Настоящее изобретение создано при государственной поддержке в силу договора номер СА 130911, заключенного с Национальным институтом здравоохранения. Государство обладает определенными правами на данное изобретение.
Перекрестные ссылки на родственные заявки
Настоящая заявка является частичным продолжением заявки на Патент США №13/502 978, поданной 19 апреля 2012 г., которая является национальной заявкой, поданной на основании международной заявки на патент № PCT/US 2010/54846, поданной 29 октября 2010 г., которая испрашивает приоритет предварительной заявки на патент №61/255 980, поданной 29 октября 2009 г, раскрытие которой включено в настоящую заявку посредством ссылки.
Уровень техники
Область изобретения
Настоящее изобретение относится к Т-клеткам, дефицитным по TCR (рецептору Т-клеток), способам получения и применения лишенных TCR Т-клеток, и способам применения указанных TCR-дефицитных Т-клеток для борьбы с заболеваниями и расстройствами. Согласно одному варианту реализации настоящее изобретение в широком смысле относится к TCR-дефицитным Т-клеткам, их выделенной популяции и композициям, их содержащих. Согласно другому варианту реализации настоящего изобретения указанные TCR-дефицитные Т-клетки также изменяют для экспрессии функционального рецептора, отличного от TRC. Также настоящее изобретение относится к способам получения указанных TCR-дефицитных Т-клеток, а также способам уменьшения или облегчения, либо профилактики или лечения заболеваний и расстройств при помощи указанных TCR-дефицитных Т-клеток, их популяций или композиций, их содержащих.
Описание предшествующего уровня техники
Глобальное бремя рака с 1975 г. до 2000 г. увеличилось вдвое, и предполагается, что рак станет ведущей причиной смертности во всем мире к 2010 г. Согласно информации Американского онкологического общества, ожидается еще двукратное увеличение к 2020 г. и трехкратное увеличение к 2030 г. Таким образом, существует потребность в более эффективных средствах лечения разных форм рака. Идеально, чтобы любое средство от рака было бы эффективным (в уничтожении раковых клеток), направленным (т.е., селективным, не убивало бы здоровые клетки), перманентным (позволяло бы избежать рецидивов и метастазов) и доступным по цене. Сегодняшние стандарты лечения большинства форм рака не оправдывают ожиданий по некоторым или по всем из перечисленных критериев.
Было показано, что клеточная терапия приводит к специфическому искоренению опухоли и обладает потенциалом в отношении создания специфических и эффективных средств от рака. (Но, W.Y., et al. 2003. Cancer Cell 3:1318-1328; Morris E.C. et al. 2003. Clin. Exp. Immunol. 131:1-7; Rosenberg, S.A. 2001. Nature 411:380-384; Boon T. and P. van der Bruggen. 1996. J. Exp. Med. 183:725-729). Т-клетки часто являлись клетками-эффекторами выбора для иммунотерапии рака вследствие их селективного распознавания и мощных эффекторных механизмов. Т-клетки распознают специфические пептиды, получаемые из внутренних белков клеток, в контексте собственного главного комплекса гистосовместимости (МНС) при помощи своих рецепторов Т-клеток (TCR).
В технике считается признанным, что комплекс TCR ассоциирован с тонким способом образования димеров и соединения указанных димеров (TRC-альфа/бета, CD3-гамма/эпсилон, CD3-дельта/эпсилон и CD3-дзета-димер) в один комплекс TRC, который может экспортироваться на поверхность клетки. Неспособность любого из указанных комплексов правильно образовываться будет приводить к подавлению сборки и экспрессии TCR (Call, М.Е, et al. (2007) Nature Rev. Immunol, 7:841-850; Call, M.E, et al. (2005) Annu Rev. Immunol, 23:101-125).
Было выяснено, что определенные остатки аминокислот в соответствующих цепях TCR важны для правильного образования димеров и сборки TCR. В частности, для TCR-альфа такими ключевыми аминокислотами в трансмембранной части являются аргинин (для соединения с CD3-дзета) и лизин (для соединения с CD3 -эпсилон/дельта-димером). Для TCR-бета ключевой аминокислотой в трансмембранной части является лизин для соединения с CD3-эпсилон/гамма-димером). Для CD3-гамма ключевой аминокислотой в трансмембранной части является глутаминовая кислота. Для CD3-дельта ключевой аминокислотой в трансмембранной части является аспарагиновая кислота. Для CD3-эпсилон ключевой аминокислотой в трансмембранной части является аспарагиновая кислота. Для CD3-дзета ключевой аминокислотой в трансмембранной части является аспарагиновая кислота (Call, М.Е, et al. (2007) Nature Rev. Immunol, 7:841-850; Call, M.E, et al. (2005) Annu Rev. Immunol, 23:101-125).
Пептиды, полученные из измененных или мутированных белков в опухолях, могут распознаваться специфическими TCR. Несколько ключевых исследований привели к идентификации антигенов, связанных со специфическими опухолями, которые оказались способны вызывать эффективные цитотоксические реакции Т-клеток (CTL) у пациентов. (Ribas, A. et al. 2003, J. Clin. Oncol 21:2415-2432). Эффекторные механизмы Т-клеток включают способность к уничтожению клеток опухоли напрямую и к выработке цитокинов, которые активируют другие клетки иммунитета хозяина и изменяют локальное микроокружение опухоли. Теоретически Т-клетки могут идентифицировать и разрушать клетку опухоли, экспрессирующую один мутированный пептид. Адаптивная иммунотерапия клонами CTL, специфическими в отношении MARTI или gp100, при низких дозах IL-2 (интерлекина-2), оказалась эффективной в снижении и стабилизации опухолевой нагрузки у некоторых пациентов (Yee, С et. Al. 2002. Proc. Natl.Acad. Sci. USA 99:16168-16173). Другие способы основаны на применении Т-клеток с определенным противоопухолевым рецептором. Такие способы включают генетически модифицированные CTL с новыми антиген-специфичными рецепторами Т-клеток, которые распознают пептиды опухоли и МСН, химерные рецепторы антигенов (CARS), полученные из фрагментов одноцепочечного антитела (scFv), соединенных с соответствующим сигнальным элементом, или применение химерных рецепторов клеток NK (естественных киллеров) (Но W.Y. et al. 2003. Cancer Cell 3:431-437; Eshhar, Ζ. et al. 1993. Proc. Natl. Acad. Sci. USA 91:817-821; Zhang, T. et al. 2005. Blood 106:1544-1551).
Клеточные препараты применяют у пациентов, которым не помогла традиционная химиотерапия или лучевая терапия, или у которых развился рецидив, у которых часто был опробован более чем один тип терапии. Иммунные клетки, отобранные у пациентов с распространенным раком, которые подверглись нескольким циклам химиотерапии, не реагируют так же сильно, как клетки здоровых людей. Кроме того, пациенты с раком часто являются пожилыми и могут страдать другими заболеваниями, которые могут ограничивать способность их иммунных клеток становиться примированными эффекторными клетками, даже после активации in vivo и экспансии. Кроме того, каждый пациент с раком должен предоставить достаточное количество своих собственным иммунных клеток, чтобы они были модифицированы так, чтобы экспрессировать новый иммунный рецептор. Поскольку каждое средство должно быть сделано индивидуально для каждого пациента, данный процесс занимает несколько недель с момента решения о проведении такой терапии; тем временем рак продолжает расти. Заявка на патент США US 2002/0039576 описывает способ имодулирования активности Τ клеток, в котором используют Τ клетки с фенотипом CD3+-αβ-TcR+CD4-CD8-CD28-NK1.1-. В публикации заявки на патент США № US 2006/0166314 раскрывается применение мутированных Т-клеток для лечения рака, при котором Т-клетки представляют собой Т-клетки с рецептором αβ-Τ-клеток, специфичным для белка MDM2, опосредующим ответ Т-клеток.
Рак является не единственным заболеванием, при котором эффективным средством могут быть манипуляции с Т-клетками. Известно, что активные рецепторы Т-клеток на Т-клетках критичны для ответа организма на стимуляцию иммунной системы. Например, было показано, что широкий спектр рецепторов Т-клеток играет определенную роль в реакции «трансплантат против хозяина» (GVHD), в частности при хронической GVHD (Anderson et al. (2004) Blood 104:1565-1573). Фактически было показано, что введение антител к рецепторам Т-лимфоцитов уменьшает симптомы острой GVHD (Maeda et al. (2005) Blood 106:749-755).
Сохраняется потребность в более эффективных средствах, основанных на Т-клетках, для лечения определенных заболеваний и расстройств, и в способах лечения, основанных на разработке новых типов Т-клеток.
Сущность изобретения
Согласно одному варианту реализации настоящее изобретение в широком смысле относится к изолированным, модифицированным Т-клеткам, которые не экспрессируют функциональных рецепторов Т-клеток (TCR). Согласно данному варианту реализации указанные Т-клетки дефицитны по TCR в том, что касается экспрессии функционального TCR. Согласно еще одному варианту реализации настоящего изобретения дефицитные по TCR Т-клекти модифицированы таким образом, чтобы экспрессировать функциональный отличный от TCR рецептор, например, такой как химерный рецептор. Указанные клетки также функционируют как платформа, позволяющая экспрессировать другие специфические рецепторы, например, рецепторы, которые могут быть полезны при специфических заболеваниях, при этом сохраняющих эффекторные функции Т-клеток, хотя и без функционирующих TCR.
Настоящее изобретение предусматривает популяции TCR-дефицитных Т-клеток, и композиции, их содержащие. Также изобретение предусматривает способы получения указанных TCR-дефицитных Т-клеток, и способы уменьшения и облегчения, либо профилактики или лечения, заболеваний и расстройств с применением указанных TCR-дефицитных Т-клеток, их популяций или терапевтических композиций, их содержащих. Согласно одному варианту реализации данную композицию можно применять для лечения рака, инфекции, одного или более аутоиммунных расстройств, лучевой болезни, или для профилактики или лечения реакции «трансплантат против хозяина» (GCHD) или отторжения трансплантата у субъекта, подвергшегося операции по пересадке ткани или органа.
Краткое описание чертежей
На Фигуре 1 проиллюстрированы химерные рецепторы NK, описываемые в настоящей заявке. Показаны внеклеточные (ЕС), трансмембранные (ТМ) и цитоплазматические участки (Сур). Показаны формы рецептора дикого типа (WT) и химерного (СН), где NH2 обозначает N-конец, а СООН обозначает С-конец.
На Фигуре 2 проиллюстрировано, что TIM снижают распознавание TCR и функцию Т-лимфоцитов человека во время культивирования с РВМС (мононуклеарными клетками периферической крови). На панели (А) показано, что TIM-трансфецированные Т-клетки, культивируемые с аллогенными РВМС, демонстрируют снижение выработки IFN-γ (интерферона гамма). Показана общая выработка IFN-γ. На панели (В) показано количество IFN-γ после вычитания количества аутологичного IFN-γ . Данное значение отображает специфический IFN-γ , вырабатываемый при распознавании аллогенных РВМС.
На Фигуре 3 проиллюстрирована активация TIM-экспрессирующих Т-клеток при помощи рекомбинантного рецептора, направленного против для рака. TIM-экспрессирующие Т-клетки, которые одновременно экспрессируют химерный рецептор NKG2D (chNKG2D), который распознают специфические лиганды на клетках опухолей многих типов, вырабатывающих повышенные количества IFN-γ при совместном культивировании с клетками миеломы RPMI8226. В некоторых лунках было включено блокирующее моноклональное антитело (mAb) NKG2D, чтобы не позволить chNKG2D распознавать свои лиганды на клетках опухоли, и это демонстрирует специфический ответ рецептора chNKG2D на указанных Т-клетках.
Подробное описание предпочтительных вариантов реализации
Определения
Следует понимать, что настоящее изобретение не ограничивается конкретной описанной методологией, протоколами, линиями клеток, видами или родами животных, и реагентами, которые могут варьировать. Также следует понимать, что терминология, применяемая в настоящей заявке, предназначена только для описания конкретных вариантов реализации, но не предназначена для ограничения области настоящего изобретения, которая ограничена только прилагаемой формулой изобретения.
В настоящей заявке единственное число включает множественные объекты, если из контекста явно не следует другое. Таким образом, например, указание на «клетку» включает множество таких клеток, а указание на «белок» включает указание на один или более белков и их эквивалентов, известных специалистам в данной области техники, и так далее. Все технические и научные термины, применяемые в настоящей заявке, имеют те же значения, как и принято понимать специалистами той области техники, к которой относится данное изобретение, если явно не указано другое.
В контексте настоящего изобретения под «TCR-дефицитной Т-клеткой» или сходной фразой следует понимать выделенную Т-клетку(и), которая не экспрессирует функциональный TCR, внутренне способный к ингибированию выработки собственных TCR, а также для которых можно с определенной вероятностью ожидать, что потомство указанной Т-клетки внутренне способно к ингибированию выработки собственных TCR. Внутренняя способность является важной в контексте терапии, когда временная шкала обмена TCR (≈ часы) гораздо быстрее, чем демонстрируемая временная шкала эффективности (дни-недели), т.е., внутренняя способность требуется для сохранения желаемого фенотипа в период лечения. Этого можно достичь, например, разными способами, описанными ниже, например, путем получения измененной Т-клетки такой, чтобы она не экспрессировала никаких функциональных TCR на своей поверхности, или путем получения измененной Т-клетки такой, чтобы она не экспрессировала одну или более субъединиц, которые содержит функциональный TCR, и, следовательно, не вырабатывала функциональный TCR, или путем конструирования Т-клетки такой, чтобы она вырабатывала незначительное количество функционального TCRHa своей поверхности, или который экспрессирует по существу поврежденный TCR, например, путем получения измененной Т-клетки такой, чтобы она экспрессировала мутированные или укороченные формы одной или более субъединиц, которые содержит TCR, в результате чего указанная Т-клетка становится неспособна экспрессировать функциональный TCR, или образуется клетка, которая экспрессирует по существу поврежденный TCR. Разные субъединицы, которые составляют функциональный TCR, описаны ниже. Экспрессирует ли клетка функциональный TCR, можно определить при помощи способов анализа, таких как способы, известные в технике и описанные в настоящей заявке. Под «по существу поврежденным TCR» авторы заявки понимают, что данный TCR не будет по существу вызывать нежелательные иммунные реакции в организме хозяина, например, реакцию GVHD.
Как более подробно описано ниже, не обязательно указанные TCR-дефицитные клетки можно сконструировать так, чтобы они содержали другие мутации или трансгены, например, мутации или трансгены, которые влияют на рост или пролиферацию Т-клеток, приводят к экспрессии или отсутствию экспрессии желаемого гена или генной конструкции, например, другого рецептора или цитокина, или другого иммуномодулятора, или полипептида, или селектируемого маркера, такого как ген доминантного селектируемого маркера, например, DHFR или неомицин-трансфераза.
Термин «аллогенная Т-клетка» относится к Т-клетке, полученной от донора, обладающего тканевым типом HLA (лимфоцитарный антиген человека), который соответствует типу реципиента. Как правило, оценку соответствия проводят на основании вариабельности в трех или более локусах гена HLA, и предпочтительно полное соответствие. В ряде случаев доноры аллогенных транспланатов могут быть родственниками (обычно родные братья или сестры с близким HLA), изогенными субъектами (монозиготный «идентичный» близнец пациента) или не-родственниками (донор, который не является родственником и обнаруживает очень высокую степень соответствия HLA). Гены HLA делятся на две категории (I типа и II типа). Как правило, несоответствие генов I типа (т.е., HLA-A, HLA-B или HLA-C) повышают риск отторжения трансплантата. Несоответствие HLAII типа (т.е., HLA-DR или HLA-DQB1) повышает риск реакции «трансплантат против хозяина».
В контексте настоящего изобретения термин «банк тканесовместимых TCR-дефицитных Т-клеток» относится к разным композициям, содержащим Т-клетки конкретного аллотипа по HLA, которым придана недостаточность по TCR согласно настоящему изобретению. В идеальном случае указанный банк должен содержать композиции, содержащие Т-клетки разных типов TLA широкого спектра, которые репрезентативны для популяции людей. Такой банк сконструированных TCR-дефицитных Т-клеток может быть полезен, поскольку он облегчает доступ к Т-лимфоцитам, пригодным для применения у разных реципиентов, например, у пациентов с раком. Согласно настоящему изобретению предложены способы создания банка TCR-дефицитных Т- клеток, обладающих разными гаплотипами по HLA. Указанные способы включают получение совокупности выделенных Т- клеток, обладающих заданным гаплотипом по HLA, который определяют посредством стандартных способов типирования (например, антитела, ПЦР (полимеразная цепная реакция) или секвенирование ДНК), и экспрессирование TCR-ингибирующей молекулы (TIM) в указанных Т-клетках, которая дестабилизирует комплекс TCR путем снижения или блокирования экспрессии компонентов указанного комплекса TCR. Это выполняют для Т-клеток, полученных от широкого круга разных индивидуумов с разными гаплотипами по HLA. Такая совокупность Т- клеток от разных доноров, которые экспрессируют TIM, содержит банк TCR-дефицитных Т- клеток. Указанный банк Т-лимфоцитов включает разные совокупности Т-клеток, каждая из которых содержит TCR-дефицитные Т-лимфоциты с HLA специфического типа. Предпочтительно, чтобы такой банк Т-лимфоцитов включал разнообразные типы HLA, например, по меньшей мере 10 разных тканевых типов HLA, по меньшей мере 50 разных тканевых типов HLA, по меньшей мере 100 разных тканевых типов HLA. Согласно одному варианту реализации указанный банк Т-клеток включает Т-клетки по меньшей мере 10 разных тканевых типов HLA. Согласно другому варианту реализации указанный банк Т-лимфоцитов включает Т-клетки по меньшей мере 100 разных тканевых типов HLA.
