Область техники, к которой относится изобретение
Настоящее изобретение относится к сконструированным Т-клеткам, способу их получения и их применению в качестве лекарственного средства, прежде всего для иммунотерапии. Сконструированные Т-клетки, предлагаемые в изобретении, отличаются тем, что в них ингибируют экспрессию бета-2-микроглобулина (В2М) и/или трансактиватора главного комплекса гистосовместимости класса II (CIITA), например, посредством применения редко расщепляющих эндонуклеаз, которые обладают способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего В2М и/или CIITA, или посредством применения молекул нуклеиновых кислот, которые ингибируют экспрессию В2М и/или CIITA. Для дополнительного обеспечения отсутствия Т-клеточной аллореактивности инактивируют по меньшей мере один ген, кодирующий компонент Т-клеточного рецептора, например посредством применения редко расщепляющих эндонуклеаз, которые обладают способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего указанный компонент TCR. Кроме того, в указанных модифицированных Т-клетках можно осуществлять стадию экспрессии иммуносупрессорного полипептида, такого как гомолог вирусной молекулы ГКГСI или лиганд (рецептора) NKG2D, с целью пролонгирования выживания указанных модифицированных Т-клеток в организме-хозяине. Указанная модифицированная Т-клетка особенно пригодна для аллогенных трансплантаций, прежде всего благодаря тому, что она позволяет снижать как риск отторжения иммунной системой хозяина, так и риск развития реакции «трансплантат-против-хозяина». Настоящее изобретение открывает путь создания стандартных и доступных стратегий адоптивной иммунотерапии с использованием Т-клеток для лечения рака, инфекций и аутоиммунных заболеваний.
Предпосылки создания изобретения
Адоптивная иммунотерапия, которая включает перенос аутологичных антигенспецифических Т-клеток, созданных ex vivo, представляет собой перспективную стратегию лечения вирусных инфекций и рака. Т-клетки, применяемые для адоптивной иммунотерапии, можно создавать либо путем размножения антигенспецифических Т-клеток, либо посредством перенаправления Т-клеток с помощью методов генетической инженерии (Park, Rosenberg и др., 2011).
В Т-клетках были успешно созданы новые специфичности посредством генетического переноса трансгенных Т-клеточных рецепторов или химерных антигенных рецепторов (CAR) (Jena, Dotti и др., 2010). CAR представляют собой синтетические рецепторы, состоящие из «нацеливающего» фрагмента, ассоциированного с одним или несколькими сигнальными доменами в одной слитой молекуле. В целом, связывающий фрагмент CAR состоит из антигенсвязывающего домена одноцепочечного антитела (scFv), который содержит легкие и вариабельные фрагменты моноклонального антитела, соединенные гибким линкером. Успешно применяли также связывающие фрагменты на основе доменов рецептора или лиганда. Сигнальные домены для первого поколения CAR получали из цитоплазматической области цепей зета-субъединицы CD3 или рецептора Fc гамма. Было продемонстрировано, что первое поколение CAR позволяло успешно перенаправлять Т-клеточную цитотоксичность, однако для них не удалось обеспечить пролонгированное размножение и противоопухолевую активность in vivo. Для повышения выживаемости и усиления пролиферации модифицированных с помощью CAR Т-клеток осуществляли добавление сигнальных доменов из костимулирующих молекул, включая CD28, ОХ-40 (CD134) и 4-1ВВ (CD137), по отдельности (второе поколение) или в комбинации (третье поколение). CAR позволяли успешно перенаправлять Т-клетки на антигены, экспрессируемые на поверхности опухолевых клеток при различных злокачественных заболеваниях, включая лимфомы и солидные опухоли (Jena, Dotti и др., 2010).
Современный протокол лечения пациентов с использованием адоптивной иммунотерапии базируется на переносе аутологичных клеток. При таком подходе из организма пациентов выделяют Т-лимфоциты, генетически модифицируют или отбирают ex vivo, культивируют in vitro для увеличения при необходимости количества клеток и, наконец, вводят их пациенту путем инфузии. Помимо введения лимфоцитов путем инфузии организм хозяина можно подвергать другим манипуляциям, которые поддерживают приживление трансплантата Т-клеток или их участие в иммунном ответе, таким, например, как предварительное кондиционирование (облучением или химиотерапией) и введение факторов роста лимфоцитов (таких как IL-2). Каждого пациента подвергают индивидуально разработанной схеме лечения, используя собственные лимфоциты пациента (т.е. имеет место аутологичная терапия). Практическое применение аутологичных терапий сталкивается со значительными техническими и логистическими препятствиями, для их разработки требуются дорогостоящие специализированные устройства и опытный персонал, они должны быть разработаны в течение короткого промежутка времени после диагностирования пациента, и при этом во многих случаях сталкиваются с тем, что предшествующее лечение пациента привело к нарушению иммунной функции, в результате чего лимфоциты пациента могут плохо функционировать и присутствовать в очень незначительных количествах. Из-за указанных препятствий каждый препарат аутологичных клеток пациента фактически является новым продуктом, что приводит к значительным вариациям с позиций эффективности и безопасности.
В идеальном варианте представляется предпочтительным применять стандартизованную терапию, для осуществления которой аллогенные терапевтические клетки должны быть предварительно приготовлены, подробно охарактеризованы и доступны для немедленного введения пациентам. Под аллогенными клетками подразумеваются клетки, полученные из индивидуумов, принадлежащих к этому же виду, но генетически не одинаковых. Однако применение аллогенных клеток в настоящее время имеет ряд недостатков. У иммунокомпетентных хозяев происходит быстрое отторжение аллогенных клеток, т.е. процесс, который называют реакцией «хозяин-против-трансплантата» (HvG), и это в значительной степени ограничивает эффективность внесенных клеток. У хозяев с нарушенной иммунологической реактивностью (иммунокомпроментированных) аллогенные клетки могут приживляться, но специфичности их эндогенных Т-клеточных рецепторов (TCR) могут распознаваться тканью хозяина как «чужие», что приводит к реакции «трансплантат-против-хозяина» (GvHD), которая приводит к серьезному повреждению ткани и смерти.
Для получения аллогенных Т-клеток авторы настоящего изобретения ранее описали метод генетического конструирования Т-клеток, при осуществлении которого различные эффекторные гены, прежде всего кодирующие Т-клеточные рецепторы, инактивировали с помощью специфических TAL-нуклеаз, более хорошо известных под товарным знаком TALEN™ (фирма Cellectis, 8 rue de la Croix Jarry, 75013, Париж). Установлено, что этот метод является высокоэффективным в случае первичных клеток при применении трансфекции РНК в качестве части платформы, обеспечивающей массовое производство аллогенных Т-клеток (WO 2013/176915).
Бета-2-микроглобулин, известный также как В2М, представляет собой легкую цепь молекул ГКГС класса I, и поэтому является составной частью главного комплекса гистосовместимости. У человека В2М кодируется геном b2m, который локализован на хромосоме 15, в отличие от других генов ГКГС, которые локализованы в генном кластере на хромосоме 6. Человеческий белок состоит из 119 аминокислот (SEQ ID NO: 1) и имеет молекулярную массу 11800 Да. На моделях, созданных на мышах с дефицитом бета-2-микроглобулина, установлено, что В2М необходим на экспрессии на клеточной поверхности молекул ГКГС класса I и стабильности пептидсвязывающего желобка. Установлено также, что трансплантаты гемопоэтических клеток из мышей с дефицитом нормальной экспрессии на клеточной поверхности молекул ГКГС I отторгались NK1.1+-клетками у здоровых мышей из-за целевой мутации в гене бета-2-микроглобулина, это позволяет предположить, что дефицит экспрессии молекул ГКГС I придает клеткам костного мозга чувствительность к отторжению иммунной системой хозяина (Bix и др., 1991).
Белок CIITA (SEQ ID NO: 4 - NCBI референс-последовательность: NP_000237.2) действует в качестве позитивного регулятора транскрипции гена главного комплекса гистосовместимости класса II, включая транскрипцию гена β2m, и его часто обозначают как «главный фактор контроля» экспрессии этих генов. мРНК CIITA (SEQ ID NO: 5) можно обнаружить только в клеточных линиях и тканях, позитивных по системе человеческих лейкоцитарных антигенов (HLA) класса И. Такое строго ограниченное распределение в тканях позволяет предположить, что экспрессия генов HLA класса II в большой степени находится под контролем CIITA (Mach B. и др., 1994).
Адаптивный иммунный ответ представляет собой сложную биологическую систему, в которой происходит взаимодействие между многочисленными клеточными компонентами. «Специализированные» антигенпрезентирующие клетки (АРС) обладают способностью процессировать чужеродные субстанции и экспонировать их на Т-клетках-хелперах в контексте молекул ГКГС класса II. В свою очередь, активированные Т-клетки-хелперы стимулируют В-клеточный ответ и ответ цитотоксических Т-клеток (CTL). CTL распознают чужеродные пептиды, презентуемые молекулами ГКГС класса I, но в случае аллореактивности распознают и уничтожают клетки, несущие чужеродные молекулы ГКГС класса I. Молекулы ГКГС класса I состоят из 2 субстанций: высокополиморфной трансмембранной тяжелой цепи и малого инвариантного полипептида, бета-2-микроглобулина (бета2-m), кодируемого геном В2М. Для экспрессии тяжелой цепи ГКГС класса I на клеточной поверхности требуется ее ассоциация с бета2-m. Таким образом, элиминация экспрессии бета2-m в модифицированных CAR Т-клетках (CAR-T-клетки) должна нарушать экспрессию ГКГС класса I и делать их «невидимыми» для CTL хозяина. Однако молекулы ГКГС класса I с дефицитом CAR-T-клеток являются чувствительными к лизису NK-клетками хозяина, мишенью которых являются клетки, у которых отсутствуют молекулы ГКГС класса I (Ljunggren H.G. и др., Immunl Today. 11, 1990, сс. 237-244).
NK-клетки обладают цитотоксическими функциями в отношении клеток, с которыми они взаимодействуют, на основе баланса между активирующими и ингибирующими сигналами, которые они получают через различные мономорфные или полиморфные рецепторы. Одним из основных активирующих рецепторов на человеческих NK-клетках является NKG2D и его лиганды, которые включают такие белки как MICA, MICB, ULBP1, ULBP2, ULBP3 (Raulet D.H., Nature Reviews Immunology 3 (10), 2003, cc. 781-779). С другой стороны, ингибирующий сигнал опосредуется через взаимодействие между NK-рецепторами типа LIR-1/ILT2 и молекулами ГКГС класса I (Ljunggren H.G. и др., Immunl Today. 11, 1990, сс. 237-244). Некоторые вирусы, такие как цитомегаловирус, имеют приобретенные механизмы, позволяющие избегать опосредуемого NK-клетками иммунного «надзора». Геном HCMV кодирует белки, которые обладают способностью препятствовать поверхностной экспрессии молекул ГКГС класса I (т.е. US2, US3, US6 и US11), экспрессируя при этом белок-гомолог молекул ГКГС класса I (UL18), который действует в качестве «ловушки», блокируя опосредуемый NK клеточный лизис (Kim Y. и др., PLOS Pathogens. 4, 2008, е1000123 и Wilkinson G. и др., J Clin Virol. 41(3), 2010, сс. 206-212). Кроме того, HCMV оказывает воздействие на путь NKG2D путем секреции белка, который обладает способностью связывать лиганды NKG2D и предупреждает их поверхностную экспрессию (Welte S.A. и др., Eur J Immunol 33 (1), 2003, сс. 194-203). В опухолевых клетках присутствуют некоторые механизмы, позволяющие избегать NKG2D-ответа путем секреции лигандов NKG2D, таких как ULBP2, MICB или MICA (Waldhauer I., Steinle A. Proteolytic release of soluble UL16-binding protein 2 from tumor cells. Cancer Res 66(5), 2006, cc. 2520-2526; Salih H.R. и др., Hum Immunol.67(3), март 2006 г., сс. 188-195; Salih H.R. и др., Blood. 102(4), 15 августа 2003 г., сс. 1389-1396; Salih H.R. и др., J Immunol, 169(8), сс. 4098-4102).
При создании настоящего изобретения разработаны стратегии иммунотерапии, с помощью которых создают Т-клетки, прежде всего аллогенные Т-клетки, наиболее пригодные для аллогенных трансплантаций, снижая риск отторжений из-за реакций «хозяин-против-трансплантата» и риск развития реакции «трансплантат-против-хозяина», и которые делают Т-клетки «невидимыми», прежде всего для АРС-клеток или NK-клеток.
Краткое изложение сущности изобретения
В настоящем изобретении предложены способы получения сконструированных Т-клеток, прежде всего аллогенных Т-клеток, полученных из донора, для того, чтобы сделать возможным их применение для иммунотерапевтических целей. Более конкретно, способы, предлагаемые в настоящем изобретении, позволяют точно модулировать экспрессию определенных эффекторных молекул, важных для иммунного распознавания и гистосовместимости.
Одним из объектов настоящего изобретения является способ получения сконструированной Т-клетки, предпочтительно аллогенной Т-клетки, полученной из донора, заключающийся в том, что осуществляют стадии, на которых:
а) получают Т-клетку, предпочтительно аллогенную донорскую Т-клетку, и
б) ингибируют экспрессию бета-2-микроглобулина (В2М) и/или трансактиватора главного комплекса гистосовместимости класса II (CIITA) в указанной Т-клетке.
Согласно некоторым вариантам осуществления изобретения ингибирование экспрессии В2М достигается путем геномной модификации, более предпочтительно посредством экспрессии в Т-клетке редко расщепляющих эндонуклеаз, которые обладают способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего В2М, такого как человеческий ген β2m, представленный в SEQ ID NO: 2 (NCBI референс-последовательность: NG_012920.1), или гена, последовательность которого идентична по меньшей мере на 70%, например, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99%, последовательности человеческого гена β2m, представленной в SEQ ID NO: 2, по всей длине SEQ ID NO: 2. Указанная редко расщепляющая эндонуклеаза может представлять собой TAL-нуклеазу, мегануклеазу, нуклеазу с «цинковыми пальцами» (ZFN) или РНК-направляемую эндонуклеазу (такую как Cas9).
Согласно другим конкретным вариантам осуществления изобретения ингибирование экспрессии В2М достигается посредством применения (например, путем интродукции в Т-клетку) молекулы нуклеиновой кислоты, которая специфически гибридизуется (например, связывается) в определенных условиях в клетке с клеточной мРНК и/или геномной ДНК, кодирующей В2М, ингибируя тем самым транскрипцию и/или трансляцию гена. Согласно конкретным вариантам осуществления изобретения ингибирование экспрессии В2М достигается посредством применения (например, путем интродукции в Т-клетку) молекулы антисмыслового олигонуклеотида, рибозима или интерферирующей РНК (PHKi). Предпочтительно указанная молекула нуклеиновой кислоты содержит по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 3 (т.е. мРНК, кодирующей человеческий В2М; NCBI референс-последовательность: NM_004048).
Согласно конкретным вариантам осуществления изобретения ингибирование экспрессии CIITA путем геномной модификации, более предпочтительно посредством экспрессии в Т-клетке редко расщепляющих эндонуклеаз, которые обладают способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего CIITA, такого как ген человеческого CIITA (NCBI референс-последовательность: NG_009628.1), или гена, последовательность которого идентична по меньшей мере на 70%, например, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99%, последовательности человеческого гена CIITA, имеющего номер NG_009628.1, по всей длине человеческого гена CIITA, имеющего номер NG_009628.1. Указанная редко расщепляющая эндонуклеаза может представлять собой TAL-нуклеазу, мегануклеазу, нуклеазу с «цинковыми пальцами» (ZFN) или РНК-направляемую эндонуклеазу (такую как Cas9).
Согласно другим конкретным вариантам осуществления изобретения ингибирование экспрессии CIITA достигается посредством применения (например, путем интродукции в Т-клетку) молекулы нуклеиновой кислоты, которая специфически гибридизуется (например, связывается) в определенных условиях в клетке с клеточной мРНК и/или геномной ДНК, кодирующей CIITA, ингибируя тем самым транскрипцию и/или трансляцию гена. Согласно конкретным вариантам осуществления изобретения ингибирование экспрессии CIITA достигается посредством применения (например, путем интродукции в Т-клетку) молекулы антисмыслового олигонуклеотида, рибозима или интерферирующей РНК (PHKi). Предпочтительно указанная молекула нуклеиновой кислоты содержит по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 5 (т.е. мРНК, кодирующей изоформу 2 человеческого CIITA).
Согласно конкретным вариантам осуществления изобретения Т-клетку можно конструировать также с получением неаллореактивной клетки, прежде всего путем инактивации одного или несколько генов, участвующих в самораспознавании, таких, например, как гены, кодирующие компоненты Т-клеточных рецепторов (TCR). Для достижения этого можно осуществлять геномную модификацию, более предпочтительно посредством экспрессии в Т-клетке редко расщепляющей эндонуклеазы, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК, предпочтительно двухцепочечного разрыва, по меньшей мере одного гена, кодирующего компонент Т-клеточного рецептора (TCR), такого как ген, который кодирует TCR-альфа или TCR-бета. Указанная редко расщепляющая эндонуклеаза может представлять собой TAL-нуклеазу, мегануклеазу, нуклеазу с «цинковыми пальцами» (ZFN) или РНК-направляемую эндонуклеазу (такую как Cas9). Предпочтительно редко расщепляющая эндонуклеаза обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего TCR-альфа.
Согласно оптимальным вариантам осуществления изобретения Т-клетку можно конструировать также так, чтобы она экспрессировала химерный антигенный рецептор (CAR), направленный по меньшей мере против одного антигена, который экспрессируется на поверхности злокачественной или инфицированной клетки, такой как антиген CD19 В-лимфоцитов.
Таким образом, еще одним объектом настоящего изобретения являются сконструированные Т-клетки, прежде всего сконструированные выделенные Т-клетки, отличающиеся тем, что в них ингибируется экспрессия бета-2-микроглобулина (В2М).
Конкретными вариантами осуществления изобретения является Т-клетка, в которой происходит экспрессия редко расщепляющей эндонуклеазы, обладающей способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего В2М. Более конкретно, указанная Т-клетка содержит экзогенную молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанную редко расщепляющую эндонуклеазу, которая может представлять собой TAL-нуклеазу, мегануклеазу, нуклеазу с «цинковыми пальцами» (ZFN) или РНК-направляемую эндонуклеазу.
Другими конкретными вариантами осуществления изобретения является Т-клетка, содержащая экзогенную молекулу нуклеиновой кислоты, которая ингибирует экспрессию В2М. Согласно конкретным вариантам осуществления изобретения указанная молекула нуклеиновой кислоты представляет собой молекулу антисмыслового олигонуклеотида, рибозима или интерферирующей РНК (PHKi). Согласно предпочтительным вариантам осуществления изобретения указанная молекула нуклеиновой кислоты содержит по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 3.
В настоящем изобретении предложены также сконструированные Т-клетки, в частности, сконструированные выделенные Т-клетки, отличающиеся тем, что в них ингибируется экспрессия трансактиватора главного комплекса гистосовместимости класса II (CIITA).
Конкретными вариантами осуществления изобретения является Т-клетка, в которой происходит экспрессия редко расщепляющей эндонуклеазы, обладающей способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего CIITA. Более конкретно, указанная Т-клетка содержит экзогенную молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую редко расщепляющую эндонуклеазу, которая может представлять собой TAL-нуклеазу, мегануклеазу, нуклеазу с «цинковыми пальцами» (ZFN) или РНК-направляемую эндонуклеазу.
Другими конкретными вариантами осуществления изобретения является Т-клетка, содержащая экзогенную молекулу нуклеиновой кислоты, которая ингибирует экспрессию CIITA. Согласно конкретным вариантам осуществления изобретения указанная молекула нуклеиновой кислоты представляет собой молекулу антисмыслового олигонуклеотида, рибозима или интерферирующей РНК (PHKi). Согласно предпочтительным вариантам осуществления изобретения указанная молекула нуклеиновой кислоты содержит по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 5.
Согласно конкретным вариантам осуществления изобретения Т-клетка может нести также по меньшей мере один инактивированный ген, кодирующий компонент рецептора TCR. Более конкретно в указанной Т-клетке может экспрессироваться редко расщепляющая эндонуклеаза, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК, предпочтительно путем двухцепочечного разрыва, указанного по меньшей мере одного гена, кодирующего компонент Т-клеточного рецептора (TCR). Таким образом, указанная Т-клетка может содержать экзогенную молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую редко расщепляющую эндонуклеазу, обладающую способностью осуществлять избирательную инактивацию путем расщепления ДНК по меньшей мере одного гена, кодирующего один компонент Т-клеточного рецептора (TCR). Разрушение TCR позволяет получать неаллореактивную Т-клетку, которую можно применять в стратегиях аллогенного лечения.
Согласно оптимальным вариантам осуществления изобретения можно конструировать Т-клетку, которая экспрессирует химерный антигенный рецептор (CAR), направленный против по меньшей мере одного антигена, экспрессируемого на поверхности злокачественной или инфицированной клетки, такой как антиген CD19 В-лимфоцитов. В частности, Т-клетка содержит экзогенную молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанный CAR. Связывание антигена-мишени с помощью CAR обладает способностью запускать Т-клеточный иммунный ответ, направленный против патологической клетки, что приводит к дегрануляции различных цитокинов и расщеплению ферментов в межклеточном пространстве.
Согласно некоторым вариантам осуществления изобретения осуществляют дополнительную модификацию Т-клеток для того, чтобы делать их «невидимыми», путем экспрессии по меньшей мере одного неэндогенного иммуноспрессорного полипептида, такого как вирусный гомолог молекул ГКГС, например, UL18, или такого как лиганд NKG2D.
Согласно некоторым вариантам осуществления изобретения Т-клетка, предлагаемая в настоящем изобретении, экспрессирует по меньшей мере один неэндогенный иммуносупрессорный полипептид. Согласно более конкретным вариантам осуществления изобретения указанный неэндогенный иммуносупрессорный полипептид представляет собой вирусный гомолог молекул ГКГС, такой как UL18. Т-клетка может содержать экзогенную молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую полипептид, идентичный по меньшей мере на 80%, предпочтительно по меньшей мере на 90% и более предпочтительно по меньшей мере на 95% SEQ ID NO: 89. Согласно другим более конкретным вариантам осуществления изобретения указанный неэндогенный иммуносупрессорный полипептид представляет собой лиганд NKG2D. Т-клетка может содержать экзогенную молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую полипептид, идентичный по меньшей мере на 80%, предпочтительно по меньшей мере на 90% и более предпочтительно по меньшей мере на 95% одной из SEQ ID NO: 90-97.
Полученные в результате осуществления настоящего изобретения, сконструированные Т-клетки можно применять в качестве терапевтических продуктов, в идеальном варианте в качестве «имеющегося в продаже» продукта, предназначенного для применения для лечения или предупреждения рака, бактериальных или вирусных инфекций или аутоиммунных заболеваний.
Таким образом, в настоящем изобретении предложена также сконструированная Т-клетка или композиция, такая как фармацевтическая композиция, содержащая ее, для применения в качестве лекарственного средства. Согласно некоторым вариантам осуществления изобретения сконструированная Т-клетка или композиция предназначена для применения для лечения рака и более конкретно для применения для лечения лимфомы. Согласно некоторым другим вариантам осуществления изобретения сконструированная Т-клетка или композиция предназначена для применения для лечения вирусной инфекции. Согласно некоторым другим вариантам осуществления изобретения сконструированная Т-клетка или композиция предназначена для применения для лечения бактериальной инфекции.
Как должно быть очевидно, подробности, указанные касательно одного из объектов изобретения, применимы также к любому из других объектов изобретения.
Краткое описание чертежей
На чертежах показано:
на фиг. 1 - схематическое изображение нормальной взаимосвязи между Т-клетками донора, Т-клетками хозяина и антигенпрезентирующими клетками;
на фиг. 2 - схематическое изображение генетически модифицированных терапевтических Т-клеток, предлагаемых в изобретении, и Т-клеток и опухолевых клеток пациента;
на фиг. 3 - сравнение распределения рассеяния в прямом направлении (FSC), являющегося индикаторов размера клеток, между TCR-позитивными и TCR-негативными клетками;
на фиг. 4 - результаты анализа методом проточной цитометрии TCR-альфа/бета и экспрессии CD3 на человеческих первичных Т-клетках после введения путем электропорации мРНК TALE-нуклеазы, мишенью которой является TRAC (альфа-цепь TCR) (TALE-нуклеаза TRAC) (вверху);
на фиг. 5 - результаты анализа методом проточной цитометрии экспрессии HLA_ABC на поверхности человеческих первичных Т-клеток в: А. контрольных Т-клетках; Б. - после введения путем электропорации мРНК TALE-нуклеазы, мишенью которой является β2m (TALE-нуклеаза β2m);
на фиг. 6 - результаты анализа методом проточной цитометрии экспрессии CAR (анти-F(ab')2) после введения путем электропорации в Т-клетки мРНК, кодирующей одноцепочечный CAR, или без нее; Б. - результаты анализа методом проточной цитометрии экспрессии CD107a (маркер дегрануляции) на подвергнутых электропорации Т-клетках, которые культивировали совместно с клетками Дауди;
на фиг. 7 - схематическое изображение возможных взаимодействий между аллогенной экспрессирующей CAR Т-клеткой (CAR-T-клетка) и различными иммунными клетками хозяина (CD8+- и CD4+-Т-клетка, АРС, такая как дендритная клетка и NK-клетка), CAR-T-клеткой, ген В2М которой инактивирован путем выключения (KO). Знак (+) обозначает активацию, а знак (-) обозначает ингибирование. Возможное взаимодействие между CAR-T-клеткой с опухолевой клеткой остается неизмененным. Инактивация гена В2М, который является одним из компонентов молекул ГКГСI, делает последние нефункциональными в отношение взаимодействий с цитотоксической Т-клеткой (CD8+) и NK-клеткой хозяина. Затем NK-клетка может оказывать свое активирующее действие на аллогенную CAR-T-клетку посредством пути активатора, такого как NKG2D/лиганд NKG2D;
на фиг. 8 - схематическое изображение возможных взаимодействий между аллогенной CAR-T-клеткой и различными иммунными клетками хозяина (CD8+- и CD4+-Т-клетка, АРС, такая как дендритная клетка и NK-клетка), CAR-T-клеткой, ген В2М которой инактивирован путем KO, и экспрессирующей вирусный гомолог ГКГСI. Знак (+) обозначает активацию, а знак (-) обозначает ингибирование. Возможное взаимодействие между CAR-T-клеткой с опухолевой клеткой остается неизмененным. Также как и на приведенном выше чертеже (KO только В2М), взаимодействие между CAR-T-клеткой и хозяйской CD8+-T-клеткой ослаблялось. В этом случае экспрессия вирусного гомолога ГКГCI делало взаимодействие с NK-клеткой неоперативным через ГКГСI/ингибитор рецептора. Двойная генетическая модификация аллогенных CAR-T-клеток путем KO В2М в сочетании с экспрессией вирусного гомолога ГКГCI усиливает их иммуносупрессорную защиту;
на фиг. 9 - схематическое изображение возможных взаимодействий между аллогенной CAR-T-клеткой и различными иммунными клетками хозяина (CD8+- и CD4+-Т-клетка, АРС, такая как дендритная клетка и NK-клетка), CAR-T-клеткой, ген В2М которой инактивирован путем KO, и экспрессирующей растворимый лиганд NKG2D. Знак (+) обозначает активацию, а знак (-) обозначает ингибирование. Возможное взаимодействие между CAR-T-клеткой с опухолевой клеткой остается неизмененным. Также как и на приведенном выше чертеже (KO только В2М), взаимодействие между CAR-T-клеткой и хозяйской CD8+-Т-клеткой ослаблялось. Экспрессия растворимого лиганда NKG2D представляет собой другой путь инактивации взаимодействия с NK-клеткой. В этом случае растворимый лиганд NKG2D может связываться с рецептором NKG2D на NK-клетке, но не оказывать действия в противоположность лиганду NKG2D CAR-T-клетки, с которой он вступает в ингибирующую конкуренцию. Двойная генетическая модификация аллогенных CAR-T-клеток путем KO В2М в сочетании с экспрессией растворимого лиганда NKG2D усиливает их иммуносупрессорную защиту;
на фиг. 10 - результаты FACS-анализа экспрессии β2-m в Т-клетках. Нетрансфектированные (вверху) и трансфектированные Т-клетки (в середине и внизу) анализировали с помощью FACS в отношении жизнеспособности (слева) и экпрессии β2-m (справа).
Подробное описание изобретения
Если специально не указано иное, то все технические и научные понятия, использованные в настоящем описании, имеют значение, которое является общепринятым для специалистов в области генной терапии, биохимии, генетики и молекулярной биологии.
