Сущность изобретения
Изобретение относится к области биологии, в частности молекулярной биологии и генетической инженерии, и может быть использовано в векторных генетических конструкциях для биотехнологии растений с целью придания целым растениям или их отдельным частям (клеткам, тканям и органам) новых признаков и свойств, и для биофарминга с целью получения в растениях или их отдельных частях (клетках, тканях и органах) рекомбинантных белков. Помимо этого, настоящее изобретение может применяться в фундаментальных исследованиях в области биологии растений с использованием методов прямой и обратной генетики. В качестве настоящего изобретения предложен фрагмент нуклеотидной последовательности промоторной области гена антимикробного пропептида pro-SmAMP2 из сорного растения Stellaria media, который является сильным конститутивным промотором для экспрессии в растениях гетерологичных генов.
Краткое описание изобретения
Растительный конститутивный промотор, выделенный из генома мокрицы (Stellaria media L.) и представленный в качестве 5'-делеционного фрагмента 495 (SEC ID NO 5) п.н промоторной области от сайта начала трансляции (кодона ATG) гена антимикробного пропептида pro-SmAMP2 для обеспечения высокого уровня экспрессии гетерологичных генов в трансгенных растениях/клетках растений.
Краткое описание чертежей
Представленные чертежи являются схематическими.
Фиг. 1. Нуклеотидная последовательность фрагмента промоторной и 5'-нетранслируемой области гена pro-SmAMP2.
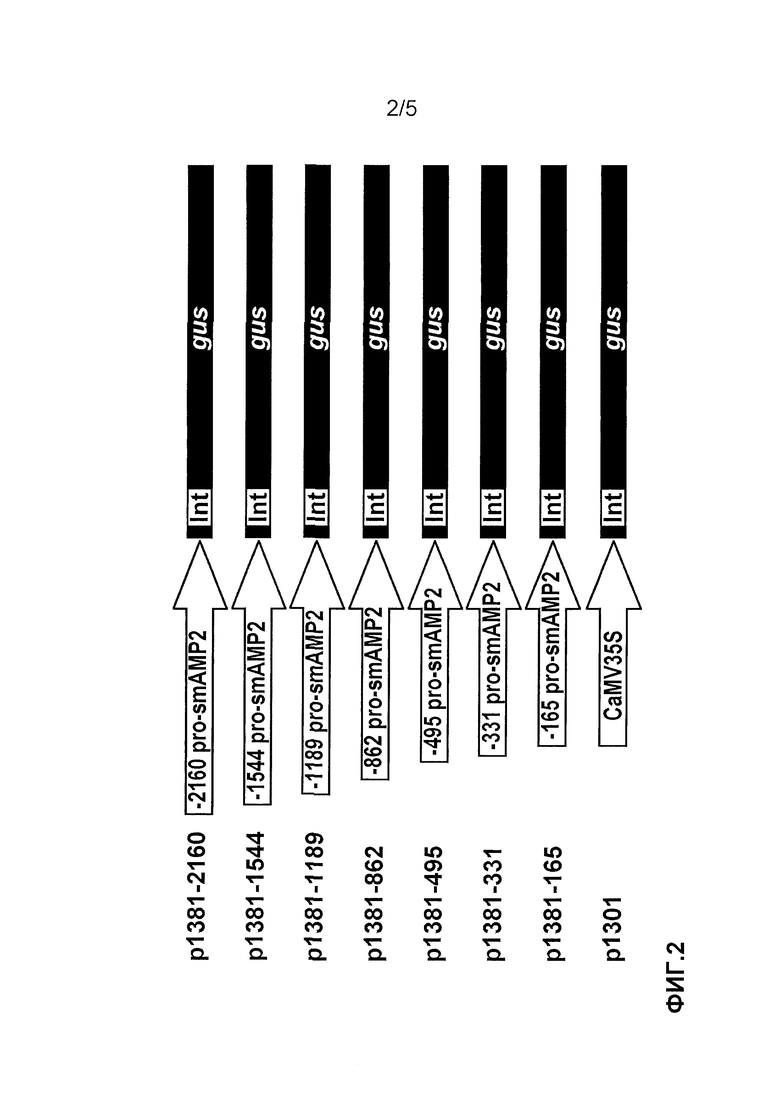
Фиг. 2. Схема конструкций с промотором pro-SmAMP2 для анализа экспрессии гена gus (бета-глюкуронидазы) в трансгенных растениях табака. 5'-делеционные фрагменты нуклеотидной последовательности промотора гена pro-SmAMP2 были слиты с репортерным геном gus в векторе pCAMBIA1381Z. В качестве конструкции с геном gus под контролем вирусного промотора CaMV35S была использована плазмида pCAMBIA1301.
