РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет предварительной заявки на патент США № 62/622468, поданной 26 января 2018 г., полное описание которой включено в настоящий документ ссылкой.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Взаимодействие антител с неонатальным Fc-рецептором (FcRn) является определяющим фактором в поддержании и продлении периода полувыведения из сыворотки крови антител и других полученных на основе Fc терапевтических средств. FcRn представляет собой гетеродимер α-домена и субъединицы β2-макроглобулина (β2-m) в MHC класса I, который распознает области тяжелой цепи Fc антитела, отличающиеся от других рецепторов Fcγ (FcγR). Несмотря на то, что FcRn экспрессируется в различных тканях, считается, что он действует главным образом на эндотелий сосудов, почки и гематоэнцефалический барьер, предотвращая деградацию IgG, экскрецию и возникновение воспалительных реакций, соответственно.
Связывание антител с FcRn сильно зависит от pH, и взаимодействие происходит с высоким сродством (от высокого наномолярного до низкого микромолярного) только при низких значениях pH (pH <6,5), но не при физиологическом pH (pH примерно 7,4). Подкисление эндосомы до рН менее 6,5 создает благоприятные условия для взаимодействия между IgG и FcRn, которое непосредственно отвечает за ингибирование деградации и стимулирование рециркуляции FcRn-связанных антител на клеточную поверхность. Увеличение рН ослабляет взаимодействие и способствует выделению антител в кровоток.
Инжиниринг Fc с использованием мутагенеза с высокой пропускной способностью активно использовался для идентификации вариантов, которые повышают способность связывания с FcRn, так как усиление связывания предположительно приводит к повышению эффективности и уменьшению частоты дозировки терапевтических антител вследствие продления периода полувыведения из сыворотки по сравнению с антителом IgG дикого типа. Однако варианты, которые повышают способность связывания с FcRn, могут иметь непредсказуемые результаты. Например, некоторые варианты IgG, такие как N434W или P257I/Q311I среди прочих, которые в исследованиях на трансгенных мышах, на обезьянах Cynomolgus и человеческих FcRn (hFcRn) демонстрируют значительное увеличение сродства к FcRn при рН 6,0, имеют периоды полувыведения из сыворотки крови, как для молекул дикого типа, или они становятся чрезвычайно короткими (см., например, Kuo et al. 2011 выше; Datta-Mannan et al. 2007, J. Biol. Chem. 282: 1709-1717; и Datta-Mannan et al. 2007, Metab. Dispos. 35. 86-94). Вариант T250Q/M428L (QL) показал специфичные для каркаса IgG результаты на животных моделях (см., например, Datta-Mannan et al. 2007, J. Biol. Chem. 282: 1709-1717; и Hinton et al. 2006, J. Immunol. 176: 346-356). Вариант M252Y/S254T/T256E (YTE, нумерация ЕС) показал 10-кратное усиление in vitro, но демонстрирует пониженную антителозависимую клеточно-опосредованную цитотоксичность (ADCC) in vivo из-за 2-кратного снижения сродства к FcγRIIIa рецептору (см., например, Dall'Acqua et al. 2002, выше).
Таким образом, по-прежнему существует потребность в альтернативных вариантах Fc, которые обладают улучшенным связыванием с FcRn и пролонгированным периодом полувыведения из кровеносной системы.
КРАТКОЕ ОПИСАНИЕ
Настоящее изобретение основано на открытии новых антител IgG, имеющих одну или несколько из следующих характеристик: более длинный период полувыведения из сыворотки крови, повышенная способность связывания с FcRn, повышенная способность связывания с FcRn при кислотном значении pH, повышенная способность связывания с FcγRIIIa по сравнению с антителом IgG дикого типа и похожая термостабильность.
Соответственно, в некоторых аспектах выделенный связывающий полипептид, содержащий модифицированный Fс-домен, содержащий аспарагиновую кислоту (D) или глутаминовую кислоту (E) в положении аминокислоты 256, и/или триптофан (W) или глутамин (Q) в положении аминокислоты 307, где положение аминокислоты 254 не является треонином (Т), и дополнительно содержит фенилаланин (F) или тирозин (Y) в положении аминокислоты 434, или тирозин (Y) в положении аминокислоты 252, где положения аминокислот соответствуют нумерации ЕС.
В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен человека. В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен IgG1.
В некоторых иллюстративных вариантах осуществления связывающий полипептид обладает способностью связывания с FcRn человека, способностью связывания с FcRn крысы или способностью связывания с FcRn и человека, и крысы.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более длинный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со способностью связывания с FcRn связывающего полипептида при повышенном, некислотном значении рН. В некоторых иллюстративных вариантах осуществления повышенная способность связывания с FcRn включает в себя сниженную скорость диссоциации с FcRn.
В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0. В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0, а некислотное значение рН составляет примерно 7,4.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет примерно такую же способность связывания с FcγRIIIa, что и связывающий полипептид, содержащий Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет примерно такую же термостабильность, что и связывающий полипептид, содержащий Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет примерно такую же термостабильность, что и связывающий полипептид, содержащий измененный Fc-домен, содержащий тройную аминокислотную замену M252Y/S254T/T256E, согласно нумерации ЕС.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид представляет собой антитело, например, моноклональное антитело. В некоторых иллюстративных вариантах осуществления выделенное антитело представляет собой химерное, гуманизированное или человеческое антитело. В некоторых иллюстративных вариантах осуществления выделенное антитело представляет собой полноразмерное антитело.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид специфически связывает одну или несколько мишеней у человека.
В других аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, содержащий комбинацию аминокислотных замен в положениях, выбранных из группы, состоящей из а) тирозина (Y) в положении аминокислоты 252 и аспарагиновой кислоты (D) в положении аминокислоты 256, b) аспарагиновой кислоты (D) в положении аминокислоты 256 и фенилаланина (F) в положении аминокислоты 434, c) аспарагиновой кислоты (D) в положении аминокислоты 256 и тирозина (Y) в положении аминокислоты 434, d) триптофана (W) в положении аминокислоты 307 и фенилаланина (F) в положении аминокислоты 434, е) тирозина (Y) в положении аминокислоты 252 и триптофана (W) в положении аминокислоты 307, где тирозин (Y) не находится в положении аминокислоты 434, f) аспарагиновой кислоты (D) в положении аминокислоты 256 и триптофана (W) в положении аминокислоты 307, где тирозин (Y) не находится в положении аминокислоты 434, g) аспарагиновой кислоты (D) в положении аминокислоты 256 и глутамина (Q) в положении аминокислоты 307, где тирозин (Y) не находится в положении аминокислоты 434, h) тирозина (Y) в положении аминокислоты 252, аспарагиновой кислоты (D) в положении аминокислоты 256 и глутамина (Q) в положении аминокислоты 307, где тирозин (Y) не находится в положении аминокислоты 434, и i) тирозина (Y) в положении аминокислоты 252, глутаминовой кислоты (E) в положении аминокислоты 256 и глутамина (Q) в положении аминокислоты 307, где треонин (T) не находится в положении аминокислоты 254, гистидин (H) не находится в положении аминокислоты 311, и тирозин (Y) не находится в положении аминокислоты 434, где аминокислотные замены представлены согласно нумерации ЕС.
В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен человека. В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен IgG1.
В некоторых иллюстративных вариантах осуществления связывающий полипептид обладает способностью связывания с FcRn человека, способностью связывания с FcRn крысы или способностью связывания с FcRn и человека, и крысы.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более длинный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со способностью связывания с FcRn связывающего полипептида при повышенном, некислотном значении рН. В некоторых иллюстративных вариантах осуществления повышенная способность связывания с FcRn включает сниженную скорость диссоциации с FcRn. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более слабую способность связывания с FcRn при некислотных значениях рН, чем связывающий полипептид, содержащий измененный Fc-домен, содержащий двойную аминокислотную замену M428L/N434S, согласно нумерации ЕС.
В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0. В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0, а некислотное значение рН составляет примерно 7,4.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет примерно такую же способность связывания с FcγRIIIa, что и связывающий полипептид, содержащий Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет примерно такую же термостабильность, что и связывающий полипептид, содержащий Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет примерно такую же термостабильность, что и связывающий полипептид, содержащий измененный Fc-домен, содержащий тройную аминокислотную замену M252Y/S254T/T256E, согласно нумерации ЕС.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид представляет собой антитело, например, моноклональное антитело. В некоторых иллюстративных вариантах осуществления выделенное антитело представляет собой химерное, гуманизированное или человеческое антитело. В некоторых иллюстративных вариантах осуществления выделенное антитело представляет собой полноразмерное антитело.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид специфически связывает одну или несколько мишеней у человека.
В других аспектах выделенный связывающий полипептид, содержащий измененный Fc-домен, содержащий а) двойную аминокислотную замену, выбранную из группы, состоящей из M252Y/T256D, M252Y/T256E, M252Y/T307Q, M252Y/T307W, T256D/T307Q, T256D/T307W, T256E/T307Q и T256E/T307W, где треонин (T) не находится в положении аминокислоты 254, гистидин (H) не находится в положении аминокислоты 311, а тирозин (Y) не находится в положении аминокислоты 434 или b) тройную аминокислотную замену, выбранную из группы, состоящей из M252Y/T256D/T307Q, M252Y/T256D/T307W, M252Y/T256E/T307Q и M252Y/T256E/T307W, где треонин (T) не является аминокислотой в положении 254 гистидин (H) не находится в положении аминокислоты 311, а тирозин (Y) не находится в положении аминокислоты 434, где аминокислотные замены соответствуют нумерации ЕС.
В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен человека. В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен IgG1.
В некоторых иллюстративных вариантах осуществления связывающий полипептид обладает способностью связывания с FcRn человека, способностью связывания с FcRn крысы или способностью связывания с FcRn и человека, и крысы.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более длинный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со способностью связывания с FcRn связывающего полипептида при повышенном, некислотном значении рН. В некоторых иллюстративных вариантах осуществления повышенная способность связывания с FcRn включает сниженную скорость диссоциации с FcRn. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более слабую способность связывания с FcRn при некислотных значениях рН, чем связывающий полипептид, содержащий измененный Fc-домен, содержащий двойную аминокислотную замену M428L/N434S, согласно нумерации ЕС.
В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0. В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0, а некислотное значение рН составляет примерно 7,4.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет примерно такую же способность связывания с FcγRIIIa, что и связывающий полипептид, содержащий Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет примерно такую же термостабильность, что и связывающий полипептид, содержащий Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет примерно такую же термостабильность, что и связывающий полипептид, содержащий измененный Fc-домен, содержащий тройную аминокислотную замену M252Y/S254T/T256E, согласно нумерации ЕС.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид представляет собой антитело, например, моноклональное антитело. В некоторых иллюстративных вариантах осуществления выделенное антитело представляет собой химерное, гуманизированное или человеческое антитело. В некоторых иллюстративных вариантах осуществления выделенное антитело представляет собой полноразмерное антитело.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид специфически связывает одну или несколько мишеней у человека.
В определенных аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, где измененный Fc-домен содержит аспарагиновую кислоту (D) в положении аминокислоты 256 и глутамин (Q) в положении аминокислоты 307, согласно нумерации ЕС.
В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен человека. В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен IgG1.
В некоторых иллюстративных вариантах осуществления связывающий полипептид обладает способностью связывания с FcRn человека, или способностью связывания с FcRn крысы, или способностью связывания с FcRn и человека, и крысы.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более длинный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со способностью связывания с FcRn связывающего полипептида при повышенном, некислотном значении рН. В некоторых иллюстративных вариантах осуществления повышенная способность связывания с FcRn включает сниженную скорость диссоциации с FcRn.
В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0. В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0, а некислотное значение рН составляет примерно 7,4.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид представляет собой моноклональное антитело. В некоторых иллюстративных вариантах осуществления антитело представляет собой химерное, гуманизированное или человеческое антитело.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид специфически связывает одну или несколько мишеней у человека.
В некоторых аспектах предложена выделенная молекула нуклеиновой кислоты, содержащая нуклеиновую кислоту, кодирующую выделенный полипептид.
В некоторых аспектах предложен вектор, содержащий выделенную молекулу нуклеиновой кислоты. В некоторых иллюстративных вариантах осуществления вектор представляет собой вектор экспрессии. В некоторых аспектах предложен вектор экспрессии, содержащий выделенную молекулу нуклеиновой кислоты.
В некоторых аспектах предложена клетка-хозяин, содержащая вектор. В некоторых аспектах предложена клетка-хозяин, содержащая вектор экспрессии.
В некоторых иллюстративных вариантах осуществления клетка-хозяин имеет эукариотическое или прокариотическое происхождение. В некоторых иллюстративных вариантах осуществления клетка-хозяин происходит от млекопитающего. В некоторых иллюстративных вариантах осуществления клетка-хозяин имеет бактериальное происхождение.
В некоторых аспектах предложен фармацевтический состав, содержащий выделенный связывающий полипептид.
В некоторых аспектах предложен фармацевтический состав, содержащий выделенное антитело.
В некоторых аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, где измененный Fc-домен содержит аспарагиновую кислоту (D) в положении аминокислоты 256 и триптофан (W) в положении аминокислоты 307, согласно нумерации ЕС.
В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен человека. В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен IgG1.
В некоторых иллюстративных вариантах осуществления связывающий полипептид обладает способностью связывания с FcRn человека, или способностью связывания с FcRn крысы, или способностью связывания с FcRn и человека, и крысы.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более длинный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со способностью связывания с FcRn связывающего полипептида при повышенном, некислотном значении рН. В некоторых иллюстративных вариантах осуществления повышенная способность связывания с FcRn включает сниженную скорость диссоциации с FcRn.
В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0. В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0, а некислотное значение рН составляет примерно 7,4.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид представляет собой моноклональное антитело. В некоторых иллюстративных вариантах осуществления антитело представляет собой химерное, гуманизированное или человеческое антитело.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид специфически связывает одну или несколько мишеней у человека.
В некоторых аспектах предложена выделенная молекула нуклеиновой кислоты, содержащая нуклеиновую кислоту, кодирующую выделенный полипептид.
В некоторых аспектах предложен вектор, содержащий выделенную молекулу нуклеиновой кислоты. В некоторых иллюстративных вариантах осуществления вектор представляет собой вектор экспрессии. В некоторых аспектах предложен вектор экспрессии, содержащий выделенную молекулу нуклеиновой кислоты.
В некоторых аспектах предложена клетка-хозяин, содержащая вектор. В некоторых аспектах предложена клетка-хозяин, содержащая вектор экспрессии.
В некоторых иллюстративных вариантах осуществления клетка-хозяин имеет эукариотическое или прокариотическое происхождение. В некоторых иллюстративных вариантах осуществления клетка-хозяин происходит от млекопитающего. В некоторых иллюстративных вариантах осуществления клетка-хозяин имеет бактериальное происхождение.
В некоторых аспектах предложен фармацевтический состав, содержащий выделенный связывающий полипептид.
В некоторых аспектах предложен фармацевтический состав, содержащий выделенное антитело.
В некоторых аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, где измененный Fc-домен содержит тирозин (Y) в положении аминокислоты 252 и аспарагиновую кислоту (D) в положении аминокислоты 256, согласно нумерации ЕС.
В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен человека. В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен IgG1.
В некоторых иллюстративных вариантах осуществления связывающий полипептид обладает способностью связывания с FcRn человека, или способностью связывания с FcRn крысы, или способностью связывания с FcRn и человека, и крысы.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более длинный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со способностью связывания с FcRn связывающего полипептида при повышенном, некислотном значении рН. В некоторых иллюстративных вариантах осуществления повышенная способность связывания с FcRn включает сниженную скорость диссоциации с FcRn.
В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0. В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0, а некислотное значение рН составляет примерно 7,4.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид представляет собой моноклональное антитело. В некоторых иллюстративных вариантах осуществления антитело представляет собой химерное, гуманизированное или человеческое антитело.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид специфически связывает одну или несколько мишеней у человека.
В некоторых аспектах предложена выделенная молекула нуклеиновой кислоты, содержащая нуклеиновую кислоту, кодирующую выделенный полипептид.
В некоторых аспектах предложен вектор, содержащий выделенную молекулу нуклеиновой кислоты. В некоторых иллюстративных вариантах осуществления вектор представляет собой вектор экспрессии. В некоторых аспектах предложен вектор экспрессии, содержащий выделенную молекулу нуклеиновой кислоты.
В некоторых аспектах предложена клетка-хозяин, содержащая вектор. В некоторых аспектах предложена клетка-хозяин, содержащая вектор экспрессии.
В некоторых иллюстративных вариантах осуществления клетка-хозяин имеет эукариотическое или прокариотическое происхождение. В некоторых иллюстративных вариантах осуществления клетка-хозяин происходит от млекопитающего. В некоторых иллюстративных вариантах осуществления клетка-хозяин имеет бактериальное происхождение.
В некоторых аспектах предложен фармацевтический состав, содержащий выделенный связывающий полипептид.
В некоторых аспектах предложен фармацевтический состав, содержащий выделенное антитело.
В некоторых аспектах выделенный связывающий полипептид, содержащий измененный Fc-домен, где измененный Fc-домен содержит комбинацию из по меньшей мере четырех аминокислотных замен, содержащих: аспарагиновую кислоту (D) или глутаминовую кислоту (E) в положении аминокислоты 256, и триптофан (W) или глутамин (Q) в положении аминокислоты 307, где положение аминокислоты 254 не является треонином (Т), и дополнительно содержит фенилаланин (F) или тирозин (Y) в положении аминокислоты 434; и тирозин (Y) в положении аминокислоты 252, где положения аминокислот соответствуют нумерации ЕС.
В некоторых аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, включающий в себя комбинацию аминокислотных замен в положениях, выбранных из группы, состоящей из: а) тирозина (Y) в положении аминокислоты 252, аспарагиновой кислоты (D) в положении аминокислоты 256, глутамина (Q) в положении аминокислоты 307 и тирозина (Y) в положении аминокислоты 434; b) тирозина (Y) в положении аминокислоты 252, глутаминовой кислоты (E) в положении аминокислоты 256, триптофана (W) в положении аминокислоты 307 и тирозина (Y) в положении аминокислоты 434; c) тирозина (Y) в положении аминокислоты 252, глутаминовой кислоты (E) в положении аминокислоты 256, глутамина (Q) в положении аминокислоты 307 и тирозина (Y) в положении аминокислоты 434; d) тирозина (Y) в положении аминокислоты 252, аспарагиновой кислоты (D) в положении аминокислоты 256, глутамина (Q) в положении аминокислоты 307 и фенилаланина (F) в положении аминокислоты 434; или е) тирозина (Y) в положении аминокислоты 252, аспарагиновой кислоты (D) в положении аминокислоты 256, триптофана (W) в положении аминокислоты 307 и тирозина (Y) в положении аминокислоты 434, где аминокислотные замены соответствуют нумерации ЕС.
В некоторых аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, содержащий: четверную аминокислотную замену, выбранную из группы, состоящей из M252Y/T256D/T307Q/N434Y, M252Y/T256E/T307W/N434Y, M252Y/T256E/T307Q/N434Y, M252Y/T256D/T307Q/N434F и M252Y/T256D/T307W/N434Y, в которых аминокислотные замены представлены в соответствии с нумерацией ЕС.
В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен человека. В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен IgG1.
В некоторых примерных вариантах осуществления связывающий полипептид обладает способностью связывания с FcRn человека. В некоторых примерных вариантах осуществления связывающий полипептид обладает способностью связывания с FcRn крысы. В некоторых примерных вариантах осуществления связывающий полипептид обладает способностью связывания с FcRn и человека, и крысы.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со связывающим полипептидом, содержащим M252Y/S254T/T256E/H433K/N434F.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при некислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при некислотных рН по сравнению со связывающим полипептидом, содержащим M252Y/S254T/T256E/H433K/N434F.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН и повышенную способность связывания с FcRn при некислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН и повышенную способность связывания с FcRn при некислотном значении рН по сравнению со связывающим полипептидом, содержащим M252Y/S254T/T256E/H433K/N434F.
В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0. В некоторых иллюстративных вариантах осуществления некислотное значение рН составляет примерно 7,4.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более короткий период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более короткий период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим M252Y/S254T/T256E/H433K/N434F.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет измененную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим M252Y/S254T/T256E/H433K/N434F.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную термостабильность по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную термостабильность по сравнению со связывающим полипептидом, содержащим M252Y/S254T/T256E/H433K/N434F.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид представляет собой антитело. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид представляет собой моноклональное антитело. В некоторых иллюстративных вариантах осуществления выделенное антитело представляет собой химерное, гуманизированное или человеческое антитело. В некоторых иллюстративных вариантах осуществления выделенное антитело представляет собой полноразмерное антитело.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид специфически связывает одну или несколько мишеней.
В некоторых аспектах предложена выделенная молекула нуклеиновой кислоты, содержащая нуклеиновую кислоту, кодирующую выделенный полипептид.
В некоторых аспектах предложен вектор, содержащий выделенную молекулу нуклеиновой кислоты.
В некоторых иллюстративных вариантах осуществления вектор представляет собой вектор экспрессии.
В некоторых аспектах предложена клетка-хозяин, содержащая вектор.
В некоторых иллюстративных вариантах осуществления клетка-хозяин имеет эукариотическое или прокариотическое происхождение. В некоторых иллюстративных вариантах осуществления клетка-хозяин происходит от млекопитающего. В некоторых иллюстративных вариантах осуществления клетка-хозяин имеет бактериальное происхождение.
В некоторых аспектах предложен фармацевтический состав, содержащий выделенный связывающий полипептид.
В некоторых аспектах предложен фармацевтический состав, содержащий выделенное антитело.
В некоторых аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, содержащий тирозин (Y) в положении аминокислоты 252, аспарагиновую кислоту (D) в положении аминокислоты 256, глутамин (Q) в положении аминокислоты 307 и тирозин (Y) в положении аминокислоты 434, согласно нумерации ЕС.
В некоторых аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, содержащий тирозин (Y) в положении аминокислоты 252, глутаминовую кислоту (Е) в положении аминокислоты 256, триптофан (W) в положении аминокислоты 307 и тирозин (Y) в положении аминокислоты 434, согласно нумерации ЕС.
В некоторых аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, содержащий тирозин (Y) в положении аминокислоты 252, глутаминовую кислоту (Е) в положении аминокислоты 256, глутамин (Q) в положении аминокислоты 307 и тирозин (Y) в положении аминокислоты 434, согласно нумерации ЕС.
В некоторых аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, содержащий тирозин (Y) в положении аминокислоты 252, аспарагиновую кислоту (D) в положении аминокислоты 256, глутамин (Q) в положении аминокислоты 307 и фенилаланин (F) в положении аминокислоты 434, согласно нумерации ЕС.
В некоторых аспектах предложен выделенный связывающий полипептид, содержащий измененный Fc-домен, содержащий тирозин (Y) в положении аминокислоты 252, аспарагиновую кислоту (D) в положении аминокислоты 256, триптофан (W) в положении аминокислоты 307 и тирозин (Y) в положении аминокислоты 434, согласно нумерации ЕС.
В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен человека. В некоторых иллюстративных вариантах осуществления измененный Fc-домен представляет собой измененный Fc-домен IgG1.
В некоторых примерных вариантах осуществления связывающий полипептид обладает способностью связывания с FcRn человека.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более короткий период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет более короткий период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим M252Y/S254T/T256E/H433K/N434F.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН и повышенную способность связывания с FcRn при некислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении рН и повышенную способность связывания с FcRn при некислотном значении рН по сравнению со связывающим полипептидом, содержащим M252Y/S254T/T256E/H433K/N434F.
В некоторых иллюстративных вариантах осуществления кислотное значение рН составляет примерно 6,0, а некислотное значение рН составляет примерно 7,4.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим домен Fc дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим M252Y/S254T/T256E/H433K/N434F.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную термостабильность, как связывающий полипептид, содержащий Fc-домен дикого типа. В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид имеет пониженную термостабильность по сравнению со связывающим полипептидом, содержащим M252Y/S254T/T256E/H433K/N434F.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид представляет собой моноклональное антитело. В некоторых иллюстративных вариантах осуществления антитело представляет собой химерное, гуманизированное или человеческое антитело.
В некоторых иллюстративных вариантах осуществления выделенный связывающий полипептид специфически связывает одну или несколько мишеней.
В некоторых аспектах предложена выделенная молекула нуклеиновой кислоты, содержащая нуклеиновую кислоту, кодирующую выделенный полипептид.
В некоторых аспектах предложен вектор экспрессии, содержащий выделенную молекулу нуклеиновой кислоты.
В некоторых аспектах предложена клетка-хозяин, содержащая вектор экспрессии.
В некоторых аспектах предложен фармацевтический состав, содержащий выделенный связывающий полипептид.
В некоторых аспектах предложен способ лечения заболевания или расстройства у субъекта, нуждающегося в этом, включающий введение субъекту терапевтически эффективного количества выделенного связывающего полипептида или введение субъекту терапевтически эффективного количества фармацевтического состава.
В некоторых иллюстративных вариантах осуществления заболевание или расстройство представляет собой онкологическое заболевание. В некоторых иллюстративных вариантах осуществления онкологическое заболевание представляет собой опухоль.
В некоторых иллюстративных вариантах осуществления заболевание или расстройство представляет собой аутоиммунное расстройство.
В некоторых аспектах предложен способ лечения онкологического заболевания у субъекта, нуждающегося в этом, включающий введение субъекту терапевтически эффективного количества выделенного связывающего полипептида или введение субъекту терапевтически эффективного количества фармацевтического состава.
В некоторых аспектах предложен способ лечения аутоиммунного расстройства у субъекта, нуждающегося в этом, включающий введение субъекту терапевтически эффективного количества выделенного связывающего полипептида или введение субъекту терапевтически эффективного количества фармацевтического состава.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Вышеизложенные и другие признаки и преимущества настоящего изобретения будут в большей степени понятны из следующего подробного описания иллюстративных вариантов осуществления во взаимосвязи с прилагаемыми графическими материалами.
Фиг. 1А - Фиг. 1В изображают структуру FcRn, взаимодействующего с Fc-участком IgG1. На Фиг. 1А изображено взаимодействие между hFcRn и Fc IgG1 (pdb: 4n0u), где показан один Fc-мономер (темно-серая лента), включая гликозилирование, показанное в виде палочек, помеченных «гликан», в комплексе с α-доменом (серым) и субъединицами β2-m (светло-серым) hFcRn. Большинство остатков антител, участвующих во взаимодействии с FcRn, расположены в петлях, непосредственно примыкающих к области контакта CH2-CH3 (пунктирная линия) и напротив сайта гликозилирования. На Фиг. 1B показано изображение поверхности кристаллической структуры Fc IgG1 (pdb: 5d4q), повернутой на 75º относительно Фиг. 1A. Область контакта при связывании с FcRn состоит из остатков в доменах CH2 и CH3. Библиотека насыщения была построена в одиннадцати положениях, показанных в виде палочек, как обозначено: M252; I253; S254; T256; K288; T307; K322; E380; L432; N434 и Y436. Все эти остатки находятся в непосредственной близости или в прямом контакте с FcRn. Поверхности критических остатков гистидина, ответственные за кластер pH-зависимости (H310, H433, H435), находятся вблизи интересующих позиций и являются такими, как обозначено.
Фиг. 2А - Фиг. 2D изображают скрининговый анализ по технологии Octet и его результаты. На Фиг. 2А схематически представлен скрининговый анализ по технологии Octet. Биосенсоры NiNTA захватывают меченый гистидином антиген, а затем - варианты антител для исследования кинетики связывания c FcRn (rFcRn) крысы. На Фиг. 2В показаны кинетические профили связывания c rFcRn при рН 6,0 варианта дикого типа (сплошная линия), варианта T307A/E380A/N434A (AAA) (короткие штрихи), LS (короткие штрихи с точкой), YTE (длинные штрихи), антитела к H435A (длинные штрихи с точкой) и антитела H310A/H435Q (длинные штрихи с двумя точками), выровненные по началу фазы ассоциирования с rFcRn. Варианты H435A и H310A/H435Q практически не связывались с FcRn. Вариант YTE имеет самую низкую скорость диссоциации с FcRn, исследованную в анализе связывания с rFcRn по технологии Octet. На Фиг. 2C графически изображена нормализация кинетики связывания c FcRn при pH 6,0 сокращенного числа мутантов, полученных из скрининга по технологии Octet. Большинство мутантов сохранили значительную степень связывания с rFcRn, но некоторые результаты были похожи на результат проверочного контроля (пунктирная линия), что указывает на потерю всего связывания с rFcRn (длинные штрихи, расположенные ниже пунктирной линии (проверка)). Два варианта (сплошные линии) имели более медленную скорость диссоциации с rFcRn, чем антитела дикого типа (жирные длинные штрихи). На Фиг. 2D изображен анализ диаграммы скоростей диссоциации с rFcRn для всех точечных мутаций с наблюдаемой кинетикой связывания c rFcRn, разделенной положением остатка. Варианты насыщения попадали в один из следующих четырех режимов скорости диссоциации с rFcRn: отсутствие связывания (не показано), более быстрое связывание (черным), связывание по типу дикого типа (белым), замедленное связывание (серым). Восемнадцать мутантов продемонстрировали значительно более медленную скорость диссоциации с rFcRn, чем антитела дикого типа (черные пунктирные линии).
На Фиг. 3 графически изображена Biacore кинетика образцов сравнения и вариантов дикого типа с FcRns человека и крысы при рН 6,0 и рН 7,4. Все кривые связывания с FcRn для серии разбавлений вариантов дикого типа (вверху слева), варианта AAA (вверху справа), варианта M428/N434S (LS) (внизу слева) и варианта M252Y/S254T/T256E (YTE) (внизу справа) показаны для FcRn человека (первый и третий столбцы) и крысы (второй и четвертый столбцы) при pH 6,0 (первый и третий ряды) и pH 7,4 (второй и четвертый ряды). Варианты AAA, LS и YTE демонстрировали более медленную диссоциацию с FcRn, чем антитела дикого типа. В целом, антитела связывают rFcRn с примерно 10-кратно увеличенной способностью по сравнению с антителом дикого типа. Вариант LS обладал наиболее сильным сродством к hFcRn при рН 7,4 и наибольшим остаточным связыванием при рН 7,4, тогда как связывание rFcRn вариантом YTE было наиболее тесным.
На Фиг. 4А графически изображена Biacore кинетика лидирующих вариантов насыщения с FcRn человека и крысы при рН 6,0. Следы кинетического связывания FcRn для серии разбавлений показаны для 18 лидирующих вариантов насыщения. Скорости диссоциации M252Y, T256D, T256E, N434F, N434P, N434Y, T307A, T307E, T307F, T307Q и T307W с FcRn человека и крысы были более медленными. Остальные варианты были специфичны только для FcRn крысы.
На Фиг. 4В графически показана кинетика связывания FcRn для WT, вариантом сравнения и лидирующих одиночных вариантов насыщения с FcRn человека при pH 6,0. Сенсограммы связывания FcRn с серийными разведениями WT, LS, YTE и 18 вариантов насыщения с FcRn человека при pH 6,0. Одиночные варианты насыщения, используемые для библиотеки комбинаций, подчеркнуты и выделены жирным шрифтом.
На Фиг. 5A - Фиг. 5D изображены данные, показывающие, что несколько вариантов имеют более медленные скорости диссоциации с FcRn человека и крысы при pH 6,0. На Фиг. 5A и 5B изображены Biacore сенсограммы различных вариантов. На рис. 5А показаны скорости диссоциации с FcRn человека при рН 6,0 для варианта YTE (длинные штрихи с точкой ), варианта LS (длинные штрихи с двумя точками), варианта дикого типа (WT; пунктирная линия) и лидирующих вариантов насыщения (лидирующие; сплошные линии разных оттенков). На Фиг. 5А показаны нормализованные сенсограммы, показывающие улучшенные скорости диссоциации с hFcRn по сравнению с WT. На Фиг. 5В показаны скорости диссоциации с FcRn крысы при рН 6,0 для варианта ААА (пунктир), варианта LS (штрихи с двумя точками), варианта YTE (штрихи с одной точкой), варианта дикого типа (сплошная линия) и лидирующих вариантов насыщения (пунктирные линии различной частоты и толщины). Репрезентативная инъекция каждого из лидирующих одиннадцати антител показана для ясности. Эти лидирующие одиночные варианты показали улучшенную кинетику диссоциации с FcRn человека и крысы по сравнению с диким типом. На Фиг. 5C и 5D показаны графики способности связывания для лидирующих вариантов насыщения антител (белые кружки) и антител дикого типа (черный кружок) с FcRn человека (Фиг. 5C) и крысы (Фиг. 5D) с использованием скоростей связывания и диссоциации, полученных из измерений кинетики Biacore. Показаны сравнительные варианты: AAA (диагональные линии, направленные вниз вправо), LS (пунктир) и YTE (диагональные линии, направленные вниз влево). Несмотря на улучшение скорости диссоциации с FcRn, большинство вариантов не обладало более сильным сродством к FcRn человека или крысы из-за более медленной кинетики ассоциации. Одиннадцать вариантов имели более медленные скорости диссоциации от обоих видов FcRn.
На Фиг. 6A - Фиг. 6D изображены данные, показывающие, что комбинации лидирующих насыщающих мутаций дополнительно улучшают скорости диссоциации с FcRn и способности связывания. На Фиг. 6А и 6В представлены типичные сенсограммы Biacore, показывающие диссоциацию с FcRn для FcRn человека и крысы, соответственно. На Фиг. 6А представлены нормализованные сенсограммы для FcRn человека типичных вариантов с одинарной (штриховая линия), двойной (сплошная светло-серая), тройной (сплошная серая линия) и четверной (сплошная черная линия) комбинациями в сравнении с вариантом дикого типа (пунктирная линия) и вариантом LS (длинные штрихи с двумя точками). На Фиг. 6В показаны нормализованные сенсограммы для FcRn крысы типичных вариантов с одинарной (длинные штрихи с двумя точками), двойной (длинные штрихи с одной точкой), тройной (длинные штрихи) и четверной (короткие штрихи) комбинациями в сравнении с вариантом дикого типа (пунктирная линия) и вариантом YTE (сплошная линия). Включение множественных мутаций уменьшало скорость диссоциации и увеличивало способность связывания с FcRn в большей степени, чем у вариантов сравнения. На Фиг. 6C и Фиг. 6D представлены графики комбинированных вариантов насыщения, показывающие скорость взаимодействия как функцию скорости диссоциации для FcRn человека (Фиг. 6C) или крысы (Фиг. 6D), из которых видно, что большинство вариантов обладало повышенным связыванием с FcRn при рН 6,0 по сравнению с вариантами сравнения. Варианты c наиболее тесным связыванием с FcRn человека и крысы представляли собой четверную и двойную комбинации, соответственно.
На Фиг.7А - Фиг.7D представлены данные, показывающие, что повышенное связывание с FcRn при рН 6,0 нарушает зависимость взаимодействия от рН. На Фиг. 7А и Фиг. 7В представлены репрезентативные Biacore сенсограммы кинетики связывания с FcRn при pH 7,4 вариантов с одинарной (длинные штрихи с двумя точками), двойной (длинные штрихи с одной точкой), тройной (длинные штрихи) и четверной (короткие штрихи) комбинациями в сравнении с вариантом дикого типа (пунктир) и вариантом LS (Фиг. 7А, сплошная линия) и вариантом YTE (Фиг. 7B, сплошная линия). Увеличение количества мутаций, повышающих связывание с FcRn, приводило к большему остаточному связыванию при физиологическом pH, при этом большинство вариантов с двойными, тройными и четверными комбинациями демонстрируют устойчивое связывание с обоими видами FcRn. На Фиг. 7C и Фиг. 7D представлено графическое изображение устойчивого состояния RU (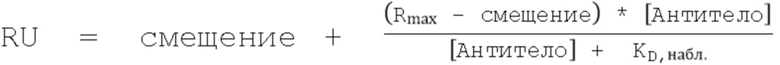 (Уравнение 2)) всех вариантов насыщения с FcRn человека (Фиг. 7C) или крысы (Фиг. 7D) при pH 7,4 как функции способности связывания при рН 6,0. На Фиг. 7С показано сравнение остаточного связывания с FcRn при рН 7,4 со способностью связывания с FcRn при рН 6,0. Лидирующие комбинации с улучшенными свойствами связывания с FcRn занимают нижнюю левую четверть изображения относительно варианта сравнения LS (ромб). На рис. 7D варианты LS (ромб) и YTE (треугольник) служат граничными условиями для подтверждения лидирования, соответственно. Эти два варианта имели способность к наиболее тесному связыванию при рН 6,0 и наибольшее остаточное связывание при рН 7,4 для FcRn человека и крысы, соответственно. На обеих Фиг. 7C и 7D показаны одинарный (белые кружки), двойной (светло-серые кружки), тройной (темно-серые кружки) и четверной (черные кружки) варианты, а также вариант YTE (треугольник).
(Уравнение 2)) всех вариантов насыщения с FcRn человека (Фиг. 7C) или крысы (Фиг. 7D) при pH 7,4 как функции способности связывания при рН 6,0. На Фиг. 7С показано сравнение остаточного связывания с FcRn при рН 7,4 со способностью связывания с FcRn при рН 6,0. Лидирующие комбинации с улучшенными свойствами связывания с FcRn занимают нижнюю левую четверть изображения относительно варианта сравнения LS (ромб). На рис. 7D варианты LS (ромб) и YTE (треугольник) служат граничными условиями для подтверждения лидирования, соответственно. Эти два варианта имели способность к наиболее тесному связыванию при рН 6,0 и наибольшее остаточное связывание при рН 7,4 для FcRn человека и крысы, соответственно. На обеих Фиг. 7C и 7D показаны одинарный (белые кружки), двойной (светло-серые кружки), тройной (темно-серые кружки) и четверной (черные кружки) варианты, а также вариант YTE (треугольник).
На 8A - фиг. 8C приведены данные, полученные из аффинной хроматографии FcRn и дифференциальной сканирующей флуориметрии (DSF) вариантов сравнения. На рис. 8А показаны нормализованные профили элюции для вариантов WT (сплошная черная линия), AAA (пунктирная линия), LS (длинные штрихи с двумя точками), YTE (длинные штрихи с одной точкой), H435A (сплошная светло-серая линия) и H310A/H435Q (AQ; сплошная темно-серая линия). рН указан в верхней части графика. Нулевые варианты связывания с FcRn (H435A, H310A/H435Q) не связываются в колонке и элюируются с потоком (<10 мл). Варианты AAA, LS и YTE элюируются при более высоком pH, чем антитело WT. На Фиг. 8В показаны профили DSF вариантов WT (черным), LS (серым) и YTE (темно-серым). YTE был дестабилизирован по сравнению с WT и LS. На Фиг. 8С показаны профили элюирования в аффинной колонке с FcRn для семи лидирующих одиночных вариантов, используемых для комбинированных вариантов, по сравнению с вариантами WT и LS (вертикальный пунктир). Два варианта (N434F/Y) элюируются при более высоком рН, чем LS, что указывает на пониженную зависимость взаимодействия с FcRn от рН у вариантов, содержащих эти мутации.
На Фиг.9А - Фиг.9D приведены данные, показывающие, что комбинированные варианты значительно нарушают зависимость от рН и термостабильность. На Фиг. 9А показаны типичные хроматограммы аффинности FcRn с одиночными (длинными штрихами с двумя точками), двойными (длинными штрихами с одной точкой), тройными (длинными штрихами) и четверными (короткими штрихами) вариантами. Увеличение количества мутаций, усиливающих связывание с FcRn, сдвигало элюирование в сторону более высоких значений pH; вариант LS (маленькая пунктирная вертикальная линия). На Фиг. 9В представлена ящичная диаграмма рН при элюировании для лидирующих вариантов насыщения и комбинированных вариантов, включая одиночные (белые кружки), двойные (горизонтальные линии), тройные (вертикальные линии) и четверные (клетчатые) мутанты, что указывает на тенденцию к повышению pH при увеличении числа мутантов, усиливающих связывание с FcRn. На Фиг. 9C показано, что высокая корреляция (R2=0,94) между pH при элюировании в аффинной хроматографии FcRn и скоростью диссоциации с hFcRn с использованием Biacore выявила потерю зависимости от pH взаимодействия антитело-FcRn при улучшении кинетики диссоциации с FcRn. Варианты AAA (диагональные линии, направленные вправо вниз), LS (пунктирные линии) и YTE (диагональные линии, направленные влево вниз) имели сходные с двойными вариантами скорости диссоциации с hFcRn и уровень pH при элюировании. На Фиг.9D представлена ящичная диаграмма для Tпл., полученная из DSF для комбинированных вариантов насыщения, из которой видно, что дополнительные мутации, усиливающие связывание с FcRn, дестабилизируют антитело по сравнению с вариантами WT, одиночными вариантами или вариантами сравнения.
