Область техники, к которой относится изобретение
Изобретение относится к области биологии, молекулярной биологии, биотехнологии и генетической инженерии растений. Изобретение может быть использовано для создания генетических конструкций с целью повышенной экспрессии рекомбинантных генов и продукции рекомбинантных белков. Изобретение может применяться для наработки рекомбинантных белков в клетках растений методом транзиентной экспрессии, для создания трансгенных растений, продуцирующих рекомбинантные белки, для отбора трансгенных клеток, каллусов, побегов и проростков растений на питательной среде с селективным агентом. Настоящее изобретение может применяться в фундаментальных исследованиях биологии растений с использованием методов прямой и обратной генетики.
Уровень техники
Промотор гена - фрагмент геномной ДНК, обеспечивающий инициацию транскрипции гена. Промоторы необходимы для координированной экспрессии генов в клетках растений в изменяющихся условиях окружающей среды. В растениях полноразмерные промоторы белок-кодирующих генов состоят из дистальной, проксимальной и «коровой» промоторных областей. «Коровая» область содержит, как правило, последовательность известного цис-элемента ТАТА-box, локализованного около - 40-20 п.н. выше (или левее) сайта инициации транскрипции (transcription start site, TSS) в котором белки основного транскрипционного механизма (RNA Pol II, TATA-binding proteins и TBP-associated factors) запускают транскрипцию (Hernandez-Garcia and Finer 2014). Изменение экспрессии генов достигается благодаря физическому взаимодействию между основным транскрипционным механизмом и регуляторными белками (факторами транскрипции), связывающими примыкающую к «коровой» проксимальную промоторную область. Дистальная промоторная область, наиболее удаленная от TSS, может влиять на эффективность экспрессии генов благодаря трехмерной подвижности ДНК и ее пространственному сближению с «коровой» областью.
Функциональную архитектуру полноразмерного промотора определяют множественные регуляторные элементы (энхансеры, сайленсеры, инсуляторы и цис-элементы) - специфические последовательности нуклеотидов для связывания регуляторными белками, распределенные вдоль всей нуклеотидной последовательности промотора. Взаимодействия между различными регуляторными элементами и связывающими их регуляторными белками играют главную роль в определении эффективности и специфичности эукариотического промотора.
Изолированные нуклеотидные последовательности промоторов и их отдельные структурные области в экспрессионных кассетах генетических конструкций имеют существенное значение для регулирования экспрессии рекомбинантных генов в трансгенных растениях. Под термином «рекомбинантный ген» обычно понимают любой искусственно созданный ген, состоящий из компонентов разных генов. Рекомбинантный ген создается путем соединения отдельных компонентов разных генов в новые комбинации с помощью методов генетической инженерии. Любой ген в составе кассеты экспрессии генетической конструкции для трансформации растений может считаться рекомбинантным.
Изолированные промоторы могут сохранять свои основные функции в составе генетических конструкций, интегрированных в геном трансгенных растений, однако качественные и количественные вариации экспрессии рекомбинантных генов под их контролем не являются редкостью и должны быть изучены для каждого промотора, который предлагается для биотехнологии растений (Hernandez-Garcia and Finer 2014).
Используемые в биотехнологии растений промоторы могут быть классифицированы как конститутивные (активны в большинстве тканей на разных стадиях развития растения), пространственно-временные (активны в определенных тканях и/или на определенных стадиях развития растения) и индуцибельные (активируются при воздействии химических или физических агентов). К отдельному классу можно отнести синтетические промоторы, состоящие из различных по происхождению регуляторных элементов (Hernandez-Garcia and Finer 2014).
Сильные конститутивные промоторы наиболее востребованы в биотехнологии растений. Промоторы этого класса наряду с эффективной экспрессией целевых генов (рекомбинантных генов предназначенных для изменения фенотипа растения) способны контролировать экспрессию селективных генов (рекомбинантных генов предназначенных для отбора трансформированных клеток растения) на уровне, достаточном для селекции трансгенных клеток, каллусов, побегов и проростков растения на питательной среде с селективным агентом.
Ранее в биотехнологии растений практически все трансгенные растения содержали два рекомбинантных гена, один из которых под контролем конститутивного промотора был необходим для селективного отбора трансформированных клеток, а другой целевой ген под контролем промотора любого типа был предназначен для изменения фенотипа растения. В настоящее время возможности мультигенной трансформации делают доступными импорт в растения целых метаболических путей, включающих экспрессию нескольких целевых генов, и разработку трансгенных культур, одновременно производящих целый спектр соединений (Zhu et al. 2007; Peremarti et al. 2010). Каждый дополнительный рекомбинантный ген нуждается в собственном промоторе, что требует применения в одной генетической конструкции различных промоторов со схожим уровнем и профилем экспрессии или одного промотора несколько раз. В первом случае ощущается дефицит доступных промоторов с необходимыми параметрами, а во втором случае введение повторяющихся последовательностей в один или разные локусы одного трансгенного растения может иметь отрицательное влияние на экспрессию и наследование входящих в них рекомбинантных генов из-за эффекта гомологозависимого замолкания (Mette et al. 1999; Lessard et al. 2002; Potenza et al. 2004; Mourrain et al. 2007). В настоящее время настоятельно рекомендуется управлять различными рекомбинантными генами, введенными в трансгенные растения, с использованием разных промоторов, чтобы избежать эффекта гомологозависимого замолкания генов из-за гомологии повторяющихся промоторов (Peremarti et al. 2010; Dutt et al. 2014; Nuccio 2017).
До настоящего времени наиболее популярным сильным конститутивным промотором для генетической инженерии растений является промотор CaMV35S, созданный на основе промоторной области 35S РНК вируса мозаики цветной капусты CaMV (Odell et al. 1985). Промотор CaMV35S обеспечивает высокий уровень экспрессии в большинстве тканей и органов растений и широко используется при разработке трансгенных растений, особенно двудольных видов, поэтому часто в исследованиях других промоторов используется в качестве сравнительного контроля (прототипа). Известны другие сильные конститутивные промоторы из геномов фитопатогенов. Например, промотор из генома вируса мозаики георгин (Kuluev et al. 2010), промотор из генома вируса мозаики норичника (Acharya et al. 2014), промотор nos гена нопалинсинтазы агробактерий (Agrobacterium tumefaciens) (Verdaguer et al. 1996) и др. Однако изолированные из патогенов растений промоторы могут приводить к нарушению развития трансгенных растений (АН and Kim 2019), они менее предпочтительны с экологической точки зрения и их эффективность может подавляться в случае заражения генетически модифицированных растений вирусами (Al-Kaff et al. 2000). Промоторы патогенов растений в кассетах экспрессии могут быть заменены промоторами растительного происхождения для получения cis-генных или intra-генных растений, свободных от нежелательных чужеродных (не растительного происхождения) последовательностей (Holme et al. 2013). Поэтому, в последние два десятилетия активно проводится поиск и тестирование новых эффективных промоторов генов растений и создание синтетических промоторов (АН and Kim 2019).
Сильных конститутивных промоторов растительного происхождения, одновременно эффективных для экспрессии целевых и селективных генов, известно крайне мало. Одним из немногих является промотор AtTCTP из арабидопсиса (Arabidopsis thaliana), конститутивной активности которого достаточно как для высокоуровневой экспрессии репортерного гена gus в различных органах трансгенных растений арабидопсиса, так и для контроля селективного гена фосфинотрицинацетилтрансферазы при отборе на питательной среде с гербицидом фосфинотрицином трансгенных каллусов и растений полевицы побегообразующей (Agvyrostis stoloniferd) (Han et al. 2015). Промотор Actinl из риса (Oryza sativa) проявил себя как сильный конститутивный при селекции на среде с антибиотиком паромомицином трансгенных каллусов и растений кукурузы (Zea mays) (Prakash et al. 2008). Относительно недавно для генетической инженерии однодольных сельскохозяйственных растений предложен сильный промотор UBI10 из коротконожки лесной (Brachypodium distachyon), который до 35 раз более эффективен, чем CaMV35S (Coussens et al. 2012). Для генетической инженерии двудольных был клонирован целый спектр сильных промоторов различных генов растений, но превосходство их над CaMV35S было не так существенно, как у однодольных. Промоторы генов UBQ1 и UBQ6 из А. thaliana активны во всех тканях табака на уровне вирусного промотора CaMV35S (Callis et al. 1990). Сильный промотор MtHP из Medicago truncatula в растениях арабидопсиса, клевера ползучего (Trifolium repens) и люцерны посевной (Medicago sativa) по эффективности примерно в полтора раза превосходил CaMV35S (Xiao et al. 2005). В сравнении с CaMV35S в трансгенных растениях табака и арабидопсиса до шести раз более эффективным был промотор гена АЦЦ-синтазы VR-ACS1 из вигны лучистой (Vigna radiata), однако его явное превосходство было результатом не столько транскрипционной, сколько трансляционной активации (Cazzonelli et al. 2005).