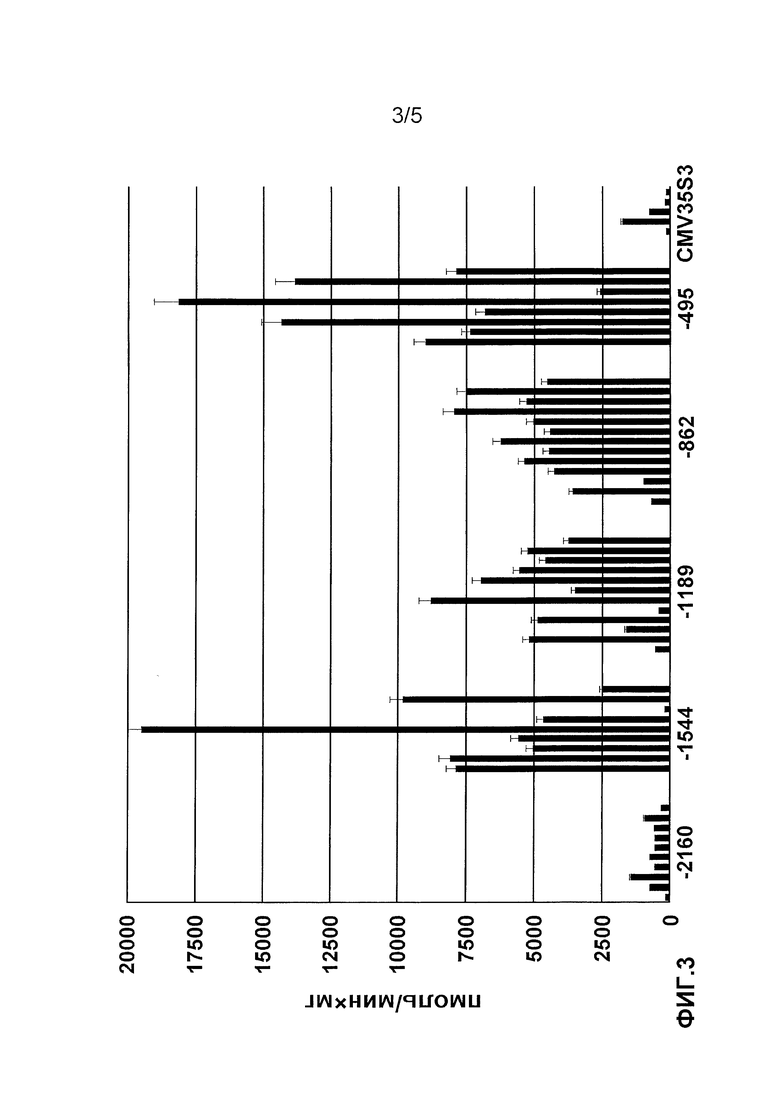
Фиг. 3. Активность репортерного белка бета-глюкуронидазы у отдельных асептически выращенных трансгенных растений N. tabacum, экспрессирующих ген gus под контролем 5'-делеционных фрагментов нуклеотидной последовательности промотора гена pro-SmAMP2 и промотора CaMV35S (приведены данные для растений с наибольшей активностью репортерного белка). Активность бета-глюкуронидазы измерена флуориметрическим способом, используя 4-метилумбеллиферил-D-глюкоранид в качестве субстрата.
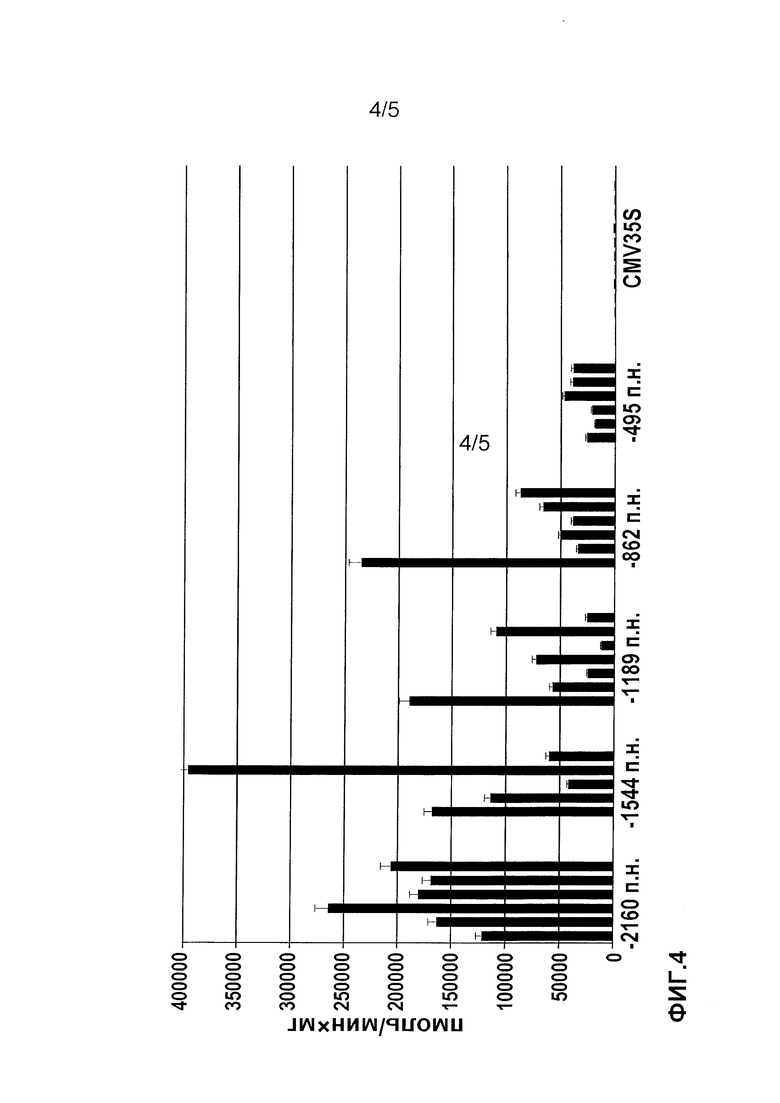
Фиг. 4. Активность репортерного белка GUS в трансгенных растениях табака N. tabacum, экспрессирующих ген gus под контролем фрагментов промоторной области гена pro-SmAMP2 и промотора CMV35S, выращенных в почве (приведены данные для растений с наибольшей активностью репортерного белка).
