмбрионный антиген (CEA), циклина, циклин A2, C-C мотив лиганда хемокина 1 (CCL-1), CD19, CD20, CD22, CD23, CD24, CD30, CD33, CD38, CD44, CD44v6, CD44v7/8, CD123, CD133, CD138, CD171, протеогликан с хондроитинсульфатами 4 (CSPG4), белок эпидермального фактора роста (EGFR), мутацию III типа рецептора эпидермального фактора роста (EGFR vIII), эпителиальный гликобелок 2 (EPG-2), эпителиальный гликобелок 40 (EPG-40), эфрин B2, рецептор эфрина A2 (EPHa2), рецептор эстрогена, Fc рецептор подобный 5 (FCRL5; также известный как Fc рецептор гомолог 5 или FCRH5), фетальный ацетилхолиновый рецептор (фетальный AchR), белок связывания фолата (FBP), фолатный рецептор альфа, ганглиозид GD2, O-ацетилированный GD2 (OGD2), ганглиозид GD3, гликобелок 100 (gp100), глипикан-3 (GPC3), член D группы 5 класса C сопряженного с G белком рецептора (GPRC5D), Her2/neu (рецептор тирозинкиназы erb-B2), Her3 (erb-B3), Her4 (erb-B4), erbB димеры, человеческий высокомолекулярный меланома-ассоциированный антиген (HMW-MAA), поверхностный антиген гепатита B, человеческий лейкоцитарный антиген A1 (HLA-A1), человеческий лейкоцитарный антиген A2 (HLA-A2), IL-22 рецептор альфа (IL-22Rα), IL-13 рецептор альфа 2 (IL-13Rα2), рецептор со встроенный киназным доменом (kdr), легкую каппа-цепь, L1 молекулу клеточной адгезии (L1-CAM), CE7 эпитоп L1-CAM, содержащий насыщенные лейцином повторы 8 член семейства A (LRRC8A), Lewis Y, меланома-ассоциированный антиген (MAGE)-A1, MAGE-A3, MAGE-A6, MAGE-A10, мезотелин (MSLN), c-Met, мышиный цитомегаловирус (CMV), муцин 1 (MUC1), MUC16, лиганды члена D группы 2 группы естественных киллеров (NKG2D), мелан A (MART-1), молекулу адгезии нервных клеток (NCAM), онкофетальный антиген, преимущественно экспрессированный антиген меланомы (PRAME), рецептор прогестерона, специфический антиген простаты, антиген стволовых клеток простаты (PSCA), специфический мембранный антиген простаты (PSMA), орфанный рецептор типа рецепторной тирозинкиназы 1 (ROR1), сурвивин, трофобластный гликопротеин (TPBG, также известный как 5T4), опухоль-ассоциированный гликопротеин 72 (TAG72), родственный тирозиназе белок 1 (TRP1, также известный как TYRP1 или gp75), родственный тирозиназе белок 2 (TRP2, также известный как допахромтаутомераза, допахром дельта-изомераза или DCT), рецептор фактора роста сосудистого эндотелия (VEGFR), рецептор 2 фактора роста сосудистого эндотелия (VEGFR2), опухоль Вильмса 1 (WT-1), патоген-специфический или патоген-экспрессированный антиген или антиген, ассоциированный с универсальной меткой, и/или биотинилированные молекулы, и/или молекулы, экспрессированные ВИЧ, HCV, HBV или другими патогенами. Антигены, на которые нацелены рецептора, в некоторых вариантах, включают антигены, ассоциированные с злокачественностью B клетки, такие как любые из множества известных маркеров B клетки. В некоторых вариантах, антиген является или включает CD20, CD19, CD22, ROR1, CD45, CD21, CD5, CD33, Ig каппа, Ig лямбда, CD79a, CD79b или CD30.
В некоторых вариантах, антиген является или включает патоген-специфический или патоген-экспрессированный антиген. В некоторых вариантах, антигеном является вирусный антиген (такой как вирусный антиген из ВИЧ, HCV, HBV и т.д.), бактериальные антигены и/или антигены паразитов.
Термин “антитело” здесь применяется в широчайшем смысле и включает поликлональные и моноклональные антитела, включая нативные антитела и функциональные (антигенсвязывающие) фрагменты антитела, включая фрагмент антигенсвязывающих (Fab) фрагментов, F(ab’)2 фрагментов, Fab’ фрагментов, Fv фрагментов, фрагментов рекомбинантного IgG (rIgG), области переменной тяжелой цепи (VH), способные специфически связывать антиген, фрагменты однонитевого антитела, включая однонитевые переменные фрагменты (scFv), и фрагменты однодоменного антитела (например, sdAb, sdFv, нанотела). Термин охватывает генетически сконструированные и/или другим образом модифицированные формы иммуноглобулинов, такие как интратела, пептитела, химерные антитела, полностью человеческие антитела, гуманизированные антитела и гетероконъюгированные антитела, мультиспецифические, например, биспецифические антитела, диатела, триатела и тетратела, тандем ди-scFv, тандем три-scFv. Если не указано иначе, термин “антитело” должно пониматься как охватывающее функциональные фрагменты антитела. Термин также охватывает нативные или полноразмерные антитела, включая антитела любого класса или подкласса, включая IgG и его подклассы, IgM, IgE, IgA и IgD.
В некоторых вариантах, антигенсвязывающие белки, антитела и их антигенсвязывающие фрагменты специфически распознают антиген полноразмерного антитела. В некоторых вариантах, тяжелые и легкие цепи антитела могут быть полноразмерными или могут быть антигенсвязывающей частью (a Fab, F(ab’)2, Fv или однонитевым Fv фрагментом (scFv)). В других вариантах, постоянную область тяжелой цепи антитела выбирают из, например, IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgD, и IgE, в частности, выбранную из, например, IgG1, IgG2, IgG3, и IgG4, более конкретно, IgG1 (например, человеческого IgG1). В другом варианте, постоянную область легкой цепи антитела выбирают из, например, каппа или лямбда, в частности, каппа.
Представленные антитела включают фрагменты антитела. “Фрагмент антитела” относится к молекуле, отличной от нативного антитела, которая содержит часть нативного антитела, которая связывает антиген, с которым связывается нативное антитело. Примеры фрагментов антитела включают, но не ограничены ими, Fv, Fab, Fab’, Fab’-SH, F(ab’)2; диатела; линейные антитела; переменные области тяжелой цепи (VH), молекулы однонитевого антитела, такие как scFv и однодоменные VH одинарные антитела; и мультиспецифические антитела, образованные из фрагментов антитела. В конкретных вариантах, антителами являются однонитевые фрагменты антитела, содержащие переменную область тяжелой цепи и/или переменную область легкой цепи, такую как scFv.
Термин “переменная область” или “переменный домен” относится к домену тяжелой или легкой цепи антитела, который вовлечен в связывание антитела с антигеном. Переменные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) природного антитела в общем имеют похожие структуры, где каждый домен содержит четыре консервативные каркасные области (FR) и три CDR. (См., например, Kindt et al. Kuby Immunology, 6th ed., W.H. Freeman и Co., page 91 (2007)). Отдельный VH или VL домен может быть достаточен для придания антигенсвязывающей специфичности. Более того, антитела которые связываются с конкретным антигеном, могут быть выделены с применением VH или VL домена из антитела, которое связывает антиген, для скрининга высвобождения комплементарных VL или VH доменов, соответственно. См, например, Portolano et al., J. Immunol. 150:880-887 (1993); Clarkson et al., Nature 352:624-628 (1991).
Однодоменные антитела включают фрагменты антитела, содержащие весь или часть переменного домена тяжелой цепи или весь или часть переменного домена легкой цепи антитела. В определенных вариантах, однодоменным антителом является человеческое однодоменное антитело. В некоторых вариантах, CAR содержит домен тяжелой цепи антитела, который специфически связывает антиген, такой как раковый маркер или антиген клеточной поверхности целевой клетки или заболевания, такой как опухолевая клетка или раковая клетка, такой как целевые антигены, описанные здесь или известные.
Фрагменты антитела могут быть получены разными методами, включая, но не ограничиваясь ими, протеолитическое расщепление нативного антитела, а также образование рекомбинантными клетками хозяина. В некоторых вариантах, антителами являются полученные рекомбинантно фрагменты, такие как фрагменты, содержащие расположения, не существующие в природе, такие как с двумя или более областями или цепями антитела, соединенными синтетическими линкерами, например, пептидными линкерами, и/или которые могут не быть произведены ферментным расщеплением существующего в природе нативного антитела. В некоторых вариантах, фрагментами антитела являются scFv.
“Гуманизированным” антителом является антитело, в котом все или по существу все аминокислотные остатки CDR получены из не человеческих CDR, и все или по существу все аминокислотные остатки FR получены из человеческих FR. Гуманизированное антитело необязательно может включать, по меньшей мере, часть постоянной области антитела, полученного из человеческого антитела. “Гуманизированная форма” не человеческого антитела относится к варианту не человеческого антитела, для которого провели гуманизацию, обычно для снижения иммуногенности у человека, при этом сохранив специфичность и аффинность исходного не человеческого антитела. В некоторых вариантах, некоторые остатки FR в гуманизированном антителе замещены соответствующими остатками из не человеческого антитела (например, антитела, из которого получены остатки CDR), например, для восстановления или улучшения специфичности или аффинности антитела.
Таким образом, в некоторых вариантах, химерный антигенный рецептор, включая TCR-подобные CAR, включает внеклеточную часть, содержащую антитело или фрагмент антитела. В некоторых вариантах, антитело или фрагмент включает scFv. В некоторых аспектах, химерный антигенный рецептор включает внеклеточную часть, содержащую антитело или фрагмент, и внутриклеточную сигнальную область. В некоторых вариантах, внутриклеточная сигнальная область содержит внутриклеточный связывающий домен. В некоторых вариантах, внутриклеточный связывающий домен является или содержит первичный сигнальный домен, сигнальный домен, который способен вызывать первичный сигнал активации в T клетке, компонент сигнального домена Т-клеточного рецептора (TCR) и/или сигнальный домен, содержащий иммунорецепторный тирозиновый активационный мотив (ITAM).
В некоторых вариантах, рекомбинантный рецептор, такой как CAR, такой как его часть антитела, дополнительно содержит спейсер, который может быть или включает, по меньшей мере, часть иммуноглобулиновой постоянной области или ее вариант или модифицированную версию, такую как шарнирная область, например, IgG4 шарнирная область, и/или CH1/CL и/или Fc область. В некоторых вариантах, рекомбинантный рецептор дополнительно содержит спейсер и/или шарнирную область. В некоторых вариантах, постоянной областью или частью является человеческий IgG, такой как IgG4 или IgG1. В некоторых аспектах, часть постоянной области служит как спейсерная область между антиген-распознающим компонентом, например, scFv, и трансмембранным доменом. Спейсер может иметь длину, которая обеспечивает повышенную отвечаемость клетки после антигенного связывания, по сравнению с отсутствием спейсера.
В некоторых примерах, спейсер имеет от около 12 аминокислот в длину или не более чем 12 аминокислот в длину. Типовые спейсеры включают такие, которые имеют, по меньшей мере, от около 10 до 229 аминокислот, от около 10 до 200 аминокислот, от около 10 до 175 аминокислот, от около 10 до 150 аминокислот, от около 10 до 125 аминокислот, от около 10 до 100 аминокислот, от около 10 до 75 аминокислот, от около 10 до 50 аминокислот, от около 10 до 40 аминокислот, от около 10 до 30 аминокислот, от около 10 до 20 аминокислот или от около 10 до 15 аминокислот, и включая любое целое число между крайними значениями любого из перечисленных интервалов. В некоторых вариантах, спейсерная область имеет около 12 аминокислот или менее, около 119 аминокислот или менее или около 229 аминокислот или менее. В некоторых вариантах, спейсер имеет менее 250 аминокислот в длину, менее 200 аминокислот в длину, менее 150 аминокислот в длину, менее 100 аминокислот в длину, менее 75 аминокислот в длину, менее 50 аминокислот в длину, менее 25 аминокислот в длину, менее 20 аминокислот в длину, менее 15 аминокислот в длину, менее 12 аминокислот в длину или менее 10 аминокислот в длину. В некоторых вариантах, спейсер имеет от или от около 10 до 250 аминокислот в длину, от 10 до 150 аминокислот в длину, от 10 до 100 аминокислот в длину, от 10 до 50 аминокислот в длину, от 10 до 25 аминокислот в длину, от 10 до 15 аминокислот в длину, от 15 до 250 аминокислот в длину, от 15 до 150 аминокислот в длину, от 15 до 100 аминокислот в длину, от 15 до 50 аминокислот в длину, от 15 до 25 аминокислот в длину, от 25 до 250 аминокислот в длину, от 25 до 100 аминокислот в длину, от 25 до 50 аминокислот в длину, от 50 до 250 аминокислот в длину, от 50 до 150 аминокислот в длину, от 50 до 100 аминокислот в длину, от 100 до 250 аминокислот в длину, от 100 до 150 аминокислот в длину или от 150 до 250 аминокислот в длину.
Типовые спейсеры включают IgG4 шарнир отдельно, IgG4 шарнир, связанный с CH2 и CH3 доменами или IgG4 шарнир, связанный с CH3 доменом. Типовые спейсеры включают, но не ограничены ими, описанные в Hudecek et al. (2013) Clin. Cancer Res., 19:3153 или публикации заявка на международный патент № WO2014031687. В некоторых вариантах, спейсер имеет последовательность, представленную в SEQ ID NO: 131, и кодирован последовательностью, представленной в SEQ ID NO: 132. В некоторых вариантах, спейсер имеет последовательность, представленную в SEQ ID NO: 133. В некоторых вариантах, спейсер имеет последовательность, представленную в SEQ ID NO: 134.
В некоторых вариантах, постоянную область или часть берут из IgD. В некоторых вариантах, спейсер имеет последовательность, представленную в SEQ ID NO: 135. В некоторых вариантах, спейсер имеет последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности с любой из SEQ ID NOS: 131, 133,134 и 135.
Домен распознавания антигена в основном связан с одним или более внутриклеточными сигнальными компонентами, такими как сигнальные компоненты, которые имитируют активацию через антигенный рецепторный комплекс, такой как TCR комплекс, в случае CAR, и/или сигнал через другой рецептор клеточной поверхности. Таким образом, в некоторых вариантах, антигенсвязывающий компонент (например, антитело) связан с одной или более трансмембранными и внутриклеточными сигнальными областями. В некоторых вариантах, трансмембранный домен слит с внеклеточным доменом. В одном варианте, применяют трансмембранный домен, который в природе ассоциирован с одним или более доменами в рецепторе, например, CAR. В некоторых случаях, трансмембранный домен выбирают или модифицируют аминокислотным замещением так, чтобы избежать связывания таких доменов с трансмембранными доменами того же или другого белков поверхностной мембраны для минимизации взаимодействий с другими членами рецепторного комплекса.
Трансмембранный домен в некоторых вариантах получают либо из природного, либо из синтетического источника. Если источник является природным, домен в некоторых аспектах получают любого связанного с мембраной или трансмембранного белка. Трансмембранная область включает такую, которая получена из (т.е. содержит, меньшей мере, трансмембранные области) альфа, бета или дзэта цепи T-клеточного рецептора, CD28, CD3 эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137, CD154. Альтернативно, трансмембранный домен в некоторых вариантах является синтетическим. В некоторых аспектах, синтетический трансмембранный домен содержит преимущественно гидрофобные остатки, такие как лейцин и валин. В некоторых аспектах, триплет из фенилаланина, триптофана и валина находят на каждом конце синтетического трансмембранного домена. В некоторых вариантах, связь происходит через линкеры, спейсеры и/или трансмембранные домены.
Внутриклеточная сигнальная область включает такую, которая имитирует или почти соответствует сигналу через природный антигенный рецептор, сигналу через такой рецептор в сочетании с костимулирующим рецептором и/или сигналу через только костимулирующий рецептор. В некоторых вариантах, присутствует короткий олиго- или полипептидный линкер, например, линкер от 2 до 10 аминокислот в длину, например, содержащий глицины и серины, например, дублет глицин-серин, и образует связь между трансмембранным доменом и цитоплазматическим сигнальным доменом CAR.
Рецептор, например, CAR, обычно включает, по меньшей мере, один внутриклеточный сигнальный компонент или компоненты. В некоторых вариантах, рецептор включает внутриклеточный компонент TCR комплекса, такой как TCR CD3 цепь, которая медиирует активацию и цитотоксичность T-клетки, например, CD3 дзэта цепь. Таким образом, в некоторых аспектах, ROR1-связывающее антитело связано с одной или более сигнальными молекулами клетки. В некоторых вариантах, сигнальная молекула клетки включает CD3 трансмембранный домен, CD3 внутриклеточные связывающие домены и/или другие CD трансмембранные домены. В некоторых вариантах, рецептор, например, CAR, дополнительно включает часть одной или более дополнительных молекул, таких как Fc рецептор γ, CD8, CD4, CD25 или CD16. Например, в некоторых аспектах, CAR включает химерную молекулу между CD3-дзэта (CD3-ζ) или Fc рецептором γ и CD8, CD4, CD25 или CD16.
В некоторых вариантах, при сшивании CAR, цитоплазматический домен или внутриклеточная сигнальная область CAR активирует, по меньшей мере, одну из нормальных эффекторных функций или ответов иммунной клетки, например, T клетки, сконструированной для экспрессии CAR. Например, в некоторых контекстах, CAR вызывает функционирование T клетки, такое как цитолитическая активность или T-хелперная активность, такая как секреция цитокинов или других факторов. В некоторых вариантах, усеченную часть внутриклеточной сигнальной области компонента антигенного рецептора или костимулирующей молекулы используют вместо нативной иммуностимулирующей цепи, например, если она передает сигнал эффекторной функции. В некоторых вариантах, внутриклеточные сигнальные области, например, содержащие внутриклеточный домен или домены, включают цитоплазматические последовательности Т-клеточного рецептора (TCR), и, в некоторых аспектах, также сорецепторов, которые в природном контексте действуют совместно с таким рецептором для инициации трансдукции сигнала после соединения с антигенным рецептором, и/или любого производного или варианта таких молекул, и/или любой синтетической последовательности, которая имеет такую функциональную возможность.
В контексте природного TCR, полная активация обычно требует не только подачи сигналов через TCR, но также костимулирующего сигнала. Таким образом, в некоторых вариантах, чтобы способствовать полной активации, компонент для создания вторичного или костимулирующего сигнала также включен в CAR. В других вариантах, CAR не включает компонент для создания костимулирующего сигнала. В некоторых аспектах, дополнительный CAR экспрессируется в той же клетке и обеспечивает компонент для создания вторичного или костимулирующего сигнала.
Активация T клетки в некоторых аспектах описана как медиированная двумя классами цитоплазматических сигнальных последовательностей: такими, которые инициируют антиген-зависимую первичную активацию через TCR (первичными цитоплазматическими сигнальными последовательностями), и такими, которые действуют антиген-независимым образом для получения вторичного или костимулирущего сигнала (вторичными цитоплазматическими сигнальными последовательностями). В некоторых аспектах, CAR включает один или оба таких сигнальных компонента.
В некоторых аспектах, CAR включает первичную цитоплазматическую сигнальную последовательность, которая регулирует первичную активацию TCR комплекса. Первичные цитоплазматические сигнальные последовательности, которые действуют стимулирующим образом, могут содержать сигнальные мотивы, которые известны как иммунорецепторные тирозиновые активационные мотивы или ITAM. Примеры содержащих ITAM первичных цитоплазматических сигнальных последовательностей включают такие, которые получены из TCR или CD3 дзэта, CD3 гамма, CD3 дельта, CD3 эпсилон, FcR гамма или FcR бета. В некоторых вариантах, цитоплазматические сигнальные молекулы в CAR содержит цитоплазматический сигнальный домен, его часть или последовательность, полученную из CD3 дзэта.
В некоторых случаях, CAR называют CAR первого, второго и/или третьего поколения. В некоторых аспектах, CAR первого поколения являются такие, которые дают только вызванный CD3-цепью сигнал при антигенном связывании; в некоторых аспектах, CAR второго поколения являются такие, которые обеспечивают такой сигнал и костимулирующий сигнал, такие, которые включают внутриклеточный связывающий домен из костимулирующего рецептора, такого как CD28 или CD137; в некоторых аспектах, CAR третьего поколения в некоторых аспектах являются такие, которые включают множество костимулирующих доменов разных костимулирующих рецепторов.
В некоторых вариантах, химерный антигенный рецептор включает внеклеточную часть, содержащую антитело или фрагмент, описанные здесь. В некоторых аспектах, химерный антигенный рецептор включает внеклеточную часть, содержащую антитело или фрагмент, описанные здесь, и внутриклеточный связывающий домен. В некоторых вариантах, антитело или фрагмент включает scFv или однодоменное VH антитело, и внутриклеточный домен содержит ITAM. В некоторых аспектах, внутриклеточный связывающий домен включает сигнальный домен дзэта цепи CD3-дзэта (CD3ζ) цепи. В некоторых вариантах, химерный антигенный рецептор включает трансмембранный домен, расположенный между внеклеточным доменом и внутриклеточной сигнальной областью.
В некоторых аспектах, трансмембранный домен содержит трансмембранную часть CD28. Внеклеточный домен и трансмембранный могут быть связаны прямо или косвенно. В некоторых вариантах, внеклеточный домен и трансмембранный связаны спейсером, таким как любой, описанный здесь. В некоторых вариантах, химерный антигенный рецептор содержит внутриклеточный домен костимулирующей молекулы T клетки, например, между трансмембранным доменом и внутриклеточным связывающим доменом. В некоторых аспектах, костимулирующей молекулой T клетки является CD28 или 4-1BB.
В некоторых вариантах, CAR содержит антитело, например, фрагмент антитела, трансмембранный домен, который является или содержит трансмембранную часть CD28 или ее функциональный вариант, и внутриклеточный связывающий домен, содержащий сигнальную часть CD28 или ее функциональный вариант, и сигнальную часть CD3 дзэта или ее функциональный вариант. В некоторых вариантах, CAR содержит антитело, например, фрагмент антитела, трансмембранный домен, который является или содержит трансмембранную часть CD28 или a ее функциональный вариант, и внутриклеточный связывающий домен, содержащий сигнальную часть 4-1BB или ее функциональный вариант, и сигнальную часть CD3 дзэта или ее функциональный вариант. В некоторых таких вариантах, рецептор дополнительно включает спейсер, содержащий часть Ig молекулы, такой как человеческая Ig молекула, такую как Ig шарнир, например, IgG4 шарнир, такой как только шарнирный спейсер.
В некоторых вариантах, трансмембранным доменом рецептора, например, CAR, является трансмембранный домен человеческого CD28 или его вариант, например, 27-аминокислотный трансмембранный домен человеческого CD28 (Образец №: P10747.1), или трансмембранный домен, который содержит последовательность аминокислот представленную в SEQ ID NO: 136, или последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности к SEQ ID NO:136; в некоторых вариантах, трансмембранный домен, содержащий часть рекомбинантного рецептора, содержит последовательность аминокислот представленную в SEQ ID NO: 137 или последовательность аминокислот, имеющую, по меньшей мере, от около 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности к ней.
В некоторых вариантах, химерный антигенный рецептор содержит внутриклеточный домен костимулирующей молекулы T клетки. В некоторых аспектах, костимулирующей молекулой T клетки является CD28 или 4-1BB.
В некоторых вариантах, внутриклеточная сигнальная область содержит внутриклеточный костимулирующий сигнальный домен человеческого CD28 или его функциональный вариант или часть, такую как его 41 аминокислотный домен и/или домен с замещением LL на GG в положениях 186-187 нативного CD28 белка. В некоторых вариантах, внутриклеточный связывающий домен может содержать последовательность аминокислот, представленную в SEQ ID NO: 138 или 139, или последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности к SEQ ID NO: 138 или 139. В некоторых вариантах, внутриклеточная область содержит внутриклеточный костимулирующий сигнальный домен 4-1BB или его функциональный вариант или часть, такую как 42-аминокислотный цитоплазматический домен человеческого 4-1BB (Образец № Q07011.1) или его функциональный вариант или часть, например, последовательность аминокислот, представленную в SEQ ID NO: 140, или последовательность аминокислот, которая демонстрирует, по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности к SEQ ID NO: 140.
В некоторых вариантах, внутриклеточная сигнальная область содержит человеческую CD3 цепь, необязательно, CD3 дзэта стимулирующий сигнальный домен или его функциональный вариант, такой как 112 AA цитоплазматический домен изоформы 3 человеческого CD3ζ (Образец №: P20963.2) или CD3 дзэта сигнальный домен, описанный в Патенте США №: 7,446,190 или Патенте США № 8,911,993. В некоторых вариантах, внутриклеточная сигнальная область содержит последовательность аминокислот, представленную в SEQ ID NO: 129, 130 или 141, или последовательность аминокислот, которая демонстрирует по меньшей мере, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичность последовательности к SEQ ID NO: 129, 130 или 141.
В некоторых аспектах, спейсер содержит только шарнирную область IgG, такую как только шарнир IgG4 или IgG1, такой как только шарнирный спейсер, представленный в SEQ ID NO:131. В других вариантах, спейсером является Ig шарнир, например, и IgG4 шарнир, связанный с CH2 и/или CH3 доменами. В некоторых вариантах, спейсером является Ig шарнир, например, IgG4 шарнир, связанный с CH2 и CH3 доменами, такой как представлен в SEQ ID NO:133. В некоторых вариантах, спейсером является Ig шарнир, например, IgG4 шарнир, связанный только с CH3 доменом, такой как представлен в SEQ ID NO:134. В некоторых вариантах, спейсер является или содержит богатую глицином-серином последовательность или другой гибкий линкер, такой как известные гибкие линкеры.
В некоторых вариантах, CAR включает анти-HPV 16 E6 или E7 антитело или фрагмент, включая sdAbs (например, содержащие только VH область) и scFv, спейсер, такой как любой из содержащих Ig-шарнир спейсеров, CD28 трансмембранный домен, CD28 внутриклеточный связывающий домен и CD3 дзэта сигнальный домен. В некоторых вариантах, CAR включает HPV 16 антитело или фрагмент, включая sdAbs и онFvs, спейсер, такой как любой из содержащих Ig-шарнир спейсеров, CD28 трансмембранный домен, CD28 внутриклеточный связывающий домен и CD3 дзэта сигнальный домен. В некоторых вариантах, такие CAR конструкты дополнительно включают T2A элемент проскока рибосомы и/или tEGFR последовательность, например, ниже CAR.
В некоторых вариантах, CAR или его антигенсвязывающий фрагмент кодирован нуклеотидной последовательностью, которая является или была кодон-оптимизирована. Типовые кодон-оптимизированные варианты описаны здесь везде.
С. TCR-подобные CAR
В некоторых вариантах, антитело или его антигенсвязывающая часть экспрессируется на клетках как часть рекомбинантного рецептора, такого как антигенный рецептор. Антигенные рецепторы включают функциональные не-TCR антигенные рецепторы, такие как химерные антигенные рецепторы (CAR). В общем, CAR, содержащий антитело или антигенсвязывающий фрагмент, который демонстрирует TCR-подобную специфичность, направленную против пептида в контексте MHC молекулы, также может называться TCR-подобный CAR.
В некоторых вариантах, CAR содержит TCR-подобное антитело, такое как антитело или антигенсвязывающий фрагмент (например, scFv), который специфически распознает внутриклеточный антиген, такой как опухоль-ассоциированный антиген, представленный на поверхности клетки как MHC-пептидный комплекс. В некоторых вариантах, антитело или его антигенсвязывающая часть, которая распознает MHC-пептидный комплекс, может экспрессироваться на клетках как часть рекомбинантного рецептора, такого как антигенный рецептор. Антигенные рецепторы включают функциональные не-TCR антигенные рецепторы, такие как химерные антигенные рецепторы (CAR). В общем, CAR, содержащий антитело или антигенсвязывающий фрагмент, который демонстрирует TCR-подобную специфичность, направленную против пептид-MHC комплексов, также может называться TCR-подобный CAR.
Ссылка на “главный комплекс гистосовместимости” (MHC) относится к белку, обычно, гликопротеину, который содержит сайт связывания полиморфного пептида или полость связывания, которая может, в некоторых случаях, образовывать комплекс с пептидными антигенами полипептидов, включая пептидные антигены, процессированные клеточным механизмом. В некоторых случаях, MHC молекулы могут быть воспроизведены или экспрессированы на клеточной поверхности, например, в виде комплекса с пептидом, т.е. MHC-пептидного комплекса, для представления антигена в конформации, распознаваемой антигенным рецептором на T клетках, таким как TCR или TCR-подобное антитело. В общем, молекулы MHC класса I являются гетеродимерами, имеющими трансмембранную α цепь, в некоторых случаях, с тремя α доменами, и не-ковалентно ассоциированный β2 микроглобулин. В общем, молекулы MHC класса II состоят из двух трансмембранных гликопротеинов, α и β, оба которых обычно охватывают мембрану. MHC молекула может включать эффективную часть MHC, которая содержит сайт или сайты антигенного связывания для связывания пептида и последовательностей, необходимых для распознавания подходящим антигенным рецептором. В некоторых вариантах, молекулы MHC класса I доставляют пептиды, зарождающиеся в цитозоле, на поверхность клетки, где MHC-пептидный комплекс распознается T клетками, такими как, в основном, CD8+ T клетки, но в некоторых случаях, CD4+ T клетки. В некоторых вариантах, молекулы MHC класса II доставляют пептиды, зарождающиеся в везикулярной системе, на поверхность клетки, где они обычно распознаются CD4+ T клетками. В общем, MHC молекулы кодированы группой связанных локусов, которые вместе называются H-2, в мышином и человеческом лейкоцитарном антигене (HLA) у человека. Следовательно, человеческий MHC также может быть назван человеческим лейкоцитарным антигеном (HLA).
Термин “MHC-пептидный комплекс” или “пептид-MHC комплекс” или его варианты относятся к комплексу или ассоциации пептидного антигена и MHC молекулы, такому как, обычно, нековалентные взаимодействия пептида в полости связывания или щели MHC молекулы. В некоторых вариантах, MHC-пептидный комплекс присутствует или воспроизводится на поверхности клетки. В некоторых вариантах, MHC-пептидный комплекс может специфически распознаваться антигенным рецептором, таким как TCR, TCR-подобный CAR или его антигенсвязывающие части.
В некоторых вариантах, пептид, такой как пептидный антиген или эпитоп, полипептида может ассоциироваться с MHC молекулой, например, для распознавания антигенным рецептором. В общем, пептид получен из или основан на фрагменте более длинной биологической молекулы, такой как полипептид или белок. В некоторых вариантах, пептид обычно имеет от около 8 до около 24 аминокислот в длину. В некоторых вариантах, пептид имеет длину от или от около 9 до 22 аминокислот для распознавания в MHC комплексе класса II. В некоторых вариантах, пептид имеет длину от или от около 8 до 13 аминокислот для распознавания в MHC комплексе класса I. В некоторых вариантах, при распознавании пептида в контексте MHC молекулы, такой как MHC-пептидный комплекс, антигенный рецептор, такой как TCR или TCR-подобный CAR, вызывает или запускает сигнал активации в T клетку, который вызывает ответ T клетки, такой как пролиферация T клетки, образование цитокина, цитотоксический ответ T клетки или другой ответ.
В некоторых вариантах, TCR-подобное антитело или его антигенсвязывающая часть, известно или может быть получено известными способами (см., например, опубликованные заявки США №№ US 2002/0150914; US 2003/0223994; US 2004/0191260; US 2006/0034850; US 2007/00992530; US20090226474; US20090304679; и международную публикацию PCT № WO 03/068201).
В некоторых вариантах, антитело или его антигенсвязывающая часть, которая специфически связывается с MHC-пептидным комплексом, может быть получено иммунизацией хозяина эффективным количеством иммуногена, содержащего определенный MHC-пептидный комплекс. В некоторых случаях, пептидом MHC-пептидного комплекса является эпитоп антигена, способного связываться с MHC, такого как опухолевый антиген, например универсальный опухолевый антиген, антиген миеломы или другой антиген, как описано здесь. В некоторых вариантах, эффективное количество иммуногена затем вводят хозяину для вызова иммунного ответа, где иммуноген сохраняет его трехмерную форму в течение периода, достаточного для вызова иммунного ответа против трехмерного представления пептида в полости связывания MHC молекулы. Сыворотку, собранную у хозяина, затем анализируют для определения того, выработались ли антитела, которые распознают трехмерное представление пептида в полости связывания MHC молекулы. В некоторых вариантах, выработанные антитела могут быть проанализированы для оценки того, что антитело может отличать MHC-пептидный комплекс от только MHC молекулы, только рассматриваемого пептида и комплекса MHC и нерелевантного пептида. Затем желаемые антитела могут быть выделены.
В некоторых вариантах, антитело или его антигенсвязывающая часть, которая специфически связывается с MHC-пептидным комплексом, может быть получена с применением способов антиген-дисплейной библиотеки, такими как фаг-антитело библиотеки. В некоторых вариантах, могут быть созданы фаг-дисплейные библиотеки мутантных Fab, scFv или других форм антител, например, в которых члены библиотеки мутированы на одном или более остатках CDR или CDR. См., например, опубликованную заявку США № US20020150914, US2014/0294841; и Cohen CJ. et al. (2003) J Mol. Recogn. 16:324-332.
Представленные варианты осуществления включают рекомбинантные рецепторы, например те, которые включают антитела, например, TCR-подобные антитела. В некоторых вариантах, антигенные рецепторы и другие химерные рецепторы специфически связываются с областью или эпитопом антигена, например, TCR-подобных антител. Антигенными рецепторами являются не-TCR антигенные рецепторы, такие как химерные антигенные рецепторы (CAR). Также представлены клетки, экспрессирующие CAR, и их применение в адоптивной клеточной терапии, такой как лечение заболеваний и расстройств, связанных с экспрессией антигена и/или эпитопа.
Таким образом, здесь представлены TCR-подобные CAR, которые содержат не-TCR молекулу, которая демонстрирует специфичность к Т-клеточному рецептору, такую как Т клеточный эпитоп или пептидный эпитоп, при воспроизведении или присутствии в контексте MHC молекулы. В некоторых вариантах, TCR-подобный CAR может содержать антитело или его антигенсвязывающую часть, например, TCR-подобное антитело, такое, как писано здесь. В некоторых вариантах, антитело или его антителосвязывающая часть реагирует на его специфический пептидный эпитоп в контексте MHC молекулы, где антитело или фрагмент антитела может отличать специфический пептид в контексте MHC молекулы от только MHC молекулы, только специфического пептида и, в некоторых случаях, нерелевантного пептида в контексте MHC молекулы. В некоторых вариантах, антитело или его антигенсвязывающая часть может демонстрировать большее сродство связывания, чем Т-клеточный рецептор.
В некоторых аспектах, трансген может включать нуклеиновые кислоты, кодирующие один или более CAR, например, первый CAR, который содержит сигнальные домены для вызова первичного сигнала, и второй CAR, который связывается с вторым антигеном и содержит компонент для создания костимулирующего сигнала. Например, первый CAR может быть активирующим CAR, и второй CAR может быть костимулирующим CAR. В некоторых аспектах, оба CAR должны быть лигированы для того, чтобы вызвать конкретную эффекторную функцию в клетке, которая может придать специфичность и селективность целевому типу клетки.
В некоторых вариантах, активирующий домен включен в один CAR, в костимулирующий компонент представлен другим химерным рецептором, распознающим другой антиген. В некоторых вариантах, CAR включают активирующие или стимулирующие CAR, и костимулирующие рецепторов, оба которых экспрессированы на одной и той же клетке (см. WO2014/055668). В некоторых аспектах, рецептор, содержащий HPV 16 E6 или E7 антитело, является стимулирующим или активирующим CAR; в других аспектах, он является костимулирующим рецептором. В некоторых вариантах, трансген дополнительно кодирует ингибирующий CAR (iCAR, см. Fedorov et al., Sci. Transl. Medicine, 5(215) (December, 2013), такой как ингибирующий рецептор, распознающий пептидный эпитоп, отличный от HPV 16 E6 или HPV16 E7, тем самым активирующий сигнал, доставленный через HPV 16-направленный CAR, ослабляется или ингибируется связыванием ингибирующего CAR с его лигандом, например, для снижения нецелевого действия.
В некоторых вариантах, трансген может включать нуклеиновые кислоты, кодирующие рекомбинантный рецептор, которые могут дополнительно кодировать дополнительный рецептор, такой как рецептор, способный доставлять костимулирующий или способствующий выживанию сигнал, такой как костимулирующий рецептор (см. WO2014/055668) и/или блокировать или изменять выход ингибирующего сигнала, такого как сигнал, обычно доставляемый через иммунную контрольную точку или другие иммуноингибирующие молекулы, такие как экспрессируются в опухолевой микросреде, например, для способствования повышенной эффективности таких сконструированных клеток. См., например, Tang et al., Am J Transl Res. 2015; 7(3): 460-473. В некоторых вариантах, клетка может дополнительно включать один или более экзогенных или рекомбинантных или сконструированных компонентов, таких как один или более экзогенных факторов и/или костимулирующих лигандов, которые экспрессированы на и в или секретированы клетками и могут способствовать функционированию, например, в микросреде. Примеры таких лигандов и компонентов включают, например, ФНОR и/или Ig семейство рецепторов или лигандов, например, 4-1BBL, CD40, CD40L, CD80, CD86, цитокинов, хемокинов и/или антител или других молекул, таких как scFv. См., например, публикации заявок на патент №№ WO2008121420 A1, WO2014134165 A1, US20140219975 A1.
D. Химерный рецептор ауто-антитела (CAAR)
В некоторых вариантах, химерным рецептором является химерный рецептор аутоантитела (CAAR). В некоторых вариантах, CAAR связывается, например, специфически связывается или распознает аутоантитело. В некоторых вариантах, клетка, экспрессирующая CAAR, такая как T клетка, сконструированная для экспрессии CAAR, может применяться для связывания с и гибели клеток, экспрессирующих аутоантитело, но не клеток, экспрессирующих нормальное антитело. В некоторых вариантах, CAAR-экспрессирующие клетки могут применяться для лечения аутоиммунного заболевания, связанного с экспрессией аутоантигенов, такие как аутоиммунные заболевания. В некоторых вариантах, CAAR-экспрессирующие клетки могут поражать B клетки, которые в конечном итоге вырабатывают аутоантитела и демонстрируют аутоантитела на поверхности их клеток, и помечают эти B клетки как болезнь-специфические цели для терапевтического вмешательства. В некоторых вариантах, CAAR-экспрессирующие клетки могут применяться для эффективного целенаправленного воздействия и уничтожения патогенных B клеток при аутоиммунных заболеваниях путем целенаправленного воздействия на вызывающие заболевание B клетки с применением рецептора антиген-специфического химерного аутоантитела. В некоторых вариантах, химерным рецептором является CAAR, такое как любое, описанное в публикации заявки на патент США № US 2017/0051035.
В некоторых вариантах, CAAR содержит связывающий аутоантитело домен, трансмембранный домен и одну или более внутриклеточных сигнальных областей или доменов (также взаимозаменяемо называемых цитоплазматический сигнальный домен или область). В некоторых вариантах, внутриклеточная сигнальная область содержит внутриклеточный связывающий домен. В некоторых вариантах, внутриклеточный связывающий домен является или содержит первичную сигнальную область, сигнальный домен, который способен стимулировать и/или вызывать первичный сигнал активации в T клетке, сигнальный домен компонента Т-клеточного рецептора (TCR) (например, внутриклеточный связывающий домен или область CD3-дзэта (CD3ζ) цепи или его функциональный вариант или сигнальная часть) и/или сигнальный домен, содержащий иммунорецепторный тирозиновый активационный мотив (ITAM).
В некоторых вариантах, аутоантитело связывающий домен содержит аутоантиген или его фрагмент. Выбор аутоантигена может зависеть от типа поражаемого аутоантитела. Например, аутоантиген может быть выбран потому, что он распознает аутоантитело на целевой клетке, такой как B клетка, ассоциированное с определенным болезненным состоянием, например, аутоиммунным заболеванием, таким как медиированное аутоантителом аутоиммунное заболевание. В некоторых вариантах, аутоиммунное заболевание включает обыкновенную пузырчатку (PV). Типовые аутоантигены включают десмоглеин 1 (Dsg1) и Dsg3.
V. КОМПОЗИЦИИ И СОСТАВЫ
Также предоставлены популяции сконструированных клеток, композиции, содержащие такие клетки и/или обогащенные такими клетками. Композиции включают фармацевтические композиции и составы для введения, например, для адоптивной клеточной терапии. Также представлены терапевтические способы введения клеток и композиций субъектам, например, пациентам.
В некоторых вариантах, предоставленная популяция клеток и/или композиция, содержащие сконструированные клетки, включают популяцию клеток, которая демонстрирует более хорошую, однородную, гомогенную и/или стабильную экспрессию и/или антигенное связывание рекомбинантного рецептора, например, демонстрируют уменьшенный коэффициент вариации, по сравнению с экспрессией и/или антигенным связыванием клеточных популяций и/или композиций, созданных с использованием обычных способов. В некоторых вариантах популяция клеток и/или композиции демонстрируют, по меньшей мере, коэффициент вариации экспрессии трансгенного и/или антигенного связывания рекомбинантного рецептора, на 100%, 95%, 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20% или 10% ниже, чем у соответствующей популяции, созданной с использованием обычных способов, например, случайной интеграция трансгена. Коэффициент вариации определен как стандартное отклонение экспрессии рассматриваемой нуклеиновой кислоты (например, трансгена, кодирующего рекомбинантный рецептор) в популяции клеток, например CD4+ и/или CD8+ T клеток, Деленное на среднее экспрессии соответствующей рассматриваемой нуклеиновой кислоты в соответствующей популяции клеток. В некоторых вариантах, популяция клеток и/или композиции демонстрируют коэффициент вариации, который ниже чем 0,70, 0,65, 0,60, 0,55, 0,50, 0,45, 0,40, 0,35 или 0,30 или менее, при измерении в популяциях CD4+ и/или CD8+ T клеток, которые сконструированы с применением представленных здесь способов.
В некоторых вариантах, представлены популяция клеток и/или композиции, которые включают клетки, которые имеют целевой нокин кодирующего рекомбинантный рецептор трансгена в одном или более локусах гена эндогенного TCR, тем самым имея нокаут одного или более локусов гена эндогенного TCR, например, нокаут целевого гена для интеграции, такого как TRAC, TRBC1 и/или TRBC2. В некоторых вариантах, все или по существу все клетки в популяции клеток, в которые интегрирован кодирующий рекомбинантный рецептор трансген, также имеют нокаут одного или более локусов гена эндогенного TCR. В некоторых вариантах, по меньшей мере, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более клеток в популяция клеток и/или композиции, которые экспрессируют рекомбинантный рецептор, содержат нокаут одного или более локусов гена эндогенного TCR, например, TRAC, TRBC1 и/или TRBC2. таким образом, в представленной популяции клеток и/или композициях, все или по существу все сконструированные клетки, которые экспрессируют рекомбинантный рецептор, также содержат нокаут эндогенного TCR, в силу направленного нокина трансгена в локусах гена эндогенного TCR.
В некоторых вариантах, представлены популяция клеток и/или композиции, которые включают множество сконструированных иммунных клеток, содержащих рекомбинантный рецептор или его антигенсвязывающий фрагмент, кодированные трансгеном, и генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), где, по меньшей мере или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции содержат генетическое разрушение в целевом положении в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC); и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или его цепь, направленный на или рядом с целевым положением через репарацию, направляемую гомологией (HDR).
В некоторых вариантах, экспрессия и/или антигенное связывание рекомбинантным рецептором может быть оценено с применением любых реагентов и/или анализов, описанных здесь, например, в разделе I.C. В некоторых вариантах, экспрессию измеряют с применением связывающей молекулы, которая распознает и/или специфически связывается с рекомбинантным рецептором или его частью. Например, в некоторых вариантах, экспрессию рекомбинантного рецептора, кодированного трансгеном, оценивают с применением анти-TCR Vβ 22 антитела, например, проточной цитометрией. В некоторых вариантах, антигенное связывание рекомбинантного рецептора, которым является TCR, может быть оценено с применением антигена, который выделен или очищен или является рекомбинантным, клеток, экспрессирующих конкретный антиген, и/или с применением TCR лиганда (MHC-пептидного комплекса).
В некоторых вариантах, представлены композиции, содержащие клетки, в которых клетки, экспрессирующие рекомбинантный рецептор и/или содержащие нокаут одного или более генов, кодирующих эндогенный TCR, составляют, по меньшей мере, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более от всех клеток в композиции или клеток определенного типа, таких как T клетки или CD8+ или CD4+ клетки.
Также представлены композиции, включающие клетки для введения, включая фармацевтические композиции и составы, такие как композиции стандартных дозированных форм, включающие количество клеток для введения в данной дозе или их часть. Фармацевтические композиции и составы обычно включают один или более необязательных фармацевтически приемлемых носителей или эксципиентов. В некоторых вариантах, композиция включает, по меньшей мере, один дополнительный терапевтический агент.
Термин «фармацевтический состав» относится к препарату, который находится в такой форме, которая позволяет эффективную биологическую активность содержащегося в ней активного ингредиента, и который не содержит дополнительных компонентов, которые являются неприемлемо токсичными для субъекта, которому вводят состав.
«Фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтическом составе, отличному от активного ингредиента, который нетоксичен для субъекта. Фармацевтически приемлемый носитель включает, но не ограничивается ими, буфер, эксципиент, стабилизатор или консервант.
В некоторых аспектах выбор носителя частично определяется конкретной клеткой и/или способом введения. Соответственно, существует множество подходящих составов. Например, фармацевтическая композиция может содержать консерванты. Подходящие консерванты могут включать, например, метилпарабен, пропилпарабен, бензоат натрия и хлорид бензалкония. В некоторых аспектах используется смесь двух или более консервантов. Консервант или их смеси обычно присутствуют в количестве от около 0,0001% до около 2% массовых от всей композиции. Носители описаны, например, в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980). Фармацевтически приемлемые носители, как правило, нетоксичны для реципиентов в используемых дозах и концентрациях, и включают, но не ограничиваются ими: буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее около 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТК; сахара, такие как сахароза, маннит, трегалоза или сорбитол; солеобразующие противоионы, такие как натрий; металлические комплексы (например, Zn-белок комплексы); и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ).
Буферные агенты, в некоторых аспектах включены в композиции. Подходящие буферные агенты включают, например, лимонную кислоту, цитрат натрия, фосфорную кислоту, фосфат калия и различные другие кислоты и соли. В некоторых аспектах используется смесь двух или более буферных агентов. Буферный агент или их смеси обычно присутствуют в количестве от около 0,001% до около 4% массовых от всей композиции. Способы вводимых приготовления фармацевтических композиций известны. Типовые способы описаны более подробно в, например, Remington: The Science и Practice of Pharmacy, Lippincott Williams & Wilkins; 21st ed. (May 1, 2005).
Составы могут включать водные растворы. Состав или композиция может также содержать более одного активного ингредиента, применимого для конкретного показания, заболевания или состояния, которые лечатся клетками, предпочтительно, такими, активность которых комплементарна к клеткам, где соответствующие активности не влияют отрицательно друг на друга. Такие активные ингредиенты обычно присутствуют в комбинации в количествах, эффективных для предполагаемой цели. Таким образом, в некоторых вариантах, фармацевтическая композиция дополнительно включает другие фармацевтически активные агенты или лекарственные средства, такие как химиотерапевтические агенты, например, аспарагиназу, бусульфан, карбоплатин, цисплатин, даунорубицин, доксорубицин, фторурацил, гемцитабин, гидроксимочевину, метотрексат, паклитабластаксел, ритуксимаб, винбластин и/или винкристин.
Фармацевтическая композиция в некоторых вариантах содержит клетки в количествах, эффективных для лечения или предотвращения заболевания или состояния, таких как терапевтически эффективное или профилактически эффективное количество. Терапевтическую или профилактическую эффективность в некоторых вариантах отслеживают путем периодической оценкой леченных субъектов. Желаемая доза может быть доставлена однократным болюсным введением клеток, многократным болюсным введением клеток или непрерывным инфузионным введением клеток.
Клетки и композиции могут быть введены с использованием стандартных методик введения, составов и/или устройств. Введение клеток может быть аутологичным или гетерологичным. В некоторых аспектах клетки выделяют из субъекта, конструируют и вводят тому же субъекту. В других аспектах их изолируют от одного субъекта, конструируют и вводят другому субъекту. Например, иммунореактивные клетки или предшественники могут быть получены от одного субъекта и введены тому же субъекту или другому совместимому субъекту. Иммунореактивные клетки, полученные из периферической крови, или их потомство (например, полученные in vivo, ex vivo или in vitro) могут быть введены локальной инъекцией, включая катетерное введение, системную инъекцию, локальную инъекцию, внутривенную инъекцию или парентеральное введение. При введении терапевтической композиции (например, фармацевтической композиции, содержащей генетически модифицированную иммунореактивную клетку) она будет изначально составляться в виде стандартной дозированной формы для инъекций (раствор, суспензия, эмульсия).
Составы включают составы для перорального, внутривенного, внутрибрюшинного, подкожного, легочного, трансдермального, внутримышечного, интраназального, буккального, сублингвального введения или введения суппозиториев. В некоторых вариантах популяции клеток вводятся парентерально. Используемый здесь термин «парентеральное» включает внутривенное, внутримышечное, подкожное, ректальное, вагинальное и внутрибрюшинное введение. В некоторых вариантах, клетки вводят субъекту с использованием периферической системной доставки внутривенной, внутрибрюшинной или подкожной инъекцией.
Композиции в некоторых вариантах представлены в виде стерильных жидких препаратов, например, изотонических водных растворов, суспензий, эмульсий, дисперсий или вязких композиций, которые в некоторых аспектах могут быть забуференными до выбранного pH. Жидкие препараты обычно легче приготовить, чем гели, другие вязкие композиции и твердые композиции. Дополнительно, жидкие композиции несколько удобнее вводить, особенно инъекцией. С другой стороны, вязкие композиции могут быть составлены с соответствующим диапазоном вязкости, чтобы обеспечить более длительные периоды контакта с конкретными тканями. Жидкие или вязкие композиции могут содержать носители, которые могут быть растворителем или диспергирующей средой, содержащей, например, воду, физиологический раствор, забуференный фосфатом физиологический раствор, полиол (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль) и их подходящие смеси.
Стерильные растворы для инъекций могут быть приготовлены путем включения клеток в растворитель, например в смесь с подходящим носителем, разбавителем или эксципиентом, таким как стерильная вода, физиологический раствор, глюкоза, декстроза или подобные. Композиции могут содержать вспомогательные вещества, такие как смачивающие, диспергирующие или эмульгирующие агенты (например, метилцеллюлозу), pH буферные агенты, желирующие или повышающие вязкость добавки, консерванты, ароматизаторы и/или красители, в зависимости от пути введения и желаемого препарата. В некоторых случаях можно сверяться со стандартными текстами для приготовления подходящих препаратов.
Могут быть добавлены различные добавки, которые повышают стабильность и стерильность композиции, включая противомикробные консерванты, антиоксиданты, хелатирующие агенты и буферы. Предотвращение действия микроорганизмов можно обеспечить с помощью различных антибактериальных и противогрибковых агентов, например парабенов, хлорбутанола, фенола и сорбиновой кислоты. Продолжительная абсорбция инъекционной фармацевтической формы может быть достигнута за счет использования агентов, замедляющих абсорбцию, например, моностеарата алюминия и желатина.
Составы, используемые для введения in vivo, в основном стерильны. Стерильность может быть легко достигнута, например, путем фильтрации через стерильные фильтрующие мембраны.
VI. СПОСОБЫ ВВЕДЕНИЯ И ПРИМЕНЕНИЕ В АДОПТИВНОЙ КЛЕТОЧНОЙ ТЕРАПИИ
Предоставлены способы введения клеток, популяций и композиций и использования таких клеток, популяций и композиций для предотвращения заболеваний, состояний и расстройств, включая рак. В некоторых вариантах клетки, популяции и композиции вводят субъекту или пациенту, имеющему конкретное заболевание или состояние, подлежащее лечению, например, с помощью адоптивной клеточной терапии, такой как адоптивная Т-клеточная терапия. В некоторых вариантах клетки и композиции, приготовленные с представленными способами, такие как сконструированные композиции и композиции на конечном этапе производственного цикла после инкубации и/или других стадий обработки, вводят субъекту, такому как субъект, имеющий или подверженный риску заболевания или состояния. В некоторых аспектах способы, таким образом, лечат, например, облегчают один или более симптомов заболевания или состояния, например, уменьшая опухолевую массу при раке, экспрессирующем антиген, распознаваемый сконструированной Т-клеткой.
Используемый здесь термин «субъект» представляет собой млекопитающее, такое как человек или другое животное, и обычно является человеком. В некоторых вариантах субъект, например, пациент, которому вводят клетки, популяции клеток или композиции, является млекопитающим, обычно приматом, таким как человек. В некоторых вариантах приматом является обезьяна или человекообразная обезьяна. Субъект может быть мужчиной или женщиной и может быть любого подходящего возраста, включая младенцев, детей, подростков, взрослых и пожилых людей. В некоторых вариантах субъектом является млекопитающее, не являющееся приматом, например грызун.
Используемый здесь термин «лечение» (и его грамматические вариации, такие как «лечить» или «обработка») относится к полному или частичному облегчению или уменьшению заболевания, или состояния, или расстройства, или симптома, побочного эффекта или результата или фенотипа, связанных с ним. Желательные эффекты лечения включают, но не ограничиваются ими, предотвращение возникновения или рецидива заболевания, облегчение симптомов, уменьшение любых прямых или косвенных патологических последствий заболевания, предотвращение метастазирования, снижение скорости прогрессирования заболевания, улучшение или паллиативное улучшение болезненного состояния, и ремиссию или улучшенный прогноз. Термины не подразумевают полного излечения заболевания или полного устранения какого-либо симптома или влияния на все симптомы или исходы.
используемый здесь термин «задержка развития заболевания» означает отсрочку, сдерживание, замедление, торможение, стабилизацию, подавление и/или отсрочку развития заболевания (например, рака). Эта задержка может иметь различную продолжительность, в зависимости от анамнеза заболевания и/или индивидуума, проходящего лечение. Достаточная или значительная отсрочка может, по сути, включать профилактику в том смысле, что у индивидуума не развивается заболевание. Например, рак на поздней стадии, такой как развитие метастазов, может быть задержан.
«Профилактика» в данном описании включает профилактику возникновения или рецидива заболевания у субъекта, который может быть предрасположен к заболеванию, но у которого еще не диагностировано заболевание. В некоторых вариантах представленные клетки и композиции используются для задержки развития заболевания или для замедления прогрессирования заболевания.
Используемый здесь термин «подавление» функции или активности означает уменьшение функции или активности по сравнению с другими такими же условиями, за исключением условия или параметра, представляющего интерес, или альтернативно, по сравнению с другим условием. Например, клетки, подавляющие рост опухоли, снижают скорость роста опухоли по сравнению со скоростью роста опухоли в отсутствие клеток.
«Эффективное количество» фармацевтического состава, клеток или композиции в контексте введения относится к эффективному количеству в дозах/количествах и в течение периодов времени, необходимых для достижения желаемого результата, такого как терапевтический или профилактический результат.
«Терапевтически эффективное количество» фармацевтического состава или клеток относится к количеству, эффективному в дозировках и в течение периодов времени, необходимых для достижения желаемого терапевтического результата, такого как лечение заболевания, состояния или расстройства и/или фармакокинетический или фармакодинамический эффект от лечения. Терапевтически эффективное количество может варьироваться в зависимости от таких факторов, как болезненное состояние, возраст, пол и вес субъекта, и популяции вводимых клеток. В некоторых вариантах представленные способы включают введение клеток и/или композиций в эффективных количествах, например, терапевтически эффективных количествах.
«Профилактически эффективное количество» относится к количеству, эффективному при дозировках и в течение периодов времени, необходимых для достижения желаемого профилактического результата. Обычно, но не обязательно, поскольку профилактическая доза используется у субъектов до или на более ранней стадии заболевания, профилактически эффективное количество будет меньше терапевтически эффективного количества. В контексте меньшей опухолевой массы профилактически эффективное количество в некоторых случаях будет выше, чем терапевтически эффективное количество.
Способы введения клеток для адоптивной клеточной терапии известны и могут применяться в связи с представленными способами и композициями. Например, способы адоптивной T клеточной терапии описаны, например, в публикации заявки на патент США № 2003/0170238 Gruenberg et al; Патенте США № 4,690,915 Rosenberg; Rosenberg (2011) Nat Rev Clin Oncol. 8(10):577-85). См., например, Themeli et al. (2013) Nat Biotechnol. 31(10): 928-933; Tsukahara et al. (2013) Biochem Biophys Res Commun 438(1): 84-9; Davila et al. (2013) PLoS ONE 8(4): e61338.
В некоторых вариантах клеточную терапию, например, адоптивную Т-клеточную терапию, проводят аутологичным переносом, при котором клетки выделяют и/или иным образом получают из субъекта, который должен получать клеточную терапию, или из образца, полученного от такого субъекта. Таким образом, в некоторых аспектах, клетки получают от субъекта, например, пациента, нуждающегося в лечении, и клетки, после выделения и обработки, вводят тому же субъекту.
В некоторых вариантах клеточную терапию, например, адоптивную Т-клеточную терапию, проводят аллогенным переносом, при котором клетки выделяют и/или иным образом получают от субъекта, отличного от субъекта, который должен получить или который, в конечном итоге, получит клеточную терапию, например, первого субъекта. В таких вариантах осуществления клетки затем вводят другому субъекту, например второму субъекту того же вида. В некоторых вариантах первый и второй субъекты генетически идентичны. В некоторых вариантах первый и второй субъекты генетически похожи. В некоторых вариантах второй субъект экспрессирует тот же класс или супертип HLA, что и первый субъект.
Клетки могут быть введены любыми подходящими средствами. Дозирование и введение могут частично зависеть от того, является ли введение кратковременным или постоянным. Различные схемы дозирования включают, но не ограничиваются ими, однократное или многократное введение в различные моменты времени, болюсное введение и импульсное вливание.
В некоторых вариантах субъекта лечат терапевтическим агентом, нацеленным на заболевание или состояние, например опухоль, до введения клетки или композиции, содержащей клетки. В некоторых аспектах субъект трудно поддается лечению или невосприимчив к другому терапевтическому агенту. В некоторых вариантах у субъекта имеется трудноизлечимое или рецидивирующее заболевание, например, после лечения другим терапевтическим вмешательством, включая химиотерапию, облучение и/или трансплантацию гемопоэтических стволовых клеток (HSCT), например, аллогенную HSCT. В некоторых вариантах введение эффективно лечит субъекта, несмотря на то, что субъект стал устойчивым к другой терапии.
В некоторых вариантах субъект чувствителен к другому терапевтическому агенту, и лечение терапевтическим агентом снижает тяжесть заболевания. В некоторых аспектах субъект изначально чувствителен к терапевтическому агенту, но со временем у него наблюдается рецидив заболевания или состояния. В некоторых вариантах у субъекта нет рецидива. В некоторых таких вариантах определено, что у субъекта имеется риск рецидива, например, высокий риск рецидива, и поэтому клетки вводят профилактически, например, для снижения вероятности или предотвращения рецидива.
В некоторых аспектах субъект не получал ранее лечения другим терапевтическим агентом.
Заболевание или состояние, которое лечат в некоторых аспектах, может быть любым, при котором экспрессия антигена, связанного с, являющегося специфическим к и/или экспрессированного на клетке или ткани заболевания, расстройства или состояния, и/или вовлеченного в этиологию заболевания, состояния или расстройства, например, вызывает, обостряет или иным образом участвует в таком заболевании, состоянии или расстройстве. Типовые заболевания и состояния могут включать заболевания или состояния, связанные со злокачественными новообразованиями или трансформацией клеток (например, рак), аутоиммунное или воспалительное заболевание или инфекционное заболевание, например, вызванное бактериальным, вирусным или другим патогеном. Типовые антигены, которые включают антигены, связанные с различными заболеваниями и состояниями, которые можно лечить, описаны здесь. В конкретных вариантах осуществления, иммуномодулирующий полипептид и/или рекомбинантный рецептор, например, химерный антигенный рецептор или TCR, специфически связывается с антигеном, связанным с заболеванием или состоянием. В некоторых вариантах у субъекта есть заболевание, расстройство или состояние, необязательно, рак, опухоль, аутоиммунное заболевание, расстройство или состояние или инфекционное заболевание.
В некоторых вариантах заболевание, расстройство или состояние включает опухоли, связанные с различными видами рака. В некоторых вариантах раком может быть любой рак, локализованный в теле субъекта, такой как, но не ограниченный ими, раки, расположенные в области головы и шеи, груди, печени, толстой кишки, яичников, простаты, поджелудочной железы, мозга, шейки матки, кости, кожи, глаза, мочевого пузыря, желудка, пищевода, брюшины или легкого. Например, противораковый агент может быть использован для лечения рака толстой кишки, рака шейки матки, рака центральной нервной системы, рака груди, рака мочевого пузыря, анальной карциномы, рака головы и шеи, рака яичников, рака эндометрия, мелкоклеточного рака легкого, немелкоклеточной карциномы легкого, нейроэндокринного рака, карциномы мягких тканей, рака полового члена, рака простаты, рака поджелудочной железы, рака желудка, рака желчного пузыря или рака пищевода. В некоторых случаях рак может быть раком крови. В некоторых вариантах заболевание, расстройство или состояние представляет собой опухоль, такую как солидная опухоль, лимфома, лейкоз, опухоль крови, метастатическая опухоль или другой тип рака или опухоли. В некоторых вариантах заболевание, расстройство или состояние выбрано из раков толстой кишки, легких, печени, груди, простаты, яичников, кожи, меланомы, рака костей, рака мозга, рака яичников, рака эпителия, почечноклеточной карциномы, аденокарциномы поджелудочной железы, карциномы шейки матки, колоректального рака, глиобластомы, нейробластомы, саркомы Юинга, медуллобластомы, остеосаркомы, синовиальной саркомы и/или мезотелиомы.
К заболеваниям, состояниям и расстройствам относятся опухоли, включающие солидные опухоли, гематологические злокачественные новообразования и меланомы, и включающие локализованные и метастатические опухоли, инфекционные заболевания, такие как инфекция вирусом или другим патогеном, например ВИЧ, HCV, HBV, CMV, HPV и паразитарные заболевания, аутоиммунные и воспалительные заболевания. В некоторых вариантах заболеванием, расстройством или состоянием является опухоль, рак, злокачественное новообразование, новообразование или другое пролиферативное заболевание или расстройство. Такие заболевания включают, но не ограничиваются ими, лейкоз, лимфому, например, острый миелоидный (или миелогенный) лейкоз (AML), хронический миелоидный (или миелогенный) лейкоз (CML), острый лимфоцитарный (или лимфобластный) лейкоз (ALL), хронический лимфоцитарный лейкоз (CML), волосатоклеточный лейкоз (HCL), малую лимфоцитарную лимфому (SLL), мантийноклеточную лимфому (MCL), лимфому маргинальной зоны, лимфому Беркитта, лимфому Ходжкина (HL), неходжкинскую лимфому (NHL), анапластическую крупноклеточную лимфому (ALCL), фолликулярную лимфому, трудно поддающуюся лечению фолликулярную лимфому, диффузную крупноклеточную B-клеточную лимфому (DLBCL) и множественную миелому (MM), В-клеточное злокачественное новообразование, выбранное из острого лимфобластного лейкоза (ALL), ALL у взрослых, хронического лимфобластного лейкоза (CML), неходжкинской лимфомы (NHL) и диффузной крупноклеточной B-лимфомы (DLBCL).
В некоторых вариантах заболеванием или состоянием является инфекционное заболевание или состояние, такое как, но не ограниченное ими, вирусные, ретровирусные, бактериальные и протозойные инфекции, иммунодефицит, цитомегаловирус (CMV), вирус Эпштейн-Барр (EBV), аденовирус, BK полиомавирус. В некоторых вариантах заболеванием или состоянием является аутоиммунное или воспалительное заболевание или состояние, такое как артрит, например, ревматоидный артрит (RA), диабет I типа, системная красная волчанка (SLE), воспалительное заболевание кишечника, псориаз, склеродермия, аутоиммунное заболевание щитовидной железы, болезнь Грейвса, болезнь Крона, рассеянный склероз, астма и/или заболевание или состояние, связанное с трансплантатом.
В некоторых вариантах, антигеном, связанным с заболеванием или расстройством, является опухолевый антиген, которым может быть глиома-ассоциированный антиген, β-человеческий хорионический гонадотропин, альфафетобелок (AFP), антиген созревания B-клетки (BCMA, BCM), рецептор фактора активации B-клетки (BAFFR, BR3) и/или трансмембранный активатор и CAML интерактор (TACI), Fc Рецептор-подобный 5 (FCRL5, FcRH5), лектин-реактивный AFP, тироглобулин, RAGE-1, MN-CA IX, обратная транскриптаза теломеразы человека, RU1, RU2 (AS), кишечная карбоксилэстераза, mut hsp70-2, M-CSF, Меланин-A/MART-1, WT-1, S-100, MBP, CD63, MUC1 (например, MUC1-8), p53, Ras, циклин B1, HER-2/neu, карциноэмбрионный антиген (CEA), gp100, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11, MAGE-A11, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-C1, BAGE, GAGE-1, GAGE-2, pl5, тирозиназа (например, родственный тирозиназе белок 1 (TRP-1) или родственный тирозиназе белок 2 (TRP-2)), β-катенин, NY-ESO-1, LAGE-1a, PP1, MDM2, MDM4, EGVFvIII, Tax, SSX2, теломераза, TARP, pp65, CDK4, виментин, S100, eIF-4A1, IFN-индуцируемый p78 и меланотрансферрин (p97), уроплакин II, специфический антиген простаты (PSA), человеческий калликреин (huK2), специфический мембранный антиген простаты (PSM) и простатическая кислая фосфатаза (PAP), нейтрофильная эластаза, эфрин B2, BA-46, бета-катенин, Bcr-abl, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, Каспаза 8 или B-Raf антиген. Другие опухолевые антигены могут включать любые, полученные из FRa, CD24, CD44, CD133, CD 166, epCAM, CA-125, HE4, Oval, рецептора эстрогена, рецептора прогестерона, uPA, PAI-1, CD19, CD20, CD22, ROR1, CD33/IL3Ra, c-Met, PSMA, Гликолипида F77, GD-2, инсулинового фактора роста (IGF)-I, IGF-II, IGF-I рецептора и мезотелина. Конкретные ассоциированные с опухолью антигены или Т клеточные эпитопы известны (см., например, van der Bruggen et al. (2013) Cancer Immun, доступно на www.cancerimmunity.org/peptide/; Cheever et al. (2009) Clin Cancer Res, 15, 5323-37).
