Область техники, к которой относится изобретение
Настоящее изобретение относится к области белковой инженерии, биотехнологии и медицины, и касается способа получения рекомбинантного белка, противовирусного агента, далее называемого Таргевир, обладающего способностью блокировать связывание с клетками и/или препятствовать размножению вируса SARS-CoV-2. Таргевир предназначен для использования в качестве терапевтического агента для лечения болезней, вызванных SARS-CoV-2, и характеризуется сниженным риском побочного воздействия за счет возможности моментальной отмены РНКазной активности при помощи природного ингибитора барстара. Изобретение может найти применение в биотехнологии и медицине при создании лекарств против КОВИД-19.
Уровень техники
Создание эффективных терапевтических агентов для таргетной терапии заболеваний, вызванных SARS-CoV-2 - одна из неотложных и важнейших задач современной биологии, биотехнологии и медицины. По состоянию на конец 2021 года зарегистрировано более 270 миллионов случаев заболевания и более 5 млн смертей в 222 странах. Коронавирусной инфекцией в активной фазе болеет более 21 млн человек, при этом в тяжелом или критическом состоянии находятся почти 87 тыс. человек. (https://www.worldometers.info/coronavirus/).
SARS-CoV-2 является членом семейства Coronavirinae и относится к роду бета-коронавирусов, который включает также MERS-CoV и SARS-CoV. Последний разделяет ~75-80% своего вирусного генома с SARS-CoV-2 (Rehman S.U. et al. // Pathogens. 2020. V. 9. P. 240; Velavan T.P., Meyer C.G. // Trop. Med. Int. Health. 2020. V. 25. 278-280; Masters P.S. // Adv. Virus Res. 2006. V. 66. P. 193-292). Вирусные частицы диаметром около 60-140 нм имеют сферическую или плеоморфную форму. Коронавирусы имеют один из самых больших геномов одноцепочечной РНК размером 27-32 килобазы (Hobman T.C. et al. // PLoS Pathog. 2020. V. 16(8). e1008762).
Бета-коронавирусы имеют три важных белка оболочки: белок шипа (S), мембранный (М) белок и белок оболочки (Е). Белок S опосредует прикрепление вируса к рецептору клеточной мембраны, слияние мембран и, в конечном итоге, проникновение вируса в клетку-хозяина. Белок М, самый многочисленный мембранный белок, вместе с белком Е отвечает за структуру мембраны коронавируса. Еще одним компонентом бета-коронавируса является белок N, который представляет собой белковый компонент спиралевидного нуклеокапсида, включающего геномную РНК [Masters P.S. // Adv. Virus Res. 2006. V. 66. P. 193-292).
Было показано, что SARS-CoV-2 использует рецептор ангиотензин-превращающего фермента 2 (ACE2) для проникновения в клетки (Zhou P. et al. // Nature. 2020. V. 579. P. 270-273). Рецептор-связывающий домен (RBD) в S-белке SARS-CoV-2 специфически распознает рецептор ACE2 своего хозяина. При исследовании образцов тканей человека рецепторы ACE2 были обнаружены в различных органах и клетках, включая носоглотку, слизистую оболочку носа и рта, тонкий кишечник, толстую кишку, почки, печень, эндотелий сосудов и эпителиальные клетки легочных альвеол (в основном пневмоциты II типа) (Hamming I. et al. // J. Pathol. 2004. V. 203. P. 631-637).
После проникновения в клетку геном вирусной +РНК высвобождается в цитоплазму и подвергается трансляции и репликации, в результате чего образуются новые +РНК и субгеномные мРНК. Последние транслируются в мембранные белки, N-белок и различные вспомогательные белки (Masters P.S. // Adv. Virus Res. 2006. V. 66. P. 193-292). Вирусные структуры и нуклеокапсид впоследствии собираются в промежуточном канале Гольджи эндоплазматического ретикулума. Новый вирион, упакованный в везикулы Гольджи, сливается с плазматической мембраной и высвобождается путем экзоцитоза. (Masters P.S. // Adv. Virus Res. 2006. V. 66. P. 193-292; Li X. et al. // J. Pharm Anal. 2020. V. 10. P. 102-108).
Инфекция SARS-CoV-2 индуцирует провоспалительные факторы, которые приводят к активации макрофагов и дендритных клеток. Презентация антигена SARS-CoV-2 через основные комплексы гистосовместимости I и II (MHC I и II) стимулирует гуморальный и клеточный иммунитет, что приводит к выработке цитокинов и антител. В тяжелых случаях COVID-19 вирус достигает нижних дыхательных путей и инфицирует пневмоциты II типа, что приводит к их апоптозу и потере сурфактанта. Приток макрофагов и нейтрофилов вызывает, так называемый, цитокиновый шторм. Негерметичные капилляры приводят к альвеолярному отеку. Все эти патологические изменения приводят к повреждению и разрушению альвеол и нарушению газообмена (Mason R.J. // Eur. Respir. J. 2020. V. 55. P. 200607).
Лекарственные препараты, применяемые для лечения КОВИД-19 можно условно разделить на несколько основных групп:
- противовирусные, подавляющие способность вируса к репликации;
- противовоспалительные и иммуномодулирующие, применяемые для восстановления иммунного баланса в результате цитокинового шторма, вызванного инфекцией;
- ингибиторы ангиотензин-превращающего фактора и блокаторы рецепторов ангиотензина;
- плазма от выздоровевших людей;
- моноклональные гуманизированные антитела, блокирующие вирусную инфекцию.
Управление по контролю за продуктами и лекарствами США (FDA) одобрило такое противовирусное средство, как Remdesivir (ингибитор вирусной РНК-зависимой РНК-полимеразы), которое, как было установлено, прерывает вирусную репликацию. Гидроксихлорохин и хлорохин привлекли значительное внимание во время пандемии КОВИД-19 с различными заявлениями о клиническом успехе, полученными, главным образом, в результате небольших испытаний. Отмечается, что хлорохин подавляет выработку/высвобождение TNF и IL-6, которые ассоциируются с цитокиновой бурей. Широкий спектр воспалительных цитокинов и хемокинов, присутствующих у пациентов с COVID-19, указывает на активацию пути JAK-STAT. Первым одобренным ингибитором JAK был руксолитиниб (Izda V. et al. // Clin. Immunol. 2021. V. 222. P. 108634; McFee R.B. // Dis. Mon. 2020. V. 66(9). P. 101058).
Микробные рибонуклеазы всегда привлекали повышенное внимание в качестве средства для борьбы с вирусами. Это объясняется несколькими причинами. Во-первых, геном многих вирусов состоит из РНК. Поэтому они - по определению - должны быть чувствительны к РНКазе. Во-вторых, микробные РНКазы способны проникать в клетки млекопитающих и создавать препятствия для размножения вируса. Более того, в определенных условиях РНКаза способна убивать клетки, зараженные вирусом, прерывая тем самым цепочку инфекции. Наиболее известными микробными РНКазами являются барназа, продуцируемая бактериями вида Bacillus amyloliquefaciens, и биназа - бактериями вида Bacillus pumilis. РНКазы не имеют дисульфидных связей и не нуждаются ни в посттрансляционных модификациях, ни в ионах двухвалентных металлов, ни в каких-либо других компонентах для проявления своей каталитической функции. В силу высокой стабильности и отсутствии восприимчивости к ингибиторам рибонуклеаз эукариот, оба фермента проявляют высокую каталитическую активность к молекулам РНК в любых клетках эукариот. Разрезание молекулы РНК происходит преимущественно рядом с остатками гуанозина с образованием гуанозин-3′-фосфатного фрагмента и свободной 5’-гидроксильной группы (Ulyanova V. et al. // FEBS J. 2011. V. 278. P. 3633-3643; Makarov A.A., Ilinskaya O.N. // FEBS Lett. 2003. V. 540(1-3). P. 15-20).
Бактерии вида Bacillus amyloliquefaciens синтезируют специальный белок барстар для защиты от барназы в случае ее попадания в цитоплазму. Барназа и барстар при взаимодействии образуют прочный комплекс и характеризуются чрезвычайно быстрой кинетикой (kon ~ 108 М-1⋅с-1) и высокой аффинностью связывания (коэффициент ассоциации Kас ~ 1014 М-1) (Fersht A.R., Schreiber G. // Nat. Struct. Biol. 1996. V. 3. P. 427 - 431.). РНКазы барназа и биназа очень похожи по структуре - гомология аминокислотных последовательностей достигает 82%, а третичные структуры практически совпадают. Ферментативные свойства также очень похожи.
Способность микробных РНКаз блокировать размножение вируса была неоднократно продемонстрирована на практике. Биназа проявила высокую активность против РНК-содержащего вируса бешенства у инфицированных морских свинок, кроликов и мышей при введении в очаг инфекции (S.V. Gribencha et al. // Vopr. Virusol. 2004. V. 49. P. 38-41; S.V. Gribencha et al. // Vopr. Virusol. 2006. V. 51. P. 41-43). Терапевтический эффект был зарегистрирован как через 2-3 ч после заражения, так и через 1 день (57-67% защиты животных). Однократное внутрибрюшинное введение биназы кроликам, зараженным вирусом ящура типа О и типа А22550, снижало смертность животных в три раза (I.I. Alekseeva et al. // Antibiot. Med. Tekhnol. 1981. V. 26. P. 527-532). Биназа была активна в отношении вирусов гриппа типов А и В (M.A. Shneider et al. // Antibiot. Khimioterap. 1990. V. 35. P. 27-33.). В нетоксичных для эпителиальных клеток концентрациях биназа проявляла противовирусную активность против пандемического вируса гриппа A/Hamburg/04/09 (H1N1). Кратковременная обработка инфекционных частиц вируса (15-30 мин) биназой в возрастающих концентрациях пропорционально снижала жизнеспособность вируса, что проявлялось в ослаблении его способности инфицировать клетки аденокарциномы легких A549 почти на порядок (R. Shah Mahmud, O.N. Ilinskaya // Acta Naturae. 2013. V. 5. P. 44-51).
Известен способ подавления биназой вирусной инфекции MERS-CoV и HCoV-229E, вирусов близкородственных SARS-CoV-2. В нетоксичных для клеток концентрациях биназа снижала существенно титры этих вирусов (Müller C. et al. // Bionanoscience. 2017. V. 7(2). P. 294-299). К недостаткам этого способа следует отнести очень высокие действующие концентрации РНКазы.
