Настоящее изобретение имеет отношение к способу идентификации кандидатных пептидов, презентируемых главным комплексом гистосовместимости (MHC) для вмешательств in vivo и/или in vitro, включая вакцинацию, индукцию иммунологической толерантности, блокирование TCRs и MHC-опосредованную доставку токсинов, для проверки иммуногенности и других in vitro тестов на реакционную способность T-клеток.
T-клетки играют центральную роль в иммунной системе. Функционирование T-клеток критически зависит от распознавания эпитопов, содержащихся в пептидах, представленных антиген-презентирующими клетками (APCs). APCs могут быть ядросодержащими клетками почти любого типа, однако, особенно эффективными в презентировании являются специализированные APCs, такие как дендритные клетки (DCs). Существуют разные группы T-клеток с различными функциями, в частности CD4+ T-клетки или CD8+ T-клетки, которые распознают MHCII-связанные эпитопы, представленные специализированными APCs, или MHCI-связанные эпитопы, представленные большинством типов клеток, соответственно. Молекулы MHC играют важную роль в презентировании клеточных антигенов или эпитопов в форме коротких линейных пептидов T-клеткам.
MHC состоит из альфа и бета цепей и пептида, закрепленного в бороздке, образованной этими цепями. Стабильность этого комплекса очень зависит от связывания пептида, вследствие чего «ненагруженные» молекулы MHC на клеточной поверхности понижающе регулируются и разрушаются. Тем не менее, «ненагруженные» молекулы MHC существуют на поверхности клеток и могут связывать внеклеточные пептиды и представлять их T-клеткам. Более того, были описаны два различных изомера «ненагруженных» MHC (Rabunowitz et al., Immunity, 9, 699–709 (1998)): менее устойчивый активный изомер и более устойчивый неактивный изомер. Набор MHC отдельного индивидуума является полигенным, и гены MHC являются высокополиморфными. Таким образом, набор MHCs, который может экспрессироваться у отдельного индивидуума, вероятнее всего очень отличается от набора другого произвольно выбранного индивидуума.
Несущие пептид MHCs взаимодействуют с T-клеточными рецепторами (TCRs), присутствующими на поверхности T-клеток, что приводит к T-клеточной активации. APCs, такие как дендритные клетки или макрофаги, могут интернализировать антигены путем фагоцитоза или рецептор-опосредованного эндоцитоза.
Для того, чтобы управлять активностью T-клеток, важно установить их специфичные пептид-лиганды и контролировать или модулировать взаимодействие MHC-пептидных комплексов с TCRs. Поэтому важно определить связывающую способность пептида с MHC. Подходящие пептиды или комплексы MHC-пептид могут использоваться в самых разных областях применения. Например, их можно использовать для идентификации или обогащения реактивных T-клеток или для анализа на иммуногенность in vitro. Также они могут использоваться для блокирования TCRs, для доставки токсинов в T-клетки in vivo или для того, чтобы перенацеливать T-клетки с помощью MHC-содержащих химерных рецепторов антигенов, MHC-CARs (Jyothi et al., Nat.Biotech, 20, 1215–1220 (2002)), чтобы вызвать иммунологическую толерантность к эпитопу, который иначе будет активировать T-клетки, специфичные для этого эпитопа. Создание вакцин, в частности противораковых вакцин на основе опухолевого специфического антигена (TSA) с целью активации T-клеток для уничтожения раковых клеток, входит в число основных целей современной медицины. Опухолеспецифичные пептиды, так называемые неоантигены, присутствуют только в опухолевых клетках и полностью отсутствуют в нормальной ткани. Такие опухолеспецифичные пептиды создаются специфичными для опухоли изменениями ДНК, которые приводят к образованию новых белковых последовательностей. Вследствие генетической гетерогенности клеток в пределах одной или между разными опухолями, даже в том случае, когда они происходят из одного и того же органа, набор неоантигенов, присутствующих в опухоли пациента, является в большинстве случаев специфичным для этого пациента. Опухолеспецифичные пептиды «выставляются» в рамках MHC на поверхности опухолевых клеток и так называемых антиген-презентирующих клеток (APCs). Как только опухолеспецифичные пептиды презентированы на клеточной поверхности, они могут быть распознаны и разрушены иммунной системой. Таким образом, вакцины, содержащие опухолеспецифичные пептиды, могут стимулировать иммунную систему к обнаружению и уничтожению раковых клеток, которые презентируют эти молекулы на своей поверхности.
Совсем недавно было показано, что TSAs, возникшие в результате соматических мутаций, накапливающихся в опухолях, зачастую распознаются лимфоцитами, инфильтрирующими опухоль (TILs) (Linnemann et al., Nat Med 1–7 (2014)). Секвенирование экзома опухолевой ДНК и сравнение с ДНК из здоровой ткани делает возможным обнаружение таких соматических мутаций, специфичных для опухолевой клетки пациента. Однако, в большинстве случаев обнаруживается множество мутаций (Vormehr et al., Curr Opin Immunol 39, 14–22 (2016)), но только некоторые из TSAs (т.е. опухоль-специфичные пептиды) должны смешиваться в одной вакцине, по техническим условиям, но также для того, чтобы свести к минимуму риск аутоиммунной реакции. Выбор этих TSAs имеет решающее значение, потому что антиген-презентирующие клетки (APCs) презентируют не все возможные пептиды на своих MHC-молекулах, а скорее набор, который соответствует конкретному аллелю MHC, который является специфичным для пациента.
Выбор пептидов, в частности, опухоль-cпецифичных пептидов, которые должны эффективно презентироваться APCs, имеет определяющее значение для многих вариантов применения, вовлекающих T-клетки, и остается ограничивающим фактором при создании противораковых вакцин. Чтобы решить эту проблему, кандидатные пептиды обычно классифицируют в соответствии с их MHC аффинностью связывания, спрогнозированной с помощью биоинформационного анализа (Sahin et al., Nature 1–19 (2017); Ott et al., Nature 547, 217–221 (2017)). Однако, новые данные показывают, что биоинформационное предсказание презентации пептида осуществляется довольно плохо, в частности для эпитопов MHC II класса (Sofron et al., Eur J Immunol 46, 319–328 (2015)). Раскрыт метод «мокрой химии», использующий антиген-презентирующую репортерную клеточную линию для измерения связывания T-клетки с комплексом MHC-пептид (через его TCR); WO2016097334 A1. Такая репортерная клеточная линия экспрессирует химерный антигенный рецептор, содержащий комплекс MHC-пептид, который активирует репортерный ген в репортерной клеточной линии после связывания с TCR T-клетки. Также с помощью такого метода опосредованно измеряется стабильность комплекса MHC-пептид, поскольку только достаточно стабильные комплексы MHC-пептид будут появляться на поверхности репортерной клеточной линии, взаимодействовать с T-клетками и, в конечном счете, активировать репортер. Однако, такой метод не обеспечивает, в частности, возможность измерения стабильности комплекса MHC-пептид, потому что активация репортерного гена также зависит от аффинности TCRs к эпитопам пептида и других иммуно-модулирующих факторов.
Следовательно, существует необходимость в обеспечении средств и методов для надежного установления пептидов, в частности TSAs, которые эффективно презентируются APCs.
Настоящее изобретение в данный момент удовлетворяет эту потребность, поскольку оно предоставляет указанные средства и методы, которые более подробно определены в пунктах формулы изобретения и следующих вариантах осуществления изобретения.
Основные аспекты изобретения имеют отношение к:
1. Способ идентификации кандидатных пептидов, презентированных главным комплексом гистосовместимости (MHC), включающий экспрессию в репортерной клеточной линии рекомбинантного комплекса MHC-пептид, содержащего ковалентно связанный кандидатный пептид, обнаружение репортерных клеток, которые демонстрируют поверхностную экспрессию комплекса MHC-пептид, и установление последовательности кандидатных пептидов, презентированных на клеточной поверхности.
2. Способ согласно 1 аспекту, включающий
(i) получение библиотек, содержащих кандидатные пептиды, клонированные впереди или в рамку считывания с рекомбинантной бета и/или альфа цепью MHC;
(ii) трансдукцию таких библиотек, вместе с соответствующей альфа и/или бета цепью MHC, в подходящие репортерные клеточные линии;
(iii) обнаружение и изолирование клеток, экспрессирующих один или более комплексов MHC-пептид на клеточной поверхности;
(iv) выделение ДНК из клеток, на клеточной поверхности которых присутствует один или более комплекс MHC-пептид;
(v) определение последовательности кандидатных пептидов, кодированных векторами, интегрированными в ДНК, выделенную из клеток, на клеточной поверхности которых присутствует комплекс(ы) MHC-пептид.
3. Способ идентификации кандидатных пептидов, презентированных главным комплексом гистосовместимости (MHC), включающий экспрессию в репортерной клеточной линии рекомбинантного комплекса MHC-пептид, содержащего ковалентно связанный кандидатный пептид, обнаружение репортерных клеток, которые демонстрируют поверхностную экспрессию комплекса MHC-пептид, и определение уровня поверхностной экспрессии комплекса MHC-пептид и последовательности кандидатных пептидов, представленных на клеточной поверхности.
4. Способ согласно 3 аспекту, включающий
(i) получение библиотек, содержащих кандидатные пептиды, клонированные впереди или в рамку считывания с рекомбинантной бета и/или альфа цепью MHC;
(ii) трансдукцию таких библиотек, вместе с соответствующей альфа и/или бета цепью MHC, в подходящие репортерные клеточные линии;
(iii) обнаружение и изолирование клеток, экспрессирующих один или более комплексов MHC-пептид на клеточной поверхности;
(iv) определение уровня экспрессии на клеточной поверхности одного или более комплексов MHC-пептид;
(v) выделение ДНК из клеток, которые презентируют один или более комплексов MHC-пептид на своей клеточной поверхности;
(vi) определение последовательности кандидатных пептидов, кодированных векторами, интегрированными в ДНК, выделенную из клеток, которые презентируют комплексы MHC-пептид на клеточной поверхности.
5. Способ согласно одному из аспектов 1 - 4, в котором молекула MHC является молекулой MHC класса II, содержащей внеклеточную альфа цепь MHC класса II и трансмембранный домен, а также внеклеточную бета цепь MHC класса II и трансмембранный домен.
6. Способ согласно одному из аспектов 1 - 4, в котором MHC молекула является молекулой MHC I класса, содержащей внеклеточную альфа цепь MHC класса I и трансмембранный домен, а также бета-2 микроглобулин.
7. Способ согласно одному из аспектов 1 - 4, в котором комплекс MHC-пептид является гибридным белком, содержащим кандидатный пептид, бета-2 макроглобулин, внеклеточную альфа цепь MHC класса I и трансмембранный домен.
8. Способ согласно аспекту 6 или 7, в котором альфа цепь MHC несет мутацию Y84A.
9. Способ любого из аспектов 1 - 8, в котором каждая цепь молекулы MHC содержит трансмембранный домен.
10. Способ любого из аспектов 1 - 9, в котором трансмембранный домен является нативным трансмембранным доменом молекулы MHC.
11. Способ любого из аспектов 1 - 9, в котором трансмембранный домен является трансмембранным доменом гетерологичной молекулы, такой как TCRб/в, TCRг/д, CD3г/д/е/ж, CD4 или CD8б/в.
12. Способ любого из аспектов 1 - 11, в котором репортерная клеточная линия является клеточной линией млекопитающего.
13. Способ любого из аспектов 1 - 12, в котором репортерная клеточная линия является клеточной линией, лишенной механизма «загрузки» пептидов MHC класса II.
14. Способ аспекта 13, в котором репортерная клеточная линия является T-клеточной гибридомой.
15. Способ любого из аспектов 1 - 14, в котором репортерная клеточная линия является клеточной линией, не имеющей функционального гена TAP1, TAP2 и/или бета-2-микроглобулина.
16. Способ аспекта 15, в котором репортерная клеточная линия является T-клеточной гибридомой с дефектным или удаленным геном TAP1, TAP2 и/или бета-2-микроглобулина.
17. Способ любого из предшествующих аспектов, в котором кандидатный пептид является опухолеспецифическим пептидом, несущим индивидуальную мутацию(и) опухолевого происхождения.
18. Способ аспекта 17, в котором указанная мутация представляет собой SNV.
19. Способ аспектов 1 - 16, в котором кандидатный пептид является антигеном, вызывающим иммунный ответ.
20. Способ согласно аспектам 1 - 16, в котором кандидатный пептид является соединением, подвергающимся тестированию на иммуногенность.
21. Способ любого из предшествующих аспектов, в котором репортерные клетки, эффективно экспрессирующие MHC на своей поверхности, обогащены посредством FACS или MACS сортировки клеток.
22. Способ любого из предшествующих аспектов, в котором последовательность кандидатных пептидов, презентированных на клеточной поверхности, определяется при помощи ПЦР и секвенирования.
23. Способ любого из аспектов 1 - 22, в котором кандидатный пептид предназначен для применения в качестве вакцины.
24. Способ аспекта 23, в котором вакцина является противораковой вакциной на основе опухолеспецифического антигена (TSA).
25. Способ любого одного из аспектов 1 - 22, в котором кандидатный пептид применяется для индукции иммунологической толерантности, по меньшей мере, в отношении одного из эпитопов, которые он содержит.
26. Способ любого из аспектов 1 - 22 или 25, в котором кандидатный пептид в случае с MHC молекулой применяется для блокировки TCRs.
27. Способ любого из аспектов 1 - 22 или 25, в котором кандидатный пептид применяется для MHC-опосредованной доставки токсина в клетки, в частности в T-клетки.
28. Способ любого из аспектов 1 - 22 или 25, в котором кандидатный пептид применяется для перенацеливания T-клеток с помощью MHC-CAR.
29. Способ любого из аспектов 1 - 22, в котором кандидатный пептид применяется в анализе на иммуногенность.
30. Способ любого из аспектов 1 - 22 или 29, в котором кандидатный пептид применяется в тестировании T-клеточной реактивности.
