Ссылка на родственные заявки
Согласно настоящей заявке испрашивается приоритет в соответствии с заявкой на патент Китая №201910462073.5 под названием «Рекомбинантный онколитический вирус, способ его получения, его применение и лекарственный препарат на его основе», поданной в Патентное ведомство Китая 30 мая 2019 года, полный объем которой включен в настоящий документ посредством ссылки.
Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к области биотехнологии, в частности, к онколитическому вирусу, способу его получения, его применению и лекарственному препарату на его основе.
Предшествующий уровень техники настоящего изобретения
Основной причиной смертности, угрожающей здоровью человека, в настоящее время являются различные формы рака. Согласно данным Global Cancer Statistics 2015, в 2015 году во всем мире было зарегистрировано приблизительно 14,1 миллиона новых случаев рака, а число смертельных исходов достигло 8,2 миллиона. В 2015 году в Китае было зарегистрировано 4,29 миллиона новых случаев рака, а число смертельных исходов достигло 2,81 миллиона. В настоящее время лечение рака преимущественно основано на традиционной хирургической резекции, лучевой терапии и химиотерапии. Хирургическая резекция позволяет устранить опухоль или по меньшей мере облегчить страдания пациентов, но не позволяет принести облегчение в случае опухолей, расположенных глубоко в теле, из-за отсутствия доступа, и не помогает при уже метастазировавших опухолях. Лучевую терапию и химиотерапию уже давно применяют в клинической практике, но область их применения сильно ограничена из-за серьезных побочных эффектов, связанных с отсутствием селективности между нормальными и опухолевыми клетками. В последние годы, особенно в последние пять лет, большое внимание при лечении различных форм рака привлекают антитела и CAR-T. Терапия антителами может замедлить прогрессирование рака, но терапевтическая эффективность далека от удовлетворительной, и широко распространено мнение, что терапия антителами больше подходит для применения в качестве вспомогательной терапии. С помощью CAR-T-терапии может быть достигнуто точное целенаправленное действие, и CAR-T-терапия может даже излечить различные формы рака, однако терапия CAR-T может в основном быть пригодной для лечения гемологических лимфом, и одна терапия предназначается специально для конкретного пациента, поэтому стоимость лечения с помощью CAR-T является чрезвычайно высокой. Более того, последствие потенциального нецелевого воздействия может быть разрушительным или даже смертельным. Для безопасного и эффективного лечения различных форм рака и облегчения страданий пациентов крайне желательно разработать новую терапию для лечения с применением совершенно новой стратегии. Среди всех возможных вариантов выделяется модифицированный методами генной инженерии онколитический вирус, поскольку было продемонстрировано, что онколитические вирусы безопасны, в то время как отсутствует такая проблема, как устойчивость к лекарственным средствам, и остается потенциал в том, что одним онколитическим вирусом можно лечить множество опухолей.
Еще 100 лет назад в клинических условиях наблюдали, что вирусная инфекция замедляет рост опухоли или даже уничтожает опухоль. И потом плазму или экстракты плазмы больных гепатитом В даже применяли для лечения болезни Ходжкина. Однако люди не верили, что вирус был возможным вариантом лечения рака, потому что не было средств придать вирусу селективность, позволяющую вирусу реплицироваться только в опухолях. В 1990-х годах прогресс в области молекулярно-биологических технологий проложил путь к осуществлению генетической инженерии вируса, специфически нацеленного на опухолевые клетки. Ранние исследования были сосредоточены на разработке дефектных по механизму репликации литических вирусов, таких как аденовирус, и недефектных по механизму репликации литических вирусов, таких как аденовирус-ассоциированный вирус, для экспрессии стимулирующих иммунитет молекул, таких как GMCSF, IL-12 и IL-17, или цитокинов, таких как TNFα и IFNα, для лечения различных форм рака путем повышения противоопухолевого иммунитета. Поскольку различные виды иммунотерапии на основе вирусов, как и обычные виды иммунотерапии, дают лишь ограниченный терапевтический эффект, считалось, что их следует применять в основном в качестве вспомогательной терапии. Для реализации потенциала вирусов при лечении рака в последние годы была сделана попытка генетической модификации литических вирусов для придания вирусу способности специфически реплицироваться в опухолевых клетках (онколитический вирус). Таким образом, вирус размножается в опухолевых клетках, распространяется на соседние клетки и уничтожает их (онколиз). Более того, клеточный дебрис, полученный из лизированных опухолевых клеток, может индуцировать опухолеспецифический иммунитет, который, в свою очередь, помогает уничтожать опухолевые клетки в первичном очаге опухоли и разрушать уже метастазированные опухолевые клетки, что дает терапевтический эффект. Из-за продемонстрированного профиля безопасности онколитических вирусов и потенциала онколитического вируса для лечения различных форм рака будущее применение онколитических вирусов в лечении рака является весьма прогнозируемым. Одобрение США, ЕС и Австралией в 2015, 2016 годах соответственно клинического применения онколитического вируса T-vec на основе вируса герпеса 1-го типа от американской компании Amgen для лечения меланомы ознаменовало новую эру в лечении различных форм рака с помощью онколитических вирусов. С тех пор исследование онколитических вирусов значительно расширилось, и только в 2017 году во всем мире было проведено 80 клинических испытаний онколитических вирусов для лечения различных опухолей. В доклинических исследованиях было хорошо продемонстрировано, что многие виды онколитических вирусов хорошо себя зарекомендовали, но в клинической практике терапевтических эффектов достигают намного меньше, чем ожидалось, даже несмотря на то, что они оказались клинически безопасными.
Краткое раскрытие настоящего изобретения
Настоящее изобретение относится к новому онколитическому вирусу, нуклеотидным последовательностям, используемым для его создания, и способу получения данного вируса, потенциальному применению онколитического вируса для лечения рака и композиции, содержащей онколитический вирус и т.п. Репликация онколитического вируса регулируется и контролируется экзогенными элементами, вставленными в его вирусный геном; посредством регуляции и контроля экспрессии вирусных генов такими экзогенными элементами онколитический вирус по-разному реплицируется в разных типах клеток, и посредством селективной репликации целевые клетки (например, опухолевые клетки) могут быть соответственно селективно уничтожены, в то время как нецелевые клетки (такие как нормальные клетки) остаются нетронутыми.
В настоящее время селективная репликация онколитических вирусов в раковых клетках достигается главным образом за счет удаления одного или нескольких несущественных вирусных генов или путем помещения экспрессии одного или нескольких существенных вирусных генов под контроль опухолеспецифического промотора. Вирусные несущественные гены являются теми генами, которые не нужны вирусу для репликации в культивируемых клетках. Для репликации вируса не нужны несущественные гены in vitro, но несущественные гены выполняют различные функции для поддержки репликации вируса in vivo, например, противодействуя противовирусным механизмам хозяина или тому подобное для облегчения репликации вируса. В случае онколитических вирусов, сконструированных с помощью опухолеспецифического промотора для управления экспрессией одного или нескольких существенных генов, хотя геном и остается интактным, нарушается скоординированная по времени экспрессия вирусных генов. Таким образом, способность вируса к репликации in vivo может быть значительно нарушена независимо от того, удален ли несущественный ген или экспрессируется существенный ген под контролем опухолеспецифического промотора. Действительно, доступные в настоящее время онколитические вирусы обычно обладают низкой эффективностью в клинической практике. Для повышения эффективности и расширения спектра применения онколитических вирусов крайне важно сохранить интактность вирусного генома, не нарушая при этом высококоординированную экспрессию вирусных генов. С помощью таких руководящих принципов был разработан и получен совершенно новый онколитический вирус по настоящему изобретению с помощью новой стратегии.
Согласно первому аспекту, настоящее изобретение относится к онколитическому вирусу с геномом онколитического вируса, содержащим следующие экзогенные элементы:
(1) первую кассету экспрессии, содержащую первый промотор и первую последовательность экспрессии интерферирующей РНК;
(2) целевую последовательность; и
(3) вторую кассету экспрессии.
В первой кассете экспрессии первая последовательность экспрессии интерферирующей РНК используется для экспрессии первой интерферирующей РНК, которая специфически связывается с целевой последовательностью; первая последовательность экспрессии интерферирующей РНК находится под управлением первого промотора с целью экспрессии первой интерферирующей РНК в первых клетках.
Целевая последовательность вставлена в 5'- или 3'-нетранслируемый участок (UTR) существенного вирусного гена в вирусном геноме.
Вторая кассета экспрессии содержит второй промотор и последовательность экспрессии ингибирующего компонента;
последовательность экспрессии ингибирующего компонента используется для экспрессии ингибирующих компонентов. Ингибирующие компоненты используются для ингибирования биосинтеза и/или биоактивности одного фермента, участвующего в биосинтезе интерферирующей РНК; и последовательность экспрессии ингибирующего компонента находится под управлением второго промотора с целью экспрессии ингибирующих компонентов во вторых клетках, но не в первых клетках.
Первая и вторая клетки являются клетками различных типов.
Что касается онколитического вируса, представленного в настоящем раскрытии, первая кассета экспрессии, целевая последовательность первой интерферирующей РНК и вторая кассета экспрессии вставлены в вирусный геном так, как показано на фиг. 1. Первая интерферирующая РНК конститутивно экспрессируется в первых клетках (нецелевые клетки, такие как нормальные клетки) под контролем первого промотора после инфицирования клеток, в то время как ингибирующие компоненты специфически экспрессируются при экспрессии под управлением второго промотора в инфицированных вторых клетках (целевых клетках, таких как опухолевые клетки). В первых клетках первая интерферирующая РНК конститутивно экспрессируется после инфицирования клеток, которая связывается с целевой последовательностью интерферирующей РНК, расположенной в 5'- или 3'-UTR относительно существенного гена вируса, что приводит в результате к расщеплению целевой мРНК или предотвращению трансляции существенного гена. В результате не вырабатывается регулируемый вирусный белок или вырабатывается гораздо меньшее его количество, что приводит к отсутствию репликации вируса незатронутыми клетками. Напротив, во вторых клетках ингибирующие компоненты после инфицирования клеток вирусом специфически экспрессируются с вирусного генома под контролем второго промотора, который ингибирует биосинтез и/или биоактивность фермента, участвующего в биосинтезе интерферирующей РНК, что приводит в результате к отсутствию или гораздо меньшему количеству интерферирующей РНК, вырабатываемой в таких клетках, что приводит к устойчивой репликации вируса и гибели клеток. Разительно отличающийся от доступных в настоящее время онколитических вирусов, онколитический вирус, представленный в настоящем раскрытии, обладает интактным геномом, сохраняя при этом способность вируса к высококоординированной экспрессии регулируемых вирусных генов во вторых клетках, для устойчивой репликации онколитического вируса в опухолевых клетках необходимы два критических признака. Следовательно, онколитический вирус обладает способностью к репликации, такой же или схожей с таковой, которую наблюдают у вируса дикого типа во вторых клетках, таким образом эффективно уничтожая опухолевые клетки. Основываясь на данной стратегии, можно ожидать, что при применении опухолеспецифического промотора, который очень активен в различных опухолевых клетках, для управления экспрессией ингибирующих компонентов со второй кассеты экспрессии онколитический вирус можно будет применять для лечения различных опухолей. Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, первая кассета экспрессии дополнительно содержит вторую последовательность экспрессии интерферирующей РНК для экспрессии второй интерферирующей РНК. Вторая интерферирующая РНК действует на открытую рамку считывания (ORF) несущественного вирусного гена онколитического вируса, препятствуя экспрессии несущественного гена, а вторая последовательность экспрессии интерферирующей РНК экспрессируется под контролем первого промотора.
Вторая интерферирующая РНК после экспрессии в первых клетках с вирусного генома связывается с ORF несущественного гена вируса, таким образом ингибируя выработку несущественного генного продукта, что дополнительно повышает безопасность вируса в первых клетках.
Кроме того, в соответствии со всеми вариантами осуществления, первая интерферирующая РНК и вторая интерферирующая РНК могут быть либо малой интерферирующей РНК (siRNA), либо микроРНК (miRNA).
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, вторые клетки являются опухолевыми клетками млекопитающего, а первые клетки являются отличными от опухолевых клетками млекопитающего.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, млекопитающее относится к человеку.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, опухолевые клетки представляют собой клетки рака легких, клетки рака печени, клетки рака молочной железы, клетки рака желудка, клетки рака предстательной железы, клетки опухоли головного мозга, клетки рака толстой кишки человека, клетки рака шейки матки, клетки рака почек, клетки рака яичников, клетки рака головы и шеи, клетки меланомы, клетки рака поджелудочной железы или клетки рака пищевода.
Следует отметить, что онколитический вирус, представленный в настоящем раскрытии, не ограничивается селективным уничтожением опухолевых клеток, но также его можно применять для уничтожения других представляющих интерес отличных от опухолевых клеток. Иными словами, в роли вторых клеток могут выступать любые представляющие интерес клетки, как упомянуто выше, то есть в роли целевых клеток, в то время как в роли первых клеток выступают не представляющие интерес клетки, то есть являются нецелевыми клетками. Например, любой один вид клеток, выбранный из нервных клеток, эритроцитов, лейкоцитов, тромбоцитов, фагоцитов, эпителиальных клеток, миокардиальных клеток, яйцеклеток, сперматозоидов и т.п., можно уничтожить в качестве вторых клеток, т.е., скажем, любая клетка, происходящая от млекопитающих, может выступать в роли вторых клеток, в то время как один вид, несколько видов или все виды клеток, которые не выбраны в качестве вторых клеток, могут выступать в роли первых клеток, и этот онколитический вирус не оказывает цитолитического действия на такие первые клетки.
Кроме того, следует также указать на онколитические вирусы, созданные с помощью настоящих идей, представленных в настоящем раскрытии, независимо оттого, что является родительским вирусом, все они подпадают под объем защиты настоящего изобретения.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, целевая последовательность выбрана из кодирующей последовательности гена отличного от млекопитающего организма.
Предпочтительно, в соответствии с определенными вариантами осуществления настоящего изобретения, целевая последовательность длиной 19-23 нуклеотида выбрана из ORF гена отличного от млекопитающего организма.
Предпочтительно, в соответствии с определенными вариантами осуществления настоящего изобретения, отличный от млекопитающего организм представляет собой дрожжи, медузу, кишечную палочку, насекомое, рыбу или растение.
Предпочтительно, в соответствии с определенными вариантами осуществления настоящего изобретения, ген отличного от млекопитающего организма можно выбрать из группы, включающей ген зеленого флуоресцентного белка, полученный из медузы, ген β-галактозидазы, полученный из Escherichia coli, и ген люциферазы, полученный из светлячка.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, нуклеотидная последовательность целевой последовательности представлена под SEQ ID NO: 1, а последовательность первой интерферирующей РНК представлена под SEQ ID NO: 2.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, целевая последовательность вставлена в 5'- или 3'-UTR одного или нескольких существенных генов рекомбинантного онколитического вируса.
Предпочтительно, количество копий целевой последовательности, вставленной в любое положение, может насчитывать одну или несколько.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, онколитический вирус выбран из ряда вирусов, включая вирус простого герпеса (HSV), аденовирус, вирус коровьей оспы, вирус болезни Ньюкасла, полиовирус, вирус Коксаки, вирус кори, вирус эпидемического паротита, вирус везикулярного стоматита (VSV) и вирус гриппа.
Предпочтительно, когда онколитический вирус представляет собой вирус простого герпеса, существенный ген выбран из группы, включающей гликопротеин L оболочки, урацил-ДНК-гликозилазу, белок капсида, субъединицу профермента геликазы, унвиндазу, связывающую точку инициации репликации ДНК, производный белок миристиновой кислоты, дезоксирибонуклеазу, серин/треониновую протеинкиназу оболочки, субъединицу 1 ДНК-упаковывающей терминазы, белок UL16 оболочки, белок UL17 упаковки ДНК, триплексную субъединицу 2 капсида, основной белок капсида, белок UL20 оболочки, нуклеопротеин UL24, белок UL25 упаковки ДНК, протеазу созревания капсида, белок капсида, гликопротеин В оболочки, связывающий одноцепочечную ДНК белок, каталитическую субъединицу ДНК-полимеразы, белок слоя выходя из ядра, белок UL32 упаковки ДНК, белок UL33 упаковки ДНК, мембранный белок выходя из ядра, большой капсидный белок, триплексную субъединицу 1 капсида, субъединицу 1 рибонуклеотидредуктазы, субъединицу 2 рибонуклеотидредуктазы, белок отключения хозяина капсулы (capsule host shutoff protein), процессирующую субъединицу ДНК-полимеразы, мембранный белок UL45, белок VP13/14 оболочки, транс-активирующий белок VP16, белок VP22 оболочки, гликопротеин N оболочки, белок UL51 оболочки, примазную субъединицу унвиндазы, гликопротеин K оболочки, ICP27, нуклеопротеин UL55, нуклеопротеин UL56, фактор регуляции транскрипции ICP4, регуляторный белок ICP22, гликопротеин оболочки D и мембранный белок US8A; а несущественный ген выбран из ICP34.5, ICP0, нуклеопротеина UL3, нуклеопротеина UL4, геликазной субъединицы профермента геликаза, кутикулярного белка UL7, гликопротеина М оболочки, белка UL14 оболочки, белка UL21 оболочки, гликопротеина Н оболочки, тимидинкиназы, субъединицы 2 фермента, завершающего упаковку ДНК, малого капсидного белка, белка UL37 оболочки, белка UL43 оболочки, гликопротеина С оболочки, белка VP11/12 оболочки, урацил-дезоксирибозидетрифосфатазы, вирусного белка US2, серин/треониновой протеинкиназы U3, мембранного гликопротеина G (гликопротеина G оболочки), гликопротеина J оболочки, гликопротеина I оболочки, гликопротеина Е оболочки, мембранного белка US9, вирусного белка US10, кутикулярного белка Us11 и ICP47.