Известен также способ блокирования активности вируса с помощью моноклональных антител. Ряд моноклональных антител находится в стадии разработки в качестве системного лечения КОВИД-19 (R. Shi et al. // Nature. 2020. V. 584. P. 120-124; D. Pinto et al. // Nature. 2020. V. 583. P. 290-295; M. Yuan et al. // Science. 2020. V. 368. P. 630-633; Y. Wu et al. // Science. 2020. V. 368. P. 1274-1278; патент CN111592595B, 11.08.2021). К недостаткам, присущим антителам, можно отнести большой размер, склонность к агрегации, а также трудности с получением. Потенциальной проблемой может также являться антитело-зависимое усиление болезни [K. L. Winarski et al. // Proc. Natl. Acad. Sci. U.S.A. 2019. V. 116. 15194-15199; A. Taylor et al. // Immunol. Rev. 2015. V. 268. P. 340-364; B. S. Graham. Science. 2020. V. 368. P. 945-946). Терапевтическое действие антител желательно усиливать. Одним из способов к усилению воздействия антител на вирусы является конъюгация антител с РНКазами, то есть создание терапевтических агентов направленного действия, которые избирательно связываются с вирусами и препятствуют их размножению.
Наиболее близким по технической сущности и достигаемому результату к предлагаемому изобретению, является способ блокирования вируса, принятый за ближайший аналог (прототип). В одной из работ (L. Cao et al. // Science. 2020. V. 370(6515). P. 426-431) при помощи компьютерных методов были сгенерированы искусственные скаффолды, связывающиеся с RBD доменом S-белком вируса SARS-CoV-2. Их структура была доработана экспериментальным путем для оптимизации связывания, сворачивания и стабильности. В результате было получено десять конструкций, которые связывались с RBD с аффинностью от 100 пМ до 10 нМ и блокировали SARS-CoV-2 инфекцию клеток Vero E6 со средним значением ингибирующей концентрации (IC50) от 24 пМ до 35 пМ. Наиболее эффективными оказались два пептида LCB1 и LCВ3 длиной 56 и 64 а.о., соответственно имеющие последовательности SEQ ID NO: 3 (L. Cao et al. // Science. 2020. V. 370(6515). P. 426-431) и SEQ ID NO: 10 (L. Cao et al. // Science. 2020. V. 370(6515). P. 426-431).
Настоящее изобретение решает техническую проблему получения следующих рекомбинантных белков:
Таргевир 1, имеющего последовательность SEQ ID NO:2, на основе белков барназа с последовательностью SEQ ID NO:5 и барстар с последовательностью SEQ ID NO:7 и адресного полипептида LCB1 с последовательностью SEQ ID NO: 3,
Таргевир 2, имеющего последовательность SEQ ID NO:9, на основе белков барназа с последовательностью SEQ ID NO:12 и барстар с последовательностью SEQ ID NO:14 и адресного полипептида LCB3 с последовательностью SEQ ID NO: 10, далее называемых также как Таргевир в качестве собирательного понятия.
Указанные рекомбинатные белки потенциально способны блокировать связывание с клетками и/или препятствовать размножению вируса SARS-CoV-2 и характеризуются сниженным риском побочного воздействия за счет возможности моментальной отмены РНКазной активности.
Поставленная техническая проблема достигается за счет:
1) рекомбинантного белка Таргевир 1, имеющего последовательность SEQ ID NO: 2, состоящего из адресного полипептида LCB1 с последовательностью SEQ ID NO: 3 и барназы с последовательностью SEQ ID NO: 5, соединенных гибким пептидным линкером с последовательностью SEQ ID NO: 4, который обладает способностью эффективно связываться с рецептор-связывающим доменом S-белка SARS-CoV-2, а также его применения в качестве вирус-нейтрализующего средства против SARS-CoV-2.
2) рекомбинантного белка Таргевир 2, имеющего последовательность SEQ ID NO: 9, состоящего из адресного полипептида LCB3 с последовательностью SEQ ID NO: 10 и барназы с последовательностью SEQ ID NO: 12, соединенных гибким пептидным линкером с последовательностью SEQ ID NO: 11, который обладает способностью высокоэффективно связываться с рецептор-связывающим доменом S-белка SARS-CoV-2, а также его применения в качестве вирус-нейтрализующего средства против SARS-CoV-2.
3) рекомбинантной плазмиды pET39b-LCB1-Ba, содержащей ген Таргевир 1 и ген барстара в составе плазмидного вектора экспрессии pET39b.
4) рекомбинантной плазмиды pET39b-LCB3-Ba, содержащей ген Таргевир 2 и ген барстара в составе плазмидного вектора экспрессии pET39b.
5) штамма E. coli BL21(DE3)/pET39b-LCB1-Ba, получаемого путем трансформации штамма BL21(DE3) плазмидой pET39b-LCB1-Ba и отбора клона трансформанта с наиболее высоким уровнем синтеза Таргевир 1;
6) штамма E. coli BL21(DE3)/pET39b-LCB3-Ba, получаемого путем трансформации штамма BL21(DE3) плазмидой pET39b-LCB3-Ba и отбора клона трансформанта с наиболее высоким уровнем синтеза Таргевир 2;
7) способа получения из биомассы штаммов Escherichia coli BL21(DE3)/pET39b-LCB1-Ba и E. coli BL21(DE3)/pET39b-LCB3-Ba рекомбинантных белков Таргевир 1 и Таргевир 2, соответственно.
В результате решения заявленной технической проблемы получают следующий технический результат:
1) высокий уровень индуцируемого синтеза (45% от суммарного клеточного белка) и стабильную продукцию Таргевир 1 и Таргевир 2, который обеспечивается штаммами E. coli BL21(DE3)/pET39b-LCB1-Ba и E. coli BL21(DE3)/pET39b-LCB3-Ba, соответственно;
2) высокую специфическую антивирусную активность Таргевир;
3) сниженный риск побочного воздействия за счет возможности моментальной отмены цитотоксичности Таргевир.
Раскрытие изобретения
Получен рекомбинантный белок Таргевир 1, имеющий последовательность SEQ ID NO: 2, состоящий из адресного полипептида LCB1 с последовательностью SEQ ID NO: 3 и барназы с последовательностью SEQ ID NO: 5, соединенных гибким пептидным линкером с последовательностью SEQ ID NO: 4, который обладает способностью эффективно связываться с рецептор-связывающим доменом S-белка SARS-CoV-2.
Получен рекомбинантный белок Таргевир 2, имеющий последовательность SEQ ID NO: 9, состоящий из адресного полипептида LCB3 с последовательностью SEQ ID NO: 10 и барназы с последовательностью SEQ ID NO: 12, соединенных гибким пептидным линкером с последовательностью SEQ ID NO: 11, который обладает способностью эффективно связываться с рецептор-связывающим доменом S-белка SARS-CoV-2.
Показано применение рекомбинантного белка Таргевир 1, имеющего последовательность SEQ ID NO:2, в качестве вируснейтрализующего средства в отношении SARS-CoV-2.
Показано применение рекомбинантного белка Таргевир 2, имеющего последовательность SEQ ID NO: 9, в качестве вируснейтрализующего средства в отношении SARS-CoV-2.
Разработан способ получения Таргевир - химерного бифункционального белка, состоящего из адресного полипептида LCB1 или LCB3 (L. Cao et al. // Science. 2020. V. 370(6515). P. 426-431), специфичного к рецептор-связывающему домену (RBD) S-белка SARS-CoV-2, и высокоактивной бактериальной рибонуклеазы барназы, соединенных гибким пептидным линкером. Таргевир обладает способностью специфически связываться с вирионами SARS-CoV-2, имеющими на своей поверхности S-белок, и блокировать их связывание с рецептором ACE-2, представленным на поверхности клеток, чувствительных к заражению вирусом. При интернализации в клетки вместе с вирусом обладает вирус статическим действием за счет гидролиза вирусной РНК и последующего прекращения биосинтеза белка, необходимого для синтеза новых вирионов. При взаимодействии Таргевир с природным ингибитором барназы - рекомбинантным белком барстар - цитотоксический эффект Таргевир моментально отменяется, что может быть использовано в случае гиперчувствительной реакции организма на РНКазную активность или передозировки лекарственного средства. Сочетание таких свойств Таргевир выгодно отличает его от всех известных и применяемых в клинической практике аналогов.
Экспрессирующие рекомбинантные плазмиды pET39b-LCB1-Ba и pET39b-LCB3-Ba получают путем клонирования последовательности, кодирующей Таргевир и барстар, в вектор pET39b (pET System Manual, 10th Edition Rev.B 0403, Novagen Inc. (2003)) по сайтам рестрикции NdeI/HindIII (фиг. 1). Для преодоления чрезвычайной токсичности рибонуклеазы барназы для бактериальной клетки ген целевого белка ставят под контроль индуцируемого промотора фага Т7, кроме того, в конструкцию включают ген ингибитора барназы, барстара, под собственным промотором. В процессе культивирования ингибитор барстар, синтезирующийся в бактериальной клетке одновременно с Таргевир, полностью ингибирует ее РНКазную активность, что позволяет проводить наработку клеток штамма-продуцента, но требует последующей очистки Таргевир от ингибитора барстара.
Путем трансформации клеток штамма Escherichia coli BL21(DE3) (Studier, F.W. and Moffatt, B.A. // J. Mol. Biol. 1986. V.189. P. 113-130) сконструированной плазмидой pET39b-LCB1-Ba или pET39b-LCB3-Ba, отбора и культивирования клонов трансформантов с высоким уровнем синтеза гибридного белка получают рекомбинантные штаммы Escherichia coli BL21(DE3)/pET39b-LCB1-Ba и Escherichia coli BL21(DE3)/pET39b-LCB3-Ba - продуценты Таргевир. Синтез Таргевир в полученном рекомбинантном штамме осуществляется при культивировании на обычных селективных средах с добавлением индуктора изопропил-D-тиогалактозида (ИПТГ) или лактозы.
В способе получения рекомбинантного белка Таргевир биомассу рекомбинантного штамма E. coli-продуцента разрушают ультразвуком в лизирующем буфере, содержащем Tris-HCl и хлорид натрия. Целевой белок, изначально снабженный олигогистидиновой последовательностью SEQ ID: 6 и SEQ ID: 13, выделяют при помощи металлохелатной аффинной хроматографии. В бактериальных клетках Таргевир образует чрезвычайно устойчивый комплекс со своим ингибитором барстаром (KD 10-13 M-1) и в стандартной процедуре аффинной хроматографии на Ni2+-сефарозе выделяется совместно с ним в виде прочного комплекса, поэтому основной проблемой при очистке является отделение ингибитора барстара. Очистку рекомбинатного белка Таргевир от барстара осуществляют в денатурирующих для комплекса Таргевир -барстар условиях с использованием 6 М гуанидингидрохлорида и 0.5 М NaCl. Затем рекомбинатный белок Таргевир дочищают при помощи анионообменной хроматографии.
