Изобретение относится к биотехнологии, в частности к генетической инженерии. Может быть использовано для получения клеток, обладающих активностью эстеразы Psychrobacter cryohalolentis K5T (цельноклеточного биокатализатора).
Цельноклеточные биокатализаторы (ЦБК) с активностью липаз и эстераз могут быть использованы для биоремедиации загрязненных территорий за счет деградации органических поллютантов, получения биотоплива на основе растительного сырья и др. [Wu С., Mulchandani A., Chen W. Versatile microbial surface-display for environmental remediation and biofuels production. Trends Microbiol. 2008. V.16. P.181-188]. ЦБК со свойствами эстеразы P. cryohalolentis K5T также может быть использован в тонком химическом синтезе в реакциях переэтерификации и гидролиза [Jaeger К.Е., Eggert Т. Lipases for biotechnology. Curr. Opin. Biotechnol. 2002. V.13. Р.390-397].
Преимуществом использования ЦБК является отсутствие необходимости разрушения клеток и очистки целевого продукта, что удешевляет процесс производства, а также повышенная стабильность препарата по сравнению с изолированным ферментом [Ishige Т., Honda К., Shimizu S. Whole organism biocatalysis. Curr. Op. Chem. Biol. 2005. V.9. P.174-180]. Преимуществом использования ЦБК со свойствами эстеразы Р. cryohalolentis K5T является высокая активность данного фермента при низких температурах [Novototskaya-Vlasova К., Petrovskaya L., Yakimov S., Gilichinsky D. Cloning, purification, and characterization of a cold-adapted esterase produced by Psychrobacter cryohalolentis K5T from Siberian cryopeg. FEMS Microbiol. Ecol. 2012. V.82. P.367-375], что позволяет использовать его для биоремедиации загрязненных территорий Севера, а также сокращение расходов на подогрев реакционных смесей и связанных с этим атмосферных выбросов углекислого газа.
Известен способ получения ЦБК с активностью липазы путем экспрессии в клетках Е. coli гибридного гена, кодирующего в единой рамке считывания с ферментом белок-транспортер. При таком способе происходит секреция рекомбинантной липазы на поверхность клеток, что обеспечивает правильный фолдинг белка, а также устраняет необходимость выделения фермента из биомассы штамма-продуцента. В качестве транспортера может быть использован белок, способствующий формированию кристаллов льда (ice nucleation protein, INP) Pseudomonas syringae [H. Jung, S. Ко, S. Ju, E. Kim, M. Kim, J. Pan. Bacterial cell surface display of lipase and its randomly mutated library facilitates high-throughput screening of mutants showing higher specific activities. Journal of Molecular Catalysis B: Enzymatic. 2003. V.26. P.177-184]. Для получения ЦБК ген липазы TliA P. fluorescens клонируют в 3′-направлении от гена inaK с образованием единой рамки считывания под контролем tac-промотора в плазмиде pKK223-3. Синтез рекомбинантного белка индуцируют добавлением 1 мМ изопропил-β-D-тиогалактопиранозида (ИПТГ). К недостаткам этого способа получения следует отнести большое количество индуктора, необходимое для достижения экспрессии, а также недостаточно строгий контроль репрессии промотора в плазмиде pKK223-3.
Известен способ получения ЦБК с активностью липазы с использованием в качестве транспортера белка FadL Е. coli [S.Н. Lee, J. Choi, S.J. Park, S.Y. Lee, В.C. Park. Display of bacterial lipase on the Escherichia coli cell surface by using FadL as an anchoring motif and use of the enzyme in enantioselective biocatalysis. Appl. Environ. Microbiol. 2004. V.70. P.5074-5080]. В этом случае ген термостабильной липазы Bacillus sp. TG43 клонируют в 3′-направлении от последовательности, кодирующей белок FadL, под контролем trc-промотора в плазмиде pTrc99A. Экспрессию гибридного гена индуцируют добавлением 0,1 мМ ИПТГ. Недостатком этого способа является низкий уровень синтеза рекомбинантного белка.
Наиболее близким к заявленному является способ получения клеток Е. coli, обладающих активностью эстеразы ApeE Salmonella enterica [Е. Schultheiss, S. Weiss, Е. Winterer, R. Maas, E. Heinzle, J. Jose. Esterase autodisplay: enzyme engineering and whole-cell activity determination in microplates with pH sensors. Applied and Environmental Microbiology. 2008. V.74. P.4782-4791]. Рекомбинантная плазмида pES02 содержит ген гибридного белка, состоящего из эстеразы АреЕ Salmonella enterica и аутотранспортера AIDA-I Е. coli под контролем промотора Т7. Синтез белка осуществляют при добавлении индуктора ИПТГ до конечной концентрации 1 мМ. Рекомбинантный штамм Е. coli BL21(DE3)/pES02 обеспечивает синтез и секрецию белка FP89 на поверхности клеток. В результате получают клетки Е. coli с активностью эстеразы АреЕ по отношению к α-нафтилацетату, составляющей 1 U на 2×107 клеток. К недостаткам описанного способа получения клеток Е. coli, обладающих эстеразной активностью, можно отнести большое количество индуктора, необходимое для достижения экспрессии, а также узкую субстратную специфичность полученного ЦБК.