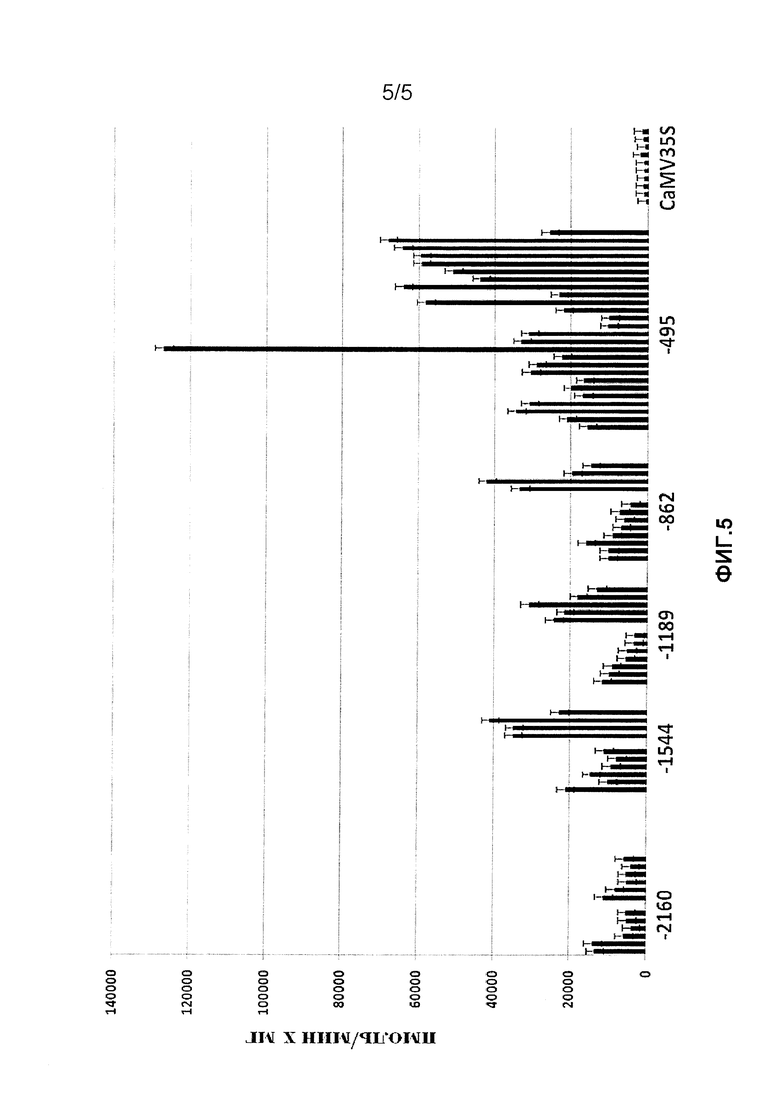
Фиг. 5. Активность репортерного белка GUS в трансгенных растениях табака N. tabacum поколения Т1, экспрессирующих ген gus под контролем фрагментов промоторной области гена pro-SmAMP2 и промотора CMV35S, выращенных в почве.
Задача изобретения
Задача изобретения заключается в создании векторов с растительным промотором, применение которых в генетической инженерии растений обеспечит высокий уровень экспрессии целевого гена с параллельным отсутствием недостатков, обозначенных ниже и характерных для промоторов из вирусов растений. В качестве растительного промотора настоящее изобретение предлагает использовать фрагмент (-495) нуклеотидной последовательности промоторной области гена pro-SmAMP2 сорного растения Stellaria media, обеспечивающий существенно более высокий уровень экспрессии репортерного гена gus (gusA) в трансгенных растениях Nicotiana tabacum по сравнению с вирусным промотором CaMV35S и известными делеционными вариантами (-2160, -1544, -1189, -862) промотора гена pro-SmAMP2.