Таким образом, настоящее изобретение включает 5 объектов:
Первый объект - рекомбинантный белок Таргевир 1, имеющий последовательность SEQ ID NO: 2, который обладает активностью к рецептор-связывающему домену S-белка SARS-CoV-2.
Второй объект - рекомбинантный белок Таргевир 2, имеющий последовательность SEQ ID NO: 9, который обладает активностью к рецептор-связывающему домену S-белка SARS-CoV-2.
Третий объект - применение указанных белков в качестве вируснейтрализующего средства в отношении SARS-CoV-2.
Четвертый объект - рекомбинантная плазмида pET39b-LCB1-Ba, обеспечивающая синтез Таргевир 1 в клетках Escherichia coli и состоящая из фрагмента ДНК с последовательностью SEQ ID NO: 1 и фрагмента плазмиды pET39b.
Пятый объект - рекомбинантная плазмида pET39b-LCB3-Ba, обеспечивающая синтез Таргевир 2 в клетках Escherichia coli и состоящая из фрагмента ДНК с последовательностью SEQ ID NO: 8 и фрагмента плазмиды pET39b.
Шестой объект - рекомбинантный штамм Escherichia coli BL21 (DE3)/pET39b-LCB1-Ba.
Седьмой объект - рекомбинантный штамм Escherichia coli BL21 (DE3)/pET39b-LCB3-Ba.
Восьмой объект - способ получения рекомбинантного белка Таргевир 1 или Таргевир 2 с использованием штамма Escherichia coli BL21 (DE3)/pET39b-LCB1-Ba или Escherichia coli BL21 (DE3)/pET39b-LCB3-Ba с высокой степенью очистки.
Краткое описание фигур.
Фиг. 1 - Физическая и генетическая карты вектора pET39b-LCB1-Ba. Обозначено положение гена Таргевир 1, других элементов вектора, уникальных сайтов рестрикции.
Фиг. 2 - Физическая и генетическая карты вектора pET39b-LCB3-Ba. Обозначено положение гена Таргевир 2, других элементов вектора, уникальных сайтов рестрикции.
Фиг. 3 - Физическая и генетическая карты вектора pMT643. Обозначено положение гена Барстар, других элементов вектора, уникальных сайтов рестрикции.
Фиг. 4 - Гель-электрофорез Таргевир 1 после осаждения сульфатом аммония. Стрелкой обозначено положение Таргевир 1. Дорожка T -образец Таргевир, дорожка МW - маркер молекулярных масс.
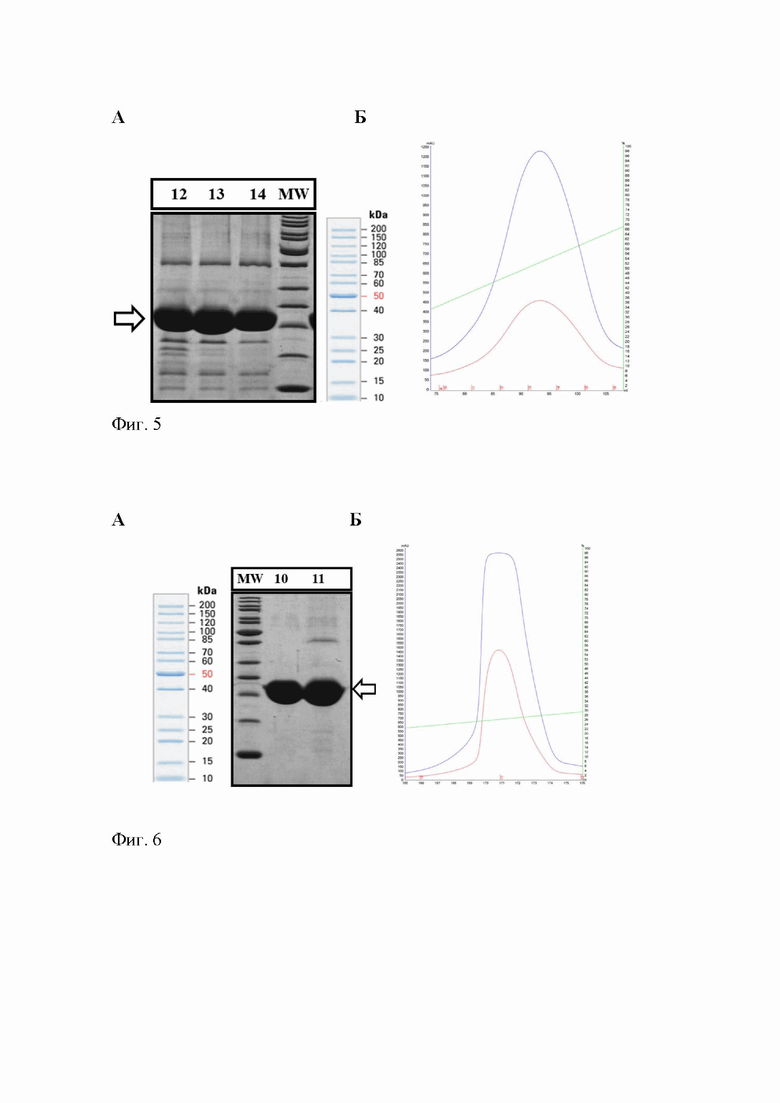
Фиг. 5 - Результаты металлохелатной аффинной хроматографии на HisTrap HP, 5 мл: (А) гель-электрофорез фракций 12, 13, 14, собранных в ходе элюции Таргевир 1 с колонки, дорожка МW - маркер молекулярных масс, стрелкой обозначено положение Таргевир1; (Б) профиль хроматографической элюции.
Фиг. 6 - Результаты финишной анионообменной хроматографии на HiTrap Q HP, 5 мл: (А) гель-электрофорез фракций 10, 11, собранных в ходе элюции Таргевир3 с колонки, дорожка МW - маркер молекулярных масс, стрелкой обозначено положение Таргевир3; (Б) профиль хроматографической элюции.
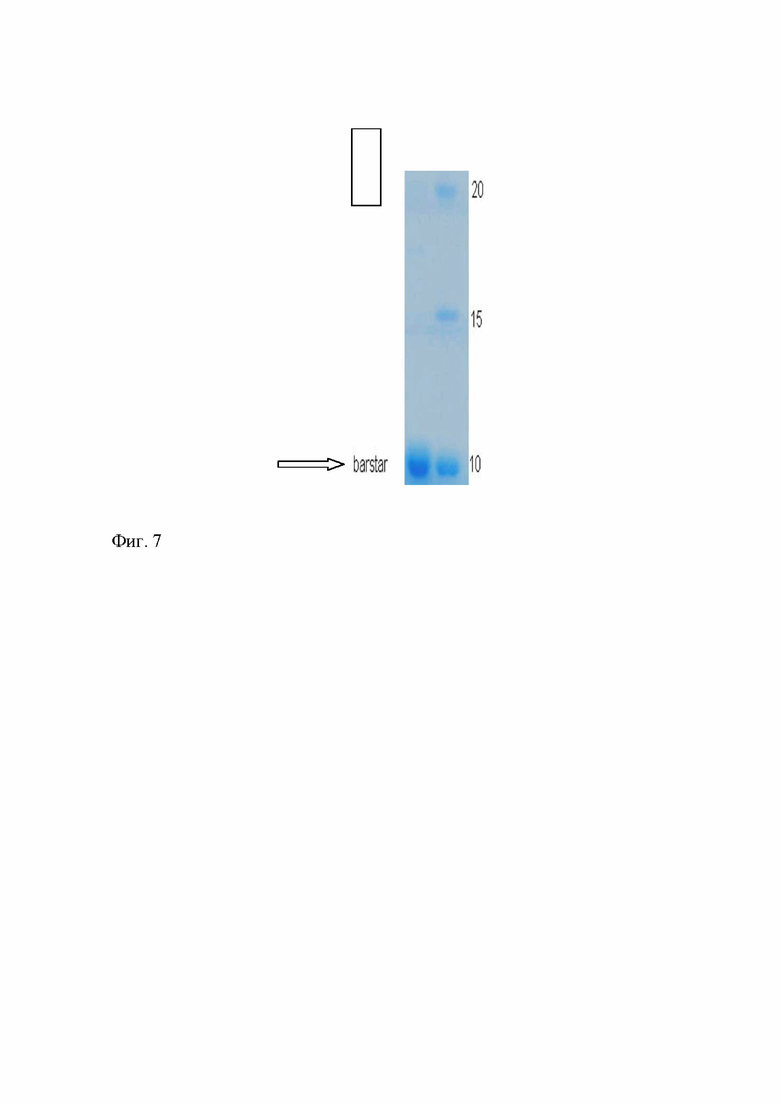
Фиг. 7 - Гель-электрофорез белка Барстар после финишной очистки анионообменной хроматографией на Сефарозе Q. Дорожка 1 - очищенный образец Барстар, дорожка М - маркер молекулярных масс.
Фиг. 8 - Определение РНКазной активности Таргевир в сравнении с барназой дикого типа методом кислотно-растворимого осадка.
Фиг. 9 - Определение способности барстар ингибировать РНКазную активность Таргевир. РНКазная активность Таргевир (пунктирная линия) и ее ингибирование барстаром (сплошная линия).