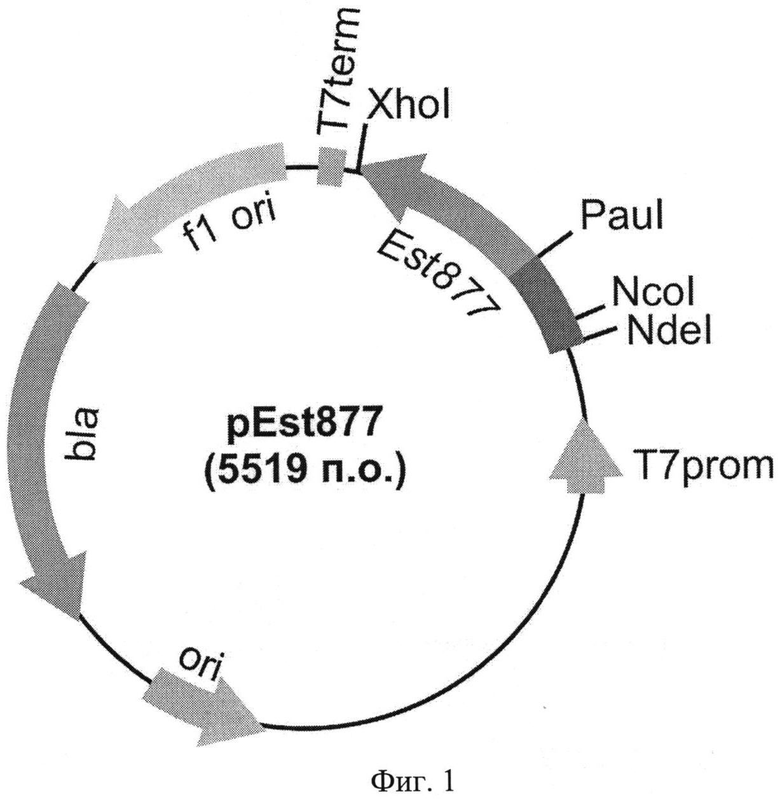
Технической задачей изобретения является получение штамма-продуцента полипептида с активностью эстеразы Psychrobacter cryohalolentis K5T на поверхности клеток, а также повышение уровня его активности по отношению к различным субстратам. Поставленная задача решается за счет конструирования рекомбинантной плазмидной ДНК pEst877, детерминирующей экспрессию полипептида с активностью эстеразы P. cryohalolentis К5Т на поверхности клеток, с мол. массой 3,64 Md (5,519 т.п.о.), состоящей из NcoI/XhoI - фрагмента ДНК плазмиды pET20b(+) длиной 3,654 т.п.о., включающего промотор T7lac, терминатор транскрипции бактериофага Т7, ген bla β-лактамазы, определяющий устойчивость трансформированных плазмидой pEst877 клеток к ампициллину, участок ori инициации репликации, сигнальную последовательность pelB пектатлиазы В Erwinia carotovora; и NcoI/XhoI - фрагмента ДНК размером 1,865 т.п.о., содержащего гибридный ген Est877, кодирующий аминокислотные последовательности эстеразы P. cryohalolentis K5T с дополнительным аминокислотным остатком аспарагиновой кислоты после сайта отщепления сигнального пептида и аутотранспортера P. cryohalolentis K5T в единой рамке считывания; содержащей уникальные сайты узнавания рестрикционными эндонуклеазами, имеющими следующие координаты: NdeI - 2089, NcoI - 2023, PauI - 1138, XhoI - 158, а также за счет штамма бактерий Е. coli BL21(DE3)pLysS/pEst877 - продуцента полипептида со свойствами эстеразы Р. cryohalolentis К5Т на клеточной поверхности.
Рекомбинантная плазмидная ДНК pEst877 кодирует индуцибельный синтез полипептида с активностью эстеразы P. cryohalolentis K5T на поверхности клеток, а штамм Е. coli BL21(DE3)pLysS/pEst877 обеспечивает синтез и секрецию этого полипептида с уровнем экспрессии не ниже 10% суммарного клеточного белка. Индуцибельный высокий уровень синтеза целевого полипептида обеспечивается тем, что плазмида pEst877 содержит промотор T7lac и усилитель трансляции гена 10 бактериофага Т7, а экспрессия на поверхности клеток - наличием в составе плазмиды кодирующих последовательностей сигнального пептида pelB и С-концевого домена аутотранспортера P. cryohalolentis K5T, а также модификацией 5′-концевой последовательности гена эстеразы P. cryohalolentis K5T.
Особенностью предложенной плазмидной конструкции является наличие в ее составе кодирующей последовательности гибридного белка, состоящего из эстеразы Р. cryohalolentis К5Т с дополнительным аминокислотным остатком аспарагиновой кислоты после сайта отщепления сигнального пептида и С-концевого домена аутотранспортера Р. cryohalolentis К5Т (SEQ ID NO 5), под контролем сильного регулируемого промотора T7lac, что в сочетании с наличием в плазмиде кодирующей последовательности сигнального пептида pelB, генов РНК-полимеразы бактериофага Т7 и компонентов системы секреции в мембране клеток в хромосоме хозяйского штамма обеспечивает индуцибельный синтез целевого белка (SEQ ID NO 6) на поверхности клеток с высоким выходом, достигаемым при малых концентрациях индуктора. В 5′-концевую последовательность гена эстеразы P. cryohalolentis К5Т добавлен кодон GAT, обеспечивающий наличие остатка аспарагиновой кислоты после сайта отщепления сигнального пептида. Известно, что суммарный отрицательный заряд N-концевой части зрелой последовательности секретируемого белка способствует более эффективной секреции через мембрану [Li P., Beckwith J., Inouye Н. Alteration of the amino terminus of the mature sequence of a periplasmic protein can severely affect protein export in Escherichia coli. Proc. Natl. Acad. Sci. USA. 1988. V.85. P.7685-7689].
Для получения штамма-продуцента полипептида с активностью эстеразы Р. cryohalolentis К5Т на поверхности клеток компетентные клетки Е. coli штамма BL21(DE3)pLysS трансформируют рекомбинантной плазмидой pEst877.
Полученный штамм Е. coli BL21(DE3)pLysS/pEst877 характеризуется следующими признаками.
Морфологические признаки: клетки мелкие, палочковидной формы, грамотрицательные, подвижные, размером 1×3-5 мкм, неспороносные.
Культуральные признаки: при росте на агаризованной LB-среде преобладают мелкие колонии диаметром 1-3 мм; круглые, гладкие, полупрозрачные, блестящие, серые, край ровный; консистенция пастообразная. Могут встречаться крупные колонии диаметром 4-5 мм; круглые, гладкие, матовые, край волнистый. Рост в жидких средах (LB, минимальная среда с глюкозой) характеризуется равномерным помутнением среды.
