Изобретение относится к области биотехнологии, в частности к генетической инженерии высших растений, и касается способа получения трансгенных растений с повышенным содержанием пролина.
Известны два способа получения трансгенных растений с модифицированным метаболизмом пролина: с помощью повышения экспрессии гена Δ1-пирролин-5-карбоксилатсинтетазы (P5CS), отвечающего за синтез пролина (Hu С-А. A., Delauney A.J., Verma D.P.S. // Proc. Natd. Acad. Sci. USA. 1992. V.89. P.9354-9358), либо снижения активности фермента пролиндегидрогеназы, определяющего скорость деградации пролина (Kiyosue Т., Yoshiba Y., Yamagichi-Shinozaki K., Shinozaki K // Plant Cell. 1996. V.8. P.1323-1335). Основным недостатком данных способов является низкая экологическая безопасность, поскольку отбор трансформантов производят на средах, содержащих антибиотик (канамицин).
Наиболее ближайшим к заявленному способу-прототипом, является способ получения трансгенных растений табака с повышенным содержанием пролина, заключающийся в клонировании кДНК гена P5CS из фасоли (Vigna aconitifolid) под контролем конститутивного промотора 35S из вируса мозаики цветной капусты, передаче полученной рекомбинантной плазмиды pBi-P5CS в штамм агробактерии (Agrobacterium tumefaciens) и получении трансгенных растений табака (Nicotiana tabacum) с помощью отбора и генерации на селективной среде с канамицином (Kavi Kishor P.B, Hong Z., Miao G. et. al. // Plant Physiol. 1995. V.108. P.1387-1394).
Основными недостатками прототипа являются:
- необходимость введения в геном растений гена устойчивости к канамицину в составе Т-ДНК рекомбинантной плазмиды, который не играет никакой другой функциональной роли, кроме возможности отбора;
- использование для отбора трансгенных растений селективной среды с добавлением антибиотика канамицина, что не позволяет сразу отбирать растения с повышенным содержание пролина, а требует дополнительной стадии отбора растений на устойчивость к канамицину.
Технической задачей настоящего изобретения является повышение экологической безопасности способа.
Поставленная техническая задача достигается предлагаемым способом, заключающимся в следующем.
Конструируют рекомбинантную плазмидную ДНК путем выделения кодирующей последовательности гена P5CS с помощью обратной транскрипции суммарной РНК, выделенной из проростков люцерны (Medicago truncatula), и последующей ПЦР-реакции со специфическими праймерами, с последующим клонированном полученной последовательности в полилинкер Т-ДНК области бинарной векторной плазмиды pBi101 (Jefferson R.A., Kavanagh T.A., Bevan M.W. // EMBO J. 1987. V.6. P.3301-3307). В результате получают рекомбинантную плазмидную ДНК pBi-P5CS, размером 14681 п.н., обеспечивающую перенос целевой последовательности ДНК в геном растений и экспрессию гена P5CS люцерны.
Плазмида состоит из следующих элементов:
- ДНК векторной плазмиды размером 13915 п.н.;
- фрагмента ДНК, содержащего кДНК гена Δ1-пирролин-5-карбоксилатсинтетазы размером 2148 п.н.;
- Промотора 35S РНК из генома вируса мозаики цветной капусты;
- 3'НТР области гена нопалинсинтазы.
Сконструированную рекомбинантную плазмиду pBi-P5CS, несущую ген Δ1-пирролин-5-карбоксилатсинтетазы, переносят в штамм агробактерий (Agrobacterium tumefaciens) AGL0 с последующей прямой трансформацией листовых дисков табака (Nicotiana tabacum) SR1 кокультивацией с агробактерией с последующим отбором и генерацией непосредственно на средах, содержащих NaCl.
Физическая карта плазмиды pBi-P5CS с указанием генетических маркеров приведена на фиг.1, где P5CS - нуклеотидная последовательность гена P5CS люцерны; 35S CaMV - промотор гена 35S РНК вируса мозаики цветной капусты; NOS PolyA - терминатор гена нопалинсинтазы из Ti-плазмиды агробактерий (Agrobacterium tumefaciens); NPTII-ген неомицинфосфотрансферазы II; LB, RB - левая и правая границы Т-ДНК области.
