Область техники
Заявленная группа изобретений относится к области молекулярной биологии и биотехнологии, а именно к молекулам РНК, способным снижать экспрессию мутантного аллеля c.607G>A гена GNAO1 человека, запуская механизм РНК-интерференции. Изобретение может быть использовано в фармацевтических и медицинских целях, в частности, при разработке генотерапевтических подходов для пациентов с GNAO1-энцефалопатией, имеющих мутацию c.607G>A (p.Gly203Arg).
Уровень техники
GNAO1-энцефалопатия является орфанным аутосомно-доминантным генетическим заболеванием и вызвана гетерозиготными мутациями в гене GNAO1, возникающими de novo. Данное заболевание поражает детей, в основном уже в период младенчества, и проявляется в виде спектра неврологических фенотипов, включая младенческую эпилептическую энцефалопатию 17 типа (OMIM 615473) и/или нарушение развития мозга с непроизвольными движениями (OMIM 617493). Ген GNAO1 (GeneID: 2775, размер около 166 кб) человека экспрессируется преимущественно в головном мозге и кодирует белок Gαo (масса около 40 кДа). Белок Gαo относится к семейству G-белков и участвует в модуляции передачи сигналов через рецепторы, сопряженные с G-белком (GPCR), в нейронах. Несмотря на то, что белок Gαo имеет огромное значение для работы нервной системы, его функция изучена не до конца.
Впервые роль мутаций в гене GNAO1 в развитии детских энцефалопатий была обнаружена в 2013 году. К настоящему времени в литературе описано около 25 различных патогенных мутаций гена GNAO1. Мутация c.607G>A в 6-ом экзоне гена GNAO1 (полиморфизм rs587777057), приводящая к аминокислотной замене Gly203Arg (G203R) в белковом продукте, встречается наиболее часто и приводит к комбинированному фенотипу с наиболее тяжелыми проявлениями, включая младенческие эпилептические припадки, тяжелую двигательную дисфункцию, задержку умственного развития и пороки развития головного мозга.
К сожалению, лечения для GNAO1-энцефалопатии на сегодняшний день не существует. Ситуация осложняется фенотипической гетерогенностью заболевания, вызванной разным влиянием конкретных мутаций на функцию Gαo и опосредованную им нейрональную передачу. Фенотипическую гетерогенность нужно принимать во внимание при подборе терапии и применять персонализированный подход с учетом конкретного патогенного варианта GNAO1 и молекулярного механизма заболевания. Данные о влиянии мутации c.607G>A (p.Gly203Arg) на функциональную активность белка Gαo до сих пор остаются противоречивыми. Аминокислотная замена G203R происходит вблизи гуанозин-трифосфат(ГТФ)-связывающего центра a-субъединицы и ранее описывалась как gain-of-function мутация [Feng et al., 2018. A Mechanistic Review on GNAO1-Associated Movement Disorder. Neurobiology of Disease 116 (August): 131-41]. Согласно последним данным, эта мутация имеет доминантно-негативный характер, что проявляется в том, что мутантный Gαo мешает активности нормального белка за счет конкурентного взаимодействия с другими субъединицами G-белкового комплекса (Gβγ), и таким образом нарушает GPCR-опосредованную передачу сигналов в нейронах [Muntean et al., 2021. Gαo Is a Major Determinant of с AMP Signaling in the Pathophysiology of Movement Disorders. Cell Reports 34 (5): 108718; Wang et al., 2022. Genetic Modeling of GNAO1 Disorder Delineates Mechanisms of Gαo Dysfunction. Human Molecular Genetics 31 (4): 510-22].
В условиях стремительного развития генетических технологий большие надежды в отношении терапии наследственных заболеваний, и в том числе GNAO1-энцефалопатии, возлагаются на генную терапию, которая позволяет исправить генетический дефект. Для лечения заболеваний, в основе которых лежат доминантно-негативные мутации или gain-of-function мутации, может быть применена генная терапия на основе РНК-интерференции, направленная на подавление экспрессии мутантного гена и соответствующего белкового продукта на стадии транскрипции или трансляции.
РНК-интерференция (РНКи) представляет собой естественный механизм регуляции экспрессии генов, а также защиты клеток эукариот от вирусов. Процесс РНК-интерференции запускают двухцепочечные молекулы РНК, где одна из цепей комплементарна участку мРНК целевого гена (мРНК-мишени) и является направляющей. Направляющая цепь включается в мультибелковый комплекс RISC (RNA-induced silencing complex), который, в свою очередь, за счет комплементарности образует комплекс с мРНК-мишенью, что приводит либо к снижению эффективности трансляции в случае неполной комплементарности, либо к деградации последовательности мРНК в случае полной комплементарности [Ramachandran P.S., Keiser M.S., Davidson B.L. Recent advances in RNA interference therapeutics for CNS diseases. Neurotherapeutics. 2013 Jul; 10(3):473-85. doi: 10.1007/s13311-013-0183-8].
РНК-интерференция с момента ее открытия широко используется для нокдауна генов-мишеней в научно-исследовательских работах, а в последние годы и для разработки новых подходов к генной терапии заболеваний. Выделяют три группы терапевтических инструментов на основе РНК-интерференции: 1) синтетические малые интерферирующие РНК (siRNA, small interfering RNA), 2) короткие шпилечные РНК (shRNA, small hairpin RNA, short hairpin RNA), которые в результате процессинга в клетке превращаются в siRNA, и 3) искусственные микроРНК (miRNA), имитирующие природные аналоги. Размер двухцепочечных молекул РНК для РНК-интерференции обычно составляет примерно 19-22 нуклеотидов. В ткани пациента такие молекулы РНК могут доставляться с помощью различных векторов, в том числе векторов на основе аденоассоциированных вирусов (ААВ), чаще всего используемых при разработке in vivo генной терапии неврологических нарушений [Hudry and Vandenberghe. 2019. Therapeutic AAV Gene Transfer to the Nervous System: A Clinical Reality. Neuron 101 (5): 839-62].
Несмотря на растущее число исследований, направленных на создание ААВ-опосредованных препаратов для генной супрессии путем РНК-интерференции с целью лечения генетических заболеваний, в настоящее время нет ни одного одобренного препарата с таким механизмом действия. В 2020 году было опубликовано пилотное исследование безопасности ААВ-опосредованного кандидатного препарата на основе miRNA для супрессии гена SOD1. Исследование было проведено на двух пациентах с SOD1-ассоциированным семейным боковым амиотрофическим склерозом [Mueller С., Berry J.D., McKenna-Yasek D.M. et al. SOD1 Suppression with AdeNo-Associated Virus and MicroRNA in Familial ALS. N Engl J Med. 2020 Jul 9; 383(2):151-158. doi: 10.1056/NEJMoa2005056]. В декабре 2021 года начались клинические испытания первого и пока единственного ААВ-опосредованного кандидатного препарата на основе miRNA (препарат АМТ-130) для лечения болезни Гентингтона (NCT05243017, при поддержке UniQure Biopharma BV).
Среди новейших биотехнологических разработок в этой области можно выделить следующие. Компания UniQure использует собственную запатентованную технологию подавления экспрессии генов miQURE для создания препаратов на основе ААВ: помимо упомянутого АМТ-130 для лечения болезни Гентингтона (патент ЕА037696), компания разрабатывает препарат АМТ-150 для лечения спиноцеребеллярной атаксии 3 типа. Компания Voyager Therapeutics разрабатывает аналогичный препарат VY-HTT01 для болезни Гентингтона, заявка на проведение клинических испытаний нового лекарственного препарата в настоящий момент находится на рассмотрении в FDA. Сразу три компании Novartis Gene Therapies, Voyager Therapeutics и ApicBio работают над созданием ААВ-опосредованной РНК-терапии для лечения семейной формы бокового амиотрофического склероза, вызванного мутациями в гене SOD1 (препараты AVXS-301, VY-SOD 101 и АРВ-102 соответственно).
Ближайшего аналога настоящему изобретению в уровне техники не было обнаружено.
Технической задачей, на решение которой направлено настоящее изобретение, является решение как минимум одной из вышеуказанных в уровне техники проблем.
Сущность изобретения
Техническим решением является использование описанного признаками в пунктах формулы изобретения.
Одной из возможных технических задач, на решение которой может быть направлено настоящее изобретение, являлось создание средства для аллель-селективного подавления экспрессии мутантного аллеля c.607G>A гена GNAO1 человека.
Задача может решаться созданием молекул двухцепочечных РНК, содержащих последовательность, комплементарную транскрипту мутантного аллеля c.607G>A гена GNAO1 человека, и способных подавлять его экспрессию на посттранскрипционном уровне через механизм РНК-интерференции.
В настоящем изобретении предлагается двухцепочечная РНК, содержащая смысловую последовательность РНК и антисмысловую последовательность РНК, которые, по существу, комплементарны, причем антисмысловая последовательность РНК содержит по меньшей мере 19 нуклеотидов и является, по существу, комплементарной последовательности мРНК-мишени, содержащей SEQ ID No: 28.
Было обнаружено, что двухцепочечная РНК по настоящему изобретению эффективно снижала экспрессию мутантного аллеля c.607G>A гена GNAO1 человека. Двухцепочечная РНК при введении в клетку, либо прямо посредством трансфекции, либо непрямо посредством доставки кодирующей ее ДНК, например, посредством трансфекции или вирусной трансдукции, при которой может экспрессироваться двухцепочечная РНК, способна селективно снижать экспрессию мутантного аллеля c.607G>A гена GNAO1 человека, сохраняя экспрессию нормального аллеля гена GNAO1 человека.
Кроме того, было показано, что двухцепочечная РНК по настоящему изобретению способна снижать экспрессию мутантного аллеля c.607G>A гена GNAO1 человека при использовании либо в форме малой интерферирующей РНК (англ. siRNA), либо в каркасе искусственной микроРНК (англ. miRNA) или короткой шпилечной РНК (англ. shRNA).
Двухцепочечная РНК по настоящему изобретению может быть получена в форме siRNA, shRNA или miRNA. Такие двухцепочечные РНК можно доставить в клетки-мишени с использованием известных из уровня техники способов, например, способов трансфекции РНК-дуплексами либо плазмидными экспрессионными векторами, содержащими ДНК, кодирующую двухцепочечные РНК по настоящему изобретению, либо трансдукции с использованием вирусного вектора, в состав которого включают кассету экспрессии для shRNA или miRNA. В случае трансдукции вирусным вектором клетки можно обеспечить постоянным поступлением двухцепочечной РНК для достижения длительной супрессии мутантного аллеля c.607G>A гена GNAO1 человека без необходимости повторного введения.
В одном из вариантов осуществления изобретения упомянутая смысловая последовательность РНК выбрана из последовательностей SEQ ID No: 1-17.
В одном из вариантов осуществления изобретения упомянутая двухцепочечная РНК представляет собой малую интерферирующую РНК. В варианте осуществления изобретения малая интерферирующая РНК содержит упомянутую смысловую последовательность РНК, выбранную из группы, состоящей из SEQ ID No: 1-4.
В другом варианте осуществления изобретения упомянутая двухцепочечная РНК является частью короткой шпилечной РНК (shRNA). В варианте осуществления изобретения короткая шпилечная РНК содержит упомянутую смысловую последовательность РНК, выбранную из группы, состоящей из SEQ ID No: 5-15. В варианте осуществления изобретения короткая шпилечная РНК содержит последовательность РНК, выбранную из группы, состоящей из SEQ ID No: 18 20, SEQ ID No: 33.
В другом варианте осуществления изобретения упомянутая двухцепочечная РНК встроена в каркас miRNA. В варианте осуществления изобретения предлагается miRNA, содержащая каркас miR-30. В варианте осуществления изобретения miRNA, содержащая каркас miR-30, содержит упомянутую смысловую последовательность РНК, выбранную из группы последовательностей SEQ ID No: 16-17.
В одном из вариантов осуществления изобретения предлагается ДНК, кодирующая любую двухцепочечную РНК по настоящему изобретению.
В одном из вариантов осуществления изобретения предлагается кассета экспрессии, кодирующая любую двухцепочечную РНК по настоящему изобретению.
В одном из вариантов осуществления изобретения предлагается вектор, содержащий упомянутую кассету экспрессии. В вариантах осуществления изобретения вектор может представлять собой плазмидный экспрессионный или вирусный вектор. В вариантах осуществления изобретения вирусный вектор может представлять собой лентивирусный вектор или вектор на основе аденоассоциированного вируса.
К объекту патентования относится любая последовательность РНК, находящаяся в «окне» длиной 36 нуклеотид, начиная с позиции 1487 и заканчивая позицией 1522 относительно начала транскрипта GNAO1 (NM 020988.2) (SEQ ID No: 28).
Изобретение иллюстрируется следующими графическими материалами:
На Фиг. 1 представлена общая схема механизма интерференции РНК в клетке.
На Фиг. 2 демонстрируется предлагаемая схема применения двухцепочечных РНК для генной терапии GNAO1-энцефалопатии, вызванной вариантом GNAO1 с. 607 G>A (p.Gly203Arg).
На Фиг. 3 представлены участки последовательности-мишени (SEQ ID No: 26), таргетируемые двухцепочечными РНК по настоящему изобретению, используемыми в форме siRNA (A), shRNA (Б) или miRNA (В) для селективного подавления мутантного варианта GNAO1 c.607G>A (p.Gly203Arg) через механизм РНК-интерференции. Схематически показан фрагмент транскрипта гена GNAO1 человека дикого типа и с мутацией c.607G>A (p.Gly203Arg). Нумерация нуклеотидов соответствует положению в транскрипте NM 020988.2. Триплет с мутацией обозначен жирным шрифтом, а мутантный нуклеотид выделен серым цветом. Номенклатура РНК по настоящему изобретению отражает положение нуклеотида в транскрипте GNAO1 (NM 020988.2), соответствующего первому нуклеотиду в смысловой цепи РНК по настоящему изобретению.
На Фиг. 4 представлены результаты скрининга кандидатных siRNA (siGNAO1, содержат SEQ ID No: 1-4), комплементарных последовательности мутантного аллеля c.607G>A гена GNAO1 человека, в гемизиготной тест-системе с использованием временной трансфекции клеток НЕК293Т методом Вестерн-блот.
Показана экспрессия белка Gαo (продукт гена GNAO1), репортерного белка GFP и референсного белка GAPDH. Колориметрический Вестерн-блот (верхние панели) и окрашивание Ponceau S (нижняя панель).
(A) Анализ лизатов клеток, экспрессирующих мутантный аллель c.607G>A гена GNAO1 человека, после добавления siRNA.
(Б) Анализ лизатов клеток, экспрессирующих нормальный аллель гена GNAO1 человека, после добавления siRNA.
(B) Серийные двукратные разведения контрольного лизата клеток (после добавления siSCR) для сравнительной оценки количества белка Gαo в экспериментальных образцах.
siSCR (от англ. Scramble), содержит SEQ ID No: 22, siEGFP-1 и siEGFP-2 (от англ. Enhanced green fluorescent protein), содержат SEQ ID No: 29 и 30 соответственно: отрицательные контроли РНКи в составе siRNA; siGNAO1-CTRL (от англ. Control), содержит SEQ ID No: 21: положительный контроль РНКи в составе siRNA.
На Фиг. 5 представлен результат количественного анализа эффективности и селективности работы siGNAO1-1488 (содержит SEQ ID No: 1) в гемизиготной тест-системе с использованием временной трансфекции клеток НЕК293Т.
Показано подавление экспрессии мутантного аллеля c.607G>A гена GNAO1 человека на уровне мРНК, метод аллель-специфической кПЦР. Данные представлены как среднее значение ± стандартное отклонение (n=4). Уровень мРНК GNAO1 в образцах, содержащих отрицательный контроль РНКи (siSCR), принят за 100%.
siSCR (содержит SEQ ID No: 22), siЕGFР-1(содержит SEQ ID No: 29) и siEGFP-2 (содержит SEQ ID No: 30): отрицательные контроли РНКи в составе siRNA; siGNAO1-CTRL (содержит SEQ ID No: 21): положительный контроль РНКи в составе siRNA; «-»: отрицательный контроль без РНКи (без добавления siRNA).
На Фиг. 6 представлены результаты анализа эффективности и селективности работы siGNAO1-1488 (содержит SEQ ID No: 1) в гетерозиготной тест-системе с временной (А-Б) и стабильной (В) экспрессией GNAO1 в клетках НЕК293Т.
(А) Показано подавление экспрессии мутантного аллеля c.607G>A гена GNAO1 человека на уровне белка, метод Вестерн-блот, в условиях временной экспрессии GNAO1. Данные демонстрируют экспрессию мутантного белка Gαo-G203R, слитого с эпитопом FLAG, экспрессию нормального белка Gαo, слитого с эпитопом туе, и экспрессию референсного белка GAPDH. На правой панели показаны серийные двукратные разведения контрольного лизата клеток, содержащих отрицательный контроль РНКи (siSCR), для сравнительной оценки количества белка Gαo в экспериментальных образцах.
(Б) Подавление экспрессии мутантного аллеля c.607G>A гена GNAO1 человека на уровне мРНК, метод аллель-специфической кПЦР, в условиях временной экспрессии GNAO1. Данные представлены как среднее значение±стандартное отклонение (n=3, технические повторности трансфекций). Уровень мРНК GNAO1 в образцах, содержащих отрицательный контроль РНКи (siSCR), принят за 100%.
(В) Показано подавление экспрессии мутантного аллеля c.607G>A гена GNAO1 человека на уровне белка, метод Вестерн-блот, в условиях стабильной экспрессии GNAO1. Данные демонстрируют экспрессию мутантного белка Gαo-G203R, слитого с эпитопом FLAG, экспрессию нормального белка Gαo, слитого с эпитопом туе, экспрессию репортерного белка EGFP, и экспрессию референсного белка GAPDH. siSCR (содержит SEQ ID No: 22) и siEGFP-1 (содержит SEQ ID No: 29): отрицательные контроли РНКи в составе siRNA; siGNAO1-CTRL (содержит SEQ ID No: 21): положительный контроль РНКи в составе siRNA; «-»: отрицательный контроль без РНКи (без добавления siRNA).
На Фиг. 7 схематично показаны основные структурные элементы двухцепочечной РНК по настоящему изобретению, заключенной в каркас короткой шпилечной РНК (shRNA) на примере shGNAO1-1489, содержащей смысловую (пассажирскую) цепь с последовательностью SEQ ID No: 7.
На Фиг. 8 показана базовая схема одного из вариантов лентивирусного экспрессионного вектора, содержащего ДНК, кодирующую РНК по настоящему изобретению в каркасе shRNA.
5' LTR 5'-длинный концевой повтор (от англ. Long Terminal Repeat), компонент лентивирусного экспрессионного вектора;
RRE - последовательность элемента, связывающего белок Rev (от англ. Rev Responsive Element), компонент лентивирусного экспрессионного вектора;
U6 promoter промотор РНК-полимеразы III;
shGNAO1 последовательность ДНК, кодирующая РНК по настоящему изобретению в каркасе shRNA;
Terminator - терминатор транскрипции;
Pol II promoter промотор РНК-полимеразы II;
PuroR ген устойчивости к пуромицину; polyA signal сигнал полиаденилирования;
sin 3' LTR - «самоинактивирующийся» 3'-длинный концевой повтор, компонент лентивирусного экспрессионного вектора.
На Фиг. 9 представлены результаты скрининга кандидатных shRNA (shGNAO1) содержащих SEQ ID No: 5-13, комплементарных последовательности мутантного аллеля c.607G>A гена GNAO1 человека, введенных в клетки с помощью лентивирусных экспрессионных векторов, содержащих ДНК, кодирующую соответствующие shRNA, в гемизиготной тест-системе с использованием временной трансфекции клеток НЕК293Т.
Показан уровень экспрессии аллеля дикого типа и мутантного аллеля c.607G>A гена GNAO1 на уровне мРНК, метод аллель-специфической кПЦР. Уровень мРНК GNAO1 в образцах, содержащих отрицательный контроль РНКи (shLuc, содержит SEQ ID No: 32), принят за 100%. Данные представлены как среднее значение±стандартное отклонение (n=3). shGNAO1-CTRL (содержит SEQ ID No: 23): положительный контроль РНКи в составе shRNA.