Ранее нами были клонированы промоторы генов антимикробных пептидов pro-SmAMP1 (MF461278) и pro-SmAMP2 (КХ196447) из растения звездчатка средняя (S. media), которые по эффективности сопоставимы или превосходят вирусный промотор CaMV35S (Komakhin et al. 2016; Vysotskii et al. 2016; Madzharova et al. 2018). Эффективность промоторов первоначально оценивали по экспрессии репортерного гена gus путем измерения активности его белкового продукта GUS. Установлено, что эти промоторы при транзиентной экспрессии в растениях Nicotiana benthamiana по эффективности до 3,4 раз превосходили вирусный промотор CaMV35S. В гомозиготных линиях трансгенных растений табака {Nicotiana tabacum) промоторы pro-SmAMP1 и pro-SmAMP2 были в 2 раза сильнее, чем вирусный промотор CaMV35S. Промотор pro-SmAMP2 по эффективности контроля гена неомицинфосфотрансферазы II (nptII) при селекции трансгенных растений табака и арабидопсиса на средах с антибиотиком канамицином в рекомендованных концентрациях не уступает дуплицированному вирусному промотору 2х CaMV35S. Но при этом выяснилось, что нуклеотидные последовательности промоторов pro-SmAMP1 и pro-SmAMP2 идентичны на 94%, что препятствует их использованию для управления целевым и селективным генами в составе одной генетической конструкции из-за возможного эффекта гомологозависимого замолкания. Поэтому в настоящее время необходим поиск новых растительных промоторов со схожим паттерном активности в клетках растений и отличающейся от промоторов pro-SmAMP1 и pro-SmAMP2 генетической основой (нуклеотидной последовательностью).
Таким образом, несмотря на существующие достижения в области молекулярной биологии и биотехнологии, необходимость высокой, конститутивной, стабильной и точной экспрессии рекомбинантных генов в трансгенных растениях обуславливает потребность поиска новых промоторов генов растений с разной генетической основой. Использование таких промоторов для экспрессии рекомбинантных генов позволит создавать стабильные трансгенные растения, эффективно экспрессирующие множество целевых генов и продуцирующие ценные вторичные метаболиты или сложные белки, состоящие из нескольких полипептидных цепей.
Раскрытие сущности изобретения
Объектом изобретения является нуклеотидная последовательность промоторной области гена дефензина SmAMP-D1 из генома растения звездчатка средняя, или мокрица, (Stellaria media). Впервые выделенный из генома промотор pro-SmAMP-D1 представлен двумя полиморфными версиями и их 5'-делеционными вариантами различной длины. Первая версия промотора pro-SmAMP-D1, обозначена как pro-SmAMP-D1-1 и представлена 5'-делеционными вариантами -303 п.н. (SEC ID NO 1) и -118 п.н. (SEC ID NO 2) относительно +1 п.н. TSS гена дефензина SmAMP-D1. Вторая версия промотора pro-SmAMP-D1, обозначена как pro-SmAMP-D1-2 и представлена 5'-делеционными вариантами -304 п.н. (SEC ID NO 3) и -118 п.н. (SEC ID NO 4) относительно +1 п.н. TSS гена дефензина SmAMP-D1. Версии промотора pro-SmAMP-D1-1 и pro-SmAMP-D1-2 идентичны на 93% и различаются заменами отдельных нуклеотидов в их последовательностях.
Изолированные нуклеотидные последовательности растительного промотора pro-SmAMP-D1 предназначены для создания генетических конструкций с целью экспрессии рекомбинантных генов на высоком и конститутивном уровнях в генетически модифицированных клетках растений и в генетически модифицированных растениях.
Генетические конструкции, несущие рекомбинантный ген (целевой или селективный) под контролем промотора pro-SmAMP-D1, также содержащие терминатор транскрипции, предназначены для генетической трансформации клеток растений, в том числе, с использованием штаммов агробактерий (Agrobacterium).
Штаммы агробактерий, содержащие генетические конструкции с целевым или селективным генами под контролем промотора pro-SmAMP-D1 в составе бинарного вектора для трансформации растений, предназначены для агробактериальной инфильтрации тканей растений или для ко-культивации с растительными эксплантами.
Инфильтрация растительных тканей с использованием штаммов агробактерий, содержащих целевой ген под контролем промотора рго-SmAMP-D1 в составе бинарного вектора для трансформации растений, обеспечивает накопление целевого белка на уровне, который сопоставим или превосходит уровень накопления целевого белка при использовании сильного вирусного промотора CaMV35S.
Агробактериальные штаммы, содержащие генетические конструкции с целевым геном под контролем промотора pro-SmAMP-D1 в составе бинарного вектора для трансформации растений, после ко-культивации с растительными эксплантами позволяют создавать трансгенные растения, продуцирующие целевой белок на уровне, который сопоставим или превосходит уровень накопления целевого белка при использовании сильного вирусного промотора CaMV35S.
Агробактериальные штаммы, содержащие генетические конструкции с селективным геном под контролем промотора pro-SmAMP-D1 в составе бинарного вектора для трансформации растений, после ко-культивации с растительными эксплантами позволяют создавать и отбирать на питательной среде с селективным агентом трансгенные клетки, каллусы, побеги и проростки растений без использования промоторов из патогенов растений.
Основной технический результат изобретения заключается в расширении арсенала новых эффективных промоторов растительного происхождения для биотехнологии растений, в том числе для экспрессии рекомбинантных генов на высоком и конститутивном уровне без использования промоторов из патогенов растений.
Краткое описание чертежей
Фиг. 1. Нуклеотидная последовательность промоторной области гена SmAMP-D1. (а) Промоторная версия pro-SmAMP-D1-1 длиною -303 п.н. относительно +1 п.н. TSS вместе с 5'-НТО гена SmAMP-D1; (б) Промоторная версия pro-SmAMP-D1-2 длиною -304 п.н. относительно +1 п.н. TSS вместе с 5'-НТО гена SmAMP-D1. 5'-НТО подчеркнута, основные цис-действующие элементы в промоторной последовательности выделены рамкой и обозначенных надписями. Сайт начала трансляции белка atg выделен красным цветом. Вертикальными красными стрелками и зелеными цифрами отмечены начальные точки нуклеотидных последовательностей 5'-делеционных вариантов промотора.
Фиг. 2. Генетические конструкции на основе растительного экспрессионного вектора pCAMBIA1381Z, в которых содержащий интрон репортерный ген gus находится под контролем 5'-делеционных вариантов -303 и -118 п.н. версии промотора pro-SmAMP-D1-1 и 5'-делеционных вариантов -304 и -118 п.н. версии промотора pro-SmAMP-D1-2. Растительный экспрессионный вектор pMOG35SintGUS с содержащим интрон геном gus под контролем вирусного промотора CaMV35S использован в качестве сравнительного контроля. Черным цветом показана кодирующая область гена gus; Int - модифицированный интрон каталазы клещевины внутри транслируемой области гена gus; PIV2 - модифицированный интрон гена ST-LS1 картофеля внутри транслируемой области гена gus. Промоторы изображены в виде стрелок с соответствующими подписями.
Фиг. 3. Генетические конструкции на основе растительного экспрессионного вектора pCAMBIA2300, в которых селективный ген nptII находится под контролем 5'-делеционных вариантов -303 и -118 п.н. версии промотора рго-SmAMP-D1-1 и 5'-делеционных вариантов -304 и -118 п.н. версии промотора pro-SmAMP-D1-2. Вектор pCAMBIA2300 с геном nptII под контролем дуплицированного промотора 2х CaMV35S использован в качестве сравнительного контроля. Кодирующая область гена nptII показана синим цветом. Промоторы изображены в виде стрелок с соответствующими подписями.
Фиг. 4. Уровни активности репортерного белка GUS в листьях растений N. benthamiana при транзиентной экспрессии гена gus под контролем разных версий и 5'-делеционных вариантов промотора pro-SmAMP-D1 и промотора CaMV35S (контроль). Горизонтальными линиями отмечены делеционные варианты на основе промоторных версий pro-SmAMP-D1-1 и pro-SmAMP-D1-2. Цифрами ниже оси абсцисс показаны размеры 5'-делеционных вариантов в п.н. относительно +1 п.н. TSS. Планки погрешностей соответствуют стандартным ошибкам. * - значения, существенно отличающиеся от контроля при р≤0.05.
