ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к области биотехнологии. Более конкретно, настоящее изобретение относится к антителам к PD-1 и их применениям.
УРОВЕНЬ ТЕХНИКИ
Утверждения в данном разделе относятся только к общей информации, связанной с настоящим изобретением, и не обязательно составляют уровень техники.
Иммунотерапия опухолей представляет собой метод лечения, в котором всецело используются и мобилизуются Т-киллеры у онкологических больных для уничтожения опухоли. В то же время уклонение опухолевых клеток является огромным препятствием для иммунотерапии опухолей. Опухолевые клетки используют свое собственное ингибирующее действие на иммунную систему, чтобы способствовать быстрому росту опухолей. Между механизмом уклонения опухолей от иммунного ответа и иммунным ответом организма на опухоли существует очень сложная взаимосвязь. На ранней стадии иммунотерапии опухолей опухолеспецифические Т-киллеры обладают своей биологической активностью, но по мере роста опухоли они теряют свою уничтожающую функцию на более поздней стадии.
Для активации Т-клеток в организме человека используются две системы сигнальных путей. В дополнение к первому сигналу, передаваемому Т-клеткам путем презентации МНС-антигенных пептидов антигенпрезентирующими клетками, для того, чтобы Т-клетки могли производить нормальные иммунные ответы, также требуется второй сигнал, передаваемый серией костимулирующих молекул. Эта двойная сигнальная система играет жизненно важную роль в балансе иммунной системы в организме. Она строго регулирует различные иммунные ответы организма на собственные и чужеродные антигены. Если второй сигнал, передаваемый костимулирующей молекулой, отсутствует, это приведет к отсутствию ответа или устойчивому специфическому иммунному ответу Т-клеток, что приведет к толерантности. Следовательно, второй сигнальный путь играет очень важную регулирующую роль во всем процессе иммунного ответа организма.
Программируемая смерть-1 (PD-1) - это белковый рецептор, экспрессируемый на поверхности Т-клеток, открытый в 1992 году и участвующий в процессе клеточного апоптоза. PD-1 принадлежит к семейству CD28. Он обладает 23% аминокислотной гомологией с антигеном-4 цитотоксических Т-лимфоцитов (CTLA-4), но его экспрессия отличается от экспрессии CTLA, и он в основном экспрессируется на активированных Т-клетках, В-клетках и миелоидных клетках. PD-1 имеет два лиганда, PD-L1 и PD-L2, соответственно. PD-L1 в основном экспрессируется на Т-клетках, В-клетках, макрофагах и дендритных клетках (ДК), и экспрессия на клетках может повышаться после активации. Экспрессия PD-L2 относительно ограничена и в основном имеет место на антигенпрезентирующих клетках, таких как активированные макрофаги и дендритные клетки.
PD-L1 подавляет иммунную систему, связываясь с PD-1 и B7-1. Многие опухолевые клетки и иммунные клетки в микроокружении опухолевой ткани экспрессируют PD-L1. Новые исследования показали, что высокая экспрессия белка PD-L1 была обнаружена при раке молочной железы, раке легкого (например, немелкоклеточном раке легкого), раке желудка, раке кишечника, раке почки, меланоме, раке ободочной кишки, раке мочевого пузыря, раке яичника, раке поджелудочной железы и раке печени, и в других опухолевых тканях человека, а уровень экспрессии PD-L1 был тесно связан с клиническими проявлениями и прогнозом у пациентов.
Моноклональное антитело к PD-1 может максимизировать ответ собственной иммунной системы пациента на опухоли, блокируя связывание PD-L1/PD-1, таким образом достигая цели уничтожения опухолевых клеток. В настоящее время антитело к PD-1 пембролизумаб (также известное как Merck keytruda, keytruda, Merck-Pemb, Merck-keytruda, Merck-PD-1, pembrolizumab) и ниволумаб (также известное как BMS Opdivo, Opdivo, BMS-Nivolumab, Nivolumab) было одобрено FDA (Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США) для лечения меланомы, пациентов с лимфомой Ходжкина, немелкоклеточного рака легкого и других опухолей. Кроме того, патентные документы, такие как WO200139722, WO2006121168, WO2010036959, WO2010089411, WO2011110604, WO2013173223, WO2013181634, US2014335093, US6803192B1, US8617546B2 и WO2015085847, также раскрывают ряд моноклональных антител к PD-1.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новому антителу к PD-1, его антигенсвязывающему фрагменту и его медицинскому применению.
В некоторых альтернативных вариантах осуществления настоящее изобретение относится к антителу к PD-1 или его антигенсвязывающему фрагменту, выбранному из любого из следующих i) - iii):
i) антитело к PD-1 или его антигенсвязывающий фрагмент, вариабельная область тяжелой цепи которого содержит HCDR1, представленную в SEQ ID NO: 8 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее, HCDR2, представленную в SEQ ID NO: 9 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее, и HCDR3, представленную в SEQ ID NO: 10 или имеющую не более 8, 3, 2 или 1 аминокислотной мутации относительно нее; вариабельная область легкой цепи которого содержит LCDR1, представленную в SEQ ID NO: 11 или имеющую не более 4, 3, 2 или 1 аминокислотной мутации относительно нее, LCDR2, представленную в SEQ ID NO: 12 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее, и LCDR3, представленную в SEQ ID NO: 13 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее;
ii) антитело к PD-1 или его антигенсвязывающий фрагмент, вариабельная область тяжелой цепи которого содержит HCDR1, представленную в SEQ ID NO: 14 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее, HCDR2, представленную в SEQ ID NO: 15 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее, и HCDR3, представленную в SEQ ID NO: 16 или имеющую не более 8, 3, 2 или 1 аминокислотной мутации относительно нее; вариабельная область легкой цепи которого содержит LCDR1, представленную в SEQ ID NO: 17 или имеющую не более 4, 3, 2 или 1 аминокислотной мутации относительно нее, LCDR2, представленную в SEQ ID NO: 12 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее, и LCDR3, представленную в SEQ ID NO: 18 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее; и
iii) антитело к PD-1 или его антигенсвязывающий фрагмент, вариабельная область тяжелой цепи которого содержит HCDR1, представленную в SEQ ID NO: 21 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее, HCDR2, представленную в SEQ ID NO: 22 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее, и HCDR3, представленную в SEQ ID NO: 23 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее; вариабельная область легкой цепи которого содержит LCDR1, представленную в SEQ ID NO: 24 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее, LCDR2, представленную в SEQ ID NO: 25 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее, и LCDR3, представленную в SEQ ID NO: 26 или имеющую не более 3, 2 или 1 аминокислотной мутации относительно нее.
В некоторых вариантах осуществления вышеупомянутое антитело к PD-1 или его антигенсвязывающий фрагмент согласно настоящему изобретению связывается с человеческим PD-1 с равновесной константой диссоциации 10-7 M или менее. В некоторых вариантах осуществления оно связывается с человеческим PD-1 с равновесной константой диссоциации, равной или менее 10-8 M, 10-9 M, 10-10 M или 10-11 M.
В некоторых альтернативных вариантах осуществления настоящее изобретение относится к антителу к PD-1 или его антигенсвязывающему фрагменту, вариабельная область тяжелой цепи которого содержит: HCDR1, представленную в SEQ ID NO: 65, HCDR2, представленную в SEQ ID NO: 66, и HCDR3, представленную в SEQ ID NO: 67; вариабельная область легкой цепи которого содержит: LCDR1, представленную в SEQ ID NO: 68, LCDR2, представленную в SEQ ID NO: 12, и LCDR3, представленную в SEQ ID NO: 69; последовательности представлены в таблице 1 ниже:
Таблица 1
(SEQ ID NO: 65)
(SEQ ID NO: 68)
(SEQ ID NO: 66)
(SEQ ID NO: 12)
(SEQ ID NO: 67)
(SEQ ID NO: 69)
В некоторых альтернативных вариантах осуществления в вышеупомянутом антителе к PD-1 или его антигенсвязывающем фрагменте вариабельная область тяжелой цепи содержит HCDR1, представленную в SEQ ID NO: 8, HCDR2, представленную в SEQ ID NO: 9, и HCDR3, представленную в SEQ ID NO: 10; вариабельная область легкой цепи содержит LCDR2, представленную в SEQ ID NO: 12, LCDR3, представленную в SEQ ID NO: 13, и LCDR1, представленную общей формулой RSSQSX13VHSX14X15X16TYLE (SEQ ID NO: 68), где X13 выбран из L, X14 выбран из N, Q, L, T или D, X15 выбран из G, A или V, и X16 выбран из N.
В некоторых альтернативных вариантах осуществления антитело к PD-1 или его антигенсвязывающий фрагмент выбраны из любого из следующих (a) - (e):
(a) антитело к PD-1 или его антигенсвязывающий фрагмент, содержащее HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10, соответственно, и LCDR2 и LCDR3, представленные в SEQ ID NO: 12 и SEQ ID NO: 13, соответственно, и LCDR1, представленную в SEQ ID NO: 11, 47, 48, 49, 50, 51 или 52;
(b) антитело к PD-1 или его антигенсвязывающий фрагмент, содержащее HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 14, SEQ ID NO: 15 и SEQ ID NO: 16, соответственно, и LCDR1, LCDR2 и LCDR3, представленные в SEQ ID NO: 17, SEQ ID NO: 12 и SEQ ID NO: 18, соответственно;
(c) антитело к PD-1 или его антигенсвязывающий фрагмент, содержащее HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 21, SEQ ID NO: 22 и SEQ ID NO: 23, соответственно, и LCDR1, LCDR2 и LCDR3, представленные в SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26, соответственно;
(d) антитело к PD-1 или его антигенсвязывающий фрагмент, содержащее HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 14, SEQ ID NO: 15 и SEQ ID NO: 16, соответственно, и LCDR2 и LCDR3, представленные в SEQ ID NO: 12 и SEQ ID NO: 13, соответственно, и LCDR1, представленную в SEQ ID NO: 11, 47, 48, 49, 50, 51 или 52; и
(e) вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, представленные в SEQ ID NO: 17, SEQ ID NO: 12 и SEQ ID NO: 18, соответственно.
В некоторых вариантах осуществления антитело к PD-1 или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, представленные в SEQ ID NO: 49, SEQ ID NO: 12 и SEQ ID NO: 13, соответственно.
В некоторых вариантах осуществления антитело к PD-1 или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 21, SEQ ID NO: 22 и SEQ ID NO: 23, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, представленные в SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26, соответственно.
В некоторых альтернативных вариантах осуществления настоящее изобретение относится к антителу к PD-1 или его антигенсвязывающему фрагменту, выбранному из любого из следующих iv) - vi):
iv) антитело к PD-1 или его антигенсвязывающий фрагмент, вариабельная область тяжелой цепи которого содержит HCDR1, HCDR2 и HCDR3 с теми же последовательностями, что и в вариабельной области тяжелой цепи, представленной в последовательности SEQ ID NO: 4, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3 с теми же последовательностями, что и в вариабельной области легкой цепи, представленной в последовательности SEQ ID NO: 5;
v) антитело к PD-1 или его антигенсвязывающий фрагмент, вариабельная область тяжелой цепи которого содержит HCDR1, HCDR2 и HCDR3 с теми же последовательностями, что и в вариабельной области тяжелой цепи, представленной в последовательности SEQ ID NO: 6, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3 с теми же последовательностями, что и в вариабельной области легкой цепи, представленной в последовательности SEQ ID NO: 7; и
vi) антитело к PD-1 или его антигенсвязывающий фрагмент, вариабельная область тяжелой цепи которого содержит HCDR1, HCDR2 и HCDR3 с теми же последовательностями, что и в вариабельной области тяжелой цепи, представленной в последовательности SEQ ID NO: 19, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3 с теми же последовательностями, что и в вариабельной области легкой цепи, представленной в последовательности SEQ ID NO: 20.
В некоторых вариантах осуществления вышеупомянутого антитела к PD-1 или его антигенсвязывающего фрагмента антитело к PD-1 или его антигенсвязывающий фрагмент представляет собой мышиное антитело или его антигенсвязывающий фрагмент, химерное антитело или его антигенсвязывающий фрагмент, полностью человеческое антитело или его антигенсвязывающий фрагмент, или гуманизированное антитело или его антигенсвязывающий фрагмент.
В некоторых вариантах осуществления вышеупомянутого антитела к PD-1 или его антигенсвязывающего фрагмента антитело к PD-1 или его антигенсвязывающий фрагмент представляет собой гуманизированное антитело или его антигенсвязывающий фрагмент.
В некоторых вариантах осуществления гуманизированное антитело содержит каркасную область, полученную из человеческого антитела, или вариант его каркасной области.
В некоторых вариантах осуществления вариант каркасной области имеет не более 11 обратных аминокислотных мутаций в каждой из каркасной области легкой цепи и/или каркасной области тяжелой цепи человеческого антитела.
В некоторых вариантах осуществления вариант каркасной области содержит мутацию(ии), выбранную из любой из следующих (f) - (h):
(f) обратная аминокислотная мутация 2G, содержащаяся в вариабельной области легкой цепи, и/или одна или более обратных аминокислотных мутаций, выбранных из 27Y, 48I, 67T, 69L, 82F и 93T, содержащихся в вариабельной области тяжелой цепи;
(g) обратная аминокислотная мутация 2V, содержащаяся в вариабельной области легкой цепи, и/или одна или более обратных аминокислотных мутаций, выбранных из 26D, 27F, 30T, 38K, 43H, 48I, 66K, 67A, 69L, 82F и 93T, содержащихся в вариабельной области тяжелой цепи; и
(h) одна или более обратных аминокислотных мутаций, выбранных из 42G, 44V и 71Y, содержащихся в вариабельной области легкой цепи, и/или обратные аминокислотные мутации 1K и/или 94S, содержащиеся в вариабельной области тяжелой цепи.
В некоторых вариантах осуществления вышеупомянутого антитела к PD-1 или его антигенсвязывающего фрагмента антитело к PD-1 или его антигенсвязывающий фрагмент содержит вариабельные области антитела, выбранные из группы, состоящей из:
(a2) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10, соответственно, и одну или более обратных аминокислотных мутаций из 27Y, 48I, 67T, 69L, 82F и 93T, содержащихся в каркасной области тяжелой цепи, и
вариабельной области легкой цепи, содержащей LCDR2 и LCDR3, представленные в SEQ ID NO: 12 и SEQ ID NO: 13, соответственно, и LCDR1, представленную в SEQ ID NO: 11, 47, 48, 49, 50, 51 или 52, и обратную аминокислотную мутацию 2G, содержащуюся в каркасной области легкой цепи;
(b2) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 14, SEQ ID NO: 15 и SEQ ID NO: 16, соответственно, и одну или более обратных аминокислотных мутаций, выбранных из 26D, 27F, 30T, 38K, 43H, 48I, 66K, 67A, 69L, 82F и 93T, содержащихся в вариабельной области тяжелой цепи; и
вариабельной области легкой цепи, содержащей LCDR1, LCDR2 и LCDR3, представленные в SEQ ID NO: 17, SEQ ID NO: 12 и SEQ ID NO: 18, соответственно, и обратную аминокислотную мутацию 2V, содержащуюся в каркасной области легкой цепи;
(c2) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 21, SEQ ID NO: 22 и SEQ ID NO: 23, соответственно, и обратные аминокислотные мутации 1K и/или 94S, содержащиеся в каркасной области тяжелой цепи, и
вариабельной области легкой цепи, содержащей LCDR1, LCDR2 и LCDR3, представленные в SEQ ID NO: 24, SEQ ID NO: 25 и SEQ ID NO: 26, соответственно, и одну или более обратных аминокислотных мутаций, выбранных из 42G, 44V и 71Y, содержащихся в каркасной области легкой цепи.
В некоторых вариантах осуществления вышеупомянутого антитела к PD-1 или его антигенсвязывающего фрагмента антитело к PD-1 или его антигенсвязывающий фрагмент содержит вариабельные области антитела, выбранные из любых из следующих (i) - (o):
(i) вариабельная область тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 4 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 4; и/или
вариабельная область легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 5 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 5;
(j) вариабельная область тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 6 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 6; и/или
вариабельная область легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 7 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 7;
(k) вариабельная область тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 19 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 19; и/или
вариабельная область легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 20 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 20;
(l) вариабельная область тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 27, 30, 31 или 32, или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 27, 30, 31 или 32, соответственно; и/или
вариабельная область легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 28, 29, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63 или 64, или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 28, 29, 34, 35, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63 или 64, соответственно;
(m) вариабельная область тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 33, 36, 37, 38, 39 или 40, или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 33, 36, 37, 38, 39 или 40, соответственно; и/или
вариабельная область легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 34, 35, 28, 29, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63 или 64, или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 34, 35, 28, 29, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63 или 64, соответственно;
(n) вариабельная область тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 41, 45 или 46, или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 41, 45 или 46, соответственно; и/или
вариабельная область легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 42, 43 или 44, или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 42, 43 или 44, соответственно;
(o) вариабельная область тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 70 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 70; и/или
вариабельная область легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 71 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 71; и
(p) вариабельная область тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 27, 30, 31 или 32, или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 27, 30, 31 или 32, соответственно; и/или
вариабельная область легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 34 или 35 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 34 или 35, соответственно;
где последовательности SEQ ID NO: 70 и SEQ ID NO: 71 представлены последовательностями общей формулы, как показано в таблице 2:
Таблица 2

В некоторых вариантах осуществления вариабельная область тяжелой цепи антитела к PD-1 соответствует представленной в SEQ ID NO: 27 или обладает по меньшей мере 90% идентичностью SEQ ID NO: 27, а последовательность вариабельной области легкой цепи антитела к PD-1 соответствует представленной в SEQ ID NO: 55 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 55.
В некоторых вариантах осуществления вариабельная область тяжелой цепи антитела к PD-1 соответствует представленной в SEQ ID NO: 46 или обладает по меньшей мере 90% идентичностью SEQ ID NO: 46, а последовательность вариабельной области легкой цепи антитела к PD-1 соответствует представленной в SEQ ID NO: 43 или обладает по меньшей мере 90% идентичностью последовательности SEQ ID NO: 43.
Вышеупомянутая «по меньшей мере 90% идентичность» включает по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность.
В некоторых других вариантах осуществления вышеупомянутого антитела к PD-1 или его антигенсвязывающего фрагмента антитело дополнительно содержит константные области антитела; в некоторых других вариантах осуществления константная область тяжелой цепи константной области антитела выбрана из константных областей человеческого IgG1, IgG2, IgG3 и IgG4 и их традиционных вариантов, константная область легкой цепи константной области антитела выбрана из константных областей κ- и λ-цепи человеческого антитела и их традиционных вариантов; в некоторых других вариантах осуществления константные области антитела содержат константную область тяжелой цепи IgG4, в которую введены одна или более мутаций из S228P, F234A и L235A, например, содержат константную область тяжелой цепи IgG4, имеющую три аминокислотные мутации из S228P, F234A и L235A; в некоторых других вариантах осуществления антитело содержит константную область тяжелой цепи, представленную в SEQ ID NO: 72 или SEQ ID NO: 79, и константную область легкой цепи, представленную в SEQ ID NO: 73.
В некоторых вариантах осуществления вышеупомянутого антитела к PD-1 или его антигенсвязывающего фрагмента антитело к PD-1 содержит легкую цепь, представленную в SEQ ID NO: 78, и тяжелую цепь, представленную в SEQ ID NO: 77 или 82; или
легкую цепь, представленную в SEQ ID NO: 75, и тяжелую цепь, представленную в SEQ ID NO: 74, 76, 80 или 81.
В некоторых вариантах осуществления антитело к PD-1 содержит:
тяжелую цепь, представленную в SEQ ID NO: 74, и легкую цепь, представленную в SEQ ID NO: 75; или
тяжелую цепь, представленную в SEQ ID NO: 77, и легкую цепь, представленную в SEQ ID NO: 78.
В некоторых вариантах осуществления предложено антитело к PD-1 или его антигенсвязывающий фрагмент, где антитело конкурирует с любым из вышеупомянутых антител к PD-1 или их антигенсвязывающих фрагментов за связывание с человеческим PD-1, или связывается с тем же эпитопом человеческого PD-1, что и любое из вышеупомянутых антител к PD-1 или их антигенсвязывающих фрагментов.
В некоторых вариантах осуществления вышеупомянутого антитела к PD-1 или его антигенсвязывающего фрагмента антитело представляет собой биспецифическое антитело или мультиспецифическое антитело.
В некоторых вариантах осуществления антитела к PD-1 или его антигенсвязывающего фрагмента антигенсвязывающий фрагмент выбран из группы, состоящей из Fab, Fab', F(ab')2, одноцепочечного антитела (scFv), димеризованной V-области (диатела) и стабилизированной дисульфидными связями V-области (dsFv).
В некоторых вариантах осуществления раскрыто выделенное моноклональное антитело или его антигенсвязывающий фрагмент, где указанное антитело конкурирует с антителом к PD-1 или его антигенсвязывающим фрагментом в соответствии с любым из вышеупомянутых за связывание с человеческим PD-1.
В некоторых вариантах осуществления настоящего изобретения также предложена фармацевтическая композиция, содержащая терапевтически эффективное количество антитела к PD-1 или его антигенсвязывающего фрагмента в соответствии с любым из вышеупомянутых, или терапевтически эффективное количество вышеупомянутого выделенного моноклонального антитела или его антигенсвязывающего фрагмента, и один или более фармацевтически приемлемых носителей, разбавителей, буферов или эксципиентов. В некоторых вариантах осуществления терапевтически эффективное количество относится к разовой дозе композиции, содержащей от 0,1 до 3000 мг вышеупомянутого антитела к PD-1 или его антигенсвязывающего фрагмента.
В некоторых вариантах осуществления настоящее изобретение также относится к молекуле нуклеиновой кислоты, кодирующей антитело к PD-1 или его антигенсвязывающий фрагмент в соответствии с любым из вышеупомянутых, или кодирующей вышеупомянутое выделенное моноклональное антитело или его антигенсвязывающий фрагмент.
В некоторых вариантах осуществления настоящее изобретение также относится к клетке-хозяину, содержащей вышеупомянутую молекулу нуклеиновой кислоты.
В некоторых вариантах осуществления настоящее изобретение также относится к способу иммунодетекции или определения PD-1, который включает стадию применения антитела к PD-1 или его антигенсвязывающего фрагмента в соответствии с любым из вышеупомянутых, или стадию применения вышеупомянутого выделенного моноклонального антитела или его антигенсвязывающего фрагмента.
В некоторых вариантах осуществления настоящее изобретение также относится к набору, содержащему вышеупомянутое антитело к PD-1 или его антигенсвязывающий фрагмент, или вышеупомянутое выделенное моноклональное антитело или его антигенсвязывающий фрагмент.
В некоторых вариантах осуществления предложено применение вышеупомянутого антитела к PD-1 или его антигенсвязывающего фрагмента для получения диагностических агентов для заболеваний, связанных с PD-1.
В некоторых вариантах осуществления настоящее изобретение также относится к способу лечения заболеваний, включающему введение субъекту терапевтически эффективного количества антитела к PD-1 или его антигенсвязывающего фрагмента в соответствии с любым из вышеупомянутых, или вышеупомянутого выделенного моноклонального антитела или его антигенсвязывающего фрагмента, или вышеупомянутой фармацевтической композиции, или вышеупомянутой молекулы нуклеиновой кислоты.
В некоторых вариантах осуществления заболевание представляет собой опухоль.
