ОБЛАСТЬ ТЕХНИКИ
[001] Настоящее изобретение относится к рекомбинантным поксвирусам и их композициям, которые содержат модифицированный Mucin 1, трансген, ассоциированный с клеточной поверхностью (MUC1), трансген человеческого карциноэмбрионального антигена (СЕА) и/или одну или более костимуляторных молекул. По меньшей мере в одном аспекте модифицированные трансгены молекул MUC1, СЕА и/или костимуляторной молекулы увеличивают стабильность поксвируса при последовательном пассировании рекомбинантного поксвируса. В дополнительных аспектах настоящее изобретение относится к рекомбинантным поксвирусам и их композициям для использования в качестве вакцин и лекарственных композиций.
УРОВЕНЬ ТЕХНИКИ
[002] Рекомбинантные поксвирусы использовались в качестве иммунотерапевтических вакцин против инфекционных организмов и, в последнее время, против опухолей. Mastrangelo et al. J Clin Invest. 2000;105(8):1031-1034. Было показано, что две из этих групп поксвирусов, авипоксвирусы и ортопоксвирусы, эффективны в борьбе с опухолями и использовались в потенциальных способах лечения рака. Id.
[003] Было показано, что один типовой вид авипоксвируса, вирус оспы птиц, является безопасным носителем для введения человеку, поскольку вирус оспы птиц проникает в клетки млекопитающих и экспрессирует белки, но не может проходить полную репликацию. Skinner et al. Expert Rev Vaccines. 2005 Feb;4(1):63-76. Кроме того, использование вируса оспы птиц в качестве средства для экспрессии оценивается в многочисленных клинических испытаниях вакцин против рака, малярии, туберкулеза и СПИДа. Id.
[004] Вирус осповакцины, наиболее известный из ортопоксвирусов, использовался при ликвидации оспы во всем мире и показал свою полезность в качестве вектора и/или вакцины. Рекомбинантный вектор вакцины был разработан для экспрессии широкого ряда встроенных генов, включая несколько генов, ассоциированных с опухолью, таких как p97, HER-2/neu, p53 и ETA (Paoletti, et al., 1993).
[005] Одним из штаммов вируса оспы, который оказался полезным в качестве иммунотерапевтической вакцины против инфекционных заболеваний и рака, является модифицированный вирус осповакцины Анкара (MVA). MVA получали путем 516 серийных пассажей на фибробластах куриных эмбрионов штамма Анкара вируса осповакцины (CVA) (для обзора см. Mayr, A., et al. Infection 3, 6-14 (1975)). В результате этих длительных пассажей в геноме результируемого вируса MVA было удалено около 31 т.н его геномной последовательности, и следовательно, он был описан как высокоограниченый по репликации к клеткам хозяевам, происходящим из птиц (Meyer, H. et al. J. Gen. Virol. 72, 1031-1038 (1991)). На различных моделях животных было показано, что полученный MVA был в значительной степени авирулентным (Mayr, A. & Danner, K., Dev. Biol. Stand. 41: 225-34 (1978)).
[006] Были описаны штаммы MVA, имеющие улучшенные профили безопасности для разработки более безопасных продуктов, таких как вакцины или фармацевтические препараты. См. международную публикацию РСТ WO2002042480 (см. также, например, патент США № 6,761,893 и 6,913,752) которые все включены в настоящее описание посредством ссылки. Такие варианты способны к репродуктивной репликации в клетках и клеточных линиях, не принадлежащих человеку, особенно в фибробластах куриных эмбрионов (CEF), но не способны к репликации в клеточных линиях человека, в частности, в том числе в клеточных линиях HeLa, HaCat и 143B. Такие штаммы также не способны к репродуктивной репликации in vivo, например, у некоторых мышиных штаммов, таких как трансгенная мышиная модель AGR 129, которая имеет серьезно нарушенный иммунитет и очень чувствительна к реплицирующемуся вирусу. Смотрите патент США № 6761893. Такие варианты MVA и его производные, включая рекомбинанты, были описаны как «MVA-BN». См. международную публикацию РСТ WO2002042480 (см. также, например, патент США № 6761893 и 6913752). Было показано, что при разработке вакцин для иммунотерапии рака опухолевый антиген MUC1 индуцирует и усиливает иммунный ответ пациента на различные виды рака при экспрессии рекомбинантными поксвирусами. Смотри, например, Mehebtash et al. Clin Cancer Res. 2011 Nov 15;17(22):7164-73.
[007] MUC1 (MUC-1, Mucin 1, ассоциированный с клеточной поверхностью) (также известный как CD227) представляет собой гликопротеин, который выстилает апикальную поверхность эпителиальных клеток в легких, желудке, кишечнике, глазах и некоторых других органах и в небольшой подгруппе неэпителиальных клеток, таких как гемопоэтические клетки и активированные Т-клетки. Его основной функцией в здоровом эпителии является обеспечение смазывающего эффекта и физического барьера к химическим и микробным агентам. Hollingsworth MA, Swanson BJ (January 2004). "Mucins in cancer: protection and control of the cell surface". Nature Reviews Cancer 4 (1): 45-60.
[008] MUC1 прикреплен к апикальной поверхности с помощью трансмембранного домена. Hattrup CL, Gendler, SJ (2008). "Structure and Function of the Cell Surface (Tethered) Mucins". Annu. Rev. Physiol. 70: 431-457. Внеклеточный домен MUC1 содержит домен с переменным числом тандемных повторов из 20 аминокислот (VNTR), который обычно сильно гликозилирован, причем количество повторов варьирует от 20 до 120 у разных индивидов. Brayman M, Thathiah A, Carson DD (January 2004). "MUC1: a multifunctional cell surface component of reproductive tissue epithelia". Reprod Biol Endocrinol 2: 4.
[009] Было продемонстрировано, что многие карциномы человека (такие как рак яичников, молочной железы, поджелудочной железы, колоректальный рак и рак простаты) и гематологические злокачественные опухоли (множественная миелома и некоторые В-клеточные неходжкинские лимфомы) аберрантно сверхэкспрессируют MUC1. Pecher et al. Anticancer Res. 2001 Jul-Aug. 21:2591-2596. В отличие от кластерной экспрессии в нормальных тканях, MUC1 равномерно распределен по всей поверхности опухолевых клеток. Correa et al. Immunology. Jan 2003; 108(1): 32-41. Кроме того, MUC1 обычно недостаточно гликозилируется в опухолях, экспонируя новые и потенциально антигенные эпитопы белкового ядра иммунной системе. Reis et al. Int J Cancer. 1998 Aug 21;79(4):402-10.
[010] Ввиду ассоциации MUC1 с карциномами человека ранее в данной области техники была предпринята попытка модифицировать MUC1 для усиления иммуногенности белка. Например, в US2006/0147458, Hamblin et al. использовался «коэффициент использования кодонов» для конструирования последовательности MUC1, имеющей пониженную гомологию с нативным MUC1, а также имеющей сегмент 7XVNTR. В US2006/0147458 Hamblin et al. был создан слитый белок HSP-70-MUC1 в попытке повысить иммуногенность. В патенте США №5744144, выданном Finn et al., белок MUC1 модифицировался путем добавления двух 20-аминокислотных тандемных повторов.
[011] Человеческий карциноэмбриональный антиген (СЕА) представляет собой гликопротеин массой 180 кДа, экспрессируемый в большинстве опухолей толстой кишки, прямой кишки, желудка и поджелудочной железы (1), в около 50% карцином молочной железы (2) и 70% карцином легких (3). СЕА также экспрессируется в ткани ЖКТ плода и в меньшей степени в нормальном эпителии толстой кишки. Иммуногенность СЕА была неоднозначной, в нескольких исследованиях сообщалось о наличии антител против СЕА у пациентов (4-7), в то время как в других исследованиях этого не обнаруживалось (8-10). CEA был впервые описан как специфичный для рака фетальный антиген в аденокарциноме пищеварительного тракта человека в 1965 г. (Gold, P. and Freeman, S. O. (1965) Exp. Med. 121:439-462). С тех пор СЕА был охарактеризован как антиген клеточной поверхности, сверхпродуцируемый почти во всех солидных опухолях желудочно-кишечного тракта человека. Ген белка СЕА человека был клонирован. (Oikawa et al (1987) Biochim. Biophys. Res. 142:511-518; European Application No. EP 0346710).
[012] Существует значительная неудовлетворенная медицинская потребность в улучшении лечения рака. Ввиду эффективности антигенов MUC1 и CEA в индукции иммунного ответа против рака, существует потребность в улучшенных вакцинах, способных эффективно вводить антигены больным раком.
[013] Кроме того, существует растущая потребность в лечении рака, которое могло бы успешно преодолеть препятствия для получения разрешения регулирующих органов. В частности, трудности, связанные с крупномасштабным производством, примесями и тому подобным, могут стать серьезным препятствием на пути получения одобрения регулирующих органов для лечения и использования этих видов лечения для получения пользы пациентами. По меньшей мере, в одном аспекте, благодаря разработке различных вариантов осуществления настоящего изобретения, трудности, связанные с крупномасштабным производством, примесями и другими проблемами, были успешно преодолены.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[014] В настоящем изобретении было определено, что различные замены нуклеиновых кислот, кодирующих MUC1, CEA и/или TRICOM, в одной или более повторяющихся нуклеотидных областях повышают стабильность трансгенов MUC1, CEA и/или TRICOM в рекомбинантных поксвирусах.
[015] Соответственно, в одном варианте осуществления изобретения, настоящее изобретение относится к рекомбинантному поксвирусу, который стабилен при последовательном пассировании рекомбинантного поксвируса. Рекомбинантный поксвирус содержит первую нуклеиновую кислоту, кодирующую пептид MUC1, имеющий по меньшей мере два домена вариабельных N-концевых повторов (VNTR), причем а) расположение по меньшей мере двух доменов VNTR перетасовано, и b) по меньшей мере два домена VNTR являются кодон-оптимизированными, причем рекомбинантный поксвирус стабилен при последовательном пассировании.
[016] В одном или более предпочтительных вариантах осуществления изобретения рекомбинантный поксвирус содержит первую нуклеиновую кислоту, по меньшей мере на 95% гомологичную SEQ ID NO:2 (336 MUC), по меньшей мере на 95% гомологичную SEQ ID NO:3 (373 MUC), по меньшей мере на 95% гомологичную SEQ ID NO:4 (399/400 MUC1) или, по меньшей мере, на 95% гомологичную SEQ ID NO:5 (420 MUC1). В более предпочтительном варианте осуществления изобретения рекомбинантный поксвирус содержит нуклеиновую кислоту, по меньшей мере на 95% гомологичную SEQ ID NO: 2 (336 MUC1). В другом более предпочтительном варианте осуществления изобретения рекомбинантный поксвирус содержит нуклеиновую кислоту, по меньшей мере на 95% гомологичную SEQ ID NO:3 (373 MUC).
[017] В еще одном предпочтительном варианте осуществления изобретения рекомбинантные поксвирусы дополнительно содержат нуклеиновую кислоту, по меньшей мере на 99% гомологичную SEQ ID NO: 13 или 14 (CEA). В предпочтительном варианте осуществления изобретения рекомбинантные поксвирусы содержат SEQ ID NO: 13 или 14.
[018] Предполагается, что рекомбинантный поксвирус может представлять собой любой тип поксвируса. В некоторых вариантах осуществления изобретения поксвирус представляет собой ортопоксвирус или авипоксвирус. В предпочтительных вариантах осуществления изобретения ортопоксвирус выбран из вируса осповакцины, вируса MVA, MVA-BN и производных MVA-BN. В других более предпочтительных вариантах осуществления изобретения ортопоксвирус представляет собой MVA, MVA-BN или производные MVA-BN. В других предпочтительных вариантах осуществления изобретения авипоксвирус представляет собой вирус оспы птиц.
[019] В других вариантах осуществления изобретения в дополнение к нуклеиновым кислотам MUC1 и/или CEA, описанным в настоящем описании, рекомбинантные поксвирусы согласно данному изобретению содержат одну или более нуклеиновых кислот, кодирующих TRICOM (триада костимулирующих молекул).
[020] В некоторых вариантах осуществления изобретения рекомбинантные поксвирусы и/или нуклеиновые кислоты согласно настоящему изобретению могут использоваться в гетерологичном режиме дозирования с прайм-бустом. В предпочтительных вариантах осуществления изобретения режим включает: а) одну или более первичных (priming) доз вируса MVA, причем вирус MVA содержит одну или более нуклеиновых кислот MUC1, CEA и/или TRICOM согласно настоящему изобретению; и b) одну или более стимулирующих доз вируса оспы птиц, содержащего одну или более нуклеиновых кислот MUC1, CEA и/или TRICOM согласно настоящему изобретению.
[021] Предполагается, что рекомбинантные поксвирусы, нуклеиновые кислоты, способы, вакцины и композиции, описанные в настоящем описании, могут быть реализованы в наборе. Соответственно, в предпочтительном варианте осуществления изобретения, настоящее изобретение относится к композиции, вакцине, набору или их применению, содержащим: рекомбинантный ортопоксвирус, такой как, но не ограничиваясь этим, MVA, рекомбинантный ортопоксвирус, содержащий одну или более из нуклеиновых кислот MUC1, CEA и/или TRICOM согласно настоящему изобретению; и рекомбинантный авипоксвирус, такой как, но не ограничиваясь этим, вирус оспы птиц, содержащий одну или более из нуклеиновых кислот MUC1, CEA и/или TRICOM согласно данному раскрытию.
[022] В других вариантах осуществления изобретения настоящее изобретение относится к одному или более способам получения рекомбинантного поксвируса, кодирующего один или более трансгенов согласно настоящему изобретению, который стабилен при последовательном пассировании рекомбинантного поксвируса.
[023] В одном варианте осуществления изобретения представлен способ получения рекомбинантного поксвируса, имеющего трансген MUC1, который стабилен при последовательном пассировании рекомбинантного поксвируса, причем способ включает: а) получение любой одной из нуклеиновых кислот или кассет экспрессии согласно настоящему изобретению; и b) вставку нуклеиновой кислоты или кассеты экспрессии в рекомбинантный поксвирус.
[024] В другом варианте осуществления изобретение относится к способу получения рекомбинантного поксвируса, который стабилен при последовательном пассировании, включающему: a) получение первой последовательности нуклеиновой кислоты, кодирующей пептид MUC1, имеющий по меньшей мере два домена вариабельных N-концевых повторов (VNTR), причем расположение по меньшей мере двух доменов VNTR перетасовано, и по меньшей мере два домена VNTR являются кодон-оптимизированными; и b) получение второй нуклеиновой кислоты, кодирующей пептид CEA, причем вторая нуклеиновая кислота содержит по меньшей мере одну нуклеотидную замену по меньшей мере в одной повторяющейся нуклеотидной области второй нуклеиновой кислоты, причем по меньшей мере одна повторяющаяся нуклеотидная область определена как а) три дополнительных последовательно повторяющихся нуклеотида G или C и/или b) три или более последовательно повторяющихся нуклеотида T; причем рекомбинантный поксвирус является стабильным при последовательном пассировании.
[025] Дополнительные цели и преимущества изобретения будут частично изложены в нижеследующем описании и частично будут очевидны из описания или могут быть изучены при практическом использовании изобретения. Цели и преимущества изобретения будут реализованы и достигнуты посредством элементов и комбинаций, конкретно указанных в прилагаемой формуле изобретения.
[026] Прилагаемые графические материалы, которые включены в данное описание и составляют его часть, иллюстрируют один или более вариантов осуществления изобретения и вместе с описанием служат для пояснения принципов изобретения.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[027] На Фиг. 1 представлен анализ ПЦР, который иллюстрирует нестабильность трансгенов PANVAC (MUC1 и CEA). Показаны результаты ампликона ПЦР используемого сайта для интеграции MUC1 и СЕА в геном TBC-FPV (IGR61/62). Отмечено положение ожидаемого фрагмента ПЦР и потенциального фрагмента дикого типа. Несколько делеционных фрагментов меньшего размера могут быть обнаружены и их может быть больше во время повторного пассирования либо при температуре 34†C, либо при температуре 37°C. Результаты показаны для пассажей PANVA-F от 0 до 7.
[028] На Фиг. 2А и 2В изображены аминокислотные последовательности MUC1 и перетасовка повторов домена VNTR в соответствии с различными вариантами осуществления изобретения. А) Аминокислотная последовательность MUC1, обнаруженная в PANVAC (SEQ ID NO:6). Проиллюстрированы 6 VNTR, обнаруженных в PANVAC MUC1. B) Аминокислотная последовательность MUC1, обнаруженная в mBN336, mBN373 и mBN420 (SEQ ID NO:30). Проиллюстрированы 3 VNTR, обнаруженные в mBN336, mBN373 и mBN420 MUC1. Подчеркнутые аминокислоты представляют собой аминокислоты, модифицированные с образованием агонистических эпитопов согласно WO 2013/103658.
[029] На Фиг. 3А-3С изображены парные выравнивания и типовая кодон-оптимизация домена VNTR MUC1 в соответствии с различными вариантами осуществления изобретения. A) Выравнивание VNTR PANVAC №2 (SEQ ID NO:7) и VNTR mBN336, mBN373, mBN420 №1 (SEQ ID NO:8), B) Выравнивание VNTR PANVAC №1 (SEQ ID NO:9) и VNTR mBN336, mBN373, mBN420 №2 (SEQ ID NO:10), C) Выравнивание VNTR PANVAC №3 (SEQ ID NO:11) и VNTR mBN336, mBN373, mBN420 №3 (SEQ ID NO:12). Подчеркнутые нуклеотиды представляют нуклеотидные области, модифицированные с образованием агонистических эпитопов согласно WO 2013/103658.
[030] На Фиг. 4А-4С изображены попарные выравнивания кодирующих последовательностей MUC1 относительно PANVAC, используемых в конструкциях на основе рекомбинантного поксвируса в соответствии с настоящим изобретением. A) PANVAC MUC1 (SEQ ID NO:1) относительно mBN336 MUC1 (SEQ ID NO:2); B) PANVAC MUC1 (SEQ ID NO:1) относительно mBN373 MUC1 (SEQ ID NO:3); C) PANVAC MUC1 (SEQ ID NO:1) относительно mBN420 MUC1 (SEQ ID NO:5). Типовые повторяющиеся области, содержащие одну или более замен, подчеркнуты.
[031] На Фиг. 5 изображено попарное выравнивание кодирующей последовательности СЕА mBN373 и mBN420 (SEQ ID NO:14) относительно СЕА PANVAC (SEQ ID NO:13), используемой в конструкциях на основе рекомбинантного поксвируса в соответствии с настоящим изобретением. Типовые повторяющиеся области, содержащие одну или более замен, подчеркнуты.
[032] На Фиг. 6 изображено попарное выравнивание кодирующей последовательности B7-1 mBN373 и mBN420 (SEQ ID NO:15) относительно B7-1 PANVAC (SEQ ID NO:16), относительно PANVAC, используемой в рекомбинантной конструкции на основе поксвируса в соответствии с настоящим изобретением. Типовые повторяющиеся области отмечены указанными заменами (области выравнивания, не отмеченные *).
[033] На Фиг. 7 изображено попарное выравнивание кодирующей последовательности ICAM-1 mBN373 и mBN420 (SEQ ID NO:18) относительно PANVAC (SEQ ID NO:19), относительно PANVAC, используемой в рекомбинантной конструкции на основе поксвируса в соответствии с настоящим изобретением. Типовые повторяющиеся области отмечены указанными заменами (области выравнивания, не отмеченные *).
[034] На Фиг. 8 изображено попарное выравнивание кодирующей последовательности LFA-3 mBN373 и mBN420 (SEQ ID NO:21) относительно PANVAC (SEQ ID NO:22), относительно PANVAC, используемой в рекомбинантной конструкции на основе поксвируса в соответствии с настоящим изобретением. Типовые повторяющиеся области отмечены указанными заменами (области выравнивания, не отмеченные *).
[035] На Фиг. 9А и 9В изображены эксперименты по исследованию стабильности трансгена MUC1 в mBN336. А) Результаты ПЦР при исследовании стабильности СЕА в течение семи пассажей, характерные для пассажей во время и после производства материалов клинического исследования (СТМ)/GMP. B) Результаты ПЦР при исследовании стабильности MUC1 в течение семи пассажей, характерные для пассажей во время и после производства материалов СТМ/GMP. C) Результаты ПЦР при исследовании стабильности TRICOM в течение 7 пассажей, характерные для пассажей во время и после производства материалов СТМ/GMP. Рекомбинационные плазмиды, использованные для получения MVA-mBN336B были использованы в качестве положительных контролей, MVA-BN® была использована в качестве отрицательного контроля (пустой векторный остов), а в качестве контроля для реакции ПЦР была использована H2O.
[036] На Фиг. 10А и 10В изображен анализ пассажей mBN336 5, 6 и 7. А) ПЦР-амплификация образцов пассажа 7, отправленных для анализа посредством секвенирования. Отдельные амплификации ПЦР проводили для каждого отдельного трансгена: CEA, MUC1 и TRICOM. B) Электроферограммы нуклеотидной последовательности MUC1, на которых изображены локусы, содержащие обнаруженную точечную мутацию, приводящую к сдвигу рамки считывания, возникшему в пассаже 5.
[037] На Фиг. 11A и 11B изображены эксперименты по исследованию стабильности трансгена MUC1 в mBN373. А) Анализ ПЦР вставленных трансгенов для каждого пассажа. Рекомбинационная плазмида, использованная для получения FPV-mBN373B, использовалась в качестве положительного контроля, FPV (штамм TBC-FPV) использовалась в качестве отрицательного контроля. B) ПЦР-анализ FPV-mBN373B при седьмом пассаже привел к получению полос ожидаемого размера 5566 п.н. (PCR1) и 5264 п.н. (PCR2), содержащих вставленные трансгены и каждую вставленную фланкирующую область. Анализ последовательности подтвердил генетическую стабильность рекомбинанта после 7 пассажей, являясь репрезентативным для пассажей во время и после производства материала CTM/GMP.
[038] На Фиг. 12 изображен анализ ПЦР, в котором проанализирована стабильность трансгенов MUC1, CEA и TRICOM в mBN420. Показан результат для ампликона ПЦР используемого сайта для интеграции всех пяти трансгенов в геном MVA-BN (IGR88/89). Отмечена высота ожидаемого фрагмента ПЦР и потенциального фрагмента дикого типа. Несколько делеционных фрагментов меньшего размера могут быть обнаружены и их может быть больше во время повторного пассирования при температуре 30°C. Результаты показаны для пассажей mBN420 от 0 до 7.
[039] Все попарные выравнивания, изображенные на графических материалах, проводились с использованием инструмента выравнивания последовательностей Clustal Omega, доступного по адресу http://www.ebi.ac.uk/Tools/msa/clustalo/.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[040] Следует понимать, что как предшествующее Краткое Описание Сущности Изобретения, так и последующее Подробное Описание Изобретения являются лишь типовыми и пояснительными и не ограничивают объем изобретения, приведенный в Формуле изобретения.
[041] В PANVAC используется гетерологичная стратегия с прайм-бустом с использованием рекомбинантных поксвирусов осповакцины (PANVAC-V) и оспы птиц (PANVAC-F), каждый из которых экспрессирует трансгены MUC1, CEA и TRICOM. Было показано, что PANVAC эффективен при лечении рака и в настоящее время проходит клинические испытания при различных видах рака, включая колоректальный рак, рак яичников, рак молочной железы и рак мочевого пузыря. MVA-CV301 представляет собой еще одну гетерологичную вакцинную комбинацию, проходящую клинические испытания (см., например, Gulley et al., Clin Cancer Res 2008; vol. 14:10, Tsang et al. Clin Cancer Res 2005; vol. 11). В MVA-CV301 используется гетерологичная стратегия с прайм-бустом с использованием MVA и оспы птиц, каждый из которых экспрессирует трансгены MUC1, CEA и TRICOM.
[042] Хотя PANVAC и MVA-CV301 эффективны при лечении рака, трансгены рекомбинантных поксвирусов PANVAC становятся менее стабильными при последовательном пассировании и продуцировании вирусов. Как указано в Таблицах 1 и 2, после последовательного пассирования PANVAC-V и PANVAC-F процент вирусов, экспрессирующих MUC1 и CEA, неуклонно снижается.
Таблица 1 Процент экспрессирующих колоний с использованием PANVAC-V MVB1, MVB2 и пассажи
*Каждое число представляет средние значения, полученные в результате трех независимых экспериментов пассирования.
Таблица 2 Процент экспрессирующих колоний с использованием PANVAC-F MVB1, MVB2 и пассажи
*Каждое число представляет средние значения, полученные в результате трех независимых экспериментов пассирования.
[043] По меньшей мере в одном аспекте снижение экспрессии MUC1 и/или CEA, по-видимому, является результатом по меньшей мере частичной потери трансгенов MUC1 и/или CEA. На Фигура 1 изображена потеря трансгенной последовательности MUC и СЕА PANVAC. На Фиг. 1, ожидаемая длина рекомбинантного продукта PANVAC-F составляет 4445 п.н. Однако, как показано, эксперименты продемонстрировали наличие нескольких фрагментов меньшего размера, которые, как было подтверждено, представляют собой фрагментированные последовательности MUC1 и CEA (данные не показаны). Потеря экспрессии и нестабильность трансгена MUC1 и предыдущих рекомбинантных поксвирусов препятствует получению и чистоте рекомбинантных поксвирусов CV301.
[044] Перед созданием различных нуклеиновых кислот и рекомбинантных поксвирусов согласно настоящему изобретению, чтобы стабилизировать трансгены, авторы изобретения предприняли многочисленные попытки адаптировать и/или модифицировать рекомбинантную осповакцину, рекомбинантный MVA и рекомбинантные вирусы оспы птиц PANVAC и MVA-CV301. В Таблицах 3 и 4 указаны модификации трансгенов и/или рекомбинантных вирусов осповакцины, рекомбинантных вирусов MVA и рекомбинантных вирусов оспы птиц: (i) изменение или модификация межгенных областей (IGR), в которые были встроены трансгены, (ii) кодон-оптимизация одного или более трансгенов, (iii) изменение промоторов трансгена и (iv) изменение количества и расположения областей VNTR в трансгене MUC1. Как описано в таблицах, многие конструкции не удалось стабильно получать из-за мутаций с потерей функции или делеций фрагментов, приводящих к потере экспрессии трансгена.
