Изобретение относится к медицине, а именно - к иммунологии, и может быть использовано для клинического применения полимер-субъединичных адъювантных вакцин против гриппа у пациентов с общей вариабельной иммунной недостаточностью (ОВИН).
Проявления первичного иммунодефицита (ПИД) варьируются от потенциально доброкачественных форм, как, например, селективный дефицит IgA, до тяжелых проявлений, требующих проведения радикальной терапии - трансплантации гемопоэтических стволовых клеток (ТГКС) для того, чтобы выжить, например, при тяжелой комбинированной иммунной недостаточности (ТКИН). Общепризнанным является тот факт, что диагностика и выявляемость ПИД значительно снижена и варьируется в зависимости от форм по данным разных источников от 1:8500 до 1:100000 среди пациентов с клиническими проявлениями. [1, 2].
В течение последних четырех десятилетий регистрация пациентов с ПИД играла важную роль в различных аспектах этих заболеваний во всем мире, включая эпидемиологические показатели, разработку политики обеспечения жизненно важными препаратами, контроль качества жизни, содействие генетическим исследованиям и клиническим испытаниям, а также улучшение понимания естественного течения болезни и функции иммунной системы. Однако из-за ограниченности устойчивых ресурсов, поддерживающих эти регистры, несоответствия диагностических критериев и отсутствия молекулярной диагностики, а также трудностей с документацией и разработкой какой-либо универсальной платформы глобальная перспектива этих заболеваний остается неясной.
В структуре ПИД иммунодефицитные состояния с нарушением синтеза антител представляют собой около 50% всех нозологий. Общая вариабельная иммунная недостаточность (ОВИН) является наиболее распространенным вариантом первичных иммунодефицитов с поражением преимущественно гуморального звена иммунитета. ОВИН характеризуется различными клиническими проявлениями, среди которых чаще всего встречается склонность к рецидивирующим инфекциям, обусловленными гипогаммаглобулинемией и нарушением выработки специфических антител в ответ на белковые и полисахаридные антигены, и проявлениям иммунной дисрегуляции [3].
Симптомы иммунной дисрегуляиции включают аутоиммунные осложнения, энтеропатию, патологическую лимфопролиферацию, повышенный в несколько сотен раз риск малигнизации, в первую очередь, лимфомы [4-6].
Основные причины ОВИН в большей степени неизвестны. Генетические мутации могут быть идентифицированы как причина заболевания примерно у 10-20% пациентов в зависимости от этнической принадлежности и популяционной структуры когорты [7]. Мутации в генах чаще всего обнаруживаются в части В-клеточного ко-рецепторного комплекса, генах трансмембранного активатора и модулятора кальция и взаимодействующего с лигандом циклофилина (transmembrane activator and calcium modulator and cyclophilin ligand interactor - TACI), рецептора фактора, активирующего B-клетки (В cell-activating factor receptor - BAFFR), индуцибельного костимулятора (inducible costimulator - ICOS) и липополисахарид-чувствительного белка (lipopolysaccharide responsive beige-like anchor - LRBA). Хотя эти новые моногенные дефекты имеют общие клинические проявления с ОВИН, их можно рассматривать как отдельную группу первичных иммунодефицитов с фенотипом, подобным ОВИН [8, 9].
Однако определенная молекулярно-генетическая диагностика не проводилась у более 80% пациентов с клинически диагностированными ОВИН, и причина их болезни пока остается неизвестной. Ожидается, что все больше и больше пациентов с ОВИН будут вскоре перенесены из большой группы с данным диагнозом в различные формы моногенных заболеваний, приводящих к иммунологическим проявлениям, подобным фенотипам ОВИН, и не исключено, что механизм формирования иммунного ответа в ответ на вакцинацию и изучение дефектов в функционировании того или иного гена будет основным в принятии решения о необходимости вакцинации больных с моногенными заболеваниями.
Остро встает вопрос о вакцинации против гриппа пациентов, находящихся на регулярной заместительной терапии препаратами внутривенных или подкожных иммуноглобулинов. У пациентов с ОВИН, относящейся к группе ПИД с нарушенным антителообразованием, несмотря на регулярное проведение заместительной терапии препаратами донорских иммуноглобулинов, ежегодная вакцинация против вируса гриппа является единственным способом формирования защиты от вируса, а также предотвращением инфекционных осложнений заболевания и показана ввиду того, что препараты ВВИГ не могут содержать антител к актуальным, циркулирующим в настоящий сезон вариантов вируса гриппа, а тот факт, что вакцины против гриппа могут индуцировать клеточный иммунитет, генерирующий специфические для гриппа CD4 и CD8 Т-лимфоциты у пациентов, не способных вырабатывать IgG-антитела против вируса гриппа, делают вакцинацию оправданной [10-12]. В то же время имеются данные о содержании перекрестно-реактивных антител A/H1N1 в ВВИГ [13], однако этот факт не подтверждается данными других научных работ (Gardulf и соавт.), где среди 48 пациентов с ОВИН, несмотря на регулярную терапию иммуноглобулинами (1 раз в неделю), антител к вирусу гриппа на довакцинацльном этапе ни у кого обнаружено не было [14].
Остается открытым вопрос о схемах вакцинации иммунокомпрометированных больных. Хотя Eib1 и Wolf считают, что вакцинация против вируса гриппа должна осуществляться по тем же схемам, что и у здоровых людей [15], данные клинических исследований показывают необходимость введения второй дозы не ранее, чем через 21 день после вакцинации, или введения двойной дозы одномоментно с целью более активной стимуляции [16, 17]. Так, например, в исследовании Hartley и соавт. было доказано, что у 5 больных с преимущественным дефектом антител по сравнению со здоровым контролем количество антиген-специфичных В-клеток памяти к гемагглютинину штамма A/Michigan/2015 H1N1, как и уровень преимущественно IgG1, было значительно ниже и не увеличивалось после повторной вакцинации [18].
На настоящий момент в мире существуют единичные исследования, которые определяли формирование поствакцинального иммунитета как гуморального, так и клеточного в ответ на вакцинацию против гриппа у ограниченного числа пациентов с ОВИН с нарушением гуморального звена иммунитета [19-21].
Моновалентная сплит-вакцина против гриппа A (H1N1) с адъювантом AS03 (Pandemrix®, GlaxoSmithKline, Бельгия) широко использовалась во время пандемии гриппа 2009-2010 гг.для формирования специфического иммунного ответа в короткие сроки как среди здорового взрослого населения, из которых 67-98,3% продуцируют защитные уровни антител против гриппа A (H1N1) через 21 день после инъекции разовой дозы (3,75 мкг) вакцины, так и среди людей с ослабленным иммунитетом, в том числе пациентов с ПИД, применение у которых показало обнадеживающие результаты. В связи с экстренной необходимостью вакцинации по эпидемическим показаниям и формирования в короткие сроки стойкого иммунитета среди всех групп граждан 2 из 4 исследований были проведены в этот период с использованием адъювантной вакцины противопандемического вируса гриппа с включением в состав А / California / 7/2009 - H1N1-подобного вируса с добавлением адъюванта AS03 (Pandemrix (GlaxoSmithKline), благодаря которому создается «депо» антигенов, обеспечивая их более медленное рассасывание, в результате чего активируются макрофаги, усиливая поглощение антигена дендритными клетками, и CD4+Т-клетки, стимулируя не только гуморальный, но и Т-клеточный иммунитет [22, 23]. В исследовании, проведенном Pedersen и др., число участников с ОВИН было всего 3, однако по результатам полученных данных оказалось, что после вакцинации двумя дозами одномоментно, что произошло в результате случайной ошибки, как отмечает сам автор, и бустеризации еще одной дозой через 3 недели (7,5 мкг + 3,75 мкг гемагглютинина) 2 из 3 пациентов ответили на вакцинацию более чем 4-кратным увеличением титра антител в реакции торможения гемагглютинации [24]. В другом и самом масштабном на данный момент исследовании, опубликованном в 2018 году, с использованием той же вакцины 48 пациентов с ОВИН были вакцинированы двухкратно против вируса гриппа с интервалом 1 месяц. Исследователями было обнаружено, что 8 (16,7%) участников достигли протективного титра специфических антител против пандемичного вируса гриппа A (H1N1) на уровне ≥1:40: 4/8 после первой вакцинации, остальные 4 - после ревакцинации через 1 месяц, при том что 7-98,3% здоровых людей формируют защитные уровни антител с 21 дня после однократного введения Pandemrix® [14]. В третьем исследовании введения одной дозы безадъювантной вакцины после иммунизации у 7 из 8 пациентов с нарушением гуморального звена иммунитета отмечался синтез антител, по крайней мере, против 1 из 3 штаммов вакцины [20]. В самом раннем же исследовании, проведенном Sander van Assen и др. в 2010 году после вакцинации трехзвалентной безадъювантной субъединичной вакциной в сезон 2006-2007 (Influvac) 18 пациентов с нарушением гуморального звена иммунитета (в том числе и ОВИН), автор посчитал, что получил неудовлетворительные результаты: у 88% и всего у 23% не было отмечено прироста титра антител в значениях ≥40 к A/H3N2 и A/H1N1, соответственно, после вакцинации. Однако во всех исследованиях было отмечено, что клеточный ответ против гриппа у пациентов с ОВИН был сопоставим с показателями групп здорового контроля.
Общая вариабельная иммунная недостаточность (ОВИН) характеризуется снижением уровня сывороточных иммуноглобулинов (Ig) из-за отсутствия нормальной дифференцировки В-клеток [4, 25]. Поскольку для дифференцировки наивных В-клеток в клетки, секретирующие антитела, требуются специфические экзогенные сигналы, многие исследователи изучали синтез Ig in vitro при ОВИН, чтобы проанализировать причины дефектов. Эти исследования показывают, что В-клетки некоторых пациентов с ОВИН сохраняли способность к синтезу Ig in vitro, в то время как В-клетки других - нет.Несмотря на то, что большинство пациентов с ОВИН имеют нормальное количество периферических В-клеток, у всех их отмечается значительно сниженное количество циркулирующих CD27 + IgD- переключенных В-клеток памяти, что связано со сниженной продукцией антител [26], и, как следствие, высокой вероятностью более неблагоприятных клинических исходов [27, 28]. Была выявлена корреляция умеренного снижения количества В-клеток с поверхностным маркером CD27 + (клетки памяти), особенно очень низкого количества В-клеток CD27 + IgD - (изотип-переключенные клетки памяти) с более низким синтезом антител как in vitro, так и in vivo [29], а также некоторыми отдельными клиническими осложнениями [28, 30, 31].
Дифференцировка CD27- наивных В-клеток человека в CD27+ В-клетки памяти и плазматические клетки обычно происходит в зародышевых центрах вторичных лимфоидных органов, где антиген-активированные наивные В-клетки проходят пролиферацию, соматическую гипермутацию генов вариабельной (V)-области Ig, переключение изотипов, и в конечном итоге дифференциацию [32, 33]. В ответ на повторное заражение антигеном В-клетки памяти подвергаются соматической гипермутации генов V-области Ig и способны генерировать Ig всех изотипов. Фактически, было показано, что CD27+ IgD+ В-клетки, которые подвергаются соматической гипермутации у здоровых субъектов, с меньшей вероятностью подвергаются таковой у субъектов с ОВИН [34]. Поскольку дифференцировка плазматических клеток происходит преимущественно из CD27+ В-клеток [35], одним из следствий является нехватка плазматических клеток при ОВИН [15, 36]. В то время как van Assen в своем исследовании из 18 пациентов с ОВИН указывает, что хоть и достижение протективного титра антител было достигнуто у 77% к A/H1N1 и у 12% к A/H3N2, адекватного увеличения IFN- гамма продукции CD4+ Т-клетками по сравнению со здоровым контролем не было отмечено.
В то время как В-клетки традиционно считаются ключевыми в адаптивном иммунитете из-за их способности продуцировать антитела, активация рецепторов врожденного иммунитета, также экспрессирующихся на В-клетках, обеспечивает ко-стимулирующий эффект, который способствует как их функционированию и выживанию [37], так и координированию врожденных и адаптивных иммунных сигналов, что приводит к широкому диапазону клеточных ответов.
В-клетки памяти обычно образуются в зародышевых центрах в ответ на Т-зависимые или Т-независимые антигены, но, как и другие антигенпрезентирующие клетки, В-клетки экспрессируют различные Toll-подобные рецепторы (TLR) [38, 39] -консервативные мембранные белки, которые обеспечивают альтернативные способы активации В-клеток [40].
Толл-подобные рецепторы (TLR) - патоген-распознающие рецепторы, которые играют ключевую роль в элиминации микробных агентов через привлечение фагоцитов или активацию дендритных клеток. Из 10 TLR, экспрессируемых у человека, на В-клетках экспрессируются TLR1, TLR2, TLR6, TLR7, TLR9 и TLR10. TLR1, 2, 4, 5 и 6 экспрессируются на внеклеточной поверхности и распознают компоненты микробной стенки, включая ЛПС, липопептиды и флагеллин. TLR3, 7, 8 и 9 в основном локализуются в эндосомальных компартментах; это расположение позволяет этим рецепторам распознавать продукты распада ДНК и РНК вирусного и бактериального происхождения. TLR экспрессируются в различных типах клеток, включая моноциты, фагоциты, дендритные клетки и субпопуляции В-клеток.
Одним из наиболее мощных стимуляторов активации и созревания В-клеток являются эндосомные Toll-подобные рецепторы (Toll-like receptor - TLR), агонистами которых являются одноцепочечные РНК или различные синтетические агонисты (TLR7) и неметилированные мотивы CpG в микробной ДНК (CpG-ДНК) (TLR9), соответственно [41]. Было показано, что связывание TLR9 с помощью CpG-ДНК активирует нормальные В-клетки, усиливает экспрессию костимулирующих молекул, запускает секрецию IL-6 и IL-10 и опосредует Т-независимое переключение изотипов и продукцию специфических антител независимо от связывания рецепторов В-клеток (BCR) [42-46]. Перекрестное же связывание В-клеточного рецептора (BCR) приводит к быстрой активации их экспрессии. Наивные В-клетки экспрессируют низкие уровни TLR, тогда как В-клетки памяти конститутивно экспрессируют TLR7, TLR8 и TLR9 на более высоких уровнях [46-49]. Предполагают, что данные различия между наивными и В-клетками памяти связаны с разными адаптивными функциями: В-клетки памяти экспрессируют повышенные уровни TLR и обладают большей способностью по сравнению с наивными В-клетками дифференцироваться в плазматические клетки через стимуляцию TLR [50].
Один из наиболее изученных сигнальных ответов TLR - ответ TLR9 на В-клетках, который запускается олигодезоксинуклеотидами (ODN), полученными из бактериальной и вирусной ДНК, содержащие неметилированные мотивы CpG (CpG-ODN) [51] Результаты передачи сигналов TLR9 на В-клетках человека разнообразны и включают клеточную активацию, регуляцию костимулирующих молекул, включая CD23, CD25, CD40, CD54, CD80, CD86, CD69 и HLA-DR, секрецию IL-6 и IL-10, Т-независимое переключение изотипов иммуноглобулинов, транскрипцию генов зародышевых линий, соматическую гипермутацию и выработку иммуноглобулина, не зависящая от В-клеточного рецептора (BCR) [52].Связывание TLR9 на В-клетках памяти дополнительно усиливает его собственную экспрессию, повышает чувствительность клеток к активации, независимо от стимулирующего антигена, при этом одновременно усиливая специфический антительный ответ. В то время как В-клетки памяти более чувствительны к сигналам с TLR, активация наивных В-клеток при определенных условиях также приводит к созреванию и секреции Ig [53].
TLR7 и TLR8, которые обладают сходной лигандной специфичностью и экспрессируются вместе с TLR9 в эндосомном компартменте В-клеток, имеют сходные сигнальные пути с TLR9. Агонисты TLR7, такие, как: продукты одноцепочечной РНК, синтетические аналоги гуанозина (имиквимод или локсорибин) и производные имидазохинолина (например, CL097), могут активировать как наивные, так и В-клетки памяти человека. Активация TLR7 приводит как к клеточной дифференцировке, так и продукции иммуноглобулинов [54]. Интересно, что для TLR7-опосредованной стимуляции, удаление плазматических дендритных клеток (пДК) снижает продукцию иммуноглобулинов, демонстрируя необходимость прямого или косвенного взаимодействия с пДК. Поскольку добавление IFN-альфа восстанавливает секрецию иммуноглобулинов, этот цитокин необходим для продукции антител в нормальных культурах В-клеток [55]. Эксперименты как in vivo, так и in vitro показали, что для переключения В-клеток на изотипы IgG необходимо одновременное присутствие, как минимум, двух сигналов наряду с вовлечением BCR: активация TLR, вовлечение CD40 и/или IFN-альфа [56]. Эти наблюдения привели к исследованиям, предполагающим, что активация TLR может обеспечивать длительный стимул, важный для поддержания пролиферации В-клеток памяти и дифференцировки в зрелые секретирующие антитела клетки, которые первоначально индуцируются BCR и посредством Т-клеток [57, 58].
Связывание и активация TLR7 и TLR9 может служить сигналом для начала дифференцировки В-клеток после антигенной стимуляции через BCR. Однако имеются данные, в которых сообщается о дефектах в TLR7 и TLR9 на В-клетках пациентов с ОВИН [41]. Поскольку активация TLR, по-видимому, является неотъемлемым механизмом активации и последующей выживаемости В-клеток памяти [59], исследование пациентов с ОВИН дефектов TLR9 выявило, что их В-клетки не активировались лигандом - CpG-ODN - даже при костимуляции BCR, как и секрецией ИЛ-6 и ИЛ-10 [41], таким образом, не происходила активация TLR, наблюдалась низкая пролиферация В-клеток, отсутствие их созревания, переключения изотипов и выработки IgG и IgA [41].