Фиг. 10 - Изучение связывания Таргевир с RBD-доменом методом ИФА
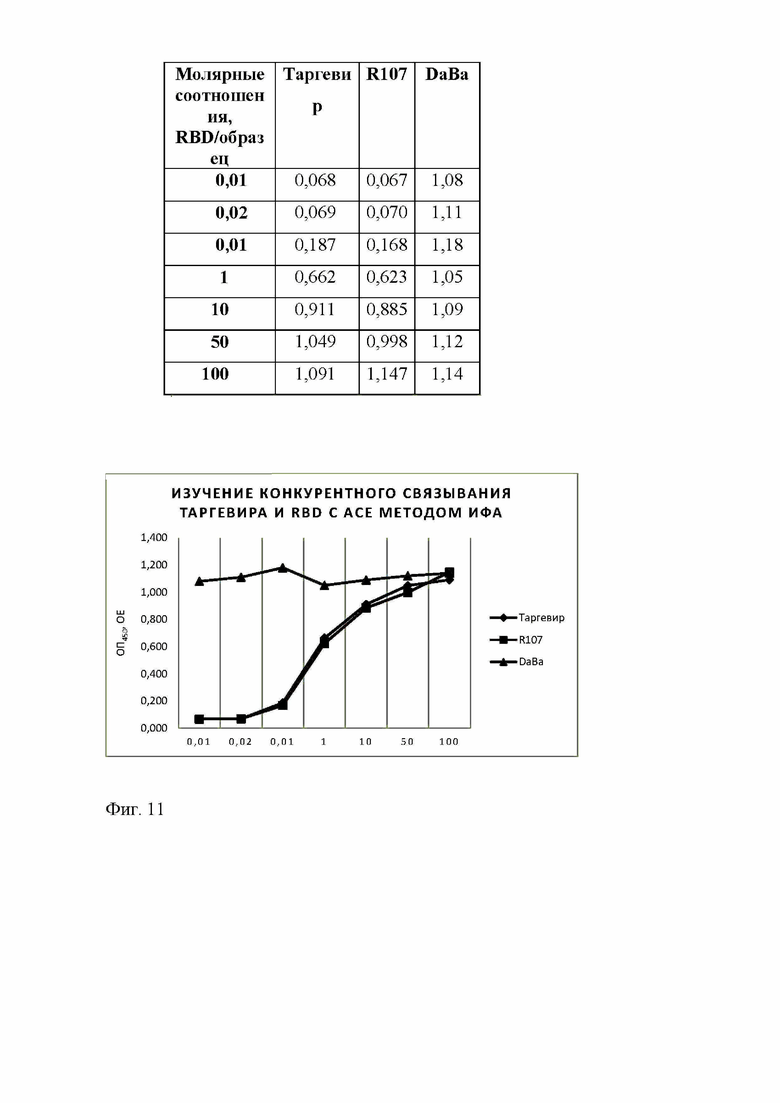
Фиг. 11 - Изучение конкурентного связывания Таргевир и RBD с рекомбинантным АСЕ - 2 методом ИФА
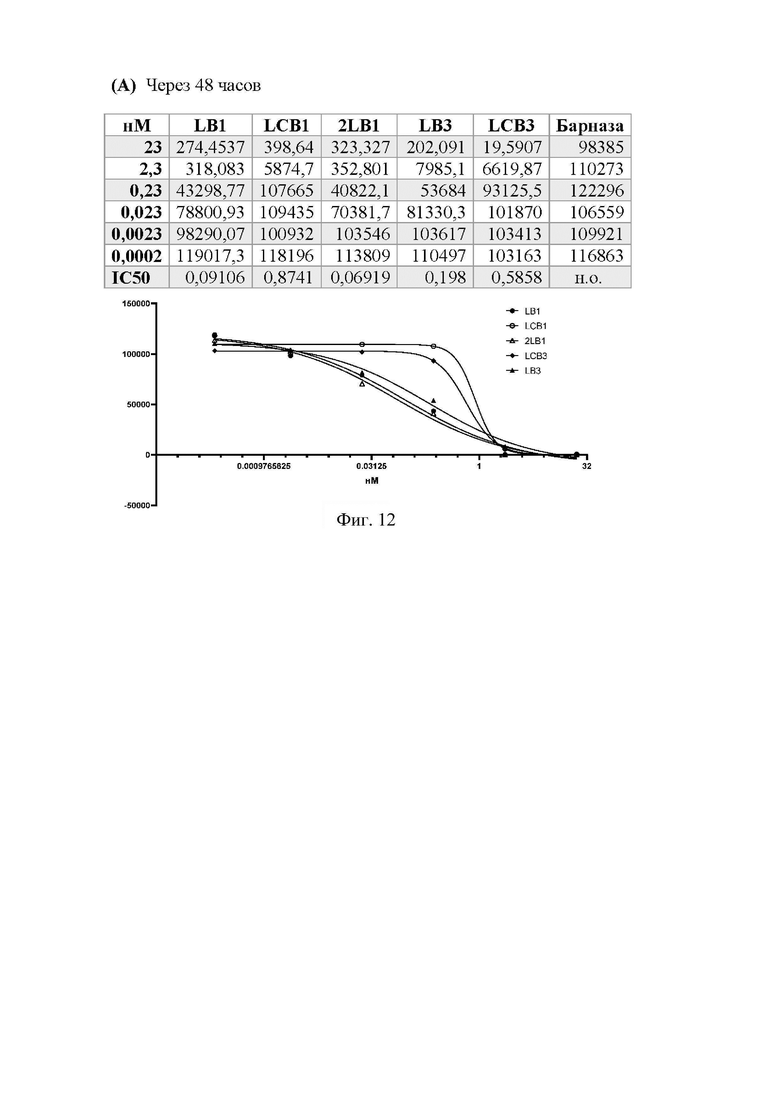
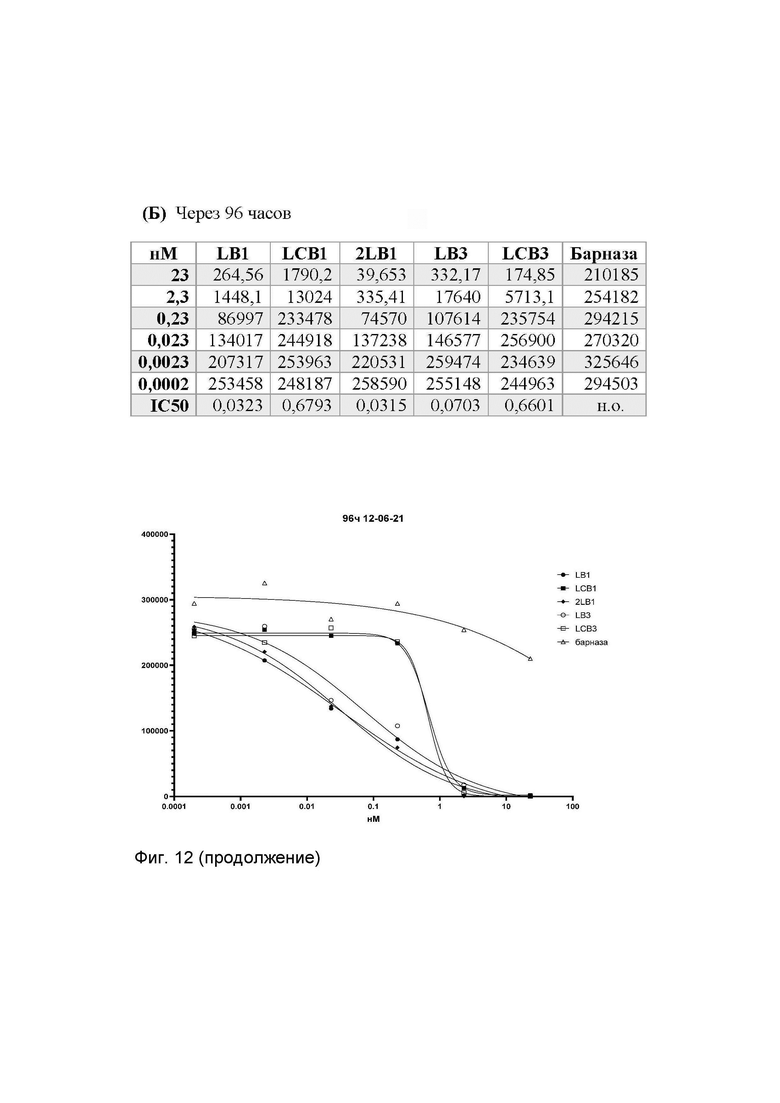
Фиг. 12 - Изучение вируснейтрализующего действия Таргевира с использованием псевдовирусной системы
Осуществление изобретения.
Пример 1. Получение рекомбинантной плазмиды pET39b-LCB1-Ba.
Плазмиду pET39b-LCB-Ba получают путем клонирования фрагмента ДНК с последовательностью SEQ ID NO: 1 в вектор pET39b (pET System Manual, 10th Edition Rev.B 0403, Novagen Inc. (2003)) по сайтам рестрикции NdeI/XhoI (Фиг. 1).
Пример 2. Получение рекомбинантной плазмиды pET39b-LCB3-Ba.
Плазмиду pET39b-LCB3-Ba получают путем клонирования фрагмента ДНК с последовательностью SEQ ID NO: 8 в вектор pET39b (pET System Manual, 10th Edition Rev.B 0403, Novagen Inc. (2003)) по сайтам рестрикции NdeI/XhoI (Фиг. 2).
Пример 3. Получение рекомбинантного штамма - продуцента Таргевир.
Рекомбинантной плазмидой pET39b-LCB1-Ba или pET39b-LCB3-Ba трансформируют штамм E. coli BL21(DE3) [F-, ompT, hsdSB (rB-, mB-), dcm, gal, λ(DE3)] (Studier, F.W. and Moffatt, B.A.//J. Mol. Biol. 1986. V.189. P. 113-130). Выбор данного штамма в качестве реципиента для продукции Таргевир обусловлен тем, что он синтезирует РНК-полимеразу фага Т7, а также обладает сниженной протеазной активностью, что способствует повышению выхода целевых белков.
Поскольку первичные трансформанты DE3 - реципиентов могу заметно отличаться по уровню экспрессии целевого белка (Vethanayagam J, Flower A. // Microbial Cell Factories. 2005. V.4. P. 3-7) для селекции наиболее активного продуцента Таргевир и параметров индукции синтеза Таргевир отдельные клоны трансформантов выращивают в условиях «автоиндукции» (Studier F.W. // Protein Expr. Purif. 2005. V.41. № 1. P. 207-234) в течение 24-48 часов при 25°С с периодическим отбором аликвот суспензии клеток. Для получения грубого экстракта осадок клеток из 100-200 мкл культуры суспендируют в 100 мкл лизирующего буфера для нанесения на ДСН-ПААГ, прогревают 3 мин при 85°С, клеточный дебрис удаляют центрифугированием (12 000 об/мин, 5 мин) и 5 мкл полученного экстракта анализируют путем электрофореза в 15% денатурирующем полиакриламидном геле.
Появление отчетливой полосы размером порядка 21,7 кДа для Таргевир 1 и порядка 22,6 кДа для Таргевир 2 в образцах анализируемых штаммов при ее отсутствии в контрольном реципиентном штамме BL21(DE3) принимают за доказательство способности штамма синтезировать Таргевир.
Клон трансформанта, отличающийся наибольшим выходом Таргевир 1, обозначают BL21(DE3)/pET39b-LCB1-Ba. Клон трансформанта, отличающийся наибольшим выходом Таргевир 2, обозначают BL21(DE3)/pET39b-LCB3-Ba.
Выход Таргевир 1 и Таргевир 2 в полученных нами штаммах E. coli BL21(DE3)/pET39b-LCB1-Ba и E. coli BL21(DE3)/pET39b-LCB3-Ba составляет 45% от суммарного белка клетки.
Клетки полученных рекомбинантных штаммов Escherichia coli BL21(DE3)/pET39b-LCB1-Ba и Escherichia coli BL21(DE3)/pET39b-LCB3-Ba характеризуются следующими признаками.