В некоторых вариантах, антигеном, связанным с заболеванием или расстройством, является вирусный антиген. Множество вирусных антигенных целей идентифицировано и известно, включая пептиды, полученные из вирусных геномов ВИЧ, HTLV и других вирусах (см., например, Addo et al. (2007) PLoS ONE, 2, e321; Tsomides et al. (1994) J Exp Med, 180, 1283-93; Utz et al. (1996) J Virol, 70, 843-51). Типовые вирусные антигены включают, но не ограничены ими, антиген из гепатита A, гепатита B (например, HBV коровые или поверхностные антигены (HBVc, HBVs)), гепатита C (HCV), вируса Эпштейн-Барр (например, EBVA), папилломавируса человека (HPV; например, E6 и E7), вируса иммунодефицита человека типа-1 (ВИЧ1), герпесвируса саркомы Капоши (KSHV), папилломавируса человека (HPV), вируса гриппа, вируса Ласса, HTLN-1, HIN-1, HIN-II, CMN, EBN или HPN. В некоторых вариантах, целевым белком является бактериальный антиген или другой патогенный антиген, такой как антигены Mycobacterium tuberculosis (MT), трипаносома, например, Tiypansoma cruzi (T. cruzi), антигены, такие как поверхностный антиген (TSA) или малярийные антигены. Конкретный вирусный антиген или эпитопы или другие патогенные антигены или Т клеточные эпитопы известны (см., например, Addo et al. (2007) PLoS ONE, 2:e321; Anikeeva et al. (2009) Clin Immunol, 130:98-109).
В некоторых вариантах, антигеном, связанным с заболеванием или расстройством, является антиген, полученный из вируса, связанного с раком, такого как онкогенный вирус. Например, онкогенным вирусом является такой, при котором известно, что заражение определенными вирусами приводят к развитию различных типов рака, например, антигены вируса гепатита A, гепатита B (например, HBV которые и поверхностные антигены (HBVc, HBVs)), гепатита C (HCV), папилломавируса человека (HPV), вирусных инфекций гепатита, вируса Эпштейн-Барр (EBV), герпесвируса человека 8 (HHV-8), вируса Т-клеточного лейкоза человека-1 (HTLV-1), вируса Т-клеточного лейкоза человека-2 (HTLV-2) или цитомегаловируса (CMV) или любые антигены, на которые нацелены рекомбинантные рецепторы, описанные здесь, например, в разделе IV.
В некоторых вариантах, антиген, связанный с заболеванием, расстройством или состоянием, выбирают из αvβ6 интегрина (avb6 интегрина), антигена созревания B клетки (BCMA), B7-H3, B7-H6, карбонангидразы 9 (CA9, также известной как CAIX или G250), раково-тестикулярного антигена, раково-тестикулярного антигена 1B (CTAG, также известного как NY-ESO-1 и LAGE-2), карциноэмбрионного антигена (CEA), циклина, циклина A2, хемокинового лиганда 1 C-C мотива 1 (CCL-1), CD19, CD20, CD22, CD23, CD24, CD30, CD33, CD38, CD44, CD44v6, CD44v7/8, CD123, CD133, CD138, CD171, хондроитинсульфатпротеогликана 4 (CSPG4), белка эпидермального фактора роста (EGFR), мутации рецептора эпидермального фактора роста III типа (EGFR vIII), эпителиального гликопротеина 2 (EPG-2), эпителиального гликопротеина 40 (EPG-40), эфрина B2, рецептора эфрина A2 (EPHa2), рецептора эстрогена, Fc рецептора подобного 5 (FCRL5; также известного как гомолог 5 Fc рецептора или FCRH5), фетального ацетилхолинового рецептора (фетального AchR), белка связывания фолата (FBP), рецептора альфа фолата, ганглиозида GD2, O-ацетилированного GD2 (OGD2), ганглиозида GD3, гликопротеина 100 (gp100), глипикана-3(GPC3), рецептора, сопряженного с G белком класса C группы 5 члена D (GPRC5D), Her2/neu (рецептора тирозинкиназы erb-B2), Her3 (erb-B3), Her4 (erb-B4), erbB димеров, антигена, связанного с человеческой высокомолекулярной меланомой (HMW-MAA), поверхностного антигена гепатита B, человеческого лейкоцитарного антигена A1 (HLA-A1), человеческого лейкоцитарного антигена A2 (HLA-A2), IL-22 рецептора альфа (IL-22Rα), IL-13 рецептора альфа 2 (IL-13Rα2), рецептора домена вставки киназы (kdr), легкой цепи каппа, L1 молекулы клеточной адгезии (L1-CAM), CE7 эпитопа L1-CAM, члена А семейства 8, содержащий насыщенные лейцином повторы (LRRC8A), Льюис Y, связанного с меланомой антигена (MAGE)-A1, MAGE-A3, MAGE-A6, MAGE-A10, мезотелина (MSLN), c-Met, мышиного цитомегаловируса (CMV), муцина 1 (MUC1), MUC16, лигандов члена D группы 2 естественного киллера (NKG2D), мелана A (MART-1), молекулы адгезии нервных клеток (NCAM), онкофетального антигена, преимущественно экспрессированного антигена меланомы (PRAME), рецептора прогестерона, специфического антигена простаты, антигена стволовых клеток простаты (PSCA), специфического мембранного антигена простаты (PSMA), рецептора тирозинкиназа подобного орфанного рецептора 1 (ROR1), сурвивина, трофобластного гликопротеина (TPBG, также известного как 5T4), связанного с опухолью гликобелка 72 (TAG72), родственного тирозиназе белка 1 (TRP1, также известного как TYRP1 или gp75), родственного тирозиназе белка 2 (TRP2, также известного как допахромтаутомераза, допахром дельта-изомераза или DCT), рецептора фактора роста сосудистого эндотелия (VEGFR), рецептора 2 фактора роста сосудистого эндотелия (VEGFR2), опухоли Вильмса 1 (WT-1), патоген-специфического или патоген-экспрессированного антигена или антигена, ассоциированного с универсальной меткой, и/или биотинилированных молекул, и/или молекул, экспрессированных ВИЧ, HCV, HBV или другими патогенами. Антигены, на которые нацелены рецепторы, в некоторых вариантах, включают антигены, ассоциированные с злокачественностью B клетки, такие как любые из множества известных маркеров B клетки. В некоторых вариантах, антиген является или включает CD20, CD19, CD22, ROR1, CD45, CD21, CD5, CD33, Ig каппа, Ig лямбда, CD79a, CD79b или CD30. В некоторых вариантах, антигеном является или включает патоген-специфический или патоген-экспрессированный антиген. В некоторых вариантах, антигеном является вирусный антиген (такой как вирусный антиген из ВИЧ, HCV, HBV и т.д.), бактериальные антигены и/или паразитарные антигены.
В некоторых вариантах, клетки вводят в желаемой дозе, которая, в некоторых аспектах, включает желаемую дозу или количество клеток или клеточных типов, и/или желаемое соотношение клеточных типов. Таким образом, доза клеток в некоторых вариантах основана на общем количестве клеток (или количестве на кг массы тела) и желательном отношении популяций или подтипов, таком как отношение CD4+ к CD8+. В некоторых вариантах, доза клеток основана на желательном общем количестве (или количестве на кг массы тела) клеток в отдельных популяциях или отдельных типах клеток. В некоторых вариантах, доза основана на сочетании таких характеристик, таких, как желаемое общее количество клеток, желаемое соотношение и желаемое общее количество клеток в отдельных популяциях.
В некоторых вариантах, популяции или подтипы клеток, таких как CD8+ и CD4+ T клетки, вводят в пределах допустимой разности желаемой дозы общего количества клеток, такой как желательная доза T клеток. В некоторых аспектах, желаемой дозой является желаемое количество клеток или желаемого количества клеток на единицу массы тела субъекта, которому вводят клетки, например, клетки/кг. В некоторых аспектах, желаемая доза превышает минимальное количество клеток или минимальное количество клеток на единицу массы тела. В некоторых аспектах, среди общего количества клеток, вводимых в желаемой дозе, отдельные популяции или суб-типы присутствуют с близким к желаемым выходным соотношением (таким как соотношение CD4+ к CD8+), например, в пределах определенной допустимой разности или ошибки такого соотношения.
В некоторых вариантах, клетки вводят в пределах допустимой разности между желаемой дозой одной или более отдельных популяций или подтипов клеток, такой как желаемая доза CD4+ клеток и/или желаемая доза CD8+ клеток. В некоторых аспектах, желаемая доза представляет собой желаемое количество клеток подтипа или популяции, или желаемое количество таких клеток на единицу массы тела субъекта, которому вводятся клетки, например, клетки/кг. В некоторых аспектах желаемая доза превышает минимальное количество клеток популяции или подтипа или минимальное количество клеток популяции или подтипа на единицу массы тела.
Таким образом, в некоторых вариантах, доза основана на желаемой фиксированной дозе общих клеток и желаемом соотношении и/или на основе желаемой фиксированной дозы одного или более, например, каждого из индивидуальных подтипов или субпопуляций. Таким образом, в некоторых вариантах, доза основана на желаемой фиксированной или минимальной дозе Т-клеток и желаемом соотношении CD4+ к CD8+ клеткам, и/или основано на желаемой фиксированной или минимальной дозе CD4+ и/или CD8+ клеток.
В определенных вариантах, клеток или отдельные субпопуляции подтипов клеток вводят субъекту в интервале от около одного миллиона до около 100 миллиардов клеток и/или такого количества клеток на килограмма массы тела, как, например, от 1 миллионов до около 50 миллиардов клеток (например, около 5 миллионов клеток, около 25 миллионов клеток, около 500 миллионов клеток, около 1 миллиарда клеток, около 5 миллиардов клеток, около 20 миллиардов клеток, около 30 миллиардов клеток, около 40 миллиардов клеток или интервала, определенного любыми двумя из вышеуказанных значений), например, от около 10 миллионов до около 100 миллиардов клеток (например, около 20 миллионов клеток, около 30 миллионов клеток, около 40 миллионов клеток, около 60 миллионов клеток, около 70 миллионов клеток, около 80 миллионов клеток, около 90 миллионов клеток, около 10 миллиардов клеток, около 25 миллиардов клеток, около 50 миллиардов клеток, около 75 миллиардов клеток, около 90 миллиардов клеток или интервала, определенного любыми двумя из вышеуказанных значений), и, в некоторых случаях, от около 100 миллионов клеток до около 50 миллиардов клеток (например, около 120 миллионов клеток, около 250 миллионов клеток, около 350 миллионов клеток, около 450 миллионов клеток, около 650 миллионов клеток, около 800 миллионов клеток, около 900 миллионов клеток, около 3 миллиардов клеток, около 30 миллиардов клеток, около 45 миллиардов клеток) или любого значения между этими интервалами, и/или на килограмм массы тела. Дозы могут варьироваться в зависимости от признаков, конкретных для заболевания или расстройства и/или пациента и/или другого лечения.
В некоторых вариантах, например, если субъектом является человек, доза включает менее чем около 5×108 общих экспрессирующих рекомбинантный рецептор (например, CAR) клеток, T клеток или мононуклеарных клеток периферической крови (PBMC), например, в интервале от около 1×106 до 5×108 таких клеток, например, 2×106, 5×106, 1×107, 5×107, 1×108 или 5×108 всего таких клеток или интервал между любыми двумя из вышеуказанных значений.
В некоторых вариантах, доза генетически сконструированных клеток содержит от или от около 1×105 до 5×108 всего CAR-экспрессирующих T клеток, от 1×105 до 2,5×108 всего CAR-экспрессирующих T клеток, от 1×105 до 1×108 всего CAR-экспрессирующих T клеток, от 1×105 до 5×107 всего CAR-экспрессирующих T клеток, от 1×105 до 2,5×107 всего CAR-экспрессирующих T клеток, от 1×105 до 1×107 всего CAR-экспрессирующих T клеток, от 1×105 до 5×106 всего CAR-экспрессирующих T клеток, от 1×105 до 2,5×106 всего CAR-экспрессирующих T клеток, от 1×105 до 1×106 всего CAR-экспрессирующих T клеток, от 1×106 до 5×108 всего CAR-экспрессирующих T клеток, от 1×106 до 2,5×108 всего CAR-экспрессирующих T клеток, от 1×106 до 1×108 всего CAR-экспрессирующих T клеток, от 1×106 до 5×107 всего CAR-экспрессирующих T клеток, от 1×106 до 2,5×107 всего CAR-экспрессирующих T клеток, от 1×106 до 1×107 всего CAR-экспрессирующих T клеток, от 1×106 до 5×106 всего CAR-экспрессирующих T клеток, от 1×106 до 2,5×106 всего CAR-экспрессирующих T клеток, от 2,5×106 до 5×108 всего CAR-экспрессирующих T клеток, от 2,5×106 до 2,5×108 всего CAR-экспрессирующих T клеток, от 2,5×106 до 1×108 всего CAR-экспрессирующих T клеток, от 2,5×106 до 5×107 всего CAR-экспрессирующих T клеток, от 2,5×106 до 2,5×107 всего CAR-экспрессирующих T клеток, от 2,5×106 до 1×107 всего CAR-экспрессирующих T клеток, от 2,5×106 до 5×106 всего CAR-экспрессирующих T клеток, от 5×106 до 5×108 всего CAR-экспрессирующих T клеток, от 5×106 до 2,5×108 всего CAR-экспрессирующих T клеток, от 5×106 до 1×108 всего CAR-экспрессирующих T клеток, от 5×106 до 5×107 всего CAR-экспрессирующих T клеток, от 5×106 до 2,5×107 всего CAR-экспрессирующих T клеток, от 5×106 до 1×107 всего CAR-экспрессирующих T клеток, от 1×107 до 5×108 всего CAR-экспрессирующих T клеток, от 1×107 до 2,5×108 всего CAR-экспрессирующих T клеток, от 1×107 до 1×108 всего CAR-экспрессирующих T клеток, от 1×107 до 5×107 всего CAR-экспрессирующих T клеток, от 1×107 до 2,5×107 всего CAR-экспрессирующих T клеток, от 2,5×107 до 5×108 всего CAR-экспрессирующих T клеток, от 2,5×107 до 2,5×108 всего CAR-экспрессирующих T клеток, от 2,5×107 до 1×108 всего CAR-экспрессирующих T клеток, от 2,5×107 до 5×107 всего CAR-экспрессирующих T клеток, от 5×107 до 5×108 всего CAR-экспрессирующих T клеток, от 5×107 до 2,5×108 всего CAR-экспрессирующих T клеток, от 5×107 до 1×108 всего CAR-экспрессирующих T клеток, от 1×108 до 5×108 всего CAR-экспрессирующих T клеток, от 1×108 до 2,5×108 всего CAR-экспрессирующих T клеток или от 2,5×108 до 5×108 всего CAR-экспрессирующих T клеток.
В некоторых вариантах, доза генетически сконструированных клеток содержит, по меньшей мере, или по меньшей мере, около или составляет или составляет около 1×105 CAR-экспрессирующих клеток, по меньшей мере, или по меньшей мере около или составляет, или составляет около 2,5×105 CAR-экспрессирующих клеток, по меньшей мере или по меньшей мере, около или составляет, или составляет около 5×105 CAR-экспрессирующих клеток, по меньшей мере или по меньшей мере, около или составляет, или составляет около 1×106 CAR-экспрессирующих клеток, по меньшей мере или по меньшей мере, около или составляет, или составляет около 2,5×106 CAR-экспрессирующих клеток, по меньшей мере или по меньшей мере, около или составляет, или составляет около 5×106 CAR-экспрессирующих клеток, по меньшей мере или по меньшей мере, около или составляет, или составляет около 1×107 CAR-экспрессирующих клеток, по меньшей мере или по меньшей мере, около или составляет, или составляет около 2,5×107 CAR-экспрессирующих клеток, по меньшей мере или по меньшей мере, около или составляет, или составляет около 5×107 CAR-экспрессирующих клеток, по меньшей мере или по меньшей мере, около или составляет, или составляет около 1×108 CAR-экспрессирующих клеток, по меньшей мере или по меньшей мере, около или составляет, или составляет около 2,5×108 CAR-экспрессирующих клеток или по меньшей мере или по меньшей мере, около или составляет, или составляет около 5×108 CAR-экспрессирующих клеток.
В некоторых вариантах, клеточная терапия включает введение дозы, содержащей количество клеток от или от около 1×105 до 5×108 всего экспрессирующих рекомбинантный рецептор клеток, всего T клеток или всего мононуклеаров периферической крови (PBMC), от или от около 5×105 до 1×107 всего экспрессирующих рекомбинантный рецептор клеток, всего T клеток или всего мононуклеаров периферической крови (PBMC) или от или от около 1×106 до 1×107 всего экспрессирующих рекомбинантный рецептор клеток, всего T клеток или всего мононуклеаров периферической крови (PBMC), все включительно. В некоторых вариантах, клеточная терапия включает введение дозы клеток, содержащей количество клеток, по меньшей мере, или по меньшей мере, около 1×105 всего экспрессирующих рекомбинантный рецептор клеток, всего T клеток или всего мононуклеаров периферической крови (PBMC), например, по меньшей мере или по меньшей мере, 1×106, по меньшей мере или по меньшей мере, около 1×107, по меньшей мере или по меньшей мере, около 1×108 таких клеток. В некоторых вариантах, количество указано со ссылкой на общее количество CD3+ или CD8+, в некоторых случаях также экспрессирующих рекомбинантный рецептор (например, CAR+) клеток. В некоторых вариантах, клеточная терапия включает введение дозы, содержащей количество клеток от или от около 1×105 до 5×108 CD3+ или CD8+ всего T клеток или CD3+ или CD8+ экспрессирующих рекомбинантный рецептор клеток, от или от около 5×105 до 1×107 CD3+ или CD8+ всего T клеток или CD3+ или CD8+ экспрессирующих рекомбинантный рецептор клеток или от или от около 1×106 до 1×107 CD3+ или CD8+ всего T клеток или CD3+ или CD8+экспрессирующих рекомбинантный рецептор клеток, все включительно. В некоторых вариантах, клеточная терапия включает введение дозы, содержащей количество клеток от или от около 1×105 до 5×108 всего CD3+/CAR+ или CD8+/CAR+ клеток, от или от около 5×105 до 1×107 всего CD3+/CAR+ или CD8+/CAR+ клеток или от или от около 1×106 до 1×107 всего CD3+/CAR+ или CD8+/CAR+ клеток, все включительно.
В некоторых вариантах, T клетки дозы включают CD4+ T клетки, CD8+ T клетки или CD4+ и CD8+ T клетки.
В некоторых вариантах, например, если субъектом является человек, CD8+ T клетки дозы, включая дозу, включающую CD4+ и CD8+ T клетки, включают от около 1×106 до 5×108 всего экспрессирующих рекомбинантный рецептор (например, CAR) CD8+ клеток, например, в интервале от около 5×106 до 1×108 таких клеток, например, 1×107, 2,5×107, 5×107, 7,5×107, 1×108 или 5×108 всего таких клеток или интервал между любыми двумя из вышеуказанных значений. В некоторых вариантах, пациенту вводят множество доз, и каждая из доз и общая доза может быть в пределах любого из вышеуказанных значений. В некоторых вариантах, доза клеток содержит введение от или от около 1×107 до 0,75×108 всего экспрессирующих рекомбинантный рецептор CD8+ T клеток, от 1×107 до 2,5×107 всего экспрессирующих рекомбинантный рецептор CD8+ T клеток, от или от около 1×107 до 0,75×108 всего экспрессирующих рекомбинантный рецептор CD8+ T клеток, все включительно. В некоторых вариантах, доза клеток содержит введение около 1×107, 2,5×107, 5×107 7,5×107, 1×108 или 5×108 всего экспрессирующих рекомбинантный рецептор CD8+ T клеток.
В некоторых вариантах дозу клеток, например, экспрессирующих рекомбинантный рецептор T клеток, вводят субъекту в виде однократной дозы или вводят только один раз в течение двух недель, одного месяца, трех месяцев, шести месяцев, 1 года или больше.
В некоторых вариантах, например, если субъектом является человек, доза включает менее около 1×108 всего экспрессирующих рекомбинантный рецептор (например, рекомбинантный TCR) клеток, T клеток или мононуклеаров периферической крови (PBMC), например, в интервале от около 1×106 до 1×108 таких клеток, например, 2×106, 5×106, 1×107, 5×107 или 1×108 или всего таких клеток или интервал между любыми двумя из вышеуказанных значений.
В некоторых вариантах, доза всего клеток и/или доза отдельных субпопуляций клеток составляет в пределах интервала от около 104 и до около 109 клеток/килограмм (кг) массы тела, например, от 105 до106 клеток/кг массы тела, например, от около 1×105 клеток/кг, 1,5×105 клеток/кг, 2×105 клеток/кг или 1×106 клеток/кг массы тела. Например, в некоторых вариантах, клетки вводят в пределах определенного интервала ошибки от около 104 и от около 109 T клетки/килограмм (кг) массы тела, например, от 105 до 106 T клеток/кг массы тела, например, от около 1×105 T клеток/кг, 1,5×105 T клеток/кг, 2×105 T клеток/кг или 1×106 T клеток/кг массы тела.
В некоторых вариантах, клетки вводят в пределах определенного интервала ошибки между от около 104 и от около 109 CD4+ и/или CD8+ клеток/килограмм (кг) массы тела, например, между 105 и 106 CD4+ и/или CD8+клетки/кг массы тела, например, от около 1×105 CD4+ и/или CD8+ клеток/кг, 1,5×105 CD4+ и/или CD8+ клеток/кг, 2×105 CD4+ и/или CD8+ клеток/кг или 1×106 CD4+ и/или CD8+ клеток/кг массы тела.
В некоторых вариантах, клетки вводят в пределах определенного интервала ошибки, более чем и/или, по меньшей мере, около 1×106, около 2,5×106, около 5×106, около 7,5×106 или около 9×106 CD4+ клетки и/или, по меньшей мере, около 1×106, около 2,5×106, около 5×106, около 7,5×106 или около 9×106 CD8+ клетки и/или, по меньшей мере, около 1×106, около 2,5×106, около 5×106, около 7,5×106 или около 9×106 T клетки. В некоторых вариантах, клетки вводят в пределах определенного интервала ошибки между около 108 и 1012 или между около 1010 и 1011 T клетки, между около 108 и 1012 или между около 1010 и 1011 CD4+ клетки и/или между около 108 и 1012 или между около 1010 и 1011 CD8+ клетки.
В некоторых вариантах, клетки вводят в пределах допустимого интервала желаемого соотношения на выходе множества популяций клеток или подтипов, таких как CD4+ и CD8+ клетки или подтипы. В некоторых аспектах, желаемое соотношение может быть удельным соотношением или может быть интервалом соотношений. Например, в некоторых вариантах, желаемое соотношение (например, соотношение CD4+ к CD8+ клеткам) находится между от около 1:5 и от около 5:1 (или более чем около 1:5 и менее около 5:1) или между от около 1:3 и от около 3:1 (или более чем около 1:3 и менее около 3:1), например, между от около 2:1 и от около 1:5 (или более чем около 1:5 и менее около 2:1, например, от около 5:1, 4,5:1, 4:1, 3,5:1, 3:1, 2,5:1, 2:1, 1,9:1, 1,8:1, 1,7:1, 1,6:1, 1,5:1, 1,4:1, 1,3:1, 1,2:1, 1,1:1, 1:1, 1:1,1, 1:1,2, 1:1,3, 1:1,4, 1:1,5, 1:1,6, 1:1,7, 1:1,8, 1:1,9: 1:2, 1:2,5, 1:3, 1:3,5, 1:4, 1:4,5 или 1:5. В некоторых аспектах, допустимая разность составляет в пределах около 1%, около 2%, около 3%, около 4% около 5%, около 10%, около 15%, около 20%, около 25%, около 30%, около 35%, около 40%, около 45%, около 50% от желаемого соотношения, включая любое значение между этими интервалами.
Для профилактики или лечения заболевания подходящая доза может зависеть от типа заболевания, которое нужно лечить, типа клеток или рекомбинантных рецепторов, тяжести и течения заболевания, от того, вводятся ли клетки в профилактических или терапевтических целях, предыдущей терапии, истории болезни субъекта и реакции на клетки, и на усмотрение лечащего врача. Композиции и клетки в некоторых вариантах подходящим образом вводят субъекту за один раз или в течение ряда курсов лечения.
В некоторых аспектах размер дозы определяется на основании одного или более критериев, таких как реакция субъекта на предшествующее лечение, например, химиотерапию, тяжесть заболевания у субъекта, такую как опухолевая масса, объем, размер или степень, распространенность или тип метастазов, стадии и/или вероятности или частоты развития токсических исходов у субъекта, например, СВЦ, синдрома активации макрофагов, синдрома лизиса опухоли, нейротоксичности и/или иммунного ответа хозяина против вводимых клеток и/или рекомбинантных рецепторов.
В некоторых аспектах размер дозы определяется тяжестью заболевания или состояния субъекта. Например, в некоторых аспектах, количество клеток, вводимых в дозе, определяется на основании опухолевой массы, которая присутствует у субъекта непосредственно перед началом введения дозы клеток. В некоторых вариантах размер первой и/или последующей дозы обратно коррелирует с тяжестью заболевания. В некоторых аспектах, например, в контексте большей тяжести заболевания, субъекту вводят небольшое количество клеток. В других вариантах, например, в контексте меньшей тяжести заболевания, субъекту вводят большее количество клеток.
Клетки могут быть введены любыми подходящими способами, например, болюсной инфузией, инъекцией, например, внутривенными или подкожными инъекциями, внутриглазной инъекцией, окологлазной инъекцией, субретинальной инъекцией, интравитреальной инъекцией, транссептальной инъекцией, субсклеральной инъекцией, интрахороидальной инъекцией, интракамеральной инъекцией, субконъектальной инъекцией, субконъюнктивальной инъекцией, субтеноновой инъекцией, ретробульбарной инъекцией, перибульбарной инъекцией или доставкой в заднюю околосклеральную область. В некоторых вариантах, их вводят парентерально, внутрилегочно и интраназально и, если желательно для местного лечения, внутриочагово. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. В некоторых вариантах, данную дозу вводят однократным болюсным введением клетки. В некоторых вариантах, ее вводят путем многократного болюсного введения клеток, например, в течение периода не более 3 дней или путем непрерывного инфузионного введения клеток.
В некоторых вариантах, клетки вводят как часть комбинированной терапии, например, одновременно или последовательно с, в любом порядке, другим терапевтическим вмешательством, таким как антитело или сконструированные клетки или рецептор или агент, такой как цитотоксический или терапевтический. Клетки в некоторых вариантах вводят совместно с одним или более дополнительными терапевтическими агентами или в сочетании с другим терапевтическим вмешательством одновременно или последовательно, в любом порядке. В некоторых контекстах клетки вводят совместно с другой терапией, достаточно близкой по времени, так что популяция клеток усиливает эффект одного или более дополнительных терапевтических агентов, или наоборот. В некоторых вариантах клетки вводят до одного или более дополнительных терапевтических агентов. В некоторых вариантах клетки вводят после одного или более дополнительных терапевтических агентов. В некоторых вариантах один или более дополнительных агентов включают цитокин, такой как IL-2, например, для повышения стойкости. В некоторых вариантах, способы включают введение химиотерапевтического агента.
После введения клеток, биологическую активность популяций сконструированных клеток, в некоторых вариантах, измеряют, например, любым из множества известных способов. Параметры оценки включают специфическое связывание сконструированной или природной T клетки или другой иммунной клетки с антигеном, in vivo, например, визуализацией, или ex vivo, например, ELISA или проточной цитометрией. В определенных вариантах, способность сконструированных клеток разрушать целевые клетки может быть измерена любыми подходящими известными способами, такими как анализы цитотоксичности, с применением любых подходящих известных способов, таких как анализы цитотоксичности, описанные в, например, Kochenderfer et al., J. Immunotherapy, 32(7): 689-702 (2009), и Herman et al. J. Immunological Methods, 285(1): 25-40 (2004). В определенных вариантах, биологическую активность клеток измеряют анализом экспрессии и/или секреции одного или более цитокинов, таких как CD 107a, IFNγ, IL-2, и TNF. В некоторых аспектах, биологическую активность измеряют оценкой клинических результатов, таких как снижение опухолевой массы или нагрузки.
В определенных вариантах, сконструированные клетки далее модифицируют любыми путями, так, чтобы повысить терапевтическую или профилактическую эффективность. Например, сконструированный CAR или TCR, экспрессированный популяцией, может быть конъюгирован либо прямо, либо косвенно через линкер с группой целенаправленного воздействия. Практика конъюгирования соединений, например, CAR или TCR, с группами целенаправленного воздействия известна. См., например, Wadwa et al., J. Drug Targeting 3: 1 1 1 (1995), и патент США 5,087,616.
VII. НАБОРЫ И ГОТОВЫЕ ИЗДЕЛИЯ
Также представлены готовые изделия, системы, аппараты и наборы, применяемые при осуществлении представленных вариантов. В некоторых вариантах, представленные готовые изделия и наборы содержат один или более компонентов одного или более агентов, способных к генетическому разрушению, и/или матричных полинуклеотидов, например, матричных полинуклеотидов, содержащих трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, или один или более вторых матричных полинуклеотидов. В некоторых вариантах, готовые изделия или наборы могут применяться в способах конструирования T клеток для экспрессирования рекомбинантного рецептора и/или других полипептидов, и оценке полученных клеток и/или популяции клеток в соответствии с представленными способами. В некоторых вариантах, готовые изделия или наборы, представленные здесь, содержат T клетки и/или T клеточные композиции, такие как любые T клетки и/или T клеточные композиции, описанные здесь.
В некоторых вариантах, готовые изделия или наборы включают полипептиды, нуклеиновые кислоты, векторы и/или полинуклеотиды, применяемые в осуществлении представленных способов. В некоторых вариантах, готовые изделия или наборы включают одну или более молекул нуклеиновой кислоты, например, плазмида или ДНК фрагмент, который содержит один или более компонентов одного или более агентов, способных вызывать генетическое разрушение, и/или матричных полинуклеотидов, например, матричных полинуклеотидов, содержащих трансген, кодирующий рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, или один или более вторых матричных полинуклеотидов. В некоторых вариантах, готовые изделия или наборы, представленные здесь, содержат контрольные векторы.
В некоторых вариантах, готовые изделия или наборы, представленные здесь, содержат один или более агентов, где каждый из одного или более агентов независимо способны вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC); и матричный полинуклеотид, содержащий трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом через репарацию, направляемую гомологией (HDR).
В некоторых вариантах, готовые изделия или наборы, представленные здесь, содержат T клетки и/или T клеточные композиции, такие как любые T клетки и/или T клеточные композиции, описанные здесь. В некоторых вариантах, T клетки, и/или T клеточные композиции любых модифицированных T клеток, применяемых в способах скрининга, описаны здесь. В некоторых вариантах, готовые изделия или наборы, представленные здесь, содержат контрольные T клетки и/или T клеточные композиции.
В некоторых вариантах, готовые изделия или наборы включают один или более компонентов, применяемых для оценки свойств популяции и/или композиции сконструированных клеток, экспрессирующих рекомбинантный рецептор. Например, готовые изделия или наборы могут включать реагенты связывания, например, антитела, MHC-пептидные тетрамеры и/или пробы, применяемые для оценки конкретных свойств введенных рекомбинантных TCR, например, экспрессию рекомбинантного TCR на поверхности клетки, распознавание пептида в контексте MHC молекулы и/или определяемый сигнал, вырабатываемый репортером, например, репортером активации T клетки. В некоторых вариантах, готовые изделия или наборы могут включать компоненты, которые применяют для определения конкретных свойств, таких как меченные компоненты, например, флуоресцентно меченные компоненты и/или компоненты, которые могут давать определяемый сигнал, например, субстраты, которые могут давать флуоресценцию или люминесценцию.
В некоторых вариантах, готовые изделия или наборы включают один или более контейнеров, обычно множество контейнеров, упаковочный материал и этикетку или вкладыш, связанные с контейнером или контейнерами и/или упаковкой, обычно включающие инструкции по применению, например, инструкции по введению компонентов в клетки для конструирования и/или для оценки популяций сконструированных клеток и/или композиции. В некоторых вариантах, готовое изделие или наборы включают одну или более инструкций для введения сконструированных клеток и/или клеточных композиций для терапии.
Готовые изделия, представленные здесь, содержат упаковочные материалы. Упаковочные материалы для применения для упаковки представленных материалов, хорошо известны. См., например, патенты США №№ 5,323,907, 5,052,558 и 5,033,252, каждый из которых включен сюда в качестве ссылки полностью. Примеры упаковочных материалов включают, но не ограничены ими, блистерные упаковки, бутылки, пробирки, ингаляторы, помпы, мешки, флаконы, контейнеры, шприцы, одноразовые лабораторные расходные материалы, например, наконечники к пипеткам и/или пластиковые тарелки или бутылки. Готовые изделия или наборы могут включать устройство, способствующее распределению материалов или способствующие применению и высокой пропускной способностью или в крупных масштабах, например, способствующие применению в роботизированном оборудовании. Обычно упаковка не реагирует с композицией, содержащейся в ней.
В некоторых вариантах, агент, способный вызывать генетическое разрушение, и/или матричные полинуклеотиды упаковывают по отдельности. В некоторых вариантах, каждый контейнер может иметь одно отделение. В некоторых вариантах, другие компоненты готового изделия или наборов упакованы по отдельности или вместе в одном отделении.
VIII. ОПРЕДЕЛЕНИЯ
Если не указано иное, все термины в данной области техники, обозначения и другие технические и научные термины или терминология, используемые здесь, имеют то же значение, которое обычно понимается специалистом в данной области техники, к которой относится заявленный предмет изобретения. В некоторых случаях термины с общепринятыми значениями определены в данном документе для ясности и/или для удобства ссылок, и включение таких определений в настоящий документ не обязательно должно толковаться как представляющее существенное различие по сравнению с тем, что обычно понимается в данной области техники.
Термины «полипептид» и «белок» используются взаимозаменяемо для обозначения полимера аминокислотных остатков, и не ограничиваются минимальной длиной. Полипептиды, включая предоставленные антитела и цепи антитела и другие пептиды, например линкеры, могут включать аминокислотные остатки, включая природные и/или не природные аминокислотные остатки. Термины также включают модификации полипептида после экспрессии, например, гликозилирование, сиалирование, ацетилирование, фосфорилирование и подобные. В некоторых аспектах, полипептиды могут содержать модификации по отношению к нативным или природным последовательностям, пока белок сохраняет желаемую активность. Эти модификации могут быть преднамеренными, например, через сайт-направленный мутагенез, или могут быть случайными, например, из-за мутаций хозяев, которые продуцируют белки, или ошибок из-за ПЦР амплификации.
«Выделенная» нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая была отделена от компонента ее естественной среды. Выделенная молекула нуклеиновой кислоты включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат молекулу нуклеиновой кислоты, но молекула нуклеиновой кислоты присутствует экстрахромосомно или в хромосомной локации, которая отличается от ее природной хромосомной локации.
«Выделенная нуклеиновая кислота, кодирующая TCR или антитело» относится к одной или более молекулам нуклеиновой кислоты, кодирующим TCR альфа или β цепи (или их фрагменты) или тяжелую и легкую цепи антитела (или их фрагменты), включая такие молекулы нуклеиновой кислоты в одинарном векторе или раздельных векторах, и такие молекулы нуклеиновой кислоты, присутствующие в одной или более локациях местах в клетке хозяина.
Термины «клетка хозяина», «колония клеток хозяина» и «культура клеток хозяина» используются взаимозаменяемо и относятся к клеткам, в которые были введены экзогенные нуклеиновые кислоты, включая потомство таких клеток. Клетки хозяина включают «трансформанты» и «трансформированные клетки», которые включают первичную трансформированную клетку и полученное от нее потомство, независимо от количества пассажей. Потомство может не быть полностью идентичным по содержанию нуклеиновых кислоты к родительской клетке, но может содержать мутации. Мутантное потомство, которое имеет ту же функцию или биологическую активность, что и проверенные или отобранные для первоначально трансформированной клетки, включены сюда.
В данном описании «процент (%) идентичности аминокислотной последовательности» и «процент идентичности» при использовании по отношению к аминокислотной последовательности (эталонной полипептидной последовательности) определяется как процент аминокислотных остатков в кандидатной последовательности (например, антителе или фрагменте), которые идентичны аминокислотным остаткам в ссылочной полипептидной последовательности, после выравнивания последовательностей и введения гэпов, если необходимо, для достижения максимального процента идентичности последовательности, и без учета каких-либо консервативных замен как части идентичности последовательности. Выравнивание для целей определения процента идентичности аминокислотной последовательности может быть достигнуто с помощью различного известного, например, с использованием общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Подходящие параметры для выравнивания последовательностей могут быть определены, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей.
В некоторых вариантах «функционально связанный» может включать сбор компонентов, таких как ДНК последовательность, например, гетерологичных нуклеиновых кислот) и регуляторных последовательностей таким образом, чтобы разрешить экспрессию гена, когда соответствующие молекулы (например, белки активаторы транскрипции) связаны с регуляторной последовательностью. Следовательно, это означает, что описанные компоненты находятся во взаимосвязи, позволяющей им функционировать предполагаемым образом.
Замещение аминокислот может включать замещение одной аминокислоты в полипептиде другой аминокислотой. Замещение может быть консервативным замещением аминокислоты или не консервативным замещением аминокислоты. Замещения аминокислот могут быть введены в рассматриваемую связывающую молекулу, например, антитело, и продукты, проверенные на желаемую активность, например, сохранение/улучшение антигенного связывания, снижение иммуногенности или улучшение АЗКЦ или КДЦ.
Аминокислоты обычно могут быть сгруппированы согласно следующим свойствам боковой цепи:
(1) гидрофобная: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральная гидрофильная: Cys, Ser, Thr, Asn, Gln;
(3) кислая: Asp, Glu;
(4) щелочная: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(6) ароматическая: Trp, Tyr, Phe.
В некоторых вариантах, консервативные замещения могут включать обмен члена одного из этих классов на другой член того же класса. В некоторых вариантах, не консервативные аминокислотные замещения могут включать обмен члена одного из этих классов на другой класс.
Используемый здесь термин «вектор» относится к молекуле нуклеиновой кислоты, способной репродуцировать другие нуклеиновые кислоты, с которыми она связана. Термин включает вектор как самовоспроизводящуюся структуру нуклеиновой кислоты, а также вектор, включенный в геном клетки хозяина, в которую он был введен. Некоторые векторы способны направлять экспрессию нуклеиновых кислот, с которыми они оперативно связаны. Такие векторы называются здесь «векторы экспрессии».
Термин «вкладыш в упаковку» используется для обозначения инструкций, обычно включаемых в коммерческие упаковки терапевтических продуктов, которые содержат информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предупреждениях относительно использования таких терапевтических продуктов.
Используемые здесь формы единственного числа включают множественное число, если контекст явно не диктует иное. Например, «a» или «an» означает «по меньшей мере, один» или «один или более». Понятно, что аспекты и варианты, описанные в данном документе, включают «состоящий» и/или «состоящий, по существу, из» аспектов и вариантов.
В этом раскрытии различные аспекты заявленного объекта изобретения представлены в формате интервала. Следует понимать, что описание в формате интервала дано просто для удобства и краткости, и не должно рассматриваться как жесткое ограничение объема заявленного объекта изобретения. Соответственно, следует считать, что описание диапазона конкретно раскрывает все возможные подинтервалы, а также отдельные числовые значения в пределах этого интервала. Например, если предоставляется интервал значений, подразумевается, что каждое промежуточное значение между верхним и нижним пределом этого интервала и любое другое заявленное или промежуточное значение в этом указанном интервале охватывается заявленным объектом изобретения. Верхние и нижние пределы этих меньших интервалов могут быть независимо включены в меньшие интервалы, а также включены в заявленный объект изобретения с учетом любого специально исключенного ограничения в указанном интервале. Если указанный интервал включает один или оба ограничения, диапазоны, исключающие один или оба включенных ограничения, также включаются в заявленный объект изобретения. Это применимо независимо от широты интервала.
Используемый здесь термин «около» относится к обычному диапазону ошибок для соответствующего значения, хорошо известному специалисту в данной области техники. Ссылка на «около» значения или параметра в данном документе включает (и описывает) варианты осуществления, которые направлены на это значение или параметр per se. Например, описание, относящееся к «около X», включает описание «X». В некоторых вариантах «около» может относиться к ±25%, ±20%, ±15%, ±10%, ±5% или ±1%.
В данном описании композиция относится к любой смеси двух или более продуктов, веществ или соединений, включая клетки. Это может быть раствор, суспензия, жидкость, порошок, паста, водная, неводная или любое их сочетание.
В данном описании утверждение, что клетка или популяция клеток является «положительной» для определенного маркера, относится к обнаруживаемому присутствию на или в клетке определенного маркера, обычно поверхностного маркера. Когда речь идет о поверхностном маркере, этот термин относится к наличию поверхностной экспрессии, обнаруживаемой с помощью проточной цитометрии, например, путем окрашивания антителом, которое специфически связывается с маркером, и обнаружением указанного антитела, где окрашивание обнаруживается проточной цитометрией на уровне по существу выше окрашивания, определенного проведением той же методики с изотипически сходным контролем в идентичных в остальном условиях и/или на уровне, по существу аналогичном уровню для клетки, заведомо положительной для маркера, и/или на уровне, по существу более высоком, чем для клетки, заведомо отрицательной для маркера.
В данном описании утверждение, что клетка или популяция клеток является «отрицательной» для определенного маркера, относится к отсутствию существенного обнаруживаемого присутствия на клетке или в клетке определенного маркера, обычно поверхностного маркера. При упоминании поверхностного маркера, этот термин относится к отсутствию поверхностной экспрессии, определенной проточной цитометрией, например, путем окрашивания антителом, которое специфически связывается с маркером, и обнаружением указанного антитела, где окрашивание не обнаруживается проточной цитометрией на уровне, по существу превышающем окрашивание, определенное проведением той же процедуры с изотипически сходным контролем в идентичных в остальном условиях и/или на уровне, по существу более низком, чем уровень для клетки, заведомо положительной для маркера, и/или на уровне, по существу аналогичном по сравнению с клеткой, заведомо отрицательной для маркера.
Все публикации, включая патентные документы, научные статьи и базы данных, упомянутые в этой заявке, включены в качестве ссылки полностью для всех целей в той же степени, как если бы каждая отдельная публикация была индивидуально включена в качестве ссылки. Если определение, изложенное здесь, противоречит или иным образом не согласуется с определением, изложенным в патентах, заявках, опубликованных заявках и других публикациях, которые включены в настоящий документ в качестве ссылки, определение, изложенное здесь, преобладает над определением, которое включено в данный документ в качестве ссылки.
Заголовки разделов, используемые здесь, предназначены только для организационных целей и не должны толковаться как ограничивающие описываемый объект.
IX. ТИПОВЫЕ ВАРИАНТЫ
Представленные варианты осуществления включают:
1. Способ получения генетически сконструированной иммунной клетки, включающий:
(a) введение в иммунную клетку, одного или более агентов, где каждый из одного или более агентов независимо способны вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), тем самым вызывая генетическое разрушение, по меньшей мере, одного целевого сайта; и
(b) введение в иммунную клетку матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
2. Способ получения генетически сконструированной иммунной клетки, включающий:
введение в иммунную клетку, имеющую генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где генетическое разрушение вызвано одним или более агентами, где каждый из одного или более агентов независимо способны вызывать генетическое разрушение, и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого положения через репарацию, направляемую гомологией (HDR).
3. Способ по варианту осуществления 1 или варианту осуществления 2, где, по меньшей мере, один из одного или более агентов способен вызывать генетическое разрушение целевого сайта в гене TRAC.
4. Способ по любому из вариантов осуществления 1-3, где, по меньшей мере, один из одного или более агентов способен вызывать генетическое разрушение целевого сайта в гене TRBC.
5. Способ по любому из вариантов осуществления 1-4, где один или более агентов содержат, по меньшей мере, один агент, который способен вызывать генетическое разрушение целевого сайта в гене TRAC и, по меньшей мере, один агент, который способен вызывать генетическое разрушение целевого сайта в гене TRBC.
6. Способ по варианту осуществления 4 или варианту осуществления 5, где TRBC геном является один или оба из генов бета константы Т-клеточного рецептора 1 (TRBC1) или бета константы Т-клеточного рецептора 2 (TRBC2).
7. Способ получения генетически сконструированной иммунной клетки, включающий:
(a) введение в иммунную клетку, одного или более агентов, где каждый из одного или более агентов независимо способны вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и гене бета константы Т-клеточного рецептора (TRBC), тем самым вызывая генетическое разрушение целевых сайтов; и
(b) введение в иммунную клетку матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом через репарацию, направляемую гомологией (HDR).
8. Способ получения генетически сконструированной иммунной клетки, включающий:
введение в иммунную клетку, имеющую генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или его цепь, где генетическое разрушение вызвано одним или более агентами, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение, и трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
9. Способ по варианту осуществления 7 или варианту осуществления 8, где TRBC геном является один или оба из генов бета константы Т-клеточного рецептора 1 (TRBC1) или бета константы Т-клеточного рецептора 2 (TRBC2).
10. Способ по любому из вариантов осуществления 1-9, где один или более агентов, способных вызывать генетическое разрушение, содержит ДНК связывающий белок или ДНК связывающую нуклеиновую кислоту, которые специфически связываются с или гибридизируются в целевой сайт.
11. Способ по варианту осуществления 10, где один или более агентов, способных вызывать генетическое разрушение, содержит (a) слитый белок, содержащий ДНК-направленный белок и нуклеазу или (b) РНК-направляемую нуклеазу.
12. Способ по варианту осуществления 11, где ДНК-направленный белок или РНК-направляемая нуклеаза содержит белок «цинковый палец» (ZFP), TAL белок или связанную с кластеризованной регулярной промежуточной короткой падлиндромной нуклеиновой кислотой (CRISPR) нуклеазу (Cas), специфическую к целевому сайту.
13. Способ по любому из вариантов осуществления 10-12, где один или более агентов содержат нуклеазу «цинковый палец» (ZFN), TAL-эффекторную нуклеазу (TALEN) или и CRISPR-Cas9 сочетание, которые специфически связываются с, распознают или гибридизируют в целевой сайт.
14. Способ по любому из вариантов осуществления 1-13, где каждый из одного или более агентов содержат направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту.
15. Способ по варианту осуществления 14, где один или более агентов вводят в виде рибонуклеопротеинового (RNP) комплекса, содержащего нРНК и Cas9 белок.
16. Способ по варианту осуществления 15, где RNP вводят через электропорацию, генную пушку, трансфекцию фосфата кальция, сгущение или сжатие клеток.
17. Способ по варианту осуществления 15 или варианту осуществления 16, где RNP вводят через электропорацию.
18. Способ по любому из вариантов осуществления 1-13, где один или более агентов вводят в виде одного или более полинуклеотидов, кодирующих нРНК и/или Cas9 белка.
19. Способ по любому из вариантов осуществления 1-18, где по меньшей мере, один целевой сайт находится внутри экзона TRAC, TRBC1 и/или TRBC2 гена.
20. Способ по любому из вариантов осуществления 14-19, где нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в TRAC гене, и содержит последовательность, выбранную из UCUCUCAGCUGGUACACGGC (SEQ ID NO:28), UGGAUUUAGAGUCUCUCAGC (SEQ ID NO:29), ACACGGCAGGGUCAGGGUUC (SEQ ID NO:30), GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31), GCUGGUACACGGCAGGGUCA (SEQ ID NO:32), CUCAGCUGGUACACGGC (SEQ ID NO:33), UGGUACACGGCAGGGUC (SEQ ID NO:34), GCUAGACAUGAGGUCUA (SEQ ID NO:35), GUCAGAUUUGUUGCUCC (SEQ ID NO:36), UCAGCUGGUACACGGCA (SEQ ID NO:37), GCAGACAGACUUGUCAC (SEQ ID NO:38), GGUACACGGCAGGGUCA (SEQ ID NO:39), CUUCAAGAGCAACAGUGCUG (SEQ ID NO:40), AGAGCAACAGUGCUGUGGCC (SEQ ID NO:41), AAAGUCAGAUUUGUUGCUCC (SEQ ID NO:42), ACAAAACUGUGCUAGACAUG (SEQ ID NO:43), AAACUGUGCUAGACAUG (SEQ ID NO:44), UGUGCUAGACAUGAGGUCUA (SEQ ID NO:45), GGCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:46), GCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:47), GGGGAAGAAGGUGUCUUC (SEQ ID NO:48), GUUUUGUCUGUGAUAUACACAU (SEQ ID NO:49), GGCAGACAGACUUGUCACUGGAUU (SEQ ID NO:50), GCAGACAGACUUGUCACUGGAUU (SEQ ID NO:51), GACAGACUUGUCACUGGAUU (SEQ ID NO:52), GUGAAUAGGCAGACAGACUUGUCA (SEQ ID NO:53), GAAUAGGCAGACAGACUUGUCA (SEQ ID NO:54), GAGUCUCUCAGCUGGUACACGG (SEQ ID NO:55), GUCUCUCAGCUGGUACACGG (SEQ ID NO:56), GGUACACGGCAGGGUCAGGGUU (SEQ ID NO:57) и GUACACGGCAGGGUCAGGGUU (SEQ ID NO:58).
21. Способ по варианту осуществления 20, где нРНК имеет домен целенаправленного воздействия, содержащий последовательность GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31).
22. Способ по любому из вариантов осуществления 14-21, где нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в одном или более из TRBC1 и TRBC2 гене и содержит последовательность, выбранную из CACCCAGAUCGUCAGCGCCG (SEQ ID NO:59), CAAACACAGCGACCUCGGGU (SEQ ID NO:60), UGACGAGUGGACCCAGGAUA (SEQ ID NO:61), GGCUCUCGGAGAAUGACGAG (SEQ ID NO:62), GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63), GAAAAACGUGUUCCCACCCG (SEQ ID NO:64), AUGACGAGUGGACCCAGGAU (SEQ ID NO:65), AGUCCAGUUCUACGGGCUCU (SEQ ID NO:66), CGCUGUCAAGUCCAGUUCUA (SEQ ID NO:67), AUCGUCAGCGCCGAGGCCUG (SEQ ID NO:68), UCAAACACAGCGACCUCGGG (SEQ ID NO:69), CGUAGAACUGGACUUGACAG (SEQ ID NO:70), AGGCCUCGGCGCUGACGAUC (SEQ ID NO:71), UGACAGCGGAAGUGGUUGCG (SEQ ID NO:72), UUGACAGCGGAAGUGGUUGC (SEQ ID NO:73), UCUCCGAGAGCCCGUAGAAC (SEQ ID NO:74), CGGGUGGGAACACGUUUUUC (SEQ ID NO:75), GACAGGUUUGGCCCUAUCCU (SEQ ID NO:76), GAUCGUCAGCGCCGAGGCCU (SEQ ID NO:77), GGCUCAAACACAGCGACCUC (SEQ ID NO:78), UGAGGGUCUCGGCCACCUUC (SEQ ID NO:79), AGGCUUCUACCCCGACCACG (SEQ ID NO:80), CCGACCACGUGGAGCUGAGC (SEQ ID NO:81), UGACAGGUUUGGCCCUAUCC (SEQ ID NO:82), CUUGACAGCGGAAGUGGUUG (SEQ ID NO:83), AGAUCGUCAGCGCCGAGGCC (SEQ ID NO:84), GCGCUGACGAUCUGGGUGAC (SEQ ID NO:85), UGAGGGCGGGCUGCUCCUUG (SEQ ID NO:86), GUUGCGGGGGUUCUGCCAGA (SEQ ID NO:87), AGCUCAGCUCCACGUGGUCG (SEQ ID NO:88), GCGGCUGCUCAGGCAGUAUC (SEQ ID NO:89), GCGGGGGUUCUGCCAGAAGG (SEQ ID NO:90), UGGCUCAAACACAGCGACCU (SEQ ID NO:91), ACUGGACUUGACAGCGGAAG (SEQ ID NO:92), GACAGCGGAAGUGGUUGCGG (SEQ ID NO:93), GCUGUCAAGUCCAGUUCUAC (SEQ ID NO:94), GUAUCUGGAGUCAUUGAGGG (SEQ ID NO:95), CUCGGCGCUGACGAUCU (SEQ ID NO:96), CCUCGGCGCUGACGAUC (SEQ ID NO:97), CCGAGAGCCCGUAGAAC (SEQ ID NO:98), CCAGAUCGUCAGCGCCG (SEQ ID NO:99), GAAUGACGAGUGGACCC (SEQ ID NO:100), GGGUGACAGGUUUGGCCCUAUC (SEQ ID NO:101), GGUGACAGGUUUGGCCCUAUC (SEQ ID NO:102), GUGACAGGUUUGGCCCUAUC (SEQ ID NO:103), GACAGGUUUGGCCCUAUC (SEQ ID NO:104), GAUACUGCCUGAGCAGCCGCCU (SEQ ID NO:105), GACCACGUGGAGCUGAGCUGGUGG (SEQ ID NO:106), GUGGAGCUGAGCUGGUGG (SEQ ID NO:107), GGGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:108), GGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:109), GCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:110), GGGCUGCUCCUUGAGGGGCU (SEQ ID NO:111), GGCUGCUCCUUGAGGGGCU (SEQ ID NO:112), GCUGCUCCUUGAGGGGCU (SEQ ID NO:113), GGUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:114), GUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:115) и GAAUGGGAAGGAGGUGCACAG (SEQ ID NO:116).
23. Способ по варианту осуществления 22, где нРНК имеет домен целенаправленного воздействия, содержащий последовательность GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63).
24. Способ по любому из вариантов осуществления 1-23, где матричный полинуклеотид содержит структуру [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии].
25. Способ по варианту осуществления 24, где 5’ плечо гомологии и 3’ плечо гомологии содержит последовательности нуклеиновых кислот, гомологичные последовательностям нуклеиновых кислот окружающим, по меньшей мере, один целевой сайт.
26. Способ по варианту осуществления 24 или варианту осуществления 25, где 5’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот 5’ целевого сайта.
27. Способ по варианту осуществления 24 или варианту осуществления 25, где 3’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот 3’ целевого сайта.
28. Способ по любому из вариантов осуществления 24-27, где 5’ плечо гомологии и 3’ плечо гомологии независимо имеет, по меньшей мере, или по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований.
29. Способ по варианту осуществления 28, где 5’ плечо гомологии и 3’ плечо гомологии независимо имеет между около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 пар оснований.
30. Способ по варианту осуществления 29, где 5’ плечо гомологии и 3’ плечо гомологии независимо имеет около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований.
31. Способ по любому из вариантов осуществления 1-30, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене.
32. Способ по любому из вариантов осуществления 1-32, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в одном или обоих TRBC1 и TRBC2 генов.
33. Способ по любому из вариантов осуществления 1-32, дополнительно включающий введение в иммунную клетку одного или более вторых матричных полинуклеотидов, содержащих один или более вторых трансгенов, где второй трансген нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
34. Способ по варианту осуществления 33, где второй матричный полинуклеотид содержит структуру [второе 5’ плечо гомологии]-[один или более вторых трансгенов]-[второе 3’ плечо гомологии].
35. Способ по варианту осуществления 34, где второе 5’ плечо гомологии и второе 3’ плечо гомологии содержат последовательности нуклеиновых кислот гомологичные последовательностям нуклеиновых кислот окружающим, по меньшей мере, один целевой сайт.
36. Способ по варианту осуществления 34 или варианту осуществления 35, где второе 5’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот второго 5’ целевого сайта.
37. Способ по варианту осуществления 34 или варианту осуществления 35, где второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот второго 3’ целевого сайта.
38. Способ по любому из вариантов осуществления 34-37, где второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет, по меньшей мере или по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований.
39. Способ по варианту осуществления 38, где второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет между около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 пар оснований.
40. Способ по варианту осуществления 39, где второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 пар оснований.
41. Способ по любому из вариантов осуществления 33-40, где один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене.
42. Способ по любому из вариантов осуществления 33-41, где один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRBC1 или TRBC2 гене.
43. Способ по любому из вариантов осуществления 33-42, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, TRBC1 гене или TRBC2 гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами, на которые не нацелен трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь.
44. Способ по любому из вариантов осуществления 33-43, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами в TRBC1 гене и/или TRBC2 гене.
45. Способ по любому из вариантов осуществления 33-44, где один или более вторых трансгенов кодирует молекулу, выбранную из костимулирующего лиганда, цитокина, растворимого однонитевого переменного фрагмента (scFv), иммуномодулирующего слитого белка, химерного рецептора переключения (CSR) или корецептора.
46. Способ по варианту осуществления 45, где кодированной молекулой является костимулирующий лиганд, необязательно выбранный из лиганда фактора некроза опухоли (TNF), выбранного из 4-1BBL, OX40L, CD70, LIGHT и CD30L или лиганда суперсемейства иммуноглобулина (Ig), выбранного из CD80 и CD86.
47. Способ по варианту осуществления 45, где кодированной молекулой является цитокин, необязательно выбранный из IL-2, IL-3, IL-6, IL-11, IL-12, IL-7, IL-15, IL-21, гранулоцитарно-моноцитарного колониестимулирующего фактора (GM-CSF), интерферона альфа (IFN-α), интерферона бета (IFN-β) или интерферона гамма (IFN-γ) и эритропоэтина.
48. Способ по варианту осуществления 45, где кодированной молекулой является растворимый однонитевой переменный фрагмент (scFv), который необязательно связывает полипептид, который обладает иммунодепрессивным действием или иммуностимулирующим действием, выбранный из CD47, PD-1, CTLA-4 и их лигандов или CD28, OX-40, 4-1BB и их лигандов.
49. Способ по варианту осуществления 45, где кодированной молекулой является иммуномодулирующий слитый белок, необязательно содержащий (a) внеклеточный связывающий домен который специфически связывает антиген, полученный из CD200R, SIRPα, CD279 (PD-1), CD2, CD95 (Fas), CD152 (CTLA4), CD223 (LAG3), CD272 (BTLA), A2aR, KIR, TIM3, CD300 или LPA5; (b) внутриклеточный связывающий домен, полученный из CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD47, CD79A, CD79B, CD134 (OX40), CD137 (4-1BB), CD150 (SLAMF1), CD278 (ICOS), CD357 (GITR), ХАРD11, DAP10, DAP12, FcRα, FcRβ, FcRγ, Fyn, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, ROR2, Ryk, Slp76, pTα, TCRα, TCRβ, TRFM, Zap70, PTCH2 или любого их сочетания; и (c) гидрофобный трансмембранный домен, полученный из CD2, CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD79A, CD79B, CD80, CD86, CD95 (Fas), CD134 (OX40), CD137 (4-1BB), CD150 (SLAMF1), CD152 (CTLA4), CD200R, CD223 (LAG3), CD270 (HVEM), CD272 (BTLA), CD273 (PD-L2), CD274 (PD-L1), CD278 (ICOS), CD279 (PD-1), CD300, CD357 (GITR), A2aR, DAP10, FcRα, FcRβ, FcRγ, Fyn, GAL9, KIR, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, PTCH2, ROR2, Ryk, Slp76, SIRPα, pTα, TCRα, TCRβ, TIM3, TRIM, LPA5 или Zap70.
50. Способ по варианту осуществления 45, где кодированной молекулой является химерный рецептор переключения (CSR) который необязательно содержит усеченный внеклеточный домен PD1 и трансмембранный и цитоплазматический сигнальные домены CD28.
51. Способ по варианту осуществления 45, где кодированной молекулой является корецептор, необязательно выбранный из CD4 или CD8.
52. Способ по любому из вариантов осуществления 33-44, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует одну цепь рекомбинантного TCR и второй трансген кодирует другую цепь рекомбинантного TCR.
53. Способ по варианту осуществления 52, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует альфа (TCRα) цепь рекомбинантного TCR и второй трансген кодирует бета (TCRβ) цепь рекомбинантного TCR.
54. Способ по любому из вариантов осуществления 1-53, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо дополнительно содержат регулирующий или контрольный элемент.
55. Способ по варианту осуществления 54, где регулирующий или контрольный элемент содержит промотор, энхансер, интрон, сигнал полиаденилирования, консенсусную последовательность Козака, внутренний участок посадки рибосомы (IRES), последовательность, кодирующую последовательности проскока рибосомы, последовательность акцептора сплайсинга или последовательность донора сплайсинга.
56. Способ по варианту осуществления 55, где регулирующий или контрольный элемент содержит промотор.
57. Способ по варианту осуществления 56, где промотор выбирают из конститутивного промотора, индуцируемого промотора, репрессируемого промотора и/или ткань-специфического промотора.
58. Способ по варианту осуществления 56 или варианту осуществления 57, где промотор выбирают из РНК pol I, pol II или pol III промотора.
59. Способ по варианту осуществления 58, где промотор выбирают из:
pol III промотора, который является U6 или H1 промотором; или
pol II промотора, который является CMV, промотором SV40 ранней области или основной поздней области аденовируса.
60. Способ по любому из вариантов осуществления 56-58, где промотор является или содержит промотор человеческого фактора элонгации 1 альфа (EF1α) или MND промотор или их вариант.
61. Способ по любому из вариантов осуществления 56-58, где промотором является индуцируемый промотор или репрессируемый промотор.
62. Способ по варианту осуществления 61, где промотор содержит Lac последовательность оператора, тетрациклиновую последовательность оператора, галактозную последовательность оператора или доксициклиновую последовательность оператора или является их аналогом или способен связываться или распознаваться Lac репрессором или тетрациклиновым репрессором или их аналогом.
63. Способ по любому из вариантов осуществления 1-62, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо содержит один или более мультицистронных элементов.
64. Способ по варианту осуществления 63, где один или более мультицистронных элементов расположены против хода транскрипции трансгена, кодирующего рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или одного или более вторых трансгенов.
65. Способ по варианту осуществления 63 или варианту осуществления 64, где мультицистронные элементы расположены между трансгеном, кодирующим рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и одним или более вторыми трансгенами.
66. Способ по любому из вариантов осуществления 63-65, где мультицистронные элементы расположены между последовательностью нуклеиновых кислот, кодирующей TCRα или его часть, и последовательностью нуклеиновых кислот, кодирующей TCRβ или его часть.
67. Способ по варианту осуществления 66, где мультицистронные элементы содержат последовательность, кодирующую элемент проскока рибосомы, выбранную из T2A, P2A, E2A или F2A или внутреннего участка посадки рибосомы (IRES).
68. Способ по варианту осуществления 67, где последовательность, кодирующая элемент проскока рибосомы нацелена на то, чтобы быть внутри рамки с геном в целевом сайте.
69. Способ по любому из вариантов осуществления 1-53, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо функционально связан с эндогенным промотором гена на целевом сайте.
70. Способ по любому из вариантов осуществления 1-69, где рекомбинантный TCR способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния.
71. Способ по варианту осуществления 70, где заболеванием, расстройством или состоянием является инфекционное заболевание, аутоиммунное заболевание, воспалительное заболевание или опухоль или рак.
72. Способ по варианту осуществления 70 или варианту осуществления 71, где антигеном является опухолевый антиген или патогенный антиген.
73. Способ по варианту осуществления 72, где патогенным антигеном является бактериальный антиген или вирусный антиген.
74. Способ по варианту осуществления 73, где антигеном является вирусный антиген, и вирусным антигеном является вирус гепатита A, гепатита B, гепатита C (HCV), папилломавирус человека (HPV), вирусные инфекции гепатита, вирус Эпштейн-Барр (EBV), герпес-вирус человека 8 (HHV-8), вирус Т-клеточного лейкоза человека-1 (HTLV-1), вирус Т-клеточного лейкоза человека-2 (HTLV-2) или цитомегаловирус (CMV).
75. Способ по варианту осуществления 74, где антигеном является антиген из HPV, выбранного из HPV-16, HPV-18, HPV-31, HPV-33 и HPV-35.
76. Способ по варианту осуществления 75, где антигеном является HPV-16 антиген, которым является HPV-16 E6 или HPV-16 E7 антиген.
77. Способ по варианту осуществления 73, где вирусным антигеном является EBV антиген, выбранный из ядерного антигена Эпштейн-Барр (EBNA)-1, EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, EBNA-лидерного белка (EBNA-LP), латентных мембранных белков LMP-1, LMP-2A и LMP-2B, EBV-EA, EBV-MA и EBV-VCA.
78. Способ по варианту осуществления 73, где вирусным антигеном является HTLV-антиген, которым является TAX.
79. Способ по варианту осуществления 73, где вирусным антигеном является HBV антиген, которым является ядерный антиген гепатита В или оболочечный антиген гепатита В.
80. Способ по любому из вариантов осуществления 70-72, где антигеном является опухолевый антиген.
81. Способ по варианту осуществления 80, где антиген выбирают из глиома-ассоциированного антигена, β-человеческого хорионического гонадотропина, альфафетобелка (AFP), лектин-реактивного AFP, тироглобулина, RAGE-1, MN-CA IX, обратной транскриптазы теломеразы человека, RU1, RU2 (AS), кишечной карбоксилэстеразы, mut hsp70-2, M-CSF, Меланина-A/MART-1, WT-1, S-100, MBP, CD63, MUC1 (например, MUC1-8), p53, Ras, циклина B1, HER-2/neu, карциноэмбрионного антигена (CEA), gp100, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11, MAGE-A11, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-C1, BAGE, GAGE-1, GAGE-2, pl5, тирозиназы, родственного тирозиназе белка 1 (TRP-1), родственного тирозиназе белка 2 (TRP-2), β-катенина, NY-ESO-1, LAGE-1a, PP1, MDM2, MDM4, EGVFvIII, Tax, SSX2, теломеразы, TARP, pp65, CDK4, виментина, S100, eIF-4A1, IFN-индуцируемого p78, меланотрансферрина (p97), Уроплакина II, специфического антигена простаты (PSA), человеческого калликреина (huK2), специфического мембранного антигена простаты (PSM) и простатической кислой фосфатазы (PAP), нейтрофильной эластазы, эфрина B2, BA-46, Bcr-abl, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, Каспазы 8, FRa, CD24, CD44, CD133, CD 166, epCAM, CA-125, HE4, Oval, рецептора эстрогена, рецептора прогестерона, uPA, PAI-1, CD19, CD20, CD22, ROR1, CD33/IL3Ra, c-Met, PSMA, Гликолипида F77, GD-2, инсулинового фактора роста (IGF)-I, IGF-II, IGF-I рецептора и мезотелина.
82. Способ по любому из вариантов осуществления 1-81, где иммунной клеткой является Т клетка.
83. Способ по варианту осуществления 82, где Т клеткой является CD8+ T клетка или ее подтипы.
84. Способ по варианту осуществления 82, где Т клеткой является CD4+ T клетка или ее подтипы.
85. Способ по любому из вариантов осуществления 1-81, где иммунную клетку получают из мультипотентной или плюрипотентной клетки, которой необязательно является иПСК.
86. Способ по любому из вариантов осуществления 1-85, где иммунная клетка содержит Т клетку, которая является аутологической к субъекту.
87. Способ по любому из вариантов осуществления 1-85, где иммунная клетка содержит Т клетку, которая является аутогенной к субъекту.
88. Способ по любому из вариантов осуществления 1-87, где первый матричный полинуклеотид, один или более вторых матричных полинуклеотидов и/или один или более полинуклеотидов, кодирующих нРНК и/или a Cas9 белок, содержатся в одном или более векторах, которые необязательно являются вирусными векторами.
89. Способ по варианту осуществления 88, где вектором является AAV вектор.
90. Способ по варианту осуществления 89, где AAV вектор выбирают из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 или AAV8 вектора.
91. Способ по варианту осуществления 90, где AAV вектором является AAV2 или AAV6 вектор.
92. Способ по варианту осуществления 88, где вирусным вектором является ретровирусный вектор.
93. Способ по варианту осуществления 92, где вирусным вектором является лентивирусный вектор.
