РОДСТВЕННЫЕ ЗАЯВКИ
[001] По настоящей заявке испрашивается приоритет по заявке США №15/648,337, названной "Универсальные донорские клетки и связанные с ними способы", поданной 12 июля 2107, целиком включенной настоящее описание посредством ссылки целиком.
ОБЛАСТЬ ТЕХНИКИ
[002] Настоящее изобретение относится к области генной экспрессии, генной инженерии и генной/клеточной терапии.
УРОВЕНЬ ТЕХНИКИ
[003] Плюрипотентные стволовые клетки человека (hPSCs) являются полезным инструментом для получения зрелых клеток любого типа для трансплантации пациентам. В принципе, терапии, основанные на клетках hPSC, имеют потенциал для лечения большинства, если не всех дегенеративных заболеваний, однако успех таких терапий может быть ограничен иммунным ответом субъекта. Иммунная система защищает организм от инфекций с многоуровневой защитой возрастающей специфичности. Проще говоря, физические барьеры препятствуют проникновению в организм болезнетворных микроорганизмов, таких как бактерии и вирусы. Если патоген нарушает эти барьеры, врожденная иммунная система обеспечивает немедленный, но неспецифический ответ.Если патогены успешно уклоняются от врожденного ответа, позвоночные обладают вторым уровнем защиты, адаптивной иммунной системой, которая активируется врожденным ответом. Адаптивная иммунная система генерирует гораздо более специфический ответ. В данном случае иммунная система адаптирует свой ответ во время инфекции, чтобы улучшить распознавание возбудителя. Этот улучшенный ответ затем сохраняется после уничтожения возбудителя в форме иммунологической памяти, что позволяет адаптивной иммунной системе ускоряться и усиливать атаки при каждом обнаружении этого патогена. Адаптивный иммунный ответ является антиген-специфическим и требует распознавание специфических «чужих» антигенов во время процесса, называемого презентацией антигена. Антигенная специфичность позволяет генерировать ответы, которые адаптированы к конкретным патогенам или патоген-инфицированным клеткам. Интерферон гамма (IFN-γ) играет важную роль в борьбе с инфекционными и неинфекционными заболеваниями. Основным источником IFN-γ при иммунном ответе человека являются Т-клетки. NK-клетки, макрофаги и интерфероны играют важную роль как во врожденном, так и в приобретенном иммунитете.
[004] Главный комплекс гистосовместимости (ГКГС, англ. - МНС) представляет собой набор белков клеточной поверхности, необходимых для регуляции иммунной системы. Основная функция молекул ГКГС - связывать антигены, полученные из патогенов, и экспонировать их на поверхности клетки для распознавания соответствующими Т-клетками. Семейство генов ГКГС подразделяется на три подгруппы: класс I, класс II и класс III. ГКГС человека также называют комплексом ЛАЧ (лейкоцитарных антигенов человека, англ. - HLA). Естественные клетки - киллеры (NK-клетки) представляют собой лимфоциты, которые функционируют на границе между врожденным и адаптивным иммунитетом. NK-клетки вносят непосредственный вклад в иммунную защиту посредством своих эффекторных функций, таких как цитотоксичность и секреция цитокинов, а также путем регуляции врожденных и адаптивных иммунных реакций. Когда мишень или клетка-хозяин сталкиваются с NK-клетками, возможны несколько результатов. Степень ответа клеток NK определяется количеством и типом активирующих и ингибирующих рецепторов на NK-клетках, а также количеством и типом активирующих и ингибирующих лигандов на клетке-мишени. См., Фиг. 1. В сценарии А, когда клетки-мишени не имеют лейкоцитарных антигенов человека (ЛАЧ) класса I и не активируют лиганды NK-клеток, NK-клетки, экспрессирующие ингибирующие рецепторы ГКГС класса I и рецепторы, активирующие лиганды, не атакуют клетки-мишени (нет ответа или ответ не разрешен). В сценарии В, когда клетки-мишени экспрессируют ЛАЧ класса I, но не имеют активирующих лигандов, NK-клетки, экспрессирующие ингибирующие рецепторы и активирующие рецепторы, не могут атаковать мишени. В сценарии С, когда клетки-мишени имеют пониженный уровень ЛАЧ класса I или не имеют ЛАЧ класса I и экспрессируют лиганды, активирующие NK-клетки, NK-клетки, экспрессирующие ингибирующие рецепторы и активирующие рецепторы, атакуют клетки-мишени. В сценарии D, когда клетки-мишени экспрессируют как лиганды, активирующие собственные белки ЛАЧ класса I, так и лиганды, активирующие NK-клетки, уровень ответных реакций NK-клеток, экспрессирующих ингибирующие рецепторы и активирующие рецепторы, определяется балансом ингибирующих и активирующих сигналов для NK-клеток. Haynes и др., THE IMMUNE SYSTEM IN HEALTH AND DISEASE, PART 15: Immune-Mediated, Inflammatory, and Rheumatotogic Disorders, 372e, Introduction to the Immune System.
[005] Исторически усилия исследователей по преодолению иммунного ответа хозяина на аллогенные клетки были сосредоточены на адаптивном иммунном ответе, то есть на вмешательстве в адгезию между Т-клетками и антигенами ГКГС класса I, представленными на чужеродных клетках. Так, для генерации генетических модификаций потери функции и, следовательно, для получения стволовых клеток, которые не экспрессируют один или несколько классических генов ГКГС / ЛАЧ, использовали системы CRISPR и TALEN. Однако эти клетки и клетки, полученные из них, все еще уязвимы для врожденного иммунного ответа хозяина (NK-клеток). Смотри, например, Parham и др. (2005) Nat Rev Immunol. 5(3):201-214. Чтобы преодолеть врожденный иммунный ответ хозяина, другие исследователи пытались ввести толерогенные факторы обратно в клетку-мишень; основное внимание было уделено "потере своего". См. публикацию WO 2016183041 A2, которая целиком включена настоящее описание посредством ссылки. Авторы настоящего изобретения неожиданно обнаружили, что ключом к уклонению от NK-опосредованного иммунного ответа хозяина является не "потеря своего", а экспрессия и уровень экспрессии лигандов, активирующих NK-клетки, на клетках-мишенях.
[006] Таким образом, остается потребность в композициях и способах для разработки клеток-мишеней, в которых отсутствует часть или вся классическая экспрессия ЛАЧ, но которые не подвергаются атакам Клеток NK с последующим лизисом.
КРАТКОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
[007] Настоящем описании раскрыты стратегии преодоления отторжения трансплантата, в частности, отторжения аллогенного иммунного трансплантата в клеточной трансплантационной терапии путем предоставления универсальных донорских клеточных линий. В одном варианте осуществления настоящего изобретения предоставлены плюрипотентные стволовые клетки человека, у которых отсутствует часть или вся классическая экспрессия белка ЛАЧ класса I на поверхности клеток и экспрессия лиганда, активирующего NK-клетки. В одном варианте осуществления настоящего изобретения предоставлена клетка, полученная из плюрипотентной стволовой клетки человека, такой как клетка поджелудочной железы, в которой отсутствует часть или вся классическая экспрессия белка ЛАЧ класса I на поверхности клеток и экспрессия NK-активирующего лиганда. В одном варианте осуществления настоящего изобретения предложен способ предотвращения отторжения клеточного трансплантата путем предоставления трансплантированных клеток поджелудочной железы, в которых по меньшей мере один ген ГКГС, такой как бета-2-микроглобулин (В2М), и по меньшей мере один ген активирующего лиганда NK, такой как Молекула 1 межклеточной адгезии (ICAM-1), разрушены, удалены, модифицированы или ингибированы. В другом варианте осуществления настоящего изобретения предложен способ предотвращения отторжения клеточного трансплантата путем предоставления трансплантированных клеток поджелудочной железы, в которых экспрессия по меньшей мере одного белка ГКГС, такого как В2М, и по меньшей мере одного белка, активирующего NK-клетки, такого как ICAM-1, разрушены, удалены, модифицированы или ингибированы. Разрушение, удаление, модификация или ингибирование В2М приводит к дефициту экспрессии и функции ЛАЧ класса I на поверхности клеток.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
[008] Фиг. 1 является репродукцией из статьи Haynes и др., см. выше, рисунок 372е-4 (которая целиком включена настоящее описание), демонстрирующий различные сценарии ответа, опосредованного NK-клетками, на клетки-мишени. В отсутствие ГКГС класса I и в отсутствие NK-активирующих лигандов на клетке-мишени, ингибирующие и активирующие рецепторы на NK-клетках не задействуются и NK-клетки остаются неактивными (Сценарий A). В присутствии ГКГС класса I, но в отсутствие NK-активирующих лигандов на клетке-мишени, задействуются ингибирующие рецепторы на NK-клетках, но активирующие рецепторы на NK-клетках не задействуются и NK-клетки остаются неактивными (Сценарий B). В отсутствие собственного ГКГС класса I, но в присутствии NK-активирующих лигандов на клетке-мишени, ингибирующие рецепторы на NK-клетках не задействованы, а активирующие рецепторы на NK-клетках задействованы и NK-клетки осуществляют атаку (Сценарий С). В присутствии ГКГС класса I и NK-активирующих лигандов на клетке-мишени, ингибирующие и активирующие рецепторы на NK-клетках задействованы и результат определяется балансом сигналов.
[009] На Фиг. 2А показан репрезентативный анализ проточной цитометрии экспрессии белка В2М поверхности клеток в клетках hES дикого типа без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Заштрихованная область обозначает базовую экспрессию без прокрашивания антителами. Обработка IFN-γ повышает экспрессию белка В2М поверхности клеток в клетках hES дикого типа.
[0010] На Фиг. 2В показан репрезентативный анализ проточной цитометрии экспрессии белка В2М поверхности клеток в клетках hES с нокаутом В2М (В2М -/-) без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Клетки hES В2М -/- обладают очень малой экспрессией белка В2М поверхности клеток, которая существенно не меняется после обработки IFN-γ.
[0011] На Фиг. 3А показан репрезентативный анализ проточной цитометрии экспрессии белка ЛАЧ-АВС поверхности клеток в клетках hES дикого типа без IFN-γ (Линия В) и после обработки IFN-γ (Линия А) с использованием антител пан-ЛАЧ класса I. Заштрихованная область обозначает базовую экспрессию без прокрашивания антителами.
[0012] На Фиг. 3В показан репрезентативный анализ проточной цитометрии экспрессии белка ЛАЧ-АВС поверхности клеток в клетках hES с нокаутом В2М без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Клетки hES В2М -/- не обладают экспрессией белка ЛАЧ-АВС поверхности клеток.
[0013] На Фиг. 4А показан репрезентативный анализ проточной цитометрии экспрессии белка В2М поверхности клеток в клетках панкреатической эндодермы (РЕС) дикого типа без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Заштрихованная область обозначает базовую экспрессию без прокрашивания антителами. Обработка IFN-γ повышает экспрессию белка В2М поверхности клеток в клетках РЕС дикого типа.
[0014] На Фиг. 4В показан репрезентативный анализ проточной цитометрии экспрессии белка В2М поверхности клеток в клетках РЕС с нокаутом В2М без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Клетки В2М -/- РЕС не обладают экспрессией белка В2М поверхности клеток.
[0015] На Фиг. 5А показан репрезентативный анализ проточной цитометрии экспрессии белка ЛАЧ-АВС поверхности клеток в клетках РЕС дикого типа без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Заштрихованная область обозначает базовую экспрессию без прокрашивания антителами. Обработка IFN-γ повышает экспрессию белка ЛАЧ-АВС поверхности клеток в клетках РЕС дикого типа.
[0016] На Фиг. 5В показан репрезентативный анализ проточной цитометрии экспрессии белка ЛАЧ-АВС поверхности клеток в клетках РЕС с нокаутом В2М без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Клетки В2М -/- РЕС не обладают экспрессией белка ЛАЧ-АВС поверхности клеток.
[0017] На Фиг. 6А показан репрезентативный анализ проточной цитометрии экспрессии белка ICAM-1 поверхности клеток в клетках hES дикого типа без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Заштрихованная область обозначает базовую экспрессию без прокрашивания антителами. Обработка IFN-γ повышает экспрессия белка ICAM-1 поверхности клеток в клетках hES дикого типа.
[0018] На Фиг. 6В показан репрезентативный анализ проточной цитометрии экспрессии белка ICAM-1 поверхности клеток в клетках hES с нокаутом В2М без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Клетки hES В2М -/- имеют экспрессию белка ICAM-1 поверхности клеток, схожую с экспрессией в клетках hES дикого типа перед обработкой IFN-γ и после нее.
[0019] На Фиг. 7А показан репрезентативный анализ проточной цитометрии экспрессии белка ICAM-1 поверхности клеток в клетках РЕС дикого типа без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Заштрихованная область обозначает базовую экспрессию без прокрашивания антителами. Обработка IFN-γ повышает экспрессию белка ICAM-1 поверхности клеток в клетках РЕС дикого типа.
[0020] На Фиг. 7В показан репрезентативный анализ проточной цитометрии экспрессии белка ICAM-1 поверхности клеток в клетках РЕС с нокаутом В2М без IFN-γ (Линия В) и после обработки IFN-γ (Линия А). После обработки IFN-γ, клетки В2М -/- РЕС имеют экспрессию белка ICAM-1 поверхности клеток, схожую с экспрессией в клетках РЕС дикого типа, которая выше, чем базовая экспрессия (заштрихованная область).
[0021] На Фиг. 8 показана гистограмма, демонстрирующая данные экспрессии мРНК ICAM-1 (анализ экспрессии на биочипе Asymetrix) в клетках hES дикого типа, В2М (-/-) клетках hES, клетках РЕС дикого типа и клетках В2М (-/-)РЕС, каждые из которых не обрабатывали IFN-γ (контроль) или обрабатывали IFN-γ. Экспрессию ICAM-1 также определяли в клетках, про которые известно, что они имеют низкую экспрессию белка ICAM поверхности клеток: раковые клетки (K562 и SKBR3), трансплантированные клетки РЕС, которые созрели и превратились в клетки to клетки, продуцирующие инсулин in vivo, островковые клетки человека и два различных образца мононуклеарные клетки периферической крови (РВМС) (без обработки IFN-γ). Экспрессия мРНК ICAM-1 повышалась после обработки клеток hES (дикого типа или В2М-/-) или РЕС (дикого типа или В2М-/-) с помощью IFN-γ.
[0022] На Фиг. 9 показан репрезентативный анализ проточной цитометрии экспрессии белков CD58 (обозначены как: LFA-3) поверхности клеток в клетках РЕС дикого типа без обработки IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Линия С обозначает базовую экспрессию без прокрашивания антителами. Обработка IFN-γ незначительно повышает экспрессию белков CD58 поверхности клеток в клетках РЕС дикого типа по сравнению с необработанным контролем. Использовали антитела компании BioLegend, кат. №330909.
[0023] На Фиг. 10 показан репрезентативный анализ проточной цитометрии экспрессии белков CD155 поверхности клеток на клетках РЕС дикого типа без обработки IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Линия С обозначает базовую экспрессию без прокрашивания антителами. После обработки IFN-γ, клетки РЕС дикого типа имели экспрессию белков CD155 поверхности клеток, сходную с экспрессией в необработанных контрольных клетках РЕС дикого типа. Обозначение гена PVR (обозначены как: CD 155, NECL-5, HVED). Использовали антитела компании Milteneyi Biotech Inc., кат. №130-105-905.
[0024] На Фиг. 11 показан репрезентативный анализ проточной цитометрии экспрессии белков СЕАСАМ1 (обозначены как: CD66a, BGP, BGP1) поверхности клеток в клетках РЕС дикого типа без обработки IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Линия С обозначает базовую экспрессию без прокрашивания антителами. Обработка IFN-γ слегка повышала экспрессия белков СЕАСАМ1 поверхности клеток в клетках РЕС дикого типа по сравнению с необработанным контролем. Использовали антитела компании Milteneyi Biotech Inc., кат. №130-098-858.
[0025] На Фиг. 12 показан репрезентативный анализ проточной цитометрии экспрессия белков ВАТ3 поверхности клеток в клетках РЕС дикого типа без обработки IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Линия С обозначает базовую экспрессию без прокрашивания антителами. Клетки РЕС дикого типа в необработанном контроле имеют экспрессию белков ВАТ3 поверхности клеток, схожую с экспрессией в РЕС дикого типа, обработанных IFN-γ. Обозначение гена BAG6 (обозначены как: ВАТ3, транскрипт 3, ассоциированный с ЛАЧ-В). Использовали антитела компании Abeam, Inc., кат. №ab210838.
[0026] На Фиг. 13 показан репрезентативный анализ проточной цитометрии экспрессии белков CADM1 (обозначены как: NECL2, TSLC1, IGSF4, RA175) поверхности клеток в клетках РЕС дикого типа без обработки IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Линия С обозначает базовую экспрессию без прокрашивания антителами. После обработки IFN-γ, клетки РЕС дикого типа имеют экспрессию белка CADM1 поверхности клеток, сходную с экспрессией в необработанном контроле. Использовали антитела компании MBL International Corp. кат. № СМ004-4.
[0027] На Фиг. 14 показан репрезентативный анализ проточной цитометрии экспрессии белков CD112 поверхности клеток в клетках РЕС дикого типа без обработки IFN-γ (Линия В) и после обработки IFN-γ (Линия А). Линия С обозначает базовую экспрессию без прокрашивания антителами. После обработки IFN-γ, клетки РЕС дикого типа имеют экспрессия белков CD 112 поверхности клеток, сходную с экспрессией в необработанном контроле. Обозначение гена PVRL2 (обозначены как; CD112, Nectin-2, PVRR2, HVEB). Использовали антитела компании Milteneyi Biotech Inc., кат. №130-109-056.
[0028] На Фиг. 15 показана гистограмма, демонстрирующая снижение лизиса клеток-мишеней NK после блокировки экспрессии ICAM-1 в клетках-мишенях с помощью антител против ICAM1 (слева направо, клетки hESC дикого типа, клетки hESC типа, обработанные IFN-γ, клетки В2М -/-hESC, клетки В2М -/-hES, обработанные IFN-γ, клетки РЕС дикого типа, клетки РЕС дикого типа, обработанные IFN-γ, клетки В2М -/- РЕС, клетки В2М -/-РЕС, обработанные IFN-γ, контрольная линия клеток K562 для анализа цитотоксичности NK). Слева направо каждый набор из трех столбцов представляет: NK-062216; NK-062216 + ICAM1 АВ (5 мкг/мл); NK-062216+ICAM1 АВ (10 мкг/мл).
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0029] Последовательности нуклеиновых кислот и аминокислот, приведенные в прилагающемся перечне последовательностей, приведены с использованием стандартных буквенных сокращений для оснований нуклеиновых кислит и трехбуквенных кодов для аминокислот, установленных 37 C.F.R. 1.822. Показана только одна цепь каждой из нуклеиновых кислот, но следует понимать, что соответствующая комплементарная цепь включена в дополнение к представленной цепи. Указанный перечень последовательностей представлен посредством текстового файла формата ASCII под названием Sequence_Listing, датированного 3 июля 2018, размером 11 KB, который включен настоящее описание посредством ссылки.
ПОДРОБНОЕ ОПИСАНИЕ
[0030] Молекулы ГКГС класса I являются молекулами одного из двух главных классов молекул главного комплекса гистосовместимости (ГКГС) (вторым классом являются молекулы ГКГС класса II). Их функция состоит в экспонировании фрагментов пептидов чужеродных белков из клеток цитотоксичным Т-клеткам; это вызывает немедленный ответ иммунной системы против конкретного чужеродного антигена, экспонированного с помощью белка ГКГС класса I. У людей молекулами ЛАЧ, соответствующим молекулам ГКГС класса I, являются молекулы ЛАЧ-А, ЛАЧ-В, ЛАЧ-С, ЛАЧ-Е, ЛАЧ-F и ЛАЧ-G. Молекулы ЛАЧ-Е, ЛАЧ-F и ЛАЧ-G человека являются неклассическими молекулами ГКГС класса I, характеризующимися ограниченным полиморфизмом и низкой экспрессией на поверхности клеток по сравнению с классическими паралогами (ЛАЧ-А, -В и -С). Все белки ГКГС класса I должны быть ассоциированы с р2-микроглобулином (В2М) для получения функционального гетеродимерного белкового комплекса ГКГС класса I перед функциональной экспрессией на поверхности клеток. Молекулы ГКГС класса I также могут выступать в качестве ингибирующих лигандов для клеток NK. Снижение нормальных уровней молекул ГКГС класса I на поверхности клеток активирует уничтожение клеток NK.
[0031] Исторически полагали, что клетки-мишени, содержащие ингибирующие лиганды ГКГС класса I, избегают атаки, когда обрабатывают NK-клетки, ввиду предполагаемой доминантной природы ингибирующего сигнала ГКГС класса I (см. на Фиг. 1 Сценарий В, воспроизведенный из "Harrison's Principles of Internal Medicine 19 E" (т. 1 и т. 2) A Major Histocompatibility Complex, часть 15, фиг. 372e-4, в этом документе). Но заявители неожиданно обнаружили, что все совсем наоборот.
[0032] Было показано, что воздействие на клетки IFN-γ повышает экспрессию мРНК молекул ГКГС-класса I, а также экспрессию белкового комплекса ГКГС класса I на поверхности клеток. Ожидалось, что это увеличение экспрессии ГКГС-1 класса ингибирует NK-клетки. Заявители обнаружили, что клетки hES дикого типа (WT) при обработке IFN-γ (которые, как было показано, увеличивают количество молекул ГКГС класса I на поверхности клеток hES, на Фиг. 2А и 3А), увеличивали цитотоксичность, опосредованную клетками NK. Смотрите Фиг. 15, показывающую опосредованную клетками NK токсичность (лизис), возрастающую с 58% до 79% (сравните первые столбцы в условиях 1 и 2 на Фиг. 15). То же самое было верно, когда клетки РЕС дикого типа обрабатывали IFN-γ, при этом указанные клетки также имели повышенную экспрессию В2М и ЛАЧ-АВС, см. Фиг. 4А и 5А, где HNK-опосредованная токсичность (лизис) увеличилась с 33% до 53% (сравните первые столбики в условиях 5 и 6 на Фиг. 15). Эти данные свидетельствуют о том, что ключевым фактором к преодолению иммунного ответа клеток NK хозяина заключается не в сверхэкспрессии ингибирующих сигналов ГКГС класса I, а в блокировании NK-активирующих сигналов лигандов.
[0033] Для дальнейшей проверки этой гипотезы в контексте ярко выраженной цитотоксичности, опосредованной клетками NK, Заявители создали клетки hES В2М -/- (нокаут) (аналогичные клеткам на Фиг. 1, Сценарий С). Как и ожидалось, клетки В2М -/- ликвидировали экспрессию на клеточной поверхности молекул ГКГС класса I на клетках hES (Фиг. 2В и 3В) и РЕС (Фиг. 4В и 5В). Также, как и ожидалось, клетки В2М -/- продемонстрировали увеличение лизиса, опосредованного клетками NK, относительно клеток WT, от 58% до 71% для клеток hES и от 33% до 54% для клеток РЕС (сравните первые столбцы в условиях 1 против 3 или 5 против 7 на Фиг. 15). Заявители обнаружили, что воздействие IFN-γ на клетки В2М -/- hES или РЕС еще больше увеличивало процент опосредованной клетками NK токсичности (лизис) (сравните первые столбцы в условиях 2 против 4 и 6 против 8 на Фиг. 15). Соответственно, Заявители обнаружили, что экспрессия на клеточной поверхности лиганда, активирующего NK-клетки (Фиг. 6В и 7В), и экспрессия мРНК (Фиг. 8) увеличиваются при обработке IFN-γ. Эти данные позволили предположить, что лиганды, активирующие NK-клетки, на клетке-мишени играют критическую роль в цитотоксичности клеток NK и это привело к гипотезе о том, что ингибирование экспрессии лиганда, активирующего NK-клетки, может защищать от опосредованной клетками NK цитотоксичности в контексте снижения экспрессии МСН класса I, например, в контексте В2М -/-.
[0034] Чтобы определить, может ли токсичность клеток NK быть уменьшена путем ингибирования действия NK-активирующих лигандов на клетках-мишенях, Заявители заблокировали экспрессию NK-активирующего лиганда в клетках WT и hES В2М -/- и РЕС, например, используя блокирующее антитело ICAM1 для блокирования белка ICAM1 на поверхности клеток-мишеней. Заявители неожиданно обнаружили, что лизис клеток-мишеней уменьшился (сравните первые столбцы со вторыми и третьими столбцами для условий 2, 4, 6 и 8 на Фиг. 15). Таким образом, Заявители обнаружили, что лизис клеток NK может быть уменьшен путем блокирования лиганда, активирующего NK. Блокирование экспрессии ICAM1 с использованием антитела против лиганда, активирующего NK, в В2М -/- клетках является доказательством концепции получения клетки, имеющей двойной нокаут (нокаут гена ЛАЧ класса I и нокаут гена лиганда, активирующего NK). При этом, Заявители могут переводить клетки-мишени (например, клетки hES и/или клетки поджелудочной железы) из Сценария С в Сценарий А на Фиг. 1. В частности, клетки, ткани и органы согласно настоящему изобретению ингибировали или не ингибировали экспрессию белков ЛАЧ класса I поверхности клеток (В2М -/-) и ингибировали или не ингибировали экспрессию лиганда, активирующего NK, экспрессию белков поверхности клеток (например, ICAM1 -/-). Ингибирование экспрессии белков поверхности клеток может быть достигнуто путем нокаута гена или блокирования экспрессии белка с использованием антител. Другие стратегии вмешательства в экспрессию белков поверхности клеток включают использование антисмысловых РНК, РНК-ложных целей, рибозимов, РНК-аптамеров, миРНК, шРНК/микроРНК, трансдоминантных негативных белков (TNP), химерных/слитых белков, нуклеаз, лигандов хемокинов, антиинфекционных клеточных белков, внутриклеточных антител (sFv), нуклеозидных аналогов (NRT1), ненуклеозидных аналогов (NNRTI), ингибиторов интеграз (олигонуклеотидов, динуклеотидов и химических агентов) и ингибиторов протеаз. Двойной или множественный нокаут гена эффективно предотвращает как опосредованную цитотоксическими Т-клетками (ЦТЛ), так и опосредованную NK-клетками токсичность, потому что ЛАЧ класса I присутствуют в малых количествах или практически отсутствуют, а белковые лиганды, активирующие NK, с которыми можно было бы связаться практически не экспрессируются на поверхности клеток CTL или NK. Кроме того, чтобы полностью исключить активацию NK, Заявители ожидают, что экспрессия множественных NK-активирующих лигандов должна быть устранена/уменьшена либо путем нокаута гена в клетке-мишени (например, клеточной терапии, основанной на клетках hES), либо с помощью блокирующих антител или других стратегий, известных в настоящее время или разработанных в будущем.
Агенты, блокирующие лиганды, активирующие NK-клетки
[0035] В соответствии с одним аспектом настоящее изобретение обеспечивает способ лечения с подавлением функции клеток NK. В соответствии с другим аспектом настоящее изобретение обеспечивает способ лечения с подавлением по меньшей мере одного иммунного ответа. Каждый из способов включает введение субъекту, нуждающемуся в лечении, агента, ингибирующего функцию клеток NK. В некоторых вариантах осуществления, указанных агентом является антитело. В некоторых вариантах осуществления указанное антитело селективно связывается с лигандами на клетках-мишенях, активирующими NK-клетки.
[0036] Подразумевается, что для предотвращения адгезии между клетками NK и лигандами на клетках-мишенях, активирующими NK-клетки, могут использоваться реагенты различных типов, включая антитела и блокирующие белки.
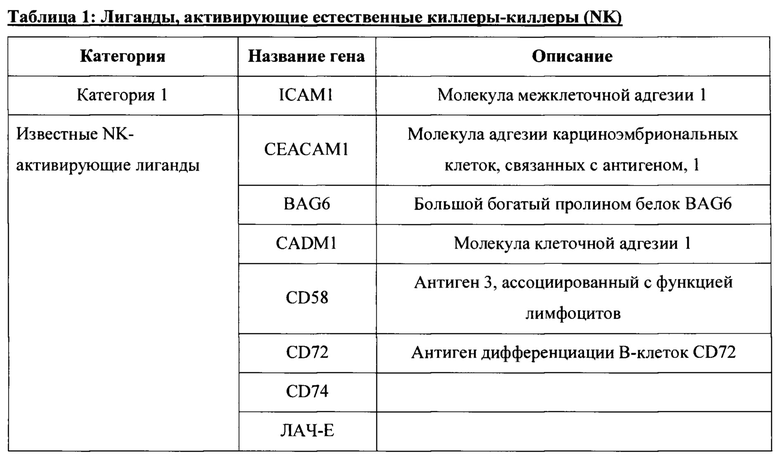
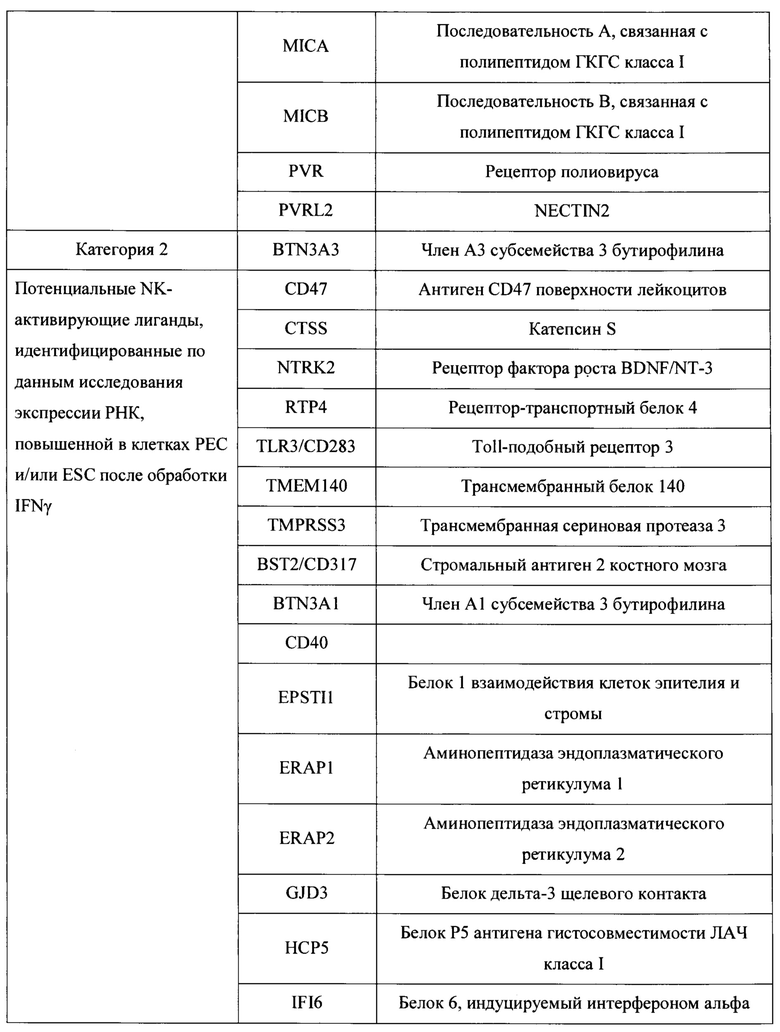
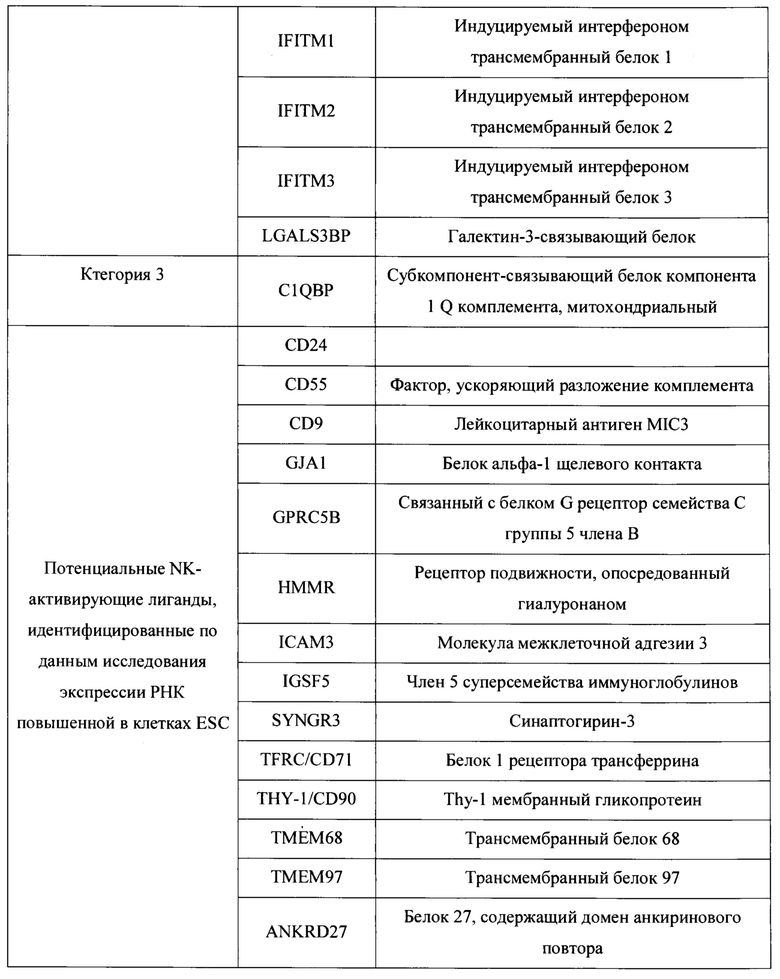
[0037] В некоторых вариантах осуществления, такие NK-активирующие лиганды могут быть выбраны из Таблицы 1.
NK-активирующие лиганды также описаны в статье Pegram и др., Activating and inhibiting receptors of NK-cells, Immunology and Cell Biology (2011) 89, 216-224, которая целиком включена в настоящее описание посредством ссылки.
[0038] Гипоиммуногенные клетки hES и их производные
[0039] ЛАЧ являются молекулами поверхности клеток, которые кодируются большим семейством генов и которые можно разделить на молекулы класса I и класса II. Молекулы ЛАЧ класса 1 находятся на поверхности каждой ядерной клетки и именно на них нацелено настоящее изобретение. Несоответствие молекул ЛАЧ между донорскими клетками (клетками-мишенями) и иммунными клетками реципиента (например, Т-клетками) в ходе трансплантации часто приводит к иммунному отторжению или отторжению трансплантата. Комплексы ЛАЧ класса I структурно состоят из полиморфных тяжелых цепей, состоящих из пептидов ЛАЧ класса I (например, ЛАЧ-А, ЛАЧ-В и ЛАЧ-С) и легких цепей бета-2-микроглобулина (β2m или В2М). В отсутствие В2М комплексы ЛАЧ класса I не могут правильно собираться и также не экспрессируются на поверхности клеток или клеточной мембране. В изобретении, описанном в настоящем описании Заявители создали линии клеток hES и клеток, являющихся их производными, посредством нарушения (добавления и удаления нескольких пар оснований, создания сдвига рамки считывания в мРНК, кодирующей белок, и внесения мутаций, вызывающих утрату активности) гена В2М, и, таким образом, сильного сокращения экспрессии ЛАЧ класса 1 на поверхности клеток hESC.
[0040] Вышеупомянутая методология также может быть использована для получения или генерирования линий клеток hES и их производных путем дополнительного разрушения генов, которые кодируют NK-активирующие лиганды, такие как ICAM1. Таким образом, в одном варианте осуществления настоящего изобретения предложены композиции и способы для получения клетки-мишени, в которой отсутствует, по меньшей мере, один антиген LAT-класса I и, по меньшей мере, один лиганд, активирующий NK, и таким образом создается гипоиммуногенная клетка. Ожидается, что такая гипоиммуногенная клетка будет менее склонна к иммунному отторжению субъектом, в который трансплантируют такие клетки. При трансплантации эта гипоиммуногенная клетка должна приживаться (и не должна отторгаться). В одном варианте осуществления такая клетка-мишень способна приживаться и выживать практически без иммунной супрессии, требуемой от реципиента.
[0041] В одном варианте осуществления ингибирование, уменьшение и/или делеция как экспрессии ЛАЧ класса I, так и экспрессии лиганда, активирующего NK, (или дефицит ЛАЧ класса I и дефицит лиганда, активирующего NK) в клетках hESC и полученных из них клетках могут служить как универсальный источник донорских клеток для трансплантационной терапии. Эти двойные нокауты (дефицит ЛАЧ класса I и дефицит лиганда, активирующего NK) могут быть трансплантированы повсеместно без совпадения с минорным комплексом гистосовместимости (MiHC), сопоставления с лейкоцитарным антигеном человека (ЛАЧ) или подавления иммунитета.