Для осуществления на практике или тестирования настоящего изобретения можно применять все методы и материалы, сходные или эквивалентные тем, которые указаны в настоящем описании, при этом в настоящей заявке описаны пригодные методы и материалы. Все публикации, заявки на патент, патенты и другие ссылки, упомянутые в настоящем описании, полностью включены в настоящее описание в качестве ссылки. В случае разночтения следует руководствоваться настоящим описанием, включая определения. Кроме того, материалы, методы и примеры представлены только для иллюстрации и не направлены на ограничение объема изобретения, если не указано иное.
При осуществлении на практике настоящего изобретения следует применять, если не указано иное, общепринятые методы клеточной биологии, культивирования клеток, молекулярной биологии, трансгенной биологии, микробиологии, рекомбинантной ДНК и иммунологии, известные специалисту в данной области. Такие методы подробно описаны в литературе, см., например, Current Protocols in Molecular Biology (Frederick M. AUSUBEL, изд-во Wiley and son Inc, Library of Congress, USA, 2000); Molecular Cloning: A Laboratory Manual, 3-е изд. (Sambrook и др., изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 2001); Oligonucleotide Synthesis, под ред. M.J. Gait, 1984; Mullis и др., U.S. №4683195; Nucleic acid Hybridization (под ред. В.D. Harries и S.J. Higgins, 1984); Transcription And Translation (под ред. В.D. Hames и S.J. Higgins, 1984); Culture Of Animal Cells (R.I. Freshney, изд-во Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (изд-во IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); серия: Methods In ENZYMOLOGY (под. ред J. Abelson и M. Simon, изд-во Academic Press, Inc., New York), прежде всего том 154 и том 155 (под ред. Wu и др.) и том 185, «Gene Expression Technology» (под ред. D. Goeddel); Gene Transfer Vectors For Mammalian Cells (под ред. J.H. Miller и M.P. Calos, 1987, изд-во Cold Spring Harbor Laboratory); Immunochemical Methods In Cell And Molecular Biology (под ред. Mayer и Walker, изд-во Academic Press, London, 1987); Handbook Of Experimental Immunology, тома I-IV (под ред. D.M. Weir и С.С. Blackwell, 1986); и Manipulating the Mouse Embryo, (изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1986).
Способы получения сконструированных Т-клеток
В целом, в настоящем изобретении предложены способы получения сконструированных Т-клеток, прежде всего аллогенных Т-клеток, полученных из донора.
Таким образом, в настоящем изобретении предложен способ получения сконструированной Т-клетки, предпочтительно аллогенной Т-клетки, полученной из донора, заключающийся в том, что осуществляют стадии, на которых:
а) получают Т-клетку, предпочтительно донорскую аллогенную Т-клетку, и
б) ингибируют экспрессию бета-2-микроглобулина (В2М) и/или трансактиватора главного комплекса гистосовместимости класса II (CIITA) в указанной Т-клетке.
Согласно конкретным вариантам осуществления изобретения способ предусматривает ингибирование экспрессии бета-2-микроглобулина (В2М). В альтернативном или дополнительном варианте способ может предусматривать ингибирование трансактиватора главного комплекса гистосовместимости класса II (CIITA).
Согласно некоторым вариантам осуществления изобретения ингибирование экспрессии В2М достигается путем геномной модификации, более конкретно посредством экспрессии в Т-клетке редко расщепляющей эндонуклеазы, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего В2М (например, человеческого гена β2m, представленного в SEQ ID NO: 2).
Согласно некоторым вариантам осуществления изобретения ингибирование экспрессии CIITA достигается путем геномной модификации, более конкретно посредством экспрессии в Т-клетке редко расщепляющей эндонуклеазы, которая обладает способность осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего CIITA (например, человеческого гена CIITA).
Под полученным путем «инактивации» или «инактивированным» геном подразумевается, что представляющий интерес ген (например, ген, кодирующий В2М или CIITA) не экспрессируется в форме функционального белка. В конкретных вариантах осуществления изобретения способ генетической модификации основан на экспрессии в предназначенной для конструирования клетке редко расщепляющей эндонуклеазы, которая катализирует расщепление в требуемом гене, инактивируя тем самым указанный требуемый ген. Разрывы нуклеотидной цепи, вызываемые эндонуклеазой, как правило репарируются с помощью различных механизмов гомологичной рекомбинации или соединения негомологичных концов (NHEJ). Однако NHEJ представляет собой несовершенный процесс репарации, который часто приводит к изменениям последовательности ДНК в сайте расщепления. Механизмы включают повторное соединение остатков двух концов ДНК посредством прямого повторного лигирования (Critchlow и Jackson, 1998) или посредством так называемого опосредуемого микрогомологией соединения концов (Betts, Brenchley и др., 2003; Ma, Kim и др., 2003). Репарация посредством соединения негомологичных концов (NHEJ) часто приводит к небольшим инсерциям или делециям и ее можно применять для создания специфических «выключений» генов. Указанная модификация может представлять собой замену, делецию или добавление по меньшей мере одного нуклеотида. Клетки, в которых произошел случай индуцированного расщеплением мутагенеза, т.е. случай мутагенеза, являющегося результатом NHEJ, можно идентифицировать и/или отбирать с помощью метода, хорошо известного в данной области.
Редко расщепляющая эндонуклеаза, которую можно применять согласно настоящему изобретению для инактивации гена β2m, может представлять собой, например, TAL-нуклеазу, мегануклеазу, нуклеазу с «цинковыми пальцами» (ZFN) или РНК-направляемую эндонуклеазу (такую как Cas9).
Согласно конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой TAL-нуклеазу.
Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой хоминг-эндонуклеазу, известную также как мегануклеаза.
Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой нуклеазу с цинковыми пальцами (ZNF).
Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой РНК-направляемую эндонуклеазу. Согласно предпочтительному варианту осуществления изобретения РНК-направляемая эндонуклеаза представляет собой комплекс Cas9/CRISPR.
Согласно конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой TAL-нуклеазу, кодируемую молекулой нуклеиновой кислоты, которая содержит нуклеотидную последовательность, представленную в SEQ ID NO: 67. Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой TAL-нуклеазу, кодируемую молекулой нуклеиновой кислоты, которая содержит нуклеотидную последовательность, представленную в SEQ ID NO: 68. Согласно еще одному конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет комбинацию TAL-нуклеазы, кодируемой молекулой нуклеиновой кислоты, которая содержит нуклеотидную последовательность, представленную в SEQ ID NO: 67, и TAL-нуклеазы, кодируемой молекулой нуклеиновой кислоты, которая содержит нуклеотидную последовательность, представленную в SEQ ID NO: 68.
Для экспрессии в Т-клетке указанную редко расщепляющую эндонуклеазу можно интродуцировать в клетку с помощью экзогенной молекулы нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанную редко расщепляющую эндонуклеазу. Согласно конкретным вариантам осуществления изобретения способ, предлагаемый в изобретении, предусматривает также интродукцию в указанную Т-клетку экзогенной молекулы нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую редко расщепляющую эндонуклеазу, предпочтительно редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего В2М (например, человеческого гена β2m, представленного в SEQ ID NO: 2). Например, экзогенная молекула нуклеиновой кислоты может содержать нуклеотидную последовательность, представленную в SEQ ID NO: 67 или SEQ ID NO: 68.
В результате получают сконструированную Т-клетку, которая содержит редко расщепляющую эндонуклеазу, предпочтительно редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего В2М. В результате, инактивация гена В2М указанной редко расщепляющей эндонуклеазой приводит к ингибированию экспрессии В2М в сконструированной Т-клетке. Таким образом, получают сконструированную Т-клетку, отличающуюся тем, что в ней ингибируется экспрессия В2М.
Редко расщепляющая эндонуклеаза, которую можно применять согласно настоящему изобретению для инактивации гена CIITA, может представлять собой, например, TAL-нуклеазу, мегануклеазу, нуклеазу с «цинковыми пальцами» (ZFN) или РНК-направляемую эндонуклеазу (такую как Cas9).
Согласно конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой TAL-нуклеазу.
Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой хоминг-эндонуклеазу, известную также как мегануклеаза.
Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой нуклеазу с «цинковыми пальцами» (ZNF).
Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой РНК-направляемую эндонуклеазу. Согласно предпочтительному варианту осуществления изобретения РНК-направляемая эндонуклеаза представляет собой комплекс Cas9/CRISPR.
Для экспрессии в Т-клетке указанную редко расщепляющую эндонуклеазу можно интродуцировать в клетку с помощью экзогенной молекулы нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанную редко расщепляющую эндонуклеазу. Согласно конкретным вариантам осуществления изобретения способ, предлагаемый в изобретении, предусматривает также интродукцию в указанную Т-клетку экзогенной молекулы нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую редко расщепляющую эндонуклеазу, предпочтительно редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего CIITA (например, человеческого гена CIITA).
В результате получают сконструированную Т-клетку, которая содержит редко расщепляющую эндонуклеазу, предпочтительно редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего CIITA. В результате, инактивация гена CIITA указанной редко расщепляющей эндонуклеазой приводит к ингибированию экспрессии CIITA в сконструированной Т-клетке. Таким образом, получают сконструированную Т-клетку, отличающуюся тем, что в ней ингибируется экспрессия CIITA.
Согласно некоторым другим вариантам осуществления изобретения ингибирование экспрессии В2М достигается путем применения (например, интродукции в Т-клетку) молекулы нуклеиновой кислоты, которая специфически гибридизуется (например, связывается) в определенных условиях в клетке с клеточной мРНК и/или геномной ДНК, кодирующей В2М, ингибируя тем самым транскрипцию и/или трансляцию гена. Согласно конкретным вариантам осуществления изобретения ингибирование экспрессии В2М достигается путем применения (например, интродукции в Т-клетку) молекулы антисмыслового олигонуклеотида, рибозима или интерферирующей РНК (PHKi).
Согласно конкретному варианту осуществления изобретения молекула нуклеиновой кислоты представляет собой антисмысловой олигонуклеотид.
Согласно конкретному варианту осуществления изобретения молекула нуклеиновой кислоты представляет собой рибозим, предпочтительно рибозим типа hammerhead (в виде головки молотка).
Согласно конкретному варианту осуществления изобретения нуклеиновая кислота представляет собой молекулу интерферирующей РНК (PHKi), такой как микроРНК (miPHK), малая интерферирующая РНК (siPHK) или короткая образующая шпильку РНК (shPHK). Таким образом, согласно предпочтительному варианту осуществления изобретения молекула нуклеиновой кислоты представляет собой микроРНК. Согласно другому предпочтительному варианту осуществления изобретения молекула нуклеиновой кислоты представляет собой малую интерферирующую РНК. Согласно другому предпочтительному варианту осуществления изобретения молекула нуклеиновой кислоты представляет собой короткую образующую шпильку РНК.
В результате получают сконструированную Т-клетку, отличающуюся тем, что в ней ингибируется экспрессия В2М.
Поскольку В2М является важным структурным компонентом главного комплекса гистосовместимости (ГКГС), ингибирование экспрессии В2М приводит к уменьшению количества или элиминации молекул ГКГС на поверхности сконструированной Т-клетки. В результате сконструированная Т-клетка перестает презентовать антигены на поверхности, которые распознаются CD8+-клетками. Прежде всего, в случае аллогенной Т-клетки, полученной из донора, уменьшение количества или элиминация «чужих» антигенов, презентуемых молекулами ГКГС на поверхности Т-клетки, препятствует распознаванию сконструированной Т-клетки, когда ее вводят путем инфузии аллогенному хозяину, хозяйскими CD8+-клетками. Это делает сконструированную Т-клетку особенно пригодной для аллогенных трансплантаций, прежде всего из-за снижения риска отторжения иммунной системой хозяина.
Согласно некоторым другим вариантам осуществления изобретения ингибирование экспрессии CIITA достигается путем применения (например, интродукции в Т-клетку) молекулы нуклеиновой кислоты, которая специфически гибридизуется (например, связывается) в определенных условиях в клетке с клеточной мРНК и/или геномной ДНК, кодирующей CIITA, ингибируя тем самым транскрипцию и/или трансляцию гена. Согласно конкретным вариантам осуществления изобретения ингибирование экспрессии CIITA достигается путем применения (например, интродукции в Т-клетку) молекулы антисмыслового олигонуклеотида, рибозима или интерферирующей РНК (PHKi).
Согласно конкретному варианту осуществления изобретения молекула нуклеиновой кислоты представляет собой антисмысловой олигонуклеотид.
Согласно конкретному варианту осуществления изобретения молекула нуклеиновой кислоты представляет собой рибозим, предпочтительно рибозим типа hammerhead.
Согласно другому конкретному варианту осуществления изобретения нуклеиновая кислота представляет собой молекулу интерферирующей РНК (PHKi), такой как микроРНК (miPHK), малая интерферирующая РНК (siPHK) или короткая образующая шпильку РНК (shPHK). Таким образом, согласно предпочтительному варианту осуществления изобретения молекула нуклеиновой кислоты представляет собой микроРНК. Согласно другому предпочтительному варианту осуществления изобретения молекула нуклеиновой кислоты представляет собой малую интерферирующую РНК. Согласно другому предпочтительному варианту осуществления изобретения молекула нуклеиновой кислоты представляет собой короткую образующую шпильку РНК.
В результате получают сконструированную Т-клетку, отличающуюся тем, что в ней ингибируется экспрессия CIITA.
Согласно настоящему изобретению сконструированная Т-клетка, предлагаемая в настоящем изобретении, не экспрессирует функциональный Т-клеточный рецептор (TCR) на ее клеточной поверхности. Т-клеточные рецепторы представляют собой расположенные на клеточной поверхности рецепторы, которые участвуют в активации Т-клеток в ответ на презентацию антигена. TCR, как правило, состоит из двух цепей, альфа и бета, которые в результате сборки образуют гетеродимер и которые ассоциированы с субъединицами, участвующими в трансдукции CD3, с образованием комплекса Т-клеточного рецептора, присутствующего на клеточной поверхности. Каждая альфа- и бета-цепь TCR состоит из иммуноглобулинподобной N-концевой вариабельной (V) и константной (С) области, гидрофобного трансмембранного домена и короткой цитоплазматической области. Также как и в молекулах иммуноглобулинов, вариабельная область альфа- и бета-цепей образуется путем рекомбинации V(D)J, что приводит к большому разнообразию антигенных специфичностей в популяции Т-клеток. Однако в отличие от иммуноглобулинов, которые распознают интактный антиген, Т-клетки активируются процессированными пептидными фрагментами в сочетании с молекулой ГКГС, вносящей дополнительное измерение в распознавание антигена Т-клетками, называемое рестрикцией по ГКГС. Распознавание несоответствий ГКГС между донором и реципиентом посредством Т клеточного рецептора приводит к пролиферации Т-клеток и потенциальному развитию реакции «трансплантат-против-хозяина» (GVHD). Установлено, что нормальная поверхностная экспрессия TCR зависит от скоординированного синтеза и сборки всех семи компонентов комплекса (Ashwell и Klusner, 1990). Инактивация TCR-альфа или TCR-бета может приводить к элиминации TCR с поверхности Т-клеток, что препятствует распознаванию аллоантигенов и, как следствие, GVHD. Таким образом, инактивация по меньшей мере одного гена, кодирующего компонент TCR, делает сконструированную Т-клетку менее аллореактивной. Под полученным путем «инактивации» или «инактивированным» геном подразумевается, что представляющий интерес ген (например, ген, кодирующий компонент TCR) не экспрессируется в форме функционального белка.
Таким образом, согласно конкретным вариантам осуществления изобретения способ, предлагаемый в настоящем изобретении, заключается также в том, что инактивируют по меньшей мере один ген, кодирующий компонент Т-клеточного рецептора. Более конкретно, инактивация достигается применением (например, интродукции в Т-клетку) редко расщепляющей эндонуклеазы, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, предпочтительно двухцепочечного разрыва, по меньшей мере одного гена, кодирующего компонент Т-клеточного рецептора (TCR). Согласно конкретным вариантам осуществления изобретения редко расщепляющая эндонуклеаза обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего TCR-альфа или TCR-бета. Согласно предпочтительному варианту осуществления изобретения редко расщепляющая эндонуклеаза обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего TCR-альфа. Прежде всего, в случае аллогенной Т-клетки, полученной из донора, инактивация по меньшей мере одного гена, кодирующего компонент TCR, прежде всего TCR-альфа, позволяет получать сконструированные Т-клетки, которые являются неаллореактивными после введения путем инфузии аллогенному хозяину. Это делает сконструированную Т-клетку особенно пригодной для аллогенных трансплантаций, прежде всего, поскольку это снижает риск возникновения реакции «трансплантат-против-хозяина».
Редко расщепляющая эндонуклеаза, которую можно применять согласно настоящему изобретению для инактивации по меньшей мере одного гена, кодирующего компонент Т-клеточного рецептора, может, например, представлять собой, например, TAL-нуклеазу, мегануклеазу, нуклеазу с «цинковыми пальцами» (ZFN) или РНК-направляемую эндонуклеазу (такую как Cas9).
Согласно конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой TAL-нуклеазу.
Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой хоминг-эндонуклеазу, известную также как мегануклеаза.
Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой нуклеазу с «цинковыми пальцами» (ZNF).
Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой РНК-направляемую эндонуклеазу. Согласно предпочтительному варианту осуществления изобретения РНК-направляемая эндонуклеаза представляет собой комплекс Cas9/CRISPR.
Для экспрессии в Т-клетке указанную редко расщепляющую эндонуклеазу можно интродуцировать в клетку с помощью экзогенной молекулы нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанную редко расщепляющую эндонуклеазу. Согласно конкретным вариантам осуществления изобретения способ, предлагаемый в изобретении, предусматривает также интродукцию в указанную Т-клетку экзогенной молекулы нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую редко расщепляющую эндонуклеазу, предпочтительно редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, предпочтительно двухцепочечного разрыва, по меньшей мере одного гена, кодирующего компонент Т-клеточного рецептора (TCR).
В результате получают сконструированную Т-клетку, которая содержит редко расщепляющую эндонуклеазу, предпочтительно редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК по меньшей мере одного гена, кодирующего компонент Т-клеточного рецептора (TCR). Таким образом, получают сконструированную Т-клетку, отличающуюся тем, что в ней инактивируется по меньшей мере один ген, кодирующий компонент Т-клеточного рецептора.
В настоящем изобретении предложена также сконструированная Т-клетка, дополнительно экспрессирующая химерный антигенный рецептор (CAR), направленный против по меньшей мере одного антигена, который экспрессируется на поверхности злокачественной или инфицированной клетки. Таким образом, согласно определенным вариантам осуществления изобретения способ, предлагаемый в изобретении, дополнительно предусматривает интродукцию в указанную Т-клетку экзогенной молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая кодирует химерный антигенный рецептор (CAR), направленный против по меньшей мере одного антигена, который экспрессируется на поверхности злокачественной или инфицированной клетки.
Т-клетка, подлежащая модификации согласно настоящему изобретению, может представлять собой любую приемлемую Т-клетку. Например, Т-клетка может представлять собой воспалительный Т-лимфоцит, цитотоксический Т-лимфоцит, регуляторную Т-клетку или хелперный Т-лимфоцит. В частности, Т-клетка представляет собой цитотоксический Т-лимфоцит. В некоторых вариантах осуществления изобретения указанную Т-клетку выбирают из CD4+-Т-лимфоцитов и CD8+-Т-лимфоцитов. Их можно экстрагировать из крови или получать из стволовых клеток. Стволовые клетки могут представлять собой зрелые стволовые клетки, эмбриональные стволовые клетки, более предпочтительно нечеловеческие стволовые клетки, стволовые клетки из пуповинной крови, клетки-предшественники, стволовые клетки костного мозга, индуцированные плюрипотентные стволовые клетки, тотипотентные стволовые клетки или гематопоэтические стволовые клетки. Репрезентативными человеческими клетками являются CD34+-клетки. В конкретных вариантах осуществления изобретения подлежащая модификации согласно изобретению Т-клетка представляет собой человеческую Т-клетку. Перед осуществлением размножения и генетической модификации клеток, предлагаемых в изобретении, из организма индивидуума, такого как пациент, можно получать источник клеток с помощью различных методов, не ограничивающих объем изобретения. Т-клетку можно получать из многочисленных источников, включая (но, не ограничиваясь только ими) мононуклеарные клетки периферической крови, костный мозг, ткань лимфатических узлов, пуповинную кровь, ткань тимуса, ткань из инфицированной области, асциты, плевральный выпот, ткань селезенки и опухоли. В некоторых вариантах осуществления настоящего изобретения можно применять любое количество Т-клеточных линий, доступных и известных специалистам в данной области. В другом варианте осуществления изобретения указанную клетку можно получать из организма здорового донора, пациента, у которого диагностирован рак, или пациента, у которого диагностирована инфекция. В другом варианте осуществления изобретения указанная клетка представляет собой часть смешанной популяции клеток, обладающих различными фенотипическими характеристиками.
Редко расщепляющая эндонуклеаза
Согласно некоторым вариантам осуществления настоящего изобретения применяют редко расщепляющие эндонуклеазы, которые обладают способностью осуществлять избирательную инактивацию путем расщепления ДНК представляющего интерес гена, такого как ген, который кодирует В2М.
Понятие «редко расщепляющая эндонуклеаза» относится к ферменту дикого типа или варианту фермента, который обладает способностью катализировать гидролиз (расщепление) связей между нуклеиновыми кислотами в молекуле ДНК или РНК, предпочтительно молекуле ДНК. В частности, указанная нуклеаза может представлять собой эндонуклеазу, более предпочтительно редко расщепляющую эндонуклеазу, которая является высокоспецифической, обладает способностью распознавать сайты-мишени нуклеиновой кислоты, состоящие из 10-45 пар оснований (bp), как правило 10-35 пар оснований, более конкретно 12-20 пар оснований. Эндонуклеаза, предлагаемая в настоящем изобретении, распознает специфические полинуклеотидные последовательности, которые обозначают также как «последовательность-мишень», и расщепляет нуклеиновую кислоту внутри указанных последовательностей-мишеней или в примыкающих к ним последовательностях в зависимости от молекулярного строения указанной эндонуклеазы. Редко расщепляющая эндонуклеаза может распознавать и создавать одно- или двухцепочечный разрыв в специфических полинуклеотидных последовательностях.
В конкретных вариантах осуществления изобретения указанная редко расщепляющая эндонуклеаза, предлагаемая в настоящем изобретении, представляет собой РНК-направляемую эндонуклеазу, такую как комплекс Cas9/CRISPR. РНК-направляемые эндонуклеазы представляют собой недавно созданный инструмент конструирования генома, в котором эндонуклеаза ассоциирована с молекулой РНК. В такой системе нуклеотидная последовательность молекулы РНК определяет специфичность мишени и активирует эндонуклеазу (Gasiunas, Barrangou и др., 2012; Jinek, Chylinski и др., 2012; Cong, Ran и др., 2013; Mali, Yang и др., 2013). Cas9, который называют также как Csnl, представляет собой крупный белок, который принимает участие как в биогенезе crРНК, так и в деструкции внедряющейся ДНК. Cas9 описан также в различных видах бактерий, таких как S. thermophiles, Listeria innocua (Gasiunas, Barrangou и др., 2012; Jinek, Chylinski и др., 2012) и S. pyogenes (Deltcheva, Chylinski и др., 2011). Крупный белок Cas9 (>1200 аминокислот) содержит два предсказанных нуклеазных домена, а именно, HNH (McrA-подобный) нуклеазный домен, который локализован в середине белка и расщепленный RuvC-подобный нуклеазный домен (Н-складка РНКазы). Вариант Cas9 может представлять собой эндонуклеазу Cas9, которая не встречается в естественных условиях и которую получают путем конструирования белка или с помощью неспецифического мутагенеза. Варианты Cas9, предлагаемые в изобретении, можно, например, получать с помощью мутаций, т.е. делеций, инсерций или замен по меньшей мере одного остатка в аминокислотной последовательности эндонуклеазы Cas9 S. pyogenes (COG3513).
В других конкретных вариантах осуществления изобретения указанная редко расщепляющая эндонуклеаза может представлять собой также хоминг-эндонуклеазу, известную также под названием мегануклеаза. Указанные хоминг-эндонуклеазы хорошо известны в данной области (Stoddard 2005). Хоминг-эндонуклеазы являются высокоспецифическими, распознают сайты-мишени в ДНК, состоящие из 12-45 пар оснований, как правило, от 14 до 40 пар оснований. Хоминг-эндонуклеаза, предлагаемая в изобретении, может, например, соответствовать эндонуклеазе LAGLIDADG, эндонуклеазе HNH или эндонуклеазе GIY-YIG. Предпочтительная хоминг-эндонуклеаза, предлагаемая в настоящем изобретении, может представлять собой вариант I-CreI. «Вариант» эндонуклеазы, т.е. эндонуклеаза, которая не встречается в естественных условиях и которую получают путем генетического конструирования или неспецифического мутагенеза, может связываться с последовательностями ДНК, отличными от тех, которые распознаются эндонуклеазами дикого типа (см. международную заявку на патент WO 2006/097854).
В других конкретных вариантах осуществления изобретения указанная редко расщепляющая эндонуклеаза может представлять собой «нуклеазы «с цинковыми пальцами»» (ZFN), которые, как правило, представляют собой слияние между доменом расщепления типа IIS рестриктазы FokI и доменом распознавания ДНК, который содержит 3 или большее количество С2Н2-мотивов «цинкового пальца». Гетеродимеризация в конкретном положении в ДНК двух индивидуальных ZFN в точной ориентации и на точном расстоянии приводит к двухцепочечному разрыву (DSB) в ДНК. Применение таких химерных эндонуклеаз широко известно в данной области, и данные обобщены у Urnov и др. (Genome editing with engineered zinc finger nucleases, Nature reviews Genetics 11, 2010, cc. 636-646). В стандартных ZFN домен расщепления слит с С-концом каждого домена «цинкового пальца». Для обеспечения димеризации двух доменов расщепления и расщепления ДНК две индивидуальные ZFN связываются с противоположными цепями ДНК с помощью их С-концов на определенном расстоянии друг от друга. Наиболее часто для применяемых линкерных последовательностей между доменом «цинковых пальцев» и доменом расщепления требуется, чтобы 5'-конец каждого сайта связывания был разделен 5-7 парами оснований. Наиболее перспективный метод создания новых массивов «цинковых пальцев» является объединение более мелких «модулей» «цинковых пальцев» известной специфичности. Наиболее распространенный процесс сборки модулей включает объединение трех различных «цинковых пальцев», каждый из которых может распознавать состоящую из трех пар оснований последовательность ДНК, с созданием массива из 3 пальцев, который может распознавать сайт-мишень, состоящий из 9 пар оснований. Можно применять различные методы селекции для создания массивов «цинковых пальцев», мишенью которых являются требуемые последовательности. В начальных процессах селекции используют фаговый дисплей для отбора белков, которые связываются с данной ДНК-мишенью, из большого пула частично рандомизированных массивов «цинковых пальцев». В более современных процессах применяли моногибридные системы на основе дрожжей, моногибридные и двугибридные системы на основе бактерий, а также клетки млекопитающих.