В некоторых других вариантах осуществления заболевание выбрано из: плоскоклеточной карциномы головы и шеи, рака головы и шеи, рака головного мозга, глиомы, мультиформной глиобластомы, нейробластомы, рака центральной нервной системы, нейроэндокринной опухоли, рака глотки, рака носоглотки, рака пищевода, рака щитовидной железы, злокачественной мезотелиомы плевры, рака легкого, рака молочной железы, рака печени, гепатомы, гепатоклеточной карциномы, гепатобилиарного рака, рака поджелудочной железы, рака желудка, рака желудочно-кишечного тракта, рака кишечника, рака ободочной кишки, колоректального рака, рака почки, светлоклеточной почечно-клеточной карциномы, рака яичника, рака эндометрия, рака шейки матки, рака мочевого пузыря, рака предстательной железы, рака яичка, рака кожи, меланомы, лейкоза, лимфомы, рака кости, хондросаркомы, миеломы, множественной миеломы, миелодиспластического синдрома, миелопролиферативного новообразования, плоскоклеточной карциномы, саркомы Юинга, системного амилоидоза легкой цепи и карциномы из клеток Меркеля; в некоторых из этих вариантов осуществления лимфома выбрана из: лимфомы Ходжкина, неходжкинской лимфомы, диффузной крупноклеточной B-клеточной лимфомы, фолликулярной лимфомы, первичной медиастинальной крупноклеточной B-клеточной лимфомы, лимфомы из мантийных клеток, малой лимфоцитарной лимфомы, крупноклеточной B-клеточной лимфомы, богатой Т-клетками/гистиоцитами, и лимфоплазматической лимфомы, рак легкого выбран из: немелкоклеточного рака легкого и мелкоклеточного рака легкого, лейкоз выбран из: хронического миелоидного лейкоза, острого миелоидного лейкоза, лимфоцитарного лейкоза, лимфобластного лейкоза, острого лимфобластного лейкоза, хронического лимфоцитарного лейкоза и миелоидного лейкоза; в некоторых других вариантах осуществления заболевание выбрано из: PD-L1-положительной меланомы, рака легкого, немелкоклеточного рака легкого, рака молочной железы, рака желудка, рака почки, рака мочевого пузыря, рака кишечника и рака ободочной кишки.
В некоторых вариантах осуществления настоящее изобретение также относится к применению антитела к PD-1 или его антигенсвязывающего фрагмента в соответствии с любым из вышеупомянутых, или вышеупомянутого выделенного моноклонального антитела или его антигенсвязывающего фрагмента, или вышеупомянутой фармацевтической композиции, или вышеупомянутой молекулы нуклеиновой кислоты для получения лекарственных средств для лечения или предупреждения заболеваний.
В некоторых вариантах осуществления заболевание представляет собой опухоль.
В некоторых других вариантах осуществления заболевание выбрано из: плоскоклеточной карциномы головы и шеи, рака головы и шеи, рака головного мозга, глиомы, мультиформной глиобластомы, нейробластомы, рака центральной нервной системы, нейроэндокринной опухоли, рака глотки, рака носоглотки, рака пищевода, рака щитовидной железы, злокачественной мезотелиомы плевры, рака легкого, рака молочной железы, рака печени, гепатомы, гепатоклеточной карциномы, гепатобилиарного рака, рака поджелудочной железы, рака желудка, рака желудочно-кишечного тракта, рака кишечника, рака ободочной кишки, колоректального рака, рака почки, светлоклеточной почечно-клеточной карциномы, рака яичника, рака эндометрия, рака шейки матки, рака мочевого пузыря, рака предстательной железы, рака яичка, рака кожи, меланомы, лейкоза, лимфомы, рака кости, хондросаркомы, миеломы, множественной миеломы, миелодиспластического синдрома, миелопролиферативного новообразования, плоскоклеточной карциномы, саркомы Юинга, системного амилоидоза легкой цепи и карциномы из клеток Меркеля; в некоторых из этих вариантов осуществления лимфома выбрана из: лимфомы Ходжкина, неходжкинской лимфомы, диффузной крупноклеточной B-клеточной лимфомы, фолликулярной лимфомы, первичной медиастинальной крупноклеточной B-клеточной лимфомы, лимфомы из мантийных клеток, малой лимфоцитарной лимфомы, крупноклеточной B-клеточной лимфомы, богатой Т-клетками/гистиоцитами, и лимфоплазматической лимфомы, рак легкого выбран из: немелкоклеточного рака легкого и мелкоклеточного рака легкого, лейкоз выбран из: хронического миелоидного лейкоза, острого миелоидного лейкоза, лимфоцитарного лейкоза, лимфобластного лейкоза, острого лимфобластного лейкоза, хронического лимфоцитарного лейкоза и миелоидного лейкоза; в некоторых других вариантах осуществления заболевание выбрано из: PD-L1-положительной меланомы, рака легкого, немелкоклеточного рака легкого, рака молочной железы, рака желудка, рака почки, рака мочевого пузыря, рака кишечника и рака ободочной кишки.
В некоторых вариантах осуществления настоящее изобретение также относится к антителу к PD-1 или его антигенсвязывающему фрагменту в соответствии с любым из вышеупомянутых, или к вышеупомянутому выделенному моноклональному антителу или его антигенсвязывающему фрагменту, или к вышеупомянутой молекуле нуклеиновой кислоты, или к вышеупомянутой фармацевтической композиции для применения в качестве лекарственных средств.
В некоторых вариантах осуществления лекарственное средство применяют для лечения или предупреждения заболеваний, связанных с PD-1.
В некоторых вариантах осуществления заболевание представляет собой опухоль.
В некоторых других вариантах осуществления заболевание выбрано из: плоскоклеточной карциномы головы и шеи, рака головы и шеи, рака головного мозга, глиомы, мультиформной глиобластомы, нейробластомы, рака центральной нервной системы, нейроэндокринной опухоли, рака глотки, рака носоглотки, рака пищевода, рака щитовидной железы, злокачественной мезотелиомы плевры, рака легкого, рака молочной железы, рака печени, гепатомы, гепатоцеллюлярной карциномы, гепатобилиарного рака, рака поджелудочной железы, рака желудка, рака желудочно-кишечного тракта, рака кишечника, рака ободочной кишки, колоректального рака, рака почки, светлоклеточной почечно-клеточной карциномы, рака яичника, рака эндометрия, рака шейки матки, рака мочевого пузыря, рака предстательной железы, рака яичка, рака кожи, меланомы, лейкоза, лимфомы, рака кости, хондросаркомы, миеломы, множественной миеломы, миелодиспластического синдрома, миелопролиферативного новообразования, плоскоклеточной карциномы, саркомы Юинга, системного амилоидоза легкой цепи и карциномы из клеток Меркеля; в некоторых из этих вариантов осуществления лимфома выбрана из: лимфомы Ходжкина, неходжкинской лимфомы, диффузной крупноклеточной B-клеточной лимфомы, фолликулярной лимфомы, первичной медиастинальной крупноклеточной B-клеточной лимфомы, лимфомы из мантийных клеток, малой лимфоцитарной лимфомы, крупноклеточной B-клеточной лимфомы, богатой Т-клетками/гистиоцитами, и лимфоплазматической лимфомы, рак легкого выбран из: немелкоклеточного рака легкого и мелкоклеточного рака легкого, лейкоз выбран из: хронического миелоидного лейкоза, острого миелоидного лейкоза, лимфоцитарного лейкоза, лимфобластного лейкоза, острого лимфобластного лейкоза, хронического лимфоцитарного лейкоза и миелоидного лейкоза; в некоторых других вариантах осуществления заболевание выбрано из: PD-L1-положительной меланомы, рака легкого, немелкоклеточного рака легкого, рака молочной железы, рака желудка, рака почки, рака мочевого пузыря, рака кишечника и рака ободочной кишки.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1: результаты теста блокирования антителами к PD-1 связывания PD-1 с его лигандом;
Фиг. 2: влияние антител к PD-1 на секрецию IFNγ из клеток МНПК;
Фиг. 3: эффективность антител к PD-1 в отношении ксенотрансплантата опухоли рака ободочной кишки MC38 у мышей;
Фиг. 4: влияние антител к PD-1 на объем опухоли рака ободочной кишки MC38 у мышей.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Обзор
Для облегчения понимания настоящего изобретения определения некоторых технических и научных терминов специально приведены ниже. Если иное не определено явным образом в настоящем документе, все технические и научные термины, используемые в настоящем документе, имеют значения, обычно понимаемые специалистами в области техники, к которой относится настоящее изобретение.
Термины «программируемая смерть 1», «программируемая клеточная смерть 1», «белок PD-1», «PD-1», «PDCD1» и «hPD-1» используются взаимозаменяемо и включают варианты, изотипы, видовые гомологи и аналоги человеческого PD-1, которые имеют по меньшей мере один общий эпитоп с PD-1. Полную последовательность PD-1 можно найти под регистрационным номером GenBank U64863.
Термин «лиганд программируемой смерти-1 (PD-L1)» представляет собой один из двух поверхностных гликопротеиновых лигандов PD-1 (второй называется PD-L2), который подавляет активацию Т-клеток и секрецию цитокинов при связывании с PD-1. В контексте настоящего документа термин «PD-L1» включает человеческий PD-L1 (hPD-L1), варианты, изотипы и межвидовые гомологи hPD-L1, а также 5 аналогов, имеющих по меньшей мере один общий эпитоп с hPD-1. Полную последовательность hPD-L1 можно найти под регистрационным номером GenBank Q9NZQ7.
Термин «цитокин» является общим термином для белков, высвобождаемых популяцией клеток и действующих на другие клетки как медиаторы межклеточных взаимодействий. Примеры таких цитокинов включают лимфокины, монокины, хемокины и традиционные полипептидные гормоны. Примеры цитокинов включают: человеческие IL-2, IFN-γ, IL-6, TNFα, IL-17 и IL-5.
Трехбуквенные коды и однобуквенные коды для аминокислот, используемые в настоящем описании, соответствуют описанным в J. biol. chem, 243, p3558 (1968).
«Антитело», описанное в настоящем описании, в общем относится к иммуноглобулину. Природное интактное антитело имеет структуру тетрапептидной цепи, состоящей из двух идентичных тяжелых цепей и двух идентичных легких цепей, связанных межцепочечными дисульфидными связями. Аминокислотный состав и последовательности константных областей тяжелой цепи иммуноглобулина различаются, поэтому их антигенность также различается. Таким образом, иммуноглобулины могут быть разделены на пять классов, иначе называемых изотипами иммуноглобулинов, а именно, IgM, IgD, IgG, IgA и IgE, и их соответствующие тяжелые цепи представляют собой μ-цепь, δ-цепь, γ-цепь, α-цепь и ε-цепь, соответственно. Один и тот же класс Ig может быть разделен на разные подклассы в соответствии с различием в аминокислотном составе шарнирной области и количеством и положением дисульфидных связей тяжелой цепи. Например, IgG можно разделить на IgG1, IgG2, IgG3 и IgG4. Легкая цепь подразделяется на κ-цепь или λ-цепь в соответствии с различием константной области. Каждый из пяти классов Ig может иметь κ-цепь или λ-цепь. Антитела, упомянутые в настоящем описании, включают антитела или их антигенсвязывающие фрагменты, включая антитела или их антигенсвязывающие фрагменты, которые были модифицированы на основе иммуноглобулина, сохраняя при этом способность связывать антиген; включая моноспецифические антитела, биспецифические антитела или мультиспецифические антитела; также включая одновалентные антитела, двухвалентные антитела или поливалентные антитела. Например, антигенсвязывающие фрагменты антител могут представлять собой антигенсвязывающие фрагменты, содержащие по меньшей мере одну структуру VH-CH1 и по меньшей мере одну структуру VL-CL, где структуры VH и VL могут сближаться друг с другом на основе межцепочечного взаимодействия и сохранять антигенсвязывающую способность. В некоторых вариантах осуществления антигенсвязывающий фрагмент антитела представляет собой одновалентный Fab-фрагмент (фрагмент Fab1), двухвалентный Fab-фрагмент (F(ab)2), трехвалентный Fab-фрагмент (F(ab)3), поливалентный (два или более) Fab-фрагмент, а также может представлять собой и другие моноспецифические, биспецифические или мультиспецифические антигенсвязывающие фрагменты, содержащие по меньшей мере один Fab-фрагмент.
«Биспецифическое антитело» относится к антителу (включая антитело или его антигенсвязывающий фрагмент, такой как одноцепочечное антитело), которое может специфически связываться с двумя разными антигенами или двумя разными эпитопами одного и того же антигена. В уровне техники раскрыты биспецифические антитела с различными структурами. По целостности молекулы IgG их можно разделить на IgG-подобные биспецифические антитела и биспецифические антитела, подобные фрагментам антител. По количеству и конфигурации антигенсвязывающих областей их можно разделить на двухвалентные, трехвалентные, четырехвалентные или имеющие более высокую валентность биспецифические антитела. В зависимости от того, является ли структура симметричной, их можно разделить на биспецифические антитела с симметричной структурой и биспецифические антитела с асимметричной структурой. Среди них биспецифические антитела на основе фрагментов антител, например, Fab-фрагменты, лишенные Fc-фрагмента, образуют биспецифические антитела путем связывания двух или более Fab-фрагментов в одной молекуле. Эти антитела имеют более низкую иммуногенность, меньшую молекулярную массу и более высокую проницаемость в опухолевую ткань. Типичными структурами антител этого типа являются биспецифические антитела, такие как F(ab)2, scFv-Fab, (scFv)2-Fab и т.д. Что же касается IgG-подобных биспецифических антител (например, с Fc-фрагментом), этот тип антител имеет большую молекулярную массу. Fc-фрагмент помогает очистке антитела на более поздних стадиях и улучшает его растворимость и стабильность. Fc-фрагмент может также связываться с рецептором FcRn и увеличивать период полужизни антитела в сыворотке. Типичными моделями структуры биспецифических антител являются KiH, CrossMAb, Triomab квадрогибридома, FcΔAdp, ART-Ig, BiMAb, Biclonics, BEAT, DuoBody, Azymetric, XmAb, 2:1 TCBs, 1Fab-IgG TDB, FynomAb, два-в-одном/DAF, scFv-Fab-IgG, DART-Fc, LP-DART, CODV-Fab-TL, HLE-BiTE, F(ab)2-CrossMAb, IgG-(scFv)2, Bs4Ab, DVD-Ig, тетравалентный-DART-Fc, (scFv)4-Fc, CODV-Ig, mAb2, F(ab)4-CrossMAb и другие биспецифические антитела (см. Aran F. Labrijn et al., Nature Reviews Drug Discovery volume 18, pages 585-608 (2019); Chen S1 et al., J Immunol Res. 2019 Feb 11; 2019:4516041).
Термин «одновалентный», «двухвалентный», «трехвалентный» или «поливалентный» относится к присутствию указанного количества антигенсвязывающих сайтов в антителе или полипептидном комплексе. Например, «одновалентное антитело» означает, что в антителе есть один антигенсвязывающий сайт, «одновалентный полипептидный комплекс» означает, что в полипептидном комплексе есть один антигенсвязывающий сайт; «двухвалентное антитело» означает, что в антителе есть два антигенсвязывающих сайта, «двухвалентный полипептидный комплекс» означает, что в полипептидном комплексе есть два антигенсвязывающих сайта; «трехвалентное антитело» означает, что в антителе есть три антигенсвязывающих сайта, «трехвалентный полипептидный комплекс» означает, что в полипептидном комплексе есть три антигенсвязывающих сайта; «поливалентное антитело» означает, что в антителе есть несколько (три или более) антигенсвязывающих сайтов, и «поливалентный полипептидный комплекс» означает, что в полипептидном комплексе есть несколько (два или более) антигенсвязывающих сайтов.
Термин «слитый белок антитела» относится к биологически активному слитому белку, образованному путем связывания представляющего интерес белка (полипептида) с иммуноглобулином. Слитый белок обладает биологической активностью как связанного белка, так и иммуноглобулина.
Последовательность из примерно 110 аминокислот около N-конца тяжелой и легкой цепи антитела сильно варьирует и представляет собой вариабельную область (Fv-область); оставшаяся аминокислотная последовательность около С-конца относительно стабильна и представляет собой константную область. Вариабельная область включает 3 гипервариабельных области (HVR) и 4 каркасных области (FR) с относительно консервативными последовательностями. 3 гипервариабельные области, которые определяют специфичность антитела, также известны как определяющие комплементарность области (CDR). Каждая из вариабельной области легкой цепи (VL) и вариабельной области тяжелой цепи (VH) состоит из 3 областей CDR и 4 областей FR. Порядок от аминоконца к карбоксильному концу представляет собой следующий: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. 3 области CDR легкой цепи относятся к LCDR1, LCDR2 и LCDR3; и 3 области CDR тяжелой цепи относятся к HCDR1, HCDR2 и HCDR3.
Антитела согласно настоящему изобретению включают мышиные антитела, химерные антитела, гуманизированные антитела и полностью человеческие антитела, предпочтительно гуманизированные антитела.
Термин «мышиное антитело» в настоящем изобретении означает моноклональное антитело к человеческому PD-1, полученное в соответствии со знаниями и навыками в данной области техники. Во время подготовки испытуемому вводят антиген PD-1, а затем выделяют гибридомы, экспрессирующие антитела с желаемой последовательностью или функциональными свойствами. В предпочтительном варианте осуществления настоящего изобретения мышиное антитело к PD-1 или его антигенсвязывающий фрагмент может дополнительно содержать константную область легкой мышиной κ-, λ-цепи или ее вариант, или дополнительно содержать константную область тяжелой цепи мышиного IgG1, IgG2, IgG3 или ее вариант.
Термин «химерное антитело» означает антитело, образованное путем слияния вариабельной области мышиного антитела с константной областью человеческого антитела, что может ослабить иммунный ответ, индуцируемый мышиным антителом. Создание химерного антитела требует сначала создания гибридомы, секретирующей мышиные специфические моноклональные антитела, затем клонирования гена вариабельной области из клеток мышиной гибридомы, а затем клонирования гена константной области человеческого антитела, если необходимо, связывания гена мышиной вариабельной области с геном человеческой константной области с образованием химерного гена, вставки химерного гена в вектор экспрессии и, наконец, экспрессии молекулы химерного антитела в эукариотической системе или прокариотической системе. В предпочтительном варианте осуществления настоящего изобретения легкая цепь антитела химерного антитела к PD-L1 дополнительно содержит константную область легкой цепи человеческой κ-, λ-цепи или ее вариант. Тяжелая цепь антитела химерного антитела к PD-1 дополнительно содержит константную область тяжелой цепи человеческого IgG1, IgG2, IgG3, IgG4 или ее вариант, предпочтительно содержит константную область тяжелой цепи человеческого IgG1, IgG2 или IgG4, или содержит варианты IgG1, IgG2 или IgG4 с аминокислотными мутациями (например, мутациями L234A и/или L235A, и/или мутациями S228P).
Термин «гуманизированное антитело», также известное как антитело с привитыми CDR, относится к антителу, полученному путем привития мышиных последовательностей CDR на каркас вариабельных областей человеческого антитела, то есть относится к антителу, получаемому из различных типов каркасных последовательностей антитела зародышевой линии человека. Оно может преодолеть гетерогенную реакцию, индуцируемую химерным антителом, поскольку оно несет большое количество компонентов мышиного белка. Такие каркасные последовательности могут быть получены из общедоступных баз данных ДНК или опубликованных ссылок, которые включают последовательности генов антител зародышевой линии. Например, последовательности ДНК зародышевой линии генов вариабельной области человеческой тяжелой и легкой цепи можно найти в базе данных последовательностей зародышевой линии человека «VBase» (доступной по ссылке www.mrccpe.com.ac.uk/vbase), а также в Kabat, E. A., et al., 1991, Sequences of Proteins of Immunological Interest, 5th edition. Чтобы избежать снижения активности, одновременно вызываемого сниженной иммуногенностью, каркасная последовательность вариабельной области человеческого антитела может быть подвергнута минимальным реверсивным мутациям или обратным мутациям для поддержания активности. Гуманизированные антитела согласно настоящему изобретению также включают гуманизированные антитела, которые были дополнительно подвергнуты созреванию аффинности CDR с помощью дрожжевого дисплея.
Вследствие того, что остатки контактируют с антигеном, привитие CDR может приводить к снижению аффинности полученного антитела или его антигенсвязывающего фрагмента к этому антигену из-за того, что каркасные остатки контактировали с антигеном. Такие взаимодействия могут быть результатом гипермутации соматических клеток. Поэтому все же может быть необходимо прививать такие донорские каркасные аминокислоты на каркас гуманизированного антитела. Аминокислотные остатки, участвующие в связывании антигена и полученные из отличных от человеческих антител или их антигенсвязывающих фрагментов, могут быть идентифицированы путем исследования последовательности и структуры вариабельной области моноклонального антитела животного. Остатки в донорском каркасе CDR, которые отличаются от зародышевой линии, можно считать родственными. Если ближайшую зародышевую линию определить невозможно, последовательность можно сравнить с консенсусной последовательностью подкласса или последовательностью антитела животного с высоким процентом сходства. Считается, что редкие каркасные остатки являются результатом гипермутации в соматических клетках и поэтому играют важную роль в связывании.
В одном из вариантов осуществления настоящего изобретения антитело или его антигенсвязывающий фрагмент может дополнительно содержать константную область легкой цепи человеческой или мышиной κ-, λ-цепи или ее вариант, или дополнительно содержит константную область тяжелой цепи человеческого или мышиного IgG1, IgG2, IgG3, IgG4 или ее вариант; предпочтительно содержит константную область тяжелой цепи человеческого IgG1, IgG2 или IgG4, или варианта IgG1, IgG2 или IgG4 с аминокислотными мутациями (например, мутацией L234A и/или L235A, и/или мутацией S228P).
«Традиционный вариант» константной области тяжелой цепи человеческого антитела и константной области легкой цепи человеческого антитела, описанный в настоящем документе, относится к варианту константной области тяжелой цепи или константной области легкой цепи, раскрытому в уровне техники, который не изменяет структуру и функцию вариабельной области антитела. Примеры вариантов включают варианты константной области тяжелой цепи IgG1, IgG2, IgG3 или IgG4 с сайт-направленными модификациями и аминокислотными заменами в константной области тяжелой цепи. Конкретные замены представляют собой такие, как мутации YTE, мутации L234A и/или L235A, мутации S228P и/или мутации для получения структуры «выступ-во-впадину» (что превращает тяжелую цепь антитела в комбинацию выступ-Fc и впадина-Fc), известные в данной области техники. Было доказано, что эти мутации наделяют антитело новыми свойствами без изменения функции вариабельной области антитела.
«HuMAb», «человеческое антитело», «полностью человеческое антитело» и «целое человеческое антитело» могут использоваться взаимозаменяемо и могут относиться к антителам, полученным от человека, или антителам, полученным от генетически модифицированного организма, который был «сконструирован» для получения определенных человеческих антител в ответ на стимуляцию антигеном, и могут быть получены любым способом, известным в данной области техники. В некоторых технологиях элементы локусов человеческой тяжелой и легкой цепей вводят в клеточные линии организмов, происходящих из линий эмбриональных стволовых клеток, в которых локусы эндогенных тяжелой цепи и легкой цепи были целевым образом нарушены агентами, нацеленными на эндогенные локусы тяжелой цепи и легкой цепи, содержащиеся в этих клеточных линиях. Трансгенные организмы могут синтезировать человеческие антитела, специфичные к человеческим антигенам, и эти организмы можно использовать для получения гибридом, секретирующих человеческие антитела. Человеческое антитело также может представлять собой антитело, в котором тяжелая и легкая цепи кодируются нуклеотидными последовательностями, полученными из одного или нескольких источников человеческой ДНК. Полностью человеческие антитела также могут быть сконструированы методами трансфекции генов или хромосом и технологией фагового дисплея, или сконструированы из B-клеток, активированных in vitro, при этом все из перечисленного известно в данной области техники.