На Фиг. 10А - Фиг. 10В приведены данные, полученные из аффинной хроматографии FcRn и DSF для семи лидирующих вариантов. На Фиг.10А приведены данные аффинной хроматографии FcRn для вариантов M252Y (сплошная линия), T256D (короткие штрихи с одной точкой), T256E (длинные штрихи), T307Q (длинные штрихи с одной точкой), T307W (длинные штрихи с двумя точками), N434F (пунктир) и N434Y (короткие штрихи). Из хроматограмм виден сдвиг рН при элюировании по сравнению с антителами дикого типа и LS (вертикальные пунктирные линии). N434F и N434Y имели более высокий pH при элюировании (pH примерно 8,3), чем вариант LS (вертикальная пунктирная линия). Значение pH при определенных объемах элюирования указано над хроматограммами для справки. На Фиг. 10B изображает профили DSF семи лидирующих вариантов, которые показали, что ни один из семи лидирующих вариантов не дестабилизировал антитела в той же степени, что и вариант YTE (вертикальная пунктирная линия). Все варианты, кроме T307Q (длинные штрихи с одной точкой), были дестабилизированы по сравнению с WT (вертикальная пунктирная линия).
На Фиг. 11A - Фиг. 11C приведены данные, показывающие, что связывание с FcγRIIIa было снижено у M252Y-содержащих комбинированных вариантов. На Фиг.11А показаны сенсограммы связывания с FcγRIIIa вариантов WT (черным), LS (серым) и YTE (темно-серым), из которых видно снижение ответа на связывание у варианта YTE. На Фиг. 11B показана ящичная диаграмма взаимодействия связывания с FcγRIIIa варианта сравнения, одинарного и комбинированного вариантов, как указано. Варианты с мутациями M252Y содержат сниженный ответ на связывание с FcγRIIIa, включая все четверные варианты. Комбинации с N434F/Y обычно демонстрируют повышенное взаимодействие с FcγRIIIa. На Фиг. 11С показаны ответы на связывание с FcγRIIIa для семи лидирующих одинарных вариантов в сравнении с вариантами WT и YTE (пунктирная горизонтальная линия). Мутация M252Y демонстрирует пониженное связывание FcγRIIIa по сравнению с WT, тогда как шесть демонстрируют сходное с WT или повышенное связывание с этим рецептором.
На Фиг. 12A - Фиг. 12D приведены данные, полученные с помощью аффинной хроматографии FcRn, DSF и связывания с FcγRIIIa для семи лидирующих комбинированных вариантов. На Фиг.12А показаны аффинные хроматограммы FcRn для семи лидирующих комбинированных вариантов в сравнении с антителом дикого типа и вариантом LS (вертикальная пунктирная линия и сплошная вертикальная линия соответственно). Каждый лидирующий вариант имел рН при элюировании, близкий к варианту LS. На Фиг.12В показаны профили DSF лидирующих комбинированных вариантов в сравнении с вариантами YTE и дикого типа (вертикальные пунктирные линии, как указано). Шесть из семи лидирующих вариантов имели Tпл., которая была аналогичной или более дестабилизированной, чем у варианта YTE: MDWN (длинные штрихи с двумя точками); YTWN (длинные штрихи); YDTN (сплошная линия); YETN (длинные штрихи с одной точкой); YDQN (пунктир); YEQN (короткие штрихи с одной точкой). Вариант MDQN имел Tпл., сходную с антителом дикого типа (короткие штрихи). На Фиг. 12С показаны Biacore сенсограммы кинетики связывания FcγRIIIa для семи лидирующих вариантов по сравнению с диким типом (большая пунктирная линия) и вариантом YTE (толстые длинные штрихи). Варианты, содержащие M252Y, YDTN (сплошная линия), YDQN (короткие штрихи с одной точкой), YTWN (длинные штрихи), YETN (длинные штрихи с одной точкой) и YEQN (меньшая пунктирная линия), каждый обладал уменьшенным устойчивым состоянием RU таким же образом, как YTE. (D) показывает устойчивое состояние RU семи лидирующих вариантов, дикого типа и варианта YTE. Только варианты MDWN и MDQN обладали сходной аффинностью к FcγRIIIa, что и антитело дикого типа.
На Фиг. 12E - Фиг. 12H приведены данные, показывающие, что три лидирующих варианта демонстрируют ряд ключевых свойств антител. На Фиг. 12E показаны профили элюирования FcRn при аффинной хроматографии для вариантов DQ (сплошным), DW (пунктиром) и YD (пунктиром) по сравнению с WT и LS (вертикальными пунктирными линиями). Каждый двойной вариант показывал рН для элюирования, который находился между WT и LS. На Фиг. 12F показаны профили флуоресценции DSF для трех вариантов в сравнении с вариантами YTE и WT (вертикальном пунктиром), по которым видно, что YD (штрихами) и DW (пунктиром) были слегка дестабилизированы по сравнению с YTE, но DQ (сплошным) была аналогична WT. На Фиг. 12G показаны сенсограммы связывания с FcγRIIIa в сравнении с WT и YTE (горизонтальным пунктиром). YD (штрихами) показал аналогичный ответ на связывание, как и YTE, тогда как DQ (сплошным) и DW (пунктиром) показали небольшое снижение по сравнению с WT. На Фиг. 12H представлены данные, показывающие, что RF-ELISA с гомогенным мостиковым соединением выявил три лидирующих варианта, а YTE продемонстрировал значительно сниженное или WT-подобное связывание RF, в отличие от LS. **p<0,001, *p<0,01.
На Фиг. 13А - Фиг. 13D приведены данные, показывающие сравнение кинетики связывания с FcRn лидирующих комбинированных вариантов при рН 6,0 и рН 7,4. На Фиг. 13A и Фиг. 13B показаны Biacore сенсограммы связывания с FcRn лидирующих комбинированных вариантов для FcRn человека (Фиг. 13A) или FcRn крысы (Фиг. 13B) по сравнению с вариантом дикого типа (пунктирной линией) и либо LS (hFcRn, Фиг. 13A, жирными длинными штрихами), либо YTE (rFcRn, Фиг. 13B, жирными длинными штрихами) при pH 6,0. Каждый комбинированный вариант имел в целом способность к более тесному связыванию с соответствующим FcRn, несмотря на изменение скорости взаимодействия и диссоциации. На Фиг. 13C и Фиг. 13D показаны Biacore сенсограммы FcRn при pH 7,4. Каждый лидирующий вариант hFcRn имел сходный или уменьшенный ответ на связывание с FcRn в устойчивом состоянии по сравнению с вариантом LS. Только варианты MDQN и MDWN показали меньшее связывание с rFcRn при рН 7,4, чем вариант YTE.
На Фиг. 14 приведена таблица, изображающая скорости диссоциации связывания с rFcRn по технологии Octet библиотеки насыщения в соответствии с некоторыми вариантами осуществления. Образцы дикого типа (WT) и подобные дикому типу (WT-подобные) обозначены белыми прямоугольниками; образцы WT такие, как указано. Варианты с небольшим или отсутствующим связыванием с rFcRn по сравнению с диким типом обозначены темно-серыми прямоугольниками. Варианты с более высокой скоростью диссоциации с rFcRn по сравнению с диким типом обозначены светло-серыми прямоугольниками, а варианты с более низкой скоростью диссоциации с rFcRn по сравнению с диким типом обозначены черными прямоугольниками.
На Фиг. 15А - Фиг. 15С показан новый анализ связывания, разработанный с использованием сенсорного чипа СМ5. На Фиг. 15А представлена схема анализа. На Фиг. 15В показана прямая иммобилизация FcRn. На Фиг. 15C показан захват стрептавидином биотинилированного FcRn.
На Фиг. 16А - Фиг. 16В изображено связывание с FcRn антитела-2 при рН 6,0. На Фиг.16А изображен FcRn человека. На Фиг. 16B изображен FcRn мыши.
На Фиг. 17А - Фиг. 17В изображено связывание с FcRn антитела-2 при рН 7,4. На Фиг.17А изображен FcRn человека. На Фиг. 17B изображен FcRn мыши.
На Фиг. 18 графически изображена pH-зависимость для различных вариантов антитела-2. Лидирующие варианты сохраняли более высокую способность связывания при рН 6 и более низкое остаточное связывание при рН 7,4, чем LS.
На Фиг. 19 показано сравнение зависимости от рН связывания с FcRn с использованием каркасов антитела-1 и антитела-2.
На Фиг. 20 показано сравнение термостабильности с использованием каркасов антитела-1 и антитела-2.
На Фиг. 21 показано сравнение связывания с FcγRIIIa с использованием каркасов антитела-1 и антитела-2.
На Фиг. 22A - Фиг. 22I изображены многочисленные графики, показывающие, что варианты DQ, DW и YD можно переносить между каркасами IgG1. Графики a-c изображают нормализованные сенсограммы связывания FcRn при pH 6,0 в трех каркасах IgG1 с вариантами WT (светло-серым), LS (темно-серым), DQ (сплошная черная), DW (пунктиром) и YD (штрихами), демонстрирующие сходную кинетику при низком pH. Эти три варианта - DQ, DW и YD - обладали немного более высокими скоростями связывания и диссоциации, чем вариант LS, но сохраняли способность к более тесному связыванию с FcRn. Графики d-f изображают сенсограммы связывания FcRn при pH 7,4; вариант сравнения LS (сплошная черная). Графики g-i изображают ответ на связывание с FcRn при pH 7,4 по сравнению со способностью связывания при pH 6,0 для каждого каркаса антитела с вариантами WT (серым), LS (темно-серым), DQ (сплошным черным), DW (незакрашено) и YD (пустой квадрат). DQ, DW и YD показывают улучшенные характеристики FcRn с повышенным связыванием при рН 6,0 и минимальным связыванием при рН 7,4.
На Фиг. 23A - Фиг. 23C показано, что три лидирующих варианта в каркасе mAb2 аналогичным образом улучшают связывание с FcRn cynomolgus. На фиг.23А показаны нормализованные сенсограммы связывания cFcRn при рН 6,0 для WT (серым), LS (темно-серым), DQ (сплошным черным), DW (пунктиром) и YD (штрихами), демонстрирующие кинетику и способность связывания, сходные hFcRn. На Фиг. 23В показано, что реакция связывания cFcRn для трех вариантов была резко снижена при физиологическом рН; однако LS (темно-серым) показал большую степень связывания, чем WT (серым), сходным образом с hFcRn. На Фиг. 23C показано сравнение остаточной реакции связывания cFcRn при pH 7,4 со способностью связывания cFcRn при pH 6,0 WT (серым), LS (темно-серым), DQ (сплошным черным), DW (незакрашено) и YD (пустой квадрат), из которого видно, что все три варианта сохраняли улучшенные свойства связывания FcRn, наблюдаемые с hFcRn.
Из Фиг. 24А-24В видно, что лидирующие варианты продлевают период полувыведения антител из сыворотки крови. Фармакокинетические профили концентрации антител в плазме крови как функции времени у обезьян cynomolgus (Фиг. 24A) и трансгенной мыши hFcRn (фиг. 24B) с антителами WT (дикого типа) (черные кружки со сплошной черной линией), LS (белые кружки с прерывистой черной линией), DQ (светло-серые кружки со сплошной светло-серой линией), DW (темно-серые кружки со сплошной темно-серой линией) и YD (черные кружки с пунктирной черной линией). Все три лидирующие варианта продлевают период полувыведения антител по сравнению с WT.
На Фиг.25 представлен график устойчивого состояния RU всех вариантов насыщения с FcRn человека при рН 7,4 как функции способности связывания при рН 6,0. Показано сравнение остаточного связывания с FcRn при рН 7,4 со способностью связывания с FcRn при рН 6,0. Четверные комбинации с улучшенными свойствами связывания с FcRn как при рН 6,0, так и при рН 7,4 показаны в рамке в правой верхней четверти изображения. Показаны одиночные (белые кружки), двойные (светло-серые кружки), тройные (темно-серые кружки) и четверные (черные кружки) варианты, а также сравнительные варианты AAA, LS и YTE (как указано).
На Фиг. 26 представлена схема метода Biotin CAPture, используемого для захвата биотинилированного FcRn.
На Фиг. 27 изображены графики кинетики связывания FcRn человека при рН 6,0 вариантом сравнения YTEKF и комбинированными вариантами, как указано.
На Фиг. 28A - Фиг. 28B показана кинетика связывания с FcRn комбинированными вариантами в сравнении с вариантом сравнения YTEKF при pH 6,0 (Фиг. 28A) и при pH 7,4 (Фиг. 28B). Дикий тип обозначен сплошной черной линией (WT), а вариант сравнения YTEKF - пунктирной линией.
На Фиг. 29 представлен график устойчивого состояния RU выбранных вариантов человеческого FcRn при рН 7,4 в зависимости от способности связывания при рН 6,0 в сравнении с вариантом сравнения YTEKF. Несколько вариантов (лидирующие четверные варианты) проявляли повышенную способность связывания с FcRn человека при рН 6,0 и рН 7,4 в сравнении с вариантом сравнения YTEKF.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение относится к связывающим полипептидам (например, антителам), имеющим измененную способность связывания с Fc-неонатальным рецептором (FcRn). В некоторых вариантах осуществления связывающие полипептиды содержат измененный домен Fc, который усиливает способность связывания с FcRn по сравнению со связывающим полипептидом, который содержит Fc-домен дикого типа (например, немодифицированный). В настоящем изобретении также предложены нуклеиновые кислоты, кодирующие связывающие полипептиды, рекомбинантные векторы экспрессии и клетки-хозяева для получения связывающих полипептидов, и фармацевтические композиции, содержащие связывающие полипептиды, раскрываемые в данном документе. Также предложены способы использования связывающих полипептидов настоящего изобретения для лечения заболеваний.
Fc-домены иммуноглобулинов участвуют в неантигенсвязывающих процессах и имеют несколько эффекторных функций, которые выполняются путем связывания с эффекторными молекулами, например, связывания с FcRn. Как показано на Фиг. 1А, Fc-домены состоят из домена СН2 и домена СН3. Большинство остатков, участвующих во взаимодействии с FcRn, расположены в петлях, непосредственно примыкающих к области контакта CH2-CH3 (Фигура 1А, пунктирная линия) и напротив сайта гликозилирования. На Фигуре 1B показано изображение поверхности кристаллической структуры Fc IgG1 (pdb: 5d4q) и показаны остатки в доменах CH2 и CH3, которые составляют область контакта при связывании с FcRn. Настоящее изобретение относится к связывающим полипептидам, содержащим модифицированный Fc-домен. Связывающие полипептиды, содержащие модифицированный Fc-домен, могут представлять собой антитела, или иммуноадгезины или, составные белки Fc.
В некоторых вариантах осуществления связывающий полипептид может содержать модифицированный Fc-домен, содержащий аминокислотную замену, которая изменяет антиген-независимые эффекторные функции антитела, в частности, период полувыведения из кровотока (например, период полувыведения из сыворотки крови) связывающего полипептида. В некоторых вариантах осуществления связывающий полипептид может содержать модифицированный Fc-домен, содержащий аминокислотную замену, которая изменяет период полувыведения связывающего полипептида из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа (то есть немодифицированный). В некоторых вариантах осуществления связывающий полипептид может содержать модифицированный Fc-домен, содержащий аминокислотную замену, которая продлевает период полувыведения связывающего полипептида из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа (то есть немодифицированный). В некоторых вариантах осуществления связывающий полипептид может содержать модифицированный Fc-домен, содержащий аминокислотную замену, которая делает более коротким период полувыведения связывающего полипептида из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа (то есть немодифицированный).
В некоторых вариантах осуществления связывающий полипептид, содержащий модифицированный Fc-домен, который изменяет (т.е. продлевает или делает более коротким) период полувыведения из кровотока (например, период полувыведения из сыворотки крови), дополнительно содержит одну или несколько мутаций в дополнение к мутации(ям), которые изменяют период полувыведения из кровотока. В некоторых вариантах осуществления одна или несколько мутаций в дополнение к мутациям, которые изменяют период полувыведения из кровотока, обеспечивают одну или несколько требуемых биохимических характеристик, таких как, например, одна или несколько ослабленных или усиленных эффекторных функций, способность к нековалентной димеризации, повышенная способность локализоваться в месте опухоли, более короткий период полувыведения из сыворотки крови, более продолжительный период полувыведения из сыворотки крови по сравнению с цельным неизмененным антителом примерно такой же иммуногенности и т.п.
Связывающие полипептиды, описанные здесь, могут проявлять либо повышенное, либо пониженное связывание с неонатальным Fc-рецептором (FcRn) по сравнению со связывающими полипептидами, в которых отсутствуют эти замены, и, следовательно, имеют более продолжительный или более короткий период полувыведения из сыворотки крови, соответственно. Fc-домены с улучшенным сродством к FcRn предположительно имеют более длительные периоды полувыведения из сыворотки крови, и такие молекулы могут с пользой применяться в способах лечения млекопитающих, где требуется продолжительный период полувыведения вводимого антитела, например, для лечения хронических заболеваний или нарушений. В противоположность этому, ожидается, что Fc-домены со сниженной способностью связывания с FcRn будут иметь более короткие периоды полувыведения, и такие молекулы также применимы, например, для введения млекопитающему, где сокращенное время циркуляции может быть предпочтительным, например, для диагностической визуализации in vivo или в ситуациях, где исходное антитело имеет токсичные побочные эффекты, если оно присутствует в кровотоке в течение продолжительных периодов времени. Fc-домены со сниженной способностью связывания с FcRn также с меньшей долей вероятности пересекают плацентарный барьер и, таким образом, также применимы в лечении заболеваний или нарушений у беременных женщин. Кроме того, другие применения, в которых может требоваться снижение способности связывания с FcRn, включают применения, локализованные в мозге, почке и/или печени.
Следует понимать, что способы, описанные в этом изобретении, не ограничиваются конкретными способами и экспериментальными условиями, раскрываемыми в данном документе, поскольку такие способы и условия могут варьироваться. Также следует понимать, что терминология, используемая в данном документе, предназначена только для целей описания конкретных вариантов осуществления и не предназначена для ограничения.
Кроме того, в описанных здесь экспериментах, если не указано иное, используются обычные молекулярные и клеточные биологические и иммунологические методы, известные специалистам в данной области. Такие методы хорошо известны квалифицированному специалисту и полностью объяснены в литературе. См., например, Ausubel, et al. , ed., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., NY, Нью-Йорк (1987-2008), включая все приложения, Molecular Cloning: A Laboratory Manual (Fourth Edition) by MR Green and J. Sambrook and Harlow et al., Antibodies: A Laboratory Manual, Chapter 14, Cold Spring Harbor Laboratory, Cold Spring Harbor (2013, 2nd edition).
Если не указано иное, научные и технические термины, используемые в данном документе, имеют значения, которые обычно понятны специалистами в данной области техники. В случае любой латентной неясности, данные в данном документе определения превалируют над любым определением, данным в словаре или используемым вне данного документа. Если контекстом не требуется иное, термины в единственном числе будут включать формы множественного числа, и термины во множественном числе будут включать форму единственного числа. Использование «или» означает «и/или», если не указано иное. Применение термина «включая», а также других форм, таких как «включать», «включает» и «включенный», не является ограничивающим.
Как правило, используемая номенклатура, связанная с культивированием клеток и тканей, молекулярной биологией, иммунологией, микробиологией, генетикой и химией и гибридизацией белков и нуклеиновых кислот, а также соответствующие методики, описанные в данном документе, хорошо известны и широко используются в данной области техники. Способы и методики, предоставленные в данном документе, обычно выполняются, если не указано иное, в соответствии с обычными способами, хорошо известными в данной области техники, и как описано в различных общих и более конкретных ссылках, которые цитируются и обсуждаются в настоящем описании. Ферментативные реакции и методики очистки выполняют в соответствии с описаниями производителя, как обычно осуществляется в данной области техники или как описано в данном документе. Терминология, а также лабораторные процедуры и их методики, используемые в контексте аналитической химии, синтетической органической химии и медицинской и фармацевтической химии и описанные в данном документе, являются общеизвестными и широко используемыми в данной области техники. Стандартные методики используются в химическом синтезе, химическом анализе, фармацевтическом получении, составлении и доставке, и у лечении пациентов.
Чтобы изобретение было более понятным, избранные термины определены ниже.
Термин «полипептид» относится к любой полимерной цепи аминокислот и охватывает природные или искусственные белки, полипептидные аналоги или варианты последовательности белка или их фрагменты, если это не противоречит контексту. Полипептид может быть мономерным или полимерным. Полипептидный фрагмент содержит, например, по меньшей мере примерно 5 граничащих друг с другом аминокислот, по меньшей мере примерно 10 граничащих друг с другом аминокислот, по меньшей мере примерно 15 граничащих друг с другом аминокислот или, по меньшей мере примерно 20 граничащих друг с другом аминокислот.
Термин «выделенный белок» или «выделенный полипептид» относится к белку или полипептиду, которые в силу своего происхождения или источника получения не связаны с естественным образом связанными с ними компонентами, сопутствующими им в их природном состоянии, по существу не содержат других белков из того же вида, экспрессируются клеткой из другого вида или не встречаются в природе. Таким образом, белок или полипептид, синтезированный химическим путем или синтезированный в клеточной системе, отличной от клетки, из которой он происходит естественным образом, будет «выделенным» из естественным образом связанных с ним компонентов. Белок или полипептид также можно сделать практически не содержащим естественным образом связанных с ним компонентов путем выделения с использованием методик очистки белка, хорошо известных из уровня техники.
Применяемое в данном документе выражение «связывающий белок» или «связывающий полипептид» относится к полипептиду (например, антителу), который содержит по меньшей мере один сайт связывания, который ответственен за избирательное связывание с представляющим интерес целевым антигеном (например, антигеном человека). Иллюстративные сайты связывания включают вариабельный домен антитела, сайт связывания лиганда рецептора или сайт связывания рецептора лиганда. В определенных аспектах связывающие белки или связывающие полипептиды содержат множество (например, два, три, четыре или более) сайтов связывания. В определенных аспектах связывающий белок или связывающий полипептид не является терапевтическим ферментом.
Термин «лиганд» относится к любому веществу, способному связываться с другим веществом или быть связанным им. Аналогично, термин «антиген» относится к любому веществу, к которому может быть создано антитело. Хотя «антиген» обычно используется в отношении субстрата, связывающего антитело, а «лиганд» часто используется при ссылке на субстраты, связывающие рецептор, эти термины не отличаются друг от друга и охватывают широкий спектр перекрывающихся химических объектов. Во избежание недоразумений, термины антиген и лиганд используются здесь взаимозаменяемо. Антигены/лиганды могут быть пептидом, полипептидом, белком, аптамером, полисахаридом, молекулой сахара, углеводом, липидом, олигонуклеотидом, полинуклеотидом, синтетической молекулой, неорганической молекулой, органической молекулой и любой другой их комбинацией.
Используемый в данном документе термин «специфически связывается» относится к способности антитела или иммуноадгезина связываться с антигеном с константой диссоциации (Kd), составляющей самое большее примерно 1×10-6 М, примерно 1×10-7 М, примерно 1×10-8 М, примерно 1×10-9 М, примерно 1×10-10 М, примерно 1×10-11 М, примерно 1×10-12 М или менее, и/или для связывания с антигеном со сродством, которое по меньшей мере примерно в два раза превышает его сродство к неспецифическому антигену.
Применяемое в данном документе выражение «антитело» относится к таким скоплениям (например, молекулам интактного антитела, иммуноадгезинам или их вариантам), которые обладают значительной известной специфической иммунореактивной активностью к представляющему интерес антигену (например, ассоциированному с опухолью антигену). Антитела и иммуноглобулины содержат легкие и тяжелые цепи с межцепочечной ковалентной связью между ними или без нее. Основные структуры иммуноглобулинов у позвоночных организмов относительно хорошо изучены.
Как будет более подробно обсуждаться ниже, общий термин «антитело» включает пять различных классов антител, которые можно различать биохимически. В то время как все пять классов антител несомненно входят в объем настоящего раскрытия, последующее обсуждение будет, в основном, направлено на молекулы иммуноглобулина класса IgG. Что касается IgG, иммуноглобулины включают две идентичные легкие цепи с молекулярной массой приблизительно 23000 дальтон и две идентичные тяжелые цепи с молекулярной массой 53000-70000 дальтон. Эти четыре цепи соединены дисульфидными связями в конфигурации «Y», где соединение легких цепей с тяжелыми цепями начинается в устье «Y» и продолжается по вариабельной области.
Легкие цепи иммуноглобулина классифицируются как каппа (κ) или лямбда (λ). Каждый класс тяжелых цепей может быть связан с легкой цепью каппа или лямбда. В общем случае, легкая и тяжелая цепи ковалентно связаны друг с другом, а «хвостовые» части двух тяжелых цепей связаны друг с другом ковалентными дисульфидными связями или нековалентными связями при создании иммуноглобулинов либо гибридомами, B клетками, либо генетически модифицированными клетками-хозяевами. В тяжелой цепи аминокислотные последовательности проходят от N-конца на разветвленных концах Y-конфигурации до С-конца в нижней части каждой цепи. Специалистам в данной области понятно, что тяжелые цепи классифицируются как гамма (γ), мю (μ), альфа (α), дельта (δ) или эпсилон (ε) с некоторыми подклассами среди них (например, γl-γ4). По природе этой цепи определяют «класс» антитела, а именно IgG, IgM, IgA IgG или IgE, соответственно. Подклассы изотипов иммуноглобулина (например, IgG1, IgG2, IgG3, IgG4, IgA1 и т.д.) хорошо охарактеризованы, и о них известно, что они придают функциональную специализацию. Специалист в данной области легко распознает модифицированные версии каждого из этих классов и изотипов в свете настоящего изобретения и, соответственно, они находятся в пределах объема настоящего изобретения.
Как легкие, так и тяжелые цепи делятся на участки структурной и функциональной гомологии. Термин «участок» относится к части или отрезку цепи иммуноглобулина или антитела и включает константный участок или вариабельные участки, а также более разрозненные части или отрезки указанных участков. Например, вариабельные участки легкой цепи включают в себя «определяющие комплементарность участки» или «CDR», расположенные между «каркасными участками» или «FR», как определено в данном документе.
Участки тяжелой или легкой цепи иммуноглобулина могут быть определены как «константный» (C) участок или «вариабельные» (V) участки на основании относительного отсутствия изменения на участках последовательности у различных членов класса в случае «константного участка», или на основании значительного изменения на участках у различных членов класса в случае «вариабельных участков». Термины «константный участок» и «вариабельный участок» также могут использоваться функционально. В связи с этим следует понимать, что вариабельные участки иммуноглобулина или антитела определяют распознавание и специфичность антигена. И наоборот, константные участки иммуноглобулина или антитела придают им важные эффекторные функции, такие как секреция, транс-плацентарная подвижность, связывание с Fc-рецептором, связывание с комплементом и тому подобное. Структуры субъединиц и трехмерные конфигурации константных участков различных классов иммуноглобулинов хорошо известны.
Константные и вариабельные участки тяжелой и легкой цепей иммуноглобулина свернуты в домены. Термин «домен» относится к глобулярной области тяжелой или легкой цепи, содержащей пептидные петли (например, содержащие от 3 до 4 пептидных петель), стабилизированные, например, β-складчатой конформацией и/или внутрицепной дисульфидной связью. Домены константного участка на легкой цепи иммуноглобулина взаимозаменяемо называют «доменами константного участка легкой цепи», «участками CL» или «доменами CL». Константные домены на тяжелой цепи (например, шарнирные, домены СН1, СН2 или СН3) взаимозаменяемо называют «доменами константного участка тяжелой цепи», «доменами участка СН» или «доменами СН». Вариабельные домены легкой цепи взаимозаменяемо называют «доменами вариабельного участка легкой цепи», «доменами участка VL» или «доменами VL». Вариабельные домены тяжелой цепи взаимозаменяемо называют «доменами вариабельного участка тяжелой цепи», «доменами участка VН» или «доменами VН».
По соглашению, нумерация аминокислот доменов вариабельного константного участка увеличивается, когда они удаляются от антигенсвязывающего сайта или аминного конца иммуноглобулина или антитела. N-конец каждой тяжелой и легкой цепи иммуноглобулина является вариабельным участком, а С-конец является константным участком. Домены CH3 и CL содержат карбокси-конец тяжелой и легкой цепей, соответственно. Соответственно, домены иммуноглобулина легкой цепи ориентированы как VL-CL, тогда как домены тяжелой цепи ориентированы как VH-CH1-шарнир-CH2-CH3.
Распределение аминокислот по каждому домену вариабельного участка соответствует определениям Kabat, Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Bethesda, MD, 1987 и 1991). Kabat также предоставляет широко используемое соглашение о нумерации (нумерация Kabat), в котором соответствующим остаткам между различными вариабельными участками тяжелой цепи или между различными вариабельными участками легкой цепи присваивается один и тот же номер. CDR 1, 2 и 3 домена VL также упоминаются в данном документе, соответственно, как CDR-L1, CDR-L2 и CDR-L3. CDR 1, 2 и 3 домена VН также упоминаются в данном документе, соответственно, как CDR-H1, CDR-H2 и CDR-H3. Если указано, распределение CDR может производиться в соответствии с IMGT® (Lefranc et al., Developmental & Comparative Immunology 27:55-77; 2003) вместо Kabat. Нумерация константного участка тяжелой цепи осуществляют с помощью индекса ЕС, как указано в Kabat (Kabat, Sequences of Proteins of Immunological Interest, National Institutes of Health, Bethesda, MD, 1987 и 1991).
Используемый здесь термин «домен VH» включает вариабельный домен аминного конца тяжелой цепи иммуноглобулина, а термин «домен VL» включает вариабельный домен аминного конца легкой цепи иммуноглобулина.
Используемый здесь термин «домен CH1» включает в себя первый (наиболее близкий к концу аминный конец) домен константного участка тяжелой цепи иммуноглобулина, который продолжается, например, примерно от положений 114-223 согласно системе нумерации Kabat (положения 118-215 по нумерации ЕС). Домен CH1 находится рядом с доменом VH и аминным концом шарнирного участка молекулы тяжелой цепи иммуноглобулина и не образует часть Fc-участка тяжелой цепи иммуноглобулина.
Используемый здесь термин «шарнирный участок» включает часть молекулы тяжелой цепи, которая присоединяет домен CH1 к домену CH2. Шарнирный участок содержит примерно 25 остатков и характеризуется гибкостью, что позволяет двум N-концевым антигенсвязывающим областям перемещаться независимо. Шарнирные участки можно подразделить на три отдельных домена: верхний, средний и нижний шарнирные домены (Roux et al. J. Immunol. 1998, 161: 4083).
Используемый здесь термин «домен CH2» включает в себя часть тяжелой цепи молекулы иммуноглобулина, которая проходит, например, примерно от положений 244-360 согласно системе нумерации Kabat (положения 231-340 по нумерации ЕС). Домен CH2 уникален тем, что он не тесно связан с другим доменом. Точнее, две N-соединенные разветвленные углеводные цепи расположены между двумя доменами CH2 интактной нативной молекулы IgG. В одном варианте осуществления связывающий полипептид настоящего изобретения содержит домен CH2, полученный из молекулы IgG1 (например, молекулы IgG1 человека).
Используемый здесь термин «домен CH3» включает в себя часть тяжелой цепи молекулы иммуноглобулина, длина которой составляет примерно 110 остатков от N-конца домена СН2 и проходит, например, примерно от положений 361-476 согласно системе нумерации Kabat (положения 341-445 по нумерации ЕС). Домен СН3 обычно образует С-концевую часть антитела. Однако в некоторых иммуноглобулинах дополнительные домены могут проходить от домена СН3, образуя С-концевую часть молекулы (например, домен СН4 в μ-цепи IgM и е-цепи IgE). В одном варианте осуществления связывающий полипептид настоящего изобретения содержит домен CH3, полученный из молекулы IgG1 (например, молекулы IgG1 человека).
Используемый здесь термин «домен CL» включает домен константной области легкой цепи иммуноглобулина, который проходит, например, от положения 107А по Kabat до положения 216 по Kabat. Домен CL граничит непосредственно с доменом VL. В одном варианте осуществления связывающий полипептид настоящего изобретения содержит домен CL, полученный из легкой цепи типа каппа (например, человеческой легкой цепи типа каппа).
Используемый здесь термин «Fc-участок» определяется как часть константного участка тяжелой цепи, начинающийся в шарнирном участке непосредственно перед сайтом расщепления папаином (т.е. остаток 216 в IgG, принимая положение первого остатка константного участка тяжелой цепи за 114) и заканчивающийся на С-конце антитела. Соответственно, полный Fc-участок содержит по меньшей мере шарнирный домен, CH2-домен и CH3-домен.
Используемый в данном документе термин «природный Fc», «нативный Fc» и «Fc дикого типа» обозначает молекулу как в мономерной, так и в мультимерной форме, содержащую последовательность, не являющуюся частью антигенсвязывающего фрагмента, которая получена в результате расщепления антитела или другими способами, и при этом она может содержать шарнирный участок. Исходный источник иммуноглобулина для природного Fc обычно имеет человеческое происхождение и может представлять собой любой из иммуноглобулинов, такой как IgG1 и IgG2. Нативные молекулы Fc составлены из мономерных полипептидов, которые могут быть связаны в димерные или мультимерные формы ковалентной (т.е. дисульфидными связями) и нековалентной связями. Количество межмолекулярных дисульфидных связей между мономерными субъединицами природных молекул Fc варьируется от 1 до 4, в зависимости от класса (например, IgG, IgA и IgE) или подкласса (например, IgG1, IgG2, IgG3, IgA1 и IgGA2). Одним примером природного Fc является димер с дисульфидной связью, образующийся в результате расщепления IgG папаином. Термин «природный Fc» или «нативный Fc», используемый в данном документе, является общим для мономерных, димерных и мультимерных форм.
Используемый здесь термин «вариант Fc» или «модифицированный Fc» относится к молекуле или последовательности, которая получена модифицированием природного Fc/Fc дикого типа, но все еще содержит сайт связывания с FcRn. Таким образом, термин «вариант Fc» может включать в себя молекулу или последовательность, полученную в результате гуманизации нативного Fc нечеловеческого происхождения. Кроме того, нативный Fc содержит участки, которые можно удалить, поскольку они обеспечивают структурные признаки или биологическое действие, которые не требуются для подобных антителу связывающих полипептидов, описываемых в данном документе. Таким образом, термин "вариант Fc" предусматривает молекулу или последовательность, в которой отсутствует один или несколько участков или остатков нативного Fc или в которой были модифицированы один или несколько участков или остатков Fc, воздействующих на или вовлеченных в: (1) образование дисульфидной связи, (2) несовместимость с выбранной клеткой-хозяином, (3) гетерогенность N-концов при экспрессии в выбранной клетке-хозяине, (4) гликозилирование, (5) взаимодействие с системой комплемента, (6) связывание с Fc-рецептором, отличным от рецептора реутилизации, или (7) антителозависимую клеточную цитотоксичность (ADCC).
В некоторых примерных вариантах осуществления вариант Fc, представленный в настоящем документе, имеет одну или несколько из следующих характеристик: более длинный период полувыведения из сыворотки крови, повышенную способность связывания с FcRn, повышенную способность связывания с FcRn при кислотном pH, повышенную способность связывания с FcγRIIIa и/или сходную термическую стабильность в сравнении с антителом IgG, содержащим Fc дикого типа.
Термин «Fc-домен» в контексте данного документа, охватывает природный Fc/Fc дикого типа, а также варианты Fc и последовательности, определяемые выше. Что касается вариантов Fc и нативных молекул Fc, термин «Fc-домен» включает молекулы в мономерной или мультимерной форме, вне зависимости от того, образованы ли они расщеплением целого антитела или получены другими способами.
Как указано выше, вариабельные участки антитела позволяют ему избирательно распознавать и специфически связываться с эпитопами на антигенах. То есть домен VL и домен VH антитела объединяются, образуя вариабельный участок (Fv), который определяет трехмерный сайт связывания антигена. Эта четвертичная структура антитела образует антигенсвязывающий сайт, присутствующий в конце каждого плеча Y. Более конкретно, антигенсвязывающий сайт определяется тремя комплементарными определяющими областями (CDR) в каждом из вариабельных участков тяжелой и легкой цепи. Используемый здесь термин «сайт связывания антигена» включает сайт, который специфически связывается (иммунореагирует) с антигеном (например, антигеном клеточной поверхности или растворимым антигеном). Сайт связывания антигена включает вариабельный участок тяжелой цепи и легкой цепи иммуноглобулина; сайт связывания, образованный этими вариабельными участками, определяет специфичность антитела. Антигенсвязывающий сайт образован вариабельными участками, которые различаются в зависимости от антитела. Измененные антитела настоящего изобретения содержат по меньшей мере один антигенсвязывающий сайт.
В некоторых вариантах осуществления связывающие полипептиды настоящего изобретения содержат по меньшей мере два антигенсвязывающих домена, которые обеспечивают взаимодействие связывающего полипептида с выбранным антигеном. Антигенсвязывающие домены не обязательно должны быть получены из одной и той же молекулы иммуноглобулина. В этом отношении вариабельный участок может быть из любого типа животного или может получен из любого типа животного, у которого можно индуцировать гуморальный ответ и генерировать иммуноглобулины против желаемого антигена. Как таковой, вариабельный участок связывающего полипептида может, например, происходить от млекопитающего, например, может быть от человека, мыши, крысы, козы, овцы, примата, не являющегося человеком (такого как обезьяны cynomolgus, макаки и т.д.), семейства волчьих или верблюжьих (например, от верблюдов, лам и родственных видов).
Во встречающихся в природе антителах шесть CDR, присутствующие в каждом мономерном антителе, представляют собой короткие, не граничащие друг с другом последовательности аминокислот, которые специально расположены так, чтобы образовывать антигенсвязывающий сайт, когда антитело приобретает трехмерную конфигурацию в водной среде. Остальные тяжелые и легкие вариабельные домены обладают меньшей межмолекулярной вариабельностью в аминокислотной последовательности и называются каркасными участками. Каркасные участки в основном принимают β-складчатую конформацию, а CDR образуют петли, которые соединяют, а в некоторых случаях являются частью β-складчатой структуры. Таким образом, под действием этих каркасных участков формируется каркасная структура, который обеспечивает позиционирование шести CDR в правильной ориентации за счет межцепочечных нековалентных взаимодействий. Антигенсвязывающий домен, сформированный позиционированными CDR, определяет поверхность, комплементарную эпитопу на иммунореактивном антигене. Эта комплементарная поверхность способствует нековалентному связыванию антитела с эпитопом иммунореактивного антигена.
Типичные связывающие полипептиды включают варианты антител. Используемый здесь термин «вариант антитела» включает синтетические и сконструированные формы антител, которые изменены таким образом, чтобы они не встречались в природе, например, антитела, которые содержат по меньшей мере две части тяжелой цепи, но не две полные тяжелые цепи (такие как антитела с удаленным доменом или миниантитело); мультиспецифичные формы антител (например, биспецифичные, триспецифичные и т.д.), измененные для связывания с двумя или более различными антигенами или с различными эпитопами на одном антигене); молекулы тяжелых цепей, соединенные с молекулами scFv и т.п. Кроме того, термин «вариант антитела» включает многовалентные формы антител (например, трехвалентные, четырехвалентные и т.д. антитела, которые связываются с тремя, четырьмя или более копиями одного и того же антигена).
Используемый здесь термин «валентность» относится к числу потенциальных сайтов связывания мишени в полипептиде. Каждый целевой сайт связывания специфически связывает одну целевую молекулу или специфический сайт на молекуле-мишени. Когда полипептид содержит более одного целевого сайта связывания, каждый целевой сайт связывания может специфически связываться с одной и той же или разными молекулами (например, может связываться с разными лигандами или разными антигенами или разными эпитопами на одном и том же антигене). Связывающие полипептиды субъекта обычно имеют по меньшей мере один сайт связывания, специфичный для молекулы человеческого антигена.
Термин «специфичность» относится к способности специфически связываться (например, вступать в иммунную реакцию) с данным целевым антигеном (например, целевым антигеном человека). Связывающий полипептид может быть моноспецифичным и содержать один или несколько сайтов связывания, которые специфически связывают мишень, или полипептид может быть мультиспецифичным и содержать два или более сайтов связывания, которые специфически связываются с одной и той же или разными мишенями. В некоторых вариантах осуществления связывающий полипептид специфичен по отношению к двум разным (например, неперекрывающихся) частям одной и той же мишени. В определенных вариантах осуществления связывающий полипептид специфичен по отношению к более чем одной мишени. Типичные связывающие полипептиды (например, антитела), которые содержат антигенсвязывающие сайты, которые связываются с антигенами, экспрессируемыми на опухолевых клетках, известны в данной области, и один или несколько CDR таких антител могут быть включены в антитело, которое описано в настоящем документе.
Термин «антиген» или «целевой антиген» в контексте настоящего описания относится к молекуле или части молекулы, с которой способен связываться сайт связывания связывающего полипептида. Целевой антиген может иметь один или несколько эпитопов.
Термин «примерно» или «приблизительно» означает в пределах 20%, например, в пределах 10%, в пределах 5% или в пределах 1% или менее от заданного значения или интервала значений.