Морфологические признаки: Клетки имеют продолговатую палочковидную форму, при делении не почкуются.
Культуральные признаки: Клетки хорошо растут на обычно используемых питательных средах. Время генерации около 30 мин в жидкой LB-среде. На 2-2,5% питательном агаре "Difco" образуются круглые, гладкие, желтоватые колонии с ровными краями. При выращивании на жидких LB- и YT-средах образуется интенсивная ровная мутность.
Физиолого-биохимические признаки: Оптимальная температура культивирования - от 25 до 37°C, оптимум рН - 7,6. Источником азота служат органические соединения (в виде триптона, дрожжевого экстракта).
Уровень синтеза Таргевир 1 и Таргевир 2 в сконструированных штаммах составляет около 50 мг/л при титре культуры 1×109 кл/мл, что следует из данных определения Таргевир и Таргевир 2 в образцах биомассы штамма-продуцента с помощью электрофореза в 15% ДСН-ПААГ.
Пример 4. Получение биомассы рекомбинантного штамма - продуцента Таргевир 1 и Таргевир 2.
Клетки E. coli BL21(DE3) трансформируют соответствующей плазмидой pET39b-LCB1-Ba или pET39b-LCB3-Ba по методу (Kurien B.T., Scofield R.H. // BioTechniques. 1995. V. 18. № 6. P. 1023-1026). Клетки из глицеринового стока, хранящегося в морозильнике, рассевают на поверхности чашки с питательной агаризованной средой. Инкубируют ночь при 37°С. Собирают около 20-30 колоний стеклянной палочкой и суспендируют в 100 мкл 0.1 М СaCl2, содержащем 1 мг PEG 8000 (pH 8.0). Инкубируют во льду в течение 5 мин. Добавляют плазмидную ДНК в количестве 5 нг и продолжают инкубацию во льду еще в течение 8 мин. Подвергают клетки тепловому шоку при 42°С в течение 30 сек и затем возвращают в лед на 2 мин. Добавляют 900 мкл среды YT (8 г бакто-триптона, 5 г дрожжевого экстракта, 5 г NaCl на 1 л воды) и высевают 100 мкл суспензии на чашки с агаризованной средой и канамицином (25 мг/л).
Для получения посевного материала собирают стеклянной палочкой около 50 колоний с чашек Петри, засеянных трансформантами, и суспендируют в 200 мкл среды YT. Суспензию клеток переносят в 200 мл жидкой автоиндукционной питательной среды ZYM-5052 (Studier F.W. // Protein Expr. Purif. 2005. V. 41. № 1. P. 207-234), содержащей канамицин (50 мг/л), и культивируют при температуре 37°С при интенсивной аэрации (скорость качалки 180 об/мин) в течение суток.
Выход биомассы рекомбинантного штамма - продуцента Таргевир 1 обычно составляет от 10.0 до 15.0 г/л культуральной среды.
Аналогичным образом получали биомассу рекомбинантного штамма - продуцента Таргевир 2, выход которой составляет от 10.0 до 15.0 г/л культуральной среды.
Пример 5. Выделение и очистка рекомбинантного белка Таргевир 1 и Таргевир 2
Биомассу ресуспендируют в 20 мл раствора 200 мM Трис-HCl, pH 8.0 и добавляют лизоцим до конечной концентрации 60 мкг/мл. Суспензию разбавляют в 2 раза дистиллированной водой, добавляют PMSF до конечной концентрации 100 мкМ и инкубируют при комнатной температуре в течение 30 мин, периодически перемешивая. Суспензию переносят в ледяную баню и обрабатывают ультразвуком при помощи Sonics Vibra-Cell VCX 130, добиваясь полного лизиса клеток (общее время обработки 5 раз по 2 мин, - циклы по 10 сек с 10 секундным интервалом, мощность 80W). Клеточный лизат центрифугируют в течение 30 мин при 18500 g и декантируют надосадочную жидкость, содержащую растворимую фракцию целевого белка. К осветленному лизату при постоянном перемешивании добавляют кристаллический сернокислый аммоний до 60% от насыщения. Суспензию оставляют на ночь в холодильнике. Типичный результат электрофоретического анализа Таргевир 1 после осаждения сульфатом аммония представлен в Фиг. 4.
Суспензию разливают по стаканам и центрифугируют в течение 30 мин при 18500 g. Надосадочную жидкость сливают, а осадок растворяют в растворе I (20 мМ NaPi, pH 7.5, 500 мМ NaCl, 30 мМ имидазол) и фильтруют через мембрану с размером пор 0.22 мкм (MilliPore).
Раствор, содержащий целевой белок, наносят на колонку HisTrap HP, 5 мл, уравновешенную 10-кратным объемом раствора I, со скоростью 5 мл/мин, затем колонку промывают от примесных белков 10 объемами раствора I с той же скоростью. Для денатурации комплекса Таргевир1:барстар, иммобилизованного на колонке, носитель промывают 25 объемами раствора I (без имидазола) с 6 М гуанидин гидрохлоридом со скоростью 1 мл/мин. Таргевир 1, освобожденный от ингибитора барстар, ренатурируют непосредственно на колонке линейным ниспадающим градиентом гуанидин гидрохлорида (60 объемов колонки) от 6 М до 0 М при скорости 1 мл/мин.
После ренатурации Таргевира 1, иммобилизованного на колонке, ее промывают 10 объемами раствора II (20 мM NaPi, pH 7.5, 100 мM NaCl) со скоростью 5 мл/мин. Таргевир 1 элюируют с колонки раствором II с 250 мМ имидазолом с той же скоростью. Типичные результаты анализа фракций, собранных в ходе элюции Таргевир1 с металлохелатного аффинного сорбента, а также профиль хроматографической элюции, представлены в Фиг. 5.
Полученный после аффинной хроматографии элюат разбавляют раствором III (20 мМ NaPi, pH 7.5) в 5 раз и наносят со скоростью 5 мл/мин на колонку HiTrap Q HP, 5 мл, уравновешенную 10-кратным объемом раствора III. Колонку промывают 10 объемами раствора III, а затем элюируют Таргевир1 со скоростью 5 мл/мин линейным градиентом NaCl от 0 до 500 мМ в растворе III. Объем градиента составляет 20 объемов колонки. Фракции, содержащие Таргевир1, объединяют и стерильно фильтруют через мембрану с диаметром пор 0,22 микрона.
По представленной методике осуществляют выделение и очистку рекомбинантного белка Таргевир 2.Концентрацию белка определяют методом УФ-спектроскопии, используя молярный коэффициент экстинкции ε280 = 34280 M-1 cм-1 для Таргевир 1 и ε280 = 28590 M-1 cм-1 для Таргевир 2. Типичные результаты анализа фракций, собранных в ходе элюции Таргевир1 с анионообменного сорбента, а также профиль хроматографической элюции, представлены в Фиг. 6.
Пример 6. Получение биомассы рекомбинантного штамма - продуцента барстар.
Для наработки биомассы, содержащей рекомбинантный барстар, клетки E. coli BL21(DE3) трансформируют плазмидой pMT643 (Hartley R.W. // Biochemistry. 1993. V. 32. P. 5978-5984), несущей ген целевого белка барстара под контролем tac-промотора (Фиг. 3). Полученные колонии трансформантов переносят в питательную среду YTPSМ (1% дрожжевой экстракт, 1% триптон, 150 мМ NaCl, 20 мМ KH2PO4, 80 мМ K2HPO4, 2 мM MgCl2, pH 7.5) и культивируют при 37°С с аэрацией до достижения стационарной фазы роста, затем клетки осаждают центрифугированием при 7000 g при 4°С. Биомассу замораживают или немедленно используют для выделения белка. Выход биомассы составляет 5.0 г/л культуральной среды.
Пример 7. Выделение и очистка препарата барстар.
Все процедуры проводят при +4°С. Клетки ресуспендируют в буфере УЗ (20 мМ Трис-HCl, 10 мМ KH2PO4, 10 мМ EDTA, 10 мМ DТТ, 100 мМ NaCl, pH 8) и разрушают с помощью ультразвука. Полученный лизат осветляют центрифугированием при 18500 g, а белки фракционируют ступенчатым высаливанием сульфатом аммония в концентрации 40 и 80 % от насыщения. Осадок, выпавший при концентрации сульфата аммония 80 % от насыщения, содержащий целевой белок, растворяют в буфере ГФ (0.1 M Трис-HCl, 10 мМ EDTA, 10 мМ DTT, pH 8.0). Исходное содержание целевого белка в сульфат-аммонийном осадке составляет около 10%. Полученный раствор целевого белка (объем прим. 2,5 мл) подвергают гель-фильтрации, а затем анионообменной хроматографии на сефарозе Q. Образец наносят на колонку C16/100 с 180 мл носителя Sephacryl S-200, уравновешенную буфером TSDT (20 мМ Трис-HCl, 20 мМ NaCl, 2 мМ DTT, 0,05% Tween-20, pH 8.5), собирают фракцию белка в объеме 115-140 мл. На этой стадии концентрация целевого белка барстара в элюате составляет 0.2-0.3 мг/мл. Содержание целевого белка барстара составляет 60-70 % по данным электрофореза. Фракцию, полученную после гель-фильтрации, содержащую целевой белок, объемом 115-140 мл наносят на колонку HiTrap c Q сефарозой FF, промывают колонку буфером TSDT, затем буфером TDG (20 мМ Трис-HСl, 4 мМ DTT, 10% глицерин, pH 8,5) и элюируют градиентом NaCl от 0 до 500 мМ в буфере TDG. Гомогенный барстар элюируют при концентрации NaCl 100 мМ. Для длительного хранения очищенный белок переводят в буфер SB2 (15 мМ трис-HCl, 1.5 мМ EDTA, 75 мМ NaCl, 40% глицерин, pH 8.0), содержащий 1 мМ DTT, и хранят при -20°С. Выход целевого белка барстара составляет 24 мг из 1 л культуральной жидкости. Содержание целевого белка барстара составляет 96 % по данным электрофореза и последующей фотометрии (Фиг. 7).
Пример 8. Проверка ферментативной (РНКазной) активности Таргевир1 и Таргевир 2.
Сохранение функций барназы как токсического агента в составе Таргевир 1 и Таргевир 2 проверяют путем определения ферментативной РНКазной активности. Для этого используют модифицированный сравнительный метод кислоторастворимого осадка. В качестве субстрата используют дрожжевую РНК.