31. Способ определения MHC аффинности связывания кандидатных пептидов, включающий экспрессию в репортерной клеточной линии рекомбинантного комплекса MHC-пептид, содержащего ковалентно связанный кандидатный пептид, обнаружение репортерных клеток, которые презентируют комплекс MHC-пептид на поверхности репортерной клетки и определение уровня такой презентации/экспрессии.
32. Способ аспекта 31, включающий
(i) получение библиотек, содержащих кандидатные пептиды, клонированные впереди рекомбинантного альфа или бета домена MHC, соединенного, по меньшей мере, с одним трансмембранным участком;
(ii) трансдукцию таких библиотек, вместе с соответствующим MHC альфа или бета доменом, в подходящие репортерные клеточные линии;
(iii) обнаружение клеток, презентирующих один или более комплексов MHC-пептид на своей клеточной поверхности;
(iv) определение уровня такой презентации/экспрессии.
Настоящее изобретение предоставляет способы идентификации кандидатных пептидов, презентированных главным комплексом гистосовместимости (MHC), включающие экспрессию в репортерной клеточной линии рекомбинантного комплекса MHC-пептид, содержащего ковалентно связанный кандидатный пептид, обнаружение репортерных клеток, которые демонстрируют поверхностную экспрессию комплекса MHC-пептид, и определение последовательности кандидатных пептидов, презентированных на клеточной поверхности.
Изобретение, по меньшей мере, отчасти, основывается на неожиданном открытии, что уровень поверхностной экспрессии рекомбинантного комплекса пептид-MHC в клеточной линии млекопитающего может быть измерен количественно напрямую. Кроме того, удивительно, что такое прямое измерение комплекса пептид-MHC было возможно с помощью стандартного лабораторного метода, основанного на окрашивании антител и проточной цитометрии, и при отсутствии сложной системы совместного культивирования при участии T-клеток и/или усовершенствованной репортерной системы. Несмотря на то, что специалистам в данной области техники может быть известно, что предположения на основе биоинформационных подходов не всегда являются очень точными, все еще неожиданно, что современный уровень метода биоинформатики оказался полностью неспособным предсказать связывание пептида со специфичными MHC вариантами, что установлено с помощью способа, раскрытого в данном описании. Далее неожиданным является то, что обнаружение MHC, включенного в рекомбинантный комплекс MCR (пептид-MHC-TCR химера, раскрытая в WO2016097334 A1), было возможно с помощью CD3e антитела независимо от конкретного пептида и/или MHC варианта, который подлежит анализу. Без ограничения какой-либо теорией, раскрытый в этом описании способ позволяет измерять прежде всего биологическую вариабельность презентации пептидов посредством MHCs без технической вариабельности, которая может вводиться, когда разные антитела используются для обнаружения различных MHC вариантов.
Пептиды могут быть ковалентно присоединены к MHC молекулам, чтобы преодолеть необходимость процессинга пептидов/механизма «загрузки». Этот подход используется для получения стабильных MHC-тетрамеров (Crawford et al., Immunity 8, 675–682 (1998)), в различных способах MHC-дисплея (Crawford et al., PLoS Biol 2, e90 (2004); Wen et al., Protein Eng Des Sel 24, 701–709 (2011)), для получения мышей, экспрессирующих отдельные пептиды на APCs (Ignatowicz et al.,Cell 84, 521–529 (1996)) или в CARs, созданных, чтобы перенацеливать T-клетки в направлении других T-клеток (Jyothi et al., Nat Biotechnol 20, 1215–1220 (2002)). Зачастую MHC молекулы должны быть мутированы, чтобы обеспечить экспрессию на поверхности клеток дрожжей или клеток насекомых (Wen et al., Protein Eng Des Sel 24, 701–709 (2011)), и в некоторых исследованиях были предприняты меры для того, чтобы стабилизировать пептиды в бороздке MHC путем встраивания дополнительного дисульфидного мостика (Truscott et al., J. Immunol. 178, 6280–6289 (2007)).
Однако, в контексте настоящего изобретения показано, что в клетках, неспособных к «загрузке» пептидов на MHC, рекомбинантные молекулы MHC без связанных пептидов, являются необнаружимыми или нестабильными (обнаружимыми на низких уровнях) на клеточной поверхности, даже в случае сверхэкспрессии (фиг. 4).
Итак, относительно стабильные изомеры «незанятых» MHC, описанные Rabunowitz (Rabunowitz et al., Immunity, 9, 699–709 (1998)), даже если присутствуют, не сохраняются очень долго на поверхности непрофессиональных антиген-презентирующих клеток. Важно отметить, в контексте настоящего изобретения показано, что не все пептиды, соединенные с рекомбинантным MHC, приводят к эффективной экспрессии на клеточной поверхности комплекса MHC. В действительности, некоторые предотвращают экспрессию на поверхности (фиг. 2a и b). Таким образом, высокая экспрессия рекомбинантных молекул MHC с соединенными кандидатными пептидами на поверхности подходящих репортерных клеточных линий в отличие от неинфицированных репортерных клеток и/или репортерных клеток, трансдуцированных MHC без ковалентно связанного пептида или нестабильными комплексами MHC-пептид, могут использоваться в качестве индикатора эффективной MHC-презентации пептида. Другими словами, репортерные клетки, демонстрирующие поверхностную экспрессию рекомбинантного комплекса MHC-пептид, могут быть обнаружены путем сравнения поверхностной экспрессии указанного комплекса MHC-пептид с уровнем исходного сигнала неинфицированных репортерных клеток и/или репортерных клеток, трансдуцированных MHC без ковалентно связанного пептида или нестабильного комплекса MHC-пептид. Более того, измерение уровня такой презентации/экспрессии (фиг. 2a и 7) обеспечивает возможность упорядочивания пептидов в соответствии с их способностью связываться с MHC. Обнаружение и/или упорядочивание пептидов, презентированных MHCs, может включать, в случае необходимости, один или более этапов нормализации, например, нормализацию по уровню исходного сигнала соответствующего MHC варианта, не имеющего ковалентно связанного кандидатного пептида или проявляющего нестабильное взаимодействие с кандидатным пептидом или случайно выбранным пептидом. Методы нормалицации и/или стандартизации, которые хорошо известны в данной области техники, cмогут использоваться, в случае необходимости, для определения аффинности связывания кандидатного пептида с MHC, исходя из измеренной поверхностной экспрессии рекомбинантного комплекса MHC-пептид.
В некоторых вариантах осуществления упорядочивание кандидатных пептидов и/или определение аффинности связывания кандидатного пептида с MHC может прямо коррелировать с поверхностной экспрессией рекомбинантного комплекса MHC-пептид, определенной с помощью CD3 окрашивания с использованием анти-CD3e, анти-CD3д или анти-CD3г антитела. Предпочтительно, MHC входит в состав MCR для указанной корреляции с указанным CD3 окрашиванием.
В данной работе впервые было показано, что соответствующим образом созданный комплекс MHC класса II-пептид может использоваться для идентификации кандидатных пептидов, эффективно презентированных APCs.
Кроме того, в рамках настоящего изобретения было обнаружено, что не все комбинации MHC-класса II пептид эффективно экспрессируются на поверхности клеток, когда используются гибридные белки MHC-TCR (MCRs), состоящие из пептидов, соединенных с нативными внеклеточными доменами MHC, слитыми с трансмембранными частями TCR. Например, наблюдалась разная поверхностная экспрессия пептида, происходящего от матриксного белка 1 гриппа (MP1), когда были проверены разные комбинации MHC-пептид. Важно отметить, что предсказательный анализ связывания пептида не показывает подобную картину в отношении аффинности связывания MHC-пептид.
В различных вариантах осуществления изобретение имеет отношение к способу идентификации кандидатных пептидов согласно предыдущему варианту осуществления, описанному в этом документе, при этом данный способ включает следующие стадии:
(i) получение библиотек, содержащих кандидатные пептиды, в частности библиотек, содержащих кандидатные пептиды, клонированные впереди или в рамку считывания с рекомбинантной бета и/или альфа цепью MHC;
(ii) трансдукцию таких библиотек, вместе с соответствующим альфа или бета доменом MHC, в подходящие репортерные клеточные линии;
(iii) обнаружение клеток, презентирующих один или более комплексов MHC-пептид на своей клеточной поверхности;
(iv) выделение ДНК из клеток, которые презентируют один или более комплексов MHC-пептид на их клеточной поверхности;
(v) определение последовательности кандидатного пептида в комплексе MHC-пептид, презентированном на клеточной поверхности.
Использованный в описании термин “главный комплекс гистосовместимости (MHC)” относится к генам MHC Класса I и MHC Класса II и белкам, кодированным этими генами. Термины MHC-I, MHC-II, MHC-1 и MHC-2 различным образом используются в этом документе, чтобы обозначать эти классы молекул.
В некоторых вариантах осуществления изобретение имеет отношение к способу идентификации кандидатных пептидов, презентированных главным комплексом гистосовместимости (MHC), включающему экспрессию в репортерной клеточной линии рекомбинантного комплекса MHC-пептид, содержащего ковалентно связанный кандидатный пептид, обнаружение репортерных клеток, которые демонстрируют поверхностную экспрессию комплекса MHC-пептид, и определение уровня поверхностной экспрессии комплекса MHC-пептид и последовательности кандидатных пептидов, презентированных на клеточной поверхности.
В различных вариантах осуществления изобретение имеет отношение к способу идентификации кандидатных пептидов согласно предшествующему варианту осуществления, описанному в данном документе, при этом указанный способ включает следующие стадии:
(i) получение библиотек, содержащих кандидатные пептиды, клонированные впереди или в рамку считывания вместе с рекомбинантной бета и/или альфа цепью MHC;
(ii) трансдукцию таких библиотек, вместе с соответствующей MHC альфа и/или бета цепью, в подходящие репортерные клеточные линии;
(iii) обнаружение и изолирование клеток, которые экспрессируют один или более комплексов MHC-пептид на клеточной поверхности;
(iv) определение уровня экспрессии на клеточной поверхности одного или более комплексов MHC-пептид;
(v) выделение ДНК из клеток, которые презентируют один или более комплексов MHC-пептид на их клеточной поверхности;
(vi) определение последовательности кандидатных пептидов, кодированных векторами, интегрированными в ДНК, выделенную из клеток, которые презентируют комплекс MHC-пептид на клеточной поверхности.
У млекопитающих существует два типа молекул MHC, обозначенных как молекулы класса I и класса II. Молекулы MHC класса I присутствуют почти на всех ядросодержащих клетках в организме и распознаются CD8+ клетками. Молекулы MHC класса II преимущественно экспрессируются на профессиональных APCs иммунной системы и распознаются CD4+ клетками. Эти молекулы дополнительно подразделяются на антигенные подтипы. Человеческие молекулы MHC класса I, также называемые человеческими лейкоцитарными антигенами (HLA), обозначаются как HLA-A, -B и -C. Человеческие молекулы MHC класса II обозначаются как HLA-DR, -DQ и -DP.
Молекулы MHC класса I обнаруживаются почти в каждой клетке организма. Молекулы MHC класса I являются гетеродимерами, состоящими из полиморфной тяжелой цепи (б), нековалентно связанной с мономорфной (у людей) не-MHC кодированной легкой (в) цепью белка около 12 kDa, называемого в2 микроглобулин (в2m). Тяжелая б цепь представляет собой полиморфный трансмембранный гликопротеин около 45kDa, состоящий из 3 внеклеточных доменов, каждый из которых содержит около 90 аминокислот (б1 на N-конце, б2 и б3), трансмембранного участка примерно из 40 аминокислот и цитоплазматического «хвоста» примерно из 30 аминокислот. Домены б1 и б2, мембранные дистальные домены, формируют пептид-связывающую бороздку или щель, имеющую достаточный размер для связывания с пептидом из 8-10 аминокислот, тогда как б3 домен является расположенным ближе (проксимально) к плазматической мембране. Будучи презентируемым, пептид удерживается пептид-связывающей бороздкой в центральном участке б1 и б2 доменов.
Молекулы MHC Класса II обнаруживаются только на некоторых типах специализированных клеток, в частности антиген-презентирующих клетках (APCs), таких как макрофаги, дендритные клетки и B-клетки. Молекулы MHC Класса II содержат две различных полипептидных цепи, 33-kD б цепь и 28-kDa в цепь, которые связаны нековалентными взаимодействиями. Подобно молекулам MHC класса I, молекулы MHC класса II являются мембрано-связанными гликопротеинами, содержащими внеклеточные домены, трансмембранный сегмент и цитоплазматический «хвост».
Каждая цепь в этих нековалентных гетеродимерных комплексах содержит два внеклеточных домена, трансмембранный домен и цитоплазматический «хвост». Внеклеточными доменами являются б1 и б2 домены и в1 и в2 домены для б цепи и в цепи, соответственно. Мембрано-дистальный домен молекулы класса II состоит из б1 и в1 доменов и образует пептид-связывающую бороздку или щель, имеющую достаточный размер для связывания с пептидом, который, как правило, состоит из 13-18 аминокислот в длину, альтернативно 10-18 аминокислот или больше. Мембрано-проксимальные пары класса II, б2 и в2, имеют структурное сходство с Ig константными (C) доменами. У людей существует три пары генов б и в цепи класса II, известные как HLA-DR, HLA-DP и HLA-DQ. Самый высокий уровень полиморфизма документально подтвержден для HLA-DR.
При использовании в описании кандидатный пептид имеет отношение к пептиду из 8-18 аминокислот или до 50 аминокислот в длину. Предпочтительная длина кандидатного пептида, связывающегося с MHC класса I, составляет 8, 9 или 10 аминокислот. Предпочтительная длина кандидатного пептида, связывающегося с MHC класса II, составляет 10, 11, 12, 13, 14, 15, 16, 17 или 18 аминокислот. В частности, кандидатный пептид входит в состав комплекса MHC-пептид, кодированного рекомбинантной нуклеиновой кислотой.
В различных вариантах осуществления изобретение имеет отношение к способу согласно любому из предшествующих вариантов осуществления, описанных в этом документе, при этом молекула MHC является молекулой MHC класса II, содержащей внеклеточную альфа цепь MHC класса II и трансмембранный домен, а также внеклеточную бета цепь MHC класса II и трансмембранный домен.