Фиг. 5. Гистохимическая локализация активности репортерного белка GUS в 2-х недельных трансгенных растениях A. thaliana поколения Т2, экспрессирующих ген gus под контролем разных версий и 5'-делеционных вариантов промотора pro-SmAMP-D1 и промотора CaMV35S (контроль). Горизонтальными линиями отмечены 5'-делеционные варианты на основе промоторных версий pro-SmAMP-D1-1 и pro-SmAMP-D1-2. Цифрами показаны размеры 5'-делеционных вариантов в п.н. относительно +1 п.н. TSS. Показана локализация активности GUS (синяя окраска тканей) в листьях, черешках, стеблях, корнях и семядолях трансгенных растений в сравнении с аналогичными органами интактных растений арабидопсиса экотипа Colambia-0.
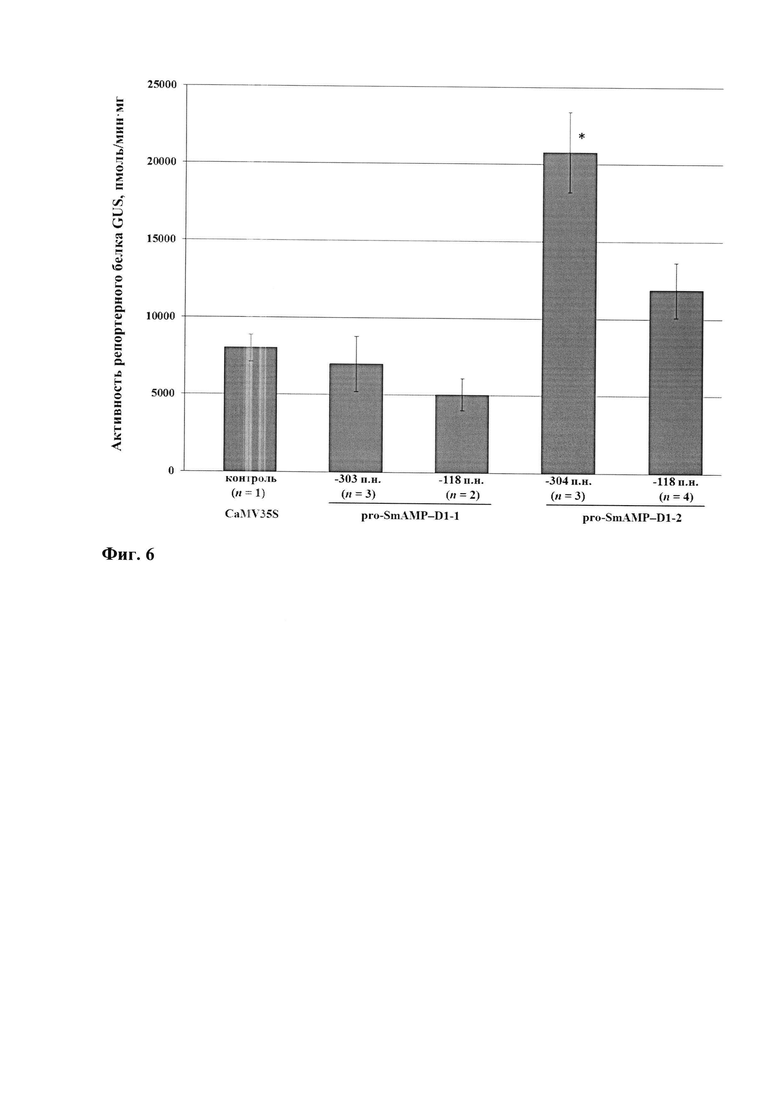
Фиг. 6. Уровни активности репортерного белка GUS в листьях 3-х недельных трансгенных растений A. thaliana поколения Т2, экспрессирующих ген gus под контролем разных версий и 5'-делеционных вариантов промотора pro-SmAMP-D1 и промотора CaMV35S (контроль). Горизонтальными линиями отмечены 5'-делеционные варианты на основе промоторных версий рго-SmAMP-D1-1 и pro-SmAMP-D1-2. Цифрами ниже оси абсцисс показаны размеры 5-делеционных вариантов в п.н. относительно +1 п.н. TSS. n-количество независимых линий трансгенных растений. Планки погрешностей соответствуют стандартным ошибкам. * - значения, существенно отличающиеся от контроля при р≤0.05.
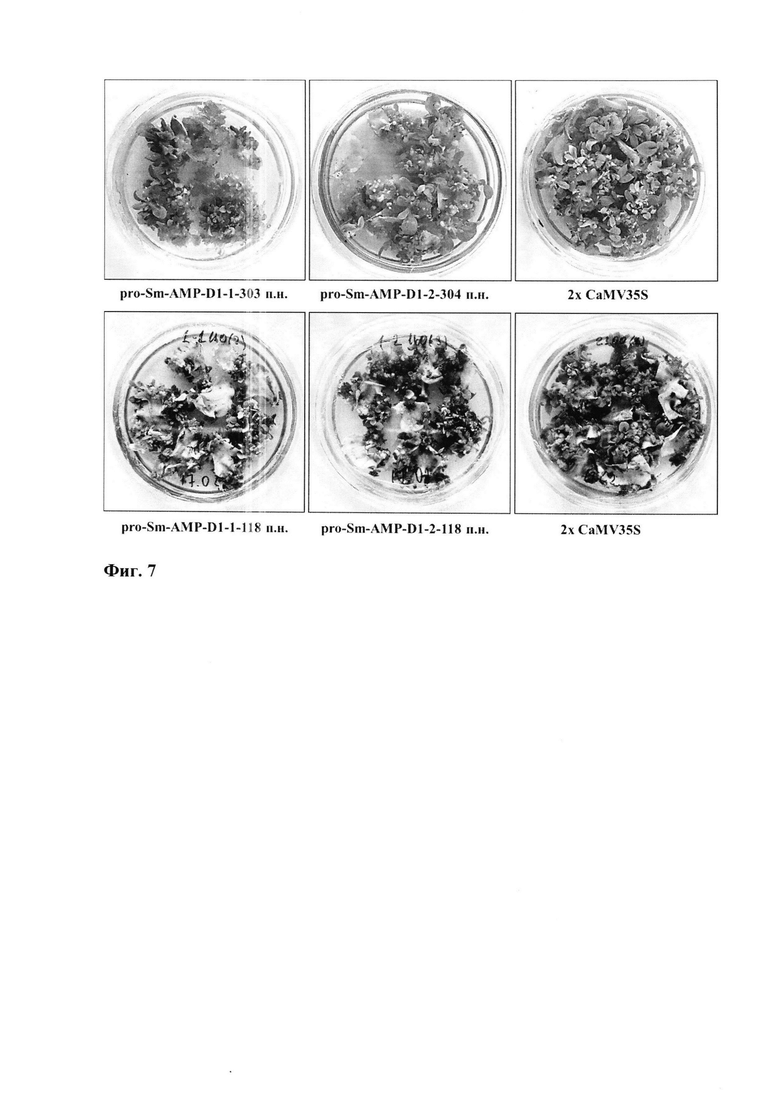
Фиг. 7. Морфогенез побегов из соматических клеток листовых эксплантов табака на питательной среде с антибиотиком канамицином в высокой концентрации 350 мг/л. Для управления экспрессией селективного гена nptII использовали разные версии и 5'-делеционные варианты промотора pro-SmAMP-D1 и промотор 2х CaMV35S (контроль). Цифрами показаны размеры 5'-делеционных вариантов промотора в п.н. относительно +1 п.н. TSS.