Физиолого-биохимические признаки. Клетки штамма продуцента могут расти в диапазоне температур 20-42°С, оптимальная температура роста составляет 37°С. Наиболее благоприятные для роста значения pH находятся в интервале 6,8-7,2. При росте в аэробных условиях, культура может усваивать азот как органических соединений (пептон, триптон, аминокислоты, дрожжевой экстракт), так и аммонийных солей. Углерод усваивается в форме углеводов, многоатомных спиртов (глицерин), аминокислот.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к ампициллину (до 200 мкг/мл), обусловленную наличием в плазмиде гена бета-лактамазы, и к хлорамфениколу (до 34 мкг/мл), обусловленную присутствием в хозяйском штамме плазмиды pLysS.
Способ, условия и состав среды для хранения штамма. LB-бульон с 15% глицерином, при температуре -70°С, в криовиалах.
Технический результат заявленного изобретения заключается в том, что полученный штамм Е. coli BL21(DE3)pLysS/pEst877 позволяет получать полипептид с активностью эстеразы P. cryohalolentis K5T на поверхности клеток в количестве не менее 10% от суммарного клеточного белка при концентрации индуктора 0,1 мМ. В отличие от прототипа в данном штамме достигается в 2 раза более высокая эстеразная активность при использовании в 10 раза меньшего количества индуктора (ИПТГ). Кроме того, в отличие от прототипа полученный штамм проявляет эстеразную активность по отношению к более широкому кругу субстратов - эфиров жирных кислот с длиной углеводородной цепи от С4 до С10. Совокупность перечисленных свойств штамма Е. coli BL21(DE3)pLysS/pEst877 обусловливает большую технологичность процесса получения ЦБК с активностью эстеразы P. cryohalolentis K5T на поверхности клеток.
Изобретение иллюстрируют графические материалы.
Фиг.1. Физическая карта рекомбинантной плазмиды pEst877. Указаны уникальные сайты эндонуклеаз рестрикции. Т7 promoter - промотор бактериофага Т7, Т7 terminator-терминатор транскрипции бактериофага Т7, Ар - ген устойчивости к ампициллину, ori - участок инициации репликации плазмиды, Est877 - рекомбинантный ген Est877, кодирующий аминокислотную последовательность полипептида с активностью эстеразы P. cryohalolentis К5Т на поверхности клеток.
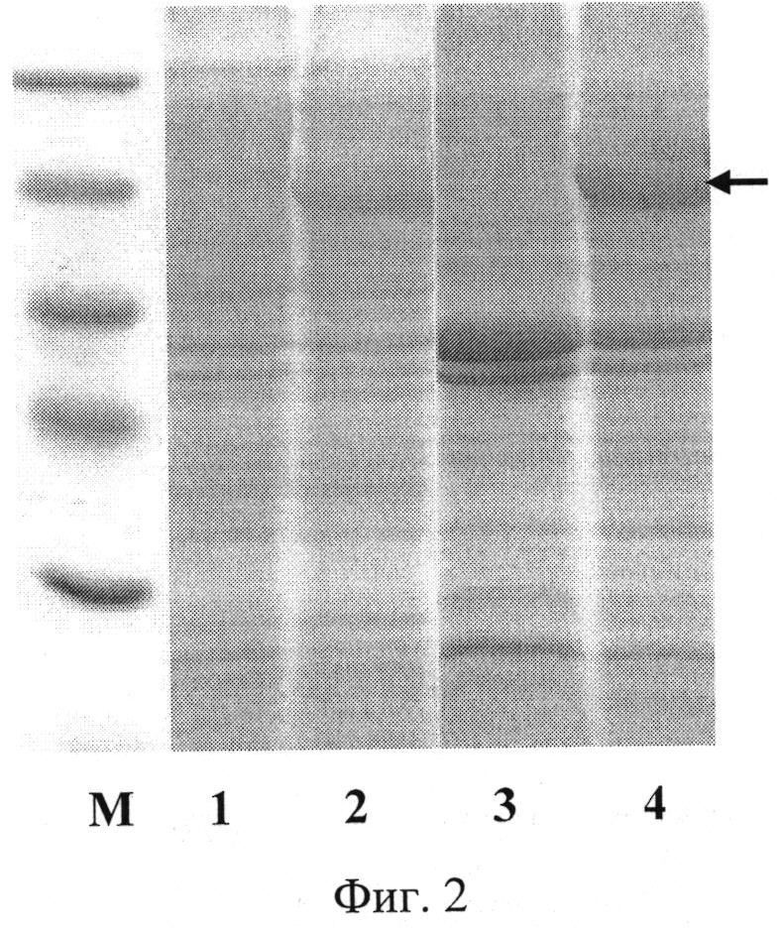
Фиг.2. Электрофореграмма лизатов клеток штамма-продуцента Е. coli BL21(DE3)pLysS/pEst877 до (дорожка 1) и после индукции (дорожки 2) и мембранных фракций клеток штамма-продуцента Е. coli BL21(DE3)pLysS/pEst877 до (дорожка 3) и после индукции ИПТГ (дорожка 4) в 13%-ном полиакриламидном геле (М - белковые маркеры молекулярной массы); стрелкой указано положение целевого полипептида.
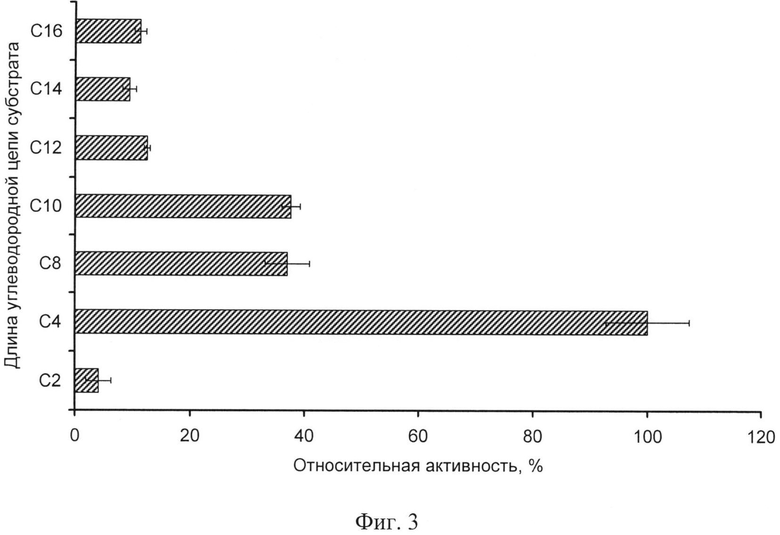
Фиг.3. Субстратная специфичность полипептида с активностью эстеразы Р. cryohalolentis К5Т на поверхности клеток.