ТАБЛИЦА 3:
Промоторы и ТГ в точности как в PANVAC-V
TRICOM без изменений в IGR148/149
TRICOM без изменений в IGR148/149
Стабильная экспрессия TG в течение 7 пассажей при температуре 30°C и 37°C (FACS с помощью BN-CVD)
CEA (как в PANVAC-V) в IGR44/45
TRICOM (как в PANVAC-V) в IGR148/149
CEA (как в PANVAC-V) в IGR44/45
TRICOM (как в PANVAC-V) в IGR148/149
ТАБЛИЦА 4:
Промоторы и ТГ в точности как в PANVAC-F
с промотором FPV-40K для MUC1-C13-opt3VNTR
(PrS)-MUC1-C13-opt3VNTR в IGR61/62 и TRICOM в BamJ
[045] После этих многочисленных попыток была создана MVA-mBN336. Как описано в настоящем описании, MVA-mBN336 представляет собой рекомбинантный поксвирус MVA-CV301, содержащий модифицированный MUC1, CEA и модифицированные трансгены TRICOM. Как изображено на Фиг. 9 и 10, MVA-mBN336 продемонстрировал стабильность трансгена по сравнению с PANVAC (см. Фигуру 1 и Таблицу 1). Как изображено на Фиг. 10, MVA-mBN336 продемонстрировал стабильность всех трансгенов (MUC1, CEA и TRICOM) при пассаже 4. Начиная с пассажа 5, мутация со сдвигом рамки считывания была обнаружена в небольшой популяции анализируемого материала. Стабильность, проиллюстрированная при пассаже 4, демонстрирует способность MVA-mBN336 преодолевать проблемы со стабильностью, связанные с PANVAC, и другими попытками получить стабильный поксвирус, содержащий MUC1. Стабильность MVA-mBN336 обеспечивает дополнительное преимущество, так как производство и более крупномасштабное производство вакцин на основе MVA обычно происходит с использованием MVA при пассаже 3 или пассаже 4. Таким образом, поскольку MVA-mBN336 является стабильным при пассаже 4, можно начать крупномасштабное производство и благодаря стабильности могут быть преодолены значительные нормативные препятствия.
[046] Для решения и исправления проблем нестабильности были синтезированы нуклеиновые кислоты согласно настоящему изобретению, которые содержат одну или более нуклеиновых кислот, которые кодируют трансген MUC1, трансаген CEA и трансгены TRICOM. Как указано в настоящем описании, нуклеиновые кислоты MUC1, CEA и TRICOM согласно настоящему изобретению приводят к увеличенной генетической стабильности рекомбинантного поксвируса и включенных в него трансгенов при последовательной пассировании рекомбинантных поксвирусов.
[047] Таким образом, в различных вариантах осуществления настоящего изобретения, настоящее изобретение относится к рекомбинантному поксвирусу, имеющему одну или более новых нуклеиновых кислот, которые кодируют антигены MUC1, CEA и/или TRICOM. Как описано более подробно в настоящем описании, по меньшей мере в одном аспекте, при включении в виде части рекомбинантного поксвируса, одна или более модифицированных последовательностей нуклеиновых кислот, кодирующих MUC1, CEA и/или TRICOM, увеличивают стабильность и присутствие трансгенов в рекомбинантном поксвирусе.
Определения
[048] При использовании в контексте настоящего изобретения, значения существительных в единственном числе включают значения существительных во множественном числе, если в контексте явно не указано иначе. Таким образом, например, ссылка на «нуклеиновую кислоту» включает одну или более нуклеиновых кислот, а ссылка на «способ» включает ссылку на эквивалентные этапы и способы, известные специалистам в данной области техники, с помощью которых могут быть модифицированы или заменены способы, описанные в настоящем описании.
[049] Если не указано иное, термин «по меньшей мере», предшествующий серии элементов, следует понимать как относящийся к каждому элементу в серии. Специалисты в данной области техники распознают или смогут определить, используя не более чем обычные эксперименты, множество эквивалентов конкретных вариантов осуществления изобретения, описанных в настоящем описании. Такие эквиваленты должны рассматриваться как входящие в объем настоящего изобретения.
[050] В настоящем описании и в следующей формуле изобретения, если контекст не требует иного, будет понятно, что слово «содержат» и такие варианты, как «содержит» и «содержащий», подразумевают включение указанного целого числа или этапа или группы целых чисел или этапов, но не исключение любого другого целого числа или этапа или группы целых чисел или этапов. При использовании в контексте данного документа термин «содержащий» может быть заменен термином «охватывающий» или «включающий», или иногда, при использовании в настоящем описании, термином «имеющий». Любой из вышеупомянутых терминов (включающий, охватывающий, содержащий, имеющий), хотя и менее предпочтительно, всякий раз, когда они используются в настоящем описании в контексте аспекта или варианта осуществления настоящего изобретения, может быть заменен термином «состоящий из». При использовании в контексте настоящего изобретения «состоящий из» исключает любой элемент, этап или компонент, не указанный в пункте формулы изобретения. При использовании в контексте настоящего изобретения «по существу состоящий из» не исключает материалы или стадии, которые не оказывают существенного влияния на основные и новые характеристики пунктов формулы изобретения.
[051] Используемый в контексте настоящего изобретения термин «и/или» между несколькими перечисленными элементами понимается как охватывающий как отдельные, так и комбинированные варианты. Например, когда два элемента объединены термином «и/или», первый вариант относится к применимости первого элемента без второго. Второй вариант относится к применимости второго элемента без первого. Третий вариант относится к применимости первого и второго элементов вместе. Предполагается, что любой из этих вариантов соответствует значению, и следовательно, удовлетворяет требованию термина «и/или», используемого в контексте настоящего изобретения. Понятно, что одновременная применимость более чем одного из вариантов соответствует значению, и следовательно, удовлетворяет требованию термина «и/или».
[052] «Мутация», описанная в настоящем описании, как определено в настоящем описании, относится к любой модификации нуклеиновой кислоты, такой как делеции, добавления, вставки и/или замены.
[053] «Костимулирующие молекулы» при использовании в контексте настоящего изобретения представляют собой молекулы, которые при связывании со своим лигандом передают второй сигнал, в результате чего Т-клетка может стать активированной. Наиболее известной костимулирующей молекулой в Т-клетке является CD28, которая связывается либо с B7-1 (также известным B7.1 или CD80), либо с B7-2 (также известным как CD86). Дополнительной костимулирующей молекулой является В7-3. Вспомогательные молекулы, которые также обеспечивают второй сигнал для активации Т-клеток, включают молекулу внутриклеточной адгезии (ICAM-1 и ICAM-2), антиген, связанный с функцией лейкоцитов (LFA-1, LFA-2 и LFA-3). Представители суперсемейства интегринов и фактора некроза опухолей (TNF) также могут выступать в качестве костимулирующих молекул.
[054] Термины «Генетическая стабильность», «стабильность», «стабильность экспрессии», «стабильность при последовательном пассировании», «стабилен при последовательном пассировании» или «стабильность экспрессии при последовательном пассировании» в отношении рекомбинантных поксвирусов при использовании в настоящем описании в сочетании с рекомбинантным поксвирусом, MUC1, CEA, TRICOM и другими трансгенами означают, что трансгенные нуклеотидные последовательности рекомбинантного поксвируса остаются материально интактными и/или материально неизменными при последовательном пассировании рекомбинантного поксвируса по меньшей мере при пассаже 3 или пассаже 4. Рекомбинантный поксвирус, обладающий стабильностью, по меньшей мере, при пассаже 3 или пассаже 4, особенно важен, поскольку конечный продукт, получаемый в результате крупномасштабного изготовления и производства поксвирусов, обычно представляет собой пассаж 3 или пассаж 4. Термин «материально интактный и/или материально неизменный» означает отсутствие единичных или фрагментных мутаций (например, замен, делеций и т. д.), которые вызывают постоянное снижение экспрессии трансгена по мере увеличения числа пассажей. Например, как показано в Таблицах 1 и 2, уровни экспрессии различных трансгенов PANVAC снижались по мере увеличения числа пассажей. В данной области техники известно множество способов, с помощью которых можно анализировать генетическую стабильность или стабильность трансгенов, включая, но не ограничиваясь этим, исследования, описанные в Примерах 2-4 данной заявки. Дополнительные известные в данной области техники способы измерения стабильности включают, но не ограничиваются этим, ПЦР, FACS, измерение коэкспрессии трансгена с помощью FACS и так далее.
[055] При использовании в контексте настоящего изобретения «клетка-хозяин» представляет собой клетку, в которую была введена чужеродная молекула, вирус или микроорганизм с целью развития и/или продуцирования чужеродной молекулы, вируса или микроорганизма. В одном неограничивающем примере, как описано в настоящем описании, клетка подходящей клеточной культуры, такой как, например, клетки CEF, может быть инфицирована поксвирусом или, в других альтернативных вариантах осуществления изобретения, плазмидным вектором, содержащим чужеродный или гетерологичный ген. Таким образом, подходящие клеточные культуры служат в качестве хозяина для поксвируса и/или чужеродного или гетерологичного гена.
[056] «Процент (%) гомологии или идентичности последовательности» по отношению к последовательностям нуклеиновой кислоты, описанным в настоящем описании, определяется как процент нуклеотидов в последовательности-кандидате, которые идентичны нуклеотидам в референтной последовательности (то есть последовательности нуклеиновой кислоты, от которой она является производной), после выравнивания последовательностей и внесения пропусков, если необходимо, для достижения максимальной процентной идентичности последовательности, и без учета каких-либо консервативных замен в качестве идентичности последовательности. Выравнивание в целях определения процента идентичности или гомологии нуклеотидной последовательности может быть достигнуто различными способами, которые известны специалисту в данной области техники, например, с использованием общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить подходящие параметры для измерения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей.
[057] Например, подходящее выравнивание для последовательностей нуклеиновых кислот обеспечивается алгоритмом локальной гомологии, как указано в Smith and Waterman, (1981), Advances in Applied Mathematics 2:482-489. Этот алгоритм может быть применен к аминокислотным последовательностям с использованием оценочной матрицы, разработанной Dayhoff, Atlas of Protein Sequences and Structure, M. O. Dayhoff ed., 5 suppl. 3:353-358, National Biomedical Research Foundation, Washington, D.C., USA, и нормализованой Gribskov (1986), Nucl. Acids Res. 14(6):6745-6763. Типовая реализация данного алгоритма для определения процента идентичности последовательности предоставляется Genetics Computer Group (Madison, WI) в приложении «BestFit». Параметры по умолчанию для этого способа описаны в Wisconsin Sequence Analysis Package Program Manual, Version 8 (1995) (доступно предоставляется Genetics Computer Group, Мадисон, Висконсин). Предпочтительным способом определения процентной идентичности в контексте настоящего изобретения является использование пакета программ MPSRCH, защищенных авторским правом Эдинбургского Университета (University of Edinburgh), разработанного авторами John F. Collins и Shane S. Sturrok и распространяемых IntelliGenetics, Inc. (Маунтин-Вью, Калифорния). Из этого набора пакетов может быть использован алгоритм Smith-Waterman, в котором для оценочной таблицы используются параметры по умолчанию (например, штраф за пропуск, вносящий разрыв, составляет 12 баллов, штраф за пропуск, продолжающий разрыв, составляет 1 балл, и штраф за пропуск составляет шесть баллов). В полученных данных «Соответствие» соответствует «идентичности последовательности». Другие подходящие программы для вычисления процента идентичности или сходства между последовательностями обычно известны в данной области техники, например, другой программой для проведения выравнивания является BLAST, используемой с параметрами по умолчанию. Например, BLASTN и BLASTP могут использоваться с использованием следующих параметров по умолчанию: генетический код=стандарт; фильтр=нет; цепь=обе; порог=60; E-значение=10; Матрица=BLOSUM62; Описания=50 последовательностей; сортировать по=HIGH SCORE; Базы данных=не избыточные (non-redundant), GenBank+EMBL+DDBJ+PDB+CDS translations+Swiss protein+Spupdate+PIR. Подробную информацию об этих программах можно найти по следующему интернет-адресу: http:// http://blast.ncbi.nlm.nih.gov/.
[058] Термин «вакцинация с прайм-бустом» или «режим с прайм-бустом» относится к стратегии или режиму вакцинации с использованием первой активирующей инъекции вакцины, нацеленной на определенный антиген, после чего через определенные промежутки времени проводится одна или более бустерных инъекций той же самой вакцины. Вакцинация с прайм-бустом может быть гомологичной или гетерологичной. В гомологичной вакцинации с прайм-бустом используется вакцина, содержащая один и тот же антиген и вектор как для активирующей инъекции, так и для одной или более бустерных инъекций. В гетерологичной вакцинации с прайм-бустом используется вакцина, содержащая один и тот же антиген как для активирующей инъекции, так и для одной или более бустерных инъекций, но разные векторы для активирующей инъекции и одной или более бустерных инъекций. Например, при гомологичной вакцинации с прайм-бустом может использовать рекомбинантный поксвирус, содержащий нуклеиновые кислоты, экспрессирующие один или более антигенов для первичной инъекции, и тот же рекомбинантный поксвирус, экспрессирующий один или более антигенов для одной или более бустерных инъекций. Напротив, при гетерологичной вакцинации с прайм-бустом может использовать рекомбинантный поксвирус, содержащий нуклеиновые кислоты, экспрессирующие один или более антигенов для первичной инъекции, и другой рекомбинантный поксвирус, экспрессирующий один или более антигенов для одной или более бустерных инъекций.
[059] Термин «рекомбинантный» означает полинуклеотид полусинтетического или синтетического происхождения, который либо не встречается в природе, либо связан с другим полинуклеотидом в расположении, не встречающемся в природе.
[060] «Последовательное пассирование» при использовании в контексте настоящего изобретения относится к получению рекомбинантных вирусов при использования клеточного пассирования. Только в качестве примера, клетки хозяина инфицируются вирусом или рекомбинантным вирусом при начальном пассаже. Вирусы реплицируются и производятся при начальном пассаже. После заражения и культивирования клеток-хозяев вирусы выделяют из клеток-хозяев и собирают в клеточной/вирусной суспензии. Эту процедуру обычно повторяют несколько раз в последующих клеточных пассажах, причем каждый пассаж продуцирует и реплицирует больше рекомбинантных вирусов.
[061] При использовании в контексте настоящего изобретения термин «трансген» или «гетерологичный» ген понимается как последовательность нуклеиновой кислоты или аминокислоты, которой нет в геноме поксвируса дикого типа (например, в вирусе осповакцины, оспы птиц или MVA). Специалист понимает, что «трансген» или «гетерологичный ген», когда он присутствует в поксвирусе, таком как вирус осповакцины, должен быть включен в поксвирусный геном таким образом, что после введения рекомбинантного поксвируса в клетку-хозяин он экспрессируется в виде соответствующего гетерологичного продукта гена, то есть как «гетерологичный антиген» и/или «гетерологичный белок». Экспрессия обычно достигается путем функционального объединения гетерологичного гена с регуляторными элементами, которые обеспечивают экспрессию в зараженной поксвирусом клетке. Предпочтительно регуляторные элементы содержат природный или синтетический поксвирусный промотор.
[062] «TRICOM» Триада костимулирующих молекул (также известная как TRICOM) включает B7-1 (также известный как B7.1 или CD80), молекулу внутриклеточной адгезии-1 (ICAM-1, также известную как CD54) и антиген-3, связанный с функцией лимфоцитов (LFA-3, также известный как CD58) и обычно вводится в рекомбинантные вирусные векторы (например, поксвирусные векторы), экспрессирующие специфический антиген для усиления антигенспецифического иммунного ответа. Отдельные компоненты TRICOM могут находиться под контролем одинаковых или разных промоторов и могут находиться в одном и том же векторе со специфическим антигеном или в отдельном векторе. Типовые векторы раскрыты, например, в Hodge et al. “A Triad of Costimulatory Molecules Synergize to Amplify T-Cell Activation,” Cancer Res. 59:5800-5807 (1999) и патенте США № 7,211,432 В2, оба из которых включены в настоящее описание посредством ссылки.
[063] «Вектор» относится к ДНК или РНК-содержащему вирусу или плазмиде, которые могут содержать гетерологичный полинуклеотид. Гетерологичный полинуклеотид может содержать представляющую интерес последовательность для целей профилактики или терапии и необязательно может быть в форме кассеты экспрессии. При использовании в контексте настоящего изобретения вектор не должен быть способен к репликации в конечной клетке-мишени или субъекте. Термин включает векторы для клонирования и вирусные векторы.
Новые последовательности нуклеиновых кислот MUC1
[064] При разработке настоящего изобретения авторы изобретения определили, что в ходе пассирования рекомбинантных поксвирусов, в частности ортопоксвирусов (например, вируса осповакцины, MVA, MVA-BN) и авипоксвирусов (например, вируса оспы птиц), один или более участков нуклеиновых кислот, кодирующих MUC1, CEA и/или TRICOM, стали мутированными (например, содержали делеции, замены и т. д.), способствуя тем самым нестабильности рекомбинантного поксвируса и трансгенов в нем.
Модификации областей VNTR
[065] Как отмечалось ранее, область VNTR представляет собой внеклеточный домен MUC1, который содержит домен с переменным числом тандемных повторов из 20 аминокислот (VNTR), причем количество повторов варьирует от 20 до 120 у разных индивидов. Смотри, Brayman et al. Хотя аминокислотная последовательность доменов VNTR обычно идентична (см., например, Фигуру 2A), нуклеотидная последовательность VNTR может варьировать. Как показано на Фиг. 2А, как часть PANVAC, MUC1 был синтезирован, чтобы иметь 6 VNTR.
[066] В одном аспекте в ходе разработки настоящего изобретения было определено, что одна или более модификаций нуклеиновых кислот, кодирующих область VNTR MUC1, улучшали стабильность трансгена MUC1 в рекомбинантном поксвирусе. Более конкретно, авторы изобретения определили, что перетасовка нуклеиновых кислот, кодирующих VNTR, дополнительно повышает стабильность трансгена MUC1 по сравнению с MUC1 PANVAC. При использовании в контексте настоящего изобретения термин «перетасовка» VNTR означает изменение порядка нуклеиновых кислот, кодирующих повторы домена VNTR. На Фиг. 2А и 2В проиллюстрирован неограничивающий пример перетасовки VNTR. На Фиг. 2А порядок доменов VNTR PANVAC отмечен как VNTR №1-6. Как изображено на Фиг. 2B, нуклеиновая кислота, кодирующая VNTR№1 из mBN336, mBN373, mBN420, соответствует той, которая представляет собой VNTR№ в PANVAC. VNTR №2 из mBN336, mBN373, mBN420 соответствует той, которая представляет собой VNTR №1 в PANVAC. Таким образом, при синтезе MUC1 mBN336, mBN373, mBN420 порядок/положение VNTR №1 и 2 PANVAC был перетасован.
[067] Согласно настоящему изобретению понятно, что домены VNTR, изображенные на Фиг. 2A и 2B, являются просто репрезентативными для доменов VNTR MUC1, и что количество доменов VNTR и схемы, в которых перетасованы VNTR, могут варьировать.
[068] В дополнение к перетасовке VNTR было установлено, что кодон-оптимизация кодонов доменов VNTR дополнительно повышает стабильность трансгена MUC1. При использовании в контексте настоящего изобретения термин «кодон-оптимизация» означает замену одного или более нуклеотидов VNTR с целью минимизации вероятности мутаций и/или делеций нуклеотидных последовательностей VNTR из-за гомологии повторяющихся нуклеотидных последовательностей.
[069] В более конкретном варианте осуществления изобретения, изображенном на Фиг. 3А-3В, одна или более нуклеиновых кислот согласно настоящему изобретению содержат различные замены в доменах VNTR. Как изображено на Фиг. 3А, VNTR1 mBN373 и mBN420 (далее mBN373/420) содержат одну или более нуклеотидных последовательностей, кодирующих эпитоп агониста согласно WO 2013/103658 (область, отмеченная подчеркиванием), в дополнение к проиллюстрированным заменам кодон-оптимизации. Как изображено на Фиг. 3В и 3С, VNTR 2 и 3 модификации mBN373/420 содержат проиллюстрированные замены кодон-оптимизации.
[070] Согласно настоящему изобретению понятно, что указанные модификации кодон-оптимизации для доменов VNTR, изображенные на Фиг. 3A-3C, представляют лишь кодон-оптимизацию домена VNTR MUC1. Только в качестве примера, согласно данному изобретения предполагается, что альтернативные нуклеотиды могут быть замещены в определенных точках модификации в доменах VNTR. Дополнительно предполагается, что конкретные точки модификации в доменах VNTR могут варьировать, таким образом что модификация является молчащей модификацией. Под молчащей модификацией в контексте настоящего изобретения подразумевается, что эта модификация не влияет на аминокислотную последовательность антигена MUC1.
[071] Таким образом, в одном варианте осуществления изобретения нуклеиновые кислоты MUC1 согласно настоящему изобретению содержат одну или более областей домена VNTR, которые 1) перетасованы и 2) являются кодон-оптимизированными.
Модификации областей MUC1, не являющихся VNTR
[072] В другом аспекте настоящего изобретения были внесены еще одни модификации в эти области за пределами доменов VNTR. В более конкретном аспекте в ходе разработки настоящего изобретения было определено, что одна или более модификаций в этих нуклеотидных областях за пределами области VNTR MUC1 (области, не являющиеся VNTR) улучшали стабильность трансгена MUC1. Типовой образец этих областей (подчеркнутые нуклеотиды) изображен ниже. Область VNTR отмечена серым фоном.
ATGACACCGG GCACCCAGTC TCCTTTCTTC CTGCTGCTGC TCCTCACAGT GCTTACAGTT 60
GTTACGGGTT CTGGTCATGC AAGCTCTACC CCAGGTGGAG AAAAGGAGAC TTCGGCTACC 120
CAGAGAAGTT CAGTGCCCAG CTCTACTGAG AAGAATGCTG TGAGTATGAC AAGCTCCGTA 180
CTCTCCAGCC ACAGCCCCGG TTCAGGCTCC TCCACCACTC AGGGACAGGA TGTCACTCTG 240
GCCCCGGCCA CGGAACCAGC TTCAGGTTCA GCTGCCTTGT GGGGACAGGA TGTCACCTCG 300
GTACCAGTTA CTAGACCAGC TTTAGGTAGC ACAGCACCTC CTGCTCATGG AGTAACTAGT 360
GCTCCTGATA CTCGTCCAGC TCCTGGCAGT ACTGCACCAC CGGCACATGG CGTAACATCA 420
GCACCTGATA CAAGACCTGC ACCTGGATCT ACAGCGCCGC CTGCGCACGG AGTGACATCG 480
GCGCCCGATA CGCGCCCCGC TCCCGGTAGC ACCGCACCGC CCGCCCACGG TGTTACAAGT 540
GCACCCGATA CCCGGCCGGC ACCCGGAAGT ACCGCTCCAC CTGCACACGG GGTCACAAGC 600
GCGCCAGACA CTCGACCTGC GCCAGGGTCG ACTGCCCCTC CGGCGCATGG TGTGACCTCA 660
GCTCCTGACA CAAGGCCAGC CCCAGCTAGC ACTCTGGTGC ACAACGGCAC CTCTGCCAGG 720
GCTACCACAA CCCCAGCCAG CAAGAGCACT CCATTCTCAA TTCCCAGCCA CCACTCTGAT 780
ACTCCTACCA CCCTTGCCAG CCATAGCACC AAGACTGATG CCAGTAGCAC TCACCATAGC 840
ACGGTACCTC CTCTCACCTC CTCCAATCAC AGCACTTCTC CCCAGTTGTC TACTGGGGTC 900
TCTTTCTTTT TCCTGTCTTT TCACATTTCA AACCTCCAGT TTAATTCCTC TCTGGAAGAT 960
CCCAGCACCG ACTACTACCA AGAGCTGCAG AGAGACATTT CTGAAATGTT TTTGCAGATT 1020
TATAAACAAG GGGGTTTTCT GGGCCTCTCC AATATTAAGT TCAGGCCAGG ATCTGTGGTG 1080
GTACAATTGA CTCTGGCCTT CCGAGAAGGT ACCATCAATG TCCACGACGT GGAGACACAG 1140
TTCAATCAGT ATAAAACGGA AGCAGCCTCT CGATATAACC TGACGATCTC AGACGTCAGC 1200
GTGAGTGATG TGCCATTTCC TTTCTCTGCC CAGTCTGGGG CTGGGGTGCC AGGCTGGGGC 1260
ATCGCGCTGC TGGTGCTGGT CTGTGTTCTG GTTGCGCTGG CCATTGTCTA TCTCATTGCC 1320
TTGGCTGTCT GTCAGTGCCG CCGAAAGAAC TACGGGCAGC TGGACATCTT TCCAGCCCGG 1380
GATACCTACC ATCCTATGAG CGAGTACCCC ACCTACCACA CCCATGGGCG CTATGTGCCC 1440
CCTAGCAGTA CCGATCGTAG CCCCTATGAG AAGGTTTCTG CAGGTAATGG TGGCAGCAGC 1500
CTCTCTTACA CAAACCCAGC AGTGGCAGCC ACTTCTGCCA ACTTGTAG 1548
[SEQ ID NO:1]
[073] Для создания рекомбинантного поксвируса, который стабилен при последовательном пассировании вируса, была синтезирована одна или более нуклеиновых кислот согласно настоящему изобретению. Более конкретно, как изображено на Фиг. 2А-2С, была выполнена одна или более замен в одной или более подчеркнутых областях за пределами областей VNTR PANVAC MUC1 (SEQ ID NO:1).
[074] Таким образом, в одном варианте осуществления изобретения представлена новая нуклеиновая кислота MUC1, которая содержит замену по меньшей мере в одной из повторяющихся нуклеотидных областей вне областей VNTR нуклеиновой кислоты MUC1. По меньшей мере, в одном аспекте одна или более повторяющихся областей определены как: (i) три или более последовательно повторяющихся нуклеотида, (ii) три или более последовательных нуклеотида G или C и/или (iii) три или более последовательных нуклеотида T или С. В более конкретных аспектах одна или более повторяющихся нуклеотидных областей дополнительно определена как (i) четыре или более последовательно повторяющихся нуклеотида, (ii) четыре или более последовательных нуклеотида G или C и/или (iii) четыре или более последовательных нуклеотида T или С. В некоторых других более конкретных аспектах последовательно повторяющиеся нуклеотиды определены как (i) последовательные нуклеотиды G, (ii) последовательные нуклеотиды C и/или (iii) последовательные нуклеотиды T.