[0042] В данном документе раскрыты новые полученные in vitro гипоиммуногенные композиции и клетки. В частности, в определенных вариантах осуществления настоящие изобретения, раскрытые в данном документе, относятся к стволовой клетке, геном которой был изменен (модифицирован) для уменьшения или удаления критических компонентов как гена(ов) ГКГС класса I, так и гена(ов) лиганда, активирующего NK. В определенных вариантах осуществления настоящие изобретения, раскрытые в данном документе, относятся к клеткам панкреатической линии, таким как клетки панкреатической эндодермы, эпителиальные клетки поджелудочной железы, клетки-предшественники поджелудочной железы, эндокринные клетки-предшественники поджелудочной железы, эндокринные клетки поджелудочной железы, пре-бета-клетки поджелудочной железы или бета-клетки поджелудочной железы, геном которых был изменен (модифицирован) для уменьшения или удаления критических компонентов как гена(ов) ГКГС класса I, так и гена(ов) лиганда, активирующего NK, тем самым генерируя гипоиммуногенные линии клеток панкреатического типа. Лиганды, активирующие естественные киллеры, включают лиганды, перечисленные в таблице 1, из категории 1, 2, 3 или их комбинации, но не ограничиваются ими. Лиганды, активирующие естественные киллеры включают, например, ICAM-1, CEACAM1, CADM1, MICA и MICB. Гены ГКГС класса I включают ЛАЧ-А, ЛАЧ-Б, ЛАЧ-С, ЛАЧ-Е, ЛАЧ-Ф, ЛАЧ-Г и В2М. В определенных аспектах такое снижение экспрессии или нокаут генов ГКГС класса I и/или ГКГС класса II осуществляется путем прямого и/или косвенного нацеливания на гены NLRC5, В2М и CIITA и другие компоненты энхансосомы ГКГС (энхансосома является беловым комплексом высшего порядка, собранный на энхансере и регулирующий экспрессию целевого гена (например, регуляторы транскрипции ГКГС класса I или ГКГС класса II).
[0043] В настоящем документе также раскрыты способы получения гипоиммуногенных клеток, причем такой способ включает модулирование экспрессии одного или нескольких NK-активирующих лигандов, экспрессируемых клеткой, и модулирование экспрессии одного или нескольких ГКГС класса I и/или ГКГС класса II посредством указанной клетки, тем самым получая гипоиммуногенную клетку. В определенных аспектах модулирование экспрессии белков поверхности клеток одного или нескольких комплексов ГКГС класса I и/или ГКГС класса II включает снижение, ингибирование и/или вмешательство в экспрессию одного или нескольких генов или белков ГКГС класса I и/или ГКГС класса II. В некоторых вариантах модулирование экспрессии одного или нескольких комплексов ГКГС класса I и/или ГКГС класса II включает удаление одного или нескольких генов, кодирующих один или несколько регуляторов транскрипции ГКГС класса I или ГКГС класса II, по меньшей мере, из одного аллеля указанной клетки. Например, в определенных вариантах осуществления такие способы включают удаление одного или нескольких генов, кодирующих один или несколько регуляторов транскрипции генов ГКГС класса I или ГКГС класса II, выбранных из группы, состоящей из LRC5, CIITA, В2М и их комбинаций. В определенных аспектах модулирование экспрессии одного или нескольких NK-активирующих лиганд включает удаление, ингибирование или уменьшение экспрессии одного или нескольких генов, кодирующих лиганды, активирующие NK. В определенных вариантах осуществления такие NK-активирующие лиганды выбирают из Таблицы 1. В определенных вариантах осуществления такие NK-активирующие лиганды выбирают из Таблицы 1, категории 1, 2, 3 или их комбинаций. В некоторых вариантах такие NK-активирующие лиганды выбирают из Таблицы 1, категории 1, 2 и 3. В некоторых вариантах такие NK-активирующие лиганды выбирают из Таблицы 1, категории 1 и 3. В некоторых вариантах осуществления такие NK-активирующие лиганды выбирают из Таблицы 1 категории 1 и 2. В определенных вариантах осуществления такие NK-активирующие лиганды выбирают из Таблицы 1, категории 2 и 3. В определенных вариантах осуществления такие NK-активирующие лиганды выбирают из группы, состоящей из ICAM-1, CEACAM1, CADM1 MICA, MICB и их комбинации.
[0044] В некоторых вариантах осуществления, указанные имплантируемые гипоиммуногенные клетки находятся в среде, свободной от продуктов животного происхождения, например, продукты "XenoFree".
[0045] Настоящее изобретение предполагает внесение изменений в целевые последовательности полинуклеотидов любыми способами, которые доступны специалисту в данной области техники, например, с использованием любых мегануклеаз или эндодезоксирибоуклеаз, нуклеаз с белковыми доменами "цинковые пальцы" (ZFN или ZNF), эффекторной нуклеазы, подобной активаторам транскрипции, (TALEN) или систем на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR/Cas или CRISPR/Cas9) или традиционных методик гомологичной рекомбинации. Такие системы CRISPR/Cas могут использовать большое разнообразие белков Cas (Haft и др. PLoS Comput Biol. 2005; 1(6)е60). В некоторых вариантах осуществления, указанной системой CRISPR/Cas является система CRISPR типа I. В некоторых вариантах осуществления, указанной системой CRISPR/Cas является система CRISPR типа II. В некоторых вариантах осуществления, указанной системой CRISPR/Cas является система CRISPR типа V. Для внесения изменений в целевые последовательности полинуклеотидов также может быть использована система NEXTGEN™ CRISPR (Transposagen Inc., Лексингтон, Кентукки), которая включает двунаправленную РНК и каталитически неактивный белок Cas9, слитой с нуклеазой FokI. Для создания указанных гипоиммуногенных клеток, описанных в настоящем документе, могут использоваться другие способы изменения последовательностей полинуклеотидов для снижения или подавления их экспрессии в клетках-мишенях, известные специалисту в данной области техники или обнаруженные позднее.
[0046] В некоторых вариантах осуществления, указанные изменения приводят к сниженной экспрессии целевой полинуклеотидной последовательности. В некоторых вариантах осуществления, указанные изменения являются гомозиготными изменениями. В некоторых вариантах осуществления, указанные изменения являются гетерозиготными изменениями.
[0047] В некоторых вариантах осуществления, указанной целевой полинуклеотидной последовательностью является последовательность генома. В некоторых вариантах осуществления, указанной целевой полинуклеотидной последовательностью является последовательность генома человека. В некоторых вариантах осуществления, указанной целевой полинуклеотидной последовательностью является последовательность генома млекопитающего. В некоторых вариантах осуществления, указанной целевой полинуклеотидной последовательностью является последовательность генома позвоночного.
[0048] В некоторых вариантах осуществления, указанными гипоиммуногенными клетками являются эмбриональные стволовые клетки. В некоторых вариантах осуществления, указанными гипоиммуногенными клетками являются плюрипотентные стволовые клетки. В некоторых вариантах осуществления, указанными гипоиммуногенными клетками являются индуцированные плюрипотентные стволовые клетки, перепрограммированные клетки, дедифференцированные или трансдифференцированные клетки. В некоторых вариантах осуществления, указанными гипоиммуногенными клетками являются мультипотентные клетки-предшественники поджелудочной железы. В некоторых вариантах осуществления, указанными гипоиммуногенными клетками являются одногормональные или полигормональные клетки. В некоторых вариантах осуществления, указанными гипоиммуногенными клетками являются клетки мезендодермы, клетки дефинитивной эндодермы, PDX1-негативные клетки энтодермы передней кишки, PDX1-позитивные клетки энтодермальные клетки передней кишки, клетки панкреатической эндодермы, эндокринные клетки-предшественники, эндокринные клетки, правильно определенные эндокринные клетки, незрелые эндокринные клетки или функциональные бета-клетки. В некоторых вариантах осуществления, указанные гипоиммуногенные клетки могут быть гомогенными или гетерогенными популяциями клеток. В некоторых вариантах осуществления, указанными гипоиммуногенными клетками являются клетки, продуцирующие одну или более биологически активную субстанцию, представляющую интерес. Гипоиммуногенные клетки изначально могут быть терапевтически неактивными при трансплантации, например, они могут быть клетками-предшественники поджелудочной железы или PDX1-позитивными клетками панкреатической энтодермы, но после трансплантации они дополнительно развиваются, созревают и оказывают терапевтическое действие.
[0049] В некоторых вариантах осуществления, указанными гипоиммуногенными клетками могут быть любые клетки, которые возможно получить из плюрипотентных стволовых клеток человека, включая любую клетку, ткань или орган, и они могут включать в себя клетки кожи, бета-клетки (т.е. клетки поджелудочной железы, расположенные в островках Лангерганса), клетки паращитовидной железы, клетки кишечника, эндокринные клетки, клетки сердца, клетки головного мозга, клетки почек, клетки печени, клетки желудочно-кишечного тракта и вспомогательных органов пищеварения, клетки слюнной железы, клетки надпочечников, клетки простаты, клетки легких, клетки поджелудочной железы, костные клетки, клетки иммунной системы, кроветворные клетки, сосудистые клетки, клетки глаза, клетки соединительной ткани, скелетно-мышечные клетки, костную ткань, костно-мышечную ткань, ткань роговицы, ткань кожи, клапаны сердца, кровеносные сосуды, иммунные клетки, соединительную ткань, легочную ткань, кожу, роговицу, почку, печень, легкое, поджелудочную железу, сердце и кишечник, но ограничиваясь ими.
[0050] В некоторых вариантах осуществления, указанными гипоиммуногенными клетками могут быть индивидуальные (единичные) клетки в суспензии или в виде агрегатов клеток. В некоторых вариантах осуществления, указанные гипоиммуногенные клетки включают тотипотентные клетки. В одном варианте осуществления, указанные гипоиммуногенные клетки включают мультипотентные клетки. В одном варианте осуществления, указанные гипоиммуногенные клетки включают унипотентные клетки.
[0051] В некоторых вариантах осуществления, указанные гипоиммуногенные клетки получают из популяции плюрипотентных клеток, лишенных функциональной экспрессии ЛАЧ класса I и экспрессии NK-активирующих лигандов. Указанные производные клетки могут быть выбраны из группы, состоящей из: любой клетки, ткани или органа, и могут включать клетки кожи, бета-клетки (т.е. клетки поджелудочной железы, расположенные в островках Лангерганса), клетки паращитовидной железы, кишечные клетки, эндокринные клетки сердца, клетки головного мозга, клетки почек, клетки печени, клетки пищеварительного тракта и вспомогательных органов пищеварения, клетки слюнных желез, клетки надпочечников, клетки предстательной железы, клетки легких, клетки поджелудочной железы, клетки кости, иммунные клетки, гемопоэтические клетки, сосудистые клетки, клетки глаза, клетки соединительной ткани, костно-мышечные клетки, костную ткань, костно-мышечную ткань, ткань роговицы, кожную ткань, клапаны сердца, кровеносные сосуды, иммунные клетки, соединительную ткань, ткань легкого, кожу, роговицу, почку, печень, легкое, поджелудочную железу, сердце и кишечник.
[0052] Гипоиммуногенная клетка, ткань и/или орган, подлежащие трансплантации, могут быть сингенными или аллогенными для субъекта, получающего трансплантат.
[0053] В одном варианте осуществления гипоиммуногенная клетка представляет собой плюрипотентную клетку человека. В одном воплощении гипоиммуногенная клетка представляет собой клетку поджелудочной железы человека. В одном воплощении гипоиммуногенная клетка представляет собой клетку эндодермы поджелудочной железы человека. В одном воплощении гипоиммуногенная клетка представляет собой клетку-предшественник поджелудочной железы человека. В одном воплощении гипоиммуногенная клетка представляет собой клетку-предшественник поджелудочной железы человека. В одном воплощении гипоиммуногенная клетка представляет собой эндокринную клетку поджелудочной железы человека. В одном воплощении гипоиммуногенная клетка представляет собой эндокринную клетку-предшественник поджелудочной железы человека. В одном воплощении гипоиммуногенная клетка представляет собой эндокринную пре-бетаклетку поджелудочной железы человека. В одном воплощении гипоиммуногенная клетка представляет собой бета-клетку поджелудочной железы человека. В одном воплощении гипоиммуногенная клетка представляет собой одногормональную или полигормональную клетку поджелудочной железы человека. В одном воплощении гипоиммуногенная клетка представляет собой клетку, экспрессирующую инсулин человека.
[0054] В одном варианте осуществления гипоиммуногенные клетки являются хорошо известными, общедоступными линиями плюрипотентных клеток. Описанное здесь изобретение полезно для всех клеток hES и линий iPSC, и, по крайней мере, для hESC, например, СуТ49, СуТ25, СуТ203 и СуТ212. Линия плюрипотентных клеток включают коммерчески доступные клетки, которые можно приобрести у WiCell в интернете по адресу: wicell.org/home/stem-cell-lines/order-stem-cell-lines/obtain-stem-cell-lines.cmsx и, в частности, включают BG01, BG02 и BG03.
[0055] В одном варианте осуществления гипоиммуногенные клетки по существу сходны с клетками, описанными в статье D'Amour и др. "Production of Pancreatic Hormone-Expressing Endocrine Cells From Human Embryonic Stem Cells" (Nov. 1, 2006) Mature Biotechnology 24, 1392-1401, которая целиком включена в настоящее описание посредством ссылки. Статья D'Amour и др. описывает 5-ступенчатый протокол дифференциации: стадия 1 (приводит в основном к продукции эндодермы), стадия 2 (приводит в основном к продукции PDX1-отрицательной эндодермы передней кишки), стадия 3 (приводит в основном к продукции PDX1-положительной эндодермы передней кишки), стадия 4 (приводит в основном к продукции панкреатической энтодермы, также называемой мультипотентными панкреатическими предшественниками или к продукции панкреатических эндокринных предшественников) и стадия 5 (приводит в основном к продукции эндокринных клеток, экспрессирующих гормоны). В одном варианте осуществления указанные гипоиммуногенные клетки, по существу, аналогичны тем, которые описаны в патентах США №№7,510,876, 7,695,965, 7,985,585, 8,586,357, 8,633,024 и 8,129,182 (которые целиком включены в настоящее описание посредством ссылки).
[0056] В одном варианте осуществления указанные гипоиммуногенные клетки по существу сходны с клетками, описанными в статье Schulz и др. A Scalable System for Production of Functional Pancreatic Progenitors from Human Embryonic Stem Cells, PLoS One 7:5 1-17 (2012), которая целиком включена в настоящее описание посредством ссылки. Статья Schulz и др. описывает способы выращивания и консервирования hESC и систему дифференциации на основе суспензии. В частности, недифференцированные плюрипотентные клетки агрегировали в кластеры в динамической ротационной суспензионной культуре с последующей тотальной дифференцировкой в течение двух недель по четырехэтапному протоколу. Вкратце, из суспензий агрегатов клеток hES монослои hESC диссоциируют с Accutase (Innovative Cell Technologies), собирают и ресуспендируют в концентрации 1×106 клеток/мл в StemPro hESC SFM (Life Technologies; комбинированная среда DMEM/F12, содержащая Glutamax, добавку StemPro hESC, БСА, и 1% (об./об.) пенициллин/стрептомицин; без FGF-2 и 2-меркаптоэтанола). Суспензии отдельных клеток распределяли по 6-луночным планшетам, не обработанным ТС (5,5 мл/лунку), и вращали при 95 об/мин на роторе Innova 2000 (New Brunswick Scientific) или распределяли по 500 мл флаконам для хранения Nalgene с фильтром (150 мл/бутылка) и вращали со скоростью 65 об/мин на роторе Sartorius Certomat RM-50 (с осью вращения 5 см). Клетки качали в течение ночи в инкубаторе при 37°С/8% СО2 и образовывали агрегаты размером приблизительно 100-200 мкм. Для агрегатов диаметром от 100 до 200 мкм могут использоваться скорости вращения от 60 до 140 об/мин для 6-луночного планшета; для бутылок объемом 500 мл можно использовать скорость вращения 5-20 об/мин. Дифференциация суспензионных агрегатов включала только несколько модификаций от D'Amour. Ингибитор TGF-βRI киназы IV добавляют во время стадии 2, а ретиноевую кислоту заменяли более стабильным аналогом ретиноида, TTNPB (3 нМ), на стадии 3. Факторы роста KGF (50 нг/мл) и EGF (50 нг/мл) добавляли на стадии 4 для сохранения клеточной массы. Ноггин (50 нг/мл) также добавляли на стадии 4. В одном варианте осуществления гипоиммуногенные клетки по существу аналогичны тем, которые описаны в патентах США №№8,008,075 и 8,895,300 (которые целиком включены в настоящее описание посредством ссылки).
[0057] В одном варианте осуществления гипоиммуногенные клетки по существу сходны с клетками, описанными в статье Agulnick и др. Insulin-Producing Endocrine Cells Differentiated In Vitro From Human Embryonic Stem Cells Function in Macroencapsulation Devices In Vivo Stem Cells. Translational Medicine, 4: 1-9(2015), которая целиком включена в настоящее описание посредством ссылки. Статья Agulnick и др. описывает модифицированный протокол для получения клеток-предшественников поджелудочной железы, так что 73-80% популяции клеток состоят из PDX1-позитивных (PDX1+) HNKX6.1 + панкреатических предшественников. Клетки-предшественники поджелудочной железы далее дифференцируют в островковые клетки (IC), которые воспроизводимо содержали 73-89% эндокринных клеток, из которых приблизительно 40-50% экспрессировали инсулин. Большая часть этих инсулин-положительных клеток была одногормон-положительной и экспрессировала факторы транскрипции PDX1 и NKX6.1. Статья Agulnick и др. описывает протокол, в котором протокол 2012 года Schulz и др. был модифицирован путем дополнительной обработки активином A, Wnt3A и херегулином β1 на стадии 3 (дни 5-7) и активином А и херегулином β1 на стадии 4 (дни 7-13). В одном варианте осуществления указанные гипоиммуногенные клетки в значительной степени сходны с клетками, описанными в патенте США №8,859,286, который целиком включен в настоящее описание посредством ссылки.
[0058] Выращивание, пассирование и пролиферация эмбриональных стволовых клеток человека может быть выполнено по существу, как описано в патентах США №№7,964,402; 8,211,699; 8,334,138; 8,008,07; и 8,153,429.
[0059] Стандартный протокол получения
[0060] Стандартный протокол для получения клетки панкреатической эндодермы (РЕС), полученной из hESC, описан ниже в Таблице 2.
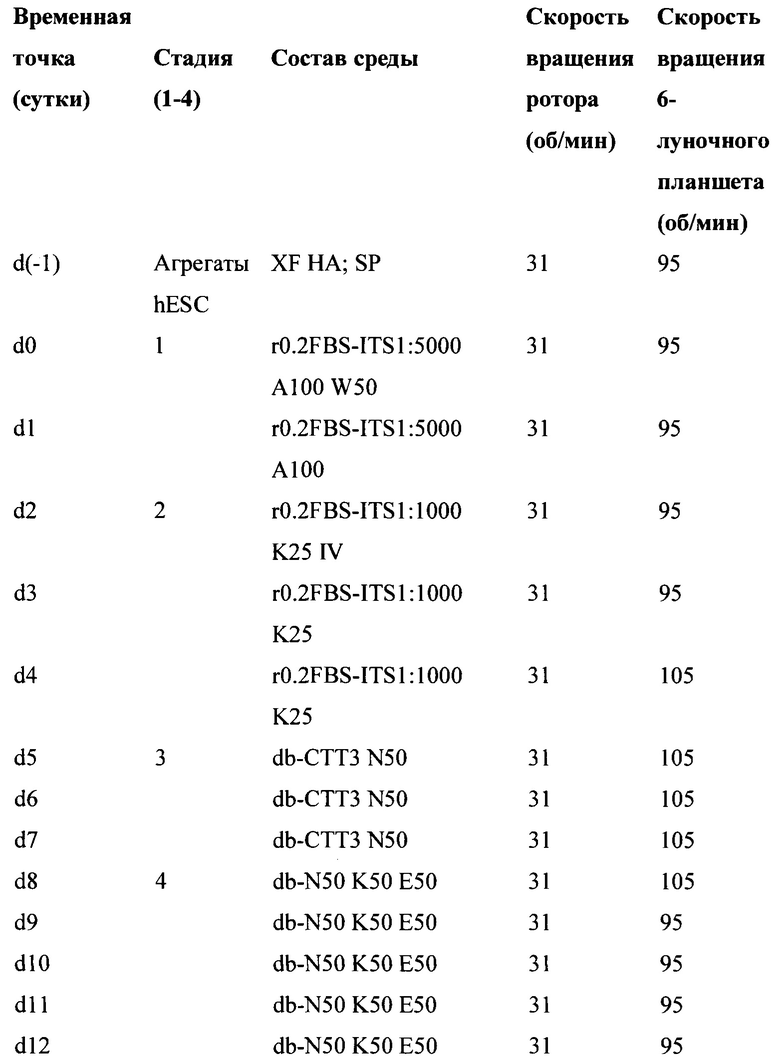
[0061] HESC Agg.: агрегаты hESC; XF НА: DMEM/F12, содержащая GlutaMAX, с добавлением 10% по объему Xeno-free замены KnockOut Serum, 1% по объему не незаменимых аминокислот, 1% по объему пенициллина/стрептомицина(все от Life Technologies), 10 нг/мл херегулин-1β (Peprotech) и 10 нг/мл активина A (R&D Systems); SP: StemPro® hESC SFM (Life Technologies); r0.2FBS: RPMI 1640 (Mediatech); 0,2% ФБС (HyClone), 1x GlutaMAX-1 (Life Technologies), 1% об./об. пенициллина/стрептомицина; ITS: инсулин-трансферрин-селен (Life Technoiogies), разведенный 1:5000 или 1:1000; A100: 100 нг/мл рекомбинантного человеческого активина A (R & D Systems); W50: 50 нг/мл рекомбинантного Wnt3A мыши (R&D Systems); K25: 25 нг/мл рекомбинантного KGF человека (R&D Systems); IV: 2,5 мкМ TGF-β RI ингибитора киназы IV (EMD Bioscience); db: DMEM HI глюкоза (HyClone), дополненная 0,5x добавкой B-27 (Life Technologies), 1x GlutaMAX и 1% по объему пенициллина/стрептомицина; СТТ3: 0,25 мкМ KAAD-циклопамин (Toronto Research Chemicals) и 3 нМ TTNPB (Sigma-Aldrich); N50: 50 нг/мл рекомбинантного человеческого Noggin (R&D Systems); K50: 50 нг/мл рекомбинантного KGF человека (R&D Systems); E50: 50 нг/мл рекомбинантного EGF человека (R&D Systems).
[0062] Анализ высвобождения кальцеина
[0063] Анализ высвобождения кальцеина является альтернативой изучения цитотоксичности клеток NK, не использующей радиоактивные изотопы. Клетки-мишени транспортируют внутрь флуоресцентную краску (кальцеин AM) и превращают его в цитоплазме в активный флуорохром, который высвобождается из клетки только в случае ее лизиса. Лизированые клетки высвобождают указанный флуорохром в супернатант, который затем собирают и количественно определяют его флуоресценцию с помощью флуориметра. Процент лизиса клеток рассчитывают по количеству флуоресценции, присутствующей в супернатанте после инкубации в присутствии или отсутствие клеток NK (эффекторов), блокирующего антитела или и того, и другого.
[0064] Удельный лизис рассчитывают по формуле, % лизиса = 100×[(средняя флуоресценция с антителом - средняя спонтанная флуоресценция)/(средняя максимальная флуоресценция - средняя спонтанная флуоресценция)]. Максимальную флуоресценцию определяли посредством лизиса клеток, инкубированных с детергентом (1% Тритон Х-100), а спонтанный лизис определяли как флуоресценцию, полученную с клетками-мишенями без каких-либо антител или эффекторных клеток.
[0065] Различные композиции клеток, полученные из плюрипотентных стволовых клеток, и способы их получения описаны в настоящем описании, а также могут быть найдены в патентных заявках США настоящего Заявителя №№10/486,408 под названием METHODS FOR CULTURE OF HESC ON FEEDER CELLS, подана 6 августа 2002; 11/021,618 под названием DEFINITIVE ENDODERM, подана 23 декабря 2004; 11/115,868 под названием PDX1 EXPRESSING ENDODERM, подана 26 апреля 2005; 11/165,305 под названием METHODS FOR IDENTIFYING FACTORS FOR DIFFERENTIATING DEFINITIVE ENDODERM, подана 23 июня 2005; 11/573,662 под названием METHODS FOR INCREASING DEFINITIVE ENDODERM DIFFERENTIATION OF PLURIPOTENT HUMAN EMBRYONIC STEM CELLS WITH PI-3 KINASE INHIBITORS, подана 15 августа 2005; 12/729, 084 под названием PDX1-EXPRESSING DORSAL AND VENTRAL FOREGUT ENDODERM, подана 27 октября 2005; 12/093,590 под названием MARKERS OF DEFINITIVE ENDODERM, подана 14 ноября 2005; 11/993,399 под названием EMBRYONIC STEM CELL CULTURE COMPOSITIONS AND METHODS OF USE THEREOF, подана 20 июня 2006; 11/588,693 под названием PDX1-EXPRESSING DORSAL AND VENTRAL FOREGUT ENDODERM, подана 27 октября 2006; 11/681,687 под названием ENDOCRINE PROGENITOR/PRECURSOR CELLS, PANCREATIC HORMONE-EXPRESSING CELLS AND METHODS OF PRODUCTION, подана 2 марта 2007; 11/807,223 под названием METHODS FOR CULTURE AND PRODUCTION OF SINGLE CELL POPULATIONS OF HESC, подана 24 марта 2007; 11/773,944 под названием METHODS OF PRODUCING PANCREATIC HORMONES, подана 5 июля 2007; 11/860,494 под названием METHODS FOR INCREASING DEFINITIVE ENDODERM PRODUCTION, подана 24 сентября 2007; 12/099,759 под названием METHODS OF PRODUCING PANCREATIC HORMONES, подана 8 апреля 2008; 12/107,020 под названием METHODS FOR PURIFYING ENDODERM AND PANCREATIC ENDODERM CELLS DERIVED FORM HUMAN EMBRYONIC STEM CELLS, подана 21 апреля 2008; 12/618,659 под названием ENCAPSULATION OF PANCREATIC LINEAGE CELLS DERIVED FROM HUMAN PLURIPOTENT STEM CELLS, подана 13 ноября 2009; 12/765,714 и 13/761,078, обе под названием CELL COMPOSITIONS FROM DEDIFFERENTIATED REPROGRAMMED CELLS, поданы 22 аперля 2010 и 6 февраля 2013; 11/838,054, под названием COMPOSITIONS AND METHODS USEFUL FOR CULTURING DIFFERENTIABLE CELLS, подана 13 августа 2007; 12/264,760 под названием STEM CELL AGGREGATE SUSPENSION COMPOSITIONS AND METHODS OF DIFFERENTIATION THEREOF, подана 4 ноября 2008; 13/259,15 под названием SMALL MOLECULES SUPPORTING PLURIPOTENT CELL GROWTH, подана 27 апреля 2010; PCT/US11/25628, под названием LOADING SYSTEM FOR AN ENCAPSULATION DEVICE, подана 21 февраля 2011; 13/992,931, под названием AGENTS AND METHODS FOR INHIBITING PLURIPOTENT STEM CELLS, подана 28 декабря 2010; и заявках США на промышленные образцы №№: 29/408,366, подана 12 декабря 2011; 29/408,368, подана 12 декабря 2011; 29/423,365, подана 31 мая 2012; и 29/447,944, подана 13 марта 2013; заявке США №14/201,630 под названием 3-DIMENSIONAL LARGE CAPACITY CELL ENCAPSULATION DEVICE ASSEMBLY, подана 7 марта 2014; и заявке США №14/106,330 под названием IN VITRO DIFFERENTIATION OF PLURIPOTENT STEM CELLS TO PANCREATIC ENDODERM CELLS (PEC) AND ENDOCRINE CELLS, подана 13 декабря 2013; PCT/US2016/061442, под названием PDX1 PANCREATIC ENDODERM CELLS IN CELL DELIVERY DEVICES AND METHODS THEREOF, подана 10 ноября 2016; каждая из которых целиком включена в настоящее описание посредством ссылки.
[0066] Различные композиции клеток, полученные из плюрипотентных стволовых клеток, и способы их получения описаны в настоящем документе и могут быть найдены в заявках, на которые настоящим Заявителем приобретены исключительные лицензии: патентная заявка США №2009/0269845 под названием Pluripotent cells, подана 24 апреля 2008; патентная заявка США №2011/0014703 под названием Differentiation of Human Embryonic Stem Cells, подана 20 июля 2010; патентная заявка США №2011/0014702 под названием Differentiation of Human Embryonic Stem Cells, подана 19 июля 2010; патентная заявка США №2011/0151561 под названием Differentiation of Human Embryonic Stem Cells, подана 16 декабря 2010; патентная заявка США №2010/0112692 под названием Differentiation of Human Embryonic Stem Cells, подана 22 октября 2009; патентная заявка США №2012/0052576 под названием Differentiation of Pluripotent Stem Cells, подана 17 августа 2011; патентная заявка США №2010/0112693 под названием Differentiation of Human Pluripotent Stem Cells, подана 23 октября 2009; патентная заявка США №2011/0151560 под названием Differentiation of Human Embryonic Stem Cells, подана 16 декабря 2010; патентная заявка США №2010/0015100 под названием Differentiation of Human Embryonic Stem Cells, подана 31 июля 2008; патентная заявка США №2009/0170198 под названием Differentiation of Human Embryonic Stem Cells, подана 25 ноября 2008; патентная заявка США №2015/0329828 под названием Use of Small Molecules to Enhance MAFA Expression in Pancreatic Endocrine Cells, подана 7 мая 2015; патентная заявка США №2013/0330823 под названием Differentiation of Human Embryonic Stem Cells into Pancreatic Endocrine Cells, подана 6 июня 2013; публикация международной заявки №WO 2013/192005 под названием Differentiation of Human Embryonic Stem Cells into Pancreatic Endocrine Cells, подана 13 июня 2013; Патентная заявка США №2014/0242693 под названием Suspension and Clustering of Human Pluripotent Stem Cells for Differentiation into Pancreatic Endocrine Cells, подана 30 декабря 2013; патентная заявка США №2014/0295552 под названием Suspension and Clustering of Human Pluripotent Stem Cells for Differentiation into Pancreatic Endocrine Cells, подана 17 июня 2014; публикация международной заявки № WO 2015/065524 под названием Suspension and Clustering of Human Pluripotent Stem Cells for Differentiation into Pancreatic Endocrine Cells, подана 21 мая 2014; патентная заявка США №2013/0330823 под названием Differentiation of Human Embryonic Stem Cells into Pancreatic Endocrine Cells, подана 6 июня 2013; патентная заявка США №2014/0186953 под названием Differentiation of Human Embryonic Stem Cells Into Pancreatic Endocrine Cells Using HB9 Regulators, подана 18 декабря 2013; патентная заявка США №14/963730, подана 9 декабря 2015; патентная заявка США №14/898,015, подана 11 декабря 2015, каждая из которых целиком включена в настоящее описание посредством ссылки.
[0067] В одном варианте осуществления, гипоиммуногенные клетки упакованы внутри устройства доставки клеток. Указанные устройства доставки клеток могут содержать нетканое полотно. Устройства доставки клеток содержат различные слои, каждый из которых выполняет одну или множество функций. В некоторых вариантах осуществления, указанные устройства доставки клеток включают как мембрану, отделяющую клетки, так и нетканое полотно. В другом варианте осуществления, указанным устройством доставки является устройство TheraCyte (ранее Baxter) (Ирвин, Калифорния). Устройства доставки клеток TheraCyte описаны в патентах США №№6,773,458; 6,156,305; 6,060,640; 5,964,804; 5,964,261; 5,882,354; 5,807,406; 5,800,529; 5,782,912; 5,741,330; 5,733,336; 5,713,888; 5,653,756; 5,593,440; 5,569,462; 5,549,675; 5,545,223; 5,453,278; 5,421,923; 5,344,454; 5,314,471; 5,324,518; 5,219,361; 5,100,392; и 5,011,494, каждый из которых целиком включен в настоящее описание посредством ссылки.
[0068] В другом варианте осуществления, указанным устройством является устройство, по существу описанное в патенте США №8,278,106 и патентной заявке США №14/201,630, поданной 7 марта 2014 и в заявке PCT/US2016/061442, поданной 10 ноября 2016, и в патентах США на промышленные образцы №29/447,944, 29/509,102, 29/484,363, 29/484,360, 29/484,359, 29/484,357, 29/484,356, 29/484,355, 29/484,362, 29/484,358, 29/408,366, 29/517,319, 29/408,368, 29/518,513, 29/518,516, 29/408,370, 29/517,144, 29/423,365, 29/530,325, 29/584,046, каждый из которых целиком включен в настоящее описание посредством ссылки. В других вариантах осуществления, устройства доставки клеток или агрегаты большой емкости включают одно или два или более уплотнений, которые дополнительно разделяют просвет устройства доставки клеток, то есть уплотнение перегородки. Смотри, например заявки США Заявителя на промышленные образцы №№29/408366, 29/408368, 29/408370, 29/423,365 и 29/584,046.
[0069] В одном варианте осуществления, гипоиммуногенные клетки имплантируют в перфорированные устройства доставки клеток, которые обеспечивают прямой межклеточный контакт между сосудистой сетью хозяина и инкапсулированными клетками. Средствами перфорации являются отверстия или поры в указанном устройстве. В некоторых вариантах осуществления не все слои устройства перфорированы. Например, смотри заявку PCT/US2016/0061442, которая целиком включена в настоящее описание посредством ссылки и которая раскрывает перфорированные устройства доставки клеток с перфорацией только в одном слое, например, в мембране, удерживающей клетки; или только в мембране, удерживающей клетки, и слое нетканого материала. В одном варианте осуществления, гипоиммуногенные клетки инкапсулированы в перфорированном устройстве и окружены нетканым материалом. В таких вариантах осуществления, указанный нетканый материал расположен на внешней стороне устройства доставки клеток. Вместо того, чтобы воздействовать на имплантированные клетки, указанный нетканый материал усиливает васкуляризацию хозяина, окружающего корпус, содержащий клетки. Смотри, например, заявку PCT/US2016/0061442 и патент США №8,278,106 (оба документа целиком включены в настоящее описание посредством ссылки), которые описывают перфорированные устройства и полимеры, использующиеся в указанном устройстве.
[0070] В одном варианте осуществления, указанные отверстия/перфорации меньше, чем клеточные агрегаты, содержащиеся в устройстве, такие как агрегаты, полученные из hPSC, например, клеточные агрегаты линии дефинитивной энтодермы, содержащиеся в нем. В одном случае перфорированное устройство для доставки клеток, имплантированное крысе или человеку, содержит перфорации только в удерживающей клетки мембране (другие слои устройства не перфорированы), причем указанные отверстия находятся на расстоянии примерно 2 мм или более друг от друга, причем диаметр указанного отверстия составляет менее чем примерно 100 мкм.
[0071] Истощение гипоиммуногенных клеток ("ген самоубийства")
[0072] Универсальность эмбриональных стволовых клеток и индуцированных плюрипотентных стволовых (iPS) клеток для замены и восстановления любой ткани в организме сопровождается повышенным риском развития рака. Повышенный риск развития рака также связан с генной терапией. Следовательно, перепрограммированные ткани, полученные из клеток ES или клеток iPS (Takahashi, K. & Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663-676, (2006), и Hanna, J.H., Saha, K. & Jaenisch, R. Pluripotency and Cellular Reprogramming: Facts, Hypotheses, Unresolved Issues. Cell 143, 508-525, (2010), каждая из статей целиком включена в настоящее описание посредством ссылки) или из других мультипотентных или прогениторных клеток, а также из клеток, обработанных векторами генной терапии, вызывают проблемы, связанные с безопасностью (Knoepfler, P.S. Deconstructing Stem Cell Tumorigenicity: A Roadmap to Safe Regenerative Medicine. Stem Cells 27, 1050-1056, (2009) целиком включена в настоящее описание посредством ссылки). Например, подкожно имплантированные клетки iPS вызывают тератомы, а у химерных животных с iPS с высокой частотой возникновения развивается примитивный злокачественный рак (Takahashi, K. & Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663-676, (2006); Knoepfler, P.S. Deconstructing Stem Cell Tumorigenicity: A Roadmap to Safe Regenerative Medicine. Stem Cells 27, 1050-1056, (2009)). В то время как доброкачественные тератомы могут быть легко удалены хирургическим путем, инвазивный рак остается риском при клеточной терапии.