В контексте настоящего изобретения под термином «терапевтически эффективное количество» специалисты в данной области техники понимают количество TCR-дефицитных Т-клеток, которое при их введении пациенту ослабляет признаки и симптомы заболевания (например, рака, инфекции или GVHD). Фактическое количество, которое следует вводить, можно определить на основании исследований, проведенных in vitro или in vivo, в которых TCR-дефицитные Т-клетки проявляли фармакологическую активность в отношении заболевания. Например, функциональные TCR-дефицитные Т-клетки могут подавлять рост in vitro или in vivo, и количество указанных функциональных TCR-дефицитных Т-клеток, которое подавляет такой рост, определяют как терапевтически эффективное количество.
Термин «фармацевтическая композиция» относится к химической или биологической композиции, пригодной для введения млекопитающему. Такие композиции можно составлять специально для введения одним или более путями, включая без ограничений буккальный, внутриартериальный, внутрисердечный, интрацеребровентрикулярный, внутрикожный, внутримышечный внутриглазной, внутрибрюшинный, интраспинальный, подоболочечный, внутривенный, пероральный, парентеральный, ректальный при помощи клизмы или суппозиториев, подкожный, субдермальный, сублингвальный, чрескожный и через слизистые оболочки. Кроме того, введение можно проводить в форме инъекции, жидкости, геля, капель или других средств введения.
В настоящей заявке под терминами «конструкция нуклеиновой кислоты» или «последовательность нуклеиновой кислоты» следует понимать молекулу ДНК, которую можно трансформировать или ввести в Т-клетку, и которая будет транскрибироваться и транслироваться с образованием продукта (например, химерного рецептора или белка суицида).
Нуклеиновые кислоты являются «функционально связанными», когда они находятся в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, ДНК для сигнальной последовательности функционально связана с ДНК для полипептида, если она экспрессируется как белок-предшественник, который участвует в секреции полипептида; промотор или энхансер функционально связаны с кодирующей последовательностью, если он влияет на транскрипцию указанной последовательности. Как правило, термин «функционально связан» означает, что последовательности ДНК, которые связаны, примыкают друг к другу и, в случае лидерной последовательности для секреции, примыкают друг к другу и находятся в рамке считывания. Однако энхансеры не обязательно должны примыкать. Связывания достигают путем сшивки в традиционных сайтах рестрикции или, в качестве альтернативы, при помощи способа ПЦР/рекомбинации, известного специалистам в данной области техники (Технология Gateway®; «Invitrogen», Карлсбад, Калифорния). Если такие сайты действительно существуют, применяют синтетические олигонуклеотиды-адаптеры или линкеры в соответствии с традиционной практикой.
Настоящее изобретение подразумевает композиции и способы уменьшения или облегчения, либо профилактики или лечения заболеваний или патологических состояний, таких как рак инфекционные заболевания, GVHD, отторжение трансплантата, одно или более аутоиммунные расстройства или лучевая болезнь. Согласно неограничивающему варианту реализации композиции основаны на концепции предоставления аллогенного источника изолированных Т- клеток человека, а именно TCR-дефицитных Т- клеток, которые можно производить до того, как пациент будет в них нуждаться, и недорого. Возможность создания единого терапевтического препарата на одном производственном участке с применением хорошо контролируемых процессов, является привлекательной с точки зрения и затрат, и качества. Переход с аутологичного источника на аллогенный источник Т- клеток дает значительные преимущества. Например, было оценено, что один здоровый донор может предоставить количество Т- клеток, достаточное для лечения двенадцати пациентов после трансдукции и экспансии.
Согласно настоящему изобретению модифицированные аллогенные Т-клетки производят таким образом, чтобы они не экспрессировали функциональных рецепторов Т-клеток (TCR). Следует понимать, что некоторые, или даже все, из субъединиц/димеров TCR могут экспрессироваться на поверхности клеток, но что указанные Т-клетки не экспрессируют достаточно функционального TCR, чтобы вызывать нежелательную реакцию у хозяина. Без функциональных TCR на своей поверхности аллогенные Т-клетки не могут вызвать нежелательный иммунный ответ на клетки в организме хозяина. Как следствие, указанные TCR-дефицитные Т-клетки не вызывают, например, CVHD, поскольку они не могут распознавать молекулы МНС хозяина. Кроме того, указанные TCR-дефицитные Т-клетки можно сконструировать так, чтобы они одновременно экспрессировали функциональные, отличный от TCR рецептор, специфичные для определенного заболевания рецепторы.
Как хорошо известно специалистам в данной области техники, есть разные способы для выделения аллогенных Т- клеток из организма субъекта. Например, применение экспрессии маркеров на поверхности клетки или применение коммерческих наборов (например, ISOCELL™ от компании «Pierce», Рокфорд, Иллинойс Rockford, Ill.).
Для терапии рака способ включает получение выделенной совокупности TCR-дефицитных Т-эффекторных клеток, например, аллотип желаемой ткани, которые не экспрессируют функциональную форму своих эндогенных TCR, или которые экспрессируют по существу пониженный уровень эндогенного TCR по сравнению с Т-клетками дикого типа, такой, что они не вызывают иммунного ответа при введении (такого как CVHD), но вместо них экспрессируют функциональный отличный от TCR рецептор, который распознает клетки опухоли, или экспрессирует другой полипептид, который не атакует ощутимо, или вообще не атакует, не связанные с заболевание клетки, например, нормальные (неонкогенные) клетки, которые не экспрессируют антиген или лиганд, распознаваемый опухоль-специфичным рецептором, или которые экспрессируют указанный антиген или лиганд на пониженном уровне по сравнению с клетками опухоли. Специалисты в данной области техники должны понимать, что определенные ассоциированные с опухолями антигены экспрессируются в нераковых тканях, но они являются возможными мишенями терапии у хозяев-носителей опухоли. В отношении изложенного среди специалистов в данной области техники принято считать, что определенные отличный от TCR рецептор, опухоль-специфичные рецепторы экспрессируются в нераковых тканях, но являются возможными мишенями терапии у хозяев-носителей опухоли, поскольку они могут экспрессироваться на значительно сниженном уровне в нормальных клетках по сравнению с клетками опухоли.
Хотя для большинства областей применения TCR-дефицитных Т- клеток субъекта и необязательно, но в некоторых случаях может быть желательно удалять некоторые или все из Т- клеток донора из организма хозяина после того, как они опосредовали свое противоопухолевое действие. Это может быть облегчено путем такого конструирования Т- клеток, чтобы они экспрессировали дополнительные рецепторы или маркеры, которые облегчают их удаление и/или идентификацию в организме хозяина, такие как GFP (зеленый флуоресцентный белок) и подобные. Хотя настоящее изобретение должно по существу исключать любую возможность GVHD или другой нежелательной иммунной реакции у реципиента, они могут быть желательны у некоторых индивидуумов. Это не должно уменьшать эффективность, поскольку уже было показано, что Т-клетки доноров не должны сохраняться долго в организме хозяина, чтобы инициировать противоопухолевое действие (Zhang, T., et al. 2007. Cancer Res. 67:11029-11036; Barber, Α. et al. 2008. J. Immunol. 180:72-78).
Согласно одному варианту реализации настоящего изобретения конструкции нуклеиновых кислот, введенные в сконструированные Т-клетки, дополнительно содержат ген суицида, такие как тимидинкиназа (ТК) вируса HSV (вируса герпеса) I типа (Bonini, et al. (1997) Science 276:1719-1724), «искусственный ген суицида» на основе Fas (Thomis, et al. (2001) Blood 97:1249-1257) или ген цитозин-диаминазы бактерии E.coli, которые активируются ганцикловиром, АР 1903 или 5-фторцитозином, соответственно. Указанный ген суицида успешно включают в конструкцию нуклеиновой кислоты согласно настоящему изобретению, чтобы обеспечить возможность удаления трансформированных Т-клеток в случае токсичности и разрушения химерной конструкции, когда опухоль будет уменьшена или удалена. Применение генов суицида для удаления трансформированных или трансфецированных клеток хорошо известно в технике. Например, Bonini, et al. ((1997) Science 276:1719-1724) показали, что лимфоциты донора, трансфецированные геном суицида HSV-TK, обеспечивают противоопухолевую активность у пациентов до одного года, а удаление трансфецированных клеток достигается при помощи ганцикловира. Далее, Gonzalez, et al. ((2004) J. Gene Med. 6:704-711) описали направленное воздействие на нейробластому при помощи клонов цитотоксических Т-клеток, генетически модифицированных так, чтобы они экспрессировали химерный иммунорецептор scFvFc:дзета на эпитоп на L1-CAM, причем указанная конструкция также экспрессирует ген суицида - гигромицин-тимидин-киназу (НуТК) для исключения трансгенных клонов.
Подразумевается, что ген суицида может экспрессироваться с того же промотора, что и кшРНК (короткая шпилечная РНК), миниген, отличный от TCR рецептор, или с другого промотора. Однако, как правило последовательности нуклеиновых кислот, кодирующие белок суицида и кшРНК, миниген или не-TCR-рецептор, находятся в одной конструкции или векторе. Экспрессии гена суицида с того же промотора, что кшРНК, миниген или He-TCR-рецептор, можно достичь при помощи любого из хорошо известных участков внутренней посадки рибосомы (1RES). Соответствующие последовательности 1RES, которые можно применять в конструкции нуклеиновой кислоты согласно настоящему изобретению, включают без ограничений 1RES из EMCV, c-myc, FGF-2, вируса полиомиелита и HTLV-1. Только в качестве иллюстрации конструкция нуклеиновой кислоты для экспрессии химерного рецептора может обладать следующей структурой: промотор -> химерный рецептор -> IRES-> ген суицида. В качестве альтернативы ген суицида может экспрессироваться с промотора, отличного от промотора химерного рецептора (например, промотор 1->химерный рецептор->промотор 2->ген суицида).
В связи с широким применением Т- клеток в клеточной терапии и усовершенствованной природой Т- клеток согласно настоящему изобретению, настоящее изобретение включает любой способ или композицию, в которых Т- клеток являются терапевтически желательными. Такие композиции и способы включают композиции и способы для снижения или облегчения, либо для профилактики или лечения рака, GVHD, отторжения трансплантата, инфекции, одного или более аутоиммунных заболеваний, лучевой болезни или других заболеваний или патологических состояний, которые основаны на применении Т- клеток, полученных из аллогенного источника, у которых не экспрессируются функциональные TCR.
Как указано, дополнительные варианты реализации настоящего изобретения включают рекомбинантную экспрессию рецепторов в указанных TCR-дефицитных Т-клетках, таких как химерный NKG2D, химерные домены Fv, NKG2D или любой другой рецептор для инициации сигналов для Т- клеток, вследствие которой создаются мощные специфические эффекторные Т-клетки. Специалист в данной области техники может выбрать соответствующий рецептор, чтобы экспрессировать его на TCR-дефицитных Т-клетках, в зависимости от заболевания, которое требуется лечить. Например, рецепторы, которые можно экспрессировать на TCR-дефицитных Т-клетках для лечения рака, могут включать любой рецептор или лиганд, который был идентифицирован в раковых клетках. Такие рецепторы включают без ограничений NKG2D, NKG2A, NKG2C, NKG2F, LLT1, AICL, CD26, NKRP1, NKp30, NKp44, NKp46, CD244 (2В4), DNAM-1 и NKp80.
Согласно другому варианту реализации настоящего изобретения такие рецепторы включают без ограничений NKG2D, NKG2A, NKG2C, NKG2F, LLT1, AICL, CD26, NKRP1, NKp30, NKp44, NKp46, CD244 ( В4), DNAM-1 и NKp80,или противоопухолевое антитело, такое как анти-Her2neu или анти-EGFR, и сигнальный домен, полученный из CD3-дзета, Dap10, СВ28, 41ВВ И CD40L. Примером химерного рецептора служит chNKG2D, в котором NKG2D соединен с цитоплазматическим доменом CD3-дзета, и ассоциирован с Dap10 для обеспечения как первичных, так и вторичных сигналов активации для Т- клеток (Zhang, T. et al. 2006. Cancer Res. 66(11): 5927-5933). Согласно одному варианту реализации настоящего изобретения химерный рецептор связывается с MIC-A, MIC-B, Her2neu, EGFR, мезотелином, CD38, CD20, CD 19, PSA, MUC1, MUC2, MUC3A, MUC3B, MUC4, MUC5AC, MUC5B, MUC6, MUC7, MUC8, MUC12, MUC13, MUC15, MUC16, MUC17, MUC19, MUC20, является рецептором эстрогена, рецептором прогестерона, RON, или одним или более членами семейства ULBP/RAET1, включая ULBP1, ULBP2, ULBP3, ULBP5 и ULBP6.
Согласно способам настоящего изобретения пациенту, страдающему раком, GVHD, отторжением трансплантата, инфекцией, одним или более аутоиммунными расстройствами или лучевой болезнью, вводят терапевтически эффективное количество композиции, содержащей указанные TCR-дефицитные Т-клетки. Согласно другому варианту настоящего изобретения терапевтически эффективное количество композиции, содержащей указанные TCR-дефицитные Т-клетки вводят для профилактики, лечения или уменьшения GVHD, отторжения трансплантата или рака.
Способы получения TCR-дефицитных Т-клеток
Т-клетки, у которых стабильно не экспрессируются функциональные TCR, согласно настоящему изобретению можно получить при помощи разных способов. Т-клетки интернализуют, сортируют и разрушают целые рецепторы Т-клеток в виде комплекса с временем полужизни приблизительно 10 часов у обычных Т- клеток, и 3 часа у стимулированных Т- клеток (von Essen, M. et al. 2004. J. Immunol. 173:384-393). Правильное функционирование комплекса TCR требует правильного стехиометрического соотношения белков, которые составляют комплекс TCR. Для функционирование TCR также требуется два функционирующих белка TCR-дзета с мотивами ITAM. Активация комплекса TCR при участии его МНС-пептидного лиганда требует участия нескольких TCR в одной Т-клетке, все из которых должны правильно передавать сигнал. Таким образом, если комплекс TCR дестабилизирован белками, которые не соединены правильным образом или не могут оптимально передавать сигнал, такая Т-клетка не активируется в достаточной степени для того, чтобы запустить клеточный ответ.
Способы согласно настоящему изобретению включают экспрессию TCR-ингибирующих молекул (TIM) в Т-клетках для дестабилизации комплекса TCR путем блокирования экспрессии основных компонентов комплекса TCR и/или приостановки экспрессии или функционирования TCR. Существуют разные классы TIM, включая без ограничений короткие шпилечные РНК (кшРНК) и доминантно-негативные белки-ингибиторы, например, процессированные белки, которые не содержат важных сигнальных мотивов; KIR-гибридные белки, которые способствуют ингибирующему сигналу; и белки с мутациями, которые нарушают правильное соединение с другими компонентами комплекса TCR и/или правильную передачу сигнала. Как правило, TIM можно применять для генерирования TCR-дефицитных Т- клеток за счет препятствования экспрессии каких-либо или очень слабо функциональных TCR на поверхности клетки, и/или для стимулирования экспрессии по существу поврежденных TCR на поверхности клетки.
Как показывают результаты в примерах экспериментов ниже, авторы настоящего изобретения продемонстрировали, что экспрессию или функцию TCR можно приостановить или устранить при помощи TIM, например, кшРНК и/или доминантно-негативных ингибиторов, получая при этом TCR-дефицитные Т-клетки. Такие линии TCR-дефицитных клеток хорошо подходят для применения в качестве средств на основе Т-клеток для лечения рака и других заболеваний и расстройств, как описано ниже.
Согласно одному варианту реализации настоящего изобретения экспрессию TCR устраняют при помощи коротких шпилечных РНК (кшРНК), которые направленно действуют на нуклеиновые кислоты, кодирующие специфические TCR (например, TCR-α и TCR-β) и/или цепи CD3 (например, CD3-дзета) в первичных Т-клетках. При блокировании экспрессии одного или более из указанных белков Т-клетка больше не сможет вырабатывать один или более из ключевых компонентов комплекса TCR, вследствие чего комплекс TCR дестабилизируется, и предотвращается экспрессия функционального TCR на поверхности клетки. Даже несмотря на то, что некоторые комплексы TCR могут возвращаться в оборот на поверхности клетки, кшРНК препятствует новой выработке белков TCR, что ведет к разрушению и удалению целого комплекса TCR, и образуются Т-лимфоциты со стабильной недостаточностью экспрессии функциональных TCR.
Экспрессии кшРНК в первичных Т-клетках можно достичь при помощи традиционной системы экспрессии, например, системы экспрессии лентивируса. Хотя лентивирусы применимы для направленного действия на большую часть первичных Т-клеток, не все Т-клетки будут экспрессировать кшРНК. Некоторые из указанных Т-клеток могут не экспрессировать достаточного количества кшРНК, чтобы достичь подавления экспрессии TCR, достаточного для изменения функциональной активности Т-клеток. Таким образом, Т-клетки, которые сохраняют умеренную или высокую экспрессию TCR после вирусной трансдукции, можно удалить, например, путем сортировки клеток или их разделения, так, чтобы оставшиеся Т-клетки были дефицитными в отношении TCR или CD3 на поверхности клеток, что позволяет провести экспансию выделенной популяции Т-клеток, дефицитных в отношении экспрессии функциональных TCR или CD3.