В других конкретных вариантах осуществления изобретения указанная редко расщепляющая эндонуклеаза представляет собой «TALE-нуклеазу» или «MBBBD-нуклеазу», полученную в результате слияния ДНК-связывающего домена, как правило, полученного из эффекторных белков, подобных активатору транскрипции (
 proteins (TALE)) или из модульного домена связывания по типу «основание-с-основанием» (связывающие домены, обладающие сходными модульными свойствами связывания нуклеиновых кислот по типу «основание-с-основанием») (
proteins (TALE)) или из модульного домена связывания по типу «основание-с-основанием» (связывающие домены, обладающие сходными модульными свойствами связывания нуклеиновых кислот по типу «основание-с-основанием») (
 (MBBBD)), с каталитическим доменом, который обладает эндонуклеазной активностью. Указанный каталитический домен, как правило, происходит из ферментов, таких, например, как I-TevI, ColE7, NucA и Fok-I. TALE-нуклеазу можно получать в мономерных или димерных формах в зависимости от выбранного каталитического домена (WO 2012/138927). Указанные сконструированные TALE-нуклеазы поступают в продажу под товарным знаком TALEN™ (фирма Cellectis, 8 rue de la Croix Jarry, 75013, Париж, Франция). Как правило, ДНК-связывающий домен получают из эффектора, подобного активатору транскрипции (TALE), в котором специфичность последовательности определяется сериями повторов, состоящих из 33-35 аминокислот, например, происходящих из белков бактерий Xanthomonas или Ralstonia, таких как (но, не ограничиваясь только ими) AvrBs3, PthXo1, AvrHah1, PthA, Tal1c. Эти повторы существенно отличаются друг от друга аминокислотными положениями, которые специфически взаимодействуют с парой оснований (Boch, Scholze и др., 2009; Moscou и Bogdanove, 2009). Каждая пара оснований в ДНК-мишени контактирует с индивидуальным повтором, при этом специфичность определяется двумя вариантами аминокислот повтора (так называемый повторяющийся вариабельный дипептид, RVD). Связывающие домены TALE могут содержать N-концевой домен транслокации, ответственный за требование в отношении первого тимидинового основания (Т0) последовательности-мишени, и С-концевой домен, который содержит сигналы ядерной локализации (NLS). Связывающий нуклеиновую кислоту домен TALE, как правило, соответствует сконструированному каркасу ядра TALE, который содержит множество повторяющихся последовательностей TALE, где каждый повтор содержит RVD, специфический для каждого из нуклеотидных оснований сайта распознавания TALE. Согласно настоящему изобретению каждая повторяющаяся последовательность TALE в указанном каркасе ядра состоит из 30-42 аминокислот, более предпочтительно 33 или 34 аминокислот, при этом две имеющие решающее значение аминокислоты (так называемый повторяющийся вариабельный дипептид, RVD) локализованы в положениях 12 и 13 и опосредуют распознавание одного нуклеотида указанной последовательности сайта связывания TALE; эквивалентно этому, две имеющие решающее значение аминокислоты могут быть локализованы в положениях, отличных от положений 12 и 13, в частности в более длинной повторяющейся последовательности TALE, состоящей более чем из 33 или 34 аминокислот. Предпочтительно, RVD, ассоциированные с распознаванием различных нуклеотидов, представляют собой HD для распознавания С, NG для распознавания Т, NI для распознавания А, NN для распознавания G или А. В другом варианте осуществления изобретения имеющие решающее значение аминокислоты в положениях 12 и 13 можно изменять в результате мутации на другие аминокислотные остатки для модуляции их специфичности в отношении нуклеотидов А, Т, С и G и, в частности, для повышения их специфичности. Связывающий нуклеиновую кислоту домен TALE, как правило, содержит от 8 до 30 повторяющихся последовательностей TALE. Более предпочтительно указанный каркас ядра, предлагаемый в настоящем изобретении, содержит от 8 до 20 повторяющихся последовательностей TALE; и еще более предпочтительно 15 повторяющихся последовательностей TALE. Он может содержать также дополнительную индивидуальную укороченную повторяющуюся последовательность TALE, состоящую из 20 аминокислот, локализованную на С-конце указанного набора повторяющихся последовательностей TALE, т.е. дополнительную С-концевую повторяющуюся половинную последовательность (полупоследовательность) TALE. Другие модульные домены связывания по типу «основание-с-основанием» специфических нуклеиновых кислот (MBBBD) описаны в WO 2014/018601. Указанные MBBBD можно конструировать, например, из вновь идентифицированных белков, а именно, белков EAV36_BURRH, E5AW43_BURRH, E5AW45_BURRH и E5AW46_BURRH из недавно секвенированного генома грибов-эндосимбионтов Burkholderia rhizoxinica. Указанные связывающие нуклеиновые кислоты полипептиды содержат модули, состоящие примерно из 31-33 аминокислот, специфических для оснований. Указанные модули характеризуются идентичностью последовательностей, составляющей менее 40%, с обычными повторами TALE Xanthomonas и характеризуются большой вариабельностью присутствующих полипептидных последовательностей. Различные домены из вышеуказанных белков (модули, N- и С-концевые) из Burkholderia и Xanthomonas можно применять для создания новых белков или каркасов, которые обладают способностью связываться со специфическими нуклеотидными последовательностями и которые можно объединять с получением химерных белков TALE-MBBBD.
(MBBBD)), с каталитическим доменом, который обладает эндонуклеазной активностью. Указанный каталитический домен, как правило, происходит из ферментов, таких, например, как I-TevI, ColE7, NucA и Fok-I. TALE-нуклеазу можно получать в мономерных или димерных формах в зависимости от выбранного каталитического домена (WO 2012/138927). Указанные сконструированные TALE-нуклеазы поступают в продажу под товарным знаком TALEN™ (фирма Cellectis, 8 rue de la Croix Jarry, 75013, Париж, Франция). Как правило, ДНК-связывающий домен получают из эффектора, подобного активатору транскрипции (TALE), в котором специфичность последовательности определяется сериями повторов, состоящих из 33-35 аминокислот, например, происходящих из белков бактерий Xanthomonas или Ralstonia, таких как (но, не ограничиваясь только ими) AvrBs3, PthXo1, AvrHah1, PthA, Tal1c. Эти повторы существенно отличаются друг от друга аминокислотными положениями, которые специфически взаимодействуют с парой оснований (Boch, Scholze и др., 2009; Moscou и Bogdanove, 2009). Каждая пара оснований в ДНК-мишени контактирует с индивидуальным повтором, при этом специфичность определяется двумя вариантами аминокислот повтора (так называемый повторяющийся вариабельный дипептид, RVD). Связывающие домены TALE могут содержать N-концевой домен транслокации, ответственный за требование в отношении первого тимидинового основания (Т0) последовательности-мишени, и С-концевой домен, который содержит сигналы ядерной локализации (NLS). Связывающий нуклеиновую кислоту домен TALE, как правило, соответствует сконструированному каркасу ядра TALE, который содержит множество повторяющихся последовательностей TALE, где каждый повтор содержит RVD, специфический для каждого из нуклеотидных оснований сайта распознавания TALE. Согласно настоящему изобретению каждая повторяющаяся последовательность TALE в указанном каркасе ядра состоит из 30-42 аминокислот, более предпочтительно 33 или 34 аминокислот, при этом две имеющие решающее значение аминокислоты (так называемый повторяющийся вариабельный дипептид, RVD) локализованы в положениях 12 и 13 и опосредуют распознавание одного нуклеотида указанной последовательности сайта связывания TALE; эквивалентно этому, две имеющие решающее значение аминокислоты могут быть локализованы в положениях, отличных от положений 12 и 13, в частности в более длинной повторяющейся последовательности TALE, состоящей более чем из 33 или 34 аминокислот. Предпочтительно, RVD, ассоциированные с распознаванием различных нуклеотидов, представляют собой HD для распознавания С, NG для распознавания Т, NI для распознавания А, NN для распознавания G или А. В другом варианте осуществления изобретения имеющие решающее значение аминокислоты в положениях 12 и 13 можно изменять в результате мутации на другие аминокислотные остатки для модуляции их специфичности в отношении нуклеотидов А, Т, С и G и, в частности, для повышения их специфичности. Связывающий нуклеиновую кислоту домен TALE, как правило, содержит от 8 до 30 повторяющихся последовательностей TALE. Более предпочтительно указанный каркас ядра, предлагаемый в настоящем изобретении, содержит от 8 до 20 повторяющихся последовательностей TALE; и еще более предпочтительно 15 повторяющихся последовательностей TALE. Он может содержать также дополнительную индивидуальную укороченную повторяющуюся последовательность TALE, состоящую из 20 аминокислот, локализованную на С-конце указанного набора повторяющихся последовательностей TALE, т.е. дополнительную С-концевую повторяющуюся половинную последовательность (полупоследовательность) TALE. Другие модульные домены связывания по типу «основание-с-основанием» специфических нуклеиновых кислот (MBBBD) описаны в WO 2014/018601. Указанные MBBBD можно конструировать, например, из вновь идентифицированных белков, а именно, белков EAV36_BURRH, E5AW43_BURRH, E5AW45_BURRH и E5AW46_BURRH из недавно секвенированного генома грибов-эндосимбионтов Burkholderia rhizoxinica. Указанные связывающие нуклеиновые кислоты полипептиды содержат модули, состоящие примерно из 31-33 аминокислот, специфических для оснований. Указанные модули характеризуются идентичностью последовательностей, составляющей менее 40%, с обычными повторами TALE Xanthomonas и характеризуются большой вариабельностью присутствующих полипептидных последовательностей. Различные домены из вышеуказанных белков (модули, N- и С-концевые) из Burkholderia и Xanthomonas можно применять для создания новых белков или каркасов, которые обладают способностью связываться со специфическими нуклеотидными последовательностями и которые можно объединять с получением химерных белков TALE-MBBBD.
Ингибирующие молекулы нуклеиновых кислот
Согласно некоторым другим вариантам осуществления настоящего изобретения применяют молекулы нуклеиновых кислот, которые ингибируют экспрессию В2М. Более конкретно, нуклеиновая кислота может представлять собой молекулу антисмыслового олигонуклеотида, рибозима или интерферирующей РНК (PHKi). Предпочтительно указанная молекула нуклеиновой кислоты содержит по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 3.
Согласно конкретным вариантам осуществления изобретения ингибирующая нуклеиновая кислота представляет собой антисмысловой олигонуклеотид, который ингибирует экспрессию В2М. Указанный антисмысловой олигонуклеотид представляет собой нуклеиновую кислоту (либо ДНК, либо РНК), которая специфически гибридизуется (например, связывается) в определенных условиях в клетке с клеточной мРНК и/или геномной ДНК, кодирующей В2М, ингибируя тем самым транскрипцию и/или трансляцию гена. Связывание может представлять собой традиционное комплементарное спаривание оснований. Альтернативно этому, связывание может, например, в случае связывания с дуплексами ДНК, происходить посредством специфических взаимодействий в главном желобке двойной спирали. При этом не требуется, хотя является предпочтительной, абсолютная комплементарность.
Кроме того, согласно настоящему изобретению применяют молекулы нуклеиновых кислот, которые ингибируют экспрессию CIITA. Более конкретно, нуклеиновая кислота может представлять собой молекулу антисмыслового олигонуклеотида, рибозима или интерферирующей РНК (PHKi). Предпочтительно указанная молекула нуклеиновой кислоты содержит по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 5.
Антисмысловые олигонуклеотиды, применяемые согласно изобретению, могут представлять собой ДНК или РНК, или их химерные смеси, или производные, или их модифицированные версии и могут быть одноцепочечными или двухцепочечными. Так, согласно предпочтительному варианту осуществления изобретения антисмысловой олигонуклеотид представляет собой одноцепочечную или двухцепочечную молекулу ДНК, более предпочтительно двухцепочечную молекулу ДНК. Согласно другому предпочтительному варианту осуществления изобретения антисмысловой олигонуклеотид представляет собой одноцепочечную или двухцепочечную молекулу РНК, более предпочтительно одноцепочечную молекулу РНК.
Согласно предпочтительным вариантам осуществления изобретения антисмысловой олигонуклеотид представляет собой модифицированный олигонуклеотид, устойчивый к эндогенным нуклеазам, например, экзонуклеазам и/или эндонуклеазам, и поэтому является стабильным in vivo и in vitro.
Антисмысловой олигонуклеотид можно модифицировать, например, во фрагменте, представляющем собой основание, фрагменте, представляющем собой сахар, или в фосфатном каркасе, для повышения стабильности молекулы. Антисмысловой олигонуклеотид может включать другие присоединенные группы, такие как пептиды (например, для «нацеливания» на клеточные рецепторы хозяина) или агенты, облегчающие транспорт через клеточную мембрану. Таким образом, антисмысловой олигонуклеотид можно конъюгировать с другой молекулой, такой как пептид или транспортирующий агент.
Согласно конкретным вариантам осуществления изобретения антисмысловой олигонуклеотид содержит по меньшей мере один модифицированный фрагмент, представляющий собой основание, который выбирают из группы, включающей (но, не ограничиваясь только ими) 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-йодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 5-(карбоксигидрокситриэтил)урацил, 5-карбоксиметиламинометил-2-тиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, бета-D-галактозилквеозин, инозин, N6-изопентиладенин, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-аденин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилквеозин, 5-метоксикарбоксиметилурацил, 5-метоксиурацил, 2-метилтио-N6-изопентиладенин, урацил-5-оксиуксусную кислоту (v), вибутоксозин, псевдоурацил, квеозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, метиловый эфир урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусную кислоту (v), 5-метил-2-тиоурацил, 3-(3-амино-3-N-2-карбоксипропил)урацил, (acp3)w и 2,6-диаминопурин.
Согласно другим конкретным вариантам осуществления изобретения антисмысловой олигонуклеотид содержит по меньшей мере один модифицированный сахарный фрагмент, выбранный из группы, включающей (но, не ограничиваясь только ими) арабинозу, 2-фторарабинозу, ксилозу и гексозу.
Согласно другим конкретным вариантам осуществления изобретения антисмысловой олигонуклеотид содержит по меньшей мере один модифицированный фосфатный каркас, выбранный из группы, включающей (но, не ограничиваясь только ими) фосфоротиоат, фосфородитиоат, фосфороамидотиоат, фосфорамидат, фосфордиамидат, метилфосфонат и алкилфосфотриэфир и формацеталь или их аналоги.
Антисмысловой олигонуклеотид можно вводить в клетку, например, в форме экспрессионного вектора, такого как плазмидный или вирусный вектор, который при транскрибировании в клетке продуцирует РНК, комплементарную по меньшей мере уникальному участку клеточной мРНК В2М. Альтернативно этому, антисмысловой олигонуклеотид можно создавать ex vivo и интродуцировать в клетку с помощью любых методов, известных в данной области. Антисмысловой олигонуклеотид можно синтезировать ex vivo стандартным методом, известным в данной области, например, с использованием автоматического синтезатора ДНК (такого как автоматический синтезатор ДНК, поступающий в продажу, например, от фирмы Applied Biosystems). Разработан ряд методов для введения антисмысловой ДНК или РНК в клетки, например, путем непосредственной инъекции или посредством модификации, созданной для направленного переноса в требуемую клетку (например, используя антисмысловые олигонуклеотиды, связанные с пептидами или антителами, которые специфически связываются с рецепторами или антигенами, экспрессируемыми на поверхности клетки-мишени).
Согласно предпочтительным вариантам осуществления изобретения применяют рекомбинантный ДНК-вектор, в котором нуклеотидную последовательность, кодирующую антисмысловой олигонуклеотид, который ингибирует экспрессию В2М или CIITA, помещают под контроль промотора, такого как сильный промотор pol III или pol II. Применение такой конструкции для трансфекции клетки-мишени, такой как Т-клетка, должно приводить к транскрипции в достаточном количестве одноцепочечной РНК, которая должна образовывать комплементарные пары оснований с эндогенным транскриптом и тем самым препятствовать трансляции мРНК В2М или CIITA. Согласно указанным вариантам осуществления изобретении ДНК-вектор, содержащий нуклеотидную последовательность, которая кодирует антисмысловой олигонуклеотид, интродуцируют в клетку, в которой происходит транскрипция антисмысловой РНК. Указанный вектор может оставаться эписомальным или может интегрироваться в хромосому, при условии, что он может транскрибироваться с образованием антисмысловой РНК. Экспрессия последовательности, кодирующей антисмысловую РНК, может находиться под контролем любого промотора, для которого в данной области известно, что он обладает активностью в клетке млекопитающих, предпочтительно человеческой клетке. Указанный промотор может представлять собой индуцибельный или конститутивный промотор. Примеры промоторов включают (но, не ограничиваясь только ими) область раннего промотора SV40, промотор, содержащий длинный 3'-концевой повтор вируса саркомы Рауса, промотор тимидина вируса герпеса и регуляторные последовательности гена металлотионеина.
Альтернативно этому, в клетку можно интродуцировать конструкции антисмысловой кДНК, которые синтезируют антисмысловую РНК конститутивно или индуцибельно в зависимости от применяемого промотора.
Согласно предпочтительным вариантам осуществления изобретения антисмысловой олигонуклеотид содержит по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 3. В случае двухцепочечной молекулы указанный двухцепочечный антисмысловой олигонуклеотид содержит первую цепь, которая содержит по меньшей мере 10 последовательных нуклеотидов SEQ ID NO: 3, и вторую цепь, комплементарную указанной первой цепи. В случае одноцепочечной молекулы указанный одноцепочечный олигонуклеотид содержит по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 3.
Согласно другим предпочтительным вариантам осуществления изобретения антисмысловой олигонуклеотид содержит по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 5. В случае двухцепочечной молекулы указанный двухцепочечный антисмысловой олигонуклеотид содержит первую цепь, содержащую по меньшей мере 10 последовательных нуклеотидов SEQ ID NO: 5, и вторую цепь, комплементарную указанной первой цепи. В случае одноцепочечной молекулы указанный одноцепочечный олигонуклеотид содержит по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 5.
Антисмысловой олигонуклеотид может содержать нуклеотидную последовательность, комплементарную некодирующей или кодирующей области мРНК В2М или CIITA. Согласно предпочтительным вариантам осуществления изобретения антисмысловой олигонуклеотид содержит нуклеотидную последовательность, комплементарную 5'-концу мРНК В2М или CIITA, например, 5'-нетранслируемую последовательность, простирающуюся вплоть до, и включая, кодон инициации AUG. Согласно другим предпочтительным вариантам осуществления изобретения антисмысловой олигонуклеотид содержит нуклеотидную последовательность, комплементарную 3'-нетранслируемой последовательности мРНК В2М или CIITA. Согласно другим предпочтительным вариантам осуществления изобретения антисмысловой олигонуклеотид содержит нуклеотидную последовательность, комплементарную кодирующей области мРНК В2М или CIITA. Если его создают для гибридизации с 5'-, 3'- или кодирующей областью мРНК В2М или CIITA, то антисмысловой олигонуклеотид должен состоять по меньшей мере из 6 нуклеотидов, предпочтительно по меньшей мере из 10 нуклеотидов и предпочтительно состоять меньшей чем из примерно 100 и более предпочтительно меньшей чем из примерно 50, 25, 20, 15 или 10 нуклеотидов. Согласно предпочтительным вариантам осуществления изобретения антисмысловой олигонуклеотид состоит из 6-25, например, 10-25 нуклеотидов.
Согласно другим конкретным вариантам осуществления изобретения молекулу рибозима, созданную для каталитического расщепления мРНК-транскрипта В2М или CIITA, применяют для предупреждения трансляции и экспрессии В2М или CIITA в Т-клетке соответственно (см., например, общее руководство в WO 90/11364 и US 5093246). Согласно предпочтительным вариантам осуществления изобретения рибозим представляет собой рибозим типа hammerhead. Рибозимы типа hammerhead расщепляют мРНК в положениях, определяемых фланкирующими областями, которые образуют комплементарные пары оснований с мРНК-мишенью, например. мРНК В2М, такой как мРНК человеческого В2М, представленная в SEQ ID NO: 3. Единственным требованием является присутствие в мРНК-мишени следующей последовательности двух оснований: 5'-UG-3'. Создание и получение рибозимов типа hammerhead хорошо известно в данной области и описано более подробно у Haseloff и Gerlach, 1988. Согласно предпочтительным вариантам осуществления изобретения рибозим конструируют так, чтобы распознаваемый ферментом сайт расщепления был локализован вблизи 5'-конца мРНК В2М. Согласно другим предпочтительным вариантам осуществления изобретения рибозим конструируют так, чтобы распознаваемый ферментом сайт расщепления был локализован вблизи 5'-конца мРНК CIITA. Это повышает эффективность и минимизирует внутриклеточное накопление нефункциональных мРНК-транскриптов.
Подобно антисмысловым олигонуклеотидам рибозим, применяемый согласно изобретению, может состоять из олигонуклеотидов, модифицированных, например, для повышения стабильности. Рибозим можно вводить в клетку любыми путями, известными в данной области. Рибозим можно вводить в Т-клетку в форме экспрессионного вектора, такого как плазмидный или вирусный вектор, которые при транскрибировании в клетках продуцирует рибозим. Согласно предпочтительным вариантам осуществления изобретения применяют рекомбинантный ДНК-вектор, в котором нуклеотидную последовательность, кодирующую рибозим, помещают под контроль промотора, такого как сильный промотор pol III или pol II, в результате чего трансфектированная клетка может продуцировать рибозим в количествах, достаточных для деструкции эндогенной мРНК и ингибирования трансляции. Поскольку рибозимы в отличие от антисмысловых олигонуклеотидов обладают каталитической активностью, для эффективного действия требуется более низкая внутриклеточная концентрация.
Согласно другим конкретным вариантам осуществления изобретения ингибирующая нуклеиновая кислота представляет собой молекулу интерферирующей РНК (PHKi). РНК-интерференция представляет собой биологический процесс, при котором молекулы РНК ингибируют экспрессию гена, как правило, в результате деструкции специфической мРНК. Примеры типов молекул PHKi включают микроРНК (miPHK), малую интерферирующую РНК (siPHK) и короткую образующую шпильку РНК (shPHK). Согласно предпочтительному варианту осуществления изобретения молекула PHKi представляет собой miPHK. Согласно другому предпочтительному варианту осуществления изобретения молекула PHKi представляет собой siPHK. Согласно еще одному предпочтительному варианту осуществления изобретения молекула PHKi представляет собой shPHK. Получение молекул PHKi in vivo и in vitro и методы их применения описаны, например, в US 6506559, WO 01/36646, WO 00/44895, US 2002/01621126, US 2002/0086356, US 2003/0108923, WO 02/44321, WO 02/055693, WO 02/055692 и WO 03/006477.
Согласно предпочтительному варианту осуществления изобретения молекула PHKi представляет собой интерферирующую РНК, комплементарную SEQ ID NO: 3. Согласно другому предпочтительному варианту осуществления изобретения молекула PHKi представляет собой молекулу рибонуклеиновой кислоты, содержащую по меньшей мере 10 последовательных нуклеотидов комплемента SEQ ID NO: 3. Согласно другому предпочтительному варианту осуществления изобретения молекула PHKi представляет собой молекулу двухцепочечной рибонуклеиновой кислоты, которая содержит первую цепь, идентичную 20-25, например, 21-23, последовательных нуклеотидов SEQ ID NO: 3, и вторую цепь, комплементарную указанной первой цепи.
Согласно предпочтительному варианту осуществления изобретения молекула PHKi представляет собой интерферирующую РНК, комплементарную SEQ ID NO: 5. Согласно другому предпочтительному варианту осуществления изобретения молекула PHKi представляет собой молекулу рибонуклеиновой кислоты, содержащую по меньшей мере 10 последовательных нуклеотид комплемента SEQ ID NO: 5. Согласно другому предпочтительному варианту осуществления изобретения молекула PHKi представляет собой молекулу двухцепочечной рибонуклеиновой кислоты, которая содержит первую цепь, идентичную 20-25, например, 21-23, последовательных нуклеотидов SEQ ID NO: 5, и вторую цепь, комплементарную указанной первой цепи.
Создание PD1/PDL1-пути Т-клеточной регуляции
В основу настоящего изобретения положена задача облегчить приживление трансплантата Т-клеток, прежде всего аллогенных Т-клеток, предпочтительно путем ингибирования экспрессии В2М и/или CIITA в сочетании с инактивацией TCR.
При создании изобретения было установлено, что в качестве альтернативы указанному подходу или в сочетании с ним в Т-клетке можно разрушать PD1 (белок запрограммированной клеточной гибели 1, который обозначают также как PD1; PD-1; CD279; SLEB2; hPD-1; hPD-1 или hSLE1), который представляет собой состоящую из 288 аминокислот молекулу поверхностного белка, кодируемого геном PDCD1 (NCBI - NC_000002.12). Указанный белок экспрессируется на Т-клетках и про-В-клетках и, как установлено, осуществляет отрицательную регуляцию Т-клеточных ответов (Carter L. и др., 2002). Образование комплекса рецептор PD-1 / лиганд PD-L1 приводит к передаче ингибирующего сигнала, который снижает пролиферацию Т-клеток.
Лиганд (рецептора) запрограммированной клеточной гибели 1 (PD-L1) представляет собой трансмембранный белок типа 1 с молекулярной массой 40 кДа, который, вероятно, играет основную роль в подавлении иммунной системы в конкретных случаях, таких как беременность, аллотрансплантации тканей, аутоиммунное заболевание и другие болезненные состояния, такие как гепатит. PDL-1 (который называют также CD274 или В7Н1) кодируется геном CD274 (NCBI - NM_014143).
Согласно конкретному объекту изобретения экспрессию как PD-1, так и TCR ингибируют в сконструированных Т-клетках, предлагаемых в изобретении, что оказывает двойное действие на активацию Т-клеток в качестве компонента аллогенной трансплантации. Однако инактивация или ингибирование PD-1 можно применять также при осуществлении аутологичной трансплантации Т-клеток, при которой не требуется ингибирование или разрушение TCR.
Согласно другому объекту изобретения ингибирование или разрушение PD1 объединяют со сверхэкспрессией его лиганда PDL-1 в трансплантированных Т-клетках. Для получения указанной сверхэкспрессии, например, можно применять трансформацию лентивирусами или ретровирусами Т-клеток, в которых PD-1 заингибирован или разрушен, или применять любые другие средства, известные в данной области. Таким образом, PDL1, для которого характерна сверхэкспрессия в Т-клетках, не должен влиять на трансплантированные [PD1-]-клетки, а только на [PD1+]-Т-клетки из организма пациента. В результате Т-клетки из организма пациента ингибируются и не обладают активностью в отношении трансплантированных клеток, что облегчает их приживление при трансплантации и сохранение в хозяине.
Согласно предпочтительному варианту осуществления изобретения в изобретении предложены сконструированные Т-клетки, которые представляют собой [PD1-][TCR-]-клетки, при этом сверхэкспрессирующие PDL1, для облегчения их трансплантации пациенту, в частности, в качестве компонента иммунотерапии.
Экспрессия по меньшей мере одного неэндогенного иммуносупрессорного полипептида
Согласно некоторым предпочтительным вариантам осуществления изобретения ингибирование экспрессии бета-2m и/или CIITA осуществляют с использованием дополнительной стадии экспрессии в указанной Т-клетке по меньшей мере одного неэндогенного иммуносупрессорного полипептида.
Под «неэндогенным» полипептидом подразумевают полипептид, который в норме не экспрессируется донорской иммунной клеткой, предпочтительно полипептид, который экспрессируется экзогенным полинуклеотидом, импортированным в геном иммунной клетки. Например, IL12 не рассматривается как неэндогенный полипептид, поскольку он экспрессируется из ранее существующего гена из донорской иммунной клетки.
Под «иммуносупрессорным полипептидом» подразумевается, что экспрессия указанного неэндогенного полипептида обладает способностью снижать иммунный ответ пациента-хозяина против донорских иммунных клеток.
Таким образом, способ, предлагаемый в настоящем изобретении, может предусматривать интродукцию в указанную Т-клетку экзогенной молекулы нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую по меньшей мере один неэндогенный иммуносупрессорный полипептид, такой как вирусный гомолог ГКГС или лиганд NKG2D.
Экспрессия вирусного гомолога ГКГС
Согласно конкретным предпочтительным вариантам осуществления изобретения указанный неэндогенный иммуносупрессорный полипептид, экспрессируемый в указанной Т-клетке, представляет собой вирусный гомолог ГКГС, такой, например, как UL18 (обозначенный как NP_044619 в базе данных белков NCBI).
Таким образом, согласно указанным вариантам осуществления изобретения способ, предлагаемый в настоящем изобретении, может предусматривать интродукцию в указанную Т-клетку молекулы экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую вирусный гомолог ГКГС, такой как UL18. Молекула экзогенной нуклеиновой кислоты может содержать нуклеотидную последовательность, кодирующую полипептид, идентичный по меньшей мере на 80%, предпочтительно по меньшей мере на 90% и более предпочтительно по меньшей мере на 95% SEQ ID NO: 89.
Взаимодействие между аллогенной Т-клеткой и иммунными клетками хозяина схематически изображено на фиг. 8 (экспрессия вирусного гомолога ГКГС) относительно ситуации, представленной на фиг. 7 (отсутствие экспрессии). На обоих чертежах молекулу ГКГС класса I предпочтительно инактивируют путем разрушения (KO) гена бета2М.
Экспрессия лиганда NKG2D
Некоторые вирусы, такие как цитомегаловирусы, имеют приобретенные механизмы, позволяющие избегать опосредуемого NK-клеткой иммунного «надзора» и взаимодействующие с путем NKG2D посредством секреции белка, который может связываться с лигандами NKG2D и предупреждать их поверхностную экспрессию (Welte S.A, Sinzger С., Lutz S.Z., Singh-Jasuja Н., Sampaio K.L., Eknigk U., Rammensee H.G., Steinle A. «Selective intracellular retention of virally induced NKG2D ligands by the human cytomegalovirus UL16 glycoprotein». Eur. J. Immunol., 33, 2003, cc. 194-203). В опухолевых клетках присутствуют некоторые механизмы, позволяющие «уклоняться» от NKG2D-ответа путем секреции лигандов NKG2D, таких как ULBP2, MICB или MICA (Salih H.R., Antropius Н., Gieseke F., Lutz S.Z., Kanz L. и др., Functional expression and release of ligands for the activating immunoreceptor NKG2D in leukemia. Blood 102, 2003, cc. 1389-1396)
Согласно другим конкретным предпочтительным вариантам осуществления изобретения неэндогенный иммуносупрессорный полипептид, подлежащий экспрессии в указанной Т-клетке, представляет собой лиганд NKG2D.