Предпочтительно, когда онколитический вирус представляет собой аденовирус, существенный ген выбран из группы, включающей ранний белок 1А, ранний белок 1B 19K, ранний белок 1B 55K, белок Iva2 инкапсидации, ДНК-полимеразу, предшественник рТР терминального белка, белок 52K инкапсидации, капсидный белок-предшественник pIIIa, пентомерную матрицу, коровый белок pVII, предшественник рХ корового белка, предшественник pVI корового белка, гексонмер, протеиназу, связывающий одноцепочечную ДНК белок, белок 100K сборки гексамера, белок 33K, белок 22K инкапсидации, предшественник капсидного белка, белок U, фибрин, открытую рамку считывания 6/7 регуляторного белка Е4, регуляторный белок Е4 34K, открытую рамку считывания 4 регуляторного белка Е4, открытую рамку считывания 3 регуляторного белка Е4, открытую рамку считывания 2 регуляторного белка Е4 и открытую рамку считывания 1 регуляторного белка Е4; а несущественный ген выбран из группы, включающей капсидный белок IX, белок 13.6K, коровый белок V, регуляторный белок Е3 12.5K, мембранный гликопротеин Е3 CR1-α, мембранный гликопротеин Е3 gp 19K, мембранный гликопротеин Е3 CR1-β, мембранный гликопротеин Е3 CR1-δ, мембранный гликопротеин Е3 RID-δ и мембранный гликопротеин Е3 14.7K.
Предпочтительно, когда онколитический вирус представляет собой вирус коровьей оспы, существенный ген выбран из группы, включающей малую субъединицу рибонуклеотидредуктазы, серин/треонинкиназу, ДНК-связывающий вирусный коровый белок, большую субъединицу полимеразы, субъединицу РНК-полимеразы, ДНК-полимеразу, сульфгидрилоксидазу, гипотетический ДНК-связывающий вирусный нуклеопротеин, ДНК-связывающий фосфопротеин, нуклеоид-цистеиновую протеиназу, РНК-геликазу NPH-II, гипотетическую металлопротеиназу, транскрипционный фактор элонгации, белок глутатионового типа, РНК-полимеразу, гипотетический вирусный нуклеопротеин, поздний транскрипционный фактор VLTF-1, ДНК-связывающий вирусный нуклеопротеин, вирусный капсидный белок, малую субъединицу полимеразы, субъединицу rpo22 РНК-полимеразы, зависящую от ДНК, субъединицу rpo147 РНК-полимеразы, зависящую от ДНК, серин/треониновую протеинфосфатазу, гепарин-связывающий поверхностный белок IMV, ДНК-зависимую РНК-полимеразу, поздний транскрипционный фактор VLTF-4, ДНК-топоизомеразу I типа, большую субъединицу фермента кэпирования мРНК, вирусный коровый белок 107, вирусный коровый белок 108, урацил-ДНК-гликозилазу, трифосфатазу, 70 кДа малую субъединицу раннего фактора транскрипции генов VETF, субъединицу rpo18 РНК-полимеразы, зависящую от ДНК, нуклеозидтрифосфатгидролазу-1, малую субъединицу фермента кэпирования мРНК, целевой сайт для рифампицина, поздний транскрипционный фактор VLTF-2, поздний транскрипционный фактор VLTF-3, путь образования дисульфидной связи, предшественник p4b корового белка 4b, 39 кДа коровый белок, субъединицу rpo19 РНК-полимеразы, зависящую от ДНК, 82 кДа большую субъединицу раннего фактора транскрипции гена VETF, 32 кДа малую субъединицу транскрипционного фактора VITF-3, мембранный белок IMV 128, предшественник Р4а корового белка 4а, мембранный белок 131 IMV, фосфорилированный мембранный белок IMV, мембранный белок IMV A17L, ДНК-унвиндазу, процессирующий фактор вирусной ДНК-полимеразы, мембранный белок IMV A21L, пальмитоиловый белок, 45 кДа большую субъединицу промежуточного фактора транскрипции гена VITF-3, субъединицу rpo132 РНК-полимеразы, зависящую от ДНК, РНК-полимеразу rpo35, зависящую от ДНК, белок A30L IMV, гипотетический АТФ-зависимый фермент, серин/треонинкиназу, белок созревания EEV, пальмитоилированный мембранный гликопротеин EEV, поверхностный белок IMV A27L, мембранный фосфатгликопротеин EEV, мембранные гликопротеины EEV и EEV, мембранный гликопротеин EEV, белок пути образования дисульфидной связи, гипотетический вирусный нуклеопротеин, мембранный белок I2LIMV, поксвирусный миристоиловый белок, мембранный белок L1R IMV, 16 кДа поздний гипотетический мембранный белок, гипотетический вирусный мембранный белок H2R, мембранный белок A21L IMV, хемокин-связывающий белок, белок, подобный эпидермальному фактору роста, и IL-18-связывающий белок; а несущественный ген выбран из группы, включающей секреторный комплемент-связывающий белок, kelch-подобный белок, факторы вирулентности, гипотетический белок, чувствительный к α-аминобелкам, белок серпинового типа, белок фосфолипазы D-типа, неохарактеризованный белок K7R, гипотетический мембранный белок типа CD47, белок алармонического типа, мембранный белок агглютинина С-типа типа II, секреторный гликопротеин, урацилдезоксирибозидетрифосфатазу, kelch-подобный белок F3L, гипотетический миристоилированный белок, большую субъединицу рибонуклеотидредуктазы, белок телец включения вируса коровьей оспы типа А, белок анкиринового типа, 6 кДа внутриклеточный вирусный белок, белок 215, подобный α-рецептору фактора некроза опухоли, белок 217, подобный α-рецептору фактора некроза опухоли, белок B4R анкиринового типа, белок 213 анкиринового типа, белок 211 анкиринового типа, белок 207 с цинковым пальцем, белок 208 с цинковым пальцем, белок 014 анкиринового типа, белок 015 анкиринового типа, белок 016 анкиринового типа, белок 017 анкиринового типа, белок 019 анкиринового типа, белок 030 анкиринового типа, гипотетическую моноглицеридную липазу 036, гипотетическую моноглицеридную липазу 037, гипотетическую моноглицеридную липазу 038, белок 199 анкиринового типа, белок 203 анкиринового типа/гипотетический белок, белок телец включения типа А, гуанилаткиназу и белок 188 анкиринового типа.
Предпочтительно, когда онколитический вирус представляет собой вирус Коксаки, существенный ген выбран из группы, включающей белок Vpg, коровый белок 2А, белок 2В, РНК-унвиндазу 2С, белок 3А, протеиназу 3С, обратную транскриптазу 3D, белок оболочки Vp4 и белок Vp1; а несущественный ген представляет собой любой из капсидных белков Vp2 или Vp3.
Предпочтительно, когда онколитический вирус представляет собой вирус кори, существенный ген выбран из группы, включающей нуклеопротеин N, фосфопротеин Р, матричный белок М, трансмембранный гликопротеин F, трансмембранный гликопротеин Н и РНК-зависимую РНК-полимеразу L; а несущественный ген представляет собой либо РНК-зависимый вспомогательный белок С РНК-полимеразы, либо РНК-зависимый вспомогательный белок V РНК-полимеразы.
Когда онколитический вирус представляет собой вирус эпидемического паротита, существенный ген выбран из группы, включающей нуклеопротеин N, фосфопротеин Р, гибридный белок F и РНК-полимеразу L; а несущественный ген выбран из группы, включающей фосфопротеин V, мембранный белок М и гемагглютинин-нейраминидазный белок HN.
Предпочтительно, когда онколитический вирус представляет собой вирус везикулярного стоматита, существенный ген выбран из группы, включающей гликопротеин G, нуклеопротеин N, фосфопротеин Р и РНК-полимеразу L; а несущественный ген представляет собой матричный белок М.
Предпочтительно, когда онколитический вирус представляет собой полиовирус, существенный ген выбран из группы, включающей капсидный белок VP1, капсидный белок VP2, капсидный белок VP3, цистеиновую протеазу 2А, белок 2В, белок 2С, белок 3А, белок 3В, протеиназу 3С, белок 3D и РНК-направленную РНК-полимеразу; а несущественный ген представляет собой капсидный белок VP4.
Предпочтительно, когда онколитический вирус представляет собой вирус гриппа, существенный ген выбран из группы, включающей гемагглютинин, нейраминидазу, нуклеопротеин, мембранный белок M1, мембранный белок М2, полимеразу РА, полимеразу PB1-F2 и полимеразу РВ2; а несущественный ген представляет собой неструктурный белок NS1 или неструктурный белок NS2.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, онколитический вирус представляет собой вирус простого герпеса 1-го типа, существенный ген представляет собой ICP27, а несущественный ген представляет собой ICP34.5.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, последовательность второй интерферирующей РНК представлена под SEQ ID NO: 3.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, первый промотор представляет собой конститутивный промотор.
Предпочтительно, если целевая последовательность вставлена в 5'- или 3'-UTR множества существенных генов, первая кассета экспрессии экспрессирует только первую интерферирующую РНК, а первый промотор представляет собой промотор Hu6 или H1 человека.
Предпочтительно, если целевая последовательность вставлена в 5'- или 3'-UTR только одного существенного гена, и ORF несущественного гена является целью для второй интерферирующей РНК, первая кассета экспрессии экспрессирует первую и вторую интерферирующую РНК одновременно, и первый промотор выбран из группы, включающей промоторы CMV, SV40 и СБА.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, второй промотор представляет собой опухолеспецифический промотор человека.
Предпочтительно, опухолеспецифический промотор человека выбран из промотора обратной транскриптазы теломеразы (hTERT), промотора рецептора-2 эпидермального фактора роста человека (HER-2), промотора E2F1, промотора остеокальцина, промотора карциноэмбрионального антигена, промотора сурвивина и промотора церулоплазмина.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, фермент представляет собой любой из Drosha, Dicer или Agonauts.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, ингибирующие компоненты содержат третью интерферирующую РНК, препятствующую экспрессии гена фермента, ингибируя биосинтез интерферирующей РНК.
Предпочтительно, ферментом является Drosha.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, последовательность оснований третьей интерферирующей РНК представлена под SEQ ID NO: 4.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, ингибирующий компонент дополнительно содержит РНК с увеличенным количеством нуклеотидных триплетных повторов для ингибирования активности Drosha или некодирующую РНК для ингибирования активности Dicer.
Предпочтительно, последовательность с увеличенным количеством нуклеотидных триплетных повторов имеет следующую общую формулу: (CGG)n, где n представляет собой целое число, равное или больше 20.
Предпочтительно, n находится в диапазоне от 60 до 150.
Предпочтительно, n равно 100.
Предпочтительно, некодирующая РНК для ингибирования активности Dicer представляет собой РНК VA1 аденовируса 5-го типа.
Предпочтительно, нуклеотидная последовательность PHKVA1 аденовируса 5-го типа имеет следующий вид (SEQ ID NO: 8):
AGCGGGCACUCUUCCGUGGUCUGGUGGAUAAAUUCGCAAGGGUAUCAUGGCGGACGACCGGGGUUCGAGCCCCGUAUCCGGCCGUCCGCCGUGAUCCAUGCGGUUACCGCCCGCGUGUCGAACCCAGGUGUGCGACGUCAGACAACGGGGGAGUGCUCCUUU.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, вторая кассета экспрессии дополнительно содержит энхансерную последовательность для усиления экспрессии ингибирующих компонентов.
Предпочтительно, энхансер представляет собой энхансер либо CMV, либо SV40.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, вышеупомянутый онколитический вирус имеет регистрационный номер ССТСС №V201919. Данный вирус был сохранен в Китайском центре сбора типовых культур (ССТСС), расположенном в Уханьском университете, Луоцзяшань, Учан, город Ухань, 24 апреля 2019 года.
Согласно второму аспекту, настоящее изобретение дополнительно относится к другому онколитическому вирусу (второй тип онколитических вирусов), геном онколитического вируса содержит следующие экзогенные элементы: целевую последовательность интерферирующей РНК и кассету экспрессии интерферирующей РНК,
причем кассета экспрессии содержит промотор и последовательность экспрессии интерферирующей РНК, и последовательность экспрессии интерферирующей РНК используется для экспрессии интерферирующей РНК, которая связывается с целевой последовательностью.
Во втором типе онколитических вирусов целевая последовательность РНК вставлена в 5'- или 3'-UTR одного или более чем одного существенного гена в геноме рекомбинантного онколитического вируса.
Экспрессия интерферирующей РНК находится под управлением промотора, специфичного для первых клеток, с целью экспрессии РНК в первых клетках, но не во вторых клетках.
Первая и вторая клетки являются клетками различных типов.
Второй вид онколитических вирусов не содержит второй кассеты экспрессии. Экспрессия интерферирующей РНК с ее кассеты экспрессии находится под управлением промотора, специфичного для первых клеток, который управляет экспрессией интерферирующей РНК только в первых клетках, но не во вторых клетках. В первых клетках интерферирующая РНК экспрессируется с вирусного генома после инфицирования клеток вирусом и связывается с целевой последовательностью интерферирующей РНК, таким образом предотвращая или ингибируя трансляцию одного или нескольких существенных генов; в результате вирус не реплицируется, и клетки остаются в безопасности. Во вторых клетках интерферирующая РНК не экспрессируется из-за отсутствия промоторной активности в клетках, что приводит в результате к устойчивой экспрессии регулируемого существенного гена, что приводит к репликации вируса и уничтожению клеток.
Онколитический вирус двух упомянутых выше видов может обеспечивать селективную репликацию во вторых клетках, при этом не затрагивая первые клетки, так чтобы нормальные клетки оставались в безопасности.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, первые клетки являются отличными от опухолевых клетками млекопитающего, а вторые клетки являются опухолевыми клетками.
Согласно третьему аспекту, настоящее изобретение относится к нуклеотидным последовательностям для создания описанного выше онколитического вируса, нуклеотидные последовательности содержат один или несколько следующих элементов:
целевую последовательность, первую кассету экспрессии и вторую кассету экспрессии.
Согласно четвертому аспекту, настоящее изобретение относится к способу конструирования плазмид для получения комплементарных клеток-хозяев, родительского вируса и онколитического вируса, которые описаны выше.
По сравнению с геномной последовательностью вируса дикого типа в геноме родительского вируса отсутствует существенный ген.
Комплементарные клетки-хозяева содержат фрагмент ДНК для экспрессии существенного гена, который отсутствует в родительском вирусе.
Согласно пятому аспекту, настоящее изобретение относится к способу получения описанного выше онколитического вируса, предусматривающему встраивание описанной выше нуклеотидной последовательности в вирусный геном.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, данный способ получения предусматривает культивирование клеток, инфицирование комплементарных клеток родительским вирусом с последующей трансфекцией клеток плазмидной ДНК, скрининг и идентификацию рекомбинантного онколитического вируса.
Согласно шестому аспекту, настоящее изобретение относится к применению описанного выше онколитического вируса для селективного уничтожения клеток.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, клетки представляют опухолевые клетки.
Согласно седьмому аспекту, настоящее изобретение относится к способу уничтожения клеток, предусматривающему инфицирование целевых для онколитического вируса клеток описанным выше онколитическим вирусом.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, целевые клетки представляют собой опухолевые клетки.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, способ направлен на лечение, не связанное с лечением заболевания.
Согласно восьмому аспекту, настоящее изобретение относится к лекарственному препарату для уничтожения клеток, который содержит описанный выше онколитический вирус и клинически приемлемое вспомогательное вещество.
Кроме того, в соответствии с определенными вариантами осуществления настоящего изобретения, клетки представляют опухолевые клетки.
Согласно девятому аспекту, настоящее изобретение относится к способу детекции описанного выше онколитического вируса с подробными стадиями, предусматривающими определение титра онколитического вируса, размножение онколитического вируса, очистку онколитического вируса, анализ экспрессии мРНК, анализ экспрессии белка и анализ экспрессии miRNA.
В настоящем описании дополнительно предложен способ лечения заболевания в клинической практике, предусматривающий введение онколитического вируса или средств комбинированной терапии, содержащих вирус, представленный в настоящем раскрытии, причем источником заболевания являются вторые клетки.
В соответствии с одним или несколькими вариантами осуществления, заболевание представляет собой рак, а вторые клетки представляют собой опухолевые клетки.
Настоящее изобретение дополнительно относится к применению онколитического вируса в соответствии с настоящим изобретением для селективного уничтожения клеток.
В соответствии с одним или несколькими вариантами осуществления клетки представляют собой опухолевые клетки.
Краткое описание чертежей
Иллюстративные признаки настоящего изобретения, его суть и различные преимущества будут очевидны из сопроводительных чертежей и последующего подробного описания различных вариантов осуществления. Неограничивающие и неисчерпывающие варианты осуществления описаны с привязкой к прилагаемым чертежам. Размеры и взаимное расположение элементов на чертежах не обязательно выполнены в масштабе. Например, формы различных элементов выбраны, увеличены и позиционированы для улучшения читаемости чертежа. Конкретные формы нарисованных элементов выбраны для облегчения распознавания на чертежах. Один или несколько вариантов осуществления описаны ниже с привязкой к прилагаемым чертежам, на которых:
Фиг. 1. Схематическое изображение экзогенных элементов, вставленных в геном онколитического вируса, представленного в настоящем раскрытии: (а): элементы, вставленные в геном, причем количество копий целевой последовательности может насчитывать одну или несколько; (b): расположение целевой последовательности, вставленной в существенный вирусный ген; (с): экспрессия первой интерферирующей РНК в первых клетках (c1) и одновременная экспрессия первой и второй интерферирующей РНК в первых клетках (с2). Первый промотор является конститутивным, который управляет непрерывной экспрессией интерферирующей РНК в первых клетках. Первая интерферирующая РНК нацелена на один или несколького существенных генов путем связывания с целевой последовательностью, вставленной в 3'-UTR одного существенного гена или генов. Вторая интерферирующая РНК нацелена на ORF несущественного гена; (d): специфическая экспрессия ингибирующих компонентов во вторых клетках, что ингибирует биосинтез интерферирующих РНК. Ингибирующие компоненты включают интерферирующую РНК и РНК с увеличенным количеством нуклеотидных триплетных повторов.
Фиг. 2. Схематическое изображение родительской плазмиды pcDNA3.1-EGFP, унифицированной для конструирования плазмиды, экспрессирующей HSV-1 ICP27. В плазмиде EGFP конститутивно экспрессируется под контролем промотора CMV, и плазмида содержит последовательность экспрессии гена устойчивости к неомицину.
Фиг. 3. Схема экзогенных элементов, вставленных в геном онколитического вируса oHSV-BJR в варианте осуществления настоящего изобретения.