Изобретение иллюстрируют следующие примеры.
Пример 1.
Конструирование рекомбинантной плазмидной ДНК рАТ877.
Культуру P. cryohalolentis К5Т выращивают в течение ночи на чашке с агаризованной средой TSB (Difco, USA) +2% NaCl. 20 мкл полученной биомассы суспендируют в 50 мкл буфера для Taq-полимеразы (75 мМ трис-HCl, pH 8,8, 20 мМ (NH4)2SO4, 0,1% Tween 20) и разрушают в течение 15 мин прогреванием на кипящей водяной бане. После центрифугирования 5 мин при скорости 13000 об/мин 5 мкл полученного супернатанта добавляют в реакционную смесь для ПЦР объемом 50 мкл, содержащую 50 pmol праймера PC877F (SEQ ID NO 3), 50 pmol праймера PC877R (SEQ ID NO 4), 2 мМ каждого из четырех dNTP, 5 мкл 10× буфера для Pfu ДНК-полимеразы (Fermentas), 2,5 е.a. Pfu ДНК-полимеразы и 0,5 е.a. Taq ДНК-полимеразы (Fermentas). Полимеразную цепную реакцию (ПЦР) проводят в следующем режиме: денатурация - 1 мин, 94°С; отжиг - 45 с, 52°С; достройка - 60 с, 72°С; количество циклов - 35. 2 мкг ПЦР-продукта обрабатывают рестриктазами NcoI и XhoI (Fermentas) в условиях, рекомендованных производителем ферментов, фрагмент длиной 1,783 т.п.о. выделяют из 1% агарозного геля. 0,5 мкг полученного фрагмента и 0,2 мкг плазмидного вектора pET20b(+), линеаризированного обработкой рестриктазами NcoI и XhoI, длиной 3,654 п.о. лигируют в течение 4 ч при 12°С в 10 мкл раствора, содержащего 40 мМ трис-HCl (pH 7,8), 10 мМ MgCl2, 10 мМ дитиотреитола, 0,5 мМ аденозинтрифосфата и 3 е.а. Т4 ДНК-лигазы (Fermentas, Литва). 5 мкл лигазной смеси используют для трансформации компетентных клеток Е. coli XL-1 Blue (Stratagene, США). Трансформанты высевают на LB-arap, содержащий 100 мкг/мл ампициллина.
Из выросших клонов выделяют плазмидную ДНК и обрабатывают рестриктазами NcoI и XhoI с последующим электрофоретическим анализом длин рестрикционных фрагментов в 1% агарозном геле. Структуру клонированного гена в отобранных клонах подтверждают определением нуклеотидной последовательности. В результате получают плазмидную ДНК рАТ877, содержащую кодирующую последовательность аутотранспортера P. cryohalolentis K5T (SEQ ID NO 1).
Пример 2.
Конструирование рекомбинантной плазмидной ДНК pEst877.
Для амплификации гена эстеразы P. cryohalolentis K5T с дополнительным аминокислотным остатком аспарагиновой кислоты после сайта отщепления сигнального пептида проводят полимеразную цепную реакцию (ПЦР) на матрице плазмидной ДНК pET32a(+)/EstPc [Novototskaya-Vlasova К., Petrovskaya L., Yakimov S., Gilichinsky D. Cloning, purification, and characterization of a cold-adapted esterase produced by Psychrobacter cryohalolentis K5T from Siberian cryopeg. FEMS Microbiol. Ecol. 2012. V.82. P.367-375] в смеси объемом 50 мкл следующего состава: 50 pmol праймера 23NcoF (SEQ ID NO 7), 50 pmol праймера 23R (SEQ ID NO 8), 2 мМ каждого из четырех dNTP, 5 мкл 10× буфера для Pfu ДНК-полимеразы (Fermentas), 2,5 е.а. Pfu ДНК-полимеразы (Fermentas). Реакцию проводят в следующем режиме: денатурация - 40 с, 94°С; отжиг - 40 с, 52°С; достройка - 60 с, 72°С; количество циклов - 25. 2 мкг ПЦР-продукта обрабатывают рестриктазами NcoI и PauI (Fermentas) в условиях, рекомендованных производителем ферментов, фрагмент длиной 0,885 т.п.о., содержащий ген эстеразы P. cryohalolentis К5Т с дополнительным аминокислотным остатком аспарагиновой кислоты после сайта отщепления сигнального пептида, выделяют из 1% агарозного геля. 0,5 мкг полученного фрагмента и 0,2 мкг плазмидного вектора рАТ877, линеаризированного обработкой рестриктазами NcoI и PauI, длиной 4,634 п.о. лигируют в течение 4 ч при 12°С в 10 мкл раствора, содержащего 40 мМ трис-HCl (pH 7,8), 10 мМ MgCl2, 10 мМ дитиотреитола, 0,5 мМ аденозинтрифосфата и 3 е.а. Т4 ДНК-лигазы (Fermentas, Литва). 5 мкл лигазной смеси используют для трансформации компетентных клеток Е. coli XL-1 Blue (Stratagene, США). Трансформанты высевают на LB-arap, содержащий 100 мкг/мл ампициллина.
Из выросших клонов выделяют плазмидную ДНК и обрабатывают рестриктазами NcoI и PauI с последующим электрофоретическим анализом длин рестрикционных фрагментов в 1% агарозном геле. Структуру клонированного гена в отобранных клонах подтверждают определением нуклеотидной последовательности. В результате получают плазмидную ДНК pEst877 (фиг.1), содержащую кодирующую последовательность гибридного полипептида с активностью эстеразы P. cryohalolentis K5T на поверхности клеток (SEQ ID NO 5).