94. Способ по любому из вариантов осуществления 1-93, где введение одного или более агентов, способных вызывать генетическое разрушение, и введение матричного полинуклеотида проводят одновременно или последовательно, в любом порядке.
95. Способ по любому из вариантов осуществления 1-94, где введение матричного полинуклеотида проводят после введения одного или более агентов, способных вызывать генетическое разрушение.
96. Способ по варианту осуществления 95, где матричный полинуклеотид вводят сразу после или в течение около 30 секунд, 1 минуты, 2 минут, 3 минут, 4 минут, 5 минут, 6 минут, 6 минут, 8 минут, 9 минут, 10 минут, 15 минут, 20 минут, 30 минут, 40 минут, 50 минут, 60 минут, 90 минут, 2 часов, 3 часов или 4 часов после введения одного или более агентов, способных вызывать генетическое разрушение.
97. Способ по любому из вариантов осуществления 33-96, где введение матричного полинуклеотида и введение одного или более вторых матричных полинуклеотидов проводят одновременно или последовательно, в любом порядке.
98. Способ по любому из вариантов осуществления 1-97, где введение одного или более агентов, способных вызывать генетическое разрушение, и введение матричного полинуклеотида проводят в одной экспериментальной реакции.
99. Способ по любому из вариантов осуществления 33-98, где введение одного или более агентов, способных вызывать генетическое разрушение, и введение матричного полинуклеотида и вторых матричных полинуклеотидов проводят в одной экспериментальной реакции.
100. Способ по любому из вариантов осуществления 1-99, где, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток во множестве сконструированных клеток содержат генетическое разрушение, по меньшей мере, одного целевого сайта в гене, кодирующем домен или область гена альфа константы Т-клеточного рецептора (TRAC) и/или гена бета константы Т-клеточного рецептора (TRBC).
101. Способ по любому из вариантов осуществления 1-99, где, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток во множестве сконструированных клеток экспрессируют рекомбинантный рецептор или его антигенсвязывающий фрагмент и/или демонстрируют антигенное связывание.
102. Способ по любому из вариантов осуществления 1-99, где коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора или его антигенсвязывающего фрагмента среди множества сконструированных клеток ниже, чем 0,70, 0,65, 0,60, 0,55, 0,50, 0,45, 0,40, 0,35 или 0,30 или менее.
103. Способ по любому из вариантов осуществления 1-99, где коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора или его антигенсвязывающего фрагмента среди множества сконструированных клеток составляет на, по меньшей мере, 100%, 95%, 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20% или 10% ниже чем коэффициент вариации экспрессии и/или антигенного связывания того же рекомбинантного рецептора, который интегрирован в геном случайной интеграцией.
104. Способ по любому из вариантов осуществления 100-103, где экспрессия и/или антигенное связывание рекомбинантного рецептора или его антигенсвязывающего фрагмента оценивают через контакт клеток в композиции со связывающим реагентом, специфическим к TCRα цепи или TCRβ цепи и оценкой связывания реагента с клетками.
105. Способ по варианту осуществления 104, где связывающим реагентом является анти-TCR Vβ антитело или является анти-TCR Vα антитело, которое специфически распознает конкретное семейство Vβ или Vα цепей.
106. Способ по варианту осуществления 104, где связывающим агентом является пептидный антиген-МНС комплекс, который необязательно является тетрамером.
107. Сконструированная клетка или множество сконструированных клеток, созданное с применением способа по любому из вариантов осуществления 1-106.
108. Композиция, содержащая сконструированные клетки или множество сконструированных клеток по варианту осуществления 107.
109. Композиция по варианту осуществления 108, где, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции содержат a генетическое разрушение, по меньшей мере, одного целевого сайта в гене, кодирующем домен или область гена альфа константы Т-клеточного рецептора (TRAC) и/или гена бета константы Т-клеточного рецептора (TRBC).
110. Композиция по варианту осуществления 108, где, по меньшей мере, или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции экспрессируют рекомбинантный рецептор или его антигенсвязывающий фрагмент и/или демонстрируют антигенное связывание.
111. Композиция по варианту осуществления 108, где коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора или его антигенсвязывающего фрагмента среди множества клеток ниже, чем 0,70, 0,65, 0,60, 0,55, 0,50, 0,45, 0,40, 0,35 или 0,30 или менее.
112. Композиция по варианту осуществления 108, где коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора или его антигенсвязывающего фрагмента среди множества клеток на, по меньшей мере, 100%, 95%, 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20% или 10% ниже, чем коэффициент вариации экспрессии и/или антигенного связывания того же рекомбинантного рецептора, который интегрирован в геном случайной интеграцией.
113. Композиция по любому из вариантов осуществления 109-112, где экспрессию и/или антигенное связывание рекомбинантного рецептора или его антигенсвязывающего фрагмента оценивают через контакт клеток в композиции со связывающим реагентом, специфическим к TCRα цепи или TCRβ цепи, и оценкой связывания реагента с клетками.
114. Композиция по варианту осуществления 113, где связывающим реагентом является анти-TCR Vβ антитело или является анти-TCR Vα антитело, которое специфически распознает конкретное семейство Vβ или Vα цепей.
115. Композиция по варианту осуществления 113, где связывающим агентом является пептидный антиген-МНС комплекс, который необязательно является тетрамером.
116. Композиция по любому из вариантов осуществления 108-115, дополнительно содержащая фармацевтически приемлемый носитель.
117. Композиция, содержащая множество сконструированных Т клеток, содержащих рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, кодированные трансгеном, и генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), где, по меньшей мере или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции содержат генетическое разрушение, по меньшей мере, одного целевого сайта в TRAC гене и/или TRBC гене; и
трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацеленный на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
118. Композиция, содержащая множество сконструированных Т клеток, содержащих рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, кодированные трансгеном, и генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), где, по меньшей мере или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток в композиции экспрессируют рекомбинантный рецептор или его антигенсвязывающий фрагмент и/или демонстрируют антигенное связывание; и
трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацеленный на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
119. Композиция, содержащая множество сконструированных Т клеток, содержащих рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, кодированные трансгеном, и генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), где коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора среди множества клеток ниже чем 0,70, 0,65, 0,60, 0,55, 0,50, 0,45, 0,40, 0,35 или 0,30 или менее.
120. Композиция, содержащая множество сконструированных Т клеток, содержащих рекомбинантный рецептор или его антигенсвязывающий фрагмент или цепь, кодированные трансгеном, и генетическое разрушение, по меньшей мере, одного целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), где коэффициент вариации экспрессии и/или антигенного связывания рекомбинантного рецептора среди множества клеток на, по меньшей мере, 100%, 95%, 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20% или 10% ниже, чем коэффициент вариации экспрессии и/или антигенного связывания того же рекомбинантного рецептора, который интегрирован в геном случайной интеграцией.
121. Композиция по любому из вариантов осуществления 117-120, где композицию получают:
(a) введением во множество T клеток одного или более агентов, где каждый из одного или более агентов независимо способны вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC), тем самым вызывая генетическое разрушение, по меньшей мере, одного целевого сайта; и
(b) введением во множество T клеток матричного полинуклеотида, содержащего трансген, кодирующий рекомбинантный Т-клеточный рецептор (TCR) или его антигенсвязывающий фрагмент или цепь, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
122. Композиция по любому из вариантов осуществления 117-121, где экспрессию и/или антигенное связывание рекомбинантного рецептора или его антигенсвязывающего фрагмента оценивают через контакт клеток в композиции со связывающим реагентом, специфическим к TCRα цепи или TCRβ цепи и оценкой связывания реагента с клетками.
123. Композиция по варианту осуществления 122, где связывающим реагентом является анти-TCR Vβ антитело или является анти-TCR Vα антитело, которое специфически распознает конкретное семейство Vβ или Vα цепей.
124. Композиция по варианту осуществления 122, где связывающим агентом является пептидный антиген-МНС комплекс, который необязательно является тетрамером.
125. Композиция по любому из вариантов осуществления 121-124, где по меньшей мере, один из одного или более агентов способен вызывать генетическое разрушение целевого сайта в гене TRAC.
126. Композиция по любому из вариантов осуществления 121-125, где по меньшей мере, один из одного или более агентов способен вызывать генетическое разрушение целевого сайта в гене TRBC.
127. Композиция по любому из вариантов осуществления 121-126, где один или более агентов содержат, по меньшей мере, один агент, который способен вызывать генетическое разрушение целевого сайта в гене TRAC и, по меньшей мере, один агент, который способен вызывать генетическое разрушение целевого сайта в гене TRBC.
128. Композиция по любому из вариантов осуществления 117-127, где TRBC геном является один или оба из генов бета константы Т-клеточного рецептора 1 (TRBC1) или бета константы Т-клеточного рецептора 2 (TRBC2).
129. Композиция по любому из вариантов осуществления 121-128, где один или более агентов, способных вызывать генетическое разрушение, содержат ДНК связывающий белок или ДНК связывающую нуклеиновую кислоту, которые специфически связываются с или гибридизируются в целевой сайт.
130. Композиция по варианту осуществления 129, где один или более агентов, способных вызывать генетическое разрушение содержат (a) слитый белок, содержащий ДНК-направленный белок и нуклеазу или (b) РНК-направляемую нуклеазу.
131. Композиция по варианту осуществления 130, где ДНК-направленный белок или РНК-направляемая нуклеаза содержит белок «цинковый палец» (ZFP), TAL белок или связанную с кластеризованной регулярной промежуточной короткой падлиндромной нуклеиновой кислотой (CRISPR) нуклеазу (Cas), специфическую к целевому сайту.
132. Композиция по любому из вариантов осуществления 129-131, где один или более агентов содержат нуклеазу «цинковый палец» (ZFN), TAL-эффекторную нуклеазу (TALEN) и/или CRISPR-Cas9 сочетание которые специфически связываются с, распознают или гибридизируют в целевой сайт.
133. Композиция по любому из вариантов осуществления 121-132, где каждый из одного или более агентов содержит направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту.
134. Композиция по варианту осуществления 133, где один или более агентов вводят в виде рибонуклеопротеинового (RNP) комплекса, содержащего нРНК и Cas9 белок.
135. Композиция по варианту осуществления 134, где RNP вводят через электропорацию, генную пушку, трансфекцию фосфата кальция, сгущение или сжатие клеток.
136. Композиция по варианту осуществления 134 или варианту осуществления 135, где RNP вводят через электропорацию.
137. Композиция по любому из вариантов осуществления 121-132, где один или более агентов вводят в виде одного или более полинуклеотидов, кодирующих нРНК и/или Cas9 белок.
138. Композиция по любому из вариантов осуществления 121-137, где по меньшей мере, один целевой сайт находится внутри экзона TRAC, TRBC1 и/или TRBC2 гена.
139. Композиция по любому из вариантов осуществления 133-138, где нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в TRAC гене и содержит последовательность, выбранную из UCUCUCAGCUGGUACACGGC (SEQ ID NO:28), UGGAUUUAGAGUCUCUCAGC (SEQ ID NO:29), ACACGGCAGGGUCAGGGUUC (SEQ ID NO:30), GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31), GCUGGUACACGGCAGGGUCA (SEQ ID NO:32), CUCAGCUGGUACACGGC (SEQ ID NO:33), UGGUACACGGCAGGGUC (SEQ ID NO:34), GCUAGACAUGAGGUCUA (SEQ ID NO:35), GUCAGAUUUGUUGCUCC (SEQ ID NO:36), UCAGCUGGUACACGGCA (SEQ ID NO:37), GCAGACAGACUUGUCAC (SEQ ID NO:38), GGUACACGGCAGGGUCA (SEQ ID NO:39), CUUCAAGAGCAACAGUGCUG (SEQ ID NO:40), AGAGCAACAGUGCUGUGGCC (SEQ ID NO:41), AAAGUCAGAUUUGUUGCUCC (SEQ ID NO:42), ACAAAACUGUGCUAGACAUG (SEQ ID NO:43), AAACUGUGCUAGACAUG (SEQ ID NO:44), UGUGCUAGACAUGAGGUCUA (SEQ ID NO:45), GGCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:46), GCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:47), GGGGAAGAAGGUGUCUUC (SEQ ID NO:48), GUUUUGUCUGUGAUAUACACAU (SEQ ID NO:49), GGCAGACAGACUUGUCACUGGAUU (SEQ ID NO:50), GCAGACAGACUUGUCACUGGAUU (SEQ ID NO:51), GACAGACUUGUCACUGGAUU (SEQ ID NO:52), GUGAAUAGGCAGACAGACUUGUCA (SEQ ID NO:53), GAAUAGGCAGACAGACUUGUCA (SEQ ID NO:54), GAGUCUCUCAGCUGGUACACGG (SEQ ID NO:55), GUCUCUCAGCUGGUACACGG (SEQ ID NO:56), GGUACACGGCAGGGUCAGGGUU (SEQ ID NO:57) и GUACACGGCAGGGUCAGGGUU (SEQ ID NO:58).
140. Композиция по варианту осуществления 139, где нРНК имеет домен целенаправленного воздействия, содержащий последовательность GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31).
141. Композиция по любому из вариантов осуществления 133-140, где нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в одном или более из TRBC1 и TRBC2 гена и содержит последовательность, выбранную из CACCCAGAUCGUCAGCGCCG (SEQ ID NO:59), CAAACACAGCGACCUCGGGU (SEQ ID NO:60), UGACGAGUGGACCCAGGAUA (SEQ ID NO:61), GGCUCUCGGAGAAUGACGAG (SEQ ID NO:62), GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63), GAAAAACGUGUUCCCACCCG (SEQ ID NO:64), AUGACGAGUGGACCCAGGAU (SEQ ID NO:65), AGUCCAGUUCUACGGGCUCU (SEQ ID NO:66), CGCUGUCAAGUCCAGUUCUA (SEQ ID NO:67), AUCGUCAGCGCCGAGGCCUG (SEQ ID NO:68), UCAAACACAGCGACCUCGGG (SEQ ID NO:69), CGUAGAACUGGACUUGACAG (SEQ ID NO:70), AGGCCUCGGCGCUGACGAUC (SEQ ID NO:71), UGACAGCGGAAGUGGUUGCG (SEQ ID NO:72), UUGACAGCGGAAGUGGUUGC (SEQ ID NO:73), UCUCCGAGAGCCCGUAGAAC (SEQ ID NO:74), CGGGUGGGAACACGUUUUUC (SEQ ID NO:75), GACAGGUUUGGCCCUAUCCU (SEQ ID NO:76), GAUCGUCAGCGCCGAGGCCU (SEQ ID NO:77), GGCUCAAACACAGCGACCUC (SEQ ID NO:78), UGAGGGUCUCGGCCACCUUC (SEQ ID NO:79), AGGCUUCUACCCCGACCACG (SEQ ID NO:80), CCGACCACGUGGAGCUGAGC (SEQ ID NO:81), UGACAGGUUUGGCCCUAUCC (SEQ ID NO:82), CUUGACAGCGGAAGUGGUUG (SEQ ID NO:83), AGAUCGUCAGCGCCGAGGCC (SEQ ID NO:84), GCGCUGACGAUCUGGGUGAC (SEQ ID NO:85), UGAGGGCGGGCUGCUCCUUG (SEQ ID NO:86), GUUGCGGGGGUUCUGCCAGA (SEQ ID NO:87), AGCUCAGCUCCACGUGGUCG (SEQ ID NO:88), GCGGCUGCUCAGGCAGUAUC (SEQ ID NO:89), GCGGGGGUUCUGCCAGAAGG (SEQ ID NO:90), UGGCUCAAACACAGCGACCU (SEQ ID NO:91), ACUGGACUUGACAGCGGAAG (SEQ ID NO:92), GACAGCGGAAGUGGUUGCGG (SEQ ID NO:93), GCUGUCAAGUCCAGUUCUAC (SEQ ID NO:94), GUAUCUGGAGUCAUUGAGGG (SEQ ID NO:95), CUCGGCGCUGACGAUCU (SEQ ID NO:96), CCUCGGCGCUGACGAUC (SEQ ID NO:97), CCGAGAGCCCGUAGAAC (SEQ ID NO:98), CCAGAUCGUCAGCGCCG (SEQ ID NO:99), GAAUGACGAGUGGACCC (SEQ ID NO:100), GGGUGACAGGUUUGGCCCUAUC (SEQ ID NO:101), GGUGACAGGUUUGGCCCUAUC (SEQ ID NO:102), GUGACAGGUUUGGCCCUAUC (SEQ ID NO:103), GACAGGUUUGGCCCUAUC (SEQ ID NO:104), GAUACUGCCUGAGCAGCCGCCU (SEQ ID NO:105), GACCACGUGGAGCUGAGCUGGUGG (SEQ ID NO:106), GUGGAGCUGAGCUGGUGG (SEQ ID NO:107), GGGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:108), GGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:109), GCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:110), GGGCUGCUCCUUGAGGGGCU (SEQ ID NO:111), GGCUGCUCCUUGAGGGGCU (SEQ ID NO:112), GCUGCUCCUUGAGGGGCU (SEQ ID NO:113), GGUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:114), GUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:115) и GAAUGGGAAGGAGGUGCACAG (SEQ ID NO:116).
142. Композиция по варианту осуществления 141, где нРНК имеет домен целенаправленного воздействия, содержащий последовательность GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63).
143. Композиция по любому из вариантов осуществления 121-142, где матричный полинуклеотид содержит структуру [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии].
144. Композиция по варианту осуществления 143, где 5’ плечо гомологии и 3’ плечо гомологии содержит последовательности нуклеиновых кислот гомологичные последовательностям нуклеиновых кислот окружающим, по меньшей мере, один целевой сайт.
145. Композиция по варианту осуществления 143 или варианту осуществления 144, где 5’ плечо гомологии содержит последовательности нуклеиновых кислот которые гомологичны последовательностям нуклеиновых кислот 5’ целевого сайта.
146. Композиция по варианту осуществления 143 или варианту осуществления 144, где 3’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот 3’ целевого сайта.
147. Композиция по любому из вариантов осуществления 143-146, где 5’ плечо гомологии и 3’ плечо гомологии независимо имеет, по меньшей мере, или по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов.
148. Композиция по варианту осуществления 147, где 5’ плечо гомологии и 3’ плечо гомологии независимо имеет между около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 нуклеотидов.
149. Композиция по варианту осуществления 148, где 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от или от около 100 до 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов.
150. Композиция по любому из вариантов осуществления 117-149, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене.
151. Композиция по любому из вариантов осуществления 117-150, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в одном или обоих TRBC1 и TRBC2 гене.
152. Композиция по любому из вариантов осуществления 117-151, где композицию получают дополнительным введением в иммунную клетку одного или более вторых матричных полинуклеотидов, содержащих один или более вторых трансгенов, где второй трансген нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
153. Композиция по варианту осуществления 152, где второй матричный полинуклеотид содержит структуру [второе 5’ плечо гомологии]-[один или более вторы трансгенов]-[второе 3’ плечо гомологии].
154. Композиция по варианту осуществления 153, где второе 5’ плечо гомологии и второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот, гомологичные последовательностям нуклеиновых кислот окружающим, по меньшей мере, один целевой сайт.
55. Композиция по варианту осуществления 153 или варианту осуществления 154, где второе 5’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот второго 5’ целевого сайта.
156. Композиция по варианту осуществления 153 или варианту осуществления 154, где второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот второго 3’ целевого сайта.
157. Композиция по любому из вариантов осуществления 153-156, где второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет, по меньшей мере, или по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов.
158. Композиция по варианту осуществления 157, где второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет между около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 нуклеотидов.
159. Композиция по варианту осуществления 158, где второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет от или от около 100 до 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов.
160. Композиция по любому из вариантов осуществления 152-159, где один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене.
161. Композиция по любому из вариантов осуществления 152-160, где один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRBC1 или TRBC2 гене.
162. Композиция по любому из вариантов осуществления 152-161, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, TRBC1 гене или TRBC2 гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами на которые не нацелен трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь.
163. Композиция по любому из вариантов осуществления 152-162, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами в TRBC1 гене и/или TRBC2 гене.
164. Композиция по любому из вариантов осуществления 152-163, где один или более вторых трансгенов кодирует молекулу, выбранную из костимулирующего лиганда, цитокина, растворимого однонитевого переменного фрагмента (scFv), иммуномодулирующего слитого белка, химерного рецептора переключения (CSR) или корецептора.
165. Композиция по варианту осуществления 164, где кодированной молекулой является костимулирующий лиганд, необязательно выбранный из лиганда фактора некроза опухоли (TNF), выбранного из 4-1BBL, OX40L, CD70, LIGHT и CD30L, или лиганда суперсемейства иммуноглобулина (Ig), выбранного из CD80 и CD86.
166. Композиция по варианту осуществления 164, где кодированной молекулой является цитокин, необязательно выбранный из IL-2, IL-3, IL-6, IL-11, IL-12, IL-7, IL-15, IL-21, гранулоцитарно-моноцитарного колониестимулирующего фактора (GM-CSF), интерферона альфа (IFN-α), интерферона бета (IFN-β) или интерферона гамма (IFN-γ) и эритропоэтина.
167. Композиция по варианту осуществления 164, где кодированной молекулой является растворимый однонитевой переменный фрагмент (scFv), который необязательно связывает полипептид, который обладает иммунодепрессивным действием или иммуностимулирующим действием, выбранный из CD47, PD-1, CTLA-4 и их лигандов или CD28, OX-40, 4-1BB и их лигандов.
168. Композиция по варианту осуществления 164, где кодированной молекулой является иммуномодулирующий слитый белок, необязательно содержащий:
(a) внеклеточный связывающий домен, который специфически связывает антиген, полученный из CD200R, SIRPα, CD279 (PD-1), CD2, CD95 (Fas), CD152 (CTLA4), CD223 (LAG3), CD272 (BTLA), A2aR, KIR, TIM3, CD300 или LPA5;
(b) внутриклеточный связывающий домен, полученный из CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD47, CD79A, CD79B, CD134 (OX40), CD137 (4-1BB), CD150 (SLAMF1), CD278 (ICOS), CD357 (GITR), ХАРD11, DAP10, DAP12, FcRα, FcRβ, FcRγ, Fyn, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, ROR2, Ryk, Slp76, pTα, TCRα, TCRβ, TRFM, Zap70, PTCH2 или любого их сочетания; и
(c) гидрофобный трансмембранный домен, полученный из CD2, CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD79A, CD79B, CD80, CD86, CD95 (Fas), CD134 (OX40), CD137 (4-1BB), CD150 (SLAMF1), CD152 (CTLA4), CD200R, CD223 (LAG3), CD270 (HVEM), CD272 (BTLA), CD273 (PD-L2), CD274 (PD-L1), CD278 (ICOS), CD279 (PD-1), CD300, CD357 (GITR), A2aR, DAP10, FcRα, FcRβ, FcRγ, Fyn, GAL9, KIR, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, PTCH2, ROR2, Ryk, Slp76, SIRPα, pTα, TCRα, TCRβ, TIM3, TRIM, LPA5 или Zap70.
169. Композиция по варианту осуществления 164, где кодированной молекулой является химерный рецептор переключения (CSR), который необязательно содержит усеченный внеклеточный домен PD1 и трансмембранный и цитоплазматический сигнальные домены CD28.
170. Композиция по варианту осуществления 164, где кодированной молекулой является корецептор, необязательно выбранный из CD4 или CD8.
171. Композиция по любому из вариантов осуществления 152-163, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует одну цепь рекомбинантного TCR и второй трансген кодирует другую цепь рекомбинантного TCR.
172. Композиция по варианту осуществления 171, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует альфа (TCRα) цепь рекомбинантного TCR и второй трансген кодирует бета (TCRβ) цепь рекомбинантного TCR.
173. Композиция по любому из вариантов осуществления 117-172, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо дополнительно содержат регулирующий или контрольный элемент.
174. Композиция по варианту осуществления 173, где регулирующий или контрольный элемент содержит промотор, энхансер, интрон, сигнал полиаденилирования, консенсусную последовательность Козака, последовательность акцептора сплайсинга или последовательность донора сплайсинга.
175. Композиция по варианту осуществления 174, где регулирующий или контрольный элемент содержит промотор.
176. Композиция по варианту осуществления 175, где промотор выбирают из конститутивного промотора, индуцируемого промотора, репрессируемого промотора и/или ткань-специфического промотора.
177. Композиция по варианту осуществления 175 или варианту осуществления 176, где промотор выбирают из РНК pol I, pol II или pol III промотора.
178. Композиция по варианту осуществления 177, где промотор выбирают из:
pol III промотора, которым является U6 или H1 промотор; или
pol II промотора, которым является CMV, промотор SV40 ранней области или основной поздней области аденовируса.
179. Композиция по любому из вариантов осуществления 175-177, где промотор является или содержит промотор человеческого фактора элонгации 1 альфа (EF1α) или MND промотор или их вариант.
180. Композиция по любому из вариантов осуществления 175-177, где промотором является индуцируемый промотор или репрессируемый промотор.
181. Композиция по варианту осуществления 180, где промотор содержит Lac последовательность оператора, тетрациклиновую последовательность оператора, галактозную последовательность оператора или доксициклиновую последовательность оператора или является их аналогом или способен связываться или распознаваться Lac репрессором или тетрациклиновым репрессором или их аналогом.
182. Композиция по любому из вариантов осуществления 108-181, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо содержит один или более мультицистронных элементов.
183. Композиция по варианту осуществления 182, где один или более мультицистронных элементов расположены против хода транскрипции трансгена, кодирующего рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или одного или более вторых трансгенов.
184. Композиция по варианту осуществления 182 или варианту осуществления 183, где мультицистронные элементы расположены между трансгеном, кодирующим рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и одним или более вторыми трансгенами.
185. Композиция по любому из вариантов осуществления 182-184, где мультицистронные элементы расположены между последовательностью нуклеиновых кислот, кодирующей TCRα или его часть, и последовательностью нуклеиновых кислот, кодирующей TCRβ или его часть.
186. Композиция по любому из вариантов осуществления 182-185, где мультицистронные элементы содержат последовательность, кодирующую элемент проскока рибосомы, выбранный из T2A, P2A, E2A или F2A, или внутренний участок посадки рибосомы (IRES).
187. Композиция по варианту осуществления 186, где последовательность, кодирующая элемент проскока рибосомы нацелена на то, чтобы быть внутри рамки с геном в целевом сайте.
188. Композиция по любому из вариантов осуществления 117-172, где при HDR, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов, независимо функционально связан с эндогенным промотором гена на целевом сайте.
189. Композиция по любому из вариантов осуществления 117-188, где рекомбинантный TCR способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния.
190. Композиция по варианту осуществления 189, где заболеванием, расстройством или состоянием является инфекционное заболевание, аутоиммунное заболевание, воспалительное заболевание или опухоль или рак.
191. Композиция по варианту осуществления 189 или варианту осуществления 190, где антигеном является опухолевый антиген или патогенный антиген.
192. Композиция по варианту осуществления 191, где патогенным антигеном является бактериальный антиген или вирусный антиген.
193. Композиция по варианту осуществления 192, где антигеном является вирусный антиген, и вирусным антигеном является вирус гепатита A, гепатита B, гепатита C (HCV), папилломавирус человека (HPV), вирусные инфекции гепатита, вирус Эпштейн-Барр (EBV), герпес-вирус человека 8 (HHV-8), вирус Т-клеточного лейкоза человека-1 (HTLV-1), вирус Т-клеточного лейкоза человека-2 (HTLV-2) или цитомегаловирус (CMV).
194. Композиция по варианту осуществления 193, где антигеном является антиген из HPV, выбранный из HPV-16, HPV-18, HPV-31, HPV-33 и HPV-35.
195. Композиция по варианту осуществления 194, где антигеном является HPV-16 антиген, который является HPV-16 E6 или HPV-16 E7 антигеном.
196. Композиция по варианту осуществления 192, где вирусным антигеном является EBV антиген, выбранный из ядерного антигена Эпштейн-Барр (EBNA)-1, EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, EBNA-лидерного белка (EBNA-LP), латентных мембранных белков LMP-1, LMP-2A и LMP-2B, EBV-EA, EBV-MA и EBV-VCA.
197. Композиция по варианту осуществления 192, где вирусным антигеном является HTLV-антиген, которым является TAX.
198. Композиция по варианту осуществления 192, где вирусным антигеном является HBV антиген, которым является ядерный антиген гепатита В или оболочечный антиген гепатита В.
199. Композиция по любому из вариантов осуществления 189-191, где антигеном является опухолевый антиген.
200. Композиция по варианту осуществления 199, где антиген выбирают из глиома-ассоциированного антигена, β-человеческого хорионического гонадотропина, альфафетобелка (AFP), лектин-реактивного AFP, тироглобулина, RAGE-1, MN-CA IX, обратной транскриптазы теломеразы человека, RU1, RU2 (AS), кишечной карбоксилэстеразы, mut hsp70-2, M-CSF, Меланин-A/MART-1, WT-1, S-100, MBP, CD63, MUC1 (например, MUC1-8), p53, Ras, циклина B1, HER-2/neu, карциноэмбрионного антигена (CEA), gp100, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11, MAGE-A11, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-C1, BAGE, GAGE-1, GAGE-2, pl5, тирозиназы, родственного тирозиназе белка 1 (TRP-1), родственного тирозиназе белка 2 (TRP-2), β-катенина, NY-ESO-1, LAGE-1a, PP1, MDM2, MDM4, EGVFvIII, Tax, SSX2, теломеразы, TARP, pp65, CDK4, виментина, S100, eIF-4A1, IFN-индуцируемого p78, меланотрансферрина (p97), Уроплакина II, специфического антигена простаты (PSA), человеческого калликреина (huK2), специфического мембранного антигена простаты (PSM), и простатической кислой фосфатазы (PAP), нейтрофильной эластазы, эфрина B2, BA-46, Bcr-abl, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, Каспазы 8, FRa, CD24, CD44, CD133, CD 166, epCAM, CA-125, HE4, Oval, рецептора эстрогена, рецептора прогестерона, uPA, PAI-1, CD19, CD20, CD22, ROR1, CD33/IL3Ra, c-Met, PSMA, Гликолипида F77, GD-2, инсулинового фактора роста (IGF)-I, IGF-II, IGF-I рецептор и мезотелина.
201. Композиция по любому из вариантов осуществления 117-200, где Т клеткой является CD8+ T клетка или ее подтипы.
202. Композиция по любому из вариантов осуществления 117-200, где Т клеткой является CD4+ T клетка или ее подтипы.
203. Композиция по любому из вариантов осуществления 117-202, где Т клетка является аутологичной к субъекту.
204. Композиция по любому из вариантов осуществления 117-202, где Т клетка является аллогенной к субъекту.
205. Композиция по любому из вариантов осуществления 117-204, где первый матричный полинуклеотид, один или более вторых матричных полинуклеотидов и/или один или более полинуклеотидов, кодирующих нРНК и/или Cas9 белок содержатся в одном или более векторах, которые необязательно являются вирусными векторами.
206. Композиция по варианту осуществления 205, где вектором является AAV вектор.
207. Композиция по варианту осуществления 206, где AAV вектор выбирают из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 или AAV8 вектора.
208. Композиция по варианту осуществления 207, где AAV вектором является AAV2 или AAV6 вектор.
209. Композиция по варианту осуществления 205, где вирусным вектором является ретровирусный вектор.
210. Композиция по варианту осуществления 209, где вирусным вектором является лентивирусный вектор.
211. Композиция по любому из вариантов осуществления 117-210, где введение одного или более агентов, способных вызывать генетическое разрушение, и введение матричного полинуклеотида проводят одновременно или последовательно, в любом порядке.
212. Композиция по любому из вариантов осуществления 117-211, где введение матричного полинуклеотида проводят после введения одного или более агентов, способных вызывать генетическое разрушение.
213. Композиция по варианту осуществления 212, где матричный полинуклеотид вводят сразу после или в течение около 30 секунд, 1 минуты, 2 минут, 3 минут, 4 минут, 5 минут, 6 минут, 6 минут, 8 минут, 9 минут, 10 минут, 15 минут, 20 минут, 30 минут, 40 минут, 50 минут, 60 минут, 90 минут, 2 часов, 3 часов или 4 часов после введения одного или более агентов, способных вызывать генетическое разрушение.
214. Композиция по любому из вариантов осуществления 152-213, где введение матричного полинуклеотида и введение одного или более вторых матричных полинуклеотидов проводят одновременно или последовательно, в любом порядке.
215. Композиция по любому из вариантов осуществления 117-214, где введение одного или более агентов, способных вызывать генетическое разрушение, и введение матричного полинуклеотида проводят в одной экспериментальной реакции.
216. Композиция по любому из вариантов осуществления 152-215, где введение одного или более агентов, способных вызывать генетическое разрушение, и введение матричного полинуклеотида и вторых матричных полинуклеотидов проводят в одной экспериментальной реакции.
217. Композиция по любому из вариантов осуществления 108-216, дополнительно содержащая фармацевтически приемлемый носитель.
218. Способ лечения, включающий введение сконструированной клетки, множества сконструированных клеток или композиции по любому из вариантов осуществления 107-216 субъекту.
219. Применение сконструированной клетки, множества сконструированных клеток или композиции по любому из вариантов осуществления 107-216 для лечения рака.
220. Применение сконструированной клетки, множества сконструированных клеток или композиции по любому из вариантов осуществления 107-216 в производстве лекарственного средства для лечения рака.
221. Сконструированная клетка, множество сконструированных клеток или композиция по любому из вариантов осуществления 107-216 для применения в лечении рака.
222. Набор, содержащий:
один или более агентов, где каждый из одного или более агентов независимо способен вызывать генетическое разрушение целевого сайта в гене альфа константы Т-клеточного рецептора (TRAC) и/или гене бета константы Т-клеточного рецептора (TRBC); и
матричный полинуклеотид, содержащий трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом через репарацию, направляемую гомологией (HDR).
223. Набор по варианту осуществления 222, где один или более агентов, способных вызывать генетическое разрушение, содержит ДНК связывающий белок или ДНК связывающую нуклеиновую кислоту, которые специфически связываются с или гибридизируются в целевой сайт.
224. Набор по варианту осуществления 223, где один или более агентов, способных вызывать генетическое разрушение содержит (a) слитый белок, содержащий ДНК-направленный белок и нуклеазу или (b) РНК-направляемую нуклеазу.
225. Набор по варианту осуществления 224, где ДНК-направленный белок или РНК-направляемая нуклеаза содержит белок «цинковый палец» (ZFP), TAL белок или связанную с кластеризованной регулярной промежуточной короткой падлиндромной нуклеиновой кислотой (CRISPR) нуклеазу (Cas), специфическую к целевому сайту.
226. Набор по любому из вариантов осуществления 222-225, где один или более агентов содержат нуклеазу «цинковый палец» (ZFN), TAL-эффекторную нуклеазу (TALEN) и/или CRISPR-Cas9 сочетание, которые специфически связываются с, распознают или гибридизируют в целевой сайт.
227. Набор по любому из вариантов осуществления 222-226, где каждый из одного или более агентов содержит направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту.
228. Набор по варианту осуществления 227, где один или более агентов вводят в виде рибонуклеопротеинового (RNP) комплекса, содержащего нРНК и Cas9 белок.
229. Набор по варианту осуществления 228, где RNP вводят через электропорацию, генную пушку, трансфекцию фосфата кальция, сгущение или сжатие клеток.
230. Набор по варианту осуществления 228 или варианту осуществления 229, где RNP вводят через электропорацию.
231. Набор по любому из вариантов осуществления 222-230, где один или более агентов вводят в виде одного или более полинуклеотидов, кодирующих нРНК и/или Cas9 белок.
232. Набор по любому из вариантов осуществления 222-231, где, по меньшей мере, один целевой сайт находится внутри экзона TRAC, TRBC1 и/или TRBC2 гена.
233. Набор по любому из вариантов осуществления 227-232, где нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в TRAC гене и содержит последовательность, выбранную из UCUCUCAGCUGGUACACGGC (SEQ ID NO:28), UGGAUUUAGAGUCUCUCAGC (SEQ ID NO:29), ACACGGCAGGGUCAGGGUUC (SEQ ID NO:30), GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31), GCUGGUACACGGCAGGGUCA (SEQ ID NO:32), CUCAGCUGGUACACGGC (SEQ ID NO:33), UGGUACACGGCAGGGUC (SEQ ID NO:34), GCUAGACAUGAGGUCUA (SEQ ID NO:35), GUCAGAUUUGUUGCUCC (SEQ ID NO:36), UCAGCUGGUACACGGCA (SEQ ID NO:37), GCAGACAGACUUGUCAC (SEQ ID NO:38), GGUACACGGCAGGGUCA (SEQ ID NO:39), CUUCAAGAGCAACAGUGCUG (SEQ ID NO:40), AGAGCAACAGUGCUGUGGCC (SEQ ID NO:41), AAAGUCAGAUUUGUUGCUCC (SEQ ID NO:42), ACAAAACUGUGCUAGACAUG (SEQ ID NO:43), AAACUGUGCUAGACAUG (SEQ ID NO:44), UGUGCUAGACAUGAGGUCUA (SEQ ID NO:45), GGCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:46), GCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:47), GGGGAAGAAGGUGUCUUC (SEQ ID NO:48), GUUUUGUCUGUGAUAUACACAU (SEQ ID NO:49), GGCAGACAGACUUGUCACUGGAUU (SEQ ID NO:50), GCAGACAGACUUGUCACUGGAUU (SEQ ID NO:51), GACAGACUUGUCACUGGAUU (SEQ ID NO:52), GUGAAUAGGCAGACAGACUUGUCA (SEQ ID NO:53), GAAUAGGCAGACAGACUUGUCA (SEQ ID NO:54), GAGUCUCUCAGCUGGUACACGG (SEQ ID NO:55), GUCUCUCAGCUGGUACACGG (SEQ ID NO:56), GGUACACGGCAGGGUCAGGGUU (SEQ ID NO:57) и GUACACGGCAGGGUCAGGGUU (SEQ ID NO:58).
234. Набор по варианту осуществления 233, где нРНК имеет домен целенаправленного воздействия, содержащий последовательность GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31).
235. Набор по любому из вариантов осуществления 227-234, где нРНК имеет домен целенаправленного воздействия, который является комплементарным к целевому сайту в одном или более из TRBC1 и TRBC2 генов и содержит последовательность, выбранную из CACCCAGAUCGUCAGCGCCG (SEQ ID NO:59), CAAACACAGCGACCUCGGGU (SEQ ID NO:60), UGACGAGUGGACCCAGGAUA (SEQ ID NO:61), GGCUCUCGGAGAAUGACGAG (SEQ ID NO:62), GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63), GAAAAACGUGUUCCCACCCG (SEQ ID NO:64), AUGACGAGUGGACCCAGGAU (SEQ ID NO:65), AGUCCAGUUCUACGGGCUCU (SEQ ID NO:66), CGCUGUCAAGUCCAGUUCUA (SEQ ID NO:67), AUCGUCAGCGCCGAGGCCUG (SEQ ID NO:68), UCAAACACAGCGACCUCGGG (SEQ ID NO:69), CGUAGAACUGGACUUGACAG (SEQ ID NO:70), AGGCCUCGGCGCUGACGAUC (SEQ ID NO:71), UGACAGCGGAAGUGGUUGCG (SEQ ID NO:72), UUGACAGCGGAAGUGGUUGC (SEQ ID NO:73), UCUCCGAGAGCCCGUAGAAC (SEQ ID NO:74), CGGGUGGGAACACGUUUUUC (SEQ ID NO:75), GACAGGUUUGGCCCUAUCCU (SEQ ID NO:76), GAUCGUCAGCGCCGAGGCCU (SEQ ID NO:77), GGCUCAAACACAGCGACCUC (SEQ ID NO:78), UGAGGGUCUCGGCCACCUUC (SEQ ID NO:79), AGGCUUCUACCCCGACCACG (SEQ ID NO:80), CCGACCACGUGGAGCUGAGC (SEQ ID NO:81), UGACAGGUUUGGCCCUAUCC (SEQ ID NO:82), CUUGACAGCGGAAGUGGUUG (SEQ ID NO:83), AGAUCGUCAGCGCCGAGGCC (SEQ ID NO:84), GCGCUGACGAUCUGGGUGAC (SEQ ID NO:85), UGAGGGCGGGCUGCUCCUUG (SEQ ID NO:86), GUUGCGGGGGUUCUGCCAGA (SEQ ID NO:87), AGCUCAGCUCCACGUGGUCG (SEQ ID NO:88), GCGGCUGCUCAGGCAGUAUC (SEQ ID NO:89), GCGGGGGUUCUGCCAGAAGG (SEQ ID NO:90), UGGCUCAAACACAGCGACCU (SEQ ID NO:91), ACUGGACUUGACAGCGGAAG (SEQ ID NO:92), GACAGCGGAAGUGGUUGCGG (SEQ ID NO:93), GCUGUCAAGUCCAGUUCUAC (SEQ ID NO:94), GUAUCUGGAGUCAUUGAGGG (SEQ ID NO:95), CUCGGCGCUGACGAUCU (SEQ ID NO:96), CCUCGGCGCUGACGAUC (SEQ ID NO:97), CCGAGAGCCCGUAGAAC (SEQ ID NO:98), CCAGAUCGUCAGCGCCG (SEQ ID NO:99), GAAUGACGAGUGGACCC (SEQ ID NO:100), GGGUGACAGGUUUGGCCCUAUC (SEQ ID NO:101), GGUGACAGGUUUGGCCCUAUC (SEQ ID NO:102), GUGACAGGUUUGGCCCUAUC (SEQ ID NO:103), GACAGGUUUGGCCCUAUC (SEQ ID NO:104), GAUACUGCCUGAGCAGCCGCCU (SEQ ID NO:105), GACCACGUGGAGCUGAGCUGGUGG (SEQ ID NO:106), GUGGAGCUGAGCUGGUGG (SEQ ID NO:107), GGGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:108), GGCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:109), GCGGGCUGCUCCUUGAGGGGCU (SEQ ID NO:110), GGGCUGCUCCUUGAGGGGCU (SEQ ID NO:111), GGCUGCUCCUUGAGGGGCU (SEQ ID NO:112), GCUGCUCCUUGAGGGGCU (SEQ ID NO:113), GGUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:114), GUGAAUGGGAAGGAGGUGCACAG (SEQ ID NO:115) и GAAUGGGAAGGAGGUGCACAG (SEQ ID NO:116).
236. Набор по варианту осуществления 235, где нРНК имеет домен целенаправленного воздействия, содержащий последовательность GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63).
237. Набор по любому из вариантов осуществления 222-236, где матричный полинуклеотид содержит структуру [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии].
238. Набор по варианту осуществления 237, где 5’ плечо гомологии и 3’ плечо гомологии содержит последовательности нуклеиновых кислот гомологичные последовательностям нуклеиновых кислот окружающим, по меньшей мере, один целевой сайт.
239. Набор по варианту осуществления 237 или варианту осуществления 238, где 5’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот 5’ целевого сайта.
240. Набор по варианту осуществления 237 или варианту осуществления 238, где 3’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот 3’ целевого сайта.
241. Набор по любому из вариантов осуществления 237-240, где 5’ плечо гомологии и 3’ плечо гомологии независимо имеет, по меньшей мере, или по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов.
242. Набор по варианту осуществления 241, где 5’ плечо гомологии и 3’ плечо гомологии независимо имеет между около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 нуклеотидов.
243. Набор по варианту осуществления 242, где 5’ плечо гомологии и 3’ плечо гомологии независимо имеет от или от около 100 до 1000 нуклеотидов, от 100 до 750 нуклеотидов, от 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов.
244. Набор по любому из вариантов осуществления 222-243, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене.
245. Набор по любому из вариантов осуществления 222-244, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в одном или обоих TRBC1 и TRBC2 генов.
246. Набор по любому из вариантов осуществления 222-245, дополнительно содержащий один или более вторых матричных полинуклеотидов, содержащих один или более вторых трансгенов, где второй трансген нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
247. Набор по варианту осуществления 246, где второй матричный полинуклеотид содержит структуру [второе 5’ плечо гомологии]-[один или более вторых трансгенов]-[второе 3’ плечо гомологии].
248. Набор по варианту осуществления 247, где второе 5’ плечо гомологии и второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот, гомологичные последовательностям нуклеиновых кислот окружающим, по меньшей мере, один целевой сайт.
249. Набор по варианту осуществления 247 или варианту осуществления 248, где второе 5’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот второго 5’ целевого сайта.
250. Набор по варианту осуществления 247 или варианту осуществления 248, где второе 3’ плечо гомологии содержит последовательности нуклеиновых кислот, которые гомологичны последовательностям нуклеиновых кислот второго 3’ целевого сайта.
251. Набор по любому из вариантов осуществления 247-250, где второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет, по меньшей мере, или по меньшей мере, около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов или менее или менее чем около 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов.
252. Набор по варианту осуществления 251, где второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет между около 50 и 100, 100 и 250, 250 и 500, 500 и 750, 750 и 1000, 1000 и 2000 нуклеотидов.
253. Набор по варианту осуществления 252, где второе 5’ плечо гомологии и второе 3’ плечо гомологии независимо имеет от или от около 100 до 1000 нуклеотидов, от 100 до 750 нуклеотидов, 100 до 600 нуклеотидов, от 100 до 400 нуклеотидов, от 100 до 300 нуклеотидов, от 100 до 200 нуклеотидов, от 200 до 1000 нуклеотидов, от 200 до 750 нуклеотидов, от 200 до 600 нуклеотидов, от 200 до 400 нуклеотидов, от 200 до 300 нуклеотидов, от 300 до 1000 нуклеотидов, от 300 до 750 нуклеотидов, от 300 до 600 нуклеотидов, от 300 до 400 нуклеотидов, от 400 до 1000 нуклеотидов, от 400 до 750 нуклеотидов, от 400 до 600 нуклеотидов, от 600 до 1000 нуклеотидов, от 600 до 750 нуклеотидов или от 750 до 1000 нуклеотидов.
254. Набор по любому из вариантов осуществления 246-253, где один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене.
255. Набор по любому из вариантов осуществления 246-253, где один или более вторых трансгенов нацелен на интеграцию в или рядом с целевым сайтом в TRBC1 или TRBC2 гене.
256. Набор по любому из вариантов осуществления 246-255, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, TRBC1 гене или TRBC2 гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами, на которые не нацелен трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь.
257. Набор по любому из вариантов осуществления 246-256, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, нацелен на интеграцию в или рядом с целевым сайтом в TRAC гене, и один или более вторых трансгенов нацелен на интеграцию в или рядом с одним или более целевыми сайтами в TRBC1 гене и/или TRBC2 гене.
258. Набор по любому из вариантов осуществления 246-257, где один или более вторых трансгенов кодирует молекулу, выбранную из костимулирующего лиганда, цитокина, растворимого однонитевого переменного фрагмента (scFv), иммуномодулирующего слитого белка, химерного рецептора переключения (CSR) или корецептора.
259. Набор по варианту осуществления 258, где кодированной молекулой является костимулирующий лиганд, необязательно выбранный из лиганда фактора некроза опухоли (TNF), выбранного из 4-1BBL, OX40L, CD70, LIGHT и CD30L, или лиганда суперсемейства иммуноглобулина (Ig), выбранного из CD80 и CD86.
260. Набор по варианту осуществления 258, где кодированной молекулой является цитокин, необязательно выбранный из IL-2, IL-3, IL-6, IL-11, IL-30, IL-7, IL-24, IL-30, гранулоцитарно-моноцитарного колониестимулирующего фактора (GM-CSF), интерферона альфа (IFN-α), интерферона бета (IFN-β) или интерферона гамма (IFN-γ) и эритропоэтина.
261. Набор по варианту осуществления 258, где кодированной молекулой является растворимый однонитевой переменный фрагмент (scFv), который необязательно связывает полипептид, который обладает иммунодепрессивным действием или иммуностимулирующим действием, выбранный из CD47, PD-1, CTLA-4 и их лигандов или CD28, OX-40, 4-1BB и их лигандов.
262. Набор по варианту осуществления 258, где кодированной молекулой является иммуномодулирующий слитой белок, необязательно содержащий:
(a) внеклеточный связывающий домен, который специфически связывает антиген, полученный из CD290R, SIRPα, CD279 (PD-1), CD2, CD95 (Fas), CD242 (CTLA4), CD223 (LAG3), CD272 (BTLA), A2aR, KIR, TIM3, CD300 или LPA5;
(b) внутриклеточный связывающий домен, полученный из CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD47, CD79A, CD79B, CD224 (OX40), CD227 (4-1BB), CD240 (SLAMF1), CD278 (ICOS), CD357 (GITR), ХАРD11, DAP10, DAP30, FcRα, FcRβ, FcRγ, Fyn, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, ROR2, Ryk, Slp76, pTα, TCRα, TCRβ, TRFM, Zap70, PTCH2 или любого их сочетания; и
(c) гидрофобный трансмембранный домен, полученный из CD2, CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD40, CD79A, CD79B, CD80, CD86, CD95 (Fas), CD224 (OX40), CD227 (4-1BB), CD240 (SLAMF1), CD242 (CTLA4), CD290R, CD223 (LAG3), CD270 (HVEM), CD272 (BTLA), CD273 (PD-L2), CD274 (PD-L1), CD278 (ICOS), CD279 (PD-1), CD300, CD357 (GITR), A2aR, DAP10, FcRα, FcRβ, FcRγ, Fyn, GAL9, KIR, Lck, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, PTCH2, ROR2, Ryk, Slp76, SIRPα, pTα, TCRα, TCRβ, TIM3, TRIM, LPA5 или Zap70.
263. Набор по варианту осуществления 258, где кодированной молекулой является химерный рецептор переключения (CSR), который необязательно содержит усеченный внеклеточный домен PD1 и трансмембранный и цитоплазматический сигнальные домены CD28.
264. Набор по варианту осуществления 258, где кодированной молекулой является корецептор, необязательно выбранный из CD4 или CD8.
265. Набор по любому из вариантов осуществления 246-257, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует одну цепь рекомбинантного TCR и второй трансген кодирует другую цепь рекомбинантного TCR.
266. Набор по варианту осуществления 265, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, кодирует альфа (TCRα) цепь рекомбинантного TCR и второй трансген кодирует бета (TCRβ) цепь рекомбинантного TCR.
267. Набор по любому из вариантов осуществления 222-266, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо дополнительно содержат регулирующий или контрольный элемент.
268. Набор по варианту осуществления 267, где регулирующий или контрольный элемент содержит промотор, энхансер, интрон, сигнал полиаденилирования, консенсусную последовательность Козака, последовательность акцептора сплайсинга или последовательность донора сплайсинга.
269. Набор по варианту осуществления 268, где регулирующий или контрольный элемент содержит промотор.
270. Набор по варианту осуществления 269, где промотор выбирают из конститутивного промотора, индуцируемого промотора, репрессируемого промотора и/или ткань-специфического промотора.
271. Набор по варианту осуществления 269 или варианту осуществления 270, где промотор выбирают из РНК pol I, pol II или pol III промотора.
272. Набор по варианту осуществления 271, где промотор выбирают из:
pol III промотора, которым является U6 или H1 промотор; или
pol II промотор, которым является CMV, промотор SV40 ранней области или основной поздней области аденовируса.
273. Набор по любому из вариантов осуществления 269-271, где промотор является или содержит промотор человеческого фактора элонгации 1 альфа (EF1α) или MND промотор или их вариант.
274. Набор по любому из вариантов осуществления 269-271, где промотором является индуцируемый промотор или репрессируемый промотор.
275. Набор по варианту осуществления 274, где промотор содержит Lac последовательность оператора, тетрациклиновую последовательность оператора, галактозную последовательность оператора или доксициклиновую последовательность оператора или является их аналогом или способен связываться или распознаваться Lac репрессором или тетрациклиновым репрессором или их аналогом.
276. Набор по любому из вариантов осуществления 222-275, где трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо содержит один или более мультицистронных элементов.
277. Набор по варианту осуществления 276, где один или более мультицистронных элементов расположены против хода транскрипции трансгена, кодирующего рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов.
278. Набор по варианту осуществления 276 или варианту осуществления 277, где мультицистронные элементы расположены между трансгеном, кодирующим рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и одним или более вторыми трансгенами.
279. Набор по любому из вариантов осуществления 276-278, где мультицистронные элементы расположены между последовательностью нуклеиновых кислот, кодирующей TCRα или его часть, и последовательностью нуклеиновых кислот, кодирующей TCRβ или его часть.
280. Набор по любому из вариантов осуществления 276-279, где мультицистронные элементы содержат последовательность, кодирующую элемент проскока рибосомы, выбранный из T2A, P2A, E2A или F2A или внутренний участок посадки рибосомы (IRES).
281. Набор по варианту осуществления 280, где последовательность, кодирующая элемент проскока рибосомы нацелена на то, чтобы быть внутри рамки с геном в целевом сайте.
282. Набор по любому из вариантов осуществления 222-266, где при HDR, трансген, кодирующий рекомбинантный TCR или его антигенсвязывающий фрагмент или цепь, и/или один или более вторых трансгенов независимо функционально связаны с эндогенным промотором гена на целевом сайте.
283. Набор по любому из вариантов осуществления 222-282, где рекомбинантный TCR способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния.
284. Набор по варианту осуществления 283, где заболеванием, расстройством или состоянием является инфекционное заболевание, аутоиммунное заболевание, воспалительное заболевание или опухоль или рак.
285. Набор по варианту осуществления 283 или варианту осуществления 284, где антигеном является опухолевый антиген или патогенный антиген.
286. Набор по варианту осуществления 285, где патогенным антигеном является бактериальный антиген или вирусный антиген.
287. Набор по варианту осуществления 286 , где антигеном является вирусный антиген, и вирусным антигеном является вирус гепатита A, гепатита B, гепатита C (HCV), папилломавирус человека (HPV), вирусные инфекции гепатита, вирус Эпштейн-Барр (EBV), герпес-вирус человека 8 (HHV-8), вирус Т-клеточного лейкоза человека-1 (HTLV-1), вирус Т-клеточного лейкоза человека-2 (HTLV-2) или цитомегаловирус (CMV).
288. Набор по варианту осуществления 287, где антигеном является антиген из HPV выбранный из HPV-25, HPV-27, HPV-31, HPV-33 и HPV-35.
289. Набор по варианту осуществления 288, где антигеном является HPV-25 антиген, который является HPV-25 E6 или HPV-25 E7 антигеном.
290. Набор по варианту осуществления 286, где вирусным антигеном является EBV антиген, выбранный из ядерного антигена Эпштейн-Барр (EBNA)-1, EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, EBNA-лидерного белка (EBNA-LP), латентных мембранных белков LMP-1, LMP-2A и LMP-2B, EBV-EA, EBV-MA и EBV-VCA.
291. Набор по варианту осуществления 286, где вирусным антигеном является HTLV-антиген, которым является TAX.
292. Набор по варианту осуществления 286, где вирусным антигеном является HBV антиген, которым является ядерный антиген гепатита В или оболочечный антиген гепатита В.
293. Набор по любому из вариантов осуществления 283-285, где антигеном является опухолевый антиген.
294. Набор по варианту осуществления 293, где антиген выбирают из глиома-ассоциированного антигена, β-человеческого хорионического гонадотропина, альфафетобелка (AFP), лектин-реактивного AFP, тироглобулина, RAGE-1, MN-CA IX, обратной транскриптазы теломеразы человека, RU1, RU2 (AS), кишечной карбоксилэстеразы, mut hsp70-2, M-CSF, Меланин-A/MART-1, WT-1, S-100, MBP, CD63, MUC1 (например, MUC1-8), p53, Ras, циклина B1, HER-2/neu, карциноэмбрионного антигена (CEA), gp100, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11, MAGE-A11, MAGE-B1, MAGE-B2, MAGE-B3, MAGE-B4, MAGE-C1, BAGE, GAGE-1, GAGE-2, pl5, тирозиназы, родственного тирозиназе белка 1 (TRP-1), родственного тирозиназе белка 2 (TRP-2), β-катенина, NY-ESO-1, LAGE-1a, PP1, MDM2, MDM4, EGVFvIII, Tax, SSX2, теломеразы, TARP, pp65, CDK4, виментина, S100, eIF-4A1, IFN-индуцируемого p78, меланотрансферрина (p97), Уроплакина II, специфического антигена простаты (PSA), человеческого калликреина (huK2), специфического мембранного антигена простаты (PSM), и простатической кислой фосфатазы (PAP), нейтрофильной эластазы, эфрина B2, BA-46, Bcr-abl, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, Каспазы 8, FRa, CD24, CD44, CD223, CD 256, epCAM, CA-224, HE4, Oval, рецептора эстрогена, рецептора прогестерона, uPA, PAI-1, CD28, CD29, CD22, ROR1, CD33/IL3Ra, c-Met, PSMA, Гликолипида F77, GD-2, инсулинового фактора роста (IGF)-I, IGF-II, IGF-I рецептора и мезотелина.
295. Набор по любому из вариантов осуществления 222-294, где первый матричный полинуклеотид, один или более вторых матричных полинуклеотидов и/или один или более полинуклеотидов, кодирующих нРНК и/или Cas9 белок, содержатся в одном или более векторах, которые необязательно являются вирусными векторами.
296. Набор по варианту осуществления 295, где вектором является AAV вектор.
297. Набор по варианту осуществления 296, где AAV вектор выбирают из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7 или AAV8 вектор.
298. Набор по варианту осуществления 297, где AAV вектором является AAV2 или AAV6 вектор.
299. Набор по варианту осуществления 295, где вирусным вектором является ретровирусный вектор.
300. Набор по варианту осуществления 299, где вирусным вектором является лентивирусный вектор.
Х. ПРИМЕРЫ
Следующие ниже примеры включены только в иллюстративных целях и не предназначены для ограничения объема изобретения.
Пример 1: Создание и оценка сконструированных Т клеток, экспрессирующих рекомбинантный Т-клеточный рецептор (TCR) нацеленным нокином или случайной интеграцией последовательностей, кодирующих TCR
Полинуклеотиды, кодирующие типовые рекомбинантные Т-клеточные рецепторы (TCR) вводят в T клетки с генетическим разрушением на локусе эндогенного гена, который кодирует цепь Т-клеточного рецептора альфа (TCRα), медиированным CRISPR/Cas9 редактированием генома и направленной интеграции на сайте генетического разрушения через репарацию, направляемую гомологией (HDR) или случайной интеграцией через лентивирусную трансдукцию.
А. Конструкты рекомбинантного TCR трансгена
Типовые матричные полинуклеотиды создают для направленной интеграцией путем HDR трансгена, содержащего последовательности нуклеиновых кислот, кодирующие один из двух типовых рекомбинантных TCR. Общая структура типовых матричных полинуклеотидов следующая: [5’ плечо гомологии]-[трансгенные последовательности]-[3’ плечо гомологии]. Плечи гомологии включают приблизительно 600 п.о. последовательностей нуклеиновых кислот, гомологичных последовательностям, окружающим целевой сайт интеграции, в экзоне 1 постоянной области человеческого TCR α (TRAC) гена (5’ плечо гомологии, последовательность, представленная в SEQ ID NO:124; 3’ плечо гомологии, последовательность, представленная в SEQ ID NO:125).
Трансген включает последовательности нуклеиновых кислот, кодирующие α и β цепи одного или более типовых рекомбинантных TCR, которые распознают эпитоп онкобелка E7 папилломавируса человека (HPV) 16 (TCR #1 и TCR #2), в котором последовательности, кодирующие TCRα и TCRβ цепи, разделены 2A элементом проскока рибосомы. Нуклеотидные последовательности, кодирующие TCR #1 и постоянную область для TCR #2, также модифицированы оптимизацией кодона и/или мутациями для того, чтобы способствовать образованию ненативной дисульфидной связи на интерфейсе между постоянными доменами TCR для повышения спаривания и стабильности TCR. Ненативные дисульфидные связи укрепляют модификацией TCR цепей на остатке 48 в постоянной области TCR альфа цепи (Cα) области от Thr до Cys, и остатке 57 постоянной области TCR бета цепи (Cβ) области от Ser до Cys (см. Kuball et al. (2007) Blood, 109:2331-2338).
Трансген также включает либо a) промотор человеческого фактора элонгации 1 альфа (EF1α) для управления экспрессией последовательностей, кодирующих рекомбинантный TCR (последовательности, представленной в SEQ ID NO:127); или b) последовательностей, кодирующих P2A элемент проскока рибосомы (последовательности, представленной в SEQ ID NO:128) выше последовательностей, кодирующих рекомбинантный TCR, для управления экспрессией рекомбинантного TCR от эндогенного TCRα локуса при HDR-медиированной направленной интеграцией внутри рамки в постоянной области гена человеческого TCR α (TRAC).
Для направленной интеграции HDR, создают конструкты вектора адено-ассоциированного вируса (AAV), содержащие матричные полинуклеотиды, описанные выше. Исходный раствор AAV получают тройной трансфекцией AAV вектора, который включает матричный полинуклеотид, серотипическую хелперную плазмиду и аденовирусную хелперную плазмиду, в колонию клеток 293T. Трансфицированные клетки собирают, лизируют, и исходный раствор AAV собирают для трансдукции клеток.
В качестве контроля, для случайной интеграции, последовательности нуклеиновых кислот, кодирующие типовые рекомбинантные конструкты TCR трансгена, описанные выше, или последовательности, кодирующие ссылочный TCR, способный связываться с HPV 16 E7, но содержащий мышиные Cα и Cβ области, под контролем EF1α промотора, вводят в типовой лентивирусный вектор, полученный из ВИЧ-1. Частицы псевдотипического лентивирусного вектора получают стандартными методами, временно трансфицируя HEK-293T клетки полученными векторами, хелперными плазмидами (содержащими gagpol плазмиды и rev плазмиды), и псевдотипирующей плазмидой, и применяют для трансдукции клеток.
В. Создание сконструированных Т клеток
Первичные человеческие CD4+ и CD8+ T клетки от 2 разных людей-доноров выделяют селекцией на основе иммуноаффинности из мононуклеаров периферической крови человека (PBMC), полученных от здоровых доноров. CD8+ клетки (для TCR #1) или CD4+ и CD8+ клетки, объединенные в соотношении 1:1 (для TCR #2), стимулируют в течение 72 часов при 37ºC культивированием с анти-CD3/анти-CD28 реагентом с соотношении 1:1 сферы:клетки в среде, содержащей человеческую сыворотку, IL-2, IL-7 и IL-15. Для введения генетического разрушения на локусе эндогенного TRAC медиированным CRISPR/Cas9 редактированием генома, анти-CD3/анти-CD28 реагент удаляют, и клетки электропорируют с 2 мкМ рибонуклеопротеиновых (RNP) комплексов, содержащих Streptococcus pyogenes Cas9 и направляющую РНК (нРНК) с доменом целенаправленного воздействия последовательности GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31), которые нацелены на генетическое разрушение в пределах экзона 1 постоянной области эндогенного TCR α (TRAC) гена. После электропорации, клетки смешивают со средой, содержащей препарат AAV, содержащий HDR матричный полинуклеотид, кодирующий типовой рекомбинантный TCR под контролем EF1α промотора для трансдукции (HDR KO). В качестве контроля, клетки обрабатывают в тех же условиях, которые применяют для электропорации, но без добавления RNP (mock KO), трансдуцируют с лентивирусным вектором, кодирующим рекомбинантный TCR (Lenti) или ссылочным TCR, способным связываться с HPV 16 E7, но содержащим мышиные Cα и Cβ области (Lenti Ref), или трансдуцируют с лентивирусным вектором, кодирующим рекомбинантный TCR, и также электропорируют с RNP комплексами, направленными на генетическое разрушение на TRAC локус (Lenti KO). После трансдукции клетки культивируют в течение приблизительно 7 дней в середе, содержащей человеческую сыворотку и IL-2, IL-7 и IL-15.
С. Экспрессия TCR
На 7 день после электропорации, клетки оценивают проточной цитометрией для окрашивания анти-CD3 антителом, анти-CD4 антителом, анти-CD8 антителом, анти-Vбета антителом, специфически для каждого рекомбинантного TCR, и пептид-MHC тетрамером в комплексе с HPV16 E7 пептидом.
Результаты для экспрессирующих TCR #1 CD8+ клеток показаны на ФИГ. 1A-1C. Как показано, клетки, экспрессирующие TCR #1 посредством HDR-медиированной направленной интеграции на TRAC локус, демонстрируют большую долю клеток, связанных тетрамером (ФИГ. 1A) и наиболее высокую среднюю интенсивность флуоресценции окрашивания тетрамера в CD8+ клетках (ФИГ. 1B). Среди CD8+ клеток, связанных тетрамером, степень связывания тетрамером обычно более однородна в клетках, которые были подвергнуты HDR-медиированной интеграции, по сравнению со случайной интеграцией лентивирусной трансдукцией, как показано более низким коэффициент вариации (стандартное отклонение сигнала в популяции клеток, деленное на среднее значение сигнала в соответствующей популяции; см. ФИГ. 1C). Результаты для TCR #2-экспрессирующих клеток показаны на ФИГ. 2A-2B. Как показано, CD4+ и CD8+ клетки, экспрессирующие TCR #2 через HDR-медиированную интеграцию на TRAC локус, демонстрируют более высокую долю клеток, связанных тетрамером (ФИГ. 2A), и наиболее высокую среднюю интенсивность флуоресценции окрашивания тетрамера (ФИГ. 2B).
D. Цитолитическая активность и образование цитокина
Цитолитическую активность и образование цитокина клеток, сконструированных для экспрессии рекомбинантного TCR #1 или рекомбинантного TCR #2 через HDR или случайной интеграцией, как описано выше, оценивают после инкубирования с целевыми клетками, экспрессирующими HPV 16 E7 in vitro.
Цитолитическую активность оценивают культивированием рекомбинантных TCR-экспрессирующих эффекторных клеток с целевыми клетками, экспрессирующими HPV 16 E7, меченный NucLight Red (NLR) при соотношении эффектора к цели (E:T) 10:1, 5:1 и 2,5:1. Способность T клеток вызывать антиген-специфический лизис целевых клеток оценивают измерением потери меченных целевых клеток каждые 2 часа вплоть до 44 часов после совместного культивирования. Цитолитическую активность определяют из среднего для 2 доноров в каждой группе, из площади под кривой (AUC) % гибели, нормализованной до Vбета экспрессии для каждого рекомбинантного TCR, и сравнивают с контрольной фиктивной трансдукцией. Образование цитокина измеряют после инкубирования рекомбинантных TCR-экспрессирующих эффекторных клеток с целевыми клетками. Секрецию интерферона-гамма (IFNγ) и интерлейкина-2 (IL-2) в надосадочной жидкости определяют ELISA, нормализуют до Vбета экспрессии и усредняют для 2 доноров в каждой группе.
Результаты для TCR #1 показаны на ФИГ. 3A-3B. Степень гибели и секреция IFNγ были выше в клетках, в которые рекомбинантный TCR #1, управляемый EF1α промотором, был введен HDR-медиированной направленной интеграцией на TRAC локус (EF1α-TCR #1 HDR KO), по сравнению со случайной интеграцией, с (TCR #1 Lenti) или без нокаута TRAC локуса (TCR #1 Lenti KO).