Термины «полноразмерное антитело», «интактное антитело», «целое антитело» и «полное антитело» используются в настоящем документе взаимозаменяемо и относятся к антителу в по существу интактной форме, в отличие от антигенсвязывающих фрагментов, определенных ниже. Эти термины относятся исключительно к антителу, в котором тяжелая цепь включает Fc-область.
Термин «антигенсвязывающий фрагмент» или «функциональный фрагмент» антитела относится к одному или более фрагментам антитела, которые сохраняют способность специфически связываться с антигеном (например, PD-1). Было показано, что фрагменты полноразмерных антител могут быть использованы для осуществления антигенсвязывающей функции антител. Примеры связывающего фрагмента, включенного в термин «антигенсвязывающий фрагмент» антитела, включают (i) Fab-фрагменты, одновалентные фрагменты, состоящие из доменов VL, VH, CL и CH1; (ii) F(ab')2-фрагменты, включая двухвалентные фрагменты из двух Fab-фрагментов, связанных дисульфидным мостиком в шарнирной области, (iii) Fd-фрагменты, состоящие из доменов VH и CH1; (iv) Fv-фрагменты, состоящие из доменов VH и VL одного плеча антитела; (V) одиночные домены или dAb-фрагменты (Ward et al., (1989) Nature 341:544-546), которые состоят из домена VH; и (vi) выделенные определяющие комплементарность области (CDR) или (vii) комбинации двух или более выделенных CDR, необязательно связанных синтетическими линкерами. Кроме того, хотя два домена VL и VH Fv-фрагмента кодируются отдельными генами, могут использоваться методы рекомбинации для их связывания с помощью синтетических линкеров, так что он может быть получен в виде единой белковой цепи, в которой области VL и VH соединяются в пары с образованием одновалентной молекулы (называемой одноцепочечным Fv (scFv); см., например, Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci USA 85: 5879-5883). Подразумевается, что такие одноцепочечные антитела также охватываются термином «антигенсвязывающий фрагмент» антитела. Такие фрагменты антител получают с использованием обычных методов, известных специалистам в данной области техники, и функции фрагментов подвергают скринингу таким же образом, как и функции интактных антител. Антигенсвязывающий фрагмент может быть получен с помощью технологии рекомбинантных ДНК или ферментативного или химического расщепления интактного иммуноглобулина. Антитела могут представлять собой антитела разных изотипов, например, антитела IgG (например, подтипы IgG1, IgG2, IgG3 или IgG4), IgA1, IgA2, IgD, IgE или IgM.
Антигенсвязывающие фрагменты согласно настоящему изобретению включают Fab, F(ab')2, Fab', одноцепочечное антитело (scFv), димеризованную V-область (диатело), стабилизированную дисульфидными связями V-область (dsFv), CDR-содержащий пептид и тому подобное.
Fab представляет собой фрагмент антитела, имеющий молекулярную массу около 50000 и обладающий антигенсвязывающей активностью, и его получают обработкой молекул антитела IgG протеазой папаином (расщепляет аминокислотный остаток в положении 224 H-цепи), в котором примерно половина N-концевой стороны H-цепи и вся L-цепь соединены вместе дисульфидной связью(ями).
Fab согласно настоящему изобретению может быть получен с использованием папаина для обработки моноклонального антитела согласно настоящему изобретению, которое специфически распознает человеческий PD-1 и связывается с аминокислотной последовательностью внеклеточной области или ее трехмерной структурой. Кроме того, Fab может быть получен путем вставки ДНК, кодирующей Fab антитела, в прокариотический вектор экспрессии или эукариотический вектор экспрессии и введения вектора в прокариотический организм или эукариотический организм для экспрессии Fab.
F(ab')2 представляет собой фрагмент антитела, который имеет молекулярную массу около 100000, обладает антигенсвязывающей активностью и содержит две области Fab, связанные в положении шарнира, и его получают путем расщепления нижней части двух дисульфидных связей в шарнирной области IgG ферментом пепсином.
F(ab')2 согласно настоящему изобретению может быть получен с использованием пепсина для обработки моноклонального антитела согласно настоящему изобретению, которое специфически распознает человеческий PD-1 и связывается с аминокислотной последовательностью внеклеточной области или ее трехмерной структурой. Кроме того, F(ab')2 может быть получен путем связывания Fab', описанного ниже, тиоэфирной связью или дисульфидной связью.
Fab' представляет собой фрагмент антитела, имеющий молекулярную массу около 50000 и обладающий антигенсвязывающей активностью, получаемый путем расщепления дисульфидной связи в шарнирной области F(ab')2. Fab' согласно настоящему изобретению может быть получен с использованием восстанавливающих агентов, например, дитиотреитола, для обработки F(ab')2 согласно настоящему изобретению, который специфически распознает PD-1 и связывается с аминокислотной последовательностью внеклеточной области или ее трехмерной структурой.
Кроме того, Fab может быть получен путем вставки ДНК, кодирующей Fab'-фрагмент антитела, в прокариотический вектор экспрессии или эукариотический вектор экспрессии, и затем введения вектора в прокариотический организм или эукариотический организм для экспрессии Fab'.
Термин «одноцепочечное антитело», «одноцепочечный Fv» или «scFv» относится к молекулам, содержащим вариабельный домен тяжелой цепи антитела (или область; VH) и вариабельный домен легкой цепи антитела (или область; VL), связанные линкером. Такие молекулы scFv могут иметь общую структуру: NH2-VL-линкер-VH-COOH или NH2-VH-линкер-VL-COOH. Подходящие линкеры в уровне техники состоят из повторяющихся аминокислотных последовательностей GGGGS или их вариантов, например, варианта с 1-4 повторами последовательности (Holliger et al. (1993), Proc. Natl. Acad. Sci. USA 90: 6444-6448). Другие линкеры, которые могут быть использованы в настоящем изобретении, описаны в Alfthan et al. (1995), Protein Eng. 8:725-731, Choi et al. (2001), Eur. J. Immunol. 31:94-106, Hu et al. (1996), Cancer Res. 56:3055-3061, Kipriyanov et al. (1999), J. Mol. Biol. 293:41-56 и Roovers et al. (2001), Cancer Immunol.
ScFv согласно настоящему изобретению может быть получен посредством следующих стадий: получение кДНК, кодирующей VH и VL моноклонального антитела согласно настоящему изобретению, которое специфически распознает человеческий PD-1 и связывается с аминокислотной последовательностью внеклеточной области или ее трехмерной структурой, конструирование ДНК, кодирующей scFv, вставка ДНК в прокариотический вектор экспрессии или эукариотический вектор экспрессии, а затем введение вектора экспрессии в прокариотический организм или эукариотический организм для экспрессии scFv.
Диатело представляет собой фрагмент антитела, в котором scFv димеризован, и представляет собой фрагмент антитела с двухвалентной антигенсвязывающей активностью. В двухвалентной антигенсвязывающей активности два антигена могут быть одинаковыми или разными.
Диатело согласно настоящему изобретению может быть получено посредством следующих стадий: получение кДНК, кодирующей VH и VL моноклонального антитела согласно настоящему изобретению, которое специфически распознает человеческий PD-1 и связывается с аминокислотной последовательностью внеклеточной области или ее трехмерной структурой, конструирование ДНК, кодирующей scFv, так что пептидный линкер имеет 8 или менее аминокислотных остатков в длину, вставка ДНК в прокариотический вектор экспрессии или эукариотический вектор экспрессии, а затем введение вектора экспрессии в прокариотический организм или эукариотический организм для экспрессии диатела.
dsFv получают путем связывания полипептидов, в которых один аминокислотный остаток в каждой из VH и VL заменен остатком цистеина, посредством дисульфидной связи между остатками цистеина. Аминокислотные остатки, замещенные остатками цистеина, могут быть выбраны согласно известному методу (Protein Engineering, 7, 697 (1994)), основанному на предсказании трехмерной структуры антитела.
dsFv согласно настоящему изобретению может быть получен посредством следующих стадий: получение кДНК, кодирующей VH и VL моноклонального антитела согласно настоящему изобретению, которое специфически распознает человеческий PD-1 и связывается с аминокислотной последовательностью внеклеточной области или ее трехмерной структурой, конструирование ДНК, кодирующей dsFv, вставка ДНК в прокариотический вектор экспрессии или эукариотический вектор экспрессии, а затем введение вектора экспрессии в прокариотический организм или эукариотический организм для экспрессии dsFv.
CDR-содержащий пептид состоит из одной или нескольких областей, включающих область (области) CDR VH или VL. Пептиды, содержащие несколько CDR, могут быть связаны напрямую или через подходящий пептидный линкер.
CDR-содержащий пептид согласно настоящему изобретению может быть получен посредством следующих стадий: конструирование последовательности(ей) кДНК, кодирующей область (области) CDR VH и VL моноклонального антитела согласно настоящему изобретению, которое специфически распознает человеческий PD-1 и связывается с аминокислотной последовательностью внеклеточной области или ее трехмерной структурой, вставка последовательности(ей) ДНК в прокариотический вектор экспрессии или эукариотический вектор экспрессии, а затем введение вектора экспрессии в прокариотический организм или эукариотический организм для экспрессии пептида. CDR-содержащий пептид также может быть получен методами химического синтеза, например, методом Fmoc или методом tBoc.
Термин «аминокислотное различие» или «аминокислотная мутация» относится к наличию аминокислотных изменений или мутаций в варианте белка или полипептида по сравнению с исходным белком или полипептидом, включая наличие 1, 2, 3 или более аминокислотных инсерций, делеций или замен относительно исходного белка или полипептида.
Термин «каркас антитела» или «FR-область» относится к фрагменту вариабельного домена VL или VH, который служит каркасом для антигенсвязывающей петли (CDR) вариабельного домена. Фактически, он представляет собой вариабельный домен без CDR.
Термин «определяющая комплементарность область», «CDR» или «гипервариабельная область» относится к одной из шести гипервариабельных областей в вариабельном домене антитела, которые в основном вносят вклад в связывание антигена. Обычно имеется три CDR (HCDR1, HCDR2 и HCDR3) в каждой вариабельной области тяжелой цепи и три CDR (LCDR1, LCDR2 и LCDR3) в каждой вариабельной области легкой цепи. Для определения границ аминокислотных последовательностей CDR может быть использована любая из хорошо известных схем, включая систему нумерации «Kabat» (см. Kabat et al. (1991), "Sequences of Proteins of Immunological Interest", 5th edition, Public Health Service, National Institutes of Health, Bethesda, MD), систему нумерации «Chothia» (см. Al-Lazikani et al., (1997) JMB 273:927-948) и систему нумерации ImmunoGenTics (IMGT) (Lefranc M. P., Immunologist, 7, 132- 136 (1999); Lefranc, M. P., et al., Dev. Comp. Immunol., 27, 55-77 (2003)), и т.п. Например, для классического формата в соответствии с системой Kabat нумерация аминокислотных остатков CDR в вариабельном домене тяжелой цепи (VH) представляет собой 31-35 (HCDR1), 50-65 (HCDR2) и 95-102 (HCDR3); нумерация аминокислотных остатков CDR в вариабельном домене легкой цепи (VL) представляет собой 24-34 (LCDR1), 50-56 (LCDR2) и 89-97 (LCDR3). В соответствии с системой Chothia нумерация аминокислотных остатков CDR в VH представляет собой 26-32 (HCDR1), 52-56 (HCDR2) и 95-102 (HCDR3); и нумерация аминокислотных остатков в VL представляет собой 26-32 (LCDR1), 50-52 (LCDR2) и 91-96 (LCDR3). В соответствии с комбинацией определений CDR согласно Kabat и Chothia, CDR состоят из аминокислотных остатков 26-35 (HCDR1), 50-65 (HCDR2) и 95-102 (HCDR3) в человеческой VH и аминокислотных остатков 24-34 (LCDR1), 50-56 (LCDR2) и 89-97 (LCDR3) в человеческой VL. В соответствии с системой IMGT нумерация аминокислотных остатков CDR в VH примерно представляет собой 26-35 (CDR1), 51-57 (CDR2) и 93-102 (CDR3), и нумерация аминокислотных остатков CDR в VL примерно представляет собой 27-32 (CDR1), 50-52 (CDR2) и 89-97 (CDR3). В соответствии с системой IMGT области CDR антитела могут быть определены с помощью программы IMGT/DomainGap Align.
Термин «эпитоп» или «антигенная детерминанта» относится к сайту на антигене, с которым специфически связывается иммуноглобулин или антитело (например, определенный сайт на молекулах PD-L1). Эпитопы обычно включают по меньшей мере 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 последовательных или непоследовательных аминокислот в уникальной пространственной конформации. См., например, Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, G.E.Morris, Ed. (1996).
Термины «специфически связывает», «селективно связывает», «связывается селективно» и «связывается специфически» относятся к связыванию антитела с эпитопом на выбранном антигене. Обычно антитела связываются с аффинностью (KD) менее приблизительно 10-8 М, например, менее приблизительно 10-9 М, 10-10 М, 10-11 М или менее.
Термин «KD» или «Kd» относится к равновесной константе диссоциации для конкретного взаимодействия антитело-антиген. Обычно антитела согласно настоящему изобретению связываются с PD-1 с равновесной константой диссоциации (KD) менее приблизительно 10-7 M, например, менее приблизительно 10-8 M или 10-9 M, например, при измерении на приборе BIACORE с использованием технологии поверхностного плазменного резонанса (SPR).
Когда термин «конкуренция» используется в контексте антигенсвязывающих белков, которые конкурируют за один и тот же эпитоп (например, нейтрализующих антигенсвязывающих белков или нейтрализующих антител), он относится к конкуренции между антигенсвязывающими белками, которая может быть определена с помощью следующего анализа: в анализе подлежащий тестированию антигенсвязывающий белок (например, антитело или его иммунологически функциональный фрагмент) предотвращает или ингибирует (например, снижает) специфическое связывание референсного антигенсвязывающего белка (например, лиганда или референсного антитела) с общим антигеном (например, PD-1 или его фрагментом). Для определения того, конкурирует ли один антигенсвязывающий белок с другим, можно использовать множество типов анализов конкурентного связывания, например: твердофазный прямой или непрямой радиоиммуноанализ (РИА), твердофазный прямой или непрямой иммуноферментный анализ (EIA), конкурентный сэндвич-анализ (см., например, Stahli et al, 1983, Methods in Enzymology 9: 242-253); твердофазный прямой биотин-авидиновый EIA (см., например, Kirkland et al, 1986, J. Immunol. 137: 3614-3619), твердофазный анализ с прямым мечением, твердофазный сэндвич-анализ с прямым мечением (см., например, Harlow and Lane, 1988, Antibodies, A Laboratory Manual, Cold Spring Harbor Press); твердофазный РИА с прямым мечением метками I-125 (см., например, Morel et al, 1988, Molec. Immunol. 25: 7-15); твердофазный прямой биотин-авидиновый EIA (см., например, Cheung, et al, 1990, Virology 176: 546-552); и РИА с прямым мечением (Moldenhauer et al, 1990, Scand. J. Immunol. 32: 77-82). Обычно анализ включает использование очищенного антигена, связанного с твердой поверхностью или клеткой, несущей либо немеченый тестируемый антигенсвязывающий белок, либо меченый референсный антигенсвязывающий белок. Конкурентное ингибирование измеряют путем измерения количества метки, связанной с твердой поверхностью или с клеткой, в присутствии тестируемого антигенсвязывающего белка. Обычно тестируемый антигенсвязывающий белок присутствует в избытке. Антигенсвязывающие белки, идентифицируемые с помощью конкурентного анализа (конкурирующие антигенсвязывающие белки), включают: антигенсвязывающие белки, которые связываются с тем же эпитопом, что и референсный антигенсвязывающий белок; и антигенсвязывающие белки, которые связываются с соседними эпитопами, расположенными достаточно близко к эпитопу, с которым связывается референсный антигенсвязывающий белок, где указанные два эпитопа пространственно затрудняют связывание друг друга. Дополнительные подробности в отношении способов определения конкуренции за связывание представлены в приведенных в настоящем документе примерах. Обычно, когда конкурирующий антигенсвязывающий белок присутствует в избытке, он будет ингибировать (например, снижать) специфическое связывание референсного антигенсвязывающего белка с общим антигеном по меньшей мере на 40-45%, 45-50%, 50-55%, 55-60%, 60-65%, 65-70%, 70%-75% или 75%, или даже более. В некоторых случаях связывание ингибируется по меньшей мере на 80-85%, 85-90%, 90-95%, 95-97% или 97%, или более.
Термин «молекула нуклеиновой кислоты» в контексте настоящего документа относится к молекулам ДНК и молекулам РНК. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной и предпочтительно представляет собой двухцепочечную ДНК или одноцепочечную мРНК или модифицированную мРНК. Когда нуклеиновая кислота находится в функциональной связи с другой последовательностью нуклеиновой кислоты, нуклеиновая кислота является «функционально связанной». Например, если промотор или энхансер влияет на транскрипцию кодирующей последовательности, то промотор или энхансер функционально связан с кодирующей последовательностью.
Термин «вектор» относится к молекуле нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, с которой она была связана. В одном из вариантов осуществления вектор представляет собой «плазмиду», которая относится к кольцевой двухцепочечной петле ДНК, с которой могут быть связаны дополнительные сегменты ДНК. В еще одном варианте осуществления вектор представляет собой вирусный вектор, в котором дополнительные сегменты ДНК могут быть связаны с вирусным геномом. Раскрытые в настоящем документе векторы могут автономно реплицироваться в клетке-хозяине, в которую они были введены (например, бактериальный вектор с бактериальной точкой начала репликации и эписомальный вектор млекопитающих), или могут быть интегрированы в геном клетки-хозяина после введения в клетку-хозяина, чтобы реплицироваться вместе с геномом хозяина (например, неэписомальный вектор млекопитающих).
Способы получения и очистки антител и антигенсвязывающих фрагментов хорошо известны в уровне техники, например, Antibodies: A Laboratory Manual, Cold Spring Harbor, главы 5-8 и 15. Например, мыши могут быть иммунизированы человеческим PD-1 или его фрагментами, и полученные антитела могут быть ренатурированы, очищены, и может быть проведено аминокислотное секвенирование обычными методами. Антигенсвязывающие фрагменты также могут быть получены обычными методами. Одну или более человеческих областей FR добавляют к нечеловеческим областям CDR антител или антигенсвязывающих фрагментов согласно изобретению с использованием методов генной инженерии. Последовательности FR зародышевой линии человека могут быть получены с веб-сайта ImmunoGeneTics (IMGT), http://imgt.cines.fr, путем сравнения базы данных IMGT генов зародышевой линии вариабельной области человеческого антитела и программного обеспечения MOE, или быть получены из The Immunoglobulin FactsBook, 2001ISBN012441351.
Термин «клетка-хозяин» относится к клетке, в которую был введен вектор экспрессии. Клетки-хозяева могут включать бактерии, микроорганизмы, клетки растений или животных. Бактерии, которые можно легко трансформировать, включают представителей Enterobacteriaceae, например, штаммы Escherichia coli или Salmonella; Bacillaceae, например, Bacillus subtilis; Pneumococcus; Streptococcus и Haemophilus influenzae. Подходящие микроорганизмы включают Saccharomyces cerevisiae и Pichia pastoris. Подходящие линии клеток-хозяев животных включают клетки СНО (линия клеток яичника китайского хомячка) и NS0.
Генетически модифицированные антитела или антигенсвязывающие фрагменты согласно настоящему изобретению могут быть получены и очищены обычными методами. Например, последовательности кДНК, кодирующие тяжелую и легкую цепи, могут быть клонированы и рекомбинированы в вектор экспрессии GS. Вектор экспрессии рекомбинантного иммуноглобулина может быть стабильно трансфицирован в клетки СНО. В качестве более рекомендуемого уровня техники системы экспрессии млекопитающих могут приводить к гликозилированию антител, особенно в высококонсервативных N-концевых сайтах Fc-области. Стабильные клоны получают путем экспрессии антител, специфически связывающихся с человеческим PD-1. Положительные клоны размножают в бессывороточной среде биореактора для получения антител. Культуральная среда, в которую секретируются антитела, может быть очищена обычными методами. Например, с использованием для очистки колонки Sepharose FF A или G с отрегулированным буфером. Неспецифически связывающиеся компоненты вымываются. Затем связывающие антитела элюируются методом градиента pH, и фрагменты антител обнаруживают с помощью SDS-PAGE и собирают. Антитела могут быть отфильтрованы и концентрированы с использованием обычных методов. Растворимые смеси и полимеры также могут быть удалены обычными методами, например, с помощью молекулярных сит и ионного обмена. Полученный продукт необходимо немедленно заморозить, например, при -70 °C, или лиофилизировать.
«Введение», «дача», «лечение» и «обработка» применительно к животным, людям, экспериментальным субъектам, клеткам, тканям, органам или биологическим жидкостям относятся к контакту экзогенного лекарственного средства, терапевтического агента, диагностического агента или композиции с животными, людьми, субъектами, клетками, тканями, органами или биологическими жидкостями. «Введение», «дача», «лечение» и «обработка» могут относиться, например, к лечению, фармакокинетике, диагностике, исследованиям и экспериментальным методам. Обработка клеток включает контакт реагентов с клетками и контакт реагентов с жидкостями, где жидкости контактируют с клетками. «Введение», «дача» и «обработка» также относятся к обработке, например, клеток реагентами, диагностическими, связывающими композициями или другой клеткой in vitro и ex vivo. «Лечение», применительно к пациенту-человеку, пациентам-животным или испытуемым, относится к терапевтическому лечению, профилактическим или превентивным мерам, исследовательским и диагностическим применениям.
«Лечение» относится к введению внутреннего или наружного терапевтического агента, например, композиции, содержащей любое из связывающих соединений согласно настоящему изобретению, пациенту, у которого есть один или более симптомов заболевания, при котором терапевтическое средство, как известно, оказывает терапевтический эффект. Обычно терапевтический агент вводят в количестве, эффективном для облегчения одного или более симптомов заболевания у получающего лечение пациента или популяции, чтобы вызвать регресс таких симптомов или подавить развитие таких симптомов в любой клинически измеренной степени. Количество терапевтического агента, эффективное для облегчения какого-либо конкретного симптома заболевания (также называемое «терапевтически эффективным количеством»), может варьироваться в зависимости от различных факторов, таких как стадия заболевания, возраст и масса тела пациента, а также способность лекарственного средства оказывать желаемый терапевтический эффект у пациента. Были ли облегчены симптомы заболевания, можно оценить с помощью любых методов клинического тестирования, обычно используемых врачами или другими специалистами в области здравоохранения для оценки степени тяжести или прогрессирования симптомов. Хотя варианты осуществления настоящего изобретения (например, способы лечения или продукты) могут быть неэффективны для облегчения каждого симптома целевого заболевания, они должны ослаблять симптом целевого заболевания у статистически значимого количества пациентов, как определено с помощью любого статистического критерия, известного в данной области техники, такого как t-критерий Стьюдента, критерий хи-квадрат, U-критерий Манна-Уитни, критерий Краскела-Уоллиса (H-критерий), критерий Джонкхира-Терпстры и критерий Уилкоксона.
