Область техники, к которой относится изобретение
Настоящее изобретение относится к антителам к миостатину и способам их применения. Настоящее изобретение относится также к полипептидам, содержащим варианты Fc-областей, и способам их применения.
Предпосылки создания изобретения
Миостатин, который называют также фактором дифференциации-8 (GDF8), представляет собой секретируемый белок и является представителем суперсемейства белков трансформирующего рост фактора-бета (TGF-бета). Представители этого суперсемейства обладают регулирующими рост и морфогенетическими свойствами (см., например, ссылки, обозначенные как Непатентная литература 1, Непатентная литература 2 и Патентная литература 1). Миостатин экспрессируется главным образом в развивающейся скелетной мускулатуре и мускулатуре взрослых и функционирует в качестве отрицательного регулятора мышечного роста. Системная сверхэкспрессия миостатина в организме взрослых мышей приводит к мышечному истощению (см., например, ссылку, обозначенную как Непатентная литература 3), в то время как в противоположном случае, мыши с «выключенным» миостатином характеризуются гипертрофией и гиперплазией скелетной мускулатуры, приводящей к 2-3-кратному увеличению мышечной массы по сравнению с их однопометниками дикого типа (см., например, ссылку, обозначенную как Непатентная литература 4).
Подобно другим представителям семейства TGF-бета миостатин синтезируется в виде крупного белка-предшественника, который содержит N-концевой пропептидный домен и С-концевой домен, представляющий собой активную молекулу (см., например, ссылки, обозначенные как Непатентная литература 5; Патентная литература 2). Две молекулы предшественника миостатина ковалентно соединены с помощью одной дисульфидной связи, присутствующей в С-концевом домене фактора роста. Активный зрелый миостатин (соединенный дисульфидной связью гомодимер, состоящий из С концевого домена фактора роста) высвобождается из предшественника миостатина посредством нескольких стадий протеолитического процессинга. На первой стадии пути активации миостатина происходит расщепление пептидной связи между N-концевым пропептидным доменом и С-концевым доменом фактора роста, т.е. Arg266-Asp267, с помощью пропротеиновой конвертазы фуринового типа в обеих цепях гомодимерного предшественника. Однако образовавшиеся три пептида (два пропептида и один зрелый миостатин (т.е. связанный дисульфидом гомодимер, состоящий из доменов фактора роста)) остаются ассоциированными, образуя нековалентный неактивный комплекс, который обозначают как «латентный миостатин». Затем зрелый миостатин может высвобождаться из латентного миостатина посредством расщепления пропептида. Представители семейства металлопротеиназ, представляющих собой костный морфогенетический белок (белок, участвующий в остеогенезе) 1 (ВМР1), расщепляют единичную пептидную связь в пропептиде, Arg98-Asp99, что сопровождается высвобождением зрелого активного миостатина в виде гомодимера (см., например, ссылку, обозначенную как Непатентная литература 6). Кроме того, латентный миостатин можно активировать также in vitro посредством диссоциации комплекса с помощью либо обработки кислотой, либо тепловой обработки (см., например, ссылку, обозначенную как Непатентная литература 7).
Миостатин проявляет свои действия через семейство трансмембранных гетеротетрамерых рецепторов серин/треониновых киназ, активация которых повышает трансфосфорилирование рецепторов, приводя к стимуляции активности серин/треониновых киназ. Установлено, что путь миостатина включает активный димер миостатина, связывающийся с рецептором активина типа IIB (ActRIIB) с высокой аффинностью, что далее приводит к возникновению и активации трансфосфорилирования низкоаффинного рецептора, активинподобной киназы 4 (ALK4) или активинподобной киназы 5 (ALK5). Установлено также, что далее активируются белки Smad 2 и Smad 3 и образуют комплекс с Smad 4, который затем транслоцируется в ядро для активации транскрипции гена-мишени. Было продемонстрировано, что ActRIIB обладает способностью опосредовать воздействие миостатина in vivo, поскольку экспрессия доминантно-негативной формы ActRIIB в организме мышей имитирует «выключение» гена миостатина (см., например, ссылку, обозначенную как Непатентная литература 8).
С мышечным истощением (т.е., с утратой или функциональным нарушением мышечной ткани) ассоциирован целый ряд нарушений или состояний, таких как мышечная дистрофия (MD; включая мышечную дистрофию Дюшенна), амиотрофический боковой склероз (ALS), мышечная атрофия, атрофия органа, состояние немощи, застойное обструктивное заболевание легких (COPD), саркопения и кахексия, являющаяся результатом рака или других нарушений, а также заболевание почек, сердечную недостаточность или болезнь и заболевание печени. На пациентов может оказывать благоприятное воздействие увеличение мышечной массы и/или мышечной силы; однако в настоящее время доступны лишь ограниченные средства лечения указанных нарушений. Таким образом, из-за его роли в качестве отрицательного регулятора роста скелетных мышц, миостатин становится важной мишенью для терапевтического или профилактического вмешательства при указанных нарушениях или состояниях или для мониторинга развития указанных нарушений или состояний. В частности, агенты, которые ингибируют активность миостатина, могут являться ценными терапевтическими средствами.
Ингибирование экспрессии миостатина приводит как к мышечной гипертрофии, так и к гиперплазии (Непатентная литература 4). Миостатин является отрицательным регулятором мышечной регенерации после повреждения, и отсутствие миостатина у «нулевых» по миостатину мышей приводит к ускорению мышечной регенерации (см., например, ссылку, обозначенную как Непатентная литература 9). Установлено, что антитела к миостатину (GDF8), которые описаны, например, в ссылках, обозначенных как Патентная литература 3, Патентная литература 4, Патентная литература 5, Патентная литература 6 и Патентная литература 7, и Патентная литература 8, Патентная литература 9 и Патентная литература 10, связываются с миостатином и ингибируют активность миостатина in vitro и in vivo, включая активность миостатина, ассоциированную с отрицательной регуляцией массы скелетных мышц. Нейтрализующие миостатин антитела увеличивают вес тела, массу скелетных мышц и размер мышц, а также силу скелетных мышц у мышей дикого типа (см., например, ссылку, обозначенную как Непатентная литература 10) и у mdx-мышей (модель мышечной дистрофии) (см., например, ссылки, обозначенные как Непатентная литература 11; Непатентная литература 12). Однако известные из существующего уровня техники антитела все являются специфическими для зрелого миостатина, но не для латентного миостатина, и в описанных стратегиях ингибирования активности миостатина использовались антитела, которые могут связываться и нейтрализовать зрелый миостатин.
Антитела привлекают внимание в качестве фармацевтических средств благодаря их высокой стабильности в крови и небольшого количества побочных действий (см., например, ссылки, обозначенные как Непатентная литература 13 и Непатентная литература 14). Почти все современные поступающие в продажу терапевтические антитела представляют собой антитела человеческого IgG1-подкласса. Одной из известных функций антител IgG-класса является антитело-обусловленная клеточнозависимая цитотоксичность (далее в контексте настоящего описания обозначена как ADCC-активность) (см., например, ссылку, обозначенную как Непатентная литература 15). Для проявления антителом ADCC-активности Fc-область антитела должна связываться с Fc-гамма-рецептором (далее в контексте настоящего описания обозначен как Fc-гамма R), который представляет собой связывающийся с антителом рецептор, присутствующий на поверхности эффекторных клеток, таких как клетки-киллеры, естественные клетки-киллеры и активированные макрофаги.
Для человека описаны изоформы Fc-гамма RIa (CD64A), Fc-гамма RIIa (CD32A), Fc-гамма RIIb (CD32B), Fc-гамма RIIIa (CD16A) и Fc-гамма RIIIb (CD16B) в качестве белков семейства Fc-гамма R, кроме того, описаны также соответствующие аллотипы (см., например, ссылку, обозначенную как Непатентная литература 16). Fc-гамма RIa, Fc-гамма RIIa и Fc-гамма RIIIa называют активирующими Fc-гамма R, поскольку они обладают иммунологически активными функциями, а Fc-гамма RIIb называют ингибирующим Fc-гамма R, поскольку он обладает иммуносупрессорными функциями (см., например, ссылку, обозначенную как Непатентная литература 17).
Установлено, что для связывания между Fc-областью и Fc-гамма R важны несколько аминокислотных остатков в шарнирной области антитела и СН2-домене и сахарная цепь, присоединенная к Asn в положении 297 (EU-нумерация), которая связана с СН2-доменом (см., например, ссылки, обозначенные как Непатентная литература 18, Непатентная литература и Непатентная литература 20). К настоящему времени получены различные варианты, обладающие способностью связываться с Fc-гамма R, прежде всего антитела с мутациями, интродуцированными в указанные сайты; и варианты Fc-области, обладающие более высокими связывающими активностями в отношении активирующего Fc-гамма R (см., например, ссылки, обозначенные как Патентная литература 11, Патентная литература 12, Патентная литература 13 и Патентная литература 14).
Когда активирующий Fc-гамма R перекрестно связывается с иммунным комплексом, он фосфорилирует активирующие мотивы на основе тирозина иммунорецептора (ITAM), входящие во внутриклеточной домен или общую гамма-цепь FcR (участвующий во взаимодействии партнер), активирует трансдуктор сигнала SYK и запускает воспалительный иммунный ответ путем инициации активации каскада сигналов (см., например, ссылку, обозначенную как Непатентная литература 21).
Fc-гамма RIIb является единственным Fc-гамма R, который экспрессируется на В-клетках (см., например, ссылку, обозначенную как Непатентная литература 22). Установлено, что взаимодействие Fc-области антитела с Fc-гамма RIIb подавляет первичный иммунный ответ В-клеток (см., например, ссылку, обозначенную как Непатентная литература 23). Кроме того, описано, что, когда Fc-гамма RIIb на В-клетках и В-клеточный рецептор (BCR) являются перекрестно связанными через иммунный комплекс в крови, то подавляется В-клеточная активация и производство антител В-клетками (см., например, ссылку, обозначенную как Непатентная литература 24). Для указанной иммуносупрессорной трансдукции сигнала, опосредуемой BCR и Fc-гамма RIIb, необходим ингибирующий мотив на основе тирозина иммунорецептора (ITIM), входящий во внутриклеточный домен Fc-гамма RIIb (см., например, ссылки, обозначенные как Непатентная литература 25 и Непатентная литература 26). Когда ITIM фосфорилируется после передачи сигнала, то SH2-содержащая инозитолполифосфат-5-фосфатаза (SHIP) вовлекается в процесс, ингибируется трансдукция каскада сигналов других Fc-гамма R и подавляется воспалительный иммунный ответ (см., например, ссылку, обозначенную как Непатентная литература 27). Кроме того, известно, что только агрегация Fc-гамма RIIb приводит к кратковременному подавлению притока кальция из-за перекрестного связывания BCR и В-клеточной пролиферации независимым от BCR образом без индукции апоптоза IgM-продуцирующих В-клеток (см., например, ссылку, обозначенную как Непатентная литература 28).
Fc-гамма RIIb экспрессируется также на дендритных клетках, макрофагах, активированных нейтрофилах, тучных клетках и базофилах. Fc-гамма RIIb ингибирует функции активирующего Fc-гамма R, такие как фагоцитоз и высвобождение провоспалительных цитокинов в этих клетках, и подавляет воспалительные иммунные ответы (см., например, ссылку, обозначенную как Непатентная литература 17).
К настоящему времени для изучения иммуносупрессорных функций Fc-гамма RIIb осуществляли исследования с использованием мышей с «выключенным» Fc-гамма RIIb. С их помощью установлено, что у мышей с «выключенным» Fc-гамма RIIb гуморальный иммунный ответ не регулируется соответствующим образом (см., например, ссылку, обозначенную как Непатентная литература 29), чувствительность к индуцированному коллагеном артриту (CIA) повышается (см., например, ссылку, обозначенную как Непатентная литература 30), присутствуют напоминающие волчанку симптомы и симптомы, напоминающие синдром Гудпасчера (см., например, ссылку, обозначенную как Непатентная литература 31).
Кроме того, известно, что с нарушением регуляции Fc-гамма RIIb связаны аутоиммунные заболевания человека. Например, описана взаимосвязь между генетическим полиморфизмом в трансмембранной области и промоторной области Fc-гамма RIIb и частотой развития системной красной волчанки (SLE) (см., например, ссылки, обозначенные как Непатентная литература 32, Непатентная литература 33, Непатентная литература 34, Непатентная литература 35 и Непатентная литература 36) и снижением экспрессии Fc-гамма RIIb на поверхности В-клеток у страдающих SLE пациентов (см., например, ссылки, обозначенные как Непатентная литература 37 и Непатентная литература 38).
С учетом данных, полученных на мышиных моделях, и самих клинических результатов считается, что Fc-гамма RIIb играет роль в контроле аутоиммунных заболеваний и воспалительных заболеваний посредством привлечения, прежде всего, В-клеток, и он является перспективной молекулой-мишенью для контроля аутоиммунных заболеваний и воспалительных заболеваний.
Известно, что IgG1, которые главным образом применяют в качестве поступающих в продажу терапевтических антител, связываются не только с Fc-гамма RIIb, но также характеризуются сильным связыванием с активирующим Fc-гамма R (см., например, ссылку, обозначенную как Непатентная литература 39). Это делает возможным разработку терапевтических антител, обладающих более высокими иммуносупрессорными свойствами по сравнению со свойствами IgG1, путем применения Fc-области с повышенной способностью связываться с Fc-гамма RIIb или с улучшенной селективностью связывания с Fc-гамма RIIb по сравнению со связыванием с активирующим Fc-гамма R. Например, высказано предположение о том, что применение антитела, имеющего вариабельную область, которая связывается с BCR, и Fc с повышенной способностью связываться с Fc-гамма RIIb, может ингибировать В-клеточную активацию (см., например, ссылку, обозначенную как Непатентная литература 40). Описано, что перекрестное сшивание Fc-гамма RIIb на В-клетках и IgE, связанного с В-клеточным рецептором, подавляет дифференцировку В-клеток в плазматические клетки, что в результате приводит к подавлению производства IgE; и у мышей с трансплантированными человеческими РВМС сохраняются концентрации человеческих IgG и IgM, в то время как концентрация человеческого IgE понижается (см., например, ссылку, обозначенную как Непатентная литература 41). Было описано что, когда Fc-гамма RIIB и CD79b, который представляет собой конститутивную молекулу комплекса В-клеточного рецептора, перекрестно сшиваются антителом, то помимо воздействия на IgE подавляется пролиферация В-клеток in vitro, и облегчаются симптомы артрита на модели индуцированного коллагеном артрита (см., например, ссылку, обозначенную как Непатентная литература 42).
Описано также, что помимо воздействия на В-клетки, перекрестное связывание Fc-эпсилон RI и Fc-гамма RIIb на тучных клетках с использованием молекул, в которых Fc-область IgG с повышенной способностью связываться с Fc-гамма RIIb слита с Fc-областью IgE, которая связывается с IgE-рецептором Fc-эпсилон RI, вызывает фосфорилирование Fc-гамма RIIb, подавляя тем самым зависящий от Fc-эпсилон RI приток кальция. Это позволяет предположить, что ингибирования дегрануляции посредством стимуляции Fc-гамма RIIb можно достигать путем повышения связывания Fc-гамма RIIb (см., например, ссылку, обозначенную как Непатентная литература 43).
Таким образом, можно предположить, что антитело, которое имеет Fc с улучшенной способностью связываться с Fc-гамма RIIb, будет перспективным в качестве терапевтического агента при воспалительных заболеваниях, таких как аутоиммунное заболевание.
Кроме того, описано, что активация макрофагов и дендритных клеток с помощью Толл-подобного рецептора 4 при стимуляции с использованием LPS, подавляется в присутствии иммунного комплекса антитело-антиген, и предполагается, что это также представляет собой действия иммунного комплекса через Fc-гамма RIIb (см., например, ссылки, обозначенные Непатентная литература 44 и Непатентная литература 45). Таким образом, ожидается, что применение антител с повышенной способностью связываться с Fc-гамма RIIb может усиливать опосредуемые TLR подавляющие действия в отношении активации сигналов; поэтому предполагается, что указанные антитела могут являться перспективными в качестве терапевтических агентов при воспалительных заболеваниях, таких как аутоиммунные заболевания.
Кроме того, ожидается, что мутанты с повышенной способностью связываться с Fc-гамма RIIb могут являться перспективными в качестве терапевтических агентов при раке, а также при воспалительных заболеваниях, таких как аутоиммунные заболевания. К настоящему времени установлено, что Fc-гамма RIIb играет важную роль в агонистической активности агонистических антител к суперсемейству TNF-рецепторов. В частности, можно предположить, что взаимодействие с Fc-гамма RIIb требуется для проявления агонистической активности антител к CD40, DR4, DR5, CD30 и CD137, которые входят в семейство TNF-рецепторов (см., например, ссылки, обозначенные как Непатентная литература 46, Непатентная литература 47, Непатентная литература 48, Непатентная литература 49, Непатентная литература 50, Непатентная литература 51 и Непатентная литература 52). В ссылке, обозначенной как Непатентная литература 46, продемонстрировано, что применение антител с повышенной способностью связываться с Fc-гамма RIIb увеличивает противоопухолевое действие антител к CD40. Таким образом, можно предположить, что антитела с повышенной способностью связываться с Fc-гамма RIIb могут влиять на увеличение агонистической активности агонистических антител, включая антитела к суперсемейству TNF-рецепторов.
Кроме того, установлено, что клеточная пролиферация подавляется при применении антитела, которое распознает Kit, тип рецепторной тирозинкиназы (RTK), для перекрестной сшивки Fc-гамма RIIb и Kit на экспрессирующих Kit клетках. Аналогичные действия описаны даже в случаях, в которых указанная Kit являлась конститутивно активированной и имела мутации, которые вызывают онкогенез (см., например, ссылку, обозначенную как Непатентная литература 53). Таким образом, можно предполагать, что применение антител с повышенной способностью связываться с Fc-гамма RIIb может увеличивать ингибирующие воздействия на клетки, которые экспрессируют RTK, имеющую конститутивно активирующие мутации.
Описаны антитела, имеющие Fc с повышенной Fc-гамма RIIb-связывающей активностью (см., например, ссылку, обозначенную как Непатентная литература 40). В этой публикации описано повышение Fc-гамма RIIb-связывающей активности путем внесения изменений, таких как S267E/L328F, G236D/S267E и S239D/S267E, в Fc-область антитела. Среди них антитело с интродуцированной мутацией S267E/L328F наиболее сильно связывалось с Fc-гамма RIIb, и у него сохранялся такой же уровень связывания с Fc-гамма RIa и Fc-гамма RIIa типа Н, в котором остаток в положении 131 Fc-гамма RIIa представляет собой His, что и во встречающемся в естественных условиях IgG1. Однако в другой публикации описано, что указанное изменение повышает связывание Fc-гамма RIIa типа R, в котором остаток в положении 131 Fc-гамма RIIa представляет собой Arg, в несколько сотен раз относительно уровня такого же связывания Fc-гамма RIIb, это означает, что избирательность Fc-гамма RIIb-связывания не повышается по сравнению с Fc-гамма RIIa типа R (см., например, ссылку, обозначенную как Патентная литература 15).
Считается, что только воздействие повышенного связывания с Fc-гамма RIIa, но не повышенного связывания с Fc-гамма RIIb, должно влиять на клетки, такие как тромбоциты, которые экспрессируют Fc-гамма RIIa, но не экспрессируют Fc-гамма RIIb (см., например, ссылку, обозначенную как Непатентная литература 17). Например, известно, что в группе пациентов, которым вводили бевацизумаб, антитело к VEGF, имели повышенный риск тромбоэмболии (см., например, ссылку, обозначенную как Непатентная литература 54). Кроме того, аналогичным образом тромбоэмболия обнаружена в клинических исследованиях разработанных антител к лиганду для CD40, и клиническое испытание было прервано (см., например, ссылку, обозначенную как Непатентная литература 55). Оба варианта указанных антител изучены в более поздних исследованиях с использованием животных моделей и было высказано предположение, что вводимые антитела вызывали агрегацию посредством связывания Fc-гамма RIIa на тромбоцитах и приводили к образованию сгустка крови (см., например, ссылки, обозначенные как Непатентная литература 56 и Непатентная литература 57). При системной красной волчанке, представляющей собой аутоиммунное заболевание, тромбоциты активируются посредством зависящего от Fc-гамма RIIa механизма и описано, что активация тромбоцитов коррелирует с серьезностью симптомов (см., например, ссылку, обозначенную как Непатентная литература 58). Введение антитела с повышенной способностью связываться с Fc-гамма RIIa тем пациентам, которые уже имели высокий риск развития тромбоэмболии, риск развития тромбоэмболии может повышаться, что является чрезвычайно опасным.
Кроме того, описано, что антитела с повышенной способностью связываться с Fc-гамма RIIa, усиливают опосредуемый макрофагами антителозависимый клеточной фагоцитоз (ADCP) (см., например, ссылку, обозначенную как Непатентная литература 59). Когда антигены, подлежащие связыванию с помощью антител, фагоцитируются макрофагами, считается, что сами антитела также одновременно фагоцитируются. Когда антитела вводят в качестве фармацевтических агентов, предполагается, что пептидные фрагменты из введенных антител, вероятно, тоже презентируются в качестве антигена, что повышает риск производства антител против терапевтических антител (антитерапевтические антитела). Более конкретно, повышенная способность связываться с Fc-гамма RIIa может повышать риск выработки антител против терапевтических антител, и это может значительно снижать их ценность в качестве фармацевтических агентов. Кроме того, предполагается, что Fc-гамма RIIb на дендритных клетках участвует в периферической толерантности путем ингибирования активации дендритных клеток, вызываемой иммунными комплексами, сформированными между антигенами и антителами, или путем подавления презентации антигена Т-клеткам посредством активации Fc-гамма-рецепторов (см., например, ссылку, обозначенную как Непатентная литература 60). Поскольку Fc-гамма RIIa экспрессируется также на дендритных клетках, то, когда антитела, которые имеют Fc с повышенной избирательной способностью связываться с Fc-гамма RIIb, применяли в качестве фармацевтических агентов, антигены не могли легко презентироваться дендритными клетками и т.п. из-за повышенной избирательной способности связываться с Fc-гамма RIIb, и риск выработки антитела к лекарственному средству мог относительно уменьшаться. Указанные антитела в этом плане также могут быть ценными.
Более конкретно, ценность в качестве фармацевтических средств может существенно снижаться при повышении способности связываться с Fc-гамма RIIa, что приводит к повышенному риску образования тромбов в результате агрегации тромбоцитов и повышенному риску выработки антитерапевтических антител из-за повышенной иммуногенности.
С этой точки зрения вышеуказанный вариант Fc с повышенной способностью связываться с Fc-гамма RIIb характеризуется в значительной степени повышенным связыванием Fc-гамма RIIa R-типа по сравнению со встречающимся в естественных условиях IgG1. Таким образом, его ценность в качестве фармацевтического средства для пациентов, которые несут Fc-гамма RIIa R-типа, существенно снижается. Типы Н и R Fc-гамма RIIa встречаются у людей европеоидной расы и афроамериканцев примерно с одинаковой частотой (см., например, ссылки, обозначенные как Непатентная литература 61 и Непатентная литература 62). Таким образом, когда указанный вариант Fc применяют для лечения аутоиммунных заболеваний, количество пациентов, для которых его можно безопасно применять с достижением его эффективности в качестве фармацевтического средства, может быть ограниченным.
Кроме того, установлено, что происходит созревание дендритных клеток, если дендритные клетки имеют дефицит Fc-гамма RIIb или в дендритных клетках происходит ингибирование взаимодействие между Fc-гамма RIIb и Fc-областью антитела антителом к Fc-гамма RIIb (см., например, ссылки, обозначенные как Непатентная литература 63 и Непатентная литература 64). В указанном исследовании высказано предположение о том, что Fc-гамма RIIb активно подавляет созревание дендритных клеток в стабильном состоянии в отсутствии воспаления или т.п.ив отсутствии активации. Fc-гамма RIIa экспрессируется на поверхности дендритных клеток в дополнение к Fc-гамма RIIb; поэтому, даже, если связывание с ингибирующим Fc-гамма RIIb повышается и если связывание с активирующим Fc-гамма R, таким как Fc-гамма RIIa, также повышается, то в результате созревание дендритных клеток может усиливаться. Более конкретно, повышение не только Fc-гамма RIIb-связывающей активности, но и также соотношения Fc-гамма RIIb-связывающей активности и Fc-гамма RIIa-связывающей активности рассматривается как важный параметр при создании антител с иммуносупрессорным действием.
Таким образом, при создании фармацевтического средства, в котором используется опосредуемое связыванием Fc-гамма RIIb иммуносупрессорное действие, существует необходимость в варианте Fc, который не только обладает повышенной Fc-гамма RIIb-связывающей активностью, но также обладает способностью связываться в обоими аллотипами Fc-гамма RIIa Н и R, которая сохраняется на сходном уровнем или ослаблена до более низкого уровня по сравнению со встречающимся в естественных условиях IgG1.
При этом, к настоящему время опубликованы данные исследований, в которых в Fc-область интродуцировали аминокислотные изменения для повышения избирательности связывания с Fc-гамма RIIb (см., например, ссылку, обозначенную как Непатентная литература 65). Однако для всех вариантов, указанных в данной публикации, которые имели улучшенную избирательность в отношении Fc-гамма RIIb, обнаружено снижение способности связываться с Fc-гамма RIIb по сравнению со встречающимся в естественных условиях IgG1. Таким образом, фактически для указанных вариантов представляется сложным индуцировать опосредуемую Fc-гамма RIIb иммуносупрессорную реакцию, более сильную по сравнению с IgG1.
Кроме того, поскольку Fc-гамма RIIb играет важную роль для указанных выше агонистических антител, ожидается, что повышение их связывающей активности приведет к повышению агонистической активности. Однако когда аналогичным образом повышается связывание с Fc-гамма RIIa, но могут непреднамеренно проявляться такие виды активности, как ADCC-активность и ADCP-активность, и это может приводить к побочным действиям. Исходя из указанной точки зрения, предпочтительно иметь возможность избирательно повышать Fc-гамма RIIb-связывающую активность.
Из этих результатов следует, что при производстве терапевтических антител, предназначенных для лечения аутоиммунных заболеваний и рака, в которых участвует Fc-гамма RIIb, важно, чтобы по сравнению по сравнению со встречающимся в естественных условиях IgG1 активности связывания с обоими аллотипами Fc-гамма RIIa сохранялась или снижалась, а связывание с Fc-гамма RIIb повышалось. Однако для Fc-гамма RIIb характерна 93%-ная идентичность последовательности внеклеточной области с Fc-гамма RIIa, который представляет собой один из активирующих Fc-гамма R, и они очень сходны структурно. Известны аллотипы Fc-гамма RIIa, Н-тип и R-тип, в которых аминокислота в положении 131 представляет собой His (Н-тип) или Arg (R-тип), и при этом каждый из них по-разному взаимодействует с антителами (см., например, ссылку, обозначенную как Непатентная литература 66). Таким образом, может оказаться трудной проблемой создание варианта Fc-области с повышенным избирательным связыванием с Fc-гамма RIIb по сравнению с каждым из аллотипов Fc-гамма RIIa, включая способность различать высокогомологичные последовательности у Fc-гамма RIIa и Fc-гамма RIIb. В свете указанных трудностей к настоящему времени было идентифицировано несколько вариантов Fc, который обладали избирательной связывающей активностью в отношении Fc-гамма RIIb по сравнению с Fc-гамма RIIa, путем осуществления широкомасштабного анализа аминокислотных модификаций в Fc-области (см., например, ссылки, обозначенные как Патентная литература 16, Патентная литература 17, Патентная литература 18, Патентная литература 19 и Патентная литература 20).
К настоящему времени описан вариант Fc-области, обладающий избирательным связыванием с Fc-гамма RIIb, среди человеческих Fc-гамма R, однако отсутствуют данные о варианте Fc-области с избирательным связыванием с Fc-гамма RIIb среди Fc-гамма R обезьян. С учетом отсутствия указанного варианта Fc, воздействия варианта Fc, избирательно связывающегося с Fc-гамма RIIb, еще не были всесторонне протестированы на обезьянах.
Помимо вышеуказанного, описано, что путем модификации заряда аминокислотных остатков, которые могут экспонироваться на поверхности антитела, приводя к повышению или понижению изоэлектрической точки (pI) антитела, можно регулировать время полужизни антитела в крови (см., например, ссылки, обозначенные как Патентная литература 21 и Патентная литература 22). В указанных патентах продемонстрировано, что можно пролонгировать время полужизни антитела в плазме путем снижения pI антитела и наоборот.
Кроме того, описано, что включение антигена в клетки можно повышать путем модификации заряда специфических аминокислотных остатков, прежде всего в СН3-домене антитела, для повышения его pI (см., например, ссылку, обозначенную как Патентная литература 23). Кроме того, описано, что путем модификации заряда аминокислотных остатков в константной области (в основном в СН1-домене) с целью снижения pI можно пролонгировать время полужизни антитела в плазме (см., например, ссылку, обозначенную как Патентная литература 24).
Перечень ссылок
Патентная литература
PTL 1: Патент США №5827733
PTL 2: WO 1994/021681
PTL 3: Патент США №6096506
PTL 4: Патент США №7261893
PTL 5: Патент США №7320789
PTL 6: Патент США №7807159
PTL 7: Патент США №7888486
PTL 8: WO 2005/094446
PTL 9: WO 2007/047112
PTL 10: WO 2010/070094
PTL 11: WO 2000/042072
PTL 12: WO 2006/019447
PTL 13: WO 2004/099249
PTL 14: WO 2004/029207
PTL 15: Публикация заявки на патент США № US 2009/0136485
PTL 16: WO 2012/115241
PTL 17: WO 2013/047752
PTL 18: WO 2013/125667
PTL 19: WO 2014/030728
PTL 20: WO 2014/163101
PTL 21: WO 2007/114319
PTL 22: WO 2009/041643
PTL 23: WO 2014/145159
PTL 24: WO 2012/016227
Непатентная литература
NPL 1: Kingsley и др., Genes Dev. 8(2), 1994, cc. 133-146.
NPL 2: Hoodless и др., Curr. Top. Microbiol. Immunol. 228, 1998, cc. 235-272.
NPL 3: Zimmers и др., Science 296(5572), 2002, cc. 1486-1488.
NPL 4: McPherron и др., Nature 387(6628), 1997, cc. 83-90.
NPL 5: McPherron и Lee, Proc. Natl. Acad. Sci. USA 94(23), 1997, cc. 12457-12461.
NPL 6: Szlama и др., FEBS J 280(16), 2013, cc. 3822-3839.
NPL 7: Lee, PloS One 3(2), 2008, c. e1628.
NPL 8: Lee, Proc. Natl. Acad. Sci. USA 98(16), 2001, cc. 9306-9311.
NPL 9: McCroskery и др., J Cell Sci. 118(15), 2005, cc. 3531-3541.
NPL 10: Whittemore и др., Biochem. Biophys. Res. Commun. 300(4), 2003, cc. 965-971.
NPL 11: Bogdanovich и др., Nature 420(6914), 2002, cc. 418-421.
NPL 12: Wagner., Ann. Neurol. 52(6), 2002, cc. 832-836.
NPL 13: Reichert и др., Nat. Biotechnol. 23, 2005, cc. 1073-1078.
NPL 14: Pavlou и др., Eur. J. Pharm. Biopharm. 59, 2005, cc. 389-396.
NPL 15: Clark и др., Chem. Immunol. 65, 1997, cc. 88-110.
NPL 16: Jefferis и др., Immunol. Lett. 82, 2002, cc. 57-65.
NPL 17: Smith и др., Nat. Rev. Immunol. 10, 2010, cc. 328-343.
NPL 18: Radaev и др., J. Biol. Chem. 276, 2001, cc. 16478-16483.
NPL 19: Greenwood и др., Eur. J. Immunol. 23, 1993, cc. 1098-1104.
NPL 20: Morgan и др., Immunology 86, 1995, cc. 319-324.
NPL 21: Nimmerjahn и др., Nat. Rev. Immunol. 8, 2008, cc. 34-47.
NPL 22: Amigorena и др., Eur. J. Immunol. 19, 1989, cc. 1379-1385.
NPL 23: Sinclair, J. Exp. Med. 129, 1969, cc. 1183-1201.
NPL 24: Heyman, Immunol. Lett. 88, 2003, cc. 157-161.
NPL 25: Amigorena и др., Science 256, 1992, cc. 1808-1812.
NPL 26: Muta и др., Nature 368, 1994, cc. 70-73.
NPL 27: Ravetch, Science 290, 2000, cc. 84-89.
NPL 28: Fournier и др., J. Immunol. 181, 2008, cc. 5350-5359.
NPL 29: J. Immunol. 163, 1999, cc. 618-622.
NPL 30: Yuasa и др., J. Exp. Med. 189, 1999, cc. 187-194.
NPL 31: Nakamura и др., J. Exp. Med. 191, 2000, cc. 899-906.
NPL 32: Blank, Hum. Genet. 117, 2005, cc. 220-227.
NPL 33: Olferiev и др., J. Biol. Chem. 282, 2007, cc. 1738-1746.
NPL 34: Chen и др., Arthritis Rheum. 54, 2006, cc. 3908-3917.
NPL 35: Floto и др., Nat. Med. 11, 2005, cc. 1056-1058.
NPL 36: Li и др., J. Immunol. 176, 2006, cc. 5321-5328.
NPL 37: Mackay и др., J. Exp. Med. 203, 2006, cc. 2157-2164.
NPL 38: Yang и др., J. Immunol. 178, 2007, cc. 3272-3280.
NPL 39: Bruhns и др., Blood 113, 2009, cc. 3716-3725.
NPL 40: Chu и др., Mol. Immunol. 45, 2008, cc. 3926-3933.
NPL 41: Chu и др., J. Allergy Clin. Immunol. 129, 2012, cc. 1102-1115.
NPL 42: Veri и др., Arthritis Rheum. 62, 20-10, cc. 1933-1943.
NPL 43: Cemerski и др., Immunol. Lett. 143, 2012, cc. 34-43.
NPL 44: Wenink и др., J. Immunol. 183, 2009, cc. 4509-4520.
NPL 45: Zhang и др., J. Immunol. 182, 2009, cc. 554-562.
NPL 46: Ravetch, Science 333, 2011, cc. 1030-1034.
NPL 47: Wilson и др., Cancer Cell 19, 2011, cc. 101-113.
NPL 48: Kohrt и др., J. Clin. Invest. 122, 2012, cc. 1066-1075.
NPL 49: Xu и др., J. Immunol. 171, 2003, cc. 562-568.
NPL 50: Zhang и др., Blood 108, 2006, cc. 705-710.
NPL 51: Chuntharapai и др., J. Immunol. 166, 2001, cc. 4891-4898.
NPL 52: Ravetch и др., Proc. Natl. Acad. Sci. USA 109, 2012, cc. 10966-10971.
NPL 53: Malbec и др., Immunol. Lett. 143, 2012, cc. 28-33.
NPL 54: Scappaticci и др., J. Natl. Cancer. Inst. 99, 2007, cc. 1232-1239.
NPL 55: Arthritis Rheum. 48, 2003, cc. 719-727.
NPL 56: Meyer и др., J. Thromb. Haemost. 7, 2008, cc. 171-181.
NPL 57: Robles-Carrillo и др., J. Immunol. 185, 2010, cc. 1577-1583.
NPL 58: Duffau и др., Sci. Transl. Med. 2:47ra63, 2010.
NPL 59: Richards и др., Mol. Cancer Ther. 7, 2008, cc. 2517-2527.
NPL 60: Desai и др., J. Immunol. 178, 2007, cc. 6217-6226.
NPL 61: Salmon и др., J. Clin. Invest. 97, 1996, cc. 1348-1354.
NPL 62: Manger и др., Arthritis Rheum. 41, 1998, cc. 1181-1189.
NPL 63: Boruchov и др., J. Clin. Invest. 115, 2005, cc. 2914-2923.
NPL 64: Dhodapkar и др., Proc. Natl. Acad. Sci. USA 102, 2005, cc. 2910-2915.
NPL 65: Armour и др., Mol. Immunol. 40, 2003, cc. 585-593.
NPL 66: Warmerdam и др., J. Exp. Med. 172, 1990, cc. 19-25.
Краткое изложение сущности изобретения
Техническая задача, положенная в основу настоящего изобретения
Задача настоящего изобретения заключалась в том, чтобы разработать антитела к миостатину, полипептиды, содержащие варианты Fc-областей, и способы их применения.
Решение задачи
В настоящем изобретении предложены антитела к миостатину и способы их применения. В изобретении предложены также белки, содержащие вариант Fc-области, и способы их применения.
В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, связывается с латентным миостатином. В других вариантах осуществления изобретения антитело связывается с эпитопом внутри фрагмента, состоящего из аминокислот 21-100 пропептида миостатина (SEQ ID NO: 78). В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, ингибирует активацию миостатина. В других вариантах осуществления изобретения антитело блокирует высвобождение зрелого миостатина из латентного миостатина. В других вариантах осуществления изобретения антитело блокирует протеолитическое высвобождение зрелого миостатина. В других вариантах осуществления изобретения антитело блокирует спонтанное высвобождение зрелого миостатина. В других вариантах осуществления изобретения антитело не связывается со зрелым миостатином. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, представленное в таблице 13. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, которое содержит пару VH и VL, указанную в таблице 13. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, представленное в таблице 2а. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, которое содержит пару VH и VL, указанную в таблице 2а. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, представленное в таблице 11а. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, которое содержит пару VH и VL, указанную в таблице 11а. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, представленное в таблице 2а, 11а или 13. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, которое содержит пару VH и VL, указанную в таблице 2а, 11а или 13.
В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, связывается с латентным миостатином с более высокой аффинностью при нейтральном рН, чем при кислом рН. В некоторых вариантах осуществления изобретения антитело к миостатину связывается с латентным миостатином с более высокой аффинностью при рН 7,4, чем при рН 5,8. В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, связывается с полипептидным фрагментом, состоящим из аминокислот 21-100 пропептида миостатина (SEQ ID NO: 78), с более высокой аффинностью при рН 7,4, чем при рН 5,8. В некоторых вариантах осуществления изобретения антитело связывается с тем же эпитопом миостатина, что и антитело, представленное в таблице 13, с более высокой аффинностью при нейтральном рН, чем при кислом рН. В дополнительных вариантах осуществления изобретения антитело к миостатину связывается с тем же эпитопом, что и антитело, представленное в таблице 13, с более высокой аффинностью при рН 7,4, чем при рН 5,8. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, которое содержит пару VH и VL, указанную в таблице 13, с более высокой аффинностью при рН 7,4, чем при рН 5,8. В некоторых вариантах осуществления изобретения антитело связывается с тем же эпитопом миостатина, что и антитело, представленное в таблице 2а, с более высокой аффинностью при нейтральном рН, чем при кислом рН. В некоторых вариантах осуществления изобретения антитело связывается с тем же эпитопом миостатина, что и антитело, представленное в таблице 2а, с более высокой аффинностью при рН 7,4, чем при рН 5,8. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, которое содержит пару VH и VL, указанную в таблице 2а, с более высокой аффинностью при рН 7,4, чем при рН 5,8. В дополнительных вариантах осуществления изобретения антитело к миостатину связывается с тем же эпитопом, что и антитело, представленное в таблице 11а, с более высокой аффинностью при нейтральном рН, чем при кислом рН. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом миостатина, что и антитело, представленное в таблице 11а, при рН 7,4, чем при рН 5,8. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, которое содержит пару VH и VL, указанную в таблице 11а, с более высокой аффинностью при рН 7,4, чем при рН 5,8. В дополнительных вариантах осуществления изобретения антитело к миостатину связывается с тем же эпитопом, что и антитело, представленное в таблице 2а, 11а или 13, с более высокой аффинностью при нейтральном рН, чем при кислом рН. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом миостатина, что и антитело, представленное в таблице 2а, 11а или 13, с более высокой аффинностью при рН 7,4, чем при рН 5,8. В других вариантах осуществления изобретения антитело связывается с тем же эпитопом, что и антитело, которое содержит пару VH и VL, указанную в таблице 2а, 11а или 13, с более высокой аффинностью при рН 7,4, чем при рН 5,8.
В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, конкурирует за связывание с латентным миостатином с антителом, указанным в настоящем описании. В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, конкурирует за связывание с латентным миостатином с антителом, представленным в таблице 13. В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, конкурирует за связыванием с латентным миостатином, с антителом, которое содержит пару VH и VL, указанную в таблице 13. В некоторых вариантах осуществления изобретения антитело конкурирует за связывание с латентным миостатином с антителом, представленным в таблице 2а. В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, конкурирует за связывание с латентным миостатином с антителом, которое содержит пару VH и VL, указанную в таблице 2а. В некоторых вариантах осуществления изобретения антитело конкурирует за связывание с латентным миостатином с антителом, представленным в таблице 11а. В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, конкурирует за связывание с латентным миостатином с антителом, которое содержит пару VH и VL, указанную в таблице 11а. В дополнительных вариантах осуществления изобретения антитело к миостатину конкурирует за связывание с латентным миостатином с антителом, представленным в таблице 2а, 11а или 13. В дополнительных вариантах осуществления изобретения антитело к миостатину конкурирует за связывание с латентным миостатином с антителом, которое содержит пару VH и VL, указанную в таблице 2а, 11а или 13. В других вариантах осуществления изобретения антитело к миостатину связывается с латентным миостатином с более высокой аффинностью при нейтральном рН, чем при кислом рН. В других вариантах осуществления изобретения антитело к миостатину связывается с латентным миостатином с более высокой аффинностью при рН 7,4, чем при рН 5,8. В других вариантах осуществления изобретения антитело к миостатину связывается с полипептидным фрагментом, состоящим из аминокислот 21-100 пропептида миостатина (SEQ ID NO: 78), с более высокой аффинностью при рН 7,4, чем при рН 5,8. Методы оценки способности антитела конкурировать с референс-антителом за связывание с латентным миостатином, указаны в настоящем описании и известны в данной области.
В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, представляет собой моноклональное антитело. В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, представляет собой человеческое, гуманизированное или химерное антитело. В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, представляет собой фрагмент антитела, который связывается с миостатином. В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, представляет собой фрагмент антитела, который связывается с латентным миостатином. В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, представляет собой фрагмент антитела, который связывается с полипептидным фрагментом, состоящим из аминокислот 21-100 пропептида миостатина (SEQ ID NO: 78). В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, представляет собой полноразмерное антитело IgG-типа.
В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в изобретении, содержит:
(а) (I) HVR-H3, содержащий аминокислотную последовательность GVPAX1SX2GGDX3, в которой X1 обозначает Y или Н, Х2 обозначает Т или Н, Х3 обозначает L или K (SEQ ID NO: 128), (II) HVR-L3, содержащий аминокислотную последовательность AGGYGGGX1YA, в которой X1 обозначает L или R (SEQ ID NO: 131), и (III) HVR-H2, содержащий аминокислотную последовательность IISX1AGX2X3YX4X5X6WAKX7, в которой X1 обозначает Y или Н, Х2 обозначает S или K, Х3 обозначает Т, М или K, Х4 обозначает Y или K, Х5 обозначает А, М или Е, Х6 обозначает S или Е, Х7 обозначает G или K (SEQ ID NO: 127);
(б) (I) HVR-H1, содержащий аминокислотную последовательность X1X2DIS, в которой X1 обозначает S или Н, Х2 обозначает Y, Т, D или Е (SEQ ID NO: 126), (II) HVR-H2, содержащий аминокислотную последовательность IISX1AGX2X3YX4X5X6WAKX7, в которой X1 обозначает Y или Н, Х2 обозначает S или K, Х3 обозначает Т, М или K, Х4 обозначает Y или K, Х5 обозначает А, М или Е, Х6 обозначает S или Е, Х7 обозначает G или K (SEQ ID NO: 127), и (III) HVR-H3, содержащий аминокислотную последовательность GVPAX1SX2GGDX3, в которой X1 обозначает Y или Н, Х2 обозначает Т или Н, Х3 обозначает L или K (SEQ ID NO: 128);
(в) (I) HVR-H1, содержащий аминокислотную последовательность X1X2DIS, в которой X1 обозначает S или Н, Х2 обозначает Y, Т, D или Е (SEQ ID NO: 126), (II) HVR-H2, содержащий аминокислотную последовательность IISX1AGX2X3YX4X5X6WAKX7, в которой X1 обозначает Y или Н, Х2 обозначает S или K, Х3 обозначает Т, М или K, Х4 обозначает Y или K, Х5 обозначает А, М или Е, Х6 обозначает S или Е, Х7 обозначает G или K (SEQ ID NO: 127), (III) HVR-H3, содержащий аминокислотную последовательность GVPAX1SX2GGDX3, в которой X1 обозначает Y или Н, Х2 обозначает Т или Н, Х3 обозначает L или K (SEQ ID NO: 128), (IV) HVR-L1, содержащий аминокислотную последовательность X1X2SQX3VX4X5X6NWLS, в которой X1 обозначает Q или Т, Х2 обозначает S или Т, Х3 обозначает S или Е, Х4 обозначает Y или F, Х5 обозначает D или Н, Х6 обозначает N, D, А или Е (SEQ ID NO: 129); (V) HVR-L2, содержащий аминокислотную последовательность WAX1TLAX2, в которой X1 обозначает S или Е, Х2 обозначает S, Y, F или W (SEQ ID NO: 130); и (VI) HVR-L3, содержащий аминокислотную последовательность AGGYGGGX1YA, в которой X1 обозначает L или R (SEQ ID NO: 131);
(г) (I) HVR-L1, содержащий аминокислотную последовательность X1X2SQX3VX4X5X6NWLS, в которой X1 обозначает Q или Т, Х2 обозначает S или Т, Х3 обозначает S или Е, Х4 обозначает Y или F, Х5 обозначает D или Н, Х6 обозначает N, D, А или Е (SEQ ID NO: 129); (II) HVR-L2, содержащий аминокислотную последовательность WAX1TLAX2, в которой X1 обозначает S или Е, Х2 обозначает S, Y, F или W (SEQ ID NO: 130); и (III) HVR-L3, содержащий аминокислотную последовательность AGGYGGGX1YA, в которой X1 обозначает L или R (SEQ ID NO: 131). В некоторых вариантах осуществления изобретения антитело, указанное в подпункте (б), дополнительно содержит каркасный участок вариабельного домена тяжелой цепи FR1, содержащий одну из аминокислотных последовательностей SEQ ID NO: 132-134; FR2, содержащий одну из аминокислотных последовательностей SEQ ID NO: 135-136; FR3, содержащий аминокислотную последовательность SEQ ID NO: 137; и FR4, содержащий аминокислотную последовательность SEQ ID NO: 138. В некоторых вариантах осуществления изобретения антитело, указанное в подпункте (г), дополнительно содержит каркасный участок вариабельного домена легкой цепи FR1, содержащий аминокислотную последовательность SEQ ID NO: 139; FR2, содержащий одну из аминокислотных последовательностей SEQ ID NO: 140-141; FR3, содержащий одну из аминокислотных последовательностей SEQ ID NO: 142-143; и FR4, содержащий аминокислотную последовательность SEQ ID NO: 144.
В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, содержит (a) HVR-H3, содержащий аминокислотную последовательность GVPAX1SX2GGDX3, в которой X1 обозначает Y или Н, Х2 обозначает Т или Н, Х3 обозначает L или K (SEQ ID NO: 128), (б) HVR-L3, содержащий аминокислотную последовательность AGGYGGGX1YA, в которой X1 обозначает L или R (SEQ ID NO: 131), и (в) HVR-H2, содержащий аминокислотную последовательность IISX1AGX2X3YX4X5X6WAKX7, в которой X1 обозначает Y или Н, Х2 обозначает S или K, Х3 обозначает Т, М или K, Х4 обозначает Y или K, Х5 обозначает А, М или Е, Х6 обозначает S или Е, Х7 обозначает G или K (SEQ ID NO: 127).
В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, содержит (a) HVR-H1, содержащий аминокислотную последовательность X1X2DIS, в которой X1 обозначает S или Н, Х2 обозначает Y, Т, D или Е (SEQ ID NO: 126), (б) HVR-H2, содержащий аминокислотную последовательность IISX1AGX2X3YX4X5X6WAKX7, в которой X1 обозначает Y или Н, Х2 обозначает S или K, Х3 обозначает Т, М или K, Х4 обозначает Y или K, Х5 обозначает А, М или Е, Х6 обозначает S или Е, Х7 обозначает G или K (SEQ ID NO: 127), и (в) HVR-H3, содержащий аминокислотную последовательность GVPAX1SX2GGDX3, в которой X1 обозначает Y или Н, Х2 обозначает Т или Н, Х3 обозначает L или K (SEQ ID NO: 128). В других вариантах осуществления изобретения антитело содержит каркасный участок вариабельного домена тяжелой цепи FR1, содержащий одну из аминокислотных последовательностей SEQ ID NO: 132-134; FR2, содержащий одну из аминокислотных последовательностей SEQ ID NO: 135-136; FR3, содержащий аминокислотную последовательность SEQ ID NO: 137; и FR4, содержащий аминокислотную последовательность SEQ ID NO: 138. В других вариантах осуществления изобретения антитело дополнительно содержит (a) HVR-L1, содержащий аминокислотную последовательность X1X2SQX3VX4X5X6NWLS, в которой X1 обозначает Q или Т, Х2 обозначает S или Т, Х3 обозначает S или Е, Х4 обозначает Y или F, Х5 обозначает D или Н, Х6 обозначает N, D, А или Е (SEQ ID NO: 129); (б) HVR-L2, содержащий аминокислотную последовательность WAX1TLAX2, в которой X1 обозначает S или Е, Х2 обозначает S, Y, F или W (SEQ ID NO: 130); и (в) HVR-L3, содержащий аминокислотную последовательность AGGYGGGX1YA, в которой X1 обозначает L или R (SEQ ID NO: 131).
В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, содержит (a) HVR-L1, содержащий аминокислотную последовательность X1X2SQX3VX4X5X6NWLS, в которой X1 обозначает Q или Т, Х2 обозначает S или Т, Х3 обозначает S или Е, Х4 обозначает Y или F, Х5 обозначает D или Н, Х6 обозначает N, D, А или Е (SEQ ID NO: 129); (б) HVR-L2, содержащий аминокислотную последовательность WAX1TLAX2, в которой X1 обозначает S или Е, Х2 обозначает S, Y, F или W (SEQ ID NO: 130); и (в) HVR-L3, содержащий аминокислотную последовательность AGGYGGGX1YA, в которой X1 обозначает L или R (SEQ ID NO: 131). В следующем варианте осуществления изобретения антитело содержит дополнительно каркасный участок вариабельного домена легкой цепи FR1, содержащий аминокислотную последовательность SEQ ID NO: 139; FR2, содержащий одну из аминокислотных последовательностей SEQ ID NO: 140-141; FR3, содержащий одну из аминокислотных последовательностей SEQ ID NO: 142-143; и FR4, содержащий аминокислотную последовательность SEQ ID NO: 144.
В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, содержит каркасный участок вариабельного домена тяжелой цепи FR1, содержащий одну из аминокислотных последовательностей SEQ ID NO: 132-134; FR2, содержащий одну из аминокислотных последовательностей SEQ ID NO: 135-136; FR3, содержащий аминокислотную последовательность SEQ ID NO: 137; и FR4, содержащий аминокислотную последовательность SEQ ID NO: 138. В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, содержит каркасный участок вариабельного домена легкой цепи FR1, содержащий аминокислотную последовательность SEQ ID NO: 139; FR2, содержащий одну из аминокислотных последовательностей SEQ ID NO: 140-141; FR3, содержащий одну из аминокислотных последовательностей SEQ ID NO: 142-143; и FR4, содержащий аминокислотную последовательность SEQ ID NO: 144.
В некоторых вариантах осуществления изобретения выделенное антитело к миостатину, предлагаемое в настоящем изобретении, содержит (а) последовательность VH, идентичную по меньшей мере на 95% одной из аминокислотных последовательностей SEQ ID NO: 13, 16-30, 32-34 и 86-95; (б) последовательность VL, идентичную по меньшей мере на 95% одной из аминокислотных последовательностей SEQ ID NO: 15, 31, 35-38 и 96-99; или (в) последовательность VH, указанную в подпункте (а), и последовательность VL, указанную в подпункте (б). В других вариантах осуществления изобретения антитело содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95. В других вариантах осуществления изобретения антитело содержит последовательность VL, представленную в одной из SEQ ID NO: 15, 31, 35-38 и 96-99. В некоторых вариантах осуществления изобретения антитело содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95. В других вариантах осуществления изобретения антитело содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95; и последовательность VL, представленную в одной из SEQ ID NO: 15, 31, 35-38 и 96-99.
Изобретение относится также к выделенным нуклеиновым кислотам, кодирующим антитело к миостатину, предлагаемое в настоящем изобретении. Изобретение относится также к клеткам-хозяевам, которые содержат нуклеиновую кислоту, предлагаемую в настоящем изобретении. Изобретение относится также к способу получения антитела, включающему культивирование клетки-хозяина, предлагаемой в настоящем изобретении, так, чтобы получать антитело.
В некоторых объектах изобретения предложен способ получения антитела к миостатину, включающий (а) культивирование клетки-хозяина, предлагаемой в настоящем изобретении, так, чтобы получать антитело; или (б) иммунизацию животного полипептидом, где полипептид содержит область, соответствующую аминокислотам в положениях 21-100 пропептида миостатина (SEQ ID NO: 78).
В изобретении предложен также способ получения антитела к миостатину. В некоторых вариантах осуществления изобретения способ включает иммунизацию животного полипептидом, где полипептид содержит область, соответствующую аминокислотам в положениях 21-100 пропептида миостатина (SEQ ID NO: 78).
Изобретение относится также к фармацевтической композиции, которая содержит антитело к миостатину, предлагаемое в настоящем изобретении, и фармацевтически приемлемый носитель.
Антитела к миостатину, предлагаемые в настоящем изобретении, можно применять в качестве лекарственного средства. В некоторых вариантах осуществления изобретения антитело применяют для приготовления лекарственного средства, предназначенного для: (а) лечения синдрома (болезни) мышечного истощения; (б) увеличения массы мышечной ткани; (в) увеличения силы мышечной ткани; или (г) снижения накопления жировой массы тела. В некоторых вариантах осуществления изобретения антитела к миостатину, предлагаемые в настоящем изобретении, можно применять для лечения синдрома мышечного истощения. Антитела к миостатину, предлагаемые в настоящем изобретении, можно применять для увеличения массы мышечной ткани. Антитела к миостатину, предлагаемые в настоящем изобретении, можно применять для увеличения силы мышечной ткани. Антитела к миостатину, предлагаемые в настоящем изобретении, можно применять для снижения накопления жировой массы тела.
В некоторых вариантах осуществления изобретения представленное в настоящем описании антитело к миостатину предназначено для применения для: (а) лечения синдрома мышечного истощения; (б) увеличения массы мышечной ткани; (в) увеличения силы мышечной ткани; или (г) снижения накопления жировой массы тела.
Антитела к миостатину, предлагаемые в настоящем изобретении, можно применять для приготовления лекарственного средства. В некоторых вариантах осуществления изобретения антитело применяют для приготовления лекарственного средства, предназначенного для лечения: (а) синдрома мышечного истощения; (б) увеличения массы мышечной ткани; (в) увеличения силы мышечной ткани; или (г) снижения накопления жировой массы тела. В некоторых вариантах осуществления изобретения лекарственное средство предназначено для лечения синдрома мышечного истощения. В некоторых вариантах осуществления изобретения лекарственное средство предназначено для увеличения массы мышечной ткани. В некоторых вариантах осуществления изобретения лекарственное средство предназначено для увеличения силы мышечной ткани. В некоторых вариантах осуществления изобретения лекарственное средство предназначено для снижения накопления жировой массы тела.
В изобретении предложен также способ лечения индивидуума, имеющего синдром мышечного истощения. В некоторых вариантах осуществления изобретения способ включает введение индивидууму в эффективном количестве антитела к миостатину, предлагаемого в настоящем изобретении. В изобретении предложен также способ увеличения массы мышечной ткани у индивидуума. В некоторых вариантах осуществления изобретения способ включает введение индивидууму в эффективном количестве антитела к миостатину, предлагаемого в настоящем изобретении, для увеличения массы мышечной ткани. В изобретении предложен также способ увеличения силы мышечной ткани у индивидуума. В некоторых вариантах осуществления изобретения способ включает введение индивидууму в эффективном количестве антитела к миостатину, предлагаемого в настоящем изобретении, для увеличения силы мышечной ткани. В изобретении предложен также способ снижения накопления жировой массы тела у индивидуума. В некоторых вариантах осуществления изобретения способ включает введение индивидууму в эффективном количестве антитела к миостатину, предлагаемого в настоящем изобретении, для снижения накопления жировой массы тела.
В частности, в изобретении предложены:
[1] антитело, которое связывается с латентным миостатином, предназначенное для применения для лечения синдрома мышечного истощения, где антитело ингибирует активацию миостатина;
[2] антитело, которое связывается с латентным миостатином, предназначенное для применения для увеличения массы мышечной ткани, где антитело ингибирует активацию миостатина;
[3] антитело, которое связывается с латентным миостатином, предназначенное для применения для увеличения силы мышечной ткани, где антитело ингибирует активацию миостатина;
[4] антитело, которое связывается с латентным миостатином, предназначенное для применения для снижения накопления жировой массы тела, где антитело ингибирует активацию миостатина;
[5] применение антитела, которое связывается с латентным миостатином, для приготовления лекарственного средства, предназначенного для лечения синдрома мышечного истощения, где антитело ингибирует активацию миостатина;
[6] применение антитела, которое связывается с латентным миостатином, для приготовления лекарственного средства, предназначенного для увеличения массы мышечной ткани, где антитело ингибирует активацию миостатина;
[7] применение антитела, которое связывается с латентным миостатином, для приготовления лекарственного средства, предназначенного для увеличения силы мышечной ткани, где антитело ингибирует активацию миостатина;
[8] применение антитела, которое связывается с латентным миостатином, для приготовления лекарственного средства, предназначенного для снижения накопления жировой массы тела, где антитело ингибирует активацию миостатина;
[9] способ лечения индивидуума, имеющего синдром мышечного истощения, включающий введение индивидууму в эффективном количестве антитела, которое связывается с латентным миостатином, где антитело ингибирует активацию миостатина;
[10] способ увеличения массы мышечной ткани у индивидуума, включающий введение индивидууму в эффективном количестве антитела, которое связывается с латентным миостатином, где антитело ингибирует активацию миостатина;
[11] способ увеличения силы мышечной ткани у индивидуума, включающий введение индивидууму в эффективном количестве антитела, которое связывается с латентным миостатином, где антитело ингибирует активацию миостатина;
[12] способ снижения накопления жировой массы тела у индивидуума, включающий введение индивидууму в эффективном количестве антитела, которое связывается с латентным миостатином, где антитело ингибирует активацию миостатина;
[13] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[12], где антитело блокирует высвобождение зрелого миостатина из латентного миостатина;
[14] антитело, предназначенное для применения, применение или способ по п. [13], где антитело блокирует протеолитическое высвобождение зрелого миостатина;
[15] антитело, предназначенное для применения, применение или способ по п. [13], где антитело блокирует спонтанное высвобождение зрелого миостатина;
[16] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[15], где антитело не связывается со зрелым миостатином;
[17] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[16], где антитело связывается с эпитопом внутри фрагмента, состоящего из аминокислот 21-100 пропептида миостатина (SEQ ID NO: 78);
[18] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[17], где антитело связывается с тем же эпитопом, что и антитело, представленное в таблицах 2а, 11а и 13;
[19] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[18], где антитело связывается с латентным миостатином с более высокой аффинностью при нейтральном рН, чем при кислом рН;
[20] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[19], где антитело представляет собой моноклональное антитело;
[21] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[20], где антитело представляет собой человеческое, гуманизированное или химерное антитело;
[22] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[21], где антитело представляет собой фрагмент антитела, который связывается с миостатином;
[23] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[22], где антитело содержит (а) HVR-H3, содержащий аминокислотную последовательность GVPAX1SX2GGDX3, в которой X1 обозначает Y или Н, Х2 обозначает Т или Н, Х3 обозначает L или K (SEQ ID NO: 128), (б) HVR-L3, содержащий аминокислотную последовательность AGGYGGGX1YA, в которой X1 обозначает L или R (SEQ ID NO: 131), и (в) HVR-H2, содержащий аминокислотную последовательность IISX1AGX2X3YX4X5X6WAKX7, в которой X1 обозначает Y или Н, Х2 обозначает S или K, Х3 обозначает Т, М или K, Х4 обозначает Y или K, Х5 обозначает А, М или Е, Х6 обозначает S или Е, Х7 обозначает G или K (SEQ ID NO: 127);
[24] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[22], где антитело содержит (a) HVR-H1, содержащий аминокислотную последовательность X1X2DIS, в которой X1 обозначает S или Н, Х2 обозначает Y, Т, D или Е (SEQ ID NO: 126), (б) HVR-H2, содержащий аминокислотную последовательность IISX1AGX2X3YX4X5X6WAKX7, в которой X1 обозначает Y или Н, Х2 обозначает S или K, Х3 обозначает Т, М или K, Х4 обозначает Y или K, Х5 обозначает А, М или Е, Х6 обозначает S или Е, Х7 обозначает G или K (SEQ ID NO: 127), и (в) HVR-H3, содержащий аминокислотную последовательность GVPAX1SX2GGDX3, в которой X1 обозначает Y или Н, Х2 обозначает Т или Н, Х3 обозначает L или K (SEQ ID NO: 128);
[25] антитело, предназначенное для применения, применение или способ по п. [24], где антитело дополнительно содержит (a) HVR-L1, содержащий аминокислотную последовательность X1X2SQX3VX4X5X6NWLS, в которой X1 обозначает Q или Т, Х2 обозначает S или Т, Х3 обозначает S или Е, Х4 обозначает Y или F, Х5 обозначает D или Н, Х6 обозначает N, D, А или Е (SEQ ID NO: 129); (б) HVR-L2, содержащий аминокислотную последовательность WAX1TLAX2, в которой X1 обозначает S или Е, Х2 обозначает S, Y, F или W (SEQ ID NO: 130); и (в) HVR-L3, содержащий аминокислотную последовательность AGGYGGGX1YA, в которой X1 обозначает L или R (SEQ ID NO: 131);
[26] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[22], где антитело содержит (a) HVR-L1, содержащий аминокислотную последовательность X1X2SQX3VX4X5X6NWLS, в которой X1 обозначает Q или Т, Х2 обозначает S или Т, Х3 обозначает S или Е, Х4 обозначает Y или F, Х5 обозначает D или Н, Х6 обозначает N, D, А или Е (SEQ ID NO: 129); (б) HVR-L2, содержащий аминокислотную последовательность WAX1TLAX2, в которой X1 обозначает S или Е, Х2 обозначает S, Y, F или W (SEQ ID NO: 130); и (в) HVR-L3, содержащий аминокислотную последовательность AGGYGGGX1YA, в которой X1 обозначает L или R (SEQ ID NO: 131);
[27] антитело, предназначенное для применения, применение или способ по п. [24], где антитело дополнительно содержит каркасный участок вариабельного домена тяжелой цепи FR1, содержащий одну из аминокислотных последовательностей SEQ ID NO: 132-134; FR2, содержащий одну из аминокислотных последовательностей SEQ ID NO: 135-136; FR3, содержащий аминокислотную последовательность SEQ ID NO: 137; и FR4, содержащий аминокислотную последовательность SEQ ID NO: 138;
[28] антитело, предназначенное для применения, применение или способ по п. [26], где антитело дополнительно содержит каркасный участок вариабельного домена легкой цепи FR1, содержащий аминокислотную последовательность SEQ ID NO: 139; FR2 содержащий одну из аминокислотных последовательностей SEQ ID NO: 140-141; FR3, содержащий одну из аминокислотных последовательностей SEQ ID NO: 142-143; и FR4, содержащий аминокислотную последовательность SEQ ID NO: 144;
[29] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[22], где антитело содержит (а) последовательность VH, идентичную по меньшей мере на 95% одной из аминокислотных последовательностей SEQ ID NO: 13, 16-30, 32-34 и 86-95; (б) последовательность VL, идентичную по меньшей мере на 95% одной из аминокислотных последовательностей SEQ ID NO: 15, 31, 35-38 и 96-99; или (в) последовательность VH, указанную в подпункте (а), и последовательность VL, указанную в подпункте (б);
[30] антитело, предназначенное для применения, применение или способ по п. [29], где антитело содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95;
[31] антитело, предназначенное для применения, применение или способ по п. [29], где антитело содержит последовательность VL, представленную в одной из SEQ ID NO: 15, 31, 35-38 и 96-99;
[32] антитело, предназначенное для лечения синдрома мышечного истощения, которое содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95, и последовательность VL, представленную в одной из SEQ ID NO: 15, 31, 35-38 и 96-99;
[33] антитело, предназначенное для увеличения массы мышечной ткани, которое содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95, и последовательность VL, представленную в одной из SEQ ID NO: 15, 31, 35-38 и 96-99;
[34] антитело, предназначенное для увеличения силы мышечной ткани, которое содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95, и последовательность VL, представленную в одной из SEQ ID NO: 15, 31, 35-38 и 96-99;
[35] антитело, предназначенное для снижения накопления жировой массы тела, которое содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95, и последовательность VL, представленную в одной из SEQ ID NO: 15, 31, 35-38 и 96-99; и
[36] антитело, предназначенное для применения, применение или способ по одному из п.п. [1]-[35], где антитело представляет собой полноразмерное антитело IgG-типа.
Изобретение относится также к полипептидам, содержащим варианты Fc-областей, и способам их получения и применения.
Одним из вариантов осуществления изобретения является Fc-гамма RIIb-связывающие полипептиды, содержащие варианты Fc-областей, и способы их применения. В некоторых вариантах осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, предлагаемый в настоящем изобретении, содержит по меньшей мере одно аминокислотное изменение в родительской Fc-области. В других вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для обезьяньего Fc-гамма RIIb]/[величина KD варианта Fc-области для обезьяньего Fc-гамма RIIb] составляет 2,0 или более. В других вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для обезьяньего Fc-гамма RIIIa]/[величина KD варианта Fc-области для обезьяньего Fc-гамма RIIIa] составляет 0,5 или менее. В других вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для человеческого Fc-гамма RIIb]/[величина KD варианта Fc-области для человеческого Fc-гамма RIIb] составляет 2,0 или более. В других вариантах осуществления изобретения соотношение [величина KD родительской Fc-области ля человеческого Fc-гамма RIIIa]/[величина KD варианта Fc-области для человеческого Fc-гамма RIIIa] составляет 0,5 или менее. В других вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для человеческого Fc-гамма RIIa (Н-типа)]/[величина KD варианта Fc-области для человеческого Fc-гамма RIIa (Н-типа)] составляет 5,0 или менее. В других вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для человеческого Fc-гамма RIIa (R)-типа]/[величина KD варианта Fc-области для человеческого Fc-гамма RIIa (R-типа)] составляет 5,0 или менее. В другом варианте осуществления изобретения величина KD варианта Fc-области для обезьяньего Fc-гамма RIIb составляет 1,0×10-6М или менее. В другом варианте осуществления изобретения величина KD варианта Fc-области для обезьяньего Fc-гамма RIIIa составляет 5,0×10-7М или более. В другом варианте осуществления изобретения величина KD варианта Fc-области для человеческого Fc-гамма RIIb составляет 2,0×10-6М или менее. В другом варианте осуществления изобретения величина KD варианта Fc-области для человеческого Fc-гамма RIIIa составляет 1,0×10-6М или более. В другом варианте осуществления изобретения величина KD варианта Fc-области для человеческого Fc-гамма RIIa (Н-типа) составляет 1,0×10-7М или более. В другом варианте осуществления изобретения величина KD варианта Fc-области для человеческого Fc-гамма RIIa (R-типа) составляет 2,0×10-7М или более.
В некоторых вариантах осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, предлагаемый в настоящем изобретении, содержит по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 233, 234, 235, 236, 237, 238, 239, 264, 266, 267, 268, 271, 295, 298, 325, 326, 327, 328, 330, 331, 332, 334 и 396, согласно EU-нумерации.
В других вариантах осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит по меньшей мере два аминокислотных изменения, включающих: (а) одно аминокислотное изменение в положении 236 и (б) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей: (I) положений: 231, 232, 233, 234, 235, 237, 238, 239, 264, 266, 267, 268, 271, 295, 298, 325, 326, 327, 328, 330, 331, 332, 334 и 396; (II) положений: 231, 232, 235, 239, 268, 295, 298, 326, 330 и 396; или (III) положений 268, 295, 326 и 330; согласно EU-нумерации.
В других вариантах осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит по меньшей мере два аминокислотных изменения, включающих: (а) одно аминокислотное изменение в положении 236 и (б) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 233, 234, 235, 237, 238, 239, 264, 266, 267, 268, 271, 295, 298, 325, 326, 327, 328, 330, 331, 332, 334 и 396, согласно EU-нумерации.
В других вариантах осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит по меньшей мере два аминокислотных изменения, включающих: (а) одно аминокислотное изменение в положении 236 и (б) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 235, 239, 268, 295, 298, 326, 330 и 396, согласно EU-нумерации.
В других вариантах осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит по меньшей мере два аминокислотных изменения, включающих: (а) одно аминокислотное изменение в положении 236 и (б) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 268, 295, 326 и 330, согласно EU-нумерации.
В некоторых вариантах осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит по меньшей мере одну аминокислоту, выбранную из группы, состоящей из: (a) Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp, Tyr в положении 231; (6) Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp, Tyr в положении 232; (в) Asp в положении 233; (г) Trp, Tyr в положении 234; (д) Trp в положении 235; (е) Ala, Asp, Glu, His, Ile, Leu, Met, Asn, Gln, Ser, Thr, Val в положении 236; (ж) Asp, Tyr в положении 237; (з) Glu, Ile, Met, Gln, Tyr в положении 238; (и) Ile, Leu, Asn, Pro, Val в положении 239; (к) Ile в положении 264; (л) Phe в положении 266; (м) Ala, His, Leu в положении 267; (н) Asp, Glu в положении 268; (о) Asp, Glu, Gly в положении 271; (п) Leu в положении 295; (р) Leu в положении 298; (с) Glu, Phe, Ile, Leu в положении 325; (т) Thr в положении 326; (у) Ile, Asn в положении 327; (ф) Thr в положении 328; (х) Lys, Arg в положении 330; (ц) Glu в положении 331; (ч) Asp в положении 332; (ш) Asp, Ile, Met, Val, Tyr в положении 334; и (щ) Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp, Tyr в положении 396; согласно EU-нумерации.
В других вариантах осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит по меньшей мере одну аминокислоту, выбранную из группы, состоящей из: (a) Gly, Thr в положении 231; (б) Asp в положении 232; (в) Trp в положении 235; (г) Asn, Thr в положении 236; (д) Val в положении 239; (е) Asp, Glu в положении 268; (ж) Leu в положении 295; (з) Leu в положении 298; (и) Thr в положении 326; (к) Lys, Arg в положении 330 и (л) Lys, Met в положении 396; согласно EU-нумерации.
Другим вариантом осуществления изобретения является полипептид, который содержит вариант Fc-области с повышенной изоэлектрической точкой (pI) и способ его применения. В некоторых вариантах осуществления изобретения полипептид, который содержит вариант Fc-области с повышенной pI, включает по меньшей мере два аминокислотных изменения в родительской Fc-области. В других вариантах осуществления изобретения каждое из аминокислотных изменений повышает изоэлектрическую точку (pI) варианта Fc-области по сравнению с родительской Fc-областью. В других вариантах осуществления изобретения аминокислота может экспонироваться на поверхности варианта Fc-области. В других вариантах осуществления изобретения полипептид содержит вариант Fc-области и антигенсвязывающий домен. В других вариантах осуществления изобретения антигенсвязывающая активность антигенсвязывающего домена изменяется в зависимости от концентрации ионов. В других вариантах осуществления изобретения вариант Fc-области с повышенной pI, предлагаемый в настоящем изобретении, содержит по меньшей мере два аминокислотных изменения по меньшей мере в двух положениях, выбранных из группы, состоящей из: 285, 311, 312, 315, 318, 333, 335, 337, 341, 342, 343, 384, 385, 388, 390, 399, 400, 401, 402, 413, 420, 422 и 431, согласно EU-нумерации. В других вариантах осуществления изобретения вариант Fc-области с повышенной pI содержит Arg или Lys в каждом из выбранных положений.
В некоторых вариантах осуществления изобретения вариант Fc-области, предлагаемый в настоящем изобретении, содержит аминокислотные изменения, которые представлены в таблицах 14-30.
В некоторых вариантах осуществления изобретения полипептид содержит вариант Fc-области, предлагаемый в настоящем изобретении. В других вариантах осуществления изобретения родительскую Fc-область получают из человеческого IgG1. В других вариантах осуществления изобретения полипептид представляет собой антитело. В других вариантах осуществления изобретения полипептид представляет собой содержащий Fc слитый белок.
В изобретении предложен полипептид, содержащий одну из аминокислотных последовательностей SEQ ID NO: 229-381.
Изобретение относится также к выделенной нуклеиновой кислоте, которая кодирует полипептид, содержащий вариант Fc-области, предлагаемый в настоящем изобретении. Изобретение относится также к клетке-хозяину, которая содержит нуклеиновую кислоту, предлагаемую в настоящем изобретении. Изобретение относится также к способу получения полипептида, который содержит вариант Fc-области, включающему культивирование хозяина, предлагаемого в настоящем изобретении, так, чтобы получать полипептид.
Изобретение относится также к фармацевтической композиции, которая содержит полипептид, содержащий вариант Fc-области, предлагаемый в настоящем изобретении, и фармацевтически приемлемый носитель.
Краткое описание чертежей
На чертежах показано:
на фиг. 1 - ингибирование протеолитической активации латентного миостатина антителом к латентному миостатину, как проиллюстрировано в примере 3. Активность активного миостатина, высвобождающегося из латентного миостатина под действием протеазы ВМР1, измеряли в присутствии антитела к латентному миостатину, используя анализ HEK Blue;
на фиг. 2 - ингибирование спонтанной активации латентного миостатина антителом к латентному миостатину, как проиллюстрировано в примере 4. Активность активного миостатина, высвобождающегося из латентного миостатина под действием инкубации при 37°С, измеряли в присутствии антитела к латентному миостатину, используя анализ HEK Blue;
на фиг. 3 - связывание антитела к латентному миостатину с пропептидным доменом, как проиллюстрировано в примере 5;
на фиг. 4 - результаты анализа методом Вестерн-блоттинга против пропептида миостатина, как проиллюстрировано в примере 6. Протеолитическое расщепление пропептида миостатина с помощью ВМР1 оценивали в присутствии и отсутствии антитела к латентному миостатину;
на фиг. 5А-5В - полученные с помощью BIACORE® сенсограммы, характеризующие связывание антитела к латентному миостатину MST1032-G1m с человеческим латентным миостатином (А), латентным миостатином обезьян циномолгус (Б) и мышиным латентным миостатином (В), как проиллюстрировано в примере 7;
на фиг. 6А - эффективность in vivo антитела к латентному миостатину и антитела к зрелому миостатину в отношении мышечной массы и жировой массы, как проиллюстрировано в примере 8. Антитело к латентному миостатину (MST1032-G1m; обозначено как MST1032 на чертежах) или антитело к зрелому миостатину (41С1Е4) вводили SCID-мышам и оценивали общую тощую (безжировую) массу тела (или тощую массу тела: LBM);
на фиг. 6Б - эффективность in vivo антитела к латентному миостатину и антитела к зрелому миостатину в отношении мышечной массы и жировой массы, как проиллюстрировано в примере 8. Антитело к латентному миостатину (MST1032-G1m; обозначено как MST1032 на чертежах) или антитело к зрелому миостатину (41С1Е4) вводили SCID-мышам и оценивали изменение общей жировой массы тела со дня 0 по день 14;
на фиг. 6В - эффективность in vivo антитела к латентному миостатину и антитела к зрелому миостатину в отношении мышечной массы и жировой массы, как проиллюстрировано в примере 8. Антитело к латентному миостатину (MST1032-G1m; обозначено как MST1032 на чертежах) или антитело к зрелому миостатину (41С1Е4) вводили SCID-мышам и оценивали мышечную массу икроножной мышцы и четырехглавой мышцы;
на фиг. 7А - сравнение эффективности in vivo нескольких антител к миостатину, как проиллюстрировано в примере 9. Антитело к латентному миостатину (MST1032-G1m; обозначено как MST1032 на чертежах) или антитело к зрелому миостатину (41С1Е4, REGN, OGD или MYO-029) вводили SCID-мышам и оценивали общую тощую массу тела;
на фиг. 7Б - сравнение эффективности in vivo нескольких антител к миостатину, как проиллюстрировано в примере 9. Антитело к латентному миостатину (MST1032-G1m; обозначено как MST1032 на чертежах) или антитело к зрелому миостатину (41С1Е4, REGN, OGD или MYO-029) вводили SCID-мышам и оценивали силу сжатия;
на фиг. 7В - сравнение эффективности in vivo нескольких антител к миостатину, как проиллюстрировано в примере 9. Антитело к латентному миостатину (MST1032-G1m; обозначено как MST1032 на чертежах) или антитело к зрелому миостатину (41С1Е4, REGN, OGD или MYO-029) вводили SCID-мышам и оценивали изменение общей жировой массы тела со дня 0 по день 14;
на фиг. 8 - ингибирование протеолитической и спонтанной активации латентного миостатина гуманизированным антителом к латентному миостатину, как проиллюстрировано в примере 10. Активность активного миостатина, высвобождающегося из латентного миостатина под действием протеазы ВМР1 (протеолитическое) или инкубации при 37°С без ВМР1 (спонтанное), измеряли в присутствии антитела к латентному миостатину, используя анализ HEK Blue;
на фиг. 9 - полученные с помощью BIACORE® сенсограммы, характеризующие связывание вариантов с замещенным гистидином антитела к латентному миостатину, как проиллюстрировано в примере 11. Комплексам антител о/антиген давали диссоциироваться при рН 7,4 с последующей дополнительной диссоциацией при рН 5,8 (обозначено стрелкой) для изучения зависящих от рН взаимодействий. В этом эксперименте тестировали следующие антитела: Ab001 (кривая, обозначенная черной сплошной линией), Ab002 (кривая, обозначенная черной пунктирной линией), Ab003 (кривая, обозначенная черной точечной линией), Ab004 (кривая, обозначенная серой пунктирной линией), Ab005 (кривая, обозначенная серой сплошной линией), Ab006 (кривая, обозначенная серыми длинными штрихами) и Ab007 (кривая, обозначенная черными длинными штрихами);
на фиг 10 - ингибирование протеолитической и спонтанной активации латентного миостатина рН-зависимыми антителам к латентному миостатину, как проиллюстрировано в примере 13. Активность активного миостатина, высвобождающегося из латентного миостатина под действием протеазы ВМР1 (протеолитическое) или инкубации при 37°С без ВМР1 (спонтанное), измеряли в присутствии антитела к латентному миостатину, используя анализ HEK Blue. На чертеже антитела MS1032LO01-SG1, MS1032LO02-SG1, MS1032LO03-SG1 и MS1032LO04-SG1 обозначены соответственно как MSLO-01, MSLO-02, MSLO-03 и MSLO-04. Ингибирование протеолитической и спонтанной активации латентного миостатина, сопоставимое с MS1032LO00-SG1, достигалось при использовании MS1032LO01-SG1, MS1032LO02-SG1, MS1032LO03-SG1 и MS1032LO04-SG1:
на фиг. 11А-11Е - полученные с помощью BIACORE® сенсограммы, характеризующие связывание рН-зависимого антитела к латентному миостатину, как проиллюстрировано в примере 14. Кинетические параметры MST1032-SG1 (A), MS1032LO00-SG1 (Б), MS1032LO01-SG1 (В), MS1032LO02-SG1 (Г), MS1032LO03-SG1 (Д) и MS1032LO04-SG1 (Е) измеряли при нейтральном рН и кислом рН;
на фиг. 12 - зависимость от времени концентрации миостатина в плазме после внутривенного введения антитела к миостатину мышам, как проиллюстрировано в примере 15. Воздействия опосредуемого Fc-гамма R поглощения клетками комплексов антител о/антиген на клиренс миостатина in vivo оценивали путем сравнения антител к миостатину, обладающих способностью связываться с Fc-гамма R (MS1032LO00-SG1), и антител к миостатину с элиминированной способностью связываться с Fc-гамма R (MS1032LO00-F760);
на фиг. 13 - зависимость от времени концентрации миостатина в плазме после внутривенного введения антитела к миостатину мышам, как проиллюстрировано в примере 16. Воздействия рН-зависимого связывания антитела к миостатину на клиренс миостатина in vivo оценивали путем сравнения рН-зависимого антитела к миостатину (MS1032LO01-SG1 или MS1032LO01-F760) и рН-независимого антитела к миостатину (MS1032LO00-SG1 или MS1032LO00-F760);
на фиг. 14А-14Д - эффективность in vivo рН-зависимых и рН-независимых антител к латентному миостатину, как проиллюстрировано в примере 17. рН-зависимое антитело к латентному миостатину (MS1032LO01-SG1; обозначено как MSLO1 на чертеже) или рН-независимое антитело к латентному миостатину (MS1032LO00-SG1; обозначено как MSLO0 на чертеже) вводили SCID-мышам и оценивали общую тощую массу тела (А), общую жировую массу тела (Б), массу четырехглавой мышцы (В); массу икроножной мышцы (Г) и силу сжатия (Д);
на фиг. 15 - активность связывания антитела к латентному миостатину MST1032 с латентным миостатином и GDF11, как проиллюстрировано в примере 19;
на фиг. 16 - ингибирующая активность антитела к латентному миостатину MST1032 в отношении протеолитической и спонтанной активации GDF11, как проиллюстрировано в примере 20. Активность активного GDF11, высвобождающегося под действием протеазы ВМР1 (протеолитическое) или инкубации при 37°С без ВМР1 (спонтанное), измеряли в присутствии антитела к латентному миостатину, используя анализ HEK Blue;
на фиг. 17 - ингибирование протеолитической активации латентного миостатина антителом к латентному миостатину, как проиллюстрировано в примере 22. Активность активного миостатина, высвобождающегося под действием протеазы ВМР1, измеряли в присутствии антитела к латентному миостатину, используя анализ HEK Blue;
на фиг. 18 - зависимость от времени концентрации миостатина в плазме после внутривенного введения антитела к латентному миостатину мышам, как проиллюстрировано в примере 23. Воздействия зависимости от pH на клиренс миостатина in vivo оценивали путем сравнения pH-независимого антитела к латентному миостатину (MS1032LO00-SG1) и различных pH-зависимых антител к латентному миостатину (MS1032LO01-SG1, MS1032LO06-SG1, MS1032LO11-SG1, MS1032LO18-SG1, MS1032LO19-SG1, MS1032LO21-SG1 и MS1032LO25-SG1);
на фиг. 19А и 19Б - зависимость от времени концентрации миостатина в плазме после внутривенного введения антитела к латентному миостатину обезьянам циномолгус, как проиллюстрировано в примере 24. (А) Воздействие зависимости от pH и конструирования Fc-области на клиренс миостатина in vivo оценивали путем сравнения pH-независимого антитела к латентному миостатину (MS1032LO00-SG1) и pH-зависимых антител к латентному миостатину со сконструированной Fc (MS1032LO06-SG1012, MS1032LO06-SG1016, MS1032LO06-SG1029, MS1032LO06-SG1031, MS1032LO06-SG1033, MS1032LO06-SG1034). (Б) Воздействие конструирования Fc на клиренс миостатина in vivo оценивали путем сравнения антител к латентному миостатину (MS1032LO19-SG1079, MS1032LO19-SG1071, MS1032LO19-SG1080, MS1032LO19-SG1074, MS1032LO19-SG1081 и MS1032LO19-SG1077);
на фиг. 20А, а также на фиг. 20Б-20И - эффективность in vivo антител к латентному миостатину (варианты MS1032) в отношении тощей массы тела (LBM), силы сжатия и жировой массы тела, как проиллюстрировано в примере 25. MS1032LO06-SG1, MS1032LO11-SG1 и MS1032LO18-SG1 вводили Scid-мышам и оценивали тощую массу тела (А);
на фиг. 20Б, а также на фиг. 20А, 20В-20И - эффективность in vivo антител к латентному миостатину (варианты MS1032) в отношении тощей массы тела (LBM), силы сжатия и жировой массы тела, как проиллюстрировано в примере 25. MS1032LO06-SG1, MS1032LO19-SG1 и MS1032LO25-SG1 вводили Scid-мышам и оценивали тощую массу тела (Б);
на фиг. 20В, а также на фиг. 20А-Б, 20Г-И - эффективность in vivo антител к латентному миостатину (варианты MS1032) в отношении тощей массы тела (LBM), силы сжатия и жировой массы тела, как проиллюстрировано в примере 25. MS1032LO01-SG, MS1032LO06-SG1 и MS1032LO11-SG1 вводили Scid-мышам и оценивали тощую массу тела (В);
на фиг. 20Г, а также на фиг. 20А-В, 20Д-И - эффективность in vivo антител к латентному миостатину (варианты MS1032) в отношении тощей массы тела (LBM), силы сжатия и жировой массы тела, как проиллюстрировано в примере 25. MS1032LO06-SG1, MS1032LO11-SG1 и MS1032LO18-SG1 вводили Scid-мышам и оценивали силу сжатия (Г);
на фиг. 20Д, а также на фиг. 20А-Г, 20Е-20И - эффективность in vivo антител к латентному миостатину (варианты MS1032) в отношении тощей массы тела (LBM), силы сжатия и жировой массы тела, как проиллюстрировано в примере 25. MS1032LO06-SG1, MS1032LO19-SG1 и MS1032LO25-SG1 вводили Scid-мышам и оценивали силу сжатия (Д);
на фиг. 20Е, а также на фиг. 20А-Д, 20Ж-И - эффективность in vivo антител к латентному миостатину (варианты MS1032) в отношении тощей массы тела (LBM), силы сжатия и жировой массы тела, как проиллюстрировано в примере 25. MS1032LO01-SG, MS1032LO06-SG1 и MS1032LO11-SG1 вводили Scid-мышам и оценивали силу сжатия (Е);
на фиг. 20Ж, а также на фиг. 20А-Е, 20З-И - эффективность in vivo антител к латентному миостатину (варианты MS1032) в отношении тощей массы тела (LBM), силы сжатия и жировой массы тела, как проиллюстрировано в примере 25. MS1032LO06-SG1, MS1032LO11-SG1 и MS1032LO18-SG1 вводили Scid-мышам и оценивали жировую массу тела (Ж);
на фиг. 20З, а также на фиг. 20А-Ж, 20И - эффективность in vivo антител к латентному миостатину (варианты MS1032) в отношении тощей массы тела (LBM), силы сжатия и жировой массы тела, как проиллюстрировано в примере 25. MS1032LO06-SG1, MS1032LO19-SG1 и MS1032LO25-SG1 вводили Scid-мышам и оценивали жировую массу тела (З);
на фиг. 20И, а также на фиг. 20А-З - эффективность in vivo антител к латентному миостатину (варианты MS1032) в отношении тощей массы тела (LBM), силы сжатия и жировой массы тела, как проиллюстрировано в примере 25. MS1032LO01-SG, MS1032LO06-SG1 и MS1032LO11-SG1 вводили Scid-мышам и оценивали жировую массу тела (И);
на фиг. 21 - ингибирующая активность в отношении активации латентного миостатина антител к латентному миостатину, как проиллюстрировано в примере 26. Количества зрелого миостатина, высвобождающегося из латентного миостатина под действием протеазы ВМР1, измеряли в присутствии антител к латентному миостатину (MST1032, MST1504, MST1538, MST1551, MST1558, MST1572 и MST1573);
на фиг. 22А - схематическая диаграмма фрагментов латентного миостатина, состоящих из 100 аминокислот каждый, созданных для эпитопного картирования антител к латентному миостатину, как проиллюстрировано в примере 26;
на фиг. 22Б - результаты анализа методом Вестерн-блоттинга против меченных GST фрагментов человеческого латентного миостатина (GST-hMSTN) с использованием антитела к GST, как проиллюстрировано в примере 26. Каждая полоса соответствует: 1, GST-hMSTN 1-100 ак; 2, GST-hMSTN 21-120 ак; 3, GST-hMSTN 41-140 ак; 4, GST-hMSTN 61-160 ак; 5, GST-hMSTN 81-180 ак; 6, GST-hMSTN 101-200 ак; 7, GST-hMSTN 121-220 ак; 8, GST-hMSTN 141-241 ак; 9, GST-контроль;
на фиг. 22В - результаты анализа методом Вестерн-блоттинга против меченных GST фрагментов человеческого латентного миостатина (GST-hMSTN) с использованием антител к латентному миостатину (MST1032, MST1538, MST1572 и MST1573), как проиллюстрировано в примере 26. Каждая полоса соответствует: 1, GST-hMSTN 1-100 ак; 2, GST-hMSTN 21-120 ак; 3, GST-hMSTN 41-140 ак; 4, GST-hMSTN 61-160 ак; 5, GST-hMSTN 81-180 ак; 6, GST-hMSTN 101-200 ак; 7, GST-hMSTN 121-220 ак; 8, GST-hMSTN 141-241 ак; 9, GST-контроль; 10, человеческий латентный миостатин (100 нг);
на фиг. 22Г - обобщение результатов анализа методом Вестерн-блоттинга D и предсказанные (выведенные) положения в эпитопе для антител к латентному миостатину (MST1032, MST1538, MST1572 и MST1573), как проиллюстрировано в примере 26;
на фиг. 23 - сравнительный анализ первичной структуры аминокислотных последовательностей обезьяньих (циномолгус, (cyno)) Fc-гамма RIIa1, Fc-гамма RIIa2, Fc-гамма RIIa3, Fc-гамма RIIb, человеческих Fc-гамма RIIaH, Fc-гамма RIIaR и Fc-гамма RIIb. Обозначенные квадратом области соответствуют предполагаем остаткам, которые взаимодействуют с Fc-доменом;
на фиг. 24 - зависимость от времени общей концентрации миостатина в плазме после внутривенного введения антитела к миостатину с вариантом Fc с повышенной способностью связываться с Fc-гамма RIIb всем трансгенным по человеческому Fc-гамма R мышам, как проиллюстрировано в примере 28. Оценивали воздействия вариантов Fc с повышенной способностью связываться с Fc-гамма RIIb на элиминацию антигена посредством человеческого Fc-гамма RIIb;
на фиг. 25 - зависимость от времени концентрации антитела в плазме после внутривенного введения антитела к миостатину с вариантом Fc с повышенной способностью связываться с Fc-гамма RIIb всем трансгенным по человеческому Fc-гамма R мышам, как проиллюстрировано в примере 28. Оценивали воздействия вариантов Fc с повышенной способностью связываться с Fc-гамма RIIb на фармакокинетику антитела;
на фиг. 26А и 26Б - зависимость от времени концентрации миостатина в плазме после внутривенного введения антител к латентному миостатину обезьяна циномолгус, как проиллюстрировано в примере 29. (А) Влияние зависимости pH и конструирования Fc на клиренс миостатина in vivo оценивали путем сравнения с pH-независимым антителом к латентному миостатину (MS1032LO00-SG1) и pH-зависимыми антителами к латентному миостатину со сконструированной Fc-областью (MS1032LO06-SG1012, MS1032LO06-SG1016, MS1032LO06-SG1029, MS1032LO06-SG1031, MS1032LO06-SG1033, MS1032LO06-SG1034). (Б) Воздействие конструирования Fc на клиренс миостатина in vivo оценивали путем сравнения антител к латентному миостатину (MS1032LO19-SG1079, MS1032LO19-SG1071, MS1032LO19-SG1080, MS1032LO19-SG1074, MS1032LO19-SG1081, and MS1032LO19-SG1077);
на фиг. 27А - зависимость от времени общей концентрации миостатина в плазме после внутривенного введения антитела к миостатину с вариантом Fc с повышенной pI трансгенным по человеческому FcRn мышам, как проиллюстрировано в примере 30. Оценивали воздействия вариантов Fc с повышенной pI на элиминацию антигена;
на фиг. 27Б - зависимость от времени концентрации антитела в плазме после внутривенного введения антитела к миостатину с вариантом Fc с повышенной pI трансгенным по человеческому FcRn мышам, как проиллюстрировано в примере 30. Оценивали воздействия вариантов Fc с повышенной pI на фармакокинетику антитела;
на фиг. 28А - зависимость от времени общей концентрации миостатина в плазме после внутривенного введения антитела к миостатину с вариантом Fc с повышенной pI трансгенным по человеческому FcRn мышам, как проиллюстрировано в примере 30. Оценивали воздействия вариантов Fc с повышенной pI на элиминацию антигена. В этом анализе вводили в избыточном количестве человеческий нормальный иммуноглобулин совместно с антителом к миостатину для имитации ситуации в человеческой плазме;
на фиг. 28Б - зависимость от времени концентрации антитела в плазме после внутривенного введения антитела к миостатину с вариантом Fc с повышенной pI трансгенным по человеческому FcRn мышам, как проиллюстрировано в примере 30. Оценивали воздействия вариантов Fc с повышенной pI на фармакокинетику антитела. В этом анализе вводили в избыточном количестве человеческий нормальный иммуноглобулин совместно с антителом к миостатину для имитации ситуации в человеческой плазме;
на фиг. 29 - зависимость от времени общей концентрации миостатина в плазме после внутривенного введения антитела к миостатину с вариантом Fc с повышенной способностью связываться с Fc-гамма RIIb трансгенным по человеческому Fc-гамма RIIb мышам, как проиллюстрировано в примере 31. Оценивали воздействия вариантов Fc с повышенной способностью связываться с Fc-гамма RIIb на элиминацию антигена посредством человеческого Fc-гамма RIIb;
на фиг. 30 - зависимость от времени концентрации антитела в плазме после внутривенного введения антитела к миостатину с вариантом Fc с повышенной способностью связываться с Fc-гамма RIIb трансгенным по человеческому Fc-гамма RIIb мышам, как проиллюстрировано в примере 31. Оценивали воздействия вариантов Fc с повышенной способностью связываться с Fc-гамма RIIb на фармакокинетику антитела;
на фиг. 31 - результаты анализа визуализации в клетках антител к миостатину с вариантом Fc с повышенной способностью связываться с Fc-гамма RIIb, как проиллюстрировано в примере 33. Каждое антитело включали в комплекс с флуоресцентно меченным миостатином и измеряли внутриклеточное поглощение комплекса антиген-антитело в клетки, экспрессирующие человеческий Fc-гамма RIIb.
Описание вариантов осуществления изобретения
Технологии и процедуры, которые описаны или на которые сделаны ссылки в настоящем описании, в целом хорошо известны специалистам в данной области, и их широко применяют с использованием общепринятых методологий, таких, например, как описанные у Sambrook и др., Molecular Cloning: А Laboratory Manual, 3-е изд., изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. 2001; Current Protocols in Molecular Biology (под ред. F.M. Ausubel и др., 2003); серии Methods in Enzymology (изд-во Academic Press, Inc.): PCR 2: A Practical Approach (под ред. M.J. MacPherson, B.D. Hames и G.R. Taylor, 1995), Antibodies, A Laboratory Manual, под ред. Harlow и Lane, 1988 и Animal Cell Culture (под ред. R.I. Freshney 1987); Oligonucleotide Synthesis (под ред. M.J. Gait, 1984); Methods in Molecular Biology, изд-во Humana Press; Cell Biology: A Laboratory Notebook (под ред. J.E. Cellis, 1998) из-во Academic Press; Animal Cell Culture (под ред. R.I. Freshney, 1987); Introduction to Cell and Tissue Culture (J.P. Mather и P.E. Roberts, 1998), изд-во Plenum Press; Cell and Tissue Culture: Laboratory Procedures (под ред. A. Doyle, J.B. Griffiths и D.G. Newell, 1993-8), изд-во J. Wiley and Sons; Handbook of Experimental Immunology (под ред. D.M. Weir и С.С. Blackwell); Gene Transfer Vectors for Mammalian Cells (под ред. J.M. Miller и M.P. Calos, 1987); PCR: The Polymerase Chain Reaction, (под ред. Mullis и др., 1994); Current Protocols in Immunology (под ред. J.E. Coligan и др., 1991); Short Protocols in Molecular Biology (изд-во Wiley and Sons, 1999); Immunobiology (C.A. Janeway и P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: A Practical Approach (под ред. D. Catty., изд-во IRL Press, 1988-1989); Monoclonal Antibodies: A Practical Approach (под ред. P. Shepherd и С. Dean, изд-во Oxford University Press, 2000); Using Antibodies: A Laboratory Manual (E. Harlow и D. Lane, изд-во Cold Spring Harbor Laboratory Press, 1999); The Antibodies (под ред. M. Zanetti и J. D. Capra, изд-во Harwood Academic Publishers, 1995); и Cancer: Principles and Practice of Oncology (под ред. V.T. DeVita и др., изд-во J.В. Lippincott Company, 1993).
I. Определения
Если не указано иное, то технические и научные понятия, применяемые в настоящем описании, имеют значения, хорошо известные обычному специалисту в области, в которой относится настоящее изобретение. У Singleton и др., Dictionary of Microbiology and Molecular Biology 2-ое изд., изд-во J. Wiley & Sons, New York, N.Y. 1994 и March, Advanced Organic Chemistry Reactions, Mechanisms and Structure 4-ое изд., изд-во John Wiley & Sons, New York, N.Y. 1992, для специалистов в данной области представлено в целом толкование многих понятий, применяемых в настоящей заявке. Все процитированные в настоящем описании ссылки, включая заявки на патент и публикации патентов, полностью включены в него в качестве ссылки.
Для интерпретации настоящего описания изобретения следует применять приведенные ниже понятия, и, если это возможно, понятия, применяемые в единственном числе, включают также их применение во множественном числе и наоборот. Должно быть очевидно, что применяемая в настоящем описании терминология представлена только для цели описания конкретных вариантов осуществления изобретения и не направлена на ограничение его объема. В случае если любое указанное ниже определение вступает в противоречие с указанным в любом документе, включенном в описание в качестве ссылки, то следует использовать указанное ниже определение.
Для целей настоящего описания «акцепторный человеческий каркасный участок» означает каркасный участок, который содержит аминокислотную последовательность каркасного участка вариабельного домена легкой цепи (VL) или каркасный участок вариабельного домена тяжелой цепи (VH), происходящую из каркасного участка человеческого иммуноглобулина или человеческого консенсусного каркасного участка, указанного ниже. Акцепторный человеческий каркасный участок, «происходящий из» каркасного участка человеческого иммуноглобулина или человеческого консенсусного каркасного участка, может иметь такую же аминокислотную последовательность или может содержать изменения в аминокислотной последовательности. В некоторых вариантах осуществления изобретения количество аминокислотных изменений составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В некоторых вариантах осуществления изобретения последовательность человеческого акцепторного каркасного участка VL идентична последовательности каркасного участка VL человеческого иммуноглобулина или последовательности человеческого консенсусного каркасного участка.
Понятие «аффинность» относится к суммарной силе всех нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). В контексте настоящего описания, если не указано иное, «аффинность связывания» относится к присущей молекуле аффинности связывания, отражающей взаимодействие по типу 1:1 между компонентами связывающейся пары (например, антителом и антигеном). Аффинность молекулы X к ее партнеру Y, как правило, можно характеризовать с помощью константы диссоциации (Kd). Аффинность можно оценивать с помощью общепринятых методов, известных в данной области, включая те, которые представлены в настоящем описании. Ниже в качестве иллюстрации представлены конкретные варианты измерения аффинности связывания.
Антитело «с созревшей аффинностью» означает антитело с одним или несколькими изменениями в одном или нескольких гипервариабельных участках (HVR) по сравнению с родительским антителом, которое не содержит указанных изменений, указанные изменения приводят к повышению аффинности антитела к антигену.
Понятия «антитело к миостатину» и «антитело, которое связывается с миостатином» относятся к антителу, которое обладает способностью связываться с миостатином с аффинностью, достаточной для того, чтобы антитело можно было применять в качестве диагностического и/или терапевтического агента, мишенью которого является миостатин. В одном из вариантов осуществления изобретения уровень связывания антитела к миостатину с неродственным не представляющим собой миостатин белком составляет менее чем примерно 10% от связывания антитела с миостатином по данным измерений, полученным, например, с помощью радиоиммуноанализа (РИА). В некоторых вариантах осуществления изобретения антитело, которое связывается с миостатином, имеет константу диссоциации (Kd) 1 мкМ или менее 100 нМ или менее, 10 нМ или менее, 1 нМ или менее, 0,1 нМ или менее, 0,01 нМ или менее или 0,001 нМ или менее (например, 10-8М или менее, например, от 10-8М до 10-13М, например, от 10-9М до 10-13М). В некоторых вариантах осуществления изобретения антитело к миостатину связывается с эпитопом миостатина, который является консервативным для миостатина из различных видов.
В контексте настоящего описания понятие «антитело» используется в его наиболее широком смысле и относится к различным структурам антител, включая (но, не ограничиваясь только ими) моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они обладают требуемой антигенсвязывающей активностью.
Понятие «фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, связывающуюся с антигеном, с которым связывается интактное антитело. Примеры фрагментов антитела включают (но, не ограничиваясь только ими) Fv, Fab, Fab', Fab'-SH, F(ab')2; димерные антитела (диабоди); линейные антитела; молекулы одноцепочечных антител (например, scFv) и мультиспецифические антитела, образованные из фрагментов антител.
Понятие «антитело, которое связывается с тем же эпитопом», что референс-антитело, относится к антителу, которое блокирует связывание референс-антитела с его антигеном при оценке с помощью анализа в условиях конкуренции (конкурентный анализ) и/или, наоборот, референс-антитело блокирует связывание антитела с его антигеном при оценке с помощью конкурентного анализа. Пример конкурентного анализа представлен в настоящем описании.
Понятие «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи происходит из конкретного источника или конкретных видов, а остальная часть тяжелой и/или легкой цепи происходит из другого источника или других видов.
Понятие «класс» антитела относится к типу константного домена или константной области, который/которая входит в его тяжелую цепь. Известно пять основных классов антител: IgA, IgD, IgE, IgG и IgM и некоторые из них можно подразделять дополнительно на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулина, обозначают как альфа, дельта, эпсилон, гамма и мю (α, δ, ε, γ и μ) соответственно.
Понятие "цитотоксический агент» в контексте настоящего описания относится к субстанции, которая ингибирует клеточную функцию или препятствует ей и/или вызывает клеточную гибель или деструкцию. Цитотоксические агенты включают (но, не ограничиваясь только ими) радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu); химиотерапевтические агенты или лекарственные средства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты); ингибирующие рост агенты; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибного, растительного или животного происхождения, включая их фрагменты и/или варианты; и различные противоопухолевые или противораковые агенты, указанные ниже.
Понятие «эффекторные функции» относится к тем видам биологической активности, которые связаны с Fc-областью антитела, которые варьируются в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплементзависимую цитотоксичность (CDC); связывание Fc-рецептора; антитело-обусловленную клеточнозависимую цитотоксичность (ADCC); фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, В-клеточного рецептора) и активацию В-клеток.
Понятие «эффективное количество» агента, например, фармацевтической композиции, означает количество, эффективное в дозах и в течение периода времени, необходимых для достижения требуемого терапевтического или профилактического результата.
Понятие «эпитоп» включает любую детерминанту, обладающую способностью связываться антителом. Эпитоп представляет собой область антигена, которая связывается с антителом, мишенью которого является антиген, и включает специфические аминокислоты, которые непосредственно контактируют с антителом. Эпитопные детерминанты могут включать химически активные поверхностные группы молекул, таких как аминокислоты, боковые сахарные цепи, фосфорильные или сульфонильные группы, и могут иметь специфически характеристики трехмерной структуры и/или специфические характеристики зарядов. Как правило, антитела, специфические в отношении конкретного антигена-мишени, должны предпочтительно распознавать эпитоп на антигене-мишени в сложной смеси белков и/или макромолекул.
«Fc-рецептор» или «FcR» означает рецептор, который связывается с Fc-областью антитела. В некоторых вариантах осуществления изобретения FcR представляет собой нативный человеческий FcR. В некоторых вариантах осуществления изобретения FcR представляет собой рецептор, который связывается с антителом IgG-типа (гамма-рецептор) и включает рецепторы подкласса Fc-гамма RI, Fc-гамма RII и Fc-гамма RIII, в том числе аллельные варианты и полученные в результате альтернативного сплайсинга формы этих рецепторов. Рецепторы Fc-гамма RII включают Fc-гамма RIIA («активирующий рецептор») и Fc-гамма RIIB («ингибирующий рецептор»), которые имеют сходные аминокислотные последовательности, отличающиеся прежде всего их цитоплазматическими доменами. Активирующий рецептор Fc-гамма RIIA содержит активирующие мотивы на основе тирозина иммунорецептора (ITAM) в его цитоплазматическом домене. Ингибирующий рецептор Fc-гамма RIIB содержит ингибирующий мотив на основе рецептора тирозина иммунорецептора (ITIM) в его цитоплазматическом домене (см., например, Daeron, Annu. Rev. Immunol. 15, 1997, cc. 203-234). Обзор сведений о FcR представлен, например, у Ravetch и Kinet, Annu. Rev. Immunol. 9, 1991, cc. 457-492; Capel и др., Immunomethods 4, 1994, cc. 25-34 и de Haas и др., J. Lab. Clin. Med. 126, 1995, cc. 330-341. Под понятие «FcR», указанное в настоящем описании, подпадают и другие FcR, включая те, которые будут идентифицированы в будущем.
Понятие «Fc-рецептор» или «FcR» включает также неонатальный рецептор, FcRn, который ответствен за перенос материнских IgG эмбриону (Guyer и др., J. Immunol. 117, 1976, сс. 587-593 и Kim и др., J. Immunol. 24, 1994, сс. 2429-2434) и регулирует гомеостаз иммуноглобулинов. Известны методы измерения связывания с FcRn (см., например, Ghetie и Ward., Immunol. Today 18(12), 1997, сс. 592-598; Ghetie и др., Nature Biotechnology 15(7), 1997, сс. 637-640; Hinton и др., J. Biol. Chem. 279(8), 2004, сс. 6213-6216; WO 2004/92219 (Hinton и др.). Связывание с человеческим FcRn in vivo и время полужизни в сыворотке человеческого FcRn, отличающегося высокой аффинностью связывания с полипептидами, можно оценивать, например, в трансгенных мышах или в трансфектированных человеческих клеточных линиях, экспрессирующих человеческий FcRn, или в приматах, которым вводят полипептиды с вариантом Fc-области. В WO 2000/42072 (Presta) описаны варианты антител с повышенной или пониженной способностью связываться с FcR (см., например, также Shields и др., J. Biol. Chem. 9(2), 2001, сс. 6591-6604).
Понятие «Fc-область» в контексте настоящего описания относится к C-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Понятие включает нативную последовательность Fc-областей и вариант Fc-областей. В одном из вариантов осуществления изобретения Fc-область тяжелой цепи человеческого IgG простирается с Cys226 или с Pro230 до карбоксильного конца тяжелой цепи. Однако C-концевой лизин (Lys447) или глицин-лизин (остатки 446-447) Fc-области может присутствовать или отсутствовать. Если специально не указано иное, то нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU, которую обозначают также как EU-индекс, описанной у Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
Понятие «содержащее Fc-область антитело» относится к антителу, которое содержит Fc-область. C-концевой лизин (остаток 447 согласно системе EU-нумерации) или C-концевой глицин-лизин (остатки 446-447) Fc-области может быть удален, например, в процессе очистки антитела или путем рекомбинантного конструирования нуклеиновой кислоты, кодирующей антитело. Таким образом, композиция, которая содержит антитело, имеющее Fc-область, предлагаемое в настоящем изобретении, может содержать антитело с G446-K447, с G446 и без K447, со всеми удаленными G446-K447 или смесь из трех описанных выше типов антител.
«Каркасный участок» или «FR», означает остатки вариабельного домена, отличные от остатков гипервариабельного участка (HVR). FR вариабельного домена, как правило, состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Таким образом, последовательности HVR и FR, как правило, имеют следующее расположение в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Понятия «полноразмерное антитело», «интактное антитело» и «цельное антитело» в контексте настоящего описания используют взаимозаменяемо для обозначения антитела, имеющего структуру, практически сходную с нативной структурой антитела, или имеющего тяжелые цепи, которые содержат представленную в настоящем описании Fc-область.
«Функциональная Fc-область» обладает «эффекторной функцией», присущей нативной последовательности Fc-области. Примеры «эффекторных функций» включают связывание C1q; (CDC); связывание Fc-рецептора; ADCC; фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, В-клеточного рецептора; BCR) и т.д. Для проявления указанных эффекторных функций, как правило, требуется, чтобы Fc-область была объединена со связывающим доменом (например, вариабельным доменом антитела), и их можно оценивать с помощью различных анализов, например, представленных в разделе «Определения» в настоящем описании.
Понятия «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используют взаимозаменяемо, и они относятся к клеткам, в которые интродуцирована экзогенная нуклеиновая кислота, включая потомство указанных клеток. К клеткам-хозяевам относятся «трансформанты» и «трансформированные клетки», которые включают первично трансформированную клетку и полученное из нее потомство безотносительно к количеству пересевов. Потомство может не быть полностью идентичным по составу нуклеиновых кислот родительской клетке, но может содержать мутации. Под объем изобретения подпадает мутантное потомство, которое обладает такой же функцией или биологической активностью, которая обнаружена в результате скрининга или отобрана у исходной трансформированной клетки.
«Человеческое антитело» представляет собой антитело, имеющее аминокислотную последовательность, которая соответствует последовательности антитела, продуцируемого в организме человека или человеческими клетками, или происходящее из нечеловеческого источника, в котором использован спектр человеческих антител или другие кодирующие человеческое антитело последовательности. Из указанного определения человеческого антитела специально исключено гуманизированное антитело, содержащее нечеловеческие антигенсвязывающие остатки.
«Человеческий консенсусный каркасный участок» представляет собой каркасный участок, в который входят наиболее часто встречающиеся аминокислотные остатки в выбранных последовательностях каркасных участков VL или VH человеческого иммуноглобулина. Как правило, выбор последовательностей VL или VH человеческого иммуноглобулина осуществляют из подгруппы последовательностей вариабельных доменов. Как правило, подгруппа последовательностей представляет собой подгруппу, описанную у Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд., публикация NIH 91-3242, Bethesda MD, т.т. 1-3, 1991. В одном из вариантов осуществления изобретения касательно VL подгруппа представляет собой подгруппу каппа I согласно Kabat и др., выше. В одном из вариантов осуществления изобретения касательно VH подгруппа представляет собой подгруппу III согласно Kabat и др., выше.
Понятие «гуманизированное» антитело относится к химерному антителу, которое содержит аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из человеческих FR. В некоторых вариантах осуществления изобретения гуманизированное антитело может содержать практически все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или практически все HVR (например, CDR) соответствуют участкам нечеловеческого антитела, а все или практически все FR соответствуют участкам человеческого антитела. Гуманизированное антитело необязательно может содержать по меньшей мере часть константной области антитела, происходящей из человеческого антитела. Понятие «гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, которое подвергали гуманизации.
Понятие «гипервариабельный участок» или «HVR» в контексте настоящего описания относится к каждому из участков вариабельного домена антитела, последовательности которых являются гипервариабельными («определяющие комплементарность участки» или «CDR») и/или формируют петли определенной структуры («гипервариабельные петли»), и/или содержат контактирующие с антигеном остатки («контакты с антигеном»). Как правило, антитела содержат шесть HVR; три в VH (H1, Н2, Н3) и три в VL (L1, L2, L3). Согласно настоящему описанию примеры HVR включают (а) гипервариабельные петли, включающие аминокислотные остатки 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia и Lesk, J. Mol. Biol. 196, 1987, cc. 901-917); (6) CDR, включающие аминокислотные остатки 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (Н3) (Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991, публикация NIH 91-3242); (в) области контакта с антигеном, включающие аминокислотные остатки 27с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (Н3) (MacCallum и др., J. Mol. Biol. 262, 1996, cc. 732-745); и (г) комбинации остатков, указанных в подпунктах (а), (б) и/или (в), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3).
Если специально не указано иное, то остатки в HVR и другие остатки в вариабельном домене (например, остатки в FR) нумеруют согласно Kabat и др., выше.
«Иммуноконъюгат» представляет собой антитело, конъюгированное с одной или несколькими гетерологичной(ыми) молекулой(ами), включая (но, не ограничиваясь только им) цитотоксический агент.
«Индивидуум» или «субъект» представляет собой млекопитающее. К млекопитающим относятся (но, не ограничиваясь только ими) одомашненные животные (например, коровы, овцы, кошки, собаки и лошади), приматы (например, человек и приматы кроме человека, например, обезьяны), кролики и грызуны (например, мыши и крысы). В некоторых вариантах осуществления изобретения индивидуум или субъект представляет собой человека.
«Выделенное» антитело представляет собой антитело, которое отделено от компонента его естественного окружения. В некоторых вариантах осуществления изобретения антитело очищают до чистоты, превышающей 95% или 99% по данным, например, электрофоретических анализов (таких, например, как ДСН-ПААГ, изоэлектрическое фокусирование (ИЭФ), капиллярный электрофорез) или хроматографических анализов (таких, например, как ионообменная хроматография или ЖХВР с обращенной фазой). Обзор методов оценки чистоты антител см., например, у Flatman и др., J. Chrom. В 848, 2007, сс. 79-87.
«Выделенная» нуклеиновая кислота представляет собой молекулу нуклеиновой кислоты, которая отделена от компонента ее естественного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетке, которая в норме включает молекулу нуклеиновой кислоты, но в которой молекула нуклеиновой кислоты присутствует вне хромосомы или в которой ее локализация на хромосоме отличается от ее встречающейся в естественных условиях локализации на хромосоме.
Понятие «выделенная нуклеиновая кислота, кодирующая антитело к миостатину» относится к одной или нескольким молекулам нуклеиновых кислот, кодирующих тяжелые и легкие цепи антитела (или их фрагменты), включая указанную(ые) молекулу(ы) в одном векторе или в различных векторах, и указанную(ые) молекулу(ы) нуклеиновой(ых) кислоты(т), присутствующую(ие) в одном или нескольких положениях в клетке-хозяине.
Понятие «моноклональное антитело» в контексте настоящего описания относится к антителу, полученному из популяции практически гомогенных антител, т.е. индивидуальные антитела, входящие в популяцию, идентичны и/или связываются с одним и тем же эпитопом, за исключением возможных вариантов антител, например, содержащих мутации, встречающиеся в естественных условиях или возникающие в процессе производства препарата моноклонального антитела, указанные варианты, как правило, присутствуют в минорных количествах. В отличие от препаратов поликлональных антител, которые, как правило, включают различные антитела к различным детерминантам (эпитопам), каждое моноклональное антитело из препарата моноклонального антитела направлено против одной детерминанты антигена. Таким образом, прилагательное «моноклональный» определяет особенность антитела, характеризуя его как полученное из практически гомогенной популяции антител, а не сконструированное в соответствии с требованиями к получению антитела с помощью какого-либо конкретного метода. Например, моноклональные антитела, которые можно применять согласно настоящему изобретению, можно создавать с помощью различных технологий, включая (но, не ограничиваясь только ими) метод гибридом, методы рекомбинантной ДНК, методы фагового дисплея и методы, основанные на использовании трансгенных животных, которые содержат все или часть локусов человеческого иммуноглобулина, указанные методы и другие, приведенные в качестве примера методы получения моноклональных антител, представлены в настоящем описании.
Понятие «голое антитело» относится к антителу, неконъюгированному с гетерологичным фрагментом (например, цитотоксическим фрагментом) или радиоактивной меткой. Голое антитело может присутствовать в фармацевтической композиции.
Понятие «нативные антитела» относится к встречающимся в естественных условиях молекулам иммуноглобулинов с различными структурами. Например, нативные антитела в виде IgG представляют собой гетеротетрамерные гликопротеины с молекулярной массой примерно 150000 Да, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, связанных дисульфидными мостиками. В направлении от N-конца к С-концу, каждая тяжелая цепь имеет вариабельную область (VH), которую называют также вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которой следует три константных домена (CH1, СН2 и СН3). Аналогично этому, в направлении от N-конца к С-концу, каждая легкая цепь имеет вариабельную область (VL), которую называют также вариабельным легким доменом или вариабельным доменом легкой цепи, за которой следует константный домен легкой цепи (CL). Легкая цепь антитела в зависимости от аминокислотной последовательности ее константного домена может принадлежать к одному из двух типов, которые обозначают каппа (κ) и лямбда (λ).
«Имеющая нативную последовательность Fc-область» содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, которая встречается в природе. Нативная последовательность человеческих Fc-областей включает нативную последовательность Fc-области человеческого IgG1 (не-А- и А-аллотипов); нативную последовательность Fc-области человеческого IgG2; нативную последовательность Fc-области человеческого IgG3 и нативную последовательность Fc-области человеческого IgG4, а также их встречающиеся в естественных условиях варианты.
Понятие «листовка-вкладыш в упаковку» относится к инструкциям, которые обычно входят в поступающие в продажу упаковки терапевтических продуктов, содержащим информацию о показаниях, применении, дозе, пути введения, комбинированной терапии, противопоказаниях и/или мерах предосторожности, которые связаны с применением указанных терапевтических продуктов.
«Процент (%) идентичности аминокислотной последовательности» относительно полипептидной референс-последовательности определяют как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в полипептидной референс-последовательности, после выравнивая последовательностей и при необходимости интродукции брешей для достижения максимального процента идентичности последовательностей, и не рассматривая какие-либо консервативные замены в качестве компонента при оценке идентичности последовательностей. Сравнительный анализ для целей определения процента идентичности аминокислотных последовательностей можно осуществлять различными путями, известными в данной области, используя, например, публично доступные компьютерные программы, такие как программа BLAST, BLAST-2, ALIGN или Megalign (DNASTAR) или GENETYX® (фирма Genetyx Co., Ltd.) Специалисты в данной области могут определять соответствующие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Авторство компьютерной программы для сравнения последовательностей ALIGN-2 принадлежит фирме Genentech, Inc., и исходный код был представлен в комплекте с документацией для пользователей в U.S. Copyright Office, Washington D.C., 20559, где он зарегистрирован как U.S. Copyright Registration № TXU510087. Программа ALIGN-2 публично доступна от фирмы Genentech, Inc., Южный Сан-Франциско, шт. Калифорния, или ее можно компилировать на основе исходного кода. Программу ALIGN-2 можно компилировать для применения в операционной системе UNIX, включая цифровую систему UNIX V4.0D. Все параметры, требуемые для сравнения последовательностей, устанавливаются программой ALIGN-2 и не должны изменяться.
В ситуациях, в которых ALIGN-2 применяют для сравнения аминокислотных последовательностей, % идентичности аминокислотных последовательностей данной аминокислотной последовательности А по сравнению, с или относительно данной аминокислотной последовательности Б (что в альтернативном варианте можно обозначать как данная аминокислотная последовательность А, которая имеет или содержит определенный % идентичности аминокислотной последовательности по сравнению, с или относительно данной аминокислотной последовательности Б), рассчитывают следующим образом: 100 × дробь X/Y, где X обозначает количество аминокислотных остатков, определенных как идентичные совпадения при оценке с помощью программы для сравнительного анализа последовательностей ALIGN-2, где с помощью программы осуществляли сравнение А и Б, и где Y обозначает общее количество аминокислотных остатков в Б. Должно быть очевидно, что, если длина аминокислотной последовательности А не равна длине аминокислотной последовательности Б, то % идентичности аминокислотных последовательностей А относительно Б не может быть равен % идентичности аминокислотной последовательности Б относительно А. Если специально не указано иное, то все величины % идентичности аминокислотных последовательностей, указанные в настоящем описании, получали с использованием описанной в предыдущем параграфе компьютерной программы ALIGN-2.
Понятие «фармацевтическая композиция» относится к препарату, который находится в такой форме, что он обеспечивает биологическую активность входящего в его состав действующего вещества, которое должно обладать эффективностью, и который не содержит дополнительных компонентов, обладающих неприемлемой токсичностью для индивидуума, которому следует вводить композицию.
«Фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтической композиции, отличному от действующего вещества, который является нетоксичным для индивидуума. Фармацевтически приемлемые носители включают (но, не ограничиваясь только ими) буфер, эксципиент, стабилизатор или консервант.
Понятие «миостатин» в контексте настоящего описания может относиться к любому нативному миостатину из любого позвоночного животного, включая млекопитающих, таких как приматы (например, человек) и грызуны (например, мыши и крысы). Если специально не указано иное, понятие «миостатин» относится к человеческому белку миостатина, имеющему аминокислотную последовательность, представленную в SEQ ID NO: 1, и содержащему концевой пропептидный домен человеческого миостатина, представленный в SEQ ID NO: 75 или 78. Понятие относится к «полноразмерному» непроцессированному миостатину, а также к форме миостатина, которая образуется в результате процессинга в клетке. Под понятие подпадают также встречающиеся в естественных условиях варианты человеческого миостатина, например, сплайсинговые варианты или аллельные варианты. Аминокислотная последовательность человеческого миостатина (промиостатина) представлена в SEQ ID NO: 1. Аминокислотная последовательность приведенного в качестве примера C-концевого домена фактора роста человеческого миостатина представлена в SEQ ID NO: 2. Аминокислотная последовательность приведенного в качестве примера N-концевого пропептидного домена человеческого миостатина представлена в SEQ ID NO: 75 или 78. Активный зрелый миостатин представляет собой связанный дисульфидом гомодимер, состоящий из двух С-концевых доменов фактора роста. Неактивный латентный миостатин представляет собой нековалентно ассоциированный комплекс из двух пропептидов и зрелого миостатина. Как указано в настоящем описании, антитела, предлагаемые в изобретении, связываются с неактивным латентным миостатином, но не связываются со зрелым активным гомодимером миостатина. В некоторых вариантах осуществления изобретения антитела, предлагаемые в изобретении, связываются с эпитопом внутри фрагмента, состоящего из аминокислот 21-100 пропептида миостатина (SEQ ID NO: 78), но не связываются со зрелым активным гомодимером миостатина. Аминокислотная последовательность приведенного в качестве примера миостатина обезьян циномолгус и мышиного миостатина (промиостатина) представлены в SEQ ID NO: 3 и 5 соответственно. Аминокислотная последовательность приведенного в качестве примера C-концевого домена фактора роста миостатина обезьян циномолгус и мышиного миостатина представлены в SEQ ID NO: 4 и 6 соответственно. Аминокислотная последовательность приведенного в качестве примера N-концевого пропептидного домена миостатина обезьян циномолгус и мышиного миостатина представлены в SEQ ID NO: 76 или 79 и 77 или 80 соответственно. GDF-11 (ВМР-11) представляет собой молекулу, близко родственную миостатину, они обе являются представителями суперсемейства TGF-бета. Аналогично миостатину, GDF11 синтезируется сначала в виде полипептида-предшественника, а затем расщепляется на N-концевой продомен и C-концевой зрелый GDF11. Аминокислотная последовательность человеческого GDF11 (предшественник) представлена в SEQ ID NO: 81. Аминокислотная последовательность C-концевого зрелого человеческого GDF11 представлена в SEQ ID NO: 82. Аминокислотная последовательность N-концевого продомена человеческого GDF11 представлена в SEQ ID NO: 83 или 84. Аминокислотные последовательности SEQ ID NO: 1, 3, 5, 78, 79, 80, 81 и 84 включают сигнальную последовательность. Ей соответствуют аминокислоты 1-24, и они удаляются во время процессинга в клетке.
В контексте настоящего описания понятие «лечение» (и его грамматические вариации, такие как «лечить» или «процесс лечения») относится к клиническому вмешательству с целью изменения естественного течения болезни у индивидуума, подлежащего лечению, и его можно осуществлять либо для профилактики, либо в процессе развития клинической патологии. Требуемыми действиями лечения являются (но, не ограничиваясь только ими) предупреждение возникновения или рецидива болезни, облегчение симптомов, уменьшение любых прямых или косвенных патологических последствий болезни, предупреждение метастазов, снижение скорости развития болезни, облегчение или временное ослабление болезненного состояния и ремиссия или улучшение прогноза. В некоторых вариантах осуществления изобретения антитела, предлагаемые в изобретении, применяют для задержки развития болезни или замедления прогрессирования болезни.
Понятие «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL соответственно) нативного антитела, как правило, имеют сходные структуры, при этом каждый домен содержит четыре консервативных каркасных участка (FR) и три гипервариабельных участка (HVR) (см., например, Kindt и др., Kuby Immunology, 6-ое изд., изд-во W.H. Freeman and Co., 2007, с. 91). Одного VH- или VL-домена может быть достаточно для обеспечения специфичности связывания антигена. Кроме того, антитела, которые связываются с конкретным антигеном, можно выделять, используя VH- или VL-домен из антитела, которое связывается с антигеном, для скрининга библиотеки комплементарных VL- или VH-доменов соответственно (см., например, Portolano и др., J. Immunol. 150, 1993, сс. 880-887; Clarkson и др., Nature 352, 1991, сс. 624-628).
«Вариант Fc-области» содержит аминокислотную последовательность, которая отличается от нативной последовательности Fc-области благодаря наличию по меньшей мере одной аминокислотной модификации (изменения), предпочтительно одной или нескольких аминокислотной(ых) замены(замен). Предпочтительно вариант Fc-области имеет по меньшей мере одну аминокислотную замену по сравнению с нативной последовательностью Fc-области родительского полипептида, например, от примерно одной до примерно десяти аминокислотных замен, и предпочтительно от примерно одной до примерно пяти аминокислотных замен в нативной последовательности Fc-области или в Fc-области родительского полипептида. В контексте настоящего описания вариант Fc-области должен обладать предпочтительно по меньшей мере примерно 80%-ной гомологией с нативной последовательностью Fc-области и/или Fc-области родительского полипептида и наиболее предпочтительно по меньшей мере примерно 90%-ной гомологией с ней, более предпочтительно по меньшей мере примерно 95%-ной гомологией с ней.
Понятие «вектор» в контексте настоящего описания относится к молекуле нуклеиновой кислоты, которая обладает способностью к размножению другой нуклеиновой кислоты, с которой она связана. Понятие включает вектор в виде самореплицирующейся структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую он интродуцирован. Некоторые векторы обладают способностью контролировать экспрессию нуклеиновых кислот, с которыми они функционально связаны. Указанные векторы в контексте настоящего описания обозначают как «экспрессионные векторы».
II. Композиции и способы
Одним из объектов изобретения являются, в частности, антитела к миостатину и их применения. Некоторыми вариантами осуществления изобретения являются антитела, которые связываются с миостатином. Антитела, предлагаемые в изобретении, можно применять, например, для диагностирования или лечения синдрома мышечного истощения.
Одним из объектов изобретения являются, в частности С5, полипептиды, содержащие варианты Fc-областей, и их применения. Одним из вариантов осуществления изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью. Другим вариантом осуществления изобретения являются содержащие варианты Fc-областей полипептиды с повышенной pI. В конкретных вариантах осуществления изобретения полипептиды, предлагаемые в изобретении, представляют собой антитела. Полипептиды, содержащие вариант Fc-области, предлагаемые в изобретении, можно применять, например, для диагностирования или лечения заболевания.
А. Примеры антител к миостатину и полипептидов, содержащих варианты Fc-областей
Одним из объектов изобретения являются выделенные антитела, которые связываются с миостатином. В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, связывается с латентным миостатином. В других вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, связывается с пропептидом миостатина (человеческий: SEQ ID NO: 75 или 78; обезьян циномолгус: SEQ ID NO: 76 или 79; мышиный: SEQ ID NO: 77 или 80). В других вариантах осуществления изобретения антитело связывается с эпитопом во фрагменте, состоящем из аминокислот 21-100 пропептида миостатина (SEQ ID NO: 78). Пропептид входит в латентный миостатин в качестве одного из компонентов, как описано выше. В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, ингибирует активацию миостатина. В некоторых вариантах осуществления изобретения антитело к миостатину блокирует высвобождение зрелого миостатина из латентного миостатина. Известно, что зрелый миостатин высвобождается посредством протеолитических и непротеолитических процессов из латентного миостатина. Антитело к миостатину, предлагаемое в настоящем изобретении, может блокировать протеолитическое и/или непротеолитическое высвобождение зрелого миостатина из латентного миостатина. В некоторых вариантах осуществления изобретения антитело к миостатину блокирует протеолитическое расщепление латентного миостатина. В некоторых вариантах осуществления изобретения антитело к миостатину блокирует доступ протеазы к латентному миостатину (прежде всего к сайту протеолитического расщепления (Arg98-Asp99) латентного миостатина). В других вариантах осуществления изобретения протеаза может представлять собой представителя семейства металлопротеаз BMP1/TLD, такую как ВМР1, TED, толлоидподобный белок-1 (TLL-1) или толлоидподбный белок-2 (TLL-2). В другом варианте осуществления изобретения антитело к миостатину блокирует непротеолитическое высвобождение зрелого миостатина из латентного миостатина. В контексте настоящего описания под непротеолитическим высвобождением подразумевают спонтанное высвобождение зрелого миостатина из латентного миостатина, которое не сопровождается протеолитическим расщеплением латентного миостатина. Непротеолитическое высвобождение включает, например, высвобождение зрелого миостатина в результате инкубации латентного миостатина, например, при 37°С в отсутствии протеазы, которая расщепляет латентный миостатин. В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, не связывается со зрелым миостатином. В некоторых вариантах осуществления изобретения антитело к миостатину связывается с тем же эпитопом, что и антитело, представленное в таблице 2а. В некоторых вариантах осуществления изобретения антитело к миостатину конкурирует за связывание с латентным миостатином с антителом, представленным в таблице 2а. В дополнительных вариантах осуществления изобретения антитело к миостатину конкурирует за связывание с латентным миостатином, с антителом, которое содержит пару VH и VL, указанную в таблице 2а. В некоторых вариантах осуществления изобретения антитело к миостатину конкурирует за связывание с фрагментом, состоящим из аминокислот 21-100, пропептида миостатина (SEQ ID NO: 78) с антителом, представленным в таблице 2а. В других вариантах осуществления изобретения антитело к миостатину связывается с тем же эпитопом, что и антитело, представленное в таблице 11а или 13. В некоторых вариантах осуществления изобретения антитело к миостатину конкурирует за связывание с латентным миостатином с антителом, представленным в таблице 11а или 13. В некоторых вариантах осуществления изобретения антитело к миостатину конкурирует за связывание с фрагментом, состоящим из аминокислот 21-100, пропептида миостатина (SEQ ID NO: 78) с антителом, представленным в таблице 11а или 13.
В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, связывается с латентным миостатином и ингибирует активацию миостатина. В других вариантах осуществления изобретения антитело: (а) блокирует высвобождение зрелого миостатина из латентного миостатина; (б) блокирует протеолитическое высвобождение зрелого миостатина; (в) блокирует спонтанное высвобождение зрелого миостатина или (г) не связывается со зрелым миостатином; или связывается с эпитопом внутри фрагмента, состоящего из аминокислот 21-100 пропептида миостатина (SEQ ID NO: 78). В других вариантах осуществления изобретения антитело конкурирует за связывание с латентным миостатином или связывается с тем же эпитопом, что и антитело, которое содержит пару VH и VL, указанную в таблицах 2а, 11а или 13. В других вариантах осуществления изобретения антитело связывается с латентным миостатин с более высокой аффинностью при нейтральном pH (например, pH 7,4), чем при кислом pH (например, pH 5,8). В других вариантах осуществления изобретения антитело представляет собой (а) моноклональное антитело, (б) человеческое, гуманизированное или химерное антитело; (в) полноразмерное антитело IgG-типа или (г) фрагмент антитела, который связывается с латентным миостатином или пропептидом миостатина.
В другом варианте осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, не связывается с GDF11 (дифференцировочный фактор роста 11). В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, не ингибирует активацию GDF11. В некоторых вариантах осуществления изобретения антитело к миостатину не блокирует высвобождение зрелого GDF11 из латентного GDF11. Антитело к миостатину, предлагаемое в настоящем изобретении, не блокирует ни протеолитическое, ни непротеолитическое высвобождение зрелого GDF11 из латентного GDF11. В некоторых вариантах осуществления изобретения антитело к миостатину не блокирует протеолитическое расщепление латентного GDF11. В некоторых вариантах осуществления изобретения антитело к миостатину не блокирует доступ протеазы к латентному GDF11 (прежде всего к сайту протеолитического расщепления латентного GDF11). В других вариантах осуществления изобретения протеаза может представлять собой металлопротеазу из семейства BMP1/TLD, такую как ВМР1, TED, толлоидподобный белок-1 (TLL-1) или толлоидподобный белок-2 (TLL-2). Под непротеолитическим высвобождением в контексте настоящего описания подразумевается спонтанное высвобождение зрелого GDF11 из латентного GDF11, которое не осуществляется посредством протеолитического расщепления латентного GDF11. Непротеолитическое высвобождение включает, например, высвобождение зрелого GDF11 посредством инкубации латентного GDF11, например, при 37°С в отсутствии протеазы, которая расщепляет латентный GDF11. Наибольшее количество известных к настоящему времени антител к миостатину не являются специфическими в отношении миостатина. Эти антитела обладают высокой аффинностью к другим представителям суперсемейства TGF-бета, таким как GDF11, и нейтрализуют их биологическую активность. GDF11 играет важную роль в процессе эмбриогенеза и ответствен за гомеозисную трансформацию аксиального скелета. Гомозиготные по «выключенному» GDF11 мыши погибали в перинатальном периоде, мыши с одной копией дикого типа гена GDF11 были жизнеспособными, но имели дефекты скелета. Поскольку GDF11 играет важную роль в процессе эмбриогенеза, то антагонист, который ингибирует GDF11, теоретически несет риски, связанные с безопасностью, которые могут проявляться либо в виде токсичности для обработанных пациентов, или в виде репродуктивной токсичности, например, для женщин с детородным потенциалом. Таким образом, существует необходимость в специфическом ингибирования активности миостатина при лечении ассоциированных с миостатином нарушений, при которых желательно увеличение массы, размера, силы и т.д. мышц, прежде всего у женщин с детородным потенциалом.
Другим объектом изобретения являются антитела к миостатину, обладающие pH-зависимыми характеристиками связывания. В контексте настоящего описания понятие «pH-зависимое связывание» означает, что антитело обладает «пониженной способностью связываться с миостатином при кислом pH по сравнению с его связыванием при нейтральном pH» (для целей настоящего описания оба понятия можно использовать взаимозаменяемо). Например, антитела с «с pH-зависимыми характеристиками связывания» включают антитела, которые связываются с миостатином с более высокой аффинностью при нейтральном pH, чем при кислом pH. В некоторых вариантах осуществления изобретения антитела, предлагаемые в настоящем изобретении, связываются с миостатином с аффинностью, по меньшей мере в 2, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 или более раз более высокой при нейтральном pH, чем при кислом pH. В некоторых вариантах осуществления изобретения антитела связываются с миостатином (например, латентным миостатином или пропептидом миостатина) с более высокой аффинностью при pH 7,4, чем при pH 5,8. В других вариантах осуществления изобретения антитела связываются с миостатином с аффинностью, по меньшей мере в 2, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 или более раз более высокой при pH 7,4, чем при pH 5,8.
Когда антиген представляет собой растворимый белок, то связывание антитела с антигеном может приводить к удлиненному времени полужизни антигена в плазме (т.е. к пониженному клиренсу антигена из плазмы), поскольку антитело может иметь более продолжительное время полужизни в плазме, чем сам антиген, и может служить в качестве носителя для антигена. Это является следствием рециклинга комплекса антиген-антитело с помощью FcRn через эндосомальный путь в клетке (Roopenian, Nat. Rev. Immunol. 7(9), 2007, cc. 715-725). Однако антитело с pH-зависимыми характеристиками связывания, которое связывается с его антигеном в нейтральном внеклеточном окружении, высвобождая при этом антиген в кислые эндосомальные компартменты после проникновения в клетки, как ожидается, должно обладать предпочтительными свойствами касательно нейтрализации и клиренса антигена относительно его копии, которая связывается pH-независимым образом (Igawa и др., Nature Biotechnol. 28(11), 2010, cc. 1203-1207; Devanaboyina и др., mAbs 5(6), 2013, cc. 851-859; WO 2009/125825).
Для целей настоящего описания «аффинность» антитела к миостатину выражают в понятиях KD антитела. KD антитела означает константу равновесия диссоциации взаимодействия антитело-антиген. Чем выше величина KD, характеризующая связывание антитела с его антигеном, тем слабее его аффинность связывания с указанным конкретным антигеном. Таким образом, в контексте настоящего описания понятие «более высокая аффинность при нейтральном pH, чем при кислом pH» (или эквивалентное понятие «pH-зависимое связывание») означает, что KD, характеризующая связывание антитела с миостатином при кислом pH, выше, чем KD, характеризующая связывание антитела с миостатином при нейтральном pH. Например, в контексте настоящего изобретения считается, что антитело связывается с миостатином с более высокой аффинностью при нейтральном pH, чем при кислом pH, если KD, характеризующая связывание антитела с миостатином при кислом pH, по меньшей мере в 2 раза выше, чем KD, характеризующая связывание антитела с миостатином при нейтральном pH. Таким образом, настоящее изобретение включает антитела, связывание которых с миостатином при кислом pH характеризуется величиной KD, которая по меньшей мере в 2, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 или более раз превышает KD, характеризующую связывание с миостатином при нейтральном pH. В другом варианте осуществления изобретения величина KD при нейтральном pH может составлять 10-7, 10-8, 10-9, 10-10, 10-11, 10-12М или менее. В другом варианте осуществления изобретения величина KD при кислом pH может составлять 10-9М, 10-8М, 10-7М, 10-6М или более.
В других вариантах осуществления изобретения считается, что антитело связывается с миостатином (например, латентным миостатином или пропептидом миостатин) с более высокой аффинностью при нейтральном pH, чем при кислом pH, если величина KD, характеризующая связывание антитела с миостатином при pH 5,8, по меньшей мере в 2 раза выше, чем KD, характеризующая связывание антитела с миостатином при pH 7,4. В некоторых вариантах осуществления изобретения связывание предложенных антител с миостатином при pH 5,8 характеризуются величиной KD, которая по меньшей мере в 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 или более раз превышает KD, характеризующую связывание антитела с миостатином при pH 7,4. В другом варианте осуществления изобретения величина KD антитела при pH 7,4 может составлять 10-7, 10-8, 10-9, 10-10, 10-11, 10-12М или менее. В другом варианте осуществления изобретения величина KD антитела при pH 5,8 может составлять 10-9, 10-8, 10-7, 10-6М или более.
Характеристики связывания антитела с конкретным антигеном можно выражать также в виде kd антитела. Величина kd антитела означает константу скорости диссоциации антитела относительно конкретного антигена и ее выражают в виде величин, обратных секунде (т.е., с-1). Повышенная величина kd означает более слабое связывание антитела с его антигеном. Таким образом, настоящее изобретение включает антитела, связывание которых с миостатином характеризуется более высокой величиной kd при кислом pH, чем при нейтральном pH. Настоящее изобретение включает антитела, связывание которых с миостатином при кислом pH характеризуется величиной kd, которая по меньшей мере в 2, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 или более раз превышает kd, характеризующую связывание антитела с миостатином при нейтральном pH. В другом варианте осуществления изобретения величина kd антитела при нейтральном pH может составлять 10-2, 10-3, 10-4, 10-5, 10-6 1/с или менее. В другом варианте осуществления изобретения величина kd антитела при кислом pH может составлять 10-3, 10-2, 10-1 1/с или более. Изобретение включает также антитела, связывание которых с миостатином (например, латентным миостатином или пропептидом миостатина) характеризуется более высокой величиной kd при pH 5,8, чем при pH 7,4. Изобретение включает антитела, связывание которых с миостатином при pH 5,8 характеризуется величиной kd, которая по меньшей мере в 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 или более раз превышает kd, характеризующую связывание антитела с миостатином при pH 7,4. В другом варианте осуществления изобретения величина kd антитела при pH 7,4 может составлять 10-2, 10-3, 10-4, 10-5, 10-6 1/с или менее. В другом варианте осуществления изобретения величина kd антитела при pH 5,8 может составлять 10-3, 10-2, 10-1 1/с или более.
В некоторых случаях «пониженную способность связываться с миостатином при кислом pH по сравнению со способностью связываться при нейтральном pH» выражают в виде соотношения величины KD, характеризующей связывание антитела с миостатином при кислом pH, и величины KD, характеризующей связывание антитела с миостатином при нейтральном pH (или наоборот). Например, антитело может рассматриваться как обладающее «пониженной способностью связываться с миостатином при кислом pH по сравнению со способностью связываться при нейтральном pH» для целей настоящего изобретения, если для антитела соотношение KD при кислом/нейтральном pH составляет 2 или более. В некоторых вариантах осуществления изобретения соотношение KD при pH 5,8/pH 7,4 для антитела к миостатину, предлагаемого в настоящем изобретении, составляет 2 или более. В некоторых вариантах осуществления изобретения соотношение KD при кислом/нейтральном pH для антитела, предлагаемого в настоящем изобретении, может составлять 2, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 или более. В другом варианте осуществления изобретения величина KD антитела при нейтральном pH может составлять 10-7, 10-8, 10-9, 10-10, 10-11, 10-12М или менее. В другом варианте осуществления изобретения величина KD антитела при кислом pH может составлять 10-9, 10-8, 10-7, 10-6М или более. В других случаях антитело может рассматриваться как обладающее «пониженной способностью связываться с миостатином (например, латентным миостатином) при кислом pH по сравнению со способностью связываться при нейтральном pH», если для антитела характерно соотношение KD при pH 5,8/pH 7,4, составляющее 2 или более. В некоторых приведенных в качестве примеров вариантах осуществления изобретения соотношение KD при pH 5,8/pH 7,4 для антитела может составлять 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 или более. В другом варианте осуществления изобретения величина KD антитела при pH 7,4 может составлять 10-7, 10-8, 10-9, 10-10, 10-11, 10-12М или менее. В другом варианте осуществления изобретения величина KD антитела при pH 5,8 может составлять 10-9, 10-8, 10-7, 10-6М или более.
В некоторых случаях «пониженную способность связываться с миостатином при кислом pH по сравнению со способностью связываться при нейтральном pH» выражают в виде соотношения величины kd, характеризующей связывание антитела с миостатином при кислом pH, и величины kd, характеризующей связывание антитела с миостатином при нейтральном pH (или наоборот). Например, антитело может рассматриваться как обладающее «пониженной способностью связываться с миостатином при кислом pH по сравнению со способностью связываться при нейтральном pH» для целей настоящего изобретения, если для антитела соотношение kd при кислом/нейтральном pH составляет 2 или более. В некоторых вариантах осуществления изобретения соотношение kd при pH 5,8/pH 7,4 для антитела к миостатину, предлагаемого в настоящем изобретении, составляет 2 или более. В некоторых вариантах осуществления изобретения соотношение kd при кислом/нейтральном pH для антитела, предлагаемого в настоящем изобретении, может составлять 2, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 или более. В другом варианте осуществления изобретения величина kd антитела при нейтральном pH может составлять 10-2, 10-3, 10-4, 10-5, 10-6 1/с или менее. В другом варианте осуществления изобретения величина kd антитела при кислом pH может составлять 10-3, 10-2, 10-1 1/с или более. В некоторых приведенных в качестве примеров вариантах осуществления изобретения соотношение kd при pH 5,8/pH 7,4 для антитела может составлять 2, 3, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 200, 400, 1000, 10000 или более. В другом варианте осуществления изобретения величина kd антитела при pH 7,4 может составлять 10-2, 10-3, 10-4, 10-5, 10-6 1/с или менее. В другом варианте осуществления изобретения величина kd антитела при pH 5,8 может составлять 10-3, 10-2, 10-1 1/с или более.
В контексте настоящего описании понятие «кислый pH» означает pH от 4,0 до 6,5. Понятие «кислый pH» включает одно из следующих значений pH 4,0, 4,1, 4,2, 4,3, 4,4, 4,5, 4,6, 4,7, 4,8, 4,9, 5,0, 5,1, 5,2, 5,3, 5,4, 5,5, 5,6, 5,7, 5,8, 5,9, 6,0, 6,1, 6,2, 6,3, 6,4 и 6,5. В конкретных объектах изобретения «кислый pH» соответствует 5,8.
В контексте настоящего описании понятие «нейтральный pH» означает pH от 6,7 до примерно 10,0. Понятие «нейтральный pH» включает одно из следующих значений pH 6,7, 6,8, 6,9, 7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9, 8,0, 8,1, 8,2, 8,3, 8,4, 8,5, 8,6, 8,7, 8,8, 8,9, 9,0, 9,1, 9,2, 9,3, 9,4, 9,5, 9,6, 9,7, 9,8, 9,9, и 10,0. В конкретных объектах изобретения «нейтральный pH» соответствует 7,4.
Указанные в настоящем описании величины KD и kd можно определять с помощью биосенсора на основе метода поверхностного плазмонного резонанса для характеризации взаимодействий антитело-антиген (см., например, пример 7 в настоящем описании). Величины KD и величины kd можно определять при 25°С или 37°С.
В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, связывается с миостатином из более чем одного вида. В других вариантах осуществления изобретения антитело к миостатину связывается с миостатином человека и животного кроме человека. В других вариантах осуществления изобретения антитело к миостатину связывается с миостатином человека, мышей и обезьян (например, обезьяны циномолгус (яванский макак-крабоед), макаки резус, мармозетки, шимпанзе или бабуины).
В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, связывается с латентным миостатином из более чем одного вида. В других вариантах осуществления изобретения антитело к миостатину связывается с латентным миостатином человека и животного кроме человека. В других вариантах осуществления изобретения антитело к миостатину связывается с латентным миостатином человека, мышей и обезьян.
В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, связывается с пропептидом миостатина из более чем одного вида. В других вариантах осуществления изобретения антитело к миостатину связывается с пропептидом миостатина человека и животного кроме человека. В других вариантах осуществления изобретения антитело к миостатину связывается с пропептидом миостатина человека, мышей и обезьян.
Другим объектом изобретения является антитело к миостатину, которое образует иммунный комплекс (т.е. комплекс антиген-антитело) с миостатином. В некоторых вариантах осуществления изобретения два или большее количество антител к миостатину связывается с двумя или большим количеством молекул миостатина с образованием иммунного комплекса. Это является возможным, поскольку миостатин существует в виде гомодимера, который содержит две молекулы миостатина, а антитело имеет два антигенсвязывающих сайта. Антитело к миостатину может связываться с одним и тем же эпитопом на молекуле миостатина или может связываться с различными эпитопами на молекуле миостатина, наиболее вероятно такое антитело представляет собой биспецифическое антитело. В целом, когда два или большее количество антител образуют иммунный комплекс с двумя или большим количеством антигенов, то образовавшийся иммунный комплекс может сильно связываться с Fc-рецепторами, присутствующими на клеточных поверхностях, благодаря эффектам авидности, через Fc-области антител в комплексе и затем могут с высокой эффективностью поглощаться клетками. Таким образом, вышеуказанное антитело к миостатину, которое обладает способностью образовывать иммунный комплекс, содержащий два или большее количество антител к миостатину и две или большее количество молекул миостатина, может приводить к быстрому клиренсу миостатина из плазмы в живом организме в результате сильного связывания с Fc-рецепторами благодаря эффектам авидности.
Кроме того, предполагается, что антитело с pH-зависимыми характеристиками связывания может обладать предпочтительными свойствами касательно нейтрализации клиренса антигена по сравнению с его копией, которая связывается pH-независимым образом (Igawa и др., Nature Biotech. 28(11), 2010, сс. 1203-1207; Devanaboyina и др., mAbs 5(6), 2013, сс. 851-859; WO 2009/125825). Таким образом, ожидается, что антитело, обладающее двумя указанными выше свойствами, т.е. антитело, имеющее pH-зависимые характеристики связывания и образующее иммунный комплекс, который содержит два или большее количество антител с двумя или большим количеством антигенов, должно иметь еще более предпочтительные свойства, касательно ускоренной элиминации антигенов из плазмы (WO 2013/081143).
Другим объектом изобретения является антитело к миостатину, которое содержит по меньшей мере один, два, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57, 114-115, 126; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, 116-120, 127; (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, 121, 128; (г) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69, 122-124, 129; (д) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72, 125, 130; и (е) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74, 131.
Другим объектом изобретения является антитело к миостатину, которое содержит по меньшей мере один, два, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60; (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64; (г) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69; (д) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72; и (е) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74.
Одним из объектов изобретения является антитело к миостатину, которое содержит по меньшей мере один, два, три, четыре, пять или шесть гипервариабельных участков (HVR), выбранных из (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 114-115; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 116-120; (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 121; (г) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 122-124; (д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 125; и (е) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74.
Другим объектом изобретения является антитело к миостатину, которое содержит по меньшей мере один, два, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 114; (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58; (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63; (г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 122; (д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71; и (е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74. Другим объектом изобретения является антитело к миостатину, которое содержит по меньшей мере один, два, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 114; (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58; (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63; (г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 123; (д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71; и (е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74.
Другим объектом изобретения является антитело к миостатину, которое содержит по меньшей мере один, два, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 126; (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 127; (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 128; (г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 129; (д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 130; и (е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 131.
Одним из объектов изобретения является антитело, которое содержит по меньшей мере одну, по меньшей мере две или все три HVR-последовательности VH, выбранные из (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57, 114-115, 126; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, 116-120, 127; (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, 121, 128. В одном из вариантов осуществления изобретения антитело содержит HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, 121, 128. В другом варианте осуществления изобретения антитело содержит HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, 121, 128, и HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74, 131. В следующем варианте осуществления изобретения антитело содержит HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, 121, 128, HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74, 131, и HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, 116-120, 127. В следующем варианте осуществления изобретения антитело содержит (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57, 114-115, 126; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, 116-120, 127; и (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, 121, 128.
Одним из объектов изобретения является антитело, которое содержит по меньшей мере одну, по меньшей мере две или все три HVR-последовательности VH, выбранные из (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60; и (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64. В одном из вариантов осуществления изобретения антитело содержит HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64. В другом варианте осуществления изобретения антитело содержит HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, и HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74. В следующем варианте осуществления изобретения антитело содержит HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74, и HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60. В следующем варианте осуществления изобретения антитело содержит (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60; и (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64.
Другим объектом изобретения является антитело, которое содержит по меньшей мере одну, по меньшей мере две или все три HVR-последовательности VH, выбранные из (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 114-115; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 116-120; и (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 121. В одном из вариантов осуществления изобретения антитело содержит HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 121. В другом варианте осуществления изобретения антитело содержит HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 121, и HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74. В следующем варианте осуществления изобретения антитело содержит HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 121, HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74, и HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 116-120. В следующем варианте осуществления изобретения антитело содержит (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 114-115; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 116-120; и (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 121.
Другим объектом изобретения является антитело, которое содержит по меньшей мере одну, по меньшей мере две или все три HVR-последовательности VH, выбранные из (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 114; (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58; и (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63. В одном из вариантов осуществления изобретения антитело содержит HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63. В другом варианте осуществления изобретения антитело содержит HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63, и HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74. В следующем варианте осуществления изобретения антитело содержит HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63, HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74, и HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58. В другом варианте осуществления изобретения антитело содержит (a) HVR-Н1, который содержит аминокислотную последовательность SEQ ID NO: 114; (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58; и (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63.
Другим объектом изобретения является антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 126; (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 127; и (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 128. В одном из вариантов осуществления изобретения антитело содержит HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 128. В другом варианте осуществления изобретения антитело содержит HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 128, и HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 131. В следующем варианте осуществления изобретения антитело содержит HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 128, HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 131, и HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 127. В другом варианте осуществления изобретения антитело содержит (a) HVR-Н1, который содержит аминокислотную последовательность SEQ ID NO: 126; (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 127; и (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 128.
Другим объектом изобретения является антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69,122-124, 129; (б) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72, 125, 130; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74, 131. В одном из вариантов осуществления изобретения антитело содержит (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69,122-124, 129; (б) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72, 125, 130; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74, 131.
Другим объектом изобретения является антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69; (б) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74. В одном из вариантов осуществления изобретения антитело содержит (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69; (б) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74.
Другим объектом изобретения является антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 122-124; (б) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 125; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74. В одном из вариантов осуществления изобретения антитело содержит (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 122-124; (б) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 125; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74.
Другим объектом изобретения является антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 122; (б) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71; и (в) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74. В одном из вариантов осуществления изобретения антитело содержит (a) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 122; (б) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71; и (в) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74. Другим объектом изобретения является антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 123; (б) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71; и (в) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74. В одном из вариантов осуществления изобретения антитело содержит (a) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 123; (б) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71; и (в) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74.
Другим объектом изобретения является антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 129; (б) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 130; и (в) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 131. В одном из вариантов осуществления изобретения антитело содержит (a) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 129; (б) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 130; и (в) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO 131.
Согласно другому объекту изобретения антитело, предлагаемое в изобретении, содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (I) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57, 114, 115, 126, (II) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, 116-120, 127, и (III) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, 121, 128; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (I) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69, 122-124, 129, (II) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72, 125, 130, и (III) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74, 131.
Согласно другому объекту изобретения антитело, предлагаемое в изобретении, содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (I) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57, (II) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, и (III) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (I) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69, (II) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72, и (III) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74.
Согласно другому объекту изобретения антитело, предлагаемое в изобретении, содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (I) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 114-115, (II) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 116-120, и (III) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 121; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (I) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 122-124, (II) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 125, и (III) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74.
Согласно другому объекту изобретения антитело, предлагаемое в изобретении, содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (I) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 114, (II) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58, и (III) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (I) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 122, (II) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71, и (III) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74. Согласно другому объекту изобретения антитело, предлагаемое в изобретении, содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (I) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 114, (II) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58, и (III) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (I) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 123, (II) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71, и (III) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74.
Согласно другому объекту изобретения антитело, предлагаемое в изобретении, содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (I) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 126, (II) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 127, и (III) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 128; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (I) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 129, (II) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 1301, и (III) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 131.
Другим объектом изобретения является антитело, содержащее (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57, 114-115, 126; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, 116-120, 127; (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, 121, 128; (г) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69, 122-124, 129; (д) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72, 125, 130; и (е) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74, 131.
Другим объектом изобретения является антитело, содержащее (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57, 114-115; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, 116-120; (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, 121; (г) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69, 122-124; (д) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72, 125; и (е) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74.
Другим объектом изобретения является антитело, содержащее (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 114-115; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 116-120; (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 121; (г) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 122-124; (д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 125; и (е) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74.
Другим объектом изобретения является антитело, содержащее (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 114; (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58; (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63; (г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 122; (д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71; и (е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74. Другим объектом изобретения является антитело, содержащее (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 114; (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58; (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63; (г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 123; (д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71; и (e) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74.
Другим объектом изобретения является антитело, содержащее (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 126; (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 127; (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 128; (г) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 129; (д) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 130; и (е) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 131.
В некоторых вариантах осуществления изобретения любая одна или несколько аминокислот в антитело к миостатину, представленном в настоящем описании, заменена в следующих положениях HV: (а) в HVR-H1 (SEQ ID NO: 55) в положениях 1 и 2; (б) в HVR-H2 (SEQ ID NO: 58) в положениях 4, 7, 8, 10, 11, 12 и 16; (в) в HVR-H3 (SEQ ID NO: 61) в положениях 5, 7 и 11; (г) в HVR-L1 (SEQ ID NO: 65) в положениях 1, 2, 5, 7, 8 и 9; (д) в HVR-L2 (SEQ ID NO:70) в положениях 3 и 7; и (е) в HVR-L3 (SEQ ID NO:73) в положении 8.
В некоторых вариантах осуществления изобретения одна или несколько аминокислотная(ых) замена(н) в антителе к миостатину представляет(ют) собой консервативную(ые) замену(ы), представленные в настоящем описании. В некоторых вариантах осуществления изобретения может быть сделана одна или несколько и следующих аминокислотных замен в любой комбинации: (а) в HVR-Н1 (SEQ ID NO: 55) S1H; Y2T, D или E; (б) в HVR-H2 (SEQ ID NO: 58) Y4H; S7K; T8M или K; Y10K; A11M или E; S12E; G16K; (в) в HVR-H3 (SEQ ID NO: 61) Y5H; T7H; L11K; (г) в HVR-L1 (SEQ ID NO: 65) Q1T; S2T; S5E; Y7F; D8H; N9D или А, или E; (д) в HVR-L2 (SEQ ID NO: 70) S3E; S7Y, F или W; и (e) в HVR-L3 (SEQ ID NO: 73) L8R.
Все возможные комбинации указанных выше замен входят в консенсусные последовательности SEQ ID NO: 126, 127, 128, 129, 130 и 131 HVR-H1, HVR-H2, HVR-H3, HVR-L1, HVR-L2 и HVR-L3 соответственно.
В любом из указанных выше вариантов осуществления изобретения антитело к миостатину может быть гуманизированным. В одном из вариантов осуществления изобретения антитело к миостатину содержит HVR, указанные в любом из приведенных выше вариантов осуществления изобретения, и дополнительно содержит акцепторный человеческий каркасный участок, например, каркасный участок человеческого иммуноглобулина или человеческий консенсусный каркасный участок. В другом варианте осуществления изобретения антитело к миостатину содержит HVR, указанные в любом из приведенных выше вариантов осуществления изобретения, и дополнительно содержит VH или VL, содержащие последовательность FR. В следующем варианте осуществления изобретения антитело к миостатину содержит следующие последовательности FR вариабельного домена тяжелой цепи и/или легкой цепи: в вариабельном домене тяжелой цепи FR1 содержит одну из аминокислотных последовательностей SEQ ID NO: 132-134, FR2 содержит одну из аминокислотных последовательностей SEQ ID NO: 135-136, FR3 содержит одну из аминокислотных последовательностей SEQ ID NO: 137, FR4 содержит одну из аминокислотных последовательностей SEQ ID NO: 138. В вариабельном домене легкой цепи FR1 содержит аминокислотную последовательность SEQ ID NO: 139, FR2 содержит одну из аминокислотных последовательностей SEQ ID NO: 140-141, FR3 содержит одну из аминокислотных последовательностей SEQ ID NO: 142-143, FR4 содержит аминокислотную последовательность SEQ ID NO: 144.
Одним из объектов изобретения является антитело к миостатину, содержащее по меньшей мере один, два, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 157-162; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 163-168; (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 169-174; (г) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 175-180; (д) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 181-186; и (е) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 187-192.
Одним из объектов изобретения является антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 157-162; (6) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 163-168; и (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 169-174. В одном из вариантов осуществления изобретения антитело содержит HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 169-174. В другом варианте осуществления изобретения антитело содержит HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 169-174, и HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 187-192. В другом варианте осуществления изобретения антитело содержит HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 169-174, HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 187-192, и HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 163-168. В следующем варианте осуществления изобретения антитело содержит (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 157-162; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 163-168; и (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 169-174.
Другим объектом изобретения является антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 175-180; (б) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 181-186; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 187-192. В одном из вариантов осуществления изобретения антитело содержит (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 175-180; (б) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 181-186; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 187-192.
Другим объектом изобретения является антитело, предлагаемое в изобретении, которое содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH HVR, выбранные из (I) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 157-162, (II) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 163-168, и (III) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 169-174; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL HVR, выбранные из (I) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 175-180, (II) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 181-186, и (III) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 187-192.
Другим объектом изобретения является антитело, содержащее (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 157-162; (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 163-168; (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 169-174; (г) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 175-180; (д) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 181-186; и (е) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 187-192.
В другом объекте изобретения антитело к миостатину содержит последовательность вариабельного домена тяжелой цепи (VH), которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична одной из аминокислотной последовательностей SEQ ID NO: 13, 16-30, 32-34 и 86-95. В некоторых вариантах осуществления изобретения последовательность VH, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в одной из SEQ ID NO: 13, 16-30 и 32-34. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30 и 32-34, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три HVR, выбранных из: (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57, (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, и (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64.
В другом объекте изобретения антитело к миостатину содержит последовательность вариабельного домена тяжелой цепи (VH), которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична одной из аминокислотной последовательностей SEQ ID NO: 13, 16-30, 32-34 и 86-95. В некоторых вариантах осуществления изобретения последовательность VH, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три HVR, выбранных из: (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57, 114-115, 126, (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, 116-120, 127, и (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64, 121, 128. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к миостатину содержит последовательность VH, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична одной из аминокислотной последовательностей SEQ ID NO: 13, 16-30, 32, 33 и 34. В некоторых вариантах осуществления изобретения последовательность VH, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в одной из SEQ ID NO: 13, 16-30, 32, 33 и 34. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VH, представленную в одной из SEQ ID NO: 13, 16-30, 32, 33 и 34, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три HVR, выбранных из: (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55-57, (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58-60, и (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61-64. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к миостатину содержит последовательность VH, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична одной из аминокислотной последовательностей SEQ ID NO: 86-95. В некоторых вариантах осуществления изобретения последовательность VH, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в одной из SEQ ID NO: 86-95. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VH, представленную в одной из SEQ ID NO: 86-95, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три HVR, выбранных из: (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 57, 114-115, 126, (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58, 116-120, 127, и (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 63, 121, 128. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к миостатину содержит последовательность VH, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 86. В некоторых вариантах осуществления изобретения последовательность VH, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в SEQ ID NO: 86. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VH, представленную в SEQ ID NO: 86, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три HVR, выбранных из: (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 114, (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58, и (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к миостатину содержит последовательность VH, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 92. В некоторых вариантах осуществления изобретения последовательность VH, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в SEQ ID NO: 92. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VH, представленную в SEQ ID NO: 92, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три HVR, выбранных из: (a) HVR-H1, который содержит аминокислотную последовательность SEQ ID NO: 114, (б) HVR-H2, который содержит аминокислотную последовательность SEQ ID NO: 58, и (в) HVR-H3, который содержит аминокислотную последовательность SEQ ID NO: 63. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к миостатину содержит последовательность VL, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична одной из аминокислотных последовательностей SEQ ID NO: 15, 31, 35-38 и 96-99. В некоторых вариантах осуществления изобретения последовательность VL, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в одной из SEQ ID NO: 15, 31, 35-38 и 96-99. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VL, представленную в одной из SEQ ID NO: 15, 31, 35-38 и 96-99, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VL содержит один, два или три HVR, выбранных из: (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69, 122-124, 129; (б) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72, 125, 130; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74, 131. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к миостатину содержит последовательность VL, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична одной из аминокислотных последовательностей SEQ ID NO: 15, 31, 35, 36, 37 и 38. В некоторых вариантах осуществления изобретения последовательность VL, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в одной из SEQ ID NO: 15, 31, 35, 36, 37 и 38. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VL, представленную в одной из SEQ ID NO: 15, 31, 35, 36, 37 и 38, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VL содержит один, два или три HVR, выбранных из: (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65-69; (б) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70-72; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73-74. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к миостатину содержит последовательность VL, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична одной из аминокислотных последовательностей SEQ ID NO: 96-99. В некоторых вариантах осуществления изобретения последовательность VL, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в одной из SEQ ID NO: 96-99. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VL, представленную в одной из SEQ ID NO: 96-99, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VL содержит один, два или три HVR, выбранных из: (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 122-124, 129; (б) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 71, 125, 130; и (в) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74, 131. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к миостатину содержит последовательность VL, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 96. В некоторых вариантах осуществления изобретения последовательность VL, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в SEQ ID NO: 96. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VL, представленную в SEQ ID NO: 96, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VL содержит один, два или три HVR, выбранных из: (a) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 122; (б) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71; и (в) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к миостатину содержит последовательность VL, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 97. В некоторых вариантах осуществления изобретения последовательность VL, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в SEQ ID NO: 97. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VL, представленную в SEQ ID NO: 97, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VL содержит один, два или три HVR, выбранных из: (a) HVR-L1, который содержит аминокислотную последовательность SEQ ID NO: 123; (б) HVR-L2, который содержит аминокислотную последовательность SEQ ID NO: 71; и (в) HVR-L3, который содержит аминокислотную последовательность SEQ ID NO: 74. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
Другим объектом изобретения является антитело к миостатину, где антитело содержит VH, представленную в одном из указанных выше вариантов осуществления изобретения, и VL, представленную в одном из указанных выше вариантов осуществления изобретения. В одном из вариантов осуществления изобретения антитело содержит последовательности VH и VL, представленные в одной из SEQ ID NO: 13, 16-30, 32-34 и 86-95 и в одной из SEQ ID NO: 15, 31, 35-38 и 96-99 соответственно, включая пост-трансляционные модификации этих последовательностей. В одном из вариантов осуществления изобретения антитело содержит последовательности VH и VL, указанные в одной из SEQ ID NO: 13, 16-30 и 32-34 и в одной из SEQ ID NO: 15, 31 и 35-38 соответственно, включая пост-трансляционные модификации этих последовательностей. В одном из вариантов осуществления изобретения антитело содержит последовательности VH и VL, указанные в одной из SEQ ID NO: 86-95 и одной из SEQ ID NO: 96-99 соответственно, включая пост-трансляционные модификации этих последовательностей. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
Другим объектом изобретения является антитело к миостатину, где антитело содержит VH, представленную в любом из указанных выше вариантов осуществления изобретения, и VL, представленную в любом из указанных выше вариантов осуществления изобретения. В одном из вариантов осуществления изобретения антитело содержит последовательности VH и VL SEQ ID NO: 86 и SEQ ID NO: 96 соответственно, включая пост-трансляционные модификации этих последовательностей. В одном из вариантов осуществления изобретения антитело содержит последовательности VH и VL SEQ ID NO: 92 и SEQ ID NO: 97 соответственно, включая пост-трансляционные модификации этих последовательностей. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
Другим объектом изобретения является антитело к миостатину, содержащее последовательность VH, идентичную по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% одной из аминокислотных последовательностей SEQ ID NO: 12, 145-150. В некоторых вариантах осуществления изобретения последовательность VH, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в одной из SEQ ID NO: 12, 145-150. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VH, представленную в одной из SEQ ID NO: 12, 145-150, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три HVR, выбранных из: (a) HVR-H1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 55, 157-162, (б) HVR-H2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 58, 163-168, и (в) HVR-H3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 61, 169-174. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В другом объекте изобретения антитело к миостатину содержит последовательность VL, которая по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична одной из аминокислотных последовательностей SEQ ID NO: 14, 151-156. В некоторых вариантах осуществления изобретения последовательность VL, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело к миостатину, содержащее указанную последовательность, сохраняет способность связываться с миостатином. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменено, встроено и/или удалено путем делеции в одной из SEQ ID NO: 14, 151-156. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антитело к миостатину содержит последовательность VL, представленную в одной из SEQ ID NO: 14, 151-156, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VL содержит один, два или три HVR, выбранных из: (a) HVR-L1, который содержит одну из аминокислотных последовательностей SEQ ID NO: 65, 175-180; (б) HVR-L2, который содержит одну из аминокислотных последовательностей SEQ ID NO: 70, 181-186; и (в) HVR-L3, который содержит одну из аминокислотных последовательностей SEQ ID NO: 73, 187-192. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
Другим объектом изобретения является антитело к миостатину, где антитело содержит VH, представленную в любом из указанных выше вариантов осуществления изобретения, и VL, представленную в любом из указанных выше вариантов осуществления изобретения. В одном из вариантов осуществления изобретения антитело содержит последовательности VH и VL, указанные в одной из SEQ ID NO: 12, 145-150 и в одной из SEQ ID NO: 14, 151-156, соответственно, включая пост-трансляционные модификации этих последовательностей. Пост-трансляционные модификации включают (но, не ограничиваясь только ими) модификацию путем пироглутамилирования глутамина или глутамата в N-концевой области тяжелой цепи или легкой цепи в пироглутамовую кислоту.
В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, содержит VH, представленную в любом из указанных выше вариантов осуществления изобретения, и константную область тяжелой цепи, содержащую одну из аминокислотных последовательностей SEQ ID NO: 7, 9, 11, 193, 195-198, 227, 228, 229-381. В некоторых вариантах осуществления изобретения антитело к миостатину, предлагаемое в настоящем изобретении, содержит VL, представленную в любом из указанных выше вариантов осуществления изобретения, и константную область легкой цепи, содержащую одну из аминокислотных последовательностей SEQ ID NO: 8 и 10.
Следующим объектом изобретения является антитело, которое связывается с тем же эпитопом, что и антитело к миостатину, представленное в настоящем описании. Следующим объектом изобретения является антитело, которое связывается с тем же эпитопом, что и антитело, указанное в таблице 2а. Следующим объектом изобретения является антитело, которое связывается с тем же эпитопом, что и антитело, указанное в таблицах 11а или 13. Некоторыми вариантами осуществления изобретения является антитело, которое связывается с эпитопом внутри фрагмента пропептида миостатина, состоящего из аминокислот 21-100 SEQ ID NO: 78. Альтернативно этому, антитело связывается с фрагментом пропептида миостатина, который состоит из аминокислот 21-80, 41-100, 21-60, 41-80, 61-100, 21-40, 41-60, 61-80 или 81-100 SEQ ID NO: 78.
В другом варианте осуществления изобретения антитело к миостатину, указанное в любом из приведенных выше вариантов осуществления изобретения, представляет собой моноклональное антитело, включая химерное, гуманизированное или человеческое антитело. В одном из вариантов осуществления изобретения антитело к миостатину представляет собой фрагмент антитела, например, Fv, Fab, Fab', димерное антитело или F(ab')2-фрагмент. В другом варианте осуществления изобретения антитело представляет собой полноразмерное антитело, например, интактное антитело изотипа IgG1 или IgG4 или другого класса или изотипа антител, указанных в настоящем описании.
В дополнительном объекте изобретения антитело к миостатину, указанное в любом из приведенных выше вариантов осуществления изобретения, может обладать любыми особенностями, индивидуально или в комбинации, описанными ниже в разделах 1-7:
1. Аффинность антител
В некоторых вариантах осуществления изобретения антитело, представленное в настоящем описании, имеет величину константы диссоциации (Kd), составляющую 1 мкМ или менее, 100 нМ или менее, 10 нМ или менее, 1 нМ или менее, 0,1 нМ или менее, 0,01 нМ или менее, или 0,001 нМ или менее (например 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М).
В одном из вариантов осуществления величину Kd измеряют с помощью анализа связывания несущего радиоактивную метку антигена (РИА). В одном из вариантов осуществления изобретения РИА осуществляют с использованием Fab-версии представляющего интерес антитела и его антигена. Например, измеряют в растворе аффинность связывания Fab с антигеном путем уравновешивания Fab минимальной концентрацией 125I-меченного антигена в присутствии полученных титрованием разведений немеченого антигена, осуществляя затем «захват» связанного антигена на сенсибилизированном антителом к Fab планшете (см., например, Chen и др., J. Mol Biol 293, 1999, сс. 865-881). Для обеспечения условий, необходимых для проведения анализа, многолуночные планшеты MICROTITER® (фирма Thermo Scientific) сенсибилизируют в течение ночи захватывающим антителом к Fab (фирма Cappel Labs) в концентрации 5 мкг/мл в 50 мМ карбонате натрия (рН 9,6) и затем блокируют с помощью 2% (мае./об) бычьего сывороточного альбумина в ЗФР в течение 2-5 ч при комнатной температуре (примерно 23°С). В неадсорбирующем планшете (фирма Nunc, №269620) смешивают взятый в концентрации 100 или 26пМ меченный с помощью 125I антиген с серийными разведениями представляющего интерес Fab (например, пригодного для оценки антитела к VEGF, Fab-12, см. Presta и др., Cancer Res. 57, 1997, сс. 4593-4599). Затем представляющий интерес Fab инкубируют в течение ночи; однако инкубацию можно осуществлять в течение более продолжительного периода времени (например, в течение 65 ч) для того, чтобы гарантировать достижение равновесия. После этого смеси переносят в подготовленный для захвата планшет и инкубируют при комнатной температуре (например, в течение 1 ч). Затем раствор удаляют и планшет промывают восемь раз 0,1% полисорбатом 20 (TWEEN-20®) в ЗФР. После просушки планшетов в них добавляют по 150 мкл/лунку сцинтилляционной жидкости ((MICROSCINT-20TM; фирма Packard) и осуществляют считывание планшетов с помощью гамма-счетчика TOPCOUNT TM (фирма Packard) в течение 10 мин. Для осуществления анализа связывания в условиях конкуренции выбирают концентрации каждого Fab, обеспечивающие уровень связывания, составляющий менее или равный 20% от максимального.
В другом варианте осуществления изобретения Kd измеряют методом поверхностного плазмонного резонанса с помощью устройства BIACORE®. Например, анализ осуществляют с помощью устройства BIACORE®-2000 или BIACORE®-3000 (фирма GE Healthcare Inc., Пискатавей, шт. Нью-Джерси) при 25°С с использованием антигена, иммобилизованного на СМ5-чипах с уровнем иммобилизации, соответствующим ~10 единицам ответа (RU). В одном из вариантов осуществления изобретения биосенсорные чипы из карбоксиметилированного декстрана (СМ5, фирма GE Healthcare Inc.) активируют с помощью гидрохлорида N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимида (NHS) согласно инструкциям поставщика. Антиген разводят 10 мМ ацетатом натрия, рН 4,8 до концентрации 5 мкг/мл (~0,2 мкМ) перед инъекцией со скоростью потока 5 мкл/мин для достижения уровня иммобилизации, соответствующего примерно 10 единицам ответа (RU) сшитого белка. После инъекции антигена инъецируют 1 М этаноламин для блокады непрореагировавших групп. Для кинетических измерений инъецируют двукратные серийные разведения Fab (от 0,78 до 500 нМ) в ЗФР, дополненном 0,05% полисорбата 20 (TWEEN-20TM) в качестве сурфактанта (ЗФРТ), при 25°С со скоростью потока примерно 25 мкл/мин. Скорость реакции ассоциации (kon) и реакции диссоциации (koff) рассчитывают с использованием простой модели связывания Ленгмюра 1:1 (программное обеспечение Evaluation версия 3.2, BIACORE®) путем одновременной аппроксимации сенсограмм ассоциации и диссоциации. Константу равновесия реакции диссоциации (Kd) рассчитывают как соотношение koff/kon (см., например, Chen и др., J Mol Biol 293, 1999, сс. 865-881). Если скорость реакции ассоциации превышает 106 М-1 с-1 при оценке с помощью описанного выше метода поверхностного плазмонного резонанса, то скорость реакции ассоциации можно определять с помощью метода гашения флуоресценции, который позволяет измерять повышение или снижение интенсивности излучения флуоресценции (длина волны возбуждения 295 нм; длина волны испускания 340 нм, полоса пропускания 16 нм) при 25°С применяемого в концентрации 20 нМ антитела к антигену (Fab-формат) в ЗФР, рН 7,2, в присутствии возрастающих концентраций антигена, осуществляя измерения с помощью спектрометра, такого как спектрофотометр, снабженный блоком для остановки потока (фирма Aviv Instruments), или спектрофотометр SLM-AMINCOTM серии 8000 (фирма ThermoSpectronic), при перемешивании содержимого кюветы.
2. Фрагменты антител
В некоторых вариантах осуществления изобретения антитело, предлагаемое в настоящем изобретении, представляет собой фрагмент антитела. Фрагменты антител включают (но, не ограничиваясь только ими) такие фрагменты как Fab, Fab', Fab'-SH, F(ab')2, Fv и scFv и другие описанные ниже фрагменты. Обзор некоторых фрагментов антител см., у Hudson и др., Nat. Med. 9, 2003, сс. 129-134. Обзор scFv-фрагментов см., например, у Plueckthun в: The Pharmacology of Monoclonal Antibodies, под ред. Rosenburg и Moore, изд-во Springer, New York, т. 113, 1994, сс. 269-315; см. также WO 93/16185; и US №№5571894 и 5587458. Обсуждение, касающееся Fab- и F(ab')2-фрагментов, которые содержат остатки эпитопа, связывающегося с рецептором спасения, и обладают удлиненным временем полужизни in vivo, см. в US №5869046.
Димерные антитела представляют собой фрагменты антител с двумя антигенсвязывающими центрами, которые могут быть двухвалентными или биспецифическими (см., например, ЕР 0404097; WO 1993/01161; Hudson и др., Nat. Med. 9, 2003, сс. 129-134; и Holliger и др., Proc. Natl. Acad. Sci. USA 90, 1993, сс. 6444-6448). Тримерные и тетрамерные антитела описаны также у Hudson и др., Nat. Med. 9, 2003, сс. 129-134).
Однодоменные антитела представляют собой фрагменты антител, содержащие весь вариабельный домен тяжелой цепи или его часть, или весь вариабельный домен легкой цепи антитела или его часть. В некоторых вариантах осуществления изобретения однодоменное антитело представляет собой человеческое однодоменное антитело (фирма Domantis, Inc., Уолтхэм, шт. Массачусетс; см., например, US №6248516 В1).
Фрагменты антител можно создавать различными методиками, включая (но, не ограничиваясь только ими) протеолитическое расщепление интактного антитела, а также получать с помощью рекомбинантных клеток-хозяев (например, Е. coli или фаг), указанных в настоящем описании.
3. Химерные и гуманизированные антитела
В некоторых вариантах осуществления изобретения антитело, указанное в настоящем описании, представляет собой химерное антитело. Некоторые химерные антитела описаны, например, в US №4816567; и у Morrison и др., Proc. Natl. Acad. Sci. USA 81, 1984, сс. 6851-6855. В одном из примеров химерное антитело содержит нечеловеческую вариабельную область (например, вариабельную область из антитела мышей, крыс, хомяков, кроликов или приматов кроме человека, таких как обезьяны) и человеческую константную область. В другом примере химерное антитело представляет собой антитело «переключенного класса», класс или подкласс которого был изменен относительно родительского антитела. Химерные антитела включают их антигенсвязывающие фрагменты.
В некоторых вариантах осуществления изобретения химерное антитело представляет собой гуманизированное антитело. Как правило, нечеловеческое антитело гуманизируют для снижения иммуногенности для людей, сохраняя при этом специфичность и аффинность родительского нечеловеческого антитела. Как правило, гуманизированное антитело содержит один или несколько вариабельных доменов, в которых HVR, например, CDR (или их участки), получают из нечеловеческого антитела, a FR (или их участки) получают из последовательностей человеческого антитела. Гуманизированное антитело необязательно может содержать по меньшей мере часть человеческой константной области. В некоторых вариантах осуществления изобретения некоторые остатки FR в гуманизированном антителе заменены на соответствующие остатки из нечеловеческого антитела (например, антитела, из которого происходят остатки HVR), например, для сохранения или улучшения специфичности или аффинности антитела.
Сведения о гуманизированных антителах и методах их создания обобщены, например, у Almagro, Front. Biosci. 13, 2008, сс. 1619-1633, и описаны также у Riechmann и др., Nature 332, 1988, сс. 323-329; Queen и др., Proc. Natl. Acad. Sci. USA 86, 1989, сс. 10029-10033; US №№5821337, 7527791, 6982321 и 7087409; Kashmiri и др., Methods 36, 2005, сс. 25-34 (описание трансплантации SDR (а-CDR)); Padlan, Mol. Immunol. 28, 1991, сс. 489-498 (описание «нанесения нового покрытия»);  и др., Methods 36, 2005, сс. 43-60 (описание «перестановки FR»); и Osbourn и др., Methods 36, 2005, сс. 61-68 и Klimka и др., Br. J. Cancer 83, 2000, сс. 252-260 (описание подхода «направленной селекции» для перестановки FR).
и др., Methods 36, 2005, сс. 43-60 (описание «перестановки FR»); и Osbourn и др., Methods 36, 2005, сс. 61-68 и Klimka и др., Br. J. Cancer 83, 2000, сс. 252-260 (описание подхода «направленной селекции» для перестановки FR).
Человеческие каркасные участки, которые можно применять для гуманизации, включают (но, не ограничиваясь только ими): каркасные участки, выбранные с использованием методов «наилучшего подбора» (см., например, Sims и др., J. Immunol. 151, 1993, сс. 2296-2308; каркасные участки, происходящие из консенсусной последовательности конкретной подгруппы вариабельных областей легких и тяжелых цепей человеческих антител (см., например, Carter и др., Proc. Natl. Acad. Sci. USA 89, 1992, сс. 4285-4289 и Presta и др., J. Immunol. 151, 1993, сс. 2623-2632); человеческие зрелые (с соматическими мутациями) каркасные области или каркасные области человеческой зародышевой линии (см., например, Almagro и Fransson, Front. Biosci. 13, 2008, с.1619-1633); и каркасные участки, полученные в результате скрининга FR-библиотек (см., например, Васа и др., J. Biol. Chem. 272, 1997, сс. 10678-10684 и Rosok и др., J. Biol. Chem. 271, 1996, сс. 22611-22618).
4. Человеческие антитела
В некоторых вариантах осуществления изобретения антитело, предлагаемое в настоящем изобретении, представляет собой человеческое антитело. Человеческие антитела можно получать, используя различные методики, известные в данной области. Человеческие антитела описаны в целом у van Dijk и van de Winkel, Curr. Opin. Pharmacol. 5, 2001, cc. 368-374 и Lonberg, Curr. Opin. Immunol. 20, 2008, cc. 450-459.
Человеческие антитела можно получать путем введения иммуногена трансгенному животному, модифицированному таким образом, чтобы оно продуцировало интактные человеческие антитела или интактные антитела с человеческими вариабельными областями в ответ на контрольное заражение антигеном. Указанные животные, как правило, содержат все или часть локусов человеческого иммуноглобулина вместо эндогенных иммуноглобулиновых локусов, которые либо присутствуют вне хромосом, либо интегрированы произвольно в хромосомы животного. У таких трансгенных мышей эндогенные иммуноглобулиновые локусы, как правило, инактивированы. Обзор методов получения человеческих антител в трансгенных животных см. у Lonberg, Nat. Biotech. 23, 2005, сс. 1117-1125 (см., например, также в US №6075181 и US №6150584 описание технологии XENOMOUSETM; в US №5770429 описание технологии HUMAB®; в US №7041870 описание технологии K-М MOUSE® и в публикации заявки на патент США 2007/0061900 описание технологии VELOCIMOUSE®). Человеческие вариабельные области из интактных антител, полученных с помощью указанных животных, можно дополнительно модифицировать, например, путем объединения с другой человеческой константной областью.
Человеческие антитела можно создавать с помощью методов на основе гибридом. Описаны клеточные линии человеческой миеломы и мышиной-человеческой гетеромиеломы, предназначенные для получения человеческих моноклональных антител (см., например, Kozbor, J. Immunol. 133, 1984, сс. 3001-3005; Brodeur и др., Monoclonal Antibody Production Techniques and Applications, изд-во Marcel Dekker, Inc., New York, 1987, cc. 51-63 и Boerner и др., J. Immunol. 147, 1991, сс. 86-95). Человеческие антитела, созданные с использованием технологии на основе человеческих В-клеточных гибридом, описаны также у Li и др., Proc. Natl. Acad. Sci. USA 103, 2006, сс. 3557-3562. Дополнительные методы включают методы, описанные, например, в US №7189826 (описание получения моноклональных человеческих антител типа IgM из клеточных линий гибридом), и у Ni, Xiandai Mianyixue 26, 2006, сс. 265-268 (описание человеческих-человеческих гибридом). Технология человеческих гибридом (технология Trioma) описана также у Vollmers и Brandlein, Histology and Histopathology 20, 2005, сс. 927-937 и Vollmers и Brandlein, Methods and Findings in Experimental and Clinical Pharmacology 27, 2005, cc. 185-191.
Человеческие антитела можно создавать также путем выделения последовательностей вариабельных доменов Fv-клонов, выбранных из фаговых дисплейных библиотек, для создания которых использовали человеческие антитела. Указанные последовательности вариабельных доменов затем можно объединять с требуемым человеческим константным доменом. Методики отбора человеческих антител из библиотек антител описаны ниже.
5. Антитела, полученные из библиотек
Антитела, предлагаемые в изобретении, можно выделять путем скрининга комбинаторных библиотек антител с требуемой активностью или видами активности. Например, в данной области известны разнообразные методы для создания фаговых дисплейных библиотек и скрининга указанных библиотек в отношении антител, обладающих требуемыми характеристиками связывания. Указанные методы обобщены, например, у Hoogenboom и др., Methods in Molecular Biology 178, 2001, сс. 1-37, и описаны также, например, у McCafferty и др., Nature 348, 1990, сс. 552-554; Clackson и др., Nature 352, 1991, сс. 624-628; Marks и др., J. Mol. Biol. 222, 1992, сс. 581-597; Marks и Bradbury, Methods in Molecular Biology 248, 2003, cc. 161-175; Sidhu и др., J. Mol. Biol. 338, 2004, cc. 299-310; Lee и др., J. Mol. Biol. 340, 2004, cc. 1073-1093; Fellouse, Proc. Natl. Acad. Sci. USA 101, 2004, cc. 12467-12472 и Lee и др., J. Immunol. Methods 284, 2004, cc. 119-132.
При осуществлении некоторых методов фагового дисплея популяции VH- и VL-генов клонируют по отдельности с помощью полимеразной цепной реакции (ПЦР) и рекомбинируют произвольно в фаговых библиотеках, которые затем подвергают скринингу в отношении антигенсвязывающего фага согласно методу, описанному у Winter и др., Ann. Rev. Immunol. 12, 1994, сс. 433-455. Фаг, как правило, экспонирует фрагменты антител либо в виде одноцепочечных Fv-фрагментов (scFv), либо в виде Fab-фрагментов. Библиотеки, полученные из иммунизированных источников, включают антитела, обладающие высокой аффинностью к иммуногену, при этом отсутствует необходимость в создании гибридом. Альтернативно этому, можно клонировать необработанную («наивную») популяцию (например, из организма человека), получая один источник антител к широкому спектру чужих, а также своих антигенов без какой-либо иммунизации, что описано у Griffiths и др., EMBO J, 12, 1993, сс. 725-734. И, наконец, «наивные» библиотеки можно получать также методами синтеза путем клонирования неперегруппированных сегментов V-гена из стволовых клеток и с помощью ПЦР-праймеров, содержащих случайную последовательность, для кодирования гипервариабельных CDR3-участков и для осуществления перегруппировки in vitro согласно методу, описанному у Hoogenboom и Winter, J. Mol. Biol. 227, 1992, сс. 381-388. Фаговые библиотеки человеческих антител описаны, например, в следующих патентных публикациях: US №5750373 и публикациях заявок на патент США 2005/0079574, 2005/0119455, 2005/0266000, 2007/0117126, 2007/0160598, 2007/0237764, 2007/0292936 и 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек человеческих антител, рассматриваются как человеческие антитела или фрагменты человеческих антител.
6. Мультиспецифические антитела
В некоторых вариантах осуществления изобретения антитело, указанное в настоящем описании, представляет собой мультиспецифическое антитело, например, биспецифическое антитело. Мультиспецифические антитела представляют собой моноклональные антитела, которые имеют специфичности, связывающиеся по меньше мере с двумя различными сайтами. В некоторых вариантах осуществления изобретения одна из связывающих специфичностей обеспечивает связывание с миостатином, а другая с любым другим антигеном. В некоторых вариантах осуществления изобретения биспецифические антитела могут связываться с двумя различными эпитопами миостатина. Биспецифические антитела можно применять также для локализации цитотоксических агентов в клетках, которые экспрессируют миостатин. Биспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител.
Методики создания мультиспецифических антител включают (но, не ограничиваясь только ими) рекомбинантную совместную экспрессию двух пар тяжелых цепей-легких цепей иммуноглобулинов, обладающих различными специфичностями (см. Milstein и Cuello, Nature 305, 1983, сс. 537-540, WO 93/08829 и Traunecker и др., EMBO J. 10,1991, сс. 3655-3659), и технологию «knob-in-hole» (обеспечение взаимодействия по типу «выступ-во впадину», см., например, US №5731168). Мультиспецифические антитела можно получать также путем создания регулируемых электростатических воздействий для получения молекул антитела с гетеродимерными Fc (WO 2009/089004); перекрестного связывания двух или большего количества антител или фрагментов (см., например, US №4676980 и Brennan и др., Science 229, 1985, сс. 81-83); применения лейциновых молний для получения биспецифических антител (см., например, Kostelny и др., J. Immunol. 148, 1992, сс. 1547-1553); применения технологии «димерных антител (диабоди)» для создания фрагментов биспецифических антител (см., например, Holliger и др., Proc. Natl. Acad. Sci. USA 90, 1993, сс. 6444-6448); и применения димеров одноцепочечных Fv (sFv) (см., например, Gruber и др., J. Immunol. 152, 1994, сс. 5368-5374); и получения триспецифических антител согласно методу, описанному, например, у Tutt и др., J. Immunol. 147, 1991, сс. 60-69).
Под объем настоящего изобретения подпадают также сконструированные антитела с тремя или большим количеством функциональных антигенсвязывающих сайтов, включая «антитела-осьминоги» (см., например, US 2006/0025576 А1).
Антитело или его фрагмент может включать также «Fab двойного действия» или «DAF», содержащий антигенсвязывающий сайт, который связывается с миостатином, а также с другим, отличным от него антигеном (см., например, US 2008/0069820).
7. Варианты антител
Некоторые варианты осуществления изобретения относятся к вариантам аминокислотных последовательностей антител, представленных в настоящем описании. Например, может требоваться повышать аффинность связывания и/или другие биологические свойства антитела. Варианты аминокислотной последовательности антитела можно получать путем интродукции соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или путем синтеза пептидов. Указанные модификации включают, например, делеции и/или инсерции, и/или замены остатков в аминокислотных последовательностях антитела. Для получения конечной конструкции можно использовать любую комбинацию делеции, инсерции и замен, при условии, что конечная конструкция обладает требуемыми характеристиками, например, способностью связываться с антигеном.
а) Полученные путем замены, инсерции и делеции варианты
Некоторыми вариантами осуществления изобретения являются варианты антител, которые имеют одну или несколько аминокислотных замен. Сайты, представляющие интерес для замещающего мутагенеза, включают HVR и FR. Консервативные замены представлены ниже в таблице 1 под заголовком «предпочтительные замены», и они описаны ниже со ссылкой на классы боковых цепей аминокислот. Более важные замены представлены ниже в таблице 1 под заголовком «приведенные в качестве примера замены», и они описаны ниже со ссылкой на классы боковых цепей аминокислот. Аминокислотные замены можно интродуцировать в представляющее интерес антитело и продукты подвергать скринингу в отношении требуемой активности, например, сохранения/повышения способности к связыванию антигена, пониженной иммуногенности или повышенной ADCC или CDC.
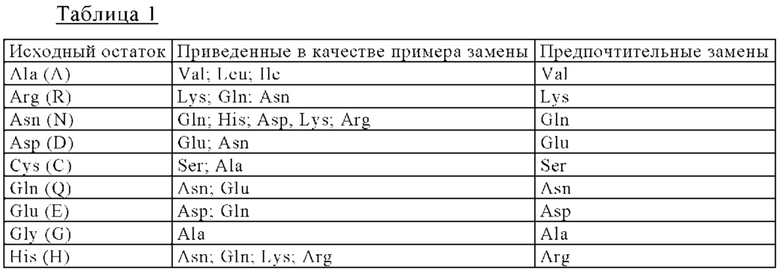
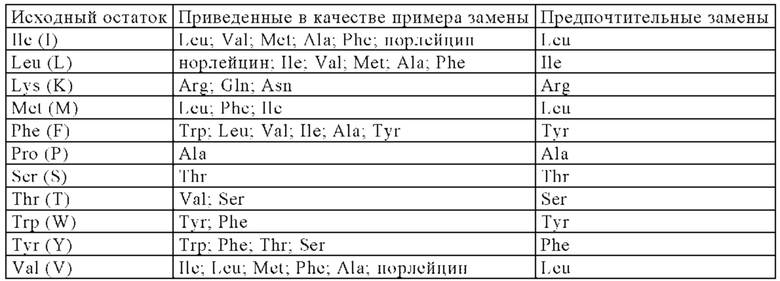
Аминокислоты можно группировать на основе общих свойств боковых цепей следующим образом: (1) гидрофобные: норлейцин, Met, Ala, Val, Leu, He; (2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln; (3) кислотные: Asp, Glu; (4)  : His, Lys, Arg; (5) остатки, которые влияют на ориентацию цепи: Gly, Pro; (6) ароматические: Trp, Tyr, Phe. Под неконсервативными заменами подразумевают замену представителя одного из указанных классов на представителя из другого класса.
: His, Lys, Arg; (5) остатки, которые влияют на ориентацию цепи: Gly, Pro; (6) ароматические: Trp, Tyr, Phe. Под неконсервативными заменами подразумевают замену представителя одного из указанных классов на представителя из другого класса.
Один из типов полученного в результате замены варианта включает замену одного или нескольких остатков гипервариабельного участка родительского антитела (например, гуманизированного или человеческого антитела). Как правило, полученный(ые) в результате вариант(ы), отобранный(ые) для дополнительного исследования, должен(ы) иметь модификации (например, улучшения) некоторых биологических свойств (например, повышенную аффинность, пониженную иммуногенность) относительно родительского антитела, и/или должен(ы) иметь практически сохраненные определенные биологические свойства родительского антитела. Примером полученного в результате замены варианта является антитело с созревшей аффинностью, которое можно создавать, например, с использованием технологий созревания аффинности на основе фагового дисплея, например, указанных в настоящем описании. В целом, метод состоит в следующем: один или несколько остатков HVR подвергают мутации и варианты антител экспонируют на фаге и подвергают скринингу в отношении конкретной биологической активности (например, аффинности связывания).
Изменения (например, замены) можно осуществлять в HVR, например, для повышения аффинности антитела. Указанные изменения можно делать в «горячих (активных) точках» в HVR, т.е. остатках, кодируемых кодонами, которые с высокой частотой подвергаются мутации в процессе соматического созревания (см., например, Chowdhury, Methods Mol. Biol. 207, 2008, сс. 179-196), и/или в остатках, которые контактируют с антигеном, с последующим тестированием варианта VH или VL в отношении аффинности связывания. Созревание аффинности путем создания и повторной селекции из вторичных библиотек описано, например, у Hoogenboom и др., Methods in Molecular Biology 178, 2002, сс. 1-37. В некоторых вариантах осуществления процесса созревания аффинности интродуцируют разнообразие в гены вариабельной области, выбранные для созревания, с помощью любого из широкого разнообразия методов (например, ПЦР пониженной точности, перестановка цепи или сайтнаправленный мутагенез с использованием олигонуклеотидов). Затем создают вторичную библиотеку. Затем библиотеку подвергают скринингу для идентификации любых вариантов антител с требуемой аффинностью. Другой метод, применяемый для интродукции разнообразия, включает подходы, мишенью которых является HVR, при которых рандомизируют несколько остатков HVR (например, одновременно 4-6 остатков). Остатки HVR, участвующие в связывании антигена, можно специфически идентифицировать, например, используя аланин-сканирующий мутагенез или моделирование. Часто мишенями являются, прежде всего, CDR-H3 и CDR-L3.
В некоторых вариантах осуществления изобретения замены, инсерции или делеции могут затрагивать один или несколько HVR, если указанные изменения не снижают существенно способность антитела связываться с антигеном. Например, в HVR могут быть сделаны консервативные изменения (например, консервативные замены, указанные в настоящем описании), которые не снижают существенно аффинность связывания. Указанные изменения, например, могут находиться вне принимающих участие в контактировании с антигеном остатков HVR. В некоторых вариантах осуществления изобретения в последовательностях вариантов VH и VL, указанных выше, каждый HVR либо является неизмененным, либо содержит не более одной, двух или трех аминокислотных замен.
Ценным методом идентификации остатков или областей антитела, которые можно подвергать мутагенезу, является так называемый «аланин-сканирующий мутагенез», описанный у Cunningham и Wells, Science 244, 1989, сс. 1081-1085. При осуществлении этого метода остаток или группу остатков-мишеней (например, заряженные остатки, такие как arg, asp, his, lys и glu) идентифицируют и заменяют на нейтральную или отрицательно заряженную аминокислоту (например, аланин или полиаланин) для решения вопроса о том, будет ли это влиять на взаимодействие антитела с антигеном. Дополнительные замены можно интродуцировать в те положения аминокислот, для которых продемонстрирована функциональная чувствительность к начальным заменам. В альтернативном или дополнительном варианте можно анализировать кристаллическую структуру комплекса антиген-антитело для идентификации точек контакта между антителом и антигеном. Указанные контактирующие остатки и соседние остатки можно рассматривать в качестве мишеней или исключать из рассмотрения в качестве кандидатов для замены. Варианты можно подвергать скринингу для решения вопроса о том, обладают ли они требуемыми свойствами.
Инсерции в аминокислотные последовательности включают амино- и/или карбоксиконцевые слияния, варьирующие по длине от одного остатка до полипептидов, содержащих сто или большее количество остатков, а также инсерции одного или нескольких аминокислотных остатков внутрь последовательности. Примером результата осуществления концевых инсерции является антитело с N-концевым метионильным остатком. Другие инсерционные варианты молекул антител включают слияние N- или С-концов антитела с ферментом (например, для ADEPT (антитело-направляемый фермент пролекарственной терапии) или полипептидом, который удлиняет время полужизни антитела в сыворотке.
б) Варианты гликозилирования
В некоторых вариантах осуществления изобретения антитело, предлагаемое в изобретении, изменяют с целью повышения или понижения степени гликозилирования антитела. Добавление или делецию сайтов гликозилирования в антителе можно легко осуществлять путем такого изменения аминокислотной последовательности, которое позволяет создавать или удалять один или несколько сайтов гликозилирования.
Если антитело содержит Fc-область, то можно изменять присоединенный к ней углевод. Нативные антитела, которые продуцируются клетками млекопитающих, как правило, содержат разветвленный биантенный олигосахарид, который, как правило, присоединен с помощью N-связи к Asn297 СН2-домена Fc-области (см., например, Wright и др., TIBTECH 15, 1997, сс. 26-32). Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стебле» биантенной олигосахаридной структуры. В некоторых вариантах осуществления изобретения модификации олигосахарида в антителе, предлагаемом в изобретении, можно осуществлять для создания вариантов антитела с определенными улучшенными свойствами.
Одним из вариантов осуществления изобретения являются варианты антитела, имеющие углеводную структуру, в которой снижено содержание фукозы, присоединенной (прямо или косвенно) к Fc-области. Например, содержание фукозы в указанной Fc-области может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Содержание фукозы определяют путем расчета среднего содержания фукозы в сахарной цепи на Asn297 относительно суммы всех гликоструктур, присоединенных к Asn297 (например, комплексных, гибридных структур и структур с высоким содержанием маннозы), которое измеряют с помощью MALDI-TOF-масс-спектрометрии согласно методу, описанному, например, в WO 2008/077546. Asn297 обозначает остаток аспарагина, присутствующий примерно в положении 297 в Fc-области (EU-нумерация остатков Fc-области); однако вследствие минорных вариаций последовательности антитела Asn297 может располагаться также в положении, находящемся на расстоянии примерно ±3 аминокислоты в прямом или обратном направлении от положения 297, т.е. между положениями 294 и 300. Указанные фукозилированные варианты могут обладать улучшенной ADCC-функцией (см., например, US 2003/0157108 (Presta L.); US 2004/0093621 (фирма Kyowa Hakko Kogyo Co., Ltd). Примерами публикаций, касающихся «дефукозилированных» вариантов антител или вариантов антител «с дефицитом фукозы» являются: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2002/031140; Okazaki и др., J. Mol. Biol. 336, 2004, cc. 1239-1249; Yamane-Ohnuki и др., Biotech. Bioeng. 87, 2004, cc. 614-622). Примерами клеточных линий, которые обладают способностью продуцировать дефукозилированные антитела, являются Lec13 СНО-клетки с дефицитом белкового фукозилирования (Ripka и др., Arch. Biochem. Biophys. 249, 1986, сс. 533-545; US 2003/0157108 (Presta L.) и WO 2004/056312 (Adams и др., см., прежде всего, пример 11), и клеточные линии с «выключенным» геном, например, СНО-клетки с «выключенным» геном альфа-1,6-фукозилтрансферазы, FUT8 (см., например, Yamane-Ohnuki и др., Biotech. Bioeng. 87, 2004, сс. 614-622); Kanda и др., Biotechnol. Bioeng., 94 (4), 2006, сс. 680-688; и WO 2003/085107).
Кроме того, описаны варианты антител с бисекционными олигосахаридами, например, в которых биантенный олигосахарид, присоединенный к Fc-области антитела, бисекционнируется с помощью GlcNAc. Указанные варианты антител могут отличаться пониженным фукозилированием и/или повышенной ADCC-функцией. Примеры указанных вариантов антител описаны, например, в WO 2003/011878 (Jean-Mairet и др.); US №6602684 (Umana и др.) и US 2005/0123546 (Umana и др.). Предложены также варианты антител по меньшей мере с одним остатком галактозы в олигосахариде, присоединенном к Fc-области. Указанные варианты антител могут обладать повышенной CDC-функцией. Указанные варианты антител описаны, например, в WO 1997/30087 (Patel и др.); WO 1998/58964 (Raju S.); и WO 1999/22764 (Raju S.).
в) Варианты Fc-области
Согласно конкретным вариантам осуществления изобретения можно интродуцировать в Fc-область антитела, представленного в настоящем описании, одну или несколько аминокислотных модификаций, создавая тем самым вариант Fc-области. Вариант Fc-области может содержать последовательность человеческой Fc-области (например, Fc-области человеческого IgG1, IgG2, IgG3 или IgG4), включающую аминокислотную модификацию (например, замену) в одном или нескольких аминокислотных положениях.
Некоторые варианты осуществления изобретения относятся к варианту антитела, который обладает некоторыми, но не всеми эффекторными функциями, что делает его перспективным кандидатом для путей применения, для которых является важным время полужизни антитела in vivo, однако некоторые эффекторные функции (такие как CDC и ADCC) не являются необходимыми или являются вредными. Можно осуществлять анализы цитотоксичности in vitro и/или in vivo для подтверждения снижения/истощения CDC- и/или ADCC-активности. Например, можно осуществлять анализы связывания Fc-рецептора (FcR) для гарантии того, что у антитела отсутствует способность к связыванию с Fc-гамма R (и поэтому, вероятно, отсутствует ADCC-активность), но сохраняется способность связываться с FcRn. Первичные клетки, опосредующие ADCC, т.е. NK-клетки, экспрессируют только Fc-гамма RIII, в то время как моноциты экспрессируют Fc-гамма RI, Fc-гамма RII и Fc-гамма RIII. Данные об экспрессии FcR на гематопоэтических клетках обобщены в таблице 3 на с. 464 у Ravetch и Kinet, Annu. Rev. Immunol. 9, 1991, сс. 457-492. Примерами анализов in vitro ADCC-активности представляющей интерес молекулы являются (но, не ограничиваясь только ими) анализы, описанные в US №5500362 (см., например, Hellstrom и др., Proc. Natl. Acad. Sci. USA 83, 1986, сс. 7059-7063 и Hellstrom и др., Proc.  Acad. Sci. USA 82, 1985, cc. 1499-1502); US №5821337 (см. Bruggemann и др., J. Exp.Med. 166, 1987, cc. 1351-1361). В альтернативном варианте можно применять нерадиоактивные методы анализа (см., например, нерадиоактивный анализ цитотоксичности АСТ1™ на основе проточной цитометрии (фирма CellTechnology, Inc. Маунтин-Вью, шт. Калифорния); и нерадиоактивный анализ цитотоксичности CytoTox 96® (фирма Promega, Мэдисон, шт. Висконсин). Пригодными для таких анализов эффекторными клетками являются мононуклеарные клетки периферической крови (РВМС) и естественные клетки-киллеры (NK). В альтернативном или дополнительном варианте ADCC-активность представляющей интерес молекулы можно оценивать in vivo, например, на животной модели, например, как описано у Clynes и др., Proc. Natl. Acad. Sci. USA 95, 1998, cc. 652-656. Можно осуществлять также анализы связывания C1q для подтверждения того, что антитело не может связывать C1q и поэтому у него отсутствует CDC-активность (см., например, описание ELISA для оценки связывания C1q и С3 с в WO 2006/029879 и WO 2005/100402). Для оценки активации комплемента можно осуществлять анализ CDC (см., например, Gazzano-Santoro и др., J. Immunol. Methods 202, 1996, сс. 163-171; Cragg и др., Blood 101, 2003, сс. 1045-1052; и Cragg, Blood 103, 2004, сс. 2738-2743). Определение FcRn-связывания и клиренса/времени полужизни in vivo можно осуществлять также с использованием методов, известных в данной области (см., например, Petkova и др.,
Acad. Sci. USA 82, 1985, cc. 1499-1502); US №5821337 (см. Bruggemann и др., J. Exp.Med. 166, 1987, cc. 1351-1361). В альтернативном варианте можно применять нерадиоактивные методы анализа (см., например, нерадиоактивный анализ цитотоксичности АСТ1™ на основе проточной цитометрии (фирма CellTechnology, Inc. Маунтин-Вью, шт. Калифорния); и нерадиоактивный анализ цитотоксичности CytoTox 96® (фирма Promega, Мэдисон, шт. Висконсин). Пригодными для таких анализов эффекторными клетками являются мононуклеарные клетки периферической крови (РВМС) и естественные клетки-киллеры (NK). В альтернативном или дополнительном варианте ADCC-активность представляющей интерес молекулы можно оценивать in vivo, например, на животной модели, например, как описано у Clynes и др., Proc. Natl. Acad. Sci. USA 95, 1998, cc. 652-656. Можно осуществлять также анализы связывания C1q для подтверждения того, что антитело не может связывать C1q и поэтому у него отсутствует CDC-активность (см., например, описание ELISA для оценки связывания C1q и С3 с в WO 2006/029879 и WO 2005/100402). Для оценки активации комплемента можно осуществлять анализ CDC (см., например, Gazzano-Santoro и др., J. Immunol. Methods 202, 1996, сс. 163-171; Cragg и др., Blood 101, 2003, сс. 1045-1052; и Cragg, Blood 103, 2004, сс. 2738-2743). Определение FcRn-связывания и клиренса/времени полужизни in vivo можно осуществлять также с использованием методов, известных в данной области (см., например, Petkova и др.,  Immunol. 18 (12), 2006, сс. 1759-1769).
Immunol. 18 (12), 2006, сс. 1759-1769).
Антитела с пониженной эффекторной функцией включают антитела с заменой одного или нескольких следующих остатков Fc-области, таких как 238, 265, 269, 270, 297, 327 и 329 (см., например, US №6737056). К указанным мутантам Fc относятся мутанты Fc с заменами в двух или большем количестве из следующих аминокислотных положений: 265, 269, 270, 297 и 327, включая так называемый мутант «DANA» Fc-области, несущий замену остатков 265 и 297 на аланин (US №7332581).
Описаны некоторые варианты антител с повышенной или пониженной способностью связываться с FcR (см., например, US №6737056; WO 2004/056312 и Shields и др., J. Biol. Chem. 276, 2001, сс. 6591-6604).
Согласно конкретным вариантам осуществления изобретения вариант антитела содержит Fc-область с одной или несколькими аминокислотными заменами, которые повышают ADCC, например, замены в положениях 298, 333, и/или 334 Fc-области (EU-нумерация остатков).
В некоторых вариантах осуществления изобретения в Fc-области осуществляют изменения, которые приводят к измененной (т.е. либо повышенной, либо пониженной) способности связывать C1q и/или комплементзависимой цитотоксичности (CDC), что описано, например, в US №6194551, WO 1999/51642 и у Idusogie и др., J. Immunol. 164, 2000, сс. 4178-4184.
Антитела с удлиненным временем полужизни и повышенной способностью к связыванию с неонатальным Fc-рецептором (FcRn), который ответствен за перенос материнских IgG эмбриону (Guyer и др., J. Immunol. 117, 1976, сс. 587-593 и Kim и др., J. Immunol. 24, 1994, сс. 2429-2434), описаны в US 2005/0014934А1 (Hinton и др.). Эти антитела содержат Fc-область с одной или несколькими заменами, которые повышают связывание Fc-области с FcRn. Указанные варианты Fc-области включают варианты с заменами одного или нескольких следующих остатков Fc-области: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, замену остатка 434 в Fc-области (US №7371826). Дополнительные примеры, касающиеся других вариантов Fc-области, описаны также у Duncan, Nature 322, 1988, сс. 738-740; в US №№5648260 и 5624821 и в WO 1994/29351.
г) Варианты антител, сконструированные с помощью цистеина
В некоторых вариантах осуществления изобретения может оказаться желательным создавать антитела, сконструированные с помощью цистеина, например, «тиоМАт», в которых один или несколько остатков в антителе заменены на остатки цистеина. В конкретных вариантах осуществления изобретения замененные остатки находятся в доступных сайтах антитела. Путем замены этих остатков на цистеин реакционноспособные тиольные группы помещаются в доступные сайты антитела и могут использоваться для конъюгации антитела с другими фрагментами, такими как фрагменты, представляющие собой лекарственное средство, или фрагменты, представляющие собой линкер, связанный с лекарственным средством, с созданием иммуноконъюгата, указанного ниже в настоящем описании. В некоторых вариантах осуществления изобретения любой(ые) один или несколько следующих остатков может(гут) быть заменен(ы) на цистеин: V205 (нумерация по Кэботу) легкой цепи; A118 (EU-нумерация) тяжелой цепи и S400 (EU-нумерация) Fc-области тяжелой цепи. Антитела, сконструированные с помощью цистеина, можно создавать согласно методу, например, описанному в US №7521541.
д) Производные антител
В некоторых вариантах осуществления изобретения антитело, представленное в настоящем описании, можно дополнительно модифицировать таким образом, чтобы оно содержало дополнительные небелковые фрагменты, известные в данной области и легко доступные. Фрагменты, пригодные для дериватизации антитела, включают (но, не ограничиваясь только ими) водорастворимые полимеры. Примерами водорастворимых полимеров являются (но, не ограничиваясь только ими) полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлоза, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (либо гомополимеры, либо статистические сополимеры) и декстран- или поли(н- винилпиролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры пропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси.
Полиэтиленгликольпропиональдегид может иметь преимущество при производстве благодаря его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Количество полимеров, присоединенных к антителу, может варьироваться и, если присоединено более одного полимера, то они могут представлять собой одинаковые или различные молекулы. В целом, количество и/или тип полимеров, применяемых для дериватизации, можно определять с учетом таких особенностей (но, не ограничиваясь только ими), как конкретные свойства или функции антитела, подлежащие усовершенствованию, предполагается ли применение производного антитела в терапии в определенных условиях, и т.д.
Другим вариантом осуществления изобретения являются конъюгаты антитела и небелкового фрагмента, которые можно избирательно нагревать, воздействуя на него излучением. В одном из вариантов осуществления изобретения небелковый фрагмент представляет собой углеродную нанотрубку (Kam и др., Proc. Natl. Acad. Sci. USA 102, 2005, cc. 11600-11605). Излучение может иметь любую длину волны и включает (но, не ограничиваясь только ими) длины волн, которые не повреждают обычные клетки, но которые нагревают небелковый фрагмент до температуры, при которой уничтожаются клетки, ближайшие к конъюгату: антитело-небелковый фрагмент.
8. Варианты Fc-областей
Одним из объектов изобретения является выделенный полипептид, содержащий вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В некоторых вариантах осуществления изобретения вариант Fc-области содержит по меньшей мере одно изменение аминокислотного остатка (например, замену) по сравнению с соответствующей нативной последовательностью Fc-области или последовательностью референс-варианта (в некоторых случаях обозначена в целом как «родительская» Fc-область). В некоторых вариантах осуществления изобретения вариант Fc- области, предлагаемый в изобретении, обладает повышенной связывающей активностью в отношении обезьяньего Fc-гамма RIIb по сравнению с родительской Fc-областью. В конкретных вариантах осуществления изобретения обезьяний Fc-гамма RIIb представляет собой Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223).
В некоторых вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для обезьяньего Fc-гамма RIIb)]/[величина KD варианта Fc-области для обезьяньего Fc-гамма RIIb] может составлять 2,0 или более, 3,0 или более, 4,0 или более, 5,0 или более, 6,0 или более, 7,0 или более, 8,0 или более, 9,0 или более, 10 или более, 15 или более, 20 или более, 25 или более, 30 или более, 40 или более или 50 или более. В других вариантах осуществления изобретения вариант Fc-области обладает пониженной активностью связывания с обезьяньим Fc-гамма RIIIa. В некоторых вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для обезьяньего Fc-гамма RIIIa]/[величина KD варианта Fc-области для обезьяньего Fc-гамма RIIIa] может составлять 0,50 или менее, 0,40 или менее, 0,30 или менее, 0,20 или менее, 0,10 или менее, 0,09 или менее, 0,08 или менее, 0,07 или менее, 0,06 или менее, 0,05 или менее, 0,04 или менее, 0,03 или менее, 0,02 или менее или 0,01 или менее. В некоторых вариантах осуществления изобретения обезьяний Fc-гамма RIIb имеет последовательность SEQ ID NO: 223 (обезьяны циномолгус). В некоторых вариантах осуществления изобретения обезьяний Fc-гамма RIIIa имеет последовательность SEQ ID NO:224 (обезьяны циномолгус).
В других вариантах осуществления изобретения вариант Fc-области обладает повышенной активностью связывания с человеческим Fc-гамма RIIb. В некоторых вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для человеческого Fc-гамма RIIb]/[величина KD варианта Fc-области для человеческого Fc-гамма RIIb] может составлять 2,0 или более, 3,0 или более, 4,0 или более, 5,0 или более, 6,0 или более, 7,0 или более, 8,0 или более, 9,0 или более, 10 или более, 15 или более, 20 или более, 25 или более, 30 или более, 40 или более или 50 или более. В других вариантах осуществления изобретения вариант Fc-области обладает пониженной активностью связывания с человеческим Fc-гамма RIIIa. В некоторых вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для человеческого Fc-гамма RIIIa]/[величина KD варианта Fc-области для человеческого Fc-гамма RIIIa] может составлять 0,50 или менее, 0,40 или менее, 0,30 или менее, 0,20 или менее, 0,10 или менее, 0,09 или менее, 0,08 или менее, 0,07 или менее, 0,06 или менее, 0,05 или менее, 0,04 или менее, 0,03 или менее, 0,02 или менее или 0,01 или менее. В некоторых вариантах осуществления изобретения человеческий Fc-гамма RIIb имеет последовательность SEQ ID NO: 212, 213 или 214. В некоторых вариантах осуществления изобретения человеческий Fc-гамма RIIIa имеет последовательность SEQ ID NO: 215, 216, 217 или 218.
В других вариантах осуществления изобретения вариант Fc-области обладает пониженной активностью связывания с человеческим Fc-гамма RIIa (Н-типа), чем с человеческим Fc-гамма RIIb. В некоторых вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для человеческого Fc-гамма RIIa (Н-типа)]/[величина KD варианта Fc-области для человеческого Fc-гамма RIIa (H-типа)] может составлять 5,0 или менее, 4,0 или менее, 3,0 или менее, 2,0 или менее, 1,0 или менее, 0,9 или менее, 0,8 или менее, 0,7 или менее, 0,6 или менее, 0,5 или менее, 0,4 или менее, 0,3 или менее, 0,2 или менее или 0,1 или менее. В других вариантах осуществления изобретения вариант Fc-области обладает пониженной активностью связывания с человеческим Fc-гамма RIIa (R-типа), чем с человеческим Fc-гамма RIIb. В некоторых вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для человеческого Fc-гамма RIIa (R-типа)]/[величина KD варианта Fc-области для человеческого Fc-гамма RIIa (R-типа)] может составлять 5,0 или менее, 4,0 или менее, 3,0 или менее, 2,0 или менее, 1,0 или менее, 0,9 или менее, 0,8 или менее, 0,7 или менее, 0,6 или менее, 0,5 или менее, 0,4 или менее, 0,3 или менее, 0,2 или менее или 0,1 или менее. В некоторых вариантах осуществления изобретения человеческий Fc-гамма RIIa (Н-типа) имеет последовательность SEQ ID NO: 211. В некоторых вариантах осуществления изобретения человеческий Fc-гамма RIIa (R-типа) имеет последовательность SEQ ID NO: 210.
В некоторых вариантах осуществления изобретения соотношение [величина KD родительской Fc-области для обезьяньего Fc-гамма RIIa]/[величина KD варианта Fc-области для обезьяньего Fc-гамма RIIa] может составлять 2,0 или более, 3,0 или более, 4,0 или более, 5,0 или более, 6,0 или более, 7,0 или более, 8,0 или более, 9,0 или более, 10 или более, 15 или более, 20 или более, 25 или более, 30 или более, 40 или более или 50 или более. В некоторых вариантах осуществления изобретения обезьяний Fc-гамма RIIa выбирают из обезьяньего Fc-гамма RIIa1 (Fc-гамма RIIa1 циномолгус (SEQ ID NO: 220)), обезьяньего Fc-гамма RIIa2 (например, Fc-гамма RIIa2 циномолгус (SEQ ID NO: 221)) и обезьяньего Fc-гамма RIIa3 (например, Fc-гамма RIIa3 циномолгус (SEQ ID NO: 222).
В другом варианте осуществления изобретения величина KD варианта Fc-области для обезьяньего Fc-гамма RIIb может составлять 1,0×10-6 М или менее, 9,0×10-7 М или менее, 8,0×10-7 М или менее, 7,0×10-7 М или менее, 6,0×10-7 М или менее, 5,0×10-7 М или менее, 4,0×10-7 М или менее, 3,0×10-7 М или менее, 2,0×10-7 М или менее или 1,0×10-7 М или менее. В другом варианте осуществления изобретения величина KD варианта Fc-области для обезьяньего Fc-гамма RIIIa может составлять 5,0×10-7 М или более, 6,0×10-7 М или более, 7,0×10-7 М или более, 8,0×10-7 М или более, 9,0×10-7 М или более, 1,0×10-6 М или более, 2,0×10-6 М или более, 3,0×10-6 М или более, 4,0×10-6 М или более, 5,0×10-6 М или более, 6,0×10-6 М или более, 7,0×10-6 М или более, 8,0×10-6 М или более, 9,0×10-6 М или более или 1,0×10-5 М или более. В другом варианте осуществления изобретения величина KD варианта Fc-области для человеческого Fc-гамма RIIb может составлять 2,0×10-6 М или менее, 1,0×10-6 М или менее, 9,0×10-7 М или менее, 8,0×10-7 М или менее, 7,0×10-7 М или менее, 6,0×10-7 М или менее, 5,0×10-7 М или менее, 4,0×10-7 М или менее, 3,0×10-7 М или менее, 2,0×10-7 М или менее или 1,0×10-7 М или менее. В другом варианте осуществления изобретения величина KD варианта Fc-области для человеческого Fc-гамма RIIIa может составлять 1,0×10-6 М или более, 2,0×10-6 М или более, 3,0×10-6 М или более, 4,0×10-6 М или более, 5,0×10-6 М или более, 6,0×10-6 М или более, 7,0×10-6 М или более, 8,0×10-6 М или более, 9,0×10-6 М или более, 1,0×10-5 М или более, 2,0×10-5 М или более, 3,0×10-5 М или более, 4,0×10-5 М или более или 5,0×10-5 М или более. В другом варианте осуществления изобретения величина KD варианта Fc-области для человеческого Fc-гамма RIIa (Н-типа) может составлять 1,0×10-7 М или более, 2,0×10-7 М или более, 3,0×10-7 М или более, 4,0×10-7 М или более, 5,0×10-7 М или более, 6,0×10-7 М или более, 7,0×10-7 М или более, 8,0×10-7 М или более, 9,0×10-7 М или более, 1,0×10-6 М или более, 2,0×10-6 М или более, 3,0×10-6 М или более, 4,0×10-6 М или более или 5,0×10-6 М или более. В другом варианте осуществления изобретения величина KD варианта Fc-области для человеческого Fc-гамма RIIa (R-типа) может составлять 2,0×10-7 М или более, 3,0 ×10-7 М или более, 4,0×10-7 М или более, 5,0×10-7 М или более, 6,0×10-7 М или более, 7,0×10-7 М или более, 8,0×10-7 М или более, 9,0×10-7 М или более, 1,0×10-6 М или более, 2,0×10-6 М или более, 3,0×10-6 М или более, 4,0×10-6 М или более или 5,0×10-6 М или более.
В другом варианте осуществления изобретения величина KD варианта Fc-области для обезьяньего Fc-гамма RIIa может составлять 1,0×10-6 М или менее, 9,0×10-7 М или менее, 8,0×10-7 М или менее, 7,0×10-7 М или менее, 6,0×10-7 М или менее, 5,0×10-7 М или менее, 4,0×10-7 М или менее, 3,0×10-7 М или менее, 2,0×10-7 М или менее или 1,0×10-7 М или менее. В некоторых вариантах осуществления изобретения обезьяний Fc-гамма RIIa можно выбирать из одного из следующих рецепторов: обезьяний Fc-гамма RIIa1, обезьяний Fc-гамма RIIa2 и обезьяний Fc-гамма RIIa3.
При разработке фармацевтического продукта для лечения болезней у человека оценка его эффективности и безопасности на обезьянах является важной из-за их биологического сходства с человеком. С учетом этого, фармацевтический продукт, подлежащий разработке, предпочтительно должен обладать перекрестной реактивностью и для человека, и для обезьян касательно активности связывания с мишенью.
«Fc-гамма-рецепторы» (в контексте настоящего описания обозначают как Fc-гамма рецепторы, Fc-гамма R или FcgR) представляют собой рецепторы, которые могут связываться с Fc-областью моноклональных антител изотипа IgG1, IgG2, IgG3 и IgG4, и прежде всего представляет собой любого представителя семейства белков, кодируемых генами Fc-гамма рецептора. У человека это семейство включает (но, не ограничиваясь только ими) Fc-гамма RI (CD64), в том числе изоформы Fc-гамма RIa, Fc-гамма Rib и Fc-гамма RIc; Fc-гамма RII (CD32), в том числе изоформы Fc-гамма RIIa (включая аллотипы Н131 (тип Н) и R131 (тип R)), Fc-гамма RIIb (в том числе Fc-гамма RIIb-1 и Fc-гамма RIIb-2), и Fc-гамма RIIc; и Fc-гамма RIII (CD16), в том числе изоформы Fc-гамма RIIIa (включая аллотипы V158 и F158), и Fc-гамма RIIIb (включая аллотипы Fc-гамма RIIIb-NA1 и Fc-гамма RIIIb-NA2), и любые человеческие Fc-гамма R, изоформы или аллотипы Fc-гамма R, которые пока еще не открыты. Fc-гамма RIIb1 и Fc-гамма RIIb2 описаны в виде сплайсинговых вариантов человеческого Fc-гамма RIIb. Кроме того, описан сплайсинговый вариант, обозначенный как Fc-гамма RIIb3 (J Exp Med, 170, 1989, сс. 1369-1385). Помимо указанных сплайсинговых вариантов человеческий Fc-гамма RIIb включает все сплайсинговые варианты, зарегистрированные в NCBI, которые представляют собой NP_001002273.1, NP_001002274.1, NP_001002275.1, NP_001177757.1 и NP_003992.3. Кроме того, человеческий Fc-гамма RIIb включает каждый из описанных ранее генных полиморфизмов, а также Fc-гамма RIIb (Arthritis Rheum. 48, 2003, сс. 3242-3252; Kono и др., Hum. Mol. Genet. 14, 2005, сс. 2881-2892 и Kyogoju и др., Arthritis Rheum. 46, 2002, сс. 1242-1254), и каждый генный полиморфизм, который будет описан в будущем.
У Fc-гамма RIIa существует два аллотипа, один, у которого аминокислота в положении 131 Fc-гамма RIIa представляет собой гистидин (тип Н), а другой, у которого аминокислота в положении 131 заменена на аргинин (тип R) (Warrmerdam, J. Exp.Med. 172, 1990, сс. 19-25).
Fc-гамма R включает Fc-гамма R, полученные из человека, мышей, крыс, кроликов и обезьян (но не ограничен указанными), и он может иметь происхождение из любого организма. Мышиные Fc-гамма R включают (но, не ограничиваясь только ими) Fc-гамма RI (CD64), Fc-гамма RII (CD32), Fc-гамма RIII (CD16) и Fc-гамма RIII-2 (CD16-2) и любые мышиные Fc-гамма R или изоформы Fc-гамма R. Если специально не указано иное, то понятие «обезьяний Fc-гамма R» или его вариация относится к Fc-гамма RIIa1 (SEQ ID NO: 220), Fc-гамма RIIa2 (SEQ ID NO: 221), Fc-гамма RIIa3 (SEQ ID NO: 222), Fc-гамма RIIb (SEQ ID NO: 223) или Fc-гамма RIIIaS (SEQ ID NO: 224) обезьян циномолгус.
Полинуклеотидная последовательность человеческого Fc-гамма RI представлена в SEQ ID NO: 199 (NM_000566.3); полинуклеотидная последовательность человеческого Fc-гамма RIIa представлена в SEQ ID NO: 200 (ВС020823.1) или SEQ ID NO: 201 (NM_001136219.1); полинуклеотидная последовательность человеческого Fc-гамма RIIb представлена в SEQ ID NO: 202 (ВС146678.1) или SEQ ID NO: 203 (NM_004001.3); полинуклеотидная последовательность человеческого Fc-гамма RIIIa представлена в SEQ ID NO: 204 (ВС033678.1) или SEQ ID NO: 205 (NM_001127593.1) и полинуклеотидная последовательность человеческого Fc-гамма RIIIb представлена в SEQ ID NO: 206 (ВС128562.1).
Аминокислотная последовательность человеческого Fc-гамма RI представлена в SEQ ID NO: 207 (NP_000557.1); аминокислотная последовательность человеческого Fc-гамма RIIa представлена в SEQ ID NO: 208 (ААН20823.1), SEQ ID NO: 209, SEQ ID NO: 210 или SEQ ID NO: 211; аминокислотная последовательность человеческого Fc-гамма RIIb представлена в SEQ ID NO: 212 (AAI46679.1), SEQ ID NO: 213 или SEQ ID NO: 214; аминокислотная последовательность человеческого Fc-гамма RIIIa представлена в SEQ ID NO: 215 (AAH33678.1), SEQ ID NO: 216, SEQ ID NO: 217 или SEQ ID NO: 218 и аминокислотная последовательность человеческого Fc-гамма RIIIb представлена в SEQ ID NO: 219 (AAI28563.1).
Аминокислотная последовательность обезьяньего (циномолгус) Fc-гамма RIIa представлена в SEQ ID NO: 220 (Fc-гамма RIIa1), SEQ ID NO: 221 (Fc-гамма RIIa2) или SEQ ID NO: 222 (Fc-гамма RIIa3); аминокислотная последовательность обезьяньего (циномолгус) Fc-гамма RIIb представлена в SEQ ID NO: 223 и аминокислотная последовательность обезьяньего (циномолгус) Fc-гамма RIIIa представлена в SEQ ID NO: 224.
Одним из объектов изобретения являются полипептиды, которые содержат варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью по сравнению с соответствующей Fc-гамма RIIb-связывающим референс-полипептидом. В других объектах изобретения полипептид, предлагаемый в изобретении, содержит по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 233, 234, 235, 236, 237, 238, 239, 264, 266, 267, 268, 271, 295, 298, 325, 326, 327, 328, 330, 331, 332, 334 и 396, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
Одним из объектов изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью, которые содержат по меньшей мере два аминокислотных изменения, включающие: (а) одно аминокислотное изменение в положении 236 и (б) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 233, 234, 235, 237, 238, 239, 264, 266, 267, 268, 271, 295, 298, 325, 326, 327, 328, 330, 331, 332, 334, и 396, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
Одним из объектов изобретения являются полипептиды, содержащие вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, который содержит аминокислотное изменение в положении 236 согласно EU-нумерации.
Одним из объектов изобретения являются полипептиды, содержащие вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, который содержит по меньшей мере два аминокислотных изменения, включающих: (а) одно аминокислотное изменение в положении 236 и (б) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 235, 239, 268, 295, 298, 326, 330 и 396, согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области содержит аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 235, 239, 268, 295, 298, 326, 330 и 396, согласно EU-нумерации. В следующем варианте осуществления изобретения вариант Fc-область содержит аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 268, 295, 326 и 330, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность обезьян циномолгус Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
Другим объектом изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью, которые содержат аминокислотные изменения в одном из следующих положений (1)-(37): (1) положения 231, 236, 239, 268 и 330; (2) положения 231, 236, 239, 268, 295 и 330; (3) положения 231, 236, 268 и 330; (4) положения 231, 236, 268, 295 и 330; (5) положения 232, 236, 239, 268, 295 и 330; (6) положения 232, 236, 268, 295 и 330; (7) положения 232, 236, 268 и 330; (8) положения 235, 236, 268, 295, 326 and 330; (9) положения 235, 236, 268, 295 и 330; (10) положения 235, 236, 268 и 330; (11) положения 235, 236, 268, 330 и 396; (12) положения 235, 236, 268 и 396; (13) положения 236, 239, 268, 295, 298 и 330; (14) положения 236, 239, 268, 295, 326 и 330; (15) положения 236, 239, 268, 295 и 330; (16) положения 236, 239, 268, 298 и 330; (17) положения 236, 239, 268, 326 и 330; (18) положения 236, 239, 268 и 330; (19) положения 236, 239, 268, 330 и 396; (20) положения 236, 239, 268 и 396; (21) положения 236 и 268; (22) положения 236, 268 и 295; (23) положения 236, 268, 295, 298 и 330; (24) положения 236, 268, 295, 326 и 330; (25) положения 236, 268, 295, 326, 330 и 396; (26) положения 236, 268, 295 и 330; (27) положения 236, 268, 295, 330 и 396; (28) положения 236, 268, 298 и 330; (29) положения 236, 268, 298 и 396; (30) положения 236, 268, 326 и 330; (31) положения 236, 268, 326, 330 и 396; (32) положения 236, 268 и 330; (33) положения 236, 268, 330 и 396; (34) положения 236, 268 и 396; (35) положения 236 и 295; (36) положения 236, 330 и 396; и (37) положения 236 и 396, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В следующем варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит по меньшей мере одну аминокислоту, выбранную из группы, состоящей из: (a) Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp, Tyr в положении 231; (б) Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp, Tyr в положении 232; (в) Asp в положении 233; (г) Trp, Tyr в положении 234; (д) Trp в положении 235; (е) Ala, Asp, Glu, His, Ile, Leu, Met, Asn, Gln, Ser, Thr, Val в положении 236; (ж) Asp, Tyr в положении 237; (з) Glu, Ile, Met, Gln, Tyr в положении 238; (и) Ile, Leu, Asn, Pro, Val в положении 239; (к) Ile в положении 264; (л) Phe в положении 266; (м) Ala, His, Leu в положении 267; (н) Asp, Glu в положении 268; (о) Asp, Glu, Gly в положении 271; (п) Leu в положении 295; (р) Leu в положении 298; (с) Glu, Phe, Ile, Leu в положении 325; (т) Thr в положении 326; (у) Ile, Asn в положении 327; (ф) Thr в положении 328; (х) Lys, Arg в положении 330; (ц) Glu в положении 331; (ч) Asp в положении 332; (ш) Asp, Ile, Met, Val, Tyr в положении 334; и (щ) Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp, Tyr в положении 396; согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В следующем варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит по меньшей мере одно аминокислотное изменение (например, замену), выбранное из группы, состоящей из: (а) Gly, Thr в положении 231; (б) Asp в положении 232; (в) Trp в положении 235; (г) Asn, Thr в положении 236; (д) Val в положении 239; (е) Asp, Glu в положении 268; (ж) Leu в положении 295; (з) Leu в положении 298; (и) Thr в положении 326; (к) Lys, Arg в положении 330 и (л) Lys, Met в положении 396 согласно EU-нумерации. В следующем варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Asn в положении 236, Glu в положении 268, Lys в положении 330 и Met в положении 396 согласно EU-нумерации. В следующем варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Asn в положении 236, Asp в положении 268 и Lys в положении 330 согласно EU-нумерации. В следующем варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Asn в положении 236, Asp в положении 268, Leu в положении 295 и Lys в положении 330 согласно EU-нумерации. В следующем варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Thr в положении 236, Asp в положении 268 и Lys в положении 330 согласно EU-нумерации. В следующем варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Asn в положении 236, Asp в положении 268, Leu в положении 295, Thr в положении 326 и Lys в положении 330 согласно EU-нумерации. В следующем варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Trp в положении 235, Asn в положении 236, Asp в положении 268, Leu в положении 295, Thr в положении 326, и Lys в положении 330 согласно EU-нумерации.
Одним из объектов изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью, которые содержат аминокислотное изменение в положении 238 согласно EU-нумерации.
Одним из объектов изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью, которые содержат по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 234, 238, 250, 264, 267, 307, и 330, согласно EU-нумерации. В других вариантах осуществления изобретения полипептид содержит по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 234, 250, 264, 267, 307 и 330, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
Другим объектом изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью, которые содержат аминокислотные изменения в одном из следующих положений (1)-(9): (1) положения 234, 238, 250, 307 и 330; (2) положения 234, 238, 250, 264, 307 и 330; (3) положения 234, 238, 250, 264, 267, 307 и 330; (4) положения 234, 238, 250, 267, 307 и 330; (5) положения 238, 250, 264, 307 и 330; (6) положения 238, 250, 264, 267, 307 и 330; (7) положения 238, 250, 267, 307 и 330; (8) положения 238, 250 и 307; и (9) положения 238, 250, 307 и 330 согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В следующем варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит по меньшей мере одно аминокислотное изменение (например, замену), выбранное из группы, состоящей из: (а) Tyr в положении 234; (б) Asp в положении 238; (в) Val в положении 250, (г) Ile в положении 264; (д) Ala в положении 267; (е) Pro в положении 307 и (ж) Lys в положении 330; согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Asp в положении 238 согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Asp в положении 238, Val в положении 250 и Pro в положении 307; согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Asp в положении 238, Val в положении 250, Pro в положении 307 и Lys в положении 330; согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Asp в положении 238, Val в положении 250, Ile в положении 264, Pro в положении 307 и Lys в положении 330; согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Asp в положении 238, Val в положении 250, Ala в положении 267, Pro в положении 307, и Lys в положении 330; согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Tyr в положении 234, Asp в положении 238, Val в положении 250, Pro в положении 307 и Lys в положении 330; согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Tyr в положении 234, Asp в положении 238, Val в положении 250, Ala в положении 267, Pro в положении 307 и Lys в положении 330; согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Asp в положении 238, Val в положении 250, Ile в положении 264, Ala в положении 267, Pro в положении 307 и Lys в положении 330; согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Tyr в положении 234, Asp в положении 238, Val в положении 250, Ile в положении 264, Pro в положении 307 и Lys в положении 330; согласно EU-нумерации. В другом варианте осуществления изобретения вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью содержит аминокислотные изменения (например, замены): Tyr в положении 234, Asp в положении 238, Val в положении 250, Ile в положении 264, Ala в положении 267, Pro в положении 307 и Lys в положении 330; согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
Другим объектом изобретения являются выделенные полипептиды, которые содержат варианты Fc-областей с повышенной изоэлектрической точкой (pI). В некоторых вариантах осуществления изобретения вариант Fc-области, указанный в настоящем описании, содержит по меньшей мере два аминокислотных изменения в родительской Fc-области. В некоторых вариантах осуществления изобретения каждое из аминокислотных изменений повышает изоэлектрическую точку (pI) варианта Fc-области по сравнению с родительской Fc-областью. В основу создания указанных полипептидов положены сведения о том, что элиминацию антигена из плазмы можно ускорять с помощью антитела, у которого pI повышена путем модификации по меньшей мере двух аминокислотных остатков, например, когда антитело вводят in vivo.
Согласно настоящему изобретению pI может представлять собой либо теоретическую или определенную экспериментальным путем pI. Величину pI можно определять, например, с помощью изоэлектрического фокусирования, известного специалистам в данной области. Величину теоретической pI можно рассчитывать, например, с помощью программного обеспечения для анализа генной и аминокислотной последовательности (фирмы Genetyx и др.).
В одном из вариантов осуществления изобретения величина pI может быть повышена, например, по меньшей мере на 0,01, 0,03, 0,05, 0,1, 0,2, 0,3, 0,4, 0,5 или более, по меньшей мере на 0,6, 0,7, 0,8, 0,9 или более, по меньшей мере на 1,0, 1,1, 1,2, 1,3, 1,4, 1,5 или более или по меньшей мере на 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 3,0 или более, по сравнению с вариантом до модификации.
В некоторых вариантах осуществления изобретения аминокислота, приводящая к увеличению pI, может экспонироваться на поверхности варианта Fc-области. Согласно настоящему изобретению аминокислота, которая может экспонироваться на поверхности, как правило, представляет собой аминокислотный остаток, локализованный на поверхности полипептида, образующего вариант Fc-области. Аминокислотный остаток, локализованный на поверхности полипептида, означает аминокислотный остаток, боковая цепь которого может иметь контакт с молекулами растворителя (наиболее часто молекулами воды). Однако боковая цепь необязательно должна полностью находиться в контакте с молекулами растворителя, и даже, если только часть боковой цепи находится в контакте с молекулами растворителя, то аминокислоту рассматривают как «аминокислотный остаток, локализованный на поверхности». Аминокислотные остатки, локализованные на поверхности полипептида, включают также аминокислотные остатки, локализованные вблизи поверхности, и которые в результате могут влиять на электрический заряд другого аминокислотного остатка, даже если боковая цепь которого только частично, имеет контакт с молекулами растворителя. Специалисты в данной области могут создать гомологичную модель полипептида, например, используя поступающее в продажу программное обеспечение. Альтернативно этому, можно применять методы, известные специалистам в данной области, такие, например, как рентгеновская кристаллография. Аминокислотные остатки, которые могут экспонироваться на поверхности, определяют, например, используя координаты трехмерной модели с применением компьютерной программы, такой как программа InsightII (фирма Accelrys). Сайты, которые могут экспонироваться на поверхности, можно определять с помощью алгоритмов, известных в данной области (например, см. Lee и Richards, J. Mol. Biol. 55, 1971, сс. 379-400; Connolly, J. Appl. Cryst. 16, 1983, сс. 548-558). Сайты, которые могут экспонироваться на поверхности, можно определять с помощью программного обеспечения, пригодного для моделирования белка, и информации о трехмерной структуре. Пригодное для этой цели программное обеспечение включает, например, программное обеспечение SYBYL Biopolymer Module (фирма Tripos Associates). Когда в алгоритме требуется ввести задаваемый пользователем параметр «размер», то «размер» образца, используемый при расчетах, можно задавать как радиус, равный примерно 1,4  или менее. Кроме того, методы определения областей, которые могут экспонироваться на поверхности, описаны у Pacios (Comput. Chem. 18(4), 1994, сс. 377-386; J. Mol. Model. 1, 1995, сс. 46-53). На основе указанной выше информации можно отбирать пригодные аминокислотные остатки, локализованные на поверхности полипептида, который образует вариант Fc-области.
или менее. Кроме того, методы определения областей, которые могут экспонироваться на поверхности, описаны у Pacios (Comput. Chem. 18(4), 1994, сс. 377-386; J. Mol. Model. 1, 1995, сс. 46-53). На основе указанной выше информации можно отбирать пригодные аминокислотные остатки, локализованные на поверхности полипептида, который образует вариант Fc-области.
В некоторых вариантах осуществления изобретения полипептид содержит как вариант Fc-области, так и антигенсвязывающий домен. В других вариантах осуществления изобретения антиген представляет собой растворимый антиген. В одном из вариантов осуществления изобретения антиген присутствует в биологических жидкостях (например, плазме, интерстициальной жидкости, лимфатической жидкости, асцитной жидкости и плевральной жидкости) индивидуумов. Антиген может представлять собой также мембранный антиген.
В других вариантах осуществления изобретения антигенсвязывающая активность антигенсвязывающего домена изменяется в зависимости от уровня концентрации ионов. В одном из вариантов осуществления изобретения на концентрацию ионов не накладывается конкретное ограничение, и она означает концентрацию ионов водорода (рН) или концентрацию ионов металла. В контексте настоящего описания ионы металлов представляют собой ионы элементов группы I за исключением водорода, такие как щелочные металлы и элементы группы меди, элементы группы II, такие как щелочноземельные металлы и элементы группы цинка, элементы группы III за исключением бора, элементы группы IV за исключением углерода и кремния, элементы группы VIII, такие как элементы группы железа и группы платины, элементы, принадлежащие к подгруппе А групп V, VI и VII, и элементы металлов, таких как сурьма, висмут и полоний. Согласно настоящему изобретению ионы металлов включают, например, ионы кальция, что описано в WO 2012/073992 и WO 2013/125667. В одном из вариантов осуществления изобретения «уровень концентрации ионов» может представлять собой влияющий на биологическое поведение антигенсвязывающего домена уровень, находящийся между низкой концентрацией ионов и высокой концентрацией ионов. Кроме того, понятие «антигенсвязывающая активность антигенсвязывающих доменов изменяется в зависимости от уровня концентрации ионов» означает, что антигенсвязывающая активность антигенсвязывающих доменов изменяется в зависимости от того, является ли концентрация ионов низкой или высокой (при этом антигенсвязывающий домен обозначают как «зависящий от концентрации ионов антигенсвязывающий домен»). Антигенсвязывающая активность антигенсвязывающих доменов при высоком уровне концентрации ионов может быть выше (сильнее) или ниже (слабее), чем при низком уровне концентрации ионов. В одном из вариантов осуществления изобретения зависящие от концентрации ионов антигенсвязывающие домены (такие как рН-зависимые антигенсвязывающие домены или зависящие от концентрации ионов кальция антигенсвязывающие домены) можно получать с помощью известных методов, например, описанных в WO 2009/125825, WO 2012/073992 и WO 2013/046722.
Согласно настоящему изобретению антигенсвязывающая активность антигенсвязывающего домена при высоком уровне концентрации ионов кальция может быть выше, чем при низком уровне концентрации ионов кальция. Высокая концентрация ионов кальция не ограничена конкретными величинами, но может представлять собой концентрацию, выбранную из диапазона от 100 мкМ до 10 мМ, от 200 мкМ до 5 мМ, от 400 мкМ до 3 мМ, от 200 мкМ до 2 мМ, от 400 мкМ до 1 мМ или от 500 мкМ до 2,5 мМ, что является предпочтительным из-за близости к концентрации ионов кальция в плазме (крови) in vivo. Кроме того, низкая концентрация ионов кальция не ограничена конкретными величинами, но может представлять собой концентрацию, выбранную из диапазона от 0,1 до 30, от 0,2 до 20, от 0,5 до 10, от 1 до 5 или от 2 до 4 мкМ, что является предпочтительным из-за близости к концентрации ионов кальция в ранних эндосомах in vivo.
В одном из вариантов осуществления изобретения соотношение между антигенсвязывающими активностями при низком уровне концентрации ионов кальция и высоком уровне концентрации ионов кальция не ограничено конкретными значениями, но соотношение константы диссоциации (KD) при низком уровне концентрации ионов кальция и KD при высоком уровне концентрации ионов кальция, т.е. KD (при низком уровне концентрации ионов кальция)/KD (при высоком уровне концентрации ионов кальция) составляет 2 или выше, 10 или выше или 40 или выше. Верхний предел соотношения может составлять 400, 1000 или 10000, если указанный антигенсвязывающий домен можно получать с помощью технологий, известных специалистам в данной области. Альтернативно этому, например, можно применять константу скорости реакции диссоциации (kd) вместо KD. В этом случае соотношение kd при низком уровне концентрации ионов кальция и kd при высоком уровне концентрации ионов кальция, т.е. kd (при низком уровне концентрации ионов кальция)/ kd (при высоком уровне концентрации ионов кальция) составляет 2 или выше, 5 или выше, 10 или выше или 30 или выше. Верхний предел соотношения может составлять 50, 100 или 200, если указанный антигенсвязывающий домен можно получать с помощью технологий, известных специалистам в данной области.
Согласно настоящему изобретению антигенсвязывающая активность антигенсвязывающего домена при низкой концентрации ионов водорода (при нейтральном рН) может быть выше, чем при высокой концентрации ионов водорода (при кислом рН). Кислый рН может представлять собой, например, рН, выбранный из диапазона рН от 4,0 до 6,5, от 4,5 до 6,5, от 5,0 до 6,5 или от 5,5 до 6,5, который является предпочтительным из-за близости к величине рН in vivo в ранних эндосомах. Кислый рН может представлять собой также, например, рН 5,8 или рН 6,0. В конкретных вариантах осуществления изобретения кислый рН представляет собой рН 5,8. Кроме того, нейтральный рН может представлять собой, например, рН, выбранный из диапазона рН от 6,7 до 10,0, от 6,7 до 9,5, от 7,0 до 9,0 или от 7,0 до 8,0, который является предпочтительным из-за близости к величине рН in vivo в плазме (крови). Нейтральный рН может представлять собой также, например, рН 7,4 или рН 7,0. В конкретном варианте осуществления изобретения нейтральный рН представляет собой рН 7,4.
В одном из вариантов осуществления изобретения соотношение между антигенсвязывающими активностями в условиях кислого рН и в условиях нейтрального рН не ограничено конкретными значениями, но соотношение константны диссоциации (KD) в условиях кислого рН и KD в условиях нейтрального рН, т.е. KD (условия кислого pH)/KD (условия нейтрального рН), составляет 2 или выше, 10 или выше или 40 или выше. Верхний предел соотношения может составлять 400, 1000 или 10000, если указанный антигенсвязывающий домен можно получать с помощью технологий, известных специалистам в данной области. Альтернативно этому, например, можно применять константу скорости реакции диссоциации (kd) вместо KD. В этом случае соотношение kd в условиях кислого рН и kd в условиях нейтрального рН, т.е., kd (условия кислого pH)/kd (условия нейтрального рН) составляет 2 или выше, 5 ли выше, 10 или выше или 30 или выше. Верхний предел соотношения может составлять 50, 100 или 200, если указанный антигенсвязывающий домен можно получать с помощью технологий, известных специалистам в данной области.
В одном из вариантов осуществления изобретения, например, по меньшей мере один аминокислотный остаток заменяют аминокислотным остатком с pKa боковой цепи 4,0-8,0 и/или по меньшей мере один аминокислотный остаток с pKa боковой цепи 4,0-8,0 встраивают в антигенсвязывающий домен, как описано в WO 2009/125825. Аминокислота может быть заменена и/или встроена в любой сайт, если антигенсвязывающая активность антигенсвязывающего домена становится слабее в условиях кислого рН по сравнению с условиями нейтрального рН относительно варианта до замены или инсерции. Когда антигенсвязывающий домен имеет вариабельную область или CDR, сайт может находиться внутри вариабельной области или CDR. Специалисты в данной области могут соответствующим образом определять количество замененных или встроенных аминокислот; и это количество может быть 1 или более. Аминокислоты с pKa боковой цепи 4,0-8,0 можно применять для изменения антигенсвязывающей активности антигенсвязывающего домена в зависимости от уровня концентрации ионов водорода. Указанные аминокислоты включают, например, встречающиеся в естественных условиях аминокислоты, такие как His (Н) и Glu (E), и не встречающиеся в естественных условиях аминокислоты, такие как аналоги гистидина (US 2009/0035836), м-NO2-Tyr (pKa 7,45), 3,5-Br2-Tyr (pKa 7,21), и 3,5-I2-Tyr (pKa 7,38) (Heyl и др., Bioorg. Med. Chem. 11(17), 2003, сс. 3761-3768). Можно применять также аминокислоты с pKa боковой цепи 6,0-7,0, к которым относится, например, His (Н).
В другом варианте осуществления изобретения предпочтительные антигенсвязывающие домены для варианта Fc-области с повышенной pI являются известными и их можно получать с помощью методов, описанных в заявках на японский патент JP2015-021371 и JP2015-185254.
В некоторых вариантах осуществления изобретения вариант Fc-области с повышенной pI содержит по меньшей мере два аминокислотных изменения по меньшей мере в двух положениях, выбранных из группы, состоящей из: 285, 311, 312, 315, 318, 333, 335, 337, 341, 342, 343, 384, 385, 388, 390, 399, 400, 401, 402, 413, 420, 422, и 431, согласно EU-нумерации.
В других вариантах осуществления изобретения вариант Fc-области с повышенной pI содержит по меньшей мере два аминокислотных изменения по меньшей мере в двух положениях, выбранных из группы, состоящей из: 311, 341, 343, 384, 399, 400, 401, 402, и 413, согласно EU-нумерации.
Другим объектом изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной pI, которые содержат аминокислотные изменения в одном из следующих положений (1)-(10): (1) положения 311 и 341; (2) положения 311 и 343; (3) положения 311, 343 и 413; (4) положения 311, 384 и 413; (5) положения 311 и 399; (6) положения 311 и 401; (7) положения 311 и 413; (8) положения 400 и 413; (9) положения 401 и 413; и (10) положения 402 и 413; согласно EU-нумерации.
Метод снижения pI белка предусматривает, например, уменьшение количества аминокислот с отрицательно заряженной боковой цепью (например, аспарагиновая кислота и глутаминовая кислота) и/или увеличение количества аминокислот с положительно заряженной боковой цепью (например, аргинин, лизин и гистидин) в условиях нейтрального рН. Аминокислоты с отрицательно заряженной боковой цепью имеют отрицательный заряд, соответствующий -1, в условиях рН, что в достаточной степени превышает величину pKa боковой цепи, согласно теории, хорошо известной специалистам в данной области. Например, теоретическая величина pKa для боковой цепи аспарагиновой кислоты составляет 3,9, и боковая цепь имеет отрицательный заряд, соответствующий -1, в условиях нейтрального рН (например, в растворе с рН 7,0). И, наоборот, аминокислоты с положительно заряженной боковой цепью имеют положительный заряд, соответствующий +1, в условиях рН, которые в достаточной степени ниже, чем величина pKa их боковой цепи. Например, теоретическая величина pKa для боковой цепи аргинина составляет 12,5, и боковая цепь имеет положительный заряд, соответствующий +1, в условиях нейтрального рН (например, в растворе с рН 7,0). При этом, как известно, аминокислоты, боковые цепи которых не имеют заряда в условиях нейтрального рН (например, в растворе с рН 7,0), включают 15 типов встречающихся в естественных условиях аминокислот, т.е. аланин, цистеин, фенилаланин, глицин, изолейцин, лейцин, метионин, аспарагин, пролин, глутамин, серин, треонин, валин, триптофан и тирозин. Само собой разумеется, что аминокислоты, которые можно применять для повышения pI, могут представлять собой не встречающиеся в естественных условиях аминокислоты.
Исходя из вышеуказанного, с помощью метода повышения pI белка в условиях нейтрального рН (например, в растворе с рН 7,0) можно осуществлять изменение заряда представляющего интерес белка на +1, например, путем замены аминокислотами с незаряженными боковыми цепями аспарагиновой кислоты или глутаминовой кислоты (боковая цепь которых имеет отрицательный заряд -1) в аминокислотной последовательности белка. Кроме того, изменение заряда на +1 можно осуществлять в белке, например, путем замены аргинином или лизином (боковая цепь которых имеет положительный заряд +1) аминокислот с незаряженными боковыми цепями. Кроме того, одновременно можно осуществлять изменение заряда белка на +2 путем замены аргинином или лизином (боковая цепь которых имеет положительный заряд +1) аспарагиновой кислоты или глутаминовой кислоты (боковая цепь которых имеет отрицательный заряд -1). Альтернативно этому, для повышения pI белка аминокислоты с незаряженными боковыми цепями и/или предпочтительно аминокислоты с положительно заряженными боковыми цепями можно добавлять или встраивать в аминокислотную последовательность белка, или аминокислоты с незаряженными боковыми цепями и/или предпочтительно аминокислоты с отрицательно заряженными боковыми цепями, присутствующие в аминокислотной последовательности белка, можно изымать путем делеции. Очевидно, что, например, N-концевые и С-концевые аминокислотные остатки белка имеют заряд, присущий их основной цепи (NH3+ аминогруппы на N-конце и COO- карбонильной группы на С-конце) помимо зарядов, присущих их боковой цепи. Так, pI белка можно повышать, осуществляя в функциональных группах основной цепи определенное добавление, делецию, замену или инсерцию.
Замена аминокислоты для повышения pI включает, например, замену аминокислотой с незаряженной боковой цепью аминокислоты с отрицательно заряженной боковой цепью, замену аминокислотой с положительно заряженной боковой цепью аминокислоты с незаряженной боковой цепью и замену аминокислотой с положительно заряженной боковой цепью аминокислоты с отрицательно заряженной боковой цепью в аминокислотной последовательности родительской Fc-области, что осуществляют индивидуально или в соответствующей комбинации.
Инсерция или добавление аминокислоты для повышения pI включает, например, инсерцию или добавление аминокислоты с незаряженной боковой цепью и/или инсерцию или добавление аминокислоты с положительно заряженной боковой цепью в аминокислотную последовательность родительской Fc-области, что осуществляют индивидуально или в соответствующей комбинации.
Делеция аминокислоты для повышения pI включает, например, делецию аминокислоты с незаряженной боковой цепью и/или делецию аминокислоты с отрицательно заряженной боковой цепью в аминокислотной последовательности родительской Fc-области, что осуществляют индивидуально или в соответствующей комбинации.
В одном из вариантов осуществления изобретения встречающиеся в естественных условиях аминокислоты, которые применяют для повышения pI, можно классифицировать следующим образом: (а) аминокислота с отрицательно заряженной боковой цепью может представлять собой Glu (Е) или Asp (D); (б) аминокислота с незаряженной боковой цепью может представлять собой Ala (А), Asn (N), Cys (С), Gln (Q), Gly (G), His (H), Ile (I), Leu (L), Met (M), Phe (F), Pro (P), Ser (S), Thr (T), Trp (W), Tyr (Y) или Val (V); и (в) аминокислота с положительно заряженной боковой цепью может представлять собой His (H), Lys (K) или Arg (R). В одном из вариантов осуществления изобретения после модификации путем инсерции или замены аминокислота представляет собой Lys (K) или Arg (R).
Другим объектом изобретения являются выделенные полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью и повышенной pI. В некоторых вариантах осуществления изобретения вариант Fc-области, указанный в настоящем описании, содержит по меньшей мере два аминокислотных изменения в родительской Fc-области.
Одним из объектов изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью и повышенной pI, которые содержат по меньшей мере три аминокислотных изменения, представляющих собой: (а) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 233, 234, 235, 236, 237, 238, 239, 264, 266, 267, 268, 271, 295, 298, 325, 326, 327, 328, 330, 331, 332, 334 и 396, согласно EU-нумерации, и (б) по меньшей мере два аминокислотных изменения по меньшей мере в двух положениях, выбранных из группы, состоящей из: 285, 311, 312, 315, 318, 333, 335, 337, 341, 342, 343, 384, 385, 388, 390, 399, 400, 401, 402, 413, 420, 422 и 431, согласно EU-нумерации.
Одним из объектов изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью и повышенной pI, которые содержат по меньшей мере три аминокислотных изменения, представляющих собой: (а) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 235, 236, 239, 268, 295, 298, 326, 330, и 396, согласно EU-нумерации, и (б) по меньшей мере два аминокислотных изменения по меньшей мере в двух положениях, выбранных из группы, состоящей из: 311, 341, 343, 384, 399, 400, 401, 402 и 413, согласно EU-нумерации.
Другим объектом изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью и повышенной pI, которые содержат аминокислотные изменения в одном из следующих положений (1)-(9): (1) положения 235, 236, 268, 295, 311, 326, 330 и 343; (2) положения 236, 268, 295, 311, 326, 330 и 343; (3) положения 236, 268, 295, 311, 330 и 413; (4) положения 236, 268, 311, 330, 396 и 399; (5) положения 236, 268, 311, 330 и 343;(6) положения 236, 268, 311, 330, 343 и 413; (7) положения 236, 268, 311, 330, 384 и 413; (8) положения 236, 268, 311, 330 и 413; и (9) положения 236, 268, 330, 396, 400 и 413; согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
Одним из объектов изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью и повышенной pI, которые содержат по меньшей мере три аминокислотных изменения, представляющих собой: (а) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 234, 238, 250, 264, 267, 307 и 330, и (б) по меньшей мере два аминокислотных изменения по меньшей мере в двух положениях, выбранных из группы, состоящей из: 285, 311, 312, 315, 318, 333, 335, 337, 341, 342, 343, 384, 385, 388, 390, 399, 400, 401, 402, 413, 420, 422, и 431, согласно EU-нумерации. В других вариантах осуществления изобретения полипептиды содержат по меньшей мере два аминокислотных изменения по меньшей мере в двух положениях, выбранных из группы, состоящей из: 311, 341, 343, 384, 399, 400, 401, 402 и 413, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
Другим объектом изобретения являются полипептиды, содержащие варианты Fc-областей с повышенной Fc-гамма RIIb-связывающей активностью и повышенной pI, которые содержат аминокислотные изменения в одном из следующих положений (1)-(16): (1) положения 234, 238, 250, 264, 307, 311, 330 и 343; (2) положения 234, 238, 250, 264, 307, 311, 330 и 413; (3) положения 234, 238, 250, 264, 267, 307, 311, 330 и 343; (4) положения 234, 238, 250, 264, 267, 307, 311, 330 и 413; (5) положения 234, 238, 250, 267, 307, 311, 330 и 343; (6) положения 234, 238, 250, 267, 307, 311, 330 и 413; (7) положения 234, 238, 250, 307, 311, 330 и 343; (8) положения 234, 238, 250, 307, 311, 330 и 413; (9) положения 238, 250, 264, 267, 307, 311, 330 и 343; (10) положения 238, 250, 264, 267, 307, 311, 330 и 413; (11) положения 238, 250, 264, 307, 311, 330 и 343; (12) положения 238, 250, 264, 307, 311, 330 и 413; (13) положения 238, 250, 267, 307, 311, 330 и 343; (14) положения 238, 250, 267, 307, 311, 330 и 413; (15) положения 238, 250, 307, 311, 330 и 343; и (16) положения 238, 250, 307, 311, 330 и 413 согласно EU-нумерации.
В следующем варианте осуществления изобретения вариант Fc-области содержит аминокислотные изменения, выбранные из любых единичных изменений, комбинации единичных изменений или комбинаций изменений, указанных в таблицах 14-30.
В некоторых вариантах осуществления изобретения полипептид содержит вариант Fc-области, предлагаемый в настоящем изобретении. В следующем варианте осуществления изобретения полипептид представляет собой константную область тяжелой цепи антитела. В следующем варианте осуществления изобретения полипептид представляет собой тяжелую цепь антитела. В следующем варианте осуществления изобретения полипептид представляет собой антитело. В следующем варианте осуществления изобретения полипептид представляет собой содержащий Fc слитый белок.
Другим вариантом осуществления изобретения является полипептид, содержащий одну из аминокислотных последовательностей SEQ ID NO: 229-381.
В контексте настоящего описания понятие «родительская Fc-область» относится к Fc-области до интродукции в нее аминокислотного(ых) изменения(й), указанного(ых) в настоящем описании. Предпочтительными примерами родительской Fc-области являются Fc-области из нативных антител. Антитела включают, например, IgA (IgA1, IgA2), IgD, IgE, IgG (IgG1, IgG2, IgG3, IgG4) и IgM или т.п. Антитела могут иметь происхождение из организма человека или обезьян (например, циномолгус, макаки резус, мармозетки, шимпанзе или бабуины). Нативные антитела могут включать также встречающиеся в естественных условиях мутации. Множество аллотипических последовательностей IgG, связанных с генетическим полиморфизмом, описаны в «Sequences proteins of immunological interest», публикация NIH №91-3242, и любую из них можно применять в настоящем изобретении. В частности, в случае человеческого IgG1 аминокислотная последовательность в положениях 356-358 (EU-нумерация) может представлять собой либо DEL, либо ЕЕМ. Предпочтительные примеры родительской Fc-области включают Fc-области из константной области тяжелой цепи человеческого IgG1 (SEQ ID NO: 195), человеческого IgG2 (SEQ ID NO: 196), человеческого IgG3 (SEQ ID NO: 197) и человеческого IgG4 (SEQ ID NO: 198). Другим предпочтительным примером родительской Fc-области является Fc-область из константной области тяжелой цепи SG1 (SEQ ID NO: 9). Кроме того, родительская Fc-область может представлять собой Fc-область, полученную путем добавления аминокислотного(ых) изменения(й), отличного(ых) от аминокислотного(ых) изменения(й), указанного(ых) в настоящем описании для Fc-области из нативного антитела.
Кроме того, аминокислотные изменения, которые осуществляют для другой(их) цели(ей), можно объединять в варианте Fc-области, указанном в настоящем описании. Например, можно добавлять аминокислотные замены, которые повышают FcRn-связывающую активность (Hinton и др., J. Immunol. 176(1), 2006, сс. 346-356; Dall'Acqua и др, J. Biol. Chem. 281(33), 2006, сс. 23514-23524; Petkova и др., Intl. Immunol. 18(12), 2006, сс. 1759-1769; Zalevsky и др., Nat. Biotechnol. 28(2), 2010, сс. 157-159; WO 2006/019447; WO 2006/053301; и WO 2009/086320), и аминокислотные замены, которые повышают гетерогенность или стабильность антитела (WO 2009/041613). Альтернативно этому, полипептиды, обладающие способностью усиливать клиренс антигена, которые описаны в WO 2011/122011, WO 2012/132067, WO 2013/046704 или WO 2013/180201, полипептиды, обладающие способностью специфически связываться с тканью-мишенью, которые описаны в WO 2013/180200, полипептиды, обладающие способностью повторно связываться с множеством молекул антигенов, которые описаны в WO 2009/125825, WO 2012/073992 или WO 2013/047752, можно объединять с вариантом Fc-области, указанном в настоящем описании. Альтернативно этому, с целью придания способности связываться с другими антигенами аминокислотные изменения, описанные в ЕР 1752471 и ЕР 1772465, можно объединять в СН3 варианта Fc-области, указанного в настоящем описании. Альтернативно этому, с целью повышения удерживания в плазме можно объединять аминокислотные изменения, которые снижают pI константной области (WO 2012/016227), в варианте Fc-области, указанном в настоящем описании. Альтернативно этому, с целью повышения поглощения клеткой можно объединять аминокислотные изменения, которые повышают pI константной области (WO 2014/145159), в варианте Fc-области, указанном в настоящем описании. Альтернативно этому, с целью усиления элиминации молекулы-мишени из плазмы можно объединять аминокислотные изменения, которые повышают pI константной области (заявки на японский патент JP2015-021371 и JP2015-185254), в варианте Fc-области, указанном в настоящем описании. В одном из вариантов осуществления изобретения указанное изменение может включать, например, замену по меньшей мере одного положения, выбранного из группы, состоящей из 311, 343, 384, 399, 400 и 413 согласно EU-нумерации. В другом варианте осуществления изобретения указанная замена может представлять собой замену аминокислоты на Lys или Arg в каждом положении.
Аминокислотные изменения, приводящие к повышению способности связываться с человеческим FcRn при кислом рН, можно объединять также в варианте Fc-области, указанным в настоящем описании. В частности, такие изменения могут включать, например, замену на Leu Met в положении 428 и замену на Ser Asn в положении 434, согласно EU-нумерации (Zalevsky и др., Nat. Biotechnol. 28, 2010, сс. 157-159); замену на Ala Asn в положении 434 (Deng и др., Metab. Dispos. 38(4), 2010, сс. 600-605); замену на Tyr Met в положении 252, замену на Thr Ser в положении 254 и замену на Glu Thr в положении 256 (Dall'Acqua и др., J. Biol. Chem. 281, 2006, сс. 23514-23524); замену на Gln Thr в положении 250 и замену на Leu Met в положении 428 (Hinton и др., J. Immunol. 176(1), 2006, сс. 346-356); замену на His Asn в положении 434 (Zheng и др., Clin. Pharmacol. Ther. 89(2), 2011, сс. 283-290), и изменения, описанные в WO 2010/106180, WO 2010/045193, WO 2009/058492, WO 2008/022152, WO 2006/050166, WO 2006/053301, WO 2006/031370, WO 2005/123780, WO 2005/047327, WO 2005/037867, WO 2004/035752 или WO 2002/060919. Такие изменения могут включать, например, по меньшей мере одно изменение, выбранное из группы, состоящей из замены на Leu Met в положении 428, замены на Ala Asn в положении 434 и замены на Thr Tyr в положении 436. Указанные изменения включают также замену на Arg Gln в положении 438 и/или замену на Glu Ser в положении 440 (заявки на японский патент JP2015-021371 и JP2015-185254).
Два или большее количество полипептидов, содержащих вариант Fc-области, указанный в настоящем описании, можно включать в одну молекулу, в которой два полипептида, содержащие варианты Fc-областей, находятся в ассоциации, максимально напоминающей ассоциацию в антителе. Тип антитела не ограничен и можно применять IgA (IgA1, IgA2), IgD, IgE, IgG (IgG1, IgG2, IgG3, IgG4) и IgM или т.п.
Два ассоциированных полипептида, содержащих варианты Fc-областей, могут представлять собой полипептиды, которые содержат варианты Fc-областей, в которые интродуцировано(ы) одинаковое(ые) аминокислотное(ые) изменение(я) (далее в контексте настоящего описания обозначены как гомологичные варианты Fc-областей), или полипептиды, которые содержат варианты Fc-областей, в которые интродуцировано(ы) различные аминокислотное(ые) изменение(я), или альтернативно этому, полипептиды, которые содержат варианты Fc-областей, отличающиеся тем, что аминокислотное(ые) изменение(я) интродуцировано(ы) только в одну из Fc-областей ((далее в контексте настоящего описания обозначены как гетерологичные полипептиды вариантов Fc-областей). Одним из предпочтительных аминокислотных изменений является изменение в петлевой структуре от положения 233 до 239 (EU-нумерация) в СН2-домене Fc-области, который участвует в связывании с Fc-гамма RIIb и Fc-гамма RIIa. Предпочтительно изменение, которое повышает Fc-гамма RIIb-связывающую активность и/или селективность, интродуцируют в петлевую структуру СН2-домена одной из Fc-областей, а в петлевую структуру СН2-домена другой Fc-области интродуцируют другое изменение, которое дестабилизирует ее. Примеры аминокислотных изменений, которые могут дестабилизировать петлевую структуру СН2-домена, могут представлять собой замену по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 235, 236, 237, 238 и 239, на другую аминокислоту. В частности, можно осуществлять дестабилизацию, например, путем изменения аминокислоты в положении 235 на Asp, Gln, Glu или Thr, изменения аминокислоты в положении 236 на Asn, изменения аминокислоты в положении 237 на Phe или Trp, изменения аминокислоты в положении 238 на Glu, Gly или Asn и изменения аминокислоты в положении 239 на Asp или Glu, согласно EU-нумерации.
Для ассоциации гетерологичных полипептидов, содержащих варианты Fc-областей, можно применять методику подавления нежелательной ассоциации гомологичных полипептидов, содержащих варианты Fc-областей, путем интродукции электростатического отталкивания в поверхность раздела СН2- или СН3-домена Fc-области, описанную в WO 2006/106905.
Примеры аминокислотных остатков, контактирующих на поверхности раздела СН2- или СН3-домена Fc-области, включают остаток в положении 356 (EU-нумерация), остаток в положении 439 (EU-нумерация), остаток в положении 357 (EU-нумерация), остаток в положении 370 (EU-нумерация), остаток в положении 399 (EU-нумерация) и остаток в положении 409 (EU-нумерация) в СН3-домене.
Более конкретно, например, можно получать Fc-область, в которой 1-3 пары аминокислотных остатков, выбранных из указанных ниже в подпунктах (1)-(3), имеют одинаковый заряд: (1) аминокислотные остатки в положениях 356 и 439 (EU-нумерация) в СН3-домене; (2) аминокислотные остатки в положениях 357 и 370 (EU-нумерация) в СН3-домене и (3) аминокислотные остатки в положениях 399 и 409 (EU-нумерация) в СН3-домене.
Кроме того, можно получать гетерологичные полипептиды, содержащие варианты Fc-областей, в которых 1-3 пары аминокислотных остатков, выбранных из указанных выше в подпунктах (1)-(3), имеют одинаковый заряд в CH3-домене первой Fc-области, и пары аминокислотных остатков, выбранных в указанной выше первой Fc-области, имеют также одинаковый заряд в CH3-домене второй Fc-области, при условии, что заряды в первой и второй Fc-областях являются противоположными.
В вышеуказанных Fc-областях, например, отрицательно заряженные аминокислотные остатки предпочтительно выбирают из глутаминовой кислоты (Е) и аспарагиновой кислоты (D), а положительно заряженные аминокислотные остатки предпочтительно выбирают из лизина (K), аргинина (R) и гистидина (Н).
Для ассоциации гетерологичных полипептидов, содержащих варианты Fc-областей, можно применять также другие известные методики. В частности, указанную методику осуществляют путем замены боковой цепи аминокислоты в одной из Fc-областей на более крупную боковую цепь (выступ; который обозначают как «выпуклость») и путем замены боковой цепи аминокислоты, присутствующей в Fc-области, на меньшую боковую цепь (впадина; которую обозначают как «полость»), помещая выступ во впадину. Это может способствовать эффективной ассоциации между содержащими Fc-области полипептидами, которые имеют отличные друг от друга аминокислотные последовательности (WO 1996/027011; Ridgway и др., Prot. Eng. 9, 1996, сс. 617-621; Merchant и др., Nat. Biotech. 16, 1998, сс. 677-681).
Кроме того, другие известные методики можно применять также для ассоциации гетерологичных полипептидов, содержащих варианты Fc-областей. Ассоциацию полипептидов, содержащих Fc-область, можно эффективно индуцировать, используя созданный на основе обмена цепи CH3-домен гетеродимеров (Davis и др., Prot. Eng. Des. & Sel., 23, 2010, сс. 195-202). Указанную методику можно применять также для эффективной индукции ассоциации между содержащими Fc-области полипептидами с различными аминокислотными последовательностями.
Кроме того, можно применять также методы получения гетеродимеризированных антител, в которых используют ассоциацию СН1 и CL антитела и ассоциацию VH и VL, описанные в WO 2011/028952.
Аналогично методу, описанному в WO 2008/119353 и WO 2011/131746, можно также применять методику получения гетеродимеризированных антител путем предварительного создания двух типов гомодимеризованных антител, инкубации антител в восстанавливающих условиях для их диссоциации и давая им возможность вновь ассоциироваться.
Аналогично методу, описанному у Strop, J. Mol. Biol. 420, 2012, сс. 204-219, можно также применять методику получения гетеродимеризированных антител путем интродукции заряженных остатков, таких как Lys, Arg, Glu и Asp, для интродукции электростатического отталкивания в CH3-домены.
Кроме того, аналогично методу, описанному в WO 2012/058768, можно применять также методику получения гетеродимеризированных антител путем введения изменений в СН2- и CH3-домены.
При одновременной экспрессии двух полипептидов, содержащих вариант Fc-области, которые имеют различные аминокислотные последовательности, для получения полипептидов, содержащих гетерологичные варианты Fc-областей, полипептиды, содержащие гомологичные варианты Fc-областей, как правило, образуются в качестве примесей. В таких случаях полипептиды, содержащие гетерологичные варианты Fc-областей, можно эффективно получать путем отделения их и очистки от полипептидов, содержащих гомологичные варианты Fc-областей, используя известные методики. Описан метод эффективного разделения и очистки гетеродимеризованных антител от гомодимеризованных антител, основанный на применении ионообменной хроматографии, путем интродукции аминокислотных изменений в вариабельные области двух типов тяжелых цепей антител для создания различия в изоэлектрических точках между гомодимеризованными антителами и гетеродимеризованными антителами (WO 2007/114325). Описан другой метод очистки гетеродимеризированных антител с использованием хроматографии на белке А путем конструирования гетеродимеризованного антитела, содержащего два типа тяжелых цепей из мышиного IgG2a, которые связываются с белком А и крысиным IgG2b, который не связывается с белком A (WO 1998/050431 и WO 1995/033844).
Кроме того, гетеродимеризированное антитело можно эффективно очищать с помощью хроматографии на белке А путем замены аминокислотных остатков в положениях 435 и 436 (EU-нумерация), которые локализованы в сайте связывания с белком А в тяжелой цепи антитела, на аминокислоты, такие как Tyr или His, с получением различных аффинностей связывания с белком А.
В настоящем изобретении аминокислотное изменение означает любое из таких изменений, как замена, делеция, добавление, инсерция и модификация, или их комбинацию. В настоящем изобретении аминокислотное изменение можно обозначать как аминокислотная мутация.
При замене аминокислотных остатков замену на другой аминокислотный остаток можно осуществлять с целью изменения характеристик, которые указаны ниже в подпунктах (а)-(в): (а) структура полипептидного каркаса области складчатой структуры или спиралеобразной структуры; (б) электрический заряд или гидрофобность в сайте-мишени; или (в) размер боковой цепи.
Аминокислотные остатки классифицируют в следующие группы на основе общих свойств боковой цепи: (а) гидрофобные: норлейцин, Met, Ala, Val, Leu и Ile; (б) нейтральные гидрофильные: Cys, Ser, Thr, Asn и Gln; (в) кислотные: Asp и Glu; (г)  : His, Lys и Arg; (д) остатки, которые влияют на ориентацию цепи: Gly и Pro; и (е) ароматические: Trp, Tyr и Phe.
: His, Lys и Arg; (д) остатки, которые влияют на ориентацию цепи: Gly и Pro; и (е) ароматические: Trp, Tyr и Phe.
Аминокислотные изменения получают с помощью различных методов, известных специалистам в данной области. Указанные методы включают (но, не ограничиваясь только ими) метод сайтнаправленного мутагенеза (Hashimoto-Gotoh и др., Gene 152, 1995, сс. 271-275; Zoller, Meth. Enzymol. 100, 1983, сс. 468-500; Kramer и др., Nucleic Acids Res. 12, 1984, сс. 9441-9456; Kramer и Fritz, Methods Enzymol. 154, 1987, сс. 350-367 и Kunkel, Proc. Natl. Acad. Sci. USA 82, 1985, сс. 488-492), метод на основе ПЦР-мутагенеза и кассетного мутагенеза.
Количество аминокислотных изменений, интродуцированных с Fc-область, не ограничено. В некоторых вариантах осуществления изобретения оно может составлять 1, 2 или менее, 3 или менее, 4 или менее, 5 или менее, 6 или менее, 8 или менее, 10 или менее, 12 или менее, 14 или менее, 16 или менее, 18 или менее или 20 или менее.
Аминокислотная модификация включает пост-трансляционную модификацию. Специфическая пост-трансляционная модификация может представлять собой добавление или делецию сахарной цепи. Например, аминокислотный остаток в положении 297 (EU-нумерации) в константной области IgG1 может быть модифицирован сахарной цепью. Структура сахарной цепи не ограничена конкретной структурой. Например, можно добавлять сиаловую кислоту в сахарную цепь Fc-области (MAbs 2(5), сентябрь-октябрь 2010 г., сс. 519-527). Как правило, антитела, которые экспрессируются в эукариотических клетках, содержат сайт гликозилирования в константной области. Например, известно, что несколько типов сахарных цепей в норме добавляются в антитела, экспрессируемые в клетках, таких как встречающиеся в естественных условиях продуцирующие антитела клетки млекопитающих или эукариотические клетки, трансформированные экспрессионным вектором, который содержит ДНК, кодирующую антитело.
В контексте настоящего описания эукариотические клетки включают клетки дрожжей и животных. Например, СНО-клетки и HEK293-клетки являются репрезентативными клетками животных, которые применяют для трансформации экспрессионным вектором, содержащим ДНК, которая кодирует антитело. С другой стороны, под объем настоящего изобретения подпадают также константные области без гликозилирования. Антитела, константные области которых не являются гликозилированными, можно получать путем экспрессии кодирующего антитело гена в прокариотических клетках, таких как Escherichia coli.
Кроме того, полипептид, содержащий вариант Fc-области, предлагаемый в настоящем изобретении, можно химически модифицировать с помощью различных молекул, таких как полиэтиленгликоль (ПЭГ) и цитотоксические субстанции. Методы указанной химической модификации полипептидов разработаны в данной области.
Одним из объектов изобретения является выделенный полипептид, содержащий вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В некоторых объектах изобретения антитело представляет собой химерное антитело или гуманизированное антитело. Происхождение антитела не ограничено конкретным происхождением, но примеры включают человеческое антитело, мышиное антитело, крысиное антитело, и кроличье антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок.
Вариабельные области антител, которые содержат вариант Fc-области, указанный в настоящем описании, и связывающие белок мотивы содержащий Fc слитых белков, которые содержат вариант Fc-области, могут распознавать любой антиген. Примеры антигенов, которые могут связываться такими антителами и слитыми белками, включают (но, не ограничиваясь только ими) лиганды (цитокины, хемокины и т.п.), рецепторы, раковые антигены, антигены ГКГС, дифференцировочные антигены, иммуноглобулины и иммунные комплексы, частично включающие иммуноглобулины.
Примеры цитокинов, которые могут связываться антителом или слитым белком, содержащим вариант Fc-области, предлагаемый в изобретении, и/или рекомбинантно слитым с полипептидом, который содержит указанный вариант Fc-области, включают (но, не ограничиваясь только ими) интерлейкины 1-18, колониестимулирующие факторы (G-CSF, M-CSF, GM-CSF и т.д.), интерфероны (IFN-альфа, IFN-бета, IFN-гамма и т.д.), факторы роста (EGF, FGF, IGF, NGF, PDGF, TGF, HGF и т.д.), факторы некроза опухоли (TNF-альфа и TNF-бета), лимфотоксин, эритропоэтин, лептин, SCF, ТРО, MCAF и BMP.
Примеры хемокинов, которые могут связываться антителом или слитым белком, содержащим вариант Fc-области, предлагаемый в изобретении, и/или рекомбинантно слитым с полипептидом, который содержит указанный вариант Fc-области, включают (но, не ограничиваясь только ими) СС-хемокины, такие как CCL1-CCL28, СХС-хемокины, такие как CXCL1-CXCL17, С-хемокины, такие как XCL1-XCL2, и СХ3С-хемокины, такие как CX3CL1.
Примеры рецепторов, которые могут связываться антителом или слитым белком, содержащим вариант Fc-области, предлагаемый в изобретении, и/или рекомбинантно слитым с полипептидом, который содержит указанный вариант Fc-области, включают (но, не ограничиваясь только ими) рецепторы, принадлежащие к семействам рецепторов, таких как семейство рецепторов гематопоэтического фактора роста, семейство цитокиновых рецепторов, семейство рецепторов тирозинкиназного-типа, семейство рецепторов серин/треонинкиназного типа, семейство рецепторов TNF, семейство рецепторов, сопряженных с белком G, семейство GPI-заякоренных рецепторов, семейство рецепторов тирозинфосфатазного типа, семейство факторов адгезии и семейство рецепторов гормонов. Рецепторы, принадлежащие к указанным семействам рецепторов, и их характеристики описаны во многих документах, таких как New Comprehesive Biochemistry под ред. Cooke, т. 18В «Hormones и their Actions Part II», 1988, сс. 1-46, изд-во Elsevier Science Publishers BV; Patthy, Cell 61(1), 1990, сс. 13-14; Ullrich, Cell 61(2), 1990, сс. 203-212; Massague, Cell 69(6), 1992, сс. 1067-1070; Miyajima и др., Annu. Rev. Immunol. 10, 1992, сс. 295-331; Taga и др., FASEB J. 6, 1992, сс. 3387-3396; Fantl и др., Annu. Rev. Biochem. 62, 1993, сс. 453-481; Smith и др., Cell 76(6), 1994, сс. 959-962 и Flower, Biochim. Biophys. Acta 1422(3), 1999, сс. 207-234.
Примеры специфических рецепторов, принадлежащих к вышеуказанным семействам рецепторов, включают человеческие или мышиные рецепторы эритропоэтина (ЕРО) (Jones и др., Blood 76(1), 1990, сс. 31-35; D'Andrea и др., Cell 57(2), 1989, сс. 277-285), человеческие или мышиные рецепторы гранулоцитарного колониестимулирующего фактора (G-CSF) (Fukunaga и др., Proc. Natl. Acad. Sci. USA 87(22), 1990, сс. 8702-8706), mG-CSFR (Fukunaga и др., Cell 61(2), 1990, сс. 341-350), человеческие или мышиные рецепторы тромбопоэтина (ТРО) (Vigon и др., Proc. Natl. Acad. Sci. USA. 89(12), 1992, сс. 5640-5644; Skoda и др., ЕМВО J. 12(7), 1993, сс. 2645-2653), человеческие или мышиные рецепторы инсулина (Ullrich и др., Nature 313(6005), 1985, сс. 756-761), человеческие или мышиные рецепторы для лиганда Flt-3 (Small и др., Proc. Natl. Acad. Sci. USA. 91(2), 1994, сс. 459-463), человеческие или мышиные рецепторы тромбоцитарного фактора роста (PDGF) (Gronwald и др., Proc. Natl. Acad. Sci. USA. 85(10), 1988, сс. 3435-3439), человеческие или мышиные рецепторы интерферона (IFN)-альфа и -бета (Uze и др., Cell 60(2), 1990, сс. 225-234 (1990); Novick и др, Cell 77(3), 1994, сс. 391-400), человеческие или мышиные рецепторы лептина, человеческие или мышиные рецепторы гормонов роста (GH), человеческие или мышиные рецепторы интерлейкина (IL)-10, человеческие или мышиные рецепторы инсулиноподобного фактора роста (IGF)-I, человеческие или мышиные рецепторы ингибирующего лейкоз фактора (LIF), и человеческие или мышиные рецепторы цилиарного нейротрофического фактора (CNTF).
Раковые антигены представляют собой антигены, которые экспрессируются на клетках, которые становятся злокачественными, и их называют также опухольспецифическими антигенами. Аномальные сахарные цепи, которые появляются на клеточной поверхности или на белковых молекулах, когда клетки становятся раковыми, относятся также к раковым антигенам, и их называют также раковыми антигенами с сахарными цепями. Примеры раковых антигенов, которые могут связываться антителом или слитым белком, содержащим вариант Fc-области, предлагаемый в изобретении, включают (но, не ограничиваясь только ими) GPC3, который является рецептором, принадлежащим к указанному выше семейству GPI-заякоренных рецепторов, и экспрессируется также при некоторых видах рака, включая рак печени (Midorikawa и др., Int. J. Cancer 103(4), 2003, сс. 455-465), а также ЕрСАМ, который экспрессируется при некоторых видах рака, включая рак легкого (Linnenbach и др., Proc. Natl. Acad. Sci. USA 86(1), 1989, сс. 27-31), СА19-9, СА15-3 и сиалил-SSEA-1 (SLX).
Антигены ГКГС грубо классифицируют на антигены ГКГС класса I и антигены ГКГС класса II. Антигены ГКГС класса I включают HLA-A, -В, -С, -Е, -F, -G и -Н, а антигены ГКГС класса II включают HLA-DR, -DQ и -DP.
Примеры дифференцировочных антигенов, которые могут связываться антителом или слитым белком, содержащим вариант Fc-области, предлагаемый в изобретении, и/или рекомбинантно слитым с полипептидом, который содержит указанный вариант Fc-области, включают (но, не ограничиваясь только ими) CD1, CD2, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15s, CD16, CD18, CD19, CD20, CD21, CD23, CD25, CD28, CD29, CD30, CD32, CD33, CD34, CD35, CD38, CD40, CD41a, CD41b, CD42a, CD42b, CD43, CD44, CD45, CD45RO, CD48, CD49a, CD49b, CD49c, CD49d, CD49e, CD49f, CD51, CD54, CD55, CD56, CD57, CD58, CD61, CD62E, CD62L, CD62P, CD64, CD69, CD71, CD73, CD95, CD102, CD106, CD122, CD126 и CDw130.
Иммуноглобулины включают IgA, IgM, IgD, IgG и IgE. Иммунные комплексы включают компонент по меньшей мере любого из иммуноглобулинов.
Другие примеры антигенов, которые могут связываться антителом или слитым белком, содержащим вариант Fc-области, предлагаемый в изобретении, и/или рекомбинантно слитым с полипептидом, который содержит указанный вариант Fc-области, включают (но, не ограничиваясь только ими) 17-IA, 4-1BB, 4Dc, 6-кето-PGF1a, 8-изо-PGF2a, 8-оксо-dG, A1-рецептор аденозина, А33, АСЕ, АСЕ-2, активин, активин А, активин АВ, активин В, активин С, активин RIA, активин RIA ALK-2, активин RIB ALK-4, активин RIIA, активин RIIB, ADAM, ADAM10, ADAM12, ADAM15, ADAM17/TACE, ADAM8, ADAM9, ADAMTS, ADAMTS4, ADAMTS5, адрессин, aFGF, ALCAM, ALK, ALK-1, ALK-7, альфа-1-антитрипсин, антагонист альфа-V/бета, ANG, Ang, APAF-1, APE, APJ, АРР, APRIL, AR, ARC, ART, артемин, анти-Id, ASPARTIC, предсердный натрийуретический пептид, av/b3-интегрин, Ax1, b2M, B7-1, В7-2, В7-Н, стимулирующий В-лимфоциты фактор (BlyS), ВАСЕ, ВАСЕ-1, Bad, BAFF, BAFF-R, Bag-1, BAK, Вах, ВСА-1, ВСАМ, Bcl, BCMA, BDNF, b-ECGF, bFGF, BID, Bik, BIM, BLC, BL-CAM, BLK, BMP, BMP-2 BMP-2a, остеогенин BMP-3, BMP-4, BMP-2b, BMP-5, BMP-6 Vgr-1, BMP-7 (OP-1), BMP-8 (BMP-8a, OP-2), BMPR, BMPR-IA (ALK-3), BMPR-IB (ALK-6), BRK-2, RPK-1, BMPR-II (BRK-3), BMP, b-NGF, BOK, бомбезин, костный нейротропный фактор, BPDE, BPDE-DNA, BTC, фактор системы комплемента 3 (С3), С3а, С4, С5, С5а, С10, СА125, CAD-8, кальцитонин, цАМФ, карциноэмбриональный антиген (СЕА), ассоциированный с раком антиген, катепсин А, катепсин В, катепсин C/DPPI, катепсин D, катепсин Е, катепсин Н, катепсин L, катепсин О, катепсин S, катепсин V, катепсин X/Z/P, CBL, CCI, CCK2, CCL, CCL1, CCL11, CCL12, CCL13, CCL14, CCL15, CCL16, CCL17, CCL18, CCL19, CCL2, CCL20, CCL21, CCL22, CCL23, CCL24, CCL25, CCL26, CCL27, CCL28, CCL3, CCL4, CCL5, CCL6, CCL7, CCL8, CCL9/10, CCR, CCR1, CCR10, CCR11, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CD1, CD2, CD3, CD3E, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15, CD16, CD18, CD19, CD20, CD21, CD22, CD23, CD25, CD27L, CD28, CD29, CD30, CD30L, CD32, CD33 (белок р67), CD34, CD38, CD40, CD40L, CD44, CD45, CD46, CD49a, CD52, CD54, CD55, CD56, CD61, CD64, CD66e, CD74, CD80 (В7-1), CD89, CD95, CD123, CD137, CD138, CD140a, CD146, CD147, CD148, CD152, CD164, CEACAM5, CFTR, цГМФ, CINC, токсин ботулотоксин, токсин Clostridium perfringens, CKb8-1, CLC, CMV, CMV UL, CNTF, CNTN-1, COX, C-Ret, CRG-2, CT-1, CTACK, CTGF, CTLA-4, CX3CL1, CX3CR1, CXCL, CXCL1, CXCL2, CXCL3, CXCL4, CXCL5, CXCL6, CXCL7, CXCL8, CXCL9, CXCL10, CXCL11, CXCL12, CXCL13, CXCL14, CXCL15, CXCL16, CXCR, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6, антиген, ассоциированный с цитокератином опухоли, DAN, DCC, DcR3, DC-SIGN, регулирующий комплемент фактор (фактор ускорения диссоциации), des(1-3)-IGF-I (IGF-1 головного мозга), Dhh, дигоксин, DNAM-1, ДКазу, Dpp, DPPIV/CD26, Dtk, ECAD, EDA, EDA-A1, EDA-A2, EDAR, EGF, EGFR (ErbB-1), EMA, EMMPRIN, ENA, рецептор эндотелина, энкефалиназу, eNOS, Eot, эотаксин 1, EpCAM, эфрин B2/EphB4, EPO, ERCC, E-селектин, ET-1, фактор IIa, фактор VII, фактор VIIIc, фактор IX, фибробласт-активирующий белок (FAP), Fas, FcR1, FEN-1, ферритин, FGF, FGF-19, FGF-2, FGF3, FGF-8, FGFR, FGFR-3, фибрин, FL, FLIP, Flt-3, Flt-4, фолликулостимулирующий гормон, фракталкин, FZD1, FZD2, FZD3, FZD4, FZD5, FZD6, FZD7, FZD8, FZD9, FZD10, G250, Gas6, GCP-2, GCSF, GD2, GD3, GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF8 (миостатин), GDF-9, GDF-15 (MIC-1), GDNF, GFAP, GFRa-1, GFR-альфа1, GFR-альфа2, GFR-альфа3, GITR, глюкагон, Glut4, гликопротеин IIb/IIIa (GPIIb/IIIa), GM-CSF, gp130, gp72, GRO, рилизинг-фактор ростового гормона, гаптен (NP-cap или NIP-cap), HB-EGF, HCC, HCMV оболочечный гликопротеин gB, оболочечный гликопротеин HCMV gH, HCMV UL, гематопоэтический фактор роста (HGF), Нер В gp120, гепараназу, Her2, Her2/neu (ErbB-2), Her3 (ErbB-3), Her4 (ErbB-4), оболочечный гликопротеин gB вируса герпеса простого (HSV), гликопротеин HSV gD, HGFA, высокомолекулярный ассоциированный с меланомой антиген (HMW-MAA), HIV gp120, HIV IIIB gp 120 V3-петли, HLA, HLA-DR, HM1.24, HMFG РЕМ, HRG, Hrk, человеческий сердечный миозин, человеческий цитомегаловирус (HCMV), человеческий гормон роста (HGH), HVEM, I-309, IAP, ICAM, ICAM-1, ICAM-3, ICE, ICOS, IFNg, Ig, IgA-рецептор, IgE, IGF, IGF-связывающий белок, IGF-1R, IGFBP, IGF-I, IGF-II, IL, IL-1, IL-1R, IL-2, IL-2R, IL-4, IL-4R, IL-5, IL-5R, IL-6, IL-6R, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-18, IL-18R, IL-23, интерферон (IFN)-альфа, IFN-бета, IFN-гамма, ингибин, iNOS, цепь А инсулина, цепь В инсулина, инсулиноподобный фактор роста1, интегрин альфа2, интегрин альфа3, интегрин альфа4, интегрин альфа4/бета1, интегрин альфа4/бета7, интегрин альфа5 (альфа V), интегрин альфа5/бета1, интегрин альфа5/бета3, интегрин альфа6, интегрин бета1, интегрин бета2, интерферон гамма, IP-10, I-TAC, JE, калликреин 2, калликреин 5, калликреин 6, калликреин 11, калликреин 12, калликреин 14, калликреин 15, калликреин L1, калликреин L2, калликреин L3, калликреин L4, KC, KDR, фактор роста кератиноцитов (KGF), ламинин 5, LAMP, LAP, LAP (TGF-1), латентный TGF-1, латентный TGF-1 bp1, LBP, LDGF, LECT2, фактор Lefty, антиген Льюис-Y, антиген, ассоциированный с системой Льюис, LFA-1, LFA-3, Lfo, LIF, LIGHT, липопротеин, LIX, LKN, Lptn, L-селектин, LT-a, LT-b, LTB4, LTBP-1, антиген поверхности легкого, лютеинизирующий гормон, рецептор лимфотоксина бета, Mac-1, MAdCAM, MAG, MAP2, MARC, MCAM, MCK-2, МСР, M-CSF, MDC, Mer, металлопротеазы, MGDF-рецептор, MGMT, МНС (HLA-DR), MIF, MIG, MIP, MIP-1-альфа, MK, ММАС1, ММР, ММР-1, ММР-10, ММР-11, ММР-12, ММР-13, ММР-14, ММР-15, ММР-2, ММР-24, ММР-3, ММР-7, ММР-8, ММР-9, MPIF, Mpo, MSK, MSP, муцин (Muc1), MUC18, ингибирующее вещество Мюллера, Mug, MuSK, NAIP, NAP, NCAD, N-кадхерин, NCA 90, NCAM, неприлизин, нейротрофин-3, -4 или -6, нейротурин, фактор роста нерва (NGF), NGFR, NGF-бета, nNOS, NO, NOS, Npn, NRG-3, NT, NTN, OB, OGG1, OPG, OPN, OSM, OX40L, OX40R, p150, p95, PADPr, паратиреоидный гормон, PARC, PARP, PBR, PBSF, PCAD, Р-кадхерин, PCNA, PDGF, PDK-1, PECAM, PEM, PF4, PGE, PGF, PGI2, PGJ2, PIN, PLA2, плацентарную щелочную фосфатазу (PLAP), PIGF, PLP, PP14, проинсулин, прорелаксин, белок С, PS, PSA, PSCA, простатспецифический мембранный антиген (PSMA), PTEN, PTHrp, Ptk, PTN, R51, RANK, RANKL, RANTES, цепь А релаксина, цепь В релаксина, ренин, респираторно-синцитиальный вирус (RSV) F, RSV Fgp, Ret, ревматоидный фактор, RLIP76, RPA2, RSK, S100, SCF/KL, SDF-1, серин, сывороточный альбумин, sFRP-3, Shh, SIGIRR, SK-1, SLAM, SLPI, SMAC, SMDF, SMOH, SOD, SPARC, Stat, STEAP, STEAP-II, TACE, TACI, TAG-72 (ассоциированный с опухолью гликопротеин-72), TARC, ТСА-3, Т-клеточный рецептор (например, Т-клеточный рецептор альфа/бета), TdT, TECK, TEM1, ТЕМ5, ТЕМ7, ТЕМ8, TERT, PLAP-подобную щелочную фосфатазу яичек, TfR, TGF, TGF-альфа, TGF-бета, TGF-бета Pan-специфический, TGF-бетаRI (ALK-5), TGF-бетаRII, TGF-бетаRIIb, TGF-бетаRIII, TGF-бета1, TGF-бета2, TGF-бета3, TGF-бета4, TGF-бета5, тромбин, Ck-1 тимуса, тиреотропный гормон, Tie, TIMP, TIQ, тканевый фактор, TMEFF2, Tmpo, TMPRSS2, TNF, TNF-альфа, TNF-альфа,бета, TNF-бета2, TNFc, TNF-RI, TNF-RII, TNFRSF10A (TRAIL R1 Apo-2, DR4), TNFRSF10B (TRAIL R2 DR5, KILLER, TRICK-2A, TRICK-B), TNFRSF10C (TRAIL R3 DcR1, LIT, TRID), TNFRSF10D (TRAIL R4 DcR2, TRUNDD), TNFRSF11A (RANK ODF R, TRANCE R), TNFRSF11B (OPG OCIF, TR1), TNFRSF12 (TWEAK R FN14), TNFRSF13B (TACI), TNFRSF13C (BAFF R), TNFRSF14 (HVEM ATAR, HveA, LIGHT R, TR2), TNFRSF16 (NGFR p75NTR), TNFRSF17 (BCMA), TNFRSF18 (GITR AITR), TNFRSF19 (TROY TAJ, TRADE), TNFRSF19L (RELT), TNFRSF1A (TNF RI CD120a, p55-60), TNFRSF1B (TNF RII CD120b, p75-80), TNFRSF26 (TNFRH3), TNFRSF3 (LTbR TNF RIII, TNFC R), TNFRSF4 (OX40 ACT35, TXGP1 R), TNFRSF5 (CD40 p50), TNFRSF6 (Fas Apo-1, APT1, CD95), TNFRSF6B (DcR3 M68, TR6), TNFRSF7 (CD27), TNFRSF8 (CD30), TNFRSF9 (4-1BB CD137, ILA), TNFRSF21 (DR6), TNFRSF22 (DcTRAIL R2 TNFRH2), TNFRST23 (DcTRAIL R1 TNFRH1), TNFRSF25 (DR3 Apo-3, LARD, TR-3, TRAMP, WSL-1), TNFSF10 (TRAIL Аро-2-лиганд, TL2), TNFSF11 (TRANCE/RANK-лиганд ODF, OPG- лиганд), TNFSF12 (TWEAK Apo-3-лиганд, DR3-лиганд), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (LIGHT HVEM-лиганд, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (GITR-лиганд AITR-лиганд, TL6), TNFSF1A (TNF-a конектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (ОХ40-лиганд gp34, TXGP1), TNFSF5 (CD40-лиганд CD154, gp39, HIGM1, IMD3, TRAP), TNFSF6 (Fas-лиганд Apo-1-лиганд, АРТ1-лиганд), TNFSF7 (CD27-лиганд CD70), TNFSF8 (CD30- лиганд CD153), TNFSF9 (4-1ВВ-лиганд CD137-лиганд), ТР-1, t-PA, Tpo, TRAIL, TRAIL R, TRAIL-R1, TRAIL-R2, TRANCE, рецептор трансферрина, TRF, Trk, TROP-2, TSG, TSLP, ассоциированный с опухолью антиген СА125, ассоциированный с опухолью антиген, экспрессирующий ассоциированные с антигеном Льюис-Y углеводы, TWEAK, TXB2, Ung, uPAR, uPAR-1, урокиназу, VCAM, VCAM-1, VECAD, VE-кадхерин, VE-кадхерин-2, VEFGR-1 (flt-1), VEGF, VEGFR, VEGFR-3 (flt-4), VEGI, VIM, вирусный антиген, VLA, VLA-1, VLA-4, VNR интегрин, фактор фон Виллебранда, WIF-1, WNT1, WNT2, WNT2B/13, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16, XCL1, XCL2, XCR1, XCR1, XEDAR, XIAP, XPD, HMGB1, IgA, А бета, CD81, CD97, CD98, DDR1, DKK1, EREG, Hsp90, IL-17/IL-17R, IL-20/IL-20R, окисленный LDL, PCSK9, прекалликреин, RON, TMEM16F, SOD1, хромогранин А, хромогранин В, tau, VAP1, высокомолекулярный кининоген, IL-31, IL-31R, Nav1.1, Nav1.2, Nav1.3, Nav1.4, Nav1.5, Nav1.6, Nav1.7, Nav1.8, Nav1.9, EPCR, C1, C1q, C1r, C1s, C2, C2a, C2b, C3, C3a, C3b, C4, C4a, C4b, C5, C5a, C5b, C6, C7, C8, C9, фактор В, фактор D, фактор Н, пропердин, склеростин, фибриноген, фибрин, протромбин, тромбин, тканевый фактор, фактор V, фактор Va, фактор VII, фактор VIIa, фактор VIII, фактор VIIIa, фактор IX, фактор IXa, фактор X, фактор Ха, фактор XI, фактор XIa, фактор XII, фактор XIIa, фактор XIII, фактор XIIIa, TFPI, антитромбин III, EPCR, тромбомодулин, TAPI, tPA, плазминоген, плазмин, PAI-1, PAI-2, GPC3, синдекан-1, синдекан-2, синдекан-3, синдекан-4, LPA и S1P; и рецепторы для гормонов и факторов роста.
Как указано в настоящем описании, допустимо наличие изменений одного или нескольких аминокислотных остатков в аминокислотных последовательностях, которые образуют вариабельные области, если сохраняется их антигенсвязывающая активность. Изменения, затрагивающие аминокислотную последовательность вариабельной области, не ограничены сайтом, в котором осуществляют изменение, и количеством измененных аминокислот. Например, аминокислоты, присутствующие в CDR и/или FR, можно изменять соответствующим образом. Когда изменяют аминокислоты в вариабельной области, то связывающая активность предпочтительно поддерживается без конкретного ограничения; и, например, по сравнению с активностью до изменения, связывающая активность может составлять 50% или более, 80% или более и 100% или более. Кроме того, связывающая активность может повышаться благодаря аминокислотным изменениям. Например, связывающая активность может быть выше в 2-, 5-, 10-раз или т.п. по сравнению с активностью до изменения. Изменение аминокислотной последовательности может представлять собой замену, добавление, делецию или модификацию по меньшей мере одного аминокислотного остатка.
Например, модификация N-концевого глутамина вариабельной области, приводящая к образованию пироглутаминовой кислоты в результате пироглутамилирования, хорошо известна специалистам в данной области. Так, когда N-конец тяжелой цепи представляет собой глутамин, то антитела, указанные в настоящем описании, могут содержать вариабельные области, в которых глутамин модифицирован с образованием пироглутаминовой кислоты.
Вариабельные области антител, указанных в настоящем описании, могут иметь любые последовательности, и они могут представлять собой вариабельные области антител любого происхождения, например, мышиных антител, крысиных антител, кроличьих антител, козьих антител, антител верблюдов, гуманизированных антител, полученных путем гуманизации указанных нечеловеческих антител, и человеческих антител. Кроме того, эти антитела могут иметь различные аминокислотные замены, интродуцированные в их вариабельные области, для улучшения их связывания с антигеном, фармакокинетических особенностей, стабильности и иммуногенности. Вариабельные области могут обладать способностью повторно связываться с антигенами благодаря их зависимого от рН связывания антигена (WO 2009/125825).
В константных областях легких цепей антител присутствуют каппа-цепь и лямбда-цепь, любая из которых является приемлемой. Кроме того, они могут иметь аминокислотные изменения, такие как замены, делеции, добавления и/или инсерции.
Кроме того, полипептиды, которые содержат варианты Fc-областей, указанные в настоящем описании, могут образовывать содержащие Fc слитые белки в результате связывания с другими белками, такими как физиологически активные пептиды. Указанные слитые белки могут представлять собой мультимеры из по меньшей мере двух полипептидов, содержащих варианты Fc-областей. Примерами других белков являются (но не ограничены ими) рецепторы, молекулы адгезии, лиганды и ферменты.
Примеры содержащих Fc слитых белков включают белки, слитые с помощью Fc-области с рецептором, который связывается с молекулой-мишенью, включая слитый белок TNFR-Fc, слитый белок IL1R-Fc, слитый белок VEGFR-Fc и слитый белок CTLA4-Fc (Economides и др,. Nat. Med. 9(1), 2003, сс. 47-52; Dumont и др., BioDrugs. 20(3), 2006, сс. 151-160). Кроме того, белки, подлежащие слиянию, могут представлять собой другие молекулы, которые обладают способностью связываться с мишенью, например, scFv (WO 2005/037989), однодоменные антитела (WO 2004/058821; WO 2003/002609), антителоподобные молекулы (Davinder, Curr. Op. Biotech. 17, 2006, сс. 653-658; Current Opinion in Biotechnology 18, 2007, сс. 1-10; Nygren и др., Curr. Op. Struct. Biol. 7, 1997, сс. 463-469 и Hoss. Protein Science 15, 2006, сс. 14-27 (2006), такие как DARPin (WO 2002/020565), аффибоди (Affibody) (WO 1995/001937), авимер (Avimer) (WO 2004/044011; WO 2005/040229) и аднектин (Adnectin) (WO 2002/032925). Кроме того, антитела и содержащие Fc слитые белки могут быть мультиспецифическими и могут связываться с несколькими типами молекул-мишеней или эпитопов.
Б. Методы рекомбинации и композиции
Антитела можно получать, используя методы рекомбинации и композиции, например, описанные в US №4816567. Одним из вариантов осуществления изобретения является выделенная нуклеиновая кислота, кодирующая антитело к миостатину, представленное в настоящем описании. Другим вариантом осуществления изобретения является выделенная нуклеиновая кислота, которая кодирует полипептид, содержащий вариант Fc-области или родительскую Fc-область, указанную в настоящем описании. Указанная нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкие и/или тяжелые цепи антитела). Следующим вариантом осуществления изобретения является(ются) один или несколько векторов (например, экспрессионных векторов), который(е) содержит(ат) указанную нуклеиновую кислоту. Еще одним вариантом осуществления изобретения является клетка-хозяин, которая содержит указанную нуклеиновую кислоту. В одном из указанных вариантов осуществления изобретения клетка-хозяин содержит (например, в результате трансформации указанными векторами): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном из вариантов осуществления изобретения клетка-хозяин представляет собой эукариотическую клетку, например, клетку яичника китайского хомячка (СНО) или лимфоидную клетку (например, Y0-, NS0-, Sp20-клетку). Одним из вариантов осуществления изобретения является способ получения антитела к миостатину, включающий культивирование клетки-хозяина, содержащей нуклеиновую кислоту, которая кодирует антитело, описанное выше, в условиях, пригодных для экспрессии антитела, и необязательно выделение антитела из клетки-хозяина (или среды для культивирования клетки-хозяина). Другим вариантом осуществления изобретения является способ получения полипептида, который содержит вариант Fc-области или родительскую Fc-область, включающий культивирование клетки-хозяина, которая содержит нуклеиновую(ые) кислоту(ы), кодирующую(ие) полипептид, такой как антитело, Fc-область или вариант Fc-области, указанный выше, в условиях, пригодных для экспрессии полипептида, и необязательно выделение полипептида из клетки-хозяина (или среды для культивирования клетки-хозяина).
Для рекомбинантного получения антитела к миостатину нуклеиновые кислоты, кодирующие антитело, например, описанные выше, выделяют и встраивают в один или несколько векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Для рекомбинантного получения Fc-области нуклеиновую кислоту, кодирующую Fc-область, выделяют и встраивают в один или несколько векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Указанную нуклеиновую кислоту легко можно выделять и секвенировать с использованием общепринятых процедур (например, путем использования олигонуклеотидных зондов, которые обладают способностью специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела).
Приемлемые клетки-хозяева для клонирования или экспрессии кодирующих антитело векторов включают прокариотические или эукариотические клетки, представленные в настоящем описании. Например, антитела можно получать в бактериях, в частности, когда не требуется гликозилирование и связанная с Fc эффекторная функция. Сведения об экспрессии фрагментов антитела и полипептидов в бактериях см. например, в US №№5648237, 5789199 и 5840523 (см. также у Charlton в: Methods in Molecular Biology, под ред. Lo В.K.С., изд-во Humana Press, Totowa, NJ, т. 248, 2003, cc. 245-254 описание экспрессии фрагментов антител в Е. coli.) После экспрессии антитело можно выделять из пасты бактериальных клеток в виде растворимой фракции и можно дополнительно очищать.
Помимо прокариотических организмов в качестве хозяев, пригодных для клонирования или экспрессии плазмид, которые кодируют антитела, можно использовать эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи, включая штаммы грибов и дрожжей, пути гликозилирования которых были «гуманизированы», что позволяет получать антитело с частично или полностью человеческой схемой гликозилирования (см. Gerngross, Nat. Biotech. 22, 2004, cc. 1409-1414 и Li и др., Nat. Biotech. 24, 2006, cc. 210-215).
Клетки-хозяева, которые можно использовать для экспрессии гликозилированного антитела, получают также из многоклеточных организмов (беспозвоночных и позвоночных животных). Примерами клеток беспозвоночных являются клетки насекомых, а также можно применять клетки растений. Были выявлены многочисленные штаммы бакуловирусов и соответствующие пригодные для них в качестве хозяев клетки насекомых, прежде всего для трансфекции клеток Spodoptera frugiperda.
В качестве хозяев можно применять также культуры растительных клеток (см., например, US №№5959177, 6040498, 6420548, 7125978 и 6417429 (описание технологии PLANTIBODIES™ для получения антител в трансгенных растениях).
В качестве хозяев можно применять также клетки позвоночных. Например, можно использовать клеточные линии млекопитающих, которые адаптированы к росту в суспензии. Другими примерами приемлемых линий клеток-хозяев млекопитающих являются линия клеток почки обезьяны CV1, трансформированная с помощью ОВ40 (COS-7); линия клеток почки эмбриона человека (HEK 293-клетки или клетки линии 293, описанные, например, у Graham и др., J. Gen. Virol., 36, 1977, сс. 59-74); клетки почки детеныша хомяка (ВНК); клетки Сертоли мыши (ТМ4-клетки, описанные, например, у Mather, Biol. Reprod., 23, 1980, сс. 243-251); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76,); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK); клетки печени бычьей крысы (BRL 3А); клетки легкого человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI, описанные, например, у Mather и др., Annals N.Y. Acad. Sci., 383, 1982, сс. 44-68); клетки MRC 5 и клетки FS4. Другими ценными линиями клеток-хозяев млекопитающих являются клетки яичника китайского хомячка (СНО), включая DHFR--СНО-клетки (Urlaub и др., Proc. Natl. Acad. Sci. USA, 77, 1980, сс. 4216-4220); и клеточные линии миеломы, такие как Y0, NS0 и Sp2/0. Обзор конкретных линий клеток-хозяев млекопитающих, которые можно применять для производства антител, см., например, у Yazaki и Wu, в: Methods in Molecular Biology под ред. В.K.С. Lo, изд-во Humana Press, Totowa, NJ, т. 248, 2003, сс. 255-268.
Антитела с рН-зависимыми характеристиками можно получать с помощью методов скрининга и/или методов мутагенеза, например, описанных в WO 2009/125825. Методы скрининга могут включать любой процесс, с помощью которого идентифицируют антитело с рН-зависимыми характеристиками связывания в популяции антител, специфических для конкретного антигена. В некоторых вариантах осуществления изобретения методы скрининга могут предусматривать измерение одного или нескольких параметров связывания (например, KD или kd) индивидуальных антител в исходной популяции антител, как при кислом, так и при нейтральном рН. Параметры связывания антител можно измерять, например, с помощью поверхностного плазмонного резонанса или любого другого аналитического метода, который позволяет количественно или качественно оценивать характеристики связывания антитела с конкретным антигеном. В некоторых вариантах осуществления изобретения методы скрининга могут предусматривать идентификацию антитела, связывание которого с антигеном характеризуется соотношением KD при кислом/нейтральном рН, составляющим 2 или более. В других вариантах осуществления изобретения методы скрининга могут предусматривать идентификацию антитела, связывание которого с антигеном характеризуется соотношением KD при рН 5,8/рН 7,4, составляющим 2 или более. Альтернативно этому, методы скрининга могут предусматривать идентификацию антитела, связывание которого с антигеном характеризуется соотношением kd при кислом/нейтральном рН, составляющим 2 или более. В других вариантах осуществления изобретения методы скрининга могут предусматривать идентификацию антитела, связывание которого с антигеном характеризуется соотношением kd при рН 5,8/рН 7,4, составляющим 2 или более.
В другом варианте осуществления изобретения методы мутагенеза могут предусматривать делецию, замену или добавление аминокислоты в тяжелую и/или легкую цепь антитела для усиления рН-зависимого связывания антитела с антигеном. В некоторых вариантах осуществления изобретения мутагенезу можно подвергать один или несколько вариабельных доменов антитела, например, один или несколько HVR (например, CDR). Например, мутагенез может приводить к замене аминокислоты в одном или нескольких HVR (например, CDR) антитела на другую аминокислоту. В некоторых вариантах осуществления изобретения мутагенез может приводить к замене аминокислоты по меньшей мере в одном HVR (например, CDR) антитела на гистидин. В некоторых вариантах осуществления изобретения «повышенное рН-зависимое связывание» означает, что мутантная версия антитела характеризуется более высоким соотношением KD при кислом/нейтральном рН, или более высоким соотношением kd при кислом/нейтральном рН, чем исходная «родительская» (т.е. в меньшей степени зависящая от рН) версия антитела до мутагенеза. В некоторых вариантах осуществления изобретения мутантная версия антитела характеризуется соотношением KD при кислом/нейтральном рН, составляющим 2 или более. В некоторых вариантах осуществления изобретения мутантная версия антитела характеризуется соотношением KD при рН 5,8/рН 7,4, составляющим 2 или более. Альтернативно этому, мутантная версия антитела характеризуется соотношением kd при кислом/нейтральном рН, составляющим 2 или более. В некоторых вариантах осуществления изобретения мутантная версия антитела характеризуется соотношением kd при рН 5,8/рН 7,4, составляющим 2 или более.
Поликлональные антитела предпочтительно вырабатываются у животных после нескольких подкожных (sc) или внутрибрюшинных (ip) инъекций релевантного антитела и адъюванта. Можно конъюгировать релевантный антиген с белком, иммуногенным для подлежащего иммунизации вида, например, гемоцианином лимфы улитки, сывороточным альбумином, бычьим тироглобулином или соевым ингибитором трипсина, с использованием бифункционального или дериватизирующего агента, например, сложного сульфосукцинимидного эфира малеимидобензоила (конъюгация через остатки цистеина), N-гидроксисукцинимида (конъюгация через остатки лизина), глутарового альдегида, янтарного ангидрида, SOCl2 или R1N=C=NR, где R и R1 обозначают различные алкильные группы.
Животных (как правило, млекопитающих кроме человека) иммунизируют с использованием антигена, иммуногенных конъюгатов или производных путем объединения, например, 100 или 5 мкг белка или конъюгата (для кроликов и мышей соответственно) с 3 объемами полного адъюванта Фрейнда, и осуществляют введение раствора внутрикожно в несколько мест. Через 1 месяц животных подвергают бустерной вакцинации (ревакцинации) с использованием от 1/5 до 1/10 исходного количества пептида или конъюгата в полном адъюванте Фрейнда путем подкожной инъекции в несколько мест. Через 7-14 дней получают образцы крови животных и оценивают в сыворотке титр антитела. Животных подвергают ревакцинации вплоть до выхода титра на плато. Предпочтительно животных ревакцинируют с использованием конъюгата, в который входит тот же антиген, но конъюгированный с другим белком и/или через другой перекрестносшивающий реагент. Конъюгаты можно получать также в рекомбинантной клеточной культуре в виде белковых слияний. Кроме того, агрегирующие агенты, такие как квасцы, можно применять для усиления иммунного ответа.
Моноклональные антитела получают из популяции практически гомогенных антител, т.е. индивидуальные антитела, входящие в популяцию, идентичны за исключением возможных встречающихся в естественных условиях мутаций и/или пост-трансляционных модификаций (например, изомеризации, амидирования), которые могут присутствовать в минорных количествах. Прилагательное «моноклональный» свидетельствует о том, что антитело не присутствует в смеси различных антител.
Например, моноклональные антитела можно получать, используя метод гибридом, впервые описанный Kohler и др., Nature 256(5517), 1975, сс. 495-497. При осуществления метода гибрид мышь или другое пригодное в качестве хозяина животное, такое как хомяк, иммунизируют согласно описанному выше методу для выработки лимфоцитов, которые продуцируют или обладают способностью продуцировать антитела, специфически связывающиеся с белком, применяемым для иммунизации. Альтернативно этому, лимфоциты можно иммунизировать in vitro.
Иммунизирующий агент должен, как правило, включать антигенный белок или его слитый вариант. Как правило, применяют либо лимфоциты периферической крови (PBL), если требуются клетки человеческого происхождения, либо применяют селезеночные клетки или клетки лимфатических узлов, если источниками являются млекопитающие кроме человека. Затем лимфоциты сливают с иммортализованной клеточной линией с помощью приемлемого агента для слияния, такого как полиэтиленгликоль, с получением клетки гибридомы (Goding, Monoclonal Antibodies: Principles and Practice, изд-во Academic Press, 1986, cc. 59-103).
Иммортализованные клеточные линии, как правило, представляют собой трансформированные клетки млекопитающих, прежде всего клетки миеломы грызунов, быков и человека. Как правило, применяют крысиные или мышиные клеточные линии миеломы. Полученные таким образом клетки гибридомы высевают и выращивают в приемлемой культуральной среде, которая предпочтительно содержит одну или несколько субстанций, которые ингибируют рост или выживание неслитых родительских клеток миеломы. Например, если в родительских клетках миеломы отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (HGPRT или HPRT), то культуральная среда для гибридом, как правило, включает гипоксантин, аминоптерин и тимидин (НАТ-среда), т.е. субстанции, которые препятствуют росту клеток с дефицитом HGPRT.
Предпочтительными иммортализованными клетками миеломы являются клетки, которые можно эффективно сливать, которые поддерживают стабильный высокий уровень производства антитела отобранными антителопродуцирующими клетками и обладают чувствительностью к среде, такой как НАТ-среда. Среди них предпочтительными являются мышиные линии миеломы, например, полученные из мышиных опухолей МОРС-21 и МРС-11, которые можно получать от фирмы Salk Institute Cell Distribution Center, Сан-Диего, шт. Калифорния, США, и SP-2-клетки (и их производные, например, Х63-Ag8-653), которые можно получать из Американской коллекции типовых культур, Манассас, шт. Вирджиния, США. Также описаны клеточные линии человеческой миеломы и мышиной-человеческой гетеромиеломы, которые можно применять для производства человеческих моноклональных антител (Kozbor и др., J. Immunol. 133(6), 1984, сс. 3001-3005; Brodeur и др., Monoclonal Antibody Production Techniques and Applications, изд-во Marcel Dekker, Inc., New York, 1987, cc. 51-63).
Культуральную среду, в которой выращивают клетки гибридомы, оценивают в отношении производства моноклональных антител против антигена. Предпочтительно специфичность связывания моноклональных антител, продуцируемых клетками гибридомы, определяют с помощью иммнопреципитации или анализа связывания in vitro, такого как радиоиммуноанализ (РИА) или твердофазный иммуноферментный анализ (ELISA). Указанные методики и анализы известны в данной области. Например, аффинность связывания можно определять с помощью анализа Скэтчарда, описанного у Munson, Anal. Biochem. 107(1), 1980, сс. 220-239.
После того, как установлено, что клетки гибридом продуцируют антитела требуемой специфичности, аффинности и/или активности, клоны можно субклонировать, используя процедуры серийного разведения, и выращивать с помощью стандартных методов (Goding, выше). Приемлемые для этой цели культуральные среды включают, например, среду D-MEM или RPMI-1640. Кроме того, клетки гибридомы можно выращивать in vivo в виде опухолей в млекопитающих.
Моноклональные антитела, секретируемые субклонами, можно отделять от культуральной среды, асцитной жидкости или сыворотки с помощью общепринятых процедур очистки иммуноглобулинов, таких, например, как хроматография на белок А-сефарозе, гидроксиапатите, гель-электрофорез, диализ или аффинная хроматография.
Антитела можно получать путем иммунизации приемлемого животного-хозяина антигеном. В одном из вариантов осуществления изобретения антиген представляет собой полипептид, содержащий полноразмерный миостатин. В одном из вариантов осуществления изобретения антиген представляет собой полипептид, содержащий латентный миостатин. В одном из вариантов осуществления изобретения антиген представляет собой полипептид, содержащий пропептид миостатина. В одном из вариантов осуществления изобретения антиген представляет собой полипептид, содержащий область, соответствующую аминокислотам в положениях 21-100 пропептида миостатина (SEQ ID NO: 78). В одном из вариантов осуществления изобретения антиген представляет собой полипептид, содержащий аминокислоты в положениях 21-80, 41-100, 21-60, 41-80, 61-100, 21-40, 41-60, 61-80 или 81-100 пропептида миостатина (SEQ ID NO: 78). Настоящее изобретение относится также к антителам, полученным путем иммунизации животного антигеном. Антитела могут обладать индивидуально или в комбинации любой из особенностей, которые описаны выше в разделе «Примеры антител к миостатину».
Fc-область можно получать путем повторной элюции фракции, адсорбированной на белке А, после частичного расщепления моноклональных антител в виде IgG1, IgG2, IgG3, IgG4 или т.п. с помощью протеазы, такой как пепсин. Протеаза не ограничена конкретной протеазой, если она может расщеплять полноразмерное антитело с образованием Fab и F(ab')2 ограниченным образом при создании соответствующих условий ферментативной реакции, таких как рН, и ее примеры включают пепсин и папаин.
Кроме того, в настоящем изобретении предложен способ получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью по сравнению с полипептидом, содержащим родительскую Fc-область, который включает интродукцию по меньшей мере одного аминокислотного изменения в родительскую Fc-область. В некоторых объектах изобретения полученный полипептид представляет собой антитело. В некоторых вариантах осуществления изобретения антитело представляет собой химерное антитело или гуманизированное антитело. В некоторых объектах изобретения полученный полипептид представляет собой содержащий Fc слитый белок.
В некоторых вариантах осуществления изобретения способ получения заключается в том, что осуществляют следующие стадии: (а) получают полипептид, содержащий родительскую Fc-область; (б) интродуцируют по меньшей мере одно аминокислотное изменение в родительскую Fc-область в полипептиде; (в) измеряют активность связывания с обезьяньим Fc-гамма RIIb полипептида, который содержит родительскую Fc-область, и полипептида, который содержит Fc-область, полученную на стадии (б); и (г) отбирают полипептид, содержащий вариант Fc-области с повышенной активностью связывания с обезьяньим Fc-гамма RIIb по сравнению с полипептидом, содержащим родительскую Fc-область.
В некоторых вариантах осуществления изобретения способ получения включает следующие стадии, на которых: (а) получают полипептид, содержащий родительскую Fc-область; (б) интродуцируют по меньшей мере одно аминокислотное изменение в родительскую Fc-область в полипептиде; (в) измеряют активность связывания с обезьяньим Fc-гамма RIIIa полипептида, который содержит родительскую Fc-область, и полипептида, который содержит Fc-область, полученную на стадии (б); и (г) отбирают полипептид, содержащий вариант Fc-области с пониженной активностью связывания с обезьяньим Fc-гамма RIIIa, по сравнению с полипептидом, содержащим родительскую Fc-область.
В некоторых вариантах осуществления изобретения способ получения заключается в том, что осуществляют следующие стадии: (а) получают полипептид, содержащий родительскую Fc-область; (б) интродуцируют по меньшей мере одно аминокислотное изменение в родительскую Fc-область в полипептиде; (в) измеряют активность связывания с человеческим Fc-гамма RIIb полипептида, который содержит родительскую Fc-область, и полипептида, который содержит Fc-область, полученную на стадии (б); и (г) отбирают полипептид, содержащий вариант Fc-области с повышенной активностью связывания с человеческим Fc-гамма RIIb, по сравнению с полипептидом, содержащим родительскую Fc-область.
В некоторых вариантах осуществления изобретения способ получения включает следующие стадии, на которых: (а) получают полипептид, содержащий родительскую Fc-область; (б) интродуцируют по меньшей мере одно аминокислотное изменение в родительскую Fc-область в полипептиде; (в) измеряют активность связывания с человеческим Fc-гамма RIIIa полипептида, который содержит родительскую Fc-область, и полипептида, который содержит Fc-область, полученную на стадии (б); и (г) отбирают полипептид, содержащий вариант Fc-области с пониженной активностью связывания с человеческим Fc-гамма RIIIa, по сравнению с полипептидом, содержащим родительскую Fc-область.
В некоторых вариантах осуществления изобретения способ получения заключается в том, что осуществляют следующие стадии: (а) получают полипептид, содержащий родительскую Fc-область; (б) интродуцируют по меньшей мере одно аминокислотное изменение в родительскую Fc-область в полипептиде; (в) измеряют активность связывания с человеческим Fc-гамма RIIa (Н-типа) полипептида, который содержит родительскую Fc-область, и полипептида, который содержит Fc-область, полученную на стадии (б); и (г) отбирают полипептид, содержащий вариант Fc-области с пониженной активностью связывания с человеческим Fc-гамма RIIa (Н-типа), по сравнению с полипептидом, содержащим родительскую Fc-область.
В некоторых вариантах осуществления изобретения способ получения заключается в том, что осуществляют следующие стадии: (а) получают полипептид, содержащий родительскую Fc-область; (б) интродуцируют по меньшей мере одно аминокислотное изменение в родительскую Fc-область в полипептиде; (в) измеряют активность связывания с человеческим Fc-гамма RIIa (R-типа) полипептида, который содержит родительскую Fc-область, и полипептида, который содержит Fc-область, полученную на стадии (б); и (г) отбирают полипептид, содержащий вариант Fc-области с пониженной активностью связывания с человеческим Fc-гамма RIIa (R-типа), по сравнению с полипептидом, содержащим родительскую Fc-область.
В одном из объектов изобретения по меньшей мере одну аминокислоту изменяют в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 233, 234, 235, 236, 237, 238, 239, 264, 266, 267, 268, 271, 295, 298, 325, 326, 327, 328, 330, 331, 332, 334 и 396, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В другом объекте изобретения изменяют одну аминокислоту в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, в положении 236.
В другом объекте изобретения по меньшей мере две аминокислоты изменяют в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, где изменения представляют собой: (а) одно аминокислотное изменение в положении 236 и (б) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 233, 234, 235, 237, 238, 239, 264, 266, 267, 268, 271, 295, 298, 325, 326, 327, 328, 330, 331, 332, 334 и 396, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В другом объекте изобретения по меньшей мере две аминокислоты изменяют в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, где изменения представляют собой: (а) одно аминокислотное изменение в положении 236, и (б) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 231, 232, 235, 239, 268, 295, 298, 326, 330 и 396, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В другом объекте изобретения по меньшей мере две аминокислоты изменяют в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, где изменения представляют собой: (а) одно аминокислотное изменение в положении 236 и (б) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 268, 295, 326, и 330, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В другом объекте изобретения аминокислоты изменяют в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, в одном из следующих положений (1)-(37): (1) в положениях 231, 236, 239, 268 и 330; (2) в положениях 231, 236, 239, 268, 295 и 330; (3) в положениях 231, 236, 268 и 330; (4) в положениях 231, 236, 268, 295 и 330; (5) в положениях 232, 236, 239, 268, 295 и 330; (6) в положениях 232, 236, 268, 295 и 330; (7) в положениях 232, 236, 268 и 330; (8) в положениях 235, 236, 268, 295, 326 и 330; (9) в положениях 235, 236, 268, 295 и 330; (10) в положениях 235, 236, 268 и 330; (11) в положениях 235, 236, 268, 330 и 396; (12) в положениях 235, 236, 268 и 396; (13) в положениях 236, 239, 268, 295, 298 и 330; (14) в положениях 236, 239, 268, 295, 326 и 330; (15) в положениях 236, 239, 268, 295 и 330; (16) в положениях 236, 239, 268, 298 и 330; (17) в положениях 236, 239, 268, 326 и 330; (18) в положениях 236, 239, 268 и 330; (19) в положениях 236, 239, 268, 330 и 396; (20) в положениях 236, 239, 268 и 396; (21) в положениях 236 и 268; (22) в положениях 236, 268 и 295; (23) в положениях 236, 268, 295, 298 и 330; (24) в положениях 236, 268, 295, 326 и 330; (25) в положениях 236, 268, 295, 326, 330 и 396; (26) в положениях 236, 268, 295 и 330; (27) в положениях 236, 268, 295, 330 и 396; (28) в положениях 236, 268, 298 и 330; (29) в положениях 236, 268, 298 и 396; (30) в положениях 236, 268, 326 и 330; (31) в положениях 23 6, 268, 326, 330 и 396; (32) в положениях 236, 268 и 330; (33) в положениях 236, 268, 330 и 396; (34) в положениях 236, 268 и 396; (35) в положениях 236 и 295; (36) в положениях 236, 330 и 396; и (37) в положениях 236 и 396 согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В следующем объекте изобретения аминокислотное изменение в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, выбирают в каждом из положений, выбранных из группы, состоящей из: (a) Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp, Tyr в положении 231; (б) Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp, Tyr в положении 232; (в) Asp в положении 233; (г) Trp, Tyr в положении 234; (д) Trp в положении 235; (е) Ala, Asp, Glu, His, Ile, Leu, Met, Asn, Gln, Ser, Thr, Val в положении 236; (ж) Asp, Tyr в положении 237; (з) Glu, Ile, Met, Gln, Tyr в положении 238; (и) Ile, Leu, Asn, Pro, Val в положении 239; (к) Ile в положении 264; (л) Phe в положении 266; (м) Ala, His, Leu в положении 267; (н) Asp, Glu в положении 268; (о) Asp, Glu, Gly в положении 271; (п) Leu в положении 295; (р) Leu в положении 298; (с) Glu, Phe, Ile, Leu в положении 325; (т) Thr в положении 326; (у) Ile, Asn в положении 327; (ф) Thr в положении 328; (х) Lys, Arg в положении 330; (ц) Glu в положении 331; (ч) Asp в положении 332; (ш) Asp, Ile, Met, Val, Tyr в положении 334 и (щ) Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp, Tyr в положении 396; согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В следующем объекте изобретения аминокислотное изменение в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, выбирают в каждом из положений, выбранных из группы, состоящей из: (a) Gly, Thr в положении 231; (б) Asp в положении 232; (в) Trp в положении 235; (г) Asn, Thr в положении 236; (д) Val в положении 239; (е) Asp, Glu в положении 268; (ж) Leu в положении 295; (з) Leu в положении 298; (и) Thr в положении 326; (к) Lys, Arg в положении 330 и (л) Lys, Met в положении 396; согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Asn в положении 236, Glu в положении 268, Lys в положении 330 и Met в положении 396 согласно EU-нумерации. В другом объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Asn в положении 236, Asp в положении 268 и Lys в положении 330 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Asn в положении 236, Asp в положении 268, Leu в положении 295 и Lys в положении 330 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Thr в положении 236, Asp в положении 268 и Lys в положении 330 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Asn в положении 236, Asp в положении 268, Leu в положении 295, Thr в положении 326 и Lys в положении 330 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Trp в положении 235, Asn в положении 236, Asp в положении 268, Leu в положении 295, Thr в положении 326 и Lys в положении 330 согласно EU-нумерации.
В одном из объектов изобретения по меньшей мере одну аминокислоту изменяют в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, по меньшей мере в одном положении, выбранном из группы, состоящей из: 234, 238, 250, 264, 267, 307 и 330, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В другом объекте изобретения одну аминокислоту изменяют в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, в положении 238.
В другом объекте изобретения по меньшей мере две аминокислоты изменяют в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, где изменения представляют собой: (I) одно аминокислотное изменение в положении 238 и (II) по меньшей мере одно аминокислотное изменение по меньшей мере в одном положении, выбранном из группы, состоящей из: 234, 250, 264, 267, 307 и 330, согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
В другом объекте изобретения аминокислоты изменяют в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, в одном из следующих положений (1)-(9): (1) в положениях 234, 238, 250, 307 и 330; (2) в положениях 234, 238, 250, 264, 307 и 330; (3) в положениях 234, 238, 250, 264, 267, 307 и 330; (4) в положениях 234, 238, 250, 267, 307 и 330; (5) в положениях 238, 250, 264, 307 и 330; (6) в положениях 238, 250, 264, 267, 307 и 330; (7) в положениях 238, 250, 267, 307 и 330; (8) в положениях 238, 250 и 307; и (9) в положениях 238, 250, 307 и 330 согласно EU-нумерации.
В следующем объекте изобретения аминокислотное изменение в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, выбирают в каждом из положений, выбранных из группы, состоящей из: (а) Tyr в положении 234; (б) Asp в положении 238; (в) Val в положении 250; (г) Ilе в положении 264; (д) Ala в положении 267; (е) Pro в положении 307; и (ж) Lys в положении 330; согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Asp в положении 238 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Asp в положении 238, Val в положении 250 и Pro в положении 307 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Asp в положении 238, Val в положении 250, Pro в положении 307 и Lys в положении 330 согласно EU-нумерации. согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Asp в положении 238, Val в положении 250, Ilе в положении 264, Pro в положении 307 и Lys в положении 330 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Asp в положении 238, Val в положении 250, Ala в положении 267, Pro в положении 307 и Lys в положении 330 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Tyr в положении 234, Asp в положении 238, Val в положении 250, Pro в положении 307 и Lys в положении 330 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Tyr в положении 234, Asp в положении 238, Val в положении 250, Ala в положении 267, Pro в положении 307 и Lys в положении 330 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Asp в положении 238, Val в положении 250, Ile в положении 264, Ala в положении 267, Pro в положении 307 и Lys в положении 330 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Tyr в положении 234, Asp в положении 238, Val в положении 250, Ile в положении 264, Pro в положении 307 и Lys в положении 330 согласно EU-нумерации. В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом получения полипептида, содержащего вариант Fc-области с повышенной Fc-гамма RIIb-связывающей активностью, представляют собой: Tyr в положении 234, Asp в положении 238, Val в положении 250, Ile в положении 264, Ala в положении 267, Pro в положении 307 и Lys в положении 330 согласно EU-нумерации. В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность Fc-гамма RIIb обезьян циномолгус (SEQ ID NO: 223). В некоторых вариантах осуществления изобретения Fc-гамма RIIb имеет последовательность человеческого Fc-гамма RIIb (например, SEQ ID NO: 212, 213 или 214).
Кроме того, в настоящем изобретении предложен способ получения полипептида, содержащего вариант Fc-области с повышенной pI по сравнению с полипептидом, содержащим родительскую Fc-область, который заключается в том, что интродуцируют по меньшей мере два аминокислотных изменения в родительскую Fc-область.
В некоторых вариантах осуществления изобретения способ получения полипептида, содержащего вариант Fc-области, представленный в настоящем описании, включает осуществление следующих стадий, на которых: (а) получают полипептид, содержащий родительскую Fc-область; (б) интродуцируют по меньшей мере два аминокислотных изменения в родительскую Fc-область в полипептиде; (в) измеряют pI полипептида, который содержит родительскую Fc-область, и полипептида, который содержит Fc-область, полученную на стадии (б); и (г) отбирают полипептид, содержащий вариант Fc-области с повышенной pI по сравнению с полипептидом, содержащим родительскую Fc-область.
В одном из объектов изобретения по меньшей мере две аминокислоты изменяют в соответствии с описанным выше способом для получения полипептида, содержащего вариант Fc-области с повышенной pI, по меньшей мере в двух положениях, выбранных группы, состоящей из: 285, 311, 312, 315, 318, 333, 335, 337, 341, 342, 343, 384, 385, 388, 390, 399, 400, 401, 402, 413, 420, 422 и 431, согласно EU-нумерации. В другом объекте изобретения полученный полипептид содержит вариант Fc-области с повышенной pI, содержащий изменения по меньшей мере в двух аминокислотных положениях, выбранных из группы, состоящей из: 311, 341, 343, 384, 399, 400, 401, 402 и 413, согласно EU-нумерации.
В другом объекте изобретения по меньшей мере две аминокислоты изменяют в соответствии с описанным выше способом для получения полипептида, содержащего вариант Fc-области с повышенной pI, в одном из следующих положений (1)-(10): (1) в положениях 311 и 341; (2) в положениях 311 и 343; (3) в положениях 311, 343 и 413; (4) в положениях 311, 384 и 413;(5) в положениях 311 и 399; (6) в положениях 311 и 401; (7) в положениях 311 и 413; (8) в положениях 400 и 413; (9) в положениях 401 и 413 и (10) в положениях 402 и 413 согласно EU-нумерации.
В другом объекте изобретения аминокислотные изменения в соответствии с описанным выше способом для получения полипептида, содержащего вариант Fc-области с повышенной pI, представляют собой: Arg в положении 400 и Lys в положении 413 согласно EU-нумерации. В другом объекте изобретения аминокислотные изменения в соответствии с описанным выше способом для получения полипептида, содержащего вариант Fc-области с повышенной pI, представляют собой: Arg в положении 311 и Lys в положении 413 согласно EU-нумерации. В другом объекте изобретения аминокислотные изменения в соответствии с описанным выше способом для получения полипептида, содержащего вариант Fc-области с повышенной pI, представляют собой: Arg в положении 311 и Arg в положении 399 согласно EU-нумерации. В другом объекте изобретения аминокислотные изменения в соответствии с описанным выше способом для получения полипептида, содержащего вариант Fc-области с повышенной pI, представляют собой: Arg в положении 311 и Arg в положении 343 согласно EU-нумерации. В другом объекте изобретения аминокислотные изменения в соответствии с описанным выше способом для получения полипептида, содержащего вариант Fc-области с повышенной pI, представляют собой: Arg в положении 311 и Arg в положении 413 согласно EU-нумерации. В другом объекте изобретения аминокислотные изменения в соответствии с описанным выше способом для получения полипептида, содержащего вариант Fc-области с повышенной pI, представляют собой: Arg в положении 311, Arg в положении 343 и Arg в положении 413 согласно EU-нумерации. В другом объекте изобретения аминокислотные изменения в соответствии с описанным выше способом для получения полипептида, содержащего вариант Fc-области с повышенной pI, представляют собой: Arg в положении 311, Arg в положении 384 и Arg в положении 413 согласно EU-нумерации.
В следующем объекте изобретения аминокислотные изменения в соответствии с описанным выше способом для получения полипептида, содержащего вариант Fc-области с повышенной pI, выбирают из Lys или Arg в каждом положении.
В следующем объекте изобретения аминокислотные изменения в соответствии с описанными выше способами получения выбирают из любых из: единичного изменения, комбинации единичных изменений или комбинации изменений, описанных в таблицах 14-30.
Необязательно следующие дополнительные стадии можно включать в вышеописанные способы для получения полипептида, содержащего вариант Fc-области, когда полипептид дополнительно содержит антигенсвязывающий домен: (д) осуществление оценки фармакокинетики антигена в плазме после введения полипептида, содержащего родительскую Fc-область, и полипептида, содержащего вариант Fc-области, в организме животных, таких как мыши, крысы, кролики, собаки, обезьяны и человек; и (е) отбор полипептида, содержащего вариант Fc-области с повышенной способностью к элиминации антигена из плазмы, по сравнению с полипептидом, содержащим родительскую Fc-область.
Полипептиды, содержащие вариант Fc-области, полученные с помощью любого из указанных выше способов или других методов, известных в данной области, включены в настоящее изобретение.
В. Анализы
Представленные в настоящем описании антитела к миостатину можно идентифицировать, осуществлять их скрининг или характеризовать их физические/химические свойства и/или виды биологической активности с помощью различных анализов, известных в данной области.
Варианты Fc-областей, представленные в настоящем описании, можно идентифицировать, осуществлять их скрининг или характеризовать их физические/химические свойства и/или виды биологической активности с помощью различных анализов, известных в данной области.
1. Анализы связывания и другие анализы
Согласно одному из объектов изобретения антитело, предлагаемое в изобретении, тестируют в отношении антигенсвязывающей активности с использованием известных методов, таких как ELISA, Вестерн-блоттинг, BIACORE® и т.д. В одном из объектов изобретения полипептид, содержащий вариант Fc-области, предлагаемый в изобретении, тестируют в отношении активности связывания с его Fc-рецептором с помощью известных методов, таких как BIACORE® и т.д.
Согласно другому объекту изобретения можно использовать анализы в условиях конкуренции для идентификации антитела, которое конкурирует за связывание с миостатином с любым антителом к миостатину, представленным в настоящем описании. В некоторых вариантах осуществления изобретения, когда указанное конкурирующее антитело присутствует в избытке, оно блокирует (например, снижает) связывание референс-антитела с миостатином по меньшей мере на 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75% или более. В некоторых случаях связывание ингибируется по меньшей мере на 80%, 85%, 90%, 95% или более. В некоторых вариантах осуществления изобретения указанное конкурирующее антитело связывается с таким же эпитопом (например, линейным или конформационным эпитопом), с которым связывается антитело к миостатину, представленное в настоящем описании (например, антитело к миостатину, описанное в таблицах 2а, 11а или 13). Согласно другим объектам изобретения референс-антитело имеет пару VH и VL, описанную в таблице 2а, 11а или 13. Подробное описание приведенных в качестве примера методов картирования эпитопа, с которым связывается антитело, представлено у Morris, Epitope Mapping Protocols, в: Methods in Molecular Biology, т. 66, изд-во Humana Press, Totowa, NJ, 1996.
В качестве примера анализа в условиях конкуренции иммобилизованный латентный миостатин или пропептид миостатина инкубируют в растворе, содержащем первое меченое антитело, которое связывается с миостатином, и второе немеченое антитело, подлежащее тестированию с отношении способности конкурировать с первым антителом за связывание с миостатином. Второе антитело может присутствовать в супернатанте гибридомы. В качестве контроля иммобилизованный миостатин инкубируют в растворе, содержащем первое меченое антитело, но не второе немеченое антитело. После инкубации в условиях, приемлемых для связывания первого антитела с миостатином, избыток несвязанного антитела удаляют и измеряют количество метки, ассоциированной с иммобилизованным миостатином. Если количество метки, ассоциированной с иммобилизованным миостатином, существенно снижается в тестируемом образце по сравнению с контрольным образцом, то это свидетельствует о том, что второе антитело конкурирует с первым антителом за связывание с миостатином (см., например, Harlow и Lane, Antibodies: A Laboratory Manual, глава 14, изд-во Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1988).
Анализы по определению активности связывания полипептида, содержащего вариант Fc-области, с одним или несколькими представителями Fc-гамма R представлены в настоящем описании или известны в данной области. Такие анализы связывания включают (но, не ограничиваясь только ими) BIACORE®-анализ, в котором используется явление поверхностного плазмонного резонанса (SPR), скрининг на основе гомогенного анализа усиленной за счет эффекта близости люминесценции (Amplified Luminescent Proximity Homogeneous Assay (ALPHA), ELISA, и сортинг клеток по интенсивности флуоресценции (FACS) (Lazar и др., Proc. Natl. Acad. Sci. USA 103(11), 2006, cc. 4005-4010).
В одном из вариантов осуществления изобретения BIACORE®-анализ можно применять для решения вопроса о том, является ли активность связывания полипептида, содержащего вариант Fc-области, повышенной, или сохраняется на том же уровне, или снижается в отношении конкретного представителя семейства Fc-гамма R. Например, для этой цели определяют снижается ли или повышается величина константны диссоциации (KD), для определения которой применяют анализ сенсограмм, когда различные Fc-гамма R подвергают взаимодействию в качестве аналита с полипептидами, содержащими вариант Fc-области, иммобилизованными или «захваченными» на сенсорном чипе, с использованием известных методов и реагентов, таких как белок А, белок L, белок A/G, белок G, антитела к лямбда-цепи, антитела к каппа-цепи, антигенные пептиды, антигенные белки. Изменения в активности связывания можно определять также путем сравнения изменений в количестве резонансных единиц (RU) на сенсограмме до и после того, как один или несколько типов Fc-гамма R подвергают взаимодействию в качестве аналитов с «захваченными» полипептидами, содержащими вариант Fc-области. Альтернативно этому, Fc-гамма R можно иммобилизовать или «захватывать» на сенсорных чипах, и полипептиды, содержащие вариант Fc-области, применять в качестве аналита.
При осуществлении BIACORE®-анализов одну из субстанций (лиганд), предназначенных для исследования взаимодействия, иммобилизуют на тонком слое золота на сенсорном чипе и при освещении светом с задней поверхности сенсорного чипа таким образом, чтобы имело место полное отражение на границе раздела между тонким слоем золота и стеклом, в части отраженного света формируется область с пониженной интенсивностью (SPR-сигнал). Подготавливают другую субстанцию (аналит), предназначенную для исследования взаимодействия, для инъекции на поверхность сенсорного чипа; и когда лиганд связывается с аналитом, масса иммобилизованной молекулы-лиганда возрастает и показатель преломления растворителя на поверхности сенсорного чипа изменяется. В результате указанного изменения показателя преломления положение SPR-сигнала сдвигается (и наоборот, положение сигнала возвращается в исходное, если происходит диссоциация указанного связывания). С помощью BIACORE®-системы определяют уровень описанного выше сдвига, или более конкретно изменение массы в зависимости от времени, откладывая изменение массы на поверхности сенсорного чипа по вертикальной оси, и таким образом получают количественные данные (сенсограмма).
Количество аналита, связанного с лигандом, иммобилизованном на поверхности сенсорного чипа, определяют из сенсограммы. Кинетические параметры, такие как константа скорости ассоциации (ka) и константа скорости диссоциации (kd), определяют из представленных в виде кривых сенсограмм, и определяют аффинность (KD) как соотношение указанных констант. В качестве метода для анализа ингибирования предпочтительно применяют BIACORE®-метод. Примеры такого метода для анализа ингибирования описаны у Lazar и др., Ргос. Natl. Acad. Sci. USA 103(11), 2006, cc. 4005-4010.
Скрининг на основе ALPHA осуществляют на основе технологии ALPHA, которая основана на описанном ниже принципе, с использованием двух типов гранул, доноров и акцепторов. Люминесцентные сигналы поддаются обнаружению только тогда, когда молекулы, связанные с гранулами-донорами, физически взаимодействуют с молекулами, связанными с гранулами-акцепторами, и когда обе гранулы находятся в непосредственной близости друг от друга. Возбужденный лазерным пучком фотосенсибилизатор в гранулах-донорах превращает кислород окружающей среды в возбужденный синглетный кислород. Когда синглетный кислород диффундирует из гранул-доноров и достигает гранул-акцепторов, локализованных в непосредственной близости, то индуцируется хемилюминесцентная реакция в гранулах, что в итоге приводит к испусканию света. Если молекулы, связанные с гранулами-донорами, не взаимодействуют с молекулами, связанными с гранулами-акцепторами, то хемилюминесцентная реакция не происходит, поскольку синглетный кислород, который продуцируется гранулами-донорами, не достигает гранул-акцепторов.
Например, биотинилированный полипептидный комплекс связывают с гранулами-донорами и Fc-гамма-рецептор, меченный глутатион-S-трансферазой (GST), связывают с гранулами-акцепторами. В отсутствии конкурирующего полипептидного комплекса, содержащего вариант Fc-области, полипептидный комплекс, содержащий родительскую Fc-область, взаимодействует с Fc-гамма-рецептором и образует сигналы с длиной волны от 520 до 620 нм. Полипептидный комплекс, содержащий немеченый вариант Fc-области, конкурирует с полипептидным комплексом, содержащим родительскую Fc-область, за связывание с Fc-гамма-рецептором Относительную аффинность связывания можно оценивать, определяя количественно снижение флуоресценции в результате конкуренции. Методы биотинилирования полипептидных комплексов, таких как антитела, с помощью сульфо-NHS-биотина или подобных агентов являются хорошо известными. Метод экспрессии Fc-гамма-рецептора и GST в клетке, несущей слитый ген, полученный путем слияния полинуклеотида, который кодирует Fc-гамма-рецептор, в рамке считывания с полинуклеотидом, который кодирует GST, в экспрессионном векторе и осуществления очистки с помощью содержащей глутатион колонки, соответствующим образом адаптируют, получая метод мечения Fc-гамма-рецептора с помощью GST. Индуцированные сигналы можно анализировать, например, посредством подгонки к односайтовой модели конкуренции на основе нелинейного регрессионного анализа с использованием такого программного обеспечения, как GRAPHPAD PRISM (фирма GraphPad; Сан-Диего).
Понятие «вариант Fc-области с пониженной Fc-гамма R-связывающей активностью» относится к Fc-области, которая связывается с Fc-гамма R с существенно более слабой связывающей активностью, чем родительская Fc-область, при осуществлении анализов с использованием практически одинаковых количеств соответствующей родительской Fc-области и варианта Fc-области. Кроме того, понятие «вариант Fc-области с повышенной Fc-гамма R-связывающей активностью» относится к Fc-области, которая связывается с Fc-гамма R с существенно более сильной связывающей активностью, чем родительская Fc-область, при осуществлении анализов с использованием практически одинаковых количеств соответствующей родительской Fc-области и варианта Fc-области. Понятие «вариант Fc-области с сохраненной Fc-гамма R-связывающей активностью» относится к Fc-области, которая связывается с Fc-гамма R с активностью, эквивалентной или практически такой же, что и активность родительской Fc-области, при осуществлении анализов с использованием практически одинаковых количеств соответствующей родительской Fc-области и полипептида, содержащего вариант Fc-области.
Повышается ли или понижается активность связывания Fc-области с различными Fc-гамма R, можно определять по увеличению или понижению уровня связывания различных Fc-гамма R с Fc-областью при определении с использованием вышеописанного метода измерения. В контексте настоящего описания уровень связывания различных Fc-гамма R с Fc-областью можно оценивать в виде величины, полученной делением разницы в уровнях RU на сенсограммах до и после взаимодействия различных Fc-гамма R, которые применяют в качестве аналита, с Fc-областью, на разницу в уровнях RU на сенсограммах, которая изменилась до и после «захвата» Fc-областей на сенсорных чипах.
Согласно настоящему изобретению повышенная Fc-гамма RIIb-связывающая активность предпочтительно означает, например, что соотношение [величина KD родительской Fc-области для Fc-гамма RIIb]/[величина KD варианта Fc-области для Fc-гамма RIIb] при измерении в виде величин KD, измеренных с использованием вышеописанного метода, предпочтительно составляет 2,0 или более, 3,0 или более, 4,0 или более, 5,0 или более, 6,0 или более, 7,0 или более, 8,0 или более, 9.0 или более, 10 или более, 15 или более, 20 или более, 25 или более, 30 или более, 35 или более, 40 или более, 45 или более, or even 50 или более, 55 или более, 60 или более, 65 или более, 70 или более, 75 или более, 80 или более, 85 или более, 90 или более, 95 или более или 100 или более.
Величина KD (моль/л) варианта Fc-области, указанного в настоящем описании, для Fc-гамма RIIb предпочтительно ниже, чем родительской Fc-области для Fc-гамма RIIb, и может составлять, например, 2,0×10-6 М или менее, 1,0×10-6 М или менее, 9,0×10-7 М или менее, 8,0×10-7 М или менее, 7,0×10-7 М или менее, 6,0×10-7 М или менее, 5,0×10-7 М или менее, 4,0×10-7 М или менее, 3,0×10-7 М или менее, 2,0×10-7 М или менее, 1,0×10-7 М или менее, 9,0×10-8 М или менее, 8,0×10-8 М или менее, 7,0×10-8 М или менее, 6,0×10-8 М или менее, 5,0×10-8 М или менее.
Кроме того, вариант Fc-области с повышенной избирательностью связывания с Fc-гамма RIIb по сравнению с Fc-гамма RIIa означает Fc-область, у которой: (а) Fc-гамма RIIb-связывающая активность повышается, а Fc-гамма RIIa-связывающая активность сохраняется на таком уровне или снижается, (б) Fc-гамма RIIb-связывающая активность повышается и Fc-гамма RIIa-связывающая активность повышается также, но степень повышения Fc-гамма RIIa-связывающей активности ниже, чем степень повышения Fc-гамма RIIb-связывающей активности; или (в) Fc-гамма RIIb-связывающая активность снижается и Fc-гамма RIIa-связывающая активность снижается также, но степень снижения Fc-гамма RIIb-связывающей активности ниже, чем степень снижения Fc-гамма RIIa-связывающей активности. Обладает ли вариант Fc-области повышенной избирательностью связывания в отношении Fc-гамма RIIb по сравнению с Fc-гамма RIIa, можно определять, например, путем сравнения соотношения величины KD для Fc-гамма RIIa и величины KD для Fc-гамма RIIb варианта Fc-области (соотношение [величина KD варианта Fc-области для Fc-гамма RIIa]/[величина KD варианта Fc-области для Fc-гамма RIIb]), с соотношением величины KD для Fc-гамма RIIa и величины KD для Fc-гамма RIIb родительской Fc-области (соотношение [величина KD родительской Fc-области для Fc-гамма RIIa]/[величина KD родительской Fc-области для Fc-гамма RIIb]), которую определяют согласно вышеуказанным примерам. В частности, когда соотношение KD для варианта Fc-области выше, чем для родительской Fc-области, то можно считать, что вариант Fc-области обладает повышенной избирательностью связывания с Fc-гамма RIIb по сравнению с Fc-гамма RIIa относительно родительской Fc-области. Касательно человеческих рецепторов, Fc-гамма RIIb-связывающая активность, вероятно, коррелирует с активностью связывания с Fc-гамма RIIa (R-типа), а не Fc-гамма RIIa (Н-типа), поскольку аминокислотная последовательность Fc-гамма RIIb характеризуется большей идентичностью с Fc-гамма RIIa (R-типа), чем с Fc-гамма RIIa (Н-типа). Таким образом, выявленное(ые) аминокислотное(ые) изменение(я), которое(ые) может(гут) повышать избирательность связывания с человеческим Fc-гамма RIIb по сравнению с человеческим Fc-гамма RIIa (R-типа), является(ются) важной(ыми) для повышения избирательности связывания с Fc-гамма RIIb по сравнению с Fc-гамма RIIa у человека.
Когда вариант Fc-области, предлагаемый в настоящем изобретении, обладает повышенной связывающей активностью с Fc-гамма RIIb и пониженной связывающей активностью с Fc-гамма RIII (таким как Fc-гамма RIIIa или Fc-гамма RIIIb) по сравнению с родительской Fc-областью, то можно считать, что вариант Fc-области является Fc-гамма RIIb-специфическим.
2. Анализы активности
Одним из объектов изобретения являются анализы, предназначенные для идентификации антител к миостатину, которые обладают биологической активностью. Биологическая активность может включать, например, ингибирование активации миостатина, блокаду высвобождения зрелого миостатина из латентного миостатина, ингибирование протеолитического расщепления латентного миостатина, блокаду доступности латентного миостатина для протеазы и т.д. Предложены также антитела, обладающие указанной биологической активностью in vivo и/или in vitro. В некоторых вариантах осуществления изобретения антитело, предлагаемое в изобретении, тестируют в отношении указанной биологической активности.
В некоторых вариантах осуществления изобретения обладает ли тестируемое антитело способностью ингибировать расщепление латентного миостатина, определяют путем детекции продукта расщепления латентного миостатина (миостатина) с использованием метода, известного в данной области, такого как электрофорез, хроматография, анализ методом иммуноблоттинга, твердофазный иммуноферментный анализ (ELISA) или масс-спектрометрия, после контакта протеазы, которая может расщеплять латентный миостатин, с латентным миостатином с присутствии тестируемого антитела или без него (см. например, Thies и др., Growth Factors 18(4), 2001, cc. 251-259 (2001)). При обнаружении пониженного количества продукта расщепления латентного миостатина (например, человеческий пропептид миостатина) в присутствии тестируемого антитела (или после контакта с ним), тестируемое антитело идентифицируют как антитело, которое может ингибировать расщепление латентного миостатина. В некоторых вариантах осуществления изобретения обладает ли тестируемое антитело способностью блокировать доступ к латентному миостатину, определяют с помощью методов оценки белковых взаимодействий между протеазой и латентным миостатином, например, с помощью ELISA или BIACORE®. При обнаружении снижения взаимодействия между протеазой и латентным миостатином в присутствии тестируемого антитела (или после контакта с ним), тестируемое антитело идентифицируют как антитело, которое может блокировать доступ протеазы к латентному миостатину.
В некоторых вариантах осуществления изобретения обладает ли тестируемое антитело способность блокировать высвобождение зрелого миостатина из латентного миостатина, определяют путем определения активности зрелого миостатина, например, активности связывания с рецептором для миостатина, или способности опосредовать трансдукцию сигнала в клетке, экспрессирующей рецептор для миостатина (например, ActRIIb). Клетки, которые можно применять в таком анализе, могут представлять собой клетки, которые экспрессируют эндогенный рецептор для миостатина, например, миоциты линии L6, или могут представлять собой генетически модифицированные, кратковременно или стабильно клетки, так, что они экспрессируют трансген, кодирующий рецептор для миостатина, рецептор для активина, такой как рецептор для активина типа II (Thies и др., выше). Связывание миостатина с рецептором для миостатина можно оценивать, используя анализ связывания с рецептором. Опосредуемую миостатином трансдукцию сигнала можно определять на любом уровне в пути трансдукции сигнала, например, путем оценки фосфорилирования Smad-полипептида, оценки экспрессии регулируемого миостатином гена, включая репортерный ген, или измерения пролиферации миостатин-зависимых клеток. При снижении активности зрелого миостатина в присутствии тестируемого антитела (или после контакта с ним), тестируемое антитело идентифицируют как антитело, которое может блокировать высвобождение зрелого миостатина из латентного миостатина.
Ингибирование активации миостатина можно выявлять и/или измерять с использованием также методов, изложенных в качестве примера в описанных примерах осуществления изобретения. С использованием этих анализов или других пригодных типов анализов тестируемые антитела можно подвергать скринингу в отношении их способности ингибировать активацию миостатина. В некоторых вариантах осуществления изобретения ингибирование активации миостатина включает снижение активации миостатина по меньшей мере на 5%, 10%, 15%, 20%, 25%, 30%, 35% или 40% или более в этом анализе по сравнению с отрицательным контролем в одинаковых условиях. В некоторых вариантах осуществления изобретения оно включает ингибирование активации миостатина по меньшей мере на 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% или более.
Другим объектом изобретения являются анализы, предназначенные для идентификации антител к миостатину, которые образуют иммунный комплекс (т.е. комплекс антиген-антитело) с миостатином. Предложены также антитела, обладающие указанной биологической активностью in vivo и/или in vitro.
В некоторых вариантах осуществления изобретения антитело, предлагаемое в изобретении, тестируют в отношении указанной биологической активности.
В некоторых вариантах осуществления изобретения образование иммунного комплекса оценивают с помощью такого метода, как хроматография методом гель-фильтрации, ультрацентрифугирование, светорассеяние, электронная микроскопия или масс-спектрометрия (Mol. Immunol. 39, 2002, сс. 77-84, Mol. Immunol. 47, 2009, сс. 357-364). Возможность осуществления этих методов основана на том, что иммунный комплекс является более крупной молекулой, чем индивидуальное антитело или индивидуальный антиген. Если крупный комплекс, содержащий два или большее количество антител и два или большее количество антигенов (например, молекул миостатина), выявляют в присутствии тестируемого антитела и антигена, то тестируемое антитело идентифицируют как антитело, которое может образовывать иммунный комплекс, содержащий два или большее количество антител и две или большее количество молекул миостатина. В другом варианте осуществления изобретения образование иммунного комплекса оценивают с помощью такого метода, как ELISA, FACS или SPR (анализ методом поверхностного плазмонного резонанса; например, с помощью BIACORE™) (Shields и др., J. Biol. Chem. 276(9), 2001, cc. 6591-6604; Singh и др., J. Immunol. Methods 50, 1982, cc. 109-114; Suzuki и др., J. Immunol. 184(4), 2010, cc. 1968-1976; Luo и др., mAbs 1(5), 2009, cc. 491-504). Возможность применения этих методов основана на том, что иммунный комплекс, содержащий два или большее количество антител, и два или большее количество антигенов, может связываться сильнее с Fc-рецептором или компонентом системы комплемента, чем антитело или антиген при их индивидуальном применении. Если обнаружено повышенное связывание с Fc-рецептором или компонентом системы комплемента в присутствии и тестируемого антитела, и антигена по сравнению с присутствием только антитела, то тестируемое антитело идентифицируют как антитело, которое может образовывать иммунный комплекс, содержащий два или большее количество антител и две или большее количество молекул миостатина. В другом варианте осуществления изобретения образование иммунного комплекса оценивают путем введения тестируемого антитела животному (например, мыши) и измерения клиренса антигена из плазмы. Как описано выше, антитело, которое образует иммунный комплекс, содержащий два или большее количество антител с двумя или большим количеством антигеном, как ожидается, должно ускорять элиминацию антигенов из плазмы. Таким образом, если обнаружена ускоренная элиминация миостатина из плазмы у мышей, которым вводили тестируемое антитело, по сравнению с мышами, которым вводили референс-антитело, то тестируемое антитело идентифицируют как антитело, которое может образовывать иммунный комплекс, содержащий два или большее количество антител и две или большее количество молекул миостатина, более эффективно, чем референс-антитело.
Г. Иммуноконъюгаты
Некоторыми вариантами осуществления изобретения являются иммуноконъюгаты, содержащие антитело миостатину, представленное в настоящем описании, конъюгированное с одним или несколькими цитотоксическим агентами, такими как химиотерапевтические агенты или лекарственные средства, ингибирующие рост агенты, токсины (например, белковые токсины, обладающие ферментативной активностью токсины бактериального, грибного, растительного или животного происхождения или их фрагменты) или радиоактивные изотопы.
В одном из вариантов осуществления изобретения иммуноконъюгат представляет собой конъюгат антитело-лекарственное средство (ADC), в котором антитело конъюгировано с одним или несколькими лекарственными средствами, включая (но, не ограничиваясь только ими) майтансиноид (см. US №№5208020, 5416064 и ЕР 0425235 В1); ауристатин, например, фрагменты DE и DF монометилауристатина (ММАЕ и MMAF) (см. US №№5635483, 5780588 и 7498298); доластатин; калихеамицин или его производные (см. US №№5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001 и 5877296; Hinman и др., Cancer Res. 53, 1993, сс. 3336-3342; и Lode и др., Cancer Res. 58, 1998, сс. 2925-2928); антрациклин, такой как дауномицин или доксорубицин (см. Kratz и др., Curr. Med. Chem. 13, 2006, сс. 477-523; Jeffrey и др., Bioorg. Med. Chem. Lett. 16, 2006, сс. 358-362; Torgov и др., Bioconjug. Chem. 16, 2005, сс. 717-721; Nagy и др., Proc. Natl. Acad. Sci. USA 97, 2000, cc. 829-834; Dubowchik и др., Bioorg. & Med. Chem. Letters 12, 2002, cc. 1529-1532; King и др., J. Med. Chem. 45, 2002, cc. 4336-4343; и US №6630579); метотрексат; виндесин; таксан, такой как доцетаксел, паклитаксел, ларотаксел, тесетаксел и ортатаксел; трихотецен и СС1065.
В другом варианте осуществления изобретения иммуноконъюгат содержит антитело, представленное в настоящем описании, конъюгированное с обладающим ферментативной активностью токсином или его фрагментом, включая (но, не ограничиваясь только ими) А-цепь дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, А-цепь экзотоксина (из Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модецина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-5), ингибитор из Momordica charantia, курцин, кротин, ингибитор из Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трихотецены.
В другом варианте осуществления изобретения иммуноконъюгат содержит антитело, представленное в настоящем описании, конъюгированное с радиоактивным атомом с образованием радиоконъюгата. Широкое разнообразие радиоактивных изотопов можно применять для получения радиоконъюгатов. Примеры включают 211At, 131I, 125I, 90Y, 186Re, 188Re, 153Sm, 212Bi, 32P, 212Pb и радиоактивные изотопы Lu. Когда радиоконъюгат применяют для детекции, то он может содержать радиоактивный атом для сцинтиграфических исследований, например, TC99m или I123, или спиновую метку для визуализации методом ядерного магнитного резонанса (ЯМР) (известный также как магнитно-резонансная визуализация, МРВ), такую как йод-123, иод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического агента можно получать с использованием широкого разнообразия бифункциональных связывающих белки агентов, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат × HCL), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бисазидопроизводные (такие как бис(пара-азидобензоил)гександиамин), производные бисдиазония (такие как бис(пара-диазониумбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат), и обладающие двойной активностью фторсодержащие соединения (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получать согласно методу, описанному у Vitetta и др., Science 238, 1987, сс. 1098-1104. Меченная с помощью С14 1-изотиоцианатбензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является примером хелатирующего агента, применяемого для конъюгации радионуклеотида с антителом (см. WO 94/11026). Линкер может представлять собой «расщепляемый линкер», облегчающий высвобождение цитотоксического лекарственного средства. Например, можно использовать неустойчивый в кислой среде линкер, чувствительный к действию пептидаз линкер, фотолабильный линкер, диметильный линкер или содержащий дисульфид линкер (Chari и др., Cancer Res. 52, 1992, сс. 127-131; US №5208020).
В контексте настоящего описания под иммуноконъюгатами или ADC подразумеваются (но, не ограничиваясь только ими) указанные конъюгаты, полученные с использованием перекрестносшивающих реагентов, которые включают (но, не ограничиваясь только ими) BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB, а также SVSB (сукцинимидил(4-винилсульфон)бензоат), которые поступают в продажу (например, от фирмы Pierce Biotechnology, Inc., Рокфорд, шт. Иллинойс, США).
Д. Способы и композиции для диагностирования и обнаружения
В некоторых вариантах осуществления изобретения любое из антител к миостатину, представленных в настоящем описании, можно применять для обнаружения присутствия миостатина в биологическом образце. Понятие «обнаружение (детекция)» в контексте настоящего описания предусматривает количественное или качественное обнаружение. В одном из вариантов осуществления изобретения биологический образец содержит клетку или ткань, например, сыворотку, цельную кровь, плазму, полученный с помощью биопсии образец, образец ткани, клеточную суспензию, слюну, мокроту, оральную жидкость, спинномозговую жидкость, амниотическую жидкость, асцитную жидкость, молоко, молозиво, секрет молочных желез, лимфу, мочу, пот, слезную жидкость, желудочный сок, синовиальную жидкость, перитонеальную жидкость, жидкость хрусталика глаза или слизь.
Одним из вариантов осуществления изобретения является антитело к миостатину, представленное в настоящем описании, для применения в способе диагностирования или обнаружения. Другой объект изобретения относится к способу обнаружения присутствия латентного миостатина или пропептида миостатина в биологическом образце. В другом варианте осуществления изобретения способ заключается в том, что приводят в контакт биологический образец с антителом к миостатину, представленным в настоящем описании, в условиях, обеспечивающих связывание антитела к миостатину с миостатином, и выявляют образование комплекса между антителом к миостатину и миостатином. Указанный способ может представлять собой способ in vitro или in vivo. В одном из вариантов осуществления изобретения антитело к миостатину применяют для отбора индивидуумов, которых можно лечить с помощью антитела к миостатину, например, когда миостатин является биомаркером для отбора пациентов.
Примеры нарушений, которые можно диагностировать с использованием антитела, предлагаемого в изобретении, включают (но, не ограничиваясь только ими) мышечную дистрофию (MD; включая мышечную дистрофию Дюшенна), амиотрофический боковой склероз (ALS), мышечную атрофию, атрофию органа, синдром канала запястья, состояние немощи, застойное обструктивное заболевание легких (COPD), саркопению, кахексию, синдромы мышечного истощения, индуцируемое ВИЧ мышечное истощение, диабет типа 2, нарушенную толерантность к глюкозе, метаболический синдром (включая синдром X), устойчивость к инсулину (включая устойчивость, индуцированную травмой, например, например, ожогом или нарушением азотистого равновесия), нарушения жировой ткани (например, ожирение, дислипидемия, неалкогольная жировая болезнь печени и т.д.), остеопороз, остеопению, остеоартрит и метаболические нарушения костной ткани (включая низкую массу костной ткани, преждевременную овариальную недостаточность, подавление андрогена, дефицит витамина Д, вторичный гиперпаратиреоз, дефицит питательных веществ и нервную анорексию).
Некоторыми вариантами осуществления изобретения являются меченые антитела к миостатину. Метки включают (но, не ограничиваясь только ими) метки или фрагменты, которые можно обнаруживать непосредственно (такие как флуоресцентные, хромофорные, электронноплотные, хемилюминесцентные и радиоактивные метки), а также фрагменты, такие как ферменты или лиганды, которые подлежат косвенному обнаружению, например, посредством ферментативной реакции или молекулярного взаимодействия. Примеры меток включают (но, не ограничиваясь только ими) радиоизотопы 32Р, 14С, 125I, 3Н и 131I, флуорофоры, такие как хелаты редкоземельных металлов или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люцифераза светляка и бактериальная люцифераза (US №4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидаза из хрена (HRP), щелочная фосфатаза, β-галактозидаза, глюкоамилаза, лизоцим, оксидазы сахаридов, например, глюкозооксидаза, галактозооксидаза и глюкозо-6-фосфатдегидрогеназа, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, сшитые с ферментом, который использует пероксид водорода для окисления предшественника красителя, такого как HRP, лактопероксидаза или микропероксидаза, биотин/авидин, спиновые метки, метки в виде бактериофагов, стабильные свободные радикалы и т.п.
Е. Фармацевтические композиции
Фармацевтические композиции антитела к миостатину, представленного в настоящем описании, получают путем смешения указанного антитела, имеющего необходимую степень чистоты, с одним или несколькими необязательными фармацевтически приемлемыми носителями ( Pharmaceutical Sciences, 16-е изд. под ред. Osol А., 1980) в форме лиофилизированных композиций или водных растворов.
Pharmaceutical Sciences, 16-е изд. под ред. Osol А., 1980) в форме лиофилизированных композиций или водных растворов.
Фармацевтические композиции полипептида, содержащего вариант Fc-области, представленный в настоящем описании, получают путем смешения указанного полипептида, имеющего необходимую степень чистоты, с одним или несколькими необязательными фармацевтически приемлемыми носителями в форме лиофилизированных композиций или водных растворов.
Фармацевтически приемлемые носители, как правило, являются нетоксичными для реципиентов в используемых дозах и концентрациях, они включают (но, не ограничиваясь только ими): буферы, такие как фосфатный, цитратный буферы, а также буферы на основе других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол и мета-крезол); полипептиды с низкой молекулярной массой (содержащие менее приблизительно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поли(винилпирролидон); аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТК; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; содержащие металл комплексы (например, комплексы Zn-белок); и/или неионогенные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). В контексте настоящего описания примеры фармацевтически приемлемых носителей включают также диспергирующие агенты интерстициальных лекарственных средств, такие как растворимые активные в нейтральной среде гликопротеины гиалуронидаз (sHASEGP), например, человеческие растворимые гликопротеины гиалуронидазы РН-20, такие как rHuPH20 (HYLENEX®, фирма Baxter International, Inc.). Некоторые примеры sHASEGP и методы их применения, в том числе rHuPH20, описаны в публикациях патентов США №№2005/0260186 и 2006/0104968. Согласно одному из объектов изобретения sHASEGP объединяют с одним или несколькими дополнительными гликозаминогликаназами, такими как хондроитиназы.
Примеры лиофилизированных композиций антител описаны в US №6267958. Водные композиции антител включают композиции, описанные в US №6171586 и WO 2006/044908, последние композиции включают гистидин-ацетатный буфер.
Композиция, предлагаемая в изобретении, может содержать также более одного действующего вещества, если это необходимо для конкретного подлежащего лечению показания, предпочтительно вещества с дополнительными видами активности, которые не оказывают отрицательного воздействия друг на друга. Такие действующие вещества должны присутствовать в комбинации в количествах, эффективных для достижения поставленной цели.
Действующие вещества можно включать также в микрокапсулы, например, полученные с помощью методов коацервации или межфазной полимеризации, например в гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли(метилметакрилатные) микрокапсулы соответственно, в коллоидные системы введения лекарственного средства (например в липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методы описаны в Pharmaceutical Sciences, 16-е изд., под ред. Osol А., 1980.
Можно приготавливать препараты с замедленным высвобождением. Приемлемыми примерами препаратов с замедленным высвобождением являются полупроницаемые матрицы из твердых гидрофобных полимеров, включающие антитело или конъюгат, такие матрицы представляют собой изделия определенной формы, например, пленки или микрокапсулы.
Композиции, предназначенные для применения in vivo, как правило, должны быть стерильными. Стерильность можно легко обеспечивать, например, путем фильтрации через стерильные фильтрующие мембраны.
Ж. Терапевтические способы и композиции
Любое из антител к миостатину, представленных в настоящем описании, можно применять в терапевтических способах. Аналогично этому, любой из полипептидов, содержащих вариант Fc-области, представленный в настоящем описании, можно применять в терапевтических способах.
Одним из объектов изобретения является антитело к миостатину, представленное в настоящем описании, для применения в качестве лекарственного средства. Другими объектами изобретения является антитело к миостатину, представленное в настоящем описании, для применения для лечения синдрома мышечного истощения. Некоторыми вариантами осуществления изобретения является антитело к миостатину для применения в способе лечения. Некоторыми вариантами осуществления изобретения является антитело к миостатину для применения в способе лечения индивидуума, имеющего синдром мышечного истощения, включающем введение индивидууму в эффективном количестве антитела к миостатину. В одном из указанных вариантов осуществления изобретения способ включает также введение индивидууму в эффективном количестве по меньшей мере одного дополнительного терапевтического средства. В других вариантах осуществления изобретения антитело к миостатину предназначено для применения для увеличения массы мышечной ткани. В некоторых вариантах осуществления изобретения антитело к миостатину предназначено для применения в способе увеличения массы мышечной ткани у индивидуума, включающем введение индивидууму в эффективном количестве антитела к миостатину для увеличения массы мышечной ткани. В следующих вариантах осуществления изобретения антитело к миостатину предназначено для применения для увеличения силы мышечной ткани. В некоторых вариантах осуществления изобретения антитело к миостатину предназначено для применения в способе увеличения силы мышечной ткани у индивидуума, включающем введение индивидууму в эффективном количестве антитела к миостатину для увеличения силы мышечной ткани. В следующих вариантах осуществления изобретения антитело к миостатину предназначено для применения для снижения накопления жировой массы тела. В некоторых вариантах осуществления изобретения антитело к миостатину предназначено для применения в способе снижения накопления жировой массы тела у индивидуума, включающем введение индивидууму в эффективном количестве антитела к миостатину для снижения накопления жировой массы тела. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Антитело к миостатину, предлагаемое в настоящем изобретении, может обладать рН-зависимыми характеристиками связывания. Другими вариантами осуществления изобретения является антитело к миостатину, предназначенное для применения для повышения клиренса миостатина из плазмы. Некоторыми вариантами осуществления изобретения является антитело к миостатину, предназначенное для применения в способе повышения клиренса миостатина из плазмы у индивидуума, включающем введение индивидууму в эффективном количестве антитела к миостатину для повышения клиренса миостатина из плазмы. В одном из вариантов осуществления изобретения антитело к миостатину с рН-зависимыми характеристиками связывания повышает клиренс миостатина из плазмы по сравнению с каноническим антителом к миостатину, которое не обладает рН-зависимыми характеристиками связывания. В следующем варианте осуществления изобретения антитело к миостатину с рН-зависимыми характеристиками связывания, обладающее способностью к связыванию при рН между рН 5,8 и рН 7,4, повышает клиренс миостатина из плазмы по сравнению с каноническим антителом к миостатину, которое не обладает рН-зависимыми характеристиками связывания. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Следующим объектом изобретения является применение антитела к миостатину для приготовления или получения лекарственного средства. В одном из вариантов осуществления изобретения лекарственное средство предназначено для лечения синдрома мышечного истощения. В другом варианте осуществления изобретения лекарственное средство применяют в способе лечения синдрома мышечного истощения, включающем введение индивидууму, имеющему синдром мышечного истощения, в эффективном количестве лекарственного средства. В одном из указанных вариантов осуществления изобретения способ включает также введение индивидууму в эффективном количестве по меньшей мере одного дополнительного терапевтического средства. В следующем варианте осуществления изобретения лекарственное средство предназначено для увеличения массы мышечной ткани. В другом варианте осуществления изобретения лекарственное средство предназначено для применения в способе увеличения массы мышечной ткани у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для увеличения массы мышечной ткани. В следующем варианте осуществления изобретения лекарственное средство предназначено для увеличения силы мышечной ткани. В другом варианте осуществления изобретения лекарственное средство предназначено для применения в способе увеличения силы мышечной ткани у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для увеличения силы мышечной ткани. В следующем варианте осуществления изобретения лекарственное средство предназначено для снижения накопления жировой массы тела. В другом варианте осуществления изобретения лекарственное средство предназначено для применения в способе снижения накопления жировой массы тела у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для снижения накопления жировой массы тела. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Антитело к миостатину, предлагаемое в настоящем изобретении, может обладать рН-зависимыми характеристиками связывания. В следующем варианте осуществления изобретения лекарственное средство предназначено для повышения клиренса миостатина из плазмы. В другом варианте осуществления изобретения лекарственное средство предназначено для применения в способе повышения клиренса миостатина из плазмы у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для повышения клиренса миостатина из плазмы. В одном из вариантов осуществления изобретения антитело к миостатину с рН-зависимыми характеристиками связывания повышает клиренс миостатина из плазмы по сравнению с каноническим антителом к миостатину, которое не обладает рН-зависимыми характеристиками связывания. В другом варианте осуществления изобретения антитело к миостатину имеет разные рН-зависимые характеристики связывания между рН 5,8 и рН 7,4. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Другим объектом изобретения является способ лечения синдрома мышечного истощения. В одном из вариантов осуществления изобретения способ включает введение индивидууму, имеющему указанный синдром мышечного истощения, в эффективном количестве антитела к миостатину, представленного в настоящем описании. В одном из вариантов осуществления изобретения способ включает также введение индивидууму в эффективном количестве по меньшей мере одного дополнительного терапевтического средства. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Другим объектом изобретения является способ увеличения массы мышечной ткани у индивидуума. В одном из вариантов осуществления изобретения способ включает введение индивидууму в эффективном количестве антитела к миостатину, представленного в настоящем описании, для увеличения массы мышечной ткани. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Другим объектом изобретения является способ увеличения силы мышечной ткани у индивидуума. В одном из вариантов осуществления изобретения способ включает введение индивидууму в эффективном количестве антитела к миостатину, представленного в настоящем описании, для увеличения силы мышечной ткани. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Другим объектом изобретения является способ снижения накопления жировой массы тела у индивидуума. В одном из вариантов осуществления изобретения способ включает введение индивидууму в эффективном количестве антитела к миостатину, представленного в настоящем описании, для снижения накопления жировой массы тела. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Антитело к миостатину, предлагаемое в настоящем изобретении, может обладать рН-зависимыми характеристиками связывания. Следующим вариантом осуществления изобретения является способ повышения клиренса миостатина из плазмы у индивидуума. В одном из вариантов осуществления изобретения способ заключается в том, что вводят индивидууму в эффективном количестве антитело к миостатину, представленное в настоящем описании, для повышения клиренса миостатина из плазмы. В одном из вариантов осуществления изобретения антитело к миостатину с рН-зависимыми характеристиками связывания повышает клиренс миостатина из плазмы по сравнению с каноническим антителом к миостатину, которое не обладает рН-зависимыми характеристиками связывания. В другом варианте осуществления изобретения антитело к миостатину имеет разные рН-зависимые характеристики связывания между рН 5,8 и рН 7,4. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Другим объектом изобретения являются фармацевтические композиции, содержащие любое из антител к миостатину, представленных в настоящем описании, которые предназначены, например, для применения в любом из указанных выше терапевтических способов. В одном из вариантов осуществления изобретения фармацевтическая композиция содержит любое из антител к миостатину, представленных в настоящем описании, и фармацевтически приемлемый носитель. В другом варианте осуществления изобретения фармацевтическая композиция содержит любое из антител к миостатину, представленных в настоящем описании, и по меньшей мере одно дополнительное терапевтическое средство.
Следующим объектом изобретения является фармацевтическая композиция, предназначенная для лечения синдрома мышечного истощения. В другом варианте осуществления изобретения фармацевтическая композиция предназначена для увеличения массы мышечной ткани. В другом варианте осуществления изобретения фармацевтическая композиция предназначена для увеличения силы мышечной ткани. В следующем варианте осуществления изобретения фармацевтическая композиция предназначена для снижения накопления жировой массы тела. Антитело к миостатину, предлагаемое в настоящем изобретении, может обладать рН-зависимыми характеристиками связывания. В следующем варианте осуществления изобретения фармацевтическая композиция предназначена для повышения клиренса миостатина из плазмы. В одном из вариантов осуществления изобретения фармацевтическую композицию вводят индивидууму, который имеет синдром мышечного истощения. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Следующим объектом изобретения являются способы приготовления лекарственного средства или фармацевтической композиции, включающие смешение любых антител к миостатину, представленных в настоящем описании, с фармацевтически приемлемым носителем, например, для применения в любом из указанных выше терапевтических способов. В одном из вариантов осуществления изобретения способы приготовления лекарственного средства или фармацевтической композиции дополнительно включают добавление по меньшей мере одного дополнительного терапевтического средства к лекарственному средству или фармацевтической композиции.
В некоторых вариантах осуществления мышечное истощение выбирают из группы, включающей мышечную дистрофию (MD; включая мышечную дистрофию Дюшенна), амиотрофический боковой склероз (ALS), мышечную атрофию, атрофию органа, синдром канала запястья, состояние немощи, застойное обструктивное заболевание легких (COPD), саркопению, кахексию, синдромы мышечного истощения, индуцируемое ВИЧ мышечное истощение, диабет типа 2, нарушенную толерантность к глюкозе, метаболический синдром (включая синдром X), устойчивость к инсулину (включая устойчивость, индуцированную травмой, например, например, ожогом или нарушением азотистого равновесия), нарушения жировой ткани (например, ожирение, дислипидемия, неалкогольная жировая болезнь печени и т.д.), остеопороз, остеопению, остеоартрит и метаболические нарушения костной ткани (включая низкую массу костной ткани, преждевременную овариальную недостаточность, подавление андрогена, дефицит витамина Д, вторичный гиперпаратиреоз, дефицит питательных веществ и нервную анорексию).
Любой из полипептидов, содержащих вариант Fc-области, представленный в настоящем описании, можно применять в терапевтических способах. Следующим объектом изобретения являются фармацевтические композиции, содержащие полипептид, который содержит любой из полипептидов, содержащих варианты Fc-областей, представленных в настоящем описании, например, для применения в терапевтических способах. В одном из вариантов осуществления изобретения фармацевтическая композиция включает полипептид, который содержит любой из полипептидов, содержащих варианты Fc-областей, представленные в настоящем описании, и фармацевтически приемлемый носитель. В другом варианте осуществления изобретения фармацевтическая композиция содержит полипептид, который содержит любой из вариантов Fc-областей, представленных в настоящем описании, и по меньшей мере одно дополнительное терапевтическое средство.
Одним из объектов изобретения является полипептид, содержащий вариант Fc-области, предназначенный для применения в качестве лекарственного средства. Другими объектами изобретения является полипептид, содержащий вариант Fc-области, предназначенный для лечения нарушения. Некоторыми вариантами осуществления изобретения является полипептид, содержащий вариант Fc-области, предназначенный для применения в способе лечения. Некоторыми вариантами осуществления изобретения является полипептид, содержащий вариант Fc-области, предназначенный для применения в способе лечения индивидуума, который имеет нарушение, включающем введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, представленный в настоящем описании. В одном из вариантов осуществления изобретения способ дополнительно включает введение индивидууму в эффективном количестве по меньшей мере одного дополнительного терапевтического средства. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения является применение полипептида, содержащего вариант Fc-области, для приготовления или получения лекарственного средства. В одном из вариантов осуществления изобретения лекарственное средство предназначено для лечения нарушения. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В другом варианте осуществления изобретения лекарственное средство предназначено для применения в способе лечения нарушения, включающем введение индивидууму, который имеет нарушение, подлежащее лечению, в эффективном количестве лекарственного средства. В одном из вариантов осуществления изобретения способ дополнительно включает введение индивидууму в эффективном количестве по меньшей мере одного дополнительного терапевтического средства. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения является способ лечения нарушения. В одном из вариантов осуществления изобретения способ включает введение индивидууму, который имеет указанное нарушение, в эффективном количестве полипептида, содержащего вариант Fc-области. В одном из вариантов осуществления изобретения способ дополнительно включает введение индивидууму в эффективном количестве по меньшей мере одного дополнительного терапевтического средства. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения являются фармацевтические композиции, которые содержат полипептид, содержащий вариант Fc-области, представленный в настоящем описании, предназначенные для применения в терапевтическом способе, таком как любой из терапевтических способов, представленных в настоящем описании. В одном из вариантов осуществления изобретения фармацевтическая композиция содержит полипептид, который содержит вариант Fc-области, представленный в настоящем описании, и фармацевтически приемлемый носитель. В другом варианте осуществления изобретения фармацевтическая композиция содержит полипептид, который содержит вариант Fc-области, представленный в настоящем описании, и по меньшей мере одно дополнительное терапевтическое средство.
Согласно следующему объекту изобретения фармацевтическая композиция предназначена для лечения нарушения. В одном из вариантов осуществления изобретения фармацевтическую композицию вводят индивидууму, который имеет нарушение. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
В других вариантах осуществления изобретения полипептид, содержащий вариант Fc-области, предназначен для применения для подавления активации В-клеток, тучных клеток, дендритных клеток и/или базофилов. В некоторых вариантах осуществления изобретения полипептид, содержащий вариант Fc-области, предназначен для применения в способе подавления активации В-клеток, тучных клеток, дендритных клеток и/или базофилов у индивидуума, включающем введение индивидууму в эффективном количестве полипептида, который содержит вариант Fc-области, для подавления активации В-клеток, тучных клеток, дендритных клеток и/или базофилов. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Другим объектом изобретения является применение полипептида, содержащего вариант Fc-области, для приготовления или получения лекарственного средства. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В другом варианте осуществления изобретения лекарственное средство предназначено для подавления активации В-клеток, тучных клеток, дендритных клеток и/или базофилов. В следующем варианте осуществления изобретения лекарственное средство предназначено для применения в способе подавления активации В-клеток, тучных клеток, дендритных клеток и/или базофилов у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для подавления активации В-клеток, тучных клеток, дендритных клеток и/или базофилов. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Другим объектом изобретения является способ подавления активации В-клеток, тучных клеток, дендритных клеток и/или базофилов у индивидуума. В одном из вариантов осуществления изобретения способ включает введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для подавления активации В-клеток, тучных клеток, дендритных клеток и/или базофилов. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения являются фармацевтические композиции, которые содержат полипептид, содержащий вариант Fc-области. В другом варианте осуществления изобретения фармацевтическая композиция предназначена для подавления активации В-клеток, тучных клеток, дендритных клеток и/или базофилов.
Полипептиды, содержащие вариант Fc-области, предлагаемый в настоящем изобретении, могут подавлять активацию В-клеток, тучных клеток, дендритных клеток и/или базофилов. Не вдаваясь в теорию, можно предположить, что указанное подавление активации является результатом избирательного связывания предлагаемых в изобретении вариантов Fc-области с Fc-гамма RIIb без активации активирующего Fc-гамма R. В контексте настоящего описания «активация В-клеток» включает пролиферацию, производство IgE, производство IgM и производство IgA. Не вдаваясь в теорию, можно предположить, что подавление активации В-клеток является результатом способности предлагаемых в изобретении вариантов Fc-области в полипептидах, содержащих вариант Fc-области, предлагаемый в настоящем изобретении, к перекрестному сшиванию Fc-гамма RIIb с IgE для подавлении производства IgE В-клетками, с IgM для подавления производства IgM В-клетками и с IgA для подавления производства IgA. Помимо вышесказанного, подавляющие действия, сходные с указанными выше, являются результатом прямого или косвенного перекрестного сшивания Fc-гамма RIIb с молекулами, которые экспрессируются на В-клетках и содержат домен ITAM внутри клетки, или взаимодействия с доменом ITAM, таким как BCR, CD19 и CD79b. В контексте настоящего описания «активация тучных клеток» включает пролиферацию, активацию с помощью IgE и дегрануляцию. Не вдаваясь в теорию, можно предположить, что подавление активации тучных клеток является результатом способности предлагаемых в изобретении вариантов Fc-области в полипептидах, содержащих вариант Fc-области, предлагаемый в настоящем изобретении, к прямому или косвенному перекрестному сшиванию Fc-гамма RIIb с молекулами IgE-рецептора, которые экспрессируются на тучных клетках и содержат домен ITAM, или к взаимодействию с доменом ITAM, таким как Fc-эпсилон RI, DAP12 и CD200R3. В контексте настоящего описания «активация базофилов» включает пролиферацию и дегрануляцию базофилов. Не вдаваясь в теорию, можно предположить, что подавление активации базофилов является результатом способности предлагаемых в изобретении вариантов Fc-области в полипептидах, содержащих вариант Fc-области, предлагаемый в настоящем изобретении, к прямому или косвенному перекрестному сшиванию Fc-гамма RIIb с молекулами на клеточной мембране, которые содержат домен ITAM внутри клетки, или к взаимодействию в доменом ITAM. В контексте настоящего описания «активация дендритных клеток» включает пролиферацию и дегрануляцию дендритных клеток. Не вдаваясь в теорию, можно предположить, что подавление активации дендритных клеток является результатом способности предлагаемых в изобретении вариантов Fc-области в полипептидах, содержащих вариант Fc-области, предлагаемый в настоящем изобретении, к прямому или косвенному перекрестному сшиванию Fc-гамма RIIb с молекулами на клеточной мембране, которые содержат домен ITAM внутри клетки, или к взаимодействию в доменом ITAM.
Следующими вариантами осуществления изобретения является полипептид, содержащий вариант Fc-области, представленный в настоящем описании, предназначенный для применения для лечения иммунологического воспалительного заболевания. В некоторых вариантах осуществления изобретения в изобретении предложен полипептид, содержащий вариант Fc-области, который предназначен для применения в способе лечения иммунологического воспалительного заболевания у индивидуума, включающем введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для лечения иммунологического воспалительного заболевания. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения является применение полипептида, содержащего вариант Fc-области, представленный в настоящем описании, для приготовления или получения лекарственного средства. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В другом варианте осуществления изобретения лекарственное средство предназначено для лечения иммунологического воспалительного заболевания. В следующем варианте осуществления изобретения лекарственное средство предназначено для применения в способе лечения иммунологического воспалительного заболевания у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для лечения иммунологического воспалительного заболевания. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Следующим объектом изобретения является способ лечения иммунологического воспалительного заболевания у индивидуума. В одном из вариантов осуществления изобретения способ включает введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для лечения иммунологических воспалительных заболеваний. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения являются фармацевтические композиции, содержащие любой из полипептидов, содержащих вариант Fc-области, представленный в настоящем описании. В другом варианте осуществления изобретения фармацевтическая композиция предназначена для лечения иммунологического воспалительного заболевания.
Как описано выше, поскольку полипептиды, содержащие вариант Fc-области, предлагаемый в настоящем изобретении, могут подавлять активацию В-клеток, тучных клеток, дендритных клеток и/или базофилов, то введение полипептидов, содержащих вариант Fc-области, предлагаемый в настоящем изобретении, в результате может лечить или предупреждать иммунологические воспалительные заболевания.
В некоторых вариантах осуществления изобретения подлежащее лечению иммунологическое воспалительное заболевание выбирают из группы, включающей ревматоидный артрит, аутоиммунный гепатит, аутоиммунный тиреоидит, аутоиммунные буллезные заболевания, аутоиммунное адренокортикальное заболевание, аутоиммунную гемолитическую анемию, аутоиммунную тромбоцитопеническую пурпуру, макроцитарную анемию, аутоиммунный атрофический гастрит, аутоиммунную нейтропению, аутоиммунный орхит, аутоиммунный энцефаломиелит, аутоиммунную рецепторную болезнь, аутоиммунное бесплодие, хронический активный гепатит, гломерулонефрит, интерстициальный легочный фиброз, рассеянный склероз, болезнь Педжета, остеопороз, множественную миелому, увеит, острый и хронический спондилит, подагрический артрит, воспалительное заболевание кишечника, респираторный дистресс-синдром взрослых (ARDS), псориаз, болезнь Крона, базедову болезнь, юношеский диабет, болезнь Аддисона, миастению тяжелую псевдопаралитическую (гравис), факоантигенный увеит, системную красную волчанку, аллергический ринит, аллергический дерматит, неспецифический язвенный колит, гиперчувствительность, мышечную дегенерацию, кахексию, системную склеродермию, локализованную склеродермию, синдром Шегрена, болезнь Бехчета, синдром Рейтера, диабет типа I и типа II, нарушение костной резорбции, реакцию «трансплантат-против-хозяина», повреждение, вызываемое ишемией-реперфузией, атеросклероз, травму головного мозга, церебральную малярию, сепсис, септический шок, синдром токсического шока, лихорадку, миалгия, связанную с малоподвижным образом жизни (malgias due to staining), апластическую анемию, гемолитическую анемию, идиопатическую тромбоцитопению, синдром Гудпасчера, синдром Гийена-Барре, тиреоидит Хашимото, пемфигус, IgA-нефропатию, поллиноз, синдром антифосфолипидных антител, полимиозит, грануломатоз Вегенера, узелковый артериит, смешанное заболевание соединительной ткани, фибромиалгию, астму, атопический дерматит, хронический атрофический гастрит, первичный билиарный цирроз, первичный склерозирующий холангит, аутоиммунный панкреатит, синдром воспаления аорты, быстро прогрессирующий гломерулонефрит, мегалобластную анемию, идиопатическую тромбоцитопеническую пурпуру, первичный гипотиреоидит, идиопатическую болезнь Аддисона, инсулинозависимый сахарный диабет, хроническую дискоидную красную волчанку, пемфигоид, герпес беременных, линейный IgA-зависимый буллезный дерматоз, приобретенный буллезный эпидермолиз, гнездную алопецию, витилиго обыкновенное, центробежную приобретенную лейкодерму Саттона, болезнь Харады, аутоиммунную нейропатию зрительного нерва, идиопатическую азооспермию, привычный выкидыш, гипогликемию, хроническую крапивницу, анкилозирующий спондилит, псориатический артрит, энтеропатический артрит, реактивный артрит, спондилоатропатию, энтезопатию, синдром воспаленного кишечника, синдром хронической усталости, дерматомиозит, миозит с тельцами включений, синдром Шмидта, болезнь Грейвса, пернициозную анемию, люпоидный гепатит, возрастную деменцию, болезнь Альцгеймера, демиелинизирующее нарушение, амиотрофический боковой склероз, гипопаратиреоидит, синдром Дресслера, синдром Итона-Лэмберта, герпетиформный дерматит, алопецию, прогрессирующий системный склероз, CREST-синдром (кальциноз, синдром Рейно, нарушение перистальтики пищевода, склеродактилия и телеангиэктазия), саркоидоз, ревматическую атаку, мультиформную эритему, синдром Кушинга, реакцию трансфузии, болезнь Хансена, артериит Такаясу, ревматическую полимиалгию, темпоральный артериит, гигантоклеточный артрит, экзему, лимфоматоидный грануломатоз, болезнь Кавасаки, эндокардит, эндомиокардиальный фиброз, эндофтальмит, эритробластоз плода, эозинофильный фасциит, синдром Фелти, пурпуру Геноха-Шенлейна, отторжение трансплантата, свинку, кардиомиопатию, гнойный артрит, семейную средиземноморскую лихорадку, синдром Макла-Уэллса и гипер-IgD-синдром.
Другие варианты осуществления изобретения относятся к полипептиду, содержащему вариант Fc-области, предназначенному для лечения или предупреждения аутоиммунных заболеваний, которые могут вызываться или могут быть ассоциированы с производством антител против аутоантигенов (аутоантитела). Некоторые варианты осуществления изобретения относятся к полипептиду, содержащему вариант Fc-области, предназначенному для применения в способе лечения или предупреждения аутоиммунного заболевания, которое может вызываться или может быть ассоциировано с производством антител против аутоантигенов (аутоантитела), у индивидуума, включающем введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для лечения или предупреждения аутоиммунного заболевания. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Следующим объектом изобретения является применение полипептида, содержащего вариант Fc-области, для приготовления или получения лекарственного средства. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В другом варианте осуществления изобретения лекарственное средство предназначено для лечения или предупреждения аутоиммунного заболевания, которое может вызываться или может быть ассоциировано с производством антител против аутоантигенов (аутоантитела). В другом варианте осуществления изобретения лекарственное средство предназначено для применения в способе лечения или предупреждения аутоиммунного заболевания, которое может вызываться или может быть ассоциировано с производством антител против аутоантигенов (аутоантитела), у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для лечения или предупреждения аутоиммунного заболевания. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Следующим объектом изобретение является способ лечения или предупреждения аутоиммунного заболевания, которое может вызываться или может быть ассоциировано с производством антител против аутоантигенов (аутоантитела), у индивидуума. В одном из вариантов осуществления изобретения способ включает введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для лечения или предупреждения аутоиммунного заболевания. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения являются фармацевтические композиции, которые содержат любой из полипептидов, содержащих вариант Fc-области, представленный в настоящем описании. Другим вариантом осуществления изобретения является фармацевтическая композиция, предназначенная для лечения или предупреждения аутоиммунного заболевания, которое может вызываться или может быть ассоциировано с производством антител против аутоантигенов (аутоантитела).
Полипептиды, содержащие вариант Fc-области, предлагаемый в настоящем изобретении, могут лечить или предупреждать аутоиммунное заболевание, которое может вызываться или может быть ассоциировано с производством антител против аутоантигенов (аутоантитела), путем подавления производства аутоантител. Установлено, что применение слитой молекулы Fc-области антитела с AchR (аутоантиген тяжелой псевдопаралитической миастении) подавляет пролиферацию В-клеток, которые экспрессируют распознающий AchR BCR, и индуцируют апоптоз (J. Neuroimmunol 227, 2010, сс. 35-43). Применение слитого белка, образованного путем слияния варианта Fc-области, предлагаемого в настоящем изобретении, и антигена, распознаваемого аутоантителом, обеспечивает перекрестное сшивание Fc-гамма RIIb с BCR указанного аутоантигена на В-клетке, что приводит к подавлению пролиферации В-клеток и/или приводит к индукции апоптоза В-клеток.
В некоторых вариантах осуществления изобретения аутоиммунное заболевание, которое можно лечить или предупреждать, выбирают из группы, включающей синдром Гийена-Барре, тяжелую псевдопаралитическую миастению, хронический атрофический гастрит, аутоиммунный гепатит, первичный билиарный цирроз, первичный склерозирующий холангит, аутоиммунный панкреатит, синдром воспаления аорты, синдром Гудпасчера, быстро прогрессирующий гломерулонефрит, мегалобластную анемию, аутоиммунную гемолитическую анемию, аутоиммунную нейтропению, идиопатическую тромбоцитопеническую пурпуру, базедову болезнь, тиреоидит Хашимото, первичный гипотиреоидит, идиопатическую болезнь Аддисона, инсулинозависимый сахарный диабет, хроническую дискоидную красную волчанку, локализованную склеродермию, пемфигус, пемфигоид, герпес беременных, линейный IgA-зависимый буллезный дерматоз, приобретенный буллезный эпидермолиз, гнездную алопецию, витилиго обыкновенное, центробежную приобретенную лейкодерму Саттона, болезнь Харады, аутоиммунную нейропатию зрительного нерва, идиопатическую азооспермию, привычный выкидыш, диабет типа II, гипогликемию и хроническую крапивницу.
Следующими вариантами осуществления изобретения является полипептид, содержащий вариант Fc-области, предназначенный для лечения заболевания, ассоциированного с дефицитом биологически важного белка. Некоторыми вариантами осуществления изобретения является полипептид, содержащий вариант Fc-области, предназначенный для применения в способе лечения заболевания, ассоциированного с дефицитом биологически важного белка у индивидуума, включающем введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для лечения заболевания. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Следующим объектом изобретения является применение полипептида, содержащего вариант Fc-области, для приготовления или получения лекарственного средства. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В другом варианте осуществления изобретения лекарственное средство предназначено для лечения заболевания, ассоциированного с дефицитом биологически важного белка. В другом варианте осуществления изобретения лекарственное средство предназначено для применения в способе лечения заболевания, ассоциированного с дефицитом биологически важного белка, у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для лечения заболевания. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Следующим объектом изобретения является способ лечения заболевания, ассоциированного с дефицитом биологически важного белка, у индивидуума. В одном варианте осуществления изобретения способ включает введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для лечения заболевания. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения являются фармацевтические композиции, которые содержат любой из полипептидов, содержащих вариант Fc-области, представленный в настоящем описании. В следующем варианте осуществления изобретения фармацевтическая композиция предназначена для лечения заболевания, ассоциированного с дефицитом биологически важного белка.
В случае заболеваний, ассоциированных с дефицитом биологически важного белка, применяют терапевтические методы, при осуществлении которых вводят и добавляют белок в качестве фармацевтического агента. Однако поскольку у пациента отсутствует белок с самого начала, то добавляемый извне белок распознается как чужеродная субстанция и продуцируются антитела против белка. В результате белок легко удаляется и фармацевтическое действие снижается. Применение слитого белка, содержащего указанный белок и вариант Fc-области, предлагаемый в настоящем изобретении, обеспечивает перекрестное сшивание Fc-гамма RIIb с BCR, который распознает белок на В-клетках, что приводит к подавлению производства антитела против белка.
В некоторых вариантах осуществления изобретения подлежащий добавлению белок выбирают из группы, состоящей из фактора VIII, фактора IX, ТРО, ЕРО, альфа-идуронидазы, идуронатсульфатазы, гепаран-N-сульфатазы А-типа, альфа-N-ацетилглюкозаминидазы В-типа, ацетилСоА: альфа-глюкозаминид-ацетилтрансферазы С-типа, N-ацетилглюкозамин-6-сульфатазы D-типа, галактозо-6-сульфатазы, N-ацетилгалактозамин-4-сульфатазы, бета-глюкуронидазы, альфа-галактозидазы, кислой альфа-галактозидазы и глюкоцереброзидазы. Эти белки можно добавлять при таких заболеваниях как гемофилия, идиопатическая тромбоцитопеническая пурпура, почечная анемия и лизосомная болезнь (мукополисахаридоз, болезнь Фабри, болезнь Помпе и болезнь Гоше) (но, не ограничиваясь только ими).
Следующими вариантами осуществления изобретения является полипептид, содержащий вариант Fc-области, предназначенный для применения для лечения вирусной инфекции. Некоторыми вариантами осуществления изобретения является полипептид, содержащий вариант Fc-области, предназначенный для применения в способе лечения вирусной инфекции у индивидуума, включающем введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для лечения вирусной инфекции. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Следующим объектом изобретения является применение полипептида, содержащего вариант Fc-области, представленный в настоящем описании, для приготовления или получения лекарственного средства. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В следующем варианте осуществления изобретения лекарственное средство предназначено для лечения вирусной инфекции. В другом варианте осуществления изобретения лекарственное средство предназначено для применения в способе лечения вирусной инфекции у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для лечении вирусной инфекции. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Следующим объектом изобретения является способ лечения вирусной инфекции у индивидуума. В одном из вариантов осуществления изобретения способ включает введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для лечения вирусной инфекции. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения являются фармацевтические композиции, которые содержат любой из полипептидов, содержащих вариант Fc-области, представленный в настоящем описании. В следующем варианте осуществления изобретения фармацевтическая композиция предназначена для лечения вирусной инфекции.
Противовирусные антитела, которые содержат вариант Fc-области, предлагаемый в настоящем изобретении, могут подавлять антителозависимое усиление (инфекции), характерное для канонических противовирусных антител. Антителозависимое усиление представляет собой явление, при котором вирус, связанный с антителом, подвергается фагоцитозу посредством активирующих Fc-гамма R, в результате чего заражение вирусом клетки усиливается. Установлено, что связывание антител против вируса Денге с Fc-гамма RIIb играет важную роль в подавлении антителозависимого усиления (Proc. Natl. Acad. Sci. USA 108, 2011, cc. 12479-12484). Перекрестное сшивание молекул Fc-гамма RIIb с помощью иммунного комплекса, включающего антитела против вируса Денге и вирус Денге, ингибирует опосредуемый Fc-гамма R фагоцитоз, что приводит к подавлению антителозависимого усиления. Примеры указанных вирусов включают (но, не ограничиваясь только ими) вирус Денге (DENV1, DENV2, DENV3 и DENV4) и ВИЧ.
Следующими вариантами осуществления изобретения является полипептид, содержащий вариант Fc-области, предназначенный для применения для предупреждения или лечения артериосклероза. В некоторых вариантах осуществления изобретения полипептид, содержащий вариант Fc-области, предназначен для применения в способе предупреждения или лечения артериосклероза у индивидуума, включающем введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для предупреждения или лечения артериосклероза. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Следующим объектом изобретения является применение полипептида, содержащего вариант Fc-области, для приготовления или получения лекарственного средства. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В другом варианте осуществления изобретения лекарственное средство предназначено для предупреждения или лечения артериосклероза. В следующем варианте осуществления изобретения лекарственное средство предназначено для применения в способе предупреждения или лечения артериосклероза у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для предупреждения или лечения артериосклероза. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Следующим объектом изобретения является способ предупреждения или лечения артериосклероза у индивидуума. В одном из вариантов осуществления изобретения способ включает введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для предупреждения или лечения артериосклероза. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения являются фармацевтические композиции, которые содержат любой из полипептидов, содержащих вариант Fc-области, представленный в настоящем описании. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В другом варианте осуществления изобретения фармацевтическая композиция предназначена для предупреждения или лечения артериосклероза.
Антитела против окисленного ЛПНП, т.е. причинного фактора артериосклероза, содержащие вариант Fc-области, предлагаемый в настоящем изобретении, могут предупреждать зависимую от Fc-гамма RIIa адгезию воспалительных клеток. Установлено, что хотя антитела против окисленного ЛПНП ингибируют взаимодействие между окисленным ЛПНП и CD36, антитела против окисленного ЛПНП связываются с эндотелиальными клетками и моноциты распознают их Fc-область Fc-гамма RIIa-зависимым или Fc-гамма RI-зависимым образом (Immunol. Lett. 108, 2007, сс. 52-61). Применение антител, содержащих вариант Fc-области, предлагаемый в настоящем изобретении, может ингибировать Fc-гамма RIIa-зависимое связывание и подавлять адгезию моноцитов с помощью опосредуемых Fc-гамма ингибирующих сигналов.
Следующим вариантом осуществления изобретения является полипептид, содержащий вариант Fc-области, предназначенный для применения для предупреждения или лечения рака. В некоторых вариантах осуществления изобретения полипептид, содержащий вариант Fc-области, предназначен для применения в способе предупреждения или лечения рака у индивидуума, включающем введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для предупреждения или лечения рака. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Следующим объектом является применение полипептида, содержащего вариант Fc-области, представленный в настоящем описании, для приготовления или получения лекарственного средства. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В другом варианте осуществления изобретения лекарственное средство предназначено для предупреждения или лечения рака. В следующем варианте осуществления изобретения лекарственное средство предназначено для применения в способе предупреждения или лечения рака у индивидуума, включающем введение индивидууму в эффективном количестве лекарственного средства для предупреждения или лечения рака. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Следующим объектом изобретения является способ предупреждения или лечения рака у индивидуума. В одном варианте осуществления изобретения способ включает введение индивидууму в эффективном количестве полипептида, содержащего вариант Fc-области, для предупреждения или лечения рака. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В одном варианте осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения являются фармацевтические композиции, которые содержат любой из полипептидов, содержащих вариант Fc-области, представленный в настоящем описании. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В другом варианте осуществления изобретения фармацевтическая композиция предназначена для предупреждения или лечения рака.
Как описано выше, известно, что усиление связывания Fc-гамма RIIb повышает агонистическую активность агонистического антитела и повышает противоопухолевое действие антитела. Таким образом, агонистические антитела, содержащие вариант Fc-области, предлагаемый в настоящем изобретении, можно применять для лечения или предупреждения рака. В частности, вариант Fc-области, предлагаемый в настоящем изобретении, повышает агонистическую активность антител, направленных, например, против рецепторов семейства TNF-рецептора, таких как CD120a, CD120b, рецептор лимфотоксина бета, CD134, CD40, FAS, TNFRSF6B, CD27, CD30, CD137, TNFRSF10A, TNFRSF10B, TNFRSF10C, TNFRSF10D, RANK, остеопротегерин, TNFRSF12A, TNFRSF13B, TNFRSF13C, TNFRSF14, рецептор фактора роста нервов, TNFRSF17, TNFRSF18, TNFRSF19, TNFRSF21, TNFRSF25 и рецептор эктодисплазина А2, и их можно применять для лечения или предупреждения рака. Кроме того, повышается агонистическая активность также антител против других молекул, которые обладают агонистической активностью в результате взаимодействия с Fc-гамма RIIb. Кроме того, путем включения варианта Fc-области, предлагаемого в настоящем изобретении, в антитело против рецепторной тирозинкиназы (RTK), такой как Kit, которая подавляет клеточную пролиферацию посредством перекрестного сшивания с Fc-гамма RIIb, можно повышать ингибирующее действие антител против клеточной пролиферации.
В некоторых вариантах осуществления изобретения предложен способ предупреждения или лечения рака, включая (но, не ограничиваясь только ими) вид рака, выбранный из: мелкоклеточного рака легкого, немелкоклеточного рака легкого, легочной аденокарциномы и плоскоклеточной карциномы легкого, крупноклеточного рака кишечника, ректального рака, рака ободочной кишки, рака молочной железы, рака желудка, рака поджелудочной железы, рака почки, рака предстательной железы, рака яичника, рака щитовидной железы, холангиокарциномы, перитонеального рака, мезотелиомы, плоскоклеточной карциномы, рака шейки матки, рака эндометрия, рака мочевого пузыря, рака пищевода, рака головы и шеи, носоглоточного рака, рака слюнных желез, тимомы, рака кожи, базальноклеточного рака, злокачественной меланомы, рака ануса, рака полового члена, рака яичек, опухоли Вильмса, острого миелоидного лейкоза (включая острый миелолейкоз, острый миелобластный лейкоз, острый промиелоцитарный лейкоз, острый миеломоноцитраный лейкоз и острый моноцитарный лейкоз), хронического миелогенного лейкоза, острого лимфобластного лейкоза, хронического лимфатического лейкоза, лимфомы Ходжкина, неходжскинской лимфомы (лимфома Беркитта, хронический лимфоцитарный лейкоз, мукозный фугоид, лимфома из клеток мантии, фолликулярная лимфома, диффузная крупноклеточная лимфома, лимфома из маргинальной зоны, пилоидная лейкемоидная плазмоцитома, лимфома из периферических Т-клеток и Т-клеточный лейкоз/Т-клеточная лимфома взрослых), гистоцитоза клеток Лангерганса, множественной миеломы, миелодисплатического синдрома, опухоли головного мозга (включая глиому, астроглиому, глиобластому, менингиому и эпендимому), нейробластомы, ретинобластомы, остеосаркомы, саркомы Капоши, саркомы Юинга, ангиосаркомы и гемангиоперицитомы.
Антитела, которые были модифицированы так, чтобы повышать активность связывания с Fc-гамма R, включая Fc-гамма RIIb, путем модификации по меньшей мере одного аминокислотного остатка, могут усиливать элиминацию антигена из плазмы, что описано или высказано в качестве предположения, например, в WO 2013/047752, WO 2013/125667, WO 2014/030728 или WO 2014/163101.
Не вдаваясь в конкретную теорию, антитело, обладающее повышенной Fc-гамма R-связывающей активностью в условиях нейтрального рН, быстро проникают в клетки вместе с антигеном, образующим комплекс с антителом, и в результате элиминация антигена из плазмы может усиливаться при введении антитела in vivo.
Антитела, которые были модифицированы так, чтобы обладать повышенной pI, путем модификации по меньшей мере одного аминокислотного остатка, который может экспонироваться на поверхности антитела, могут более быстро включаться в клетки или могут усиливать элиминацию антигена из плазмы, что описано или высказано в качестве предположения, например, в WO 2007/114319, WO 2009/041643, WO 2014/145159 или WO 2012/016227.
Не вдаваясь в конкретную теорию, можно предположить, что рН биологических жидкостей (например, плазмы) находится в диапазоне нейтральных значений рН. В биологических жидкостях чистый положительный заряд антитела с увеличенной pI возрастает вследствие увеличения pI, и в результате антитело привлекается с большей силой посредством физико-химического кулоновского взаимодействия к поверхности эндотелиальной клетки, которая имеет чистый отрицательный заряд, по сравнению с антителом, у которого не повышена pI. Это означает, что посредством такого неспецифического связывания проникновение в клетки антитела в комплексе с его антигеном повышается, что приводит к повышенной элиминации антигена из плазмы. Предполагается, что это явление широко распространено in vivo вне зависимости от типа клеток, типа тканей, типа органа и т.д.
В контексте настоящего описания усиление элиминации антигена из плазмы означает, например, что скорость элиминации антигена из плазмы возрастает по сравнению со скоростью при введении полипептида, который содержит соответствующую, но не модифицированную таким же образом Fc-область. Повышение элиминации антигена из плазмы можно оценивать, например, измеряя концентрацию антигена в плазме. В данной области известны различные методы измерения концентрации антигена в плазме.
Другими вариантами осуществления изобретения является полипептид, содержащий вариант Fc-области, предназначенный для применения для усиления элиминации антигена из плазмы. Некоторыми вариантами осуществления изобретения является полипептид, который содержит вариант Fc-области, предназначенный для применения в способе усиления элиминации антигена из плазмы у индивидуума, заключающемся в том, что вводят индивидууму в эффективном количестве полипептид, который содержит вариант Fc-области, для усиления элиминации антигена из плазмы. В указанных вариантах осуществления изобретения предпочтительно, чтобы полипептид дополнительно содержал антигенсвязывающий домен. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения предпочтительно представляет собой человека.
Следующим объектом изобретения является применение полипептида, который содержит вариант Fc-области, для изготовления или получения лекарственного средства. В некоторых объектах изобретения полипептид представляет собой антитело. В некоторых объектах изобретения полипептид представляет собой содержащий Fc слитый белок. В другом варианте осуществления изобретения лекарственное средство предназначено для усиления элиминации антигена из плазмы. В следующем варианте осуществления изобретения лекарственное средство предназначено для применения в способе усиления элиминации антигена из плазмы у индивидуума, заключающемся в том, что вводят индивидууму в эффективном количестве лекарственное средство для усиления элиминации антигена из плазмы. В указанных вариантах осуществления изобретения предпочтительно, чтобы полипептид дополнительно содержал антигенсвязывающий домен. «Индивидуум» в контексте любого из указанных выше вариантов осуществления изобретения может представлять собой человека.
Другим вариантом осуществления изобретения является способ усиления элиминации антигена из плазмы у индивидуума. В одном из вариантов осуществления изобретения способ заключается в том, что вводят индивидууму в эффективном количестве полипептид, который содержит вариант Fc-области, для усиления элиминации антигена из плазмы. В указанных вариантах осуществления изобретения предпочтительно, чтобы полипептид дополнительно содержал антигенсвязывающий домен. В одном из вариантов осуществления изобретения «индивидуум» представляет собой человека.
Следующим объектом изобретения являются фармацевтические композиции, которые содержат любой из полипептидов, содержащих вариант Fc-области, представленный в настоящем описании. Следующим вариантом осуществления изобретения является фармацевтическая композиция, предназначенная для усиления элиминации антигена из плазмы. В указанном варианте осуществления изобретения предпочтительно, чтобы полипептид дополнительно содержал антигенсвязывающий домен.
В одном из вариантов осуществления изобретения по сравнению с вариантом до аминокислотной модификации антитело, которое имеет вариант Fc-области, предлагаемый в настоящем изобретении, повышает элиминацию антигена из плазма, например, по меньшей мере в 1,1, 1,25, 1,5, 1,75, 2,0, 2,25, 2,5, 2,75, 3,0, 3,25, 3,5, 3,75, 4,0, 4,25, 4,5, 4,75, 5,0, 5,5, 6,0, 6,5, 7,0, 7,5, 8,0, 8,5, 9,0, 9,5 или 10,0 раз или более при введении антитела in vivo.
Следующим объектом изобретения являются способы приготовления лекарственного средства или фармацевтической композиции, заключающиеся в том, что смешивают любые полипептиды, содержащие варианты Fc-области, представленные в настоящем описании, с фармацевтически приемлемым носителем, например, для применения в любом из указанных выше терапевтических способов. В одном из вариантов осуществления изобретения способы приготовления лекарственного средства или фармацевтической композиции дополнительно включают добавление по меньшей мере одного дополнительного терапевтического средства к лекарственному средству или фармацевтической композиции.
Антитела, предлагаемые в изобретении, для лечения можно применять либо индивидуально, либо в сочетании с другими агентами. Например, антитело, предлагаемое в изобретении, можно вводить совместно по меньшей мере с одним дополнительным терапевтическим средством.
Аналогично этому, полипептиды, содержащие вариант Fc-области, предлагаемый в изобретении, для лечения можно применять либо индивидуально, либо в сочетании с другими агентами. Например, полипептид, содержащий вариант Fc-области, предлагаемый в изобретении, можно вводить совместно по меньшей мере с одним дополнительным терапевтическим средством.
Такие комбинированные терапии, указанные выше, предусматривают совместное введение (когда два или большее количество терапевтических средств включены в одну и ту же или различные композиции) и раздельное введение, в каждом случае введение антитела или полипептида, содержащего вариант Fc-области, предлагаемый в изобретении, можно осуществлять до, одновременно и/или после введения дополнительного терапевтического средства или средств. В одном из вариантов осуществления изобретения введение антитела к миостатину и введение дополнительного терапевтического средства можно осуществлять в пределах примерно месяца или в пределах примерно 1, 2 или 3 недель, или в пределах примерно 1, 2, 3, 4, 5 или 6 дней друг относительно друга.
В другом варианте осуществления изобретения введение полипептида, содержащего вариант Fc-области, и введение дополнительного терапевтического средства можно осуществлять в пределах примерно месяца или в пределах примерно 1, 2 или 3 недель, или в пределах примерно 1, 2, 3, 4, 5 или 6 дней друг относительно друга.
Антитело или полипептид, содержащий вариант Fc-области, предлагаемый в изобретении (и необязательно любое дополнительное терапевтическое средство), можно вводить любыми приемлемыми путями, включая парентеральный, внутрилегочный и интраназальный и при необходимости применять для местной обработки, введения в повреждение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Дозирование можно осуществлять любым приемлемым путем, например, путем инъекций, таких как внутривенные или подкожные инъекции, в зависимости, в том числе, от того, является ли лечение кратковременным или хроническим. Можно применять различные схемы дозирования, включая (но, не ограничиваясь только ими) однократное введение или несколько введений в различные моменты времени, болюсное введение и пульсирующую инфузию.
Антитела или полипептиды, содержащие вариант Fc-области, предлагаемый в изобретении, можно включать в состав композиций, дозировать и вводить в соответствии с надлежащей клинической практикой. Рассматриваемые в этом контексте факторы включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние индивидуального пациента, причину нарушения, область введения агента, метод введения, схему введения и другие факторы, известные практикующим медикам. Антитело можно, но это не является обязательным, включать в композицию в сочетании с одним или несколькими другими агентами, которые в настоящее время применяют для профилактики и лечения рассматриваемого нарушения. Эффективное количество указанных других агентов зависит от количества антитела, присутствующего в композиции, типа нарушения или лечения и других указанных выше факторов. Их, как правило, применяют в таких же дозах и с использованием таких же путей введения, которые указаны в настоящем описании, или в количестве, составляющем от 1 до 99% от дозы, указанной в настоящем описании, или в любой дозе и с помощью любого пути, которые по данным эмпирической/клинической оценки являются пригодными.
Для предупреждения или лечения заболевания приемлемая доза антитела, предлагаемого в изобретении (при его применении индивидуально или в сочетании с одним или несколькими другими дополнительными терапевтическими средствами), должна зависеть от типа заболевания, подлежащего лечению, типа антитела, типа полипептида, содержащего вариант Fc-области, серьезности и течения болезни, применяют ли антитело или полипептид, содержащий вариант Fc-области, для превентивных или терапевтических целей, предшествующей терапии, истории болезни пациента и ответа на антитело или полипептид, содержащий вариант Fc-области, а также предписания лечащего врача. Антитело или полипептид, содержащий вариант Fc-области, можно вводить пациенту однократно или в виде серий обработок. В зависимости от типа и серьезности заболевания примерно от 1 мкг/кг до 15 мг/кг (например, 0,5-10 мг/кг) антитела может представлять собой начальную возможную дозу для введения пациенту, например, с помощью одной или нескольких отдельных обработок или с помощью непрерывной инфузии. Одним из примеров типичных суточных доз может являться доза, составляющая от примерно 1 мкг/кг до 100 мг/кг или более в зависимости от указанных выше факторов. При использовании повторных введений в течение нескольких дней или более длительного периода времени, в зависимости от состояния, лечение, как правило, следует продолжать до достижения подавления симптомов заболевания. Одним из примеров доз антитела или полипептида, содержащего вариант Fc-области, является доза, составляющая от примерно 0,05 до примерно 10 мг/кг. Так, пациенту можно вводить одну или несколько следующих доз: примерно 0,5, 2,0, 4,0 или 10 мг/кг (или любую их комбинацию). Указанные дозы можно вводить дробно, например, каждую неделю или каждые три недели (например, так, чтобы пациент получал от примерно двух до примерно двадцати, например, примерно шесть доз антитела или полипептида, содержащего вариант Fc-области). Можно применять начальную повышенную ударную дозу с последующим введением одной или нескольких более низких доз. С помощью общепринятых методов и анализов можно легко осуществлять мониторинг действия указанной терапии.
Очевидно, что любые из вышеуказанных композиций или терапевтических способов можно применять с использованием иммуноконъюгата, предлагаемого в изобретении, вместо или в дополнение к антителу к миостатину.
Очевидно, что любые из вышеуказанных композиций или терапевтических способов можно применять с использованием иммуноконъюгата, предлагаемого в изобретении, вместо или в дополнение к полипептиду, содержащему вариант Fc-области, представленный в настоящем описании.
З. Изделия
Другим объектом изобретения является изделие, которое содержит продукты, применяемые для лечения, предупреждения и/или диагностирования указанных выше нарушений. Изделие представляет собой контейнер и этикетку или листовку-вкладыш в упаковку, которые размещены на контейнере или вложены в него. Приемлемыми контейнерами являются, например банки, пузырьки, шприцы, пакеты для внутривенного раствора и т.д. Контейнеры можно изготавливать из различных материалов, таких как стекло или пластмасса. Контейнер содержит композицию, которая сама или в сочетании с другой композицией является эффективной для лечения, предупреждения и/или диагностирования состояния, и может иметь стерильный порт доступа (например, контейнер может представлять собой пакет для внутривенного раствора или пузырек, снабженный пробкой, которую можно прокалывать с помощью иглы для подкожных инъекций). По меньшей мере одно действующее вещество в композиции представляет собой антитело, предлагаемое в изобретении. На этикетке или листовке-вкладыше в упаковку указано, что композицию применяют для лечения выбранного состояния. Кроме того, изделие может включать (а) первый контейнер с находящейся в нем композицией, где композиция содержит антитело, предлагаемое в настоящем изобретении; и (б) второй контейнер с находящейся в нем композицией, где композиция содержит дополнительное цитотоксическое или иное терапевтическое средство. Согласно этому варианту осуществления изобретения изделие может содержать листовку-вкладыш в упаковку, которая содержит информацию о том, что композиции можно использовать для лечения конкретного состояния. В альтернативном или дополнительном варианте изделие может дополнительно включать второй (или третий) контейнер с фармацевтически приемлемым буфером, таким как бактериостатическая вода для инъекций (БСВИ), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Кроме того, оно может включать другие продукты, необходимые с коммерческой точки зрения и с точки зрения потребителя, в частности, другие буферы, разбавители, фильтры, иглы и шприцы.
Как должно быть очевидно, любое из указанных выше изделий может включать иммуноконъюгат, предлагаемый в изобретении, вместо или в дополнение к антителу к миостатину.
Примеры
Ниже приведены примеры методов и композиций, предлагаемых в изобретении. Как должно быть очевидно, различные другие варианты осуществления изобретения можно применять на практике с учетом общего описания изобретения, представленного выше.
Пример 1
Экспрессия и очистка латентной и зрелой форм миостатина человека, обезьян циномолгус и мышей
Человеческий латентный миостатин (обозначенный в настоящем описании также как латентная форма человеческого миостатина) (SEQ ID NO: 1) кратковременно экспрессировали, используя клетки FreeStyle293-F (FS293-F-клетки) (фирма Thermo Fisher, Карлсбад, шт. Калифорния, США). Кондиционированные среды, содержащие экспрессированную латентную форму человеческого миостатина, подкисляли до рН 6,8 и разводили, используя 1/2 объема, водой milliQ, после чего вносили в ионообменную колонку с Q-сефарозой FF (фирма GE healthcare, Уппсала, Швеция). Значение рН прошедшей через колонку фракции доводили до рН 5,0 и вносили в катионообменную колонку с SP-сефарозой HP (фирма GE healthcare, Уппсала, Швеция), а затем элюировали с использованием градиента NaCl. Фракции, содержащие латентную форму человеческого миостатина, собирали и затем подвергали гель-фильтрации на колонке с Супердекс 200 (фирма GE healthcare, Уппсала, Швеция) уравновешенной с помощью 1×ЗФР. Затем фракции, содержащие латентную форму человеческого миостатина, объединяли и хранили при -80°C.
Человеческий зрелый миостатин (обозначенный в настоящем описании также как зрелая форма человеческого миостатина) (SEQ ID NO: 2) очищали из очищенной латентной формы человеческого миостатина. Латентную форму человеческого миостатина подкисляли путем добавления 0,1% трифторуксусной кислоты (ТФК) и вносили в колонку с обращенной фазой Vydac 214ТР С4 (фирма Grace, Дирфилд, шт. Иллинойс, США) и элюировали с использованием градиента ТФК/CH3CN. Фракции, содержащие человеческий зрелый миостатин, объединяли, сушили и хранили при -80°С. Для восстановления человеческий зрелый миостатин растворяли в 4 мМ HCl.
Экспрессию и очистку латентной и зрелой формы миостатина обезьян циномолгус (циномолгус или cyno) (SEQ ID NO: 3 и 4 соответственно) и мышей (SEQ ID NO: 5 и 6 соответственно) осуществляли во всех случаях совершенно таким же образом, что и в случае человеческой копии. Касательно гомологии последовательностей зрелая форма человеческого, cyno и мышиного миостатина были идентичны на 100%, поэтому любой зрелый миостатин вне зависимости от вида применяли в качестве зрелого миостатина во всех проводимых экспериментах.
Пример 2
Идентификация антитела к латентному миостатину
Антитела к латентному миостатину получали, отбирали и анализировали следующим образом.
Новозеландских белых кроликов (NZW) возрастом 12-16 недель иммунизировали внутрикожно мышиным латентным миостатином и/или человеческим латентным миостатином (50-100 мкг/дозу/кролика). Обработку указанной дозой повторяли 3-4 раз в течение 1 месяца. Через 1 неделю после последней иммунизации собирали селезенки и образцы крови иммунизированного кролика. Антигенспецифические В-клетки окрашивали меченым антигеном, сортировали с использованием клеточного FCM-сортера (FACS aria III, фирма BD) и высевали в 96-луночные планшеты с плотностью 1 клетка/лунку вместе с 25000 EL4-клеток/лунку (клетки из Европейской коллекции клеточных культур) и вместе с кондиционированной средой для кроличьих Т-клеток, разведенной в 20 раз, и культивировали в течение 7-12 дней. EL4-клетки предварительно обрабатывали митомицином С (фирма Sigma) в течение 2 ч и промывали 3 раза. Кондиционированную среду для кроличьих Т-клеток получали путем культивирования кроличьих тимоцитов в среде RPMI-1640, содержащей фитогемагглютинин-М (фирма Roche), форбол-12-миристат-13-ацетат (фирма Sigma) и 2% FBS. После культивирования супернатанты В-клеточных культур собирали для дополнительного анализа и дебрис криоконсервировали.
ELISA-анализ применяли для оценки специфичности антител в супернатанте В-клеточной культуры. 384-луночные планшеты MAXISorp (фирма Nunc) сенсибилизировали стрептивидином (фирма GeneScript) в концентрации 50 нМ в ЗФР в течение 1 ч при комнатной температуре. Затем планшеты блокировали с помощью реагента Blocking One (фирма Nacalai Tesque), разведенного в 5 раз. Человеческий или мышиный латентный миостатин метили с помощью NHS-ПЭГ4-биотина (фирма PIERCE) и добавляли в заблокированные планшеты для ELISA, инкубировали в течение 1 ч и промывали забуференным Трис физиологическим раствором с 0,05% Твин-20 (TBS-T). Супернатанты В-клеточных культур вносили в планшеты для ELISA, инкубировали в течение 1 ч и промывали TBS-T. Связывание выявляли с помощью козьего антикроличьего IgG, конъюгированного с пероксидазой из хрена (фирма BETHYL), для чего добавляли ABTS (фирма KPL).
В целом 17818 В-клеточных линий подвергали скринингу в отношении специфического связывания с мышиным и/или человеческим латентным миостатином, отбирали 299 линий и обозначали их как MST0255-287, 630-632, 677-759, 910, 932-1048, 1050-1055, 1057-1066, 1068, 1070-1073, 1075-1110, 1113-1119. РНК очищали из соответствующих клеточных дебрисов с помощью наборов ZR-96 Quick-RNA (фирма ZYMO RESEARCH).
ДНК, кодирующие вариабельные области тяжелой и легкой цепей каждой отобранной клеточной линии, амплифицировали с помощью ПЦР с обратной транскрипцией и клонировали в экспрессионных векторах с последовательностью константной области тяжелой цепи G1m (SEQ ID NO: 50 (аминокислотная последовательность представлена в SEQ ID NO: 7)) и с последовательностью константной области легкой цепи k0MTC или k0MC (SEQ ID NO: 53 или 194 (аминокислотная последовательность (в обоих случаях одна и та же) представлена в SEQ ID NO: 8)) соответственно. Рекомбинантные антитела кратковременно экспрессировали, используя клетки FreeStyle FS293-F и 293фектин (фирма Life technologies), согласно инструкции производителя. Для скрининга использовали супернатант культуры или рекомбинантные антитела. Рекомбинантные антитела очищали с помощью белка А (фирма GE Healthcare) и элюировали в D-ЗФР или His-буфере (20 мМ гистидин, 150 мМ NaCl, рН 6,0). Дополнительно при необходимости проводили гель-фильтрацию для удаления высокомолекулярного и/или низкомолекулярного компонента.
Пример 3
Характеризация антитела к латентному миостатину (HEK-Blue-анализ (BMP1-активация))
Анализ репортерного гена применяли для оценки биологической активности активного миостатина in vitro. Клетки HEK-Blue™ TGF-бета (фирма Invivogen), которые экспрессируют индицируемые Smad3/4-связывающими элементами (SBE) репортерные гены SEAP (секретируемая щелочная фосфатаза), позволяют осуществлять детекцию биоактивного миостатина, осуществляя мониторинг активации рецепторов для активина типа 1 и типа 2. Активный миостатин стимулирует производство SEAP, которая секретируется в клеточный супернатант. Затем определяют количество секретируемой SEAP с помощью QUANTIBlue™ (фирма Invivogen).
Клетки HEK-Blue™ TGF-бета поддерживали в среде DMEM (фирма Gibco), дополненной 10% фетальной бычьей сыворотки, 50 мкг/мл стрептомицина, 50 ед./мл пенициллина, 100 мкг/мл Normocin™, 30 мкг/мл бластицидина, 200 мкг/мл HygroGold™ и 100 мкг/мл Zeocin™. При проведении функционального анализа среду для клеток заменяли на среду для анализа (DMEM, дополненная 0,1% бычьего сывороточного альбумина, стрептомицином, пенициллином и Normocin™) и высевали в 96-луночный планшет. Латентный миостатин человека, циномолгус или мышей инкубировали с рекомбинантным человеческим ВМР1 (фирма R&D Systems) и антителом к латентному миостатину при 37°С в течение ночи. Смеси образцов переносили в клетки. После 24-часовой инкубации клеточные супернатанты смешивали с QUANTIBlue™ и измеряли оптическую плотность при 620 нм в колориметрическом планшет-ридере. Как продемонстрировано на фиг. 1, МАт MST1032-G1m (см. в таблице 2а аминокислотную и нуклеотидную последовательности) препятствовало опосредуемой протеазой активации латентного миостатина человека, циномолгус и мышей, о чем свидетельствуют пониженные концентрации секретируемой SEAP. Продемонстрировано, что MST1032-G1m обладал практически сопоставимой ингибирующей активностью в отношении латентного миостатина человека, циномолгус и мышей.
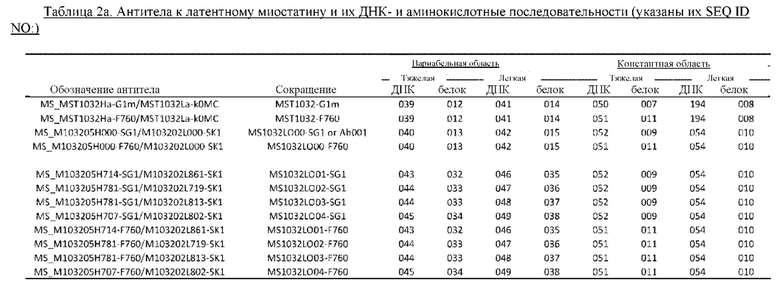