Фиг. 4. Экспрессия miRNA EGFP и ICP34.5 с онколитического вируса oHSV-BJR в нормальных клетках и ингибирование с их помощью биосинтеза белков целевых генов ICP27 и ICP34.5. Клетки Vero инфицировали посредством 0,25 MOI (вирус/клетка) вируса KOS HSV-1 дикого типа или онколитического вируса oHSV-BJR. Часть клеток собирали через одни сутки, выделяли малые РНК и детектировали miRNA с помощью нозерн-блоттинга (А). Остальные клетки собирали через двое суток после инфицирования, выделяли белки и определяли уровни белков ICP27 и ICP34.5 с помощью вестерн-блоттинга (В).
Фиг. 5. Функциональный путь биосинтеза малых интерферирующих РНК, наблюдаемый в опухолевых клетках; значительное снижение экспрессии Drosha и биосинтез малых интерферирующих РНК, а также устойчивая экспрессия ICP27 и ICP34.5, наблюдаемая в опухолевых клетках, инфицированных онколитическим вирусом. Для детекции того, обладают ли опухолевые клетки функциональным путем биосинтеза малой интерферирующей РНК, клетки линии Hela опухоли шейки матки, клетки линии siHA плоскоклеточного рака шейки матки, клетки линии SK-BR3 рака молочной железы и клетки линии МЕ-180 рака молочной железы трансфицировали плазмидой экспрессии ICP27 или плазмидой коэкспрессии ICP27+ целевой последовательности и miRNA. Клетки собирали через двое суток после трансфекции, выделяли белки и детектировали белок существенного гена ICP27 HSV-1 с помощью вестерн-блоттинга (А). Для определения того, влияет ли экспрессия ингибирующих триплетных повторов и siRNA Drosha из онколитического вируса oHSV-BJR в раковых клетках на экспрессию Drosha, ингибируется или блокируется путь биосинтеза малых интерферирующих РНК в раковых клетках, инфицированных онколитическим вирусом, и могут ли ICP27 и ICP34.5 устойчиво экспрессироваться с онколитического вируса в раковых клетках, раковые клетки Hela, SiHA, SK-BR3 и МЕ-180 инфицировали посредством KOS или онколитическим вирусом oHSV-BJR (0,5 MOI) соответственно. Клетки собирали через одни сутки после инфицирования и выделяли малые РНК и белки. Триплетные повторы и siRNA Drosha (В), а также miRNA EGFP и ICP34.5 (D) детектировали с помощью нозерн-блоттинга, тогда как белки Drosha (С), ICP27 и ICP34.5 (Е) детектировали с помощью вестерн-блоттинга.
Фиг. 6. В раковых клетках наблюдали сходную кинетику репликации онколитического вируса oHSV-BJR и вируса дикого типа KOS. Различные раковые клетки инфицировали посредством 0,1 MOI KOS или oHSV-BJR. Клетки в среде собирали в разные сутки после инфицирования и оставшиеся в клетках вирусы высвобождали в культуральную среду с помощью трех циклов замораживания и оттаивания. Комплементарные клетки инфицировали вирусами и титры вирусов определяли с помощью анализа бляшкообразования (бляшкообразующая единица/миллилитр, БОЕ/мл). А: клетка линии Hela опухоли шейки матки; В: клетка линии siHA плоскоклеточного рака шейки матки; С: клетка линии SK-BR3 рака молочной железы; и D: клетка линии МЕ-180 рака молочной железы.
Фиг. 7. Значительное ингибирование роста опухоли онколитическим вирусом oHSV-BJR в животных моделях опухолей. Культивированные клетки линии А549 немелкоклеточного рака легкого человека, клетки линии NCI-N87 рака желудка и клетки линии SK-HEP-1 рака печени вводили подкожно мышам BALB/c (рак легкого и рак желудка) или мышам NPG (рак печени); в тот день, когда опухоли достигали размера 1000 мм3, опухоли иссекали, разрезали на мелкие кусочки и подкожно имплантировали мышам; когда опухоль достигала размера 40-120 мм3, начинали внутриопухолевое введение вируса. Модель рака прямой кишки создавали путем подкожной инъекции клеток линии НСТ-8 ректальной аденокарциномы мышам BALB/c, и когда опухоль достигала размера 40-120 мм3, начинали внутриопухолевую инъекцию онколитического вируса. Внутриопухолевую инъекцию онколитического вируса производили путем многоточечной инъекции один раз каждые 3 суток, всего 3 раза, каждый раз дозой 2×107 инфекционных единиц (суспендированных в 40 мкл PBS). Инъекция PBS служила отрицательным контролем. После первой инъекции онколитического вируса размер опухоли измеряли два раза в неделю, и исследование завершалось через 25-32 суток после первой инъекции вируса в зависимости от того, когда необходимо было умерщвлять животных в группе отрицательного контроля. Строили кривую роста опухоли на основе размера опухоли в течение нескольких суток после первой инъекции вируса (А: рак легких; В: рак печени; С: рак желудка; и D: рак прямой кишки), и относительную степень ингибирования (Е) рассчитывали путем сравнения размеров опухолей между тестируемой группой и отрицательным контролем в конце исследования.
Подробное раскрытие настоящего изобретения
Для демонстрации признаков настоящего изобретения, его сути и различных преимуществ, были реализованы иллюстративные варианты осуществления, которые подробно описаны ниже. Все эксперименты были проведены с применением стандартных способов, описанных в литературе. Реагенты были приобретены у коммерческих поставщиков и были использованы в соответствии с инструкциями производителя.
Применяемые в контексте настоящего документа термины «последовательность оснований» и «нуклеотидная последовательность» можно применять взаимозаменяемо, и обычно они относятся к составу и порядку расположения нуклеотидов в ДНК или РНК.
Термин «праймер» относится к синтетическому олигонуклеотиду, который необходим для синтеза нуклеиновой кислоты de novo. После связывания с полинуклеотидной матрицей праймер удлиняется в направлении от 5' к 3' по матрице под каталитическим действием ДНК-полимеразы, тем самым образуя удлиненный дуплекс. Добавление нуклеотидов во время удлинения определяется последовательностью матрицы. Длина праймера обычно составляет 18-23 нуклеотида. Однако длина праймера определяется несколькими факторами, включая нуклеотидный состав и точку плавления праймера, а также последующее применение ПЦР-продукта после амплификации.
Термин «промотор» обычно относится к последовательности ДНК, которая расположена выше 5'-UTR гена, может быть специфически идентифицирована и связываться РНК-полимеразой и необходима для транскрипции.
Термин «энхансер» относится к последовательности ДНК, которая увеличивает частоту транскрипции связанного с ней гена. Энхансер усиливает транскрипцию за счет увеличения активности промотора. Энхансер может быть расположен либо на 5'-конце, либо на 3'-конце гена и даже может существовать в форме интрона в пределах гена. Энхансер может значительно влиять на экспрессию гена, что может увеличивать транскрипцию гена в 10-200 раз или даже в тысячу раз.
Применяемый в контексте настоящего документа термин «интерферирующая РНК» относится к молекуле РНК, которая может связываться со своей целевой последовательностью, таким образом ингибируя экспрессию целевого гена. К молекулам интерферирующей РНК относятся без ограничения короткая шпилечная РНК (shRNA), siRNA, микроРНК (miRNA), синтезированный дуплекс РНК длиной 21-23 нуклеотида.
Термины «субъект», «индивидуум» и «пациент» можно применять в настоящем документе взаимозаменяемо, и они относятся к позвоночному, предпочтительно млекопитающему, наиболее предпочтительно человеку. К млекопитающему относятся без ограничения мышь, обезьяна, человек, домашнее животное или выращиваемый на ферме домашний скот.
Ниже подробно на основе примеров описаны признаки и суть настоящего изобретения.
Пример 1
Получение рекомбинантного онколитического вируса
Онколитический вирус, представленный в настоящем примере, был разработан путем вставки экзогенных элементов в геном вируса герпеса 1-го типа (HSV-1) KOS дикого типа путем гомологичной рекомбинации. Геном данного онколитического вируса имеет следующие структурные признаки.
(1) Состоит из трех вставленных элементов: целевая последовательность miRNA EGFP, вставленная в 3'-UTR существенного гена HSV-1 ICP27, за которым следует поли(А) SV 40, первая кассета экспрессии и вторая кассета экспрессии. Целевая последовательность miRNA EGFP представляет собой небольшую часть кодирующей последовательности EGFP, которая в дальнейшем называется целевой последовательностью miRNA EGFP. Первая и вторая кассеты экспрессии расположены между поли(А) SV40 и 3'-UTR ICP27. Первая кассета экспрессии экспрессирует как miRNA EGFP, так и miRNA ICP34.5. MiRNA ICP34.5 связывается с ORF ICP34.5. Вторая кассета экспрессии экспрессирует siRNA для целенаправленного воздействия на ORF Dorsa и повторы CGG для ингибирования активности Dorsa.
(2) Первая кассета экспрессии включала: промотор CMV, последовательность экспрессии miRNA EGFP (первой miRNA) и последовательность экспрессии miRNA ICP34.5 (второй интерферирующей РНК, за которой следовала последовательность 1 поли(А) SV40.
(3) Вторая кассета экспрессии, включающая гибридный промотор, состоящий из опухолеспецифического промотора hTERT, слитого с энхансером CMV, последовательность экспрессии ингибирующего компонента для одновременной экспрессии siRNA к Drosha и триплетных повторов CGG и последовательность поли(А), расположенную ниже последовательности экспрессии ингибирующих компонентов.
Данный онколитический вирус oHSV-BJR получали в соответствии с описанными далее способами. Во-первых, комплементарные клетки, экспрессирующие ICP27, получали с использованием клеток почек африканской зеленой мартышки (клеток Vero) в качестве исходного материала с целью подтверждения получения, идентификации и размножения рекомбинантных вирусов. Во-вторых, родительский вирус HSV-EGP, в котором ICP27 HSV-1 заменена на EGFP, получали путем гомологичной рекомбинации между плазмидой и KOS дикого типа с комплементарными клетками в качестве хозяина. В-третьих, oHSV-BJR получали путем гомологичной рекомбинации между плазмидой и родительским вирусом HSV- с комплементарными клетками в качестве хозяина. Далее описаны подробные стадии эксперимента.
(1) Получение комплементарных клеток, экспрессирующих ICP27 ДНК вируса герпеса KOS дикого типа применяли в качестве матрицы, кодирующий участок гена ICP27 амплифицировали с помощью ПЦР и амплифицированный фрагмент вставляли в сайты HindIII и XbaI плазмиды pcDNA3.1-EGFP (см. фиг. 2) для замены ORF EGFP. Полученная в результате плазмида была названа pcDNA3.1-ICP27. В плазмиде pcDNA3.1-ICP27 ген ICP27 HSV-1 экспрессировался под контролем промотора CMV. Кроме того, для облегчения скрининга комплементарных клеток в клетках млекопитающих с плазмиды pcDNA3.1-ICP27 экспрессировался ген устойчивости к неомицину.
Клетки Vero обрабатывали посредством G418 в разных концентрациях, культуральную среду, содержащую G418, заменяли каждые трое суток средой, содержащей G418 в разных концентрациях, и каждые сутки отслеживали гибель клеток. Была определена минимальная концентрация G418, необходимая для гибели всех клеток спустя 6 суток после обработки посредством G418. Такую концентрацию G418 (500 мкг/мл) использовали для получения комплементарных клеток.
3,5×105 клеток Vero высевали в лунки 6-луночного планшета для культивирования клеток и культивировали в течение ночи в не содержащей антибиотики среде. Клетки трансфицировали плазмидой, экспрессирующей ICP-27, с применением липофектамина 2000 в качестве реагента для трансфекции (4 мкг ДНК/лунка) и собирали через 24 часа после трансфекции. Клетки разводили с применением среды, содержащей 500 мкг/мл G418, в 5, 10, 20, 40, 60 раз. 3 мл клеток каждого разведения высевали в лунки 6-луночных планшетов. Среду меняли каждые трое суток, всего от 6 до 7 раз. После достижения клонами клеток диаметра 3-4 мм клоны клеток отбирали с помощью цилиндра для отбора клонов. Клетки от каждого клона постепенно размножали из лунок 24-луночного планшета до объема культуральных колб Т150. Выделяли белки и экспрессию ICP27 из клеток, полученных из каждого клона, анализировали с помощью вестерн-блоттинга. Клетки с наивысшим уровнем экспрессии ICP27 отбирали в качестве комплементарных клеток для поддержки роста и репликации дефектных по механизму репликации вирусов, у которых не экспрессируется ICP27; клетки были названы CICP27. Комплементарная клетка была сохранена в Китайском центре сбора типовых культур (ССТСС), расположенном в Уханьском университете, Луоцзяшань, Учан, город Ухань, 24 апреля 2019 года, с регистрационным номером ССТСС №С201974. (2) Получение родительского вируса
Родительский вирус HSV-EGFP, в геноме которого ICP27 HSV-1 заменен геном EGFP, был разработан путем гомологичной рекомбинации между плазмидой и вирусом герпеса KOS дикого типа. Конструкция родительского вируса должна была облегчить скрининг онколитического вируса на последующих стадиях.
Фрагмент А ДНК, включающий 5'-UTR гена ICP27, промотор CMV, ORF EGFP, поли(А) гена бычьего гормона роста (поли(А) BGH) и 3'-UTR гена ICP27, синтезировали с нуклеотидной последовательностью, представленной под SEQ ID NO: 5. Ниже представлено подробное описание фрагмента А:
сайт 1-6: нерелевантная последовательность для увеличения конечной длины с целью облегчения ферментативного расщепления;
сайт 7-12: сайт Xho1, C/TCGAG;
сайт 13-575: 5'-UTRICP27;
сайт 576-1163: промотор CMV;
сайт 1164-1174: спейсер;
сайт 1175-1180: последовательность Козака для усиления трансляции белков;
сайт 1181-1900: ORF EGFP;
сайт 1901-2144: поли(А) BGH;
сайт 2145-2667: 3'-UTR ICP27;
сайт 2668-2773: сайт HindIII, A/AGCTT; и
сайт 2774-2779: нерелевантная последовательность для увеличения конечной длины для облегчения ферментативного расщепления.
Фрагмент А ДНК был расщеплен и лигирован с сайтами HindIII и Xho1 плазмиды pcDNA3.1-EGFP. Полученная плазмида была названа плазмидой экспрессии EGFP.
3,5×105 комплементарных клеток CICP27 высевали в каждую лунку 6-луночного планшета и культивировали в течение ночи в среде, не содержащей антибиотиков. Клетки из каждой лунки инфицировали посредством 0,1, 0,5, 1 или 3 MOI (вирус/клетка) вируса KOS дикого типа и трансфицировали плазмидой экспрессии EGFP, полученной на предыдущей стадии (4 мкг ДНК/лунка), с применением липофектамина 2000 в качестве реагента для трансфекции через 1 час после заражения.
Через 4 часа после трансфекции смесь для трансфекции заменяли полной средой в 6-луночном планшете. После того, как у всех клеток была обнаружена цитопатическая экспрессия и они стали округлыми, собирали клетки в среде. Центрифугировали клеточные смеси после трех циклов замораживания и оттаивания и собирали супернатанты.
Разводили супернатанты и комплементарные клетки CIcp27 инфицировали вирусом в различных разведениях, а вирусы отделяли с помощью анализа бляшкообразования с наложенной полутвердой метилцеллюлозой в качестве среды-носителя. Через 4-5 суток после инфицирования бляшки подвергали скрининговому исследованию под флуоресцентным микроскопом и отбирали зеленые бляшки. Проводили еще 2-3 рабочих цикла скрининга до получения чистых зеленых бляшек под флуоресцентным микроскопом. Брали бляшку с наиболее яркой зеленой флуоресценцией и размножали с применением комплементарных клеток CIcp27 в качестве хозяина. Полученный вирус являлся родительским вирусом HSV-EGFP.
(3) Конструирование плазмиды, содержащей ICP27 и регуляторные компоненты
Плазмиду для клонирования ТА модифицировали так, чтобы сайт множественного клонирования в плазмиде содержал сайт Xho1. Полученная в результате плазмида была названа плазмидой TA-Xho1.
Синтезировали фрагмент В ДНК, содержащий 5'-UTR ICP27 с включенным эндогенным промотором ICP27, ORF ICP27, две копии целевой последовательности, вставленные тандемно (единичная копия целевой последовательности miRNA EGFP показана под SEQ ID NO: 1 и в таблице 1), и последовательность поли(А) SV40, за которой следовала последовательность 3'-UTR ICP27 (5'- и 3'-фрагмента ДНК, оба из которых содержали один сайт XhoI; и один сайт HindIII был вставлен между поли(А) SV40 и последовательностью 3'-UTR ICP27); и нуклеотидная последовательность фрагмента В ДНК представлена под SEQ ID NO: 6. Ниже представлена подробная информация о фрагменте В ДНК.
сайт 1-6: нерелевантная дополнительная последовательность для увеличения конечной длины с целью облегчения ферментативного расщепления;
сайт 7-12: сайт Xho1, C/TCGAG;
сайт 13-683: 5'-UTRICP27, включая промотор ICP27;
сайт 684-2222: ORF ICP27;
сайт 2223-2227: спейсерная последовательность;
сайт 2228-2249 и 2253-2274: целевая последовательность miRNA EGFP (SEQ ID NO: 1);
сайт 2250-2252: спейсер;
сайт 2275-2800: поли(А) SV40;
сайт 2801-2806: сайт HindIII, A/AGCTT;
сайт 2807-3326: 3'-UTR ICP27;
сайт 3327-3332: сайт Xho1; и
сайт 3333-3338: нерелевантная последовательность для увеличения конечной длины с целью облегчения ферментативного расщепления.
Фрагмент В ДНК расщепляли с помощью XhoI и вставляли в сайт XhoI плазмиды TA-XhoI. Полученная в результате плазмида была названа плазмидой TA-XhoI-mICP27.