Исследуемую пробу соответствующего белка растворяют в концентрации 1,25 мкМ в буферном растворе 0,125 М Трис-HCl, pH 8.5 и затем получают серию последовательных 5-кратных разведений образца в том же буфере. В качестве отрицательного контрольного образца используют буферный раствор 0,125 М Трис-HCl, pH 8.5. В качестве положительных контрольных образцов используют образцы барназы дикого типа с известной молярной концентрацией. К 40 мкл каждого образца добавляют 160 мкл раствора дрожжевой РНК (в концентрации 2 г/л) в 0,125 М Трис-HCl, pH 8.5 на льду. Реакционную смесь инкубируют 15 минут при +37°С. Реакцию останавливают добавлением 200 мкл 6% хлорной кислоты и инкубируют смесь 15 мин при 0°С. После чего центрифугируют при 16 000 g в течение 10 минут. Супернатанты разбавляют в 10 раз и измеряют оптическую плотность при 260 нм (OD260) относительно контрольного образца. Большим значениям оптической плотности соответствует большая рибонуклеазная активность. 1 ед. акт. рибонуклеазы в исследуемой пробе соответствует приращению OD260 на 0,05 единиц.
Величина РНКазной активности Таргевир 1 и Таргевир 2 обычно колеблется в диапазоне от 4.8 × 106 до 11,3 × 106 ед. акт./нмоль, что составляет от 30 до 70% рибонуклеазной активности рекомбинантной барназы из E. coli (Фиг. 8).
Пример 9. Проверка ингибирования барстаром РНКазной активности Таргевир 1 и Таргевир 2.
Способность барстар связываться с Таргевир 1 и Таргевир 2 оценивают по его способности ингибировать рибонуклеазную активность Таргевир. Серию разведений образцов барстар в диапазоне концентраций от 125 до 0,2 мкМ в 0,125 М Трис-HCl (рН 8,5) предварительно инкубируют с 24 мкМ раствором Таргевира 1 и Таргевира 2, а затем используют эти смеси для определения активности РНКазы методом осаждения нерастворимой в кислоте дрожжевой РНК, как описано в примере 7. Барстар полностью ингибирует РНКазную активность Таргевир 1 и Таргевир 2 при добавлении в 1,5-кратном молярном избытке (Фиг. 9).
Пример 10. Изучение связывания Таргевира с RBD-доменом методом ИФА
Исследования связывания Таргевира 1 и Таргевира 2 с рекомбинантным RBD-доменом проводили вместе со следующими контролями - рекомбинантный внеклеточный домен ангеотензинпревращающего фермента-2 человека (АСЕ2) в качестве положительного контроля и Darpin9.29-Barnase (DaBa) (Патент RU 2610179) в качестве отрицательного контроля. Последний контроль был выбран в силу схожести его структуры с Таргевиром 1 и Таргевир 2.
В лунки 96-луночных планшетов Nunc MaxiSorp (Nunc, США) вносили по 100 мкл раствора каждого из исследуемых рекомбинантных белков (Таргевир, DaBa и ACE-2) в концентрации 2 мкг/мл в 50 мМ гидрокарбонате натрия, рН 10.6 из расчета по 6 лунок для каждого исследуемого белка и инкубировали в течение 16 ч при 4°С без перемешивания. По окончании вносили блокирующий раствор в объеме 100 мкл (фосфатно-солевой буфер (PBS), 0.1% Tween 20, 3% БСА), и инкубировали в течение 1 ч при 37°С без перемешивания. Лунки промывали 5 раз промывочным раствором (PBS, 0.1% Tween 20) и добавляли по 100 мкл растворов рекомбинантного RBD-домена в PBS в концентрациях: 1000, 300, 100, 30, 10, и 0 нг/мл для каждого из исследуемых белков. Планшет инкубировали в течение 1 ч при 37°С и перемешивании; промывали 5 раз промывочным раствором. В лунки, содержащие раствор RBD-домена, добавляли моноклональные антитела мыши специфичные к RBD-домену (кат. RBD5305, HyTest) в разведении 1/10 000 в растворе для разведения конъюгатов (PBS, 0.1% Tween 20, 0.1% БСА) и инкубировали в течение 1 ч при 37°С и перемешивании. После промывки планшетов промывочным раствором (5 раз) добавляли антитела козы к Fc-фрагменту иммуноглобулина класса G мыши, конъюгированные с пероксидазой хрена (кат. A2554 Sigma-Aldrich), в разведении 1/30 000 в растворе для разведения конъюгатов и инкубировали в течение 1 ч при 37°С и перемешивании. Планшеты промывали 5 раз промывочным раствором, добавляли 100 мкл раствора субстрата ТМБ и инкубировали в темноте в течение 10 мин. Реакцию останавливали внесением в лунки планшета по 100 мкл 10% раствора фосфорной кислоты и измеряли абсорбцию при 450 нм на планшетном спектрофотометре.
Таргевир 1 и Таргевир 2 связываются с RBD так же эффективно, как и рекомбинантный внеклеточный домен ангеотензинпревращающего фермента-2 человека (АСЕ2) (Фиг. 10). В тех же условиях Darpin9.29-Barnase (DaBa), выступавший в качестве отрицательного контроля, не проявляет способности связываться с RBD.
Пример 11. Изучение связывания RBD с АСЕ-2 в присутствии Таргевир методом ИФА
Исследования связывания рекомбинантного RBD с рекомбинантным внеклеточным доменом ангеотензинпревращающего фермента-2 человека (АСЕ-2) в присутствии Таргевир 1 и Таргевир 2 проводили методом ИФА. При этом были использованы следующие контроли:
(1) моноклональное антитело мыши R107 (кат. R107, HyTest), специфичное к RBD-домену, в качестве положительного контроля. Это антитело, как было заявлено производителем, блокирует связывание RBD с ACE-2;
(2) рекомбинантный белок АСЕ-2;
(3) рекомбинантный белок Darpin9-29-Barnase (DaBa) (Патент РФ№ 2610179) в качестве отрицательного контроля. Этот контроль был выбран в силу схожести его структуры с Таргевиром.
В лунки 96-луночных планшетов Nunc MaxiSorp (Nunc, США) вносили по 100 мкл раствора рекомбинантного белка ACE-2 в концентрации 2 мкг/мл в 50 мМ гидрокарбонате натрия, рН 10.6 и инкубировали в течение 16 ч при 4°С без перемешивания. По окончании вносили блокирующий раствор в объеме 100 мкл (фосфатно-солевой буфер (PBS), 0.1% Tween 20, 3% БСА), и инкубировали в течение 1 ч при 37°С без перемешивания. Лунки промывали 5 раз промывочным раствором (PBS, 0.1% Tween 20) и вносили по 100 мкл растворов, содержащих рекомбинантный RBD-домен в концентрации 300 нг/мл, и один из исследуемых белков (LB1, DaBa и R107) в концентрациях, соответствующих следующим молярным соотношениям RBD-домен/исследуемый белок: 1:100, 1:10, 1:1, 1:0.1 и 1:0.01 в PBS для каждого из исследуемых белков. Планшет инкубировали в течение 1 ч при 37°С, промывали 5 раз промывочным раствором, добавляли моноклональные антитела мыши, специфичные к RBD-домену (кат. RBD5305, HyTest) в разведении 1/10 000 в растворе для разведения конъюгатов (PBS, 0.1% Tween 20, 0.1% БСА) и инкубировали в течение 1 ч при 37°С и перемешивании. После промывки планшетов промывочным раствором (5 раз) добавляли антитела козы к Fc-фрагменту иммуноглобулина класса G мыши, конъюгированные с пероксидазой хрена (кат. A2554 Sigma-Aldrich), в разведении 1/30 000 в растворе для разведения конъюгатов и инкубировали в течение 1 ч при 37°С и перемешивании. По окончании инкубации планшеты промывали 5 раз промывочным раствором, добавляли 100 мкл раствора субстрата ТМБ и инкубировали в темноте 10 мин. Реакцию останавливали внесением в лунки планшета по 100 мкл 10% раствора фосфорной кислоты и измеряли абсорбцию при 450 нм на планшетном спектрофотометре.
Таргевир 1 и Таргевир 2 блокирует связывание RBD с внеклеточным доменом ангеотензинпревращающего фермента-2 человека (АСЕ2), иммобилизованном на стенках лунок планшета, также эффективно, как и коммерческое моноклональное антитело мыши R107 (кат. R107, HyTest), специфичное к RBD-домену, которое как было заявлено производителем, блокирует связывание RBD с ACE-2 (Фиг. 11). В тех же условиях Darpin9.29-Barnase (DaBa), выступавший в качестве отрицательного контроля, не влияет на связывание RBD c ACE-2.
Пример 12. Изучение вируснейтрализующего действия Таргевира с использованием псевдовирусной системы
Вируснейтрализующее действие Таргевира 1 и Таргевира 2 изучали в так называемой псевдовирусной системе. Для этого была создана линия клеток HEK293T-ACE-2, несущая на своей поверхности ACE-2, а также получены рекомбинантные лентивирусные частицы, псевдотипированные S-белком SARS-CoV-2 и содержащие в качестве репортерного гена ген люциферазы светлячка (Kruglova N. et al. // Viruses. 2021. V. 13(6). P. 1133). Детекция вируснейтрализующей активности происходит за счет измерения люминесценции клеток, несущих на своей поверхности АСЕ2, при добавлении к ним псевдовирусных частиц, прединкубированных с испытуемыми веществами. При этом чем больше вируснейтрализующая активность испытуемого вещества, тем меньше уровень детектируемой люминесценции.
Линию клеток HEK293T-ACE-2, гиперэкспрессирующую на поверхности рецептор ACE-2 человека, получали путем трансдукции HEK293T псевдовирусными частицами с мембранным гликопротеином VSV-g (гликопротеин G вируса везикулярного стоматита), содержащими в геноме гены рецептора ACE-2 человека и зеленого флуоресцентного белка GFP. После трех последовательных обогащений трансдуцированных клеток, чередующихся пассажированием, методом проточной цитофлуориметрии были отобраны клетки, стабильно экспрессирующие GFP. Присутствие ACE2 на поверхности пула клеток подтверждали проточной цитофлуориметрией. Клетки инкубировали с рекомбинантным RBD-Fc с дальнейшей визуализацией поликлональными антителами козы, кроссреактивными с Fc-доменом человека, коньюгированными с флуоресцентной меткой DL650, испускание которой детектируется в канале APC. Согласно полученным с помощью проточной цитофлуориметрии данным 96% клеток несут на своей поверхности АСЕ2. Полученные клетки ACE2-HEK293T используют далее в тестах.