Таким образом, согласно указанным вариантам осуществления изобретения способ, предлагаемый в настоящем изобретении, может предусматривать интродукцию в указанную Т-клетку молекулы экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую лиганд NKG2D. Молекула нуклеиновой кислоты может содержать нуклеотидную последовательность, кодирующую полипептид, идентичный по меньшей мере на 80%, предпочтительно по меньшей мере на 90% и более предпочтительно по меньшей мере на 95% любой из SEQ ID NO: 90-97.
Взаимодействие между аллогенной Т-клеткой и иммунными клетками хозяина схематически изображено на фиг. 9 (экспрессия растворимого лиганда NKG2D) относительно ситуации, представленной на фиг. 7 (отсутствие экспрессии). На обоих чертежах молекулу ГКГС класса I инактивировали путем разрушения (KO) гена бета2М.
В таблице 10, которая приведена ниже в настоящем описании, представлены вирусный гомолог ГКГС (UL18) и панель лигандов NKG2D и их полипептидные последовательности, которые можно экспрессировать согласно настоящему изобретению.
Химерные антигенные рецепторы (CAR)
Адоптивная иммунотерапия, в которой применяют перенос аутологичных антигенспецифических Т-клеток, созданных ex vivo, представляет собой перспективную стратегию для лечения вирусных инфекций и рака. Т-клетки, применяемые для адоптивной иммунотерапии, можно создавать либо путем размножения антигенспецифических Т-клеток, либо посредством перенаправления Т-клеток с помощью методов генетической инженерии (Park, Rosenberg и др., 2011). Перенос Т-клеток, специфических в отношении вирусных антигенов, является хорошо зарекомендовавшей себя процедурой, которую применяют для лечения ассоциированных с трансплантацией вирусных инфекций и редких связанных с вирусами злокачественных заболеваний. Продемонстрировано также, что выделение и перенос специфических в отношении опухоли Т-клеток приводит к успеху при лечении меланомы.
В Т-клетках были успешно созданы новые специфичности посредством генетического переноса трансгенных Т-клеточных рецепторов или химерных антигенных рецепторов (CAR) (Jena, Dotti и др., 2010). CAR представляют собой синтетические рецепторы, состоящие из «нацеливающего» фрагмента, ассоциированного с одним или несколькими сигнальными доменами в одной слитой молекуле. В целом, связывающий фрагмент CAR состоит из антигенсвязывающего домена одноцепочечного антитела (scFv), который содержит легкие и вариабельные фрагменты моноклонального антитела, соединенные гибким линкером. Успешно применяли также связывающие фрагменты на основе доменов рецептора или лиганда. Сигнальные домены для первого поколения CAR получали из цитоплазматической области цепей зета CD3 или гамма Fc-рецептора. Было продемонстрировано, что первое поколение CAR позволяло успешно перенаправлять Т-клеточную цитотоксичность, однако для них не удалось обеспечить пролонгированное размножение и противоопухолевую активность in vivo. Для повышения выживаемости и усиления пролиферации модифицированных с помощью CAR Т-клеток осуществляли добавление сигнальных доменов из костимулирующих молекул, включая CD28, ОХ-40 (CD134) и 4-1ВВ (CD137), по отдельности (второе поколение) или в комбинации (третье поколение). CAR позволяли успешно перенаправлять Т-клетки на антигены, экспрессируемые на поверхности опухолевых клеток при различных злокачественных заболеваниях, включая лимфомы и солидные опухоли (Jena, Dotti и др., 2010).
CD19 представляет собой привлекательную мишень для иммунотерапии, поскольку огромное большинство клеток острого В-клеточного лимфобластного лейкоза (В-ALL) постоянно экспрессируют CD19, в то время как экспрессия отсутствует на негематопоэтических клетках, а также на миелоидных, эритроидных и Т-клетках и стволовых клетках костного мозга. В настоящее время проводятся клинические опыты по оценке направленного воздействия на CD19 при В-клеточных злокачественных заболеваниях, в которых получены обнадеживающие противоопухолевые ответы. В большинстве случаев вводимые путем инфузии Т-клетки генетически модифицировали таким образом, чтобы они экспрессировали химерный антигенный рецептор (CAR) со специфичностью, полученный из scFv-области CD19-специфического мышиного моноклонального антитела FMC63 (WO 2013/126712).
Таким образом, согласно некоторым вариантам осуществления изобретения химерный антигенный рецептор, экспрессируемый сконструированной Т-клеткой, направлен против антигена CD19 В-лимфоцитов.
Согласно некоторым вариантам осуществления изобретения химерный антигенный рецептор представляет собой одноцепочечный химерный антигенный рецептор. Например, одноцепочечный химерный антигенный рецептор, подлежащий экспрессии в сконструированных Т-клетках, предлагаемых в настоящем изобретении, представляет собой единичный полипептид, который содержит по меньшей мере один внеклеточный лигандсвязывающий домен, трансмембранный домен и по меньшей мере один домен трансдукции сигнала, в котором указанный внеклеточный лигандсвязывающий домен содержит scFV, происходящий из специфического моноклонального антитела к CD19, такого как 4G7. После трансдукции в Т-клетку, например, посредством трансдукции ретровирусом или лентивирусом, указанный CAR принимает участие в распознавании антигена CD19, присутствующего на поверхности злокачественных В-клеток при лимфоме или лейкозе.
Согласно конкретным вариантам осуществления изобретения химерный антигенный рецептор представляет собой полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 6, или ее вариант, который содержит аминокислотную последовательность, идентичную по меньшей мере на 70%, например, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере 99% аминокислотной последовательности, представленной в SEQ ID NO: 6 по всей длине SEQ ID NO: 6. Предпочтительно вариант обладает способностью связываться с CD19.
Наиболее предпочтительный химерный антигенный рецептор представляет собой полипептид, который содержит аминокислотную последовательность, представленную в SEQ ID NO: 7 или ее вариант, который содержит аминокислотную последовательность, идентичную по меньшей мере на 70%, например, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере 99% аминокислотной последовательности, представленной в SEQ ID NO: 7 по всей длине SEQ ID NO: 7. Указанный вариант может отличаться от полипептида, представленного в SEQ ID NO: 7, заменой по меньшей мере одного, по меньшей мере двух или по меньшей мере трех аминокислотного(ых) остатка(ов). Предпочтительно указанный вариант обладает способностью связываться с CD19.
Согласно другим конкретным вариантам осуществления изобретения химерный антигенный рецептор может быть направлен против другого антигена, экспрессируемого на поверхности злокачественной или инфицированной клетки, такой как молекула из кластера дифференцировки, такая как CD16, CD64, CD78, CD96, CLL1, CD116, CD117, CD71, CD45, CD71, CD123 и CD138, ассоциированный в опухолью поверхностный антиген, такой как ErbB2 (HER2/neu), карциноэмбриональный антиген (СЕА), молекула клеточной адгезии эпителия (ЕрСАМ), рецептор эпидермального фактора роста (EGFR), вариант EGFR III (EGFRvIII), CD19, CD20, CD30, CD40, дисиалоганглиозид GD2, муцин протоков-эпителия, gp36, TAG-72, гликосфинголипиды, ассоциированный с глиомой антиген, человеческий хорионический гонадотропин β, альфафетопротеин (AFP), лектин-реактивный AFP, тироглобулин, RAGE-1, MN-СА IX, человеческая обратная транскриптаза теломеразы, RU1, RU2 (AS), кишечная карбоксилэстераза, mut hsp70-2, M-CSF, простаза, простатспецифический антиген (PSA), PAP, NY-ESO-1, LAGA-1a, р53, простеин, PSMA, сурвивин и теломераза, опухолевый антиген-1 карциномы предстательной железы (РСТА-1), MAGE, ELF2M, эластаза нейтрофилов, эфрин В2, CD22, инсулиновый фактор роста (IGF1)-I, IGF-II, рецептор IGFI, мезотелин, молекула главного комплекса гистосовместимости (ГКГС), презентующая опухольспецифический пептидный эпитоп, 5Т4, ROR1, Nkp30, NKG2D, антигены стромы опухоли, экстра-домен A (EDA) и экстра-домен В (EDB) фибронектина и А1-домен тенасцина-С (TnC А1) и фибробласт-ассоциированный белок (fap); специфический для линии дифференцировки или тканеспецифический антиген, такой как CD3, CD4, CD8, CD24, CD25, CD33, CD34, CD133, CD138, CTLA-4, В7-1 (CD80), В7-2 (CD86), GM-CSF, цитокиновые рецепторы, эндоглин, молекула главного комплекса гистосовместимости (ГКГС), ВСМА (CD269, TNFRSF 17), антиген множественной миеломы или лимфобластного лейкоза, например, выбранный из TNFRSF17 (UNIPROT Q02223), SLAMF7 (UNIPROT Q9NQ25), GPRC5D (UNIPROT Q9NZD1), FKBP11 (UNIPROT Q9NYL4), KAMP3, ITGA8 (UNIPROT Р53708) и FCRL5 (UNIPROT Q68SN8), вирус-специфический поверхностный антиген, такой как ВИЧ-специфический антиген (такой как gp120 ВИЧ), EBV-специфический антиген, CMV-специфический антиген, HPV-специфический антиген, антиген, специфический для вируса Ласа, антиген, специфический для вируса гриппа, а также любое производное или любой вариант указанных поверхностных антигенов.
В некоторых других вариантах осуществления изобретения химерный антигенный рецептор представляет собой многоцепочечный химерный антигенный рецептор. Известные из существующего уровня техники химерные антигенные рецепторы, интродуцированные в Т-клетки, образовывали одноцепочечные полипептиды обязательно с присоединенными сериями сигнальных доменов. Однако путем сдвига сигнальных доменов с их естественного положения вблизи мембраны можно влиять на их функцию. Для преодоления этого недостатка при создании настоящего изобретения конструировали многоцепочечный CAR, происходящий из FcεRI, для обеспечения нормального положения вблизи мембраны всех представляющих интерес сигнальных доменов. В указанной новой архитектуре высокоаффинный IgE-связывающий домен альфа-цепи FcεRI заменяли на внеклеточный лигандсвязывающий домен, такой как scFv, для того, чтобы перенаправлять Т-клеточную специфичность против клеток-мишеней, и N- и/или С-концевые хвосты бета-цепи FcεRI применяли для помещения костимулирующих сигналов в нормальные положения вблизи мембраны, как описано в WO 2013/176916.
Таким образом, CAR, экспрессируемый сконструированной Т-клеткой, согласно изобретению может представлять собой многоцепочечный химерный антигенный рецептор, наиболее адаптированный к производству и размножению сконструированных Т-клеток, предлагаемых в настоящем изобретении. Такие многоцепочечные CAR содержат по меньшей мере два следующих компонента:
а) один полипептид, содержащий трансмембранный домен альфа-цепи FcεRI и внеклеточный лигандсвязывающий домен,
б) один полипептид, содержащий часть N- и С-концевого цитоплазматического хвоста и трансмембранный домен бета-цепи FcεRI и/или
в) по меньшей мере два полипептида, каждый из которых содержит часть внутрицитоплазматического хвоста и трансмембранный домен гамма-цепи FcεRI, при этом различные полипептиды спонтанно образуют мультимеры, такие как димерный, тримерный или четырехмерный CAR.
При такой архитектуре лигандсвязывающие домены и сигнальные домены находятся на различных полипептидах. Различные полипептиды закрепляются в мембране в непосредственной близости, что обеспечивает их взаимодействие друг с другом. При такой архитектуре сигнальные и костимулирующие домены могут находиться в положениях вблизи мембраны (т.е. примыкать к клеточной мембране с ее внутренней стороны), что, по-видимому, позволяет улучшать функцию костимулирующих доменов. Кроме того, включающая несколько субъединиц архитектура является более гибкой и дает возможность создавать CAR, которые в большей степени контролируют Т-клеточную активацию. Например, можно включать несколько внеклеточных антигенраспознающих доменов, имеющих различную специфичность, с получением архитектуры CAR с несколькими специфичностями. Можно также контролировать относительное соотношение между различными субъединицами в многоцепочечном CAR. Такой тип архитектуры в настоящее время подробно описан авторами настоящего изобретения в PCT/US2013/058005.
Сборка различных цепей в виде части одного многоцепочечного CAR делает возможным, например, применять различные альфа-, бета- и гамма-цепи высокоаффинного рецептора для IgE (FcεRI) (Metzger, Alcaraz и др., 1986), которые сливают с сигнальными и костимулирующими доменами. Гамма-цепь содержит трансмембранную область и цитоплазматический хвост, содержащий один мотив активации иммунных рецепторов на основе тирозина (ITAM) (Cambier, 1995).
Многоцепочечный CAR может содержать несколько внеклеточных лигандсвязывающих доменов для одновременного связывания различных элементов мишени, тем самым усиливая активацию и функцию иммунных клеток. В одном из вариантов осуществления изобретения внеклеточные лигандсвязывающие домены могут находиться в виде тандема на одном и том же трансмембранном полипептиде и необязательно могут разделяться линкером. В другом варианте осуществления изобретения указанные различные внеклеточные лигандсвязывающие домены могут находиться на различных трансмембранных полипептидах, из которых состоит многоцепочечный CAR.
Домен трансдукции сигналов или внутриклеточный сигнальный домен многоцепочечного(ых) CAR, предлагаемого(ых) в изобретении, ответствен за внутриклеточную передачу сигналов после связывания внеклеточного лигандсвязывающего домена с мишенью, что приводит к активации иммунной клетки и иммунного ответа. Другими словами, домен трансдукции сигналов ответствен за активацию по меньшей мере одной из нормальных эффекторных функций иммунной клетки, в которой экспрессируется многоцепочечный CAR. Например, эффекторная функция Т-клетки может представлять собой цитолитическую активность или хелперную активность, включая секрецию цитокинов.
В контексте настоящего описания понятие «домен трансдукции сигналов» относится к участку белка, который трансдуцирует эффекторную сигнальную функцию сигнала и принуждает клетку осуществлять специализированную функцию.
Предпочтительными примерами домена трансдукции сигналов, которые можно применять в одноцепочечном или многоцепочечном CAR, могут являться цитоплазматические последовательности Fc-рецептора или Т-клеточного рецептора и корецепторов, совместное действие которых состоит в инициации трансдукции сигналов после контакта с антигенным рецептором, а также любые производные или варианты указанных последовательностей и любые синтетические последовательности, имеющие такие функциональные особенности. Домен трансдукции сигналов содержит два различных класса цитоплазматических сигнальных последовательностей, те, которые инициируют антиген-зависимую первичную активацию, и те, которые действуют вне зависимости от антигена, обеспечивая вторичный или костимулирующий сигнал. Первичная цитоплазматическая сигнальная последовательность может содержать сигнальные мотивы, которые известны как мотивы активации иммунных рецепторов на основе тирозина, ITAM. ITAM представляют собой хорошо известные сигнальные мотивы, присутствующие во внутрицитоплазматическом хвосте различных рецепторов, которые служат в качестве сайтов связывания для тирозинкиназ класса syk/zap70. Примерами ITAM, применяемых согласно изобретению, могут служить (но, не ограничиваясь только ими) ITAM, происходящие из TCRзета, FcRгамма, FcRбета, FcRэпcилoн, CD3гамма, CD3дельта, CD3эпсилон, CD5, CD22, CD79a, CD79b и CD66d. В предпочтительном варианте осуществления изобретения домен трансдукции сигнала многоцепочечного CAR может содержать происходящий из CD3зета сигнальный домен или внутрицитоплазматический домен бета- или гамма цепей FcεRI.
Согласно конкретным вариантам осуществления изобретения домен трансдукции сигналов многоцепочечных CAR, предлагаемых в настоящем изобретении, содержит костимулирующую сигнальную молекулу. Костимулирующая молекула представляет собой молекулу на клеточной поверхности, отличную от антигенного рецептора или его лигандов, которая требуется для эффективного иммунного ответа.
Лигандсвязывающие домены могут представлять собой любой применявшийся и описанный ранее антигенный рецептор касательно одноцепочечного CAR, описанного в литературе, в частности, scFv из моноклональных антител.
Сконструированные Т-клетки
При осуществлении настоящего изобретения можно получать сконструированные Т-клетки с улучшенными характеристиками. В частности, в настоящем изобретении предложена сконструированная, предпочтительно выделенная Т-клетка, отличающаяся тем, что в ней ингибируется экспрессия В2М и/или CIITA.
Согласно некоторым вариантам осуществления изобретения в настоящем изобретении предложена сконструированная, предпочтительно выделенная Т-клетка, которая экспрессирует редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК, предпочтительно двухцепочечного разрыва, гена, кодирующего В2М. Согласно конкретным вариантам осуществления изобретения указанная Т-клетка содержит молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанную редко расщепляющую эндонуклеазу. Согласно более конкретным вариантам осуществления изобретения указанная редко расщепляющая эндонуклеаза представляет собой TAL-нуклеазу, мегануклеазу, нуклеазу с «цинковыми пальцами» (ZFN) или РНК-направляемую эндонуклеазу. Таким образом, согласно конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой TAL-нуклеазу. Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой мегануклеазу. Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой нуклеазу с «цинковыми пальцами». Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой РНК-направляемую эндонуклеазу, такую как Cas9.
Согласно некоторым другим вариантам осуществления изобретения в настоящем изобретении предложена сконструированная, предпочтительно выделенная Т-клетка, которая содержит молекулу экзогенной нуклеиновой кислоты, которая ингибирует экспрессию В2М. Согласно конкретным вариантам осуществления изобретения указанная Т-клетка содержит молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую молекулу нуклеиновой кислоты, которая ингибирует экспрессию В2М. Согласно более конкретным вариантам осуществления изобретения молекула нуклеиновой кислоты, которая ингибирует экспрессию В2М, представляет собой молекулу антисмыслового олигонуклеотида, рибозима или интерферирующей РНК (PHKi). Таким образом, согласно конкретному варианту осуществления изобретения молекула нуклеиновой кислоты, которая ингибирует экспрессию В2М, представляет собой антисмысловый олигонуклеотид. Согласно другому конкретному варианту осуществления изобретения молекула нуклеиновой кислоты, которая ингибирует экспрессию В2М, представляет собой рибозим и предпочтительно рибозим hammerhead-типа. Согласно другому конкретному варианту осуществления изобретения молекула нуклеиновой кислоты, которая ингибирует экспрессию В2М, представляет собой молекулу интерферирующей РНК.
Согласно некоторым вариантам осуществления изобретения в настоящем изобретении предложена сконструированная, предпочтительно выделенная Т-клетка, которая экспрессирует редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК, предпочтительно двухцепочечного разрыва, гена, кодирующего CIITA. Согласно конкретным вариантам осуществления изобретения указанная Т-клетка содержит молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанную редко расщепляющую эндонуклеазу. Согласно более конкретным вариантам осуществления изобретения указанная редко расщепляющая эндонуклеаза представляет собой TAL-нуклеазу, мегануклеазу, нуклеазу с «цинковыми пальцами» (ZFN) или РНК-направляемую эндонуклеазу. Таким образом, согласно конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой TAL-нуклеазу. Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой мегануклеазу. Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой нуклеазу с «цинковыми пальцами». Согласно другому конкретному варианту осуществления изобретения редко расщепляющая эндонуклеаза представляет собой РНК- или ДНК-направляемую эндонуклеазу, такую как Cas9 или эндонуклеазу семейства Argonaute.
Согласно некоторым другим вариантам осуществления изобретения в настоящем изобретении предложена сконструированная, предпочтительно выделенная Т-клетка, которая содержит молекулу экзогенной нуклеиновой кислоты, которая ингибирует экспрессию CIITA. Согласно конкретным вариантам осуществления изобретения указанная Т-клетка содержит молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую молекулу нуклеиновой кислоты, которая ингибирует экспрессию CIITA. Согласно более конкретным вариантам осуществления изобретения молекула нуклеиновой кислоты, которая ингибирует экспрессию CIITA, представляет собой молекулу антисмыслового олигонуклеотида, рибозима или интерферирующей РНК (PHKi). Таким образом, согласно конкретному варианту осуществления изобретения молекула нуклеиновой кислоты, которая ингибирует экспрессию CIITA, представляет собой антисмысловый олигонуклеотид. Согласно другому конкретному варианту осуществления изобретения молекула нуклеиновой кислоты, которая ингибирует экспрессию CIITA, представляет собой рибозим и предпочтительно рибозим hammerhead-типа. Согласно другому конкретному варианту осуществления изобретения молекула нуклеиновой кислоты, которая ингибирует экспрессию CIITA, представляет собой молекулу интерферирующей РНК.
Согласно некоторым вариантам осуществления изобретения сконструированная Т-клетка экспрессирует также редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК, предпочтительно двухцепочечного разрыва, по меньшей мере одного гена, кодирующего компонент Т-клеточного рецептора (TCR), такой как TCR-альфа. Согласно конкретным вариантам осуществления изобретения указанная Т-клетка содержит молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанную редко расщепляющую эндонуклеазу.
Согласно некоторым вариантам осуществления изобретения сконструированная Т-клетка дополнительно экспрессирует химерный антигенный рецептор (CAR), направленный против по меньшей мере одного антигена, экспрессируемого на поверхности злокачественной или инфицированной клетки. Согласно конкретным вариантам осуществления изобретения указанная Т-клетка содержит молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанный CAR.
Согласно некоторым другим вариантам осуществления изобретения в настоящем изобретении предложена сконструированная, предпочтительно выделенная Т-клетка, которая экспрессирует по меньшей мере один неэндогенный иммуносупрессорный полипептид. Согласно конкретным вариантам осуществления изобретения указанный неэндогенный иммуносупрессорный полипептид представляет собой вирусный гомолог ГКГС, такой как UL18. Таким образом, Т-клетка может содержать молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую полипептид, идентичный по меньшей мере на 80%, предпочтительно по меньшей мере на 90% и более предпочтительно по меньшей мере на 95% SEQ ID NO: 89. Согласно другим конкретным вариантам осуществления изобретения указанный неэндогенный иммуносупрессорный полипептид представляет собой лиганд NKG2D. Таким образом, Т-клетка может содержать молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую полипептид, идентичный по меньшей мере на 80%, предпочтительно по меньшей мере на 90% и более предпочтительно по меньшей мере на 95% любой одной из SEQ ID NO: 90-97.
Следует понимать, что все детали, касающиеся конкретно редко расщепляющей эндонуклеазы, которая обладает способностью осуществлять избирательную активацию путем расщепления ДНК гена, кодирующего В2М, молекулы нуклеиновой кислоты, которая ингибирует экспрессию В2М, редко расщепляющей эндонуклеазы, которая обладает способностью осуществлять избирательную активацию путем расщепления ДНК гена, кодирующего компонент Т клеточного рецептора (TCR), и химерный антигенный рецептор, также включены в указанный объект изобретения.
Кроме того, под объем настоящего изобретения подпадает также клетка или клеточная линия, полученная из сконструированной согласно изобретению Т-клетки, предпочтительно характеризующаяся одним из следующих фенотипов:
[b2m]-[TCR]-
[TCR]- [PD1]- [PDL-1]+
[b2m]-[TCR]- [PD1]-
[b2m]-[TCR]- [PD1]- [PDL-1]+
[b2m]- [вирусный гомолог ГКГС]+
[b2m]- [TCR]- [вирусный гомолог ГКГС]+
[b2m]- [лиганд NKG2D]+
[b2m]- [TCR]- [лиганд NKG2D]+
Т-клетки, предлагаемые в настоящем изобретении, предпочтительно представляют собой [CAR]+-клетки, т.е. несут химерный антигенный рецептор, предназначенный для специфического распознавания опухолевых клеток.
Методы доставки
При создании настоящего изобретения рассматривались любые средства, известные в данной области, которые позволяют осуществлять доставку внутрь клеток или в субклеточные компартменты указанных клеток молекул нуклеиновых кислот, применяемых согласно изобретению. Эти средства включают (но, не ограничиваясь только ими) вирусную трансдукцию, электропорацию, а также средства доставки, основанные на применении липосом, полимерных носителей, химических носителей, липоплексов, полиплексов, дендримеров, наночастиц, эмульсий, естественного пути эндоцитоза или фагоцитоза.
Согласно настоящему изобретению молекулы нуклеиновых кислот, указанные конкретно в контексте настоящего описания, можно интродуцировать в Т-клетку с помощью любых приемлемых методов, известных в данной области. Приемлемыми методами являются (но не ограничиваясь только ими) интродукция молекулы нуклеиновой кислоты в Т-клетку согласно методам стабильной трансформации, при осуществлении которых молекулу нуклеиновой кислоты интегрируют в геном клетки, методам кратковременной трансформации, при осуществлении которых молекулу нуклеиновой кислоты не интегрируют в геном клетки, и опосредуемые вирусными векторами методы. Указанную молекулу нуклеиновой кислоты можно интродуцировать в клетку с помощью, например, рекомбинантного вирусного вектора (например, ретровирусного, аденовирусного), липосомы и т.п. Методы кратковременной трансформации включают, например, микроинъекцию, электропорацию или бомбардировку частицами. В некоторых вариантах осуществления изобретения молекула нуклеиновой кислоты представляет собой вектор, такой как вирусный вектор или вирусная плазмида. Предпочтительно указанный вектор представляет собой экспрессионный вектор, который может обеспечивать экспрессию соответствующего(их) полипептида(ов) или белка(ов), конкретно указанных в настоящем описании, в Т-клетке.
Молекула нуклеиновой кислоты, интродуцированная в Т-клетку, может представлять собой ДНК или РНК. В некоторых вариантах осуществления изобретения молекула нуклеиновой кислоты, интродуцированная в Т-клетку, представляет собой ДНК. В некоторых вариантах осуществления изобретения молекула нуклеиновой кислоты, интродуцированная в Т-клетку, представляет собой РНК, и конкретно мРНК, кодирующую полипептид или белок, подробно описанный в настоящем описании, указанную мРНК интродуцируют непосредственно в Т-клетку, например, с помощью электропорации. Приемлемый метод электропорации описан, например, в международной публикации WO 2013/176915 (в частности, в разделе, озаглавленном «Электропорация», на сс. 29-30). Конкретная молекула нуклеиновой кислоты, которая может представлять собой мРНК, представляет собой молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять инактивацию путем расщепления ДНК гена, кодирующего В2М. Другая конкретная молекула нуклеиновой кислоты, которая может представлять собой мРНК, представляет собой молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять инактивацию путем расщепления ДНК гена, кодирующего CIITA. Еще одна конкретная молекула нуклеиновой кислоты, которая может представлять собой мРНК, представляет собой молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую редко расщепляющую эндонуклеазу, которая обладает способностью осуществлять инактивацию путем расщепления ДНК по меньшей мере одного гена, кодирующего один компонент Т-клеточного рецептора (TCR).
В качестве предпочтительного варианта осуществления изобретения молекулами нуклеиновых кислот, кодирующими эндонуклеазы, предлагаемые в настоящем изобретении, осуществляют трансфекции в форме мРНК для получения кратковременной экспрессии во избежание хромосомной интеграции чужеродной ДНК, например, путем электропорации. При создании изобретения были определены различные оптимальные условия для введения мРНК путем электропорации в Т-клетку, которые представлены в таблице 1. При создании изобретения использовали технологию CytoPulse, которая позволяет путем применения пульсирующих электрических полей кратковременно повышать проницаемость живых клеток для введения материалов в клетки (US 6010613 и WO 2004/083379). Продолжительность, интенсивность импульса, а также интервал между импульсами можно модифицировать для достижения наилучших условий для высокоэффективной трансфекции с минимальной смертностью. По существу, первые электрические импульсы высокого напряжения обеспечивают образование пор, а последующие электрические импульсы более низкого напряжения обеспечивают проникновение полинуклеотида в клетку. В одном из объектов настоящего изобретения описаны стадии, которые позволяют достигать эффективности трансфекции мРНК в Т-клетки, составляющей >95%, и применение протокола электропорации для кратковременной экспрессии различных типов белков в Т-клетки. В частности, в изобретении описан способ трансформации Т-клетки, заключающийся в том, что приводят в контакт указанную Т-клетку с РНК и прилагают к Т-клетке последовательность быстрых импульсов, состоящую из:
а) одного электрического импульса, имеющего напряжение в диапазоне от 2250 до 3000 В на см, ширину импульса 0,1 мс и период повторения импульсов (импульсный интервал между электрическими импульсами) на стадии (а) и (б), составляющий от 0,2 до 10 мс;
б) одного электрического импульса, имеющего напряжение в диапазоне от 2250 до 3000 В, ширину импульса 100 мс и импульсный интервал между электрическим импульсом на стадии (б) и первым электрическим импульсом на стадии (в), составляющий 100 мс; и
в) 4 электрических импульсов, имеющих напряжение 325 В, ширину импульса 0,2 мс и импульсный интервал между каждым из 4 электрических импульсов, составляющий 2 мс.