Синтезировали фрагмент С ДНК, включающий промотор CMV, последовательность экспрессии miRNA EGFP, последовательность экспрессии miRNA ICP34.5, поли(А) BGH, гибридный промотор hTERT-CMV, последовательность экспрессии siRNA Drosha и последовательность экспрессии триплетного повтора CGG с последующим поли(А) SV40. Фрагмент С ДНК содержал один сайт HindIII на 5'- и 3'-концах соответственно. Последовательность оснований фрагмента С ДНК показана под SEQ ID NO: 7 со следующими подробностями:
сайт 1-8: нерелевантная последовательность для увеличения конечной длины с целью облегчения ферментативного расщепления;
сайт 9-14: сайт HindIII, A/AGCTT;
сайт 15-629: промотор CMV;
сайт 630-706: последовательность экспрессии miRNA EGFP (последовательность, представленная под SEQ ID NO: 2 и в таблице 2);
сайт 707-762: нерелевантная последовательность, служащая спейсером;
сайт 763-830: последовательность экспрессии miRNA ICP34.5 (последовательность, представленная под SEQ ID NO: 3 и в таблице 1);
сайт 831-989: нерелевантная последовательность, служащая спейсером;
сайт 990-1213: поли(А) BGH
где последовательность с нуклеотида 15 по нуклеотид 1213 представляет собой первую кассету экспрессии;
сайт 1214-1660 (обратно-комплементарная): поли(А) SV40;
сайт 1661-1667 (обратно-комплементарная): нерелевантная последовательность, служащая спейсером;
сайт 1668-1967 (обратно-комплементарная): триплетные повторы CGG (общая формула триплетных повторов имеет вид (CGG) 100;
сайт 1968-1976 (обратно-комплементарная): нерелевантная последовательность, служащая спейсером;
сайт 1977-2026 (обратно-комплементарная): последовательность экспрессии siRNA Drosha (последовательность, представленная под SEQ ID NO: 4 и в таблице 1 f);
сайт 2027-2044 (обратно-комплементарная): нерелевантная последовательность, служащая спейсером;
сайт 2045-2152 (обратно-комплементарная): энхансер CMV;
сайт 2153-2608 (обратный комплементарная): промотор hTERT;
последовательность с нуклеотида 214 по нуклеотид 2608 представляет собой вторую кассету экспрессии (обратно-комплементарную);
сайт 2609-2614: сайт HindIII, A/AGCTT;
сайт 2615-2622: нерелевантная последовательность для увеличения конечной длины с целью облегчения ферментативного расщепления.
Фрагмент С ДНК расщепляли с помощью HindIII и вставляли в сайт HindIII плазмиды TA-XhoI-mICP27, полученной на предыдущей стадии, с получением плазмиды TA-XhoI-mICP27-R.EG-R.NA для получения рекомбинантного онколитического вируса.
(4) Конструирование онколитического вируса герпеса oHSV-BJR
Онколитический вирус герпеса конструировали путем гомологичной рекомбинации между плазмидой TA-XhoI-mICP27-REG-RNA и родительским вирусом HSV-EGFP в комплементарных клетках CICP27.
Манипуляции заключались в следующем.
3,5×105 комплементарных клеток CICP27 высевали в 6-луночный планшет и культивировали в течение ночи в среде, не содержащей антибиотиков. Клетки из каждой лунки инфицировали посредством 0,1, 0,5, 1 или 3 MOI родительского вируса HSV-EGFP соответственно и через 1 час трансфицировали рекомбинантной плазмидой TA-XhoI-mICP27-REG-PHK (4 мкг ДНК/лунка) с применением липофектамина 2000 в качестве реагента для трансфекции. Через 4 часа после трансфекции смесь для трансфекции заменяли полной средой. После того, как у всех клеток была обнаружена цитопатическая экспрессия и они стали округлыми, собирали клетки в среде. Центрифугировали клеточные смеси после трех циклов замораживания и оттаивания и собирали супернатанты. Разводили супернатанты и инфицировали комплементарные клетки CICP27 разведенными вирусами. Вирусы разделяли с помощью анализа бляшкообразования с наложенной полутвердой метилцеллюлозой в качестве среды-носителя. Через 4-5 суток инкубации бляшки подвергали скрининговому исследованию под флуоресцентным микроскопом и отбирали черные бляшки. Проводили еще 2-3 рабочих цикла скрининга до получения чистых бляшек под флуоресцентным микроскопом. Вирусы из нескольких чистых бляшек размножали с применением комплементарных клеток CiCp27 в качестве хозяина. Выделяли ДНК инфицированных клеток. Рекомбинантный вирус идентифицировали с помощью ПЦР-амплификации с применением специфических праймеров и секвенирования. Рекомбинантный вирус был назван oHSV-BJR.
Онколитический вирус oHSV-BJR был сохранен в Китайском центре сбора типовых культур (ССТСС), расположенном в Уханьском университете в Луоцзяшане, Учан, город Ухань, 24 апреля 2019 года под регистрационным номером ССТСС №V201919.
Вставленные экзогенные элементы и их местоположения в геноме показаны на фиг. 3:
целевую последовательность miRNA EGFP и последовательность поли(А) SV40 вставляли в 3'-UTR существенного гена ICP27;
первая кассета экспрессии располагалась ниже последовательности поли(А) SV40, включая промотор CMV, последовательность экспрессии miRNA EGFP, последовательность экспрессии miRNA ICP34.5 и поли(А) BGH; и
вторая кассета экспрессии располагалась ниже первой кассеты экспрессии, включая гибридный промотор hTERT-CMV, последовательность экспрессии siRNA Drosha, последовательность экспрессии триплетных повторов CGG и последовательность поли(А) SV40 (вторая кассета экспрессии является обратно-комплементарной по отношению к первой кассете экспрессии).
Пример 2
Анализ титра, размножение и очистка онколитического вируса; анализ экспрессии miRNA, мРНК и белков; и тест на уничтожение опухолевых клеток
(1) Определение титра онколитического вируса oHSV-BJR
3,5×105 комплементарных клеток CICP27 высевали в 6-луночный планшет и культивировали в течение ночи в полной среде. Выполняли серию 10-кратных разведений исходной биомассы вируса, полученного в примере 1, и клетки в лунках инфицировали посредством 0,1 мл вируса каждого разведения соответственно. Среду в лунках аспирировали через 1 час и в каждую лунку добавляли 3 мл полной среды, содержащей 1,25% метил целлюлозы. Клетки инкубировали при 37°С в инкубаторе с 5% СО2 в течение 4-5 суток. В лунки вносили 0,1% кристаллического фиолетового, приготовленного в 50% метаноле и 50% этаноле, и промывали для удаления красителя водопроводной водой, и подсчитывали бляшки. Рассчитывали титр вируса (БОЕ/мл).
(2) Размножение онколитического вируса oHSV-BJR
5,5×106 комплементарных клеток CIcp27 высевали в 150-мл колбу для культивирования и культивировали в течение ночи. Клетки инфицировали посредством 0,03 MOI (количество вирусов на клетку) онколитического вируса oHSV-BJR, полученного из примера 1, и инкубировали при 37°С в инкубаторе с СО2 до тех пор, пока по меньшей мере у 90% клеток не наблюдали цитопатической экспрессии. Собирали клетки в среде. Клеточная смесь клеток представляла собой неочищенный исходную биомассу вирусов.
(3) Очистка онколитического вируса oHSV-BJR
Неочищенную биомассу онколитического вируса oHSV-BJR трижды подвергали циклу замораживания и оттаивания при -80°С/37°С, и очищали при 4°С с помощью низкоскоростного центрифугирования, и собирали супернатант. Супернатант фильтровали и концентрировали с помощью 0,6-мкМ половолоконного фильтра с последующей ультрафильтрацией и концентрированием с применением 0,1-мкМ половолоконного фильтра. Затем исходную биомассу вируса дополнительно очищали с помощью аффинной хроматографии с применением гепарина. Чистый вирус концентрировали с применением дополнительного 0,1-мкМ половолоконного фильтра.
(4) Анализ экспрессии мРНК
Собирали клетки и выделяли РНК с применением набора для очистки РНК Qiagen. Синтезировали кДНК с применением реагента для обратной транскрипции Thermofisher. Уровни мРНК ICP27 и ICP34.5 анализировали с помощью полуколичественной ПЦР (20 циклов ПЦР) с применением праймеров, специфичных к ICP27 или ICP34.5. В качестве контрольной загрузки служил β-актин.
(5) Анализ экспрессии белков
Клетки собирали, промывали посредством 1×PBS и собирали центрифугированием. Белки выделяли с применением буферного раствора RIPA. Концентрацию белка измеряли с помощью ВСА с применением BSA в качестве стандарта для построения стандартной кривой. Белки разделяли на геле SDS-PAGE с градиентом 4-20% и переносили на PVDF-мембрану. Мембрану блокировали с помощью 5% сухого молока, приготовленного в PBS, содержащем 0,05% Твин 20, затем инкубировали с первичными антителами, приготовленными в PBST, содержащем 2,5% сухое молоко, при комнатной температуре в течение 2 часов. Иммуноблот 3 раза промывали посредством PBST и инкубировали со вторичными антителами, приготовленными в PBST, содержащем 2,5% сухое молоко, при комнатной температуре в течение 1 часа. Мембрану инкубировали с хемилюминесцентными субстратами от Piece, а бэнды белка визуализировали с помощью ChemiDoc (Bio-Rad). В качестве контрольной загрузки использовали β-актин.
(6) Анализ экспрессии miRNA
MiRNA выделяли с применением набора для выделения чистой miRNA от Thermofisher, РНК-зонды метили с применением набора для мечения РНК DIG от Roche и miRNA анализировали с помощью нозерн-блоттинга.
(7) Культивирование опухолевых клеток
Клетки линии Hela рака шейки матки, клетки линии siHA плоскоклеточного рака шейки матки, клетки линии SK-BR3 рака молочной железы и клетки линии МЕ-180 рака молочной железы приобретали у АТСС, США. Hela, siHA и МЕ-180 культивировали в среде DMEM с добавлением 7,5% фетальной бычьей сыворотки (FBS) и 1× смеси пенициллина/стрептомицина. SK-BR3 культивировали в среде McCoy с добавлением 7,5% FBS и 1× смеси пенициллина/стрептомицина. Для поддержания клетки пересевали каждые три суток.
Пример 3
Введенные молекулы miRNA экспрессировались с oHSV-BJ и значительно влияли на экспрессию целевого гена
Для изучения, экспрессируются ли miRNA с онколитического вируса oHSV-BJR в нормальных клетках, как и ожидалось, и влияют ли они на экспрессию целевых вирусных генов, клетки Vero инфицировали посредством 3 MOI онколитического вируса oHSV-BJR или вируса KOS дикого типа. Клетки собирали через 1 сутки после инфицирования, выделяли малые РНК и белки. MiRNA EGFP и ICP34.5 анализировали с помощью нозерн-блоттинга соответственно, тогда как белки ICP27 и ICP34.5 HSV-1 анализировали с помощью вестерн-блоттинга с применением антител, специфичных к ICP27 и ICP34.5.
Первичная miRNA EGFP (pri-miRNA), miRNA-предшественница EGFP (pre-miRNA) и зрелая miRNA EGFP не были детектированы в клетках, инфицированных посредством KOS дикого типа. Тем не менее, все из ICP34.5-специфической pri-miRNA, pre-miRNA и зрелой miRNA экспрессировались до легко детектируемого уровня, при этом наблюдали больше pre-miRNA и зрелой miRNA по сравнению с pri-miRNA, явление, которое согласовывалось с публикациями в литературе о том, что клетки-хозяева кодируют miRNA к ICP34.5 для ограничения репликации HSV-1. Все из pri-miRNA, pre-miRNA и зрелой miRNA как EGFP, так и ICP34.5 экспрессировались до детектируемого уровня в клетках, инфицированных посредством oHSV-BJR, с гораздо большим количеством pre-miRNA и зрелой miRNA EGFP и ICP34.5, чем наблюдаемое количество pri-miRNA. Более того, уровни pre-miRNA и зрелой miRNA ICP34.5 в инфицированных онколитическим вирусом клетках были намного выше, чем уровни, наблюдаемые в KOS-инфицированных клетках (фиг. 4А). В KOS-инфицированных клетках белки ICP27 и ICP34.5 продуцировались до легко детектируемого уровня. Однако в клетках, инфицированных посредством oHSV-BJR, данные два белка были ниже предела детекции (фиг. 4В). Такие результаты свидетельствовали, что введенные miRNA EGFP и ICP34.5 надежно экспрессировались с онколитического вируса oHSV-BJR в нормальных клетках и значительно ингибировали экспрессию целевых вирусных генов.
Пример 4
Опухолевые клетки обладают функциональным путем биосинтеза интерферирующей РНК; продуцировалось следовое количество Drosha, в то время как в опухолевых клетках, инфицированных онколитическим вирусом oHSV-BJR, биосинтез интерферирующей РНК был значительно ингибирован или полностью заблокирован. В результате целевые гены устойчиво экспрессировались в клетках, инфицированных онколитическим вирусом oHSV-BJR, при этом уровни экспрессии были аналогичны уровням, наблюдаемым в клетках, инфицированных вирусом KOS дикого типа.
Для проверки, имеют ли опухолевые клетки функциональный путь биосинтеза малой интерферирующей РНК, опухолевые клетки линии Hela, siHA SK-BR3 или МЕ-180 трансфицировали плазмидой, экспрессирующей ICP27, или плазмидой коэкспрессии ICP27 с целевой последовательностью и miRNA соответственно. Клетки собирали через двое суток после трансфекции и выделяли белки. Белок ICP27 анализировали с помощью вестерн-блоттинга с применением антител, специфических к ICP27.
Для определения, влияет ли экспрессия siRNA Drosha и триплетных повторов онколитического вируса oHSV-BJR в опухолевых клетках на экспрессию Drosha, ингибирует или блокирует ли синтез интерферирующей РНК в клетках, и могут ли ICP27 и ICP34.5 устойчиво экспрессироваться с онколитического вируса в опухолевых клетках, опухолевые клетки линии Hela, siHA, SK-BR3 или МЕ-180 инфицировали посредством 0,5 MOI KOS или онколитического вируса oHSV-BJR соответственно. Клетки собирали через 2 суток после инфицирования, выделяли малые РНК и белки.
Триплетные повторы и siRNA Drosha, а также miRNA EGFP и ICP34.5 детектировали с помощью нозерн-блоттинга, тогда как белки Drosha, ICP27 и ICP34.5 анализировали с помощью вестерн-блоттинга.
Экспрессия miRNA EGFP ингибировала экспрессию ICP27 из ICP27 с помощью плазмиды коэкспрессии целевой последовательности и miRNA (фиг. 5А). Pri-PHK Drosha и триплетные повторы CGG экспрессировались до легко детектируемого уровня (фиг. 5Б) во всех четырех опухолевых клетках, инфицированных онколитическим вирусом, но количество как pre-siRNA, так и зрелой siRNA Drosha было ниже уровня детекции (результаты не показаны). Белок Drosha достигал детектируемого уровня во всех четырех опухолевых клетках, инфицированных вирусом KOS дикого типа, но во всех четырех опухолевых клетках, инфицированных онколитическим вирусом, уровень белка Drosha был очень низким (фиг. 5С). Pri-miRNA EGFP и ICP34.5 достигала детектируемого уровня, тогда как уровни pre-miRNA и зрелой miRNA EGFP и ICP34.5 были очень низкими или не поддавались детекции (фиг. 5D). Соответственно, белки ICP27 и ICP34.5 в опухолевых клетках, инфицированных посредством oHSV-BJR, экспрессировались до уровня, в основном идентичного тому, который наблюдается в клетках, инфицированных KOS (фиг. 5Е). Из результатов видно, что в опухолевых клетках, инфицированных онколитическим вирусом oHSV-BJR, путь синтеза интерферирующей РНК был ингибирован или полностью заблокирован, а целевые вирусные гены oHSV-BJR могли устойчиво экспрессироваться с эффективностью, аналогичной эффективности, наблюдаемой с вирусом KOS дикого типа.
Пример 5
Онколитический вирус oHSV-BJR обладает способностью к репликации, аналогичной той, что наблюдается с вирусом KOS дикого типа в раковых клетках
Для того, чтобы понять механизм, с помощью которого онколитический вирус уничтожает раковые клетки, оценивали репликационную способность вируса в опухолевых клетках. 4 раковых клетки, в том числе клетки линии Hela опухоли шейки матки, клетки линии siHA плоскоклеточного рака шейки матки, клетки линии SK-BR3 рака молочной железы и клетки линии МЕ-180 рака молочной железы инфицировали посредством 0,1 MOI KOS или oHSV-BJR соответственно. Клетки в среде собирали в разные сутки после инфицирования. Вирусы, оставшиеся в клетках, высвобождали в среду посредством трех циклов замораживания и оттаивания при -80/37°С и исходные биомассы вирусов очищали с помощью низкоскоростного центрифугирования. Комплементарные клетки инфицировали вирусами и титры вирусов определяли с помощью анализа бляшкообразования (бляшкообразующая единица/миллилитр, БОЕ/мл). Хотя рекомбинантный вирус oHSV-BJR размножался с разной скоростью в Hela (фиг. 6А), siHA (фиг. 6В), SK-BR3 (фиг. 6С) и МЕ-180 (фиг. 6D), кинетика репликации рекомбинантного вируса в каждом типе клеток была в основном идентичной таковой, наблюдаемой у KOS.
Пример 6
Онколитический вирус oHSV-BJR, как и вирус KOS дикого типа, эффективно уничтожал опухолевые клетки.
Для анализа активности онколитического вируса oHSV-BJR по уничтожению опухолевых клеток опухолевые клетки шейки матки линии Hela, клетки линии siHA плоско клеточного рака шейки матки, клетки линии SK-BR3 рака молочной железы или клетки МЕ-180 линии рака молочной железы инфицировали посредством 0,25 или 0,5 MOI KOS дикого типа или oHSV-BJR соответственно. Жизнеспособность клеток анализировали в разные сутки после инфицирования и рассчитывали уровни гибели клеток. Больше клеток погибало в случае MOI, используемым для инфицирования онколитическим вирусом, которое увеличивалось в любые сутки после инфицирования у всех четырех опухолевых клеток. Уровень гибели клеток одного типа клеток отличался от другого в определенные сутки после инфицирования онколитическим вирусом, но общий профиль уничтожения опухолевых клеток онколитическим вирусом был схож или даже идентичен таковому, наблюдаемому у клеток, инфицированных KOS, для данного типа клеток (таблицы 2-5).