Получение рекомбинантных лентивирусных частиц, псевдотипированных S-белком SARS-CoV-2 и содержащих в качестве репортерного гена ген люциферазы светлячка, проводили как описано в работе (Kruglova N. et al. // Viruses. 2021. V. 13(6). P. 1133). Псевдовирус способен проникать в клетки HEK293T-ACE-2, несущие на своей поверхности АСЕ2, где на рекомбинантном РНК-геноме синтезируется ДНК провируса, содержащая маркерный ген люциферазы светлячка, встраивание которого в геном клетки-мишени придает ей способность люминесцировать.
В тесте на вируснейтрализующую активность в качестве контролей выступали белковые компоненты, из которых построен Таргевир, а именно, LCB1 и LCB3, как направляющие модули, и барназа, как цитотоксичный и вирус-интерферирующий компонент. LB1, LB3, 2LB1 являются условными обозначениями для Таргевир 1, Таргевир 2 и одного из мутантных вариантов Таргевир, соответственно.
Тест на нейтрализующую активность проводили следующим образом. Клетки ACE2-HEK293T рассевали в лунки 96-луночной планшета в количестве 5000 клеток на 100 мкл среды DMEM с 10% FBS (Gibco) и инкубировали в течение 24 часов при 37°C, 5% CO2. Псевдовирусы SARS-CoV-2 вносили по 50 мкл в лунки 96-луночной плашки, добавляли к ним тестируемые вещества (LB1, LCB1, 2LCB1, LB3, LCB3 и рекомбинантную барназу) в концентрациях 46, 4,6, 0,46, 0,046, 0,0046, 0,00046 нМ и инкубировали при 37°C, 5% CO2 в течение 1часа. Из лунок 96-луночной плашки с клетками ACE2-HEK293T удаляли по 50 мкл культуральной среды, добавляли прединкубированные смеси псевдовирусных частиц SARS-CoV-2 с указанными концентрациями для каждого из тестируемых веществ и инкубировали при 37°C, 5% CO2. Через 24 часа проводили смену среды, и еще через 48 и 96 часов культуральную среду удаляли, клетки лизировали с помощью GLO lysis buffer (кат. E2661, Promega), смешивали с субстратом Bright-Glo™ Luciferase Assay System (кат. E2620, Promega), измеряли уровень люминесценции и строили график уровня люминисценции в зависимости от концентрации тестируемого вещества. Обработку данных и расчет значений IC50 проводили с помощью программного обеспечения GraphPad Prism 8. Увеличение продолжительности инкубирования клеток, зараженных вирусом, до 96 ч было необходимо для того, чтобы позволить барназе, которая предположительно будет попадать внутрь клетки вместе с вирусом, проявить свою РНКазную активность.
Таргевир 1 и Таргевир 2, а также направляющие модули LCB1 и LCB3, которые содержатся в их составе, эффективно нейтрализуют вирус (Фиг. 12). Однако концентрации полумаксимального ингибирования (IC50) для Таргевир существенно меньше. Наиболее активным является Таргевир 1, у которого IC50 в 10 раз меньше, чем у LCB1, и составляет 0,09 нМ против 0,87 нМ у LCB1 через 48 ч (Фиг. 12А). Через 96 ч IC50 для этой пары белков различаются уже в 20 раз (0,03 нМ против 0,68 нМ) (Фиг. 12Б). Таргевир 2 демонстрирует в целом меньшую противовирусную активность. IC50 для Таргевир 2 равняется 0,07 нМ против 0,2 нМ для LCB3 (Фиг. 12А). Также, как и в случае Таргевир 1 увеличение продолжительности инкубирования усиливает эффект от применения этого белка (Фиг. 12Б). В исследуемом диапазоне концентраций барназа не проявила никакой противовирусной активности (Фиг. 12).
--->
Перечень последовательностей
Sequence Listing Information:
DTD Version: V1_3
File Name: Targevir.xml
Software Name: WIPO Sequence
Software Version: 1.1.0
Production Date: 2021-12-23
General Information:
Current application / IP Office: RU
Current application / Application number: 1
Current application / Filing date: 2021-12-23
Current application / Applicant file reference: 1
Earliest priority application / IP Office: RU
Earliest priority application / Application number: 1
Earliest priority application / Filing date: 2021-12-23
Applicant name: ИБХ РАН
Applicant name / Language: en
Applicant name / Name Latin: IBCh RAS
Invention title: СПОСОБ ПОЛУЧЕНИЯ ЛЕКАРСТВЕННОГО СРЕДСТВА ПРОТИВ
КОВИД НА ОСНОВЕ БЕЛКОВ БАРНАЗА-БАРСТАР И АДРЕСНОГО ПЕПТИДА С ЭФФЕКТОМ
МОМЕНТАЛЬНОЙ ОТМЕНЫ ЦИТОТОКСИЧЕСКОГО ДЕЙСТВИЯ ( ru )
Sequence Total Quantity: 14
Sequences:
Sequence Number (ID): 1
Length: 1110
Molecule Type: DNA
Features Location/Qualifiers:
- source, 1..1110
> mol_type, other DNA
> organism, Escherichia coli
- CDS, 4..577
> protein_id, 2
> translation, MDKEWILQKIYEIMRLLDELGHAEASMRVSDLIYEFMKKGD
ERLLEEAERLLEEVEREFPKPSTPPGSSGGAPAQVINTFDGVADYLQTYHKLPDNYITKSEAQALGWVASKGNLADV
APGKSIGGDIFSNREGKLPGKSGRTWREADINYTSGFRNSDRILYSSDWLIYKTTDHYQTFTKIRGAPHHHHH
- CDS, 4..174
> protein_id, 3
> translation, MDKEWILQKIYEIMRLLDELGHAEASMRVSDLIYEFMKKG
DERLLEEAERLLEEVER
- CDS, 175..222
> protein_id, 4
> translation, EFPKPSTPPGSSGGAP
- CDS, 223..552
> protein_id, 5
> translation, AQVINTFDGVADYLQTYHKLPDNYITKSEAQALGWVASKGNL
ADVAPGKSIGGDIFSNREGKLPGKSGRTWREADINYTSGFRNSDRILYSSDWLIYKTTDHYQTFTKIR
- CDS, 830..1099
> protein_id, 7
> translation, MKKAVINGEQIRSISDLHQTLKKELALPEYYGENLDALWDCL
TGWVEYPLVLEWRQFEQSKQLTENGAESVLQVFREAKAEGCDITIILS
- CDS, 562..577
> protein_id, 6
> translation, HHHHH
Residues:
catatggata aagaatggat tctgcaaaaa atttacgaga tcatgcgtct gctggacgaa 60
ctgggccacg cggaggcttc catgcgtgtg agcgatctga tctatgagtt catgaagaaa 120
ggtgacgaac gtctgctgga agaagcagaa cgcctgctgg aagaagttga acgcgaattc 180
cccaaacctt ctactccacc cgggagcagt ggtggcgcgc ctgcacaggt tatcaacacg 240
tttgacgggg ttgcggatta tcttcagaca tatcataagc tacctgataa ttacattaca 300
aaatcagaag cacaagccct cggctgggtg gcatcaaaag ggaaccttgc agacgtcgct 360
ccggggaaaa gcatcggcgg agacatcttc tcaaacaggg aaggcaaact cccgggcaaa 420
agcggacgaa catggcgtga agcggatatt aactatacat caggcttcag aaattcagac 480
cggattcttt actcaagcga ctggctgatt tacaaaacaa cggaccatta tcagaccttt 540
acaaaaatca gaggcgcgcc gcaccatcat caccattgac gtctaagctt ataacgaaaa 600
aaacggcttc ctgcggaggc cgtttttttc agctttacat aaagtgtgta ataaattttt 660
cttcaaactc tgatcggtca atttcacttt ccggctctag agtccggtcc aatctgcagc 720
cgtccgagac aggaggacat cgtccagctg aaaccggggc agaatccggc catttctgaa 780
gagaaaaatg gtaaactgat agaataaaat cataagaaag gagccgcaca tgaaaaaagc 840
agtcattaac ggggaacaaa tcagaagtat cagcgacctc caccagacat tgaaaaagga 900
gcttgccctt ccggaatact acggtgaaaa cctggacgct ttatgggatt gtctgaccgg 960
atgggtggag tacccgctcg ttttggaatg gaggcagttt gaacaaagca agcagctgac 1020
tgaaaatggc gccgagagtg tgcttcaggt tttccgtgaa gcgaaagcgg aaggctgcga 1080
catcaccatc atactttctt aatactcgag 1110
Sequence Number (ID): 2
Length: 191
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..191
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
MDKEWILQKI YEIMRLLDEL GHAEASMRVS DLIYEFMKKG DERLLEEAER LLEEVEREFP 60
KPSTPPGSSG GAPAQVINTF DGVADYLQTY HKLPDNYITK SEAQALGWVA SKGNLADVAP 120
GKSIGGDIFS NREGKLPGKS GRTWREADIN YTSGFRNSDR ILYSSDWLIY KTTDHYQTFT 180
KIRGAPHHHH H 191
Sequence Number (ID): 3
Length: 57
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..57
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
MDKEWILQKI YEIMRLLDEL GHAEASMRVS DLIYEFMKKG DERLLEEAER LLEEVER 57
Sequence Number (ID): 4
Length: 16
Molecule Type: AA
Residues:
EFPKPSTPPG SSGGAP 16
Sequence Number (ID): 5
Length: 110
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..110
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
AQVINTFDGV ADYLQTYHKL PDNYITKSEA QALGWVASKG NLADVAPGKS IGGDIFSNRE 60
GKLPGKSGRT WREADINYTS GFRNSDRILY SSDWLIYKTT DHYQTFTKIR 110
Sequence Number (ID): 6
Length: 5
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..5
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
HHHHH 5
Sequence Number (ID): 7
Length: 90
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..90
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
MKKAVINGEQ IRSISDLHQT LKKELALPEY YGENLDALWD CLTGWVEYPL VLEWRQFEQS 60
KQLTENGAES VLQVFREAKA EGCDITIILS 90
Sequence Number (ID): 8
Length: 1134
Molecule Type: DNA
Features Location/Qualifiers:
- source, 1..1134
> mol_type, other DNA
> organism, Escherichia coli
- CDS, 4..600
> protein_id, 9
> translation, MNDDELHMLMTDLVYEALHFAKDEEIKKRVFQLFELADKA
YKNNDRQKLEKVVEELKELLERLLSEFPKPSTPPGSSGGAPAQVINTFDGVADYLQTYHKLPDNYITKSEAQALGW
VASKGNLADVAPGKSIGGDIFSNREGKLPGKSGRTWREADINYTSGFRNSDRILYSSDWLIYKTTDHYQTFTKIRGA