Поставленная задача решается тем, что в генетических конструкциях для трансформации растений (клеток растений) используется нуклеотидная последовательность промоторной области гена антимикробного пептида pro-SmAMP2, вместо нуклеотидной последовательности промотора CaMV35S.
Описание изобретения
В настоящее время в биотехнологии растений и в биофарминге широко используется конститутивный промотор CaMV35S, созданный на основе промоторной области гена белка оболочки вируса мозаики цветной капусты (Patents US 5352605, US 5530196). CaMV35S, как правило, обеспечивает относительно высокий уровень экспрессии гетерологичных генов в растениях, однако CaMV35S имеет ряд существенных недостатков.
Промотор CaMV35S является вирусным по происхождению, и инфицирование трансгенных растений вирусом CaMV может приводить к сайленсингу перенесенного гена (Al-Kaff et al., 2000). Широкое использование промотора CaMV35S увеличивает также вероятность транскрипционной инактивации генов (Matzke and Matzke, 1995; STAM et al., 1997). Множество трансгенных растений проявляют феномен гомологично-зависимого генного сайленсинга, который возникает при взаимодействии между собой тесно связанных повторяющихся элементов на одной молекуле ДНК или на гомологичных последовательностях ДНК молекулы, в обеих аллельных и неаллельных позициях (Jakowitsch et al., 1999; Park et al., 1996). Кроме этого, сайленсинг возникает при использовании промотора CaMV35S в качестве одного и того же промотора двух и более гетерологичных генов в векторах для трансформации растений (Matzke and Matzke, 1995; McCabe et al., 1999; Park et al., 1996). В других случаях, таких как внедрение дополнительного гена в трансгенное растение, которое уже содержит ген с промотором CaMV35S, введение добавочных копий CaMV35S может усиливать метилирование и сайленсинг несвязанных гомологичных копий (Dong and von Arnim, 2003; Mette et al., 2000).
В биофарминге основные усилия направлены на повышение содержания рекомбинантного белка в трансгенном растении, которое в свою очередь зависит от величины экспрессии в растениях целевого гена (Medrano et al., 2009). В настоящее время в биофарминге относительно низкий уровень экспрессии целевого гена в растениях является одним из основных препятствий для коммерциализации конечного продукта.
Настоящим изобретением является фрагмент ДНК, обеспечивающий высокий уровень экспрессии генов в растениях и представляющий собой промотор гена рго-SmAMP2, соединенный с его 5'-нетранслируемой областью, где указанный фрагмент имеет нуклеотидную последовательность SEC ID NO 5 или ее фрагмент, содержащий по меньшей мере участок от 1829 до 2160 нуклеотида последовательности SEC ID NO 1.
Другим объектом настоящего изобретения является экспрессионный вектор, содержащий указанный фрагмент ДНК и по меньшей мере одну рекомбинантную последовательность ДНК.
Объектом настоящего изобретения также является способ экспрессии по меньшей мере одной рекомбинантной последовательности ДНК в растении или его части, включающий трансформацию клеток растения указанным вектором и экспрессию по меньшей мере одной рекомбинантной последовательности ДНК.
Вариантом указанного способа является способ, где частью растения является клетка.
Другим вариантом указанного способа является способ, где частью растения является ткань или орган.
Еще одним вариантом указанного способа является способ, включающий регенерацию трансгенного растения из трансформированных клеток растений.
Примеры
Приведенное выше раскрытие описывает изобретение в общих чертах. Более полное понимание может быть достигнуто за счет приведения соответствующих специфических примеров. Представленные здесь примеры приводятся исключительно с целью иллюстрации и никоим образом не претендуют на то, чтобы сузить область данного изобретения в рамках этих примеров. Изменение формы или замещение эквивалентов допускается на основании его целесообразности в соответствующих условиях. Хотя здесь и будут применяться специфические термины, они используются исключительно с описательной целью.
Пример 1
Этот пример раскрывает особенности установления нуклеотидной последовательности промотора гена pro-SmAMP2.
Геномная ДНК была выделена из тканей листьев Stellaria media с помощью GenElute Plant Genomic DNA Miniprep Kit ("Sigma", США) согласно инструкции производителя.
Нуклеотидная последовательность промотора гена pro-SmAMP2 была определена с помощью метода «прогулка по геному». Для этого был использован набор Genome Walker Universal Kit (Clontech Laboratories Inc., Palo Alto, CA, USA) и два антисмысловых праймера, специфичных к последовательности гена pro-SmAMP2: 5'- CTG TGT CTG AGC CTG GTA ATA AGC CT -3' и 5'- GCC TGC GGC AGT AGG CTC AAT CTC AGA AA -3'. Искомая нуклеотидная последовательность была амплифицирована путем двухступенчатой ПЦР согласно инструкции фирмы-производителя. Полученный ПЦР-продукт длиной 2400 п.н. был клонирован в вектор pGEM-T («Promega»). Его секвенирование показало, что он содержит фрагмент кодирующей нуклеотидной последовательности гена pro-SmAMP2 длиной 240 п.н. и фрагмент длиной 2160 п.н. влево (5'-upstream gene sequence) от сайта начала трансляции - ко дона ATG и потому относится к промоторной области этого гена. Нуклеотидная последовательность секвенированного фрагмента ДНК показана на фиг. 1.
