Изобретение относится к области биохимии и касается способа определения активности ферментов репарации: апурин/апиримидин-эндонуклеазы человека (АП-эндонуклеаза, фермент АРЕ1), селективной к одноцепочечным участкам монофункциональной урацил ДНК-гликозилазы (фермент SMUG1), алкиладенин-ДНК-гликозилазы (фермент AAG), NEI-подобной гликозилазы 1 (фермент NEIL1) и 8-оксогуанин ДНК-гликозилазы (фермент OGG1).
Клеточная ДНК во время своего функционирования постоянно подвергается воздействию различных эндо- и экзогенных факторов: высокореактивные клеточные метаболиты, алкилирующие соединения, ультрафиолетовое и ионизирующее излучение могут приводить к модификации нуклеотидов. Такие повреждения геномной ДНК могут вызывать сердечно-сосудистые, нейродегенеративные и онкологические заболевания [S. Maynard, S.H. Schurman, С. Harboe, N.C. de Souza-Pinto, V.A. Bohr. Carcinogenesis (2009)]. Кроме того, окислительный стресс, вызывающий накопление повреждений ДНК, приводит к ускоренному развитию дегенеративных процессов и преждевременному старению [F. Coppede, L. Migliore Mutat. Res. Mol. Mech. Mutagen. (2015)].
Распознавание и удаление необъемных поврежденных азотистых оснований реализуется по пути эксцизионной репарации оснований (BER), включающем в себя последовательные действия ДНК-гликозилазы, АП-эндонуклеазы, ДНК-полимеразы и ДНК-лигазы [Е.С. Friedberg, G.C. Walker, W. Siede, R.D. Wood, R.A. Schultz, T. Ellenberger, DNA Repair and Mutagenesis, ASM Press, Washington, 2006].
В процессе BER, восстановление поврежденного участка ДНК начинается с распознавания поврежденного нуклеотида ДНК-гликозилазой и гидролиза N-гликозидной связи с модифицированным азотистым основанием. У млекопитающих обнаружено 11 различных ДНК-гликозилаз, задействованных в данном процессе, которые обладают специфичностью к различным типам модифицированных нуклеотидов. Последующие этапы BER: формирование одноцепочечного разрыва за счет разрыва фосфодиэфирной связи по механизму гидролиза или β-элиминирования, удаление блокирующих групп на концах разрыва, заполнение бреши путем присоединения на 3'-конец разрыва нового нуклеотида и лигирование - называют «общими стадиями BER». Однако эти стадии могут реализоваться по различным механизмам (коротко-заплаточный и длинно-заплаточный пути), зависящим от типа инициирующей ДНК-гликозилазы, активности других участников BER и белок-белковой координации всех белков, принимающих участие в репарационном процессе.
Помимо изучения функционирования отдельных ферментов BER, белок-белковые взаимодействия активно исследуются в последнее время [Moor N.A.; Lavrik O.I. Biochemistry 2018, Howard M.; Horton J.; Zhao M.-L.; Wilson S.J. Biol. Chem. 2020]. Полученные данные свидетельствуют о том, что эффективная репарация поврежденной ДНК происходит через хорошо скоординированное действие ферментов, при котором предшествующий фермент остается связанным со своим ДНК-продуктом до тех пор, пока не будет вытеснен следующим ферментом в цепи реакции.
На сегодняшний день известно о существовании в человеческой популяции большого количества полиморфных вариантов (SNP) ферментов репарации, которые могут влиять на эффективность всего пути BER. Литературные данные свидетельствуют, что наличие аминокислотных замен, связанных с SNP, может приводить не только к ухудшению функциональной активности конкретного фермента, но и быть причиной нарушения белок-белковых взаимодействий с другими участниками BER, что может вызвать развитие различных заболеваний, в том числе и онкологических [Wallace S.S.; Murphy D.L.; Sweasy J.B. Cancer Lett. 2012, Czarny P.; Kwiatkowski D.; Toma M.; Kubiak J.; Sliwinska A.; Talarowska M.; Szemraj J.; Maes M.; Galecki P.; Sliwinski T. Mol. Neurobiol. 2017, Marsden C.G.; Dragon J.A.; Wallace S.S.; Sweasy J.B. Methods Enzym. 2017].