Определяющими отличительными признаками заявляемого способа, по сравнению с прототипом, являются:
- в качестве векторной плазмиды используют вектор pBi101, в котором ген β-глюкуронидазы был заменен на кДНК гена Δ1-пирролин-5-карбоксилатсинтетазы, что позволило получить конструкцию меньшего размера, более оптимальную для трансформации, в то время как в прототипе был использован вектор pBi121, где целевой ген с промотором и терминатором был встроен за геном β-глюкуронидазы;
- отбор трансгенных растений осуществляют непосредственно на стрессовом фоне (на среде, содержащей 200 mM NaCl), что позволяет получать стрессоустойчивые формы растений и повысить экологическую безопасность способа за счет возможности исключения из сконструированной рекомбинантной плазмиды pBi-P5CS гена устойчивости к антибиотику канамицину.
Изобретение поясняется следующими примерами конкретного выполнения способа.
Пример 1.
Конструирование генетической конструкции pBi-P5CS.
Клонирование кДНК гена Δ1-пирролин-5-карбоксилатсинтетазы (P5CS) в составе вектора pBi101 осуществляли в один этап посредством тройного лигирования. В конструкцию собирали три фрагмента: промотор 35S РНК из плазмиды pRT104, кДНК P5CS и часть плазмиды pBi101, содержащую в Т-области ген NPTII и 3'-нетранслируемый участок гена нопалинсинтазы, включающий сигнал полиаденилирования.
Кодирующая последовательность гена P5CS была выделена с помощью обратной транскрипции суммарной РНК, выделенной из проростков люцерны (Medicago truncatula), и последующей ПЦР-реакции со специфическими праймерами. В качестве праймеров использовались олигонуклеотиды, гомологичные 5'- и 3'- районам кодирующей последовательности гена P5CS (праймеры: 5'-gttcatctcatagctgtactatcat-3'; 5'-caagtgaacttccttgcctgccgtc-3'). Полученная кДНК гена P5CS длиной 2252 п.н. с включенными сайтами SalI и Есl136II была обработана рестриктазами SalI и Есl136II. Плазмида pRT104 была обработана рестриктазами HindIII и Sfr274I (XhoI). Плазмида pBi101 была обработана рестриктазами HindIII и Есl136II. Продукты рестрикции разделяли в 1% агарозном геле, гель окрашивали раствором бромистого этидия в воде (1 мкг/мл) и методом сорбции ДНК на силикагеле выделяли фрагменты: ДНК P5CS длиной 2148 п.н., фрагмент pRT104 длиной 441 п.н., соответствующий 35S промотору, фрагмент pBi101 длиной 11996 п.н. Выделенные фрагменты были обработаны ДНК-лигазой, лигазной смесью трансформировали компетентные клетки E.coli штамма XL1 Blue. Трансформанты высевали на чашку с LB-агаром, содержащим 50 мкг/мл канамицина. Клоны, несущие рекомбинантные плазмиды со встроенным геном, отбирали методом ПЦР анализа ДНК отдельных колоний E.coli с праймерами на нуклеотидную последовательность P5CS люцерны.
Пример 2
Получение трансгенных растений, несущих конструкцию pBi-P5CS.