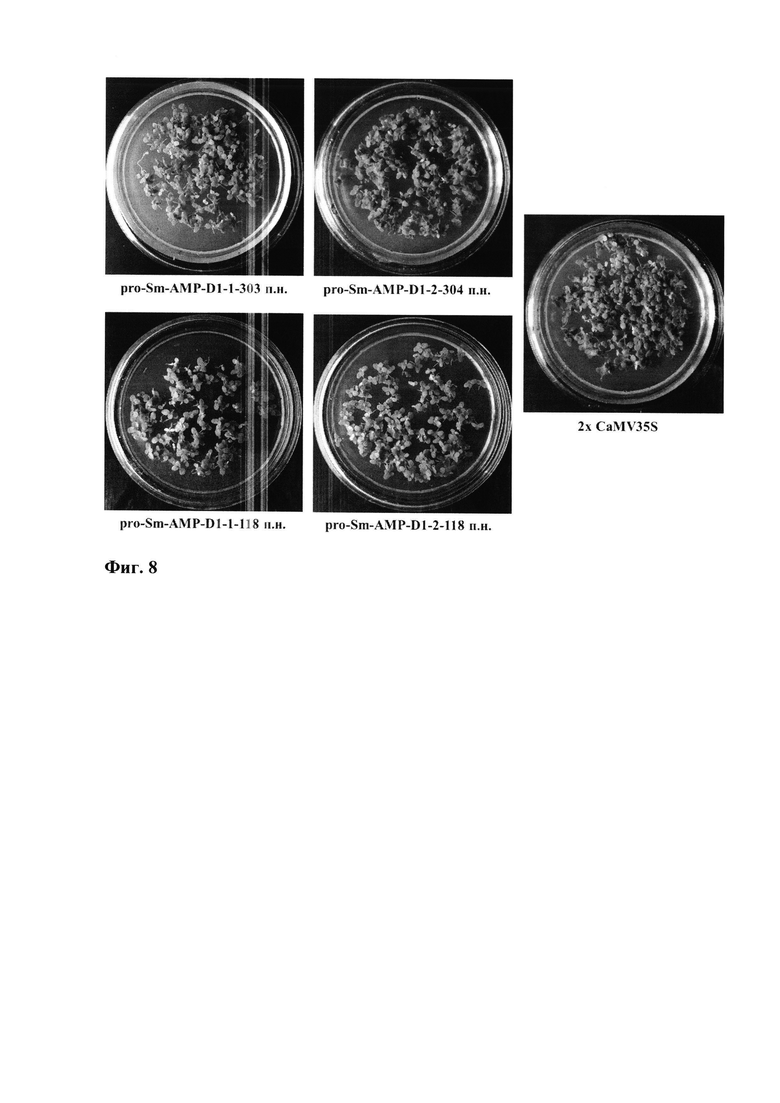
Фиг. 8. Сегрегация проростков табака поколения T1 на селективной питательной среде с антибиотиком канамицином. Зеленые растения устойчивы к канамицину. Чувствительные к канамицину растения - белые. Для контроля экспрессии селективного гена nptII использовали разные версии и 5'-делеционные варианты промотора pro-SmAMP-D1 и промотор 2х CaMV35S (контроль). Цифрами показаны размеры 5'-делеционных вариантов промотора в п.н. относительно +1 п.н. TSS.
Фиг. 9. Анализ с помощью ПЦР геномной ДНК растений табака поколения T1 на наличие ДНК-сегмента, состоящего из нуклеотидных последовательностей гена nptII и 5'-делеционных вариантов версии промотора рго-SmAMP-D1-1, версии промотора pro-SmAMP-D1-2 и промотора 2х CaMV35S. М - ДНК-маркер. «К+» - положительный контроль. «K-» - отрицательный контроль. Цифрами показаны индивидуальные образцы растений.
Фиг. 10. Гистохимическая локализация активности репортерного белка GUS в трансгенных растениях картофеля, экспрессирующих ген gus под контролем 5'-делеционного варианта -304 п.н. промотора pro-SmAMP-D1-2 и промотора CaMV35S. Показана локализация активности GUS в листьях, черешках, стеблях, цветках и клубнях трансгенных растений в сравнении с аналогичными органами интактных растений сорта Вектор (К). Слово «нет» свидетельствует об отсутствие цветков у растения.
Осуществление изобретения
Приведенное выше раскрытие описывает изобретение в общих чертах. Более полное понимание может быть достигнуто за счет представления соответствующих примеров. Представленные здесь примеры приводятся исключительно с целью иллюстрации и никоим образом не претендуют на то, чтобы сузить область данного изобретения в рамках этих примеров. Изменение формы или замещение эквивалентов допускается на основании его целесообразности в соответствующих условиях. Хотя здесь и будут применяться специфические термины, они используются исключительно с описательной целью.
Пример 1. Клонирование промотора гена дефензина SmAMP-D1 из генома растения звездчатка средняя (Stellaria media)
Геном растения звездчатки средней в настоящее время не секвенирован, поэтому не представляется возможным непосредственно амплифицировать нуклеотидную последовательность промотора гена дефензина SmAMP-D1 из интактной ДНК растения S. media. Однако ранее клонировали нуклеотидную последовательность гена SmAMP-D1, включающую 5'-нетранслируемую (5'-НТО) и кодирующую белок области (Slavokhotova et al. 2011). Поэтому в данной работе для клонирования промотора вместе с 5'-НТО гена SmAMP-D1 использовали метод, который ранее разработали для обнаружения локусов интеграции Т-ДНК в геноме растений арабидопсиса (Pogorelko and Fursova 2008). Для этого выделили геномную ДНК растений звездчатки и гидролизовали ее с использованием рестриктазы HinfI, для которой обнаружили сайт рестрикции в прилежащей кодирующей области гена SmAMP-D1. Затем в соответствии с методикой лигировали ДНК и использовали в качестве матрицы для последовательной амплификации с двумя парами ориентированных противоположно праймеров. Для этого разработали праймеры комплементарные 5'-НТО и прилежащей кодирующей области гена SmAMP-D1: «внешние» smDa1 (5'-aaatccagtgcattattttctgtgtcga-3') и smDa2 (5'-aggaaagtaacatggatacccaacaca-3') и «внутренние» smDb3 (5'-aacttacatattcctcttaggctaat-3') и smDb4 (5'-tcttttttgtttagtttaagtcgatca-3'). В результате последовательных раундов амплификации, сначала с «внешними», а потом с «внутренними» праймерами получили единственный ампликон размером около 750 п.н. При его секвенировании установили, что между последовательностями для отжига «внутренних» праймеров находятся известные нуклеотидные последовательности гена SmAMP-D1, между которыми располагается предполагаемая промоторная область. С целью проверки существования данной области в интактном геноме звездчатки средней, а также для клонирования промотора, разработали два новых праймера. Первый праймер smD4.3(E) (5'-gaattcaactataacattttgtattctttt-3') комплементарен 5'-концу предполагаемой нуклеотидной последовательности промотора и содержит сайт рестрикции EcoRI. Второй - smD11(N) (5'-ccatggtttcttttttgtttagtttaagtc-3') комплементарен 5-НТО вплоть до инициирующего кодона atg гена SmAMP-D1 и содержит сайт рестрикции NcoI В результате амплификации интактной ДНК звездчатки с использованием праймеров smD4.3(E) и smD11(N) получили ампликон размером около 350 п.н. При его секвенировании в составе Т-вектора установили, что он состоит из известной 5-НТО гена SmAMP-D1 размером 42 п.н. и примыкающей к ней проксимальной и «коровой» промоторных областей гена SmAMP-D1 размером около 300 п.н. С помощью сравнения десяти клонов установили, что в геноме звездчатки средней присутствуют как минимум две идентичные на 93% версии промотора гена SmAMP-D1, обозначенные как pro-SmAMP-D1-1 (размером -303 п.н. относительно +1 п.н. TSS) и pro-SmAMP-D1-2 (размером -304 п.н. относительно +1 п.н. TSS), различающиеся полиморфизмом отдельных нуклеотидов (фиг. 1).
С использование программы Blast не обнаружили в GenBank (https://www.ncbi.nlm.nih.gov/) других нуклеотидных последовательностей, идентичных последовательностям нового промотора pro-SmAMP-D1.
Анализ in silico нуклеотидных последовательностей обеих версий промотора рrо-SmAMP-D1-1 и pro-SmAMP-D1-2 выполнили с помощью программы «New place A Database of Plant Cis-acting Regulatory DNA Elements» (https://www.dna.affrc.go.jp/PLACE/?action=newplace). Установили, что обе версии промотора содержат консервативный цис-элемент ТАТА-box в ожидаемом положении -34 п.н. влево относительно +1 п.н. TSS. В области минимального промотора в положении -107 п.н. относительно +1 п.н. TSS обнаружили консервативный цис-элемент СААТ-box (фиг. 1). Кроме этого, идентифицировали и другие потенциальные цис-элементы в промоторе pro-SmAMP-D1, способные обеспечить ткане-специфичную или индуцибельную экспрессию гена SmAMP-D1 или выступить в качестве энхансеров или супрессоров его экспрессии. Они включали цис-элементы реагирующие на свет (GATABOX, GT1CONSENSUS) и на фитогормоны (PYRIMIDINEBOXHVEPB1), необходимые для ткане- и органспецифической экспрессии (CACTFTPPCA1, WRKY710S, RAV1AAT), для ответа на абиотический стресс (ССААТВОХ1, CBFHV и MYCCONSENSUSAT), для метаболизма углеводов (DOFCOREZM, WBOXHVISO1), для ответа на атаку патогенов (BIHD10S, SEBFCONSSTPR10A) и др.