В конкретном варианте осуществления изобретения способ трансформации Т-клетки заключается в том, что приводят в контакт указанную Т-клетку с РНК и прилагают к Т-клетке последовательность быстрых импульсов, состоящую из:
а) одного электрического импульса, имеющего напряжение 2250, 2300, 2350, 2400, 2450, 2500, 2550, 2400, 2450, 2500, 2600, 2700, 2800, 2900 или 3000 В на см, ширину импульса 0,1 мс и импульсный интервал между электрическими импульсами на стадии (а) и (б), составляющий 0,2, 0,5, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 мс;
б) одного электрического импульса, имеющего напряжение 2250, 2300, 2350, 2400, 2450, 2500, 2550, 2400, 2450, 2500, 2600, 2700, 2800, 2900 или 3000 В, ширину импульса 100 мс и импульсный интервал между электрическим импульсом на стадии (б) и первым электрическим импульсом на стадии (в), составляющий 100 мс; и
в) 4 электрических импульсов, имеющих напряжение 325 В, ширину импульса 0,2 мс и импульсный интервал между каждым из 4 электрических импульсов, составляющий 2 мс.
Под объем изобретения подпадают все величины, включенные в диапазон, указанный в настоящем описании. Среда для осуществления электропорации может представлять собой любую приемлемую среду, известную в данной области. Предпочтительно среда для осуществления электропорации имеет проводимость в диапазоне, составляющем от 0,01 до 1,0 мСм.
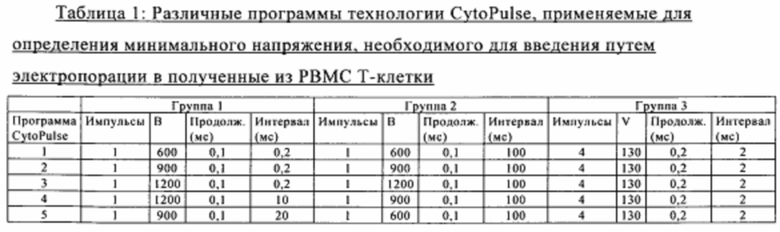
Неаллореактивные Т-клетки:
Хотя способ, предлагаемый в изобретении, можно осуществлять in vivo, например, в качестве компонента генной терапии, путем применения вирусных векторов, мишенью которых являются Т-клетки в кровотоке, которые должны включать генные последовательности, экспрессирующие специфическую редко расщепляющую эндонуклеазу, наряду с другими генными последовательностями, экспрессирующими, например, CAR, способ, предлагаемый в изобретении, в большей степени предназначен для практического применения ex vivo с использованием культивируемых Т-клеток, которые можно получать из организма пациентов или доноров. Сконструированные Т-клетки для применения ex vivo можно либо повторно имплантировать пациенту, из организма которого они получены, в качестве компонента аутологичного лечения, либо применять в качестве компонента аллогенного лечения. В указанном последнем случае предпочтительно также создавать неаллореактивные клетки для гарантии соответствующего приживления трансплантата. Таким образом, способ, предлагаемый в изобретении, может включать дополнительные стадии получения Т-клеток донора и инактивации их генов, участвующих в распознавании ГКГС или являющихся мишенями иммуносупрессорных лекарственных средств, которые описаны, например, в WO 2013/176915.
Т-клеточные рецепторы (TCR) представляют собой рецепторы клеточной поверхности, которые участвуют в активации Т-клеток в ответ на презентацию антигена. TCR, как правило, состоит из двух цепей, альфа и бета, которые в результате сборки образуют гетеродимер и которые ассоциированы с субъединицами, участвующими в трансдукции CD3, с образованием комплекса Т-клеточного рецептора, присутствующего на клеточной поверхности. Каждая альфа- и бета-цепь TCR состоит из иммуноглобулинподобной N-концевой вариабельной (V) и константной (С) области, гидрофобного трансмембранного домена и короткой цитоплазматической области. Также как и в молекулах иммуноглобулинов, вариабельная область альфа- и бета-цепей образуется путем рекомбинации V(D)J, что приводит к большому разнообразию антигенных специфичностей в популяции Т-клеток. Однако в отличие от иммуноглобулинов, которые распознают интактный антиген, Т-клетки активируются процессированными пептидными фрагментами в сочетании с молекулой ГКГС, вносящей дополнительное измерение в распознавание антигена Т-клетками, называемое рестрикцией по ГКГС. Распознавание несоответствий ГКГС между донором и реципиентом посредством Т-клеточного рецептора приводит к пролиферации Т-клеток и потенциальному развитию реакции «трансплантат-против-хозяина» (GVHD). Установлено, что нормальная поверхностная экспрессия TCR зависит от скоординированного синтеза и сборки всех семи компонентов комплекса (Ashwell и Klusner, 1990). Инактивация TCR-альфа или TCR-бета может приводить к элиминации TCR с поверхности Т-клеток, что препятствует распознаванию аллоантигенов и, как следствие, GVHD.
Кроме того, согласно изобретению приживление трансплантата Т-клеток можно улучшать путем инактивации по меньшей мере одного гена, кодирующего компонент TCR. TCR становится нефункциональным в клетках в результате инактивации гена TCR-альфа и/или гена(ов) TCR-бета.
Касательно применения системы Cas9/CRISPR при создании изобретения были установлены соответствующие последовательности-мишени в 3 экзонах, кодирующих TCR, что позволяло снижать токсичность в отношении живых клеток, сохраняя при этом эффективность расщепления. Предпочтительные последовательности-мишени представлены в таблице 2 (+ обозначает более низкое относительное содержание TCR-негативных клеток, ++ обозначает промежуточное относительное содержание, +++ обозначает более высокое относительное содержание).
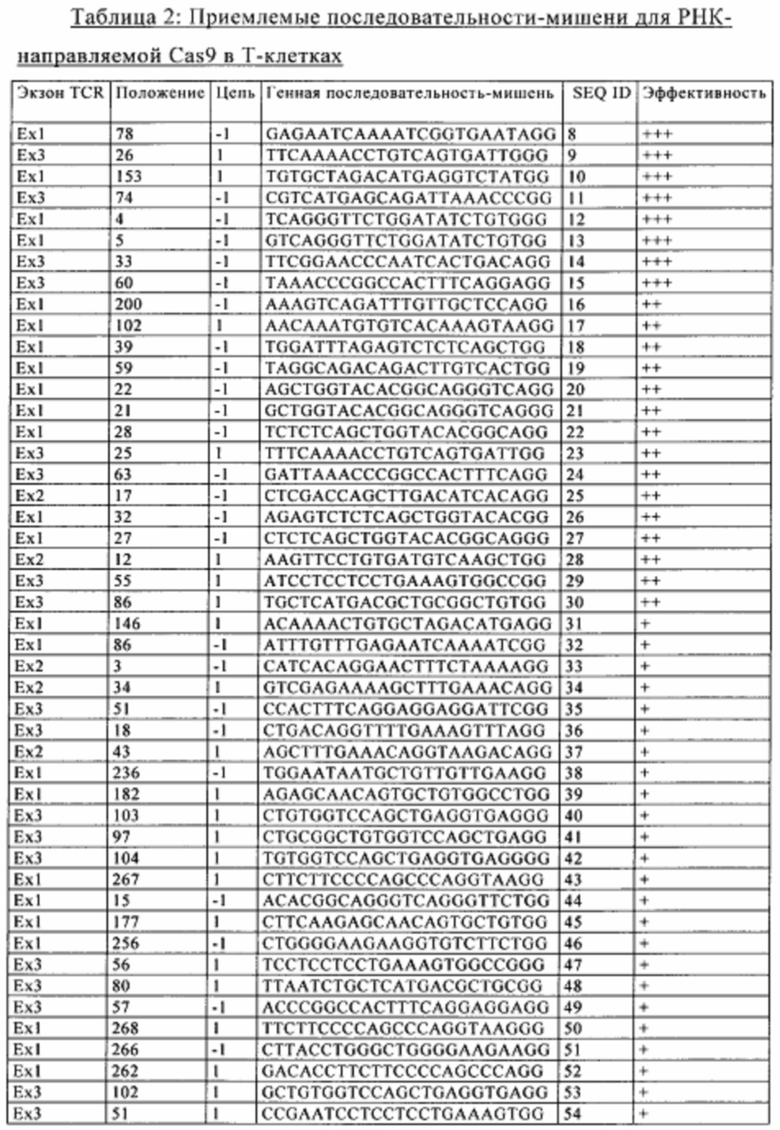
Антигены ГКГС также представляют собой белки, которые играют основную роль при реакциях на трансплантацию. Отторжение опосредуется Т-клетками, взаимодействующими с антигенами гистосовместимости на поверхности имплантируемых тканей, и самую большую группу указанных антигенов представляют собой антигены главного комплекса гистосовместимости (ГКГС). Эти белки экспрессируются на поверхности клеток всех высших позвоночных животных и в человеческих клетках их обозначают как HLA-антигены (человеческие лейкоцитарные антигены). Подобно TCR, белки ГКГС играют ключевую роль в стимуляции Т-клеток. Антигенпрезентирующие клетки (часто дендритные клетки) экспонируют пептиды, которые являются продуктами расщепления чужеродных белков, на клеточной поверхности на ГКГС. В присутствии костимулирующего сигнала Т-клетка становится активированной и может воздействовать на клетку-мишень, которая также экспонирует комплекс, включающий такой же пептид/ГКГС. Например, мишенью стимулированной Т-клетки-хелпера может являться макрофаг, экспонирующий антиген в сочетании с его ГКГС, или цитотоксическая Т-клетка (CTL) может воздействовать на инфицированную клетку, экспонирующую чужеродные вирусные пептиды.
Таким образом, для получения Т-клеток, обладающих меньшей аллореактивностью, способ, предлагаемый в изобретении, может дополнительно включать стадию инактивации или мутации гена HLA.
Кластер генов HLA класса I у человека включает три основных локуса, В, С и А, а также несколько минорных локусов. Кластер генов HLA класса II включает также три основных локуса, DP, DQ и DR, и кластеры генов как класса I, так и класса II являются полиморфными, в том плане, что в популяции присутствуют несколько различных аллелей генов как класса I, так и класса II. Существует также несколько вспомогательных белков, которые также играют роль в функционировании HLA. Субъединицы Tap1 и Тар2 являются компонентами транспортного комплекса ТАР, который играет важную роль для «загрузки» пептидных антигенов в комплекс HLA класса I, а протеосомные субъединицы LMP2 и LMP7 играют роль в протеолитическом расщеплении антигенов с образованием пептидов для экспонирования на HLA. Было установлено, что уменьшение количества LMP7 приводит к уменьшению количества молекул ГКГС класса I на клеточной поверхности, вероятно из-за отсутствия стабилизации (Fehling и др., Science 265, 1999, сс. 1234-1237). Помимо ТАР и LMP известен ген тапасина, продукт которого формирует мостик между комплексом ТАР и цепями HLA класса I и повышает «загрузку» пептидами. Уменьшение уровней тапасина приводит к возникновению клеток с нарушенной сборкой ГКГС класса I, сниженным уровнем экспрессии на клеточной поверхности ГКГС класса I и нарушенным иммунным ответом (Grandea и др., Immunity 13, 2000, сс. 213-222 и Garbi и др., Nat. Immunol. 1, 2000, сс. 234-238). Любой из указанных выше генов может быть инактивирован в контексте настоящего изобретения, что описано, например, в WO 2012/012667.
Таким образом, согласно некоторым вариантам осуществления изобретения способ, предлагаемый в изобретении, предусматривает также инактивацию по меньшей мере одного гена, выбранного из группы, состоящей из RFXANK, RFX5, RFXAP, ТАР1, ТАР2, ZXDA, ZXDB и ZXDC. Для достижения инактивации можно, например, использовать геномную модификацию, более конкретно, включающую экспрессию в Т клетке редко расщепляющей эндонуклеазы, обладающей способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, выбранного из группы, состоящей из RFXANK, RFX5, RFXAP, ТАР1, ТАР2, ZXDA, ZXDB и ZXDC.
Активация и размножение Т-клеток
Способ, предлагаемый в изобретении, может включать дополнительную стадию активации и/или размножения Т-клетки(ок). Это можно осуществлять до или после генетической модификации Т-клетки(ок) с использованием методов, описанных, например, в US 6352694; 6534055; 6905680; 6692964; 5858358; 6887466; 6905681; 7144575; 7067318; 7172869; 7232566; 7175843; 5883223; 6905874; 6797514; 6867041; и публикации заявки на патент США №20060121005. Согласно указанным методам Т-клетки, предлагаемые в изобретении, можно размножать путем приведения их в контакт с поверхностью, к которой прикреплен агент, который стимулирует сигнал, ассоциированный с комплексом CD3/TCR, и лиганд, который стимулирует костимулирующую молекулу на поверхности Т-клеток.
В частности, Т-клеточные популяции можно стимулировать in vitro, например, путем приведения их в контакт с антителом к CD3 или его антигенсвязывающим фрагментом или с антителом к CD2, иммобилизованном на поверхности, или путем приведения в контакт с активатором протеинкиназы С (например, бриостатином) в сочетании с кальциевым ионофором. Для костимуляции вспомогательной молекулы на поверхности Т-клеток применяют лиганд, который связывается с вспомогательной молекулой. Например, популяцию Т-клеток можно приводить в контакт с антителом к CD3 и антителом к CD28 в условиях, пригодных для стимуляции пролиферации Т-клеток. Для стимуляции пролиферации либо CD4+-Т-клеток, либо CD8+-Т-клеток применяют антитело к CD3 и антитело к CD28. Например, агенты, обеспечивающие каждый сигнал, могут присутствовать в растворе или быть сшитыми с поверхностью. Как должно быть очевидно обычным специалистам в данной области, соотношение частиц и клеток может зависеть от размера частиц относительно клетки-мишени. В других вариантах осуществления настоящего изобретения клетки, такие как Т-клетки, объединяют с покрытыми агентом гранулами, затем разделяют гранулы и клетки, после чего клетки культивируют. В альтернативном варианте осуществления изобретения перед культивированием покрытые агентом гранулы и клетки не разделяют, а культивируют вместе. Белки клеточной поверхности можно лигировать, давая возможность контактировать парамагнитным гранулам, с которыми связаны антитела к CD3 и к CD28 (3×28 гранул), с Т-клетками. В одном из вариантов осуществления изобретения клетки (например, 4-10 Т-клеток) и гранулы (например, парамагнитные гранулы DYNABEADS® М-450 CD3/CD28 Т в соотношении 1:1) объединяют в буфере, предпочтительно ЗФР (без двухвалентных катионов, таких как кальций и магний). И в этом случае обычным специалистам в данной области должно быть очевидно, что можно применять любые концентрации клеток. Смесь можно культивировать в течение промежутка времени, составляющего от нескольких часов (примерно 3 ч) до примерно 14 дней, или в течение любых целых величин часов между указанными значениями. В другом варианте осуществления изобретения смесь можно культивировать в течение 21 дня. Условия, пригодные для культивирования Т-клеток, включают соответствующие среды (например, минимальная поддерживающая среда или среда RPMI 1640, или X-vivo 5, (фирма Lonza)), которые могут содержать факторы, необходимые для пролиферации и жизнеспособности, включая сыворотку (например, фетальную бычью или человеческую сыворотку), интерлейкин-2 (IL-2), инсулин, IFN-g, 1L-4, 1L-7, GM-CSF, -10, -2, 1L-15, TGFp и TNF, или любые другие добавки для роста клеток, известные специалисту в данной области. Другие добавки для роста клеток включают (но, не ограничиваясь только ими) поверхностно-активное вещество, плазманат и восстановители, такие как N-ацетилцистеин и 2-меркаптоэтанол. Среды могут включать RPMI 1640, A1M-V, DMEM, MEM, а-МЕМ, F-12, Х-Vivo 1 и Х-Vivo 20, Optimizer с добавлением аминокислот, пирувата натрия и витаминов, представлять собой бессывороточные среды или среды, дополненные сывороткой (или плазмой) в соответствующем количестве или определенным набором гормонов и/или цитокином(ами) в количестве, достаточном для роста и размножения Т-клеток. Антибиотики, например, пенициллин и стрептомицин, включают только в экспериментальные культуры, но не включают в культуры клеток, предназначенные для вливания индивидууму. Указанные требуемые клетки выращивают в условиях, необходимых для поддержания роста, например, при соответствующей температуре (например, 37°С) и атмосфере (например, воздух плюс 5% СО2). Т-клетки, которых стимулировали в течение различных промежутков времени, могут иметь различные характеристики.
В другом конкретном варианте осуществления изобретения указанные клетки можно размножать путем совместного культивирования с тканью или клетками. Указанные клетки можно размножать также in vivo, например, в крови индивидуума после введения указанной клетки индивидууму.
Терапевтические применения
Т-клетки, которые можно получать согласно настоящему изобретению, предназначены для применения в качестве лекарственного средства и, в частности, для лечения среди прочего рака, инфекций (таких как вирусные инфекции) или иммунных заболеваний у пациента, который нуждается в этом. Таким образом, в настоящем изобретении предложены сконструированные Т-клетки для применения в качестве лекарственного средства. В частности, в настоящем изобретении предложены сконструированные Т-клетки для применения для лечения рака, такого как лимфома, или вирусной инфекции. Предложены также композиции, в частности, фармацевтические композиции, которые содержат по меньшей мере одну сконструированную Т-клетку, предлагаемую в настоящем изобретении. В некоторых вариантах осуществления изобретения композиция может содержать популяцию сконструированных Т-клеток, предлагаемых в настоящем изобретении.
Лечение может быть облегчающим симптомы, излечивающим или профилактическим. Оно может представлять собой либо компонент аутологичной иммунотерапии, либо компонент аллогенной иммунотерапии. Под аутологичным лечением подразумевается, что клетки, клеточная линия или популяция клеток, которые применяют для лечения пациентов, имеют происхождение из указанного пациента или из человеческого лейкоцитарного антигена (HLA) совместимого донора. Под аллогенным лечением подразумевают, что клетки или популяция клеток, которые применяют для лечения пациентов, имеют происхождение не из указанного пациента, а из донора.
Изобретение наиболее пригодно для аллогенной иммунотерапии, поскольку оно позволяет превращать Т-клетки, как правило, полученные из доноров, в неаллореактивные клетки. Это можно осуществлять с использованием стандартных протоколов и повторять столько раз, сколько является необходимым. Полученные в результате модифицированные Т-клетки можно объединять и вводить одному или нескольким пациентам, получая поступающий в продажу терапевтический продукт.
Лечение предназначено, прежде всего, для пациентов с диагностированным раком. Рак предпочтительно включает лейкозы и лимфомы, представляющие собой жидкие опухоли, но могут рассматриваться также и солидные опухоли. Типы рака, подлежащие лечению с помощью генетически сконструированных Т-клеток, предлагаемых в изобретении, включают (но, не ограничиваясь только ими) карциному, бластому и саркому и некоторые лейкозы или лимфоидные злокачественные заболевания, доброкачественные и злокачественные опухоли и злокачественные заболевания, такие как саркомы, карциномы и меланомы. Включены также опухоли/типы рака взрослых и детские опухоли/типы рака.
Лечение можно осуществлять в комбинации с одной или несколькими терапиями, выбранными из группы, включающей терапию с использованием антител, химиотерапию, лечение цитокинами, лечение дендритными клетками, генную терапию, гормональную терапию, лечение лазерным излучением и лучевую терапию.
Согласно некоторым вариантам осуществления изобретения Т-клетки, предлагаемые в изобретении, могут подвергаться существенному размножению in vivo после введения пациенту и могут сохраняться в общей воде организма в течение пролонгированных периодов времени, предпочтительно в течение недели, более предпочтительно 2 недель, еще более предпочтительно по меньшей мере одного месяца. Хотя ожидается, что Т-клетки, предлагаемые в изобретении, могут сохраняться в течение указанных периодов, требуемая продолжительность их жизни в организме пациента не превышает одного года, предпочтительно 6 месяцев, более предпочтительно 2 месяцев и еще более предпочтительно одного месяца.
Введение клеток или популяции клеток, предлагаемых в настоящем изобретении, можно осуществлять любым приемлемым путем, например, с помощью аэрозольной ингаляции, инъекции, приема внутрь, трансфузии, имплантации или трансплантации. Композиции, указанные в настоящем описании, можно вводить пациенту подкожно, внутрикожно, внутрь опухоли, интранодально, интрамедуллярно, внутримышечно, путем внутривенной или внутрилимфатической инъекции или внутрибрюшинно. В одном из вариантов осуществления изобретения содержащие клетки композиции, предлагаемые в настоящем изобретении, предпочтительно вводят путем внутривенной инъекции.
Введение клеток или популяции клеток может предусматривать введение 104-109 клеток на кг веса тела, предпочтительно 105-106 клеток/кг веса тела, включая все целые величины количеств клеток в указанных диапазонах. Клетки или популяцию клеток можно вводить в виде одной или нескольких доз. В другом варианте осуществления изобретения указанное эффективное количество клеток вводят в виде однократной дозы. В другом варианте осуществления изобретения указанное эффективное количество клеток вводят в виде нескольких доз в течение определенного периода времени. Режим введения регламентируется лечащим врачом и зависит от клинического состояния пациента. Клетки или популяцию клеток можно получать из любого источника, такого как банк крови или донор. Поскольку индивидуальные потребности варьируются, определение оптимальных диапазонов эффективных количеств данного типа клеток для конкретного заболевания или конкретных состояний осуществляет специалист в данной области. Эффективное количество означает количество, которое обладает терапевтической или профилактической пользой. Введенная доза должна зависеть от возраста, состояния здоровья и веса реципиента, типа сопутствующего лечения, если оно проводится, частоты обработок и природы требуемого воздействия.
В других вариантах осуществления изобретения клетки или композицию, содержащую указанные клетки, в указанном эффективном количестве вводят парентерально. Указанное введение может представлять собой внутривенное введение. Указанное введение может представлять собой непосредственную инъекцию в опухоль.
В некоторых вариантах осуществления изобретения клетки вводят пациенту в сочетании с (например, до, одновременно или после) любым количеством соответствующих лечебных подходов, включая (но, не ограничиваясь только ими) лечение агентами, такими как средства противовирусной терапии цидофовир и интерлейкин-2, цитарабин (который обозначают также как ARA-C) или натализтимаб в случае лечения страдающих MS пациентов, или лечение эфализтимабом в случае страдающих псориазом пациентов или другие пути лечения пациентов, страдающих PML. В других вариантах осуществления изобретения Т-клетки, предлагаемые в изобретении, можно применять в комбинации с химиотерапией, облучением, иммуносупрессорными агентами, такими как циклоспорин, азатиоприн, метотрексат, микофенолят и FK506, антитела, или другими средствами для иммуноабляции, такими как CAMPATH, антитела к CD3, или другими терапиями на основе антител, цитоксина, флударабина, циклоспорина, FK506, рапамицина, микофеноловой кислоты, стероидов, FR901228, цитокинов и облучения. Указанные лекарственные средства ингибируют либо кальцийзависимую фосфатазу кальцийнеурин (циклоспорин и FK506), либо ингбируют киназу p70S6, важную для индуцируемой фактором роста передачи сигналов (рапамицин) (Liu и др., Cell 66, 1991, сс. 807-815; Henderson и др., Immun. 73, 1991, сс. 316-321; Bierer и др., Citrr. Opin.mm n. 5, 1993, сс. 763-773). В другом варианте осуществления изобретения содержащую клетки композицию, предлагаемую в настоящем изобретении, вводят пациенту в сочетании с (например, до, одновременно или после) трансплантацией костного мозга, Т-клеточной аблятивной терапией с использованием либо химиотерапевтических средств, таких как флударабин, либо наружной дистанционной лучевой терапии (XRT), циклофосфамида, либо антител, таких как OKT3 или CAMPATH. В другом варианте осуществления изобретения содержащие клетки композиции, предлагаемые в настоящем изобретении, вводят после В-клеточной аблятивной терапии, например, с использованием агентов, которые взаимодействуют с CD20, например, ритуксана. Например, в одном из вариантов осуществления изобретения индивидуумов можно подвергать стандартному лечению с использованием химиотерапевтических средств в высоких дозах с последующей трансплантацией стволовых клеток периферической крови. В некоторых вариантах осуществления изобретения после трансплантации индивидуумам вводят путем инфузии размноженные генетически сконструированные Т-клетки, предлагаемые в настоящем изобретении. В дополнительном варианте осуществления изобретения размноженные клетки вводят до или после хирургического вмешательства.
Объектами изобретения являются также способы лечения пациента, который нуждается в этом, заключающиеся в том, что а) получают по меньшей мере одну сконструированную Т-клетку, предлагаемую в настоящем изобретении, предпочтительно популяцию указанных Т-клеток; и б) вводят указанную Т-клетку или популяцию указанному пациенту.
Объектами изобретения являются также способы приготовления лекарственного средства с использованием по меньшей мере одной сконструированной Т-клетки, предлагаемой в настоящем изобретении, и предпочтительно популяции указанных Т-клеток. Таким образом, в настоящем изобретении предложено применение по меньшей мере одной сконструированной Т-клетки, предлагаемой в настоящем изобретении, и предпочтительно популяции указанных Т-клеток, для производства лекарственного средства. Предпочтительно указанное лекарственное средство предназначено для применения для лечения рака, такого как лимфома, или вирусной инфекции.
Другие определения
- Аминокислотные остатки в полипептидной последовательности обозначают в настоящем описании с помощью однобуквенного кода, согласно которому, например, Q обозначает Gln или остаток глутамина, R обозначает Arg или остаток аргинина и D обозначает Asp или остаток аспарагиновой кислоты.
- Аминокислотная замена обозначает замену одного аминокислотного остатка на другой, например, замена остатка аргинина на остаток глутамина в пептидной последовательности представляет собой аминокислотную замену.
- Нуклеотиды обозначают следующим образом: однобуквенный код используют для обозначения основания нуклеозида: а обозначает аденин, t обозначает тимин, с обозначает цитозин и g обозначает гуанин. Для вырожденных нуклеотидов r обозначает g или а (пуриновые нуклеотиды), k обозначает g или t, s обозначает g или с, w обозначает а или t, m обозначает а или с, у обозначает t или с (пиримидиновые нуклеотиды), d обозначает g, а или t, v обозначает g, а или с, b обозначает g, t или с, h обозначает a, t или с и n обозначает g, a, t или с.
- В контексте настоящего описания понятия «нуклеиновая кислота» или «полинуклеотиды» относятся к нуклеотидам и/или полинуклеотидам, таким как дезоксирибонуклеиновая кислота (ДНК) или рибонуклеиновая кислота (РНК), олигонуклеотидам, фрагментам, созданным с помощью полимеразной цепной реакции (ПЦР), и фрагментам, созданным путем лигирования, расщепления, обработки эндонуклеазами и обработки экзонуклеазами. Молекулы нуклеиновой кислоты могут состоять из мономеров, которые являются встречающимися в естественных условиях нуклеотидами (такими как ДНК или РНК) или аналогами встречающихся в естественных условиях нуклеотидов (например, энантиомерными формами встречающихся в естественных условиях нуклеотидов), или представляют собой их комбинацию. Модифицированные нуклеотиды могут иметь изменения в сахарных фрагментах и/или во фрагментах, содержащих пиримидиновые или пуриновые основания. Модификации сахаров включают, например, замену одной или нескольких гидроксильных групп на галогены, алкильные группы, амины и азидогруппы, или сахара могут быть функционализированы до простых или сложных эфиров. Кроме того, весь сахарный фрагмент может быть заменен стерически и электронно сходными структурами, такими как аза-сахара и карбоциклические аналоги сахаров. Примерами модификаций во фрагменте, представляющем собой основание, могут служить алкилированные пурины и пиримидины, ацилированные пурины или пиримидины, или другие хорошо известные гетероциклические заместители. Мономеры нуклеиновой кислоты могут быть сшиты фосфодиэфирными связями или аналогами таких связей. Нуклеиновые кислоты могут быть одноцепочечными или двухцепочечными.