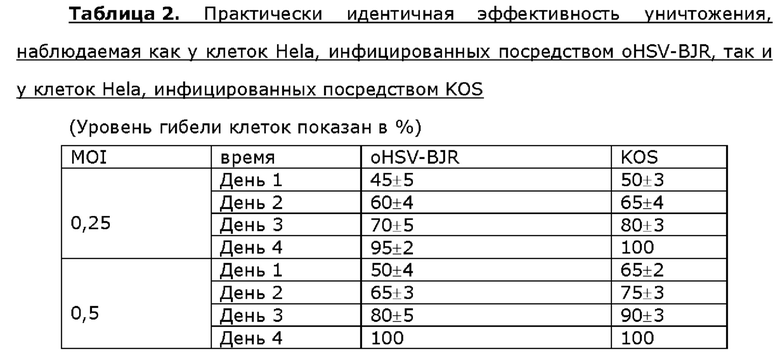
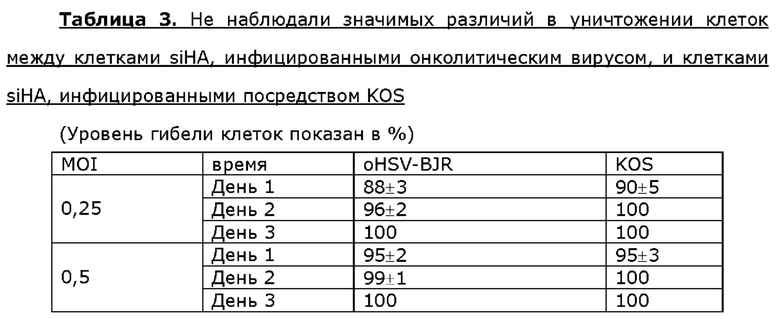
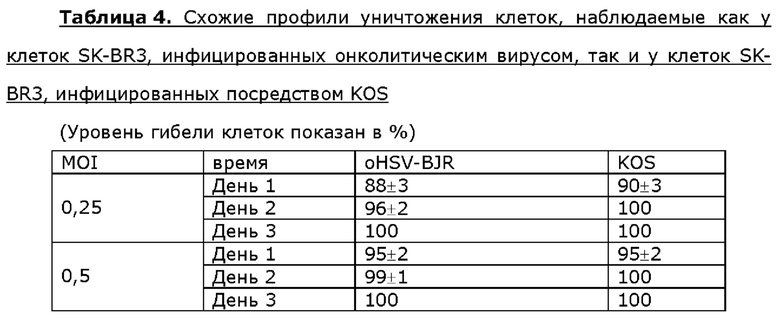
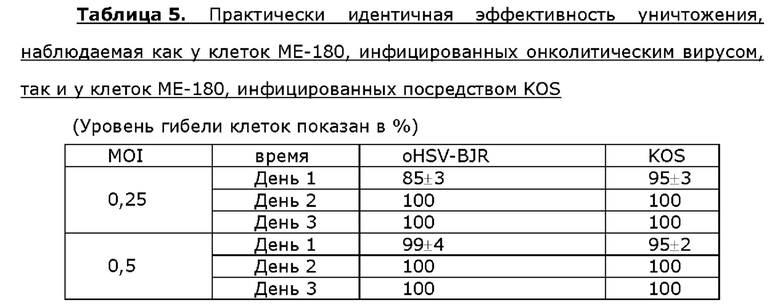
Пример 7
Онколитический вирус oHSV-BJR безопасен для нормальных клеток Для оценки, является ли онколитический вирус безопасным для нормальных клеток, клетки Vero или первичные эпидермальные клетки роговицы человека инфицировали посредством 2 MOI онколитического вируса oHSV-BJR (2 MOI) или посредством 0,5 MOI KOS. Жизнеспособность клеток, инфицированных посредством oHSV-BJR и ложно инфицированных (без обработки) клеток, исследовали через 3 суток после инфицирования, а жизнеспособность клеток, инфицированных вирусом KOS дикого типа, оценивали через 2 суток после инфицирования.
Все клетки Vero и первичные эпидермальные клетки роговицы человека погибали через 2 суток после инфицирования посредством KOS (таблица 6). Лишь незначительная часть клеток Vero и первичных эпидермальных клеток роговицы человека погибала на любые сутки после инфицирования онколитическим вирусом (таблица 6), а выживаемость клеток все еще составляла 95%, что в основном было идентично наблюдаемому у клеток без обработки. Из данных результатов видно, что онколитический вирус oHSV-BJR, полученный в примере 1, относительно безопасен для нормальных клеток.


Пример 8
Онколитический вирус oHSV-BJR значимо ингибировал рост опухоли легких, желудка, печени и прямой кишки у животных
Для оценки эффективности и широкого спектра онколитического вируса oHSV-BJR при лечении опухолей создавали модели опухолей у мышей для опухолей легких, желудка, печени и толстой кишки человека. Культивируемые in vitro клетки немелкоклеточной опухоли человека А549, опухоли желудка NCI-N87 и рака печени SK-HEP-1 вводили подкожно мышам BALB/c (опухоли легких и желудка) или NPG (опухоль печени). При достижении опухолями размера 800-1000 мм3 опухоли иссекали, разрезали на части по 30 мм3, а затем имплантировали мышам. При достижении опухолями размера 40-120 мм3 начинали внутриопухолевое введение онколитического вируса oHSV-BJR.
Модель опухоли прямой кишки создавали путем прямой подкожной инъекции мышам BALB/c культивированных клеток аденокарциномы прямой кишки НСТ-8. При достижении опухолью размера 40-120 мм3 начинали внутриопухолевое введение онколитического вируса oHSV-BJR.
Внутриопухолевую инъекцию онколитического вируса oHSV-BJR производили путем многоточечной инъекции один раз каждые трое суток дня, всего три раза, каждый раз инъекцией дозой 2×107 инфекционных единиц, суспендированных в 40 мкл PBS. Каждая группа в каждой модели включала 8 животных, а инъекция 40 мкл PBS служила отрицательным контролем. Объем опухоли измеряли два раза в неделю после первой инъекции вируса, а исследование продолжалось в течение 25-32 суток после первой инъекции вируса, в зависимости от того, когда необходимо было умерщвлять животных в контрольной группе. Строили кривую роста по объему опухоли в течение нескольких суток после первой инъекции вируса и рассчитывали относительную степень ингибирования путем сравнения объема опухоли в тестируемой группе с объемом опухоли в контрольной группе.
Объем опухоли у животных в тестируемой группе был меньше, чем в контрольной группе, начиная с 8-х суток или около того после первой инъекции вируса (фиг. 7А: опухоль легкого; В: опухоль печени; С: опухоль желудка; и D: опухоль прямой кишки). Разница в объемах опухолей между группой с инъекцией вируса и контрольной группой увеличивалась в течение нескольких суток после первой инъекции вируса. Показатели степени ингибирования онколитическим вирусом роста опухолей легких, печени, желудка и прямой кишки в конце исследования составляли 52,2, 19,5, 45,2, 64,6% соответственно.
Таким образом, автор настоящего изобретения полагает, что основные недостатки доступного в настоящее время онколитического вируса, т.е. низкую эффективность лечения опухолей для данного типа опухоли и небольшую группу пациентов, подходящих для данного вируса, можно объяснить предостережениями, связанными со стратегиями его разработки: селективная репликация доступных в настоящее время онколитических вирусов в опухолевых клетках достигается за счет репликации вируса.
В настоящее время онколитические вирусы разрабатываются в основном с применением следующих трех стратегий:
1) Онколитические вирусы получают путем удаления одного или нескольких несущественных вирусных генов, которые необходимы для репликации вируса в нормальных клетках, но не нужны вирусу в опухолевых клетках, что позволяет вирусу селективно реплицироваться в опухолевых клетках и, в конечном итоге, уничтожать их. Нормальные клетки кодируют несколько механизмов ограничения вирусной инфекции, в то время как каждый вирус кодирует один или несколько генных продуктов для противодействия противовирусной защите хозяина, что позволяет вирусу реплицироваться в нормальных клетках. Удаление такого одного или нескольких вирусных генов из вирусного генома делает вирусные мутанты (онколитические вирусы) неспособными размножаться в нормальных клетках, в то время как такие мутанты по-прежнему сохраняют способность реплицироваться в опухолевых клетках, поскольку во многих опухолевых клетках по разным причинам нарушены противовирусные функции. Для того, чтобы гарантировать безопасность онколитического вируса для нормальных клеток, необходимо удалить более одного гена. Но такой мутантный вирус обычно плохо реплицируется в различных опухолевых клетках, поскольку отсутствуют или нарушены лишь одна или две противовирусные функции среди множества защитных механизмов для любого данного типа опухолевых клеток. В исследованиях было продемонстрировано, что онколитический вирус, полученный с помощью такой стратегии, реплицируется со скоростью от нескольких раз до одного порядка медленнее, чем наблюдаемая скорость репликации у вируса дикого типа в опухолевых клетках in vitro, и скорость репликации такого онколитического вируса может быть ниже, чем у вируса дикого типа, на 1-3 порядка в опухолях у животных. Также, по результатам клинических исследований было видно, что терапевтическая польза от такого онколитического вируса, как правило, неудовлетворительна. Большинство доступных в настоящее время онколитических вирусов было получено с применением такой стратегии с незначительными модификациями или без них.
2) Онколитические вирусы получают путем помещения экспрессии одного или двух существенных вирусных генов под контроль опухолеспецифического промотора. Онколитический вирус, полученный с применением такой стратегии, несомненно, безопасен для нормальных клеток, поскольку опухолеспецифический промотор не активен в нормальных клетках, что не приводит к репликации вируса из-за отсутствия продуцируемого продукта регулируемого гена. Онколитические вирусы такого типа обладают преимуществом перед онколитическими вирусами, полученными с применением первой стратегии, т.е. онколитические вирусы такого типа обладают интактным геномом. Но такие онколитические вирусы по своей природе ассоциированы с проблемой, связанной с репликацией вируса, т.е. с нарушением высококоординированного порядка экспрессии вирусных генов. Вирусные гены делятся на несколько категорий в соответствии с временным порядком экспрессии генов после инфицирования. Гены каждой категории экспрессируются с экспрессией, продолжающейся только в течение короткого периода времени, после чего следует экспрессия генов следующей категории, обеспечивая временную экспрессию вирусных генов. Опухолеспецифический промотор обеспечивает непрерывную экспрессию одного или нескольких регулируемых вирусных генов из онколитического вирусного генома в опухолевых клетках, что определенно препятствует своевременному инициированию экспрессии одного или нескольких генов следующего класса, что приводит к нарушению вирусной репликации онколитического вируса в опухолевых клетках. Можно ожидать, что онколитический вирус, получаемый с помощью такой стратегии, не должен хорошо реплицироваться во многих опухолевых клетках, и по результатам исследования in vitro и in vivo было видно, что это имело место как в условиях in vitro, так и в условиях in vivo. К настоящему времени с помощью такой стратегии было получено несколько онколитических вирусов.
3) Онколитические вирусы получают путем вставки тканеспецифической целевой последовательности miRNA в 3'-UTR существенного вирусного гена. Тканеспецифическая miRNA может быть сверхэкспрессирована в нормальных клетках, в то время как такая miRNA не продуцируется или такое продуцирование сильно нарушено в опухолевых клетках. Следовательно, вставка целевой последовательности miRNA в 3'-UTR существенного вирусного гена может привести к отсутствию или нарушению продуцирования регулируемого вирусного генного продукта у вируса, что приведет к отсутствию или серьезному нарушению репликации вируса в нормальных клетках, в то время как в опухолевых клетках вирус может реплицироваться. С онколитическим вирусом, разработанным с применением такой стратегии, могут быть неразрывно ассоциированы следующие две проблемы: 1) такой онколитический вирус можно было бы применять только для лечения определенного типа опухоли с ограниченной группой пациентов; 2) терапевтические преимущества могут быть ограничены, потому что для любого заданного типа опухоли продуцирование miRNA может быть нарушено, но не исключено, что приведет к ограниченной репликации вируса в опухолевых клетках. С применением такой стратегии было разработано лишь ограниченное количество онколитических вирусов.
Было показано, что онколитические вирусы, полученные с применением вышеупомянутых стратегий, безопасны в клинической практике, и каждый вирус потенциально можно применять для лечения различных опухолей. Но доступные в настоящее время онколитические вирусы обычно обладают низкой эффективностью. Для преодоления недостатков, связанных с доступными в настоящее время онколитическими вирусами, и реализации потенциала онколитического вируса в лечении опухолей, авторы настоящего изобретения использовали новую стратегию, описанную в настоящем раскрытии, для разработки эффективных онколитических вирусов широкого спектра действия для лечения опухолей. Основная идея стратегии состоит в том, чтобы вставить экзогенные элементы в вирусный геном и использовать встроенные элементы для регулирования выработки белка одного или нескольких вирусных генов. Онколитические вирусы, полученные с помощью такой стратегии, обладают двумя важными характеристиками, необходимыми для надежной вирусной репликации, которые, как правило, отсутствуют у большинства доступных в настоящее время онколитических вирусов: 1) интактный вирусный геном без удаления или замены одного нуклеотида из генома, что критически важно для надежной репликации вируса, поскольку каждый генный продукт прямо или косвенно участвует или способствует репликации вируса; 2) экспрессия вирусных генов скоординированным по времени образом. Вирусные гены делятся на несколько категорий в соответствии с порядком экспрессии генов после инфицирования. Гены каждой категории экспрессируются с экспрессией, продолжающейся только в течение короткого периода времени, после чего следует экспрессия генов следующей категории. Нарушение такого порядка, например, за счет использования клеточного или опухолеспецифического промотора для управления экспрессией вирусного гена, может привести к серьезному нарушению репликации вируса. В случае онколитических вирусов, полученных с применением стратегии авторов настоящего изобретения, при экспрессии регулируемых генов их экспрессия управляется их эндогенными промоторами, таким образом поддерживая временной порядок экспрессии всех вирусных генов для обеспечения эффективной репликации вируса. Вставленные последовательности, используемые для получения онколитических вирусов с применением стратегии авторов настоящего изобретения, в основном включают три элемента: 1) целевую последовательность интерферирующей РНК, вставленную в 5'- или 3'-UTR одного, двух или более вирусных существенных генов; 2) первую кассету экспрессии, которая экспрессирует первую интерферирующую РНК в первых клетках; и 3) вторую кассету экспрессии, которая специфически экспрессирует ингибирующие компоненты во вторых клетках, что препятствует или блокирует путь биосинтеза интерферирующей РНК. В первых клетках первая интерферирующая РНК, которая может быть либо siRNA, либо miRNA, конститутивно экспрессируется с первой кассеты экспрессии, таким образом целенаправленно воздействуя на целевую последовательность интерферирующей РНК, расположенную в 5'- или 3'-UTR мРНК регулируемого вирусного гена, что приводит к отсутствию белка или к продуцированию следовых количеств белка. В результате в таких клетках вирус не реплицируется или плохо реплицируется. Во вторых клетках слитый промотор, который состоит из опухолеспецифического промотора, слитого с энхансером, управляет экспрессией ингибирующих компонентов со второй кассеты экспрессии, чтобы как препятствовать биосинтезу, так и ингибировать активность одного или нескольких ферментов, участвующих в пути биосинтеза интерферирующей РНК, что приводит в результате к отсутствию интерферирующей РНК или образованию следовых количеств интерферирующей РНК, продуцируемой в таких клетках, что приводит к устойчивой экспрессии одного или нескольких регулируемых генов и репликации вируса. Для повышения безопасности онколитических вирусов, в соответствии с определенными вариантами осуществления, более одной копии целевой последовательности интерферирующей РНК вставляют в 5'- или 3'-UTR одного существенного вирусного гена; в соответствии с определенными вариантами осуществления, целевую последовательность интерферирующей РНК вставляют в 5'- или 3'-UTR более чем одного вирусного существенного гена; или, альтернативно, в соответствии с определенными вариантами осуществления, вторая интерферирующая РНК, которая нацелена на ORF несущественного вирусного гена, экспрессируется с первой кассеты экспрессии, когда целевая последовательность первой интерферирующей РНК вставлена в 5'- или 3'-UTR одного вирусного существенного гена для обеспечения отсутствия репликации вируса в первых клетках. Для расширения сферы клинического применения онколитических вирусов слитый промотор, состоящий из опухолеспецифического промотора, активного в различных опухолевых клетках, и энхансера, выбирают так, чтобы он управлял экспрессией ингибирующих компонентов со второй кассеты экспрессии, чтобы как препятствовать биосинтезу, так и ингибировать активность одного или нескольких ферментов, участвующих в пути биосинтеза интерферирующей РНК. Опухолеспецифический промотор обуславливает экспрессию ингибирующих компонентов, в частности, в опухолевых клетках. Опухолеспецифический промотор, активный в ряде опухолевых клеток, такой как промотор hTERT, который, как было показано, активен у 85-90% протестированных опухолевых клеток, выбирают так, чтобы он обеспечивал, что в большинстве опухолевых клеток с вирусов могли экспрессироваться ингибирующие компоненты. Добавление энхансера для образования слитого промотора должно обеспечивать достаточную экспрессию ингибирующих компонентов даже в таких опухолевых клетках, в которых активность опухолеспецифического промотора низкая. По сравнению со всеми стратегиями, применяемыми для получения онколитических вирусов, стратегия авторов настоящего изобретения, описанная в настоящем раскрытии, полностью отличается и является совершенно новой. И безопасность, и эффективность онколитического вируса, полученного с применением такой стратегии, была продемонстрирована в условиях in vitro и на животных с помощью иллюстративных вариантов осуществления.
Вышеупомянутые варианты осуществления были реализованы лишь для демонстрации концепций и идеи, представленных в настоящем раскрытии, которое не предназначалось для идентификации ключевых или существенных признаков заявляемого предмета изобретения и не предназначалось для ограничения объема заявляемого предмета изобретения. Настоящее раскрытие можно модифицировать и изменить с помощью различных способов, известных специалисту в настоящей области техники. Модификации, замены и комбинации, сделанные на основе настоящего раскрытия, будут охватываться объемом защиты настоящего изобретения.
Промышленная применимость
Онколитический вирус, представленный в настоящем раскрытии, может селективно уничтожать целевые клетки (такие как опухолевые клетки) посредством селективной репликации, но не затрагивает нецелевые клетки (такие как нормальные клетки). В частности, онколитический вирус, полученный с применением стратегии, представленной в настоящем раскрытии, можно применять для лечения рака.