PHHHHH
- CDS, 4..198
> protein_id, 10
> translation, MNDDELHMLMTDLVYEALHFAKDEEIKKRVFQLFELADKAY
KNNDRQKLEKVVEELKELLERLLS
- CDS, 199..246
> protein_id, 11
> translation, EFPKPSTPPGSSGGAP
- CDS, 247..576
> protein_id, 12
> translation, AQVINTFDGVADYLQTYHKLPDNYITKSEAQALGWVASKGN
LADVAPGKSIGGDIFSNREGKLPGKSGRTWREADINYTSGFRNSDRILYSSDWLIYKTTDHYQTFTKIR
- CDS, 586..600
> protein_id, 13
> translation, HHHHH
- CDS, 854..1123
> protein_id, 14
> translation, MKKAVINGEQIRSISDLHQTLKKELALPEYYGENLDALWDCL
TGWVEYPLVLEWRQFEQSKQLTENGAESVLQVFREAKAEGCDITIILS
Residues:
catatgaatg atgatgaact gcacatgctg atgactgacc tggtttatga agcactgcat 60
ttcgctaaag acgaggaaat caagaaacgt gtgttccagc tgtttgaact ggcggacaaa 120
gcctacaaaa acaacgatcg tcagaagctg gaaaaagttg tagaagaact gaaagaactg 180
ctggagcgcc tgctgtctga attccccaaa ccttctactc cacccgggag cagtggtggc 240
gcgcctgcac aggttatcaa cacgtttgac ggggttgcgg attatcttca gacatatcat 300
aagctacctg ataattacat tacaaaatca gaagcacaag ccctcggctg ggtggcatca 360
aaagggaacc ttgcagacgt cgctccgggg aaaagcatcg gcggagacat cttctcaaac 420
agggaaggca aactcccggg caaaagcgga cgaacatggc gtgaagcgga tattaactat 480
acatcaggct tcagaaattc agaccggatt ctttactcaa gcgactggct gatttacaaa 540
acaacggacc attatcagac ctttacaaaa atcagaggcg cgccgcacca tcatcaccat 600
tgacgtctaa gcttataacg aaaaaaacgg cttcctgcgg aggccgtttt tttcagcttt 660
acataaagtg tgtaataaat ttttcttcaa actctgatcg gtcaatttca ctttccggct 720
ctagagtccg gtccaatctg cagccgtccg agacaggagg acatcgtcca gctgaaaccg 780
gggcagaatc cggccatttc tgaagagaaa aatggtaaac tgatagaata aaatcataag 840
aaaggagccg cacatgaaaa aagcagtcat taacggggaa caaatcagaa gtatcagcga 900
cctccaccag acattgaaaa aggagcttgc ccttccggaa tactacggtg aaaacctgga 960
cgctttatgg gattgtctga ccggatgggt ggagtacccg ctcgttttgg aatggaggca 1020
gtttgaacaa agcaagcagc tgactgaaaa tggcgccgag agtgtgcttc aggttttccg 1080
tgaagcgaaa gcggaaggct gcgacatcac catcatactt tcttaatact cgag 1134
Sequence Number (ID): 9
Length: 199
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..199
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
MNDDELHMLM TDLVYEALHF AKDEEIKKRV FQLFELADKA YKNNDRQKLE KVVEELKELL 60
ERLLSEFPKP STPPGSSGGA PAQVINTFDG VADYLQTYHK LPDNYITKSE AQALGWVASK 120
GNLADVAPGK SIGGDIFSNR EGKLPGKSGR TWREADINYT SGFRNSDRIL YSSDWLIYKT 180
TDHYQTFTKI RGAPHHHHH 199
Sequence Number (ID): 10
Length: 65
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..65
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
MNDDELHMLM TDLVYEALHF AKDEEIKKRV FQLFELADKA YKNNDRQKLE KVVEELKELL 60
ERLLS 65
Sequence Number (ID): 11
Length: 16
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..16
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
EFPKPSTPPG SSGGAP 16
Sequence Number (ID): 12
Length: 110
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..110
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
AQVINTFDGV ADYLQTYHKL PDNYITKSEA QALGWVASKG NLADVAPGKS IGGDIFSNRE 60
GKLPGKSGRT WREADINYTS GFRNSDRILY SSDWLIYKTT DHYQTFTKIR 110
Sequence Number (ID): 13
Length: 5
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..5
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
HHHHH 5
Sequence Number (ID): 14
Length: 90
Molecule Type: AA
Features Location/Qualifiers:
- SOURCE, 1..90
> MOL_TYPE, protein
> ORGANISM, Escherichia coli
Residues:
MKKAVINGEQ IRSISDLHQT LKKELALPEY YGENLDALWD CLTGWVEYPL VLEWRQFEQS 60
KQLTENGAES VLQVFREAKA EGCDITIILS 90
END
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИПЕПТИД С АНТИ-SARS-CoV-2 АКТИВНОСТЬЮ И ПРОЛОНГИРОВАННЫМ ВРЕМЕНЕМ ЦИРКУЛЯЦИИ В КРОВОТОКЕ | 2023 |
|
RU2822355C1 |
| ИММУНОЛИПОСОМА, СВЯЗЫВАЮЩАЯСЯ С RBD S-БЕЛКА SARS-COV-2 | 2023 |
|
RU2827165C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ БАРНАЗЫ В КЛЕТКАХ ESCHERICHIA COLI, ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ БАРНАЗЫ И СПОСОБ ПОЛУЧЕНИЯ БАРНАЗЫ. | 2017 |
|
RU2650871C1 |
| Способ получения рекомбинантного противоопухолевого токсина на основе белков барназа-барстар и адресного полипептида дарпина с эффектом моментальной отмены цитотоксического действия | 2015 |
|
RU2610179C1 |
| Рекомбинантный RBD S-белок коронавируса SARS-CoV-2 и способ его получения | 2023 |
|
RU2833839C1 |
| Плазмидная генетическая конструкция pET21a_SKP, обеспечивающая экспрессию в прокариотической системе E.coli рекомбинантного белка SKP, и рекомбинантный белок SKP, обладающий широконейтрализующими свойствами однодоменного наноантитела против SARS-CoV-2 | 2024 |
|
RU2839376C1 |
| Плазмидная генетическая конструкция pET21_Ab_CoV-2_1.3, обеспечивающая экспрессию рекомбинантного белка AB_COV-2_1.3 в прокариотической системе E. coli, и рекомбинантный белок AB_COV-2_1.3, обладающий свойствами однодоменного наноантитела против SARS-CoV-2 | 2022 |
|
RU2798508C1 |
| Интегративный плазмидный вектор pVEAL3-RBDdel, обеспечивающий синтез и секрецию рекомбинантного белка рецепторсвязывающего домена RBDdelta коронавируса SARS-CoV-2 в клетках млекопитающих, рекомбинантный штамм клеточной линии CHO-K1- RBDdelta и рекомбинантный белок RBDdelta SARS-CoV-2, продуцируемый штаммом клеточной линии | 2023 |
|
RU2816175C1 |
| СПОСОБ ИСПОЛЬЗОВАНИЯ РЕКОМБИНАНТНЫХ БЕЛКОВ SARS-COV-2 В СОСТАВЕ ТЕСТ-СИСТЕМЫ ДЛЯ ИММУНОФЕРМЕНТНОГО АНАЛИЗА С ОПРЕДЕЛЕНИЕМ УРОВНЕЙ АНТИТЕЛ КЛАССОВ IgM, IgG, IgA В СЫВОРОТКЕ/ПЛАЗМЕ КРОВИ БОЛЬНЫХ COVID-19 | 2020 |
|
RU2730897C1 |
| Моноклональное антитело к RBD фрагменту в составе S белка вируса SARS-CoV-2 | 2020 |
|
RU2744274C1 |




Изобретение относится к области белковой инженерии, биотехнологии и медицины. Описан рекомбинантный белок, имеющий последовательность SEQ ID NO: 2 или SEQ ID NO: 9, связывающийся с рецептор-связывающим доменом S-белка SARS-CoV-2, а также его применение в качестве средства, обладающего способностью связываться с рецептор-связывающим доменом S-белка SARS-CoV-2. Раскрыта рекомбинантная плазмида, обеспечивающая синтез в клетках Escherichia coli указанного рекомбинантного белка, причем указанная плазмида получена путем клонирования фрагмента ДНК с нуклеотидной последовательностью SEQ ID NO: 8 или SEQ ID NO: 1 в плазмидный вектор pET39b по сайтам рестрикции NdeI/XhoI. Представлен рекомбинантный штамм Escherichia coli BL21(DE3), содержащий указанную рекомбинантную плазмиду – продуцент описанного рекомбинантного белка. Кроме того, представлен способ получения описанного рекомбинантного белка из биомассы указанного рекомбинантного штамма Escherichia coli BL21(DE3), при котором осуществляют разрушение клеток дезинтеграцией, удаляют клеточный дебрис, выделяют белок при помощи двухстадийной хроматографии, включающей стадию белковой денатурации/ренатурации на металлохелатном аффинном сорбенте, и очистку при помощи анионообменной хроматографии. Изобретение может найти применение в биотехнологии и медицине при создании лекарств против КОВИД-19. 5 н.п. ф-лы, 12 ил., 12 пр.
1. Рекомбинантный белок, имеющий последовательность SEQ ID NO: 2 или SEQ ID NO: 9, связывающийся с рецептор-связывающим доменом S-белка SARS-CoV-2.
2. Применение рекомбинантного белка по п. 1 в качестве средства, обладающего способностью связываться с рецептор-связывающим доменом S-белка SARS-CoV-2.
3. Рекомбинантная плазмида, обеспечивающая синтез в клетках Escherichia coli рекомбинантного белка по п. 1, причем указанная плазмида получена путем клонирования фрагмента ДНК с нуклеотидной последовательностью SEQ ID NO: 8 или SEQ ID NO: 1 в плазмидный вектор pET39b по сайтам рестрикции NdeI/XhoI.
4. Рекомбинантный штамм Escherichia coli BL21(DE3), содержащий рекомбинантную плазмиду по п. 3 – продуцент рекомбинантного белка по п. 1.
5. Способ получения рекомбинантного белка по п. 1 из биомассы рекомбинантного штамма Escherichia coli BL21(DE3) по п. 4, при котором осуществляют разрушение клеток дезинтеграцией, удаляют клеточный дебрис, выделяют белок по п. 1 при помощи двухстадийной хроматографии, включающей стадию белковой денатурации/ренатурации на металлохелатном аффинном сорбенте, и очистку при помощи анионообменной хроматографии.
| RU 2020126021 A, 31.01.2022 | |||
| RU 2020130238 A, 14.03.2022 | |||
| WO 2021181355 A1, 16.09.2021 | |||
| WO 2021206636 A1, 14.10.2021. |