Таким образом, определение эффективности репарации ДНК у конкретного организма является важным направлением исследований для практического внедрения. В связи с этим, разработка и оптимизация новых способов определения активности ферментов репарации в клетках человека является актуальной задачей и чрезвычайно важна для определения уровня репарационного статуса людей. Понимание эффективности репарационного процесса позволит проводить оптимальный подбор или корректировку противоракового лечения.
Активность фермента выражается в скорости накопления продукта или скорости убытия субстрата в пересчете на количество материала, содержащего фермент. За международную единицу активности фермента (Е) принимается количество фермента, способного превратить 1 мкмоль субстрата за 1 минуту в стандартных условиях [Номенклатурный комитет Международного союза биохимии (NC-IUB) (1979) "Единицы активности ферментов". Eur. J. Биохимия].
Известен способ флуоресцентного определения активности генно-инженерных очищенных препаратов ферментов OGG1 и NEIL1 путем взаимодействия с субстратом, содержащим в составе самокомплементарной FRET-шпильки повреждение, специфичное для данного типа ДНК-гликозилазы (7,8-дигидро-8-оксогуанин (oxoG) для OGG1 и 5,6-дигидрокси-5,6-дигидротимин (Tg) для NEIL1) Реакционную смесь инкубируют при 37°С в течение 40 мин (OGG1) или 5 мин (NEIL1), а затем регистрируют FRET-сигнал, характеризующий активность данных ферментов [Donley N., Jaruga P., Coskun E., Dizdaroglu M., McCullough A.K., Lloyd R.S. ACS Chem. Biol. 2015].
Также известен способ одновременного определения активности ферментов OGG1 и AAG в ядерном экстракте клеток аденокарциномы легкого А549, основанный на использовании смеси двух ДНК-зондов, содержащих разные сочетания повреждение-флуорофор (oxoG-Су3 и Нх-Су5) для OGG1 и AAG, соответственно [J. Hu, М. Liu, Y. Li, В. Tang, С. Zhang Chem. Sci., 2018]. Для этого 10 мкл реакционной смеси распределяли на покровном стекле для визуализации изображения отдельных молекул с помощью микроскопа TIRF (Nikon, Ti-E, Япония). Для одновременного возбуждения флуоресценции Су3 и Су5 использовались лазеры с длиной волны 561 нм и 640 нм, соответственно. Фотоны собирали с использованием иммерсионного объектива (CFIApochrom TIRF 100). Флуоресценция разделялась на канал Су3 (фильтр 573-613 нм) и канал Су5 (фильтр 661,5-690,5 нм) с помощью дихроичного зеркала и выводилась на камеру EMCCD (Photometries, Evolve 512). Для анализа данных была выбрана интересующая область размером 600-600 пикселей для подсчета молекул Су3 и Су5 с использованием программного обеспечения Image J. Средние значения счетов Су3 и Су5 были получены путем усреднения десяти кадров. Используя результаты, полученные для клеточного экстракта и данные калибровки, определяют концентрацию гликозилаз OGG1 и AAG в клетках аденокарциномы легкого А549.
Известен способ определения активности урацил ДНК-гликозилазы (UDG) в экстракте клеток HeLa с использованием других ферментов, например, терминальной дезоксинуклеотидилтрансферазы (TdT), которая необходима для удлинения, расщепленного ДНК-зонда [G. Liu, W. Не, С. Liu Talanta, 2019].
Известен способ определения активности фермента АП-эндонуклеазы (АРЕ1) в сыворотке крови человека путем взаимодействия с субстратом, содержащим остаток 2-оксиметил-3-окси-тетрагидрофурана (F), где в качестве субстрата используют ДНК-дуплекс, содержащий FRET-пару ROX/BHQ2 [S. Fang, L. Chen, M. Zhao Anal. Chem. 2015]. Интенсивность флуоресценции регистрировали в режиме реального времени на приборе Rotor-Gene Q 5plex (QIAGEN, Hilden, Germany]. Программа была установлена на 250 циклов при 37°С, 5 сек на цикл, и флуоресценция измерялась в конце каждого цикла с коэффициентом усиления 10. Длины волн возбуждения/испускания 470 нм/510 нм для FAM и 585 нм/610 нм для ROX, соответственно. Скорость накопления продукта определяли по наклону линейного участка флуоресцентной кривой. Используя данные калибровки, которые были получены на чистом препарате АРЕ1, были рассчитаны концентрации фермента в образце сыворотки, которые составили 2.1±0.2 U/мл (1.2±0.1 нг/мл).