[075] Как изображено на Фиг. 2А-2С, новая нуклеиновая кислота MUC1 может содержать замену по меньшей мере в 2, 3, 4 или 5 повторяющихся нуклеотидных областях вне областей VNTR нуклеиновой кислоты MUC1. В дополнительных аспектах новая нуклеиновая кислота MUC1 может содержать замену по меньшей мере в 10, 15, 20 или 25 повторяющихся нуклеотидных областях вне областей VNTR.
[076] В других дополнительных аспектах новая нуклеиновая кислота MUC1 может содержать по меньшей мере одну замену в тех областях за пределами областей VNTR, которые более подвержены мутированию при последовательном пассировании рекомбинантного поксвируса. В типовом аспекте новая нуклеиновая кислота MUC1 может содержать по меньшей мере одну замену в одной или более из таких повторяющихся областей нуклеотидов MUC1 вне областей VNTR, выбранных из областей нуклеотидов PANVAC MUC1 (SEQ ID NO:1) и/или их комбинаций, указанных в Таблице 5.
ТАБЛИЦА 5:
Более предпочтительно, новая нуклеиновая кислота MUC1 может содержать по меньшей мере одну замену в этих повторяющихся областях нуклеотидов MUC1 вне областей VNTR, выбранных из областей нуклеотидов PANVAC MUC1 (SEQ ID NO:1) и/или их комбинаций, указанных в Таблице 6.
ТАБЛИЦА 6:
Согласно настоящему изобретению понятно, что положения нуклеотидов, перечисленные в Таблицах 5 и 6, являются лишь репрезентативными для повторяющихся областей нуклеиновой кислоты MUC1, обнаруженных в областях MUC1, не являющихся VNTR. Таким образом, хотя описанная в настоящем описании область повторов имеет определенное нуклеотидное положение в SEQ ID NO:1 (например, 240-253), эта конкретная область может соответствовать другому нуклеотидному положению в другой нуклеиновой кислоте MUC1.
[077] В дополнительных вариантах осуществления изобретения модификации повторяющихся областей за пределами VNTR и/или модификации областей VNTR представляют собой молчащую модификацию, что означает, что эта модификация не влияет на аминокислотную последовательность антигена MUC1. По меньшей мере, в одном аспекте повышение стабильности трансгена MUC1 путем модификации одной или более повторяющихся областей было затруднительным из-за того, что лишь некоторые нуклеотиды и/или повторяющиеся области могли быть модифицированы без влияния на аминокислотную последовательность MUC1.
[078] С учетом вышеизложенного, в одном или более вариантах осуществления настоящее изобретение включает одну или более нуклеиновых кислот MUC1, содержащих 1) одну или более модификаций повторов домена VNTR, выбранных из а) перетасовки и b) кодон-оптимизации; и 2) одну или более модификации повторяющихся областей за пределами VNTR.
[079] В другом аспекте нуклеиновые кислоты MUC1 согласно настоящему изобретению могут содержать одну или более модификаций, сконфигурированных для усиления иммуногенности трансгена MUC1 у субъекта. В одном неограничивающем примере нуклеиновые кислоты MUC1 могут быть модифицированы для включения одного или более агонистических эпитопов, описанных в WO 2013/103658, который включен в настоящее описание посредством ссылки. В более конкретном варианте осуществления изобретения нуклеиновые кислоты MUC1 согласно настоящему изобретению содержат агонистические эпитопы, выбранные из группы, состоящей из: YLAPPAHGV [SEQ ID NO: 24], YLDTRPAPV [SEQ ID NO: 25], YLAIVYLIAL [SEQ ID NO: 26], YLIALAVCQV [SEQ ID NO: 27], YLSYTNPAV [SEQ ID NO: 28], и SLFRSPYEK [SEQ ID NO: 29] (подчеркнутые участки представляют собой замещенные аминокислоты).
[080] В предпочтительных вариантах осуществления изобретения нуклеиновая кислота MUC1 содержит нуклеотидную последовательность, по меньшей мере на 95% гомологичную SEQ ID NO: 2 (336 MUC), SEQ ID NO:3 (373 MUC), SEQ ID NO:4 (399/400 MUC1), или SEQ ID NO:5 (420 MUC1). В дополнительных предпочтительных вариантах осуществления изобретения нуклеиновая кислота MUC1 содержит нуклеотидную последовательность, по меньшей мере, 96%, 97% или 98%, гомологичную SEQ ID NO: 2 (336 MUC), SEQ ID NO:3 (373 MUC), SEQ ID NO:4 (399/400 MUC1), или SEQ ID NO:5 (420 MUC1). В более предпочтительном варианте осуществления изобретения нуклеиновая кислота MUC1 содержит нуклеотидную последовательность, выбранную из SEQ ID NO: 2 (336 MUC), SEQ ID NO:3 (373 MUC), SEQ ID NO:4 (399/400 MUC1), или SEQ ID NO:5 (420 MUC1).
[081] В других предпочтительных вариантах осуществления изобретения нуклеиновая кислота MUC1 содержит нуклеотидную последовательность, по меньшей мере на 95% гомологичную SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33; или SEQ ID NO: 34. В дополнительных предпочтительных вариантах осуществления изобретения нуклеиновая кислота MUC1 содержит нуклеотидную последовательность, по меньшей мере, 96%, 97% или 98%, гомологичную SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33; или SEQ ID NO: 34. В более предпочтительном варианте осуществления изобретения нуклеиновая кислота MUC1 содержит нуклеотидную последовательность, выбранную из SEQ ID NO: 31, SEQ ID NO: 32, SEQ ID NO: 33; или SEQ ID NO: 34.
Новые последовательности нуклеиновых кислот CEA
[082] В другом аспекте настоящего изобретения было определено, что одна или более модификаций в повторяющихся областях нуклеиновых кислот СЕА увеличивали стабильность трансгена СЕА. Типовой образец этих областей изображен в парном выравнивании на Фиг. 5. Эти типовые повторяющиеся области отмечены указанными заменами (области выравнивания, не отмеченные *).
[083] По меньшей мере, в одном аспекте замена дополнительно повышает стабильность рекомбинантного поксвируса. Это демонстрируется, по меньшей мере частично, данными стабильности mBN336, указанными на Фиг. 9 и 10. Как отмечалось ранее, mBN336 содержит MUC1, CEA и TRICOM. В то время как mBN336 содержит модифицированную нуклеиновую кислоту MUC1, mBN336 не содержит каких-либо дополнительных модификаций CEA и содержит только промежуточные модификации в костимулирующих молекулах TRICOM. Таким образом, хотя раскрытые в настоящем описании модификации в MUC1 увеличивают стабильность рекомбинантного поксвируса, нестабильность, начиная с пассажа 5, сохраняется. Как только модифицированный СЕА был включен в состав вируса оспы птиц в mBN373, стабильность трансгена и вируса оспы птиц была продемонстрирована после 5-го и 7-го пассажа (см., например, Фигуру 11).
[084] Соответственно, в различных вариантах осуществления изобретения, настоящее изобретение включает нуклеиновую кислоту, кодирующую пептид CEA (нуклеиновую кислоту CEA), содержащую по меньшей мере одну нуклеотидную замену по меньшей мере в одной повторяющейся нуклеотидной области нуклеиновой кислоты CEA, причем по меньшей мере одна повторяющаяся нуклеотидная область определена как а) три дополнительных последовательно повторяющихся нуклеотида G или C и/или b) три или более последовательно повторяющихся нуклеотида T. В дополнительных вариантах осуществления изобретения повторяющиеся нуклеотидные области дополнительно определены как а) три или более последовательно повторяющихся нуклеотида G и/или b) три или более последовательно повторяющихся нуклеотида C.
[085] В предпочтительных вариантах осуществления изобретения повторяющиеся нуклеотидные области нуклеиновой кислоты CEA определены как (i) четыре или более последовательно повторяющихся нуклеотида, (ii) четыре или более последовательных нуклеотида G или C и/или (iii) четыре или более последовательных нуклеотида T. В дополнительных предпочтительных вариантах осуществления изобретения повторяющаяся нуклеотидная область дополнительно определена как (i) четыре или более последовательных нуклеотида G, (ii) четыре или более последовательных нуклеотида C и/или (iii) четыре или более последовательных нуклеотида T.
[086] В одном или более вариантах осуществления изобретения нуклеиновая кислота СЕА содержит, по меньшей мере, одну замену, по меньшей мере, в 2, 3, 4, 5 или 10 повторяющихся нуклеотидных областях второй нуклеиновой кислоты. В предпочтительном варианте осуществления изобретения нуклеиновая кислота СЕА содержит, по меньшей мере, одну нуклеотидную замену, по меньшей мере, в 10, по меньшей мере, в 12, по меньшей мере, в 15 и/или, по меньшей мере, в 19 повторяющихся нуклеотидных областях. В более предпочтительном варианте осуществления изобретения нуклеиновая кислота СЕА содержит, по меньшей мере, одну нуклеотидную замену в 19 областях второй нуклеиновой кислоты.
[087] В более предпочтительных вариантах осуществления изобретения нуклеиновая кислота СЕА содержит SEQ ID NO: 14 (mBN373/420 CEA).
Новые последовательности нуклеиновых кислот TRICOM
[088] В другом аспекте настоящего изобретения была произведена еще одна модификация одной или более нуклеиновых кислот, кодирующих костимулирующие молекулы TRICOM. В более конкретном аспекте в ходе разработки настоящего изобретения было определено, что одна или более модификаций в повторяющихся областях нуклеиновых кислот TRICOM увеличивают стабильность трансгенов TRICOM. Типовой образец этих областей изображен в парном выравнивании на Фиг. 6-8. Эти типовые повторяющиеся области отмечены указанными заменами (области выравнивания, не отмеченные *).
[089] По меньшей мере, в одном аспекте, одна или более замен дополнительно повышают стабильность рекомбинантного поксвируса. Это демонстрируется, по меньшей мере частично, данными стабильности mBN336, указанными на Фиг. 9 и 10. Как отмечалось ранее, mBN336 содержит модифицированный MUC1. Однако mBN336 не содержит каких-либо дополнительных модификаций CEA и содержит только промежуточные модификации в костимулирующих молекулах TRICOM. Таким образом, хотя раскрытые в настоящем описании модификации MUC1 увеличивают стабильность mBN336, нестабильность после пассажа 5 сохраняется. Как только модифицированные трансгены были включены в состав вируса оспы птиц, стабильность трансгена и поксвируса была продемонстрирована после пассажа 5 и при пассаже 7 (см., например, Фигуру 11).
[090] В одном варианте осуществления изобретения новые костимулирующие молекулы TRICOM содержат нуклеотидную последовательность, по меньшей мере на 80% гомологичную SEQ ID NO: 15 или 17 (для B7-1), нуклеотидную последовательность, по меньшей мере на 80% гомологичную SEQ ID NO:18 или 20 (для ICAM-1), и нуклеотидную последовательность, по меньшей мере на 80% гомологичную SEQ ID NO:21 или 23 (для LFA-3). В других дополнительных предпочтительных вариантах осуществления изобретения нуклеиновые кислоты TRICOM содержат нуклеотидную последовательность, по меньшей мере на 85%, 90% или 95%, гомологичную SEQ ID NO:15 или 17 (для B7-1), SEQ ID NO:18 или 20 (для ICAM-1) и/или SEQ ID NO:21 или 23 (для LFA-3). В других более предпочтительных вариантах осуществления изобретения нуклеиновые кислоты TRICOM содержат нуклеотидную последовательность, по меньшей мере на 85%, 90% или 95%, гомологичную SEQ ID NO:17 (для B7-1), SEQ ID NO:20 (для ICAM-1) и/или SEQ ID NO:23 (для LFA-3)
[091] В другом варианте осуществления изобретения костимулирующие молекулы TRICOM содержат SEQ ID NO: 15 или 17 (для B7-1), SEQ ID NO:18 или 20 (для ICAM-1) и/или SEQ ID NO:21 или 23 (для LFA-3).
[092] В другом варианте осуществления изобретения костимулирующие молекулы TRICOM содержат SEQ ID NO: 17 (для B7-1), SEQ ID NO:20 (для ICAM-1) и/или SEQ ID NO:23 (для LFA-3).
[093] В одном предпочтительном варианте осуществления изобретения новые костимулирующие молекулы TRICOM содержат нуклеотидную последовательность, по меньшей мере на 80% гомологичную SEQ ID NO: 15 (для B7-1), нуклеотидую последовательность, по меньшей мере на 80% гомологичную SEQ ID NO:18 (для ICAM-1), и нуклеотидную последовательность, по меньшей мере на 80% гомологичную SEQ ID NO:21 (для LFA-3). В других дополнительных предпочтительных вариантах осуществления изобретения нуклеиновые кислоты TRICOM содержат нуклеотидную последовательность, по меньшей мере на 85%, 90% или 95%, гомологичную SEQ ID NO:15 (для B7-1), SEQ ID NO:18 (для ICAM-1) и/или SEQ ID NO:21 (для LFA-3).
[094] В другом предпочтительном варианте осуществления изобретения новые костимулирующие молекулы TRICOM содержат нуклеотидную последовательность, по меньшей мере на 80%, 90% или 95%, гомологичную SEQ ID NO: 17 (для B7-1), нуклеотидную последовательность, по меньшей мере на 80%, 90% или 95%, гомологичную SEQ ID NO:20 (для ICAM-1), и нуклеотидную последовательность, по меньшей мере на 80%, 90% или 95% гомологичную SEQ ID NO:23 (для LFA-3).
[095] В другом варианте осуществления изобретения костимулирующие молекулы TRICOM содержат SEQ ID NO: 15 (для B7-1), SEQ ID NO:18 (для ICAM-1) и/или SEQ ID NO:21 (для LFA-3).
[096] В другом варианте осуществления изобретения костимулирующие молекулы TRICOM содержат SEQ ID NO: 17 (для B7-1), SEQ ID NO:20 (для ICAM-1) и/или SEQ ID NO:23 (для LFA-3).
[097] Предполагается, что данное раскрытие охватывает те последовательности нуклеиновых кислот, которые являются комплементарными новым последовательностям нуклеиновых кислот, представленным в настоящем описании.
Рекомбинантные поксвирусы
[098] В одном или более вариантах осуществления изобретения, изобретение включает рекомбинантный поксвирус, содержащий одну или более нуклеиновых кислот MUC1, описанных в настоящем описании. В более предпочтительных вариантах осуществления изобретения рекомбинантный поксвирус содержит последовательность нуклеиновой кислоты MUC1 и последовательность нуклеиновой кислоты CEA, описанные в настоящем описании.
[099] В предпочтительных вариантах осуществления изобретения нуклеиновая кислота MUC1 содержит нуклеотидную последовательность, по меньшей мере на 95% гомологичную SEQ ID NO:2, SEQ ID NO: 3 (373 MUC), SEQ ID NO:5 (420 MUC1) или SEQ ID NO:4 (399/400 MUC1), а последовательность нуклеиновой кислоты CEA содержит SEQ ID NO: 13 или 14.
[0100] В других дополнительных вариантах осуществления изобретения рекомбинантные поксвирусы согласно настоящему изобретению содержат одну или более костимулирующих молекул, таких как, но не ограничиваясь этим, молекулы, описанные в настоящем описании. В одном предпочтительном варианте осуществления изобретения костимулирующие молекулы включают TRICOM (B7-1, ICAM-1 и LFA-3). В более предпочтительном варианте осуществления изобретения костимулирующие молекулы B7-1 выбраны из последовательности нуклеиновой кислоты, содержащей SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17. В более предпочтительном варианте осуществления изобретения костимулирующая молекула ICAM-1 выбрана из последовательности нуклеиновой кислоты, содержащей SEQ ID NO:18, SEQ ID NO: 19 и SEQ ID NO: 20. В более предпочтительном варианте осуществления изобретения костимулирующая молекула LFA-3 выбрана из последовательности нуклеиновой кислоты, содержащей SEQ ID NO: 21, SEQ ID NO: 22 и SEQ ID NO: 23. В более предпочтительном варианте осуществления изобретения B7-1, ICAM-1 и LFA-3 выбраны из последовательности нуклеиновой кислоты, содержащей SEQ ID NO: 15, SEQ ID NO: 18 и SEQ ID NO: 21 соответственно. В другом более предпочтительном варианте осуществления изобретения B7-1, ICAM-1 и LFA-3 выбраны из последовательности нуклеиновой кислоты, содержащей SEQ ID NO: 17, SEQ ID NO: 20 и SEQ ID NO: 23 соответственно.
[0101] В различных вариантах осуществления настоящего изобретения рекомбинантный поксвирус представляет собой, предпочтительно, ортопоксвирус, такой как, но не ограничиваясь этим, вирус осповакцины, модифицированный вирус осповакцины Анкара (MVA), MVA-BN или производные от MVA-BN.
[0102] Примерами штаммов вируса осповакцины являются штаммы Temple of Heaven, Copenhagen, Paris, Budapest, Dairen, Gam, MRIVP, Per, Tashkent, TBK, Tom, Bern, Patwadangar, BIEM, B-15, Lister, EM-63, New York City Board of Health, Elstree, Ikeda и WR. Предпочтительным штаммом вируса осповакцины (VV) является штамм Wyeth (DRYVAX) (патент США 7,410,644).
[0103] Другим предпочтительным штаммом VV является модифицированный вирус осповакцины Анкара (MVA) (Sutter, G. et al. [1994], Vaccine 12: 1032-40). Примерами штаммов вируса MVA, которые могут быть использованы в практике настоящего изобретения и которые были депонированы в соответствии с требованиями Будапештского договора, являются штаммы MVA 572, депонированные в Европейской Коллекции Культур Клеток Животных (ECACC), Лаборатории Изучения и Производства Вакцин, Лаборатории Службы Общественного Здравоохранения, Центре Прикладной Микробиологии и Исследований, Портон-Даун, Солсбери, Уилтшир SP4 0JG, Великобритания, с номером депонирования ECACC 94012707, депонированный 27 января 1994 года и MVA 575, депонированный под номером ECACC 00120707 7 декабря 2000 года, MVA-BN, депонированный 30 августа 2000 года в Европейской Коллекции Клеточных Культур (ECACC) под номером V00083008, и производные MVA-BN, все являются дополнительными типовыми штаммами.
[0104] «Производные» MVA-BN относятся к вирусам, проявляющим по существу те же характеристики репликации, что и MVA-BN, как описано в настоящем описании, но проявляющим различия в одной или более частях их геномов. MVA-BN, а также его производные являются неспособными к репликации, что означает неспособность к репродуктивной репликации in vivo и in vitro. Более конкретно, in vitro, MVA-BN или его производные были описаны как способные к репродуктивной репликации в фибробластах куриных эмбрионов (CEF), но не способные к репродуктивной репликации в клеточной линии кератиноцитов человека HaCat (Boukamp et al. (1988), J. Cell Biol. 106:761-771), клеточной линии остеосаркомы кости человека 143B (номер депонирования ECACC 91112502), клеточной линии почки эмбриона человека 293 (номер депонирования ECACC 85120602) и клеточной линии аденокарциномы шейки матки человека HeLa (номер депонирования ATCC CCL-2). Кроме того, MVA-BN или его производные имеют скорость амплификации вируса, по меньшей мере, в два раза меньшую, более предпочтительно, в три раза меньшую, чем у MVA-575 в клетках Hela и клеточных линиях HaCaT. Испытания и исследование этих свойств MVA-BN и его производных описаны в WO 02/42480 (заявка на патент США № 2003/0206926) и WO 03/048184 (заявка на патент США № 2006/0159699).
[0105] Термин «не способен к репродуктивной репликации» или «неспособность к репродуктивной репликации» в клеточных линиях человека in vitro, как описано ранее, описан, например, в WO 02/42480, в котором также описано как получить MVA, имеющий желаемые свойства, как указано выше. Термин применяется к вирусу, который имеет скорость амплификации вируса in vitro через 4 дня после заражения менее 1 при использовании исследований, описанных в WO 02/42480 или в патенте США № 6,761,893.
[0106] Термин «неспособность к репродуктивной репликации» относится к вирусу, который имеет скорость амплификации вируса в клеточных линиях человека in vitro, как описано ранее, через 4 дня после заражения менее 1. Исследования, описанные в WO 02/42480 или в патенте США № 6,761,893, применимы для определения скорости амплификации вируса.
[0107] Амплификация или репликация вируса в клеточных линиях человека in vitro, как описано ранее, обычно выражается как отношение вируса, продуцируемого клеткой (выход), к количеству, первоначально использовавшемуся для заражения клетки вначале (стартовое к-во) и называется «скорость амплификации». Коэффициент амплификации «1» означает статус амплификации, когда количество вируса, продуцируемого инфицированными клетками, совпадает с количеством, первоначально использованным для инфицирования клеток, что означает, что инфицированные клетки допускают вирусную инфекцию и размножение. Напротив, коэффициент амплификации менее 1, то есть уменьшение выхода по сравнению со стартовым количеством, указывает на отсутствие репродуктивной репликации и, следовательно, аттенуацию вируса.
[0108] В другом варианте осуществления изобретения рекомбинантный поксвирус, содержащий MUC1 и/или другие нуклеиновые кислоты, раскрытые в настоящем описании, представляет собой авипоксвирус, такой как, но не ограничиваясь этим, вирус оспы птиц.
[0109] Термин «авипоксвирус» относится к любому авипоксвирусу, такому как Fowlpoxvirus, Canarypoxvirus, Uncopoxvirus, Mynahpoxvirus, Pigeonpoxvirus, Psittacinepoxvirus, Quailpoxvirus, Peacockpoxvirus, Penguinpoxvirus, Sparrowpoxvirus, Starlingpoxvirus и Turkeypoxvirus. Предпочтительными авипоксвирусами являются Canarypoxvirus и Fowlpoxvirus.
[0110] Примерами вируса оспы птиц являются штаммы FP-1, FP-5, TROVAC (патент США № 5,766,598), POXVAC-TC (патент США № 7,410,644), TBC-FPV (Therion Biologics- FPV), FP-1 представляет собой штамм Duvette, модифицированный для использования в качестве вакцины у однодневных цыплят. Штамм представляет собой коммерческий вакцинный штамм вируса оспы птиц, обозначенный O DCEP 25/CEP67/2309 October 1980, и доступен в Institute Merieux, Inc. FP-5 представляет собой коммерческий вакцинный штамм вируса оспы птиц, полученный из эмбрионов кур, доступный в American Scientific Laboratories (Division of Schering Corp.) Мэдисон, Висконсин, Ветеринарная Лицензия США № 165, серийный № 30321.
[0111] В некоторых предпочтительных вариантах осуществления изобретения представлен рекомбинантный ортопоксвирус, такой как вирус осповакцины, MVA, MVA-BN или производные от MVA-BN, содержащий последовательность нуклеиновой кислоты MUC1, выбранную из SEQ ID NO: 5 (420 MUC1), SEQ ID NO:4 (399/400 MUC1), SEQ ID NO:3 (373 MUC1), ИЛИ SEQ ID NO:2 (336 MUC1). В некоторых более предпочтительных вариантах осуществления изобретения рекомбинантный ортопоксвирус представляет собой вирус MVA, содержащий последовательность нуклеиновой кислоты MUC1, выбранную из SEQ ID NO: 2 (420 MUC1), нуклеиновую кислоту CEA, выбранную из SEQ ID NO: 13 или 14 и TRICOM. В наиболее предпочтительном варианте осуществления изобретения представлен рекомбинантный MVA, содержащий последовательность нуклеиновой кислоты MUC1, содержащую SEQ ID NO: 2 (336 MUC1), нуклеиновую кислоту СЕА, содержащую SEQ ID NO: 13 и TRICOM. В другом наиболее предпочтительном варианте осуществления изобретения TRICOM содержит одну или более нуклеиновых кислот, содержащих SEQ ID NO: 17, (B7-1), SEQ ID NO:20 (ICAM-1) и SEQ ID NO:23 (LFA-3).
[0112] В некоторых других предпочтительных вариантах осуществления изобретения представлен рекомбинантный авипоксвирус, такой как вирус оспы птиц, содержащий последовательность нуклеиновой кислоты MUC1, содержащую SEQ ID NO: 3 (373 MUC1). В некоторых более предпочтительных вариантах осуществления изобретения рекомбинантный авипоксвирус представляет собой вирус оспы птиц, содержащий нуклеиновую кислоту MUC1, содержащую SEQ ID NO: 3 (373), нуклеиновую кислоту CEA, выбранную из SEQ ID NO: 13 или 14 и TRICOM. В наиболее предпочтительном варианте осуществления изобретения представлен рекомбинантный вирус оспы птиц, содержащий последовательность нуклеиновой кислоты MUC1, содержащую SEQ ID NO: 3 (373 MUC1), нуклеиновую кислоту СЕА, содержащую SEQ ID NO: 14 и TRICOM. В другом наиболее предпочтительном варианте осуществления изобретения TRICOM содержит одну или более нуклеиновых кислот, содержащих SEQ ID NO: 15 (B7-1), SEQ ID NO:18 (ICAM-1) и SEQ ID NO:21 (LFA-3).
Кассеты экспрессии/Контрольные последовательности
[0113] В различных аспектах одна или более нуклеиновых кислот, описанных в настоящем описании, заключены в одну или более кассет экспрессии, в которых одна или более нуклеиновых кислот функционально связаны с последовательностями, контролирующими экспрессию. «Функционально связанный» означает, что описанные компоненты находятся во взаимосвязи, позволяющей им функционировать согласно назначению, например, промотору для транскрипции экспрессируемой нуклеиновой кислоты. Последовательность, контролирующая экспрессию, функционально связанная с кодирующей последовательностью, объединяется таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с последовательностями, контролирующими экспрессию. Последовательности, контролирующие экспрессию включают, но не ограничиваются этим, соответствующие промоторы, энхансеры, терминаторы транскрипции, стартовый кодон в начале открытой рамки считывания, кодирующей белок, сигналы сплайсинга интронов и стоп-кодоны в рамке считывания. Подходящие промоторы включают, но не ограничиваются этим, ранний промотор SV40, промотор RSV, ретровирусный LTR, главный поздний промотор аденовируса, немедленный ранний промотор I CMV человека и различные промоторы поксвируса, включая, но не ограничиваясь этим, следующие промоторы осповакцины или промоторы, полученные от MVA и FPV: промотор 30K, промотор I3, промотор PrS, промотор PrS5E, Pr7.5K, длинный промотор Pr13.5, промотор 40K, промотор MVA-40K, промотор FPV 40K, промотор 30k, промотор PrSynIIm и промотор PrLE1. Дополнительные промоторы дополнительно описаны в WO 2010/060632, WO 2010/102822, WO 2013/189611 и WO 2014/063832, которые включены в настоящее описание в полном объеме посредством ссылки.