Используемый здесь термин «вводить» или «введение» относится к выполнению инъекции или иным образом выполняемой физической доставки вещества в том виде, в котором оно существует вне организма (например, выделенного связывающего полипептида, представленного в настоящем документе), пациенту, например, но не ограничиваясь этим, путем легочной (например, ингаляционной), слизистой (например, интраназальной), внутрикожной, внутривенной, внутримышечной доставки и/или любым другим способом физической доставки, описанным здесь или известным в данной области. При ведении заболевания или его симптома или их лечения введение вещества, как правило, происходит после выявления заболевания или появления его симптомов. При предотвращении заболевания или его симптомов введение вещества обычно происходит до начала заболевания или появления его симптомов и может быть продолжено постоянно для отсрочки или уменьшения появления или тяжести связанных с заболеванием симптомов.
Используемый здесь термин «состав» предназначен охватывать продукт, содержащий указанные ингредиенты (например, выделенный связывающий полипептид, представленный в настоящем документе), необязательно, в указанных количествах, а также любой продукт, который прямо или косвенно получается при комбинировании указанных ингредиентов, при необходимости, в указанных количествах.
«Эффективное количество» означает количество активной фармацевтической субстанции (например, выделенного связывающего полипептида настоящего изобретения), достаточное для достижения желаемого физиологического результата у лица, нуждающегося в этой активной субстанции. Эффективное количество может варьироваться среди людей в зависимости от состояния здоровья и физического состояния человека, нуждающегося в лечении, таксономической группы людей, нуждающихся в лечении, композиции состава, оценки состояния здоровья человека и других имеющих к этому отношение факторов.
Используемые здесь термины «субъект» и «пациент» используются взаимозаменяемо. Субъект в контексте данного документа может быть млекопитающим, отличающимся от примата (например, коровы, свиньи, лошади, кошки, собаки, крысы и т.д.), или приматом (например, обезьяна и человек). В определенных вариантах осуществления термин «субъект», используемый в данном документе, относится к позвоночному животному, такому как млекопитающее. Млекопитающие включают, без ограничения, людей, нечеловекоподобных приматов, диких животных, одичавших животных, сельскохозяйственных животных, животных, используемых в спорте, и домашних животных.
Используемый здесь термин «терапия» относится к любому протоколу, способу и/или средству, которые можно использовать для профилактики, ведения, лечения и/или улучшения заболевания или связанного с ним симптома. В некоторых вариантах осуществления термин «терапия» относится к любому протоколу, способу и/или средству, которые можно использовать для модулирования у субъекта иммунного ответа на инфекцию или связанный с ней симптом. В некоторых вариантах осуществления термины «виды терапии» и «терапия» относятся к биологической терапии, поддерживающей терапии и/или другим видам терапии, применимым в предупреждении, контроле, лечении и/или уменьшении интенсивности заболевания или связанного с ним симптома, известным специалисту в данной области, такому как медицинский персонал. В других вариантах осуществления термины «виды терапии» и «терапия» относятся к биологической терапии, поддерживающей терапии и/или другим видам терапии, применимым для модулирования у субъекта иммунного ответа на инфекцию или связанный с ней симптом, известным специалисту в данной области, такому как медицинский персонал.
Используемые в настоящем документе термины «лечить», «лечение» и «лечащий» относятся к снижению или ослаблению прогрессирования, тяжести и/или продолжительности заболевания или связанного с ним симптома, в результате введения одного или более видов терапии (включая, но не ограничиваясь этим, введение одного или нескольких профилактических или терапевтических средств, таких как выделенный связывающий полипептид, предложенный в данном документе). Используемый здесь термин «лечение» может также относиться к изменению течения заболевания у субъекта, получающего лечение. Терапевтическое действие лечения включает, без ограничения, предотвращение возникновения или рецидива заболевания, ослабление симптома(ов), уменьшение прямых или косвенных патологических последствий заболевания, снижение скорости прогрессирования заболевания, улучшение или смягчение тяжести заболевания и ремиссия или улучшение прогноза.
Связывающие полипептиды
В одном аспекте настоящего раскрытия предложено связывание полипептидов (например, антител, иммуноадгезинов, вариантов антител и составных белков), содержащих модифицированный Fc-домен. Связывающие полипептиды, раскрываемые в данном документе, охватывают любой связывающий полипептид, который содержит модифицированный Fc-домен. В определенных вариантах осуществления связывающий полипептид представляет собой антитело, или иммуноадгезин, или его производное. Любое антитело из любого источника или вида можно использовать в связывающих полипептидах, раскрываемых здесь. Подходящие антитела включают, без ограничения, человеческие антитела, гуманизированные антитела или химерные антитела. Подходящие антитела включают, без ограничения, моноклональные антитела, поликлональные антитела, полноразмерные антитела или одноцепочечные антитела.
Fc-домены из любого класса иммуноглобулинов (например, IgM, IgG, IgD, IgA и IgE) и их видов можно использовать в связывающих полипептидах, раскрываемых здесь. Также можно использовать химерные Fc-домены, содержащие части Fc-доменов из разных видов или классов Ig. В некоторых вариантах осуществления Fc-домен представляет собой Fc-домен человека. В некоторых вариантах осуществления Fc-домен представляет собой Fc-домен IgG1. В других вариантах осуществления Fc-домен представляет собой Fc-домен IgG4. В некоторых вариантах осуществления Fc-домен представляет собой Fc-домен человеческого IgG1 или IgG4. В некоторых вариантах осуществления Fc-домен представляет собой Fc-домен человеческого IgG1. В случае Fc-доменов других видов и/или классов или изотипов Ig специалист в данной области поймет, что любая из описанных здесь аминокислотных замен может быть адаптирована соответствующим образом. В некоторых вариантах осуществления модифицированный Fc-домен может содержать аминокислотную замену, выбранную из M252, I253, S254, T256, K288, T307, K322, E380, L432, N434 или Y436 и любых их комбинаций, в соответствии с нумерацией ЕС. В некоторых вариантах осуществления модифицированный Fc-домен может содержать двойную аминокислотную замену в любых двух положениях аминокислот, выбранных из M252, I253, S254, T256, K288, T307, K322, E380, L432, N434 и Y436, в соответствии с нумерацией ЕС. В некоторых вариантах осуществления модифицированный Fc-домен может содержать тройную аминокислотную замену в любых трех положениях аминокислот, выбранных из M252, I253, S254, T256, K288, T307, K322, E380, L432, N434 и Y436, в соответствии с нумерацией ЕС. В некоторых вариантах осуществления модифицированный Fc-домен может содержать четверную аминокислотную замену в любых четырех положениях аминокислот, выбранных из M252, I253, S254, T256, K288, T307, K322, E380, L432, N434 и Y436, в соответствии с нумерацией ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену в любом из положений аминокислот, выбранных из M252, I253, S254, T256, K288, T307, K322, E380, L432 или Y436 и любых их комбинаций, где положение аминокислоты N434 не замещено (то есть позиция аминокислоты N434 дикого типа), согласно нумерации ЕС.
В некоторых вариантах осуществления модифицированный Fc-домен может содержать аминокислотную замену, выбранную из M252Y (т.е. тирозин в позиции аминокислоты 252), T256D, T256E, K288D, K288N, T307A, T307E, T307F, T307M, T307Q, T307W, E380C, N434F, N434P, N434Y, Y436H, Y436N или Y436W и любых их комбинаций, в соответствии с нумерацией ЕС. В некоторых вариантах осуществления модифицированный Fc-домен может содержать двойную аминокислотную замену, выбранную из M252, где замена представляет собой M252Y; T256, где замена представляет собой T256D или T256E; K288, где замена представляет собой K288D или K288N; T307, где замена представляет собой T307A, T307E, T307F, T307M, T307Q или T307W; E380, где замена представляет собой E380C; N434, где замена представляет собой N434F, N434P или N434Y; Y436, где замена представляет собой Y436H, Y436N или Y436W, в соответствии с нумерацией ЕС. В некоторых вариантах осуществления модифицированный Fc-домен может содержать тройную аминокислотную замену, выбранную из M252, где замена представляет собой M252Y; T256, где замена представляет собой T256D или T256E; K288, где замена представляет собой K288D или K288N; T307, где замена представляет собой T307A, T307E, T307F, T307M, T307Q или T307W; E380, где замена представляет собой E380C; N434, где замена представляет собой N434F, N434P или N434Y; Y436, где замена представляет собой Y436H, Y436N или Y436W, в соответствии с нумерацией ЕС. В некоторых вариантах осуществления модифицированный Fc-домен может содержать четверную аминокислотную замену, выбранную из M252, где замена представляет собой M252Y; T256, где замена представляет собой T256D или T256E; K288, где замена представляет собой K288D или K288N; T307, где замена представляет собой T307A, T307E, T307F, T307M, T307Q или T307W; E380, где замена представляет собой E380C; N434, где замена представляет собой N434F, N434P или N434Y; Y436, где замена представляет собой Y436H, Y436N или Y436W, в соответствии с нумерацией ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену в любом из положений аминокислот, выбранных из M252Y, T256D, T256E, K288D, K288N, T307A, T307E, T307F, T307M, T307Q, T307W, E380C, Y436H, Y436N или Y436W и любых их комбинаций, где положение аминокислоты N434 не замещено фенилаланином (F) или тирозином (Y), согласно нумерации ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену в любом из положений аминокислот, выбранных из M252Y, T256D, T256E, K288D, K288N, T307A, T307E, T307F, T307M, T307Q, T307W, E380C, Y436H, Y436N или Y436W и любых их комбинаций, где положение аминокислоты N434 не замещено тирозином (Y), согласно нумерации ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену в любом из положений аминокислот, выбранных из M252Y, T256D, T256E, K288D, K288N, T307A, T307E, T307M, T307Q, T307W, E380C, Y436H, Y436N или Y436W и любых их комбинаций, где положение аминокислоты N434 не замещено (то есть позиция аминокислоты N434 дикого типа), согласно нумерации ЕС.
В определенных вариантах осуществления модифицированный Fc-домен может содержать аминокислотную замену, выбранную из M252, T256, T307 или N434 и любых их комбинаций, в соответствии с нумерацией ЕС. В определенных вариантах осуществления модифицированный Fc-домен может содержать двойную аминокислотную замену в любых двух положениях аминокислот, выбранных из M252, T256, T307 и N434, в соответствии с нумерацией ЕС. В определенных вариантах осуществления модифицированный Fc-домен может содержать тройную аминокислотную замену в любых трех положениях аминокислот, выбранных из M252, T256, T307 и N434, в соответствии с нумерацией ЕС. В определенных вариантах осуществления модифицированный Fc-домен может содержать четверную аминокислотную замену в положениях аминокислот M252, T256, T307 и N434, в соответствии с нумерацией ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену, выбранную из M252, T256 или T307 и любых их комбинаций, где положение аминокислоты N434 не замещено (то есть позиция аминокислоты N434 дикого типа), согласно нумерации ЕС.
В иллюстративных вариантах осуществления модифицированный Fc-домен может содержать аминокислотную замену, выбранную из M252, где замена представляет собой M252Y; T256, где замена представляет собой T256D или T256E; T307, где замена представляет собой T307Q или T307W; или N434, где замена представляет собой N434F или N434Y, и любые их комбинации в соответствии с нумерацией ЕС. В иллюстративных вариантах осуществления модифицированный Fc-домен может содержать двойную аминокислотную замену в любых двух положениях аминокислоты, выбранных из M252, где замена представляет собой M252Y; T256, где замена представляет собой T256D или T256E; T307, где замена представляет собой T307Q или T307W; или N434, где замена представляет собой N434F или N434Y, согласно нумерации ЕС. В иллюстративных вариантах осуществления модифицированный Fc-домен может содержать тройную аминокислотную замену в любых трех положениях аминокислоты, выбранных из M252, где замена представляет собой M252Y; T256, где замена представляет собой T256D или T256E; T307, где замена представляет собой T307Q или T307W; или N434, где замена представляет собой N434F или N434Y, согласно нумерации ЕС. В иллюстративных вариантах осуществления модифицированный Fc-домен может содержать четверную аминокислотную замену в положениях аминокислоты, выбранных из M252, где замена представляет собой M252Y; T256, где замена представляет собой T256D или T256E; T307, где замена представляет собой T307Q или T307W; или N434, где замена представляет собой N434F или N434Y, согласно нумерации ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену, выбранную из M252Y, T256D, T256E, T307Q или T307W и любых их комбинаций, где положение аминокислоты N434 не замещено фенилаланином (F) или тирозином (Y), согласно нумерации ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену, выбранную из M252Y, T256D, T256E, T307Q или T307W и любых их комбинаций, где положение аминокислоты N434 не замещено тирозином (Y), согласно нумерации ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену, выбранную из M252Y, T256D, T256E, T307Q или T307W и любых их комбинаций, где положение аминокислоты N434 не замещено (то есть позиция аминокислоты N434 дикого типа), согласно нумерации ЕС.
В некоторых вариантах осуществления модифицированный Fc-домен может содержать аминокислотную замену, выбранную из T256D, или T256E, и/или T307W, или T307Q, и дополнительно содержит аминокислотную замену, выбранную из N434F, или N434Y, или M252Y, согласно нумерации ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену, выбранную из T256D, или T256E, и/или T307W, или T307Q, и дополнительно содержит аминокислотную замену M252Y, где положение аминокислоты N434 не замещено фенилаланином (F) или тирозином (Y), согласно нумерации ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену, выбранную из T256D, или T256E, и/или T307W, или T307Q, и дополнительно содержит аминокислотную замену M252Y, где положение аминокислоты N434 не замещено тирозином (Y), согласно нумерации ЕС. В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислотную замену, выбранную из T256D, или T256E, и/или T307W, или T307Q, и дополнительно содержит аминокислотную замену M252Y, где положение аминокислоты N434 не содержит замены (т.е. это аминокислотное положение N434 дикого типа), согласно нумерации ЕС.
В некоторых вариантах осуществления модифицированный Fc-домен может содержать двойную аминокислотную замену, выбранную из M252Y/T256D, M252Y/T256E, M252Y/T307Q, M252Y/T307W, M252Y/N434F, M252Y/N434Y, T256D/T307Q, T256D/N434F, T256D/N434Y, T256E/T307Q, T256E/T307W, T256E/N434F, T256E/N434Y, T307Q/N434F, T307Q/N434Y, T307W/N434F и T307W/N434Y, согласно нумерации ЕС. В некоторых вариантах осуществления модифицированный Fc-домен может содержать тройную аминокислотную замену, выбранную из M252Y/T256D/T307Q, M252Y/T256D/T307W, M252Y/T256D/N434F, M252Y/T256D/N434Y, M252Y/T256E/T307Q, M252Y/T256E/T307W, M252Y/T256E/N434F, M252Y/T256E/N434Y, M252Y/T307Q/N434F, M252Y/T307Q/N434Y, M252Y/T307W/N434F, M252T/T307W/N434Y, T256D/307Q/N434F, T256D/307W/N434F, T256D/307Q/N434Y, T256D/307W/N434Y, T256E/307Q/N434F, T256E/307W/N434F, T256E/307Q/N434Y и T256E/307W/N434Y, согласно нумерации ЕС.
В некоторых вариантах осуществления модифицированный Fc-домен может содержать четверную аминокислотную замену, выбранную из M252Y/T256D/T307Q/N434F, M252Y/T256E/T307Q/N434F, M252Y/T256D/T307W/N434F, M252Y/T256E/T307W/N434F, M252Y/T256D/T307Q/N434Y, M252Y/T256E/T307Q/N434Y, M252Y/T256D/T307W/N434Y и M252Y/T256E/T307W/N434Y, согласно нумерации ЕС.
В некоторых вариантах осуществления может быть желательно, чтобы модифицированный Fc-домен содержал аминокислоту дикого типа в положении аминокислоты N434 согласно нумерации ЕС. В некоторых вариантах осуществления может быть желательно, чтобы Fc-домен не содержал фенилаланин (F) или тирозин (Y) в положении аминокислоты N434, согласно нумерации ЕС. В некоторых вариантах осуществления может быть желательно, чтобы Fc-домен не содержал тирозин (Y) в положении аминокислоты N434, согласно нумерации ЕС. В некоторых вариантах осуществления модифицированный Fc-домен может содержать двойную аминокислотную замену, выбранную из M252Y/T256D, M252Y/T256E, M252Y/T307Q, M252Y/T307W, T256D/T307Q, T256D/T307W, T256E/T307Q и T256E/T307W, в соответствии с нумерацией ЕС. В некоторых вариантах осуществления модифицированный Fc-домен может содержать тройную аминокислотную замену, выбранную из M252Y/T256D/T307Q, M252Y/T256D/T307W, M252Y/T256E/T307Q и M252Y/T256E/T307W, в соответствии с нумерацией ЕС.
В одном варианте осуществления связывающий полипептид с измененным связыванием с FcRn содержит Fc-домен, имеющий одну или несколько аминокислотных замен, как раскрывается в настоящем документе. В одном варианте осуществления связывающий полипептид с повышенной способностью связывания с FcRn содержит Fc-домен, имеющий одну или несколько аминокислотных замен, как раскрывается в настоящем документе. В одном варианте осуществления связывающий полипептид с повышенной способностью связывания с FcRn содержит Fc-домен, имеющий две или более аминокислотные замены, как раскрывается в настоящем документе. В одном варианте осуществления связывающий полипептид с повышенной способностью связывания с FcRn содержит Fc-домен, имеющий три или более аминокислотные замены, как раскрывается в настоящем документе.
В некоторых вариантах осуществления связывающий полипептид может проявлять видоспецифическую способность связывания с FcRn. В одном варианте осуществления связывающий полипептид может проявлять способность связывания с FcRn человека. В одном варианте осуществления связывающий полипептид может проявлять способность связывания с FcRn крысы. В некоторых вариантах осуществления связывающий полипептид может проявлять межвидовую способность связывания с FcRn. О таких связывающих полипептидах говорят, что они являются перекрестно-реактивными по отношению к одному или нескольким различным видам. В одном варианте осуществления связывающий полипептид может проявлять способность связывания с FcRn и человека, и крысы.
Неонатальный рецептор Fc (FcRn) взаимодействует с Fc-участком антител, чтобы способствовать рециркуляции за счет восстановления нормальной лизосомальной деградации. Этот процесс является рН-зависимым процессом, который происходит в эндосомах при кислотном значении рН (например, рН менее 6,5), но не в физиологических условиях рН кровотока (например, некислотного рН). В некоторых вариантах осуществления связывающий полипептид настоящего изобретения, содержащий измененный Fc-домен, имеет повышенную способность связывания с FcRn при кислотном значении рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых вариантах осуществления связывающий полипептид, содержащий измененный Fc-домен, обладает повышенной способностью связывания с FcRn при pH ниже 7, например, при pH примерно 6,5, при pH примерно 6,0, при pH примерно 5,5, при pH примерно 5,0 по сравнению со связывающим полипептидом содержащий Fc-домен дикого типа. В некоторых вариантах осуществления связывающий полипептид, содержащий измененный Fc-домен, обладает повышенной способностью связывания с FcRn при рН ниже 7, например, при рН примерно 6,5, при рН примерно 6,0, при рН примерно 5,5, при рН примерно 5,0 по сравнению со способностью связывания с FcRn связывающего полипептида при повышенном некислотном значении рН. Повышенный некислотный рН может быть, например, рН выше 7, рН примерно 7, рН примерно 7,4, рН примерно 7,6, рН примерно 7,8, рН примерно 8,0, рН примерно 8,5, рН примерно 9,0.
В некоторых вариантах осуществления может быть желательным, чтобы связывающий полипептид, содержащий измененный Fc-домен, проявлял примерно такую же способность связывания с FcRn при некислом pH, что и связывающий полипептид, содержащий Fc-домен дикого типа. В некоторых вариантах осуществления может быть желательным, чтобы связывающий полипептид, содержащий измененный Fc-домен, проявлял более слабую способность связывания с FcRn при некислотных значениях рН, чем связывающий полипептид, содержащий измененный Fc-домен, содержащий двойную аминокислотную замену M428L/N434S, согласно нумерации ЕС. Соответственно, может быть желательным, чтобы связывающий полипептид, содержащий измененный Fc-домен, проявлял минимальное нарушение рН-зависимого связывания FcRn.
В некоторых вариантах осуществления связывающий полипептид, содержащий измененный Fc-домен, имеющий повышенную способность связывания с FcRn при кислотном значении рН, имел сниженную (т.е. более медленную) скорость диссоциации с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых вариантах осуществления связывающий полипептид, содержащий измененный Fc-домен, имеющий повышенную способность связывания с FcRn при кислотном значении рН по сравнению со способностью связывания с FcRn связывающего полипептида при повышенном некислотном значении рН, обладает более медленной скоростью диссоциации с FcRn при кислотном значении рН по сравнению со скоростью диссоциации с FcRn связывающего полипептида при повышенном некислотном значении рН.
В некоторых вариантах осуществления предложен связывающий полипептид, содержащий измененный Fc-домен, который проявляет более сильную способность связывания с FcRn при некислотных рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых вариантах осуществления предложен связывающий полипептид, содержащий измененный Fc-домен, который проявляет более сильную способность связывания с FcRn при кислотных рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых вариантах осуществления предложен связывающий полипептид, содержащий измененный Fc-домен, который проявляет более сильную способность связывания с FcRn при некислотных рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, проявляет более сильную способность связывания с FcRn при кислотных рН по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. Соответственно, в определенных вариантах осуществления предложен связывающий полипептид, содержащий измененный Fc-домен, который демонстрирует потерю рН-зависимого связывания с FcRn.
Определенные варианты осуществления включают антитела, которые, в дополнение к описанным здесь мутациям Fc, которые изменяют способность связывания с FcRn, содержат по меньшей мере одну аминокислоту в одном или нескольких доменах константного участка и/или по меньшей мере одну аминокислоту в одном или нескольких доменах вариабельных участков, которые были удалены или иным образом изменены, чтобы обеспечить желаемые биохимические характеристики, такие как, например, пониженные или усиленные эффекторные функции, способность нековалентно димеризоваться, повышенную способность локализоваться в месте опухоли, более короткий период полувыведения из сыворотки крови, более длинный период полувыведения из сыворотки крови по сравнению с цельным неизмененным антителом примерно такой же иммуногенности и т.п.
В некоторых других вариантах осуществления связывающие полипептиды содержат константные участки, полученные из разных изотипов антител (например, константные участки из двух или более человеческих IgG1, IgG2, IgG3 или IgG4). В других вариантах осуществления антитела к PAI-1 содержат химерный шарнирный участок (т.е. шарнирный участок, содержащий части шарнирного участка, происходящие из шарнирных доменов различных изотипов антител, например, верхний шарнирный домен из молекулы IgG4 и средний шарнирный домен IgG1).
В определенных вариантах осуществления домен Fc может быть подвергнут мутации для увеличения или уменьшения эффекторной функции с использованием методик, известных в данной области техники. В некоторых вариантах осуществления связывающий полипептид по настоящему изобретению, содержащий измененный Fc-домен, имеет измененную способность связывания с Fc-рецептором. Существует несколько разных типов Fc-рецепторов, которых классифицируют по типу антител, которые они распознают. Например, Fc-гамма-рецепторы (FcγR) связываются с антителами класса IgG, Fc-альфа-рецепторы (FcαR) связываются с антителами класса IgA, а Fc-эпсилон-рецепторы (FcεR) связываются с антителами класса IgE. FcγR относятся к семейству, которое включает членов, например, FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa и FcγRIIIb. В некоторых вариантах осуществления связывающий полипептид, содержащий измененный Fc-домен, имеет измененную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых вариантах осуществления связывающий полипептид, содержащий измененный Fc-домен, имеет пониженную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых вариантах осуществления связывающий полипептид, содержащий измененный Fc-домен, имеет повышенную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. В некоторых вариантах осуществления связывающий полипептид, содержащий измененный Fc-домен, имеет примерно такую же способность связывания с FcγRIIIa, что и связывающий полипептид, содержащий Fc-домен дикого типа.
В других вариантах осуществления связывающие полипептиды для применения в способах диагностики и лечения, описываемых в данном документе, имеют константный участок, например, константный участок тяжелой цепи IgG1, измененный так, чтобы уменьшать или устранять гликозилирование. Например, связывающие полипептиды (например, антитела или иммуноадгезины), содержащие измененный Fc-домен, могут дополнительно содержать аминокислотную замену, которая изменяет гликозилирование Fc антитела. Например, указанный измененный Fc-домен может характеризоваться сниженным гликозилированием (например, N- или O-сцепленным гликозилированием).
Иллюстративные аминокислотные замены, придающие сниженное или измененное гликозилирование, раскрыты в международной публикации согласно PCT № WO05/018572, полностью включенной в данный документ ссылкой. В некоторых вариантах осуществления связывающие полипептиды изменены так, чтобы устранить гликозилирование. Такие связывающие полипептиды могут упоминаться как «agly»-связывающие полипептиды (например, «agly»-антитела). Не будучи связанными какой-либо теорией, полагают, что «agly»-связывающие полипептиды могут иметь улучшенный профиль безопасности и стабильности in vivo. Agly-связывающие полипептиды могут иметь любой изотип или их подкласс, например, IgG1, IgG2, IgG3 или IgG4. Для получения "agly"-антител или антител с измененными гликанами применяются многочисленные признанные в данной области способы. Например, для получения таких антител можно использовать генетически сконструированные клетки-хозяева (например, модифицированные дрожжи, например клетки Picchia или клетки СНО) с модифицированными сигнальными путями гликозилирования (например, с делецией гликозилтрансферазы).
В некоторых вариантах осуществления связывающие полипептиды могут содержать константный участок антитела (например, константный участок IgG, например, константный участок IgG человека, например, константный участок IgG1 человека), который участвует в одной или нескольких эффекторных функциях. Например, связывание C1-комплекса системы с константной областью антитела может активировать систему комплемента. Активация системы комплемента важна для опсонизации и лизиса клеточных патогенов. Активация системы комплемента также стимулирует воспалительный ответ и может также быть вовлечена в аутоиммунную гиперчувствительность. Кроме того, антитела связываются с рецепторами на различных клетках через Fc-домен (сайты связывания с Fc-рецептором в Fc-участке антитела связываются с Fc-рецептором (FcR) на поверхности клетки). Существует несколько Fc-рецепторов, специфичных для различных классов антител, в том числе IgG (гамма-рецепторы), IgE (эпсилон-рецепторы), IgA (альфа-рецепторы) и IgM (мю-рецепторы). Связывание антитела с Fc-рецепторами на поверхности клеток запускает несколько важных и разнообразных биологических реакций, включающих поглощение и разрушение частиц, покрытых антителами, выведение иммунных комплексов, лизис клеток-мишеней, покрытых антителами, клетками-киллерами (называемый антителозависимой клеточноопосредованной цитотоксичностью или ADCC), высвобождение медиаторов воспаления, трансплацентарный перенос и регуляцию выработки иммуноглобулинов. В некоторых вариантах осуществления связывающие полипептиды (например, антитела или иммуноадгезины) связываются с Fc-гамма-рецептором. В альтернативных вариантах осуществления связывающие полипептиды могут содержать константный участок, лишенный одной или нескольких эффекторных функций (например, активности ADCC) и/или способности связывания с Fcγ-рецептором.
Белки, включая антитела, с низкой термодинамической стабильностью, имеют повышенную склонность к неправильному сворачиванию и агрегации и могут ограничивать или препятствовать активности, эффективности и действенности белка в качестве полезного терапевтического средства. В некоторых вариантах осуществления связывающий полипептид, содержащий измененный Fc-домен, имеет примерно такую же термостабильность, что и связывающий полипептид, содержащий Fc-домен дикого типа. В некоторых вариантах осуществления связывающий полипептид, содержащий измененный Fc-домен, имеет примерно такую же термическую стабильность, что и связывающий полипептид, содержащий измененный Fc-домен, имеющий тройную аминокислотную замену M252Y/S254T/T256E (YTE).
Обусловленные этим физиологический профиль, биодоступность и другие биохимические эффекты этих изменений, такие как локализация в опухоли, биораспределение и период полувыведения из сыворотки крови, можно легко измерить и определить количественно с помощью хорошо известных иммунологических методик без проведения излишних экспериментов.
В некоторых вариантах осуществления связывающий полипептид настоящего изобретения может содержать антигенсвязывающий фрагмент антитела. Термин «антигенсвязывающий фрагмент» относится к полипептидному фрагменту иммуноглобулина или антитела, которое связывает антиген или конкурирует с интактным антителом (т.е. с интактным антителом, из которого они были получены) за связывание антигена (т.е. специфическое связывание). Антигенсвязывающие фрагменты можно получать рекомбинантными или биохимическими методами, которые хорошо известны в данной области. Примеры антигенсвязывающих фрагментов включают Fv, Fab, Fab'и (Fab')2. В иллюстративных вариантах осуществления связывающий полипептид настоящего изобретения содержит антигенсвязывающий фрагмент и измененный Fc-домен.
В некоторых вариантах осуществления связывающий полипептид содержит одноцепочечную последовательность вариабельного участка (ScFv). Одноцепочечная последовательность вариабельного участка содержат одиночный полипептид, имеющий один или несколько антигенсвязывающих сайтов, например домен VL, связанный гибким линкером с доменом VH. Молекулы ScFv могут быть сконструированы в ориентации VH-линкер-VL или ориентации VL-линкер-VH. Гибкий шарнир, который связывает домены VL и VH, образующие сайт связывания антигена, включает от около 10 до примерно 50 аминокислотных остатков. Связывающие пептиды известны в данной области. Связывающие полипептиды могут содержать по меньшей мере один scFv и/или по меньшей мере один константный участок. В одном варианте осуществления связывающий полипептид настоящего изобретения может содержать по меньшей мере один scFv, присоединенный или слитый с измененным Fc-доменом.
В некоторых вариантах осуществления связывающий полипептид настоящего изобретения представляет собой многовалентное (например, четырехвалентное) антитело, которое получают слиянием последовательности ДНК, кодирующей антитело, с молекулой ScFv (например, измененной молекулой ScFv). Например, в одном варианте осуществления эти последовательности объединяют так, что молекула ScFv (например, измененная молекула ScFv) связана на своем N-конце или С-конце с фрагментом Fc антитела через гибкий линкер (например, гли-/сер-линкер). В другом варианте осуществления четырехвалентное антитело настоящего изобретения может быть получено путем слияния молекулы ScFv с соединяющим пептидом, который соединен с измененным Fc-доменом для конструирования четырехвалентной молекулы ScFv-Fab.
В другом варианте осуществления связывающий полипептид настоящего изобретения представляет собой измененное миниантитело. Измененное миниантитело настоящего описания представляет собой димерную молекулу, состоящую из двух полипептидных цепей, каждая из которых содержит молекулу ScFv, которая соединена с измененным Fc-доменом через соединяющий пептид. Миниантитела можно получать, конструируя компонент ScFv и соединяя пептидные компоненты с использованием способов, описанных в данной области (см., например, патент US 5837821 или WO 94/09817A1). В другом варианте осуществления может быть сконструировано четырехвалентное миниантитело. Четырехвалентные миниантитела могут быть сконструированы так же, как и миниантитела, за исключением того, что две молекулы ScFv связаны гибким линкером. Связанную конструкцию scFv-scFv затем присоединяют к измененному Fc-домену.
В другом варианте осуществления связывающий полипептид настоящего изобретения содержит димерное антитело. Димерные антитела представляют собой димерные, четырехвалентные молекулы, каждая из которых имеет полипептид, подобный молекулам scFv, но обычно содержит короткий линкер из (менее 10, например, от около 1 до около 5) аминокислотных остатков, соединяющий оба вариабельных домена, таким образом, что домены VL и VH на одной и той же полипептидной цепи не могут взаимодействовать друг с другом. Вместо этого домен VL и VH одной полипептидной цепи взаимодействуют с доменом VH и VL (соответственно) на второй полипептидной цепи (см., например, WO 02/02781). Димерные антитела настоящего изобретения содержат scFv-подобную молекулу, соединенную с измененным Fc-доменом.
В других вариантах осуществления связывающие полипептиды содержат мультиспецифические или многовалентные антитела, включающие один или несколько вариабельных доменов последовательно на одной и той же полипептидной цепи, например полипептиды тандемного вариабельного домена (TVD). Типичные полипептиды TVD включают в себя конфигурацию «двойная головка» или «Dual-Fv», описанную в патенте США № 5989830. В конфигурации Dual-Fv вариабельные домены двух разных антител экспрессируются в тандемной ориентации на двух отдельных цепях (одной тяжелой цепи и одной легкой цепи), где одна полипептидная цепь имеет два последовательных домена VH, разделенных пептидным линкером (VH1-линкер-VH2), а другая полипептидная цепь состоит из комплементарных доменов VL, соединенных последовательно пептидным линкером (VL1-линкер-VL2). В конфигурации перекрестной двойной головки вариабельные домены двух разных антител экспрессируются в тандемной ориентации на двух отдельных полипептидных цепях (одной тяжелой цепи и одной легкой цепи), где одна полипептидная цепь имеет два последовательных домена VH, разделенных пептидным линкером (VH1-линкер-VH2), а другая полипептидная цепь состоит из комплементарных доменов VL, соединенных последовательно пептидным линкером в противоположной ориентации (VL2-линкер-VL1). Дополнительные варианты антител в формате «Dual-Fv» включают биспецифическое антитело с двумя вариабельными доменами IgG (DVD-IgG) (см. патент США № 7612181) и формат TBTI (см. US 2010/0226923 A1). В некоторых вариантах осуществления связывающие полипептиды содержат мультиспецифические или многовалентные антитела, включающие один или несколько вариабельных доменов последовательно на одной и той же полипептидной цепи, соединенной с измененным Fc-доменом.
В другом типичном варианте осуществления связывающий полипептид содержит биспецифическое антитело с двумя перекрестными вариабельными доменами IgG (CODV-IgG), основанное на конфигурации «двойной головки» (см. US20120251541 А1, который полностью включен в настоящее описание ссылкой).
В другом иллюстративном варианте осуществления связывающий полипептид представляет собой иммуноадгезин. Используемый здесь термин «иммуноадгезин» относится к связывающему полипептиду, содержащему один или несколько связывающих доменов (например, из рецептора, лиганда или молекулы клеточной адгезии), связанных с константным доменом иммуноглобулина (то есть Fc-участком) (см., например, Ashkenazi et al. 1995, Methods 8 (2): 104-115, и Isaacs (1997) Brit. J. Rheum. 36:305, которые в полном объеме включены в данный документ ссылкой). Иммуноадгезины обозначаются суффиксом «-цепт» в их международных непатентованных названиях (INN). Как и антитела, иммуноадгезины имеют длительный период полувыведения, их легко очищать с помощью методов, основанных на аффинности, и обладают преимуществами авидности благодаря их бивалентности. Примеры поступающих в продажу терапевтических иммуноадгезинов включают этанерцепт (ENBREL®), абатацепт (ORENCIA®), рилонацепт (ARCALYST®), афлиберцепт (ZALTRAP® / EYLEA®) и белатацепт (NULOJIX®).
В определенных вариантах осуществления связывающий полипептид включает иммуноглобулиноподобные домены. Подходящие иммуноглобулиноподобные домены включают, без ограничения, фибронектиновые домены (см., например, Koide et al. (2007), Methods Mol. Biol. 352 : 95-109, которая полностью включена в настоящее описание ссылкой), DARPin (см., например, Stumpp et al. (2008) Drug Discov. Today 13 (15-16): 695-701, которая полностью включена в настоящее описание ссылкой), Z-доменов белка A (см. Nygren et al. (2008) FEBS J. 275 (11): 2668-76, которая полностью включена в настоящее описание ссылкой), липокалины (см., например, Skerra et al. (2008) FEBS J. 275 (11): 2677-83, которая в полном объеме включена в настоящее описание ссылкой), аффилины (см., например, Ebersbach et al. (2007) J. Mol. Biol. 372 (1): 172-85, которая полностью включена в данный документ ссылкой), аффитины (см., например, Krehenbrink et al. (2008). J. Mol. Biol. 383 (5): 1058-68 , которая в полном объеме включена в настоящее описание ссылкой), авимеры (см., например, Silverman et al. (2005) Nat. Biotechnol. 23 (12): 1556-61 , которая в полном объеме включена в настоящее описание ссылкой), финомеры (см., например, Grabulovski et al. (2007) J Biol Chem 282 (5): 3196-3204 , которая полностью включена в настоящее описание ссылкой), и пептиды домена Kunitz (см., например, N ixon et al. (2006) Curr Opin Drug Discov Devel 9 (2): 261-8, которая полностью включена в настоящее описание ссылкой).
Для связывающих полипептидов и иммуноадгезинов настоящего изобретения связывающие полипептиды могут быть нацелены практически на любой антиген, включая, но не ограничиваясь ими, белки, субъединицы, домены, мотивы и/или эпитопы антигенов-мишеней, которые включают оба растворимых фактора, такие как цитокины и связанные с мембраной факторы и трансмембранные рецепторы.
Связывающий полипептид настоящего изобретения, содержащий измененный Fc-домен, описываемый в настоящем документе, может включать последовательности CDR или последовательности вариабельного домена известного "родительского" антитела. В некоторых вариантах осуществления родительское антитело и антитело данного изобретения могут иметь сходные или идентичные последовательности, за исключением измененного Fc-домена, как описано в настоящем документе.
Нуклеиновые кислоты и векторы экспрессии
В одном аспекте изобретение относится к полинуклеотидам, кодирующим связывающие полипептиды, раскрываемые в данном документе. Также предложены способы получения связывающего полипептида, включающие экспрессию этих полинуклеотидов.
Полинуклеотиды, кодирующие связывающие полипептиды, раскрываемые в данном документе, обычно вставляют в вектор экспрессии для введения в клетки-хозяева, которые можно использовать для получения желаемого количества заявляемых антител или иммуноадгезинов. Соответственно, в определенных аспектах изобретения предложены векторы экспрессии, содержащие полинуклеотиды, раскрываемые в данном документе, и клетки-хозяева, содержащие эти векторы и полинуклеотиды.
Термин «вектор» или «вектор экспрессии» используется в данном описании и формуле изобретения для обозначения векторов, используемых для введения и экспрессии желаемого гена в клетке. Как известно специалистам в данной области, такие векторы можно легко выбрать из группы, состоящей из плазмид, фагов, вирусов и ретровирусов. Как правило, вектор будет содержать селективный маркер, соответствующие сайты рестрикции для облегчения клонирования желаемого гена и способность проникать и/или реплицироваться в эукариотических или прокариотических клетках.
Могут быть использованы многочисленные системы векторов экспрессии. Например, в одном классе векторов используются элементы ДНК, происходящие из вирусов животных, таких как вирус папилломы крупного рогатого скота, вирус полиомы, аденовирус, вирус коровьей оспы, бакуловирус, ретровирусы (RSV, MMTV или MOMLV) или вирус SV40. В других предусматривается применение полицистронных систем с внутренними сайтами связывания рибосом. Дополнительно, клетки с интегрированной ДНК в их хромосомах можно подвергать отбору путем введения одного или нескольких маркеров, позволяющих проводить отбор трансфицированных клеток-хозяев. Маркер может обеспечивать прототрофность ауксотрофному хозяину, устойчивость к биоцидам (например, антибиотикам) или устойчивость к тяжелым металлам, таким как медь. Селектируемый маркерный ген может быть непосредственно связан с последовательностями ДНК, подлежащими экспрессии, либо введен в ту же клетку путем котрансформации. Для оптимального синтеза мРНК также могут быть необходимы дополнительные элементы. Эти элементы могут включать в себя сигнальные последовательности, сигналы сплайсинга, а также транскрипционные промоторы, энхансеры и сигналы терминации. В некоторых вариантах осуществления клонированные гены вариабельного участка встраивают в вектор экспрессии вместе с генами константного участка тяжелой и легкой цепи (такими, как гены человека), синтезированными, как обсуждалось выше.
В других вариантах осуществления связывающий полипептид, как описано в настоящем документе, может быть экспрессирован с использованием полицистронных конструкций. В таких системах экспрессии несколько продуктов генов, представляющих интерес, таких как тяжелая и легкая цепи антител, можно получать из одной полицистронной конструкции. В этих системах преимущественно используют сайт внутренней посадки рибосомы (IRES) для обеспечения относительно высоких уровней полипептидов в эукариотических клетках-хозяевах. Совместимые последовательности IRES раскрыты в патенте США № 6193980, включенном в данный документ ссылкой. Специалистам в данной области будет понятно, что такие системы экспрессии можно применять для эффективного получения полного спектра полипептидов, раскрытых в настоящей заявке.
В более общем смысле, после того как был получен вектор или последовательность ДНК, кодирующая связывающий полипептид настоящего изобретения, вектор экспрессии можно вводить в соответствующую клетку-хозяина. Иными словами, клетку-хозяина можно трансформировать. Введение плазмиды в клетку-хозяина можно выполнить с помощью различных технологий, хорошо известных специалистам в данной области. Они включают, без ограничения, трансфекцию (в том числе электрофорез и электропорацию), слияние протопластов, осаждение фосфатом кальция, слияние клеток с ДНК в оболочке, микроинъекцию и инфицирование интактным вирусом. См. Ridgway, A. A. G. "Mammalian Expression Vectors" Chapter 24.2, pp. 470-472, Vectors, Rodriguez and Denhardt, Eds. (Butterworths, Boston, MА 1988). Трансформированные клетки выращивают в условиях, подходящих для продуцирования легких цепей и тяжелых цепей, и анализируют в отношении синтеза белков тяжелой и/или легкой цепи. Иллюстративные аналитические технологии включают твердофазный иммуноферментный анализ (ELISA), радиоиммунологический анализ (RIA) или анализ на клеточном сортере с активацией флуоресценции (FACS), иммуногистохимический анализ и т.п.