--->
Перечень последовательностей
<110> ЦЗЕТАНГ ВУ
<120> Рекомбинантный онколитический вирус, способ его получения, его
применение и лекарственный препарат на его основе
<130> OPI21301130RU
<150> CN201910462073.5
<151> 2019-5-30
<150> PCT/CN2020/093006
<151> 2020-5-28
<160> 8
<170> PatentIn version 3.5
<210> 1
<211> 22
<212> DNA
<213> Искусственная последовательность
<220>
<223> Последовательность оснований целевой последовательности
<400> 1
caagctgacc ctgaagttca ta 22
<210> 2
<211> 21
<212> RNA
<213> Искусственная последовательность
<220>
<223> Последовательность оснований первой интерферирующей РНК
<400> 2
augaacuuca gggucagcuu g 21
<210> 3
<211> 22
<212> RNA
<213> Искусственная последовательность
<220>
<223> Последовательность второй интерферирующей РНК
<400> 3
cuugccuguc uaacucgcua gu 22
<210> 4
<211> 25
<212> RNA
<213> Искусственная последовательность
<220>
<223> Последовательность оснований третьей интерферирующей РНК
<400> 4
cuugcugaau acuugguccu uggug 25
<210> 5
<211> 2679
<212> DNA
<213> Искусственная последовательность
<220>
<223> Последовательность оснований фрагмента A ДНК
<400> 5
aaataactcg agctgcgggc gctgttgttc catcatcctg tcgggcatcg caatgcgatt 60
gtgttatatc gccgtggtgg ccggggtggt gctcgtggcg cttcactacg agcaggagat 120
ccagaggcgc ctgtttgatg tatgacgtca catccaggcc ggcggaaacc ggaacggcat 180
atgcaaactg gaaactgtcc tgtcttgggg cccacccacc cgacgcgtca tatgtaaatg 240
aaaatcgttc ccccgaggcc atgtgtagcc tggatcccaa cgaccccgcc catgggtccc 300
aattggccgt cccgttacca agaccaaccc agccagcgta tccacccccg cccgggtccc 360
cgcggaagcg gaacggtgta tgtgatatgc taattaaata catgccacgt acttatggtg 420
tctgattggt ccttgtctgt gccggaggtg gggcgggggc cccgcccggg gggcggaact 480
aggaggggtt tgggagagcc ggccccggca ccacgggtat aaggacatcc accacccggc 540
cggtggtggt gtgcagccgt gttccaacca cggtcgttga cattgattat tgactagtta 600
ttaatagtaa tcaattacgg ggtcattagt tcatagccca tatatggagt tccgcgttac 660
ataacttacg gtaaatggcc cgcctggctg accgcccaac gacccccgcc cattgacgtc 720
aataatgacg tatgttccca tagtaacgcc aatagggact ttccattgac gtcaatgggt 780
ggactattta cggtaaactg cccacttggc agtacatcaa gtgtatcata tgccaagtac 840
gccccctatt gacgtcaatg acggtaaatg gcccgcctgg cattatgccc agtacatgac 900
cttatgggac tttcctactt ggcagtacat ctacgtatta gtcatcgcta ttaccatggt 960
gatgcggttt tggcagtaca tcaatgggcg tggatagcgg tttgactcac ggggatttcc 1020
aagtctccac cccattgacg tcaatgggag tttgttttgg caccaaaatc aacgggactt 1080
tccaaaatgt cgtaacaact ccgccccatt gacgcaaatg ggcggtaggc gtgtacggtg 1140
ggaggtctat ataagcagag ctctctggct aactgccacc atggtgagca agggcgagga 1200
gctgttcacc ggggtggtgc ccatcctggt cgagctggac ggcgacgtaa acggccacaa 1260
gttcagcgtg tccggcgagg gcgagggcga tgccacctac ggcaagctga ccctgaagtt 1320
catctgcacc accggcaagc tgcccgtgcc ctggcccacc ctcgtgacca ccctgaccta 1380
cggcgtgcag tgcttcagcc gctaccccga ccacatgaag cagcacgact tcttcaagtc 1440
cgccatgccc gaaggctacg tccaggagcg caccatcttc ttcaaggacg acggcaacta 1500
caagacccgc gccgaggtga agttcgaggg cgacaccctg gtgaaccgca tcgagctgaa 1560
gggcatcgac ttcaaggagg acggcaacat cctggggcac aagctggagt acaactacaa 1620
cagccacaac gtctatatca tggccgacaa gcagaagaac ggcatcaagg tgaacttcaa 1680
gatccgccac aacatcgagg acggcagcgt gcagctcgcc gaccactacc agcagaacac 1740
ccccatcggc gacggccccg tgctgctgcc cgacaaccac tacctgagca cccagtccgc 1800
cctgagcaaa gaccccaacg agaagcgcga tcacatggtc ctgctggagt tcgtgaccgc 1860
cgccgggatc actctcggca tggacgagct gtacaagtaa cgggcctcga ctgtgccttc 1920
tagttgccag ccatctgttg tttgcccctc ccccgtgcct tccttgaccc tggaaggtgc 1980
cactcccact gtcctttcct aataaaatga ggaaattgca tcgcattgtc tgagtaggtg 2040
tcattctatt ctggggggtg gggtggggca ggacagcaag ggggaggatt gggaagacaa 2100
tagcaggcat gctggggatg cggtgggctc tatggcttct gaggtacaat aaaaacaaaa 2160
catttcaaac aaatcgcccc acgtgttgtc cttctttgct catggccggc ggggcgtggg 2220
tcacggcaga tggcgggggt gggcccggcg tacggcctgg gtgggcggag ggaactaacc 2280
caacgtataa atccgtcccc gctccaaggc cggtgtcata gtgcccttag gagcttcccg 2340
cccgggcgca tccccccttt tgcactatga cagcgacccc cctcaccaac ctgttcttac 2400
gggccccgga cataacccac gtggcccccc cttactgcct caacgccacc tggcaggccg 2460
aaacggccat gcacaccagc aaaacggact ccgcttgcgt ggccgtgcgg agttacctgg 2520
tccgcgcctc ctgtgagacc agcggcacaa tccactgctt tttctttgcg gtatacaagg 2580
acacccacca tacccctccg ctgattaccg agctccgcaa ctttgcggac ctggttaacc 2640
acccgccggt cctacgcgaa ctggtcgaag ctttttata 2679
<210> 6
<211> 3338
<212> DNA
<213> Искусственная последовательность
<220>
<223> Последовательность оснований фрагмента B ДНК
<400> 6
cccaaactcg agctgcgggc gctgttgttc catcatcctg tcgggcatcg caatgcgatt 60
gtgttatatc gccgtggtgg ccggggtggt gctcgtggcg cttcactacg agcaggagat 120
ccagaggcgc ctgtttgatg tatgacgtca catccaggcc ggcggaaacc ggaacggcat 180
atgcaaactg gaaactgtcc tgtcttgggg cccacccacc cgacgcgtca tatgtaaatg 240
aaaatcgttc ccccgaggcc atgtgtagcc tggatcccaa cgaccccgcc catgggtccc 300
aattggccgt cccgttacca agaccaaccc agccagcgta tccacccccg cccgggtccc 360
cgcggaagcg gaacggtgta tgtgatatgc taattaaata catgccacgt acttatggtg 420
tctgattggt ccttgtctgt gccggaggtg gggcgggggc cccgcccggg gggcggaact 480
aggaggggtt tgggagagcc ggccccggca ccacgggtat aaggacatcc accacccggc 540
cggtggtggt gtgcagccgt gttccaacca cggtcacgct tcggtgcctc tccccgattc 600
gggcccggtc gcttgctacc ggtgcgccac caccagaggc catatccgac accccagccc 660
cgacggcagc cgacagcccg gtcatggcga ctgacattga tatgctaatt gacctcggcc 720
tggacctctc cgacagcgat ctggacgagg acccccccga gccggcggag agccgccgcg 780
acgacctgga atcggacagc aacggggagt gttcctcgtc ggacgaggac atggaagacc 840
cccacggaga ggacggaccg gagccgatac tcgacgccgc tcgcccggcg gtccgcccgt 900
ctcgtccaga agaccccggc gtacccagca cccagacgcc tcgtccgacg gagcggcagg 960
gccccaacga tcctcaacca gcgccccaca gtgtgtggtc gcgcctcggg gcccggcgac 1020
cgtcttgctc ccccgagcgg cacgggggca aggtggcccg cctccaaccc ccaccgacca 1080
aagcccagcc tgcccgcggc ggacgccgtg ggcgtcgcag gggtcggggt cgcggtggtc 1140
ccggggccgc cgatggtttg tcggaccccc gccggcgtgc ccccagaacc aatcgcaacc 1200
cggggggacc ccgccccggg gcggggtgga cggacggccc cggcgccccc catggcgagg 1260
cgtggcgcgg aagtgagcag cccgacccac ccggaggccc gcggacacgg agcgtgcgcc 1320
aagcaccccc cccgctaatg acgctggcga ttgccccccc gcccgcggac ccccgcgccc 1380
cggccccgga gcgaaaggcg cccgccgccg acaccatcga cgccaccacg cggttggtcc 1440
tgcgctccat ctccgagcgc gcggcggtcg accgcatcag cgagagcttc ggccgcagcg 1500
cacaggtcat gcacgacccc tttggggggc agccgtttcc cgccgcgaat agcccctggg 1560
ccccggtgct ggcgggccaa ggagggccct ttgacgccga gaccagacgg gtctcctggg 1620
aaaccttggt cgcccacggc ccgagcctct atcgcacttt tgccggcaat cctcgggccg 1680
catcgaccgc caaggccatg cgcgactgcg tgctgcgcca agaaaatttc atcgaggcgc 1740
tggcctccgc cgacgagacg ctggcgtggt gcaagatgtg catccaccac aacctgccgc 1800
tgcgccccca ggaccccatt atcgggacgg ccgcggcggt gctggataac ctcgccacgc 1860
gcctgcggcc ctttctccag tgctacctga aggcgcgagg cctgtgcggc ctggacgaac 1920
tgtgttcgcg gcggcgtctg gcggacatta aggacattgc atccttcgtg tttgtcattc 1980
tggccaggct cgccaaccgc gtcgagcgtg gcgtcgcgga gatcgactac gcgacccttg 2040
gtgtcggggt cggagagaag atgcatttct acctccccgg ggcctgcatg gcgggcctga 2100
tcgaaatcct agacacgcac cgccaggagt gttcgagtcg tgtctgcgag ttgacggcca 2160
gtcacatcgt cgcccccccg tacgtgcacg gcaaatattt ttattgcaac tccctgtttt 2220
aggatcgcaa gctgaccctg aagttcatag tccaagctga ccctgaagtt catacgtcca 2280
catgaagcag cacgacttct gaccacatga agcagcacga cttgatccag acatgataag 2340
atacattgat gagtttggac aaaccacaac tagaatgcag tgaaaaaaat gctttatttg 2400
tgaaatttgt gatgctattg ctttatttgt aaccattata agctgcaata aacaagttaa 2460
caacaacaat tgcattcatt ttatgtttca ggttcagggg gaggtgtggg aggtttttta 2520
aagcaagtaa aacctctaca aatgtggtat ggctgattat gatcctgcaa gcctcgtcgt 2580
cctggccgga ccacgctatc tgtgcaaggt ccccggcccc ggacgcgcgc tccatgagca 2640
gagcgcccgc cgccgaggcg aagactcggg cggcgccctg cccgtcccac caggtcaaca 2700
ggcggtaacc ggcctcttca tcgggaatgc gcgcgacctt cagcatcgcc ggcatgtccc 2760
cctggcggac gggaagtatc cagctcgacc aagctgtttt aagcttgtac aataaaaaca 2820
aaacatttca aacaaatcgc cccacgtgtt gtccttcttt gctcatggcc ggcggggcgt 2880
gggtcacggc agatggcggg ggtgggcccg gcgtacggcc tgggtgggcg gagggaacta 2940
acccaacgta taaatccgtc cccgctccaa ggccggtgtc atagtgccct taggagcttc 3000
ccgcccgggc gcatcccccc ttttgcacta tgacagcgac ccccctcacc aacctgttct 3060
tacgggcccc ggacataacc cacgtggccc ccccttactg cctcaacgcc acctggcagg 3120
ccgaaacggc catgcacacc agcaaaacgg actccgcttg cgtggccgtg cggagttacc 3180
tggtccgcgc ctcctgtgag accagcggca caatccactg ctttttcttt gcggtataca 3240
aggacaccca ccatacccct ccgctgatta ccgagctccg caactttgcg gacctggtta 3300
accacccgcc ggtcctacgc gaactgctcg aggggaaa 3338
<210> 7
<211> 2622
<212> DNA
<213> Искусственная последовательность
<220>
<223> Последовательность оснований фрагмента C ДНК
<400> 7
aataatataa gcttgttgac attgattatt gactagttat taatagtaat caattacggg 60
gtcattagtt catagcccat atatggagtt ccgcgttaca taacttacgg taaatggccc 120
gcctggctga ccgcccaacg acccccgccc attgacgtca ataatgacgt atgttcccat 180
agtaacgcca atagggactt tccattgacg tcaatgggtg gagtatttac ggtaaactgc 240
ccacttggca gtacatcaag tgtatcatat gccaagtacg ccccctattg acgtcaatga 300
cggtaaatgg cccgcctggc attatgccca gtacatgacc ttatgggact ttcctacttg 360
gcagtacatc tacgtattag tcatcgctat taccatggtg atgcggtttt ggcagtacat 420
caatgggcgt ggatagcggt ttgactcacg gggatttcca agtctccacc ccattgacgt 480
caatgggagt ttgttttggc accaaaatca acgggacttt ccaaaatgtc gtaacaactc 540
cgccccattg acgcaaatgg gcggtaggcg tgtacggtgg gaggtctata taagcagagc 600
tcgtttagtg aaccgtcaga tcgcctggat gctgatgaac ttcagggtca gcttggtttt 660
ggccactgac tgaccaagct gactgaagtt catcaggctg gctcagcact gctatgttgc 720
ctgctcttac tggctggagt gaagaccagc aaaggccatt cctgctgatg aacttcaggg 780
tcagcttggt tttggccact gactgaccaa gctgactgaa gttcatcagg cctgctgagt 840
gacagcaccc ctttggagcc ccctccctta tatctaatgg aagattatgt gggcaacccg 900
gtggtagccg gggtggtaga gtttgacagg caagcatgtg cgtgcagagg cgagtagtgc 960
ttgcctgtct aactcgctag tctcggcgac tgtgccttct agttgccagc catctgttgt 1020