В других вариантах осуществления молекула MHC является молекулой MHC класса I, содержащей внеклеточную альфа цепь MHC класса I и трансмембранный домен, а также бета-2 микроглобулин.
В еще одном варианте осуществления комплекс MHC-пептид является гибридным белком, содержащим кандидатный пептид, бета-2 макроглобулин, внеклеточную альфа цепь MHC класса I и трансмембранный домен.
В отношении молекул MHC класса I было показано, что Y84A мутация в альфа цепи открывает связывающий пептид «карман», дающий возможность лучшего размещения связанного пептида (Mitaksov, V. et al. Chem Biol 14, 909–922 (2007)).
Соответственно, в отдельном варианте осуществления MHC альфа цепь молекулы MHC класса I несет мутацию Y84A. Например, мутация Y84A может быть введена с помощью сайт-направленного мутагенеза.
В некоторых вариантах осуществления изобретение имеет отношение к способу согласно любому из предшествующих вариантов осуществления, описанных в этом документе, при этом молекула MHC содержит один или два трансмембранных домена.
В различных вариантах осуществления изобретение имеет отношение к способу согласно любому из предшествующих вариантов осуществления, описанных в этом документе, при этом трансмембранный домен является нативным трансмембранным доменом молекулы MHC.
Использованный в описании термин “нативный трансмембранный домен” относится к трансмембранному домену MHC, который сохраняет конструктивную особенность трансмембранного участка молекулы MHC в его нативном состоянии.
В конкретных вариантах осуществления трансмембранный домен является трансмембранным доменом гетерологичной молекулы, включая, но не ограничиваясь этим, TCRб/в, TCRг/д, CD3г/д/е/ж, CD4 или CD8б/в.
Репортерная клеточная линия для осуществления способа изобретения может быть любой клеточной линией или производной от клеточной линии, которая может использоваться в биологических вариантах применения. Подходящими линиями клеток или их производными являются, например, но без ограничения, клетки, происходящие от человека, мыши, крысы, обезьяны, хомяка, рыбы, лягушки или насекомого, или дрожжевые клетки, например, но не ограничиваясь этим, HEK, CHO, 3T3, гибридомы, HeLa, Sf9, Шнайдер 2 клетки, Jurkat, HUVEC, HL-60, MCF-7, Saos-2 клетки, IMR-90, Raw 264.7, PC3, Vero, Cos, GH3, PC12, Dog MDCK, Xenopus A6 или Zebrafish AB9.
В отдельном варианте осуществления указанная репортерная клеточная линия является клеточной линией млекопитающего.
Подходящими клеточными линиями млекопитающего являются, например, но без ограничения, T-клеточные гибридомы, T-клеточные клоны, Jurkat, BW5147б-в-сливающиеся партнеры, миелома, HEK, CHO, 3T3.
В частности, репортерная клеточная линия, подходящая для использования в настоящем изобретении, предпочтительно клеточная линия млекопитающего, является клеточной линией, утратившей способность к экспонированию пептидов, происходящих из эндогенных белков, исходно существующих в клетке. Таким образом, клетка должна презентировать исключительно комплексы MHC-пептид, которые несут кандидатные пептиды, полученные из представляющей интерес библиотеки пептидов. Процессинг пептидов и их «загрузка» на молекулы MHC для экспонирования является многоэтапным процессом, в котором используется множество видов молекул, которые определяют механизм процессинга и презентирования. Молекулы MHC класса I «нагружаются» пептидами посредством транспортера, называемого TAP (транспортер, связанный с процессированием антигена), который перемещает пептиды в ER, где они связываются с молекулами MHC класса I. «Загрузка» молекулы MHC класса II происходит посредством фагоцитоза или аутофагии; молекулы MHC класса II доставляются в фаголизосомы и нагружаются пептидами до их перемещения на клеточную поверхность. Этот механизм процессирования пептида/»загрузки» для молекул MHC класса II присутствует исключительно в APCs.
В соответствии со сказанным выше, в одном варианте осуществления репортерная клеточная линия, подходящая для осуществления способа изобретения, является линией, не имеющей механизма для «загрузки» пептидов на MHC класса II. Предпочтительно, репортерная клеточная линия, не имеющая механизма для «загрузки» пептидов на MHC класса II пептид, является линией клеток млекопитающего.
В отдельном варианте осуществления репортерная клеточная линия является T-клеточной гибридомой.
В другом варианте осуществления репортерная клеточная линия, подходящая для осуществления способа изобретения, является клеточной линией, не имеющей функционального гена TAP1, TAP2 и/или бета-2-микроглобулина.
В отдельном варианте осуществления репортерная клеточная линия является T-клеточной гибридомой с дефектным или удаленным геном TAP1, TAP2 и/или геном бета-2-микроглобулина. Другие примеры репортерных клеточных линий с дефектным или удаленным геном TAP1, TAP2 и/или бета-2-микроглобулина включают, но не ограничиваются этим, T- и B-клеточные клоны или гибридомы, HEK клетки или 3T3 клетки.
Секвенирование экзома может использоваться для идентификации TSAs, которые присутствуют только лишь в опухоли. Например, TSAs могут быть идентифицированы путем секвенирования опухолевой ДНК, полученной от пациентов, и сравнения ее с ДНК, полученной от здоровых субъектов. TSAs также могут быть идентифицированы для отдельных пациентов путем секвенирования опухолевой и нормальной ДНК каждого пациента. На основе этого анализа, множество кандидатных пептидов, каждый из которых имеет мутацию(и) опухолевого происхождения, может использоваться для получения комплексов MHC-пептид. Таким образом, в одном варианте осуществления изобретения используется библиотека пептидов. Эти пептиды могут нести мутацию(и) опухолевого происхождения или быть нативными пептидами.
Соответственно, в различных вариантах осуществления изобретение имеет отношение к способу согласно любому из предшествующих вариантов осуществления, описанных в данном документе, при этом кандидатный пептид является опухолеспецифическим пептидом, несущим индивидуальную мутацию(и) опухолевого происхождения.
Термин «мутация опухолевого происхождения», использованный в описании, имеет отношение к мутациям в последовательностях нуклеиновых кислот, которые являются специфическими для опухоли и отсутствуют в ДНК из здоровой ткани.
В отдельном варианте осуществления мутация опухолевого происхождения является однонуклеотидным вариантом (SNV). SNV включает, но не ограничивается этим, различные мутации p53, KRAS и BRAF.
В другом варианте осуществления кандидатный пептид является антигеном, который вызывает иммунный ответ. Например, пептиды могут происходить из патогенов.
В еще одном варианте осуществления кандидатный пептид является соединением, подвергающимся тестированию на иммуногенность.
В одном варианте осуществления изобретение имеет отношение к способу, использующему библиотеку кандидатных пептидов, при этом библиотека содержит мутантные формы нативного пептида(ов). Например, известно, что нативный пептид(ы) может неэффективно презентироваться данной молекулой MHC. Таким образом, для того, чтобы подвергнуть скринингу мутантные пептиды, которые более эффективно презентируются данной молекулой MHC, может использоваться библиотека, содержащая мутантные пептиды.
В другом варианте осуществления изобретение имеет отношение к способу, использующему библиотеку кандидатных пептидов, при этом данная библиотека содержит пептиды, образованные в результате случайного отклонения или расщепления кДНК или ДНК, происходящей из представляющих интерес клеток или патогенов. Такая библиотека может охватить все пептиды, присутствующие в таких клетках. Например, создание MHC-дисплейных библиотек из раковой ткани или ткани, подвергающейся аутоиммунной атаке, может обеспечить возможность установления всех тканевых пептидов, потенциально представленных данным MHC. Таким образом, в будущем не будет необходимости в проведении тестирования, и эффективно ли презентируется отдельный пептид данным MHC, можно будет посмотреть в таблице.
В одном варианте осуществления библиотека комплексов MHC-пептид создается при помощи рекомбинантных векторов экспрессии. Рекомбинантные векторы экспрессии представляют собой реплицируемые ДНК конструкции, содержащие комплект из (1) агента(ов), обладающего регуляторной ролью в экспрессии гена, например, промоторов, операторов или энхансеров, функционально связанных с (2) нуклеотидной последовательностью, кодирующей желательный белок (такой как комплекс MHC-пептид), которая транскрибируется в мРНК и транслируется в белок, и (3)
соответствующих последовательностей транскрипции, инициации трансляции и терминации. Выбор промотора и других регуляторных элементов, как правило, варьирует в соответствии с предполагаемой для использования репортерной клеточной линией. Векторы экспрессии часто имеют форму “плазмид”, под которыми имеются в виду кольцевые двухцепочечные петли ДНК, и которые, в их векторной форме, не являются связанными с хромосомой. Подходящие прокариотические векторы экспрессии включают плазмиды из E. coli, например, Col E1, pCR1, pBR322, pMB9 и их производные, плазмиды более широкого круга хозяев, например, RP4, ДНК фага, такого как фаг лямбда и его различные производные, M13 и тому подобное. Дополнительные векторы E. coli описаны, например, в Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, NY (1982) и Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, NY (1989). Однако, изобретение имеет целью включать другие подобные формы векторов экспрессии, которые исполняют эквивалентные функции и которые станут известными в данной области техники в дальнейшем. Эукариотические векторы экспрессии, реплицирующиеся эписомально, такие как pCEP4 или BKV, или другие векторы, происходящие от вирусов, таких как ретровирусы, например, pMY, pMX, pSIR, аденовирусы, например, pAd, и тому подобное, могут использоваться. В большинстве случаев в векторах экспрессии регуляторные элементы, контролирующие транскрипцию или трансляцию, могут происходить из генов млекопитающего, микроорганизма, вирусных генов или генов насекомого. Способность к репликации, обычно обеспечиваемая точкой начала репликации, и выбор гена для облегчения распознавания трансформантов дополнительно могут быть включены. Векторы экспрессии, содержащие регуляторные элементы из эукариотических вирусов, как правило, используются в эукариотических векторах экспрессии, например, SV40 векторах, векторах папиллома вируса и векторах, происходящих от вируса Эпштейна-Барр. Другие характерные эукариотические векторы включают pMSG, pAV009/A+, pMTO10/A+, pMAMneo-5, бакуловирус pDSVE, и любой другой вектор, обеспечивающий экспрессию белков под контролем CMV промотора, SV40 раннего промотора, SV40 позднего промотора, промотора металлотионеина, промотора вируса опухоли молочной железы мышей, промотора вируса саркомы Рауса, промотора гена полиэдрина или других промоторов, продемонстрировавших эффективность в отношении экспрессии в эукариотических клетках.
“Промотор” определяется как набор нуклеиновокислотных регуляторных последовательностей, который управляет транскрипцией нуклеиновой кислоты. Использованный в данном документе промотор включает необходимые нуклеиновокислотные последовательности вблизи сайта начала транскрипции, такой как, например, в случае промотора полимеразы II типа, TATA элемент. Промотор также необязательно включает дистальный энхансерный или репрессорный элементы, которые могут располагаться даже за несколько тысяч пар оснований от сайта начала транскрипции. Промоторы, которые можно использовать в эукариотических клетках-хозяевах, известны специалистам в данной области техники. Иллюстративные примеры таких промоторов включают, но не ограничиваются этим, промоторы вируса обезьян 40 (SV40), промотор вируса опухоли молочной железы мышей (MMTV), промоторы вируса иммунодефицита человека (HIV), такие как промотор длинного концевого повтора (LTR) HIV, промоторы вируса Молони, ALV промоторы, промоторы цитомегаловируса (CMV), такой как CMV промотор раннего немедленного ответа, промотор вируса Эпштейна-Барр (EBV), промотор вируса саркомы Рауса (RSV), а также промоторы из таких человеческих генов, как человеческий актин, человеческий миозин, человеческий гемоглобин, человеческий мышечный креатин и человеческий металлотионеин. Другие примеры подходящих промоторов включают CAG промотор (гибридный промотор, содержащий
CMV энхансер, промотор куриного в-актина и акцепторный участок сплайсинга кроличьего в-глобина и поли(A) последовательность).
Термин “функционально связанный” имеет отношение к функциональной связи между последовательностью контроля экспрессии нуклеиновой кислоты (такой как промотор или набор сайтов соединения фактора транскрипции) и второй нуклеиновокислотной последовательностью, при этом данная последовательность контроля экспрессии направляет транскрипцию нуклеиновой кислоты, соответствующей второй последовательности.
Библиотека MHC-пептидов вводится в подходящие репортерные клеточные линии путем трансдукции вектора экспрессии в клетку-хозяина с помощью стандартных методов, известных в данной области техники. Подходящие методы описаны, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, NY (1989). Чтобы получить псевдо-ретровирусы для трансдукции, пакующие клеточные линии, постоянно экспрессирующие ретровирусные белки GAG, POL и ENV (подобно например клеточной линии Phoenix), временно трансфицируются конструкциями, содержащими вирусный геном, состоящий из LTRs, сигналов упаковки и генов, представляющих интерес (в этом случае цепей MHC, несущих пептид). Альтернативно, подходящие клеточные линии, подобные HEK, 3T3 или другим, временно трансфицируются смесью векторов, кодирующих отдельно ретровирусные белки GAG, POL и ENV и вирусный геном, состоящий из LTRs, сигналов упаковки и генов, представляющих интерес. Эти широко используемые стратегии обеспечивают получение дефектных псевдоретровирусов, которые способны инфицировать клетки-мишени и ввести интересующие гены в их геномную ДНК. Однако, инфицированные клетки-мишени не способны продуцировать ретровирусы, потому что псевдоретровирусы не несут gag, pol и env гены в своем геноме. Альтернативно, MHC-пептидные библиотеки вводятся в подходящие репортерные клеточные линии путем трансфекции с помощью реагентов на основе липидов, фосфата кальция, катионных полимеров или DEAE-декстрана, или путем электропорации.
Термины “инфекция” и “трансдукция” репортерной клеточной линии используются в описании взаимозаменяемым образом.