Пример 4
Характеризация антитела к латентному миостатину (HEK Blue-анализ (спонтанная активация))
Латентный миостатин человека, циномолгус или мышей инкубировали с антителом к латентному миостатину при 37°С в течение ночи, и смесь образцов вносили в клетки HEK-Blue™ TGF-бета. После 24-часовой инкубация клеточный супернатант смешивали с QUANTIBlue™ и измеряли оптическую плотность при 620 нм в колориметрическом планшет-ридере. Как продемонстрировано на фиг. 2, после инкубации при 37°С обнаружено высвобождение активного миостатина из латентной формы. В присутствии МАт MST1032-G1m происходило ингибирование активации миостатина, и в результате в клеточном супернатанте был выявлен более низкий уровень SEAP. MST1032-G1m ингибировало спонтанную активацию латентного миостатина, и для него продемонстрирована практически сопоставимая ингибирующая активность в отношении латентного миостатина человека, циномолгус и мышей.
Пример 5
Характеризация антитела к латентному миостатину (ELISA)
Несенсибилизированные планшеты для ELISA (планшет NUNC-IMMUNO, поверхность MAXISORP, фирма Nalge Nunc International) покрывали 20 мкл 50 нМ стрептивидина (фирма GenScript) в течение 1 ч при комнатной температуре. Затем планшеты трижды промывали ЗФРТ и блокировали в течение ночи, используя 50 мкл 20% Blocking One (фирма Nacalai Tesque). На следующий день каждую лунку каждого планшета инкубировали с биотинилированным зрелым миостатином, человеческим или мышиным биотинилированным латентным миостатином или биотинилированным рекомбинантным химерным белком мышиный пропептид миостатина-Fc (фирма R&D Systems) из расчета 4 нМ/лунку/20 мкл в течение 2 ч. После промывки добавляли в лунки 20 мкл образца антитела и планшеты выдерживали в течение 1 ч. Планшеты промывали и добавляли 20 мкл античеловеческого IgG, конъюгированного с пероксидазой из хрена (HRP) (фирма Abcam), разведенного в забуференном HEPES физиологическом растворе, и планшеты выдерживали в течение еще 1 ч. Затем планшеты вновь промывали и после этого добавляли в каждую лунку по 50 мкл ABTS (фирма KPL), и планшеты инкубировали в течение 1 ч. Обнаружение сигнала осуществляли при 405 нм в колориметрическом планшет-ридере. Результаты эксперимента по связыванию представлены на фиг. 3. MST1032-G1m связывалось с латентным миостатином (т.е. нековалентным комплексом зрелого миостатина и пропептидов) и пропептидами, но не связывалось с зрелым миостатином. Эти результаты свидетельствуют о том, что MST1032-G1m специфически связывается с пропептидом миостатина (например, пропептидной частью), но не связывается с активным миостатином (например, зрелой областью).
Пример 6
Характеризация антитела к латентному миостатину (Вестерн-блоттинг)
Мышиный латентный миостатин инкубировали с рекомбинантным человеческим ВМР1 (фирма R&D Systems) с MST1032-G1m или без антитела при 37°С в течение ночи. Затем образцы смешивали с 4× восстанавливающим буфером для образца, применяемым для ДСН-ПААГ (фирма Wako), и выдерживали при 95°С в течение 5 мин и затем вносили в систему для гель-электрофореза в присутствии ДСН. Белки переносили на мембрану с помощью системы для переноса Trans-Blot Turbo™ (фирма Bio-rad). Пропептид миостатина выявляли с использованием овечьего антитела к мышиному пропептиду GDF8 (фирма R&D Systems), для обнаружения которого затем применяли антитело к овечьему IgG-HRP (фирма Santa Cruz). Мембрану инкубировали с субстратом для ECL и визуализировали с использованием устройства ImageQuant LAS 4000 (фирма GE Healthcare). Как продемонстрировано на фиг. 4, расщепление пропептида с помощью ВМР1 ингибировалось MST1032-G1m.
Пример 7
Характеризация антитела к латентному миостатину (BIACORE®)
Кинетические параметры антител к латентному миостатину в отношении латентного миостатина человека, обезьян циномолгус (cyno) и мышей оценивали при 37°С при рН 7,4, используя устройство BIACORE® Т200 (фирма GE Healthcare). ProA/G (белок A/G) (фирма Pierce) иммобилизовывали на всех проточных ячейках СМ4-чипа с использованием набора для аминного сочетания (фирма GE Healthcare). Антитело к латентному миостатину и аналиты приготавливали в ACES, рН 7,4 (20 мМ ACES, 150 мМ NaCl, 1,2 мМ CaCl2, 0,05% Твин 20, 0,005% NaN3). Антитело «захватывали» на поверхности сенсорного чипа с помощью ProA/G. Уровни иммобилизации антител, как правило, соответствовали 150-220 резонансным единицам (RU). Затем латентный миостатин человека, cyno или мышей инъецировали в концентрациях от 3,125 до 50 нМ, которые получали путем двукратных серийных разведений, после чего давали пройти диссоциации. Поверхность сенсора регенерировали с помощью 25 мМ NaOH. Кинетические параметры определяли путем обработки и подгонки данных к модели связывания 1:1, используя программное обеспечение BIACORE® Т200 Evaluation, версия 2.0 (фирма GE Healthcare). Сенсограммы приведены на фиг. 5. Данные о скорости ассоциации (ka), скорости диссоциации (kd) и аффинности связывания (KD) представлены в таблице 3. Кинетические параметры MST1032-G1m, характеризующие связывание с латентным миостатином человека, cyno и мышей, оказались сопоставимыми.