На Фиг. 10 представлены результаты тестирования shGNAO1-1489 (содержит SEQ ID No: 7), и shGNAO1-1500 (содержит SEQ ID No: 11), введенных в клетки с помощью лентивирусных экспрессионных векторов, содержащих кодирующую их ДНК, в гетерозиготной тест-системе со стабильной экспрессией GNAO1 в клетках НЕК293Т.
Показана экспрессия белка Gαo с мутантного аллеля c.607G>A GNAO1 человека, слитого с FLAG-эпитопом, экспрессия белка Gαo с аллеля дикого типа GNAO1, слитого с myc-эпитопом, и экспрессия референсного белка GAPDH, метод Вестерн-блот.shEGFP (содержит SEQ ID No: 31) и shSCR (содержит SEQ ID No: 24) - отрицательные контроли РНКи в составе shRNA; shGNAO 1 -CTRL (содержит SEQ ID No: 23) - положительный контроль РНКи в составе shRNA; «-»: отрицательный контроль без РНКи (без добавления shRNA).
На Фиг. 11 представлена базовая схема одного из вариантов экспрессионного ААВ-вектора, кодирующего РНК по настоящему изобретению в каркасе shRNA.
5' ITR - 5'-инвертированный концевой повтор (от англ. Inverted Terminal Repeat), компонент аденоассоциированного вирусного экспрессионного вектора;
U6 promoter РНК-полимераза III-зависимый промотор U6;
shGNAO1 - последовательность ДНК, кодирующая РНК по настоящему изобретению в каркасе shRNA;
Terminator терминатор транскрипции;
CMV promoter РНК-полимераза II-зависимый промотор цитомегаловируса человека.
Reporter - ген репортерного белка, например, флуоресцентного белка EGFP;
polyA signal - сигнал полиаденилирования;
3' ITR 3'-инвертированный концевой повтор, компонент аденоассоциированного вирусного экспрессионного вектора.
На Фиг. 12 представлены результаты тестирования shGNAO1-1489 (содержит SEQ ID No: 7) и shGNAO1-1500 (содержит SEQ ID No: 11) в экспрессионных ААВ-векторах при временной трансфекции в гетерозиготной системе в клетках НЕК293Т.
(А): анализ эффективности и селективности подавления экспрессии мутантного аллеля c.607G>A гена GNAO1 человека на уровне белка методом Вестерн-блота. Показана экспрессия белка Gαo с мутантного аллеля GNAO1, слитого с FLAG-эпитопом, экспрессия белка Gαo с аллеля дикого типа GNAO1, слитого с myc-эпитопом, и экспрессия референсного белка GAPDH.
(Б): анализ эффективности и селективности подавления экспрессии мутантного аллеля c.607G>A гена GNAO1 на уровне транскрипта GNAO1 методом аллель-специфической кПЦР. Данные представлены как среднее значение±стандартное отклонение (n=3, независимые эксперименты с двумя техническими повторностями трансфекций каждый). shSCR (содержит SEQ ID No: 24): отрицательный контроль РНКи в составе shRNA; shGNAO1-CTRL (содержит SEQ ID No: 23): положительный контроль РНКи в составе shRNA.
На Фиг. 13 демонстрируется результат тестирования shGNAO1-1489-transcript (содержит SEQ ID No: 33) и модифицированных вариантов shGNAO1-1489-v2-transcript (содержит SEQ ID No: 18), shGNAO1-1489-v3-transcript (содержит SEQ ID No: 19) и shGNAO1-1489-v4-transcript (содержит SEQ ID No: 20), введенных в клетки в составе аденоассоциированных экспрессионных плазмид, содержащих кодирующую их ДНК, в гетерозиготной тест-системе с использованием временной трансфекции клеток НЕК293Т. shGNAO1-1489-transcript (содержит SEQ ID No: 33), shGNAO1- 1489-v2-transcript (содержит SEQ ID No: 18), shGNAO1-1489-v3-transcript (содержит SEQ ID No: 19) и shGNAO1-1489-v4-transcript (содержит SEQ ID No: 20) обозначены как shGNAO1-1489, shGNAO1-1489-v2, shGNAO1-1489-v3 и shGNAO1-1489-v4 соответственно.
(А): анализ эффективности и селективности подавления экспрессии мутантного аллеля c.607G>A гена GNAO1 на уровне белка методом Вестерн-блота. Показана экспрессия белка Gαo с мутантного аллеля GNAO1, слитого с FLAG-эпитопом, экспрессия белка Gαo с аллеля дикого типа GNAO1, слитого с myc-эпитопом, и экспрессия референсного белка GAPDH.
(Б): анализ эффективности и селективности подавления экспрессии мутантного аллеля c.607G>A гена GNAO1 на уровне транскрипта GNAO1 методом аллель-специфической кПЦР. Данные представлены как среднее значение±стандартное отклонение (n=2).
shSCR-1 и shSCR-2: отрицательные контроли РНКи в составе shRNA; «No shRNA»: отрицательный контроль без РНКи (без добавления shRNA).
На Фиг. 14 схематично показаны основные структурные элементы двухцепочечной РНК по настоящему изобретению, заключенной в каркас искусственной микроРНК (miRNA) на примере miR30-GNAO1-1500(-1), содержащей смысловую (пассажирскую) цепь с последовательностью SEQ ID No: 17.
На Фиг. 15 представлены результаты тестирования miR30-GNAO1-1489(-1) (содержит SEQ ID No: 16), введенной в клетки в составе лентивирусной экспрессионной плазмиды, в гетерозиготной тест-системе с использованием временной трансфекции клеток НЕК293Т.
(А): анализ эффективности и селективности подавления экспрессии мутантного аллеля c.607G>A гена GNAO1 на уровне белка методом Вестерн-блота. Показана экспрессия белка Gαo с мутантного аллеля GNAO1, слитого с FLAG-эпитопом, экспрессия белка Gαo с аллеля дикого типа GNAO1, слитого с myc-эпитопом, экспрессия репортерного белка EGFP и референсного белка GAPDH.
(Б): анализ эффективности и селективности подавления экспрессии мутантного аллеля c.607G>A гена GNAO1 на уровне транскрипта GNAO1 методом аллель-специфической кПЦР. Данные представлены как среднее значение±стандартное отклонение (n=2).
shEGFP, shSCR-1 и shSCR-2: отрицательные контроли РНКи в составе shRNA; miR30-GNAO1-CTRL (содержит SEQ ID No: 25): положительный контроль РНКи в составе miRNA.
На Фиг. 16 представлены результаты тестирования работы miR30-GNAO1-1489(-1) (содержит SEQ ID No: 16) и miR30-GNAO1-1500(-1) (содержит SEQ ID No: 17), введенных в клетки в составе аденоассоциированной вирусной плазмиды, содержащей кодирующую их ДНК под контролем промотора U6, либо промотора РНК-полимеразы II, в гетерозиготной тест-системе с использованием временной трансфекции клеток НЕК293Т.
(А) Анализ эффективности и селективности подавления экспрессии мутантного аллеля c.607G>A гена GNAO1 на уровне белка методом Вестерн-блота. Показана экспрессия белка Gαo с мутантного аллеля GNAO1, слитого с FLAG-эпитопом, экспрессия белка Gαo с аллеля дикого типа GNAO1, слитого с myc-эпитопом, и экспрессия референсного белка GAPDH.
(Б) Анализ эффективности и селективности подавления экспрессии мутантного аллеля c.607G>A гена GNAO1 на уровне транскрипта GNAO1 методом аллель-специфической кПЦР. Данные представлены как среднее значение±стандартное отклонение (n=3).
«No miRNA»: отрицательный контроль без РНКи (без добавления miRNA); miR30-SCR: отрицательный контроль РНКи в составе miRNA; miR30-GNAO1-CTRL (содержит SEQ ID No: 25) под контролем промотора U6, либо промотора РНК-полимеразы II: положительные контроли РНКи в составе miRNA.
На Фиг. 17 представлены результаты тестирования shGNAO1-1489 (содержит SEQ ID No: 7) и shGNAO1-1500 (содержит SEQ ID No: 11), доставленных в пациент-специфические нейроны с гетерозиготным вариантом GNAO1 c.607G>A посредством трансдукции ААВ серотипа DJ.
(A) Характеристика нейрональной культуры. Иммуноцитохимическое исследование экспрессии белка Gαo и белка β-тубулин III класса (маркера зрелых нейронов) в дифференцированных в нейрональном направлении иПСК пациента с мутацией GNAO1 c.607G>A. DAPI - окраска ядер; Merge - наложение всех 3-х изображений друг на друга.
(Б) Анализ эффективности подавления экспрессии гена GNAO1 на уровне транскрипта GNAO1 в культуре пациент-специфических нейронов методом аллель-специфической кПЦР. Данные кПЦР представлены как среднее±стандартная ошибка среднего (n=5).
(B) Анализ эффективности и селективности подавления экспрессии мутантного аллеля c.607G>A гена GNAO1 на уровне транскрипта GNAO1 методом аллель-специфической кПЦР. Данные представлены как соотношение уровней экспрессии аллеля дикого типа и мутантного аллеля c.607G>A гена GNAO1 (кратность изменения соотношения).
shSCR (содержит SEQ ID No: 24): отрицательный контроль РНКи в составе shRNA; shGNAO1-CTRL (содержит SEQ ID No: 23): положительный контроль РНКи в составе shRNA.
Детальное описание изобретения
Если не указано иначе, предполагается, что все термины, обозначения и другие научные термины, используемые в данной заявке, имеют значения, которые обычно понимают специалисты в области, к которой относится настоящее изобретение. В некоторых случаях определения терминов с общепринятыми значениями приведены в данной заявке для ясности и/или для быстрой справки и понимания, и включение таких определений в настоящее описание не должно истолковываться как наличие существенного отличия значения термина от обычно подразумеваемого в данной области.
Кроме того, если по контексту не требуется иное, термины в единственном числе включают в себя термины во множественном числе, и термины во множественном числе включают в себя термины в единственном числе. Как правило, используемая классификация и методы культивирования клеток, трансфекции и трансдукции клеток эукариот, методы молекулярной биологии, аналитические методы определения специфических белков в образце, описанные в настоящем документе, хорошо известны специалистам и широко применяются в данной области. Реакция количественной ПЦР и методы работы с нуклеиновыми кислотами и белками осуществляют в соответствии с инструкциями производителя, как это обычно осуществляется в данной области, или как описано в настоящем документе.
Определения
Термин «комплементарность» в контексте описания настоящего изобретения относится к молекуле нуклеиновой кислоты (РНК или ДНК), которая образует водородные связи с другой молекулой нуклеиновой кислоты или с самой собой при спаривании оснований по Уотсону-Крику. Спаривание оснований по Уотсону-Крику относится к следующим парам оснований (нуклеотидов), образующих водородную связь: А (аденин): Т (тимин) и С (цитозин): G (гуанин) (для ДНК); и А (аденин): U (урацил) и С (цитозин): G (гуанин) (для РНК). Например, две или более комплементарных цепей молекулы нуклеиновой кислоты могут иметь одинаковое количество нуклеотидов (т.е. иметь одинаковую длину и образовывать одну двухцепочечную область с выступом (англ. overhang) или без него) или иметь разное количество нуклеотидов (например, одна цепь может быть короче, но полностью содержаться внутри другой цепи).
Термин «смысловая последовательность» или «смысловая цепь» в контексте описания настоящего изобретения относится к последовательности РНК в составе двухцепочечной РНК по настоящему изобретению, которая может обладать значительной идентичностью последовательности мРНК-мишени или быть идентичной последовательности мРНК-мишени.
Термин «антисмысловая последовательность» или «антисмысловая цепь» в контексте описания настоящего изобретения относится к последовательности РНК в составе двухцепочечной РНК по настоящему изобретению, которая по существу или полностью комплементарна последовательности мРНК-мишени, способна связываться с последовательностью мРНК-мишени и запускать механизм РНК-интерференции.
Термин «кассета экспрессии» в контексте описания настоящего изобретения относится к фрагменту ДНК, который может быть частью плазмидного или вирусного вектора, в который может быть вставлена целевая ДНК, а также регуляторные элементы для обеспечения ее экспрессии в клетке. Кассеты экспрессии для двухцепочечных РНК хорошо известны в данной области техники, и может быть пригодна кассета экспрессии любого типа, например, в которой может быть использован промотор pol III, промотор pol II или промотор pol I [среди прочего, ter Brake О, 't Hooft К., Liu Y.P. et al. Lentiviral vector design for multiple shRNA expression and durable HIV-1 inhibition. Mol Ther. 2008 Mar; 16(3):557-64; Maczuga P., Koornneef A., Borel F. et al. Optimization and comparison of knockdown efficacy between polymerase II expressed shRNA and artificial miRNA targeting luciferase and Apolipoprotein B100. BMC BiotechNol. 2012 Jul 24; 12:42]. Кассеты экспрессии по настоящему изобретению можно поместить в клетку с использованием, например, способов трансфекции. Любые подходящие средства могут быть достаточными для переноса кассеты экспрессии по настоящему изобретению. Предпочтительно используют вирусные векторы, которые обеспечивают в клетке-мишени стабильную экспрессию двухцепочечных РНК по настоящему изобретению. Подходящими векторами могут быть лентивирусные векторы или векторы на основе аденоассоциированного вируса (ААВ-векторы). Очевидно, что, например, лентивирусные векторы содержат РНК-геном, РНК-геном будет кодировать кассету экспрессии, так что после трансдукции в клетке образуются последовательность ДНК и кассета экспрессии.
Термины «мутантный аллель», или «мутантный аллель GNAOh> или «мутантный аллель гена GNAOh> в контексте описания настоящего изобретения относятся к мутантному аллелю c.607G>A гена GNAO1 человека. Термины «нормальный аллель», или «здоровый аллель», или «аллель дикого типа» в контексте описания настоящего изобретения относятся к нормальному аллелю гена GNAO1 человека, в котором отсутствует мутация c.607G>A.
Термин «аллель-селективное подавление» означает подавление экспрессии гена, при котором подавление экспрессии мутантного аллеля выше, чем подавление экспрессии аллеля дикого типа.
Термин «трансфекция» в контексте настоящего описания относится к процессу доставки в эукариотические клетки нуклеиновой кислоты (ДНК или РНК) невирусным методом. В качестве невирусного метода доставки нуклеиновой кислоты может быть использована кальций-фосфатная трансфекция, когда нуклеиновая кислота адсорбируется на кристаллах фосфата кальция, которые поглощаются клеткой путем фагоцитоза. В качестве невирусного метода доставки нуклеиновой кислоты также может быть использован метод липофекции, когда ДНК или РНК инкапсулированы в липосомы липидные структуры, которые могут беспрепятственно сливаться с мембраной эукариотической клетки. Для невирусного метода доставки нуклеиновой кислоты также могут быть использованы катионные полимеры, такие как линейный полиэтиленимин (linear PEI, LPEI). Трансфекция обеспечивает временную экспрессию доставляемой нуклеиновой кислоты в клетке.
Термин «трансдукция» в контексте настоящего описания относится к процессу доставки в эукариотические клетки нуклеиновой кислоты с помощью вирусных векторов. В качестве вирусных векторов могут быть использованы ретровирусы, в частности лентивирусы, а также аденоассоциированные вирусы. Трансдукция может обеспечивать стабильную экспрессию доставляемой нуклеиновой кислоты в клетке в течение длительного времени.
Термин «вектор» в контексте настоящего описания относится к плазмидной ДНК или частицам лентивируса/аденоассоциированного вируса, которые используются в качестве носителя нуклеиновой кислоты, содержащей последовательность ДНК, кодирующую РНК по настоящему изобретению, для ее доставки в эукариотические клетки посредством трансфекции или трансдукции соответственно. Используемые плазмидные и/или вирусные векторы, приведенные ниже, используются для примера и не ограничивают объем прав настоящего изобретения.
Термин «гемизиготная тест-система» в контексте настоящего описания относится к культуре клеток НЕК293Т (от англ. Human Embryonic Kidney 293, клетки, полученные из эмбриональных почек человека), в которые с помощью временной трансфекции или вирусной трансдукции доставлена ДНК, кодирующая один из аллелей гена GNAO1 человека: либо аллель дикого типа, либо мутантный аллель.
Термин «гетерозиготная тест-система» в контексте настоящего описания относится к культуре клеток НЕК293Т, в которые с помощью временной трансфекции или вирусной трансдукции одновременно доставлена ДНК, кодирующая аллель дикого типа гена GNAO1 человека, а также ДНК, кодирующая мутантный аллель гена GNAO1 человека.
Термин «отрицательный контроль РНКи» в контексте настоящего описания относится к РНК, которая не содержит последовательности, комплементарные последовательностям генома человека и не должна вызывать процесс РНК-интерференции эндогенных транскриптов генов человека. В качестве отрицательного контроля могут быть использованы рандомизированные неспецифические РНК, так называемые scramble, а также РНК, способные подавлять экспрессию экзогенных генов, например, репортерных генов, таких как ген зеленого флуоресцентного белка (GFP/EGFP) или ген люциферазы (Luc). Отрицательный контроль РНКи может быть представлен в форме малой интерферирующей РНК (siRNA), короткой шпилечной РНК (shRNA) или искусственной микроРНК (miRNA). Отрицательный контроль РНКи в форме siRNA может быть доставлен в клетки с помощью трансфекции. Отрицательный контроль РНКи в форме shRNA или miRNA может быть вставлен в экспрессионный вектор в виде ДНК, кодирующей соответствующую shRNA или miRNA, и доставлен в клетки с помощью трансфекции или трансдукции.
Термин «положительный контроль РНКи» в контексте настоящего описания относится к РНК, которая комплементарна последовательности гена GNAO1 человека на участке транскрипта, не содержащего мутацию c.607G>A. Положительный контроль РНКи инициирует процесс РНК-интерференции гена GNAO1 человека, что приводит к полному, или «тотальному» подавлению экспрессии обоих аллелей гена GNAO1 человека: как аллеля дикого типа, так и мутантного аллеля. Положительный контроль РНКи может быть представлен в форме малой интерферирующей РНК (siRNA), короткой шпилечной РНК (shRNA) или искусственной микроРНК (miRNA). Положительный контроль РНКи в форме siRNA может быть доставлен в клетки с помощью трансфекции. Положительный контроль РНКи в форме shRNA или miRNA может быть вставлен в экспрессионный вектор в виде ДНК, кодирующей соответствующую shRNA или miRNA, и доставлен в клетки с помощью трансфекции или трансдукции.
Термин «отрицательный контроль без РНКи» в контексте настоящего описания относится к образцу, не содержащему РНК, способную индуцировать РНК-интерференцию, в том числе не содержащему отрицательный контроль РНКи. Это могут быть клетки без добавления РНК по настоящему изобретению, представленной в форме siRNA, либо клетки без добавления плазмидного или вирусного вектора, содержащего РНК по настоящему изобретению, представленную в форме shRNA или miRNA.
В настоящем изобретении предлагается двухцепочечная РНК, содержащая смысловую последовательность РНК и антисмысловую последовательность РНК, которые, по существу, комплементарны, причем антисмысловая последовательность РНК имеет длину, равную по меньшей мере 19 нуклеотидам, и является, по существу, комплементарной последовательности SEQ ID No: 28. При этом следует понимать, что термин «по существу, комплементарные» означает, что нет необходимости в том, чтобы все нуклеотиды смысловой и антисмысловой последовательностей РНК были парными, то есть полностью комплементарными, или чтобы все нуклеотиды антисмысловой последовательности РНК и SEQ ID No: 28 были парными. Если двухцепочечная РНК способна индуцировать РНК-интерференцию транскрипта гена GNAO1, содержащего мутацию c.607G>A, за счет последовательности, которая специфически нацелена на последовательность SEQ ID No: 28, то такая, по существу, комплементарность является предметом настоящего изобретения.
SEQ ID No: 28 (5'-TCAGGCTGTTTGACGTCAGAGGCCAGCGATCTGAAC-3') соответствует 36-нуклеотидной последовательности мутантного аллеля гена GNAO1, содержащей мутацию c.607G>A. SEQ ID No: 28 соответствует нуклеотидам с номерами с 1487 по 1522 последовательности SEQ ID No: 26.