Способы обнаружения клеток, которые презентируют находящиеся на клеточной поверхности комплексы MHC-пептид, известны в данной области техники. Подходящие способы включают методы клеточной сортировки, такие как проточная цитометрия, сортировка флуоресцентно-активированных клеток (FACS) и магнитная сортировка клеток (MACS).
Соответственно, в различных вариантах осуществления изобретение имеет отношение к способу согласно любому из предшествующих вариантов осуществления, описанных в этом документе, в котором репортерные клетки, эффективно экспрессирующие MHC на их клеточной поверхности, обогащаются путем сортировки флуоресцентно-активированных клеток (FACS).
FACS имеет отношение к способу разделения популяции клеток на одну или более субпопуляций, исходя из наличия, отсутствия или уровня одного или более специфических полипептидов, экспрессированных данными клетками. FACS основывается на оптических свойствах, включая флуоресценцию, индивидуальных клеток, для того, чтобы рассортировать клетки на субпопуляции. Сортировщики клеток, подходящие для осуществления способа, описанного в данном документе, хорошо известны в данной области техники и коммерчески доступны. Примеры клеточных сортеров включают сортер MoFlo (DakoCytomation, Fori Collins, Colo.), FACSAria™, FACSArray™, FACS Vantage™, BD™ LSR II и FACSCaiibur™ (BD Biosciences, San Jose, Calif.) и другие равноценные клеточные сортеры, производимые другими коммерческими поставщиками, такими как Sony, Bio-Rad и Beckman Coulter.
В другом варианте осуществления изобретение имеет отношение к способу согласно любому из предшествующих вариантов осуществления, описанных в этом документе, в котором репортерные клетки, эффективно экспрессирующие MHC на их клеточной поверхности, обогащаются с помощью магнитной сортировки MACS.
“MACS” относится к способу разделения популяции клеток на одну или более субпопуляций, основываясь на присутствии, отсутствии или уровне одного или более отбираемых с помощью MACS полипептидов, экспрессированных клетками. MACS основывается на свойствах магнитной чувствительности помеченных индивидуальных клеток, для того, чтобы рассортировать клетки на субпопуляции. В качестве меток для MACS могут использоваться магнитные частицы (например, доступные от компании Miltenyi Biotec Bergisch Gladbach, Германия; 130-048-402). MACS клеточные сортеры, подходящие для осуществления способа, описанного в этом документе, хорошо известны в данной области техники и коммерчески доступны. Примеры MACS клеточных сортеров включают autoMACS Pro Separator (Miltenyi Biotec).
В различных вариантах осуществления клетки могут контактировать с антителом, специфическим в отношении MHC-I или MHC-II или с антителами, обнаруживающими белки, связанные с гибридами. Например, антитела, обнаруживающие CD3г, CD3д или CD3е, могут использоваться, если MHC внеклеточные домены слиты с TCR трансмембранными участками. Антитело может быть напрямую конъюгировано с обнаружимой меткой. Альтернативно, вторичное антитело, конъюгированное с обнаружимой меткой и специфическое в отношении первого антитела, может контактировать с клетками. Обнаружимые метки, подходящие для использования, включают любое вещество, обнаружимое спектроскопическими, фотохимическими, биохимическими, иммунохимическими, оптическими или химическими способами. Подходящие для использования в настоящем изобретении метки включают биотин, магнитные частицы (например, Dynabeads™), флюоресцентные метки (например, флюоресцеин, техасский красный, родамин, зеленый флуоресцирующий белок, дансил, умбеллиферон, PE, APC, CY5, Cy7, PerCP, красители Alexa и тому подобное), радиометки (например, 3H, 1251, 35S, 1 C или 32P), ферментные (например, лошадиная пероксидаза хрена, щелочная фосфатаза и другие) и колориметрические метки, такие как коллоидное золото или зерна из цветного стекла или пластика (например, полистирола, полипропилена, латекса и т.д.). Ряд подходящих флуоресцентных меток дополнительно описан, например, в руководстве The Molecular Probes Handbook: A Guide to Fluorescent Probes and Labeling Technologies, 11th Edition.
Сортировка в результате дает популяцию нефлуоресцирующих клеток и, по меньшей мере, одну популяцию флуоресцирующих клеток, в зависимости от того, как много флуоресцентных меток использовалось. Присутствие, по меньшей мере, одной клеточной популяции с флуоресцирующими клетками служит признаком того, что, по меньшей мере, один кандидатный пептид эффективно презентируется APCs. Таким образом, FACS обеспечивает сортировку популяции клеток, чтобы получить популяцию клеток, обогащенных клетками, содержащими экспонированный на поверхности MHC-I или MHC-II.
Варианты осуществления настоящего изобретения используют способы выделения ДНК, известные специалистам в данной области техники. В общем, цель заключается в том, чтобы отделить ДНК, присутствующую в ядре клетки от других клеточных компонентов. Выделение ДНК обычно начинается с лизиса или разрушения клеток. Этот процесс имеет большое значение для разрушения белковых структур и обеспечивает высвобождение нуклеиновых кислот из ядра. Лизис проводится в растворе соли, содержащем поверхностно-активные вещества для денатурации белков или протеазы (расщепляющие белки ферменты), такие как протеиназа K, или в некоторых случаях и то и другое. Это заканчивается распадом клеток и растворением мембран. Способы выделения ДНК включают, но не ограничиваются этим, фенольно-хлороформную экстракцию, высокосолевое осаждение, щелочную денатурацию, ионообменную колоночную хроматографию, связывание со смолой и связывание парамагнитными гранулами.
Варианты осуществления настоящего изобретения используют способы получения кДНК, известные специалистам в данной области техники. В общем, цель заключается в том, чтобы преобразовать выделенную РНК, присутствующую в клетках, в ДНК, так называемую копи-ДНК, для того, чтобы использовать ее в качестве матрицы для полимеразной цепной реакции (ПЦР). Выделение РНК обычно начинается с лизиса или разрушения клеток. Этот процесс необходим для разрушения белковых структур и обеспечивает высвобождение из них нуклеиновых кислот. Лизис обычно проводится в растворе, содержащем фенол (например, TRIzolTM). Это заканчивается распадом клеток и растворением мембран и обеспечивает отделение РНК от других клеточных компонентов. Затем выделенная РНК преобразуется в кДНК с помощью обратной транскриптазы (например, SuperscriptTM, GoscriptTM).
Затем последовательность кандидатных пептидов в комплексе MHC-пептид, представленном на клеточной поверхности, амплифицируется с помощью ПЦР и может быть установлена с помощью любого из методов, известных в данной области техники.
В разных вариантах осуществления изобретение имеет отношение к способу согласно любому из предшествующих вариантов осуществления, описанных в этом документе, при этом последовательность кандидатных пептидов определяется с помощью ПЦР и секвенирования.
В одном варианте осуществления последовательность кандидатных пептидов определяется с помощью цифровой ПЦР. Цифровая полимеразная цепная реакция (цифровая ПЦР, dPCR или dePCR) является результатом усовершенствования методов обычной полимеразной цепной реакции, которая может использоваться для прямого определения количества и клональной амплификации нуклеиновых кислот, включая ДНК, кДНК или РНК.
Секвенирование также может осуществляться при помощи микроструйной техники (микрофлюидики). Микрофлюидика задействует микромасштабные устройства, которые оперируют маленькими объемами жидкостей. Поскольку микрофлюидика может точно и воспроизводимо контролировать и отмерять маленькие объемы жидкости, в частности объемы меньше 1 мкл, использование микроструйной техники обеспечивает значительную экономию затрат. Применение микрофлюидной технологии уменьшает время цикла, сокращает время до результата и увеличивает производительность. Более того, внедрение микрофлюидной технологии повышает системную интеграцию и автоматизацию. Микрожидкостные реакции обычно проводятся в микрокаплях.
В некоторых вариантах осуществления секвенирование проводится с использованием технологии секвенирования второго поколения (или следующего поколения или Next-Gen), третьего поколения (или Next-Next-Gen) или секвенирования четвертого поколения (или N3-Gen), включая, но не ограничиваясь этим, пиросеквенирование, секвенирование лигированием, одномолекулярное секвенирование, секвенирование методом синтеза (SBS), массовое параллельное секвенирование, массовое параллельное одномолекулярное SBS, массовое параллельное одномолекулярное секвенирование в реальном времени, массовое параллельное одномолекулярное секвенирование в реальном времени с использованием технологии нанопор. Morozova и Marra предоставляют обзор некоторых таких технологий в Genomics, 92: 255 (2008).
В некоторых вариантах осуществления до, после или одновременно с секвенированием нуклеиновые кислоты амплифицируются. Иллюстративные неограничивающие примеры методов амплификации нуклеиновых кислот включают, но не ограничиваются этим, полимеразную цепную реакцию (ПЦР), полимеразную цепную реакцию с обратной транскрипцией (RT-PCR), транскрипционно-опосредованную амплификацию (TMA), лигазную цепную реакцию (LCR), амплификацию с замещением цепей (SDA) и амплификацию, основанную на последовательности нуклеиновых кислот (NASBA). Специалисту в данной области техники понятно, что в некоторых методах амплификации (например, ПЦР) требуется, чтобы РНК была обратно транскрибирована в ДНК до амплификации (например, RT-PCR), тогда как другие методы амплификации напрямую амплифицируют РНК (например, TMA и NASBA).
В одном варианте осуществления изобретение имеет отношение к способу согласно любому из предшествующих вариантов осуществления, описанных в этом документе, согласно которому кандидатный пептид используется в качестве вакцины.
В отдельном варианте осуществления вакцина представляет собой противораковую вакцину на основе опухоль-специфического антигена (TSA).
Термин “вакцинация” или его эквиваленты хорошо известны в данной области техники. Например, понятно, что термин вакцинация обозначает процесс, который увеличивает иммунную реакцию субъекта на антиген и, следовательно, способность противостоять или побеждать болезнь. Термин “вакцина” обозначает композицию для получения иммунитета с целью профилактики и/или лечения болезней (например, рака). Соответственно, вакцины являются медикаментами, которые содержат антигены и предназначаются для использования среди людей или животных с целью создания специфической защиты и защитного фактора путем вакцинации. Термин “противораковая вакцина на основе TSA” имеет отношение к вакцине, содержащей смешанный образец опухоль-специфических антигенов, например, по меньшей мере, один, по меньшей мере, два, по меньшей мере, три, по меньшей мере, четыре, по меньшей мере, пять, или более опухоль-специфических пептидов. Для того, чтобы вакцина была эффективной, короткие линейные пептиды, содержащие антиген или эпитоп, должны презентироваться MHC на поверхности APC. Набор экспрессированных вариантов MHCs сильно различается между разными индивидуумами.
Повторяющиеся мутации опухолевого происхождения могут служить в качестве общих опухоль-специфических антигенов, предоставляя возможность разработки противораковых вакцин на основе TSA, применимых к более широким когортам пациентов. Соответственно, способ настоящего изобретения может использоваться для идентификации пациентов, содержащих определенные MHC варианты и эффективно презентирующих эти типичные/общие TSAs иммунной системе. Однако, многие мутации опухолевого происхождения, по-видимому, происходят в результате пациент-специфичных изменений, и любые из этих изменений могут влиять на связывание TSA пептида с MHCs. Таким образом, способ настоящего изобретения также может использоваться для идентификации специфических для пациента кандидатных пептидов для создания персонализированных вакцин.
Более того, опухоль пациента часто содержит много TSAs, каждый из которых может содержаться в подгруппе опухолевых клеток. В некоторых вариантах осуществления набор кандидатных TSAs может вследствие этого состоять из TSAs, происходящих из разных групп опухолевых клеток. Поэтому важным этапом при создании противораковой вакцины является выбор из TSAs кандидатов, которые содержатся и предпочтительно хорошо экспрессируются в опухолевых клетках пациента, кандидатов с высокой аффинностью к MHC, которые экспрессируются у пациента. Для получения оптимального иммунного ответа набор TSAs, содержащихся в вакцине, должен дополнительно активировать и CD4+ и CD8+ T-клетки, которые распознают комплексы MHCII или MHCI-пептид, соответственно. Таким образом, способ настоящего изобретения может использоваться для идентификации связывания TSAs со специфическими вариантами MHCII и MHCI пациента.
В одном варианте осуществления кандидатный пептид используется для индукции иммунологической толерантности, по меньшей мере, к одному из эпитопов, которые он содержит. При использовании в описании «иммунологическая толерантность» имеет отношение к уменьшению иммунологической реактивности хозяина к специфическому антигену или антигенам. Антигены содержат иммунные детерминанты/эпитопы, которые, при отсутствии толерантности, вызывают нежелательный иммунный ответ. Иммунологическую толерантность можно индуцировать для того, чтобы предотвратить или устранить отторжение трансплантата, аутоиммунный ответ, аллергическую реакцию или другой нежелательный иммунный ответ. Без ограничения какой-либо теорией, иммунологическая толерантность может быть достигнута путем выработки регуляторных T-клеток, которые действуют как отрицательные регуляторы иммунного ответа. Баланс различных типов T-клеток также может зависеть от взаимодействия TCRs и комплексов MHC-пептид. Следовательно, способ настоящего изобретения может использоваться для выбора комплексов MHC-пептид, подходящих для получения иммунологической толерантности к данному пептиду. Другие возможные механизмы для достижения иммунологической толерантности включают блокирование TCRs специфических T-клеток и/или уничтожение специфических T-клеток.
Таким образом, в одном варианте осуществления кандидатный пептид используется для блокировки TCRs в случае с MHC молекулой. “Блокировка TCRs” относится к любому агенту, который включает комплекс пептид-MHC, блокирующий естественное взаимодействие TCR-MHC. Для предсказания эффективности TCR-блокировки комплекса MHC-пептид важной является стабильность этого комплекса.
В другом варианте осуществления кандидатный пептид используется для MHC-опосредованной доставки токсина в клетки, в частности в T-клетки.