Пример 3.
Получение штамма-продуцента полипептида с активностью эстеразы Р. cryohalolentis К5Т на поверхности клеток.
Рекомбинантной плазмидной ДНК pEst877 трансформируют компетентные клетки Е. coli BL21(DE3)pLysS (Novagen) и после выращивания рекомбинантных клонов на LB-агаре с ампициллином (100 мкг/мл) и хлорамфениколом (34 мкг/мл) при 37°С получают штамм-продуцент полипептида с активностью эстеразы P. cryohalolentis K5T на поверхности клеток.
Пример 4.
Определение продуктивности штамма-продуцента полипептида с активностью эстеразы P. cryohalolentis K5T на поверхности клеток.
Для определения продуктивности клетки E.coli BL21(DE3)pLysS/pEst877 выращивают при 37°С в 5 мл жидкой среды LB, содержащей 100 мкг/мл ампициллина, в течение 16 ч на качалке при 250 об/мин. Полученную ночную культуру в объеме 200 мкл переносят в 5 мл жидкой среды LB (разбавление до оптической плотности 0,15-0,20 при длине волны 560 нм), содержащей 100 мкг/мл ампициллина, и выращивают до оптической плотности 0,8 (длина волны 560 нм) при 37°С и перемешивании 250 об/мин. Добавляют ИПТГ до конечной концентрации 0,1 мМ, после чего клетки выращивают в течение 3 ч при 27°С и перемешивании 200 об/мин. Отбирают пробу культуры в количестве 1 оптическая единица (длина волны 560 нм) и центрифугируют 5 мин при скорости 7000 об/мин. Осажденные клетки суспендируют в 100 мкл буфера, содержащего 125 мМ трис-HCl, pH 6,8, 20% глицерин, 3% додецилсульфат натрия, 3% меркаптоэтанол, 0,005% бромфеноловый синий, инкубируют 10 мин в кипящей водяной бане, образцы объемом 10 мкл анализируют электрофорезом в 13% полиакриламидном геле с додецилсульфатом натрия. По окончании электрофореза гель прокрашивают при помощи кумасси R-250. После отмывки красителя гель сканируют и проводят математическую обработку результатов с помощью программы Scion Image (Scion Corp., США). По данным сканирования содержание гибридного белка, состоящего из эстеразы и С-концевого домена аутотранспортера P. cryohalolentis К5Т, составляет 20% от общего клеточного белка. На фиг.2 представлена электрофореграмма лизатов клеток штамма-продуцента Е. coli BL21(DE3)pLysS/pEst877 (дорожки 1 и 2) в 13%-ном полиакриламидном геле до (дорожка 1) и после индукции ИПТГ (дорожки 2) (М - белковые маркеры молекулярной массы; стрелкой указано положение целевого полипептида).
Пример 5.
Анализ локализации полипептида с активностью эстеразы P. cryohalolentis K5T на поверхности клеток.
Проводят индукцию биосинтеза целевого белка в клетках штамма-продуцента Е. coli BL21(DE3)pLysS/pEst877 ИПТГ (конечная концентрация 0,1 мМ) в течение 3 ч при 27°С. Клетки центрифугируют 15 мин при 7000 об/мин. 1 г влажной биомассы индуцированных клеток ресуспендируют в 10 мл буфера (50 мМ Трис-HCl, 200 мМ NaCl, pH 8,0, после чего разрушают клетки обработкой в ультразвуковом дезинтеграторе Branson Sonifier 450, не допуская нагрева выше 10°С. Полученную суспензию центрифугируют 30 мин при 6000 об/мин. Полученный супернатант повторно центрифугируют 1 час при 100000 g. Осадок суспендируют в 1 мл буфера 50 мМ Трис-HCl pH 8,0; 5 мкл полученной суспензии смешивают с 5 мкл буфера, содержащего 125 мМ трис-HCl, pH 6,8, 20% глицерин, 3% додецилсульфат натрия, 3% меркаптоэтанол, 0,005% бромфеноловый синий, инкубируют 10 мин в кипящей водяной бане, образцы объемом 10 мкл анализируют электрофорезом в 13% полиакриламидном геле с додецилсульфатом натрия. По окончании электрофореза гель прокрашивают при помощи кумасси R-250. Гибридный белок, состоящий из эстеразы P. cryohalolentis K5T с дополнительным аминокислотным остатком аспарагиновой кислоты после сайта отщепления сигнального пептида и С-концевого домена аутотранспортера P. cryohalolentis K5T, обнаруживается в мембранной фракции клеток штамма-продуцента. На фиг.2 представлена электрофореграмма мембранных фракций клеток штамма-продуцента Е. coli BL21(DE3)pLysS/pEst877 в 13%-ном полиакриламидном геле до (дорожка 3) и после индукции ИПТГ (дорожка 4) (стрелкой указано положение полипептида с активностью эстеразы P. cryohalolentis К5Т на поверхности клеток).
Пример 6.
Определение активности полипептида со свойствами эстеразы P. cryohalolentis K5T на поверхности клеток.
Проводят индукцию биосинтеза гибридного полипептида со свойствами эстеразы P. cryohalolentis К5Т на поверхности клеток, кодируемого плазмидой pEst877, в клетках штамма-продуцента Е. coli BL21(DE3)pLysS/pEst877 ИПТГ (конечная концентрация 0,1 мМ) в течение 3 ч при 27°С. В качестве контроля используют культуру клеток Е. coli BL21(DE3)pLysS, трансформированную плазмидным вектором pET20b(+) и индуцированную в тех же условиях. Отбирают пробы обеих культур в количестве 1 оптической единицы (длина волны 560 нм) и центрифугируют 5 мин при скорости 7000 об/мин. Осадки промывают 200 мкл буфера PBS и центрифугируют в тех же условиях. Осадки суспендируют в 50 мкл буфера 50 мМ Трис-HCl (рН 8,0), 100 мМ NaCl. Активность эстеразы на поверхности клеток определяют, используя паранитрофенилбутират (Sigma) в качестве субстрата. К реакционной смеси объемом 1 мл, содержащей 50 мМ Трис-HCl (pH 8,0), 100 мМ NaCl, 0,25 мМ паранитрофенилбутирата (Sigma) добавляют 50 мкл клеточной суспензии, смесь инкубируют 15 мин при 30°С. По 200 мкл реакционной смеси после центрифугирования при 13000 об/мин в течение 5 мин раскапывают в лунки 96-луночного планшета (Costar). Измеряют величину абсорбции при 415 нм на ридере Model680 (BioRad). Измерения проводят в трех повторностях, затем вычисляют среднее значение. За единицу эстеразной активности принимают количество фермента, необходимое для высвобождения 1 мкмоль паранитрофенола за 1 мин. Общая эстеразная активность ЦБК со свойствами эстеразы Р. cryohalolentis К5Т составляет 1 U на 107 клеток, что в 2 раза больше, чем активность прототипа.