Пример 2. Создание 5'-делеционных вариантов промотора pro-SmAMP-D1 и генетических конструкций для агробактериальной трансформации растений
На основе проведенного анализа нуклеотидных последовательностей версий pro-SmAMP-D1-1 и pro-SmAMP-D1-2 подобрали праймеры для создания 5'-делеционных вариантов с целью последующей оценки промоторной эффективности при управлении экспрессией рекомбинантных генов. Создали один новый «прямой» праймер SmD12(E) (5'-gaattcgcatgtcacaatt-3'), содержащий сайт рестрикции EcoRI, ограничивающий области обеих версий промотора до -118 п.н. относительно +1 п.н. TSS (фиг. 1). В качестве «обратного» использовали ранее созданный праймер smD11(N).
В качестве матриц для амплификации новых 5'-делеционных вариантов обеих версий рго-SmAMP-D1-1 и pro-SmAMP-D1-2 размером -118 п.н. использовали плазмидные ДНК с их более длинными 5'-делеционными вариантами.
С применением новых праймеров амплифицировали и клонировали по сайтам рестрикции EcoRI и NcoI в растительный экспрессионный (бинарный) вектор pCAMBIA1381Z (Cambia, Australia) два новых коротких 5'-делеционных варианта промоторных версий, обозначенные, как pro-SmAMP-D1-1-118 и pro-SmAMP-D1-2-118, а также созданные ранее их более длинные 5'-делеционные варианты pro-SmAMP-D1-1-303 и pro-SmAMP-D1-2-304. Схема полученных генетических конструкций, обозначенных, соответственно, pC1381Z-pro-SmAMP-D1-1-303, pC1381Z-pro-SmAMP-D1-1-118, pC1381Z-pro-SmAMP-D1-2-304 и pC1381Z-pro-SmAMP-D1-2-118 отображена на фиг. 2.
Созданные генетические конструкции предназначены для экспрессии в растениях целевого гена, находящегося под контролем промотора pro-SmAMP-D1. В качестве целевого гена использовали содержащий интрон репортерный ген gus (gusA или uidA), позволяющий качественно и количественно оценить промоторную эффективность по активности его белкового продукта - фермента β-глюкуронидазы или GUS. Растительный экспрессионный вектор pMOG35SintGUS, содержащий ген gus под контролем известного сильного и конститутивного вирусного промотора CaMV35S применили в качестве сравнительного контроля. Перечисленные выше генетические конструкции использовали для создания агробактериальных штаммов на основе клеток GV3101 Agrobacterium tumefaciens.
При создании трансгенных растений для оптимизации селекции трансформированных клеток и проростков на питательной среде с относительно дешевым антибиотиком канамицином использовали растительный (бинарный) вектор pCAMBIA2301 (Cambia, Australia). Для этого по сайтам рестрикции Eco91I и EcoRI перенесли в вектор pCAMBIA2301 фрагменты из генетических конструкций pC1381Z-pro-SmAMP-D1-1-303, pC1381Z-pro-SmAMP-D1-1-118, pC1381Z-pro-SmAMP-D1-2-304 и pC1381Z-pro-SmAMP-D1-2-118, содержащие нуклеотидные последовательности соответствующих 5'-делеционных вариантов промотора и гена gus, и предварительно вырезанные по сайтам Eco91I и EcoRI. В результате создали генетические конструкции pC2301-pro-SmAMP-D1-1-303, pC2301-pro-SmAMP-D1-1-118, pC2301-pro-SmAMP-D1-2-304 и pC2301-pro-SmAMP-D1-2-118.
Для оценки возможности использования промотора pro-SmAMP-D1 для контроля экспрессии селективного гена неомицинфосфотрансферазы II (nptII), придающего клеткам растений устойчивость к антибиотику канамицину, создали генетические конструкции на основе бинарного вектора pCAMBIA2300 (Cambia, Australia). Для этого клонировали по сайтам рестрикции EcoRI и NcoI в вектор pCAMBIA2300 нуклеотидные последовательности 5'-делеционных вариантов -303 и -118 п.н. версии промотора рго-SmAMP-D1-1 и 5-делеционных вариантов -304 и -118 п.н. версии промотора pro-SmAMP-D1-2. Созданные генетические конструкции, обозначенные как pC2300-pro-SmAMP-D1-1-303, pC2300-pro-SmAMP-D1-1-118, pC2300-pro-SmAMP-D1-2-304 и pC2300-pro-SmAMP-D1-2-118, содержат ген nptII под контролем различных 5'-делеционных вариантов промотора pro-SmAMP-D1 (фиг. 3). В качестве сравнительного контроля использовали интактный вектор pCAMBIA2300 с геном nptII под контролем дуплицированного вирусного промотора 2х CaMV35S.
Генетические конструкции на основе плазмид pCAMBIA2300 и pCAMBIA2301 использовали для получения агробактериальных штаммов с применением клеток AGLO A. tumefaciens.
Пример 3. Транзиентная экспрессия репортеного гена gus под контролем различных версий и 5'-делеционных вариантов промотора pro-SmAMP-D1 в агроинфильтрированных растениях Nicotiana benthamiana
Инфильтрацию листьев растений N. benthamiana проводили пятью вариантами агробактерий штамма GV3101 с плазмидными генетическими конструкциями pC1381Z-pro-SmAMP-D1-1-303, pC1381Z-pro-SmAMP-D1-1-118, pC1381Z-pro-SmAMP-D1-2-304 и pC1381Z-pro-SmAMP-D1-2-118 и pMOG35SintGUS в качестве сравнительного контроля. Первоначально штаммы агробактерий культивировали на агаризованной питательной среде LB с антибиотиками канамицином (100 мг/л), рифампицином (100 мг/л) и гентамицином (25 мг/л). Затем, для выполнения агроинфильтрации листьев растений штаммы выращивали в жидкой питательной среде LB при тех же концентрациях перечисленных выше антибиотиков в течение суток при температуре 28°С и перемешивании (160 об/мин). Для подавления РНК-интерференции в клетках растений использовали штамм A. tumifaciens GV2260/C58C1 pLH7000 р19, который выращивали в жидкой питательной среде LB с антибиотиками рифампицином (100 мг/л) и стрептомицином (50 мг/л) в течении суток при температуре 28°С и перемешивании (160 об/мин).
Суспензии клеток агробактерий разбавляли до оптической плотности 0.6 при длине волны 600 нм. Затем штаммы GV3101 pMOG35SintGUS, GV3101 pC1381Z-pro-SmAMP-D1-1-303, GV3101 pC1381Z-pro-SmAMP-D1-1-118, GV3101 pC1381Z-pro-SmAMP-D1-2-304 и GV3101 pC1381Z-pro-SmAMP-D1-2-118 смешивали в соотношение 1:1 с суспензией агробактерий GV2260/C58C1 pLH7000 р19. Готовую смесь агробактерий вводили в листья растений N. benthamiana шприцом без иглы с внутренней стороны. После инфильтрации растения содержали при температуре 21°С и 16/8 часовом искусственном освещении. На 7 день из места инокуляции брали высечки массой около 10 мг и замораживали при -70°С.
Затем из каждой высечки выделяли белковые экстракты в которых измеряли активность рекомбинантного фермента β-глюкуронидазы с использованием субстрата 4-methylumbelliferyl-β-D-glucuronide Trihydrate (MUG, PhytoTechnology Laboratories) и концентрацию водорастворимых белков по методу Бредфорд; на основе полученных результатов рассчитывали активность репортерного белка GUS в пмоль 4MU/мг⋅мин или пмоль/мг мин. Для каждой генетической конструкции проанализировали от 60 до 100 высечек, полученных за три повторения эксперимента. Средний уровень активности GUS при использовании каждого делеционного варианта промотора pro-SmAMP-D1 и вирусного промотора CaMV35S представлен на фиг. 4.
Как следует из фиг. 4, новый промотор pro-SmAMP-D1 функционален для транзиентной экспрессии репортерного гена gus и продукции его белкового продукта GUS в клетках гетерологичных растений. Средний уровень активности GUS в листьях N. benthamiana, агроинфильтрированных генетическими конструкциями pCambia1381Z, несущими различные 5'-делеционные варианты промоторов рго-SmAMP-D1-1 и pro-SmAMP-D1-2, варьировал от 29914 до 98338 пмоль/мин⋅мг, что как ниже, так и выше активности, стимулируемой промотором CaMV35S (53161 пмоль/мин⋅мг). В частности, длинный 5'-делеционный вариант -304 п.н. версии промотора pro-SmAMP-D1-2 был в 1,9 раз достоверно сильнее, чем вирусный промотор CaMV35S. Короткий 5'-делеционный вариант -118 п.н. версии pro-SmAMP-D1-2 и длинный 5'-делеционный вариант -303 п.н. версии pro-SmAMP-D1-1 были сравнимы по эффективности с промотором CaMV35S. Короткий 5'-делеционный вариант -118 п.н. версии рго-SmAMP-D1-1 был в 1,7 раз достоверно слабее, чем CaMV35S. В целом, короткие 5'-делеционные варианты pro-SmAMP-D1 ожидаемо, менее эффективны, чем их более длинные 5'-делеционные варианты.