“MHC-опосредованная доставка токсинов” относится к способам ковалентного связывания токсических агентов (белков или других) с тетрамерами пептид-MHC или другими мультимерами MHC, для того чтобы передать указанный токсин в клетку, в частности, T-клетку, чтобы вызвать смерть клетки.
Раскрытый в описании способ предоставляет возможность измерения стабильности специфических пар MHC-пептид и, таким образом, может использоваться для создания комплексов MHC-пептид с целью блокировки специфических TCRs или уничтожения специфических T-клеток, в частности для применения in vivo.
В еще одном варианте осуществления кандидатный пептид используется для перенацеливания T-клеток в направлении других пептид-MHC-специфических T-клеток посредством MHC-CARs.
Часто используется перенацеливание T-клеток в направлении других клеток с CARs, состоящих из антител, связанных с внутриклеточной цепью CD247(CD3дзэта) и даже в клинике. T-клетки, в частности цитотоксические T-клетки, «оснащенные» CARs, состоящими из пептид-MHC, связанного с внутриклеточной цепью CD247(CD3дзэта), или T-клетки, «оснащенные» MCRs, могут использовать эти рецепторы для распознавания T-клеток, специфических для этих комплексов пептид-MHC, и уничтожать их (Jyothi et al., Nat.Biotech, 20, 1215–1220 (2002)). Раскрытый в этом документе способ дает возможность измерения стабильности специфических пар MHC-пептид и, таким образом, может использоваться для создания комплексов MHC-пептид для использования в CARs или MCRs для перенацеливания уничтожения специфических T-клеток, в частности для вариантов применения in vivo.
При использовании многих видов терапии требуется модулирование иммунного ответа, идентификация TCRs, вовлеченных в этот иммунный ответ. Существуют различные способы, хорошо известные в данной области техники, которые позволяют идентифицировать такие TCRs и основываются на взаимодействии TCRs с эпитопом, содержащимся в комплексе MHC-пептид. Решающим предварительным условием является информация о том, насколько хорошо комплекс MHC-пептид презентируется на поверхности APC. Если эпитоп не презентируется на MHCs APC, использованных в экспериментальном анализе in vitro, соответствующее TCR связывание с этим эпитопом может быть неидентифицировано. Более того, если в библиотеке или комплексах MHC-пептид, представленных APCs в таком анализе, один из пептидов презентируется преимущественно, это может привести к выбору субоптимального TCR. Для процесса оптимизированной идентификации и выбора TCR важно, следовательно, преодолеть неопределенность, связанную с презентацией комплекса MHC-пептид. Следовательно, способ настоящего изобретения может использоваться для предоставления количественной информации об аффинности специфического пептида к специфическому MHC и выбора подходящих комплексов для анализов in vitro, используемых для идентификации TCRs, распознающих эпитоп или для обогащения T-клеток, экспрессирующих такой TCR.
В одном варианте осуществления кандидатный пептид используется для проведения теста на реакционную способность T-клеток.
Термин «T-клеточная реактивность», использованный в описании, имеет отношение к способности вещества вызывать T-клеточную активацию. Конкретнее, «T-клеточная реактивность» означает способность пептида индуцировать пролиферацию, дифференцировку и/или выработку цитокинов T-клеток. Способы анализа T-клеточной реактивности хорошо известны в данной области техники и часто включают совместное культивирование APCs, презентирующих комплексы MHC-пептид, и T-клеток. Такие методы анализа могут использоваться для наращивания T-клеток для клеточной терапии или для установления TCRs связывания со специфическим эпитопом, например, с целью адаптивной T-клеточной терапии или T-клеточной терапии с использованием химерного антигенного рецептора (CAR). Тесты на T-клеточную реактивность могут дополнительно использоваться для тестирования иммуногенности.
В некотором варианте осуществления кандидатный пептид используется для анализа на иммуногенность.
Термин “анализ (тест) на иммуногенность”, использованный в описании, имеет отношение к измерению возможных иммунных ответов на биотерапевтические средства. Биотерапевтические средства могут вызывать иммунный ответ, который может оказывать влияние на их безопасность и эффективность. Тестирование иммуногенности используется для контролирования и оценки гуморального (антитела) или клеточного (T-клетки) ответов в ходе клинических и доклинических исследований. Обычно тестирование иммуногенности биотерапевтических средств включает измерение антител, специфически выработанных против биотерапевтических средств. С помощью способа настоящего изобретения стало возможным идентифицировать пептиды, которые эффективно презентируются MHC молекулами, в частности MHCs, характерными для индивидуума, и таким образом, потенциально вызывают опосредованный Т-клетками иммунный ответ, в методе in vitro. Это может помочь в предоставлении более полной и точной картины общего иммуногенного профиля соединения и в то же время может уменьшить нагрузку на пациента или участников клинических испытаний.
В следующем варианте осуществления изобретение предоставляет комплекс MHC-пептид, содержащий кандидатный пептид, ковалентно связанный с внеклеточной частью альфа или бета домена MHC молекулы, внеклеточной частью бета домена MHC и, по меньшей мере, одним трансмембранным доменом гетерологичной молекулы.
В одном варианте осуществления изобретение предоставляет комплекс MHC-пептид в соответствии с предшествующим вариантом осуществления, в котором трансмембранный домен является трансмембранным доменом гетерологичной молекулы, такой как TCRб/в, TCRг/д, CD3г/д/е/ж, CD4 или CD8б/в.
В одном варианте осуществления MHC молекула является молекулой MHC класса II, содержащей внеклеточную альфа цепь MHC класса II, внеклеточную бета цепь MHC класса II и, по меньшей мере, два трансмембранных домена.
В другом варианте осуществления молекула MHC является молекулой MHC класса I, содержащей внеклеточную альфа цепь MHC класса I, бета-2 макроглобулин и, по меньшей мере, один трансмембранный домен.
В еще одном варианте осуществления MHC молекула является MHC гибридной молекулой, содержащей гибрид пептида, в2-микроглобулин и внеклеточную альфа цепь MHC класса I и, по меньшей мере, один трансмембранный домен.
В конкретном варианте осуществления MHC альфа цепь молекулы MHC класса I несет мутацию Y84A.
В различных вариантах осуществления изобретение предоставляет комплекс MHC-пептид, в котором кандидатный пептид является опухолеспецифическим пептидом, несущим индивидуальную мутацию(и), происходящую из опухоли.
В конкретном варианте осуществления происходящая из опухоли мутация является однонуклеотидным вариантом (SNV).
В следующем варианте осуществления изобретение предоставляет рекомбинантную конструкцию, содержащую нуклеотидную последовательность, кодирующую комплекс MHC-пептид, как определено в любом из предшествующих вариантов осуществления.
В следующем варианте осуществления изобретение предоставляет кассету экспрессии, содержащую промоторную последовательность, соединенную с рекомбинантной конструкцией предшествующего варианта осуществления.
В другом варианте осуществления изобретение предоставляет вектор, содержащий кассету экспрессии предшествующего варианта осуществления.
В еще одном варианте осуществления изобретение предоставляет клетку, содержащую комплекс MHC-пептид изобретения.
В одном варианте осуществления изобретение имеет отношение к клетке согласно предшествующему варианту осуществления, содержащей комплекс пептид-MHC класса II, при этом клетка является клеткой млекопитающего, лишенной механизма «загрузки» пептида MHC класса II.
В другом варианте осуществления клетка содержит комплекс пептид-MHC класса I, при этом клетка является клеткой млекопитающего, лишенной функционального гена TAP1, TAP2 и/или бета-2-микроглобулина.
В еще одном варианте осуществления клетка содержит комплекс пептид-MHC класса I, при этом клетка является T-клеточной гибридомой с дефектным или удаленным геном TAP1, TAP2 и/илибета-2-микроглобулина.
Изобретение дополнительно содержит способ определения MHC аффинности связывания кандидатных пептидов, включающий экспрессию в репортерной клеточной линии рекомбинантного комплекса MHC-пептид, содержащего ковалентно связанный кандидатный пептид, и обнаружение репортерных клеток, которые презентируют комплекс MHC-пептид на поверхности репортерной клетки, включая измерение уровня такой презентации/экспрессии. Репортерная клеточная линия может быть клеточной линией млекопитающего. В одном варианте осуществления использованная клеточная линия млекопитающего демонстрирует измененные характеристики в отношении нативной клеточной линии, например, она несет делеции ферментов, мутации и/или сверхэкспрессию ферментов и/или MHC молекул.
В одном варианте осуществления изобретение имеет отношение к способу, согласно предшествующему варианту осуществления, включающему:
(i) получение библиотек, содержащих кандидатные пептиды, клонированные впереди рекомбинантного альфа или бета домена, соединенного с трансмембранным участком;
(ii) трансдукцию таких библиотек, вместе с соответствующим MHC альфа или бета доменом, в подходящие репортерные клеточные линии;
(iii) обнаружение клеток, презентирующих один или более комплексов MHC-пептид на своей клеточной поверхности;
(iv) определение уровня такой презентации/экспрессии.
Способы настоящего изобретения также могут использоваться в высокопроизводительном скрининге. Технология высокопроизводительного скрининга (HTS) обычно используется для обозначения быстрой обработки клеток в большом масштабе. В некоторых вариантах осуществления различные библиотеки кандидатных пептидов могут подвергаться скринингу параллельно. Системы высокопроизводительного скрининга коммерчески доступны и, как правило, обеспечивают полную автоматизацию всех операций, включая дозирование всех образцов и реагентов, подачу жидких компонентов, регулировку по времени периодов инкубации и снятие окончательных данных с микропланшетов в детекторе(ах), подходящем для анализа. Такие предусматривающие различные конфигурации системы обеспечивают высокую пропускную способность и быстрый запуск, а также высокую степень вариативности и специфицирования.
Под термином «пептид», использованным в данном документе, подразумевается, по меньшей мере, две ковалентно-связанные аминокислоты. Как правило, пептиды MHC класса I имеют 8 или 9 аминокислот в длину, но могут варьировать в пределах от 7 до 10 аминокислот в длину. Пептиды MHC класса II могут варьировать в пределах от 15 до 24 аминокислот в длину. Необязательно, они могут варьировать в пределах от 10 до 30 аминокислот или более в длину.
При использовании в описании связывание или “аффинность связывания” пептида с MHC или взаимодействие пептида с MHC имеет отношение к взаимодействию пептид-MHC, которое происходит в пептид-связывающей бороздке или щели. Такое связывание или взаимодействие также может происходить в естественных условиях.
“Ковалентное связывание” или “прикрепление” пептида к MHC при использовании в описании имеет отношение к рекомбинантному комплексу MHC-пептид, в котором ковалентная связь создается при помощи генной инженерии.
Комплекс MHC-пептид при использовании в описании имеет отношение к комплексу, в котором пептид связан с MHC. Это связывание может происходить посредством пептид-связывающей бороздки или щели и/или путем ковалентного связывания пептида с MHC.
В рекомбинантном комплексе MHC-пептид, используемом в данном документе, кандидатный пептид всегда ковалентно связывается с MHC, однако пептид может или может неэффективно связываться со щелью MHC.
Стабильность комплекса MHC-пептид имеет отношение к устойчивости взаимодействия пептид-MHC, происходящего в пептид-связывающей бороздке, которая коррелирует с поверхностной экспрессией указанного комплекса MHC-пептид. Это не имеет отношения к стабильности ковалентной связи пептид-MHC.
Термин “антиген”, использованный в описании, относится к всему или частям, пептида или белка, способным вызывать иммунный ответ к нему самому или его частям. Этот иммунный ответ может включать в себя или выработку антител или активацию специфических иммунологически компетентных клеток, или и то и другое.
Термин “механизм загрузки пептида”, использованный в описании, относится ко всем требуемым компонентам, необходимым для загрузки пептидов на MHC молекулу. Например, механизм загрузки пептида на MHC класса I присутствует почти во всех клетках и включает среди прочего TAP транспортер, тапасин и кальретикулин. Механизм загрузки пептида на MHC класса I постоянно присутствует только в APCs, но он может быть индуцирован в других клетках, подобных T-клеткам.
Использованный в описании термин “трансмембранный домен” определяется как преимущественно гидрофобная последовательность аминокислот, которая способна располагаться в клеточной мембране. Термин “трансмембранный участок” используется в описании как синоним “трансмембранного домена”.
Термин «библиотека» или его эквиваленты, использованные в описании, означает множество молекул. В случае комплексов MHC-пептид библиотека предоставляет в достаточной степени структурно различную популяцию пептидов, чтобы обеспечить с достаточным уровнем вероятности соответствующую линейку клеточных ответов, чтобы предоставить одну или более клеток, демонстрирующих желательный ответ. В предпочтительном варианте осуществления, по меньшей мере, 10, предпочтительно, по меньшей мере, 50, более предпочтительно, по меньшей мере, 200 и наиболее предпочтительно, по меньшей мере, 1000 пептидов одновременно анализируются в способе изобретения. Библиотеки могут создаваться с целью максимального увеличения размера и разнообразия библиотеки.
Термин “рекомбинантный” при использовании со ссылкой, например, на клетку или нуклеиновую кислоту, белок или вектор, указывает, что клетка, нуклеиновая кислота, белок или вектор, модифицируется путем введения гетерологичной нуклеиновой кислоты или белка или изменения нативной нуклеиновой кислоты или белка, или что клетка происходит из клетки модифицированной таким образом. Таким образом, например, рекомбинантные клетки экспрессируют гены, которые не обнаруживаются в нативной (нерекомбинантной) форме клетки или экспрессируют нативные гены, которые аномально экспрессируются, экспрессируются в меньших количествах меньше или не экспрессируются совсем. Рекомбинантная нуклеиновая кислота изначально создается in vitro, в большинстве случаев, путем манипуляции с нуклеиновой кислотой, например, с использованием полимераз и эндонуклеаз, в форме, которая при обычных условиях не обнаруживается в природе. Таким образом, достигается функциональная связь различных последовательностей. Таким образом, выделенная нуклеиновая кислота в линейной форме или вектор экспрессии, образованный in vitro путем лигирования ДНК молекул, которые не соединяются в обычных условиях, оба считаются рекомбинантными для целей данного изобретения. Понятно, что как только рекомбинантная нуклеиновая кислота получена и вновь «вставлена» в клетку-хозяина или организм, она будет реплицироваться нерекомбинантным путем, т.е. с использованием in vivo клеточных механизмов клетки-хозяина, а не in vitro манипуляции; однако, такие нуклеиновые кислоты, ранее полученные рекомбинантно, хотя впоследствии реплицируются нерекомбинантно, по-прежнему считаются рекомбинантными для целей изобретения. Аналогичным образом, рекомбинантный белок, такой как комплекс MHC-пептид изобретения, является белком, полученным с использованием рекомбинантных методов, т.е. посредством экспрессии рекомбинантной нуклеиновой кислоты, как описано выше.