[0114] Дополнительные последовательности, контролирующие экспрессию включают, но не ограничиваются этим, лидерные последовательности, кодоны терминации, сигналы полиаденилирования и любые другие последовательности, необходимые для соответствующей транскрипции и последующей трансляции последовательности нуклеиновой кислоты, кодирующей желаемый рекомбинантный белок (например, MUC1, CEA и/или TRICOM) в желаемой системе-хозяине. Вектор поксвируса может также содержать дополнительные элементы, необходимые для переноса и последующей репликации вектора экспрессии, содержащего последовательность нуклеиновой кислоты в желаемой системе-хозяине. Кроме того, специалист в данной области поймет, что такие векторы легко конструировать с использованием обычных способов (Ausubel et al., (1987) в “Current Protocols in Molecular Biology,” John Wiley and Sons, Нью-Йорк) и они коммерчески доступны.
В некоторых вариантах осуществления изобретения рекомбинантный ортопоксвирус и/или авипоксвирус согласно настоящему изобретению содержит один или более цитокинов, таких как IL-2, IL-6, IL-12, RANTES, GM-CSF, TNF-α или IFN-γ, один или более факторов роста, таких как GM-CSF или G-CSF, одну или более костимулирующих молекул, таких как ICAM-1, LFA-3, CD72, B7-1, B7-2 или другие связанные с B7 молекулы; одну или более молекул, таких как OX-40L или 41 BBL, или комбинации этих молекул, которые могут быть использованы в качестве биологических адъювантов (см., например, Salgaller et al., 1998, J. Surg. Oncol. 68(2):122-38; Lotze et al., 2000, Cancer J. Sci. Am. 6(Suppl 1):S61-6; Cao et al., 1998, Stem Cells 16(Suppl 1):251-60; Kuiper et al., 2000, Adv. Exp. Med. Biol. 465:381-90). Эти молекулы могут быть введены хозяину системно (или локально). В нескольких примерах вводят IL-2, RANTES, GM-CSF, TNF-α, IFN-γ, G-CSF, LFA-3, CD72, B7-1, B7-2, B7-1, B7-2, OX-40L. 41 BBL и ICAM-1.
Получение рекомбинантных поксвирусов, содержащих трансгены
[0115] Рекомбинантные поксвирусы, предоставленные в настоящем описании, могут быть получены обычными способами, известными в данной области техники. Способы получения рекомбинантных поксвирусов или встраивания экзогенных кодирующих последовательностей в поксвирусный геном хорошо известны специалисту в данной области техники. Например, способы с использованием стандартных методик молекулярной биологии, таких как клонирование ДНК, выделение ДНК и РНК, вестерн-блоттинг, способы амплификации ОТ-ПЦР и ПЦР, описаны в Molecular Cloning, A Laboratory Manual (2nd Ed.) [J. Sambrook et al., Cold Spring Harbor Laboratory Press (1989)], а также способы работы с вирусами и манипуляции с ними описаны в Virology Methods Manual [B.W.J. Mahy et al. (eds.), Academic Press (1996)]. Точно так же способы и знания для обработки, манипуляции и генной инженерии MVA описаны в Molecular Virology: A Practical Approach [A.J. Davison & R.M. Elliott (Eds.), The Practical Approach Series, IRL Press at Oxford University Press, Оксфорд, Великобритания (1993) (см., например, Главу 9: Expression of genes by Vaccinia virus vectors)] и Current Protocols in Molecular Biology [John Wiley & Son, Inc. (1998) (см., например, Главу 16, Раздел IV: Expression of proteins in mammalian cells using vaccinia viral vector)].
[0116] Для получения различных рекомбинантных поксвирусов, описанных в настоящем описании, могут быть применимы разные способы. Последовательность ДНК, которая должна быть вставлена в вирус, может быть помещена в плазмидную конструкцию E.coli, в которую была вставлена ДНК, гомологичная части ДНК поксвируса. Отдельно, вставляемая последовательность ДНК может быть лигирована с промотором. Связь промотор-ген может быть расположена в плазмидной конструкции таким образом, чтобы связь промотор-ген фланкировалась на обоих концах ДНК, гомологичной последовательности ДНК, фланкирующей область поксвирусной ДНК, содержащей несущественный локус. Полученная плазмидная конструкция может быть амплифицирована путем размножения в бактериях E.coli и выделена. Выделенная плазмида, содержащая вставляемую последовательность гена ДНК, может быть трансфицирована в клеточную культуру, например фибробласты куриных эмбрионов (CEF), в то же время культура инфицируется поксвирусом. Рекомбинация между гомологичной поксвирусной ДНК в плазмиде и вирусным геномом, соответственно, может привести к получению поксвируса, модифицированного добавлением чужеродных последовательностей ДНК.
[0117] В соответствии с предпочтительным вариантом осуществления изобретения клетка подходящей клеточной культуры, такой как, например, клетки CEF, может быть инфицирована поксвирусом. Затем инфицированная клетка может быть трансфицирована первой плазмидой, содержащей чужеродный или гетерологичный ген или гены, такие как одна или более нуклеиновых кислот MUC1, CEA и/или TRICOM, представленных в данном раскрытии; предпочтительно под транскрипционным контролем элемента контроля экспрессии поксвируса. Как объяснено выше, плазмида также содержит последовательности, способные направлять вставку экзогенной последовательности в выбранную часть поксвирусного генома. Необязательно, плазмидный вектор также содержит кассету, содержащую маркерный и/или селективный ген, функционально связанный с поксвирусным промотором. Подходящими маркерными или селективными генами являются, например, гены, кодирующие зеленый флуоресцентный белок, β-галактозидазу, неомицин-фосфорибозилтрансферазу или другие маркеры. Использование селективных или маркерных кассет упрощает идентификацию и выделение полученного рекомбинантного поксвируса. Однако рекомбинантный поксвирус также может быть идентифицирован с помощью технологии ПЦР. Далее, следующая клетка может быть инфицирована рекомбинантным поксвирусом, полученным, как описано выше, и трансфицирована вторым вектором, содержащим второй чужеродный или гетерологичный ген или гены. В случае, если этот ген должен быть введен в другой сайт встраивания поксвирусного генома, второй вектор также отличается поксвирус-гомологичными последовательностями, направляющими интеграцию второго чужеродного гена или генов в геном поксвируса. После того, как произошла гомологичная рекомбинация, рекомбинантный вирус, содержащий два или более чужеродных или гетерологичных гена, может быть выделен. Для введения дополнительных чужеродных генов в рекомбинантный вирус этапы заражения и трансфекции можно повторять, используя рекомбинантный вирус, выделенный на предыдущих этапах для инфекции, и используя дополнительный вектор, содержащий дополнительный чужеродный ген или гены для трансфекции.
[0118] Альтернативно, этапы заражения и трансфекции, как описано выше, являются взаимозаменяемыми, то есть подходящая клетка может сначала трансфицироваться плазмидой, содержащей чужеродный ген, а затем инфицироваться поксвирусом. В качестве дополнительной альтернативы также возможно ввести каждый чужеродный ген в разные вирусы, коинфицировать клетку всеми полученными рекомбинантными вирусами и провести скрининг рекомбинантов, содержащих все чужеродные гены. Третья альтернатива представляет собой лигирование генома ДНК и чужеродных последовательностей in vitro и восстановление ДНК генома рекомбинированного вируса осповакцины с использованием вируса-помощника. Четвертая альтернатива представляет собой гомологичную рекомбинацию в E.coli или других видах бактерий между геномом поксвируса, клонированным в виде бактериальной искусственной хромосомы (BAC) и линейной чужеродной последовательностью, фланкированной последовательностями ДНК, гомологичными последовательностям, фланкирующим желаемый сайт интеграции в геном вируса осповакцины.
[0119] Одна или более нуклеиновых кислот согласно настоящему изобретению могут быть вставлены в любую подходящую часть поксвируса. В предпочтительном аспекте поксвирус, используемый для настоящего изобретения, включает вирус MVA и/или вирус оспы птиц. Подходящими частями MVA и вируса оспы птиц являются несущественные части генома вирусов MVA и оспы птицы.
[0120] Для MVA несущественные части генома MVA могут представлять собой межгенные области или известные сайтами делеции 1-6 генома MVA. Альтернативно или дополнительно, несущественные части рекомбинантного MVA могут представлять собой кодирующую область генома MVA, которая не является необходимой для роста вируса. Однако сайты встраивания не ограничены этими предпочтительными сайтами встраивания в геноме MVA, поскольку в объем настоящего изобретения входят нуклеиновые кислоты согласно настоящему изобретению (например, MUC1, CEA и TRICOM) и любые сопутствующие промоторы как описано в настоящем описании, которые могут быть вставлены в любом месте вирусного генома в тех случаях, пока возможно получить рекомбинанты, которые можно амплифицировать и размножать по меньшей мере в одной системе культуры клеток, такой как фибробласты куриных эмбрионов (клетки CEF).
[0121] Предпочтительно, нуклеиновые кислоты согласно настоящему изобретению могут быть встроены в одну или более межгенных областей (IGR) вируса MVA и/или вируса оспы птиц. Термин «межгенная область» относится предпочтительно к тем частям вирусного генома, которые расположены между двумя смежными открытыми рамками считывания (ОРС) генома вируса MVA и/или вируса оспы птиц, предпочтительно между двумя существенными ОРС генома вируса MVA и/или вируса оспы птиц. Для MVA в некоторых вариантах осуществления изобретения IGR выбрана из IGR 07/08, IGR 44/45, IGR 64/65, IGR 88/89, IGR 136/137 и IGR 148/149. Для вируса оспы птиц IGR выбрана из BamH1,
[0122] Для вируса MVA нуклеотидные последовательности могут быть дополнительно или альтернативно встроены в один или более известных сайтов делеции, то есть сайтов делеции I, II, III, IV, V или VI генома MVA. Термин «известный сайт делеции» относится к тем частям генома MVA, которые были удалены при непрерывном пассировании на клетках CEF, охарактеризованным в 20 пассаже 516, в отношении генома родительского вируса, от которого происходит MVA, в частности родительского хориоаллантоисного вируса осповакцины Анкара (CVA), например, как описано в Meisinger-Henschel et al. (2007), Journal of General Virology 88:3249-3259.
Вакцины
[0123] В некоторых вариантах осуществления изобретения рекомбинантные поксвирусы согласно настоящему изобретению могут быть приготовлены в качестве части вакцины. Для приготовления вакцин поксвирус может быть преобразован в физиологически приемлемую форму. В некоторых вариантах осуществления изобретения такое приготовление основано на опыте приготовления вакцин против вируса оспы, используемых для вакцинации против натуральной оспы, как описано, например, в Stickl, H. et al., Dtsch. med. Wschr. 99, 2386-2392 (1974).
[0124] Типовое приготовление описано далее. Очищенный вирус хранится при температуре -80°С с титром 5×108 TCID50/мл, приготовленным в 10 мМ Трис, 140 мМ NaCl, рН 7,4. Для приготовления инъекций вакцины, например, 102-108 частиц вируса могут быть лиофилизированы в фосфатно-солевом буфере (ФСБ) в присутствии 2% пептона и 1% человеческого альбумина в ампуле, предпочтительно в стеклянной ампуле. Альтернативно, инъекции вакцины могут быть получены путем поэтапной лиофилизации вируса в приготовлении. В некоторых вариантах осуществления изобретения приготовление содержит дополнительные компоненты, такие как маннит, декстран, сахар, глицин, лактоза, поливинилпирролидон или другие добавки, такие как, включая, но не ограничиваясь этим, антиоксиданты или инертный газ, стабилизаторы или рекомбинантные белки (например, сывороточный альбумин человека), пригодные для введения in vivo. Затем ампула запечатывается и может храниться при подходящей температуре, например, при температуре между 4°С и комнатной температурой, в течение нескольких месяцев. Однако до тех пор, пока в этом нет необходимости, ампулу хранят предпочтительно при температуре ниже -20°С.
[0125] В различных вариантах осуществления изобретения, включающих вакцинацию или терапию, лиофилизат растворяют в 0,1-0,5 мл водного раствора, предпочтительно физиологического солевого раствора или трис-буфера, и вводят системно или локально, то есть парентерально, подкожно, внутривенно, внутримышечно, интраназально, внутрикожно или любым другим путем введения, известным квалифицированному специалисту. Оптимизация способа введения, дозы и количества введений известна специалисту в данной области техники.
[0126] В некоторых вариантах осуществления изобретения аттенуированные штаммы вируса осповакцины полезны для индукции иммунного ответа у животных с ослабленным иммунитетом, например у обезьян (CD4 <400/мкл крови), инфицированных SIV, или у людей с ослабленным иммунитетом. Термин «ослабленный иммунитет» описывает состояние иммунной системы индивидуума, которая проявляет лишь неполные иммунные ответы или имеет пониженную эффективность в защите от инфекционных агентов.
Наборы, Композиции и Способы Применения
[0127] В одном из различных вариантов осуществления изобретение охватывает наборы и/или композиции, содержащие рекомбинантный поксвирус, который содержит нуклеиновые кислоты, описанные в настоящем описании. Предпочтительно композиция представляет собой фармацевтическую или иммуногенную композицию.
[0128] В одном варианте осуществления изобретения представлен набор и/или композиция, содержащая комбинацию двух или более рекомбинантных поксвирусов, каждый из рекомбинантных поксвирусов содержит нуклеиновые кислоты MUC1, CEA и/или TRICOM согласно настоящему изобретению. Комбинация содержит а) ортопоксвирус, такой как вирус осповакцины, MVA, MVA-BN или производные от MVA-BN, содержащий нуклеиновые кислоты MUC1, CEA и/или TRICOM согласно настоящему изобретению, и b) авипоксвирус, такой как вирус оспы птиц, содержащий нуклеиновые кислоты MUC1, CEA и/или TRICOM согласно настоящему изобретению. Предполагается, что комбинация ортопоксвируса и вируса оспы птиц может быть введена в соответствии с гомологичным или гетерологичным режимом введения с прайм-бустом.
[0129] В другом варианте осуществления изобретения набор и/или композиция, содержащая комбинацию двух или более рекомбинантных поксвирусов, содержит а) вирус MVA, содержащий нуклеиновые кислоты MUC1, СЕА и/или TRICOM согласно настоящему изобретению, и b) авипоксвирус, такой как вирус оспы птиц, содержащий нуклеиновые кислоты MUC1, CEA и/или TRICOM согласно настоящему изобретению. Предполагается, что комбинация вируса MVA и вируса оспы птиц может быть введена в соответствии с гомологичным или гетерологичным режимом введения с прайм-бустом.
[0130] В дополнительном варианте осуществления изобретения каждый из одного или более рекомбинантных поксвирусов дополнительно содержит одну или более костимулирующих молекул согласно настоящему изобретению. В предпочтительном варианте осуществления изобретения одна или более костимулирующих молекул представляют собой одну или более молекул TRICOM согласно настоящему изобретению.
[0131] Предполагается, что набор и/или композиция могут содержать одну или более емкостей или флаконов рекомбинантных поксвирусов согласно настоящему изобретению вместе с инструкциями по введению рекомбинантных поксвирусов.
[0132] Наборы и/или композиции, представленные в настоящем описании, обычно могут содержать один или более фармацевтически приемлемых и/или утвержденных носителей, добавок, антибиотиков, консервантов, адъювантов, разбавителей и/или стабилизаторов. Такими вспомогательные вещества могут представлять собой воду, физиологический раствор, глицерин, этанол, смачивающие или эмульгирующие агенты, буферизующее вещество для поддержания рН или тому подобное. Подходящими носителями обычно являются крупные медленно метаболизируемые молекулы, такие как белки, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, аминокислотные сополимеры, липидные агрегаты или тому подобное.
[0133] Для приготовления композиций (например, фармацевтических и/или иммуногенных композиций) рекомбинантные поксвирусы, представленные в настоящем описании, могут быть преобразованы в физиологически приемлемую форму. Это можно сделать, основываясь на опыте приготовления вакцин против вируса оспы, используемых для вакцинации против натуральной оспы, как описано в H. Stickl et al., Dtsch. med. Wschr. 99:2386-2392 (1974).
[0134] Например, очищенные вирусы могут храниться при температуре -80°C с титром 5×108 TCID50/мл, приготовленными в около 10 мМ Трис, 140 мМ NaCl, рН 7,4. Для приготовления инъекций вакцины, например, 102-108 или 102-109 частиц вируса могут быть лиофилизированы в 100 мл фосфатно-солевого буфера (ФСБ) в присутствии 2% пептона и 1% человеческого альбумина в ампуле, предпочтительно в стеклянной ампуле. Альтернативно, инъекции вакцины могут быть получены путем поэтапной лиофилизации вируса в приготовлении. Эта композиция может содержать дополнительные добавки, такие как маннит, декстран, сахар, глицин, лактоза или поливинилпирролидон или другие вспомогательные вещества, такие как антиоксиданты или инертный газ, стабилизаторы или рекомбинантные белки (например, сывороточный альбумин человека), подходящие для введения in vivo. Типовая вирусосодержащая композиция, подходящая для лиофилизации, содержит 10 мМ трис-буфера, 140 мМ NaCl, 18,9 г/л декстрана (MW 36 000-40 000), 45 г/л сахарозы, 0,108 г/л моногидрата монокалиевой соли L-глутаминовой кислоты, pH 7,4. Стеклянная ампула затем запечатывается и может храниться при температуре от 4°C до комнатной температуры в течение нескольких месяцев. Однако до тех пор, пока в этом нет необходимости, ампулу хранят предпочтительно при температуре -20°С или ниже.
[0135] Для вакцинации или терапии лиофилизат может быть растворен в водном растворе (например, 0,1-0,5 мл), предпочтительно в воде для инъекций, физиологическом растворе или трис-буфере, и вводиться системно или локально, то есть парентерально, подкожно, внутривенно, внутримышечно, интраназально или любым другим путем введения, известным квалифицированному специалисту. Способ введения, доза и количество введений могут быть оптимизированы специалистами в данной области техники известным способом.
[0136] В различных других вариантах осуществления изобретения представлен один или более способов, связанных с обеспечением и/или увеличением стабильности рекомбинантного поксвируса и/или содержащихся в нем трансгенов при последовательном пассировании рекомбинантного поксвируса. В более конкретном варианте осуществления изобретения рекомбинантный поксвирус является стабильным в течение по меньшей мере до пассажа 3 или 4 включительно.
[0137] Наличие стабильного рекомбинантного поксвируса при проведении множественных пассажей особенно важно по многим причинам, некоторые из которых включают крупномасштабное производство рекомбинантного вируса и его использование в качестве лекарственного средства, а также государственную политику обеспечения стабильности вакцины при проведении множественных пассажей. Для рекомбинантных поксвирусов согласно настоящему изобретению важным является получение стабильного рекомбинантного поксвируса при меньшей мере 3 или 4 пассажей, так как PANVAC-V и PANVAC-F начали демонстрировать нестабильность и/или потерю жизнеспособности трансгена при пассаже 1 (см., например, Таблицы 1 и 2 и Фигуры 1А и 1В).
[0138] В одном варианте осуществления изобретения представлен способ получения поксвируса, имеющего трансген MUC1, который стабилен при последовательном пассировании рекомбинантного поксвируса, причем способ включает: a) получение любой одной из нуклеиновых кислот или кассет экспрессии согласно настоящему изобретению; и b) вставку нуклеиновой кислоты или кассеты экспрессии в рекомбинантный поксвирус, причем рекомбинантный поксвирус стабилен при последовательном пассировании.
Типовые Способы согласно данному раскрытию
1. В другом варианте осуществления изобретения представлен способ получения рекомбинантного поксвируса, который стабилен при последовательном пассировании рекомбинантного поксвируса, причем способ включает:
а) получение первой нуклеиновой кислоты, кодирующей пептид MUC1, имеющий по меньшей мере два домена вариабельных N-концевых повторов (VNTR), причем а) расположение по меньшей мере двух доменов VNTR перетасовано, и b) по меньшей мере два домена VNTR являются кодон-оптимизированными, причем рекомбинантный поксвирус стабилен при последовательном пассировании рекомбинантного поксвируса.
2. В другом варианте осуществления изобретения представлен способ получения стабильного рекомбинантного поксвируса, который стабилен при последовательном пассировании рекомбинантного поксвируса, причем способ включает: получение первой нуклеиновой кислоты, кодирующей белок MUC1, причем белок MUC1 содержит по меньшей мере два домена VNTR; перестановку или перестановку порядка по меньшей мере двух повторов домена VNTR; кодон-оптимизацию по меньшей мере двух повторов домена VNTR; вставку первой последовательности нуклеиновой кислоты в поксвирус для получения рекомбинантного поксвируса, который является стабильным при последовательном пассировании рекомбинантного поксвируса.
3. Способ по любому из пп.1 и 2, в котором первая нуклеиновая кислота по меньшей мере на 95% гомологична SEQ ID NO:2, на 95% гомологична SEQ ID NO: 4, на 95% гомологична SEQ ID NO: 3, или на 95% гомологична SEQ ID NO: 5.
4. Способ по любому из пп.1-3, в котором нуклеиновая кислота, по меньшей мере на 95% гомологична SEQ ID NO: 2.
5. Способ по любому из пп.1-4, в котором нуклеиновая кислота, по меньшей мере на 95% гомологична SEQ ID NO: 3.
6. Способ по любому из пп.1-5, в котором нуклеиновая кислота содержит SEQ ID NO: 2.
7. Способ по любому из пп.1-6, в котором нуклеиновая кислота содержит SEQ ID NO: 5.
8. Способ по любому из пп.1-7, в котором способ дополнительно включает замену по меньшей мере одного нуклеотида в повторяющейся нуклеотидной области второй нуклеиновой кислоты, кодирующей пептид СЕА, причем повторяющаяся нуклеотидная область определена как: (i) три или более последовательно повторяющиеся нуклеотида, (ii) три или более последовательных нуклеотида G или C и/или (iii) три или более последовательных нуклеотида T или C; и вставку второй нуклеиновой кислоты в рекомбинантный поксвирус.
9. Способ по п.8, в котором область повторов второй нуклеиновой кислоты дополнительно определена как (i) три или более последовательных нуклеотида G, (ii) три или более последовательных нуклеотида C и/или (iii) три или более последовательных нуклеотида T.
10. Способ по любому из пп.8-9, в котором повторяющаяся нуклеотидная область второй нуклеиновой кислоты дополнительно определена как (i) четыре или более последовательно повторяющихся нуклеотида, (ii) четыре или более последовательных нуклеотида G или C и/или (iii) четыре или более последовательных нуклеотида Т или С.
11. В одном аспекте способов 1-10 нуклеотидная область СЕА дополнительно определена как (i) четыре или более последовательно повторяющихся нуклеотида, (ii) четыре или более последовательных нуклеотида G или C и/или (iii) четыре или более последовательных нуклеотида Т или С.
12. В одном аспекте способов 1-11 область повторов СЕА дополнительно определена как (i) четыре или более последовательных нуклеотида G, (ii) четыре или более последовательных нуклеотида C и/или (iii) четыре или более последовательных нуклеотида T.
13. В одном аспекте способов 1-12 замена представляет собой по меньшей мере 2, 3, 4, 5 или 10 повторяющихся нуклеотидных областей нуклеиновой кислоты СЕА.
14. В одном аспекте способов 1-13 нуклеиновая кислота СЕА содержит SEQ ID NO: 14.
15. В одном аспекте способов 1-14 способ дополнительно включает замену по меньшей мере одного нуклеотида в повторяющейся нуклеотидной области нуклеиновой кислоты, кодирующей костимуляторную молекулу, выбранную из B7-1, ICAM-1 и/или LFA-3, СЕА, причем повторяющаяся нуклеотидная область определена как: (i) три или более последовательно повторяющихся нуклеотида, (ii) три или более последовательных нуклеотида G или C и/или (iii) три или более последовательных нуклеотида T или C; и вставку в рекомбинантный поксвирус нуклеиновой кислоты, кодирующей костимуляторную молекулу.
16. В одном аспекте способов 1-15 повторяющаяся область костимуляторной молекулы дополнительно определена как (i) три или более последовательных нуклеотида G, (ii) три или более последовательных нуклеотида C и/или (iii) три или более последовательных нуклеотида T.
17. В одном аспекте способов 1-16 повторяющаяся нуклеотидная область костимуляторной молекулы дополнительно определена как (i) четыре или более последовательно повторяющихся нуклеотида, (ii) четыре или более последовательных нуклеотида G или C и/или (iii) четыре или более последовательных нуклеотида Т или С.
18. В одном аспекте способов 1-17 область повторов костимуляторной молекулы дополнительно определена как (i) четыре или более последовательных нуклеотида G, (ii) четыре или более последовательных нуклеотида C и/или (iii) четыре или более последовательных нуклеотида T.
19. В одном аспекте способов 1-18 замена представляет собой по меньшей мере 2, 3, 4, 5 или 10 повторяющихся нуклеотидных областей нуклеиновой кислоты костимуляторной молекулы.
20. В одном аспекте способов 1-19 нуклеиновая кислота, кодирующая костимуляторную молекулу, выбрана из B7-1 (SEQ ID NO:15-17); ICAM-1 (SEQ ID NO:18-20) и LFA-3 (SEQ ID NO:21-23).
21. В одном аспекте способов 1-20 нуклеиновая кислота, кодирующая костимуляторную молекулу, по меньшей мере на 80%, на 85%, на 90% или на 95% гомологична по меньшей мере одному из B7-1 (SEQ ID NO:15; ICAM-1 (SEQ ID NO:18) и LFA-3 (SEQ ID NO:21).
22. В одном аспекте способов 1-21 нуклеиновая кислота, кодирующая костимуляторную молекулу, по меньшей мере на 80%, на 85%, на 90% или на 95% гомологична по меньшей мере одному из B7-1 (SEQ ID NO:17; ICAM-1 (SEQ ID NO:20) и LFA-3 (SEQ ID NO:23).