[0073] Были рассмотрены стратегии преодоления туморогенности стволовых клеток, включая стратегию гена самоубийства (Knoepfler, P.S. Deconstructing Stem Cell Tumorigenicity: A Roadmap to Safe Regenerative Medicine. Stem Cells 27, 1050-1056, (2009)). В частности, некий ген может быть избирательно введен в указанные имплантируемые клетки, который кодирует фермент, который локально метаболизирует системно доступные пролекарство в активный противоопухолевый агент. Например, обработка ганцикловиром, который превращается тимидинкиназой в соединения, которые становятся токсичными после трифосфорилирования клеточными киназами, приводила к разрушению опухолевых клеток in vitro. Таким образом, имплантированные клетки могут быть модифицированы так, чтобы искусственно создать биохимические различия между тканями хозяина и имплантированными клетками. Нацеливание на указанные имплантируемые клетки достигается путем выбора вектора, используемого для доставки "гена самоубийства", а также с помощью биологии системы гена самоубийства/пролекарства. В результате, высокие дозы препарата, генерируемого только в той среде, где имплантируют клетки, ограничивают побочные эффекты в других тканях.
[0074] Истощение гипоиммуногенных клеток может быть достигнуто путем избирательного введения гена в гипоиммуногенную клетку, экспрессия которого либо непосредственно приводит к гибели гипоиммуногенной клетки, либо делает гипоиммуногенную клетку специфически восприимчивой к другим фармакологическим агентам. В соответствии с этим способом истощение гипоиммуногенной клетки in vivo или ex vivo может быть достигнуто путем доставки нужного гена в гипоиммуногенную клетку с использованием вирусных систем доставки генов, таких как система доставки гена ретровируса, аденовируса или аденоассоциированного вируса, но не ограничивается ими. Желаемая система доставки вируса может содержать вирус, геном которого кодирует белок, который, например, непосредственно вызывает гибель клетки, например, индуцируя апоптоз гипоиммуногенной клетки. В качестве альтернативы, система доставки вируса может содержать вирус, геном которого кодирует, например, ген тимидинкиназы вируса простого герпеса. Экспрессия гена тимидинкиназы вируса простого герпеса в гипоиммуногенной клетке делает гипоиммуногенную клетку чувствительной к фармакологическим дозам ганцикловира. Таким образом, последующий контакт вирусно-трансдуцированной гипоиммуногенной клетки с ганцикловиром приводит к гибели гипоиммуногенной клетки. Истощение гипоиммуногенных клеток может быть достигнуто путем введения так называемого "гена самоубийства" посредством систем редактирования генома, например, системы ZFN, CRISPR/cas и TALEN.
[0075] Такие агенты, как ганцикловир, которые вызывают гибель клетки при экспрессии гена, такого как тимидинкиназа, в настоящем документе называются "агентом, индуцирующим гибель клеток".
[0076] Гены, которые можно использовать для уничтожения гипоиммуногенных клеток, включают ген тимидинкиназы вируса простого герпеса и цитозиндеаминазы или любой ген, который вызывает гибель клетки, который может быть помещен под контроль индуцируемого промотора/регуляторной последовательности (упоминаемых в настоящем как "промотор/регуляторная последовательность" или "промотор"), но не ограничиваются ими. Указанный ген переносят в гипоиммуногенную клетку, клетки отбирают при соответствующем селективном давлении, затем указанные клетки переносят пациенту и дают возможность прижиться в нем. Затем пациенту вводят агент, который индуцирует активность промотора, тем самым индуцируя экспрессию гена, продукт которого вызывает гибель гипоиммуногенных клеток. В случае тимидинкиназы также могут быть использованы другие агенты, которые облегчают гибель клетки с помощью этого фермента, такие как, например, ганцикловир (Bonini и др., 1997, Science 276:1719-1724; Bordignon и др., 1995, Human Gene Therapy 6:813-819; Minasi и др., 1993, J. Exp. Med. 177:1451-1459; Braun и др., 1990, Biology of Reproduction 43:684-693). Другие гены, пригодные для этой цели, включают гены конститутивно активных форм каспаз 3, 8, и 9, ген bax, ген гранзимов, гены дифтерийного токсина, токсина Pseudomonas А, рицина и других токсинов, которые раскрыты в другом месте настоящего документа, но не ограничиваются ими. Создание соответствующих конструкций для доставки таких генов человеку будет очевидно специалистам в данной области техники и описано, например, в Sambrook и др. (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York) и в Ausubel и др. (1997, Current Protocols in Molecular Biology, John Wiley & Sons, New York).
[0077] Важно, чтобы ген, который переносится в гипоиммуногенные клетки с целью уничтожения клеток, находился под контролем соответствующей последовательности промотора, так что индукция экспрессии указанного гена может осуществляться при добавлении к клеткам (введение млекопитающему) соответствующего индуктора. Такие индуцируемые промоторные последовательности включают промоторы, которые индуцируются при добавлении к клеткам металла, стероид-индуцируемые промоторы и подобные им, но не ограничиваются ими. В одном предпочтительном варианте осуществления можно использовать промоторную систему экдизона. В этом варианте осуществления промотор экдизона клонируют перед последовательностью белка рецептора экдизона, которая расположена выше последовательности второго промотора, который управляет экспрессией сайта связывания экдизона, функционально связанного с желаемым геном, например, желаемым токсином. Индукция промотора индуцирует экспрессию токсина, тем самым вызывая уничтожение гибель клетки, в которой находится ген токсина.
[0078] Клетки, в которые трансдуцировали ген для уничтожения клеток, когда такие клетки трансдуцируют ex vivo, могут быть отобраны (т.е. отделены от клеток, которые не содержат указанный ген) путем предоставления клеткам маркера селекции в дополнение к трансдуцированному гену. Маркеры селекции хорошо известны в данной области техники и описаны, например, в Sambrook и др. (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, N.Y.).
[0079] Кроме того, истощение гипоиммуногенных клеток может быть достигнуто путем введения в популяцию гипоиммуногенных клеток олигонуклеотида (например, антисмысловой молекулы, но не ограничиваясь ей) или рибозима, которые способны вызывать гибель гипоиммуногенной клетки, или индукции нарушения функции гипоиммуногенной клеточной. Такие олигонуклеотиды включают олигонуклеотиды, которые нацелены на существенную функцию гипоиммуногенной клетки, называемые в настоящем документе как олигонуклеотиды, которые либо убивают гипоиммуногенную клетку, либо нарушают функцию гипоиммуногенной клетки в отношении стимуляции Т-клеток. Такие функции гипоиммуногенной клетки включают среди других совместную стимуляторную функцию В71 и В72, CD40, но не ограничиваются ими. Таким образом, олигонуклеотиды и рибозимы, которые пригодны в способах согласно настоящему изобретению, включают олигонуклеотиды и рибозимы, которые направлены против этих мишеней, но не ограничиваются ими.
[0080] Как отмечено в настоящем документе, истощение гипоиммуногенных клеток включает нарушение функции гипоиммуногенных клеток. Нарушение функции гипоиммуногенных клеток включает все формы нарушения в гипоиммуногенных клетках с физическим удалением или истощением гипоиммуногенных клеток или без них. Так, нарушение функции гипоиммуногенных клеток включает использование антитела, которое блокирует функцию молекул гипоиммуногенной поверхности клетки, которые являются критическими для гипоиммуногенной функции клетки.
[0081] В качестве альтернативы, пептиды, которые блокируют функцию молекул поверхности гипоиммуногенной клеточной, блокирование которых приводит к нарушению функции гипоиммуногенных клеток, могут быть использованы для эффективного истощения гипоиммуногенных клеток в организме хозяина. Такие пептиды включают пептиды, которые предназначены для специфического связывания рецепторных молекул на поверхности гипоиммуногенных клеток, и пептиды, которые предназначены, например, для ингибирования важных ферментативных функций в этих клетках, но не ограничиваются ими.
[0082] Аналогично, гены и олигонуклеотиды, которые предназначены для той же цели, что и гены и олигонуклеотиды описанные в настоящем документе, также включены в качестве инструментов в способы согласно настоящему изобретению. Таким образом, пептиды, олигонуклеотиды и гены, которые нарушают биологическую функцию гипоиммуногенной клетки, как этот термин определен в настоящем документе, также предполагаются для использования в способах согласно настоящему изобретению, раскрытых в данном документе.
[0083] Настоящее изобретение также охватывает применение фармацевтических композиций, подходящих в качестве композиций, истощающих гипоиммуногенные клетки, для практического осуществления способов согласно настоящему изобретению, причем указанные композиции содержат пригодную композицию, истощающую гипоиммуногенные клетки, и фармацевтически приемлемый носитель. В некоторых вариантах осуществления указанная композиция, истощающая клетки, представляет собой химерную композицию, содержащую антитело и токсин.
[0084] Используемый в настоящем описании термин "фармацевтически приемлемый носитель" означает химическую композицию, с которой можно комбинировать подходящую композицию, истощающую гипоиммуногенные клетки, и которую после комбинации можно использовать для введения подходящей композиции, истощающей гипоиммуногенные клетки, в млекопитающее.
[0085] Фармацевтические композиции, пригодные для использования в способах согласно настоящему изобретению, можно вводить системно в виде пероральных твердых составов, офтальмологических, суппозиторных, аэрозольных, местных или других подобных составов. В дополнение к композиции, истощающей гипоиммуногенные клетки, такие фармацевтические композиции могут содержать фармацевтически приемлемые носители и другие ингредиенты, которые, как известно, усиливают и облегчают введение лекарственного средства. Другие возможные составы, такие как наночастицы, липосомы, повторно запечатанные эритроциты и системы, основанные на иммунологических принципах, также могут быть использованы для введения соответствующей композиции, истощающей гипоиммуногенные клетки, в соответствии со способами настоящего изобретения.
[0086] Способы введения "генов самоубийства" в клетки раскрыты в заявке US 20060222633, которая целиком включена в настоящее описание посредством ссылки.
[0087] Настоящее изобретение включает способ истощения гипоиммуногенных клеток у хозяина млекопитающего. После того, как указанные гипоиммуногенные клетки трансплантировали хозяину, указанный способ включает контакт гипоиммуногенных клеток с композицией, истощающей клетки, для воздействия на функцию гипоиммуногенных клеток или уничтожение гипоиммуногенных клеток, тем самым истощая гипоиммуногенные клетки у хозяина млекопитающего.
[0088] В другом аспекте указанная композиция, разрушающая гипоиммуногенные клетки, выбрана из группы, состоящей из токсина, антитела, радиоактивной молекулы, нуклеиновой кислоты, пептида, пептидомиметика и рибозима.
[0089] В одном аспекте указанным токсином является иммунотоксин. Указанный токсин выбран из группы, состоящей из рицина, дифтерийного токсина и псевдомонадного экзотоксина А.
[0090] В другом варианте осуществления, указанное антитело выбрано из группы, состоящей из антитела, специфичного к CD1a, антитела, специфичного к CD11c, антитела, специфичного к ГKГCII, антитела, специфичного к CD11b, антитела, специфичного к DEC205, антитела, специфичного к В71, антитела, специфичного к В72, антитела, специфичного к CD40, антитела, специфичного к лектинам типа 1, и антитела, специфичного к лектинам типа II.
[0091] В еще одном варианте осуществления молекула нуклеиновой кислоты выбрана из группы, состоящей из гена и олигонуклеотида.
[0092] В следующем варианте осуществления указанная радиоактивная молекула представляет собой радиоактивно меченное антитело.
[0093] В другом варианте осуществления, указанная антигенная истощающая композиция представляет собой химерную композицию, содержащую антитело и токсин. Указанный токсин может быть выбран из группы, состоящей из рицина, дифтерийного токсина и псевдомонадного экзотоксина А.
[0094] В другом варианте осуществления, указанное антитело выбрано из группы, состоящей из антитела, специфичного к CD1a, антитела, специфичного к CD11c, антитела, специфичного к ГKГCII, антитела, специфичного к CD11b, антитела, специфичного к DEC205, антитела, специфичного к В71, антитела, специфичного к В72, антитела, специфичного к CD40, антитела, специфичного к лектинам типа I, и антитела, специфичного к лектинам типа II.
Комбинированный продукт
[0095] Варианты осуществления, описанные в данном документе, также раскрывают комбинированный продукт, который относится к устройству, нагруженному гипоиммуногенными клетками или терапевтическим агентом, то есть каждый из них может быть подходящим медицинским устройством или продуктом клеток, но при совместном использовании они образуют комбинированный продукт. В одном случае, указанный комбинированный продукт относится к перфорированному устройству, нагруженному гипоиммуногенными клетками. Это называется "комбинированный продукт" или "перфорированный комбинированный продукт". Указанное устройство (перфорированное или нет) может представлять собой любое макроустройство доставки клеток, описанное в настоящем документе, включая устройства для инкапсуляции клеток, которые описаны в патентах США №№8,278,106 и 9,526,880, заявке PCT/US2016/0061442 и патентах США на промышленные образцы №№ D714956, D718472, D718467, D718466, D718468, D718469, D718470, D718471, D720469, D726306, D726307, D728095, D734166, D734847, D747467, D747468, D747798, D750769, D750770, D755986, D760399, D761423, D761424 (целиком включенные в настоящее описание посредством ссылки), но не ограничиваются ими. Клетки, загруженные в указанное устройство (перфорированное или нет), могут быть любыми гипоиммуногенными клетками, обсуждаемыми выше, включая клетки дефинитивной эндодермы, PDX1-позитивной энтодермы, PDX1-позитивной эндодермы передней кишки, панкреатической эндодермы, клетки панкреатической эндодермы, экспрессирующие PDX1 nNKX6.1, эндокринные предшественники, эндокринные предшественники, экспрессирующие NKX6.1 и INS, незрелые бета-клетки, незрелые бета-клетки, экспрессирующие NKX6.1, INS и MAFB, зрелые эндокринные клетки, зрелые эндокринные клетки, экспрессирующие INS, GCG, SST и РР, и зрелые бета-клетки и зрелые бета-клетки, экспрессирующие INS и MAFA, но не ограничиваются ими.
[0096] Перфорированные устройства доставки, нагруженные клетками панкреатической энтодермы или панкреатическими предшественниками гипоиммуногенных клеток, которые созревают при имплантации in vivo, предназначены для уменьшения инсулиновой зависимости и/или уменьшения гипогликемии у пациентов с диабетом. Это включает пациентов с высоким риском диабетом I типа, которые не знают о гипогликемии, лабильны или получили трансплантацию органов и которые могут переносить или уже находятся на иммуносупрессивной терапии, но не ограничивается ими. Как по существу описано в заявке PCT/US2016/0061442 (целиком включена посредством ссылки), основным методом действия является использование клеток панкреатической эндодермы (РЕС) человека или гипоиммуногенных клеток предшественников поджелудочной железы, содержащихся в проницаемом, долговечном имплантируемом медицинском устройстве, что облегчает прямую васкуляризацию хозяина. Указанные гипоиммуногенные клетки РЕС после имплантации дифференцируются и созревают в терапевтические, чувствительные к глюкозе, высвобождающие инсулин гипоиммуногенные клетки. Таким образом, перфорированный комбинированный продукт поддерживает секрецию инсулина человека. Перфорированный комбинированный продукт ограничивает распространение (выход) РЕС гипоиммуногенных клеток in vivo. Перфорированный комбинированный продукт имплантируют в место, в котором обеспечивается достаточная степень прорастания сосудов для поддержания популяции терапевтических гипоиммуногенных клеток в устройстве и облегчения распределения инсулина и других панкреатических продуктов в кровоток. Перфорированный комбинированный продукт предназначен для имплантации и извлечения с помощью обычных хирургических инструментов, а также для обеспечения терапевтической дозы в течение двух и более лет. Указанное устройство предназначено для сохранения адекватной дозы продукта гипоиммуногенных клеток РЕС во время создания, срока годности, обработки и хирургической имплантации для достижения клинической эффективности и обеспечения того, чтобы клеточный продукт находился внутри капсулы ткани в соответствии с требованиями безопасности.
Нокин
[0097] В некоторых вариантах осуществления, толерогенные факторы могут быть введены или повторно введены в линии стволовых клеток с отредактированным геномом, чтобы создать иммунно привилегированные линии универсальных донорских стволовых клеток. В некоторых случаях заявленные в настоящем описании универсальные стволовые клетки были дополнительно модифицированы для экспрессии одного или нескольких толерогенных факторов. Типичные толерогенные факторы включают один или несколько ЛАЧ-С, ЛАЧ-Е, ЛАЧ-F, ЛАЧ-G, PD-L1, CTLA-4-Ig, CD47, CI-ингибитор и IL-35, но не ограничиваются ими. Для облегчения введения толерогенных факторов, таких как указанные выше толерогенные факторы, может быть использован любой метод редактирования генов в локус AAVS1 для активного ингибирования иммунного отторжения.
[0098] В частности, в некоторых вариантах осуществления, настоящие изобретения, раскрытые в настоящем документе, относятся к стволовым клеткам, геном которых был изменен для ослабления или удаления критических компонентов, по меньшей мере, одного гена ГКГС-класса I и, по меньшей мере, одного гена лиганда, активирующего NK, и который был дополнительно изменен для увеличения экспрессии одного или нескольких толерогенных факторов. В некоторых вариантах осуществления, изобретения, раскрытые в настоящем документе, относятся к стволовой клетке, геном которой был изменен для ослабления или удаления критических компонентов, по меньшей мере, одного гена ГКГС-класса I и, по меньшей мере, одного гена лиганда, активирующего NK, и который был дополнительно изменен для увеличения экспрессии одного или нескольких из белков ЛАЧ-С, ЛАЧ-Е, ЛАЧ-Ф, ЛАЧ-Г, PD-L1, CTLA-4-Ig, CD47, CI-ингибитора и IL-35.
Варианты осуществления
[0099] Другие варианты осуществления настоящего изобретения описаны со ссылкой на пронумерованные параграфы ниже.
[00100] Относящиеся к блокирующим антителам: композиция
[00101] Параграф 1: Композиция, содержащая плюрипотентную клетку, в которой отсутствует по меньшей мере один ген человеческого лейкоцитарного антигена (ЛАЧ) класса I, и по меньшей мере один агент, который связывается с лигандом, активирующим естественные клетки-киллеры (NK).
[00102] Параграф 2: Композиция согласно параграфу 1, в которой указанным агентом является антитело.
[00103] Параграф 3: Композиция согласно параграфу 1, в которой указанным геном ЛАЧ класса I является В2М.
[00104] Параграф 4: Композиция согласно параграфам 1-3, в которой указанным лигандом, активирующим NK-клетки, являются ICAM-1, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00105] Параграф 5: Композиция согласно параграфам 1-2, в которой указанным лигандом, активирующим NK-клетки, являются ICAM-1 и CEACAM1.
[00106] Параграф 6: Композиция согласно параграфам 1-2, в которой указанным лигандом, активирующим NK-клетки, являются ICAM-1, CEACAM1, CADM1, MICA и MICB.
[00107] Параграф 7: Композиция согласно параграфам 1-2, в которой указанные клетки, полученные из плюрипотентных клеток, дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных плюрипотентных клеток.
[00108] Параграф 8: Композиция согласно параграфу 7, в которой указанным агентом, способным вызывать гибель клеток, является ганцикловир.
[00109] Параграф 9: Композиция согласно любому параграфу 1-8, в которой клетки, полученные из плюрипотентных клеток, дополнительно сверхэкспрессируют один или несколько толерогенных факторов.
[00110] Параграф 10: Композиция согласно параграфу 9, в которой указанными толерогенными факторами являются ЛАЧ-С, ЛАЧ-Е, ЛАЧ-G, PD-L1, CTLA-4-Ig, CD47, CI-ингибитор или IL-35.
[00111] Параграф 11: Композиция согласно параграфу 9, в которой указанными толерогенными факторами являются ЛАЧ-С, ЛАЧ-Е и ЛАЧ-G.
[00112] Относящиеся к блокирующим антителам: Способ
[00113] Параграф 1: Способ предотвращения отторжения трансплантата клеток, полученных из плюрипотентных клеток человека, включающий введение субъекту, нуждающемуся в лечении, композиции, включающей популяцию клеток-мишеней, в которой отсутствует по меньшей мере один ген ЛАЧ класса I, и по меньшей мере один агент, связывающий лиганд, активирующий NK-клетку, на клетке-мишени в количестве, эффективном для подавления атаки NK-клетками субъекта, тем самым предотвращая отторжение трансплантата клеток, полученных из плюрипотентных клеток человека.
[00114] Параграф 2: Способ согласно параграфу 1, в котором указанным агентом является антитело.
[00115] Параграф 3: Способ согласно параграфу 1, в котором иммунный ответ субъекта на лиганд, активирующий NK-клетку, подавляется.
[00116] Параграф 4: Способ согласно параграфу 1, в котором указанным лигандом, активирующим NK-клетку, является ICAM-1, CEACAM1, CADM1, MICA или MICB.
[00117] Параграф 5: Способ согласно параграфу 1, в котором указанным лигандом, активирующим NK-клетку, является ICAM-1, CEACAM1, CADM1, MICA и MICB.
[00118] Параграф 6: Способ согласно параграфу 1, в котором указанным субъектом является человек.
[00119] Параграф 7: Способ согласно параграфу 2, в котором указанным антителом является антитело человека.
[00120] Относящиеся к двойному нокауту hES-клеток: композиции [00121] Параграф 1: Популяция клеток in vitro, включающая клетки, полученные из плюрипотентных клеток, в которой клетки, полученные из плюрипотентных клеток, не имеют по меньшей мере одного гена ЛАЧ класса I и по меньшей мере одного гена лиганда, активирующего клетку естественного киллера (NK).
[00122] Параграф 2: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанным геном ЛАЧ класса является В2М.
[00123] Параграф 3: Указанная популяция клеток in vitro согласно параграфу 1-2, в которой указанным лигандом, активирующим NK-клетку, является ICAM-1, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00124] Параграф 4: Указанная популяция клеток in vitro согласно параграфу 1-2, в которой указанным лигандом, активирующим NK-клетку, является ICAM-1 и CEACAM1.
[00125] Параграф 5: Указанная популяция клеток in vitro согласно параграфу 1-2, в которой указанным лигандом, активирующим NK-клетку, является ICAM-1, CEACAM1, CADM1, MICA и MICB.
[00126] Параграф 6: Указанная популяция клеток in vitro согласно параграфу 1-5, в которой указанные клетки, полученные из плюрипотентных клеток, дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных клеток, полученных из плюрипотентных клеток.
[00127] Параграф 7: Указанная популяция клеток in vitro согласно параграфу 6, в которой указанным агентом, вызывающим гибель клеток, является ганцикловир.
[00128] Параграф 8: Указанная популяция клеток in vitro согласно любому из параграфов 1-7, в которой указанные клетки, полученные из плюрипотентных клеток, дополнительно суперэкспрессируют один или более толерогенных факторов.
[00129] Параграф 9: Указанная популяция клеток in vitro согласно параграфу 8, в которой указанными толерогенными факторами являются ЛАЧ-С, ЛАЧ-Е, ЛАЧ-G, PD-L1, CTLA-4-Ig, CD47, CI-ингибитор, IL-35 или их комбинация.
[00130] Параграф 10: Указанная популяция клеток in vitro согласно параграфу 9, в которой указанными толерогенными факторами являются ЛАЧ-С, ЛАЧ-Е и ЛАЧ-G.
[00131] Относящиеся к тройному нокауту плюрипотентных стволовых клеток: композиции
[00132] Плюрипотентная стволовая клетка человека, содержащая модифицированный геном, содержащий: (а) первую модификацию генома, посредством которой ген В2М был отредактирован для уменьшения или устранения экспрессии В2М на поверхности клетки и/или активности В2М в клетке; (b) вторую модификацию генома, в которой ген ICAM-1 был отредактирован для уменьшения или устранения экспрессии ICAM-1 на поверхности клетки и/или активности ICAM-1 в клетке; и (с) третью модификацию генома, в которой ген CEACAM1 был отредактирован для уменьшения или устранения экспрессии CEACAM1 на поверхности клетки и/или активности CEACAM1 в клетке.
[00133] Относящиеся к нокауту регуляторов транскрипции: композиции
[00134] Параграф 12: Клетка, полученная из плюрипотентных клеток, содержащая пониженную экспрессию одного или нескольких генов ГКГС класса I или ГКГС класса II или комплексов белок и одного или нескольких лигандов, активирующих NK-клетку, по сравнению с плюрипотентной стволовой клетки дикого типа, в которой плюрипотентная стволовая клетка имеет один или более генов, кодирующих один или несколько регуляторов транскрипции генов-активирующих лигандов клеток ГКГС-класса I или ГКГС-класса II и NK, удаленных по меньшей мере из одного аллеля клетки.
[00135] Параграф 13: Плюрипотентная стволовая клетка, обладающая пониженной экспрессией одного или нескольких лигандов, активирующих NK-клетки, по сравнению с плюрипотентными стволовыми человеческими клетками дикого типа.
[00136] Параграф 14: Плюрипотентная стволовая клетка человека, которая не экспрессирует В2М или ICAM-1.
[00137] Параграф 15: Плюрипотентная стволовая клетка человека, которая не экспрессирует CIITA или ICAM-1.
[00138] Параграф 16: Плюрипотентная стволовая клетка человека, которая не экспрессирует LRC5 или ICAM-1.
[00139] Параграф 17: Плюрипотентная стволовая клетка человека, которая не экспрессирует один или более из NLRC5, CIITA и В2М, и дополнительно не экспрессирует один или более из ICAM-1, CEACAM1, CADM1, MICA и MICB.
[00140] Параграф 18: Плюрипотентная стволовая клетка человека, которая не экспрессирует один или более из ЛАЧ-А, ЛАЧ-В и ЛАЧ-С и дополнительно не экспрессирует один или более из ICAM-1, CEACAM1, CADM1, MICA и MICB.
[00141] Параграф 19: Плюрипотентная стволовая клетка человека, которая не экспрессирует один или более из антигенов ГКГС класса I и один или более лигандов, активирующих NK-клетки, и дополнительно один или более толерогенных факторов, вставлен в локус, являющийся безопасным местом, по крайней мере одного аллеля клетки.
[00142] Параграф 20: Плюрипотентная стволовая клетка человека, содержащая модифицированный геном, содержащий первую геномную модификацию, в которой ген В2М был отредактирован для уменьшения или устранения экспрессии В2М на поверхности клетки и/или активности В2М в клетке, и в котором ген ICAM-1 был отредактирован для уменьшения или устранения экспрессии ICAM-1 на поверхности клетки и/или активности ICAM-1 в клетке.
[00143] Параграф 21: Плюрипотентная стволовая клетка человека, содержащая модифицированный геном, содержащий первую геномную модификацию, в которой ген В2М был отредактирован для уменьшения или устранения экспрессии В2М на поверхности клетки и/или активности В2М в клетке, и в котором ген ICAM-1 был отредактирован для уменьшения или устранения экспрессии ICAM-1 на поверхности клетки и/или активности ICAM-1 в клетке.
[00144] Относящиеся к двойному нокауту клеток hES: композиции
[00145] Параграф 1: Популяция клеток in vitro, включающая плюрипотентные клетки, в которых у указанных плюрипотентных клеток отсутствует по меньшей мере один функциональный белок поверхности ГКГС класса I и по крайней мере один функциональный белок поверхности клеток, активирующий естественные клетки-киллеры (NK).
[00146] Параграф 2: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанным белком поверхности клеток ГКГС класса I являются ЛАЧ-А, ЛАЧ-В, ЛАЧ-С или их комбинация.
[00147] Параграф 3: Указанная популяция клеток in vitro согласно параграфам 1-2, в которой указанным белком поверхности клеток ГКГС класса I является белок В2М.
[00148] Параграф 4: Указанная популяция клеток in vitro по любому из параграфов 1-3, в которой у указанных плюрипотентных клеток отсутствуют по меньшей мере два функциональных белка поверхности клеток, активирующих NK-клетки.
[00149] Параграф 5: Указанная популяция клеток in vitro по любому из параграфов 1-4, в которой у указанных плюрипотентных клеток отсутствует по меньшей мере три функциональных белка поверхности клеток, активирующих NK-клетки.
[00150] Параграф 6: Указанная популяция клеток in vitro по любому из параграфов 1-5, в которой указанным белком поверхности клеток, активирующим NK-клетки, является ICAM-1, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00151] Параграф 7: Указанная популяция клеток in vitro по любому из параграфов 1-5, в которой указанным белком поверхности клеток, активирующим NK-клетки, является ICAM-1 и CEACAM1.
[00152] Параграф 8: Указанная популяция клеток in vitro по любому из параграфов 1-7, в которой указанные плюрипотентные клетки дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных плюрипотентных клеток.
[00153] Параграф 9: Указанная популяция клеток in vitro по любому из параграфов 1-8, в которой указанным белком, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанным агентом, способным вызывать гибель указанных плюрипотентных клеток, является белок вируса простого герпеса, тимидинкиназа или цитозиндеаминаза.
[00154] Параграф 10: Указанная популяция клеток in vitro по любому из параграфов 8-9, в которой указанным агентом, вызывающим гибель клеток, является ганцикловир.
[00155] Относящиеся к двойному нокауту клеток hES: композиции
[00156] Параграф 1: Популяция клеток in vitro, включающая плюрипотентные клетки, в которых указанные плюрипотентные клетки имеют пониженную экспрессию по меньшей мере одного белка поверхности клеток ГКГС класса I и пониженную функцию и/или экспрессию по меньшей мере одного белкового лиганда клеточной поверхности, активирующего NK-клетки, по сравнению с исходным генотипом или по сравнению с человеческой клеткой дикого типа.
[00157] Параграф 2: Популяция клеток in vitro, включающая плюрипотентные клетки, в которой указанные плюрипотентные клетки имеют пониженную экспрессию одного или более из белков поверхности клеток ЛАЧ-А, ЛАЧ-В и ЛАЧ-С и пониженную функцию и/или экспрессию по меньшей мере одного белкового лиганда клеточной поверхности, активирующего NK-клетки, по сравнению с исходным генотипом или по сравнению с человеческой клеткой дикого типа.
[00158] Параграф 3: Популяция клеток in vitro, включающая плюрипотентные клетки, в которой у указанных плюрипотентных клеток отсутствует экспрессия функциональных белков поверхности клеток ЛАЧ, экспрессия лигандов белков поверхности клеток, активирующих NK-клетки, и имеется белок, который экспрессируется в указанных плюрипотентных клетках в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных плюрипотентных клеток.
[00159] Параграф 4: Стволовая клетка, в которой экспрессия одного или более белков поверхности клеток ЛАЧ класса I и одного или более белкового лиганда поверхности клеток, активирующего NK-клетки, снижена по сравнению со стволовой клеткой дикого типа.
[00160] Параграф 5: Плюрипотентная клетка, в которой экспрессия одного или более белка поверхности клеток ЛАЧ класса I, одного или более белкового лиганда поверхности клеток, активирующего NK-клетки, и одного или более толерогенного белкового фактора поверхности клеток снижена по сравнению с плюрипотентной клеткой дикого типа и в которой указанные плюрипотентные клетки дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных плюрипотентных клеток.
[00161] Параграф 6: Популяция клеток in vitro, включающая плюрипотентные клетки, в которой у указанных плюрипотентных клеток отсутствует функциональные гены ГКГС класса I и гены лиганда, активирующего естественные клетки-киллеры (NK), и в которой указанные плюрипотентные клетки обладают повышенной экспрессией толерогенного белкового фактора поверхности клеток по сравнению с плюрипотентной клеткой дикого типа и в которой указанные плюрипотентные клетки дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных плюрипотентных клеток.
[00162] Относящиеся к двойному нокауту клеток РЕС: композиция
[00163] Параграф 1: Популяция клеток in vitro, содержащая клетки панкреатической эндодермы (РЕС), в которой в указанных клетках РЕС отсутствует по меньшей мере один функциональный ген ЛАЧ класса I и по меньшей мере один ген лиганда, активирующего естественные клетки-киллеры (NK).
[00164] Параграф 2: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанным по меньшей мере одним геном ЛАЧ класса I является ген В2М.
[00165] Параграф 3: Указанная популяция клеток in vitro согласно параграфу 1-2, в которой указанным по меньшей мере одним лигандом, активирующим NK-клетки, является ICAM-1, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00166] Параграф 4: Указанная популяция клеток in vitro согласно параграфу 1-2, в которой указанным по меньшей мере одним лигандом, активирующим NK-клетки, является ICAM-1 и CEACAM1.
[00167] Параграф 5: Указанная популяция клеток in vitro согласно параграфу 1-2, в которой указанным по меньшей мере одним лигандом, активирующим NK-клетки, является ICAM-1, CEACAM1, CADM1, MICA и MICB.
[00168] Параграф 6: Указанная популяция клеток in vitro согласно параграфу 1-5, в которой указанные клетки РЕС дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных клеток РЕС.
[00169] Параграф 7: Указанная популяция клеток in vitro согласно параграфу 6, в которой указанным агентом, вызывающим гибель клеток, является ганцикловир.
[00170] Параграф 8: Указанная популяция клеток in vitro согласно параграфу 6, в которой указанным геном, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных клеток РЕС, является ген простого вируса герпеса, тимидинкиназы или цитозиндеаминазы.
[00171] Параграф 9: Указанная популяция клеток in vitro по любому из параграфов 1-8, в которой указанные клетки РЕС дополнительно суперэкспрессируют один или более толерогенный белок поверхности клеток.
[00172] Параграф 10: Указанная популяция клеток in vitro согласно параграфам 1-9, в которой указанными толерогенными факторами являются ЛАЧ-С, ЛАЧ-Е, ЛАЧ-G, PD-L1, CTLA-4-Ig, CD47, CI-ингибитор, IL-35 или их комбинация.
[00173] Параграф 11: Указанная популяция клеток in vitro согласно параграфам 1-9, в которой указанными толерогенными факторами являются ЛАЧ-С, ЛАЧ-Е и ЛАЧ-G.
[00174] Параграф 12: Популяция клеток in vitro, содержащая клетки панкреатической эндодермы (РЕС), в которой в указанных клетках РЕС отсутствует по меньшей мере один функциональный ген ГКГС класса I, ген ГКГС класса II и ген лиганда, активирующего естественные клетки-киллеры (NK).
[00175] Параграф 13: Популяция клеток in vitro, содержащая клетки панкреатической эндодермы (РЕС), в которой в указанных клетках РЕС отсутствует по меньшей мере один функциональный ген ГКГС класса I и ген ГКГС класса II, а также отсутствуют по меньшей мере два гена лиганда, активирующего естественные клетки-киллеры (NK).
[00176] Параграф 14: Клетка панкреатической эндодермы (РЕС), в которой экспрессия одного или более генов ЛАЧ класса I и одного или более генов лиганда, активирующего NK-клетки, снижена по сравнению с клетками РЕС дикого типа.
[00177] Параграф 15: Клетка панкреатической эндодермы (РЕС), в которой экспрессия одного или более белков поверхности клеток ЛАЧ класса I, одного или более лиганда, активирующего NK-клетки, и одного или более толерогенных факторов снижена по сравнению с клетками РЕС дикого типа и в которой указанная клетка панкреатической эндодермы дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных клеток РЕС.
[00178] Параграф 16: Клетка панкреатической эндодермы (РЕС), которая не экспрессирует В2М или ICAM-1.
[00179] Параграф 17: Клетка панкреатической эндодермы (РЕС), которая не экспрессирует CIITA или ICAM-1.
[00180] Параграф 18: Клетка панкреатической эндодермы (РЕС), которая не экспрессирует LRC5 или ICAM-1.
[00181] Параграф 19: Клетка панкреатической эндодермы (РЕС), которая не экспрессирует один или более из NLRC5, CIITA и В2М и дополнительно не экспрессирует один или более из ICAM-1, CEACAM1, CADM1, MICa и MICB.
[00182] Параграф 20: Клетка панкреатической эндодермы (РЕС), которая не экспрессирует один или более из ЛАЧ-А, ЛАЧ-В и ЛАЧ-С и дополнительно не экспрессирует один или более из ICAM-1, CEACAM1, CADM1, MICa и MICB.
[00183] Параграф 21: Клетка панкреатической эндодермы (РЕС), которая не экспрессирует один или более белков поверхности клеток ГКГС класса I и один или более лигандов, активирующих NK-клетки, и дополнительно имеет один или более толерогенных факторов, введенных в безопасный локус по меньшей мере одного аллеля указанной клетки.
[00184] Параграф 22: Клетка панкреатической эндодермы (РЕС), содержащая модифицированный геном, содержащий первую модификацию генома, в которой указанный ген В2М был изменен для снижения или упразднения экспрессии В2М на поверхности указанной клетки и/или активности в указанной клетке и, в которой указанный ген ICAM-1 был изменен для снижения или упразднения Экспрессии ICAM-1 на поверхности указанной клетки и/или активности в указанной клетке.
[00185] Параграф 23: Клетка панкреатической эндодермы (РЕС), содержащая модифицированный геном, содержащий: первую модификацию генома, в которой указанный ген В2М был изменен для снижения или упразднения экспрессии В2М на поверхности указанной клетки и/или активности в указанной клетке; и (b) вторую модификацию генома, в которой указанный ген ICAM-1 был изменен для снижения или упразднения экспрессии ICAM-1 на поверхности указанной клетки и/или активности в указанной клетке.