Согласно неограничивающему варианту реализации настоящего изобретения были разработаны примерные кшРНК для ключевых компонентов комплекса TCR, как указано ниже (Таблица 1).


мРНК TCR-альфа, TCR-бета, TCR-гамма, TCR-дельта, CD3-гамма, CD3-дельта, CD3-эпсилон или CD3-дзета могут обладать направленным действием по отдельности или вместе с применением разных направляющих кшРНК. Цепи TCR-β и TCR-α состоят из вариабельных и константных частей. Было разработано несколько направляющих кшРНК для константных частей указанных последовательностей TCR/CD3. Одну из комбинаций кшРНК можно применять для каждой из молекулярных мишеней для идентификации наиболее эффективного ингибитора экспрессии TCR. Применяя установленные протоколы каждую конструкцию кшРНК клонируют, например, в плазмиду pLko.1 или вектор pSMc2, и экспрессия контролируется промотором, стандартно применяемым в технике, например, промотором U6p.Полученный в результате конструкцию можно подвергнуть скринингу и подтвердить точность секвенирования. Плазмиду экспрессии кшРНК затем можно трансфецировать в любую подходящую клетку-хозяин (например, 293Т), вместе с упаковывающей плазмидой и плазмидой оболочки для упаковки. Первичные мононуклеарные клетки крови человека (PNMC) выделяют у здоровых доноров и активируют низкой дозой растворимого анти-CD3 и, например, от 25 Е/мл до 50 Е/мл, rhuIL-2 в течение 72 часов. Хотя для ретровирусной трансдукции не требуется активировать Т-лимофциты, трасндукция проходит более эффективно и позволяет клеткам продолжать развиваться в IL-2. Активированные клетки отмывают и проводят их трансдукцию, например, путем 1-часовой спин-фекции при 30°С, после которой наступает 7-часовой период покоя.
Согласно другому варианту реализации настоящего изобретения гиперэкспрессия доминантно-негативного белка-ингибитора способна привести к приостановке экспрессии или функции TCR. Согласно данному варианту реализации настоящего изобретения готовят миниген, который включает часть или весь полинуклеотид, кодирующий один из компонентов TCR (например, TCR-альфа, TCR-бета, CD3-гамма, CD3-дельта, CD3-эпсилон или CD3-дзета), но модифицируют его так, чтобы: (1) он не содержал основного сигнального мотива (например, ITAM), нужного для функции белка; (2) модифицирован так, чтобы он не соединялся правильно с другими своими природными компонентами TCR; или (3) мог правильно соединяться, но не связывал лигандов (например, процессированный миниген TCr-бета). Кроме того, миниген можно изменить так, чтобы он включал ингибиторный сигнальный мотив, например, цитоплазматический домен из белка KIR, который изменяет передачу сигнала в клетке и способствует ингибиторным сигналам посредством рекрутинга фосфатаз, например, SHP1 И SHP2.
Указанные минигены могут также кодировать часть белка, который служит как средство идентификации минигена, который гиперэкспрессируется. Например, полинуклеотиды, кодирующие процессированный белок CD 19, который содержит связывающий сайт для анти-СТ)19 mAb, могут быть функционально связаны с минигеном так, чтобы получаемая в результате клетка, которая экспрессирует указанный миниген, экспрессировала кодируемый белок и могла быть идентифицирована при помощи анти-CD19 mAb. Такая идентификация позволяет определять степень экспрессии минигена и выделять клетки, экспрессирующие указанный белок (и которые не содержат функционального TCR).
Согласно одному варианту реализации настоящего изобретения гиперэкспрессия минигена, не содержащего сигнального мотива(ов), приводит к образованию комплекса TCR, который не может правильно передавать сигнал, когда TCR контактирует со своим МСН-пептидом-лигандом на антагонистической клетке. Согласно неограничивающему варианту реализации настоящего изобретения высокая степень экспрессии указанного минигена (и кодируемого полипептида) приводит к вытеснению природного полного белка, когда компоненты TCR связываются, что ведет к образованию комплекса TCR, который не может передавать сигнал. Согласно другому варианту реализации указанный миниген включает, или в качестве альтернативы состоит из полинуклеотида, кодирующего полные полипептиды или часть полипептидов CD3-дзета, CD3-гамма, CD3-дельта или CD3-эпсилон, не содержащие мотивы ITAM, которые нужны для передачи сигнала. Белок CD3-дзета содержит три мотива IT AM в цитоплазматической части, и согласно одному варианту реализации настоящего изобретения, удаление всех из указанных мотивов посредством процессинга приводит к подавлению правильной передачи сигнала с TCR в любых комплексах, в которые включены указанные модифицированные белки. См., например, TIM5-8 в Таблице 2, который соответствует SEQ ID №: 72-79. Указанный конструкцию может включать ITIM или другой мотив передачи сигнала, о котором известно, что он изменяет передачу сигнала в клетке и способствуют ингибиторным сигналам посредством рекрутинга таких фосфатаз, как SHP1 и SHP2. См., например, TIM9-13 в Таблице 2, который соответствует SEQ ID №: 80-89.
Согласно одному варианту реализации настоящего изобретения указанный миниген содержит полинуклеотид, кодирующий полные полипептиды или часть полипептидов CD3-дзета, CD3-гамма, CD3-дельта или CD3-эпсилон с мутациями, например, изменением одного нуклеотида, которая приводит к изменению аминокислоты, кодируемой указанным полинуклеотидом. См., например, TIM14-19 в Таблице 2, которые соответствуют SEQ ID №: 90-101.
Согласно еще одному варианту реализации настоящего изобретения гиперэкспрессия минигена модифицирована так, чтобы кодируемый полипептид мог соединяться с некоторыми, но не со всеми из своих природных партнеров, создавая конкуренцию с нормальным белком за указанные соединяющиеся белки. Согласно еще одной неограничивающей гипотезе в настоящем изобретении высокий уровень экспрессии указанного минигена (и кодируемого полипептида) приводит к вытеснению природных белков-партнеров и препятствует сборке функционального комплекса TCR, для которой требуются все компоненты для соединения в правильных соотношениях и с правильным белок-белковым взаимодействием. Согласно другому варианту реализации настоящего изобретения, минигены содержат, или в качестве альтернативы состоят из всех или части полинуклеотидов, кодирующих полноразмерный белок (например, TCR-альфа, TCR-бета, CD3-гамма, CD3-дельта, CD3-эпсилон или CD3-дзета), но содержащих избранные замены в последовательности, кодирующей аминокислоты в трансмембранной части белка, которая, как известно, нужна для сборки с другими белками TCR/CD3.
Согласно одному предпочтительному варианту реализации настоящего изобретения избранные замены в последовательности, кодирующей аминокислоты в трансмембранной части белка, которая, как известно нужна для сборки с другими белками TCR/CD3, включают без ограничений остаток аргинина в положении 5 в трансмембранной области TCR-альфа; остаток лизина в положении 10 в трансмембранной области TCR-альфа; остаток лизина в положении 9 в трансмембранной области TCR-бета; остаток глутаминовой кислоты в трансмембранной области CD3-гамма; остаток аспарагиновой кислоты в трансмембранной области CD3-эпсилон; и остаток аспарагиновой кислоты в трансмембранной области CD3-дзета.
Гиперэкспрессия процессированного белка TCR-альфа, TCR-бета, CD3 -гамма или CD3-дельта приводит к образованию комплекса TCR, который не может связываться с МСН пептидами-лигандами, и таким образом, не функционирует, активируя Т-клетки. См., Фигуру 2, панели (А) и (В). Согласно другому варианту реализации настоящего изобретения минигены содержат, или в качестве альтернативы состоят из полинуклеотидов, кодирующих полные трансмембранные или цитоплазматические части указанных белков и части внеклеточной области, но не содержат полинуклеотидов, кодирующих весь или часть первого внеклеточного домена (т.е., самый наружный домен, содержащий лиганд-связывающий сайт). Согласно предпочтительному варианту реализации полинуклеотиды указанных минигенов не кодируют полипептиды V-альфа и V-бета цепей TCR-альфа и TCR-бета. Согласно одному варианту реализации полинуклеотиды указанных минигенов могут быть функционально связаны с полинуклеотидами, кодирующими белок-эпитопную метку (например, CD 19), что позволяет проводить идентификацию клеток, экспрессирующих указанные гены, при помощи mAb.
Согласно другому варианту реализации указанные минигены можно экспрессировать при помощи сильного вирусного промотора, такого как 5ʹLTR ретровируса, CMV или промотора SV40. Обычно указанный промотор находится непосредственно перед указанным минигеном и управляет высоко интенсивной экспрессией мРНК минигена. Слогласно другому варианту реализации указанная конструкция кодирует последовательность второго полинуклеотида под контролем того же промотора (например, с применением последовательности ДНК 1RES между ними) или под контролем другого промотора. Последовательность указанного второго полинуклеотида может кодировать функциональный отличный от TCR рецептор, обеспечивающий специфичность в отношении Т-клеток. Примеры указанного полинуклеотида включают без ограничений химерный NKG2D, химерный NKp30, химерный NKp46 или химерный анти-Her2neu. Согласно дополнительному варианту реализации комплекс промотор-минигены конструируют в плазмиде ретровируса или другой подходящей плазмиде экспрессии и проводят трансдукцию непосредственно Т-клеток с применением стандартных способов (Zhang, T. et al., (2006) Cancer Res., 66(11) 5927-5933; Barber, A. et al, (2007) Cancer Res., 67(10):5003-5008).
После трансдукции вирусом и экспансии с применением любого из способов, обсуждаемых ранее, любые Т-клетки, которые все еще экспрессируют TCR/CD3, удаляют при помощи анти-CD3 mAb, и магнитных гранул с применением колонок разделения Miltenyi, как было описано ранее (Barber, A. et al., (2007) Cancer Res., 67(10):5003-5008). Впоследствии Т-клетки отмывают и культивируют с IL-2 (25 Е/мл) в течение 3-7 дней, чтобы произошла экспансия эффекторных клеток, аналогично применению клеток in vivo.
Экспрессию TCR βα и CD3 можно оценить при помощи проточной цитометрии и количественной ПЦР в режиме реального времени (кРВ-ПЦР). Экспрессию мРНК TCR-α, TCR-β, CD3ε, CD3-ζ и GAPDH (в качестве контроля) можно анализировать при помощи кРВ-ПЦР с применением инструмента для ПЦР в режиме реального времени ABI7300 и геноспецифичных праймеров TAQMAN® при помощи способов, сходных со способами, описанными у Sentman, CL. et al. ((2004) J. Immunol. 173:6760-6766). Изменения экспрессии на поверхности клетки можно определять при помощи антител, специфичных в отношении TCR-α, TCR-β, CD3ε, CD8, CD4, CD5 и CD45.
Возможно, что одного вида кшРНК может быть не достаточно для подавления экспрессии TCR на поверхности клетки. В данном случае можно одновременно применять несколько кшРНК TCR, направленных на разные компоненты комплекса TCR. Для сборки комплекса TCR на поверхности клетки нужен каждый компонент комплекса TCR, поэтому утрата одного из указанных белков может приводить к утрате экспрессии TCR на поверхности клетки. Хотя может сохраняться некоторая часть или даже вся экспрессия TCR, будет ли рецептор вызывать иммунный ответ, определяется функцией рецептора. Критическим показателем является функциональная недостаточность, а не полное отсутствие на поверхности. Как правило, чем ниже уровень экспрессии TCR, тем меньше вероятность того, что возникнет достаточная перекрестная сшивка TCR, приводящая к активации Т-клеток через комплекс TCR. Хотя частные варианты реализации охватывают направленное действие на TCR-альфа, TCR-бета и CD3-эпсилон, можно направленно воздействовать также и на другие компоненты комплекса TCR, такие как CD3-гамма, CD3-дельта или CD3-дзета.
Основная цель удаления TCR с поверхности клетки состоит в предотвращении активации Т-клеток несовместимыми аллелями МНС. Чтобы определить, достаточно ли снижения экспрессии TCR при помощи каждой конструкции кшРНК или минигена для изменения функции Т-клетки, можно проводить исследование Т-клеток по параметрам: (1) выживания клеток in vitro; (2) пролиферации в присутствии митомицин С-обработанных аллогенных РВМС; и (3) выработки цитокинов в ответ на аллогенные РВМС, анти-CD3 mAb или анти-TCR mAb.
Чтобы оценить выживание клеток Т-лимфоциты, подвергнутые трансдукции, распределяют в полной среде RPMI (Онкологический институт Розуэлла Парка) с rhuIL-2 (например. 25 Е/мл - 50 Е/мл). Клетки помещают в чашки Петри при одинаковой плотности в начале культивирования, и можно отбирать пробы для подсчета клеток и оценки жизнеспособности раз в день в течение 7 или более дней. Чтобы определить, экспрессируют ли Т-клетки достаточно TCR, чтобы индуцировать ответ на аллогенные клетки, трансдуцированные или контрольные Т-клетки культивируют с обработанными митомицином С аллогенными или синергичными РВМС, например, в отношении 4:1. Указанные Т-клетки заранее нагружают CFSE, который является проникающим красителем клеток, который равномерно распределяется между дочерними клетками после деления. Степень деления клеток можно легко определить посредством проточной цитометрии. Другой отличительной особенностью активации Т- клеток является выработка цитокинов. Чтобы определить, подавляет ли каждый конструкция кшРНК функцию Т- клеток, подвергнутые трансдукции Т-клетки культивируют с разными дозами анти-CD3 mAb (от 1,6 до 5000 нг/мл). Через 24 часа отбирают надосадочную жидкость, не содержащую клеток, и определяют количество выработанных IL-2 и/или INF-γ при помощи ELISA (твердофазного иммуноферментного анализа). В качестве положительного контроля для стимуляции Т- клеток применяли РМА/иономицин, и Т-клетки одни применяют в качестве отрицательного контроля.
Оценивали действие примерных TIM, например, кшРНК, процессированных доминантно-негативных белков, KIR-гибридных доминантно-негативных белков и доминантно-негативных белков с измененной последовательностью аминокислот в следствие однонуклеотидной замены, разработанных для воздействия на ключевые компоненты комплекса TCR, например, CD3-эпсилон или CD3-дзета, на эффекторные Т-клетки, и результаты такой оценки представлены ниже в Таблице 2. Указанные результаты демонстрируют, что при применении TIM можно получать TCR-дефицитные Т-клетки.
Возможно, что удаление компонентов TCR-альфа или TCR-бета может позволить добиться предпочтительной экспансии TCR-гамма/дельта Т-лимфоцитов. Такие Т-клетки весьма редко встречаются в крови, однако присутствие указанных клеток можно определить при помощи антител на TCR-гамма/дельта. Если происходит естественное развитие данных клеток, можно применять направленное воздействие на CD3-эпсилон, который нужен для экспрессии на поверхности клетки TCR-альфа/бета и TCR-гамма/дельта. И Il-2, и INF-γ являются основными эффекторными цитокинами, которые запускают экспансию Т-клеток и активацию макрофагов. Следовательно, отсутствие выработки указанных цитокинов является признаком функциональной инактивации. Также можно измерить изменения уровня других цитокинов, таких как TNF-α (фактор некроза опухоли-α). Любое снижение выживаемости Т-клеток при исключении экспрессии TCR можно определить посредством культивирования TCR-дефицитных Т-клеток с РВМС, которые лучше соответствуют окружению in vivo и обеспечивают поддержку для выживания Т-клеток.
Способы получения TCR-дефицитных Т-клеток, экспрессирующих функциональный рецептор, не свойственный Т-клеткам
Согласно другому варианту реализации настоящего изобретения Т-клетки, стабильно дефектные в отношении экспрессии функционального TCR, экспрессируют функциональный отличный от TCR рецептор. Согласно данному варианту реализации устранение функции TCR (как было описано ранее) также сочетают с экспрессией одного или более экзогенных He-TCR-направленных рецепторов (например, таких как химерные молекулы NKG2D (chNKG2D) или Fv). Согласно данному варианту реализации предложены «универсальные» клеточные продукты, которые можно хранить для будущей терапии любого пациента с любым типом раком, при условии, что применяется подходящий направляющий рецептор.
Дополнительные варианты реализации настоящего изобретения включают рекомбинантную экспрессию рецепторов в указанных TCR-дефицитных Т-клетках, таких как chNKG2D, химерные домены Fv, NKG2D или любой другой рецептор для инициации сигналов в Т-лимфоцитах, в результате чего создаются мощные, специфические эффекторные Т-клетки. Специалисты в данной области техники могут выбрать соответствующий рецептор, который следует экспрессировать в TCR-дефицитных Т-клетках, в зависимости от заболевания, которое требуется лечить. Например, рецепторы, которые можно экспрессировать в TCR-дефицитных Т-клетках для лечения рака, могут включать любой рецептор к лиганду, который был идентифицирован в раковых клетках. Такие рецепторы включают без ограничений NKG2D (номер доступа в системе GENBANK ВС039836), NKG2A (номер доступа в системе GENBANK AF461812), NKG2C (номер доступа в системе GENBANK AJ001684), NKG2F, LLT1, AICL, CD26, NKRP1, NKp30 (например, номер доступа в системе GENBANK АВ055881), NKp44 (например, номер доступа в системе GENBANK AJ225109), NKp46 (например, номер доступа в системе GENBANK AJ001383), CD244 (2В4), DNAM-1 и NKp80.