23. В одном аспекте способов 1-22 нуклеиновая кислота, кодирующая костимуляторную молекулу, содержит по меньшей мере одно из B7-1 (SEQ ID NO:17; ICAM-1 (SEQ ID NO:20) и LFA-3 (SEQ ID NO:23)
24. В одном аспекте способов 1-23 нуклеиновая кислота, кодирующая костимуляторную молекулу, содержит: B7-1 (SEQ ID NO:15; ICAM-1 (SEQ ID NO:18) и LFA-3 (SEQ ID NO:21).
25. В одном аспекте способов 1-24 первая нуклеиновая кислота, кодирующая MUC1, выбрана из SEQ ID NO: 31, 32, 33 и 34.
26. Согласно данному раскрытию, рекомбинантный поксвирус способов 1-26 может быть выбран из ортопоксвируса или авипоксвируса. В предпочтительных вариантах осуществления изобретения ортопоксвирус выбран из вируса осповакцины, вируса MVA, MVA-BN и производных MVA-BN. В более предпочтительном варианте осуществления изобретения ортопоксвирус представляет собой либо MVA, MVA-BN, либо производное от MVA-BN. В еще одном более предпочтительном варианте осуществления изобретения авипоксвирус представляет собой вирус оспы птиц.
[0139] В других вариантах осуществления изобретения предложено использование a) нуклеиновой кислоты, b) кассеты экспрессии, c) композиции, d) клетки-хозяина или e) вектора согласно данному раскрытию в способе получения рекомбинантного поксвируса который стабилен при последовательном пассировании поксвируса.
[0140] В других вариантах осуществления изобретения предложено использование а) рекомбинантного поксвируса, b) нуклеиновой кислоты, c) кассеты экспрессии, d) композиции, d) клетки-хозяина или е) вектора согласно данному раскрытию в приготовлении лекарственного средства, предпочтительно вакцины.
[0141] В других вариантах осуществления изобретения предложен рекомбинантный поксвирус, b) нуклеиновая кислота, b) кассета экспрессии, c) композиция, d) клетка-хозяин или e) вектор согласно настоящему изобретению для применения в качестве лекарственного средства, предпочтительно вакцины.
[0142] В других дополнительных вариантах осуществления изобретения предложен рекомбинантный поксвирус, b) нуклеиновая кислота, b) кассета экспрессии, c) композиция, d) клетка-хозяин или e) вектор согласно данного раскрытия для использования в способе для введение кодирующей последовательности в клетку-мишень.
ПРИМЕРЫ
[0143] Следующие примеры иллюстрируют изобретение, но, конечно, не должны рассматриваться как ограничивающие объем формулы изобретения.
Пример 1: Создание Рекомбинантных Поксвирусов
[0144] Создание поксвирусов, кодирующих MUC1 (например, mBN399, mBN400, mBN336, mBN373 и mBN420), осуществляли путем вставки указанных последовательностей нуклеиновых кислот MUC1 и CEA с их промоторами посредством одновременной инфекции и трансфекции культур CEF с последующей допустимой гомологичной рекомбинацией между вирусным геномом и рекомбинационной плазмидой pBN146. Вирус, несущий вставку, был выделен, охарактеризован и был подготовлен пул вируса.
[0145] Для конструирования mBN398 и mBN400 использовали рекомбинационную плазмиду MVA, содержащую гомологичные последовательности, которые также присутствуют в вирусе осповакцины в IGR88/89. Нуклеотидная последовательность MUC1 и СЕА была встроена между последовательностями вируса осповакцины в IGR 88/89 для обеспечения возможности рекомбинации в вирусный геном осповакцины. Таким образом, была сконструирована плазмида, которая содержала нуклеотидную последовательность MUC1 и СЕА после промотора поксвируса. Для mBN398 и mBN400 были использованы SEQ ID NO: 1 (MUC1) и SEQ ID NO:13 (CEA). В качестве промоторов для MUC1 и CEA в mBN398 были использованы промотор PrS (MUC1) и промотор 40k-MVA1 (CEA) соответственно. В качестве промоторов для MUC1 и CEA в mBN400 были использованы промотор Pr13.5long (MUC1) и промотор PrS5E (CEA), соответственно. Костимулирующие молекулы TRICOM были включены в состав mBN398 и mBN400. Эти последовательности включали: B7-1, ICAM-1 и LFA-3 и содержат SEQ ID NO: 16, 19 и 21 соответственно.
[0146] Для конструирования mBN336 три рекомбинационные плазмиды были использованы для трех трансгенов pBN 374 (для TRICOM), pBN 515 (для CEA SEQ ID NO:13), pBN 525 (для MUC1 SEQ ID NO:2), встроенных последовательностей, которые также присутствуют в MVA (IGR88/89 (MUC1), IGR 44/45 (CEA), IGR 148/149 (TRICOM). Нуклеотидная последовательность MUC1 и CEA была встроена между вирусными последовательностями MVA для обеспечения возможности рекомбинации в вирусный геном MVA. Таким образом, была сконструирована плазмида, которая содержала нуклеотидную последовательность MUC1 и СЕА после промотора поксвируса. Для mBN336 были использованы SEQ ID NO: 2 (MUC1) и SEQ ID NO:13 (CEA). В качестве промоторов были использованы промотор PrS (для MUC1) и промотор 40k (для CEA). Костимулирующие молекулы TRICOM были включены в состав mBN336. Эти последовательности включали: B7-1, ICAM-1 и LFA-3 и содержат SEQ ID NO: 17, 20 и 23 соответственно. pBN632 содержит последовательности, которые также присутствуют в MVA (в IGR 88/89). Нуклеотидная последовательность MUC1 и CEA была встроена между вирусными последовательностями MVA для обеспечения возможности рекомбинации в вирусный геном MVA. Таким образом, была сконструирована плазмида, которая содержала нуклеотидную последовательность MUC1 и СЕА после промотора поксвируса. Для mBN420 были использованы SEQ ID NO: 5 (MUC1) и SEQ ID NO:14 (CEA). В качестве промоторов для MUC1 и CEA были использованы промотор Pr13.5 (см. публикацию патента США 2015/0299267) (MUC1) и промотор 40k MVA1 (CEA), соответственно. Костимулирующие молекулы TRICOM были включены в состав mBN420 и интегрированы в IGR 88/89. Эти последовательности включали: B7-1, ICAM-1 и LFA-3 и содержат SEQ ID NO: 15, 18 и 21 соответственно.
[0147] Для конструирования mBN373 рекомбинационная плазмида pBN563 содержит последовательности, которые также присутствуют в вирусе оспы птиц. Нуклеотидная последовательность MUC1 и СЕА была встроена между последовательностями вируса оспы птиц в область BamH1 для обеспечения возможности рекомбинации в геном вируса оспы птиц. Таким образом, была сконструирована плазмида, которая содержала нуклеотидную последовательность MUC1 и СЕА после промотора поксвируса. Для mBN373 были использованы SEQ ID NO: 3 (MUC1) и SEQ ID NO:14 (CEA). В качестве промоторов для MUC1 и CEA были использованы промотор 40K FPV-1 PrS (MUC1) и промотор 40k-MVA1 (CEA) соответственно. Костимулирующие молекулы TRICOM были включены в состав mBN373. Эти последовательности включали: B7-1, ICAM-1 и LFA-3 и содержат SEQ ID NO: 15, 18 и 21 соответственно.
[0148] Вышеуказанные рекомбинационные плазмиды также содержали селекционную кассету, содержащую синтетический промотор вируса осповакцины (Ps), ген устойчивости к лекарственному средству GPT, внутренний сайт связывания рибосомы (IRES) и усиленный зеленый флуоресцентный белок (EGFP) и ген устойчивости к лекарственному средству гуанин-ксантинфосфорибозилтрансферазы (Ecogpt) в сочетании с мономерным красным флуоресцентным белком. Все селекционные гены (GFP, NPTII и mRFP1) кодировались в составе одного бицистронного транскрипта.
[0149] Культуры CEF инокулировали вирусом осповакцины для mBN399/400, MVA-BN для mBN336, mBN420 или FPV для mBN373, и каждую культуру CEF также трансфицировали плазмидной ДНК. В свою очередь, образцы из этих клеточных культур инокулировали в культуры CEF в среде, содержащей лекарственные средства для проведения отбора, и экспрессирующие EGFP вирусные клоны выделяли путем очистки колоний. Пулы вирусов, которые росли в присутствии лекарственных средств для проведения отбора и экспрессировали EGFP, были обозначены как одно из следующего: mBN399, mBN400 (вирусы осповакцины), mBN336, mBN420 (вирус MVA) и mBN373 (вирус оспы птиц). Создание рекомбинантных вирусов и подготовка вирусного пула включали от 5 до 12 последовательных пассажей, в том числе от одной (1) до пяти (5) очисток колоний.
[0150] Рекомбинантные поксвирусы пассировали в клеточных культурах CEF в отсутствие лекарственных средств для проведения отбора. Отсутствие лекарственных средств для проведения отбора обеспечило возможность потери области, кодирующей гены для проведения отбора, gpt и EGFP и связанный с ней промотор (кассету для экспрессии) из вставленной последовательности. Рекомбинация, приводящая к потере селекционной кассеты, опосредуется областью F1 I4L и участком этой области, F1-повтором (F1 rpt), которые фланкируют селекционную кассету в плазмиде каждой конструкции. Эти дублированные последовательности были включены для обеспечения рекомбинации, что приводит к утрате селекционной кассеты, оставляя только последовательности MUC1 и CEA, встроенные в описанные межгенные области конструкций, описанных в настоящем описании.
[0151] Был приготовлен очищенный из колоний вирус, в котором отсутствует селекционная кассета. Такая подготовка включала пятнадцать (15) пассажей, включая пять (5) очисток из колоний.
[0152] Присутствие последовательности MUC1 и CEA и отсутствие родительского вируса MVA-BN в пулах mBN336, mBN420 и mBN373 было подтверждено с помощью анализа ПЦР, и гнездовая ПЦР была использована для подтверждения отсутствия селекционной кассеты (гены gpt и EGFP/NPTII и mRFP1).
[0153] Экспрессия белков MUC1 и CEA была продемонстрирована в клетках, инокулированных MVA-BN-MUC1-CEA-TRICOM in vitro.
Пример 2: ПЦР-анализ пассажей 1-7 MVA-mBN336
[0154] Генетическую стабильность MVA-mBN336B оценивали путем культивирования в течение семи пассажей. MVA-mBN336B кодирует 5 человеческих трансгенов, при этом муцин 1 человека (MUC-1) и карциноэмбриональный антиген человека (CEA) представляют собой антигены-мишени данной вакцины-кандидата, и 3 гена, кодирующих иммунные костимулирующие молекулы человека (обозначенные как триада костимулирующих молекул или TRICOM) в качестве поддержки для индукции надежного и направленного иммунного ответа: связанный с функцией лейкоцитов антиген-3 (LFA-3), молекула внутриклеточной адгезии 1 (ICAM-1) и B7-1. Трансгены были встроены в три межгенных области (IGR) MVA-BN®: IGR 44/45, содержащая CEA, IGR 88/89, содержащая hMUC1, и IGR 148/149, содержащая гены TRICOM. Экспрессия трансгена управляется промоторами вируса оспы 40k-MVA1, 30k, I3L и PrS.
[0155] Первичные клетки фибробластов куриных эмбрионов (CEF) готовили, высевали в роллерные флаконы (RB) (7×107 клеток) в среде VP-SFM и инкубировали в течение 4 дней при температуре 37°C. Среду VP-SFM заменяли 100 мл среды RPMI и клетки инфицировали при MOI приблизительно 0,3-00,1, что соответствует числу клеток 1×108 клеток/RB, и культивировали в течение 3 дней при температуре 30°C. После инкубации образцы вируса отбирали путем замораживания RB при температуре -20°C в течение по меньшей мере 16 часов с последующим оттаиванием RB для отбора суспензии клеточного вируса. Определяли точный объем клеточной суспензии, образцы вируса обрабатывали ультразвуком, а затем разделяли на аликвоты и хранили при температуре -80°С. Эту процедуру повторяли шесть раз, что привело к семи пассажам.
[0156] Анализ ПЦР встроенных трансгенов проводили для каждого пассажа после культивирования при температуре 30°С. На Фиг. 9А изображены результаты ПЦР для оценки стабильности СЕА в течение семи пассажей. На Фиг. 9В изображены результаты ПЦР для оценки стабильности MUC1 в течение семи пассажей. На Фиг. 9C изображены результаты ПЦР для оценки стабильности TRICOM в течение 7 пассажей. Рекомбинационные плазмиды, использованные для получения MVA-mBN336B были использованы в качестве положительных контролей, MVA-BN® была использована в качестве отрицательного контроля (пустой векторный остов), а в качестве контроля для реакции ПЦР была использована H2O.
[0157] На Фиг. 10А и 10В изображен анализ образца пассажа 7. На Фиг. 10А изображен результат ПЦР-амплификации образцов пассажа 7, отправленных для исследования с помощью секвенирования. Отдельные амплификации ПЦР проводили для каждого отдельного трансгена: CEA, MUC1 и TRICOM. B) Электроферограммы нуклеотидной последовательности MUC1, на которых изображены локусы, содержащие обнаруженную точечную мутацию, приводящую к сдвигу рамки считывания. Точечная мутация была обнаружена в пассаже 5 впервые при ПЦР-амплификации и электрофореграммах нуклеотидной последовательности MUC1, соответствующих локусам, содержащим обнаруженную точечную мутацию, приводящую к сдвигу рамки считывания. Точечная мутация, обнаруженная впервые в пассаже 5, представляет собой мутации, обнаруженные при анализе электрофореграммы пассажей 5, 6 и 7.
[0158] На Фиг. 9 и 10 изображено, что комбинация MUC1, CEA и TRICOM в mBN336 продемонстрировала улучшенную и повышенную стабильность по сравнению с трансгенами MUC1, CEA и TRICOM в PANVAC-V и PANVAC-F (сравните, например, Фигуру 1 и Таблицы 1 и 2 с Фигурами 3 и 4). Начиная с пассажа 5, мутация со сдвигом рамки считывания была обнаружена в небольшой популяции анализируемого материала.
[0159] Стабильность, проиллюстрированная при пассаже 4, демонстрирует способность MVA-mBN336 преодолевать проблемы со стабильностью, связанные с PANVAC, и другими попытками получить стабильный поксвирус, содержащий MUC1. Стабильность MVA-mBN336 обеспечивает дополнительное преимущество, так как производство и более крупномасштабное производство вакцин на основе MVA обычно происходит с использованием MVA при пассаже 3 или пассаже 4. Таким образом, поскольку MVA-mBN336 является стабильным при пассаже 4, можно начать крупномасштабное производство и благодаря стабильности могут быть преодолены значительные нормативные препятствия.
Пример 3: Увеличенная Стабильность FPV-mBN373
[0160] Генетическую стабильность FPV-mBN373B оценивали в течение семи пассажей. Культивирование проводили в роллерных флаконах (RB), применяемых во время крупномасштабного производства, используемого для изготовления материала для клинического исследования. Каждый пассаж анализировали на предмет титра вируса с помощью проточной цитометрии и на предмет правильного размера трансгенной вставки с помощью ПЦР. Кроме того, последний пассаж (P7) анализировали с помощью секвенирования трансгенов.
[0161] Первичные клетки фибробластов куриных эмбрионов (CEF) готовили, высевали в RB (7×107 клеток/RB) в среду VP-SFM и инкубировали в течение 3 дней при температуре 37°C. Среду VP-SFM заменяли 100 мл среды RPMI, и клетки инфицировали при MOI 0,1, что соответствует количеству клеток 1×108/RB, и культивировали в течение 4 дней при температуре 37°C. После инкубации образцы вируса отбирали путем замораживания RB при температуре -20°C в течение по меньшей мере 16 часов с последующим оттаиванием RB для отбора суспензии клеточного вируса. Определяли точный объем клеточной суспензии, образцы вируса обрабатывали ультразвуком, а затем разделяли на аликвоты и хранили при температуре -80°С. Титр патогенного вируса определяли после каждого пассажа для мониторинга титров вируса и для обеспечения возможности заражения следующего пассажа при определенной MOI. Эту процедуру повторяли шесть раз, что привело к семи пассажам.
[0162] Как изображено на Фиг. 11А, анализ ПЦР встроенных трансгенов проводили для каждого пассажа после культивирования при температуре 37°С. Рекомбинационная плазмида, используемая для получения FPV-mBN373B, использовалась в качестве положительного контроля, FPV использовалась в качестве отрицательного контроля (пустой векторный остов), а H2O использовалась в качестве контроля реакции ПЦР.
[0163] Как изображено на Фиг. 11B, секвенирование седьмого пассажа проводили после амплификации J-сайта BamHI, содержащего трансгены и по меньшей мере 600 п.н. каждой фланкирующей области. Ампликон ПЦР FPV-mBN373B, проанализированный при седьмом пассаже (37°C), соответствовал ожидаемому размеру продукта 5566 п.н. (PCR1) и 5264 п.н. (PCR2), охватывая встроенные трансгены и по меньшей мере 600 п.н. каждой фланкирующей области. Результаты продемонстрировали 100% идентичность сконструированной последовательности по сравнению с теоретической последовательностью, подтверждая генетическую стабильность FPV-mBN373B в течение 7 пассажей при температуре 37°C.
[0164] По меньшей мере, в одном аспекте результирующая стабильность трансгена MUC1, SEQ ID NO: 3, в mBN373 была неожиданной, поскольку и mBN373, и mBN336 содержат SEQ ID NO:3. Соответственно, хотя MUC1 SEQ ID NO:3 начинает проявлять нестабильность при пассаже 5 в mBN336 (вирус MVA), тот же SEQ ID NO:3 стабилен в mBN373 (вирус оспы птиц), по крайней мере, до пассажа 7 включительно.
Пример 4: Стабильность MVA-mBN420
[0165] Генетическую стабильность MVA-mBN 420 оценивали в течение семи пассажей. Культивирование проводили в роллерных флаконах (RB), применяемых во время крупномасштабного производства, используемого для изготовления материала для клинического исследования. Исследование проводилось при температурах 30°C и 34°C с использованием MOI приблизительно от 0,05 до 0,1 и периода инкубации вируса 4 дня, поскольку эти условия характерны для типичного крупномасштабного производства, используемого для изготовления материала для клинического исследования. Каждый пассаж анализировали на предмет титра вируса с помощью проточной цитометрии и на предмет правильного размера трансгенной вставки с помощью ПЦР.
[0166] Первичные клетки фибробластов куриных эмбрионов (CEF) готовили, высевали в RB (7×107 клеток/RB) в среду VP-SFM и инкубировали в течение 3 дней при температуре 37°C. Среду VP-SFM заменяли 100 мл среды RPMI, и клетки инфицировали при MOI от 0,05 до 0,1, что соответствует количеству клеток 1×108/RB, и культивировали в течение 4 дней при температурах 30°C и 34°C. После инкубации образцы вируса отбирали путем замораживания RB при температуре -20°C в течение по меньшей мере 16 часов с последующим оттаиванием RB для отбора суспензии клеточного вируса. Образцы вируса обрабатывали ультразвуком, а затем разделяли на аликвоты и хранили при температуре -80°С. Титр патогенного вируса определяли после каждого пассажа для мониторинга титров вируса и для обеспечения возможности заражения следующего пассажа при определенной MOI. Эту процедуру повторяли шесть раз, что привело к семи пассажам.
[0167] Анализ ПЦР встроенных трансгенов проводили для каждого пассажа после культивирования при температуре 30°С. Результаты пассирования, выполненного при температуре 30°С, изображены на Фиг. 12. Рекомбинационная плазмида, используемая для создания mBN420, использовалась в качестве положительного контроля, MVA-BN использовалась в качестве отрицательного контроля (пустой векторный остов), а H2O использовалась в качестве контроля реакции ПЦР.
[0168] Как изображено на Фиг. 12, стабильность MVA в mBN420 была снижена по сравнению с MVA в mBN336 и вирусом оспы птиц в mBN373.
Пример 5: Увеличенная Стабильность дополнительных рекомбинантных вирусов MVA и рекомбинантных вирусов оспы птиц, кодирующих MUC1 и CEA
[0169] Получение дополнительных рекомбинантных вирусов MVA и рекомбинантных вирусов оспы птиц согласно настоящему изобретению проводят, как описано в Примере 1. Нуклеиновые кислоты, кодирующие трансгены MUC1, CEA и TRICOM, содержащие SEQ ID NO: 31, 32, 33 или 34 (для MUC1) и SEQ ID NO:13 или 14 (для CEA), встраивают в MVA-BN, как описано в Примере 1. Кроме того, TRICOM встраивают в MVA, последовательности TRICOM, включая SEQ ID NO:15 или 17 (для B7.1), SEQ ID NO:18 или 20 (для ICAM-1) и SEQ ID NO:21 или 23 (для LFA-3) встраивают в MVA, как описано в Примере 1.
[0170] Кроме того, нуклеиновые кислоты, кодирующие трансгены MUC1 и CEA, содержащие SEQ ID NO: 31, 32, 33 или 34 (для MUC1) и SEQ ID NO:13 или 14 (для CEA), встраивают в MVA-BN, как описано в Примере 1. Кроме того, TRICOM встраивают в вирус оспы птиц, последовательности TRICOM, включающие SEQ ID NO:15 или 17 (для B7.1), SEQ ID NO:18 или 20 (для ICAM-1) и SEQ ID NO:21 или 23 (для LFA-3) встраивают в вирус оспы птиц, как описано в Примере 1.
[0171] SEQ ID NO:31, 32, 33 или 34, каждый, кодируют пептид MUC1, содержащий SEQ ID NO:35.
[0172] Новые нуклеиновые кислоты MUC1 SEQ ID NO: 31, 32, 33 и 34 каждая кодируют вариации нуклеиновых кислот согласно настоящему изобретению без агонистических эпитопов согласно WO 2013/103658. В некоторых аспектах замена и/или удаление агонистических эпитопов не влияет на стабильность рекомбинантных поксвирусов согласно настоящему изобретению, так как присутствие агонистических эпитопов функционирует для усиления иммуногенности MUC1, а не стабильности или нестабильности.
[0173] Экспрессия белков MUC1, CEA и TRICOM демонстрируется в клетках, инокулированных MVA-BN-MUC1-CEA-TRICOM in vitro, как описано в Примере 1.
[0174] Увеличенная генетическая стабильность трансгенов в вирусах MVA и/или оспы птиц оценивается в течение семи пассажей. Культивирование проводится в роллерных флаконах (RB), применяемых во время крупномасштабного производства, используемого для изготовления материала для клинического исследования. Исследование проводится при температурах 30°C, 34°C или 37°C (в зависимости от используемой векторной системы) с использованием MOI приблизительно 00.05-00.1 и периода инкубации вируса 2, 3, 4, 5, 6 или 7 дней, поскольку эти условия соответствуют типичному крупномасштабному производству, используемому для изготовления материала для клинического исследования. Каждый пассаж анализируется на предмет титра вируса с помощью проточной цитометрии и на предмет правильного размера трансгенной вставки с помощью ПЦР. Кроме того, последний пассаж (P7) анализируется с помощью секвенирования трансгенов.
[0175] Первичные клетки фибробластов куриных эмбрионов (CEF) готовят, высевают в RB (7×107 клеток/RB) в среду VP-SFM и инкубируют в течение 3 дней при температуре 37°C. Среду VP-SFM заменяют 100 мл среды RPMI, и клетки инфицируют при MOI от 0,005 до 0,1 и культивируют в течение 4 дней при температурах 30°C, 34°C или 37°C (в зависимости от используемой векторной системы). После инкубации образцы вируса отбирают путем замораживания RB при температуре -20°C в течение по меньшей мере 16 часов с последующим оттаиванием RB для отбора суспензии клеточного вируса. Образцы вируса обрабатываются ультразвуком, а затем разделяют на аликвоты и хранят при температуре -80°С. Титр патогенного вируса определяют после каждого пассажа для мониторинга титров вируса и для обеспечения возможности заражения следующего пассажа при определенной MOI. Эту процедуру повторяют шесть раз, что приводит к семи пассажам.
[0176] Анализ ПЦР встроенных трансгенов проводят для каждого пассажа после культивирования при температурах 30°С, 34°С или 37°С (в зависимости от векторной системы). Рекомбинационная плазмида, используемая для получения каждого соответствующего поксвируса (например, вируса MVA-BN или вируса оспы птиц), используется в качестве положительного контроля, MVA-BN или вирус оспы птиц используется в качестве отрицательного контроля (пустой векторный остов), а H2O используется в качестве контроля для реакции ПЦР.
[0177] Секвенирование седьмого пассажа проводят после амплификации сайта IGR, содержащего трансгены и по меньшей мере 600 п.н. каждой фланкирующей области. Ампликон ПЦР каждой конструкции анализируют при седьмом пассаже. Секвенирование нуклеиновых кислот MUC1, CEA и/или TRICOM проводят для подтверждения того, что вирус MVA и/или вирус оспы птиц стабилен в отношении трансгенов.