«Консервативная модификация» или «консервативное замещение или замена» относится к замене аминокислот в белке другими аминокислотами, имеющими схожие характеристики (например, заряд, размер боковой цепи, гидрофобность/гидрофильность, конформацию и жесткость основной цепи и т.д.), чтобы можно было часто вносить изменения без изменения биологической активности белка. Специалистам в данной области техники известно, что в общем случае замена одной аминокислоты в несущественных областях полипептида по существу не изменяет биологическую активность (см., например, Watson et al. (1987) Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., Page 224, (4th edition)). Кроме того, маловероятно, что замена аминокислотами с аналогичной структурой или функцией повлияет на биологическую активность. Иллюстративные консервативные аминокислотные замены приведены в таблице «Иллюстративные консервативные аминокислотные замены» ниже.
Таблица 3. Иллюстративные консервативные аминокислотные замены
«Эффективное количество» или «эффективная доза» относится к количеству лекарственного средства, соединения или фармацевтической композиции, необходимому для получения любого одного или более полезных или желаемых терапевтических результатов. Для профилактических целей полезные или желаемые результаты включают устранение или снижение риска, снижение степени тяжести или отсрочку начала заболевания, включая биохимические, гистологические и поведенческие проявления заболевания, его осложнения и промежуточные патологические фенотипы, которые имеют место во время развития процесса заболевания. Для терапевтических целей полезные или желаемые результаты включают клинические результаты, такие как снижение частоты различных расстройств согласно настоящему изобретению, связанных с антигеном-мишенью, или улучшение одного или более симптомов расстройства, снижение дозы других агентов, необходимых для лечения расстройства, повышение терапевтической эффективности другого агента и/или задержка прогрессирования расстройства согласно настоящему изобретению, связанного с антигеном-мишенью, у пациента.
«Экзогенный» относится к веществам, продуцируемым вне организмов, клеток или человеческих тел, в зависимости от обстоятельств. «Эндогенный» относится к веществам, продуцируемым внутри клеток, организмов или человеческих тел, в зависимости от обстоятельств.
«Гомология» относится к сходству последовательностей между двумя полинуклеотидными последовательностями или между двумя полипептидами. Когда положения в двух сравниваемых последовательностях заняты одним и тем же основанием или субъединицей мономера аминокислоты, например, если положение в каждой из двух молекул ДНК занято аденином, то молекулы гомологичны в этом положении. Процент гомологии между двумя последовательностями является функцией количества совпадающих или гомологичных положений, общих для двух последовательностей, деленного на количество сравниваемых положений, а затем умноженного на 100. Например, когда две последовательности оптимально выровнены, если 6 из 10 положений в двух последовательностях совпадают или гомологичны, то две последовательности гомологичны на 60%; если 95 из 100 положений в двух последовательностях совпадают или гомологичны, то две последовательности гомологичны на 95%. Обычно, когда две последовательности выровнены, сравнение проводят для получения максимального процента гомологии. Например, сравнение может проводиться алгоритмом BLAST, в котором параметры алгоритма выбирают так, чтобы обеспечить максимальное совпадение для каждой последовательности по всей длине каждой референсной последовательности. Следующие ссылки относятся к алгоритму BLAST, часто используемому для анализа последовательностей: АЛГОРИТМЫ BLAST: Altschul, S. F. et al., (1990) J. Mol. Biol. 215:403-410; Gish, W. et al., (1993) Nature Genet. 3:266-272; Madden, T. L. et al., (1996) Meth. Enzymol. 266:131-141; Altschul, S. F. et al., (1997) Nucleic Acids Res. 25:3389-3402; Zhang, J. et al., (1997) Genome Res. 7:649-656. Другие традиционные алгоритмы BLAST, такие как доступные от NCBI BLAST, также хорошо известны специалистам в данной области техники.
Выражения «клетка», «клеточная линия» и «культура клеток», используемые в настоящем документе, могут использоваться взаимозаменяемо, и все такие названия включают потомство. Следовательно, слова «трансформант» и «трансформированная клетка» включают первичные тестируемые клетки и полученные из них культуры, независимо от количества пассажей. Также следует иметь в виду, что из-за преднамеренных или случайных мутаций все потомство не может быть абсолютно одинаковым с точки зрения содержания ДНК. Включено мутантное потомство с той же функцией или биологической активностью, что и обнаруженная при скрининге исходных трансформированных клеток. Когда имеется в виду другое понятие, это будет ясно из контекста.
«Полимеразная цепная реакция» или «ПЦР» в контексте настоящего документа относится к процедуре или методике, в которой следовое количество определенной части нуклеиновой кислоты, РНК и/или ДНК, амплифицируют, как описано, например, в патенте США № 4683195. В общем случае информацию о последовательности на конце или вне целевой области необходимо получать для того, чтобы можно было конструировать олигонуклеотидные праймеры; эти праймеры являются такими же или подобными с точки зрения последовательности соответствующим цепям матрицы, подлежащей амплификации. 5'-концевые нуклеотиды двух праймеров могут быть идентичны концам вещества, подлежащего амплификации. ПЦР может быть использована для амплификации конкретных последовательностей РНК, конкретных последовательностей ДНК из тотальной геномной ДНК, а также последовательностей кДНК, фагов или плазмид, транскрибированных из тотальной клеточной РНК, и т.д. См. в общем Mullis et al. (1987) Cold Spring Harbor, Symp. Ouant. Biol. 51:263; Erlich ed., (1989) PCR TECHNOLOGY (Stockton Press, N.Y.). Используемая в настоящем документе ПЦР рассматривается как пример, но не единственный пример метода полимеразной реакции нуклеиновых кислот для амплификации тестируемого образца нуклеиновой кислоты, и этот метод включает использование известных нуклеиновых кислот в качестве праймеров и полимераз нуклеиновых кислот для амплификации или получения определенной части нуклеиновой кислоты.
«Выделенный» относится к очищенному состоянию и в этом случае означает, что обозначенная молекула по существу не содержит других биомолекул, например, нуклеиновых кислот, белков, липидов, углеводов или других веществ, например, клеточного детрита и питательной среды. В целом, термин «выделенный» не подразумевает полного отсутствия этих веществ или отсутствия воды, буферов или солей, если только они не присутствуют в количестве, которое существенно препятствует экспериментальному или терапевтическому применению соединения, как описано в настоящем документе.
«Необязательный» или «необязательно» означает, что событие или среда, описанные далее, могут, но не обязательно должны, иметь место, и описание включает случаи, когда событие или среда имеют или не имеют места. Например, «необязательно содержащий от 1 до 3 вариабельных областей тяжелой цепи антитела» означает, что вариабельные области тяжелой цепи антитела с определенными последовательностями могут, но не обязательно должны, присутствовать.
«Фармацевтическая композиция» означает смесь, содержащую одно или более соединений, описанных в настоящем документе, или их физиологически/фармацевтически приемлемую соль или пролекарство, и другие химические компоненты, например, физиологические/фармацевтически приемлемые носители и эксципиенты. Назначение фармацевтической композиции состоит в том, чтобы облегчить введение соединения в организм, что способствует абсорбции активного ингредиента, в результате чего он проявляет биологическую активность.
Термин «фармацевтически приемлемый носитель» относится к любому неактивному веществу, подходящему для использования в составе для доставки антител или антигенсвязывающих фрагментов. Носитель может представлять собой антиадгезивный агент, связующее, оболочку, разрыхлитель, наполнитель или разбавитель, консервант (такой как антиоксидант, антибактериальный или противогрибковый агент), подсластитель, агент, замедляющий абсорбцию, смачивающий агент, эмульгатор, буфер и т.д. Примеры подходящих фармацевтически приемлемых носителей включают воду, этанол, многоатомные спирты (например, глицерин, пропиленгликоль, полиэтиленгликоль и т.д.), декстрозу, растительные масла (например, оливковое масло), физиологический раствор, буфер, забуференный физиологический раствор и изотонические агенты, например, сахара, многоатомные спирты, сорбит и хлорид натрия.
Кроме того, настоящее изобретение включает агенты для лечения заболеваний, связанных с клетками, положительными на антиген-мишень (например, PD-1), где указанные агенты содержат антитело к PD-1 или его антигенсвязывающий фрагмент согласно настоящему изобретению в качестве активного ингредиента.
Заболевание, связанное с PD-1, не ограничено в настоящем изобретении, при условии, что оно представляет собой заболевание, связанное с PD-1. Например, терапевтический ответ, индуцированный молекулой согласно настоящему изобретению, может быть достигнут путем связывания с человеческим PD-1, а затем ингибирования связывания PD-1 и его лигандов PD-L1 и PD-L2, или уничтожения опухолевых клеток, сверхэкспрессирующих PD-1. Следовательно, будучи включенными в препараты и составы, подходящие для терапевтического применения, молекулы согласно настоящему изобретению очень эффективны для людей, имеющих опухоль или рак, предпочтительно меланому, рак ободочной кишки, рак молочной железы, рак легкого, рак желудка, рак кишечника, рак почки, немелкоклеточный рак легкого, рак мочевого пузыря и т.д.
Кроме того, настоящее изобретение относится к способам иммунодетекции или определения антигенов-мишеней (например, PD-1), реагентам для иммунодетекции или определения антигенов-мишеней (например, PD-1), способам иммунодетекции или определения клеток, экспрессирующих антигены-мишени (например, PD-1), и диагностическим агентам для диагностики заболеваний, связанных с клетками, положительными на антиген-мишень (например, PD-1), которые включают антитело или фрагмент антитела согласно настоящему изобретению, которые специфически распознают антиген-мишень (например, человеческий PD-1) и связываются с аминокислотной последовательностью внеклеточной области или ее трехмерной структурой, будучи используемыми в качестве активного ингредиента.
В настоящем изобретении способ, используемый для обнаружения или измерения количества антигена-мишени (например, PD-1) может представлять собой любой известный способ. Например, он включает методы иммунодетекции или измерения.
Методы иммунодетекции или измерения представляют собой методы обнаружения или измерения количества антитела или антигена с использованием меченых антигенов или антител. Примеры методов иммунодетекции или измерения включают радиоиммуноанализ (РИА), иммуноферментный анализ (EIA или ELISA), флуоресцентный иммуноанализ (FIA), люминесцентный иммуноанализ, вестерн-блоттинг, физико-химические методы и т.д.
Вышеупомянутые заболевания, связанные с PD-1-положительными клетками-мишенями, могут быть диагностированы путем обнаружения или измерения клеток, экспрессирующих PD-1, с использованием антител или фрагментов антител согласно настоящему изобретению.
Для обнаружения клеток, экспрессирующих полипептид, могут быть использованы известные методы иммунодетекции, предпочтительно с использованием иммунопреципитации, флуоресцентного окрашивания клеток, иммуногистохимического окрашивания и т.д. Кроме того, может быть использован метод флуоресцентного окрашивания антителами с помощью системы FMAT8100HTS (Applied Biosystem).
В настоящем изобретении образца in vivo, используемого для обнаружения или измерения антигена-мишени (например, PD-1), нет конкретных ограничений при условии, что он может содержать клетки, экспрессирующие антиген-мишень (например, PD-1), например, гистоциты, кровь, плазма, сыворотка, панкреатический сок, моча, кал, интерстициальная жидкость или культуральная жидкость.
В зависимости от требуемого способа диагностики диагностический агент, содержащий моноклональное антитело или его фрагмент согласно настоящему изобретению, может также содержать реагенты для проведения реакции антиген-антитело или реагенты для обнаружения такой реакции. Реагенты, используемые для проведения реакции антиген-антитело, включают буферы, соли и т.д. Реагенты, используемые для обнаружения, включают агенты, обычно используемые в методах иммунодетекции измерения, например, меченые вторичные антитела, распознающие моноклональное антитело, его фрагмент или конъюгат, и субстрат, соответствующий метке, и т.д.
Получение антигенов
1. Конструирование антигенов:
Человеческий слитый белок PD-1-IgG1Fc сконструирован и синтезирован со 150 аминокислотами внеклеточной области человеческого PD-1 на N-конце и Fc-фрагментом человеческого IgG1 (hIgG1Fc) на C-конце. Рекомбинантный белок PD-1-Fc высокой чистоты может быть получен после очистки с помощью аффинной колонки с белком А и использован для обнаружения связывания антитела к PD-1 с антигеном.
Человеческий PD-1-IgG1Fc (SEQ ID NO: 1):

Примечание: подчеркнутая часть представляет собой сигнальный пептид, не выделенная часть представляет собой внеклеточную область человеческого PD-1, а выделенная курсивом часть представляет собой hIgG1Fc (сигнальный пептид + внеклеточная область + hIgG1Fc).
Человеческий PD-1-his (SEQ ID NO: 2):
MEFGLSWLFLVAILKGVQCPGWFLDSPDRPWNPPTFSPALLVVTEGDNATFTCSFSNTSESFVLNWYRMSPSNQTDKLAAFPEDRSQPGQDCRFRVTQLPNGRDFHMSVVRARRNDSGTYLCGAISLAPKAQIKESLRAELRVTERRAEVPTAHPSPSPRPAGQFQTLVGSSDYKDDDDKHHHHHH.
Антиген PD-1, кодируемый нуклеиновыми кислотами, трансфицированными в клетки (SEQ ID NO: 3):
MQIPQAPWPVVWAVLQLGWRPGWFLDSPDRPWNPPTFSPALLVVTEGDNATFTCSFSNTSESFVLNWYRMSPSNQTDKLAAFPEDRSQPGQDCRFRVTQLPNGRDFHMSVVRARRNDSGTYLCGAISLAPKAQIKESLRAELRVTERRAEVPTAHPSPSPRPAGQFQTLVVGVVGGLLGSLVLLVWVLAVICSRAARGTIGARRTGQPLKEDPSAVPVFSVDYGELDFQWREKTPEPPVPCVPEQTEYATIVFPSGMGTSSPARRGSADGPRSAQPLRPEDGHCSWPL.
Получение антител
Антитела к человеческому PD-1 могут быть получены путем иммунизации мышей, а также могут быть получены из фаговой библиотеки против человеческого PD-1, полученной путем иммунизации мышей.
Способ получения антитела к человеческому PD-1 путем иммунизации мышей представляет собой следующий:
1. Иммунизация: лабораторные белые мыши SJL, самки в возрасте 6-8 недель, и белые мыши Balb/c, самки в возрасте 6-8 недель. Условия содержания: уровень СПФ (свободный от патогенной микрофлоры). После приобретения мышей содержали в лабораторных условиях в течение 1 недели для адаптации, с циклом свет/темнота 12/12 часов при температуре 20-25 °C; влажность 40-60%. Мышей, адаптированных к окружающей среде, иммунизировали по разным протоколам, по 6-10 мышей в каждой группе. Иммунантиген мог представлять собой очищенный рекомбинантный белок PD-1-IgG1Fc (см. SEQ ID NO: 1), PD-1-his (см. SEQ ID NO: 2) или клетки Jurkat/CHO-PD-1, трансфицированные PD-1, в качестве антигена (см. SEQ ID NO: 3). Один антиген в сочетании с разными иммунными адъювантами или разными типами иммуногенов мог использоваться для перекрестной иммунизации. Сайт иммунизации мог представлять собой брюшную полость или область под кожей на спине, или же иммунизация могла проводиться поочередно в двух сайтах. Для перекрестной иммунизации использовали иммуноадъювант TiterMax® Gold Adjuvant (далее называемый Titermax, приобретен у Sigma, кат. № T2684) и адъювант квасцов Imject Alum Adjuvant (далее называемый Alum, приобретен у Pierce, кат. № 77161). Отношение антигена к адъюванту (Titermax) составляло 1:1, отношение антигена к адъюванту (Alum) составляло 3:1, 25-50 мкг/животное (первая иммунизация), 50 мкг/животное (бустер-иммунизация) или 1×107 клеток Jurkat/CHO-PD-1/животное. В день 0 25-50 мкг/животное эмульгированного антигена вводили внутрибрюшинно, один раз в неделю или раз в две недели после первой иммунизации, поочередно использовали Titermax и Alum, в общей сложности 5-8 раз.
2. Слияние клеток: мышей с высокими титрами антител в сыворотке отбирали для слияния клеток селезенки. Из глаз мышей брали кровь через 72 ч после бустер-иммунизации. Мышей умерщвляли путем сворачивания шеи и помещали в 75% этанол для дезинфекции. Лимфоциты селезенки были слиты с клетками миеломы Sp2/0 (Китайская академия наук) с получением гибридомных клеток с применением оптимизированной процедуры слияния, опосредованной ПЭГ. Слитые гибридомные клетки ресуспендировали в полной среде HAT (среда RPMI-1640, содержащая 20% FBS, 1×HAT и 1×OPI), и отмеряли аликвоты в 96-луночные планшеты для культивирования клеток (1×105/150 мкл/лунку), инкубировали при 37 °C, 5% CO2 и высевали в общей сложности в примерно 10-30 планшетов. На 5 день после слияния добавляли полную среду HAT в количестве 50 мкл/лунку и инкубировали при 37 °C, 5% CO2. С 7-8 дня после слияния, в зависимости от плотности роста клеток, среду полностью меняли при 200 мкл/лунку и инкубировали при 37 °C, 5% CO2.
3. Скрининг гибридомных клеток: через 7-9 дней после слияния, в зависимости от плотности роста клеток, осуществляли анализ методом ELISA для обнаружения связывания антитела с PD-1, и клетки в положительных лунках дополнительно определяли с помощью ELISA для обнаружения блокирования связывания PD-1/PDL1. Среду в положительных лунках меняли, и клетки переносили в 24-луночные планшеты после достижения соответствующей плотности клеток. Линии клеток, перенесенные в 24-луночные планшеты, были повторно протестированы, а затем сохранены и впервые субклонированы. Клетки, положительные в скрининге первого субклонирования, сохраняли, и проводили второе или третье субклонирование до получения клонов потомства одной клетки. Путем многократных слияний получали гибридомные клетки, обладавшие эффектом блокирования связывания PD-1 и PDL1.
Способ получения антител к человеческому PD-1 с помощью фаговой мышиной иммунной библиотеки к человеческому PD-1 представлял собой следующий:
1. Конструирование фаговой мышиной иммунной библиотеки к человеческому PD-1: отбирали селезенки мышей с высоким титром антител в сыворотке и экстрагировали тотальную РНК из ткани с использованием Trizol (Invitrogen, кат. № 15596-018). кДНК получали с использованием набора PrimeScript™ II 1st Strand cDNA Synthesis Kit (Takara, кат. № 6210A) для обратной транскрипции. Праймеры для конструирования библиотеки были разработаны и синтезированы согласно базе данных IMGT. Одноцепочечные фрагменты антител получали с помощью трех раундов реакций ПЦР. Одноцепочечные фрагменты антител и модифицированный вектор для построения библиотеки pCantab5E (Amersham Biosciences/GE, кат. № 27-9400-01) расщепляли с помощью Sfi1 (NEB, кат. № R0123L), и очищали и выделяли с помощью набора E. Z. N. A.® Gel Extraction Kit (Omega, кат. № D2500-02) после электрофореза. Затем Т4 ДНК-лигазу (NEB, кат. № M0202L) использовали для лигирования при 16 °C в течение 16-18 часов, и указанный выше набор использовали для очистки и выделения, и, наконец, проводили элюирование деионизированной водой. Брали 1 мкг продукта лигирования и смешивали с 1 флаконом компетентного TG1 (Lucigen, кат. № 60502-2) для электротрансформации, и проводили электротрансформацию с помощью электропоратора (Bio Rad Micropulser) с параметрами, установленными на 2,5 кВ, 200 Ом и 25 мкФ. Трансформацию повторяли 10 раз. Продукт распределяли по планшетам и культивировали в перевернутом состоянии при 37 °C в течение 16-18 часов. Затем все колонии отделяли и смешивали вместе, добавляли глицерин до конечной концентрации 15% и хранили при -80 °C для дальнейшего использования.
2. Скрининг фаговой мышиной иммунной библиотеки к человеческому PD-1: упакованная фаговая иммунная библиотека к человеческому PD-1 (1×1012 - 1×1013) и 100 мкл микрогранул со стрептомицином (Miltenyi Biotec, Оберн, Калифорния) добавляли к 1 мл натрий-фосфатного буфера, содержащего 2% обезжиренного молока (MPBS), и инкубировали при комнатной температуре в течение 1 ч, помещали на магнитную подставку и собирали супернатант. К супернатанту добавляли 10 мкг/мл биотинилированного белка человеческого PD-1-ECD-his (приобретенного у Sino Biological) и инкубировали в течение 1 ч при комнатной температуре. Затем добавляли 100 мкл покрытых стрептавидином магнитных гранул (предварительно инкубированных с 1 мл MPBS) и инкубировали в течение 1 ч при комнатной температуре. Продукт загружали на систему магнитного штатива для сортировки и аспирировали супернатант. Добавляли 1 мл PBST (фосфатный буфер, содержащий 0,1% Tween-20) и несколько раз переворачивали. После полной аспирации добавляли свежий промывочный раствор, повторяли 11 раз для удаления несвязанных фрагментов антител и добавляли 0,5 мл элюирующего раствора (50 мкл 10 мг/мл исходного раствора трипсина, добавленного в 450 мкл PBS). Его встряхивали в течение 15 мин при комнатной температуре, помещали на магнитный штатив и аспирировали супернатант в новую пробирку Эппендорфа. TG1 высевали в среду 2YT и размножали до плотности бактериальной культуры OD600 = 0,4. В каждую пробирку добавляли 1,75 мл TG1 (OD600 = 0,4) и добавляли 250 мкл элюированного фага (фага), инкубировали на водяной бане при 37 °C в течение 30 минут и распределяли по планшетам с градиентным разведением для проверки титра. Оставшийся раствор TG1 центрифугировали, распределяли по планшетам и инкубировали в течение ночи при 37 °C.
Фаговую мышиную иммунную библиотеку подвергали 2-3 раундам скрининга с использованием биотинилированного антигена человеческого PD-1-ECD-his, со скринингом MACS (магнитные гранулы со стрептомицином, Invitrogen), и, наконец, были получены и подтверждены секвенированием моноклональные антитела, способные связываться с PD-1 и способные блокировать связывание PD-1 с PD-L1. Были получены последовательности вариабельной области этих антител.
Очистка рекомбинантных антигенных белков/антител
1. Отделение и очистка супернатанта гибридомы/аффинная хроматография с белком G:
Для очистки супернатанта мышиной гибридомы наиболее предпочтительной была аффинная хроматография с белком G. Культуру гибридомы центрифугировали для сбора супернатанта и, исходя из объема супернатанта, добавляли 10-15% объема 1 М трис-HCl (pH 8,0-8,5) для регулирования pH супернатанта. Для колонки с белком G использовали 6 M гидрохлорид гуанидина для промывки 3-5 колоночными объемами, а затем использовали чистую воду для промывки 3-5 колоночными объемами; буферную систему, такую как 1×PBS (pH 7,4) использовали в качестве уравновешивающего буфера для уравновешивания колонки 3-5 колоночными объемами. Клеточный супернатант загружали и связывали с использованием низкой скорости потока, которую контролировали так, чтобы время удерживания составляло около 1 мин или более; 1×PBS (pH 7,4) использовали для промывки хроматографической колонки 3-5 колоночными объемами до тех пор, пока УФ-поглощение не снижалось до исходного уровня. Для элюирования образца использовали буфер 0,1 М уксусная кислота/ацетат натрия (pH 3,0), собирали пики элюирования в соответствии с УФ-детекцией, и использовали 1 М трис-HCl (pH 8,0) для быстрого доведения pH элюированного продукта до 5-6 для временного хранения. Для элюированного продукта замена раствора может быть осуществлена методами, хорошо известными специалистам в данной области техники, такими как использование трубок для ультрафильтрации и концентрирование и замена раствора на требуемую буферную систему, или использование молекулярной эксклюзии, например, G-25, для обессаливания и замены на требуемую буферную систему, или использование колонок для молекулярной эксклюзии с высоким разрешением, таких как Superdex 200, для удаления компонентов, агрегированных в элюируемом продукте, для повышения чистоты образца.