[00186] Параграф 24: Клетка панкреатической эндодермы (РЕС), содержащая модифицированный геном, содержащий: первую модификацию генома, в которой указанный ген В2М был изменен для снижения или упразднения экспрессии В2М на поверхности указанной клетки и/или активности в указанной клетке; (b) вторую модификацию генома, в которой указанный ген ICAM-1 был изменен для снижения или упразднения экспрессии ICAM-1 на поверхности указанной клетки и/или активности в указанной клетке; и (с) третью модификацию генома, в которой указанный ген CEACAM1 был изменен для снижения или упразднения экспрессии CEACAM1 на поверхности указанной клетки и/или активности в указанной клетке.
Относящиеся к гипоиммуногенные клетки
[00187] Параграф 1: Популяция клеток in vitro, содержащая гипоиммуногенные клетки, в которой в указанных гипоиммуногенных клетках отсутствует по меньшей мере один функциональный белок поверхности клеток ЛАЧ класса I и по меньшей мере один функциональный Белок лиганда, активирующего NK-клетки, поверхности клеток.
[00188] Параграф 2: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанным белком поверхности клеток ЛАЧ класса I является В2М.
[00189] Параграф 3: Указанная популяция клеток in vitro согласно параграфу 1-2, в которой указанным лигандом, активирующим NK-клетку, является ICAM-1, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00190] Параграф 4: Указанная популяция клеток in vitro согласно параграфу 1-2, в которой указанным лигандом, активирующим NK-клетку, является ICAM-1 и CEACAM1.
[00191] Параграф 5: Указанная популяция клеток in vitro согласно параграфу 1-2, в которой указанным лигандом, активирующим NK-клетку, является ICAM-1, CEACAM1, CADM1, MICA и MICB.
[00192] Параграф 6: Указанная популяция клеток in vitro согласно параграфу 1-5, в которой указанные гипоиммуногенные клетки дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных гипоиммуногенных клеток.
[00193] Параграф 7: Указанная популяция клеток in vitro по любому из параграфов 1-6, в которой указанные гипоиммуногенные клетки дополнительно суперэкспрессируют один или более толерогенных факторов.
[00194] Параграф 8: Указанная популяция клеток in vitro согласно параграфам 1-7, в которой указанными толерогенными факторами являются ЛАЧ-С, ЛАЧ-Е, ЛАЧ-G, PD-L1, CTLA-4-Ig, CD47, CI-ингибитор, IL-35 и их комбинации.
[00195] Параграф 9: Указанная популяция клеток in vitro согласно параграфам 1-7, в которой указанными толерогенными факторами являются ЛАЧ-С, ЛАЧ-Е и ЛАЧ-G.
[00196] Параграф 10: Популяция клеток in vitro, содержащая гипоиммуногенные клетки, в которой в указанных гипоиммуногенных клетках отсутствует по меньшей мере один функциональный ген ГКГС класса I и по меньшей мере один Ген лиганда, активирующего NK-клетки.
[00197] Параграф 11: Популяция клеток in vitro, содержащая гипоиммуногенные клетки, в которой в указанных гипоиммуногенных клетках отсутствует по меньшей мере один функциональный ген ГКГС класса I, ген ГКГС класса II и ген лиганда, активирующего NK-клетки.
[00198] Параграф 12: Указанная популяция клеток in vitro согласно параграфам 10 или 11, в которой указанным геном ГКГС класса I является ЛАЧ-А, ЛАЧ-В, ЛАЧ-С или их комбинация.
[00199] Параграф 13: Указанная популяция клеток in vitro согласно параграфам 10, 11 или 12, в которой указанным геном ГКГС класса I является В2М.
[00200] Параграф 14: Указанная популяция клеток in vitro по любому из параграфов 10-13, в которой в указанных гипоиммуногенных клетках отсутствует по меньшей мере два функциональных гена лиганда, активирующего NK-клетки.
[00201] Параграф 15: Указанная популяция клеток in vitro по любому из параграфов 10-13, в которой в указанных гипоиммуногенных клетках отсутствует по меньшей мере три функциональных гена лиганда, активирующего NK-клетки.
[00202] Параграф 16: Указанная популяция клеток in vitro по любому из параграфов 10-13, в которой лигандом, активирующим NK-клетки, является ICAM-1, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00203] Параграф 17: Указанная популяция клеток in vitro по любому из параграфов 10-13, в которой указанным лигандом, активирующим NK-клетку, является ICAM-1 и CEACAM1.
[00204] Параграф 18: Указанная популяция клеток in vitro по любому из параграфов 10-13, в которой указанным лигандом, активирующим NK-клетку, является ICAM-1, CEACAM1, CADM1, MICA и MICB.
[00205] Параграф 19: Указанная популяция клеток in vitro по любому из параграфов 10-18, в которой указанные гипоиммуногенные клетки дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных гипоиммуногенных клеток.
[00206] Параграф 20: Указанная популяция клеток in vitro по любому из параграфов 7-15, в которой указанными гипоиммуногенными клетками являются клетки hES или линия панкреотических клеток.
[00207] Параграф 21: Гипоиммуногенная клетка, в которой экспрессия одного или более белков поверхности клеток ЛАЧ класса 1 и одного или более белок лиганда, активирующего NK-клетки, поверхности клеток снижена по сравнению с гипоиммуногенной клеткой дикого типа.
[00208] Параграф 22: Гипоиммуногенная клетка, в которой экспрессия одного или более белка поверхности клеток ЛАЧ класса I, одного или более лиганда, активирующего NK-клетки, и одного или более толерогенного белкового фактора поверхности клеток снижена по сравнению с гипоиммуногенной клеткой дикого типа и в которой указанные плюрипотентные клетки дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных гипоиммуногенных клеток.
[00209] Параграф 23: Гипоиммуногенная стволовая клетка, в которой снижена экспрессия одного или более белков поверхности клеток ГКГС класса I или ГКГС класса II и одного или более лигандов, активирующих NK-клетки, по сравнению с плюрипотентной стволовой клеткой, в которой в указанной плюрипотентной стволовой клетке один или более генов, кодирующих один или более регуляторов транскрипции ГКГС класса I или ГКГС класса II и один или более генов лиганда, активирующего NK-клетки, удалены из по меньшей мере одного аллеля указанной клетки.
[00210] Параграф 24: Гипоиммуногенная клетка, в которой снижена экспрессия одного или более лигандов, активирующих NK-клетки, по сравнению с человеческими гипоиммуногенными клетками дикого типа.
[00211] Параграф 25: Гипоиммуногенная клетка человека, которая не экспрессирует В2М или ICAM-1.
[00212] Параграф 26: Гипоиммуногенная клетка человека, которая не экспрессирует СИТА или ICAM-1.
[00213] Параграф 27: Гипоиммуногенная клетка человека, которая не экспрессирует LRC5 или ICAM-1.
[00214] Параграф 28: Гипоиммуногенная клетка человека, которая не экспрессирует один или более из NLRC5, СИТА и В2М и дополнительно не экспрессирует один или более из ICAM-1, CEACAM1, CADM1, MICa и MICB.
[00215] Параграф 29: Гипоиммуногенная клетка человека, которая не экспрессирует один или более из ЛАЧ-А, ЛАЧ-В и ЛАЧ-С и дополнительно не экспрессирует один или более из ICAM-1, CEACAM1, CADM1, MICa и MICB.
[00216] Параграф 30: Гипоиммуногенная клетка человека, которая не экспрессирует один или более из одного или более белков поверхности клеток ГКГС класса I и одного или более лигандов, активирующих NK-клетки, и дополнительно содержит один или более толерогенных факторов, введенных в безопасный локус по меньшей мере одного аллеля указанной клетки.
[00217] Параграф 31: Гипоиммуногенная клетка, содержащая модифицированный геном, содержащий первую модификацию генома, в которой указанный ген В2М был изменен для снижения или упразднения экспрессии В2М на поверхности указанной клетки и/или активности в указанной клетке и в которой указанный ген ICAM-1 был изменен для снижения или упразднения экспрессии ICAM-1 на поверхности указанной клетки и/или активности в указанной клетке.
[00218] Параграф 32: Гипоиммуногенная стволовая клетка, содержащая модифицированный геном содержащий: первую модификацию генома, в которой указанный ген В2М был изменен для снижения или упразднения экспрессии В2М на поверхности указанной клетки и/или активности в указанной клетке; и (b) вторую модификацию генома, в которой указанный ген ICAM-1 был изменен для снижения или упразднения экспрессии ICAM-1 на поверхности указанной клетки и/или активности в указанной клетке.
[00219] Параграф 33: Гипоиммуногенная стволовая клетка, содержащая модифицированный геном, содержащий: первую модификацию генома, в которой указанный ген В2М был изменен для снижения или упразднения экспрессии В2М на поверхности указанной клетки и/или активности в указанной клетке; (b) вторую модификацию генома, в которой указанный ген ICAM-1 был изменен для снижения или упразднения экспрессии ICAM-1 на поверхности указанной клетки и/или активности в указанной клетке; и (с) третью модификацию генома, в которой указанный ген CEACAM1 был изменен для снижения или упразднения экспрессии CEACAM1 на поверхности указанной клетки и/или активности в указанной клетке.
Способы получения двойного нокаута
[00220] Параграф 1: Способ снижения вероятности отторжения трансплантата, включающий:
а) введение субъекту, нуждающемуся в трансплантате, эффективного количества трансплантата, содержащего популяцию клеток панкреатической эндодермы, в которой функция по меньшей мере один белок поверхности клеток ЛАЧ класса I и по меньшей мере один белок лиганда, активирующего NK-клетки, поверхности клеток нарушена.
[00221] Параграф 2: Способ согласно параграфу 1, в которой указанным белком поверхности клеток ЛАЧ класса I является В2М.
[00222] Параграф 3: Способ согласно параграфу 1-2, в которой указанным белком лиганда, активирующего NK-клетки, поверхности клеток является ICAM-1, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00223] Параграф 4: Способ согласно параграфу 1-2, в которой указанным лигандом, активирующим NK-клетку, является ICAM-1 и CEACAM1.
[00224] Параграф 5: Способ согласно параграфу 1-2, в которой указанным лигандом, активирующим NK-клетку, является ICAM-1, CEACAM1, CADM1, MICA и MICB.
[00225] Параграф 6: Способ снижения количества гипоиммуногенных клеток в популяции клеток, включающий приведение в контакт указанных гипоиммуногенных клеток с композицией, снижающей количество гипоиммуногенных клеток, для ослабления функции гипоиммуногенных клеток или приводящей к гибели указанных гипоиммуногенных клеток, снижая таким образом количество указанных гипоиммуногенных клеток в указанной популяции клеток.
[00226] Параграф 7: Способ по параграфу 6, в котором в указанных гипоиммуногенных клетках отсутствует экспрессия по меньшей мере одного функционального белка поверхности клеток ЛАЧ класса I и по меньшей мере одного лиганда, активирующего NK-клетки.
[00227] Параграф 8: Способ удаления гипоиммуногенных клеток из хозяина-млекопитающего, включающий: (а) перенос гипоиммуногенных клеток указанному хозяину-млекопитающему; и (b) приведение хозяина в контакт с композицией, уменьшающей количество гипоиммуногенных клеток, для ослабления функции указанных гипоиммуногенных клеток или вызывающей гибель указанных гипоиммуногенных клеток, что приводит к удалению указанных гипоиммуногенных клеток в указанном хозяине-млекопитающем.
[00228] Параграф 9: Способ согласно параграфу 8, в которой указанная функция по меньшей мере одного белка поверхности клеток ЛАЧ класса I и по меньшей мере одного лиганда, активирующего NK-клетки, уменьшена.
[00229] Параграф 10: Способ повышения количества NK-активирующих лигандов в популяции клеток-мишеней, включающий экспонирование популяции клеток-мишеней с IFN-γ, повышая таким образом количество NK-активирующих лигандов в клеток-мишени по сравнению с клеткой дикого типа.
Относящиеся к плюрипотентным клеткам, в которых функция как гена ГКГС класса I, так и гена лиганда, активирующего NK-клетки, нарушена или ингибирована: композиции
[00230] Параграф 1: Популяция клеток in vitro, содержащая плюрипотентные клетки, в которых функция по меньшей мере одного гена главного комплекса гистосовместимости (ГКГС) класса I и по меньшей мере одного гена лиганда, активирующего естественные клетки-киллеры (NK), нарушена или заингибирована.
[00231] Параграф 2: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанный ген ГКГС кодирует бета-2 микроглобулин (В2М).
[00232] Параграф 3: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанным лигандом, активирующим NK-клетку, является ICAM1, CD58, PVR, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00233] Параграф 4: Указанная популяция клеток in vitro согласно параграфу I, в которой указанным лигандом, активирующим NK-клетку, является ICAM1 и CD58.
[00234] Параграф 5: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанным лигандом, активирующим NK-клетку, является ICAM1, CD58, CD155, CEACAM1, CADM1, MICA и MICB.
[00235] Параграф 6: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанными плюрипотентными клетками являются эмбриональные стволовые клетки человека.
[00236] Параграф 7: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанные плюрипотентные клетки дифференцированы в клетки панкреатической эндодермы.
[00237] Параграф 8: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанные плюрипотентные клетки дополнительно содержат белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных клеток.
[00238] Параграф 9: Указанная популяция клеток in vitro согласно параграфу 1, в которой указанный ген ГКГС класса I разрушен с использованием средств для редактирования генома.
[00239] Параграф 10: Указанная популяция клеток in vitro согласно параграфу 9, в которой указанным средством для редактирования генома являются нуклеаза белкового домена "цинковые пальцы" (ZFN), система кластерных, регулярно перемежающихся, коротких, палиндромных повторов (CRISPR)/cas или система эффекторной нуклеазы, подобной активаторам транскрипции, (TALEN).
[00240] Параграф 11: Указанная популяция клеток in vitro согласно параграфу 1, в которой ген указанного лиганда, активирующего NK-клетку, разрушен с использованием средств для редактирования генома.
[00241] Параграф 12: Указанная популяция клеток in vitro согласно параграфу 1, в которой функция указанного лиганда, активирующего NK-клетку, нарушена с использованием агента против лиганда, активирующего NK-клетку.
[00242] Параграф 13: Указанная популяция клеток in vitro согласно параграфу 8, в которой указанным агентом является антитело.
Относящиеся к линии панкреотических клеток, в которых функции как гена ГКГС класса I, так и гена лиганда, активирующего NK-клетки, нарушены или ингибированы: композиции
[00243] Параграф 14: Популяция клеток in vitro, содержащая линию панкреотических клеток, в которой функция по меньшей мере одного гена главного комплекса гистосовместимости (ГКГС) класса I и по меньшей мере одного гена лиганда, активирующего естественные клетки-киллеры (NK), нарушен или ингибирован.
[00244] Параграф 15: Указанная популяция клеток in vitro согласно параграфу 14, в которой указанный ген ГКГС класса I кодирует В2М.
[00245] Параграф 16: Указанная популяция клеток in vitro согласно параграфу 14, в которой указанным лигандом, активирующим NK-клетку, является ICAM1, CD58, CD155, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00246] Параграф 17: Указанная популяция клеток in vitro согласно параграфу 14, в которой указанным лигандом, активирующим NK-клетку, является ICAM1 и CD58.
[00247] Параграф 18: Указанная популяция клеток in vitro согласно параграфу 14, в которой указанным лигандом, активирующим NK-клетку, является ICAM1, CD58, CD 155, CEACAM1, CADM1, MICA и MICB.
[00248] Параграф 19: Указанная популяция клеток in virto согласно параграфу 14, в которой указанной линией панкреотических клеток являются клетки дефинитивной эндодермы, эндодермы передней части пищеварительного тракта, клетки панкреатической эндодермы, предшественники эндокринных клеток или клетки, продуцирующие инсулин.
[00249] Параграф 20: Указанная популяция клеток in vitro согласно параграфу 14, в которой в указанной линии панкреотических клеток дополнительно экспрессируют белок, который экспрессируется в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанных клеток.
Способы предотвращения отторжения трансплантата клеток поджелудочной железы
[00250] Параграф 21: Способ предотвращения отторжения трансплантата клеток поджелудочной железы человека, включающий:
[00251] а. обеспечение популяции клеток поджелудочной железы человека, не экспрессирующих по меньшей мере один белок поверхности клеток главного комплекса гистосовместимости (ГКГС) класса I и не экспрессирующих по меньшей мере один лиганд, активирующий естественные клетки-киллеры (NK); и
[00252] б. трансплантацию указанной популяции клеток поджелудочной железы человека в субъект, представляющий собой млекопитающее, в которой отсутствие экспрессии белка поверхности клеток ГКГС класса I и лиганда, активирующего NK-клетки, предотвращает отторжение трансплантата клеток.
[00253] Параграф 22: Способ согласно параграфу 21, в которой указанным белком поверхности клеток главного комплекса гистосовместимости (ГКГС) класса I является бета-2 микроглобулин (В2М) или белок поверхности клеток ЛАЧ-АВС.
[00254] Параграф 23: Способ согласно параграфу 21, в которой указанным лигандом, активирующим NK-клетку, является ICAM1, CD58, CD155, PVR, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00255] Параграф 24: Способ согласно параграфу 21, в которой указанной популяцией клеток поджелудочной железы человека являются клетки панкреатической эндодермы (РЕС).
[00256] Параграф 25. Способ согласно параграфу 21, в которой указанная популяция клеток поджелудочной железы человека дополнительно содержит ген белка, который экспрессируется в указанной популяции клеток человека в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанной популяции клеток поджелудочной железы человека.
Способы получения инсулина
[00257] Параграф 26. Способ получения инсулина, включающий:
[00258] а) обеспечение популяции клеток панкреатической эндодермы человека, не экспрессирующих по меньшей мере один белок поверхности клеток главного комплекса гистосовместимости (ГКГС) класса I и не экспрессирующих по меньшей мере один лиганд, активирующий естественные клетки-киллеры (NK); и
[00259] б) трансплантацию указанной популяции клеток панкреатической эндодермы человека в субъект, представляющий собой млекопитающее, в которой указанные клетки панкреатической эндодермы созревают в субъекте, представляющем собой млекопитающее, и производят инсулин в ответ на стимуляцию глюкозой.
[00260] Параграф 27: Способ согласно параграфу 26, в которой указанным белком поверхности клеток главного комплекса гистосовместимости (ГКГС) класса I является бета-2 микроглобулин (В2М) или белок поверхности клеток ЛАЧ-АВС.
[00261] Параграф 28. Способ согласно параграфу 26, в которой указанным лигандом, активирующим NK-клетку, является ICAM1, CD58, CD155, PVR, CEACAM1, CADM1, MICA, MICB или их комбинация.
[00262] Параграф 29: Способ согласно параграфу 26, в которой указанная популяция клеток поджелудочной железы человека дополнительно содержит ген белка, который экспрессируется в указанной популяции клеток поджелудочной железы человека в присутствии агента, вызывающего гибель клеток, при этом указанный агент способен вызывать гибель указанной популяции клеток поджелудочной железы человека.
[00263] Параграф 30. Способ согласно параграфу 29, в которой указанным белком является тимидинкиназа простого вируса герпеса, а указанным агентом является ганцикловир.
[00264] Способ получения гипоиммуногенной стволовой клетки, включающий снижение экспрессии одного или более белков поверхности клеток ГКГС класса I и одного или более NK-активирующих лигандов в указанной гипоиммуногенной стволовой клетке и получение таким образом указанной гипоиммуногенной стволовой клетки.
[00265] Способ получения гипоиммуногенной стволовой клетки, включающий снижение экспрессии одного или более белков поверхности клеток ГКГС класса I, одного или более лиганда, активирующего NK-клетки, и снижение экспрессии одного или более толерогенных факторов в указанной стволовой клетке и получение таким образом указанной гипоиммуногенной стволовой клетки.
[00266] Способ снижения экспрессии одного или более белков поверхности клеток ГКГС класса I и лиганда, активирующего NK-клетки, белка поверхности клеток в стволовой клетке, включающий делецию одного или более генов, кодирующих один или более регуляторов транскрипции генов ГКГС класса I и лигандов, активирующих NK-клетки, из по меньшей мере одного аллеля указанной клетки и снижение таким образом экспрессии указанного одного или более белка поверхности клеток ГКГС класса I и лиганда, активирующего NK-клетки, белка поверхности клеток.
[00267] Гипоиммуногенная стволовая клетка, содержащая модифицированный геном, содержащий (а) первую модификацию генома, в которой указанный ген В2М был изменен для снижения или элиминирования экспрессии В2М на поверхности указанной клетки и/или активности в указанной клетке посредством приведения в контакт указанной клетки либо с белком Cas или нуклеиновой кислотой, кодирующей белок Cas, в комбинации с либо плазмидой, кодирующей рибонуклеиновую кислоту, которая гомологична любой из последовательностей SEQ IDNO: 1-3, либо с рибонуклеиновой кислотой, которая гомологична любой из последовательностей SEQ ID NO: 1-3; и (б) вторую модификацию генома, в которой указанный ген ICAM-1 был изменен для снижения или элиминирования экспрессии ICAM-1 на поверхности указанной клетки и/или активности в указанной клетке посредством приведения в контакт указанной клетки либо с белком Cas, либо с нуклеиновой кислотой, кодирующей белок Cas, в комбинации либо с плазмидой, кодирующей рибонуклеиновую кислоту, которая гомологична любой из последовательностей SEQ ID NO: 4-6, либо с рибонуклеиновой кислотой, которая гомологична любой из последовательностей SEQ ID NO: 4-6.
[00268] Гипоиммуногенная стволовая клетка, содержащая модифицированный геном, содержащий (а) первую модификацию генома, в которой указанный ген В2М был изменен для снижения или элиминирования экспрессии В2М на поверхности указанной клетки и/или активности в указанной клетке посредством приведения в контакт указанной клетки либо с белком Cas, либо с нуклеиновой кислотой, кодирующей белок Cas, в комбинации либо с плазмидой, кодирующей рибонуклеиновую кислоту, которая гомологична любой из последовательностей SEQ ID NO: 1-3, либо с рибонуклеиновой кислотой, которая гомологична любой из последовательностей SEQ ID NO: 1-3; (б) вторую модификацию генома, в которой указанный ген ICAM-1 был изменен для снижения или элиминирования экспрессии ICAM-1 на поверхности указанной клетки и/или активности в указанной клетке посредством приведения в контакт указанной клетки либо с белком Cas, либо с нуклеиновой кислотой, кодирующей белок Cas, в комбинации либо с плазмидой, кодирующей рибонуклеиновую кислоту, которая гомологична любой из последовательностей SEQ ID NO: 4-6, либо с рибонуклеиновой кислотой, которая гомологична любой из последовательностей SEQ ID NO: 4-6; и (с) третью модификацию генома, в которой указанный ген CEACAM1 был изменен для снижения или элиминирования экспрессии CEACAM1 на поверхности указанной клетки и/или активности в указанной клетке посредством приведения в контакт указанной клетки либо с белком Cas, либо с нуклеиновой кислотой, кодирующей белок Cas, в комбинации либо с плазмидой, кодирующей рибонуклеиновую кислоту, которая гомологична любой из последовательностей SEQ ID No: 7-9, либо с рибонуклеиновой кислотой, которая гомологична любой из последовательностей SEQ ID NO: 7-9.
[00269] Плюрипотентная стволовая клетка, содержащая модифицированный геном, содержащий (а) первую модификацию генома, в которой указанный ген В2М был изменен для снижения или элиминирования экспрессии В2М на поверхности указанной клетки и/или активности в указанной клетке посредством приведения в контакт указанной клетки либо с белком Cas, либо с нуклеиновой кислотой, кодирующей белок Cas, в комбинации либо с плазмидой, кодирующей рибонуклеиновую кислоту, которая гомологична любой из последовательностей SEQ ID NO: 1-3, либо с рибонуклеиновой кислотой, которая гомологична любой из последовательностей SEQ ID NO: 1-3; и (б) вторую модификацию генома, в которой указанный ген ICAM-1 был изменен для снижения или элиминирования экспрессии ICAM-1 на поверхности указанной клетки и/или активности в указанной клетке посредством приведения в контакт указанной клетки либо с белком Cas, либо с нуклеиновой кислотой, кодирующей белок Cas, в комбинации либо с плазмидой, кодирующей рибонуклеиновую кислоту, которая гомологична любой из последовательностей SEQ ID NO: 4-6, либо с рибонуклеиновой кислотой, которая гомологична любой из последовательностей SEQ ID NO: 4-6.
[00270] Указанные последовательности SEQ ID No: 1-9 приведены ниже в Таблице 4, а также эти и дополнительные последовательности описаны ниже.
[00271] SEQ ID NO. 1: Экзон 1, отрицательная цепь.
[00272] SEQ ID NO. 2: Экзон 2, отрицательная цепь.
[00273] SEQ ID NO. 3: Экзон 1, отрицательная цепь.
[00274] SEQ ID NO. 4: Экзон 2, положительная цепь.
[00275] SEQ ID NO. 5: Экзон 2, отрицательная цепь.
[00276] SEQ ID NO. 6: Экзон 1, положительная цепь.
[00277] SEQ ID NO. 7: Экзон 1, отрицательная цепь.
[00278] SEQ ID NO. 8: Экзон 1, положительная цепь.
[00279] SEQ ID NO. 9: Экзон 1, положительная цепь.
[00280] SEQ ID NO. 10: кодирует последовательность ICAM1 человека.
[00281] SEQ ID NO. 11: кодирует последовательность CEACAM1 человека.
[00282] SEQ ID NO. 12: кодирует последовательность В2М человека.
[00283] SEQ ID NO. 13: кодирует последовательность CADM1 человека.
[00284] SEQ ID NO. 14: кодирует последовательность CD58 человека.
[00285] SEQ ID NO. 15: кодирует последовательность CD155 человека.
[00286] Целевые последовательности для расщепления CRISPR7Cas9, включающие РАМ (NGG), приведены в таблице ниже.
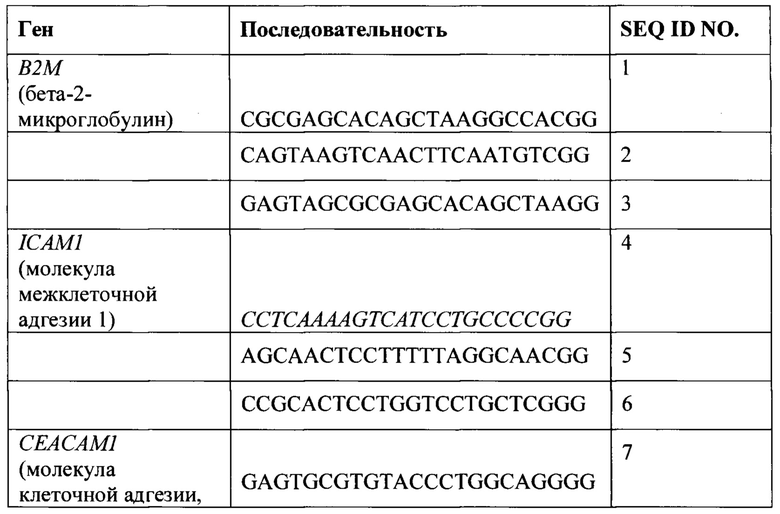
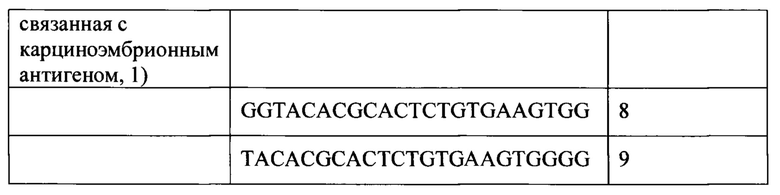
[00287] Клетка панкреатической эндодермы (РЕС), содержащая модифицированный геном, содержащий (а) первую модификацию генома, в которой указанный ген В2М был изменен для снижения или упразднения экспрессии В2М на поверхности указанной клетки и/или активности в указанной клетке посредством приведения в контакт указанной клетки с белком Cas или нуклеиновой кислотой, кодирующей белок Cas, и рибонуклеиновой кислотой, содержащей любую последовательность из SEQ ID NO: 1-3; (б) вторую модификацию генома, в которой указанный ген ICAM-1 был изменен для снижения или упразднения экспрессии ICAM-1 на поверхности указанной клетки и/или активности в указанной клетке посредством приведения в контакт указанной клетки с белком Cas или нуклеиновой кислотой, кодирующей белок Cas, и рибонуклеиновой кислотой, содержащей любую последовательность из SEQ ID NO: 4-6.; и (в) третью модификацию генома, в которой указанный ген CEACAM1 был изменен для снижения или упразднения экспрессии CEACAM1 на поверхности указанной клетки и/или активности в указанной клетке посредством приведения в контакт указанной клетки с белком Cas или нуклеиновой кислотой, кодирующей белок Cas, и рибонуклеиновой кислотой, содержащей любую последовательность из SEQ ID NO: 7-9.
[00288] Способ снижения гипогликемии, включающий:
а) введение субъекту, нуждающемуся в трансплантате, эффективного количества трансплантата, содержащего популяцию клеток панкреатической эндодермы, в которой нарушена функция по меньшей мере одного белка поверхности клеток ЛАЧ класса I и по меньшей мере одного белка лиганда поверхности клеток, активирующего NK-клетки, в которой указанная популяция клеток панкреатической эндодермы созревает in vivo и производит инсулин в ответ на стимуляцию глюкозой in vivo, снижая таким образом гипогликемию у пациента.
[00289] Способ снижения зависимости от инсулина, включающий: а) введение субъекту, нуждающемуся в трансплантате, эффективного количества трансплантата, содержащего популяцию клеток панкреатической эндодермы, в которой нарушена функция по меньшей мере одного белка поверхности клеток ЛАЧ класса I и по меньшей мере одного белка лиганда поверхности клеток, активирующего NK-клетки, в которой указанная популяция клеток панкреатической эндодермы созревает in vivo и производит инсулин в ответ на стимуляцию глюкозой in vivo, снижая таким образом зависимость от инсулина у пациента.
ОПРЕДЕЛЕНИЯ
[00290] "Гипоиммуногенные" или "универсальные донорские клетки" или "мутантные клетки" или их эквиваленты означают клетки с пониженной или устраненной экспрессией по меньшей мере одного белка поверхности клеток ЛАЧ класса I и по меньшей мере одного лиганда, активирующего NK-клетки. Ожидается, что такая клетка будет менее подвержена иммунному отторжению или отторжению трансплантата субъектом, которому трансплантируются такие клетки или трансплантат. Например, по сравнению с неизмененными клетками дикого типа, такая гипоиммуногенная клетка может быть примерно на 2,5%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97,5%, 99% или более менее подвержена иммунному отторжению субъектом, в который трансплантировали такие клетки.
[00291] Термин "лечение" или "вылечивание" или их эквиваленты относится к терапевтическому вмешательству, которое уменьшает (улучшает) признак или симптом.
[00292] Термин "пациент", "хозяин", "хозяин-млекопитающее", "субъект" или их эквиваленты относится к живым многоклеточным позвоночным организмам, категории, которая включает как млекопитающих, так и людей. В некоторых вариантах осуществления указанным субъектом является человек. Предпочтительным пациентом для лечения является человек. Целевые группы пациентов могут меняться со временем клинического использования / опыта способами, которые не зависят от самого комбинированного продукта, а скорее связаны с характером режима иммуносупрессии или его отсутствием. Например, комбинированный продукт может использоваться в популяции T1D с использованием терапии с помощью гипоиммуногенных клеток в комбинации с режимом ведения иммунодепрессанта (ISD), который обеспечивает операционную переносимость или низкий профиль токсичности и побочных эффектов.
[00293] Термин "блокирующий агент" в данном документе относится к любому агенту, способному связываться с лигандом, активирующим NK-клетки, на поверхности клетки-мишени, включая антитело, но не ограничиваясь им; или относится к агенту, который предотвращает или ингибирует экспрессию белка лиганда, активирующего NK-клетки, включая белок, фермент или химическое вещество, известное в настоящее время или разработанное позднее, но не ограничивается ими.
[00294] Термин "антитело" в данном документе используется в самом широком смысле и конкретно охватывает интактные моноклональные антитела, поликлональные антитела, полиспецифические антитела (например, биспецифичные антитела), образованные из по меньшей мере двух интактных антител, и фрагменты антител, если они проявляют желаемую биологическую блокирующую активность.
[00295] Используемый в настоящем описании термин "фрагменты антител" относится к части интактного антитела, предпочтительно содержащей его антигенсвязывающую или вариабельную области. Примеры фрагментов антител включают фрагменты Fab, Fab', F(ab')2 и Fv, диатела, линейные антитела, молекулы одноцепочечных антител и мультиспецифичные антитела, образованные из фрагментов антител.
[00296] Термин "блокирующее антитело", используемый в настоящем документе, относится к антителу, которое, когда оно связывается с лигандом, активирующим NK-клетки, на клетке-мишени in vivo или in vitro, приводит к предупреждению или снижению способности NK-клеток вызывать лизис клетки-мишени.
[00297] Используемый в настоящем описании термин "сингенный" относится к клеткам, тканям или органам, которые являются генетически идентичными или получены из генетически идентичного источника для реципиента трансплантата (например, идентичного близнеца), особенно в отношении антигенов или иммунологических реакций. Такие клетки, ткани или органы называют изографтами. Используемый в настоящем описании термин "аллогенный" относится к клеткам, тканям или органам, которые не являются генетически идентичными или получены из не генетически идентичного источника для реципиента трансплантата (например, неродственного донора), особенно в отношении антигенов или иммунологических реакций. Такие клетки, ткани или органы называются аллотрансплантатами, аллогенными трансплантатами, гомотрансплантатами или аллотрансплантатами.
[00298] Используемый в настоящем описании термин "промотор/регуляторная последовательность" означает последовательность нуклеиновой кислоты, которая необходима для экспрессии продукта гена, функционально связанного с промотором/регуляторной последовательностью. В некоторых случаях эта последовательность может быть основной последовательностью промотора, а в других случаях эта последовательность может также включать последовательность энхансера и другие регуляторные элементы, которые необходимы для экспрессии генного продукта. Указанный промотор/регуляторная последовательность может, например, представлять собой последовательность, которая экспрессирует генный продукт тканеспецифическим образом.
[00299] Термин "эффективное количество" или "терапевтически эффективное количество" или их эквиваленты относится к количеству агента, достаточному для достижения желаемого эффекта у субъекта или клетки, подвергаемой лечению. Например, это может быть количество клеток, необходимое для ингибирования или для значительного снижения уровня глюкозы в крови и, в конечном итоге, для достижения гомеостатического гликемического контроля. Это также может означать эффективное количество агента для изменения функции или структуры клетки или субъекта. Терапевтически эффективное количество агента может быть введено в одной дозе или в нескольких дозах. Тем не менее, эффективное количество будет зависеть от конкретного применяемого средства, субъекта, которого лечат, тяжести и типа заболевания, а также от способа введения.
[00300] Термины "уменьшение", "разрушение", "сниженный", "снижение" и "ингибирование" используются в настоящем описании, как правило, для обозначения уменьшения, в частности уменьшения на статистически значимое значение. Однако во избежание сомнений "уменьшенный", "сниженный", "снижение", "заингибированный" включает уменьшение по меньшей мере на 10% по сравнению с контрольным уровнем, например, уменьшение по меньшей мере примерно на 20% или по меньшей мере примерно на 30%, или, по меньшей мере, примерно на 40%, или, по меньшей мере, примерно на 50%, или, по меньшей мере, примерно на 60%, или, по меньшей мере, примерно на 70%, или, по меньшей мере, примерно на 80%, или, по меньшей мере, примерно на 90% или вплоть до в том числе снижение на 100% (т.е. отсутствует по сравнению с контрольным образцом) или любое снижение на 10-100% по сравнению с контрольным уровнем, или, по крайней мере, примерно в 2 раза, или, по крайней мере, примерно в 3 раза, или по меньшей мере, примерно в 4 раза, или, по меньшей мере, примерно в 5 раз, или, по меньшей мере, примерно в 10 раз, или любое уменьшение в 2-10 раз или более по сравнению с контрольным уровнем.
[00301] Термины "увеличенный", "увеличивать" или "усиливать" или "активировать" используются в настоящем описании для обозначения увеличения на статически значимую величину; во избежание каких-либо сомнений термины "увеличенный", "увеличивать" или "усиливать" или "активировать" означают увеличение по меньшей мере на 10% по сравнению с контрольным уровнем, например, увеличение по меньшей мере примерно на 20%, или по меньшей мере, примерно на 30%, или, по меньшей мере, примерно на 40%, или, по меньшей мере, примерно на 50%, или, по меньшей мере, примерно на 60%, или, по меньшей мере, примерно на 70%, или, по меньшей мере, примерно на 80%, или, по меньшей мере, примерно на 90%, или вплоть до увеличения на 100%, или любое увеличение на 10-100% по сравнению с контрольным уровнем, или, по меньшей мере, примерно в 2 раза, или, по меньшей мере, примерно в 3 раза, или, по меньшей мере, примерно в 4 раза, или, по крайней мере, примерно в 5 раз, или, по крайней мере, примерно в 10 раз, или любое увеличение в 2-10 раз или больше по сравнению с контрольным уровнем.