[0178] Будет очевидно, что точные детали способов или композиций, описанных в настоящем описании, могут быть изменены или модифицированы без отклонения от сущности описанного изобретения. Мы заявляем о всех таких модификациях и вариациях, которые входят в объем и сущность приведенной ниже формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> BAVARIAN NORDIC A/S
<120> КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ПОВЫШЕНИЯ СТАБИЛЬНОСТИ ТРАНСГЕНОВ В
ПОКСВИРУСАХ
<130> BNIT0011PCT
<140>
<141>
<160> 35
<170> PatentIn версия 3.5
<210> 1
<211> 1548
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность MUC1 из PANVAC
<400> 1
atgacaccgg gcacccagtc tcctttcttc ctgctgctgc tcctcacagt gcttacagtt 60
gttacgggtt ctggtcatgc aagctctacc ccaggtggag aaaaggagac ttcggctacc 120
cagagaagtt cagtgcccag ctctactgag aagaatgctg tgagtatgac aagctccgta 180
ctctccagcc acagccccgg ttcaggctcc tccaccactc agggacagga tgtcactctg 240
gccccggcca cggaaccagc ttcaggttca gctgccttgt ggggacagga tgtcacctcg 300
gtaccagtta ctagaccagc tttaggtagc acagcacctc ctgctcatgg agtaactagt 360
gctcctgata ctcgtccagc tcctggcagt actgcaccac cggcacatgg cgtaacatca 420
gcacctgata caagacctgc acctggatct acagcgccgc ctgcgcacgg agtgacatcg 480
gcgcccgata cgcgccccgc tcccggtagc accgcaccgc ccgcccacgg tgttacaagt 540
gcacccgata cccggccggc acccggaagt accgctccac ctgcacacgg ggtcacaagc 600
gcgccagaca ctcgacctgc gccagggtcg actgcccctc cggcgcatgg tgtgacctca 660
gctcctgaca caaggccagc cccagctagc actctggtgc acaacggcac ctctgccagg 720
gctaccacaa ccccagccag caagagcact ccattctcaa ttcccagcca ccactctgat 780
actcctacca cccttgccag ccatagcacc aagactgatg ccagtagcac tcaccatagc 840
acggtacctc ctctcacctc ctccaatcac agcacttctc cccagttgtc tactggggtc 900
tctttctttt tcctgtcttt tcacatttca aacctccagt ttaattcctc tctggaagat 960
cccagcaccg actactacca agagctgcag agagacattt ctgaaatgtt tttgcagatt 1020
tataaacaag ggggttttct gggcctctcc aatattaagt tcaggccagg atctgtggtg 1080
gtacaattga ctctggcctt ccgagaaggt accatcaatg tccacgacgt ggagacacag 1140
ttcaatcagt ataaaacgga agcagcctct cgatataacc tgacgatctc agacgtcagc 1200
gtgagtgatg tgccatttcc tttctctgcc cagtctgggg ctggggtgcc aggctggggc 1260
atcgcgctgc tggtgctggt ctgtgttctg gttgcgctgg ccattgtcta tctcattgcc 1320
ttggctgtct gtcagtgccg ccgaaagaac tacgggcagc tggacatctt tccagcccgg 1380
gatacctacc atcctatgag cgagtacccc acctaccaca cccatgggcg ctatgtgccc 1440
cctagcagta ccgatcgtag cccctatgag aaggtttctg caggtaatgg tggcagcagc 1500
ctctcttaca caaacccagc agtggcagcc acttctgcca acttgtag 1548
<210> 2
<211> 1365
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность MUC1 из mBN336
<400> 2
atgacaccgg gcacccagtc tcctttcttc ctgctgctgc tcctcacagt gcttacagtt 60
gttacgggtt ctggtcatgc aagctctacc ccaggtggag aaaaggagac ttcggctacc 120
cagagaagtt cagtgcccag ctctactgag aagaatgctg tgagtatgac aagctccgta 180
ctctccagcc acagcccagg ttcaggctcc tccaccactc agggacagga tgtcactctg 240
gcaccggcca cggaaccagc ttcaggttca gctgccttgt ggggacagga tgtcacctcg 300
gtaccagtta ctagaccagc tttaggctac ctggcgccac cggctcatgg cgttacatcg 360
tatttggaca ctcgaccggc accagttagc acagcacctc ccgcacacgg tgtaactagc 420
gcgcctgata cacgtcccgc tcccggatct accgctccgc cagcgcacgg agtgacgtca 480
gcaccagata cgaggccagc gcctgctagc actctggtgc acaacggcac ctctgccagg 540
gctaccacaa ccccagccag caagagcact ccattctcaa ttcccagcca ccactctgat 600
actcctacca cccttgccag ccatagcacc aagactgatg ccagtagcac tcaccatagc 660
acggtacctc ctctcacctc ctccaatcac agcacttctc cccagttgtc tactggggtc 720
tctttctttt tcctgtcttt tcacatttca aacctccagt ttaattcctc tctggaagat 780
cccagcaccg actactacca agagctgcag agagacattt ctgaaatgtt tttgcagatt 840
tataaacaag ggggttttct gggcctctcc aatattaagt tcaggccagg atctgtggtg 900
gtacagttga ctctggcctt ccgagaaggt accatcaatg tccacgacgt ggagacacag 960
ttcaatcagt ataaaacgga agcagcctct cgatataacc tgacgatctc agacgtcagc 1020
gtgagtgatg tgccatttcc tttctctgcc cagtctgggg ctggggtgcc aggctggggc 1080
atcgcgctgc tggtgctggt ctgtgttctg gtttacctgg ccattgtcta tctcattgcc 1140
ttggctgtct gtcaggtccg ccgaaagaac tacgggcagc tggacatctt tccagcccgg 1200
gatacctacc atcctatgag cgagtacccc acctaccaca cccatgggcg ctatgtgccc 1260
cctagcagtc tgttccgtag cccctatgag aaggtttctg caggtaatgg tggcagctac 1320
ctctcttaca caaacccagc agtggcagcc acttctgcca acttg 1365
<210> 3
<211> 1365
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность MUC1 из mBN373
<400> 3
atgacaccgg gcacccagtc tcctttcttc ctgctgctgc tcctcacagt gcttacagtt 60
gttacgggtt ctggtcatgc aagctctacc ccaggtggag aaaaggagac ttcggctacc 120
cagagaagtt cagtgcccag ctctactgag aagaatgctg tgagtatgac aagctccgta 180
ctctccagcc acagcccagg ttcaggctcc tccaccactc agggacagga tgtcactctg 240
gcaccggcca cggaaccagc ttcaggttca gctgccttgt ggggacagga tgtcacctcg 300
gtaccagtta ctagaccagc tttaggctac ctggcgccac cggctcatgg cgttacatcg 360
tatttggaca ctcgaccggc accagttagc acagcacctc ccgcacacgg tgtaactagc 420
gcgcctgata cacgtcccgc tcccggatct accgctccgc cagcgcacgg agtgacgtca 480
gcaccagata cgaggccagc gcctgctagc actctggtgc acaacggcac ctctgccagg 540
gctaccacaa ccccagccag caagagcact ccattctcaa ttcccagcca ccactctgat 600
actcctacca cccttgccag ccatagcacc aagactgatg ccagtagcac tcaccatagc 660
acggtacctc ctctcacctc ctccaatcac agcacttctc cccagttgtc tactggggtc 720
tctttctttt tcctgtcttt tcacatttca aacctccagt ttaattcctc tctggaagat 780
cccagcaccg actactacca agagctgcag agagacattt ctgaaatgtt tttgcagatt 840
tataaacaag ggggttttct gggcctctcc aatattaagt tcaggccagg atctgtggtg 900
gtacagttga ctctggcctt ccgagaaggt accatcaatg tccacgacgt ggagacacag 960
ttcaatcagt ataaaacgga agcagcctct cgatataacc tgacgatctc agacgtcagc 1020
gtgagtgatg tgccatttcc tttctctgcc cagtctgggg ctggggtgcc aggctggggc 1080
atcgcgctgc tggtgctggt ctgtgttctg gtttacctgg ccattgtcta tctcattgcc 1140
ttggctgtct gtcaggtccg ccgaaagaac tacgggcagc tggacatctt tccagcccgg 1200
gatacctacc atcctatgag cgagtacccc acctaccaca cccatgggcg ctatgtgccc 1260
cctagcagtc tgttccgtag cccctatgag aaggtttctg caggtaatgg tggcagctac 1320
ctctcttaca caaacccagc agtggcagcc acttctgcca acttg 1365
<210> 4
<211> 1365
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность MUC1 из mBN398 и mBN400
<400> 4
atgacacctg gcactcagtc accattcttc ctgctgttac tcttgacagt gcttacagtt 60
gttacaggtt ctggtcatgc aagctctacc ccaggtggag aaaaggagac ttcggctacc 120
cagcggagtt cagtgcctag ctctactgag aagaatgctg tgagtatgac aagctccgta 180
ctctccagcc acagcccagg ttcaggctcc agcaccactc aaggacagga tgtcactctg 240
gcaccggcca cggaaccagc ttcaggttca gctgccttgt ggggacagga tgtcacatcg 300
gtaccagtta ctagaccagc tttaggcagt actgcgccac cggctcatgg cgttacatcg 360
gcacctgaca ctcgaccggc accaggtagc acagcacctc ccgcacacgg tgtaactagc 420
gcgcctgata cacgtcccgc tcccggatct accgctccgc cagcgcacgg agtgacgtca 480
gcaccagata cgaggccagc gcctgctagc actctggtgc acaatggcac atctgccagg 540
gctaccacaa ctccagccag caagagcact ccattctcaa ttccaagcca tcactctgat 600
actcctacca cacttgccag ccatagcacc aagactgatg ccagtagcac tcaccatagc 660
acggtacctc cactcacctc atccaatcac agcacttctc ctcagttgtc tactggagtc 720
tccttctttt tcctgtcctt tcacatttca aacttgcagt tcaattcttc cctggaagat 780
cccagcaccg actactacca agagctgcag agagacattt ctgagatgtt cttgcagatt 840
tataaacaag gtggattcct tggcctctct aatattaagt tcaggccagg atctgtggtc 900
gtacagttga ctctggcctt cagagaaggt accatcaatg tccacgacgt ggagacacag 960
ttcaatcagt ataagacgga agcagcctca cgatataacc tgacgatctc agacgtcagc 1020
gttagtgatg tgccatttcc tttctctgcc cagtctggag ctggtgtgcc aggctggggc 1080
atcgcgctgc tcgtgttggt ctgtgttctg gttgcgctgg ccattgtcta tctcattgcc 1140
ttggctgttt gtcagtgcag acgcaagaac tacggacagc tggacatctt tccagctcgg 1200
gatacctacc atcctatgag cgagtaccct acctaccaca cacatggtcg ctatgtgcca 1260
cctagcagta ccgatcgtag tccctatgag aaagtttctg caggtaatgg tggcagcagc 1320
ctctcttaca caaacccagc agtggcagcc acttctgcca acttg 1365
<210> 5
<211> 1365
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Нуклеотидная последовательность MUC1 из mBN420
<400> 5
atgacacctg gcactcagtc accattcttc ctgctgttac tcttgacagt gcttacagtt 60
gttacaggtt ctggtcatgc aagctctacc ccaggtggag aaaaggagac ttcggctacc 120
cagcggagtt cagtgcctag ctctactgag aagaatgctg tgagtatgac aagctccgta 180
ctctccagcc acagcccagg ttcaggctcc agcaccactc aaggacagga tgtcactctg 240
gcaccggcca cggaaccagc ttcaggttca gctgccttgt ggggacagga tgtcacatcg 300
gtaccagtta ctagaccagc tttaggctac ctggcgccac cggctcatgg cgttacatcg 360
tatttggaca ctcgaccggc accagttagc acagcacctc ccgcacacgg tgtaactagc 420
gcgcctgata cacgtcccgc tcccggatct accgctccgc cagcgcacgg agtgacgtca 480
gcaccagata cgaggccagc gcctgctagc actctggtgc acaatggcac atctgccagg 540
gctaccacaa ctccagccag caagagcact ccattctcaa ttccaagcca tcactctgat 600
actcctacca cacttgccag ccatagcacc aagactgatg ccagtagcac tcaccatagc 660
acggtacctc cactcacctc atccaatcac agcacttctc ctcagttgtc tactggagtc 720
tccttctttt tcctgtcctt tcacatttca aacttgcagt tcaattcttc cctggaagat 780
cccagcaccg actactacca agagctgcag agagacattt ctgagatgtt cttgcagatt 840
tataaacaag gtggattcct tggcctctct aatattaagt tcaggccagg atctgtggtc 900
gtacagttga ctctggcctt cagagaaggt accatcaatg tccacgacgt ggagacacag 960
ttcaatcagt ataagacgga agcagcctca cgatataacc tgacgatctc agacgtcagc 1020
gttagtgatg tgccatttcc tttctctgcc cagtctggag ctggtgtgcc aggctggggc 1080
atcgcgctgc tcgtgttggt ctgtgttctg gtttacctgg ccattgtcta tctcattgcc 1140
ttggctgttt gtcaggtcag acgcaagaac tacggacagc tggacatctt tccagctcgg 1200
gatacctacc atcctatgag cgagtaccct acctaccaca cacatggtcg ctatgtgcca 1260
cctagcagtc tgttccgtag tccctatgag aaagtttctg caggtaatgg tggcagctac 1320
ctctcttaca caaacccagc agtggcagcc acttctgcca acttg 1365
<210> 6
<211> 515
<212> PRT
<213> Искусственная Последовательность
<220>
<223> Аминокислотная последовательность MUC 1, обнаруженная в PANVAC
<400> 6
Met Thr Pro Gly Thr Gln Ser Pro Phe Phe Leu Leu Leu Leu Leu Thr
1. 5 10 15
Val Leu Thr Val Val Thr Gly Ser Gly His Ala Ser Ser Thr Pro Gly
20 25 30
Gly Glu Lys Glu Thr Ser Ala Thr Gln Arg Ser Ser Val Pro Ser Ser
35 40 45
Thr Glu Lys Asn Ala Val Ser Met Thr Ser Ser Val Leu Ser Ser His
50 55 60
Ser Pro Gly Ser Gly Ser Ser Thr Thr Gln Gly Gln Asp Val Thr Leu
65 70 75 80
Ala Pro Ala Thr Glu Pro Ala Ser Gly Ser Ala Ala Leu Trp Gly Gln
85 90 95
Asp Val Thr Ser Val Pro Val Thr Arg Pro Ala Leu Gly Ser Thr Ala
100 105 110
Pro Pro Ala His Gly Val Thr Ser Ala Pro Asp Thr Arg Pro Ala Pro
115 120 125
Gly Ser Thr Ala Pro Pro Ala His Gly Val Thr Ser Ala Pro Asp Thr
130 135 140
Arg Pro Ala Pro Gly Ser Thr Ala Pro Pro Ala His Gly Val Thr Ser
145 150 155 160
Ala Pro Asp Thr Arg Pro Ala Pro Gly Ser Thr Ala Pro Pro Ala His
165 170 175
Gly Val Thr Ser Ala Pro Asp Thr Arg Pro Ala Pro Gly Ser Thr Ala
180 185 190
Pro Pro Ala His Gly Val Thr Ser Ala Pro Asp Thr Arg Pro Ala Pro
195 200 205
Gly Ser Thr Ala Pro Pro Ala His Gly Val Thr Ser Ala Pro Asp Thr
210 215 220
Arg Pro Ala Pro Ala Ser Thr Leu Val His Asn Gly Thr Ser Ala Arg
225 230 235 240
Ala Thr Thr Thr Pro Ala Ser Lys Ser Thr Pro Phe Ser Ile Pro Ser
245 250 255
His His Ser Asp Thr Pro Thr Thr Leu Ala Ser His Ser Thr Lys Thr
260 265 270
Asp Ala Ser Ser Thr His His Ser Thr Val Pro Pro Leu Thr Ser Ser
275 280 285
Asn His Ser Thr Ser Pro Gln Leu Ser Thr Gly Val Ser Phe Phe Phe
290 295 300
Leu Ser Phe His Ile Ser Asn Leu Gln Phe Asn Ser Ser Leu Glu Asp
305 310 315 320
Pro Ser Thr Asp Tyr Tyr Gln Glu Leu Gln Arg Asp Ile Ser Glu Met
325 330 335
Phe Leu Gln Ile Tyr Lys Gln Gly Gly Phe Leu Gly Leu Ser Asn Ile
340 345 350
Lys Phe Arg Pro Gly Ser Val Val Val Gln Leu Thr Leu Ala Phe Arg
355 360 365
Glu Gly Thr Ile Asn Val His Asp Val Glu Thr Gln Phe Asn Gln Tyr
370 375 380
Lys Thr Glu Ala Ala Ser Arg Tyr Asn Leu Thr Ile Ser Asp Val Ser
385 390 395 400
Val Ser Asp Val Pro Phe Pro Phe Ser Ala Gln Ser Gly Ala Gly Val
405 410 415
Pro Gly Trp Gly Ile Ala Leu Leu Val Leu Val Cys Val Leu Val Ala
420 425 430
Leu Ala Ile Val Tyr Leu Ile Ala Leu Ala Val Cys Gln Cys Arg Arg
435 440 445
Lys Asn Tyr Gly Gln Leu Asp Ile Phe Pro Ala Arg Asp Thr Tyr His
450 455 460
Pro Met Ser Glu Tyr Pro Thr Tyr His Thr His Gly Arg Tyr Val Pro
465 470 475 480
Pro Ser Ser Thr Asp Arg Ser Pro Tyr Glu Lys Val Ser Ala Gly Asn
485 490 495
Gly Gly Ser Ser Leu Ser Tyr Thr Asn Pro Ala Val Ala Ala Thr Ser
500 505 510
Ala Asn Leu
515
<210> 7
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность VNTR #2 из PANVAC
<400> 7
ggcagtactg caccaccggc acatggcgta acatcagcac ctgatacaag acctgcacct 60
<210> 8
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность VNTR #1 из mBN420 и mBN336
<400> 8
ggctacctgg cgccaccggc tcatggcgtt acatcgtatt tggacactcg accggcacca 60
<210> 9
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность VNTR #1 из PANVAC
<400> 9
ggtagcacag cacctcctgc tcatggagta actagtgctc ctgatactcg tccagctcct 60
<210> 10
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность VNTR #2 из mBN420 и mBN336
<400> 10
gttagcacag cacctcccgc acacggtgta actagcgcgc ctgatacacg tcccgctccc 60
<210> 11
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность VNTR #3 из PANVAC
<400> 11
ggatctacag cgccgcctgc gcacggagtg acatcggcgc ccgatacgcg ccccgctccc 60
<210> 12
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность VNTR #3 из mBN420 и mBN336
<400> 12
ggatctaccg ctccgccagc gcacggagtg acgtcagcac cagatacgag gccagcgcct 60
<210> 13
<211> 2103
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность CEA из PANVAC
<400> 13
atggagtctc cctcggcccc tccccacaga tggtgcatcc cctggcagag gctcctgctc 60
acagcctcac ttctaacctt ctggaacccg cccaccactg ccaagctcac tattgaatcc 120
acgccgttca atgtcgcaga ggggaaggag gtgcttctac ttgtccacaa tctgccccag 180
catctttttg gctacagctg gtacaaaggt gaaagagtgg atggcaaccg tcaaattata 240
ggatatgtaa taggaactca acaagctacc ccagggcccg catacagtgg tcgagagata 300
atatacccca atgcatccct gctgatccag aacatcatcc agaatgacac aggattctac 360
accctacacg tcataaagtc agatcttgtg aatgaagaag caactggcca gttccgggta 420
tacccggaac tccctaagcc ttctattagc tccaataata gtaagcctgt cgaagacaaa 480
gatgccgtcg cttttacatg cgagcccgaa actcaagacg caacatatct ctggtgggtg 540
aacaaccagt ccctgcctgt gtcccctaga ctccaactca gcaacggaaa tagaactctg 600
accctgttta acgtgaccag gaacgacaca gcaagctaca aatgcgaaac ccaaaatcca 660
gtcagcgcca ggaggtctga ttcagtgatt ctcaacgtgc tttacggacc cgatgctcct 720
acaatcagcc ctctaaacac aagctataga tcaggggaaa atctgaatct gagctgtcat 780
gccgctagca atcctcccgc ccaatacagc tggtttgtca atggcacttt ccaacagtcc 840
acccaggaac tgttcattcc caatattacc gtgaacaata gtggatccta cacgtgccaa 900
gctcacaata gcgacaccgg actcaaccgc acaaccgtga cgacgattac cgtgtatgag 960
ccaccaaaac cattcataac tagtaacaat tctaacccag ttgaggatga ggacgcagtt 1020
gcattaactt gtgagccaga gattcaaaat accacttatt tatggtgggt caataaccaa 1080
agtttgccgg ttagcccacg cttgcagttg tctaatgata accgcacatt gacactcctg 1140
tccgttactc gcaatgatgt aggaccttat gagtgtggca ttcagaatga attatccgtt 1200
gatcactccg accctgttat ccttaatgtt ttgtatggcc cagacgaccc aactatatct 1260
ccatcataca cctactaccg tcccggcgtg aacttgagcc tttcttgcca tgcagcatcc 1320
aacccccctg cacagtactc ctggctgatt gatggaaaca ttcagcagca tactcaagag 1380
ttatttataa gcaacataac tgagaagaac agcggactct atacttgcca ggccaataac 1440
tcagccagtg gtcacagcag gactacagtt aaaacaataa ctgtttccgc ggagctgccc 1500
aagccctcca tctccagcaa caactccaaa cccgtggagg acaaggatgc tgtggccttc 1560
acctgtgaac ctgaggctca gaacacaacc tacctgtggt gggtaaatgg tcagagcctc 1620
ccagtcagtc ccaggctgca gctgtccaat ggcaacagga ccctcactct attcaatgtc 1680
acaagaaatg acgcaagagc ctatgtatgt ggaatccaga actcagtgag tgcaaaccgc 1740
agtgacccag tcaccctgga tgtcctctat gggccggaca cccccatcat ttccccccca 1800
gactcgtctt acctttcggg agcggacctc aacctctcct gccactcggc ctctaaccca 1860
tccccgcagt attcttggcg tatcaatggg ataccgcagc aacacacaca agttctcttt 1920
atcgccaaaa tcacgccaaa taataacggg acctatgcct gttttgtctc taacttggct 1980
actggccgca ataattccat agtcaagagc atcacagtct ctgcatctgg aacttctcct 2040
ggtctctcag ctggggccac tgtcggcatc atgattggag tgctggttgg ggttgctctg 2100
ata 2103
<210> 14
<211> 2103
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность CEA из mBN373 и mBN420
<400> 14
atggagtctc cctcggctcc tccacacaga tggtgcatcc cttggcagag gctcctgctc 60
acagcctcac ttctaacctt ctggaacccg cccaccactg ccaagctcac tattgaatcc 120
acgccgttca atgtcgcaga ggggaaggag gtgcttctac ttgtccacaa tctgcctcag 180
catctctttg gctacagctg gtacaaaggt gaaagagtgg atggcaaccg tcaaattata 240
ggatatgtaa taggaactca acaagctact ccagggcccg catacagtgg tcgagagata 300
atatacccta atgcatccct gctgatccag aacatcatcc agaatgacac aggattctac 360
accctacacg tcataaagtc agatcttgtg aatgaagaag caactggcca gttccgggta 420
taccctgaac tccctaagcc ttctattagc tccaataata gtaagcctgt cgaagacaaa 480
gatgccgtcg ctttcacatg cgagcccgaa actcaagacg caacatatct ctggtgggtg 540
aacaaccagt ccctgcctgt gtctcctaga ctccaactca gcaacggaaa tagaactctg 600
accctgttta acgtgaccag gaacgacaca gcaagctaca aatgcgaaac ccaaaatcca 660
gtcagcgcca ggaggtctga ttcagtgatt ctcaacgtgc tttacggacc cgatgctcct 720
acaatcagcc ctctaaacac aagctataga tcaggagaaa atctgaatct gagctgtcat 780