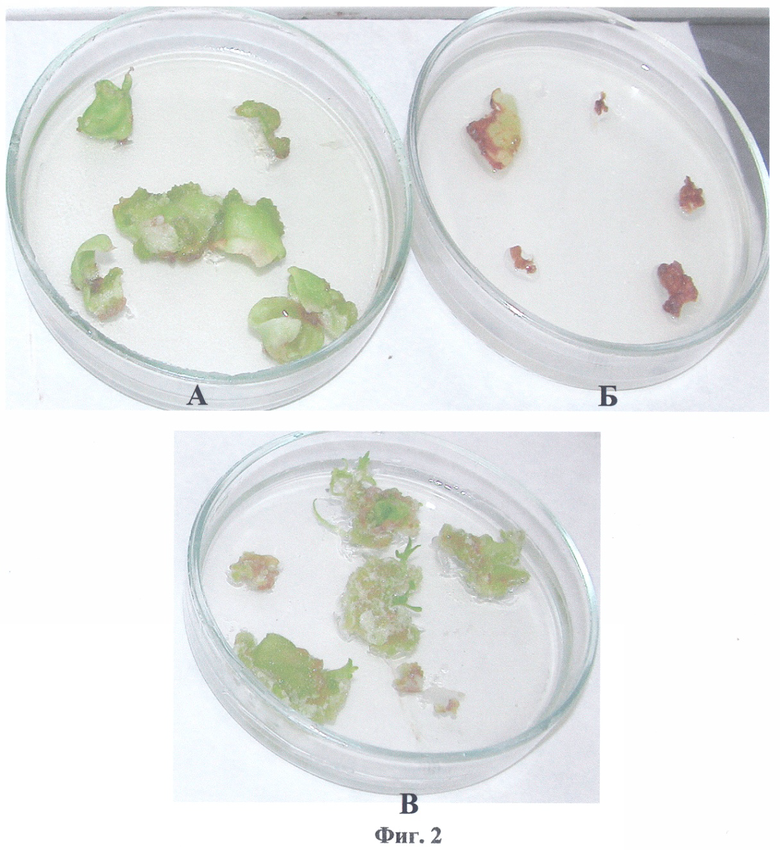
Полученная по примеру 1 конструкция pBi-P5CS, несущая кДНК гена Δ1-пирролин-5-карбоксилатсинтетазы, была использована для прямой трансформации клеток агробактерий (Agrobacterium tumefaciens) штамма AGL0 известным способом, описанным в (Horsch R., Fry J., Hoffman N. et al. // Plant Molecular Biology Manual. Dordrecht. Kluwer Academic Publishers. 1988, P.A5/1-A5/9). Для агробактериальной трансформации в качестве исходных эксплантов использовали листья трехнедельных растений табака (Nicotiana tabacum) SR1. Отбор трансформантов проводили на питательной среде Мурашиге и Скуга (МС) (Murasige Т., Skoog F. // Physiol. Plant. 1962. V.15, P.473-497) с добавлением 200 mM NaCl. На фиг.2 представлены результаты селекции листовых дисков табака, трансформированных конструкцией pBi-P5CS и контрольной конструкцией pBi101, которая представляет базовый вектор, без целевого гена, где
А, Б - трансформация конструкциями pBi101 и pBi-P5CS, отбор на среде, содержащей 200 mM NaCl;
В - трансформация конструкцией pBi101, отбор на канамицине.
На фиг.2А видно, что каллусообразование на листовых дисках с конструкцией pBi-P5CS на среде с 200 mM NaCl происходило нормально, тогда как диски с контрольной конструкцией pBi101 (фиг.2Б) некротизировались. Листовые диски с контрольной конструкцией на среде с канамицином развивались нормально и давали побеги (фиг.2В).
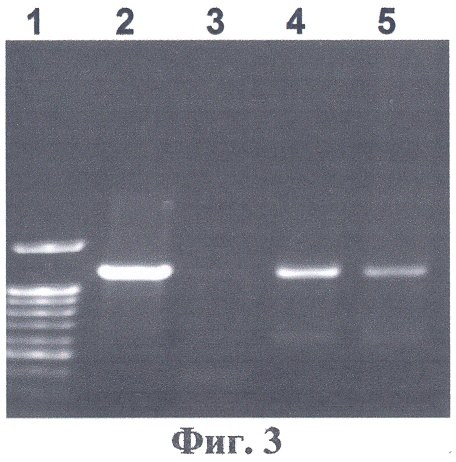
На фиг.3 представлена электрофореграмма ПЦР-анализа геномной ДНК проростков трансгенных растений на наличие встройки кДНК гена Δ1-пирролин-5-карбоксилатсинтетазы. Нанесения по дорожкам: 1-2 - трансгенные табаки SR1, экспрессирующие ген Δ1-пирролин-5-карбоксилатсинтетазы; 3 - нетрансгенные табаки SR1 (отрицательный контроль): 4, 5 - трансгенные табаки SR1, экспрессирующие ген Δ1-пирролин-5-карбоксилатсинтетазы. Из фиг.3 видно, что ПЦР-анализ с праймерами на нуклеотидную последовательность P5CS подтверждает наличие нуклеотидной последовательности P5CS люцерны в геномах отобранных растений.