Еще в 2010 году Danilo Escobar и др. исследовали активацию В-клеток у 14 пациентов с ОВИН, индуцированную Т1Л9-специфическим лигандом (CpG-ODN) и бактериальными лизатами S. pneumoniae и Н. influenzae, которые, как известно, стимулируют несколько типов TLR. Было обнаружено, что у пациентов с ОВИН В-клетки как при коактивации анти-IgM антителами, так и без экспрессируют более низкие уровни CD86 после стимуляции TLR9 лигандами CpG-ODN, S.pneumoniae и Н. influenzae, а также демонстрируют более низкий индекс пролиферации при стимуляции бактериальными экстрактами. Ученые пришли к выводу о дефектах в передаче сигналов TLR В-лимфоцитов пациентов с ОВИН [60].
Как следует из данных приведенных исследований, результаты весьма противоречивы. Расхождение в них обусловлено не только разными схемами вакцинации, но и выбором вакцинного препарата. Учитывая, что у больных с ПИД возникает необходимость повышения эффективности вакцин, которые должны активировать все звенья иммунной системы.
В настоящее время нами была исследована иммуногенность четырехвалентной инактивированной адъювантной вакцины против гриппа (adjuvanted quadrivalent inactivated vaccine - aQIV) у здоровых людей и пациентов с первичным иммунодефицитом - общим вариабельным иммунодефицитом (ОВИН) (KOSTINOVA A.M. et al. Assessment of Immunogenicity of Adjuvanted Quadrivalent Inactivated Influenza Vaccine in Healthy People and Patients With Common Variable Immune Deficiency // FRONTIERS IN IMMUNOLOGY, 2020, Volume 11, Article 1876). Данное исследование является прототипом изобретения. Всем больным вводили инактивированную субъединичную адьювантную противогриппозную вакцину. Однократную вакцинацию проводили согласно инструкции производителя. В данной работе для оценки титров антител через 21 день после вакцинации против штаммов гриппа А и В использовали реакцию ингибирования гемагглютинации (РТГ). Были выявлены статистически значимые различия уровня всех критериев между группами здоровых и больных ОВИН независимо от штамма вируса. Однако у большинства пациентов с ОВИН выявлено повышение титра поствакцинальных антител без достижения условно-защитного уровня антител.
Таким образом, у больных ОВИН поиск новых схем вакцинации является предметом дальнейших исследований. В связи с этим целью изобретения было сравнение эффективности разных режимов и схем вакцинации против гриппа пациентов с общей вариабельной иммунной недостаточностью.
В основу изобретения положено создание способа формирования поствакцинального иммунитета против вируса гриппа у пациентов с общей вариабельной иммунной недостаточностью, включающего одномоментное введение субъекту двух доз по 0,5 мл инактивированной полимер-субъединичной адъювантной вакцины, содержащей по 5 мкг протективных антигенов эпидемически актуальных штаммов вирусов гриппа типа А подтипов A(H1N1), A(H3N2), типа В и 500 мкг азоксимера бромида.
Техническим результатом заявленного изобретения является развитие высокой иммуногенности у пациентов с ОВИН после введения одномоментно двух доз субъединичной адъювантной вакцины против гриппа, при этом доля пациентов с 2-х кратным и более приростом титра антител через 3-4 недели после одномоментной вакцинации двумя дозами вакцинации после в 9,3 раз выше по сравнению с вакцинацией одной дозой. Разработанный способ вакцинации также активирует клеточный иммунитет, а именно, одномоментная вакцинация двумя дозами приводит к увеличению доли гранулоцитов, лимфоцитов, экспрессирующих TLR9, а на моноцитах - TLR3 и TLR9, которые играют ключевую роль в элиминации микробных агентов через привлечение фагоцитов или активацию дендритных клеток. Доказан высокий профиль безопасности трех- и четырехвалентных адъювантных вакцин у больных ОВИН. Иммунизация со сниженным количеством белка гемагглютинина (5 мкг) ко всем штаммам вируса за счет применения азоксимера бромида формирует протективный иммунитет.
Разработанный способ формирования поствакцинального иммунитета активирует синтез противогриппозных антител и запускает каскада иммунных реакций по распознаванию патогенов в поствакцинальном периоде при ежегодной вакцинации или первичном введении двух доз адъювантной вакцины против гриппа у пациентов с ОВИН.
Краткое описание графических материалов.
ФИГ. 1. Уровень серопротекции к штаммам вируса гриппа с учетом изначального уровня AT через 3-4 недели после введения aQIV у здоровых.
ФИГ. 2. Уровень сероконверсии к штаммам вируса гриппа с учетом изначального уровня AT через 3-4 недели после введения aQIV у здоровых.
ФИГ. 3. Фактор сероконверсии к штаммам вируса гриппа с учетом изначального уровня AT через 3-4 недели после введения aQIV у здоровых.
ФИГ. 4. Средние геометрические концентрации титров AT (GMT) к штаммам вируса гриппа с учетом изначального уровня AT через 3-4 недели после введения aQIV у здоровых.
ФИГ. 5. Доля серопозитивных к штамму A/H1N1 пациентов с ПИД на различных этапах вакцинации, приведены процентные значения и 95% доверительный интервал
(• - различия на пограничном уровне значимости р<0,10, применялся точный критерий Мак-Немара).
ФИГ. 6. Титр антител у пациентов с ПИД в зависимости от этапа вакцинации одной дозой препарата исходно и через 3-4 недели, приведены индивидуальные значения, средний геометрический титр (GMT) и его 95% доверительный интервал.
ФИГ. 7. Доля пациентов с 2-х кратным и более приростом титра AT через 3-4 недели после вакцинации в зависимости от этапа, приведены процентные показатели и 95% доверительный интервал. Применялся критерий Кохрана-Мантеля-Хенселя с обобщением отношения шансов по штаммам.
ФИГ. 8. Титр антител к штамму A/H1N1 у пациентов с ПИД в зависимости от схемы первичной вакцинации исходно и через 3-4 недели, приведены индивидуальные значения, средний геометрический титр (GMT) и его 95% доверительный интервал (* - статистически значимые различия до и после вакцинации на уровне р<0,05, для расчета использовалась линейная модель смешанных эффектов).
ФИГ. 9. Фактор сероконверсии к штамму A/H1N1 у пациентов с ПИД в зависимости от схемы первичной вакцинации, приведены индивидуальные значения, средний геометрический прирост (GMR) и его 95% доверительный интервал (* - статистически значимые различия между группами исследования на уровне р<0,05,. - тенденция к различиям между группами исследования на уровне р<0,10. Применялся критерий Манна-Уитни).
ФИГ. 10. Титр антител к штамму A/H3N2 у пациентов с ПИД в зависимости от схемы первичной вакцинации исходно и через 3-4 недели, приведены индивидуальные значения, средний геометрический титр (GMT) и его 95% доверительный интервал (* -статистически значимые различия до и после вакцинации на уровне р<0,05, для расчета использовалась линейная модель смешанных эффектов).
ФИГ. 11. Фактор сероконверсии к штамму A/H3N2 у пациентов с ПИД в зависимости от схемы первичной вакцинации, приведены индивидуальные значения, средний геометрический прирост (GMR) и его 95% доверительный интервал (* - статистически значимые различия между группами исследования на уровне р<0,05,. - тенденция к различиям между группами исследования на уровне р<0,10. Применялся критерий Манна-Уитни).
ФИГ. 12. Титр антител к штамму В/Колорадо у пациентов с ПИД в зависимости от схемы первичной вакцинации исходно и через 3-4 недели, приведены индивидуальные значения, средний геометрический титр (GMT) и его 95% доверительный интервал (* - статистически значимые различия до и после вакцинации на уровне р<0,05, для расчета использовалась линейная модель смешанных эффектов).
ФИГ. 13. Фактор сероконверсии к штамму В/Колорадо у пациентов с ПИД в зависимости от схемы первичной вакцинации, приведены индивидуальные значения, средний геометрический прирост (GMR) и его 95% доверительный интервал (* - статистически значимые различия между группами исследования на уровне р<0,05,. - тенденция к различиям между группами исследования на уровне р<0,10. Применялся критерий Манна-Уитни)
ФИГ. 14. Доля пациентов с 2-х кратным и более приростом титра AT через 3-4 недели после вакцинации в зависимости от схемы вакцинации, приведены процентные показатели и 95% доверительный интервал. Применялся критерий Кохрана-Мантеля-Хенселя с обобщением отношения шансов по штаммам.
ФИГ. 15. Процентное содержание гранулоцитов, экспрессирующих TLR3 и TLR9 (а, б), и лимфоцитов, экспрессирующих TLR9 (в), у пациентов с ПИД в зависимости от схемы первичной вакцинации исходно и через 3-4 недели, приведены индивидуальные значения, медианы и интерквартильный размах.
ФИГ. 16. Дельта (Δ - разница между показателем через 3-4 недели и исходным) процентного содержания гранулоцитов, экспрессирующих TLR3 и TLR9 (а, б), и лимфоцитов, экспрессирующих TLR9 (в), у пациентов с ПИД, вакцинированных первично одной и двумя дозами, приведены индивидуальные значения, медиана и ее 95% доверительный интервал.
ФИГ. 17. Процентное содержание моноцитов, экспрессирующих TLR3 и TLR9, у пациентов с ПИД в зависимости от схемы первичной вакцинации исходно и через 3-4 недели, приведены индивидуальные значения, медианы и интерквартильный размах (t(13)=3,8, р=0,002 для TLR3, t(13)-3,4, р=0,005 для TLR9).
ФИГ. 18. Дельта (Δ - разница между показателем через 3-4 недели и исходным) процентного содержания моноцитов, экспрессирующих TLR3 и TLR9, у пациентов с ПИД, вакцинированных первично одной и двумя дозами, приведены индивидуальные значения, медиана и ее 95% доверительный интервал (различается статистически значимо (р=0,01) в обоих случаях).
ФИГ. 19. Процентное содержание (а) и дельта (Δ - разница между показателем через 3-4 недели и исходным) процентного содержания (б) моноцитов, экспрессирующих TLR8, у пациентов с ПИД в зависимости от схемы первичной вакцинации исходно и через 3-4 недели, приведены индивидуальные значения, медианы и интерквартильный размах.
Осуществление изобретения.
Участники исследования
Для проведения исследования из регистра Института иммунологии были проанализированы 297 амбулаторных карт пациентов с ПИД и отобраны 203 пациента с диагнозом ОВИН, который был установлен в соответствии с критериями European Society on Immunodeficiency Disorders (ESID). Однако в связи с коморбидностью больных, необходимостью купирования обострения при поступлении пациентов в стационар, в соответствии со строгими критериями включениями в исследование, а также по причине ежегодной смены штаммового состава вакцин против гриппа, ограниченными сроками проведения вакцинации против гриппа (осенне-зимний период), осуществить иммунизацию оказалось возможным только 15 пациентам с ОВИН.
Таким образом, в сезон гриппа 2018-2019 на базе отделения «Иммунопатологии взрослых» клиники ФГБУ «ГНЦ Институт иммунологии» ФМБА России и ФГБНУ «Научно-исследовательский институт вакцин и сывороток им. И.И. Мечникова» МЗ РФ в исследовании приняли участие: первая группа - 6 пациентов с диагнозом общей вариабельной иммунной недостаточности, которым однократно вводилась 1 доза (0,5 мл) четырехвалентной субъединичной адъювантной вакцины против вируса гриппа Гриппол Квадривалент (ООО "НПО Петровакс Фарм", Россия). Ровно через год 5 из 6 испытуемых были повторно вакцинированы трехвалентной субъединичной адъювантной вакциной против вируса гриппа Гриппол Плюс (ООО "НПО Петровакс Фарм", Россия) из семейства вакцин Гриппол. Средний возраст пациентов с ОВИН составил 36,6±2,03 лет. Вторая группа пациентов с ОВИН: в сезон гриппа 2019-2020 после получения информированного согласия были иммунизированы 9 человек одномоментно двумя дозами по 0,5 мл трехвалентной субъединичной адъювантной вакцины против вируса гриппа Гриппол Плюс.
В качестве группы контроля для определения рабочих концентраций иммунологических показателей выступали здоровые добровольцы, которые были вакцинированы по аналогичным схемам, что и пациенты с ОВИН. 32 здоровых добровольца возрастом от 19 до 48 лет (32,56±1,67 лет) в сезон 2018-2019 были также вакцинированы одной дозой четырехвалентной вакцины, 10 из которых были повторно вакцинированы через 1 год (в сезон 2019-2020) трехвалентной адъювантной вакциной против гриппа Гриппол Плюс.
Между двумя группами не было значительных различий в отношении возраста и пола. Все участники в предшествующие два сезона (2016-2017 гг., 2017-2018 гг.) не были вакцинированы против гриппа так же, как и не было зарегистрированных случаев подтвержденной перенесенной гриппозной инфекции.
Критерии включения и не включения
• Подтвержденный диагноз ОВИН в соответствии с диагностическими критериями, установленными Европейским обществом по Иммунодефицитным заболеваниям (http://esid.orR/WorkingParties/Registry/Diagnosis-criteria) и Американской академией по Аллергии, Астме и Иммунологии для диагностики и лечения ПИД [61].
• Проведение заместительной иммунотерапии препаратами ВВИГ не позднее, чем за 28 дней до вакцинации и не ранее, чем через 21 день после нее, то есть перерыв между двумя последующими введениями иммуноглобулинов не менее 7 недель
• Исключены причины вторичной гипогаммаглобулинемии
• Исключены лица с когнитивными или поведенческими нарушениями, или психическими расстройствами, или злоупотребляющие алкоголем, что могло помешать участию в исследовании
• Добровольное подписание информированного согласия на участие в исследовании
• Отсутствие симптомов гриппа и гриппоподобных заболеваний в течение последних 6 месяцев
• Отсутствие симптомов острой инфекции на момент вакцинации
• Отсутствие приема на момент исследования и за 3 месяца до его начала глюкокортикостероидных препаратов или иной иммуносупрессивной терапии
• Отсутствие симптомов энтеропатии с потерей белка, а также подозрения на онкологическое или лимфопролиферативное заболевание у пациентов с ОВИН на момент исследования
• Отсутствие уровня специфических противовирусных антител в защитных титрах (>1:40) в довакцинальных образцах крови
• Отсутствие в анамнезе аллергии на яичные белки или какой-либо компонент исследуемой вакцины
• Отсутствие вакцинации против любых иных инфекций в течение 1,5-2 мес.до включения в исследование
Вакцины и вакцинация
Вакцины Гриппол Квадривалент и Гриппол Плюс (ООО "НПО Петровакс Фарм", Россия) были доступны для иммунизации лиц от 18 до 60 лет в сезон гриппа 2018-2019 и 2019-2020, соответственно, и включали штаммы, соответствующие рекомендациям ВОЗ в северном полушарии 2018-2019 годов для четырехвалентных вакцин (A/Michigan/45/2015(H1N1)pdm09-подобный вирус; A/Singapore/INFIMH-16-0019/2016 (H3N2)-подобный вирус; B/Colorado/06/2017-подобный вирус (B/Victoria/2/87 линия); B/Phuket/3073/2013-подобный вирус (B/Yamagata/16/88 линия)) и 2019-2020 годов для трехвалентных вакцин (А/Brisbane/02/2018(H1N1)pdm09-подобный вирус; А/Kansas/14/2017(H3N2)-подобный вирус; B/Colorado/06/2017-подобный вирус (линия B/Victoria/2/87)). Вакцины гриппозные инактивированные полимер-субъединичные представляет собой протективные антигены (гемагглютинин и нейраминидаза), выделенные из очищенных вирусов гриппа типа А и В, выращенных на куриных эмбрионах, связанные с водорастворимым высокомолекулярным иммуноадъювантом N-оксидированным производным поли-1,4-этиленпиперазина (азоксимера бромид). Вакцины содержали по 5 мкг гемагглютинина каждого включенного штамма гриппа и азоксимера бромида (полиоксидоний) - 500 мкг. Штаммы антигенов вируса гриппа - в соответствии с рекомендациями ВОЗ на текущий эпидемический сезон. Включение в вакцинный препарат иммуноадъюванта (Полиоксидоний®), обладающего широким спектром иммунофармакологического действия, обеспечивает увеличение иммуногенности и стабильности антигенов, позволяет повысить иммунологическую память, существенно снизить прививочную дозу антигенов.
Вакцинация проводилась в отделении иммунопатологии Института иммунологии ФМБА России с соблюдением всех правил асептики и антисептики, а также в соответствии с инструкцией, прилагаемой к препаратам.
Безопасность
Добровольцев наблюдали в течение 30-45 мин после каждой иммунизации для мониторинга немедленных побочных реакций. Участникам были предоставлены дневники наблюдений, а также постоянно поддерживалась связь путем ежедневного телефонного опроса с регистрацией частоты развития местных и системных реакций в течение 14 дней после вакцинации. Местные реакции оценивались по частоте возникновения боли в месте введения, эритемы, экхимозов, уплотнения и отека. К системным реакциям были отнесены: озноб, недомогание, миалгия, артралгия, головная боль, тошнота, потливость, кашель и повышение температуры тела (температура в подмышечной области фебрильная ≥38,0°С; пиретическая ≥39,0°С), а также использование анальгетиков или жаропонижающих препаратов. В зависимости от диаметра местные реакции расценивались как «нет реакции» - от 1 до 10 мм, легкие - от 11 до 25 мм, умеренные - от 26 до 50 мм, а от 51 до 100 мм или >100 мм - тяжелые. Клиническое наблюдение проводилось за всеми здоровыми и пациентами с ОВИН с регистрацией необычных явлений с 1-го по 30-й день.
Методы исследования
Лабораторная часть исследования осуществлялась в лаборатории механизмов иммунной регуляции ФГБНУ «Научно-исследовательский институт вакцин и сывороток им. И.И. Мечникова» МЗ РФ в Москве.