Пример 8
Эффективность in vivo антитела к латентному миостатину в отношении мышечной массы и жировой массы
Эффективность in vivo МАт MST1032-G1m изучали в опытах на мышах. В этом опыте антитело к зрелому миостатину 41С1Е4 (описанное в US №7632499) применяли в качестве положительного контроля. Во избежание потенциальной иммуномодуляции вследствие ответа у мышей на человеческое антитело опыты in vivo осуществляли для мышах с тяжелым комбинированным иммунодефицитом (SCID). SCID (С.В-17 SCID)-мышей возрастом 5 недель (фирма Charles River Laboratories Japan, Inc. (Канагава, Япония)) обрабатывали моноклональными антителами в различных дозах или наполнителем (ЗФР), которые вводили внутривенно один раз в неделю в течение 2 недель. В дни 0, 7 и 14 определяли общую тощую массу и жировую массу тела мышей с помощью ядерного магнитного резонанса (ЯМР) (устройство minispec LF-50, фирма Bruker Bio Spin, (Канагава, Япония)). Животных умерщвляли в день 14 и изымали и взвешивали икроножные и четырехглавые мышцы.
Статистическую значимость определяли с помощью дисперсионного анализа, t-критерия Стьюдента и критерия Дюннетта с использованием программного обеспечения JMP 9 (фирма SAS, Inc.). Величину р ниже 0,05 рассматривали как значимую.
Результаты указанного эксперимента представлены на фиг. 6А-В. Оба антитела (MST1032-G1m и 41С1Е4) повышали тощую массу тела в зависимости от дозы, и MST1032-G1m, при его введение в дозе 40 мг/кг, существенно снижало жировую массу по сравнению с обработанной наполнителем (ЗФР) группой в день 14.
После двухнедельной обработки антитела, которые вводили в дозах 10 и 40 мг/кг, приводили к значимому приросту массы во влажном состоянии четырехглавых и икроножных мышц относительно обработанной наполнителем группы.
Пример 9
Сравнение in vivo различных родственных антител к миостатину
Все применяемые экспериментальные схемы соответствовали описанным в примере 8. Различные родственные антитела к миостатину 41С1Е4, REGN, OGD, и MYO-029 тестировали на 5-недельных SCID-мышах в течение 2 недель. REGN, OGD и MYO-029 представляют собой антитела к миостатину, описанные в WO 2011/150008 как H4H1657N2, в WO 2013/186719 как OGD1.0.0. и в WO 2004/037861 как MYO-029 соответственно. Указанные антитела вводили внутривенно один раз в неделю. Тощую массу и жировую массу тела определяли путем ЯМР-сканирования всего тела в дни 0 и 14. Силу сжатия измеряли с помощью устройства для осуществления теста силы сжатия (например, GPM-100В, фирма MELQUEST Ltd., (Тояма, Япония)) в день 14. Как продемонстрировано в результатах, представленных на фиг. 7А-В, тощая масса тела увеличивалась при использовании указанных антител кроме MYO-029 (Р>0,05). Сила сжатия в группе, которой вводили MST1032-G1m, значимо повышалась по сравнению с обработанной наполнителем (ЗФР) группой при применении в дозе 10 мг/кг (Р<0,05). 41С1Е4 и REGN также значимо повышали силу сжатия по сравнению с обработкой наполнителем (ЗФР). Общая жировая масса тела имела тенденцию к снижению при применении MST1032-G1m в дозе 10 мг/кг. Эффективность в отношении снижения жировой массы у MST1032-G1m оказалась более высокой по сравнению с другими антителами к зрелому миостатину.
Пример 10
Гуманизация антитела к латентному миостатину
Аминокислотные остатки вариабельной области нумеровали согласно Кэботу (Kabat и др., Sequence of proteins of immunological interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991).
Создавали вариабельные области тяжелых и легких цепей для гуманизированных антител MST1032. Некоторые из гуманизированных антител MST1032 содержали обратную мутацию в каркасном участке. Полинуклеотиды созданных вариабельных областей тяжелых и легких цепей синтезировали на фирме GenScript Inc. и клонировали в экспрессионных векторах, содержащих последовательность константной области тяжелой цепи SG1 (SEQ ID NO: 52 (аминокислотная последовательность представлена в SEQ ID NO: 9)) и последовательность константной области легкой цепи SK1 (SEQ ID NO: 54 (аминокислотная последовательность представлена в SEQ ID NO: 10)) соответственно. Гуманизированные антитела кратковременно экспрессировали в клетках FS293-F и осуществляли анализ HEK Blue и BIACORE® согласно описанной выше процедуре. Как продемонстрировано на фиг. 8 и в таблице 4, и при сравнении с фиг. 1 и 2, установлено, что гуманизированное антитело (MS1032LO00-SG1) характеризуется ингибирующей активностью и аффинностью, сопоставимой с химерным антителом (MST1032-G1m).