Согласно другому варианту реализации настоящего изобретения такие рецепторы включают без ограничений химерные рецепторы, содержащие лиганд-связывающий домен, полученный из NKG2D, NKG2A, NKG2C, NKG2F, LLT1, AICL, CD26, NKRP1, NKp30, NKp44, NKp46, CD244 (2 В4), DNAM-1 и NKp80, или противоопухолевое антитело, такое как анти-Her2neu и анти-EGFR, и сигнальный домен, полученный из CD3-дзета (CD3ζ) (например, номер доступа в системе GENBANK NM_198053) (SEQ ID №: 62), Dap 10 (например, номер доступа в системе GENBANK AF072845), CD28, 41BB и/или CD40L.
Согласно дополнительному варианту реализации настоящего изобретения указанный химерный рецептор связывается с MIC-A, MIC-B, Her2neu, EGFR, мезотелином, CD38, CD20, CD 19, PSA, MUC1, MUC2, MUC3A, MUC3B, MUC4, MUC5AC, MUC5B, MUC6, MUC7, MUC8, MUC12, MUC13, MUC15, MUC16, MUC17, MUC19, MUC20, рецептором эстрогена, рецептором прогестерона, RON, или одним или более членами семейства ULBP/RAET1, включая ULBP1, ULBP2, ULBP3, ULBP4, ULBP5 и ULBP6.
Только с целью иллюстрации кшРНК или минигены, для которых было показано устранение экспрессии на поверхности клетки комплекса TCR, экспрессируют совместно с рецептором chNKG2D при помощи одного или более вирусных векторов. Для достижения совместной экспрессии в одном векторе кшРНК можно запускать с промотора U6, а рецептор chNKG2D с промотора PGK. Согласно другому варианту реализации, если для разделения генетических элементов применяют последовательность 1RES, то применяют только один промотор.
Белок-рецептор лектин-подобного домена С-типа клеток NK, особенно подходящий для применения в химерном рецепторе, включает рецептор, экспрессируемый на поверхности клеток-естественных киллеров, причем при связывании с его родственным лигандом(ами), он изменяет активацию клетки NK. Указанный рецептор может функционировать один или совместно с другими молекулами. Лиганды к указанным рецепторам, как правило, экспрессируются на поверхности клеток одного или более типов опухолей, например, опухолей, ассоциированных с раком ободочной кишки, легких, молочной железы, почек, яичников, шейки матки и предстательной железы; меланом; миелом; лейкозов и лимфом Wu, et al. (2004) J. Clin. Invest. 114:60-568; Groh, et al. (1999) Proc. Natl. Acad. Sci. USA 96:6879-6884; Pende, et al. (2001) Eur. J. Immunol. 31:1076-1086), но не экпрессируются широко на поверхности клеток в нормальных тканях.
Примеры таких лигандов включают без ограничений MIC-A, MIC-B, белки теплового шока, ULBP-связывающие белки (например, ULBP 1-4) и неклассические молекулы HLA, такие как HLA-E И HLA-G, тогда как классические молекулы МНС, такие как HLA-A, HLA-B или HLA-C и их аллели не считаются сильными лигандами белка-рецептора лектин-подобного домена С-типа клеток NK согласно настоящему изобретению. Рецепторы лектин-подобного домена С-типа клеток NK, которые связываются с указанными лигандами, как правило, обладают структурой белка II типа, в которой N-конец белка является внеклеточным. Кроме рецепторов клеток NK, перечисленных выше, дополнительные примеры рецепторов клеток NK включают без ограничений дектин-1 (номер доступа в системе GENBANK AJ312373 или AJ312372) антиген, ассоциированный с функцией тучных клеток (номер доступа в системе GENBANK AF097358), HNKR-P1A (номер доступа в системе GENBANK U11276), LLT1 (номер доступа в системе GENBANK ΦΑΙ 33299), CD69 (номер доступа в системе GENBANK NM_001781), гомолог СВ69, CD72 (номер доступа в системе GENBANK NM_001782), CD94 (номер доступа в системе GENBANK NM_00262 или NM_007334), KLRF1 (Номер доступа в системе GENBANK NM_016523), рецептор окисленной LDL (номер доступа в системе GENBANK NM_002543), CLEC-1, CLEC-2 (номер доступа в системе GENBANK NM_016509), NKG2D (номер доступа в системе GENBANK ВС039836), NKG2C (номер доступа в системе GENBANK AJ001684), NKG2A (номер доступа в системе GENBANK AF461812), NKG2E (номер доступа в системе GENBANK AF461157), WUGSC:H_DJ0701016.2 или миелоид ОАР12-ассоциированный лектин (MDL-1; номер доступа в системе GENBANK AJ271684). Согласно предпочтительным вариантам реализации настоящего изобретения рецептор клеток NK является NKG2D человека (SEQ ID №: 58) или NKG2C человека (SEQ ID №: 59).
Аналогичные рецепторы I типа, которые могут быть полезны в составе химерного рецептора, включают NKp46 (номер доступа в системе GENBANK AJ001383), NKp30 (номер доступа в системе GENBANK АВ055881) или NKp44 (номер доступа в системе GENBANK AJ225109).
В качестве альтернативы белку-рецептору лектин-подобного домена С-типа клеток NK в составе химерного белка-рецептора можно применять белок, ассоциированный с белком-рецептором лектин-подобного домена С-типа клеток NK. Как правило, под белками, ассоциированными с белком-рецептором лектин-подобного домена С-типа клеток NK, понимают белки, которые взаимодействуют с рецептором и посредством этого индуцируют сигнал. Подходящие белки человека, которые функционируют таким образом, дополнительно включают без ограничений DAP 10 (например, номер доступа в системе GENBANK AF072845)(SEQ ID №: 60), DAP 12 (например, номер доступа в системе GENBANK AF019562)(SEQ ID №: 61) и FcR гамма.
К N-концу рецептора лектин-подобного домена С-типа клеток NK присоединяют рецептор иммунной сигнализации, обладающий мотивом активации иммунорецепторов на основе тирозина (ITAM), (Asp/Glu)-Xaa-Xaa-Tyr*-Xaa-Xaa-(Ile/Leu)-Xaa6-8-Tyr*-Xaa-Xaa-(Ile/Leu) (SEQ ID №: 55-57), который участвует в активации ответов клетки через иммунные рецепторы. Аналогично, когда применяют белок, ассоциированный с рецептором лектин-подобного домена С-типа клеток NK, рецептор иммунной сигнализации можно соединять с С-концом указанного белка (ФИГ. 1). Подходящие рецепторы иммунной сигнализации для применения в составе химерного рецептора согласно настоящему изобретению включают без ограничений дзета-цепь рецептора Т-клеток, эта-цепь, которая отличается от дзета-цепи только по самому крайнему экзону на С-конце вследствие альтернативного сплайсинга мРНК дзета, дельта-, гамма- и эпсилон-цепи рецептора Т-клеток (цепи CD3) и гамма-субъединицу рецептора FcR1. Согласно конкретным вариантам реализации наряду с рецепторами иммунной сигнализации, описанными выше, рецептором иммунной сигнализации является также CD3-дзета (CD3ζ) (например, номер доступа в системе GENBANK для генов человека NM_198053 и NM-000734) (SEQ ID №:62 и SEQ ID №:64, соответственно), или гамма-цепь рецептора Fc-эпсилон (например, номер доступа в системе GENBANK М33195) (SEQ ID №: 63) или цитоплазматический домен, или его вариант сплайсинга. В частности, например, CD3-дзета обладает 2 альтернативными транскриптами-вариантами сплайсинга, кодирующими разные изоформы, т.е., вариант транскрипта 1 (SEQ ID №: 62) и вариант транскрипта 2 (SEQ ID №: 64). В кодируемой изоформе варианта 2 (SEQ ID №: 65) пропущена внутренняя аминокислота по сравнению с вариантом 1.
Согласно конкретным вариантам реализации химерным рецептором согласно настоящему изобретению является гибрид между NKG2D и CD3-дзета, или Dap10 и CD3-дзета.
В конструкции нуклеиновой кислоты согласно настоящему изобретению промотор функционально связан с последовательностью нуклеиновой кислоты, кодирующей химерный рецептор согласно настоящему изобретению, т.е., он расположен так, чтобы способствовать транскрипции информационной РНК с ДНК, кодирующей указанный химерный рецептор. Указанный промотор может быть геномного происхождения или может быть сгенерирован путем синтеза. В технике хорошо известен целый ряд промоторов для Т-клеток (например, С04-промотор, раскрываемый у Marodon, et al. (2003) Blood 101(9):3416-23). Такой промотор может быть конститутивным или индуцибельным, причем индукция ассоциирована со специфическим типом клеток или специфическим уровнем зрелости. В качестве альтернативы подходит также целый ряд хорошо известных вирусных промоторов. Промоторы, представляющие интерес, включают промотор β-актина, ранний и поздний промоторы SV40, промотор иммуноглобулина, промотор цитомегаловируса человека, промотор ретровируса и промотор вируса некроза селезенки Фрейнда. Указанные промоторы могут быть ассоциированы или не ассоциированы с энхансерами, причем указанные энхансеры могут быть от природы ассоциированы с определенным промотором или с разными промоторами.
Последовательность открытой рамки считывания, кодирующую химерный рецептор, можно получить из ДНК геномного источника, источника кДНК или можно синтезировать (например, посредством ПЦР) или комбинируя данный источники. В зависимости от размера геномной ДНК и количества интронов может быть желательно применять кДНК или их сочетание, поскольку было показано, что интроны стабилизируют мРНК или обеспечивают специфическую для Т-клеток экспансию (Barthel and Goldfeld (2003) J. Immunol. 171(7):3612-9). Также дополнительную пользу может принести применение эндогенных или экзогенных некодирующих областей для стабилизации мРНК.
Для экспрессии химерного рецептора согласно настоящему изобретению можно применять существующую в природе или эндогенную область инициации транскрипции или последовательность нуклеиновой кислоты, кодирующую N-концевой компонент указанного химерного рецептора, чтобы сгенерировать химерный рецептор в целевом хозяине. В качестве альтернативы можно применять экзогенную область инициации транскрипции, которая позволяет проводить конститутивную или индуцибельную экспрессию, причем экспрессию можно контролировать в зависимости от целевого хозяина, желаемого уровня экспрессии, природы целевого хозяина и т.п.
Аналогично, сигнальная последовательность, направляющая химерный рецептор на поверхность мембраны, может быть эндогенной сигнальной последовательностью N-концевого компонента химерного рецептора. Необязательно, в некоторых случаях, может быть желательно заменить данную последовательность на другую сигнальную последовательность. Однако выбираемая сигнальная последовательность должна быть совместима с секреторным путем Т-клеток, чтобы химерный рецептор присутствовал на поверхности Т-клетки.
Аналогично, область терминации может быть существующей в природе или эндогенной областью терминации транскрипции последовательности нуклеиновой кислоты, кодирующей С-концевой компонент химерного рецептора. В качестве альтернативы область терминации можно получить из другого источника. В большинстве случаев считается, что источник получения области терминации не является критичным для экспрессии рекомбинантного белка, и можно применять широкой спектр областей терминации, что не должно отрицательно сказаться на экспрессии.
Как должно быть понятно специалистам в данной области техники, в некоторых случаях несколько аминокислот на концах рецептора лектин-подобного домена С-типа естественных киллеров (или белка, ассоциированного с ним) или рецептора иммунной сигнализации можно удалить, обычно не более 10, еще чаще не более 5 остатков. Также может быть желательно ввести небольшое число аминокислот на границах, обычно не более 10, еще чаще не более 5 остатков. Делеция или вставка аминокислот обычно являются следствием необходимости получить конструкцию, предусматривающую традиционные сайты рестрикции, простоту манипулирования, повышенный уровень экспрессии, или тому подобное. Кроме того, по сходным причинам может происходить замена одной или более аминокислот на другие аминокислоты, но обычно не заменяют более пяти аминокислот в любом одном из доменов.
Химерную конструкцию, которая кодирует химерный рецептор, можно получить традиционными способами. Поскольку в большинстве случаев применяют природные последовательности, природные гены выделяют и манипулируют с ними надлежащим образом (например, когда применяют рецептор II типа, рецептор иммунной сигнализации может потребоваться инвертировать), чтобы было возможно правильное соединение разных компонентов. Таким образом, последовательность нуклеиновой кислоты, кодирующая белки на N-конце и С-конце химерного рецептора можно выделить при помощи полимеразной цепной реакции (ПЦР) с применением соответствующих праймеров, которая приводит к делеции нежелательных частей гена. В качестве альтернативы, для создания химерной конструкции можно применять расщепление рестриктазами клонированных генов. В любом случае, последовательности можно выбирать так, чтобы были сайты рестрикции, которые содержат «тупые» концы, или содержат комплементарное перекрывание.
Разные манипуляции при получении химерной конструкции можно проводить in vitro, и согласно конкретным вариантам реализации указанную химерную конструкцию вводят в векторы для клонирования и экспрессии у соответствующего хозяина с применением стандартных способов трансформации или трансфекции. Таким образом, после каждой манипуляции конструкцию, полученную в результате соединения последовательностей ДНК, клонируют, вектор выделяют, и последовательность подвергают скринингу, чтобы убедиться, что указанная последовательность кодирует желаемый химерный рецептор. Можно проводить скрининг последовательности посредством рестрикционного анализа, секвенирования или т.п.
Подразумевается, что химерную конструкцию можно вводить в Т-клетки в виде оголенной ДНК или в составе подходящего вектора. Способы стабильной трансфекции Т-клеток при помощи электропорации с применением оголенной ДНК известны в технике. См., например, Патент США №6410319. Термин «оголенная ДНК», как правило, относится к ДНК, кодирующей химерный рецептор согласно настоящему изобретению, содержащейся в векторе экспрессии на основе плазмиды в правильной ориентации для экспрессии. Предпочтительно, применение оголенной ДНК уменьшает время, которое требуется для получения Т-лимфоцитов, экспрессирующих химерный рецептор согласно настоящему изобретению.
В качестве альтернативы, для введения химерного конструкции в Т-клетки можно применять вирусный вектор (например, вектор на основе ретровируса, вектор на основе аденовируса, вектор на основе аденоассоциированного вируса или лентивирусный вектор). Подходящие векторы для применения при реализации способа согласно настоящему изобретению не реплицируются в Т-клетках субъекта. Известен широкий спектр векторов, которые основаны на вирусах, когда количество копий вируса в клетке поддерживается на низком уровне, достаточном, чтобы сохранить жизнеспособность клетки. Иллюстрационные векторы включают векторы pFB-neo (STRATAGENE™), а также векторы на основе ВИЧ, SV40, EBV, HSV или BPV. Как только установлено, что Т-клетка, подвергнутая трансфекции или трансдукции, способна экспрессировать химерный рецептор как поверхностный мембранный белок с желаемой регуляцией и на желаемом уровне, можно определить, является ли указанный химерный рецептор функциональным в клетке-хозяине, чтобы обеспечить желаемую индукцию сигнала (например, выработку Rantes, Mipl -alpha, GM-CSF при стимуляции соответствующим лигандом).
Как было описано выше, первичные РВМС человека выделяют у здоровых доноров и активируют низкими дозами малорастворимого анти-CD3 (например, 40 нг/мл) и rhuIL-2 (например, 50 Е/мл), гранулами с анти-CD3/анти-CD28 и rhuIL-2, или облученными антиген-презентирующими клетками и rhuIL-2. Затем активированные Т-клетки отмывают и подвергают трансдукции с применением ретровируса, например, обработка ультразвуковом в течение 1 часа при 32°С с последующим периодом покоя 7 часов. Хотя для лентивирусной трансдукции не требуется активировать Т-клетки, трансдукция проходит более эффективно и позволяет клеткам продолжать развиваться в IL-2. Активированные клетки отмывают и проводят их трансдукцию, как описано выше, после которой следует период покоя, а затем их передают на этап отбора, например, с G418 в течение 3 дней. После отбора клетки отмывают и культивируют в IL-2 в течение 2-7 дней, чтобы стала возможна экспансия эффекторных клеток аналогично тому, как это происходит при применении клеток in vivo. Изменение экспрессии рецепторов на поверхности клетки анализируют при помощи антител, специфичных в отношении CD3, CD4, NKG2D или CD5. Ожидается, что экспрессия экзогенного, отличного от TCR рецептора будет повышена в клетках, которые подверглись трансдукции с целью экспрессии такого конкретного рецептора, например, ожидается, что в Т-клетках, подвергнутых трансдукции chNKG2D-экспрессирующим ретровирусом, будет повышен уровень экспрессии chNKG2D на поверхности клетки.
Экспрессию TCRαβ, CD3 и NKG2D можно оценить при поморщи проточной цитометрии и количественной кРВ-ПЦР, как обсуждается в настоящей заявке. Также можно определять количество CD4+ и CD8+-Т-лимфоцитов. Общее количество клеток и процент Т-лимфоцитов, дефицитных по комплексу TCR, TCR-компетентных и экспрессирующих chNKG2D, можно определять при помощи проточной цитометрии. Указанное количество можно сравнивать с РВМС, которые подвергли трансдукции только кшРНК или генами chNKG2D (в качестве контроля). В виде контроля можно также включить клетки, подвергнутые трансдукции только вектором.