Трансформанты, несущие конструкцию pBi-P5CS, были проанализированы на содержание свободного пролина в листовой ткани. Для измерения использовали растения одного возраста (2-3 недели), выращенные стерильно в условиях климатокамеры, листья отбирали приблизительно одного размера с одного уровня растений в каждом эксперименте. Содержание пролина в листовых экстрактах оценивали по методу Бэйтса (Bates L.S., Waldren R.P., Teare I.D. // Plants and Soil. 1973. V.39. P.205-207) в микрограммах (мкг) свободного на грамм сырой массы (гсм) образца. Результаты измерений приведены в таблице.
Из представленных в таблице данных видно, что полученные трансгенные растения табака, содержащие рекомбинантную плазмиду pBi-P5CS, имеют повышенный в несколько раз уровень содержания пролина, по сравнению с контрольными растениями.
Таким образом, предлагаемый способ отвечает требованиям экологической безопасности и позволяет получать трансгенные растения, несущие кДНК гена P5CS и эффективно повышать уровень содержания пролина в трансгенных растениях, что увеличивает их стрессоустойчивость.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324737C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ОБЕСПЕЧИВАЮЩАЯ ЭКСПРЕССИЮ ГЕНА ЭКСТРАКЛЕТОЧНОЙ РИБОНУКЛЕАЗЫ Zinnia elegans ZRNaseII В ТРАНСГЕННЫХ РАСТЕНИЯХ (ВАРИАНТЫ), И СПОСОБ ПОЛУЧЕНИЯ ВИРУСОУСТОЙЧИВЫХ ФОРМ РАСТЕНИЙ | 2009 |
|
RU2393226C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ МОРКОВИ, ПРОДУЦИРУЮЩИХ ИНТЕРЛЕЙКИН-10 ЧЕЛОВЕКА | 2007 |
|
RU2374321C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBi101-IL18, КОДИРУЮЩАЯ СИНТЕЗ ИНТЕРЛЕЙКИНА-18 ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ | 2005 |
|
RU2302460C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, УСТОЙЧИВЫХ К ВИРУСНОЙ ИНФЕКЦИИ | 2005 |
|
RU2300567C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pART27INT6 И СПОСОБ ПОЛУЧЕНИЯ НА ЕЕ ОСНОВЕ ИНБРЕДНОЙ ЛИНИИ РАСТЕНИЙ ТАБАКА, СИНТЕЗИРУЮЩЕГО ВНУТРИКЛЕТОЧНЫЙ ГАММА-ИНТЕРФЕРОН БЫКА | 2014 |
|
RU2564115C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PGII, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ БЕТА-ИНТЕРФЕРОНА ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ, И СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, УСТОЙЧИВЫХ К ВИРУСНОЙ ИНФЕКЦИИ | 1994 |
|
RU2103361C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pBM И СПОСОБ ПОЛУЧЕНИЯ С ЕЕ ИСПОЛЬЗОВАНИЕМ БЕЗМАРКЕРНЫХ ТРАНСГЕННЫХ РАСТЕНИЙ, СИНТЕЗИРУЮЩИХ ЦЕЛЕВЫЕ ПРОДУКТЫ | 2009 |
|
RU2410433C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pBi121-ESAT6-CFP10-gIFN, КОДИРУЮЩАЯ БЕЛКИ ESAT6 И CFP10 MYCOBACTERIUM TUBERCULOSIS И ГАММА-ИНТЕРФЕРОН ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ | 2011 |