ttgcccctcc cccgtgcctt ccttgaccct ggaaggtgcc actcccactg tcctttccta 1080
ataaaatgag gaaattgcat cgcattgtct gagtaggtgt cattctattc tggggggtgg 1140
ggtggggcag gacagcaagg gggaggattg ggaagacaat agcaggcatg ctggggatgc 1200
ggtgggctct atgggatact tcccgtccgc cagggggaca tgccggcgat gctgaaggtc 1260
gcgcgcattc ccgatgaaga ggccggttac cgcctgttga cctggtggga cgggcagggc 1320
gccgcccgag tcttcgcctc ggcggcgggc gctctgctca tggagcgcgc gtccggggcc 1380
ggggaccttg cacagatagc gtggtccggc caggacgacg aggcttgcag gatcataatc 1440
agccatacca catttgtaga ggttttactt gctttaaaaa acctcccaca cctccccctg 1500
aacctgaaac ataaaatgaa tgcaattgtt gttgttaact tgtttattgc agcttataat 1560
ggttacaaat aaagcaatag catcacaaat ttcacaaata aagcattttt ttcactgcat 1620
tctagttgtg gtttgtccaa actcatcaat gtatcttatc aggatccccg ccgccgccgc 1680
cgccgccgcc gccgccgccg ccgccgccgc cgccgccgcc gccgccgccg ccgccgccgc 1740
cgccgccgcc gccgccgccg ccgccgccgc cgccgccgcc gccgccgccg ccgccgccgc 1800
cgccgccgcc gccgccgccg ccgccgccgc cgccgccgcc gccgccgccg ccgccgccgc 1860
cgccgccgcc gccgccgccg ccgccgccgc cgccgccgcc gccgccgccg ccgccgccgc 1920
cgccgccgcc gccgccgccg ccgccgccgc cgccgccgcc gccgccgtct agagtaaagg 1980
accaagtatt cagcaagttc gcttgctgaa tacttggtcc ttgctgccta gttcaaagtc 2040
gccgtccagg cgccgcgctg ttccgtttct tgctgcctca agatcctcca ggcgatctga 2100
cggttcacta aacgagctct gcttatatag gcctcccacc gtacacgcct accgcggggg 2160
tggccggggc cagggcttcc cacgtgcgca gcaggacgca gcgctgcctg aaactcgcgc 2220
cgcgaggaga gggcggggcc gcggaaagga aggggagggg ctgggagggc ccggaggggg 2280
ctgggccggg gacccgggag gggtcgggac ggggcggggt ccgcgcggag gaggcggagc 2340
tggaaggtga aggggcagga cgggtgcccg ggtccccagt ccctccgcca cgtggggagc 2400
gcggtcctgg gcgtctgtgc ccgcgaatcc actgggagcc cggcctggcc ccgacagcgc 2460
agctgctccg ggcggacccg ggggtctggg ccgcgcttcc ccgcccgcgc gccgctcgcg 2520
ctcccagggt gcagggacgc cagcgagggc cccagcggag agaggtcgaa tcggcctagg 2580
ctgtggggta acccgaggga ggggccataa gcttatatat aa 2622
<210> 8
<211> 162
<212> RNA
<213> Искусственная последовательность
<220>
<223> Последовательность оснований РНК VA1 аденовируса 5-го типа
<400> 8
agcgggcacu cuuccguggu cugguggaua aauucgcaag gguaucaugg cggacgaccg 60
ggguucgagc cccguauccg gccguccgcc gugauccaug cgguuaccgc ccgcgugucg 120
aacccaggug ugcgacguca gacaacgggg gagugcuccu uu 162
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ОНКОЛИТИЧЕСКИЙ HSV-ВЕКТОР | 2014 |
|
RU2719190C2 |
| КОНСТРУИРОВАНИЕ ОБЛИГАТНОГО ВЕКТОРА НА ОСНОВЕ ОНКОЛИТИЧЕСКИХ ВИРУСОВ ПРОСТОГО ГЕРПЕСА (oHSV) И КОНСТРУКЦИИ ДЛЯ ТЕРАПИИ РАКА | 2016 |
|
RU2831084C2 |
| ОНКОЛИТИЧЕСКИЕ ВИРУСЫ ПРОСТОГО ГЕРПЕСА 1 ТИПА ДЛЯ ЛЕЧЕНИЯ ОПУХОЛЕЙ ГОЛОВНОГО МОЗГА | 2021 |
|
RU2824514C1 |
| КОНСТРУИРОВАНИЕ ОБЛИГАТНОГО ВЕКТОРА НА ОСНОВЕ ОНКОЛИТИЧЕСКИХ ВИРУСОВ ПРОСТОГО ГЕРПЕСА (oHSV) И КОНСТРУКЦИИ ДЛЯ ТЕРАПИИ РАКА | 2016 |
|
RU2751236C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ, НАБОРЫ И СПОСОБЫ ЛЕЧЕНИЯ ОПУХОЛЕЙ | 2019 |
|
RU2810906C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЙ ОНКОЛИТИЧЕСКИЙ ВИРУС ПРОСТОГО ГЕРПЕСА, ДОСТАВЛЯЮЩИЙ ХЕМОКИН И ОПУХОЛЬ-АССОЦИИРОВАННЫЙ/СПЕЦИФИЧЕСКИЙ АНТИГЕН | 2022 |
|
RU2827338C1 |
| ТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ РАКА | 2016 |
|
RU2720984C2 |
| АДЕНОВИРУС, СОДЕРЖАЩИЙ АЛЬБУМИН-СВЯЗЫВАЮЩИЙ УЧАСТОК | 2015 |
|
RU2711371C2 |
| Двухцепочечная РНК, способная снижать экспрессию мутантного аллеля с.607GA гена GNAO1 человека | 2022 |
|
RU2816137C1 |
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ АНТИВИЧ-ТЕРАПИИ | 2010 |
|
RU2426788C1 |
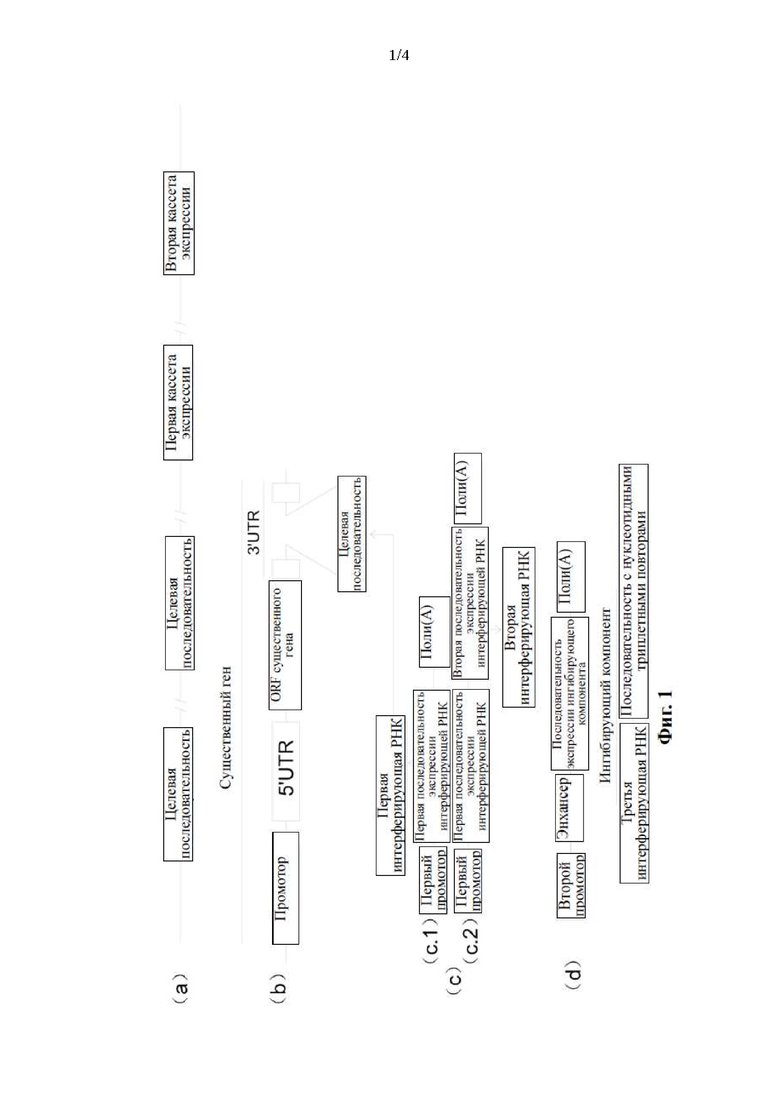
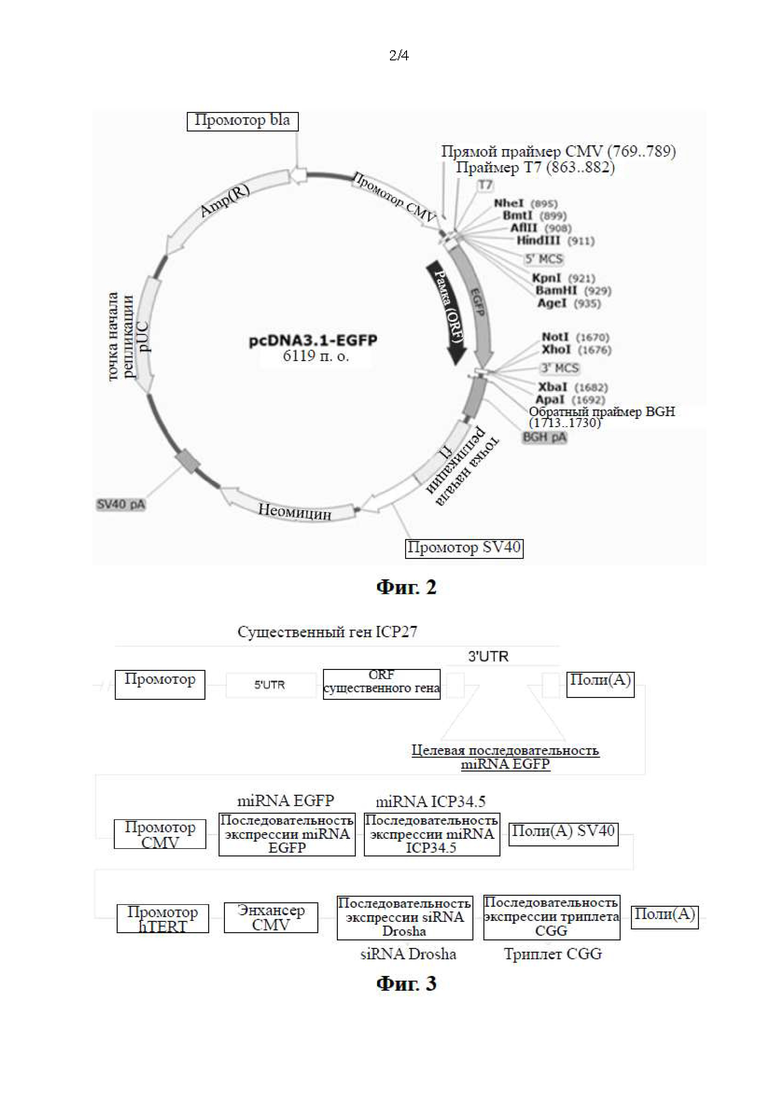
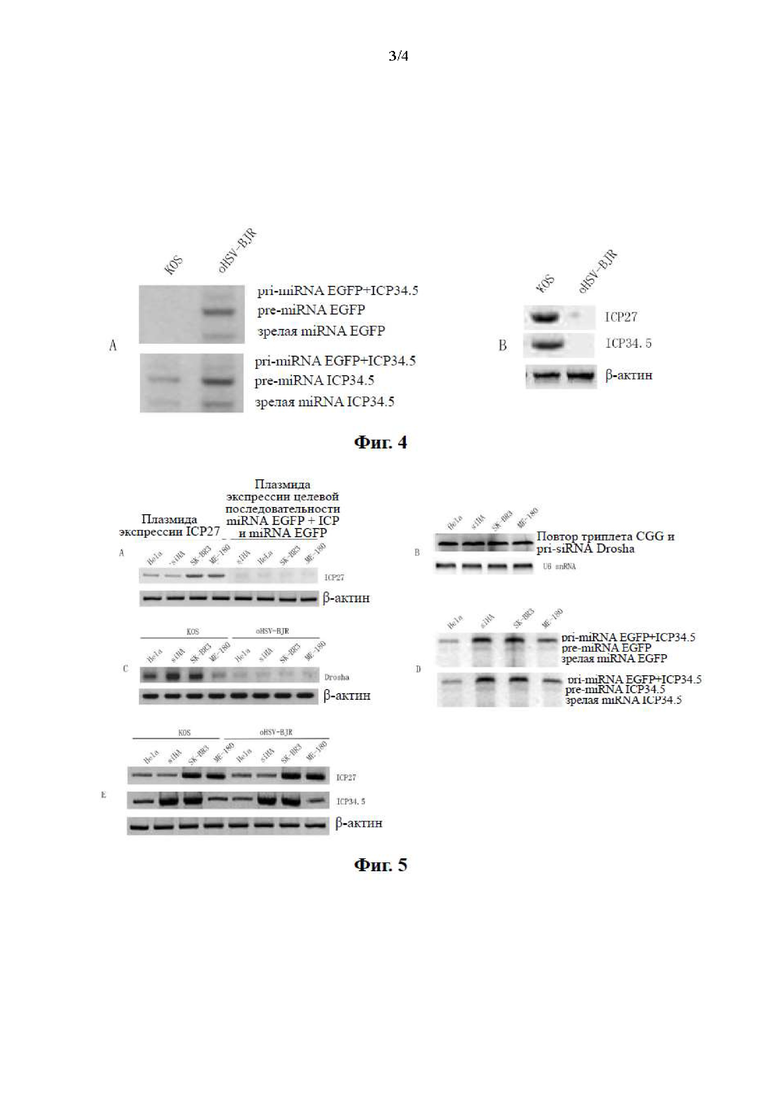
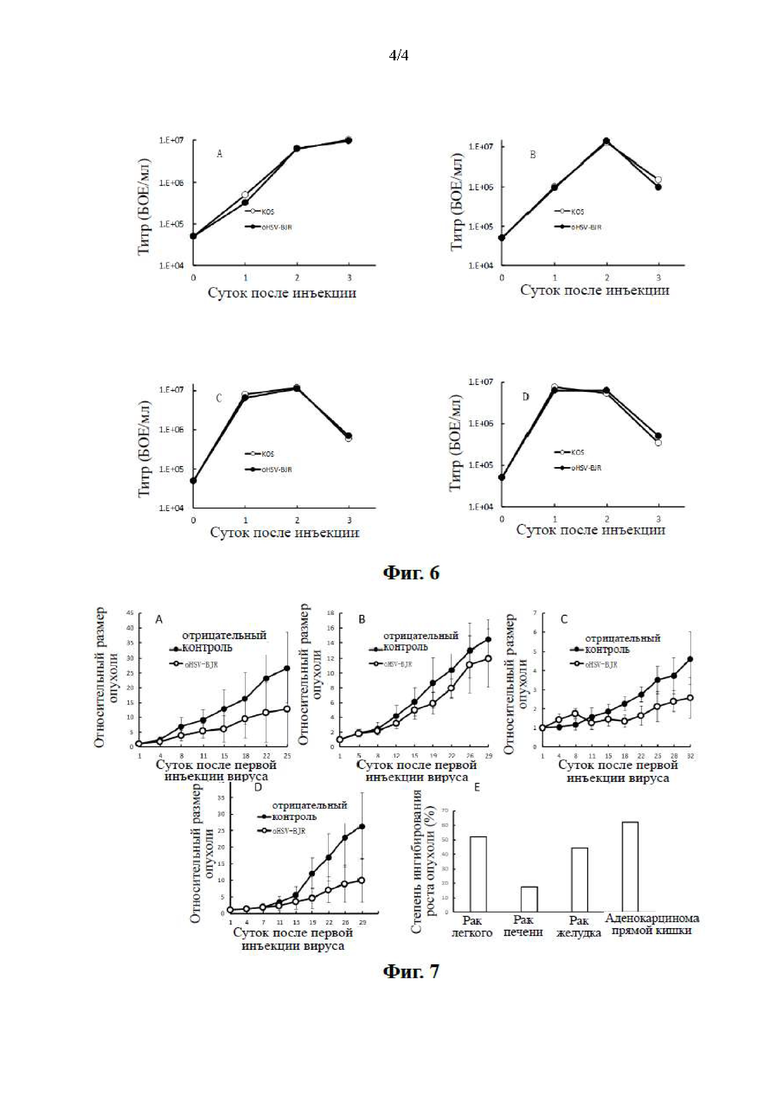
Настоящее изобретение относится к биотехнологии, вирусологии и медицине. Представлен онколитический вирус, который содержит (1) первую кассету экспрессии, содержащую первый промотор и первую последовательность экспрессии интерферирующей РНК; (2) целевую последовательность; и (3) вторую кассету экспрессии. Первая последовательность экспрессии интерферирующей РНК используется для экспрессии первой интерферирующей РНК, которая связывается с целевой последовательностью. Первая последовательность экспрессии интерферирующей РНК находится под управлением первого промотора с целью экспрессии первой интерферирующей РНК в первых клетках. Целевая последовательность расположена в 5’- или 3’-некодирующем участке существенного гена, необходимого для репликации онколитического вируса. Вторая кассета экспрессии содержит второй промотор и последовательность экспрессии ингибирующего компонента. Последовательность экспрессии ингибирующего компонента используется для экспрессии ингибирующих компонентов. Ингибирующие компоненты используются для ингибирования биосинтеза и/или биоактивности ферментов, участвующих в биосинтезе интерферирующей РНК. Последовательность экспрессии ингибирующего компонента находится под управлением второго промотора с целью экспрессии ингибирующих компонентов во вторых клетках, но не в первых клетках. Первая и вторая клетки являются клетками различных типов, и первые клетки являются отличными от опухолевых клетками млекопитающего, а вторые клетки являются опухолевыми клетками млекопитающего. Представлены: молекула нуклеиновой кислоты для применения в качестве генома онколитического вируса; композиция для получения онколитического вируса; способ получения онколитического вируса; применение онколитического вируса для приготовления лекарственного препарата, селективно уничтожающего клетки; способ уничтожения клеток; лекарственный препарат для уничтожения клеток; способ лечения заболевания у субъекта; применение онколитического вируса для селективного уничтожения клеток. Онколитический вирус может селективно реплицироваться в различных типах клеток, и, таким образом, могут быть селективно уничтожены опухолевые клетки, а нормальные клетки остаются без повреждений. 9 н. и 24 з.п. ф-лы, 7 ил., 6 табл., 8 пр.
1. Онколитический вирус, причем геном онколитического вируса содержит следующие экзогенные элементы:
(1) первую кассету экспрессии, содержащую первый промотор и первую последовательность экспрессии интерферирующей РНК;
(2) целевую последовательность; и
(3) вторую кассету экспрессии,
причем первая последовательность экспрессии интерферирующей РНК используется для экспрессии первой интерферирующей РНК, которая связывается с целевой последовательностью; первая последовательность экспрессии интерферирующей РНК находится под управлением первого промотора с целью экспрессии первой интерферирующей РНК в первых клетках;
целевая последовательность расположена в 5'- или 3'-некодирующем участке существенного гена, необходимого для репликации онколитического вируса;
вторая кассета экспрессии содержит второй промотор и последовательность экспрессии ингибирующего компонента, причем последовательность экспрессии ингибирующего компонента используется для экспрессии ингибирующих компонентов, ингибирующие компоненты используются для ингибирования биосинтеза и/или биоактивности ферментов, участвующих в биосинтезе интерферирующей РНК; и последовательность экспрессии ингибирующего компонента находится под управлением второго промотора с целью экспрессии ингибирующих компонентов во вторых клетках, но не в первых клетках; и
первая и вторая клетки являются клетками различных типов, и первые клетки являются отличными от опухолевых клетками млекопитающего, а вторые клетки являются опухолевыми клетками млекопитающего.
2. Онколитический вирус по п. 1, причем первая кассета экспрессии дополнительно содержит вторую последовательность экспрессии интерферирующей РНК для экспрессии второй интерферирующей РНК, вторая интерферирующая РНК действует на открытую рамку считывания несущественного гена, который не требуется для репликации вируса in vitro, препятствуя экспрессии несущественного гена, а вторая последовательность экспрессии интерферирующей РНК экспрессируется под контролем первого промотора.
3. Онколитический вирус по п. 2, причем первая интерферирующая РНК и вторая интерферирующая РНК могут быть либо малой интерферирующей РНК, либо микроРНК.
4. Онколитический вирус по п. 2, причем
вторые клетки являются опухолевыми клетками млекопитающего, а первые клетки являются отличными от опухолевых клетками млекопитающего;
предпочтительно, млекопитающее является человеком; и
предпочтительно, опухолевые клетки представляют собой клетки рака легких, клетки рака печени, клетки рака молочной железы, клетки рака желудка, клетки рака предстательной железы, клетки опухоли головного мозга, клетки рака толстой кишки человека, клетки рака шейки матки, клетки рака почек, клетки рака яичников, клетки рака головы и шеи, клетки меланомы, клетки рака поджелудочной железы или клетки рака пищевода.
5. Онколитический вирус по п. 2, причем целевая последовательность представляет собой короткую последовательность, выбранную из гена отличного от млекопитающего организма;
предпочтительно, целевая последовательность длиной 19-23 нуклеотида выбрана из открытой рамки считывания гена отличного от млекопитающего организма;
предпочтительно, рыбы или растения; и
предпочтительно, ген отличного от млекопитающего организма выбран из группы, состоящей из гена зеленого флуоресцентного белка, полученного из медузы, гена β-галактозидазы, полученного из Escherichia coli, и гена люциферазы, полученного из светлячка.
6. Онколитический вирус по п. 5, причем целевая последовательность представлена под SEQ ID NO: 1, а последовательность для первой интерферирующей РНК представлена под SEQ ID NO: 2.
7. Онколитический вирус по пп. 1-6, причем целевая последовательность вставлена в 5'- или 3'-некодирующие участки одного или нескольких существенных генов рекомбинантного онколитического вируса; и
предпочтительно, одна или несколько копий целевой последовательности вставлены в любой сайт вставки.