Пример 7.
Определение субстратной специфичности полипептида с активностью эстеразы Р. cryohalolentis K5T на поверхности клеток.
Субстратную специфичность полипептида с активностью эстеразы P. cryohalolentis К5Т на поверхности клеток определяют в реакциях с добавлением p-нитрофенильных производных эфирных субстратов с различной длиной углеводородной цепи (С2-С16). К реакционной смеси объемом 1 мл, содержащей 50 мМ Трис-HCl (pH 8,0), 100 мМ NaCl, 50 мкл клеточной суспензии добавляют по 0,25 мМ одного из субстратов (Sigma) - паранитрофенилацетата (С2), паранитрофенилбутирата (С4), паранитрофенилоктаноата (С8), паранитрофенилдеканоата (С10) паранитрофенилдодеканоата (С12), паранитрофенилмиристата (С14), паранитрофенилпальмитата (С16), смесь инкубируют 15 мин при 30°С. По 200 мкл реакционной смеси после центрифугирования при 13000 об/мин в течение 5 мин раскапывают в лунки 96-луночного планшета (Costar). Измеряют величину абсорбции при 415 нм на ридере Model680 (BioRad). Вычисляют активность по отношению к различным субстратам, принимая за 100% максимальное значение активности. На фиг.3 представлена диаграмма субстратной специфичности полипептида с активностью эстеразы P. cryohalolentis K5T на поверхности клеток. Максимальная активность наблюдается при использовании паранитрофенилбутирата (С4) в качестве субстрата, активность в отношении паранитрофенилоктаноата (С8) и паранитрофенилдеканоата (С10) составляет около 37% от максимальной.
Перечень последовательностей, представленных в описании изобретения
SEQ ID NO 1.
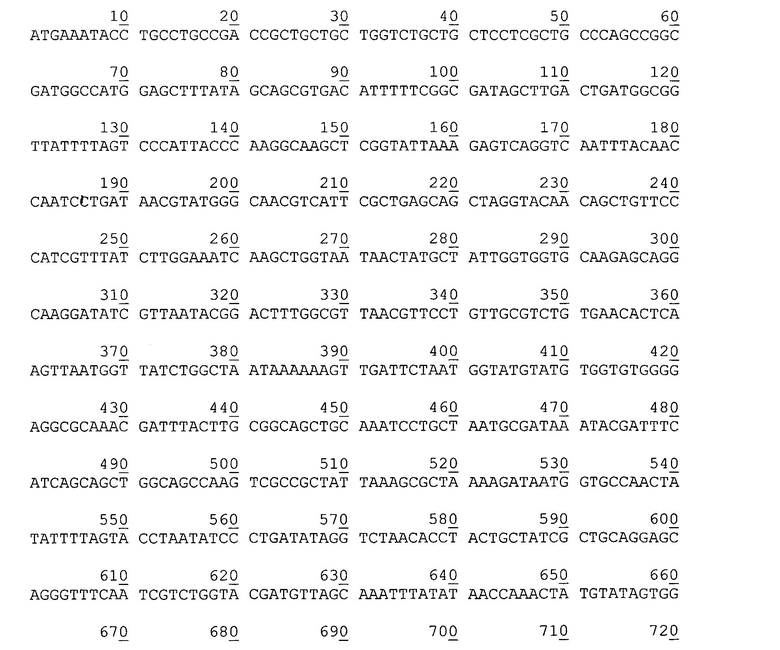
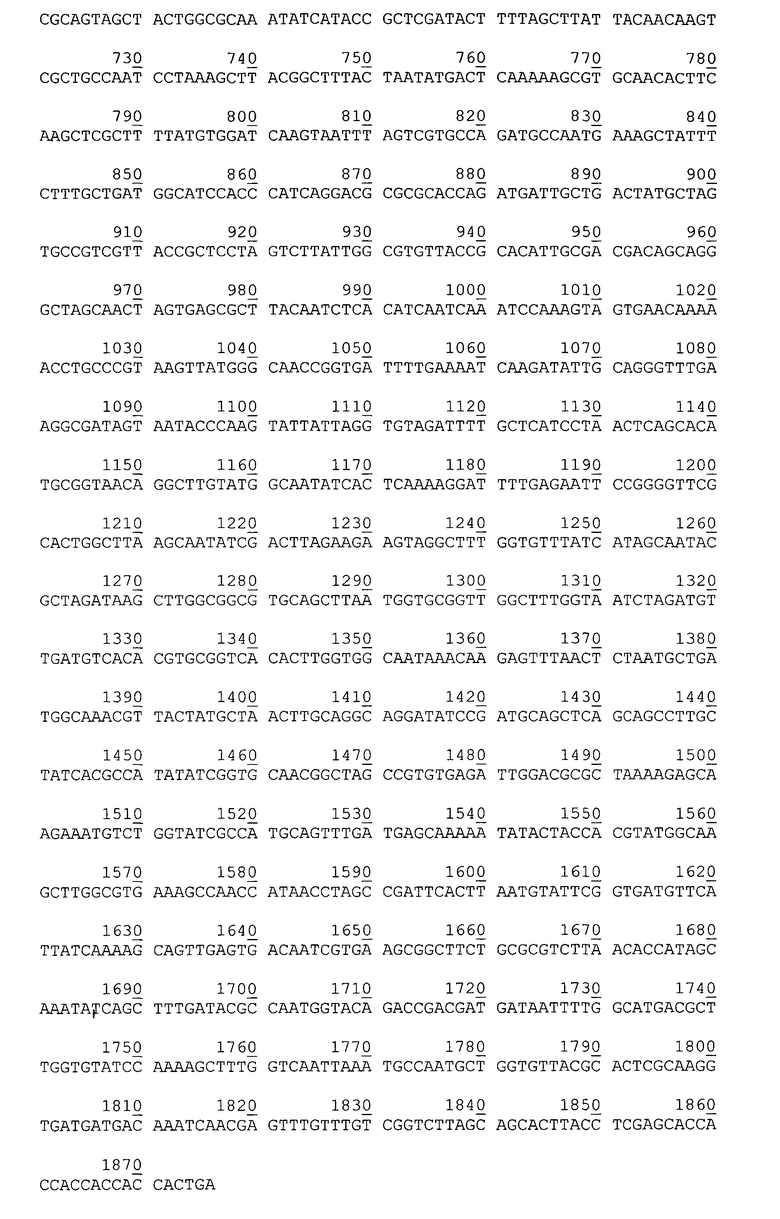
Нуклеотидная последовательность гена аутотранспортера P. cryohalolentis К5Т в плазмиде рАТ877.