gccgctagca atcctccagc tcaatacagc tggtttgtca atggcacttt ccaacagtcc 840
acccaggaac tgttcattcc caatattacc gtgaacaata gtggatccta cacgtgccaa 900
gctcacaata gcgacaccgg actcaaccgc acaaccgtga cgacgattac cgtgtatgag 960
ccaccaaaac cattcataac tagtaacaat tctaacccag ttgaggatga ggacgcagtt 1020
gcattaactt gtgagccaga gattcaaaat accacttatt tatggtgggt caataaccaa 1080
agtttgccgg ttagcccacg cttgcagttg tctaatgata accgcacatt gacactcctg 1140
tccgttactc gcaatgatgt aggaccttat gagtgtggca ttcagaatga attatccgtt 1200
gatcactccg accctgttat ccttaatgtt ttgtatggcc cagacgaccc aactatatct 1260
ccatcataca cctactaccg tcccggcgtg aacttgagcc tttcttgcca tgcagcatct 1320
aatccacctg cacagtactc ctggctgatt gatggaaaca ttcagcagca tactcaagag 1380
ttatttataa gcaacataac tgagaagaac agcggactct atacttgcca ggccaataac 1440
tcagccagtg gtcacagcag gactacagtt aaaacaataa ctgtttccgc ggagctgccc 1500
aagccctcca tctccagcaa caactccaaa cccgtggagg acaaggatgc tgtggccttc 1560
acctgtgaac ctgaggctca gaacacaacc tacctgtggt gggtaaatgg tcagagcctc 1620
ccagtcagtc ccaggctgca gctgtccaat ggcaacagga ccctcactct attcaatgtc 1680
acaagaaatg acgcaagagc ctatgtatgt ggaatccaga actcagtgag tgcaaaccgc 1740
agtgacccag tcaccctgga tgtcctctat ggaccggaca cacccatcat ttcacctcca 1800
gactcgtctt acctttcggg agcggacctc aacctctcct gccactcggc ctctaaccca 1860
tctccgcagt attcttggcg tatcaatggg ataccgcagc aacacacaca agttctcttt 1920
atcgccaaaa tcacgccaaa taataacggg acctatgcct gttttgtctc taacttggct 1980
actggccgca ataattccat agtcaagagc atcacagtct ctgcatctgg aacttctcct 2040
ggtctctcag ctggagccac tgtcggcatc atgattggag tgctggttgg ggttgctctg 2100
ata 2103
<210> 15
<211> 864
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Оптимизированная кодирующая последовательность B7.1 из mBN373 и
BN420
<400> 15
atgggacaca ccagaaggca gggcacaagc ccatccaagt gtccctacct gaacttcttt 60
cagctcctgg tgctggctgg cctgtcccac ttctgctccg gagtgatcca cgtgaccaag 120
gaggtcaaag aagtcgccac actgagctgc gggcacaatg tgtccgtgga ggaactggct 180
cagacacgga tctactggca gaaagagaag aaaatggtgc tgaccatgat gtccggcgac 240
atgaacatct ggcctgagta caagaaccgc accatcttcg acatcaccaa caatctgagc 300
atcgtgatcc tcgctctgag gccctccgac gagggaacat acgagtgcgt ggtgctgaag 360
tacgagaagg acgccttcaa acgcgagcac ctggccgagg tcaccctgtc cgtgaaggca 420
gacttcccaa cacccagcat cagcgacttc gagatcccta ccagcaacat ccggcggatt 480
atctgcagca cctccggagg cttcccagag cctcacctga gctggctcga gaacggcgaa 540
gagctcaacg ccatcaacac taccgtgtcc caggaccctg agacagagct gtacgctgtg 600
agcagcaagc tggacttcaa catgaccaca aatcacagct ttatgtgcct catcaagtac 660
ggccacctga gagtgaatca gaccttcaac tggaatacaa ccaagcagga acacttccca 720
gacaatctcc tgccctcctg ggctatcaca ctgattagcg tgaatggcat cttcgtgatc 780
tgctgtctga cctactgctt cgctcccaga tgccgggagc gcaggagaaa cgagaggctg 840
agacgggaat ccgtgaggcc cgtg 864
<210> 16
<211> 864
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность B7.1 из PANVAC
<400> 16
atgggccaca cacggaggca gggaacatca ccatccaagt gtccatacct caatttcttt 60
cagctcttgg tgctggctgg tctttctcac ttctgttcag gtgttatcca cgtgaccaag 120
gaagtgaaag aagtggcaac gctgtcctgt ggtcacaatg tttctgttga agagctggca 180
caaactcgca tctactggca aaaggagaag aaaatggtgc tgactatgat gtctggagac 240
atgaatatat ggcccgagta caagaaccgg accatctttg atatcactaa taacctctcc 300
attgtgatcc tggctctgcg cccatctgac gagggcacat acgagtgtgt tgttctgaag 360
tatgaaaaag acgctttcaa gcgggaacac ctggctgaag tgacgttatc agtcaaagct 420
gacttcccta cacctagtat atctgacttt gaaattccaa cttctaatat tagaaggata 480
atttgctcaa cctctggagg ttttccagag cctcacctct cctggttgga aaatggagaa 540
gaattaaatg ccatcaacac aacagtttcc caagatcctg aaactgagct ctatgctgtt 600
agcagcaaac tggatttcaa tatgacaacc aaccacagct tcatgtgtct catcaagtat 660
ggacatttaa gagtgaatca gaccttcaac tggaatacaa ccaagcaaga gcattttcct 720
gataacctgc tcccatcctg ggccattacc ttaatctcag taaatggaat tttcgtgata 780
tgctgcctga cctactgctt tgccccacgc tgcagagaga gaaggaggaa tgagagattg 840
agaagggaaa gtgtacgccc tgta 864
<210> 17
<211> 864
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Оптимизированная кодирующая последовательность B7.1 из mBN336
<400> 17
atgggccaca ccagaaggca gggcaccagc ccctccaagt gcccctacct gaacttcttc 60
cagctcctgg tgctggccgg cctgtcccac ttctgctccg gcgtgatcca cgtgaccaaa 120
gaggtcaaag aagtcgccac actgagctgc ggccacaatg tgtccgtgga ggaactggct 180
cagacccgga tctactggca gaaagaaaag aaaatggtgc tgaccatgat gtccggcgac 240
atgaacatct ggcctgagta caagaaccgc accatcttcg acatcaccaa caacctgagc 300
atcgtgatcc tcgccctgag gccctccgac gagggcacct acgagtgcgt ggtgctgaag 360
tacgagaagg acgccttcaa gcgcgagcac ctggccgagg tcaccctgtc cgtgaaggcc 420
gacttcccaa cccccagcat cagcgacttc gagatcccaa ccagcaacat ccggcggatc 480
atctgcagca cctccggcgg cttccccgag cctcacctga gctggctcga gaacggcgaa 540
gaactcaacg ccatcaacac taccgtgtcc caggaccccg agacagagct gtacgccgtg 600
agcagcaagc tggacttcaa catgaccaca aaccacagct ttatgtgcct catcaagtac 660
ggccacctga gagtgaatca gaccttcaac tggaacacca ccaagcagga acacttcccc 720
gacaatctgc tgccctcctg ggctatcacc ctgattagcg tgaatggcat cttcgtgatc 780
tgctgtctga cctactgctt cgcccccaga tgccgggagc ggcggagaaa cgagcggctg 840
cggcgggaat ccgtgaggcc cgtg 864
<210> 18
<211> 1596
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Оптимизированная кодирующая последовательность ICAM-1 из mBN373 и
mBN420
<400> 18
atggctccta gctcacctag accagctctg cctgccctgc tcgtgctgct cggagctctg 60
ttccctggac caggcaacgc ccagaccagc gtgtcaccta gcaaagtgat tctgcccaga 120
ggaggctccg tgctggtcac atgtagcacc agctgcgacc agcccaagct cctcgggatc 180
gagacacctc tgcccaagaa agagctgctc ctgccaggca acaatcggaa agtgtacgag 240
ctgtccaatg tgcaggaaga tagccagccc atgtgctact ccaactgtcc cgacggccag 300
agcaccgcca agacctttct gaccgtgtac tggacacctg agcgggtgga actggctcca 360
ctgcccagct ggcagccagt gggcaagaat ctgaccctgc ggtgccaggt ggaaggcgga 420
gctcccagag ccaacctgac agtggtgctc ctgagaggcg agaaagagct gaagcgggaa 480
cctgccgtgg gcgagccagc cgaagtgacc acaaccgtgc tcgtgcggag ggaccaccac 540
ggagccaact tcagctgcag aaccgagctg gacctcaggc cacagggcct ggaactgttc 600
gagaacacca gcgctcccta ccagctccag accttcgtgc tcccagcaac accacctcag 660
ctggtgtcac ctcgggtgct ggaagtggac acccagggca cagtcgtgtg cagcctggac 720
ggcctgtttc ccgtgtccga agctcaggtc cacctggctc tcggagacca gagactgaac 780
cctaccgtga cctacggcaa tgacagcttc agcgccaagg cctccgtgtc cgtgaccgcc 840
gaggatgaag gcacccagag gctgacatgc gccgtgattc tgggcaacca gagccaggaa 900
accctgcaga ccgtcaccat ctatagcttc cctgcaccta atgtgatcct gacaaagccc 960
gaggtgtccg agggcactga agtgaccgtg aaatgcgagg cccaccctag agccaaagtg 1020
accctgaacg gcgtgccagc ccagccactc ggaccaagag cacagctcct gctgaaagcc 1080
acacccgagg ataacggccg gtccttctcc tgcagcgcta ccctcgaagt ggccggacag 1140
ctgatccaca agaaccagac cagagagctg agagtgctgt acggccctag actggacgag 1200
agagactgcc caggcaactg gacctggccc gagaactccc agcagacacc catgtgccag 1260
gcttggggca acccactgcc agagctgaag tgcctgaagg acggcacctt ccctctgccc 1320
atcggcgagt ccgtgacagt gaccagggac ctggaaggca cctacctgtg cagagccaga 1380
tccacacagg gcgaagtgac acgggaggtc accgtgaatg tgctgtcacc tcgctacgag 1440
atcgtgatca tcaccgtggt cgctgcagct gtgatcatgg gcacagccgg actgagcaca 1500
tacctgtaca accggcagcg gaagatcaag aagtacaggc tgcagcaggc ccagaaaggc 1560
acacccatga agcccaacac ccaggccact cctccc 1596
<210> 19
<211> 1596
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность ICAM-1 из PANVAC
<400> 19
atggctccca gcagcccccg gcccgcgctg cccgcactcc tggtcctgct cggggctctg 60
ttcccaggac ctggcaatgc ccagacatct gtgtccccct caaaagtcat cctgccccgg 120
ggaggctccg tgctggtgac atgcagcacc tcctgtgacc agcccaagtt gttgggcata 180
gagaccccgt tgcctaaaaa ggagttgctc ctgcctggga acaaccggaa ggtgtatgaa 240
ctgagcaatg tgcaagaaga tagccaacca atgtgctatt caaactgccc tgatgggcag 300
tcaacagcta aaaccttcct caccgtgtac tggactccag aacgggtgga actggcaccc 360
ctcccctctt ggcagccagt gggcaagaac cttaccctac gctgccaggt ggagggtggg 420
gcaccccggg ccaacctcac cgtggtgctg ctccgtgggg agaaggagct gaaacgggag 480
ccagctgtgg gggagcccgc tgaggtcacg accacggtgc tggtgaggag agatcaccat 540
ggagccaatt tctcgtgccg cactgaactg gacctgcggc cccaagggct ggagctgttt 600
gagaacacct cggcccccta ccagctccag acctttgtcc tgccagcgac tcccccacaa 660
cttgtcagcc cccgggtcct agaggtggac acgcagggga ccgtggtctg ttccctggac 720
gggctgttcc cagtctcgga ggcccaggtc cacctggcac tgggggacca gaggttgaac 780
cccacagtca cctatggcaa cgactccttc tcggccaagg cctcagtcag tgtgaccgca 840
gaggacgagg gcacccagcg gctgacgtgt gcagtaatac tggggaacca gagccaggag 900
acactgcaga cagtgaccat ctacagcttt ccggcgccca acgtgattct gacgaagcca 960
gaggtctcag aagggaccga ggtgacagtg aagtgtgagg cccaccctag agccaaggtg 1020
acgctgaatg gggttccagc ccagccactg ggcccgaggg cccagctcct gctgaaggcc 1080
accccagagg acaacgggcg cagcttctcc tgctctgcaa ccctggaggt ggccggccag 1140
cttatacaca agaaccagac ccgggagctt cgtgtcctgt atggcccccg actggacgag 1200
agggattgtc cgggaaactg gacgtggcca gaaaattccc agcagactcc aatgtgccag 1260
gcttggggga acccattgcc cgagctcaag tgtctaaagg atggcacttt cccactgccc 1320
atcggggaat cagtgactgt cactcgagat cttgagggca cctacctctg tcgggccagg 1380
agcactcaag gggaggtcac ccgcgaggtg accgtgaatg tgctctcccc ccggtatgag 1440
attgtcatca tcactgtggt agcagccgca gtcataatgg gcactgcagg cctcagcacg 1500
tacctctata accgccagcg gaagatcaag aaatacagac tacaacaggc ccaaaaaggg 1560
acccccatga aaccgaacac acaagccacg cctccc 1596
<210> 20
<211> 1596
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Оптимизированная кодирующая последовательность ICAM-1 из mBN336
<400> 20
atggccccta gcagccctag accagccctg cctgccctgc tggtgctgct gggcgctctg 60
ttccccggac ccggcaacgc ccagaccagc gtgtccccca gcaaagtgat tctgcccaga 120
ggcggctccg tgctggtcac atgtagcacc agctgcgacc agcccaagct cctcgggatc 180
gagacacccc tgcccaagaa agagctgctg ctgcccggca acaaccggaa agtgtacgag 240
ctgtccaatg tgcaggaaga tagccagccc atgtgctact ccaactgccc cgacggccag 300
agcaccgcca agacctttct gaccgtgtac tggacccccg agcgggtgga actggcccca 360
ctgcccagct ggcagcccgt gggcaagaat ctgaccctgc ggtgccaggt ggaaggcgga 420
gcccccagag ccaacctgac agtggtgctc ctgcggggcg aaaaagagct gaagcgggag 480
cctgccgtgg gcgagccagc cgaagtgacc acaaccgtgc tcgtgcggag ggaccaccac 540
ggcgccaact tcagctgcag aaccgagctg gacctcaggc cacagggcct ggaactgttc 600
gagaacacca gcgcccccta ccagctccag accttcgtgc tcccagcaac cccccctcag 660
ctggtgtccc ctcgggtgct ggaagtggac acccagggca cagtcgtgtg cagcctggac 720
ggcctgtttc ccgtgtccga agctcaggtc cacctggctc tcggggacca gagactgaac 780
cctaccgtga cctacggcaa tgacagcttc agcgccaagg cctccgtgtc cgtgaccgcc 840
gaggatgagg gcacccagag gctgacatgc gccgtgattc tgggcaacca gagccaggaa 900
accctgcaga ccgtcaccat ctatagcttc cctgccccca atgtgatcct gacaaagccc 960
gaggtgtccg agggcactga agtgaccgtg aaatgcgagg cccaccccag ggccaaagtg 1020
accctgaacg gcgtgccagc ccagccactc ggaccaagag cacagctcct gctgaaagcc 1080
acccccgagg ataacggccg gtccttctcc tgcagcgcta ccctcgaagt ggccgggcag 1140
ctgatccaca agaaccagac ccgggagctg agagtgctgt acggccccag actggacgag 1200
agagactgcc ccggcaactg gacctggccc gagaactccc agcagacccc catgtgccag 1260
gcttggggca acccactgcc agagctgaag tgcctgaagg acggcacctt ccctctgccc 1320
atcggcgagt ccgtgacagt gacccgggac ctggaaggca cctacctgtg ccgggccaga 1380
tccacacagg gcgaagtgac acgggaggtc accgtgaatg tgctgtcccc ccgctacgag 1440
atcgtgatca tcaccgtggt cgctgcagct gtgatcatgg gcacagccgg cctgagcaca 1500
tacctgtaca accggcagcg gaagatcaag aagtacaggc tgcagcaggc ccagaaaggc 1560
acccccatga agcccaacac ccaggccacc cctccc 1596
<210> 21
<211> 750
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Оптимизированная кодирующая последовательность LFA-3 из mBN373 и
mBN420
<400> 21
atggtggctg gctctgatgc agggagagcc ctgggagtgc tgtctgtcgt gtgcctgctg 60
cactgcttcg gcttcatcag ctgcttcagc cagcagatct acggagtggt ctacggcaac 120
gtgaccttcc acgtgcccag caacgtgcct ctgaaagagg tgctctggaa gaaacagaag 180
gacaaggtcg cagagctgga gaacagcgag ttccgggcct tcagcagctt caagaaccgg 240
gtgtacctgg acaccgtgtc cggcagcctg accatctaca acctgaccag cagcgacgag 300
gacgagtacg agatggaaag ccctaacatc accgacacca tgaagttctt tctgtacgtg 360
ctggaaagcc tgcccagccc aacactgacc tgtgccctga ccaacggctc catcgaggtg 420
cagtgcatga ttcccgagca ctacaactcc cacagaggcc tgatcatgta ctcttgggac 480
tgccctatgg aacagtgcaa gcgcaacagc accagcatct acttcaagat ggagaacgac 540
ctccctcaga agatccagtg cacactgagc aatccactgt tcaacaccac atccagcatc 600
atcctgacaa cctgtattcc cagcagtggc cacagcagac acagatacgc cctgatccct 660
attccactgg ccgtgatcac cacatgcatc gtgctgtaca tgaacggcat cctgaagtgc 720
gaccggaagc ccgaccggac caacagcaac 750
<210> 22
<211> 750
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность LFA-3 из PANVAC
<400> 22
atggttgctg ggagcgacgc ggggcgggcc ctgggggtcc tcagcgtggt ctgcctgctg 60
cactgctttg gtttcatcag ctgtttttcc caacaaatat atggtgttgt gtatgggaat 120
gtaactttcc atgtaccaag caatgtgcct ttaaaagagg tcctatggaa aaaacaaaag 180
gataaagttg cagaactgga aaattctgaa ttcagagctt tctcatcttt taaaaatagg 240
gtttatttag acactgtgtc aggtagcctc actatctaca acttaacatc atcagatgaa 300
gatgagtatg aaatggaatc gccaaatatt actgatacca tgaagttctt tctttatgtg 360
cttgagtctc ttccatctcc cacactaact tgtgcattga ctaatggaag cattgaagtc 420
caatgcatga taccagagca ttacaacagc catcgaggac ttataatgta ctcatgggat 480
tgtcctatgg agcaatgtaa acgtaactca accagtatat attttaagat ggaaaatgat 540
cttccacaaa aaatacagtg tactcttagc aatccattat ttaatacaac atcatcaatc 600
attttgacaa cctgtatccc aagcagcggt cattcaagac acagatatgc acttataccc 660
ataccattag cagtaattac aacatgtatt gtgctgtata tgaatggtat tctgaaatgt 720
gacagaaaac cagacagaac caactccaat 750
<210> 23
<211> 750
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Оптимизированная кодирующая последовательность LFA-3 из mBN336
<400> 23
atggtggctg gctctgatgc aggcagagcc ctgggcgtgc tgtctgtcgt gtgcctgctg 60
cactgcttcg gcttcatcag ctgcttcagc cagcagatct acggcgtggt gtacggcaac 120
gtgaccttcc acgtgcccag caacgtgcct ctgaaagagg tgctctggaa gaagcagaag 180
gacaaggtcg cagagctgga aaacagcgag ttccgggcct tcagcagctt caagaaccgg 240
gtgtacctgg acaccgtgtc cggcagcctg accatctaca acctgaccag cagcgacgag 300
gacgagtacg agatggaaag ccccaacatc accgacacca tgaagttctt tctgtacgtg 360
ctggaaagcc tgcccagccc caccctgacc tgtgccctga ccaacggctc catcgaggtg 420
cagtgcatga tccccgagca ctacaactcc caccggggcc tgatcatgta ctcttgggac 480
tgccctatgg aaacgtgcaa gcgcaacagc accagcatct acttcaagat ggaaaacgac 540
ctcccccaga aaatccagtg caccctgagc aaccccctgt tcaacaccac ctccagcatc 600
atcctgacca cctgtatccc cagcagcggc cacagcagac acagatacgc cctgatcccc 660
atccccctgg ccgtgatcac cacatgcatc gtgctgtaca tgaacggcat cctgaagtgc 720
gaccggaagc ccgaccggac caacagcaac 750
<210> 24
<211> 9
<212> PRT
<213> Искусственная Последовательность
<220>
<223> Агонистический эпитоп MUC1
<400> 24
Tyr Leu Ala Pro Pro Ala His Gly Val
1. 5
<210> 25
<211> 9
<212> PRT
<213> Искусственная Последовательность
<220>
<223> Агонистический эпитоп MUC1
<400> 25
Tyr Leu Asp Thr Arg Pro Ala Pro Val
1. 5
<210> 26
<211> 10
<212> PRT
<213> Искусственная Последовательность
<220>
<223> Агонистический эпитоп MUC1
<400> 26
Tyr Leu Ala Ile Val Tyr Leu Ile Ala Leu
1. 5 10
<210> 27
<211> 10
<212> PRT
<213> Искусственная Последовательность
<220>
<223> Агонистический эпитоп MUC1
<400> 27
Tyr Leu Ile Ala Leu Ala Val Cys Gln Val
1. 5 10
<210> 28
<211> 9
<212> PRT
<213> Искусственная Последовательность
<220>
<223> Агонистический эпитоп MUC1
<400> 28
Tyr Leu Ser Tyr Thr Asn Pro Ala Val
1. 5
<210> 29
<211> 9
<212> PRT
<213> Искусственная Последовательность
<220>
<223> Агонистический эпитоп MUC1
<400> 29
Ser Leu Phe Arg Ser Pro Tyr Glu Lys
1. 5
<210> 30
<211> 455
<212> PRT
<213> Искусственная Последовательность
<220>
<223> Аминокислотная последовательность MUC1, обнаруженная в mBN336,
mBN373, and mBN420
<400> 30
Met Thr Pro Gly Thr Gln Ser Pro Phe Phe Leu Leu Leu Leu Leu Thr
1. 5 10 15
Val Leu Thr Val Val Thr Gly Ser Gly His Ala Ser Ser Thr Pro Gly
20 25 30
Gly Glu Lys Glu Thr Ser Ala Thr Gln Arg Ser Ser Val Pro Ser Ser
35 40 45
Thr Glu Lys Asn Ala Val Ser Met Thr Ser Ser Val Leu Ser Ser His
50 55 60
Ser Pro Gly Ser Gly Ser Ser Thr Thr Gln Gly Gln Asp Val Thr Leu
65 70 75 80
Ala Pro Ala Thr Glu Pro Ala Ser Gly Ser Ala Ala Leu Trp Gly Gln
85 90 95
Asp Val Thr Ser Val Pro Val Thr Arg Pro Ala Leu Gly Tyr Leu Ala
100 105 110
Pro Pro Ala His Gly Val Thr Ser Tyr Leu Asp Thr Arg Pro Ala Pro
115 120 125
Val Ser Thr Ala Pro Pro Ala His Gly Val Thr Ser Ala Pro Asp Thr
130 135 140
Arg Pro Ala Pro Gly Ser Thr Ala Pro Pro Ala His Gly Val Thr Ser
145 150 155 160
Ala Pro Asp Thr Arg Pro Ala Pro Ala Ser Thr Leu Val His Asn Gly
165 170 175
Thr Ser Ala Arg Ala Thr Thr Thr Pro Ala Ser Lys Ser Thr Pro Phe
180 185 190
Ser Ile Pro Ser His His Ser Asp Thr Pro Thr Thr Leu Ala Ser His
195 200 205
Ser Thr Lys Thr Asp Ala Ser Ser Thr His His Ser Thr Val Pro Pro
210 215 220
Leu Thr Ser Ser Asn His Ser Thr Ser Pro Gln Leu Ser Thr Gly Val
225 230 235 240
Ser Phe Phe Phe Leu Ser Phe His Ile Ser Asn Leu Gln Phe Asn Ser
245 250 255
Ser Leu Glu Asp Pro Ser Thr Asp Tyr Tyr Gln Glu Leu Gln Arg Asp
260 265 270