Применяемый в данном документе термин "трансформация" следует употреблять в широком смысле для обозначения введения ДНК в реципиентную клетку-хозяина, которое изменяет генотип и вследствие этого приводит к изменению в реципиентной клетке.
Таким же образом «клетки-хозяева» относятся к клеткам, которые были трансформированы векторами, сконструированными с использованием методов рекомбинантной ДНК и кодирующими по меньшей мере один гетерологичный ген. В описаниях способов выделения полипептидов из рекомбинантных хозяев термины "клетка" и "культура клеток" применяются взаимозаменяемо для обозначения источника антитела, если четко не определено иное. Иными словами, выделение полипептида из "клеток" может означать выделение из целых клеток, осажденных центрифугированием, или из культуры клеток, содержащей как среду, так и суспендированные клетки.
В одном варианте осуществления линия клеток-хозяев, используемая для экспрессии связывающего полипептида, имеет эукариотическое или прокариотическое происхождение. В одном варианте осуществления линия клеток-хозяев, используемая для экспрессии связывающего полипептида, имеет бактериальное происхождение. В одном варианте осуществления линия клеток-хозяев для экспрессии связывающего полипептида происходит от млекопитающих; специалисты в данной области могут определить конкретные линии клеток-хозяев, которые лучше всего подходят для экспрессии в них желаемого генного продукта. Иллюстративные линии клеток-хозяев включают, без ограничения, DG44 и DUXB11 (линии клеток яичника китайского хомячка, DHFR-отрицательные), HELA (клетки карциномы шейки матки человека), CVI (линию клеток почки обезьяны), COS (производную CVI с T-антигеном SV40), R1610 (фибробласты китайского хомячка), BALBC/3T3 (фибробласты мыши), HAK (линию клеток почки хомяка), SP2/O (клетки миеломы мыши), BFA-1c1BPT (бычьи эндотелиальные клетки), RAJI (лимфоциты человека), 293 (клетки почки человека). В одном варианте осуществления линия клеток предусматривает измененное гликозилирование, например, афукозилирование, антитела, экспрессируемого в ней (например, линии клеток PER.C6.RTM. (Crucell) или FUT8-нокаутированные клеточные линии CHO (клетки POTELLIGENTT ) (Biowa, Princeton, Нью Джерси)). В одном варианте осуществления можно применять клетки NS0. Линии клеток-хозяев обычно доступны от коммерческих служб, Американской коллекции типовых культур или из опубликованных литературных источников.
Продуцирование in vitro позволяет увеличить масштаб для получения больших количеств требуемых связывающих полипептидов. Методики культивирования клеток млекопитающих в условиях культуры ткани известны из уровня техники и включают культивирование в однородной суспензии, например, в аэролифтном реакторе или в реакторе с непрерывным перемешиванием, или культивирование иммобилизованных или захваченных клеток, например, в полых волокнах, микрокапсулах, на микрогранулах агарозы или в керамических картриджах. При необходимости и/или по желанию растворы полипептидов можно очищать с помощью традиционных хроматографических способов, например, гель-фильтрации, ионообменной хроматографии, хроматографии на DEAE-целлюлозе и/или (иммуно-)аффинной хроматографии.
Один или несколько генов, кодирующих связывающие полипептиды, также можно экспрессировать в клетках, отличных от клеток млекопитающих, таких как бактерии, дрожжи или клетки растений. В этом отношении будет понятно, что различные одноклеточные микроорганизмы, отличные от млекопитающих, такие как бактерии, т.е. способные к выращиванию в культурах или брожению, также можно трансформировать. Бактерии, подверженные трансформации, включают представителей Enterobacteriaceae, таких как штаммы Escherichia coli или Salmonella; Bacillaceae, таких как Bacillus subtilis; Pneumococcus; Streptococcus и Haemophilus influenzae. Дополнительно будет понятно, что при экспрессии у бактерий полипептиды могут стать частью телец-включений. Полипептиды должны быть выделены, очищены и затем собраны в функциональные молекулы.
В дополнение к прокариотам, также можно использовать эукариотические микроорганизмы. Saccharomyces cerevisiae, или обыкновенные пекарские дрожжи, являются наиболее широко используемыми среди эукариотических микроорганизмов, хотя ряд других штаммов является общедоступным. Для экспрессии у Saccharomyces обычно применяют, например, плазмиду YRp7 (Stinchcomb et al., Nature, 282:39 (1979); Kingsman et al., Gene, 7:141 (1979); Tschemper et al., Gene, 10:157 (1980)). Эта плазмида уже содержит ген TRP1, представляющий собой маркер отбора для мутантного штамма дрожжей, у которого отсутствует способность к росту на триптофане, например, ATCC № 44076 или PEP4-1 (Jones, Genetics, 85:12 (1977)). Наличие повреждения TRP1 в качестве характеристики генома дрожжевой клетки-хозяина в этом случае обеспечивает эффективную среду для обнаружения трансформации по росту в отсутствие триптофана.
Способы лечения
В одном аспекте изобретение относится к способам лечения или диагностики нуждающегося в этом пациента, включающим введение эффективного количества связывающего полипептида, раскрываемого в данном документе. В определенных вариантах осуществления в настоящем раскрытии предложены наборы и способы для диагностики и/или лечения расстройств, например, опухолевых новообразований у млекопитающего, нуждающегося в таком лечении. В некоторых иллюстративных вариантах осуществления субъект является человеком.
Связывающие полипептиды настоящего изобретения полезны для ряда различных применений. Например, в одном варианте осуществления связывающие полипептиды настоящего изобретения применимы для уменьшения или устранения клеток, несущих в себе эпитоп, распознаваемый связывающим доменом связывающего полипептида. В другом варианте осуществления упомянутые связывающие полипептиды эффективны для снижения концентрации или удаления растворимого антигена из кровотока. В другом варианте осуществления связывающие полипептиды данного изобретения эффективны в качестве активаторов Т-клеток. В одном варианте осуществления связывающие полипептиды могут уменьшать размер опухоли, подавлять рост опухоли и/или продлевать время выживания животных, несущих в себе эту опухоль. Соответственно, это изобретение также относится к способу лечения опухолей у человека или другого животного путем введения такому человеку или животному эффективного, нетоксичного количества модифицированного антитела.
В одном варианте осуществления связывающие полипептиды данного изобретения применимы для лечения заболевания или расстройства. Например, связывающие полипептиды данного изобретения применимы для лечения расстройства, связанного с антителом, или расстройства, состояния или заболевания, чувствительного к антителам. Используемые в настоящем документе термины «расстройство, связанное с антителами» или «расстройство, реагирующее на антитела», или «состояние» или «заболевание» относятся или описывают заболевание или расстройство, которое может быть ослаблено введением фармацевтического состава, содержащего антитело или связывающий полипептид настоящего изобретения.
В одном варианте осуществления связывающие полипептиды данного изобретения полезны для лечения рака. Используемые здесь термины «онкологическое заболевание», «рак», «онкологический» или «раковый» относятся или описывают физиологическое состояние, которое обычно характеризуется нерегулируемым ростом клеток. Примеры рака включают, но не ограничиваются ими, карциному, лимфому, бластому, саркому (включая липосаркому), нейроэндокринные опухоли, мезотелиому, шванному, менингиому, аденокарциному, меланому и лейкемию или лимфоидные злокачественные новообразования. Более конкретные примеры таких видов рака включают плоскоклеточный рак (например, плоскоклеточный рак эпителия), рак легкого, включая мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого и плоскоклеточный рак легкого, рак брюшины, гепатоцеллюлярный рак, рак ЖКТ или желудка, включая рак желудочно-кишечного тракта, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, гепатому, рак молочной железы, рак толстой кишки, рак прямой кишки, колоректальный рак, рак эндометрия или матки, рак слюнной железы, рак почки, рак простаты, рак влагалища, рак щитовидной железы, рак печени, рак анального канала, рак полового члена, рак яичка, рак пищевода, опухоли желчных путей, а также рак головы и шеи.
В другом варианте осуществления изобретения связывающие полипептиды полезны для лечения других расстройств, включая, без ограничения, инфекционные заболевания, аутоиммунные расстройства, воспалительные расстройства, заболевания легких, нейрональные или нейродегенеративные заболевания, заболевания печени, заболевания позвоночника, заболевания матки, депрессивные расстройства и т.п. Неограничивающие примеры инфекционных заболеваний включают заболевания, вызываемые РНК-вирусами (например, ортомиксовирусами (например, вирусом гриппа), парамиксовирусами (например, респираторно-синцитиальным вирусом, вирусом парагриппа, метапневмовирусом), рабдовирусами (например, вирусом бешенства), коронавирусами, альфа-вирусами (например, вирусом чикунгуньи), лентивирусами (например, ВИЧ) и т.п.) или ДНК-вирусами. Примеры инфекционных заболеваний также включают, без ограничения, бактериальные инфекционные заболевания, вызываемые, например, Staphylococcus aureus, Staphylococcus epidermidis, Enterococcus, Streptococcus, Escherichia coli, и другие инфекционные заболевания, включая, например, вызываемые Candida albicans. Другие инфекционные заболевания включают, без ограничения, малярию, атипичную пневмонию, желтую лихорадку, боррелиоз Лайма, лейшманиоз, сибирскую язву и менингит. Типичные аутоиммунные расстройства включают, но не ограничиваются ими, псориаз, ревматоидный артрит, синдром Шегрена, отторжение трансплантата, болезнь Грейвса, миастению и волчанку (например, системную красную волчанку). Соответственно, это изобретение относится к способу лечения различных состояний, на которые использование связывающего полипептида данного изобретения имело бы благоприятное воздействие, например, повышался бы период полувыведения.
Специалист в данной области сможет путем обычных экспериментов определить, какое эффективное нетоксичное количество модифицированного связывающего полипептида может быть использовано для лечения злокачественных новообразований. Например, терапевтически активное количество связывающего полипептида настоящего изобретения может варьироваться в зависимости от таких факторов, как стадия заболевания (например, стадия I в сравнении со стадией IV), возраст, пол, медицинские осложнения (например, иммунодепрессивные состояния или заболевания) и вес тела субъекта, а также от способности антитела вызывать желаемый ответ у субъекта. Режим дозирования можно корректировать для получения оптимального терапевтического эффекта. Например, ежедневно можно вводить несколько разделенных доз, или дозу можно пропорционально уменьшить, как указывают требования терапевтической ситуации.
В общем, составы, предложенные в настоящем изобретении, можно использовать для профилактического или терапевтического лечения любого новообразования, содержащего антигенный маркер, с помощью который можно нацеливать модифицированное антитело на раковые клетки.
Фармацевтические составы и их применение
Способы получения и введения связывающих полипептидов настоящего изобретения субъекту хорошо известны, или специалисты в данной области могут их легко определить. Путь введения связывающих полипептидов настоящего изобретения может быть пероральным, парентеральным, ингаляционным или местным. Используемый в данном документе термин "парентеральный" включает внутривенное, внутриартериальное, внутрибрюшинное, внутримышечное, подкожное, ректальное или вагинальное введение. Хотя все эти формы введения явным образом находятся в пределах объема настоящего изобретения, формой для введения будет раствор для инъекций, в частности, для внутривенной или внутриартериальной инъекции или капельного вливания. Обычно подходящий фармацевтический состав для инъекций может содержать буфер (например, ацетатный, фосфатный или цитратный буфер), поверхностно-активное вещество (например, полисорбат), необязательно, стабилизирующий агент (например, человеческий альбумин) и т.д. В некоторых вариантах осуществления связывающие полипептиды могут быть доставлены непосредственно к месту неблагоприятной клеточной популяции, тем самым увеличивая воздействие на пораженную ткань терапевтического агента.
Препараты для парентерального введения включают стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъекционные органические сложные эфиры, такие как этилолеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, в том числе физиологический раствор и буферные среды. В композициях и способах настоящего изобретения фармацевтически приемлемые носители включают, но не ограничиваются ими, 0,01-0,1 М, например, 0,05 М фосфатный буфер или 0,8%-ный физиологический раствор. Другие распространенные среды-носители для парентерального введения включают растворы фосфата натрия, раствор Рингера с декстрозой, раствор декстрозы и хлорида натрия, лактатный раствор Рингера или нелетучие масла. Среды-носители для внутривенного введения включают растворы с добавками жидкости и питательных веществ, растворы с добавками электролитов, как, например, на основе раствора Рингера с декстрозой, и т. п. Также могут присутствовать консерванты и другие добавки, такие как, например, противомикробные средства, антиоксиданты, хелатообразователи и инертные газы и т.п. Более конкретно, фармацевтические составы, подходящие для инъекционного применения, включают стерильные водные растворы (если они являются водорастворимыми) или дисперсии и стерильные порошки для приготовления стерильных инъекционных растворов или дисперсий для немедленного приема. В таких случаях состав должен быть стерильным и должен быть жидким в такой степени, чтобы сохранялась возможность для его легкого введения через шприц. Он должен быть стабильным в условиях производства и хранения и предпочтительно защищен от загрязняющего действия микроорганизмов, таких как бактерии и грибы. Среда-носитель может представлять собой растворитель или дисперсионную среду, содержащие, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Надлежащую текучесть можно поддерживать, например, путем применения покрытия, такого как лецитин, путем поддержания необходимого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ.
Нежелательное действие микроорганизмов можно предотвратить с помощью различных антибактериальных и противогрибковых средств, например, парабенов, хлорбутанола, фенола, аскорбиновой кислоты, тимерозала и т.п. Во многих случаях в композицию будут включены изотонические агенты, например сахара, полиспирты, такие как маннит, сорбит или хлорид натрия. Длительного всасывания инъекционных композиций можно добиться путем включения в композицию средства, задерживающего всасывание, например, моностеарата алюминия и желатина.
В любом случае стерильные инъекционные растворы можно получить путем включения действующей субстанции (например, модифицированного связывающего полипептида самого по себе или в комбинации с другими действующими началами) в требуемом количестве в соответствующем растворителе вместе с одним ингредиентом или их комбинацией, перечисленных в данном документе, в случае необходимости, с последующей стерилизующей фильтрацией. Как правило, дисперсии получают путем включения активного соединения в стерильное инертное вещество, которое содержит основную дисперсионную среду и другие необходимые ингредиенты из перечисленных выше. В случае стерильных порошков для получения стерильных инъекционных растворов иллюстративные способы получения включают вакуумную сушку и сублимационную сушку, в результате которых получают порошковую форму активного ингредиента с любым дополнительным требуемым ингредиентом из его раствора, предварительно прошедшего стерилизующую фильтрацию. Препараты для инъекций обрабатывают, ими наполняют контейнеры, такие как ампулы, пакеты, колбы, шприцы или флаконы, которые при этом герметично закрывают в асептических условиях согласно способам, известным из уровня техники. Кроме того, препараты могут быть упакованы и поступать в продажу в форме набора. Такие изделия, как правило, будут иметь этикетки или листки-вкладыши в упаковке, на которых указано, что соответствующие композиции применимы для лечения субъекта, страдающего от аутоиммунных или неопластических нарушений или предрасположенного к ним.
Эффективные дозы композиций по настоящему изобретению для лечения описанных выше состояний варьируются в зависимости от множества различных факторов, включающих способ введения, участок-мишень, физиологическое состояние пациента, того, является ли пациент человеком или животным, других вводимых лекарственных препаратов и того, является ли лечение профилактическим или терапевтическим. Обычно пациент является человеком, однако также можно лечить млекопитающих, отличных от человека, в том числе трансгенных млекопитающих. Лечебные дозы можно подобрать с помощью стандартных способов, известных специалистам в данной области, для оптимизации безопасности и эффективности.
Связывающие полипептиды настоящего изобретения можно вводить многократно. Интервалы между однократными дозами могут быть еженедельными, ежемесячными или ежегодными. Интервалы также могут быть нерегулярными и определяться по измерению уровней модифицированного связывающего полипептида или антигена в крови пациента. В некоторых способах дозировку корректируют так, чтобы достичь концентрации модифицированного связывающего полипептида в плазме крови примерно 1-1000 мкг/мл, а в некоторых способах - примерно 25-300 мкг/мл. Альтернативно, связывающие полипептиды можно вводить в виде рецептуры с замедленным высвобождением, в этом случае требуется менее частое введение. В случае антител доза и частота варьируются в зависимости от периода полувыведения антитела у пациента. Как правило, гуманизированные антитела имеют наиболее продолжительный период полувыведения, за ними следуют химерные антитела и антитела, отличные от человеческих.
Доза и частота введения могут варьироваться в зависимости от того, является ли лечение профилактическим или терапевтическим. При профилактическом применении составы, содержащие антитела настоящего изобретения или коктейль из них, вводят пациенту, который еще не находится в состоянии болезни, для повышения устойчивости пациента. Такое количество определяется как "профилактическая эффективная доза". При таком применении точные количества опять-таки зависят от состояния здоровья и общего иммунитета пациента, но обычно находятся в пределах от примерно 0,1 до примерно 25 мг на дозу, особенно от примерно 0,5 до примерно 2,5 мг на дозу. Относительно низкую дозу вводят с относительно длительными интервалами в течение длительного периода времени. Некоторые пациенты продолжают получать лечение до конца жизни. При терапевтических применениях иногда необходимо вводить относительно высокую дозу (например, от примерно 1 до 400 мг/кг антитела на дозу, при этом дозы от примерно 5 до 25 мг более широко применяются для радиоиммуноконъюгатов, а более высокие дозы - для молекул, конъюгированных с цитотоксическими лекарственными средствами) с относительно короткими интервалами до уменьшения или прекращения прогрессирования заболевания и, в конкретных вариантах осуществления - до тех пор, пока у пациента не будет наблюдаться частичное или полное облегчение симптомов заболевания. После этого пациенту можно производить введение в профилактическом режиме.
Связывающие полипептиды настоящего изобретения по выбору можно вводить в комбинации с другими средствами, эффективными при лечении ими расстройства или состояния, требующего лечения (например, профилактического или терапевтического). Эффективные однократные лечебные дозы (т.е. терапевтически эффективные количества) модифицированных антител, меченых 90Y, согласно настоящему изобретению находятся в пределах от примерно 5 до примерно 75 мКи, например, от примерно 10 до примерно 40 мКи. Эффективные однократные лечебные дозы, не разрушающие костный мозг, 131I-модифицированных антител находятся в пределах от примерно 5 до примерно 70 мКи и от примерно 5 до примерно 40 мКи. Эффективные однократные лечебные разрушающие дозы (т.е. после которых может потребоваться аутологичная трансплантация костного мозга) 131I-меченых антител находятся в пределах от примерно 30 до примерно 600 мКи, например от примерно 50 до менее чем примерно 500 мКи. В случае химерного антитела, в связи с более длительным периодом полувыведения из крови по сравнению с антителами мыши, эффективные одноразовые лечебные дозы, не разрушающие костный мозг, химерных антител, меченых йодом-131, находятся в пределах от примерно 5 до примерно 40 мКи, как, например, менее чем примерно 30 мКи. Критерии визуализации, например, для метки 111In, обычно составляют менее чем примерно 5 мКи.
Хотя связывающие полипептиды можно вводить, как описано выше, следует подчеркнуть, что в других вариантах осуществления связывающий полипептид можно вводить здоровым в других отношениях пациентам в качестве терапии первой линии. В таких вариантах осуществления связывающие полипептиды можно вводить пациентам, имеющим нормальный или средний краснокостномозговой резерв, и/или пациентам, которые еще не проходили и в данный момент не проходят лечение. В данном контексте введение модифицированных антител или иммуноадгезинов вместе или в сочетании с дополнительной терапией означает последовательное, одновременное, совпадающее по времени, сопровождающее или сопутствующее введение или применение терапии и раскрываемых антител. Специалистам в данной области будет понятно, что введение или применение различных компонентов схемы комбинированной терапии можно спланировать по времени так, чтобы повысить общую эффективность лечения.
Как обсуждалось ранее, связывающие полипептиды настоящего изобретения, иммуноадгезины или их рекомбинантные молекулы можно вводить в фармацевтически эффективном количестве для лечения заболеваний млекопитающих in vivo. В связи с этим будет понятно, что рецептуры раскрываемых связывающих полипептидов будут составлены так, чтобы облегчать введение и обеспечивать стабильность активного начала.
Фармацевтический состав в соответствии с настоящим изобретением может содержать фармацевтически приемлемый, нетоксичный, стерильный носитель, такой как физиологический раствор, нетоксичные буферы, консерванты и т.п. Для целей настоящего изобретения фармацевтически эффективное количество связывающего полипептида, иммуноадгезина или его рекомбинантной молекулы, конъюгированного или неконъюгированного с терапевтическим средством, означает количество, достаточное для достижения эффективного связывания с мишенью и для достижения благоприятного эффекта, например, для уменьшения тяжести симптомов заболевания или нарушения или для обнаружения вещества или клетки. В случае опухолевых клеток модифицированный связывающий полипептид может взаимодействовать с выбранными иммунореактивными антигенами на неопластических или иммунореактивных клетках и приводить к увеличению гибели этих клеток. Конечно, фармацевтические составы настоящего изобретения можно вводить в однократной или многократных дозах для обеспечения фармацевтически эффективного количества модифицированного связывающего полипептида.
В объеме настоящего изобретения, связывающие полипептиды данного изобретения можно вводить человеку или другому животному в соответствии с вышеупомянутыми способами лечения в количестве, достаточном для того, чтобы вызвать терапевтический или профилактический эффект. Связывающие полипептиды данного изобретения можно вводить такому человеку или другому животному в традиционной лекарственной форме, получаемой путем объединения антитела данного изобретения с традиционным фармацевтически приемлемым носителем или разбавителем согласно известным методикам. Специалисту в данной области будет понятно, что форма и тип фармацевтически приемлемого носителя или разбавителя определяются количеством активного ингредиента, с которым его следует объединить, путем введения и другими хорошо известными переменными факторами. Специалистам в данной области также должно быть понятно, что коктейль, содержащий один или несколько видов связывающих полипептидов, описываемых в настоящем изобретении, может оказаться особенно эффективным.
Содержание статей, патентов и патентных заявок, а также всех других документов и информации, доступной в электронном виде, упомянутых или цитируемых в настоящем документе, ссылкой в полном объеме включено в данный документ в той же степени, как если бы каждая отдельная публикация была специально и индивидуально указана для включения ссылкой. Заявители оставляют за собой право физически включать в эту заявку любые материалы и информацию из любых таких статей, патентов, патентных заявок или других физических и электронных документов.
Хотя настоящее изобретение описано со ссылкой на его конкретные варианты осуществления, специалистам в данной области техники должно быть понятно, что могут быть сделаны различные изменения и эквивалентные заменены без отклонения от истинной сущности и объема изобретения. Специалистам в данной области техники будет очевидно, что другие подходящие изменения и переделки описанных здесь способов могут быть выполнены с использованием подходящих эквивалентов, не выходя за рамки раскрытых здесь вариантов осуществления. Кроме того, может быть сделано много изменений для адаптации конкретных ситуации, материала, состава вещества, процесса, стадии или стадий процесса к объекту, сущности и объему настоящего изобретения. Предполагается, что все такие изменения находятся в пределах объема прилагаемой формулы изобретения. Теперь, после подробного описания некоторых вариантов осуществления, они будут более понятны при рассмотрении следующих примеров, которые включены только в целях иллюстрации и не предназначены для ограничения.
ПРИМЕРЫ
Настоящее изобретение дополнительно иллюстрируют следующие примеры, которые не должны рассматриваться как дополнительное ограничение.
Пример 1: Материалы и способы
Белковые реагенты:
Следующие белки были экспрессированы и выделены: антиген с С-концевой меткой 8xHistidine; rFcRn (UniProt: P1359, субъединица p51: остатки 23-298; UniProt: P07151, β2-m: остатки 21-119); биотинилированный FcRn от cynomolgus (UniProt: Q8SPV9, субъединица p51: остатки 24-297 с С-концевой меткой Avi; UniProt: Q8SPW0, β2-m: остатки 21-119); биотинилированный hFcRn (UniProt: P55899, субъединица p51: остатки 24-297 с С-концевой меткой Avi; UniProt: P61769, β2-m: остатки 21-119); человеческий CD16a (UniProt: P08637, FcγRIIIa: остатки 17-208 с С-концевой меткой HPC4 и валин в положении 158 (V158)). Варианты тяжелых цепей H435A и H310A/H435Q были получены из кондиционированной среды HEK293. Варианты mAb2 клонировали с помощью Evitria и очищали из суспензионной кондиционированной среды CHO K1, используя аффинные колонки mAbSelect SuRe (GE Healthcare), и буфер заменяли в фосфатно-солевом буфере (PBS) pH 7,4 для последующих экспериментов.
Построение библиотеки насыщения:
Тяжелые и легкие цепи WT IgG1 антитела mAb1 с лидерными последовательностями ДНК были включены в плазмиды экспрессии млекопитающих pBH6414 и pBH6368, соответственно, с использованием сайтов рестриктаз NcoI и HindIII. Библиотека насыщения была создана с использованием набора для сайт-направленного мутагенеза (Lightning Site Directed Mutagenesis Kit, Agilent) и NNK (N= A/C/G/T, K=G/T) и праймеров WWC (W=A/T) (IDT Technologies), чтобы вставить все возможные аминокислоты в следующих положениях: M252, I253, S254, T256, K288, T307, K322, E380, L432, N434 и Y436 (нумерация ЕС). Последовательности ДНК тяжелой цепи трех контрольных вариантов в каркасе mAb1, AAA (T307A/E380A/N434A), LS (M428L/N434S) и YTE (M252Y/S254T/T256E), были встроены в вектор pBH6414, производство LakePharma.
Комбинаторную библиотеку насыщения получали посредством сайт-направленного мутагенеза тяжелой цепи mAb1 с помощью набора для мутагенеза Q5 (Q5 Mutagenesis Kit, NEBiolabs) и праймеров T256D, T256E, T307Q, T307W, N434F и N434Y с матрицами WT и M252Y в реакции ПЦР. Включение мутаций в каркас Ab3 проводили с использованием набора для мутагенеза Q5 (Q5 Mutagenesis Kit, NEBiolabs) с праймерами M252Y, T256D, T307Q и T307W. Создание всех вариантов Fc было подтверждено с помощью Sanger Sequencing (Genewiz, Inc.).
Экспрессия и очистка рекомбинантных антител:
Для скрининга в кондиционированных средах ДНК, содержащую мутантную тяжелую цепь и легкую цепь дикого типа mAb1, трансфицировали в 1 мл клеток млекопитающих Expi293 (Invitrogen) для экспрессии в соответствии с инструкциями производителя. Клетки инкубировали при 37°С, 5% диоксида углерода и 80% влажности при встряхивании со скоростью 900 оборотов в минуту (об/мин) в 2-мл 96-луночном планшете (Greiner Bio-One) и плотно закрывали аэрированной мембраной. Кондиционированные среды собирали через пять дней после трансфекции и хранили при -80ºC до использования. Лидирующие варианты в каркасах mAb1 и Ab3 экспрессировали в масштабе 30 мл в 125-миллилитровых колбах с вентилируемыми крышками по 0,2 мкм (Corning). 125-миллилитровые колбы для культуры встряхивали при 125 об/мин в течение всей продолжительности экспрессии. Кондиционированную среду собирали через пять дней после трансфекции, отфильтровывали через 0,22 мкм конические фильтры по 50 мл (Corning) и хранили при 4ºC до очистки.
Выделение mAb1 и Ab3 проводили с использованием 1 мл колонок mAbSelect SuRe HiTrap (GE Healthcare). После стадии промывания, используя PBS pH 7,4 в размере десяти объемов колонки, антитела элюировали пятью объемами колонки 0,1 M лимонной кислоты pH 3,0 (Sigma) и нейтрализовали 0,5 мл 1 M основания трис pH 9,0 (Sigma). В элюированных антителах заменяли буфер на PBS pH 7,4 и концентрировали до > 1 мг мл-1, используя 30 кДа MWCO Amicon Concentrators (Millipore) для последующих исследований. Концентрацию очищенных антител определяли по их УФ-поглощению при 280 нм (УФ280) с соответствующим коэффициентом экстинкции.
Отбор и анализ кондиционированных сред Octet:
Скрининг кондиционированной среды, содержащей варианты mAb1, проводили на Octet QK 384 с Ni-NTA биосенсорами (PALL Life Sciences). Антиген с меткой His выдерживали при концентрации 15 мкг/мл в течение 300 сек в PBS, 0,1% бычьего сывороточного альбумина (BSA, Sigma) и 0,01% Tween-20 (Sigma), рН 7,4 (PBST-BSA 7,4), а затем 20 сек промывали в PBST-BSA pH 7,4. Антитела выдерживали в течение 200 сек в кондиционированной среде, разбавленной 1:1 PBST-BSA, pH 7,4. После стадий промывки буфером с pH 6,0 кинетику связывания с FcRn получали с использованием 200 нМ rFcRn для времени ассоциации и диссоциации, равного 150 и 200 сек, соответственно, при pH 6,0. На всех стадиях во время скрининга Octet температура была 30ºC при скорости встряхивания 1000 об/мин. Кинетические профили связывания rFcRn скорректировали до начала фазы ассоциации FcRn и смоделировали в соответствии с моделью связывания 1:1, используя программное обеспечение для анализа Octet Analysis Software 7.1.
Кинетика связывания с FcRn:
Кинетику связывания FcRn при рН 6,0 и рН 7,4 измеряли на приборе Biacore T200 (GE Healthcare) с использованием измененных протоколов либо с прямой иммобилизацией FcRn, либо с набором для улавливания биотином CAPture (GE Healthcare) (см., например, Abdiche et al., MAbs (2015) 7: 331-343; Karlsson et al., Anal. Biochem. (2016) 502: 53-63). Для прямой иммобилизации биотинилированный FcRn при концентрации 20 мкг/мл иммобилизовали в течение 180 сек при 10 мкл/мин в 10 мМ ацетате натрия pH 4,5 (GE Healthcare) до ~20 RU на поверхности сенсорного чипа C1 с помощью реакции сочетания по аминогруппе (GE Healthcare). При использовании набора с биотином CAPture реагент CAPture был захвачен на поверхности чипа CAP до RU связывания >2000 RU, с последующим добавлением 0,1 мкг/мл FcRn в соответствующих каналах в течение 24 сек при 30 мкл/мин до окончательного связывания RU на уровне ~2 RU. Рабочий буфер для экспериментов по кинетике связывания FcRn представлял собой PBS с 0,05% ПАВ P-20 (PBS-P +, GE Healthcare) при pH 6,0 или 7,4. Серии концентраций 4-кратного серийного разведения, начиная с 1000 нМ антитела, проводили в четырех повторностях для каждого варианта, включая 0 нМ контроль. Кинетические измерения получали для времен ассоциации и диссоциации 180 и 300 сек, соответственно, при расходе 10 мкл/мин. Сенсорные чипы C1 и CAP регенерировали с помощью 10 мМ тетрабората натрия, 1 М NaCl, рН 8,5 (GE Healthcare) в течение 30 сек при 50 мкл/мин или 6 М гидрохлорида гуанидина, 250 мМ гидроксида натрия (GE Healthcare) в течение 120 сек при 50 мкл/мин, соответственно, с последующей дополнительной стадией стабилизации в течение 60-90 сек в PBS-P+ pH 6,0. Измерения RU в устойчивом состоянии при pH 7,4 получали для всех вариантов при 1000 нМ в трех повторностях с использованием того же сенсорного чипа C1 или CAP и кинетических параметров, как описано выше, за исключением того, что уровень захвата FcRn был увеличен в 10-20 раз для обоих методов.
Кинетические параметры для серии концентраций при рН 6,0 были согласованы с двухвалентной моделью с использованием оценочного программного обеспечения Biacore T200 за счет эффекта авидности. См., например, Suzuki et al., J. Immunol. (2010) 184: 1968-1976. Каждую серию разведений подбирали независимо, чтобы получить средние значения скоростей связывания и диссоциации и способности связывания. Наблюдаемую способность связывания рассчитывали по первым скоростям связывания и диссоциации, полученным из двухвалентной модели. Остаточное связывание при рН 7,4 измеряли, используя 1000 нМ каждого антитела в трех экземплярах одновременно для сравнения ответа. Ответ в устойчивом состоянии в каждой повторности усредняли для получения среднего значения и стандартного отклонения.
Аффинная хроматография FcRn:
В одном эксперименте аффинную колонку для FcRn выполняли согласно протоколам, адаптированным из Schlothauer et al. 2013, mAbs 5: 576-586. Колонку Streptavidin HP HiTrap объемом 1 мл (GE Healthcare) уравновешивали буфером для связывания (20 мМ фосфат натрия (Sigma) pH 7,4, 150 мМ хлорид натрия (NaCl; Sigma)) при 1 мл/мин в размере пяти объемов колонки с последующим впрыском 4 миллиграммов биотинилированного cynoFcRn. Колонку промывали буфером для связывания и хранили при 4ºC до использования.
Аффинную колонку для FcRn уравновешивали буфером с низким рН (20 мМ 2-(N-морфолино)этансульфокислота (MES; Sigma), рН 5,5; 150 мМ NaCl) в размере пяти объемов колонки перед впрыском 300 мкг каждого антитела. рН растворов антител доводили до 5,5 с помощью буфера с низким рН. После промывки в размере десяти объемов колонки буфером с низким pH антитела элюировали линейным градиентом pH с помощью буфера с высоким pH (20 мМ 1,3-бис (трис(гидроксиметил)метиламино)пропан (бис трис пропан; Sigma) pH 9,5; 150 мМ NaCl) в размере 30 объемов колонки при 1 мл/мин фракциями по 1 мл, осуществляя наблюдение с помощью УФ280. Аффинную колонку для FcRn повторно уравновешивали буфером с низким pH в размере десяти объемов колонки для последующих прогонов или буфером для связывания случае подготовки к хранению. Все варианты были выполнены в трех повторностях.
Профиль элюирования аффинной колонки с FcRn для каждого варианта был смоделирован по одному Гауссовому распределению, используя уравнение 1 в Sigmaplot 11 (Systat Software, Inc.) для определения объема элюирования на максимуме УФ280.
 (Уравнение 1)
(Уравнение 1)
где x0 - объем элюирования на максимуме пика UV280, y0 - исходный уровень абсорбции UV280, a и b относятся к полной ширине на половине максимума распределения. рH каждой фракции измеряли с помощью pH-метра Corning Pinnacle 540 и соотносили с объемом элюирования, используя линейную регрессию.
В другом эксперименте аффинную колонку для FcRn приспосабливали из Schlothauer et al. 2013, mAbs 5: 576-586 с биотинилированным hFcRn на колонке с 1 мл Streptavidin HP HiTrap (GE Healthcare). В колонку впрыскивали 300 мкг каждого антитела в буфере с низким pH (20 мМ 2-(N-морфолино)этансульфокислота (MES; Sigma) pH 5,5; 150 мМ NaCl) в системе AKTA Pure (AKTA). Антитела элюировали с помощью линейного градиента рН, создаваемого буфером с низким и высоким рН (20 мМ 1,3-бис(трис(гидроксиметил метиламино)пропан (бис трис пропан; Sigma), рН 9,5; 150 мМ NaCl) в размере 30 объемов колонки при 0,5 мл/мин, наблюдая за поглощением и рН. Колонку повторно уравновешивали буфером с низким рН для последующих опытов. Все варианты были выполнены в трех повторностях. Профили элюирования аффинной колонки с FcRn подгоняли к одному Гауссовому распределению в Sigmaplot 11 (Systat Software, Inc.) для определения объема элюирования и рН на максимуме УФ280.
Дифференциальная сканирующая флуориметрия:
Эксперименты по дифференциальной сканирующей флуориметрии (DSF) проводили на системном термоциклере BioRad CFX96 в реальном времени (BioRad) для реакций объемом 20 мкл. Образцы антител и 5000-кратный запас красителя Sypro Orange (Invitrogen) разводили до 0,4 мг/мл и 10x, соответственно, в PBS pH 7,4. Антитела и Sypro Orange смешивали в соотношении 1:1 в 96-луночных планшетах для ПЦР и запечатывали с помощью липкого microseal (BioRad) до конечной концентрации 0,2 мг/мл каждого антитела и 5x красителя Sypro Orange. Все варианты антител были выполнены в трех повторностях. Программа термоциклирования состояла из 2-минутного стадии уравновешивания при 20ºC с последующей постоянной скоростью изменения температуры 0,5ºC/5 сек до конечной температуры 100ºC. Измерения флуоресценции каждой лунки были получены с использованием детекторов длины волны с возбуждением FAM (485 нм) и эмиссиией ROX (625 нм), подходящих для флуоресценции Sypro Orange (см., например, Biggar et al. 2012, Biotechniques 53: 231-238). Профиль интенсивности флуоресценции DSF и первая производная были взяты из BioRad CFX Manager и проанализированы в Sigmaplot 11. Tпл. определяли как середину первого перехода в профиле интенсивности флуоресценции.
Кинетика связывания с FcγRIIIa:
Кинетику и способность связывания измеряли на приборе Biacore T200 (GE Healthcare) (Zhou et al., 2008 Biotechnol. Bioeng. 99. 652-665). Антитело к HPC4 (Roche), при 50 мкг/мл в ацетате, рН 4,5, связывали с поверхностью сенсорного чипа CM5 в течение 600 сек при 10 мкл/мин реакцией по аминогруппе до конечной плотности >20000 RU. Рабочий буфер для экспериментов по кинетике связывания с FcγRIIIa представлял собой забуференный физиологический раствор HEPES с 0,05% ПАВ P-20 (HBS-P+, GE Healthcare) и 2 мМ хлорида кальция (CaCl2, Fluka) при pH 7,4. Каждый кинетический след инициализировали захватом 1,25 мкг/мл FcγRIIIa-V158 с HPC4-меткой в течение 30 сек при 5 мкл/мин. Кинетику ассоциации и диссоциации в 300 нМ каждого варианта измеряли для каждого варианта в течение 120-180 сек для каждой стадии при 5 мкл/мин. После завершения кинетических измерений чип CM5 регенерировали буфером HBS-P+ с добавлением 10 мМ EDTA (Ambion). Перед следующими кинетическими измерениями чип CM5 промывали в течение 120 сек, используя HBS-P+ с CaCl2.
В одном эксперименте кинетические эксперименты FcγRIIIa анализировали аналогично тому, как описано для связывания с FcRn при pH 7,4. Для WT, сравнительного, лидирующих одинарного и комбинированного вариантов была получена кинетика в ряду 3-кратного серийного разведения от 1000 нМ для определения способности связывания с FcγRIIIa. RU в устойчивом состоянии при каждой концентрации и повторности определяли, наносили на график как функцию концентрации антител и приводили в соответствие с моделью устойчивого состояния, как показано в уравнении 2.
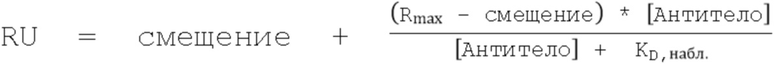 (Уравнение 2)
(Уравнение 2)
где смещение - это исходный уровень RU при 0 нМ антитела, Rmax - это плато RU при высоких концентрациях антитела, [Антитело] - концентрация антитела и KD,набл. - наблюдаемая способность связывания при взаимодействии между вариантами и FcγRIIIa.
В другом эксперименте кинетические эксперименты с FcγRIIIa анализировали аналогично тому, как описано для связывания с FcRn при рН 7,4, используя средний ответ на связывание в устойчивом состоянии. Для всех вариантов устойчивое состояние RU 300 нМ антитела определяли в трех повторностях и усредняли. Кратность изменения изменения ответа относительно WT (кратность изменения ответа) определяли для сравнения вариантов в каждом каркасе.
Изоэлектрическая фокусировка
Изоэлектрическую точку (pI) лидирующих вариантов определяли с помощью капиллярного электрофореза на приборе Maurice С (Protein Simple). Каждый образец объемом 200 мкл содержал 0,35% метилцеллюлозы (Protein Simple), 4% фармалита 3-10 (GE Healthcare), 10 мМ аргинина (Protein Simple), 0,2 мг/мл антитела и 4,05 и 9,99 pI маркеров (Protein Simple). Образец загружали в капилляр в течение 1 мин при 1500 В с последующей фазой разделения в течение 6 мин при 3000 В и контролировали по флуоресценции триптофана. рI для каждого варианта определяли с использованием программного обеспечения Maurice C и определяли как pH при максимуме флуоресценции для основных видов.