SEQ ID NO 2.
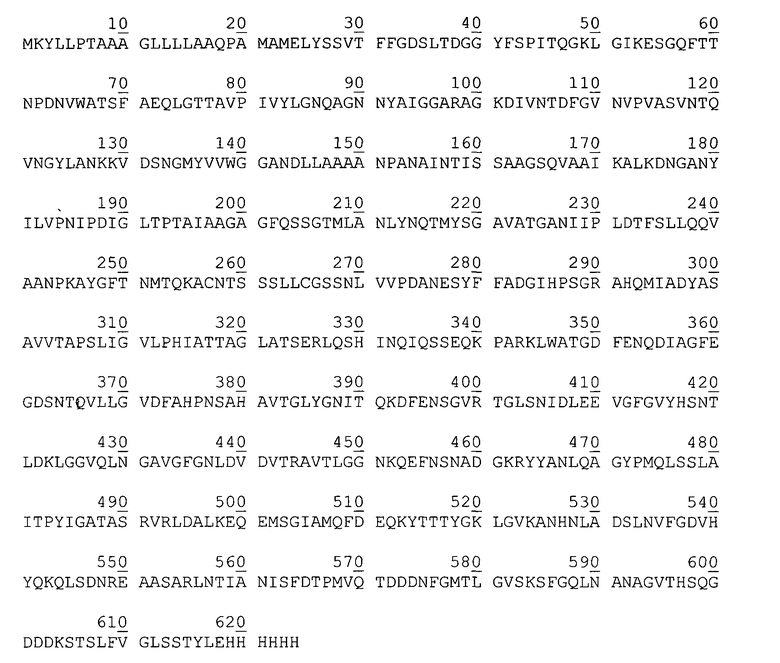
Аминокислотная последовательность аутотранспортера P. cryohalolentis К5Т, кодируемого рекомбинантной плазмидой рАТ877.
SEQ ID NO 3.
PC877F CATATCCATGGAGCTTTATAGCAGCGTGAC
Нуклеотидная последовательность праймера, использованного для амплификации гена аутотранспортера P. cryohalolentis K5T.
SEQ ID NO 4.
PC877R ACATCTCGAGGTAAGTGCTGCTAAGACCGA
Нуклеотидная последовательность праймера, использованного для амплификации гена аутотранспортера P. cryohalolentis K5T.
SEQ ID NO 5.
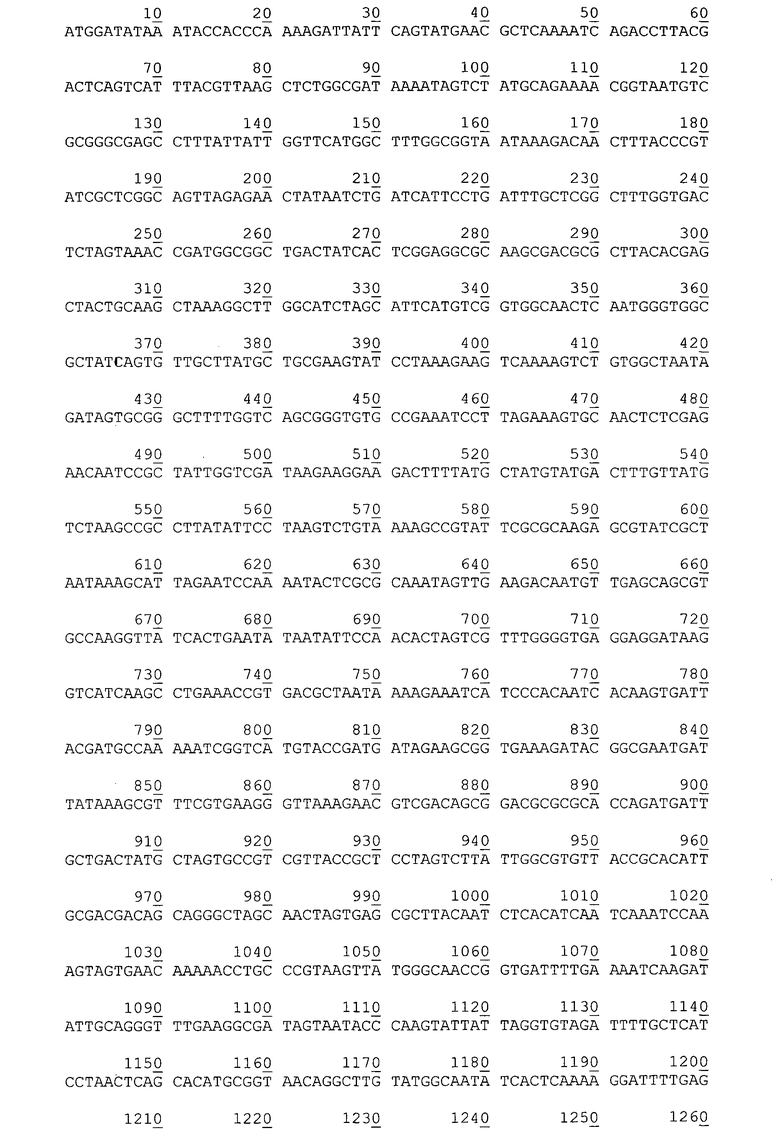
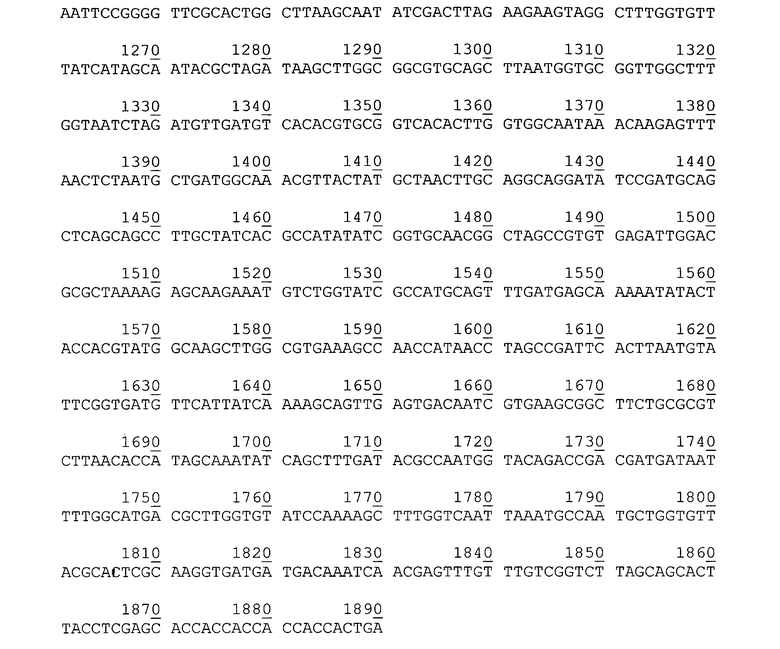
Нуклеотидная последовательность гена гибридного полипептида с активностью эстеразы P. cryohalolentis К5Т на поверхности клеток, в плазмиде pEst877.
SEQ ID NO 6.


Аминокислотная последовательность полипептида с активностью эстеразы Р. cryohalolentis К5Т на поверхности клеток, кодируемого рекомбинантной плазмидой pEst877.
SEQ ID NO 7.
23NcoF CATATCCATGGATATAAATACCACCCAAAAGATTATTC
Нуклеотидная последовательность праймера, использованного для амплификации гена эстеразы P. cryohalolentis K5T.
SEQ ID NO 8.
23R AGTATGCGCGCGTCCGCTGTCGACGTTCTTTAACCCTTCACGAAAC Нуклеотидная последовательность праймера, использованного для амплификации гена эстеразы P. cryohalolentis К5Т.
Изобретение относится к микробиологии и биотехнологии и касается рекомбинантной плазмидной ДНК pEst877, детерминирующей экспрессию полипептида с активностью эстеразы P. cryohalolentis К5Т на поверхности клеток, с мол. массой 3,64 Md (5,519 т.п.о.), состоящей из NcoI/XhoI - фрагмента ДНК плазмиды pET20b(+) длиной 3,654 т.п.о., включающего промотор T7lac, терминатор транскрипции бактериофага Т7, ген bla β-лактамазы, определяющий устойчивость трансформированных плазмидой pEst877 клеток к ампициллину, участок ori инициации репликации, сигнальную последовательность pelB пектатлиазы В Erwinia carotovora; и NcoI/XhoI - фрагмента ДНК размером 1,865 т.п.