После трансдукции вирусом и экспансии TCR+ и TCR- клетки можно разделить при помощи mAb с магнитными гранулами на колонках разделения Miltenyi, и TCR-дефецитные Т-клетки, экспрессирующие рецептор chNKG2D, выявляют и выделяют. Например, экспрессию chNKG2D можно проверить при помощи количественной ПЦР в режиме реального времени с применением специфических праймеров на рецептор chNKG2D (Zhang, T. et al. (2007) Cancer Res. 67:11029-11036; Barber, A. et al. (2008) J. Immunol. 180:72-78). Функцию указанных TCR-дефицитных chNKG2D+ клеток можно определить путем культивирования указанных клеток с PNMC или клетками опухоли, которые экспрессируют лиганды chNKG2D. Пролиферацию Т-клеток и выработку ими цитокинов (например, IFN-γ и/или IL-2) можно определять при помощи проточной цитометрии и ELISA, соответственно. Для того, чтобы идентифицировать Τ клетки, которые потеряли функцию TCR и сохранили функцию chNKG2D, трансдуцированные или контрольные Т-клетки культивируют с антителом против-CD3 (1.6 - 5000 нг/мл), аллогенными мононукеарами периферической крови (МНПК), обработанными митомицином С, или сингенными МНПК. Отбирают супернатанты культур, и определяют уровень продукции цитокинов (IFN-gamma и/или IL-2) с помощью ELISA. Указанные Т-клетки можно заранее нагрузить CFSE, который является проникающим красителем клеток, который равномерно распределяется между дочерними клетками после деления. Степень деления клеток можно легко определить посредством проточной цитометрии.
Другой отличительной особенностью активации Т-клеток является выработка цитокинов. Чтобы определить, вызывают ли TCR-дефецитные chNKG2D+-клетки активацию Т-клетки, указанные Т-клетки культивируют совместно с аллогенными РВМС, обработанными митомицином С, или клетками опухоли: P815-MICA (опухоль мыши, экспрессирующая MICA - лиганд для NKG2D), Р815, А2008 (опухоль яичников человека - лиганд+ NKG2D) и U266 (линия клеток миеломы человека, лиганд+ NKG2D). Через 24 часа отбирают надосадочную жидкость, не содержащую клеток, и при помощи ELISA определяют количество выработанных IL-2 и INF-γ . В качестве отрицательного контроля применяли Т-клетки одни и в культуре с РМВС. В Т-клетках, экспресировавших TIM7 и TIM8, которые также одновременно экспрессировали chNKG2D, наблюдали более чем 40% снижение выработки IFN-γ (результаты не показаны на Фигуре 3).
Впоследствии подвергнутые трансдукции Т-клетки повторно вводят субъекту для активации противоопухолевых реакций у указанного субъекта. Для облегчения введения подвергнутые трансдукции Т-клетки согласно настоящему изобретению можно преобразовать в фармацевтическую композицию или выполнить из них имплантат, подходящие для введения in vivo с соответствующими основами и растворителями, которые также могут быть фармацевтически приемлемыми. Способы получения такой композиции или имплантата были описаны в технике (см, например, Remingtonʹs Pharmaceutical Sciences, 16th Ed., Mack, ed. (1980)). Если применимо, подвергнутые трансдукции Т-клетки можно преобразовать в препарат в полутвердой или жидкой форме, например, капсулы, раствор, препарат для инъекций, ингаляций или аэрозоль, обычными способами для соответствующего им пути введения. Для предупреждения или минимизации всасывания композиции до того, как она не достигнет ткани или органа-мишени, или для обеспечения своевременного высвобождения композиции, можно применять способы, известные в технике. Однако желательно применять фармацевтически приемлемую форму, которая не будет неэффективной в отношении клеток, экспрессирующих химерный рецептор. Таким образом, желательно, чтобы подвергнутые трансдукции Т-клетки можно было сделать в виде фармацевтической композиции, содержащей сбалансированный раствор солей, предпочтительно сбалансированный солевой раствор Хенкса или нормальный физиологический раствор.
Способы облегчения или уменьшения симптомов, лечения или профилактики заболеваний или расстройств с применением TCR-дефицитных клеток
Также настоящее изобретение направлено на способы уменьшения или облегчения, либо профилактики или лечения заболеваний и расстройств с применением TCR-дефицитных Т-клеток, описанных в настоящей заявке, их выделенных популяций или терапевтических композиций, их содержащих. Согласно одному варианту реализации TCR-дефицитные Т-клетки, описанные в настоящей заявке, их выделенные популяции или терапевтические композиции, их содержащие, применяют для уменьшения или облегчения, либо для профилактики или лечения рака, инфекции, одного или более аутоиммунных расстройств, лучевой болезни, или для профилактики или лечения реакции «трансплантат против хозяина» (GVHD) или отторжения трансплантата у субъекта, подвергшегося операции по пересадке органа или ткани.
TCR-дефицитные Т-клетки, описанные в настоящей заявке, их выделенные популяции или терапевтические композиции, их содержащие, применимы для изменения аутоиммунных реакций или отторжения трансплантата, поскольку данные эффекторные клетки можно вырастить в TGF-β во время развития, и они будут дифференцироваться, становясь индуцированными Т-регуляторными лимфоцитами. Согласно одному варианту реализации функциональные отличные от TCR рецепторы применяют для придания указанным индуцированным Т-регуляторным лимфоцитам функциональной специфичности, которая нужна им для осуществления их ингибиторной функции в участке ткани, пораженном болезнью. Таким образом, удается вырастить большое количество антиген-специфичных регуляторных Т-клеток для применения у пациентов. Экспрессию FoxP3, которая важна для дифференцировки Т-регуляторных лимфоцитов, можно анализировать при помощи проточной цитометрии, и функциональное подавление пролиферации Т-клеток указанными Т-регуляторными лимфоцитами можно анализировать, изучая снижение пролиферации Т-клеток после стимуляции анти-СБЗ при совместном культивировании.
Другой вариант реализации настоящего изобретения направлен на применение TCR-дефицитных Т-клеток, описанных в настоящей заявке, их выделенных популяций или терапевтических композиций, их содержащих, для профилактики или лечения лучевой болезни. Одной из проблем после воздействия излучения (например, взрыв «грязной» ядерной бомбы, утечка радиации) или другого патологического состояния, которое поражает костный мозг (определенные виды медикаментозного лечения) является восстановление системы гемапоэза. У пациентов, подвергшихся трансплантации костного мозга, абсолютное количество лимфоцитов на 15-ый день после трансплантации коррелирует с успешным исходом. Такие пациенты с большим количеством лимфоцитов хорошо восстанавливаются, поскольку важно, чтобы было хорошее восстановление лимфоцитов. Причина такого действия не ясна, но это может быть связано с защитой лимфоцитов от инфекции и/или выработкой факторов роста, которые благоприятствуют восстановлению гемапоэза.
Согласно данному варианту реализации TCR-дефицитные Т-клетки, описанные в настоящей заявке, их выделенные популяции или терапевтические композиции, их содержащие, приводят к выработке большого количества Т-клеток, которые неспособны отвечать на аллогенные антигены МНС. Следовательно, указанные Т-клетки можно применять для восстановления людей, и они могут обеспечивать защиту от инфекции, что ведет к более быстрому восстановлению людей, страдающих полным или частичным поражением костного мозга в связи с действием излучения. В случаях катастрофического или непредвиденного воздействия высоких доз излучения TCR-дефицитные Т-клетки, описанные в настоящей заявке, обладающие другим функциональным рецептором, их выделенные популяции или терапевтические композиции, их содержащие, можно быстро ввести путем инфузии пациентам, чтобы обеспечить некоторую степень восстановления их иммунного ответа и выработки факторов роста в течение первых дней - недель, пока собственные гемопоэтические клетки сами не восстановятся, или пока человек не получит в качестве лечения дополнительный источник гемопоэтических стволовых клеток (например, трансплантация костного мозга).
Специалисты в данной области техники должны понимать, как лечить рак, инфекцию, отторжение трансплантата, одно или более аутоиммунные расстройства, лучевую болезнь или GVHD на основании их опыта применения других типов Т-клеток.
В дополнение к иллюстративным TCR-дефицитным chNKG2D+ Т-клеткам, описываемым в настоящей заявке, подразумевается, что TCR-дефицитные Т-клетки можно модифицировать или разработать так, чтобы они экспрессировали другие функциональные рецепторы, применимые при лечении таких заболеваний, как рак или инфекция, как было описано ранее. Если кратко, способы лечения согласно настоящему изобретению подразумевают применение TCR-дефицитных Т-клеток, экспрессирующих функциональные отличные от TCR рецепторы, такие как chNKG2D, химерные домены Fv, NKG2D, или любой другой рецептор для инициации сигнала к Т-клеткам, вследствие чего удается создать мощные, специфические эффекторные Т-клетки. Специалисты в данной области техники должны быть способны выбрать соответствующий рецептор для его экспрессии TCR-дефицитными Т-клетками в зависимости от заболевания, которое требуется лечить. Например, рецепторы, которые могут экспрессироваться TCR дефицитными Т-клетками для лечения рака, могут включать любой рецептор, который связывается с лигандом, который был идентифицирован на раковых клетках. Такие рецепторы включают без ограничений NKG2D, NKG2A, NKG2C, NKG2F, LLT1, AICL, CD26, NKRP1, NKp30, NKp44, NKp46, CD244 (2В4), DNAM-1 и NKp80.
Согласно другому варианту реализации настоящего изобретения такие рецепторы включают без ограничений химерные рецепторы, содержащие лиганд-связывающий домен из NKG2D, NKG2A, NKG2C, NKG2F, LLT1, AICL, CD26, NKRP1, NKp30, NKp44, NKp46, CD244 (2В4), DNAM-1 и NKp80, или из противоопухолевого антитела, такого как анти-Нег2пеи и анти-EGFR, и сигнальный домен, полученный из CD3-дзета, Dap10, CD28, 41NN и CD40L.
Согласно дополнительному варианту реализации указанный химерный рецептор связывается с MIC-A, MIC-B, Her2neu, EGFR, мезотелином, CD38, CD20, CD19, PSA, MUC1, MUC2, MUC3A, MUC3B, MUC4, MUC5AC, MUC5B, MUC6, MUC7, MUC8, MUC12, MUC13, MUC15, MUC16, MUC17, MUC19, MUC20, рецептором эстрогена, рецептором прогестерона, RON, или одним или более членами семейства ULBP/RAET1, включая ULBP1, ULBP2, ULBP3, ULBP5 и ULBP6.
Также настоящее изобретение охватывает TCR-дефицитные Т-клетки, которые экспрессируют отличный от TCR патоген-ассоциированный рецептор, и применение указанных TCR-дефицитных Т-клеток, экспрессирующих рецептор к патогену, для лечения или профилактики инфекционных заболеваний. Согласно данному варианту реализации указанный отличный от TCR рецептор связывается с антигеном вируса или вирус-индуцированным антигеном, обнаруживаемом на поверхности инфицированной клетки. Инфекция, которую требуется лечить или предупредить, может быть вызвана, например, вирусом, бактерией, простейшими или паразитами. Вирусы, с которыми можно бороться, включают без ограничений HCMV, EBV (вирус Эпштейна-Барр), гепатит С (HCV), вирус Эбола, VSV (вирус везикулярного стоматита), грипп, ветряную оспу, аденовирус, вирус простого герпеса 1 (ВПГ-1), вирус простого герпеса 2 (ВПГ-2), вирус чумы КРС, риновирус, эховирус, ротавирус, респираторно-синцитиальный вирус, вирус папиломы, цитомегаловирус (CMV), эхиновирус, арбовирус, хантавирус, вирус коксаки, вирус свинки, вирус кори, вирус краснухи, вирус полиомиелита и/или вирус иммунодефицита человека 1 или 2 типа (ВИЧ-1, ВИЧ-2). Невирусные инфекции, которые можно лечить при помощи TCR-дефицитных Т-клеток, включают без ограничений инфекции, вызываемые видами Staphylococcus, видами Enterococcus, Bacillus anthracis, видами Lactobacillus, видами Listeria, Cotynebacterium diphtheriae, видами Nocardia, видами Streptococcus, видами Pseudomonas, видами Gardnerella, видами Streptomyces, Thermoactinomyces vulgaris, видами Treponema, видами Camplyobacter, видами Raeruginosa, видами Legionella, N. gonorrhoeae, Ν. meningitides, F. meningosepticum, F. odoraturn, видами Brucella, В. pertussis, В. bronchiseptica, Ε. coli, Klebsiella, Enterobacter, S. marcescens, S. liquefaciens, Edwardsiella, P. mirabilis, P. vulgaris, Streptobacillus, R. fickettsfi, C. psittaci, C. trachomatis, M. tuberculosis, M. intracellulare M. folluiturn, M. laprae, M. avium, M. bovis, M. africanum, M. kansasii, M. lepraernurium, трипаносомой, хламидией или рикетсией.
Эффективность композиций согласно настоящему изобретению можно продемонстрировать в наиболее подходящей модельной системе in vivo, в зависимости от типа лекарственного препарата, который требуется разработать. В медицинской литературе приведено подробное раскрытие преимуществ и способов применения широкого спектра таких моделей. Например, существует много моделей рака разных типов, которые стандартно применяют для изучения фармацевтической активности лекарственных препаратов от рака, таких как модель ксенотрансплантата у мышей (например, Не, S. et al. 2008. J. Immunol. 181:7581-7592) и модель хронической GVHD (e.g., Xiao, Ζ.Y. et al. 2007. Life Sci. 81:1403-1410).
Когда удалось показать, что композиции согласно настоящему изобретению эффективны у животных in vivo, можно разрабатывать программу клинических исследований на основании таких доз, которые показали свою безопасность и эффективность на животных. Специалисты в данной области техники могут разработать такие клинические исследования на основании стандартных протоколов, которые описаны в таких учебных пособиях, как Spilker (2000. Guide to Clinical Trials. Lippincott Williams & Wilkins: Philadelphia).
Введение
Согласно одному варианту реализации настоящего изобретения TCR-дефицитные Т-клетки вводят реципиенту в количестве от приблизительно 106 до приблизительно 1011 клеток. Согласно предпочтительному варианту реализации настоящего изобретения TCR-дефицитные Т-клетки вводят реципиенту в количестве от 108 до 109 клеток. Согласно предпочтительному варианту реализации настоящего изобретения TCR-дефицитные Т-клетки вводят реципиенту с частотой раз в двадцать шесть недель или менее, раз в шестнадцать недель или менее, раз в восемь недель или менее, или раз в четыре недели или менее.
Указанные значения представляют собой общие указания в отношении диапазона подвергнутых трансдукции Т-клеток, которое следует применять практикующему врачу при оптимизации способа согласно настоящему изобретению для реализации изобретения. Изложение в настоящей заявке таких диапазонов никоим образом не препятствует применению большего или меньшего количества компонента, что может быть оправдано в рамках конкретной области применения. Например, реальная доза и режим введения могут варьировать в зависимости от того, вводят ли композиции в сочетании с другими фармацевтическими композициями, или в зависимости от межиндивидуальных различий фармакокинетики, распределения лекарственного препарата и его метаболизма. Специалисты в данной области техники могут легко внести любые необходимые корректировки в зависимости от потребностей конкретной ситуации.
Специалист в данной области техники должен быть способен определить эффективную дозу и частоту введения на основании сведений, имеющихся в технике, или путем стандартного экспериментирования, например, руководствуясь раскрытием в настоящей заявке и сведениями, приведенными в Goodman, L.S., Gilman, Α., Brunton, L.L., Lazo, J.S., & Parker, К.L. (2006). Goodman & Gilmanʹs the pharmacological basis of therapeutics. New York: McGraw-Hill; Howland, R.D., Mycek, M.J., Harvey, R.Α., Champe, P.С, & Mycek, M.J. (2006). Pharmacology. Lippincottʹs illustrated reviews. Philadelphia: Lippincott Williams & Wilkins; and Golan, D.E. (2008). Principles of pharmacology: the pathophysiologic basis of drug therapy. Philadelphia, Pa., [etc.]: Lippincott Williams & Wilkins. Режим введения можно выбирать на основании общепринятых способов клеточной терапии (см., например, Topalian and Rosenberg (1987) Acta Haematol. 78 Suppl 1:75-6; Патент США №4690915), или можно применять альтернативную стратегию непрерывной инфузии.
Согласно другому варианту реализации настоящего изобретения TCR-дефицитные Т-клетки вводят субъекту в виде фармацевтической формы.
Согласно одному варианту реализации настоящего изобретения TCR-дефицитные Т-клетки можно необязательно вводить в сочетании с одним или более активными агентами. Такие активные агенты включают обезболивающие, жаропонижающие, противовоспалительные, антимикробные, противовирусные и противоцитокинные агенты. Активные агенты включают агонисты, антагонисты и модуляторы TNF-α, IL-2, IL-4, IL-6, IL-10, IL-12, IL-13, IL-18, IFN-α, IFN-γ , BAFF, CXCL13, IP-10, VEGF, EPO, EGF, HRG, фактора роста гепатоцитов (HGF), гепцидина, включая антитела, направленные против любых из перечисленных выше веществ, и антитела, направленные против любого из их рецепторов. Активные агенты также включают 2-арилпропионовые кислоты, ацеклофенак, ацеметацин, ацетилсалициловую кислоту (аспирин), алклофенак, алминопрофен, амоксипирин, ампирон, арилалконовые кислоты, азапропазон, бенорилат/беноорилят, беноксапрофиен, бромфенак, капрофен, холин магния салицилат, клофезон, ингибиторы ЦОГ-2, дексибупрофен, декскетопрофен, диклофенак, дифлунизал, дроксикам, этензамид, этодолак, эторикоксиб, файсламин, фенаминовую кислоту, фенбуфен, фенопрофен, флуфенаминовую кислоту, флуноксапрофен, флубипрофен, флупрофен, ибупроксам, индометацин, индопрофен, кебузон, кетопрофен, кетолак, лорноксикам, локсопрофен, лумиракоксиб, салицилат магния, меклофенаминовую кислоту, мефенаминовую кислоту, мелоксикам, метамизол, метилсалицилат, мофебутазон, набуметон, напроксен, N-арилантранильные кислоты, фактор роста нервов (NGF), оксаметацин, оксапрозин, оксикамс, оксифенбутазон, парекоксиб, феназон, фенилбутазон, фенилбутазон, пироксикам, пирофен, профены, проглуметацин, производные пиразолидина, рофекоксиб, салицил-салицилат, салицикламид, салицилаты, сульфинпиразон, сулиндак, супрофен, теноксикам, тиапрофеновую кислоту, толфенаминовую кислоту, толметим и вальдекоксиб.