ИФА с гомогенным мостиковым ревматоидным фактором (RF)
Антитела биотинилировали и метили дигоксигеном, используя наборы EZ-Link для мечения Sulfo-NHS-LC-Biotin и Mix-n-Stain™ Digoxigenin Antibody Labeling (Biotium) в соответствии с инструкциями производителя. Исходный раствор, содержащий 4 мкг/мл биотинилированных и дигоксигенин-меченых антител, готовили для каждого варианта и смешивали в соотношении 1:1 с 300 ед/мл RF (Abcam). После инкубировании при комнатной температуре в течение 20 часов 100 мкл каждой смеси антитело-RF добавляли в планшеты Streptawell (Sigma-Aldrich) и инкубировали при комнатной температуре в течение 2 часов. Планшет трижды промывали PBS, pH 7,4, с 0,05% Tween-20 и 100 мкл разведенного 1:2000 HRP-конъюгированного вторичного антитела к дигоксигенину (Abcam) добавляли в каждую лунку. После 2-часового инкубирования при комнатной температуре лунки промывали и обрабатывали 100 мкл субстрата TMB (Abcam) в течение 15 минут при комнатной температуре. Реакцию останавливали с помощью 100 мкл стоп-раствора (Abcam) и измеряли оптическую плотность при 450 нм на считывающем устройстве для планшетов SpectraMax. Лунка, не содержащая смесь антитело-RF, обеспечивала вычитание холостых проб, и эксперимент повторяли три раза. P-значения определяли с помощью t-критерия Стьюдента.
Фармакокинетика in vivo
Фармакокинетические исследования проводили на обезьянах cynomolgus и трансгенных мышах hFcRn (штамм Tg32, Jackson Laboratory, Бар-Харбор, Мэн). В исследованиях на обезьянах варианты с WT, LS, DQ, DW и YD в каркасе mAb2 вводили в виде однократной внутривенной дозы 2,5 мг/кг в брахиоцефальную вену при объеме дозы 1,5 мл/кг трем самцам cynomolgus, не подвергавшимся лечению. Пробы крови (0,5 мл) отбирали венопункцией подкожной вены, отбирая пробы 8 раз после введения дозы на: 0,0035, 0,17, 1, 3, 7, 14, 21 и 28 дни. После отбора проб крови центрифугировали при 4°С в течение 10 минут при 1500 g и хранили при -80°С.
Мышам hFcRn варианты антител вводили в виде однократной внутривенной дозы 2,5 мг/кг в хвостовую вену при объеме дозы 5 мл/кг. В каждый из моментов времени отбирали 20 мкл крови из подкожной вены, используя предварительно заполненные гепарином капилляры. Собранные пробы крови переносили в микропробирки и центрифугировали при 1500 g в течение 10 минут при 4°C. Образцы плазмы собирали, объединяли для каждой временной точки (по 6 мышей на образец) и хранили при -80°C до анализа.
Все исследования in vivo проводили в соответствии с политикой Sanofi по институциональному содержанию животных. Исследования на обезьянах и мышах были одобрены французским «Ministère de l'Enseignement Superieur et de la Recherche» и немецким «Regierungspraesidium Darmstadt».
Концентрацию каждого варианта mAb2 в каждый момент времени определяли с помощью восходящего анализа ЖХ-МС/МС. После осаждения аликвоты плазмы осадок плазмы подвергали денатурации, восстановлению, алкилированию, расщеплению трипсином и твердофазной экстракции белка перед анализом суррогатных пептидов. Калибровочные стандарты готовили путем вливания варианта mAb2 в плазму при 1,00, 2,00, 5,00, 10,0, 20,0, 50,0, 100, 200 и 400 мкг/мл. Разделение пептидов проводили на системе Waters Acquity UPLC с обращенно-фазовой колонкой XBridge BEH C18 (2,1×150 мм, 3,5 мкМ, 300Å, Waters) с расходом 300 мкл/мин со ступенчатым градиентом 0,1% муравьиной кислоты в воде и 0,1% муравьиной кислоты в ацетонитриле. Для обнаружения использовали масс-спектрометр Sciex API5500 в режиме положительных ионов с температурой источника 700°С, напряжением ионного распыления 5500 В, газами занавесы и распылителя при 40°С и газом столкновения в середине. Время выдержки составляло 20 мсек, а входной потенциал составлял 10 В для каждого перехода. Для определения концентрации относительно стандартов и контрольных образцов использовали преобразования мониторинга множественных реакций для двух уникальных суррогатных пептидов каркаса mAb2 с использованием площади пика из алгоритма интеграции MQIII программного обеспечения Analyst. Скорость клиренса и период полувыведения из сыворотки крови получали из некомпартментной модели концентрации антител как функции времени с использованием программного обеспечения Phoenix Software (Certara). Все моменты времени, когда наблюдалось резкое снижение концентрации, были исключены из средней концентрации в плазме из-за предположения, не связанного с какой-либо теорией, о распределении лекарственного средства, обусловленного мишенью (target-mediated drug disposition, TMDD), и/или взаимодействии антител с лекарственным средством (anti-drug antibody, ADA).
Пример 2: Скрининг Octet насыщающих точечных мутаций в кондиционированных средах
FcRn представляет собой гетеродимер α-домена и субъединицы β2-макроглобулина (β2-m) MHC класса I (Фигура 1А), общий для большинства рецепторов Fc, который распознает участки тяжелой цепи антитела Fc, отличные из других FcγR (см., например, Oganesyan et al. 2014 J. Biol. Chem. 289: 7812-7824; и Shields et al. 2001 г., выше).
Чтобы идентифицировать варианты с более медленными скоростями диссоциации с FcRn, чем у антитела WT, был разработан анализ на основе бислойной интерферометрии (BLI) для скрининга вариантов антител в кондиционированных средах с высокой пропускной способностью (Фигура 2А). Этот анализ был разработан с использованием нескольких вариантов сравнения, которые усиливают (AAA, LS и YTE) или уменьшают (H435A, H310A/H435Q) сродство к FcRn при pH 6,0 по сравнению с антителом WT. Биосенсоры NiNTA улавливали антиген с his-меткой, а затем для каждого варианта антитела при рН 7,4 имитировали кондиционированную среду (Фигура 2А). Кинетику связывания с крысиным FcRn (rFcRn) при рН 6,0, скорость диссоциации которого в ~25 раз более медленная по сравнению с человеческим IgG1, и который легче исследовать методом Octet, чем человеческий FcRn (hFcRn), измеряли для каждого из шести вариантов (Фигура 2B). Варианты H435A (Фигура 2B, длинные штрихи с вкраплениями одной точки) и H310A/H435Q (Фигура 2B, длинные штрихи с двумя точками) демонстрируют практически полное отсутствие какой-либо кинетики связывания с FcRn (также см., например, Shields et al. 2001, выше; Medesan et al., 1997, выше, и Raghavan et al., 1995, выше). Варианты AAA (Фигура 2B, короткие штрихи), LS (Фигура 2B, короткие штрихи с одной точкой) и YTE (Фигура 2B, длинные штрихи) демонстрируют более медленную кинетику диссоциации по сравнению с WT (Фигура 2B, сплошная линия) со снижением скорости диссоциации с FcRn в 2-7,3 раза. Это продемонстрировало то, что скрининг Octet подходит для распознавания вариантов с использованием нарушенной кинетики диссоциации с rFcRn.
Антитело IgG1 - mAb1, служило модельной системой для создания библиотеки насыщающего мутагенеза для скрининга мутантов с пониженной скоростью диссоциации с FcRn. Одиннадцать позиций в Fc-участке mAb1 были выбраны на основе их близости или прямого участии в области контакта с FcRn (Фигуры 1A и 1B) (см., например, Oganesyan et al. 2014, выше; и Shields et al. 2001, выше). Все точечные мутации в этих положениях были сконструированы с использованием сайт-направленного мутагенеза и трансфицированы в клетки Expi293 для экспрессии. Проводили скрининг кондиционированных сред для библиотеки насыщающего мутагенеза, как описано выше. Нормализованные FcRn-связывающие сенсограммы Octet для подмножества вариантов показаны на Фигуре 2C (длинные штрихи) с диким типом (Фигура 2C, толстые длинные штрихи) и имитационным отрицательным контролем (Фигура 2C, пунктирная линия). Имитационный контроль показал отсутствие заметного связывания с FcRn. Несколько мутантов явно нарушали связывание с rFcRn, так как в кинетических профилях не наблюдалось практически никакого изменения сигнала (Фигура 2C, длинные штрихи, расположенные ниже пунктирной линии (имитационный контроль)). Граница для вариантов с улучшенной скоростью диссоциации с FcRn была определена как три стандартных отклонения ниже среднего значения для антитела WT. В подмножестве мутаций, показанных на Фигуре 2C, две (Фигура 2C, сплошные линии) имели значительно сниженные скорости диссоциации по сравнению с антителами дикого типа (Фигура 2C, толстые длинные штрихи), тогда как остальные варианты имели значения, сходные (Фигура 2C, короткие штрихи с одной точкой) или быстрее (Фигура 2C, длинные штрихи выше пунктирной линии (имитационный контроль)) скорости диссоциации с rFcRn.
Скорости диссоциации с rFcRn для всех одноточечных мутаций показаны на Фигуре 2D и Фигуре 14 по позициям и мутациям. На Фигуре 14 данные отсортированы по четырем категориям в зависимости от кратности изменения скорости диссоциации с rFcRn по сравнению с диким типом, а типы дикого типа обозначены черными квадратами.
На Фигуре 14 кратное изменение скорости диссоциации с rFcRn для всех возможных замен в одиннадцати положениях библиотеки насыщения было нормализовано по среднему значению для антитела WT и кодировано по цвету. Все мутанты подпадают под одну из четырех категорий: практически не связывающийся (темно-серым), более высокая скорость диссоциации с rFcRn (серым), WT-подобная скорость диссоциации с rFcRn (горизонтальными линиями) и более медленная скорость диссоциации с rFcRn (сеткой). Множественные варианты обладали более медленными скоростями диссоциации с rFcRn, чем антитела WT (сеткой).
Мутанты, обозначенные темно-серым на Фигуре 14, практически не связывались с rFcRn так же, как и имитационный контроль (Фигура 2C, пунктирная линия), и локализованы в петле M252, I253 и S254. Единственными мутациями в I253 были метионин и валин, и обе значительно увеличивали скорость диссоциации с rFcRn, дополнительно подтверждая важность I253 для взаимодействия с FcRn. Еще 120 вариантов (Фигура 2D и Фигура 14, светло-серые прямоугольники) дестабилизируют взаимодействие с rFcRn, причем примерно 50% из них находятся в каждом из доменов CH2 и CH3. Двадцать пять мутантов имеют сходную с WT скорость диссоциации (Фигура 2D и Фигура 14, белые прямоугольники), причем восемь из 11 позиций имеют по крайней мере одну WT-подобную мутацию (Фигура 14, белые прямоугольники). Следующие мутации имели значительно сниженную скорость диссоциации с rFcRn по сравнению с диким типом (Фигура 2D и Фигура 14, черные прямоугольники): M252Y, T256D/E, K288D/N, T307A/E/F/M/Q/W, E380C, N434F/P/Y и Y436H/N/W. Мутации M252Y, N434F и N434Y имели скорость диссоциации более чем в два раза медленнее, чем у антитела WT (Фигура 2D). Эти мутации экспрессировали и очищали с помощью хроматографии на белке А для дальнейшего изучения кинетически с FcRn in vitro.
Пример 3: Кинетика связывания Biacore с FcRn при рН 6,0
Варианты AAA, LS и YTE служили в качестве положительного контроля при измерениях кинетики связывания с FcRn с использованием Biacore с FcRn и человека, и крысы при pH 6,0. Зависимое от концентрации связывание с FcRn наблюдали для всех вариантов, включая дикий тип, образец сравнения (Фигура 3) и лидирующие (Фигуры 4А и 4В), а профиль связывания одной инъекции с FcRn человека и крысы показан на Фигурах 5А и 5B, соответственно. Антитело дикого типа имело способность связывания с FcRn человека и крысы, равную 2380 ± 470 нМ и 207 ± 43 нМ, соответственно (таблица 1).
Таблица 1. Характеристические параметры, полученные in vitro для очищенных лидирующих антител к mAb1.
pH 6,0
Скорость диссоциации
Показатель pH
Все данные, приведенные в таблице 1, были получены экспериментальными методами, указанными в верхней части каждого столбца.
Скорость диссоциации с rFcRn по Octet с использованием очищенных белков измеряли в качестве сравнения с кинетическими константами, полученными при скрининге в кондиционированных средах. рH элюирования определяли с помощью аффинной хроматографии FcRn в трех повторностях (n=3) и с помощью DSF исследовали термостабильность в трех повторностях (n=3). Кинетики связывания с FcRn для FcRn человека и крысы были получены из Biacore с серийным разведением антител в двух повторностях (n=2) и подгоняли независимо. Ответ на связывание в устойчивом состоянии (RU) каждого варианта с FcRn человека и крысы при рН 7,4 измеряли с помощью 1000 нМ антитела в трех повторностях (n=3) с использованием Biacore. Единицы измерения для каждого измерения: Octet, рH 6,0, скорость диссоциации с rFcRn (x10-3 сек-1); рН элюирования (безразмерная величина); DSF Tпл. (°C); Biacore, pH 6,0, скорость связывания с hFcRn (х104 М-1 сек-1), скорость диссоциации (х10-1 сек-1) и KD,набл. (х109 М); Biacore, pH 6,0, скорость связывания с rFcRn (х104 М-1 сек-1), скорость диссоциации (х10-3 сек-1) и KD,набл. (х109 М); Biacore, pH 7.4, ответ на связывание в устойчивом состоянии (RU).
На Фиг. 5B варианты AAA (пунктиром), LS (штрихами с двумя точками) и YTE (штрихами с одной точкой) имели улучшенную в 1,6-10,4 раза способность связывания по сравнению с WT. Характер сравнительного варианта с самым сильным сродством к FcRn был видоспецифичным, поскольку LS обладал самым сильным сродством к hFcRn, в то время как rFcRn обладал более сильным сродством к YTE (таблица 2A).
Таблица 2A. Характеристические параметры, полученные in vitro для очищенных антител с двойной комбинацией для mAb1.
Подавляющее большинство лидирующих вариантов как для человеческого, так и для крысиного FcRn (Фигуры 5А и 5В, сплошные линии разных оттенков) имели значительно более низкие скорости связывания, чем WT или варианты сравнения (>2 раз) (таблица 1). Мутации N434F и N434Y были единственными вариантами, которые продемонстрировали повышенную скорость связывания у обоих видов FcRn. Не будучи связанными какой-либо теорией, в результате более медленной кинетики ассоциации с hFcRn, наблюдаемые способности связывания у лидирующих вариантов, как правило, были слабее, чем у WT, в отличие от rFcRn (Фигуры 5C и 5D, Таблица 1). Сродство к rFcRn было слабее, чем у YTE (Фигура 5D, диагональные линии обращены внизу слева, таблица 2A). Без связи с какой-либо теорией, эти результаты показали, что одной мутации недостаточно для повышения сродства, которое было бы лучше, чем у вариантов LS и YTE. Ранжирование скоростей диссоциации с FcRn (из-за слабой способности связывания вариантов с hFcRn) выявило подмножество со сниженными скоростями диссоциации с FcRn и человека, и крысы: M252Y, N434F/P/Y, T256D/E и T307A/E/F/Q/W (Таблица 2A). Эти варианты в комбинации представляют дополнительный интерес для дальнейшего улучшения способности связывания с FcRn в Fc-участка, чтобы превзойти варианты сравнения.
Характеризующие параметры in vitro для лидирующих вариантов приведены в таблице 2В.
Таблица 2B. Характеристики in vitro лидирующих вариантов.
V158
Все данные, приведенные в таблице 2B, были получены с использованием экспериментальных методик, указанных в верхней части каждого столбца. Аффинную хроматографию с FcRn, DSF и связывание с FcγRIIIa проводили трижды (n=3). Кинетику связывания FcRn с FcRn человека и крысы получали в четырех повторностях и подгоняли независимо. Единицы: DSF Tпл. (°C); связывание с FcγRIIIa (кратность изменения относительно WT), Biacore, pH 6,0, скорость связывания с hFcRn (х104 М-1 сек-1), скорость диссоциации (х10-1 сек-1) и KD,набл. (х109 М); Biacore, pH 6,0, скорость связывания с rFcRn KD,набл. (х109 М); Biacore, pH 7,4, hFcRn и rFcRn в устойчивом состоянии RU (RU).
Пример 4: Комбинированные варианты дополнительно снижают скорость диссоциации связывания с FcRn
Множественные лидирующие мутации были локализованы в одном положении (Фигура 14, черные прямоугольники), таком как T307 и N434, где были идентифицированы шесть и три мутации, соответственно, которые показали более медленную кинетику диссоциации с FcRn. Только мутации с самыми медленными скоростями диссоциации с FcRn к hFcRn в этих положениях были использованы для создания комбинированных вариантов. В этом случае T307Q, T307W, N434F и N434Y были смешаны с M252Y, T256D и T256E для получения двойных, тройных и четверных вариантов с использованием ПЦР со смешанным праймером и сайт-направленного мутагенеза. В целом, комбинаторная библиотека состояла из 54 вариантов, включая семь лидирующих одинарных, 18 двойных, 20 тройных, 8 четверных вариантов и антитело WT. Номенклатура этих вариантов следующая: фон дикого типа содержит M252, T256, T307 и N434 и обозначен как MTTN. Таким образом, тройной вариант - YTQY - содержит мутации M252Y, T307Q и N434Y , сохраняя при этом треонин WT в положении 256.
Как и в случае одинарных мутаций, кинетику связывания с FcRn при рН 6,0 с использованием Biacore использовали для определения того, какие комбинированные варианты имеют улучшенное сродство. Типичный кинетический след FcRn-связывания каждого из одиночного (длинные штрихи с двумя точками), двойного (длинные штрихи с одной точкой), тройного (длинные штрихи) и четверного (короткие штрихи) показан на Фигурах 6А и 6В в сравнении с WT (пунктирная линия) и вариантом сравнения с наиболее сильным сродством к соответствующим им видам FcRn (hFcRn: LS (длинные штрихи с двумя точками); rFcRn: YTE (сплошная линия)). Из скоростей связывания и диссоциации с hFcRn (Фигура 6C) видно, что два одинарных, 15 двойных, 18 тройных и восемь четверных вариантов имели повышенную способность связывания, чем вариант LS (Фигура 6C, пунктир). Таким же образом все комбинации, кроме одного тройного варианта, имели более сильное сродство к rFcRn, чем YTE (Фигура 6D, диагональные линии, направленные вниз влево). В случае hFcRn, дополнительные FcRn-усиливающие мутации дополнительно увеличивали способность связывания (Фигура 6C). Все пять комбинаций с наиболее сильным сродством к hFcRn представляли собой четверные варианты (Фигура 6С, заштриховано в клетку) со способностью связывания, примерно в 500 раз превышающими дикий тип. Подобное явление не происходило с rFcRn (Фигура 6D), так как варианты с наибольшим сродством были двойными вариантами (Фигура 6D, горизонтальные линии). Тройной (Фигура 6D, вертикальные линии) и четверной (Фигура 6D, заштриховано в клетку) варианты, как правило, демонстрировали лишь незначительное снижение скорости диссоциации (менее чем в 2 раза), а также проявляли сниженные скорости ассоциации (Фигура 6D). Без связи с какой-либо теорией, из этих результатов можно предположить, что возможно существует более низкий предел в отношении наблюдаемой способности связывания с FcRn (примерно 0,5 нМ), который был достигнут с rFcRn, но не с hFcRn (Фигура 6B). В целом, более 40 комбинированных вариантов имели более сильное сродство, чем варианты сравнения, и требуется дополнительное исследование для выбора комбинаций с благоприятными свойствами для исследований in vivo.
Пример 5: Комбинированные варианты сохраняют значительное связывание при физиологическом pH
В результате значительно улучшенного сродства к FcRn при рН 6,0 было исследовано влияние на зависимость от рН, используя аффинную хроматографию с FcRn и Biacore измерения стационарного состояния при рН 7,4. Аффинная хроматография для FcRn использует линейный градиент рН для прямого измерения нарушения рН-зависимости в результате мутаций. H435A и H310A/H435Q - варианты со слабым связыванием с FcRn - не связывались с колонкой независимо от pH (Фигура 8A). Элюирование WT происходит при примерно физиологическом pH (pH 7,37 ± 0,05), в то время как AAA, LS и YTE требуют более высокого pH (таблица 2B). Для всех комбинированных вариантов и семи лидирующих вариантов требовалось более высокое значение pH для элюирования из аффинной колонки, чем для WT (Фигуры 8A и 8C). Элюирование вариантов N434F/Y происходило при более высоком pH, чем LS (таблица 2B), что, без связи с научной теорией, указывает на то, что эти варианты, как по отдельности, так и в сочетании, нарушали зависимость от pH. Характерные хроматограммы показали отчетливый сдвиг в сторону элюирования при более высоком pH с увеличением числа мутаций (Фигуры 9A и 9B). Сильная корреляция (R2=0,94) между pH элюирования и скоростями диссоциации с hFcRn (Фигура 9C) указывает на то, что более медленная скорость диссоциации с FcRn при pH 6,0 напрямую способствовала повышению pH элюирования для вариантов FcRn.
Кинетические эксперименты по связыванию с FcRn проводили при pH 7,4 с использованием Biacore для измерения остаточной активности связывания в физиологических условиях. RU устойчивого состояния использовали в качестве меры остаточной способности связывания с FcRn, так как некоторые варианты демонстрировали ненадежную кинетику и практически не связывались при этом рН. Характерные кинетические следы одинарного (длинные штрихи с двумя точками), двойного (длинные штрихи с одной точкой), тройного (длинные штрихи) и четверного (короткие штрихи) вариантов показаны на Фигурах 7А и 7В в сравнении с LS (Фигура 7A, сплошным) и YTE (Фигура 7B, сплошным). Эти два варианта показали наибольшее остаточное связывание с FcRn человека и крысы при рН, 7,4 соответственно. Большинство лидирующих одиночных вариантов имели слегка повышенное связывание с FcRn по сравнению с WT (4,3 ± 1,0 RU), но менее, чем AAA (13,1 ± 1,7 RU), LS (18,5 ± 2,6 RU) и YTE (13,1 ± 1,6 RU), за исключением мутаций N434F/Y (таблицы 2А и 2В). Комбинированные варианты также обладали значительным остаточным связыванием с обоими видами FcRn при pH 7,4 (Фигуры 7A и 7B) в еще большей степени, чем N434F/Y. Не ограничиваясь какой-либо теорией, идеальным кандидатом для исследований in vivo являются варианты с повышенным связыванием с FcRn при низком pH (такие как варианты AAA, LS и YTE), но поддерживающие низкий уровень связывания при повышенном pH, аналогичным образом с WT. На графике, показанном на Фигурах 7C и 7D, эти комбинации будут занимать нижнюю левую четверть изображения, обозначенную сродством вариантов LS и YTE при каждом pH к FcRn человека и крысы, соответственно.
Пример 6: Аффинная хроматография FcRn:
Комбинированные варианты показали умеренную положительную корреляцию (hFcRn: R2=0,69, rFcRn: R2=0,71) между наблюдаемой способностью связывания при рН 6,0 и устойчивым состоянием RU при рН 7,4 (Фигуры 7С и 7D). Без связи с какой-либо теорией, эти результаты показывают, что более высокое сродство при рН 6,0 обычно приводит к большему остаточному связыванию с FcRn при рН 7,4. Эти варианты могут оставаться связанными с FcRn в кровотоке и иметь короткие периоды полувыведения из сыворотки крови и/или способствовать их клиренсу, аналогично мутациям abdeg с высокой аффинностью к FcRn (см., например, Swiercz et al. 2014, выше; и Vaccaro et al. 2005, выше). Поскольку взаимодействие антитело-FcRn зависит от pH и происходит только при низком pH (<pH 6,5), мутации насыщения могут усиливать взаимодействие между вкладами, обусловленными гидрофобными свойствами или зарядом, это может нарушать депротонирование особенно важных остатков гистидина (Фигура 1B, как указано) и ослаблять это взаимодействие при физиологических значениях pH.
Аффинная хроматография FcRn использует линейный градиент рН для прямого измерения нарушения рН-зависимости взаимодействия с FcRn (см., например, Schlothauer et al. 2013, выше). Аффинная хроматография FcRn с вариантами AAA, LS, YTE, H435A и H310A/H435Q показала, что H435A (Фигура 8A, сплошная светло-серая линия) и H310A/H435Q (Фигура 8A, AQ, сплошная темно-серая линия) не связываются с FcRn даже при рН 5,5 и элюируются потоком. Антитело дикого типа элюировалось при физиологическом pH (pH 7,37±0,05), в то время как AAA, LS и YTE, которые имеют более медленную скорость диссоциации и способность к более тесному связыванию с FcRn, чем дикий тип, по данным Octet (Фигура 2) и Biacore (Фигура 3), требуется значительно более высокий рН для диссоциации от колонки (ААА: 7,94 ± 0,06; LS: 8,29 ± 0,03; YTE: 8,14 ± 0,03). Из профилей элюирования видно, что все варианты из комбинаторной библиотеки требуют более высокого pH для элюирования из аффинной колонки, чем вариант дикого типа. Характерные хроматограммы при среднем pH элюирования для одинарного (длинные штрихи с двумя точками), двойного (длинные штрихи с одной точкой), тройного (длинные штрихи) и четверного (короткие штрихи) вариантов показаны на Фигуре 9А. Для семи лидирующих вариантов требовалось более высокое значение рН для диссоциации из колонки по сравнению с WT (Фигура 10А, таблица 3), в то время как варианты, имеющие кинетику к hFcRn, как у варианта дикого типа (K288D/N, Y436H/H/W), элюировались при рН, сходном с рН для варианта дикого типа.
Таблица 3. Характеристические параметры лидирующих вариантов антител, полученные in vitro
(х104 М-1 сек-1)
(х10-1 сек-1)
(x109 M)
(х10-3 сек-1)
Все данные были получены с использованием экспериментальных методик, указанных в верхней части каждого столбца. рH элюирования определяли с помощью аффинной хроматографии FcRn в трех повторностях (n=3) и с помощью DSF исследовали термостабильность в трех повторностях (n=3). Кинетики связывания с FcRn для FcRn человека и крысы были получены из Biacore с серийным разведением антител (n=4) и подгоняли независимо. Единицы измерения для каждого измерения: рН элюирования (безразмерная величина); DSF Tпл. (°C); Biacore, pH 6,0, скорость связывания с hFcRn (х104 М-1 сек-1), скорость диссоциации (х10-1 сек-1) и KD,набл. (х109 М); Biacore, pH 6,0, скорость связывания с rFcRn (х104 М-1 сек-1), скорость диссоциации (х10-3 сек-1) и KD,набл. (х109 М).
Оба варианта N434F/Y элюируются при более высоком pH, чем вариант LS (N434F: 8,30 ± 0,05; N434Y: 8,46 ± 0,02) и показали значительное связывание с FcRn при рН 7,4 (таблица 4). Эти результаты показывают, что только эти варианты могут нарушить зависимость от рН. В целом, средние значение pH элюирования увеличивалось с увеличением количества мутаций, усиливающих связывание с FcRn (Фигура 9B). Появилась сильная корреляция (R2=0,94) с pH элюирования по сравнению со скоростями диссоциации hFcRn (Фигура 9C); без привязки к какой-либо теории, это указывает на то, что нарушение зависимости взаимодействия от рН прямо способствует более медленным скоростям диссоциации FcRn, наблюдаемым для комбинаторной библиотеки при рН 6,0.
Пример 7: Термостабильность
Большинство белков, включая антитела, с низкой термодинамической стабильностью, имеют повышенную склонность к неправильному сворачиванию и агрегации и могут ограничивать или препятствовать их активности, эффективности и действенности в качестве новых терапевтических средств. Термостабильность каждого варианта определяли, используя DSF, и регистрируемую температуру плавления (Tпл.) определяли как середину первого перехода в профиле интенсивности флуоресценции красителя Sypro Orange. По сравнению с WT, Tпл. которого составляет 69,0 ± 0,2°C, вариант LS является WT-подобным (68,5 ± 0,3°C), а AAA и YTE термически дестабилизированы на ~8°C (AAA: 61,3 ± 0,6°С; YTE: 61,2 ± 0,3°C) (Фигуры 8B, 9B и 10B; и таблицы 2B, 3 и 4). По сравнению с WT и LS, Tпл. которого составляет 69,0 ± 0,2°C, варианты AAA и YTE имели термостабильность ниже на ~8°C согласно данным DSF.
Таблица 4. Характеристические параметры сравнительных и лидирующих комбинаций, полученные in vitro
V158
(ºC)
(x104
М-1 сек-1)
(x10-1
сек-1)
(x109 M)
М-1 сек-1)
(x10-3
сек-1)
Мутации, введенные в каркас дикого типа, выделены жирным шрифтом и подчеркнуты. Все данные были получены с использованием экспериментальных методик, указанных в верхней части каждого столбца. рH элюирования и Tпл. определяли в трех повторностях (n=3). Кинетику связывания с FcRn при pH 6,0 с FcRn человека и крысы получали с использованием Biacore (n=4) и подгоняли независимо. Ответ на связывание c FcRn в устойчивом состоянии при рН 7,4 измеряли, используя Biacore, при одной концентрации антител в трех повторностях. Способность связывания с FcγRIIIa определяли по серии разведения антител в двух повторностях, используя Biacore. Единицы измерения для каждого измерения: рН элюирования (безразмерная величина); DSF Tпл. (°C); Biacore, pH 6,0, скорость связывания с hFcRn (х105 М-1 сек-1), скорость диссоциации (х10-2 сек-1) и KD,набл. (х109 М); Biacore, pH 6,0, скорость связывания с rFcRn (х105 М-1 сек-1), скорость диссоциации (х10-3 сек-1) и KD,набл. (х109 М); Biacore, pH 7,4, ответ на связывание в устойчивом состоянии (RU) и FcγRIIIa KD,набл. (x109 M).
Двенадцать из 18 лидирующих вариантов насыщения имели пониженную Tпл. по сравнению с диким типом, а у некоторых из мутантов T307 (T307E/F/M/Q) наблюдалась небольшая стабилизация (таблица 4). Ни один из семи одинарных вариантов, использованных для комбинаций (Фигура 10B и таблица 4), не был значительно дестабилизирован по сравнению с YTE (Фигура 8B и таблица 5). Добавление двойного (Фигура 9D, горизонтальные линии), тройного (Фигура 9D, вертикальные линии) и четверного (Фигура 9D, заштрихован в клетку) вариантов привело к дальнейшему снижению общей термостабильности по сравнению с одинарными вариантами (Фигура 9D, белые кружки). У множественных вариантов Tпл. была ниже, чем AAA или YTE (61,2 ± 0ºC), при этом >60% этих вариантов содержали T307W. Четверные варианты (Фигура 9D, заштрихованы в клетку) показали четкое бимодальное распределение температур плавления, при этом комбинации, содержащие T307Q, имели термостабильность примерно на 6ºC выше, чем у вариантов, несущих T307W (Фигура 9D).
Пример 8: Варианты Fc изменяют связывающее взаимодействие с FcγRIIIa
Помимо взаимодействия с FcRn, шарнирные участки Fc и домены CH2 отвечают за взаимодействие с другими рецепторами Fc, включая FcγRIIIa. Поскольку пять из семи отдельных вариантов, используемых для создания комбинаторной библиотеки насыщения, расположены в домене CH2, способность взаимодействовать с этими рецепторами может быть нарушена в сравнении с диким типом, несмотря на то, что они расположены далеко от области контакта при взаимодействии. Использование Biacore для измерения связывания с FcγRIIIa таким же образом, как и связывание с FcRn при pH 7,4, выявило, что вариант YTE (Фигура 11A, темно-серым) показал примерно 50%-ное снижение ответа на связывание по сравнению с диким типом (Фигура 11A, черным). Без связи с какой-либо теорией, пониженное связывание FcγRIIIa с YTE является результатом мутации M252Y (Фигура 11B, самый нижний белый кружок), так как только этот вариант значительно снижает сродство к этому рецептору. Другие одиночные мутации не имели этого сниженного сродства (Фигура 11B, белые кружки), и только варианты N434F/Y усиливали связывание на 16-40%. Эти эффекты были переданы большинству, но не всем, соответствующим им комбинациям. Например, комбинации, содержащие M252Y, имели снижение связывания с FcγRIIIa на 17-72% (таблица 5).
Таблица 5. Концентрации вариантов библиотеки насыщения в кондиционированных средах
Один из вариантов - MDQF (Фигура 11B, самый высокий в категории с тройным вариантом), показал значительное увеличение связывания с FcγRIIIa на 140%. Таким образом, комбинаторная библиотека насыщения предлагает варианты с широким спектром функциональных возможностей Fc-рецепторов, которые можно использовать для адаптации терапевтических антител с конкретными эффекторными функциями.
На Фигуре 11C показана ящичная диаграмма ответов на связывание с FcγRIIIa для семи лидирующих вариантов в сравнении с вариантами WT и YTE.
Пример 9: Семь лидирующих комбинаций уравновешивают рН-зависимость взаимодействия с FcRn
Без связи с какой-либо теорией, варианты-кандидаты для дальнейшего изучения in vivo занимали нижнюю левую часть графиков, показанных на Фигурах 7C и 7D. Семь вариантов удовлетворяли этим критериям для hFcRn и включали пять двойных и две тройные комбинации (MDQN, MDWN, YDTN, YETN, YTWN, YDQN и YEQN) и не содержали мутации в положении N434 (таблица 3). Каждую из этих комбинаций элюировали из аффинной колонки с FcRn между AAA (pH 7,94 ± 0,06) и LS (pH 8,29 ± 0,03), причем YDQN, элюировался при самом высоком pH 8,51 ± 0,14 (Фигура 12A, таблица 5), что указывает только на небольшое нарушение зависимости от рН и большее остаточное связывание при рН 7,4 (таблица 2А). Один из вариантов (MDQN) обладал термостабильностью, подобной дикому типу, и шесть имели сходную или сниженную Tпл. по сравнению с вариантом YTE (Фигура 12B, таблица 4). В анализе связывания FcγRIIIa пять комбинированных вариантов показали такое же снижение, что и YTE (таблица 4). Дальнейшее исследование с отдельными мутациями показало, что M252Y значительно влияет на связывание FcγRIIIa и, не будучи связанным какой-либо теорией, транслирует этот эффект в комбинации с этой мутацией. Остальные шесть одиночных мутаций были WT-подобными или обладали слегка улучшенным связыванием с этим рецептором.
Три комбинированных варианта были выбраны для дальнейших исследований на основе их свойств связывания с FcRn, термической стабильности и связывания FcγRIIIa. DQ (T256D/T307Q), DW (T256D/T307W) и YD (M252Y/T256D) обеспечили оптимальные свойства связывания с FcRn (таблица 2B) в качестве варианта LS (Фигура 12E). Каждый вариант обладает разнообразным диапазоном термостабильности и связывающих свойств FcγRIIIa (Фигуры 12F и 12G, таблица 2B), что обеспечивает ряд функциональных возможностей. Фигура 12H представляет собой диаграмму гомогенного мостикового соединения RF.
Повышение наблюдаемой способности связывания для FcRn и человека, и крысы при pH 6,0 по сравнению с вариантами LS (Фигура 13A, толстые длинные штрихи) и вариантами YTE (Фигура 13B, толстые длинные штрихи), соответственно, было компромиссом между скоростями связывания и диссоциации (Фигуры 13А и 13В, таблица 4). Как правило, комбинации с более быстрыми скоростями диссоциации также обладают более быстрыми скоростями связывания и наоборот. Это наблюдалось для FcRn и человека, и крысы (таблица 4). Кроме того, у всех этих вариантов был более слабый ответ в устойчивом состоянии, чем у варианта LS (Фигура 13C, толстые длинные штрихи), на hFcRn при pH 7,4. Эти результаты не согласовывались с rFcRn, так как пять вариантов, содержащих M252Y, YDTN, YETN, YTWN, YDQN и YEQN, имели повышенное связывание с FcRn при pH 7,4 (Фигура 13, таблица 5) по сравнению с YTE. Варианты MDQN и MDWN были единственными комбинациями, которые были перекрестно реактивными по отношению к FcRn человека и крысы. Кроме того, эти два варианта не нарушали взаимодействие с FcγRIIIa в той же степени, что и M252Y-содержащие варианты (Фигуры 12C и 12D и таблица 5; MDQN: 600 ± 4 нМ; MDWN: 512 ± 30 нМ; WT: 467 ± (99 нМ). Таким образом, мутагенез насыщения и комбинации в ключевых положениях взаимодействия с FcRn привел к идентификации лидирующих вариантов, которые уравновешивали pH-зависимость взаимодействия, сохраняли функциональность с Fc-рецептором, могли усиливать функциональность FcRn in vivo и могли увеличивать период полувыведения из сыворотки крови терапевтических антител.
Пример 10: Характеристики связывания ревматоидного фактора лидирующих комбинированных вариантов
Была исследована изоэлектрическая точка и RF-связывание лидирующих вариантов, поскольку эти мутации могут изменять поверхностный заряд антитела и иммуногенность. Считается, что более кислотные антитела продлевают фармакокинетику антител. По сравнению с контрольными WT и LS все три лидера приводили к снижению рН на ~0,2 единицы в pI в результате замещения T256D. Мутации, усиливающие FcRn, могут одновременно изменять связывание с антителами хозяина, такими как ревматоидный фактор (RF), из-за перекрывающихся областей контакта. Гомогенный мостиковый ИФА был адаптирован для измерения изменения RF-связывания у лидирующих вариантов. Интересно, что LS и YTE продемонстрировали совершенно противоположные сдвиги при RF-связывании по сравнению с WT (Фигура 12H). LS значительно увеличивал RF-связывание, в то время как YTE показывал значительное снижение (p <0,001). YD (p<0,001) и DW (p<0,01) также значительно снижали RF-связывание, в то время как DQ вызывал ответ, аналогичный WT. Без связи с какой-либо теорией, эти результаты показывают, что DQ, DW и YD могут обеспечить иммуногенное преимущество по сравнению с LS. Варианты YD, DW и DQ имеют ряд ключевых характеристик антител, которые можно использовать в сочетании с улучшенными свойствами связывания с FcRn по сравнению с вариантами сравнения YTE и LS.
Пример 11: Лидирующие комбинированные варианты можно переносить на другие антитела
Новый анализ связывания был разработан с использованием сенсорного чипа CM5, как показано на Фигуре 15А. Анализ связывания включает стадию иммобилизации стрептавидина на сенсорном чипе CM5 для захвата биотинилированного FcRn до примерно 30 RU, который можно пополнять по необходимости. Кинетику связывания антител измеряли при рН 6,0 и 7,4 и рН 8,5 для регенерации. На Фигурах 15B и 15C показана прямая иммобилизация FcRn и захвата стрептавидином биотинилированного FcRn, соответственно, с использованием нового анализа связывания.
Связывание с FcRn антитела-2 при рН 6,0: В случае мышиного FcRn лидирующие варианты антитела-2 демонстрируют более медленные скорости диссоциации, чем у варианта LS (штрихи) и дикого типа (черным) (Фигура 16А). В случае FcRn человека все лидирующие варианты имеют более высокие скорости связывания, но такие же скорости диссоциации, что и у LS (штрихи) (Фигура 16B).
Связывание с FcRn антитела-2 при рН 7,4: Все лидирующие варианты показали пониженное связывание с FcRn человека при рН 7,4 по сравнению с LS (штрихи) (Фигура 17А). Как и в случае антитела-1, варианты DW (MDWN) и DQ (MDQN) также демонстрировали более низкое остаточное связывание с FcRn мыши (крысы) при pH 7,4 (Фигура 17B).
Лидирующие варианты поддерживали более высокую способность связывания при рН 6,0 и более низкое остаточное связывание при рН 7,4 по сравнению с LS (Фигура 18). Важное наблюдение было сделано о том, что варианты можно переносить между различными базовыми IgG1 без сколько-нибудь значимого влияния на связывание с FcRn. Как показано на Фигуре 19, LS имел одинаковый рН элюирования независимо от базы. WT, DQ и DW на базе антитела-2 показали более высокий рН элюирования, чем на базе антитела-1, возможно, в результате более тесного связывания при рН 6,0 с базой антитела-2.
Все варианты на базе антитела-2 показали слегка повышенную термостабильность, как показано на Фигуре 20.
Как показано на Фигуре 21, аналогично базе антитела-1, YD (YDTN) показал снижение ответа на связывание FcγRIIIa (слева) и сродства (справа). DQ (светло-серым) и DW (темно-серым) показали свойства связывания FcγRIIIa, сходные с WT (черным) на базе антитела-2. Эффект на связывание FcγRIIIa для LS согласуется между антителом-1 и антителом-2.
Так, связывание с FcRn, зависимость от рН, термостабильность или связывание FcγRIIIa лидирующих вариантов на базе антитела-2 значительно не отличаются от этих показателей для тех же лидирующих вариантов на базе антитела-1.