2. Очистка белков или антител с помощью аффинной хроматографии с белком A:
Сначала супернатант культуры клеток, экспрессирующей антигенный белок или антитело, центрифугировали при высокой скорости для сбора супернатанта. Для аффинной колонки с белком А использовали 6 М гидрохлорид гуанидина для промывки 3-5 колоночными объемами, а затем использовали чистую воду для промывки 3-5 колоночными объемами. Буферную систему, такую как 1×PBS (pH 7,4), использовали в качестве уравновешивающего буфера для уравновешивания хроматографической колонки 3-5 колоночными объемами. Клеточный супернатант загружали и связывали с использованием низкой скорости потока, которую контролировали так, чтобы время удерживания составляло около 1 мин или более. После завершения связывания использовали 1×PBS (pH 7,4) для промывки хроматографической колонки 3-5 колоночными объемами до тех пор, пока УФ-поглощение не снижалось до исходного уровня. Для элюирования образца использовали буфер 0,1 М уксусная кислота/ацетат натрия (pH 3,0-3,5), собирали пики элюирования в соответствии с УФ-детекцией, и использовали 1 М трис-HCl (pH 8,0) для быстрого доведения pH элюированного продукта до 5-6 для временного хранения. Для элюированного продукта замена раствора может быть осуществлена методами, хорошо известными специалистам в данной области техники, такими как использование трубок для ультрафильтрации и концентрирование и замена раствора на требуемую буферную систему, или использование молекулярной эксклюзии, например, G-25, для обессаливания и замены на требуемую буферную систему, или использование колонок для молекулярной эксклюзии с высоким разрешением, таких как Superdex 200, для удаления компонентов, агрегированных в элюируемом продукте, для повышения чистоты образца.
ПРИМЕРЫ
Настоящее изобретение дополнительно описано ниже в сочетании с примерами, но эти примеры не ограничивают объем настоящего изобретения. Экспериментальные методы, условия для которых конкретно не указаны в примерах настоящего изобретения, обычно проводились при обычных условиях, таких как описанные в Antibodies: A Laboratory Manual and Molecular Cloning: A Laboratory Manual, Cold Spring Harbor; или при условиях, рекомендованных производителем сырья или продукта. Реагенты, источники которых конкретно не указаны, являются обычными реагентами, доступными на рынке.
Пример 1. Получение мышиных антител к человеческому PD-1
Мышиные антитела к человеческому PD-1, полученные вышеуказанным способом, подвергали экспериментам в отношении связывания антигена и проводили скрининг, чтобы получить несколько штаммов антител с хорошей активностью, куда были включены M23, M32 и M33. Клоны потомства одной клетки размножали и культивировали, РНК экстрагировали, и вырожденные праймеры мышиного Ig использовали для проведения обратной транскрипции-амплификации (ОТ-ПЦР) для получения последовательности вариабельной области антитела. Последовательность вариабельной области мышиного антитела связывали с последовательностью константной области человеческого антитела. Химерное антитело из мышиного моноклонального антитела было клонировано и рекомбинантно экспрессировано, и были проведены эксперименты по определению активности in vitro для подтверждения того, что полученная последовательность вариабельной области моноклонального антитела являлась правильной.
Последовательности вариабельных областей мышиных антител M23, M32 и M33 определены следующим образом:
Вариабельная область тяжелой цепи мышиного антитела M23 (SEQ ID NO: 4):
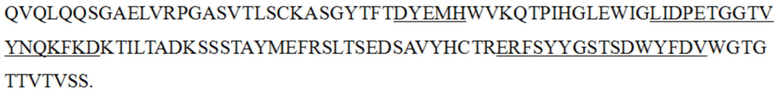
Вариабельная область легкой цепи мышиного антитела M23 (SEQ ID NO: 5):

Вариабельная область тяжелой цепи мышиного антитела M32 (SEQ ID NO: 6):
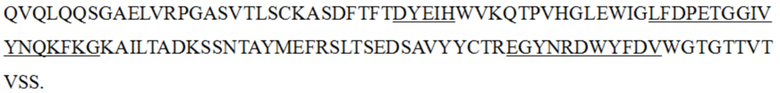
Вариабельная область легкой цепи мышиного антитела M32 (SEQ ID NO: 7):

Вариабельная область тяжелой цепи мышиного антитела M33 (SEQ ID NO: 19):

Вариабельная область легкой цепи мышиного антитела M33 (SEQ ID NO: 20):

Примечание: в последовательностях вариабельной области тяжелой цепи и вариабельной области легкой цепи вышеуказанных антител подчеркнуты последовательности CDR, определенные в соответствии с системой нумерации Kabat, в следующем порядке: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4.
Таблица 4. Последовательности областей CDR тяжелой и легкой цепи мышиного антитела M23, M32 и M33
(SEQ ID NO: 8)
(SEQ ID NO: 11)
(SEQ ID NO: 9)
(SEQ ID NO: 12)
(SEQ ID NO: 10)
(SEQ ID NO: 13)
(SEQ ID NO: 14)
(SEQ ID NO: 17)
(SEQ ID NO: 15)
(SEQ ID NO: 12)
(SEQ ID NO: 16)
(SEQ ID NO: 18)
(SEQ ID NO: 21)
(SEQ ID NO: 24)
(SEQ ID NO: 22)
(SEQ ID NO: 25)
(SEQ ID NO: 23)
(SEQ ID NO: 26)
Примечание: последовательности CDR антител в таблице определены в соответствии с системой нумерации Kabat.
Пример 2. Гуманизация моноклональных антител к человеческому PD-1
Путем выравнивания с базой данных IMGT генов зародышевой линии вариабельной области тяжелой и легкой цепи человеческого антитела и анализа в программном обеспечении MOE в качестве каркасов были выбраны гены вариабельной области тяжелой и легкой цепей зародышевой линии человека с высокой идентичностью последовательностям легкой и тяжелой цепи M23, M32 и M33. CDR из этих 3 мышиных антител были привиты на соответствующие каркасы человеческого антитела для построения на их основе соответствующих гуманизированных антител, соответственно.
1. Гуманизация мышиного антитела M23
1.1 Выбор каркаса для гуманизации мышиного антитела M23
Каркасами легких цепей для гуманизации мышиного антитела M23 являются IGKV2-40*01 и IGKJ4*01, а каркасами тяжелых цепей для гуманизации являются IGHV1-69*02 и IGHJ6*01. Последовательности вариабельных областей после гуманизации представляли собой следующие (подчеркнуты последовательности CDR):
Hu23VH-CDR-привитая: (SEQ ID NO: 27)

Hu23VL-CDR-привитая: (SEQ ID NO: 28)
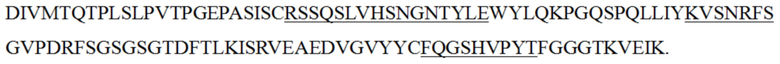
1.2 Выбор каркаса для гуманизации и дизайн обратных мутаций мышиного антитела M23
Таблица 5. Обратные мутации гуманизированных антител на основе мышиного антитела M23
Примечание: «привитая» означает, что CDR мышиного антитела пересажены в последовательности области FR зародышевой линии человека. Аминокислотные остатки определены и аннотированы в соответствии с системой нумерации Kabat, например, I2G означает, что I в положении 2 нумерации Kabat мутирован обратно в G в соответствии с системой нумерации Kabat.
Последовательности вариабельных областей легкой/тяжелой цепи гуманизированных антител на основе M23 представляют собой следующие:
> Hu23VL1 (такая же, как Hu23VL-CDR-привитая): (SEQ ID NO: 28)
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSNGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK.
> Hu23VL2 (SEQ ID NO: 29)
DGVMTQTPLSLPVTPGEPASISCRSSQSLVHSNGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK.
> Hu23VH1 (такая же, как Hu23VH-CDR-привитая): (SEQ ID NO: 27)
EVQLVQSGAEVKKPGSSVKVSCKASGGTFSDYEMHWVRQAPGQGLEWMGLIDPETGGTVYNQKFKDRVTITADKSTSTAYMELSSLRSEDTAVYYCARERFSYYGSTSDWYFDVWGQGTTVTVSS.
> Hu23VH2 (SEQ ID NO: 30)
EVQLVQSGAEVKKPGSSVKVSCKASGYTFSDYEMHWVRQAPGQGLEWMGLIDPETGGTVYNQKFKDRVTLTADKSTSTAYMELSSLRSEDTAVYYCARERFSYYGSTSDWYFDVWGQGTTVTVSS.
> Hu23VH3 (SEQ ID NO: 31)
EVQLVQSGAEVKKPGSSVKVSCKASGYTFSDYEMHWVRQAPGQGLEWIGLIDPETGGTVYNQKFKDRTTLTADKSTSTAYMEFSSLRSEDTAVYYCARERFSYYGSTSDWYFDVWGQGTTVTVSS.
> Hu23VH4 (SEQ ID NO: 32)
EVQLVQSGAEVKKPGSSVKVSCKASGYTFSDYEMHWVRQAPGQGLEWIGLIDPETGGTVYNQKFKDRTTLTADKSTSTAYMEFSSLRSEDTAVYYCTRERFSYYGSTSDWYFDVWGQGTTVTVSS.
1.3 Комбинация гуманизированных последовательностей мышиного антитела M23
Антитела и комбинации их вариабельных областей, полученные путем гуманизации мышиного антитела M23, приведены в таблице ниже.
Таблица 6. Комбинации вариабельных областей гуманизированного антитела Hu23
VH
Примечание: «Hu23-1» относится к антителу, в котором вариабельная область легкой цепи представляет собой Hu23VL1, а вариабельная область тяжелой цепи представляет собой Hu23VH1, и так далее.
Комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu23-1), представленные в приведенной выше таблице, могут быть связаны с константными областями легкой/тяжелой цепи антитела с образованием полноразмерных антител, соответственно; если в настоящем описании не указано иное, при образовании полноразмерного антитела вариабельная область легкой цепи связывается с константной областью каппа-цепи, представленной в SEQ ID NO: 73, с образованием легкой цепи антитела, а вариабельная область тяжелой цепи связывается с константной областью тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, или с константной областью тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, с образованием тяжелой цепи антитела, и название в таблице, относящееся к комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu23-1), с суффиксом «. IgG4AA» означает полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4AA, с суффиксом «. IgG4P» - полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4-P. Например, «Hu23-1.IgG4AA» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu23VH1 и константной области тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, и легкой цепи, образованной путем связывания вариабельной области легкой цепи Hu23VL1 и константной области каппа-цепи, представленной в SEQ ID NO: 73. «Hu23-1.IgG4P» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu23VH1 и константной области тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, и легкой цепи, образованный путем связывания вариабельной области легкой цепи Hu23VL1 и константной области каппа-цепи, представленной в SEQ ID NO: 73.
2. Гуманизация мышиного антитела M32
2.1 Выбор каркаса для гуманизации мышиного антитела M32
Каркасами легких цепей для гуманизации мышиного антитела M32 являются IGKV2-40*01 и IGKJ4*01, а каркасами тяжелых цепей для гуманизации являются IGHV1-69*02 и IGHJ6*01. Последовательности гуманизированных вариабельных областей представляли собой следующие (подчеркнуты последовательности CDR):
Hu32VH-CDR-привитая: (SEQ ID NO: 33) IGHV1-69*02 и IGHJ6*01

Hu32VL-CDR-привитая: (SEQ ID NO: 34)

2.2 Выбор каркаса для гуманизации и дизайн обратных мутаций мышиного антитела M32
Таблица 7. Обратные мутации гуманизированных антител на основе мышиного антитела M32
Примечание: «привитая» означает, что CDR мышиного антитела пересажены в последовательности области FR зародышевой линии человека. Аминокислотные остатки определены и аннотированы в соответствии с системой нумерации Kabat, например, I2V означает, что I в положении 2 нумерации Kabat мутирован обратно в V в соответствии с системой нумерации Kabat.
Последовательности вариабельных областей легкой и тяжелой цепи гуманизированных антител на основе мышиного антитела M32 представляют собой следующие:
> Hu32VL1 (такая же, как Hu32VL-CDR-привитая): (SEQ ID NO: 34)
DIVMTQTPLSLPVTPGEPASISCRSSQSIVHSNGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYAFGGGTKVEIK.
> Hu32VL2 (SEQ ID NO: 35)
DVVMTQTPLSLPVTPGEPASISCRSSQSIVHSNGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYAFGGGTKVEIK.
> Hu32VH1 (такая же, как Hu23VH-CDR-привитая): (SEQ ID NO: 33)
EVQLVQSGAEVKKPGSSVKVSCKASGGTFSDYEIHWVRQAPGQGLEWMGLFDPETGGIVYNQKFKGRVTITADKSTSTAYMELSSLRSEDTAVYYCAREGYNRDWYFDVWGQGTTVTVSS.
> Hu32VH2 (SEQ ID NO: 36)
EVQLVQSGAEVKKPGSSVKVSCKASGFTFSDYEIHWVRQAPGQGLEWMGLFDPETGGIVYNQKFKGRVTLTADKSTSTAYMELSSLRSEDTAVYYCTREGYNRDWYFDVWGQGTTVTVSS.
> Hu32VH3 (SEQ ID NO: 37)
EVQLVQSGAEVKKPGSSVKVSCKASDFTFSDYEIHWVRQAPGQGLEWMGLFDPETGGIVYNQKFKGRVTLTADKSTSTAYMELSSLRSEDTAVYYCTREGYNRDWYFDVWGQGTTVTVSS.
> Hu32VH4 (SEQ ID NO: 38)
EVQLVQSGAEVKKPGSSVKVSCKASGFTFSDYEIHWVRQAPGQGLEWIGLFDPETGGIVYNQKFKGRATLTADKSTSTAYMEFSSLRSEDTAVYYCTREGYNRDWYFDVWGQGTTVTVSS.
> Hu32VH5 (SEQ ID NO: 39)
EVQLVQSGAEVKKPGSSVKVSCKASDFTFTDYEIHWVRQAPGQGLEWIGLFDPETGGIVYNQKFKGRATLTADKSTSTAYMEFSSLRSEDTAVYYCTREGYNRDWYFDVWGQGTTVTVSS.
> Hu32VH6 (SEQ ID NO: 40)
EVQLVQSGAEVKKPGSSVKVSCKASDFTFTDYEIHWVKQAPGHGLEWIGLFDPETGGIVYNQKFKGKATLTADKSTSTAYMEFSSLRSEDTAVYYCTREGYNRDWYFDVWGQGTTVTVSS.
2.3 Комбинация гуманизированных последовательностей мышиного антитела M32
Антитела и комбинации их вариабельных областей, полученные путем гуманизации мышиного антитела M32.
Таблица 8. Комбинации вариабельных областей легкой/тяжелой цепи гуманизированного антитела Hu32
VH
Примечание: например, в таблице «Hu32-1» относится к антителу с комбинацией вариабельной области легкой цепи Hu32VL1 и вариабельной области тяжелой цепи Hu32VH1, и так далее.
Комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu32-1), представленные в приведенной выше таблице, могут быть связаны с константными областями легкой/тяжелой цепи антитела с образованием полноразмерных антител, соответственно; если в настоящем описании не указано иное, при образовании полноразмерного антитела вариабельная область легкой цепи связывается с константной областью каппа-цепи, представленной в SEQ ID NO: 73, с образованием легкой цепи антитела, а вариабельная область тяжелой цепи связывается с константной областью тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, или с константной областью тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, с образованием тяжелой цепи антитела, и название в таблице, относящееся к комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu32-1), с суффиксом «. IgG4AA» означает полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4AA, с суффиксом «. IgG4P» - полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4-P. Например, «Hu32-1.IgG4AA» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu32VH1 и константной области тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, и легкой цепи, образованной путем связывания вариабельной области легкой цепи Hu32VL1 и константной области каппа-цепи, представленной в SEQ ID NO: 73. «Hu32-1.IgG4P» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu32VH1 и константной области тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, и легкой цепи, образованный путем связывания вариабельной области легкой цепи Hu32VL1 и константной области каппа-цепи, представленной в SEQ ID NO: 73.
3. Гуманизация мышиного антитела M33
3.1 Выбор каркаса для гуманизации мышиного антитела M33
Каркасами легких цепей для гуманизации мышиного антитела M33 являются IGKV1-39*01 и IGKJ4*01, а каркасами тяжелых цепей для гуманизации являются IGHV3-7 и IGHJ6*01. Последовательности гуманизированных вариабельных областей представляли собой следующие (подчеркнуты последовательности CDR):
Hu33VH-CDR-привитая (SEQ ID NO: 41):

Hu33VL-CDR-привитая (SEQ ID NO: 42):

3.2 Выбор каркаса для гуманизации и дизайн обратных мутаций мышиного антитела M33
Таблица 9. Обратные мутации гуманизированных антител на основе мышиного антитела M33
Примечание: «привитая» означает, что CDR мышиного антитела пересажены в последовательности области FR зародышевой линии человека. Аминокислотные остатки определены и аннотированы в соответствии с системой нумерации Kabat, например, F71Y означает, что F в положении 71 нумерации Kabat мутирован обратно в Y в соответствии с системой нумерации Kabat.
Последовательности вариабельной области легкой цепи и вариабельной области тяжелой цепи гуманизированных антител на основе мышиного антитела M33 представляют собой следующие:
> Hu33VL1 (такая же, как Hu33VL-CDR-привитая): (SEQ ID NO: 42)
DIQMTQSPSSLSASVGDRVTITCRASQDINNFLNWYQQKPGKAPKLLIYYTSSLHSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQGNTLPWTFGGGTKVEIK.
> Hu33VL2 (SEQ ID NO: 43)
DIQMTQSPSSLSASVGDRVTITCRASQDINNFLNWYQQKPGKAPKLLIYYTSSLHSGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGGGTKVEIK.
> Hu33VL3 (SEQ ID NO: 44)
DIQMTQSPSSLSASVGDRVTITCRASQDINNFLNWYQQKPGGAVKLLIYYTSSLHSGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGGGTKVEIK.
> Hu33VH1 (такая же, как Hu33VH-CDR-привитая): (SEQ ID NO: 41)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVATISGGGVDTYYQDNVQGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCARPYGHGYFDVWGQGTTVTVSS.
> Hu33VH2 (SEQ ID NO: 45)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVATISGGGVDTYYQDNVQGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCASPYGHGYFDVWGQGTTVTVSS.
> Hu33VH3 (SEQ ID NO: 46)
KVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVATISGGGVDTYYQDNVQGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCASPYGHGYFDVWGQGTTVTVSS.
3.3 Комбинация гуманизированных последовательностей мышиного антитела M33
Таблица 10. Комбинации вариабельных областей легкой и тяжелой цепи гуманизированного антитела
VH
Примечание: например, в таблице «Hu33-6» относится к антителу с комбинацией вариабельной области легкой цепи Hu33VL2 и вариабельной области тяжелой цепи Hu33VH3, и так далее.
Комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu33-6), представленные в приведенной выше таблице, могут быть связаны с константными областями легкой/тяжелой цепи антитела с образованием полноразмерных антител, соответственно; если в настоящем описании не указано иное, при образовании полноразмерного антитела вариабельная область легкой цепи связывается с константной областью каппа-цепи, представленной в SEQ ID NO: 73, с образованием легкой цепи антитела, а вариабельная область тяжелой цепи связывается с константной областью тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, или с константной областью тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, с образованием тяжелой цепи антитела, и название в таблице, относящееся к комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu33-6), с суффиксом «. IgG4AA» означает полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4AA, с суффиксом «.IgG4P» - полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4-P. Например, «Hu32-6.IgG4AA» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu33VH3 и константной области тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, и легкой цепи, образованной путем связывания вариабельной области легкой цепи Hu33VL2 и константной области каппа-цепи, представленной в SEQ ID NO: 73. «Hu33-6.IgG4P» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu33VH3 и константной области тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, и легкой цепи, образованный путем связывания вариабельной области легкой цепи Hu33VL2 и константной области каппа-цепи, представленной в SEQ ID NO: 73.
4. Мутанты гуманизированных антител
4.1 Мутантные антитела на основе гуманизированного антитела Hu23
Посредством компьютерного моделирования аминокислоты в определенных сайтах LCDR1 легкой цепи (SEQ ID NO: 11) гуманизированного антитела Hu23 были подвергнуты сайт-направленным мутациям, и конкретные мутации приведены в таблице 11:
Таблица 11. Мутантные последовательности LCDR1 легкой цепи Hu23:
(SEQ ID NO: 47)
(SEQ ID NO: 48)
(SEQ ID NO: 49)
(SEQ ID NO: 50)
(SEQ ID NO: 51)
(SEQ ID NO: 52)
Примечание: Hu23LCDR1 (N28Q) означает последовательность с мутацией LCDR1, в которой N в положении 28 в соответствии с системой нумерации Kabat вариабельной области легкой цепи Hu23VL1 или Hu23VL2 гуманизированного антитела Hu23 мутирован в Q, Hu23LCDR1 (G29A) означает последовательность с мутацией LCDR1, в которой G в положении 29 в соответствии с системой нумерации Kabat вариабельной области легкой цепи Hu23VL1 или Hu23VL2 гуманизированного антитела Hu23 мутирован в A (CDR определены с помощью системы нумерации Kabat).
Последовательности вариабельной области легкой цепи гуманизированного антитела Hu23 после введения мутаций в LCDR1 представляют собой следующие:
> Последовательность Hu23VL1(N28Q) представляет собой:
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSQGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 53)
> Последовательность Hu23VL1(N28L) представляет собой:
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSLGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 54)
> Последовательность Hu23VL1(N28T) представляет собой:
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSTGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 55)
> Последовательность Hu23VL1(N28D) представляет собой:
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSDGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 56)
> Последовательность Hu23VL1(G29A) представляет собой:
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSNANTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 57)
> Последовательность Hu23VL1(G29V) представляет собой:
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSNVNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 58)
> Последовательность Hu23VL2(N28Q) представляет собой:
DGVMTQTPLSLPVTPGEPASISCRSSQSLVHSQGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 59)
> Последовательность Hu23VL2(N28L) представляет собой:
DGVMTQTPLSLPVTPGEPASISCRSSQSLVHSLGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 60)
> Последовательность Hu23VL2(N28T) представляет собой:
DGVMTQTPLSLPVTPGEPASISCRSSQSLVHSTGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 61)
> Последовательность Hu23VL2(N28D) представляет собой:
DGVMTQTPLSLPVTPGEPASISCRSSQSLVHSDGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 62)
> Последовательность Hu23VL2(G29A) представляет собой:
DGVMTQTPLSLPVTPGEPASISCRSSQSLVHSNANTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 63)
> Последовательность Hu23VL2(G29V) представляет собой:
DGVMTQTPLSLPVTPGEPASISCRSSQSLVHSNVNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIK (SEQ ID NO: 64)
Таблица 12. Комбинации вариабельных областей легкой и тяжелой цепи гуманизированного антитела Hu23
VL
Примечание: например, в таблице «Hu23-11» относится к антителу с комбинацией вариабельной области легкой цепи Hu23VL1(N28T) и вариабельной области тяжелой цепи Hu23VH1, и так далее.
Комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu23-11), представленные в приведенной выше таблице, могут быть связаны с константными областями легкой/тяжелой цепи антитела с образованием полноразмерных антител, соответственно; если в настоящем описании не указано иное, при образовании полноразмерного антитела вариабельная область легкой цепи связывается с константной областью каппа-цепи, представленной в SEQ ID NO: 73, с образованием легкой цепи антитела, а вариабельная область тяжелой цепи связывается с константной областью тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, или с константной областью тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, с образованием тяжелой цепи антитела, и название в таблице, относящееся к комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu23-11), с суффиксом «. IgG4AA» означает полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4AA, с суффиксом «. IgG4P» - полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4-P. Например, «Hu23-11.IgG4AA» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu23VH1 и константной области тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, и легкой цепи, образованной путем связывания вариабельной области легкой цепи Hu23VL1(N28T) и константной области каппа-цепи, представленной в SEQ ID NO: 73. «Hu23-11.IgG4P» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu23VH1 и константной области тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, и легкой цепи, образованный путем связывания вариабельной области легкой цепи Hu23VL1(N28T) и константной области каппа-цепи, представленной в SEQ ID NO: 73.
Результаты экспериментов показали, что после сайт-направленных мутаций все гуманизированные антитела Hu23LCDR1 (N28Q), Hu23LCDR1 (N28L), Hu23LCDR1 (N28T), Hu23LCDR1 (N28D), Hu23LCDR1 (G29A) и Hu23LCDR1 (G29V) сохранили способность связываться с PD-1 (таблица 16).
4.2 Мутантные антитела на основе гуманизированного антитела Hu32
Серия гуманизированных антител Hu23, полученных из M23, и серия гуманизированных антител Hu32, полученных из M32, имели высокую идентичность последовательностей при анализе последовательностей. Вариабельную область легкой цепи Hu23 и вариабельную область тяжелой цепи Hu32 объединяли в новые комбинации вариабельных областей легкой и тяжелой цепи. Результаты экспериментов показали, что гуманизированные антитела, содержащие новые комбинации вариабельных областей легкой и тяжелой цепей, сохраняют способность связываться с антигеном PD-1 (таблица 16).
Таблица 13. Общие формулы консенсусных последовательностей вариабельной области антител Hu32 и Hu23
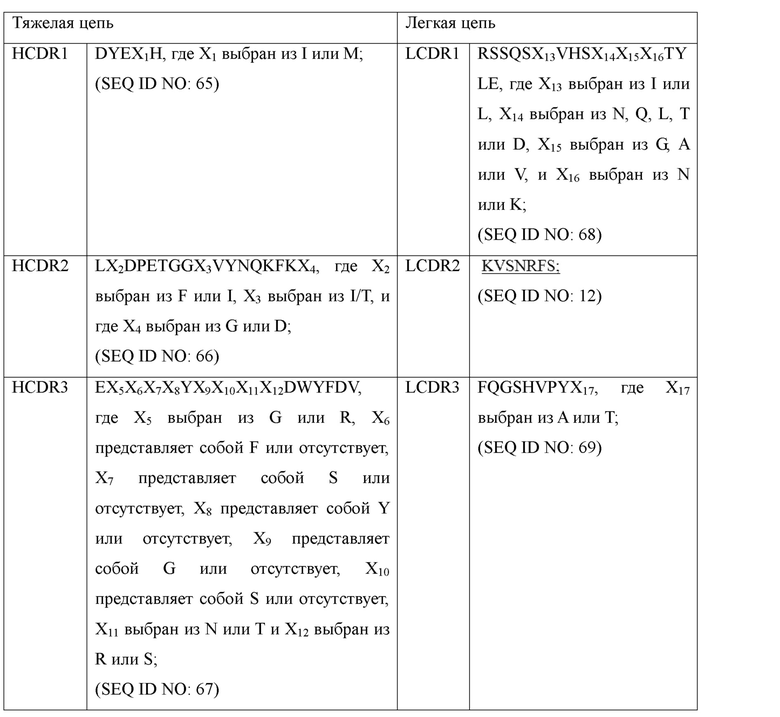

Таблица 14. Комбинации вариабельных областей тяжелой цепи Hu32 и вариабельных областей легкой цепи Hu23
VL
Примечание: например, в таблице «Hu32a-85» относится к антителу с вариабельной областью легкой цепи Hu23VL1(N28T) и вариабельной областью тяжелой цепи Hu32VH6, и так далее.
Комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu32a-85), представленные в приведенной выше таблице, могут быть связаны с константными областями легкой/тяжелой цепи антитела с образованием полноразмерных антител, соответственно; если в настоящем описании не указано иное, при образовании полноразмерного антитела вариабельная область легкой цепи связывается с константной областью каппа-цепи, представленной в SEQ ID NO: 73, с образованием легкой цепи антитела, а вариабельная область тяжелой цепи связывается с константной областью тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, или с константной областью тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, с образованием тяжелой цепи антитела, и название в таблице, относящееся к комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu32a-85), с суффиксом «. IgG4AA» означает полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4AA, с суффиксом «. IgG4P» - полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4-P. Например, «Hu32a-85.IgG4AA» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu32VH6 и константной области тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, и легкой цепи, образованной путем связывания вариабельной области легкой цепи Hu23VL1(N28T) и константной области каппа-цепи, представленной в SEQ ID NO: 73. «Hu32a-85.IgG4P» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu32VH6 и константной области тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, и легкой цепи, образованный путем связывания вариабельной области легкой цепи Hu23VL1(N28T) и константной области каппа-цепи, представленной в SEQ ID NO: 73.
Таблица 15. Комбинации вариабельных областей тяжелой цепи Hu32 и вариабельных областей легкой цепи Hu23
VL
Примечание: например, в таблице «Hu23a-57» относится к антителу с комбинацией вариабельной области легкой цепи Hu32VL1 и вариабельной области тяжелой цепи Hu23VH1, и так далее.
Комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu23a-57), представленные в приведенной выше таблице, могут быть связаны с константными областями легкой/тяжелой цепи антитела с образованием полноразмерных антител, соответственно; если в настоящем описании не указано иное, при образовании полноразмерного антитела вариабельная область легкой цепи связывается с константной областью каппа-цепи, представленной в SEQ ID NO: 73, с образованием легкой цепи антитела, а вариабельная область тяжелой цепи связывается с константной областью тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, или с константной областью тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, с образованием тяжелой цепи антитела, и название в таблице, относящееся к комбинации вариабельных областей легкой/тяжелой цепи антитела (например, Hu32a-85), с суффиксом «. IgG4AA» означает полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4AA, с суффиксом «. IgG4P» - полноразмерное антитело, образованное путем лигирования с константной областью тяжелой цепи IgG4-P. Например, «Hu23a-57.IgG4AA» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu23VH1 и константной области тяжелой цепи IgG4-AA, представленной в SEQ ID NO: 72, и легкой цепи, образованной путем связывания вариабельной области легкой цепи Hu32VL1 и константной области каппа-цепи, представленной в SEQ ID NO: 73. «Hu23a-57.IgG4P» означает полноразмерное антитело, образованное путем связывания тяжелой цепи, образованной путем связывания вариабельной области тяжелой цепи Hu23VH1 и константной области тяжелой цепи IgG4-P, представленной в SEQ ID NO: 79, и легкой цепи, образованный путем связывания вариабельной области легкой цепи Hu32VL1 и константной области каппа-цепи, представленной в SEQ ID NO: 73.
5. Скрининг гуманизированных антител
Определение аффинности различных гуманизированных антител было выполнено с помощью Biacore (см. метод в тестовом примере 3), и результаты приведены в таблице 16. Результаты показали, что различные гуманизированные антитела сохраняют способность связываться с PD-1, а некоторые гуманизированные антитела имеют аффинность даже в значительной степени близкую к аффинности мышиных антител, из которых они получены.
Таблица 16. Аффинность гуманизированного антитела Hu23 к человеческому PD-1
Пример 3. Конструирование и экспрессия гуманизированных антител к PD-1
Каждый фрагмент гена VH/VK гуманизированного антитела конструировали с помощью сконструированных праймеров посредством ПЦР, а затем использовали для конструирования вектора VH-CH1-Fc-pHr/VK-CL-pHr, экспрессирующего полноразмерное антитело, путем гомологичной рекомбинации с вектором экспрессии pHr (с сигнальным пептидом и фрагментом гена константной области (CH1-Fc/CL)). IgG4-P представляет собой мутацию S228P (соответствует положению 108 SEQ ID NO: 72 или SEQ ID NO: 79), а IgG4-AA представляет собой мутации F234A (соответствует положению 114 последовательности SEQ ID NO: 72 или SEQ ID NO: 79), L235A (соответствует положению 115 SEQ ID NO: 72 или SEQ ID NO: 79) и S228P (соответствует положению 108 SEQ ID NO: 72 или SEQ ID NO: 79). Форматы антител IgG4-AA и IgG4-P могут быть получены путем простых точечных мутаций в формате антитела IgG4.
Последовательность константной области тяжелой цепи IgG4-AA представляет собой следующую (SEQ ID NO: 72):

Последовательность константной области легкой цепи (каппа-цепи) антитела представляет собой следующую (SEQ ID NO: 73):
RTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC.
Примеры последовательностей сконструированных полноразмерных антител формы IgG4AA представляют собой следующие:
Тяжелая цепь антитела Hu23-11.IgG4AA (SEQ ID NO: 74):
EVQLVQSGAEVKKPGSSVKVSCKASGGTFSDYEMHWVRQAPGQGLEWMGLIDPETGGTVYNQKFKDRVTITADKSTSTAYMELSSLRSEDTAVYYCARERFSYYGSTSDWYFDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK.
Легкая цепь Hu23-11.IgG4AA (SEQ ID NO: 75):
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSTGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC.
Тяжелая цепь Hu32a-85.IgG4AA (SEQ ID NO: 76):
EVQLVQSGAEVKKPGSSVKVSCKASDFTFTDYEIHWVKQAPGHGLEWIGLFDPETGGIVYNQKFKGKATLTADKSTSTAYMEFSSLRSEDTAVYYCTREGYNRDWYFDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK.
Легкая цепь Hu32a-85.IgG4AA (такая же, как легкая цепь Hu23-11.IgG4AA, SEQ ID NO: 75):
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSTGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC.
Тяжелая цепь Hu33-6.IgG4AA (SEQ ID NO: 77):
KVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVATISGGGVDTYYQDNVQGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCASPYGHGYFDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK.
Легкая цепь Hu33-6.IgG4AA (SEQ ID NO: 78):
DIQMTQSPSSLSASVGDRVTITCRASQDINNFLNWYQQKPGKAPKLLIYYTSSLHSGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC.
Последовательность константной области тяжелой цепи IgG4-P представляет собой следующую (SEQ ID NO: 79):
ASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK.
Примеры последовательностей сконструированных полноразмерных антител формы IgG4-P представляют собой следующие:
Тяжелая цепь антитела Hu23-11.IgG4P (SEQ ID NO: 80):
EVQLVQSGAEVKKPGSSVKVSCKASGGTFSDYEMHWVRQAPGQGLEWMGLIDPETGGTVYNQKFKDRVTITADKSTSTAYMELSSLRSEDTAVYYCARERFSYYGSTSDWYFDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK.
Легкая цепь Hu23-11.IgG4P (такая же, как легкая цепь Hu23-11.IgG4AA, SEQ ID NO: 75):
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSTGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC.
Тяжелая цепь Hu32a-85.IgG4P (SEQ ID NO: 81):
EVQLVQSGAEVKKPGSSVKVSCKASDFTFTDYEIHWVKQAPGHGLEWIGLFDPETGGIVYNQKFKGKATLTADKSTSTAYMEFSSLRSEDTAVYYCTREGYNRDWYFDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK.
Легкая цепь Hu32a-85.IgG4P (такая же, как легкая цепь Hu23-11.IgG4AA, SEQ ID NO: 75):
DIVMTQTPLSLPVTPGEPASISCRSSQSLVHSTGNTYLEWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCFQGSHVPYTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC.
Тяжелая цепь Hu33-6.IgG4P (SEQ ID NO: 82):
KVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVATISGGGVDTYYQDNVQGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCASPYGHGYFDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK.
Легкая цепь Hu33-6.IgG4P (такая же, как легкая цепь Hu33-6.IgG4AA, SEQ ID NO: 78):
DIQMTQSPSSLSASVGDRVTITCRASQDINNFLNWYQQKPGKAPKLLIYYTSSLHSGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCQQGNTLPWTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC.
Тестовые примеры
Тестовый пример 1. ELISA-исследование связывания антител к PD-1 с лигандом PD-1 и блокирования связывания in vitro
PD-L1 на поверхности опухолевых клеток связывается с PD-1 на поверхности Т-клеток, тем самым подавляя пролиферацию Т-клеток. Антитела к PD-1 могут блокировать сигнальный путь PD-L1/PD-1 путем связывания с PD-1, тем самым стимулируя пролиферацию Т-клеток. Эксперимент по блокированию связывания PD-1/PD-L1 использовали для обнаружения блокирующей активности антител к PD-1 в отношении сигнального пути.
В этом эксперименте после нанесения на 96-луночные планшеты покрытия из белка PD-1-His (кат. № 10377H08H, Sino Biological) по отдельности добавляли тестируемые антитела к PD-1 (включая антитела: Hu23-11.IgG4AA, Hu32a-85.IgG4AA и Hu33-6.IgG4AA, антитело положительного контроля: H005-1 (см. антитело H005-1 в WO2015085847)) и инкубировали реакционную смесь; затем добавляли меченое HRP козье антитело к человеческому IgG (H+L) (кат. № 109-035-003, Jackson ImmunoResearch) и инкубировали. После промывки планшетов определяли связывающие количества меченого HRP козьего антитела к человеческому IgG (H+L) и рассчитывали значения EC50 связывания антител к PD-1 с лигандом PD-1.
В этом эксперименте после нанесения покрытия из белка PD-1, слитого с Fc во внеклеточной области (PD-1-Fc, последовательность см. в SEQ ID NO: 1), на 96-луночные планшеты, по отдельности добавляли тестируемые антитела к PD-1 (включая антитела: Hu23-11.IgG4AA, Hu32a-85.IgG4AA и Hu33-6.IgG4AA, антитело положительного контроля: H005-1 (см. антитело H005-1 в WO2015085847)) и инкубировали; затем добавляли меченые биотином PD-L1/PD-L2 и инкубировали. После промывки планшетов определяли связывающие количества меченного биотином PD-L1/PD-L2 и рассчитывали значения IC50 антител к PD-1 в отношении блокирования связывания лиганда PD-L1/PD-L2.
Буфер CB с pH 9,6 (1,59 г Na2CO3 и 2,93 г NaHCO3, растворенные в 1 л дистиллированной воды) использовали для разведения PD-1-Fc до 1 мкг/мл, который добавляли в 96-луночные планшеты в объеме 100 мкл/лунку и выдерживали при 4 °C в течение 16-20 часов. Буфер PBS аспирировали из 96-луночных планшетов, и планшеты однократно промывали буфером PBST (pH 7,4, PBS, содержащий 0,05% tween 20). Добавляли 120 мкл/лунку PBST/1% молока и инкубировали при комнатной температуре в течение 1 ч для блокирования. Блокирующий раствор удаляли, и планшеты однократно промывали буфером PBST. Добавляли 90 мкл тестируемого антитела к PD-1, разведенного до соответствующей концентрации разбавителем для образцов (pH 7,4 PBS, содержащий 5% BSA, 0,05% Tween20), и предварительно инкубировали при 4 °C в течение 1 часа. Добавляли 10-кратную концентрацию меченого биотином PD-L1/PD-L2 (Beijing Sino Biological Co., Ltd.) (10 мкг/мл) в объеме 10 мкл/лунку, встряхивали и хорошо перемешивали на шейкере, и затем инкубировали при 37 °C в течение 1 ч. Реакционную систему удаляли, и планшеты 6 раз промывали PBST. Добавляли 100 мкл/лунку полимера стрептавидин-пероксидазы, разведенного 1:400 буфером PBST, и инкубировали при встряхивании при комнатной температуре в течение 50 мин. Планшеты 6 раз промывали PBST. Добавляли 100 мкл/лунку TMB и инкубировали при комнатной температуре в течение 5-10 мин. Добавляли 100 мкл/лунку 1M H2SO4 для остановки реакции. Значения поглощения при 450 нм считывали с использованием считывающего устройства для микропланшетов и рассчитывали значения IC50 антител к PD-1 в отношении блокирования связывания лиганда PD-L1/PD-L2. Данные подробно представлены ниже в таблице 17.
Таблица 17. ELISA-анализ антител к PD-1 согласно настоящему изобретению в отношении связывания с PD-1 и блокирования связывания лиганда PD-L1/PD-L2
Все иллюстративные антитела к PD-1 Hu23-11.IgG4AA, Hu32a-85.IgG4AA и Hu33-6.IgG4AA согласно настоящему изобретению способны эффективно блокировать связывание PD-1 с PD-L1/PD-L2, и их блокирующая активность аналогична активности антитела положительного контроля.
Тестовый пример 2. Исследование блокирования лиганда иллюстративными антителами
Было изучено блокирующее действие антител на связывание PD-1 и PD-L1. Процесс проведения эксперимента кратко описывается следующим образом:
Клетки CHOK1/PD-L1 (Promega) расщепляли, добавляли в 96-луночные планшеты в количестве 100 мкл/лунку и помещали в инкубатор при 37 °C, 5% CO2 для инкубации в течение 24 ч. Контроль и образцы разводили до желаемых концентраций с помощью PBS. Клетки Jurkat/PD-1 (клетки Jurkat, стабильно трансфицированные PD-1) подсчитывали и высевали в определенной пропорции (90 мкл/лунку) в планшеты для культивирования клеток с клетками CHOK1/PD-L1, и добавляли 10 мкл/лунку разведенного антитела (антитело: Hu23-11.IgG4AA, Hu32a-85.IgG4AA и Hu33-6.IgG4AA, антитело положительного контроля: H005-1, отрицательный контроль: белок IgG4, концентрация градиентного разведения антител: 0,3 мг/мл, 3 мг/мл, 30 мг/мл) и помещали в инкубатор при 37 °C, 5% CO2 для инкубации в течение 5 ч. Планшеты для культивирования клеток вынимали и выдерживали при комнатной температуре в течение 5 мин. Затем в каждую лунку добавляли 50 мкл реагента Bio-Glo™ и инкубировали при комнатной температуре в течение 5 минут перед считыванием планшета. Результаты эксперимента показаны на фиг. 1.
Результаты показали, что иллюстративные антитела к PD-1 Hu23-11.IgG4AA, Hu32a-85.IgG4AA и Hu33-6.IgG4AA согласно настоящему изобретению могут эффективно блокировать связывание PD-1 и PD-L1.
Тестовый пример 3. Эксперимент по определению аффинности антител с использованием BIAcore в отношении иллюстративных антител
Осуществляли аффинный захват IgG с использованием биосенсорного чипа с белком А (кат. № 29127556, GE). Антиген человеческого PD-1 (кат. № 10377H08H, Sino Biological) и антиген PD-1 яванского макака (приобретенный у Sino Biological) пропускали по поверхности чипа и получали кривые связывания и диссоциации с помощью детекции в режиме реального времени сигналов реакции антитела к PD-1 и антигена PD-1 с помощью прибора Biacore T200. После того, как диссоциация каждого экспериментального цикла была завершена, биосенсорный чип промывали и регенерировали 10 мМ буфером глицин-HCl с рН 1,5. Экспериментальная буферная система представляла собой буферный раствор 1×HBS-EP (кат. № BR-1001-88, GE). После эксперимента использовали программное обеспечение GE Biacore T200 Evaluation версии 3.0 для аппроксимации данных моделью Ленгмюра (1:1), и было получено значение аффинности. Результаты приведены в таблице 18.
Таблица 18. Аффинность антител к PD-1 к человеческому PD1 и PD-1 яванского макака
Результаты показали, что все иллюстративные антитела к PD-1 Hu23-11.IgG4AA, Hu32a-85.IgG4AA и Hu33-6.IgG4AA согласно настоящему изобретению могут связываться с человеческим PD-1 и PD-1 яванского макака.
Тестовый пример 4. Влияние антител на секрецию IFNγ клетками в эксперименте по активации МНПК-T-лимфоцитов
Для изучения влияния антител к PD-1 на функцию первичных человеческих Т-лимфоцитов собирали и очищали мононуклеарные клетки периферической крови (МНПК), стимулировали туберкулином (ТБ) в течение 5 дней и определяли уровень секреции цитокина IFNγ. Процесс проведения эксперимента кратко описывается следующим образом:
МНПК получали из свежей крови с использованием Ficoll-Hypaque (17-5442-02, GE) для центрифугирования в градиенте плотности (Stem Cell Technologies), культивировали в среде RPMI 1640 (SH30809.01, GE) с добавлением 10% (об./об.) FBS (10099-141, Gibco) при 37 °C и 5% CO2.
Свежевыделенные и очищенные МНПК доводили до плотности 2×106 клеток/мл с помощью среды RPMI 1640. 40 мкл туберкулина (97-8800, Synbiotics) добавляли к 20 мл клеточной суспензии и культивировали в инкубаторе при 37 °C, 5% CO2 в течение 5 дней. На 5 день вышеупомянутые культивированные клетки собирали центрифугированием, ресуспендировали в свежей среде RPMI 1640, доводили до плотности 1,1×106 клеток/мл и высевали в 96-луночные планшеты для культивирования клеток по 90 мкл на лунку. Одновременно добавляли образцы антител (включая антитела согласно настоящему изобретению: Hu23-11.IgG4AA, Hu32a-85.IgG4AA и Hu33-6.IgG4AA, антитело положительного контроля H005-1 и белок IgG4 отрицательного контроля, концентрация градиентного разведения антител 0,3 мг/мл, 3 мг/мл и 30 мг/мл), градиент разводили в PBS (B320, Shanghai BasalMedia Technologies Co., Ltd.), 10 мкл/лунку. Планшеты для культивирования клеток помещали в инкубатор при 37 °C, 5% CO2 для инкубации в течение 3 дней. Вынимали планшеты для культивирования клеток и собирали супернатант клеточных культур центрифугированием (4000 об/мин, 10 мин). Уровни IFN-γ определяли с помощью метода ELISA (набор для определения человеческого IFN-γ (EHC102g.96, Neobioscience)). Для конкретных операций использовали инструкции, прилагаемые к реагентам.
Результаты теста показаны на фиг. 2. Результаты показали, что все антитела к PD-1 Hu23-11.IgG4AA, Hu32a-85.IgG4AA и Hu33-6.IgG4AA согласно настоящему изобретению могут эффективно активировать секрецию IFN-γ.
Тестовый пример 5. Влияние антител к PD-1 на модель рака ободочной кишки MC38 у трансгенных по PD-1 мышей
Клетки MC38 инокулировали в дозе 5×105 клеток/мышь/100 мкл 90 мышам hPD-1 TG (Biocytogen) подкожно в область правых ребер. Через 10 дней животные со слишком большими или слишком маленькими опухолями были исключены. На основании среднего объема опухоли около 120 мм3 мышей случайным образом делили на: группу холостого контроля носителем (PBS), положительного контроля H005-1 3 мг/кг, Hu32a-85.IgG4AA 1 мг/кг, Hu32a-85.IgG4AA 3 мг/кг, Hu23-11.IgG4AA 1 мг/кг, Hu23-11.IgG4AA 3 мг/кг и Hu33-6.IgG4AA 3 мг/кг, в общей сложности 7 групп по 8 животных в каждой. Антитела каждой группы вводили внутрибрюшинно три раза в неделю, начиная с дня 0. По прошествии первой недели введения было обнаружено, что опухоли были значительно ингибированы. Во вторую и третью недели частота введения была скорректирована до одного раза в неделю, в общей сложности 5 введений. Объем опухолей и массу животных контролировали два раза в неделю, и данные записывали. Когда объем опухоли превысил 2000 мм3, или большинство опухолей демонстрировали изъязвления, или масса тела снижалась на 20%, животных с опухолями умерщвляли в качестве конечной точки эксперимента.