Основными недостатками известных способов являются низкая точность в случае определения активности по одной временной точке [Donley N., Jaruga P., Coskun E., Dizdaroglu M., McCullough A.K., Lloyd R.S. ACS Chem. Biol. 2015]; трудоемкость и использование дорогостоящего оборудования [J. Hu, М. Liu, Y. Li, В. Tang, С. Zhang Chem. Sci., 2018 и S. Fang, L. Chen, M. Zhao Anal. Chem. 2015]; многостадийность и многокомпонетность [G. Liu, W. He, C. Liu Talanta, 2019].
Наиболее ближайшим к заявленному способу - прототипом, является способ определения активности фермента АП-эндонуклеазы (АРЕ1), заключающийся в следующем Реакцию запускают путем смешивания растворов субстрата и фермента в соотношении 1:1 с помощью спектрофотометра остановленного потока (Applied Photophysics Ltd., Великобритания) и регистрируют изменение интенсивности флуоресценции субстрата в интервале времени 1-2000 с. Для этого, в кювете флуориметра смешивают растворы субстрата (1,0×10-6 М), буфера (50 мМ Tris-HCl (рН 7,5), 50 мМ KCl, 5 мМ MgCl2, 1 мМ дитиотреит, 9% глицерин) и образца, содержащего рекомбинантный фермент АП-эндонуклеазы человека в концентрациях 1,0×10-7 М, 1,0×10-8 М, 1,0×10-9 М, 1,0×10-10 М [Кузнецов Н.А., Коваль В.В., Федорова О.С. Способ определения активности АП-эндонуклеазы человека. [Патент RU 2389026 С1, опубл. 10.05.2010].
В качестве субстрата используют двуспиральный комплекс комплементарных олигодезоксирибонуклеотидов длиной 13 или более звеньев, содержащий в центральном положении остаток 2-оксиметил-3-окси-тетрагидрофурана (F), например
5'-FLU-С-С-Т-С-Т-C-F-С-С-Т-Т-С-С-3'
3'-DABCYL-G-G-A-G-A-G-C-G-G-A-A-G-G-5',
где FLU - остаток флуоресцеина, DABCYL - остаток дабцила.
Взаимодействие субстрата и фермента АРЕ1 приводит к расщеплению рибозо-фосфатного остова ДНК по специфическому сайту F. В результате чего в продукте реакции цепи олигонуклеотидов расходятся, что приводит к пространственному разделению флуорофора и тушителя. Интенсивность флуоресценции увеличивается по мере накопления продукта реакции.
Регистрируют изменение интенсивности флуоресценции субстрата в реакционной смеси в спектральном диапазоне 500-550 нм, возбуждение флуоресценции проводят на длине волны 494 нм. Кинетические кривые обрабатывают методом нелинейной регрессии.
Недостатками прототипа являются недостаточная точность и достоверность результатов определения активности фермента, использование устаревшего типа флуорофора (DABCYL), невозможность использования ДНК-дуплекса в качестве ДНК-зонда в клеточных системах, а также большой расход реагентов, необходимый для метода остановленного потока.
Задачей изобретения является повышение точности и достоверности результатов определения активности ферментов репарации ДНК у человека, расширение функциональных возможностей известного способа, и применение этого способа для анализа активности в клетках человека.
Поставленная задача достигается предлагаемым способом, заключающимся в регистрации во времени изменения интенсивности флуоресценции субстрата в реакционной смеси при взаимодействии с клеточным экстрактом, полученным из клеточных линий рака яичника человека (SCOV-3, 79, OVCAR-3, TOV-21G, TOV112D, MES-OV).
Запускают реакцию путем смешивания растворов субстрата и клеточного экстракта в соотношении 1:1 и регистрируют изменение интенсивности флуоресценции субстрата в интервале времени 15-3600 с. Для этого, в 96-луночном планшете для флуориметра смешивают растворы субстрата (1,0×10-6 М), буфера (50 мМ Tris-HCl рН 7.5, 50 мМ KCl, 1 мМ ЭДТА, 5 мМ MgCl2, 1 мМ ДТТ, 7% глицерин) и образца экстракта клеточных линий рака яичника человека (25 мкг общего белка на один анализ).