В одном варианте осуществления варианты DQ (T256D/T307Q), DW (T256D/T307W) и YD (M252Y/T256D) были включены в дополнительное антитело IgG1 и рекомбинантный фрагмент Fc: mAb2 распознает антиген, отличный от mAb1, и Ab3 представляет собой Fc фрагмент. В каждом случае pH-зависимая кинетика связывания с FcRn (Фигура 22) была очень похожа наряду с pH элюирования, термостабильностью и способностью связывания с FcγRIIIa (таблица 2B и таблица 6). Не ограничиваясь какой-либо теорией, эти результаты показывают, что варианты DQ, DW и YD придают свои улучшенные свойства связывания с FcRn белкам, состоящим из Fc-домена.
Таблица 6. Концентрации вариантов библиотеки насыщения в кондиционированных средах
V158
97
360
Пример 12: Лидирующие варианты продлевают период полувыведения антител из плазмы крови in vivo
Фармакокинетику (ФК) вариантов DQ, DW и YD исследовали с позиции влияния на период полувыведения антител у обезьян cynomolgus и трансгенных мышей hFcRn (штамм Tg32) (см., например, Avery et al. Mabs (2016) 8: 1064-1078) по сравнению с контрольными WT и LS. Исследования связывания с FcRn на Cynomolgus FcRn выявили сходные с hFcRn способности связывания (Фигуры 23A-23B; таблица 6). Каждому животному внутривенно вводили варианты WT, LS, DQ, DW или YD, и концентрацию антител определяли количественно с помощью масс-спектрометрического подхода для определения скорости клиренса и периода полувыведения из сыворотки крови у обезьян (Фигура 24А) и трансгенных мышей hFcRn (Фигура 24B). Скорости клиренса и период полувыведения из сыворотки крови были получены из некомпартментной модели концентрации антител как функции времени. Все три лидирующих варианта и LS показали значительно сниженную скорость клиренса по сравнению с WT как у обезьян, так и у мышей (р <0,001). Период полувыведения из плазмы крови антитела WT для обезьян и мышей составлял 9,9 ± 0,5 и 11,7 дня, соответственно. Кроме того, сравнительный LS и выявленные варианты показали значительное увеличение периода полувыведения по сравнению с диким типом у обоих видов (увеличение в 2,5 и 1,7 раза соответственно у обезьян и мышей) (таблица 7). DQ, DW и YD показали аналогичное продление периода полувыведения по сравнению со сравнительным LS (таблица 7). Мутации DQ, DW и YD, идентифицированные здесь с помощью насыщающего мутагенеза, продемонстрировали значительно более длительный период полувыведения из плазмы крови, чем их аналоги WT, на животных моделях с использованием нечеловекоподобных приматов и мышей.
Таблица 7. Скорость клиренса и период полувыведения из сыворотки крови сравнительного и лидирующих вариантов
(мл/
день/
кг)
(мл/
день/
кг)
В таблице 7 скорость клиренса и период полувыведения из плазмы крови определяли, используя mAb2. Каждое значение скорости клиренса и периода полувыведения представляли собой среднее значение n=3 для обезьяны cynomolgus и единичную оценку из пула n=6 трансгенных мышей hFcRn. Кратность по отношению к WT и Кратность по отношению к LS показывают относительное улучшение периода полувыведения из сыворотки крови по сравнению с WT и LS, соответственно. *n=2 вследствие взаимодействия антител с лекарственным средством, ** n=2 из-за частичного подкожного пути введения
Пример 13: Комбинированные варианты с повышенным связыванием с FcRn при рН 6,0 и рН 7,4
На основании Octet скрининга (скрининга на основе BLI), как описано в примере 2, были получены различные одинарные, двойные, тройные и четверные варианты и оценено их связывание с FcRn при рН 6,0 и рН 7,4 (таблица 8).
Таблица 8. Способность связывания (рН 6,0) и связывание в устойчивом состоянии (рН 7,4) вариантов
В таблице 8 показана способность связывания с FcRn при рН 6,0 и связывание с FcRn в устойчивом состоянии при рН 7,4 для различных одинарных, двойных, тройных и четверных мутантов, а также сравнительные варианты (AAA, LS, YTE).
Эти значения приведены на Фигуре 25. На Фигуре 25 показано сравнение способности связывания при рН 6,0 и RU при рН 7,4. Как видно, вариант сравнения LS имеет самую высокую способность связывания при рН 6,0 и наибольшее остаточное связывание при рН 7,4 из протестированных сравнительных вариантов (AAA, LS, YTE).
Было установлено, что несколько комбинированных вариантов, показанных на Фигуре 25, проявляют повышенную способность связывания с FcRn при рН 6,0 и при рН 7,4. Чтобы выяснить, показал ли какой-либо из комбинированных вариантов более тесное связывание, чем вариант MST-HN (называемый здесь «сравнительным вариантом YTEKF»), содержащий мутации в Met252, Ser254, Thr256, His433 и Asn434 в Tyr252, Thr254, Glu256, Lys433 и Phe434) при pH 6,0 и pH 7,4 была выполнено исследование по следующей методологии. Захват биотинилированного FcRn человека, Cynomolgus и мыши проводили с помощью метода Biotin CAPture (см. схему на Фигуре 26). В случае рН 6,0 выполняли серии разведений (5 ед.) от 1000 нМ в двух повторностях. В случае рН 7,4 впрыск с одной концентрацией (1000 нМ) проводили в трех повторностях (уровень захвата каждого FcRn увеличивали в 10 раз, чтобы наблюдать связывание при этом рН). Ассоциация: 180 сек; Диссоциация: 300 сек.
Кинетику связывания с FcRn человека при рН 6,0 сравнительного варианта YTEKF и различных комбинированных вариантов показаны на Фигуре 27. Как видно из Фигуры 27, все исследованные варианты продемонстрировали на два порядка более тесное сродство к человеческому FcRn по сравнению с диким типом (WT).
На Фигурах 28А и 28В показана кинетика связывания с FcRn комбинированных вариантов в сравнении со сравнительным вариантом YTEKF при рН 6,0 (Фигура 28А) и при рН 7,4 (Фигура 28В). Из Фигуры 28А видно, что большинство вариантов имели более медленные скорости диссоциации, чем сравнительный вариант YTEKF, и имели аналогичные или более медленные скорости связывания. Из фигуры 28B видно значительное связывание у YTEKF при pH 7,4, а четыре варианта показывают более высокое остаточное связывание.
Таблица 9. Способность связывания (рН 6,0) и связывание в устойчивом состоянии (рН 7,4) отобранных вариантов
В таблице 9 приведена способность связывания с FcRn при рН 6,0 и связывание с FcRn в устойчивом состоянии при рН 7,4 для отобранных комбинированных вариантов, а также сравнительного варианта YTEKF и WT.
На Фигуре 29 показано сравнение способности связывания при рН 6,0 и RU при рН 7,4 для отобранных комбинированных вариантов, как указано в таблице 9. Как показано в Таблице 9 и на Фигуре 29, было обнаружено, что четыре четверных варианта имеют более высокое сродство при рН 6,0 и рН 7,4 к FcRn в сравнении со сравнительным вариантом YTEKF. Мутации T256D, T307Q и N434Y в четырех четверных вариантах оказали благоприятное действие. Эти четверные варианты демонстрируют примерно 500-кратное и 3-кратное улучшение сродства (при рН 6,0) в сравнении с WT и YTEKF, соответственно.
Другие характеристические параметры, например термостабильность, связывание с FcγRIIIa и pH элюирования, были определены и приведены в таблице 10.
Таблица 10. Другие характеристические параметры четверных лидирующих вариантов
* Без намерения быть связанным теорией, из-за присутствия "YTE" в YTEKF, термическая стабильность и связывание с FcγRIIIa, как ожидается, будут аналогичны четверным лидирующим вариантам
Как показано в таблице 10, было обнаружено, что все четверные лидирующие варианты термически дестабилизированы и демонстрируют пониженные способности связывания с FcγRIIIa.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕННО-ИНЖЕНЕРНЫЕ ПОЛИПЕПТИДЫ И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2822498C2 |
| СПОСОБ ИЗМЕНЕНИЯ УДЕРЖАНИЯ В ПЛАЗМЕ И ИММУНОГЕННОСТИ АНТИГЕНСВЯЗЫВАЮЩЕЙ МОЛЕКУЛЫ | 2012 |
|
RU2799423C1 |
| АНТИТЕЛА С МОДИФИЦИРОВАННОЙ АФФИННОСТЬЮ К FcRn, КОТОРЫЕ УВЕЛИЧИВАЮТ КЛИРЕНС АНТИГЕНОВ | 2011 |
|