[00302] Термин "статистически значимый" или "значительно" относится к статистической значимости и обычно означает два стандартных отклонения (2SD) ниже нормальной или более низкой концентрации эталона. Термин также может означать два стандартных отклонения (2SD) выше нормальной или более высокой концентрации эталона. Термин относится к статистическому свидетельству того, что обнаруживается разница. Он определяется как вероятность принятия решения отклонить нулевую гипотезу, когда нулевая гипотеза действительно верна. Решение часто принимается с использованием значения р.
[00303] Используемый в настоящем описании термин "пониженная гипогликемия" или его эквиваленты означает уменьшение числа эпизодов гипогликемии вместе с отсутствием ухудшения гликемического контроля, определяемое увеличением HbA1c на 0,2%.
[00304] Используемый в настоящем описании термин "уменьшенная зависимость от инсулина" или его эквиваленты означает уменьшение количества и/или дозы экзогенных инъекций инсулина вместе с отсутствием ухудшения гликемического контроля, определяемое увеличением HbA1c на 0,2%.
[00305] Используемый в настоящем описании термин "тканевая капсула" или его эквиваленты означает капсулу инородного тела, которую формируют вокруг имплантата или трансплантата. Комбинированный продукт и/или устройство или перфорированное устройство, содержащее определенные клетки, предназначены для удержания в капсуле в течение периода имплантации.
[00306] "Трансплантация" или его эквиваленты относится к дифференциации популяции предшественников или созревания клеточной популяции в зрелые клеточные типы. Например, трансплантация PDX1-позитивной популяции клеток панкреатической эндодермы, созревающей в популяцию эндокринных клеток поджелудочной железы.
[00307] "Трансплантат" относится к дифференцированной клеточной популяции, инкапсулированной или доставленной в устройствах, описанных в настоящем документе. Например, клеточные популяции, включая панкреатическую эндодерму, предшественник поджелудочной железы, PDX-1-позитивную панкреатическую эндодерму, эндокринный предшественник поджелудочной железы, панкреатический эндокринный трансплантат, однократно или полигормональный эндокринный трансплантат, пре-бета, бета, и/или инсулин секретирующие трансплантаты, но не ограничиваются ими.
[00308] Термин "по существу" или его эквиваленты означает в основном или de minimus или уменьшенное количество компонента или клетки, присутствующей в любой клеточной популяции или культуре, например, зрелой культуре бета-клеток, которые "по существу зрелые бета-клетки, экспрессирующие INS, NKX6.1 и PDX1 и по существу не экспрессирующие NGN3". Другие примеры включают "по существу клетки hES", "по существу клетки дефинитивной эндодермы", "по существу клетки эндодермы передней части пищеварительного тракта", "по существу PDX1-негативные клетки эндодермы передней части пищеварительного тракта", "по существу PDX1-позитивные клетки панкреатической эндодермы», "по существу предшественники эндокринных клеток поджелудочной железы", "по существу эндокринные клетки поджелудочной железы" и тому подобное, но не ограничиваются ими.
[00309] Что касается клеток в клеточных культурах или в клеточных популяциях, термин "по существу не содержит" или его эквиваленты означает, что указанный тип клеток, от которых свободны клетки культуры или клеточная популяция, присутствует в количестве менее чем примерно 10%, менее примерно 9%, менее примерно 8%, менее примерно 7%, менее примерно 6%, менее примерно 5%, менее примерно 4%, менее примерно 3%, менее примерно 2% или менее чем около 1% от общего числа клеток, присутствующих в указанных культурах клеток или клеточной популяции.
[00310] Термин "нетканый материал" или его эквивалент включает в себя укрепленные ткани, формованные ткани или искусственные ткани, которые производятся способами, отличными от ткачества или вязания, но не ограничивается ими.
[00311] Следует понимать, что изобретения, раскрытые в данном документе, не ограничиваются в своем применении деталями, изложенными в настоящем описании или в качестве примеров. Настоящее изобретение охватывает другие варианты осуществления и может быть практически воплощено или осуществлено различными способами. Кроме того, следует понимать, что фразеология и терминология, используемые в настоящем документе, предназначены для целей описания и не должны рассматриваться как ограничивающие каким-либо образом рамки настоящего изобретения.
[00312] Хотя некоторые композиции, способы и анализы согласно настоящему изобретению были конкретно описаны в соответствии с определенными вариантами осуществления, следующие примеры служат только для иллюстрации способов и композиций настоящего изобретения, которые не ограничивают каким-либо образом рамки настоящего изобретения.
[00313] Артикли единственного числа (единственное число), используемые в настоящем описании и в формуле изобретения, если не указано иное, следует понимать, как включающие множественное число. Пункты формулы или описания, которые включают союз "или" между одним или более членами группы, считаются выполненными, если один, более одного или все члены группы присутствуют, используются или иным образом относятся к данному продукту или процессу, если не указано обратное или это очевидно из контекста. Настоящее изобретение включает варианты осуществления, в которых один член группы точно присутствует, используется или иным образом относится к данному продукту или процессу. Настоящее изобретение также включает варианты осуществления, в которых более одного члена группы или все члены группы присутствуют, используются или иным образом относятся к данному продукту или процессу.
[00314] Кроме того, следует понимать, что настоящее изобретение охватывает все варианты, комбинации и перестановки, в которых одно или более из ограничений, элементов, пунктов, описательных терминов и т.д. из одного или более из перечисленных пунктов формулы изобретения включаются в другой пункт формулы, зависимый оттого же независимого пункта формулы (или, в зависимости от обстоятельств, от любого другого пункта формулы), если не указано иное, или если специалисту в данной области очевидно, что может возникнуть противоречие или несоответствие. Если элементы представлены в виде списков (например, в форме структуры Маркуша или в аналогичном формате), следует понимать, что каждая подгруппа элементов также раскрывается, и любой элемент(ы) может быть удален из группы. Следует понимать, что, в общем случае, когда настоящее изобретение или аспекты настоящего изобретения упоминаются как содержащие конкретные элементы, признаки и т.д., некоторые варианты осуществления настоящего изобретения или аспекты настоящего изобретения состоят или по существу состоят из таких элементов, признаки и т.д. В целях упрощения эти варианты осуществления не во всех случаях конкретно изложены в настоящем описании подробно с использованием такого количества слов. Также следует понимать, что любой вариант осуществления или аспект настоящего изобретения может быть явно исключен из формулы изобретения, независимо от того, указано ли конкретное исключение в описании. Публикации и другие справочные материалы, на которые даны ссылки в настоящем документе для описания предыстории настоящего изобретения и для предоставления дополнительных подробностей относительно его практики, полностью включены в настоящее описание посредством ссылок. Настоящее описание иллюстрируется следующими примерами, не ограничивающими рамки настоящего изобретения.
ПРИМЕРЫ
ПРИМЕР 1: ПОЛУЧЕНИЕ КЛЕТОК HES, ДЕФИЦИТНЫХ ПО В2М
[00315] Клетки hES, дефицитные по В2М, получали с использованием линии клеток СуТ49, однако может использоваться любая линия плюрипотентных стволовых клеток человека. Целевое нарушение гена В2М образует клетки, которые не экспрессируют любые белки ЛАЧ класса I на поверхности этих клеток. Оба аллеля локуса В2М в линии СуТ49 hESC нарушали с использованием технологии CRISPR/Cas9 с использованием известных методов, описанных в публикации заявки РСТ WO2016183041A (которая целиком включена в настоящее описание). Однако, для редактирования генов могут использоваться другие нуклеазы, включающие цинк-пальцевую нуклеазу (ZFN) и эффекторную нуклеазу, подобную активаторам транскрипции (TALEN), а также традиционная гомологичная рекомбинация и подобные им методы. Примеры опубликованных последовательностей гена В2М приведены в SEQ ID NO: 1, 2, и 3. Для редактирования гена использовали NEXTGEN™ CRISPR (Transposagen Inc., Лексингтон, Кентукки), которая включает двойную направляющую РНК и каталитически неактивный белок Cas9, соединенный с нуклеазой FokI. Плазмиды, содержащие направляющие РНК и Cas9, вводили в СуТ49 hESC посредством электропорации, и полученные клетки высевали на чашки с тканевой культурой. Через 12 суток после электропорации клетки сортировали по отрицательной активности в отношении антител к В2М (BioLegend, кат. №316306) посредством сортировки флуоресцентно-активированных клеток (FACS). Отсортированные клетки высевали с плотностью, необходимой для клонирования. Индивидуальные клоны собирали и высевали за 25 суток. Клоны наращивали и криоконсервировали. Выращенные клоны, которые демонстрировали нормальный кариотип при G-окрашивании и у которых отсутствовала экспрессия белка В2М и экспрессия белков ЛАЧ класса I на поверхности этих клеток по данным проточной цитометрии и/или иммунофлуоресценции, выбирали для дальнейших экспериментов.
[00316] Экспрессию В2М на поверхности указанной клетки дикого типа (WT) и нокаутированных клеток оценивали посредством проточной цитометрии в нормальных условиях и условиях воспаления (после обработки интерфероном (IFN)-γ). Смотри на Фиг. 2А: Нормальные условия: необработанная среда для роста (Линия В); условия воспаления: обработанные 100 нг/мл IFN-γ в течение 18-24 часов (Линия А). Воспалительный ответ происходил в соответствии с травмой ткани. Он приводил к высвобождению провоспалительных цитокинов, некоторыми из которых являются IL-1-α, IL-1-β, TNF-α, IL-6, IL-8 и IFN-γ. Хотя клетки WT и В2М-/- ESC и РЕС обрабатывали с помощью IFN-γ, эти наблюдения также можно расширить на другие цитокины.
[00317] На Фиг. 2А показана экспрессия В2М в клетках hES дикого типа (Линия В) без IFN-γ и экспрессия В2М после обработки клеток hES дикого типа с помощью IFN-γ (Линия А). Изменение (повышение) интенсивности флюоресценции в необработанных клетках hES дикого типа (Линия В) по сравнению с базовым уровнем (заштрихованная область) указывает на то, что клетки hES дикого типа экспрессируют В2М. Дальнейшее изменение (повышение) интенсивности флуоресценции выше экспрессии В2М в WT (Линия В) после обработки клеток hES дикого типа с помощью IFN-γ указывает на то, что экспрессия В2М повышается в клетках hES дикого типа после обработки IFN-γ (Линия А). Как таковая экспрессия белков ЛАЧ класса I поверхности клеток может повышаться при клеточном стрессе и воспалении, таком которое возникает при, по меньшей мере, обработке IFN-γ.
[00318] На Фиг. 2В показана экспрессия В2М в нокаутированных по В2М клетках hES, полученных с использованием системы CRISPR/Cas. По существу не наблюдается изменение (повышение) интенсивности флуоресценции по сравнению с базовым уровнем (заштрихованная область) при обработке IFN-γ или без нее, позволяющее предположить, что нокаутированные по В2М клетки hES имели сниженную или отсутствующую экспрессию В2М на поверхности указанной клетки и что экспрессия В2М не индуцировалась посредством обработки IFN-γ. В таких нокаутах по В2М, экспрессия белков поверхности клеток ЛАЧ класса I не повышается при клеточном стрессе и воспалении, обусловленном обработкой IFN-γ.
[00319] Настоящий Пример демонстрирует, что нокаутированные по В2М клетки hES имели сниженную или отсутствующую экспрессию В2М на поверхности указанной клетки как это было показано с использованием антител к В2М.
ПРИМЕР 2: АНАЛИЗ ЭКСПРЕССИИ БЕЛКОВ ПОВЕРХНОСТИ КЛЕТОК ЛАЧ КЛАССА I В КЛЕТКАХ WT И ДЕФИЦИТНЫХ ПО В2М
[00320] Затем, клетки hES дикого типа и нокаутированные по В2М клетки анализировали с использованием моноклонального антитела Пан-ЛАЧ-АВС (BD Pharmingen, кат. №560169), чтобы подтвердить, что эти нокаутированные клетки не экспрессируют белки ЛАЧ класса I на поверхности клеток. Указанное антитело Пан-ЛАЧ-АВС реагирует с белками класса I главного комплекса гистосовместимости (ГКГС) класса ЛАЧ-А, -В, и -С. Экспрессию с использованием антитела Пан-ЛАЧ-АВС оценивали в клетках дикого типа и нокаутных клетках с использованием проточной цитометрии при нормальных и воспалительных условиях после обработки IFN-γ. Нормальные условия: без ИФН-γ (Линия В); воспалительные условия: обработка 100 нг/мл IFN -γ в течение 18-24 часов (Линия А).
[00321] На Фиг. 3А показана пан-экспрессия белков ЛАЧ-АВС поверхности клеток в клетках hES дикого типа (Линия В) и пан-экспрессия белков ЛАЧ-АВС поверхности клеток после обработки клеток hES дикого типа с помощью IFN-γ (Линия А). Изменение (увеличение) интенсивности флуоресценции в указанных необработанных клетках hES дикого типа (Линия В) по сравнению с базовым уровнем (заштрихованная область) указывает на то, что клетки hES дикого типа экспрессируют пан-ЛАЧ-АВС. Изменение (увеличение) интенсивности флуоресценции по отношению к экспрессии hES клеток WT после обработки IFN-γ указывает на то, что пан-ЛАЧ-АВС экспрессия повышается в клетках hES дикого типа после обработки IFN-γ.
[00322] На Фиг. 3В показана пан-экспрессия белка поверхности клеток ЛАЧ-АВС в нокаутированных по В2М клетках hES с использованием системы CRISPR/Cas. Сдвига (увеличения) интенсивности флуоресценции по сравнению с базовым уровнем (заштрихованная область) после обработки IFN-γ или без нее не наблюдалось, что может свидетельствовать о том, что нокаутированные клетки имели сниженную или отсутствующую экспрессию белков поверхности клеток ЛАЧ класса I и что экспрессия белков ЛАЧ класса I не индуцировалась посредством обработки IFN-γ.
[00323] Настоящий Пример демонстрирует, что нокаутированные по В2М клетки hES имели сниженную или отсутствующую экспрессию белков поверхности клеток ЛАЧ класса I как это было показано с использованием антител к пан-ЛАЧ-АВС.
ПРИМЕР 3: ДИФФЕРЕНПИРОВКА ДЕФИЦИТНЫХ ПО В2М КЛЕТОК В ЛИНИЮ ПАНКРЕОТИЧЕСКИХ КЛЕТОК
[00324] Нокаутированные по В2М клетки hES выращивали, пассировали и размножали в тех же условиях, что и клетки hES дикого типа, описано в Schulz м dp. A Scalable System for Production of Functional Pancreatic Progenitors from Human Embryonic Stem Cells PLoS One 7:5 1-17 (2012) и как описано в патенте США №8,895,300, оба документа целиком включены в настоящее описание посредством ссылки. В частности, Schulz и др. описывают адгезивное распространение hESC и их дифференцировку в суспензии.
[00325] Вкратце, клетки hES дикого типа и В2М-/- клетки hES дифференцировали суспензионных агрегатах с использованием четырех (4) стадийной процедуры в течение примерно 2 недель (или 14 дней) для получения популяции клеток панкреатического типа, включая предшественники панкреатических клеток, предшественники эндокринных клеток и клеток, экспрессирующих гормоны, совместно обозначаемых как клетки панкреатической эндодермы (РЕС). Клетки ES человека диссоциировали с использованием аккутазы, а отдельные клетки собирали в роллерные флаконы. Чтобы инициировать дифференцировку, агрегаты объединяли в коническую(ые) трубу(ы) и оставляли оседать под действием силы тяжести с последующей промывкой с использованием среды Stage-1 без факторов роста (RPMI + 0,2% об./об. ФБС, содержащая 1:5000 разведение инсулин-трансферрин-селен (ITS)). Указанные агрегаты повторно собирали, затем ресуспендировали в однодневной среде, которая содержала RPMI + 0,2% об./об. ФБС, содержащей 1:5000 разведение инсулин-трансферрин-селен (ITS), активин А (100 нг/мл) и wnt3a (50 нг/мл), и распределяли в роллерные флаконы при плотности 2 мкл/мл. Указанные роллерные флаконы помещали в цифровой клеточный роллер FlexiRoll (Argos Technologies), работающий со скоростью 31 об/мин. Культуры вращали со скоростью примерно 31 об/мин в течение всего оставшегося процесса дифференцировки с ежедневной заменой среды, так это описано в Таблице 2 ниже, адаптированного по сравнению с Schulz и др., (2012), см. выше. Выращивание, пассирование и размножение клеток hES проводили, по существу, как описано в патентах США №№7,964,402; 8,211,699; 8,334,138; 8,008,07 и 8,153,429. Стандартный способ получения клеток панкреатической эндодермы (РЕС), выведенных из эмбриональных стволовых клеток человека, описан ниже в Таблице 2.


[00327] hESC Agg.: агрегаты hESC; XF HA: DMEM/F12, содержащая GlutaMAX, дополненная 10% об./об. Xeno-free заменителем сыворотки для нокаута, 1% об./об. заменимых аминокислот, 1% об./об. пенициллин/стрептомицин (все из Life Technologies), 10 нг/мл херегулина-1β (Peprotech) и 10 нг/мл активина A (R&D Systems); SP: StemPro® hESC SFM (Life Technologies); r0.2FBS: RPMI 1640 (Mediatech); 0,2% FBS (HyCione), 1x GlutaMAX-1 (Life Technologies), 1% об./об. пенициллин/стрептомицин; ITS: инсулин-трансферрин-селен (Life Technologies) разведения 1:5000 или 1:1000; A100: 100 нг/мл рекомбинантного Активина А человека (R&D Systems); W50: 50 нг/мл рекомбинантного Wnt3A человека (R&D Systems); K25: 25 нг/мл рекомбинантного KGF человека (R&D Systems); IV: 2,5 мкМ ингибитора IV TGF-β RI киназы (EMD Bioscience); db: DMEM HI Glucose (HyClone), дополненной 0,5x B-27 Supplement (Life Technologies), 1x GlutaMAX, и 1% об./об. пенициллин/стрептомицин; СТТ3: 0,25 мкМ KAAD-Cyclopamine (Toronto Research Chemicals) и 3 нМ TTNPB (Sigma-Aldrich); N50: 50 нг/мл рекомбинантного Ногина человека (R&D Systems); K50: 50 нг/мл рекомбинантного KGF человека (R&D Systems); E50: 50 нг/мл рекомбинантного EGF человека (R&D Systems).
[00328] Дифференцированные B2M -/- и РЕС дикого типа анализировали с использованием проточной цитометрии для определения относительного количества предшественников эндокринных и панкреатических клеток в популяции на стадии 4 как показано в Таблице 3.
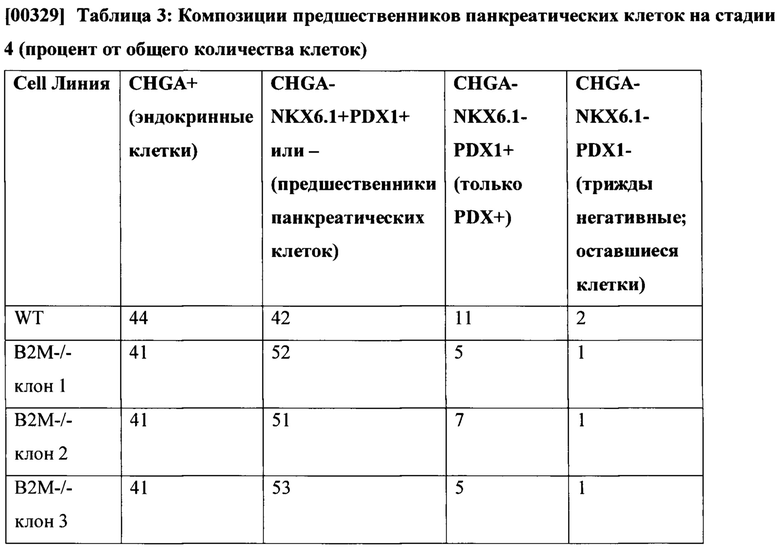
[00330] Относительные уровни эндокринных клеток поджелудочной железы, предшественников, только клеток PDX-1 и тройных отрицательных клеток в дифференцированных В2М -/- клетках во всех 3 клонах, по существу, аналогичны уровням, наблюдаемым в клетках WT (верхний ряд).
[00331] Настоящий Пример демонстрирует, что указанные В2М-/- клетки hES могут дифференцировать в линии панкреатических клеток также как клетки hES дикого типа.
ПРИМЕР 4: АНАЛИЗ ЭКСПРЕССИИ В2М В WT И В2М ДЕФИЦИТНЫХ КЛЕТКАХ ПАНКРЕАТИЧЕСКОЙ ЭНДОДЕРМЫ
[00332] Затем клетки дикого типа и нокаутированные по В2М клетки панкреатической эндодермы (РЕС) из Примера 3 анализировали с использованием проточной цитометрии с обработкой IFN-γ и без нее: без IFN-γ (Линия В); обработанные 100 нг/мл IFN-γ в течение 18-24 часов (Линия А).
[00333] На Фиг. 4А показана экспрессия В2М в клетках РЕС дикого типа без обработки (Линия В) и после обработки с помощью IFN-γ (Линия А). Изменение (увеличение) интенсивности флуоресценции в указанных необработанных клетках РЕС дикого типа (Линия В) по сравнению с базовым уровнем (заштрихованная область) указывает на то, что клетки РЕС дикого типа экспрессируют В2М. Изменение (увеличение) интенсивности флуоресценции выше экспрессию WT после обработки РЕС дикого типа с помощью IFN-γ (Линия А) указывает на то, что В2М экспрессия повышается в клетках РЕС дикого типа после обработки IFN-γ. То есть, обработка IFN-γ повышает экспрессию В2М в клетках РЕС дикого типа.
[00334] На Фиг. 4 В показана экспрессия В2М в клетках РЕС, полученных из нокаутированных по В2М клетках hES. Смещения (увеличения) интенсивности флуоресценции по сравнению с базовым уровнем (заштрихованная область) с обработкой IFN-γ или без нее не наблюдалось, что указывает на то, что клетки РЕС с нокаутом В2М имели сниженную или отсутствующую экспрессию В2М на поверхности клеток и что экспрессия В2М не может быть индуцирована в клетках РЕС, полученных из нокаутированных по В2М клеток, посредством обработки IFN-γ.
[00335] Настоящий Пример показывает, что клетки РЕС, полученные из клеток hES, в которых были снижена/элиминирована экспрессия В2М, имели сниженную или отсутствующую экспрессии В2М на поверхности указанной клетки.
ПРИМЕР 5: АНАЛИЗ ЭКСПРЕССИИ БЕЛКОВ ПОВЕРХНОСТИ КЛЕТОК ЛАЧ КЛАССА I В КЛЕТКАХ WT И НОКАУТИРОВАННЫХ ПО В2М КЛЕТКАХ ПАНКРЕАТИЧЕСКОЙ ЭНДОДЕРМЫ
[00336] Аналогично Примеру 2, клетки дикого типа и клетки РЕС с нокаутом по В2М анализировали с использованием пан-ЛАЧ-АВС моноклонального антитела (BD Pharmingen, кат. №560169) для определения экспрессии ЛАЧ класса I поверхности клеток. Экспрессию измеряли посредством проточной цитометрии в клетках дикого типа и нокаутных клетках в двух условиях: (1) необработанные (Линия В) и (2) обработанные с помощью 100 нг/мл IFN-γ в течение 18-24 часов (Линия А).
[00337] На Фиг. 5А показана экспрессия белков поверхности клеток ЛАЧ класса I в клетках РЕС дикого типа в условиях без обработки (Линия В) и экспрессия после обработки с помощью IFN-γ (Линия А). Изменение (увеличение) интенсивности флуоресценции в указанных необработанных РЕС дикого типа (Линия В) по сравнению с базовым уровнем (заштрихованная область) указывает на то, что РЕС дикого типа экспрессируют ЛАЧ класса I на поверхности клеток. Изменение (увеличение) интенсивности флуоресценции выше уровня экспрессии РЕС дикого типа после обработки IFN-γ указывает на то, что экспрессия ЛАЧ класса I на поверхности клеток повышается в клетках РЕС дикого типа после обработки IFN-γ.
[00338] На Фиг. 5 В показана экспрессия белков поверхности клеток ЛАЧ класса I в клетках РЕС, полученных из нокаутированных по В2М клеток hES. Изменение (увеличение) интенсивности флуоресценции по сравнению с базовым уровнем (заштрихованная область) в клетках РЕС с обработкой IFN-γ или без нее отсутствовало, что позволяет утверждать, что указанные нокаутированные клетки имели сниженную или отсутствующую экспрессия ЛАЧ на поверхности клеток и что экспрессия ЛАЧ класса I в клетках В2М-/- РЕС не может быть индуцирована посредством обработки IFN-γ.
[00339] Как таковая, уменьшенная или отсутствующая экспрессия белков поверхности клеток ЛАЧ класса I наблюдалась в тех клетках РЕС, в которых была уменьшена/элиминирована экспрессия В2М.
ПРИМЕР 6: АНАЛИЗ ЭКСПРЕССИИ БЕЛКА ICAM-1 ПОВЕРХНОСТИ КЛЕТОК В КЛЕТКАХ WT И НОКАУТИРОВАННЫХ ПО В2М КЛЕТКАХ HES
[00340] Для дальнейшего определения влияния обработки IFN-γ на клетки-мишени, о посредством проточной цитометрии оценивали экспрессию ICAM-1 в клетках WT и нокаутированных по В2М клетках hES в двух условиях: (1) без обработки (Линия В) и (2) обработка 100 нг/мл IFN-γ в течение 18-24 часов (Линия А). Белку ICAM-1 приписывают несколько иммунологических функций, включая презентацию антигена в клетках-мишенях, и этот белок известен в качестве лиганда, активирующего NK-клетки. In vivo, возможно получить иммунологические преимущества, нарушая межклеточное связывающее взаимодействие ICAM / LFA путем применения специфических моноклональных антител ("мАт"), то есть антител против ICAM-1 или против LFA-1. Смотри Isobe и др., Specific Acceptance of Cardiac Allograft After Treatment With Antibodies to ICAM-1 и LFA-1, 255 SCIENCE 1125-1127 (Feb. 1992). Заявитель использовал антитела против ICAM-1, полученные от Milteney Biotec Inc., кат. №130-103-909.
[00341] На Фиг. 6А показана экспрессия белка поверхности клеток ICAM-1 в необработанных клетках hES дикого типа (Линия В) и после их обработки с помощью IFN-γ (Линия А). Изменение (увеличение) интенсивности флуоресценции по сравнению с базовым уровнем (заштрихованная область) в необработанных клетках WT hES (Линия В) указывает на то, что клетки hES дикого типа экспрессируют белок ICAM-1 на своей поверхности. Изменение (увеличение) интенсивности флуоресценции выше экспрессии в WT после обработки клеток hES дикого типа с помощью IFN-γ указывает на то, что экспрессия ICAM-1 повышается после обработки IFN-γ.
[00342] На Фиг. 6В показана экспрессия белка поверхности клеток ICAM-1 в необработанных нокаутированных по В2М клетках hES (Линия В) и после их обработки с помощью IFN-γ (Линия А). На Фиг. 6В показано, что экспрессия белка поверхности клеток ICAM-1 в указанных нокаутированных по В2М клетках hES была сходной с экспрессией в клетках hES дикого типа.
[00343] Настоящий Пример демонстрирует, что обработка клеток WT и нокаутированных по В2М клетках hES с помощью IFN-γ повышает экспрессию белков поверхности клеток ICAM-1.
ПРИМЕР 7: АНАЛИЗ ЭКСПРЕССИИ БЕЛКА ПОВЕРХНОСТИ КЛЕТОК ICAM-1 В КЛЕТКАХ WT И НОКАУТИРОВАННЫХ ПО В2М КЛЕТКАХ ПАНКРЕАТИЧЕСКОЙ ЭНДОДЕРМЫ
[00344] По аналогии с Примерами 4 и 5, клетки дикого типа и клетки РЕС с нокаутом по В2М анализировали с использованием антитела к известному лиганду, активирующему NK-клетки, например ICAM-1. Экспрессию белка поверхности клеток ICAM-1 оценивали посредством проточной цитометрии в клетках WT и в нокаутированных клетках РЕС в двух условиях: (1) без обработки и (2) обработка с помощью 100 нг/мл IFN-γ в течение 18-24 часов.
[00345] На Фиг. 7А показана экспрессия белка поверхности клеток ICAM-1 в клетках РЕС дикого типа (Линия В) и экспрессия белка поверхности клеток ICAM-1 после обработки клеток РЕС дикого типа с помощью IFN-γ (Линия А). Изменение (увеличение) интенсивности флуоресценции в указанных необработанных РЕС дикого типа (Линия В) по сравнению с базовым уровнем (заштрихованная область) указывает на то, что РЕС дикого типа экспрессируют белок ICAM-1 на своей поверхности. Дальнейшее изменение (увеличение) интенсивности флуоресценции выше экспрессию в РЕС дикого типа после обработки IFN-γ указывает на то, что экспрессия ICAM-1 повышается в клетках РЕС дикого типа после обработки IFN-γ. Это демонстрирует, что NK-активирующие лиганды, и в частности ICAM-1, являются сильно индуцируемыми посредством стимуляции с помощью IFN-γ.
[00346] На Фиг. 7 В показана экспрессия белка поверхности клеток ICAM-1 в клетках РЕС с нокаутом В2М. Экспрессия белка поверхности клеток ICAM-1 в указанных клетках РЕС с нокаутом В2М была сходной с экспрессией в РЕС дикого типа, с обработкой IFN-γ и без нее, с незначительным снижением флуоресценции. Таким образом, обработка клеток РЕС с нокаутом В2М с помощью IFN-γ повышает экспрессию белка поверхности клеток ICAM-1.
[00347] На Фиг. 8 показаны данные экспрессии РНК на биочипе (Affymetrix), демонстрирующие, что на уровне мРНК, обработка IFN-γ повышает экспрессию ICAM-1 в клетках WT hESC, В2М-/- hESC, РЕС дикого типа и В2М-/- РЕС. Авторы настоящего изобретения обнаружили, что экспрессия белков поверхности клеток лиганда ICAM-1, активирующего NK-клетки, увеличивалась в дифференцированных типах клеток (РЕС дикого типа и В2М-/- РЕС) после обработки IFN-γ. Таким образом, ICAM-1 сильно индуцируется посредством стимулирования IFN-γ в РЕС.
ПРИМЕР 8: ВЛИЯНИЕ ОБРАБОТКИ IFN-GAMMA НА ДОПОЛНИТЕЛЬНЫЕ ЛИГАНДЫ, АКТИВИРУЮЩИЕ NK
[00348] Для того, чтобы далее охарактеризовать влияние обработки IFN-γ на клетки-мишени, клетки РЕС дикого типа изучали посредством проточной цитометрии в двух условиях: (1) без обработки (Линия В) и (2) обработка с помощью 100 нг/мл IFN-γ в течение 18-24 часов (Линия А), и анализировали экспрессию белков поверхности клеток других известных лигандов, активирующих NK.
[00349] На Фиг. 9 показана экспрессия белка поверхности клеток CD58 (также известного как LFA-3) с использованием антитела от BioLegend, кат. №330909, в необработанных клетках РЕС дикого типа (Линия В) и после их обработки с помощью IFN-γ (Линия А). Большой сдвиг (увеличения) интенсивности флуоресценции по сравнению с базовым уровнем (Линия С) в указанных необработанных клетках hES дикого типа (Линия В) указывает на то, что большинство из указанных клеток экспрессируют белок CD58 белок на своей поверхности. Наблюдалось небольшое дополнительное изменение (увеличение) интенсивности флуоресценции после обработки IFN-γ.
[00350] На Фиг. 10 показана экспрессия белка поверхности клеток CD155 (также известного как PVR, NECL-5, HVED) с использованием антитела от Milteneyi Biotech Inc., кат. №130-105-905 в необработанных клетках РЕС дикого типа (Линия В) и после их обработки с помощью IFN-γ (Линия А). Изменение (увеличение) интенсивности флуоресценции по сравнению с базовым уровнем (Линия С) в указанных необработанных РЕС дикого типа (Линия В) указывает на то, что РЕС дикого типа экспрессируют CD155. Дополнительного изменения (увеличения) интенсивности флуоресценции по сравнению с необработанными клетками после обработки IFN-γ не наблюдалось, что говорит о том, что экспрессия CD 155 не увеличивалась в клетках РЕС дикого типа после обработки IFN-γ.
[00351] На Фиг. 11 показана экспрессия белка поверхности клеток CEACAM1 (также известного как CD66a, BGP, и BGP1) с использованием антитела от Milteneyi Biotech Inc., кат. №130-098-858 в необработанных клетках РЕС дикого типа (Линия В) и после их обработки с помощью IFN-γ (Линия А). Изменение (увеличение) интенсивности флуоресценции по сравнению с базовым уровнем (Линия С) в указанных необработанных РЕС дикого типа (Линия В) указывает на то, что РЕС дикого типа экспрессируют белок CEACAM1 на поверхности клеток. Изменение (увеличение) интенсивности флуоресценции по отношению к условиям без обработки после обработки РЕС дикого типа IFN-γ указывает на то, что экспрессия белка CEACAM1 на поверхности клеток повышается в клетках РЕС дикого типа после обработки IFN-γ.
[00352] На Фиг. 12 показана экспрессия белка поверхности клеток ВАТ3 (также известного как BAG6) с использованием антитела от Abeam Inc., кат. № ab210838 в необработанных клетках РЕС дикого типа (Линия В) и после их обработки с помощью IFN-γ (Линия А). Изменение (увеличение) интенсивности флуоресценции по сравнению с базовым уровнем (Линия С) в указанных необработанных РЕС (Линия В) указывает на то, что указанные клетки экспрессируют CEACAM1. Дополнительного изменения (увеличения) интенсивности флуоресценции по сравнению с необработанными клетками после обработки РЕС дикого типа IFN-γ не наблюдалось, что указывает на то, что экспрессия ВАТ3 не увеличивалась в клетках РЕС дикого типа после обработки IFN-γ.
[00353] На Фиг. 13 показана экспрессия белка поверхности клеток CADM1 (также известного как NECL2, TSLC1, IGSF4, RA175) с использованием антитела от MBL international Corporation, кат. № СМ004-4 в необработанных клетках РЕС дикого типа (Линия В) и после их обработки с помощью IFN-γ (Линия А). Изменение (увеличение) интенсивности флуоресценции по сравнению с базовым уровнем (Линия С) в указанных необработанных условиях (Линия В) указывает на то, что РЕС дикого типа экспрессируют белок CADM1 на своей поверхности. Дополнительного изменения (увеличения) интенсивности флуоресценции по отношению к условиям без обработки после обработки РЕС с помощью IFN-γ не наблюдалось, что указывает на то, что экспрессия CADM1 не увеличивается после обработки IFN-γ.
[00354] На Фиг. 14 показана экспрессия CD112 (также известного как Нектин-2, PVRR2, HVEB) с использованием антитела от Milteneyi Biotech Inc., кат. №130-109-056 в необработанных клетках РЕС дикого типа (Линия В) и после их обработки с помощью IFN-γ (Линия А). Изменение (увеличение) интенсивности флуоресценции по сравнению с базовым уровнем (Линия С) в указанных необработанных условиях (Линия В) указывает на то, что РЕС дикого типа экспрессируют белок CD112 на их поверхности. Дополнительного изменения (увеличения) интенсивности флуоресценции по отношению к условиям без обработки после обработки РЕС с помощью IFN-γ не наблюдалось, что свидетельствует о том, что экспрессия CD112 не увеличивается после обработки IFN-γ.
ПРИМЕР 9: КЛЕТКИ, ДЕФИЦИТНЫЕ ПО ГКГС КЛАССА I, ДЕФИЦИТНЫЕ ПО ЛИГАНДАМ, АКТИВИРУЮЩИМ NK-КЛЕТКИ, ПРЕДОТВРАЩАЮТ ЛИЗИС, ОПОСРЕДОВАННЫЙ NK-КЛЕТКАМИ
[00355] Чтобы проверить, является ли комбинация пониженной или элиминированной экспрессии белка ЛАЧ класса I и пониженной или элиминированной экспрессии лиганда, активирующего NK-клетки, достаточной для предотвращения лизиса, опосредованного NK-клетками, блокировали экспрессию ICAM-1 на клетках-мишенях с использованием антитела, блокирующего ICAM-1 в концентрации 5 и 10 мкг/мл. Добавление антитела, блокирующего ICAM-1, к клеткам WT или В2М -/- ES или РЕС вызывало снижение лизиса NK клеток-мишеней после обработки IFN-γ (см. Фиг. 15).