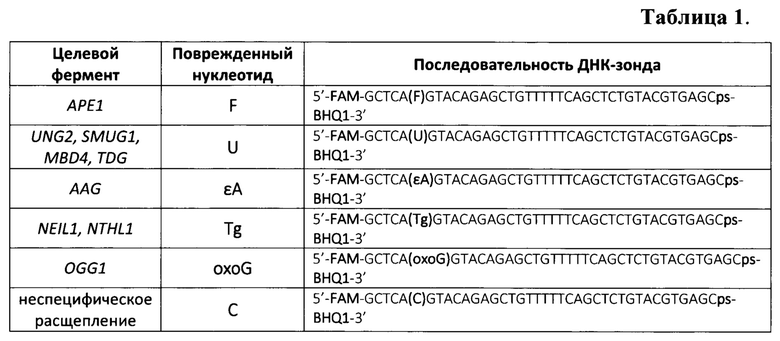
В качестве субстрата используют олигодезоксирибонуклеотиды, формирующие шпильку и несущие FRET-пару FAM/BHQ1 на концах цепи (где FAM - остаток флуоресцеина, BHQ1 - остаток тушителя black hole quencher 1), а также содержащие в своем составе повреждение, специфичное для определяемого фермента. Структура и специфичность использованных ДНК-зондов представлена в таблице 1. Для предотвращения неспецифической 3'-5'-экзонуклеазной деградации ДНК-зонда в клеточном экстракте 3'-концевая межнуклеотидная фосфатная группа заменена на тиофосфатную (ps).
Взаимодействие фермента с ДНК-зондом приводит к узнаванию специфического сайта и сопровождается расщеплением 2'-дезоксирибозо-фосфатного остова ДНК. В результате расщепления одной из цепей в продукте реакции происходит диссоциация укороченных фрагментов олигонуклеотидов, что сопровождается пространственным отдалением флуорофора и тушителя. Интенсивность флуоресценции увеличивается по мере накопления продукта реакции.
Возбуждение флуоресценции проводят на длине волны 494 нм, изменение интенсивности флуоресценции ДНК-зонда регистрируют на длине волны 500-550 нм.
Начальные линейные участки полученных кинетических кривых обрабатывают с помощью программы OriginPro8.1 (OriginLab Corp., США), используя для обработки данных уравнение прямой: F=F0+kobs×t,
где F0 - амплитуда, kobs - наблюдаемая константа скорости образования продукта. В результате обработки получают зависимость значений наблюдаемой константы скорости от концентрации фермента для каждого типа зонда.
Определяющими отличиями заявленного способа от прототипа являются:
- использование в качестве ДНК-зонда одного олигонуклеотида, несущего комбинацию флуоресцентных красителей флуоресцеин и BHQ1, способного гибридизоваться с образованием самокомплементарной шпильки;
- регистрацию интенсивности флуоресценции субстрата проводят в планшете, а не в кювете, что позволяет значительно сократить объем биологического материала для проведения анализа;
- кинетические кривые обрабатывают с помощью программы OriginPro8.1 (OriginLab Corp., США), которая позволяет минимизировать время обработки кинетических кривых и достоверно рассчитать kobs - наблюдаемую константу скорости образования продукта.
Использование способа позволяет повысить точность и достоверность определения активности ферментов BER, и, наряду с этим, ускорить и упростить проведение анализа.
Изобретение иллюстрируется следующим примером конкретного выполнения.
Пример 1
Определение активности АП-эндонуклеазы 1 (АРЕ1) в экстракте клеточной линии рака яичников человека TOV112 с использованием заявленного способа.
Для определения активности АП-эндонуклеазы 1 готовят буфер следующего состава: 50 мМ Tris-HCl рН 7.5, 50 мМ KCl, 1 мМ ЭДТА, 5 мМ MgCl2, 1 мМ ДТТ, 7% глицерин. Приготавливают в буфере 1,0×10-6 М раствор субстрата, представляющий собой самокомплементарную последовательность, формирующую шпильку: 5'-FAM-GCTCA(F)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3', где FAM - остаток флуоресцеина, BHQ1 - остаток тушителя black hole quencher 1, F - остаток 2-оксиметил-3-окси-тетрагидрофурана, ps - межнуклеотидная тиофосфатная группа.