К объекту патентования относится любая последовательность РНК длиной по меньшей мере 19 нуклеотидов, находящаяся в «окне» длиной 36 нуклеотидов, начиная с позиции 1487 и заканчивая позицией 1522 относительно начала транскрипта GNAO1 (NM 020988.2).
Одна из последовательностей РНК по настоящему изобретению включена в «смысловую» цепь двухцепочечной РНК, которую также называют «пассажирской» цепью, поскольку она может обладать значительной идентичностью последовательности-мишени мРНК или быть идентичной последовательности мРНК-мишени. Другая последовательность РНК включена в «антисмысловую» цепь, также называемую «направляющей» цепью, поскольку она, по существу или полностью комплементарна последовательности мРНК-мишени. Смысловая (пассажирская) и антисмысловая (направляющая) последовательности РНК включены в двухцепочечную РНК и являются, по существу или полностью, комплементарными.
Двухцепочечная РНК по настоящему изобретению предназначена для запуска РНК-интерференции, с помощью которой снижается экспрессия мутантного аллеля гена GNAO1, при этом уровень экспрессии нормального аллеля гена GNAO1 становится выше, чем уровень экспрессии мутантного аллеля. В одном из вариантов осуществления изобретения двухцепочечная РНК по настоящему изобретению содержит смысловую последовательность РНК и антисмысловую последовательность РНК, которые являются, по существу, комплементарными, причем антисмысловая последовательность РНК имеет длину последовательности, равную по меньшей мере 19 нуклеотидам, и является, по существу, комплементарной последовательности SEQ ID No: 28, способна вызывать РНК-интерференцию, специфически снижающую экспрессию РНК-транскрипта, содержащего SEQ ID No: 28, а также белка, кодируемого мутантным аллелем c.607G>A гена GNAO1.
Снижение экспрессии мутантного аллеля гена GNAO1 можно определить с использованием стандартных тестов, известных из уровня техники, например, методом аллель-специфической кПЦР и Вестерн-блот, которые описаны в примерах и известны в данной области техники [патент РФ 2777663].
В данной области техники хорошо известны двухцепочечные структуры РНК, способные индуцировать РНК-интерференцию. Например, малая интерферирующая РНК (siRNA) содержит две раздельные РНК-цепи, причем одна цепь содержит направляющую (антисмысловую) последовательность РНК, которая связывается с, по существу, комплементарным целевым транскриптом, а другая цепь содержит пассажирскую (смысловую) последовательность РНК. Часто используемая конструкция siRNA состоит из 19-нуклеотидного двухцепочечного участка и двух выступающих нуклеотидов на 3'-конце каждой цепи (например, dTdT или UU). Каждая из двух цепей РНК имеет фосфатную группу на 5'-конце и гидроксильную группу на 3'-конце [Elbashir, et al. (2001) Functional anatomy of siRNA for mediating efficient RNAi in Drosophila melaNogaster embryo lysate. EMBO J 20: 6877-6888]. Длина двухцепочечного, по существу, комплементарного участка может быть от 19 до 21 нуклеотидов и более, вплоть до 30 нуклеотидов.
Двухцепочечная РНК по настоящему изобретению не требует, чтобы смысловая (пассажирская) и антисмысловая (направляющая) последовательности РНК были включены в две раздельные цепи. Пассажирская и направляющая последовательности РНК также могут быть включены в одну цепь РНК, например, в каркас shRNA (короткую шпилечную РНК). Такой подход позволяет доставлять в клетку siRNA для супрессии целевого гена посредством экспрессирующих векторов. Молекулы shRNA представляют собой одноцепочечную РНК и благодаря инвертированным повторам нуклеотидной последовательности, соответствующим последовательностям смысловой и антисмысловой цепи в siRNA, образуют внутримолекулярное спаривание оснований, формируя так называемый «стебель». Инвертированные повторы в составе shRNA обычно имеют длину от 21 до 29 нуклеотидов и разделены спейсерной последовательностью, которая формирует так называемую «петлю». Длина и нуклеотидный состав «петли» могут быть различными и не ограничиваются вариантами, используемыми в примерах осуществления настоящего изобретения. Подходящие петлевые последовательности хорошо известны в данной области техники [Jensen S.M., Schmitz A., Pedersen F.S. et al. Functional selection of shRNA loops from randomized retroviral libraries. PLoS One. 2012; 7(8):e43095; Schopman N.C., Liu Y.P., KonstantiNova P. et al. Optimization of shRNA inhibitors by variation of the terminal loop sequence. Antiviral Res. 2010May; 86(2):204-11].
В качестве «петель» в составе shRNA могут быть использованы, например, 6-нуклеотидная петля (CTCGAG) и 9-нуклеотидная петля miR-22 (CCTGACCCA). Одноцепочечные молекулы shRNA могут кодироваться соответствующей последовательностью ДНК и таким образом быть клонированы в экспрессирующие векторы. Последовательность ДНК, кодирующая shRNA, содержит на 3'-конце от 4 до 6 тиминов, которые служат сайтом терминации транскрипции. На 5'-конце последовательность ДНК, кодирующая shRNA, может содержать или не содержать выступающие нуклеотиды. ДНК, кодирующая shRNA, транскрибируется под контролем промотора РНК-полимеразы III (например, U6 или HI). В клетке транскрибируемая с экспрессирующего вектора молекула РНК сворачивается в шпильку, экспортируется в цитоплазму и используется в качестве субстрата для эндонуклеазы Dicer с последующим образованием соответствующей двухцепочечной siRNA, включением направляющей цепи в комплекс RISC и запуском процесса РНК-интерференции.
Таким образом, shRNA по настоящему изобретению имеет одноцепочечную структуру по типу «стебель-петля» и может содержать, начиная с 5'-конца:
- выступающие нуклеотиды (необязательно);
- пассажирскую (смысловую) последовательность РНК;
- последовательность петли;
- направляющую (антисмысловую) последовательность РНК;
- короткий поли (U) хвост (Фиг. 7). Альтернативно, shRNA может содержать, начиная с 5'-конца:
- выступающие нуклеотиды (необязательно);
- направляющую (антисмысловую) последовательность РНК;
- последовательность петли;
- пассажирскую (смысловую) последовательность РНК;
- короткий поли (U) хвост.
Такая молекула РНК образует внутримолекулярные пары оснований за счет, по существу, комплементарных пассажирской и направляющей последовательностей РНК.
Двухцепочечная РНК по настоящему изобретению также может быть включена в каркас искусственной miRNA. Искусственные miRNA являются аналогами эндогенных miRNA, регулирующих генную экспрессию в клетке. Молекула miRNA, как и shRNA, представляет собой одноцепочечную РНК и содержит инвертированные повторы, соответствующие последовательностям смысловой (пассажирской) и антисмысловой (направляющей) цепи в siRNA. Молекула miRNA образует внутримолекулярные пары оснований за счет, по существу, комплементарных пассажирской и направляющей последовательностей РНК, формируя так называемый «стебель». Длина пассажирской и, соответственно, направляющей последовательности обычно составляет 22 нуклеотида. Последовательность, формирующая «петлю», может различаться в зависимости от типа каркаса miRNA. Так, часто используются каркасы miR-30, miR-155, и miR-451 [Kotowska-Zimmer A., Pewinska М., Olejniczak М. Artificial miRNAs as therapeutic tools: Challenges and opportunities. Wiley Interdiscip Rev RNA. 2021 Jul; 12(4):el640. doi: 10.1002/wrna.1640. Epub 2021 Jan 1. PMID: 33386705], и другие. Длина и нуклеотидный состав «петли» могут быть различными и не ограничиваются вариантами, используемыми в примерах осуществления настоящего изобретения. В одном из примеров осуществления настоящего изобретения в составе miRNA используется 15-нуклеотидная петля каркаса miR-30 (GUGAAGCCACAGAUG).
РНК по настоящему изобретению в каркасе miRNA имеет одноцепочечную структуру по типу «стебель-петля» и может содержать:
- пассажирскую (смысловую) последовательность РНК длиной 22 нуклеотида;
- последовательность петли;
- направляющую (антисмысловую) последовательность РНК длиной 22 нуклеотида;
- нуклеотиды апикального и базального соединений, служащие сайтами расщепления ферментами Drosha и Dicer;
- дополнительные фланкирующие последовательности (Фиг. 14).
Альтернативно miRNA может содержать:
- направляющую (антисмысловую) последовательность РНК длиной 22 нуклеотида;
- последовательность петли;
- пассажирскую (смысловую) последовательность РНК длиной 22 нуклеотида;
- нуклеотиды апикального и базального соединений, служащие сайтами расщепления ферментами Drosha и Dicer;
- дополнительные фланкирующие последовательности.
Одноцепочечные молекулы miRNA могут кодироваться соответствующей последовательностью ДНК и таким образом быть клонированы в экспрессирующие векторы. ДНК, кодирующая miRNA, может транскрибироваться как под контролем промотора РНК-полимеразы III, так и под контролем промотора РНК-полимеразы II. В последнем случае, это позволяет использовать тканеспецифические промоторы, например, нейроспецифические, для экспрессии miRNA. Процессинг miRNA начинается с формирования длинных первичных транскриптов (pri-miRNA), которые кэпируются на 5'-конце и полиаденилируются на 3'-конце. Затем, с помощью фермент-белкового комплекса DGCR8/Drosha в ядре pri-miRNA разрезается до предшественников со шпилечной структурой по типу shRNA - так называемые pre-miRNA. Далее pre-miRNA экспортируется в цитоплазму и используется в качестве субстрата для эндонуклеазы Dicer с последующим образованием соответствующей двухцепочечной siRNA длиной до 22 нуклеотидов. Далее направляющая цепь включается в комплекс RISC и запускается процесс РНК-интерференции [Matsuyama Н., Suzuki H.I. Systems and Synthetic microRNA Biology: From Biogenesis to Disease Pathogenesis. Int J Mol Sci. 2019 Dec 24; 21(1): 132].
В составе miRNA обычно присутствуют нуклеотиды апикального и базального соединений, служащие сайтами расщепления ферментами Drosha и Dicer, а также дополнительные фланкирующие последовательности.
В одном из вариантов осуществления изобретения предлагается молекула РНК, способная снижать экспрессию мутантного аллеля гена GNAO1 человека, содержащего мутацию c.607G>A (p.Gly203Arg). Предлагаемая молекула РНК содержит смысловую (пассажирскую) последовательность РНК и антисмысловую (направляющую) последовательность РНК, в которой смысловая и антисмысловая последовательности РНК являются, по существу, комплементарными, причем смысловая последовательность РНК может быть выбрана из группы, состоящей из SEQ ID No: 1-17.
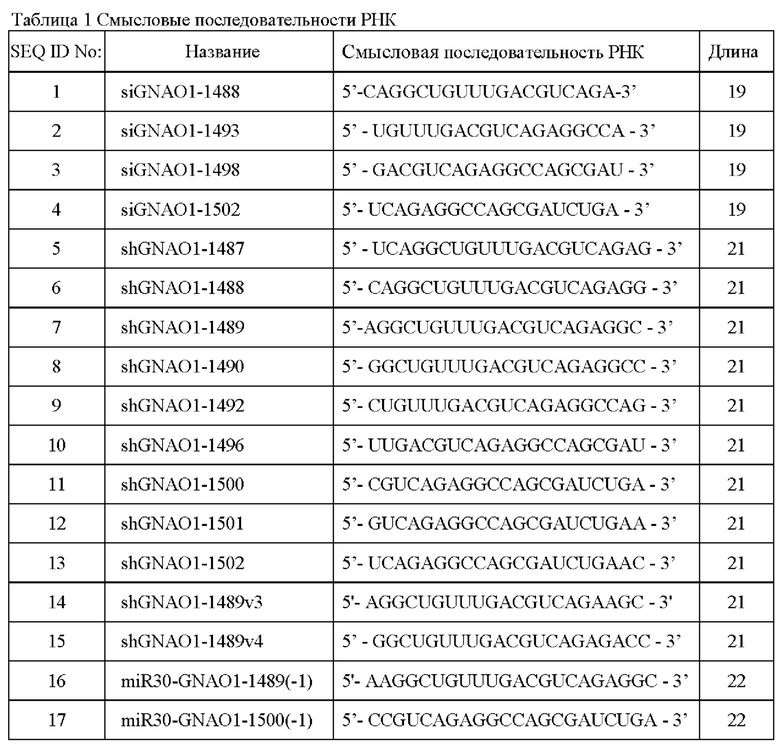
В табл.1 показаны смысловые (пассажирские) последовательности РНК, которые входят в состав двухцепочечных РНК по настоящему изобретению, способных специфически ингибировать экспрессию транскриптов, содержащих SEQ ID No: 28, как описано в разделе с примерами осуществления настоящего изобретения.
РНК по настоящему изобретению были протестированы для аллель-селективного подавления экспрессии мутантного аллеля гена GNAO1, содержащего мутацию c.607G>A (p.Gly203Arg). В результате были отобраны последовательности РНК, показавшие наибольшую эффективность аллель-селективного подавления мутантного аллеля гена GNAO1. РНК по настоящему изобретению могут быть представлены различными по структуре молекулами: siRNA, shRNA и miRNA.
В одном из вариантов осуществления изобретения РНК представляют собой малые интерферирующие РНК (siRNA). siRNA по настоящему изобретению может содержать последовательность нуклеотидов, выбранную из SEQ ID No: 14. Последовательности siRNA могут доставляться в клетку в виде РНК-дуплексов методом трансфекции с использованием трансфицирующего агента. Введение в клетки siRNA-дуплекса, содержащего последовательность SEQ ID No: 14 привело к снижению экспрессии мутантного аллеля гена GNAO1 на уровне РНК (до 37% в гемизиготной и 43% в гетерозиготной тест-системе при использовании РНК с последовательностью SEQ ID No: 1, siGNAO1-1488) при сохранении экспрессии аллеля дикого типа на нормальном уровне (до 84% в гемизиготной и 113% в гетерозиготной тест-системе при использовании РНК с последовательностью SEQ ID No: 1, siGNAO1-1488). Экспрессия мутантного аллеля гена GNAO1 на уровне белка также была снижена при сохранении экспрессии аллеля дикого типа на нормальном уровне.
В одном из вариантов осуществления изобретения РНК включена в каркас шпилечной РНК (shRNA). shRNA по настоящему изобретению может содержать последовательность нуклеотидов, выбранную из SEQ ID No: 5-15. Для доставки в клетки shRNA могут быть использованы векторы как на основе экспрессионных плазмид, в том числе вирусных, так и на основе вирусных частиц. Введение в клетки плазмидного вектора, экспрессирующего ДНК, кодирующую shRNA, которая содержит последовательность, выбранную из SEQ ID No: 5-15, привело к снижению экспрессии мутантного аллеля гена GNAO1 на уровне РНК и белка при сохранении экспрессии аллеля дикого типа на нормальном уровне. Так, экспрессия мутантного аллеля гена GNAO1 на уровне РНК снизилась до 21% и 40% в гемизиготной и 32% и 31% в гетерозиготной тест-системе при использовании РНК с последовательностью SEQ ID No: 7, shGNAO1-1489 и SEQ ID No: 11, shGNAO1-1500 соответственно при сохранении экспрессии аллеля дикого типа на нормальном уровне: до 50% и 110% в гемизиготной и 125% и 102% в гетерозиготной тест-системе при использовании РНК с последовательностью SEQ ID No: 7, shGNAO1-1489 и SEQ ID No: 11, shGNAO1-1500 соответственно. Экспрессия мутантного аллеля гена GNAO1 на уровне белка также была снижена при сохранении экспрессии аллеля дикого типа на нормальном уровне. Трансдукция культуры нейронов, полученных с помощью направленной дифференцировки индуцированных плюрипотентных стволовых клеток (иПСК) пациента с GNAO1-энцефалопатией, вирусным вектором, экспрессирующим shRNA по настоящему изобретению, привело к увеличению соотношения уровней мРНК GNAO1 дикого типа и мутантного варианта в 1,7 и 5,7 раз относительно отрицательного контроля РНКи, принятого за 1, при использовании РНК с последовательностью SEQ ID No: 7 (shGNAO1-1489) и SEQ ID No: 11 (shGNAO1-1500) соответственно.
В другом варианте осуществления изобретения предлагаются модифицированные варианты shRNA, созданные на основе нуклеотидной последовательности транскрипта shRNA (каркаса) shGNAO1-1489-transcript (SEQ ID No: 33):
shGNAO1-1489-v2-transcript (SEQ ID No: 18), каркас которой содержит пассажирскую последовательность SEQ ID No: 7 и представлен последовательностью, в которой удалены два гуанина (GG), фланкирующие 5'-конец пассажирской цепи shRNA, и использована 9-нуклеотидная петля CCUGACCCA подобно петле miR-22;
shGNAO1-1489-v3-transcript (SEQ ID No: 19), каркас которой содержит пассажирскую последовательность SEQ ID No: 14, которая содержит аденин в позиции 19, начиная от 5'-конца, и комплементарную ей направляющую цепь, и каркас представлен последовательностью, в которой удалены два гуанина (GG), фланкирующие 5'-конец пассажирской цепи shRNA, и использована 9-нуклеотидная петля CCUGACCCA подобно петле miR-22;
shGNAO1- 1489-v4-transcript (SEQ ID No: 20), каркас которой содержит пассажирскую последовательность SEQ ID No: 15, которая содержит аденин в позиции 19, начиная от 5'-конца, и комплементарную ей направляющую цепь, и каркас представлен последовательностью, в которой удалены два гуанина (GG), фланкирующие 5'-конец пассажирской цепи shRNA, и использована 9-нуклеотидная петля CCUGACCCA подобно петле miR-22, а также введен дополнительный аденин, фланкирующий 5'-конец пассажирской цепи.
Введение в клетки плазмидного вектора, экспрессирующего shRNA по настоящему изобретению, привело к снижению экспрессии мутантного аллеля гена GNAO1 на уровне РНК до 50%, 40% и 27% при использовании РНК с последовательностью SEQ ID No: 18 (shGNAO1-1489-v2-transcript), SEQ ID No: 19 (shGNAO1-1489-v3-transcript) и SEQ ID No: 20 (shGNAO1-1489-v4-transcript) соответственно при сохранении экспрессии аллеля дикого типа на нормальном уровне: 100%, 100% и 76% в гетерозиготной тест-системе. Экспрессия мутантного аллеля гена GNAO1 на уровне белка также была снижена при сохранении экспрессии аллеля дикого типа на нормальном уровне.
В одном из вариантов осуществления изобретения РНК включена в каркас miRNA. В качестве каркаса miRNA по настоящему изобретению может быть использован каркас, производный от miR-30. miRNA по настоящему изобретению может содержать последовательность нуклеотидов, выбранную из SEQ ID No: 16-17. Для доставки в клетки miRNA могут быть использованы векторы как на основе экспрессионных плазмид, в том числе вирусных, так и на основе вирусных частиц. Введение в клетки плазмидного вектора, экспрессирующего ДНК, кодирующую miRNA, которая содержит последовательность, выбранную из SEQ ID No: 16-17, в гетерозиготной тест-системе привело к снижению экспрессии мутантного аллеля гена GNAO1 на уровне РНК до 17% и 47% при сохранении высокой экспрессии аллеля дикого типа: 161% и 249% в случае, когда miRNA находится под контролем промотора U6 при использовании РНК с последовательностью SEQ ID No: 16 (miR30-GNAO1-1489(-1)) и SEQ ID No: 17 (miR30-GNAO1-1500(-1)) соответственно. Экспрессия мутантного аллеля гена GNAO1 на уровне белка также была снижена при сохранении экспрессии аллеля дикого типа на нормальном уровне.
Двухцепочечные РНК по настоящему изобретению, включенные в siPHK, либо в shPHK или miRNA каркас, могут быть доставлены в клетку с использованием способов, известных в данной области техники, таких как липофекция, трансфекция, трансдукция, или с использованием любых других подходящих способов. siPHK могут быть доставлены в клетки напрямую посредством трансфекции РНК-дуплексов. shRNA и miRNA могут быть доставлены в клетки-мишени в составе кассет экспрессии плазмидных экспрессионных векторов или вирусных векторов с помощью трансфекции и трансдукции соответственно, путем введения в векторы ДНК-вставок, кодирующих соответствующие молекулы РНК, известными из уровня техники способами.