Пример 2
Этот пример иллюстрирует приготовление конструкций pro-SmAMP2-GUS (1-7) с различными участками нуклеотидной последовательности промотора pro-SmAMP2, слитых с репортерным геном gusA для делеционного анализа промотора в трансгенных растениях табака.
Конструкции pro-SmAMP2-GUS (1-7) были получены слиянием отдельных фрагментов промотора pro-SmAMP2 (с -2160 (SEC ID NO 1), -1544 (SEC ID NO 2), -1189 (SEC ID NO 3), -862 (SEC ID NO 4), -495 (SEC ID NO 5), -331 (SEC ID NO 6), -165 (SEC ID NO 7) п.н.) до - 1 п.н., где первый нуклеотид сайта трансляции ATG был обозначен как +1) с кодирующей областью репортерного гена gusA в плазмиде pCambial381Z (Cambia, Австралия) (фиг. 2). Выбор фрагментов был осуществлен на основании анализа нуклеотидной последовательности промоторной области и расположения cis-регуляторных элементов на нем. Для конструкций pro-SmAMP2-GUS (1-7) были синтезированы следующие праймеры: один обратный, специфичный к pro-SmAMP2: 5'-AGC CCA TGG ТТТ САС TTG ATT ТТТ AGT GTG ACT AGT -3' с добавленным рестриктным сайтом Nco1 на 5'-конце и 4 прямые:

каждый с добавленным рестриктным сайтом EcoR1 на их 5'-концы (обозначены выделением). ПЦР-амплификация с вышеперечисленными праймерами была проведена смесью Pfu ДНК полимеразы (СибЭнзим, Россия)/7ад ДНК полимеразы (Синтол, Россия) в соотношении 1:10 в стандартном реакционном буфере для термостабильной полимеразы (60 мМ Tris-HCl (рН 8,5 при 25°С); 1.5 мМ MgCl2; 25 мМ KCl; 10 мМ 2-меркаптоэтанол; 0,1% Тритон Х-100) с dNTP. В качестве ДНК-матрицы был использован клонированный в вектор pGEM-T участок промоторной области гена pro-SmAMP2. Условия амплификации для конечного ПЦР продукта 2178, 1582 и 1207 п.н. были следующими: денатурация - 94°С, 30 с; отжиг праймеров - 60°С, 40 с; элонгация - 72°С, 2 мин; всего 25 циклов; для ПЦР продукта 880, 513, 348 и 183 п.н.: денатурация - 94°С, 30 с; отжиг праймеров - 60°С, 30 с; элонгация - 72°С, 1 мин; всего 20 циклов. Полученные ампликоны очищали из ПЦР-смеси с помощью колонки «Omnix» (Россия) и после рестрикции с помощью Nco1 и EcoR1 (Fermentas) в двухкратном Tango буфере (Fermentas) лигировали в плазмиду pCambia1381Z. Плазмиду pCambia1381Z подвергали действию тех же рестриктаз. Продукты рестрикции разделяли в 1%-ном агарозном геле, вырезали из геля, очищали на колонках «Omnix», определяли концентрацию и смешивали с ПЦР-ампликоном в соотношении вставка/вектор 15:1. Для лигирования использовали Т4 ДНК-лигазу (2000000 е.а./мл) бактериофага лямбда (СибЭнзим, Россия). Реакцию лигирования проводили при 16°С 2 часа в реакционном буфере (50 мМ Tris-HCl (рН 7.8 при 25°С); 10 мМ MgCl2; 10 мМ DTT; 1 мМ АТР) и затем оставляли на ночь при 4°С.
Далее лигазной смесью (объемом 25 мкл) трансформировали компетентные клетки Е. coli (штамм DH5α), высевали на чашки Петри с питательной низкосолевой средой LB (Бакто-триптон 1 г, Бакто-дрож. экст.0,5 г, NaCl 0,5 г на 100 мл среды) с канамицином (50 мг/мл) и инкубировали при 37°С в течение ночи. Отбирали с газона клеток отдельные клоны и рассевали по секторам на другой чашке. Присутствие вставки в клонах определяли методом ПЦР. Далее клон клеток, несущий рекомбинантную плазмиду, переносили из чашек Петри в 10 мл среды LB с канамицином. Инкубировали в течение ночи при 37°С на шейкере со скоростью 180 об/мин. Из ночной культуры выделяли плазмиду и проверяли наличие в ней искомой нуклеотидной последовательности рестрикцией. Кроме этого, для контроля над возможными ошибками при ПЦР амплификации все полученные конструкции: p1381Z-2160, p1381Z-1544, p1381Z-1189, p1381Z-862, p1381Z-495, p1381Z-331, p1381Z-165 были проверены секвенированием. Схематически полученные конструкции изображены на фиг. 2.