Типовые результаты для TCR #2 показаны на ФИГ. 4A-4G. Секреция целевыми клетками AUC и IFNγ и IL-2 через 24 часа была такой же или более высокой в клетках, в которые рекомбинантный TCR #2, управляемый EF1α промотором, вводят HDR-медиированной направленной интеграцией (EF1α-TCR #2 HDR KO), по сравнению со случайной интеграцией, с (TCR #2 Lenti) или без нокаута TRAC локуса (TCR #2 Lenti KO), и ссылочным TCR, введенным случайной интеграцией (Lenti Ref) (ФИГ. 4A-4C). CD8+ и CD4+ клетки, экспрессирующие TCR #2, введенный HDR на TRAC локусе демонстрируют более высокую гибель целевых клеток по сравнению со случайной интеграцией, с или без нокаута TRAC локуса, и ссылочным TCR, введенным случайной интеграцией (ФИГ. 4D-4E). Наблюдается образование IFNγ CD8+ клетками при соотношении E:T 2,5:1, или CD4+ клетками при соотношении E:T 10:1, такое же как для клеток, экспрессирующих TCR #2, введенный HDR на TRAC локус или случайной интеграцией (ФИГ. 4F-4G).
Е. Жизнеспособность клеток, экспрессирующих рекомбинантные TCR
Жизнеспособность CD4+ и CD8+ клеток, сконструированных для экспрессии TCR #2, введенного разными способами, оценивают при криоконсервации и после оттаивания законсервированных клеток, окрашиванием акридиновым оранжевым (AO) и йодидом пропидия (PI). Как показано на ФИГ. 5A-5B, на жизнеспособность клеток по существу не влияло введение TCR #2-кодирующих последовательностей посредством HDR, по сравнению с обработанными пробой клетками или клетками, экспрессирущими ссылочный TCR.
F. Заключение
В общем, результаты соответствуют наблюдению, что направленная интеграция посредством HDR последовательностей нуклеиновых кислот, кодирующих типовые рекомбинантные TCR, дает более высокую экспрессию рекомбинантного TCR человеческого TCR в человеческих T клетках по сравнению с введением TCR случайной интеграцией, тем самым приводя к более высокой функциональной активности клеток, экспрессирующих рекомбинантный TCR.
Пример 2. Создание и оценка экспрессии рекомбинантных Т-клеточных рецепторов (TCR) направленным нокином или случайной интеграцией последовательностей, кодирующих TCR, в T клетки с нокаутом эндогенных TCRα и TCRβ цепей
Полинуклеотид, кодирующий один из типовых рекомбинантных TCR, нацеливают на интеграцию на один из сайтов генетического разрушения через репарацию, направляемую гомологией (HDR), или вводят случайной интеграцией через лентивирусную трансдукцию, в клетки, сконструированные для генетического разрушения локусов эндогенного гена, которые кодируют цепи Т-клеточного рецептора альфа (TCRα) и бета (TCRβ) или обе, посредством CRISPR/Cas9 медиированного редактирования генома.
Для направленной интеграции посредством HDR, препараты AAV, содержащие конструкты матричного полинуклеотида, кодирующего типовой рекомбинантный TCR #1, создают по существу как описано в примерах 1, за исключением следующих отличий: создают дополнительный AAV конструкт, содержащий MND промотор (последовательность, представленную в SEQ ID NO:126), который является синтетическим промотором, который содержит U3 область модифицированного MoMuLV LTR с энхансером вируса миелопролиферативной саркомы, для контроля экспрессии последовательностей, кодирующих рекомбинантный TCR.
Для случайной интеграции, создают лентивирусные препараты, содержащие последовательности нуклеиновых кислот, кодирующие типовой рекомбинантный TCR #1, в общем, как описано в примере 1. Для этих исследований, лентивирусный конструкт трансдукции дополнительно содержит полинуклеотид, кодирующий усеченный рецептор, выделенный из рекомбинантного TCR трансгена, последовательностью, кодирующей T2A последовательности проскока рибосомы для экспрессии и рекомбинантного TCR и усеченного рецептора из одного и того же конструкта; усеченный рецептор служит в качестве суррогатного маркера для трансдукции. В качестве контроля, создают полинуклеотид, кодирующий химерный TCR, где Cα и Cβ рекомбинантного TCR замещены постоянными областями из мышиного TCR (мышиной Cα последовательностью, представленной в SEQ ID NO:122; мышиной Cβ последовательностью, представленной в SEQ ID NO:123), или клетки подвергают фиктивной трансдукции.
Первичные человеческие CD4+ и CD8+ T клетки выделяют и конструируют для введения генетического разрушения на локусах эндогенного TRAC и TRBC посредством CRISPR/Cas9-медиированной редактирование генома, и трансдуцируют с AAV препаратами, содержащими матричными полинуклеотидами для HDR или лентивирусных препаратов для случайной интеграции, обычно как описано в примере 1B выше. Клетки электропорируют с рибонуклеопротеиновыми (RNP) комплексами, содержащими TRAC-нацеленную направляющую РНК (нРНК), описанную в примере 1B, и RNP, содержащими нРНК, направленную на целевой сайт консенсусной последовательности, общей для экзона 1 или обеих TCR β постоянных областей 1 и 2 (с доменом целенаправленного воздействия последовательности GGCCUCGGCGCUGACGAUCU (SEQ ID NO:63)) (TCRαβ KO). В качестве контроля, клетки обрабатывают в тех же условиях, которые применяют для электропорации, но без добавления RNP (фиктивная KO; также обозначенная как TCRαβ WT).
Клетки затем культивируют в течение четырех (4) дней, затем оценивают проточной цитометрией для окрашивания анти-CD3 антителом, анти-Vбета антителом, которое распознает рекомбинантный TCR #1, и с пептид-MHC тетрамером в комплексе с антигеном, распознанным рекомбинантным TCR (HPV16 E7 пептид). Клетки также совместно окрашивают для CD8 или CD4.
Результаты показаны на ФИГ. 6A-6E. Как показано на ФИГ. 6A и 6B, CRISPR/Cas9 медиированный нокаут (KO) TRAC и TRBC (панель, обозначенная “TCRαβ KO” на чертежах) дает почти полное прекращение экспрессии TCR в CD8+ клетках, как видно по отсутствию CD3 окрашивания в клетках, подвергнутых KO и фиктивной трансдукции (панель, обозначенная TCRαβ KO/mock transd.). Экспрессия рекомбинантного TCR (как показано клетками, окрашенными специфическим Vбета антителом или клетками, положительными для окрашивания тетрамера среди CD8+ клеток) незначительно улучшалась после лентивирусной трансдукции в клетки, который имели KO для эндогенного TCR (TCRαβ KO/lenti человек) по сравнению с клетками, которые сохраняли экспрессию эндогенного TCR (TCRαβ WT/lenti человек). В клетках, сохраняющих эндогенный TCR, экспрессия рекомбинантного TCR улучшалась при лентивирусной трансдукции рекомбинантного TCR, содержащего мышиный постоянный домен, по сравнению с лентивирусной трансдукцией полностью человеческого рекомбинантного TCR (сравнение TCRαβ WT/lenti человек и TCRαβ WT/lenti мышь).
HDR-медиированный направленный нокин рекомбинантного TCR и KO эндогенного TCR дают по существу большую долю клеток, экспрессирующих рекомбинантный TCR, чем наблюдается после лентивирусной трансдукции (первые две левые панели, обозначенные “HDR” по сравнению с TCRαβ KO/lenti человек на ФИГ. 6A и 6B). Геометрическая средняя флуоресценция (gMFI) экспрессии рекомбинантного TCR, оцененная Vбета или окрашиванием тетрамера в CD8+ клетках (ФИГ. 6C) или окрашиванием Vбета в CD4+ клетках (ФИГ. 6D), также была значительно выше в клетках, подвергнутых HDR по сравнению с лентивирусной трансдукцией. Степень экспрессии рекомбинантного TCR посредством HDR была аналогичной независимо от того, был ли рекомбинантный TCR под контролем EF1α или MND промотора.
Среди клеток, которые были положительными для экспрессии рекомбинантного TCR, степень экспрессии была обычно более однородной или плотнее в клетках, которые были подвергнуты HDR-медиированной направленной интеграции, по сравнению со случайной интеграцией лентивирусной трансдукцией (см. ФИГ. 6A и 6B). Как показано на ФИГ. 6E и ФИГ. 6F, меньший коэффициент вариации (стандартное отклонение сигнала в популяции клеток, деленное на среднее значение сигнала в соответствующей популяции) экспрессии рекомбинантного TCR, определенный связыванием пептид-MHC тетрамера, и экспрессии Vбета, соответственно, наблюдали в CD8+ клетках, которые были подвергнуты HDR-медиированной интеграции, по сравнению со случайной интеграцией. Результаты соответствуют открытию, что направленный нокин рекомбинантного TCR в локус эндогенного TCRα, в сочетании с нокаутом цепей эндогенного TCRαβ, дает более высокий и однородный уровень экспрессии в популяции клеток, сконструированных для экспрессии рекомбинантного TCR, по сравнению с другими способами.
Пример 3. Оценка HDR-медиированного нокина последовательностей, кодирующих рекомбинантный Т-клеточный рецептор (TCR) в T клетках с нокаутом цепей эндогенного TCRα или TCRβ или обоих
Для дальнейшей оценки экспрессии рекомбинантного TCR посредством HDR, последовательности нуклеиновых кислот, кодирующие типовой рекомбинантный TCR в TRAC локусе, нацеливают для интеграции в клетки, содержащие двойной нокаут TRAC и TRBC локусов, нокаут только TRAC локуса или нокаут только TRBC локуса.
А. Конструкты трансгена рекомбинантного TCR и создание сконструированных клеток
Эти исследования проводят с применением AAV (для HDR) и лентивирусных конструктов (для случайной интеграции), кодирующих типовой TCR #1 по существу так, как описано в примерах 1 и 2, за исключением следующих различий: лентивирусные конструкты создают содержащими полинуклеотид, кодирующий рекомбинантный TCR под функциональным контролем EF1α или MND промотора. Лентивирусный конструкт, содержащий полинуклеотид, кодирующий рекомбинантный TCR #1 под функциональным контролем EF1α промотора, и усеченный рецептор, отделенный от трансгена рекомбинантного TCR последовательностью, кодирующей T2A последовательности проскока рибосомы, также создают для сравнения.
Для направленной интеграции посредством HDR, первичные человеческие CD4+ и CD8+ T клетки стимулируют, культивируют и подвергают электропорации с рибонуклеопротеиновыми (RNP) комплексами, содержащими только TRAC-направленную нРНК, только TRBC-направленную нРНК или обе TRAC- и TRBC-направленные нРНК как в общем описано в примерах 1 и 2. После электропорации, клетки трансдуцируют AAV препаратами, содержащими полинуклеотиды, которые кодируют рекомбинантный TCR для целенаправленного воздействия на локус эндогенного TRAC, в общем, как описано выше.
Для случайной интеграции, первичные человеческие CD4+ и CD8+ T клетки оттаивают, стимулируют и культивируют по существу как описано в примерах 1 и 2, с последующей трансдукцей с лентивирусным препаратом, который кодирует рекомбинантный TCR. В этом исследовании, лентивирусные препараты трансдуцируют в первичные T клетки, которые сохраняют эндогенный TCR. В качестве контроля, клетки обрабатывают в тех же условиях, которые применяют для лентивирусной трансдукции, но без добавления лентивируса (фиктивная трансдукция (mock transd.)).
В. Экспрессия TCR
Клетки затем культивируют в течение 4-10 дополнительных дней и оценивают проточной цитометрией после окрашивания анти-CD3 антителом, анти-Vбета антителом, специфическим для рекомбинантного TCR #1, и пептид-MHC тетрамера в комплексе с антигеном, распознанным TCR (HPV16 E7 пептид). Клетки также совместно окрашивают для CD8 или CD4.
Результаты CD3 окрашивания показаны на ФИГ. 7A-7C, для окрашивания тетрамера показаны на ФИГ. 8A-8C, и для Vбета окрашивания показаны на ФИГ. 9A-9D.
Как показано на ФИГ. 7A и 7C, электропорация с RNP в комплексе с нРНК, направленными на TRAC и TRBC, дает эффективный нокаут эндогенного TCR, что подтверждается отсутствием поверхностной экспрессии CD3 (см. панель, обозначенную KO/mock transd. на ФИГ. 7A; см. также группу, обозначенную KO mock на ФИГ. 7C, которая показывает долю CD3+CD8+ клеток среди CD8+ клеток). Степень KO с нРНК, направленными на оба TRAC и TRBC, была больше, чем для клеток, электропорированных с RNP в комплексе с нРНК, направленными только на TRAC или только на TRBC, что соответствует наблюдению, что двойное целенаправленное воздействие обоих постоянных доменов TCR цепей α и β улучшает эффективность прерывания экспрессии эндогенного TCR. В клетках, трансдуцированных лентивирусными векторами, в которых не проводится разрушение эндогенного TCR, экспрессия CD3 была такая же, как во всех тестированных условиях (ФИГ. 7B и 7C). Как показано на ФИГ. 7A и 7C, экспрессия CD3 также была одинаковой среди клеток, в которые рекомбинантный TCR был введен посредством HDR, что соответствует TCR/CD3 поверхностной экспрессии в клетках, в которые введен рекомбинантный TCR.
Как показано на ФИГ. 8A и 8C, доля CD8+ клеток, которые связывают пептид-MHC тетрамер, показывают, что экспрессия рекомбинантного TCR была выше в условиях, в которых HDR проводят в клетках, нокаутированных для обоих TRAC и TRBC, по сравнению с только TRAC (сравнение верхнего и среднего рядов на ФИГ. 8A; сравнение TRAC&TRBC с только TRAC на ФИГ. 8C). Как показано на ФИГ. 8C, подобные результаты наблюдали в дни 7 и 13. Похожую экспрессию рекомбинантного TCR наблюдают в клетках, где HDR проводят с конструктом для интеграции под контролем экзогенного EF1α или MND промотора или под контролем эндогенного TCR промотора (P2A-содержащим конструктом). Как показано на ФИГ. 8B и ФИГ. 8C, меньшее количество клеток экспрессирует рекомбинантный TCR, по оценке окрашивания тетрамера, после медиированной лентивирусом трансдукции, независимо от присутствия усеченного рецептора в лентивирусных конструктах.
Как показано на ФИГ. 9A-9C, подобно указанным выше результатам, экспрессию рекомбинантного TCR на CD8+ T клетках наблюдают при прямом окрашивании для рекомбинантного TCR антителом, которое специфически распознает Vбета цепь рекомбинантного TCR. Окрашивание анти-Vбета, которое также способно распознавать рекомбинантный TCR на CD4+ T клетках (CD8 отрицательная популяция), также показало, что экспрессия рекомбинантного TCR наблюдается в CD4+ клетках (ФИГ. 9A и ФИГ. 9D).
Для всех способов оценки экспрессии рекомбинантного TCR, показанных выше (анти-CD3, тетрамер и анти-Vбета), приведенные выше результаты также показывают, что направленная интеграция рекомбинантного TCR в TRAC посредством HDR является специфической для вызванного нуклеазой разрыва ДНК на TRAC локусе, так как клетки, электропорированные TRBC-направленной RNP, не экспрессируют рекомбинантный TCR (см., например, условие “только TRBC” на ФИГ. 8A, 8C, 9A, 9C и 9D).
С. Цитолитическая активность и образование цитокина
Цитолитическую активность и образование цитокина CD8+ клетками, сконструированными для экспрессии рекомбинантного TCR #1 посредством HDR или случайной интеграцией, как описано выше, оценивают после инкубирования с целевыми клетками, экспрессирующими HPV 16 E7 in vitro. В дополнение к клеткам, описанным выше, также оценивают первичные человеческие CD8+ клетки, трансдуцированные лентивирусом, кодирующим ссылочный TCR, способный связываться с HPV 16 E7, но содержащие мышиные Cα и Cβ области.
Цитолитическую активность оценивают инкубированием экспрессирующих рекомбинантный TCR клеток с целевыми клетками, экспрессирующими HPV 16 E7, при соотношении эффектора к цели (E:T) 10:1, 5:1 и 2,5:1. Способность T клеток вызывать антиген-специфический лизис целевых клеток оценивают через 4 часа после совместного культивирования. Цитолитическую активность определяют из площади под кривой (AUC) % гибели, нормализованной до Vбета экспрессии для каждого рекомбинантного TCR, и сравнивают с контрольной фиктивной трансдукцией. Результаты показаны на ФИГ. 10. Степень гибели была выше в клетках, в которых и рекомбинантный TCR вводят HDR-медиированной направленной интеграцией, по сравнению со случайной интеграцией, что соответствует открытию, что более высокая экспрессия рекомбинантного TCR в клетках дает более высокую функциональную активность.
Образование цитокина также отслеживают после инкубации экспрессирующих рекомбинантный TCR CD8+ эффекторных клеток с целевыми клетками, экспрессирующими HPV 16 E7 при соотношении E:T 10:1 и 2,5:1 в течение 48 часов. Секрецию IFNγ в надосадочной жидкости определяют ELISA и нормализуют до Vбета экспрессии для каждой группы. Результаты показаны на ФИГ. 11. Так же как и представленные выше результаты для цитолитической активности, большее образование IFNγ наблюдается в клетках, подвергнутых HDR-медиированной интеграции, по сравнению со случайной интеграцией. В анализе цитолитической активности и оценке секреции IFNγ, функциональная активность клеток, экспрессирующих рекомбинантный TCR, посредством HDR-медиированной интеграции, была такой же, как активность клеток, экспрессирующих, посредством лентивирусной трансдукции, ссылочный TCR, содержащий мышиные постоянные домены.
Пролиферацию экспрессирующих рекомбинантный TCR клеток оценивают после инкубирования с SCC152 целевыми клетками или T2 целевыми клетками, в которые вводили антигенный пептид. Клетки метят CellTrace™ фиолетовым (ThermoFisher) красителем. На деление живых T клеток указывает разбавление CellTrace™ фиолетового красителя, по оценке проточной цитометрией.
Результаты разных функциональных анализов показаны на ФИГ. 12. Как показано в тепловой карте, изображающей относительную активность экспрессирующей рекомбинантный TCR популяции клеток в различных функциональных активностях (AUC % гибели при соотношениях E:T 10:1, 5:1 и 2,5:1 (обозначенная “AUC”), связывание тетрамера в CD8+ клетках в дни 7 и 13 (обозначенное “тетрамер CD8”), анализ пролиферации (обозначенный “CTV импульс”) с применением SCC152 клеток или T2 целевых клеток, в которые вводили антигенный пептид, и секреция IFNγ из CD8+ клеток (обозначенная “CD8 секретирующие IFNγ”)), функциональная активность клеток с рекомбинантным TCR, направленным на нокин на локусе постоянного домена цепи эндогенного TCRα, и нокаут эндогенного TCRαβ гена или TCRα гена, обычно наблюдают как более высокую, по сравнению с клетками, где полинуклеотид, кодирующий рекомбинантный TCR, был интегрирован случайно.
В общем, результаты соответствуют наблюдению, что направленная интеграция посредством HDR дает более высокую экспрессию рекомбинантного TCR человеческого TCR в человеческих T клетках, по сравнению с введением TCR случайной интеграцией, тем самым приводят к более высокой функциональной активности клеток, экспрессирующих рекомбинантный TCR.
Пример 4. Оценка in vivo противоопухолевых эффектов у мышей T клеток, сконструированных для экспрессии рекомбинантного Т-клеточного рецептора (TCR), созданного HDR-медиированным нокином
Противоопухолевую активность и фармакокинетику CD4+ и CD8+ клеток, экспрессирующих типовой TCR, созданных HDR-медиированным нокином или случайной интеграцией через лентивирусную трансдукцию последовательностей, кодирующих рекомбинантный TCR, оценивают введением сконструированных клеток в мышиную модель опухоли.
А. Масса опухоли и выживание
Мышиную модель опухоли создают подкожной инъекцией 4×106 колонии клеток плоскоклеточной карциномы UPCI:SCC152 (ATCC® CRL-3240™) самкам мышей NOD/SCID/IL-2Rγnull (NSG). Опухоль приживается в течение приблизительно 25 дней, и ее стадию определяют на основе объема опухоли (P >0,95) до введения сконструированных клеток.
Первичные CD4+ клетки и CD8+ клетки, сконструированные для экспрессии типового рекомбинантного TCR #2 посредством направленного нокина или случайной интеграцией последовательности, кодирующей TCR, создают по существу так, как описано выше в примере 1, водят через 26 дней после инъекции опухолевых клеток. Вводят две разные суммарные дозы клеток, экспрессирующих рекомбинантный TCR (3×106 или 6×106 экспрессирующих TCR клеток), где доза основана на доле клеток, положительных к окрашиванию анти-Vбета антителом, которое распознает рекомбинантный TCR #2. Сравнивают следующие группы: TCR#2 контролируемый промотором человеческого фактора элонгации 1 альфа (EF1α), нацеленный на интеграцию на TRAC локусе посредством HDR (TCR #2 HDR KO EF1α); TCR#2, контролируемый промотором эндогенного TRAC (расположенным выше внутри рамки P2A элементом проскока рибосомы), нацеленный на интеграцию на TRAC локусе посредством HDR (TCR #2 HDR KO P2A); TCR#2, случайно интегрированный с применением лентивирусного конструкта (TCR #2 Lenti); TCR#2, случайно интегрированный с применением лентивирусного конструкта в клетки, содержащие нокаут эндогенного TRAC (TCR #2 Lenti KO); и ссылочный TCR, способный связываться с HPV 16 E7, но содержащий мышиные Cα и Cβ области, случайно интегрированный с применением лентивирусного конструкта (Lenti Ref). В качестве контроля применяют мышей, которые не получают сконструированные клетки (только опухоль), или которым вводят клетки, обработанные в тех же условиях, которые применяют для электропорации, но без добавления RNP (mock KO). В таблице E1 также указаны номера мышей и общее количество введенных T клеток для каждой дозы в каждой группе.
Таблица E1. План исследования для оценки in vivo противоопухолевого действия в модели опухоли у мышей.
(Vбета)
Средний размер опухоли оценивают два раза в неделю в течение вплоть до 58 дней после введения сконструированных клеток. Изменения массы тела, выживание мышей и количество мышей, не имеющих опухоль на 58 день также отслеживают.
Результаты противоопухолевого действия (снижение массы опухоли) и выживания мышей, которым вводили экспрессирующие TCR #2 клетки в двух разных дозах показаны на ФИГ. 13A-13B и ФИГ. 14A-14B. Как показано на ФИГ. 13A-13B, снижение объема опухоли у мышей, которым вводили экспрессирующие TCR #2 клетки посредством HDR-медиированной направленной интеграции, под контролем либо EF1α (TCR #2HDR KO EF1α), либо промотора эндогенного TRAC (TCR #2HDR KO P2A), было больше, чем у мышей, которым вводили клетки, экспрессирующие TCR #2, случайной интеграцией посредством лентивирусной трансдукции (TCR #2 Lenti или TCR #2 Lenti KO). В обеих дозах, снижение объема опухоли у мышей, которым вводили экспрессирующие TCR #2 клетки посредством HDR-медиированной направленной интеграции последовательностей, кодирующих TCR, под контролем EF1α промотора (TCR #2HDR KO EF1α), было сравнимо или больше по сравнению с мышами, которым вводили клетки, экспрессирующие ссылочный TCR (Lenti Ref). Как показано на ФИГ. 14A-14B, % выживания мышей, которым вводили экспрессирующие TCR #2 клетки посредством HDR-медиированной направленной интеграции, был выше, чем у мышей, которым вводили клетки, экспрессирующие TCR, созданные случайной интеграцией (TCR #2 Lenti или TCR #2 Lenti KO). Как показано в таблице E2, количество мышей, которые не имели опухоль на 58 день после введения клеток, также было больше в группах, которым вводили экспрессирующие TCR #2 клетки посредством HDR-медиированной направленной интеграции, под контролем либо EF1α (TCR #2HDR KO EF1α), либо промотора эндогенного TRAC (TCR #2HDR KO P2A), по сравнению с другими группами. Изменение массы тела в курсе исследования также отслеживали, как показано на ФИГ. 15A-15B.
Таблица E2. Количество мышей без опухоли на 58 день после введения сконструированных клеток.
Эти результаты согласуются с наблюдением, что введение клеток, созданных направленной интеграцией последовательностей, кодирующих типовые рекомбинантные TCR посредством HDR, дает большую in vivo противоопухолевую активность по сравнению с клетками, созданным введением кодирующих TCR последовательностей случайной интеграцией.
В. Оценка фармакокинетики (ФК)
Жизнестойкость и размножение клеток оценивают в мышиной модели, описанной выше в примере 4A. Мышей в каждой группе, перечисленной в таблице E1, отбирают альтернативно каждые 2 недели (первая когорта из 4 мышей в дни 7 и 21; вторая когорта из 3 мышей в дни 14 и 28), и каждую леченную группу оценивают один раз в неделю (в дни 7, 14, 21 и 28). Для оценки фармакокинетики введенных клеток, клетки считают, и экспрессию разных маркеров (CD3, CD4, CD8, CD45, CD45RA, CCR7, PD-1) и рекомбинантного TCR (с применением анти-Vбета, специфического для рекомбинантного TCR), и связывание пептид-MHC тетрамера определяют проточной цитометрией в каждый момент времени.
Клетки, экспрессирующие TCR #2 посредством HDR-медиированной интеграции под контролем либо EF1α, либо P2 промотора, показали раннее начальное повышение с последующим быстрым снижением в анализе крови экспрессирующих TCR клеток между днем 14 и днем 21. Клетки, экспрессирующие ссылочный TCR, показали другую кинетику размножения и жизнестойкости in vivo, меньшее снижение циркулирующих, TCR-экспрессирующих клеток. Из анализа доли CD4 или CD8 клетки в общем количестве TCR+ клеток в течение времени, видно, что клетки сохраняющие экспрессию эндогенного TCR, демонстрируют большую долю CD4 в ранние моменты времени, по сравнению с клетками, который содержат нокаут локуса эндогенного TRAC. В общем, повышение доли CD4 наблюдается с течение времени для большинства групп.
Окрашивание клеточными фенотипическими маркерами, такими как CD45RA и CCR7, показал, что фенотип большинства циркулирующих клеток, экспрессирующих рекомбинантный TCR, изменился с CCR7+CD45RA+ (связанного с более интактным фенотипом) на CCR7-CD45RA- (связанный с более зрелым фенотипом) между днем 7 и днем 14. Доля экспрессирующих PD-1 TCR+ клеток также повышается в течение времени, от практически 0% окрашивания до около 60% клеток, демонстрирующих окрашивание PD-1, в большинстве групп.
В общем, фармакокинетический анализ демонстрирует увеличение периферического размножения и повышение численности клеток, экспрессирующих рекомбинантный TCR #2 посредством HDR-медиированной интеграции под контролем EF1α промотора, согласующееся с наблюдением высокой противоопухолевой активности в низкой дозе. Кроме того, клетки, экспрессирующие рекомбинантный TCR #2, обычно демонстрируют боле зрелый фенотип с течением времени, и видно, что общая кинетика размножения и жизнеспособности отличается от клеток, экспрессирующих ссылочный TCR.
Пример 5. Оценка длины плеч гомологии для эффективной HDR-медиированной интеграции трансгенных последовательностей
Полинуклеотиды, содержащие типовой трансген, окруженные плечом гомологии разной длины, вводят в T клетки для направленной интеграции трансгена через репарацию, направляемую гомологией (HDR), и оценивают эффективность интеграции.
А. Рекомбинантные трансгенные конструкты и создание конструированных Т клеток
Типовые HDR матричные полинуклеотиды, содержащие трансгенную последовательность, кодирующую зеленый флуоресцентный белок (GFP), окруженный плечами гомологии различной длины, для направленной интеграции в локус постоянной области человеческого TCRα (TRAC), вводят в T клетки. Более конкретно, для интеграции посредством HDR, препараты AAV, содержащие конструкты матричного нуклеотида, кодирующие GFP, создают по существу как описано в примерах 1-3, за исключением следующих отличий: создают конструкты AAV, содержащие полинуклеотид, кодирующий GFP под функциональным контролем MND промотора, и связывают с SV40 поли(A) последовательностью, окруженной 50, 100, 200, 300, 400, 500 или 600 парами оснований 5’ и 3’ плеч гомологии (SEQ ID NOS: 227-233 и 234-240, соответственно), гомологичных последовательностям, окружающим целевой сайт интеграции в гене постоянной области человеческого TCRα (TRAC). Полную длину AAV конструкта делают постоянной с применением филлер-ДНК последовательностей между SV40 поли(A) последовательностью и 3’ плечом гомологии.
Для направленной интеграции посредством HDR, первичные человеческие CD4+ и CD8+ T клетки от четырех доноров-людей (доноры 1-4) объединяют в соотношении 1:1, стимулируют и подвергают электропорации с рибонуклеопротеиновыми (RNP) комплексами, содержащими TRAC-направленную нРНК, в основном как описано в примерах 1-3. После электропорации, клетки трансдуцируют с AAV препаратами, содержащими HDR матричные полинуклеотиды, содержащие разные длины плеч гомологии, описанные выше. Экспрессию GFP измеряют проточной цитометрией через 24, 48, 72, 96 часов и 7 дней после трансдукции с AAV, для определения коэффициента интеграции (представляющего долю всех AAV внутри клетки, которая интегрирована в геном) на основе следующей формулы:
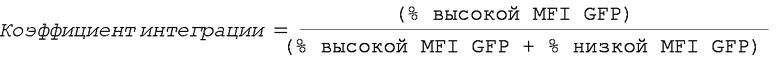
% высокой MFI и % низкой MFI показывает долю клеток, которая выше или ниже порога MFI согласно проточной цитометрии, представляющую клетки, содержащие интегрированный GFP трансген или не интегрированный AAV конструкт, соответственно. Изменения коэффициента интеграции при повышении длины плеча гомологии оценивают вычитанием коэффициента интеграции следующей наиболее короткой длины плеча из коэффициента интеграции конкретной длины плеча.
А. Оценка коэффициента интеграции
Как показано на ФИГ. 16A-16B, постоянный или повышенный коэффициент интеграции, наблюдаемый в различные моменты времени, оценивают для каждой длины плеча гомологии, сто согласуется с не интегрированными AAV конструктами, ослабевающими в течение времени. Оценка изменений в шаблонах экспрессии GFP в течение времени показала, что доля клеток с интегрированным GFP трансгеном (высокая MFI) обычно остается постоянной через 72 часа, но доля клеток, содержащих только не интегрированный AAV (низкая MFI), продолжает снижаться.
Как показано на ФИГ. 17A-17B, похоже, что наиболее значительный прирост коэффициента интеграции возникает при 200 п.о. плеча гомологии, с незначительным приростом при 300 п.о. Никакого значительного прироста не наблюдается между 300 п.о. и 500 п.о., и повышенный коэффициент интеграции наблюдается между 500 п.о. и 600 п.о., у всех доноров. Результаты поддерживают применение минимум 300 п.о. плеч гомологии для высокой эффективности интеграции, где 600 п.о. плеч гомологии, дают даже большую эффективность интеграции.
Пример 6. Создание и оценка сконструированных Т клеток, экспрессирующих химерный антигенный рецептор (CAR), направленным нокином или случайной интеграцией последовательностей, кодирующих TCR
Полинуклеотиды, кодирующие типовые химерные антигенные рецептор (CAR) вводят в T клетки с генетическим разрушением на локусе эндогенного гена, который кодирует цепь Т-клеточного рецептора альфа (TCRα), посредством CRISPR/Cas9 медиированного редактирования генома и направленной интеграции на сайте генетического разрушения, через репарацию, направляемую гомологией (HDR) или случайной интеграцией.
А. Типовые анти-CD19 CAR
а. Экспрессия типовых анти-CD19 CAR
Последовательности нуклеиновых кислот, кодирующие типовой CAR, специфический для антигенного кластера дифференциации 19 (анти-CD19 CAR) нацеливают для интеграции в один из сайтов генетического разрушения через репарацию, направляемую гомологией (HDR), или вводят случайной интеграцией посредством ретровирусной трансдукции, в клетки, сконструированные для генетического разрушения локуса эндогенного гена, который кодирует цепь Т-клеточного рецептора альфа (TCRα), посредством CRISPR/Cas9 медиированного редактирования генома, в общем как описано в примерах 1 и 2.
Проводят три исследования с применением AAV (для HDR) и ретровирусных конструктов (для случайной интеграции) по существу как описано в примерах 1 и 2, за исключением следующих отличий: трансгенные последовательности включают последовательности нуклеиновых кислот, кодирующие типовой анти-CD19 CAR и EF1α промотор для управления экспрессией типовых анти-CD19 CAR последовательностей (TRAC HDR EF1α промотор) или последовательностей, кодирующих P2A элемент проскока рибосомы выше типовых анти-CD19 CAR последовательностей (TRAC HDR P2A), для управления экспрессией типового анти-CD19 CAR из локуса эндогенного TCRα при HDR-медиированной направленной интеграции внутри рамки в гене человеческой TCR α постоянной области (TRAC).
Для направленной интеграции посредством HDR, первичные человеческие CD4+ и CD8+ T клетки стимулируют, культивируют и подвергают электропорации с рибонуклеопротеиновыми (RNP) комплексами, содержащими TRAC-направленную нРНК, в общем как описано в примерах 1 и 2. После электропорации, клетки трансдуцируют с AAV препаратами, содержащими полинуклеотиды, которые кодированы типовыми анти-CD19 CAR для целенаправленного воздействия на локус эндогенного TRAC, в общем как описано выше. Для случайной интеграции, первичные человеческие CD4+ и CD8+ T клетки трансдуцируют с ретровирусным препаратом, который кодирован типовым анти-CD19 CAR, с (ретровирус TRAC RNP или ретровирус TCR KO) или без (только ретровирус) электропорации RNP комплексов, содержащих TRAC-направленную нРНК. В качестве контроля, клетки подвергают HDR, направленной на TRAC локус после электропорации с RNP комплексами, содержащими TRBC-направленную нРНК, или фиктивно обрабатывают (Mock). На 9 день после оттаивания клеток, сконструированные клетки оценивают проточной цитометрией для окрашивания анти-CD3 антителом или анти-идиотипическим антителом (анти-ID), которое специфически распознает анти-CD19 CAR.
Как показано на ФИГ. 18A, экспрессирующие анти-CD19 CAR T клетки, созданные HDR-медиированной направленной интеграцией на TRAC локусе, демонстрируют наивысшую долю клеток, связанных анти-ID антителом. Как показано на ФИГ. 18B, эффективность интеграции и экспрессии типового анти-CD19 CAR, созданного HDR-медиированной направленной интеграцией на TRAC локусе, сравнима с или выше чем эффективность, наблюдаемая в клетках, сконструированных с применением случайной интеграции, с или без редактирования генома локуса эндогенного TRAC. Результаты согласуются с наблюдением подобной или улучшенной экспрессии типового анти-CD19 CAR в клетках, сконструированных HDR-медиированной направленной интеграции на TRAC локус, по сравнению с клетками, сконструированными случайной интеграцией.
b. Экспрессия типовых анти-CD19 CAR после серийной повторной стимуляцией
Клеточную поверхностную экспрессию CAR оценивают после множественных раундов обработки целевым антигеном. Способность CAR T клеток размножаться и демонстрировать антиген-специфическую функцию ex vivo после повторных раундов антигенного стимулирования, может коррелировать с in vivo функцией и/или способностью клетки выживать in vivo (например, после введения и исходной активации в ответ на столкновение с антигеном) (Zhao et al. (2015) Cancer Cell, 28:415-28).
Первичные человеческие T клетки, экспрессирующие типовой анти-CD19 CAR, созданные HDR-медиированной направленной интеграцией или случайной интеграцией, как описано выше, инкубируют с K562 клетками человеческого хронического миелогенного лейкоза (CML), сконструированными для экспрессии CD19 (K562-CD19). Облученные K562-CD19 целевые клетки добавляют в отношении эффектора к цели (E:T) 2,5:1. Каждые 4 дня (начало каждого нового раунда), CAR T клетки считают, собирают и повторно высевают с исходной плотностью посева со свежей средой со свежеоттаявшими, свежеоблученными целевыми клетками в количестве всего 3 раундов. На каждом раунде долю CAR-экспрессирующих клеток, среднюю интенсивность флуоресценции (MFI), коэффициент вариации (КВ) экспрессии CAR оценивают проточной цитометрией.
Как показано на ФИГ. 19A, доля клеток, экспрессирующих анти-CD19 CAR, обычно повышается или является стабильной во всех тестированных группах. Как показано на ФИГ. 19B и 19C, в течение повторяющихся стимуляций, типовые анти-CD19 CAR-экспрессирующие клетки, сконструированные HDR-медиированной направленной интеграцией последовательности нуклеиновых кислот на локусе эндогенного TRAC, под контролем EF1α промотора или P2A (эндогенного TRAC промотора), демонстрируют более однородную и менее переменную экспрессию CAR, по сравнению с клетками, сконструированными случайной интеграцией с применением ретровирусного вектора. MFI обычно выше в клетках, сконструированных случайной интеграцией (ФИГ. 19B). Анти-CD19 CAR-экспрессирующие клетки, сконструированные с применением HDR-медиированного целенаправленного воздействия на TRAC локус, демонстрируют более низкий коэффициент вариации (стандартное отклонение сигнала в популяции клеток, деленное на среднее значение сигнала в соответствующей популяции; ФИГ. 19C), показывающий меньшую флуктуацию в экспрессии в течение повторяющегося стимулирования.
с. Цитолитическая активность и образование цитокина
Образование цитокина (интерферон-гамма; IFNγ) и цитолитическую активность отслеживают после инкубации анти-CD19 CAR+ T эффекторных клеток с K562 целевыми клетками, сконструированными для экспрессии CD19 (K562-CD19) или не сконструированным контролем (K562 исходные), при E:T отношении 2:1 в течение 48 часов, в общем как описано в примере 1.D выше.