Объем опухоли (TV) = 1/2 × Lдлинный × Lкороткий2
Скорость роста опухоли (T/C%) = (T-T0)/(C-C0) × 100%
Степень ингибирования роста опухоли (TGI%) = 1- T/C %
Где T и T0 представляют собой объемы опухоли в конце теста и в начале теста в группе введения антитела, соответственно, а C и C0 представляют собой объемы опухоли в конце теста и начале теста в холостой контрольной группе, соответственно.
Результаты теста показаны в таблице 19 и на фиг. 3. Результаты теста показали, что по сравнению с холостым контролем все антитела согласно настоящему изобретению способны значительно ингибировать рост опухолей ксенотрансплантата рака ободочной кишки MC38 у мышей. Среди них группа Hu32a-85.IgG4AA-3 мг/кг имела самую высокую степень ингибирования опухоли, и степень ингибирования опухоли составляла 77,64% при последнем измерении. Когда частота дозирования составляла три раза в неделю для 3 введений, при измерении на седьмой день результаты показали, что все степени ингибирования опухоли у антител согласно настоящему изобретению были значительно лучше, чем у антитела положительного контроля H005-1; после этого частота дозирования была уменьшена до одного раза в неделю, после двух введений (день 21) разница в эффективности антител согласно настоящему изобретению постепенно увеличивалась и проявлялась дозозависимым образом, среди которых Hu32a-85.IgG4AA было значительно лучше, чем та же доза H005-1 (p < 0,05). Кроме того, мыши с опухолями могут хорошо переносить все антитела к PD-1, при этом масса тела постоянно увеличивалась на протяжении всего процесса введения и не наблюдалось явной потери массы тела, вызванной лекарственным средством, и других симптомов.
Таблица 19. Влияние антител к PD-1 на степень ингибирования роста опухоли ободочной кишки MC38 у мышей (мм3)
Тестовый пример 6. Влияние антител к PD-1 на модель рака ободочной кишки MC38 у трансгенной по PD-1 мыши
Трансгенные по PD-1 мыши были приобретены у ISIS INNOVATION LIMITED, University Offices, Wellington Square, Оксфорд OX1 2JD, Англия, и выведены в Cephrim Biosciences, Inc. для получения мышей пятого поколения. Клетки MC38 инокулировали в дозе 5×105 клеток/100 мкл/животное трансгенным по hPD-1 мышам (половина самок и половина самцов) подкожно в заднюю область правых ребер. Когда средний объем опухоли у мышей достиг 80-100 мм3, животные со слишком большой или слишком малой массой тела или опухолями были исключены. В соответствии с объемом опухоли мышей с опухолями случайным образом делили на 5 групп (по 8 животных в каждой): отрицательного контроля hIgG 30 мг/кг, H005-1 10 мг/кг, H005-1 30 мг/кг, Hu33-6.IgG4AA 10 мг/кг и Hu33-6.IgG4AA 30 мг/кг. Дата разделения на группы и введения была установлена как день 0. После разделения на группы каждое лекарственное средство вводили внутрибрюшинно в течение 22 дней, один раз в два дня, в общей сложности 11 раз. Объем опухолей измеряли 2 раза в неделю, измеряли массу тела и записывали данные. Масса тела животных и объем опухолей в каждой группе были представлены в виде среднего значения ± стандартное отклонение (среднее значение ± SEM), и для построения графиков использовали Graphpad Prism 5 и программное обеспечение Excel, а для статистического анализа использовали t-критерий Стьюдента.
Объем опухоли (TV) = 0,5236 × Lдлинный × Lкороткий2
Скорость роста опухоли T/C% = (T-T0)/(C-C0) × 100%
Степень ингибирования роста опухоли %TGI = 1- T/C %
Где T и T0 представляют собой объемы опухоли в конце теста и в начале теста в группе введения антитела, соответственно, а C и C0 представляют собой объемы опухоли в конце теста и начале теста в холостой контрольной группе, соответственно.
Результаты теста показаны в таблице 20 и на фиг. 4. Результаты теста показали, что по сравнению с контрольной группой антитела согласно настоящему изобретению способны значительно ингибировать рост ксенотрансплантатных опухолей рака ободочной кишки MC38 у мышей. Среди них группа Hu33-6.IgG4AA-30 мг/кг имела самую высокую степень ингибирования опухоли, и степень ингибирования опухоли составляла 80,4% на 20 день. В группе с низкой дозой (10 мг/кг) эффективность Hu33-6.IgG4AA-10 мг/кг была лучше, чем в группе положительного контроля H005-1-10 мг/кг.
Таблица 20. Влияние антител к PD-1 на объем опухоли ободочной кишки MC38 у мышей
Примечание: единица измерения среднего объема опухоли для каждой группы в таблице представляет собой мм3.
Тестовый пример 7. Фармакокинетический тест антител к PD-1 на яванских макаках
Яванские макаки, использованные в эксперименте, представляли собой 6 самцов, 2-5 лет, 2-5 кг, приобретенных у Guangdong Frontier Biotechnology Co., Ltd., номер лицензии: SCXK (Guangdong) 2015-0037, номер сертификата животного: 44613900000219.
Условия содержания: температуру в помещении поддерживали на уровне 18°C-26°C, относительная влажность составляла 40%-70%, и освещение представляло собой поочередно свет и темноту по 12 ч. Мышам предоставляли свободный доступ к пище и воде, за исключением случаев, когда было необходимо голодание.
Перед введением животных взвешивали, и их масса составляла от 2,81 до 3,52 кг. Введение осуществляли с помощью инъекционной помпы для подкожной внутривенной инфузии в передние или задние конечности. Доза для каждой группы составляла 1 мг/кг, ее вводили путем однократной внутривенной инъекции со скоростью 0,1 мл/кг/мин в течение периода введения около 30 минут. До введения и через 5 минут, 0,25 часа, 0,5 часа (сразу после завершения введения), 1 час, 2 часа, 4 часа, 8 часов, 1 день, 2 дня, 3 дня, 4 дня, 5 дней, 7 дней, 10 дней, 13 дней, 14 дней, 21 день и 28 дней после начала внутривенной инфузии собирали цельную кровь животного из вен задних конечностей и отделяли сыворотку. Из них около 2 мл цельной крови было собрано до введения и через 14 дней, 21 день и 28 дней после начала внутривенной инфузии, и около 1 мл цельной крови было собрано в остальные моменты забора крови. Концентрацию лекарственного средства в крови в сыворотке определяли с помощью ELISA и проводили ФК анализ. Результаты приведены в таблице 21.
Таблица 21. Фармакокинетика гуманизированных антител к PD1 у яванских макаков
Результаты показали, что Hu23-11.IgG4AA и Hu33-6.IgG4AA имеют хорошие фармакокинетические активности.
Хотя для ясного понимания вышеупомянутое изобретение было подробно описано с помощью фигур и примеров, описания и примеры не следует интерпретировать как ограничивающие объем настоящего изобретения. Раскрытие всех патентов и научных документов, цитируемых в данном документе, полностью и явным образом включено посредством ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЦЗЯНСУ ХЭНЖУЙ МЕДСИН КО., ЛТД.
ШАНХАЙ ХЭНЖУЙ ФАРМАСЬЮТИКАЛ КО., ЛТД.
<120> АНТИТЕЛО К PD-1, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ
<130> 2019
<160> 82
<170> SIPOSequenceListing 1.0
<210> 1
<211> 401
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность человеческого PD-1-IgG1Fc
<400> 1
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp Asn
20 25 30
Pro Pro Thr Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp Asn
35 40 45
Ala Thr Phe Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val Leu
50 55 60
Asn Trp Tyr Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala Ala
65 70 75 80
Phe Pro Glu Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg Val
85 90 95
Thr Gln Leu Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg Ala
100 105 110
Arg Arg Asn Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu Ala
115 120 125
Pro Lys Ala Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val Thr
130 135 140
Glu Arg Arg Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro Arg
145 150 155 160
Pro Ala Gly Gln Phe Gln Thr Leu Val Glu Pro Lys Ser Ser Asp Lys
165 170 175
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
180 185 190
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
195 200 205
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
210 215 220
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
225 230 235 240
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
245 250 255
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
260 265 270
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
275 280 285
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
290 295 300
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
305 310 315 320
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
325 330 335
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
340 345 350
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
355 360 365
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
370 375 380
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
385 390 395 400
Lys
<210> 2
<211> 186
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность человеческого PD-1-his
<400> 2
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp Asn
20 25 30
Pro Pro Thr Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp Asn
35 40 45
Ala Thr Phe Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val Leu
50 55 60
Asn Trp Tyr Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala Ala
65 70 75 80
Phe Pro Glu Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg Val
85 90 95
Thr Gln Leu Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg Ala
100 105 110
Arg Arg Asn Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu Ala
115 120 125
Pro Lys Ala Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val Thr
130 135 140
Glu Arg Arg Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro Arg
145 150 155 160
Pro Ala Gly Gln Phe Gln Thr Leu Val Gly Ser Ser Asp Tyr Lys Asp
165 170 175
Asp Asp Asp Lys His His His His His His
180 185
<210> 3
<211> 288
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность антигена PD-1 для трансфекции клеток
<400> 3
Met Gln Ile Pro Gln Ala Pro Trp Pro Val Val Trp Ala Val Leu Gln
1 5 10 15
Leu Gly Trp Arg Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp
20 25 30
Asn Pro Pro Thr Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp
35 40 45
Asn Ala Thr Phe Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val
50 55 60
Leu Asn Trp Tyr Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala
65 70 75 80
Ala Phe Pro Glu Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg
85 90 95
Val Thr Gln Leu Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg
100 105 110
Ala Arg Arg Asn Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu
115 120 125
Ala Pro Lys Ala Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val
130 135 140
Thr Glu Arg Arg Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro
145 150 155 160
Arg Pro Ala Gly Gln Phe Gln Thr Leu Val Val Gly Val Val Gly Gly
165 170 175
Leu Leu Gly Ser Leu Val Leu Leu Val Trp Val Leu Ala Val Ile Cys
180 185 190
Ser Arg Ala Ala Arg Gly Thr Ile Gly Ala Arg Arg Thr Gly Gln Pro
195 200 205
Leu Lys Glu Asp Pro Ser Ala Val Pro Val Phe Ser Val Asp Tyr Gly
210 215 220
Glu Leu Asp Phe Gln Trp Arg Glu Lys Thr Pro Glu Pro Pro Val Pro
225 230 235 240
Cys Val Pro Glu Gln Thr Glu Tyr Ala Thr Ile Val Phe Pro Ser Gly
245 250 255
Met Gly Thr Ser Ser Pro Ala Arg Arg Gly Ser Ala Asp Gly Pro Arg
260 265 270
Ser Ala Gln Pro Leu Arg Pro Glu Asp Gly His Cys Ser Trp Pro Leu
275 280 285
<210> 4
<211> 125
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность вариабельной области тяжелой цепи M23
<400> 4
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Lys Gln Thr Pro Ile His Gly Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asp Pro Glu Thr Gly Gly Thr Val Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Thr Ile Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr His Cys
85 90 95
Thr Arg Glu Arg Phe Ser Tyr Tyr Gly Ser Thr Ser Asp Trp Tyr Phe
100 105 110
Asp Val Trp Gly Thr Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 5
<211> 112
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность вариабельной области легкой цепи M23
<400> 5
Asp Gly Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp His Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 6
<211> 120
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность вариабельной области тяжелой цепи M32
<400> 6
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Asp Phe Thr Phe Thr Asp Tyr
20 25 30
Glu Ile His Trp Val Lys Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Leu Phe Asp Pro Glu Thr Gly Gly Ile Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Ile Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Glu Phe Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Glu Gly Tyr Asn Arg Asp Trp Tyr Phe Asp Val Trp Gly Thr
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 7
<211> 112
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность вариабельной области легкой цепи M32
<400> 7
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Ala Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 8
<211> 5
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность HCDR1 M23
<400> 8
Asp Tyr Glu Met His
1 5
<210> 9
<211> 17
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность HCDR2 M23
<400> 9
Leu Ile Asp Pro Glu Thr Gly Gly Thr Val Tyr Asn Gln Lys Phe Lys
1 5 10 15
Asp
<210> 10
<211> 16
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность HCDR3 M23
<400> 10
Glu Arg Phe Ser Tyr Tyr Gly Ser Thr Ser Asp Trp Tyr Phe Asp Val
1 5 10 15
<210> 11
<211> 16
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность LCDR1 M23
<400> 11
Arg Ser Ser Gln Ser Leu Val His Ser Asn Gly Lys Thr Tyr Leu Glu
1 5 10 15
<210> 12
<211> 7
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность LCDR2 M23/M32
<400> 12
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 13
<211> 9
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность LCDR3 M23
<400> 13
Phe Gln Gly Ser His Val Pro Tyr Thr
1 5
<210> 14
<211> 5
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность HCDR1 M32
<400> 14
Asp Tyr Glu Ile His
1 5
<210> 15
<211> 17
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность HCDR2 M32
<400> 15
Leu Phe Asp Pro Glu Thr Gly Gly Ile Val Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 16
<211> 11
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность HCDR3 M32
<400> 16
Glu Gly Tyr Asn Arg Asp Trp Tyr Phe Asp Val
1 5 10
<210> 17
<211> 16
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность LCDR1 M32
<400> 17
Arg Ser Ser Gln Ser Ile Val His Ser Asn Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 18
<211> 9
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность LCDR3 M32
<400> 18
Phe Gln Gly Ser His Val Pro Tyr Ala
1 5
<210> 19
<211> 118
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность вариабельной области тяжелой цепи M33
<400> 19
Lys Val Met Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Val Asp Thr Tyr Tyr Gln Asp Asn Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ser Pro Tyr Gly His Gly Tyr Phe Asp Val Trp Gly Thr Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 20
<211> 107
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность вариабельной области легкой цепи M33
<400> 20
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Asn Asn Phe
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 21
<211> 5
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность HCDR1 M33
<400> 21
Ser Tyr Ala Met Ser
1 5
<210> 22
<211> 17
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность HCDR2 M33
<400> 22
Thr Ile Ser Gly Gly Gly Val Asp Thr Tyr Tyr Gln Asp Asn Val Gln
1 5 10 15
Gly
<210> 23
<211> 9
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность HCDR3 M33
<400> 23
Pro Tyr Gly His Gly Tyr Phe Asp Val
1 5
<210> 24
<211> 11
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность LCDR1 M33
<400> 24
Arg Ala Ser Gln Asp Ile Asn Asn Phe Leu Asn
1 5 10
<210> 25
<211> 7
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность LCDR2 M33
<400> 25
Tyr Thr Ser Ser Leu His Ser
1 5
<210> 26
<211> 9
<212> Белок
<213> Mus musculus
<220>
<221> DOMAIN
<223> Последовательность LCDR3 M33
<400> 26
Gln Gln Gly Asn Thr Leu Pro Trp Thr
1 5
<210> 27
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VH1 /Hu23VH-CDR-привитой
<400> 27
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Leu Ile Asp Pro Glu Thr Gly Gly Thr Val Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Phe Ser Tyr Tyr Gly Ser Thr Ser Asp Trp Tyr Phe
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 28
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL1 /Hu23VL-CDR-привитой
<400> 28
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 29
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL2
<400> 29
Asp Gly Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 30
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VH2
<400> 30
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Leu Ile Asp Pro Glu Thr Gly Gly Thr Val Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Phe Ser Tyr Tyr Gly Ser Thr Ser Asp Trp Tyr Phe
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 31
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VH3
<400> 31
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asp Pro Glu Thr Gly Gly Thr Val Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Thr Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Phe Ser Tyr Tyr Gly Ser Thr Ser Asp Trp Tyr Phe
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 32
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VH4
<400> 32
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asp Pro Glu Thr Gly Gly Thr Val Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Thr Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Glu Arg Phe Ser Tyr Tyr Gly Ser Thr Ser Asp Trp Tyr Phe
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 33
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu32VH1 /Hu32VH-CDR-привитой
<400> 33
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Asp Tyr
20 25 30
Glu Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Leu Phe Asp Pro Glu Thr Gly Gly Ile Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Tyr Asn Arg Asp Trp Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 34
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu32VL1 /Hu32VL-CDR-привитой
<400> 34
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 35
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu32VL2
<400> 35
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 36
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu32VH2
<400> 36
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Glu Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Leu Phe Asp Pro Glu Thr Gly Gly Ile Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Glu Gly Tyr Asn Arg Asp Trp Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 37
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu32VH3
<400> 37
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Asp Phe Thr Phe Ser Asp Tyr
20 25 30
Glu Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Leu Phe Asp Pro Glu Thr Gly Gly Ile Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Glu Gly Tyr Asn Arg Asp Trp Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 38
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu32VH4
<400> 38
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Glu Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Leu Phe Asp Pro Glu Thr Gly Gly Ile Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Glu Gly Tyr Asn Arg Asp Trp Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 39
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu32VH5
<400> 39
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Asp Phe Thr Phe Thr Asp Tyr
20 25 30
Glu Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Leu Phe Asp Pro Glu Thr Gly Gly Ile Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Glu Gly Tyr Asn Arg Asp Trp Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 40
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu32VH6
<400> 40
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Asp Phe Thr Phe Thr Asp Tyr
20 25 30
Glu Ile His Trp Val Lys Gln Ala Pro Gly His Gly Leu Glu Trp Ile
35 40 45
Gly Leu Phe Asp Pro Glu Thr Gly Gly Ile Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Glu Gly Tyr Asn Arg Asp Trp Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 41
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu33VH1/Hu33VH-CDR-привитой
<400> 41
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Val Asp Thr Tyr Tyr Gln Asp Asn Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Tyr Gly His Gly Tyr Phe Asp Val Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 42
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu33VL1/Hu33VL-CDR-привитой
<400> 42
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Asn Asn Phe
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 43
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu33VL2
<400> 43
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Asn Asn Phe
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 44
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu33VL3
<400> 44
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Asn Asn Phe
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gly Ala Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 45
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu33VH2
<400> 45
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Val Asp Thr Tyr Tyr Gln Asp Asn Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Pro Tyr Gly His Gly Tyr Phe Asp Val Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 46