Ile Ser Glu Met Phe Leu Gln Ile Tyr Lys Gln Gly Gly Phe Leu Gly
275 280 285
Leu Ser Asn Ile Lys Phe Arg Pro Gly Ser Val Val Val Gln Leu Thr
290 295 300
Leu Ala Phe Arg Glu Gly Thr Ile Asn Val His Asp Val Glu Thr Gln
305 310 315 320
Phe Asn Gln Tyr Lys Thr Glu Ala Ala Ser Arg Tyr Asn Leu Thr Ile
325 330 335
Ser Asp Val Ser Val Ser Asp Val Pro Phe Pro Phe Ser Ala Gln Ser
340 345 350
Gly Ala Gly Val Pro Gly Trp Gly Ile Ala Leu Leu Val Leu Val Cys
355 360 365
Val Leu Val Tyr Leu Ala Ile Val Tyr Leu Ile Ala Leu Ala Val Cys
370 375 380
Gln Val Arg Arg Lys Asn Tyr Gly Gln Leu Asp Ile Phe Pro Ala Arg
385 390 395 400
Asp Thr Tyr His Pro Met Ser Glu Tyr Pro Thr Tyr His Thr His Gly
405 410 415
Arg Tyr Val Pro Pro Ser Ser Leu Phe Arg Ser Pro Tyr Glu Lys Val
420 425 430
Ser Ala Gly Asn Gly Gly Ser Tyr Leu Ser Tyr Thr Asn Pro Ala Val
435 440 445
Ala Ala Thr Ser Ala Asn Leu
450 455
<210> 31
<211> 1365
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность MUC1 mBN336 без агонистических эпитопов
согласно WO 2013/103658
<400> 31
atgacaccgg gcacccagtc tcctttcttc ctgctgctgc tcctcacagt gcttacagtt 60
gttacgggtt ctggtcatgc aagctctacc ccaggtggag aaaaggagac ttcggctacc 120
cagagaagtt cagtgcccag ctctactgag aagaatgctg tgagtatgac aagctccgta 180
ctctccagcc acagcccagg ttcaggctcc tccaccactc agggacagga tgtcactctg 240
gcaccggcca cggaaccagc ttcaggttca gctgccttgt ggggacagga tgtcacctcg 300
gtaccagtta ctagaccagc tttaggcagc acagcgccac cggctcatgg cgttacatcg 360
gctcctgaca ctcgaccggc accaggcagc acagcacctc ccgcacacgg tgtaactagc 420
gcgcctgata cacgtcccgc tcccggatct accgctccgc cagcgcacgg agtgacgtca 480
gcaccagata cgaggccagc gcctgctagc actctggtgc acaacggcac ctctgccagg 540
gctaccacaa ccccagccag caagagcact ccattctcaa ttcccagcca ccactctgat 600
actcctacca cccttgccag ccatagcacc aagactgatg ccagtagcac tcaccatagc 660
acggtacctc ctctcacctc ctccaatcac agcacttctc cccagttgtc tactggggtc 720
tctttctttt tcctgtcttt tcacatttca aacctccagt ttaattcctc tctggaagat 780
cccagcaccg actactacca agagctgcag agagacattt ctgaaatgtt tttgcagatt 840
tataaacaag ggggttttct gggcctctcc aatattaagt tcaggccagg atctgtggtg 900
gtacagttga ctctggcctt ccgagaaggt accatcaatg tccacgacgt ggagacacag 960
ttcaatcagt ataaaacgga agcagcctct cgatataacc tgacgatctc agacgtcagc 1020
gtgagtgatg tgccatttcc tttctctgcc cagtctgggg ctggggtgcc aggctggggc 1080
atcgcgctgc tggtgctggt ctgtgttctg gttgcactgg ccattgtcta tctcattgcc 1140
ttggctgtct gtcagtgccg ccgaaagaac tacgggcagc tggacatctt tccagcccgg 1200
gatacctacc atcctatgag cgagtacccc acctaccaca cccatgggcg ctatgtgccc 1260
cctagcagta ccgatcgtag cccctatgag aaggtttctg caggtaatgg tggcagcagt 1320
ctctcttaca caaacccagc agtggcagcc acttctgcca acttg 1365
<210> 32
<211> 1365
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность MUC1 mBN373 без агонистических эпитопов
согласно WO 2013/103658
<400> 32
atgacaccgg gcacccagtc tcctttcttc ctgctgctgc tcctcacagt gcttacagtt 60
gttacgggtt ctggtcatgc aagctctacc ccaggtggag aaaaggagac ttcggctacc 120
cagagaagtt cagtgcccag ctctactgag aagaatgctg tgagtatgac aagctccgta 180
ctctccagcc acagcccagg ttcaggctcc tccaccactc agggacagga tgtcactctg 240
gcaccggcca cggaaccagc ttcaggttca gctgccttgt ggggacagga tgtcacctcg 300
gtaccagtta ctagaccagc tttaggcagc acagcgccac cggctcatgg cgttacatcg 360
gctcctgaca ctcgaccggc accaggcagc acagcacctc ccgcacacgg tgtaactagc 420
gcgcctgata cacgtcccgc tcccggatct accgctccgc cagcgcacgg agtgacgtca 480
gcaccagata cgaggccagc gcctgctagc actctggtgc acaacggcac ctctgccagg 540
gctaccacaa ccccagccag caagagcact ccattctcaa ttcccagcca ccactctgat 600
actcctacca cccttgccag ccatagcacc aagactgatg ccagtagcac tcaccatagc 660
acggtacctc ctctcacctc ctccaatcac agcacttctc cccagttgtc tactggggtc 720
tctttctttt tcctgtcttt tcacatttca aacctccagt ttaattcctc tctggaagat 780
cccagcaccg actactacca agagctgcag agagacattt ctgaaatgtt tttgcagatt 840
tataaacaag ggggttttct gggcctctcc aatattaagt tcaggccagg atctgtggtg 900
gtacagttga ctctggcctt ccgagaaggt accatcaatg tccacgacgt ggagacacag 960
ttcaatcagt ataaaacgga agcagcctct cgatataacc tgacgatctc agacgtcagc 1020
gtgagtgatg tgccatttcc tttctctgcc cagtctgggg ctggggtgcc aggctggggc 1080
atcgcgctgc tggtgctggt ctgtgttctg gttgcactgg ccattgtcta tctcattgcc 1140
ttggctgtct gtcagtgccg ccgaaagaac tacgggcagc tggacatctt tccagcccgg 1200
gatacctacc atcctatgag cgagtacccc acctaccaca cccatgggcg ctatgtgccc 1260
cctagcagta ccgatcgtag cccctatgag aaggtttctg caggtaatgg tggcagcagt 1320
ctctcttaca caaacccagc agtggcagcc acttctgcca acttg 1365
<210> 33
<211> 1365
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность MUC1 mBN420 без агонистических эпитопов
согласно WO 2013/103658
<400> 33
atgacacctg gcactcagtc accattcttc ctgctgttac tcttgacagt gcttacagtt 60
gttacaggtt ctggtcatgc aagctctacc ccaggtggag aaaaggagac ttcggctacc 120
cagcggagtt cagtgcctag ctctactgag aagaatgctg tgagtatgac aagctccgta 180
ctctccagcc acagcccagg ttcaggctcc agcaccactc aaggacagga tgtcactctg 240
gcaccggcca cggaaccagc ttcaggttca gctgccttgt ggggacagga tgtcacatcg 300
gtaccagtta ctagaccagc tttaggcagc acagcgccac cggctcatgg cgttacatcg 360
gctcctgaca ctcgaccggc accaggcagc acagcacctc ccgcacacgg tgtaactagc 420
gcgcctgata cacgtcccgc tcccggatct accgctccgc cagcgcacgg agtgacgtca 480
gcaccagata cgaggccagc gcctgctagc actctggtgc acaatggcac atctgccagg 540
gctaccacaa ctccagccag caagagcact ccattctcaa ttccaagcca tcactctgat 600
actcctacca cacttgccag ccatagcacc aagactgatg ccagtagcac tcaccatagc 660
acggtacctc cactcacctc atccaatcac agcacttctc ctcagttgtc tactggagtc 720
tccttctttt tcctgtcctt tcacatttca aacttgcagt tcaattcttc cctggaagat 780
cccagcaccg actactacca agagctgcag agagacattt ctgagatgtt cttgcagatt 840
tataaacaag gtggattcct tggcctctct aatattaagt tcaggccagg atctgtggtc 900
gtacagttga ctctggcctt cagagaaggt accatcaatg tccacgacgt ggagacacag 960
ttcaatcagt ataagacgga agcagcctca cgatataacc tgacgatctc agacgtcagc 1020
gttagtgatg tgccatttcc tttctctgcc cagtctggag ctggtgtgcc aggctggggc 1080
atcgcgctgc tcgtgttggt ctgtgttctg gttgcactgg ccattgtcta tctcattgcc 1140
ttggctgttt gtcagtgcag acgcaagaac tacggacagc tggacatctt tccagctcgg 1200
gatacctacc atcctatgag cgagtaccct acctaccaca cacatggtcg ctatgtgcca 1260
cctagcagta ccgatcgtag tccctatgag aaagtttctg caggtaatgg tggcagcagt 1320
ctctcttaca caaacccagc agtggcagcc acttctgcca acttg 1365
<210> 34
<211> 1365
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Кодирующая последовательность MUC1 оптимизированного mBN без
агонистических эпитопов согласно WO 2013/103658
<400> 34
atgacacctg gcactcagtc accattcttc ctgctgttac tcttgacagt gcttacagtt 60
gttacaggtt ctggtcatgc aagctctacc ccaggtggag aaaaggagac ttcggctacc 120
cagcggagtt cagtgcctag ctctactgag aagaatgctg tgagtatgac aagctccgta 180
ctctccagcc acagcccagg ttcaggctcc agcaccactc aaggacagga cgtcactctg 240
gcaccggcca cggaaccagc ttcaggttca gctgccttgt ggggacagga tgtgacatcg 300
gtaccagtta ctagaccagc tttaggcagc acagcgccac cggctcatgg cgttacatcg 360
gctcctgaca ctcgaccggc accaggcagc acagcacctc ccgcacacgg tgtaactagc 420
gcgcctgata cacgtcccgc tcccggatct accgctccgc cagcgcacgg agtgacgtca 480
gcaccagata cgaggccagc gcctgctagc actctggtgc acaatggcac atctgccagg 540
gctaccacaa ctccagccag caagagcact ccattctcaa ttccaagcca tcactctgat 600
actcctacca cacttgccag ccatagcacc aagactgatg ccagtagcac tcaccatagc 660
acggtacctc cactcacctc atccaatcac agcacttctc ctcagttgtc tactggagtc 720
tccttctttt tcctgtcctt tcacatttca aacttgcagt tcaattcttc cctggaagat 780
cccagcaccg actactacca agagctgcag agagacattt ctgagatgtt cttgcagatt 840
tataaacaag gtggattcct tggcctctct aatattaagt tcaggccagg atctgtggtc 900
gtacagttga ctctggcctt cagagaaggt accatcaatg tccacgacgt ggagacacag 960
ttcaatcagt ataagacgga agcagcctca cgatataacc tgacgatctc agacgtcagc 1020
gttagtgatg tgccatttcc tttctctgcc cagtctggag ctggtgtgcc aggctggggc 1080
atcgcgctgc tcgtgttggt ctgtgttctg gttgcactgg ccattgtcta tctcattgcc 1140
ttggctgttt gtcagtgcag acgcaagaac tacggacagc tggacatctt tccagctcgg 1200
gatacctacc atcctatgag cgagtaccct acctaccaca cacatggtcg ctatgtgcca 1260
cctagcagta ccgatcgtag tccctatgag aaagtttctg caggtaatgg tggcagcagt 1320
ctctcttaca caaacccagc agtggcagcc acttctgcca acttg 1365
<210> 35
<211> 455
<212> PRT
<213> Искусственная Последовательность
<220>
<223> Аминокислотная последовательность MUC1 из mBN336/mBN373/mBN420 без
агонистических эпитопов согласно WO 2013/103658
<400> 35
Met Thr Pro Gly Thr Gln Ser Pro Phe Phe Leu Leu Leu Leu Leu Thr
1. 5 10 15
Val Leu Thr Val Val Thr Gly Ser Gly His Ala Ser Ser Thr Pro Gly
20 25 30
Gly Glu Lys Glu Thr Ser Ala Thr Gln Arg Ser Ser Val Pro Ser Ser
35 40 45
Thr Glu Lys Asn Ala Val Ser Met Thr Ser Ser Val Leu Ser Ser His
50 55 60
Ser Pro Gly Ser Gly Ser Ser Thr Thr Gln Gly Gln Asp Val Thr Leu
65 70 75 80
Ala Pro Ala Thr Glu Pro Ala Ser Gly Ser Ala Ala Leu Trp Gly Gln
85 90 95
Asp Val Thr Ser Val Pro Val Thr Arg Pro Ala Leu Gly Ser Thr Ala
100 105 110
Pro Pro Ala His Gly Val Thr Ser Ala Pro Asp Thr Arg Pro Ala Pro
115 120 125
Gly Ser Thr Ala Pro Pro Ala His Gly Val Thr Ser Ala Pro Asp Thr
130 135 140
Arg Pro Ala Pro Gly Ser Thr Ala Pro Pro Ala His Gly Val Thr Ser
145 150 155 160
Ala Pro Asp Thr Arg Pro Ala Pro Ala Ser Thr Leu Val His Asn Gly
165 170 175
Thr Ser Ala Arg Ala Thr Thr Thr Pro Ala Ser Lys Ser Thr Pro Phe
180 185 190
Ser Ile Pro Ser His His Ser Asp Thr Pro Thr Thr Leu Ala Ser His
195 200 205
Ser Thr Lys Thr Asp Ala Ser Ser Thr His His Ser Thr Val Pro Pro
210 215 220
Leu Thr Ser Ser Asn His Ser Thr Ser Pro Gln Leu Ser Thr Gly Val
225 230 235 240
Ser Phe Phe Phe Leu Ser Phe His Ile Ser Asn Leu Gln Phe Asn Ser
245 250 255
Ser Leu Glu Asp Pro Ser Thr Asp Tyr Tyr Gln Glu Leu Gln Arg Asp
260 265 270
Ile Ser Glu Met Phe Leu Gln Ile Tyr Lys Gln Gly Gly Phe Leu Gly
275 280 285
Leu Ser Asn Ile Lys Phe Arg Pro Gly Ser Val Val Val Gln Leu Thr
290 295 300
Leu Ala Phe Arg Glu Gly Thr Ile Asn Val His Asp Val Glu Thr Gln
305 310 315 320
Phe Asn Gln Tyr Lys Thr Glu Ala Ala Ser Arg Tyr Asn Leu Thr Ile
325 330 335
Ser Asp Val Ser Val Ser Asp Val Pro Phe Pro Phe Ser Ala Gln Ser
340 345 350
Gly Ala Gly Val Pro Gly Trp Gly Ile Ala Leu Leu Val Leu Val Cys
355 360 365
Val Leu Val Ala Leu Ala Ile Val Tyr Leu Ile Ala Leu Ala Val Cys
370 375 380
Gln Cys Arg Arg Lys Asn Tyr Gly Gln Leu Asp Ile Phe Pro Ala Arg
385 390 395 400
Asp Thr Tyr His Pro Met Ser Glu Tyr Pro Thr Tyr His Thr His Gly
405 410 415
Arg Tyr Val Pro Pro Ser Ser Thr Asp Arg Ser Pro Tyr Glu Lys Val
420 425 430
Ser Ala Gly Asn Gly Gly Ser Ser Leu Ser Tyr Thr Asn Pro Ala Val
435 440 445
Ala Ala Thr Ser Ala Asn Leu
450 455
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА ПУТЕМ ВНУТРИВЕННОГО ВВЕДЕНИЯ РЕКОМБИНАНТНОГО MVA И АНТИТЕЛА | 2018 |
|
RU2795103C2 |
| ПРОМОТОРЫ ДЛЯ УСИЛЕНИЯ ЭКСПРЕССИИ В ПОКСВИРУСАХ | 2016 |
|
RU2753884C2 |
| ПЕРСОНАЛИЗИРОВАННАЯ ВАКЦИНА | 2018 |
|
RU2779987C2 |
| ВАКЦИНА ПРОТИВ АФРИКАНСКОЙ ЧУМЫ СВИНЕЙ | 2020 |
|
RU2819672C2 |
| ДИСПЛЕЙ ИНТЕГРАЛЬНОГО МЕМБРАННОГО БЕЛКА НА ВНЕКЛЕТОЧНЫХ ОБОЛОЧЕЧНЫХ ВИРИОНАХ ПОКСВИРУСА | 2017 |
|
RU2759846C2 |
| Выделенный рекомбинантный вирус на основе вируса гриппа для индукции специфического иммунитета к вирусу гриппа и/или профилактики заболеваний, вызванных вирусом гриппа | 2021 |
|
RU2813150C2 |
| ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА С ПОМОЩЬЮ ВНУТРИОПУХОЛЕВОГО И/ИЛИ ВНУТРИВЕННОГО ВВЕДЕНИЯ РЕКОМБИНАНТНОГО MVA, КОДИРУЮЩЕГО 4-1BBL (CD137L) И/ИЛИ CD40L | 2019 |
|
RU2830601C2 |
| РЕКОМБИНАНТНЫЕ КЛЕТКИ, ПРОДУЦИРУЮЩИЕ ГИАЛУРОНОВУЮ КИСЛОТУ | 2022 |
|
RU2839983C2 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2020 |
|
RU2760301C1 |
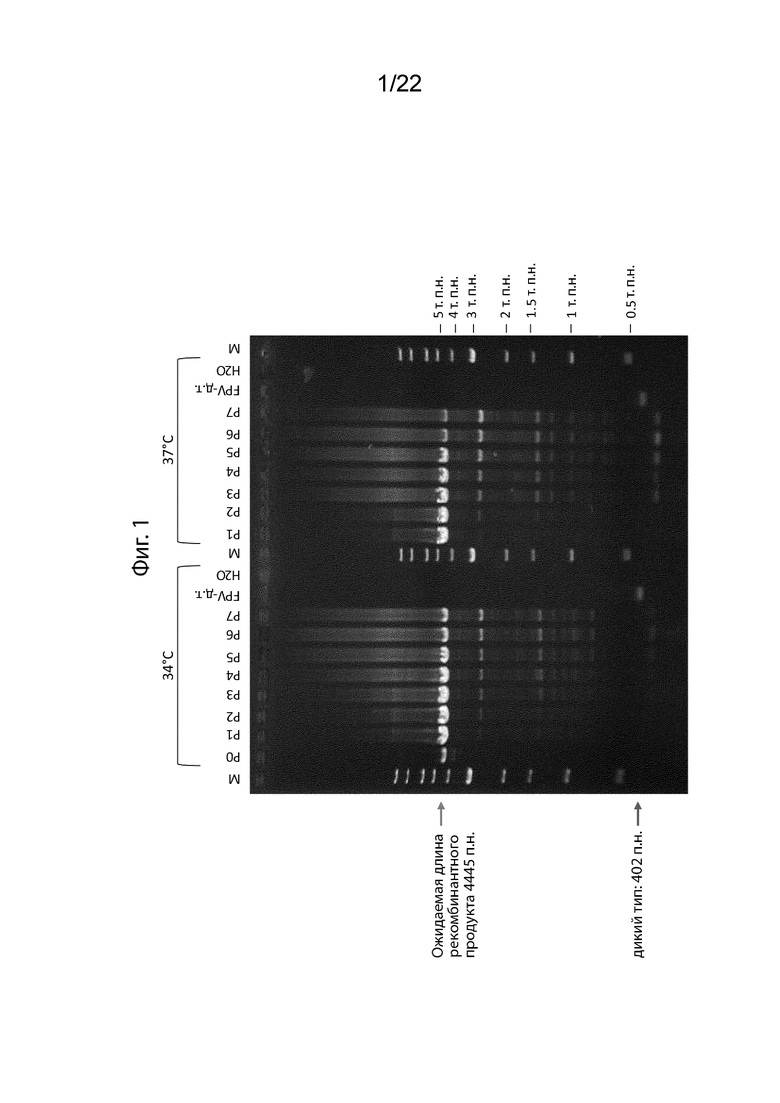



















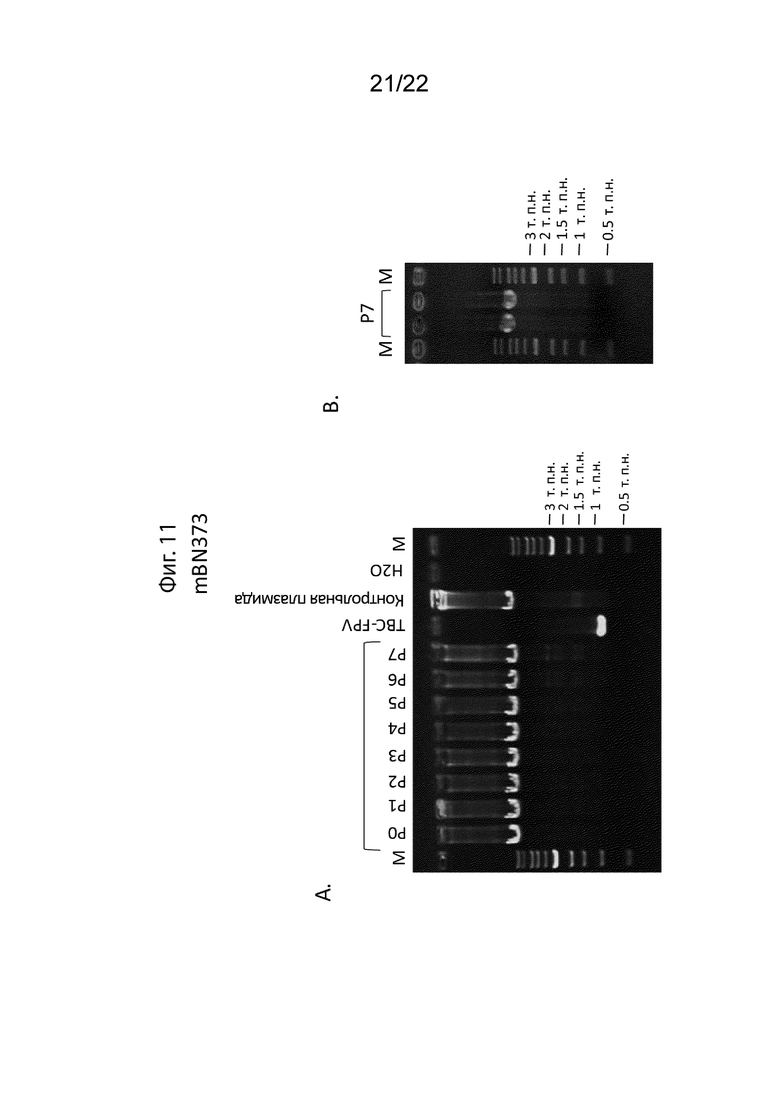
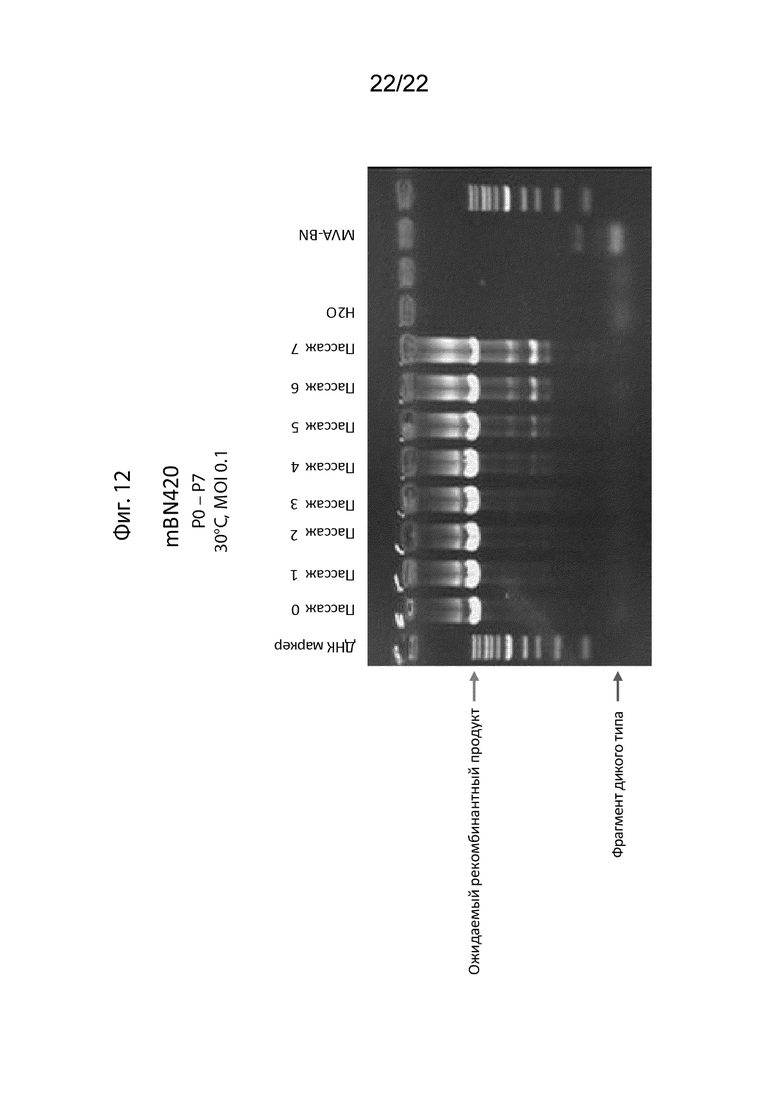
Изобретение относится к биотехнологии. Описан рекомбинантный поксвирус для лечения рака. Указанный рекомбинантный поксвирус включает нуклеиновую кислоту, кодирующую пептид MUC1, имеющий три вариабельных домена N-концевых повторов (VNTR), где (a) порядок расположения нуклеиновых кислот, кодирующих по меньшей мере два домена VNTR MUC1, отличается от их порядка расположения в PANVAC, и (b) домены VNTR были оптимизированы по кодонам. При этом рекомбинантный поксвирус является стабильным при последовательном пассировании рекомбинантного поксвируса. Также описан способ получения указанного поксвируса. Изобретение расширяет арсенал средств для борьбы с раком. 2 н. и 14 з.п. ф-лы, 12 ил., 6 табл., 5 пр.
1. Рекомбинантный поксвирус для лечения рака, где указанный рекомбинантный поксвирус включает нуклеиновую кислоту, кодирующую пептид MUC1, имеющий три вариабельных домена N-концевых повторов (VNTR), где (a) порядок расположения нуклеиновых кислот, кодирующих по меньшей мере два домена VNTR MUC1, отличается от их порядка расположения в PANVAC, и (b) домены VNTR были оптимизированы по кодонам, и где рекомбинантный поксвирус является стабильным при последовательном пассировании рекомбинантного поксвируса.
2. Рекомбинантный поксвирус по п. 1, где последовательность нуклеиновой кислоты содержит SEQ ID NO: 2.
3. Рекомбинантный поксвирус по п. 1, дополнительно содержащий нуклеиновую кислоту, кодирующую карциноэмбриональный антиген (CEA).
4. Рекомбинантный поксвирус по п. 3, где нуклеиновая кислота, кодирующая карциноэмбриональный антиген (CEA), содержит по меньшей мере одну нуклеотидную замену по меньшей мере в одной повторяющейся нуклеотидной области, где по меньшей мере одна повторяющаяся нуклеотидная область определена как (a) три или более последовательно повторяющихся нуклеотида G или C и/или (b) три или более последовательно повторяющихся нуклеотида T.
5. Рекомбинантный поксвирус по п. 4, где нуклеиновая кислота, кодирующая карциноэмбриональный антиген (CEA), содержит SEQ ID NO: 13 или SEQ ID NO: 14.
6. Рекомбинантный поксвирус по п. 1, где рекомбинантный поксвирус выбран из ортопоксвируса и авипоксвируса.
7. Рекомбинантный поксвирус по п. 6, где ортопоксвирус представляет собой модифицированный вирус осповакцины Ankara (MVA), который представляет собой MVA-BN.
8. Рекомбинантный поксвирус по п. 1, где нуклеиновая кислота, кодирующая пептид MUC1, дополнительно содержит по меньшей мере одну нуклеотидную последовательность, кодирующую пептидный фрагмент, выбранный из группы, состоящей из: YLAPPAHGV (SEQ ID NO: 24), YLDTRPAPV (SEQ ID NO: 25), YLAIVYLIAL (SEQ ID NO: 26), YLIALAVCQV (SEQ ID NO: 27), YLSYTNPAV (SEQ ID NO: 28) и SLFRSPYEK (SEQ ID NO: 29).
9. Рекомбинантный поксвирус по п. 1, где поксвирус также содержит нуклеиновую кислоту, кодирующую одну или более костимулирующих молекул, выбранных из B7-1, ICAM-1, LFA-3 и их комбинаций.
10. Способ получения рекомбинантного поксвируса, который является стабильным при последовательном пассировании рекомбинантного поксвируса, где указанный способ включает:
(1) получение нуклеиновой кислоты, кодирующей белок MUC1, имеющий три вариабельных домена N-концевых повторов (VNTR), где (a) порядок расположения по меньшей мере двух доменов VNTR отличается от их порядка расположения в PANVAC и (b) домены VNTR были оптимизированы по кодонам,
(2) вставку первой последовательности нуклеиновой кислоты в поксвирус для получения рекомбинантного поксвируса; и
(3) амплификацию рекомбинантного поксвируса в культуре клеток;
где рекомбинантный поксвирус является стабильным при последовательном пассировании.
11. Способ по п. 10, дополнительно включающий получение нуклеиновой кислоты, кодирующей карциноэмбриональный антиген (CEA), и ее вставку в поксвирус.
12. Способ по п. 11, где нуклеиновая кислота, кодирующая карциноэмбриональный антиген (CEA), содержит по меньшей мере одну нуклеотидную замену по меньшей мере в одной повторяющейся нуклеотидной области, где по меньшей мере одна повторяющаяся нуклеотидная область определена как (a) три или более последовательно повторяющихся нуклеотида G или C и/или (b) три или более последовательно повторяющихся нуклеотида T.
13. Способ по п. 10, где рекомбинантный поксвирус выбран из ортопоксвируса и авипоксвируса.
14. Способ по п. 13, где ортопоксвирус представляет собой вирус осповакцины, являющийся модифицированным вирусом осповакцины Ankara (MVA), который представляет собой MVA-BN.
15. Способ по п. 10, где первая нуклеиновая кислота дополнительно содержит по меньшей мере одну нуклеотидную последовательность, кодирующую пептидный фрагмент, выбранный из группы, состоящей из: YLAPPAHGV (SEQ ID NO: 24), YLDTRPAPV (SEQ ID NO: 25), YLAIVYLIAL (SEQ ID NO: 26), YLIALAVCQV (SEQ ID NO: 27), YLSYTNPAV (SEQ ID NO: 28) и SLFRSPYEK (SEQ ID NO: 29).
16. Способ по п. 10, где поксвирус дополнительно содержит нуклеиновую кислоту, кодирующую одну или более костимулирующих молекул, выбранных из B7-1, ICAM-1, LFA-3 и их комбинаций.
| WO 2005046614 A3, 26.05.2005 | |||
| КОЛЬЦЕВАЯ ХЛЕБОПЕКАРНАЯ ПЕЧЬ НЕПРЕРЫВНОГО ДЕЙСТВИЯ | 1927 |
|
SU21064A1 |