Как показано на ФИГ. 20A, анти-CD19 CAR-экспрессирующие клетки, сконструированные с применением HDR-медиированного целенаправленного воздействия на TRAC локус, демонстрируют более высокое образование антиген-специфического IFNγ, где клетки, сконструированные посредством HDR с EF1α промотором, демонстрируют наивысшее образование антиген-специфического IFNγ (левая панель), и более низкое образование не специфического или нецелевого цитокина (правая панель). Как показано на ФИГ. 20B, степень уничтожения целевой клетки была одинаковой во всех группах анти-CD19 CAR-экспрессирующих клеток (левая панель). Не специфическое уничтожение клеток варьируется, где клетки, сконструированные HDR с P2A (эндогенным TRAC промотором), демонстрируют наименьшее неспецифическое уничтожение клеток (правая панель).
В. Типовые анти-BCMA CAR
а. Экспрессия типового Анти-BCMA CAR
Последовательности нуклеиновых кислот, кодирующие разные типовые CAR, специфические для антигена созревания B клетки (анти-BCMA CAR) вводят направленной интеграцией, медиированной HDR на TRAC локусе под контролем EF1α или промотора эндогенного TRAC (интеграцией P2A последовательности), или случайной интеграцией с применением лентивирусного вектора, с (LV-TRAC или LV-KO) или без (только LV) введения RNP комплексов, содержащих TRAC-направленную нРНК, в общем как описано в примерах 1, 2 и 6A выше. В качестве контроля, клетки подвергают HDR нацеливанию на TRAC локус после электропорации с RNP комплексами, содержащими TRBC-направленную нРНК, или фиктивно обрабатывают (Фиктивная). На 9 день после оттаивания клеток, сконструированные клетки оценивают проточной цитометрией для определения экспрессии анти-BCMA CAR оттаиванием с BCMA-Fc слитым полипептидом, содержащим растворимый человеческий BCMA, слитый на его C-окончании с Fc областью IgG, который специфически связывается с анти-BCMA CAR, и экспрессии CD3.
Как показано на ФИГ. 21A, экспрессирующие анти-BCMA CAR- T клетки, созданные HDR-медиированной направленной интеграцией на TRAC локус, демонстрирует сравнимую долю клеток, связанных BCMA-Fc, с клетками, сконструированными с применением случайной интеграции. Как показано на ФИГ. 21B, доля клеток, экспрессирующих типовой анти-BCMA CAR, созданных HDR-медиированной направленной интеграцией на TRAC локусе, была сравнима с долей, наблюдаемой в клетках, сконструированных с применением случайной интеграции, с или без редактирования генома локуса эндогенного TRAC. Результаты согласуются с наблюдением подобной или улучшенной экспрессии типового анти-BCMAХАР в клетках, сконструированных HDR-медиированной направленной интеграцией на TRAC локусе, по сравнению с клетками, сконструированными с применением случайной интеграции.
b. Экспрессия и активность типового BCMA CAR после серийного повторного стимулирования
Экспрессию CAR на поверхности клеток и антиген-специфическую активность сконструированных клеток оценивают после множества раундов обработки целевым антигеном. Экспрессирующие анти-BCMA CAR клетки, созданные как описано выше, инкубируют с облученными экспрессирующими BCMA RPMI-8226 (колония клеток множественной миеломы BCMA+) целевые клетки, при отношении эффектора к цели (E:T) 1:1. Каждые 3-7 дней после начала каждого нового раунда, CAR T клетки считают, собирают и повторно высевают при исходной плотности посева со свежей средой и свежеоттаявшими, свежеоблученными целевыми клетками в течение всего 4 раундов. Доля клеток, экспрессирующих анти-BCMA CAR, определяют проточной цитометрией. Для оценки образования цитокина, клетки на первом, втором или третьем раунде серийного стимулирования собирают через 24 часа после стимулирования, и оценивают образование интерферона гамма (IFNγ) и интерлейкина-2 (IL-2).
Как показано на ФИГ. 22A, обычно наблюдают, что доля экспрессирующих анти-BCMA CAR клеток является одинаковой в течение множества раундов стимулирования, в клетках, сконструированных разными способами. Как показано на ФИГ. 22B, наблюдают, что образование IFNγ (верхняя панель) является одинаковым в клетках, сконструированных с применением разных способов. Клетки, сконструированные направленной интеграцией на TRAC локусе посредством HDR, под функциональным контролем EF1α промотора, демонстрируют наивысшее образование IL-2 после стимулирования с целевыми клетками.
С. Заключение
Как показано, клетки, сконструированные для экспрессии типовых CAR посредством направленной интеграции последовательности нуклеиновых кислот в локус эндогенного TRAC, демонстрируют похожую или улучшенную экспрессию и антиген-специфическую активность и более однородную экспрессию, в том числе в контексте множественных раундов стимулирования.
Настоящее изобретение не предназначено для ограничения объема конкретными раскрытыми вариантами осуществления, которые предоставляются, например, для иллюстрации различных аспектов изобретения. Различные модификации описанных композиций и способов станут очевидными из приведенного здесь описания и идей. Такие изменения могут быть применены на практике без отклонения от истинного объема и сути раскрытия, и предназначены для того, чтобы входить в объем настоящего раскрытия.
ПОСЛЕДОВАТЕЛЬНОСТИ
NCBI Ссылочная последовательность: NG_001333,2, TRBC1
NCBI Ссылочная последовательность: NG_001333,2, TRBC2
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> JUNO THERAPEUTICS, INC.
EDITAS MEDICINE, INC.
SATHER, Blythe D.
BORGES, Christopher
BURLEIGH, Stephen Michael
NYE, Christopher Heath
VONG, Queenie
WELSTEAD, G. Grant
<120> СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ РЕКОМБИНАНТНЫЙ
РЕЦЕПТОР, И РОДСТВЕННЫЕ КОМПОЗИЦИИ
<130> 735042012740
<140> Еще не передано
<141> Одновременно с этим
<150> 62/653,522
<151> 2018-04-05
<160> 256
<170> FastSEQ for Windows Version 4.0
<210> 1
<211> 4627
<212> ДНК
<213> Homo sapiens
<220>
<223> Человеческий альфа константа TCR (TRAC)
<300>
<308> NG_001332.3
<309> 2019-02-26
<400> 1
atatccagaa ccctgaccct gccgtgtacc agctgagaga ctctaaatcc agtgacaagt 60
ctgtctgcct attcaccgat tttgattctc aaacaaatgt gtcacaaagt aaggattctg 120
atgtgtatat cacagacaaa actgtgctag acatgaggtc tatggacttc aagagcaaca 180
gtgctgtggc ctggagcaac aaatctgact ttgcatgtgc aaacgccttc aacaacagca 240
ttattccaga agacaccttc ttccccagcc caggtaaggg cagctttggt gccttcgcag 300
gctgtttcct tgcttcagga atggccaggt tctgcccaga gctctggtca atgatgtcta 360
aaactcctct gattggtggt ctcggcctta tccattgcca ccaaaaccct ctttttacta 420
agaaacagtg agccttgttc tggcagtcca gagaatgaca cgggaaaaaa gcagatgaag 480
agaaggtggc aggagagggc acgtggccca gcctcagtct ctccaactga gttcctgcct 540
gcctgccttt gctcagactg tttgcccctt actgctcttc taggcctcat tctaagcccc 600
ttctccaagt tgcctctcct tatttctccc tgtctgccaa aaaatctttc ccagctcact 660
aagtcagtct cacgcagtca ctcattaacc caccaatcac tgattgtgcc ggcacatgaa 720
tgcaccaggt gttgaagtgg aggaattaaa aagtcagatg aggggtgtgc ccagaggaag 780
caccattcta gttgggggag cccatctgtc agctgggaaa agtccaaata acttcagatt 840
ggaatgtgtt ttaactcagg gttgagaaaa cagctacctt caggacaaaa gtcagggaag 900
ggctctctga agaaatgcta cttgaagata ccagccctac caagggcagg gagaggaccc 960
tatagaggcc tgggacagga gctcaatgag aaaggagaag agcagcaggc atgagttgaa 1020
tgaaggaggc agggccgggt cacagggcct tctaggccat gagagggtag acagtattct 1080
aaggacgcca gaaagctgtt gatcggcttc aagcagggga gggacaccta atttgctttt 1140
cttttttttt tttttttttt tttttttttt tgagatggag ttttgctctt gttgcccagg 1200
ctggagtgca atggtgcatc ttggctcact gcaacctccg cctcccaggt tcaagtgatt 1260
ctcctgcctc agcctcccga gtagctgaga ttacaggcac ccgccaccat gcctggctaa 1320
ttttttgtat ttttagtaga gacagggttt cactatgttg gccaggctgg tctcgaactc 1380
ctgacctcag gtgatccacc cgcttcagcc tcccaaagtg ctgggattac aggcgtgagc 1440
caccacaccc ggcctgcttt tcttaaagat caatctgagt gctgtacgga gagtgggttg 1500
taagccaaga gtagaagcag aaagggagca gttgcagcag agagatgatg gaggcctggg 1560
cagggtggtg gcagggaggt aaccaacacc attcaggttt caaaggtaga accatgcagg 1620
gatgagaaag caaagagggg atcaaggaag gcagctggat tttggcctga gcagctgagt 1680
caatgatagt gccgtttact aagaagaaac caaggaaaaa atttggggtg cagggatcaa 1740
aactttttgg aacatatgaa agtacgtgtt tatactcttt atggcccttg tcactatgta 1800
tgcctcgctg cctccattgg actctagaat gaagccaggc aagagcaggg tctatgtgtg 1860
atggcacatg tggccagggt catgcaacat gtactttgta caaacagtgt atattgagta 1920
aatagaaatg gtgtccagga gccgaggtat cggtcctgcc agggccaggg gctctcccta 1980
gcaggtgctc atatgctgta agttccctcc agatctctcc acaaggaggc atggaaaggc 2040
tgtagttgtt cacctgccca agaactagga ggtctggggt gggagagtca gcctgctctg 2100
gatgctgaaa gaatgtctgt ttttcctttt agaaagttcc tgtgatgtca agctggtcga 2160
gaaaagcttt gaaacaggta agacaggggt ctagcctggg tttgcacagg attgcggaag 2220
tgatgaaccc gcaataaccc tgcctggatg agggagtggg aagaaattag tagatgtggg 2280
aatgaatgat gaggaatgga aacagcggtt caagacctgc ccagagctgg gtggggtctc 2340
tcctgaatcc ctctcaccat ctctgacttt ccattctaag cactttgagg atgagtttct 2400
agcttcaata gaccaaggac tctctcctag gcctctgtat tcctttcaac agctccactg 2460
tcaagagagc cagagagagc ttctgggtgg cccagctgtg aaatttctga gtcccttagg 2520
gatagcccta aacgaaccag atcatcctga ggacagccaa gaggttttgc cttctttcaa 2580
gacaagcaac agtactcaca taggctgtgg gcaatggtcc tgtctctcaa gaatcccctg 2640
ccactcctca cacccaccct gggcccatat tcatttccat ttgagttgtt cttattgagt 2700
catccttcct gtggtagcgg aactcactaa ggggcccatc tggacccgag gtattgtgat 2760
gataaattct gagcacctac cccatcccca gaagggctca gaaataaaat aagagccaag 2820
tctagtcggt gtttcctgtc ttgaaacaca atactgttgg ccctggaaga atgcacagaa 2880
tctgtttgta aggggatatg cacagaagct gcaagggaca ggaggtgcag gagctgcagg 2940
cctcccccac ccagcctgct ctgccttggg gaaaaccgtg ggtgtgtcct gcaggccatg 3000
caggcctggg acatgcaagc ccataaccgc tgtggcctct tggttttaca gatacgaacc 3060
taaactttca aaacctgtca gtgattgggt tccgaatcct cctcctgaaa gtggccgggt 3120
ttaatctgct catgacgctg cggctgtggt ccagctgagg tgaggggcct tgaagctggg 3180
agtggggttt agggacgcgg gtctctgggt gcatcctaag ctctgagagc aaacctccct 3240
gcagggtctt gcttttaagt ccaaagcctg agcccaccaa actctcctac ttcttcctgt 3300
tacaaattcc tcttgtgcaa taataatggc ctgaaacgct gtaaaatatc ctcatttcag 3360
ccgcctcagt tgcacttctc ccctatgagg taggaagaac agttgtttag aaacgaagaa 3420
actgaggccc cacagctaat gagtggagga agagagacac ttgtgtacac cacatgcctt 3480
gtgttgtact tctctcaccg tgtaacctcc tcatgtcctc tctccccagt acggctctct 3540
tagctcagta gaaagaagac attacactca tattacaccc caatcctggc tagagtctcc 3600
gcaccctcct cccccagggt ccccagtcgt cttgctgaca actgcatcct gttccatcac 3660
catcaaaaaa aaactccagg ctgggtgcgg gggctcacac ctgtaatccc agcactttgg 3720
gaggcagagg caggaggagc acaggagctg gagaccagcc tgggcaacac agggagaccc 3780
cgcctctaca aaaagtgaaa aaattaacca ggtgtggtgc tgcacacctg tagtcccagc 3840
tacttaagag gctgagatgg gaggatcgct tgagccctgg aatgttgagg ctacaatgag 3900
ctgtgattgc gtcactgcac tccagcctgg aagacaaagc aagatcctgt ctcaaataat 3960
aaaaaaaata agaactccag ggtacatttg ctcctagaac tctaccacat agccccaaac 4020
agagccatca ccatcacatc cctaacagtc ctgggtcttc ctcagtgtcc agcctgactt 4080
ctgttcttcc tcattccaga tctgcaagat tgtaagacag cctgtgctcc ctcgctcctt 4140
cctctgcatt gcccctcttc tccctctcca aacagaggga actctcctac ccccaaggag 4200
gtgaaagctg ctaccacctc tgtgcccccc cggcaatgcc accaactgga tcctacccga 4260
atttatgatt aagattgctg aagagctgcc aaacactgct gccaccccct ctgttccctt 4320
attgctgctt gtcactgcct gacattcacg gcagaggcaa ggctgctgca gcctcccctg 4380
gctgtgcaca ttccctcctg ctccccagag actgcctccg ccatcccaca gatgatggat 4440
cttcagtggg ttctcttggg ctctaggtcc tgcagaatgt tgtgaggggt ttattttttt 4500
ttaatagtgt tcataaagaa atacatagta ttcttcttct caagacgtgg ggggaaatta 4560
tctcattatc gaggccctgc tatgctgtgt atctgggcgt gttgtatgtc ctgctgccga 4620
tgccttc 4627
<210> 2
<211> 1448
<212> ДНК
<213> Homo sapiens
<220>
<223> Человеческий бета константа TCR 1 (TRBC1)
<300>
<308> NG_001333.2
<309> 2018-09-23
<400> 2
aggacctgaa caaggtgttc ccacccgagg tcgctgtgtt tgagccatca gaagcagaga 60
tctcccacac ccaaaaggcc acactggtgt gcctggccac aggcttcttc cccgaccacg 120
tggagctgag ctggtgggtg aatgggaagg aggtgcacag tggggtcagc acagacccgc 180
agcccctcaa ggagcagccc gccctcaatg actccagata ctgcctgagc agccgcctga 240
gggtctcggc caccttctgg cagaaccccc gcaaccactt ccgctgtcaa gtccagttct 300
acgggctctc ggagaatgac gagtggaccc aggatagggc caaacccgtc acccagatcg 360
tcagcgccga ggcctggggt agagcaggtg agtggggcct ggggagatgc ctggaggaga 420
ttaggtgaga ccagctacca gggaaaatgg aaagatccag gtagcagaca agactagatc 480
caaaaagaaa ggaaccagcg cacaccatga aggagaattg ggcacctgtg gttcattctt 540
ctcccagatt ctcagcccaa cagagccaag cagctgggtc ccctttctat gtggcctgtg 600
taactctcat ctgggtggtg ccccccatcc ccctcagtgc tgccacatgc catggattgc 660
aaggacaatg tggctgacat ctgcatggca gaagaaagga ggtgctgggc tgtcagagga 720
agctggtctg ggcctgggag tctgtgccaa ctgcaaatct gactttactt ttaattgcct 780
atgaaaataa ggtctctcat ttattttcct ctccctgctt tctttcagac tgtggcttta 840
cctcgggtaa gtaagccctt ccttttcctc tccctctctc atggttcttg acctagaacc 900
aaggcatgaa gaactcacag acactggagg gtggagggtg ggagagacca gagctacctg 960
tgcacaggta cccacctgtc cttcctccgt gccaacagtg tcctaccagc aaggggtcct 1020
gtctgccacc atcctctatg agatcctgct agggaaggcc accctgtatg ctgtgctggt 1080
cagcgccctt gtgttgatgg ccatggtaag caggagggca ggatggggcc agcaggctgg 1140
aggtgacaca ctgacaccaa gcacccagaa gtatagagtc cctgccagga ttggagctgg 1200
gcagtaggga gggaagagat ttcattcagg tgcctcagaa gataacttgc acctctgtag 1260
gatcacagtg gaagggtcat gctgggaagg agaagctgga gtcaccagaa aacccaatgg 1320
atgttgtgat gagccttact atttgtgtgg tcaatgggcc ctactacttt ctctcaatcc 1380
tcacaactcc tggctcttaa taacccccaa aactttctct tctgcaggtc aagagaaagg 1440
atttctga 1448
<210> 3
<211> 1489
<212> ДНК
<213> Homo sapiens
<220>
<223> Человеческий бета константа TCR 2 (TRBC2)
<300>
<308> NG_001333.2
<309> 2018-09-23
<400> 3
aggacctgaa aaacgtgttc ccacccgagg tcgctgtgtt tgagccatca gaagcagaga 60
tctcccacac ccaaaaggcc acactggtat gcctggccac aggcttctac cccgaccacg 120
tggagctgag ctggtgggtg aatgggaagg aggtgcacag tggggtcagc acagacccgc 180
agcccctcaa ggagcagccc gccctcaatg actccagata ctgcctgagc agccgcctga 240
gggtctcggc caccttctgg cagaaccccc gcaaccactt ccgctgtcaa gtccagttct 300
acgggctctc ggagaatgac gagtggaccc aggatagggc caaacccgtc acccagatcg 360
tcagcgccga ggcctggggt agagcaggtg agtggggcct ggggagatgc ctggaggaga 420
ttaggtgaga ccagctacca gggaaaatgg aaagatccag gtagcggaca agactagatc 480
cagaagaaag ccagagtgga caaggtggga tgatcaaggt tcacagggtc agcaaagcac 540
ggtgtgcact tcccccacca agaagcatag aggctgaatg gagcacctca agctcattct 600
tccttcagat cctgacacct tagagctaag ctttcaagtc tccctgagga ccagccatac 660
agctcagcat ctgagtggtg tgcatcccat tctcttctgg ggtcctggtt tcctaagatc 720
atagtgacca cttcgctggc actggagcag catgagggag acagaaccag ggctatcaaa 780
ggaggctgac tttgtactat ctgatatgca tgtgtttgtg gcctgtgagt ctgtgatgta 840
aggctcaatg tccttacaaa gcagcattct ctcatccatt tttcttcccc tgttttcttt 900
cagactgtgg cttcacctcc ggtaagtgag tctctccttt ttctctctat ctttcgccgt 960
ctctgctctc gaaccagggc atggagaatc cacggacaca ggggcgtgag ggaggccaga 1020
gccacctgtg cacaggtgcc tacatgctct gttcttgtca acagagtctt accagcaagg 1080
ggtcctgtct gccaccatcc tctatgagat cttgctaggg aaggccacct tgtatgccgt 1140
gctggtcagt gccctcgtgc tgatggccat ggtaaggagg agggtgggat agggcagatg 1200
atgggggcag gggatggaac atcacacatg ggcataaagg aatctcagag ccagagcaca 1260
gcctaatata tcctatcacc tcaatgaaac cataatgaag ccagactggg gagaaaatgc 1320
agggaatatc acagaatgca tcatgggagg atggagacaa ccagcgagcc ctactcaaat 1380
taggcctcag agcccgcctc ccctgcccta ctcctgctgt gccatagccc ctgaaaccct 1440
gaaaatgttc tctcttccac aggtcaagag aaaggattcc agaggctag 1489
<210> 4
<211> 1189
<212> ДНК
<213> Искусственная последовательность
<220>
<223> EF1 альфа промотор
<300>
<308> GenBank: J04617.1
<309> 1994-11-07
<400> 4
cgtgaggctc cggtgcccgt cagtgggcag agcgcacatc gcccacagtc cccgagaagt 60
tggggggagg ggtcggcaat tgaaccggtg cctagagaag gtggcgcggg gtaaactggg 120
aaagtgatgt cgtgtactgg ctccgccttt ttcccgaggg tgggggagaa ccgtatataa 180
gtgcagtagt cgccgtgaac gttctttttc gcaacgggtt tgccgccaga acacaggtaa 240
gtgccgtgtg tggttcccgc gggcctggcc tctttacggg ttatggccct tgcgtgcctt 300
gaattacttc cacgcccctg gctgcagtac gtgattcttg atcccgagct tcgggttgga 360
agtgggtggg agagttcgag gccttgcgct taaggagccc cttcgcctcg tgcttgagtt 420
gaggcctggc ctgggcgctg gggccgccgc gtgcgaatct ggtggcacct tcgcgcctgt 480
ctcgctgctt tcgataagtc tctagccatt taaaattttt gatgacctgc tgcgacgctt 540
tttttctggc aagatagtct tgtaaatgcg ggccaagatc tgcacactgg tatttcggtt 600
tttggggccg cgggcggcga cggggcccgt gcgtcccagc gcacatgttc ggcgaggcgg 660
ggcctgcgag cgcggccacc gagaatcgga cgggggtagt ctcaagctgg ccggcctgct 720
ctggtgcctg gcctcgcgcc gccgtgtatc gccccgccct gggcggcaag gctggcccgg 780
tcggcaccag ttgcgtgagc ggaaagatgg ccgcttcccg gccctgctgc agggagctca 840
aaatggagga cgcggcgctc gggagagcgg gcgggtgagt cacccacaca aaggaaaagg 900
gcctttccgt cctcagccgt cgcttcatgt gactccacgg agtaccgggc gccgtccagg 960
cacctcgatt agttctcgag cttttggagt acgtcgtctt taggttgggg ggaggggttt 1020
tatgcgatgg agtttcccca cactgagtgg gtggagactg aagttaggcc agcttggcac 1080
ttgatgtaat tctccttgga atttgccctt tttgagtttg gatcttggtt cattctcaag 1140
cctcagacag tggttcaaag tttttttctt ccatttcagg tgtcgtgaa 1189
<210> 5
<211> 1205
<212> ДНК
<213> Искусственная последовательность
<220>
<223> EF1 альфа промотор
<400> 5
cgtgaggctc cggtgcccgt cagtgggcag agcgcacatc gcccacagtc cccgagaagt 60
tggggggagg ggtcggcaat tgaaccggtg cctagagaag gtggcgcggg gtaaactggg 120
aaagtgatgt cgtgtactgg ctccgccttt ttcccgaggg tgggggagaa ccgtatataa 180
gtgcactagt cgccgtgaac gttctttttc gcaacgggtt tgccgccaga acacaggtaa 240
gtgccgtgtg tggttcccgc gggcctggcc tctttacggg ttatggccct tgcgtgcctt 300
gaattacttc cacctggctg cagtacgtga ttcttgatcc cgagcttcgg gttggaagtg 360
ggtgggagag ttcgtggcct tgcgcttaag gagccccttc gcctcgtgct tgagttgtgg 420
cctggcctgg gcgctggggc cgccgcgtgc gaatctggtg gcaccttcgc gcctgtctcg 480
ctgctttcga taagtctcta gccatttaaa atttttgatg acctgctgcg acgctttttt 540
tctggcaaga tagtcttgta aatgcgggcc aagatcagca cactggtatt tcggtttttg 600
gggccgcggg cggcgacggg gcccgtgcgt cccagcgcac atgttcggcg aggcggggcc 660
tgcgagcgcg gccaccgaga atcggacggg ggtagtctca agctgcccgg cctgctctgg 720
tgcctggcct cgcgccgccg tgtatcgccc cgccctgggc ggcaaggctg gcccggtcgg 780
caccagttgc gtgagcggaa agatggccgc ttcccggccc tgctgcaggg agcacaaaat 840
ggaggacgcg gcgctcggga gagcgggcgg gtgagtcacc cacacaaagg aaaagggcct 900
ttccgtcctc agccgtcgct tcatgtgact ccacggagta ccgggcgccg tccaggcacc 960
tcgattagtt ctccagcttt tggagtacgt cgtctttagg ttggggggag gggttttatg 1020
cgatggagtt tccccacact gagtgggtgg agactgaagt taggccagct tggcacttga 1080
tgtaattctc cttggaattt gccctttttg agtttggatc ttggttcatt ctcaagcctc 1140
agacagtggt tcaaagtttt tttcttccat ttcaggtgtc gtgaaaacta cccctaaaag 1200
ccaaa 1205
<210> 6
<211> 24
<212> PRT
<213> Искусственная последовательность
<220>
<223> T2A
<400> 6
Leu Glu Gly Gly Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp
1 5 10 15
Val Glu Glu Asn Pro Gly Pro Arg
20
<210> 7
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> T2A
<400> 7
Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro
1 5 10 15
Gly Pro
<210> 8
<211> 22
<212> PRT
<213> Искусственная последовательность
<220>
<223> P2A
<400> 8
Gly Ser Gly Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val
1 5 10 15
Glu Glu Asn Pro Gly Pro
20
<210> 9
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> P2A
<400> 9
Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val Glu Glu Asn
1 5 10 15
Pro Gly Pro
<210> 10
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> E2A
<400> 10
Gln Cys Thr Asn Tyr Ala Leu Leu Lys Leu Ala Gly Asp Val Glu Ser
1 5 10 15
Asn Pro Gly Pro
20
<210> 11
<211> 22
<212> PRT
<213> Искусственная последовательность
<220>
<223> F2A
<400> 11
Val Lys Gln Thr Leu Asn Phe Asp Leu Leu Lys Leu Ala Gly Asp Val
1 5 10 15
Glu Ser Asn Pro Gly Pro
20
<210> 12
<211> 357
<212> PRT
<213> Искусственная последовательность
<220>
<223> EGFRt
<400> 12
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Arg Lys Val Cys Asn Gly Ile Gly Ile Gly
20 25 30
Glu Phe Lys Asp Ser Leu Ser Ile Asn Ala Thr Asn Ile Lys His Phe
35 40 45
Lys Asn Cys Thr Ser Ile Ser Gly Asp Leu His Ile Leu Pro Val Ala
50 55 60
Phe Arg Gly Asp Ser Phe Thr His Thr Pro Pro Leu Asp Pro Gln Glu
65 70 75 80
Leu Asp Ile Leu Lys Thr Val Lys Glu Ile Thr Gly Phe Leu Leu Ile
85 90 95
Gln Ala Trp Pro Glu Asn Arg Thr Asp Leu His Ala Phe Glu Asn Leu
100 105 110
Glu Ile Ile Arg Gly Arg Thr Lys Gln His Gly Gln Phe Ser Leu Ala
115 120 125
Val Val Ser Leu Asn Ile Thr Ser Leu Gly Leu Arg Ser Leu Lys Glu
130 135 140
Ile Ser Asp Gly Asp Val Ile Ile Ser Gly Asn Lys Asn Leu Cys Tyr
145 150 155 160
Ala Asn Thr Ile Asn Trp Lys Lys Leu Phe Gly Thr Ser Gly Gln Lys
165 170 175
Thr Lys Ile Ile Ser Asn Arg Gly Glu Asn Ser Cys Lys Ala Thr Gly
180 185 190
Gln Val Cys His Ala Leu Cys Ser Pro Glu Gly Cys Trp Gly Pro Glu
195 200 205
Pro Arg Asp Cys Val Ser Cys Arg Asn Val Ser Arg Gly Arg Glu Cys
210 215 220
Val Asp Lys Cys Asn Leu Leu Glu Gly Glu Pro Arg Glu Phe Val Glu
225 230 235 240
Asn Ser Glu Cys Ile Gln Cys His Pro Glu Cys Leu Pro Gln Ala Met
245 250 255
Asn Ile Thr Cys Thr Gly Arg Gly Pro Asp Asn Cys Ile Gln Cys Ala
260 265 270
His Tyr Ile Asp Gly Pro His Cys Val Lys Thr Cys Pro Ala Gly Val
275 280 285
Met Gly Glu Asn Asn Thr Leu Val Trp Lys Tyr Ala Asp Ala Gly His
290 295 300
Val Cys His Leu Cys His Pro Asn Cys Thr Tyr Gly Cys Thr Gly Pro
305 310 315 320
Gly Leu Glu Gly Cys Pro Thr Asn Gly Pro Lys Ile Pro Ser Ile Ala
325 330 335
Thr Gly Met Val Gly Ala Leu Leu Leu Leu Leu Val Val Ala Leu Gly
340 345 350
Ile Gly Leu Phe Met
355
<210> 13
<211> 335
<212> PRT
<213> Искусственная последовательность
<220>
<223> EGFRt
<400> 13
Arg Lys Val Cys Asn Gly Ile Gly Ile Gly Glu Phe Lys Asp Ser Leu
1 5 10 15
Ser Ile Asn Ala Thr Asn Ile Lys His Phe Lys Asn Cys Thr Ser Ile
20 25 30
Ser Gly Asp Leu His Ile Leu Pro Val Ala Phe Arg Gly Asp Ser Phe
35 40 45
Thr His Thr Pro Pro Leu Asp Pro Gln Glu Leu Asp Ile Leu Lys Thr
50 55 60
Val Lys Glu Ile Thr Gly Phe Leu Leu Ile Gln Ala Trp Pro Glu Asn
65 70 75 80
Arg Thr Asp Leu His Ala Phe Glu Asn Leu Glu Ile Ile Arg Gly Arg
85 90 95
Thr Lys Gln His Gly Gln Phe Ser Leu Ala Val Val Ser Leu Asn Ile
100 105 110
Thr Ser Leu Gly Leu Arg Ser Leu Lys Glu Ile Ser Asp Gly Asp Val
115 120 125
Ile Ile Ser Gly Asn Lys Asn Leu Cys Tyr Ala Asn Thr Ile Asn Trp
130 135 140
Lys Lys Leu Phe Gly Thr Ser Gly Gln Lys Thr Lys Ile Ile Ser Asn
145 150 155 160
Arg Gly Glu Asn Ser Cys Lys Ala Thr Gly Gln Val Cys His Ala Leu
165 170 175
Cys Ser Pro Glu Gly Cys Trp Gly Pro Glu Pro Arg Asp Cys Val Ser
180 185 190
Cys Arg Asn Val Ser Arg Gly Arg Glu Cys Val Asp Lys Cys Asn Leu
195 200 205
Leu Glu Gly Glu Pro Arg Glu Phe Val Glu Asn Ser Glu Cys Ile Gln
210 215 220
Cys His Pro Glu Cys Leu Pro Gln Ala Met Asn Ile Thr Cys Thr Gly
225 230 235 240
Arg Gly Pro Asp Asn Cys Ile Gln Cys Ala His Tyr Ile Asp Gly Pro
245 250 255
His Cys Val Lys Thr Cys Pro Ala Gly Val Met Gly Glu Asn Asn Thr
260 265 270
Leu Val Trp Lys Tyr Ala Asp Ala Gly His Val Cys His Leu Cys His
275 280 285
Pro Asn Cys Thr Tyr Gly Cys Thr Gly Pro Gly Leu Glu Gly Cys Pro
290 295 300
Thr Asn Gly Pro Lys Ile Pro Ser Ile Ala Thr Gly Met Val Gly Ala
305 310 315 320
Leu Leu Leu Leu Leu Val Val Ala Leu Gly Ile Gly Leu Phe Met
325 330 335
<210> 14
<211> 137
<212> PRT
<213> Mus musculus
<220>
<223> Мышиный альфа константа TCR
<400> 14
Asp Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro Arg
1 5 10 15
Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile
20 25 30
Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala
50 55 60
Trp Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr
65 70 75 80
Asn Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr
85 90 95
Glu Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Ser
100 105 110
Val Met Gly Leu Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu
115 120 125
Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135
<210> 15
<211> 137
<212> PRT
<213> Mus musculus
<220>
<223> Мышиный альфа константа TCR
<400> 15
Asn Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro Arg
1 5 10 15
Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile
20 25 30
Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala
50 55 60
Trp Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr
65 70 75 80
Asn Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr
85 90 95
Glu Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Ser
100 105 110
Val Met Gly Leu Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu
115 120 125
Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135
<210> 16
<211> 173
<212> PRT
<213> Mus musculus
<220>
<223> Мышиный бета константа TCR
<300>
<308> Uniprot P01852
<309> 1986-07-21
<400> 16
Glu Asp Leu Arg Asn Val Thr Pro Pro Lys Val Ser Leu Phe Glu Pro
1 5 10 15
Ser Lys Ala Glu Ile Ala Asn Lys Gln Lys Ala Thr Leu Val Cys Leu
20 25 30
Ala Arg Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
35 40 45
Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Ala Tyr Lys
50 55 60
Glu Ser Asn Tyr Ser Tyr Cys Leu Ser Ser Arg Leu Arg Val Ser Ala
65 70 75 80
Thr Phe Trp His Asn Pro Arg Asn His Phe Arg Cys Gln Val Gln Phe
85 90 95
His Gly Leu Ser Glu Glu Asp Lys Trp Pro Glu Gly Ser Pro Lys Pro
100 105 110
Val Thr Gln Asn Ile Ser Ala Glu Ala Trp Gly Arg Ala Asp Cys Gly
115 120 125
Ile Thr Ser Ala Ser Tyr Gln Gln Gly Val Leu Ser Ala Thr Ile Leu
130 135 140
Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala Val Leu Val Ser
145 150 155 160
Thr Leu Val Val Met Ala Met Val Lys Arg Lys Asn Ser
165 170
<210> 17
<211> 172
<212> PRT
<213> Mus musculus
<220>
<223> Мышиный бета константа TCR
<400> 17
Asp Leu Arg Asn Val Thr Pro Pro Lys Val Ser Leu Phe Glu Pro Ser
1 5 10 15
Lys Ala Glu Ile Ala Asn Lys Gln Lys Ala Thr Leu Val Cys Leu Ala
20 25 30
Arg Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn Gly
35 40 45
Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Ala Tyr Lys Glu
50 55 60
Ser Asn Tyr Ser Tyr Cys Leu Ser Ser Arg Leu Arg Val Ser Ala Thr
65 70 75 80
Phe Trp His Asn Pro Arg Asn His Phe Arg Cys Gln Val Gln Phe His
85 90 95
Gly Leu Ser Glu Glu Asp Lys Trp Pro Glu Gly Ser Pro Lys Pro Val
100 105 110
Thr Gln Asn Ile Ser Ala Glu Ala Trp Gly Arg Ala Asp Cys Gly Ile
115 120 125
Thr Ser Ala Ser Tyr His Gln Gly Val Leu Ser Ala Thr Ile Leu Tyr
130 135 140
Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala Val Leu Val Ser Gly
145 150 155 160
Leu Val Leu Met Ala Met Val Lys Arg Lys Asn Ser
165 170
<210> 18
<211> 181
<212> ДНК
<213> Искусственная последовательность
<220>
<223> MND промотор
<400> 18
gggtctctct ggttagacca gatctgagcc tgggagctct ctggctaact agggaaccca 60
ctgcttaagc ctcaataaag cttgccttga gtgcttcaag tagtgtgtgc ccgtctgttg 120
tgtgactctg gtaactagag atccctcaga cccttttagt cagtgtggaa aatctctagc 180
a 181
<210> 19
<211> 142
<212> PRT
<213> Homo sapiens
<220>
<223> Человеческий альфа константа TCR
<300>
<308> Uniprot P01848
<309> 2018-07-18
<400> 19
Pro Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
1 5 10 15
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
20 25 30
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
35 40 45
Thr Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
50 55 60
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
65 70 75 80
Ser Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys
85 90 95
Asp Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn
100 105 110
Phe Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val
115 120 125
Ala Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 20
<211> 177
<212> PRT
<213> Homo sapiens
<220>
<223> Человеческий бета константа TCR 1
<300>
<308> Uniprot P01850
<309> 2018-07-18
<400> 20
Glu Asp Leu Asn Lys Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro
1 5 10 15
Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu
20 25 30
Ala Thr Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
35 40 45
Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys
50 55 60
Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu
65 70 75 80
Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys
85 90 95
Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp
100 105 110
Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg
115 120 125
Ala Asp Cys Gly Phe Thr Ser Val Ser Tyr Gln Gln Gly Val Leu Ser
130 135 140
Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala
145 150 155 160
Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp
165 170 175
Phe
<210> 21
<211> 178
<212> PRT
<213> Homo sapiens
<220>
<223> Человеческий бета константа TCR 2
<300>
<308> Uniprot A0A5B9
<309> 2018-07-18
<400> 21
Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro Ser
1 5 10 15
Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu Ala
20 25 30
Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val Asn Gly
35 40 45
Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys Glu
50 55 60
Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu Arg
65 70 75 80
Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys Gln
85 90 95
Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp Arg
100 105 110
Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg Ala
115 120 125
Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu Ser Ala
130 135 140
Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala Val
145 150 155 160
Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp Ser
165 170 175
Arg Gly
<210> 22
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер
<220>
<221> ПОВТОР
<222> (0)…(0)
<223> SGGGG повторяется 5 или 6 раз
<400> 22
Pro Gly Gly Gly Ser Gly Gly Gly Gly Pro
1 5 10
<210> 23
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 23
Gly Ser Ala Asp Asp Ala Lys Lys Asp Ala Ala Lys Lys Asp Gly Lys
1 5 10 15
Ser
<210> 24
<211> 141
<212> PRT
<213> Homo sapiens
<220>
<223> Человеческий альфа константа TCR
<300>
<308> CAA26636.1
<309> 2016-07-25
<400> 24
Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
1 5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 25
<211> 179
<212> PRT
<213> Homo sapiens
<220>
<223> Человеческий бета константа TCR
<300>
<308> A0A0G2JNG9
<400> 25
Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro
1 5 10 15
Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu
20 25 30
Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
35 40 45
Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys
50 55 60
Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu
65 70 75 80
Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys
85 90 95
Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp
100 105 110
Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg
115 120 125
Ala Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu Ser
130 135 140
Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala
145 150 155 160
Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp
165 170 175
Ser Arg Gly
<210> 26
<211> 149
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC гРНК транскрипционная последовательность
<400> 26
agcgctctcg tacagagttg gcattataat acgactcact ataggggaga atcaaaatcg 60
gtgaatgttt tagagctaga aatagcaagt taaaataagg ctagtccgtt atcaacttga 120
aaaagtggca ccgagtcggt gcttttttt 149
<210> 27
<211> 100
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC гРНК последовательность
<400> 27
gagaaucaaa aucggugaau guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc 60
cguuaucaac uugaaaaagu ggcaccgagu cggugcuuuu 100
<210> 28
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-10 гРНК домен целенаправленного воздействия
<400> 28
ucucucagcu gguacacggc 20
<210> 29
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-110 гРНК домен целенаправленного воздействия
<400> 29
uggauuuaga gucucucagc 20
<210> 30
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-116 гРНК домен целенаправленного воздействия
<400> 30
acacggcagg gucaggguuc 20
<210> 31
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-16 гРНК домен целенаправленного воздействия
<400> 31
gagaaucaaa aucggugaau 20
<210> 32
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-4 гРНК домен целенаправленного воздействия
<400> 32
gcugguacac ggcaggguca 20
<210> 33
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-49 гРНК домен целенаправленного воздействия
<400> 33
cucagcuggu acacggc 17
<210> 34
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-2 гРНК домен целенаправленного воздействия
<400> 34
ugguacacgg caggguc 17
<210> 35
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-30 гРНК домен целенаправленного воздействия
<400> 35
gcuagacaug aggucua 17
<210> 36
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-43 гРНК домен целенаправленного воздействия
<400> 36
gucagauuug uugcucc 17
<210> 37
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-23 гРНК домен целенаправленного воздействия
<400> 37
ucagcuggua cacggca 17
<210> 38
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-34 гРНК домен целенаправленного воздействия
<400> 38
gcagacagac uugucac 17
<210> 39
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-25 гРНК домен целенаправленного воздействия
<400> 39
gguacacggc aggguca 17
<210> 40
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-128 гРНК домен целенаправленного воздействия
<400> 40
cuucaagagc aacagugcug 20
<210> 41
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-105 гРНК домен целенаправленного воздействия
<400> 41
agagcaacag ugcuguggcc 20
<210> 42
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-106 гРНК домен целенаправленного воздействия
<400> 42
aaagucagau uuguugcucc 20
<210> 43
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-123 гРНК домен целенаправленного воздействия
<400> 43
acaaaacugu gcuagacaug 20
<210> 44
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-64 гРНК домен целенаправленного воздействия
<400> 44
aaacugugcu agacaug 17
<210> 45
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-97 гРНК домен целенаправленного воздействия
<400> 45
ugugcuagac augaggucua 20
<210> 46
<211> 22
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-148 гРНК домен целенаправленного воздействия
<400> 46
ggcuggggaa gaaggugucu uc 22
<210> 47
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-147 гРНК домен целенаправленного воздействия
<400> 47
gcuggggaag aaggugucuu c 21
<210> 48
<211> 18
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-234 гРНК домен целенаправленного воздействия
<400> 48
ggggaagaag gugucuuc 18
<210> 49
<211> 22
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-167 гРНК домен целенаправленного воздействия
<400> 49
guuuugucug ugauauacac au 22
<210> 50
<211> 24
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-177 гРНК домен целенаправленного воздействия
<400> 50
ggcagacaga cuugucacug gauu 24
<210> 51
<211> 23
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-176 гРНК домен целенаправленного воздействия
<400> 51
gcagacagac uugucacugg auu 23
<210> 52
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-257 гРНК домен целенаправленного воздействия
<400> 52
gacagacuug ucacuggauu 20
<210> 53
<211> 24
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-233 гРНК домен целенаправленного воздействия
<400> 53
gugaauaggc agacagacuu guca 24
<210> 54
<211> 22
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-231 гРНК домен целенаправленного воздействия
<400> 54
gaauaggcag acagacuugu ca 22
<210> 55
<211> 22
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-163 гРНК домен целенаправленного воздействия
<400> 55
gagucucuca gcugguacac gg 22
<210> 56
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-241 гРНК домен целенаправленного воздействия
<400> 56
gucucucagc ugguacacgg 20
<210> 57
<211> 22
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-179 гРНК домен целенаправленного воздействия
<400> 57
gguacacggc agggucaggg uu 22
<210> 58
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRAC-178 гРНК домен целенаправленного воздействия
<400> 58
guacacggca gggucagggu u 21
<210> 59
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-40 гРНК домен целенаправленного воздействия
<400> 59
cacccagauc gucagcgccg 20
<210> 60
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-52 гРНК домен целенаправленного воздействия
<400> 60
caaacacagc gaccucgggu 20
<210> 61
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-25 гРНК домен целенаправленного воздействия
<400> 61
ugacgagugg acccaggaua 20
<210> 62
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-35 гРНК домен целенаправленного воздействия
<400> 62
ggcucucgga gaaugacgag 20
<210> 63
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-50 гРНК домен целенаправленного воздействия
<400> 63
ggccucggcg cugacgaucu 20
<210> 64
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-39 гРНК домен целенаправленного воздействия
<400> 64
gaaaaacgug uucccacccg 20
<210> 65
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-49 гРНК домен целенаправленного воздействия
<400> 65
augacgagug gacccaggau 20
<210> 66
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-51 гРНК домен целенаправленного воздействия
<400> 66
aguccaguuc uacgggcucu 20
<210> 67