- Под «полинуклеотидом, который содержит расположенные последовательно первую область, гомологичную последовательностям, расположенным в обратном направлении относительно указанного двухцепочечного разрыва, последовательность, подлежащую встраиванию в геном указанной клетки, и вторую область, гомологичную последовательностям, расположенным в прямом направлении относительно указанного двухцепочечного разрыва» подразумевают ДНК-конструкцию или матрикс, содержащую/содержащий первый и второй участки, гомологичные 5'- и 3'-областям в ДНК-мишени in situ. ДНК-конструкция содержит также третий участок, расположенный между первым и вторым участком, который обладает некоторой гомологией с соответствующей ДНК-последовательностью in situ или в другом варианте не обладает никакой гомологией с 5'- и 3'-областями ДНК-мишени in situ. После расщепления ДНК-мишени стимулируется случай гомологичной рекомбинации между геномом, который содержит ген-мишень, входящий в представляющий интерес локус, и указанным матриксом, в котором геномная последовательность, содержащая ДНК-мишень, заменена на третий участок матрикса и вариабельный участок первого и второго участка указанного матрикса.
- Под «ДНК-мишенью» «последовательностью ДНК-мишенью», «последовательностью нуклеиновой кислоты-мишенью, «последовательностью-мишенью» или «сайтом процессирования» подразумевают полинуклеотидную последовательность, которая может являться мишенью и процессироваться редко расщепляющей эндонуклеазой согласно настоящему изобретению. Эти понятия относятся к специфической локализации ДНК, предпочтительно геномной локализации в клетке, но также и к участку генетического материала, который может существовать независимо от основной массы генетического материала, такому как плазмиды, эписомы, вирус, транспозоны, или находиться в органеллах, таких как митохондрии (но, не ограничиваясь только указанным). Примерами РНК-направляемых последовательностей-мишеней являются (но, не ограничиваясь только ими) такие геномные последовательности, которые могут гибридизоваться с гидовой РНК, которая направляет РНК-направляемую эндонуклеазу к требуемому локусу.
- Под «вектором для доставки» или «векторами для доставки» подразумевают любой вектор для доставки, который можно применять согласно настоящему изобретению для приведения в контакт с клеткой (т.е. для «контактирования») или доставки внутрь клетки или в субклеточные компартменты (т.е. для «интродукции») агентов/химических веществ и молекул (белков или нуклеиновых кислот), требующихся для осуществления настоящего изобретения. К ним относятся (но, не ограничиваясь только ими) липосомальные векторы для доставки, вирусные векторы для доставки, векторы для доставки лекарственного средства, химические носители, полимерные носители, липоплексы, полиплексы, дендримеры, микропузырьки (ультразвуковые контрастные агенты), наночастицы, эмульсии или другие пригодные векторы для переноса. Такие векторы для доставки позволяют доставлять молекулы, химические вещества, макромолекулы (гены, белки) или другие векторы, такие как плазмиды, или проникающие пептиды. В этих случаях векторы для доставки представляют собой молекулы-носители.
- Понятия «вектор» или «векторы» относятся к молекуле нуклеиновой кислоты, обладающей способностью транспортировать другую нуклеиновую кислоту, с которой она сцеплена. К «векторам», применяемым согласно настоящему изобретению, относятся (но, не ограничиваясь только ими) вирусный вектор, плазмида, РНКовый вектор или молекула линейной или кольцевой ДНК или РНК, которые могут состоять из хромосомальных, нехромосомальных, полусинтетических или синтетических нуклеиновых кислот. Предпочтительными векторами являются векторы, которые способны автономно реплицировать (эписомальный вектор) и/или экспрессировать нуклеиновые кислоты, с которыми они сцеплены (экспрессионные векторы). Специалистам в данной области известно большое количество пригодных векторов, которые поступают в продажу.
К вирусным векторам относятся ретровирус, аденовирус, парвовирус (например, аденоассоциированные вирусы), коронавирус, РНКовые вирусы с негативной цепью, такие как ортомиксовирус (например, вирус гриппа), рабдовирус (например, вирус бешенства и вирус везикулярного стоматита), парамиксовирус (например, вирус кори и вирус Сендай), РНКовые вирусы с позитивной цепью, такие как пикорнавирус и альфавирус, и вирусы с двухцепочечной ДНК, включая аденовирус, вирус герпеса (например, вирус герпеса простого типов 1 и 2, вирус Эпшейна-Барра, цитомегаловирус) и вирус оспы (например, вирус коровьей оспы, вирус оспы птиц и вирус оспы канарейки). Другие вирусы включают, например, вирус Норвалк, тогавирус, флавивирус, реовирусы, паповирус, гепаднавирус и вирус гепатита. Примерами ретровирусов являются: вирус птичьего лейкоза-саркомы, вирусы млекопитающих С-типа, В-типа, вирусы D-типа, группа HTLV-BLV, лентивирус, спумавирус (Coffin J.М., Retroviridae: The viruses and their replication, в: Fundamental Virology, 3-е изд., под ред. В.N. Fields и др., изд-во Lippincott-Raven Publishers, Philadelphia, 1996).
- Под «лентивирусным вектором» подразумевают лентивирусные векторы на основе ВИЧ, которые являются очень перспективными с точки зрения доставки гена благодаря их относительно большой способности к упаковке, пониженной иммуногенности и их способности стабильно трансдуцировать с высокой эффективностью большой спектр различных типов клеток. Лентивирусные векторы, как правило, создают посредством кратковременной трансфекции клеток-продуцентов тремя (упаковывающая плазмида, оболочечная плазмида и плазмида переноса) или большим количеством плазмид. Подобно ВИЧ, лентивирусные векторы проникают в клетку-мишень посредством взаимодействия гликопротеинов вирусной поверхности с рецепторами на клеточной поверхности. После проникновения вирусная РНК подвергается обратной транскрипции, опосредуемой комплексом вирусной обратной транскриптазы. Продуктом обратной транскрипции является двухцепочечная линейная вирусная ДНК, которая служит субстратом для вирусной интеграции в ДНК инфицированных клеток. Понятие «интегрирующие лентивирусные векторы (или LV)» обозначает, например (но, не ограничиваясь только ими), такие векторы, которые способны интегрироваться в геном клетки-мишени. В противоположность этому, понятие «неинтегрирующие лентивирусные векторы (или NILV)» обозначает эффективные векторы для доставки генов, которые не интегрируются в геном клетки-мишени под действием вирусной интегразы.
- Векторы для доставки и векторы можно объединять или комбинировать с любыми методами клеточной пермеабилизации, такими как сонопорация или электропорация, или вариантами этих методов.
- Под клеткой или клетками подразумевают любые эукариотические живые клетки, первичные клетки и клеточные линии, выведенные из указанных организмов для получения культур in vitro.
- Под «первичной клеткой» или «первичными клетками» подразумевают клетки, взятые непосредственно из живой ткани (т.е. материал, полученный путем биопсии) и подготовленные для выращивания in vitro, в популяции которых имело место лишь небольшое количество удвоений, и, следовательно, они являются более репрезентативными с точки зрения основных функциональных компонентов и характеристик тканей, из которых они выведены, по сравнению с непрерывно поддерживаемыми или искусственно иммортализованными онкогенными клеточными линиями.
Примерами таких клеточных линий могут являться (но, не ограничиваясь только ими) клеточные линии, выбранные из группы, состоящей из клеток СНО-K1, клеток HEK293; клеток Сасо2; клеток U2-OS; клеток NIH 3Т3; клеток NSO; клеток SP2; клеток CHO-S; клеток DG44; клеток K-562, клеток U-937; клеток MRC5; клеток IMR90; клеток Jurkat; клеток HepG2; клеток HeLa; клеток НТ-1080; клеток НСТ-116; клеток Hu-h7; клеток Huvec; клеток Molt.
Все эти клеточные линии можно модифицировать с помощью способа, предлагаемого в настоящем изобретении, создавая модели клеточных линий для получения, экспрессии, количественной оценки, обнаружения, изучения представляющего интерес гена или белка; эти модели можно применять также скрининга представляющих интерес биологически активных молекул для исследовательских и промышленных целей в различных областях, например, таких как (но, не ограничиваясь только ими) химия, биотопливо, терапевтические средства и агрономия.
- Под «мутацией» подразумевают замену, делецию или инсерцию одного/одной, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати, двенадцати, тринадцати, четырнадцати, пятнадцати, двадцати, двадцати пяти, тридцати, сорока, пятидесяти или большего количества нуклеотидов/аминокислот в полинуклеотидной (кДНК, ген) или полипептидной последовательности. Мутация может влиять на кодирующую последовательность гена или его регуляторную последовательность. Она может влиять также на структуру геномной последовательности или структуру/стабильность кодируемой мРНК.
- Под «вариантом(ами)» подразумевают вариант повтора, вариант, ДНК-связывающий вариант, вариант TALE-нуклеазы, вариант полипептида, полученный в результате мутации или замены по меньшей мере одного остатка в аминокислотной последовательности родительской молекулы.
- Под «функциональным вариантом» подразумевают обладающий каталитической активностью мутант белка или домена белка; такой мутант может обладать такой же активностью, что и родительский белок или домен белка, или дополнительными свойствами, или более высокой или более низкой активностью.
- Под «геном» подразумевают основную единицу наследственности, состоящую из сегмента ДНК, расположенную линейно вдоль хромосомы, которая кодирует специфический белок или сегмент белка. Ген, как правило, включает промотор, 5'-нетранслируемую область, одну или несколько кодирующих последовательностей (экзоны), необязательно интроны, 3'-нетранслируемую область. Ген может содержать также терминатор, энхансеры и/или элементы, вызывающие «молчание».
- В контексте настоящего описания понятие «локус» означает специфическую физическую локализацию последовательности ДНК (например, гена) на хромосоме. Понятие «локус» может относиться к специфической физической локализации последовательности-мишени редко расщепляющей эндонуклеазы на хромосоме. Такой локус может содержать последовательность-мишень, которая распознается и/или расщепляется редко расщепляющей эндонуклеазой, предлагаемой в изобретении. Следует понимать, что представляющий интерес согласно настоящему изобретению локус может относиться не только к последовательности нуклеиновой кислоты, которая существует в основой массе генетического материала (т.е. в хромосоме) клетки, но также и к участку генетического материала, который может существовать независимо от основной массы генетического материала, такому как плазмиды, эписомы, вирус, транспозоны, или находиться в органеллах, таких как митохондрии (но не ограничиваясь только указанным).
- Понятие «расщепление» относится к разрыву ковалентно связанного каркаса полинуклеотида. Расщепление можно инициировать различными методами, включая (но, не ограничиваясь только ими) ферментативный или химический гидролиз фосфодиэфирной связи. Возможно как одноцепочечное расщепление, так и двухцепочечное расщепление, и двухцепочечное расщепление может иметь место в результате двух различных случаев одноцепочечного расщепления. Расщепление двухцепочечной ДНК, РНК или гибрида ДНК/РНК может приводить к образованию либо «тупых» концов, либо «шатающихся» концов.
- Под «слитым белком» подразумевают результат хорошо известного в данной области процесса, включающего соединение двух или большего количества генов, которые исходно кодируют разные белки или их части, трансляцию указанного «слитого гена», которая приводит к образованию одного полипептида с функциональными свойствами каждого из исходных белков.
- Понятие «идентичность» относится к идентичности последовательностей двух молекул нуклеиновых кислот или полипептидов. Идентичность можно оценивать путем сравнения положений в каждой из последовательностей, которые можно выравнивать для целей сравнения. Если положение в сравниваемой последовательности занято тем же самым основанием или той же самой аминокислотой, то молекулы являются идентичными в этом положении. Степень сходства или идентичности между двумя нуклеотидными или аминокислотными последовательностями является функцией количества идентичных или совпадающих нуклеотидов или аминокислот в соответствующих положениях нуклеотидных или аминокислотных последовательностей. Для расчета идентичности двух последовательностей можно использовать различные алгоритмы и/или программы сравнительного анализа первичной структуры последовательностей, включая FASTA или BLAST, которые доступны в виде компонента пакета программ анализа последовательностей GCG (Университет Висконсина, Мэдисон, шт. Висконсин), и их можно применять, например, с использованием задаваемых по умолчанию параметров. Например, можно рассматривать полипептиды, обладающие идентичностью, составляющей по меньшей мере 70, 85, 90, 95, 98 или 99%, с конкретными полипептидами, представленными в настоящем описании, и предпочтительно обладающие практически такими же функциями, а также полинуклеотиды, кодирующие такие полипептиды.
- Понятие «ингибирование» или «процесс ингибирования» экспрессии В2М означает, что экспрессия В2М в клетке снижена по меньшей мере на 1%, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50% по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 99% или 100%. Более конкретно, «ингибирование» или «процесс ингибирования» экспрессии В2М означает, что количество В2М в клетке снижено по меньшей мере на 1%, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50% по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 99% или 100%. Экспрессию или количество белка в клетке можно определять любыми приемлемыми методами, известными в данной области, такими как ELISA, иммуногистохимия, Вестерн-блоттинг или проточная цитометрия, используя специфические для В2М антитела. Указанные антитела поступают в продажу от различных поставщиков, таких как фирма Merck Millipore, Биллерика, шт. Массачусетс, США или фирма Abeam plc, Кембридж, Великобритания.
- Понятие «ингибирование» или «процесс ингибирования» экспрессии CIITA означает, что экспрессия CIITA в клетке снижена по меньшей мере на 1%, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50% по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 99% или 100%. Более конкретно, «ингибирование» или «процесс ингибирования» экспрессии CIITA означает, что количество CIITA в клетке снижено по меньшей мере на 1%, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50% по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 99% или 100%. Экспрессию или количество белка в клетке можно определять любыми приемлемыми методами, известными в данной области, такими как ELISA, иммуногистохимия, Вестерн-блоттинг или проточная цитометрия, используя специфические для CIITA антитела. Указанные антитела поступают в продажу от различных поставщиков, таких как фирма Abcam plc, Кембридж, Великобритания или фирма Santa Cruz Biotechnology, Inc., Санта-Круз, шт. Калифорния, США.
- Понятия «домен трансдукции сигнала» или «костимулирующий лиганд» относятся к молекуле на антигенпрезентирующей клетке, которая специфически связывается с когнатной костимулирующей молекулой на Т-клетке, обеспечивая тем самым сигнал, который в дополнение к первичному сигналу, обеспечиваемому, например, связыванием комплекса TCR/CD3 с молекулой ГКГС, загруженной пептидом, опосредует Т-клеточный ответ, включая (но, не ограничиваясь только ими) активацию пролиферации, дифференцировку и т.п. Костимулирующий лиганд может включать (но, не ограничиваясь только ими) CD7, В7-1 (CD80), В7-2 (CD86), PD-L1, PD-L2, 4-1BBL, OX40L, индуцибельный костимулирующий лиганд (ICOS-L), молекулу межклеточной адгезии (ICAM, CD30L, CD40, CD70, CD83, HLA-G, MICA, MICB, HVEM, рецептор лимфотоксина бета, 3/TR6, ILT3, ILT4, агонист или антитело, который/которое связывается с лигандом Толл-рецептора, и лиганд, который специфически связывается с В7-Н3. К костимулирующему лиганду относятся также среди прочего антитело, которое специфически связывается с костимулирующей молекулой, присутствующей на Т-клетке, такой как (но, не ограничиваясь только ими) CD27, CD28, 4-IBB, ОХ40, CD30, CD40, PD-1, ICOS, ассоциированный с функцией лимфоцитов антиген-1 (LFA-1), CD2, CD7, LTGHT, NKG2C, В7-Н3, лиганд, который специфически связывается с CD83.
Понятие «костимулирующая молекула» относится к когнатному связывающему партнеру на Т-клетке, который специфически связывается с костимулирующим лигандом, опосредуя тем самым костимулирующий ответ клетки, такой как (но, не ограничиваясь только им) пролиферация. Костимулирующие молекулы включают (но, не ограничиваясь только ими) молекулу ГКГС класса I, BTLA и лиганд Толл-рецептора. В контексте настоящего описания понятие «костимулирующий сигнал» относится к сигналу, который в сочетании с первичным сигналом, таким как сигнал, обусловленный сцеплением TCR/CD3, приводит к пролиферации Т-клеток и/или повышающей или понижающей регуляции имеющих решающее значение молекул.
- Понятие «биспецифическое антитело» относится к антителу, которое имеет сайты связывания для двух различных антигенов в одной молекуле антитела. Специалистам в данной области должно быть очевидно, что можно создавать другие молекулы, которые помимо структур канонического антитела имеют две специфичности связывания. Также очевидно, что связывание антигенов биспецифическими антителами может быть одновременным или последовательным. Биспецифические антитела можно получать с помощью химических методов (см., например, Kranz и др., Proc. Natl. Acad. Sci. USA 78, 1981Ю с. 5807), методов «полидом» (см. US 4474893) или методами рекомбинантной ДНК, которые все являются хорошо известными. В качестве примера, но не ограничивающего объем изобретения, каждый связывающий домен содержит по меньшей мере одну вариабельную область из тяжелой цепи антитела («VH» или» Н-область»), при этом VH-область первого связывающего домена специфически связывается с маркером лимфоцитов, таким как CD3, а VH-область второго связывающего домена специфически связывается с опухолевым антигеном.
- В контексте настоящего описания понятие «внеклеточный лигандсвязывающий домен» обозначает олиго- или полипептид, который обладает способностью связываться с лигандом. Предпочтительно домен должен обладать способностью взаимодействовать с молекулой клеточной поверхности. Например, можно выбирать внеклеточный лигандсвязывающий домен для распознавания лиганда, который функционирует в качестве маркера клеточной поверхности на клетках-мишенях, ассоциированных с конкретным болезненным состоянием. Так, примерами маркеров клеточной поверхности, которые могут функционировать в качестве лигандов, являются маркеры, ассоциированные с вирусными, бактериальными и паразитарными инфекциями, аутоиммунным заболеванием и раковыми клетками.
В контексте настоящего описания понятие «индивидуум» или «пациент» включает всех представителей царства животных, в том числе приматов кроме человека и человека.
В представленном выше описании изобретения изложены подход и способ его осуществления и применения, так что любой специалист в данной области способен осуществлять и применять его, эта возможность относится, прежде всего, к сущности изобретения, изложенной в прилагаемой формуле изобретения, которая представляет собой составную часть исходного описания.
Если в настоящем описании указаны численные пределы или численный диапазон, то они включают их границы. Кроме того, считается, что включены все значения и поддиапазоны, находящиеся в указанных пределах или в указанном численном диапазоне, как если бы они были специально указаны.
Имея общее описание настоящего изобретения, можно достигать более полного понимания после ознакомления с некоторыми конкретными примерами, которые представлены в настоящем описании только для целей иллюстрации и не направлены на ограничение объема, если специально не указано иное.
Примеры
Расщепление TALE-нуклеазами человеческого CIITA
мРНК, кодирующую TALE-нуклеазы, мишенью которых являются экзоны человеческого гена CIITA, заказывали на фирме Cellectis Bioresearch (8, rue de la Croix Jarry, 75013, Париж). Ниже в таблице 3 приведены последовательности-мишени, расщепляемые каждым из двух независимых элементов (обозначенные как полу-TALЕ-нуклеазы), каждый из которых содержит последовательность-повтор, созданную для связывания и расщепления в области между последовательностями-мишенями, состоящими из двух последовательностей длиной 17 пар оснований (обозначенные как полупоследовательности-мишени), которые разделены спейсером длиной 15 пар оснований. Поскольку экзоны 2 и 3 характерны для всех транскрипционных вариантов CIITA, то для экзонов 2 и 3 создавали две пары TALEN. Не предполагалось наличие никаких очевидных посторонних сайтов-мишеней в человеческом геноме для TALE-нуклеаз, для которых эти последовательности являлись мишенью.
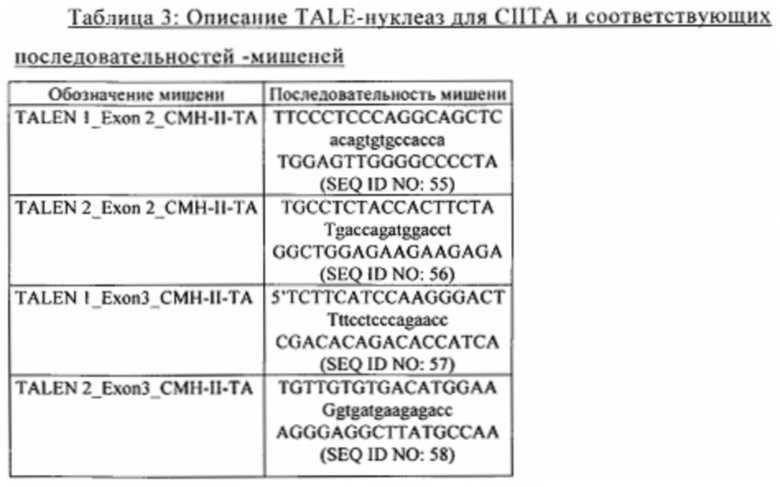
TALE-нуклеазы, расщепляющие человеческий β2m
мРНК, кодирующую TALE-нуклеазы, мишенью которых являются экзоны человеческого гена β2m, заказывали на фирме Cellectis Bioresearch (8, rue de la Croix Jarry, 75013, Париж). Ниже в таблице 4 приведены последовательности-мишени, расщепляемые каждым из двух независимых элементов (обозначенные как полу-TALE-нуклеазы), каждый из которых содержит последовательность-повтор, созданную для связывания и расщепления в области между последовательностями-мишенями, состоящими из двух последовательностей длиной 17 пар оснований (обозначенные как полупоследовательности-мишени), которые разделены спейсером длиной 15 пар оснований.

TALE-нуклеазы, расщепляющие гены человеческого TCR (TRAC и TRBC)
В человеческом геноме содержатся две функциональные бета-цепи Т-клеточного рецептора (TRBC1 и TRBC2). В процессе развития альфа/бета Т-лимфоцитов одна из указанных двух константных цепей отбирается в каждой клетке для сращивания (сплайсинга) с вариабельной областью TCR-бета и образованием функциональной полноразмерной бета-цепи. Ниже в таблице 5 представлены последовательности-мишени TRAC и двух TRBC и соответствующие им последовательности TALEN. Две TRBC-мишени выбраны в последовательностях, консервативных для TRBC1 и TRBC2, в результате чего соответствующая TALE-нуклеаза может одновременно расщеплять как TRBC1, так и TRBC2.
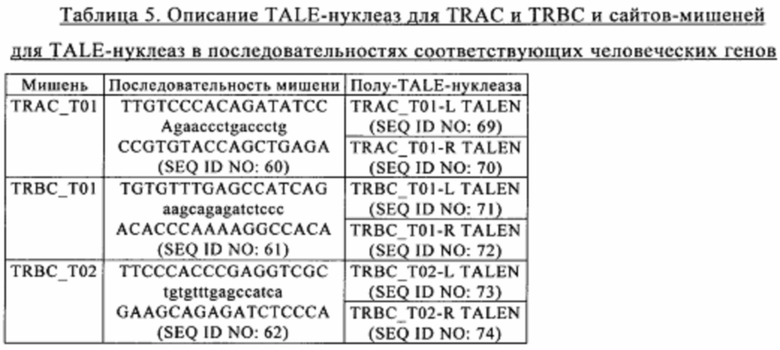
Создавали другие последовательности-мишени в генах TRAC и CD52, которые представлены в таблице 6.

Введение путем электропорации мРНК в очищенные Т-клетки, активированные с помощью технологии CytoPulse
После определения наиболее оптимальной программы CytoPulse, которая обеспечивала эффективное введение путем электропорации в Т-клетки ДНК, при создании изобретения оценивали возможность применения этого метода для введения путем электропорации мРНК.
5×106 очищенных Т-клеток, предварительно активированных в течение 6 дней с помощью PHA/IL2, ресуспендировали в буфере Т для электропорации клеток с помощью технологии CytoPulse (устройство BTX-Harvard) и осуществляли введение путем электропорации в 0,4-сантиметровых кюветах 10 мкг мРНК, кодирующей GFP, или 20 мкг плазмид, кодирующих GFP или pUC, используя предпочтительную программу CytoPulse, описанную в таблице 7.
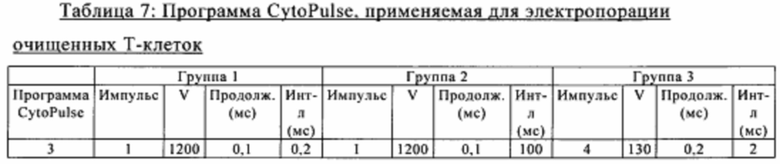
Через 48 ч после трансфекции клетки окрашивали с целью определения жизнеспособности (краситель eFluor-450) и определяли с помощью проточной цитометрии жизнеспособность клеток и процент живых GFP+-клеток.
Введение путем электропорации РНК в оптимальных условиях, определенных при создании изобретении, оказалось нетоксичным и позволяло осуществлять трансфекцию более чем 95% живых клеток.
В целом, исходя из всех полученных данных, установлено, что Т-клетки можно эффективно трансфецировать либо ДНК, либо РНК. В частности, трансфекция РНК не оказывала влияния на жизнеспособность клеток и позволяла получать однородные уровни экспрессии представляющего интерес применяемого для трансфекии гена в клеточной популяции.
Эффективную трансфекцию можно обеспечивать сразу после клеточной активации вне зависимости от применяемого метода активации (PHA/IL-2 или покрытие CD3/CD28 гранулы). При создании изобретения удалось достичь успешной трансфекции клеток через 72 ч после активации с эффективностью >95%. Кроме того, при использовании такого же протокола электропорации можно обеспечивать эффективную трансфекцию Т-клеток также и после оттаивания и активации.
Введение путем электропорации мРНК в первичные человеческие Т-клетки для функциональной экспрессии TALE-нуклеазы
После установления того, что введение мРНК путем электропорации позволяет получать эффективную экспрессию GFP в первичных человеческих Т-клетках, при создании изобретения оценивали пригодность этого метода для экспрессии других представляющих интерес белков. Нуклеазы эффектора, подобного активатору транскрипции (TALE-нуклеаза), представляют собой сайтспецифические нуклеазы, полученные путем слияния ДНК-связывающего домена TAL с доменом расщепления ДНК. Они представляют собой мощные инструменты редактирования генома, поскольку они индуцируют двухцепочечные разрывы практически любой требуемой последовательности ДНК. Указанные двухцепочечные разрывы активируют соединение негомологичных концов (NHEJ), подверженный ошибкам механизм репарации ДНК, который может приводить к инактивации любого представляющего интерес требуемого гена. Альтернативно этому, если в клетку в то же самое время интродуцируют соответствующую матрицу для репарации, то индуцируемые TALE-нуклеазой разрывы ДНК можно репарировать путем гомологичной рекомбинации, получая тем самым возможность модифицировать при желании генную последовательность.
При создании изобретения применяли введение путем электропорации мРНК для экспрессии TALE-нуклеазы, созданной для специфического расщепления последовательности в человеческом гене, кодирующей альфа-цепь Т-клеточного рецептора антигена (TRAC). Индуцированные в этой последовательности мутации приводили в результате к инактивации гена и утрате комплекса TCRαβ на клеточной поверхности. РНК, предназначенную для расщепления TRAC TALE-нуклеазы (TRAC-TALE-нуклеаза), или некодирующую РНК, используемую в качестве контроля, применяли для трансфекции активированных первичных Т-лимфоцитов с использованием технологии CytoPulse. Последовательность осуществления электропорации включала 2 импульса по 1200 В, за которыми следовали четыре импульса по 130 В, что представлено в таблице 7.
С помощью анализа методом проточной цитометрии экспрессии TCR на поверхности клеток через 7 дней после электропорации (фиг. 4, верхняя панель) установлено, что у 44% Т-клеток отсутствовала экспрессия TCRαβ. Осуществляли анализ геномной ДНК трансфецированных клеток путем ПЦР-амплификации локуса TRAC, после чего осуществляли высокопроизводительное секвенированием с помощью системы 454. 33% секвенированных аллелей (727 из 2153) одержали инсерцию или делецию в сайта расщепления TALE-нуклеазой.
Указанные данные свидетельствуют о том, что введение путем электропорации мРНК с использованием технологии CytoPulse приводит к функциональной экспрессии TRAC-TALE-нуклеазы.