Для построения калибровочной линии, в кювете флуориметра смешивают растворы ДНК-зонда (1,0×10-6 М), буфера (50 мМ Tris-HCl (рН 7,5), 50 мМ KCl, 5 мМ MgCl2, 1 мМ дитиотреит, 9% глицерин) и рекомбинантного фермента АП-эндонуклеазы человека (в концентрациях 1,0×10-7 М, 1,0×10-8 М, 1,0×10-9 М, 1,0×10-10 М).
Регистрируют изменение интенсивности флуоресценции субстрата в интервале времени 15-3600 с при температуре 37°С. Возбуждение флуоресценции проводят на длине волны 494 нм, регистрацию флуоресценции проводят на длине волны 500-550 нм.
Начальные линейные участки полученных кинетических кривых обрабатывают с помощью программы OriginPro8.1 (OriginLab Corp., США), используя для обработки данных уравнение прямой:
F=F0+kobs×t,
где F0 - амплитуда, kobs - наблюдаемая константа скорости образования продукта при данной концентрации рекомбинантного фермента.
В результате обработки получают зависимость значения наблюдаемой константы скорости от концентрации АП-эндонуклеазы 1, которую в дальнейшем используют как калибровочную линию.
Лизис клеток линии TOV112 проводят в буфере состава 10 мМ Tris-HCl, рН 7.5, 1 мМ MgCl2, 1 мМ EDTA, 0,5% CHAPS, 10% глицерин, 0,1 мМ PMSF, 0,5 мМ β-меркаптоэтанол. К клеточному осадку добавляют 150 мкл лизис-буфера, выдерживают на льду в течение 30 минут, затем центрифугируют (14500 об/мин, 10 мин). Общую концентрацию белка в полученном супернатанте определяют по методу Бредфорда (для анализа в каждую пробу берут по 25 мкг общего белка).
Запускают реакцию путем смешивания растворов субстрата и экстракта в соотношении 1:1 и регистрируют изменение интенсивности флуоресценции субстрата в интервале времени 15-3600 с при температуре 37°С. Возбуждение флуоресценции проводят на длине волны 494 нм, регистрацию флуоресценции проводят на длине волны 500-550 нм.
Начальный линейный участок кинетических кривых, полученных при взаимодействии с ДНК-зонда с экстрактом, обрабатывают аналогично калибровочным кривым.
Полученное значение наблюдаемой константы скорости образования продукта сравниваю с калибровочной линией и определяют концентрацию фермента в тестируемом клеточном экстракте. На фиг. 1 представлены результаты обработки и приведены значения наблюдаемой константы скорости расщепления ДНК-зонда и концентрации АП-эндонуклеазы 1, количественно определенные для данной линии клеток. Концентрация АП-эндонуклеазы 1 в экстракте клеток линии TOV112 составила 12 нМ.
Пример 2
Определение активности урацил-ДНК-гликозилазы (SMUG1) в экстракте клеточной линии рака яичников человека TOV112 с использованием заявленного способа.
Определение активности SMUG1 проводят аналогично примеру 1, за исключением того, что в качестве субстрата используют самокомплементарный олигонуклеотид, формирующий шпильку: 5'-FAM-GCTCA(U)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3', где FAM - остаток флуоресцеина, BHQ1 - остаток тушителя black hole quencher 1, U - уридин, ps - межнуклеотидная тиофосфатная группа.
На фиг. 2 представлены результаты обработки и приведены значения наблюдаемой константы скорости расщепления ДНК-зонда и концентрации SMUG1, количественно определенные для данной линии клеток. Концентрация SMUG 1 в экстракте клеток линии TOV112 составила 2,4 нМ.
Пример 3
Определение активности алкиладенин-ДНК-гликозилазы (AAG) в экстракте клеточной линии рака яичников человека TOV112 с использованием заявленного способа.
Определение активности AAG проводят аналогично примеру 1, за исключением того, что в качестве субстрата используют самокомплементарный олигонуклеотид, формирующий шпильку: 5'-FAM-GCTCA(εA)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3', где FAM - остаток флуоресцеина, BHQ1 - остаток тушителя black hole quencher 1, εА - 1,N6-этеноаденозин, ps - межнуклеотидная тиофосфатная группа.