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-26 гРНК домен целенаправленного воздействия
<400> 67
cgcugucaag uccaguucua 20
<210> 68
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-47 гРНК домен целенаправленного воздействия
<400> 68
aucgucagcg ccgaggccug 20
<210> 69
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-45 гРНК домен целенаправленного воздействия
<400> 69
ucaaacacag cgaccucggg 20
<210> 70
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-34 гРНК домен целенаправленного воздействия
<400> 70
cguagaacug gacuugacag 20
<210> 71
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-227 гРНК домен целенаправленного воздействия
<400> 71
aggccucggc gcugacgauc 20
<210> 72
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-41 гРНК домен целенаправленного воздействия
<400> 72
ugacagcgga agugguugcg 20
<210> 73
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-30 гРНК домен целенаправленного воздействия
<400> 73
uugacagcgg aagugguugc 20
<210> 74
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-206 гРНК домен целенаправленного воздействия
<400> 74
ucuccgagag cccguagaac 20
<210> 75
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-32 гРНК домен целенаправленного воздействия
<400> 75
cgggugggaa cacguuuuuc 20
<210> 76
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-276 гРНК домен целенаправленного воздействия
<400> 76
gacagguuug gcccuauccu 20
<210> 77
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-274 гРНК домен целенаправленного воздействия
<400> 77
gaucgucagc gccgaggccu 20
<210> 78
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-230 гРНК домен целенаправленного воздействия
<400> 78
ggcucaaaca cagcgaccuc 20
<210> 79
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-235 гРНК домен целенаправленного воздействия
<400> 79
ugagggucuc ggccaccuuc 20
<210> 80
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-38 гРНК домен целенаправленного воздействия
<400> 80
aggcuucuac cccgaccacg 20
<210> 81
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-223 гРНК домен целенаправленного воздействия
<400> 81
ccgaccacgu ggagcugagc 20
<210> 82
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-221 гРНК домен целенаправленного воздействия
<400> 82
ugacagguuu ggcccuaucc 20
<210> 83
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-48 гРНК домен целенаправленного воздействия
<400> 83
cuugacagcg gaagugguug 20
<210> 84
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-216 гРНК домен целенаправленного воздействия
<400> 84
agaucgucag cgccgaggcc 20
<210> 85
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-210 гРНК домен целенаправленного воздействия
<400> 85
gcgcugacga ucugggugac 20
<210> 86
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-268 гРНК домен целенаправленного воздействия
<400> 86
ugagggcggg cugcuccuug 20
<210> 87
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-193 гРНК домен целенаправленного воздействия
<400> 87
guugcggggg uucugccaga 20
<210> 88
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-246 гРНК домен целенаправленного воздействия
<400> 88
agcucagcuc cacguggucg 20
<210> 89
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-228 гРНК домен целенаправленного воздействия
<400> 89
gcggcugcuc aggcaguauc 20
<210> 90
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-43 гРНК домен целенаправленного воздействия
<400> 90
gcggggguuc ugccagaagg 20
<210> 91
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-272 гРНК домен целенаправленного воздействия
<400> 91
uggcucaaac acagcgaccu 20
<210> 92
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-33 гРНК домен целенаправленного воздействия
<400> 92
acuggacuug acagcggaag 20
<210> 93
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-44 гРНК домен целенаправленного воздействия
<400> 93
gacagcggaa gugguugcgg 20
<210> 94
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-211 гРНК домен целенаправленного воздействия
<400> 94
gcugucaagu ccaguucuac 20
<210> 95
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-253 гРНК домен целенаправленного воздействия
<400> 95
guaucuggag ucauugaggg 20
<210> 96
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-18 гРНК домен целенаправленного воздействия
<400> 96
cucggcgcug acgaucu 17
<210> 97
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-6 гРНК домен целенаправленного воздействия
<400> 97
ccucggcgcu gacgauc 17
<210> 98
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-85 гРНК домен целенаправленного воздействия
<400> 98
ccgagagccc guagaac 17
<210> 99
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-129 гРНК домен целенаправленного воздействия
<400> 99
ccagaucguc agcgccg 17
<210> 100
<211> 17
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-93 гРНК домен целенаправленного воздействия
<400> 100
gaaugacgag uggaccc 17
<210> 101
<211> 22
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-415 гРНК домен целенаправленного воздействия
<400> 101
gggugacagg uuuggcccua uc 22
<210> 102
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-414 гРНК домен целенаправленного воздействия
<400> 102
ggugacaggu uuggcccuau c 21
<210> 103
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-310 гРНК домен целенаправленного воздействия
<400> 103
gugacagguu uggcccuauc 20
<210> 104
<211> 18
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-308 гРНК домен целенаправленного воздействия
<400> 104
gacagguuug gcccuauc 18
<210> 105
<211> 22
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-401 гРНК домен целенаправленного воздействия
<400> 105
gauacugccu gagcagccgc cu 22
<210> 106
<211> 24
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-468 гРНК домен целенаправленного воздействия
<400> 106
gaccacgugg agcugagcug gugg 24
<210> 107
<211> 18
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-462 гРНК домен целенаправленного воздействия
<400> 107
guggagcuga gcuggugg 18
<210> 108
<211> 24
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-424 гРНК домен целенаправленного воздействия
<400> 108
gggcgggcug cuccuugagg ggcu 24
<210> 109
<211> 23
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-423 гРНК домен целенаправленного воздействия
<400> 109
ggcgggcugc uccuugaggg gcu 23
<210> 110
<211> 22
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-422 гРНК домен целенаправленного воздействия
<400> 110
gcgggcugcu ccuugagggg cu 22
<210> 111
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-420 гРНК домен целенаправленного воздействия
<400> 111
gggcugcucc uugaggggcu 20
<210> 112
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-419 гРНК домен целенаправленного воздействия
<400> 112
ggcugcuccu ugaggggcu 19
<210> 113
<211> 18
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-418 гРНК домен целенаправленного воздействия
<400> 113
gcugcuccuu gaggggcu 18
<210> 114
<211> 24
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-445 гРНК домен целенаправленного воздействия
<400> 114
ggugaauggg aaggaggugc acag 24
<210> 115
<211> 23
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-444 гРНК домен целенаправленного воздействия
<400> 115
gugaauggga aggaggugca cag 23
<210> 116
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC-442 гРНК домен целенаправленного воздействия
<400> 116
gaaugggaag gaggugcaca g 21
<210> 117
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность 1
<400> 117
attcaccgat tttgattctc 20
<210> 118
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> TRBC целевая последовательность 2
<400> 118
agatcgtcag cgccgaggcc 20
<210> 119
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> HBB акцептор сайта сплайсинга
<400> 119
ctgacctctt ctcttcctcc cacag 25
<210> 120
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> IgG акцептор сайта сплайсинга
<400> 120
tttctctcca cag 13
<210> 121
<211> 138
<212> PRT
<213> mus musculus
<220>
<223> Мышиный альфа константа TCR
<300>
<308> Uniprot P01849
<309> 1986-07-21
<400> 121
Pro Tyr Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro
1 5 10 15
Arg Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln
20 25 30
Ile Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys
35 40 45
Thr Val Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile
50 55 60
Ala Trp Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu
65 70 75 80
Thr Asn Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu
85 90 95
Thr Glu Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu
100 105 110
Ser Val Met Gly Leu Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn
115 120 125
Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135
<210> 122
<211> 136
<212> PRT
<213> Искусственная последовательность
<220>
<223> Модифицированный мышиный альфа константа TCR
<400> 122
Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp Pro Arg Ser
1 5 10 15
Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile Asn
20 25 30
Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Cys Val
35 40 45
Leu Asp Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala Trp
50 55 60
Ser Asn Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr Asn
65 70 75 80
Ala Thr Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr Leu Thr Glu
85 90 95
Lys Ser Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn Leu Leu Val
100 105 110
Ile Val Leu Arg Ile Leu Leu Leu Lys Val Ala Gly Phe Asn Leu Leu
115 120 125
Met Thr Leu Arg Leu Trp Ser Ser
130 135
<210> 123
<211> 173
<212> PRT
<213> Искусственная последовательность
<220>
<223> Модифицированный мышиный бета константа TCR
<400> 123
Glu Asp Leu Arg Asn Val Thr Pro Pro Lys Val Ser Leu Phe Glu Pro
1 5 10 15
Ser Lys Ala Glu Ile Ala Asn Lys Gln Lys Ala Thr Leu Val Cys Leu
20 25 30
Ala Arg Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
35 40 45
Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Ala Tyr Lys
50 55 60
Glu Ser Asn Tyr Ser Tyr Cys Leu Ser Ser Arg Leu Arg Val Ser Ala
65 70 75 80
Thr Phe Trp His Asn Pro Arg Asn His Phe Arg Cys Gln Val Gln Phe
85 90 95
His Gly Leu Ser Glu Glu Asp Lys Trp Pro Glu Gly Ser Pro Lys Pro
100 105 110
Val Thr Gln Asn Ile Ser Ala Glu Ala Trp Gly Arg Ala Asp Cys Gly
115 120 125
Ile Thr Ser Ala Ser Tyr Gln Gln Gly Val Leu Ser Ala Thr Ile Leu
130 135 140
Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala Val Leu Val Ser
145 150 155 160
Thr Leu Val Val Met Ala Met Val Lys Arg Lys Asn Ser
165 170
<210> 124
<211> 611
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 5' плечо гомологии
<400> 124
ctctatcaat gagagagcaa tctcctggta atgtgataga tttcccaact taatgccaac 60
ataccataaa cctcccattc tgctaatgcc cagcctaagt tggggagacc actccagatt 120
ccaagatgta cagtttgctt tgctgggcct ttttcccatg cctgccttta ctctgccaga 180
gttatattgc tggggttttg aagaagatcc tattaaataa aagaataagc agtattatta 240
agtagccctg catttcaggt ttccttgagt ggcaggccag gcctggccgt gaacgttcac 300
tgaaatcatg gcctcttggc caagattgat agcttgtgcc tgtccctgag tcccagtcca 360
tcacgagcag ctggtttcta agatgctatt tcccgtataa agcatgagac cgtgacttgc 420
cagccccaca gagccccgcc cttgtccatc actggcatct ggactccagc ctgggttggg 480
gcaaagaggg aaatgagatc atgtcctaac cctgatcctc ttgtcccaca gatatccaga 540
accctgaccc tgccgtgtac cagctgagag actctaaatc cagtgacaag tctgtctgcc 600
tattcaccga t 611
<210> 125
<211> 628
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 3' плечо гомологии
<400> 125
tttgattctc aaacaaatgt gtcacaaagt aaggattctg atgtgtatat cacagacaaa 60
actgtgctag acatgaggtc tatggacttc aagagcaaca gtgctgtggc ctggagcaac 120
aaatctgact ttgcatgtgc aaacgccttc aacaacagca ttattccaga agacaccttc 180
ttccccagcc caggtaaggg cagctttggt gccttcgcag gctgtttcct tgcttcagga 240
atggccaggt tctgcccaga gctctggtca atgatgtcta aaactcctct gattggtggt 300
ctcggcctta tccattgcca ccaaaaccct ctttttacta agaaacagtg agccttgttc 360
tggcagtcca gagaatgaca cgggaaaaaa gcagatgaag agaaggtggc aggagagggc 420
acgtggccca gcctcagtct ctccaactga gttcctgcct gcctgccttt gctcagactg 480
tttgcccctt actgctcttc taggcctcat tctaagcccc ttctccaagt tgcctctcct 540
tatttctccc tgtctgccaa aaaatctttc ccagctcact aagtcagtct cacgcagtca 600
ctcattaacc caccaatcac tgattgtg 628
<210> 126
<211> 347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> MND промотор
<400> 126
gaacagagaa acaggagaat atgggccaaa caggatatct gtggtaagca gttcctgccc 60
cggctcaggg ccaagaacag ttggaacagc agaatatggg ccaaacagga tatctgtggt 120
aagcagttcc tgccccggct cagggccaag aacagatggt ccccagatgc ggtcccgccc 180
tcagcagttt ctagagaacc atcagatgtt tccagggtgc cccaaggacc tgaaatgacc 240
ctgtgcctta tttgaactaa ccaatcagtt cgcttctcgc ttctgttcgc gcgcttctgc 300
tccccgagct ctatataagc agagctcgtt tagtgaaccg tcagatc 347
<210> 127
<211> 544
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ef1 альфа промотор с HTLV1 энхансером
<400> 127
ggatctgcga tcgctccggt gcccgtcagt gggcagagcg cacatcgccc acagtccccg 60
agaagttggg gggaggggtc ggcaattgaa ccggtgccta gagaaggtgg cgcggggtaa 120
actgggaaag tgatgtcgtg tactggctcc gcctttttcc cgagggtggg ggagaaccgt 180
atataagtgc agtagtcgcc gtgaacgttc tttttcgcaa cgggtttgcc gccagaacac 240
agctgaagct tcgaggggct cgcatctctc cttcacgcgc ccgccgccct acctgaggcc 300
gccatccacg ccggttgagt cgcgttctgc cgcctcccgc ctgtggtgcc tcctgaactg 360
cgtccgccgt ctaggtaagt ttaaagctca ggtcgagacc gggcctttgt ccggcgctcc 420
cttggagcct acctagactc agccggctct ccacgctttg cctgaccctg cttgctcaac 480
tctacgtctt tgtttcgttt tctgttctgc gccgttacag atccaagctg tgaccggcgc 540
ctac 544
<210> 128
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<223> P2A нуклеотидная последовательность
<400> 128
ggatctggag cgacgaattt tagtctactg aaacaagcgg gagacgtgga ggaaaaccct 60
ggacct 66
<210> 129
<211> 112
<212> PRT
<213> homo sapiens
<220>
<223> CD3 дзэта
<400> 129
Arg Val Lys Phe Ser Arg Ser Ala Glu Pro Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 130
<211> 112
<212> PRT
<213> homo sapiens
<220>
<223> CD3 дзэта
<400> 130
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 131
<211> 12
<212> PRT
<213> homo sapiens
<220>
<223> спейсер (IgG4 шарнир)
<400> 131
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 132
<211> 36
<212> ДНК
<213> homo sapiens
<220>
<223> спейсер (IgG4 шарнир)
<400> 132
gaatctaagt acggaccgcc ctgcccccct tgccct 36
<210> 133
<211> 119
<212> PRT
<213> homo sapiens
<220>
<223> Шарнир-CH3 спейсер
<400> 133
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Gly Gln Pro Arg
1 5 10 15
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
20 25 30
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
35 40 45
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
50 55 60
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
65 70 75 80
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
85 90 95
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
100 105 110
Leu Ser Leu Ser Leu Gly Lys
115
<210> 134
<211> 229
<212> PRT
<213> homo sapiens
<220>
<223> Шарнир-CH2-CH3 спейсер
<400> 134
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 135
<211> 282
<212> PRT
<213> homo sapiens
<220>
<223> IgD-шарнир-Fc
<400> 135
Arg Trp Pro Glu Ser Pro Lys Ala Gln Ala Ser Ser Val Pro Thr Ala
1 5 10 15
Gln Pro Gln Ala Glu Gly Ser Leu Ala Lys Ala Thr Thr Ala Pro Ala
20 25 30
Thr Thr Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Lys Glu Lys
35 40 45
Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro
50 55 60
Ser His Thr Gln Pro Leu Gly Val Tyr Leu Leu Thr Pro Ala Val Gln
65 70 75 80
Asp Leu Trp Leu Arg Asp Lys Ala Thr Phe Thr Cys Phe Val Val Gly
85 90 95
Ser Asp Leu Lys Asp Ala His Leu Thr Trp Glu Val Ala Gly Lys Val
100 105 110
Pro Thr Gly Gly Val Glu Glu Gly Leu Leu Glu Arg His Ser Asn Gly
115 120 125
Ser Gln Ser Gln His Ser Arg Leu Thr Leu Pro Arg Ser Leu Trp Asn
130 135 140
Ala Gly Thr Ser Val Thr Cys Thr Leu Asn His Pro Ser Leu Pro Pro
145 150 155 160
Gln Arg Leu Met Ala Leu Arg Glu Pro Ala Ala Gln Ala Pro Val Lys
165 170 175
Leu Ser Leu Asn Leu Leu Ala Ser Ser Asp Pro Pro Glu Ala Ala Ser
180 185 190
Trp Leu Leu Cys Glu Val Ser Gly Phe Ser Pro Pro Asn Ile Leu Leu
195 200 205
Met Trp Leu Glu Asp Gln Arg Glu Val Asn Thr Ser Gly Phe Ala Pro
210 215 220
Ala Arg Pro Pro Pro Gln Pro Gly Ser Thr Thr Phe Trp Ala Trp Ser
225 230 235 240
Val Leu Arg Val Pro Ala Pro Pro Ser Pro Gln Pro Ala Thr Tyr Thr
245 250 255
Cys Val Val Ser His Glu Asp Ser Arg Thr Leu Leu Asn Ala Ser Arg
260 265 270
Ser Leu Glu Val Ser Tyr Val Thr Asp His
275 280
<210> 136
<211> 27
<212> PRT
<213> homo sapiens
<220>
<223> CD28
<300>
<308> No. P10747
<309> 1989-07-01
<400> 136
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
20 25
<210> 137
<211> 66
<212> PRT
<213> homo sapiens
<220>
<223> CD28
<300>
<308> No. P10747
<309> 1989-07-01
<400> 137
Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu Lys Ser Asn
1 5 10 15
Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro Ser Pro Leu
20 25 30
Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val Val Gly Gly
35 40 45
Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe
50 55 60
Trp Val
65
<210> 138
<211> 41
<212> PRT
<213> homo sapiens
<220>
<223> CD28
<300>
<308> P10747
<309> 1989-07-01
<400> 138
Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr
1 5 10 15
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
20 25 30
Pro Arg Asp Phe Ala Ala Tyr Arg Ser
35 40
<210> 139
<211> 41
<212> PRT
<213> homo sapiens
<220>
<223> CD28 (LL до GG)
<400> 139
Arg Ser Lys Arg Ser Arg Gly Gly His Ser Asp Tyr Met Asn Met Thr
1 5 10 15
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
20 25 30
Pro Arg Asp Phe Ala Ala Tyr Arg Ser
35 40
<210> 140
<211> 42
<212> PRT
<213> homo sapiens
<220>
<223> 4-1BB
<300>
<308> Q07011.1
<309> 1995-02-01
<400> 140
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
35 40
<210> 141
<211> 112
<212> PRT
<213> homo sapiens
<220>
<223> CD3 дзэта
<400> 141
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 142
<211> 96
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовой гРНК комплементарный домен
<220>
<221> мдифицированное_основание
<222> (1)...(20)
<223> a, c, u, g, неизвестный или другой
<220>
<221> misc_признак
<222> (1)...(20)
<223> n is a, c, g, или u
<400> 142
nnnnnnnnnn nnnnnnnnnn guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc 60
cguuaucaac uugaaaaagu ggcaccgagu cggugc 96
<210> 143
<211> 104
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовой гРНК комплементарный домен
<220>
<221> модифицированное_основание
<222> (1)...(20)
<223> a, c, u, g, неизвестный или другой
<220>
<221> misc_признак
<222> (1)...(20)
<223> n is a, c, g, или u
<400> 143
nnnnnnnnnn nnnnnnnnnn guuuuagagc uaugcugaaa agcauagcaa guuaaaauaa 60
ggcuaguccg uuaucaacuu gaaaaagugg caccgagucg gugc 104
<210> 144
<211> 106
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовой гРНК комплементарный домен
<220>
<221> модифицированное_основание
<222> (1)...(20)
<223> a, c, u, g, неизвестный или другой
<220>
<221> misc_признак
<222> (1)...(20)
<223> n is a, c, g, или u
<400> 144
nnnnnnnnnn nnnnnnnnnn guuuuagagc uaugcuggaa acagcauagc aaguuaaaau 60
aaggcuaguc cguuaucaac uugaaaaagu ggcaccgagu cggugc 106
<210> 145
<211> 116
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовой гРНК комплементарный домен
<220>
<221> модифицированное_основание
<222> (1)...(20)
<223> a, c, u, g, неизвестный или другой
<220>
<221> misc_признак
<222> (1)...(20)
<223> n is a, c, g, или u
<400> 145
nnnnnnnnnn nnnnnnnnnn guuuuagagc uaugcuguuu uggaaacaaa acagcauagc 60
aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu ggcaccgagu cggugc 116
<210> 146
<211> 96
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовая гРНК
<220>
<221> модифицированное_основание
<222> (1)...(20)
<223> a, c, u, g, неизвестный или другой
<220>
<221> misc_признак
<222> (1)...(20)
<223> n is a, c, g, или u
<400> 146
nnnnnnnnnn nnnnnnnnnn guauuagagc uagaaauagc aaguuaauau aaggcuaguc 60
cguuaucaac uugaaaaagu ggcaccgagu cggugc 96
<210> 147
<211> 96
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовая гРНК
<220>
<221> модифицированное_основание
<222> (1)...(20)
<223> a, c, u, g, неизвестный или другой
<220>
<221> misc_признак
<222> (1)...(20)
<223> n is a, c, g, или u
<400> 147
nnnnnnnnnn nnnnnnnnnn guuuaagagc uagaaauagc aaguuuaaau aaggcuaguc 60
cguuaucaac uugaaaaagu ggcaccgagu cggugc 96
<210> 148
<211> 116
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовая гРНК
<220>
<221> модифицированное_основание
<222> (1)...(20)
<223> a, c, u, g, неизвестный или другой
<220>
<221> misc_признак
<222> (1)...(20)
<223> n is a, c, g, или u
<400> 148
nnnnnnnnnn nnnnnnnnnn guauuagagc uaugcuguau uggaaacaau acagcauagc 60
aaguuaauau aaggcuaguc cguuaucaac uugaaaaagu ggcaccgagu cggugc 116
<210> 149
<211> 47
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовой проксимальный и хвостовой домен
<400> 149
aaggcuaguc cguuaucaac uugaaaaagu ggcaccgagu cggugcu 47
<210> 150
<211> 49
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовой проксимальный и хвостовой домен
<400> 150
aaggcuaguc cguuaucaac uugaaaaagu ggcaccgagu cgguggugc 49
<210> 151
<211> 51
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовой проксимальный и хвостовой домен
<400> 151
aaggcuaguc cguuaucaac uugaaaaagu ggcaccgagu cggugcggau c 51
<210> 152
<211> 31
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовой проксимальный и хвостовой домен
<400> 152
aaggcuaguc cguuaucaac uugaaaaagu g 31
<210> 153
<211> 18
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовой проксимальный и хвостовой домен
<400> 153
aaggcuaguc cguuauca 18
<210> 154
<211> 12
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовой проксимальный и хвостовой домен
<400> 154
aaggcuaguc cg 12
<210> 155
<211> 102
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовая химерная гРНК
<220>
<221> модифицированное основание
<222> (1)...(20)
<223> a, c, u, g, неизвестный или другой
<220>
<221> misc_признак
<222> (1)...(20)
<223> n is a, c, g, или u
<400> 155
nnnnnnnnnn nnnnnnnnnn guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc 60
cguuaucaac uugaaaaagu ggcaccgagu cggugcuuuu uu 102
<210> 156
<211> 102
<212> РНК
<213> Искусственная последовательность
<220>
<223> типовая химерная гРНК
<220>
<221> модифицированное основание
<222> (1)...(20)
<223> a, c, u, g, неизвестный или другой
<220>
<221> misc_признак
<222> (1)...(20)
<223> n is a, c, g, или u
<400> 156
nnnnnnnnnn nnnnnnnnnn guuuuaguac ucuggaaaca gaaucuacua aaacaaggca 60
aaaugccgug uuuaucucgu caacuuguug gcgagauuuu uu 102
<210> 157
<211> 1344
<212> PRT
<213> Искусственная последовательность
<220>
<223> Streptococcus mutans Cas9
<400> 157
Lys Lys Pro Tyr Ser Ile Gly Leu Asp Ile Gly Thr Asn Ser Val Gly
1. 5 10 15
Trp Ala Val Val Thr Asp Asp Tyr Lys Val Pro Ala Lys Lys Met Lys
20 25 30
Val Leu Gly Asn Thr Asp Lys Ser His Ile Glu Lys Asn Leu Leu Gly
35 40 45
Ala Leu Leu Phe Asp Ser Gly Asn Thr Ala Glu Asp Arg Arg Leu Lys
50 55 60
Arg Thr Ala Arg Arg Arg Tyr Thr Arg Arg Arg Asn Arg Ile Leu Tyr
65 70 75 80
Leu Gln Glu Ile Phe Ser Glu Glu Met Gly Lys Val Asp Asp Ser Phe
85 90 95
Phe His Arg Leu Glu Asp Ser Phe Leu Val Thr Glu Asp Lys Arg Gly
100 105 110
Glu Arg His Pro Ile Phe Gly Asn Leu Glu Glu Glu Val Lys Tyr His
115 120 125
Glu Asn Phe Pro Thr Ile Tyr His Leu Arg Gln Tyr Leu Ala Asp Asn
130 135 140
Pro Glu Lys Val Asp Leu Arg Leu Val Tyr Leu Ala Leu Ala His Ile
145 150 155 160
Ile Lys Phe Arg Gly His Phe Leu Ile Glu Gly Lys Phe Asp Thr Arg
165 170 175
Asn Asn Asp Val Gln Arg Leu Phe Gln Glu Phe Leu Ala Val Tyr Asp
180 185 190
Asn Thr Phe Glu Asn Ser Ser Leu Gln Glu Gln Asn Val Gln Val Glu
195 200 205
Glu Ile Leu Thr Asp Lys Ile Ser Lys Ser Ala Lys Lys Asp Arg Val
210 215 220
Leu Lys Leu Phe Pro Asn Glu Lys Ser Asn Gly Arg Phe Ala Glu Phe
225 230 235 240
Leu Lys Leu Ile Val Gly Asn Gln Ala Asp Phe Lys Lys His Phe Glu
245 250 255
Leu Glu Glu Lys Ala Pro Leu Gln Phe Ser Lys Asp Thr Tyr Glu Glu
260 265 270
Glu Leu Glu Val Leu Leu Ala Gln Ile Gly Asp Asn Tyr Ala Glu Leu
275 280 285
Phe Leu Ser Ala Lys Lys Leu Tyr Asp Ser Ile Leu Leu Ser Gly Ile
290 295 300
Leu Thr Val Thr Asp Val Gly Thr Lys Ala Pro Leu Ser Ala Ser Met
305 310 315 320
Ile Gln Arg Tyr Asn Glu His Gln Met Asp Leu Ala Gln Leu Lys Gln
325 330 335
Phe Ile Arg Gln Lys Leu Ser Asp Lys Tyr Asn Glu Val Phe Ser Asp
340 345 350
Val Ser Lys Asp Gly Tyr Ala Gly Tyr Ile Asp Gly Lys Thr Asn Gln
355 360 365
Glu Ala Phe Tyr Lys Tyr Leu Lys Gly Leu Leu Asn Lys Ile Glu Gly
370 375 380
Ser Gly Tyr Phe Leu Asp Lys Ile Glu Arg Glu Asp Phe Leu Arg Lys
385 390 395 400
Gln Arg Thr Phe Asp Asn Gly Ser Ile Pro His Gln Ile His Leu Gln
405 410 415
Glu Met Arg Ala Ile Ile Arg Arg Gln Ala Glu Phe Tyr Pro Phe Leu
420 425 430
Ala Asp Asn Gln Asp Arg Ile Glu Lys Leu Leu Thr Phe Arg Ile Pro
435 440 445
Tyr Tyr Val Gly Pro Leu Ala Arg Gly Lys Ser Asp Phe Ala Trp Leu
450 455 460
Ser Arg Lys Ser Ala Asp Lys Ile Thr Pro Trp Asn Phe Asp Glu Ile
465 470 475 480
Val Asp Lys Glu Ser Ser Ala Glu Ala Phe Ile Asn Arg Met Thr Asn
485 490 495
Tyr Asp Leu Tyr Leu Pro Asn Gln Lys Val Leu Pro Lys His Ser Leu
500 505 510
Leu Tyr Glu Lys Phe Thr Val Tyr Asn Glu Leu Thr Lys Val Lys Tyr
515 520 525
Lys Thr Glu Gln Gly Lys Thr Ala Phe Phe Asp Ala Asn Met Lys Gln
530 535 540
Glu Ile Phe Asp Gly Val Phe Lys Val Tyr Arg Lys Val Thr Lys Asp
545 550 555 560
Lys Leu Met Asp Phe Leu Glu Lys Glu Phe Asp Glu Phe Arg Ile Val
565 570 575
Asp Leu Thr Gly Leu Asp Lys Glu Asn Lys Val Phe Asn Ala Ser Tyr
580 585 590
Gly Thr Tyr His Asp Leu Cys Lys Ile Leu Asp Lys Asp Phe Leu Asp
595 600 605
Asn Ser Lys Asn Glu Lys Ile Leu Glu Asp Ile Val Leu Thr Leu Thr
610 615 620
Leu Phe Glu Asp Arg Glu Met Ile Arg Lys Arg Leu Glu Asn Tyr Ser
625 630 635 640
Asp Leu Leu Thr Lys Glu Gln Val Lys Lys Leu Glu Arg Arg His Tyr
645 650 655
Thr Gly Trp Gly Arg Leu Ser Ala Glu Leu Ile His Gly Ile Arg Asn
660 665 670
Lys Glu Ser Arg Lys Thr Ile Leu Asp Tyr Leu Ile Asp Asp Gly Asn
675 680 685
Ser Asn Arg Asn Phe Met Gln Leu Ile Asn Asp Asp Ala Leu Ser Phe
690 695 700
Lys Glu Glu Ile Ala Lys Ala Gln Val Ile Gly Glu Thr Asp Asn Leu
705 710 715 720
Asn Gln Val Val Ser Asp Ile Ala Gly Ser Pro Ala Ile Lys Lys Gly
725 730 735
Ile Leu Gln Ser Leu Lys Ile Val Asp Glu Leu Val Lys Ile Met Gly
740 745 750
His Gln Pro Glu Asn Ile Val Val Glu Met Ala Arg Glu Asn Gln Phe
755 760 765
Thr Asn Gln Gly Arg Arg Asn Ser Gln Gln Arg Leu Lys Gly Leu Thr
770 775 780
Asp Ser Ile Lys Glu Phe Gly Ser Gln Ile Leu Lys Glu His Pro Val
785 790 795 800
Glu Asn Ser Gln Leu Gln Asn Asp Arg Leu Phe Leu Tyr Tyr Leu Gln
805 810 815
Asn Gly Arg Asp Met Tyr Thr Gly Glu Glu Leu Asp Ile Asp Tyr Leu
820 825 830
Ser Gln Tyr Asp Ile Asp His Ile Ile Pro Gln Ala Phe Ile Lys Asp
835 840 845
Asn Ser Ile Asp Asn Arg Val Leu Thr Ser Ser Lys Glu Asn Arg Gly
850 855 860
Lys Ser Asp Asp Val Pro Ser Lys Asp Val Val Arg Lys Met Lys Ser
865 870 875 880
Tyr Trp Ser Lys Leu Leu Ser Ala Lys Leu Ile Thr Gln Arg Lys Phe
885 890 895
Asp Asn Leu Thr Lys Ala Glu Arg Gly Gly Leu Thr Asp Asp Asp Lys
900 905 910
Ala Gly Phe Ile Lys Arg Gln Leu Val Glu Thr Arg Gln Ile Thr Lys
915 920 925
His Val Ala Arg Ile Leu Asp Glu Arg Phe Asn Thr Glu Thr Asp Glu
930 935 940
Asn Asn Lys Lys Ile Arg Gln Val Lys Ile Val Thr Leu Lys Ser Asn
945 950 955 960
Leu Val Ser Asn Phe Arg Lys Glu Phe Glu Leu Tyr Lys Val Arg Glu
965 970 975
Ile Asn Asp Tyr His His Ala His Asp Ala Tyr Leu Asn Ala Val Ile
980 985 990
Gly Lys Ala Leu Leu Gly Val Tyr Pro Gln Leu Glu Pro Glu Phe Val
995 1000 1005
Tyr Gly Asp Tyr Pro His Phe His Gly His Lys Glu Asn Lys Ala Thr
1010 1015 1020
Ala Lys Lys Phe Phe Tyr Ser Asn Ile Met Asn Phe Phe Lys Lys Asp
1025 1030 1035 1040
Asp Val Arg Thr Asp Lys Asn Gly Glu Ile Ile Trp Lys Lys Asp Glu
1045 1050 1055
His Ile Ser Asn Ile Lys Lys Val Leu Ser Tyr Pro Gln Val Asn Ile
1060 1065 1070
Val Lys Lys Val Glu Glu Gln Thr Gly Gly Phe Ser Lys Glu Ser Ile
1075 1080 1085
Leu Pro Lys Gly Asn Ser Asp Lys Leu Ile Pro Arg Lys Thr Lys Lys
1090 1095 1100
Phe Tyr Trp Asp Thr Lys Lys Tyr Gly Gly Phe Asp Ser Pro Ile Val
1105 1110 1115 1120
Ala Tyr Ser Ile Leu Val Ile Ala Asp Ile Glu Lys Gly Lys Ser Lys
1125 1130 1135
Lys Leu Lys Thr Val Lys Ala Leu Val Gly Val Thr Ile Met Glu Lys
1140 1145 1150
Met Thr Phe Glu Arg Asp Pro Val Ala Phe Leu Glu Arg Lys Gly Tyr
1155 1160 1165
Arg Asn Val Gln Glu Glu Asn Ile Ile Lys Leu Pro Lys Tyr Ser Leu
1170 1175 1180
Phe Lys Leu Glu Asn Gly Arg Lys Arg Leu Leu Ala Ser Ala Arg Glu
1185 1190 1195 1200
Leu Gln Lys Gly Asn Glu Ile Val Leu Pro Asn His Leu Gly Thr Leu
1205 1210 1215
Leu Tyr His Ala Lys Asn Ile His Lys Val Asp Glu Pro Lys His Leu
1220 1225 1230
Asp Tyr Val Asp Lys His Lys Asp Glu Phe Lys Glu Leu Leu Asp Val
1235 1240 1245
Val Ser Asn Phe Ser Lys Lys Tyr Thr Leu Ala Glu Gly Asn Leu Glu
1250 1255 1260
Lys Ile Lys Glu Leu Tyr Ala Gln Asn Asn Gly Glu Asp Leu Lys Glu
1265 1270 1275 1280
Leu Ala Ser Ser Phe Ile Asn Leu Leu Thr Phe Thr Ala Ile Gly Ala
1285 1290 1295
Pro Ala Thr Phe Lys Phe Phe Asp Lys Asn Ile Asp Arg Lys Arg Tyr
1300 1305 1310
Thr Ser Thr Thr Glu Ile Leu Asn Ala Thr Leu Ile His Gln Ser Ile
1315 1320 1325
Thr Gly Leu Tyr Glu Thr Arg Ile Asp Leu Asn Lys Leu Gly Gly Asp
1330 1335 1340
<210> 158
<211> 1367
<212> PRT
<213> Искусственная последовательность
<220>
<223> Streptococcus pyogenes Cas9
<400> 158
Asp Lys Lys Tyr Ser Ile Gly Leu Asp Ile Gly Thr Asn Ser Val Gly
1 5 10 15
Trp Ala Val Ile Thr Asp Glu Tyr Lys Val Pro Ser Lys Lys Phe Lys
20 25 30
Val Leu Gly Asn Thr Asp Arg His Ser Ile Lys Lys Asn Leu Ile Gly
35 40 45
Ala Leu Leu Phe Asp Ser Gly Glu Thr Ala Glu Ala Thr Arg Leu Lys
50 55 60
Arg Thr Ala Arg Arg Arg Tyr Thr Arg Arg Lys Asn Arg Ile Cys Tyr
65 70 75 80
Leu Gln Glu Ile Phe Ser Asn Glu Met Ala Lys Val Asp Asp Ser Phe
85 90 95
Phe His Arg Leu Glu Glu Ser Phe Leu Val Glu Glu Asp Lys Lys His
100 105 110
Glu Arg His Pro Ile Phe Gly Asn Ile Val Asp Glu Val Ala Tyr His
115 120 125
Glu Lys Tyr Pro Thr Ile Tyr His Leu Arg Lys Lys Leu Val Asp Ser
130 135 140
Thr Asp Lys Ala Asp Leu Arg Leu Ile Tyr Leu Ala Leu Ala His Met
145 150 155 160
Ile Lys Phe Arg Gly His Phe Leu Ile Glu Gly Asp Leu Asn Pro Asp
165 170 175
Asn Ser Asp Val Asp Lys Leu Phe Ile Gln Leu Val Gln Thr Tyr Asn
180 185 190
Gln Leu Phe Glu Glu Asn Pro Ile Asn Ala Ser Gly Val Asp Ala Lys
195 200 205
Ala Ile Leu Ser Ala Arg Leu Ser Lys Ser Arg Arg Leu Glu Asn Leu
210 215 220
Ile Ala Gln Leu Pro Gly Glu Lys Lys Asn Gly Leu Phe Gly Asn Leu
225 230 235 240
Ile Ala Leu Ser Leu Gly Leu Thr Pro Asn Phe Lys Ser Asn Phe Asp
245 250 255
Leu Ala Glu Asp Ala Lys Leu Gln Leu Ser Lys Asp Thr Tyr Asp Asp
260 265 270
Asp Leu Asp Asn Leu Leu Ala Gln Ile Gly Asp Gln Tyr Ala Asp Leu
275 280 285
Phe Leu Ala Ala Lys Asn Leu Ser Asp Ala Ile Leu Leu Ser Asp Ile
290 295 300
Leu Arg Val Asn Thr Glu Ile Thr Lys Ala Pro Leu Ser Ala Ser Met
305 310 315 320
Ile Lys Arg Tyr Asp Glu His His Gln Asp Leu Thr Leu Leu Lys Ala
325 330 335
Leu Val Arg Gln Gln Leu Pro Glu Lys Tyr Lys Glu Ile Phe Phe Asp
340 345 350
Gln Ser Lys Asn Gly Tyr Ala Gly Tyr Ile Asp Gly Gly Ala Ser Gln
355 360 365
Glu Glu Phe Tyr Lys Phe Ile Lys Pro Ile Leu Glu Lys Met Asp Gly
370 375 380
Thr Glu Glu Leu Leu Val Lys Leu Asn Arg Glu Asp Leu Leu Arg Lys
385 390 395 400
Gln Arg Thr Phe Asp Asn Gly Ser Ile Pro His Gln Ile His Leu Gly
405 410 415
Glu Leu His Ala Ile Leu Arg Arg Gln Glu Asp Phe Tyr Pro Phe Leu
420 425 430
Lys Asp Asn Arg Glu Lys Ile Glu Lys Ile Leu Thr Phe Arg Ile Pro
435 440 445
Tyr Tyr Val Gly Pro Leu Ala Arg Gly Asn Ser Arg Phe Ala Trp Met
450 455 460
Thr Arg Lys Ser Glu Glu Thr Ile Thr Pro Trp Asn Phe Glu Glu Val
465 470 475 480
Val Asp Lys Gly Ala Ser Ala Gln Ser Phe Ile Glu Arg Met Thr Asn
485 490 495
Phe Asp Lys Asn Leu Pro Asn Glu Lys Val Leu Pro Lys His Ser Leu
500 505 510
Leu Tyr Glu Tyr Phe Thr Val Tyr Asn Glu Leu Thr Lys Val Lys Tyr
515 520 525
Val Thr Glu Gly Met Arg Lys Pro Ala Phe Leu Ser Gly Glu Gln Lys
530 535 540
Lys Ala Ile Val Asp Leu Leu Phe Lys Thr Asn Arg Lys Val Thr Val
545 550 555 560
Lys Gln Leu Lys Glu Asp Tyr Phe Lys Lys Ile Glu Cys Phe Asp Ser
565 570 575
Val Glu Ile Ser Gly Val Glu Asp Arg Phe Asn Ala Ser Leu Gly Thr
580 585 590
Tyr His Asp Leu Leu Lys Ile Ile Lys Asp Lys Asp Phe Leu Asp Asn
595 600 605
Glu Glu Asn Glu Asp Ile Leu Glu Asp Ile Val Leu Thr Leu Thr Leu
610 615 620
Phe Glu Asp Arg Glu Met Ile Glu Glu Arg Leu Lys Thr Tyr Ala His
625 630 635 640
Leu Phe Asp Asp Lys Val Met Lys Gln Leu Lys Arg Arg Arg Tyr Thr
645 650 655
Gly Trp Gly Arg Leu Ser Arg Lys Leu Ile Asn Gly Ile Arg Asp Lys
660 665 670
Gln Ser Gly Lys Thr Ile Leu Asp Phe Leu Lys Ser Asp Gly Phe Ala
675 680 685
Asn Arg Asn Phe Met Gln Leu Ile His Asp Asp Ser Leu Thr Phe Lys
690 695 700
Glu Asp Ile Gln Lys Ala Gln Val Ser Gly Gln Gly Asp Ser Leu His
705 710 715 720
Glu His Ile Ala Asn Leu Ala Gly Ser Pro Ala Ile Lys Lys Gly Ile
725 730 735
Leu Gln Thr Val Lys Val Val Asp Glu Leu Val Lys Val Met Gly Arg
740 745 750
His Lys Pro Glu Asn Ile Val Ile Glu Met Ala Arg Glu Asn Gln Thr
755 760 765
Thr Gln Lys Gly Gln Lys Asn Ser Arg Glu Arg Met Lys Arg Ile Glu
770 775 780
Glu Gly Ile Lys Glu Leu Gly Ser Gln Ile Leu Lys Glu His Pro Val
785 790 795 800
Glu Asn Thr Gln Leu Gln Asn Glu Lys Leu Tyr Leu Tyr Tyr Leu Gln
805 810 815
Asn Gly Arg Asp Met Tyr Val Asp Gln Glu Leu Asp Ile Asn Arg Leu
820 825 830
Ser Asp Tyr Asp Val Asp His Ile Val Pro Gln Ser Phe Leu Lys Asp
835 840 845
Asp Ser Ile Asp Asn Lys Val Leu Thr Arg Ser Asp Lys Asn Arg Gly
850 855 860
Lys Ser Asp Asn Val Pro Ser Glu Glu Val Val Lys Lys Met Lys Asn
865 870 875 880
Tyr Trp Arg Gln Leu Leu Asn Ala Lys Leu Ile Thr Gln Arg Lys Phe
885 890 895
Asp Asn Leu Thr Lys Ala Glu Arg Gly Gly Leu Ser Glu Leu Asp Lys
900 905 910
Ala Gly Phe Ile Lys Arg Gln Leu Val Glu Thr Arg Gln Ile Thr Lys
915 920 925
His Val Ala Gln Ile Leu Asp Ser Arg Met Asn Thr Lys Tyr Asp Glu
930 935 940
Asn Asp Lys Leu Ile Arg Glu Val Lys Val Ile Thr Leu Lys Ser Lys
945 950 955 960
Leu Val Ser Asp Phe Arg Lys Asp Phe Gln Phe Tyr Lys Val Arg Glu
965 970 975
Ile Asn Asn Tyr His His Ala His Asp Ala Tyr Leu Asn Ala Val Val
980 985 990
Gly Thr Ala Leu Ile Lys Lys Tyr Pro Lys Leu Glu Ser Glu Phe Val
995 1000 1005
Tyr Gly Asp Tyr Lys Val Tyr Asp Val Arg Lys Met Ile Ala Lys Ser
1010 1015 1020
Glu Gln Glu Ile Gly Lys Ala Thr Ala Lys Tyr Phe Phe Tyr Ser Asn
1025 1030 1035 1040
Ile Met Asn Phe Phe Lys Thr Glu Ile Thr Leu Ala Asn Gly Glu Ile
1045 1050 1055
Arg Lys Arg Pro Leu Ile Glu Thr Asn Gly Glu Thr Gly Glu Ile Val
1060 1065 1070
Trp Asp Lys Gly Arg Asp Phe Ala Thr Val Arg Lys Val Leu Ser Met
1075 1080 1085
Pro Gln Val Asn Ile Val Lys Lys Thr Glu Val Gln Thr Gly Gly Phe
1090 1095 1100
Ser Lys Glu Ser Ile Leu Pro Lys Arg Asn Ser Asp Lys Leu Ile Ala
1105 1110 1115 1120
Arg Lys Lys Asp Trp Asp Pro Lys Lys Tyr Gly Gly Phe Asp Ser Pro
1125 1130 1135
Thr Val Ala Tyr Ser Val Leu Val Val Ala Lys Val Glu Lys Gly Lys
1140 1145 1150
Ser Lys Lys Leu Lys Ser Val Lys Glu Leu Leu Gly Ile Thr Ile Met
1155 1160 1165
Glu Arg Ser Ser Phe Glu Lys Asn Pro Ile Asp Phe Leu Glu Ala Lys
1170 1175 1180
Gly Tyr Lys Glu Val Lys Lys Asp Leu Ile Ile Lys Leu Pro Lys Tyr
1185 1190 1195 1200
Ser Leu Phe Glu Leu Glu Asn Gly Arg Lys Arg Met Leu Ala Ser Ala
1205 1210 1215
Gly Glu Leu Gln Lys Gly Asn Glu Leu Ala Leu Pro Ser Lys Tyr Val
1220 1225 1230
Asn Phe Leu Tyr Leu Ala Ser His Tyr Glu Lys Leu Lys Gly Ser Pro
1235 1240 1245
Glu Asp Asn Glu Gln Lys Gln Leu Phe Val Glu Gln His Lys His Tyr
1250 1255 1260
Leu Asp Glu Ile Ile Glu Gln Ile Ser Glu Phe Ser Lys Arg Val Ile
1265 1270 1275 1280
Leu Ala Asp Ala Asn Leu Asp Lys Val Leu Ser Ala Tyr Asn Lys His
1285 1290 1295
Arg Asp Lys Pro Ile Arg Glu Gln Ala Glu Asn Ile Ile His Leu Phe
1300 1305 1310
Thr Leu Thr Asn Leu Gly Ala Pro Ala Ala Phe Lys Tyr Phe Asp Thr
1315 1320 1325
Thr Ile Asp Arg Lys Arg Tyr Thr Ser Thr Lys Glu Val Leu Asp Ala
1330 1335 1340
Thr Leu Ile His Gln Ser Ile Thr Gly Leu Tyr Glu Thr Arg Ile Asp
1345 1350 1355 1360
Leu Ser Gln Leu Gly Gly Asp
1365
<210> 159
<211> 1387
<212> PRT
<213> Искусственная последовательность
<220>
<223> Streptococcus thermophilus Cas9
<400> 159
Thr Lys Pro Tyr Ser Ile Gly Leu Asp Ile Gly Thr Asn Ser Val Gly
1 5 10 15
Trp Ala Val Thr Thr Asp Asn Tyr Lys Val Pro Ser Lys Lys Met Lys
20 25 30
Val Leu Gly Asn Thr Ser Lys Lys Tyr Ile Lys Lys Asn Leu Leu Gly
35 40 45
Val Leu Leu Phe Asp Ser Gly Ile Thr Ala Glu Gly Arg Arg Leu Lys
50 55 60
Arg Thr Ala Arg Arg Arg Tyr Thr Arg Arg Arg Asn Arg Ile Leu Tyr
65 70 75 80
Leu Gln Glu Ile Phe Ser Thr Glu Met Ala Thr Leu Asp Asp Ala Phe
85 90 95
Phe Gln Arg Leu Asp Asp Ser Phe Leu Val Pro Asp Asp Lys Arg Asp
100 105 110
Ser Lys Tyr Pro Ile Phe Gly Asn Leu Val Glu Glu Lys Ala Tyr His
115 120 125