В одном из вариантов осуществления настоящего изобретения двухцепочечные РНК по настоящему изобретению закодированы в последовательности ДНК. Последовательность ДНК, кодирующая РНК, например, заключенную в shPHK или miRNA каркас, включена в кассету экспрессии. Кассеты экспрессии для двухцепочечных РНК хорошо известны в данной области техники. Для экспрессии shRNA могут быть пригодны кассеты экспрессии, в которых экспрессия shRNA контролируется промотором РНК-полимеразы III (такими как U6, HI или 7SK). Для экспрессии miRNA могут быть пригодны различные кассеты экспрессии, в которых экспрессия miRNA контролируется либо промотором РНК-полимеразы III, либо промотором РНК-полимеразы II. В качестве промоторов РНК-полимеразы II могут использоваться как конститутивные, так и тканеспецифические промоторы. В случае последних может быть снижен потенциальный токсический эффект РНК-интерференции на организм [ter Brake О, 't Hooft К., Liu Y.P. et al. Lentiviral vector design for multiple shRNA expression and durable HIV-1 inhibition. Mol Ther. 2008 Mar;16(3):557-64. doi: 10.1038/sj.mt.6300382; Maczuga P., Koornneef A., Borel F. et al. Optimization and comparison of knockdown efficacy between polymerase II expressed shRNA and artificial miRNA targeting luciferase and Apolipoprotein B100. BMC BiotechNol. 2012 Jul 24; 12:42. doi: 10.1186/1472-6750-12-42]. Примеры кассет экспрессии, экспрессирующих двухцепочечную РНК по настоящему изобретению, изображены на Фиг. 8 и Фиг. 11.
Кассета экспрессии, экспрессирующая двухцепочечную РНК по настоящему изобретению, может быть введена в плазмидный или вирусный вектор. В качестве плазмидных векторов могут быть использованы любые экспрессионные векторы, в том числе на основе лентивирусной или аденоассоциированной экспрессионной плазмиды.
В качестве вирусных векторов могут быть использованы лентивирусные или аденоассоциированные вирусные, либо другие вирусные векторы, содержащие кассету экспрессии shRNA или miRNA. Таким образом, при вирусной доставке (трансдукции) в клетках-мишенях обеспечивается стабильная экспрессия РНК по настоящему изобретению, что позволяет однократно вводить вектор. В случае доставки РНК по настоящему изобретению в центральную нервную систему предпочтительно использование аденоассоциированных вирусных векторов [Kimura S., Harashima Н. Current Status and Challenges Associated with CNS-Targeted Gene Delivery across the BBB. Pharmaceutics. 2020 Dec 15; 12(12): 1216. https://doi.org/10.3390/pharmaceuticsl2121216].
Выбор вектора определяется задачами, для которых вектор будет в дальнейшем использован и не ограничивает объем настоящего изобретения. Способы получения векторов известны из уровня техники. Векторы, содержащие ДНК с последовательностью нуклеотидов, кодирующей любую из РНК по настоящему изобретению, выбранную из shRNA и miRNA, могут быть получены специалистами в области генетической инженерии.
В одном из вариантов осуществления изобретения предлагается плазмидный экспрессионный вектор на основе трансферной плазмиды лентивируса pLKO. 1 - TRC cloning vector [Addgene, #10878; Moffat J., Grueneberg D.A., Yang X. et al. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell. 2006 Mar 24;124(6):1283-98. doi: 10.1016/j.cell.2006.01.040. PMID: 16564017], содержащий ДНК с последовательностью нуклеотидов, кодирующей любую молекулу РНК по настоящему изобретению, выбранную из shRNA и miRNA (SEQ ID No: 5-20).
В другом варианте осуществления изобретения предлагается плазмидный экспрессионный вектор на основе трансферной плазмиды аденоассоциированного вируса pAAV-LacZ (Agilent), содержащий ДНК с последовательностью нуклеотидов, кодирующей любую молекулу РНК по настоящему изобретению, выбранную из shRNA и miRNA, содержащую SEQ ID No: 5-20.
В другом варианте осуществления изобретения предлагается вирусный вектор на основе аденоассоциированного вируса, содержащий ДНК, кодирующую РНК по настоящему изобретению.
Было показано, что РНК по настоящему изобретению в форме siRNA, shRNA и каркасе miRNA эффективно и селективно снижают экспрессию мутантного аллеля c.607G>A при сохранении экспрессии аллеля дикого типа гена GNAO1 на уровне РНК и белкового продукта в клеточных моделях in vitro.
Таким образом, одна из технических задач настоящего изобретения, например, создание средства для аллель-селективного подавления экспрессии мутантного аллеля гена GNAO1 с мутацией c.607G>A (p.Gly203Arg), может быть решена с помощью любого изобретения из заявленной группы изобретений.
Селективное связывание молекул РНК по настоящему изобретению с мРНК, транскрибируемой с мутантного аллеля c.607G>A гена GNAO1, приводит к подавлению экспрессии мутантного аллеля гена GNAO1 при сохранении экспрессии аллеля дикого типа GNAO1.
Изобретение может быть использовано для разработки генотерапевтического препарата для лечения пациентов с GNAO1-энцефалопатией, имеющих подтвержденную молекулярно-генетически мутацию c.607G>A (p.Gly203Arg). Применительно к GNAO 1 -энцефалопатии с вариантом c.607G>A (p.Gly203Arg) ожидается, что РНК по настоящему изобретению, специфически связывающиеся с мРНК мутантного аллеля гена GNAO1, будут снижать содержание мутантного белкового продукта, вероятно обладающего токсическим действием на нейроны пациента. При этом работа нормального аллеля, производящего функциональный белок, затронута не будет.
Сущность и промышленная применимость заявленных изобретений поясняется следующими примерами осуществления:
Пример 1.1. Получение плазмидных экспрессионных конструкций
Для создания геми- и гетерозиготной клеточной тест-системы использовали две плазмидные конструкции, содержащие кодирующую рамку GNAO1 дикого типа, слитую с эпитопом myc, и кодирующую рамку GNAO1 с мутацией с. 607 G>A, слитую с эпитопом FLAG: pGNAO1-Myc и pGNAO 1-G203R-FLAG соответственно.
Данные конструкции были созданы на основе плазмид pGNAO1 и pGNAO1-G203R [Лунев Е.А., Шмидт А.А., Васильева С.Г. и др. Эффективная вирусная доставка в нейрональную культуру генетических конструкций для моделирования и генной терапии GNAO1-энцефалопатии. Молекулярная биология, 2022;56(4):604-618]. Для этого на N-конец белкового продукта GNAO1 дикого типа и с мутацией с. 607 G>A (p.G203R) были добавлены myc- и FLAG-эпитопы соответственно. Для получения соответствующих плазмидных конструкций использовали метод безлигазного клонирования. Для линеаризации вектора pGNAO1 методом ПЦР использовали пару праймеров: mycGNAO-F: AAACTCATCTCAGAAGAGGATCTGGGATGTACTCTGAGCGCAGAGGAG и GNAOmyc-R: TCCTCTTCTGAGATGAGTTTTTGTTCCATGGTGGCGACCGGTAGCG, для линеаризации вектора pGNAO1-G203R - праймеры flagGNAO-F: ACTACAAAGACGATGACGACAAGGGATGTACTCTGAGCGCAGAGGAG и GNAOflag-R: TCGTCATCGTCTTTGTAGTCCATGGTGGCGACCGGTAGCG. В результате были получены финальные плазмидные конструкции pGNAO1-Мус и pGNAO1-G203R-FLAG.
Пример 1.2. Основные экспериментальные методы, используемые в работе
Для анализа использовали методы кПЦР и Вестерн-блот. Метод кПЦР позволяет количественно оценивать подавление экспрессии мутантного и дикого аллелей гена GNAO1 на уровне мРНК. Нами ранее был разработан метод аллель-специфической кПЦР с использованием специфических праймеров и зондов для определения уровня экспрессии транскриптов GNAO1 дикого типа и с мутацией c.607G>A, подробно описанный в патенте РФ 2777663. Метод Вестерн-блот позволяет проводить качественную и полуколичественную (относительно экспрессии белка домашнего хозяйства, например, глицеральдегид-3-фосфатдегидрогеназы GAPDH) оценку подавления экспрессии мутантного и дикого аллелей гена GNAO1 на уровне экспрессии белкового продукта Gαo. В работе использовали стандартный протокол, подробно описанный в статье: Лунев Е.А., Шмидт А.А., Васильева С.Г. и др. Эффективная вирусная доставка в нейрональную культуру генетических конструкций для моделирования и генной терапии GNAO1-энцефалопатии. Молекулярная биология, 2022; 56(4):604-618. DOI: 10.31857/S0026898422040061. Для окрашивания мембраны использовали антитела, специфичные к GNAO1, а также к FLAG и туе эпитопам для детекции мутантного аллеля и аллеля дикого типа GNAO1 соответственно. В качестве контроля нагрузки для нормализации уровня экспрессии целевых белков использовали GAPDH и/или EGFP.
Пример 1.3. Номенклатура наименований последовательностей РНК Наименование последовательности РНК по настоящему изобретению базируется на следующем принципе: номер в названии соответствует положению первого нуклеотида смысловой последовательности РНК по настоящему изобретению относительно первого нуклеотида мРНК гена GNAO1 человека (NM 020988.2).
Пример 2. Дизайн и тестирование РНК по настоящему изобретению в форме siRNA Пример 2.1. Дизайн siRNA
Основной подход к дизайну siRNA базируется на знаниях о структуре эндогенных siRNA, которые образуются в результате разрезания двухцепочечных РНК рибонуклеазой Dicer. Такие эндогенные siRNA имеют размер 21-23 нуклеотида и состоят из смысловой и антисмысловой цепи, которые комплементарны друг другу, за исключением двух выступающих нуклеотидов на 3'-конце, так называемых «оверхенгов» (от англ. overhang выступ).
Последовательность каждой РНК по настоящему изобретению, представленной в форме siRNA, включала в себя 19 целевых нуклеотидов, в составе направляющей цепи полностью комплементарных последовательности транскрипта гена GNAO1 с мутацией c.607G>A (p.Gly203Arg), (SEQ ID No: 28), а также 2 выступающих нуклеотида (оверхенг).
Для дизайна последовательности РНК по настоящему изобретению, представленной в форме siRNA, использовали последовательность матричной РНК гена GNAO1, имеющуюся в свободном доступе в базе данных NCBI (идентификационный номер последовательности: NM 020988.2). Был выбран участок размером 41 нуклеотид, содержащий сайт мутации (c.607G>A, p.G203R), фланкированный 20-ю нуклеотидами с каждой стороны. Далее с помощью программного обеспечения siDESIGN Center (https://horizondiscovery.com/en/ordering-and-calculation-tools/sidesign-center, дата обращения 25.10.2022), находящегося в свободном доступе и предназначенного для дизайна и оценки siRNA, были получены несколько последовательностей-кандидатов размером 19 нуклеотидов. Из полученных вариантов siRNA выбрали 4 последовательности-кандидата, основываясь на наиболее низком содержании GC нуклеотидов (в диапазоне 53-63%), а также учитывая различное расположение последовательности относительно сайта мутации.
Участки последовательности-мишени (SEQ ID No: 26), таргетируемые двухцепочечными РНК по настоящему изобретению в форме siRNA, показаны на Фиг. 3А. В качестве положительного контроля РНКи в форме siRNA (siGNAO 1-CTRL, содержит SEQ ID No: 21) использовали ранее опубликованную последовательность siRNA, для которой было показано эффективное подавление экспрессии гена GNAO1 на уровне белка [Liu Z., Zhang J., Wu L. et al. Overexpression of GNAO1 correlates with poor progNosis in patients with gastric cancer and plays a role in gastric cancer cell proliferation and apoptosis. Int J Mol Med. 2014 Mar; 33(3):589-96. doi: 10.3892/ijmm.2013.1598].
В качестве отрицательного контроля РНКи в форме siRNA использовали РНК, способные вызывать РНК-интерференцию и направленные на подавление экспрессии экзогенных генов, например EGFP (siEGFP-1, содержит SEQ ID No: 29 и siEGFP-2, содержит SEQ ID No: 30), либо РНК с рандомизированной последовательностью (scramble), которые потенциально не должны влиять на экспрессию эндогенных генов в клетке, в том числе гена GNAO1. Scramble siRNA имеет тот же нуклеотидный состав, что и целевая последовательность, но порядок нуклеотидов в ней другой. Для дизайна таких scramble siRNA использовали программное обеспечение InvivoGen Scramble siRNA (https://www.invivogen.com/sirnawizard/scrambled.php, дата обращения 25.10.2022), находящееся в свободном доступе. За основу брали последовательность положительного контроля РНКи. В результате была получена последовательность scramble siRNA (siSCR, SEQ ID No: 22).
Ha 3'-конец смысловой и антисмысловой последовательностей каждой siRNA были добавлены 2 дополнительных нуклеотида для обеспечения большей стабильности синтезируемого РНК-дуплекса. В качестве выступающих динуклеотидов использовали либо два уридина (UU), либо два дезокситимидина (dTdT).
Описанные выше последовательности (SEQ ID No: 14) были синтезированы в виде одноцепочечных олигонуклеотидов РНК без дополнительных модификаций (ГенТерра, Россия). РНК-дуплексы получали путем отжига смысловой и антисмысловой пары олигонуклеотидов по стандартной технологии, описанной, например, в работе Elbashir, S., Harborth, J., Lendeckel, W. et al. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 411, 494 498 (2001). https://doi.org/10.1038/35078107.
Пример 2.2. Скрининг siRNA в гемизиготной тест-системе
Скрининг 4-х выбранных последовательностей siRNA проводили в гемизиготной тест-системе с использованием временной трансфекции клеток НЕК293Т. Для этого на первом этапе клетки НЕК293Т были трансфицированы плазмидами, кодирующими нормальный или мутантный аллель гена GNAO1 (pGNAO1-Myc и pGNAO1-G203R-FLAG соответственно). Трансфекцию проводили с помощью трансфицирующего агента LPEI, в соотношении LPEI к плазмидной ДНК 4:1. Через 24 часа после этого проводили второй этап трансфекции: к клеткам, рассеянным в 24-луночные планшеты, добавляли РНК-дуплексы, полученные путем отжига олигонуклеотидов с последовательностью, выбранной из SEQ ID No: 14, в концентрации 50 пмоль на 1 мкл трансфицирующего агента. В качестве трансфицирующего агента использовали Lipofectamine 2000 (Thermo Fisher). Наибольший эффект РНК-интерференции обычно наблюдается через 48-72 часа после попадания в клетку синтетических немодифицированных siRNA [https://www.thermofisher.com/ru/ru/home/references/ambion-tech-support/rnai-sirna/general-articles/top-ten- ways-to-optimize-sirna-delivery-in-cultured-cells.html, дата обращения 25.10.2022], поэтому через 48 часов после трансфекции проводили сбор образцов для дальнейшего анализа. Оценку аллель-селективности и эффективности работы РНК по настоящему изобретению в форме siRNA проводили методом Вестерн-блот (Фиг. 4).
Было обнаружено, что все исследованные siRNA подавляли экспрессию мутантного аллеля, причем siGNAO1-1488 (содержит SEQ ID No:l) демонстрировала наиболее выраженное подавление синтеза мутантного белка Gαo-G203R, сопоставимое с уровнем подавления мутантного аллеля положительным контролем РНКи. При этом все четыре кандидатные молекулы не затронули экспрессию аллеля дикого типа.
Эффективность и аллель-селективность работы siGNAO1-1488 (содержит SEQ ID No:l) были также подтверждены с помощью количественного анализа методом аллель-специфической кПЦР. Было показано, что siGNAO1-1488 снижает экспрессию мутантного аллеля GNAO1 до 37% относительно отрицательного контроля РНКи (siSCR), что сопоставимо с уровнем подавления при добавлении положительного контроля РНКи (34%). При этом экспрессия аллеля дикого типа сохраняется на высоком уровне (84%), таким образом siGNAO1-1488 демонстрирует высокую степень аллель-селективности и эффективности (Фиг. 5).
В итоге, на основе данных кПЦР и Вестерн-блота, для дальнейшего тестирования была выбрана кандидатная последовательность РНК siGNAO1-1488 (содержит SEQ ID No:1).
Пример 2.3. Тестирование эффективности работы siGNAO1-1488 в гетерозиготной тест-системе
Далее эффективность работы siGNAO1-1488 (содержит SEQ ID No: 1) проверяли в гетерозиготной тест-системе, которая более полно воспроизводит гетерозиготное состояние пациента с GNAO1-энцефалопатией. Для создания гетерозиготной тест-системы клетки НЕК293Т были ко-трансфицированы плазмидами, кодирующими как нормальный, так и мутантный аллель гена GNAO1 (плазмиды pGNAO1-Мус и pGNAO1-G203R-FLAG соответственно). Соотношение плазмид pGNAO1-Мус и pGNAO1-G203R-FLAG было 1:1.
Трансфекцию проводили в два этапа, с интервалом в 24 часа, как описано ранее. По результатам Вестерн-блота siGNAO1-1488 в гетерозиготной тест-системе демонстрировала аллель-селективное подавление экспрессии мутантного белка Gαo (Фиг. 6А). Анализ кПЦР показал, что экспрессия мутантного аллеля GNAO1 при воздействии siGNAO1-1488 снижается до 43%, что сопоставимо с работой положительного контроля (38%), при этом сохраняется высокий уровень экспрессии аллеля дикого типа (113%) (Фиг. 6Б).
Эффективность работы siGNAO1-1488 также была продемонстрирована при стабильно высоком уровне экспрессии GNAO1, что отражает динамику синтеза белка Gαo в нейрональных тканях. Для этого с помощью лентивирусной трансдукции с последующей селекцией зараженных клеточных клонов были получены линии клеток НЕК293Т, стабильно экспрессирующие нормальную или мутантную форму GNAO1. Далее полученные клетки, стабильно экспрессирующие мутантную или нормальную копию GNAO1, смешивали в соотношении Пик клеткам добавляли siRNA, как описано в примере 2.2. Методом Вестерн-блота было показано, что уровень экспрессии мутантного белка значительно снижался, в то время как уровень экспрессии белка Gαo дикого типа оставался высоким (Фиг. 6 В).
Пример 3. Дизайн и тестирование последовательностей РНК в форме shRNA
Пример 3.1. Дизайн shRNA
Последовательность каждой РНК (SEQ ID No: 5-15), представленной в форме коротких шпилечных РНК (shRNA), включала в себя 21 целевой нуклеотид, в составе направляющей цепи полностью комплементарных последовательности транскрипта гена GNAO1 с мутацией c.607G>A (SEQ ID No: 28).
Для дизайна последовательности РНК по настоящему изобретению, представленной в форме shRNA, использовали последовательность матричной РНК гена GNAO1, имеющуюся в свободном доступе в базе данных NCBI (идентификационный номер последовательности: NM 020988.2). Был выбран участок, размером 35 нуклеотидов, содержащий сайт мутации (c.607G>A, p.Gly203Arg), фланкированный 17-го нуклеотидами с каждой стороны. Далее с помощью программного обеспечения GPP Web Portal (https://portals.broadinstitute.org/gpp/public/seq/search, дата обращения 25.10.2022), находящегося в свободном доступе и предназначенного для дизайна и ранжирования последовательностей шпилечных РНК, были получены 15 последовательностей-кандидатов, размером 21 нуклеотид. Из представленных 15-ти выбрали 5 последовательностей, основываясь на наиболее высоком показателе рейтинга (intrinsic score) и учитывая различное расположение последовательности относительно сайта мутации. Дополнительно были сконструированы еще 4 последовательности. Участки последовательности-мишени (SEQ ID No: 26), таргетируемые двухцепочечными РНК по настоящему изобретению в каркасе shRNA показаны на Фиг. 3Б.
В качестве положительного контроля РНКи в форме shRNA (shGNAO1-CTRL, содержит SEQ ID No: 23) использовали последовательность shRNA, которая была предоставлена компанией Dharmacon в виде плазмидного вектора из набора TRC Lentiviral Human GNAO1 shRNA kit (Horizon Dharmacon, США, RHS4533-EG2775) и которая показала наибольшую эффективность в отношении подавления экспрессии гена GNAO1 человека.