Исследование иммуногенности
Иммуногенность вакцин оценивали с помощью стандартной методики (МУ 3.3.2 1758-03) в реакции торможения гемагглютинации (РТГА), проводимой на образцах сыворотки, собранных до, через 24±3 дня и 3 месяца после вакцинации против гриппа, путем измерения титров антител к штаммам гриппа, входящих в состав вакцин. РТГА проводили с агглютинирующими единицами антигенов и 0,5% куриными эритроцитами с предварительным нагреванием исследуемых сывороток до 56°С в течение 1 часа (для удаления неспецифических ингибиторов и повышения чувствительности реакции). Первичный анализ оценки иммуногенности у пациентов с ОВИН проводили через 24±3 дня после введения препарата, а вторичный анализ - через 3 месяца только в группе больных, вакцинированных в сезон гриппа 2018-2019 годов, в соответствии с протоколом и был обоснован с медицинской точки зрения как потенциально влияющий на результаты иммуногенности.
Иммунологическая эффективность вакцинации против гриппа оценивалась по следующим критериям: уровень сероконверсии, серопротекции и фактор сероконверсии в соответствии с критериями последней редакцией Руководства по клинической оценке вакцин от 26 апреля 2018 г. Комитета по лекарственным препаратам для человека (Committee on Human Medicinal Products - CHMP) для взрослых пациентов.
Уровень серопротекции - доля исследуемых с титром антител (AT) ≥1:40 на 21-й день после вакцинации (референсные значения для здоровых - выше 70%).
Уровень сероконверсии - доля исследуемых, у которых титр AT на 21-й день после вакцинации повысился более, чем в 4 раза по сравнению с исходным уровнем при уровне AT≥10 или при довакцинальном титре АТ≤10 до значений ≤40 (референсные значения для здоровых - выше 40%).
Фактор сероконверсии (GMR) - кратность нарастания среднего геометрического значения титра антител (GMT) на 21-й день после вакцинации в сравнении с исходным (референсные значения для здоровых - более чем в 2,5 раза).
В зависимости от исходных титров антител здоровые добровольцы были разделены на две группы - серонегативные (≤1:20) и серопозитивные (≥1:40) к разным штаммам, входящим в состав четырехвалентной адъювантной вакцины против гриппа.
Однако в соответствии с методическими рекомендациями Европейского агентства по лекарственным средствам от 2018 года по клинической оценке вакцин помимо оценки иммуногенности вакцин по указанным выше трем критериям (средние геометрические титры AT (GMT) и их соотношение до и после вакцинации (GMR), уровень сероконверсии и серопротекции), были впервые выделены еще 4 критерия, в том числе и по оценке клеточного и клеточно-опосредованного иммунитета, которые в меньшей степени проводятся в рутинной практике, а некоторые - и вовсе недоступны в данном исследовании впервые был проведен анализ иммуногенности четырехвалентной полимер-субъединичной (адъювантной) вакцины против гриппа с оценкой поствакцинальных уровней серопротекции и сероконверсии отдельно для серопозитивных и серонегативных участников на исходном уровне.
Оценка субпопуляционной структуры лимфоцитов
Субпопуляционную структуру лимфоцитов периферической крови у пациентов с ОВИН под воздействием вакцин против гриппа исследовали in vitro методом проточной цитометрии на приборе FC-500 (Beckman Coulter, США) с использованием моноклональных антител (мАТ) к CD45-FITC/CD3-ECD/CD19-PE, CD45-FITC/CD3-ECD/CD4-PE, CD45-FITC/CD3-ECD/CD8-PE, CD45-FITC/CD3-ECD/CD16CD56-РЕ,
Оценка внутриклеточных Толл-подобных рецепторов иммунокомпетентных клеток
Содержание TLRs-экспрессирующих гранулоцитов, лимфоцитов и моноцитов периферической крови у пациентов с ОВИН под воздействием вакцин против гриппа исследовали in vitro методом проточной цитометрии с применением мАТ к TLR3-PE, TLR7-FITC, TLR8-FITC, TLR9-PE (e-bioscience, США) с использованием проточного цитометра Cytomix FC-500 (Beckman Coulter, США) по методике, описанной в инструкции производителя.
Статистическая обработка данных
Обработка данных по антителам
Проверка нормальности распределения признаков проводилась критерием Шапиро-Уилкса (Shapiro-Wilk normality test). В результате были выявлены значительные отклонения распределения признаков от нормального. Описательная статистика уровня AT представлена средним геометрическим титром и его 95% доверительным интервалом (GMT, 95% CI). Также было рассчитано геометрическое среднее кратное увеличение по сравнению с исходным уровнем (фактор сероконверсии) и его 95% доверительный интервал (GMR, 95% CI).
Анализ динамики признаков и сравнение между группами исследования проводился с помощью построения робастной линейной модель смешанных эффектов (RLMEM) [62]. Статистическая значимость коэффициентов модели была определена с использованием аппроксимации степеней свободы Саттертуэйта [63]. Апостериорные сравнения (между группами в контрольных точках и между контрольными точками для каждой группы) проводились с помощью построения соответствующих контрастов на основе рассчитанной модели с помощью пакета emmeans [64]. Построение модели проводилось на преобразованных исходных данных, применялась логарифмическая трансформация [65]. При сравнении формирования поствакцинального иммунитета после первичной вакцинации одной и двумя дозами в качестве фиксированных факторов были определены время после введения вакцинного препарата (24±3 дня) и одна или две дозы вакцины, отдельные пациенты были заданы как случайные факторы. При сравнении формирования поствакцинального иммунитета после первичной и после повторной вакцинации одной дозой препарата в качестве фиксированных факторов были определены время после введения вакцинного препарата (24±3 дня) и этап вакцинации (первичная/повторная), отдельные пациенты (учитывался связанный характер выборок как во времени, так и этапами вакцинации) были заданы как случайные факторы.
Сравнение фактора сероконверсии между группами исследования проводилось с помощью критерия Манна-Уитни в случае несвязанных выборок (одна и две дозы вакцины) и с помощью парного критерия Вилкоксона в случае связанных выборок (первичная и повторная вакцинация одной дозой).
Для качественных признаков находили абсолютные и относительные (в %) частоты, для относительных показателей рассчитывался 95% доверительный интервал, рассчитанный по методу Вилсона. Сравнения двух групп по качественным номинальным показателям проводили в ходе анализа таблиц сопряженности точным критерием Фишера. Для связанных выборок (до - после вакцинации) применялся точный тест Мак-Немара [66]. В случае более 2-х измерений применялся омнибусный тест Q Кохонена, с последующим проведением апостериорных сравнений точным тестом Мак-Немара.
Различия считали статистически значимыми при р≤0,05, незначимыми - при р≥0,10, в промежуточных случаях (0,05<р<0,1) обсуждали тенденции к различиям [67]. Расчеты и графические построения производились с использованием программы GraphPad Prism (v.9.3.0 лицензия GPS-1963924) и статистической среды R (v.3.6, лицензия GNU GPL2).
Обработка результатов оценки клеточного звена иммунитета (TLR) Проверка нормальности распределения признаков проводилась критерием Шапиро-Уилкса (Shapiro-Wilk normality test). В результате были выявлены значительные отклонения распределения признаков от нормального.
Анализ динамики признаков и сравнение между группами исследования проводился с помощью построения робастной линейной модель смешанных эффектов (RLMEM) [62]. Статистическая значимость коэффициентов модели была определена с использованием аппроксимации степеней свободы Саттертуэйта [63]. Апостериорные сравнения (между группами в контрольных точках и между контрольными точками для каждой группы) проводились с помощью построения соответствующих контрастов на основе рассчитанной модели с помощью пакета emmeans [64]. Построение модели проводилось на преобразованных исходных данных, применялось арксинусное преобразование и логарифмическая трансформация [68]. При сравнении формирования поствакцинального иммунитета после первичной вакцинации одной и двумя дозами в качестве фиксированных факторов были определены время после введения вакцинного препарата (24±3 дня) и одна или две дозы вакцины, отдельные пациенты были заданы как случайные факторы. При сравнении формирования поствакцинального иммунитета после первичной и после повторной вакцинации одной дозой препарата в качестве фиксированных факторов были определены время после введения вакцинного препарата (24±3 дня) и этап вакцинации (первичная/повторная), отдельные пациенты (учитывался связанный характер выборок как во времени, так и этапами вакцинации) были заданы как случайные факторы.
Сравнение дельты изменений числа клеток между группами исследования проводилось с помощью критерия Манна-Уитни в случае несвязанных выборок (одна и две дозы вакцины) и с помощью парного критерия Вилкоксона в случае связанных выборок (первичная и повторная вакцинация одной дозой).
Различия считали статистически значимыми при р≤0,05, незначимыми - при р≥0,10, в промежуточных случаях (0,05<р<0,1) обсуждали тенденции к различиям [67]. Расчеты и графические построения производились с использованием программы GraphPad Prism (v.9.3.0 лицензия GPS-1963924) и статистической среды R (v.3.6, лицензия GNU GPL2).
ПРИМЕРЫ
Пример 1. Клиническая оценка течения поствакцинального периода у пациентов с ОВИН и здоровых при введении 3- и 4-валентных полимер-субъединичных (адъювантных) препаратов.
Все участники завершили исследование без развития серьезных поствакцинальных реакций вне зависимости от примененного препарата так же, как и схемы введения. Среди 32 здоровых добровольцев, которым вводилась четырехвалентная вакцина против гриппа, местные реакции были зарегистрированы у 9% (3 человека) и характеризовались локальной умеренной гиперемией, сохранявшейся в течение 2-х суток. Системная реакция - недомогание - наблюдалась у 3% (одного человека) через 3 ч после вакцинации и сохранялась в течение суток. Наблюдаемые поствакцинальные явления не требовали назначения каких-либо лекарственных средств. В течение года, следующим за проведением вакцинации, проводился анализ возникновения гриппа в исследуемой группе пациентов, а также сбор информации о болезни контактных лиц.
В ходе исследования по наблюдению за течением поствакцинального периода у пациентов с ОВИН (15 пациентов), вакцинированных против гриппа с использованием четырех- или трехвалентных вакцин как при первичной, так и повторной (через 1 год) вакцинации, одной или двумя одновременно дозами, лишь у 6% (1 больного) местные реакции в виде умеренной гиперемии (диаметром до 50 мм) и локального отека появились через 2 ч после инъекции и сохранялись в течение 3 суток после вакцинации. Только одна (6%) системная реакция (озноб) была зарегистрирована в день вакцинации без последующего повышения температуры и регрессировала через 12 часов. Следует отметить, что несмотря на длительный период без проведения базисной терапии ВВИГ в течение 7 недель в связи с проведением вакцинации, ни одному больных ОВИН не потребовалось его срочное введение до окончания выше указанного срока в связи с отсутствием обострения сопутствующей хронической патологии так же, как и присоединением ОРИ.
2 (22%) из 9 пациентов с ОВИН, вакцинированных одновременно двумя дозами вакцины, и одна пациентка (16%) из 6 больных, вакцинированная одной дозой препарата, отметили в течение 1-2 месяцев после иммунизации значительное улучшение состояния, усиление активности, а также то, что при контакте с членами семей, проживавшими с ними на одной территории и заболевшими лабораторно подтвержденным вирусом гриппа, не отметили у себя симптомов заболевания.
Пример 2. Исследование показателей иммуногенности вакцины против гриппа у здоровых.
У здоровых пациентов, иммунизированных четырехвалентной адъювантной вакциной со сниженным количеством антигенов против всех 4 штамм-специфических поверхностных антигенов до 5 мкг, вакцина доказала свою иммуногенность по таким критериям, как уровень серопротекции (≥70%), сероконверсии (≥40%), фактор сероконверсии (≥2,5) как спустя 24±3 дня после вакцинации, так и через 3 месяца, что проводилось на первом году (2018-2019 гг.) проводимого исследования для оценки динамики формирования гуморального иммунитета у пациентов с ОВИН (n=6), вакцинированных одной дозой четырехвалентной субъединичной адъювантной вакциной. Учитывая ограниченное число пациентов с ОВИН (6 человек), вначале приведена оценка иммуногенности четырехвалентной адъювантной вакцины в группе здоровых, далее индивидуально по каждому больному.
В табл.1, фиг. 1 представлен уровень серопротекции (HI≥40, должен составлять>70%) через 24±3 дня после введения aQIV в группе здоровых исследуемых с учетом изначального уровня AT.
До вакцинации в группе изначально серопозитивных участников уровень серопротекции был 100% ко всем рассматриваемым штаммам. В поствакцинальном периоде он практически не менялся, оставаясь на уровне 95-100%. В группе изначально серонегативных исследуемых уровень серопротекции до вакцинации составил 0% ко всем штаммам. Через 24±3 дня после вакцинации отмечается статистически значимый прирост уровня серопротекции до 70% к штамму A/H1N1, 100% к штамму A/H3N2, 54% к штамму В/Колорадо и 75% к штамму В/Пхукет.
В табл.2, фиг. 2 представлен уровень сероконверсии (критерий сероконверсии >40% с 4-х кратным нарастанием титров AT против исходного) в группе здоровых исследуемых с учетом изначального уровня AT.
В группе здоровых добровольцев изначально серонегативных уровень сероконверсии соответствует критерию эффективности по всем рассматриваемым штаммам и составляет 70% к штамму A/H1N1, 100% к штамму A/H3N2, 46% к штамму В/Колорадо и 63% к штамму В/Пхукет. В группе изначально серопозитивных пациентов уровень сероконверсии через 24±3 дня после иммунизации составил 40% к штамму A/H1N1, также 40% к штамму A/H3N2, 43% к штамму В/Колорадо и 39% к штамму В/Пхукет.
В табл. 3, фиг. 3 представлен фактор сероконверсии (критерий GMR до и после вакцинации должен быть >2,5) в группе здоровых исследуемых с учетом изначального уровня AT.
В группе здоровых, изначально являющихся серонегативными, через 24±3 дня фактор сероконверсии составил для штаммов A/H1N1, A/H3N2, В/Колорадо, В/Пхукет соответственно 9,8, 34,3, 3,8, 5,9, а для исходно серопозитивных лиц в те же сроки исследования он составил 4,8, 2,8, 2,5 и 2,8, соответственно.
В табл. 4, фиг. 4 представлены средние геометрические концентрации титров AT (GMT) в группе здоровых исследуемых с учетом изначального уровня AT. В поствакцинальном периоде у здоровых добровольцев, относящихся к серонегативным, через 24±3 дня GMT AT к штаммам вируса гриппа достоверно повышалась против исходного до 65,0, 226,3, 27,4 и 51,9 соответственно для штаммов A/H1N1, A/H3N2, В/Колорадо, В/Пхукет, а у серопозитивных лиц к исследуемым разновидностям линий вируса гриппа - также значимое увеличение GMT AT до 452,5, 288,4, 214,9 и 198,0, соответственно.
Индивидуальный анализ содержания антител к различным штаммам вируса гриппа показал, что 2 из 6 пациентов с ОВИН исходно были серопозитивны, у одного выявлялись титры AT 1:40 к штаммам A/H1N1, A/H3N2, а у другого пациента к этим же штаммам 1:80 (табл. 5). Остальные 4 пациента были серонегативными ко всем 4 вакцинным штаммам вируса гриппа. В поствакцинальном периоде через 24±3 дня дополнительно один пациент стал серопозитивным (1:80) к штамму A/H3N2.
Спустя 3 мес.число серопозитивных увеличилось еще на одного человека к штаммам A/H1N1, A/H3N2 - титр AT равен 1:40. Выявлена тенденция к росту титров AT через 3 мес.к штамму A/H3N2 с 1:5 до 1:20, у 3 пациентов - к штамму В/Колорадо с 1:5 до 1:20; с 1:5 до 1:10; с 1:10 до 1:20. К штамму В/Пхукет лишь в одном случае выявлен прирост AT с 1:5 до 1:10 через 3 мес.
Пример 3. Оценка критериев иммуногенности четырехвалентной адъювантной вакцины против гриппа у пациентов с ОВИН, иммунизированных одной дозой.
На основании индивидуального анализа титра специфических антител в течении 3 месяцев у каждого пациента с ОВИН, вакцинированных одной дозой четырехвалентной субъединичной адъювантной вакциной против гриппа, можно отметить, что у 3 из 6 пациентов отмечалось увеличение титра антител более, чем в 2 раза к линии гриппа B/Victoria (более того, у одного пациента рост титра антител составил 4 раза) через 3 месяца после вакцинации, чего не наблюдалось при анализе крови через 24±3 дня. Однако защитный титр антител ≥1: 40 к штаммам двух линий (Yamagata и Victoria) не был достигнут в целом и составлял не более 1:20. Следует отметить, что для пациентов с ОВИН условным показателем эффективности вакцинации является повышение поствакцинального уровня антител в 2-4 раза по сравнению с уровнем до вакцинации.
Что касается уровня антител к A/H3N2, то у 3 пациентов отмечалось повышение титра антител до 1:20, 1:40 и 1:80 (в 2, 4 и 8 раз соответственно); они также были обнаружены в 2 из 6 проб крови через 3 месяца и одной из /6 - через 3 недели после иммунизации. Для штамма A/H1N1 только у одного пациента в поствакцинальном периоде отмечалось увеличение титра антител в 2 раза через 3 месяца по сравнению с исходным (1:20) и достигало защитного уровня (1:40) - таблица 5.
Пример 4. Оценка критериев иммуногенности четырехвалентной адъювантной вакцины против гриппа у пациентов с ОВИН, иммунизированных одной дозой первично и повторно через 1 год.
В таблице 6 приведен уровень серопротекции в группе пациентов с ПИД, вакцинированных одной дозой вакцины первично и повторно через год после первой вакцинации.
Анализ динамики уровня серопротекции к штамму A/H1N1 на разных этапах вакцинации выявил наличие изменений на пограничном уровне статистической значимости (р=0,07). В частности, через 24±3 дня после повторной вакцинации по сравнению с исходным уровнем (до первичной вакцинации) доля серопозитивных пациентов выросла с 20[4;62]% до 80[38;96]% (р=0,08) (фиг. 5). Уровень серопротекции к остальным штаммам оставался без изменений в течение всего периода исследования.