Термин “гетерологичный” при использовании в описании в отношении участков нуклеиновой кислоты показывает, что нуклеиновая кислота содержит две или более последовательностей, которые не обнаруживаются в обычных условиях в тех же самых взаимоотношениях друг с другом в природе. Например, произведенной рекомбинантно является нуклеиновая кислота, имеющая две или более последовательностей, например, из неродственных генов, расположенных в определенном порядке, полученная с целью создания новой функциональной нуклеиновой кислоты, например, имеющая промотор из одного источника и кодирующую область из другого источника. Аналогичным образом, термин гетерологичный белок будет зачастую относиться к двум или более последовательностям, которые не обнаруживаются в аналогичных взаимоотношениях друг с другом в природе (например, гибридный белок).
Термин «рак», использованный в описании, определяется как болезнь, характеризующаяся быстрым и неконтролируемым ростом аномальных клеток. Раковые клетки могут распространяться локально или посредством кровообращения и лимфатической системы к другим частям организма. Примеры различных видов рака включают, но не ограничиваются этим, рак молочной железы, рак предстательной железы, рак яичников, рак шейки матки, рак кожи, рак поджелудочной железы, колоректальный рак, рак почки, рак печени, рак мозга, лимфому, лейкемию, рак легких и тому подобное.
Кроме того, в пунктах формулы изобретения слово «содержащий» не исключает другие элементы или стадии, а форма единственного числа включает множественное число обозначаемого объекта, если контекст явно не диктует иное.
Если не указано иное, все технические и научные термины, использованные в этом документе, имеют то же самое значение, которое обычно понятно среднему специалисту в области техники, к которой относится данное изобретение. Хотя сходные или эквивалентные описанным в этом документе методы и материалы могут использоваться при осуществлении на практике или тестировании настоящего изобретения, подходящие методы и материалы описаны ниже. В случае противоречий, настоящее подробное описание, включая определения, будет обеспечивать контроль. Кроме того, материалы, методы и примеры предназначаются только для иллюстрации, но не являются ограничивающими.
Отдельные варианты осуществления настоящего изобретения дополнительно иллюстрируются следующими примерами. Однако, следует понимать, что изобретение не ограничивается конкретными подробностями этих примеров. Следующие примеры включены с целью демонстрации предпочтительных вариантов осуществления изобретения. Специалистам в данной области техники должно быть ясно, что раскрытые в изложенных ниже примерах приемы представляют способы, использованные в настоящем изобретении для успешного осуществления изобретения на практике, и таким образом, могут считаться предпочтительными способами для осуществления на практике. Однако, специалистам в данной области техники должно быть ясно, в свете настоящего раскрытия, что может быть сделано множество изменений в конкретных раскрываемых вариантах осуществления, и при этом могут быть получены подобные или сходные результаты без отступления от существа и объема изобретения.
Фиг. 1 показывает структуру гибридной молекулы MHC-TCR (MCR2) и нативного MHC класса II.
Фиг. 2. Были изготовлены конструкции, кодирующие MCRs, несущие альфа и бета цепи человеческих MHC молекул (HLA-DRB1_4, DRB1_15 и DRB5_1) с ковалентно связанным пептидом матриксного белка 1 гриппа MP1103-120, (a), MP162-72 (b) или библиотека пептидов, происходящих из кДНК гриппа (c). После трансдукции клеточной линии, лишенной MHC класса II, анти-HLA-DR APC антитело было использовано для измерения MCR поверхностной экспрессии с помощью FACS. Графики показывают эффективную поверхностную экспрессию MCR, содержащего комплекс HLA-DRB1_4-MP1103-120, в то время как DRB1_15-MP1103-120, содержащие MCRs, обнаруживались на очень низких уровнях (a). Мы с трудом обнаружили MCRs, содержащие DRB5_1-MP1103-120(a), но не обнаружили эффективную экспрессию DRB5_1-MP162-72, содержащих MCRs (b). Важно отметить, что когда библиотека пептидов, происходящих из кДНК гриппа, была привязана к HLA DBR 1_4 или HLA DRB 1_15, наблюдались сходные уровни MCR поверхностной экспрессии (c). “UI” означает неинфицированные клетки.
Фиг. 3 показывает пример анализа клеток, трансфицированных MCRs, содержащими различные HLA бета цепи, несущие различные пептиды (pep1, pep2 и pep3), окрашенные анти-HLA-DR и анти-CD3e. Двойное положительное окрашивание показывает, что оба антитела являются одинаково эффективными при обнаружении MCR поверхностной экспрессии, и диагональная форма популяции подтверждает эффективную связь MCR с CD3 цепями.
Фиг. 4 показывает низкую поверхностную экспрессию MHC и MCR молекул без ковалентно связанных пептидов. Клетки, трансфектированные конструкциями HLA-DRB1.4 (слева) или MCR2-DRB1.4 (справа), были окрашены или αHLA-DR (вверху) или αCD3 (внизу).
Фиг. 5 показывает предсказание связывания пептида для MP1103-120 с использованием анализа Базы данных иммунных эпитопов (IEDB).
Фиг. 6 показывает FACS анализ клеток, трансдуцированных MCRs, содержащими различные HLA-DRB цепи, несущие различные пептиды, содержащие коровый эпитоп MP1107-117. Анти-CD3e или анти-HLA-DR PE антитела использовали для измерения MCR поверхностной экспрессии. Графики показывают, что коровый MP1107-117 пептид может эффективно презентироваться только на HLA-DRB1_4 и также не HLA-DRB1_15 не HLA-DRB5_1, и что фланкирующие участки пептидов не играют главной роли. Наличие MCRs во всех образцах было подтверждено с помощью ПЦР и секвенирования (не показано).
Фиг. 7. Библиотека случайных пептидов, связанных с альфа цепью мышиного MCR, была трансфектирована в репортерные клетки, отдельные клетки, экспрессирующие MCR на поверхности, были отсортированы, и была определена последовательность иллюстративных клонов. График представляет FACS анализ характерных клонов, несущих MCRs с такими пептидами, окрашенными анти-мышиными MHC класса II и соответствующие последовательности пептидов.
Пример 1. Характерная поверхностная экспрессия MCRs, содержащих пептид матриксного белка 1 гриппа
Изобретатели сконструировали MHC-TCR гибридные белки (MCRs), состоящие из пептидов, связанных с нативными внеклеточными доменами MHC, слитыми с трансмембранными участками TCR (фиг. 1; см. также WO 2016/097334 A1). В частности, были созданы конструкции, кодирующие MCR, несущие альфа и бета цепи человеческих MHC молекул (HLA-DRB1_4, DRB1_15 и DRB5_1), с бета цепями, несущими ковалентно связанный пептид матриксного белка 1 гриппа (MP1103-120), MP162-72 или библиотека пептидов, происходящих от кДНК гриппа. Супернатанты, содержащие ретровирус, получили согласно стандартному протоколу в экотропной Phoenix пакующей клеточной линии и использовали для инфицирования репортерных клеточных линий, и сортировали клетки. После трансдукции клеточной линии, лишенной MHC класса II (T клеточной гибридомы), анти-HLA-DR APC антитело использовали для измерения MCR поверхностной экспрессии с помощью FACS (фиг. 2). MP1-происходящий пептид, содержащий аминокислоты 103-120, связанные к HLA DRB 1_4, приводит к эффективной экспрессии MCR на поверхности репортерной клеточной линии млекопитающего. Тот же самый пептид, связанный с HLA DRB 1_15, приводит к очень низкой MCR экспрессии, а в случае HLA DRB 5_1 не экспрессируется совсем (фиг. 2a). Однако, другой MP1- происходящий пептид, содержащий аминокислоты 62-72, приводит к эффективной экспрессии MCR, содержащего HLA DRB 5-1, но не других протестированных HLAs (фиг. 2b). Важно отметить, что когда библиотека пептидов, происходящих от кДНК гриппа, была связана с HLA DBR 1_4 или HLA DRB 1_15, наблюдались сходные уровни поверхностной экспрессии, показывая, что оба HLA 1_4 и HLA 1_15 могут эффективно экспрессироваться на клеточной поверхности, как части MCR (фиг. 2c). Из опубликованных исследований (Schmid et al Immunity 2007) известно, что MP1 пептиды, содержащие аминокислоты 103-117 и 107-121, эффективно презентируются HLA DRB 1-4, указывая на то, что только если ковалентно присоединенный пептид эффективно связывается с MHC пептид-связывающей бороздкой, MHC комплекс является достаточно стабильным для поверхностной экспрессии, которая может быть обнаружена путем окрашивания антителами. Фиг. 6 дополнительно показывает, что коровый MP1107-117 пептид может эффективно презентироваться только на HLA-DRB1_4 вне зависимости от фланкирующих участков пептида.
Пример 2. Обнаружение поверхностной экспрессии MCR с помощью различных антител
Поверхностная экспрессия MCRs, содержащих различные HLA бета цепи, несущие различные пептиды (pep1, pep2 и pep3), была обнаружена путем окрашивания антителами с использованием анти-HLA-DR и анти-CD3e и проанализирована на BDFortessa проточном цитометре (фиг. 3). Двойное позитивное окрашивание показывает, что оба антитела являются эффективными в обнаружении MCR поверхностной экспрессии и что анти-CD3e окрашивание может использоваться универсально для обнаружения MCRs, несущих различные HLA аллели. Диагональная форма популяции дополнительно показывает эффективную связь MCR с CD3 цепями.
Пример 3. Предсказание MHC аффинности связывания для MP1103-120
MHC аффинность связывания для MP1103-120 была предсказана с использованием анализа Базы данных иммунных эпитопов (IEDB) (Wang, P. et al. A Systematic Assessment of MHC class II Peptide Binding Predictions and Evaluation of a Consensus Approach. PLoS Comput Biol 4, e1000048 (2008); Wang, P. et al. Peptide binding predictions for HLA DR, DP и DQ molecules. BMC Bioinformatics 11, 568 (2010)). В отношении MP1103-121 способ настоящего изобретения показал, что HLA DRB 4_1 должен связываться наилучшим образом и приводить к высокой поверхностной экспрессии HLA, HLA DRB 1_15 должен связываться умеренно и приводить к низкой поверхностной экспрессии, а HLA DRB 5_1 не связывается вовсе. Однако, с использованием анализа IEDB предсказательный анализ пептид-связывания показал, что HLA DRB 5_1 должен быть лучшим MP1103-120 связующим компонентом (фиг. 5).
Пример 4. Обнаружение поверхностной экспрессии MCRs, содержащих пептиды, слитые с альфа цепью
Изобретатели сконструировали библиотеку мышиных MCRs, несущих случайные пептиды, соединенные с альфа цепью. После трансдукции клеточной линии, лишенной MHC класса II (T-клеточная гибридома), использовали анти-MHC антитело для измерения MCR поверхностной экспрессии с помощью FACS (фиг. 7). Эффективная экспрессия MCRs, несущих множество различных пептидов была обнаружена, показывая, что пептиды, связанные с альфа цепью MCR могут также стабилизировать MHC комплекс достаточно для поверхностной экспрессии.