Антибиотики включают амикацин, аминогликозиды, амоксициллин, ампициллин, ансамицины, арсфенамины, азитромицин, азлоциллин, азтреонам, бацитрацин, карбацефем, карбапенемы, карбециллин, цефаклор, цефадроксил, цефалексин, цефалотин, цефалотин, цефамандол, цефазолин, цефдирин, цефдиторен, цефепим, цефиксим, цефоперазон, цефатоксим, цефокситин, цефодоксим, цефпрозил, цефтазидим, цефбутен, цефтизозим, цефтобипрол, цефтриаксон, цефуроксим, цефалоспорины, хлорамфеникол, циластатин, ципрофлоксацин, кларитромицин, клиндамицин, клоксациллин, колистин, котримоксазол, дальфопристин, демкелоциклин, диклоксациллин, диритромицин, дорипенем, доксициклин, эноксацин, эртапенем, эритромицин, этрамбутол, флуклоксациллин, фосфомицин, фурозолидон, фусидовая кислота, гатифлоксацин, гелдамицин, гентамицин, циклопептиды, Хербимицин, имипенем, изониазид, канамицин, левофлоксацин, линкомицин, линезолид, ломефлоксацин, лоракарбеф, макролиды, мафенид, меропенем, метициллин, метронидазол, мезлоциллин, миноциклин, монобактамы, моксифлоксацин, мупироцин, нафциллин, нетилмицин, нгитрофурантоин, норфлоксацин, офлоксацин, оксациллин, окситетрациклин, паромицин, пенициллин, пенициллины, пиперациклин, платенсимицин, полимиксин В, поипептиды, пронтозил, пиразинамид, хинолоны, квинпиристин, рифампицин, рокситромицин, спектиномицин, стрептомицин, сульфацетамид, сульфаметизол, сульфанилимид, сульфазалазин, сульфисоксазол, сульфонамиды, тейкопланин, телитромицин, тетрациклин, тетрациклины, тикарциллин, тинидазол, тобрамицин, триметоприм, триметоприм-сульфаметоксазол, тролеандомицин, тровафлоксацин и ванкомицин.
Активные агенты также включают альдостерон, беклометазон, метаметазон, кортикостероиды, кортизол, кортизон ацетат, дезоксикаортикостерон ацетат, дексаметазон, флудрокортизон ацетат, глюкокортикоиды, гидрокортизон, метилпреднизолон, преднизолон, преднизон, стероиды и триамцинолон. Также подразумевается любое подходящее сочетание указанных активных агентов.
«Фармацевтическое вспомогательное вещество» или фармацевтически приемлемое вспомогательное вещество» представляет собой основу, как правило, жидкую, в которой находится активный терапевтический агент. Согласно одному варианту реализации настоящего изобретения указанным активным терапевтическим агентом является популяция TCR-дефицитных Т-клеток. Согласно одному варианту реализации настоящего изобретения указанным активным терапевтическим агентом является популяция TCR-дефицитных Т-клеток, экспрессирующих функциональный не TCR-рецептор. Как правило, вспомогательное вещество не придает какой-либо фармакологической активности лекарственной форме, хотя оно может обеспечивать химическую и/или биологическую стабильность. Примеры лекарственных форм можно найти, например, в Remingtonʹs Pharmaceutical Sciences, 19th Ed., Grennaro, Α., Ed., 1995, который включен в настоящую заявку посредством ссылки.
В настоящей заявке термины «фармацевтически приемлемая основа» или «вспомогательное вещество» включают любые растворители, диспергенты, покрытия, противомикробные и противогрибковые агенты, изотоничные агенты и агенты, замедляющие всасывание, которые являются физиологически совместимыми. Согласно одному варианту реализации указанная основа может подходить для парентерального введения. В качестве альтернативы указанная основа может подходить для внутривенного, внутрибрюшинного, внутримышечного или сублингвального введения. Фармацевтически приемлемые основы включают стерильные водные растворы или дисперсии для приготовления стерильных растворов или дисперсий для инъекций непосредственно перед применением. Применение таких сред и агентов для фармацевтически активных веществ хорошо известно в технике. Кроме случаев, когда какая-либо традиционная среда или агент несовместимы с активным соединением, подразумевается их применение в фармацевтических композициях согласно настоящему изобретению. Также в композиции можно включать дополнительные активные соединения.
Согласно конкретному предпочтительному варианту реализации настоящего изобретения подходящие основы включают без ограничения сбалансированный солевой раствор Хенкса (HNSS), фосфатно-солевой буфер (ФСБ) или любую замороженную среду, содержащую, например, 10% DMSO (диметилсульфоксид) и 90% сыворотки человека.
Как правило, фармацевтические композиции дожны быть стерильными и стабильными при условиях производства и хранения. Изобретение подразумевает, что фармацевтическая композиция присутствует в лиофилизированной форме. Композиция может быть преобразована в форму раствора. Основой может быть диспергент, содержащий, например, воду.
Для каждого из перечисленных вариантов реализации соединения можно вводить в разных лекарственных формах. Подразумевается любая биологически приемлемая лекарственная форма, известная специалистам в данной области техники, и их сочетания. Примеры таких лекарственных форм включают без ограничений жидкости, растворы, суспензии, эмульсии, формы для инъекций (включая подкожные, внутримышечные, внутривенные и внутрикожные), инфузии и их сочетания.
Описание, которое дано выше, разных проиллюстрированных вариантов реализации настоящего изобретения не должно рассматриваться как исчерпывающее или ограничивающее изобретение точной формой, которая раскрывается. Хотя в настоящей заявке описаны специфические варианты реализации и примеры настоящего изобретения с целью иллюстрации, в пределах области настоящего изобретения возможны разные эквивалентные модификации, которые должны понимать специалисты в соответствующей области техники. Сведения, представленные в настоящей заявке, можно применять в других целях, отличных от примеров, описанных выше.
В изобретение могут быть внесены указанные и другие изменения в свете приведенного выше подробного описания. Как правило, в следующей формуле изобретения применяемые термины не предназначены для ограничения изобретения специфическими вариантами реализации, раскрываемыми в описании и формуле изобретении. Следовательно, настоящее изобретение не ограничивается раскрытием, а, напротив, область изобретения должна определяться полностью нижеследующими пунктами формулы изобретения.
Настоящее изобретение можно реализовывать способами, отличными от тех, что конкретно описаны в приведенном выше описании и примерах. Целый ряд модификаций и вариантов возможен в свете приведенных выше сведений и, следовательно, попадает в область прилагаемой формулы изобретения.
Определенные сведения, связанные с композициями с Т-клетками, дефицитными по рецептору Т-клеток, и способами их применения, были раскрыты в Предварительном описании изобретения на патент США №61/255 980, поданном 29 октября 2009 г., раскрытие, которое включено в настоящую заявку посредством ссылки на его полную версию.
Определенные сведения, связанные с получением Т-клеток, экспрессирующих химерные рецепторы, и способами их применения, были раскрыты в публикации заявки на патент США №2010/0029749, опубликованной 4 февраля 2010 г., раскрытие, которое включено в настоящую заявку посредством ссылки на его полную версию.
Определенные последовательности полинуклеотидов, применимые при получении Т-клеток, дефицитных по рецептору Т-клеток, согласно настоящему изобретению раскрываются в перечне последовательностей, прилагаемом к данной заявке на патент, и раскрытие указанного перечня последовательностей включено в настоящую заявку посредством ссылки на ее полную версию.
Полное раскрытие каждого цитируемого документа (включая патенты, заявки на патенты, статьи в журналах, рефераты, руководства, монографии или другие раскрытия) в разделе «Уровень техники», «Подробное описание» и «Примеры» включены в настоящую заявку посредством ссылок на ее полную версию.
Следующие примеры приведены, чтобы предоставить специалистам в данной области техники полное раскрытие и описание того, как выполнено и применяется настоящее изобретение, и не предназначены для ограничения области, которая закреплена за изобретением. Были предприняты все усилия для обеспечения точности в том, что касается применяемых числовых значений (например, количества, температура, концентрации и т.д.), но некоторые экспериментальные погрешности и отклонения могут быть допустимы. Если не указано другого, части являются массовыми частями, молекулярная масса является средней молекулярной массой, температура выражена в градусах Цельсия, а давление близко к атмосферному.
ПРИМЕРЫ
Пример 1: Получение Т-клеток, дефицитных по рецептору Т-клеток (TCR)
Минигены кодируются плазмидой экспрессии на основе ретровируса (например, pFB-neo или pSFG), содержащей на 3ʹ и 5ʹ-конце последовательности LTR. Указанные плазмиды упаковывают в линию клеток, упаковывающую ретровирус, например, РТ67 и PG13, и вирусные частицы отбирают, когда упаковывающие клетки разрастаются до смыкания монослоя. Затем Т-клетки активируют РИА (фетогемагглютинином), моноклональными антителами анти-СВ3 или анти-СВ3/2 8 в течение 1-3 дней в полной среде (или среде без сыворотки) с добавлением rIL-2 (25 Е/мл), и проводят трансдукцию Т-клеток посредством спинокуляции при 32°С в присутствии ретронектина или полибрена. После периода покоя от 5 до 7 часов клетки промывают и помещают в свежую среду с добавлением IL-2 на 2-7 дней. Периодически проводят подсчет клеток, чтобы избежать избыточных концентраций клеток (т.е., >2×106 клеток/мл) и повторно пересевают с плотностью 7×105 клеток/мл. Необязательно для удаления не подвергшихся трансдукции Т-клеток применяют среду для селекции спустя 2 дня в течение периода 3-5 дней. Живые клетки собирают при помощи градиента Lymphoprep™ («Sentinel», Милан, Италия) и снова рассевают на 1-3 дня.
После инкубации клетки анализируют с целью оценки экспрессии и функции TCR. Также одновременно можно анализировать экспрессию функционального отличного от TCR рецептора, если нужно. Для оценки экспрессии TCR/CD3 применяют проточную цитометрию с применением антител, меченных флуорохромом. Живые клетки окрашивают антителами на CD5, CD8 и CD4 в сочетании с антителом на CD3ε, TCRα, TCRβ, TCRγ или TCRδ. Если применяют экспрессию генов CD3 или TCR, экспрессия и белков TCR, и белков CD3 должна быть значительно снижена по сравнению с Т-клетками, обработанными контрольным вектором. Для контроля фоновой флуоресценции применяют антитела для контроля изотипа. Для идентификации Т-лимфоцитов клетки запускают на CD5, а затем определяют экспрессию CD4, CD8, CD3 и TCR. Для каждого варианта обработки применяют несколько проб, и применяют соответствующую компенсацию спектра испускания флуорохрома. Экспрессию другого рецептора (например, chNKG2D) определяют при помощи специфических антител и проточной цитометрии, как было ранее описано в технике (Zhang, T. et al., (2006) Cancer Res., 66(11) 5927-5933; Barber, A. et al., (2007) Cancer Res., 67(10):5003-5008).
Для оценки функциональной недостаточности TCR, в конце процесса культивирования применяют анти-CD3 стимуляцию эффекторных клеток, чтобы измерить выработку интерферона (IFN)-гамма через 24 часа. Т-клетки (2×105) культивируют с растворимым анти-CD3 (ОКТ3) mAb в полной среде. Через 24 часа кондиционированную среду, не содержащую клеток, отбирают и проводят анализ посредством ELISA, определяя IFN-гамма. Изменения экспрессии или функции TCR должно отразиться на выработке IFN-гамма.
Для оценки функции функциональных отличных от TCR рецепторов применяют определение выработки специфических цитокинов Т-клетками, инкубируемыми с клетками опухоли, которые экспрессируют или не экспрессируют свои специфические лиганды. Например, для оценки функции chNKG2D 105 Т-клеток инкубируют с 105 клетками опухоли P815-MICA (лиганд+), 105 клетками Р815 (лиганд-), 105 клетками RPMI8226 (лиганд+) или только с Т-клетками. Через 24 часа кондиционированную среду, не содержащую клеток, отбирают и определяют IFN-g посредством ELISA. Химерные Т-клетки NKG2D вырабатывают INF-γ после культивирования с лиганд-экспрессирующими клетками опухоли (Zhang, T. et al., (2006) Cancer Res., 66(11) 5927-5933; Barber, A. et al., (2007) Cancer Res., 67(10):5003-5008). Также можно оценить клеточную цитотоксичность в отношении лиганд+-клеток опухоли, как ранее было описано в технике (Zhang, T. et al., (2006) Cancer Res., 66(11) 5927-5933). Специфичность показывают при помощи лиганд- клеток опухоли или специфических mAb, блокирующих рецепторы.
Пример 2: Получение Т-клеток, дефицитных по рецептору Т-клеток (TCR), экспрессирующих chNKG2D
В данном примере проводят одновременную экспрессию рецептора chNKG2D и подавление экспрессии эндогенного TCR. В данном примере применяют рецептор chNKG2D мыши, состоящий из NKG2D в сочетании с присоединенным на N-конце CD3-дзета. Рецептор chNKG2D генерируют и экспрессируют в Т-клетках мыши. NKG2D является белком II типа, в котором N-конец расположен во внеклеточном пространстве Raulet (2003) Nat. Rev. Immunol. 3:781-790), тогда как цепь CD3-дзета является белком I типа с С-концом, расположенным в цитоплазме (Weissman, et al. (1988) Proc. Natl. Acad. Sci. USA 85:9709-9713). Для генерирования химерного белка NKG2D-CD3-дзета кодон инициации ATG помещают перед кодирующей последовательностью цитоплазматической области CD3-дзета цепи (без стоп-кодона ТАА), после которой идет ген NKG2D дикого типа. При экспрессии ориентация дзета-части переворачиваются внутрь клетки. Внеклеточные и трансмембранные домены происходят из NKG2D. Также конструируют второй химерный ген, кодирующий Dap 10, за которым идет фрагмент, кодирующий цитоплазматический домен CD3-дзета. На Фигуре 1 представлены структуры химерного рецептора и рецептора дикого типа.
кшРНК функционально связана в лентивирусном векторе с рецептором chNKG2D. Для достижения экспрессии обоих генов кшРНК запускают с промотора U6, а рецептор chNKG2D с промотора PGK. Первичные РВМС человека выделяют у здоровых доноров и активируют низкими дозами малорастворимого анти-CD3 и 25 Е/мл rhuIL-2 в течение 48 часов. Хотя для лентивирусной трансдукции не требуется активировать Т-лимофциты, трасндукция проходит более эффективно и позволяет клеткам продолжать развиваться в IL-2. Активированные клетки отмывают и проводят их трансдукцию посредством спин-фекции в течение 1 часа при 30°С, после которой идет период покоя 7 ч. Клетки отмывают и культивируют с 25 Е/мл IL-2 в течение 3-7 дней, чтобы стала возможна экспансия эффекторных клеток аналогично тому, как мы делаем при применении клеток in vivo. Экспрессию TCRαβ, CD3 и NKG2D оценивают посредством проточной цитометрии и количественной ПЦР в режиме реального времени (кРВ-ПЦР). Количество CD4+и CD8+-Т-клеток определяют посредством проточной цитометрии. Полученные значения сравнивают с РВМС, которые подвергли трансдукции одними кшРНК или геном chNKG2D (в качестве контроля). Также в качестве контроля включают клетки, которые подвергли трансдукции одним только вектором.
Ожидается, что указанные клетки, не экспрессирующие TCR, или экспрессирующие его в малом количестве на своей поверхности, будут экспрессировать большее количество NKG2D на поверхности вследствие совместной экспрессии рецептора chNKG2D.
В качестве альтернативы трансдукцию можно проводить двумя вирусами одновременно, один из которых содержит конструкцию кшРНК, а другой - конструкцию chNKG2D. Чтобы обеспечить высокий уровень экспрессии chNKG2D в Т-клетках, не экспрессирующих TCR, применяют большее количество вируса с chNKG2D. TCR+T-клетки, которые могут оставаться, удаляют, чтобы получить TCR-, chNKG2D+Т-клетки.
После вирусной трансдукции и экспансии TCR+ и TCR- клетки разделяют при помощи mAb с применением магнитных гранул на колонках Miltenyi. Проверку экспрессии chNKG2D проводят при помощи количественной ПЦР в режиме реального времени с применением специфических праймеров на рецептор chNKG2D.
Чтобы определить, утратили ли Т-клетки функцию TCR и сохранили ли они функцию chNKG2D, подвергнутые трансдукции или контрольные Т-клктеи культивируют с аллогенными РВМС, обработанными митомицином С, или синергичными РВМС. Указанные Т-клетки предварительно нагружают CFSE, который является проникающим красителем клеток, который равномерно распределяется между дочерними клетками после деления. Степень деления клеток можно легко определить посредством проточной цитометрии.