|
RU2468082C1 |
| РАСТИТЕЛЬНЫЕ КЛЕТКИ И ОРГАНИЗМЫ РАСТЕНИЙ С МОДИФИЦИРОВАННЫМ КЛЕТОЧНЫМ РОСТОМ, РАЗВИТИЕМ И ДИФФЕРЕНЦИРОВКОЙ | 2004 |
|
RU2349642C2 |
Изобретение относится к генетической инженерии растений. Сущность изобретения: конструируют рекомбинантную плазмидную ДНК pBi-P5CS, размером 14681 п.н., обеспечивающую перенос целевой последовательности ДНК в геном растений и экспрессию гена P5CS люцерны, состоящую из следующих элементов:
- ДНК векторной плазмиды размером 13915 п.н.; - фрагмента ДНК, содержащего кДНК гена Δ1-пирролин-5-карбоксилатсинтетазы размером 2148 п.н.; - промотора 35S РНК из генома вируса мозаики цветной капусты; - 3'НТР области гена нопалинсинтазы. Сконструированную рекомбинантную плазмиду pBi-P5CS, несущую ген Δ1-пирролин-5-карбоксилатсинтетазы, переносят в штамм Agrobacterittm tumefaciens AGL0 с последующей прямой трансформацией листовых дисков Nicotiana tabacum SR1 сокультивацией с агробактерией. Отбор и генерацию производят непосредственно на средах, содержащих NaCl. Способ позволяет получать стрессоустойчивые растения табака с повышенным уровнем содержания пролина без использования антибиотиков. 3 ил., 1 табл.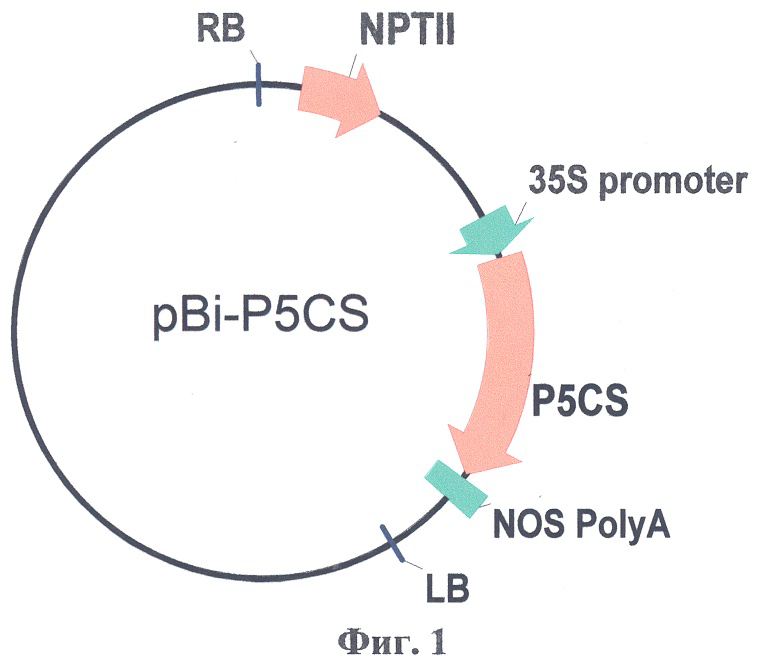
Способ получения трансгенных растений табака с повышенным содержанием пролина, включающий конструирование рекомбинантной плазмидной ДНК, содержащей ген Δ1-пирролин-5-карбоксилатсинтетазы, передачу полученной плазмидной ДНК в штамм Agrobacterium tumefaciens, инфицирование последним растительного материала, отбор трансгенных растений на селективной питательной среде с последующим тестированием отобранных трансформантов на повышенное содержание пролина, отличающийся тем, что конструируют и передают в штамм указанной бактерии рекомбинантную плазмидную ДНК pBi-P5CS, состоящую из следующих элементов:
ДНК векторной плазмиды pBI101, не содержащей ген бета-глюкуронидазы;
фрагмента ДНК, содержащего кДНК гена Δ1-пирролин-5-карбоксилатсинтетазы из Medicago truncatula размером 2148 п.н.;
промотора 35S РНК из генома вируса мозаики цветной капусты;
3'НТР области гена нопалинсинтазы, при этом отбор трансгенных растений осуществляют на питательной среде, содержащей 200 mM NaCl.
| KISHOR P | |||
| et al | |||
| PLANT PHISIOLOGY, 1995, v.108(4):1387-1394 | |||
| HU C.A | |||
| et al | |||
| PROC NATL ACAD SCI USA, 1992, v.89(19):9354-8 | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| JP 2001054385, 27.02.2001 | |||
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| JP 10057069, 03.03.1998 | |||
| US 5639950 A, 17.06.1997 | |||
| US 5344923 A, 06.09.1997. | |||