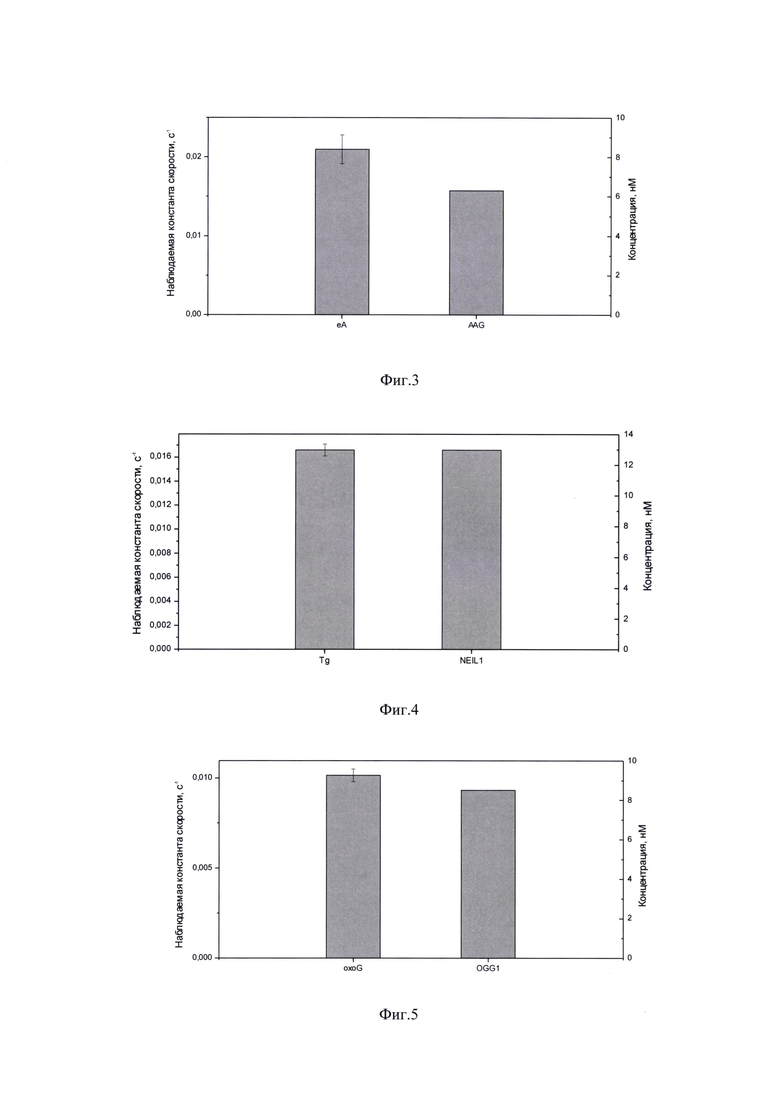
На фиг. 3 представлены результаты обработки и приведены значения наблюдаемой константы скорости расщепления ДНК-зонда и концентрации AAG, количественно определенные для данной линии клеток. Концентрация AAG в экстракте клеток линии TOV112 составила 6,3 нМ.
Пример 4
Определение активности ДНК-гликозилазы (NEIL1) в экстракте клеточной линии рака яичников человека MES-OV с использованием заявленного способа.
Определение активности NEIL1 проводят аналогично примеру 1, за исключением того, что в качестве субстрата используют самокомплементарный олигонуклеотид, формирующий шпильку: 5'-FAM-GCTCA(Tg)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3', где FAM - остаток флуоресцеина, BHQ1 - остаток тушителя black hole quencher 1, Tg-5,6-дигидрокси-5,6-дигидротимидин, ps - межнуклеотидная тиофосфатная группа.
На фиг. 4 представлены результаты обработки и приведены значения наблюдаемой константы скорости расщепления ДНК-зонда и концентрации NEIL1, количественно определенные для данной линии клеток. Концентрация NEIL1 в экстракте клеток линии MES-OV составила 13 нМ.
Пример 5
Определение активности 8-оксогуанин-ДНК-гликозилазы OGG1 в экстракте клеточной линии рака яичников человека MES-OV с использованием заявленного способа.