В качестве отрицательного контроля РНКи в форме shRNA использовали РНК, способные вызывать РНК-интерференцию и направленные на подавление экспрессии экзогенных генов, например EGFP (shEGFP, содержит SEQ ID No: 31) [Horizon Dharmacon, RHS4459] и Luc (shLuc, содержит SEQ ID No: 32) [TRCN0000072256], либо РНК с рандомизированной последовательностью (scramble), которые потенциально не должны влиять на экспрессию эндогенных генов в клетке, в том числе гена GNAO1. РНК с рандомизированной последовательностью в форме shRNA (shSCR, содержит SEQ ID No: 24) была предоставлена в виде плазмидного вектора профессором David Sabatini [плазмида Addgene, #1864, Sarbassov D.D., Guertin D.A., Ali S.M., Sabatini D.M. Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex. Science 2005 Feb 18; 307(5712): 1098-101. 10.1126/science. 1106148, PubMed 15718470].
РНК в форме shRNA доставляли в клетки в виде кодирующей их ДНК в экспрессионных кассетах в составе плазмидных экспрессионных векторов или вирусных векторов. Для клонирования последовательностей РНК в форме shRNA был проведен дизайн олигонуклеотидов согласно протоколу Addgene pLKO. 1 protocol, находящемуся в открытом доступе [http://www.addgene.org/protocols/plko, дата обращения 25.10.2022]. Для клонирования ДНК, кодирующей РНК по настоящему изобретению в форме shRNA, использовали экспрессионный плазмидный вектор pLKO.1-TRC Cloning Vector (Addgene, #10878), как описано, например, в: Moffat J., Grueneberg D.A., Yang X. et al. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell. 2006 Mar 24; 124(6): 1283-98 или в Лунев Е.А., Шмидт А.А., Васильева С.Г. и др. Эффективная вирусная доставка в нейрональную культуру генетических конструкций для моделирования и генной терапии GNAO1 - энцефалопатии. Молекулярная биология, 2022; 56(4):604-618. Также для клонирования использовали плазмидный экспрессионный вектор на основе pAAV-LacZ (Agilent).
Для получения плазмидных экспрессионных конструкций на основе pAAV-LacZ (Agilent) в векторе pAAV-LacZ заменяли последовательность гена LacZ на последовательность гена EGFP методом безлигазного клонирования. Для этого вектор линеаризовали методом ПЦР с использованием пары праймеров:
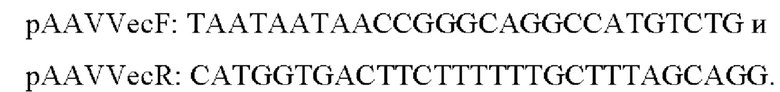
Последовательность гена зеленого флуоресцентного белка EGFP нарабатывали с матрицы pEGFP-C1 (Clontech) с использованием пары праймеров:
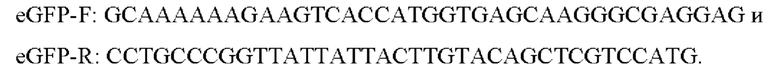
В результате была получена плазмида pAAV-eGFP. На основе данной плазмиды собирали конструкции, содержащие ДНК, кодирующую РНК по настоящему изобретению. Для создания векторов для экспрессии РНК в составе shRNA в вектор pAAV-eGFP вносили экспрессионную кассету, содержащую промотор U6 и последовательность ДНК, кодирующую соответствующую последовательность РНК. Субклонирование данной кассеты проводили с использованием вектора pLKO.1-TRC Cloning Vector (Addgene, #10878) согласно протоколу, описанному в статье: Лунев ЕА, Шмидт АА, Васильева СГ, и др. Эффективная вирусная доставка в нейрональную культуру генетических конструкций для моделирования и генной терапии GNAO1-энцефалопатии. Молекулярная биология, 2022; 56(4):604-618.
Схемы финальных плазмидных экспрессионных векторов, содержащих кассету экспрессии РНК в форме shRNA, приведены на Фиг. 8 и Фиг. 11.
ДНК, кодирующие РНК по настоящему изобретению в форме shRNA, имели одинаковый дизайн: пассажирская цепь длиной 21 нуклеотид, шестинуклеотидная петля с сайтом рестрикции XhoI и направляющая цепь длиной 21 нуклеотид.
Пример 3.2. Скрининг shRNA
На первом этапе эффективность и аллель-специфичность shRNA оценивали при введении плазмидных лентивирусных векторов, полученных как описано в примере 3.1., в клетки НЕК293Т в гемизиготной тест-системе. Клетки были ко-трансфицированы плазмидами, кодирующими нормальный или мутантный аллель гена GNAO1 (pGNAO1-Мус или pGNAO1-G203R-FLAG соответственно), плазмидой pEGFP-C1 для контроля трансфекции, а также лентивирусными плазмидами, экспрессирующими ДНК, кодирующую shRNA. Параллельно проводили трансфекцию клеток лентивирусными плазмидами, экспрессирующими ДНК, кодирующую последовательности положительного или отрицательного контролей РНКи. Трансфекцию осуществляли с помощью трансфицирующего агента LPEI, в соотношении LPEI к плазмидной ДНК 4:1, в 24-луночном формате. Через 72 часа после трансфекции проводили сбор образцов для количественного анализа аллель-селективности и эффективности работы shRNA методом аллель-специфической кПЦР (Фиг. 9). Все кандидатные молекулы продемонстрировали снижение уровня экспрессии мутантного аллеля гена GNAO1 по сравнению с аллелем дикого типа. По результатам анализа 9-ти shRNA были выбраны наиболее перспективные кандидаты: shGNAO1-1489 (содержит SEQ ID No: 7) и shGNAO1-1500 (содержит SEQ ID No: 11). Эти shRNA демонстрировали оптимальное соотношение эффективности и аллель-селективности, а именно подавляли экспрессию мутантного аллеля GNAO1 (до 21% относительно отрицательного контроля РНКи (shSCR) в случае shGNAO1-1489 и до 40% относительно shSCR в случае shGNAO1-1500), при этом сохранялся высокий уровень экспрессии аллеля дикого типа GNAO1 (50% и 110% относительно shSCR для shGNAO1-1489 и shGNAO1-1500 соответственно).
Пример 3.3. Тестирование эффективности работы shGNAO1-1489 и shGNAO1-1500 в гетерозиготной тест-системе
С выбранными shRNA были собраны лентивирусные частицы согласно стандартному протоколу с использованием описанных в Примере 3.1 плазмидных экспрессионных векторов, и проведена оценка их работы в гетерозиготной тест-системе со стабильно высоким уровнем экспрессии гена GNAO1 человека. Для этого были получены линии НЕК293Т, стабильно экспрессирующие как нормальную, так и мутантную форму GNAO1. Анализ работы лентивирусов, экспрессирующих shGNAO1-1489 (содержит SEQ ID No:7) и shGNAO1-1500 (содержит SEQ ID No: 11), проводили методом Вестерн-блот через 7 дней после заражения стабильных линий НЕК293Т. Было показано, что shGNAO1-1489 отличается высокой эффективностью, а именно подавляет экспрессию мутантного аллеля гена GNAO1 на уровне, сопоставимом с положительным контролем РНКи (shGNAO1-CTRL). При этом селективность выше у кандидата shGNAO1-1500, который не затрагивает работу аллеля дикого типа, но обладает менее выраженным эффектом подавления мутантного аллеля (Фиг. 10).
Далее работу shGNAO1-1489 и shGNAO1-1500 оценивали при введении в клетки кодирующей их ДНК с помощью описанных в Примере 3.1. плазмидных экспрессионных ААВ векторов, в гетерозиготной тест-системе на основе клеток НЕК293Т. Схема плазмидного экспрессионного ААВ вектора, используемого для клонирования и тестирования shRNA, показана на Фиг. 11. Клетки НЕК293Т были ко-трансфицированы плазмидами pGNAO1-Myc и pGNAO1-G203R-FLAG, а также плазмидными ААВ векторами, экспрессирующими shGNAO1-1489 или shGNAO1-1500, как описано в Примере 3.2. Параллельно проводили трансфекцию клеток плазмидными ААВ векторами, экспрессирующими последовательности положительного или отрицательного контролей РНКи в форме shRNA. Сбор образцов для анализа проводили через 72 часа. Методом Вестерн-блот было показано, что shGNAO1-1489 и shGNAO1-1500 при введении с помощью плазмидных экспрессионных ААВ векторов характеризуются высоким уровнем эффективности и селективности (Фиг. 12А). По результатам аллель-специфической кПЦР было показано, что shGNAO1-1489 и shGNAO1-1500 снижают экспрессию мутантного аллеля GNAO1 до 32% и 31% соответственно относительно отрицательного контроля РНКи (shSCR). При этом уровень экспрессии аллеля дикого типа GNAO1 остается максимально высоким (125% и 102% относительно shSCR) (Фиг. 12Б).
Для исследования возможности повышения эффективности и аллель-селективности shRNA были проведены модификации как собственно смысловой (пассажирской) последовательности shGNAO1-1489 (SEQ ID No: 7), так и других элементов содержащего ее каркаса shRNA (SEQ ID No: 33). Исходная последовательность пассажирской цепи shGNAO1-1489 (SEQ ID No: 7) была включена либо в состав исходного каркаса с образованием последовательности shGNAO1-1489-transcript (SEQ ID No: 33), либо в модифицированный каркас с образованием последовательности shGNAO1-1489-v2-transcript (SEQ ID No: 18). Модифицированный вариант пассажирской последовательности shGNAO1- 1489-v3 (SEQ ID No: 14) был включен в состав модифицированного каркаса с образованием последовательности shGNAO1-1489-v3-transcript (SEQ ID No: 19). Модифицированный вариант пассажирской последовательности shGNAO1-1489-v4 (SEQ ID No: 15) был включен в состав модифицированного каркаса с образованием последовательности shGNAO1-1489-v4-transcript (SEQ ID No: 20). Оценку работы новых модифицированных shRNA проводили при введении в клетки кодирующей их ДНК с помощью плазмидных экспрессионных ААВ векторов, в гетерозиготной тест-системе на основе клеток НЕК293Т, как описано в Примере 3.2. По результатам Вестерн-блот анализа было показано, что пассажирская цепь shGNAO1-1489-v4 (SEQ ID No: 15) в составе каркаса shGNAO1-1489-v4-transcript (SEQ ID No: 20) характеризуется наибольшей эффективностью, приводя к практически полному исчезновению мутантного белка Gαo-G203R (Фиг. 13А). По результатам аллель-специфической кПЦР shRNA с каркасом shGNAO1-1489-v2-transcript (SEQ ID No: 18), shGNAO1-1489-v3-transcript (SEQ ID No: 19) и shGNAO1-1489-v4-transcript (SEQ ID No: 20) продемонстрировали высокую эффективность работы, снижая экспрессию мутантного аллеля GNAO1 до 50%, 40% и 27% соответственно, относительно отрицательного контроля РНКи (shSCR). При этом в случае с shGNAO1-1489-v2-transcript (SEQ ID No: 18) и shGNAO1-1489-v3-transcript (SEQ ID No: 19) экспрессия аллеля дикого типа GNAO1 сохранялась на максимальном уровне (100%) (Фиг. 13Б).
Пример 3.4. Тестирование эффективности работы shGNAO1-1489 и shGNAO1-1500, доставленных с помощью вирусных частиц ААВ в культуру пациент-специфических нейронов
На следующем этапе тестировали эффективность работы shGNAO1-1489 (содержит SEQ ID No:7) и shGNAO1-1500 (содержит SEQ ID No: 11) в условиях, имитирующих генную терапию. В качестве модели GNAO1-энцефалопатии использовали нейроны, полученные с помощью направленной дифференцировки индуцированных плюрипотентных стволовых клеток, полученных в свою очередь из биоптата кожи пациента с de novo гетерозиготным вариантом GNAO1 c.607G>A. Такие нейроны характеризуются высокой эндогенной экспрессией GNAO1, что приводит к образованию белка Gαo на уровне, хорошо детектируемом методами иммунофлуоресценции (Фиг. 17А). Ранее нами было показано, что здоровый и мутантный аллели гена GNAO1 в пациент-специфических нейронах экспрессируются на одинаковом уровне, в соотношении, близком к 1:1 [патент РФ 2777663]. Экспрессия маркера β-III-тубулина в дифференцированных клетках подтверждает их нейрональный статус (Фиг. 17А). Для доставки shGNAO1-1489 и shGNAO1-1500 в культуру пациент-специфических нейронов использовали вирусную трансдукцию посредством аденоассоциированных вирусов. Для наработки вирусных частиц ААВ использовали экспрессионные ААВ-векторы, кодирующие shGNAO1-1489 и shGNAO1-1500. Продукцию и очистку вирусных частиц ААВ проводили согласно стандартным методикам, описанным, например, в: Лунев ЕА, Шмидт АА, Васильева СГ и др. Эффективная вирусная доставка в нейрональную культуру генетических конструкций для моделирования и генной терапии GNAO1-энцефалопатии. Молекулярная биология, 2022;56(4):604-618. Трансдукцию проводили аналогично описанным ранее методам [Там же]. Анализ экспрессии аллелей GNAO1 проводили методом аллель-специфической кПЦР (Фиг. 17Б и 17 В). Было показано, что положительный контроль РНКи (CTRL) в нейронах снижает экспрессию GNAO1 до 28% (Фиг. 17Б) относительно отрицательного контроля РНКи (shSCR); при этом положительный контроль РНКи (shGNAO1-CTRL) не меняет соотношение здорового и мутантного транскриптов GNAO1 (Фиг. 17 В). Несмотря на то, что аллель-селективные shGNAO1-1489 и shGNAO1-1500 в меньшей степени влияют на общий уровень мРНК GNAO1 (Фиг. 17Б), их экспрессия в нейронах приводит к увеличению соотношения уровней мРНК дикого типа и мутантного варианта в 1,7 и 5,7 раз соответственно относительно отрицательного контроля РНКи, принятого за 1(Фиг. 17 В).
Пример 4. Дизайн и тестирование последовательностей РНК по настоящему изобретению в каркасе miRNA
Пример 4.1. Дизайн miRNA
Перспективные кандидатные последовательности РНК shGNAO1-1489 (SEQ ID No:7) и shGNAO1-1500 (SEQ ID No: 11) были также протестированы в каркасе искусственной микроРНК (miRNA). Подход на основе miRNA обладает несколькими преимуществами. Во-первых, кодирующая miRNA ДНК может экспрессироваться с промотора РНК-полимеразы II, что позволяет проводить исследования тканеспецифичной супрессии через РНК-интерференцию, используя тканеспецифичные промоторы. Во-вторых, ДНК, кодирующая РНК, способную вызывать РНК-интерференцию, может быть помещена в 3'-нетранслируемую область репортерного гена для непосредственного мониторинга ее экспрессии. И в-третьих, РНК в каркасе miRNA менее склонны вызывать побочные токсичные эффекты в организме, так как не нарушают процессов эндогенной РНК-интерференции [Fellmann С, Hoffmann Т, Sridhar V, et al. An optimized microRNA backbone for effective single-copy RNAi. Cell Rep.2013 Dec 26; 5(6): 1704-13. doi: 10.1016/j.celrep.2013.11.020].
В качестве каркаса искусственной miRNA была выбрана miR-30, конфигурация петель которой соответствует недавно открытому «правилу подсчета петель» (от англ. «loop-counting rule»), обеспечивая точное разрезание pre-miRNA ферментом Dicer и уменьшая нецелевые эффекты [Gu S., Jin L., Zhang Y. et al. The loop position of shRNAs and pre-miRNAs is critical for the accuracy of dicer processing in vivo. Cell. 2012 Nov 9; 151(4):900-911. doi: 10.1016/j.cell.2012.09.042].
Дизайн miR-30 предполагает удлинение пассажирской и направляющей последовательностей shRNA на один нуклеотид, как описано, например, в [Chang К., Marran К., Valentine A. et al. Creating an miR30-based shRNA vector. Cold Spring Harb Protoc. 2013 Jul 1; 2013(7):631-5. doi: 10.1101/pdb.prot075853]. Поэтому последовательность РНК для РНК-интерференции в составе miR-30 имеет длину 22 нуклеотида. Для простоты понимания номенклатура, базирующаяся на положении первого нуклеотида пассажирской последовательности РНК относительно первого нуклеотида мРНК гена GNAO1, была сохранена, и последовательности РНК в составе miRNA имеют следующие названия: miR30-GNAO1-1489(-1) (SEQ ID No: 16) и miR30-GNAO1-1500(-1) (SEQ ID No: 17). Дизайн последовательности положительного контроля РНКи в форме miRNA (miR30-GNAO 1 -CTRL, SEQ ID No: 25) проводили аналогичным образом на основе соответствующего положительного контроля РНКи в форме shRNA (shGNAO1-CTRL, SEQ ID No: 23). В качестве отрицательного контроля РНКи в форме miRNA использовали РНК с рандомизированной последовательностью (scramble), которые потенциально не должны влиять на экспрессию эндогенных генов в клетке, в том числе гена GNAO1.
Для клонирования последовательностей РНК miR30-GNAO1-1489(-1) и miR30-GNAO1-1500(-1) в виде кодирующих эти последовательности ДНК-вставок был проведен дизайн олигонуклеотидов. Транскрибируемые с ДНК-вставки miRNA имели одинаковый дизайн типа «стебель-петля» (Фиг. 14) и включали в себя:
- пассажирскую (смысловую) последовательность РНК длиной 22 нуклеотида;
- последовательность петли miR-30 (GUGAAGCCACAGAUG);
- направляющую (антисмысловую) последовательность РНК длиной 22 нуклеотида;
- нуклеотиды апикального и базального соединений, служащие сайтами расщепления ферментами Drosha и Dicer;
- дополнительные фланкирующие последовательности.
Участки последовательности-мишени (SEQ ID No: 26), таргетируемые двухцепочечными РНК по настоящему изобретению в каркасе miRNA показаны на Фиг. 3В.
Клонирование в плазмидные экспрессионные векторы проводили по сайтам рестрикции Agel и EcoRI, поэтому при дизайне олигонуклеотидов на 5'- и 3'-конец соответственно каждой ДНК-вставки были добавлены указанные сайты рестрикции. То, что РНК с выбранной последовательностью формирует правильную вторичную структуру со шпилькой, было предварительно подтверждено in silico с помощью программного обеспечения uFold, находящегося в открытом доступе [https://ufold.ics.uci.edu/, дата обращения 25.10.2022].
Для клонирования использовали два типа конфигурации вектора на основе плазмидного экспрессионного ААВ вектора: #1 вектор, содержащий ДНК, кодирующую РНК по настоящему изобретению, под контролем промотора РНК-полимеразы III (U6), и #2 - вектор, содержащий ДНК, кодирующую РНК по настоящему изобретению, в 3'-нетранслируемой области репортерного гена eGFP под контролем промотора РНК-полимеразы II (CMV).
Плазмидные векторы конфигурации #1 были созданы на основе вектора pAAV-eGFP, описанного в Примере 3.1. Для создания векторов конфигурации #2 в вектор pAAV-eGFP в 3'-нетранслируемую область гена eGFP вносили последовательность полилинкера, содержащую сайты эндонуклеаз рестрикции AgeI и EcoRI, для удобства клонирования ДНК, кодирующей РНК по настоящему изобретению в каркасе miRNA.
Пример 4.1. Тестирование эффективности работы miR30-GNAO1-1489(-1) в гетерозиготной тест-системе при введении в составе лентивирусного плазмидного вектора
Эффективность и селективность работы miR30-GNAO1-1489(-1) (содержит SEQ ID No: 16) оценивали при введении его в клетки с помощью плазмидного лентивирусного экспрессионного вектора, в гетерозиготной тест-системе посредством временной трансфекции клеток НЕК293Т. Трансфекцию клеток проводили как описано в Примере 3.2. Образцы собирали на анализ через 72 часа после трансфекции. С помощью метода Вестерн-блот было показано, что miR30-GNAO1-1489(-1) эффективно и селективно подавляет экспрессию мутантного белка Gαo-G203R (Фиг. 15А).
По данным количественного анализа методом аллель-специфической кПЦР miR30-GNAO1-1489(-1) демонстрировал подавление экспрессии мутантного аллеля GNAO1 на таком же уровне, как и положительной контроль РНКи - miR30-GNAO1-CTRL (6%). При этом экспрессия нормального аллеля сохранялась на высоком уровне относительно отрицательного контроля РНКи (57%) (Фиг. 15Б).