RU2745989C2 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМ СВЯЗЫВАНИЕМ FcRn И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714116C2 |
| Fc-ВАРИАНТЫ С ИЗМЕНЕННЫМ СВЯЗЫВАНИЕМ С FcRn | 2008 |
|
RU2517621C2 |
| ВАРИАНТЫ ИММУНОГЛОБУЛИНА И ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2536937C2 |
| АФФИННАЯ ХРОМАТОГРАФИЯ С ПРИМЕНЕНИЕМ FC-РЕЦЕПТОРОВ | 2013 |
|
RU2624128C2 |
| ГИПЕРГЛИКОЗИЛИРОВАННЫЕ СВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2708314C2 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМИ СВОЙСТВАМИ СВЯЗЫВАНИЯ FcRn И БЕЛКА А | 2015 |
|
RU2713131C1 |
| КОМПОЗИЦИИ ДЛЯ УВЕЛИЧЕНИЯ ПЕРИОДА ПОЛУЖИЗНИ ТЕРАПЕВТИЧЕСКОГО СРЕДСТВА У СОБАК И СПОСОБЫ ПРИМЕНЕНИЯ | 2020 |
|
RU2828219C2 |
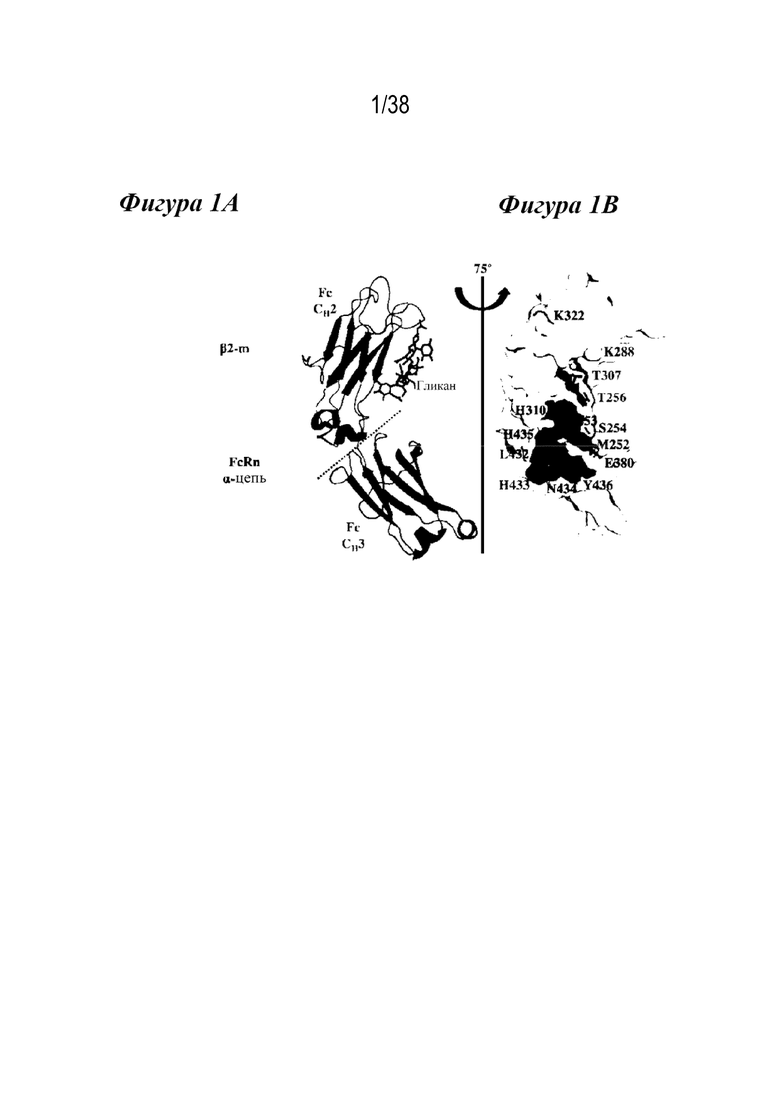
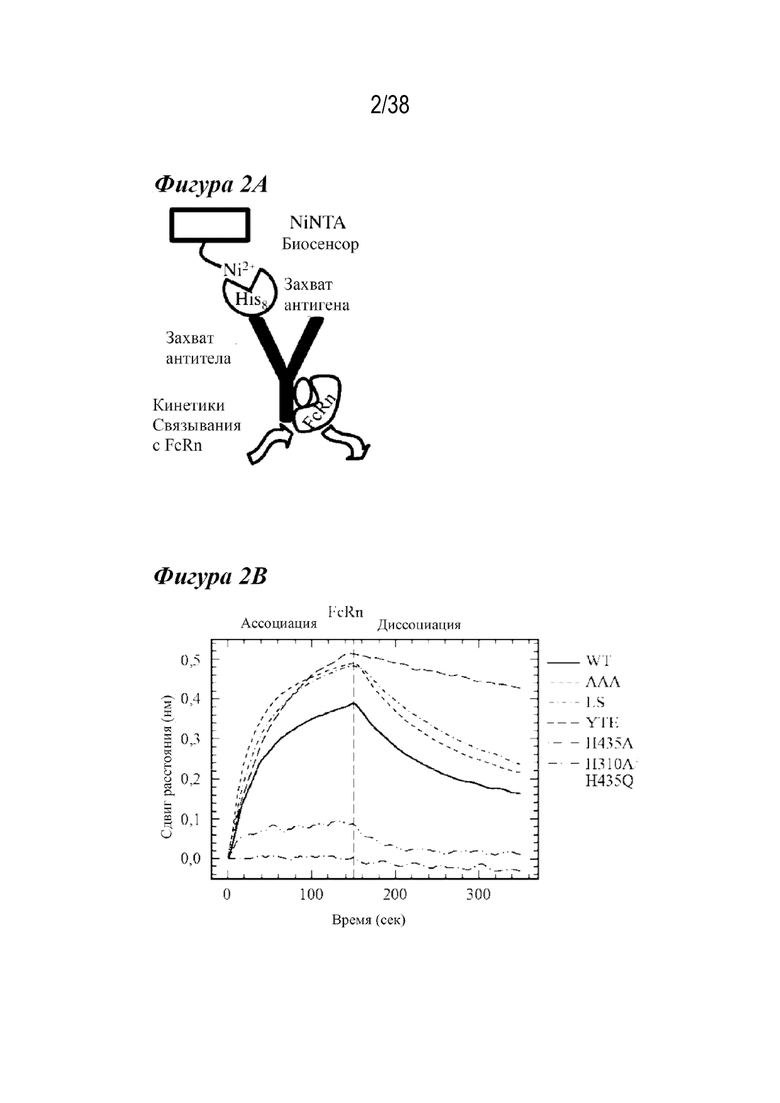
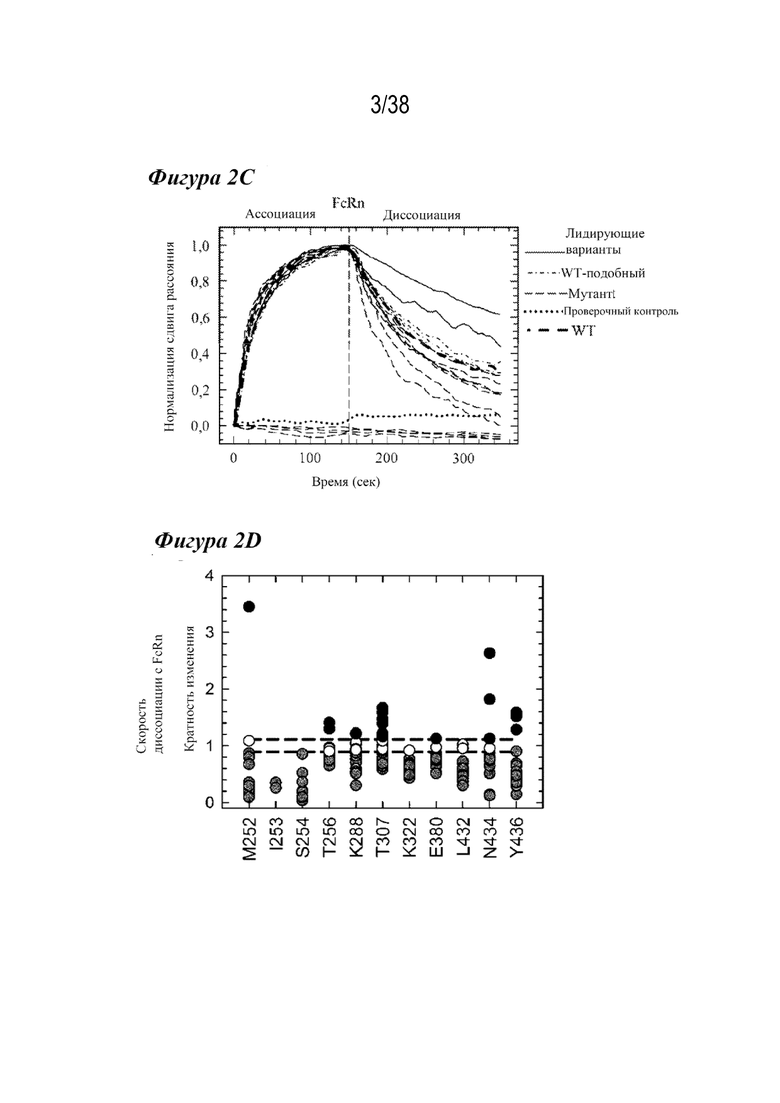
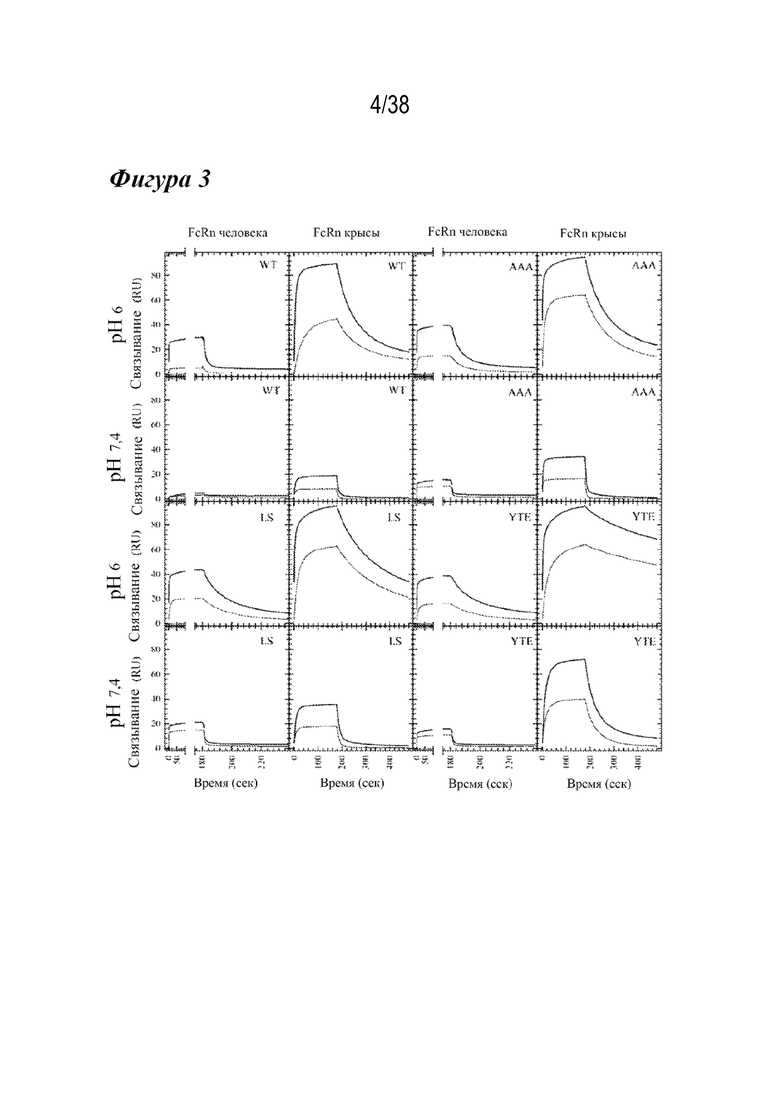
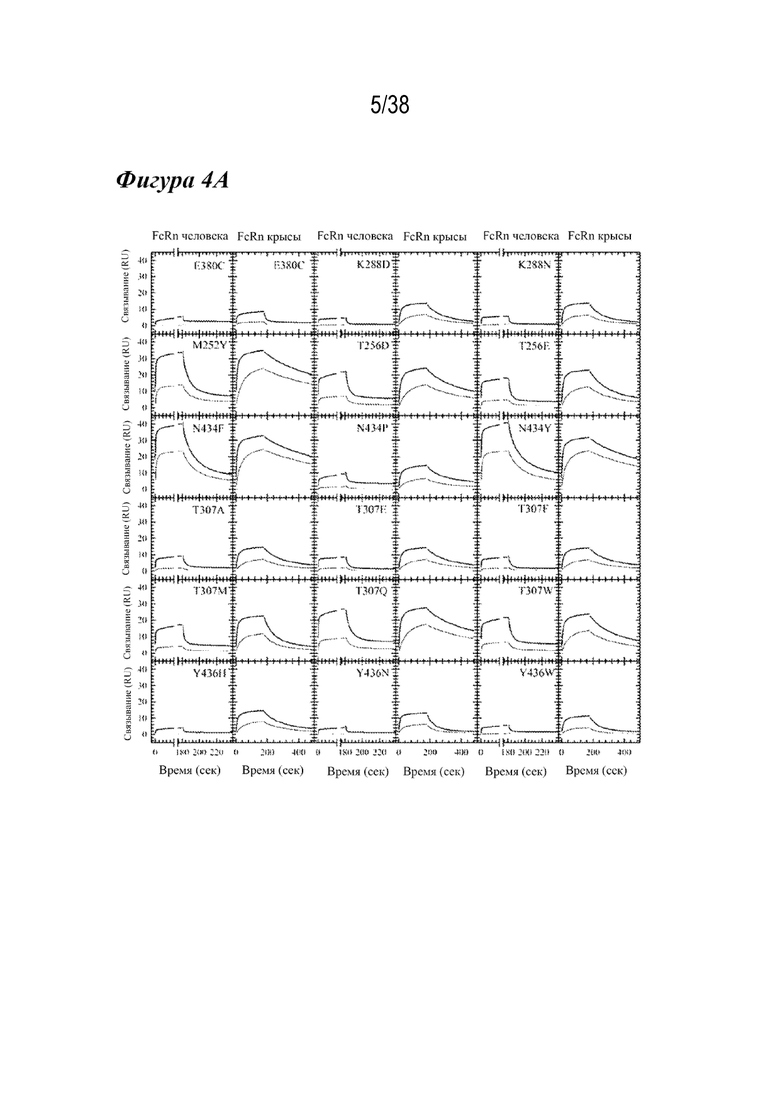
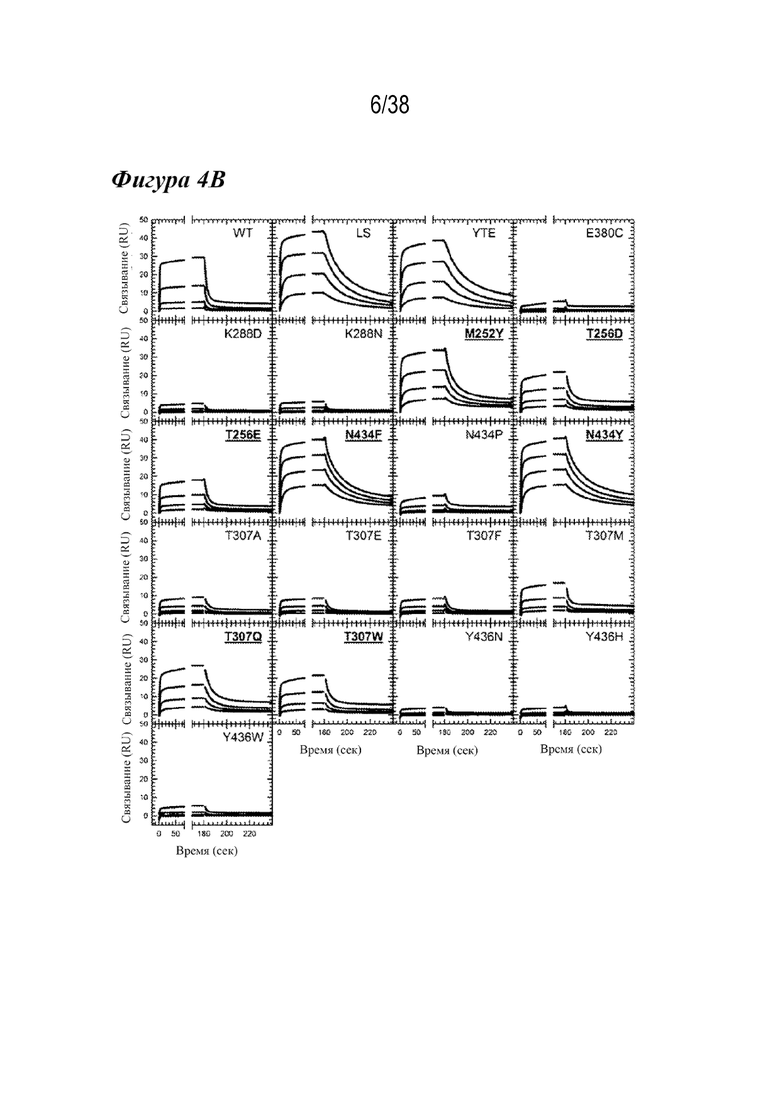
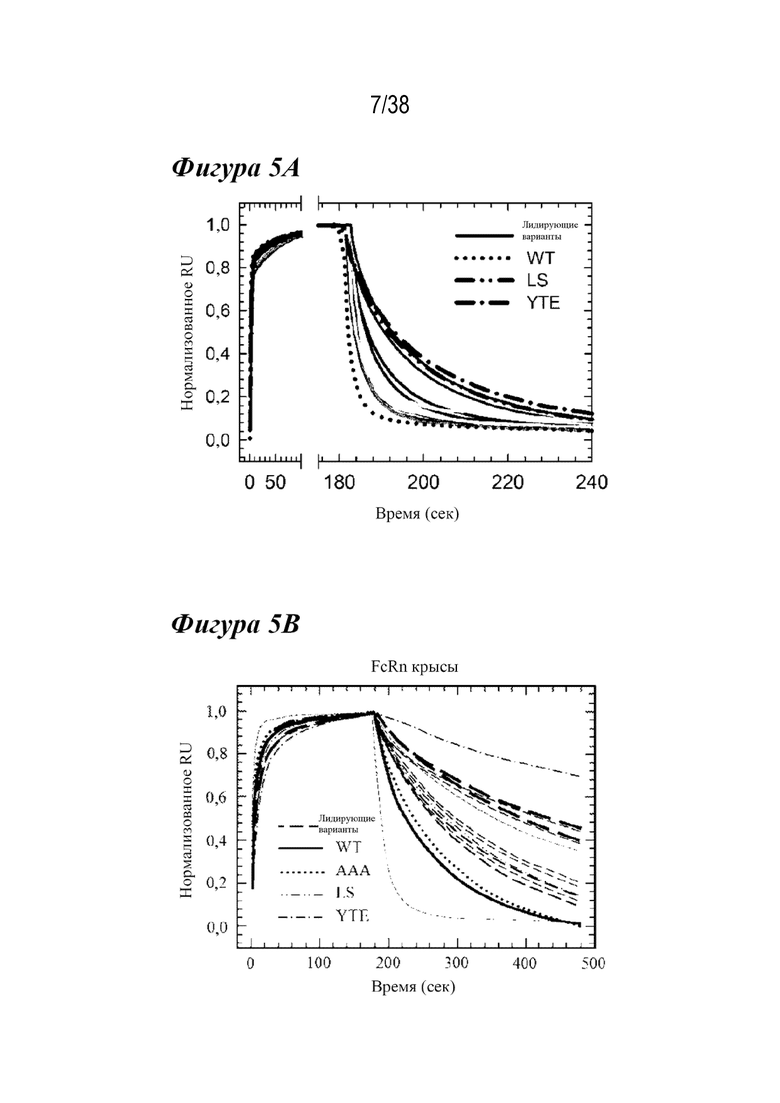
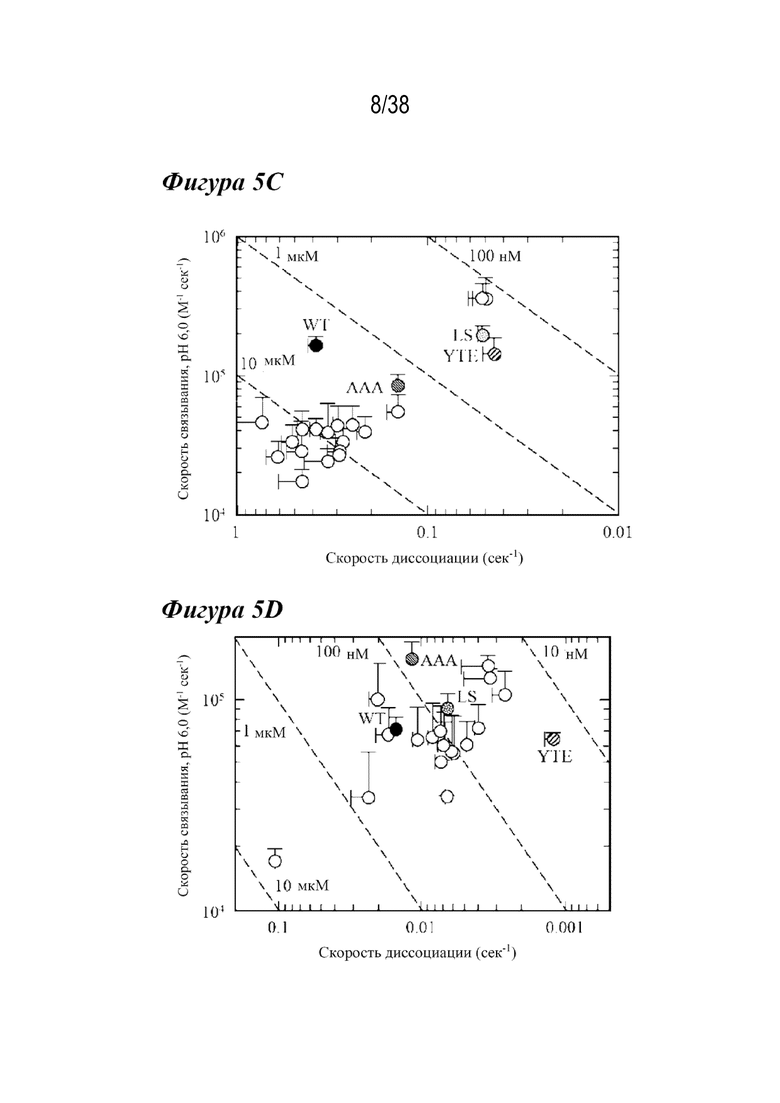
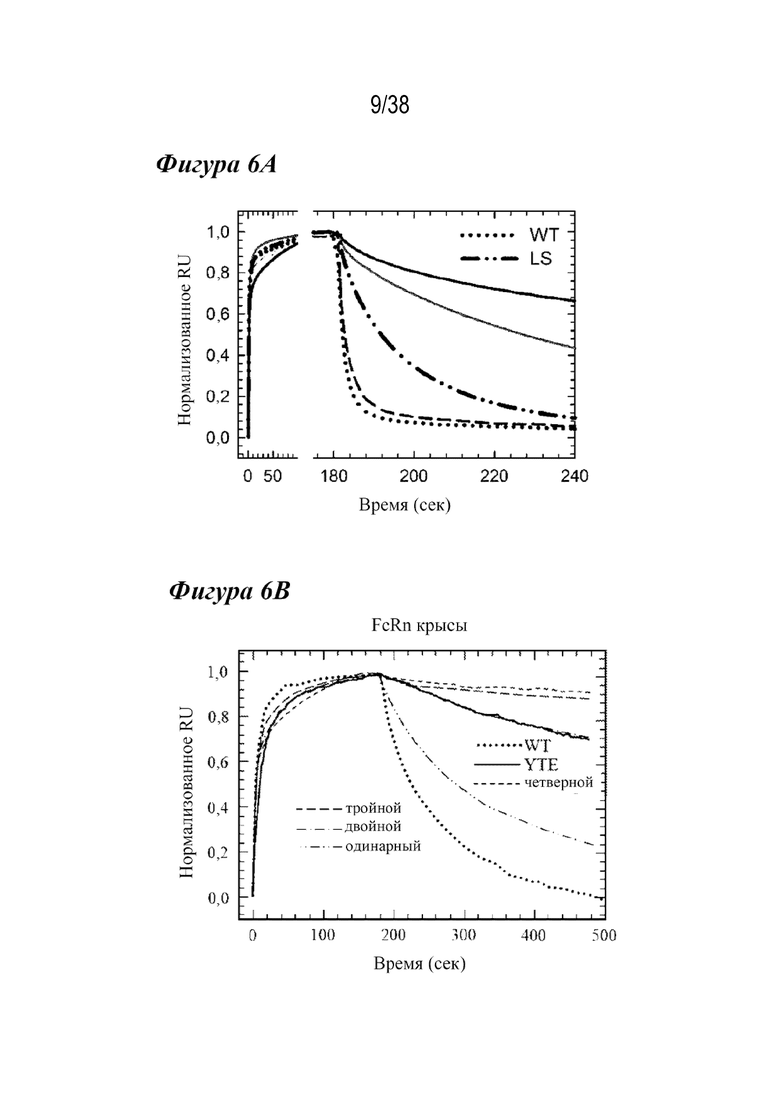
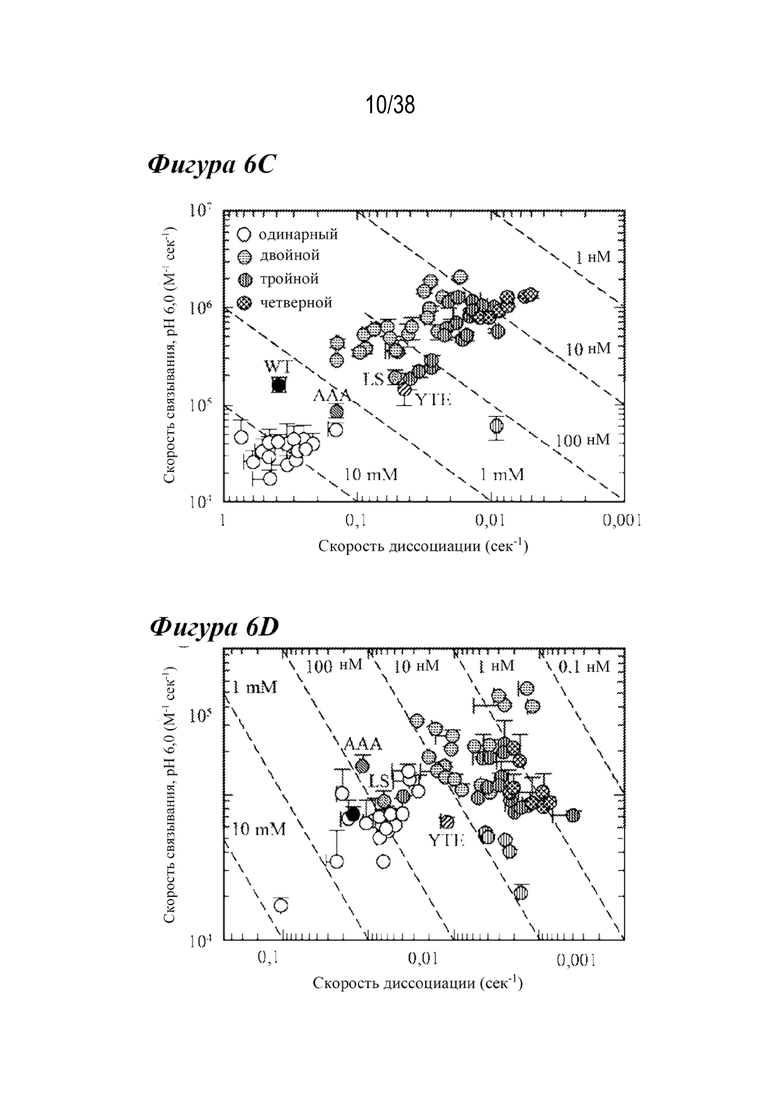
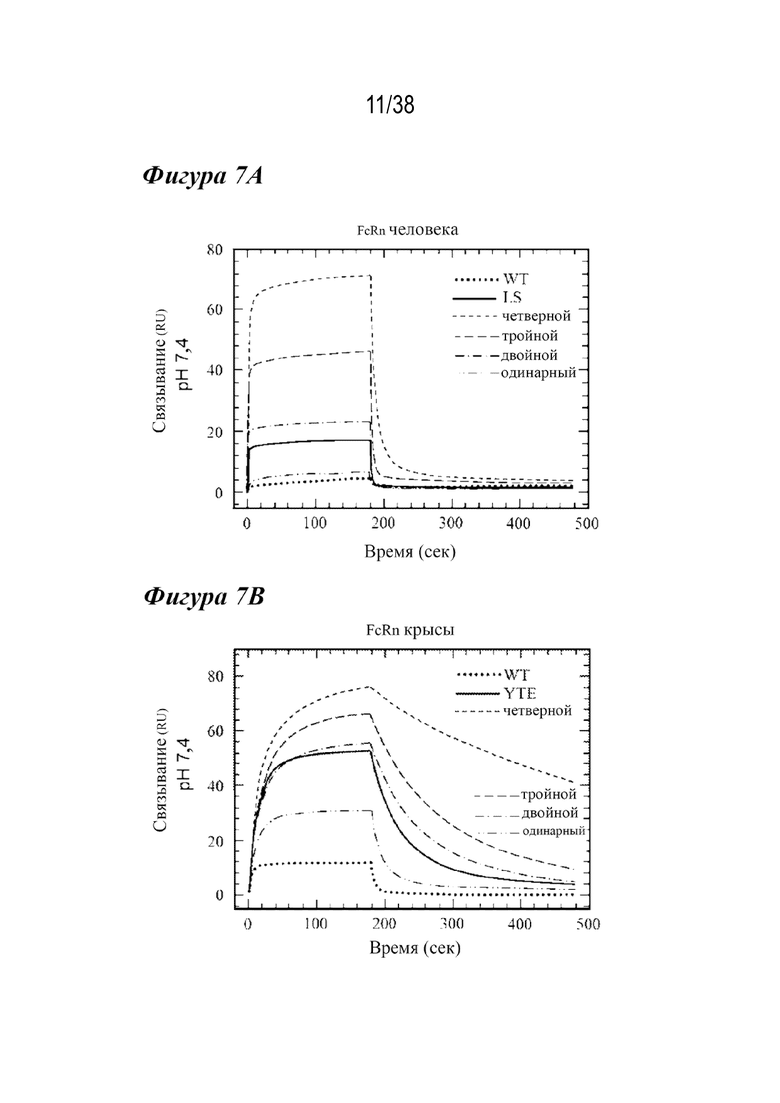
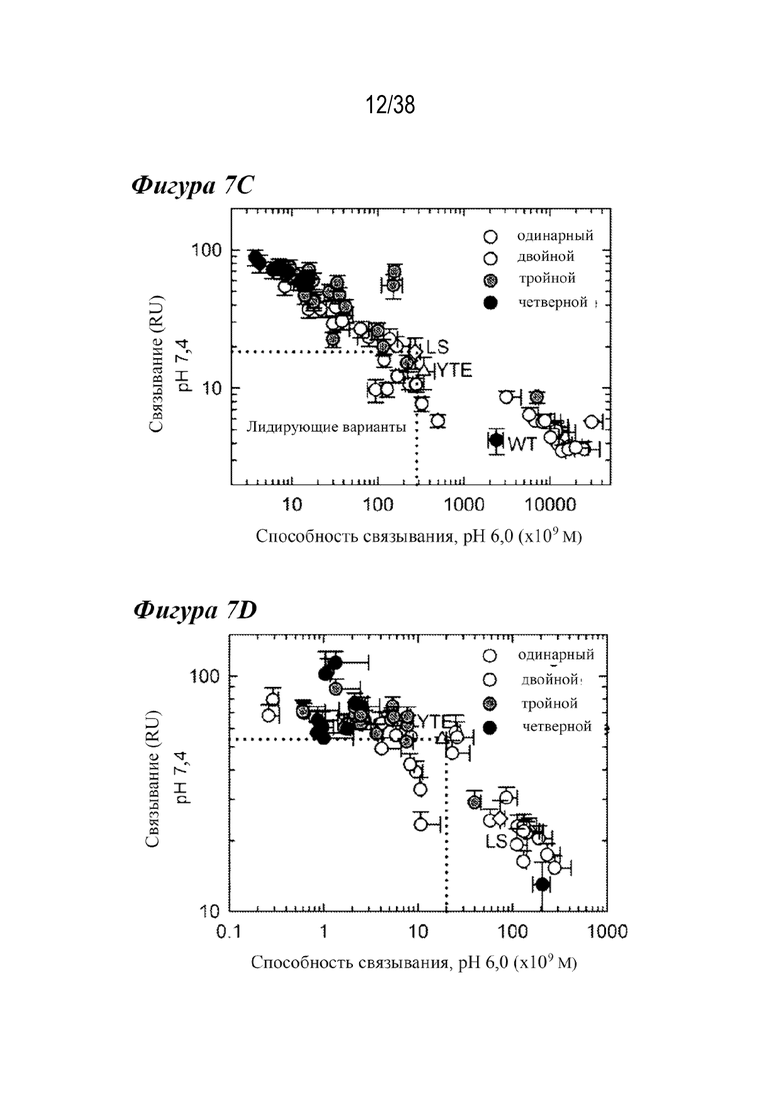
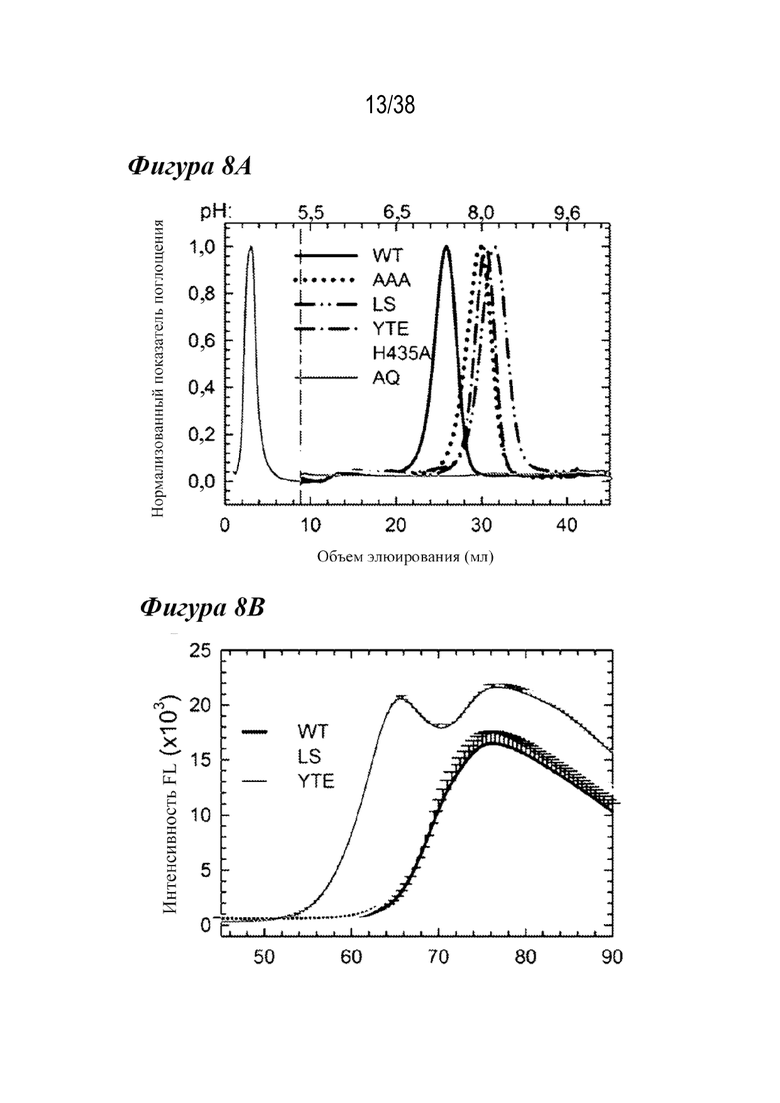
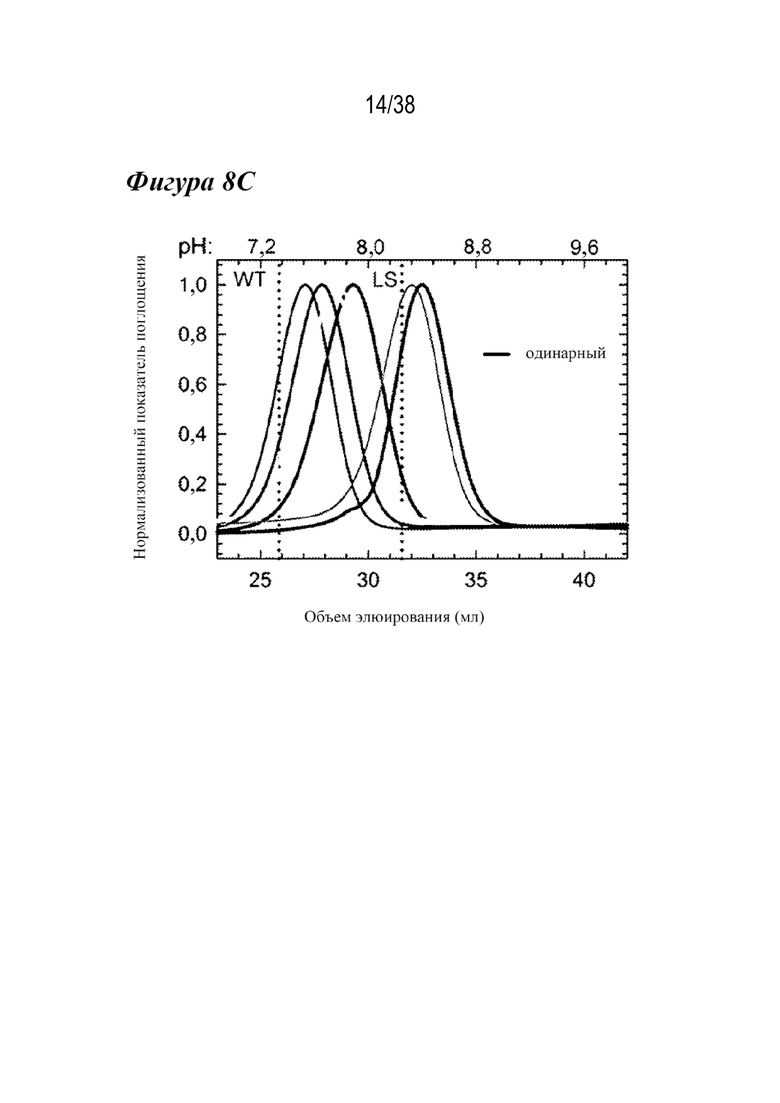
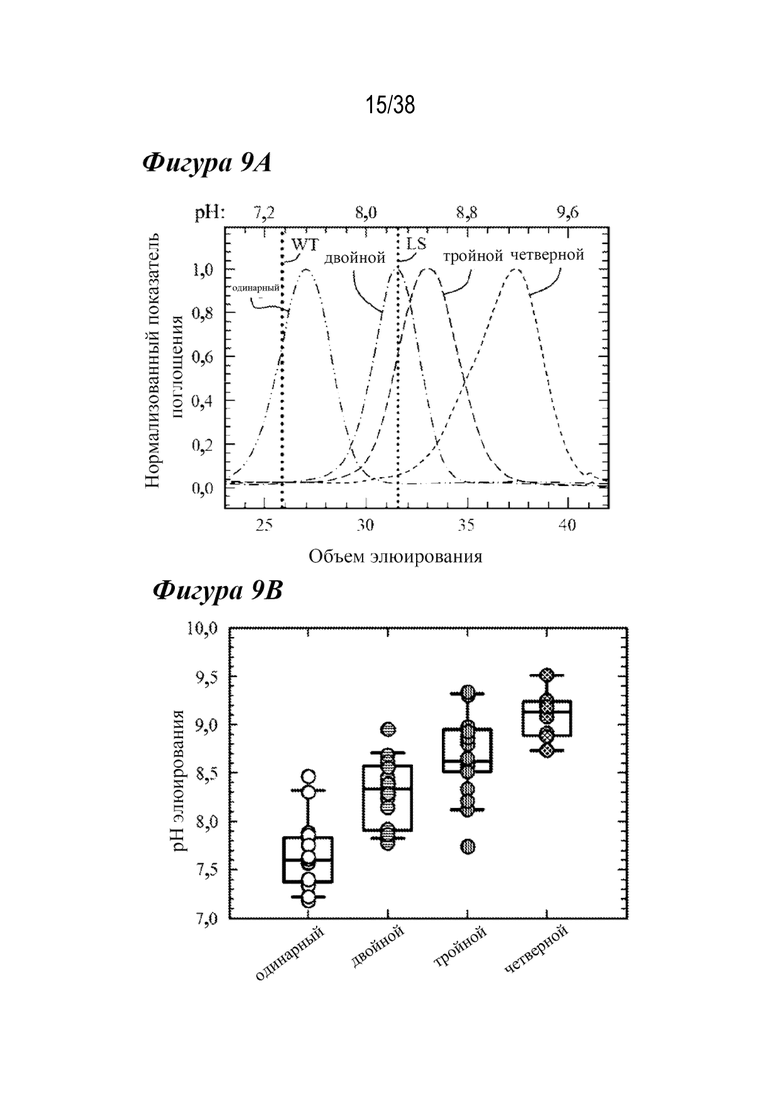
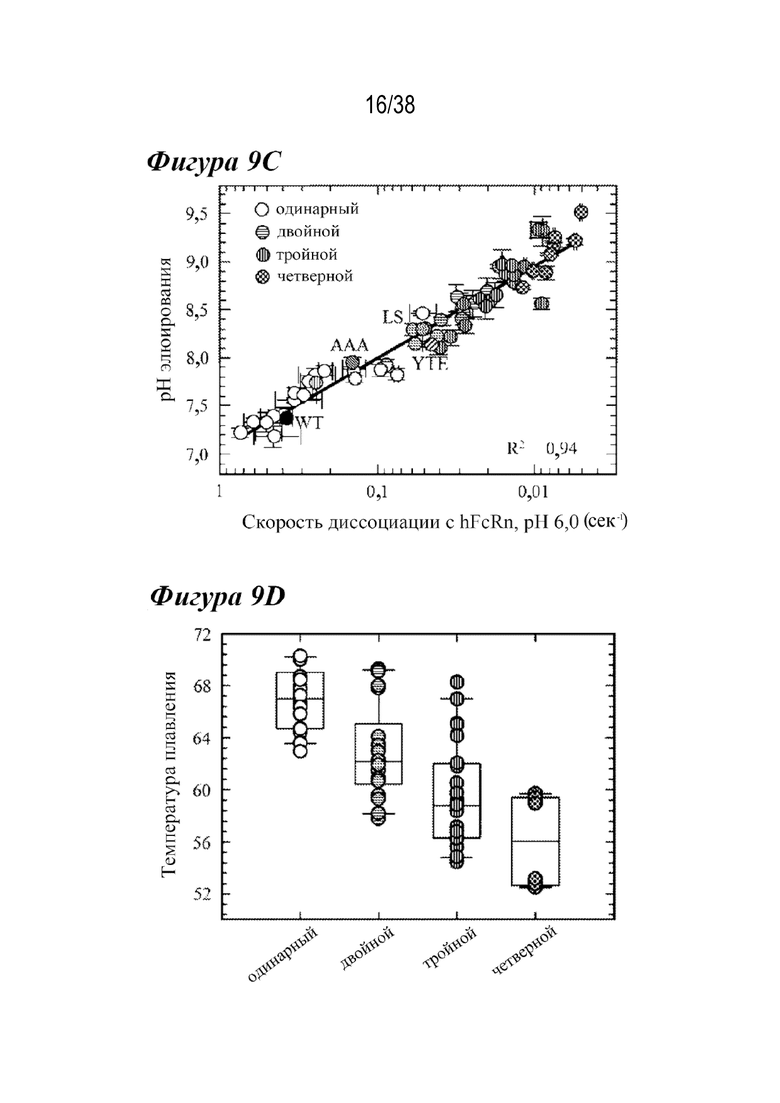
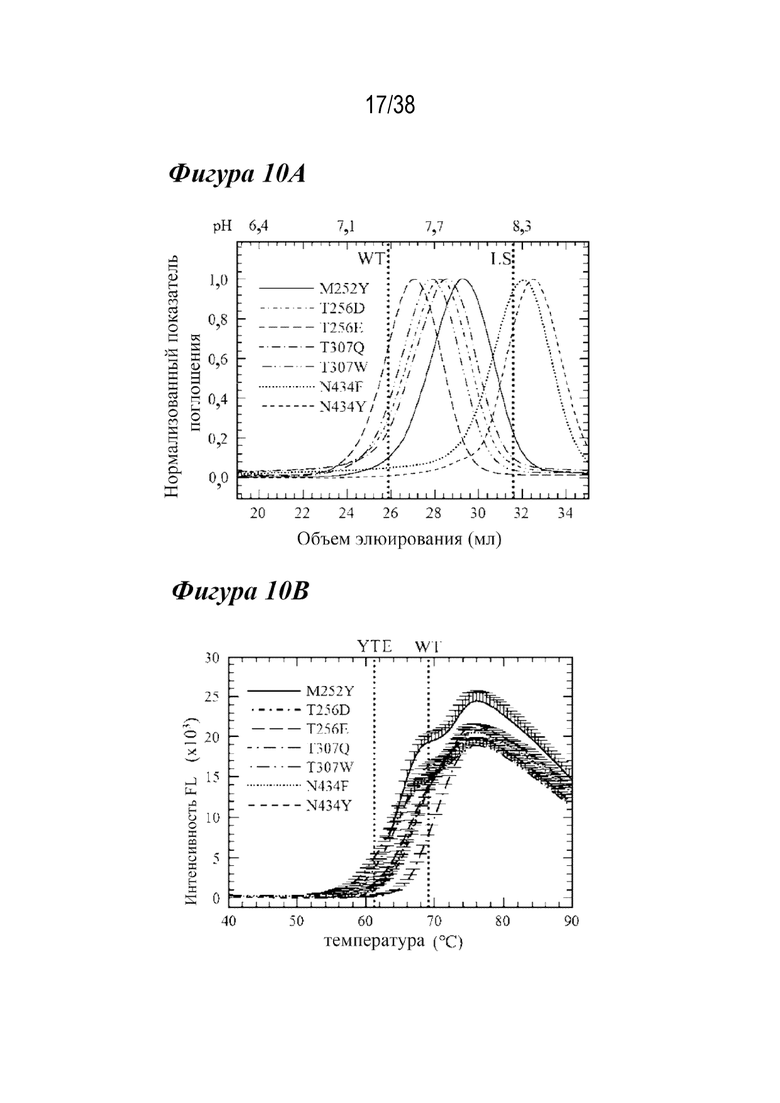
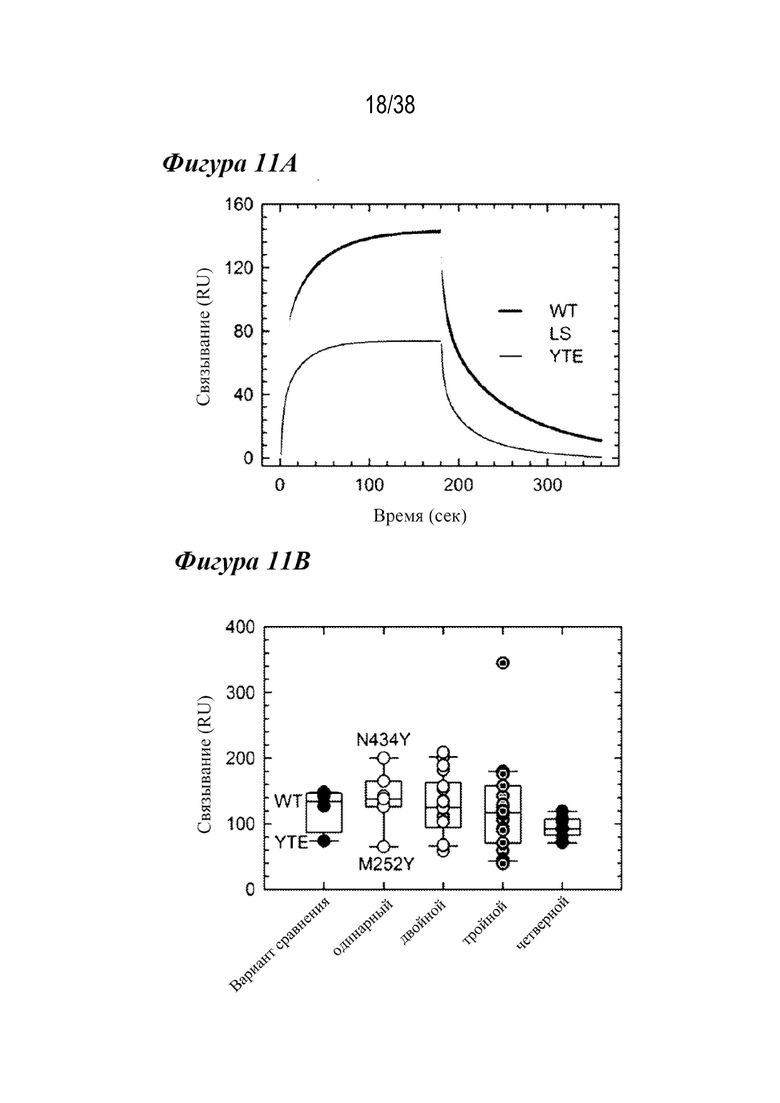
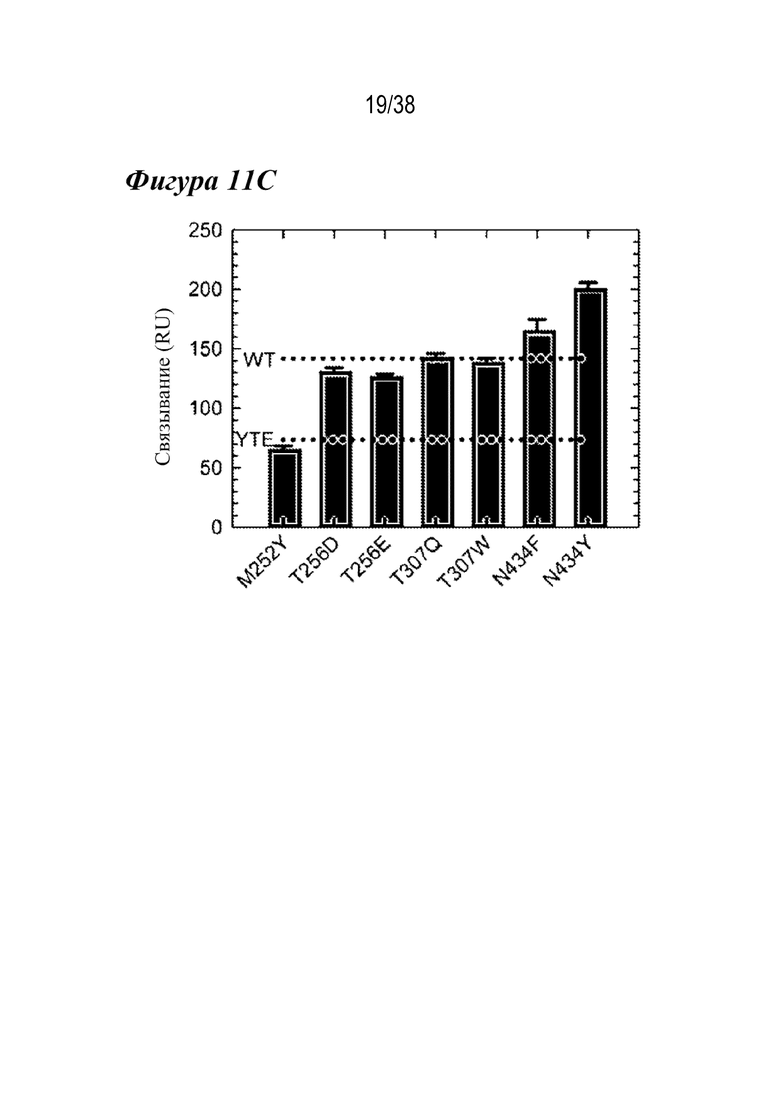
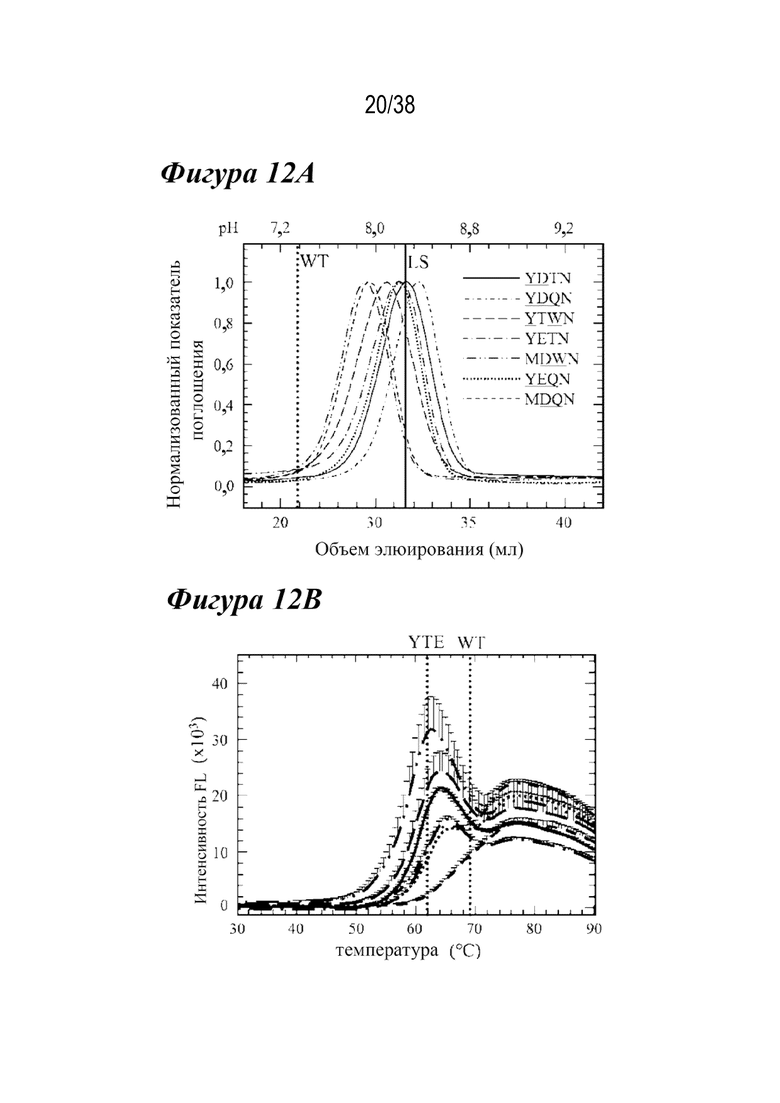
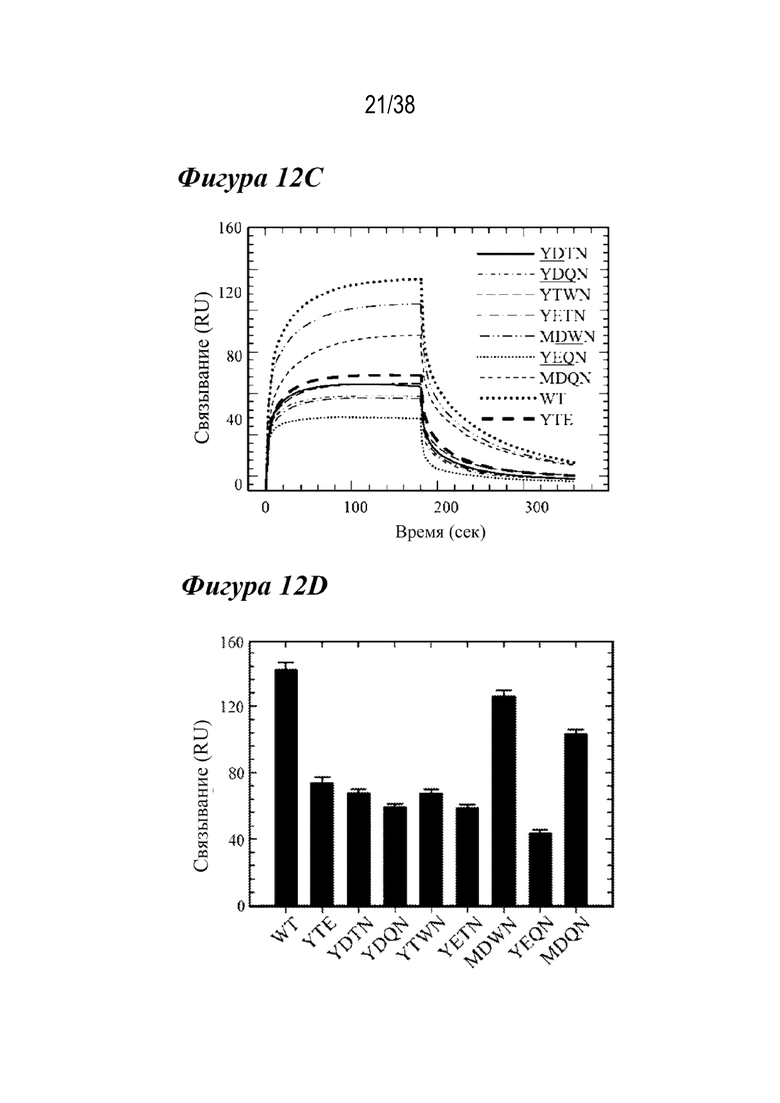
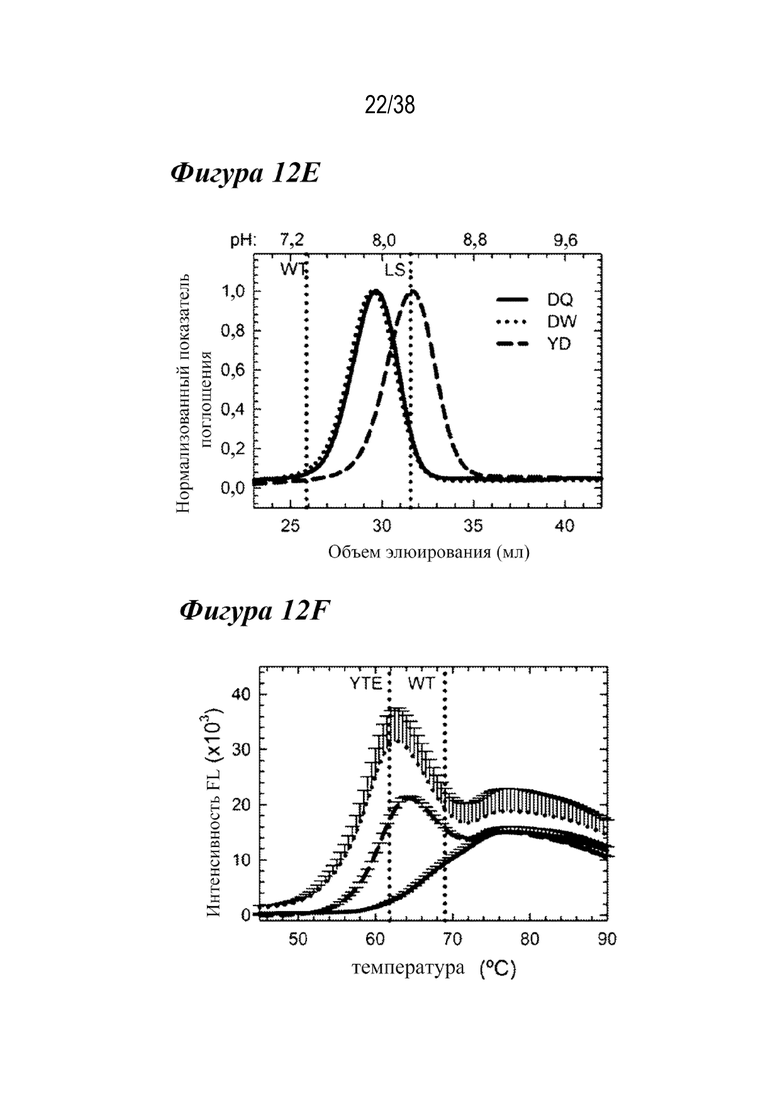
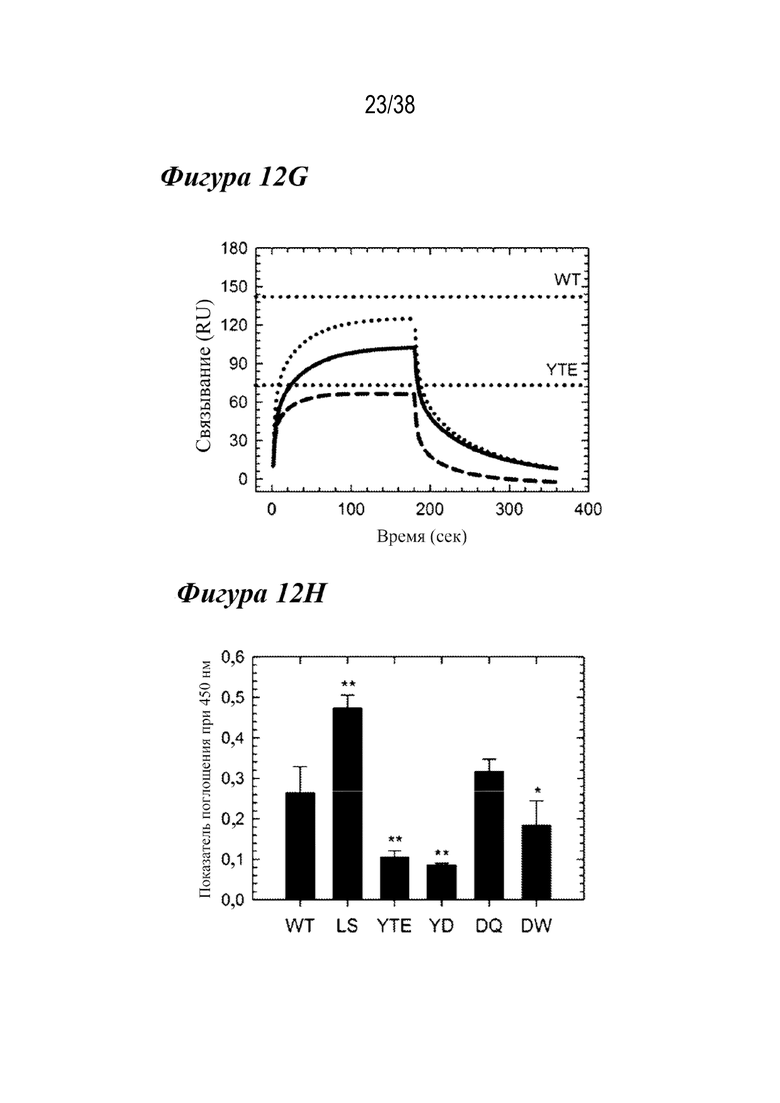
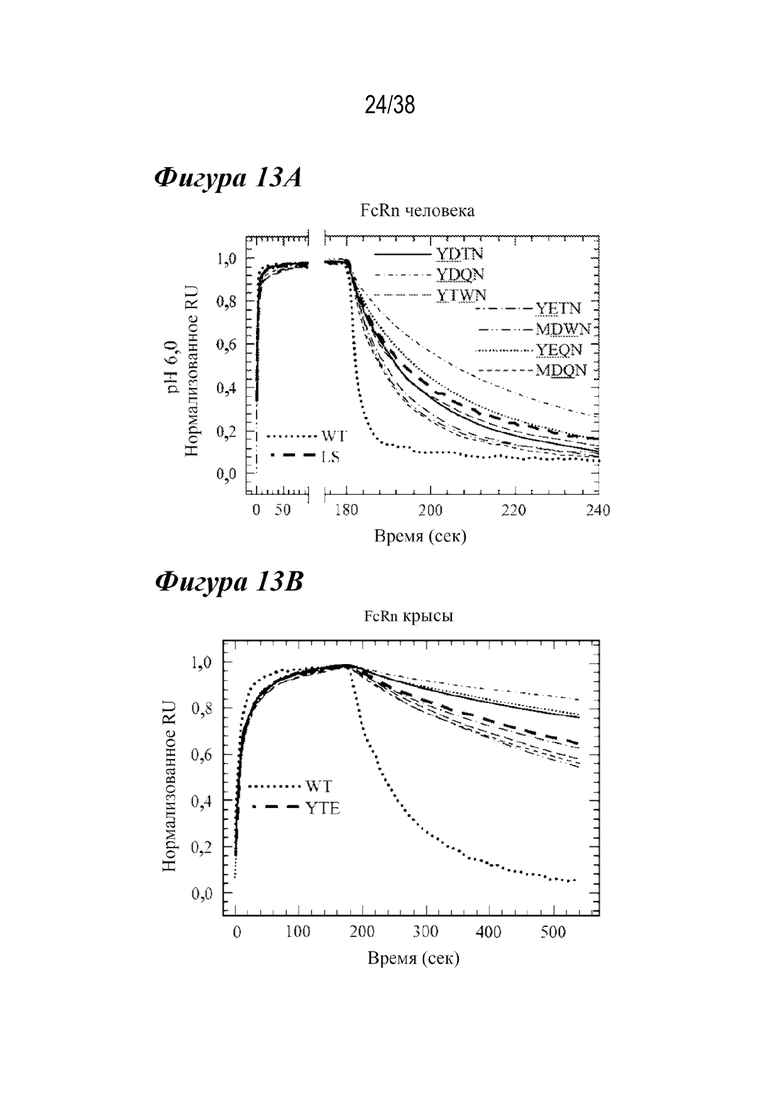
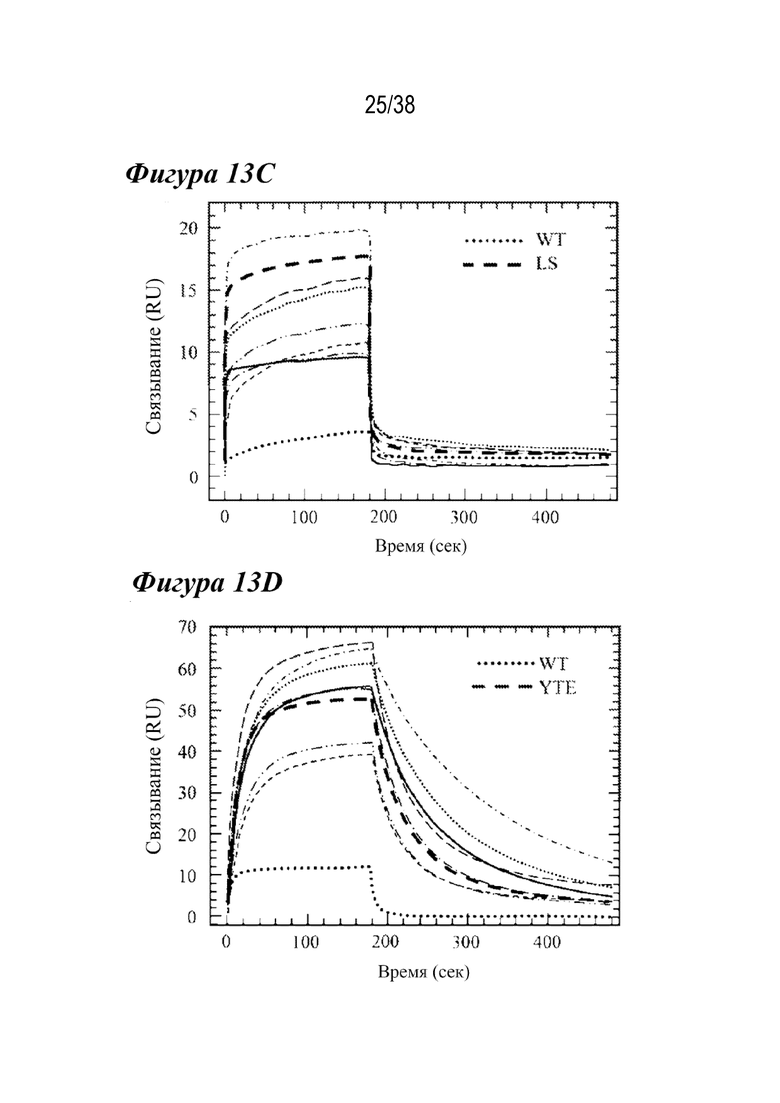

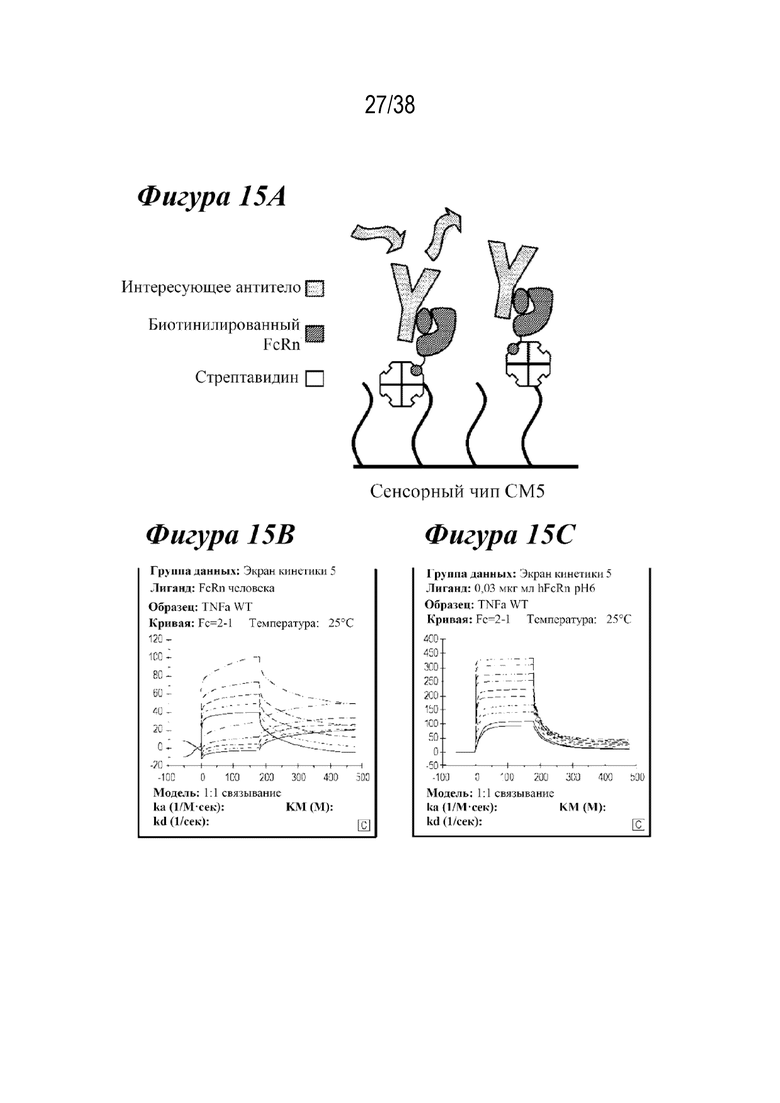
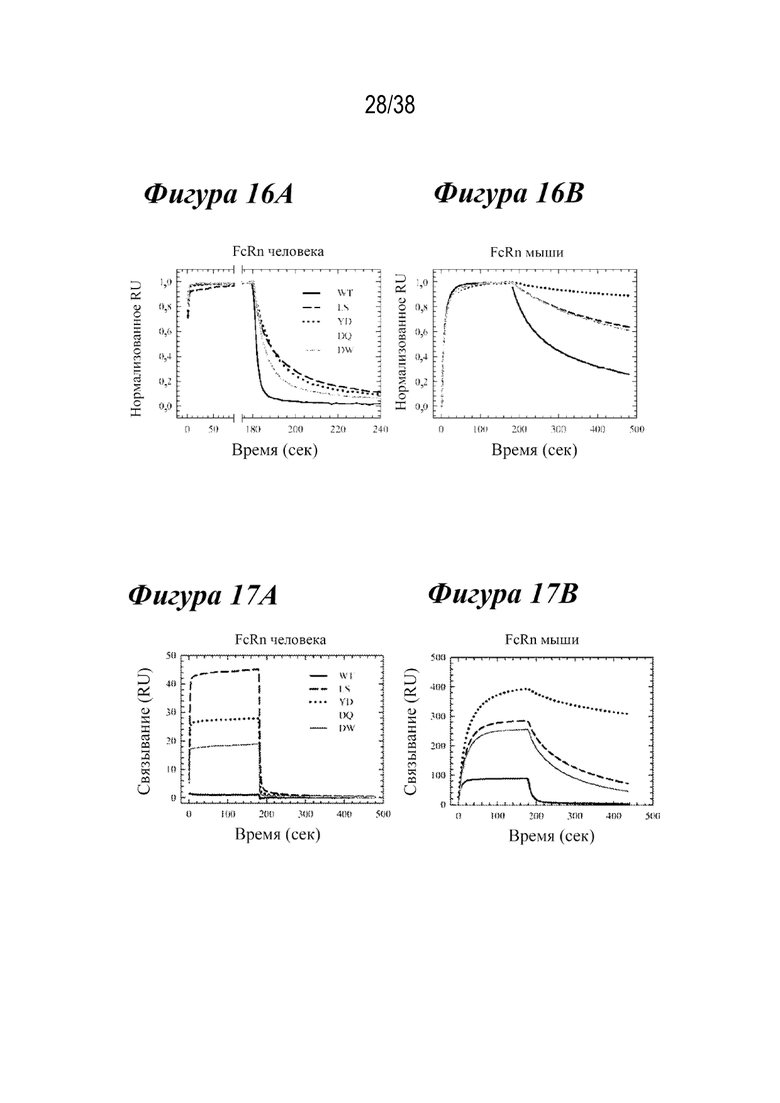
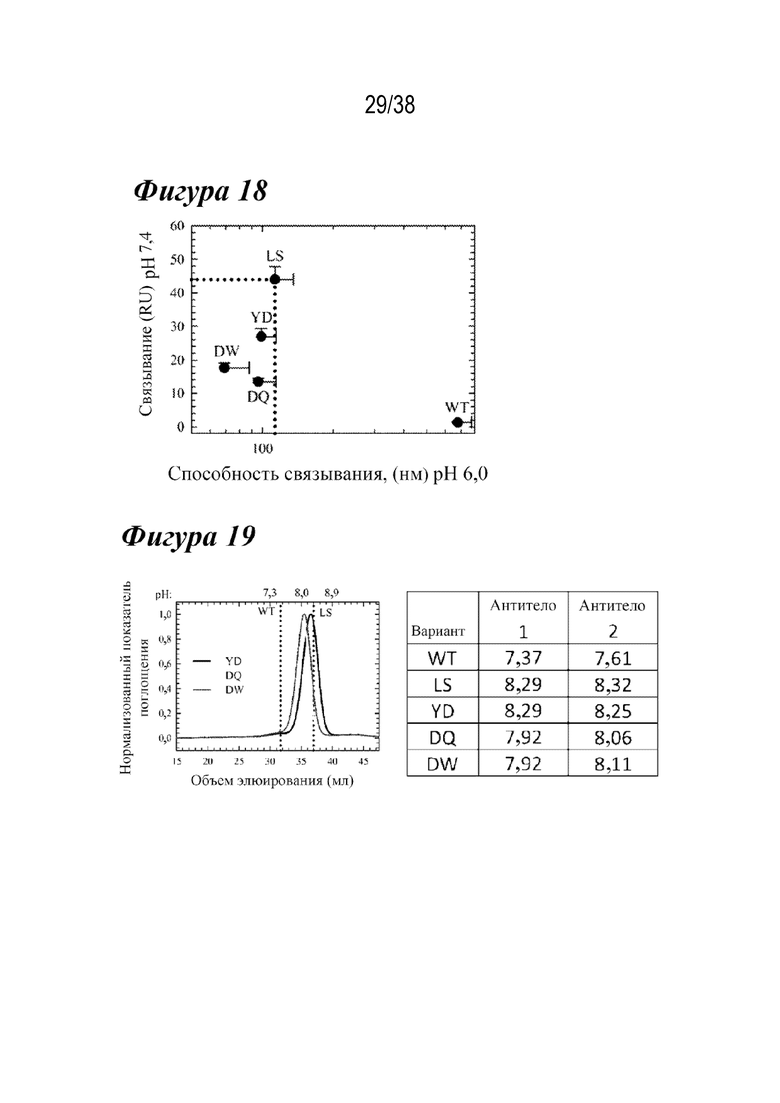
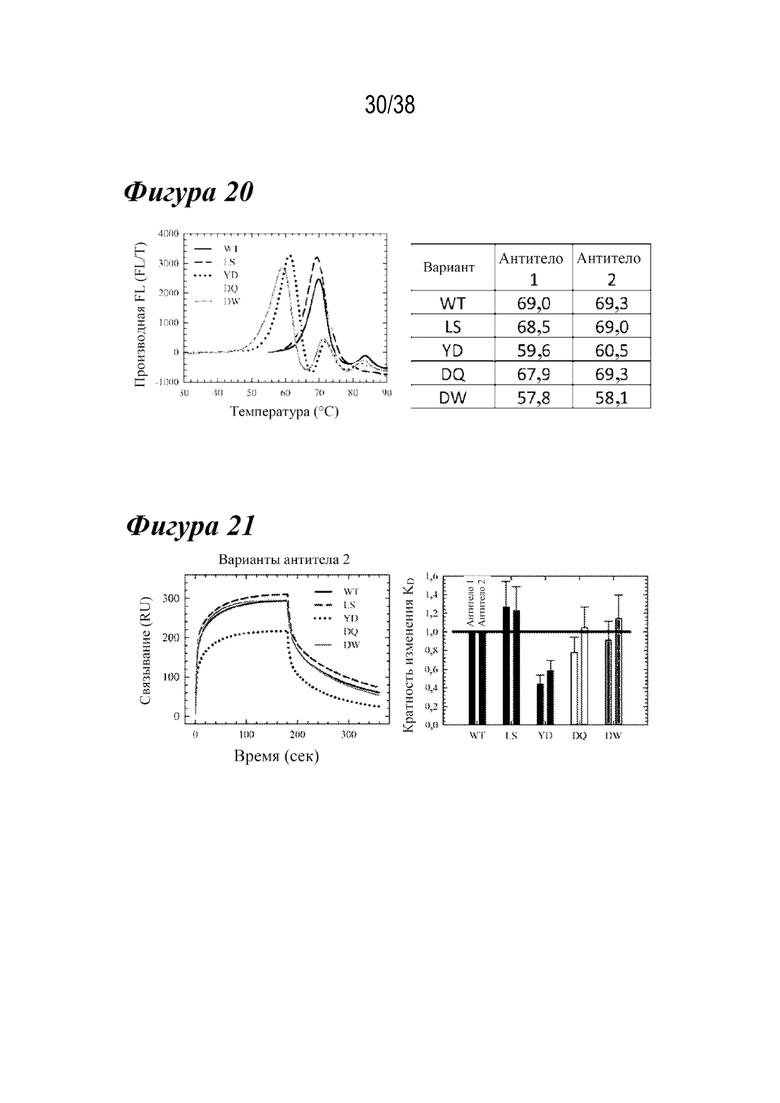
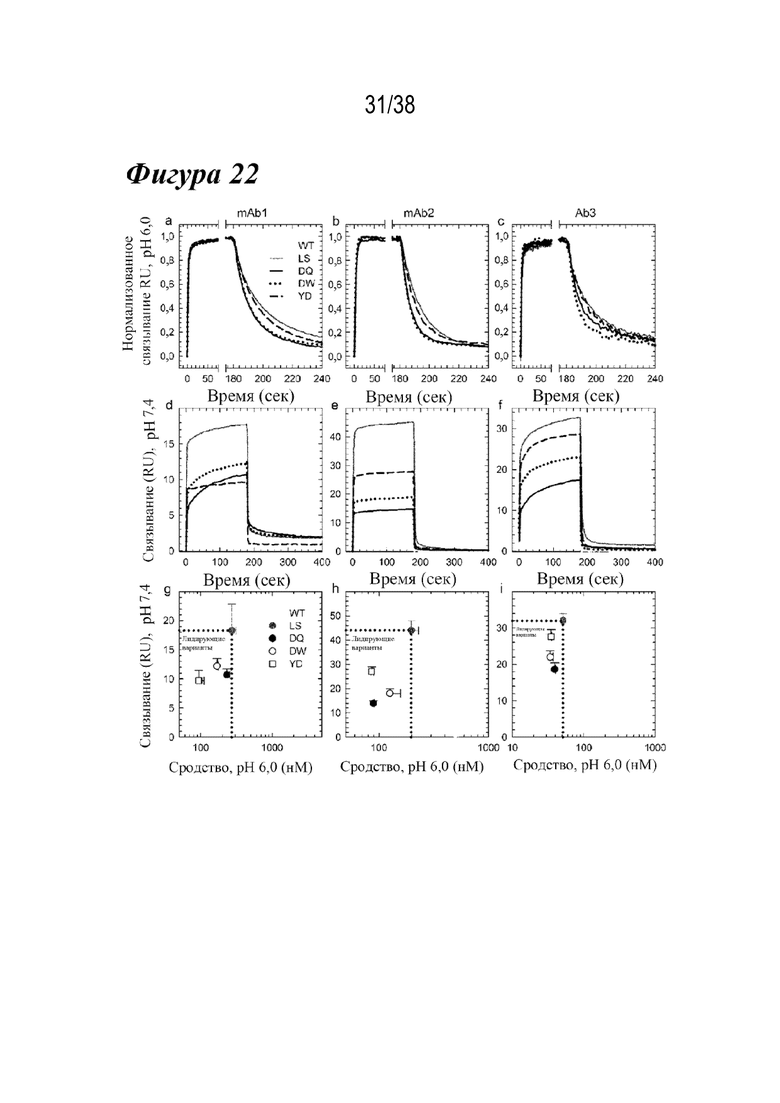
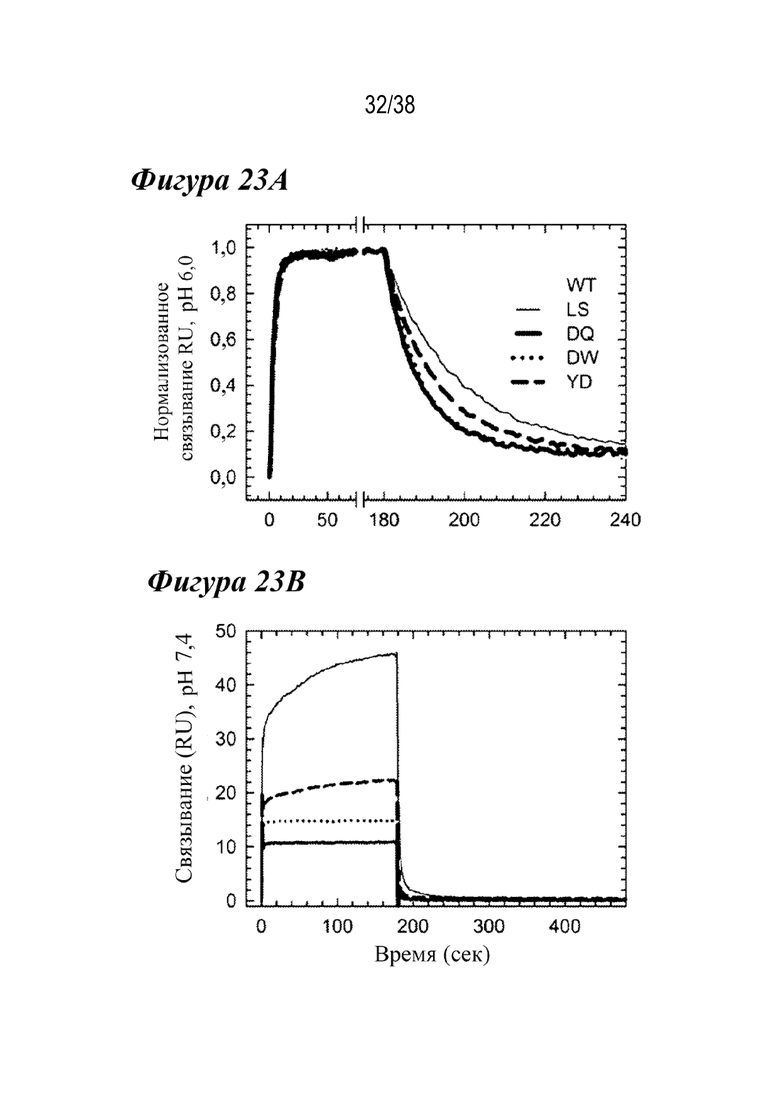
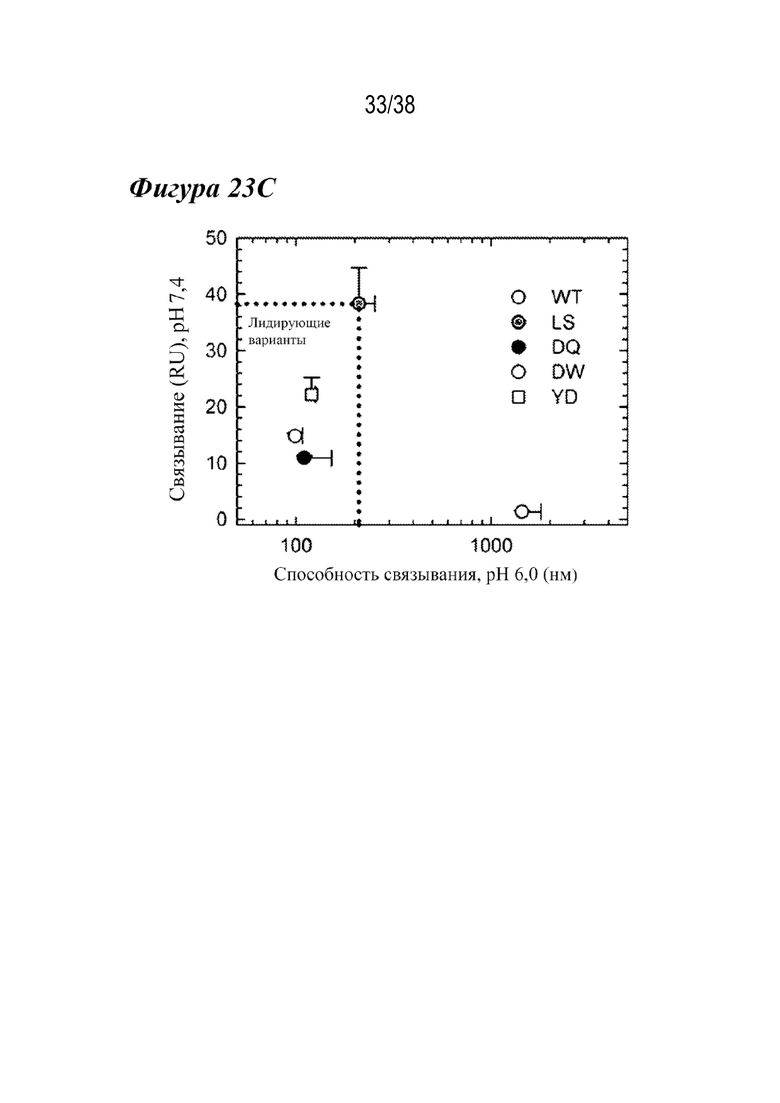
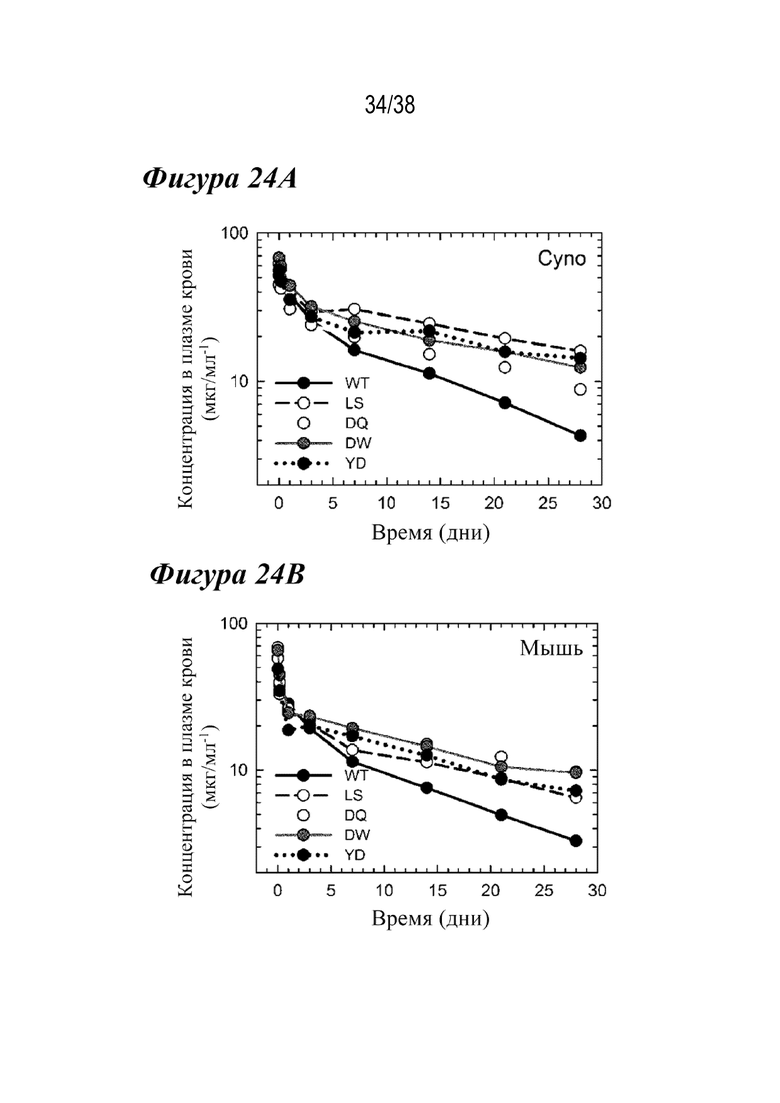
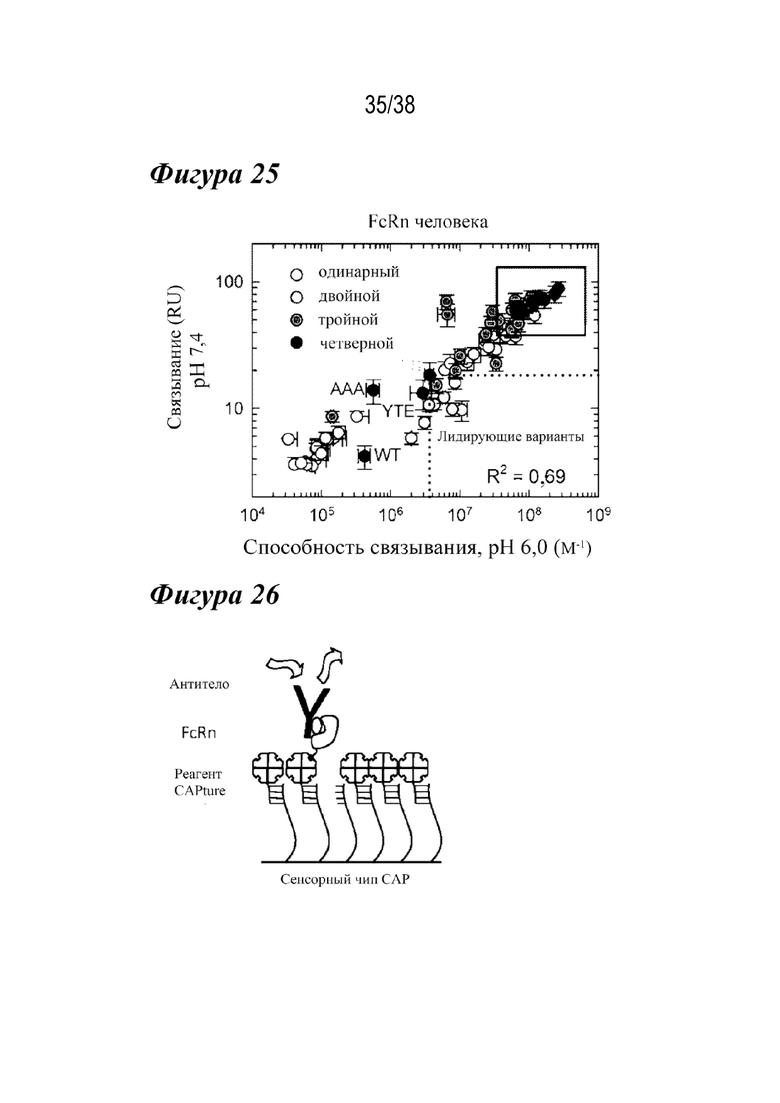
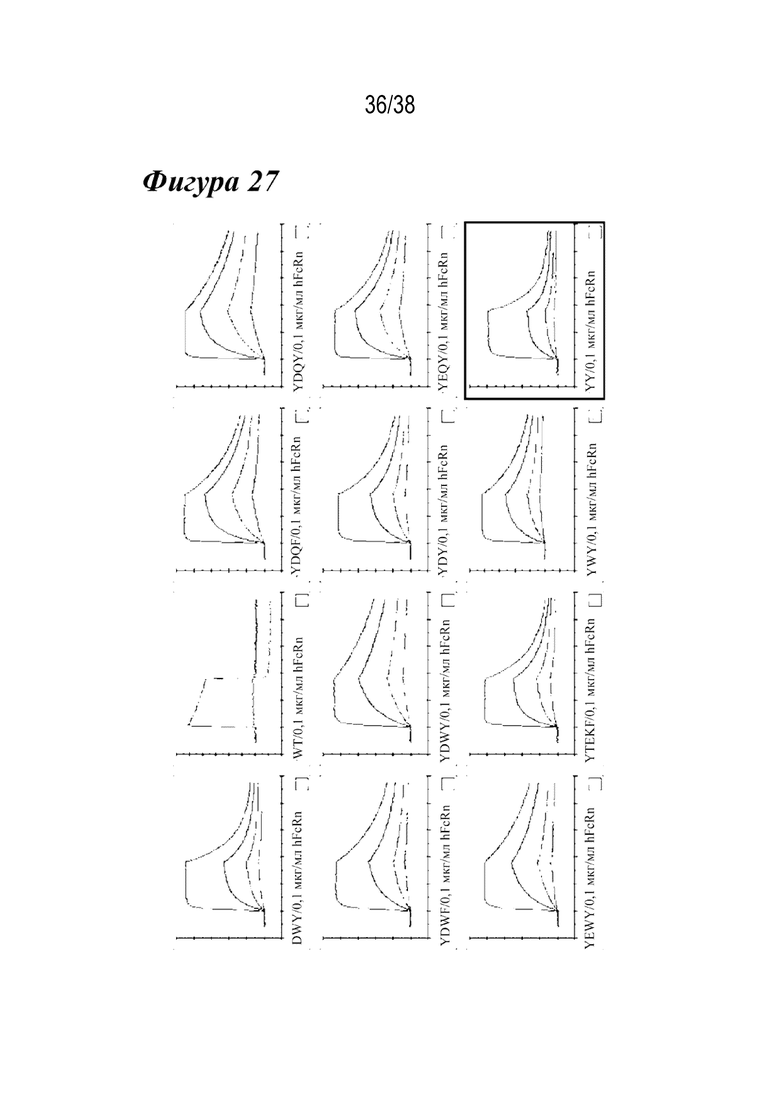
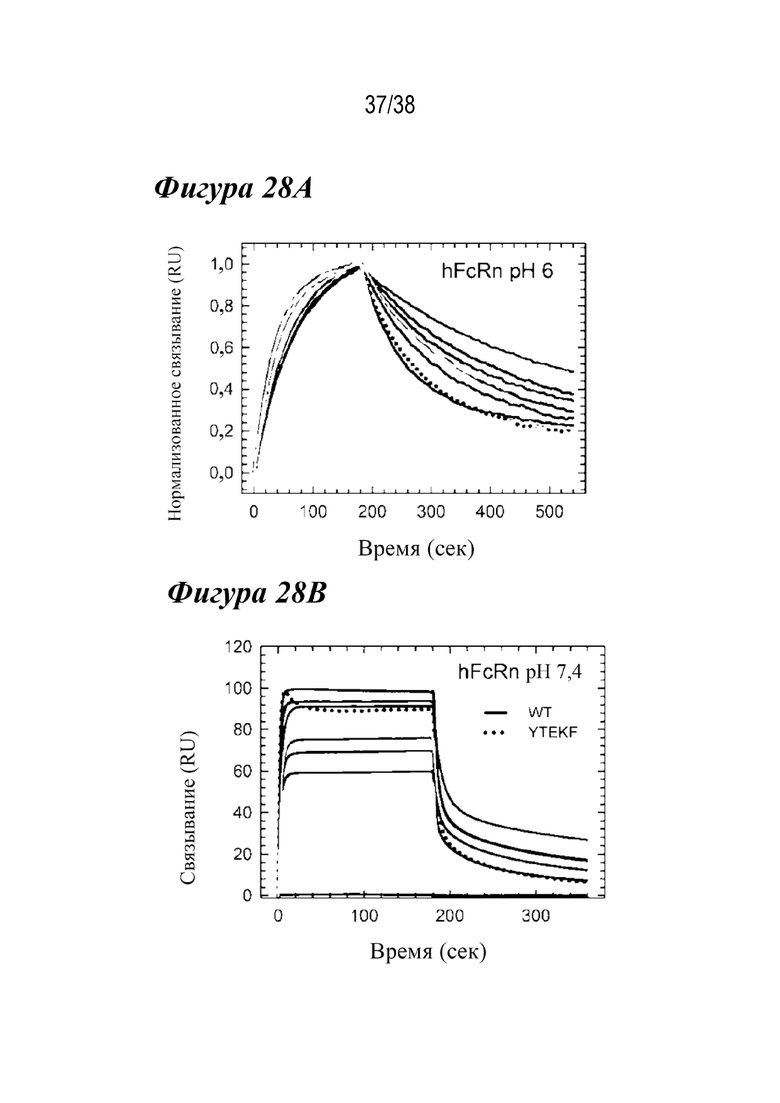
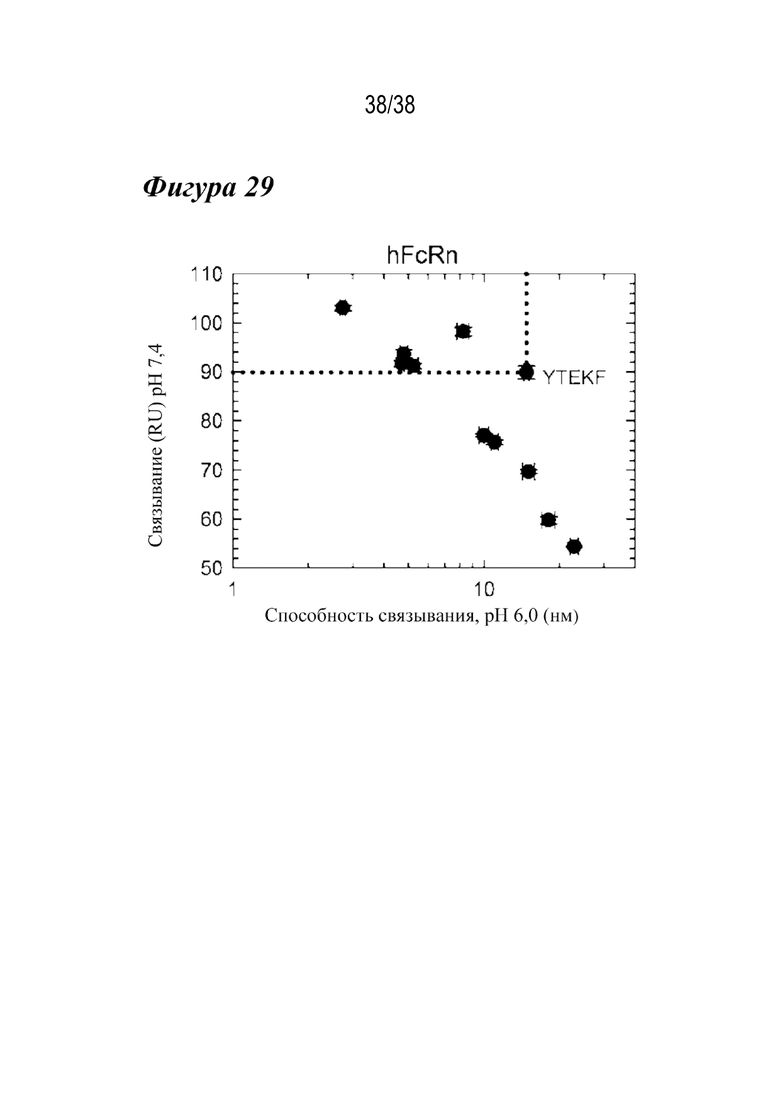
Настоящее изобретение относится к области биотехнологии, в частности к новым связывающим полипептидам, и может быть применимо в медицине. Изобретение раскрывает полипептиды с модифицированным Fc-доменом, в который были введены точечные мутации для повышения его способности связываться с FcRn и FcγRIIIa. Полипептиды, содержащие такой модифицированный Fc-домен, характеризуются увеличенным периодом полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа. 20 н. и 14 з.п. ф-лы, 29 ил., 10 табл., 13 пр.
1. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают:
аспарагиновую кислоту (D) или глутаминовую кислоту (E) в положении аминокислоты 256 и/или триптофан (W) или глутамин (Q) в положении аминокислоты 307, где положение аминокислоты 254 не является треонином (T), и дополнительно:
фенилаланин (F) или тирозин (Y) в положении аминокислоты 434; или
тирозин (Y) в положении аминокислоты 252,
где положения аминокислот соответствуют нумерации ЕС.
2. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека
(I) включают комбинацию аминокислотных остатков, выбранных из группы, состоящей из:
а) тирозина (Y) в положении аминокислоты 252 и аспарагиновой кислоты (D) в положении аминокислоты 256;
b) аспарагиновой кислоты (D) в положении аминокислоты 256 и фенилаланина (F) в положении аминокислоты 434;
c) аспарагиновой кислоты (D) в положении аминокислоты 256 и тирозина (Y) в положении аминокислоты 434;
d) триптофана (W) в положении аминокислоты 307 и фенилаланина (F) в положении аминокислоты 434;
е) тирозина (Y) в положении аминокислоты 252 и триптофана (W) в положении аминокислоты 307, где тирозин (Y) не находится в положении аминокислоты 434;
f) аспарагиновой кислоты (D) в положении аминокислоты 256 и триптофана (W) в положении аминокислоты 307, где тирозин (Y) не находится в положении аминокислоты 434;
g)тирозина (Y) в положении аминокислоты 252, аспарагиновой кислоты (D) в положении аминокислоты 256 и глутамина (Q) в положении аминокислоты 307, где тирозин (Y) не находится в положении аминокислоты 434; и
h) тирозина (Y) в положении аминокислоты 252, глутаминовой кислоты (E) в положении аминокислоты 256 и глутамина (Q) в положении аминокислоты 307, где треонин (T) не находится в положении аминокислоты 254, гистидин (H) не находится в положении аминокислоты 311, а тирозин (Y) не находится в положении аминокислоты 434; или
(II) состоит из аспарагиновой кислоты (D) в положении аминокислоты 256 и глутамина (Q) в положении аминокислоты 307, где тирозин (Y) не находится в положении аминокислоты 434;
где аминокислотные замены соответствуют нумерации ЕС.
3. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека:
a) включают двойную аминокислотную замену, выбранную из группы, состоящей из M252Y/T256D, M252Y/T256E, M252Y/T307Q, M252Y/T307W, T256D/T307W, T256E/T307Q и T256E/T307W, где треонин (T) не находится в положении аминокислоты 254, гистидин (H) не находится в положении аминокислоты 311, а тирозин (Y) не находится в положении аминокислоты 434;
b) состоят из двойной аминокислотной замены T256D/T307Q; или
с) включают тройную аминокислотную замену, выбранную из группы, состоящей из M252Y/T256D/T307Q, M252Y/T256D/T307W, M252Y/T256E/T307Q и M252Y/T256E/T307W, где треонин (T) не находится в положении аминокислоты 254, гистидин (H) не находится в положении аминокислоты 311, а тирозин (Y) не находится в положении аминокислоты 434;
где аминокислотные замены соответствуют нумерации ЕС.
4. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека состоят из аспарагиновой кислоты (D) в положении аминокислоты 256 и глутамина (Q) в положении аминокислоты 307, согласно нумерации ЕС.
5. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают аспарагиновую кислоту (D) в положении аминокислоты 256 и триптофан (W) в положении аминокислоты 307, согласно нумерации ЕС.
6. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают тирозин (Y) в положении аминокислоты 252 и аспарагиновую кислоту (D) в положении аминокислоты 256, согласно нумерации ЕС.
7. Выделенный связывающий полипептид по любому из пп. 1-6, где выделенный связывающий полипептид имеет более длинный период полувыведения из сыворотки крови по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа.
8. Выделенный связывающий полипептид по любому из пп. 1-7, где выделенный связывающий полипептид имеет повышенную способность связывания с FcRn при кислотном значении pH по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где необязательно кислотное значение pH составляет примерно 6,0.
9. Выделенный связывающий полипептид по любому из пп. 1-8, где выделенный связывающий полипептид имеет более высокую способность связывания с FcRn при кислотном значении pH по сравнению со способностью связывания с FcRn связывающего полипептида при некислотном значении pH, где необязательно кислотное значение pH составляет примерно 6,0, и где необязательно некислотное значение pH составляет примерно 7,4.
10. Выделенный связывающий полипептид по любому из пп. 1-9, где выделенный связывающий полипептид имеет измененную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где необязательно FcγRIIIa представляет собой FcγRIIIa человека.
11. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают:
тирозин (Y) в положении аминокислоты 252,
аспарагиновую кислоту (D) или глутаминовую кислоту (E) в положении аминокислоты 256,
триптофан (W) или глутамин (Q) в положении аминокислоты 307, и
фенилаланин (F) или тирозин (Y) в положении аминокислоты 434,