Пример 4. Трансгенные растения Arabidopsis thaliana, экспрессирующие репортерный ген gus под контролем различных версий и 5'-делеционных вариантов промотора pro-SmAMP-D1
С использованием конструкций pC2301-pro-SmAMP-D1-1-303, рС2301-pro-SmAMP-D1-1-118, pC2301-pro-SmAMP-D1-2-304 и pC2301-pro-SmAMP-D1-2-118 выполнили генетическую трансформацию растений арабидопсиса (A. thaliana) экотипа Colambia-0 методом погружения соцветий в суспензию агробактерий. При использовании всех генетических конструкций на селективной питательной среде с антибиотиком канамицином в концентрации 50 мг/л отобрали устойчивые растения поколения Т1, продуцирующие репортерный белок GUS. В результате самоопыления трансгенных растений получили семена поколения Т2. С помощью их селекции на питательной среде с антибиотиком канамицином отобрали популяции растений с моногенным наследованием Т-ДНК, из которых выбрали индивидуальные гомозиготные растения (по уровню активности GUS и отсутствию сегрегации на селективном агенте проростков поколения Т3), которые использовали для исследования (фиг. 5).
Установили, что все версии и 5'-делеционные варианты промотора pro-SmAMP-D1 способны управлять экспрессией гена gus в разных тканях и органах трансгенных растений A. thaliana (фиг. 5). Активность репортера GUS, визуализированная с использованием субстрата 5-bromo-4-chloro-3-indolyl-β-D-glucuronic acid sodium salt (X-Gluc, PhytoTechnology Laboratories) была представлена в корнях, стеблях, семядолях и листьях, хотя интенсивность окрашивания тканей визуально различалась в зависимости от использованного варианта генетической конструкции.
Для ясной дифференциации уровней экспрессии gus среди версий и 5'-делеционных вариантов промотора pro-SmAMP-D1 выполнили количественное определение активности GUS в листьях 3-х недельных трансгенных растений (фиг. 6). Средний уровень активности GUS в листьях трансгенных растений A. thaliana варьировал от 5054 до 20770 пмоль/мин⋅мг, что как ниже, так и выше активности, стимулируемой промотором CaMV35S (около 8050 пмоль/мин⋅мг). В частности, длинный 5'-делеционный вариант -304 п.н. версии pro-SmAMP-D1-2 в 2,1 раза достоверно сильнее, чем вирусный промотор CaMV35S. Короткий 5'-делеционный вариант -118 п.н. версии pro-SmAMP-D1-2, длинный 5'-делеционный вариант -303 п.н. и короткий 5'-делеционный вариант -118 п.н. версии pro-SmAMP-D1-1 сравнимы по эффективности с промотором CaMV35S. Картина распределения уровней активности GUS в зависимости от использованной промоторной версии и 5'-делеционного варианта в трансгенных растениях арабидопсиса (фиг. 6) в целом соответствовала распределению активностей при транзиентной экспрессии гена gus в растениях N. benthamiana (фиг. 4).
Пример 5. Использование селективного гена nptll под контролем различных версий и 5 -делеционных вариантов промотора pro-SmAMP-D1 для селекции трансгенных клеток и растений табака
Методом агробактериальной трансформации листовых эксплантов (дисков) растений табака (Nicotiana tabacum) оценили эффективность селекции трансгенных клеток, каллусов и побегов на питательной среде с антибиотиком канамицином. Для этого выполнили агробактериальную трансформацию растений табака с помощью генетических конструкций pC2300-pro-SmAMP-D1-1-303, pC2300-pro-SmAMP-D1-1-118, pC2300-pro-SmAMP-D1-2-304, pC2300-pro-SmAMP-D1-2-118 и pC2300 "газонным" методом, использованным нами ранее (Komakhin et al. 2010; Madzharova et al. 2018). Установили, что все версии и 5'-делеционные варианты промотора pro-SmAMP-D1 достаточны для конститутивного уровня экспрессии селективного гена nptII, поскольку позволяют селекцию трансформированных клеток, каллусов и побегов табака на питательной среде с канамицином (фиг. 7).
В частности, на питательной среде с канамицином в рекомендованной концентрации 100 мг/л (Дрейпер и др. 1991) за три месяца селекции обе версии и все 5'-делеционные варианты промотора pro-SmAMP-Dl и промотор 2х CaMV35S позволили сформировать устойчивые к канамицину каллусы и побеги у всех листовых эксплантов табака. На питательной среде с канамицином в избыточной концентрации 350 мг/л за четыре месяца селекции обе версии и все 5'-делеционные варианты промотора pro-SmAMP-D1 позволили сформировать устойчивые к канамицину каллусы и побеги у 35-55% листовых эксплантов табака (табл. 1, столбец «В/А × 100%»).


В этих же условиях при использовании промотора 2х CaMV35S морфогенез наблюдали у 95-100% эксплантов (табл. 1, столбец «В/А × 100%»).
Регенерацию побегов табака, устойчивых к антибиотику канамицину, оценивали в течение трех месяцев культивирования на селективной питательной среде с рекомендованной 100 мг/л и избыточной 350 мг/л концентрацией. Сформированные побеги отделяли от каллуса и селектировали на питательной среде с канамицином в концентрации 100 мг/л в течение 1,5 месяца.
Укореняющиеся побеги из расчета 1,5-1,8 побега на один эксплант взятый изначально для трансформации получили при использовании обеих версий и всех 5'-делеционных вариантов промотора pro-SmAMP-D1 и промотора 2х CaMV35S на питательной среде с канамицином в рекомендованной концентрации 100 мг/л. При этих условиях селекции побегов достоверных различий между вариантами генетических конструкций не обнаружили.
Укореняющиеся побеги с эффективностью 0,3-0,6 побега на один эксплант получили при использовании обеих версий и всех 5'-делеционных вариантов промотора pro-SmAMP-D1 на питательной среде с канамицином в избыточной концентрации 350 мг/л (табл. 1, столбец «D/А, шт.»). При использовании промотора 2х CaMV35S эффективность образования побегов составила 1,2-1,3 побега на один эксплант. Более высокая эффективность промотора 2х CaMV35S в условиях избытка канамицина была ожидаема, поскольку в интактной плазмиде рС2300 он содержит две промоторные копии CaMV35S, которые заведомо определяют его более высокую эффективность для экспрессии nptII (фиг. 3). Эти результаты призваны дополнительно продемонстрировать способность промотора pro-SmAMP-D1 для эффективной работы в условиях кратного избытка селективного агента. Ранее в растениях табака только промотор pro-SmAMP2 из звездчатки средней был сопоставимым по эффективности с промотором 2х CaMV35S для продукции побегов на питательной среде с канамицином в концентрации 350 мг/л (Madzharova et al. 2018). В этих же условиях промотор pro-SmAMP1 из звездчатки средней оказался в три раза менее эффективным.
С использованием метода ПЦР и ранее созданных праймеров virE2F и virE2R (Komakhin et al. 2010) к последовательности гена VirE2 агробактерий установили, что укоренившиеся на питательной среде с канамицином растения табака не имели агробактериальной контаминации. Для проверки присутствия гена nptII под контролем разных промоторов в укоренившихся побегах использовали метод ПЦР и первоначально созданные праймеры к последовательностям длинных и коротких 5'-делеционных вариантов промотора pro-SmAMP-D1, соответственно, smD4.3(E) или SmD12(E). Для анализа растений, созданных с использованием промотора 2х CaMV35S использовали разработанный ранее праймер 35Splus (Madzharova et al. 2018). В качестве «обратного» во всех анализах использовали первоначально созданный праймер olgminus, который комплементарен участку в последовательности гена nptII (Madzharova et al. 2018). С использованием образцов ДНК из десяти укоренившихся растений каждого варианта генетических конструкций установили, что они все содержали нуклеотидные последовательности, соответствующие гибридной области, состоящей из последовательности соответствующего 5'-делеционного варианта промотора и гена nptII. В частности, для длинных 5'-делеционных вариантов промотора pro-SmAMP-D1 и фрагмента гена nptII были получены ампликоны ожидаемого размера около 400 п.н., для коротких 5'-делеционных вариантов промотора pro-SmAMP-D1 и фрагмента гена nptII - около 300 п.н. При использовании образцов ДНК из трансгенных растений с дуплицированным промотором 2х CaMV35S ожидаемо получили два ампликона около 500 и 800 п.н., соответственно, для каждой из копий CaMV35S и фрагмента гена nptII. Таким образом, создали первичные трансформанты табака, которые обозначили как T02300-pro-SmAMP-D1-1-303, T02300-pro-SmAMP-D1-1-118, T02300-pro-SmAMP-D1-2-304, T02300-pro-SmAMP-D1-2-118 и T02300.