Чтобы определить, обеспечила ли конструкция кшРНК подавление функции TCR и оставляет ли он возможность для функционирования рецептора chNKG2D, подвергнутые трансдукции или контрольные Т-клктеи культивируют с аллогенными РВМС, обработанными митомицином С, синергичными РВМС или клетками опухоли: Р815-MICA (опухоль мыши, экспрессирующая MICA человека - лиганд для NKG2D), Р815, А2008 (клетки опухоли яичника человека, NKG2D-лиганд+) и U266 (линия клеток миеломы человека, NKG2D-лиганд+). Через 48 часов отбирают надосадочную жидкость, не содержащую клеток, и оценивают количество выработанных IL-2 и IFN-γ посредством ELISA. В качестве отрицательного контроля применяют одни Т-клетки.
Пример 3: Введение Т-клеток, дефицитных по рецептору Т-клеток (TCR), экспрессирующих chNKGID
В данном примере TCR-дефицитные Т-клетки, экспрессирующие рецептор chNKG2D мыши, полученные в Примере 2, вводят мышам, чтобы оценить терапевтический потенциал указанных Т-клеток in vivo при определенных типах рака. Т-клетки - носители химерного chNKG2D (106) вводят подкожно мышам линии C57BL/6, совместно с клетками опухоли RMA/Rae-1β (105). На терапевтическую противоопухолевую активность у таких мышей указывает отсутствие опухоли или угнетение роста опухоли RMA/Rae-1β у мышей, которым ввели TCR-дефицитные Т-клетки - носители химерного chNKG2D, через 30 дней.
Во второй и более строгой модели подвергнутые трансдукции Т-клетки (107) адаптивно вводят в/в мышам В6 за один день до п/к прививки опухоли RMA/Rae-1β в правый бок. Подавление роста опухоли RMA/Rae-1β (п/к) по сравнению с контрольными модифицированными вектором Т-клетками указывает на терапевтическую противоопухолевую активность у данных мышей. Что касается токсичности лечения Т-клетками, модифицированными химерным chNKG2D, ожидается, что животные не будут проявлять каких-то явных признаков воспалительного поражения (т.е., взъерошенность шерсти, горб или диарея, и т.п.) при их лечении Т-клетками - носителями химерного chNKG2D, что может указывать на отсутствие явной токсичности.
В более строгой модели установленных опухолей яичника (ID8) трансдуцированные chNKG2D Т-клетки (5×106 Т-клеток в/б) вводят мышам - носителям опухоли в течение 5 недель. Впоследствии мышам вводят Т-клетки через 7 и 9 недель после провокации опухолью. При указанных условиях мыши, которым ввели chNKG2D-содержащие Т-лимфоциты, должны оставаться без опухолей в течение более чем 250 дней, тогда как мыши, которым в аналогичном режиме вводили контрольные Т-лимфоциты, должны погибать от роста опухоли в течение 100 дней. Что касается токсичности лечения Т-клетками, модифицированными химерным chNKG2D, ожидается, что животные не будут проявлять каких-то явных признаков воспалительного поражения (т.е. взъерошенность шерсти, горб или диарея, и т.п.) при их обработке Т-клетками -носителями химерного chNKG2D, что может указывать на отсутствие явной токсичности.
В модели множественной миеломы мышам-носителям клеток опухоли 5Т33ММ на 12-ый день после инфузии клеток опухоли вводят chNKG2D-содержащие Т-клетки (5×106, в/в). Указанное воздействие должно приводить к увеличенной продолжительности жизни всех мышей, и приблизительно половина из указанных мышей должны быть долгожителями без опухолей. Мыши, которым ввели контрольные Т-клетки, погибают от опухолей в первые 30 дней. При лечении chNKG2D-содержащими Т-клетками никаких явных признаков токсичности не наблюдается.
Поскольку иммунная система может отбирать варианты опухолей, наиболее эффективными средствами иммунотерапии при раке, вероятно, должны быть те, которые вызывают иммунитет против нескольких антигенов опухоли. В третьем эксперименте проверяется, может ли лечение Т-клетками, содержащими химерный NKG2D, вызывать иммунитет против клеток опухоли дикого типа в организме хозяина. Мыши, которым вводили Т-клетки - носители химерного NKG2D и клетки опухоли 5Т33ММ, и у которых не было опухолей затем в течение 80 дней, подвергались провокации клетками опухоли 5Т33ММ. Мыши, выживающие без развития опухоли, устойчивы к последующей провокации клетками 5Т33ММ (3×105), по сравнению с контрольными не подвергнутыми лечению мышами, которые погибают от опухоли в среднем в первые 27 дней. Однако мыши, выживающие без развития опухоли, не устойчивы к последующей провокации клетками опухоли RMA-Rael (3×105), и погибают от опухоли с приблизительно той же продолжительностью жизни, что и у мышей, не подвергшихся лечению (20 дней). Это указывает на то, что адаптивный перенос Т-клеток - носителей химерного NKG2D может позволить хозяину генерировать опухолеспецифичную память Т-клеток.
Согласно настоящему изобретению предложено четыре класса TCR-ингибирующих молекул (TIM), которые влияют на функцию Т-клеток. В Таблице 2 приведены обобщенные данные о действии 19 разных TIM на функцию эффекторных Т-клеток, либо в ответ на стимуляцию растворимым антителом против-CD3 (ОКТ3 200 нг/мл) (CD3), либо культурой аллогенных РВМС (Allo). Обозначение слева показывает класс TIM. Численные значения указывают процент снижения при ингибировании TCR по сравнению с контрольными Т-клетками, подвергнутыми трансдукции контрольным вектором pFB. N1: нет ингибирования. ND: не сделано.
Таблица 2: краткий обзор действия TCR-ингибирующих молекул (TIM) на функцию Т- клеток.
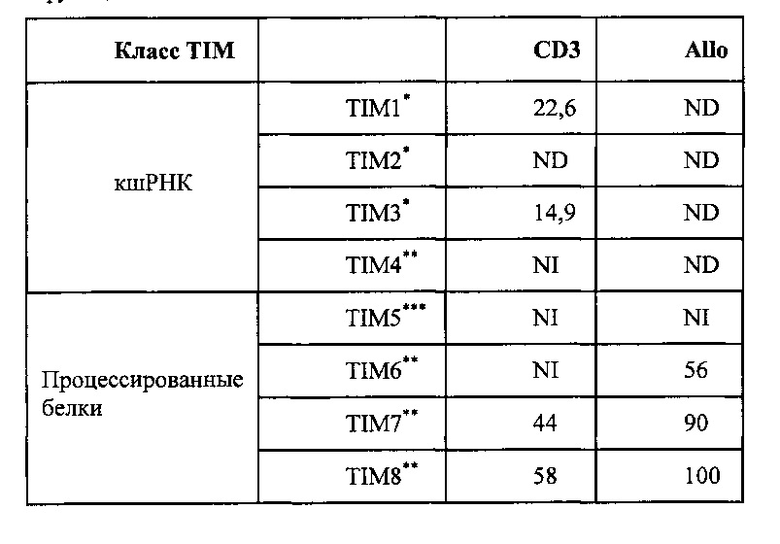
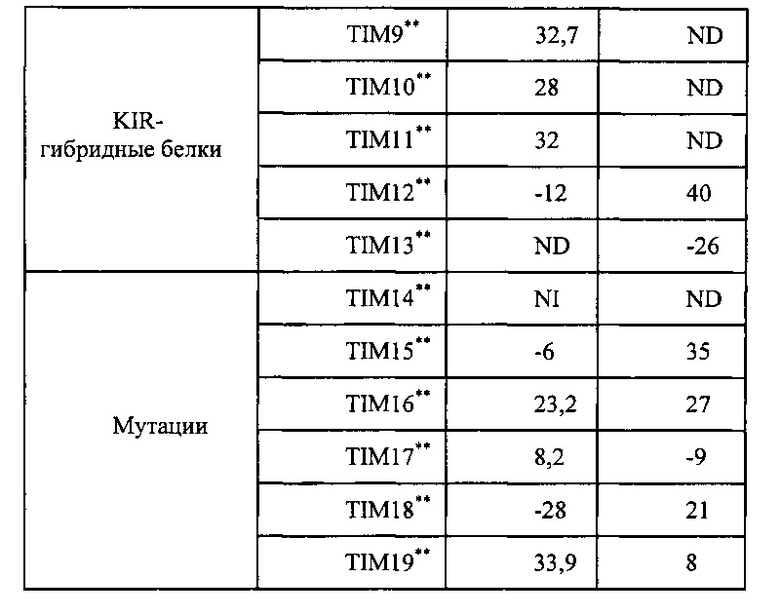
Пример 4: Получение Т-клеток, дефицитных по рецептору Т-клеток (TCR), с применением кшРНК, направленных на нуклеиновые кислоты, кодирующие CD3-эпсилон и CD3-дзета
В данном примере экспрессию эндогенного TCR ингибировали при помощи последовательностей кшРНК, которые направлены на нуклеиновые кислоты, кодирующие CD3-эпсилон или CD3-дзета.
Применяли последовательности кшРНК, клонированные в ретровирусный вектор pSM2c («Ореп Biosystems»), в котором экспрессия контролировалась промотором U6. Указанные конструкции кшРНК применяли для блокирования экспрессии CD3-эпсилон и/или CD3-дзета так, чтобы Т-клетка больше не вырабатывала один из ключевых компонентов комплекса TCR. Впоследствии комплекс TCR дестабилизировался, и нарушалась экспрессия функционального TCR на поверхности клетки, что вело к снижению функции Т-клеток, связанной с TCR-комплексом. Последовательность кшРНК, направленных против CD3-эпсилон или CD3-дзета, описана в Таблице 1, и она соответствует SEQ ID №: 9-26 и 68-71, соответственно.
Чтобы определить, изменяют ли указанные кшРНК функцию TCR, измеряли выработку IFN-гамма в ответ на (i) стимуляцию растворимым анти-CD3 (CD3) или (ii) в ответ на культивирование с аллогенными РВМС (Allo). В частности, Т-лимфоциты, обработанные TIM1 и TIM3, демонстрировали снижение на 22,6% или 14,9% при ингибировании TCR, соответственно, после стимуляции 200 нг/мл анти-CD3 моноклонального антитела. См. Таблицу 2 выше.
Пример 4: Получение Т-клеток, дефицитных по рецептору Т-клеток (TCR), с применением доминантного негативного ингибитора CD3-дзета
В данном примере гиперэкспрессия доминантно-негативного белка-ингибитора, т.е. TIM, приводила к приостановке экспрессии и функции TCR. Экспрессию эндогенного TCR ингибировали при помощи доминантного негативнбого белка-ингибитора, содержащего CD3-дзета, измененный так, чтобы он включал ингибиторный сигнал от KIR2DL1, и полученные в результате Т-клетки не активировались в ответ на стимуляцию TCR.
Конструкции минигенов, которые включали весь или часть модифицированного полинуклеотида, кодирующего CD3-дзета, генерировали посредством ПЦР с применением кДНК-матриц CD3-дзета и KIR2DL1, соответствующих SEQ ID №: 64 и SEQ ID №: 66, соответственно, приобретенных у компании «Open Biosystems» (Хантсвилл, Алабама). Все ПЦР проводили с применением высокоточного слияния ДНК-полимеразы (New England Biolabs, Ипсвич, Массачусетс), а праймеры синтезировала компания «Integrated DNA Technologies» (Коралвилл, Айова). При применении установленных протоколов каждую конструкцию клонировали в ретровирусный вектор pFB-neo ("Stratagene»), и экспрессия контролировалась промотором 5ʹ LTR. Полученные в результате конструкции подвергали скринингу и подтверждали точность секвенирования, а также анализировали при помощи DNA dynamo («Blue Tractor Software Ltd»). Последовательности ДНК и прогнозируемые для них последовательности белков соответствуют SEQ ID №: 68-101.
TIM экспрессировали в первичных Т-клетках при помощи системы экспрессии на основе ретровируса. Для получения вирусов с низким или высоким титром применяли две разных линии упаковывающих клеток. Вирус с низким титром получали при помощи клеток GP2-293T, которые временно трансфецировали упаковывающей плазмидой и плазмидой оболочки. Через 72 часа вирусную надосадочную жидкость отбирали, и титры измеряли путем инфицирования клеток NIH-3T3, после чего проводили селекцию с G418. Титры вирусов, произведенных данной системой составляли от 5×105 до 1×107 КОЕ/мл. Чтобы получить вирусы с высоким титром, вирус вырабатывали в системе GP2-293T для трансдукции упаковывающих клеток РТ67. Клетки РТ67, инфицированные вирусными частицами, отбирали путем обработки G418 в течение 5 дней. Т1М-экспрессирующие клетки РТ67 рассевали и применяли их для получения вируса. Через 72 часа после того, как клетки достигали слияния монослоя, вирусную надосадочную жидкость отбирали, и измеряли титры в NIH-3T3. Титры вирусов, полученных в данной системе, были в диапазоне от 7×107 до 2×108 КОЕ/мл.
Для трансдукции Т-клеток человека первичные РВМС человека выделяли у здоровых доноров и активировали 40 нг/мл растворимого анти-CD3 и 50 Е/мл rhuIL-2 в течение 72 часов. Активированные Т-клетки отмывали и проводили их трансдукцию ретровирусом, полученным либо в виде вируса с низким титром, либо с высоким, путем 1-часовой спин-инфекции при 32°С, после которой наступал 6-часовой период покоя. Клетки отмывали и культивировали в 50 Е/мл IL-2 в течение 48 часов, а затем передавали для отбора в течение 3 дней. После отбора живые клетки выделяли при помощи Lymphoprep (Mediatech), и эффекторные клетки распределяли в 50 Е/мл IL-2 на 48 часов, после истечения которых клетки применяли для функционального анализа. Эндогенную экспрессию CD3-эпсилон и CD3-дзета в клетках, подвергнутых трансдукции кшРНК, и снижение экспрессии указанных генов из-за кшРНК анализировали при помощи количественной ПЦР в режиме реального времени (кРВ-ПЦР). Если кратко, РНК экстрагировали из подвергнутых трансдукции Т-клеток и 0,5-1 мкг тотальной РНК подвергали обратной транскрипции при помощи QuantiTect Rev, набор для транскрипции (Qiagen). Полученную кДНК применяли с SYBR зеленым («Applied Biosystems») для количественной ПЦР в режиме реального времени, и данные нормировали на уровень глицеральдегид-3-фосфат-дегидрогеназы (GAPDH). Изменения экспрессии на поверхности клетки анализировали с применением антител, специфических в отношении CD3, CD8, CD4 и CD5, и не было выявлено никаких различий в экспрессии молекул на поверхности данных клеток у TIM-экспрессирующих Т-клеток по сравнению с контрольным вектором.
Чтобы определить, было ли уменьшение экспрессии TCR при применении каждой конструкции кшРНК или минигена (которые устраняют или разрушают TCR на поверхности клетки) достаточным для предупреждения активации Т-клеток при стимуляции TCR, проводили оценку Т-клеток на предмет: (1) выживания клеток in vitro; и (2) выработки цитокинов в ответ на аллогенные РВМС и/или анти-CD3 mAb.
Для оценки выживаемости клеток подвергнутые трансдукции Т-клетки размножали в среде RPMI с rhuIL-2 (50 Е/мл). Клетки помещали в чашки Петри при одинаковой плотности в начале культивирования, и отбирали пробы для подсчета клеток и оценки жизнеспособности раз в день в течение 7 или более дней. Не было выявлено никаких различий в росте TIM-экспрессирующих клеток и соответствующих Т-клеток, экспрессирующих контрольный вектор. Чтобы определить, экспрессируют ли Т-клетки достаточно TCR, чтобы индуцировать ответ на аллогенные клетки, трансдуцированные или контрольные Т-клетки культивировали с аллогенными аутологичными РВМС в отношении 4:1. Через 24 часа надосадочную жидкость, не содержащую клеток, отбирали и в ней определяли количество выработанного IFN-γ при помощи ELISA. В качестве отрицательного контроля применяли одни Т-клетки, включая РВМС, и клетки, подвергнутые трансдукции. Среди проанализированных TCR-ингибирующих молекул были определены два минигена (TIM7 и TIM8), которые могли значительно снижать функцию TCR в Т-клетках. См., Фигуру 2. Аллогенный анализ проводили с 19 разными донорами, экспрессирующими TIM7 и TIM8, где клетки каждого донора культивировали с 3 разными аллогенными РВМС. Среднее уменьшение выработки IFN-γ в TIM7-экспрессирующих Т-клетках составило 49%, а среднее уменьшение в TIM8-экспрессирующих Т-клетках составило 60%.
Чтобы определить, подавляет ли каждый TIM функцию Т-клеток при прямой стимуляции антителом комплекса TCR, подвергнутые трансдукции TIM Т-клетки обрабатывали разными дозами анти-CD3 mAb (of 1,6 до 5000 нг/мл). Через 24 часа отбирали надосадочную жидкость, не содержащую клеток, и определяли количество выработанных IL-2 и/или INF-γ при помощи ELISA. В качестве отрицательного контроля применяли одни Т-клетки. Когда клетки стимулировали 200 нг/мл анти-CD3 mAb в течение 24 ч, максимальное снижение выработки IFN-γ 44% и 58% наблюдалось в Т-клетках, экспрессирующих TIM7 и TIM8, соответственно. В совокупности указанные данные свидетельствуют о том, что снижения экспрессии TCR, например, при применении TIM для удаления или разрушения TCR, достаточно для изменения функции Т-клеток.