Пример 11
Создание рН-зависимого антитела к латентному миостатину
Для создания рН-зависимых антител к латентному миостатину осуществляли гистидин-сканирующий мутагенез всех CDR МАт MS1032LO00-SG1. Каждую аминокислоту в CDR индивидуально заменяли на гистидин, используя набор для клонирования In-Fusion HD (фирма Clontech Inc. или Takara Bio company) согласно инструкциям производителя. После подтверждения с помощью секвенирования правильности мутации в каждом варианте варианты кратковременно экспрессировали и очищали согласно описанному выше методу. Все варианты с замещением на гистидин оценивали с помощью BIACORE®-анализа, модифицированного по сравнению с описанным выше анализом. В целом, метод состоял в следующем: при проведении BIACORE®-анализа осуществляли дополнительную фазу диссоциации при рН 5,8 непосредственно после фазы диссоциации при рН 7,4. Это осуществляли для оценки рН-зависимой диссоциации антитела (Ат) и антигена (Аг) из комплексов, полученных при рН 7,4, в сравнении с соответствующей диссоциацией при рН 5,8. Скорость диссоциации в буфере с рН 5,8 определяли путем обработки и подгонки данных с использованием программного обеспечения для аппроксимации кривых Scrubber 2.0 (фирма BioLogic Software).
Как продемонстрировано на фиг. 9, для родительского антитела (Ab001) не обнаружено уменьшение ответа в виде связывания при рН 5,8 по сравнению с фазой диссоциации при рН 7,4. Несколько единичных замен на гистидин приводили к снижению от умеренного до сильного ответа в виде связывания при рН 5,8 по сравнению с фазой диссоциации при рН 7,4. Скорость диссоциации при рН 5,8 для каждого варианта с единичной заменой на гистидин представлена в таблице 5. Как продемонстрировано в таблице 5, для Ab002 обнаружена наиболее быстрая скорость диссоциации из комплекса Ат/Аг при рН 5,8, превышающая более чем в 200 раз скорость для родительского антитела (Ab001). Затем создавали антитела с комбинацией указанных мутаций в CDR. Последовательности CDR антител, содержащих указанные различные мутации, приведены в таблице 6.
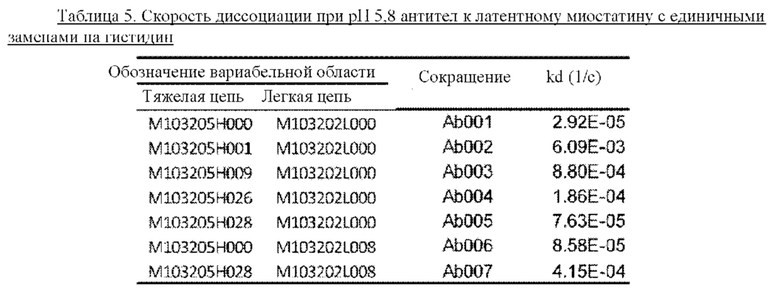
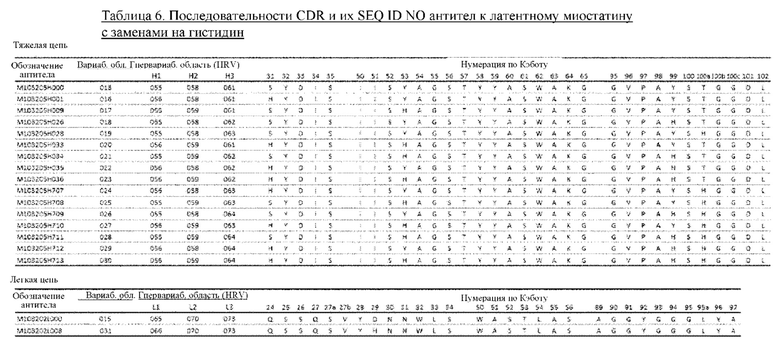
Пример 12
Повышение аффинности рН-зависимого антитела к латентному миостатину
Для идентификации мутаций, повышающих аффинность при рН 7,4 и/или in vitro ингибирующую активность, создавали более 500 вариантов тяжелых и легких цепей соответственно, используя по меньшей мере один вариант, созданный в примере 11, в качестве матрицы. В этих вариантах каждую аминокислоту в CDR заменяли на 18 других аминокислот, за исключением исходной аминокислоты и цистеина. Способность к связыванию вариантов с человеческим латентным миостатином оценивали при 37°С при рН 7,4 с помощью устройства BIACORE® 4000 (фирма GE Healthcare). Супернатант культуры, содержащей варианты, экспрессируемые в клетках FS293-F, получали в ACES-буфере, рН 7,4, содержащем 10 мг/мл БСА и 0,1 мг/мл карбоксиметилдекстрана (CMD). Каждое антитело «захватывали» на проточных ячейках до достижения уровня иммобилизации, составлявшего примерно 200 RU. Затем 25 нМ человеческий латентный миостатин инъецировали на проточные ячейки. При проведении BIACORE®-анализа осуществляли дополнительную фазу диссоциации при рН 5,8 сразу после фазы диссоциации при рН 7,4. На указанной дополнительной фазе диссоциации оценивали рН-зависимую диссоциацию антитела (Ат) и антигена (Аг) из комплексов, полученных при рН 7,4. Поверхность проточных ячеек регенерировали с помощью 25 мМ NaOH. Кинетические параметры определяли с помощью программного обеспечения BIACORE® 4000 Evaluation, версия 2.0 (фирма GE Healthcare). Анализ дополнительной диссоциации при рН 5,8 осуществляли с помощью Scrubber2 (программное обеспечение фирмы BioLogic).
Отбирали варианты с повышенной аффинностью при рН 7,4 и/или in vitro ингибирующей активностью и объединяли с мутациями, повышающими зависимость от рН, идентифицированными в примере 11. После объединения указанных мутаций отбирали 4 варианта (MS1032LO01, 02, 03 и 04) и кратковременно экспрессировали с SG1 или F760 (SEQ ID NO: 11 и 51) для кинетических анализов связывания in vitro и/или in vivo с помощью BIACORE®. И SG1, и F760 представляют собой человеческие константные области тяжелой цепи. Аффинности связывания SG1 с человеческими Fc-гамма R сопоставимы с аффиностью константной области встречающегося в естественных условиях IgG1, но аффинность F760 элиминирована из-за модификации Fc (что обозначено в настоящем описании также как молчание Fc). Аминокислотные и нуклеотидные последовательности четырех вариантов антител к миостатину представлены в таблице 2а.
Пример 13
Характеризация рН-зависимых вариантов MS1032
(HEK Blue-анализ (ВМР1 и спонтанная активация))
Все условия экспериментов соответствовали описанным в примерах 3 и 4. Как продемонстрировано на фиг. 10, у всех вариантов обнаружена сопоставимая с MS1032LO00-SG1 ингибирующая активность в отношении человеческого латентного миостатина.
Пример 14
Характеризация рН-зависимых вариантов MS1032 (BIACORE®)
Все условия экспериментов соответствовали описанным в примере 7, за исключением того, что измерения осуществляли также в ACES-буфере, рН 5,8 в дополнение к условиям ACES, рН 7,4. Активность некоторых антител оценивали только в отношении человеческого латентного миостатина. Уровень иммобилизации антител должен соответствовать 185 RU и 18,5 RU для анализа авидности и аффинности соответственно. Кинетические параметры определяли путем подгонки данных к модели связывания 1:1, используя программное обеспечение BIACORE® Т200 Evaluation, версия 2.0 (фирма GE Healthcare). Сенсограммы для всех антител к человеческому латентному миостатину, полученные в условиях, применявшихся для оценки авидности, представлены на фиг. 11, a ka, kd, и KD, рассчитанные для различных условий, представлены в таблицах 7-10. В условиях, применявшихся для оценки авидности, для всех антител за исключением MS1032LO00-SG1 обнаружена более быстрая скорость диссоциации при кислом рН, чем при нейтральном рН. В условиях, применявшихся для оценки аффинности, для всех антител за исключением MS1032LO00-SG1 обнаружено слабое взаимодействие с латентным миостатином при кислом рН и поэтому кинетические параметры не удалось определить.