о., содержащего гибридный ген Est877, кодирующий аминокислотные последовательности эстеразы P. cryohalolentis К5Т с дополнительным аминокислотным остатком аспарагиновой кислоты после сайта отщепления сигнального пептида и аутотранспортера P. cryohalolentis K5T в единой рамке считывания. Изобретение также касается штамма бактерий Е. coli BL21(DE3)pLysS/pEst877 - продуцента полипептида с активностью эстеразы P. cryohalolentis К5Т на поверхности клеток. Изобретение позволяет получать высокоактивную эстеразу с широкой субстратной специфичностью. 2 н.п. ф-лы, 3 ил., 7 пр.
1. Рекомбинантная плазмидная ДНК pEst877, детерминирующая экспрессию полипептида с активностью эстеразы P. cryohalolentis K5T на поверхности клеток, с мол. массой 3,64 Md (5,519 т.п.о.), состоящая из NcoI/XhoI - фрагмента ДНК плазмиды pET20b(+) длиной 3,654 т.п.о., включающего промотор T7lac, терминатор транскрипции бактериофага Т7, ген bla β-лактамазы, определяющий устойчивость трансформированных плазмидой pEst877 клеток к ампициллину, участок ori инициации репликации, сигнальную последовательность pelB пектатлиазы В Erwinia carotovora; и NcoI/XhoI - фрагмента ДНК размером 1,865 т.п.о., содержащего гибридный ген Est877, кодирующий аминокислотные последовательности эстеразы P. cryohalolentis K5T с дополнительным аминокислотным остатком аспарагиновой кислоты после сайта отщепления сигнального пептида и аутотранспортера P. cryohalolentis K5T в единой рамке считывания.
2. Штамм бактерий Е. coli BL21(DE3)pLysS/pEst877 - продуцент полипептида с активностью эстеразы P. cryohalolentis K5T на поверхности клеток.
| SCHULTHEISS E et.al.Esterase autodisplay: enzyme engineering and whole-cell activity determination in microplates with pH sensors.Appl Environ Microbiol | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| LEE SH et.al.Display of bacterial lipase on the Escherichia coli cell surface by using FadL as an anchoring motif and use of | |||