где положения аминокислот соответствуют нумерации ЕС.
12. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают комбинацию аминокислотных остатков, выбранных из группы, состоящей из:
a) тирозина (Y) в положении аминокислоты 252, аспарагиновой кислоты (D) в положении аминокислоты 256, глутамина (Q) в положении аминокислоты 307 и тирозина (Y) в положении аминокислоты 434;
b) тирозина (Y) в положении аминокислоты 252, глутаминовой кислоты (E) в положении аминокислоты 256, триптофана (W) в положении аминокислоты 307 и тирозина (Y) в положении аминокислоты 434;
с) тирозина (Y) в положении аминокислоты 252, глутаминовой кислоты (E) в положении аминокислоты 256, глутамина (Q) в положении аминокислоты 307 и тирозина (Y) в положении аминокислоты 434;
d) тирозина (Y) в положении аминокислоты 252, аспарагиновой кислоты (D) в положении аминокислоты 256, глутамина (Q) в положении аминокислоты 307 и фенилаланина (F) в положении аминокислоты 434;
е) тирозина (Y) в положении аминокислоты 252, аспарагиновой кислоты (D) в положении аминокислоты 256, триптофана (W) в положении аминокислоты 307 и тирозина (Y) в положении аминокислоты 434; и
f) тирозина (Y) в положении аминокислоты 252, аспарагиновой кислоты (D) в положении аминокислоты 256, триптофана (W) в положении аминокислоты 307 и фенилаланина (F) в положении аминокислоты 434,
где аминокислотные замены соответствуют нумерации ЕС.
13. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают четверную аминокислотную замену, выбранную из группы, состоящей из
M252Y/T256D/T307Q/N434Y,
M252Y/T256E/T307W/N434Y,
M252Y/T256E/T307Q/N434Y,
M252Y/T256D/T307Q/N434F,
M252Y/T256D/T307W/N434Y и
M252Y/T256D/T307W/N434F,
где аминокислотные замены соответствуют нумерации ЕС.
14. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают тирозин (Y) в положении аминокислоты 252, аспарагиновую кислоту (D) в положении аминокислоты 256, глутамин (Q) в положении аминокислоты 307 и тирозин (Y) в положении аминокислоты 434, согласно нумерации ЕС.
15. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают тирозин (Y) в положении аминокислоты 252, глутаминовую кислоту (Е) в положении аминокислоты 256, триптофан (W) в положении аминокислоты 307 и тирозин (Y) в положении аминокислоты 434, согласно нумерации ЕС.
16. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают тирозин (Y) в положении аминокислоты 252, глутаминовую кислоту (Е) в положении аминокислоты 256, глутамин (Q) в положении аминокислоты 307 и тирозин (Y) в положении аминокислоты 434, согласно нумерации ЕС.
17. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают тирозин (Y) в положении аминокислоты 252, аспарагиновую кислоту (D) в положении аминокислоты 256, глутамин (Q) в положении аминокислоты 307 и фенилаланин (F) в положении аминокислоты 434, согласно нумерации ЕС.
18. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают тирозин (Y) в положении аминокислоты 252, аспарагиновую кислоту (D) в положении аминокислоты 256, триптофан (W) в положении аминокислоты 307 и тирозин (Y) в положении аминокислоты 434, согласно нумерации ЕС.
19. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают:
a) тирозин (Y) в положении аминокислоты 252 и тирозин (Y) в положении аминокислоты 434,
b) тирозин (Y) в положении аминокислоты 252, триптофан (W) в положении аминокислоты 307 и тирозин (Y) в положении аминокислоты 434,
c) тирозин (Y) в положении аминокислоты 252, аспарагиновую кислоту (D) в положении аминокислоты 256 и тирозин (Y) в положении аминокислоты 434, или
d) аспарагиновую кислоту (D) в положении аминокислоты 256, триптофан (W) в положении аминокислоты 307 и тирозин (Y) в положении аминокислоты 434,
где положения аминокислот соответствуют нумерации ЕС.
20. Выделенный связывающий полипептид, содержащий измененный Fc-домен IgG человека, имеющий повышенную способность связывания с FcRn по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где модификации Fc-домена IgG человека включают комбинацию аминокислотных замен, выбранных из группы, состоящей из
M252Y/N434Y,
M252Y/T307W/N434Y,
M252Y/T256D/N434Y и
T256D/307W/N434Y,
где аминокислотные замены соответствуют нумерации ЕС.
21. Выделенный связывающий полипептид по любому из пп. 11-20, где измененный Fc-домен IgG человека имеет повышенную способность связывания с FcRn при кислотном значении pH и повышенную способность связывания с FcRn при некислотном значении pH по сравнению с Fc-доменом дикого типа, где необязательно кислотное значение pH составляет примерно 6,0, и где необязательно некислотное значение pH составляет примерно 7,4, где дополнительно необязательно FcRn представляет собой FcRn человека.
22. Выделенный связывающий полипептид по любому из пп. 11-21, где измененный Fc-домен IgG человека имеет повышенную способность связывания с FcRn при кислотном значении pH и повышенную способность связывания с FcRn при некислотном значении pH по сравнению с Fc-доменом, содержащим пять аминокислотных замен M252Y/S254T/T256E/H433K/N434F, где необязательно кислотное значение pH составляет примерно 6,0, и где необязательно некислотное значение pH составляет примерно 7,4, где дополнительно необязательно FcRn представляет собой FcRn человека.
23. Выделенный связывающий полипептид по любому из пп. 11-22, где измененный Fc-домен IgG человека имеет пониженную способность связывания с FcγRIIIa по сравнению со связывающим полипептидом, содержащим Fc-домен дикого типа, где дополнительно необязательно FcγRIIIa представляет собой FcγRIIIa человека.
24. Выделенный связывающий полипептид по любому из пп. 11-23, где измененный Fc-домен IgG человека имеет пониженную способность связывания с FcγRIIIa по сравнению с Fc-доменом, содержащим пять аминокислотных замен M252Y/S254T/T256E/H433K/N434F, где дополнительно необязательно FcγRIIIa представляет собой FcγRIIIa человека.
25. Выделенный связывающий полипептид по любому из пп. 1-24, где измененный Fc-домен IgG человека представляет собой измененный Fc-домен IgG1 человека.
26. Выделенный связывающий полипептид по любому из пп. 1-25, где связывающий полипептид обладает способностью связывания с FcRn человека, и, необязательно, способностью связывания с FcRn крысы.
27. Выделенный связывающий полипептид по любому из пп. 1-26, где выделенный связывающий полипептид специфически связывает одну или несколько мишеней у человека.
28. Выделенный связывающий полипептид по любому из пп. 1-27, где выделенный связывающий полипептид представляет собой моноклональное антитело.
29. Выделенный связывающий полипептид по п.28, где антитело представляет собой химерное, гуманизированное или человеческое антитело.
30. Выделенная молекула нуклеиновой кислоты для кодирования выделенного связывающего полипептида по любому из пп. 1-29, содержащая нуклеотидную последовательность, кодирующую выделенный связывающий полипептид по любому из пп. 1-29.
31. Вектор экспрессии, содержащий выделенную молекулу нуклеиновой кислоты по п.30.
32. Клетка-хозяин для получения выделенного связывающего полипептида по любому из пп. 1-29, содержащая вектор экспрессии по п.31.
33. Клетка-хозяин по п.32, где клетка-хозяин представляет собой клетку млекопитающего.
34. Способ получения выделенного связывающего полипептида, включающий
культивирование клетки-хозяина по п.33 в условиях, которые обеспечивают экспрессию связывающего полипептида, и
выделение связывающего полипептида из культуры.
| WO 2014177459 A2, 06.11.2014 | |||
| Способ получения синтетических кислот путем окисления минеральных масел | 1934 |
|
SU42072A1 |
| ACQUA W.F.D | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Универсальный двойной гаечный ключ | 1920 |
|
SU169A1 |
| WO 2013011076 A2, 24.01.2013 | |||
| WO 2011089211 A1, 28.07.2011 | |||
| US 9023996 B2, 05.05.2015 | |||
| WO 2016071376 | |||