Пример 3
Этот пример описывает получение трансгенных растений табака с геном gus А, находящегося под контролем различных участков нуклеотидной последовательности промотора pro-SmAMP2 (SEC ID NO 1, SEC ID NO 2, SEC ID NO 3, SEC ID NO 4, SEC ID NO 5, SEC ID NO 6 и SEC ID NO 7) и CaMV35S, при помощи агробактериальной трансформации.
Трансформацию Agrobacterium tumefaciens полученными конструкциями для GUS-делеционного анализа промотора pro-SmAMP2 проводили, используя так называемый «трехродительский» метод трансформации агробактерий (Wise et al., 2006).
1. Штамм AGL0 Agrobacterium tumefaciens выращивали в 2 мл среды LB с рифампицином (100 мкг/мл) при 28°С 12 часов при 180 об/мин.
2. "Хелперный" штамм НВ101 Е. coli, содержащий плазмиду pRK2013, а также "донорные" штаммы Е. coli, содержащие плазмиды pCAMBIA 1381Z с фрагментами промотора и pCAMBIA 1301 (в качестве контроля использована плазмида, в составе которой ген gus находится под контролем стандартного конститутивного промотора CaMV35S) выращивали в 2 мл среды LB с канамицином (100 мкг/мл) при 37°С в течении 12 часов при 180 об/мин.
3. Далее смешивали по 100 мкл каждого донорного и хелперного штамма с 200 мкл штамма агробактерий AGL0.
4. Смесь высевали на твердую среду LB без антибиотиков, инкубировали 12 часов при 28°С и затем пересевали на твердую среду с соответствующими антибиотиками: рифампицином (100 мкг/мл) и канамицином (100 мкг/мл) и выращивали 2 дня при 28°С.
Полученные клоны агробактерий проверяли методом ПЦР на наличие соответствующих конструкций и в дальнейшем использовали для трансформации Nicotiana tabacum.
Трансгенные растения Nicotiana tabacum, трансформированные конструкциями p1381Z-2160, p1381Z-1544, p1381Z-1189, p1381Z-862, p1381Z-495, p1381Z-331, p1381Z-165 и pCAMBIA 1301, получали в соответствии с методом (Horsch et al., 1985) с некоторыми модификациями. В частности, кокультивацию эксплантов табака с агробактериями проводили газонным методом в темноте в течение 48 ч при температуре 20°С. Для селекции трансгенного каллуса, получения регенерантов и отбора трансформированных побегов использовали канамицин в концентрации 100 мг/л. Молекулярно-биологический анализ первичных трансформантов табака То на отсутствие агробактериальной контаминации и наличия целевых генов проводили методом ПЦР.
Пример 4
Данный пример иллюстрирует сравнение активности репортерного белка GUS в асептически выращенных трансгенных растениях табака, трансформированных конструкциями p1381Z-2160, p1381Z-1544, p1381Z-1189, p1381Z-862, p1381Z-495, p1381Z-331, p1381Z-165 и pCAMBIA 1301.
Измерение активности репортерного белка GUS в экстрактах трансгенных растений табака проводили в соответствии с методом Jefferson (Jefferson et al., 1987). Для этого образцы тканей листьев табака (около 50 мг) гомогенизировали с 150 мкл экстракционного буфера (50 мМ NaH2PO4, рН 7.0, 10 мМ ЭДТА, 0.1% Triton Х-100, 0.1% (масс/об.) лаурилсаркозин натрия, 10 мМ бета-меркаптоэтанола). Гомогенат центрифугировали 10 минут при 15000 об/мин. и 4°С, затем отбирали 100 мкл супернатанта и повторно центрифугировали при аналогичных условиях. К полученному супернатанту в количестве 50 мкл добавляли 250 мкл экстракционного буфера, хранили при 4°С в течение 1-2 дней. Для длительного хранения образцы помещали в морозильную камеру при -70°С.
Для измерения активности репортерного белка GUS 0,1 мкл исходного раствора образца вносили в 100 мкл среды, содержащей экстракционный буфер с добавлением 4-метилумбеллиферил-D-глюкоранида (4MUG) (Sigma-Aldrich, США) в конечной концентрации 1 мМ и инкубировали 30 мин при 37°С. Реакцию останавливали добавлением 900 мкл 0,2 M Na2CO3. Флуоресценцию образцов измеряли при 455 нм и длине волны возбуждения 365 им (флуориметр LS55 Perkin Elmer, США). Калибровку флуоресценции проводили с помощью раствора 4MU в 0,2 M Na2CO3. Удельную активность GUS рассчитывали после определения концентрации белка в образцах, измеренной методом Bradford (1976) с использованием раствора BSA в качестве стандарта.