Пример 4.2. Тестирование эффективности работы miR30-GNAO1-1489(-1) и miR30-GNAO1-1500(-1) в гетерозиготной тест-системе при введении в составе ААВ плазмидного вектора.
В данном примере использовали два типа конфигурации плазмидного экспрессионного ААВ вектора, описанные в Примере 4.1. Тестирование проводили в гетерозиготной тест-системе посредством временной трансфекции клеток НЕК293Т, как описано в Примере 3.2. Через 72 часа после трансфекции проводили сбор образцов. Результаты Вестерн-блота показали, что в лизатах клеток, обработанных плазмидными экспрессионными векторами обоих вышеупомянутых типов конфигурации, несущих ДНК, кодирующую либо miR30-GNAO1-1489(-1) (содержит SEQ ID No: 16), либо miR30-GNAO1-1500(-1) (содержит SEQ ID No: 17), не наблюдается экспрессии мутантного белка Gαo-G203R, тогда как экспрессия Gαo дикого типа отчетливо детектируется, что говорит об эффективной и аллель-селективной работе miR30-GNAO1-1489(-1) и miR30-GNAO1-1500(-1) (Фиг. 16А).
Эффективность и аллель-селективность работы miR30-GNAO1-1489(-1) и miR30-GNAO1-1500(-1) были также подтверждены с помощью количественного анализа методом аллель-специфической кПЦР (Фиг. 16Б). Было показано, что miR30-GNAO1-1489(-1) под контролем промотора РНК-полимеразы III (U6) снижает экспрессию мутантного аллеля GNAO1 до 17% относительно отрицательного контроля РНКи (miR30-SCR), при этом экспрессия аллеля дикого типа сохраняется на очень высоком уровне (161%). Для miR30-GNAO1-1500(-1), находящегося под контролем промотора РНК-полимеразы III (U6), было показано снижение экспрессии мутантного аллеля GNAO1 до 47% относительно отрицательного контроля РНКи (shSCR), при этом детектировался очень высокий уровень экспрессии аллеля дикого типа (249%). В случае, когда miR30-GNAO1-1489(-1) и miR30-GNAO1-1500(-1) находились в 3'-нетранслируемой области репортерного гена EGFP под контролем промотора РНК-полимеразы II (CMV) наблюдался более выраженный эффект подавления экспрессии мутантного аллеля гена GNAO1 (до 13% и 20% относительно отрицательного контроля РНКи (miR30-SCR) для miR30-GNAO1-1489(-1) и miR30-GNAO1-1500(-1) соответственно). При этом экспрессия аллеля дикого типа GNAO1 оставалась на высоком уровне (104% и 122% для miR30-GNAO1-1489(-1) и miR30-GNAO1-1500(-1) соответственно).
Все публикации, патенты и заявки на патенты включены в настоящий документ посредством ссылки. Хотя в вышеприведенном описании это изобретение было описано в отношении некоторых предпочтительных вариантов его осуществления, и многие детали были изложены в целях иллюстрации, для специалистов в данной области техники будет очевидно, что изобретение допускает дополнительные варианты осуществления и что некоторые детали, описанные в данном документе, могут значительно изменяться без отклонения от сущности изобретения.
Использование терминов в единственном числе в контексте описания изобретения должно толковаться как охватывающее как единственное, так и множественное число, если иное не указано в данном документе или явно не противоречит контексту. Термины «состоящий из», «имеющий», «включающий» и «содержащий» следует толковать как неограничивающие термины, т.е. означающие «включая, но не ограничиваясь», если не указано иное. Все способы, описанные в данном документе, могут выполняться в любом подходящем порядке, если иное не указано в данном документе или иным образом явно не противоречит контексту. Использование любых и всех примеров или иллюстративного языка (например, «такой как»), представленных в данном документе, предназначено просто для лучшего описания изобретения и не налагает ограничения на объем изобретения, если иное не заявлено. Никакие формулировки в описании не следует истолковывать как указывающие на какой-либо не заявленный элемент как существенный для практического применения изобретения.
Здесь описаны варианты осуществления этого изобретения, включая лучший из известных изобретателям способов осуществления изобретения. Разновидности этих вариантов осуществления могут стать очевидными для специалистов в данной области техники после прочтения предшествующего описания. Авторы ожидают, что квалифицированные специалисты будут использовать такие варианты в зависимости от обстоятельств, и авторы предполагают, что изобретение будет реализовано на практике иначе, чем конкретно описано в данном документе. Соответственно, это изобретение включает в себя все модификации и эквиваленты признаков, изложенных в прилагаемой формуле изобретения, как это разрешено действующим законодательством. Более того, любая комбинация вышеописанных признаков во всех их возможных вариациях охватывается изобретением, если иное не указано в данном документе или иным образом явно не противоречит контексту.
Заявитель просит рассмотреть представленные материалы заявки «Двухцепочечная РНК, способная снижать экспрессию мутантного аллеля c.607G>A гена GNAO1 человека» на предмет выдачи патента на изобретение.
--->
<ST26SequenceListing dtdVersion="V1_3" fileName="Марлин Биотех
GNAO1_перечень.xml" softwareName="WIPO Sequence"
softwareVersion="2.2.0" productionDate="2022-11-16">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2022129743</ApplicationNumberText>
<FilingDate>2022-11-16</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference> </ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2022129743</ApplicationNumberText>
<FilingDate>2022-11-16</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Общество с ограниченной
ответственностью «Марлин Биотех»</ApplicantName>
<ApplicantNameLatin>Marlin Biotech LLC</ApplicantNameLatin>
<InventionTitle languageCode="ru">Двухцепочечная РНК, способная
снижать экспрессию мутантного аллеля с.607G>A гена GNAO1
человека</InventionTitle>
<SequenceTotalQuantity>33</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>caggctgtttgacgtcaga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q3">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tgtttgacgtcagaggcca</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gacgtcagaggccagcgat</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q5">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tcagaggccagcgatctga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="5">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tcaggctgtttgacgtcagag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="6">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q7">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>caggctgtttgacgtcagagg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="7">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q55">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aggctgtttgacgtcagaggc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="8">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q57">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ggctgtttgacgtcagaggcc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="9">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q59">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ctgtttgacgtcagaggccag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="10">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q61">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ttgacgtcagaggccagcgat</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="11">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q63">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cgtcagaggccagcgatctga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="12">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q65">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gtcagaggccagcgatctgaa</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="13">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q67">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tcagaggccagcgatctgaac</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="14">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q69">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aggctgtttgacgtcagaagc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="15">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q71">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ggctgtttgacgtcagagacc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="16">
<INSDSeq>
<INSDSeq_length>22</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..22</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q73">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aaggctgtttgacgtcagaggc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="17">