В следующем поколении Т1 проростки табака, несущие nptll под контролем обеих версий и всех 5'-делеционных вариантов промотора pro-SmAMP-D1, сегрегировали в зависимости от резистентности к канамицину на устойчивые и чувствительные растения, в некоторых популяциях в соотношении 3:1 (устойчивые:чувствительные), предполагающем моногенное наследование трансгена (фиг. 8). Все растения табака, устойчивые к канамицину (от 8 до 14 шт. для каждой конструкции) были свободны от агробактериальной контаминации и в соответствии с результатами ПЦР все содержали ожидаемого размера гибридные нуклеотидные последовательности между промоторными вариантами и геном nptII (фиг. 9).
Таким образом, новый промотор pro-SmAMP-D1 является конститутивным и заведомо достаточным для селекции трансформированных клеток, каллусов, побегов и проростков на среде с рекомендованной для создания трансгенных растений табака концентрацией антибиотика канамицина от 50 до 100 мг/л (Дрейпер и др. 1991). Промотор pro-SmAMP-D1-2 не уступает в эффективности конститутивному вирусному промотору CaMV35S при селекции на питательной среде с канамицином в рекомендованной концентрации 100 мг/л. С экологической точки зрения и с позиции современной науки в области биологии растительной клетки растительный промотор pro-SmAMP-D1-2 предпочтительнее для генетической модификации растений, чем вирусный промотор CaMV35S.
Пример 6. Трансгенные растения картофеля, экспрессирующие репортерный ген gus под контролем промотора pro-SmAMP-D1
Для проверки эффективности промотора pro-SmAMP-D1 для генетической модификации сельскохозяйственных растений, выполнили агробактериальную трансформацию растений картофеля (Solanum tuberosum) сорта Вектор (оригинатор сорта Федеральное государственное бюджетное научное учреждение «Всероссийский научно-исследовательский институт картофельного хозяйства имени А.Г. Лорха»). Генетическую трансформацию выполнили в соответствие с методикой опубликованной нами ранее (Vetchinkina et al. 2016). В результате исследований создали девять независимых трансформантов картофеля с использованием каждой генетической конструкции pC2301-pro-SmAMP-D1-2-304 и pMOG35SintGUS, которые были устойчивы к антибиотику канамицину в концентрации 75 мг/л и продуцировали репортерный белок GUS.
Установили, что 5'-делеционный вариант -304 п.н. версии промотора pro-SmAMP-D1-2, наиболее перспективный для биотехнологии растений, способен управлять экспрессией гена gus в разных тканях и органах трансгенных растений картофеля (фиг. 10). Активность репортера GUS, визуализированная окрашиванием тканей с использованием субстрата X-Gluc, была представлена в стеблях, черешках, листьях, клубнях и цветках трансгенных растений на высоком уровне при использовании обоих промоторов pro-SmAMP-D1-2 и CaMV35S. Заметные различия между паттернами промоторов наблюдали только при окрашивании листьев. В частности, промотор pro-SmAMP-D1-2 способствовал более интенсивной окраске проводящих тканей листа, в то время как промотор CaMV35S способствовал более равномерной окраске паренхимы листа.
Таким образом, новый промотор pro-SmAMP-D1-2 может быть использован для биотехнологии растений картофеля и по эффективности экспрессии рекомбинантного гена в разных органах растений не уступает сильному вирусному промотору CaMV35S. С экологической точки зрения и с позиции современной науки в области биологии растительной клетки растительный промотор pro-SmAMP-D1-2 предпочтительнее для генетической модификации растений, чем вирусный промотор CaMV35S.
Заключение
Промотор pro-SmAMP-D1 может быть рекомендован для биотехнологии в качестве регуляторного элемента, эффективно управляющего экспрессией рекомбинантных генов в клетках генетически модифицированных растений. Промотор pro-SmAMP-D1 может быть рекомендован для управления целевыми или селективными генами в составе кассет экспрессии генетических конструкций. Применение разных версий и 5'-делеционных вариантов промотора pro-SmAMP-D1 может быть использовано для оптимизации экспрессии рекомбинантных генов на разных уровнях, в том числе превышающем уровень экспрессии при использовании сильного вирусного промотора CaMV35S. Преимуществом промотора pro-SmAMP-D1 является то, что он имеет растительное происхождение и представлен нуклеотидной последовательностью, отличающейся от нуклеотидных последовательностей других известных растительных промоторов, и поэтому может быть использован в составе одной генетической конструкции с другими промоторами генов растений.
Список литературы
Acharya S, Ranjan R, Pattanaik S et al. (2014) Efficient chimeric plant promoters derived from plant infecting viral promoter sequences. Planta 239:381-396. https://doi.org/10.1007/s00425-013-1973-2
Ali S, Kim W-C (2019) A Fruitful Decade Using Synthetic Promoters in the Improvement of Transgenic Plants. Frontiers in Plant Science https://doi.org/10.3389/fpls.2019.01433
Al-Kaff NS, Kreike MM, Covey SN et al. (2000) Plants rendered herbicide-susceptible by cauliflower mosaic virus-elicited suppression of a 35S promoter-regulated transgene. Nature Biotechnol. 18:995-999. https://doi.org/10.1038/79501
Cazzonelli CI, McCallum EJ, Lee R, Botella JR. (2005) Characterization of a strong, constitutive mung bean (Vigna radiata L.) promoter with a complex mode of regulation in planta. Transgenic Res. 14(6):941-67. doi: 10.1007/s11248-005-2539-2
Coussens G, Aesaert S, Verelst W, Demeulenaere M et al. Brachypodium distachyon promoters as efficient building blocks for transgenic research in maize, J. Exp.Bot., 2012, vol. 63, pp. 4263-4273.
Dutt M, Dhekney S, Soriano L, Kandel R, Grosser JW (2014) Temporal and spatial control of gene expression in horticultural crops. Hortic Res. 1:14047. https://doi.org/10.1038/hortres.2014.47
Callis J, Raasch JA, Vierstra RD (1990) Ubiquitin extension proteins of Arabidopsis thaliana. Structure, localization, and expression of their promoters in transgenic tobacco. J Biol Chem. 265:12486-12493.
Han Y, Kim Y, Hwang О et al. (2015) Characterization of a small constitutive promoter from Arabidopsis translationally controlled tumor protein (AtTCTP) gene for plant transformation. Plant Cell Rep 34:265-275. https://doi.org/10.1007/s00299-014-1705-5
Hernandez-Garcia CM, Finer JJ (2014) Identification and validation of promoters and cis-acting regulatory elements. Plant Science 217:109-119. https://doi.org/10.1016/j.plantsci.2013.12.007
Holme IB, Wendt T, Holm PB (2013) Intragenesis and cisgenesis as alternatives to transgenic crop development. J Plant. Biotechnol. 11:395-407. https://doi.org/10.1111/pbi.12055
Komakhin RA, Komakhina VV, Milyukova NA et al. (2010) Transgenic tomato plants expressing recA and NLS-recA-licBM3 genes as a model for studying meiotic recombination. Russ. J. Genet. 46:1440-1448. https://doi.org/10.1134/S1022795410120069
Komakhin RA, Vysotskii DA, Shukurov RR et al. (2016) Novel strong promoter of antimicrobial peptides gene pro-SmAMP2 from chickweed (Stellaria media). BMC Biotechnol. 16(1):43. doi: 10.1186/s12896-016-0273-x.
KuluevBR, Knyazev AV, Chemeris, AV and Vakhitov VA. Morphological features of transgenic tobacco plants expressing the AINTEGUMENTA gene of rape under control of the dahlia mosaic virus promoter. Russ. J. Dev. Biol., 2013, vol. 44, pp. 86-89.
Lessard PA et al. (2002) Manipulating gene expression for the metabolic engineering of plants. Metabolic Engineering. 4(1):67-79.
Madzharova NV, Kazakova KA, Strelnikova SR et al. (2018) Promoters pro-SmAMPl and pro-SmAMP2 from Wild Plant Stellaria media for the Biotechnology of Dicotyledons. Russ J Plant Physiol 65:750-761. https://doi.org/10.1134/S1021443718040040
Mette MF et al. (1999) Production of aberrant promoter transcripts contributes to methylation and silencing of unlinked homologous promoters in trans // The EMBO journal. 18(1):241-248.