Всего было получено 19 трансгенных растений То табака Nicotiana tabaccum для конструкции (-2160), 22 растения для (-1544), 17 растений для (-1189), 20 растений для (-862), 18 растений для (-495), 20 растений для (-331), 19 растений для (-165) и 10 растений для CaMV35S:GUS. Измерение активности репортерного белка GUS в экстрактах из листьев асептических трансгенных растений показало, что все делеционные варианты нового промотора, за исключением двух наиболее коротких (-330 и -150 п.н.), экспрессировали ген gus. Уменьшение длины нуклеотидной последовательности промотора до -330 п.н. и более приводит к нарушению его активности. Уровень активности репортерного белка GUS был наименьшим в экстрактах из трансгенных растений с наиболее длинным (-2160 п.н.) фрагментом промотора pro-SmAMP2 (фиг. 3). Абсолютные значения активности репортерного белка GUS снижались по мере укорочения фрагмента, начиная с -1544 делеционного фрагмента (фиг. 3). Уровень активности репортерного белка GUS в большинстве растений с 5'-делеционными вариантами промотора pro-SmAMP2 был существенно выше, чем в растениях, содержащих ген gus под контролем промотора CaMV35S.
Пример 5
Данный пример иллюстрирует сравнение активности репортерного белка GUS в трансгенных растениях табака, трансформированных конструкциями p1381Z-2160, p1381Z-1544, p1381Z-1189, p1381Z-862, p1381Z-495, pCAMBIA 1301 и выращенных в почве в теплице.
Трансгенные растения табака Т0, выращенные в асептических условиях и имеющие высокий уровень активности репортерного белка GUS, были пересажены в почву и помещены в теплицу: 6 растений для конструкции (-2160), 5 растения для (-1544), 6 растений для (-1189), 6 растений для (-862), 6 растений для (-495) и 10 растений для CaMV35S:GUS. Растения выращивали при температуре 24-26°С и 16-часовом световом периоде. Измерение активности репортерного белка GUS проводили у трансгенных растений табака через два месяца после пересадки в почву по методике, приведенной в примере 4. Для анализа использовали листья средних ярусов. Уровень активности репортерного белка GUS у всех растений, экспрессирующих gus под контролем 5'-делеционных вариантов промотора pro-SmAMP2, был на порядок выше, чем в растениях, содержащих ген gus под контролем промотора CaMV35S (фиг. 4). Из сравнения данных, приведенных на фиг. 3 и фиг. 4, видно, что трансгенные растения, выращенные в почве, имеют гораздо более высокий уровень активности репортерного белка по сравнению с растениями, выращенными в асептических условиях. Интересно, что в отличие от примера 4 наиболее высокую активность репортерного белка имели растения, экспрессирующие gus под контролем самого длинного фрагмента промотора (-2160) pro-SmAMP2. От всех трансгенных растений, выращенных в почве, были получены семена T1. Измерения активности репортерного белка GUS в трансгенных растениях поколения Τ1 показали, что высокая активность промотора pro-SmAMP2 сохранилась у потомства, при этом она по-прежнему существенно выше, чем у потомства трансгенных растений с промотором CaMV 35S (фиг. 5).
У большинства растений Т], экспрессирующих ген gus под контролем самого короткого и активного фрагмента промотора (-495), уровень активности GUS был не ниже, чем у их родительских растений Т0 (см. фиг. 4 и фиг. 5). Во всех образцах Т1, экспрессирующих ген gus под контролем остальных фрагментов промотора (-2160, -1544, -1189 и -862), уровень активности GUS существенно понизился в сравнении с их родительскими растениями Т0. Наиболее сильно, более чем в 25-36 раз, падение наблюдалось у растений Τ1 с фрагментом промотора -2160. Примерно в 8-11 раз активность GUS уменьшилась у растений Τ1 (-1544) и в 3-10 раз у растений Τ1 (-1189 и -862). Таким образом, самая высокая и стабильная экспрессия гена gus наблюдалась у растений табака при использовании фрагмента промотора -495, который по нашим данным можно считать наиболее перспективным делеционным вариантом промотора рго-SmAMP2 для биотехнологии растений.
Список литературы
Al-Kaff, N.S. et al., 2000. Plants rendered herbicide-susceptible by cauliflower mosaic virus-elicited suppression of a 35S promoter-regulated transgene. Nat Biotech, 18(9): 995-999.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72(1-2): 248-254.
Dong, Y. and von Arnim, Α., 2003. Novel plant activation-tagging vectors designed to minimize 35S enhancer-mediated gene silencing. Plant Molecular Biology Reporter, 21(4): 349-358.
Horsch, R.B., Rogers, S.G. and Fraley, R.T., 1985. Transgenic plants. Cold Spring Harb Symp Quant Biol, 50: 433-7.
Jakowitsch, J. et al., 1999. Molecular and cytogenetic characterization of a transgene locus that induces silencing and methylation of homologous promoters intrans. The Plant Journal, 17(2): 131-140.
Jefferson, R.A., Kavanagh, T.A. and Bevan, M.W., 1987. GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J, 6(13): 3901-7.
Matzke, M.A. and Matzke, Α., 1995. How and Why Do Plants Inactivate Homologous (Trans)genes? Plant Physiol., 107(3): 679-685.
McCabe, M.S. et al., 1999. Increased stable inheritance of herbicide resistance in transgenic lettuce carrying a petE promoter-bar gene compared with a CaMV 35S-bar gene. TAG Theoretical and Applied Genetics, 99(3): 587-592.