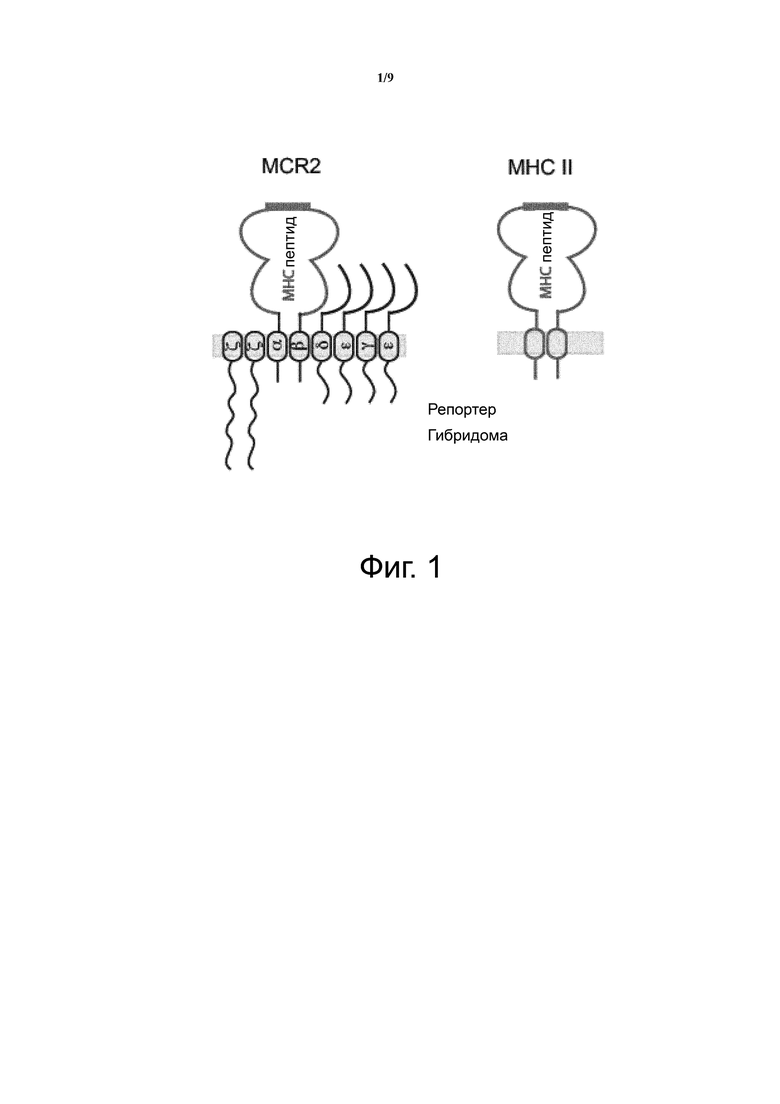
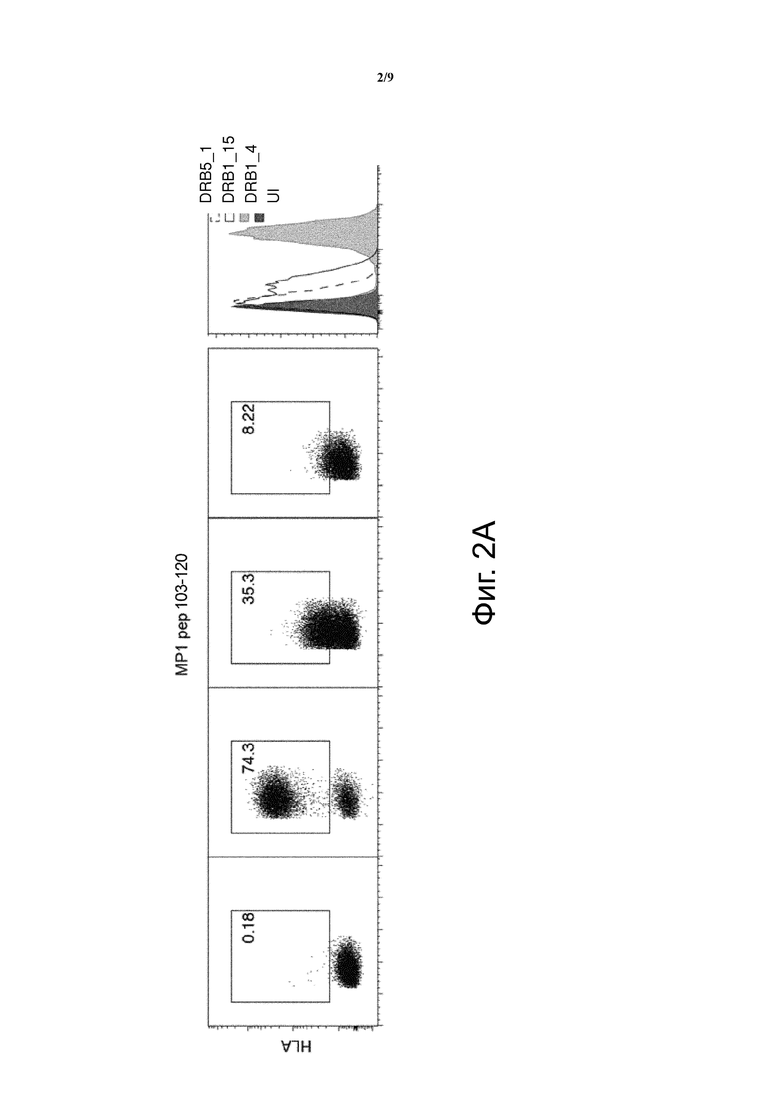
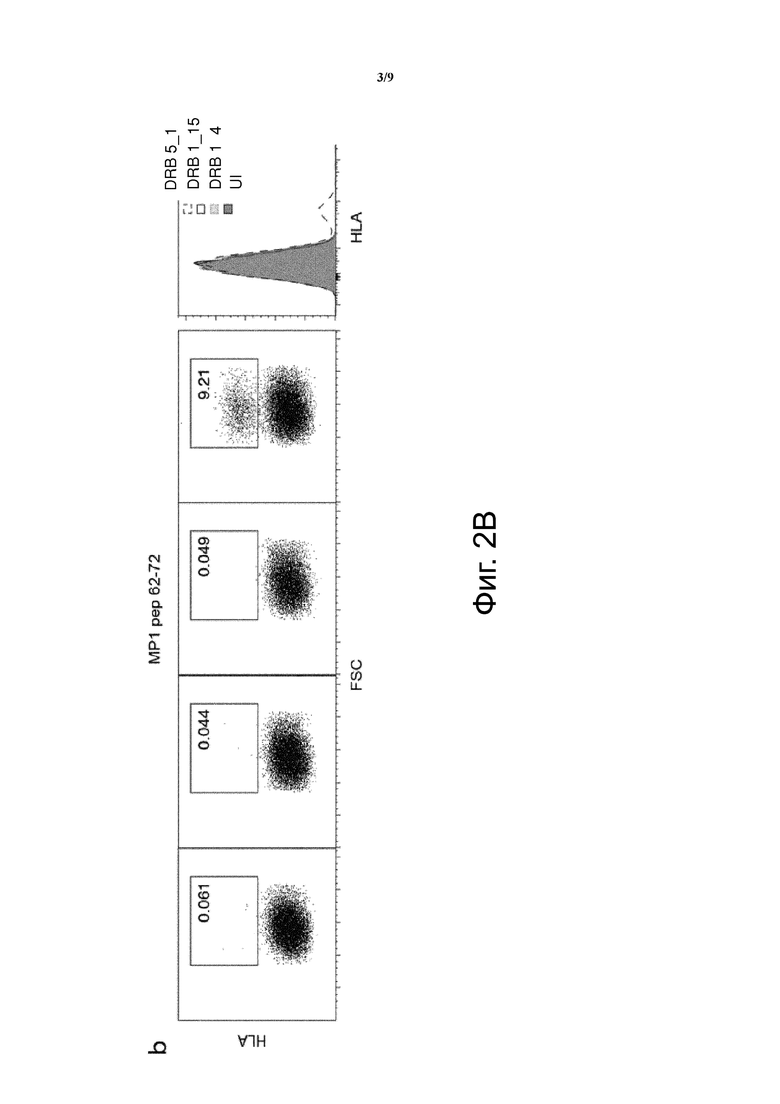
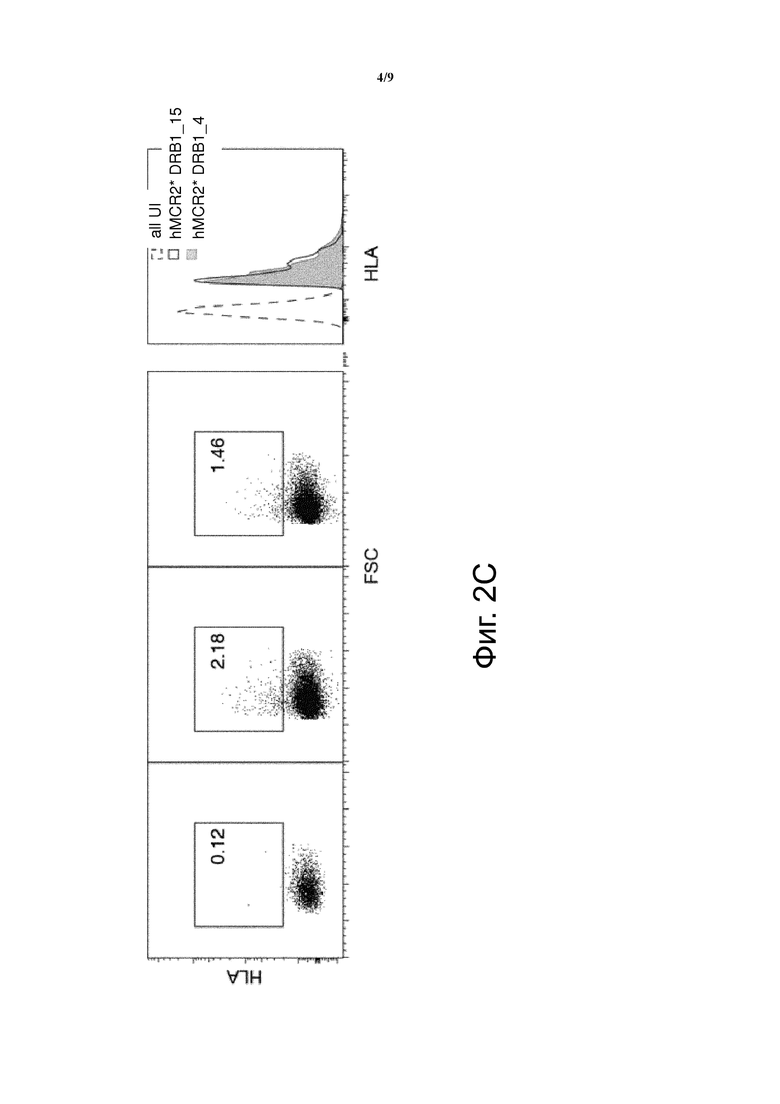
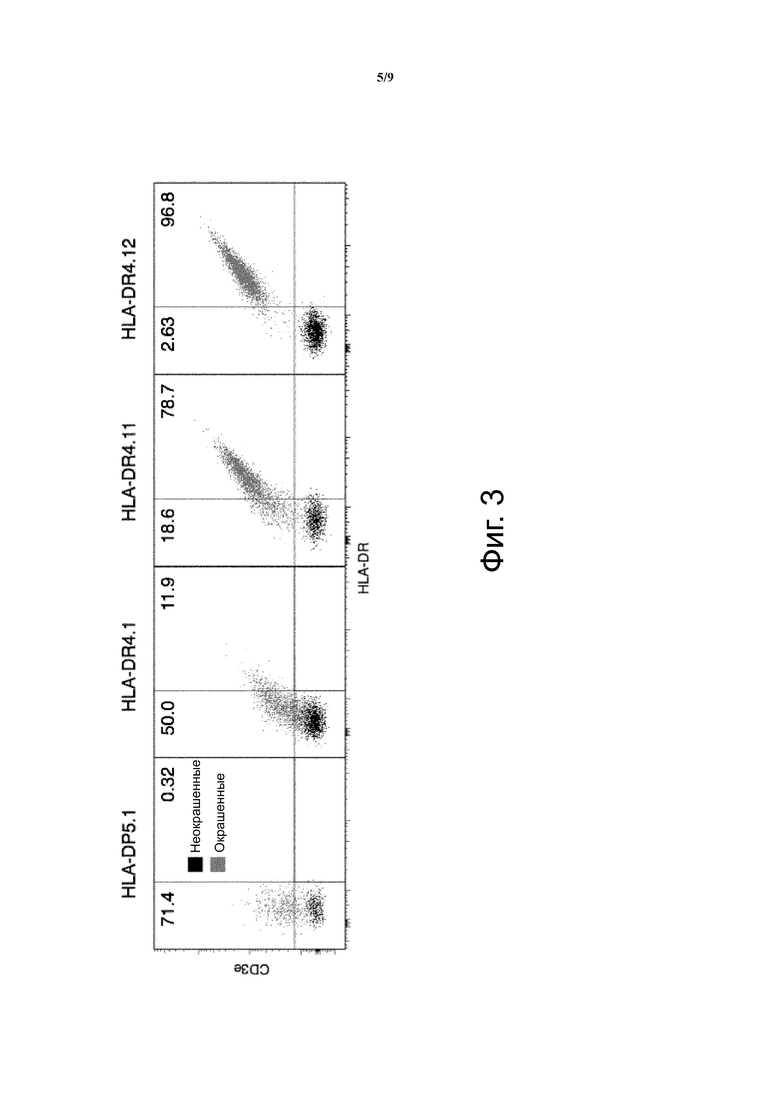
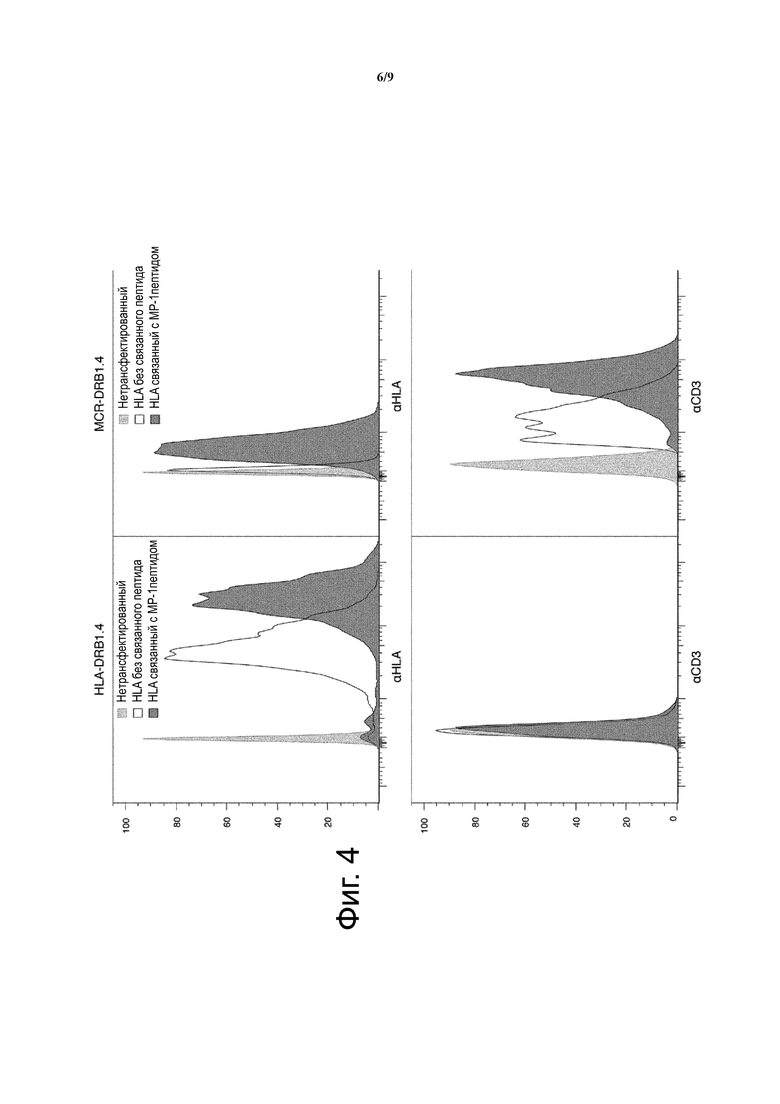

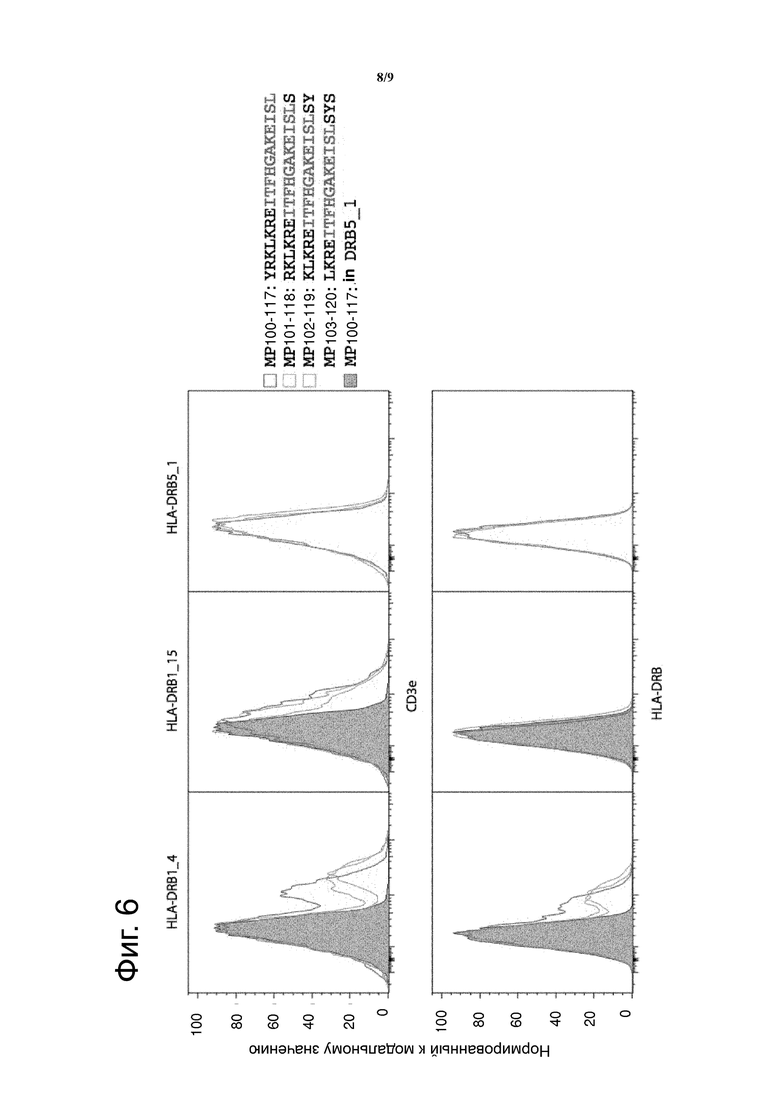
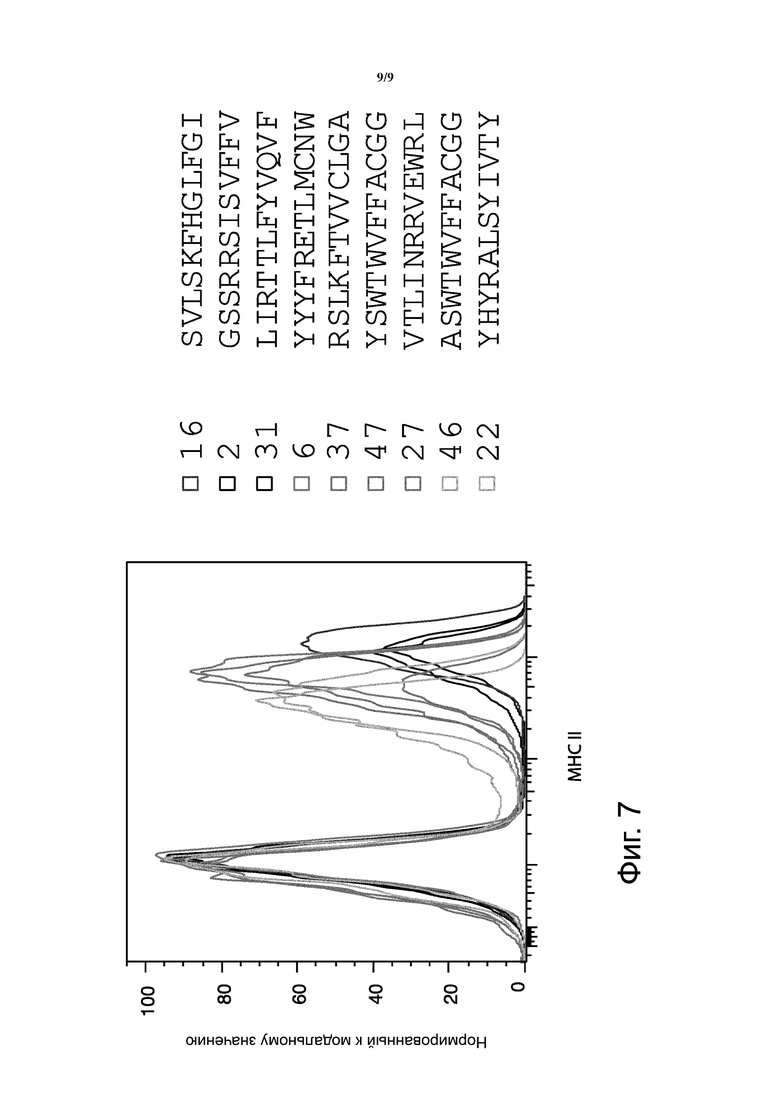
Настоящее изобретение относится к области биотехнологии. Описан способ идентификации кандидатного пептида, презентированного главным комплексом гистосовместимости (MHC). Способ включает стадии: экспрессии рекомбинантного комплекса MHC-пептид в репортерной клеточной линии; обнаружения и выделения клеток, которые экспрессируют один или несколько рекомбинантный(ых) комплекс(ов) MHC-пептид на поверхности клетки; определения аминокислотной последовательности того кандидатного пептида, который презентирован на клеточной поверхности клетки, выделенной на предыдущей стадии; идентификации кандидатного пептида как презентированного MHC. Также описан способ определения аффинности связывания MHC с кандидатным пептидом. Технический результат заключается в обеспечении средств и методов для надежного установления пептидов, в частности TSAs, которые эффективно презентируются APCs. 2 н. и 37 з.п. ф-лы, 9 ил., 4 пр.
1. Способ идентификации кандидатного пептида, презентированного главным комплексом гистосовместимости (MHC), где способ включает стадии:
(а) экспрессии рекомбинантного комплекса MHC-пептид в репортерной клеточной линии, где кандидатный пептид ковалентно связан с МНС и где репортерная клеточная линия является линией клеток млекопитающих,
(b) обнаружения и выделения клеток, которые экспрессируют один или несколько рекомбинантный(ых) комплекс(ов) MHC-пептид на поверхности клетки, где обнаружение и выделение клетки содержит стадию контакта репортерных клеток с антителом, направленным против рекомбинантного комплекса MHC-пептид,
(с) определения аминокислотной последовательности того кандидатного пептида, который презентирован на клеточной поверхности клетки, выделенной на стадии (b),
(d) идентификации кандидатного пептида стадии (с) как презентированного MHC.
2. Способ по п. 1, где стадия определения аминокислотной последовательности кандидатного пептида включает стадии выделения ДНК из клеток, которые выделены на стадии (b), и определения последовательности ДНК, кодирующей кандидатный пептид.
3. Способ по п. 2, где определение последовательности кандидатного пептида включает стадию ПЦР и секвенирования.
4. Способ по любому из пп. 1-3, дополнительно включающий стадию определения уровня поверхностной экспрессии комплекса MHC-пептид.
5. Способ определения аффинности связывания MHC с кандидатным пептидом, где способ включает стадии:
a) экспрессии рекомбинантного комплекса MHC-пептид в репортерной клеточной линии, где кандидатный пептид ковалентно связан с МНС и где репортерная клеточная линия является линией клеток млекопитающих,
b) обнаружения репортерных клеток, которые презентируют комплекс MHC-пептид на поверхности репортерной клетки, антителом, направленным против комплекса MHC-пептид,
c) определения уровня такого презентирования/экспрессии, и
d) определения аффинности связывания MHC с кандидатным пептидом.
6. Способ по любому из пп. 1-5, где кандидатный пептид клонирован выше рекомбинантной бета- и/или альфа-цепи MHC внутри одной рамки считывания.
7. Способ по любому из пп. 1-6, где кандидатный пептид закодирован в библиотеке кандидатных пептидов.
8. Способ по п. 7, где библиотека кандидатных пептидов получена путем клонирования множества полинуклеотидов, кодирующих кандидатные пептиды выше в рамке считывания с полинуклеотидом, кодирующим рекомбинантную альфа- и/или бета-цепь MHC.
9. Способ по любому из пп. 1-8, в котором молекула MHC молекула является
a) молекулой MHC класса II, содержащей внеклеточную альфа-цепь MHC класса II и трансмембранный домен, и внеклеточную бета-цепь MHC класса II и трансмембранный домен, или
b) молекулой MHC класса I, содержащей внеклеточную альфа-цепь MHC класса I и трансмембранный домен, а также бета-2-микроглобулин.
10. Способ по любому из пп. 1-8, в котором комплекс MHC-пептид является гибридным белком, содержащим кандидатный пептид, бета-2-макроглобулин, внеклеточную альфа-цепь MHC класса I и трансмембранный домен.
11. Способ по п. 9 или 10, согласно которому альфа-цепь MHC класса I несет Y84A мутацию.