<INSDSeq>
<INSDSeq_length>22</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..22</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q75">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ccgtcagaggccagcgatctga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="18">
<INSDSeq>
<INSDSeq_length>56</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..56</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q35">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aggctgtttgacgtcagaggccctgacccagcctctgacgtcaaacagcctt
tttt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="19">
<INSDSeq>
<INSDSeq_length>56</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..56</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q37">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aggctgtttgacgtcagaagccctgacccagcttctgacgtcaaacagcctt
tttt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="20">
<INSDSeq>
<INSDSeq_length>57</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..57</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q78">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aggctgtttgacgtcagagacccctgacccaggtctctgacgtcaaacagcc
ttttt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="21">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q39">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cccacttcacattcaagaa</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="22">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q41">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gcacaatacctatcaccta</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="23">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q43">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cgctcacccaacaaagaaata</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="24">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q45">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cctaaggttaagtcgccctcg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="25">
<INSDSeq>
<INSDSeq_length>22</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..22</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q47">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>acgctcacccaacaaagaaata</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="26">
<INSDSeq>
<INSDSeq_length>3332</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..3332</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>mRNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q81">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Homo sapiens</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gctgacgtcagctggcggcccgcggagccgggtgcgcagagccgctggcgca
ctcgcgcgctgcgctggcctcctccccgccccgggcccaggcgcgcgcgcgcttgttcgctctcccattt
tttttttcccctccctccctgcctctctctctctccctctccctcgagctcccggctggctgcggctccc
tggcgctctccctctctctccggtaggctcaccgagcgatgcgagctctgggagacagcgacgccgcctc
ccgctagagacctgcccctcggcccggccccctgcccaaccctgcccagcgcgcgggggtcggcgaaggc
gccgcggacgcaccgacggctgaagagcggcgatgcacatgcactagcagcaccccctaactcactccct
ccacatcccgcgccgccgccgccgcctcctccacctcctcctccgccgccgccgcctcctcctcctccgg
cagccgcggcagcaggacccaccctgccccccaccccaccctctgtcggctccggctgcggctccagcct
cgactattattttatttattttgggtcgtgcacaagcctcagtgcctgcagtccgcgcctcctcggcccg
cgggcgcctcctcccttggctccggagccccagaccccggccaccctcgattcgacaaccccagacccct
gccagctgccgcgagtctccgctgctggaatcttgttagcggctgtctttttggagggttctggtttccc
gacatttttgtttccagcccaggagaggatatcgtgattttccccccttgagcccaggctctgctctctg
ggggggtggggggcgctccaagccggggagccgtgccagccgagtcgtgcgggctgtggcagggaagggg
ccaccatgggatgtactctgagcgcagaggagagagccgccctcgagcggagcaaggcgattgagaaaaa
cctcaaagaggatggcatcagcgccgccaaagacgtgaaattactcctgctcggggctggagaatcagga
aaaagcaccattgtgaagcagatgaagatcatccatgaagatggcttctccggagaagacgtgaaacagt
acaagcctgttgtctacagcaacactatccagtccctggcagccatcgtccgggccatggacactttggg
catcgaatatggtgataaggagagaaaggctgacgccaagatggtgtgtgatgtggtgagtcggatggaa
gacaccgagcccttctctgcagagctgctttctgccatgatgcggctctggggcgactcaggaatccaag
agtgcttcaaccggtcccgggagtatcagctcaacgactctgccaaatactacctggacagcctggatcg
gattggggccgccgactaccagcccaccgagcaggacatcctccgaaccagggtcaaaaccactggcatc
gtagaaacccacttcacattcaagaacctccacttcaggctgtttgacgtcagaggccagcgatctgaac
gcaagaagtggatccattgcttcgaggacgtcacggccatcattttctgtgtcgcgctcagcggctatga
ccaggtgctccacgaagacgaaaccacgaaccgcatgcacgagtctctcatgctcttcgactccatctgt
aacaacaagttcttcatcgatacctccatcattctcttcctcaacaagaaagatctctttggcgagaaga
tcaagaagtcacctttgaccatctgctttcctgaatacacaggccccaatacctatgaagacgcagccgc
ctacatccaagcacaatttgaaagcaaaaaccgctcacccaacaaagaaatatattgtcacatgacttgt
gccacagacacgaataacatccaggtggtgttcgacgccgtcaccgacatcatcattgccaacaacctcc
ggggctgcggcttgtactgacctcttgtcctgtatagcaacctatttgactgcttcacggactctttgct
gttgacgttgatctcctggtagcatgaccttttggcctttgtaagacacacagcctttctgtatcaagcc
cctgtctaacctacgaccccagagtgactgacggctgtgtatttctgtagaatgctgtagaatccggttt
tagttgagtcttcacatttagaatttgaaaggaaaaacagaacatttctcatgtgctttgtagcttaaaa
aaaggaaaactcaccatttcatccatatttctttttcttgcaaaacaaacatacccatctgcaccgacgc
ctgcccccgtcccacctcgcggcgcagcccctggagctcctgctggggggcagggcccatctgccgcctg
gggctccggtgcggtgcactggtccgaggagtgtgctcaggccgggctttccagaaggccacaggccact
gcaaaccctgctgcctgccggccgcccagacaccacagacgccacctggcggccctaagccactcccgcc
acacgctcactccaggttcgtatagaaaaagcacagctctcactggccagtagctgccaaaaagaacacc
catcgtataaggaaatccagccccatagaaaacaatctgcccaccgaccccctttctaaggatagccttt
gtaggggttttttggttccttttttttttttttcagtttttgtttttacttttgtcccgccgagtactat
gtattaccagccccccaccccacagatcagttttcctagtctttgggaaacgggttgtgttttttttcct
ttgccgtgttggatttccggtcccggctgacacctcccttcgtgtgtgcgcccctccccacccctgcctt
tgtttcggtgccctgtgtccgctctctcatgtggggatgtttgtgctgcagacaaaaagaacaaaaaaaa
atttttaaataccacaagatggaaaaaaaaaacaaaaaaatttaaaaagatggaatgtagtgtttacatt
aaagccgccgttttcatgaccaatcctatgtcatttctactttgactaatacccaaaccctaaacctctt
cacggtttcacttttaagacttaatctttgctgaagaagaccagtcacagccatttcaaataaagaggag
gcaaaaagacaaaacacacaaaaaattaaaaaaaaaaacttaaaaaaaaaaggaaaaaagaaaaaaaagc
ccacgggtctttgtaagcctttctattcccaatcggtgaggtttctgtgatatcttacacactgccagtc
tacttctctgactaatggaaaattcctgtgtagctcaataaaagagaatgtttgtctgta</INSDSeq_
sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="27">
<INSDSeq>
<INSDSeq_length>3332</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..3332</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>mRNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q83">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Homo sapiens</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gctgacgtcagctggcggcccgcggagccgggtgcgcagagccgctggcgca
ctcgcgcgctgcgctggcctcctccccgccccgggcccaggcgcgcgcgcgcttgttcgctctcccattt
tttttttcccctccctccctgcctctctctctctccctctccctcgagctcccggctggctgcggctccc
tggcgctctccctctctctccggtaggctcaccgagcgatgcgagctctgggagacagcgacgccgcctc
ccgctagagacctgcccctcggcccggccccctgcccaaccctgcccagcgcgcgggggtcggcgaaggc
gccgcggacgcaccgacggctgaagagcggcgatgcacatgcactagcagcaccccctaactcactccct
ccacatcccgcgccgccgccgccgcctcctccacctcctcctccgccgccgccgcctcctcctcctccgg
cagccgcggcagcaggacccaccctgccccccaccccaccctctgtcggctccggctgcggctccagcct
cgactattattttatttattttgggtcgtgcacaagcctcagtgcctgcagtccgcgcctcctcggcccg
cgggcgcctcctcccttggctccggagccccagaccccggccaccctcgattcgacaaccccagacccct
gccagctgccgcgagtctccgctgctggaatcttgttagcggctgtctttttggagggttctggtttccc
gacatttttgtttccagcccaggagaggatatcgtgattttccccccttgagcccaggctctgctctctg
ggggggtggggggcgctccaagccggggagccgtgccagccgagtcgtgcgggctgtggcagggaagggg
ccaccatgggatgtactctgagcgcagaggagagagccgccctcgagcggagcaaggcgattgagaaaaa
cctcaaagaggatggcatcagcgccgccaaagacgtgaaattactcctgctcggggctggagaatcagga
aaaagcaccattgtgaagcagatgaagatcatccatgaagatggcttctccggagaagacgtgaaacagt
acaagcctgttgtctacagcaacactatccagtccctggcagccatcgtccgggccatggacactttggg
catcgaatatggtgataaggagagaaaggctgacgccaagatggtgtgtgatgtggtgagtcggatggaa
gacaccgagcccttctctgcagagctgctttctgccatgatgcggctctggggcgactcaggaatccaag
agtgcttcaaccggtcccgggagtatcagctcaacgactctgccaaatactacctggacagcctggatcg
gattggggccgccgactaccagcccaccgagcaggacatcctccgaaccagggtcaaaaccactggcatc
gtagaaacccacttcacattcaagaacctccacttcaggctgtttgacgtcggaggccagcgatctgaac
gcaagaagtggatccattgcttcgaggacgtcacggccatcattttctgtgtcgcgctcagcggctatga
ccaggtgctccacgaagacgaaaccacgaaccgcatgcacgagtctctcatgctcttcgactccatctgt
aacaacaagttcttcatcgatacctccatcattctcttcctcaacaagaaagatctctttggcgagaaga
tcaagaagtcacctttgaccatctgctttcctgaatacacaggccccaatacctatgaagacgcagccgc
ctacatccaagcacaatttgaaagcaaaaaccgctcacccaacaaagaaatatattgtcacatgacttgt