Окрашивание клеток-мишеней с помощью Кальцеина-АМ
[00356] Анализ высвобождения кальцеина является нерадиоактивной альтернативой для изучения цитотоксичности NK-клеток. Клетки-мишени поглощают флуоресцентный краситель (кальцеин AM) и цитоплазматически превращают его в активный флуорохром, который выделяется из клеток после лизиса. Лизированные клетки высвобождают флуорохром в супернатант, который затем собирают, и количество флуоресценции определяют количественно в флуориметре. Процент лизиса клеток рассчитывают по количеству флуоресценции, присутствующей в супернатанте после инкубации в присутствии или отсутствии клеток NK (эффекторов), блокирующего антитела, или обоих компонентов.
[00357] Клетки-мишени содержали либо WT ESC, В2М-/- ESC, РЕС дикого типа или В2М-/-РЕС, обработанные 100 нг/мл IFN-γ или не обработанные им перед мечением. Для получения указанных клеток-мишеней, популяции указанных клеток-мишеней окрашивали с помощью среды для окрашивания с 2 мкг/мл кальцеина AM (Enzo biosciences, 1 мг/мл концентрированного раствора (кат. № С3100МР)). Указанные клетки-мишени инкубировали в течение 1 часа при 37°С в инкубаторе с 8% СО2 с перемешиванием с перерывами. Указанные клетки-мишени дважды промывали удаления всего свободного кальцеина AM и ресуспендировали до концентрации 1×105 клеток/мл в полной среде RPMI (RPMI, 10% инактивированной теплом ФБС и 1% антибиотиков).
Совместно культивируемые клетки, NK-клетки и блокирующее антитело
[00358] Меченые кальцеином AM клетки-мишени инкубировали с клетками NK (эффекторные клетки) в соотношении эффектор: клетки-мишени (соотношение Е:Т) - 10:1. В частности, 100 мкл клеток NK при плотности 1×106 клеток/мл добавляли в каждую лунку 96-луночного планшета с V-образным дном, а затем добавляли 100 мкл окрашенных кальцеином клеток ESC или РЕС (1×105 клеток на лунку). Там, где указано, в лунки добавляли блокирующее антитело к поверхностному антигену ICAM-1 человека (R&D Systems, Inc., кат. № ВВА3) в концентрации 5 и 10 мкг/мл для определения будет ли снижаться уровень лизиса клеток, опосредованный NK-клетками.
[00359] Планшеты инкубировали в течение 4 часов при 37°С в инкубаторе с 8% СО2. После инкубации, планшеты центрифугировали при 200×g в течение 2 минут. 100 мкл супернатанта осторожно удаляли и переносили в черный пигментированный 96-луночный планшет и измеряли флуоресценцию с использованием считывателя планшетов Molecular Device (фильтр возбуждения: 485 нм/фильтр испускания: 530 нм). Специфический лизис рассчитывали с использованием следующей формулы, % лизис = 100×[(средняя флуоресценция с антителом - средняя спонтанная флуоресценция)/(средняя максимальная флуоресценция - средняя спонтанная флуоресценция)]. Максимальную флуоресценцию определяли посредством лизиса клеток, инкубированных с детергентом (1% Тритон Х-100), а спонтанным лизисом считали флуоресценцию, полученную с клетками-мишенями без какого-либо антитела или эффекторных клеток.
Результаты
[00360] Как показано на Фиг. 1, целью является переход от Сценария С (NK-клетки атакуют клетки-мишени) к Сценарию А (отсутствие ответа или пониженный ответ). На Фиг. 15, этот сценарий показан в условиях 4 и 8. В условиях 4 и 8 у клеток-мишеней (В2М-/- клетки hES или РЕС) отсутствует экспрессия функционального поверхностного белка ЛАЧ класса I и они обработаны IFN-γ. Как обсуждалось выше и видно в условиях 4 и 8 обработка IFN-γ вызывает увеличение количества NK-активирующих лигандов (ICAM-1), которое приводит к повышенному лизису клеток по сравнению с клетками с нокаутом ЛАЧ класса I без обработки IFN-γ (сравнивайте первые столбики в условии 3 по сравнению с условием 4 и условии 7 по сравнению с условием 8). Однако, при обработке антителом, ингибирующим ICAM-1, которое служит для блокирования экспрессии NK-активирующих сигналов на клетке-мишени, NK-опосредованный лизис клеток падает с 83% до 73% в В2М-/- клетках hES и с 72% до 61% в В2М-/- клетках РЕС, обработанных с помощью IFN-γ. Процент NK-опосредованного лизиса клеток не падает до нуля, потому что экспрессия белка поверхности клеток ICAM-1 не может быть полностью заблокирована с использованием указанного блокирующего антитела и, как описано выше, указанные клетки-мишени экспрессируют более одного лиганда, активирующего NK-клетки. Ожидается, что процент NK-опосредованного лизиса клеток упадет в большей степени, когда экспрессия ICAM-1 будет заингибирована еще сильнее и другие NK-активирующие лиганды также заблокированы в указанных клетках-мишенях. Поэтому, указанное антитело, ингибирующее ICAM-1, может быть объединено с дополнительными антителами, ингибирующими лиганды, активирующие NK-клетки, включая ингибирующие антитела к любому из лигандов, перечисленных в Таблице 1, и предпочтительно к лигандам категории 1 (известные активирующие лиганды) и 2 (потенциальные кандидаты в активирующие лиганды, идентифицированные на основе данных, полученных на ДНК-чипе, лиганды, регуляция которых повышается в клетках РЕС и/или ESC после обработки IFNγ). В одном варианте осуществления, указанный ген ICAM1 и гены других лигандов, активирующих NK-клетки, разрушены для полной блокировки их активности.
[00361] Сценарий D на Фиг. 1 представлен в первых столбиках условий 2 и 6 на Фиг. 15. В условиях 2 и 6, у указанных клеток hES дикого типа и РЕС наблюдается повышенная экспрессия белков поверхности клеток как антигенов ЛАЧ класса I, так и NK-активирующих лигандов в результате их обработки IFN-γ. Когда указанные клетки инкубируют с антителом, ингибирующим ICAM-1, это демонстрирует ситуацию с движением от Сценария D к Сценарию В на Фиг. 1. После инкубации с антителом, ингибирующим ICAM1, гибель клеток снижается: с 79% до 60% для клеток hES дикого типа и с 53% до 27% для клеток РЕС дикого типа, обработанных IFN-γ. Действительно, когда РЕС дикого типа обрабатывают IFN-γ и инкубируют с антителом, ингибирующим ICAM-1, NK-опосредованный лизис клеток снижается по сравнению с необработанными РЕС дикого типа: 27% для РЕС дикого типа, IFN-γ и антитела, ингибирующего ICAM-1, по сравнению с 37% для РЕС дикого типа, антитела, ингибирующего ICAM-1, без обработки IFN-γ.
[00362] Сценарий В на Фиг. 1 представлен столбиками с обработкой антителом, ингибирующим ICAM-1, в условиях 1, 2, 5 и 6 на Фиг. 15. Здесь, указанные клетки-мишени hES и PEC имеют ЛАЧ класса I и NK-активирующие лиганды, но когда указанные клетки-мишени не обработаны IFN-γ, не происходит увеличения экспрессии белков поверхности клеток ICAM-1. В результате, указанное антитело, ингибирующее ICAM-1, имеет меньший эффект. Поэтому, лизис клеток остается примерно одинаковым: 58% и 57% у клеток hES и 33% и 37% в клетках РЕС.
[00363] Сценарий С на Фиг. 1 сходен с первыми столбцами в условиях 3, 4, 7 и 8 на Фиг. 15. В этих условиях, указанные клетки не имеют экспрессии ЛАЧ класса I на поверхности клеток как результат генотипа В2М-/-. Поэтому, когда NK-активирующие лиганды не активированы посредством IFN-γ, обработка указанных клеток указанным антителом, ингибирующим ICAM-1, имеет небольшой эффект, 71% и 70% у клеток hES и 54% и 57% у клеток РЕС.
[00364] В качестве общего наблюдения, NK-опосредованный лизис клеток менее выражен в популяциях дифференцированных клеток (РЕС дикого типа и В2М-/- РЕС) по сравнению с популяциями недифференцированных клеток (WT hES и В2М-/- hES). NK-опосредованный лизис клеток повышается в большей степени, когда клетки (WT или В2М -/- нокауты, hES или РЕС) активированы IFN-γ.
ПРИМЕР 10: ПОЛУЧЕНИЕ В2М-/- НОКАУТНЫХ КЛЕТОК HES, ДЕФИЦИТНЫХ ПО ЛИГАНДУ, АКТИВИРУЮЩЕМУ NK-КЛЕТКИ
[00365] В Примере 9 описано, что ингибирование или ослабление экспрессии ICAM-1 защищает обработанные IFN-γ В2М-/- клетки РЕС от NK-опосредованной гибели клеток. Поэтому, для защиты В2М-/- клеток РЕС от гибели, опосредованной клетками NK, после трансплантации, желательно нарушить ген лиганда, активирующего NK-клетки. Например, на основании информации в вышеприведенных Примерах, ICAM-1, известный ген лиганда, активирующего NK-клетки, может быть нарушен или "нокаутирован". Предпочтительно, чтобы были нарушены оба аллеля локуса ICAM-1 в линии клеток В2М-/- СуТ49 hESC с использованием CRTSPR/Cas9 или любой другой технологии редактирования генов, известной в настоящее время или которая станет известной в будущем, см. публикацию РСТ WO2016183041A, которая целиком включена в настоящее описание посредством ссылки. Примеры опубликованных последовательностей ICAM-1 приведены в SEQ ID NO: 4, 5, и 6. Например, в одном варианте осуществления настоящего изобретения, для редактирования генов указанных клеток используют NEXTGEN™ CRISPR (Transposagen Inc., Лексингтон, Кентукки), которая включает двойную направляющую РНК и каталитически неактивный белок Cas9, соединенный с нуклеазой FokI.
[00366] Плазмиды, содержащие направляющие РНК и Cas9, могут быть введены в клетки В2М -/- СуТ49 hESC посредством электропорации, после чего полученные клетки высевают на чашки с тканевой культурой. После электропорации указанные клетки могут быть отсортированы по отрицательной реактивности к антителу ICAM-1 путем проточной цитометрии. Отсортированные клетки могут быть рассеяны с плотностью, необходимой для клонирования. Индивидуальные клоны могут быть собраны и пересеяны. Клоны могут быть размножены и криоконсервированы. Размноженные клоны, которые показали нормальный кариотип посредством G-окрашивания, и являются отрицательными в отношении экспрессии белка ICAM-1 и поверхностной экспрессии белков ICAM-1 могут быть собраны для дальнейших экспериментов с помощью проточной цитометрии и/или иммунофлуоресценции.
[00367] Как описано выше, клетки, дефицитные по ICAM1 и В2М, не могут экспрессировать по меньшей мере один лиганд, активирующий NK-клетки, и по меньшей мере один или все белки ГКГС класса I на поверхности указанных клеток и, поэтому, не могут связываться с рецепторами, активирующими NK клетки, и защищены от NK-опосредованной клеточной гибели.
ПРИМЕР 11: ПОЛУЧЕНИЕ В2М-/- НОКАУТНЫХ КЛЕТОК HES, ДЕФИЦИТНЫХ ПО МНОЖЕСТВУ ЛИГАНДОВ, АКТИВИРУЮЩИХ NK-КЛЕТКИ
[00368] В Примерах 9 и 10 продемонстрировано, что ингибирование функциональной экспрессии на поверхности клеток (анти-лиганда, активирующего NK-клетки) и нарушение гена лиганда, активирующего NK-клетки, (например, ICAM1-/-) в комбинации с нарушением экспрессии ГКГС класса I на поверхности клеток (например, В2М-/-) может обеспечивать клеткам-мишеням защиту от NK-опосредованной клеточной гибели. Трансплантация клеток, дефицитных по более чем одному лиганду, активирующему NK-клетки, может быть произведена и это обеспечит дальнейшую защиту от NK-опосредованной клеточной гибели.
[00369] Ген CD58 был выбран для нокаута в качестве гена лиганда, активирующего NK-клетки. Оба аллеля локуса CD58 могут быть нарушены с использованием технологии CRISPR/Cas9 с использованием известных методик, описанных в WO2016183041A, в указанной линии клеток СуТ49 hESC с двойным нокаутом В2М-/-:ICAM-/-. Примеры опубликованных последовательностей CEACAM1 представлены в SEQ ID NO: 7, 8 и 9. Еще раз, указанной версией для редактирования может быть NEXTGEN™ CRISPR (Transposagen Inc., Лексингтон, Кентукки).
[00370] Плазмиды, содержащие направляющие РНК и Cas9, могут быть введены в В2М -/-, ICAM -/- нокаут СуТ49 hESC посредством электропорации, а полученные клетки высевают на чашки с тканевой культурой. После электропорации клетки могут быть отсортированы по отрицательной реактивности к антителу CD58 посредством проточной цитометрии. Отсортированные клетки могут быть высеяны с плотностью, необходимой для клонирования. Индивидуальные клоны могут быть собраны и пересеяны. Клоны могут быть размножены и криоконсервированы. Размноженные клоны, которые показали нормальный кариотип посредством G-окрашивания, и являются отрицательными по экспрессии белка CD58 и для поверхностной экспрессии белков CD58 могут быть выбраны для дальнейших экспериментов посредством проточной цитометрии и/или иммунофлуоресценции.
[00371] Как описано выше, клетки с нарушенными, делетированными или модифицированными ICAM1, CD58 и В2М не могут экспрессировать ни ICAM1 и CD58, ни белки ГКГС класса I на поверхности клеток и, поэтому, не должны связываться с рецепторами, активирующими NK клетки, и защищены от NK-опосредованной клеточной гибели.
[00372] Ген CD155 (также называемый PVR) также может быть выбран для нокаута в качестве гена лиганда, активирующего NK-клетки. Ген CD155 может быть выбран в дополнение к CD58 или вместо него. Также как описано выше, ген CD28 может быть нарушен с использованием технологии CRISPR/Cas9. Ген CD155 также может быть выбран для нокаута в качестве гена лиганда, активирующего NK-клетки. Ген CD155 может быть выбран в дополнение к CD58 или вместо него. Указанный ген САЕСАМ1 также может быть выбран для нокаута в качестве гена лиганда, активирующего NK-клетки. Ген САЕСАМ1 может быть выбран в дополнение к CD58 и/или CD155 или вместо них.
[00373] ПРИМЕР 12: ИНАКТИВАЦИЯ NK-АКТИВИРУЮШИХ ЛИГАНДОВ ICAM1 И CD58
[00374] Для того, чтобы полностью элиминировать активацию NK, может потребоваться элиминирование/снижение активности множества NK-активирующих лигандов либо посредством нокаута в клетках-мишенях (например, в клеточной терапии, основанной на использовании клеток hES), либо с использованием блокирующего антитела или других стратегий, известных в настоящее время или разработанных в будущем. Чтобы определить, может ли токсичность NK-клеток быть уменьшена путем ингибирования влияния NK-активирующих лиганд на клетки-мишени, экспрессия NK-активирующих лигандов в WT и Клетки hES В2М -/- и РЕС может быть заблокирована с использованием ICAM1 и CD58 блокирующего антитела, чтобы заблокировать белки ICAM1 и CD58 на поверхности клеток-мишеней. Можно ожидать, что клеточный лизис клеток-мишеней будет снижен (аналогично Фиг. 15). Следовательно, лизис клеток с помощью NK-клеток можно уменьшить, блокируя NK-активирующие лиганды, такие как ICAM1 и CD58. Блокирование экспрессии ICAM1 и CD58 с использованием антител против NK-активирующих лигандов в В2М -/- клетках является доказательством идеи создания клетки, имеющей 3 нокаута (нокаут гена класса 1 ЛАЧ и нокаут гена лиганда, активирующего NK-клетки). При этом клетки-мишени (например, линия hES и/или линия панкреотических клеток) переходят из Сценария С в Сценарий А на Фиг. 1. В частности, у указанных клеток, тканей и органов согласно настоящему изобретению заингибированы или не экспрессируются белки поверхности клеток ЛАЧ класса I (В2М -/-) и заингибированы или не экспрессируются лиганды белков поверхности клеток, активирующие экспрессию NK-клеток (например, ICAM1 -/- и CD58 -/-). Ингибирование экспрессии белков поверхности клеток может быть достигнуто путем нокаута генов или блокирования экспрессии белков с использованием антител. Другие стратегии вмешательства в экспрессию белков поверхности клеток включают использование антисмысловой РНК, РНК-ловушек, рибозимов, РНК-аптамеров, миРНК, шРНК/микроРНК, трансдоминантных негативных белков (TNP), химерных/слитых белков, нуклеаз, лигандов хемокинов, анти-инфекционных клеточных белков, внутриклеточных антител (sFv), нуклеозидных аналогов (NRTI), ненуклеозидных аналогов (NNRTI), ингибиторов интегразы (олигонуклеотиды, динуклеотиды и химические агенты) и ингибиторов протеаз. Множественные нокауты эффективно предотвращали бы как опосредованную цитотоксическими Т-клетками (ЦТЛ) токсичность, так и токсичность, опосредованную NK-клетками, потому что практически не было бы и почти не было белков ЛАЧ класса I и лигандов, активирующих NK-клеток, экспрессируемых на поверхности клеток для связывания с клетками ЦТЛ или NK-клетками.
[00375] ПРИМЕР 13: ИНАКТИВАЦИЯ NK-АКТИВИРУЮЩИХ ЛИГАНДОВ CADM1 и CD58
[00376] Для того, чтобы полностью инактивировать активацию NK, необходимо элиминировать/снизить экспрессию множества NK-активирующих лигандов либо посредством нокаута гена в клетке-мишени (например, клеточная терапия на основе клеток hES), или с использованием блокирующих антител или других стратегий, известных в настоящее время, или стратегий, которые будут разработаны в будущем. Чтобы определить, может ли токсичность NK-клеток быть снижена путем ингибирования воздействия NK-активирующих лигандов на клетки-мишени, экспрессия NK-активирующих лигандов в клетках hES WT и клетках hES В2М -/- и РЕС может быть заблокирована с использованием антитела, блокирующего CADM1 и CD58, для блокирования CADM1 и CD58 белков на поверхности клеток. Можно ожидать, что клеточный лизис клеток-мишеней будет уменьшен (аналогично тому, как показано на Фиг. 15). Следовательно, лизис клеток NK-клетками может быть уменьшен посредством блокирования NK-активирующих лигандов, таких как CADM1 и CD58. Блокирование экспрессии CADM1 и CD58 антителами против NK-активирующих лигандов в В2М -/- клетках является доказательством идеи создания клетки, имеющей 3 нокаута (нокаут гена ЛАЧ класса I и нокаут гена лиганда, активирующего NK-клетки). При этом клетки-мишени (например, hES и/или линия панкреотических клеток) переходят из Сценарий С в Сценарий А на Фиг 1. Конкретно, у клеток, тканей и органов согласно настоящему изобретению заингибированы или не экспрессируются белки поверхности клеток ЛАЧ класса I (В2М -/-) и заингибировали или не экспрессируются лиганды, активирующие экспрессию NK-клеток в поверхностных клетках (например, CADM1 -/-) и CD58 -/-). Ингибирование экспрессии белков поверхности клеток может быть достигнуто путем нокаута генов или блокирования экспрессии белков с использованием антител. Другие стратегии по подавлению экспрессии белков поверхностей клеток включают использование антисмысловой РНК, РНК-ловушек, рибозимов, РНК-аптамеров, миРНК, шРНК/микроРНК, трансдоминантных негативных белков (TNP), химерных/слитых белков, нуклеаз, хемокиновых лигандов, антиинфекционных клеточных белков, внутриклеточных антител (sFvs), нуклеозидных аналогов (NRT1), ненуклеозидных аналогов (NNRT1), ингибиторов интеграз (олигонуклеотиды, динуклеотиды и химические агенты) и ингибиторов протеаз. Многократные нокауты генов эффективно предотвращали бы как токсичность, опосредованную цитотоксическими Т-клетками (CTL), так и токсичность, опосредованную NK-клетками, потому что практически отсутствовали бы белки ЛАЧ класса I и белки лигандов, активирующих NK-клетки, экспрессирующихся на поверхности клеток для связывания с клетками CTL или NK.
[00377] ПРИМЕР 14: ИНАКТИВАЦИЯ NK-АКТИВИРУЮЩИХ ЛИГАНДОВ CD155 и CD58
[00378] Для того, чтобы полностью инактивировать активацию NK, необходимо элиминировать/снизить экспрессию множества NK-активирующих лигандов либо посредством нокаута гена в клетке-мишени (например, клеточная терапия на основе клеток hES), или с использованием блокирующих антител или других стратегий, известных в настоящее время, или стратегий, которые будут разработаны в будущем. Чтобы определить, может ли токсичность NK-клеток быть снижена путем ингибирования воздействия NK-активирующих лигандов на клетки-мишени, экспрессия NK-активирующих лигандов в клетках hES WT и клетках hES В2М -/- и РЕС может быть заблокирована с использованием антитела, блокирующего CD155 и CD58, для блокирования белков CD155 и CD58 на поверхности клеток-мишеней. Можно ожидать, что клеточный лизис клеток-мишеней будет уменьшен (аналогично тому, как показано на Фиг. 15). Следовательно, лизис клеток NK-клетками может быть уменьшен посредством блокирования NK-активирующих лигандов, таких как CD155 и CD58. Блокирование экспрессии CD155 и CD58 антителами против NK-активирующих лигандов в В2М -/- клетках является доказательством идеи создания клетки, имеющей 3 нокаута (нокаут гена ЛАЧ класса I и нокаут гена лиганда, активирующего NK-клетки). При этом клетки-мишени (например, hES и/или линия панкреотических клеток) переходят из Сценарий С в Сценарий А на Фиг 1. Конкретно, у клеток, тканей и органов согласно настоящему изобретению заингибированы или не экспрессируются белки поверхности клеток ЛАЧ класса I (В2М -/-) и заингибировали или не экспрессируются лиганды, активирующие экспрессию NK-клеток в поверхностных клетках (например, CD155 -/- и CD58-/-). Ингибирование экспрессии белков поверхности клеток может быть достигнуто путем нокаута генов или блокирования экспрессии белков с использованием антител. Другие стратегии по подавлению экспрессии белков поверхностей клеток включают использование антисмысловой РНК, РНК-ловушек, рибозимов, РНК-аптамеров, миРНК, шРНК/микроРНК, трансдоминантных негативных белков (TNP), химерных/слитых белков, нуклеаз, хемокиновых лигандов, антиинфекционных клеточных белков, внутриклеточных антител (sFvs), нуклеозидньгх аналогов (NRTI), ненуклеозидных аналогов (NNRTI), ингибиторов интеграз (олигонуклеотиды, динуклеотиды и химические агенты) и ингибиторов протеаз. Многократные нокауты генов эффективно предотвращали бы как токсичность, опосредованную цитотоксическими Т-клетками (CTL), так и токсичность, опосредованную NK-клетками, потому что практически отсутствовали бы белки ЛАЧ класса I и белки лигандов, активирующих NK-клетки, экспрессирующихся на поверхности клеток для связывания с клетками CTL или NK.
[00379] ПРИМЕР 15: ИНАКТИВАЦИЯ NK-АКТИВИРУЮЩИХ ЛИГАНДОВ ICAM1, CD155, и CD58
[00380] Для того, чтобы полностью инактивировать активацию NK, необходимо элиминировать/снизить экспрессию множества NK-активирующих лигандов либо посредством нокаута гена в клетке-мишени (например, клеточная терапия на основе клеток hES), или с использованием блокирующих антител или других стратегий, известных в настоящее время, или стратегий, которые будут разработаны в будущем. Чтобы определить, может ли токсичность NK-клеток быть снижена путем ингибирования воздействия NK-активирующих лигандов на клетки-мишени, экспрессия NK-активирующих лигандов в клетках hES WT и клетках hES В2М -/- и РЕС может быть заблокирована с использованием антитела, блокирующего ICAM1, CD155 и CD58, для блокирования белков ICAM1, CD155 и CD58 на поверхности клетки-мишени. Можно ожидать, что клеточный лизис клеток-мишеней будет уменьшен (аналогично тому, как показано на Фиг. 15). Следовательно, лизис клеток NK-клетками может быть уменьшен посредством блокирования NK-активирующих лигандов, таких как ICAM1, CD58 и CD155. Блокирование экспрессии ICAM1, CD58 и CD 155 антителами против NK-активирующих лигандов в В2М -/- клетках является доказательством идеи создания клетки, имеющей 3 нокаута (нокаут гена ЛАЧ класса I и нокаут гена лиганда, активирующего NK-клетки). При этом клетки-мишени (например, hES и/или линия панкреотических клеток) переходят из Сценарий С в Сценарий А на Фиг 1. Конкретно, у клеток, тканей и органов согласно настоящему изобретению заингибированы или не экспрессируются белки поверхности клеток ЛАЧ класса I (В2М -/-) и заингибировали или не экспрессируются лиганды, активирующие экспрессию NK-клеток в поверхностных клетках (например, ICAM1 -/-, CD58-/- и CD 155-/-). Ингибирование экспрессии белков поверхности клеток может быть достигнуто путем нокаута генов или блокирования экспрессии белков с использованием антител. Другие стратегии по подавлению экспрессии белков поверхностей клеток включают использование антисмысловой РНК, РНК-ловушек, рибозимов, РНК-аптамеров, миРНК, шРНК/микроРНК, трансдоминантных негативных белков (TNP), химерных/слитых белков, нуклеаз, хемокиновых лигандов, антиинфекционных клеточных белков, внутриклеточных антител (sFvs), нуклеозидных аналогов (NRTI), ненуклеозидных аналогов (NNRTI), ингибиторов интеграз (олигонуклеотиды, динуклеотиды и химические агенты) и ингибиторов протеаз. Многократные нокауты генов эффективно предотвращали бы как токсичность, опосредованную цитотоксическими Т-клетками (CTL), так и токсичность, опосредованную NK-клетками, потому что практически отсутствовали бы белки ЛАЧ класса 1 и белки лигандов, активирующих NK-клетки, экспрессирующихся на поверхности клеток для связывания с клетками CTL или NK.
[00381] ПРИМЕР 16: ИНАКТИВАЦИЯ NK-АКТИВИРУЮЩИХ ЛИГАНДОВ ICAM1, CD155, CD58, и CADM1
[00382] Для того, чтобы полностью инактивировать активацию NK, необходимо элиминировать/снизить экспрессию множества NK-активирующих лигандов либо посредством нокаута гена в клетке-мишени (например, клеточная терапия на основе клеток hES), или с использованием блокирующих антител или других стратегий, известных в настоящее время, или стратегий, которые будут разработаны в будущем. Чтобы определить, может ли токсичность NK-клеток быть снижена путем ингибирования воздействия NK-активирующих лигандов на клетки-мишени, экспрессия NK-активирующих лигандов в клетках hES WT и клетках hES В2М -/- и РЕС может быть заблокирована с использованием антитела, блокирующего ICAM1, CD155, CD58 и CADM1, для блокирования белков ICAM1, CD155, CD58 и CADM1 на поверхности клетки-мишени. Можно ожидать, что клеточный лизис клеток-мишеней будет уменьшен (аналогично тому, как показано на Фиг. 15). Следовательно, лизис клеток NK-клетками может быть уменьшен посредством блокирования NK-активирующих лигандов, таких как ICAM1, CD58, CD155 и CADM1. Блокирование экспрессии ICAM1, CD58, CD155 и CADM1 антителами против NK-активирующих лигандов в В2М -/- клетках является доказательством идеи создания клетки, имеющей 3 нокаута (нокаут гена ЛАЧ класса I и нокаут гена лиганда, активирующего NK-клетки). При этом клетки-мишени (например, hES и/или линия панкреотических клеток) переходят из Сценарий С в Сценарий А на Фиг 1. Конкретно, у клеток, тканей и органов согласно настоящему изобретению заингибированы или не экспрессируются белки поверхности клеток ЛАЧ класса 1 (В2М -/-) и заингибировали или не экспрессируются лиганды, активирующие экспрессию NK-клеток в поверхностных клетках (например, ICAM1 -/-, CD58-/-, CD155-/- и CADM1-/-). Ингибирование экспрессии белков поверхности клеток может быть достигнуто путем нокаута генов или блокирования экспрессии белков с использованием антител. Другие стратегии по подавлению экспрессии белков поверхностей клеток включают использование антисмысловой РНК, РНК-ловушек, рибозимов, РНК-аптамеров, миРНК, шРНК/микроРНК, трансдоминантных негативных белков (TNP), химерных/слитых белков, нуклеаз, хемокиновых лигандов, антиинфекционных клеточных белков, внутриклеточных антител (sFvs), нуклеозидных аналогов (NRTI), ненуклеозидных аналогов (NNRT1), ингибиторов интеграз (олигонуклеотиды, динуклеотиды и химические агенты) и ингибиторов протеаз. Многократные нокауты генов эффективно предотвращали бы как токсичность, опосредованную цитотоксическими Т-клетками (CTL), так и токсичность, опосредованную NK-клетками, потому что практически отсутствовали бы белки ЛАЧ класса I и белки лигандов, активирующих NK-клетки, экспрессирующихся на поверхности клеток для связывания с клетками CTL или NK.
[00383] ПРИМЕР 17: ИНАКТИВАЦИЯ NK-АКТИВИРУЮЩИХ ЛИГАНДОВ ICAM1, CD155, и CADM1
[00384] Для того, чтобы полностью инактивировать активацию NK, необходимо элиминировать/снизить экспрессию множества NK-активирующих лигандов либо посредством нокаута гена в клетке-мишени (например, клеточная терапия на основе клеток hES), или с использованием блокирующих антител или других стратегий, известных в настоящее время, или стратегий, которые будут разработаны в будущем. Чтобы определить, может ли токсичность NK-клеток быть снижена путем ингибирования воздействия NK-активирующих лигандов на клетки-мишени, экспрессия NK-активирующих лигандов в клетках hES WT и клетках hES В2М -/- и РЕС может быть заблокирована с использованием антитела, блокирующего ICAM1, CD 155, и CADM1, для блокирования белков ICAM1, CD155 и CADM1 на поверхности клетки-мишени. Можно ожидать, что клеточный лизис клеток-мишеней будет уменьшен (аналогично тому, как показано на Фиг. 15). Следовательно, лизис клеток NK-клетками может быть уменьшен посредством блокирования NK-активирующих лигандов, таких как ICAM1, CD155 и CADM1. Блокирование экспрессии ICAM1, CD155 и CADM1 антителами против NK-активирующих лигандов в В2М -/- клетках является доказательством идеи создания клетки, имеющей 3 нокаута (нокаут гена ЛАЧ класса I и нокаут гена лиганда, активирующего NK-клетки). При этом клетки-мишени (например, hES и/или линия панкреотических клеток) переходят из Сценарий С в Сценарий А на Фиг 1. Конкретно, у клеток, тканей и органов согласно настоящему изобретению заингибированы или не экспрессируются белки поверхности клеток ЛАЧ класса 1 (В2М -/-) и заингибировали или не экспрессируются лиганды, активирующие экспрессию NK-клеток в поверхностных клетках (например, ICAM1 -/-, CD155-/- и CADM1-/-). Ингибирование экспрессии белков поверхности клеток может быть достигнуто путем нокаута генов или блокирования экспрессии белков с использованием антител. Другие стратегии по подавлению экспрессии белков поверхностей клеток включают использование антисмысловой РНК, РНК-ловушек, рибозимов, РНК-аптамеров, миРНК, шРНК/микроРНК, трансдоминантных негативных белков (TNP), химерных/слитых белков, нуклеаз, хемокиновых лигандов, антиинфекционных клеточных белков, внутриклеточных антител (sFvs), нуклеозидных аналогов (NRTI), ненуклеозидных аналогов (NNRTI), ингибиторов интеграз (олигонуклеотиды, динуклеотиды и химические агенты) и ингибиторов протеаз. Многократные нокауты генов эффективно предотвращали бы как токсичность, опосредованную цитотоксическими Т-клетками (CTL), так и токсичность, опосредованную NK-клетками, потому что практически отсутствовали бы белки ЛАЧ класса I и белки лигандов, активирующих NK-клетки, экспрессирующихся на поверхности клеток для связывания с клетками CTL или NK.
[00385] Блокирующими антитела, пригодными для настоящего изобретения, являются: антитело CADM1, клон 9D2, Creative Biolabs, Кат. № BRD-0007MZ; антитело CD155, клон SKII.4, Biolegend, Кат. №337602; и антитело CD58, клон TS2/9, Biolegend, Кат. №330911.
--->
SEQUENCE LISTING
<110> ViaCyte, Inc.