12. Способ по любому из пп. 1-11, согласно которому каждая цепь молекулы MHC содержит трансмембранный домен.
13. Способ по п. 12, согласно которому трансмембранный домен является нативным трансмембранным доменом молекулы MHC или
где трансмембранный домен является трансмембранным доменом гетерологичной молекулы, такой как TCRα/β, TCRγ/δ, CD3γ/δ/ε/ζ, CD4 или CD8α/β.
14. Способ по любому из пп. 1-13, где антитело направлено против MHC-I или MHC-II или где антитело направлено против CD3γ, CD3δ или CD3ε.
15. Способ по любому из пп. 1-14, где клетки эффективно экспрессирующие рекомбинантный комплекс МНС-пептид на своей клеточной поверхности выделяются с помощью FACS или MACS.
16. Способ по любому из пп. 1-15, согласно которому репортерная клеточная линия является клеточной линией, лишенной механизма загрузки пептида на MHC класса II.
17. Способ по п. 16, согласно которому репортерная клеточная линия является T-клеточной гибридомой.
18. Способ по любому из пп. 1-17, согласно которому репортерная клеточная линия является клеточной линией, лишенной функционального гена TAP1, TAP2 и/или бета-2-микроглобулина.
19. Способ по п. 18, согласно которому репортерная клеточная линия является T-клеточной гибридомой с дефектным или удаленным геном TAP1, TAP2 и/или бета-2-микроглобулина.
20. Способ по любому из предшествующих пунктов, согласно которому кандидатный пептид является опухоль-специфическим пептидом, несущим индивидуальную мутацию(и) опухолевого происхождения.
21. Способ по п. 20, согласно которому указанная мутация представляет собой SNV.
22. Способ по пп. 1-19, согласно которому кандидатный пептид является антигеном, вызывающим иммунный ответ.
23. Способ по п. 22, согласно которому кандидатный пептид используется в качестве вакцины.
24. Способ по п. 23, согласно которому вакцина является противораковой вакциной на основе опухоль-специфического антигена (TSA).
25. Способ по п. 22, согласно которому кандидатный пептид используется для индукции иммунологической толерантности, по меньшей мере, против одного эпитопа, который он содержит.
26. Способ по п. 22 или 25, согласно которому кандидатный пептид в контексте молекулы MHC используется с целью блокирования TCR.
27. Способ по п. 22 или 25, согласно которому кандидатный пептид используется для MHC-опосредованной доставки токсина в клетки, в частности в T-клетки.
28. Способ по п. 22 или 25, согласно которому кандидатный пептид используется для перенацеливания T-клеток с помощью MHC-CAR.
29. Способ по п. 22, согласно которому кандидатный пептид используется для тестирования иммуногенности.
30. Способ по п. 22 или 29, согласно которому кандидатный пептид используется для тестирования T-клеточной реактивности.
31. Способ по пп. 1-19, согласно которому кандидатный пептид является веществом, подвергающимся тестированию на иммуногенность.
32. Способ по п. 31, согласно которому кандидатный пептид используется в качестве вакцины.
33. Способ по п. 32, согласно которому вакцина является противораковой вакциной на основе опухоль-специфического антигена (TSA).
34. Способ по п. 31, согласно которому кандидатный пептид используется для индукции иммунологической толерантности, по меньшей мере, против одного эпитопа, который он содержит.
35. Способ по любому из пп. 31 или 34, согласно которому кандидатный пептид в контексте молекулы MHC используется с целью блокирования TCR.
36. Способ по любому из пп. 31 или 34, согласно которому кандидатный пептид используется для MHC-опосредованной доставки токсина в клетки, в частности в T-клетки.
37. Способ по любому из пп. 31 или 34, согласно которому кандидатный пептид используется для перенацеливания T-клеток с помощью MHC-CAR.
38. Способ по п. 31, согласно которому кандидатный пептид используется для тестирования иммуногенности.
39. Способ по пп. 31 или 38, согласно которому кандидатный пептид используется для тестирования T-клеточной реактивности.
| WO 2016097334, 23.06.2016 | |||
| FEI WEN et al | |||
| Cell surface display of functional human MHC class II proteins: yeast display versus insect cell display | |||
| Protein Eng Des Sel | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| 701-709 | |||
| WO 2004015395, 19.02.2004 | |||
| WO 2015143558, 01.10.2015 | |||
| RU 2015126651 A, 16.01.2017. | |||