N/T: не тестировали
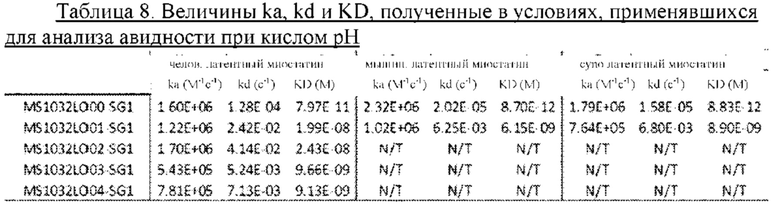
N/T: не тестировали

N/T: не тестировали

n.d.: не определяли, N/T: не тестировали
Пример 15
Сравнение концентрации общего миостатина в плазме мышей при применении антител, обладающих способностью связываться с Fc-гамма R, и с элиминированной способностью связываться с Fc-гамма R
Опыт in vivo с использованием мышей С.В-17 SCID
Накопление эндогенного миостатина оценивали in vivo после введения антитела к латентному миостатину мышам С.В-17 SCID (фирма In Vivos, Сингапур). Антитело к латентному миостатину (3 мг/мл) вводили в виде одной дозы 10 мл/кг в каудальную вену. Образцы крови собирали через 5 мин, 7 ч, 1 день, 2 дня, 3 дня, 7 дней, 14 дней, 21 день и 28 дней после введения. Собранную кровь немедленно центрифугировали при 14000 об/мин при 4°С в течение 10 мин для отделения плазмы. Отделенную плазму хранили при температуре -80°С или ниже вплоть до анализа. В качестве антител к латентному миостатину применяли MS1032LO00-SG1 и MS1032LO00-F760.
Измерение концентрации общего миостатина в плазме с помощью электрохемилюминесценции (ECL)
Концентрацию общего миостатина в плазме мышей измеряли с помощью ECL. Планшеты с иммобилизованным антителом к зрелому миостатину получали путем нанесения антитела к зрелому миостатину RK35 (описанного в WO 2009/058346) на 96-луночный планшет MULTI-ARRAY (фирма Meso Scale Discovery) и инкубации в течение ночи при 4°С. Приготавливали образцы зрелого миостатина для получения калибровочной кривой и образцы плазмы, разведенные в 40 или более раз. Образцы смешивали в кислом растворе (0,2 М глицин-HCl, рН 2,5) для отделения путем диссоциации зрелого миостатина от связывающего его белка (такого как пропептид). Затем образцы вносили в иммобилизованный антителом к зрелому миостатину планшет и давали связываться в течение 1 ч при комнатной температуре, после чего осуществляли промывку. Затем добавляли меченное SULFO-меткой антитело к зрелому миостатину RK22 (описанное в WO 2009/058346) и планшет инкубировали в течение 1 ч при комнатной температуре, после чего осуществляли промывку. В планшет немедленно добавляли буфер для считывания (Read Buffer) Т (×4) (фирма Meso Scale Discovery) и детекцию сигнала осуществляли с помощью устройства SECTOR Imager 2400 (фирма Meso Scale Discovery). Концентрацию зрелого миостатина рассчитывали на основе ответа относительно калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (фирма Molecular Devices). Зависимость от времени концентрации общего миостатина в плазме после внутривенного введения антитела к латентному миостатину, определенная с помощью указанного метода, представлена на фиг. 12.
Воздействие связывания с Fc-гамма R на накопление миостатина in vivo
После введения антитела MS1032LO00-F760 концентрация в плазме общего миостатина, накопленного ко дню 28, в 248 раз превышала концентрацию в плазме общего миостатина через 5 мин. В противоположность этому, после введения MS1032LO00-SG1 концентрация в плазме общего миостатина, накопленного ко дню 28, в 37 раз превышала концентрацию в плазме общего миостатина через 5 мин. Примерно 7-кратное различие концентрации в плазме общего миостатина в день 28 обнаружено между MS1032LO00-F760 (молчащая Fc) и MS1032LO00-SG1 в результате связывания с Fc-гамма R. В случае человеческого растворимого IL-6R (hsIL-6R) не выявлено существенного различия концентрации в плазме hsIL-6R при применении антитела к hsIL-6R F760 (молчащая Fc) и SG1, что описано в WO 2013/125667. Поскольку hsIL-6R является мономерным антигеном, комплекс антитело-hsIL-6R содержит только одну Fc. Таким образом, результаты, полученные в WO 2013/125667, позволяют предположить, что комплекс антитело-антиген с одной Fc не связывается в такой степени с Fc-гамма R in vivo, чтобы иммунный комплекс поглощался клетками. С другой стороны, с мультимерным антигеном (таким как миостатин) антитело может образовывать крупный иммунный комплекс, и комплекс антитело-антиген содержит более двух Fc. Таким образом, иметь место существенное различие в концентрации антигена в плазме может между F760 и SG1 из-за сильной авидности связывания с Fc-гамма R. Этот результат позволяет предположить, что MST1032 может образовывать крупный иммунный комплекс с миостатином, который содержит более двух антител.
Пример 16
Сравнение концентрации в плазме общего миостатина при применении рН-независимого антитела к латентному миостатину и рН-зависимого антитела к латентному миостатину у мышей
Опыт in vivo с использованием мышей С.В-17 SCID
Накопление эндогенного миостатина оценивали in vivo после введения антитела к латентному миостатину мышам С.В-17 SCID (фирма In Vivos, Сингапур) согласно методу, описанному в примере 15. Применяли антитела к латентному миостатину MS1032LO01-SG1 и MS1032LO01-F760.
Измерение концентрации общего миостатина в плазме с помощью ECL
Концентрацию общего миостатина в плазме мышей измеряли с помощью ECL согласно методу, описанному в примере 15. Зависимость от времени концентрации общего миостатина в плазме после внутривенного введения антитела к латентному миостатину, определенная с помощью указанного метода, представлена на фиг. 13.
Воздействие рН-зависимого связывания миостатина на накопление миостатина in vivo
рН-зависимые антитела к латентному миостатину (MS1032LO01-SG1 и MS1032LO01-F760) тестировали in vivo и осуществляли сравнение концентрации общего миостатина в плазме. Данные о концентрации общего миостатина после введения MS1032LO00-SG1 и MS1032LO00-F760 представлены в примере 15, а также на фиг. 13. MS1032LO00 представляет собой рН-независимое антитело, а MS1032LO01 представляет собой рН-зависимое антитело. Как продемонстрировано на фиг. 13, концентрации общего миостатина после введения MS1032LO01-F760 снижалась по сравнению с MS1032LO00-F760 из-за рН-зависимого связывания. Кроме того, концентрации общего миостатина после введения MS1032LO01-SG1 резко снижалась по сравнению с MS1032LO00-SG1 из-за рН-зависимого связывания и повышенного проникновения в клетки в результате связывания с Fc-гамма R. Можно ожидать, что MS1032LO01-SG1 обладает очень высокой способностью повышать клиренс миостатина из плазмы в качестве «Sweeping®-антитела» (антитела-уборщика) (т.е. антитела, обладающего способностью активно элиминировать растворимые антигены из кровотока»).
Пример 17
Эффективность in vivo рН-зависимого антитела к латентному миостатину
Все применяемые экспериментальные схемы соответствовали описанным в примере 8. Как продемонстрировано на фиг. 14, после внутривенного введения SCID-мышам в дозах 0,2, 1 и 5 мг/кг MS1032LO00-SG1 (не относящееся к антителам-уборщикам) и MS1032LO01-SG1 (антитело-уборщик) повышали массу скелетных мышц в зависимости от дозы через 2 недели. Антитело MS1032LO01-SG1 значимо повышало массу во влажном состоянии четырехглавой мышцы и силу сжатия при применении в дозе 1 и 5 мг/кг и значимо повышало при применении в дозе 5 мг/кг тощую массу тела и массу во влажном состоянии икроножной мышцы по сравнению с обработанной наполнителем (ЗФР) группой. MS1032LO01-SG1 также значимо снижало жировую массу при применении в дозах 1 и 5 мг/кг относительно обработанной наполнителем (ЗФР) группы. При применении MS1032LO00-SG1 в дозе 0,2 мг/кг не обнаружено никакого повышения тощей массы тела. С другой стороны, MS1032LO01-SG1 при применении в дозе 0,2 мг/кг резко повышало тощую массу тела, массу во влажном состоянии четырехглавой мышцы, массу во влажном состоянии и силу сжатия икроножной мышцы. Таким образом, антитело с рН-зависимыми характеристиками связывания в большей степени усиливало рост мышечной массы и мышечную силу по сравнению с антителом с независимым от рН связыванием.
Пример 18
Экспрессия и очистка человеческого и мышиного латентного GDF11
Человеческий GDF11 с Flag-меткой на N-конце (обозначенный также в настоящем описании как Flag-hGDF11, человеческий латентный GDF11, hGDF11 или человеческий GDF11, SEQ ID NO: 85) кратковременно экспрессировали с использованием клеток FreeStyle293-F (фирма Thermo Fisher). Применяли кондиционированные среды, экспрессирующие Flag-hGDF11, для набивки колонки с обладающей аффинностью к антителу к Flag М2 смолой (фирма Sigma) и элюировали с использованием пептида Flag (фирма Sigma). Фракции, содержащие Flag-hGDF11, собирали и затем подвергали гель-фильтрации на колонке Супердекс 200 (фирма GE healthcare), уравновешенной 1 × ЗФР. Затем фракции, содержащие Flag-hGDF11, объединяли и хранили при -80°С.
Пример 19
Характеризация антитела к латентному миостатину (ELISA)
Несенсибилизированные планшеты для ELISA (планшет NUNC-IMMUNO, поверхность MAXISORP, фирма Nalge Nunc International) покрывали 20 мкл 50 нМ стрептивидина (фирма GenScript) в течение 2 ч при комнатной температуре. Затем планшеты трижды промывали ЗФРТ и блокировали в течение ночи, используя 50 мкл 20% Blocking One (фирма Nacalai Tesque). На следующий день каждую лунку планшета инкубировали с биотинилированным человеческим латентным миостатином из расчета 2 нМ/лунку/20 мкл или биотинилированным человеческим латентным GDF11 из расчета 20 нМ/лунку/20 мкл в течение 2 ч. После промывки добавляли в лунки 20 мкл образца антитела и планшеты выдерживали в течение 1 ч. Планшеты промывали и добавляли 20 мкл античеловеческого IgG, конъюгированного с пероксидазой из хрена (HRP) (фирма Abcam), разведенного в забуференном HEPES физиологическом растворе, и планшеты выдерживали в течение еще 1 ч. Затем планшеты вновь промывали и после этого добавляли в каждую лунку по 50 мкл ABTS (фирма KPL), и планшеты инкубировали в течение 1 ч. Обнаружение сигнала осуществляли при 405 нм в колориметрическом планшет-ридере. Результаты эксперимента по связыванию представлены на фиг. 15. Установлено, что MST1032-G1m связывалось с латентным миостатином (т.е. нековалентным комплексом зрелого миостатина и пропептидов), но не связывалось с hGDF11. Эти результаты демонстрируют, что MST1032-G1m специфически связывается с миостатином, но не связывается с hGDF11.
Пример 20
Характеризация антитела к латентному миостатину (HEK Blue-анализ (ВМР1 и спонтанная активация))
Анализ репортерного гена применяли для оценки биологической активности активного GDF11 in vitro. Клетки HEK-Blue™ TGF-бета (фирма Invivogen), которые экспрессируют индицируемые Smad3/4-связывающими элементами (SBE) репортерные гены SEAP, позволяют осуществлять детекцию биоактивного GDF11, осуществляя мониторинг активации рецепторов для активина типа 1 и типа 2. Активный GDF11 стимулирует производство и секрецию SEAP в клеточный супернатант. Количество секретируемой SEAP затем определяют с помощью QUANTIBlue™ (фирма Invivogen).
Клетки HEK-Blue™ TGF-бета поддерживали в среде DMEM (фирма Gibco), дополненной 10% фетальной бычьей сыворотки, 50 мкг/мл стрептомицина, 50 ед./мл пенициллина, 100 мкг/мл Normocin™, 30 мкг/мл бластицидина, 200 мкг /мл HygroGold™ и 100 мкг/мл Zeocin™. При проведении функционального анализа среду для клеток заменяли на среду для анализа (DMEM, дополненная 0,1% бычьего сывороточного альбумина, стрептомицином, пенициллином и Normocin™) и высевали в 96-луночный планшет. Человеческий GDF11 инкубировали с рекомбинантным человеческим ВМР1 (фирма Calbiochem) или без него и антителом к латентному миостатину (MST1032-G1m) при 37°С в течение ночи. Смеси образцов переносили в клетки. После 24-часовой инкубации клеточные супернатанты смешивали с QUANTIBlue™ и оптическую плотность при 620 нм измеряли в колориметрическом планшет-ридере. Как продемонстрировано на фиг. 16, MST1032-G1m не препятствовало опосредуемой протеазой активации или спонтанной активации человеческого GDF11 и поэтому не ингибировало секрецию SEAP.
Пример 21
Дополнительная оптимизация вариантов MS1032 для повышения «sweeping-эффекта» (эффекта уборщика)
Поскольку рН-зависимое антитело, MS1032LO01-SG1, как продемонстрировано, обладает очень высокой эффективностью, при изучении на мышах, осуществляли дополнительную оптимизацию для повышения зависимости от рН, повышения поглощения клетями, повышения стабильности и т.д., путем интродукции мутаций в CDR антитела или путем изменения каркасных участков. Оценивали более тысячи вариантов с помощью BIACORE®- и/или HEK Blue-анализов, которые описаны выше, и создавали MS1032LO06-SG1, MS1032LO07-SG1, MS1032LO10-SG1, MS1032LO11-SG1, MS1032LO12-SG1, MS1032LO18-SG1, MS1032LO19-SG1, MS1032LO21-SG1, MS1032LO23-SG1, MS1032LO24-SG1, MS1032LO25-SG1 и MS1032LO26-SG1. Аминокислотные и нуклеотидные последовательности указанных созданных антител представлены в таблице 11.
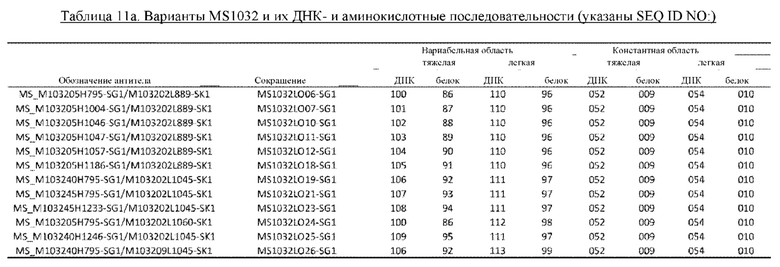
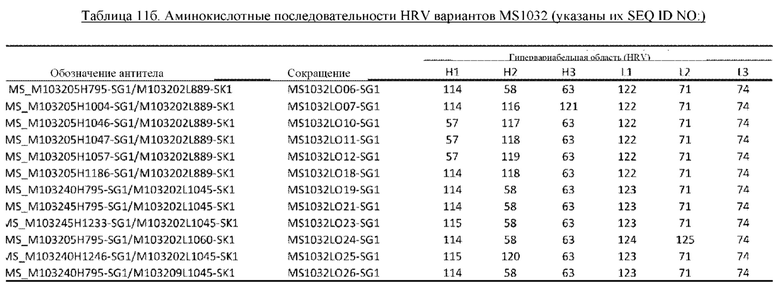
Аффинность связывания вариантов MS1032 с латентным миостатином человека, обезьян циномолгус (cyno) и мышей оценивали при 37°С при рН 7,4 и рН 5,8, используя устройство BIACORE® Т200 (фирма GE Healthcare), для оценки воздействия рН на связывание антигена. ProA/G (фирма Pierce) иммобилизовали на всех проточных ячейках СМ4-чипа с использованием набора для аминного сочетания (фирма GE Healthcare). Все антитела и аналиты приготавливали в ACES-буфере, рН 7,4 или рН 5,8, содержащем 20 мМ ACES, 150 мМ NaCl, 1,2 мМ CaCl2, 0,05% Твин 20, 0,005% NaN3. Каждое антитело «захватывали» на поверхности сенсорного чипа с помощью ProA/G. Уровни иммобилизации антител, как правило, соответствовали 130-240 резонансных единиц (RU). Латентный миостатин человека, cyno или мышей инъецировали в концентрациях от 3,125 до 50 нМ, которые получали путем двукратных серийных разведений, после чего давали пройти диссоциации. Поверхность сенсора регенерировали после каждого цикла с помощью 10 мМ глицина-HCl, рН 1,5. Аффинность связывания определяли путем обработки и подгонки данных к модели связывания 1:1, используя программное обеспечение BIACORE® Т200 Evaluation, версия 2.0 (фирма GE Healthcare).
Данные об аффинности связывания (KD) вариантов MS1032 с латентным миостатином человека, cyno и мышей при рН 7,4 и рН 5,8 представлены в таблице 12. Для всех вариантов получено соотношение величин KD ((KD при рН 5,8)/(KD при рН 7,4)), составляющее более 5, что свидетельствует о рН-зависимом связывании с латентным миостатином.
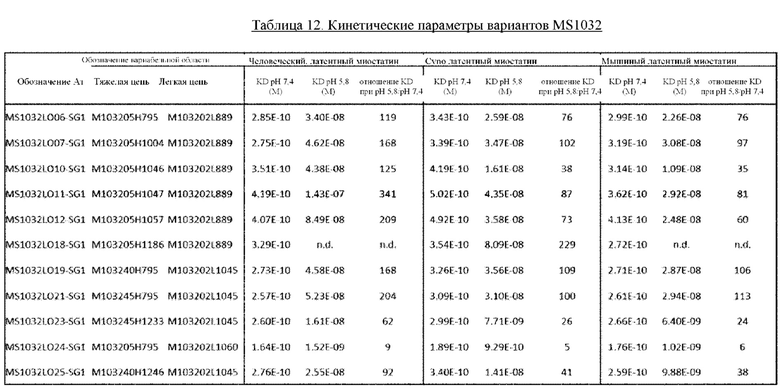
n.d.: не определяли
Пример 22
Оценка нейтрализующей активности полученных в результате дополнительной оптимизации вариантов с помощью HEK Blue-анализа
При создании изобретения оценивали нейтрализующую активность MS1032LO06-SG1, MS1032LO07-SG1, MS1032LO10-SG1, MS1032LO11-SG1, MS1032LO12-SG1, MS1032LO18-SG1, MS1032LO19-SG1, MS1032LO21-SG1, MS1032LO23-SG1 и MS1032LO25-SG1 в отношении человеческого латентного миостатина с помощью метода, описанного в примере 3. Как продемонстрировано на фиг. 17, для всех вариантов обнаружена активность, сопоставимая с активностью MS1032LO01-SG1.
Пример 23
Сравнение концентрации общего миостатина в плазме мышей при применении рН-независимого антитела к латентному миостатину и различных рН-зависимых антител к латентному миостатину
Опыт in vivo с использованием мышей С.В-17 SCID
Накопление эндогенного миостатина оценивали in vivo после введения антитела к латентному миостатину мышам С.В-17 SCID (фирма In Vivos, Сингапур). Антитело к латентному миостатину (3 мг/мл) вводили в виде одной дозы 10 мл/кг в каудальную вену. Образцы крови собирали через 5 мин, 7 ч, 1 день, 2 дня, 3 дня, 7 дней, 14 дней, 21 день и 28 дней после введения. Собранную кровь немедленно центрифугировали при 14000 об/мин при 4°С в течение 10 мин для отделения плазмы. Отделенную плазму хранили при температуре -80°С или ниже вплоть до анализа. В качестве антител к латентному миостатину применяли MS1032LO00-SG1, MS1032LO01-SG1, MS1032LO06-SG1, MS1032LO11-SG1, MS1032LO18-SG1, MS1032LO19-SG1, MS1032LO21-SG1 и MS1032LO25-SG1.
Измерение концентрации общего миостатина в плазме с помощью электрохемилюминесценции (ECL)
Концентрацию общего миостатина в плазме мышей измеряли с помощью ECL. Планшеты с иммобилизованным антителом к зрелому миостатину получали путем нанесения биотинилированного антитела к зрелому миостатину RK35 (описанного в WO 2009/058346) на 96-луночный покрытый стрептавидином планшет MULTI-ARRAY (фирма Meso Scale Discovery) и инкубации в блокирующем буфере в течение 2 ч при комнатной температуре. Приготавливали образцы зрелого миостатина для получения калибровочной кривой и образцы плазмы, разведенные в 40 или более раз. Образцы смешивали в кислом растворе (0,2 М глицин-HCl, рН 2,5) для отделения путем диссоциации зрелого миостатина от связывающего его белка (такого как пропептид). Затем образцы вносили в иммобилизованный антителом к зрелому миостатину планшет и давали связываться в течение 1 ч при комнатной температуре, после чего осуществляли промывку. Затем добавляли меченное SULFO-меткой антитело к зрелому миостатину RK22 (описанное в WO 2009/058346) и планшет инкубировали в течение 1 ч при комнатной температуре, после чего осуществляли промывку. В планшет немедленно добавляли буфер для считывания (Read Buffer) Т (×4) (фирма Meso Scale Discovery) и осуществляли детекцию сигнала с помощью устройства SECTOR Imager 2400 (фирма Meso Scale Discovery). Концентрацию зрелого миостатина рассчитывали на основе ответа относительно калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (фирма Molecular Devices). Зависимость от времени концентрации общего миостатина в плазме после внутривенного введения антитела к латентному миостатину, определенная с помощью указанного метода, представлена на фиг. 18.
Воздействие рН-зависимого связывания с миостатином на накопление миостатина у мышей в опыте in vivo
Воздействие зависимости от рН на накопление миостатина у мышей сравнивали, используя рН-независимое антитело к латентному миостатину (MS1032LO00-SG1) и различные рН-зависимые антитела к латентному миостатину (MS1032LO01-SG1, MS1032LO06-SG1, MS1032LO11-SG1, MS1032LO18-SG1, MS1032LO19-SG1, MS1032LO21-SG1 и MS1032LO25-SG1). Интродукция рН-зависимости в значительной степени ускоряла клиренс миостатина из плазмы SCID-мышей. Как продемонстрировано на фиг. 18, рН-зависимые антитела к латентному миостатину (MS1032LO01-SG1, MS1032LO06-SG1, MS1032LO11-SG1, MS1032LO18-SG1, MS1032LO19-SG1, MS1032LO21-SG1 и MS1032LO25-SG1) могли снижать накопление миостатина по сравнению с рН-независимым антителом к латентному миостатину (MS1032LO00-SG1) в день 28.
Пример 24
Сравнение концентрации общего миостатина в плазме обезьян циномолгус при применении рН-независимого антитела к латентному миостатину и рН-зависимых антител к латентному миостатину, имеющих сконструированную Fc-область
Опыт in vivo с использованием обезьян циномолгус Накопление эндогенного миостатина оценивали in vivo после введения антитела к латентному миостатину 2-4-летним Масаса fascicularis (яванский макак-крабоед, обезьяны циномолгус) из Камбоджи (фирма Shin Nippon Biomedical Laboratories Ltd., Япония). Дозу 30 мг/кг инъецировали в цефалическую вену предплечья, используя одноразовый шприц, удлиняющую трубку, находящуюся внутри иглу и инфузионный насос. Скорость дозирования составляла 30 мин на организм животного. Образцы крови собирали перед началом дозирования и либо через 5 мин, 7 ч и 1, 2, 3, 7, 14, 21, 28, 35, 42, 49 и 56 дней после окончания дозирования, либо через 5 мин и 2, 4 и 7 ч и 1, 2, 3, 7, 14, 21, 28, 35, 42, 49 и 56 дней после окончания дозирования. Кровь брали из бедренной вены с помощью шприца, содержащего гепарин натрия. Кровь немедленно охлаждали на льду и плазму получали центрифугированием при 4°С С, 1700 × g в течение 10 мин. Образцы плазмы хранили в холодильнике глубокой заморозки (приемлемый диапазон: -70°С или ниже) вплоть до анализа. В качестве антител к латентному миостатину применяли MS1032LO00-SG1, MS1032LO06-SG1012, MS1032LO06-SG1016, MS1032LO06-SG1029, MS1032LO06-SG1031, MS1032LO06-SG1033 и MS1032LO06-SG1034 (в контексте настоящего описания SG1012, SG1016, SG1029, SG1031, SG1033 и SG1034 обозначают константные области тяжелых цепей, сконструированные на основе SG1 согласно описанному ниже методу).
В цефалическую вену предплечья или подкожную вену ноги с использованием одноразового шприца и находящейся внутри иглы вводили в дозе 2 мг/кг антитела к латентному миостатину MS1032LO19-SG1079, MS1032LO19-SG1071, MS1032LO19-SG1080, MS1032LO19-SG1074, MS1032LO19-SG1081 и MS1032LO19-SG1077 (в контексте настоящего описания SG1079, SG1071, SG1080, SG1074, SG1081 и SG1077 обозначают константные области тяжелых цепей, сконструированные на основе SG1 согласно описанному ниже методу). Образцы крови собирали перед началом дозирования и через 5 мин и через 2, 4 и 7 ч, и через 1, 2, 3, 7, 14 дней после окончания дозирования. Кровь обрабатывали согласно описанной выше процедуре. Образцы плазмы хранили в холодильнике глубокой заморозки (приемлемый диапазон: -70°С или ниже) вплоть до анализа.
Измерение концентрации общего миостатина в плазме с помощью электрохемилюминесценции (ECL)
Концентрацию общего миостатина в плазме обезьян измеряли с помощью ECL согласно методу, описанному в примере 23. Зависимость от времени концентрации общего миостатина в плазме после внутривенного введения антитела к латентному миостатину, определенная с помощью указанного метода, представлена на фиг. 19.
Измерение ADA в плазме обезьян с помощью электрохемилюминесценции (ECL)
Биотинилированное лекарственное средство применяли для сенсибилизации 96-луночного стрептавидинового планшета MULTI-ARRAY (фирма Meso Scale Discovery) и инкубировали в LowCross®-буфере (фирма Candor) в течение 2 ч при комнатной температуре. Образцы обезьяньей плазмы разводили в 20 раз в LowCross®-буфере перед внесением в планшет. Образцы инкубировали в течение ночи при 4°С. На следующий день планшет промывали трижды буфером для промывки перед добавлением меченного с помощью SULFO-метки вторичного антитела к обезьяньему IgG (фирма Thermo Fisher Scientific). После инкубации в течение 1 ч при комнатной температуре планшет промывали трижды буфером для промывки. В планшет немедленно добавляли буфер для считывания (Read Buffer Т) (×4) (фирма Meso Scale Discovery) и детекцию сигнала осуществляли с помощью устройства SECTOR Imager 2400 (фирма Meso Scale Discovery).
Влияние зависимости от рН и конструирования Fc на накопление миостатина у обезьян in vivo
Введение обезьянам циномолгус рН-независимого антитела (MS1032LO00-SG1) приводило по меньшей мере к 60-кратному повышению концентрации миостатина относительно исходного уровня в день 28. В день 28 рН-зависимые антитела к латентному миостатину MS1032LO06-SG1012 и MS1032LO06-SG1033 обеспечивали 3- и 8-кратное повышение относительно исходного уровня соответственно. Сильный swipping-эффект главным образом связан с повышением аффинности к Fc-гамма RIIb обезьян циномолгус. В день 28, рН-зависимые антитела к латентному миостатину MS1032LO06-SG1029, MS1032LO06-SG1031 и MS1032LO06-SG1034 смогли элиминировать антиген до уровня ниже исходного. Причиной сильного swipping-эффекта в случае MS1032LO06-SG1029, MS1032LO06-SG1031 и MS1032LO06-SG1034 является повышение неспецифического поглощения клеткой из-за повышения положительно заряженного кластера антитела и повышение опосредуемого Fc-гамма R клеточного поглощения из-за усиления связывания с Fc-гамма R.
Введение рН-зависимых антител к латентному миостатину MS1032LO19-SG1079, MS1032LO19-SG1071, MS1032LO19-SG1080, MS1032LO19-SG1074, MS1032LO19-SG1081 и MS1032LO19-SG1077 снижали концентрацию миостатина до уровня ниже предела обнаружения (<0,25 нг/мл), начиная со дня 1 у обезьян циномолгус. В день 14 концентрация миостатина превышала предел обнаружения при применении MS1032LO19-SG1079 и MS1032LO19-SG1071, хотя концентрация миостатина оставалась на уровне ниже порога обнаружения при применении MS1032LO19-SG1080, MS1032LO19-SG1074, MS1032LO19-SG1081 и MS1032LO19-SG1077. Более слабое подавление, обнаруженное для MS1032LO19-SG1079 и MS1032LO19-SG1071, может являться результатом различных мутаций, приводящих к изменению pI.
Результаты позволяют предположить, что сильной элиминации миостатина из плазмы можно достигать с помощью мутаций, которые повышают связывание с Fc-гамма RIIb или комбинации мутаций, которые повышают положительный заряд антитела и повышают связывание с Fc-гамма RIIb. Можно ожидать, что у человека можно достигать сильной элиминации миостатина путем объединения мутаций, которые повышают положительный заряд антитела и повышают связывание с Fc-гамма R.
Пример 25
Эффективность in vivo дополнительно оптимизированных вариантов в опытах на мышах
Как описано в примере 8, эффективность in vivo оценивали на Scid-мышах, используя MS1032LO06-SG1, MS1032LO11-SG1, MS1032LO18-SG1, MS1032LO19-SG1 и MS1032LO25-SG1. Осуществляли три независимых опыта и MS1032LO01 применяли в качестве контроля.
Результаты этих экспериментов представлены на фиг. 20А-20И. Для всех антител (MS1032LO06-SG1, MS1032LO11-SG1, MS1032LO18-SG1, MS1032LO19-SG1 и MS1032LO25-SG1) обнаружена способность повышать тощую массу тела (LBM), а также силу сжатия аппендикулярных мышц. Для всех антител продемонстрировано также снижение жировой массы тела в зависимости от дозы.
Пример 26
Эпитопное картирование антител к латентному миостатину
Дополнительные антитела к человеческому латентному миостатину создавали для картирования соответствующих им эпитопов, с которыми они связываются. Двух NZW-кроликов иммунизировали и получали антитела согласно методу, описанному в примере 2. 1760 идентифицированных продуцирующих антитела В-клеточных линий дополнительно подвергали скринингу в отношении их способности блокировать опосредуемую ВМР1 активацию человеческого латентного миостатина. В целом, метод состоял в следующем: супернатант В-клеток, содержащий секретированные антитела, инкубировали с человеческим латентным миостатином в присутствии рекомбинантного человеческого ВМР1 (фирма R&D Systems) при 37°С в течение ночи. Затем 50 мкл реакционной смеси переносили в 96-луночный планшет MULTI-ARRAY фирмы Meso Scale Discovery, сенсибилизированный антителом к зрелому миостатину RK35 (описано в WO 2009/058346). После инкубации в течение 1 ч при комнатной температуре со встряхиванием планшеты инкубировали с биотинилированным антителом к зрелому миостатину RK22 (описано WO 2009/058346), а затем с меченным SULFO-меткой стрептавидином. Затем в планшет добавляли буфер для считывания (Read Buffer Т) (×4) (фирма Meso Scale Discovery) и детекцию сигнала осуществляли с помощью устройства SECTOR Imager 2400 (фирма Meso Scale Discovery). Для последующего анализа отбирали 94 линии, характеризующиеся различными уровнями нейтрализующей активности (MST1495-MST1588). Вариабельные области указанных отобранных линий клонировали согласно методу, описанному в примере 2, за исключением того, что применяли экспрессионный вектор с последовательностью константной области тяжелой цепи F1332m (SEQ ID NO: 193).
Ингибирующую активность в отношении активации человеческого латентного миостатина дополнительно оценивали для панели из 7 антител к латентному миостатину (MST1032, MST1504, MST1538, MST1551, MST1558, MST1572 и MST1573; идентифицирующие номера последовательности аминокислотных последовательностей этих антител приведены в таблице 13). Как продемонстрировано на фиг. 21, все антитела обладали способностью ингибировать опосредуемую ВМР1 активацию человеческого латентного миостатина в зависимости от дозы.
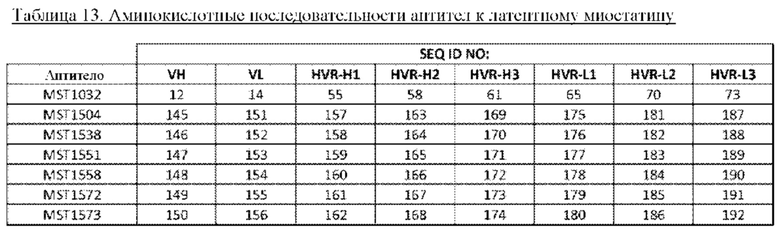
Четыре антитела, которые оказались перспективными по результатам анализа методом Вестерн-блоттинга, отбирали для эпитопного картирования. Фрагменты кодирующей N-концевой пропептид области человеческого латентного миостатина клонировали в векторе pGEX4.1 так, чтобы получать меченные GST фрагменты пропептида, каждый состоящей из 100 аминокислот с 80 перекрывающимися аминокислотами (фиг. 22А). Экспрессию белка индуцировали в трансформированных BL21 компетентных клетках с использованием системы для самоиндукции экспрессии в течение ночи (Overnight Express Autoinduction System) (фирма Merck Millipore) и белок экстрагировали с помощью реагента для экстракции белков BugBuster (фирма Novagen). Экспрессию требуемых белковых фрагментов размером 37 кДа подтверждали с помощью анализа методом Вестерн-блоттинга, который описан в примере 6 (антитело к GST (фирма Abcam)) (фиг. 22Б). При изучении антител к человеческому латентному миостатину, указанных на фиг. 22В, установлено, что хотя все 4 антитела могли ингибировать активацию латентного миостатина, они распознавали разные эпитопы на человеческом латентном миостатине. Антитело MST1032 могло обнаруживать первые пять фрагментов, не выявлены полосы после удаления аминокислот 81-100 (SEQ ID NO: 78). Оба антитела MST1538 и MST1572 связывались только с первыми тремя фрагментами, не выявлены полосы в отсутствии аминокислот 41-60 SEQ ID NO: 78. Антитело MST1573 сильно связывалось только с первыми двумя фрагментами, что позволяет предположить, что его эпитоп лежит в аминокислотах 21-40 SEQ ID NO: 78 (фиг. 22Г).
Пример 27
Создание новых вариантов Fc с повышенной активностью в отношении Fc-гамма RIIb
В этом примере проиллюстрировано конструирование Fc для повышения клиренса миостатина.
В WO 2013/125667 продемонстрировано, что клиренс растворимого антигена можно повышать путем введения связывающих указанный антиген молекул, содержащих Fc-домен, характеризующийся повышенной аффинностью в отношении Fc-гамма RIIb. Кроме того, Fc-варианты, для которых установлена повышенная способность связываться с человеческим Fc-гамма RIIb, проиллюстрированы в WO 2012/115241 и WO 2014/030728. Кроме того, продемонстрировано, что указанные Fc-варианты могут обладать избирательно повышенной способностью связываться с человеческим Fc-гамма RIIb и пониженной способностью связываться с другими активными Fc-гамма R. Указанное избирательное повышение связывания с Fc-гамма RIIb может оказывать благоприятное воздействие не только на клиренс растворимого антигена, но также на снижение риска нежелательных эффекторных функций и иммунного ответа.
При разработке антитела в качестве лекарственного средства следует изучать эффективность, фармакокинетику и безопасность на животных кроме человека, в отношении которых лекарственное средство обладает фармакологической активностью. Если оно активно только для человека, следует применять альтернативные подходы, такие как использование суррогатного антитела (Int. J. Тох. 28, 2009, сс. 230-253). Однако с его помощью трудно прогнозировать воздействия взаимодействия между Fc-областью и Fc-гамма R в организме человека при применении суррогатного антитела, поскольку схемы экспрессии и/или функции Fc-гамма R у животных кроме человека не всегда такие же, как у человека. Предпочтительно, чтобы Fc-области антител, применяемых в качестве лекарственных средств, обладали перекрестной реактивностью с областями животных кроме человека, прежде всего обезьян циномолгус, которые имеют схемы экспрессии и функции Fc-гамма R, близкие к человеческим, поэтому результаты, полученные на животных кроме человека, можно экстраполировать на человека.
Поэтому при создании настоящего изобретения создавали Fc-варианты, обладающие перекрестной реактивностью для Fc-гамма R человека и cyno.
Измерение аффинности существующих вариантов Fc с повышенной активностью в отношении Fc-гамма RIIb для человеческих и обезьяньих Fc-гамма R
Ген тяжелой цепи варианта с повышенной активностью в отношении человеческого Fc-гамма RIIb (в настоящем описании понятие «с повышенной активностью в отношении Fc-гамма RIIb» означает «с повышенной Fc-гамма RIIb-связывающей активностью»), описанного в WO 2012/115241, создавали путем замены Pro в положении 238 согласно EU-нумерации на Asp в тяжелой цепи MS1032LO06-SG1, который обозначили как M103205H795-SG1 (VH, SEQ ID NO: 86; СН, SEQ ID NO: 9). Полученную тяжелую цепь обозначили как M103205H795-MY009 (VH, SEQ ID NO: 86; СН, SEQ ID NO: 252). В качестве легкой цепи антитела применяли M103202L889-SK1 (VL, SEQ ID NO: 96; CL, SEQ ID NO: 10). Рекомбинантное антитело экспрессировали согласно методу, описанному в примере 34.
Внеклеточные домены Fc-гамма R получали следующим образом. Синтез генов внеклеточного домена человеческих Fc-гамма R осуществляли с помощью методов, известных специалистам в данной области, на основе информации, зарегистрированной в NCBI. В частности, основой для создания Fc-гамма RIIa была последовательность NCBI, имеющая код доступа № NM_001136219.1, Fc-гамма RIIb - NM_004001.3, Fc-гамма RIIIa - NM_001127593.1 соответственно. Аллотипы Fc-гамма RIIa и Fc-гамма RIIIa получали на основе данных о полиморфизме Fc-гамма RIIa (J. Exp. Med. 172, 1990, cc. 19-25) и Fc-гамма RIIIa (J. Clin. Invest. 100(5), 1997, cc. 1059-1070) соответственно. Синтез гена внеклеточного домена Fc-гамма R обезьян циномолгус осуществляли путем клонирования кДНК каждого Fc-гамма R из обезьян циномолгус, используя методы, известные специалистам в данной области. Аминокислотные последовательности сконструированных внеклеточных доменов Fc-гамма R представлены в перечне последовательностей: (SEQ ID NO: 210 для человеческого Fc-гамма RIIaR, SEQ ID NO: 211 для человеческого Fc-гамма RIIaH, SEQ ID NO: 214 для человеческого Fc-гамма RIIb, SEQ ID NO: 217 для человеческого Fc-гамма RIIIaF, SEQ ID NO: 218 для человеческого Fc-гамма RIIIaV, SEQ ID NO: 220 для cyno Fc-гамма RIIa1, SEQ ID NO: 221 для cyno Fc-гамма RIIa2, SEQ ID NO: 222 для cyno Fc-гамма RIIa3, SEQ ID NO: 223 для cyno Fc-гамма RIIb, SEQ ID NO: 224 для cyno Fc-гамма RIIIaS). Затем добавляли His-метку на их С-конец и каждый полученный ген встраивали в экспрессионный вектор, созданный для экспрессии в клетках млекопитающих. Экспрессионный вектор интродуцировали в клетки, полученные из почки человеческого эмбриона линии FreeStyle293 (фирма Invitrogen), для экспрессии требуемого белка. После культивирования полученный супернатант культуры фильтровали и очищали в основном с помощью указанных ниже четырех стадий очистки. Катионообменную хроматографию с использованием SP Сефарозы FF проводили в качестве первой стадии, аффинную хроматографию против His-метки (HisTrap HP) осуществляли в качестве второй стадии, гель фильтрацию на колонках (Супердекс200) применяли в качестве третьей стадии, и стерилизацию фильтрацией применяли в качестве четвертой стадии. Абсорбцию при 280 нм очищенных белков измеряли с помощью спектрофотометра и концентрацию очищенного белка определяли, используя коэффициент экстинкции, рассчитанный методом РАСЕ (Protein Science 4, 1995, сс. 2411-2423).
Кинетический анализ взаимодействий между этими антителами и Fc-гамма R осуществляли, используя BIACORE® Т200 или BIACORE® 4000 (фирма GE Healthcare). В качестве подвижного буфера применяли HBS-EP+ (фирма GE Healthcare) и температуру измерения устанавливали на 25°С. Чип получали путем иммобилизации белка А, белка A/G или мышиной легкой каппа-цепи к человеческому IgG (фирма BD Biosciences) на сенсорном СМ4- или СМ5-чипе серий S (фирма GE Healthcare), применяя метод аминного сочетания. Представляющее интерес антитело «захватывали» на поверхности чипа для взаимодействий с каждым Fc-гамма R, который разводили в подвижном буфере, и измеряли связывание с антителом. После измерения отмывали антитело, иммобилизованное на чипе, давая пройти реакции с 10 мМ глицином-HCl, рН 1,5 и 25 мМ NaOH, в результате чего происходила регенерация чипа и его использовали повторно. Полученные сенсограммы, являющиеся результатами измерения, анализировали с помощью модели связывания 1:1 Ленгмюра, используя программное обеспечение BIACORE® Evaluation для расчета константы скорости связывания ka (л/моль/с) и константы скорости реакции диссоциации kd (1/с), и на основе этих величин рассчитывали константу диссоциация KD (моль/л). Поскольку связывание MS1032LO06-MY009 с человеческим Fc-гамма RIIaH, человеческим Fc-гамма RIIIaV и cynoFc-гамма RIIIaS было слабым, кинетические параметры, такие как KD, не удалось рассчитать с помощью вышеуказанного аналитического метода. Описывающие эти взаимодействия величины KD рассчитывали, используя модель связывания 1:1, описанную ниже в программном обеспечении BIACORE® Т100 Handbook BR1006-48 Edition АЕ.
Поведение взаимодействующих молекул на основе модели связывания по типу 1:1 BIACORE® может быть описано уравнением 1: Req = С × Rmax/(KD+C)+RI, в котором, Req обозначает функциональную зависимость уровней стационарного связывания от концентрации аналита, С обозначает концентрацию, RI обозначает общий вклад коэффициента преломления для образца и Rmax обозначает способность поверхности к связыванию аналита. Если преобразовать указанное уравнение, то можно выразить KD в виде уравнения 2: KD = С × Rmax/(Req-RI)-C. KD можно рассчитывать, заменяя соответствующими значениями параметры Rmax, RI и С в уравнении 1 или уравнении 2. Для рассматриваемых условий измерений можно использовать значения RI=0, С=2 мкмоля/л. Кроме того, величину Rmax полученную путем глобальной аппроксимации сенсограммы, измеренной при анализе взаимодействия каждого Fc-гамма R с IgG1, с использованием ленгмюровской модели связывания 1:1, делили на количество захваченного SG1, результат умножали на количество захваченного MY009 и полученную в итоге величину использовали в качестве Rmax. Это расчет основан на гипотезе о том, что предельное количество каждого Fc-гамма R, которое может быть связано с SG1, остается неизменным для всех вариантов, полученных путем интродукции мутаций в SG1, и что величина Rmax в момент осуществления измерения пропорциональна количеству антитела, связанному с чипом в момент осуществления измерения. Req определял как количество связываний каждого Fc-гамма R с каждым вариантом на сенсорном чипе, выявленном в момент осуществления измерения.
В таблице 14 представлены результаты кинетического анализа связывания SG1 и MY009 с человеческим и cynoFc-гамма R. Величины KD в клетках, обозначенных жирным шрифтом, рассчитывали с помощью уравнения 2, поскольку их аффинность была слишком низкой для точного ее определения с помощью кинетического анализа. Кратность величин KD рассчитывали путем деления величины KD для SG1 на величину KD для варианта для каждого Fc-гамма R.
Как продемонстрировано в таблице 14, у варианта MY009 с повышенной активностью в отношении человеческого Fc-гамма RIIb не обнаружено повышенное связывание с cynoFc-гамма RIIb, но обнаружено повышенное связывание с человеческим Fc-гамма RIIb. Его аффинность к cynoFc-гамма RIIb даже оказалась пониженной в 0,4 раза по сравнению SG1, что свидетельствует об отсутствии у варианта MY009 с повышенной активностью в отношении человеческого Fc-гамма RIIb перекрестной реактивности к cynoFc-гамма R.
Разработка нового Fc-варианта, характеризующегося повышенным связыванием и с человеческим, и с cynoFc-гамма RIIb
В идеальном случае новый вариант Fc должен обладать повышенным избирательным связыванием и с человеческим, и с cynoFc-гамма RIIb и пониженным связыванием с другими активными Fc-гамма R. Однако, поскольку предполагаемые ответственные за Fc-связывание остатки в cynoFc-гамма RIIb полностью соответствуют остаткам в любом аллотипе cynoFc-гамма RIIa (фиг. 23), то теоретически невозможно достигать избирательного повышения связывания с cynoFc-гамма RIIb относительно связывания cynoFc-гамма RIIa. Таким образом, новый вариант Fc должен обладать избирательно повышенной способностью к связыванию с человеческим Fc-гамма RIIb, cynoFc-гамма RIIb и cynoFc-гамма RIIa.
Для получения нового варианта Fc, характеризующегося избирательно повышенным связыванием и с человеческим, и с cyno-Fc-гамма RIIb, применяли опыт с использованием всеохватывающего мутагенеза (полный перебор аминокислотных замен), описанного в WO 2012/11524. В опыте с использованием всеохватывающего мутагенеза всеохватывающие мутации интродуцировали во все положения в области связывания Fc-гамма R в антителах IgG1-изотипа, и осуществляли полный анализ связывания с каждым Fc-гамма R согласно изложенным ниже процедурам.