Asp Glu Phe Pro Thr Ile Tyr His Leu Arg Lys Tyr Leu Ala Asp Ser
130 135 140
Thr Lys Lys Ala Asp Leu Arg Leu Val Tyr Leu Ala Leu Ala His Met
145 150 155 160
Ile Lys Tyr Arg Gly His Phe Leu Ile Glu Gly Glu Phe Asn Ser Lys
165 170 175
Asn Asn Asp Ile Gln Lys Asn Phe Gln Asp Phe Leu Asp Thr Tyr Asn
180 185 190
Ala Ile Phe Glu Ser Asp Leu Ser Leu Glu Asn Ser Lys Gln Leu Glu
195 200 205
Glu Ile Val Lys Asp Lys Ile Ser Lys Leu Glu Lys Lys Asp Arg Ile
210 215 220
Leu Lys Leu Phe Pro Gly Glu Lys Asn Ser Gly Ile Phe Ser Glu Phe
225 230 235 240
Leu Lys Leu Ile Val Gly Asn Gln Ala Asp Phe Arg Lys Cys Phe Asn
245 250 255
Leu Asp Glu Lys Ala Ser Leu His Phe Ser Lys Glu Ser Tyr Asp Glu
260 265 270
Asp Leu Glu Thr Leu Leu Gly Tyr Ile Gly Asp Asp Tyr Ser Asp Val
275 280 285
Phe Leu Lys Ala Lys Lys Leu Tyr Asp Ala Ile Leu Leu Ser Gly Phe
290 295 300
Leu Thr Val Thr Asp Asn Glu Thr Glu Ala Pro Leu Ser Ser Ala Met
305 310 315 320
Ile Lys Arg Tyr Asn Glu His Lys Glu Asp Leu Ala Leu Leu Lys Glu
325 330 335
Tyr Ile Arg Asn Ile Ser Leu Lys Thr Tyr Asn Glu Val Phe Lys Asp
340 345 350
Asp Thr Lys Asn Gly Tyr Ala Gly Tyr Ile Asp Gly Lys Thr Asn Gln
355 360 365
Glu Asp Phe Tyr Val Tyr Leu Lys Lys Leu Leu Ala Glu Phe Glu Gly
370 375 380
Ala Asp Tyr Phe Leu Glu Lys Ile Asp Arg Glu Asp Phe Leu Arg Lys
385 390 395 400
Gln Arg Thr Phe Asp Asn Gly Ser Ile Pro Tyr Gln Ile His Leu Gln
405 410 415
Glu Met Arg Ala Ile Leu Asp Lys Gln Ala Lys Phe Tyr Pro Phe Leu
420 425 430
Ala Lys Asn Lys Glu Arg Ile Glu Lys Ile Leu Thr Phe Arg Ile Pro
435 440 445
Tyr Tyr Val Gly Pro Leu Ala Arg Gly Asn Ser Asp Phe Ala Trp Ser
450 455 460
Ile Arg Lys Arg Asn Glu Lys Ile Thr Pro Trp Asn Phe Glu Asp Val
465 470 475 480
Ile Asp Lys Glu Ser Ser Ala Glu Ala Phe Ile Asn Arg Met Thr Ser
485 490 495
Phe Asp Leu Tyr Leu Pro Glu Glu Lys Val Leu Pro Lys His Ser Leu
500 505 510
Leu Tyr Glu Thr Phe Asn Val Tyr Asn Glu Leu Thr Lys Val Arg Phe
515 520 525
Ile Ala Glu Ser Met Arg Asp Tyr Gln Phe Leu Asp Ser Lys Gln Lys
530 535 540
Lys Asp Ile Val Arg Leu Tyr Phe Lys Asp Lys Arg Lys Val Thr Asp
545 550 555 560
Lys Asp Ile Ile Glu Tyr Leu His Ala Ile Tyr Gly Tyr Asp Gly Ile
565 570 575
Glu Leu Lys Gly Ile Glu Lys Gln Phe Asn Ser Ser Leu Ser Thr Tyr
580 585 590
His Asp Leu Leu Asn Ile Ile Asn Asp Lys Glu Phe Leu Asp Asp Ser
595 600 605
Ser Asn Glu Ala Ile Ile Glu Glu Ile Ile His Thr Leu Thr Ile Phe
610 615 620
Glu Asp Arg Glu Met Ile Lys Gln Arg Leu Ser Lys Phe Glu Asn Ile
625 630 635 640
Phe Asp Lys Ser Val Leu Lys Lys Leu Ser Arg Arg His Tyr Thr Gly
645 650 655
Trp Gly Lys Leu Ser Ala Lys Leu Ile Asn Gly Ile Arg Asp Glu Lys
660 665 670
Ser Gly Asn Thr Ile Leu Asp Tyr Leu Ile Asp Asp Gly Ile Ser Asn
675 680 685
Arg Asn Phe Met Gln Leu Ile His Asp Asp Ala Leu Ser Phe Lys Lys
690 695 700
Lys Ile Gln Lys Ala Gln Ile Ile Gly Asp Glu Asp Lys Gly Asn Ile
705 710 715 720
Lys Glu Val Val Lys Ser Leu Pro Gly Ser Pro Ala Ile Lys Lys Gly
725 730 735
Ile Leu Gln Ser Ile Lys Ile Val Asp Glu Leu Val Lys Val Met Gly
740 745 750
Gly Arg Lys Pro Glu Ser Ile Val Val Glu Met Ala Arg Glu Asn Gln
755 760 765
Tyr Thr Asn Gln Gly Lys Ser Asn Ser Gln Gln Arg Leu Lys Arg Leu
770 775 780
Glu Lys Ser Leu Lys Glu Leu Gly Ser Lys Ile Leu Lys Glu Asn Ile
785 790 795 800
Pro Ala Lys Leu Ser Lys Ile Asp Asn Asn Ala Leu Gln Asn Asp Arg
805 810 815
Leu Tyr Leu Tyr Tyr Leu Gln Asn Gly Lys Asp Met Tyr Thr Gly Asp
820 825 830
Asp Leu Asp Ile Asp Arg Leu Ser Asn Tyr Asp Ile Asp His Ile Ile
835 840 845
Pro Gln Ala Phe Leu Lys Asp Asn Ser Ile Asp Asn Lys Val Leu Val
850 855 860
Ser Ser Ala Ser Asn Arg Gly Lys Ser Asp Asp Val Pro Ser Leu Glu
865 870 875 880
Val Val Lys Lys Arg Lys Thr Phe Trp Tyr Gln Leu Leu Lys Ser Lys
885 890 895
Leu Ile Ser Gln Arg Lys Phe Asp Asn Leu Thr Lys Ala Glu Arg Gly
900 905 910
Gly Leu Ser Pro Glu Asp Lys Ala Gly Phe Ile Gln Arg Gln Leu Val
915 920 925
Glu Thr Arg Gln Ile Thr Lys His Val Ala Arg Leu Leu Asp Glu Lys
930 935 940
Phe Asn Asn Lys Lys Asp Glu Asn Asn Arg Ala Val Arg Thr Val Lys
945 950 955 960
Ile Ile Thr Leu Lys Ser Thr Leu Val Ser Gln Phe Arg Lys Asp Phe
965 970 975
Glu Leu Tyr Lys Val Arg Glu Ile Asn Asp Phe His His Ala His Asp
980 985 990
Ala Tyr Leu Asn Ala Val Val Ala Ser Ala Leu Leu Lys Lys Tyr Pro
995 1000 1005
Lys Leu Glu Pro Glu Phe Val Tyr Gly Asp Tyr Pro Lys Tyr Asn Ser
1010 1015 1020
Phe Arg Glu Arg Lys Ser Ala Thr Glu Lys Val Tyr Phe Tyr Ser Asn
1025 1030 1035 1040
Ile Met Asn Ile Phe Lys Lys Ser Ile Ser Leu Ala Asp Gly Arg Val
1045 1050 1055
Ile Glu Arg Pro Leu Ile Glu Val Asn Glu Glu Thr Gly Glu Ser Val
1060 1065 1070
Trp Asn Lys Glu Ser Asp Leu Ala Thr Val Arg Arg Val Leu Ser Tyr
1075 1080 1085
Pro Gln Val Asn Val Val Lys Lys Val Glu Glu Gln Asn His Gly Leu
1090 1095 1100
Asp Arg Gly Lys Pro Lys Gly Leu Phe Asn Ala Asn Leu Ser Ser Lys
1105 1110 1115 1120
Pro Lys Pro Asn Ser Asn Glu Asn Leu Val Gly Ala Lys Glu Tyr Leu
1125 1130 1135
Asp Pro Lys Lys Tyr Gly Gly Tyr Ala Gly Ile Ser Asn Ser Phe Thr
1140 1145 1150
Val Leu Val Lys Gly Thr Ile Glu Lys Gly Ala Lys Lys Lys Ile Thr
1155 1160 1165
Asn Val Leu Glu Phe Gln Gly Ile Ser Ile Leu Asp Arg Ile Asn Tyr
1170 1175 1180
Arg Lys Asp Lys Leu Asn Phe Leu Leu Glu Lys Gly Tyr Lys Asp Ile
1185 1190 1195 1200
Glu Leu Ile Ile Glu Leu Pro Lys Tyr Ser Leu Phe Glu Leu Ser Asp
1205 1210 1215
Gly Ser Arg Arg Met Leu Ala Ser Ile Leu Ser Thr Asn Asn Lys Arg
1220 1225 1230
Gly Glu Ile His Lys Gly Asn Gln Ile Phe Leu Ser Gln Lys Phe Val
1235 1240 1245
Lys Leu Leu Tyr His Ala Lys Arg Ile Ser Asn Thr Ile Asn Glu Asn
1250 1255 1260
His Arg Lys Tyr Val Glu Asn His Lys Lys Glu Phe Glu Glu Leu Phe
1265 1270 1275 1280
Tyr Tyr Ile Leu Glu Phe Asn Glu Asn Tyr Val Gly Ala Lys Lys Asn
1285 1290 1295
Gly Lys Leu Leu Asn Ser Ala Phe Gln Ser Trp Gln Asn His Ser Ile
1300 1305 1310
Asp Glu Leu Cys Ser Ser Phe Ile Gly Pro Thr Gly Ser Glu Arg Lys
1315 1320 1325
Gly Leu Phe Glu Leu Thr Ser Arg Gly Ser Ala Ala Asp Phe Glu Phe
1330 1335 1340
Leu Gly Val Lys Ile Pro Arg Tyr Arg Asp Tyr Thr Pro Ser Ser Leu
1345 1350 1355 1360
Leu Lys Asp Ala Thr Leu Ile His Gln Ser Val Thr Gly Leu Tyr Glu
1365 1370 1375
Thr Arg Ile Asp Leu Ala Lys Leu Gly Glu Gly
1380 1385
<210> 160
<211> 1333
<212> PRT
<213> Искусственная последовательность
<220>
<223> Listeria innocua Cas9
<400> 160
Lys Lys Pro Tyr Thr Ile Gly Leu Asp Ile Gly Thr Asn Ser Val Gly
1 5 10 15
Trp Ala Val Leu Thr Asp Gln Tyr Asp Leu Val Lys Arg Lys Met Lys
20 25 30
Ile Ala Gly Asp Ser Glu Lys Lys Gln Ile Lys Lys Asn Phe Trp Gly
35 40 45
Val Arg Leu Phe Asp Glu Gly Gln Thr Ala Ala Asp Arg Arg Met Ala
50 55 60
Arg Thr Ala Arg Arg Arg Ile Glu Arg Arg Arg Asn Arg Ile Ser Tyr
65 70 75 80
Leu Gln Gly Ile Phe Ala Glu Glu Met Ser Lys Thr Asp Ala Asn Phe
85 90 95
Phe Cys Arg Leu Ser Asp Ser Phe Tyr Val Asp Asn Glu Lys Arg Asn
100 105 110
Ser Arg His Pro Phe Phe Ala Thr Ile Glu Glu Glu Val Glu Tyr His
115 120 125
Lys Asn Tyr Pro Thr Ile Tyr His Leu Arg Glu Glu Leu Val Asn Ser
130 135 140
Ser Glu Lys Ala Asp Leu Arg Leu Val Tyr Leu Ala Leu Ala His Ile
145 150 155 160
Ile Lys Tyr Arg Gly Asn Phe Leu Ile Glu Gly Ala Leu Asp Thr Gln
165 170 175
Asn Thr Ser Val Asp Gly Ile Tyr Lys Gln Phe Ile Gln Thr Tyr Asn
180 185 190
Gln Val Phe Ala Ser Gly Ile Glu Asp Gly Ser Leu Lys Lys Leu Glu
195 200 205
Asp Asn Lys Asp Val Ala Lys Ile Leu Val Glu Lys Val Thr Arg Lys
210 215 220
Glu Lys Leu Glu Arg Ile Leu Lys Leu Tyr Pro Gly Glu Lys Ser Ala
225 230 235 240
Gly Met Phe Ala Gln Phe Ile Ser Leu Ile Val Gly Ser Lys Gly Asn
245 250 255
Phe Gln Lys Pro Phe Asp Leu Ile Glu Lys Ser Asp Ile Glu Cys Ala
260 265 270
Lys Asp Ser Tyr Glu Glu Asp Leu Glu Ser Leu Leu Ala Leu Ile Gly
275 280 285
Asp Glu Tyr Ala Glu Leu Phe Val Ala Ala Lys Asn Ala Tyr Ser Ala
290 295 300
Val Val Leu Ser Ser Ile Ile Thr Val Ala Glu Thr Glu Thr Asn Ala
305 310 315 320
Lys Leu Ser Ala Ser Met Ile Glu Arg Phe Asp Thr His Glu Glu Asp
325 330 335
Leu Gly Glu Leu Lys Ala Phe Ile Lys Leu His Leu Pro Lys His Tyr
340 345 350
Glu Glu Ile Phe Ser Asn Thr Glu Lys His Gly Tyr Ala Gly Tyr Ile
355 360 365
Asp Gly Lys Thr Lys Gln Ala Asp Phe Tyr Lys Tyr Met Lys Met Thr
370 375 380
Leu Glu Asn Ile Glu Gly Ala Asp Tyr Phe Ile Ala Lys Ile Glu Lys
385 390 395 400
Glu Asn Phe Leu Arg Lys Gln Arg Thr Phe Asp Asn Gly Ala Ile Pro
405 410 415
His Gln Leu His Leu Glu Glu Leu Glu Ala Ile Leu His Gln Gln Ala
420 425 430
Lys Tyr Tyr Pro Phe Leu Lys Glu Asn Tyr Asp Lys Ile Lys Ser Leu
435 440 445
Val Thr Phe Arg Ile Pro Tyr Phe Val Gly Pro Leu Ala Asn Gly Gln
450 455 460
Ser Glu Phe Ala Trp Leu Thr Arg Lys Ala Asp Gly Glu Ile Arg Pro
465 470 475 480
Trp Asn Ile Glu Glu Lys Val Asp Phe Gly Lys Ser Ala Val Asp Phe
485 490 495
Ile Glu Lys Met Thr Asn Lys Asp Thr Tyr Leu Pro Lys Glu Asn Val
500 505 510
Leu Pro Lys His Ser Leu Cys Tyr Gln Lys Tyr Leu Val Tyr Asn Glu
515 520 525
Leu Thr Lys Val Arg Tyr Ile Asn Asp Gln Gly Lys Thr Ser Tyr Phe
530 535 540
Ser Gly Gln Glu Lys Glu Gln Ile Phe Asn Asp Leu Phe Lys Gln Lys
545 550 555 560
Arg Lys Val Lys Lys Lys Asp Leu Glu Leu Phe Leu Arg Asn Met Ser
565 570 575
His Val Glu Ser Pro Thr Ile Glu Gly Leu Glu Asp Ser Phe Asn Ser
580 585 590
Ser Tyr Ser Thr Tyr His Asp Leu Leu Lys Val Gly Ile Lys Gln Glu
595 600 605
Ile Leu Asp Asn Pro Val Asn Thr Glu Met Leu Glu Asn Ile Val Lys
610 615 620
Ile Leu Thr Val Phe Glu Asp Lys Arg Met Ile Lys Glu Gln Leu Gln
625 630 635 640
Gln Phe Ser Asp Val Leu Asp Gly Val Val Leu Lys Lys Leu Glu Arg
645 650 655
Arg His Tyr Thr Gly Trp Gly Arg Leu Ser Ala Lys Leu Leu Met Gly
660 665 670
Ile Arg Asp Lys Gln Ser His Leu Thr Ile Leu Asp Tyr Leu Met Asn
675 680 685
Asp Asp Gly Leu Asn Arg Asn Leu Met Gln Leu Ile Asn Asp Ser Asn
690 695 700
Leu Ser Phe Lys Ser Ile Ile Glu Lys Glu Gln Val Thr Thr Ala Asp
705 710 715 720
Lys Asp Ile Gln Ser Ile Val Ala Asp Leu Ala Gly Ser Pro Ala Ile
725 730 735
Lys Lys Gly Ile Leu Gln Ser Leu Lys Ile Val Asp Glu Leu Val Ser
740 745 750
Val Met Gly Tyr Pro Pro Gln Thr Ile Val Val Glu Met Ala Arg Glu
755 760 765
Asn Gln Thr Thr Gly Lys Gly Lys Asn Asn Ser Arg Pro Arg Tyr Lys
770 775 780
Ser Leu Glu Lys Ala Ile Lys Glu Phe Gly Ser Gln Ile Leu Lys Glu
785 790 795 800
His Pro Thr Asp Asn Gln Glu Leu Arg Asn Asn Arg Leu Tyr Leu Tyr
805 810 815
Tyr Leu Gln Asn Gly Lys Asp Met Tyr Thr Gly Gln Asp Leu Asp Ile
820 825 830
His Asn Leu Ser Asn Tyr Asp Ile Asp His Ile Val Pro Gln Ser Phe
835 840 845
Ile Thr Asp Asn Ser Ile Asp Asn Leu Val Leu Thr Ser Ser Ala Gly
850 855 860
Asn Arg Glu Lys Gly Asp Asp Val Pro Pro Leu Glu Ile Val Arg Lys
865 870 875 880
Arg Lys Val Phe Trp Glu Lys Leu Tyr Gln Gly Asn Leu Met Ser Lys
885 890 895
Arg Lys Phe Asp Tyr Leu Thr Lys Ala Glu Arg Gly Gly Leu Thr Glu
900 905 910
Ala Asp Lys Ala Arg Phe Ile His Arg Gln Leu Val Glu Thr Arg Gln
915 920 925
Ile Thr Lys Asn Val Ala Asn Ile Leu His Gln Arg Phe Asn Tyr Glu
930 935 940
Lys Asp Asp His Gly Asn Thr Met Lys Gln Val Arg Ile Val Thr Leu
945 950 955 960
Lys Ser Ala Leu Val Ser Gln Phe Arg Lys Gln Phe Gln Leu Tyr Lys
965 970 975
Val Arg Asp Val Asn Asp Tyr His His Ala His Asp Ala Tyr Leu Asn
980 985 990
Gly Val Val Ala Asn Thr Leu Leu Lys Val Tyr Pro Gln Leu Glu Pro
995 1000 1005
Glu Phe Val Tyr Gly Asp Tyr His Gln Phe Asp Trp Phe Lys Ala Asn
1010 1015 1020
Lys Ala Thr Ala Lys Lys Gln Phe Tyr Thr Asn Ile Met Leu Phe Phe
1025 1030 1035 1040
Ala Gln Lys Asp Arg Ile Ile Asp Glu Asn Gly Glu Ile Leu Trp Asp
1045 1050 1055
Lys Lys Tyr Leu Asp Thr Val Lys Lys Val Met Ser Tyr Arg Gln Met
1060 1065 1070
Asn Ile Val Lys Lys Thr Glu Ile Gln Lys Gly Glu Phe Ser Lys Ala
1075 1080 1085
Thr Ile Lys Pro Lys Gly Asn Ser Ser Lys Leu Ile Pro Arg Lys Thr
1090 1095 1100
Asn Trp Asp Pro Met Lys Tyr Gly Gly Leu Asp Ser Pro Asn Met Ala
1105 1110 1115 1120
Tyr Ala Val Val Ile Glu Tyr Ala Lys Gly Lys Asn Lys Leu Val Phe
1125 1130 1135
Glu Lys Lys Ile Ile Arg Val Thr Ile Met Glu Arg Lys Ala Phe Glu
1140 1145 1150
Lys Asp Glu Lys Ala Phe Leu Glu Glu Gln Gly Tyr Arg Gln Pro Lys
1155 1160 1165
Val Leu Ala Lys Leu Pro Lys Tyr Thr Leu Tyr Glu Cys Glu Glu Gly
1170 1175 1180
Arg Arg Arg Met Leu Ala Ser Ala Asn Glu Ala Gln Lys Gly Asn Gln
1185 1190 1195 1200
Gln Val Leu Pro Asn His Leu Val Thr Leu Leu His His Ala Ala Asn
1205 1210 1215
Cys Glu Val Ser Asp Gly Lys Ser Leu Asp Tyr Ile Glu Ser Asn Arg
1220 1225 1230
Glu Met Phe Ala Glu Leu Leu Ala His Val Ser Glu Phe Ala Lys Arg
1235 1240 1245
Tyr Thr Leu Ala Glu Ala Asn Leu Asn Lys Ile Asn Gln Leu Phe Glu
1250 1255 1260
Gln Asn Lys Glu Gly Asp Ile Lys Ala Ile Ala Gln Ser Phe Val Asp
1265 1270 1275 1280
Leu Met Ala Phe Asn Ala Met Gly Ala Pro Ala Ser Phe Lys Phe Phe
1285 1290 1295
Glu Thr Thr Ile Glu Arg Lys Arg Tyr Asn Asn Leu Lys Glu Leu Leu
1300 1305 1310
Asn Ser Thr Ile Ile Tyr Gln Ser Ile Thr Gly Leu Tyr Glu Ser Arg
1315 1320 1325
Lys Arg Leu Asp Asp
1330
<210> 161
<211> 1082
<212> PRT
<213> Искусственная последовательность
<220>
<223> Neisseria meningitidis Cas9
<400> 161
Met Ala Ala Phe Lys Pro Asn Ser Ile Asn Tyr Ile Leu Gly Leu Asp
1 5 10 15
Ile Gly Ile Ala Ser Val Gly Trp Ala Met Val Glu Ile Asp Glu Glu
20 25 30
Glu Asn Pro Ile Arg Leu Ile Asp Leu Gly Val Arg Val Phe Glu Arg
35 40 45
Ala Glu Val Pro Lys Thr Gly Asp Ser Leu Ala Met Ala Arg Arg Leu
50 55 60
Ala Arg Ser Val Arg Arg Leu Thr Arg Arg Arg Ala His Arg Leu Leu
65 70 75 80
Arg Thr Arg Arg Leu Leu Lys Arg Glu Gly Val Leu Gln Ala Ala Asn
85 90 95
Phe Asp Glu Asn Gly Leu Ile Lys Ser Leu Pro Asn Thr Pro Trp Gln
100 105 110
Leu Arg Ala Ala Ala Leu Asp Arg Lys Leu Thr Pro Leu Glu Trp Ser
115 120 125
Ala Val Leu Leu His Leu Ile Lys His Arg Gly Tyr Leu Ser Gln Arg
130 135 140
Lys Asn Glu Gly Glu Thr Ala Asp Lys Glu Leu Gly Ala Leu Leu Lys
145 150 155 160
Gly Val Ala Gly Asn Ala His Ala Leu Gln Thr Gly Asp Phe Arg Thr
165 170 175
Pro Ala Glu Leu Ala Leu Asn Lys Phe Glu Lys Glu Ser Gly His Ile
180 185 190
Arg Asn Gln Arg Ser Asp Tyr Ser His Thr Phe Ser Arg Lys Asp Leu
195 200 205
Gln Ala Glu Leu Ile Leu Leu Phe Glu Lys Gln Lys Glu Phe Gly Asn
210 215 220
Pro His Val Ser Gly Gly Leu Lys Glu Gly Ile Glu Thr Leu Leu Met
225 230 235 240
Thr Gln Arg Pro Ala Leu Ser Gly Asp Ala Val Gln Lys Met Leu Gly
245 250 255
His Cys Thr Phe Glu Pro Ala Glu Pro Lys Ala Ala Lys Asn Thr Tyr
260 265 270
Thr Ala Glu Arg Phe Ile Trp Leu Thr Lys Leu Asn Asn Leu Arg Ile
275 280 285
Leu Glu Gln Gly Ser Glu Arg Pro Leu Thr Asp Thr Glu Arg Ala Thr
290 295 300
Leu Met Asp Glu Pro Tyr Arg Lys Ser Lys Leu Thr Tyr Ala Gln Ala
305 310 315 320
Arg Lys Leu Leu Gly Leu Glu Asp Thr Ala Phe Phe Lys Gly Leu Arg
325 330 335
Tyr Gly Lys Asp Asn Ala Glu Ala Ser Thr Leu Met Glu Met Lys Ala
340 345 350
Tyr His Ala Ile Ser Arg Ala Leu Glu Lys Glu Gly Leu Lys Asp Lys
355 360 365
Lys Ser Pro Leu Asn Leu Ser Pro Glu Leu Gln Asp Glu Ile Gly Thr
370 375 380
Ala Phe Ser Leu Phe Lys Thr Asp Glu Asp Ile Thr Gly Arg Leu Lys
385 390 395 400
Asp Arg Ile Gln Pro Glu Ile Leu Glu Ala Leu Leu Lys His Ile Ser
405 410 415
Phe Asp Lys Phe Val Gln Ile Ser Leu Lys Ala Leu Arg Arg Ile Val
420 425 430
Pro Leu Met Glu Gln Gly Lys Arg Tyr Asp Glu Ala Cys Ala Glu Ile
435 440 445
Tyr Gly Asp His Tyr Gly Lys Lys Asn Thr Glu Glu Lys Ile Tyr Leu
450 455 460
Pro Pro Ile Pro Ala Asp Glu Ile Arg Asn Pro Val Val Leu Arg Ala
465 470 475 480
Leu Ser Gln Ala Arg Lys Val Ile Asn Gly Val Val Arg Arg Tyr Gly
485 490 495
Ser Pro Ala Arg Ile His Ile Glu Thr Ala Arg Glu Val Gly Lys Ser
500 505 510
Phe Lys Asp Arg Lys Glu Ile Glu Lys Arg Gln Glu Glu Asn Arg Lys
515 520 525
Asp Arg Glu Lys Ala Ala Ala Lys Phe Arg Glu Tyr Phe Pro Asn Phe
530 535 540
Val Gly Glu Pro Lys Ser Lys Asp Ile Leu Lys Leu Arg Leu Tyr Glu
545 550 555 560
Gln Gln His Gly Lys Cys Leu Tyr Ser Gly Lys Glu Ile Asn Leu Gly
565 570 575
Arg Leu Asn Glu Lys Gly Tyr Val Glu Ile Asp His Ala Leu Pro Phe
580 585 590
Ser Arg Thr Trp Asp Asp Ser Phe Asn Asn Lys Val Leu Val Leu Gly
595 600 605
Ser Glu Asn Gln Asn Lys Gly Asn Gln Thr Pro Tyr Glu Tyr Phe Asn
610 615 620
Gly Lys Asp Asn Ser Arg Glu Trp Gln Glu Phe Lys Ala Arg Val Glu
625 630 635 640
Thr Ser Arg Phe Pro Arg Ser Lys Lys Gln Arg Ile Leu Leu Gln Lys
645 650 655
Phe Asp Glu Asp Gly Phe Lys Glu Arg Asn Leu Asn Asp Thr Arg Tyr
660 665 670
Val Asn Arg Phe Leu Cys Gln Phe Val Ala Asp Arg Met Arg Leu Thr
675 680 685
Gly Lys Gly Lys Lys Arg Val Phe Ala Ser Asn Gly Gln Ile Thr Asn
690 695 700
Leu Leu Arg Gly Phe Trp Gly Leu Arg Lys Val Arg Ala Glu Asn Asp
705 710 715 720
Arg His His Ala Leu Asp Ala Val Val Val Ala Cys Ser Thr Val Ala
725 730 735
Met Gln Gln Lys Ile Thr Arg Phe Val Arg Tyr Lys Glu Met Asn Ala
740 745 750
Phe Asp Gly Lys Thr Ile Asp Lys Glu Thr Gly Glu Val Leu His Gln
755 760 765
Lys Thr His Phe Pro Gln Pro Trp Glu Phe Phe Ala Gln Glu Val Met
770 775 780
Ile Arg Val Phe Gly Lys Pro Asp Gly Lys Pro Glu Phe Glu Glu Ala
785 790 795 800
Asp Thr Leu Glu Lys Leu Arg Thr Leu Leu Ala Glu Lys Leu Ser Ser
805 810 815
Arg Pro Glu Ala Val His Glu Tyr Val Thr Pro Leu Phe Val Ser Arg
820 825 830
Ala Pro Asn Arg Lys Met Ser Gly Gln Gly His Met Glu Thr Val Lys
835 840 845
Ser Ala Lys Arg Leu Asp Glu Gly Val Ser Val Leu Arg Val Pro Leu
850 855 860
Thr Gln Leu Lys Leu Lys Asp Leu Glu Lys Met Val Asn Arg Glu Arg
865 870 875 880
Glu Pro Lys Leu Tyr Glu Ala Leu Lys Ala Arg Leu Glu Ala His Lys
885 890 895
Asp Asp Pro Ala Lys Ala Phe Ala Glu Pro Phe Tyr Lys Tyr Asp Lys
900 905 910
Ala Gly Asn Arg Thr Gln Gln Val Lys Ala Val Arg Val Glu Gln Val
915 920 925
Gln Lys Thr Gly Val Trp Val Arg Asn His Asn Gly Ile Ala Asp Asn
930 935 940
Ala Thr Met Val Arg Val Asp Val Phe Glu Lys Gly Asp Lys Tyr Tyr
945 950 955 960
Leu Val Pro Ile Tyr Ser Trp Gln Val Ala Lys Gly Ile Leu Pro Asp
965 970 975
Arg Ala Val Val Gln Gly Lys Asp Glu Glu Asp Trp Gln Leu Ile Asp
980 985 990
Asp Ser Phe Asn Phe Lys Phe Ser Leu His Pro Asn Asp Leu Val Glu
995 1000 1005
Val Ile Thr Lys Lys Ala Arg Met Phe Gly Tyr Phe Ala Ser Cys His
1010 1015 1020
Arg Gly Thr Gly Asn Ile Asn Ile Arg Ile His Asp Leu Asp His Lys
1025 1030 1035 1040
Ile Gly Lys Asn Gly Ile Leu Glu Gly Ile Gly Val Lys Thr Ala Leu
1045 1050 1055
Ser Phe Gln Lys Tyr Gln Ile Asp Glu Leu Gly Lys Glu Ile Arg Pro
1060 1065 1070
Cys Arg Leu Lys Lys Arg Pro Pro Val Arg
1075 1080
<210> 162
<211> 1368
<212> PRT
<213> Искусственная последовательность
<220>
<223> Streptococcus pyogenes Cas9
<400> 162
Met Asp Lys Lys Tyr Ser Ile Gly Leu Asp Ile Gly Thr Asn Ser Val
1 5 10 15
Gly Trp Ala Val Ile Thr Asp Glu Tyr Lys Val Pro Ser Lys Lys Phe
20 25 30
Lys Val Leu Gly Asn Thr Asp Arg His Ser Ile Lys Lys Asn Leu Ile
35 40 45
Gly Ala Leu Leu Phe Asp Ser Gly Glu Thr Ala Glu Ala Thr Arg Leu
50 55 60
Lys Arg Thr Ala Arg Arg Arg Tyr Thr Arg Arg Lys Asn Arg Ile Cys
65 70 75 80
Tyr Leu Gln Glu Ile Phe Ser Asn Glu Met Ala Lys Val Asp Asp Ser
85 90 95
Phe Phe His Arg Leu Glu Glu Ser Phe Leu Val Glu Glu Asp Lys Lys
100 105 110
His Glu Arg His Pro Ile Phe Gly Asn Ile Val Asp Glu Val Ala Tyr
115 120 125
His Glu Lys Tyr Pro Thr Ile Tyr His Leu Arg Lys Lys Leu Val Asp
130 135 140
Ser Thr Asp Lys Ala Asp Leu Arg Leu Ile Tyr Leu Ala Leu Ala His
145 150 155 160
Met Ile Lys Phe Arg Gly His Phe Leu Ile Glu Gly Asp Leu Asn Pro
165 170 175
Asp Asn Ser Asp Val Asp Lys Leu Phe Ile Gln Leu Val Gln Thr Tyr
180 185 190
Asn Gln Leu Phe Glu Glu Asn Pro Ile Asn Ala Ser Gly Val Asp Ala
195 200 205
Lys Ala Ile Leu Ser Ala Arg Leu Ser Lys Ser Arg Arg Leu Glu Asn
210 215 220
Leu Ile Ala Gln Leu Pro Gly Glu Lys Lys Asn Gly Leu Phe Gly Asn
225 230 235 240
Leu Ile Ala Leu Ser Leu Gly Leu Thr Pro Asn Phe Lys Ser Asn Phe
245 250 255
Asp Leu Ala Glu Asp Ala Lys Leu Gln Leu Ser Lys Asp Thr Tyr Asp
260 265 270
Asp Asp Leu Asp Asn Leu Leu Ala Gln Ile Gly Asp Gln Tyr Ala Asp
275 280 285
Leu Phe Leu Ala Ala Lys Asn Leu Ser Asp Ala Ile Leu Leu Ser Asp
290 295 300
Ile Leu Arg Val Asn Thr Glu Ile Thr Lys Ala Pro Leu Ser Ala Ser
305 310 315 320
Met Ile Lys Arg Tyr Asp Glu His His Gln Asp Leu Thr Leu Leu Lys
325 330 335
Ala Leu Val Arg Gln Gln Leu Pro Glu Lys Tyr Lys Glu Ile Phe Phe
340 345 350
Asp Gln Ser Lys Asn Gly Tyr Ala Gly Tyr Ile Asp Gly Gly Ala Ser
355 360 365
Gln Glu Glu Phe Tyr Lys Phe Ile Lys Pro Ile Leu Glu Lys Met Asp
370 375 380
Gly Thr Glu Glu Leu Leu Val Lys Leu Asn Arg Glu Asp Leu Leu Arg
385 390 395 400
Lys Gln Arg Thr Phe Asp Asn Gly Ser Ile Pro His Gln Ile His Leu
405 410 415
Gly Glu Leu His Ala Ile Leu Arg Arg Gln Glu Asp Phe Tyr Pro Phe
420 425 430
Leu Lys Asp Asn Arg Glu Lys Ile Glu Lys Ile Leu Thr Phe Arg Ile
435 440 445
Pro Tyr Tyr Val Gly Pro Leu Ala Arg Gly Asn Ser Arg Phe Ala Trp
450 455 460
Met Thr Arg Lys Ser Glu Glu Thr Ile Thr Pro Trp Asn Phe Glu Glu
465 470 475 480
Val Val Asp Lys Gly Ala Ser Ala Gln Ser Phe Ile Glu Arg Met Thr
485 490 495
Asn Phe Asp Lys Asn Leu Pro Asn Glu Lys Val Leu Pro Lys His Ser
500 505 510
Leu Leu Tyr Glu Tyr Phe Thr Val Tyr Asn Glu Leu Thr Lys Val Lys
515 520 525
Tyr Val Thr Glu Gly Met Arg Lys Pro Ala Phe Leu Ser Gly Glu Gln
530 535 540
Lys Lys Ala Ile Val Asp Leu Leu Phe Lys Thr Asn Arg Lys Val Thr
545 550 555 560
Val Lys Gln Leu Lys Glu Asp Tyr Phe Lys Lys Ile Glu Cys Phe Asp
565 570 575
Ser Val Glu Ile Ser Gly Val Glu Asp Arg Phe Asn Ala Ser Leu Gly
580 585 590
Thr Tyr His Asp Leu Leu Lys Ile Ile Lys Asp Lys Asp Phe Leu Asp
595 600 605
Asn Glu Glu Asn Glu Asp Ile Leu Glu Asp Ile Val Leu Thr Leu Thr
610 615 620
Leu Phe Glu Asp Arg Glu Met Ile Glu Glu Arg Leu Lys Thr Tyr Ala
625 630 635 640
His Leu Phe Asp Asp Lys Val Met Lys Gln Leu Lys Arg Arg Arg Tyr
645 650 655
Thr Gly Trp Gly Arg Leu Ser Arg Lys Leu Ile Asn Gly Ile Arg Asp
660 665 670
Lys Gln Ser Gly Lys Thr Ile Leu Asp Phe Leu Lys Ser Asp Gly Phe
675 680 685
Ala Asn Arg Asn Phe Met Gln Leu Ile His Asp Asp Ser Leu Thr Phe
690 695 700
Lys Glu Asp Ile Gln Lys Ala Gln Val Ser Gly Gln Gly Asp Ser Leu
705 710 715 720
His Glu His Ile Ala Asn Leu Ala Gly Ser Pro Ala Ile Lys Lys Gly
725 730 735
Ile Leu Gln Thr Val Lys Val Val Asp Glu Leu Val Lys Val Met Gly
740 745 750
Arg His Lys Pro Glu Asn Ile Val Ile Glu Met Ala Arg Glu Asn Gln
755 760 765
Thr Thr Gln Lys Gly Gln Lys Asn Ser Arg Glu Arg Met Lys Arg Ile
770 775 780
Glu Glu Gly Ile Lys Glu Leu Gly Ser Gln Ile Leu Lys Glu His Pro
785 790 795 800
Val Glu Asn Thr Gln Leu Gln Asn Glu Lys Leu Tyr Leu Tyr Tyr Leu
805 810 815
Gln Asn Gly Arg Asp Met Tyr Val Asp Gln Glu Leu Asp Ile Asn Arg
820 825 830
Leu Ser Asp Tyr Asp Val Asp His Ile Val Pro Gln Ser Phe Leu Lys
835 840 845
Asp Asp Ser Ile Asp Asn Lys Val Leu Thr Arg Ser Asp Lys Asn Arg
850 855 860
Gly Lys Ser Asp Asn Val Pro Ser Glu Glu Val Val Lys Lys Met Lys
865 870 875 880
Asn Tyr Trp Arg Gln Leu Leu Asn Ala Lys Leu Ile Thr Gln Arg Lys
885 890 895
Phe Asp Asn Leu Thr Lys Ala Glu Arg Gly Gly Leu Ser Glu Leu Asp
900 905 910
Lys Ala Gly Phe Ile Lys Arg Gln Leu Val Glu Thr Arg Gln Ile Thr
915 920 925
Lys His Val Ala Gln Ile Leu Asp Ser Arg Met Asn Thr Lys Tyr Asp
930 935 940
Glu Asn Asp Lys Leu Ile Arg Glu Val Lys Val Ile Thr Leu Lys Ser
945 950 955 960
Lys Leu Val Ser Asp Phe Arg Lys Asp Phe Gln Phe Tyr Lys Val Arg
965 970 975
Glu Ile Asn Asn Tyr His His Ala His Asp Ala Tyr Leu Asn Ala Val
980 985 990
Val Gly Thr Ala Leu Ile Lys Lys Tyr Pro Lys Leu Glu Ser Glu Phe
995 1000 1005
Val Tyr Gly Asp Tyr Lys Val Tyr Asp Val Arg Lys Met Ile Ala Lys
1010 1015 1020
Ser Glu Gln Glu Ile Gly Lys Ala Thr Ala Lys Tyr Phe Phe Tyr Ser
1025 1030 1035 1040
Asn Ile Met Asn Phe Phe Lys Thr Glu Ile Thr Leu Ala Asn Gly Glu
1045 1050 1055
Ile Arg Lys Arg Pro Leu Ile Glu Thr Asn Gly Glu Thr Gly Glu Ile
1060 1065 1070
Val Trp Asp Lys Gly Arg Asp Phe Ala Thr Val Arg Lys Val Leu Ser
1075 1080 1085
Met Pro Gln Val Asn Ile Val Lys Lys Thr Glu Val Gln Thr Gly Gly
1090 1095 1100
Phe Ser Lys Glu Ser Ile Leu Pro Lys Arg Asn Ser Asp Lys Leu Ile
1105 1110 1115 1120
Ala Arg Lys Lys Asp Trp Asp Pro Lys Lys Tyr Gly Gly Phe Asp Ser
1125 1130 1135
Pro Thr Val Ala Tyr Ser Val Leu Val Val Ala Lys Val Glu Lys Gly
1140 1145 1150
Lys Ser Lys Lys Leu Lys Ser Val Lys Glu Leu Leu Gly Ile Thr Ile
1155 1160 1165
Met Glu Arg Ser Ser Phe Glu Lys Asn Pro Ile Asp Phe Leu Glu Ala
1170 1175 1180
Lys Gly Tyr Lys Glu Val Lys Lys Asp Leu Ile Ile Lys Leu Pro Lys
1185 1190 1195 1200
Tyr Ser Leu Phe Glu Leu Glu Asn Gly Arg Lys Arg Met Leu Ala Ser
1205 1210 1215
Ala Gly Glu Leu Gln Lys Gly Asn Glu Leu Ala Leu Pro Ser Lys Tyr
1220 1225 1230
Val Asn Phe Leu Tyr Leu Ala Ser His Tyr Glu Lys Leu Lys Gly Ser
1235 1240 1245
Pro Glu Asp Asn Glu Gln Lys Gln Leu Phe Val Glu Gln His Lys His
1250 1255 1260
Tyr Leu Asp Glu Ile Ile Glu Gln Ile Ser Glu Phe Ser Lys Arg Val
1265 1270 1275 1280
Ile Leu Ala Asp Ala Asn Leu Asp Lys Val Leu Ser Ala Tyr Asn Lys
1285 1290 1295
His Arg Asp Lys Pro Ile Arg Glu Gln Ala Glu Asn Ile Ile His Leu
1300 1305 1310
Phe Thr Leu Thr Asn Leu Gly Ala Pro Ala Ala Phe Lys Tyr Phe Asp
1315 1320 1325
Thr Thr Ile Asp Arg Lys Arg Tyr Thr Ser Thr Lys Glu Val Leu Asp
1330 1335 1340
Ala Thr Leu Ile His Gln Ser Ile Thr Gly Leu Tyr Glu Thr Arg Ile
1345 1350 1355 1360
Asp Leu Ser Gln Leu Gly Gly Asp
1365
<210> 163
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность 2
<400> 163
gagaatcaaa atcggtgaat 20
<210> 164
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRBC целевая последовательность 2
<400> 164
ggcctcggcg ctgacgatct 20
<210> 165
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 3
<400> 165
gagaatcaaa atcggtgaat agg 23
<210> 166
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 4
<400> 166
ttcaaaacct gtcagtgatt ggg 23
<210> 167
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 5
<400> 167
tgtgctagac atgaggtcta tgg 23
<210> 168
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 6
<400> 168
cgtcatgagc agattaaacc cgg 23
<210> 169
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 7
<400> 169
tcagggttct ggatatctgt ggg 23
<210> 170
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 8
<400> 170
gtcagggttc tggatatctg tgg 23
<210> 171
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 9
<400> 171
ttcggaaccc aatcactgac agg 23
<210> 172
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 10
<400> 172
taaacccggc cactttcagg agg 23
<210> 173
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 11
<400> 173
aaagtcagat ttgttgctcc agg 23
<210> 174
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 12
<400> 174
aacaaatgtg tcacaaagta agg 23
<210> 175
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 13
<400> 175
tggatttaga gtctctcagc tgg 23
<210> 176
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 14
<400> 176
taggcagaca gacttgtcac tgg 23
<210> 177
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 15
<400> 177
agctggtaca cggcagggtc agg 23
<210> 178
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 16
<400> 178
gctggtacac ggcagggtca ggg 23
<210> 179
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 17
<400> 179
tctctcagct ggtacacggc agg 23
<210> 180
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 18
<400> 180
tttcaaaacc tgtcagtgat tgg 23
<210> 181
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 19
<400> 181
gattaaaccc ggccactttc agg 23
<210> 182
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 20
<400> 182
ctcgaccagc ttgacatcac agg 23
<210> 183
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 21
<400> 183
agagtctctc agctggtaca cgg 23
<210> 184
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 22
<400> 184
ctctcagctg gtacacggca ggg 23
<210> 185
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 23
<400> 185
aagttcctgt gatgtcaagc tgg 23
<210> 186
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 24
<400> 186
atcctcctcc tgaaagtggc cgg 23
<210> 187
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 25
<400> 187
tgctcatgac gctgcggctg tgg 23
<210> 188
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 26
<400> 188
acaaaactgt gctagacatg agg 23
<210> 189
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 27
<400> 189
atttgtttga gaatcaaaat cgg 23
<210> 190
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 28
<400> 190
catcacagga actttctaaa agg 23
<210> 191
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 29
<400> 191
gtcgagaaaa gctttgaaac agg 23
<210> 192
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 30
<400> 192
ccactttcag gaggaggatt cgg 23
<210> 193
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 31
<400> 193
ctgacaggtt ttgaaagttt agg 23
<210> 194
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 32
<400> 194
agctttgaaa caggtaagac agg 23
<210> 195
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 33
<400> 195
tggaataatg ctgttgttga agg 23
<210> 196
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 34
<400> 196
agagcaacag tgctgtggcc tgg 23
<210> 197
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 35
<400> 197
ctgtggtcca gctgaggtga ggg 23
<210> 198
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 36
<400> 198
ctgcggctgt ggtccagctg agg 23
<210> 199
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 37
<400> 199
tgtggtccag ctgaggtgag ggg 23
<210> 200
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 38
<400> 200
cttcttcccc agcccaggta agg 23
<210> 201
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 39
<400> 201
acacggcagg gtcagggttc tgg 23
<210> 202
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 40
<400> 202
cttcaagagc aacagtgctg tgg 23
<210> 203
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 41
<400> 203
ctggggaaga aggtgtcttc tgg 23
<210> 204
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 42
<400> 204
tcctcctcct gaaagtggcc ggg 23
<210> 205
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 43
<400> 205
ttaatctgct catgacgctg cgg 23
<210> 206
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 44
<400> 206
acccggccac tttcaggagg agg 23
<210> 207
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 45
<400> 207
ttcttcccca gcccaggtaa ggg 23
<210> 208
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 46
<400> 208
cttacctggg ctggggaaga agg 23
<210> 209
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 47
<400> 209
gacaccttct tccccagccc agg 23
<210> 210
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 48
<400> 210
gctgtggtcc agctgaggtg agg 23
<210> 211
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC целевая последовательность с PAM 49
<400> 211
ccgaatcctc ctcctgaaag tgg 23
<210> 212
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRBC целевая последовательность с PAM 3
<400> 212
gctgtcaagt ccagttctac ggg 23
<210> 213
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 1
<400> 213
ctatggactt caagagcaac agtgctgt 28
<210> 214
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 2
<400> 214
ctcatgtcta gcacagtttt gtctgtga 28
<210> 215
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 3
<400> 215
gtgctgtggc ctggagcaac aaatctga 28
<210> 216
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 4
<400> 216
ttgctcttga agtccataga cctcatgt 28
<210> 217
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 5
<400> 217
ctgtggcctg gagcaacaaa tctgact 27
<210> 218
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 6
<400> 218
ctgttgctct tgaagtccat agacctca 28
<210> 219
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 7
<400> 219
ctgtggcctg gagcaacaaa tctgactt 28
<210> 220
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 8
<400> 220
ctgactttgc atgtgcaaac gccttcaa 28
<210> 221
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 9
<400> 221
ttgttgctcc aggccacagc actgttgc 28
<210> 222
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 10
<400> 222
tgaaagtggc cgggtttaat ctgctcat 28
<210> 223
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 11
<400> 223
aggaggattc ggaacccaat cactgaca 28
<210> 224
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC ZFN целевая последовательность 12
<400> 224
gaggaggatt cggaacccaa tcactgac 28
<210> 225
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRBC ZFN целевая последовательность 1
<400> 225
ccgtagaact ggacttgaca gcggaagt 28
<210> 226
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRBC ZFN целевая последовательность 2
<400> 226
tctcggagaa tgacgagtgg acccagga 28
<210> 227
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 50 п.о. 5' плечо гомологии
<400> 227
agctgagaga ctctaaatcc agtgacaagt ctgtctgcct attcaccgat 50
<210> 228
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 100 п.о. 5' плечо гомологии
<400> 228
ctgatcctct tgtcccacag atatccagaa ccctgaccct gccgtgtacc agctgagaga 60
ctctaaatcc agtgacaagt ctgtctgcct attcaccgat 100
<210> 229
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 200 п.о. 5' плечо гомологии
<400> 229
gtgacttgcc agccccacag agccccgccc ttgtccatca ctggcatctg gactccagcc 60
tgggttgggg caaagaggga aatgagatca tgtcctaacc ctgatcctct tgtcccacag 120
atatccagaa ccctgaccct gccgtgtacc agctgagaga ctctaaatcc agtgacaagt 180
ctgtctgcct attcaccgat 200
<210> 230
<211> 300
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 300 п.о. 5' плечо гомологии
<400> 230
cctcttggcc aagattgata gcttgtgcct gtccctgagt cccagtccat cacgagcagc 60
tggtttctaa gatgctattt cccgtataaa gcatgagacc gtgacttgcc agccccacag 120
agccccgccc ttgtccatca ctggcatctg gactccagcc tgggttgggg caaagaggga 180
aatgagatca tgtcctaacc ctgatcctct tgtcccacag atatccagaa ccctgaccct 240
gccgtgtacc agctgagaga ctctaaatcc agtgacaagt ctgtctgcct attcaccgat 300
<210> 231
<211> 400
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 400 п.о. 5' плечо гомологии
<400> 231
attaaataaa agaataagca gtattattaa gtagccctgc atttcaggtt tccttgagtg 60
gcaggccagg cctggccgtg aacgttcact gaaatcatgg cctcttggcc aagattgata 120
gcttgtgcct gtccctgagt cccagtccat cacgagcagc tggtttctaa gatgctattt 180
cccgtataaa gcatgagacc gtgacttgcc agccccacag agccccgccc ttgtccatca 240
ctggcatctg gactccagcc tgggttgggg caaagaggga aatgagatca tgtcctaacc 300
ctgatcctct tgtcccacag atatccagaa ccctgaccct gccgtgtacc agctgagaga 360
ctctaaatcc agtgacaagt ctgtctgcct attcaccgat 400
<210> 232
<211> 500
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 500 п.о. 5' плечо гомологии
<400> 232
ctccagattc caagatgtac agtttgcttt gctgggcctt tttcccatgc ctgcctttac 60
tctgccagag ttatattgct ggggttttga agaagatcct attaaataaa agaataagca 120
gtattattaa gtagccctgc atttcaggtt tccttgagtg gcaggccagg cctggccgtg 180
aacgttcact gaaatcatgg cctcttggcc aagattgata gcttgtgcct gtccctgagt 240
cccagtccat cacgagcagc tggtttctaa gatgctattt cccgtataaa gcatgagacc 300
gtgacttgcc agccccacag agccccgccc ttgtccatca ctggcatctg gactccagcc 360
tgggttgggg caaagaggga aatgagatca tgtcctaacc ctgatcctct tgtcccacag 420
atatccagaa ccctgaccct gccgtgtacc agctgagaga ctctaaatcc agtgacaagt 480
ctgtctgcct attcaccgat 500
<210> 233
<211> 600
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 600 п.о. 5' плечо гомологии
<400> 233
agagagcaat ctcctggtaa tgtgatagat ttcccaactt aatgccaaca taccataaac 60
ctcccattct gctaatgccc agcctaagtt ggggagacca ctccagattc caagatgtac 120
agtttgcttt gctgggcctt tttcccatgc ctgcctttac tctgccagag ttatattgct 180
ggggttttga agaagatcct attaaataaa agaataagca gtattattaa gtagccctgc 240
atttcaggtt tccttgagtg gcaggccagg cctggccgtg aacgttcact gaaatcatgg 300
cctcttggcc aagattgata gcttgtgcct gtccctgagt cccagtccat cacgagcagc 360
tggtttctaa gatgctattt cccgtataaa gcatgagacc gtgacttgcc agccccacag 420
agccccgccc ttgtccatca ctggcatctg gactccagcc tgggttgggg caaagaggga 480
aatgagatca tgtcctaacc ctgatcctct tgtcccacag atatccagaa ccctgaccct 540
gccgtgtacc agctgagaga ctctaaatcc agtgacaagt ctgtctgcct attcaccgat 600
<210> 234
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 50 п.о. 3' плечо гомологии
<400> 234
tttgattctc aaacaaatgt gtcacaaagt aaggattctg atgtgtatat 50
<210> 235
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 100 п.о. 3' плечо гомологии
<400> 235
tttgattctc aaacaaatgt gtcacaaagt aaggattctg atgtgtatat cacagacaaa 60
actgtgctag acatgaggtc tatggacttc aagagcaaca 100
<210> 236
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 200 п.о. 3' плечо гомологии
<400> 236
tttgattctc aaacaaatgt gtcacaaagt aaggattctg atgtgtatat cacagacaaa 60
actgtgctag acatgaggtc tatggacttc aagagcaaca gtgctgtggc ctggagcaac 120
aaatctgact ttgcatgtgc aaacgccttc aacaacagca ttattccaga agacaccttc 180
ttccccagcc caggtaaggg 200
<210> 237
<211> 300
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 300 п.о. 3' плечо гомологии
<400> 237
tttgattctc aaacaaatgt gtcacaaagt aaggattctg atgtgtatat cacagacaaa 60
actgtgctag acatgaggtc tatggacttc aagagcaaca gtgctgtggc ctggagcaac 120
aaatctgact ttgcatgtgc aaacgccttc aacaacagca ttattccaga agacaccttc 180
ttccccagcc caggtaaggg cagctttggt gccttcgcag gctgtttcct tgcttcagga 240