Анализ уровня сероконверсии показал, что в результате первичной вакцинации 4-х кратный и более прирост AT был зафиксирован только у одного пациента (с 20 до 80 к штамму A/H3N2). После повторной вакцинации относительно уровня до повторной вакцинации (через год после первичной вакцинации) ни у одного пациента не было зарегистрировано 4-х кратного увеличения AT, относительно исходного уровня (до первичной вакцинации) только у одного участника исследования наблюдалось 4-х кратное увеличение уровня AT (с 20 до 80 к штаммам A/H1N1 и A/H3N2 у одного и того же пациента).
В таблице 7 приведены средние геометрические титры AT (GMT) в контрольных точках исследования (до первичной и повторной вакцинации и через 24±3 дня после) в группах пациентов с ПИД, вакцинированных одной дозой вакцины. Там же приведены результаты построения линейных моделей смешанных эффектов (RLMEM) зависимости уровня AT от этапа вакцинации (первично/повторно) и времени после введения вакцинного препарата отдельно для каждого штамма.
Выявлено статистически значимое разнонаправленное изменение уровня AT в зависимости от этапа вакцинации (первичная/повторная) к штамму A/H1N1 (t=2,5, р=0,05, df=13) - фигура 6. Если через 24±3 дня после первичной вакцинации уровень AT остается без значительных изменений, то после повторной вакцинации отмечается статистически значимый прирост уровня AT до 34,8[8,47; 143,2] по сравнению с уровнем до ревакцинации (р=0,05, проводилась через год после первичной вакцинации) и с уровнем после первичной вакцинации (р=0,02).
Фактор сероконверсии (средний геометрический прирост - GMR) через 24±3 дня после первичной и повторной вакцинации, а также за весь период наблюдения (через 24±3 дня после повторной вакцинации относительно уровня до первичной вакцинации) представлен в таблице 8.
По фактору сероконверсии (среднему геометрическому приросту уровня AT) между первичной и повторной вакцинацией не было выявлено статистически значимых различий ни для одного из рассматриваемых штаммов.
Отдельно рассмотрен такой показатель, как доля случаев с 2-х кратным и более приростом AT - фигура 7. После первичной вакцинации было зарегистрировано по одному случаю на 5 пациентов к штаммам A/H3N2 и В/Колорадо. После повторной вакцинации 2-х кратный и более прирост наблюдался у 4-х человек к штамму A/H1N1, у 2-х пациентов к штамму A/H3N2, у одного - к штамму В/Колорадо и у 3-х - к штамму В/Пхукет. По каждому отдельному штамму статистически значимых различий выявлено не было, но связанный характер выборок и гомогенность отношения шансов прироста AT по исследуемым штаммам (р=0,17, проверялась критерием Берслоу-Доу) позволяет применить критерий Кохрана-Мантеля-Хенселя, который показал, что обобщенный по всем рассматриваемым штаммам шанс 2-х кратного прироста AT, в результате повторной вакцинации одной дозой в 6,7[1,4; 34,5] раз выше (отношение шансов Мантеля-Хенселя), чем при первичной вакцинации одной дозой (р=0,02).
Пример 5. Оценка критериев иммуногенности вакцин против гриппа у пациентов с ОВИН, иммунизированных одной и двумя (одномоментно) дозами.
В таблице 9 приведен уровень серопротекции и уровень сероконверсии в группах пациентов с ПИД, вакцинированных первично одной и двумя дозами вакцины.
Ни по одному из показателей не выявлено статистически значимых различий ни в динамике у каждого пациента, ни между группами. Но стоит отметить, что данные показатели (уровень серопротекции и сероконверсии) изначально предназначены для оценки иммунного ответа здоровых пациентов, для пациентов с ПИД они является слишком «грубыми», поэтому далее рассмотрены непосредственно титры AT и их изменение.
Средние геометрические титры AT (GMT) в группах пациентов с ПИД, вакцинированных первично одной и двумя дозами вакцины, исходно и через 24±3 дня после вакцинации приведены в таблице 10. Там же приведены результаты построения линейных моделей смешанных эффектов (RLMEM) зависимости уровня AT от схемы первичной вакцинации (одна/две дозы) и времени после вакцинации отдельно для каждого штамма.
Фактор сероконверсии через 24±3 дня после первичной вакцинации представлен в таблице 11.
Построение и анализ линейной модели смешанных эффектов зависимости уровня AT к штамму A/H1N1 от количества доз и времени после вакцинации позволил выявить, что в среднем за рассматриваемый период титр AT к A/H1N1 в группах пациентов с ПИД, первично вакцинированных одной и двумя дозами, статистически значимо не различался (t=-0,1, p=0,93, df=14). Также стоит отметить отсутствие согласованной динамики в группах исследования (t=-1,4, р=0,19, df=13), однако была выявлена статистически значимая разнонаправленная динамика (t=2,5, р=0,02, df=13) - фигура 8.
В частности, в группе пациентов, вакцинированных одной дозой, через 24±3 дня после вакцинации отмечается статистически не значимое снижение титра AT, объясняемое случайными флуктуациями, в то время как в группе пациентов, вакцинированных двумя дозами, выявлен статистически значимый прирост относительно исходного уровня с 25 [12; 54] до 34[16; 75] (р=0,02). Средний геометрический прирост титра также статистически значимо выше в группе пациентов, вакцинированных двумя дозами: 1,36[1,03; 1,80) против 0,79[0,55; 1,16] (р=0,03) - фигура 9.
Анализ поствакцинального ответа к штамму A/H3N2 показал, что через 24±3 дня после вакцинации титр AT в группах исследования изменяется по-разному (t=2,l, р=0,05, df=13), и если в группе пациентов, вакцинированных одной дозой, прирост статистически не значимый (р=0,89), то в группе пациентов, вакцинированных двумя дозами, отмечается статистически значимое увеличение титра AT: с 11 [6; 19] до 17[10; 29] (р=0,01) - фигура 10.
Интенсивность изменения титра AT (средний геометрический прирост) выше в группе пациентов, вакцинированных двумя дозами: 1,59 [1,00; 2,52] против 1,00 [0,63; 1,58] (р=0,07 - на уровне тенденции) - фигура 11.
Анализ поствакцинального ответа к штамму В/Колорадо выявил, что в среднем за период уровень AT не зависит от схемы вакцинации (t=-0,1l, р=0,91, df=21) и не имеет выраженного направления изменений в результате вакцинации (t=0,04, р=0,96, df=13), но при этом в зависимости от схемы вакцинации средний титр AT меняется по-разному (t=2,5, р=0,02, df=13) - фигура 12. Так, если в группе пациентов, вакцинированных одной дозой вакцины, средний геометрический титр через 24±3 дня после вакцинации остается без изменений относительно исходного уровня, то в группе пациентов, вакцинированных двумя дозами, отмечается статистически значимый прирост титра - с 7 [5; 10] до 11 [7; 18] (р=0,02) - фигура 13.
В целом, если рассматривать все штаммы в совокупности, можно утверждать, что в группе пациентов, вакцинированных одной дозой, случаи прироста титра AT у отдельных пациентов являются единичными, в то время как в группе пациентов, вакцинированных двумя дозами, доля таких случаев значительно выше - фигура 14. По каждому отдельному штамму статистически значимых различий выявлено не было, но связанный характер выборок (каждый из 3-х штаммов рассматривался на одном и том же пациенте) и гомогенность отношения шансов прироста AT по исследуемым штаммам (р=0,62, проверялась критерием Берслоу-Доу) позволяет применить более мощный критерий (критерий Кохрана-Мантеля-Хенселя), который показал, что шанс прироста AT в результате вакцинации двумя дозами в 9,3 [1,6; 51,4] раз выше (отношение шансов Мантеля-Хенселя), чем при вакцинации одной дозой (р=0,02), независимо от штамма.
Пример 6. Влияние вакцин против гриппа на содержание TLR3, TLR8, TLR9 у пациентов с ОВИН, привитых одной и двумя (одномоментно) дозами.
Гранулоциты
После вакцинации одной дозой отмечается статистически значимое снижение доли гранулоцитов, экспрессирующих TLR9 - с 93,6(92,4-96,9)% до 86,0(81,0-91,4)% (р=0,05) -и на пограничном уровне значимости экспрессирующих TLR3 - с 93,8(91,3-95,0)% до 89,5(81,9-91,1)% (р=0,08). После вакцинации двумя дозами подобных изменений выявлено не было - фигура - 15 а, б.
Вследствие описанных выше изменений, через 24±3 дня после вакцинации двумя дозами доля гранулоцитов, экспрессирующих TLR9, была статистически значимо выше, чем при вакцинации одной дозой - 96,7(90,4-98,1)% против 86,0(81,0-91,4)% (р=0,02). Дельта изменения доли гранулоцитов (относительно исходного уровня), экспрессирующих TLR3 и TLR9 составила соответственно -5,6[-9,8; -1,0]% и -8,8[-12,2; -3,2]% при вакцинации одной дозой и +3,5[-2,9;+19,4]% и +4,5[-0,7;+26,5]% при вакцинации двумя дозами (р=0,11для TLR3 и р=0,03 для TLR9 при сравнении с одной дозой) - фигура 16 а, б.
Процентный уровень гранулоцитов, экспрессирующих TLR8, статистически значимо не меняется вследствие вакцинации (t(13)=-1,2, р=0,25) и не зависит от количества доз (t(20)=0,9, р=0,36), показывая сопоставимую динамику для одной и двух доз (t(13)=0,5, р=0,66). Данный показатель составлял (до и после вакцинации соответственно) 93,7(85,2-96,0)% - 88,9(85,7-91,0)% при вакцинации одной дозой и 94,5(82,7-99,1)% - 89,8(84,8-97,9)% при вакцинации двумя дозами.
Лимфоциты
После вакцинации двумя дозами отмечался статистически значимый прирост доли лимфоцитов, экспрессирующих TLR9 - с 2,70(0,40-6,60)% до 15,40(1,90-26,75)% (р=0,02). После вакцинации одной дозой подобных изменений выявлено не было. В результате данного прироста через 24±3 дня после вакцинации двумя дозами доля лимфоцитов, экспрессирующих TLR9, была выше, чем после вакцинации одной дозой - 15,40(1,90-26,75)% против 0,30(0,05-1,28)% (р<0,001) - рисунок 15 в.
Изменение доли лимфоцитов, экспрессирующих TLR9, в результате вакцинации двумя дозами составило+11[+1,5;+29,4]%, а в результате вакцинации одной дозой - -0,1[-0,3;+0,3]%, различия статистически значимы (р=0,05) - рисунок 16в.
Процентный уровень лимфоцитов, экспрессирующих TLR3 и TLR8, в результате вакцинации статистически значимо не изменился (t(13)=-l,0, р=0,32 и t(13)=-0,7, р=0,50 соответственно), не зависел от дозы (t(24)=0,5, р=0,60 и t(26)=0,8, р=0,42) и показывал сопоставимую динамику для одной и двух доз (t(13)=l,5, р=0,16 и t(13)=0,2, р=0,84). Доля лимфоцитов, экспрессирующих TLR3, составляла (для одной и двух доз соответственно) -0,25(0,09-1,73)% и 0,60(0,06-2,95)% до вакцинации и 0,20(0,09-0,43)% и 0,80(0,35-3,45)% после вакцинации. Доля лимфоцитов, экспрессирующих TLR8, составляла (для одной и двух доз соответственно) - 0,25(0,10-1,85)% и 1,30(0,45-5,60)% до вакцинации и 0,33(0,08-0,43)% и 0,20(0,10-2,60)% после вакцинации.
Моноциты
После вакцинации отмечалось статистически значимое разнонаправленное изменение доли моноцитов, экспрессирующих TLR3 и TLR9, в зависимости от числа доз (t(13)=3,8, р=0,002 для TLR3, t(13)=3,4, р=0,005 для TLR9) - фигура 17а, б.
Если в результате вакцинации двумя дозами отмечался прирост данных показателей, то в результате вакцинации одной дозой, напротив, отмечалось статистически значимое снижение. В результате вакцинации двумя дозами исходной уровень лимфоцитов, экспрессирующих TLR3 и TLR9, вырос на +10,7[-2,7; +29,2]% с 16,2(9,3-27,8)% до 31,2(17,3-39,1)% (р=0,05) и на 21,4[+7,0;+51,3]% с 38,4(16,0-42,6)% до 49,2(30,8-73,0)% (р=0.04), соответственно. В то время как после вакцинации одной дозой, снижение данных показателей составило -25,1 [-45,6;+0,2]% с 53,1(22,8-75,6)% до 7,3(2,7-44,0)% (р<0,001 для TLR3) и на -23,3[-57,6; -7,8]% с 66,1(35,3-80,8)% до 6,3(1,6-73,0)% (р=0,01 для TLR9). Интенсивность изменения доли лимфоцитов, экспрессирующих TLR3 и TLR9, в результате вакцинации между группами одной и двумя дозами различается статистически значимо (р=0,01 в обеих случаях) - фигура 18а, б.
При этом стоит отметить, что в группе пациентов с введением двух доз вакцины исходный уровень лимфоцитов, экспрессирующих TLR3, был ниже, чем в группе с одной дозой - 16,2(9,3-27,8)% против 53,1(22,8-75,6)% (р=0,01). Аналогичная картина наблюдалась и для TLR9 - 38,4(16,0-42,6)% против 66,1(35,3-80,8)% (р=0,07 -пограничный уровень значимости). Но в результате вакцинации двумя дозами доля лимфоцитов, экспрессирующих TLR3, стала сопоставимой с уровнем после вакцинации одной дозой - 31,2(17,3-39,1)% и 7,3(2,7-44,0)%, соответственно (р=0,16), а доля лимфоцитов, экспрессирующих TLR9, после вакцинации стала выше в группе с двумя дозами по сравнению с группой с одной дозой препарата - 49,2(30,8-73,0)% против 6,3(1,6-73,0)% (р=0,05).
Изменение уровня моноцитов, экспрессирующих TLR8, не зависит от числа доз (t(25)=-0,8, р=0,41) и показывает согласованную динамику для одной и двух доз (t(13)=0,7, р=0,52), но при этом зависит от самого факта вакцинации (t(13)=-3,5, р=0,004). В результате вакцинации как одной, так и двумя дозами доля моноцитов, экспрессирующих TLR8, снизилась статистически значимо: на 34,9[-67,0; -15,4]% с 67,5(13,9-87,4)% до 7,4(4,6-24,7)% (р<0,001) в группе с одной дозой и на 39,0[-58,3; -10,3]% с 59,4(26,2-62,2)% до 7,2(1,5-23,1)% в группе с двумя дозами - фигура 19а.
Интенсивность снижения между группами с одной и двумя дозами статистически значимо не различалась (р=0,96) - фигура 19б.
Ели при первичной вакцинации одной дозой отмечалось снижение большинства показателей (доли гранулоцитов, экспрессирующих TLR3, TLR9, и доли моноцитов, экспрессирующих TLR3, TLR8, TLR9), то при вакцинации двумя дозами, наоборот, наблюдался прирост части показателей (доли лимфоцитов, экспрессирующих TLR9, и доли моноцитов, экспрессирующих TLR3, TLR9), кроме доли моноцитов, экспрессирующих TLR8, которые снизились.
Таким образом, в проведенном нами исследовании обнаружена значительная экспрессия TLR9 после повторной вакцинации через год и при одновременном введении 2 доз адъювантных вакцин против гриппа, что сопоставимо с достоверным нарастанием уровней антител ко всем штаммам вируса гриппа, что не было отмечено при однократной вакцинации больных ОВИН одной дозой.
В результате проведенных исследований показана целесообразность изменения существующей схемы вакцинации пациентов с ОВИН против гриппа: более высокая иммуногенность отмечается после введения одномоментно двух доз полимер-субъединичной (адъювантной) вакцины или повторной вакцинации через 1 год одной дозой вакцинного препарата. Полученные результаты свидетельствуют об активации синтеза противогриппозных антител и запуска каскада иммунных реакций по распознаванию патогенов в поствакцинальном периоде при ежегодной вакцинации или первичном введении двух доз адъювантной вакцины против гриппа у пациентов с ОВИН. Полученные данные могут послужить основой для формирования рекомендаций по вакцинации пациентов с ОВИН против гриппа.
Список литературы:
1. В Gathmann, В Grimbacher, J Beaute, Y Dudoit, N Mahlaoui, A Fischer, V Knerr G.K. The European internet-based patient and research database for primary immunodeficiencies: results 2006-2008 / В Gathmann, В Grimbacher, J Beaute, Y Dudoit, N Mahlaoui, A Fischer, V Knerr G.K. // Clinical and Experimental Immunology - 2009. - Т. 157 - №Supplement_1.
2. Wen-I Lee, Jing-Long Huang, Tang-Her Jaing, Shyh-Dar Shyur, Kunder D Yang Y.-H.C. et al. Distribution, clinical features and treatment in Taiwanese patients with symptomatic primary immunodeficiency diseases (PIDs) in a nationwide population-based study during 1985-2010 / Wen-I Lee 1, Jing-Long Huang, Tang-Her Jaing, Shyh-Dar Shyur, Kunder D Yang Y.-H.C. et al. // Immunobiology - 2011. - T. 216 - №12 - C. 1286-1294.
3. Azizi G. Circulating Helper T-Cell Subsets and Regulatory T Cells in Patients With Common Variable Immunodeficiency Without Known Monogenic Disease / Azizi G., Mirshafiey A., Abolhassani H., Yazdani R., Jafarnezhad-Ansariha F., Shaghaghi M., Mortazavi-Jahromi S., Noorbakhsh F., Rezaei N., Aghamohammadi A. // Journal of Investigational Allergology and Clinical Immunology - 2018. - T. 28 - №3 - C.172-181.
4. Chapel H. Common variable immunodeficiency disorders: division into distinct clinical phenotypes / Chapel H., Lucas M., Lee M., Bjorkander J., Webster D., Grimbacher В., Fieschi C, Thon V., Abedi M.R., Hammarstrom L. // Blood - 2008. - Т. 112 - №2 - C. 277-286.
5. Azizi G. Autoimmunity in Primary Antibody Deficiencies / Azizi G., Ahmadi M., Abolhassani H., Yazdani R., Mohammadi H., Mirshafiey A., Rezaei N., Aghamohammadi A. // International Archives of Allergy and Immunology - 2016. - T. 171 - №3 - 4- C. 180-193.