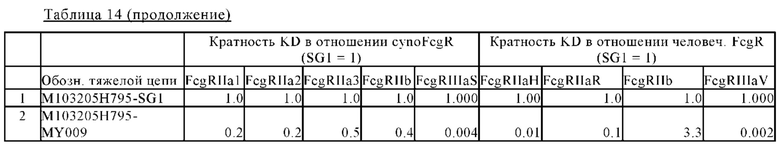
Вариабельную область антитела к глипикану 3, содержащую CDR GpH 7, представляющего собой антитело к глипикану 3 с повышенными кинетическими характеристиками в плазме, которое описано в WO 2009/041062, применяли в качестве вариабельной области тяжелой цепи антитела (GpH 7: SEQ ID NO: 225). Аналогично этому, в качестве легкой цепи антитела, GpL16-k0 (SEQ ID NO: 226), применяли антитело к глипикану 3 с повышенными кинетическими характеристиками в плазме, которое описано WO 2009/041062. Кроме того, последовательность антитела В3 (SEQ ID NO: 228), в которой мутация K439E была интродуцирована в G1d (SEQ ID NO: 227), полученную путем удаления С-концевых Gly и Lys IgG1, применяли в качестве константной области Н-цепи. Указанную тяжелую цепь, созданную путем слияния GpH 7 и В3, обозначали как GpH 7-В3 (VH, SEQ ID NO: 225; СН, SEQ ID NO: 228).
В GpH 7-В3 аминокислотные остатки, которые, как предполагается, могут участвовать в связывании с Fc-гамма R, и окружающие их аминокислотные остатки (положения 234-239, 265-271, 295, 296, 298, 300 и 324-337 согласно EU-нумерации) заменяли соответственно 18 аминокислотными остатками, исключая исходный аминокислотный остаток и Cys. Эти варианты Fc обозначали как В3-варианты. В3-варианты экспрессировали и связывание очищенных на белке А антител с каждым из Fc-гамма R (Fc-гамма RIIa типа Н, Fc-гамма RIIa типа R, Fc-гамма RIIb и Fc-гамма RIIIaF) всесторонне оценивали следующим образом. Анализ взаимодействия между каждым измененным антителом и Fc-гамма-рецептором, полученным согласно описанному выше методу, осуществляли с помощью BIACORE® Т100 (фирма GE Healthcare), BIACORE® Т200 (фирма GE Healthcare), BIACORE® А100 или BIACORE® 4000. В качестве подвижного буфера применяли HBS-EP+ (фирма GE Healthcare) и температуру измерения устанавливали на 25°С. Чип получали путем иммобилизации белка А (фирма Thermo Scientific), белка A/G (фирма Thermo Scientific) или белка L (фирма ACTIGEN или BioVision) на сенсорном СМ4- или СМ5-чипе серий S (фирма GE Healthcare), применяя метод аминного сочетания. После «захвата» представляющих интерес антител на этих сенсорных чипах давали прореагировать Fc-гамма-рецептору, разведенному в подвижном буфере, и измеряли количество связанных с антителом рецепторов. Однако, поскольку количество связанных Fc-гамма-рецепторов зависело от количества «захваченных» антител, количество связанных Fc-гамма-рецепторов делили на количество каждого «захваченного» антитела для получения скорректированных величин, и сравнивали эти величины. Кроме того, антитела, иммобилизованные на чипах, отмывали с помощью 10 мМ глицина-HCl, рН 1,5, в результате чего происходила регенерация чипа и его использовали повторно. Уровень связывания Fc-гамма R с каждым В3-вариантом делили на уровень связывания с Fc-гамма R родительского антитела В3 (антитело, имеющее последовательность встречающегося в естественных условиях человеческого IgG1 в положениях 234-239, 265-271, 295, 296, 298, 300, и 324-337 согласно EU-нумерации). Величину, полученную умножением этого уровня на 100, применяли в качестве показателя относительной Fc-гамма R-связывающей активности каждого варианта.
В таблице 15 представлен профиль связывания перспективных замен, выбранных на основе следующих критериев: превышение более чем на 40% связывания с человеческим Fc-гамма RIIb, понижение более чем на 100% связывания с человеческим Fc-гамма RIIaR и Fc-гамма RIIaH, понижение более чем на 10% связывания с человеческим Fc-гамма RIIIaF по сравнению с родительским антителом В3.

На следующей стадии оценивали перекрестную реактивность этих замен в отношении cynoFc-гамма R. 16 указанных мутаций интродуцировали в M103205H795-SG1. Варианты экспрессировали с использованием M103202L889-SK1 в качестве легкой цепи и анализировали их аффинность к cynoFc-гамма RIIa1, IIa2, IIa3, IIb, IIIaS.
В таблице 16 представлен профиль связывания указанных 16 вариантов с cynoFc-гамма R. Уровень связывания Fc-гамма R получали путем деления количества связанных Fc-гамма R на количество каждого иммобилизованного антитела. Эту величину стандартизовали, принимая величину, полученную для SG1, за 100.

Из отобранных 16 вариантов Fc только у MY001 сохранилось связывание с cynoFc-гамма RIIb, составляющее 85% относительно связывания SG1. Кроме того, у MY001 обнаружено пониженное связывание с cynoFc-гамма RIIIaS, а его связывание с cynoFc-гамма RIIa1, IIa2 и IIa3 сохранялось. В результате установлено, что мутация G236N представляет собой уникальную замену, которая обеспечивает сходный профиль связывания и с человеческими, и с cynoFc-гамма R.
Хотя замена G236N характеризуется перекрестной реактивностью с и человеческими, и cynoFc-гамма R, при ее введении аффинность к Fc-гамма RIIb ниже по сравнению с SG1 (75% для человеческого Fc-гамма RIIb и 85% для cynoFc-гамма RIIb соответственно). Для повышения аффинности к Fc-гамма RIIb оценивали дополнительные замены. В частности, на основе результате анализа на основе всеохватывающего мутагенеза (таблица 17) отбирали замены, для которых характерно повышенное связывание с человеческим Fc-гамма RIIb, пониженное связывание с человеческим Fc-гамма RIIIa и повышенная избирательность в отношении человеческого Fc-гамма RIIb по сравнению с человеческим Fc-гамма RIIa.
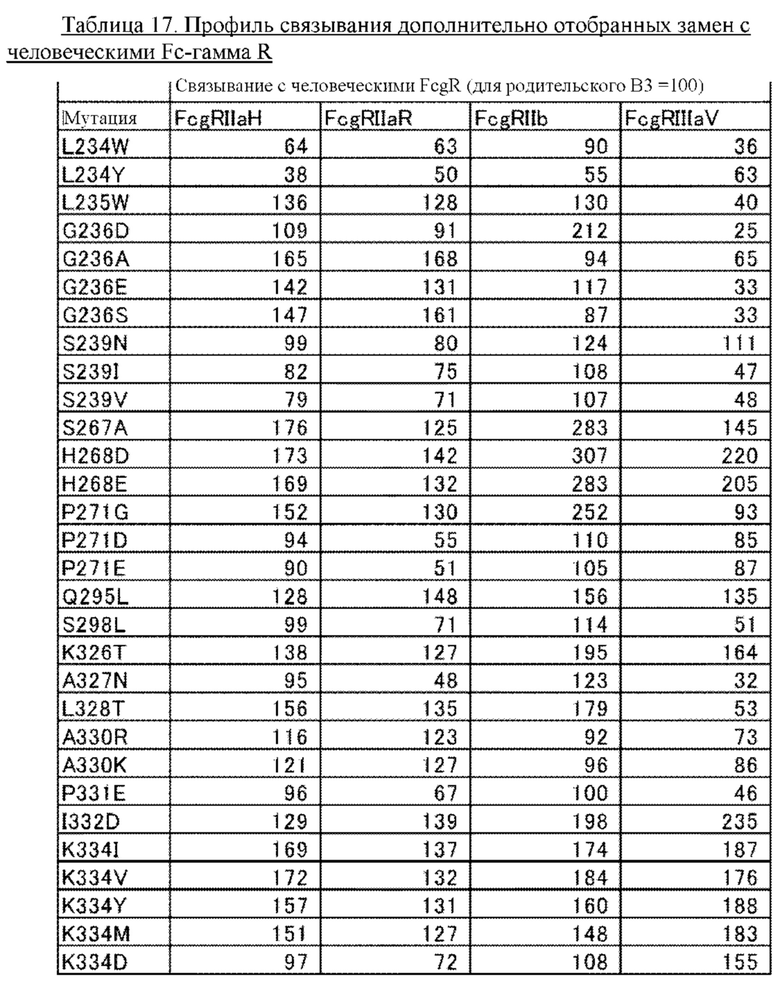
Среди этих замен L234W и L234Y отбирали для повышения избирательности к человеческому Fc-гамма RIIb по сравнению с человеческим Fc-гамма RIIa. Кроме того, отбирали G236A, G236S, A330R, A330K и Р331Е для снижения связывания с человеческим Fc-гамма RIIIa. Все другие замены отбирали для повышения связывания с человеческим Fc-гамма RIIb. Оценивали перекрестную реактивность отобранных замен к cynoFc-гамма R. Кроме того, оценивали также три замены Р396М, V264I и E233D. Замены интродуцировали в M103205H795-SG1 и варианты экспрессировали, используя M103202L889-SK1 в качестве легкой цепи.
В таблице 18 приведены результаты кинетического анализа отобранных замен, включая G236N, в отношении как человеческих, таких и cynoFc-гамма R. В частности, замены интродуцировали в M103205H795-SG1. Варианты экспрессировали, используя M103202L889-SK1 в качестве легкой цепи, и анализировали их аффинность к cynoFc-гамма RIIa1, IIa2, IIa3, IIb, IIIaS.
Величины KD в клетках, обозначенных жирным шрифтом, рассчитывали с помощью уравнения 2, поскольку их аффинность была слишком низкой для точного ее определения с помощью кинетического анализа. Кратность величин KD рассчитывали путем деления величины KD для SG1 на величину KD для варианта для каждого Fc-гамма R.
Для G236N обнаружена сопоставимая с SG1 аффинность связывания с cynoFc-гамма RIIa1, cynoFc-гамма RIIa2, cynoFc-гамма RIIa3 и cynoFc-гамма RIIb. Хотя аффинность к человеческому Fc-гамма RIIb снижалась до 0,6, аффинность к человеческому Fc-гамма RIIaH и аффинность к человеческому Fc-гамма RIIaR снижались до 0,2 и 0,1, соответственно, что свидетельствует о том, что эта замена обладает высокой избирательностью в отношении человеческого Fc-гамма RIIb по сравнению с человеческим Fc-гамма RIIa. Кроме того, аффинности к cynoFc-гамма RIIIaS и человеческому Fc-гамма RIIIaV составляли 0,03 и 0,04 соответственно, что является предпочтительным для элиминации ADCC-активности. На основе этого результата установлено, что G236N характеризуется практически идеальной перекрестной реактивностью с человеческими и cynoFc-гамма R, хотя при ее введении аффинность и к человеческому и к cynoFc-гамма RIIb может повышаться.
Среди изученных дополнительных замен S298L, G236A, Р331Е, E233D, K334Y, K334M, L235W, S239V, K334I, L234W, K328T, Q295L, K334V, K326T, Р396М, I332D, Н268Е, P271G, S267A и H268D приводили к повышенному связыванию и с человеческими, и с cynoFc-гамма RIIb. В частности, для H268D продемонстрировано самое высокое действие (7-кратное для человеческого Fc-гамма RIIb и 5,3-кратное для cynoFc-гамма RIIb). Касательно эффекта снижения связывания с Fc-гамма RIIIa, для G236S, A330K и G236D установлено менее, чем 0,5-кратная аффинность по сравнению с SG1.
Для повышения аффинности MY001 и к человеческому, и к cynoFc-гамма RIIb оценивали комбинации замен, перечисленных в таблице 18. В частности, замены интродуцировали в M103205H795-SG1. Варианты экспрессировали, применяя M103202L889-SK1 в качестве легкой цепи, и анализировали их аффинность. В таблице 19 представлены результаты кинетического анализа связывания с человеческими и cynoFc-гамма R. Величины KD в клетках, обозначенных жирным шрифтом, рассчитывали с помощью уравнения 2, поскольку их аффинность была слишком низкой для точного ее определения с помощью кинетического анализа. Кратность величин KD рассчитывали путем деления величины KD для SG1 на величину KD для варианта для каждого Fc-гамма R.
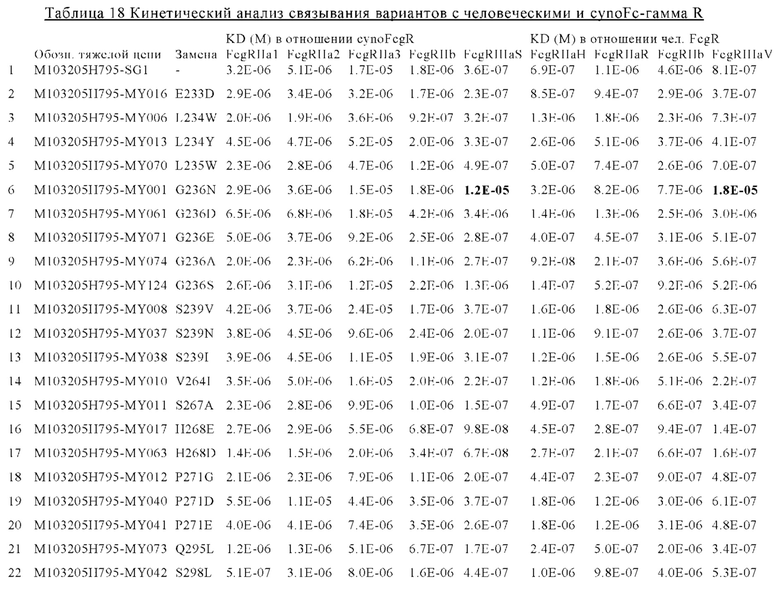
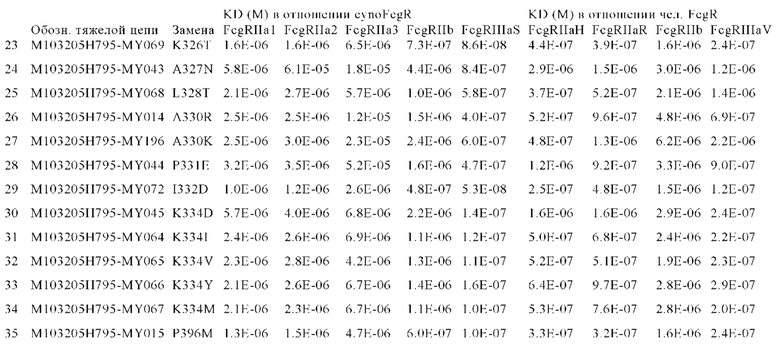
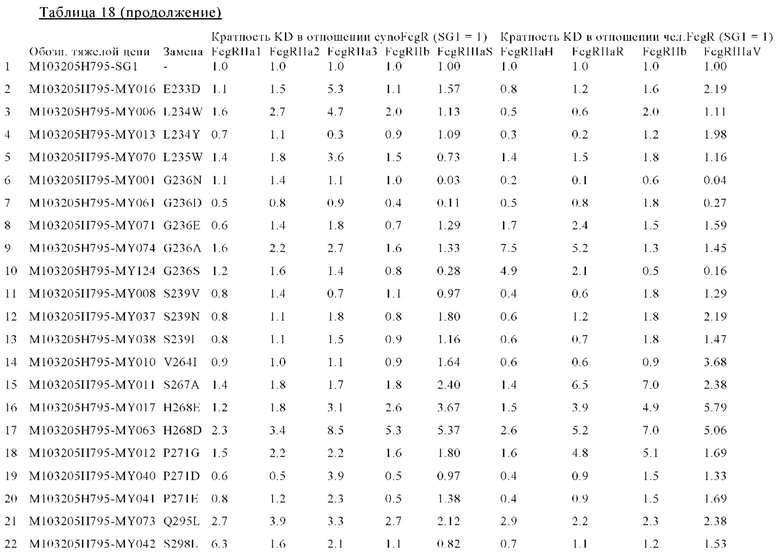
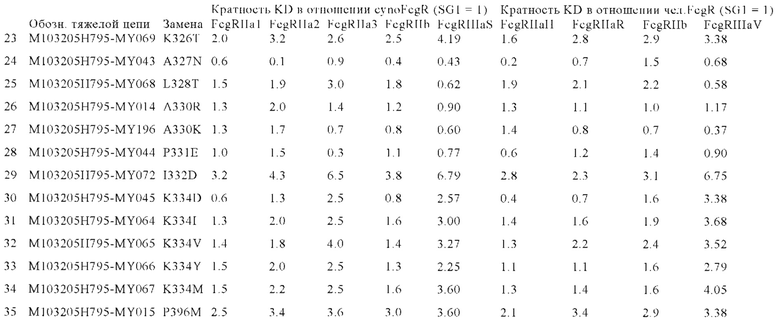
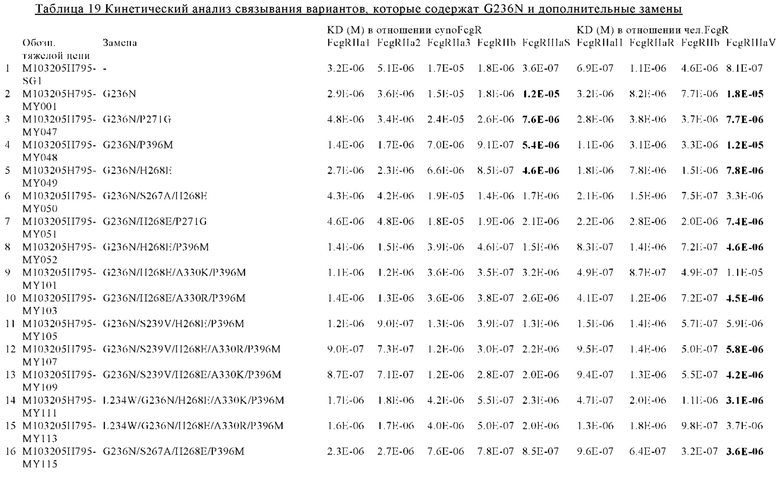
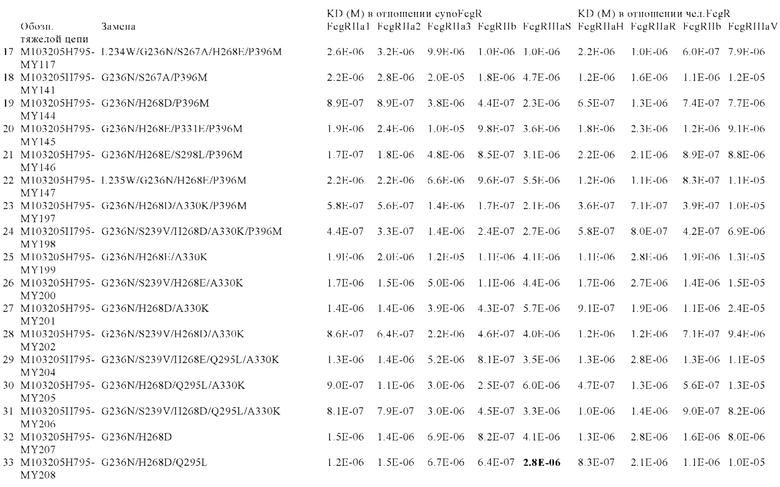
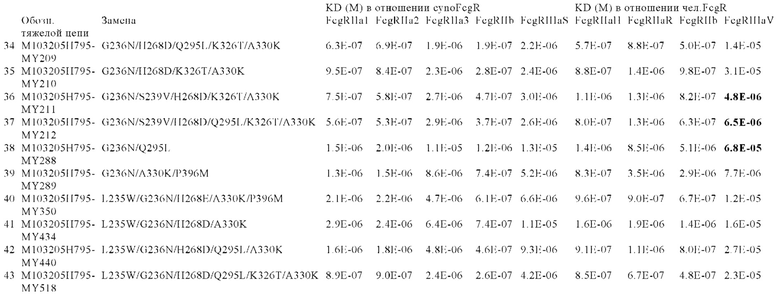
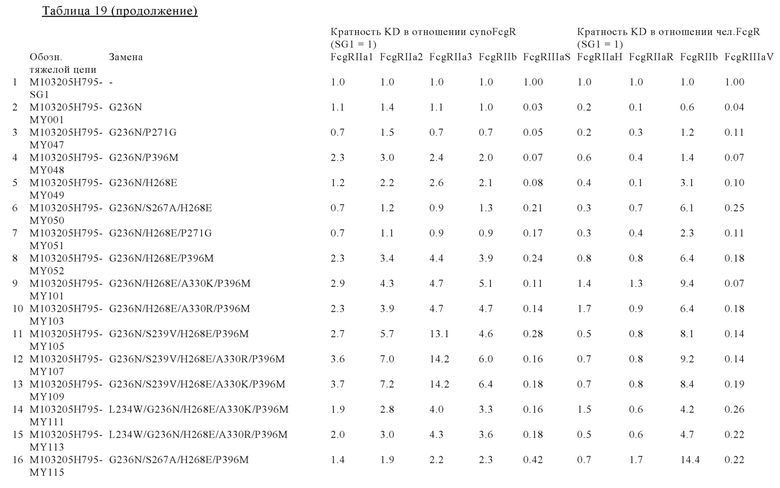
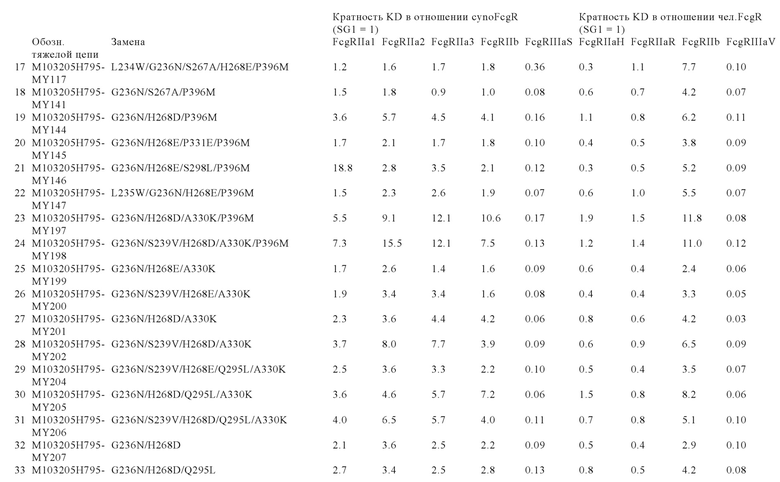
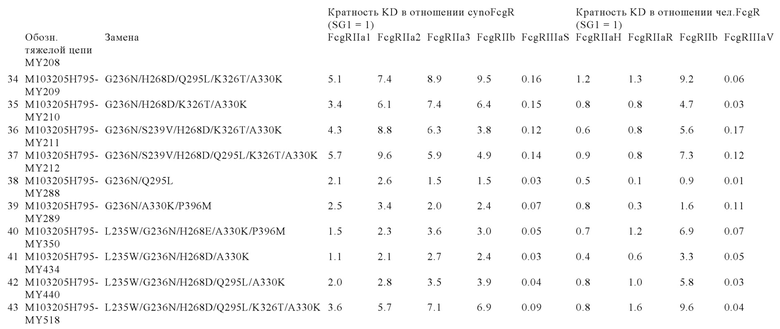
Все варианты подавляли аффинность к человеческому Fc-гамма RIIIaV менее чем в 0,26 раза и к cynoFc-гамма RIIIaS менее чем в 0,42 раза по сравнению с SG1. Кроме того, для всех вариантов за исключением MY047, MY051 и MY141, успешно продемонстрировано повышенное связывание и с человеческим, и с cynoFc-гамма RIIb по сравнению с MY001, их связывание с человеческим Fc-гамма RIIa сохранялось на менее чем 2-кратном уровне по сравнению с SG1. Среди них MY201, MY210, MY206, MY144, MY103, MY212, MY105, MY205, MY109, MY107, MY209, MY101, MY518, MY198 и MY197 характеризовались повышенным связыванием и с человеческим, и с cynoFc-гамма RIIb, превышающем более чем в 4 раза связывание SG1. Кроме того, для MY205, MY209, MY198 и MY197 продемонстрировано повышенное более чем в 7 раз связывание и с человеческим, и с cynoFc-гамма RIIb.
В WO 2014030728 проиллюстрировано, что замены в положении 396 в СН3-домене повышали аффинность к человеческому Fc-гамма RIIb. Полный перебор аминокислотных замен интродуцировали в положение 396 М103205Н795-MY052, которое содержит замены G236N/H268E/P396M. Полученные варианты экспрессировали, используя M103202L889-SK1 в качестве легкой цепи, и оценивали их аффинность к человеческим и cynoFc-гамма R (таблица 20). Кратность величин KD рассчитывали путем деления величины KD для SG1 на величину KD для варианта для каждого Fc-гамма R.
Среди изученных замен P396I, P396K и P396L поддерживали связывание с человеческим Fc-гамма RIIb на более чем 5-кратном уровне по сравнению с SG1, и только замена P396L приводила к повышенной аффинности к человеческому Fc-гамма RIIb по сравнению с MY052. Касательно связывания с cynoFc-гамма RIIb у родительского MY052 обнаружена наиболее высокая аффинность, в 3,9 раз превышающая аффинность SG1.
Поскольку G236N характеризуется идеальной перекрестной реактивностью с человеческими и cynoFc-гамма R, изучали другие замены в положении 236, которые не были изучены в предыдущих примерах. В частности, Asn в M103205H795-MY201 заменяли на Met, His, Val, Gln, Leu, Thr и Ile. Полученные варианты экспрессировали, используя M103202L889-SK1 в качестве легкой цепи, и оценивали их аффинность к человеческим и cynoFc-гамма R (таблица 21). Величины KD в клетках, обозначенных жирным шрифтом, рассчитывали с помощью уравнения 2, поскольку их аффинность была слишком низкой для точного ее определения с помощью кинетического анализа. Кратность величин KD рассчитывали путем деления величины KD для SG1 на величину KD для варианта для каждого Fc-гамма R.
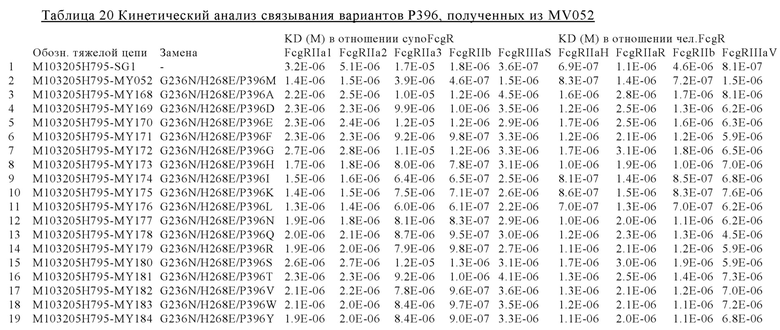
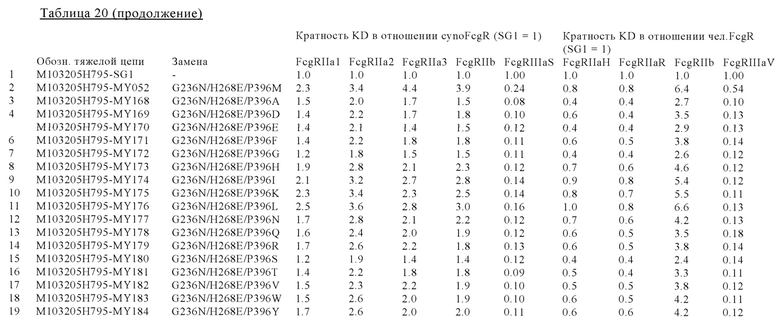

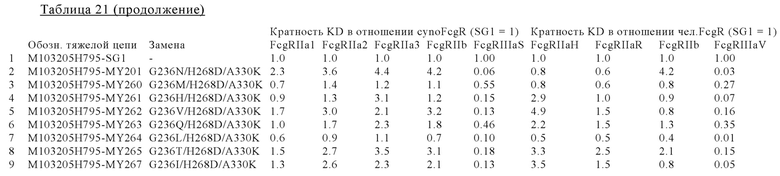


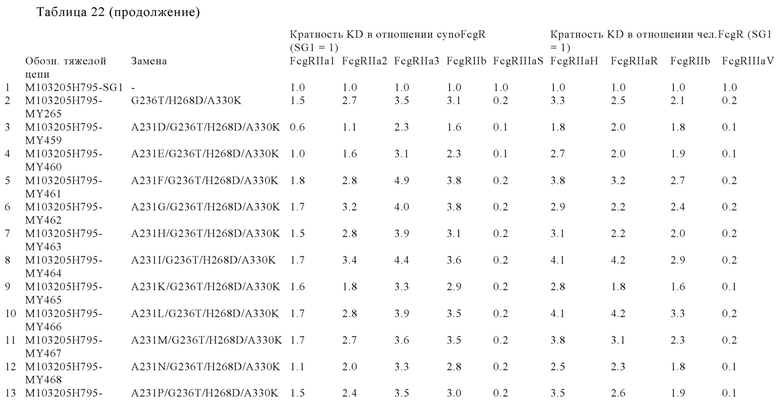
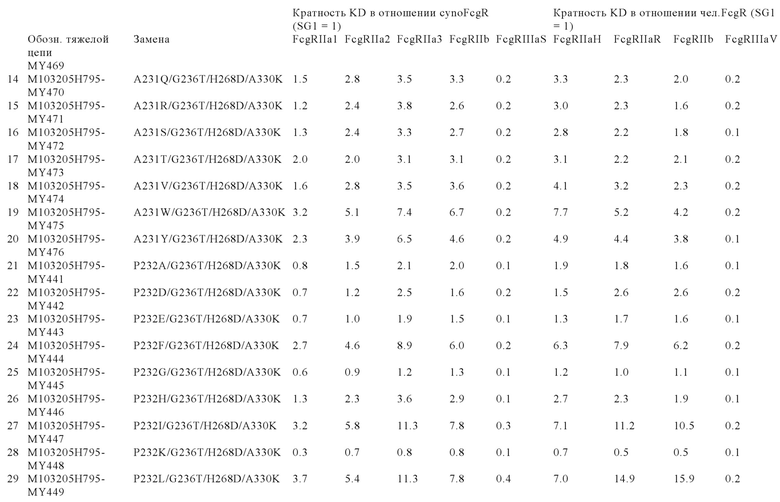
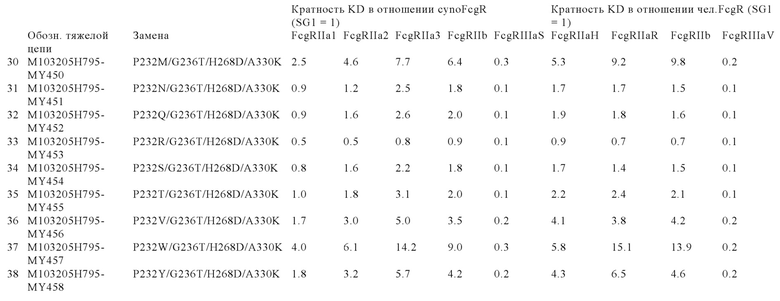
Хотя все варианты обладали пониженной аффинностью и к человеческому, и к cynoFc-гамма RIIb по сравнению с родительской молекулой MY201, MY265, который получали путем замены Asn на Thr в положении 236 MY201, сохранял повышенный 2,1-кратный уровень связывания с человеческим Fc-гамма RIIb и 3,1-кратный уровень связывания с cynoFc-гамма RIIb соответственно по сравнению с SG1. Вариант MY265 также сохранял пониженный уровень связывания и с человеческим, и с cynoFc-гамма RIIIa. Хотя аффинность к человеческим Fc-гамма RIIaH и Fc-гамма RIIaR повышалась более чем в 2,5 раза по сравнению с SG1, G236T представляла собой вторую предпочтительную замену в положении 236.
Для улучшения избирательности и аффинности MY265 к человеческому Fc-гамма RIIb осуществляли полный перебор аминокислотных замен в положениях 231 и 232. В частности, аминокислоту в положение 231 и 232 М103205Н795-MY265 заменяли одним из 18 аминокислотных остатков, исключая исходную аминокислоту и Cys. Полученные варианты экспрессировали, используя M103202L889-SK1 в качестве легкой цепи, и оценивали их аффинность к человеческим и cynoFc-гамма R (таблица 22). Кратность величин KD рассчитывали путем деления величины KD для SG1 на величину KD для варианта для каждого Fc-гамма R.
Касательно аффинности к Fc-гамма RIIb добавление А231Т, А231М, A231V, A231G, A231F, A231I, A231L, А231, A231W, P232V, P232Y, P232F, Р232М, P232I, P232W, P232L повышало связывание и с человеческим, и с cynoFc-гамма RIIb по сравнению с родительской молекулой MY265. Более того, добавление А231Т и A231G снижало связывание с человеческим Fc-гамма RIIaR по сравнению с MY265.
Изучали комбинации замен, которые не исследовали в предыдущих примерах. Протестированные и предположительно перспективные замены интродуцировали в M103205H795-SG1 в комбинации. Полученные варианты экспрессировали, используя M103202L889-SK1 в качестве легкой цепи, и оценивали их аффинность к человеческим и cynoFc-гамма R. Результаты представлены в таблице 23А «Кинетический анализ связывания других комбинаций» (см. Приложение) и в таблице 23Б «Кинетический анализ связывания других комбинаций» (см. Приложение) при этом величины KD в клетках, обозначенных жирным шрифтом, рассчитывали с помощью уравнения 2, поскольку их аффинность была слишком низкой для точного ее определения с помощью кинетического анализа. Кратность величин KD рассчитывали путем деления величины KD для SG1 на величину KD для варианта для каждого Fc-гамма R. Для сравнения вновь приведены величины для MY209, выбранного в качестве перспективных вариантов по результатам, представленным в таблице 19.
Среди протестированных вариантов только у MY213 обнаружена более высокая аффинность связывания и с человеческим, и с cynoFc-гамма RIIb по сравнению с MY209. Однако аффинность MY213 к человеческому Fc-гамма RIIaH и человеческому Fc-гамма RIIaR оказалась выше, чем у MY209.
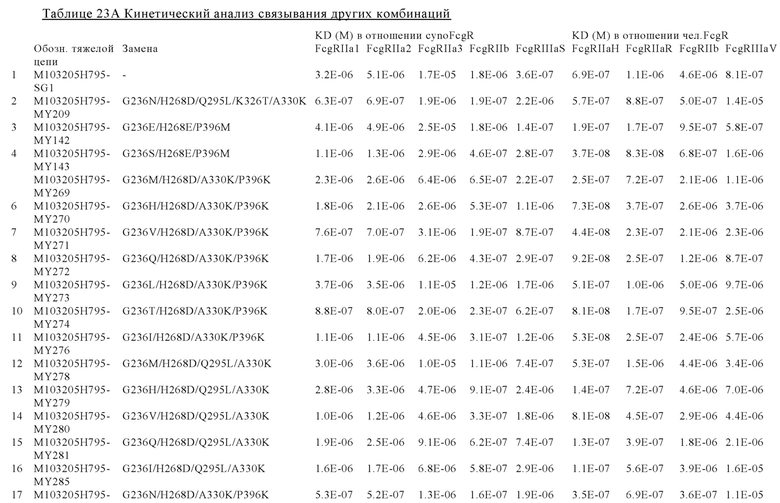
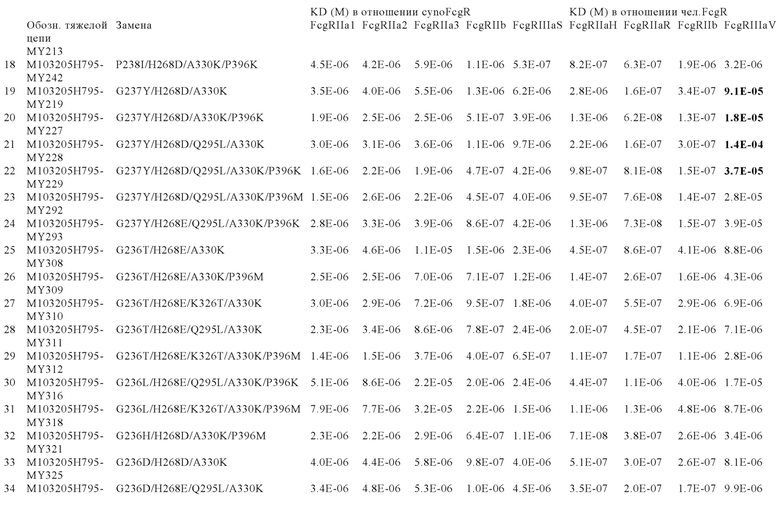

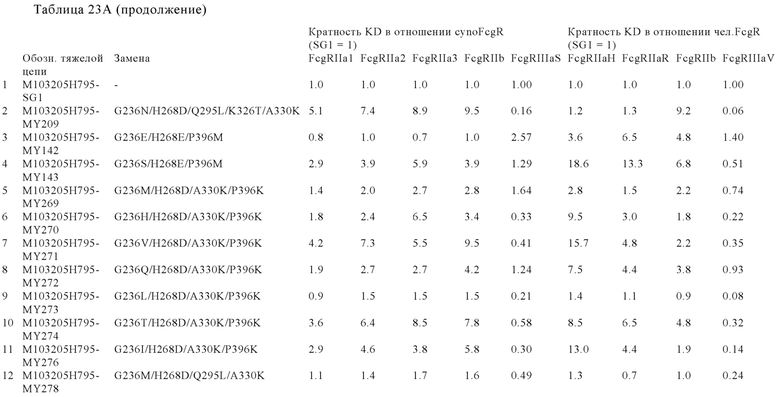
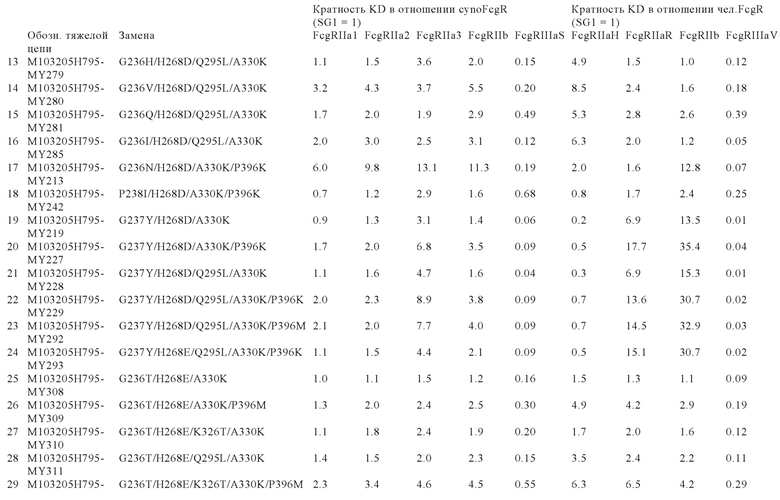
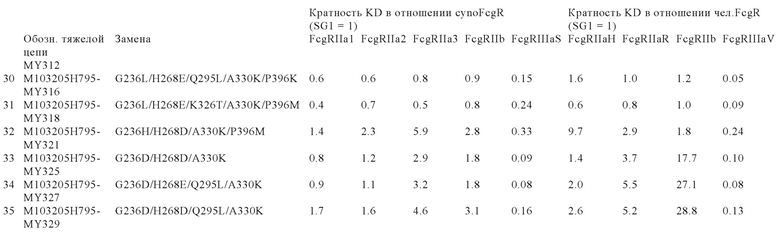


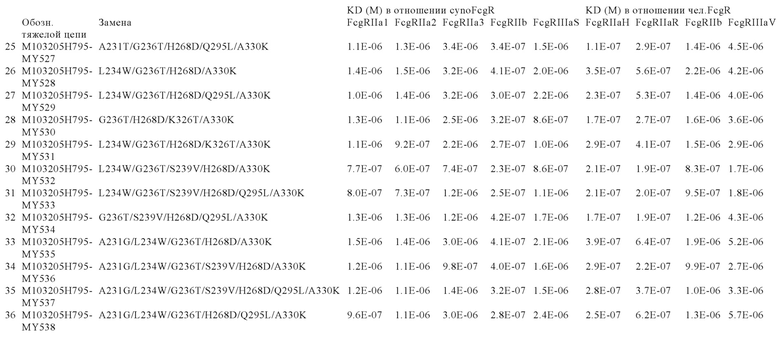

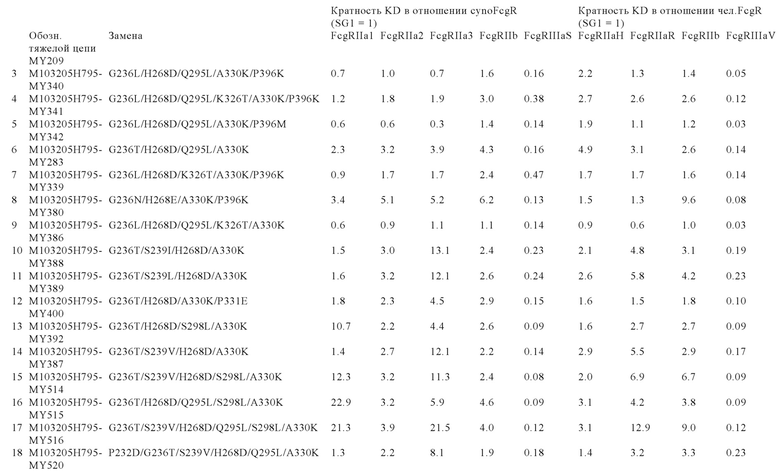
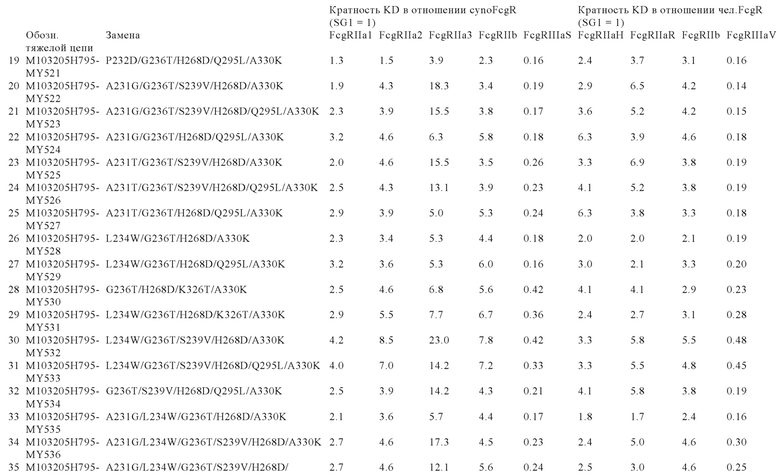

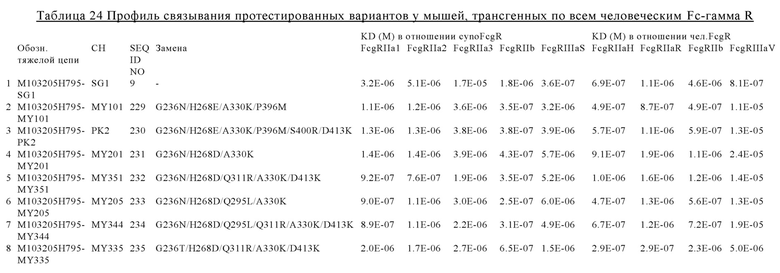
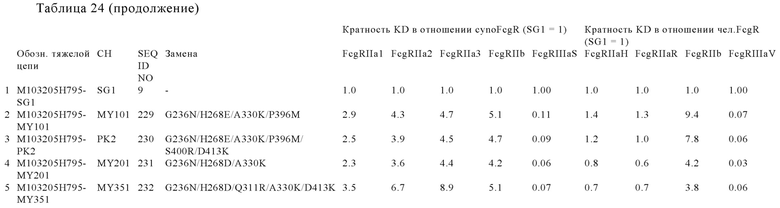

Пример 28
Оценка клиренса миостатина при применении вариантов Fc с повышенной активностью в отношении Fc-гамма RIIb у мышей, трансгенных по всем человеческим Fc-гамма R
Воздействие на клиренс миостатина варианта Fc с повышенной активностью в отношении Fc-гамма RIIb, сконструированного согласно методу, описанному в примере 27, оценивали на мышах, у которых все мышиные Fc-гамма R были удавлены путем делеции и человеческие Fc-гамма R, кодируемые в виде трансгенов, были встроены в мышиный геном (Proc. Natl. Acad. Sci., 109, 2012, с. 6181). У этих мышей все мышиные Fc-гамма R заменены человеческими рецепторами, поэтому на таких мышах можно оценивать воздействие повышенной аффинности к человеческому Fc-гамма RIIb на клиренс растворимого антигена. Кроме того, оценивали приводящие к увеличению pI замены в комбинации с вариантами Fc с повышенной активностью в отношении Fc-гамма RIIb, которые конструировали согласно методу, описанному в примере 27.
Получение и профиль тестируемых вариантов
Сведения о 8 тестируемых антителах и их профилях связывания обобщены в таблице 24. Тяжелую цепь, MS103205H795-PK2, получали путем интродукции замен, приводящих к повышению pI (S400R/D413K) в MS103205H795-MY101. MS103205H795-MY351, MS103205H795-MY344, MS103205H795-MY335 получали путем интродукции других приводящих к повышению pI замен (Q311R/D413K) в MS103205H795-MY201, MS103205H795-MY205, MS103205H795-MY265 соответственно. Все варианты MS1032LO06 экспрессировали с M103202L889-SK1 в качестве легкой цепи согласно методу, описанному в примере 27, и их аффинность к человеческим и cynoFc-гамма R оценивали, используя метод, описанный в примере 27.
Основываясь на SPR-анализе, результаты которого обобщены в таблице 24, было подтверждено, что приводящие к повышению pI замены не влияют на связывание с Fc-гамма R вариантов Fc с повышенной активностью в отношении Fc-гамма RIIb. У MY101 и PK2, которые получали путем интродукции S400R/D413K в MY101, обнаружен повышенный в 9 и 8 раз уровень связывания с человеческим Fc-гамма RIIb соответственно. Аффинность MY101 и PK2 к человеческому Fc-гамма RIIa оставалась сопоставимой с SG1. Для других человеческих и cynoFc-гамма R обнаружен практически сходный профиль связывания. Аналогично этому, для МУ351 установлен сходный профиль связывания с родительской молекулой MY201, для MY344 с родительской молекулой MY205, и для MY335 с родительской молекулой MY265 (таблица 21) соответственно. Эти результаты свидетельствуют о том, что любая из пар замен, повышающих pI, S400R/D413K или Q311R/D413K, не влияет на аффинность к человеческим и cynoFc-гамма R.
ФК-исследование, проведенное на трансгенных по всем человеческим Fc-гамма R мышах
Тестирование in vivo с использованием трансгенных по всем человеческим Fc-гамма R мышей
Элиминацию миостатина и антитела к латентному миостатину оценивали in vivo после совместного введения антитела к латентному миостатину и человеческого латентного миостатина трансгенным по всем человеческим Fc-гамма R мышам (фиг. 24 и 25). Антитело к латентному миостатину (0,3 мг/мл) и латентный миостатин (0,05 мг/мл) вводили в одной дозе 10 мл/кг в каудальную вену. Антитело к CD4 антитело (1 мг/мл) вводили три раза (каждые 10 дней) в дозе 10 мл/кг в каудальную вену для подавления антител к лекарственному средству. Образцы крови собирали через 5 мин, 15 мин, 1 ч, 4 ч, 7 ч, 1 день, 2 дня, 7 дней, 14 дней, 21 день и 28 дней после введения. Собранную кровь немедленно центрифугировали при 15000 об/мин при 4°С в течение 5 мин для отделения плазмы. Отделенную плазму хранили при температуре -20°С или ниже до анализа. В качестве антител к латентному миостатину применяли MS1032LO06-SG1, MS1032LO06-MY101, MS1032LO06-PK2, MS1032LO06-MY201, MS1032LO06-MY351, MS1032LO06-MY205, MS1032LO06-MY344 и MS1032LO06-MY335.
Измерение концентрации общего миостатина в плазме с помощью электрохемилюминесценции (ECL)
Концентрацию общего миостатина в плазме мышей измеряли с помощью ECL. Планшеты с иммобилизованным антителом к зрелому миостатину получали путем нанесения антитела к зрелому миостатину RK35 (описанного в WO 2009/058346) на 96-луночный планшет MULTI-ARRAY (фирма Meso Scale Discovery) и инкубации в блокирующем буфере в течение 2 ч при комнатной температуре. Приготавливали образцы зрелого миостатина для получения калибровочной кривой и образцы плазмы, разведенные в 40 или более раз. Образцы смешивали в кислом растворе (0,2М глицин-HCl, pH 2,5) для отделения путем диссоциации зрелого миостатина от связывающего его белка (такого как пропептид). Затем образцы вносили в иммобилизованный антителом к зрелому миостатину планшет и давали связываться в течение 1 ч при комнатной температуре, после чего осуществляли промывку. Затем добавляли меченное биотином антитело к зрелому миостатину RK22 (описанное в WO 2009/058346) и планшет инкубировали в течение 1 ч при комнатной температуре, после чего осуществляли промывку. Затем, добавляли меченный SULFO-меткой стрептавидином (фирма Meso Scale Discovery) и планшет инкубировали в течение 1 ч при комнатной температуре, после чего осуществляли промывку. В планшет немедленно добавляли буфер для считывания (Read Buffer) Т (×4) (фирма Meso Scale Discovery) и детекцию сигнала осуществляли с помощью устройства SECTOR Imager 2400 (фирма Meso Scale Discovery). Концентрацию зрелого миостатина рассчитывали на основе ответа относительно калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (фирма Molecular Devices). Зависимость от времени концентрации общего миостатина в плазме после внутривенного введения антитела к латентному миостатину и латентного миостатина, определенная с помощью указанного метода, представлена на фиг. 24.
Измерение концентрации антитела к латентному миостатину в плазме с помощью высокоэффективной жидкостной хроматографии-тандемной спектрометрии с ионизацией электроспреем (ЖХ/ESI-MC/MC)
Концентрацию антитела к латентному миостатину в плазме мышей измеряли с помощью ЖХ/ESI-MC/MC. Концентрации калибровочных стандартов в плазме мышей составляли 1,5625, 3,125, 6,25, 12,5, 25, 50, 100 и 200 мкг /мл. По 3 мкл калибровочных стандартов и образцов плазмы добавляли к 50 мкл Ab-Capture Mag (фирма ProteNova) и давали инкубироваться в течение 2 ч при комнатной температуре. Затем магнитные гранулы выделяли из образцов и промывали дважды 0,2 мл (10 ммолей/л) ЗФР с 0,05% Твин 20. Затем магнитные гранулы промывали ЗФР (10 ммолей/л) для гарантии удаления Твин 20. После промывки магнитные гранулы суспендировали в 25 мкл смеси, содержащей мочевину (7,5 моля/л), дитиотреитол (8 ммолей/л) и лизоцим (белок куриных яиц) (1 мкг/мл), 50 ммолей/л бикарбоната аммония, и суспендированные образцы инкубировали в течение 45 мин при 56°С. Затем добавляли 2 мкл йодацетамида (500 ммолей/л) и образцы инкубировали в течение 30 мин при 37°С в темноте. Затем осуществляли расщепление лизилэндопептидазой, добавляя 150 мкл раствора, содержащего 0,67 мкг/мл лизилэндопептидазы, пригодной для биохимических анализов (фирма Wako), в бикарбонате аммония (50 ммолей/л), и образцы инкубировали в течение 3 ч при 37°С. Затем осуществляли расщепление трипсином, добавляя 10 мкл (10 мкг/мл) модифицированного трипсина, имеющего чистоту, пригодную для секвенирования (фирма Promega), в бикарбонате аммония (50 ммолей/л). Давали пройти расщеплению в образцах путем перемешивания в течение ночи при 37°С и для прекращения реакции добавляли 5 мкл 10%-ной трифторуксусной кислоты. 50 мкл расщепленных образцов подвергали анализу с помощью ЖХ /ESI-MC/MC. ЖХ/ESI-MC/MC осуществляли, используя тройное квадрупольное устройство Xevo TQ-S (фирма Waters), снабженное 2D-UPLC I-класса (фирма Waters). Осуществляли мониторинг специфического для антитела к латентному миостатину пептида YAFGQGTK и специфического для лизоцима пептида GTDVQAWIR, применяемого в качестве внутреннего стандарта, используя режим мониторинга выбранных реакций (SRM). SRM-переход представлял собой [M+2H]2+ (m/z 436,2) до иона у8 (m/z 637,3) для антитела к латентному миостатину и [M+2H]2+ (m/z 523,3) до иона у8 (m/z 545,3) для лизоцима. Внутреннюю калибровочную кривую создавали с помощью взвешенной (1/× или 1/×2) линейной регрессии, используя зависимость площади пика от концентраций. Концентрацию в плазме мышей рассчитывали с помощью калибровочной кривой, используя аналитическое программное обеспечение Masslynx, версия 4.1 (фирма Waters). Зависимость от времени концентрации антитела в плазме после внутривенного введения антитела к латентному миостатину и латентного миостатина, измеренная с помощью указанного метода, представлена на фиг. 25.
Воздействие pI и связывания Fc-гамма R на концентрацию миостатина in vivo
После введения MS1032LO06-SG1 концентрация общего миостатина в плазме через 7 ч снижалась в 5 раз по сравнению с концентрацией общего миостатина в плазме через 5 мин. В противоположность этому, после введения MS1032LO06-MY101, MS1032LO06-MY201 и MS1032LO06-MY205 концентрация общего миостатина в плазме через 7 ч снижалась в 28-200 раз по сравнению с концентрацией общего миостатина в плазме через 5 мин. Кроме того, после введения MS1032LO06-PK2, MS1032LO06-MY351, MS1032LO06-MY344 и MS1032LO06-MY335 концентрация общего миостатина в плазме через 7 ч снижалась в 361-419 раз по сравнению с концентрацией общего миостатина в плазме через 5 мин. С другой стороны, различие в концентрации каждого антитела в каждый тестируемый момент времени по сравнению с MS1032LO06-SG1 оказалось примерно 2-кратным, и pI-варианты не повышали элиминацию антитела из плазмы. Замены, приводящие и к повышению связывания с человеческим Fc-гамма RIIb, и к увеличению pI, повышали элиминацию миостатина, но не антител.
Варианты с высокой pI имели больший положительный заряд в плазме. Поэтому указанный положительный заряд взаимодействовал с отрицательно заряженной клеточной поверхностью, иммунный комплекс антиген-антитело вариантов с высокой pI сильнее контактировал с клеточной поверхностью, что приводило к повышенному поглощению клеткой иммунного комплекса антиген-антитело вариантов с высокой pI.
Пример 29
Оценка клиренса миостатина при применении вариантов Fc с повышенной активностью в отношении Fc-гамма RIIb у обезьян
Воздействие повышенной способности к связыванию вариантов Fc, созданных согласно методу, описанному в примере 27, с cynoFc-гамма RIIb на элиминацию миостатина оценивали у обезьян циномолгус. Кроме того, оценивали также комбинированное действие с заменами, приводящими к повышению pI.
Получение и профиль оцененных вариантов
Сведения о 14 протестированных антителах и профилях их связывания обобщены в таблице 25. Описано, что присутствие варианта Fc с повышенной способностью связываться с FcRn при кислом pH повышает время полужизни антитела in vivo (J. Biol. Chem. 281, 2006, cc. 23514-23524; Nat. Biotechnol. 28, 2010, cc. 157-159, Clin Pharm. & Thera. 89(2), 2011, cc. 283-290). Для повышения времени полужизни антитела без связывания с ревматоидным фактором при создании изобретения объединяли эти замены с вариантами Fc с повышенной активностью в отношении Fc-гамма RIIb и повышенной pI.
Тяжелые цепи MS103205H795-SG1012, MS103205H795-SG1029, MS103205 H795-SG1031, MS103205H795-SG1033, MS103205H795-SG1034 получали путем интродукции замены N434A в MS103205H795-MY101, MS103205 H795-MY344, MS103205H795-MY351, MS103205H795-MY201, MS103205H795-MY335, соответственно. MS103205H795-SG1016 получали путем интродукции приводящих к повышению pI замен (Q311R/D399K) в MS 103205H795-SG1012. MS103240H795-SG1071 и MS103240H795-SG1079 получали путем интродукции замен M428L/N434A/Y436T/ Q438R/S440E и замен N434A/Q438R/S440E в MS103240 H795-MY344 соответственно (MS103240H795: VH, SEQ ID NO: 92).
MS103240H795-SG1074 и MS103240H795-SG1077 получали путем интродукции приводящих к повышению pI замен Q311R/P343R и замен M428L/N434A/Y436T/Q438R/S440E в MS103240H795-MY209 и MS103240H795-MY518 соответственно. MS103240H795-SG1080 и MS103240H795-SG1081 получали путем интродукции приводящих к повышению pI замен Q311R/P343R и замен N434A/Q438R/S440E в MS103240H795-MY209 и MS103240H795-MY518 соответственно. MS103240H795-SG1071 получали путем интродукции приводящих к повышению pI замен Q311R/D413K и замен M428L/N434A/Y436T/Q438R/S440E в MS103240H795-MY205. MS103240H795-SG1079 получали путем интродукции приводящих к повышению pI замен Q311R/D413K и замен N434A/Q438R/S440E в MS103240H795-MY205. Указанные варианты MS1032LO06 и варианты MS1032LO19 экспрессировали с M103202L889-SK1 и M103202L1045-SK1 соответственно (M103202L1045: VL, SEQ ID NO: 97) в качестве легкой цепи согласно методу, описанному в примере 34, и их аффинность к человеческим и cynoFc-гамма R оценивали с помощью метода, описанного в примере 27. В этом примере кратность величины KD для каждого варианта Fc рассчитывали путем деления величины KD для родительской молекулы SG1 на величину KD для варианта для каждого Fc-гамма R.
Например, кратность величин KD для MS1032LO06-SG1012 и MS1032LO19-SG1071 рассчитывали путем деления величин KD для MS1032LO06-SG1 и MS1032LO19-SG1 на величины KD для MS1032LO06-SG1012 и MS1032LO19-SG1071 соответственно.
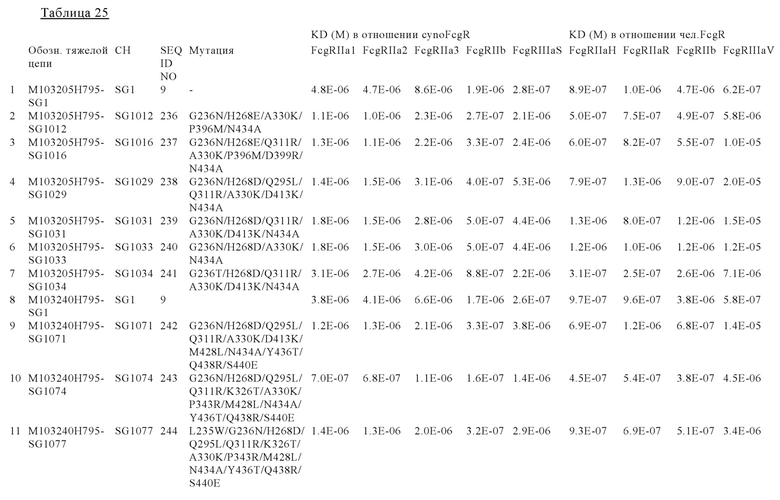

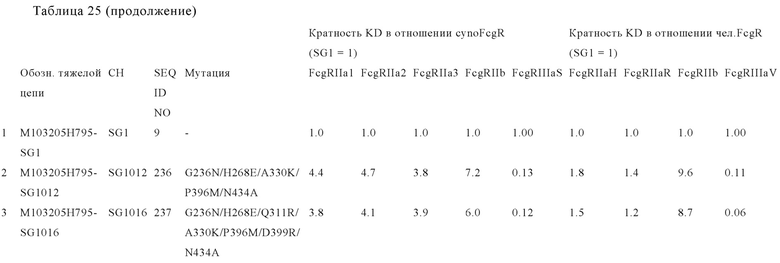
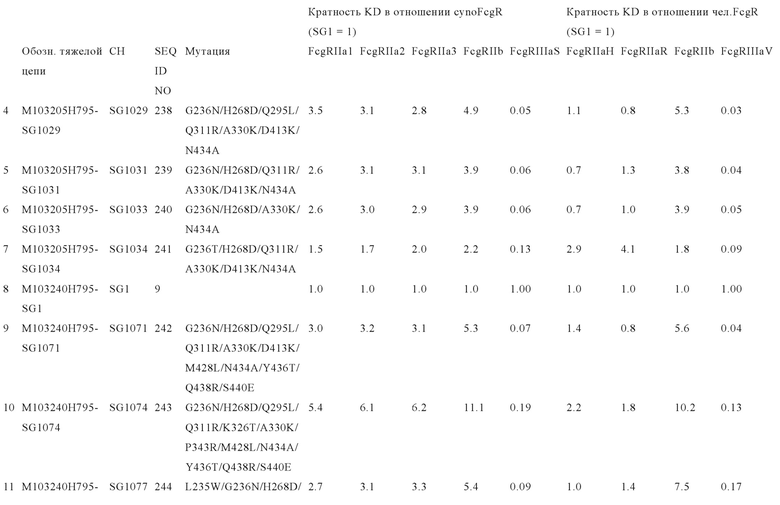
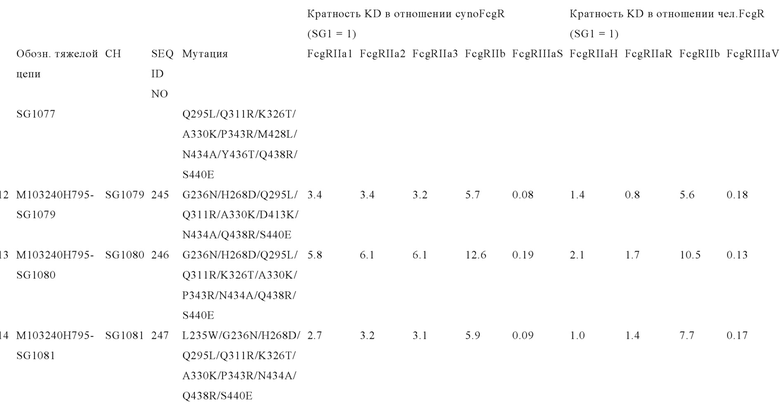
В таблице 25 обобщены результаты SPR-анализа. Из указанных вариантов для SG1012, SG1016, SG1074 и SG1080 установлена наиболее сильная аффинность к человеческому Fc-гамма RIIb, которая в 10 раз превышала аффинность к человеческому Fc-гамма RIIb по сравнению с SG1.
29.2. ФК-исследование, проведенное на обезьянах
Тестирование in vivo с использованием обезьян циномолгус
Накопление эндогенного миостатина оценивали in vivo после введения антитела к латентному миостатину 2-4-летним Масаса fascicularis (яванский макак-крабоед, обезьяны циномолгус) из Камбоджи (фирма Shin Nippon Biomedical Laboratories Ltd., Япония). Дозу 30 мг/кг инъецировали в цефалическую вену предплечья, используя одноразовый шприц, удлиняющую трубку, находящуюся внутри иглу и инфузионный насос. Скорость дозирования составляла 30 мин на организм животного. Образцы крови собирали перед началом дозирования и либо через 5 мин, 7 ч и 1, 2, 3, 7, 14, 21, 28, 35, 42, 49 и 56 дней после окончания дозирования, либо через 5 мин и через 2, 4 и 7 ч, и через 1, 2, 3, 7, 14, 21, 28, 35, 42, 49 и 56 дней после окончания дозирования. Кровь брали из бедренной вены с помощью шприца, содержащего гепарин натрия. Кровь немедленно охлаждали на льду и плазму получали центрифугированием при 4°С С, 1700×g в течение 10 мин. Образцы плазмы хранили в холодильнике глубокой заморозки (приемлемый диапазон: -70°С или ниже) вплоть до анализа. В качестве антител к латентному миостатину применяли MS1032LO00-SG1, MS1032LO06-SG1012, MS1032LO06-SG1016, MS1032LO06-SG1029, MS1032LO06-SG1031, MS1032LO06-SG1033 и MS1032LO06-SG1034 (в контексте настоящего описания SG1012, SG1016, SG1029, SG1031, SG1033 и SG1034 обозначают константные области тяжелых цепей, сконструированные на основе SG1 согласно описанному ниже методу).
В цефалическую вену предплечья или подкожную вену ноги с использованием одноразового шприца и находящейся внутри иглы вводили в дозе 2 мг/кг антитела к латентному миостатину MS1032LO19-SG1079, MS1032LO19-SG1071, MS1032LO19-SG1080, MS1032LO19-SG1074, MS1032LO19-SG1081 и MS1032LO19-SG1077 (в контексте настоящего описания SG1079, SG1071, SG1080, SG1074, SG1081 и SG1077 обозначают константные области тяжелых цепей, сконструированные на основе SG1 согласно описанному ниже методу). Образцы крови собирали перед началом дозирования и через 5 мин и 2, 4 и 7 ч, и 1, 2, 3, 7, 14 дней после окончания дозирования. Кровь обрабатывали согласно описанной выше процедуре. Образцы плазмы хранили в холодильнике глубокой заморозки (приемлемый диапазон: -70°С или ниже) вплоть до анализа.
Измерение концентрации общего миостатина в плазме с помощью электрохемилюминесценции (ECL)
Концентрацию общего миостатина в плазме обезьян измеряли с помощью ECL согласно методу, описанному в примере 23. Зависимость от времени концентрации общего миостатина в плазме после внутривенного введения антитела к латентному миостатину, определенная с помощью указанного метода, представлена на фиг. 26.
Измерение ADA в плазме обезьян с помощью электрохемилюминесценции (ECL)
Биотинилированное лекарственное средство применяли для сенсибилизации 96-луночного стрептавидинового планшета MULTI-ARRAY (фирма Meso Scale Discovery) и инкубировали в LowCross®-буфере (фирма Candor) в течение 2 ч при комнатной температуре. Образцы обезьяньей плазмы образцы разводили в 20 раз в LowCross®-буфере перед внесением в планшет. Образцы инкубировали в течение ночи при 4°С. На следующий день планшет промывали трижды буфером для промывки перед добавлением меченного с помощью SULFO-метки вторичного антитела к обезьяньему IgG (фирма Thermo Fisher Scientific). После инкубации в течение 1 ч при комнатной температуре планшет трижды промывали буфером для промывки. В планшет немедленно добавляли буфер для считывания (Read Buffer Т) (×4) (фирма Meso Scale Discovery) и детекцию сигнала осуществляли с помощью устройства SECTOR Imager 2400 (фирма Meso Scale Discovery).
Влияние зависимости от pH и конструирования Fc на накопление миостатина у обезьян in vivo
Введение обезьянам циномолгус pH-независимого антитела (MS1032LO00-SG1) приводило по меньшей мере к 60-кратному повышению концентрации миостатина в день 28 относительно исходного уровня. В день 28 pH-зависимые антитела к латентному миостатину MS1032LO06-SG1012 и MS1032LO06-SG1033 обеспечивали 3-й 8-кратное повышение относительно исходного уровня соответственно. Сильный sweeping-эффект главным образом связан с повышением аффинности к Fc-гамма RIIb обезьян циномолгус. pH-зависимые антитела к латентному миостатину MS1032LO06-SG1029, MS1032LO06-SG1031 и MS1032LO06-SG1034 смогли элиминировать антиген ко дню 28 до уровня ниже исходного. Причиной сильного sweeping-эффекта, обусловленного MS1032LO06-SG1029, MS1032LO06-SG1031 и MS1032LO06-SG1034, является повышение неспецифического поглощения клеткой из-за повышения положительно заряженного кластера антител и повышение опосредуемого Fc-гамма R клеточного поглощения из-за усиление связывания с Fc-гамма R.
Введение pH-зависимых антител к латентному миостатину MS1032LO19-SG1079, MS1032LO19-SG1071, MS1032LO19-SG1080, MS1032LO19-SG1074, MS1032LO19-SG1081 и MS1032LO19-SG1077 снижали концентрацию миостатина до уровня ниже предела обнаружения (<0,25 нг/мл), начиная со дня 1 у обезьян циномолгус. В день 14 концентрация миостатина превышала предел обнаружения при применении MS1032LO19-SG1079 и MS1032LO19-SG1071, хотя концентрация миостатина оставалась на уровне ниже порога обнаружения при применении MS1032LO19-SG1080, MS1032LO19-SG1074, MS1032LO19-SG1081 и MS1032LO19-SG1077. Более слабое подавление, обнаруженное для MS1032LO19-SG1079 и MS1032LO19-SG1071, может являться результатом различных мутаций, приводящих к изменению pI.
Результаты позволяют предположить, что можно достигать сильной элиминации миостатина из плазмы с помощью мутаций, которые усиливают связывание с Fc-гамма RIIb или комбинации мутаций, которые повышают положительный заряд антитела и усиливают связывание с Fc-гамма RIIb. Можно ожидать, что у человека можно достигать сильной элиминации миостатина путем объединения мутаций, которые повышают положительный заряд антитела и повышают связывание с Fc-гамма R.
Пример 30
Скрининг приводящих к увеличению pI замен для повышения клиренса миостатина
В этом примере оценивали роль приводящих к увеличению pI замен в Fc-области антитела в повышении клиренса миостатина. Метод введения аминокислотных замен в константную область антитела для повышения pI не ограничен указанным методом, но его, например, можно осуществлять согласно методу, описанному в WO 2014/145159. Также как и в случае вариабельной области, аминокислотные замены, которые интродуцировали в константную область, предпочтительно представляли собой замены, которые снижают количество отрицательно заряженных аминокислот (таких как аспарагиновая кислота и глутаминовая кислота), повышая при этом количество положительно заряженных аминокислот (таких как аргинин и лизин). Кроме того, аминокислотные замены можно интродуцировать в любое положение в константной области антитела, и они могут представлять собой единичную аминокислотную замену или комбинации нескольких аминокислотных замен. Сайты интродукции аминокислотных замен предпочтительно представляют собой положения (но, не ограничиваясь только ими), в которых боковые цепи аминокислот могут экспонироваться на поверхности молекулы антитела. Наиболее предпочтительные примеры включают метод интродукции комбинации нескольких аминокислотных замен в такие положения, которые могут экспонировать на поверхности молекулы антитела. Альтернативно этому, несколько аминокислотных замен, интродуцированных при создании настоящего изобретения, предпочтительно располагали так, чтобы они находились в структурной близости друг к другу. Кроме того (но, не ограничиваясь указанным) несколько аминокислотных замен, интродуцированных при создании настоящего изобретения, предпочтительно представляли собой замены на положительно заряженные аминокислоты, чтобы это предпочтительно приводило к состоянию, при котором множество положительных зарядов присутствуют в структурно близких положениях.
Получение и профиль изученных вариантов
Сведения об изученных антителах обобщены в таблице 26. Тяжелую цепь MS103205H795-SG141 получали путем интродукции приводящих к увеличению pI замен Q311R/D399R в MS103205H795-SG1. Другие варианты тяжелых цепей получали также путем интродукции соответствующих замен, представленных в таблице 26, в MS103205H795-SG1. Все варианты MS1032LO06 экспрессировали с M103202L889-SK1 в качестве легкой цепи согласно методу, описанному в примере 34.
Оценка связывающей активности с мышиным Fc-гамма RII вариантов Fc с повышенной pI с помощью BIACORE®
Вне зависимости от получения содержащих вариант Fc-области антител, анализы связывания между растворимым мышиным Fc-гамма RII и комплексами антиген-антитело осуществляли с помощью BIACORE®T200 (фирма GE Healthcare). Растворимый мышиный Fc-гамма RII получали в форме меченной His молекулы с использованием метода, известного специалистам в данной области. Соответствующее количество антитела к His фиксировали на сенсорном чипе СМ5 (фирма GE Healthcare) с помощью метода аминного сочетания, используя набор для «захвата» His (фирма GE Healthcare) для иммобилизации мышиного Fc-гамма RII. Затем инъецировали комплекс антитело-антиген и подвижный буфер (в качестве референс-раствора) и давали пройти взаимодействию с мышиным Fc-гамма RII, иммобилизованном на сенсорном чипе. Добавляли 20 мМ N-(2-ацетамидо)-2-аминоэтансульфоновую кислоту, 150 мМ NaCl, 1,2 мМ CaCl2 и 0,05% (мас./об.) Твин 20, рН 7,4 в качестве подвижного буфера, и соответствующий буфер применяли также для разведения растворимого мышиного Fc-гамма RII. Для регенерации сенсорного чипа использовали 10 мМ глицин-HCl, рН 1,5. Все измерения проводили при 25°С. Анализы осуществляли на основе связывания (RU), рассчитанного из сенсограмм, полученных с помощью измерений, и представлены относительные величины, для получения которых уровень связывания SG1 принимали за 1,00. Для расчета параметров применяли программное обеспечение Evaluation BIACORE® Т100 (фирма GE Healthcare).
Результаты SPR-анализа обобщены в таблице 26. Установлено, что несколько вариантов Fc обладали повышенной аффинностью в отношении мышиного Fc-гамма RII, фиксированного на сенсорном чипе BIACORE®.
Не ограничиваясь какой-либо конкретной теорией, этой результат можно объяснить следующем образом. Как известно, сенсорный чип BIACORE® является отрицательно заряженным, и можно предполагать, что такое заряженное состояние напоминает состояние поверхности клеточной мембраны. Более конкретно, аффинность комплекса антиген-антитело к мышиному Fc-гамма RII, фиксированному на отрицательно заряженном сенсорном чипе BIACORE®, вероятно, может напоминать связывание комплекса антиген антитело с мышиным Fc-гамма RII, присутствующим на аналогично отрицательно заряженной поверхности клеточной мембраны.
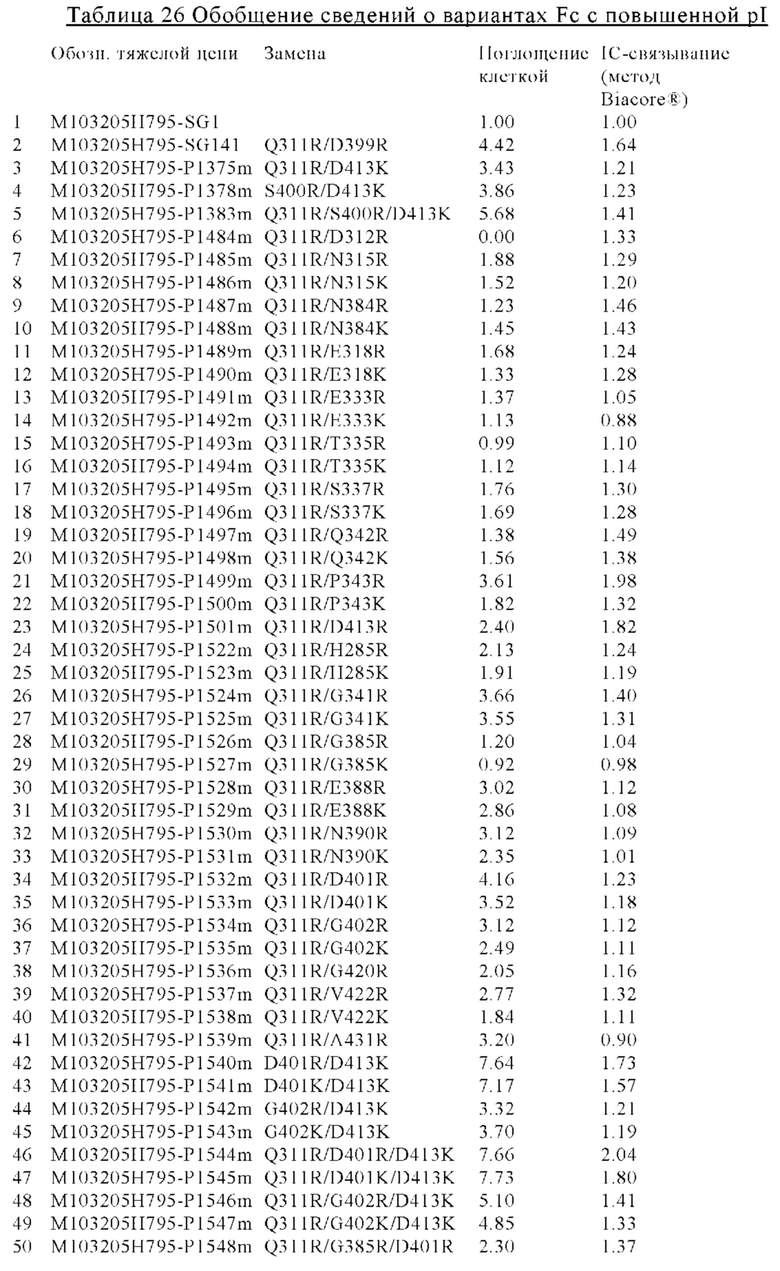
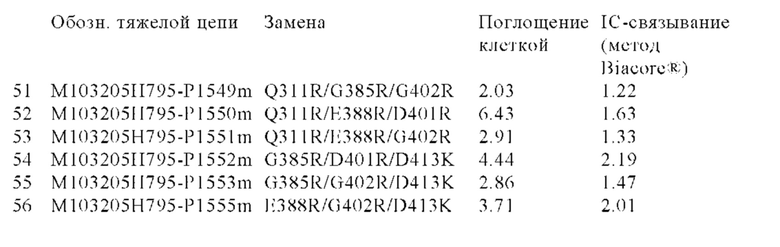
Согласно настоящему описанию антитела, полученные путем интродукции повышающих pI модификаций в Fc-область, представляют собой антитела, в которых заряд Fc-области является в большей степени положительным по сравнению с зарядом до интродукции модификаций. Таким образом, можно предполагать, что кулоновское взаимодействие между Fc-областью (положительный заряд) и поверхностью сенсорного чипа (отрицательный заряд) будет усиливаться при использовании аминокислотных модификаций, повышающих pI. Кроме того, ожидается, что аналогичные явления имеют место на такой же отрицательно заряженной поверхности клеточной мембраны; поэтому можно ожидать также, что они отражают явления увеличения скорости поглощения клеткой in vivo.
Из вариантов Fc с повышенной pI с двумя аминокислотными заменами относительно SG1 для комплекса антиген-антитело, образованного с помощью SG141, P1499m, P1501m и P1540m, обнаружено наиболее высокий уровень связывания с человеческим Fc-гамма RIIb. Подтверждено, что аминокислотные замены Q311R/D399R, Q311R/P343R, Q311R/D413R и D401R/D413K оказывают сильное обусловленное зарядом воздействие на связывание с человеческим Fc-гамма RIIb на сенсорном чипе.
Клеточное поглощение вариантов Fc с повышенной pI
Для оценки скорости внутриклеточного поглощения экспрессирующей человеческий Fc-гамма RIIb клеточной линией осуществляли описанный ниже анализ. Клетки линии MDCK (линия клеток почки собаки Майдин-Дерби), которые конститутивно экспрессируют человеческий Fc-гамма RIIb, получали известными методами. С помощью этих клеток оценивали внутриклеточное поглощение комплексов антиген-антитело.
В частности, pHrodoRed (фирма Life Technlogies) применяли для мечения человеческого латентного миостатина (антиген) согласно известному протоколу, и получали комплексы антиген-антитело в культуральном растворе с концентрацией антитела 10 мг/мл и концентрацией антигена 2,5 мг/мл. Культуральный раствор, содержащий комплексы антиген-антитело, вносили в культуральные планшеты с вышеуказанными MDCK-клетками, которые конститутивно экспрессируют человеческий Fc-гамма RIIb, и инкубировали в течение 1 ч, а затем количественно определяли интенсивность флуоресценции антигена, поглощенного клетками, с помощью анализатора InCell 6000 (фирма GE healthcare). Количество поглощенного антигена выражали в виде относительных величин, при этом величину для SG1 принимали за 1,00.
Количественные результаты клеточного поглощения обобщены в таблице 26.
При использовании некоторых вариантов Fc обнаружена сильная флуоресценция в клетках, присущая антигену.
Не ограничиваясь какой-либо конкретной теорией, этой результат можно объяснить следующем образом.
Антиген и антитела, добавленные в раствор для культивирования клеток, образуют комплексы антиген-антитело в культуральном растворе. Комплексы антиген-антитело связываются с человеческим Fc-гамма RIIb, экспрессированом на клеточной мембране, через Fc-область антитела и поглощаются клетками рецептор-зависимым образом. Антитела, применяемые в этом эксперименте, связываются с антигеном рН-зависимым образом; таким образом, антитело может отделяться в результате диссоциации от антигена в эндосомах (кислые условия рН) внутри клеток. Поскольку диссоциированный антиген, как описано ранее, метили pHrodoRed, то в эндосомах происходит его флуоресценция. Таким образом, сильная интенсивность флуоресценции внутри клетки, вероятно, свидетельствует о том, что поглощение комплексов антиген-антитело клетками происходило с большей скоростью или в больших количествах.
Среди вариантов Fc с повышенной pI с двумя аминокислотными заменами относительно SG1 для комплексов антиген-антитело, образованных из SG141, P1375m, P1378m, P1499m, P1524m, P1525m, P1532m, P1533m, P1540m, P154m и P1543m, обнаружено более сильное поглощение антигена клетками. Подтверждено, что аминокислотные замены Q311R/D399R, Q311R/D413K, S400R/D413K, Q311R/P343R, Q311R/G341R, Q311R/G341K, Q311R/D401R, Q311R/D401K, D401R/D413K, D401K/D413K и G402K/D413K оказывали сильное опосредованное зарядом действие на поглощение клетками комплекса антиген-антитело.
ФК-исследование, проведенное на трансгенных по человеческому FcRn мышах
Тестирование in vivo с использованием трансгенных по человеческому FcRn мышей
Элиминацию миостатина и антитела к латентному миостатину оценивали in vivo после совместного введения антитела к латентному миостатину и латентного миостатина трансгенным по человеческому FcRn мышам. Антитело к латентному миостатину (0,1 мг/мл) и мышиный латентный миостатин (0,05 мг/мл) вводили в одной дозе 10 мл/кг в каудальную вену в эксперименте PK-2 (описан на фиг. 27). Антитело к латентному миостатину (0,3 мг/мл), человеческий латентный миостатин (0,05 мг/мл) и человеческий нормальный иммуноглобулин (фирма CSL Behring AG) (100 мг/мл) вводили в одной дозе 10 мл/кг в каудальную вену в эксперименте PK-4 (описан на фиг. 28). Образцы крови собирали через 5 мин, 15 мин, 1 ч, 4 ч, 7 ч, 1 день, 2 дня, 7 дней, 14 дней, 21 день и 28 дней после введения в эксперименте PK-2. Образцы крови собирали через 5 мин, 1 ч, 4 ч, 7 ч, 1 день, 7 дней, 14 дней, 21 день и 30 дней после введения в эксперименте PK-4. Собранную кровь немедленно центрифугировали при 15000 об/мин при 4°С в течение 5 мин для отделения плазмы. Отделенную плазму хранили при температуре -20°С или ниже до анализа. Антитела к латентному миостатину MS1032LO06-SG1, MS1032LO06-P1375m, MS1032LO06-P1378m, MS1032LO06-P1383m использовали в эксперименте PK-2, а MS1032LO06-P1375m, MS 1032LO06-P1499m использовали в эксперименте PK-4.
Измерение концентрации общего миостатина в плазме с помощью электрохемилюминесценции (ECL)
Концентрацию общего миостатина в плазме мышей измеряли с помощью ECL согласно методу, описанному в примере 28 (Ravetch PK). Данные о зависимости от времени концентрации общего миостатина в плазме после внутривенного введения антитела к латентному миостатину и латентного миостатина, полученные с помощью указанного метода, представлены на фиг. 27А и 28А.
Измерение концентрации антитела к латентному миостатину в плазме с помощью твердофазного иммуноферментного анализа (ELISA)
Концентрацию антитела к латентному миостатину в плазме мышей измеряли с помощью ELISA в эксперименте PK-2. F(ab')2-фрагмент антитела к человеческому IgG (специфический для гамма-цепи) (фирма Sigma), наносили на Nunc-Immuno-планшет MaxiSorp (фирма Nalge Nunc International) и давали выстояться в течение ночи при 4°С для приготовления планшетов с иммобилизованным античеловеческим IgG. Приготавливали образцы для создания калибровочной кривой с концентрацией в плазме 2,5, 1,25, 0,625, 0,313, 0,156, 0,078 и 0,039 мкг/мл и образцы мышиной плазмы, разведенные в 100 или более раз. Затем образцы вносили на иммобилизованные античеловеческим IgG планшеты и давали выстояться в течение 1 ч при комнатной температуре. Затем добавляли конъюгат козий античеловеческий IgG (специфический для гамма-цепи)-биотин (фирма Southern Biotech) и давали прореагировать в течение 1 ч при комнатной температуре. Затем добавляли стрептавидин-Po1yHRP80 (фирма Stereospecific Detection Technologies) и давали прореагировать в течение 1 ч при комнатной температуре и осуществляли хромогенную реакцию с использованием в качестве субстрата ТМВ One Component HRP Microwell (фирма BioFX Laboratories). После прекращения реакции с помощью 1н. серной кислоты (фирма Showa Chemical) измеряли абсорбцию при 450 нм с применением ридера для микропланшетов. Концентрацию в плазме мышей рассчитывали на основе абсорбции калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (фирма Molecular Devices). Данные о зависимости от времени концентрации антитела в плазме после внутривенного введения антитела к латентному миостатину и латентного миостатина, полученные указанным методом, представлены на фиг. 27Б.
В эксперименте PK-4 концентрацию антитела к латентному миостатину в плазме мышей измеряли с помощью метода Gyrolab Bioaffy™ CD (фирма Gyros). Биотинилированное антитело к человеческому IgG-Fc пропускали через колонку со стрептавидиновыми гранулами в микроструктуре CD. Приготавливали образцы для калибровочной кривой с концентрациями в плазме 0,5, 1, 2, 4, 8, 16 и 32 мкг/мл, впрыскивая в плазму в концентрации 5 мг/мл человеческий нормальный иммуноглобулин (фирма CSL Behring AG) и в концентрации 200 мкг/мл мышиный латентный миостатин, и образцы мышиной плазмы разводили в 25 или более раз, впрыскивая в плазму в концентрации 200 мкг/мл мышиный латентный миостатин. Затем образцы добавляли в CD и пропускали через колонку с гранулами. Затем добавляли в CD меченное с помощью Alexa козье поликлональное антитело к человеческому IgG (фирма BETHYL) и пропускали через колонку с гранулами. Концентрацию в мышиной плазме рассчитывали на основе калибровочной кривой, используя программу Gyrolab Evaluator. Данные о зависимости от времени концентрации антитела в плазме после внутривенного введения антитела к латентному миостатину и латентного миостатина, полученные указанным методом, представлены на фиг. 28Б.
Воздействие pI на концентрацию миостатина in vivo
После введения MS1032LO06-SG1 концентрация в плазме общего миостатина через 7 ч снижалась в 12 раз по сравнению с концентрацией в плазме общего миостатина через 5 мин в эксперименте PK-2. В противоположность этому, после введения MS1032LO06-P1375m, MS1032LO06-P1378m и MS1032LO06-P1383m концентрация в плазме общего миостатина через 7 ч снижалась 26-67 раз по сравнению с концентрацией в плазме общего миостатина через 5 мин в эксперименте PK-2. Концентрация антител MS1032LO06-P1378m и MS1032LO06-P1383m снижалась более чем в 3 раза по сравнению с MS1032LO06-SG1, хотя различия в концентрации MS1032LO06-P1375m в каждый тестируемый момент времени по сравнению с MS1032LO06-SG1 оказались примерно 2-кратным в эксперименте PK-2. В присутствии pI-вариантов, включающих аминокислотные замены Q311R/D413K, обнаружена повышенная элиминация миостатина из плазмы без повышения элиминации антитела из плазмы.
Кроме того, определяли концентрацию общего миостатина и концентрацию антитела после введения MS1032LO06-P1499m при совместном введении человеческого нормального иммуноглобулина для имитации человеческой плазмы. После введения MS1032LO06-P1375m и MS1032LO06-P1499m концентрация общего миостатина в плазме через 7 ч снижалась в 3,4 и 5,3 раз по сравнению с концентрацией общего миостатина в плазме через 5 мин, и концентрация общего миостатина в плазме и концентрация антитела в каждый тестируемый момент времени при применении MS1032LO06-P1499m в 1,5 раза превышали установленные для MS1032LO06-P1375m в эксперименте PK-4. В присутствии pI-вариантов, включающих аминокислотные замены Q311R/P343R, обнаружена повышенная элиминация миостатина из плазмы без повышения элиминации антитела из плазмы.
Варианты с высокой pI имели больший положительный заряд в плазме. Поэтому указанный положительный заряд взаимодействовал с отрицательно заряженной клеточной поверхностью, иммунный комплекс антиген-антитело вариантов с высокой pI сильнее контактировал с клеточной поверхностью, что приводило к повышенному поглощению клеткой иммунного комплекса антиген-антитело вариантов с высокой pI.
Пример 31
Замены, приводящие к увеличению pI, в комбинации с вариантом Fc с повышенной активностью в отношении Fc-гамма RIIb
Воздействие на клиренс миостатина варианта Fc с повышенной активностью в отношении Fc-гамма RIIb в комбинации с другими приводящими к увеличению pI заменами оценивали на трансгенных по человеческому Fc-гамма RIIb мышах. Трансгенных по человеческому Fc-гамма RIIb мышей создавали путем микроинъекции ВАС-(бактериальная искусственная хромосома) вектора, содержащего экзоны человеческого гена FCGR2B в пронуклеусе оплодотворенных яиц мышей линии C57BL/6N с помощью стандартных методик (см., например, J. Immunol., 195(7), 1 октября 2015 г., сс. 3198-3205). Мышей с дефицитом мышиного Fc-гамма RIIb создавали, используя нуклеазы с «цинковыми пальцами» (ZFN), которые создавали для воздействия на экзон 1. Мышей с гуманизированным Fc-гамма RIIb создавали путем скрещивания трансгенных по человеческому Fc-гамма RIIb мышей с мышами с «выключенным» (КО) мышиным Fc-гамма RIIb. С помощью этих мышей можно изучать комбинированное воздействие повышенной аффинности к человеческому Fc-гамма RIIb и увеличения pI на клиренс растворимого антигена.
Получение и профиль тестируемых вариантов
Сведения о 7 протестированных антителах и их профилях связывания обобщены в таблице 27. Тяжелую цепь, MS103205H795-MY352, MS103205H795-PK55, MS103205H795-PK56, MS103205H795-PK57, получали путем интродукции приводящих к увеличению pI замен Q311R/D413R, Q311R/P343R, Q311R/P343R/D413R, Q311R/N384R/D413R, соответственно в MS103205H795-MY201. Все варианты MS1032LO06 экспрессировали с использованием M103202L889-SK1 в качестве легкой цепи согласно методу, описанному в примере 30, и определяли их аффинность к человеческим и cynoFc-гамма R с помощью метода, описанного в примере 27.



На основе результатов SPR-анализа, которые обобщены в таблице 27, подтверждено, что замены, приводящие к увеличению pI, не влияли на связывание с Fc-гамма R вариантов Fc с повышенной активностью в отношении Fc-гамма RIIb. У MY201 и MY351, которые получали путем интродукции Q311R/D413K в MY201, обнаружено повышенная в 6 раз и 5 раз способность связываться с человеческим Fc-гамма RIIb соответственно. Они имели практически одинаковый профиль связывания с другими человеческими и cynoFc-гамма R. Аналогично этому, у MY352, PK55, PK56 и PK57 обнаружен профиль связывания, сходный с профилем родительской молекулы MY201 (таблица 27). Эти результаты свидетельствуют о том, что любая пара замен, приводящих к увеличению pI, не влияла на аффинность к человеческим и cynoFc-гамма R.
ФК-исследование, проведенное на трансгенных по человеческому FcRn мышах
Тестирование in vivo с использованием трансгенных по человеческому FcRn мышах
Элиминацию миостатина и антитела к латентному миостатину оценивали in vivo после совместного введения антитела к латентному миостатину и человеческого латентного миостатина трансгенным по человеческому Fc-гамма RIIb мышам, у которых мышиный Fc-гамма RII был заменен человеческим Fc-гамма RIIb. Антитело к латентному миостатину (0,3 мг/мл) и мышиный латентный миостатин (0,05 мг/мл) вводили в одной дозе 10 мл/кг в каудальную вену. Образцы крови собирали через 5 мин, 1 ч, 4 ч, 7 ч, 1 день, 7 дней, 14 дней, 21 день и 28 дней после введения. Собранную кровь немедленно центрифугировали при 15000 об/мин при 4°С в течение 5 мин для отделения плазмы. Отделенную плазму хранили при температуре -20°С или ниже до анализа. Использовали следующие антитела к латентному миостатину MS1032LO06-SG1, MS1032LO06-MY201, MS1032LO06-MY351, MS1032LO06-MY352, MS1032LO06-PK55, MS1032LO06-PK56 и MS1032LO06-PK57.
Измерение концентрации общего миостатина в плазме с помощью электрохемилюминесценции (ECL)
Концентрацию общего миостатина в плазме мышей измеряли с помощью ECL согласно методу, описанному в примере 28. Данные о зависимости от времени концентрации общего миостатина в плазме после внутривенного введения антитела к латентному миостатину и латентного миостатина, полученные с помощью указанного метода, представлены на фиг. 29.
Измерение концентрации антитела к латентному миостатину в плазме с помощью твердофазного иммуноферментного анализа (ELISA)
Концентрацию антитела к латентному миостатину в плазме мышей измеряли с помощью ELISA. F(ab')2-фрагмент антитела к человеческому IgG (специфический для гамма-цепи) (фирма Sigma), наносили на Nunc-Immuno-планшет MaxiSorp (фирма Nalge Nunc International) и давали выстояться в течение ночи при 4°С для получения планшетов с иммобилизованным античеловеческим IgG. Приготавливали образцы для создания калибровочной кривой с концентрацией в плазме 2,5, 1,25, 0,625, 0,313, 0,156, 0,078 и 0,039 мкг/мл и образцы мышиной плазмы, разведенные в 100 или более раз. Затем образцы вносили на иммобилизованные античеловеческим IgG планшеты и давали выстояться в течение 1 ч при комнатной температуре. Затем добавляли конъюгат козий античеловеческий IgG (специфический для гамма-цепи)-биотин (фирма Southern Biotech) и давали прореагировать в течение 1 ч при комнатной температуре. Затем добавляли стрептавидин-PolyHRP80 (фирма Stereospecific Detection Technologies) и давали прореагировать в течение 1 ч при комнатной температуре и осуществляли хромогенную реакцию с использованием в качестве субстрата ТМВ One Component HRP Microwell (фирма BioFX Laboratories). После прекращения реакции с помощью 1н. серной кислоты (фирма Showa Chemical) измеряли абсорбцию при 450 нм с применением ридера для микропланшетов. Концентрацию в плазме мышей рассчитывали на основе абсорбции калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (фирма Molecular Devices). Данные о зависимости от времени концентрации антитела в плазме после внутривенного введения антитела к латентному миостатину и латентного миостатина, полученные указанным методом, представлены на фиг. 30.
Воздействие pI и Fc-гамма R-связывания на концентрацию миостатина in vivo
После введения MS1032LO06-MY201 концентрация в плазме общего миостатина через 7 ч снижалась в 8 раз по сравнению с концентрацией в плазме общего миостатина через 5 мин. В противоположность этому, после введения MS1032LO06-PK57 концентрация в плазме общего миостатина через 7 ч снижалась в 376 раз по сравнению с концентрацией в плазме общего миостатина через 5 мин. Для других вариантов с высокой pI обнаружено сходное повышение элиминации миостатина из плазмы. Концентрация вариантов антител с высокой pI через 28 дней снижалась в 2-3 раза по сравнению с концентрацией антитела MS1032LO06-SG1, и при анализе для pi-вариантов обнаружено небольшое повышение элиминации антител из плазмы.
Варианты с высокой pI имели больший положительный заряд в плазме. Поэтому указанный положительный заряд взаимодействовал с отрицательно заряженной клеточной поверхностью, иммунный комплекс антиген-антитело вариантов с высокой pI сильнее контактировал с клеточной поверхностью, что приводило к повышенному поглощению клеткой иммунного комплекса антиген-антитело вариантов с высокой pI.
Пример 32
Создание вариантов Fc с повышенной активностью в отношении человеческого Fc-гамма RIIb
Конструировали варианты Fc с повышенной активностью в отношении человеческого Fc-гамма RIIb, имеющие вариабельную область антитела к миостатину, и оценивали их аффинность к человеческим Fc-гамма R. В частности, конструировали варианты Fc с повышенной активностью в отношении человеческого Fc-гамма RIIb путем интродукции замен в MS103240H795-SG1 (VH, SEQ ID NO: 92; СН, SEQ ID NO: 9) в сочетании с заменами T250V/T307P. Кроме того, интродуцировали также замены (Q311R/D413K или Q311R/P343R). Варианты экспрессировали, используя M103202L1045-SK1 в качестве легкой цепи, и их аффинность к человеческим Fc-гамма R анализировали согласно методу, описанному в примере 34. В таблице 28 «Профиль связывания вариантов Fc с повышенной активностью в отношении человеческого Fc-гамма RIIb» (см. Приложение) представлены результаты кинетических анализов в отношении человеческих и cynoFc-гамма R. Величины KD, которые указаны в клетках таблицы, выделенных жирным шрифтом, рассчитывали с помощью уравнения 2, поскольку их аффинность была слишком низкой для точного ее определения с помощью кинетического анализа.
У всех изученных вариантов обнаружена повышенная аффинность к человеческому Fc-гамма RIIb и пониженная способность связываться с человеческими Fc-гамма RIIaR, Fc-гамма RIIaH, Fc-гамма RIIIaV по сравнению с SG1. Аффинность к человеческому Fc-гамма RIIb варьировалась от 4,1-кратного до 30,5-кратного уровня по сравнению с SG1. Аффинность к человеческому Fc-гамма RIIaR находилась между 0,02-кратным и 0,22-кратным уровнем. Аффинность к человеческому Fc-гамма RIIaH и человеческому Fc-гамма RIIIaV оказалась менее чем 0,04-кратной и 0,012-кратной соответственно. Установлено, что замены (T250V/T307P) не влияли на профиль связывания с человеческими Fc-гамма R по сравнению с аффинностью MY009 (P238D) и MY214 (P238D/T250V/T307P). С другой стороны, обе пары приводящих к увеличению pI замен (Q311R/D413K и Q311R/P343R) снижали аффинность связывания примерно в 0,5 раза по сравнению с вариантами, которые не содержали приводящие к увеличению pI замены. Например, хотя для ТТ14 обнаружена в 30,5 раз более высокая аффинность к человеческому Fc-гамма RIIb, установлено лишь 16,1-кратное и 145-кратное повышение при использовании в комбинации с Q311R/D413K (ТТ33) и Q311R/P343R (ТТ32) соответственно.
Кроме того, замены, применение которых описано в примере 29, которые удлиняли время полужизни антитела и снижали связывание с ревматоидным фактором, интродуцировали в варианты Fc с повышенной активностью в отношении человеческого Fc-гамма RIIb и анализировали их аффинность к человеческим Fc-гамма R (таблица 29). В указанных SPR-анализах все данные получали с использованием мышиного античеловеческого IgG с легкой каппа-цепью для «захвата» представляющих интерес антител. В таблице 30 «Профиль связывания вариантов Fc с повышенной активностью в отношении человеческого Fc-гамма RIIb» (см. Приложение) представлены результаты и при этом, величины KD в клетках, обозначенных жирным шрифтом, рассчитывали с помощью уравнения 2, поскольку их аффинность была слишком низкой для точного ее определения с помощью кинетического анализа.