Для определения активности OGG1 проводят аналогично примеру 1, за исключением того, что в качестве субстрата используют самокомплементарный олигонуклеотид, формирующий шпильку: 5'-FAM-GCTCA(oxoG)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3', где FAM - остаток флуоресцеина, BHQ1 - остаток тушителя black hole quencher 1, oxoG - 7,8-дигидро-8-оксогуанозин, ps - межнуклеотидная тиофосфатная группа.
На фиг. 5 представлены результаты обработки и приведены значения наблюдаемой константы скорости расщепления ДНК-зонда и концентрации OGG1, количественно определенные для данной линии клеток. Концентрация OGG1 в экстракте клеток линии MES-OV составила 8,5 нМ.
Использование предлагаемого способа позволит повысить точность и достоверность результатов определения активности ферментов BER, ускорить и упростить проведение анализа, обеспечить возможность использования разработанных ДНК-зондов в клеточных системах, а также снизить расход реагентов, необходимый для метода остановленного потока.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ определения активности эндонуклеаз рестрикции в клеточных экстрактах микроорганизмов | 2023 |
|
RU2810742C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АКТИВНОСТИ АПУРИН/АПИРИМИДИН-ЭНДОНУКЛЕАЗЫ ЧЕЛОВЕКА | 2009 |
|
RU2389026C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ МОДИФИЦИРОВАННЫХ НУКЛЕОТИДОВ РНК | 2012 |
|
RU2522863C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АКТИВНОСТИ 8-ОКСОГУАНИН-ДНК-ГЛИКОЗИЛАЗЫ ЧЕЛОВЕКА | 2006 |
|
RU2321637C1 |
| МОЛЕКУЛА СУБСТРАТА | 2016 |
|
RU2755495C2 |
| КОМПОЗИЦИИ ДЛЯ ВОССТАНОВЛЕНИЯ КОЖИ, СОДЕРЖАЩИЕ АКТИВАТОРЫ ЦИРКАДНЫХ ГЕНОВ И СИНЕРГИЧЕСКУЮ КОМБИНАЦИЮ АКТИВАТОРОВ ГЕНА sirt1 | 2010 |
|
RU2494756C1 |
| СПОСОБ ОДНОВРЕМЕННОГО ДНК-ТЕСТИРОВАНИЯ НА НАЛИЧИЕ ПОЛИМОРФИЗМОВ Н63D И C282Y В ГЕНЕ HFE, СВЯЗАННЫХ С НАСЛЕДСТВЕННЫМ ГЕМОХРОМАТОЗОМ | 2006 |
|
RU2304170C1 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ РИСКА НЕВЫНАШИВАНИЯ БЕРЕМЕННОСТИ | 2014 |
|
RU2552302C1 |
| 2-Ацетил-6-(2-(2-(4-бромбензилиден)гидразинил) тиазол-4-ил)-3, 7, 9-тригидрокси-8, 9b-диметилдибензо[b, d]фуран-1(9bH)-он, проявляющий ингибирующее действие в отношении фермента тирозил-ДНК-фосфодиэстеразы 1 человека | 2016 |
|
RU2627764C1 |
| СПОСОБ АМПЛИФИКАЦИИ И ДЕТЕКЦИИ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ В РЕАКЦИОННОЙ СМЕСИ И ИСПОЛЬЗУЕМЫЙ В НЕМ НАБОР | 2009 |
|
RU2523589C2 |

Изобретение относится к области биохимии. Описан способ определения активности фермента эксцизионной репарации оснований ДНК группы: АРЕ1, UNG2, SMUG1, MBD4, TDG, AAG, NEIL1, NTHL1 или OGG1 - в клетках человека. Он включает смешивание субстрата, буфера и клеточного экстракта в соотношении 1:1 и регистрацию изменения интенсивности флуоресценции субстрата с помощью флуориметра с последующей обработкой полученных кинетических кривых с помощью программы. Способ отличается тем, что в качестве субстрата используют ДНК-зонды: 5-FAM-GCTCA(F)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3, 5-FAM-GCTCA(U)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3, 5-FAM-GCTCA(εA)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3, 5-FAM-GCTCA(Tg)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3 или 5-FAM-GCTCA(oxoG)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3 - представляющие собой олигодезоксирибонуклеотиды, формирующие шпильку и несущие FRET-пару FAM/BHQ1 на концах цепи. Зонды также содержат в своем составе повреждение, специфичное для определяемого фермента. При этом для предотвращения неспецифической 3-5-экзонуклеазной деградации ДНК-зонда в клеточном экстракте 3-концевую межнуклеотидную фосфатную группу заменяют на тиофосфатную. Технический результат заключается в повышении точности и достоверности результатов определения активности ферментов репарации ДНК у человека, расширении функциональных возможностей известного способа, и применении этого способа для анализа активности в клетках человека. 4 з.п. ф-лы, 5 ил., 1 табл., 5 пр.