8. Онколитический вирус по п. 7, причем онколитический вирус выбран из ряда вирусов, включая вирус простого герпеса, аденовирус, вирус коровьей оспы, вирус болезни Ньюкасла, полиовирус, вирус Коксаки, вирус кори, вирус эпидемического паротита, вирус коровьей оспы, вирус везикулярного стоматита и вирус гриппа,
предпочтительно, когда онколитический вирус представляет собой вирус простого герпеса, существенный ген выбран из группы, состоящей из гликопротеина L оболочки, урацил-ДНК-гликозилазы, белка капсида, субъединицы профермента геликазы, унвиндазы, связывающей точки инициации репликации ДНК, производного белка миристиновой кислоты, дезоксирибонуклеазы, серин/треониновой протеинкиназы оболочки, субъединицы 1 ДНК-упаковывающей терминазы, белка UL16 оболочки, белка UL17 упаковки ДНК, триплексной субъединицы 2 капсида, основного белка капсида, белка UL20 оболочки, нуклеопротеина UL24, белка UL25 упаковки ДНК, протеазы созревания капсида, белка капсида, гликопротеина В оболочки, связывающего одноцепочечную ДНК белка, каталитической субъединицы ДНК-полимеразы, белка слоя выходя из ядра, белка UL32 упаковки ДНК, белка UL33 упаковки ДНК, мембранного белка выходя из ядра, большого капсидного белка, триплексной субъединицы 1 капсида, субъединицы 1 рибонуклеотидредуктазы, субъединицы 2 рибонуклеотидредуктазы, белка отключения хозяина капсулы (capsule host shutoff protein), процессирующей субъединицы ДНК-полимеразы, мембранного белка UL45, белка VP13/14 оболочки, трансактивирующего белка VP16, белка VP22 оболочки, гликопротеина N оболочки, белка UL51 оболочки, примазной субъединицы унвиндазы, гликопротеина K оболочки, ICP27, нуклеопротеина UL55, нуклеопротеина UL56, фактора регуляции транскрипции ICP4, регуляторного белка ICP22, гликопротеина оболочки D и мембранного белка US8A; а несущественный ген выбран из группы, состоящей из ICP34.5, ICP0, нуклеопротеина UL3, нуклеопротеина UL4, геликазной субъединицы профермента геликаза, кутикулярного белка UL7, гликопротеина M оболочки, белка UL14 оболочки, белка UL21 оболочки, гликопротеина H оболочки, тимидинкиназы, субъединицы 2 фермента, завершающего упаковку ДНК, малого капсидного белка, белка UL37 оболочки, белка UL43 оболочки, гликопротеина С оболочки, белка VP11/12 оболочки, урацил-дезоксирибозидетрифосфатазы, ICP0, вирусного белка US2, серин/треониновой протеинкиназы U3, мембранного гликопротеина G, гликопротеина J оболочки, гликопротеина I оболочки, гликопротеина E оболочки, мембранного белка US9, вирусного белка US10, кутикулярного белка Us11 и ICP47;
предпочтительно, когда онколитический вирус представляет собой аденовирус, существенный ген выбран из группы, состоящей из раннего белка 1А, раннего белка 1В 19K, раннего белка 1B 55K, белка Iva2 инкапсидации, ДНК-полимеразы, предшественника рТР терминального белка, белка 52K инкапсидации, капсидного белка-предшественника pIIIa, пентомерной матрицы, корового белка pVII, предшественника рХ корового белка, предшественника pVI корового белка, гексонмера, протеиназы, связывающего одноцепочечную ДНК белка, белка 100K сборки гексамера, белка 33K, белка 22K инкапсидации, предшественника капсидного белка, белка U, фибрина, открытой рамки считывания 6/7 регуляторного белка Е4, регуляторного белка Е4 34K, открытой рамки считывания 4 регуляторного белка Е4, открытой рамки считывания 3 регуляторного белка Е4, открытой рамки считывания 2 регуляторного белка Е4 и открытой рамки считывания 1 регуляторного белка Е4; а несущественный ген выбран из группы, состоящей из капсидного белка IX, белка 13.6K, корового белка V, регуляторного белка Е3 12.5K, мембранного гликопротеина Е3 CR1-α, мембранного гликопротеина Е3 gp19K, мембранного гликопротеина Е3 CR1-β, мембранного гликопротеина Е3 CR1-δ, мембранного гликопротеина Е3 RID-δ и мембранного гликопротеина Е3 14.7K;
предпочтительно, когда онколитический вирус представляет собой вирус коровьей оспы, существенный ген выбран из группы, состоящей из малой субъединицы рибонуклеотидредуктазы, серин/треонинкиназы, ДНК-связывающего вирусного корового белка, большой субъединицы полимеразы, субъединицы РНК-полимеразы, ДНК-полимеразы, сульфгидрилоксидазы, гипотетического ДНК-связывающего вирусного нуклеопротеина, ДНК-связывающего фосфопротеина, нуклеоид-цистеиновой протеиназы, РНК-геликазы NPH-II, гипотетической металлопротеиназы, транскрипционного фактора элонгации, белка глутатионового типа, РНК-полимеразы, гипотетического вирусного нуклеопротеина, позднего транскрипционного фактора VLTF-1, ДНК-связывающего вирусного нуклеопротеина, вирусного капсидного белка, малой субъединицы полимеразы, субъединицы rpo22 РНК-полимеразы, зависящей от ДНК, субъединицы rpo147 РНК-полимеразы, зависящей от ДНК, серин/треониновой протеинфосфатазы, гепарин-связывающего поверхностного белка IMV, ДНК-зависимой РНК-полимеразы, позднего транскрипционного фактора VLTF-4, ДНК-топоизомеразы I типа, большой субъединицы фермента кэпирования мРНК, вирусного корового белка 107, вирусного корового белка 108, урацил-ДНК-гликозилазы, трифосфатазы, 70 кДа малой субъединицы раннего фактора транскрипции генов VETF, субъединицы rpo18 РНК-полимеразы, зависящей от ДНК, нуклеозидтрифосфатгидролазы-1, малой субъединицы фермента кэпирования мРНК, целевого сайта для рифампицина, позднего транскрипционного фактора VLTF-2, позднего транскрипционного фактора VLTF-3, пути образования дисульфидной связи, предшественника p4b корового белка 4b, 39 кДа корового белка, субъединицы rpo19 РНК-полимеразы, зависящей от ДНК, 82 кДа большой субъединицы раннего фактора транскрипции гена VETF, 32 кДа малой субъединицы транскрипционного фактора VITF-3, мембранного белка IMV 128, предшественника Р4а корового белка 4а, мембранного белка 131 IMV, фосфорилированного мембранного белка IMV, мембранного белка IMV A17L, ДНК-унвиндазы, процессирующего фактора вирусной ДНК-полимеразы, мембранного белка IMV A21L, пальмитоилового белка, 45 кДа большой субъединицы промежуточного фактора транскрипции гена VITF-3, субъединицы rpo132 РНК-полимеразы, зависящей от ДНК, РНК-полимеразы rpo35, зависящей от ДНК, белка A30L IMV, гипотетического АТФ-зависимого фермента, серин/треонинкиназы, белка созревания EEV, пальмитоилированного мембранного гликопротеина EEV, поверхностного белка IMV A27L, мембранного фосфатгликопротеина EEV, мембранных гликопротеинов EEV и EEV, мембранного гликопротеина EEV, белка пути образования дисульфидной связи, гипотетического вирусного нуклеопротеина, мембранного белка I2L IMV, поксвирусного миристоилового белка, мембранного белка L1R IMV, 16 кДа позднего гипотетического мембранного белка, гипотетического вирусного мембранного белка H2R, мембранного белка A21L IMV, хемокин-связывающего белка, белка, подобного эпидермальному фактору роста, и IL-18-связывающего белка; а несущественный ген выбран из группы, состоящей из секреторного комплемент-связывающего белка, kelch-подобного белка, факторов вирулентности, гипотетического белка, чувствительного к α-аминобелкам, белка серпинового типа, белка фосфолипазы D-типа, неохарактеризованного белка K7R, гипотетического мембранного белка типа CD47, белка алармонического типа, мембранного белка агглютинина С-типа типа II, секреторного гликопротеина, урацилдезоксирибозидетрифосфатазы, kelch-подобного белка F3L, гипотетического миристоилированного белка, большой субъединицы рибонуклеотидредуктазы, белка телец включения вируса коровьей оспы типа А, белка анкиринового типа, 6 кДа внутриклеточного вирусного белка, белка 215, подобного α-рецептору фактора некроза опухоли, белка 217, подобного α-рецептору фактора некроза опухоли, белка B4R анкиринового типа, белка 213 анкиринового типа, белка 211 анкиринового типа, белка 207 с цинковым пальцем, белка 208 с цинковым пальцем, белка 014 анкиринового типа, белка 015 анкиринового типа, белка 016 анкиринового типа, белка 017 анкиринового типа, белка 019 анкиринового типа, белка 030 анкиринового типа, гипотетической моноглицеридной липазы 036, гипотетической моноглицеридной липазы 037, гипотетической моноглицеридной липазы 038, белка 199 анкиринового типа, белка 203 анкиринового типа/гипотетического белка, белка телец включения типа А, гуанилаткиназы и белка 188 анкиринового типа;
предпочтительно, когда онколитический вирус представляет собой вирус Коксаки, существенный ген выбран из группы, состоящей из белка Vpg, корового белка 2А, белка 2В, РНК-унвиндазы 2С, белка 3А, протеиназы 3С, обратной транскриптазы 3D, белка оболочки Vp4 и белка Vp1; а несущественный ген выбран из группы, состоящей из капсидных белков Vp2 и Vp3;
предпочтительно, когда онколитический вирус представляет собой вирус кори, существенный ген выбран из группы, состоящей из нуклеопротеина N, фосфопротеина Р, матричного белка М, трансмембранного гликопротеина F, трансмембранного гликопротеина H и РНК-зависимой РНК-полимеразы L; а несущественный ген выбран из группы, состоящей из РНК-зависимого вспомогательного белка С РНК-полимеразы и РНК-зависимого вспомогательного белка V РНК-полимеразы;
когда онколитический вирус представляет собой вирус эпидемического паротита, существенный ген выбран из группы, состоящей из нуклеопротеина N, фосфопротеина Р, гибридного белка F и РНК-полимеразы L; а несущественный ген выбран из группы, состоящей из фосфопротеина V, мембранного белка M и гемагглютинин-нейраминидазного белка HN;
предпочтительно, когда онколитический вирус представляет собой вирус везикулярного стоматита, существенный ген выбран из группы, состоящей из гликопротеина G, нуклеопротеина N, фосфопротеина P и РНК-полимеразы L; а несущественный ген представляет собой матричный белок М;
предпочтительно, когда онколитический вирус представляет собой полиовирус, существенный ген выбран из группы, состоящей из капсидного белка VP1, капсидного белка VP2, капсидного белка VP3, цистеиновой протеазы 2А, белка 2В, белка 2С, белка 3А, белка 3В, протеиназы 3С, белка 3D и РНК-направленной РНК-полимеразы; а несущественный ген представляет собой капсидный белок VP4; и
предпочтительно, когда онколитический вирус представляет собой вирус гриппа, существенный ген выбран из группы, состоящей из гемагглютинина, нейраминидазы, нуклеопротеина, мембранного белка M1, мембранного белка М2, полимеразы РА, полимеразы PB1-F2 и полимеразы РВ2; а несущественный ген представляет собой неструктурный белок NS1 или неструктурный белок NS2.
9. Онколитический вирус по п. 8, причем онколитический вирус представляет собой вирус простого герпеса, существенный ген представляет собой ICP27, а несущественный ген представляет собой ICP34.5.
10. Онколитический вирус по п. 2, причем последовательность второй интерферирующей РНК представлена под SEQ ID NO: 3.
11. Онколитический вирус по п. 2, причем
первый промотор представляет собой конститутивный промотор;
предпочтительно, если целевая последовательность вставлена в 5'-некодирующий участок или 3'-некодирующий участок множества существенных генов, первая кассета экспрессии экспрессирует только первую интерферирующую РНК, а первый промотор представляет собой промотор Hu6 или H1 человека; и
предпочтительно, если целевая последовательность вставлена в 5'-некодирующий участок или 3'-некодирующий участок только одного существенного гена, первая кассета экспрессии экспрессирует первую интерферирующую РНК и вторую интерферирующую РНК, а первый промотор выбран из группы, состоящей из промоторов CMV, SV40 и СВА.
12. Онколитический вирус по п. 1, причем второй промотор представляет собой опухолеспецифический промотор человека; и
предпочтительно, ракоспецифический промотор человека выбран из группы, состоящей из промотора обратной транскриптазы теломеразы (hTERT), промотора рецептора-2 эпидермального фактора роста человека (HER-2), промотора E2F1, промотора остеокальцина, промотора карциноэмбрионального антигена, промотора сурвивина и промотора церулоплазмина.
13. Онколитический вирус по п. 1, причем фермент может быть любым из ферментов, участвующих в биосинтезе интерферирующей РНК, в том числе Dicer, Drosha и Agonauts.
14. Онколитический вирус по п. 1, причем ингибирующий компонент содержит третью интерферирующую РНК для препятствования экспрессии гена фермента, ингибируя биосинтез фермента; и
предпочтительно, ферментом является Drosha.
15. Онколитический вирус по п. 14, причем последовательность третьей интерферирующей РНК представлена под SEQ ID NO: 4.
16. Онколитический вирус по п. 14, причем ингибирующий компонент дополнительно содержит увеличенное количество нуклеотидных триплетных повторов для ингибирования биоактивности Drosha или некодирующую РНК для ингибирования активности Dicer;
предпочтительно, увеличенное количество нуклеотидных триплетных повторов имеют следующую общую формулу: (CGG)n, где n представляет собой целое число, равное или больше 20;
предпочтительно, n находится в диапазоне от 60 до 150;
предпочтительно, n равно 100;
предпочтительно, некодирующая РНК для ингибирования активности Dicer представляет собой РНК VA1 аденовируса 5-го типа; и
предпочтительно, последовательность оснований РНК VA1 аденовируса 5-го типа представлена под SEQ ID NO: 8.
17. Онколитический вирус по пп. 12-16, причем вторая кассета экспрессии дополнительно содержит энхансерную последовательность для усиления экспрессии последовательности экспрессии ингибирующего компонента; и
предпочтительно, энхансерная последовательность представляет собой энхансер CMV или SV40.
18. Онколитический вирус по п. 17, причем регистрационным номером онколитического вируса является ССТСС № V201919.
19. Молекула нуклеиновой кислоты для применения в качестве генома онколитического вируса по пп. 1-18, причем молекула нуклеиновой кислоты содержит следующие элементы:
целевую последовательность, первую кассету экспрессии и вторую кассету экспрессии,
причем первая кассета экспрессии содержит первый промотор и первую последовательность экспрессии интерферирующей РНК; и
вторая кассета экспрессии содержит второй промотор и последовательность экспрессии ингибирующего компонента, причем последовательность экспрессии ингибирующего компонента используется для экспрессии ингибирующих компонентов, ингибирующие компоненты используются для ингибирования биосинтеза и/или биоактивности ферментов, участвующих в биосинтезе интерферирующей РНК; и последовательность экспрессии ингибирующего компонента находится под управлением второго промотора с целью экспрессии ингибирующих компонентов во вторых клетках, но не в первых клетках;
причем первые клетки являются отличными от опухолевых клетками млекопитающего, а вторые клетки являются опухолевыми клетками млекопитающего.
20. Композиция для получения онколитического вируса по пп. 1-18, содержащая: молекулу нуклеиновой кислоты по п. 19 и родительский вирус и/или комплементарные клетки-хозяева, причем
по сравнению с геномом вируса дикого типа в геноме родительского вируса отсутствует существенный ген; и
комплементарные клетки-хозяева содержат фрагмент ДНК для экспрессии существенного гена.
21. Способ получения онколитического вируса по пп. 1-18, предусматривающий встраивание молекулы нуклеиновой кислоты по п. 19 в геном онколитического вируса дикого типа.
22. Способ по п. 21, предусматривающий совместное культивирование молекулы нуклеиновой кислоты, родительского вируса и комплементарных клеток-хозяев и сбор онколитического вируса из культуры, причем
по сравнению с геномной последовательностью онколитического вируса дикого типа в геноме родительского вируса отсутствует существенный ген; и
комплементарные клетки-хозяева содержат фрагмент ДНК для экспрессии существенного гена.
23. Способ по п. 22, причем конкретные рабочие стадии способа предусматривают получение линии клеток Vero для экспрессии ICP27, получение родительского вируса, конструирование модифицированной методами генной инженерии плазмиды экспрессии ICP27 и регуляторного компонента и конструирование рекомбинантного онколитического вируса герпеса.
24. Применение онколитического вируса по пп. 1-18 для приготовления лекарственного препарата, селективно уничтожающего клетки.
25. Применение по п. 24, причем клетки представляют собой опухолевые клетки.
26. Способ уничтожения клеток, предусматривающий инфицирование целевых клеток онколитическим вирусом по пп. 1-18.
27. Способ по п. 26, причем целевые клетки представляют собой опухолевые клетки.
28. Способ по п. 26, причем способ направлен на лечение, не связанное с лечением заболевания.
29. Лекарственный препарат для уничтожения клеток, содержащий рекомбинантные онколитические вирусы по пп. 1-18 и фармацевтически приемлемое вспомогательное вещество.
30. Лекарственный препарат по п. 29, причем клетки представляют собой опухолевые клетки.
31. Способ лечения заболевания у субъекта, предусматривающий:
введение субъекту онколитического вируса по пп. 1-18 или композиции, содержащей данный онколитический вирус,
причем источником заболевания являются вторые клетки,
причем вторые клетки представляют собой опухолевые клетки.
32. Способ по п. 31, причем заболевание представляет собой рак.
33. Применение онколитического вируса по пп. 1-18 для селективного уничтожения клеток, причем предпочтительно клетки представляют собой опухолевые клетки.
| CN 108884448 A, 23.11.2018 | |||
| СПИРИН П.В | |||
| и др., Сравнение эффективности подавления функциональной активности целевых генов с помощью синтетических модифицированных siPHK и shPHK, экспрессируемых рекомбинантными лентивирусными векторами, Acta naturae, N 2, 2009 с | |||
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| стр | |||
| Облицовка комнатных печей | 1918 |
|
SU100A1 |