6. Yazdani R. Asthma and Allergic Diseases in a Selected Group of Patients With Common Variable Immunodeficiency / Yazdani R., Heydari A., Azizi G., Abolhassani H., Aghamohammadi A. // Journal of Investigational Allergology and Clinical Immunology - 2016. - T. 26 - №3 - C. 209-211.
7. Yazdani R. Evaluation of Known Defective Signaling-Associated Molecules in Patients Who Primarily Diagnosed as Common Variable Immunodeficiency / Yazdani R., Abolhassani H., Rezaei N., Azizi G., Hammarstrom L., Aghamohammadi A. // International Reviews of Immunology - 2016. - T. 35 - №1 - C. 7-24.
8. Yazdani R. Impaired Akt phosphorylation in B-cells of patients with common variable immunodeficiency / Yazdani R., Ganjalikhani-Hakemi M., Esmaeili M., Abolhassani H., Vaeli S., Rezaei A., Sharifi Z., Azizi G., Rezaei N., Aghamohammadi A. // Clinical Immunology - 2017. - T. 175 - C. 124-132.
9. Azizi G. Clinical, immunologic, molecular analyses and outcomes of iranian patients with LRBA deficiency: A longitudinal study / Azizi G., Abolhassani H., Mahdaviani S.A., Chavoshzadeh Z., Eshghi P., Yazdani R., Kiaee F., Shaghaghi M., Mohammadi J., Rezaei N., Hammarstrom L., Aghamohammadi A. // Pediatric Allergy and Immunology - 2017. - T. 28 - №5 - C. 478-484.
10. Shearer W.T. Recommendations for live viral and bacterial vaccines in immunodeficient patients and their close contacts / Shearer W.T., Fleisher T.A., Buckley R.H., Ballas Z., Ballow M., Blaese R.M., Bonilla F.A., Conley M.E., Cunningham-Rundles C, Filipovich A.H., Fuleihan R., Gelfand E.W., Hemandez-Trujillo V., Holland S.M., Hong R., Lederman H.M., Malech H.L., Miles S., Notarangelo L.D., Ochs H.D., Orange J.S., Puck J.M., Routes J.M., Stiehm E.R., Sullivan K., Torgerson Т., Winkelstein J. // Journal of Allergy and Clinical Immunology - 2014. - T. 133 - №4 - C. 961-966.
11. Bonilla F.A. Update: Vaccines in primary immunodeficiency / Bonilla F.A. // Journal of Allergy and Clinical Immunology - 2018. - T. 141 - №2 - C. 474-481.
12. Rubin L.G. Executive Summary: 2013 IDS A Clinical Practice Guideline for Vaccination of the Immunocompromised Host / Rubin L.G., Levin M.J., Ljungman P., Davies E.G., Avery R., Tomblyn M., Bousvaros A., Dhanireddy S., Sung L., Keyserling H., Kang I. // Clinical Infectious Diseases - 2014. - T. 58 - №3 - C. 309-318.
13. Hong D.K. CROSS-REACTIVE NEUTRALIZING ANTIBODY AGAINST PANDEMIC 2009 H1N1 INFLUENZA A VIRUS IN INTRAVENOUS IMMUNOGLOBULIN PREPARATIONS / Hong D.K., Tremoulet A.H., Burns J.C., Lewis D.B. // Pediatric Infectious Disease Journal - 2011. - T. 30 - №1 - C. 67-69.
14. Gardulf A. Predictive markers for humoral influenza vaccine response in patients with common variable immunodeficiency / Gardulf A., Abolhassani H., Gustafson R., Eriksson L.E., Hammarstrom L. // Journal of Allergy and Clinical Immunology - 2018. -T. 142-№6-C1922- 1931.e2.
15. W.E. P.Fundamental Immunology, 6th edition / P. W.E. - Philadelphia: Wolters Kluwer/ Lippincott Williams & Wilkins, 2008.- 1603c.
16. Hanitsch L.G. Cellular and humoral influenza-specific immune response upon vaccination in patients with common variable immunodeficiency and unclassified antibody deficiency / Hanitsch L.G., Lobel M., Mieves J.F., Bauer S., Babel N., Schweiger В., Wittke K., Grabowski P., Volk H.D., Scheibenbogen C. // Vaccine - 2016. -T. 34-№21 - C. 2417-2423.
17. Assen S. van Patients with humoral primary immunodeficiency do not develop protective anti-influenza antibody titers after vaccination with trivalent subunit influenza vaccine / Assen S. van, Holvast A., Telgt D.S.C., Benne C.A., Haan A. de, Westra J., Kallenberg C.G.M., Bijl M. // Clinical Immunology - 2010. - T. 136 - №2 - C. 228-235.
18. Hartley G.E. Influenza-specific IgG1+ memory B-cell numbers increase upon booster vaccination in healthy adults but not in patients with predominantly antibody deficiency / Hartley G.E., Edwards E.S.J., Bosco J.J., Ojaimi S., Stirling R.G., Cameron P.U., Flanagan K., Plebanski M., Hogarth P.M., O'Hehir R.E., Zelm M.C. // Clinical & Translational Immunology - 2020. - T. 9 - №10.
19. Alexia C. Polyoxidonium® Activates Cytotoxic Lymphocyte Responses Through Dendritic Cell Maturation: Clinical Effects in Breast Cancer / Alexia C, Cren M., Louis-Plence P., Vo D.-N., Ahmadi Y. El, Dufourcq-Lopez E., Lu Z.-Y., Hernandez J., Shamilov F., Chernysheva O., Vasilieva M., Vorotnikov I., Vishnevskay Y., Tupitsyn N., Rossi J.-F., Villalba M. // Frontiers in Immunology - 2019. - T. 10.
20. Kostinov M.P. Prospective randomized open-label comparative study of immunogenicity after subunit and polymeric subunit influenza vaccines administration among mothers and infants / Kostinov M.P., Cherdantsev A.P., Kuselman A.I., Akhmatova N.K., Kostinova A.M., Deryabina E.V., Demina E.O., Kostinov A.M. // Human Vaccines & Immunotherapeutics - 2018. - T. 14 - №12 - C. 2971-2978.
21. Society E. ESID Registry - Working definitions for clinical diagnosis of PID / Society E., Immunodeficiencies F. // ESID Registry - Working definitions for clinical diagnosis of PID - 2019. - №Cid - C. 1-205.
22. Roman F. AS03 A -Adjuvanted Influenza A (H1N1) 2009 Vaccine for Adults up to 85 Years of Age / Roman F., Vaman Т., Kafeja F., Hanon E., Damme P. Van // Clinical Infectious Diseases - 2010. - T. 51 - №6 - C.668-677.
23. Кузнецова T.A., Персиянова E.B., Запорожец T.C. Б.Н.Н. Адъюванты гриппозных вакцин: новые возможности применения сульфатированных полисахаридов из морских бурых водорослей. / Кузнецова Т.А., Персиянова Е.В., Запорожец Т.С.Б.Н.Н. // Вопросы вирусологии. - 2019. - Т. 64 - №1 - С. 5-11.
24. Pedersen G. Pandemic Influenza Vaccination Elicits Influenza-Specific CD4+Thl-cell Responses in Hypogammaglobulinaemic Patients: Four case reports / Pedersen G., Halstensen A., Sjursen H., Naess A., Kristoffersen E.K., Cox R.J. // Scandinavian Journal of Immunology - 2011. - T. 74 - №2 - C. 210-218.
25. Notarangelo L.D. Primary immunodeficiencies: 2009 update / Notarangelo L.D., Fischer A., Geha R.S., Casanova J.-L., Chapel H., Conley M.E., Cunningham-Rundles C, Etzioni A., Hammartrom L., Nonoyama S., Ochs H.D., Puck J., Roifman C, Seger R., Wedgwood J. // Journal of Allergy and Clinical Immunology - 2009. - T. 124 - №6 - C .1161-1178.
26. Ко J. Immune competence and switched memory В cells in common variable immunodeficiency / Ко J., Radigan L., Cunningham-Rundles C. // Clinical Immunology - 2005.- T. 116 - №1 - C. 37-41.
27. Warnatz K. Severe deficiency of switched memory В cells (CD27+IgM-IgD-) in subgroups of patients with common variable immunodeficiency: a new approach to classify a heterogeneous disease / Warnatz K., Denz A., Drager R., Braun M., Groth C, Wolff-Vorbeck G., Eibel H., Schlesier M., Peter H.H. // Blood - 2002. - T. 99 - №5 - C. 1544-1551.
28. Wehr C. The EUROclass trial: defining subgroups in common variable immunodeficiency / Wehr C., Kivioja Т., Schmitt C., Ferry В., Witte Т., Eren E., Vlkova M., Hernandez M., Detkova D., Bos P.R., Poerksen G., Bernuth H. von, Baumann U., Goldacker S., Gutenberger S., Schlesier M., Bergeron-van der Cruyssen F., Garff M. Le, Debre P., Jacobs R., Jones J., Bateman E., Litzman J., Hagen P.M. van, Plebani A., Schmidt R.E., Thon V., Quinti I., Espanol Т., Webster A.D., Chapel H., Vihinen M., Oksenhendler E., Peter H.H., Warnatz K. // Blood - 2008. - Т. 111 - №1 - C. 77-85.
29. Taubenheim N. Defined Blocks in Terminal Plasma Cell Differentiation of Common Variable Immunodeficiency Patients / Taubenheim N., Hornung M. von, Durandy A., Warnatz K., Corcoran L., Peter H.-H., Eibel H. // The Journal of Immunology - 2005. - T. 175 - №8 - C .5498-5503.
30. Sanchez-Ramon S. Memory В cells in common variable immunodeficiency: Clinical associations and sex differences / Sanchez-Ramon S., Radigan L., Yu J.E., Bard S., Cunningham-Rundles C. // Clinical Immunology - 2008. - T. 128 - №3 - C. 314-321.
31. Yu J.E. TLR-mediated В cell defects and IFN-a in common variable immunodeficiency / Yu J.E., Zhang L., Radigan L., Sanchez-Ramon S., Cunningham-Rundles C. // Journal of Clinical Immunology - 2012. - T. 32 - №1 - C. 50-60.
32. Bonilla F.A. International Consensus Document (ICON): Common Variable Immunodeficiency Disorders / Bonilla F.A., Barlan I., Chapel H., Costa-Carvalho B.T., Cunningham-Rundles C., la Morena M.T. de, Espinosa-Rosales F.J., Hammarstrom L Nonoyama S., Quinti I., Routes J.M., Tang M.L.K., Wamatz K. // Journal of Allergy and Clinical Immunology: In Practice - 2016. - T. 4 - №1 - C. 38-59.
33. Sobh A. Vaccination in Primary Immunodeficiency Disorders / Sobh A., Bonilla F.A. // The Journal of Allergy and Clinical Immunology: In Practice - 2016. - T. 4 - №6 - C. 1066-1075.
34. World Health OrganizationGlobal influenza strategy 2019-2030. Geneva; 2019. Licence: CC BY-NC-SA 3.0 IGO.Cataloguing-in-Publication (CIP) data. / World Health Organization, Centers for Disease Control and Prevention. CDC -, 2019. - 34 c.
35. Meyts I. Exome and genome sequencing for inborn errors of immunity / Meyts I., Bosch B., Bolze A., Boisson В., Itan Y., Belkadi A., Pedergnana V., Moens L., Picard C., Cobat A., Bossuyt X., Abel L., Casanova J.-L. // Journal of Allergy and Clinical Immunology - 2016. - T. 138 - №4 - C. 957-969.
36. Krammer F. Influenza / Krammer F., Smith G.J.D., Fouchier R.A.M., Peiris M., Kedzierska K., Doherty P.C., Palese P., Shaw M.L., Treanor J., Webster R.G., Garcla-Sastre A. // Nature Reviews Disease Primers - 2018. - T. 4 - №1 - C. 3.
37. 93. Booth J. Modulation of В cell responses by Toll-like receptors / Booth J., Wilson H., Jimbo S., Mutwiri G. // Cell and Tissue Research - 2011. - T. 343 - №1 - C. 131-140.
38. 94. Bernasconi N.L. A role for Toll-like receptors in acquired immunity: up-regulation of TLR9 by BCR triggering in naive В cells and constitutive expression in memory В cells / Bernasconi N.L., Onai N., Lanzavecchia A. // Blood - 2003. - T. 101 - №11 - C. 4500-4504.
39. 95. Bourke E. The toll-like receptor repertoire of human В lymphocytes: inducible and selective expression of TLR9 and TLR10 in normal and transformed cells / Bourke E., Bosisio D., Golay J., Polentarutti N., Mantovani A. // Blood - 2003. - T. 102 - №3 - C. 956-963.
40. 96. Trinchieri G. Cooperation of Toll-like receptor signals in innate immune defence / Trinchieri G., Sher A. // Nature Reviews Immunology - 2007. - T. 7 - №3 - C. 179-190.
41. 97. Yu J.E. Toll-like receptor 7 and 9 defects in common variable immunodeficiency / Yu J.E., Knight A.K., Radigan L., Marron T.U., Zhang L., Sanchez-Ramon S., Cunningham-Rundles C. // Journal of Allergy and Clinical Immunology - 2009. - T. 124 - №2 - C. 15-22.
42. 98. Crampton S.P. Innate pathways to B-cell activation and tolerance / Crampton S.P., Voynova E., Bolland S. // Annals of the New York Academy of Sciences - 2010. - T. 1183 - №1 - C 58-68.
43. 15. Hannoun C. The evolving history of influenza viruses and influenza vaccines. / Hannoun C. // Expert review of vaccines - 2013. - T. 12 - №9 - C. 1085-1094.
44.16. Europe I. Recommended composition of influenza virus vaccines for use in the 2018-2019 northern hemisphere influenza season / Europe I. // Releve epidemiologique hebdomadaire - 2018. - T. 93 -№12 - C. 133-141.
45. 17. Ambrose C.S. The rationale for quadrivalent influenza vaccines / Ambrose C.S., Levin M.J. // Human vaccines immunotherapeutics - 2012. - T. 8 - №1 - C. 1-8.
46.18. Caini S. Epidemiological and virological characteristics of influenza B: Results of the Global Influenza В Study / Caini S., Huang Q.S., Ciblak M.A., Badur S. // Influenza and other Respiratory Viruses - 2015. - T. 9 - №SI - C. 3-12.
47. 19. С.М. Харит, А.В. Рудакова, А.Н. Усков, Л.Н. Коновалова Ю.В.Л. Предотвращенный ущерб при вакцинации против гриппа 3-й 4-валентными вакцинами / С.М. Харит, А.В. Рудакова, А.Н. Усков, Л.Н. Коновалова Ю.В.Л. // Журнал Инфектологии - 2017. - Т. 9 - №2 - С. 17-22.
48. 20. Eichner М, Schwehm М, Hain J, Uphoff Н, Salzberger В, Knuf М S. - O.R. Flu - an individual based simulation tool to study the effects of quadrivalent vaccination on seasonal influenza in Germany. / Eichner M, Schwehm M, Hain J, Uphoff H, Salzberger B, Knuf M S.-O.R. // BMC Infectious Diseases - 2014. - T. 14 - C 365.
49. 21.  P. Retrospective public health impact of a quadrivalent influenza vaccine in the United States / P., Boer P.T., Postma M.J., Pitman R. // Influenza and Other Respiratory Viruses - 2015. - T. 9 - №S1 - C. 39-46.
P. Retrospective public health impact of a quadrivalent influenza vaccine in the United States / P., Boer P.T., Postma M.J., Pitman R. // Influenza and Other Respiratory Viruses - 2015. - T. 9 - №S1 - C. 39-46.
50. 22. Moa A.M. Immunogenicity and safety of inactivated quadrivalent influenza vaccine in adults: A systematic review and meta-analysis of randomised controlled trials / Moa A.M., Chughtai A.A., Muscatello D.J., Turner R.M., Maclntyre CR. // Vaccine - 2016. -T. 34 - №35 - C. 4092-4102.98. Crampton S.P. Innate pathways to B-cell activation and tolerance / Crampton S.P., Voynova E., Bolland S. // Annals of the New York Academy ofSciences - 2010. - T. 1183 - №1 - C. 58-68.
51. 99. Krieg A.M. CpG motifs in bacterial DNA and their immune effects / Krieg A.M. // Annual Review of Immunology - 2002. - T. 20 - №1 - C. 709-760.
52. 100. Aranburu A. TLR Ligation Triggers Somatic Hypermutation in Transitional В Cells Inducing the Generation of IgM Memory В Cells / Aranburu A., Ceccarelli S., Giorda E., Lasorella R., Ballatore G., Carsetti R. // The Journal of Immunology - 2010. - T. 185 - №12 - C .7293-7301.
53. 101. Traggiai E. Antigen dependent and independent mechanisms that sustain serum antibody levels / Traggiai E., Puzone R., Lanzavecchia A. // Vaccine - 2003. - T. 21 - C. S35-S37.
54.102. Glaum M.C Toll-like receptor 7-induced naive human B-cell differentiation and immunoglobulin production / Glaum M.C, Narula S., Song D., Zheng Y., Anderson A.L., Pletcher C.H., Levinson A.I. // Journal of Allergy and Clinical Immunology - 2009. - T. 123 - №1 - C. 224- 230.e4.
55. 103. Bekeredjian-Ding LB. Plasmacytoid Dendritic Cells Control TLR7 Sensitivity of Naive В Cells via Type IIFN / Bekeredjian-Ding I.В., Wagner M., Hornung V., Giese Т., Schnurr M., Endres S., Hartmann G. // The Journal of Immunology - 2005. - T. 174 - №7 - C .4043-4050.
56. 104. Heer A.K. TLR Signaling Fine-Tunes Anti-Influenza В Cell Responses without Regulating Effector T Cell Responses / Heer A.K., Shamshiev A., Donda A., Uematsu S., Akira S., Kopf M., Marsland B.J. // The Journal of Immunology - 2007. - T. 178 - №4 - C. 2182-2191.