<120> UNIVERSAL DONOR CELLS AND RELATED METHODS
<130> P-191628F
<150> US 15/648,337<151> 2017-07-21
<160> 15
<170> PatentIn version 3.5
<210> 1<211> 23<212> DNA<213> Homo sapiens
<400> 1cgcgagcaca gctaaggcca cgg 23
<210> 2<211> 23<212> DNA<213> Homo sapiens
<400> 2cagtaagtca acttcaatgt cgg 23
<210> 3<211> 23<212> DNA<213> Homo sapiens
<400> 3gagtagcgcg agcacagcta agg 23
<210> 4<211> 23<212> DNA<213> Homo sapiens
<400> 4cctcaaaagt catcctgccc cgg 23
<210> 5<211> 23<212> DNA<213> Homo sapiens
<400> 5agcaactcct ttttaggcaa cgg 23
<210> 6<211> 23<212> DNA<213> Homo sapiens
<400> 6ccgcactcct ggtcctgctc ggg 23
<210> 7<211> 23<212> DNA<213> Homo sapiens
<400> 7gagtgcgtgt accctggcag ggg 23
<210> 8<211> 23<212> DNA<213> Homo sapiens
<400> 8ggtacacgca ctctgtgaag tgg 23
<210> 9<211> 23<212> DNA<213> Homo sapiens
<400> 9tacacgcact ctgtgaagtg ggg 23
<210> 10<211> 1599<212> DNA<213> Homo sapiens
<400> 10atggctccca gcagcccccg gcccgcgctg cccgcactcc tggtcctgct cggggctctg 60
Ttcccaggac ctggcaatgc ccagacatct gtgtccccct caaaagtcat cctgccccgg 120
Ggaggctccg tgctggtgac atgcagcacc tcctgtgacc agcccaagtt gttgggcata 180
Gagaccccgt tgcctaaaaa ggagttgctc ctgcctggga acaaccggaa ggtgtatgaa 240
Ctgagcaatg tgcaagaaga tagccaacca atgtgctatt caaactgccc tgatgggcag 300
Tcaacagcta aaaccttcct caccgtgtac tggactccag aacgggtgga actggcaccc 360
Ctcccctctt ggcagccagt gggcaagaac cttaccctac gctgccaggt ggagggtggg 420
Gcaccccggg ccaacctcac cgtggtgctg ctccgtgggg agaaggagct gaaacgggag 480
Ccagctgtgg gggagcccgc tgaggtcacg accacggtgc tggtgaggag agatcaccat 540
Ggagccaatt tctcgtgccg cactgaactg gacctgcggc cccaagggct ggagctgttt 600
Gagaacacct cggcccccta ccagctccag acctttgtcc tgccagcgac tcccccacaa 660
Cttgtcagcc cccgggtcct agaggtggac acgcagggga ccgtggtctg ttccctggac 720
Gggctgttcc cagtctcgga ggcccaggtc cacctggcac tgggggacca gaggttgaac 780
Cccacagtca cctatggcaa cgactccttc tcggccaagg cctcagtcag tgtgaccgca 840
Gaggacgagg gcacccagcg gctgacgtgt gcagtaatac tggggaacca gagccaggag 900
Acactgcaga cagtgaccat ctacagcttt ccggcgccca acgtgattct gacgaagcca 960
Gaggtctcag aagggaccga ggtgacagtg aagtgtgagg cccaccctag agccaaggtg 1020
Acgctgaatg gggttccagc ccagccactg ggcccgaggg cccagctcct gctgaaggcc 1080
Accccagagg acaacgggcg cagcttctcc tgctctgcaa ccctggaggt ggccggccag 1140
Cttatacaca agaaccagac ccgggagctt cgtgtcctgt atggcccccg actggacgag 1200
Agggattgtc cgggaaactg gacgtggcca gaaaattccc agcagactcc aatgtgccag 1260
Gcttggggga acccattgcc cgagctcaag tgtctaaagg atggcacttt cccactgccc 1320
Atcggggaat cagtgactgt cactcgagat cttgagggca cctacctctg tcgggccagg 1380
Agcactcaag gggaggtcac ccgcaaggtg accgtgaatg tgctctcccc ccggtatgag 1440
Attgtcatca tcactgtggt agcagccgca gtcataatgg gcactgcagg cctcagcacg 1500
Tacctctata accgccagcg gaagatcaag aaatacagac tacaacaggc ccaaaaaggg 1560
Acccccatga aaccgaacac acaagccacg cctccctga 1599
<210> 11<211> 1407<212> DNA<213> Homo sapiens
<400> 11atggggcacc tctcagcccc acttcacaga gtgcgtgtac cctggcaggg gcttctgctc 60
Acagcctcac ttctaacctt ctggaacccg cccaccactg cccagctcac tactgaatcc 120
Atgccattca atgttgcaga ggggaaggag gttcttctcc ttgtccacaa tctgccccag 180
Caactttttg gctacagctg gtacaaaggg gaaagagtgg atggcaaccg tcaaattgta 240
Ggatatgcaa taggaactca acaagctacc ccagggcccg caaacagcgg tcgagagaca 300
Atatacccca atgcatccct gctgatccag aacgtcaccc agaatgacac aggattctac 360
Accctacaag tcataaagtc agatcttgtg aatgaagaag caactggaca gttccatgta 420
Tacccggagc tgcccaagcc ctccatctcc agcaacaact ccaaccctgt ggaggacaag 480
Gatgctgtgg ccttcacctg tgaacctgag actcaggaca caacctacct gtggtggata 540
Aacaatcaga gcctcccggt cagtcccagg ctgcagctgt ccaatggcaa caggaccctc 600
Actctactca gtgtcacaag gaatgacaca ggaccctatg agtgtgaaat acagaaccca 660
Gtgagtgcga accgcagtga cccagtcacc ttgaatgtca cctatggccc ggacaccccc 720
Accatttccc cttcagacac ctattaccgt ccaggggcaa acctcagcct ctcctgctat 780
Gcagcctcta acccacctgc acagtactcc tggcttatca atggaacatt ccagcaaagc 840
Acacaagagc tctttatccc taacatcact gtgaataata gtggatccta tacctgccac 900
Gccaataact cagtcactgg ctgcaacagg accacagtca agacgatcat agtcactgag 960
Ctaagtccag tagtagcaaa gccccaaatc aaagccagca agaccacagt cacaggagat 1020
Aaggactctg tgaacctgac ctgctccaca aatgacactg gaatctccat ccgttggttc 1080
Ttcaaaaacc agagtctccc gtcctcggag aggatgaagc tgtcccaggg caacaccacc 1140
Ctcagcataa accctgtcaa gagggaggat gctgggacgt attggtgtga ggtcttcaac 1200
Ccaatcagta agaaccaaag cgaccccatc atgctgaacg taaactataa tgctctacca 1260
Caagaaaatg gcctctcacc tggggccatt gctggcattg tgattggagt agtggccctg 1320
Gttgctctga tagcagtagc cctggcatgt tttctgcatt tcgggaagac cggcaggacc 1380
Actccaatga cccacctaac aagatga 1407
<210> 12<211> 360<212> DNA<213> Homo sapiens
<400> 12atgtctcgct ccgtggcctt agctgtgctc gcgctactct ctctttctgg cctggaggct 60
Atccagcgta ctccaaagat tcaggtttac tcacgtcatc cagcagagaa tggaaagtca 120
Aatttcctga attgctatgt gtctgggttt catccatccg acattgaagt tgacttactg 180
Aagaatggag agagaattga aaaagtggag cattcagact tgtctttcag caaggactgg 240
Tctttctatc tcttgtacta cactgaattc acccccactg aaaaagatga gtatgcctgc 300
Cgtgtgaacc atgtgacttt gtcacagccc aagatagtta agtgggatcg agacatgtaa 360
<210> 13<211> 1329<212> DNA<213> Homo sapiens
<400> 13atggcgagtg tagtgctgcc gagcggatcc cagtgtgcgg cggcagcggc ggcggcggcg 60
Cctcccgggc tccggctccg gcttctgctg ttgctcttct ccgccgcggc actgatcccc 120
Acaggtgatg ggcagaatct gtttacgaaa gacgtgacag tgatcgaggg agaggttgcg 180
Accatcagtt gccaagtcaa taagagtgac gactctgtga ttcagctact gaatcccaac 240
Aggcagacca tttatttcag ggacttcagg cctttgaagg acagcaggtt tcagttgctg 300
Aatttttcta gcagtgaact caaagtatca ttgacaaacg tctcaatttc tgatgaagga 360
Agatactttt gccagctcta taccgatccc ccacaggaaa gttacaccac catcacagtc 420
Ctggtcccac cacgtaatct gatgatcgat atccagaaag acactgcggt ggaaggtgag 480
Gagattgaag tcaactgcac tgctatggcc agcaagccag ccacgactat caggtggttc 540
Aaagggaaca cagagctaaa aggcaaatcg gaggtggaag agtggtcaga catgtacact 600
Gtgaccagtc agctgatgct gaaggtgcac aaggaggacg atggggtccc agtgatctgc 660
Caggtggagc accctgcggt cactggaaac ctgcagaccc agcggtatct agaagtacag 720
Tataagcctc aagtgcacat tcagatgact tatcctctac aaggcttaac ccgggaaggg 780
Gacgcgcttg agttaacatg tgaagccatc gggaagcccc agcctgtgat ggtaacttgg 840
Gtgagagtcg atgatgaaat gcctcaacac gccgtactgt ctgggcccaa cctgttcatc 900
Aataacctaa acaaaacaga taatggtaca taccgctgtg aagcttcaaa catagtgggg 960
Aaagctcact cggattatat gctgtatgta tacgatcccc ccacaactat ccctcctccc 1020
Acaacaacca ccaccaccac caccaccacc accaccacca tccttaccat catcacagat 1080
Tcccgagcag gtgaagaagg ctcgatcagg gcagtggatc atgccgtgat cggtggcgtc 1140
Gtggcggtgg tggtgttcgc catgctgtgc ttgctcatca ttctggggcg ctattttgcc 1200
Agacataaag gtacatactt cactcatgaa gccaaaggag ccgatgacgc agcagacgca 1260
Gacacagcta taatcaatgc agaaggagga cagaacaact ccgaagaaaa gaaagagtac 1320
Ttcatctag 1329
<210> 14<211> 753<212> DNA<213> Homo sapiens
<400> 14atggttgctg ggagcgacgc ggggcgggcc ctgggggtcc tcagcgtggt ctgcctgctg 60
Cactgctttg gtttcatcag ctgtttttcc caacaaatat atggtgttgt gtatgggaat 120
Gtaactttcc atgtaccaag caatgtgcct ttaaaagagg tcctatggaa aaaacaaaag 180
Gataaagttg cagaactgga aaattctgaa ttcagagctt tctcatcttt taaaaatagg 240
Gtttatttag acactgtgtc aggtagcctc actatctaca acttaacatc atcagatgaa 300
Gatgagtatg aaatggaatc gccaaatatt actgatacca tgaagttctt tctttatgtg 360
Cttgagtctc ttccatctcc cacactaact tgtgcattga ctaatggaag cattgaagtc 420
Caatgcatga taccagagca ttacaacagc catcgaggac ttataatgta ctcatgggat 480
Tgtcctatgg agcaatgtaa acgtaactca accagtatat attttaagat ggaaaatgat 540
Cttccacaaa aaatacagtg tactcttagc aatccattat ttaatacaac atcatcaatc 600
Attttgacaa cctgtatccc aagcagcggt cattcaagac acagatatgc acttataccc 660
Ataccattag cagtaattac aacatgtatt gtgctgtata tgaatggtat tctgaaatgt 720
Gacagaaaac cagacagaac caactccaat tga 753
<210> 15<211> 1254<212> DNA<213> Homo sapiens
<400> 15atggcccgag ccatggccgc cgcgtggccg ctgctgctgg tggcgctact ggtgctgtcc 60
Tggccacccc caggaaccgg ggacgtcgtc gtgcaggcgc ccacccaggt gcccggcttc 120
Ttgggcgact ccgtgacgct gccctgctac ctacaggtgc ccaacatgga ggtgacgcat 180
Gtgtcacagc tgacttgggc gcggcatggt gaatctggca gcatggccgt cttccaccaa 240
Acgcagggcc ccagctattc ggagtccaaa cggctggaat tcgtggcagc cagactgggc 300
Gcggagctgc ggaatgcctc gctgaggatg ttcgggttgc gcgtagagga tgaaggcaac 360
Tacacctgcc tgttcgtcac gttcccgcag ggcagcagga gcgtggatat ctggctccga 420
Gtgcttgcca agccccagaa cacagctgag gttcagaagg tccagctcac tggagagcca 480
Gtgcccatgg cccgctgcgt ctccacaggg ggtcgcccgc cagcccaaat cacctggcac 540
Tcagacctgg gcgggatgcc caatacgagc caggtgccag ggttcctgtc tggcacagtc 600
Actgtcacca gcctctggat attggtgccc tcaagccagg tggacggcaa gaatgtgacc 660
Tgcaaggtgg agcacgagag ctttgagaag cctcagctgc tgactgtgaa cctcaccgtg 720
Tactaccccc cagaggtatc catctctggc tatgataaca actggtacct tggccagaat 780
Gaggccaccc tgacctgcga tgctcgcagc aacccagagc ccacaggcta taattggagc 840
Acgaccatgg gtcccctgcc accctttgct gtggcccagg gcgcccagct cctgatccgt 900
Cctgtggaca aaccaatcaa cacaacttta atctgcaacg tcaccaatgc cctaggagct 960
Cgccaggcag aactgaccgt ccaggtcaaa gagggacctc ccagtgagca ctcaggcatg 1020
Tcccgtaacg ccatcatctt cctggttctg ggaatcctgg tttttctgat cctgctgggg 1080
Atcgggattt atttctattg gtccaaatgt tcccgtgagg tcctttggca ctgtcatctg 1140
Tgtccctcga gtacagagca tgccagcgcc tcagctaatg ggcatgtctc ctattcagct 1200
Gtgagcagag agaacagctc ttcccaggat ccacagacag agggcacaag gtga 1254
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| УНИВЕРСАЛЬНЫЕ ДОНОРСКИЕ КЛЕТКИ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2018 |
|
RU2827758C2 |
| СПОСОБ СОЗДАНИЯ Т-КЛЕТОК, ПРИГОДНЫХ ДЛЯ АЛЛОГЕННОЙ ТРАНСПЛАНТАЦИИ | 2015 |
|
RU2752933C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ГАММА ДЕЛЬТА Т-КЛЕТКИ | 2015 |
|
RU2756247C2 |
| УНИВЕРСАЛЬНЫЕ АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ И ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2809113C2 |
| ИММУНОСТИМУЛИРУЮЩАЯ КОМБИНАЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ГЕПАТИТА С | 2006 |
|
RU2431499C2 |
| СПОСОБЫ EX VIVO ЭКСПАНСИИ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2814083C2 |
| КОМПОЗИЦИИ Т-КЛЕТОК С НЕДОСТАТОЧНОСТЬЮ РЕЦЕПТОРОВ Т-КЛЕТОК | 2013 |
|
RU2653761C2 |
| ДНК-ВАКЦИНЫ ПРОТИВ ОПУХОЛЕВОГО РОСТА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2343195C2 |
| СКОНСТРУИРОВАННЫЕ КЛЕТКИ, ЭКСПРЕССИРУЮЩИЕ МНОЖЕСТВЕННЫЕ ИММУНОМОДУЛЯТОРЫ, И ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2573912C2 |
| ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА К CD33 | 2015 |
|
RU2747384C2 |
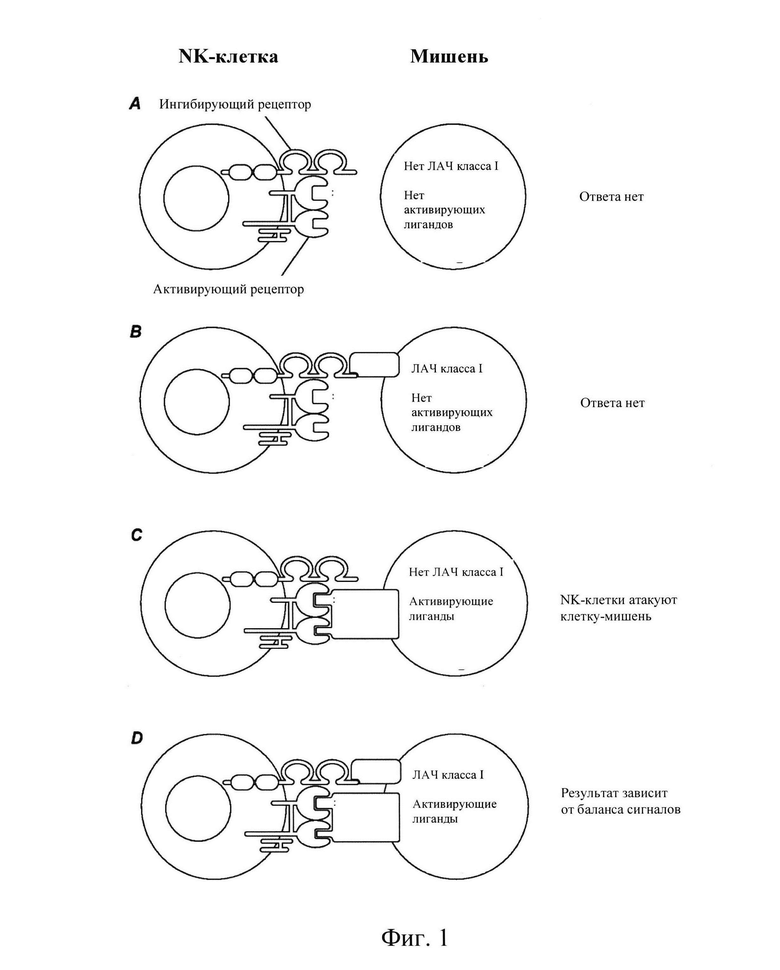
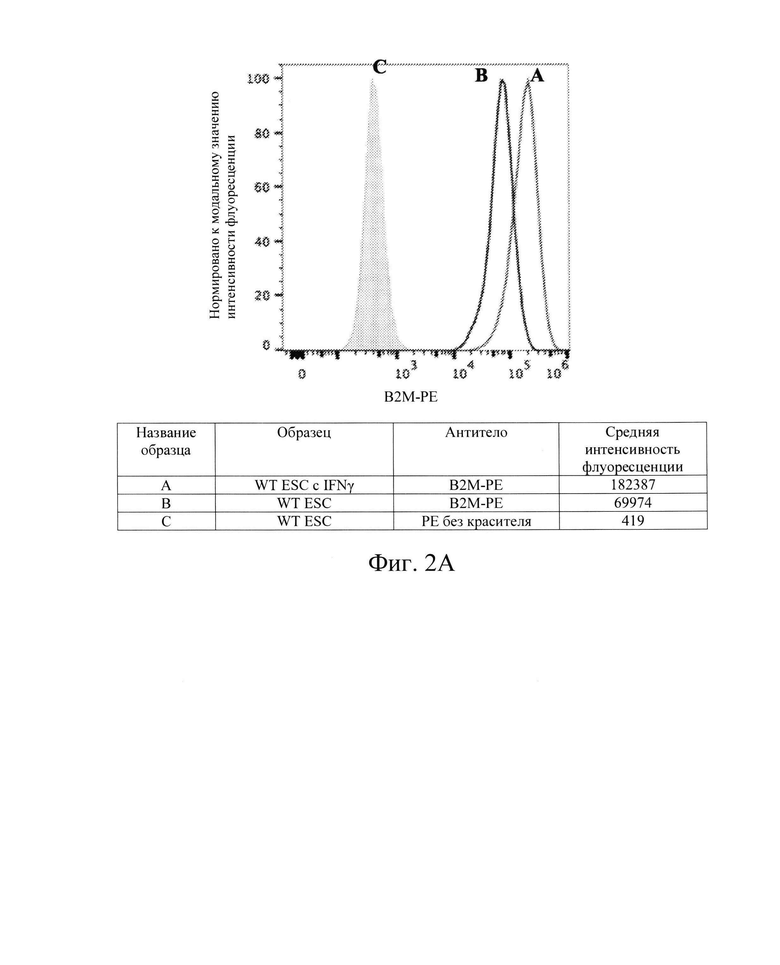
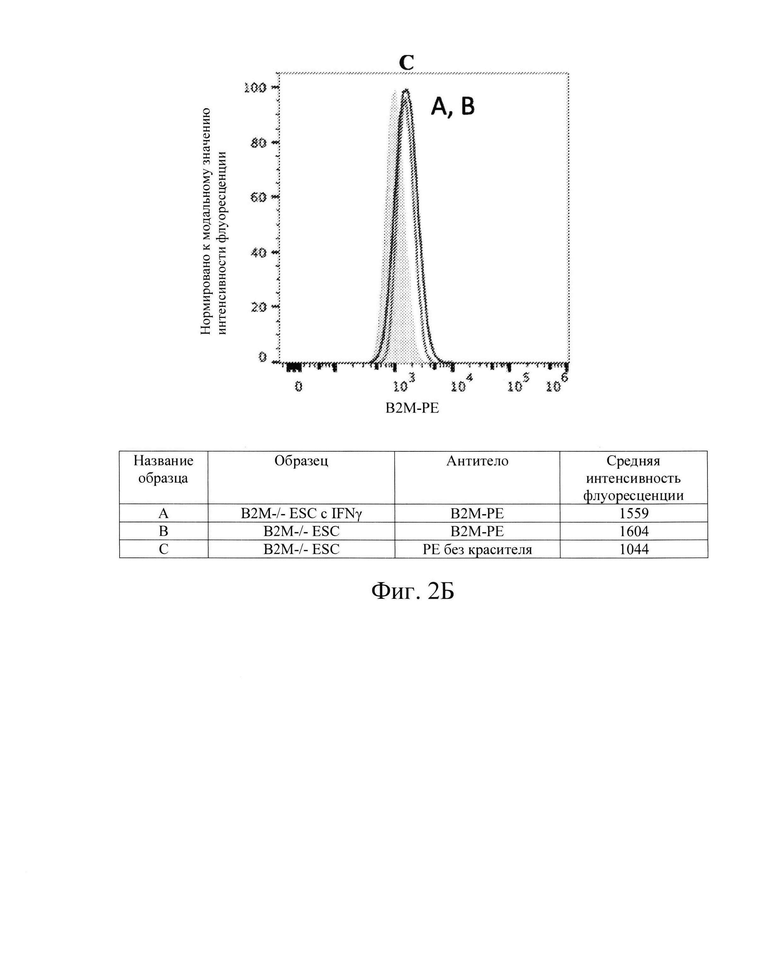
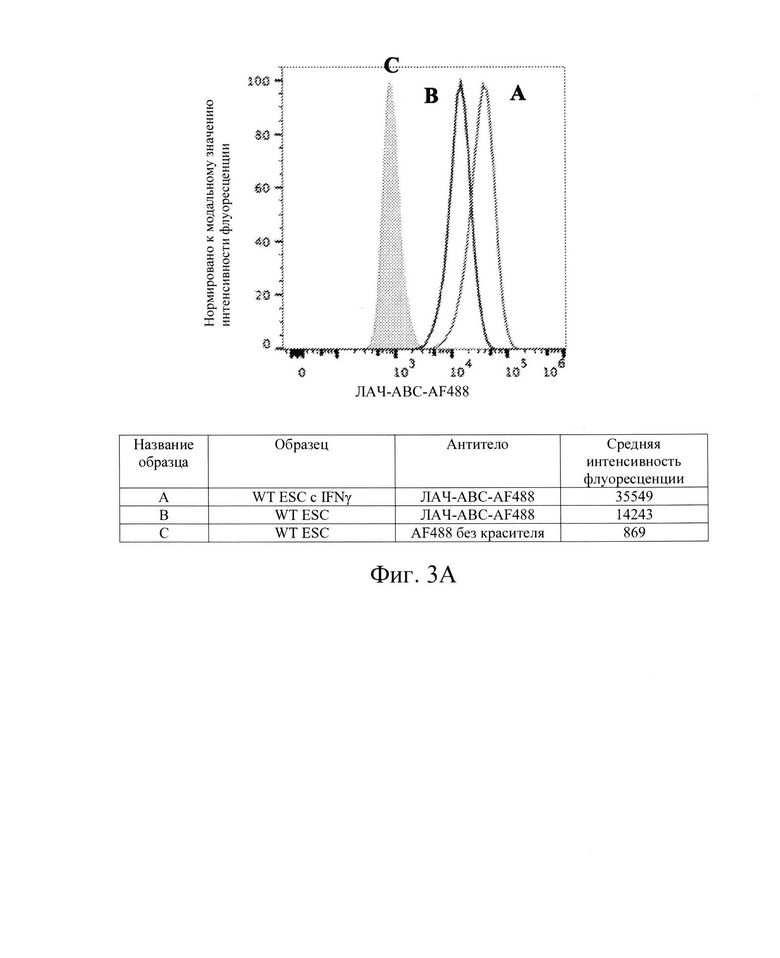
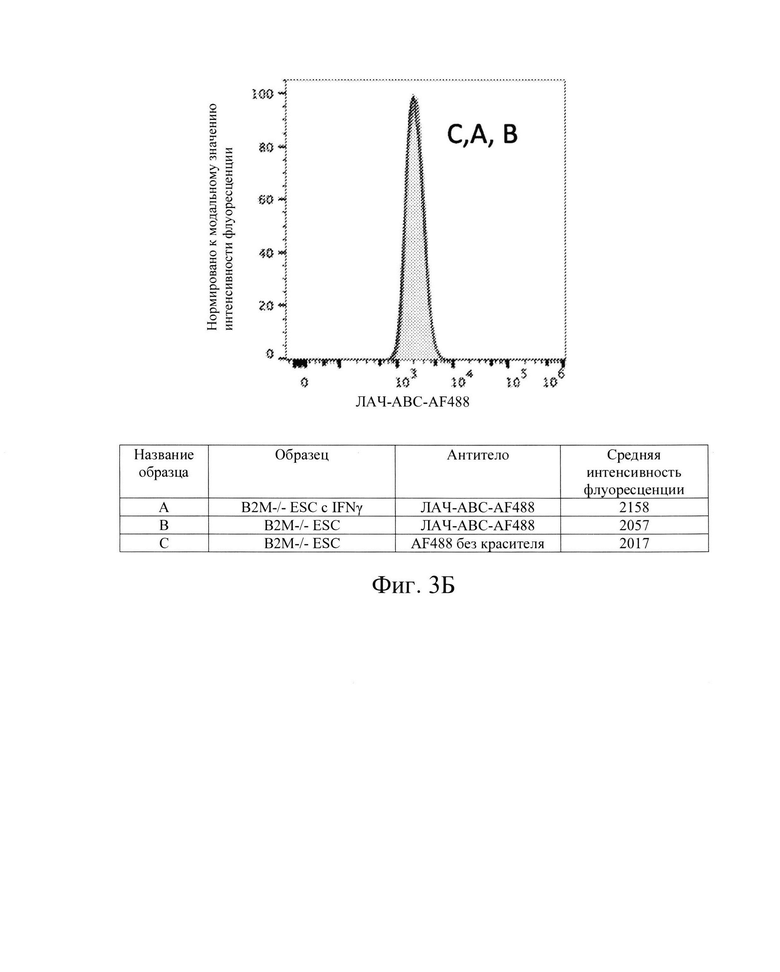
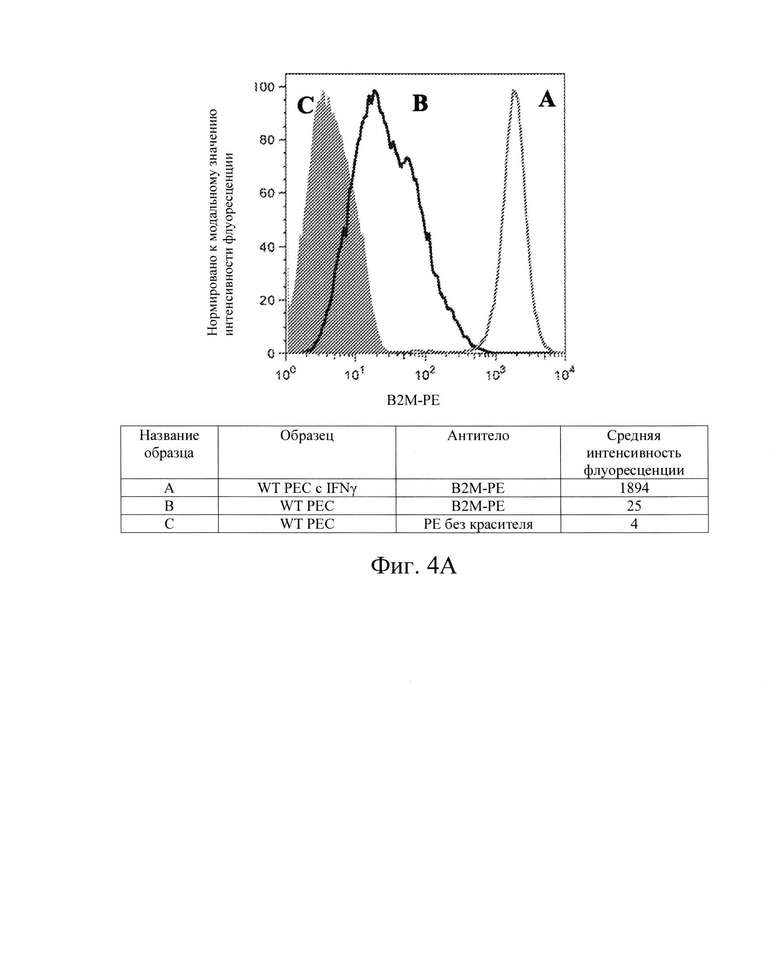
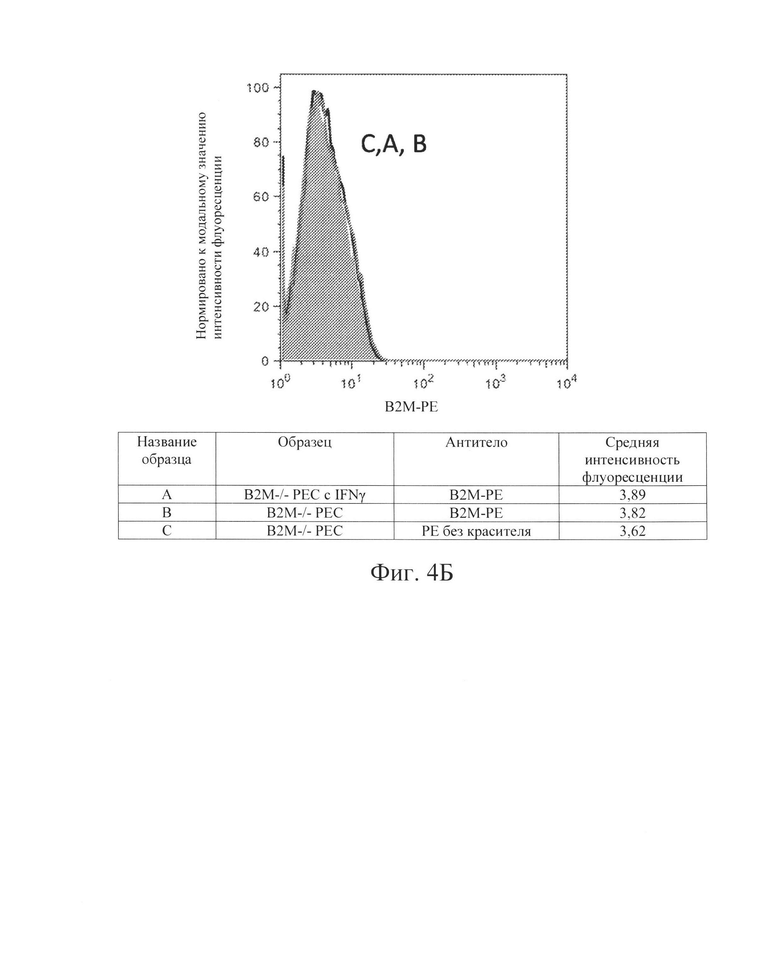
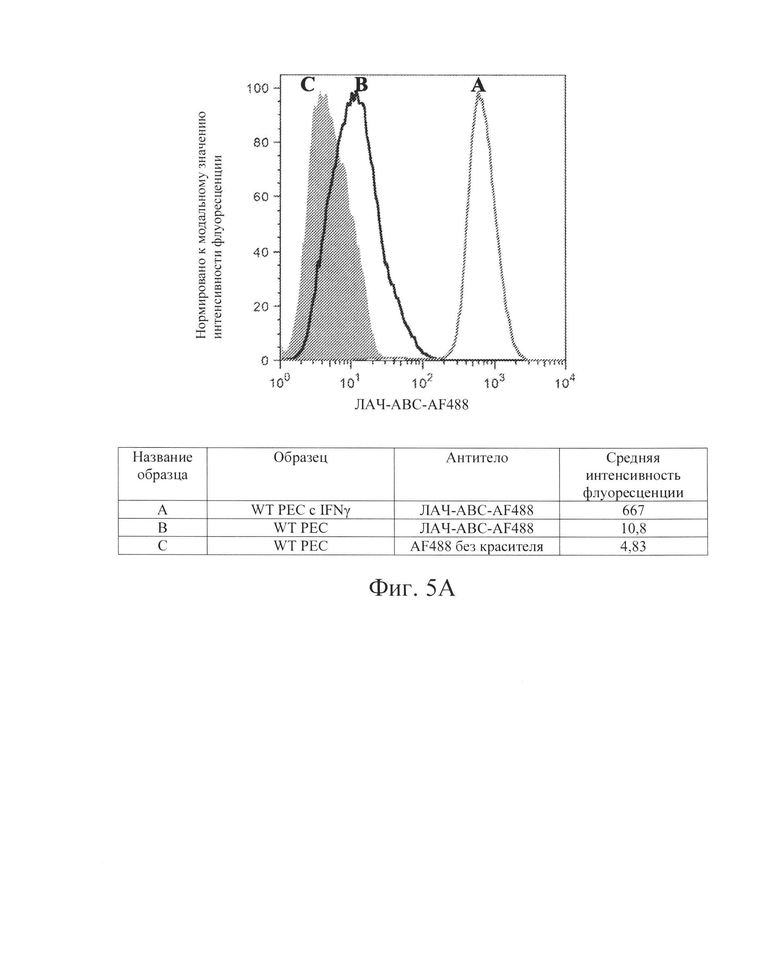
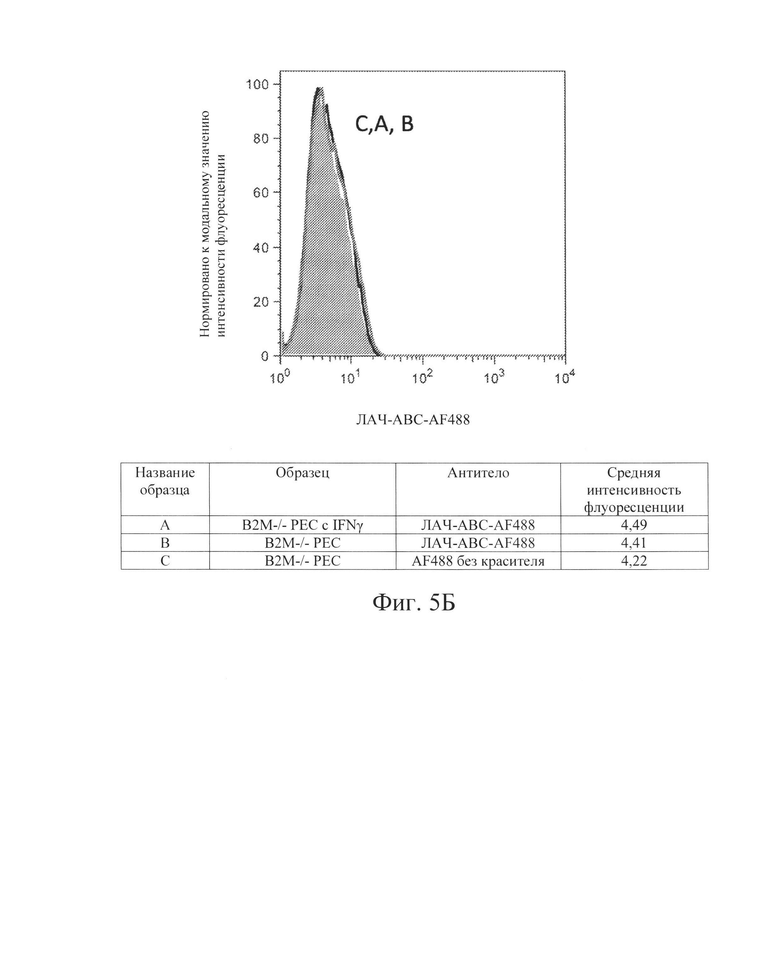
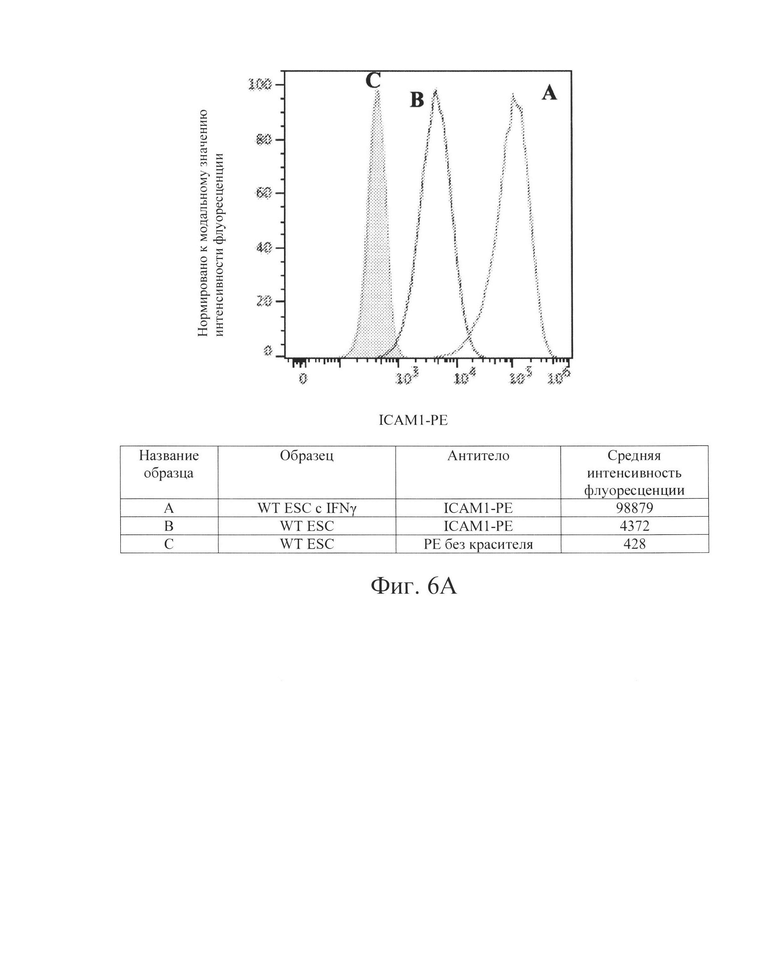
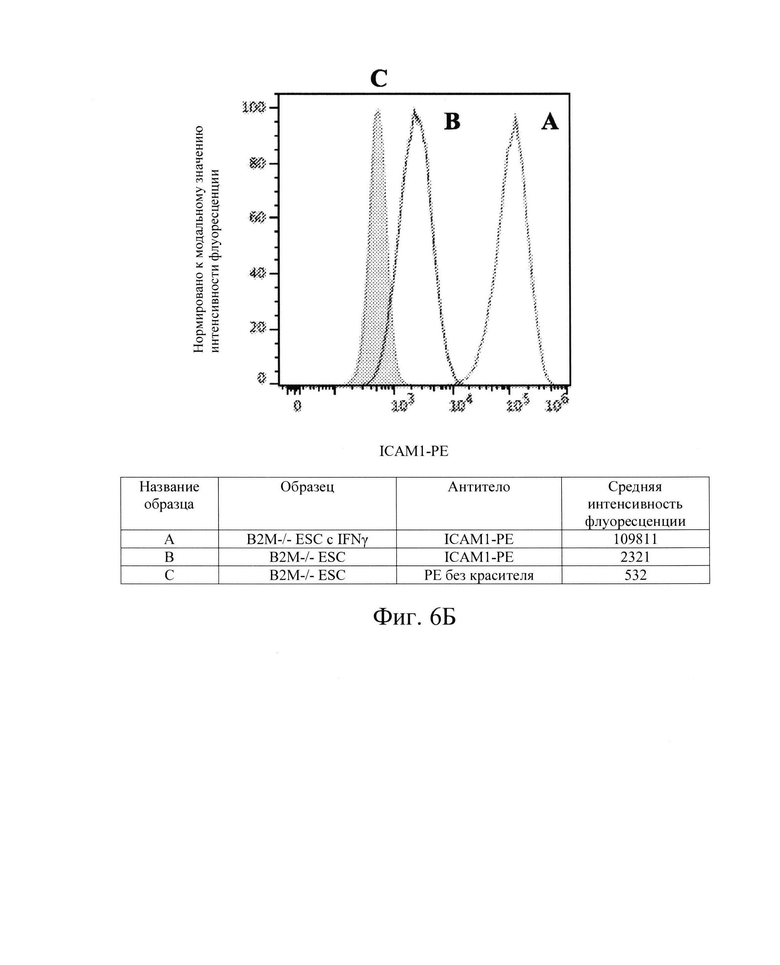
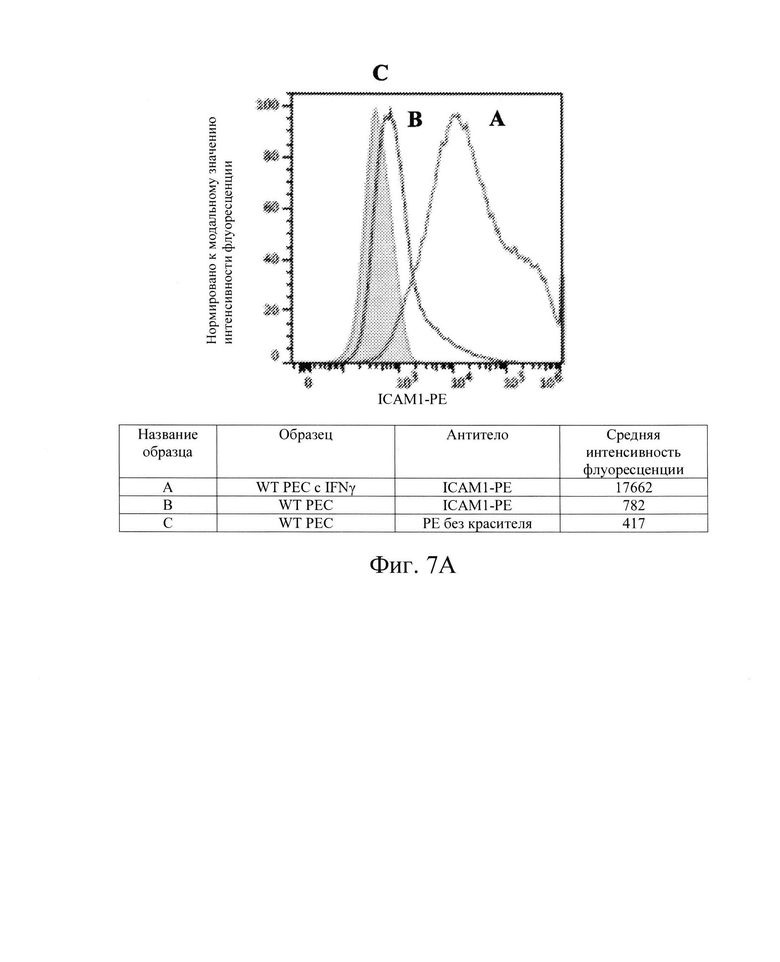
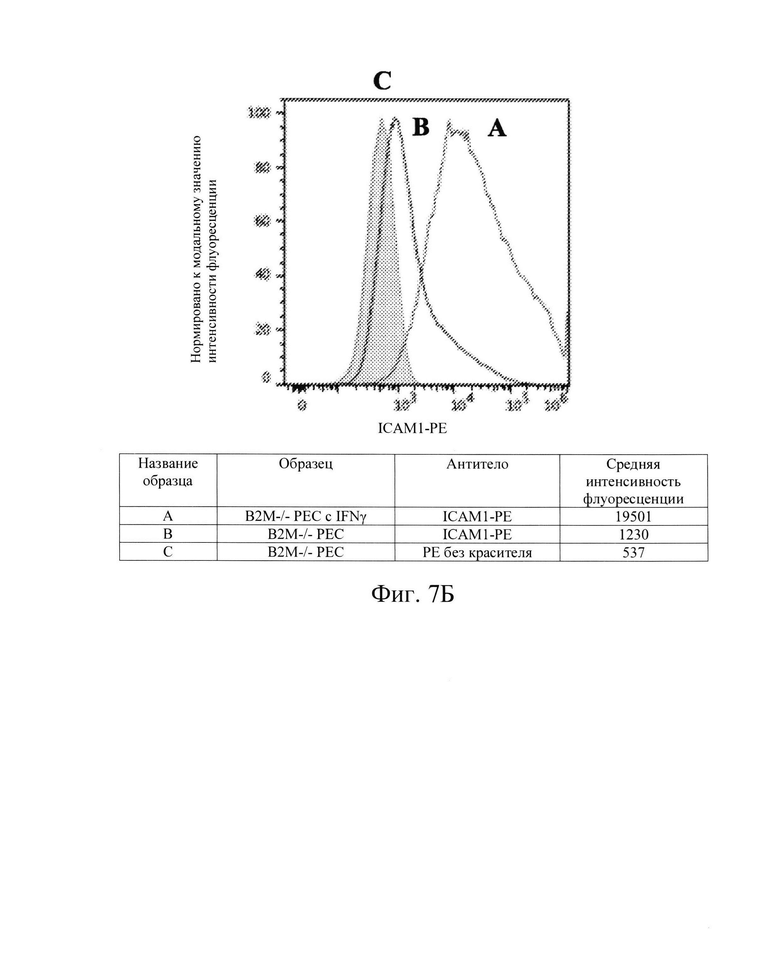
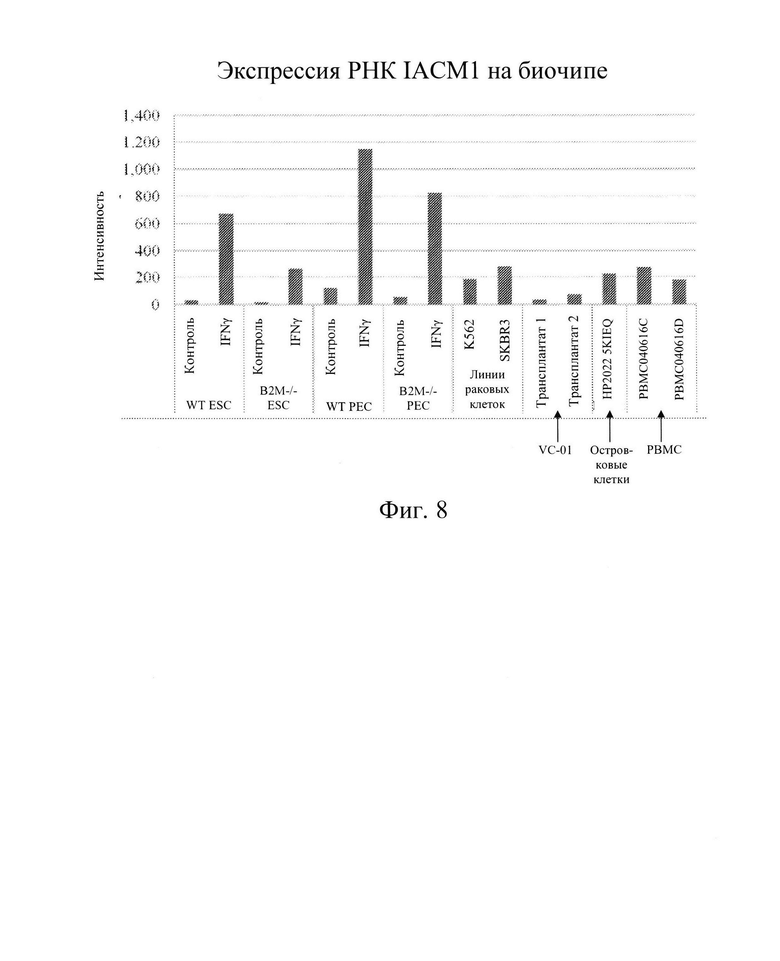
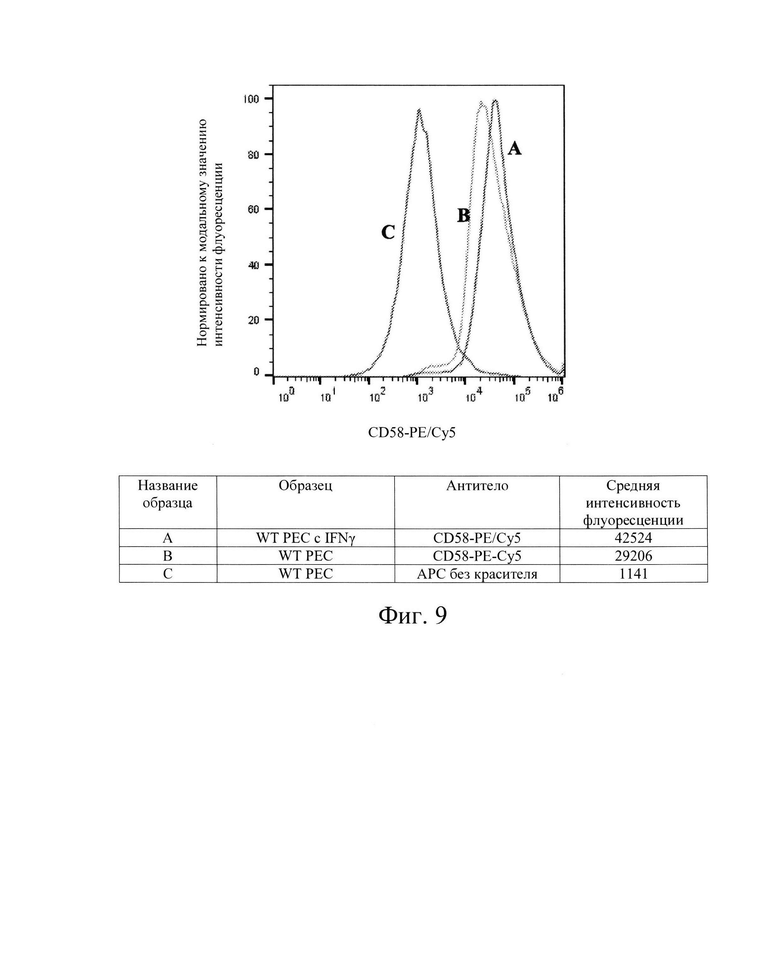
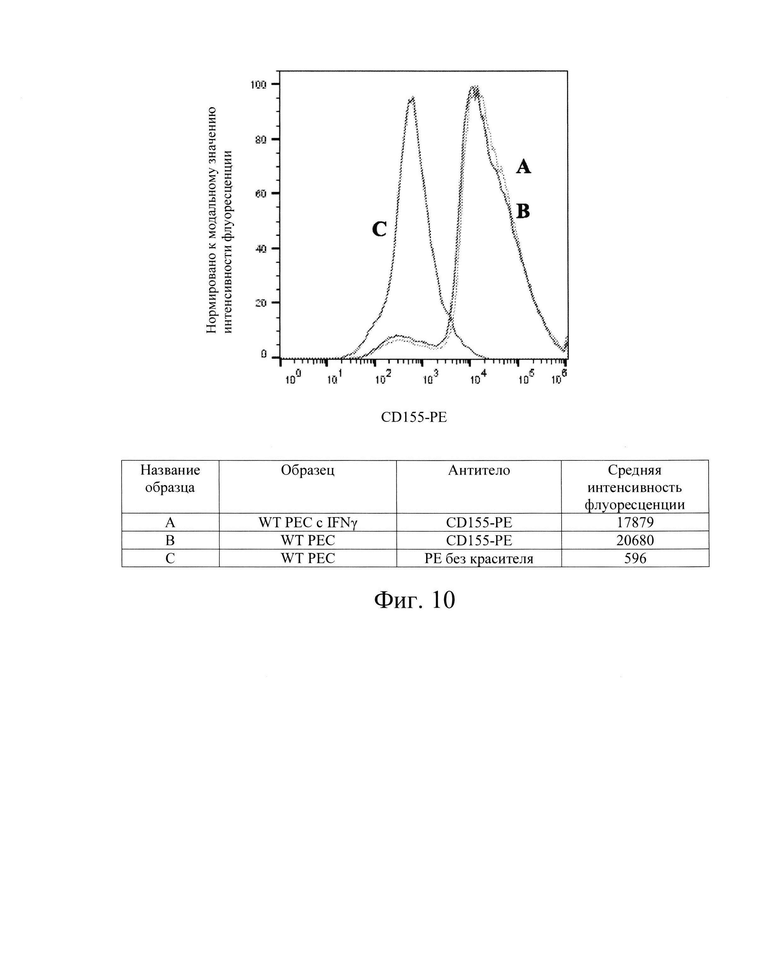
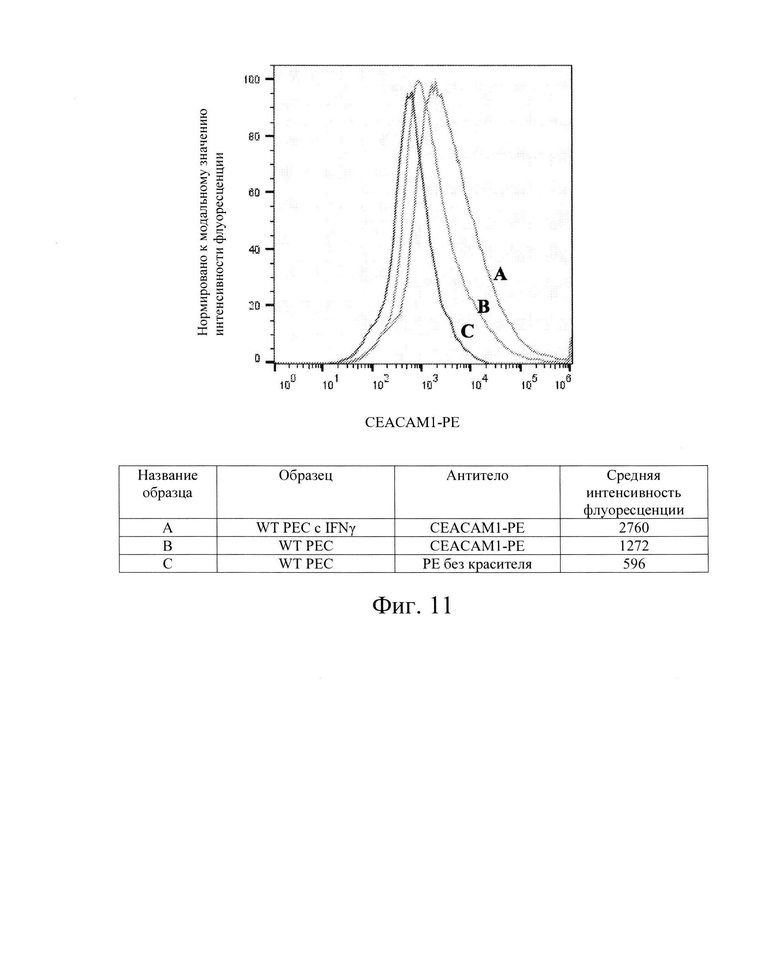
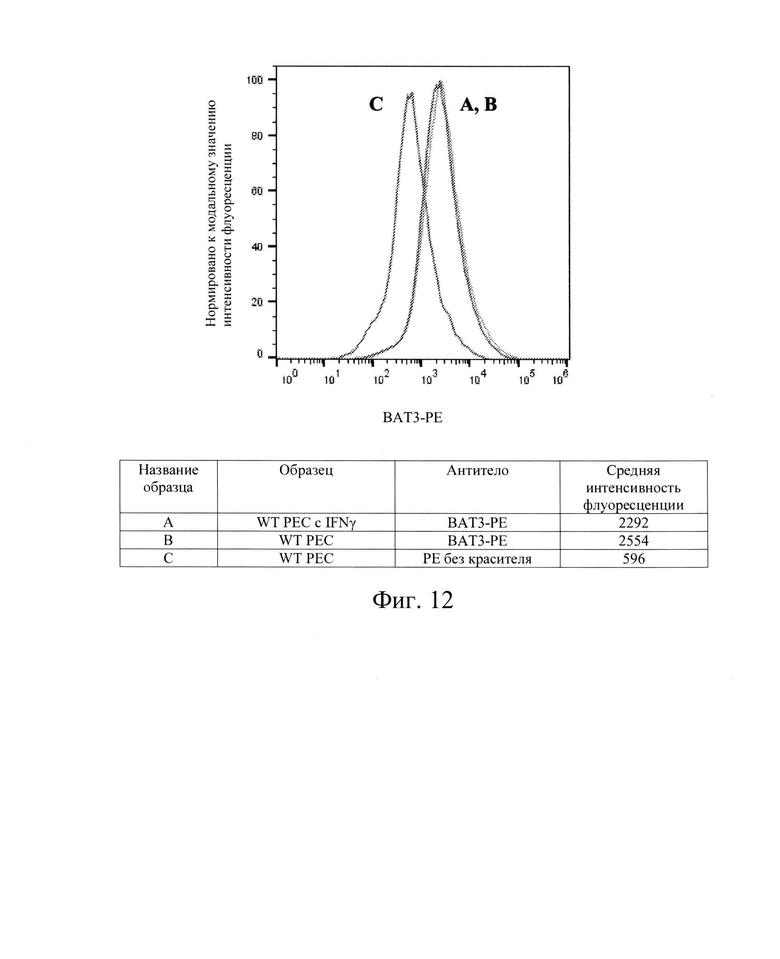
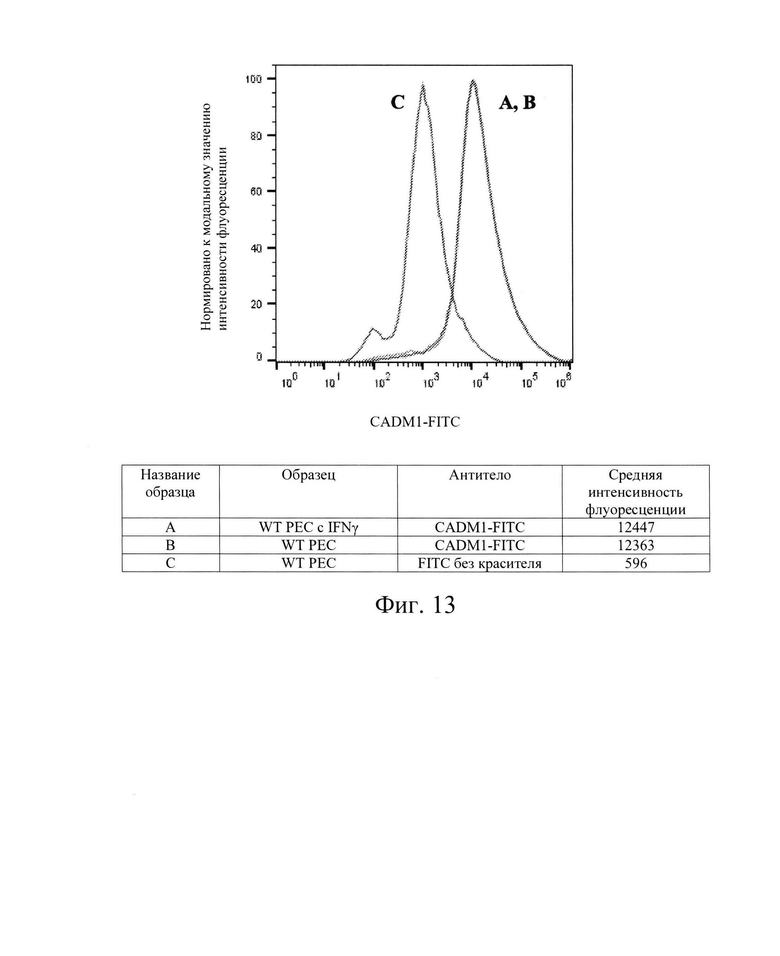
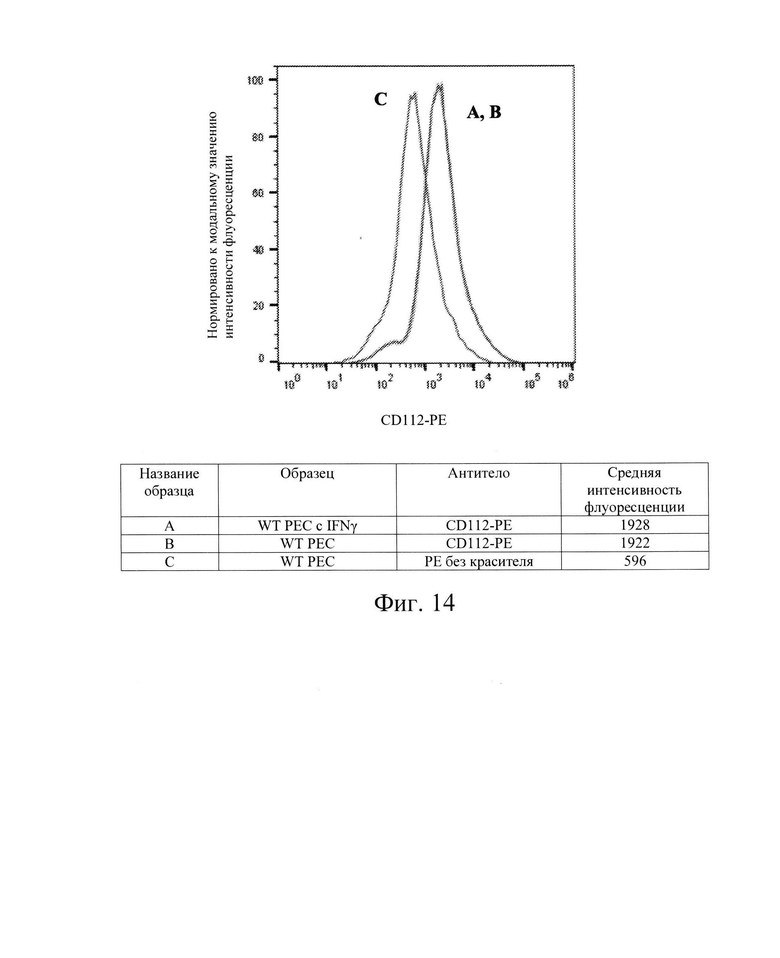
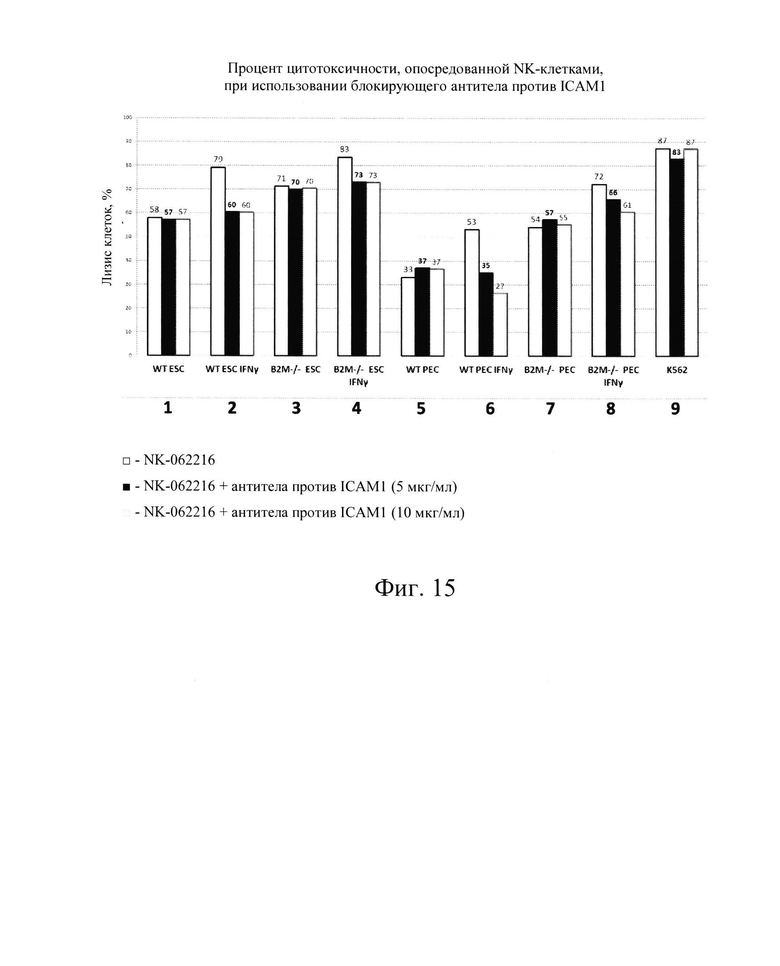