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu33VH3
<400> 46
Lys Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Val Asp Thr Tyr Tyr Gln Asp Asn Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Pro Tyr Gly His Gly Tyr Phe Asp Val Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 47
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23LCDR1(N33Q)
<400> 47
Arg Ser Ser Gln Ser Leu Val His Ser Gln Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 48
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23LCDR1(N33L)
<400> 48
Arg Ser Ser Gln Ser Leu Val His Ser Leu Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 49
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23LCDR1(N33T)
<400> 49
Arg Ser Ser Gln Ser Leu Val His Ser Thr Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 50
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23LCDR1(N33D)
<400> 50
Arg Ser Ser Gln Ser Leu Val His Ser Asp Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 51
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23LCDR1(G34A)
<400> 51
Arg Ser Ser Gln Ser Leu Val His Ser Asn Ala Asn Thr Tyr Leu Glu
1 5 10 15
<210> 52
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23LCDR1(G34V)
<400> 52
Arg Ser Ser Gln Ser Leu Val His Ser Asn Val Asn Thr Tyr Leu Glu
1 5 10 15
<210> 53
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL1(N33Q)
<400> 53
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Gln Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 54
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL1(N33L)
<400> 54
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Leu Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 55
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL1(N33T)
<400> 55
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Thr Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 56
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL1(N33D)
<400> 56
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asp Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 57
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL1(G34A)
<400> 57
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Ala Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 58
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL1(G34V)
<400> 58
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Val Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 59
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL2(N33Q)
<400> 59
Asp Gly Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Gln Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 60
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL2(N33L)
<400> 60
Asp Gly Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Leu Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 61
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL2(N33T)
<400> 61
Asp Gly Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Thr Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 62
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL2(N33D)
<400> 62
Asp Gly Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asp Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 63
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL2(G34A)
<400> 63
Asp Gly Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Ala Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 64
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность Hu23VL2(G34V)
<400> 64
Asp Gly Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Val Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 65
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Общая формула последовательности HCDR1 антитела Hu32 и Hu23
<220>
<221> DOMAIN
<222>(4)..(4)
<223> Xaa выбран из Ile или Met
<400> 65
Asp Tyr Glu Xaa His
1 5
<210> 66
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Общая формула последовательности HCDR2 антитела Hu32 и Hu23
<220>
<221> DOMAIN
<222>(2)..(2)
<223> Xaa выбран из Phe или Ile
<220>
<221> DOMAIN
<222>(9)..(9)
<223> Xaa выбран из Ile или Thr
<220>
<221> DOMAIN
<222>(17)..(17)
<223> Xaa выбран из Gly или Asp
<400> 66
Leu Xaa Asp Pro Glu Thr Gly Gly Xaa Val Tyr Asn Gln Lys Phe Lys
1 5 10 15
Xaa
<210> 67
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Общая формула последовательности HCDR3 антитела Hu32 и Hu23
<220>
<221> DOMAIN
<222>(2)..(2)
<223> Xaa выбран из Gly или Arg
<220>
<221> DOMAIN
<222>(3)..(3)
<223> Xaa выбран из Phe или отсутствует
<220>
<221> DOMAIN
<222>(4)..(4)
<223> Xaa выбран из Ser или отсутствует
<220>
<221> DOMAIN
<222>(5)..(5)
<223> Xaa выбран из Tyr или отсутствует
<220>
<221> DOMAIN
<222>(7)..(7)
<223> Xaa выбран из Gly или отсутствует
<220>
<221> DOMAIN
<222>(8)..(8)
<223> Xaa выбран из Ser или отсутствует
<220>
<221> DOMAIN
<222>(9)..(9)
<223> Xaa выбран из Asn или Thr
<220>
<221> DOMAIN
<222>(10)..(10)
<223> Xaa выбран из Arg или Ser
<400> 67
Glu Xaa Xaa Xaa Xaa Tyr Xaa Xaa Xaa Xaa Asp Trp Tyr Phe Asp Val
1 5 10 15
<210> 68
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Общая формула последовательности LCDR1 антитела Hu32 и Hu23
<220>
<221> DOMAIN
<222>(6)..(6)
<223> Xaa выбран из Ile или Leu
<220>
<221> DOMAIN
<222>(10)..(10)
<223> Xaa выбран из Asn, Gln, Leu, Thr или Asp
<220>
<221> DOMAIN
<222>(11)..(11)
<223> Xaa выбран из Gly, Ala или Val
<220>
<221> DOMAIN
<222>(12)..(12)
<223> Xaa выбран из Asn или Lys
<400> 68
Arg Ser Ser Gln Ser Xaa Val His Ser Xaa Xaa Xaa Thr Tyr Leu Glu
1 5 10 15
<210> 69
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Общая формула последовательности LCDR3 антитела Hu32 и Hu23
<220>
<221> DOMAIN
<222>(9)..(9)
<223> Xaa выбран из Ala или Thr
<400> 69
Phe Gln Gly Ser His Val Pro Tyr Xaa
1 5
<210> 70
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Общая формула последовательности вариабельной области тяжелой цепи антитела Hu32 и Hu23
<220>
<221> DOMAIN
<222>(26)..(26)
<223> Xaa выбран из Gly или Asp
<220>
<221> DOMAIN
<222>(27)..(27)
<223> Xaa выбран из Gly, Phe или Tyr
<220>
<221> DOMAIN
<222>(30)..(30)
<223> Xaa выбран из Ser или Thr
<220>
<221> DOMAIN
<222>(34)..(34)
<223> Xaa выбран из Ile или Met
<220>
<221> DOMAIN
<222>(38)..(38)
<223> Xaa выбран из Arg или Lys
<220>
<221> DOMAIN
<222>(43)..(43)
<223> Xaa выбран из Gln или His
<220>
<221> DOMAIN
<222>(48)..(48)
<223> Xaa выбран из Ile или Met
<220>
<221> DOMAIN
<222>(51)..(51)
<223> Xaa выбран из Phe или Ile
<220>
<221> DOMAIN
<222>(58)..(58)
<223> Xaa выбран из Ile или Thr
<220>
<221> DOMAIN
<222>(66)..(66)
<223> Xaa выбран из Gly или Asp
<220>
<221> DOMAIN
<222>(67)..(67)
<223> Xaa выбран из Arg или Lys
<220>
<221> DOMAIN
<222>(68)..(68)
<223> Xaa выбран из Val, Ala или Thr
<220>
<221> DOMAIN
<222>(70)..(70)
<223> Xaa выбран из Arg или Lys
<220>
<221> DOMAIN
<222>(83)..(83)
<223> Xaa выбран из Leu или Phe
<220>
<221> DOMAIN
<222>(97)..(97)
<223> Xaa выбран из Ala или Thr
<220>
<221> DOMAIN
<222>(100)..(100)
<223> Xaa выбран из Gly или Arg
<220>
<221> DOMAIN
<222>(101)..(101)
<223> Xaa выбран из Phe или отсутствует
<220>
<221> DOMAIN
<222>(102)..(102)
<223> Xaa выбран из Ser или отсутствует
<220>
<221> DOMAIN
<222>(103)..(103)
<223> Xaa выбран из Tyr или отсутствует
<220>
<221> DOMAIN
<222>(105)..(105)
<223> Xaa выбран из Gly или отсутствует
<220>
<221> DOMAIN
<222>(106)..(106)
<223> Xaa выбран из Ser или отсутствует
<220>
<221> DOMAIN
<222>(107)..(107)
<223> Xaa выбран из Asn или Thr
<220>
<221> DOMAIN
<222>(108)..(108)
<223> Xaa выбран из Arg или Ser
<400> 70
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Xaa Xaa Thr Phe Xaa Asp Tyr
20 25 30
Glu Xaa His Trp Val Xaa Gln Ala Pro Gly Xaa Gly Leu Glu Trp Xaa
35 40 45
Gly Leu Xaa Asp Pro Glu Thr Gly Gly Xaa Val Tyr Asn Gln Lys Phe
50 55 60
Lys Xaa Xaa Xaa Thr Xaa Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Xaa Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Xaa Arg Glu Xaa Xaa Xaa Xaa Tyr Xaa Xaa Xaa Xaa Asp Trp Tyr Phe
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 71
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Общая формула последовательности вариабельной области легкой цепи антитела Hu32 и Hu23
<220>
<221> DOMAIN
<222>(2)..(2)
<223> Xaa выбран из Ile, Val или Gly
<220>
<221> DOMAIN
<222>(29)..(29)
<223> Xaa выбран из Ile или Leu
<220>
<221> DOMAIN
<222>(33)..(33)
<223> Xaa выбран из Asn, Gln, Leu, Thr или Asp
<220>
<221> DOMAIN
<222>(34)..(34)
<223> Xaa выбран из Gly, Ala или Val
<220>
<221> DOMAIN
<222>(35)..(35)
<223> Xaa выбран из Asn или Lys
<220>
<221> DOMAIN
<222>(102)..(102)
<223> Xaa выбран из Ala или Thr
<400> 71
Asp Xaa Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Xaa Val His Ser
20 25 30
Xaa Xaa Xaa Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Xaa Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 72
<211> 327
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность константной области тяжелой цепи IgG4-AA
<400> 72
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 73
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность константной области легкой каппа-цепи антитела
<400> 73
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 74
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность тяжелой цепи антитела Hu23-11.IgG4AA
<400> 74
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Leu Ile Asp Pro Glu Thr Gly Gly Thr Val Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Phe Ser Tyr Tyr Gly Ser Thr Ser Asp Trp Tyr Phe
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser
130 135 140
Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys
195 200 205
Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu
210 215 220
Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Leu Gly Lys
450
<210> 75
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность легкой цепи Hu23-11.IgG4AA/Hu32a-85.IgG4AA/Hu23-11.IgG4P/Hu32a-85.IgG4P
<400> 75
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Thr Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 76
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность тяжелой цепи Hu32a-85.IgG4AA
<400> 76
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Asp Phe Thr Phe Thr Asp Tyr
20 25 30
Glu Ile His Trp Val Lys Gln Ala Pro Gly His Gly Leu Glu Trp Ile
35 40 45
Gly Leu Phe Asp Pro Glu Thr Gly Gly Ile Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Glu Gly Tyr Asn Arg Asp Trp Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 77
<211> 445
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность тяжелой цепи Hu33-6.IgG4AA
<400> 77
Lys Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Val Asp Thr Tyr Tyr Gln Asp Asn Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Pro Tyr Gly His Gly Tyr Phe Asp Val Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 78
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность легкой цепи Hu33-6.IgG4AA/Hu33-6.IgG4P
<400> 78
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Asn Asn Phe
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 79
<211> 327
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность константной области тяжелой цепи IgG4-P
<400> 79
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 80
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность тяжелой цепи антитела Hu23-11. IgG4P
<400> 80
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Asp Tyr
20 25 30
Glu Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Leu Ile Asp Pro Glu Thr Gly Gly Thr Val Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Phe Ser Tyr Tyr Gly Ser Thr Ser Asp Trp Tyr Phe
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser
130 135 140
Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys
195 200 205
Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu
210 215 220
Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Leu Gly Lys
450
<210> 81
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Hu32a-85. Последовательность тяжелой цепи Hu32a-85. IgG4P
<400> 81
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Asp Phe Thr Phe Thr Asp Tyr
20 25 30
Glu Ile His Trp Val Lys Gln Ala Pro Gly His Gly Leu Glu Trp Ile
35 40 45
Gly Leu Phe Asp Pro Glu Thr Gly Gly Ile Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Phe Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Glu Gly Tyr Asn Arg Asp Trp Tyr Phe Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 82
<211> 445
<212> Белок
<213> Искусственная последовательность
<220>
<221> DOMAIN
<223> Последовательность тяжелой цепи Hu33-6. IgG4P
<400> 82
Lys Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Val Asp Thr Tyr Tyr Gln Asp Asn Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Pro Tyr Gly His Gly Tyr Phe Asp Val Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<---
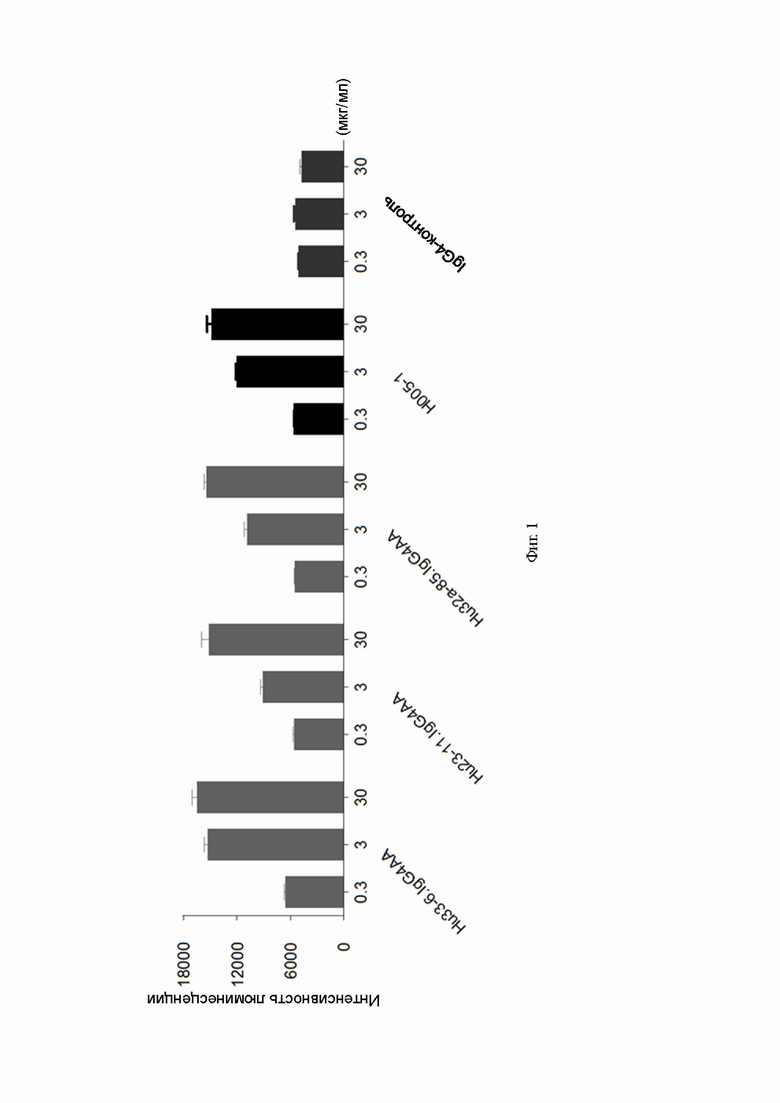
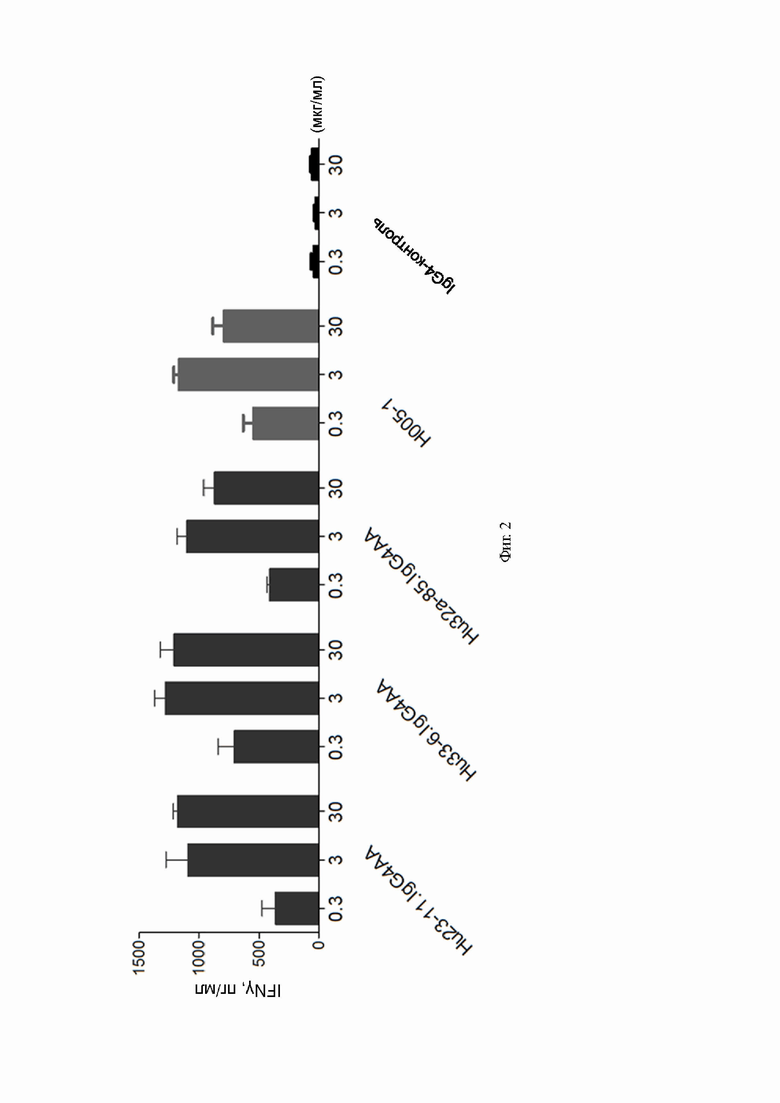
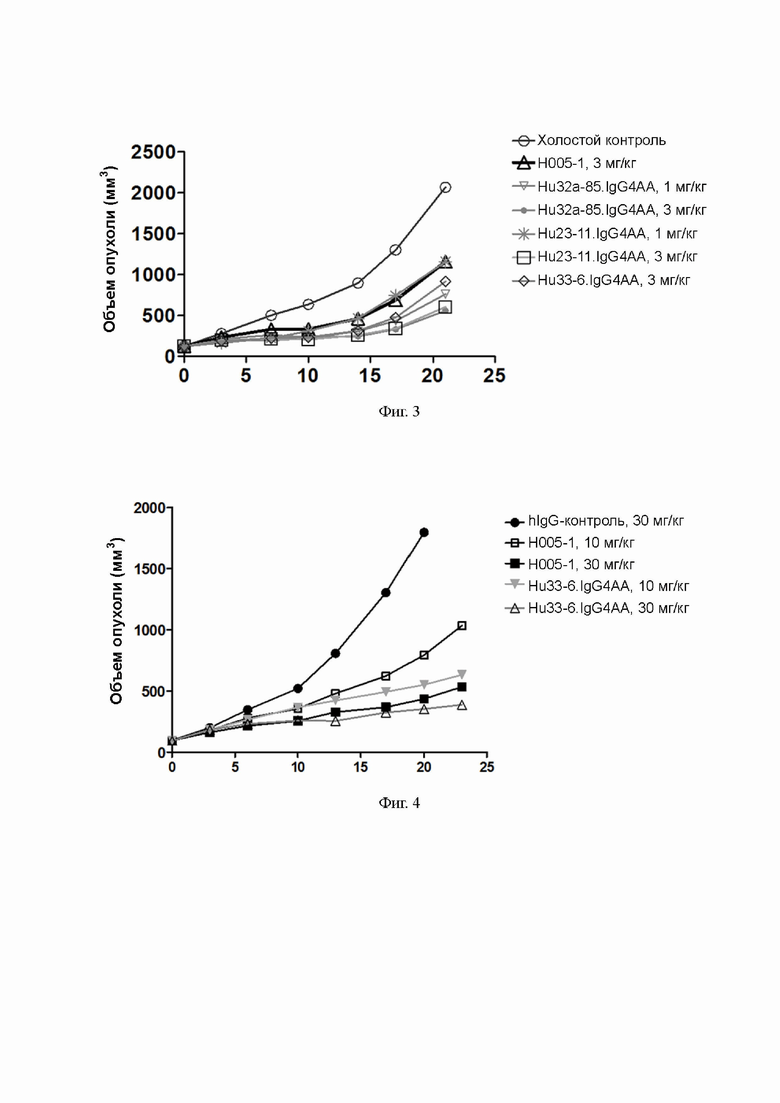
Группа изобретений относится к биотехнологии. Предложены: антитело к PD-1, его антигенсвязывающий фрагмент, фармацевтическая композиция, содержащая антитело к PD-1 и его антигенсвязывающий фрагмент, молекула нуклеиновой кислоты, кодирующая антитело к PD-1 или его антигенсвязывающий фрагмент. В частности, предложена клетка-хозяин для получения антитела к PD-1 или его антигенсвязывающего фрагмента и способ лечения заболеваний, связанных с PD-1. Изобретение применяется для лечения онкологических заболеваний. 5 н. и 15 з.п. ф-лы, 3 ил., 21 табл., 7 пр.
1. Антитело к PD-1 или его антигенсвязывающий фрагмент, содержащее вариабельную область тяжелой цепи и вариабельную область легкой цепи, где комбинация вариабельной области тяжелой цепи и вариабельной области легкой цепи является следующей:
вариабельная область тяжелой цепи, содержащая HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10 соответственно, и
вариабельная область легкой цепи, содержащая LCDR2 и LCDR3, представленные в SEQ ID NO: 12 и SEQ ID NO: 13 соответственно, и LCDR1, представленную в SEQ ID NO: 11, 47, 48, 49, 50, 51 или 52.
2. Антитело к PD-1 или его антигенсвязывающий фрагмент по п. 1, содержащее комбинацию вариабельной области тяжелой цепи и вариабельной области легкой цепи, представленную ниже:
(a1) вариабельная область тяжелой цепи, содержащая HCDR1, HCDR2 и HCDR3, представленные в SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10 соответственно, и
вариабельная область легкой цепи, содержащая LCDR1, LCDR2 и LCDR3, представленные в SEQ ID NO: 49, SEQ ID NO: 12 и SEQ ID NO: 13 соответственно.
3. Антитело к PD-1 или его антигенсвязывающий фрагмент по п. 1 или 2, где антитело к PD-1 или его антигенсвязывающий фрагмент представляет собой мышиное антитело или его антигенсвязывающий фрагмент, химерное антитело или его антигенсвязывающий фрагмент, полностью человеческое антитело или его антигенсвязывающий фрагмент, или гуманизированное антитело или его антигенсвязывающий фрагмент.
4. Антитело к PD-1 или его антигенсвязывающий фрагмент по п. 3, где антитело к PD-1 представляет собой гуманизированное антитело, содержащее каркасную область, полученную из человеческого антитела, или вариант его каркасной области, где:
вариант каркасной области содержит мутацию(ии), как указано ниже:
обратная аминокислотная мутация 2G, содержащаяся в вариабельной области легкой цепи, и/или
обратные аминокислотные мутации, содержащиеся в вариабельной области тяжелой цепи, как указано в одном из следующих случаев:
(f1) обратные аминокислотные мутации 27Y и 69L, содержащиеся в вариабельной области тяжелой цепи; или
(f2) обратные аминокислотные мутации 27Y, 48I, 67T, 69L и 82F, содержащиеся в вариабельной области тяжелой цепи; или
(f3) обратные аминокислотные мутации 27Y, 48I, 67T, 69L, 82F и 93T, содержащиеся в вариабельной области тяжелой цепи;
где положения аминокислот пронумерованы и определены в соответствии с системой Kabat.
5. Антитело к PD-1 или его антигенсвязывающий фрагмент по п. 3, где комбинация вариабельной области тяжелой цепи и вариабельной области легкой цепи антитела выбрана из (i) или (l):
(i) вариабельная область тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 4; и/или
вариабельная область легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 5;
(l) вариабельная область тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 27, 30, 31 или 32, и/или
вариабельная область легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 28, 29, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63 или 64.
6. Антитело к PD-1 или его антигенсвязывающий фрагмент по п. 5, где комбинация вариабельной области тяжелой цепи и вариабельной области легкой цепи выбрана из:
(p) вариабельной области тяжелой цепи, последовательность которой соответствует представленной в SEQ ID NO: 27; и/или
вариабельной области легкой цепи, последовательность которой соответствует представленной в SEQ ID NO: 55.
7. Антитело к PD-1 или его антигенсвязывающий фрагмент по любому из пп. 1-6, где антитело содержит константную область антитела.
8. Антитело к PD-1 или его антигенсвязывающий фрагмент по п. 7, где константная область тяжелой цепи константной области антитела выбрана из группы, состоящей из константных областей человеческого IgG1, IgG2, IgG3 и IgG4 и их традиционных вариантов, константная область легкой цепи константной области антитела выбрана из группы, состоящей из константных областей κ- и λ-цепи человеческого антитела и их традиционных вариантов.
9. Антитело к PD-1 или его антигенсвязывающий фрагмент по п. 8, где антитело содержит константную область тяжелой цепи, представленную в SEQ ID NO: 72 или 79, и константную область легкой цепи, представленную в SEQ ID NO: 73.
10. Антитело к PD-1 или его антигенсвязывающий фрагмент по любому из пп. 1-9, где антитело к PD-1 содержит легкую цепь, представленную в SEQ ID NO: 75, и тяжелую цепь, представленную в SEQ ID NO: 74 или 80.
11. Антитело к PD-1 или его антигенсвязывающий фрагмент по п. 10, где антитело к PD-1 содержит:
тяжелую цепь, представленную в SEQ ID NO: 74, и легкую цепь, представленную в SEQ ID NO: 75.
12. Антитело к PD-1 или его антигенсвязывающий фрагмент по любому из пп. 1-11, где антигенсвязывающий фрагмент выбран из группы, состоящей из Fab, Fab', F(ab')2, одноцепочечного антитела (scFv), димеризованной V-области (диатела) и стабилизированной дисульфидными связями V-области (dsFv).
13. Фармацевтическая композиция для лечения заболеваний, связанных с PD-1, содержащая:
терапевтически эффективное количество антитела к PD-1 или его антигенсвязывающего фрагмента по любому из пп. 1-12 и один или более фармацевтически приемлемых носителей, разбавителей, буферов или эксципиентов.
14. Молекула нуклеиновой кислоты, кодирующая антитело к PD-1 или его антигенсвязывающий фрагмент по любому из пп. 1-12.
15. Клетка-хозяин для получения антитела к PD-1 или его антигенсвязывающего фрагмента по любому из пп. 1-12, содержащая молекулу нуклеиновой кислоты по п. 14.
16. Способ лечения заболеваний, связанных с PD-1, включающий введение субъекту терапевтически эффективного количества антитела к PD-1 или его антигенсвязывающего фрагмента по любому из пп. 1-12 или фармацевтической композиции по п. 13, или молекулы нуклеиновой кислоты по п. 14.
17. Способ по п. 16, где заболевание представляет собой опухоль.
18. Способ по п. 17, где опухоль выбрана из группы, состоящей из: плоскоклеточной карциномы головы и шеи, рака головы и шеи, рака головного мозга, глиомы, мультиформной глиобластомы, нейробластомы, рака центральной нервной системы, нейроэндокринной опухоли, рака глотки, рака носоглотки, рака пищевода, рака щитовидной железы, злокачественной мезотелиомы плевры, рака легкого, рака молочной железы, рака печени, гепатомы, гепатоклеточной карциномы, гепатобилиарного рака, рака поджелудочной железы, рака желудка, рака желудочно-кишечного тракта, рака кишечника, рака ободочной кишки, колоректального рака, рака почки, светлоклеточной почечно-клеточной карциномы, рака яичника, рака эндометрия, рака шейки матки, рака мочевого пузыря, рака предстательной железы, рака яичка, рака кожи, меланомы, лейкоза, лимфомы, рака кости, хондросаркомы, миеломы, множественной миеломы, миелодиспластического синдрома, миелопролиферативного новообразования, плоскоклеточной карциномы, саркомы Юинга, системного амилоидоза легкой цепи и карциномы из клеток Меркеля.
19. Способ по п. 18, где лимфома выбрана из группы, состоящей из: лимфомы Ходжкина, неходжкинской лимфомы, диффузной крупноклеточной B-клеточной лимфомы, фолликулярной лимфомы, первичной медиастинальной крупноклеточной B-клеточной лимфомы, лимфомы из мантийных клеток, малой лимфоцитарной лимфомы, крупноклеточной B-клеточной лимфомы, богатой Т-клетками/гистиоцитами, и лимфоплазматической лимфомы, рак легкого выбран из группы, состоящей из: немелкоклеточного рака легкого и мелкоклеточного рака легкого, лейкоз выбран из группы, состоящей из: хронического миелоидного лейкоза, острого миелоидного лейкоза, лимфоцитарного лейкоза, лимфобластного лейкоза, острого лимфобластного лейкоза, хронического лимфоцитарного лейкоза и миелоидного лейкоза.
20. Способ по п. 17, где опухоль выбрана из группы, состоящей из: PD-L1-положительной меланомы, рака легкого, немелкоклеточного рака легкого, рака молочной железы, рака желудка, рака почки, рака мочевого пузыря, рака кишечника и рака ободочной кишки.
| CN 104250302 A, 31.12.2014 | |||
| CN 108976300 A, 11.12.2018 | |||
| CN 106573052 A, 19.04.2017 | |||
| СПОСОБ СТРОИТЕЛЬСТВА КУСТА СКВАЖИН | 2011 |
|
RU2439273C1 |
| АНТИТЕЛО К PD-1 И ЕГО ПРИМЕНЕНИЕ | 2014 |
|
RU2663795C2 |