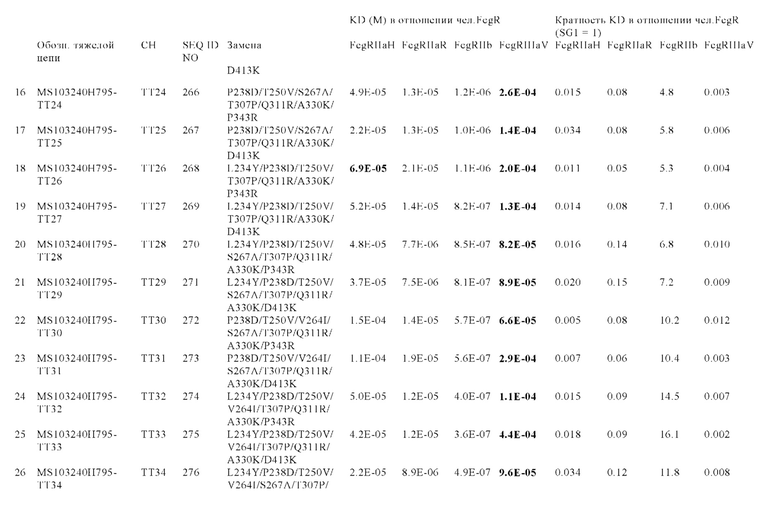

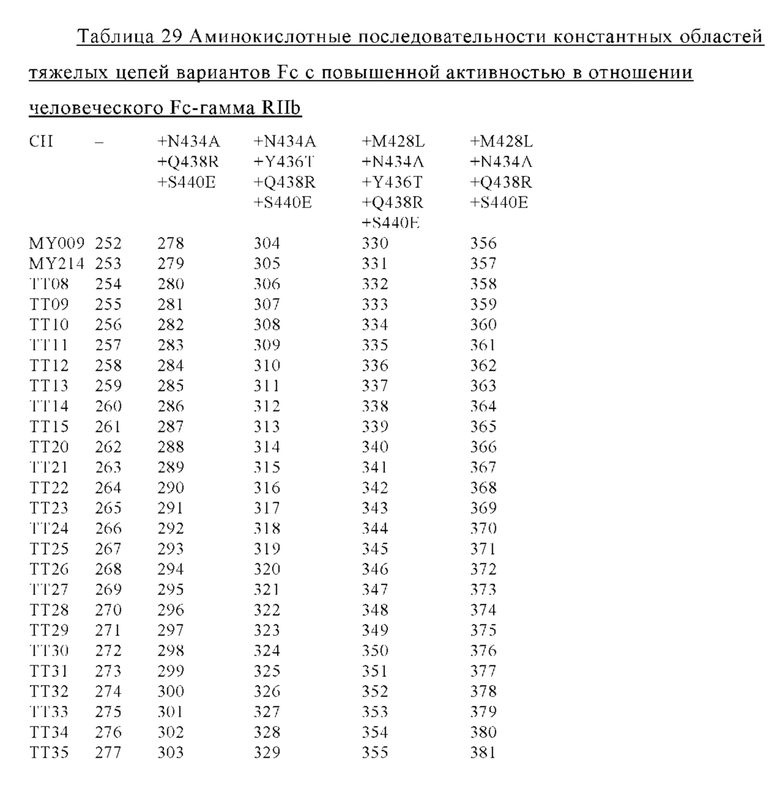
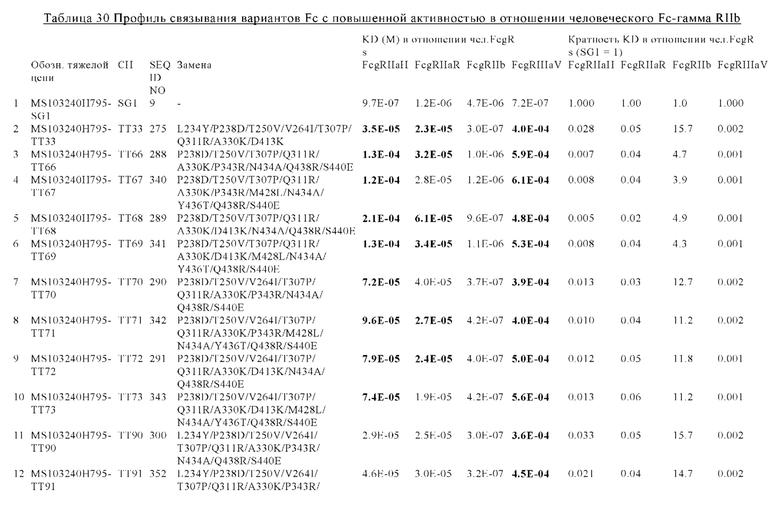

В этом опыте вновь оценивали SG1 и ТТ33, при этом метод «захвата» отличался от того, результаты которого представлены в таблице 28. В результате установлено, что величины «кратности KD» для ТТ33 соответствовали полученным при использовании метода, результаты которого приведены в таблице 28 (16,1-кратный уровень в таблице 28 и 15,7-кратный уровень в таблице 30, характеризующий связывание с человеческим Fc-гамма RIIb). Для ТТ92 и ТТ93, которые конструировали путем интродукции N434A/Q438R/S440E и M428L/N434A/Y436T/Q438R/S440E в ТТ33, обнаружен сопоставимый с ТТ33 профиль связывания. Аналогично этому, для ТТ90 и ТТ91, которые получали из ТТ32, ТТ72 и ТТ73, которые получали из ТТ31, ТТ70 и ТТ71, которые получали из ТТ30, ТТ68 и ТТ69, которые получали из ТТ21, и ТТ66 и ТТ67, которые получали из ТТ20, обнаружен профиль связывания, сопоставимый с родительскими антителами, приведенным в таблице 28. Эти результаты свидетельствуют о том, что замены, предназначенные для повышения времени полужизни антитела и снижения связывания с ревматоидным фактором, не влияют на профиль связывания с человеческими Fc-гамма R вариантов Fc с повышенной активностью в отношении человеческого Fc-гамма RIIb.
Пример 33
Анализ на основе визуализации клеток вариантов Fc с повышенной активностью в отношении человеческого Fc-гамма RIIb
Для изучения внутриклеточного поглощения комплекса антиген-антитело, образованного с антителами, указанными в примере 32, осуществляли анализ с применением визуализации клеток, описанный в примере 30. Во избежание насыщения сигнала в этом примере комплексы антиген-антитело получали в культуральном растворе, в котором концентрация антитела составляла 1,25 мг/мл и концентрация антигена составляла 0,34 мг/мл.
Результаты анализа представлены на фиг. 31. При повышении аффинности связывания с человеческим Fc-гамма RIIb поглощение клетками комплекса антиген-антитело возрастало. В случае ТТ14, отличающегося повышенной в 30 раз аффинностью связывания с человеческим Fc-гамма RIIb, по сравнению с SG1, интенсивность флуоресценции антигена внутри клеток в 7,5 раза превышала интенсивность флуоресценции SG1.
Хотя приводящие к увеличению pI замены (Q311R/D413K и Q311R/P343R) снижали аффинность связывания с человеческим Fc-гамма RIIb примерно в 0,5 раза по сравнению с вариантами, которые не содержали приводящие к увеличению pI замены, поглощение клетками комплекса антиген-антитело с вариантами Fc, содержащими замены, приводящие к увеличению pI, возрастало по сравнению с вариантами Fc без увеличивающих pI замен. Для ТТ33 обнаружено 36-кратное повышение поглощения клетками комплекса антиген-антитело по сравнению с SG1, в то время как для родительской молекулы ТТ14 обнаружено 7,5-кратное повышение поглощения клетками. Кроме того, для ТТ33 обнаружено поглощение клетками комплекса антиген-антитело, сопоставимое с поглощением клетками вариантов SG1071, SG1074, SG1077, SG1079, SG1080 и SG1081, для которых обнаружен сильный sweeping-эффект при изучении на обезьянах циномолгус. Эти результаты позволяют предположить, что объединение приводящих к увеличению pI замен с вариантами Fc с повышенной активностью в отношении Fc-гамма RIIb является эффективным инструментом для элиминации антигенов.
Пример 34
Конструирование экспрессионных векторов для антител и экспрессия, и очистка антител
Синтез полноразмерных генов, кодирующих нуклеотидные последовательности Н-цепи и L-цепи вариабельных областей антител, осуществляли с использованием методов получения, известных специалистам в данной области, с использованием применяемой для сборки ПЦР и т.п. Интродукцию аминокислотных замен осуществляли согласно методам, известным специалистам в данной области, с использованием ПЦР или т.п. Полученные фрагменты плазмид встраивали в экспрессионный вектор для клеток животных, и получали экспрессионный вектор для Н-цепи и экспрессионный вектор для L-цепи. Нуклеотидную последовательность полученного экспрессионного вектора определяли с помощью методов, известных специалистам в данной области. Полученные плазмиды кратковременно интродуцировали в клеточную линию HEK293H, полученную из клеток рака почки человеческого эмбриона (фирма Invitrogen), или клетки FreeStyle293 (фирма Invitrogen) для экспрессии антител. Полученный супернатант культуры собирали и пропускали через фильтр с размером пор 0,22 мкм MILLEX®-GV (фирма Millipore) или через фильтр с размером пор 0,45 мкм MILLEX®-GV (фирма Millipore) с получением супернатанта культуры. Антитела очищали из полученного супернатанта культуры с помощью методов, известных специалистам в данной области, применяя r(рекомбинантный) белок А-сефароза Fast Flow (фирма GE Healthcare) или белок G-сефароза 4 Fast Flow (фирма GE Healthcare). Для определения концентрации очищенных антител измеряли их абсорбцию при 280 нм с помощью спектрофотометра. Из полученных значений концентрацию антитела определяли, используя коэффициент экстинкции, рассчитанный с помощью таких методов, как РАСЕ (Protein Science 4, 1995, сс. 2411-2423).
Хотя выше изобретение описано для лучшего понимания с указанием некоторых деталей с целью иллюстрации и примеров, описание и примеры не должны рассматриваться в качестве ограничивающих объем изобретения. Описание всей патентной и научной литературы, процитированной в настоящем описании, специально полностью включено в настоящее описание в качестве ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЧУГАИ СЕЙЯКУ КАБУСИКИ КАЙСЯ
<120> АНТИТЕЛА К МИОСТАТИНУ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
<130> C1-A1607P
<150> JP 2016-120337
<151> 2016-06-17
<160> 381
<170> PatentIn version 3.5
<210> 1
<211> 375
<212> PRT
<213> Homo sapiens
<400> 1
Met Gln Lys Leu Gln Leu Cys Val Tyr Ile Tyr Leu Phe Met Leu Ile
1 5 10 15
Val Ala Gly Pro Val Asp Leu Asn Glu Asn Ser Glu Gln Lys Glu Asn
20 25 30
Val Glu Lys Glu Gly Leu Cys Asn Ala Cys Thr Trp Arg Gln Asn Thr
35 40 45
Lys Ser Ser Arg Ile Glu Ala Ile Lys Ile Gln Ile Leu Ser Lys Leu
50 55 60
Arg Leu Glu Thr Ala Pro Asn Ile Ser Lys Asp Val Ile Arg Gln Leu
65 70 75 80
Leu Pro Lys Ala Pro Pro Leu Arg Glu Leu Ile Asp Gln Tyr Asp Val
85 90 95
Gln Arg Asp Asp Ser Ser Asp Gly Ser Leu Glu Asp Asp Asp Tyr His
100 105 110
Ala Thr Thr Glu Thr Ile Ile Thr Met Pro Thr Glu Ser Asp Phe Leu
115 120 125
Met Gln Val Asp Gly Lys Pro Lys Cys Cys Phe Phe Lys Phe Ser Ser
130 135 140
Lys Ile Gln Tyr Asn Lys Val Val Lys Ala Gln Leu Trp Ile Tyr Leu
145 150 155 160
Arg Pro Val Glu Thr Pro Thr Thr Val Phe Val Gln Ile Leu Arg Leu
165 170 175
Ile Lys Pro Met Lys Asp Gly Thr Arg Tyr Thr Gly Ile Arg Ser Leu
180 185 190
Lys Leu Asp Met Asn Pro Gly Thr Gly Ile Trp Gln Ser Ile Asp Val
195 200 205
Lys Thr Val Leu Gln Asn Trp Leu Lys Gln Pro Glu Ser Asn Leu Gly
210 215 220
Ile Glu Ile Lys Ala Leu Asp Glu Asn Gly His Asp Leu Ala Val Thr
225 230 235 240
Phe Pro Gly Pro Gly Glu Asp Gly Leu Asn Pro Phe Leu Glu Val Lys
245 250 255
Val Thr Asp Thr Pro Lys Arg Ser Arg Arg Asp Phe Gly Leu Asp Cys
260 265 270
Asp Glu His Ser Thr Glu Ser Arg Cys Cys Arg Tyr Pro Leu Thr Val
275 280 285
Asp Phe Glu Ala Phe Gly Trp Asp Trp Ile Ile Ala Pro Lys Arg Tyr
290 295 300
Lys Ala Asn Tyr Cys Ser Gly Glu Cys Glu Phe Val Phe Leu Gln Lys
305 310 315 320
Tyr Pro His Thr His Leu Val His Gln Ala Asn Pro Arg Gly Ser Ala
325 330 335
Gly Pro Cys Cys Thr Pro Thr Lys Met Ser Pro Ile Asn Met Leu Tyr
340 345 350
Phe Asn Gly Lys Glu Gln Ile Ile Tyr Gly Lys Ile Pro Ala Met Val
355 360 365
Val Asp Arg Cys Gly Cys Ser
370 375
<210> 2
<211> 109
<212> PRT
<213> Homo sapiens
<400> 2
Asp Phe Gly Leu Asp Cys Asp Glu His Ser Thr Glu Ser Arg Cys Cys
1 5 10 15
Arg Tyr Pro Leu Thr Val Asp Phe Glu Ala Phe Gly Trp Asp Trp Ile
20 25 30
Ile Ala Pro Lys Arg Tyr Lys Ala Asn Tyr Cys Ser Gly Glu Cys Glu
35 40 45
Phe Val Phe Leu Gln Lys Tyr Pro His Thr His Leu Val His Gln Ala
50 55 60
Asn Pro Arg Gly Ser Ala Gly Pro Cys Cys Thr Pro Thr Lys Met Ser
65 70 75 80
Pro Ile Asn Met Leu Tyr Phe Asn Gly Lys Glu Gln Ile Ile Tyr Gly
85 90 95
Lys Ile Pro Ala Met Val Val Asp Arg Cys Gly Cys Ser
100 105
<210> 3
<211> 375
<212> PRT
<213> Macaca fascicularis
<400> 3
Met Gln Lys Leu Gln Leu Cys Val Tyr Ile Tyr Leu Phe Met Leu Ile
1 5 10 15
Val Ala Gly Pro Val Asp Leu Asn Glu Asn Ser Glu Gln Lys Glu Asn
20 25 30
Val Glu Lys Glu Gly Leu Cys Asn Ala Cys Thr Trp Arg Gln Asn Thr
35 40 45
Lys Ser Ser Arg Ile Glu Ala Ile Lys Ile Gln Ile Leu Ser Lys Leu
50 55 60
Arg Leu Glu Thr Ala Pro Asn Ile Ser Lys Asp Ala Ile Arg Gln Leu
65 70 75 80
Leu Pro Lys Ala Pro Pro Leu Arg Glu Leu Ile Asp Gln Tyr Asp Val
85 90 95
Gln Arg Asp Asp Ser Ser Asp Gly Ser Leu Glu Asp Asp Asp Tyr His
100 105 110
Ala Thr Thr Glu Thr Ile Ile Thr Met Pro Thr Glu Ser Asp Phe Leu
115 120 125
Met Gln Val Asp Gly Lys Pro Lys Cys Cys Phe Phe Lys Phe Ser Ser
130 135 140
Lys Ile Gln Tyr Asn Lys Val Val Lys Ala Gln Leu Trp Ile Tyr Leu
145 150 155 160
Arg Pro Val Glu Thr Pro Thr Thr Val Phe Val Gln Ile Leu Arg Leu
165 170 175
Ile Lys Pro Met Lys Asp Gly Thr Arg Tyr Thr Gly Ile Arg Ser Leu
180 185 190
Lys Leu Asp Met Asn Pro Gly Thr Gly Ile Trp Gln Ser Ile Asp Val
195 200 205
Lys Thr Val Leu Gln Asn Trp Leu Lys Gln Pro Glu Ser Asn Leu Gly
210 215 220
Ile Glu Ile Lys Ala Leu Asp Glu Asn Gly His Asp Leu Ala Val Thr
225 230 235 240
Phe Pro Gly Pro Gly Glu Asp Gly Leu Asn Pro Phe Leu Glu Val Lys
245 250 255
Val Thr Asp Thr Pro Lys Arg Ser Arg Arg Asp Phe Gly Leu Asp Cys
260 265 270
Asp Glu His Ser Thr Glu Ser Arg Cys Cys Arg Tyr Pro Leu Thr Val
275 280 285
Asp Phe Glu Ala Phe Gly Trp Asp Trp Ile Ile Ala Pro Lys Arg Tyr
290 295 300
Lys Ala Asn Tyr Cys Ser Gly Glu Cys Glu Phe Val Phe Leu Gln Lys
305 310 315 320
Tyr Pro His Thr His Leu Val His Gln Ala Asn Pro Arg Gly Ser Ala
325 330 335
Gly Pro Cys Cys Thr Pro Thr Lys Met Ser Pro Ile Asn Met Leu Tyr
340 345 350
Phe Asn Gly Lys Glu Gln Ile Ile Tyr Gly Lys Ile Pro Ala Met Val
355 360 365
Val Asp Arg Cys Gly Cys Ser
370 375
<210> 4
<211> 109
<212> PRT
<213> Macaca fascicularis
<400> 4
Asp Phe Gly Leu Asp Cys Asp Glu His Ser Thr Glu Ser Arg Cys Cys
1 5 10 15
Arg Tyr Pro Leu Thr Val Asp Phe Glu Ala Phe Gly Trp Asp Trp Ile
20 25 30
Ile Ala Pro Lys Arg Tyr Lys Ala Asn Tyr Cys Ser Gly Glu Cys Glu
35 40 45
Phe Val Phe Leu Gln Lys Tyr Pro His Thr His Leu Val His Gln Ala
50 55 60
Asn Pro Arg Gly Ser Ala Gly Pro Cys Cys Thr Pro Thr Lys Met Ser
65 70 75 80
Pro Ile Asn Met Leu Tyr Phe Asn Gly Lys Glu Gln Ile Ile Tyr Gly
85 90 95
Lys Ile Pro Ala Met Val Val Asp Arg Cys Gly Cys Ser
100 105
<210> 5
<211> 376
<212> PRT
<213> Mus musculus
<400> 5
Met Met Gln Lys Leu Gln Met Tyr Val Tyr Ile Tyr Leu Phe Met Leu
1 5 10 15
Ile Ala Ala Gly Pro Val Asp Leu Asn Glu Gly Ser Glu Arg Glu Glu
20 25 30
Asn Val Glu Lys Glu Gly Leu Cys Asn Ala Cys Ala Trp Arg Gln Asn
35 40 45
Thr Arg Tyr Ser Arg Ile Glu Ala Ile Lys Ile Gln Ile Leu Ser Lys
50 55 60
Leu Arg Leu Glu Thr Ala Pro Asn Ile Ser Lys Asp Ala Ile Arg Gln
65 70 75 80
Leu Leu Pro Arg Ala Pro Pro Leu Arg Glu Leu Ile Asp Gln Tyr Asp
85 90 95
Val Gln Arg Asp Asp Ser Ser Asp Gly Ser Leu Glu Asp Asp Asp Tyr
100 105 110
His Ala Thr Thr Glu Thr Ile Ile Thr Met Pro Thr Glu Ser Asp Phe
115 120 125
Leu Met Gln Ala Asp Gly Lys Pro Lys Cys Cys Phe Phe Lys Phe Ser
130 135 140
Ser Lys Ile Gln Tyr Asn Lys Val Val Lys Ala Gln Leu Trp Ile Tyr
145 150 155 160
Leu Arg Pro Val Lys Thr Pro Thr Thr Val Phe Val Gln Ile Leu Arg
165 170 175
Leu Ile Lys Pro Met Lys Asp Gly Thr Arg Tyr Thr Gly Ile Arg Ser
180 185 190
Leu Lys Leu Asp Met Ser Pro Gly Thr Gly Ile Trp Gln Ser Ile Asp
195 200 205
Val Lys Thr Val Leu Gln Asn Trp Leu Lys Gln Pro Glu Ser Asn Leu
210 215 220
Gly Ile Glu Ile Lys Ala Leu Asp Glu Asn Gly His Asp Leu Ala Val
225 230 235 240
Thr Phe Pro Gly Pro Gly Glu Asp Gly Leu Asn Pro Phe Leu Glu Val
245 250 255
Lys Val Thr Asp Thr Pro Lys Arg Ser Arg Arg Asp Phe Gly Leu Asp
260 265 270
Cys Asp Glu His Ser Thr Glu Ser Arg Cys Cys Arg Tyr Pro Leu Thr
275 280 285
Val Asp Phe Glu Ala Phe Gly Trp Asp Trp Ile Ile Ala Pro Lys Arg
290 295 300
Tyr Lys Ala Asn Tyr Cys Ser Gly Glu Cys Glu Phe Val Phe Leu Gln
305 310 315 320
Lys Tyr Pro His Thr His Leu Val His Gln Ala Asn Pro Arg Gly Ser
325 330 335
Ala Gly Pro Cys Cys Thr Pro Thr Lys Met Ser Pro Ile Asn Met Leu
340 345 350
Tyr Phe Asn Gly Lys Glu Gln Ile Ile Tyr Gly Lys Ile Pro Ala Met
355 360 365
Val Val Asp Arg Cys Gly Cys Ser
370 375
<210> 6
<211> 109
<212> PRT
<213> Mus musculus
<400> 6
Asp Phe Gly Leu Asp Cys Asp Glu His Ser Thr Glu Ser Arg Cys Cys
1 5 10 15
Arg Tyr Pro Leu Thr Val Asp Phe Glu Ala Phe Gly Trp Asp Trp Ile
20 25 30
Ile Ala Pro Lys Arg Tyr Lys Ala Asn Tyr Cys Ser Gly Glu Cys Glu
35 40 45
Phe Val Phe Leu Gln Lys Tyr Pro His Thr His Leu Val His Gln Ala
50 55 60
Asn Pro Arg Gly Ser Ala Gly Pro Cys Cys Thr Pro Thr Lys Met Ser
65 70 75 80
Pro Ile Asn Met Leu Tyr Phe Asn Gly Lys Glu Gln Ile Ile Tyr Gly
85 90 95
Lys Ile Pro Ala Met Val Val Asp Arg Cys Gly Cys Ser
100 105
<210> 7
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 7
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 8
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 8
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Cys
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 9
<211> 328
<212> PRT
<223> An artificially synthesized sequence
<400> 9
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 10
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 10
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 11
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 11
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Arg Gly Gly Pro Lys Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 12
<211> 116
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 12
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Gly Ser
1 5 10 15
Leu Thr Val Thr Cys Arg Val Ser Gly Ile Asp Leu Ser Ser Tyr Asp
20 25 30
Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Met Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly Val
85 90 95
Pro Ala Tyr Ser Thr Gly Gly Asp Leu Trp Gly Pro Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 13
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 13
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
15
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser Thr Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 14
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 14
Ala Gln Val Leu Thr Gln Thr Ala Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ser Ser Gln Ser Val Tyr Asp Asn
20 25 30
Asn Trp Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Leu
65 70 75 80
Glu Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Leu Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 15
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 15
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr Asp Asn
20 25 30
Asn Trp Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Leu Tyr Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 16
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 16
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser Thr Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 17
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 17
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser His Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser Thr Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 18
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 18
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala His Ser Thr Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 19
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 19
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 20
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 20
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser His Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser Thr Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 21
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 21
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser His Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala His Ser Thr Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 22
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 22
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala His Ser Thr Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 23
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 23
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser His Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala His Ser Thr Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 24
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 24
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 25
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 25
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser His Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 26
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 26
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala His Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 27
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 27
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser His Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 28
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 28
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
15 35 40 45
Gly Ile Ile Ser His Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala His Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 29
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 29
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala His Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 30
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 30
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser His Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala His Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 31
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 31
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr His Asn
20 25 30
Asn Trp Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Leu Tyr Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 32
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 32
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Thr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 33
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 33
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Thr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Met Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 34
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 34
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Tyr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 35
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 35
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Thr Ser Ser Gln Ser Val Tyr His Asn
20 25 30
Asn Trp Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Tyr Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Arg Tyr Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 36
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 36
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Ser Ser Gln Ser Val Tyr His Asn
20 25 30
Asn Trp Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Phe Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Leu Tyr Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 37
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 37
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gln Thr Ser Gln Ser Val Tyr His Asp
20 25 30
Asn Trp Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Arg Tyr Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 38
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 38
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Thr Ser Ser Gln Ser Val Tyr His Asp
20 25 30
Asn Trp Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Tyr Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Leu Tyr Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 39
<211> 348
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 39
cagtcgctgg aggagtccgg gggtcgcctg gtaacgcctg gaggatccct gacagtcacc 60
tgcagagtct ctggaatcga cctcagtagt tacgacatca gctgggtccg ccaggctcca 120
gggaaggggc tggagtggat cggaatcatt agttatgctg gtagcacata ctacgcgagc 180
tgggcaaaag gccgattcac catctccaaa acctcgacca cggtggatct gaaaatgacc 240
agtctgacaa ccgaggacac ggccacctat ttctgtgcca gaggtgtgcc tgcttatagt 300
actggtggtg acttgtgggg cccaggcacc ctggtcaccg tctcctcc 348
<210> 40
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 40
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcacag tgagtgggat cgacctgagc agctacgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtcggcatc attagctacg ccgggagcac atactatgcc 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactcta ccggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 41
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 41
gcgcaagtgc tgacccagac tgcatcatcc gtgtctgcag ctgtgggagg cacagtcacc 60
atcaattgcc agtccagtca gagtgtttat gataacaact ggttatcctg gtttcagcag 120
aaaccagggc agcctcccaa gctcctgatc tattgggcat ccactctggc atctggggtc 180
ccatcgcggt tcaaaggcag tggatctggg acacagttca ctctcaccat cagcgacctg 240
gagtgtgacg atgctgccac ttactactgt gcaggcggtt atggtggtgg tctgtatgct 300
ttcggcggag ggaccaaggt ggagatcaaa 330
<210> 42
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 42
gacattgtga tgactcagtc tcctgccact ctgtcactga gccctggcga gcgagcaacc 60
ctgtcttgcc agagcagcca gtccgtgtac gacaacaatt ggctgagctg gttccagcag 120
aagccagggc agccccctaa actgctgatc tattgggcat caacactggc cagcggcgtc 180
ccctccagat tctccggatc tggcagtggg actgacttta ccctgaccat cagcagcctc 240
cagcccgagg atgccgctac ctactattgt gctggcgggt acggaggcgg gctgtatgca 300
tttggacagg gcacaaaggt ggaaatcaaa 330
<210> 43
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 43
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcacag tgagtgggat cgacctgagc cacaccgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtcggcatc attagctacg ccgggagcac atactatgcc 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 44
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 44
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcacag tgagtgggat cgacctgagc cacaccgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtcggcatc attagctacg ccgggagcac atactatatg 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 45
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 45
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcacag tgagtgggat cgacctgagc cactacgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtcggcatc attagctacg ccgggagcac atactatgcc 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 46
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 46
gacattgtga tgactcagtc tcctgccact ctgtcactga gccctggcga gcgagcaacc 60
ctgtcttgca ccagcagcca gtccgtgtac cacaacaatt ggctgagctg gttccagcag 120
aagccagggc agccccctaa actgctgatc tattgggcat caacactggc ctacggcgtc 180
ccctccagat tctccggatc tggcagtggg actgacttta ccctgaccat cagcagcctc 240
cagcccgagg atgccgctac ctactattgt gctggcgggt acggaggcgg gcggtatgca 300
tttggacagg gcacaaaggt ggaaatcaaa 330
<210> 47
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 47
gacattgtga tgactcagtc tcctgccact ctgtcactga gccctggcga gcgagcaacc 60
ctgtcttgcc agagcagcca gtccgtgtac cacaacaatt ggctgagctg gttccagcag 120
aagccagggc agccccctaa actgctgatc tattgggcat caacactggc cttcggcgtc 180
ccctccagat tctccggatc tggcagtggg actgacttta ccctgaccat cagcagcctc 240
cagcccgagg atgccgctac ctactattgt gctggcgggt acggaggcgg gctgtatgca 300
tttggacagg gcacaaaggt ggaaatcaaa 330
<210> 48
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 48
gacattgtga tgactcagtc tcctgccact ctgtcactga gccctggcga gcgagcaacc 60
ctgtcttgcc agaccagcca gtccgtgtac cacgacaatt ggctgagctg gttccagcag 120
aagccagggc agccccctaa actgctgatc tattgggcat caacactggc cagcggcgtc 180
ccctccagat tctccggatc tggcagtggg actgacttta ccctgaccat cagcagcctc 240
cagcccgagg atgccgctac ctactattgt gctggcgggt acggaggcgg gcggtatgca 300
tttggacagg gcacaaaggt ggaaatcaaa 330
<210> 49
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 49
gacattgtga tgactcagtc tcctgccact ctgtcactga gccctggcga gcgagcaacc 60
ctgtcttgca ccagcagcca gtccgtgtac cacgacaatt ggctgagctg gttccagcag 120
aagccagggc agccccctaa actgctgatc tattgggcat caacactggc ctacggcgtc 180
ccctccagat tctccggatc tggcagtggg actgacttta ccctgaccat cagcagcctc 240
cagcccgagg atgccgctac ctactattgt gctggcgggt acggaggcgg gctgtatgca 300
tttggacagg gcacaaaggt ggaaatcaaa 330
<210> 50
<211> 987
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 50
gcttccacca agggcccatc ggtcttcccc ctggcaccct cctccaagtc cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac ctccggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctctcctc cgtggtgacc gtgccctcct cgtccttggg cacccagacc 240
tacatctgca acgtgaatca caagccctcc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgtcc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
tccacgtacc gtgtggtctc cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggaa 720
atgaccaaga accaggtctc cctgacctgc ctggtcaaag gcttctatcc ctccgacatc 780
gccgtggagt gggagtccaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac tccaagctca ccgtggacaa gtccaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagtcgc tctccctgtc tccgtag 987
<210> 51
<211> 984
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 51
gctagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact ccggggggga 360
ccgaaagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc tccg 984
<210> 52
<211> 987
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 52
gcttccacca agggcccatc ggtcttcccc ctggcaccct cctccaagtc gacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagtggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagtag tgtggtgacc gtgccctcca gtagtttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagt aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgagct cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagt cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agtacgtacc gtgtggtcag tgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag tctgacctgc ctggtcaaag gcttctatcc cagtgacatc 780
gccgtggagt gggagagtaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctac agtaagctca ccgtggacaa gagtaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagtc tctccctgtc tccgtga 987
<210> 53
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 53
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
tccaaggact gcacctactc cctctcctcc accctgacgc tgtccaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgtcctcgcc cgtcacaaag 300
tccttcaaca ggggagagtg ttga 324
<210> 54
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 54
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggtaccgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agtaaggaca gtacctacag tctcagtagt accctgacgc tgtccaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgtcctcgcc cgtcacaaag 300
tccttcaaca ggggagagtg ttga 324
<210> 55
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 55
Ser Tyr Asp Ile Ser
1 5
<210> 56
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 56
His Tyr Asp Ile Ser
1 5
<210> 57
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> An artificially synthesized sequence
<400> 57
His Thr Asp Ile Ser
1 5
<210> 58
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 58
Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 59
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 59
Ile Ile Ser His Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 60
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 60
Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Met Ser Trp Ala Lys Gly
1 5 10 15
<210> 61
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 61
Gly Val Pro Ala Tyr Ser Thr Gly Gly Asp Leu
1 5 10
<210> 62
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 62
Gly Val Pro Ala His Ser Thr Gly Gly Asp Leu
1 5 10
<210> 63
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 63
Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu
1 5 10
<210> 64
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 64
Gly Val Pro Ala His Ser His Gly Gly Asp Leu
1 5 10
<210> 65
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 65
Gln Ser Ser Gln Ser Val Tyr Asp Asn Asn Trp Leu Ser
1 5 10
<210> 66
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 66
Gln Ser Ser Gln Ser Val Tyr His Asn Asn Trp Leu Ser
1 5 10
<210> 67
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 67
Thr Ser Ser Gln Ser Val Tyr His Asn Asn Trp Leu Ser
1 5 10
<210> 68
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 68
Gln Thr Ser Gln Ser Val Tyr His Asp Asn Trp Leu Ser
1 5 10
<210> 69
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 69
Thr Ser Ser Gln Ser Val Tyr His Asp Asn Trp Leu Ser
1 5 10
<210> 70
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 70
Trp Ala Ser Thr Leu Ala Ser
1 5
<210> 71
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 71
Trp Ala Ser Thr Leu Ala Tyr
1 5
<210> 72
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 72
Trp Ala Ser Thr Leu Ala Phe
1 5
<210> 73
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 73
Ala Gly Gly Tyr Gly Gly Gly Leu Tyr Ala
1 5 10
<210> 74
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 74
Ala Gly Gly Tyr Gly Gly Gly Arg Tyr Ala
1 5 10
<210> 75
<211> 242
<212> PRT
<213> Homo sapiens
<400> 75
Glu Asn Ser Glu Gln Lys Glu Asn Val Glu Lys Glu Gly Leu Cys Asn
1 5 10 15
Ala Cys Thr Trp Arg Gln Asn Thr Lys Ser Ser Arg Ile Glu Ala Ile
20 25 30
Lys Ile Gln Ile Leu Ser Lys Leu Arg Leu Glu Thr Ala Pro Asn Ile
35 40 45
Ser Lys Asp Val Ile Arg Gln Leu Leu Pro Lys Ala Pro Pro Leu Arg
50 55 60
Glu Leu Ile Asp Gln Tyr Asp Val Gln Arg Asp Asp Ser Ser Asp Gly
65 70 75 80
Ser Leu Glu Asp Asp Asp Tyr His Ala Thr Thr Glu Thr Ile Ile Thr
85 90 95
Met Pro Thr Glu Ser Asp Phe Leu Met Gln Val Asp Gly Lys Pro Lys
100 105 110
Cys Cys Phe Phe Lys Phe Ser Ser Lys Ile Gln Tyr Asn Lys Val Val
115 120 125
Lys Ala Gln Leu Trp Ile Tyr Leu Arg Pro Val Glu Thr Pro Thr Thr
130 135 140
Val Phe Val Gln Ile Leu Arg Leu Ile Lys Pro Met Lys Asp Gly Thr
145 150 155 160
Arg Tyr Thr Gly Ile Arg Ser Leu Lys Leu Asp Met Asn Pro Gly Thr
165 170 175
Gly Ile Trp Gln Ser Ile Asp Val Lys Thr Val Leu Gln Asn Trp Leu
180 185 190
Lys Gln Pro Glu Ser Asn Leu Gly Ile Glu Ile Lys Ala Leu Asp Glu
195 200 205
Asn Gly His Asp Leu Ala Val Thr Phe Pro Gly Pro Gly Glu Asp Gly
210 215 220
Leu Asn Pro Phe Leu Glu Val Lys Val Thr Asp Thr Pro Lys Arg Ser
225 230 235 240
Arg Arg
<210> 76
<211> 242
<212> PRT
<213> Macaca fascicularis
<400> 76
Glu Asn Ser Glu Gln Lys Glu Asn Val Glu Lys Glu Gly Leu Cys Asn
1 5 10 15
Ala Cys Thr Trp Arg Gln Asn Thr Lys Ser Ser Arg Ile Glu Ala Ile
20 25 30
Lys Ile Gln Ile Leu Ser Lys Leu Arg Leu Glu Thr Ala Pro Asn Ile
35 40 45
Ser Lys Asp Ala Ile Arg Gln Leu Leu Pro Lys Ala Pro Pro Leu Arg
50 55 60
Glu Leu Ile Asp Gln Tyr Asp Val Gln Arg Asp Asp Ser Ser Asp Gly
65 70 75 80
Ser Leu Glu Asp Asp Asp Tyr His Ala Thr Thr Glu Thr Ile Ile Thr
85 90 95
Met Pro Thr Glu Ser Asp Phe Leu Met Gln Val Asp Gly Lys Pro Lys
100 105 110
Cys Cys Phe Phe Lys Phe Ser Ser Lys Ile Gln Tyr Asn Lys Val Val
115 120 125
Lys Ala Gln Leu Trp Ile Tyr Leu Arg Pro Val Glu Thr Pro Thr Thr
130 135 140
Val Phe Val Gln Ile Leu Arg Leu Ile Lys Pro Met Lys Asp Gly Thr
145 150 155 160
Arg Tyr Thr Gly Ile Arg Ser Leu Lys Leu Asp Met Asn Pro Gly Thr
165 170 175
Gly Ile Trp Gln Ser Ile Asp Val Lys Thr Val Leu Gln Asn Trp Leu
180 185 190
Lys Gln Pro Glu Ser Asn Leu Gly Ile Glu Ile Lys Ala Leu Asp Glu
195 200 205
Asn Gly His Asp Leu Ala Val Thr Phe Pro Gly Pro Gly Glu Asp Gly
210 215 220
Leu Asn Pro Phe Leu Glu Val Lys Val Thr Asp Thr Pro Lys Arg Ser
225 230 235 240
Arg Arg
<210> 77
<211> 242
<212> PRT
<213> Mus musculus
<400> 77
Glu Gly Ser Glu Arg Glu Glu Asn Val Glu Lys Glu Gly Leu Cys Asn
1 5 10 15
Ala Cys Ala Trp Arg Gln Asn Thr Arg Tyr Ser Arg Ile Glu Ala Ile
20 25 30
Lys Ile Gln Ile Leu Ser Lys Leu Arg Leu Glu Thr Ala Pro Asn Ile
35 40 45
Ser Lys Asp Ala Ile Arg Gln Leu Leu Pro Arg Ala Pro Pro Leu Arg
50 55 60
Glu Leu Ile Asp Gln Tyr Asp Val Gln Arg Asp Asp Ser Ser Asp Gly
65 70 75 80
Ser Leu Glu Asp Asp Asp Tyr His Ala Thr Thr Glu Thr Ile Ile Thr
85 90 95
Met Pro Thr Glu Ser Asp Phe Leu Met Gln Ala Asp Gly Lys Pro Lys
100 105 110
Cys Cys Phe Phe Lys Phe Ser Ser Lys Ile Gln Tyr Asn Lys Val Val
115 120 125
Lys Ala Gln Leu Trp Ile Tyr Leu Arg Pro Val Lys Thr Pro Thr Thr
130 135 140
Val Phe Val Gln Ile Leu Arg Leu Ile Lys Pro Met Lys Asp Gly Thr
145 150 155 160
Arg Tyr Thr Gly Ile Arg Ser Leu Lys Leu Asp Met Ser Pro Gly Thr
165 170 175
Gly Ile Trp Gln Ser Ile Asp Val Lys Thr Val Leu Gln Asn Trp Leu
180 185 190
Lys Gln Pro Glu Ser Asn Leu Gly Ile Glu Ile Lys Ala Leu Asp Glu
195 200 205
Asn Gly His Asp Leu Ala Val Thr Phe Pro Gly Pro Gly Glu Asp Gly
210 215 220
Leu Asn Pro Phe Leu Glu Val Lys Val Thr Asp Thr Pro Lys Arg Ser
225 230 235 240
Arg Arg
<210> 78
<211> 266
<212> PRT
<213> Homo sapiens
<400> 78
Met Gln Lys Leu Gln Leu Cys Val Tyr Ile Tyr Leu Phe Met Leu Ile
1 5 10 15
Val Ala Gly Pro Val Asp Leu Asn Glu Asn Ser Glu Gln Lys Glu Asn
20 25 30
Val Glu Lys Glu Gly Leu Cys Asn Ala Cys Thr Trp Arg Gln Asn Thr
35 40 45
Lys Ser Ser Arg Ile Glu Ala Ile Lys Ile Gln Ile Leu Ser Lys Leu
50 55 60
Arg Leu Glu Thr Ala Pro Asn Ile Ser Lys Asp Val Ile Arg Gln Leu
65 70 75 80
Leu Pro Lys Ala Pro Pro Leu Arg Glu Leu Ile Asp Gln Tyr Asp Val
85 90 95
Gln Arg Asp Asp Ser Ser Asp Gly Ser Leu Glu Asp Asp Asp Tyr His
100 105 110
Ala Thr Thr Glu Thr Ile Ile Thr Met Pro Thr Glu Ser Asp Phe Leu
115 120 125
Met Gln Val Asp Gly Lys Pro Lys Cys Cys Phe Phe Lys Phe Ser Ser
130 135 140
Lys Ile Gln Tyr Asn Lys Val Val Lys Ala Gln Leu Trp Ile Tyr Leu
145 150 155 160
Arg Pro Val Glu Thr Pro Thr Thr Val Phe Val Gln Ile Leu Arg Leu
165 170 175
Ile Lys Pro Met Lys Asp Gly Thr Arg Tyr Thr Gly Ile Arg Ser Leu
180 185 190
Lys Leu Asp Met Asn Pro Gly Thr Gly Ile Trp Gln Ser Ile Asp Val
195 200 205
Lys Thr Val Leu Gln Asn Trp Leu Lys Gln Pro Glu Ser Asn Leu Gly
210 215 220
Ile Glu Ile Lys Ala Leu Asp Glu Asn Gly His Asp Leu Ala Val Thr
225 230 235 240
Phe Pro Gly Pro Gly Glu Asp Gly Leu Asn Pro Phe Leu Glu Val Lys
245 250 255
Val Thr Asp Thr Pro Lys Arg Ser Arg Arg
260 265
<210> 79
<211> 266
<212> PRT
<213> Macaca fascicularis
<400> 79
Met Gln Lys Leu Gln Leu Cys Val Tyr Ile Tyr Leu Phe Met Leu Ile
1 5 10 15
Val Ala Gly Pro Val Asp Leu Asn Glu Asn Ser Glu Gln Lys Glu Asn
20 25 30
Val Glu Lys Glu Gly Leu Cys Asn Ala Cys Thr Trp Arg Gln Asn Thr
35 40 45
Lys Ser Ser Arg Ile Glu Ala Ile Lys Ile Gln Ile Leu Ser Lys Leu
50 55 60
Arg Leu Glu Thr Ala Pro Asn Ile Ser Lys Asp Ala Ile Arg Gln Leu
65 70 75 80
Leu Pro Lys Ala Pro Pro Leu Arg Glu Leu Ile Asp Gln Tyr Asp Val
85 90 95
Gln Arg Asp Asp Ser Ser Asp Gly Ser Leu Glu Asp Asp Asp Tyr His
100 105 110
Ala Thr Thr Glu Thr Ile Ile Thr Met Pro Thr Glu Ser Asp Phe Leu
115 120 125
Met Gln Val Asp Gly Lys Pro Lys Cys Cys Phe Phe Lys Phe Ser Ser
130 135 140
Lys Ile Gln Tyr Asn Lys Val Val Lys Ala Gln Leu Trp Ile Tyr Leu
145 150 155 160
Arg Pro Val Glu Thr Pro Thr Thr Val Phe Val Gln Ile Leu Arg Leu
165 170 175
Ile Lys Pro Met Lys Asp Gly Thr Arg Tyr Thr Gly Ile Arg Ser Leu
180 185 190
Lys Leu Asp Met Asn Pro Gly Thr Gly Ile Trp Gln Ser Ile Asp Val
195 200 205
Lys Thr Val Leu Gln Asn Trp Leu Lys Gln Pro Glu Ser Asn Leu Gly
210 215 220
Ile Glu Ile Lys Ala Leu Asp Glu Asn Gly His Asp Leu Ala Val Thr
225 230 235 240
Phe Pro Gly Pro Gly Glu Asp Gly Leu Asn Pro Phe Leu Glu Val Lys
245 250 255
Val Thr Asp Thr Pro Lys Arg Ser Arg Arg
260 265
<210> 80
<211> 267
<212> PRT
<213> Mus musculus
<400> 80
Met Met Gln Lys Leu Gln Met Tyr Val Tyr Ile Tyr Leu Phe Met Leu
1 5 10 15
Ile Ala Ala Gly Pro Val Asp Leu Asn Glu Gly Ser Glu Arg Glu Glu
20 25 30
Asn Val Glu Lys Glu Gly Leu Cys Asn Ala Cys Ala Trp Arg Gln Asn
35 40 45
Thr Arg Tyr Ser Arg Ile Glu Ala Ile Lys Ile Gln Ile Leu Ser Lys
50 55 60
Leu Arg Leu Glu Thr Ala Pro Asn Ile Ser Lys Asp Ala Ile Arg Gln
65 70 75 80
Leu Leu Pro Arg Ala Pro Pro Leu Arg Glu Leu Ile Asp Gln Tyr Asp
85 90 95
Val Gln Arg Asp Asp Ser Ser Asp Gly Ser Leu Glu Asp Asp Asp Tyr
100 105 110
His Ala Thr Thr Glu Thr Ile Ile Thr Met Pro Thr Glu Ser Asp Phe
115 120 125
Leu Met Gln Ala Asp Gly Lys Pro Lys Cys Cys Phe Phe Lys Phe Ser
130 135 140
Ser Lys Ile Gln Tyr Asn Lys Val Val Lys Ala Gln Leu Trp Ile Tyr
145 150 155 160
Leu Arg Pro Val Lys Thr Pro Thr Thr Val Phe Val Gln Ile Leu Arg
165 170 175
Leu Ile Lys Pro Met Lys Asp Gly Thr Arg Tyr Thr Gly Ile Arg Ser
180 185 190
Leu Lys Leu Asp Met Ser Pro Gly Thr Gly Ile Trp Gln Ser Ile Asp
195 200 205
Val Lys Thr Val Leu Gln Asn Trp Leu Lys Gln Pro Glu Ser Asn Leu
210 215 220
Gly Ile Glu Ile Lys Ala Leu Asp Glu Asn Gly His Asp Leu Ala Val
225 230 235 240
Thr Phe Pro Gly Pro Gly Glu Asp Gly Leu Asn Pro Phe Leu Glu Val
245 250 255
Lys Val Thr Asp Thr Pro Lys Arg Ser Arg Arg
260 265
<210> 81
<211> 407
<212> PRT
<213> Homo sapiens
<400> 81
Met Val Leu Ala Ala Pro Leu Leu Leu Gly Phe Leu Leu Leu Ala Leu
1 5 10 15
Glu Leu Arg Pro Arg Gly Glu Ala Ala Glu Gly Pro Ala Ala Ala Ala
20 25 30
Ala Ala Ala Ala Ala Ala Ala Ala Ala Gly Val Gly Gly Glu Arg Ser
35 40 45
Ser Arg Pro Ala Pro Ser Val Ala Pro Glu Pro Asp Gly Cys Pro Val
50 55 60
Cys Val Trp Arg Gln His Ser Arg Glu Leu Arg Leu Glu Ser Ile Lys
65 70 75 80
Ser Gln Ile Leu Ser Lys Leu Arg Leu Lys Glu Ala Pro Asn Ile Ser
85 90 95
Arg Glu Val Val Lys Gln Leu Leu Pro Lys Ala Pro Pro Leu Gln Gln
100 105 110
Ile Leu Asp Leu His Asp Phe Gln Gly Asp Ala Leu Gln Pro Glu Asp
115 120 125
Phe Leu Glu Glu Asp Glu Tyr His Ala Thr Thr Glu Thr Val Ile Ser
130 135 140
Met Ala Gln Glu Thr Asp Pro Ala Val Gln Thr Asp Gly Ser Pro Leu
145 150 155 160
Cys Cys His Phe His Phe Ser Pro Lys Val Met Phe Thr Lys Val Leu
165 170 175
Lys Ala Gln Leu Trp Val Tyr Leu Arg Pro Val Pro Arg Pro Ala Thr
180 185 190
Val Tyr Leu Gln Ile Leu Arg Leu Lys Pro Leu Thr Gly Glu Gly Thr
195 200 205
Ala Gly Gly Gly Gly Gly Gly Arg Arg His Ile Arg Ile Arg Ser Leu
210 215 220
Lys Ile Glu Leu His Ser Arg Ser Gly His Trp Gln Ser Ile Asp Phe
225 230 235 240
Lys Gln Val Leu His Ser Trp Phe Arg Gln Pro Gln Ser Asn Trp Gly
245 250 255
Ile Glu Ile Asn Ala Phe Asp Pro Ser Gly Thr Asp Leu Ala Val Thr
260 265 270
Ser Leu Gly Pro Gly Ala Glu Gly Leu His Pro Phe Met Glu Leu Arg
275 280 285
Val Leu Glu Asn Thr Lys Arg Ser Arg Arg Asn Leu Gly Leu Asp Cys
290 295 300
Asp Glu His Ser Ser Glu Ser Arg Cys Cys Arg Tyr Pro Leu Thr Val
305 310 315 320
Asp Phe Glu Ala Phe Gly Trp Asp Trp Ile Ile Ala Pro Lys Arg Tyr
325 330 335
Lys Ala Asn Tyr Cys Ser Gly Gln Cys Glu Tyr Met Phe Met Gln Lys
340 345 350
Tyr Pro His Thr His Leu Val Gln Gln Ala Asn Pro Arg Gly Ser Ala
355 360 365
Gly Pro Cys Cys Thr Pro Thr Lys Met Ser Pro Ile Asn Met Leu Tyr
370 375 380
Phe Asn Asp Lys Gln Gln Ile Ile Tyr Gly Lys Ile Pro Gly Met Val
385 390 395 400
Val Asp Arg Cys Gly Cys Ser
405
<210> 82
<211> 109
<212> PRT
<213> Homo sapiens
<400> 82
Asn Leu Gly Leu Asp Cys Asp Glu His Ser Ser Glu Ser Arg Cys Cys
1 5 10 15
Arg Tyr Pro Leu Thr Val Asp Phe Glu Ala Phe Gly Trp Asp Trp Ile
20 25 30
Ile Ala Pro Lys Arg Tyr Lys Ala Asn Tyr Cys Ser Gly Gln Cys Glu
35 40 45
Tyr Met Phe Met Gln Lys Tyr Pro His Thr His Leu Val Gln Gln Ala
50 55 60
Asn Pro Arg Gly Ser Ala Gly Pro Cys Cys Thr Pro Thr Lys Met Ser
65 70 75 80
Pro Ile Asn Met Leu Tyr Phe Asn Asp Lys Gln Gln Ile Ile Tyr Gly
85 90 95
Lys Ile Pro Gly Met Val Val Asp Arg Cys Gly Cys Ser
100 105
<210> 83
<211> 274
<212> PRT
<213> Homo sapiens
<400> 83
Ala Glu Gly Pro Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
1 5 10 15
Ala Gly Val Gly Gly Glu Arg Ser Ser Arg Pro Ala Pro Ser Val Ala
20 25 30
Pro Glu Pro Asp Gly Cys Pro Val Cys Val Trp Arg Gln His Ser Arg
35 40 45
Glu Leu Arg Leu Glu Ser Ile Lys Ser Gln Ile Leu Ser Lys Leu Arg
50 55 60
Leu Lys Glu Ala Pro Asn Ile Ser Arg Glu Val Val Lys Gln Leu Leu
65 70 75 80
Pro Lys Ala Pro Pro Leu Gln Gln Ile Leu Asp Leu His Asp Phe Gln
85 90 95
Gly Asp Ala Leu Gln Pro Glu Asp Phe Leu Glu Glu Asp Glu Tyr His
100 105 110
Ala Thr Thr Glu Thr Val Ile Ser Met Ala Gln Glu Thr Asp Pro Ala
115 120 125
Val Gln Thr Asp Gly Ser Pro Leu Cys Cys His Phe His Phe Ser Pro
130 135 140
Lys Val Met Phe Thr Lys Val Leu Lys Ala Gln Leu Trp Val Tyr Leu
145 150 155 160
Arg Pro Val Pro Arg Pro Ala Thr Val Tyr Leu Gln Ile Leu Arg Leu
165 170 175
Lys Pro Leu Thr Gly Glu Gly Thr Ala Gly Gly Gly Gly Gly Gly Arg
180 185 190
Arg His Ile Arg Ile Arg Ser Leu Lys Ile Glu Leu His Ser Arg Ser
195 200 205
Gly His Trp Gln Ser Ile Asp Phe Lys Gln Val Leu His Ser Trp Phe
210 215 220
Arg Gln Pro Gln Ser Asn Trp Gly Ile Glu Ile Asn Ala Phe Asp Pro
225 230 235 240
Ser Gly Thr Asp Leu Ala Val Thr Ser Leu Gly Pro Gly Ala Glu Gly
245 250 255
Leu His Pro Phe Met Glu Leu Arg Val Leu Glu Asn Thr Lys Arg Ser
260 265 270
Arg Arg
<210> 84
<211> 298
<212> PRT
<213> Homo sapiens
<400> 84
Met Val Leu Ala Ala Pro Leu Leu Leu Gly Phe Leu Leu Leu Ala Leu
1 5 10 15
Glu Leu Arg Pro Arg Gly Glu Ala Ala Glu Gly Pro Ala Ala Ala Ala
20 25 30
Ala Ala Ala Ala Ala Ala Ala Ala Ala Gly Val Gly Gly Glu Arg Ser
35 40 45
Ser Arg Pro Ala Pro Ser Val Ala Pro Glu Pro Asp Gly Cys Pro Val
50 55 60
Cys Val Trp Arg Gln His Ser Arg Glu Leu Arg Leu Glu Ser Ile Lys
65 70 75 80
Ser Gln Ile Leu Ser Lys Leu Arg Leu Lys Glu Ala Pro Asn Ile Ser
85 90 95
Arg Glu Val Val Lys Gln Leu Leu Pro Lys Ala Pro Pro Leu Gln Gln
100 105 110
Ile Leu Asp Leu His Asp Phe Gln Gly Asp Ala Leu Gln Pro Glu Asp
115 120 125
Phe Leu Glu Glu Asp Glu Tyr His Ala Thr Thr Glu Thr Val Ile Ser
130 135 140
Met Ala Gln Glu Thr Asp Pro Ala Val Gln Thr Asp Gly Ser Pro Leu
145 150 155 160
Cys Cys His Phe His Phe Ser Pro Lys Val Met Phe Thr Lys Val Leu
165 170 175
Lys Ala Gln Leu Trp Val Tyr Leu Arg Pro Val Pro Arg Pro Ala Thr
180 185 190
Val Tyr Leu Gln Ile Leu Arg Leu Lys Pro Leu Thr Gly Glu Gly Thr
195 200 205
Ala Gly Gly Gly Gly Gly Gly Arg Arg His Ile Arg Ile Arg Ser Leu
210 215 220
Lys Ile Glu Leu His Ser Arg Ser Gly His Trp Gln Ser Ile Asp Phe
225 230 235 240
Lys Gln Val Leu His Ser Trp Phe Arg Gln Pro Gln Ser Asn Trp Gly
245 250 255
Ile Glu Ile Asn Ala Phe Asp Pro Ser Gly Thr Asp Leu Ala Val Thr
260 265 270
Ser Leu Gly Pro Gly Ala Glu Gly Leu His Pro Phe Met Glu Leu Arg
275 280 285
Val Leu Glu Asn Thr Lys Arg Ser Arg Arg
290 295
<210> 85
<211> 409
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 85
Met Lys Trp Val Thr Phe Leu Leu Leu Leu Phe Ile Ser Gly Ser Ala
1 5 10 15
Phe Ser Asp Tyr Lys Asp Asp Asp Asp Lys Ala Glu Gly Pro Ala Ala
20 25 30
Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Gly Val Gly Gly Glu
35 40 45
Arg Ser Ser Arg Pro Ala Pro Ser Val Ala Pro Glu Pro Asp Gly Cys
50 55 60
Pro Val Cys Val Trp Arg Gln His Ser Arg Glu Leu Arg Leu Glu Ser
65 70 75 80
Ile Lys Ser Gln Ile Leu Ser Lys Leu Arg Leu Lys Glu Ala Pro Asn
85 90 95
Ile Ser Arg Glu Val Val Lys Gln Leu Leu Pro Lys Ala Pro Pro Leu
100 105 110
Gln Gln Ile Leu Asp Leu His Asp Phe Gln Gly Asp Ala Leu Gln Pro
115 120 125
Glu Asp Phe Leu Glu Glu Asp Glu Tyr His Ala Thr Thr Glu Thr Val
130 135 140
Ile Ser Met Ala Gln Glu Thr Asp Pro Ala Val Gln Thr Asp Gly Ser
145 150 155 160
Pro Leu Cys Cys His Phe His Phe Ser Pro Lys Val Met Phe Thr Lys
165 170 175
Val Leu Lys Ala Gln Leu Trp Val Tyr Leu Arg Pro Val Pro Arg Pro
180 185 190
Ala Thr Val Tyr Leu Gln Ile Leu Arg Leu Lys Pro Leu Thr Gly Glu
195 200 205
Gly Thr Ala Gly Gly Gly Gly Gly Gly Arg Arg His Ile Arg Ile Arg
210 215 220
Ser Leu Lys Ile Glu Leu His Ser Arg Ser Gly His Trp Gln Ser Ile
225 230 235 240
Asp Phe Lys Gln Val Leu His Ser Trp Phe Arg Gln Pro Gln Ser Asn
245 250 255
Trp Gly Ile Glu Ile Asn Ala Phe Asp Pro Ser Gly Thr Asp Leu Ala
260 265 270
Val Thr Ser Leu Gly Pro Gly Ala Glu Gly Leu His Pro Phe Met Glu
275 280 285
Leu Arg Val Leu Glu Asn Thr Lys Arg Ser Arg Arg Asn Leu Gly Leu
290 295 300
Asp Cys Asp Glu His Ser Ser Glu Ser Arg Cys Cys Arg Tyr Pro Leu
305 310 315 320
Thr Val Asp Phe Glu Ala Phe Gly Trp Asp Trp Ile Ile Ala Pro Lys
325 330 335
Arg Tyr Lys Ala Asn Tyr Cys Ser Gly Gln Cys Glu Tyr Met Phe Met
340 345 350
Gln Lys Tyr Pro His Thr His Leu Val Gln Gln Ala Asn Pro Arg Gly
355 360 365
Ser Ala Gly Pro Cys Cys Thr Pro Thr Lys Met Ser Pro Ile Asn Met
370 375 380
Leu Tyr Phe Asn Asp Lys Gln Gln Ile Ile Tyr Gly Lys Ile Pro Gly
385 390 395 400
Met Val Val Asp Arg Cys Gly Cys Ser
405
<210> 86
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 86
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Asp
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 87
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 87
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Asp
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Met Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Lys Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 88
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 88
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Thr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Lys Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Lys Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 89
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 89
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Thr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Lys Lys Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 90
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 90
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Thr
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Ser Lys Tyr Lys Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 91
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 91
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser His Asp
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ser Tyr Ala Gly Lys Lys Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 92
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 92
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser His Asp
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 93
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 93
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Lys Val Ser Gly Ile Asp Leu Ser His Asp
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 94
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 94
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Lys Val Ser Gly Ile Asp Leu Ser His Glu
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ile Ile Ser Tyr Ala Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 95
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 95
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser His Glu
20 25 30
Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ile Ile Ser Tyr Ala Gly Ser Lys Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Gly Val Pro Ala Tyr Ser His Gly Gly Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 96
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 96
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Thr Ser Ser Gln Ser Val Tyr His Ala
20 25 30
Asn Trp Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Tyr Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Arg Tyr Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 97
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 97
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Thr Thr Ser Gln Ser Val Tyr His Glu
20 25 30
Asn Trp Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Tyr Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Arg Tyr Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 98
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 98
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Thr Ser Ser Gln Ser Val Phe His Glu
20 25 30
Asn Trp Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Trp Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Arg Tyr Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 99
<211> 110
<212> PRT
<213> Artificial sequence
<220>
40 <223> An artificially synthesized sequence
<400> 99
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Thr Thr Ser Gln Ser Val Tyr His Glu
20 25 30
Asn Trp Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Trp Ala Ser Thr Leu Ala Tyr Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Gly Gly
85 90 95
Gly Arg Tyr Ala Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 100
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 100
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcacag tgagtgggat cgacctgagc cacgacgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtcggcatc attagctacg ccgggagcac atactatgcc 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 101
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 101
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcacag tgagtgggat cgacctgagc cacgacgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtcggcatc attagctacg ccgggagcat gtactatgcc 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga taagtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 102
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 102
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcacag tgagtgggat cgacctgagc cacaccgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtcggcatc attagctacg ccgggaagac atactatgcc 180
agctgggcta aaaaaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 103
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 103
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcacag tgagtgggat cgacctgagc cacaccgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtcggcatc attagctacg ccgggaagaa atactatgcc 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 104
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 104
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcacag tgagtgggat cgacctgagc cacaccgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtcggcatc attagctacg ccgggagcaa atacaaggcc 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 105
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 105
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcacag tgagtgggat cgacctgagc cacgacgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtcggcatc attagctacg ccgggaagaa atactatgcc 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 106
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 106
caggtgcagc tggtcgaatc tggaggagga ctggtgcagc caggagggtc actgcgactg 60
agctgtgccg tgtccgggat cgacctgtca cacgacgata ttagctgggt gagacaggca 120
cccgggaagg gactggaatg ggtctctatc attagttacg ccggcagcac ctactatgcc 180
tcttgggcta aaggcaggct gaccatttct aaggacacaa gtaaaaacca ggtggtcctg 240
accatgacaa atatggaccc cgtggatact gcaacctact attgcgcccg gggcgtccct 300
gcttatagcc acggcggcga tctgtgggga caggggacac tggtgactgt cagctcc 357
<210> 107
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 107
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcaaag tgagtgggat cgacctgagc cacgacgaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtctccatc attagctacg ccgggagcac atactatgcc 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 108
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 108
caggtgacac tgaaagaaag cggccccgtg ctggtgaagc caaccgagac actgactctg 60
acctgcaaag tgagtgggat cgacctgagc cacgaggaca tctcctgggt gcggcaggca 120
cccggaaagg gcctggaatg ggtctccatc attagctacg ccgggagcac atactatgcc 180
agctgggcta aaggaagact gactatctcc aaggacacct ctaaaaacca ggtggtcctg 240
actatgacca atatggaccc cgtggataca gcaacttact attgtgccag gggcgtccct 300
gcttactctc acggagggga tctgtgggga cagggaaccc tggtgacagt ctctagt 357
<210> 109
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 109
caggtgcagc tggtcgaatc tggaggagga ctggtgcagc caggagggtc actgcgactg 60
agctgtgccg tgtccgggat cgacctgtca cacgaggata ttagctgggt gagacaggca 120
cccgggaagg gactggaatg ggtctctatc attagttacg ccggcagcaa gtactatgcc 180
tcttgggcta aaggcaggct gaccatttct aaggacacaa gtaaaaacca ggtggtcctg 240
accatgacaa atatggaccc cgtggatact gcaacctact attgcgcccg gggcgtccct 300
gcttatagcc acggcggcga tctgtgggga caggggacac tggtgactgt cagctcc 357
<210> 110
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 110
gacattgtga tgactcagtc tcctgccact ctgtcactga gccctggcga gcgagcaacc 60
ctgtcttgca ccagcagcca gtccgtgtac cacgccaatt ggctgagctg gttccagcag 120
aagccagggc agccccctaa actgctgatc tattgggcat caacactggc ctacggcgtc 180
ccctccagat tctccggatc tggcagtggg actgacttta ccctgaccat cagcagcctc 240
cagcccgagg atgccgctac ctactattgt gctggcgggt acggaggcgg gcggtatgca 300
tttggacagg gcacaaaggt ggaaatcaaa 330
<210> 111
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 111
gacattgtga tgactcagtc tcctgccact ctgtcactga gccctggcga gcgagcaacc 60
ctgtcttgca ccaccagcca gtccgtgtac cacgagaatt ggctgagctg gttccagcag 120
aagccagggc agccccctaa actgctgatc tattgggcat caacactggc ctacggcgtc 180
ccctccagat tctccggatc tggcagtggg actgacttta ccctgaccat cagcagcctc 240
cagcccgagg atgccgctac ctactattgt gctggcgggt acggaggcgg gcggtatgca 300
tttggacagg gcacaaaggt ggaaatcaaa 330
<210> 112
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 112
gacattgtga tgactcagtc tcctgccact ctgtcactga gccctggcga gcgagcaacc 60
ctgtcttgca ccagcagcca gtccgtgttc cacgagaatt ggctgagctg gttccagcag 120
aagccagggc agccccctaa actgctgatc tattgggcat caacactggc ctggggcgtc 180
ccctccagat tctccggatc tggcagtggg actgacttta ccctgaccat cagcagcctc 240
cagcccgagg atgccgctac ctactattgt gctggcgggt acggaggcgg gcggtatgca 300
tttggacagg gcacaaaggt ggaaatcaaa 330
<210> 113
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 113
gacattgtga tgactcagtc tcctgccact ctgtcactga gccctggcga gcgagcaacc 60
ctgtcttgca ccaccagcca gtccgtgtac cacgagaatt ggctgagctg gtaccagcag 120
aagccagggc agccccctaa actgctgatc tattgggcat caacactggc ctacggcgtc 180
ccctccagat tctccggatc tggcagtggg actgacttta ccctgaccat cagcagcctc 240
cagcccgagg atgtcgctac ctactattgt gctggcgggt acggaggcgg gcggtatgca 300
tttggacagg gcacaaaggt ggaaatcaaa 330
<210> 114
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 114
His Asp Asp Ile Ser
1 5
<210> 115
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 115
His Glu Asp Ile Ser
1 5
<210> 116
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 116
Ile Ile Ser Tyr Ala Gly Ser Met Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 117
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 117
Ile Ile Ser Tyr Ala Gly Lys Thr Tyr Tyr Ala Ser Trp Ala Lys Lys
1 5 10 15
<210> 118
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 118
Ile Ile Ser Tyr Ala Gly Lys Lys Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 119
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 119
Ile Ile Ser Tyr Ala Gly Ser Lys Tyr Lys Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 120
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 120
Ile Ile Ser Tyr Ala Gly Ser Lys Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 121
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 121
Gly Val Pro Ala Tyr Ser His Gly Gly Asp Lys
1 5 10
<210> 122
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 122
Thr Ser Ser Gln Ser Val Tyr His Ala Asn Trp Leu Ser
1 5 10
<210> 123
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 123
Thr Thr Ser Gln Ser Val Tyr His Glu Asn Trp Leu Ser
1 5 10
<210> 124
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 124
Thr Ser Ser Gln Ser Val Phe His Glu Asn Trp Leu Ser
1 5 10
<210> 125
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 125
Trp Ala Ser Thr Leu Ala Trp
1 5
<210> 126
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<220>
<221> misc_feature
<222> (1)..(1)
<223> Xaa is Ser or His
<220>
<221> misc_feature
<222> (2)..(2)
<223> Xaa is Tyr, Thr, Asp or Glu
<400> 126
Xaa Xaa Asp Ile Ser
1 5
<210> 127
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa is Tyr or His
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa is Ser or Lys
<220>
<221> misc_feature
<222> (8)..(8)
<223> Xaa is Thr, Met or Lys
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa is Tyr or Lys
<220>
<221> misc_feature
<222> (11)..(11)
<223> Xaa is Ala, Met or Glu
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa is Ser or Glu
<220>
<221> misc_feature
<222> (16)..(16)
<223> Xaa is Gly or Lys
<400> 127
Ile Ile Ser Xaa Ala Gly Xaa Xaa Tyr Xaa Xaa Xaa Trp Ala Lys Xaa
1 5 10 15
<210> 128
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<220>
<221> misc_feature
<222> (5)..(5)
<223> Xaa is Tyr or His
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa is Thr or His
<220>
<221> misc_feature
<222> (11)..(11)
<223> Xaa is Leu or Lys
<400> 128
Gly Val Pro Ala Xaa Ser Xaa Gly Gly Asp Xaa
1 5 10
<210> 129
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<220>
<221> misc_feature
<222> (1)..(1)
<223> Xaa is Gln or Thr
<220>
<221> misc_feature
<222> (2)..(2)
<223> Xaa is Ser or Thr
<220>
<221> misc_feature
<222> (5)..(5)
<223> Xaa is Ser or Glu
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa is Tyr or Phe
<220>
<221> misc_feature
<222> (8)..(8)
<223> Xaa is Asp or His
<220>
<221> misc_feature
<222> (9)..(9)
<223> Xaa is Asn, Asp, Ala or Glu
<400> 129
Xaa Xaa Ser Gln Xaa Val Xaa Xaa Xaa Asn Trp Leu Ser
1 5 10
<210> 130
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<220>
<221> misc_feature
<222> (3)..(3)
<223> Xaa is Ser or Glu
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa is Ser, Tyr, Phe or Trp
<400> 130
Trp Ala Xaa Thr Leu Ala Xaa
1 5
<210> 131
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<220>
<221> misc_feature
<222> (8)..(8)
<223> Xaa is Leu or Arg
<400> 131
Ala Gly Gly Tyr Gly Gly Gly Xaa Tyr Ala
1 5 10
<210> 132
<211> 30
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 132
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser
20 25 30
<210> 133
<211> 30
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 133
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser
20 25 30
<210> 134
<211> 30
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 134
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Lys Val Ser Gly Ile Asp Leu Ser
20 25 30
<210> 135
<211> 14
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 135
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Gly
1 5 10
<210> 136
<211> 14
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 136
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 137
<211> 32
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 137
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Val Leu Thr
1 5 10 15
Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
20 25 30
<210> 138
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 138
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 139
<211> 23
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 139
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys
20
<210> 140
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 140
Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 141
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 141
Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 142
<211> 32
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 142
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Ala Ala Thr Tyr Tyr Cys
20 25 30
<210> 143
<211> 32
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 143
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys
20 25 30
<210> 144
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 144
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 145
<211> 121
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 145
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Arg Tyr Tyr
20 25 30
Val Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Tyr Thr Gly Gly Asn Thr Asn Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Ala Ser Thr Thr Val Asp Leu Lys Met Thr
65 70 75 80
Gly Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Gly Phe Leu
85 90 95
Asp Tyr Ser His Tyr Thr Thr Gly Gly Tyr Tyr Leu Asp Leu Trp Gly
100 105 110
Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 146
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 146
Gln Ser Leu Glu Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly Thr
1 5 10 15
Leu Thr Leu Thr Cys Lys Ala Ser Gly Ile Asp Phe Ser Ser Tyr Tyr
20 25 30
Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Cys Ile Tyr Ala Gly Ile Ser Gly Ser Thr Tyr Tyr Ala Ser Trp
50 55 60
Ala Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Thr
65 70 75 80
Leu Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Asp Pro Tyr His Phe Asp Tyr Gly Ser Leu Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 147
<211> 116
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 147
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Arg Tyr Trp
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Ser Ser Gly Gly Thr Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Asp Ser
85 90 95
Arg Tyr Gly Ala Ala Arg Leu Asp Leu Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 148
<211> 109
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 148
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Tyr Ile
20 25 30
Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Phe Ile Tyr Val Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asp Leu Arg Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Val Arg Thr Trp
85 90 95
Asp Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
100 105
<210> 149
<211> 115
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 149
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Tyr Ala
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Tyr Gly Val Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Ala
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Asp Asp
85 90 95
Tyr Gly Trp Gly Tyr Thr Asn Leu Trp Gly Pro Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 150
<211> 117
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 150
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Ser Ser
20 25 30
Met Ile Trp Val Arg Gln Ala Pro Gly Glu Gly Leu Lys Tyr Ile Gly
35 40 45
Phe Ile Ser Lys Ser Gly Tyr Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly Gly
85 90 95
Tyr Ser Gly Asp Trp Asp Tyr Phe Thr Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 151
<211> 112
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 151
Gln Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly Gly
1 5 10 15
Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Val Tyr Asn Asn Lys
20 25 30
Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Ala Ser Thr Leu Pro Ser Gly Val Ser Ser Arg Phe Lys
50 55 60
Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val Gln
65 70 75 80
Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Glu Phe Ser Cys Gly
85 90 95
Ser Ala Asp Cys Thr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 152
<211> 112
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 152
Ala Leu Val Met Thr Gln Thr Pro Ala Ser Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Val Tyr Gly Gly
20 25 30
Asn Glu Leu Ser Trp Tyr Gln Gln Glu Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Ser Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Ala Tyr Tyr Cys Ala Gly Tyr Arg Thr Tyr
85 90 95
Ser Asp Asp Asp Asn Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 153
<211> 112
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 153
Gln Val Leu Thr Gln Thr Pro Ala Ser Val Ser Ala Ala Val Gly Gly
1 5 10 15
Thr Val Thr Ile Asn Cys Gln Ser Ser Gln Ser Val Ala Gly Asn Asp
20 25 30
Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Ala Ser Asn Leu Ala Ser Gly Val Ser Ser Arg Phe Lys
50 55 60
Gly Ser Gly Ser Gly Thr Arg Phe Thr Leu Thr Ile Ser Asp Val Gln
65 70 75 80
Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Asn Tyr Asp Cys Ser
85 90 95
Ser Ala Asp Cys Asp Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 154
<211> 109
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 154
Gln Val Leu Thr Gln Thr Pro Ser Ser Thr Ser Ala Ala Val Gly Gly
1 5 10 15
Thr Val Thr Ile Asn Cys Gln Ser Ser Gln Ser Val Tyr Asp Asn Asn
20 25 30
Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Arg Leu
35 40 45
Ile Tyr Gly Ala Ser Thr Leu Asp Ser Gly Val Pro Ser Arg Phe Lys
50 55 60
Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Asp Leu Glu
65 70 75 80
Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Gly Tyr Ser Gly Asn
85 90 95
Ile Tyr Pro Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 155
<211> 111
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 155
Gln Val Leu Thr Gln Thr Pro Ser Ser Val Ser Ala Ala Val Gly Gly
1 5 10 15
Thr Val Thr Ile Asn Cys Gln Ser Ser Gln Ser Val Tyr Asn Asn Asn
20 25 30
Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Lys
50 55 60
Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val Gln
65 70 75 80
Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Ser Tyr Tyr Ser Ser
85 90 95
Gly Trp Tyr Arg Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 156
<211> 114
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 156
Ala Asp Ile Val Met Thr Gln Thr Pro Ala Ser Val Glu Ala Ala Val
1 5 10 15
Gly Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Glu Ser Ile Gly Asn
20 25 30
Ala Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Ala Phe Asn Leu Glu Ser Gly Val Pro Ser Arg Phe Arg
50 55 60
Gly Ser Gly Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Asp Leu Glu
65 70 75 80
Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Tyr Phe Trp Ser Ser
85 90 95
Ser Thr Ile Tyr Ala Phe Trp Ala Phe Gly Gly Gly Thr Lys Val Glu
100 105 110
Ile Lys
<210> 157
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 157
Arg Tyr Tyr Val Ser
1 5
<210> 158
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 158
Ser Tyr Tyr Tyr Met Cys
1 5
<210> 159
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 159
Arg Tyr Trp Met Ser
1 5
<210> 160
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 160
Ser Tyr Ile Met Gly
0> 161
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 161
Ser Tyr Ala Met Ser
1 5
<210> 162
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 162
Ser Ser Ser Met Ile
1 5
<210> 163
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 163
Ile Ile Tyr Thr Gly Gly Asn Thr Asn Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 164
<211> 18
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 164
Cys Ile Tyr Ala Gly Ile Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala
1 5 10 15
Lys Gly
<210> 165
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 165
Ile Ile Ser Ser Gly Gly Thr Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 166
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 166
Phe Ile Tyr Val Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 167
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 167
Ile Ile Tyr Gly Val Gly Ser Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 168
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 168
Phe Ile Ser Lys Ser Gly Tyr Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 169
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 169
Phe Leu Asp Tyr Ser His Tyr Thr Thr Gly Gly Tyr Tyr Leu Asp Leu
1 5 10 15
<210> 170
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 170
Asp Pro Tyr His Phe Asp Tyr Gly Ser Leu
1 5 10
<210> 171
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 171
Asp Ser Arg Tyr Gly Ala Ala Arg Leu Asp Leu
1 5 10
<210> 172
<211> 4
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 172
Thr Trp Asp Leu
1
<210> 173
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 173
Asp Asp Tyr Gly Trp Gly Tyr Thr Asn Leu
1 5 10
<210> 174
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 174
Gly Gly Tyr Ser Gly Asp Trp Asp Tyr Phe Thr Leu
1 5 10
<210> 175
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 175
Gln Ala Ser Gln Ser Val Tyr Asn Asn Lys Asn Leu Ala
1 5 10
<210> 176
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 176
Gln Ala Ser Gln Ser Val Tyr Gly Gly Asn Glu Leu Ser
1 5 10
<210> 177
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 177
Gln Ser Ser Gln Ser Val Ala Gly Asn Asp Asn Leu Ala
1 5 10
<210> 178
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 178
Gln Ser Ser Gln Ser Val Tyr Asp Asn Asn Trp Leu Ala
1 5 10
<210> 179
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 179
Gln Ser Ser Gln Ser Val Tyr Asn Asn Asn Tyr Leu Ala
1 5 10
<210> 180
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 180
Gln Ala Ser Glu Ser Ile Gly Asn Ala Leu Ala
1 5 10
<210> 181
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 181
Tyr Ala Ser Thr Leu Pro Ser
1 5
<210> 182
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 182
Ser Ala Ser Lys Leu Ala Ser
1 5
<210> 183
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 183
Tyr Ala Ser Asn Leu Ala Ser
1 5
<210> 184
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 184
Gly Ala Ser Thr Leu Asp Ser
1 5
<210> 185
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 185
Asp Ala Ser Lys Leu Ala Ser
1 5
<210> 186
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 186
Gly Ala Phe Asn Leu Glu Ser
1 5
<210> 187
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 187
Gln Gly Glu Phe Ser Cys Gly Ser Ala Asp Cys Thr Ala
1 5 10
<210> 188
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 188
Ala Gly Tyr Arg Thr Tyr Ser Asp Asp Asp Asn Ala
1 5 10
<210> 189
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 189
Leu Gly Asn Tyr Asp Cys Ser Ser Ala Asp Cys Asp Ala
1 5 10
<210> 190
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 190
Gln Gly Gly Tyr Ser Gly Asn Ile Tyr Pro
1 5 10
<210> 191
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 191
Gln Gly Ser Tyr Tyr Ser Ser Gly Trp Tyr Arg Ala
1 5 10
<210> 192
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 192
Gln Gly Tyr Phe Trp Ser Ser Ser Thr Ile Tyr Ala Phe Trp Ala
1 5 10 15
<210> 193
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 193
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Arg Arg Gly Pro Lys Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 194
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 194
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agtaaggact gcacctacag tctcagtagt accctgacgc tgtccaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgtcctcgcc cgtcacaaag 300
tccttcaaca ggggagagtg ttga 324
<210> 195
<211> 330
<212> PRT
<213> Homo sapiens
<400> 195
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 196
<211> 326
<212> PRT
<213> Homo sapiens
<400> 196
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 197
<211> 377
<212> PRT
<213> Homo sapiens
<400> 197
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 198
<211> 327
<212> PRT
<213> Homo sapiens
<400> 198
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 199
<211> 1125
<212> DNA
<213> Homo sapiens
<400> 199
atgtggttct tgacaactct gctcctttgg gttccagttg atgggcaagt ggacaccaca 60
aaggcagtga tcactttgca gcctccatgg gtcagcgtgt tccaagagga aaccgtaacc 120
ttgcactgtg aggtgctcca tctgcctggg agcagctcta cacagtggtt tctcaatggc 180
acagccactc agacctcgac ccccagctac agaatcacct ctgccagtgt caatgacagt 240
ggtgaataca ggtgccagag aggtctctca gggcgaagtg accccataca gctggaaatc 300
cacagaggct ggctactact gcaggtctcc agcagagtct tcacggaagg agaacctctg 360
gccttgaggt gtcatgcgtg gaaggataag ctggtgtaca atgtgcttta ctatcgaaat 420
ggcaaagcct ttaagttttt ccactggaat tctaacctca ccattctgaa aaccaacata 480
agtcacaatg gcacctacca ttgctcaggc atgggaaagc atcgctacac atcagcagga 540
atatctgtca ctgtgaaaga gctatttcca gctccagtgc tgaatgcatc tgtgacatcc 600
ccactcctgg aggggaatct ggtcaccctg agctgtgaaa caaagttgct cttgcagagg 660
cctggtttgc agctttactt ctccttctac atgggcagca agaccctgcg aggcaggaac 720
acatcctctg aataccaaat actaactgct agaagagaag actctgggtt atactggtgc 780
gaggctgcca cagaggatgg aaatgtcctt aagcgcagcc ctgagttgga gcttcaagtg 840
cttggcctcc agttaccaac tcctgtctgg tttcatgtcc ttttctatct ggcagtggga 900
ataatgtttt tagtgaacac tgttctctgg gtgacaatac gtaaagaact gaaaagaaag 960
aaaaagtggg atttagaaat ctctttggat tctggtcatg agaagaaggt aatttccagc 1020
cttcaagaag acagacattt agaagaagag ctgaaatgtc aggaacaaaa agaagaacag 1080
ctgcaggaag gggtgcaccg gaaggagccc cagggggcca cgtag 1125
<210> 200
<211> 951
<212> DNA
<213> Homo sapiens
<400> 200
atgactatgg agacccaaat gtctcagaat gtatgtccca gaaacctgtg gctgcttcaa 60
ccattgacag ttttgctgct gctggcttct gcagacagtc aagctgctcc cccaaaggct 120
gtgctgaaac ttgagccccc gtggatcaac gtgctccagg aggactctgt gactctgaca 180
tgccaggggg ctcgcagccc tgagagcgac tccattcagt ggttccacaa tgggaatctc 240
attcccaccc acacgcagcc cagctacagg ttcaaggcca acaacaatga cagcggggag 300
tacacgtgcc agactggcca gaccagcctc agcgaccctg tgcatctgac tgtgctttcc 360
gaatggctgg tgctccagac ccctcacctg gagttccagg agggagaaac catcatgctg 420
aggtgccaca gctggaagga caagcctctg gtcaaggtca cattcttcca gaatggaaaa 480
tcccagaaat tctcccattt ggatcccacc ttctccatcc cacaagcaaa ccacagtcac 540
agtggtgatt accactgcac aggaaacata ggctacacgc tgttctcatc caagcctgtg 600
accatcactg tccaagtgcc cagcatgggc agctcttcac caatgggggt cattgtggct 660
gtggtcattg cgactgctgt agcagccatt gttgctgctg tagtggcctt gatctactgc 720
aggaaaaagc ggatttcagc caattccact gatcctgtga aggctgccca atttgagcca 780
cctggacgtc aaatgattgc catcagaaag agacaacttg aagaaaccaa caatgactat 840
gaaacagctg acggcggcta catgactctg aaccccaggg cacctactga cgatgataaa 900
aacatctacc tgactcttcc tcccaacgac catgtcaaca gtaataacta a 951
<210> 201
<211> 954
<212> DNA
<213> Homo sapiens
<400> 201
atgactatgg agacccaaat gtctcagaat gtatgtccca gaaacctgtg gctgcttcaa 60
ccattgacag ttttgctgct gctggcttct gcagacagtc aagctgcagc tcccccaaag 120
gctgtgctga aacttgagcc cccgtggatc aacgtgctcc aggaggactc tgtgactctg 180
acatgccagg gggctcgcag ccctgagagc gactccattc agtggttcca caatgggaat 240
ctcattccca cccacacgca gcccagctac aggttcaagg ccaacaacaa tgacagcggg 300
gagtacacgt gccagactgg ccagaccagc ctcagcgacc ctgtgcatct gactgtgctt 360
tccgaatggc tggtgctcca gacccctcac ctggagttcc aggagggaga aaccatcatg 420
ctgaggtgcc acagctggaa ggacaagcct ctggtcaagg tcacattctt ccagaatgga 480
aaatcccaga aattctccca tttggatccc accttctcca tcccacaagc aaaccacagt 540
cacagtggtg attaccactg cacaggaaac ataggctaca cgctgttctc atccaagcct 600
gtgaccatca ctgtccaagt gcccagcatg ggcagctctt caccaatggg gatcattgtg 660
gctgtggtca ttgcgactgc tgtagcagcc attgttgctg ctgtagtggc cttgatctac 720
tgcaggaaaa agcggatttc agccaattcc actgatcctg tgaaggctgc ccaatttgag 780
ccacctggac gtcaaatgat tgccatcaga aagagacaac ttgaagaaac caacaatgac 840
tatgaaacag ctgacggcgg ctacatgact ctgaacccca gggcacctac tgacgatgat 900
aaaaacatct acctgactct tcctcccaac gaccatgtca acagtaataa ctaa 954
<210> 202
<211> 876
<212> DNA
<213> Homo sapiens
<400> 202
atgggaatcc tgtcattctt acctgtcctt gccactgaga gtgactgggc tgactgcaag 60
tccccccagc cttggggtca tatgcttctg tggacagctg tgctattcct ggctcctgtt 120
gctgggacac ctgcagctcc cccaaaggct gtgctgaaac tcgagcccca gtggatcaac 180
gtgctccagg aggactctgt gactctgaca tgccggggga ctcacagccc tgagagcgac 240
tccattcagt ggttccacaa tgggaatctc attcccaccc acacgcagcc cagctacagg 300
ttcaaggcca acaacaatga cagcggggag tacacgtgcc agactggcca gaccagcctc 360
agcgaccctg tgcatctgac tgtgctttct gagtggctgg tgctccagac ccctcacctg 420
gagttccagg agggagaaac catcgtgctg aggtgccaca gctggaagga caagcctctg 480
gtcaaggtca cattcttcca gaatggaaaa tccaagaaat tttcccgttc ggatcccaac 540
ttctccatcc cacaagcaaa ccacagtcac agtggtgatt accactgcac aggaaacata 600
ggctacacgc tgtactcatc caagcctgtg accatcactg tccaagctcc cagctcttca 660
ccgatgggga tcattgtggc tgtggtcact gggattgctg tagcggccat tgttgctgct 720
gtagtggcct tgatctactg caggaaaaag cggatttcag ccaatcccac taatcctgat 780
gaggctgaca aagttggggc tgagaacaca atcacctatt cacttctcat gcacccggat 840
gctctggaag agcctgatga ccagaaccgt atttag 876
<210> 203
<211> 933
<212> DNA
<213> Homo sapiens
<400> 203
atgggaatcc tgtcattctt acctgtcctt gccactgaga gtgactgggc tgactgcaag 60
tccccccagc cttggggtca tatgcttctg tggacagctg tgctattcct ggctcctgtt 120
gctgggacac ctgcagctcc cccaaaggct gtgctgaaac tcgagcccca gtggatcaac 180
gtgctccagg aggactctgt gactctgaca tgccggggga ctcacagccc tgagagcgac 240
tccattcagt ggttccacaa tgggaatctc attcccaccc acacgcagcc cagctacagg 300
ttcaaggcca acaacaatga cagcggggag tacacgtgcc agactggcca gaccagcctc 360
agcgaccctg tgcatctgac tgtgctttct gagtggctgg tgctccagac ccctcacctg 420
gagttccagg agggagaaac catcgtgctg aggtgccaca gctggaagga caagcctctg 480
gtcaaggtca cattcttcca gaatggaaaa tccaagaaat tttcccgttc ggatcccaac 540
ttctccatcc cacaagcaaa ccacagtcac agtggtgatt accactgcac aggaaacata 600
ggctacacgc tgtactcatc caagcctgtg accatcactg tccaagctcc cagctcttca 660
ccgatgggga tcattgtggc tgtggtcact gggattgctg tagcggccat tgttgctgct 720
gtagtggcct tgatctactg caggaaaaag cggatttcag ctctcccagg ataccctgag 780
tgcagggaaa tgggagagac cctccctgag aaaccagcca atcccactaa tcctgatgag 840
gctgacaaag ttggggctga gaacacaatc acctattcac ttctcatgca cccggatgct 900
ctggaagagc ctgatgacca gaaccgtatt tag 933
<210> 204
<211> 765
<212> DNA
<213> Homo sapiens
<400> 204
atgtggcagc tgctcctccc aactgctctg ctacttctag tttcagctgg catgcggact 60
gaagatctcc caaaggctgt ggtgttcctg gagcctcaat ggtacagggt gctcgagaag 120
gacagtgtga ctctgaagtg ccagggagcc tactcccctg aggacaattc cacacagtgg 180
tttcacaatg agagcctcat ctcaagccag gcctcgagct acttcattga cgctgccaca 240
gttgacgaca gtggagagta caggtgccag acaaacctct ccaccctcag tgacccggtg 300
cagctagaag tccatatcgg ctggctgttg ctccaggccc ctcggtgggt gttcaaggag 360
gaagacccta ttcacctgag gtgtcacagc tggaagaaca ctgctctgca taaggtcaca 420
tatttacaga atggcaaagg caggaagtat tttcatcata attctgactt ctacattcca 480
aaagccacac tcaaagacag cggctcctac ttctgcaggg ggcttgttgg gagtaaaaat 540
gtgtcttcag agactgtgaa catcaccatc actcaaggtt tgtcagtgtc aaccatctca 600
tcattctttc cacctgggta ccaagtctct ttctgcttgg tgatggtact cctttttgca 660
gtggacacag gactatattt ctctgtgaag acaaacattc gaagctcaac aagagactgg 720
aaggaccata aatttaaatg gagaaaggac cctcaagaca aatga 765
<210> 205
<211> 765
<212> DNA
<213> Homo sapiens
<400> 205
atgtggcagc tgctcctccc aactgctctg ctacttctag tttcagctgg catgcggact 60
gaagatctcc caaaggctgt ggtgttcctg gagcctcaat ggtacagggt gctcgagaag 120
gacagtgtga ctctgaagtg ccagggagcc tactcccctg aggacaattc cacacagtgg 180
tttcacaatg agagcctcat ctcaagccag gcctcgagct acttcattga cgctgccaca 240
gtcgacgaca gtggagagta caggtgccag acaaacctct ccaccctcag tgacccggtg 300
cagctagaag tccatatcgg ctggctgttg ctccaggccc ctcggtgggt gttcaaggag 360
gaagacccta ttcacctgag gtgtcacagc tggaagaaca ctgctctgca taaggtcaca 420
tatttacaga atggcaaagg caggaagtat tttcatcata attctgactt ctacattcca 480
aaagccacac tcaaagacag cggctcctac ttctgcaggg ggctttttgg gagtaaaaat 540
gtgtcttcag agactgtgaa catcaccatc actcaaggtt tggcagtgtc aaccatctca 600
tcattctttc cacctgggta ccaagtctct ttctgcttgg tgatggtact cctttttgca 660
gtggacacag gactatattt ctctgtgaag acaaacattc gaagctcaac aagagactgg 720
aaggaccata aatttaaatg gagaaaggac cctcaagaca aatga 765
<210> 206
<211> 702
<212> DNA
<213> Homo sapiens
<400> 206
atgtggcagc tgctcctccc aactgctctg ctacttctag tttcagctgg catgcggact 60
gaagatctcc caaaggctgt ggtgttcctg gagcctcaat ggtacagcgt gcttgagaag 120
gacagtgtga ctctgaagtg ccagggagcc tactcccctg aggacaattc cacacagtgg 180
tttcacaatg agagcctcat ctcaagccag gcctcgagct acttcattga cgctgccaca 240
gtcaacgaca gtggagagta caggtgccag acaaacctct ccaccctcag tgacccggtg 300
cagctagaag tccatatcgg ctggctgttg ctccaggccc ctcggtgggt gttcaaggag 360
gaagacccta ttcacctgag gtgtcacagc tggaagaaca ctgctctgca taaggtcaca 420
tatttacaga atggcaaaga caggaagtat tttcatcata attctgactt ccacattcca 480
aaagccacac tcaaagatag cggctcctac ttctgcaggg ggcttgttgg gagtaaaaat 540
gtgtcttcag agactgtgaa catcaccatc actcaaggtt tggcagtgtc aaccatctca 600
tcattctctc cacctgggta ccaagtctct ttctgcttgg tgatggtact cctttttgca 660
gtggacacag gactatattt ctctgtgaag acaaacattt ga 702
<210> 207
<211> 374
<212> PRT
<213> Homo sapiens
<400> 207
Met Trp Phe Leu Thr Thr Leu Leu Leu Trp Val Pro Val Asp Gly Gln
1 5 10 15
Val Asp Thr Thr Lys Ala Val Ile Thr Leu Gln Pro Pro Trp Val Ser
20 25 30
Val Phe Gln Glu Glu Thr Val Thr Leu His Cys Glu Val Leu His Leu
35 40 45
Pro Gly Ser Ser Ser Thr Gln Trp Phe Leu Asn Gly Thr Ala Thr Gln
50 55 60
Thr Ser Thr Pro Ser Tyr Arg Ile Thr Ser Ala Ser Val Asn Asp Ser
65 70 75 80
Gly Glu Tyr Arg Cys Gln Arg Gly Leu Ser Gly Arg Ser Asp Pro Ile
85 90 95
Gln Leu Glu Ile His Arg Gly Trp Leu Leu Leu Gln Val Ser Ser Arg
100 105 110
Val Phe Thr Glu Gly Glu Pro Leu Ala Leu Arg Cys His Ala Trp Lys
115 120 125
Asp Lys Leu Val Tyr Asn Val Leu Tyr Tyr Arg Asn Gly Lys Ala Phe
130 135 140
Lys Phe Phe His Trp Asn Ser Asn Leu Thr Ile Leu Lys Thr Asn Ile
145 150 155 160
Ser His Asn Gly Thr Tyr His Cys Ser Gly Met Gly Lys His Arg Tyr
165 170 175
Thr Ser Ala Gly Ile Ser Val Thr Val Lys Glu Leu Phe Pro Ala Pro
180 185 190
Val Leu Asn Ala Ser Val Thr Ser Pro Leu Leu Glu Gly Asn Leu Val
195 200 205
Thr Leu Ser Cys Glu Thr Lys Leu Leu Leu Gln Arg Pro Gly Leu Gln
210 215 220
Leu Tyr Phe Ser Phe Tyr Met Gly Ser Lys Thr Leu Arg Gly Arg Asn
225 230 235 240
Thr Ser Ser Glu Tyr Gln Ile Leu Thr Ala Arg Arg Glu Asp Ser Gly
245 250 255
Leu Tyr Trp Cys Glu Ala Ala Thr Glu Asp Gly Asn Val Leu Lys Arg
260 265 270
Ser Pro Glu Leu Glu Leu Gln Val Leu Gly Leu Gln Leu Pro Thr Pro
275 280 285
Val Trp Phe His Val Leu Phe Tyr Leu Ala Val Gly Ile Met Phe Leu
290 295 300
Val Asn Thr Val Leu Trp Val Thr Ile Arg Lys Glu Leu Lys Arg Lys
305 310 315 320
Lys Lys Trp Asp Leu Glu Ile Ser Leu Asp Ser Gly His Glu Lys Lys
325 330 335
Val Ile Ser Ser Leu Gln Glu Asp Arg His Leu Glu Glu Glu Leu Lys
340 345 350
Cys Gln Glu Gln Lys Glu Glu Gln Leu Gln Glu Gly Val His Arg Lys
355 360 365
Glu Pro Gln Gly Ala Thr
370
<210> 208
<211> 316
<212> PRT
<213> Homo sapiens
<400> 208
Met Thr Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu
1 5 10 15
Trp Leu Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp
20 25 30
Ser Gln Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro Trp
35 40 45
Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly Ala
50 55 60
Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn Leu
65 70 75 80
Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn
85 90 95
Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp
100 105 110
Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr Pro
115 120 125
His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His Ser
130 135 140
Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly Lys
145 150 155 160
Ser Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile Pro Gln Ala
165 170 175
Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr
180 185 190
Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro Ser
195 200 205
Met Gly Ser Ser Ser Pro Met Gly Val Ile Val Ala Val Val Ile Ala
210 215 220
Thr Ala Val Ala Ala Ile Val Ala Ala Val Val Ala Leu Ile Tyr Cys
225 230 235 240
Arg Lys Lys Arg Ile Ser Ala Asn Ser Thr Asp Pro Val Lys Ala Ala
245 250 255
Gln Phe Glu Pro Pro Gly Arg Gln Met Ile Ala Ile Arg Lys Arg Gln
260 265 270
Leu Glu Glu Thr Asn Asn Asp Tyr Glu Thr Ala Asp Gly Gly Tyr Met
275 280 285
Thr Leu Asn Pro Arg Ala Pro Thr Asp Asp Asp Lys Asn Ile Tyr Leu
290 295 300
Thr Leu Pro Pro Asn Asp His Val Asn Ser Asn Asn
305 310 315
<210> 209
<211> 317
<212> PRT
<213> Homo sapiens
<400> 209
Met Thr Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu
1 5 10 15
Trp Leu Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp
20 25 30
Ser Gln Ala Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro
35 40 45
Trp Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly
50 55 60
Ala Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn
65 70 75 80
Leu Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn
85 90 95
Asn Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser
100 105 110
Asp Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr
115 120 125
Pro His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His
130 135 140
Ser Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly
145 150 155 160
Lys Ser Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile Pro Gln
165 170 175
Ala Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly
180 185 190
Tyr Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro
195 200 205
Ser Met Gly Ser Ser Ser Pro Met Gly Ile Ile Val Ala Val Val Ile
210 215 220
Ala Thr Ala Val Ala Ala Ile Val Ala Ala Val Val Ala Leu Ile Tyr
225 230 235 240
Cys Arg Lys Lys Arg Ile Ser Ala Asn Ser Thr Asp Pro Val Lys Ala
245 250 255
Ala Gln Phe Glu Pro Pro Gly Arg Gln Met Ile Ala Ile Arg Lys Arg
260 265 270
Gln Leu Glu Glu Thr Asn Asn Asp Tyr Glu Thr Ala Asp Gly Gly Tyr
275 280 285
Met Thr Leu Asn Pro Arg Ala Pro Thr Asp Asp Asp Lys Asn Ile Tyr
290 295 300
Leu Thr Leu Pro Pro Asn Asp His Val Asn Ser Asn Asn
305 310 315
<210> 210
<211> 218
<212> PRT
<213> Homo sapiens
<400> 210
Met Thr Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu
1 5 10 15
Trp Leu Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp
20 25 30
Ser Gln Ala Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro
35 40 45
Trp Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly
50 55 60
Ala Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn
65 70 75 80
Leu Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn
85 90 95
Asn Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser
100 105 110
Asp Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr
115 120 125
Pro His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His
130 135 140
Ser Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly
145 150 155 160
Lys Ser Gln Lys Phe Ser Arg Leu Asp Pro Thr Phe Ser Ile Pro Gln
165 170 175
Ala Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly
180 185 190
Tyr Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro
195 200 205
Ser Met Gly Ser Ser Ser Pro Met Gly Ile
210 215
<210> 211
<211> 218
<212> PRT
<213> Homo sapiens
<400> 211
Met Thr Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu
1 5 10 15
Trp Leu Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp
20 25 30
Ser Gln Ala Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro
35 40 45
Trp Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly
50 55 60
Ala Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn
65 70 75 80
Leu Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn
85 90 95
Asn Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser
100 105 110
Asp Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr
115 120 125
Pro His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His
130 135 140
Ser Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly
145 150 155 160
Lys Ser Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile Pro Gln
165 170 175
Ala Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly
180 185 190
Tyr Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro
195 200 205
Ser Met Gly Ser Ser Ser Pro Met Gly Ile
210 215
<210> 212
<211> 291
<212> PRT
<213> Homo sapiens
<400> 212
Met Gly Ile Leu Ser Phe Leu Pro Val Leu Ala Thr Glu Ser Asp Trp
1 5 10 15
Ala Asp Cys Lys Ser Pro Gln Pro Trp Gly His Met Leu Leu Trp Thr
20 25 30
Ala Val Leu Phe Leu Ala Pro Val Ala Gly Thr Pro Ala Ala Pro Pro
35 40 45
Lys Ala Val Leu Lys Leu Glu Pro Gln Trp Ile Asn Val Leu Gln Glu
50 55 60
Asp Ser Val Thr Leu Thr Cys Arg Gly Thr His Ser Pro Glu Ser Asp
65 70 75 80
Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
85 90 95
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Thr
100 105 110
Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
115 120 125
Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His Leu Glu Phe Gln Glu
130 135 140
Gly Glu Thr Ile Val Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
145 150 155 160
Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser Lys Lys Phe Ser Arg
165 170 175
Ser Asp Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
180 185 190
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Leu Tyr Ser Ser Lys
195 200 205
Pro Val Thr Ile Thr Val Gln Ala Pro Ser Ser Ser Pro Met Gly Ile
210 215 220
Ile Val Ala Val Val Thr Gly Ile Ala Val Ala Ala Ile Val Ala Ala
225 230 235 240
Val Val Ala Leu Ile Tyr Cys Arg Lys Lys Arg Ile Ser Ala Asn Pro
245 250 255
Thr Asn Pro Asp Glu Ala Asp Lys Val Gly Ala Glu Asn Thr Ile Thr
260 265 270
Tyr Ser Leu Leu Met His Pro Asp Ala Leu Glu Glu Pro Asp Asp Gln
275 280 285
Asn Arg Ile
290
<210> 213
<211> 310
<212> PRT
<213> Homo sapiens
<400> 213
Met Gly Ile Leu Ser Phe Leu Pro Val Leu Ala Thr Glu Ser Asp Trp
1 5 10 15
Ala Asp Cys Lys Ser Pro Gln Pro Trp Gly His Met Leu Leu Trp Thr
20 25 30
Ala Val Leu Phe Leu Ala Pro Val Ala Gly Thr Pro Ala Ala Pro Pro
35 40 45
Lys Ala Val Leu Lys Leu Glu Pro Gln Trp Ile Asn Val Leu Gln Glu
50 55 60
Asp Ser Val Thr Leu Thr Cys Arg Gly Thr His Ser Pro Glu Ser Asp
65 70 75 80
Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
85 90 95
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Thr
100 105 110
Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
115 120 125
Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His Leu Glu Phe Gln Glu
130 135 140
Gly Glu Thr Ile Val Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
145 150 155 160
Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser Lys Lys Phe Ser Arg
165 170 175
Ser Asp Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
180 185 190
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Leu Tyr Ser Ser Lys
195 200 205
Pro Val Thr Ile Thr Val Gln Ala Pro Ser Ser Ser Pro Met Gly Ile
210 215 220
Ile Val Ala Val Val Thr Gly Ile Ala Val Ala Ala Ile Val Ala Ala
225 230 235 240
Val Val Ala Leu Ile Tyr Cys Arg Lys Lys Arg Ile Ser Ala Leu Pro
245 250 255
Gly Tyr Pro Glu Cys Arg Glu Met Gly Glu Thr Leu Pro Glu Lys Pro
260 265 270
Ala Asn Pro Thr Asn Pro Asp Glu Ala Asp Lys Val Gly Ala Glu Asn
275 280 285
Thr Ile Thr Tyr Ser Leu Leu Met His Pro Asp Ala Leu Glu Glu Pro
290 295 300
Asp Asp Gln Asn Arg Ile
305 310
<210> 214
<211> 217
<212> PRT
<213> Homo sapiens
214
Met Gly Ile Leu Ser Phe Leu Pro Val Leu Ala Thr Glu Ser Asp Trp
1 5 10 15
Ala Asp Cys Lys Ser Pro Gln Pro Trp Gly His Met Leu Leu Trp Thr
20 25 30
Ala Val Leu Phe Leu Ala Pro Val Ala Gly Thr Pro Ala Ala Pro Pro
35 40 45
Lys Ala Val Leu Lys Leu Glu Pro Gln Trp Ile Asn Val Leu Gln Glu
50 55 60
Asp Ser Val Thr Leu Thr Cys Arg Gly Thr His Ser Pro Glu Ser Asp
65 70 75 80
Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
85 90 95
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Thr
100 105 110
Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
115 120 125
Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His Leu Glu Phe Gln Glu
130 135 140
Gly Glu Thr Ile Val Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
145 150 155 160
Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser Lys Lys Phe Ser Arg
165 170 175
Ser Asp Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
180 185 190
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Leu Tyr Ser Ser Lys
195 200 205
Pro Val Thr Ile Thr Val Gln Ala Pro
210 215
<210> 215
<211> 254
<212> PRT
<213> Homo sapiens
<400> 215
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ser Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile Arg Ser Ser Thr Arg Asp Trp
225 230 235 240
Lys Asp His Lys Phe Lys Trp Arg Lys Asp Pro Gln Asp Lys
245 250
<210> 216
<211> 254
<212> PRT
<213> Homo sapiens
<400> 216
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Phe
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile Arg Ser Ser Thr Arg Asp Trp
225 230 235 240
Lys Asp His Lys Phe Lys Trp Arg Lys Asp Pro Gln Asp Lys
245 250
<210> 217
<211> 208
<212> PRT
<213> Homo sapiens
<400> 217
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Phe
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
<210> 218
<211> 208
<212> PRT
<213> Homo sapiens
<400> 218
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
<210> 219
<211> 233
<212> PRT
<213> Homo sapiens
<400> 219
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Ser Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asn Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Asp Arg Lys Tyr Phe His His Asn Ser Asp Phe His Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Ser Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile
225 230
<210> 220
<211> 211
<212> PRT
<213> Macaca fascicularis
<400> 220
Met Ser Gln Asn Val Cys Pro Gly Asn Leu Trp Leu Leu Gln Pro Leu
1 5 10 15
Thr Val Leu Leu Leu Leu Ala Ser Ala Asp Ser Gln Thr Ala Pro Pro
20 25 30
Lys Ala Val Leu Lys Leu Glu Pro Pro Trp Ile Asn Val Leu Arg Glu
35 40 45
Asp Ser Val Thr Leu Thr Cys Gly Gly Ala His Ser Pro Asp Ser Asp
50 55 60
Ser Thr Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
65 70 75 80
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Arg
85 90 95
Cys Gln Thr Gly Arg Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
100 105 110
Leu Ser Glu Trp Leu Ala Leu Gln Thr Pro His Leu Glu Phe Arg Glu
115 120 125
Gly Glu Thr Ile Met Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
130 135 140
Ile Lys Val Thr Phe Phe Gln Asn Gly Ile Ala Lys Lys Phe Ser His
145 150 155 160
Met Asp Pro Asn Phe Ser Ile Pro Arg Ala Asn His Ser His Ser Gly
165 170 175
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Pro Tyr Ser Ser Lys
180 185 190
Pro Val Thr Ile Thr Val Gln Val Pro Ser Val Gly Ser Ser Ser Pro
195 200 205
Met Gly Ile
210
<210> 221
<211> 211
<212> PRT
<213> Macaca fascicularis
<400> 221
Met Ser Gln Asn Val Cys Pro Gly Asn Leu Trp Leu Leu Gln Pro Leu
1 5 10 15
Thr Val Leu Leu Leu Leu Ala Ser Ala Asp Ser Gln Thr Ala Pro Pro
20 25 30
Lys Ala Val Leu Lys Leu Glu Pro Pro Trp Ile Asn Val Leu Arg Glu
35 40 45
Asp Ser Val Thr Leu Thr Cys Gly Gly Ala His Ser Pro Asp Ser Asp
50 55 60
Ser Thr Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
65 70 75 80
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Arg
85 90 95
Cys Gln Thr Gly Arg Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
100 105 110
Leu Ser Glu Trp Leu Ala Leu Gln Thr Thr His Leu Glu Phe Arg Glu
115 120 125
Gly Glu Thr Ile Met Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
130 135 140
Ile Lys Val Ala Phe Phe Gln Asn Gly Lys Ser Lys Asn Phe Ser His
145 150 155 160
Met Asn Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
165 170 175
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Pro Tyr Ser Ser Lys
180 185 190
Pro Val Thr Ile Thr Val Gln Val Pro Ser Val Gly Ser Ser Ser Pro
195 200 205
Met Gly Ile
210
<210> 222
<211> 211
<212> PRT
<213> Macaca fascicularis
<400> 222
Met Ser Gln Asn Val Cys Pro Gly Asn Leu Trp Leu Leu Gln Pro Leu
1 5 10 15
Thr Val Leu Leu Leu Leu Ala Ser Ala Asp Ser Gln Thr Ala Pro Pro
20 25 30
Lys Ala Val Leu Lys Leu Glu Pro Pro Trp Ile Asn Val Leu Arg Glu
35 40 45
Asp Ser Val Thr Leu Thr Cys Gly Gly Ala His Ser Pro Asp Ser Asp
50 55 60
Ser Thr Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
65 70 75 80
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Arg
85 90 95
Cys Gln Thr Gly Arg Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
100 105 110
Leu Ser Glu Trp Leu Ala Leu Gln Thr Thr His Leu Glu Phe Arg Glu
115 120 125
Gly Glu Thr Ile Met Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
130 135 140
Ile Lys Val Ala Phe Phe Gln Asn Gly Ile Ser Lys Lys Phe Ser Pro
145 150 155 160
Met Asn Pro Asn Phe Ser Ile Pro Arg Ala Asn His Ser His Ser Gly
165 170 175
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Pro Tyr Ser Ser Lys
180 185 190
Pro Val Thr Ile Thr Val Gln Val Pro Ser Val Gly Ser Ser Ser Pro
195 200 205
Met Gly Ile
210
<210> 223
<211> 217
<212> PRT
<213> Macaca fascicularis
<400> 223
Met Gly Ile Leu Ser Phe Leu Pro Val Leu Ala Thr Glu Ser Asp Trp
1 5 10 15
Ala Asp Cys Lys Ser Ser Gln Pro Trp Gly His Met Leu Leu Trp Thr
20 25 30
Ala Val Leu Phe Leu Ala Pro Val Ala Gly Thr Pro Ala Ala Pro Pro
35 40 45
Lys Ala Val Leu Lys Leu Glu Pro Pro Trp Ile Asn Val Leu Arg Glu
50 55 60
Asp Ser Val Thr Leu Thr Cys Gly Gly Ala His Ser Pro Asp Ser Asp
65 70 75 80
Ser Thr Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
85 90 95
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Arg
100 105 110
Cys Gln Thr Gly Arg Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
115 120 125
Leu Ser Glu Trp Leu Ala Leu Gln Thr Pro His Leu Glu Phe Arg Glu
130 135 140
Gly Glu Thr Ile Met Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
145 150 155 160
Ile Lys Val Thr Phe Phe Gln Asn Gly Ile Ser Lys Lys Phe Ser His
165 170 175
Met Asn Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
180 185 190
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Pro Tyr Ser Ser Lys
195 200 205
Pro Val Thr Ile Thr Val Gln Val Pro
210 215
<210> 224
<211> 208
<212> PRT
<213> Macaca fascicularis
<400> 224
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Ala Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Arg Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Thr Ser Ser Tyr Phe Ile Ala Ala Ala Arg
65 70 75 80
Val Asn Asn Ser Gly Glu Tyr Arg Cys Gln Thr Ser Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Glu Ser Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Leu Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Gly Arg Lys Tyr Phe His Gln Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Ile
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Asp Leu Ala Val Ser Ser Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
<210> 225
<211> 115
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 225
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Thr Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Ile Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Glu Ser Phe
50 55 60
Gln Asp Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 226
<211> 219
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 226
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Gln Ala Ser Glu Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 227
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 227
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 228
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 228
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 229
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 229
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Met Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 230
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 230
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Met Val Leu Asp Arg Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 231
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 231
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 232
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 232
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 233
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 233
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 234
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 234
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 235
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 235
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Thr Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 236
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 236
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Met Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 237
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 237
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Met Val Leu Arg Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 238
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 238
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 239
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 239
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 240
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 240
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 241
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 241
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Thr Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 242
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 242
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 243
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 243
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 244
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 244
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 245
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 245
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 246
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 246
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 247
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 247
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Trp Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Leu Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Thr Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 248
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 248
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Arg Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 249
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 249
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 250
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 250
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Arg Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 251
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 251
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Asn Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Asp Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Arg Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Arg Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 252
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 252
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 253
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 253
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 254
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 254
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 255
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 255
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 256
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 256
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 257
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 257
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 258
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 258
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 259
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 259
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 260
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 260
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 261
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 261
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 262
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 262
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 263
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 263
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 264
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 264
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 265
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 265
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 266
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 266
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 267
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 267
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 268
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 268
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 269
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 269
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 270
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 270
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 271
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 271
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 272
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 272
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 273
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 273
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 274
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 274
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 275
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 275
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 276
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 276
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 277
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 277
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 278
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 278
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 279
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 279
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 280
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 280
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
35 <210> 281
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 281
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 282
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 282
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 283
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 283
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 284
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 284
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 285
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 285
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 286
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 286
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 287
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 287
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 288
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 288
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 289
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 289
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 290
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 290
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 291
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 291
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 292
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 292
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 293
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 293
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 294
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 294
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 295
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 295
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 296
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 296
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 297
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 297
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 298
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 298
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 299
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 299
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 300
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 300
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 301
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 301
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 302
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 302
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 303
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 303
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 304
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 304
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 305
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 305
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 306
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 306
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 307
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 307
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
ys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 308
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 308
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 309
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 309
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
10 145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 310
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 310
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 311
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 311
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 312
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 312
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 313
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 313
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 314
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 314
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 315
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 315
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 316
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 316
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 317
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 317
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 318
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 318
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 319
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 319
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 320
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 320
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 321
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 321
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 322
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 322
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 323
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 323
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 324
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 324
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 325
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 325
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 326
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 326
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 327
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 327
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 328
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 328
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 329
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 329
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 330
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 330
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 331
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 331
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 332
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 332
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 333
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 333
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 334
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 334
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 335
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 335
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 336
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 336
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 337
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 337
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 338
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 338
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 339
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 339
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 340
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 340
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 341
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 341
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
80 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 342
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 342
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 343
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 343
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 344
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 344
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 345
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 345
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 346
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 346
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 347
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 347
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 348
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 348
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 349
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 349
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 350
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 350
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 351
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 351
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 352
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 352
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 353
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 353
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 354
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 354
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 355
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 355
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Thr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 356
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 356
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 357
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 357
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 358
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 358
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 359
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 359
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 360
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 360
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 361
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 361
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 362
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 362
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 363
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 363
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 364
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 364
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
ro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 365
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 365
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 366
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 366
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 367
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 367
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 368
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 368
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 369
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 369
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 370
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 370
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 371
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 371
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 372
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 372
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 373
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 373
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 374
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 374
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 375
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 375
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 376
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 376
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 377
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 377
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 378
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 378
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 379
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 379
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 380
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 380
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Arg Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<210> 381
<211> 328
<212> PRT
<213> Artificial sequence
<220>
<223> An artificially synthesized sequence
<400> 381
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Tyr Leu Gly Gly Asp Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Val Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Ile Asp Val Ala His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Pro Val Leu
180 185 190
His Arg Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Lys Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Lys Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Leu His Glu Ala Leu His Ala His Tyr Thr
305 310 315 320
Arg Lys Glu Leu Ser Leu Ser Pro
325
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2789884C2 |
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2799522C2 |
| АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2829536C2 |
| АНТИТЕЛА К ВИРУСУ ДЕНГЕ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2758596C2 |
| IL-12 ГЕТЕРОДИМЕРНЫЕ СЛИТЫЕ БЕЛКИ FC | 2019 |
|
RU2819097C2 |
| БИСПЕЦИФИЧЕСКИЕ 2+1 КОНТОРСТЕЛА | 2018 |
|
RU2797305C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ КОСТИМУЛЯТОРНОГО TNF-РЕЦЕПТОРА | 2016 |
|
RU2761115C1 |
| Новые гибридные ActRIIB белки-ловушки лигандов для лечения заболеваний, связанных с мышечной атрофией | 2016 |
|
RU2834707C1 |
| ГИБРИДНЫЕ БЕЛКИ ВАРИАНТА sPD-1—FC | 2019 |
|
RU2785993C2 |
| КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ СВЯЗАННЫХ С IL-8 ЗАБОЛЕВАНИЙ | 2017 |
|
RU2766112C2 |
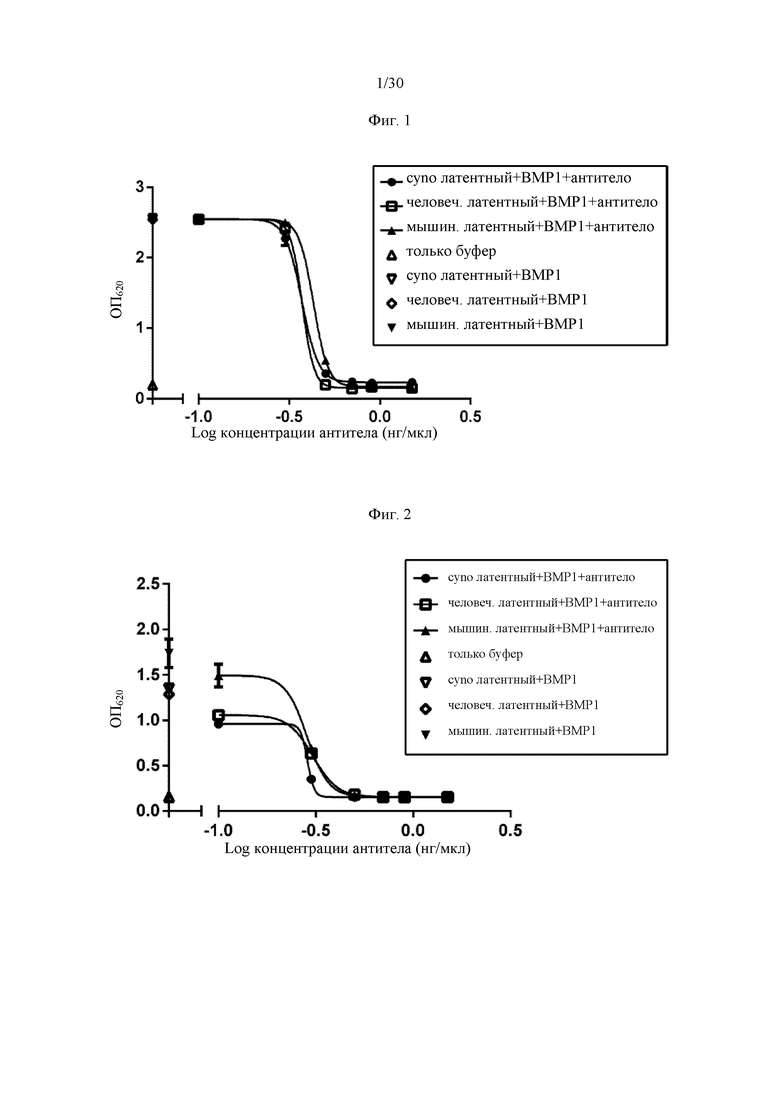
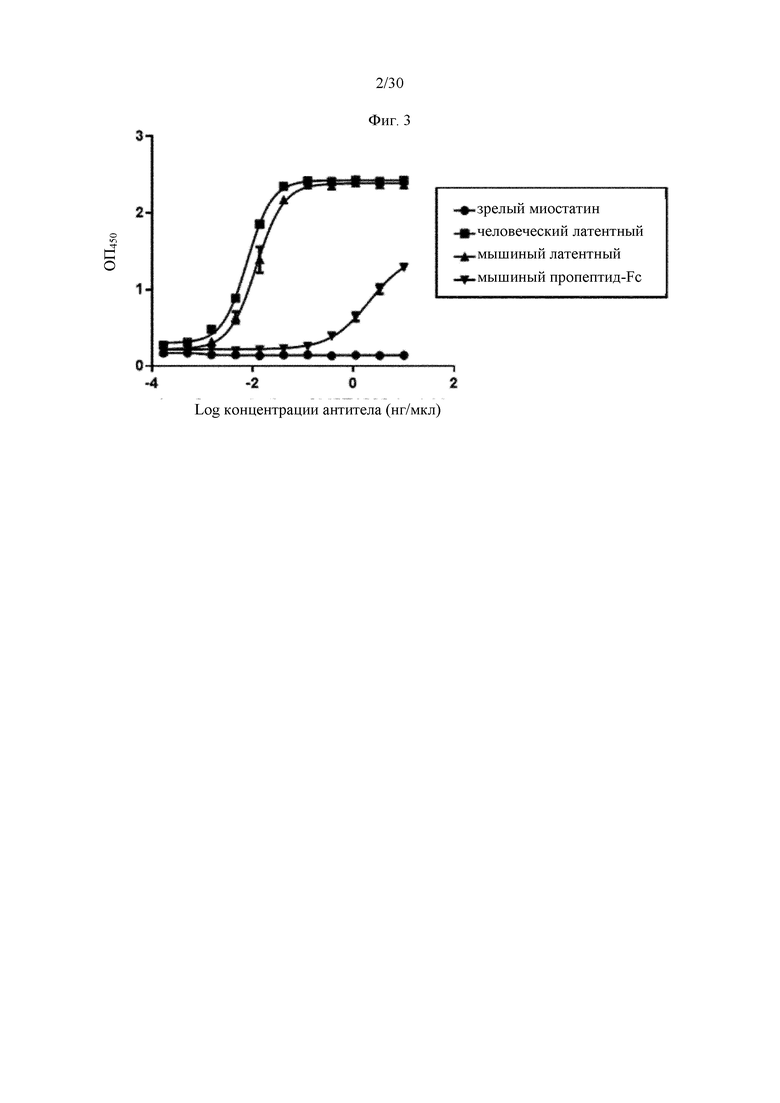
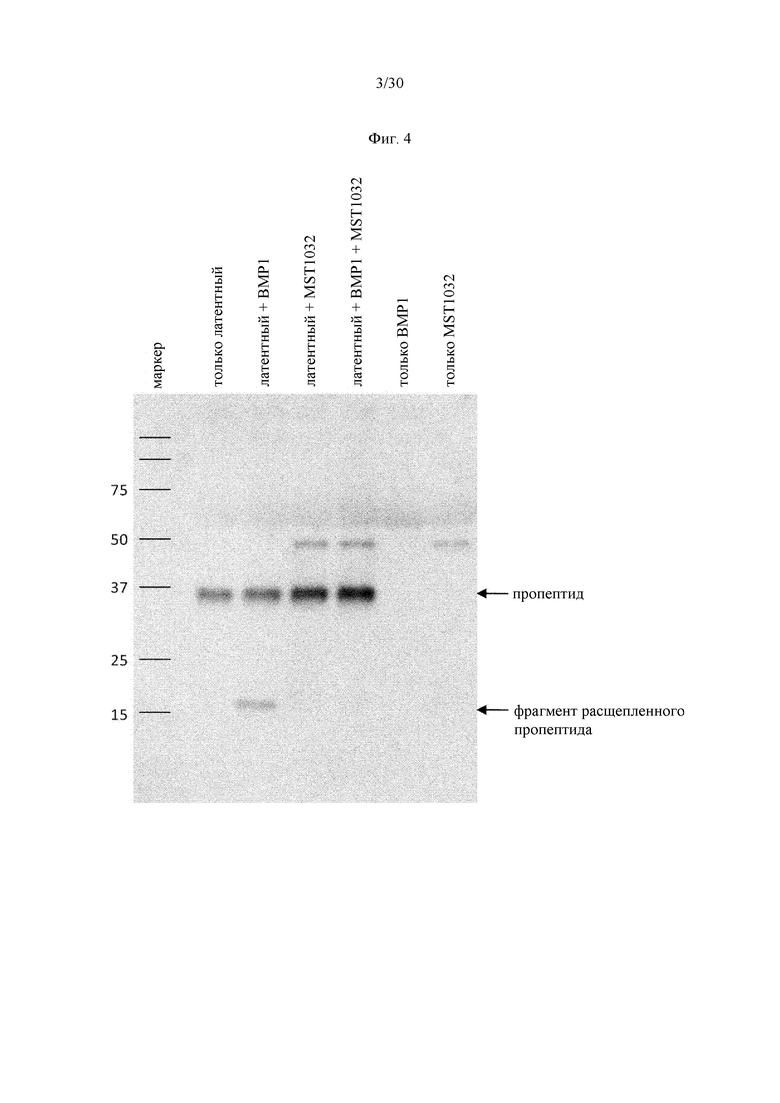
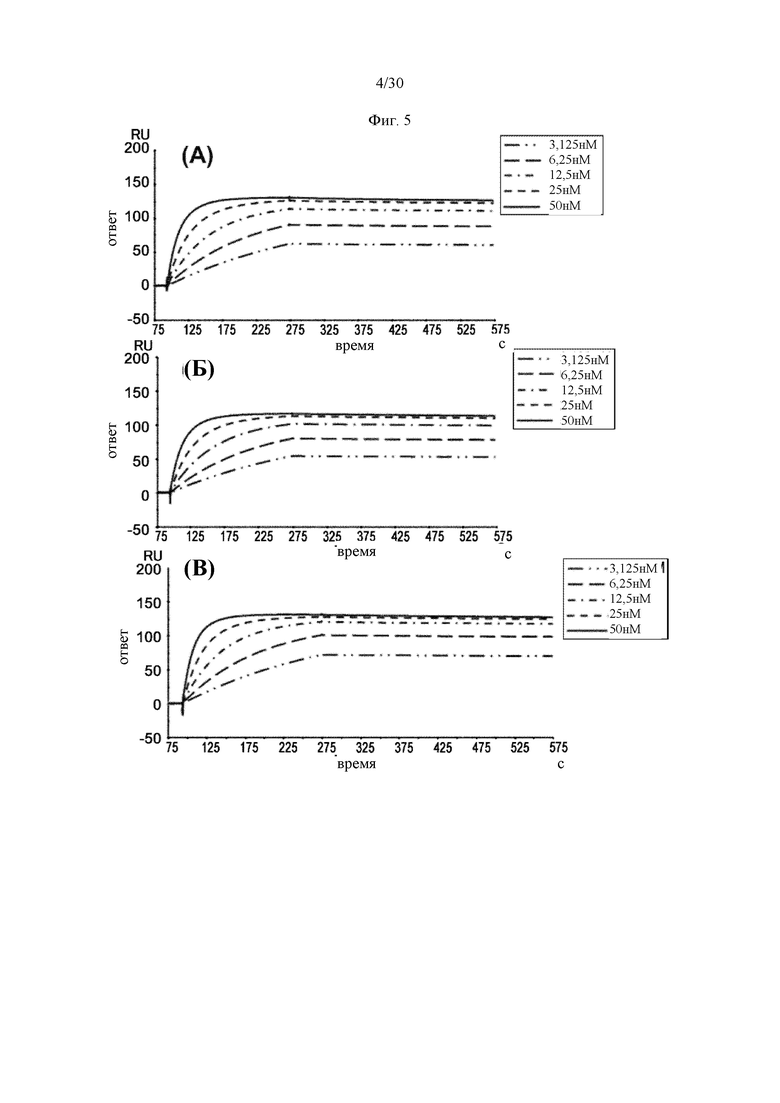
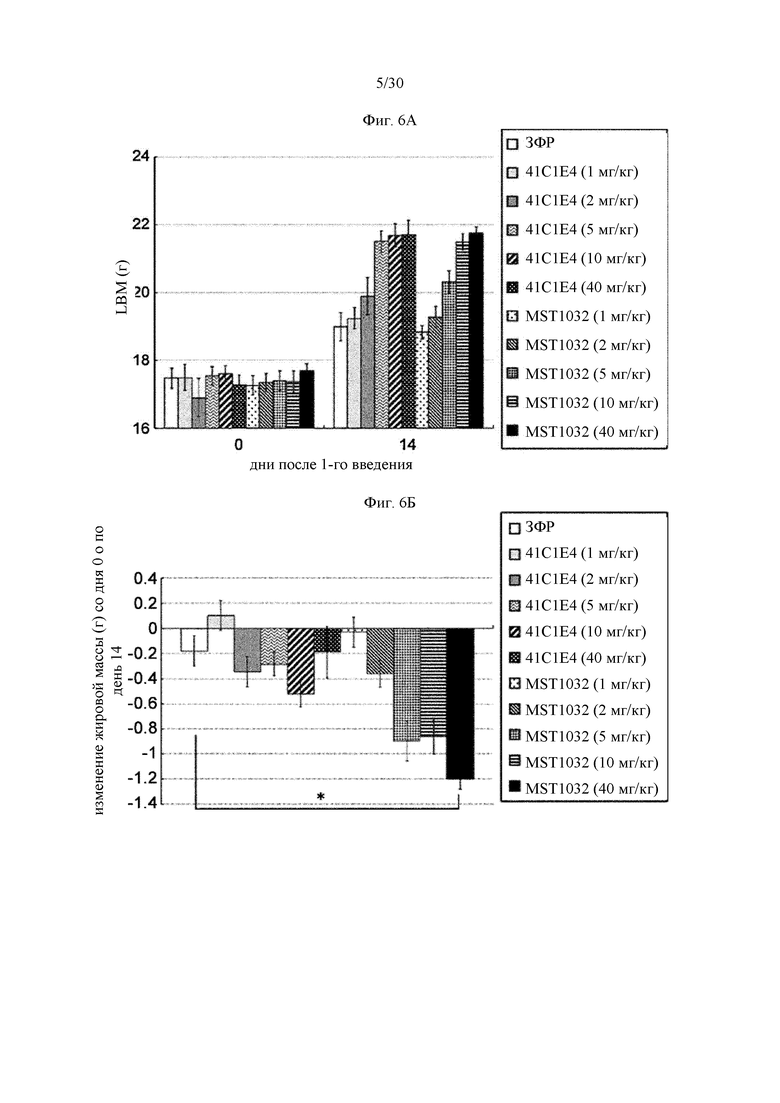