Активность TRAC-TALE-нуклеаы и TRBC-TALE-нуклеазы в HEK293-клетках
Каждую конструкцию TALE-нуклеазы субклонировали, используя расщепление рестриктазой, в экспрессионном векторе млекопитающих под контролем длинного промотора pEF1альфа. Один миллион HEK293-клеток высевали за один день до трансфекции. Для трансфекции клеток использовали по 2,5 мкг каждой из двух плазмид, кодирующих TALE-нуклеазы, которые распознают две полумишени в представляющей интерес геномной последовательности в области константной цепи Т-клеточного рецептора альфа (TRAC) или области константной цепи Т-клеточного рецептора бета (TRBC), под контролем промотора EF1-альфа, или 5 мкг контрольного вектора pUC (pCLS0003), применяя 25 мкл липофектамина (фирма Invitrogen) согласно инструкциям производителя. Двухцепочечный разрыв, созданной TALE-нуклеазами в кодирующей последовательности TRAC, репарировали в живых клетках с помощью соединения негомологичных концов (NHEJ), представляющего собой механизм пониженной точности. Активность TALE-нуклеаз в живых клетках измеряли по частоте инсерций или делеций в целевом генном локусе. Через 48 ч после трансфекции геномную ДНК выделяли из трансфецированных клеток и осуществляли локус-специфические ПЦР с применением следующих праймеров: для TRAC: 5'-ATCACTGGCATCTGGACTCCA-3' (SEQ ID NO: 75), для TRBC1: 5'-AGAGCCCCTACCAGAACCAGAC-3' (SEQ ID NO: 76 или для TRBC2: 5'-GGACCTAGTAACATAATTGTGC-3' (SEQ ID NO: 77), и обратного праймера для TRAC: 5'-CCTCATGTCTAGCACAGTTT-3' (SEQ ID NO: 78), для TRBC1 и TRBC2: 5'-ACCAGCTCAGCTCCACGTGGT-3' (SEQ ID NO: 79). ПЦР-продукты секвенировали с помощью системы 454 для секвенирования (фирма 454 Life Sciences). Получали примерно 10000 последовательностей на ПЦР-продукт и затем анализировали в отношении присутствия сайт-специфических случаев инсерции или делеции; результаты представлены в таблице 8.
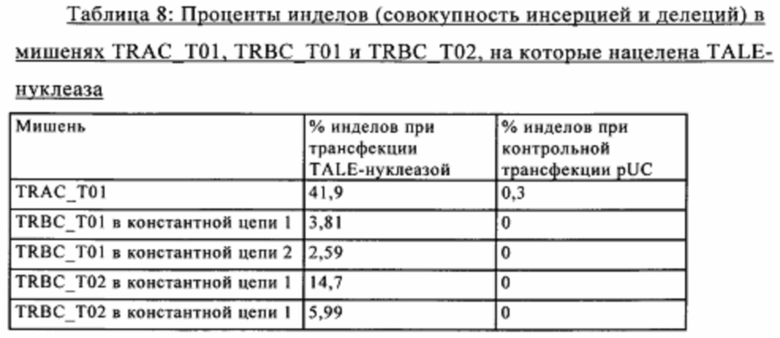
Активность β2m- и TRAC-TALE-нуклеаз в первичных Т-лимфоцитах
Каждую конструкцию TALE-нуклеазы субклонировали, используя расщепление рестриктазой, в экспрессионном векторе млекопитающих под контролем промотора Т7.
мРНК, кодирующую TALE-нуклеазу, расщепляющую геномную последовательность β2m, TRAC и TRBC, синтезировали из плазмиды, которая несла кодирующие последовательности, расположенные в прямом направлении относительно промотора Т7. Т-лимфоциты, выделенные из периферической крови, активировали в течение 5 дней, используя активирующие гранулы, покрытые антителами к CD3/CD28 (фирма Life technologies), и затем 5 миллионов клеток трансфецировали путем электропорации, используя по 10 мкг каждой из двух мРНК, кодирующих обе полу-TALE-нуклеазы (или некодирующую РНК в качестве контроля), применяя устройство CytoLVT-P. В результате инсерций и делеций, индуцированных NHEJ, кодирующая последовательность β2m и/или TRAC должна находиться вне рамки считывания во фракции клеток, несущих нефункциональные гены. Через 5 дней после электропорации клетки метили конъюгированным с флуорохромом антителом к β2m или антителом к TCR и оценивали с помощью проточной цитометрии в отношении присутствия β2m или TCR на клеточной поверхности. Поскольку все Т-лимфоциты, размноженные из периферической крови, в норме экспрессируют β2m и TCR, то относительное содержание β2m-негативных или TCR-негативных клеток является непосредственной мерой активности TALE-нуклеазы.
Функциональный анализ Т-клеток, в которых мишенью является ген TRAC
Целью инактивации гена TRAC является придание Т-лимфоцитам нечувствительности к стимуляции Т-клеточного рецептора. Согласно методу, описанному в предыдущем параграфе, Т-лимфоциты трансфецировали мРНК, кодирующей TALE-нуклеазу, которая расщепляет TRAC. Через 16 дней после трансфекции клетки обрабатывали, используя в концентрации вплоть до 5 мкг/мл фитогемагглютинин (РНА, фирма Sigma-Aldrich), который представляет собой митоген для Т-клеток, действующий через Т-клеточный рецептор. У клеток с функциональным Т-клеточным рецептором должен увеличиваться размер после обработки РНА. После инкубации в течение 3 дней клетки метили конъюгированным в флуорохромом антителом к TCR и анализировали с помощью проточной цитометрии, сравнивая распределение по размерам между TCR-позитивными и TCR-негативными клетками. На фиг. 3 продемонстрировано, что размер TCR-позитивных клеток существенно увеличивался после обработки РНА, в то время как TCR-негативные клетки имели такой же размер, что и необработанные клетки, что свидетельствует о том, что инактивация TRAC придает им нечувствительность к поступающим от TCR сигналам.
Функциональный анализ Т-клеток, в которых мишенью является ген β2m
Аналогично описанному выше, трансфецированные TALEN клетки и контрольные клетки (трансфекция без РНК) окрашивали меченным флуорохромом антителом к белку В2М, а также антителом, которое распознает все три класса молекул ГКГС-I (HLA-A, -В или-С). Трансфекция TALEN индуцировала снижение поверхностной экспрессии В2М и молекул ГКГС-I более чем у 37% Т-клеток (см. фиг. 5)
Безопасность в отношении генома β2m-TALE-нуклеазы и TRAC-TALE-нуклеазы в первичных Т-лимфоцитах
Поскольку полученные при создании изобретения конструкции включали субъединицы нуклеаз, важным является вопрос о том, может ли множественная трансфекция TALE-нуклеазой приводить к генотоксичности и расщеплению вне мишени в последовательностях «близко соответствующих» мишени или посредством ошибочного спаривания полу-TALE-нуклеаз. Для оценки воздействия TRAC-TALE-нуклеазы и β2m-TALE-нуклеазы на целостность клеточных геномов при создании изобретения исследовали перечень последовательностей в человеческом геноме, которые могут представлять собой потенциальный сайт нецелевого расщепления. Для создания указанного перечня идентифицировали все последовательности в геноме, имеющие вплоть до 4 замен по сравнению с исходными полумишенями, и затем идентифицировали пары потенциальных полумишеней в ориентации по типу «голова-к-голове» со спейсером, расположенным на расстоянии от 9 до 30 пар оснований друг от друга. Указанный анализ включал сайты, являющиеся потенциальной мишенью гомодимеров одной молекулы полу-TALE-нуклеазы или гетеродимеров, образованных одной полу-TALE-нуклеазой, мишенью которой является β2m, и одной полу-TALE-нуклеазой, мишенью которой является TRAC. При создании изобретения оценивали (в баллах) потенциальные нецелевые мишени на основе данных о специфичности, принимая в расчет «стоимость» индивидуальных замен и положения замен (при этом мисмэтчи были более приемлемыми для оснований на 3'-конце полумишени). При создании изобретения было получено 173 уникальных последовательности, которые имели балл, отражающий оценку вероятности расщепления. С помощью глубокого секвенирования было отобрано 15 последовательностей с высокими баллами и была проанализирована частота встречаемости мутаций, обнаруженных в этих локусах в Т-клетках, которые одновременно трансфецировали β2m- и TRAC-TALE-нуклеазой и очищали с помощью магнитной сепарации с получением β2m-негативных, TCRαβ-негативных клеток. Результаты продемонстрировали, что наибольшая частота встречаемости инсерций/делеций составляла 7×10-4. Эти результаты позволяют предположить, что мутация нецелевых мишеней является по меньшей мере в 600 раз менее вероятной, чем требуемых мишеней. Таким образом, реагенты, представляющие собой TALE-нуклеазы, которые применяли в этом опыте, являются чрезвычайно специфическими.
Введение путем электропорации в Т-клетки моноцистронной мРНК, кодирующей CD19-специфический одноцепочечный химерный антигенный рецептор (CAR)
5×106 Т-клеток, предварительно активированных в течение нескольких дней (3-5) с помощью покрытых антителом к CD3/CD28 гранулами и IL2, ресуспендировали в буфере Т для цитопорации и осуществляли введение путем электропорации в 0,4-сантиметровых кюветах без мРНК или с 10 мкг мРНК, кодирующей одноцепочечный CAR (SEQ ID NO: 6), применяя программу, описанную в таблице 7.
Через 24 ч после электропорации клетки окрашивали с помощью фиксирующего живые клетки красителя eFluor-780 и конъюгированного с РЕ козьего антимышиного IgG, F(ab')2-фрагмент специфического для оценки экспрессии CAR на клеточной поверхности живых клеток. Данные представлены на фиг. 6. Установлено, что огромное большинство живых Т-клеток, в которые путем электропорации вводили описанную выше моноцистронную мРНК, экспрессировали CAR на их поверхности. Через 24 ч после электропорации Т-клетки совместно культивировали с клетками Дауди (CD19+) в течение 6 ч и анализировали с помощью проточной цитометрии для определения экспрессии маркера дегрануляции CD107a на их поверхности (Betts, Brenchley и др., 2003).
Представленные на фиг. 6 данные демонстрируют, что в большинстве клеток, в которые вводили путем электропорации описанную выше моноцистронную мРНК, имела место дегрануляция в присутствии клеток-мишеней, экспрессирующих CD19. Эти результаты четко демонстрируют, что CAR, который экспрессируется на поверхности подвергнутых электропорации Т-клеток, является активным.
В следующих примерах заявители представили предназначенный для пролонгирования выживания и повышения терапевтической активности способ предупреждения опосредуемого NK-клетками отторжения терапевтических аллогенных Т-клеток, заключающийся в создании аллогенных Т-клеток посредством инактивации гена В2М с помощью специфической TALEN, объединенной либо с: I) с экспрессией химерной одноцепочечной молекулы, состоящей из UL18 и β2М B2M-UL18), либо с II) секрецией лигандов NKG2D. Конкретно способ основывался на применении к первичным Т-клеткам механизма, который обычно встречается в опухолевых клетках или инфицированных вирусом клетках. Таким образом, механизм действия был потенициально различным: в опухолевых клетках «слущивание» лигандов NKG2D приводит к снижению их присутствия на поверхности, в то время как в сконструированных клетках, секретируемый(ые) лиганд(ы) NKG2D должны служить в качестве «ловушки» для некоторых других лигандов NKG2D, которые все еще могут присутствовать на поверхности Т-клетки.
Эффективное «выключение» гена В2М с помощью специфической В2М-TALEN.
Создавали специфическую являющуюся мишенью TALEN последовательность (Т01, SEQ ID NO: 81) в первом кодирующем экзоне гена В2М (GenBank, регистрационный номер NC_000015) (левые RVD ДНК-связывающего домена: NN-NN-HD-HD-NG-NG-NI-NN-HD-NG-NN-NG-NN-HD-NG-NG с SEQ ID NO: 82 и правые RVD ДНК-связывающего домена: NI-NN-HD-HD-NG-HD-HD-NI-NN-NN-HD-HD-NI-NN-NI-NG с SEP ID NO: 83). Ниже в таблице 9 представлены последовательности нуклеаз, мишенями которых является последовательность Т01, а также две дополнительные мишени Т02 и Т03 и соответствующие им левые и правые последовательности TALE.







Для оценки способности указанной В2М-специфической TALEN увеличивать количество случаев NHEJ пониженной точности в локусе В2М вводили путем электропорации 2 или 10 мкг мРНК, кодирующей TALEN, в первичные Т-клетки, используя технологию Pulse Agile согласно протоколу производителя. Через 3 дня после трансфекции клетки выделяли и метили специфическим для β2-микроглобулина антителом, сшитым с флуорохромом фитоэритрином. Затем клетки анализировали с помощью проточной цитометрии в отношении жизнеспособности и экспрессии β2-m. Результаты представлены на фиг. 10. На верхней панели продемонстрировано, что около 100% нетрансфецированных Т-клеток экспрессировали β2-m (верхняя правая панель). В Т-клетках, трансфецированных специфической для В2М TALEN, экспрессия β2-m резко снижалась, так 38% (средняя правая панель) и 80% Т-клеток (нижняя правая панель) становились негативными по бета2-m при трансфекци 2 мкг или 10 мкг мРНК TALEN соответственно. Указанные данные свидетельствуют о том, что в Т-клетках можно с высокой эффективностью достигать «выключения» В2М.
Получение и экспрессия одноцепочечной молекулы B2M-UL18 в Т-клетках
Ген HCMV UL18 кодирует тип I трансмембранного гликопротеина, для которого характерен высокий уровень идентичности аминокислотной последовательность с молекулами ГКГС класса I, ассоциированными с бета2-m, и он связывает эндогенные пептиды. В связи с тем, что задача, положенная в основу изобретения, заключалась в том, чтобы экспрессировать эту молекулу в Т-клетках с дефектным геном В2М, стратегия настоящего изобретения включала получение химерной молекулы, в которой бета2-m и UL18 сливали с получением одноцепочечного полипептида. В SEQ ID NO: 89 представлена аминокислотная последовательность химерного белка. Лентивирусными частицами, содержащими химеру B2M-UL18, трансдуцировали Т-клетки. Мониторинг экспрессии трансгена осуществляли с помощью FACS-анализа, применяя антитело к бета2-m. Целью настоящего эксперимента была демонстрация того, что химерный белок B2M-UL18 эффективно экспрессируется в Т-клетках.
Получение и экспрессия лигандов NKG2D в Т-клетках
Встречающиеся в естественных условиях лиганды NKG2D представляют собой трансмембранные или заякоренные с помощью GPI белки. Для секреции этих молекул Т-клетками внеклеточные домены лигандов NKG2D сливали на их N-конце с формой секреторного пептида. Аминокислотные последовательности секретируемых химерных лигандов NKG2D представлены ниже (SEQ ID NO: 90 - SEQ ID NO: 97). Лентивирусными частицами, содержащими химерные лиганды NKG2D, трансдуцировали Т-клетки. Мониторинг экспрессии трансгена в супернатанте культуры осуществляли с помощью метода Вестерн-блоттинга с использованием специфических антител. Целью настоящего эксперимента была демонстрация того, что химерные белки лигандов NKG2D эффективно экспрессируются в Т-клетках.
CAR-T-клетки с дефицитом бета2-М не распознаются аллогенными Т-клетками
РВМС из здорового донора А совместно культивировали с облученными или обработанными митомицином сконструированными Т-клетками с дефицитом бета2-m, которые получали из донора Б. В качестве контроля РВМС из здорового донора А совместно культивировали с облученными или обработанными митомицином сконструированными бета2-m-позитивными Т-клетками из донора Б. Через 7 дней оценивали пролиферацию клеток из донора А с помощью включающего реагент ХТТ колориметрического анализа или по растворению CFSE (FACS-анализ). Хотя в контроле обнаружена клеточная пролиферация, отсутствие пролиферации или ограниченная клеточная пролиферация обнаружена, когда сконструированные Т-клетки не экспрессировали бета2-m. Целью настоящего эксперимента была демонстрация того, что аллореактивные Т-клетки не обладают способностью распознавать и размножаться в ответ на обработку Т-клетками с дефицитом бета2-m.
Эффективное ингибирование опосредуемого NK лизиса сконструированных Т-клеток
NK-клетки очищали из РВМС здорового донора А. В качестве мишеней получали сконструированные Т-клетки из здорового донора Б, которые перечислены ниже: а) сконструированные Т-клетки (отрицательный контроль), б) сконструированные Т-клетки с дефицитом бета2-m (положительный контроль), в) сконструированные Т-клетки с дефицитом бета2-m, экспрессирующие B2M-UL18 (SEQ ID NO: 89), г-л) сконструированные Т-клетки с дефицитом бета2-m, экспрессирующие соответственно SP-MICAed (SEQ ID NO: 90), SP-MICBed (SEQ ID NO: 91), SP-ULBP1ed (SEQ ID NO: 92), SP-ULBP2ed (SEQ ID NO: 93), SP-ULBP3ed (SEQ ID NO: 94), SP-N2DL4ed (SEQ ID NO: 95), SP-RET1Ged (SEQ ID NO: 96), SP-RAETILed (SEQ ID NO: 97). Эти последовательности предсавлены ниже в таблице 10.
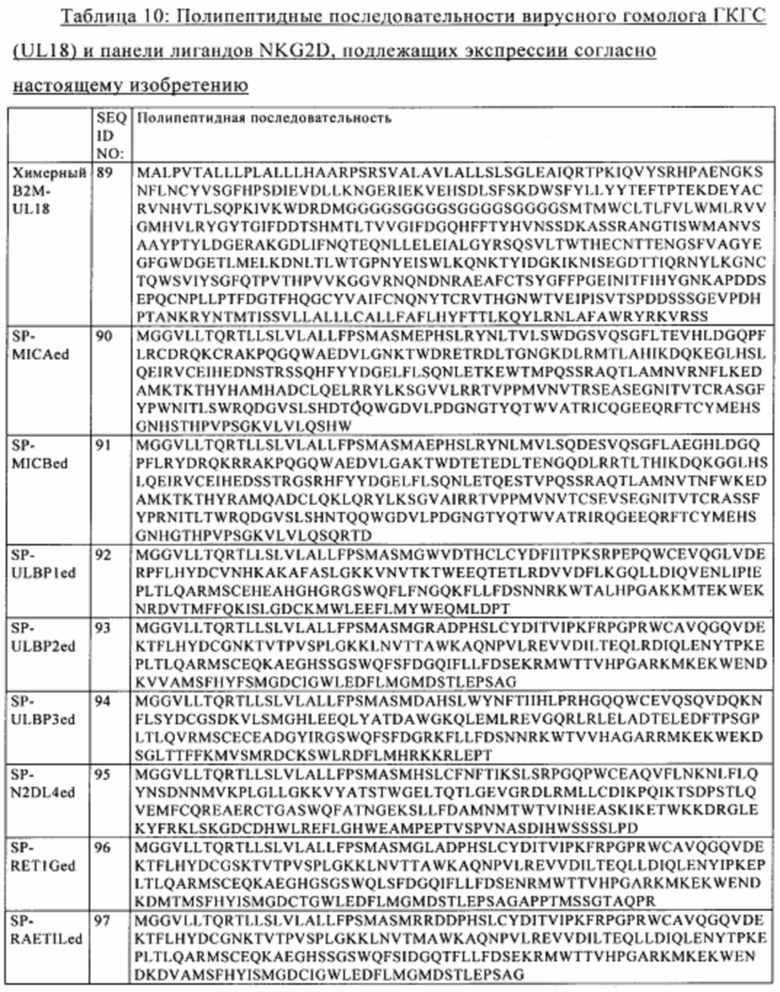
Цитотоксичность, опосредуемую NK-клетками, определяли с помощью анализа, основанного на мечении с использованием CFSE. Клетки-мишени метили с помощью CFSE, промывали в ЗФР, смешивали с NK-клетками в различных соотношениях эффекторных и Т-клеток (Е:Т) и инкубировали в течение 4 ч при 37°С. Затем клетки анализировали с помощью проточной цитометрии и определяли проценты CFSE-позитивных сконструированных Т-клеток, свидетельствующие о выживаемости сконструированных Т-клеток в присутствии NK-клеток. Установлено, что хотя опосредуемый NK клеточный лизис обнаружен в положительном контроле (сконструированные Т-клетки с дефицитом бета2-m), опосредуемый NK клеточный лизис отсутствовал или оказался ограниченным, когда сконструированные Т-клетки с дефицитом бета2-m экспрессировали B2M-UL18 (SEQ ID NO: 89) или секретировали лиганды NKG2D (SP-MICAed (SEQ ID NO: 90), SP-MICBed (SEQ ID NO: 91), SP-ULBP1ed (SEQ ID NO: 92), SP-ULBP2ed (SEQ ID NO: 93), SP-ULBP3ed (SEQ ID NO: 94), SP-N2DL4ed (SEQ ID NO: 95), SP-RET1Ged (SEQ ID NO: 96), SP-RAETILed (SEQ ID NO: 97). Целью настоящего эксперимента была демонстрация того, что опосредуемая аллогенными NK-клетками цитотоксическая активность нарушается, когда химерные молекулы, которые экспрессируются в сконструированных Т-клетках, действуют в качестве «ловушки» либо для ингибирующего сигнального рецептора (B2M-UL18), либо для стимулирующего сигнального рецептора (лиганды NKG2D).
Перечень процитированных в настоящем описании ссылок
Ashwell J.D. и Klusner R.D., Genetic and mutational analysis of the T-cell antigen receptor, Annu Rev Immunol 8, 1990, cc. 139-167;
Betts M.R., Brenchley J.M. и др., Sensitive and viable identification of antigen-specific CD8+ T cells by a flow cytometric assay for degranulation, J Immunol Methods 281(1-2), 2003, cc. 65-78;
Bierer B.E. и др., Cyclosporin A and FK506: molecular mechanisms of immunosuppression and probes for transplantation biology, Curr Opin Immunol 5(5), 1993, cc. 763-773;
Bix M. и др., Rejection of class I МНС-deficient haemopoietic cells by irradiated МНС-matched mice, Nature 349(6307), 1991, cc. 329-331;
Boch J., Scholze H. и др., Breaking the code of DNA binding specificity of TAL-type III effectors, Science 326(5959), 2009, cc. 1509-1512;
Cambier J.C., Antigen and Fc receptor signaling. The awesome power of the immunoreceptor tyrosine-based activation motif (ITAM), J Immunol 155(7), 1995, cc. 3281-3285;
Carter L. и др., PD-1 : PD-L inhibitory pathway affects both CD4(+) and CD8(+) T cells and is overcome by IL-2, Eur. J. Immunol. 32 (3), 2002, cc. 634-643;
Cong L., Ran F.А. и др., Multiplex genome engineering using CRISPR/Cas systems, Science 339(6121), 2013, cc. 819-823;
Critchlow S.E. и Jackson S.P., DNA end-joining: from yeast to man, Trends Biochem Sci 23(10), 1998, cc. 394-398;
Deltcheva E., Chylinski K. и др., CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III, Nature 471(7340), 2011, cc. 602-607;
Gasiunas G. и др., Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria, Proc Natl Acad Sci USA 109(39), 2012, cc. E2579-2586;
Haseloff и Gerlach, Simple RNA enzymes with new and highly specific endoribonuclease activities, Nature 334, 1988, cc. 585-591;
Jena В., Dotti G. и др., Redirecting T-cell specificity by introducing a tumor-specific chimeric antigen receptor, Blood 116(7), 2010, cc. 1035-1044;
Jinek M., Chylinski K. и др., A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity, Science 337(6096), 2012, cc. 816-821;
Liu L. и др., Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes, Cell 66(4), 1991, cc. 807-815;
Ma J.L., Kim E.M. и др., Yeast Mre11 and Rad1 proteins define a Ku-independent mechanism to repair double-strand breaks lacking overlapping end sequences, Mol Cell Biol 23(23), 2003, cc. 8820-8828;
Mach В., Steimle V., Reith W., MHC class II-deficient combined immunodeficiency: a disease of gene regulation, Immunol. Rev. 138 (1), 1994, cc. 207-221;
Mali P., Yang L. и др., RNA-guided human genome engineering via Cas9, Science 339(6121), 2013, cc. 823-826;
Moscou M.J. и Bogdanove A.J., A simple cipher governs DNA recognition by TAL effectors, Science 326(5959), 2009, c. 1501;
Park T.S., Rosenberg S.А. и др., Treating cancer with genetically engineered T cells, Trends Biotechnol 29(11), 2011, cc. 550-557;
Stoddard B.L., Homing endonuclease structure and function, Q Rev Biophys 38(1), 2005, cc. 49-95;
Urnov F.D. и др., Genome editing with engineered zinc finger nucleases, Nature reviews Genetics 11, 2010, cc. 636-646.