Medrano, G. et al., 2009. Rapid system for evaluating bioproduction capacity of complex pharmaceutical proteins in plants. Methods Mol Biol, 483: 51-67.
Mette, M.F., Aufsatz, W., van der Winden, J., Matzke, M.A. and Matzke, A.J.M., 2000. Transcriptional silencing and promoter methylation triggered by double-stranded RNA. EMBO J, 19(19): 5194-5201.
Park, Y.D. et al., 1996. Gene silencing mediated by promoter homology occurs at the level of transcription and results in meiotically heritable alterations in methylation and gene activity. The Plant Journal, 9(2): 183-194.
STAM, M., MOL, J.N.M. and KOOTER, J.M., 1997. Review Article: The Silence of Genes in Transgenic Plants. Annals of Botany, 79(1): 3-12.
Wise, A.A., Liu, Z. and Binns, A.N., 2006. Three methods for the introduction of foreign DNA into Agrobacterium. Methods Mol Biol, 343: 43-53.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ эффективного биосинтеза рекомбинантных белков в двудольных растениях с использованием промотора гена pro-SmAMP1 из растения Stellaria media | 2017 |
|
RU2697014C2 |
| Промотор pro-SmAMP-X из растения звездчатка белая (Stellaria media L.) для экспрессии рекомбинантных генов в клетках растений | 2020 |
|
RU2766095C1 |
| Промотор pro-SmAMP-D1 из растения звездчатка средняя (Stellaria media L.) для биотехнологии растений | 2022 |
|
RU2799014C1 |
| КОНСТРУКЦИЯ И СПОСОБ КОНСТРУИРОВАНИЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА UBI1 | 2012 |
|
RU2639538C2 |
| ПЕСТИЦИДНЫЕ КОМПОЗИЦИИ И ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ | 2013 |
|
RU2662672C2 |
| СПОСОБ СОЗДАНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ С ВЫСОКИМ УРОВНЕМ ЭКСПРЕССИИ ТРАНСГЕННОГО БЕЛКА | 2011 |
|
RU2507736C2 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |
| СИСТЕМА ДЛЯ УНИЧТОЖЕНИЯ РАСТИТЕЛЬНЫХ КЛЕТОК | 2001 |
|
RU2285045C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С НУКЛЕОТИДНОЙ ДЕЛЕЦИЕЙ В ПРОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772577C2 |
| ИСКУССТВЕННЫЙ ПРОМОТОР ДЛЯ ЭКСПРЕССИИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДНК В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2003 |
|
RU2326167C2 |





Изобретение относится к области биологии, в частности молекулярной биологии и генетической инженерии. В качестве настоящего изобретения предложен фрагмент ДНК, обеспечивающий высокий уровень экспрессии генов в растениях и представляющий собой промотор гена pro-SmAMP2, соединенный с его 5'-нетранслируемой областью, где указанный фрагмент ДНК имеет нуклеотидную последовательность SEC ID NO 5. Изобретение может быть использовано в векторных генетических конструкциях для биотехнологии растений с целью придания целым растениям или их отдельным частям (клеткам, тканям и органам) новых признаков и свойств, и для биофарминга с целью получения в растениях или их отдельных частях (клетках, тканях и органах) рекомбинантных белков. Помимо этого, настоящее изобретения может применяться в фундаментальных исследованиях в области биологии растений с использованием методов прямой и обратной генетики. 3 н. и 3 з.п. ф-лы, 5 ил., 5 пр.
1. Фрагмент ДНК, обеспечивающий высокий уровень экспрессии генов в растениях и представляющий собой промотор гена pro-SmAMP2, соединенный с его 5'-нетранслируемой областью, где указанный фрагмент ДНК имеет нуклеотидную последовательность SEC ID NO 5.
2. Экспрессионный вектор, содержащий фрагмент ДНК по п. 1 и по меньшей мере одну рекомбинантную последовательность ДНК.
3. Способ экспрессии по меньшей мере одной рекомбинантной последовательности ДНК в растении или его части, включающий трансформацию клеток растения экспрессионным вектором по п. 2 и экспрессию по меньшей мере одной рекомбинантной последовательности ДНК.
4. Способ экспрессии по п. 3, где частью растения является клетка.
5. Способ экспрессии по п. 3, где частью растения является ткань или орган.
6. Способ экспрессии по п. 3, включающий регенерацию трансгенного растения из трансформированных клеток растений.
| ШУКУРОВ Р.М., Антимикробные пептиды сорного растения Stellaria media и их гены: экспрессия и устойчивость к фитопатогенным грибам, диссертация, Москва, 2011, с.84-91 | |||
| ШУКУРОВ Р.М., Повышение устойчивости растений Arabidopsis thaliana к фитопатогенным грибам, экспрессирующих гевеиноподобные пептиды из сорного растения Stellaria media, Российская сельскохозяйственная наука, с.24-26 | |||
| US 5352605 A, 04.10.1994. |