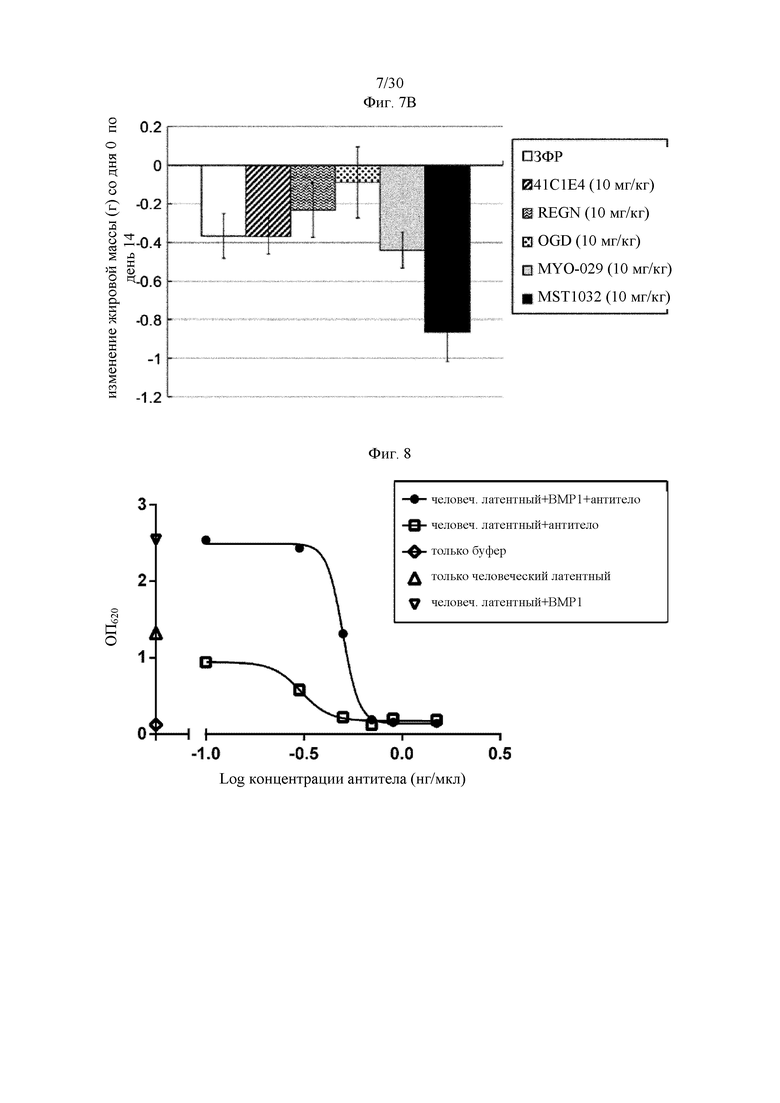
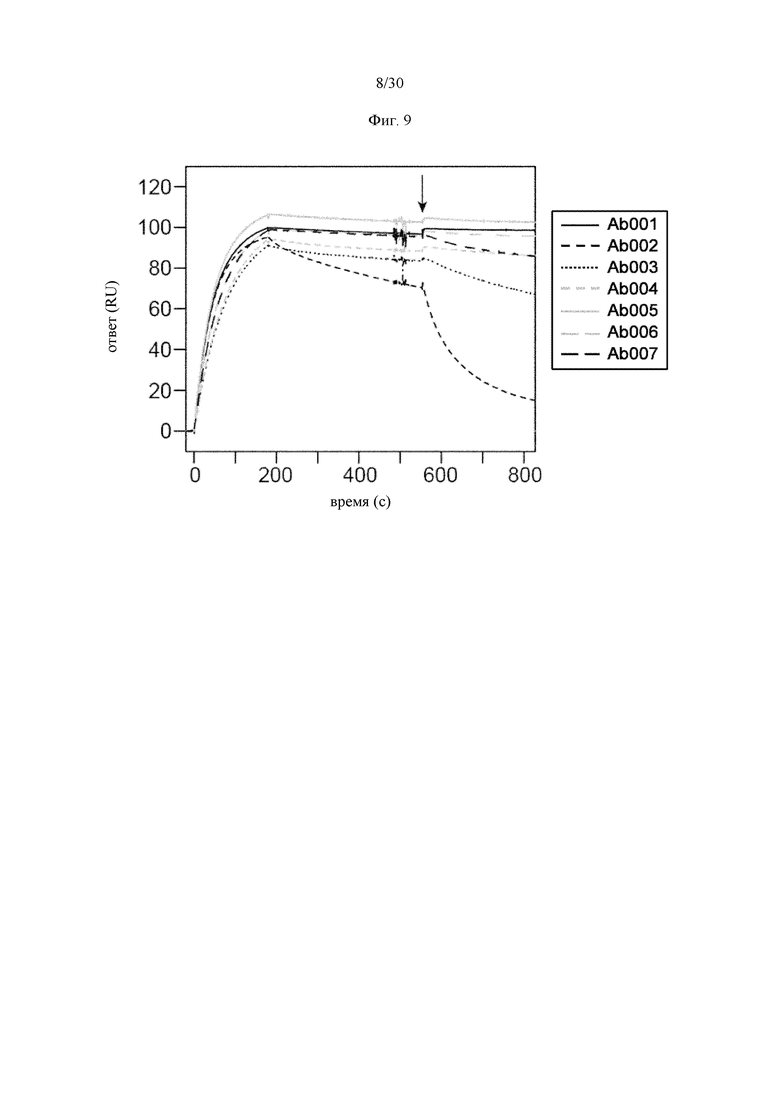
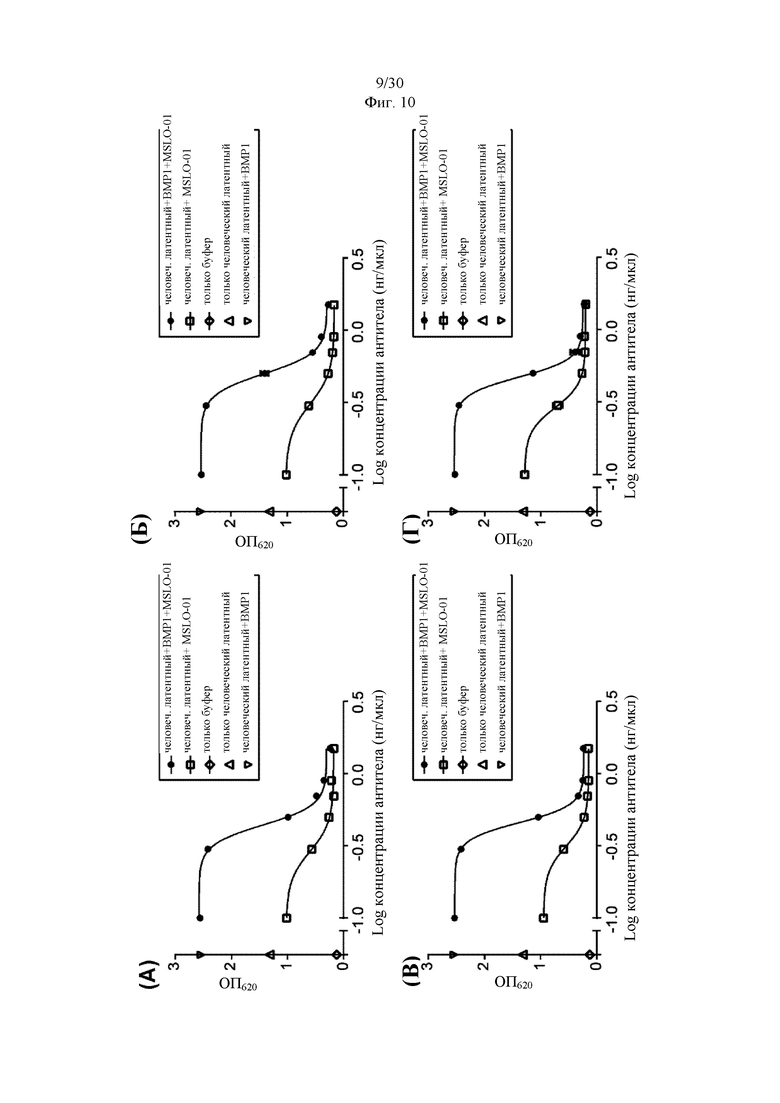
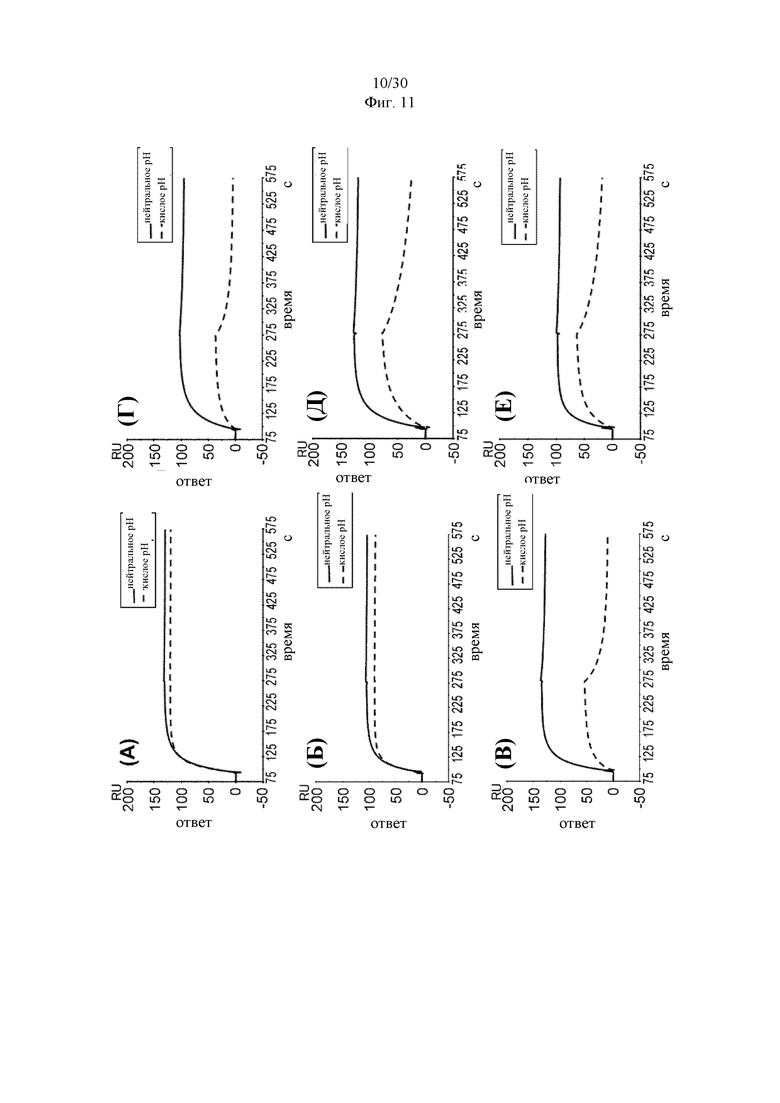
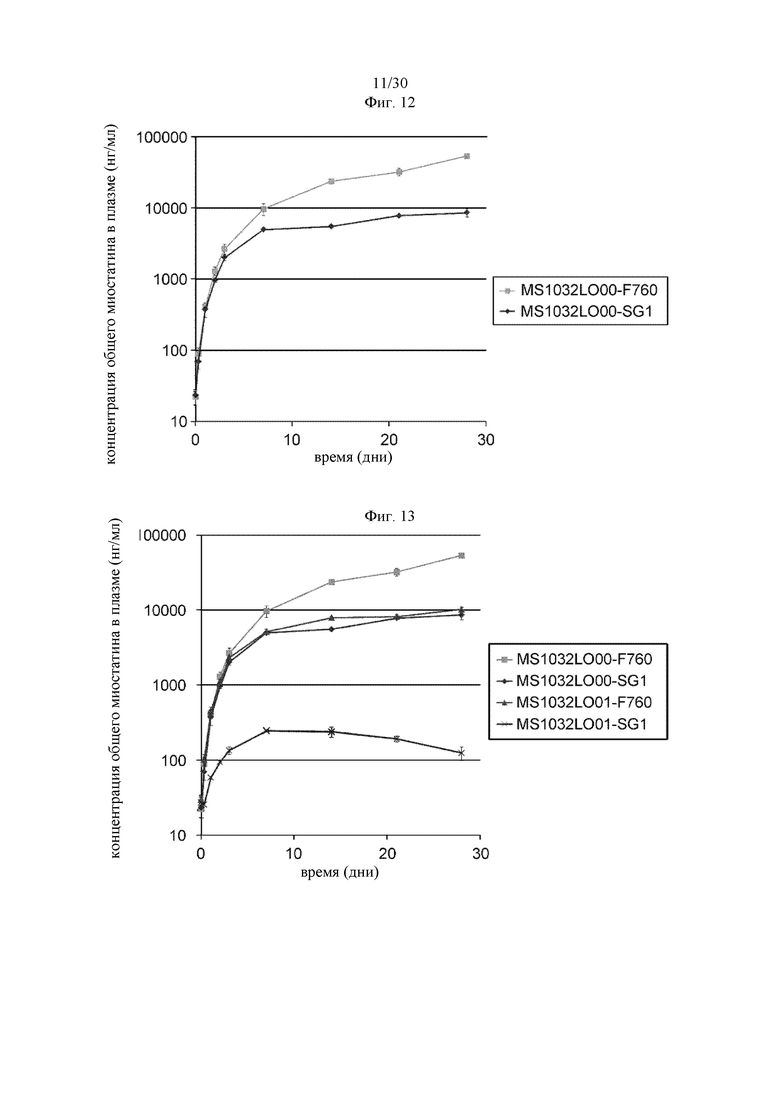
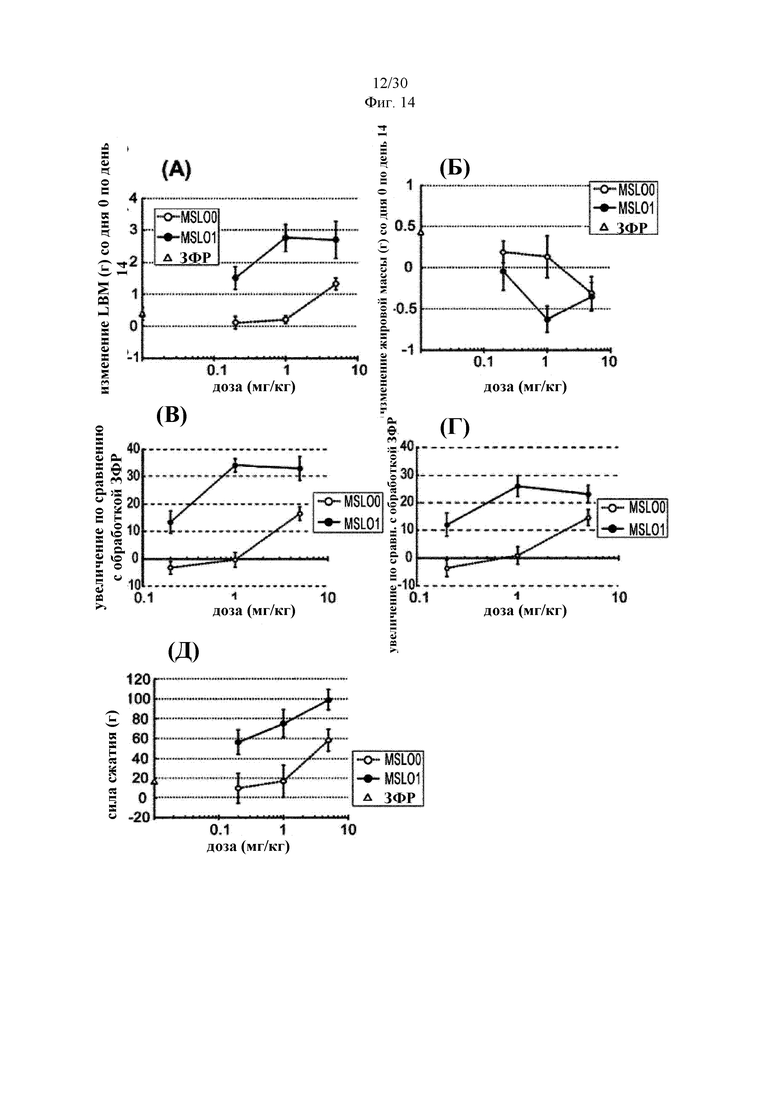
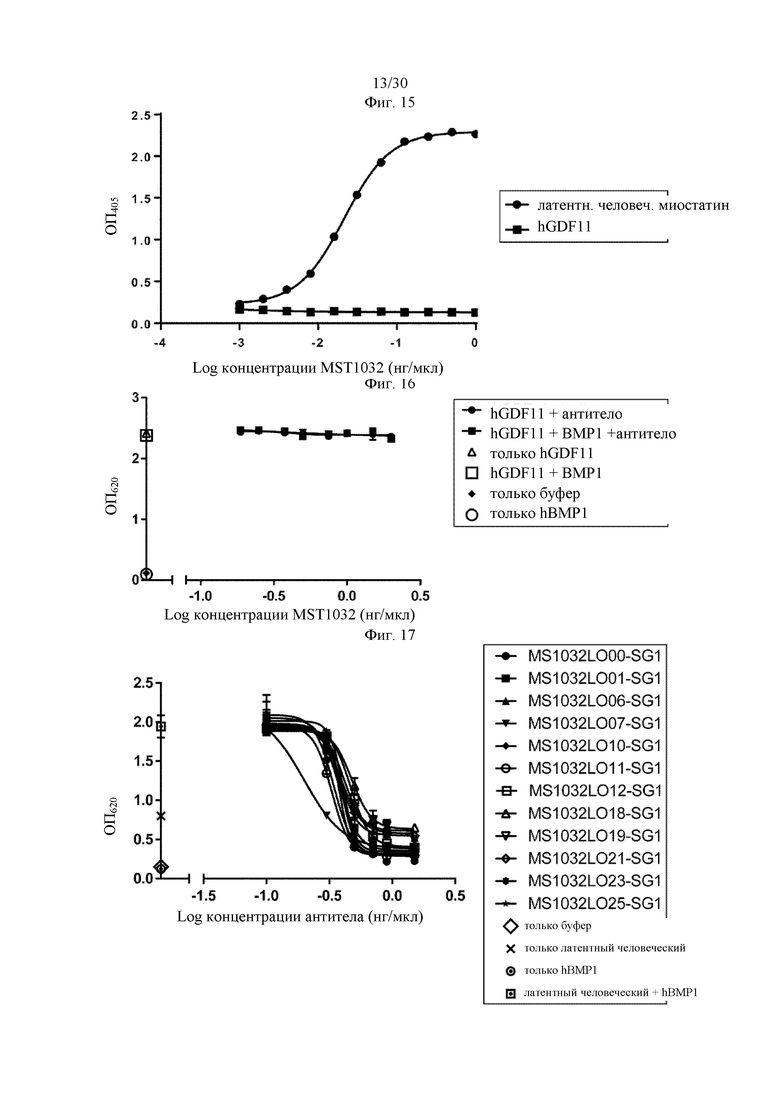
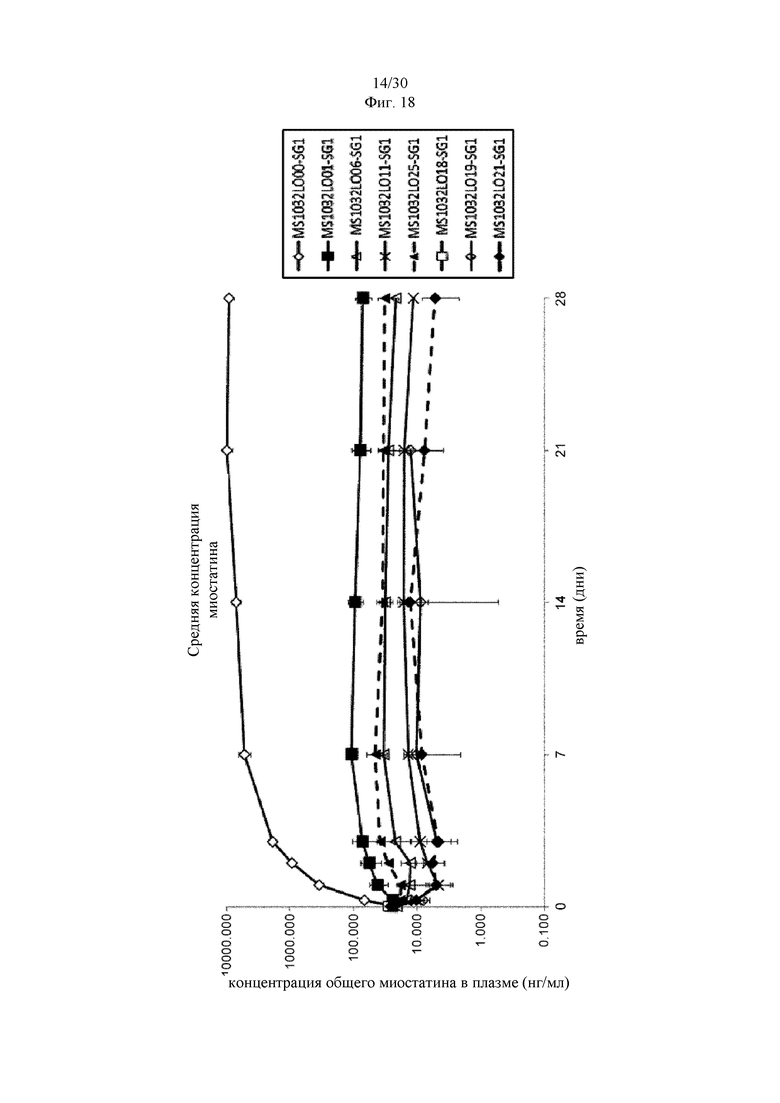
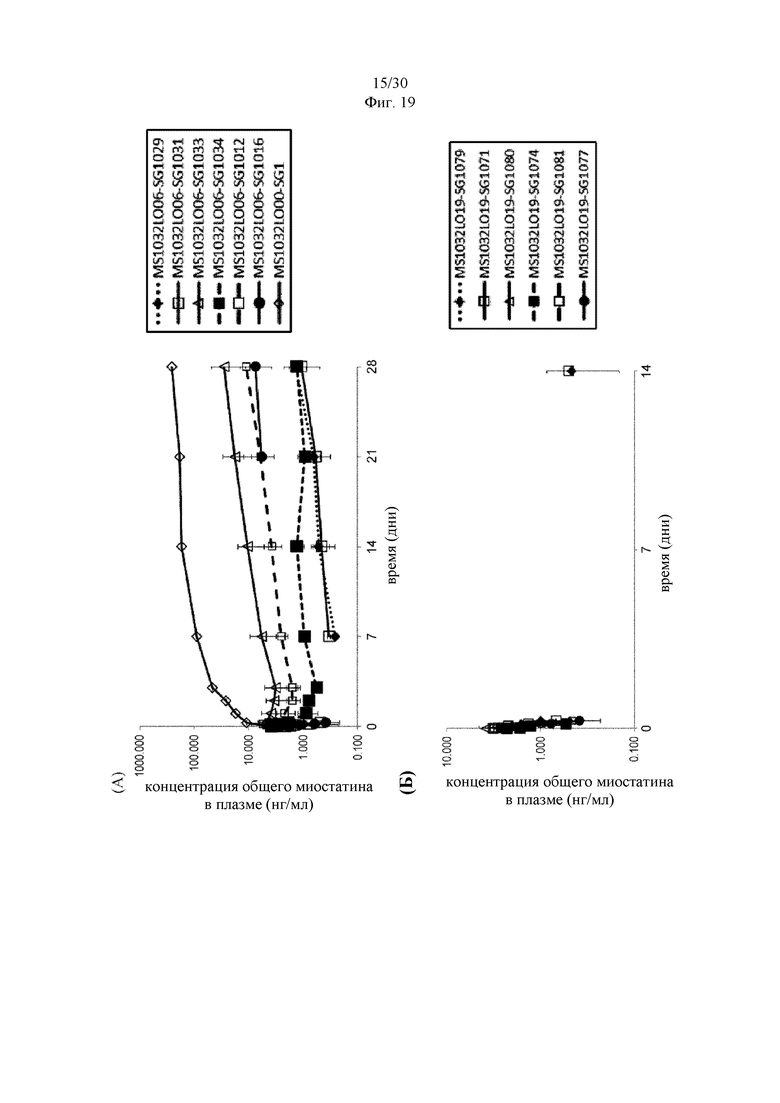
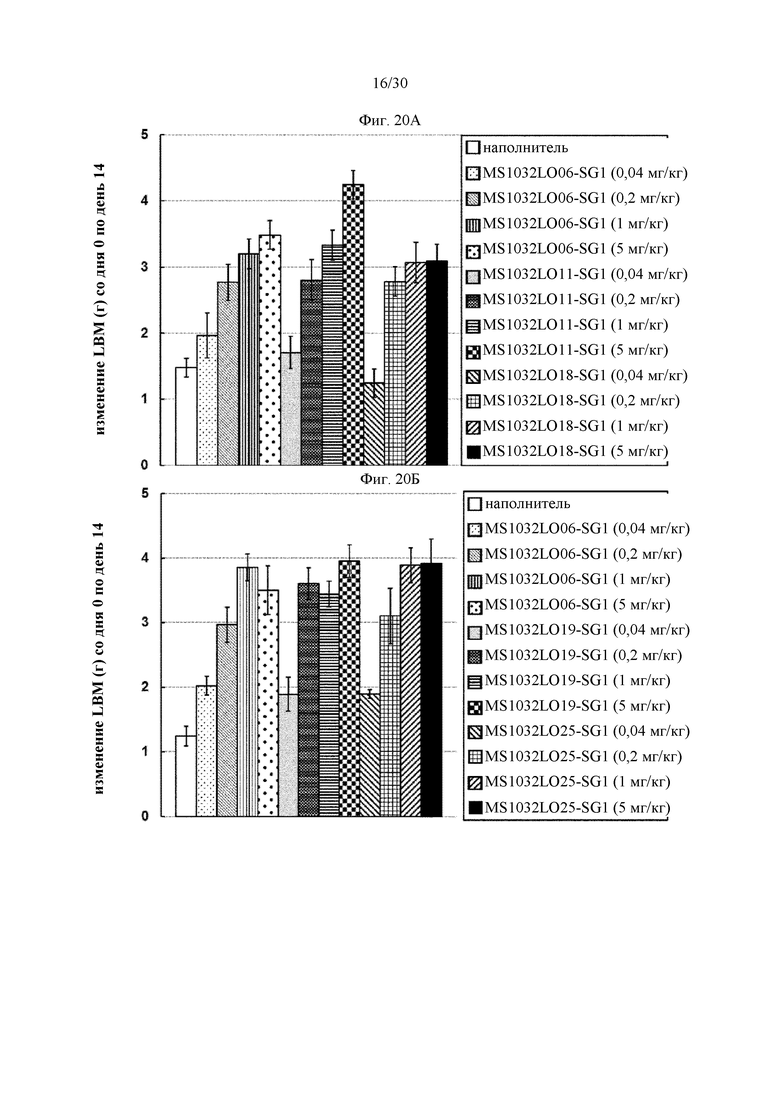
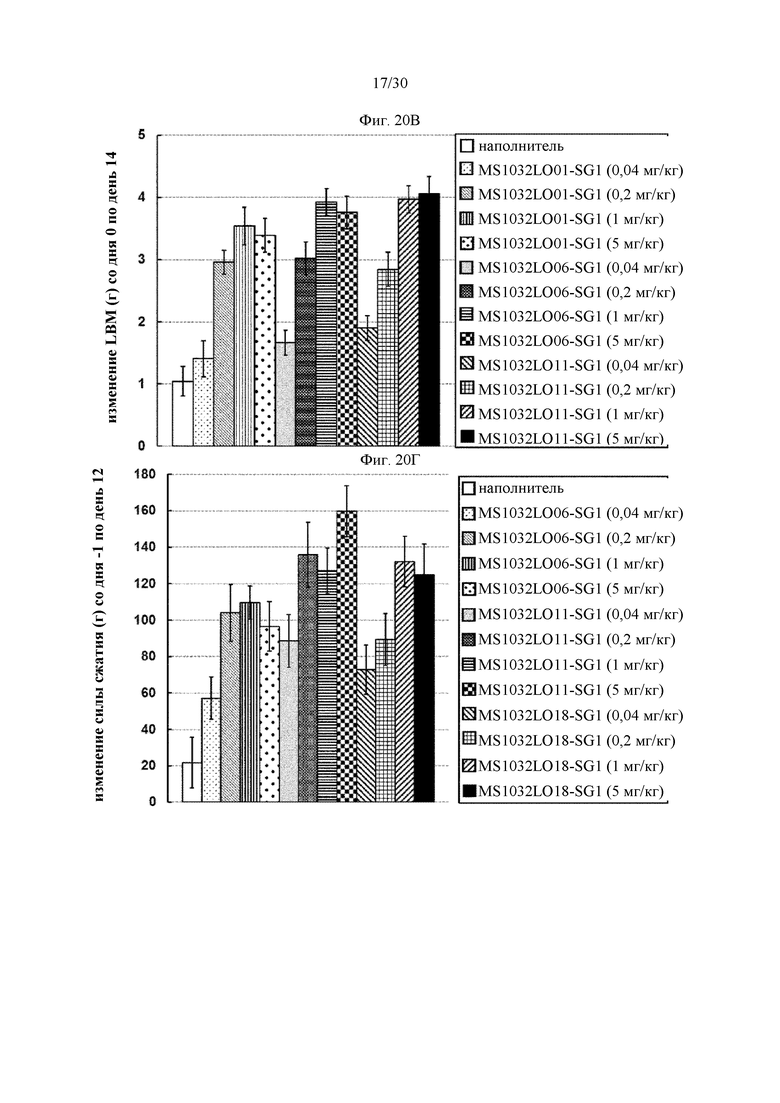
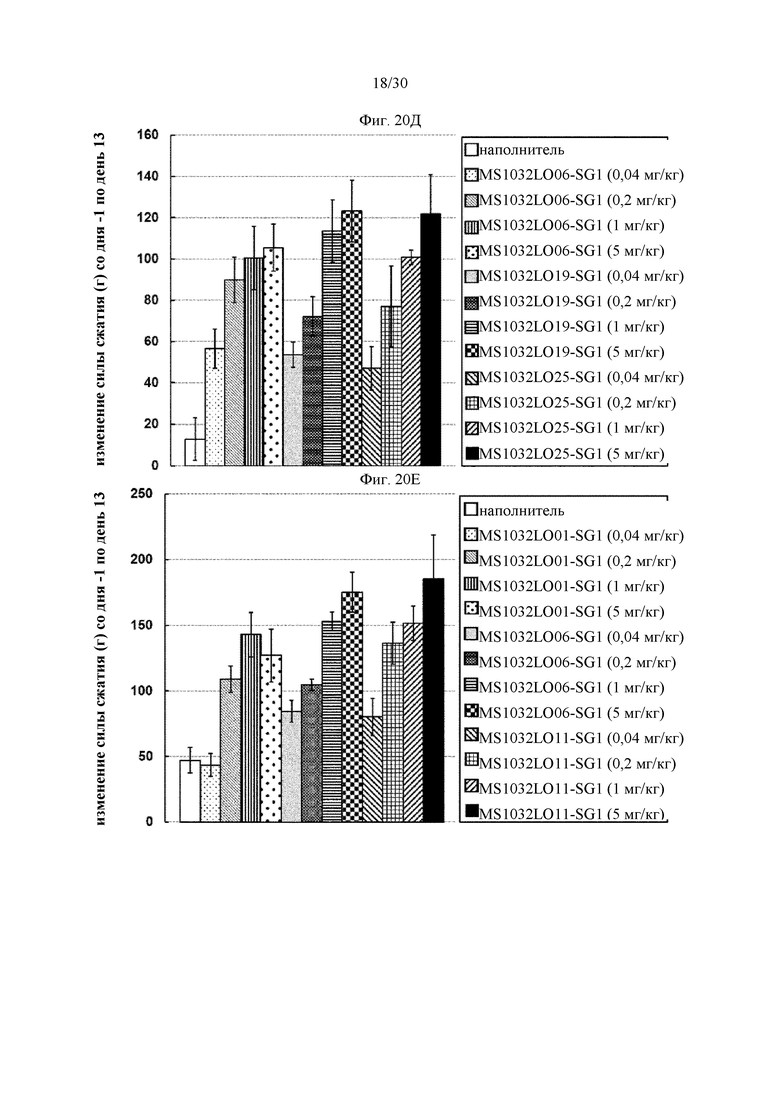

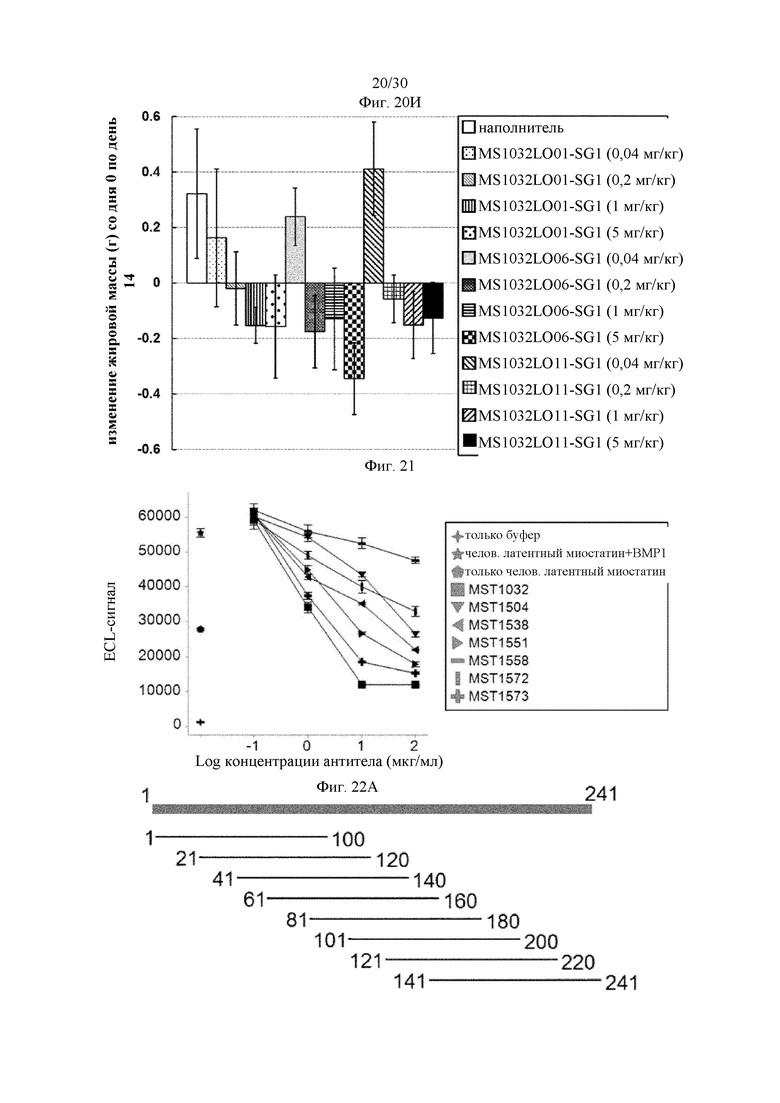
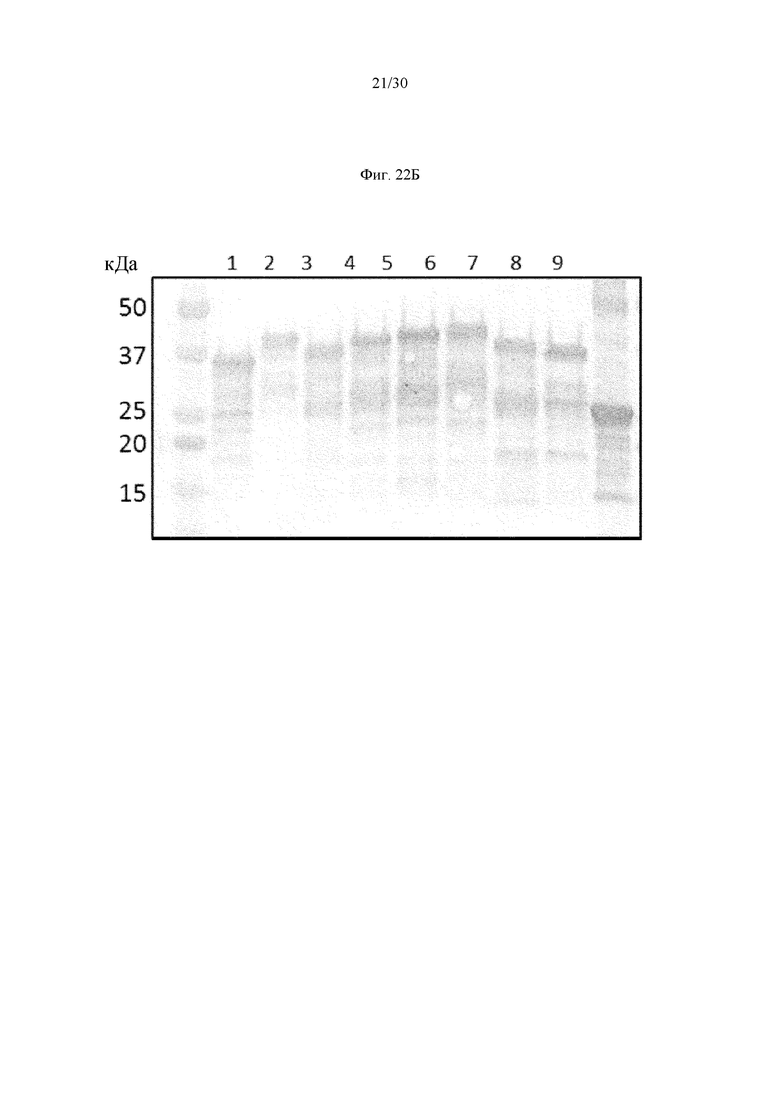

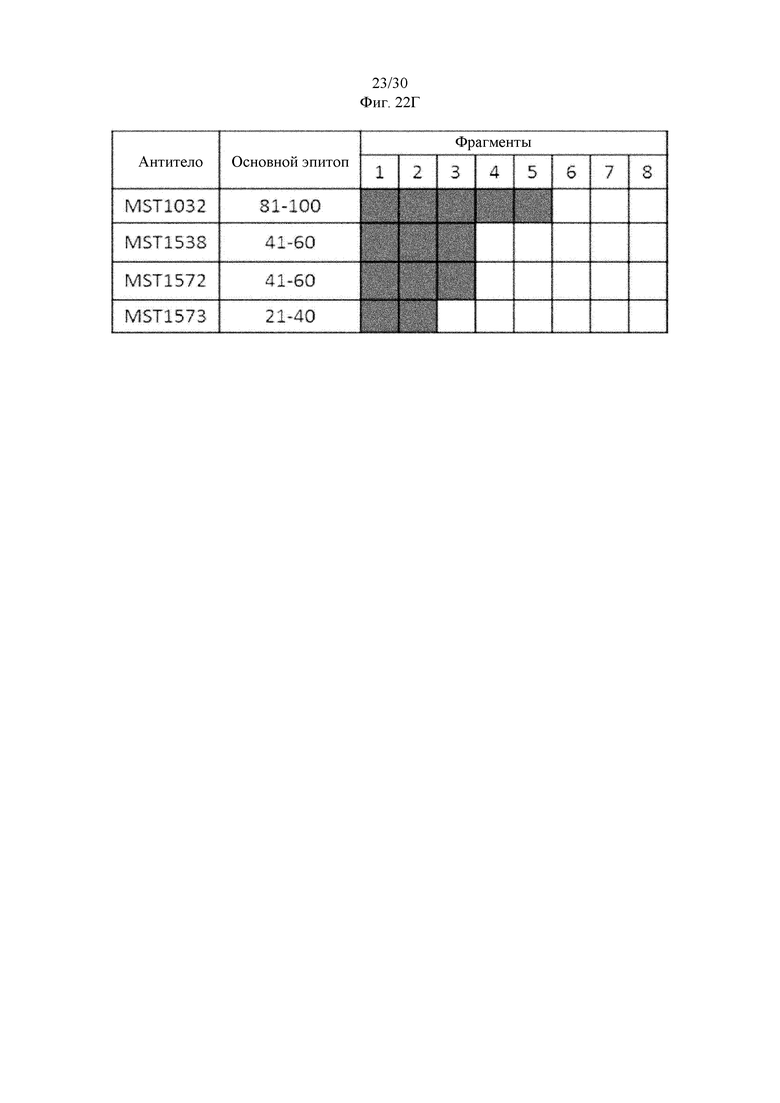
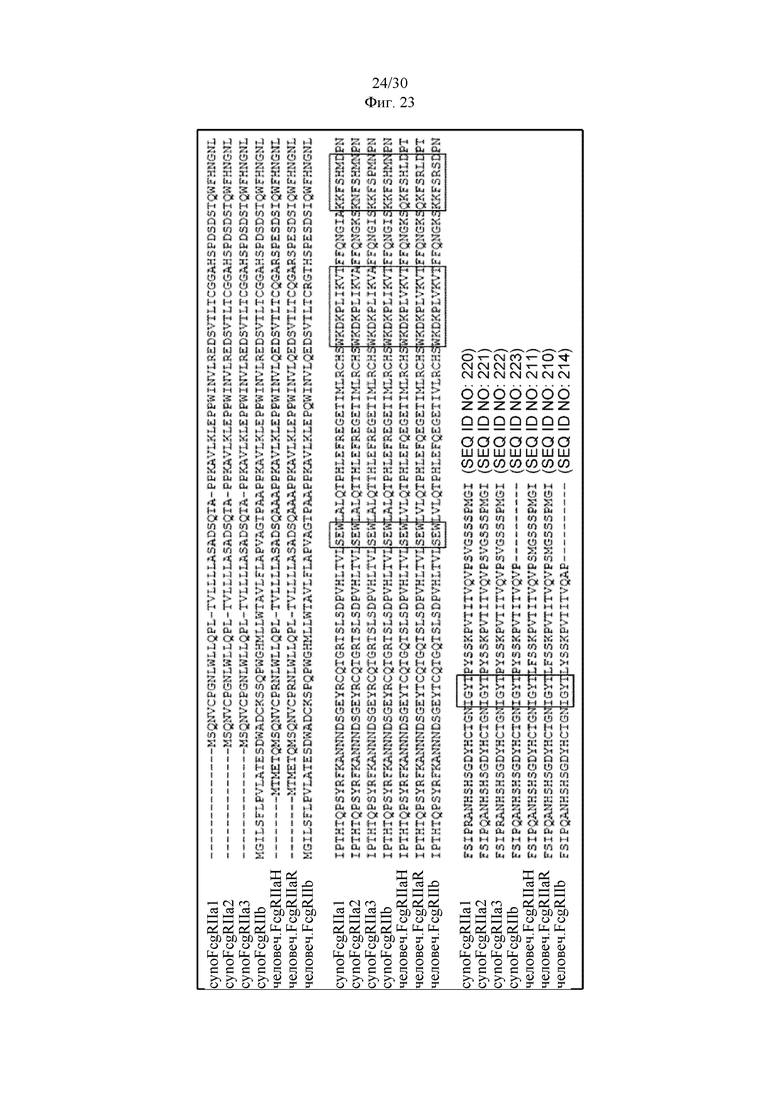
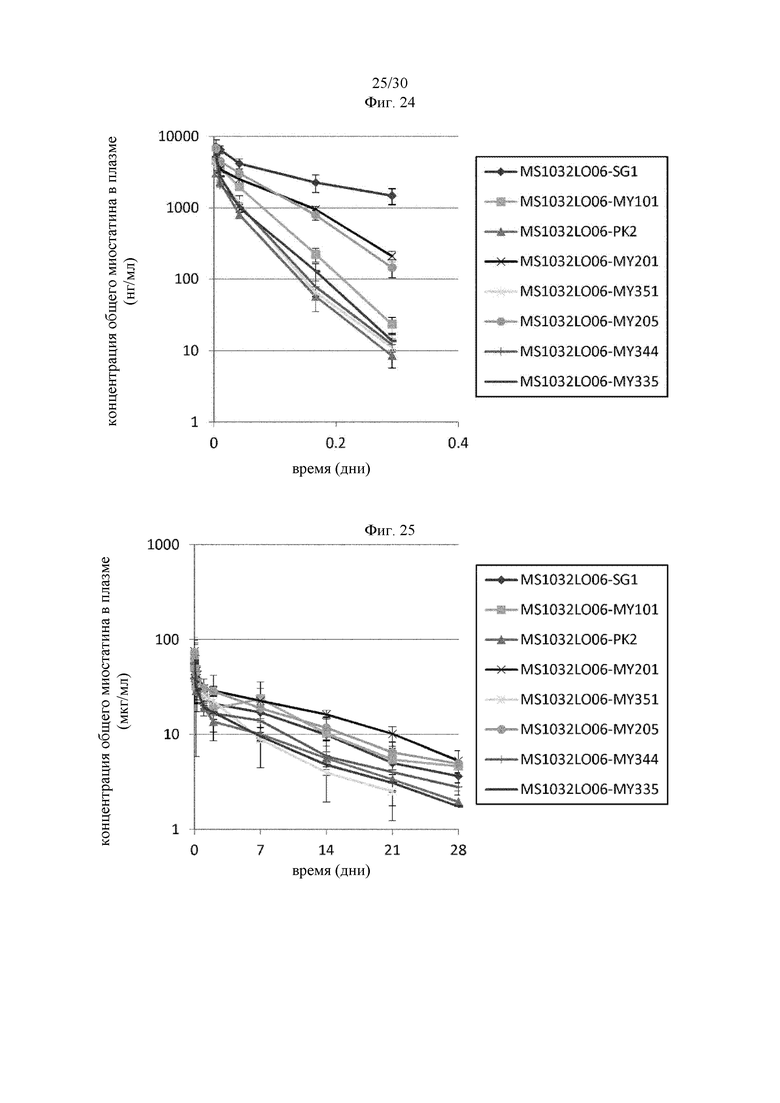
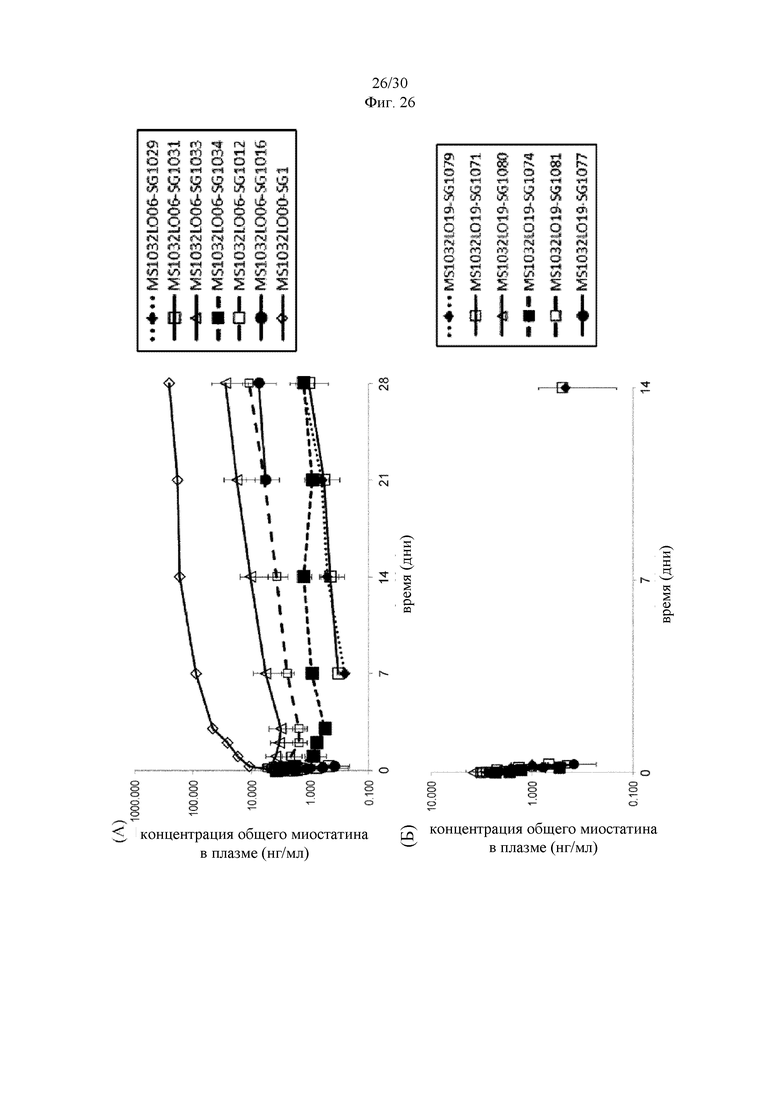
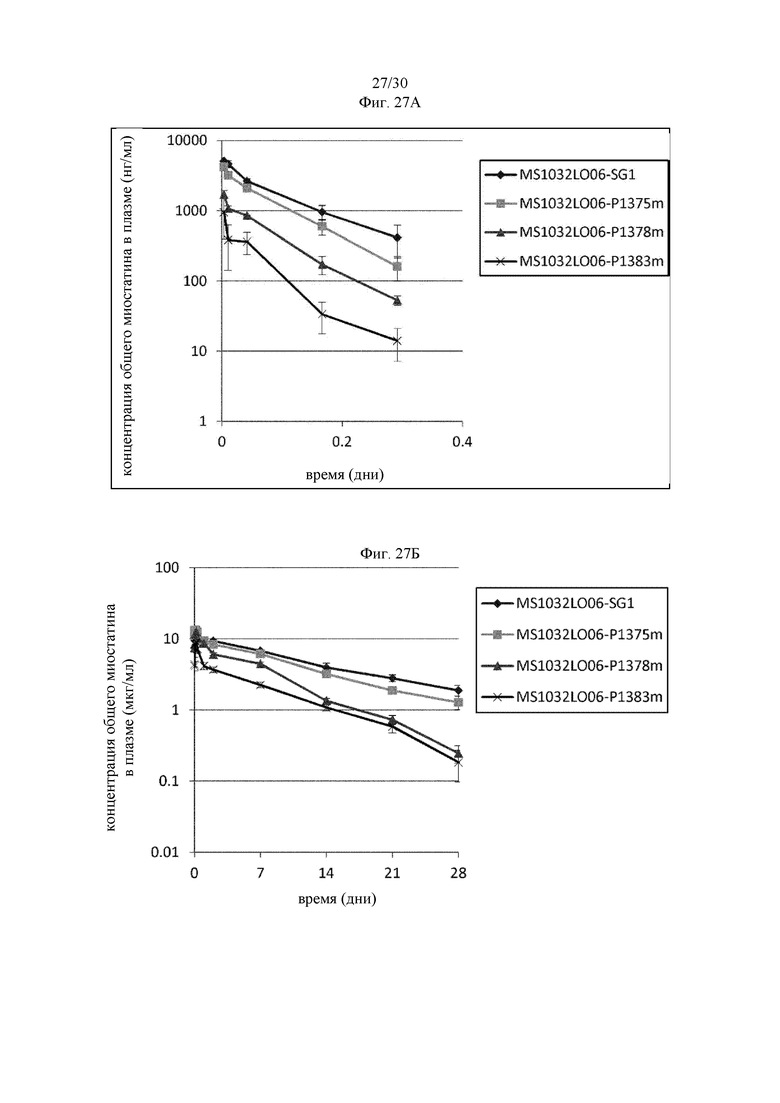
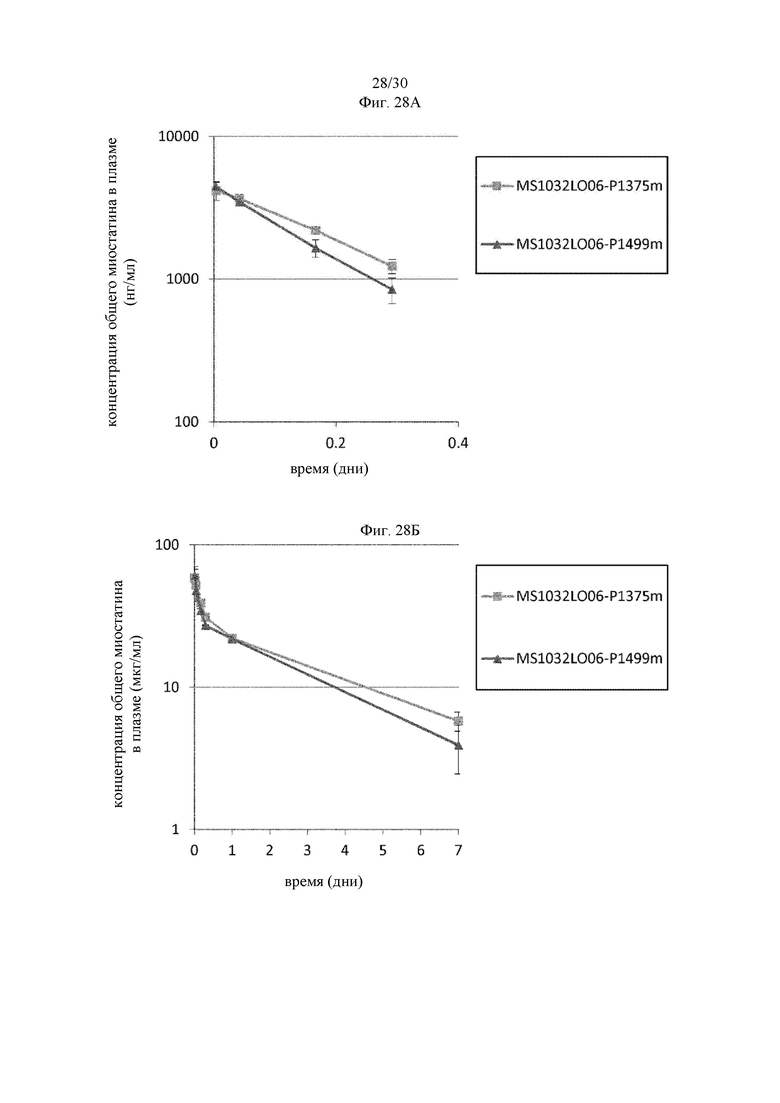
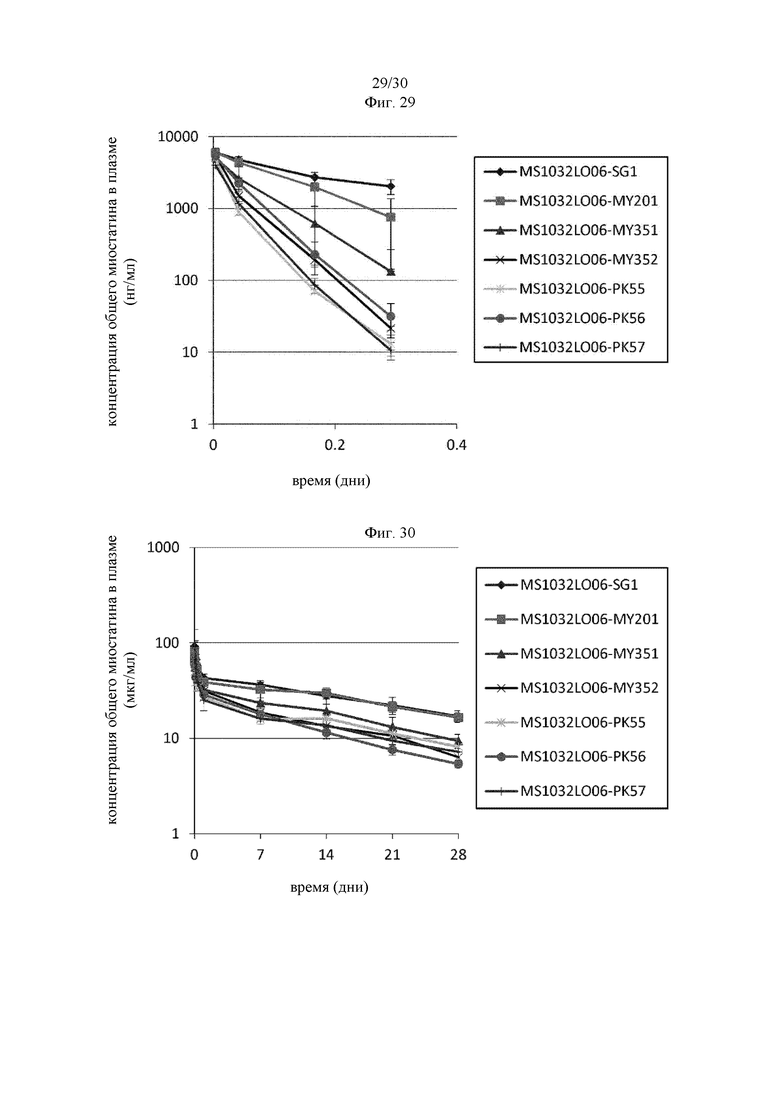
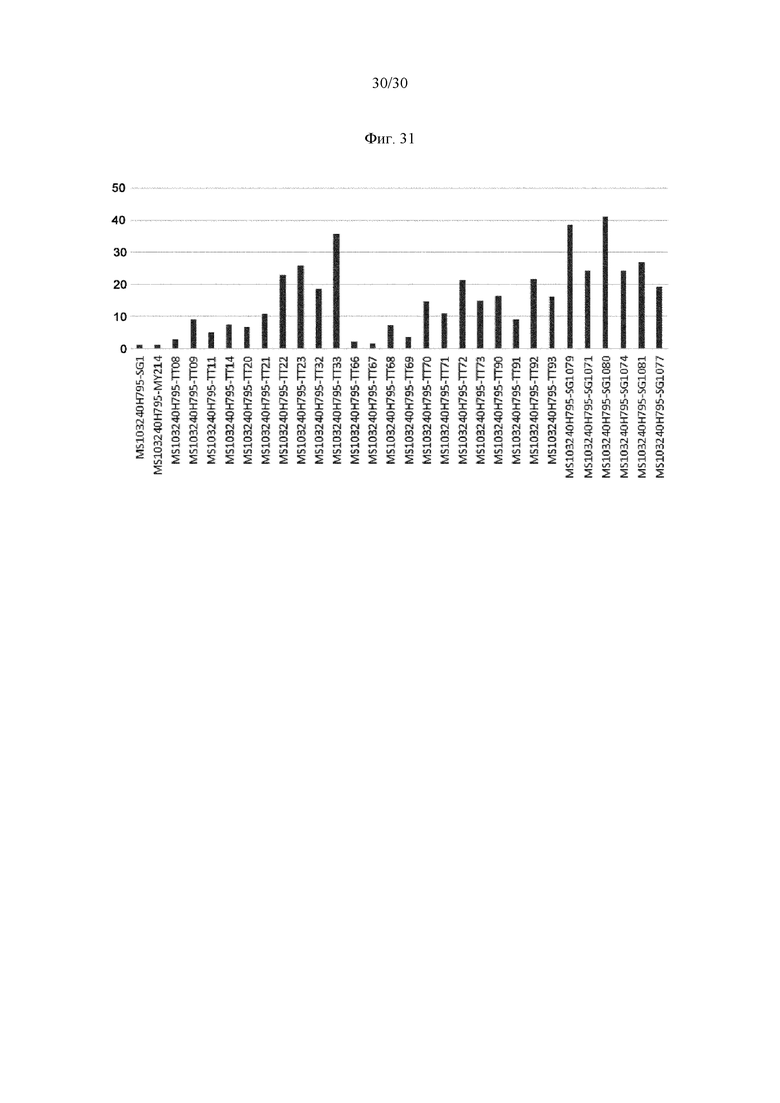
Изобретение относится к области биотехнологии. Предложены варианты применения антитела, которое связывается с латентным миостатином, для приготовления лекарственного средства. Также предложены способы применения указанного антитела у индивидуума. Лекарственные средства и способы предназначены для лечения синдрома мышечного истощения, увеличения массы мышечной ткани, увеличения силы мышечной ткани или снижения накопления жировой массы тела. Изобретение может быть использовано в терапевтических целях. 8 н. и 11 з.п. ф-лы, 48 ил., 33 табл., 34 пр.
1. Применение антитела, которое связывается с латентным миостатином, для приготовления лекарственного средства, предназначенного для лечения синдрома мышечного истощения, где антитело ингибирует активацию миостатина и где антитело содержит:
(a) последовательность VH SEQ ID NO: 86 и последовательность VL SEQ ID NO: 96;
(b) последовательность VH SEQ ID NO: 87 и последовательность VL SEQ ID NO: 96;
(c) последовательность VH SEQ ID NO: 88 и последовательность VL SEQ ID NO: 96;
(d) последовательность VH SEQ ID NO: 89 и последовательность VL SEQ ID NO: 96;
(e) последовательность VH SEQ ID NO: 90 и последовательность VL SEQ ID NO: 96;
(f) последовательность VH SEQ ID NO: 91 и последовательность VL SEQ ID NO: 96;
(g) последовательность VH SEQ ID NO: 92 и последовательность VL SEQ ID NO: 97;
(h) последовательность VH SEQ ID NO: 93 и последовательность VL SEQ ID NO: 97;
(i) последовательность VH SEQ ID NO: 94 и последовательность VL SEQ ID NO: 97; или
(j) последовательность VH SEQ ID NO: 95 и последовательность VL SEQ ID NO: 97.
2. Применение антитела, которое связывается с латентным миостатином, для приготовления лекарственного средства, предназначенного для увеличения массы мышечной ткани, где антитело ингибирует активацию миостатина и где антитело содержит:
(a) последовательность VH SEQ ID NO: 86 и последовательность VL SEQ ID NO: 96;
(b) последовательность VH SEQ ID NO: 87 и последовательность VL SEQ ID NO: 96;
(c) последовательность VH SEQ ID NO: 88 и последовательность VL SEQ ID NO: 96;
(d) последовательность VH SEQ ID NO: 89 и последовательность VL SEQ ID NO: 96;
(e) последовательность VH SEQ ID NO: 90 и последовательность VL SEQ ID NO: 96;
(f) последовательность VH SEQ ID NO: 91 и последовательность VL SEQ ID NO: 96;
(g) последовательность VH SEQ ID NO: 92 и последовательность VL SEQ ID NO: 97;
(h) последовательность VH SEQ ID NO: 93 и последовательность VL SEQ ID NO: 97;
(i) последовательность VH SEQ ID NO: 94 и последовательность VL SEQ ID NO: 97; или
(j) последовательность VH SEQ ID NO: 95 и последовательность VL SEQ ID NO: 97.
3. Применение антитела, которое связывается с латентным миостатином, для приготовления лекарственного средства, предназначенного для увеличения силы мышечной ткани, где антитело ингибирует активацию миостатина и где антитело содержит:
(a) последовательность VH SEQ ID NO: 86 и последовательность VL SEQ ID NO: 96;
(b) последовательность VH SEQ ID NO: 87 и последовательность VL SEQ ID NO: 96;
(c) последовательность VH SEQ ID NO: 88 и последовательность VL SEQ ID NO: 96;
(d) последовательность VH SEQ ID NO: 89 и последовательность VL SEQ ID NO: 96;
(e) последовательность VH SEQ ID NO: 90 и последовательность VL SEQ ID NO: 96;
(f) последовательность VH SEQ ID NO: 91 и последовательность VL SEQ ID NO: 96;
(g) последовательность VH SEQ ID NO: 92 и последовательность VL SEQ ID NO: 97;
(h) последовательность VH SEQ ID NO: 93 и последовательность VL SEQ ID NO: 97;
(i) последовательность VH SEQ ID NO: 94 и последовательность VL SEQ ID NO: 97; или
(j) последовательность VH SEQ ID NO: 95 и последовательность VL SEQ ID NO: 97.
4. Применение антитела, которое связывается с латентным миостатином, для приготовления лекарственного средства, предназначенного для снижения накопления жировой массы тела, где антитело ингибирует активацию миостатина и где антитело содержит:
(a) последовательность VH SEQ ID NO: 86 и последовательность VL SEQ ID NO: 96;
(b) последовательность VH SEQ ID NO: 87 и последовательность VL SEQ ID NO: 96;
(c) последовательность VH SEQ ID NO: 88 и последовательность VL SEQ ID NO: 96;
(d) последовательность VH SEQ ID NO: 89 и последовательность VL SEQ ID NO: 96;
(e) последовательность VH SEQ ID NO: 90 и последовательность VL SEQ ID NO: 96;
(f) последовательность VH SEQ ID NO: 91 и последовательность VL SEQ ID NO: 96;
(g) последовательность VH SEQ ID NO: 92 и последовательность VL SEQ ID NO: 97;
(h) последовательность VH SEQ ID NO: 93 и последовательность VL SEQ ID NO: 97;
(i) последовательность VH SEQ ID NO: 94 и последовательность VL SEQ ID NO: 97; или
(j) последовательность VH SEQ ID NO: 95 и последовательность VL SEQ ID NO: 97.
5. Способ лечения индивидуума, имеющего синдром мышечного истощения, включающий введение индивидууму в эффективном количестве антитела, которое связывается с латентным миостатином, где антитело ингибирует активацию миостатина и где антитело содержит:
(a) последовательность VH SEQ ID NO: 86 и последовательность VL SEQ ID NO: 96;
(b) последовательность VH SEQ ID NO: 87 и последовательность VL SEQ ID NO: 96;
(c) последовательность VH SEQ ID NO: 88 и последовательность VL SEQ ID NO: 96;
(d) последовательность VH SEQ ID NO: 89 и последовательность VL SEQ ID NO: 96;
(e) последовательность VH SEQ ID NO: 90 и последовательность VL SEQ ID NO: 96;
(f) последовательность VH SEQ ID NO: 91 и последовательность VL SEQ ID NO: 96;
(g) последовательность VH SEQ ID NO: 92 и последовательность VL SEQ ID NO: 97;
(h) последовательность VH SEQ ID NO: 93 и последовательность VL SEQ ID NO: 97;
(i) последовательность VH SEQ ID NO: 94 и последовательность VL SEQ ID NO: 97; или
(j) последовательность VH SEQ ID NO: 95 и последовательность VL SEQ ID NO: 97.
6. Способ увеличения массы мышечной ткани у индивидуума, включающий введение индивидууму в эффективном количестве антитела, которое связывается с латентным миостатином, где антитело ингибирует активацию миостатина и где антитело содержит:
(a) последовательность VH SEQ ID NO: 86 и последовательность VL SEQ ID NO: 96;
(b) последовательность VH SEQ ID NO: 87 и последовательность VL SEQ ID NO: 96;
(c) последовательность VH SEQ ID NO: 88 и последовательность VL SEQ ID NO: 96;
(d) последовательность VH SEQ ID NO: 89 и последовательность VL SEQ ID NO: 96;
(e) последовательность VH SEQ ID NO: 90 и последовательность VL SEQ ID NO: 96;
(f) последовательность VH SEQ ID NO: 91 и последовательность VL SEQ ID NO: 96;
(g) последовательность VH SEQ ID NO: 92 и последовательность VL SEQ ID NO: 97;
(h) последовательность VH SEQ ID NO: 93 и последовательность VL SEQ ID NO: 97;
(i) последовательность VH SEQ ID NO: 94 и последовательность VL SEQ ID NO: 97; или
(j) последовательность VH SEQ ID NO: 95 и последовательность VL SEQ ID NO: 97.
7. Способ увеличения силы мышечной ткани у индивидуума, включающий введение индивидууму в эффективном количестве антитела, которое связывается с латентным миостатином, где антитело ингибирует активацию миостатина и где антитело содержит:
(a) последовательность VH SEQ ID NO: 86 и последовательность VL SEQ ID NO: 96;
(b) последовательность VH SEQ ID NO: 87 и последовательность VL SEQ ID NO: 96;
(c) последовательность VH SEQ ID NO: 88 и последовательность VL SEQ ID NO: 96;
(d) последовательность VH SEQ ID NO: 89 и последовательность VL SEQ ID NO: 96;
(e) последовательность VH SEQ ID NO: 90 и последовательность VL SEQ ID NO: 96;
(f) последовательность VH SEQ ID NO: 91 и последовательность VL SEQ ID NO: 96;
(g) последовательность VH SEQ ID NO: 92 и последовательность VL SEQ ID NO: 97;
(h) последовательность VH SEQ ID NO: 93 и последовательность VL SEQ ID NO: 97;
(i) последовательность VH SEQ ID NO: 94 и последовательность VL SEQ ID NO: 97; или
(j) последовательность VH SEQ ID NO: 95 и последовательность VL SEQ ID NO: 97.
8. Способ снижения накопления жировой массы тела у индивидуума, включающий введение индивидууму в эффективном количестве антитела, которое связывается с латентным миостатином, где антитело ингибирует активацию миостатина и где антитело содержит:
(a) последовательность VH SEQ ID NO: 86 и последовательность VL SEQ ID NO: 96;
(b) последовательность VH SEQ ID NO: 87 и последовательность VL SEQ ID NO: 96;
(c) последовательность VH SEQ ID NO: 88 и последовательность VL SEQ ID NO: 96;
(d) последовательность VH SEQ ID NO: 89 и последовательность VL SEQ ID NO: 96;
(e) последовательность VH SEQ ID NO: 90 и последовательность VL SEQ ID NO: 96;
(f) последовательность VH SEQ ID NO: 91 и последовательность VL SEQ ID NO: 96;
(g) последовательность VH SEQ ID NO: 92 и последовательность VL SEQ ID NO: 97;
(h) последовательность VH SEQ ID NO: 93 и последовательность VL SEQ ID NO: 97;
(i) последовательность VH SEQ ID NO: 94 и последовательность VL SEQ ID NO: 97; или
(j) последовательность VH SEQ ID NO: 95 и последовательность VL SEQ ID NO: 97.
9. Применение или способ по одному из пп. 1-8, где антитело блокирует высвобождение зрелого миостатина из латентного миостатина.
10. Применение или способ по п. 9, где антитело блокирует протеолитическое высвобождение зрелого миостатина.
11. Применение или способ по п. 9, где антитело блокирует спонтанное высвобождение зрелого миостатина.
12. Применение или способ по одному из пп. 1-11, где антитело не связывается со зрелым миостатином.
13. Применение или способ по одному из пп. 1-12, где антитело связывается с эпитопом внутри фрагмента, состоящего из аминокислот 21-100 пропептида миостатина (SEQ ID NO: 78).
14. Применение или способ по одному из пп. 1-13, где антитело связывается с тем же эпитопом, что и антитело, представленное в таблице 2a или 11a.
15. Применение или способ по одному из пп. 1-14, где антитело связывается с латентным миостатином с более высокой аффинностью при нейтральном pH, чем при кислом pH.
16. Применение или способ по одному из пп. 1-15, где антитело представляет собой моноклональное антитело.
17. Применение или способ по одному из пп. 1-16, где антитело представляет собой человеческое, гуманизированное или химерное антитело.
18. Применение или способ по одному из пп. 1-17, где антитело представляет собой фрагмент антитела, который связывается с миостатином.
19. Применение или способ по одному из пп. 1-18, где антитело представляет собой полноразмерное антитело IgG-типа.
| WO 2016073853 A1, 12.05.2016 | |||
| WO 2010070094 A1, 24.06.2010 | |||
| WO 2011151432 A1, 08.12.2011 | |||
| НЕЙТРАЛИЗУЮЩИЕ АНТИТЕЛА ПРОТИВ GDF-8 И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2360925C2 |
| US 7731961 B1, 08.06.2010 | |||
| LATRES E | |||
| et al., Myostatin blockade with a fully human monoclonal antibody induces muscle hypertrophy and reverses muscle atrophy in young and aged mice, Skeletal Muscle, 2015, 5:34. | |||