57. 105. Lanzavecchia A. Understanding and making use of human memory В cells / Lanzavecchia A., Bernasconi N., Traggiai E., Ruprecht C.R., Corti D., Sallusto F. // Immunological Reviews - 2006. - T. 211 - №1 - C.303-309.
58.106. Ruprecht CR. Toll-like receptor stimulation as a third signal required for activation of human naive В cells / Ruprecht C.R., Lanzavecchia A. // European Journal of Immunology - 2006. - T. 36 - №4 - C. 810-816.
59. 107. Bernasconi N.L. Maintenance of Serological Memory by Polyclonal Activation of Human Memory В Cells / Bernasconi N.L., Traggiai E., Lanzavecchia A. // Science -2002. - T. 298-№5601 - C. 2199-2202.
60. 108. Escobar D. Defective В cell response to TLR9 ligand (CpG-ODN), Streptococcus pneumoniae and Haemophilus influenzae extracts in common variable immunodeficiency patients / Escobar D., Pons J., Clemente A., Iglesias J., Regueiro V., Bengoechea J.A., Matamoros N., Ferrer J.M. // Cellular Immunology - 2010. - T. 262 - №2 - C. 105-111.
61. Eibl M.M. Vaccination in patients with primary immune deficiency, secondary immune deficiency and autoimmunity with immune regulatory abnormalities / Eibl M.M., Wolf H.M. // Immunotherapy - 2015. - T. 7 - №12 - C. 1273-1292.
62. Koller M. robustlmm: An R Package for Robust Estimation of Linear Mixed-Effects Models / Koller M. // Journal of Statistical Software - 2016. - T. 75 - №6.
63. Luke S.G. Evaluating significance in linear mixed-effects models in R / Luke S.G. // Behavior Research Methods - 2017. - T. 49 - №4 - C. 1494-1502.
64. Russell V. Lenth emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.6.1.//- 2021.
65. Reverberi R. The statistical analysis of immunohaematological data / Reverberi R. // Blood transfusion = Trasfusione del sangue - 2008. - T. 6 - №1 - C. 37-45.
66. Fay M.P. Two-sided Exact Tests and Matching Confidence Intervals for Discrete Data / Fay M.P. // The R Journal - 2010. - T. 2 - №1 - C .53-58.
67. Д.Ю. НохринЛабораторный практикум по биостатистике / Д.Ю. Нохрин - Челябинск: Изд-во Челяб. гос. ун-та, 2018. - 289 с.
68. Sokal R.R., Rohlf F.J. Biometry: The principles and practice of statistics in biological research / F.J. Sokal R.R., Rohlf-New York: W.H. Freeman, 2012. Вып. 4th.
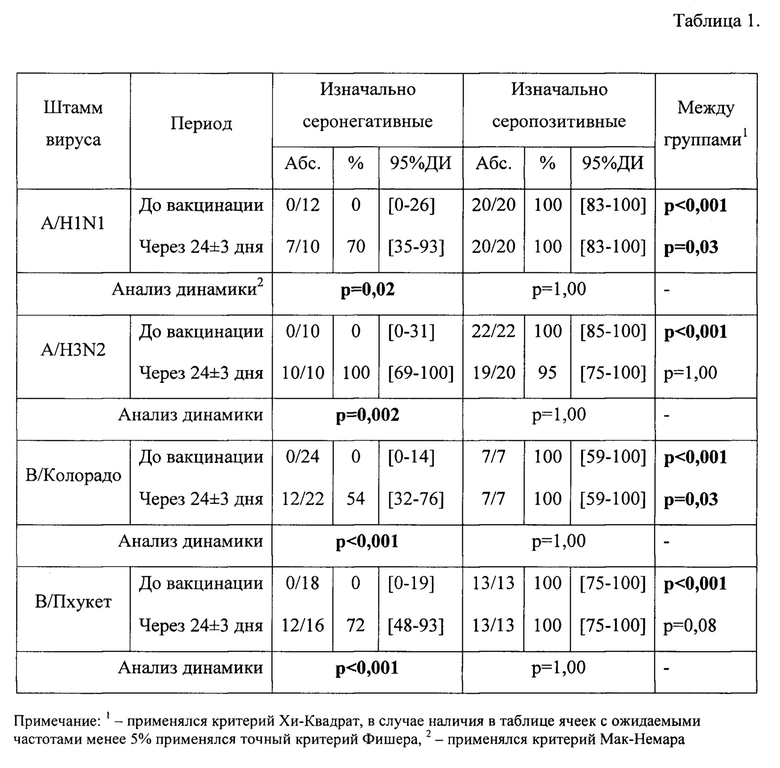