| название | год | авторы | номер документа |
|---|---|---|---|
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
| ИНСЕРЦИЯ ТАРГЕТНОГО ГЕНА ДЛЯ УЛУЧШЕННОЙ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2017 |
|
RU2824204C2 |
| МНОГОЦЕПОЧЕЧНЫЙ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР И ЕГО ПРИМЕНЕНИЯ | 2013 |
|
RU2663725C2 |
| CD33-СПЕЦИФИЧЕСКИЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ИММУНОТЕРАПИИ РАКА | 2015 |
|
RU2701341C2 |
| ИММУННЫЕ КЛЕТКИ, ДЕФЕКТНЫЕ ПО Suv39h1 | 2018 |
|
RU2784531C2 |
| CD19-СПЕЦИФИЧЕСКИЙ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР И ЕГО ПРИМЕНЕНИЯ | 2014 |
|
RU2727447C2 |
| СПОСОБ КОНСТРУИРОВАНИЯ АЛЛОГЕННЫХ И УСТОЙЧИВЫХ К ЛЕКАРСТВЕННЫМ ПРЕПАРАТАМ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2689558C1 |
| СПОСОБЫ КОНСТРУИРОВАНИЯ АЛЛОГЕННЫХ И ВЫСОКОАКТИВНЫХ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2736616C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР, И РОДСТВЕННЫЕ КОМПОЗИЦИИ | 2019 |
|
RU2835579C2 |
| СПОСОБЫ КОНСТРУИРОВАНИЯ ВЫСОКОАКТИВНЫХ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2725542C2 |
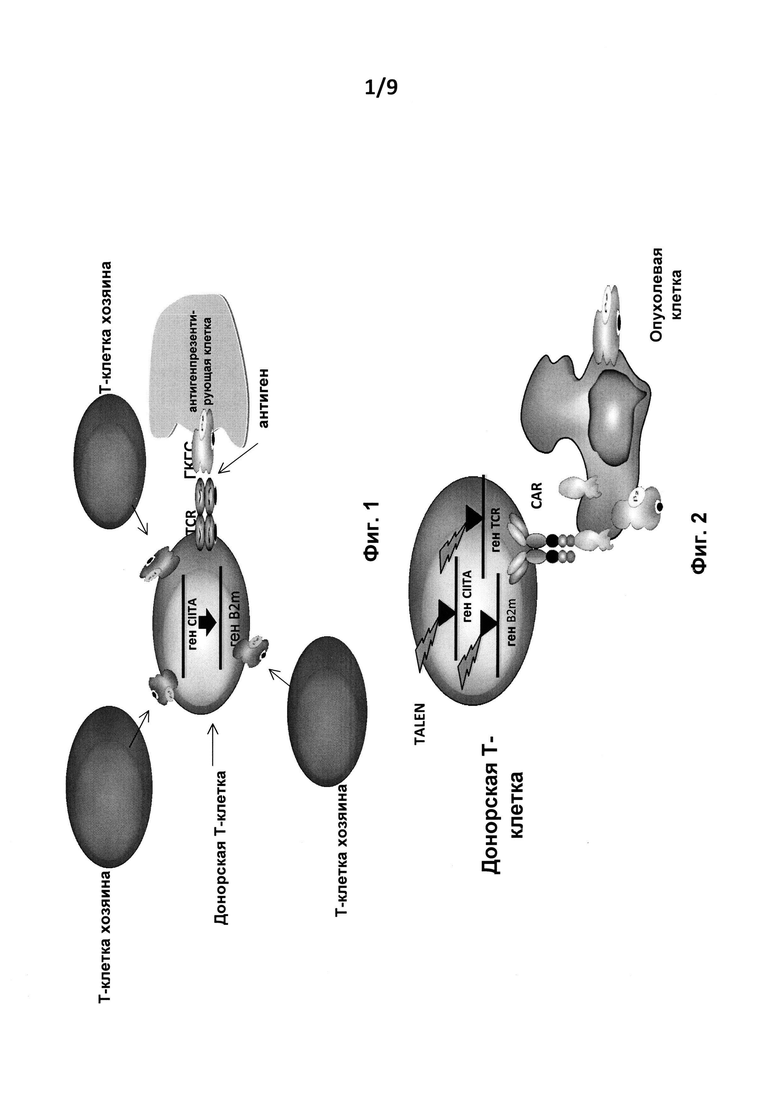
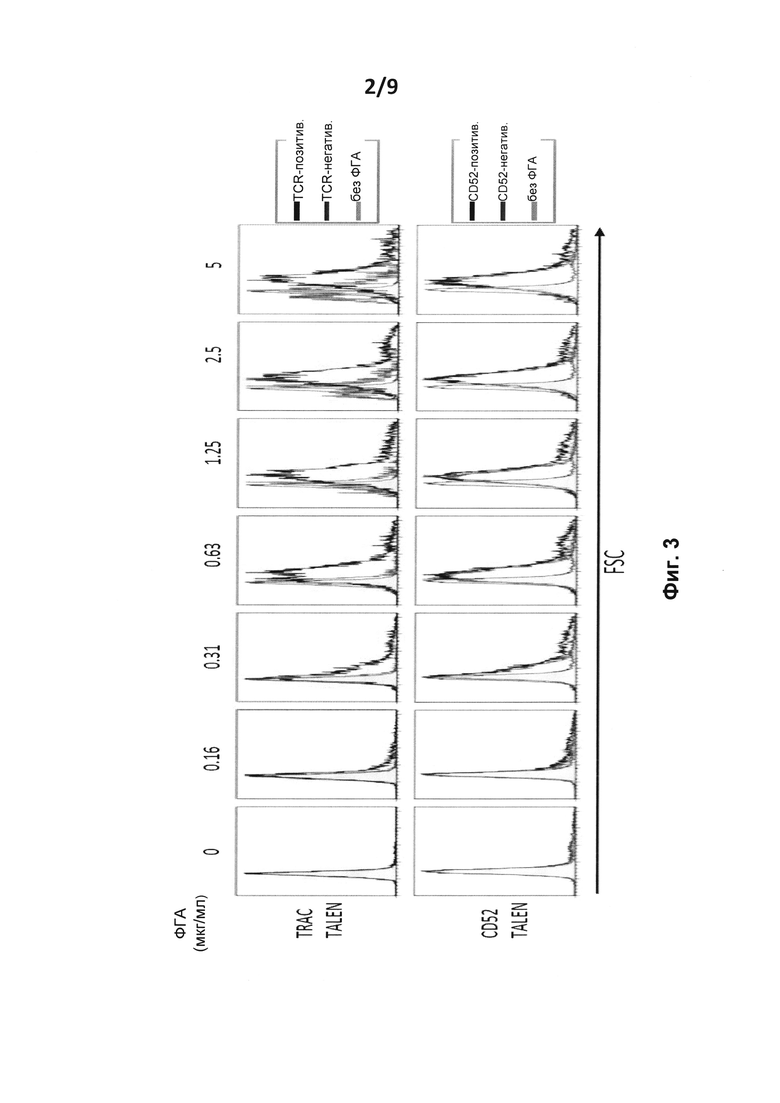
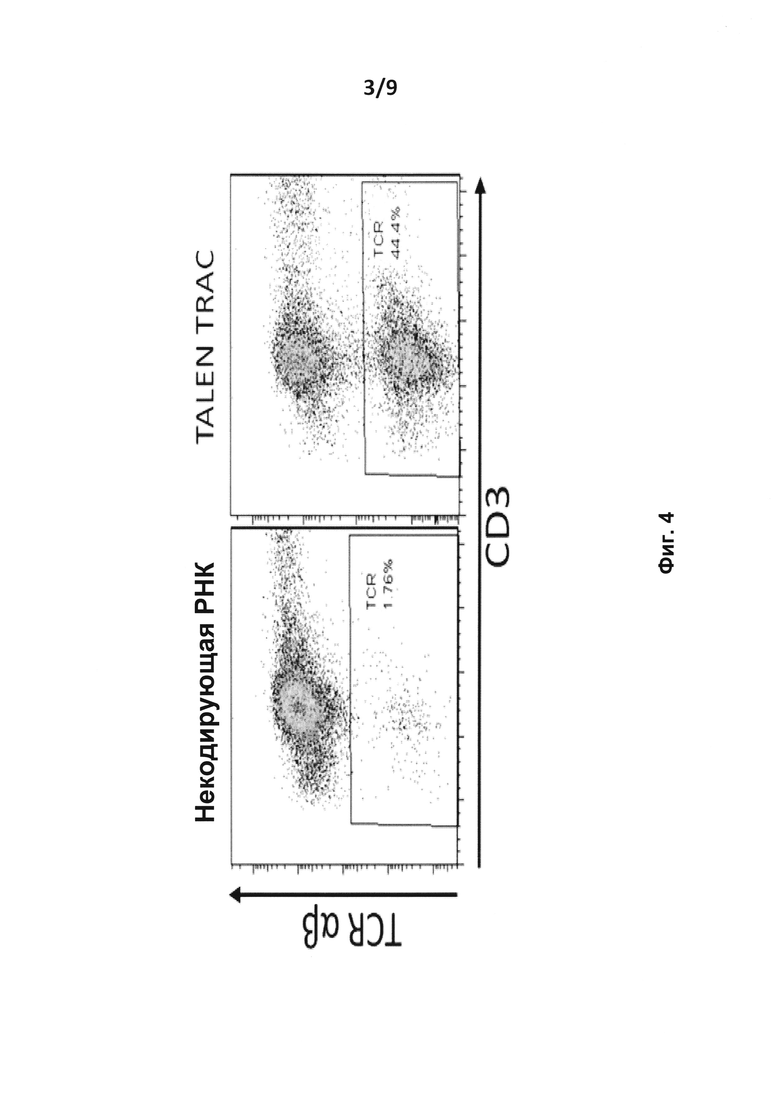
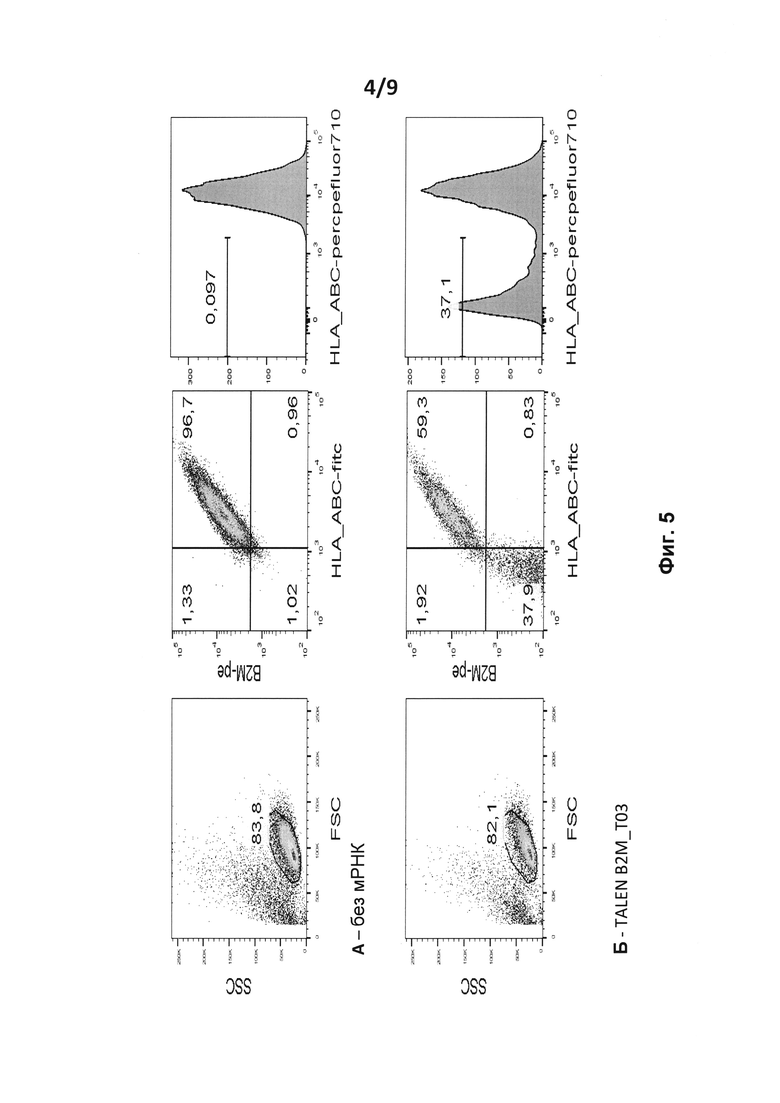
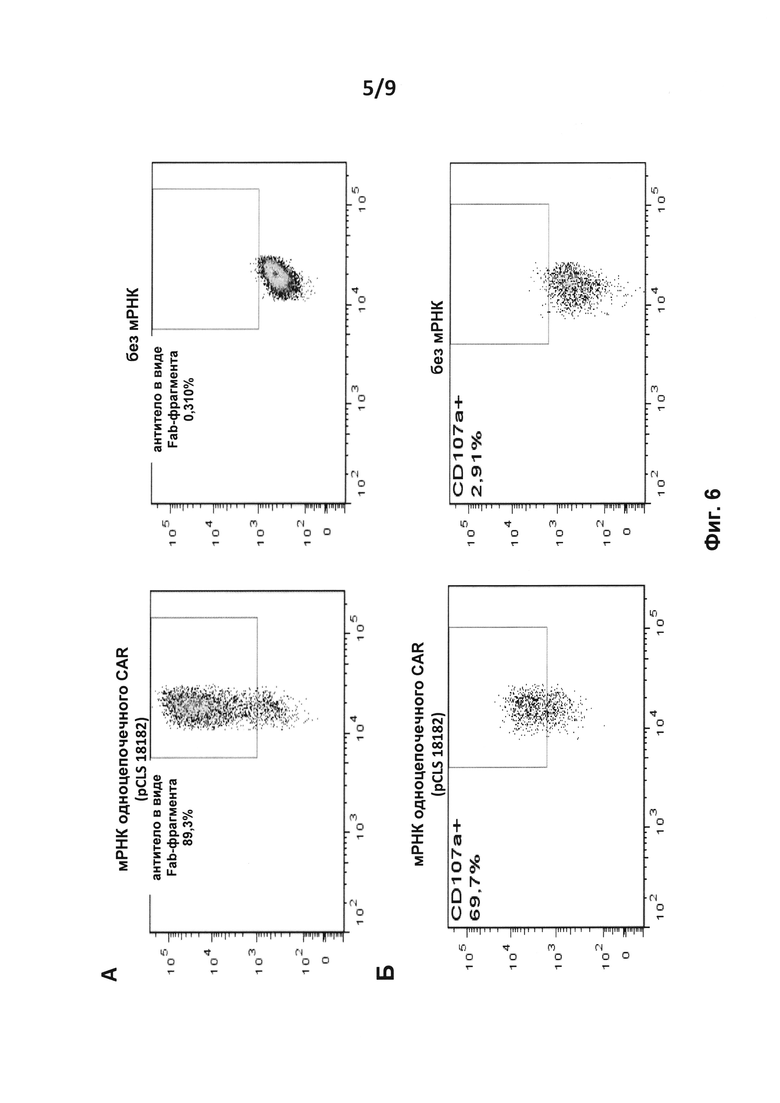
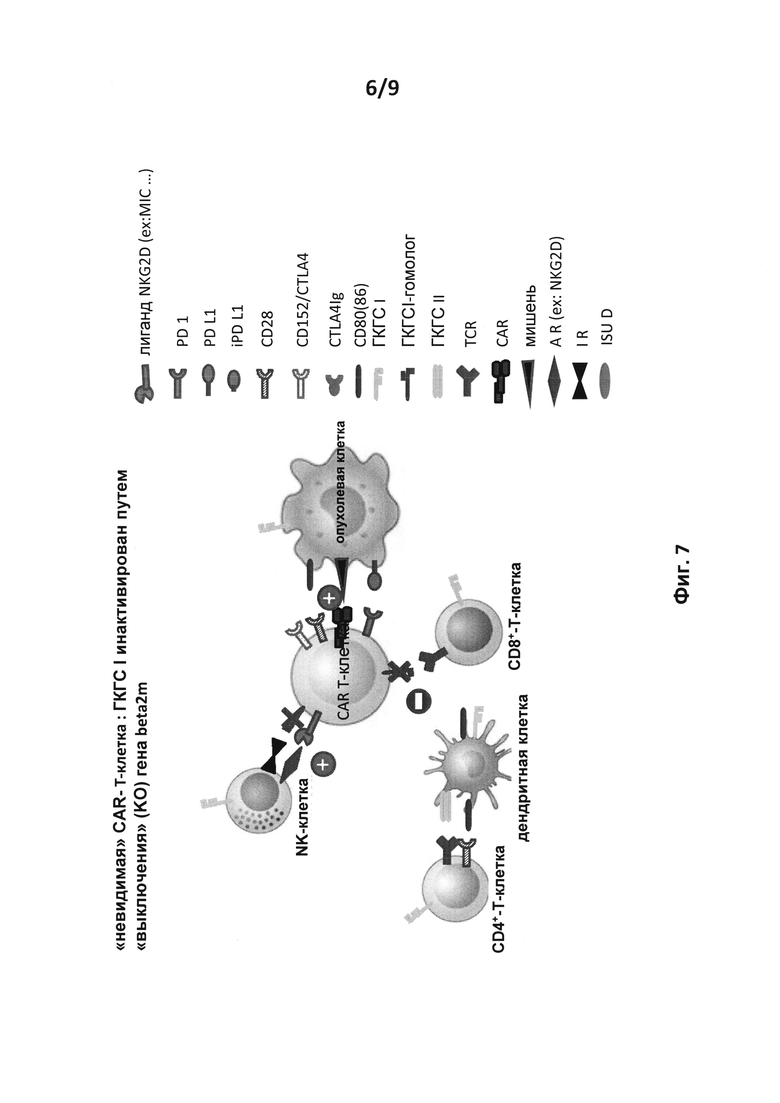
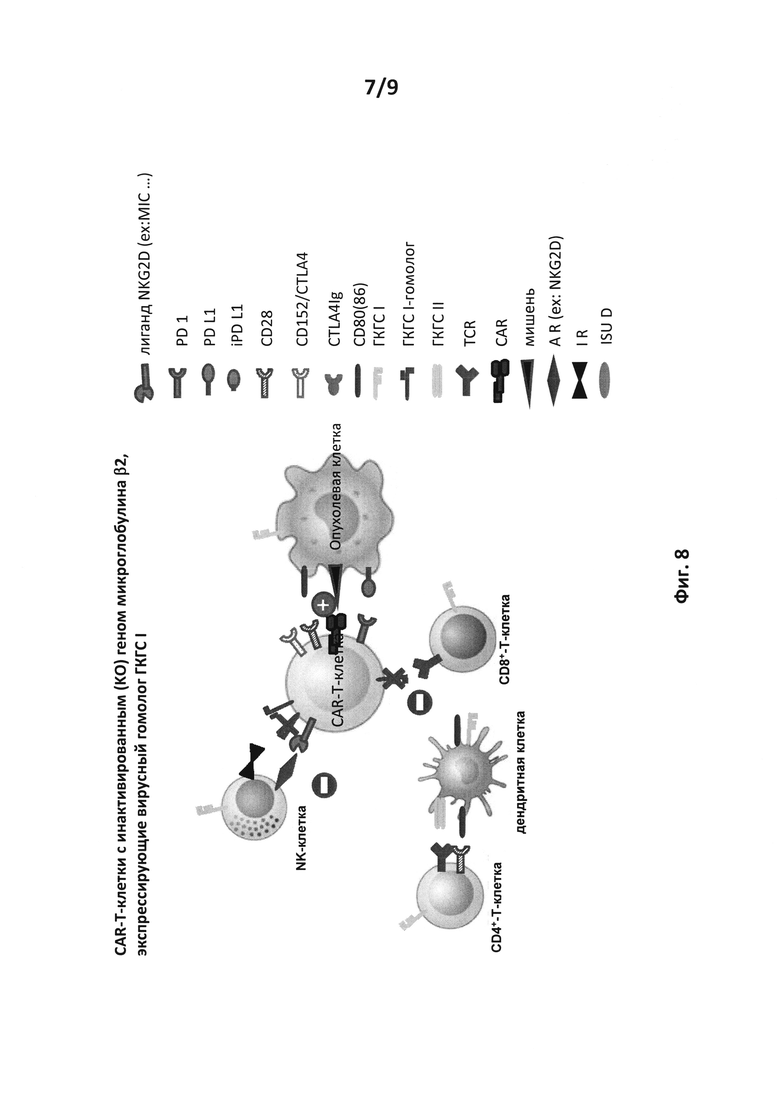
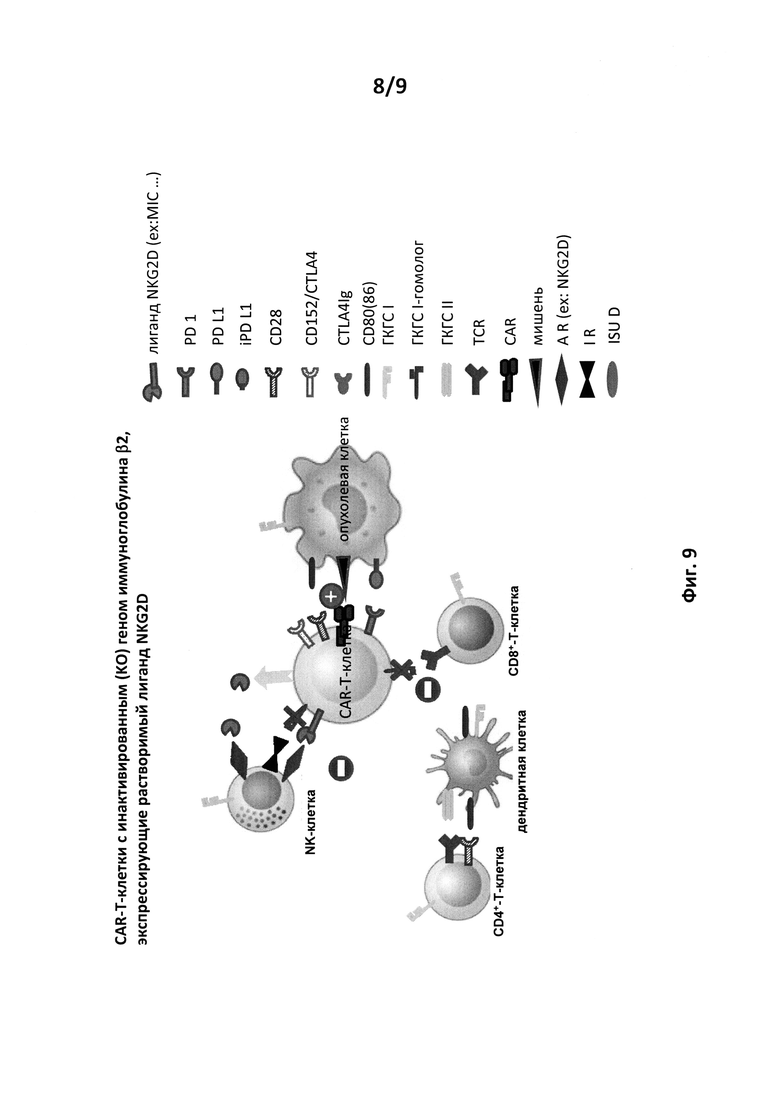
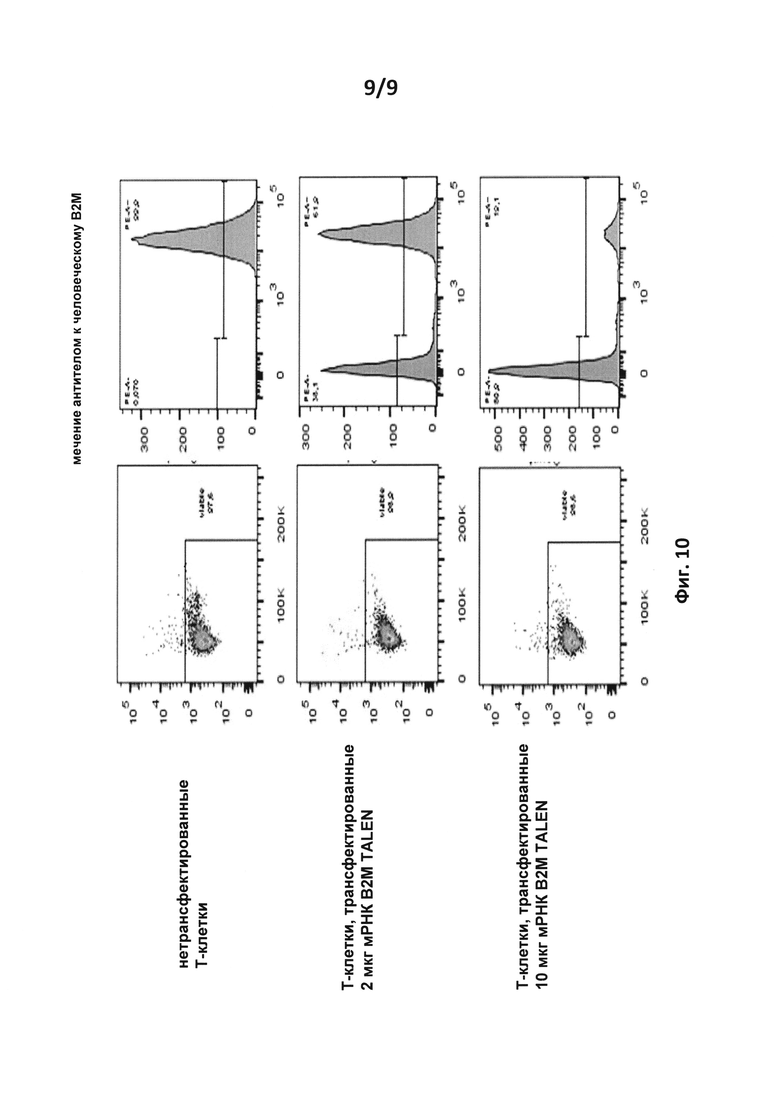
Изобретение относится к области биотехнологии, конкретно к способу получения Т-клеток для иммунотерапии, и может быть использовано в медицине. Заявленный способ, включающий инактивацию гена бета 2-микроглобулина (В2М) и по меньшей мере одного гена, кодирующего компонент Т-клеточного рецептора (TCR), путем применения направленных редкощепящих эндонуклеаз, позволяет получать Т-клетки для экспрессии химерного антигенного рецептора (CAR), пригодные для аллотрансплантации и иммунотерапии рака. 4 н. и 17 з.п. ф-лы, 10 ил., 10 табл., 1 пр.
1. Способ получения сконструированного цитотоксического Т-лимфоцита для применения в иммунотерапии, включающий стадии, на которых:
а) получают цитотоксический Т-лимфоцит от здорового донора;
б) ингибируют экспрессию бета 2-микроглобулина (В2М) в указанном Т-лимфоците путем применения редкощепящей эндонуклеазы, выбранной из TAL-нуклеазы, мегануклеазы, нуклеазы с «цинковыми пальцами» (ZFN) или РНК-направляемой эндонуклеазы, которая обладает способностью осуществлять избирательную инактивацию путем расщепления ДНК гена, кодирующего указанный В2М;
в) инактивируют по меньшей мере один ген, кодирующий компонент Т-клеточного рецептора (TCR), выбранного из TCR-альфа и TCR-бета, применяя редкощепящую эндонуклеазу, выбранную из TAL-нуклеазы, мегануклеазы, нуклеазы с «цинковыми пальцами» (ZFN) и РНК-направляемой эндонуклеазы, которая обладает способностью осуществлять избирательную инактивацию указанного гена путем расщепления ДНК, предпочтительно с помощью двухцепочечного разрыва; и
г) интродуцируют в указанный Т-лимфоцит молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую химерный антигенный рецептор (CAR), направленный против антигена, выбранного из CD19, CD20, CD22, CD30, CD33, ErbB2, CEA, EGFRvIII, GD2, MAGE, мезотелина, NKG2D, NY-ESO-1, PSMA, ROR1, 5Т4 и TAG-72, и экспрессирующий указанный CAR в указанный Т-лимфоцит.
2. Способ по п. 1, в котором стадию б) осуществляют, применяя TAL-нуклеазу.
3. Способ по п. 1, в котором стадию б) осуществляют, применяя РНК- направляемую эндонуклеазу.
4. Способ по п. 3, в котором РНК-направляемая эндонуклеаза представляет собой Cas9.
5. Способ по п. 1, где стадия в) проводится с использованием TAL-нуклеазы.
6. Способ по п. 1, где стадия в) проводится с использованием РНК-направляемой эндонуклеазы.
7. Способ по п. 1, в котором компонент TCR представляет собой TCR-альфа.
8. Способ по п. 1, в котором химерный антигенный рецептор направлен против антигена CD19 В-лимфоцита.
9. Сконструированный цитотоксический Т-лимфоцит для применения в иммунотерапии, экспрессирующий химерный антигенный рецептор (CAR), направленный против антигена, выбранного из CD19, CD20, CD22, CD30, CD33, ErbB2 (HER2/neu), CEA, EGFRvIII, GD2, MAGE, мезотелина, NKG2D, NY-ESO- 1, PSMA, ROR1, 5T4 и TAG-72, причем указанный Т-лимфоцит также отличается тем, что i) в нем ингибирована экспрессия бета 2-микроглобулина (В2М) и ii) инактивирован по меньшей мере один ген, кодирующий компонент Т-клеточного рецептора (TCR), выбранного из TCR-альфа и TCR-бета; где указанный Т-лимфоцит экспрессирует редкощепящую эндонуклеазу, выбранную из TAL-нуклеазы, мегануклеазы, нуклеазы с «цинковыми пальцами» (ZFN) и РНК-направляемой эндонуклеазы, обладающую способностью осуществлять избирательную инактивацию гена, кодирующего В2М, путем расщепления ДНК, предпочтительно с помощью двухцепочечного разрыва; и причем указанный Т-лимфоцит дополнительно экпрессирует редкощепящую эндонуклеазу, выбранную из TAL-нуклеазы, мегануклеазы, нуклеазы с «цинковыми пальцами» (ZFN) и РНК-направляемой эндонуклеазы, обладающую способностью осуществлять избирательную инактивацию по меньшей мере одного указанного гена, кодирующего компонент Т-клеточного рецептора (TCR), путем расщепления ДНК, предпочтительно с помощью двухцепочечного разрыва.
10. Сконструированный цитотоксический Т-лимфоцит по п. 9, где указанный Т-лимфоцит содержит молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанную редкощепящую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию гена, кодирующего В2М, путем расщепления ДНК, предпочтительно с помощью двухцепочечного разрыва.
11. Сконструированный цитотоксический Т-лимфоцит по п. 9, в котором указанная редкощепящая эндонуклеаза, способная осуществлять избирательную инактивацию гена, кодирующего В2М, путем расщепления ДНК, предпочтительно с помощью двухцепочечного разрыва, представляет собой TAL-нуклеазу.
12. Сконструированный цитотоксический Т-лимфоцит по п. 9, в котором указанная редкощепящая эндонуклеаза, способная осуществлять избирательную инактивацию гена, кодирующего В2М, путем расщепления ДНК, предпочтительно с помощью двухцепочечного разрыва, представляет собой РНК-направляемую эндонуклеазу.
13. Сконструированный цитотоксический Т-лимфоцит по п. 12, в которой РНК-направляемая эндонуклеаза представляет собой Cas9.
14. Сконструированный цитотоксический Т-лимфоцит по п. 10, в котором указанная нуклеиновая кислота представляет собой вектор, обеспечивающий экспрессию указанных редкощепящих эндонуклеаз указанным Т-лимфоцитом.
15. Сконструированный цитотоксический Т-лимфоцит по п. 10, в котором указанная нуклеиновая кислота представляет собой трансфецированную мРНК.
16. Сконструированный цитотоксический Т-лимфоцит по п. 9, где указанный Т-лимфоцит включает молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанную редкощепящую эндонуклеазу, которая обладает способностью осуществлять избирательную инактивацию указанного по меньшей мере одного гена, кодирующего компонент Т-клеточного рецептора (TCR), путем расщепления ДНК, предпочтительно с помощью двухцепочечного разрыва.
17. Сконструированный цитотоксический Т-лимфоцит по п. 9, где указанный Т-лимфоцит содержит молекулу экзогенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую указанный CAR.
18. Сконструированный цитотоксический Т-лимфоцит по п. 9, в котором указанный CAR направлен против антигена CD19 В-лимфоцита.
19. Сконструированный цитотоксический Т-лимфоцит по п. 9, где указанный Т-лимфоцит получают от здорового донора.
20. Применение сконструированного цитотоксического Т-лимфоцита по п. 9 в лечении лимфомы.
21. Применение сконструированного цитотоксического Т-лимфоцита по п. 9 в лечении рака.
| LU P | |||
| et al., Generating hypoimmunogenic human embryonic stem cells by the disruption of beta 2-microglobulin, Stem Cell Reviews and Reports, 2013, V | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| TORIKAI H | |||
| et al., A foundation for universal T-cell based immunotherapy: T cells engineered to express a CD19-specific chimeric-antigen-receptor and eliminate expression of | |||