Пример 5: Получение Т-клеток, дефицитных по рецептору Т-клеток (TCR), экспрессирующих chNKG2D
В данном примере добивались одновременной гиперэкспрессии доминантно-негативного белка-ингибитора TCR, т.е., TIM, и экспрессии химерного рецептора, направленного против опухоли. В частности экспрессию эндогенного TCR подавляли при помощи TIM, и экспрессировали химерный рецептор chNKG2D, т.е. NKG2D, связанный с цитоплазматическим доменом CD3-дзета. NKG2D ассоциирован с Dap 10 для обеспечения как первичной, так и вторичной активации сигналов к Т-клеткам. См., Zhang, T. et al. 2006. Cancer Res. 66(11): 5927-5933. Лиганды к NKG2D экспрессирует большинство клеток опухолей человек, но не большинство нормальных клеток.
Чтобы оценить экспрессию и TIM, и химерного рецептора, направленного против опухолей, первичные РВМС человека выделяли у здоровых доноров и активировали 40 нг/мл растворимого анти-CD3 и 50 Е/мл rhuIL-2 в течение 72 часов. Активированные Т-лимфоциты отмывали и проводили их трансдукцию ретровирусами с высоким титром путем 1-часовой спинокуляции при 32°С, после чего наступал 7-часовой период покоя. Для трансдукции применяли эквивалентное количество TIM и chNKG2D-вируса. Клетки отмывали и культивировали в 50 Е/мл IL-2 в течение 48 часов, а затем передавали для отбора с G418 в течение 3 дней. После отбора живые клетки выделяли при помощи Lymphoprep (Mediatech), и эффекторные клетки распределяли в 50 Е/мл IL-2 на 48 часов, после истечения которых клетки применяли для функционального анализа. Изменение экспрессии на поверхности клетки анализировали с применением антител, специфических в отношении CD3, CD8, CD4, NKG2D и CD5. Не было выявлено никаких различий в экспрессии молекул на поверхности данных клеток у TIM-экспрессирующих Т-клеток по сравнению с контрольным вектором, за исключением более высокого уровня экспрессии рецептора NKG2D в клетках, подвергшихся трансдукции вирусом chNKG2D, что было ожидаемо.
Чтобы определить, могут ли TIM+ chNKG2D+ клетки демонстрировать снижение ответа на аллогенные клетки, но усиление ответа на клетки опухоли, Т-клетки культивировали совместно с аллогенными РВМС, синергичными РВМС или клетками опухоли: RPMI8226 (линия клеток миеломы человека, NKG2D лиганд+), PANC-1 ((линия клеток из поджелудочной железы человека, NKG2D+) или NIH-3T3 (линия нормальных фибробластов мыши, NKG2D лиганд-), в качестве отрицательного контроля. Через 24 часа отбирали надосадочную жидкость, не содержащую клеток, и определяли количество выработанного INF-γ при помощи ELISA. В качестве отрицательного контроля применяли одни Т-клетки и культуру с синергичными РВМС.
При аллогенном анализе 45% снижение выработки INF-γ наблюдалось в TIM7-экспрессирующих Т-клетках и 44% снижение выработки INF-γ в TIM8-экспрессирующих Т-клетках, которые совместно экспрессировали chNKG2D, по сравнению с клетками, экспрессировавшими контрольный вектор. При культивировании с клетками опухоли наблюдалось значимое увеличение объема выработки INF-γ в ответ на клетки опухоли в TIM+ chNKG2D+ клетках по сравнению с клетками, экспрессирующими только TIM, когда клетки опухоли экспрессировали лиганды chNKG2D (RPMI8226 и PANC-1), но не при культивировании с лиганд-дефицитными клетками опухолей (NIH-3T3). См. Фигуру 3, показывающую репрезентативный эксперимент с применением RPMI8226. Этот же эксперимент также продемонстрировал, что более высокая выработка INF-γ зависела от NKG2D, поскольку инкубация с блокирующим mAb на NKG2D не приводила к повышению выработки INF-γ.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ Т-КЛЕТОК С НЕДОСТАТОЧНОСТЬЮ РЕЦЕПТОРОВ Т-КЛЕТОК | 2013 |
|
RU2781979C2 |
| СПОСОБ СОЗДАНИЯ Т-КЛЕТОК, ПРИГОДНЫХ ДЛЯ АЛЛОГЕННОЙ ТРАНСПЛАНТАЦИИ | 2015 |
|
RU2752933C2 |
| УЛУЧШЕННЫЕ ИММУННЫЕ КЛЕТКИ С ДВОЙНОЙ кшРНК И КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ИХ | 2019 |
|
RU2793922C2 |
| СПОСОБЫ ЭКСПАНДИРОВАНИЯ АНТИГЕНСПЕЦИФИЧЕСКИХ CAR-T-КЛЕТОК, КОМПОЗИЦИИ И ПРИМЕНЕНИЯ, СВЯЗАННЫЕ С НИМИ | 2019 |
|
RU2800920C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ГАММА ДЕЛЬТА Т-КЛЕТКИ | 2015 |
|
RU2756247C2 |
| ИММУННЫЕ КЛЕТКИ, ДЕФЕКТНЫЕ ПО Suv39h1 | 2018 |
|
RU2784531C2 |
| СПОСОБ ПРОИЗВОДСТВА T-КЛЕТОК | 2015 |
|
RU2813668C2 |
| РАЗМНОЖЕНИЕ И ПРИМЕНЕНИЕ НЕГЕМОПОЭТИЧЕСКИХ ТКАНЕРЕЗИДЕНТНЫХ γδ Т-КЛЕТОК | 2016 |
|
RU2784566C2 |
| УЛУЧШЕННЫЕ КОМПОЗИЦИИ НА ОСНОВЕ T-КЛЕТОК | 2015 |
|
RU2719030C2 |
| СПОСОБЫ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ ИСТОЩЕНИЯ TCRαβ КЛЕТОК | 2020 |
|
RU2829624C2 |







































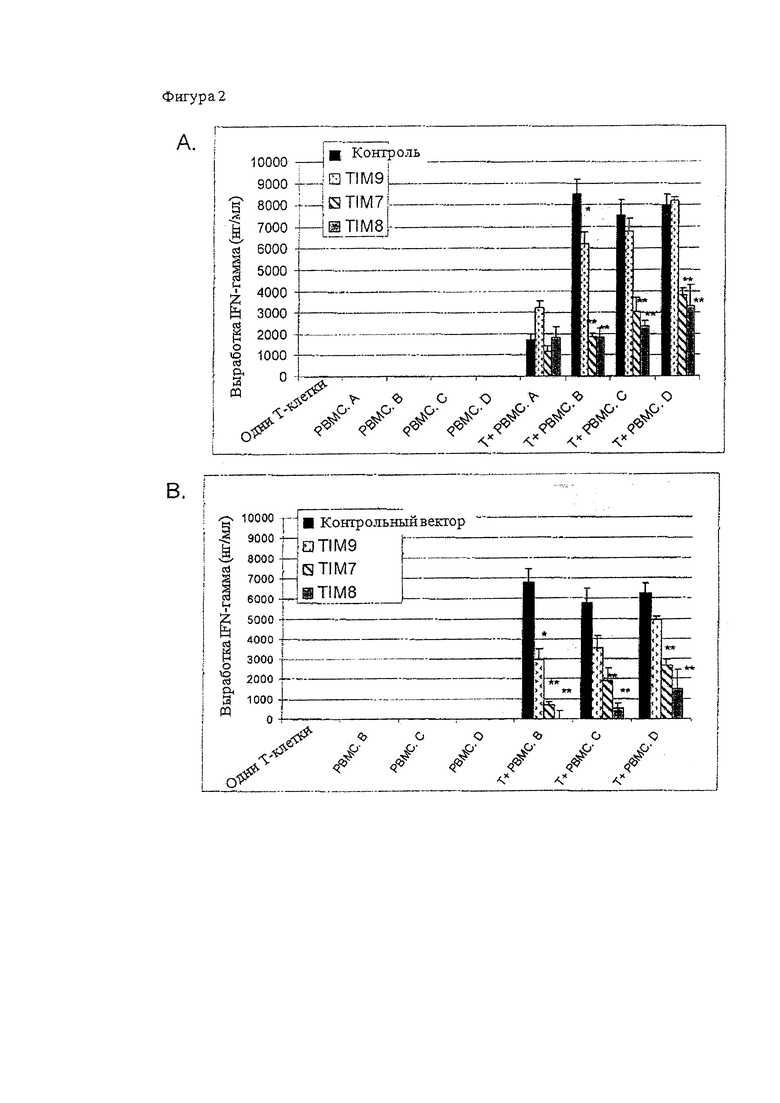
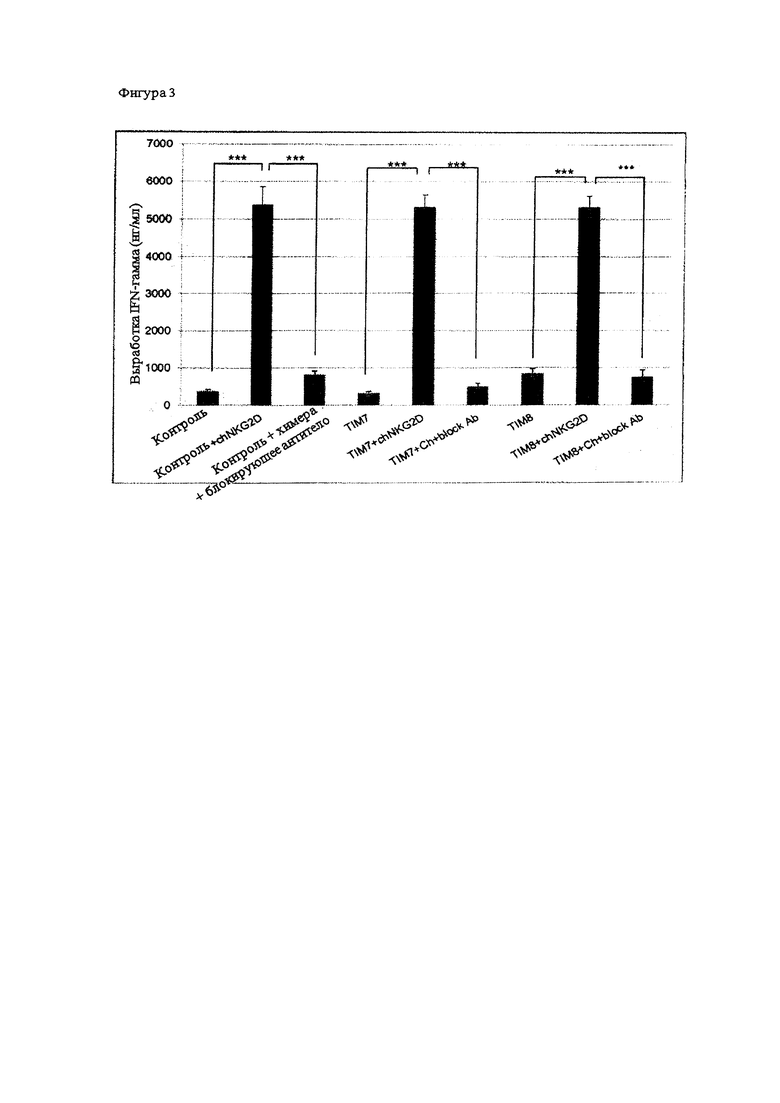
Настоящее изобретение относится к области биотехнологии, конкретно к генетически-модифицированным TCR-дефицитным Т-клеткам, и может быть использовано в медицине для лечения рака. Способ направлен на получение модифицированной первичной человеческой Т-клетки из первичной человеческой Т-клетки от донора-человека, которая модифицирована с целью уменьшения экспрессии эндогенного Т-клеточного рецептора (TCR). Способ включает введение в первичную человеческую Т-клетку, полученную от человеческого донора, по меньшей мере одного минигена, имеющего последовательность нуклеиновой кислоты, кодирующей усеченный полипептид CD3-дзета и экспрессию указанного минигена с целью уменьшения экспрессии эндогенного Т-клеточного рецептора. Изобретение позволяет уменьшить реакцию «трансплантат против хозяина» (GVHD) в гистосовместимых реципиентах-людях по сравнению с реакцией GVHD, вызываемой первичной человеческой Т-клеткой, полученной от того же донора-человека, которая не модифицирована введением по меньшей мере одного минигена. 5 н. и 13 з.п. ф-лы, 3 ил., 2 табл., 5 пр.
1. Способ получения модифицированной первичной человеческой Т-клетки, которая получена из первичной человеческой Т-клетки, полученной от донора-человека, и которая модифицирована с целью уменьшения экспрессии эндогенного Т-клеточного рецептора (TCR), где указанный способ включает введение в первичную человеческую Т-клетку, полученную от человеческого донора, по меньшей мере одного минигена, имеющего последовательность нуклеиновой кислоты, кодирующей усеченный полипептид CD3-дзета SEQ ID NO: 73, 75, 77 или 79, и экспрессию указанного минигена, где экспрессия указанного минигена уменьшает экспрессию эндогенного Т-клеточного рецептора и где указанная модифицированная первичная Т-клетка не вызывает или вызывает уменьшенную реакцию «трансплантат против хозяина» (GVHD) в гистосовместимых реципиентах-людях по сравнению с реакцией GVHD, вызываемой первичной человеческой Т-клеткой, полученной от того же донора-человека, которая не модифицирована введением по меньшей мере одного минигена.
2. Способ по п.1, где миниген имеет последовательность нуклеиновой кислоты SEQ ID NO: 72, 74, 76 или 78.
3. Способ по п. 1, отличающийся тем, что указанный миниген имеет последовательность нуклеиновой кислоты, кодирующей усечнный полипептид CD3-дзета, согласно SEQ ID №:77 или 79.
4. Способ по п.3, отличающийся тем, что указанный миниген имеет последовательность нуклеиновой кислоты SEQ ID №:76 или 78.
5. Способ по одному из пп.1-4, отличающийся тем, что способ далее включает модификацию модифицированной первичной Т-клетки, чтобы экспрессировать химерный рецептор, направленный против опухоли.
6. Способ по любому из пп.1-5, где указанный миниген дестабилизирует комплекс TCR путем уменьшения или блокирования экспрессии компонентов комплекса TCR на поверхности модифицированной первичной человеческой Т-клетки.
7. Способ по любому из пп. 1-6, в котором уменьшенную реакцию GVHD подтверждают по уменьшенной экспрессии гамма-интерферона, вызванной модифицированной первичной человеческой Т-клеткой по сравнению с первичной человеческой Т-клеткой, полученной от того же донора-человека, которая не модифицирована путем введения по меньшей мере одного минигена.
8. Способ по любому из пп. 1-6, в котором уменьшенную реакцию GVHD подтверждают по отсутствию вызывания повышения экспрессии гамма-интерферона модифицированной первичной человеческой Т-клеткой по сравнению с первичной человеческой Т-клеткой, полученной от того же донора-человека, которая не модифицирована путем введения по меньшей мере одного минигена.
9. Способ по любому из пп. 1-8, в котором модифицированная первичная человеческая Т-клетка не вызывает реакцию GVHD у человеческого субъекта.
10. Модифицированная первичная человеческая Т-клетка, полученная согласно любому из пп.1-9.
11. Модифицированная первичная человеческая Т-клетка по п.10, которая получена согласно п.5.
12. Фармацевтическая композиция, содержащая терапевтически эффективное количество модифицированной первичной человеческой Т-клетки по п. 11, для использования в лечении рака.
13. Способ лечения рака, включающий введение субъекту, нуждающемуся в лечении, терапевтически эффективного количества модифицированной первичной человеческой Т-клетки согласно п.11 или фармацевтической композиции по п.12.
14. Вектор, содержащий миниген, имеющий последовательность нуклеиновой кислоты, кодирующей усеченный полипептид CD3-дзета, согласно SEQ ID №:73, 75, 77 или 79 для экспрессии усеченного полипептида CD3-дзета согласно SEQ ID №:73, 75, 77 или 79 в модифицированной первичной человеческой Т-клетке по пп. 10 или 11, или для получения модифицированной первичной Т-клетки согласно любому из пп. 1-9.
15. Вектор по п. 14, отличающийся тем, что последовательность нуклеиновой кислоты, кодирующая усеченный полипептид CD3-дзета, представлена SEQ ID №:72, 74, 76 или 78.
16. Вектор по п.14, в котором указанный миниген кодирует усеченный полипептид CD3-дзета SEQ ID №: 77 или 79.
17. Вектор по п.16, в котором указанный миниген имеет последовательность нуклеиновой кислоты SEQ ID №: 76 или 78.
18. Вектор по любому из пп.14-17, в котором вектор является ретровирусной экспрессионной плазмидой.
| WO 2011059836 A2, 19.05.2011 | |||
| WO 2006036445 A2, 06.04.2006 | |||
| BARBER A | |||
| ET AL., Chimeric NKG2D receptor-expressing T cells as an immunotherapy for multiple myeloma, Exp | |||
| Hematol., 2008, v.36, n.10, p:1318-1328 | |||
| POLIC B | |||
| ET AL., How alpha beta T cells deal with induced TCR alpha ablation, Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| USA, 2001, v.98,n.15,p:8744-8749 | |||
| Приспособление к прессовому ящику для удержания на определенном уровне порций хлопка прессованием его в кипы | 1927 |
|
SU20963A1 |
| Самоходный конвейерный перегружатель | 1977 |
|
SU693020A1 |
| СПОСОБ НАПРАВЛЕНИЯ (ИНДУКЦИИ) ИММУННОГО ОТВЕТА, ДНК, ВЕКТОР, СПОСОБ ЛЕЧЕНИЯ | 1992 |
|
RU2161044C2 |