atggccaggt tctgcccaga gctctggtca atgatgtcta aaactcctct gattggtggt 300
<210> 238
<211> 400
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 400 п.о. 3' плечо гомологии
<400> 238
tttgattctc aaacaaatgt gtcacaaagt aaggattctg atgtgtatat cacagacaaa 60
actgtgctag acatgaggtc tatggacttc aagagcaaca gtgctgtggc ctggagcaac 120
aaatctgact ttgcatgtgc aaacgccttc aacaacagca ttattccaga agacaccttc 180
ttccccagcc caggtaaggg cagctttggt gccttcgcag gctgtttcct tgcttcagga 240
atggccaggt tctgcccaga gctctggtca atgatgtcta aaactcctct gattggtggt 300
ctcggcctta tccattgcca ccaaaaccct ctttttacta agaaacagtg agccttgttc 360
tggcagtcca gagaatgaca cgggaaaaaa gcagatgaag 400
<210> 239
<211> 500
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 500 п.о. 3' плечо гомологии
<400> 239
tttgattctc aaacaaatgt gtcacaaagt aaggattctg atgtgtatat cacagacaaa 60
actgtgctag acatgaggtc tatggacttc aagagcaaca gtgctgtggc ctggagcaac 120
aaatctgact ttgcatgtgc aaacgccttc aacaacagca ttattccaga agacaccttc 180
ttccccagcc caggtaaggg cagctttggt gccttcgcag gctgtttcct tgcttcagga 240
atggccaggt tctgcccaga gctctggtca atgatgtcta aaactcctct gattggtggt 300
ctcggcctta tccattgcca ccaaaaccct ctttttacta agaaacagtg agccttgttc 360
tggcagtcca gagaatgaca cgggaaaaaa gcagatgaag agaaggtggc aggagagggc 420
acgtggccca gcctcagtct ctccaactga gttcctgcct gcctgccttt gctcagactg 480
tttgcccctt actgctcttc 500
<210> 240
<211> 600
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TRAC 600 п.о. 3' плечо гомологии
<400> 240
tttgattctc aaacaaatgt gtcacaaagt aaggattctg atgtgtatat cacagacaaa 60
actgtgctag acatgaggtc tatggacttc aagagcaaca gtgctgtggc ctggagcaac 120
aaatctgact ttgcatgtgc aaacgccttc aacaacagca ttattccaga agacaccttc 180
ttccccagcc caggtaaggg cagctttggt gccttcgcag gctgtttcct tgcttcagga 240
atggccaggt tctgcccaga gctctggtca atgatgtcta aaactcctct gattggtggt 300
ctcggcctta tccattgcca ccaaaaccct ctttttacta agaaacagtg agccttgttc 360
tggcagtcca gagaatgaca cgggaaaaaa gcagatgaag agaaggtggc aggagagggc 420
acgtggccca gcctcagtct ctccaactga gttcctgcct gcctgccttt gctcagactg 480
tttgcccctt actgctcttc taggcctcat tctaagcccc ttctccaagt tgcctctcct 540
tatttctccc tgtctgccaa aaaatctttc ccagctcact aagtcagtct cacgcagtca 600
<210> 241
<211> 141
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий альфа константа TCR
<400> 241
Asn Ile Gln Lys Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
1 5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Ala Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 242
<211> 141
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий альфа константа TCR
<400> 242
His Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
1 5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 243
<211> 141
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий альфа константа TCR
<400> 243
Tyr Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
1 5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 244
<211> 141
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий альфа константа TCR
<400> 244
Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
1 5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 245
<211> 142
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий альфа константа TCR
<400> 245
Pro Asn Ile Gln Lys Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser
1 5 10 15
Lys Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln
20 25 30
Thr Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys
35 40 45
Thr Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val
50 55 60
Ala Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn
65 70 75 80
Ser Ile Ile Pro Ala Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys
85 90 95
Asp Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn
100 105 110
Phe Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val
115 120 125
Ala Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 246
<211> 141
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий альфа константа TCR
<400> 246
Asn Ile Gln Lys Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
1 5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Cys
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Ala Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 247
<211> 141
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий альфа константа TCR
<400> 247
His Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
1 5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Cys
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 248
<211> 140
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий альфа константа TCR
<400> 248
Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys Ser
1 5 10 15
Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr Asn
20 25 30
Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Cys Val
35 40 45
Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala Trp
50 55 60
Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser Ile
65 70 75 80
Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp Val
85 90 95
Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe Gln
100 105 110
Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala Gly
115 120 125
Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 249
<211> 141
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий альфа константа TCR
<400> 249
Asn Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Cys
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 250
<211> 141
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий альфа константа TCR
<400> 250
Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
1 5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Cys
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 251
<211> 177
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий бета константа TCR
<400> 251
Glu Asp Leu Asn Lys Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro
1 5 10 15
Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu
20 25 30
Ala Thr Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
35 40 45
Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Pro Leu Lys
50 55 60
Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu
65 70 75 80
Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys
85 90 95
Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp
100 105 110
Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg
115 120 125
Ala Asp Cys Gly Phe Thr Ser Val Ser Tyr Gln Gln Gly Val Leu Ser
130 135 140
Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala
145 150 155 160
Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp
165 170 175
Phe
<210> 252
<211> 180
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий бета константа TCR
<400> 252
Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu
1 5 10 15
Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys
20 25 30
Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val
35 40 45
Asn Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu
50 55 60
Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg
65 70 75 80
Leu Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg
85 90 95
Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln
100 105 110
Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly
115 120 125
Arg Ala Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu
130 135 140
Ser Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr
145 150 155 160
Ala Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys
165 170 175
Asp Ser Arg Gly
180
<210> 253
<211> 180
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий бета константа TCR
<400> 253
Leu Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu
1 5 10 15
Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys
20 25 30
Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val
35 40 45
Asn Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu
50 55 60
Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg
65 70 75 80
Leu Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg
85 90 95
Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln
100 105 110
Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly
115 120 125
Arg Ala Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu
130 135 140
Ser Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr
145 150 155 160
Ala Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys
165 170 175
Asp Ser Arg Gly
180
<210> 254
<211> 179
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий бета константа TCR
<400> 254
Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro
1 5 10 15
Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu
20 25 30
Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
35 40 45
Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Pro Leu Lys
50 55 60
Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu
65 70 75 80
Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys
85 90 95
Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp
100 105 110
Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg
115 120 125
Ala Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu Ser
130 135 140
Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala
145 150 155 160
Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp
165 170 175
Ser Arg Gly
<210> 255
<211> 180
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий бета константа TCR
<400> 255
Thr Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu
1 5 10 15
Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys
20 25 30
Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val
35 40 45
Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Pro Leu
50 55 60
Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg
65 70 75 80
Leu Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg
85 90 95
Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln
100 105 110
Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly
115 120 125
Arg Ala Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu
130 135 140
Ser Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr
145 150 155 160
Ala Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys
165 170 175
Asp Ser Arg Gly
180
<210> 256
<211> 180
<212> PRT
<213> Искусственная последовательность
<220>
<223> Человеческий бета константа TCR
<400> 256
Leu Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu
1 5 10 15
Pro Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys
20 25 30
Leu Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val
35 40 45
Asn Gly Lys Glu Val His Ser Gly Val Cys Thr Asp Pro Gln Pro Leu
50 55 60
Lys Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg
65 70 75 80
Leu Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg
85 90 95
Cys Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln
100 105 110
Asp Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly
115 120 125
Arg Ala Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu
130 135 140
Ser Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr
145 150 155 160
Ala Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys
165 170 175
Asp Ser Arg Gly
180
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИММУНООНКОЛОГИИ | 2016 |
|
RU2771624C2 |
| НЕ РЕСТРИКТИРОВАННЫЕ ПО HLA T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812917C2 |
| КОНСТРУКТЫ Т-КЛЕТОЧНОГО РЕЦЕПТОРА И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2785954C2 |
| Т-КЛЕТКИ С ХИМЕРНЫМИ АНТИГЕННЫМИ РЕЦЕПТОРАМИ (CAR-T) ДЛЯ ЛЕЧЕНИЯ РАКА | 2019 |
|
RU2811466C2 |
| МЫШИ, СОДЕРЖАЩИЕ МУТАЦИИ, ВСЛЕДСТВИЕ КОТОРЫХ ЭКСПРЕССИРУЕТСЯ УКОРОЧЕННЫЙ НА С-КОНЦЕ ФИБРИЛЛИН-1 | 2017 |
|
RU2721125C1 |
| ДНК-СОДЕРЖАЩИЕ ПОЛИНУКЛЕОТИДЫ И РУКОВОДЯЩИЕ ПОСЛЕДОВАТЕЛЬНОСТИ ДЛЯ СИСТЕМ CRISPR ТИПА V И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2021 |
|
RU2832314C1 |
| ВАРИАНТЫ, НА ОСНОВЕ АДЕНОАССОЦИИРОВАННОГО ВИРУСА, СОСТАВЫ И СПОСОБЫ ДЛЯ ПУЛЬМОНАЛЬНОЙ ДОСТАВКИ | 2021 |
|
RU2829874C1 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ КОМПЛЕКСА ОПУХОЛЕВЫЙ АНТИГЕН NY-ESO-1/HLA-A*02 | 2016 |
|
RU2775394C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2017 |
|
RU2775623C2 |
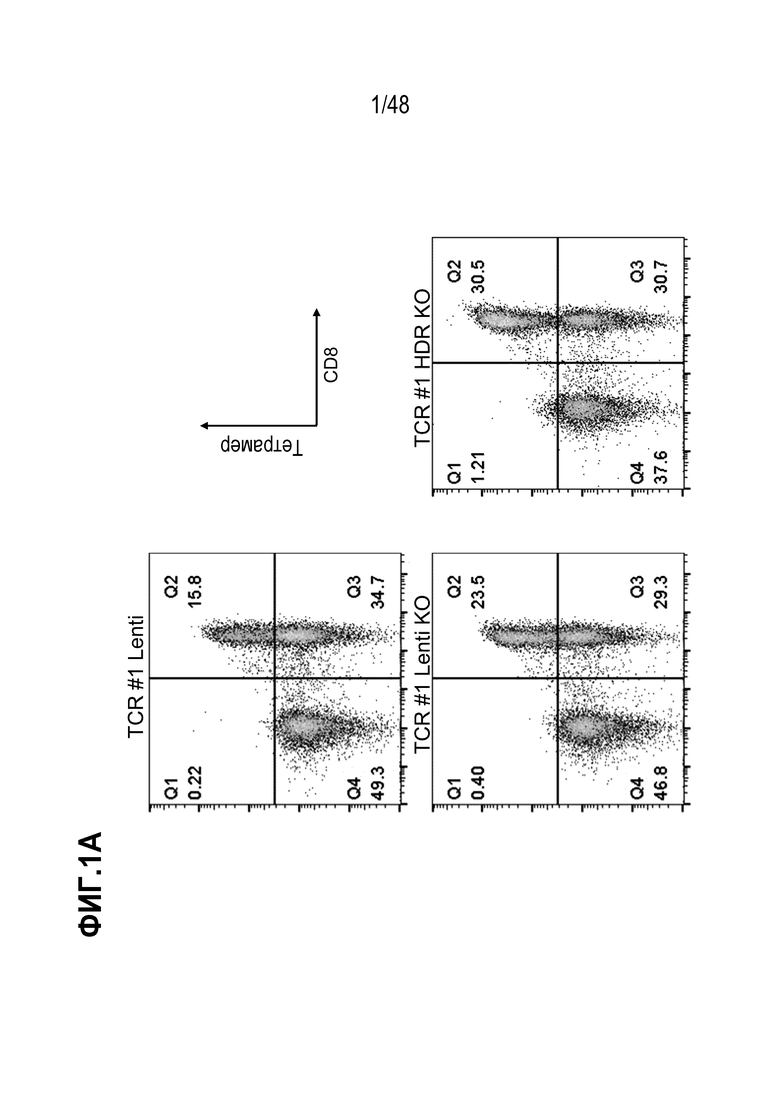
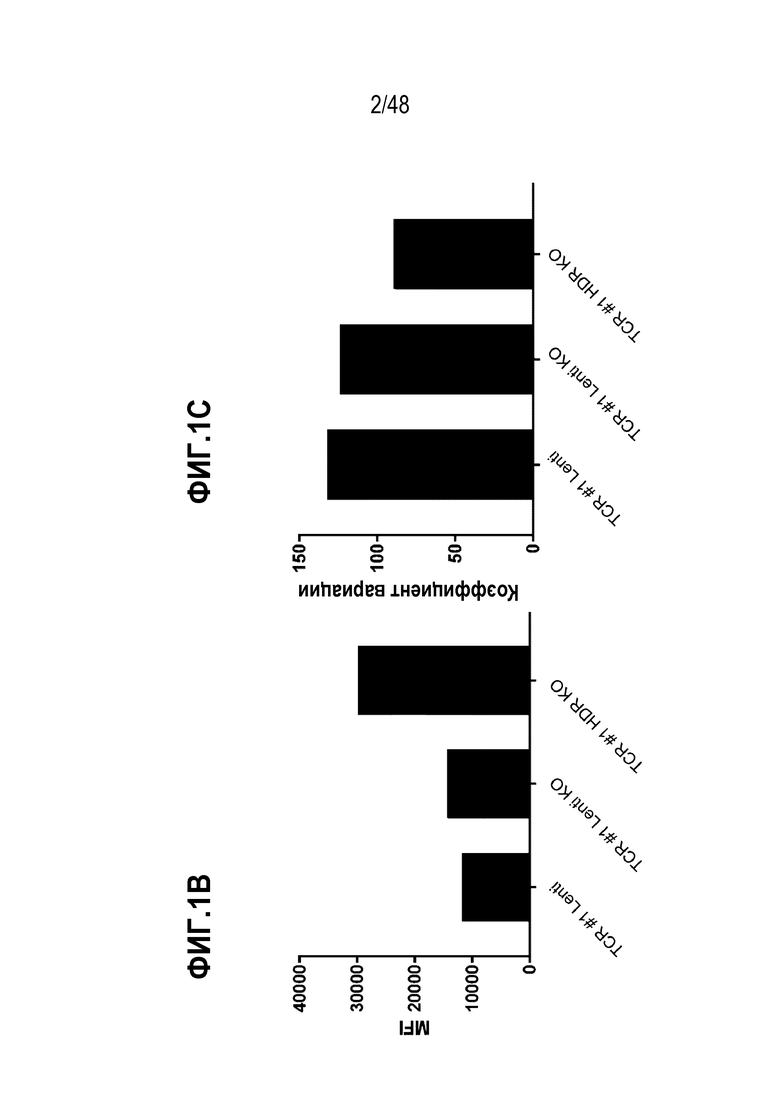
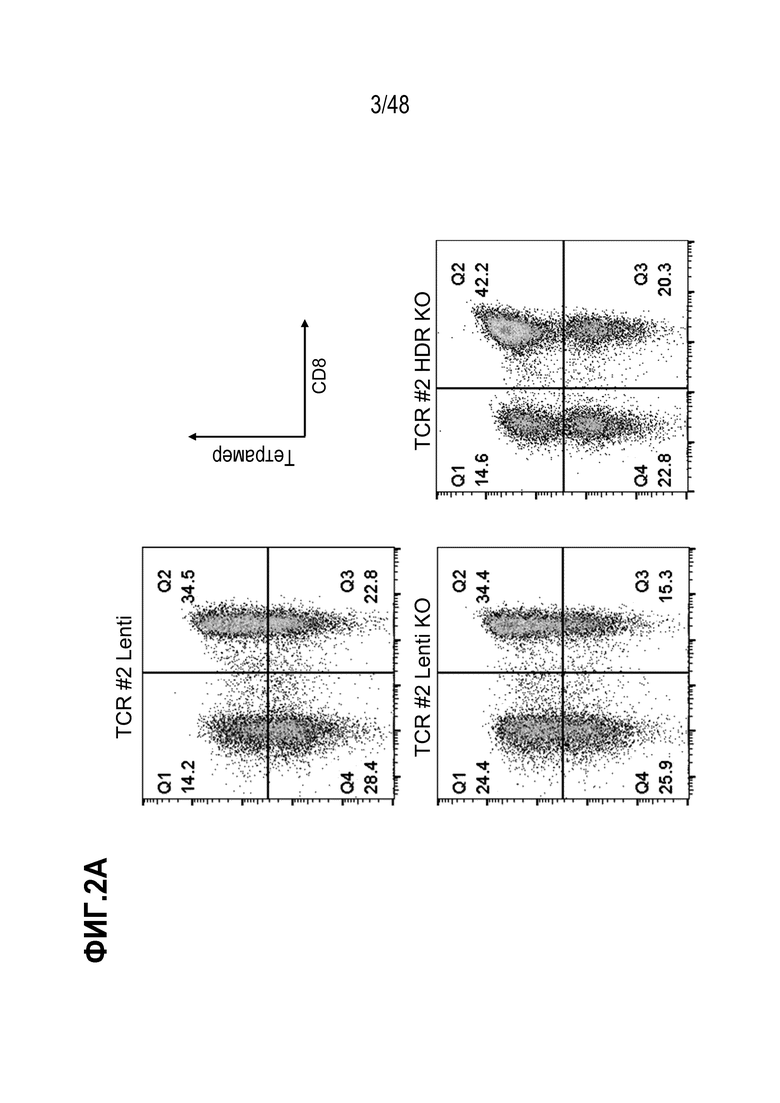
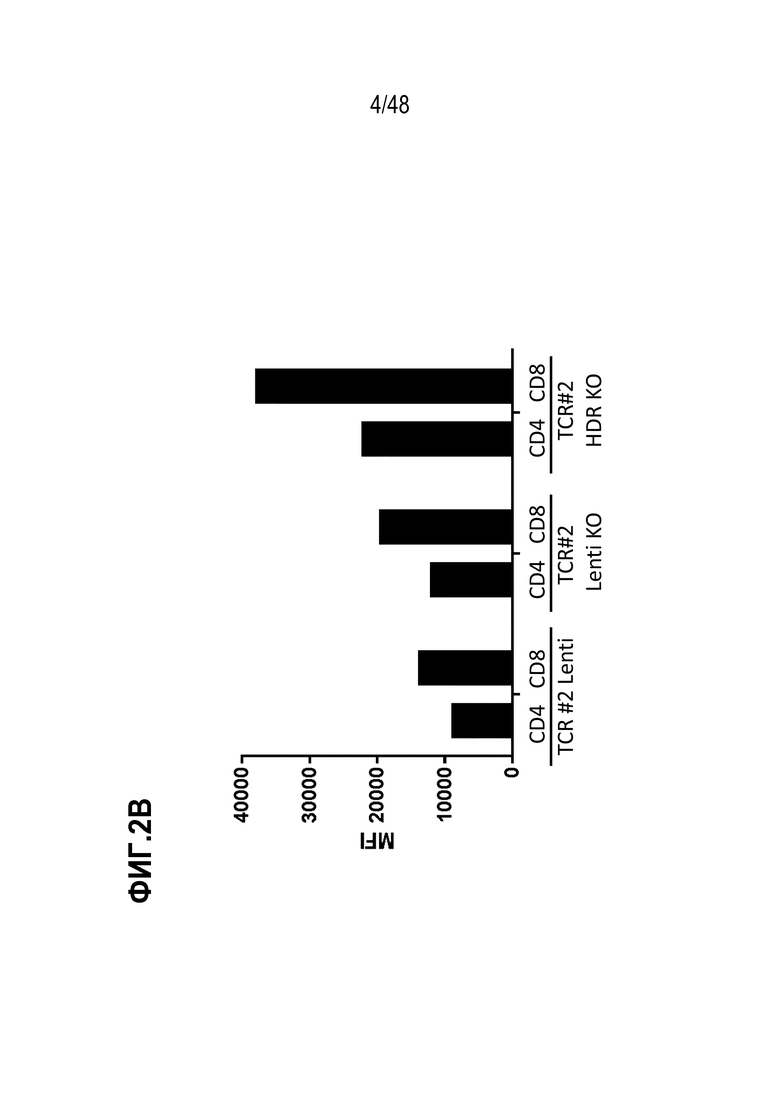
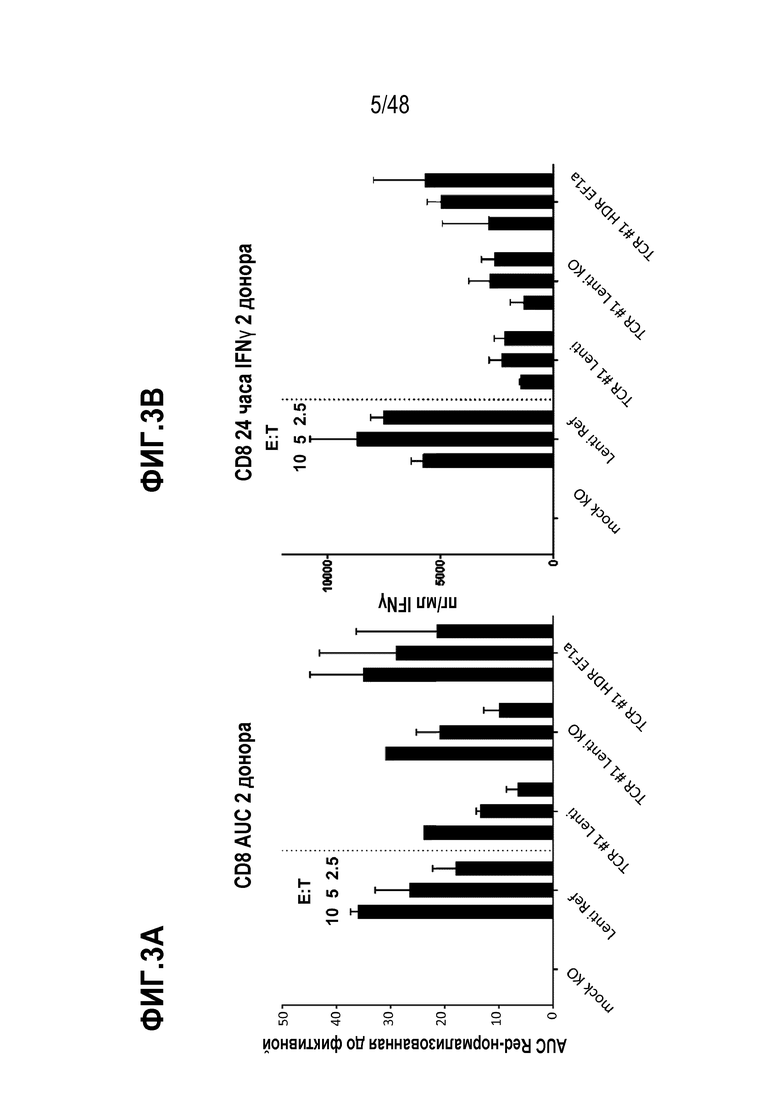
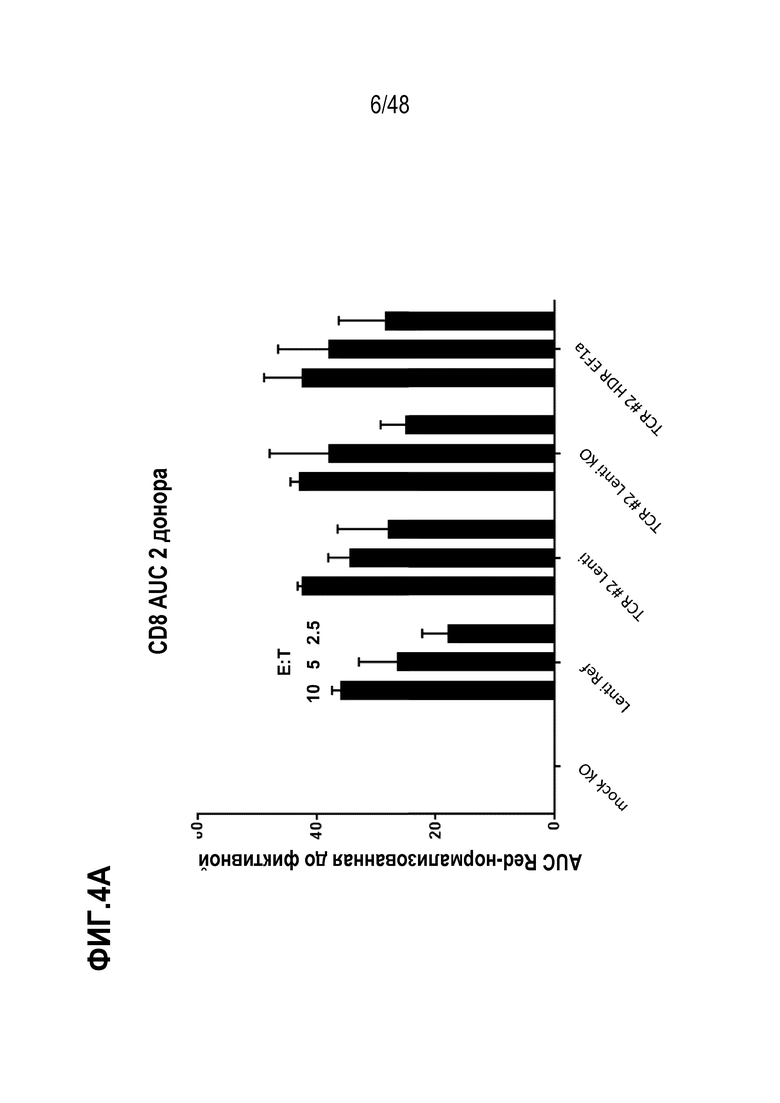
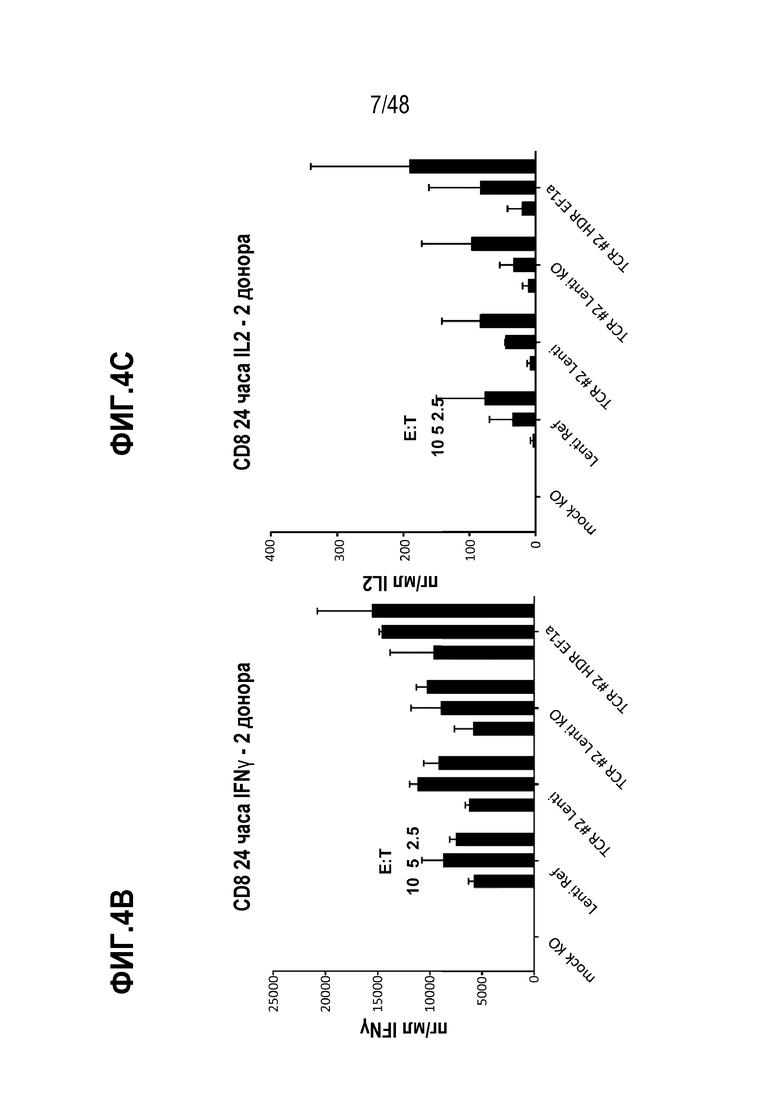
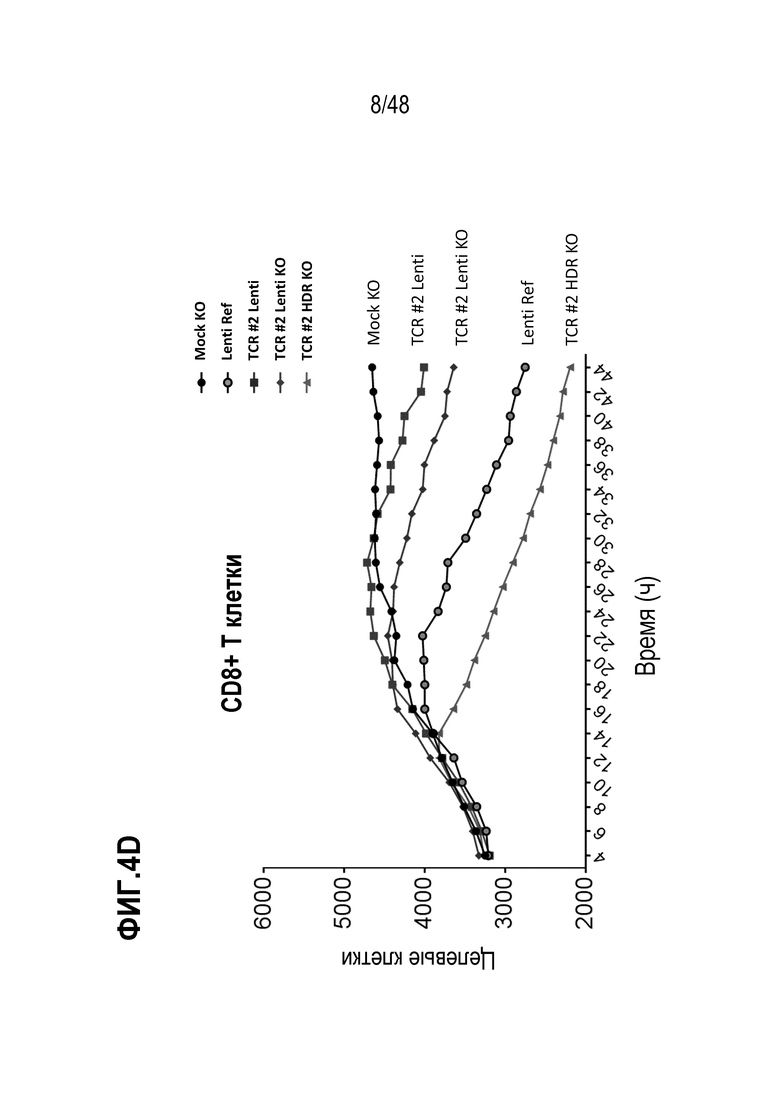
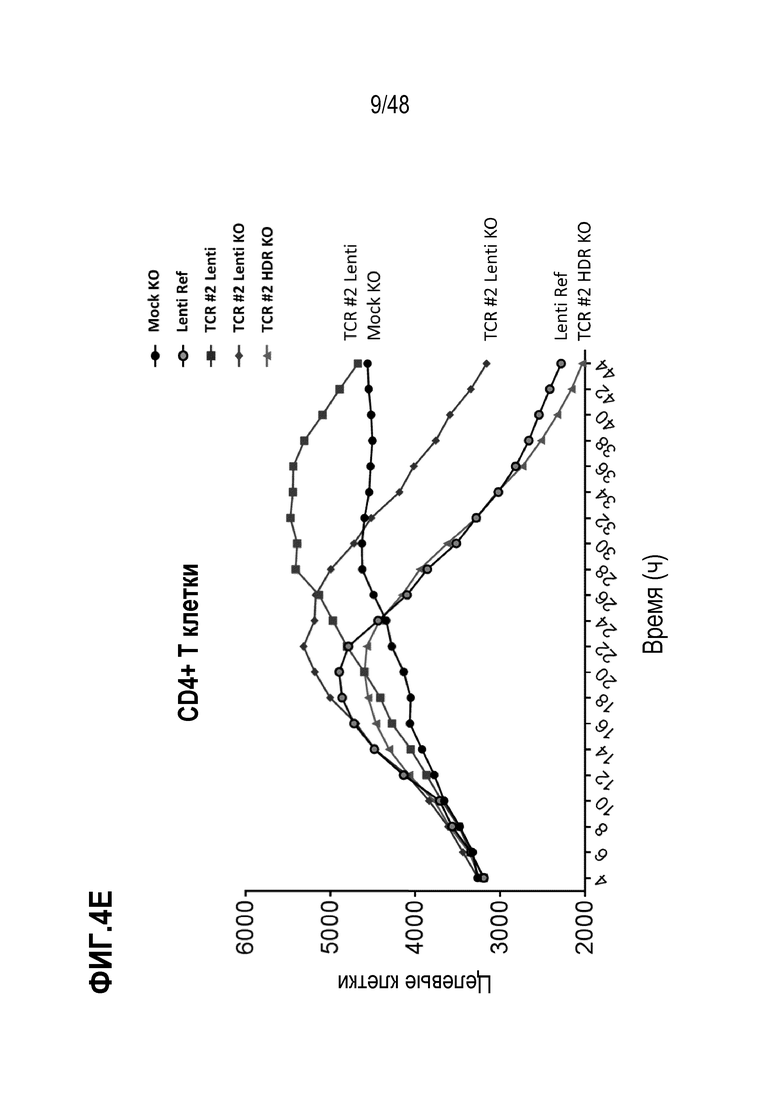
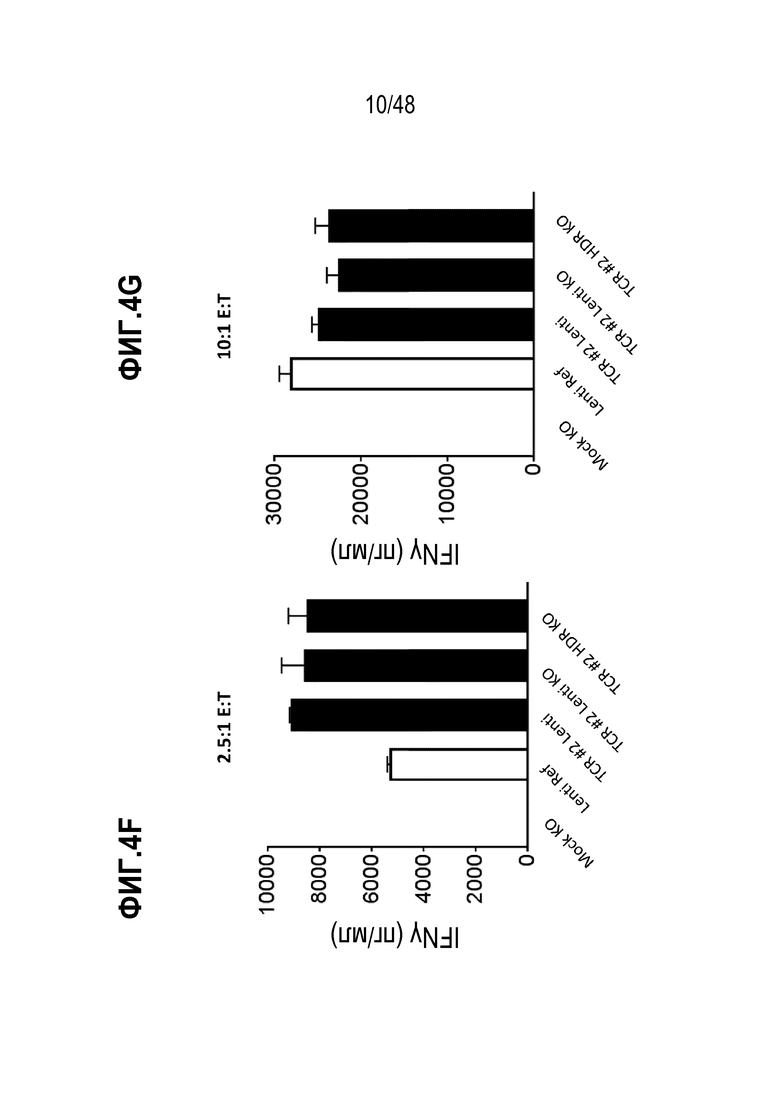
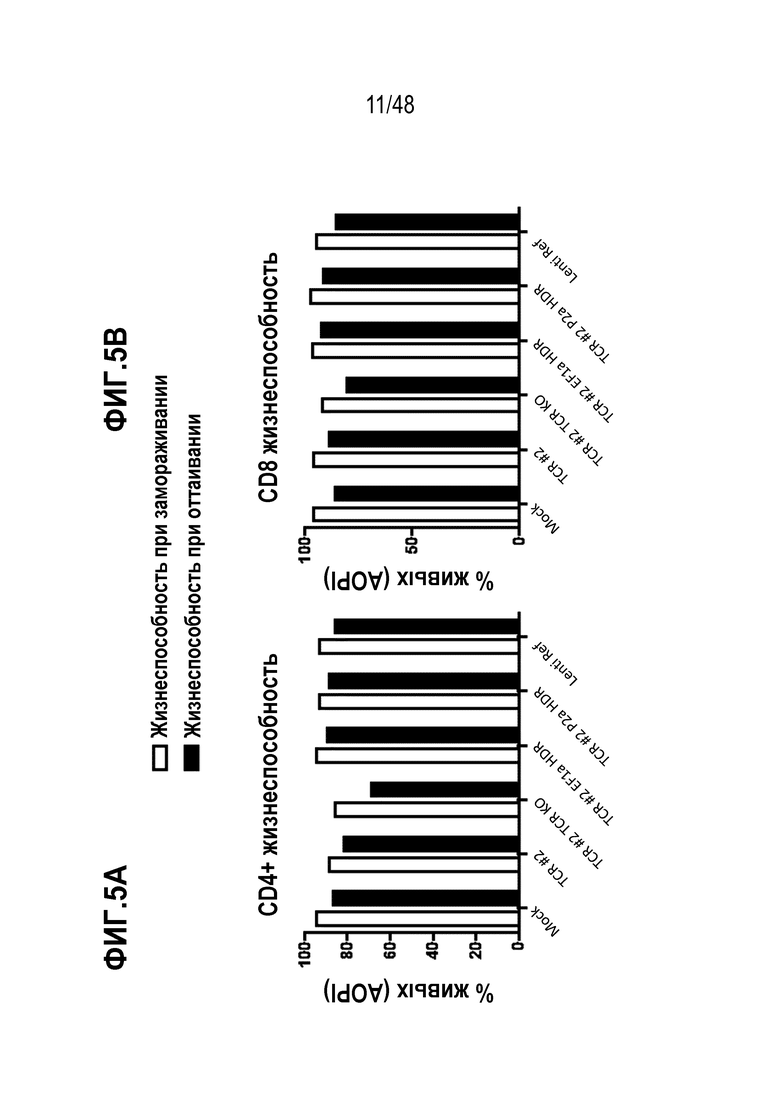
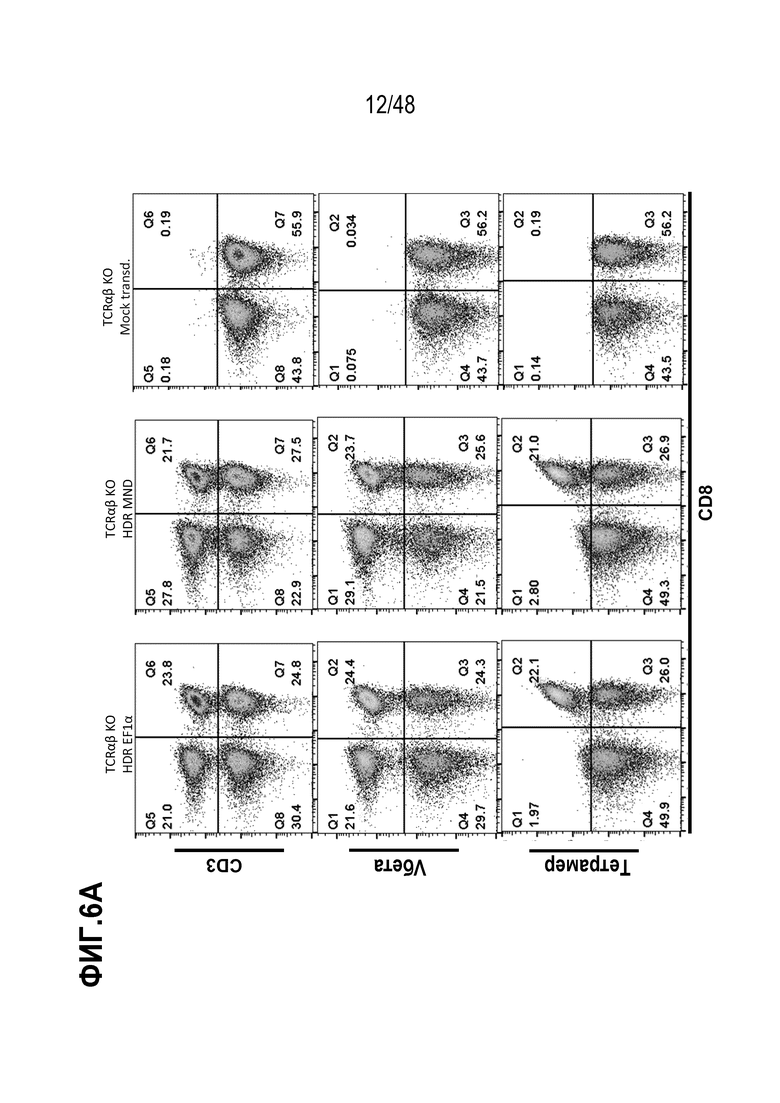
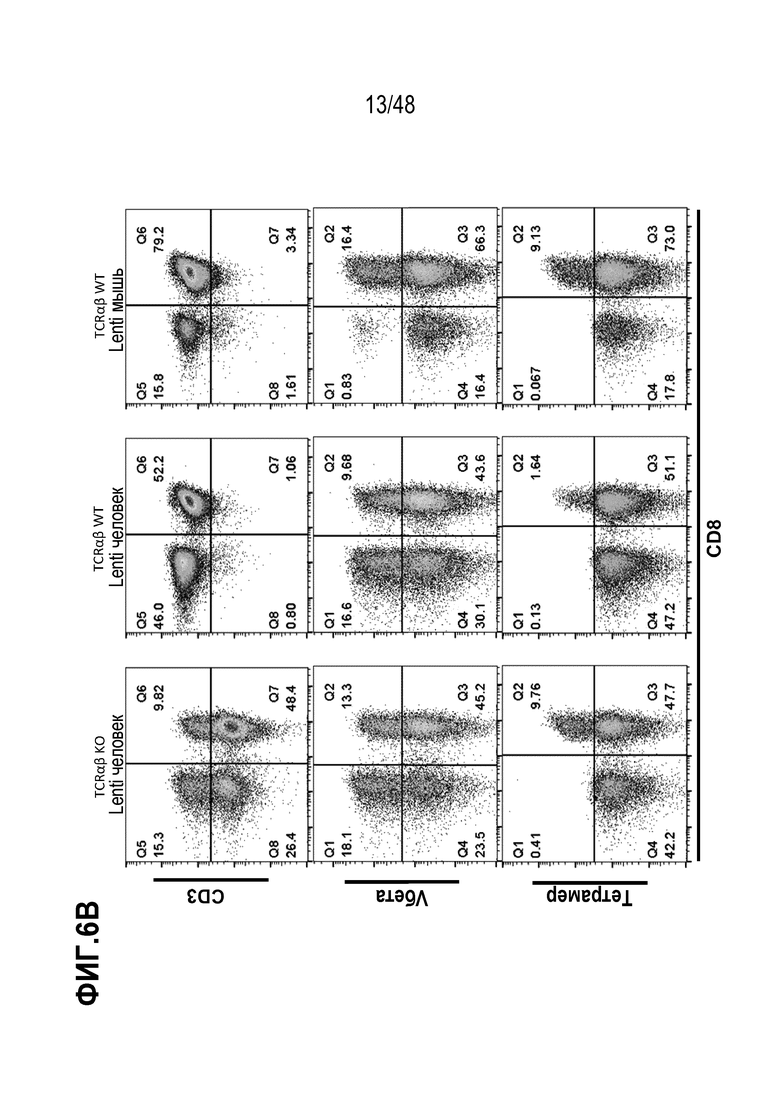
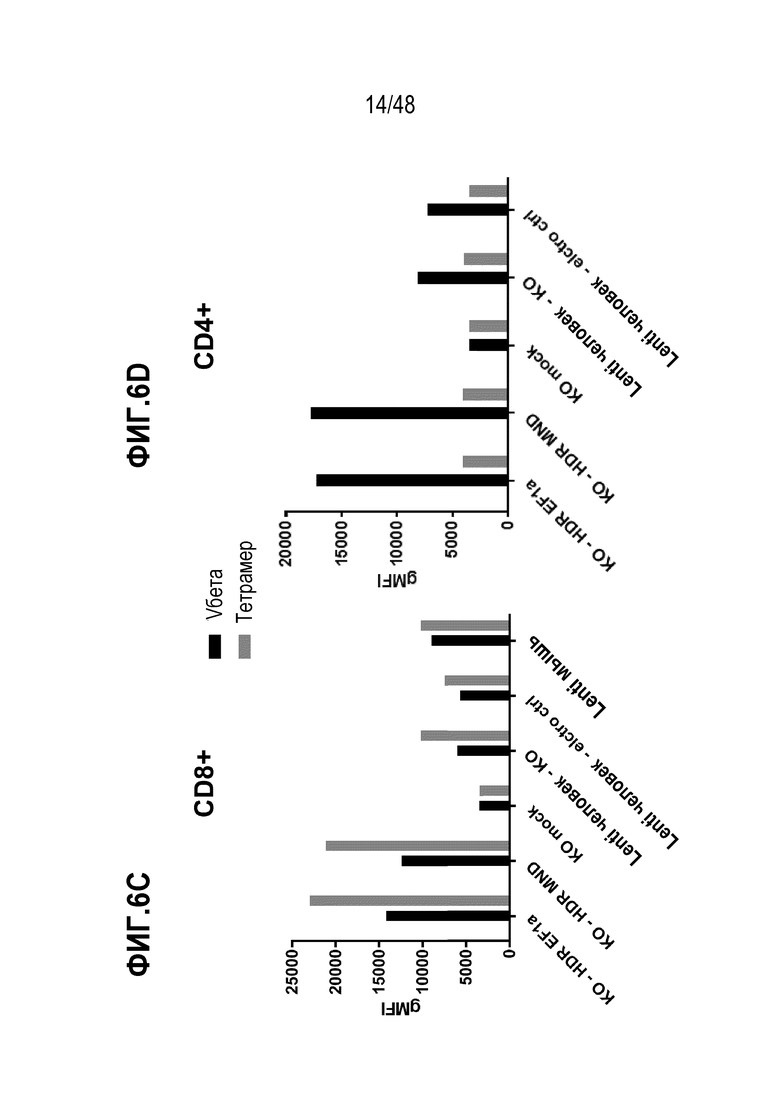
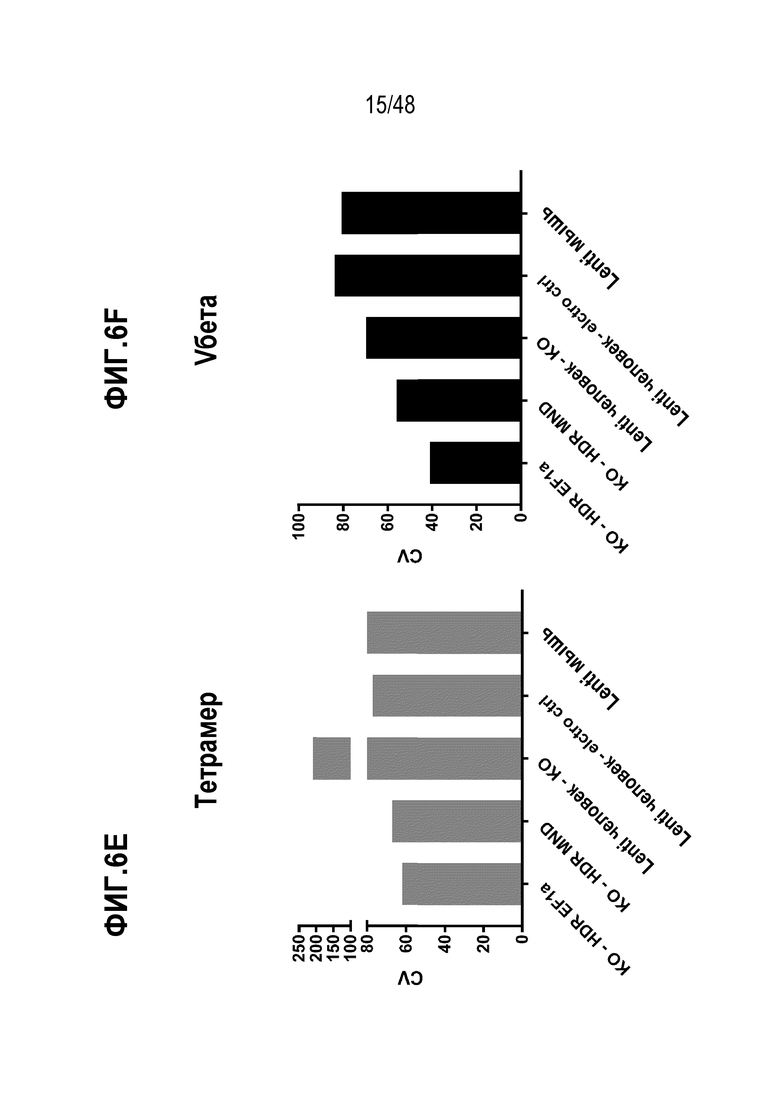
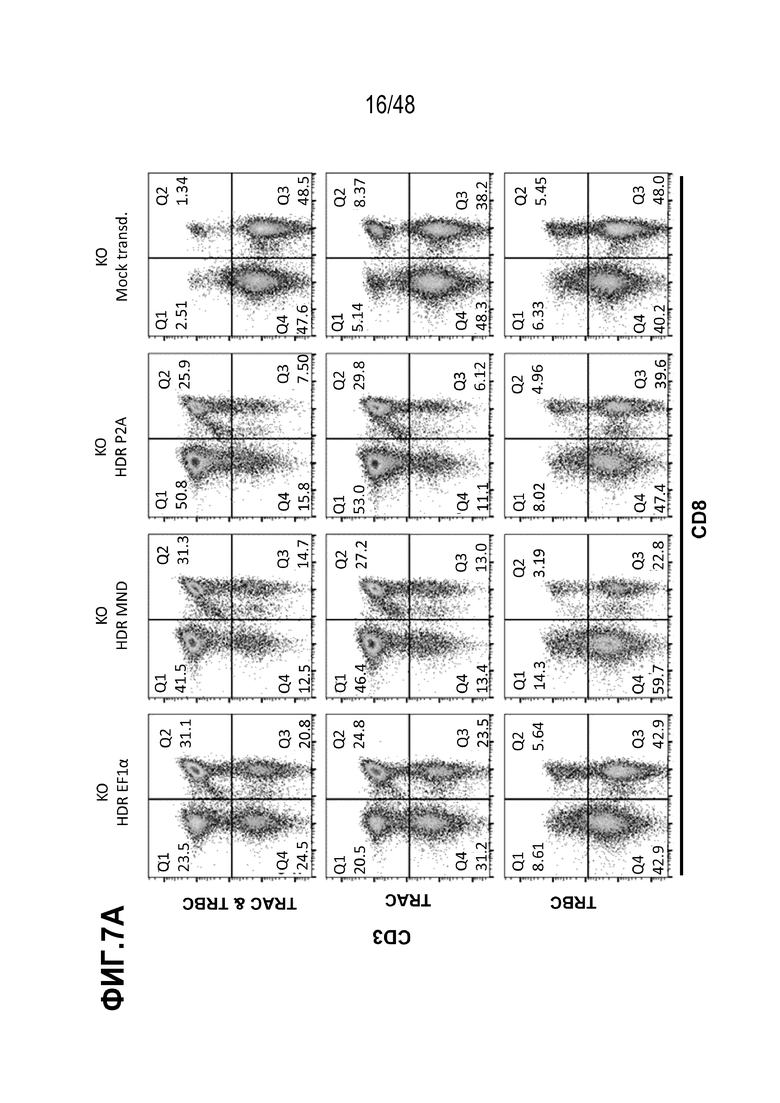
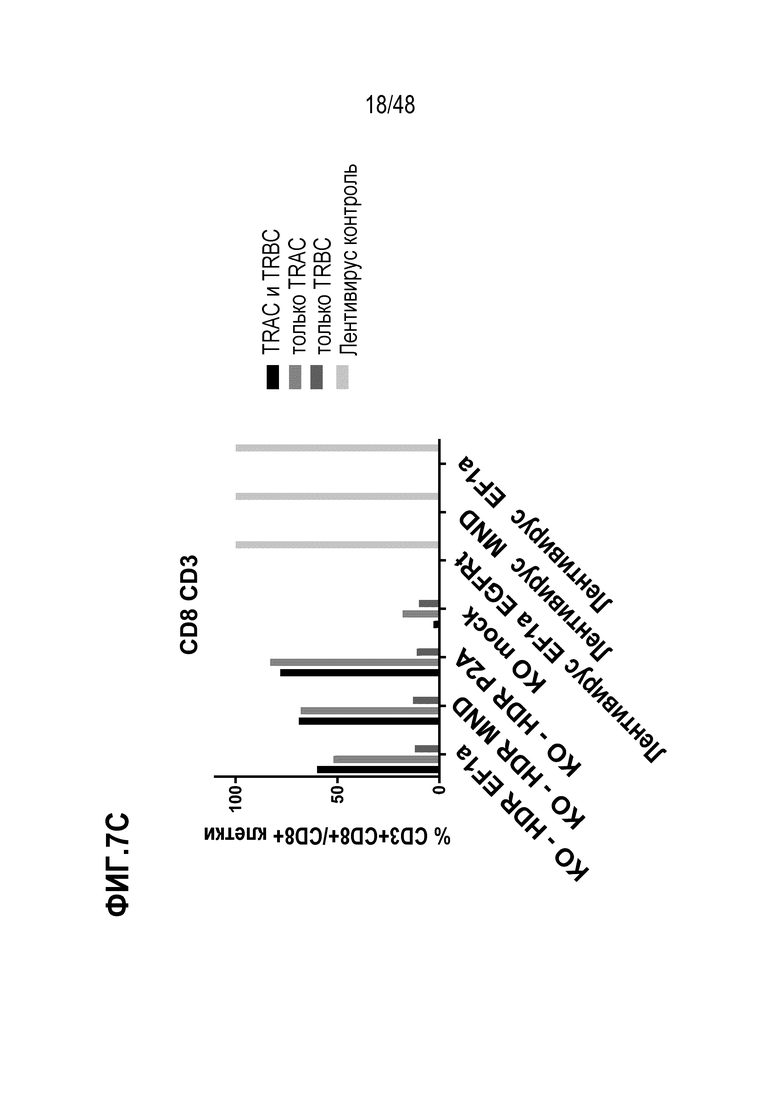
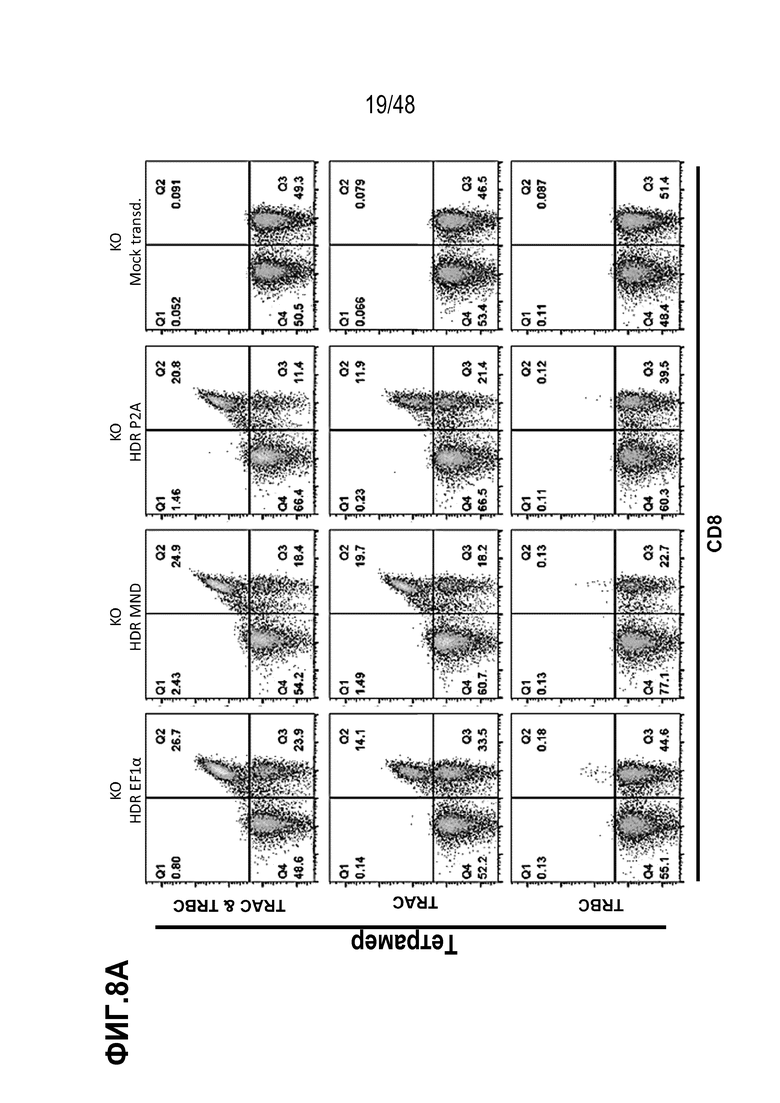
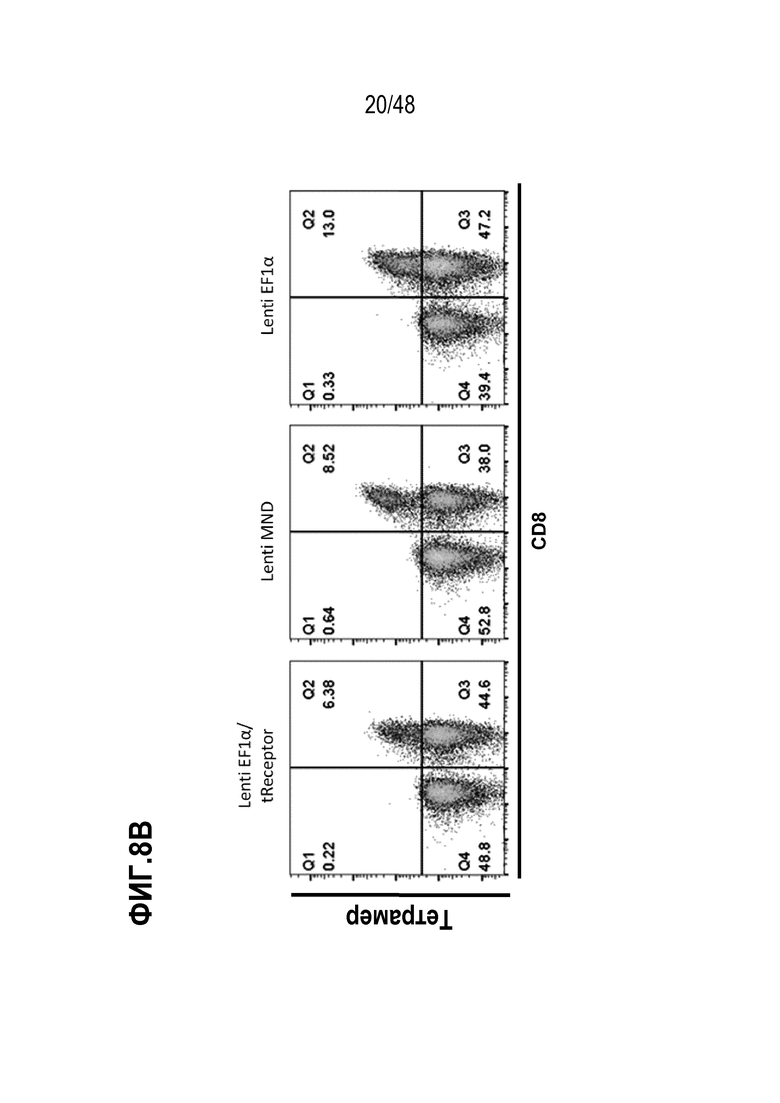
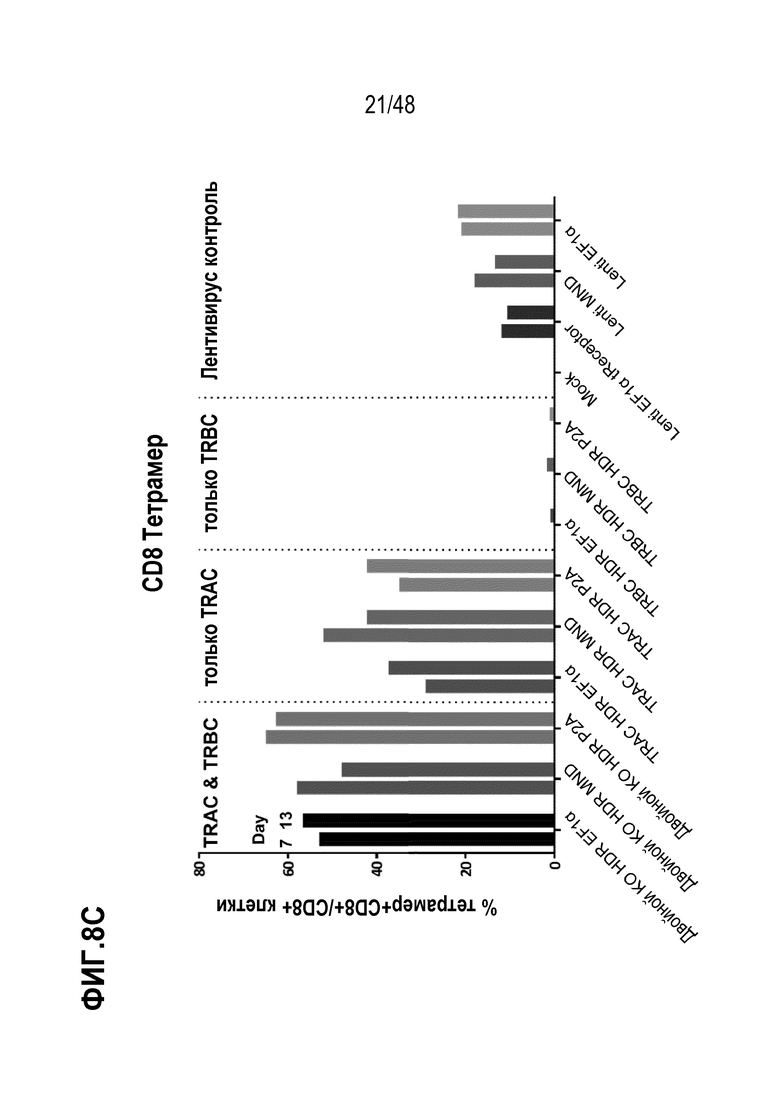

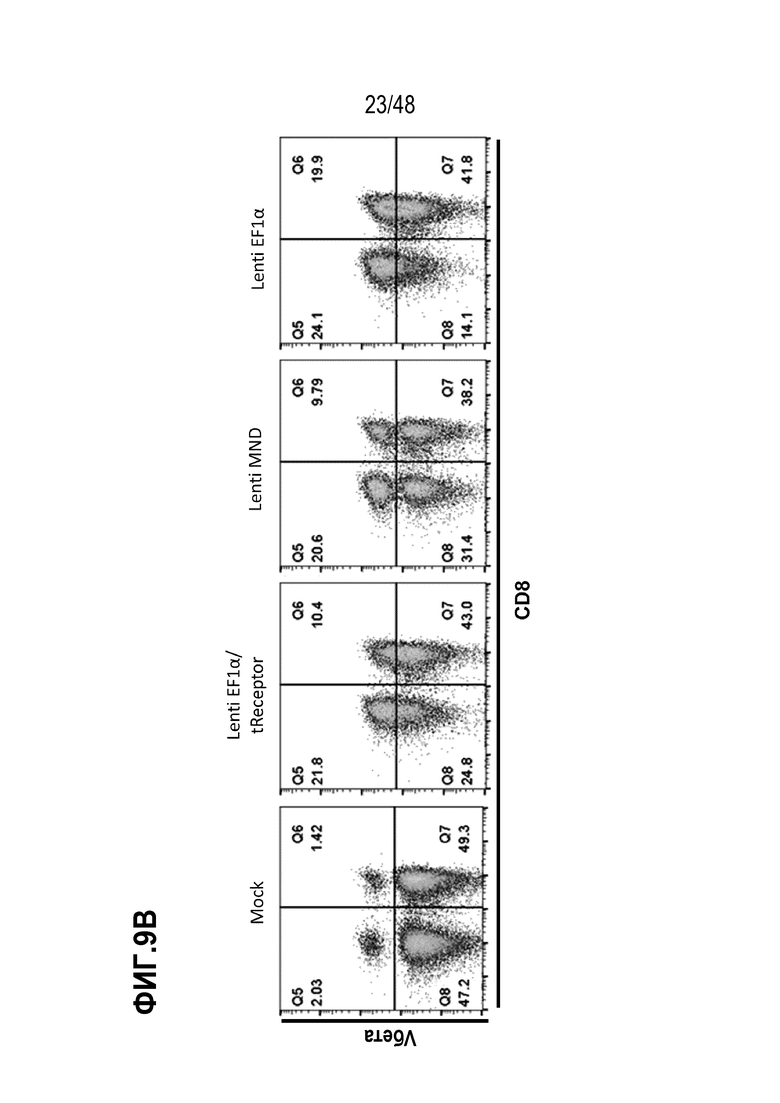
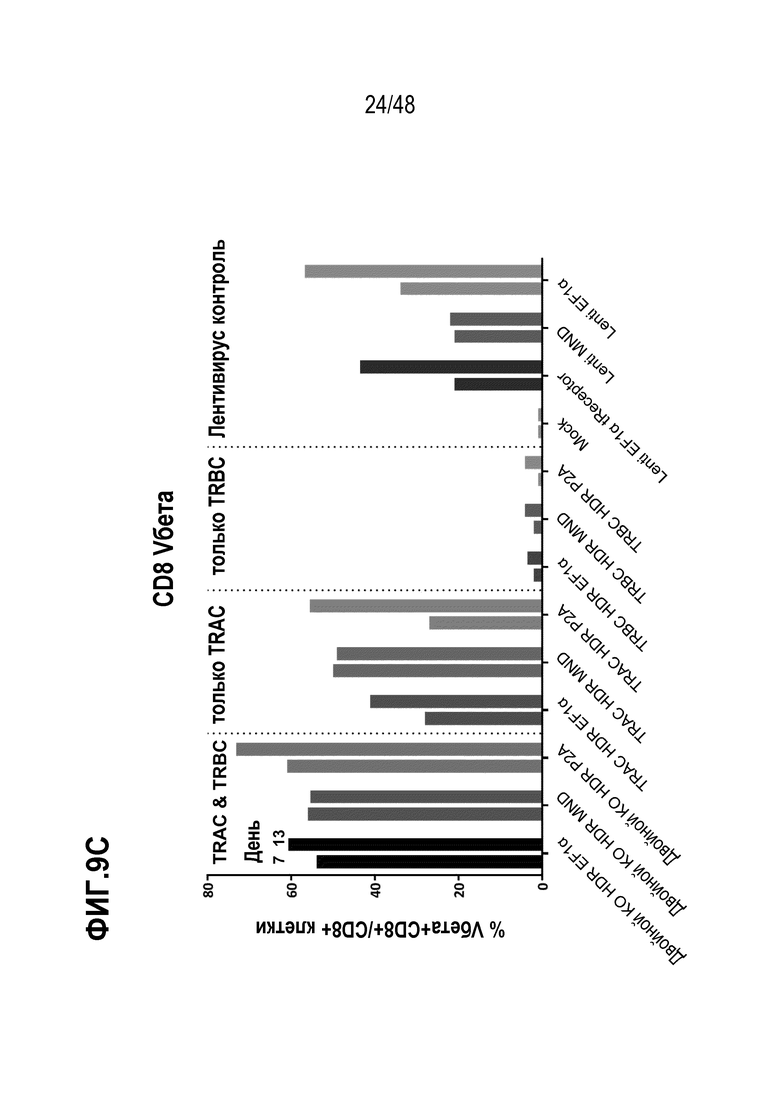
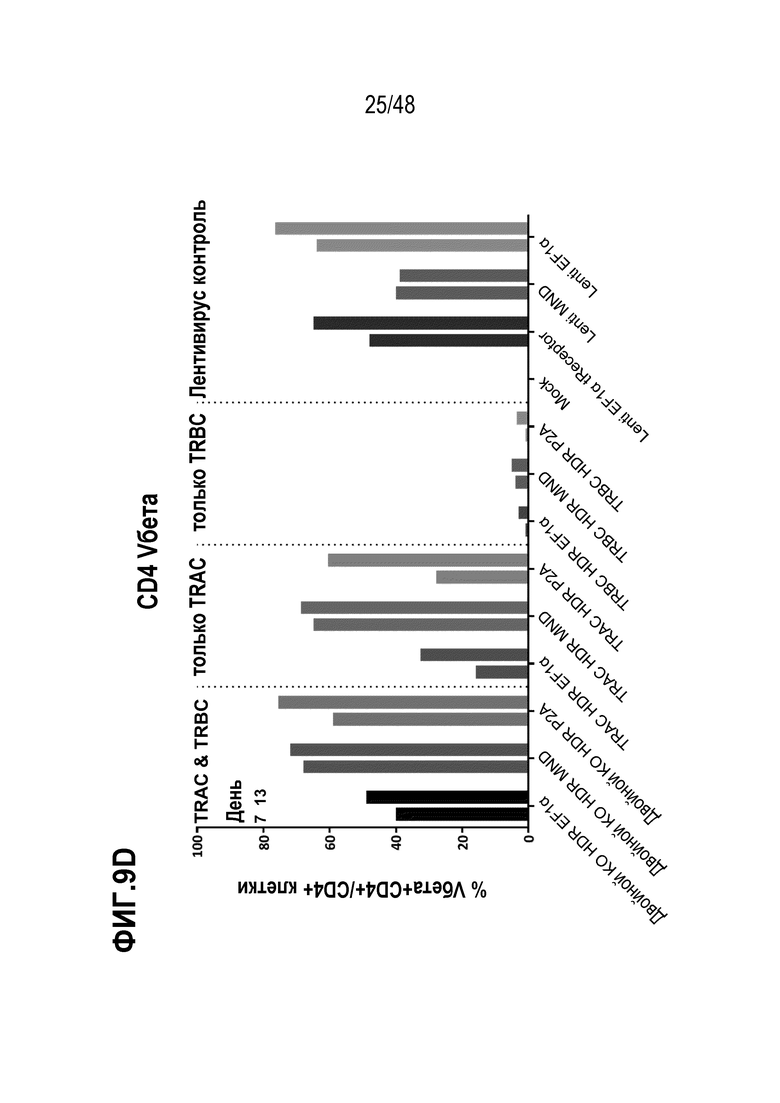
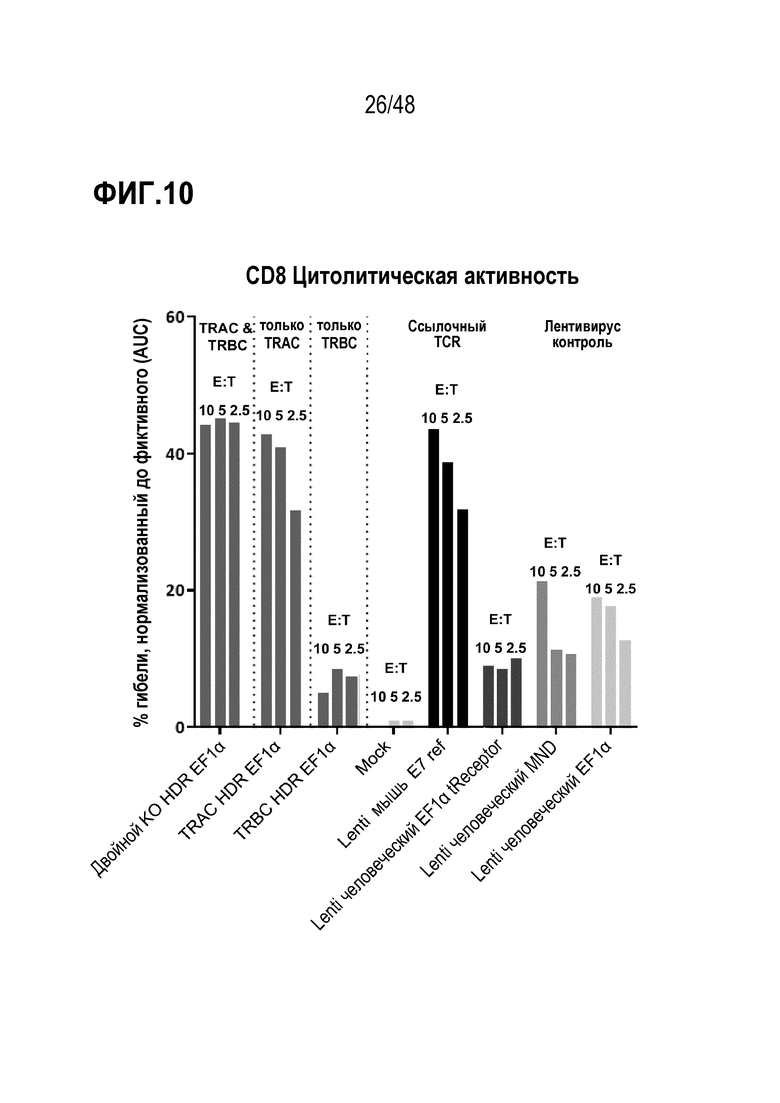
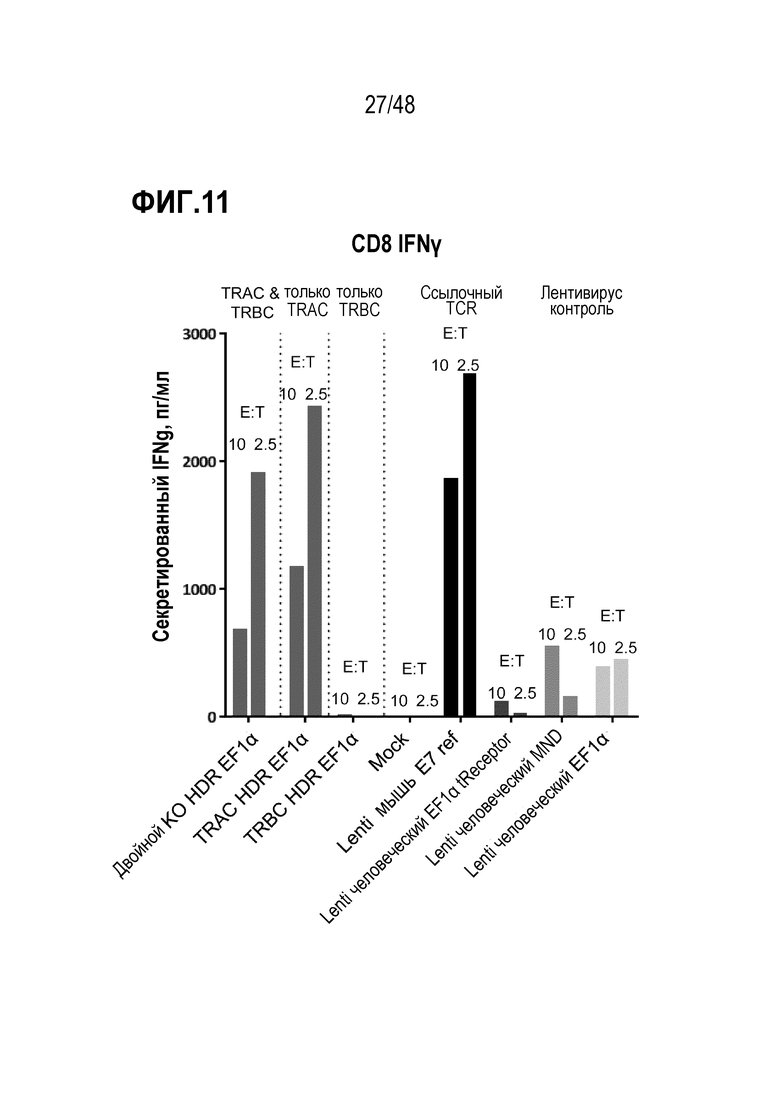

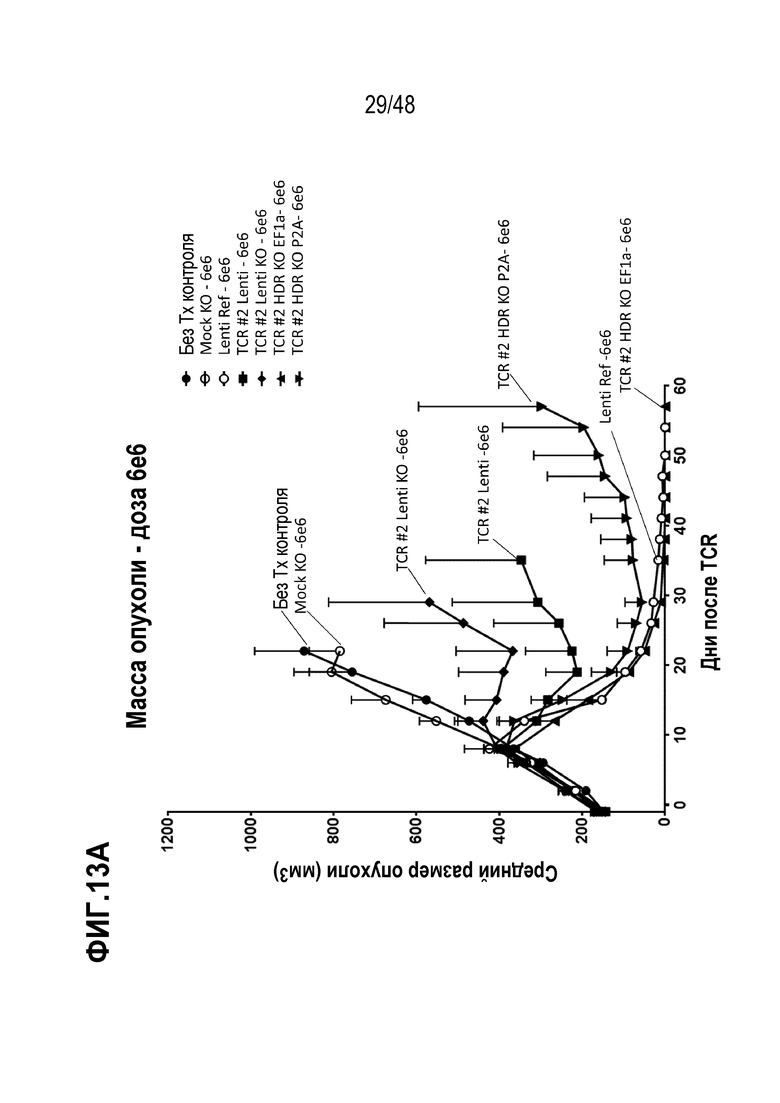
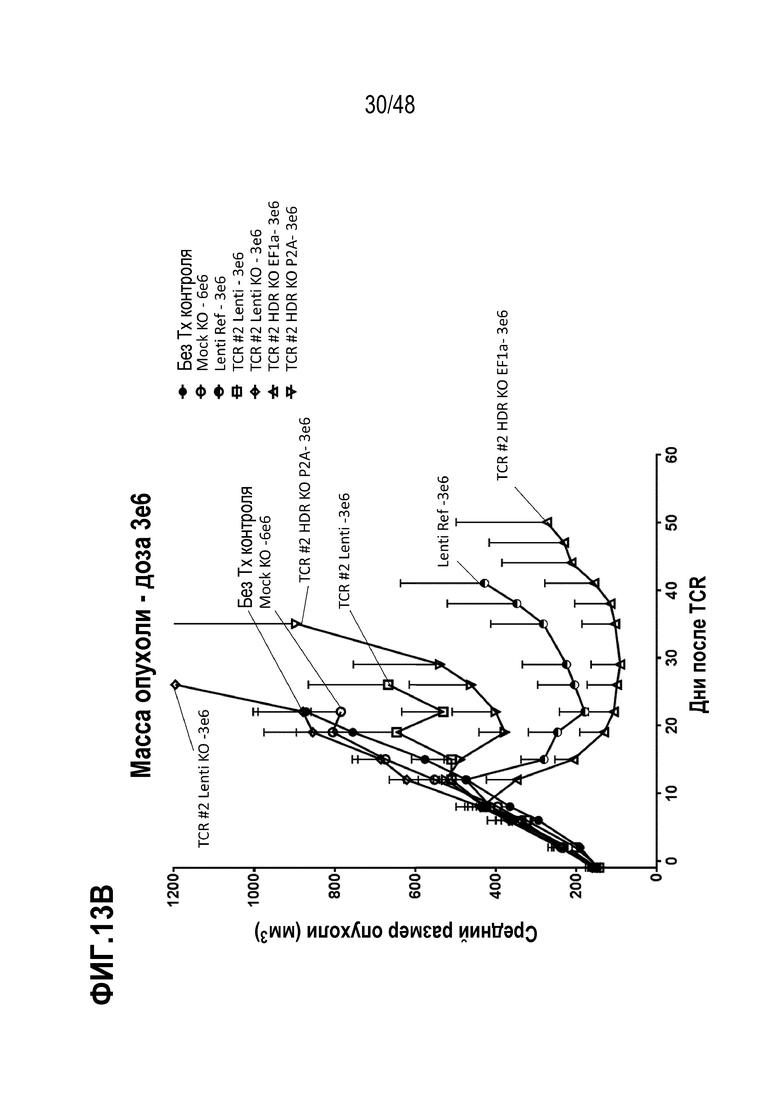
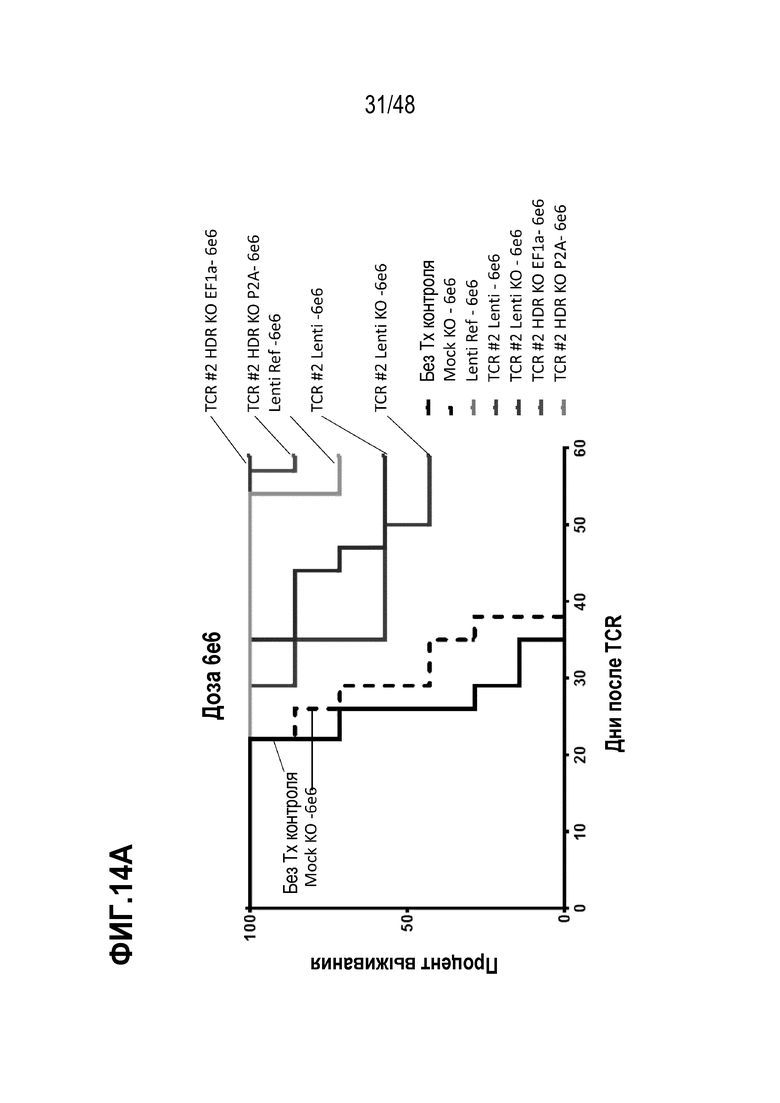
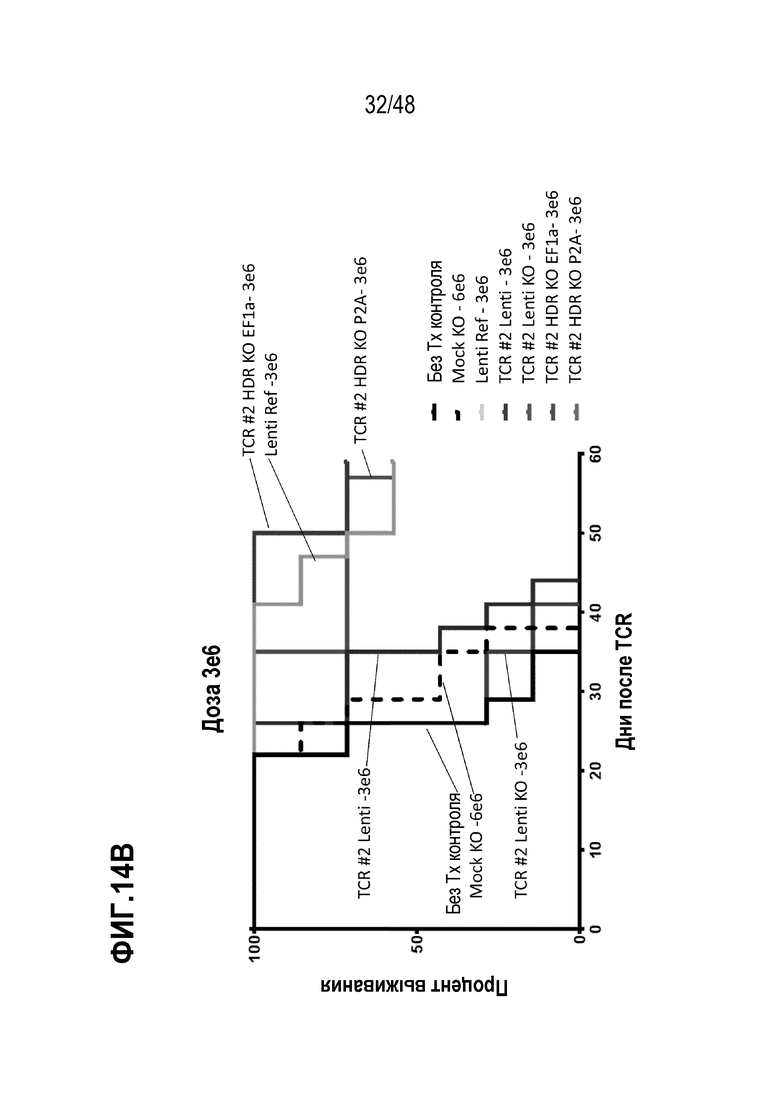
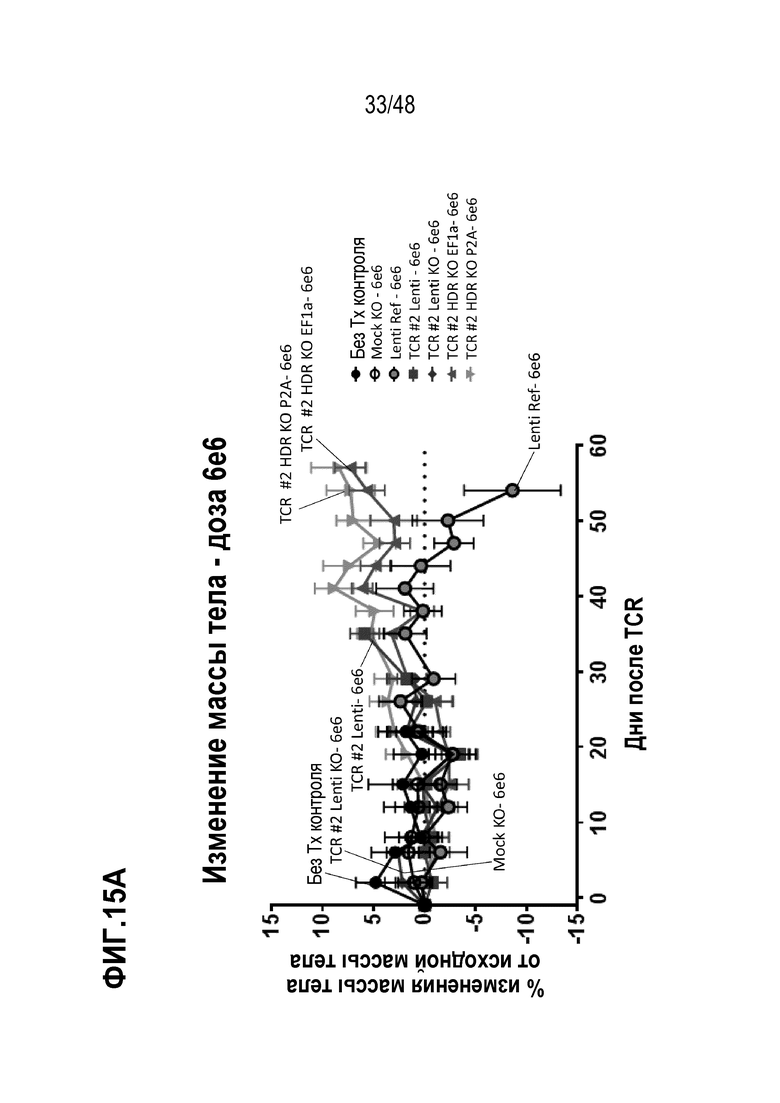
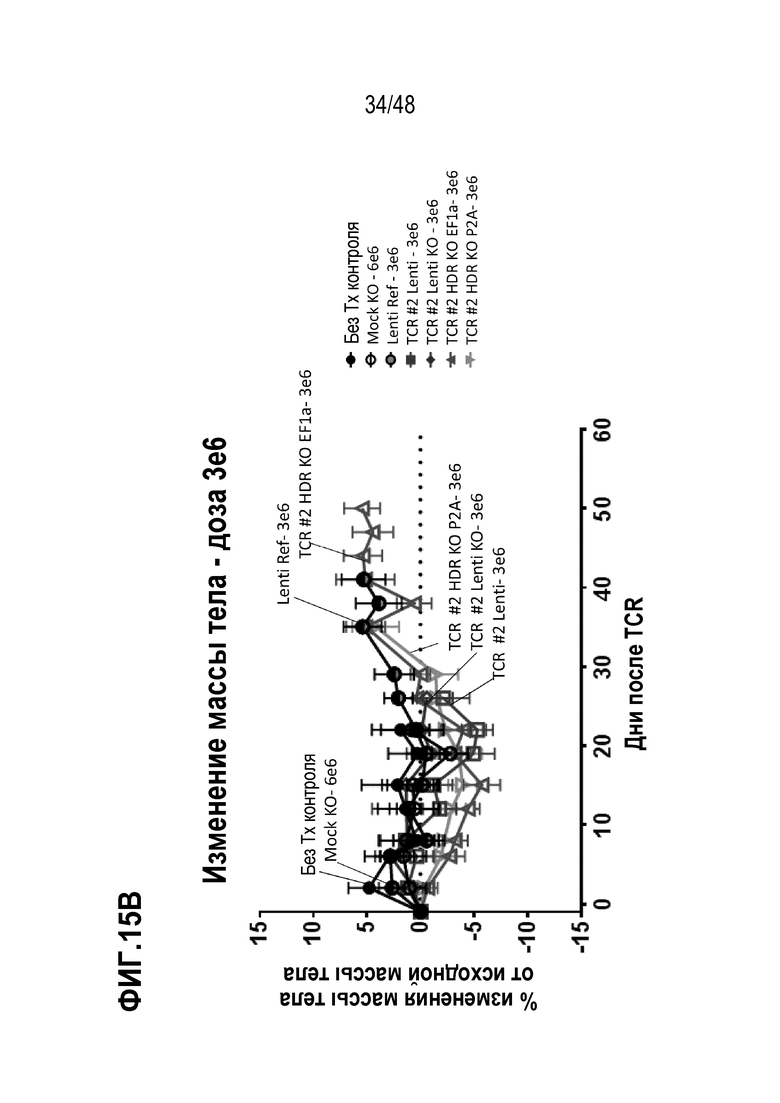
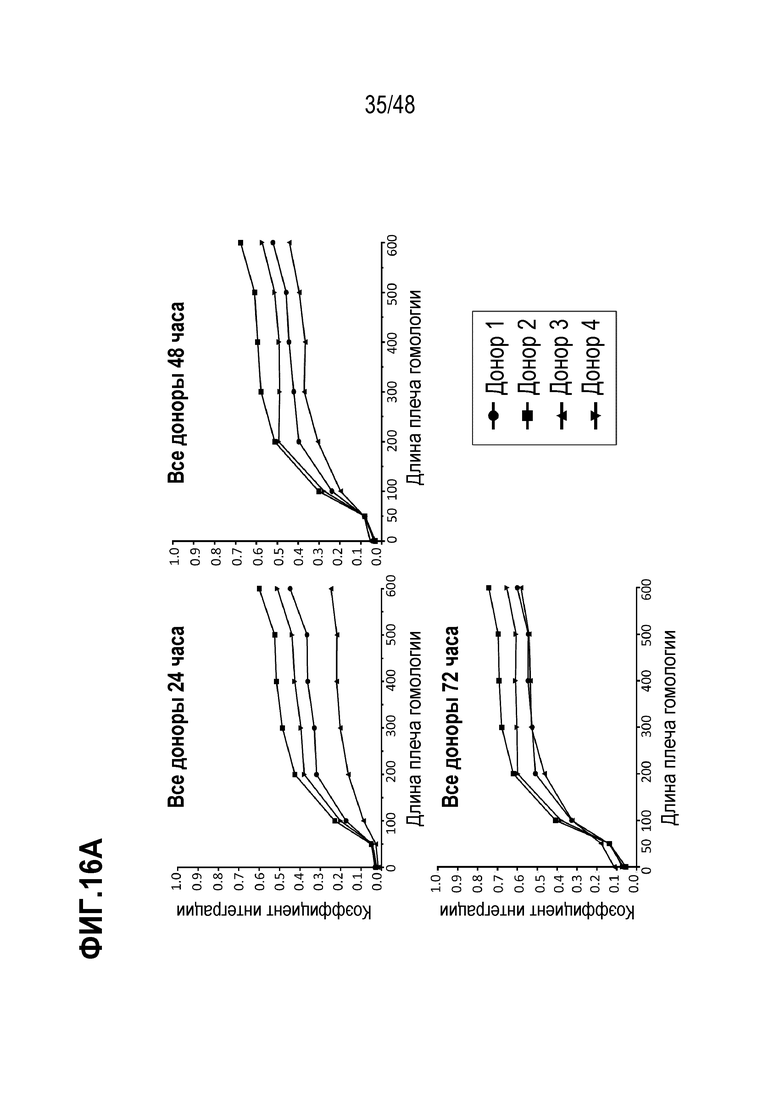
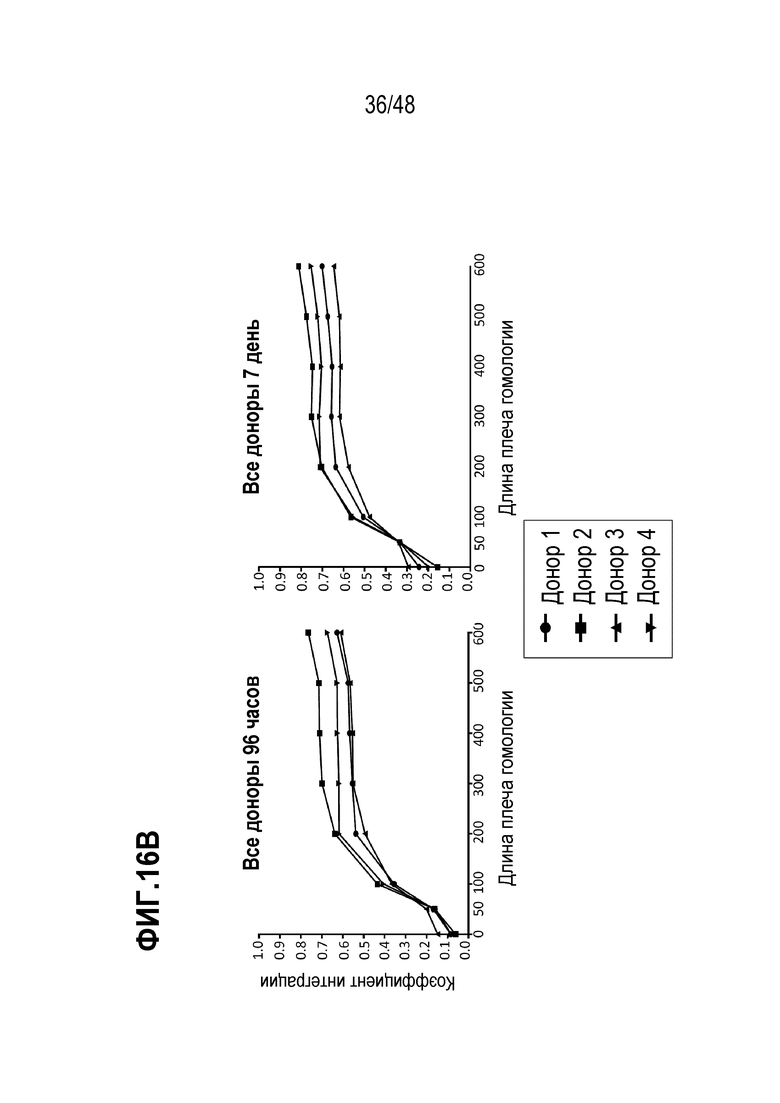
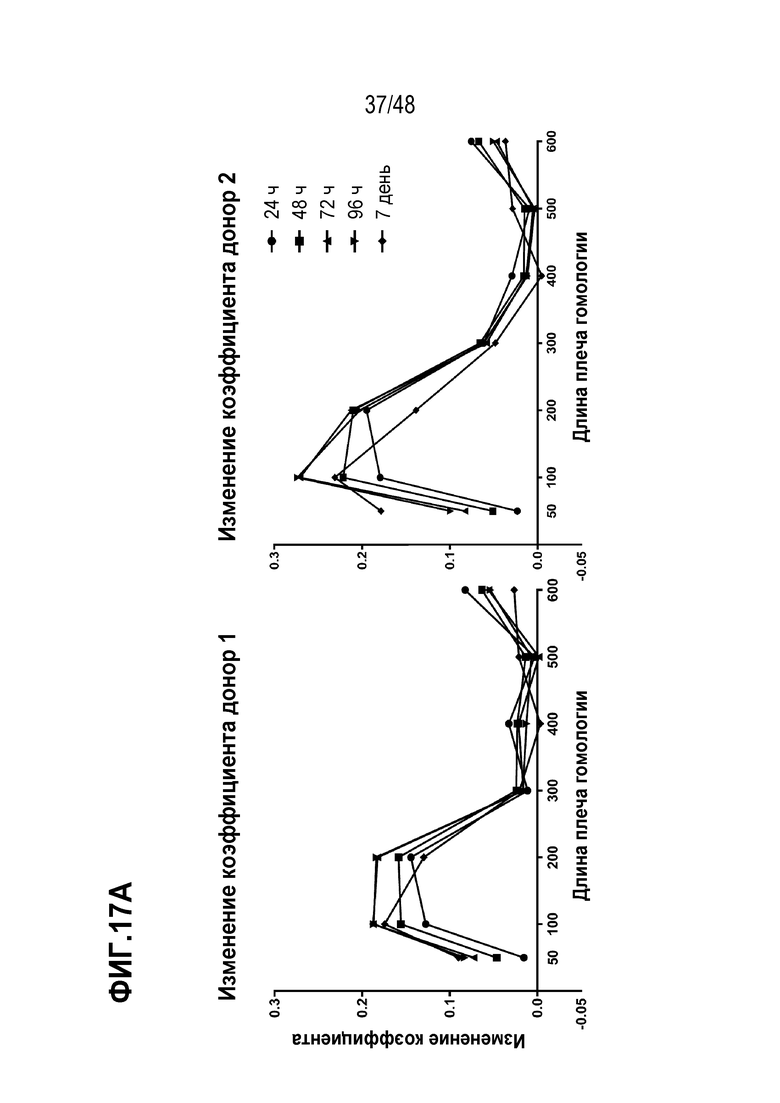
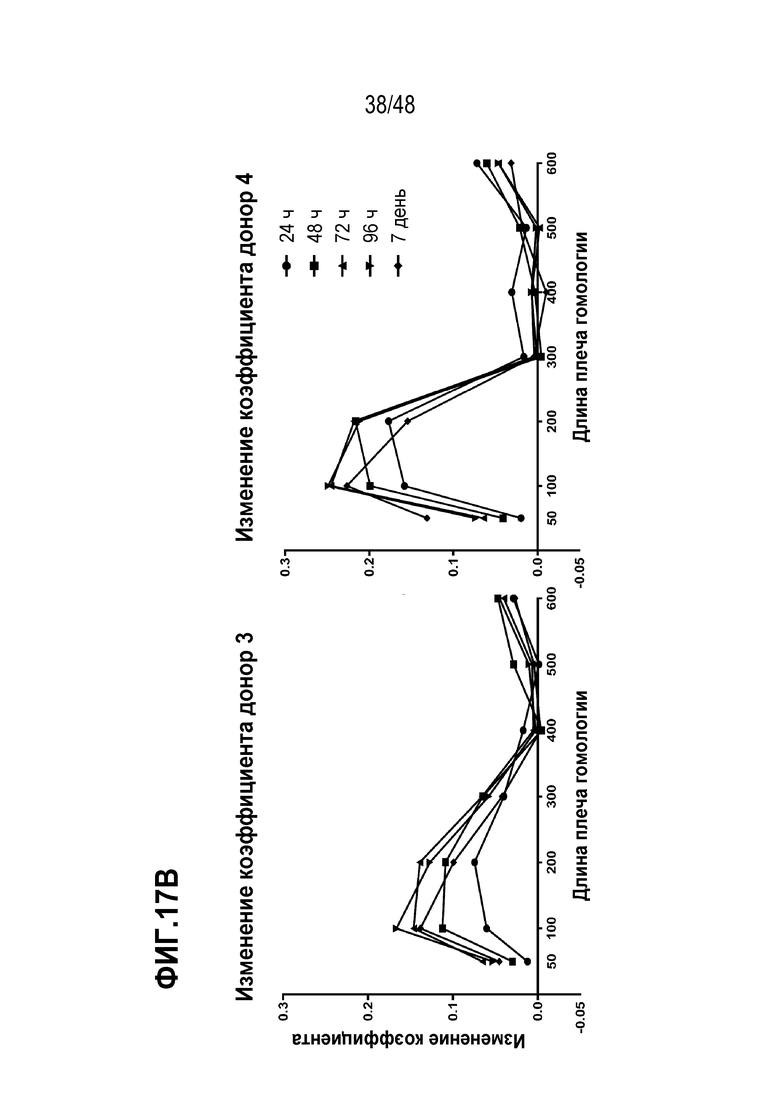
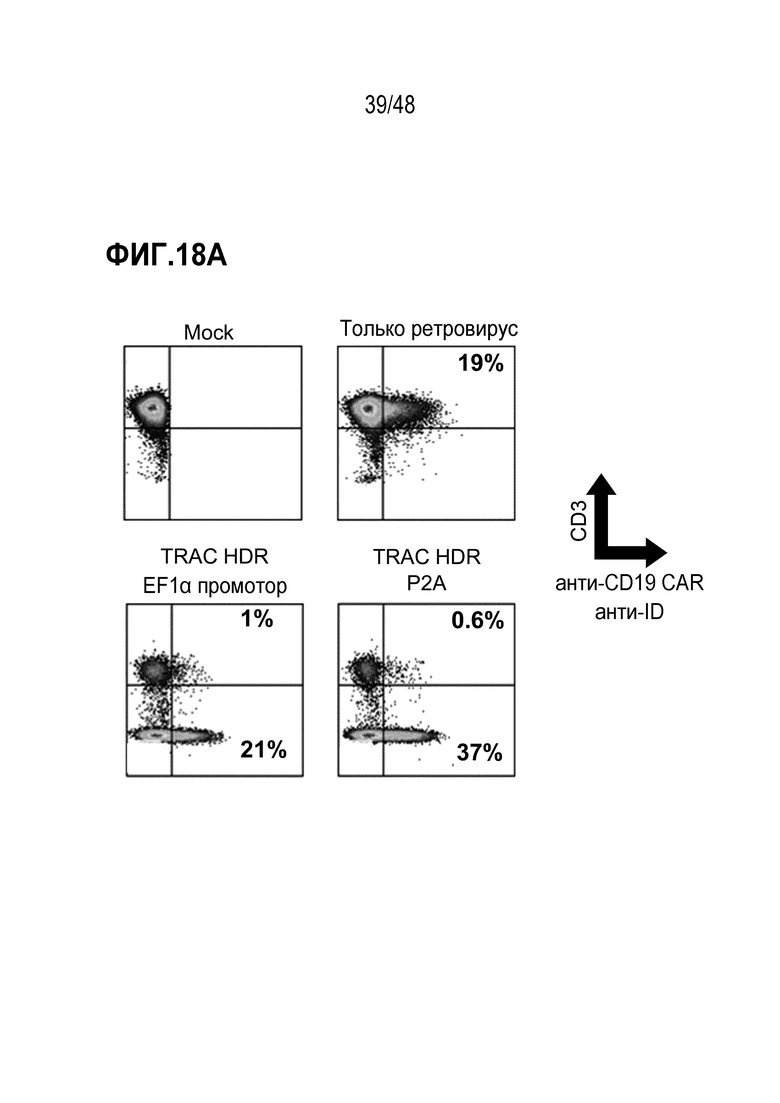
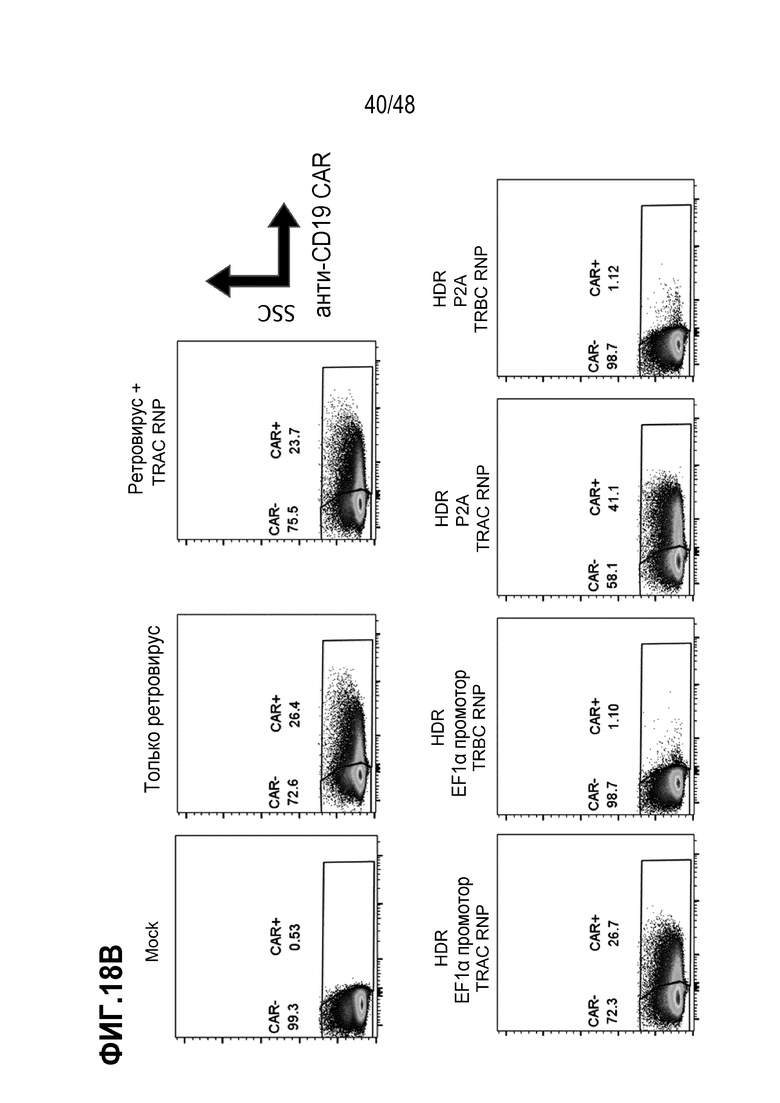
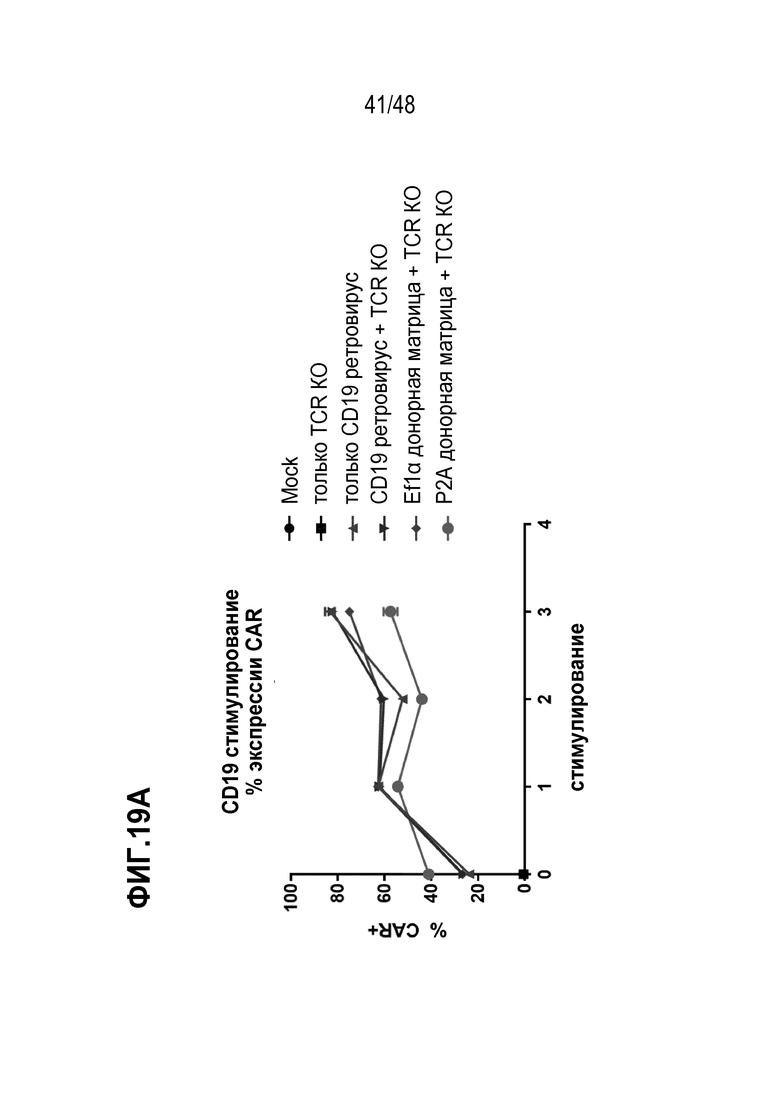
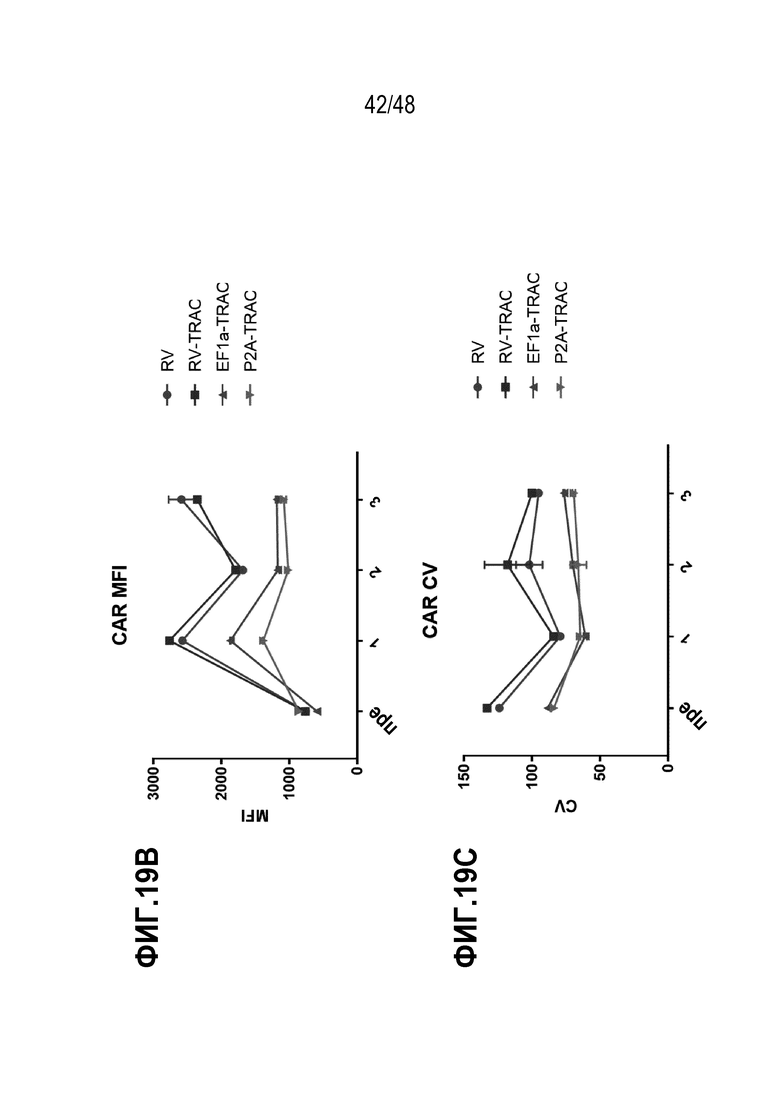
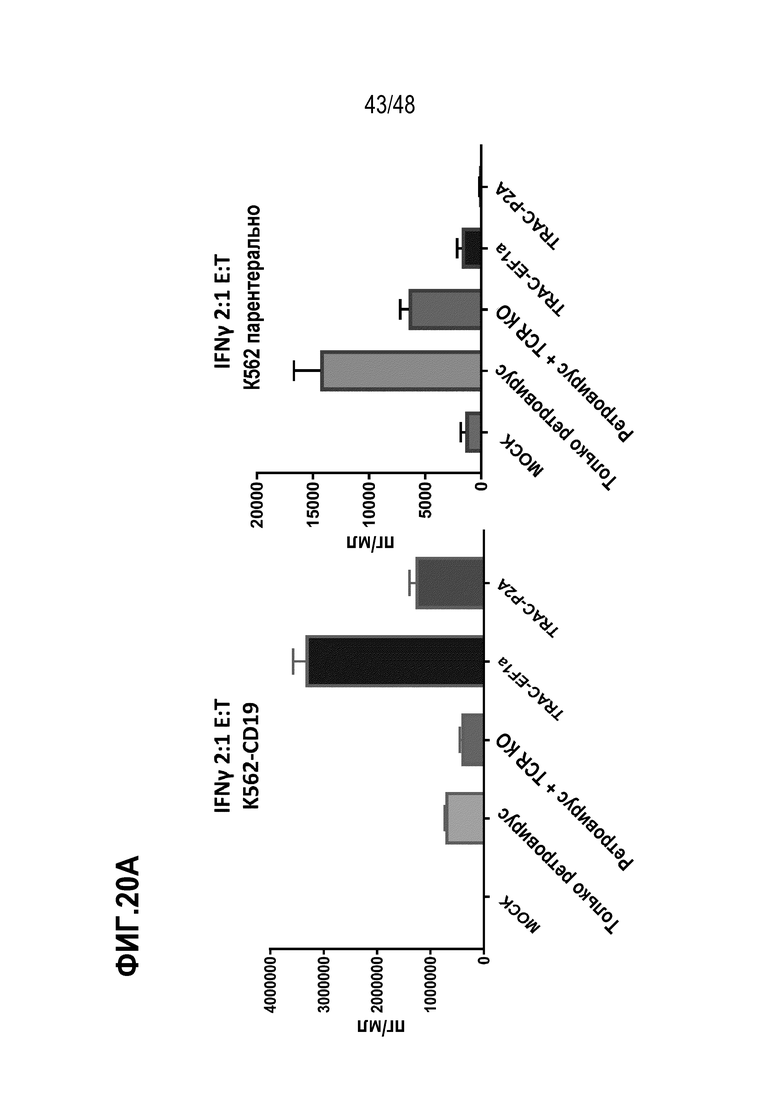
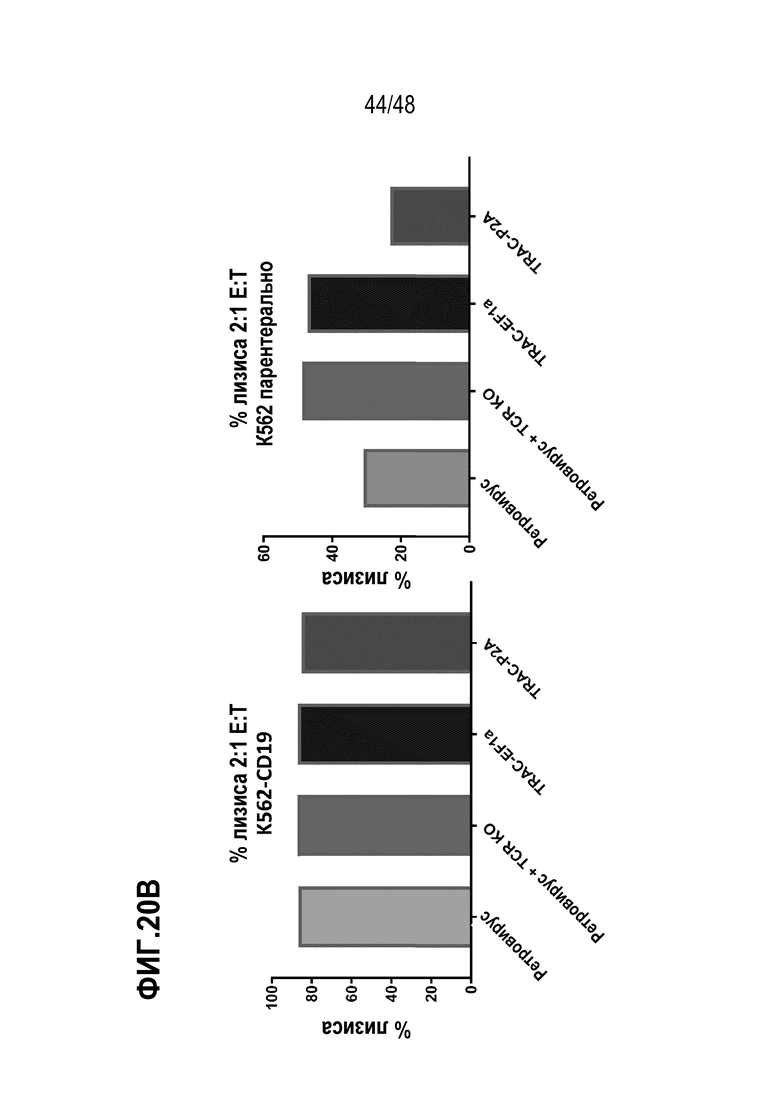
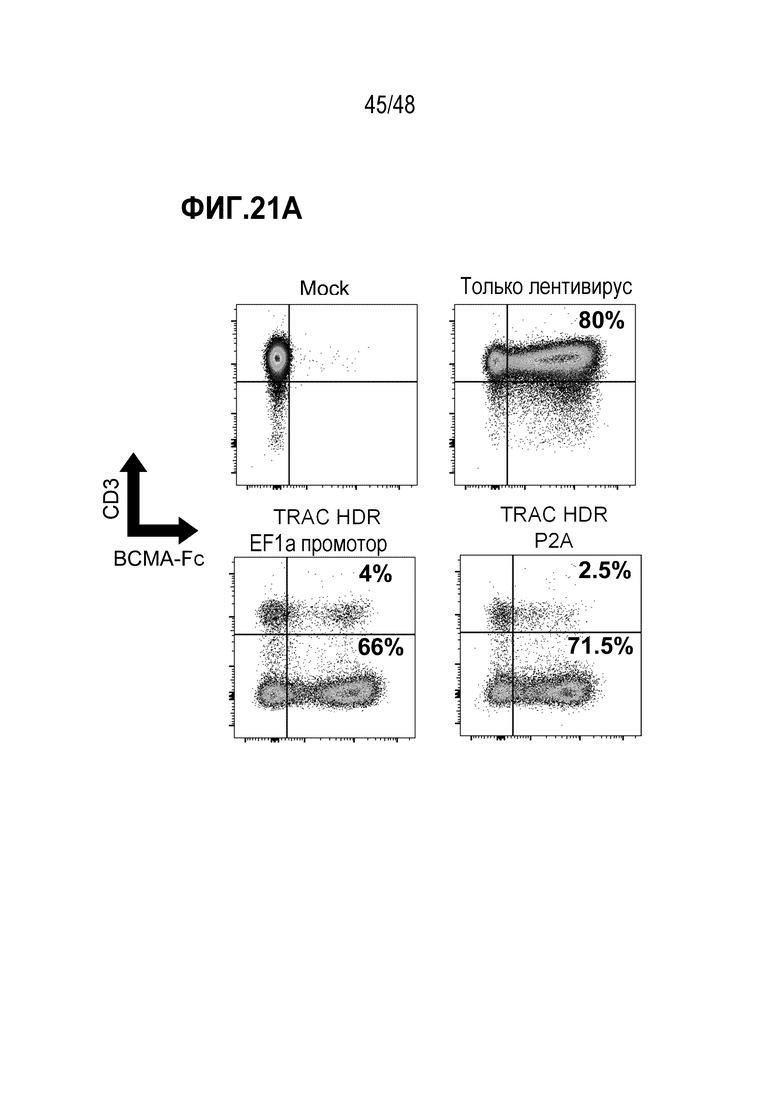
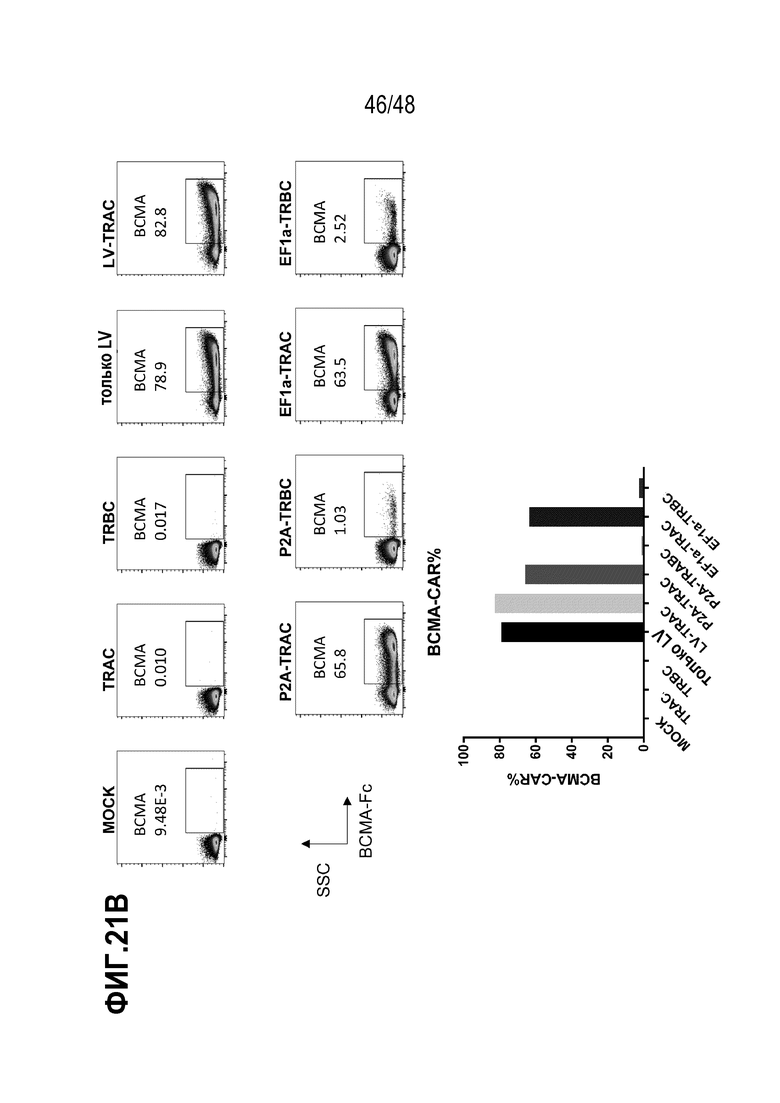
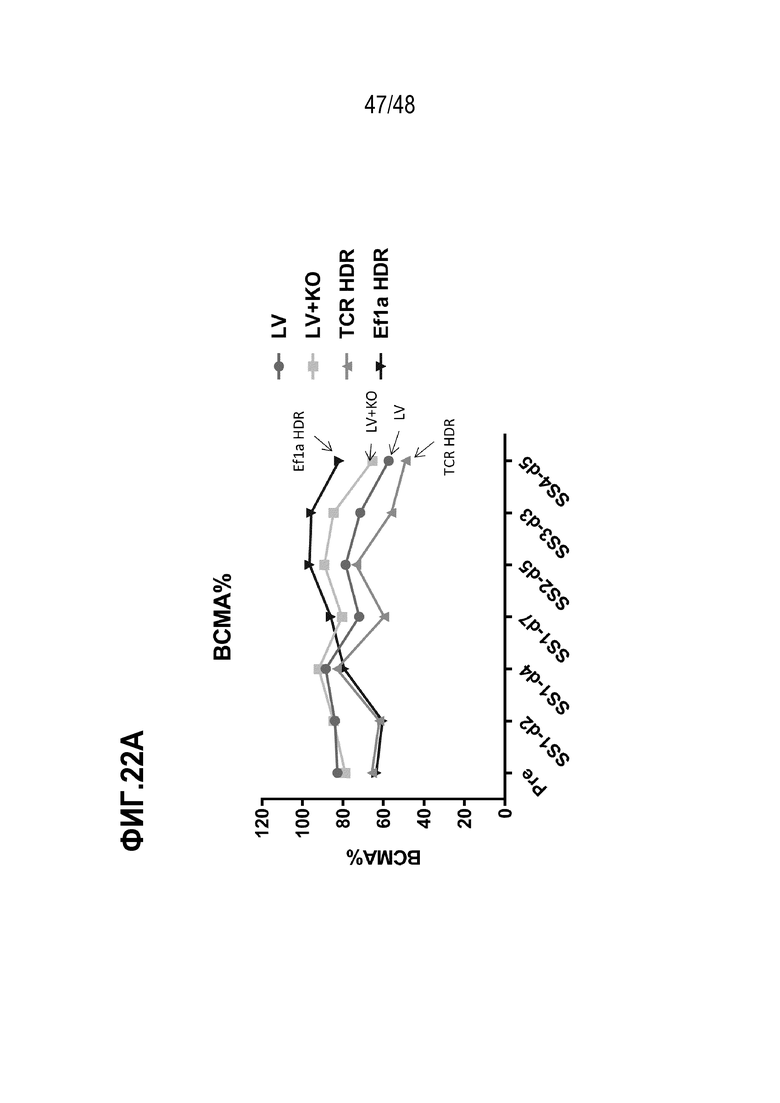
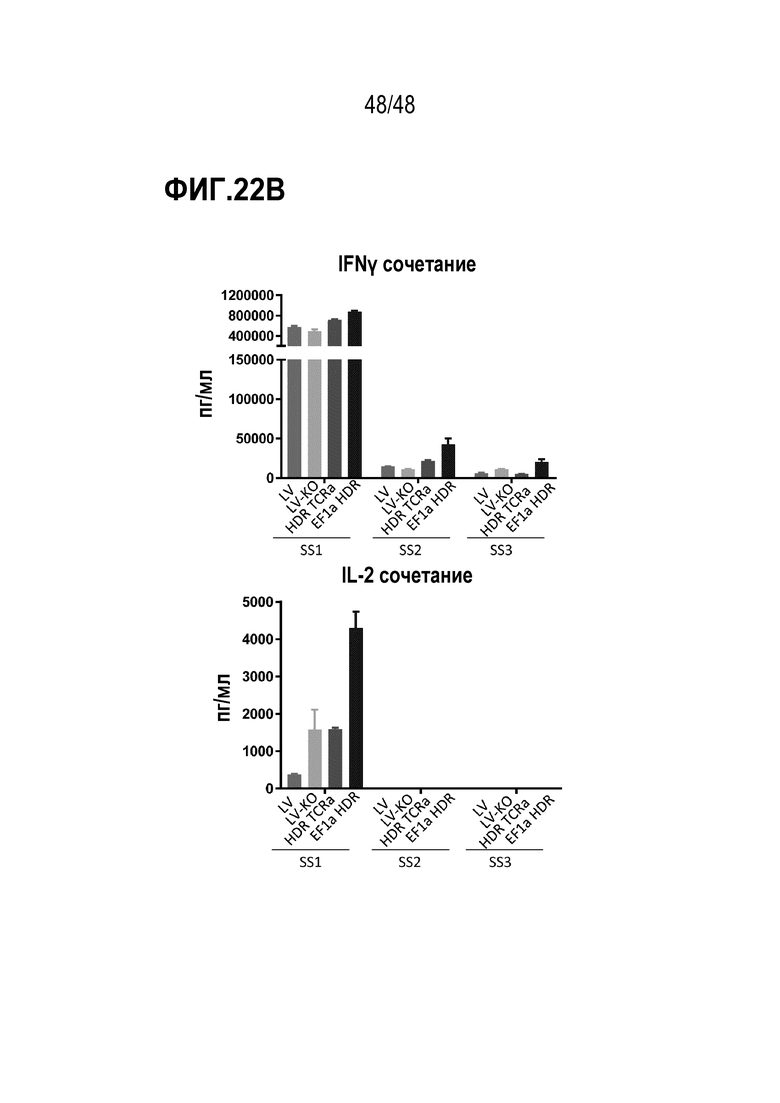
Группа изобретений относится к биотехнологии. Раскрыты способы получения генетически сконструированных T-клеток, включающие: введение в T-клетки агента CRISPR, содержащего направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту в пределах экзона 1 гена альфа константы Т-клеточного рецептора (TRAC) и/или гена бета константы Т-клеточного рецептора (TRBC), где указанный агент CRISPR способен вызвать генетическое разрушение, по меньшей мере, одного целевого сайта, что приводит к эффективному нокауту эндогенного T-клеточного рецептора (TCR) и введение в указанные T-клетки матричного полинуклеотида, где указанный матричный полинуклеотид содержит структуру [5' плечо гомологии]-[трансген]-[3' плечо гомологии]. Также представлены способ получения генетически сконструированных T-клеток, включающий введение в T-клетки, имеющие генетическое разрушение, по меньшей мере, одного целевого сайта в пределах экзона 1 гена альфа константы Т-клеточного рецептора (TRAC), приводящее к эффективному нокауту эндогенного T-клеточного рецептора (TCR), матричного полинуклеотида, где указанный матричный полинуклеотид содержит структуру [5' плечо гомологии]-[трансген]-[3' плечо гомологии], где указанный трансген кодирует рекомбинантный TCR, содержащий альфа (TCRα) цепь и бета (TCRβ) цепь, где указанный трансген содержит последовательность нуклеиновой кислоты, кодирующую TCRα цепь, и последовательность нуклеиновой кислоты, кодирующую TCRβ цепь и набор для осуществления представленных способов. Изобретение может быть использовано для инжиниринга иммунных клеток, в частности Т-клеток. 4 н. и 65 з.п. ф-лы, 48 ил., 10 табл., 6 пр.
1. Способ получения генетически сконструированных T-клеток, включающий:
(a) введение в T-клетки агента CRISPR, содержащего направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту в пределах экзона 1 гена альфа константы Т-клеточного рецептора (TRAC), где указанный агент CRISPR способен вызвать генетическое разрушение, по меньшей мере, одного целевого сайта, что приводит к эффективному нокауту эндогенного Т-клеточного рецептора (TCR); и
(b) введение в указанные T-клетки матричного полинуклеотида, где указанный матричный полинуклеотид содержит структуру [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии], где:
указанный трансген кодирует рекомбинантный TCR, содержащий альфа (TCRα) цепь и бета (TCRβ) цепь, где указанный трансген содержит последовательность нуклеиновой кислоты, кодирующую TCRα цепь, и последовательность нуклеиновой кислоты, кодирующую TCRβ цепь, и где указанный рекомбинантный TCR способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния,
и
плечи гомологии представляют собой приблизительно 600 пар оснований каждое с 5’- и 3’-стороны последовательности нуклеиновой кислоты, кодирующей указанный рекомбинантный TCR для целенаправленного воздействия в экзоне 1 эндогенного гена TRAC, где трансген, кодирующий указанный рекомбинантный TCR, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR), и
где введение матричного полинуклеотида проводят после введения указанного агента CRISPR.
2. Способ по п.1, где матричный полинуклеотид вводят сразу после или в течение 30 секунд, 1 минуты, 2 минут, 3 минут, 4 минут, 5 минут, 6 минут, 8 минут, 9 минут, 10 минут, 15 минут, 20 минут, 30 минут, 40 минут, 50 минут, 60 минут, 90 минут, 2 часов, 3 часов или 4 часов после введения указанного агента CRISPR, необязательно в течение 2 часов после введения указанного агента CRISPR.
3. Способ по п.1 или 2, где Т-клетки содержат CD4+ T клетки или их подтипы, или CD8+ T-клетки или их подтипы.
4. Способ по п.3, где Т-клетки содержат CD4+ и CD8+ T клетки, и соотношение CD4+ к CD8+ T-клеткам составляет от 1:3 до 3:1, необязательно, от 1:2 до 2:1, необязательно, 1:1.
5. Способ по любому из пп.1-4, где агент CRISPR содержит CRISPR-Cas9 сочетание.
6. Способ по п.5, где CRISPR-Cas9 сочетанием является рибонуклеопротеиновый (RNP) комплекс, содержащий нРНК и Cas9 белок.
7. Способ по п.6, где концентрация RNP составляет от 1 мкМ до 5 мкМ, необязательно, где концентрация RNP составляет 2 мкМ.
8. Способ по любому из пп.1-7, где введение указанного агента CRISPR проводят электропорацией.
9. Способ по любому из пп.1-8, где матричный полинуклеотид содержится в вирусном векторе, и введение матричного полинуклеотида проводят трансдукцией.
10. Способ по п.9, где указанным вирусным вектором является AAV вектор.
11. Способ по любому из пп.1-10, где до введения указанного агента CRISPR, способ включает инкубирование указанных T-клеток, in vitro со стимулирующим агентом (стимулирующими агентами) в условиях стимулирования или активации указанных T-клеток.
12. Способ по п.11, где указанный стимулирующий агент (указанные стимулирующие агенты) содержит (содержат) анти-CD3 антитело и/или анти-CD28 антитело, необязательно где указанный стимулирующий агент (указанные стимулирующие агенты) содержит (содержат) анти-CD3/анти-CD28 сферы, необязательно, где соотношение сфер к клеткам составляет 1:1.
13. Способ по п.11 или 12, включающий удаление указанного стимулирующего агента (указанных стимулирующих агентов) из указанных T-клеток до введения указанного агента CRISPR.
14. Способ по любому из пп.1-13, где способ дополнительно включает инкубирование клеток до, во время или после введения указанного агента CRISPR и/или введения матричного полинуклеотида с одним или более рекомбинантными цитокинами, необязательно, где один или более рекомбинантных цитокинов выбирают из группы, состоящей из IL-2, IL-7, и IL-15.
15. Способ по п.14, где указанные один или более рекомбинантных цитокинов добавляют в концентрации, выбранной из концентрации IL-2 от 10 Ед/мл до 200 Ед/мл, необязательно, от 50 МЕд/мл до 100 Ед/мл; IL-7 в концентрации от 0,5 нг/мл до 50 нг/мл, необязательно от 5 нг/мл до 10 нг/мл и/или IL-15 в концентрации от 0,1 нг/мл до 20 нг/мл, необязательно от 0,5 нг/мл до 5 нг/мл.
16. Способ по п.14 или 15, где инкубирование проводят после введения указанного агента CRISPR и введения матричного полинуклеотида в течение вплоть до приблизительно 24 часов, 36 часов, или 48 часов, или 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или 21 дней, необязательно вплоть до 7 дней.
17. Способ по любому из пп.1-16, где указанный трансген, кодирующий рекомбинантный TCR, дополнительно содержит гетерологичный регулирующий или контрольный элемент, необязательно где указанный гетерологичный регулирующий или контрольный элемент содержит гетерологичный промотор.
18. Способ получения генетически сконструированных T-клеток, включающий:
(a) введение в T-клетки агента CRISPR, содержащего направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту в пределах экзона 1 гена альфа константы Т-клеточного рецептора (TRAC) и/или гена бета константы Т-клеточного рецептора (TRBC), где указанный агент CRISPR способен вызвать генетическое разрушение, по меньшей мере, одного целевого сайта, что приводит к эффективному нокауту эндогенного T-клеточного рецептора (TCR); и
(b) введение в указанные T-клетки матричного полинуклеотида, где указанный матричный полинуклеотид содержит структуру [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии], где указанный трансген кодирует рекомбинантный TCR, содержащий альфа (TCRα) цепь и бета (TCRβ) цепь, где указанный трансген содержит последовательность нуклеиновой кислоты, кодирующую TCRα цепь, и последовательность нуклеиновой кислоты, кодирующую TCRβ цепь, и
где плечи гомологии представляют собой приблизительно 600 пар оснований каждое с 5’- и 3’-стороны последовательности нуклеиновой кислоты, кодирующей указанный рекомбинантный TCR для целенаправленного воздействия в экзоне 1 эндогенного гена TRAC, где указанный трансген содержит гетерологичный промотор, и где трансген нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
19. Способ по п.18, где введение указанного агента CRISPR и введение матричного полинуклеотида проводят одновременно или последовательно, в любом порядке.
20. Способ по п.18 или 19, где введение матричного полинуклеотида проводят после введения агента CRISPR.
21. Способ по п.20, где матричный полинуклеотид вводят сразу после или в течение 30 секунд, 1 минуты, 2 минут, 3 минут, 4 минут, 5 минут, 6 минут, 8 минут, 9 минут, 10 минут, 15 минут, 20 минут, 30 минут, 40 минут, 50 минут, 60 минут, 90 минут, 2 часов, 3 часов или 4 часов после введения указанного агента CRISPR или в течение 2 часов после введения указанного агента CRISPR.
22. Способ по п.18 или 19, где введение указанного агента CRISPR и введение матричного полинуклеотида проводят в одной экспериментальной реакции.
23. Способ по любому из пп.18-22, где до введения указанного агента CRISPR способ включает инкубирование клеток, in vitro со стимулирующим агентом (стимулирующими агентами) в условиях стимулирования или активации указанных T-клеток.
24. Способ по п.23, где указанный стимулирующий агент (указанные стимулирующие агенты) содержит (содержат) анти-CD3 антитело и/или анти-CD28 антитело, необязательно где указанный стимулирующий агент (стимулирующие агенты) содержит (содержат) анти-CD3/анти-CD28 сферы, необязательно, где соотношение сфер к клеткам составляет около 1:1.
25. Способ по п.23 или 24, включающий удаление указанного стимулирующего агента (указанных стимулирующих агентов) из указанных T-клеток до введения указанного агента CRISPR.
26. Способ по любому из пп.18-25, где способ дополнительно включает инкубирование клеток до, во время или после введения указанного агента CRISPR и/или введения матричного полинуклеотида с одним или более рекомбинантными цитокинами, необязательно, где указанные один или более рекомбинантных цитокинов выбирают из группы, состоящей из IL-2, IL-7, и IL-15.
27. Способ по п.26, где указанные один или более рекомбинантных цитокинов добавляют в концентрации, выбранной из концентрации IL-2 от 10 Ед/мл до 200 Ед/мл, необязательно от 50 МЕд/мл до 100 Ед/мл; IL-7 в концентрации от 0,5 нг/мл до 50 нг/мл, необязательно от 5 нг/мл до 10 нг/мл и/или IL-15 в концентрации от 0,1 нг/мл до 20 нг/мл, необязательно от 0,5 нг/мл до 5 нг/мл.
28. Способ по п.26 или 27, где инкубирование проводят после введения указанного агента CRISPR и введения матричного полинуклеотида в течение вплоть до приблизительно 24 часов, 36 часов, или 48 часов, или 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или 21 дней, необязательно вплоть до 7 дней.
29. Способ получения генетически сконструированных T-клеток, включающий:
введение в T-клетки, имеющие генетическое разрушение, по меньшей мере, одного целевого сайта в пределах экзона 1 гена альфа константы Т-клеточного рецептора (TRAC), приводящее к эффективному нокауту эндогенного T-клеточного рецептора (TCR), матричного полинуклеотида, где указанный матричный полинуклеотид содержит структуру [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии], где:
указанный трансген кодирует рекомбинантный TCR, содержащий альфа (TCRα) цепь и бета (TCRβ) цепь, где указанный трансген содержит последовательность нуклеиновой кислоты, кодирующую TCRα цепь, и последовательность нуклеиновой кислоты, кодирующую TCRβ цепь, и
где плечи гомологии представляют собой приблизительно 600 пар оснований каждое с 5’- и 3’-стороны последовательности нуклеиновой кислоты, кодирующей указанный рекомбинантный TCR для целенаправленного воздействия в экзоне 1 эндогенного гена TRAC,
где указанный трансген, кодирующий рекомбинантный TCR, содержит гетерологичный промотор, где генетическое разрушение вызвано агентом CRISPR, содержащим направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к указанному, по меньшей мере, одному целевому сайту, и указанный трансген, кодирующий рекомбинантный TCR, нацелен на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
30. Способ по любому из пп.18-29, где указанный агент CRISPR содержит CRISPR-Cas9 сочетание.
31. Способ по п.30, где CRISPR-Cas9 сочетанием является рибонуклеопротеиновый (RNP) комплекс, содержащий нРНК и Cas9 белок.
32. Способ по п.31, где концентрация RNP составляет от 1 мкМ до 5 мкМ, необязательно, где концентрация RNP составляет 2 мкМ.
33. Способ по п.31 или 32, где RNP вводят посредством электропорации.
34. Способ по любому из пп.18-33, где Т-клетки содержат CD8+ T-клетки или их подтипы.
35. Способ по любому из пп.18-33, где Т-клетки содержат CD4+ T-клетки или их подтипы.
36. Способ по любому из пп.18-33, где Т-клетки содержат CD4+ T-клетки или их подтипы и CD8+ T-клетки или их подтипы.
37. Способ по п.36, где соотношение CD4+ к CD8+ T-клеткам составляет от 1:3 до 3:1, необязательно от 1:2 до 2:1, необязательно 1:1.
38. Способ по любому из пп.18-37, где матричный полинуклеотид содержится в векторе, необязательно вирусном векторе.
39. Способ по п.38, где указанным вектором является вирусный вектор, и указанным вирусным вектором является AAV вектор.
40. Способ по п.39, где указанный AAV вектор выбирают из группы, состоящей из AAV1 вектора, AAV2 вектора, AAV3 вектора, AAV4 вектора, AAV5 вектора, AAV6 вектора, AAV7 вектора и AAV8 вектора.
41. Способ по п.40, где указанным AAV вектором является AAV2 вектор или AAV6 вектор.
42. Способ по п.38, где указанным вектором является вирусный вектор, и указанным вирусным вектором является ретровирусный вектор, необязательно, лентивирусный вектор.
43. Способ по любому из пп.1-42, где нРНК содержит последовательность, выбранную из группы, состоящей из UCUCUCAGCUGGUACACGGC (SEQ ID NO:28), UGGAUUUAGAGUCUCUCAGC (SEQ ID NO:29), ACACGGCAGGGUCAGGGUUC (SEQ ID NO:30), GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31), GCUGGUACACGGCAGGGUCA (SEQ ID NO:32), CUCAGCUGGUACACGGC (SEQ ID NO:33), UGGUACACGGCAGGGUC (SEQ ID NO:34), GCUAGACAUGAGGUCUA (SEQ ID NO:35), GUCAGAUUUGUUGCUCC (SEQ ID NO:36), UCAGCUGGUACACGGCA (SEQ ID NO:37), GCAGACAGACUUGUCAC (SEQ ID NO:38), GGUACACGGCAGGGUCA (SEQ ID NO:39), CUUCAAGAGCAACAGUGCUG (SEQ ID NO:40), AGAGCAACAGUGCUGUGGCC (SEQ ID NO:41), AAAGUCAGAUUUGUUGCUCC (SEQ ID NO:42), ACAAAACUGUGCUAGACAUG (SEQ ID NO:43), AAACUGUGCUAGACAUG (SEQ ID NO:44), UGUGCUAGACAUGAGGUCUA (SEQ ID NO:45), GGCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:46), GCUGGGGAAGAAGGUGUCUUC (SEQ ID NO:47), GGGGAAGAAGGUGUCUUC (SEQ ID NO:48), GUUUUGUCUGUGAUAUACACAU (SEQ ID NO:49), GGCAGACAGACUUGUCACUGGAUU (SEQ ID NO:50), GCAGACAGACUUGUCACUGGAUU (SEQ ID NO:51), GACAGACUUGUCACUGGAUU (SEQ ID NO:52), GUGAAUAGGCAGACAGACUUGUCA (SEQ ID NO:53), GAAUAGGCAGACAGACUUGUCA (SEQ ID NO:54), GAGUCUCUCAGCUGGUACACGG (SEQ ID NO:55), GUCUCUCAGCUGGUACACGG (SEQ ID NO:56), GGUACACGGCAGGGUCAGGGUU (SEQ ID NO:57) и GUACACGGCAGGGUCAGGGUU (SEQ ID NO:58).
44. Способ по любому из пп.1-43, где нРНК имеет домен целенаправленного воздействия, содержащий последовательность GAGAAUCAAAAUCGGUGAAU (SEQ ID NO:31).
45. Способ по любому из пп.1-44, где 5’ плечо гомологии и 3’ плечо гомологии содержит последовательности нуклеиновых кислот гомологичные последовательностям нуклеиновых кислот окружающим, по меньшей мере, один целевой сайт.
46. Способ по п.45, где 5’ плечо гомологии содержит последовательность, представленную в SEQ ID NO: 124, и/или 3’ плечо гомологии содержит последовательность, представленную в SEQ ID NO: 125.
47. Способ по любому из пп.1-46, где трансген дополнительно содержит один или более мультицистронных элементов, и мультицистронные элементы расположены между последовательностью нуклеиновых кислот, кодирующей TCRα или его часть, и последовательностью нуклеиновых кислот, кодирующей TCRβ или его часть.
48. Способ по п.47, где мультицистронные элементы содержат последовательность, кодирующую элемент проскока рибосомы, выбранный из T2A, P2A, E2A или F2A или внутренний участок посадки рибосомы (IRES).
49. Способ по любому из пп.1-48, где способ дополнительно включает введение в указанные T-клетки одного или более вторых матричных полинуклеотидов, содержащих один или более вторых трансгенов, где указанные один или более вторых трансгенов нацелены на интеграцию в или рядом с одним из, по меньшей мере, одного целевого сайта через репарацию, направляемую гомологией (HDR).
50. Способ по п.49, где указанные один или более вторых трансгенов кодируют молекулу, выбранную из костимулирующего лиганда, цитокина, растворимого одноцепочечного вариабельного фрагмента (scFv), иммуномодулирующего слитого белка, химерного рецептора переключения (CSR) или корецептора.
51. Способ по п.49 или 50, где указанные один или более вторых трансгенов независимо дополнительно содержат гетерологичный регулирующий или контрольный элемент, необязательно, промотор.
52. Способ по любому из пп.17-51, где указанным гетерологичным промотором является или он содержит промотор человеческого фактора элонгации 1 альфа (EF1α) или MND промотор или их вариант.
53. Способ по любому из пп.17-51, где указанным гетерологичным промотором является индуцируемый промотор или репрессируемый промотор.
54. Способ по любому из пп.1-53, где TCRα цепь содержит константную область (Cα) область, в которую введен один или более цистеиновых остатков, и/или TCRβ цепь содержит константную (Cβ) область, в которую введен один или более цистеиновых остатков, где один или более введенных цистеиновых остатков способны образовывать один или более не нативных дисульфидных мостиков между альфа цепью и бета цепью.
55. Способ по п.54, где введение одного или более цистеиновых остатков включает замещение не цистеинового остатка цистеиновым остатком.
56. Способ по п.54 или 55, где Cα область содержит цистеин в положении, соответствующем положению 48 с нумерацией, указанной в любой из SEQ ID NO: 24; и/или Cβ область содержит цистеин в положении, соответствующем положению 57 с нумерацией, указанной в SEQ ID NO: 20.
57. Способ по любому из пп.1-56, где указанные T-клетки получают из мультипотентной или плюрипотентной клетки, которой необязательно является иПСК.
58. Способ по любому из пп.1-56, где указанные T-клетки представляют собой первичные клетки от субъекта.
59. Способ по п.58, где субъект имеет или предположительно имеет заболевание, расстройство или состояние.
60. Способ по п.59, где субъект является или предположительно является здоровым.
61. Способ по п.58 или 59, где указанные T-клетки являются аутологичными к субъекту.
62. Способ по любому из пп.58-61, где указанные T-клетки являются аллогенными к субъекту.
63. Способ по любому из пп.1-62, где матричный полинуклеотид имеет, по меньшей мере, 2500, 2750, 3000, 3250, 3500, 3750, 4000, 4250, 4500, 4760, 5000, 5250, 5500, 5750, 6000, 7000, 7500, 8000, 9000 или 10000 нуклеотидов в длину или любое значение между любыми из вышеописанных.
64. Способ по любому из пп.1-63, где указанный матричный полинуклеотид имеет между 2500 и 5000 нуклеотидов, 3500 и 4500 нуклеотидов или 3750 нуклеотидов и 4250 нуклеотидов в длину.
65. Способ по любому из пп.1-64, где по меньшей мере или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% T-клеток во множестве генетически сконструированных T-клеток, полученных указанным способом, содержат генетическое разрушение указанного, по меньшей мере, одного целевого сайта в пределах экзона 1 гена TRAC.
66. Способ по любому из пп.1-65, где по меньшей мере или более чем 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80% или 90% клеток во множестве генетически сконструированных T-клеток, полученных указанным способом, экспрессируют указанный рекомбинантный TCR.
67. Способ по любому из пп.1-66, где коэффициент вариации экспрессии рекомбинантного TCR среди множества генетически сконструированных T-клеток, полученных указанным способом, ниже чем 0,70, 0,65, 0,60, 0,55, 0,50, 0,45, 0,40, 0,35 или 0,30 или менее.
68. Способ по любому из пп.1-67, где коэффициент вариации экспрессии рекомбинантного TCR среди множества генетически сконструированных T-клеток, полученных указанным способом, на, по меньшей мере, 100%, 95%, 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20% или 10% ниже, чем коэффициент вариации экспрессии того же рекомбинантного TCR среди множества генетически сконструированных T-клеток, полученных случайной интеграцией.
69. Набор для осуществления способа по любому из пп.1-68, где указанный набор содержит:
CRISPR-Cas9 сочетание, содержащее направляющую РНК (нРНК), имеющую домен целенаправленного воздействия, который является комплементарным к, по меньшей мере, одному целевому сайту в гене альфа константы Т-клеточного рецептора (TRAC), где указанное CRISPR-Cas9 сочетание представляет собой рибонуклеопротеиновый (RNP) комплекс, содержащий нРНК и Cas9 белок; и
матричный полинуклеотид, который содержится в вирусном векторе, который представляет собой AAV-вектор, где указанный матричный полинуклеотид содержит структуру [5’ плечо гомологии]-[трансген]-[3’ плечо гомологии], где указанный трансген кодирует рекомбинантный T-клеточный рецептор (TCR), содержащий альфа (TCRα) цепь и бета (TCRβ) цепь, где указанный трансген содержит последовательность нуклеиновой кислоты, кодирующую TCRα цепь, и последовательность нуклеиновой кислоты, кодирующую TCRβ цепь, и где указанный трансген находится под контролем конститутивного промотора; и
где плечи гомологии представляют собой приблизительно 600 пар оснований каждое с 5’- и 3’-стороны последовательности нуклеиновой кислоты, кодирующей указанный рекомбинантный TCR для целенаправленного воздействия в экзоне 1 эндогенного гена TRAC, где указанный трансген, кодирующий указанный рекомбинантный TCR, подвергается целенаправленному воздействию для интеграции в или рядом с указанным по меньшей мере одним целевым сайтом через репарацию, направляемую гомологией (HDR), где указанный рекомбинантный рецептор способен связываться с антигеном, который ассоциирован с, является специфическим к и/или экспрессирован на клетке или ткани заболевания, расстройства или состояния, где указанные заболевание, расстройство или состояние представляют собой рак;
и инструкции по проведению способа по любому из пп.1-68.
| WO 2013074916 A1, 23.05.2013 | |||
| EYQUEM J | |||
| et al | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Nature | |||
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| Приспособление для открывания боковых откидных стенок вагонетки | 1922 |
|
SU543A1 |
| No | |||
| Приспособление для остановки вагонов при обрыве поезда | 1927 |
|
SU7643A1 |
| Способ обработки грубых шерстей на различных аппаратах для мериносовой шерсти | 1920 |
|
SU113A1 |
| РОЙТ А | |||
| и др., Иммунология, Москва, "Мир", 2000, стр | |||
| Способ получения борнеола из пихтового или т.п. масел | 1921 |
|
SU114A1 |
| КЛЕТКИ, ПРОИЗВОДНЫЕ ОТ КАРДИАЛЬНОЙ ТКАНИ | 2010 |
|
RU2642282C1 |