1. Способ определения активности ферментов эксцизионной репарации оснований ДНК: АРЕ1, UNG2, SMUG1, MBD4, TDG, AAG, NEIL1, NTHL1 или OGG1 - в клетках человека, включающий смешивание субстрата, буфера и клеточного экстракта в соотношении 1:1 и регистрацию изменения интенсивности флуоресценции субстрата с помощью флуориметра с последующей обработкой полученных кинетических кривых с помощью программы, отличающийся тем, что в качестве субстрата используют ДНК-зонды: 5-FAM-GCTCA(F)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3, 5-FAM-GCTCA(U)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3, 5-FAM-GCTCA(εA)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3, 5-FAM-GCTCA(Tg)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3 или 5-FAM-GCTCA(oxoG)GTACAGAGCTGTTTTTCAGCTCTGTACGTGAGCps-BHQ1-3 - представляющие собой олигодезоксирибонуклеотиды, формирующие шпильку и несущие FRET-пару FAM/BHQ1 на концах цепи, а также содержащие в своем составе повреждение, специфичное для определяемого фермента, при этом для предотвращения неспецифической 3-5-экзонуклеазной деградации ДНК-зонда в клеточном экстракте 3-концевую межнуклеотидную фосфатную группу заменяют на тиофосфатную.
2. Способ по п. 1, отличающийся тем, что ДНК-зонд используют в концентрации, равной 1,0×10-6 М, ферменты для построения калибровочной линии используют в концентрации, равной 1,0×10-7 М, 1,0×10-8 М, 1,0×10-9 М, 1,0×10-10 М, а общее количество белка в тестируемом экстракте не превышает 25 мкг.
3. Способ по п. 1, отличающийся тем, что буфер содержит 50 мМ Tris-HCl рН 7.5, 50 мМ KCl, 1 мМ ЭДТА, 5 мМ MgCl2, 1 мМ ДТТ, 7% глицерин.
4. Способ по п. 1, отличающийся тем, что регистрацию изменений интенсивности флуоресценции осуществляют в спектральном диапазоне 500-550 нм в интервале времени 15-3600 с.
5. Способ по п. 1, отличающийся тем, что начальные линейные участки полученных кинетических кривых обрабатывают с помощью программы OriginPro8.1 (OriginLab Corp., США).
| СПОСОБ ОЦЕНКИ АКТИВНОСТИ СИСТЕМЫ ЭКСЦИЗИОННОЙ РЕПАРАЦИИ НУКЛЕОТИДОВ МЛЕКОПИТАЮЩИХ | 2012 |
|
RU2492242C1 |
| Способ применения БПЛА и система осуществления способа | 2024 |
|
RU2836926C1 |
| О.А.Кладова, Н.А.Кузнецов, О.С.Федорова.НАЧАЛЬНЫЕ СТАДИИ ЭКСЦИЗИОННОЙ РЕПАРАЦИИ ОСНОВАНИЙ ДНК В НУКЛЕОСОМАХ | |||
| Молекулярная биология, 2021, T | |||
| Устройство двукратного усилителя с катодными лампами | 1920 |
|
SU55A1 |
| Кран машиниста для автоматических тормозов с сжатым воздухом | 1921 |
|
SU194A1 |
| Н.И | |||
| Речкунова, Ю.С | |||
| Красикова, О.И | |||
| Лаврик | |||
| ИНТЕРАКТОМ СИСТЕМ РЕПАРАЦИИ ОСНОВАНИЙ И НУКЛЕОТИДОВ | |||
| Молекулярная биология, 2021, T | |||
| Устройство двукратного усилителя с катодными лампами | 1920 |
|
SU55A1 |