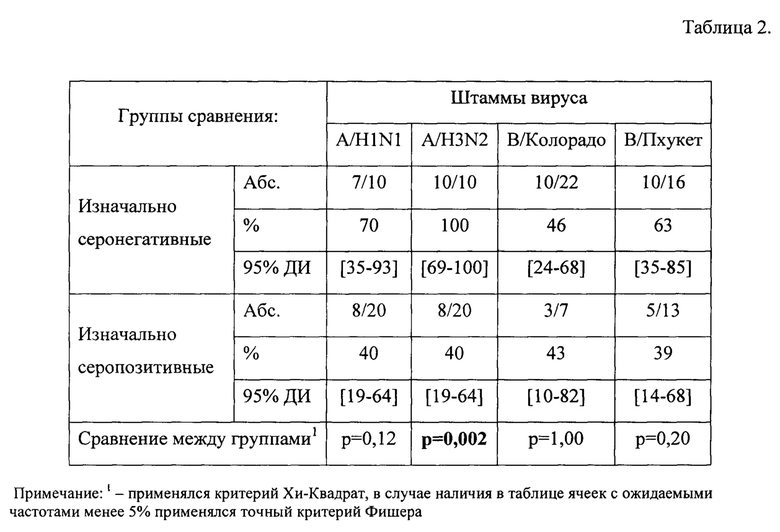
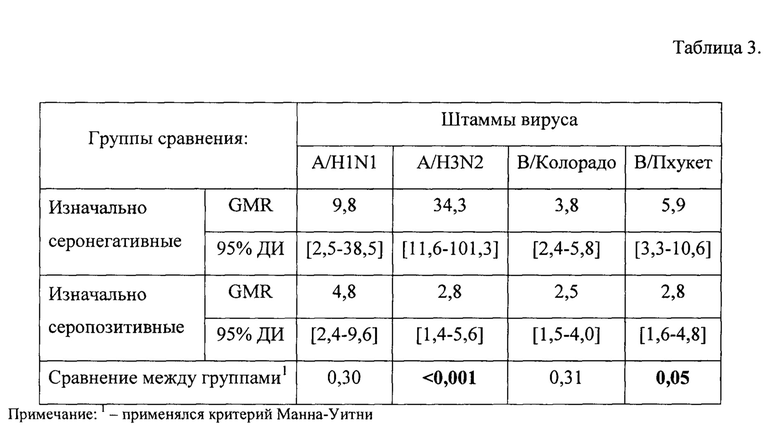
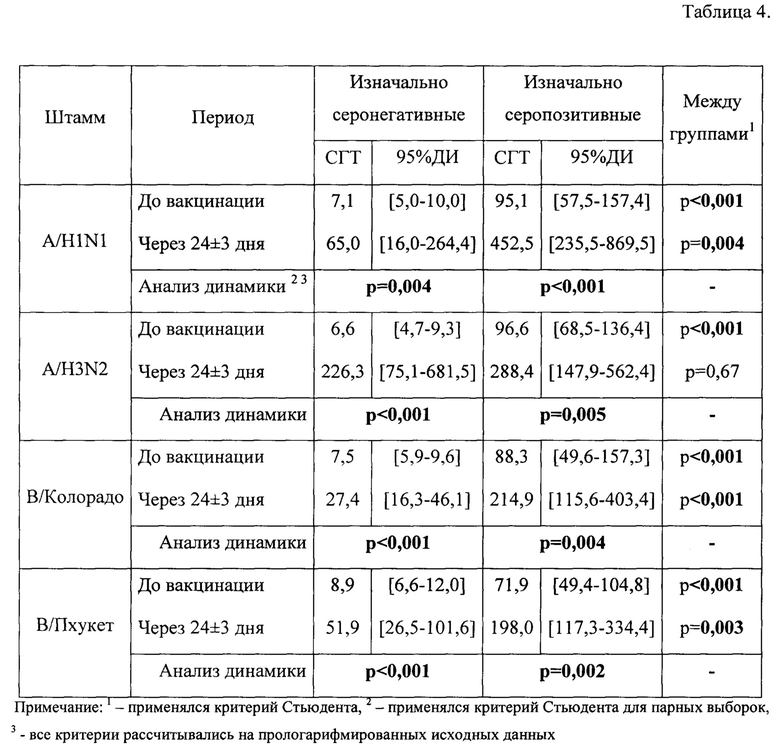
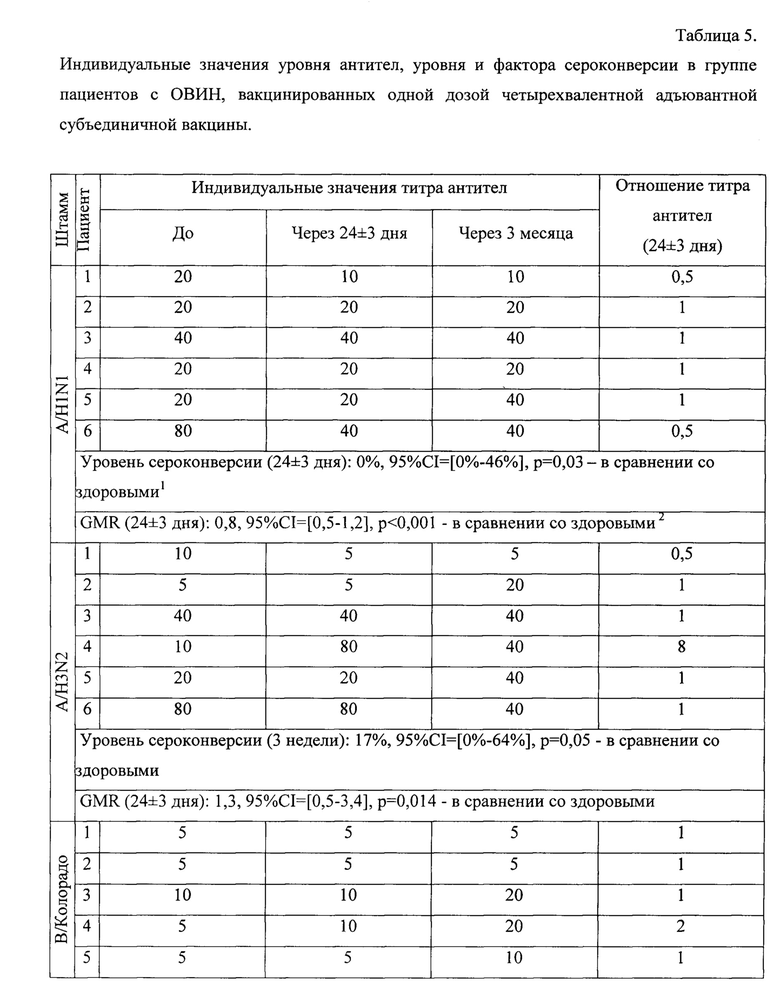
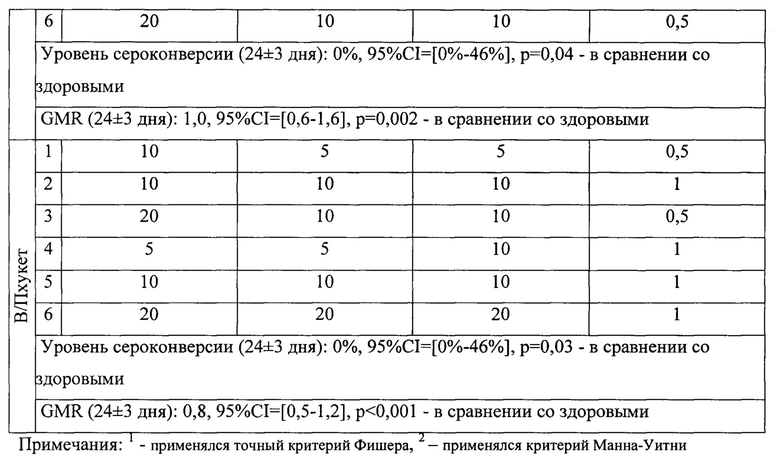
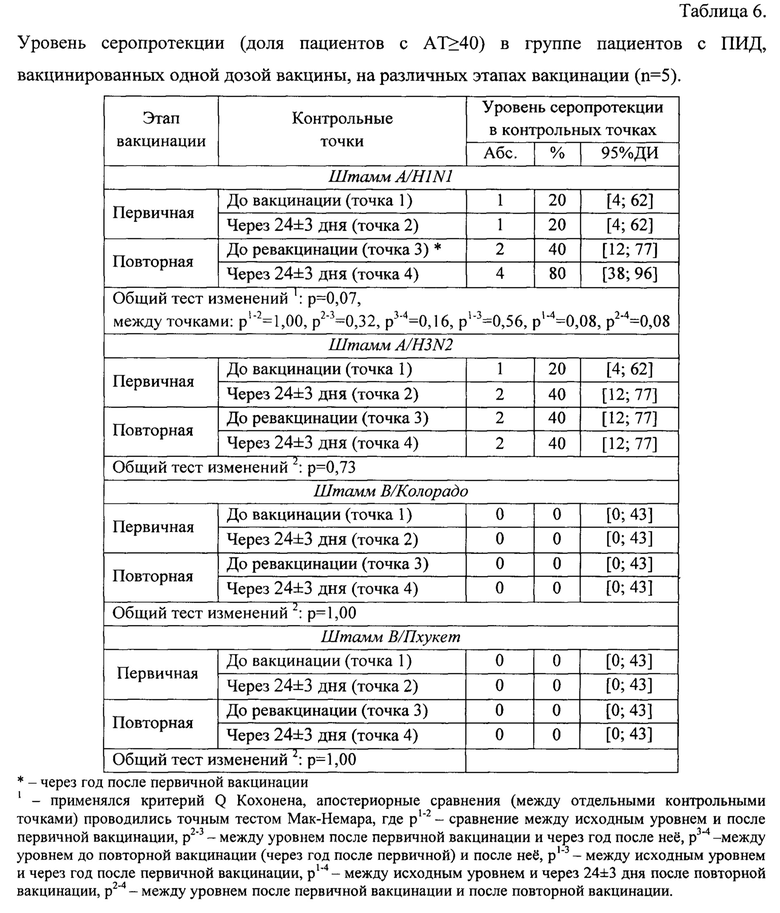


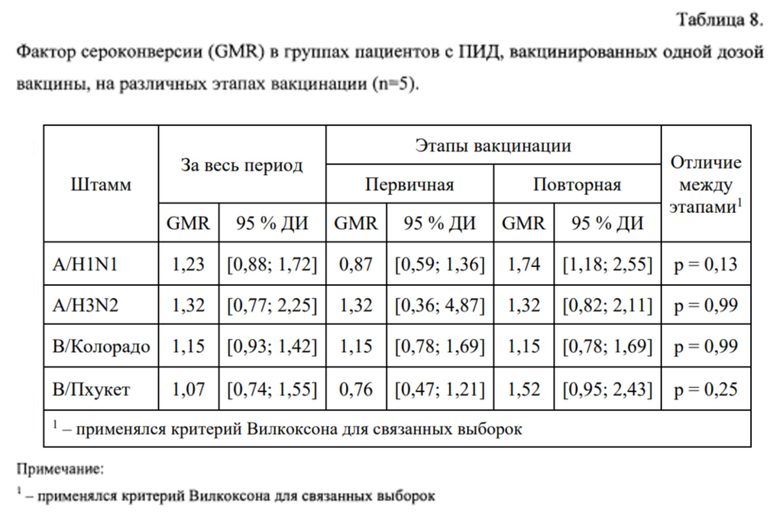
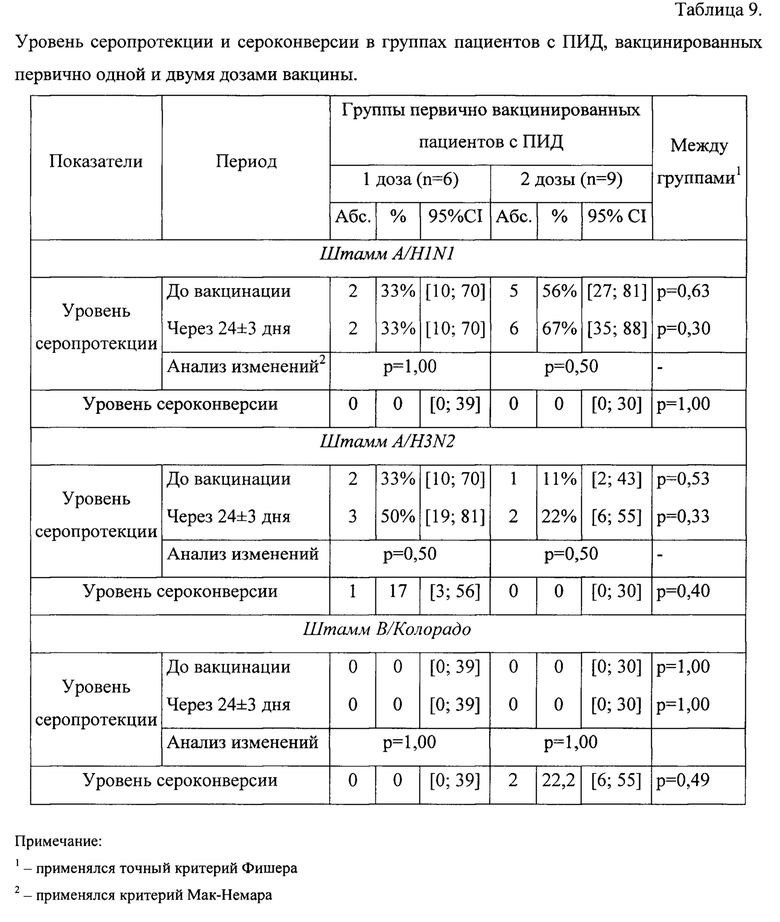
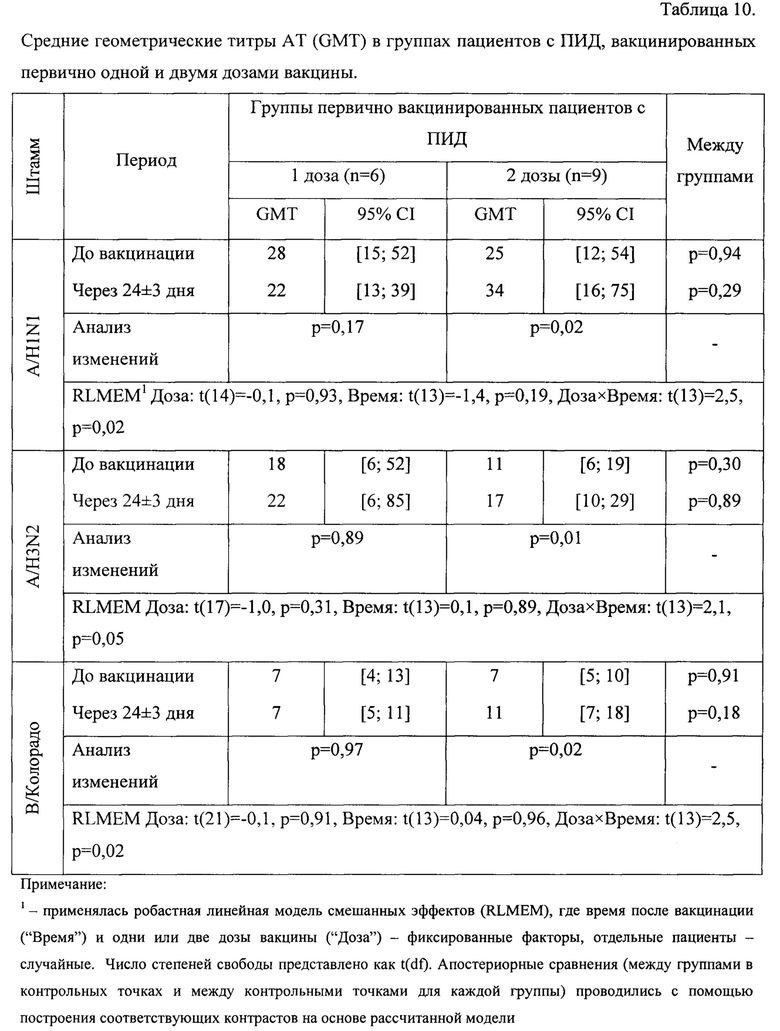
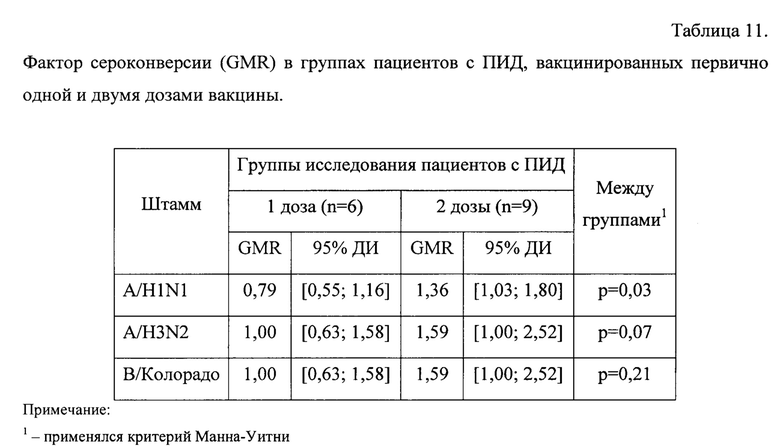
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения аттенуированных штаммов вируса гриппа а - кандидатов в живые гриппозные вакцины | 2017 |
|
RU2670514C1 |
| СПОСОБ ПОВЫШЕНИЯ ИММУНОГЕННОСТИ ИНАКТИВИРОВАННОЙ ГРИППОЗНОЙ ВАКЦИНЫ | 2007 |
|
RU2323742C1 |
| Способ оценки репродуктивной активности вирусов гриппа в составе тривалентной и квадривалентной живой гриппозной вакцины | 2019 |
|
RU2735293C1 |
| АТТЕНУИРОВАННЫЙ ХОЛОДОАДАПТИРОВАННЫЙ ШТАММ ВИРУСА ГРИППА А/КРАСНОДАР/101/59/35 (H2N2) ДЛЯ ПОЛУЧЕНИЯ ШТАММОВ ЖИВОЙ ИНТРАНАЗАЛЬНОЙ ГРИППОЗНОЙ ВАКЦИНЫ | 2007 |
|
RU2354695C1 |
| Набор консервативных эпитопов вирусных белков, способ их идентификации и отбора, вакцинная композиция и способ вакцинации | 2022 |
|
RU2814020C2 |
| ИММУНОАДЪЮВАНТНАЯ КОМПОЗИЦИЯ ДЛЯ ВАКЦИН ПРОТИВ ИНФЕКЦИОННЫХ АГЕНТОВ ВИРУСНОЙ И БАКТЕРИАЛЬНОЙ ПРИРОДЫ | 2018 |
|
RU2697527C1 |
| РЕАССОРТАНТНЫЙ ШТАММ ВИРУСА ГРИППА RN2/57-HUMAN A(H7N2) ДЛЯ ОПРЕДЕЛЕНИЯ АНТИТЕЛ К НЕЙРАМИНИДАЗЕ ПРИ ГРИППОЗНОЙ ИНФЕКЦИИ И ВАКЦИНАЦИИ | 2011 |
|
RU2464312C1 |
| СПОСОБ ПОЛУЧЕНИЯ ВИРУСА ГРИППА | 2008 |
|
RU2484136C2 |
| РЕКОМБИНАНТНАЯ ПРОТИВОГРИППОЗНАЯ ВАКЦИНА С ШИРОКИМ СПЕКТРОМ ЗАЩИТЫ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2017 |
|
RU2757013C2 |
| Рекомбинантный штамм вируса гриппа A/PR8-NS124-TB10.4-2A-HspX и способ специфической профилактики туберкулеза легких с использованием вакцины мукозального применения на его основе | 2019 |
|
RU2726106C1 |

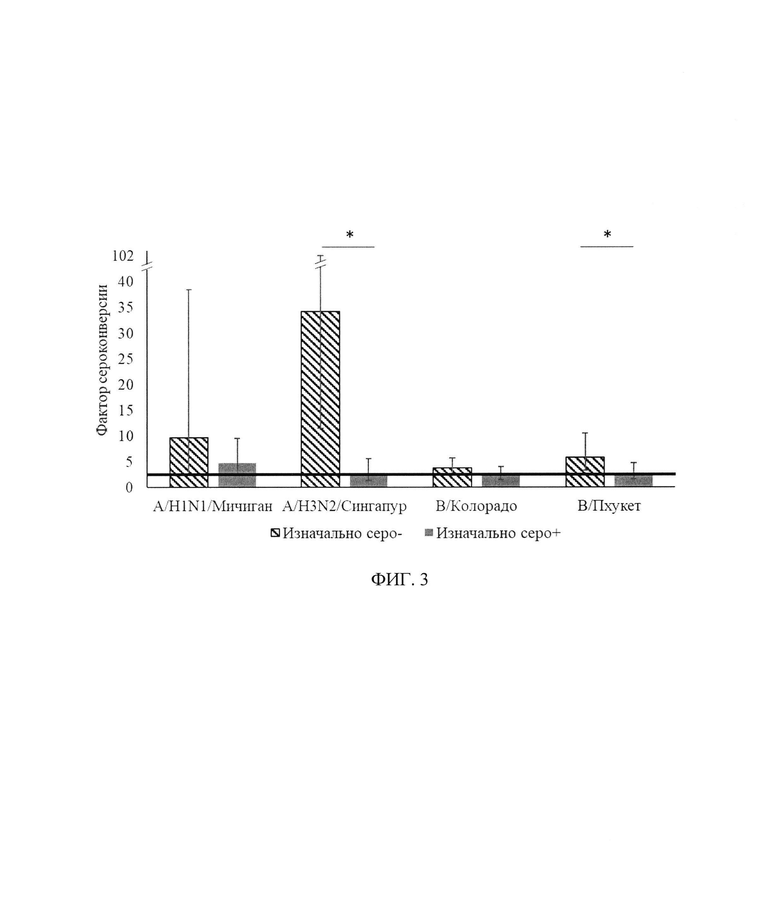
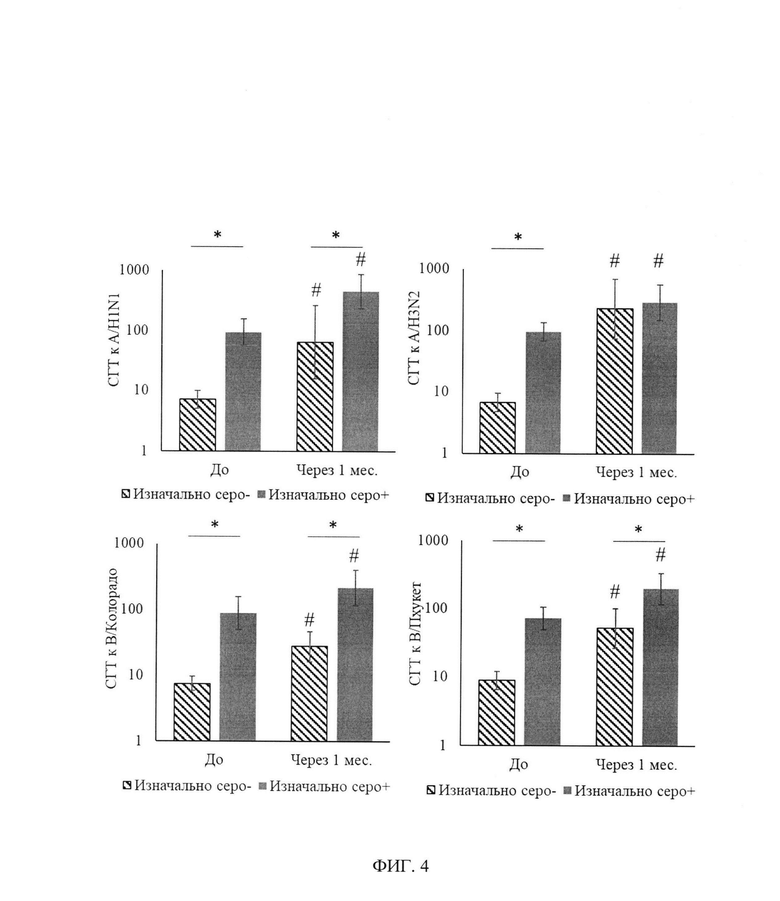
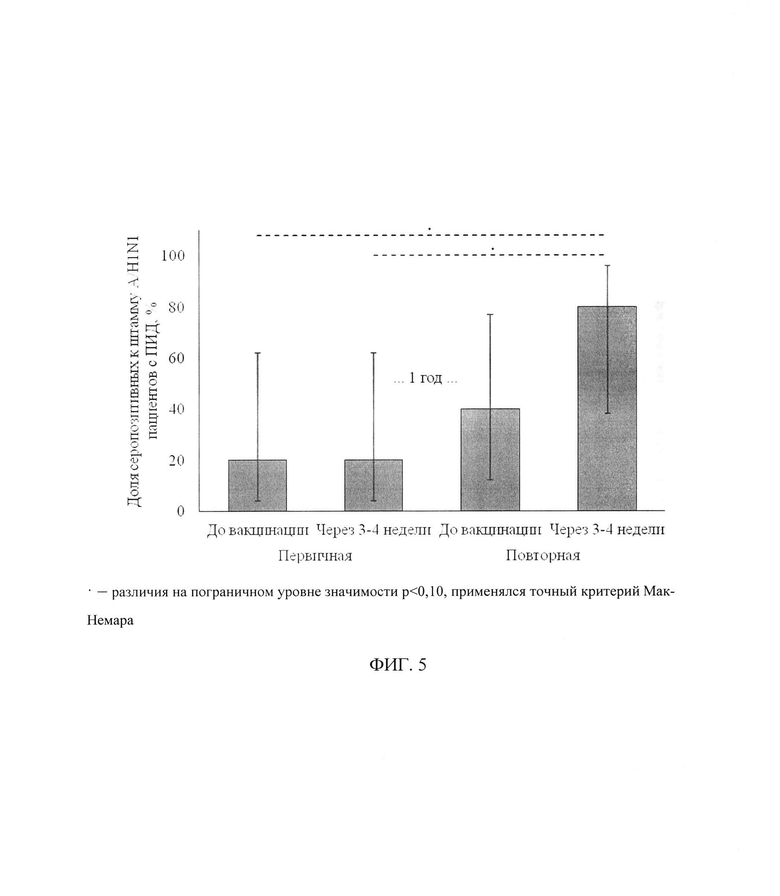
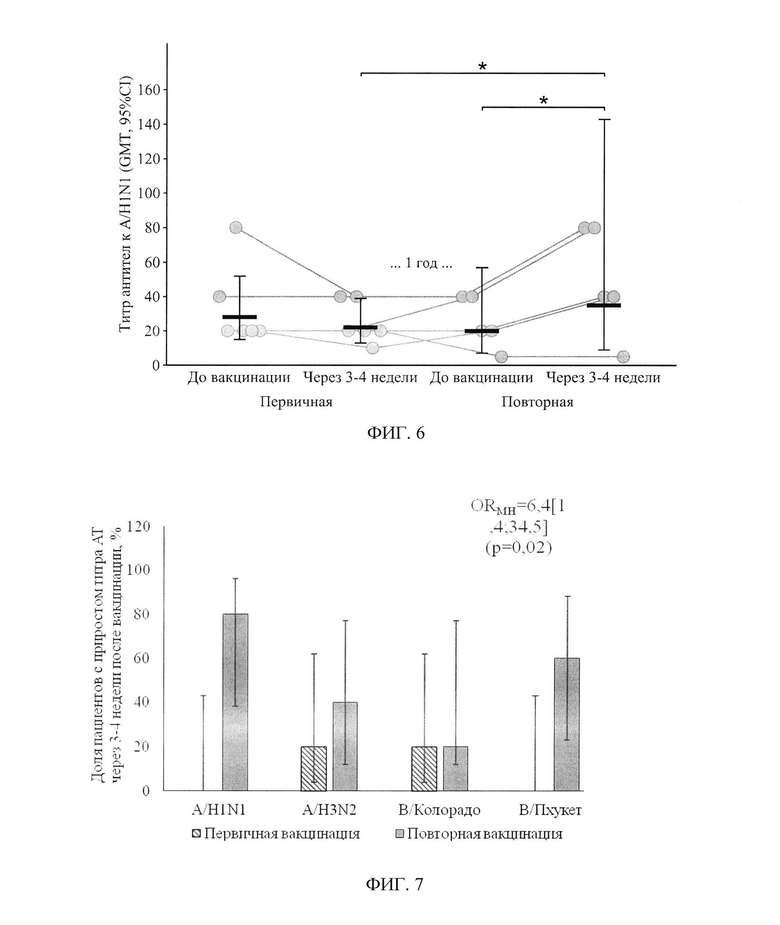



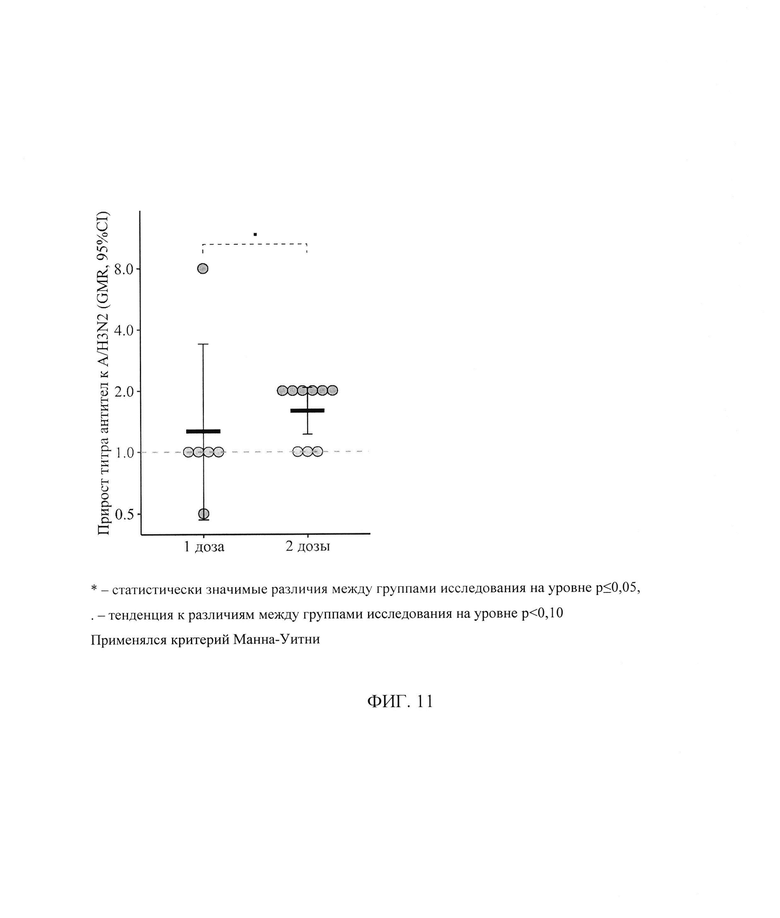
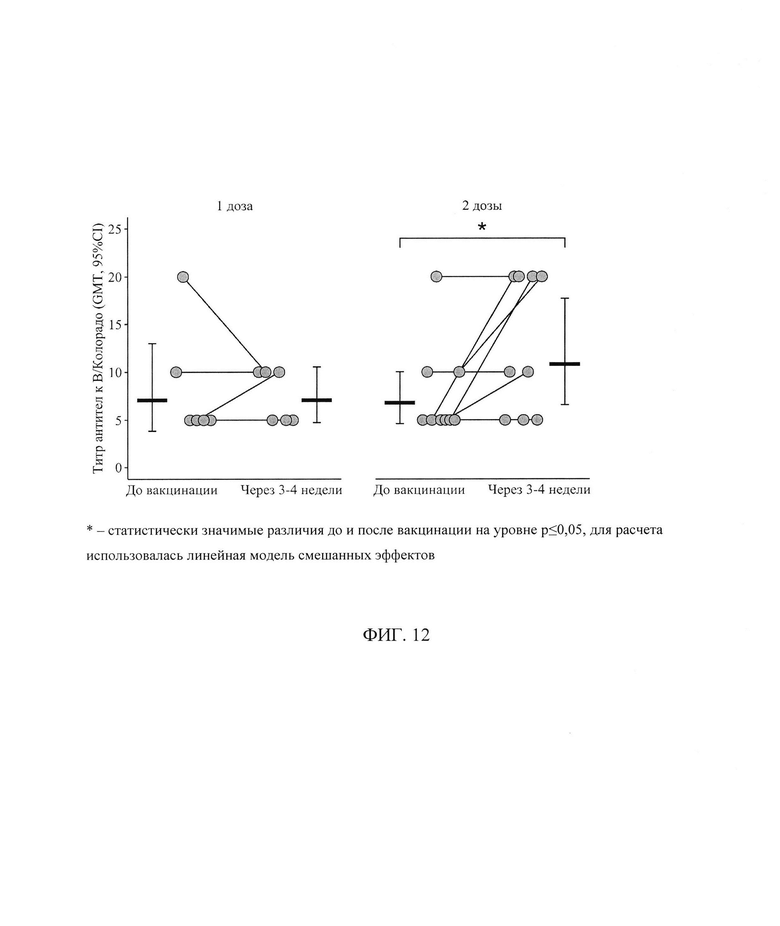
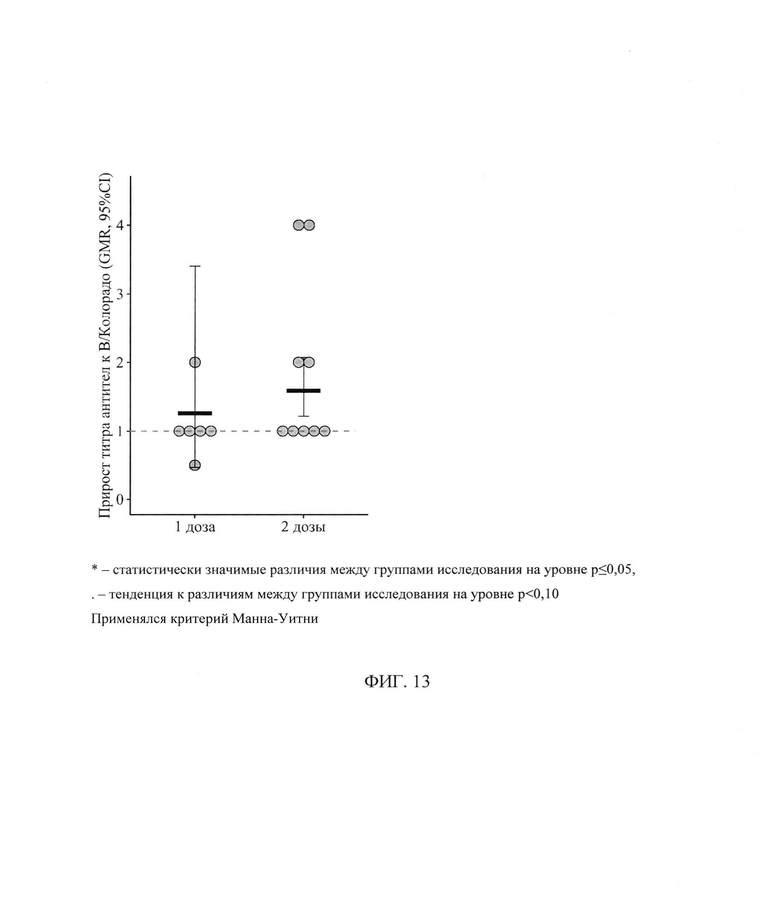
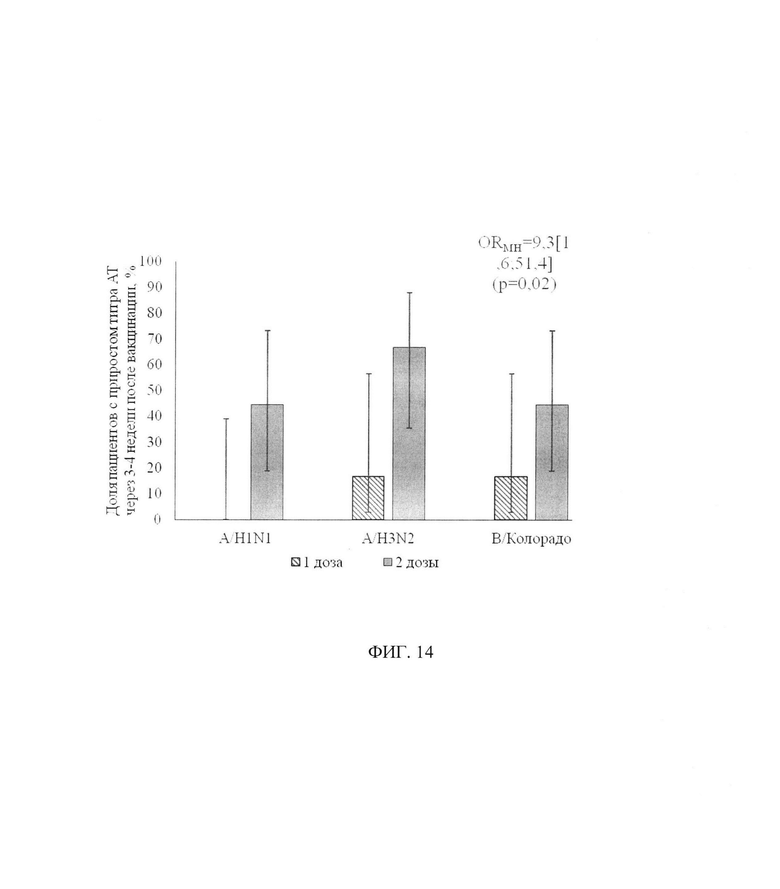
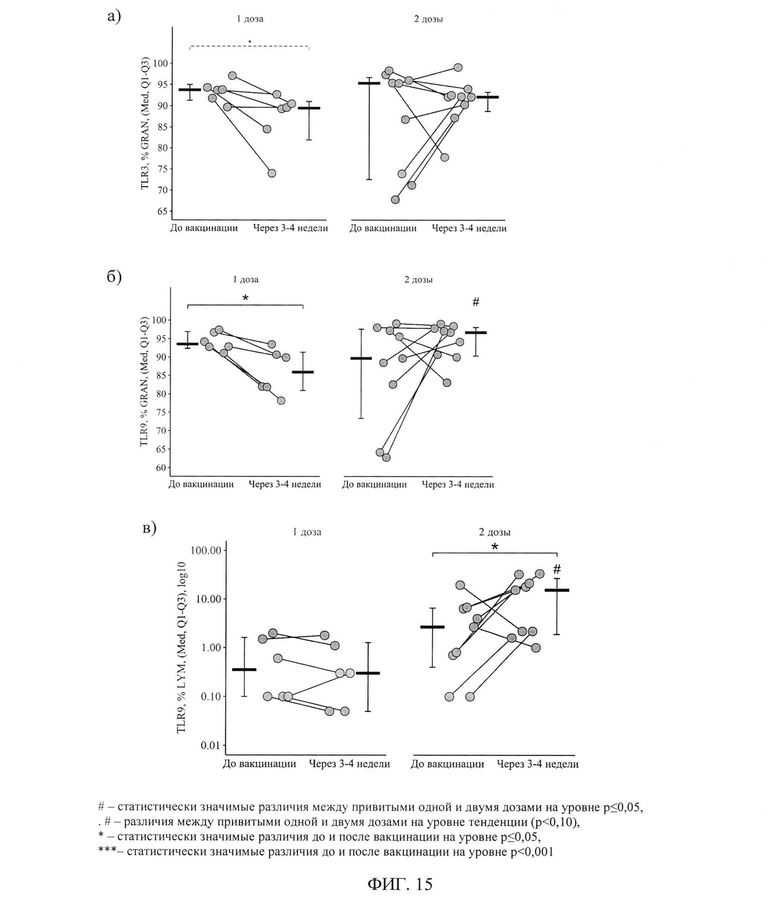
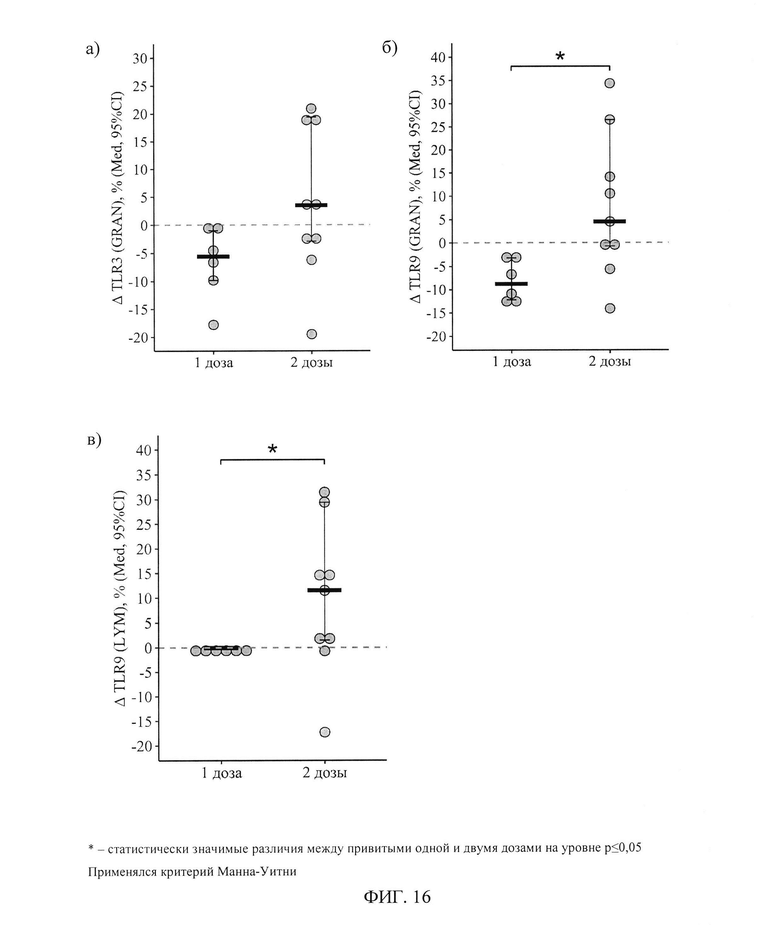