Mourrain P et al. (2007) A single transgene locus triggers both transcriptional and post-transcriptional silencing through double-stranded RNA production // Planta. 225(2):365-379.
Nuccio ML (2017) A Brief history of promoter development for use in transgenic maize applications. Methods Mol Biol. 1676:61-93. https://doi.org/10.1007/978-1-4939-7315-6_4.
Odell JT, Nagy F, Chua NH (1985) Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature. 313(6005):810-812.
Prakash NS, Prasad V, Chidambram TP et al. Effect of promoter driving selectable marker on corn transformation, Transgenic Res., 2008, vol. 17, pp. 695-704.
Peremarti A et al. (2010) Promoter diversity in multigene transformation // Plant molecular biology. 73(4-5):363-378.
Pogorelko GV and Fursova OV (2008) A highly efficient miPCR method for isolating FSTs from transgenic Arabidopsis thaliana plants. Journal of Genetics. 87(2):133-140.
Potenza C, Aleman L, Sengupta-Gopalan С (2004) Targeting transgene expression in research, agricultural, and environmental pplications: Promoters used in plant transformation. In Vitro CellDevBiol-Plant. 40(1):1-22.
Slavokhotova AA, Odintsova TI, Rogozhin EA et al. (2011) Isolation, molecular cloning and antimicrobial activity of novel defensins from common chickweed (Stellaria media L.) seeds. Biochimie 93 (2011) 450e456 doi:10.1016/j.biochi.2010.10.019.
Verdaguer B, de Kochko A, Beachy RN, and Fauquet C. Isolation and expression in transgenic tobacco and rice plants, of the cassava vein mosaic virus (CVMV) promoter. Plant Mol. Biol, 1996, vol. 31, pp. 1129-1139.
Vetchinkina EM, Komakhina VV, Vysotskii DA et al. (2016) Expression of plant antimicrobial peptide pro-SmAMP2 gene increases resistance of transgenic potato plants to Alternaria and Fusarium pathogens. Russ. J. Genet. 52(9): 1055-68.
Vysotskii DA, Strelnikova SR, Efremova LN et al. (2016) Structural and functional analysis of new plant promoter pro-SmAMPl from Stellaria media. Russ J Plant Physiol 63:663-672. https://doi.org/10.1134/S1021443716050174.
Xiao K, Zhang C, Harrison M, Wang Z-Y (2005) Isolation and characterization of a novel plant promoter that directs strong constitutive expression of transgenes in plants. Mol Breeding. 15(2):221-31. https://doi.org/10.1007/sll032-004-5679-9.
Zhu С et al. (2007) Transgenic strategies for the nutritional enhancement of plants. Trends in plant science. 12(12):548-555.
Дрейпер Д, Скотт P, Армитидж Ф и др., Генная инженерия растений. Лабораторное руководство. Учебное издание. Москва: Мир. 1991. 408 с.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<210> SEC ID NO 1
<211> Длина: 303
<212> Тип: ДНК
<213> Организм: Stellaria media
<400> Последовательность: 1-1-303
aactataaca ttttgtattc tttttttttg ctaccctatt tataaatttc ttgatcctcc 60
cctggtttga ttatcacccg atgttgctaa attttaattc ttaaaaaata atagtaaaac 120
gatagttttt tccaaaaaac gaaaatcgac gtccctgcta tccccttcct cttgttgtcg 180
cccgtgcatg tcacaattat tttatttgct cttatcatga caacacttgc gtcgccttac 240
atgaatattc ttcgaagagg cctataaaac accccattgt cactctaaaa tgatcaaaat 300
ctt 303
<210> SEC ID NO 2
<211> Длина: 118
<212> Тип: ДНК
<213> Организм: Stellaria media
<400> Последовательность: 1-1-118
gcatgtcaca attattttat ttgctcttat catgacaaca cttgcgtcgc cttacatgaa 60
tattcttcga agaggcctat aaaacacccc attgtcactc taaaatgatc aaaatctt 118
<210> SEC ID NO 3
<211> Длина: 304
<212> Тип: ДНК
<213> Организм: Stellaria media
<400> Последовательность: 1-2-304
aactataaca ttttgtattc ttttttttgc taccctattt atgaatttct ggatcctcac 60
ctgatttgat taccacccga tgtcgctaca ttttaattct taaaatataa tattaaaatg 120
atagtttttc ccaaaaaaaa gaaggatcga cgtccctgct atcccgttcc tcttgtcgtc 180
gcccgtgcat gtcacaattc tttcatttgc tcttatcatg acaacacttg tgtcgcctta 240
catgactatt cttcgaagag gcctataaaa taccccattg tcactctaaa atgatcaaaa 300
tctt 304
<210> SEC ID NO 4
<211> Длина: 118
<212> Тип: ДНК
<213> Организм: Stellaria media
<400> Последовательность: 1-2-118
gcatgtcaca attctttcat ttgctcttat catgacaaca cttgtgtcgc cttacatgac 60
tattcttcga agaggcctat aaaatacccc attgtcactc taaaatgatc aaaatctt 118
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Промотор pro-SmAMP-X из растения звездчатка белая (Stellaria media L.) для экспрессии рекомбинантных генов в клетках растений | 2020 |
|
RU2766095C1 |
| СИЛЬНЫЙ РАСТИТЕЛЬНЫЙ ПРОМОТОР PRO-SMAMP2 ИЗ СОРНОГО РАСТЕНИЯ STELLARIA MEDIA | 2014 |
|
RU2615456C1 |
| Способ эффективного биосинтеза рекомбинантных белков в двудольных растениях с использованием промотора гена pro-SmAMP1 из растения Stellaria media | 2017 |
|
RU2697014C2 |
| СПОСОБ СОЗДАНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ С ВЫСОКИМ УРОВНЕМ ЭКСПРЕССИИ ТРАНСГЕННОГО БЕЛКА | 2011 |
|
RU2507736C2 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324737C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324736C1 |
| ТОЛЕРАНТНОЕ К СТРЕССУ ТРАНСГЕННОЕ РАСТЕНИЕ ПШЕНИЦЫ | 2005 |
|
RU2376377C2 |
| СПОСОБ ИДЕНТИФИКАЦИИ ИЛИ СЕЛЕКЦИИ ЭУКАРИОТИЧЕСКИХ КЛЕТОК | 1994 |
|
RU2127759C1 |
| ЭКСПРЕССИЯ ПРОМОТОРА СИНТАЗЫ АЦЕТООКСИКИСЛОТ В ИНТРОДУЦИРОВАННЫХ ГЕНАХ РАСТЕНИЙ | 1995 |
|
RU2197527C2 |







Изобретение относится к области биохимии, в частности к фрагменту ДНК для генетической модификации растительной клетки с целью экспрессии рекомбинантного гена в растении. Также раскрыта генетическая конструкция, содержащая указанный фрагмент ДНК. Раскрыт способ экспрессии рекомбинантного гена в растительной клетке. Изобретение позволяет эффективно экспрессировать рекомбинантный ген в растении. 3 н.п. ф-лы, 10 ил., 1 табл., 6 пр.
1. Фрагмент ДНК для генетической модификации растительной клетки с целью экспрессии рекомбинантного гена в растении, имеющий нуклеотидную последовательность, выбранную из группы, включающей SEC ID NO 1, SEC ID NO 2, SEC ID NO 3 и SEC ID NO 4.
2. Генетическая конструкция, содержащая фрагмент ДНК по п. 1 и рекомбинантную последовательность ДНК, включающую ген рекомбинантного белка и терминатор транскрипции, для экспрессии рекомбинантного гена в растении.
3. Способ экспрессии рекомбинантного гена в растительной клетке, включающий использование генетической конструкции по п. 2, состоящий из этапов: создание генетической конструкции по п. 2, содержащей нуклеотидную последовательность по п. 1; введение генетической конструкций по п. 2 в растительную клетку; селекция трансформированной клетки на питательной среде и регенерация трансформированного растения; экспрессия рекомбинантного гена в растении.
| Промотор pro-SmAMP-X из растения звездчатка белая (Stellaria media L.) для экспрессии рекомбинантных генов в клетках растений | 2020 |
|
RU2766095C1 |
| WO 2014171860 A1, 23.10.2014 | |||
| WO 2014023285 A2, 13.02.2014 | |||
| МАДЖАРОВА Н.В | |||
| и др., Промоторы pro-smamp1 и pro-smamp2 из дикорастущего растения stellaria media для биотехнологии двудольных растений, ФИЗИОЛОГИЯ РАСТЕНИЙ, 2018, том 65. | |||