gccacagacacgaataacatccaggtggtgttcgacgccgtcaccgacatcatcattgccaacaacctcc
ggggctgcggcttgtactgacctcttgtcctgtatagcaacctatttgactgcttcacggactctttgct
gttgacgttgatctcctggtagcatgaccttttggcctttgtaagacacacagcctttctgtatcaagcc
cctgtctaacctacgaccccagagtgactgacggctgtgtatttctgtagaatgctgtagaatccggttt
tagttgagtcttcacatttagaatttgaaaggaaaaacagaacatttctcatgtgctttgtagcttaaaa
aaaggaaaactcaccatttcatccatatttctttttcttgcaaaacaaacatacccatctgcaccgacgc
ctgcccccgtcccacctcgcggcgcagcccctggagctcctgctggggggcagggcccatctgccgcctg
gggctccggtgcggtgcactggtccgaggagtgtgctcaggccgggctttccagaaggccacaggccact
gcaaaccctgctgcctgccggccgcccagacaccacagacgccacctggcggccctaagccactcccgcc
acacgctcactccaggttcgtatagaaaaagcacagctctcactggccagtagctgccaaaaagaacacc
catcgtataaggaaatccagccccatagaaaacaatctgcccaccgaccccctttctaaggatagccttt
gtaggggttttttggttccttttttttttttttcagtttttgtttttacttttgtcccgccgagtactat
gtattaccagccccccaccccacagatcagttttcctagtctttgggaaacgggttgtgttttttttcct
ttgccgtgttggatttccggtcccggctgacacctcccttcgtgtgtgcgcccctccccacccctgcctt
tgtttcggtgccctgtgtccgctctctcatgtggggatgtttgtgctgcagacaaaaagaacaaaaaaaa
atttttaaataccacaagatggaaaaaaaaaacaaaaaaatttaaaaagatggaatgtagtgtttacatt
aaagccgccgttttcatgaccaatcctatgtcatttctactttgactaatacccaaaccctaaacctctt
cacggtttcacttttaagacttaatctttgctgaagaagaccagtcacagccatttcaaataaagaggag
gcaaaaagacaaaacacacaaaaaattaaaaaaaaaaacttaaaaaaaaaaggaaaaaagaaaaaaaagc
ccacgggtctttgtaagcctttctattcccaatcggtgaggtttctgtgatatcttacacactgccagtc
tacttctctgactaatggaaaattcctgtgtagctcaataaaagagaatgtttgtctgta</INSDSeq_
sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="28">
<INSDSeq>
<INSDSeq_length>36</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..36</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>mRNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q85">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Homo sapiens</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tcaggctgtttgacgtcagaggccagcgatctgaac</INSDSeq_sequen
ce>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="29">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q87">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ggcacaagctggagtacaa</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="30">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q89">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>acttcaaggaggacggcaa</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="31">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q91">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tacaacagccacaacgtctat</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="32">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q93">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>acgctgagtacttcgaaatgt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="33">
<INSDSeq>
<INSDSeq_length>55</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..55</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q95">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ggaggctgtttgacgtcagaggcctcgaggcctctgacgtcaaacagccttt
ttt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕРАПЕВТИЧЕСКИЕ СОЕДИНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2015 |
|
RU2711147C2 |
| Технология получения мышей с гуманизированным участком Gnao1 в области однонуклеотидного полиморфизма rs587777057 для тестирования РНК-терапии для GNAO1 c.607 G>A пациентов | 2021 |
|
RU2757121C1 |
| ЛИНИИ СКОНСТРУИРОВАННЫХ КЛЕТОК-ПРОДУЦЕНТОВ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2823068C2 |
| МОЛЕКУЛА RNAI, НАЦЕЛИВАЮЩАЯ ТИМИДИЛАТСИНТАЗУ И ЕЕ ПРИМЕНЕНИЕ | 2010 |
|
RU2535993C2 |
| Средство для ингибирования репликации вируса SARS-CoV-2, опосредованного РНК-интерференцией | 2020 |
|
RU2733361C1 |
| УСОВЕРШЕНСТВОВАНИЕ ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИВИЧ ТЕРАПИИ | 2013 |
|
RU2533817C1 |
| ВАРИАНТ СРЕДСТВА ДЛЯ RNAi | 2018 |
|
RU2789647C2 |
| Молекула РНК-проводника sgRNA для внесения мутаций в консервативный участок промоторной области гена PPD-D1 мягкой пшеницы с применением системы редактирования генома CRISPR/Cas9 | 2024 |
|
RU2822358C1 |
| ДВУХЦЕПОЧЕЧНЫЙ ОЛИГОНУКЛЕОТИД, СПЕЦИФИЧНЫЙ К ГЕНУ CTGF, И СОДЕРЖАЩАЯ ЕГО КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ФИБРОЗНЫХ ЗАБОЛЕВАНИЙ И ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ДЫХАТЕЛЬНОЙ СИСТЕМОЙ | 2020 |
|
RU2807108C1 |
| Реагенты для лечения окулофарингеальной мышечной дистрофии (OPMD) и их применение | 2017 |
|
RU2755544C2 |
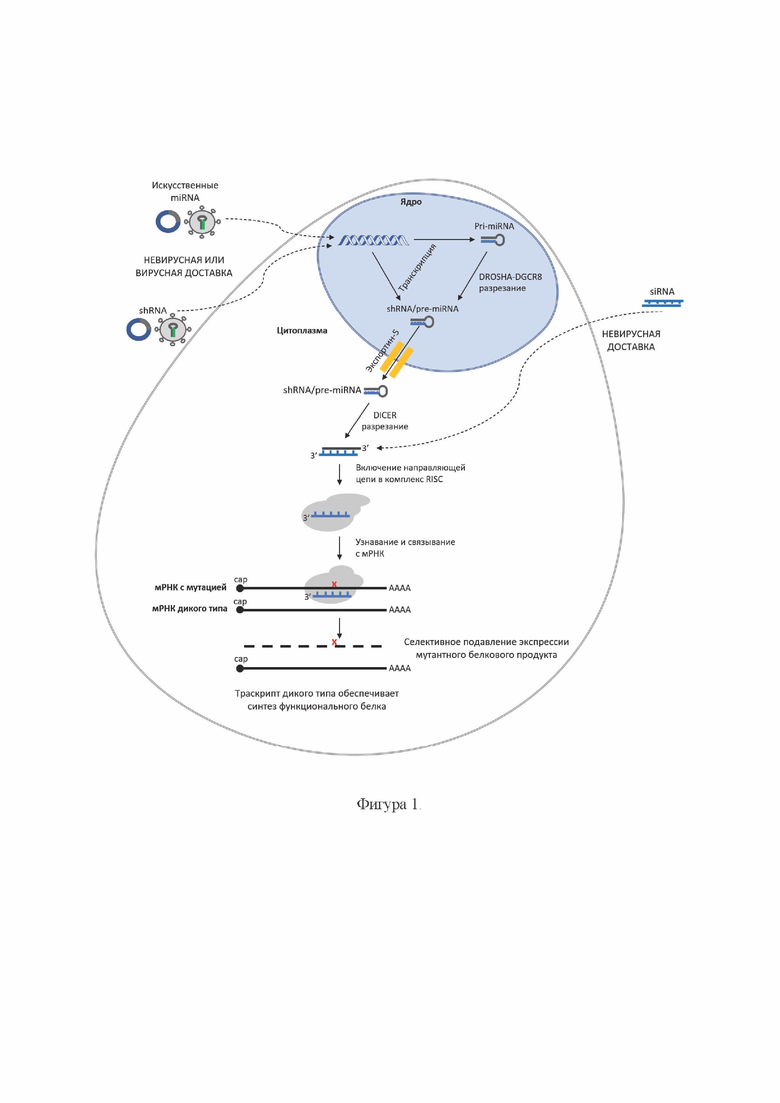
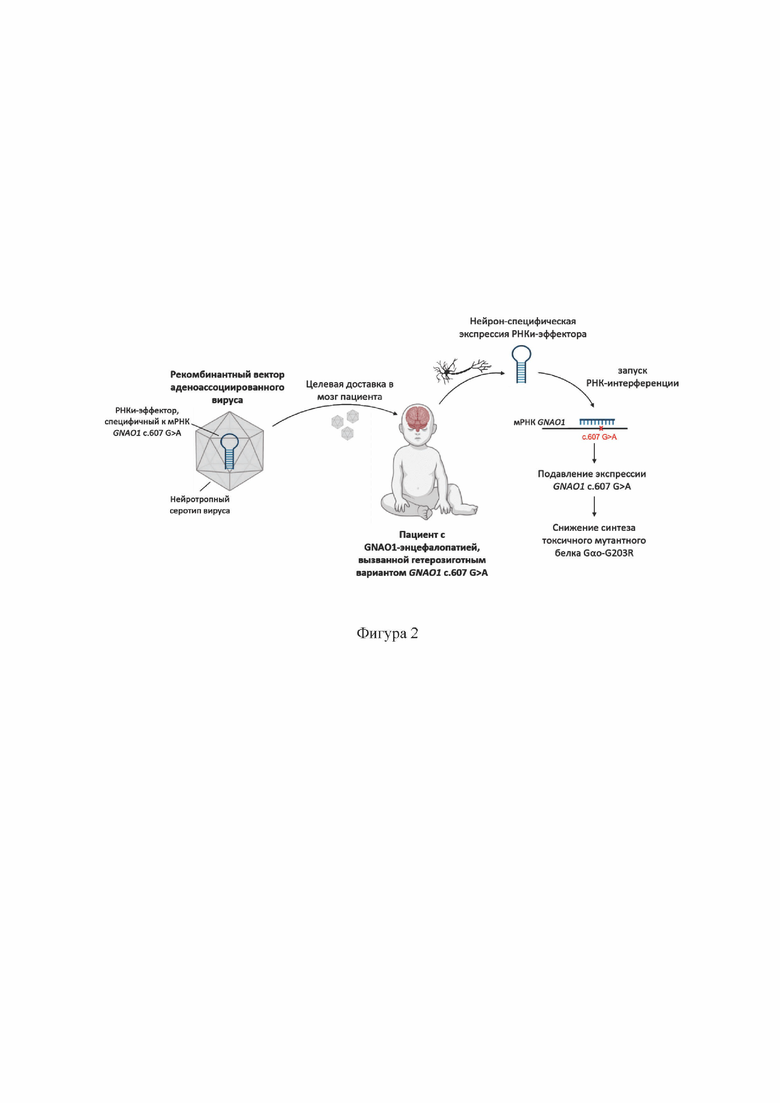

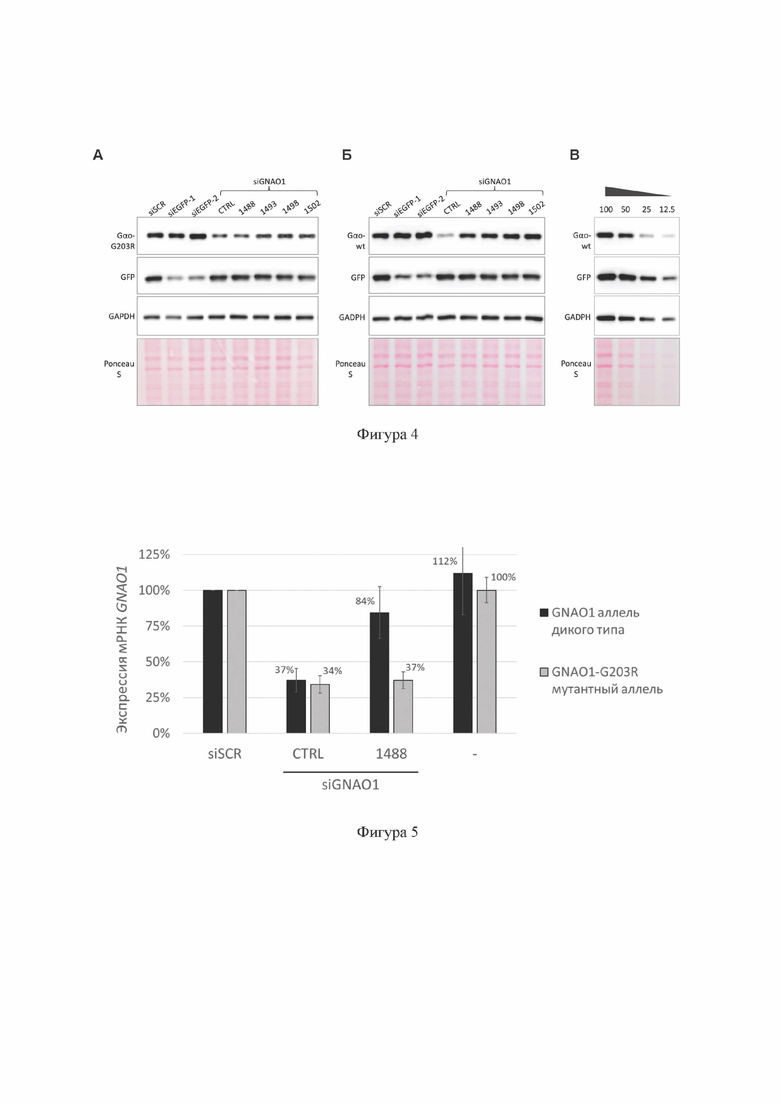
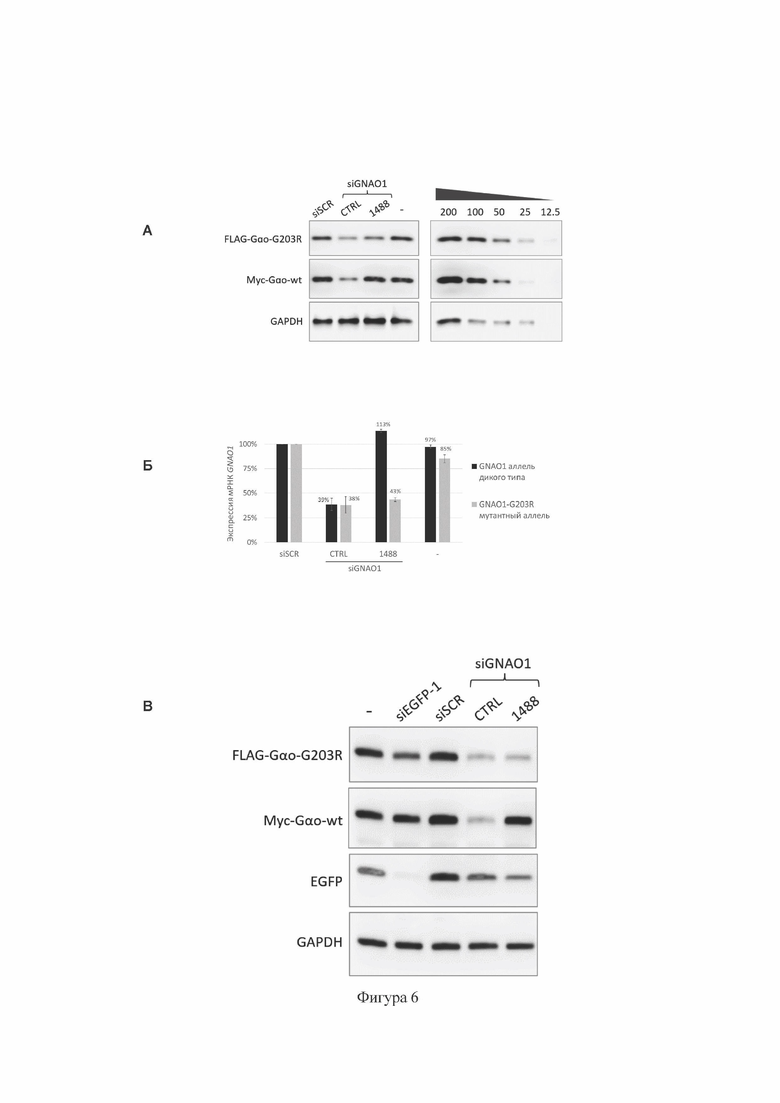
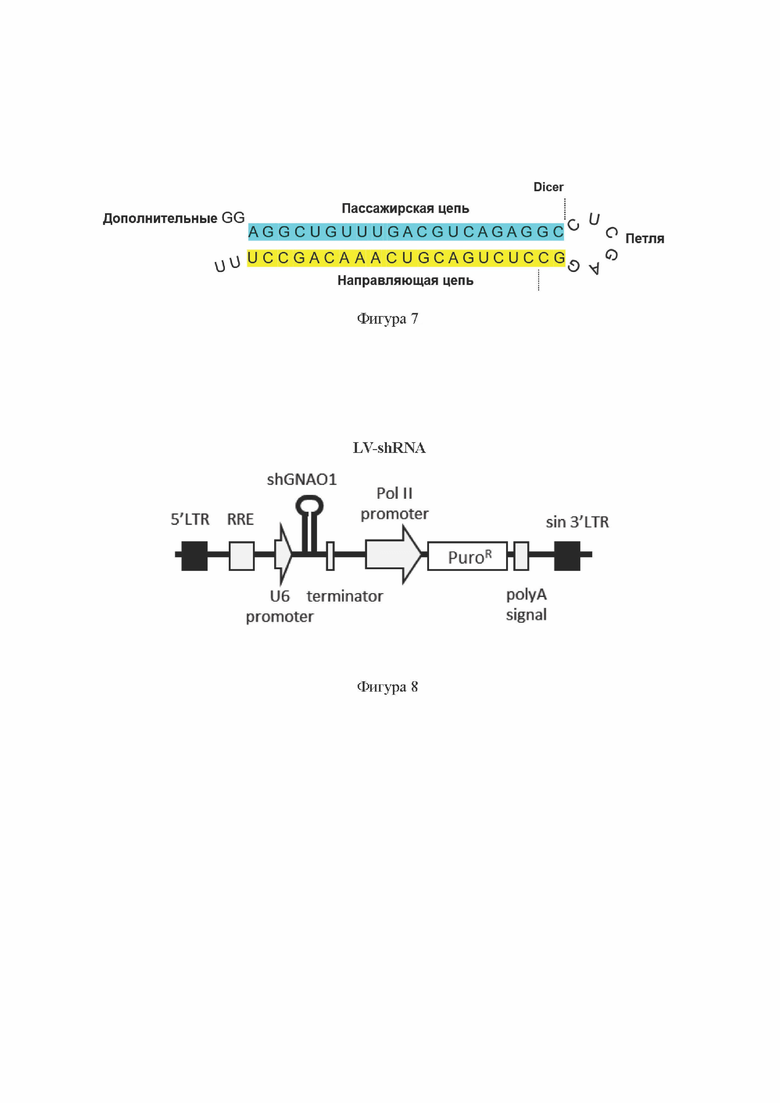
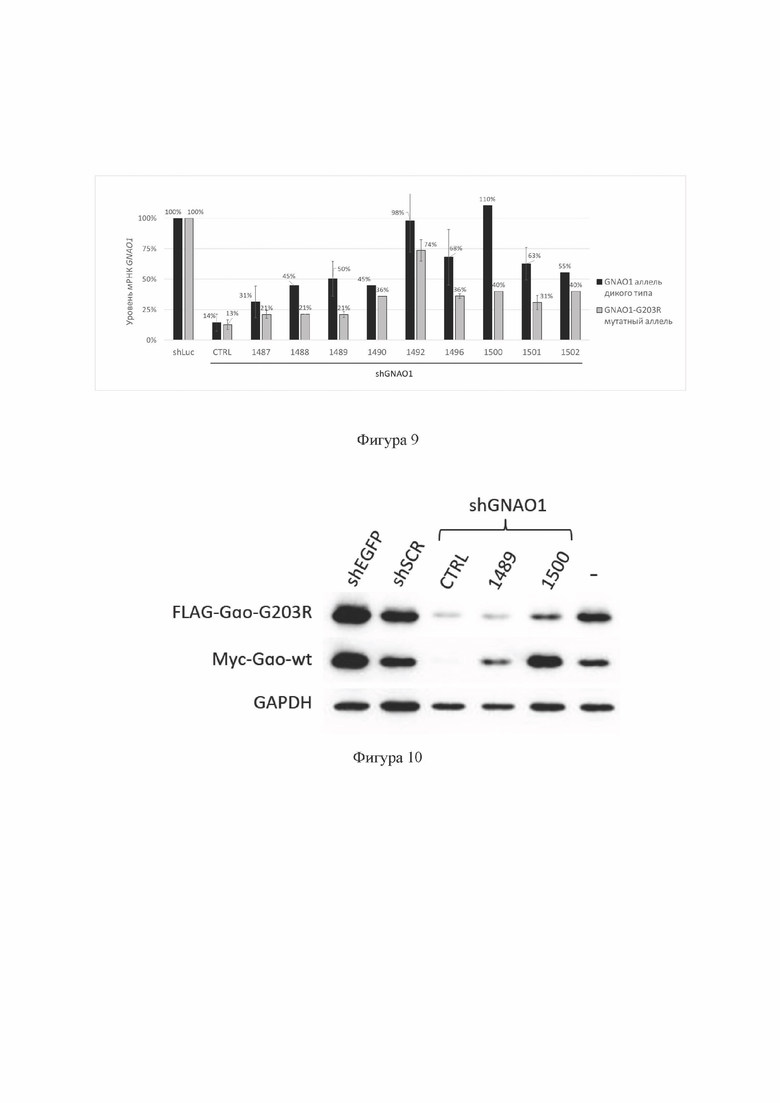
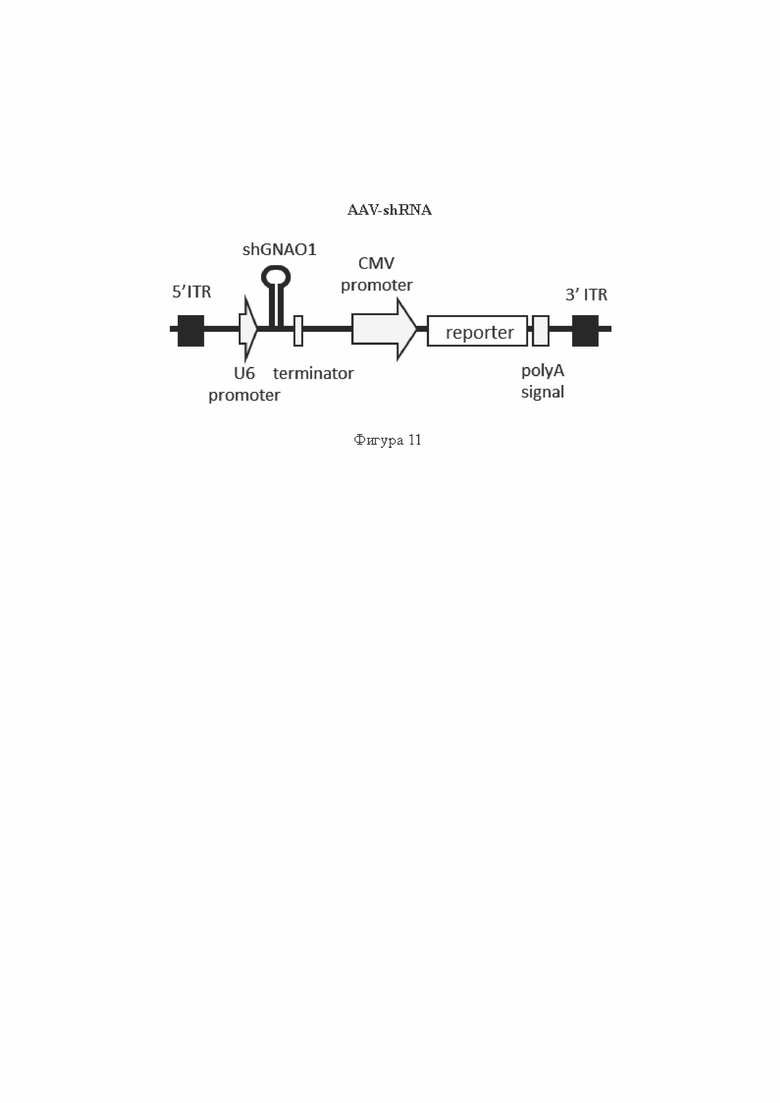
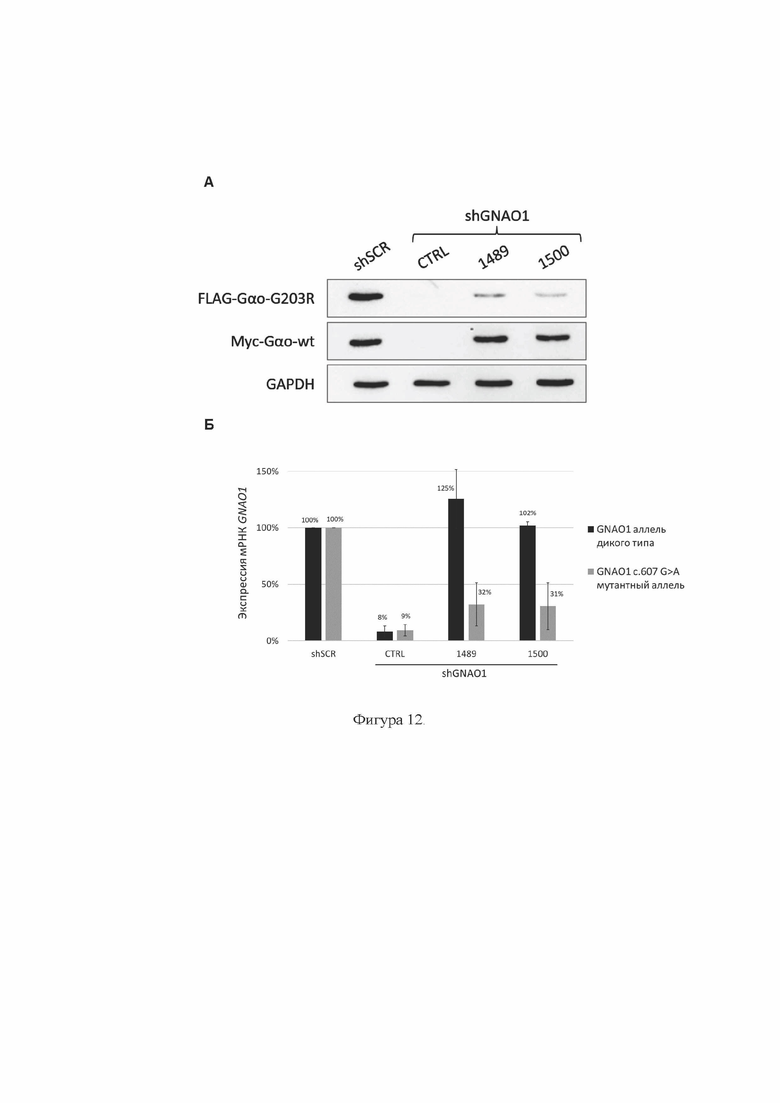
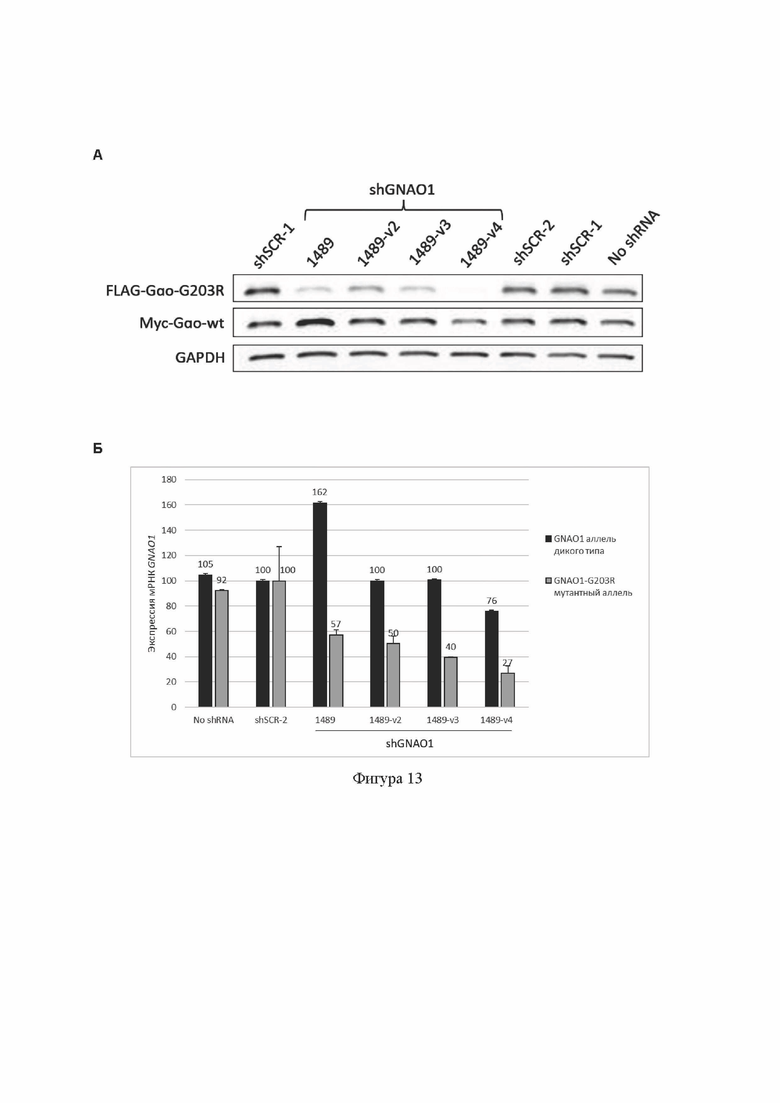
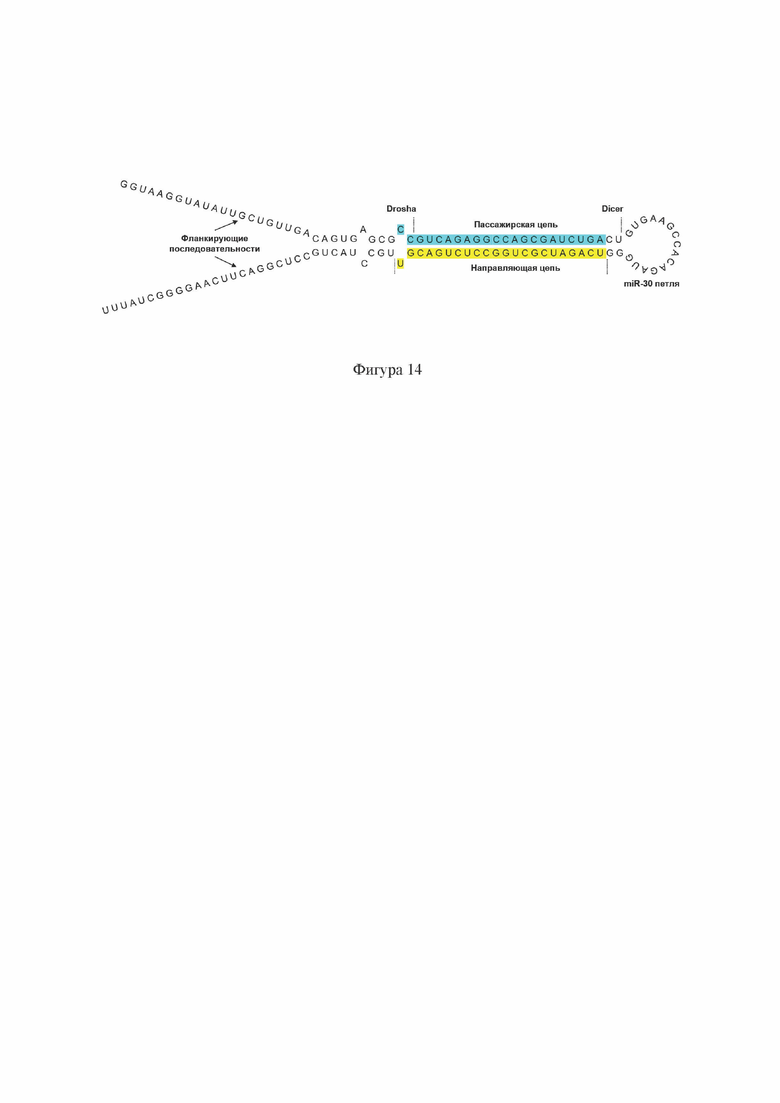
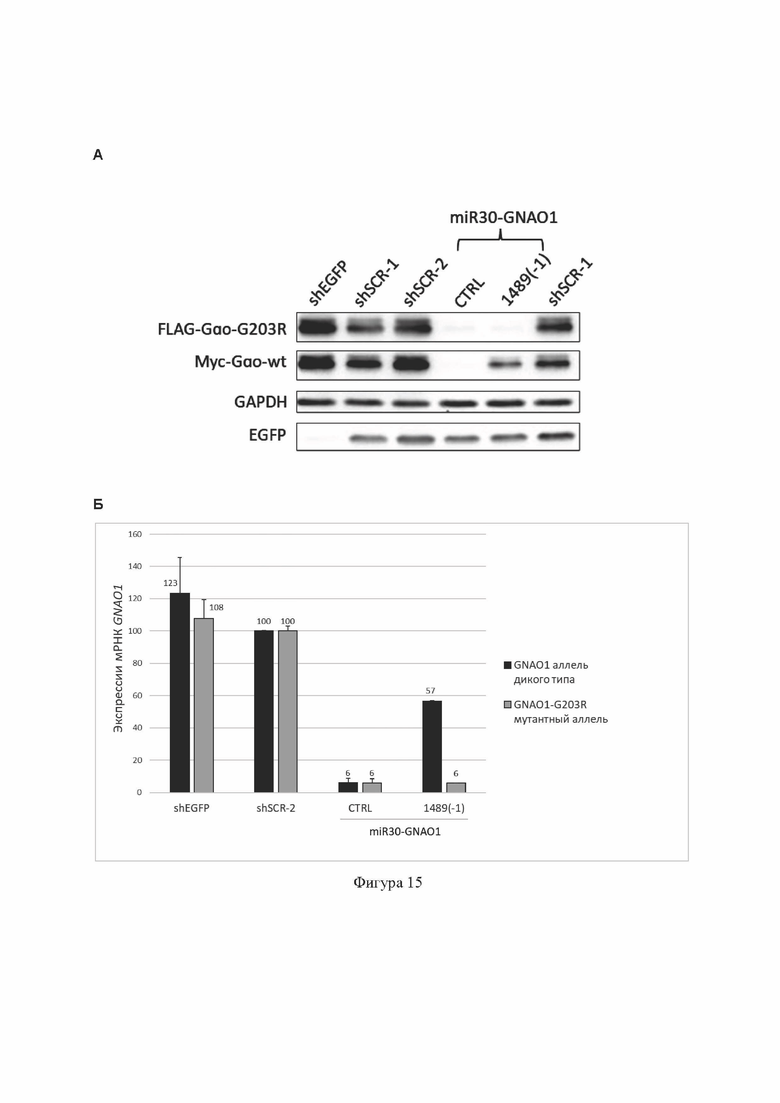
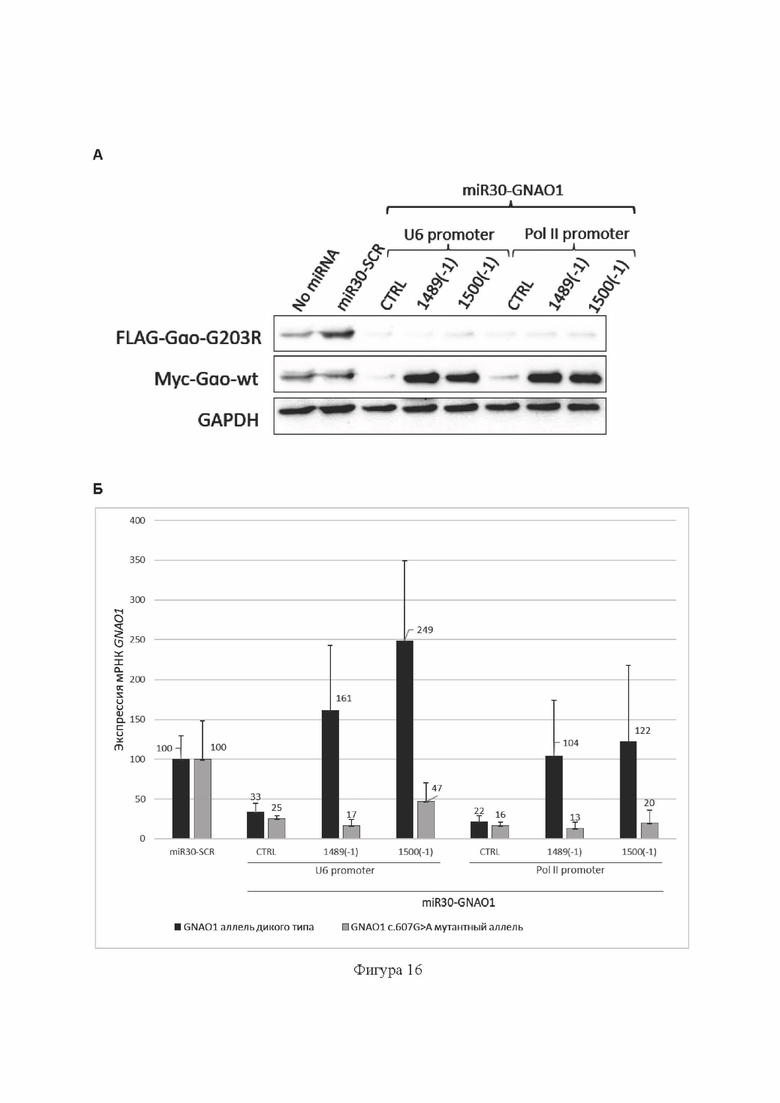
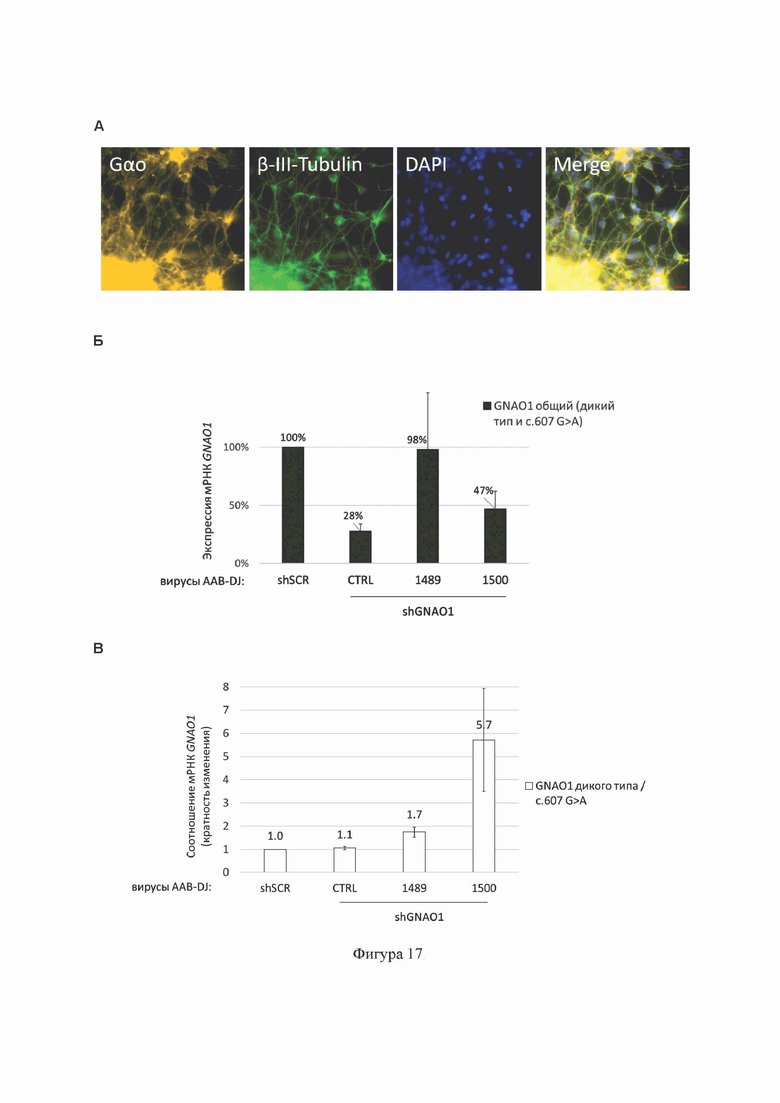
Изобретение относится к области биотехнологии, конкретно к молекулам РНК, способным снижать экспрессию мутантного аллеля c.607G>A гена GNAO1 человека, запуская механизм РНК-интерференции, и может быть использовано в медицине при разработке генотерапевтических подходов для пациентов с GNAO1-энцефалопатией, имеющих мутацию c.607G>A (p.Gly203Arg). Предложена двухцепочечная РНК, содержащая смысловую последовательность РНК и антисмысловую последовательность РНК, которые комплементарны, причем антисмысловая последовательность РНК содержит по меньшей мере 19–22 нуклеотида и является комплементарной последовательности мРНК-мишени SEQ ID NO: 28, либо смысловая последовательность РНК выбрана из группы, состоящей из SEQ ID NO: 1–17. Также предложены экспрессионные векторы, содержащие ДНК, кодирующую указанную двухцепочечную РНК. Изобретение позволяет аллель-селективно ингибировать экспрессию мутантного аллеля с.607G>A гена GNAO1 человека. 6 н. и 11 з.п. ф-лы, 17 ил., 1 табл., 4 пр.
1. Двухцепочечная РНК, способная снижать экспрессию мутантного аллеля с.607G>A гена GNAO1 человека через механизм РНК-интерференции, содержащая смысловую последовательность РНК и антисмысловую последовательность РНК, которые комплементарны, причем антисмысловая последовательность РНК содержит по меньшей мере 19-22 нуклеотида и является комплементарной последовательности мРНК-мишени SEQ ID No: 28, либо смысловая последовательность РНК выбрана из группы, состоящей из SEQ ID No: 1–17.
2. Двухцепочечная РНК по п. 1, где смысловая последовательность РНК выбрана из группы, состоящей из SEQ ID No: 1–17.
3. Двухцепочечная РНК по п. 1 или 2, представляющая собой малую интерферирующую РНК.
4. Двухцепочечная РНК по п. 3, где смысловая последовательность РНК выбрана из группы, состоящей из SEQ ID No: 1–4.
5. Двухцепочечная РНК по п. 1 или 2, где двухцепочечная РНК включена в каркас короткой шпилечной РНК.
6. Двухцепочечная РНК по п. 5, где смысловая последовательность РНК выбрана из группы, состоящей из SEQ ID No: 5–15.
7. Двухцепочечная РНК по п. 5, где каркас короткой шпилечной РНК содержит последовательность РНК, выбранную из группы, состоящей из SEQ ID No: 18–20, SEQ ID No: 33.
8. Двухцепочечная РНК по п. 1 или 2, где двухцепочечная РНК включена в каркас искусственной микроРНК.
9. Двухцепочечная РНК по п. 8, где двухцепочечная РНК включена в каркас искусственной микроРНК на основе miR-30.
10. Двухцепочечная РНК по п. 8 или 9, где смысловая последовательность выбрана из группы, состоящей из SEQ ID No: 16–17.
11. ДНК, кодирующая двухцепочечную РНК по любому из пп. 1–10.
12. Кассета экспрессии, содержащая ДНК по п. 11.
13. Экспрессионный вектор, содержащий кассету экспрессии по п. 12.
14. Плазмидный экспрессионный вектор, содержащий кассету экспрессии по п. 12.
15. Вирусный экспрессионный вектор, содержащий кассету экспрессии по п. 12.
16. Вирусный вектор по п. 15, где вектор представляет собой лентивирусный вектор.
17. Вирусный вектор по п. 15, где вектор представляет собой вектор на основе аденоассоциированного вируса.
| ЛУНЕВ Е.А | |||
| и др | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Приспособление для разматывания лент с семенами при укладке их в почву | 1922 |
|
SU56A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| ЖЕЛЕЗНЫЙ КАРКАС ДЛЯ ЭЛЕКТРОДВИГАТЕЛЯ | 1922 |
|
SU604A1 |
| БД GenBank последовательность под номером NM_020988.2, размещена 16.09.2018 | |||
| YANG S | |||
| et al | |||
| Specific | |||