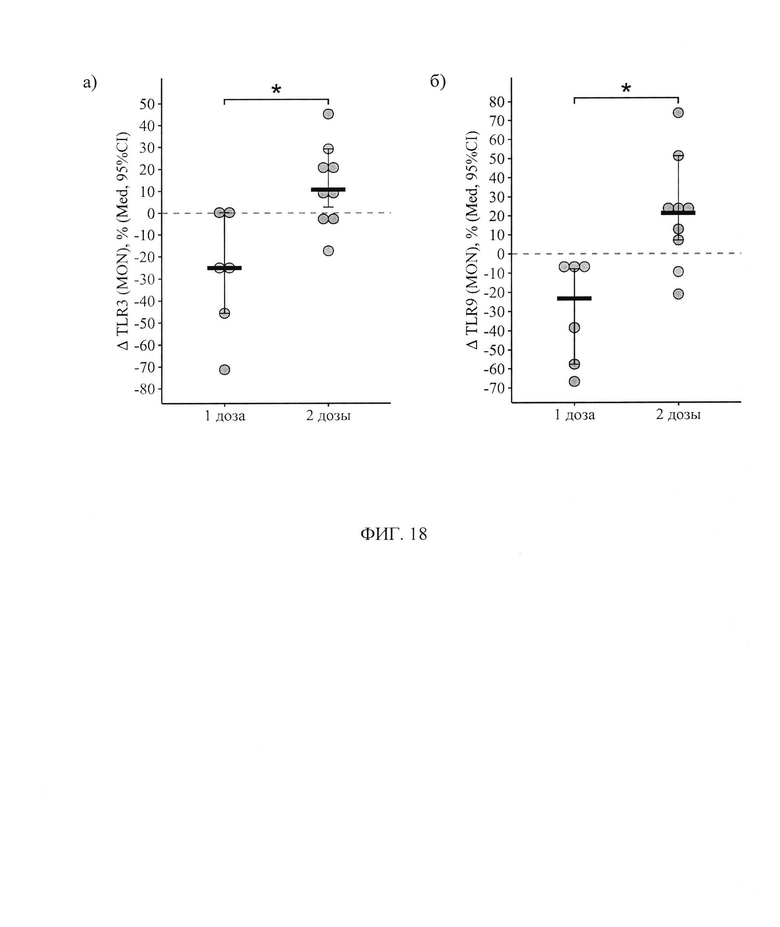
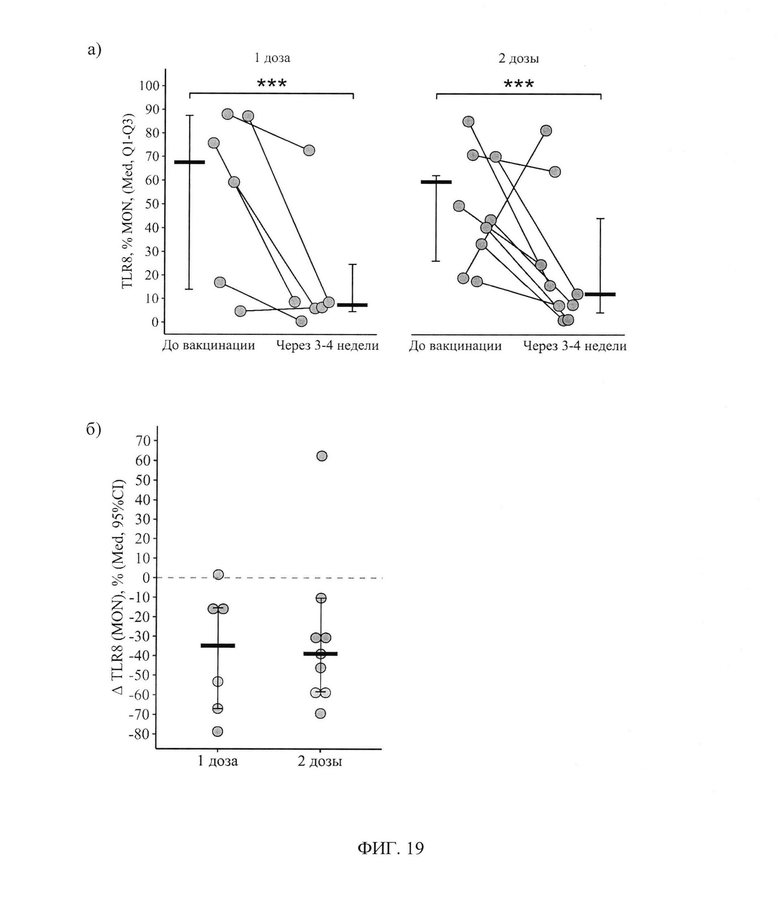
Настоящее изобретение относится к способу формирования поствакцинального иммунитета против вируса гриппа у пациентов с общей вариабельной иммунной недостаточностью, включающему одномоментное введение субъекту двух доз по 0,5 мл инактивированной полимер-субъединичной адъювантной вакцины, содержащей по 5 мкг протективных антигенов эпидемически актуальных штаммов вирусов гриппа типа А подтипов A(H1N1), A(H3N2), типа В и 500 мкг азоксимера бромида. Настоящее изобретение обеспечивает развитие высокой иммуногенности у пациентов с общей вариабельной иммунной недостаточностью (ОВИН) после введения одномоментно двух доз субъединичной адъювантной вакцины против гриппа, при этом доля пациентов с 2-кратным и более приростом титра антител через 3-4 недели после одномоментной вакцинации двумя дозами вакцинации после в 9,3 раз выше по сравнению с вакцинацией одной дозой, также обеспечивает увеличение доли гранулоцитов, лимфоцитов, экспрессирующих TLR9, а на моноцитах - TLR3 и TLR9, которые играют ключевую роль в элиминации микробных агентов через привлечение фагоцитов или активацию дендритных клеток, также обеспечивает высокий профиль безопасности трех- и четырехвалентных адъювантных вакцин у больных ОВИН. 19 ил., 6 пр., 11 табл.
Способ формирования поствакцинального иммунитета против вируса гриппа у пациентов с общей вариабельной иммунной недостаточностью, включающий одномоментное введение субъекту двух доз по 0,5 мл инактивированной полимер-субъединичной адъювантной вакцины, содержащей по 5 мкг протективных антигенов эпидемически актуальных штаммов вирусов гриппа типа А подтипов A(H1N1), A(H3N2), типа В и 500 мкг азоксимера бромида.
| Регистр лекарственных средств России РЛС Доктор: Педиатрия | |||
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| / Под ред | |||
| Г.Л | |||
| Вышковского | |||
| - М.: ВЕДАНТА, 2015 | |||
| АППАРАТ ДЛЯ РАСПРЕДЕЛЕНИЯ ТОРФА НА СУШИЛЬНОМ ПОЛЕ | 1922 |
|
SU608A1 |
| Костинова А.М | |||
| Иммуногенность квадривалентной иммуноадъювантной вакцины против гриппа у здоровых лиц и больных с общей вариабельной иммунной недостаточностью | |||
| Вопросы гематологии/онкологии и иммунопатологии в | |||