Настоящее изобретение относится к клеточной биологии и медицине, в частности к композиции клеток для трансплантации, содержащая популяцию панкреатических клеток человека in vitro, её применению для трансплантации и для получения инсулина в субъекте, а также способу снижения отторжения трансплантата клеток панкреатической эндодермы в человеке и способу получения инсулина в человеке, обладающем здоровым иммунитетом или ослабленным иммунитетом. Указанная композиция клеток содержит популяцию панкреатических клеток человека in vitro, содержащую CHGA-, NKX6.1+, PDX1+ клетки-предшественники панкреатических клеток, и CHGA+ панкреатические эндокринные клетки, в которых функция по меньшей мере одного гена главного комплекса гистосовместимости (ГКГС) класса I и по меньшей мере одного лиганда, активирующего естественные клетки-киллеры (NK), нарушена или заингибирована, что приводит к снижению связывания NK-активирующего лиганда с NK-активирующим рецептором в указанной популяции панкреатических клеток человека in vitro. Настоящее изобретение позволяет получать клетки-мишени, в которых отсутствует часть или вся классическая экспрессия клеток лейкоцитарного антигена человека, но которые не подвергаются атакам NK-клеток с последующим лизисом. 6 н. и 20 з.п. ф-лы, 21 ил., 5 табл., 17 пр.
1. Композиция клеток для трансплантации, содержащая популяцию панкреатических клеток человека in vitro, содержащая CHGA-, NKX6.1+, PDX1+ клетки-предшественники панкреатических клеток, и CHGA+ панкреатические эндокринные клетки, в которых функция по меньшей мере одного гена главного комплекса гистосовместимости (ГКГС) класса I и по меньшей мере одного лиганда, активирующего естественные клетки-киллеры (NK), нарушена или заингибирована, что приводит к снижению связывания NK-активирующего лиганда с NK-активирующим рецептором в указанной популяции панкреатических клеток человека in vitro.
2. Композиция клеток для трансплантации по п. 1, отличающаяся тем, что указанный ген ГКГС класса I кодирует бета-2 микроглобулин (В2М).
3. Композиция клеток для трансплантации по п. 1 или 2, отличающаяся тем, что указанным лигандом, активирующим NK-клетки, является:
а) молекула межклеточной адгезии 1 (ICAM1), CD58, CD155, молекула клеточной адгезии, связанная с карциноэмбриональным антигеном 1 (СЕАСАМ1), молекула клеточной адгезии 1 (CADM1), белок, связанный с полипептидом главного комплекса гистосовместимости класса I A (MICA), белок, связанный с полипептидом главного комплекса гистосовместимости класса I В (MICB), или их комбинация;
б) ICAM1 и CD58;
в) ICAM1, CD58, CD155, СЕАСАМ1, CADM1, MICA и MICB;
г) ICAM1,CD58, CD155;
д) ICAM1, CD58, CD155 и CADM1;
е) CD58 и CADM1; или
ж) ICAM1, CADM1 и CD155.
4. Композиция клеток для трансплантации по любому из пп. 1-3, отличающаяся тем, что указанная популяция панкреатических клеток экспрессирует белок, который будучи экспрессирован в присутствии агента, вызывающего гибель клеток, способен вызывать гибель клеток указанной популяции клеток.
5. Композиция клеток для трансплантации по п. 4, отличающаяся тем, что указанным белком является тимидинкиназа простого вируса герпеса, а указанным агентом, вызывающим гибель клеток, является ганцикловир.
6. Применение композиции клеток для трансплантации по любому из пп. 1-5 для снижения отторжения трансплантата популяции панкреатических клеток человека, при котором нарушение или ингибирование функции гена ГКГС класса I и лиганда, активирующего NK-клетки, приводящее к снижению связывания NK-активирующего лиганда с NK-активирующим рецептором в указанной популяции панкреатических клеток человека in vitro, снижает отторжение трансплантата клеток по сравнению с панкреатическими клетками человека без нарушения или ингибирования функции указанного гена ГКГС класса I и лиганда, активирующего NK-клетки.
7. Применение композиции клеток для трансплантации по любому из пп. 1-5 для получения инсулина в субъекте, являющемся млекопитающим, в котором указанная популяция панкреатических клеток человека in vitro созревают в субъекте, представляющем собой млекопитающее, в клетки, которые производят инсулин в ответ на стимуляцию глюкозой.
8. Способ снижения отторжения трансплантата клеток панкреатической эндодермы (РЕС) в человеке, обладающем здоровым иммунитетом или ослабленным иммунитетом, включающий:
трансплантацию устройства с клетками, упакованными в капсулу, содержащего популяцию клеток, содержащую клетки РЕС, которые являются CHGA-, NKX6.1+, PDX1+ клетками-предшественниками панкреатических клеток, и РЕС, которые являются CHGA+ панкреатическими эндокринными клетками, в локацию в человеке, обладающем здоровым иммунитетом или ослабленным иммунитетом, которая допускает васкуляризацию в реципиенте,
при этом функция по меньшей мере одного гена главного комплекса гистосовместимости (ГКГС) класса I и по меньшей мере одного лиганда, активирующего естественные клетки-киллеры (NK), нарушена или заингибирована в указанной популяции клеток, что приводит к снижению связывания NK-активирующего лиганда с NK-активирующим рецептором, а нарушение или ингибирование по меньшей мере одного гена ГКГС класса I и по меньшей мере одного лиганда, активирующего естественные NK-клетки, приводит к ингибированию отторжения трансплантата клеток указанной популяции клеток в человеке, обладающем здоровым иммунитетом или ослабленным иммунитетом.
9. Способ по п. 8, отличающийся тем, что указанной локацией в человеке, обладающем здоровым иммунитетом или ослабленным иммунитетом, является подкожная локация.
10. Способ по п. 8 или 9, отличающийся тем, что указанный ген ГКГС класса I кодирует бета-2 микроглобулин (В2М) или кодирует белок поверхности клеток лейкоцитарного антигена человека (ЛАЧ)-АВС.
11. Способ по любому из пп. 8-10, отличающийся тем, что указанным лигандом, активирующим NK-клетки, является:
а) молекула межклеточной адгезии 1 (ICAM1), CD58, CD155, молекула клеточной адгезии, связанная с карциноэмбриональным антигеном 1 (СЕАСАМ1), молекула клеточной адгезии 1 (CADM1), белок, связанный с полипептидом главного комплекса гистосовместимости класса I A (MICA), или белок, связанный с полипептидом главного комплекса гистосовместимости класса I В (М1СВ);
б) ICAM1 и CD58;
в) ICAM1, CD58, CD155;
г) ICAM1, CD58, CD155 и CADM1;
д) СD58 и CADM1; или
е) ICAM1, CADM1 и CD155.
12. Способ по любому из пп. 8-11, отличающийся тем, указанная популяция клеток экспрессирует белок, который будучи экспрессирован в присутствии агента, вызывающего гибель клеток, способен вызывать гибель клеток указанной популяции клеток.
13. Способ по п. 12, отличающийся тем, что указанным белком является тимидинкиназа простого вируса герпеса, а указанным агентом, вызывающим гибель клеток, является ганцикловир.
14. Способ по любому из пп. 8-13, отличающийся тем, что указанным человеком является человек, обладающий здоровым иммунитетом.
15. Способ по любому из пп. 8-13, отличающийся тем, что указанным человеком является человек, обладающий ослабленным иммунитетом.
16. Способ получения инсулина в человеке, обладающем здоровым иммунитетом или ослабленным иммунитетом, включающий:
трансплантацию устройства с клетками, упакованными в капсулу, содержащего популяцию клеток, содержащую клетки РЕС, которые являются CHGA-, NKX6.1+, PDX1+ клетками-предшественниками панкреатических клеток, и РЕС, которые являются CHGA+ панкреатическими эндокринными клетками, в локацию в человеке, обладающем здоровым иммунитетом или ослабленным иммунитетом, которая допускает васкуляризацию в реципиенте,
при этом функция по меньшей мере одного гена главного комплекса гистосовместимости (ГКГС) класса I и по меньшей мере одного лиганда, активирующего естественные клетки-киллеры (NK), нарушена или заингибирована в указанной популяции клеток, что приводит к снижению связывания NK-активирующего лиганда с NK-активирующим рецептором, при этом указанная популяция клеток созревает в указанном человеке, обладающем здоровым иммунитетом или ослабленным иммунитетом, в популяцию зрелых клеток, что приводит к продукции инсулина в ответ на стимуляцию глюкозой.
17. Способ по п. 16, отличающийся тем, что указанной локацией в человеке, обладающем здоровым иммунитетом или ослабленным иммунитетом, является подкожная локация.
18. Способ по п. 16 и 17, отличающийся тем, что указанный ген ГКГС класса I кодирует бета-2 микроглобулин (В2М) или кодирует белок поверхности клеток лейкоцитарного антигена человека (ЛАЧ)-АВС.
19. Способ по любому из пп. 16-18, отличающийся тем, что указанным лигандом, активирующим NK-клетки, является:
а) молекула межклеточной адгезии 1 (ICAM1), CD58, CD155, молекула клеточной адгезии, связанная с карциноэмбриональным антигеном 1 (СЕАСАМ1), молекула клеточной адгезии 1 (CADM1), белок, связанный с полипептидом главного комплекса гистосовместимости класса I A (MICA), или белок, связанный с полипептидом главного комплекса гистосовместимости класса I В (М1СВ);
б) ICAM1 и CD58;
в) ICAM1, CD58 и CD155;
г) ICAM1, CD58, CD155 и CADM1;
д) CD58 и CADM1; или
е) ICAM1, CADM1 и CD155.
20. Способ по любому из пп. 16-19, отличающийся тем, что указанная популяция клеток экспрессирует белок, который будучи экспрессирован в присутствии агента, вызывающего гибель клеток, способен вызывать гибель клеток указанной популяции клеток.
21. Способ по п. 20, отличающийся тем, что указанным белком является тимидинкиназа простого вируса герпеса, а указанным агентом, вызывающим гибель клеток, является ганцикловир.
22. Способ по любому из пп. 16-21, отличающийся тем, что указанным человеком является человек, обладающий здоровым иммунитетом.
23. Способ по любому из пп. 16-21, отличающийся тем, что указанным человеком является человек, обладающий ослабленным иммунитетом.
24. Композиция клеток для трансплантации, содержащая популяцию панкреатических клеток in vitro, содержащая CHGA-, NKX6.1+, PDX1+ клетки-предшественники панкреатических клеток, и CHGA+ панкреатические эндокринные клетки, в которых функция по меньшей мере одного гена главного комплекса гистосовместимости (ГКГС) класса I и по меньшей мере одного лиганда, активирующего естественные клетки-киллеры (NK), нарушена или заингибирована, что приводит к снижению связывания NK-активирующего лиганда с NK-активирующим рецептором в указанной популяции панкреатических клеток in vitro.
25. Композиция клеток для трансплантации по п. 24, отличающаяся тем, что указанный ген ГКГС класса I кодирует бета-2 микроглобулин (В2М).
26. Композиция клеток для трансплантации по п. 24 или 25, отличающаяся тем, что указанным лигандом, активирующим NK-клетки, является:
а) молекула межклеточной адгезии 1 (ICAM1), CD58, CD155, молекула клеточной адгезии, связанная с карциноэмбриональным антигеном 1 (СЕАСАМ1), молекула клеточной адгезии 1 (CADM1), белок, связанный с полипептидом главного комплекса гистосовместимости класса I A (MICA), белок, связанный с полипептидом главного комплекса гистосовместимости класса I В (MICB), или их комбинация;
б) ICAM1 и CD58;
в) ICAM1, CD58, CD155, СЕАСАМ1, CADM1, MICA и MICB;
г) ICAM1, CD58 и CD155;
в) ICAM1, CD58, CD155 и CADM1;
е) CD58 и CADM1; или
ж) ICAM1, CADM1 и CD155.
| WO 2016183041 A2, 11.06.2016 | |||
| GARRIDO F | |||
| et al., Generation of MHC class I diversity in primary tumors and selection of the malignant phenotype, International Journal of Cancer, Int | |||
| J | |||
| Cancer, 2016, 138, pp | |||
| Искроудержатель для паровозов | 1920 |
|
SU271A1 |
| US 2014134195 A1, 15.05.2014 | |||
| БОРИСОВ М.А | |||
| и др., Клеточные подходы к лечению инсулинзависимого диабета, Acta naturae, 2016, Т | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |

