Перекрестные ссылки на родственные заявки
Настоящая заявка испрашивает приоритет по предварительной заявке на патент США с серийным номером 62/162174, поданной 15 мая 2015 года, с серийным номером 62/252055, поданной 6 ноября 2015 года, с серийным номером 62/301980, поданной 1 марта 2016 года и с серийным номером 62/331156, поданной 3 мая 2016 года, раскрытие каждой из которых включено в настоящий документ посредством ссылки.
Заявление по вопросу прав Правительства
Данное изобретение было сделано при поддержке правительства в рамках грантов Национального института здоровья HD032652 и DK094538. Правительство имеет определенные права на изобретение.
Предпосылки создания изобретения
Мукополисахаридозы (MPS) представляют собой группу из 11 болезней накопления, обусловленных нарушениями катаболизма гликозаминогликанов (GAG), что приводит к их накоплению в лизосомах (Muenzer, 2004; Munoz-Rojas et al., 2008). Манифестации различной степени тяжести включают органомегалию, дисплазию скелета, сердечную и легочную обструкцию и неврологическое ухудшение. Для мукополисахаридоза MPS I, характеризующегося дефицитом идуронидазы (IDUA), степень тяжести изменяется от слабой (синдром Шейе) до умеренной (Гурлер-Шейе) и до тяжелой (синдром Гурлер), при этом тяжелая форма приводит к неврологическому дефициту и смерти в возрасте до 15 лет (Muenzer, 2004; Munoz-Rojas et al., 2008). Существующие методы лечения MPS по большей части являются паллиативными. Однако для некоторых заболеваний MPS, включая синдром Гурлер, аллогенная трансплантация гемопоэтических стволовых клеток (HSCT) показала эффективность (Krivit, 2004; Orchard et al., 2007; Peters et al., 2003). К тому же, ферментная заместительная терапия (ERT) становится доступной для все большего числа заболеваний MPS (Brady, 2006). Как правило, HSCT и ERT обеспечивают очистку материалов накопления и улучшение периферических состояний, хотя некоторые проблемы сохраняются после лечения (скелетные, сердечные, помутнение роговицы). Основной проблемой этих клеточных и ферментных терапий является отсутствие эффективности в отношении неврологических манифестаций, так как периферически вводимый фермент не проникает через гематоэнцефалический барьер, и было обнаружено, что HSCT является полезной для некоторых, но не для всех заболеваний MPS.
Мукополисахаридоз I типа (MPS I) является одним из наиболее широко изученных заболеваний MPS для разработки клеточных и молекулярных методов лечения. Эффективность аллогенной HSCT, скорее всего, является результатом метаболической кросс-коррекции, в результате которой отсутствующий фермент высвобождается из клеток, полученных от доноров, а затем поглощается клетками-хозяевами и переносится в лизосомы, где фермент способствует лизосомному метаболизму (Fratantoni et al., 1968). Очистка материалов накопления GAG последовательно наблюдается в периферических органах, таких как печень и селезенка, происходит облегчение сердечно-легочной обструкции и улучшение помутнения роговицы (Orchard et al., 2007). Особенно важным является эффект аллогенной трансплантации стволовых клеток на возникновение неврологических манифестаций в заболеваниях MPS. В этой связи, для нескольких заболеваний MPS имеется подтверждение того, что индивидуумы, подвергнутые аллогенной трансплантации стволовых клеток, показали улучшенный результат по сравнению с пациентами, не подвергнутыми трансплантации (Bjoraker et al., 2006; Krivit, 2004; Orchard et al., 2007; Peters et al., 2003). Главная гипотеза, объясняющая неврологическую пользу аллогенной трансплантации гемопоэтических стволовых клеток, состоит в проникновении донорских гемопоэтических клеток (наиболее вероятно микроглии) (Hess et al., 2004; Unger et al., 1993) в центральную нервную систему, где отсутствующий фермент экспрессируется приживленными клетками, после чего диффундирует в ткани CNS и участвует в очистке материалов накопления. Таким образом, уровень фермента, поступающего в ткани CNS, ограничивается таким количеством, которое экспрессируется и высвобождается из донорских клеток, приживленных в головном мозге. Хотя такое приживление является полезным для MPS I, реципиенты, тем не менее, продолжают демонстрировать IQ ниже нормального и нарушенную нейрокогнитивную способность (Ziegler and Shapiro, 2007).
Явление метаболической кросс-коррекции также объясняет эффективность ERT для нескольких лизосомных болезней накопления (Brady, 2006), в первую очередь, MPS I. Однако эффективность ферментной терапии в отношении неврологических манифестаций лизосомной болезни накопления (LSD) не наблюдалась из-за необходимости проникновения через гематоэнцефалический барьер (ВВВ) фермента, отсутствующего в конкретной лизосомной болезни накопления (LSD), для эффективного достижения CNS (Brady, 2006). Ферменты почти всегда имеют слишком большой размер и обычно слишком заряжены для эффективного пересечения ВВВ. Это послужило толчком для изучения инвазивного интратекального введения фермента (Dickson et al., 2007), эффективность которого была продемонстрирована на собачьей модели MPS I (Kakkis et al., 2004) и начались клинические исследования с участием людей с MPS I (Pastores, 2008; Munoz-Rojas et al., 2008). К числу основных недостатков ферментной терапии относятся высокие затраты (>200000 долл. США в год) и необходимость повторных вливаний рекомбинантного белка. Проводимые в настоящее время клинические исследования интратекального введения IDUA предусматривают введение фермента только раз в три месяца, поэтому эффективность этого режима дозирования остается неопределенной.
Краткое описание изобретения
Векторы AAV, используемые в способах согласно изобретению, являются полезными для доставки генов в CNS. В одном варианте осуществления изобретение обеспечивает интраназальную доставку в CNS терапевтических белков посредством AAV, например, для предупреждения, ингибирования или лечения нейрокогнитивной дисфункции или неврологического заболевания. Как описано здесь, интраназальная доставка вектора приводит к трансдукции переднего мозга (обонятельной луковицы) и экспрессии терапевтического белка. Белок диффундирует во все области головного мозга. Таким образом, использование интраназальной доставки векторов AAV для экспрессии, например, секретируемого белка, позволяет осуществлять лечение многих различных неврологических нарушений, например, MPS I, MPS II, MPS III, других метаболических заболеваний, включая болезнь Паркинсона и болезнь Альцгеймера, и т.п. Например, анализ экстрактов из всех подвергнутых микродиссекции отделов головного мозга показал широкое распределение по всему головному мозгу альфа-L-идуронидазы, доставляемой посредством rAAV.
В одном варианте осуществления rAAV доставляют млекопитающему интратекально (IT), эндоваскулярно (IV), церебровентрикулярно (ICV) или интраназально (IN) для предупреждения, ингибирования или лечения нейрокогнитивной дисфункции или неврологического заболевания. В одном варианте осуществления интраназальное введение приводит к неинвазивному прямому введению в CNS с метаболической кросс-коррекцией. В одном варианте осуществления млекопитающее подвергают иммуносупрессии. В одном варианте осуществления млекопитающего подвергают индукции иммунологической толерантности.
В одном варианте осуществления заболевание, подлежащее предупреждению, ингибированию или лечению с помощью конкретного гена, включает, но без ограничения, MPS I (IDUA), MPS II (IDS), MPS ΠΙΑ (renapaH-N-сульфатаза; сульфамидаза), MPS ШВ (альфа-М-ацетил-глюкозаминидаза), MPS ШС (ацетил-СоА: альфа-М-глюкозаминид-ацетилтрансфераза), MPS HID (N-ацетилглюкозамин-6-сульфатаза), MPS VII (бета-глюкуронидаза), болезнь Гоше (кислая бета-гликозидаза), альфа-маннозидоз (альфа-маннозидаза), бета-маннозидоз (бета-маннозидаза), альфа-фукозидоз (альфа-фукозидаза), сиалидоз (альфа-сиалидаза), галактосиалидоз (катепсин А), аспартилглюкозаминурия (аспартилглюкозаминидаза), GMl-ганглиозидоз (бета-галактозидаза), болезнь Тея-Сакса (альфа-субъединица бета-гексозаминидазы), синдром Сандхоффа (бета-субъединица бета-гексозаминидазы), ОМ2-ганглиозидоз/вариант АВ (белок-активатор GM2), болезнь Краббе (галактоцереброзидаза), метахроматическая лейкодистрофия (арилсульфатаза А) и другие неврологические нарушения, включая, но без ограничения, болезнь Альцгеймера (экспрессия антитела, такого как антитело к бета-амилоиду, или фермент, который атакует бляшки и фибриллы, связанные с болезнью Альцгеймера), или болезни Альцгеймера и Паркинсона (экспрессия нейропротекторных белков, включая, но без ограничения, GDNF или нейротурин).
Таким образом, описаны способы предупреждения, ингибирования и/или лечения, например, одного или нескольких симптомов, связанных с заболеванием центральной нервной системы (CNS), у млекопитающего, нуждающегося в этом. Способы включают доставку в CNS млекопитающего, нуждающегося в лечении, композиции, содержащей эффективное количество рекомбинантного вектора на основе аденоассоциированного вируса (rAAV), содержащего открытую рамку считывания, кодирующую генный продукт, например, продукт терапевтического гена. Целевые генные продукты, которые могут быть кодированы вектором rAAV, включают, но без ограничения, альфа-L-идуронидазу, идуронат-2-сульфатазу, гепарансульфат-сульфатазу, N-ацетил-альфа-О-глюкозаминидазу, бета-гексозаминидазу, альфа-галактозидазу, бета-галактозидазу, бета-глюкуронидазу или глюкоцереброзидазу, а также те, которые описаны выше. Заболевания, которые можно предупредить, ингибировать или лечить с использованием описанных здесь способов, включают, но без ограничения, нарушение мукополисахаридоз I типа, нарушение мукополисахаридоз II типа или нарушение мукополисахаридоз VII типа, а также нарушения, указанные выше. Вектор AAV может быть введен различными способами для того, чтобы гарантировать его доставку в CNS/головной мозг и успешную трансдукцию трансгена в CNS/головном мозге субъекта. Пути доставки в CNS/головной мозг включают, но без ограничения, интратекальное введение, интракраниальное введение, например, интрацеребровентрикулярное введение или латеральное церебровентрикулярное введение, интраназальное введение, эндоваскулярное введение и интрапаренхимальное введение.
В одном варианте осуществления способы включают доставку в CNS взрослого млекопитающего, нуждающегося в лечении, композиции, содержащей эффективное количество вектора rAAV серотипа 9 (rAAV9), содержащего открытую рамку считывания, кодирующую ген. В одном варианте осуществления способы включают доставку в CNS взрослого млекопитающего, нуждающегося в лечении, композиции, содержащей эффективное количество вектора rAAV9, содержащего открытую рамку считывания, кодирующую IDUA. Эти способы отчасти основаны на открытии того, что вектор AAV9 может эффективно трансдуцировать терапевтический трансген в головной мозг/CNS взрослых субъектов, восстанавливая уровни фермента до уровней, наблюдаемых у животных дикого типа (см. ниже фигуру 15). Результаты, полученные с использованием AAV9, являются неожиданными, принимая во внимание предыдущую работу, в которой было продемонстрировано, что интраваскулярная доставка AAV9 взрослым мышам не достигает широкого прямого нацеливания на нейроны (см. Foust et al., 2009), а также дополнительные данные, свидетельствующие о том, что прямая инъекция AAV8-IDUA в CNS взрослых ШиА-дефицитных мышей приводит к слабой экспрессии трансгена (фигура 18). В примерах, описанных в настоящем документе, использована доклиническая модель для лечения MPS1, наследственного метаболического нарушения, обусловленного дефицитом лизосомного фермента альфа-L-идуронидазы (IDUA). В этих примерах продемонстрировано, что непосредственное применение AAV9-IDUA в CNS иммунокомпетентных взрослых Ши А-дефицитных мышей приводит к экспрессии и активности фермента IDUA, которые являются такими же или более высокими, чем экспрессия и активность фермента IDUA у взрослых мышей дикого типа (см. ниже фигуру 15).
В дополнительном варианте осуществления изобретения в примерах также продемонстрировано, что совместная терапия для индукции иммуносупрессии или иммунологической толерантности, или лечение иммунодефицитных животных может обеспечивать даже более высокие уровни экспрессии и активности фермента IDUA. В одном варианте осуществления пациентов с генотипами, которые стимулируют иммунный ответ, нейтрализующий активность фермента (см., например, Barbier et al., 2013), подвергают лечению иммуносупрессантом в дополнение к вектору rAAV, содержащему открытую рамку считывания, кодирующую генный продукт, такой как как IDUA.
Новорожденные IDUA"" мыши являются иммунологически интактными. Введение AAV8-IDUA новорожденным IDUA"'" мышам привело к экспрессии IDUA (Wolf et al., 2011), таким образом, вызывая у животных толерантность к IDUA. Как описано здесь, применимость AAV-опосредованного переноса генов у взрослых (иммунокомпетентных) мышей путем прямой инфузии AAV в центральную нервную систему была продемонстрирована с использованием различных путей введения. Например, AAV9-IDUA вводили путем прямой инъекции в боковые желудочки взрослых IDAA-дефицитных мышей, которые были либо иммунокомпетентными, иммунодефицитными (NODSCID/IDUA-/-), подвергнутыми иммуносупрессии циклофосфамидом (CP), либо подвергнутыми индукции иммунологической толерантности путем еженедельной инъекции человеческого белка идуронидазы (Альдуразим), начиная с рождения. Подвергнутым иммуносупрессии циклофосфамидом (CP) животным также вводили AAV9-IDUA путем интраназальной инфузии, путем интратекальной инъекции и путем эндоваскулярной инфузии с маннитом для разрушения гематоэнцефалического барьера и без него. Животных умерщвляли через 8 недель после введения вектора, и головной мозг извлекали и подвергали микродессекции для оценки активности фермента IDUA, гликозаминогликанов ткани и векторных последовательностей IDUA по сравнению с нормальными и пораженными контрольными мышами. Результаты этих исследований показали, что могут быть использованы различные способы введения AAV-вектора непосредственно в CNS, например, для достижения более высоких уровней доставки белка и/или активности фермента в CNS. Кроме того, несмотря на то, что головной мозг является иммунологически привилегированной областью, введение иммуносупрессанта или индукция иммунологической толерантности может повысить активность в головном мозге после введения AAV. Более высокие уровни экспрессии на введение и/или менее инвазивные пути введения клинически являются более приемлемыми для пациентов.
Таким образом, изобретение включает применение рекомбинантных векторов AAV (rAAV), которые кодируют генный продукт, обладающий терапевтическими эффектами при экспрессии в CNS млекопитающего. В одном варианте осуществления млекопитающее представляет собой иммунокомпетентное млекопитающее, имеющее заболевание или нарушение CNS (неврологическое заболевание). Используемое здесь «иммунокомпетентное» млекопитающее представляет собой млекопитающее, находящееся в возрасте, когда как клеточный, так и гуморальный иммунные ответы вызываются под воздействием антигенного стимула, путем апрегуляции функций Thl или продукции IFN-γ в ответ на поликлональные стимулы, в отличие от новорожденного млекопитающего, имеющего врожденный иммунитет и иммунитет, полученный от матери, например, в период беременности или посредством грудного кормления. Взрослое млекопитающее, которое не имеет иммунодефицитное заболевание, является примером иммунокомпетентного млекопитающего. Например, иммунокомпетентный человек, как правило, находится в возрасте по меньшей мере 1, 2, 3, 4, 5 или 6 месяцев и включает взрослых людей, не имеющих иммунодефицитного заболевания. В одном варианте осуществления AAV вводят интратекально. В одном варианте осуществления AAV вводят интракраниально (например, интрацеребровентрикулярно). В одном варианте осуществления AAV вводят интраназально, с усилителем проницаемости или без него. В одном варианте осуществления AAV вводят эндоваскулярно, например, в сонную артерию с усилителем проницаемости или без него. В одном варианте осуществления млекопитающее, которому вводят AAV, является иммунодефицитным или же подвергнутым индукции иммунологической толерантности или иммуносупрессии, например, для индукции более высоких уровней экспрессии терапевтического белка по сравнению с соответствующим млекопитающим, которому вводили AAV, но не подвергали индукции иммунологической толерантности или иммуносупрессии, В одном варианте осуществления иммуносупрессорный агент вводят для индукции иммуносупрессии. В одном варианте осуществления млекопитающее, которому вводят AAV, не подвергается индукции иммунологической толерантности или иммуносупрессии (например, терапевтический эффект обеспечивается введением только AAV).
В одном варианте осуществления в изобретении предлагается способ увеличения концентрации секретируемого белка в центральной нервной системе млекопитающего, имеющего неврологическое заболевание, которое может включать нейрокогнитивную дисфункцию. Способ включает интраназальное введение млекопитающему композиции, содержащей эффективное количество рекомбинантного вектора на основе аденоассоциированного вируса (rAAV), содержащего открытую рамку считывания, кодирующую секретируемый белок, экспрессия которого у млекопитающего уменьшает невропатологию и/или усиливает нейрокогнитивные функции по всему головному мозгу по сравнению с млекопитающим, имеющим заболевание или дисфункцию, но которому не вводили rAAV. В одном варианте осуществления кодируемый белок представляет собой нейропротекторный белок, например, GDNF (глиальный нейротрофический фактор) или Neurturin (нейротурин). В одном варианте осуществления кодируемый белок представляет собой антитело, например, антитело, которое связывает бета-амилоид. В одном варианте осуществления белок представляет собой фермент, который расщепляет бляшки или фибриллы, связанные с болезнью Альцгеймера. В одном варианте осуществления млекопитающее не подвергают лечению иммуносупрессантом. В другом варианте осуществления, например, у субъектов, которые могут генерировать иммунный ответ, нейтрализующий активность терапевтического белка, млекопитающее подвергают лечению иммуносупрессантом, например, глюкокортикоидом, цитостатическими агентами, включая алкилирующий агент, антиметаболит, цитотоксический антибиотик, антитело или агент, связывающийся с иммунофилином, такой как азотистый иприт, нитрозомочевина, соединения платины, метотрексат, азатиоприн, меркаптопурин, фторурацил, дактиномицин, антрациклин, митомицин С, блеомицин, митрамицин, направленные на рецептор IL-2 (CD25-) или CD3-антитела, антитела к IL-2, циклоспорин, такролимус, сиролимус, IFN-β, IFN-γ, опиоид или связывающее TNF-α (фактор некроза опухоли-альфа) средство. В одном варианте осуществления rAAV и иммуносупрессант вводят совместно или иммуносупрессант вводят после rAAV. В одном варианте осуществления иммуносупрессант вводят интратекально. В одном варианте осуществления иммуносупрессант вводят интрацребровентрикулярно. В одном варианте осуществления вектор rAAV представляет собой вектор rAAVl, rAAV3, rAAV4, rAAV5, rAA rhlO или rAAV9. В одном варианте осуществления млекопитающее подвергают индукции иммунологической толерантности до введения композиции.
В одном варианте осуществления в изобретении предлагается способ предупреждения, ингибирования или лечения неврологического заболевания, которое может включать нейрокогнитивную дисфункцию у млекопитающего. Способ включает интраназальное введение млекопитающему композиции, содержащей эффективное количество рекомбинантного вектора на основе аденоассоциированного вируса (rAAV), содержащего открытую рамку считывания, кодирующую белок, экспрессия которого у млекопитающего предупреждает, ингибирует или лечит невропатологию и/или нейрокогнитивную дисфункцию. В одном варианте осуществления кодируемый белок представляет собой нейропротекторный белок, например, GDNF или нейротурин. В одном варианте осуществления кодируемый белок представляет собой антитело, например, антитело, которое связывает бета-амилоид. В одном варианте осуществления белок представляет собой фермент, который расщепляет бляшки или фибриллы, связанные с болезнью Альцгеймера. В одном варианте осуществления млекопитающее не подвергают лечению иммуносупрессантом. В другом варианте осуществления, например, у субъектов, которые могут генерировать иммунный ответ, нейтрализующий активность терапевтического белка, млекопитающее подвергают лечению иммуносупрессантом, например, глюкокортикоидом, цитостатическими агентами, включая алкилирующий агент, антиметаболит, цитотоксический антибиотик, антитело или агент, связывающийся с иммунофилином, такой как азотистый иприт, нитрозомочевина, соединение платины, метотрексат, азатиоприн, меркаптопурин, фторурацил, дактиномицин, антрациклин, митомицин С, блеомицин, митрамицин, направленные на рецептор IL-2 (CD25-) или CD3-антитела, антитела к IL-2, циклоспорин, такролимус, сиролимус, IFN-β, IFN-γ, опиоид или связывающее TNF-a (фактор некроза опухоли-альфа) средство. В одном варианте осуществления rAAV и иммуносупрессант вводят совместно или иммуносупрессант вводят после rAAV. В одном варианте осуществления иммуносупрессант вводят интратекально. В одном варианте осуществления иммуносупрессант вводят интрацребровентрикулярно. В одном варианте осуществления вектор rAAV представляет собой вектор rAAVl, rAAV3, rAAV4, rAAV5, rAA rh10 или rAAV9. В одном варианте осуществления млекопитающее подвергают индукции иммунологической толерантности перед введением композиции. В одном варианте осуществления млекопитающее имеет болезнь Альцгеймера или болезнь Паркинсона. В одном варианте осуществления в изобретении предлагается способ обеспечения кросс-коррекции секретируемого белка в центральной нервной системе у млекопитающего, имеющего неврологическое заболевание, которое может включать нейрокогнитивную дисфункцию. Способ включает: интраназальное, интратекальное, интрацеребровентрикулярное или внутривенное введение млекопитающему эффективного количества композиции, содержащей эффективное количество рекомбинантного вектора на основе аденоассоциированного вируса (rAAV), содержащего открытую рамку считывания, кодирующую секретируемый белок, экспрессия которого у млекопитающего обеспечивает кросс-коррекцию. В одном варианте осуществления кодируемый белок представляет собой нейропротекторный белок, например, GDNF или нейротурин. В одном варианте осуществления кодируемый белок представляет собой антитело, например, антитело, которое связывает бета-амилоид. В одном варианте осуществления белок представляет собой фермент, который расщепляет бляшки или фибриллы, связанные с болезнью Альцгеймера. В одном варианте осуществления млекопитающее не подвергают лечению иммуносупрессантом. В одном варианте осуществления, например, у субъектов, которые могут генерировать иммунный ответ, нейтрализующий активность терапевтического белка, млекопитающее подвергают лечению иммуносупрессантом, например, глюкокортикоидом, цитостатическими агентами, включая алкилирующий агент, антиметаболит, цитотоксический антибиотик, антитело или агент, связывающийся с иммунофилином, такой как азотистый иприт, нитрозомочевина, соединение платины, метотрексат, азатиоприн, меркаптопурин, фторурацил, дактиномицин, антрациклин, митомицин С, блеомицин, митрамицин, направленные на рецептор IL-2 (CD25-) или CD3-антитела, антитела к IL-2, циклоспорин, такролимус, сиролимус, IFN-β, IFN-γ, опиоид или связывающее TNF-α (фактор некроза опухоли-альфа) свредство. В одном варианте осуществления rAAV и иммуносупрессант вводят совместно или иммуносупрессант вводят после rAAV. В одном варианте осуществления иммуносупрессант вводят интратекально. В одном варианте осуществления иммуносупрессант вводят интрацребровентрикулярно. В одном варианте осуществления вектор rAAV представляет собой вектор rAAV1, rAAV3, rAAV4, rAAV5, rAA rh10 или rAAV9. В одном варианте осуществления млекопитающее подвергают индукции иммунологической толерантности перед введением композиции.
В изобретении предлагается способ предупреждения, ингибирования или лечения нейрокогнитивной дисфункции, связанной с заболеванием или нарушением центральной нервной системы у млекопитающего, нуждающегося в этом. Способ включает интратекальное, например, в поясничную область, или интрацеребровентрикулярное, например, в боковой желудочек, введение млекопитающему композиции, содержащей эффективное количество вектора rAAV, содержащего открытую рамку считывания, кодирующую генный продукт, экспрессия которого в центральной нервной системе млекопитающего предупреждает, ингибирует или лечит нейрокогнитивную дисфункцию. В одном варианте осуществления генный продукт представляет собой лизосомной фермент накопления. В одном варианте осуществления млекопитающее представляет собой иммунокомпетентное взрослое млекопитающее. В одном варианте осуществления вектор rAAV представляет собой вектор AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh10 или AAV9. В одном варианте осуществления млекопитающее представляет собой человека. В одном варианте осуществления вводят многократные дозы. В одном варианте осуществления композицию вводят раз в неделю, раз в месяц, или раз в два или более месяцев.
В одном варианте осуществления способ включает интратекальное, например, в поясничную область, введение млекопитающему композиции, содержащей эффективное количество вектора rAAV, содержащего открытую рамку считывания, кодирующую генный продукт, экспрессия которого в центральной нервной системе млекопитающего предупреждает, ингибирует или лечит нейрокогнитивную дисфункцию, и, необязательно, введение усилителя проницаемости. В одном варианте осуществления усилитель проницаемости вводят до введения композиции. В одном варианте осуществления композиция содержит усилитель проницаемости. В одном варианте осуществления усилитель проницаемости вводят после введения композиции. В одном варианте осуществления генный продукт представляет собой лизосомный фермент, связанный с лизосомной болезнью накопления. В одном варианте осуществления млекопитающее представляет собой иммунокомпетентное взрослое млекопитающее. В одном варианте осуществления вектор rAAV представляет собой вектор AAV-1, AAV-3, AAV-4, AAV-5, AAV-6, AAV-7, AAV-8, AAV rh10 или AAV-9. В одном варианте осуществления млекопитающее представляет собой человека. В одном варианте осуществления вводят многократные дозы. В одном варианте осуществления композицию вводят раз в неделю, раз в месяц, или раз в два или более месяцев. В одном варианте осуществления млекопитающее, которому интратекально вводят AAV, не подвергают индукции иммунологической толерантности или иммуносупрессии (например, терапевтический эффект обеспечивает введение только AAV). В одном варианте осуществления млекопитающее, которому интратекально вводят AAV, является иммунодефицитным или подвергается индукции иммунологической толерантности или иммуносупрессии, например, для индукции более высоких уровней экспрессии терапевтического белка по сравнению с соответствующим млекопитающим, которому интратекально вводили AAV, но не подвергали индукции иммунологической толерантности или иммуносупрессии.
В одном варианте осуществления способ включает интрацеребровентрикулярное, например, в боковой желудочек, введение иммунокомпетентному млекопитающему композиции, содержащей эффективное количество вектора rAAV, содержащего открытую рамку считывания, кодирующую генный продукт, экспрессия которого в центральной нервной системе млекопитающего предупреждает, ингибирует или лечит нейрокогнитивную дисфункцию. В одном варианте осуществления генный продукт представляет собой лизосомный фермент, связанный с лизосомной болезнью накопления. В одном варианте осуществления вектор rAAV представляет собой вектор AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh10 или AAV9. В одном варианте осуществления вектор rAAV не является вектором rAAV5. В одном варианте осуществления млекопитающее представляет собой человека. В одном варианте осуществления вводят многократные дозы. В одном варианте осуществления композицию вводят раз в неделю, раз в месяц, или раз в два или более месяцев. В одном варианте осуществления млекопитающего, которому интрацеребровентрикулярно вводят AAV, не подвергают индукции иммунологической толерантности или иммуносупрессии (например, терапевтический эффект обеспечивает введение только AAV). В одном варианте осуществления млекопитающее, которому интрацеребровентрикулярно вводят AAV, является иммунодефицитным, или подвергается индукции иммунологической толерантности или иммуносупрессии, например, для индукции более высоких уровней экспрессии терапевтического белка по сравнению с соответствующим млекопитающим, которому интрацеребровентрикулярно вводили AAV, но не подвергали индукции иммунологической толерантности или иммуносупрессии. В одном варианте осуществления млекопитающее подвергали индукции иммунологической толерантности к генному продукту до введения композиции, содержащей AAV.
Кроме того, предлагается способ предупреждения, ингибирования или лечения нейрокогнитивной дисфункции, связанной с заболеванием или нарушением центральной нервной системы, у млекопитающего, нуждающегося в этом. Способ включает эндоваскулярное введение млекопитающему композиции, содержащей эффективное количество вектора rAAV, содержащего открытую рамку считывания, кодирующую генный продукт, экспрессия которого в центральной нервной системе млекопитающего предупреждает, ингибирует или лечит дисфункцию, и эффективное количество усилителя проницаемости. В одном варианте осуществления композиция содержит усилитель проницаемости. В одном варианте осуществления усилитель проницаемости включает маннит, гликохолат натрия, таурохолат натрия, дезоксихолат натрия, салицилат натрия, каприлат натрия, капрат натрия, лаурилсульфат натрия, полиоксиэтилен-9-лауриловый эфир или ЭДТА. В одном варианте осуществления генный продукт представляет собой лизосомный фермент, связанный с лизосомной болезнью накопления. В одном варианте осуществления млекопитающее представляет собой иммунокомпетентное взрослое млекопитающее. В одном варианте осуществления вектор rAAV представляет собой вектор AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh10 или AAV9. В одном варианте осуществления вектор rAAV не является вектором rAAV5. В одном варианте осуществления млекопитающее представляет собой человека. В одном варианте осуществления вводят многократные дозы. В одном варианте осуществления композицию вводят раз в неделю. В одном варианте осуществления композицию вводят раз в неделю, раз в месяц, или раз в два или более месяцев. В одном варианте осуществления млекопитающего, которому эндоваскулярно вводят AAV, не подвергают индукции иммунологической толерантности или иммуносупрессии (например, терапевтический эффект обеспечивает введение только AAV). В одном варианте осуществления млекопитающее, которому эндоваскулярно вводят AAV, является иммунодефицитным или подвергается индукции иммунологической толерантности или иммуносупрессии, например, для индукции более высоких уровней экспрессии терапевтического белка по сравнению с соответствующим млекопитающим, которому эндоваскулярно вводили AAV, но не подвергали индукции иммунологической толерантности или иммуносупрессии.
В одном варианте осуществления способ включает интраназальное введение млекопитающему композиции, содержащей эффективное количество вектора rAAV9, содержащего открытую рамку считывания, кодирующую генный продукт, экспрессия которого в центральной нервной системе млекопитающего предупреждает, ингибирует или лечит нейрокогнитивную дисфункцию, и, необязательно, введение усилителя проницаемости. В одном варианте осуществления интраназальная доставка может быть осуществлена, как описано в патенте США 8609088, раскрытие которого включено здесь посредством ссылки. В одном варианте осуществления усилитель проницаемости вводят до введения композиции. В одном варианте осуществления композиция содержит усилитель проницаемости. В одном варианте осуществления усилитель проницаемости вводят после введения композиции. В одном варианте осуществления генный продукт представляет собой лизосомный фермент, связанный с лизосомной болезнью накопления.
В одном варианте осуществления млекопитающее представляет собой иммунокомпетентное взрослое млекопитающее. В одном варианте осуществления млекопитающее представляет собой человека. В одном варианте осуществления вводят многократные дозы. В одном варианте осуществления композицию вводят раз в неделю, раз в месяц, или раз в два или более месяцев. В одном варианте осуществления млекопитающего, которому интраназально вводят AAV, не подвергают индукции иммунологической толерантности или иммуносупрессии. В одном варианте осуществления млекопитающего, которому интраназально вводят AAV, подвергают индукции иммунологической толерантности или иммуносупрессии, например, для индукции более высоких уровней экспрессии белка IDUA по сравнению с соответствующим млекопитающим, которому интраназально вводили AAV, но не подвергали индукции иммунологической толерантности или иммуносупрессии.
Также, предлагается способ предупреждения, ингибирования или лечения нейрокогнитивной дисфункции, связанной с заболеванием центральной нервной системы, у млекопитающего, нуждающегося в этом. Способ включает введение млекопитающему композиции, содержащей эффективное количество вектора rAAV, содержащего открытую рамку считывания, кодирующую генный продукт, экспрессия которого в центральной нервной системе млекопитающего предупреждает, ингибирует или лечит, и иммуносупрессант.В одном варианте осуществления иммуносупрессант представляет собой циклофосфамид. В одном варианте осуществления иммуносупрессант представляет собой глюкокортикоид, цитостатические агенты, включая алкилирующий агент или антиметаболит, такой как метотрексат, азатиоприн, меркаптопурин или цитотоксический антибиотик, антитело или агент, связывающийся с иммунофилином. В одном варианте осуществления иммуносупрессант представляет собой азотистый иприт, нитрозомочевину, соединение платины, метотрексат, азатиоприн, меркаптопурин, фторурацил, дактиномицин, антрациклин, митомицин С, блеомицин, митрамицин, направленные на рецептор IL-2 (CD25-) или СО3-антитела, антитела к IL-2, циклоспорин, такролимус, сиролимус, IFN-β, IFN-γ, опиоид или связывающие TNF-α (фактор некроза опухоли-альфа) средства, такие как инфликсимаб (Ремикейд), этанерцепт (Энбрел) или адалимумаб (Хумира). В одном варианте осуществления rAAV и иммунодепрессант вводят совместно. В одном варианте осуществления rAAV вводят до и, необязательно, после введения иммуносупрессанта. В одном варианте осуществления иммуносупрессант вводят до введения rAAV. В одном варианте осуществления rAAV и иммунодепрессант вводят интратекально. В одном варианте осуществления rAAV и иммунодепрессант вводят интрацребровентрикулярно. В одном варианте осуществления rAAV вводят интратекально и иммунодепрессант вводят внутривенно. В одном варианте осуществления генный продукт представляет собой лизосомный фермент, связанный с лизосомной болезнью накопления. В одном варианте осуществления млекопитающее представляет собой взрослое млекопитающее. В одном варианте осуществления вектор rAAV представляет собой вектор AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrhlO или AAV9. В одном варианте осуществления млекопитающее представляет собой человека. В одном варианте осуществления вводят многократные дозы. В одном варианте осуществления композицию вводят раз в неделю. В одном варианте осуществления композицию вводят раз в неделю, раз в месяц, или раз в два или более месяцев.
В изобретении также предлагается способ предупреждения, ингибирования или лечения нейрокогнитивной дисфункции, связанной с заболеванием центральной нервной системы, у млекопитающего, нуждающегося в этом. Млекопитающему, подвергшемуся индукции иммунологической толерантности к генному продукту, который связан с заболеванием, вводили композицию, содержащую эффективное количество вектора rAAV, содержащего открытую рамку считывания, кодирующую генный продукт, экспрессия которого в центральной нервной системе млекопитающего предупреждает, ингибирует или лечит один или несколько симптомов. В одном варианте осуществления генный продукт представляет собой лизосомный фермент, связанный с лизосомной болезнью накопления. В одном варианте осуществления млекопитающее представляет собой взрослое млекопитающее. В одном варианте осуществления вектор rAAV представляет собой вектор AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrhlO или AAV9. В одном варианте осуществления млекопитающее представляет собой человека. В одном варианте осуществления вводят многократные дозы. В одном варианте осуществления композицию вводят раз в неделю.
Генные продукты, которые могут быть кодированы векторами rAAV, включают, но без ограничения, альфа-Ь-идуронидазу, идуронат-2-сульфатазу, гепарансульфат-сульфатазу, 1М-ацетил-альфа-0-глюкозаминидазу, бета-гексозаминидазу, альфа-галактозидазу, бета-галактозидазу, бета-глюкуронидазу, глюкоцереброзидазу, фактор рост фибробластов-2 (FGF-2), мозговой фактор роста (BDGF), нейротурин, глиальный фактор роста (GDGF), тирозингидроксилазу, допаминдекарбоксилазу или декарбоксилазу глутаминовой кислоты.
Заболевания, которые могут проявлять неврологические симптомы или нейрокогнитивную дисфункцию, которые можно предупреждать, ингибировать или лечить с использованием способов, описанных в настоящем документе, включают, но без ограничения, адренолейкодистрофию, болезнь Альцгеймера, боковой амиотрофический склероз, синдром Ангельмана, телеангиоэктатическую атаксию, синдром Шарко-Мари-Тута, синдром Коккейна, тугоухость, миодистрофию Дюшенна, эпилепсию, эссенциальный тремор, синдром фрагильной Х-хромосомы, наследственную атаксию Фридрейха, болезнь Гоше, болезнь Хантингтона, синдром Леша-Найхана, болезнь кленового сиропа, синдром Менкеса, миотоническую дистрофию, нарколепсию, нейрофиброматоз, болезнь Ниманна-Пика, болезнь Паркинсона, фенилкетонурию, синдром Прадера-Вилли, болезнь Рефсума, синдром Ретта, спинальную мышечную атрофию, спинально-церебеллярную атаксию, танжерскую болезнь, болезнь Тея-Сакса, туберозный склероз, синдром фон Геппеля-Линдау, болезнь Вильямса, болезнь Вильсона или синдром Цельвегера. В одном варианте осуществления заболевание представляет собой лизосомную болезнь накопления, например, отсутствие или дефицит лизосомного фермента, связанного с лизосомной болезнью накопления. Лизосомные болезни накопления включают, но без ограничения, мукополисахаридоз (MPS), например, мукополисахаридоз I типа, например, синдром Гурлер и варианты синдрома Шейе и синдром Гурлер-Шейе (дефицит альфа-Ь-идуронидазы); синдром Хантера (дефицит идуронат-2-сульфатазы); мукополисахаридоз III типа, например, болезнь Санфилиппо (А, В, С или D, дефицит гепарансульфатсульфатазы,]ЧТ-ацетил-альфа-0-глюкозаминидазы, ацетил-СоА: альфа-глюкозаминидазы N-ацетилтрансферазы или N-ацетилглюкозамин-6-сульфатсульфатазы); мукополисахаридоз IV типа, например, синдром Моркио (дефицит галактозамин-6-сульфатсульфатазы или бета-галактозидазы); мукополисахаридоз VI типа, например, синдром Марото-Лами (дефицит арилсульфатазы В); мукополисахаридоз II типа; мукополисахаридоз III типа (А, В, С или D; дефицит гепарансульфатсульфатазы, N-ацетил-альфа-О-глюкозаминидазы, ацетил-СоА: альфа-глюкозаминидазы N-ацетилтрансферазы или N-ацетилглюкозамин-6-сульфатсульфатазы); мукополисахаридоз IV типа (А или В; дефицит галактозамин-6-сульфатазы и бета-галатакозидазы); мукополисахаридоз VI типа (дефицит арилсульфатазы В); мукополисахаридоз VII типа (дефицит бета-глюкуронидазы); мукополисахаридоз VIII типа (дефицит глюкозамин-6-сульфатсульфатазы); мукополисахаридоз ГХ типа (дефицит гиалуронидазы); болезнь Тея-Сакса (дефицит альфа-субъединицы бета-гексозаминидазы); синдром Сандгоффа (дефицит альфа- и бета-субъединиц бета-гексозаминидазы); II болезнь Сандгоффа (тип I или тип II); Болезнь Фабри (дефицит альфа-галактозидазы); метахроматическую лейкодистрофию (дефицит арилсульфатазы А); болезнь Помпе (дефицит кислой мальтазы); фукозидоз (дефицит фукозидазы); альфа-маннозидоз (дефицит альфа-маннозидазы); бета-маннозидоз (дефицит бета-маннозидазы), нейронный восковидный липофусциноз и болезнь Гоше (I, II и III типов; дефицит глюкоцереброзидазы), а также расстройства, такие как синдром Германского-Пудлака; амавротическую идиотию; танжерскую болезнь; аспартилглюкозаминурию; врожденное нарушение гликозилирования, тип 1а; Синдром Чедиака-Хигаши; макулярную дистрофию роговицы, 1; нефропатический цистиноз; синдром Фанкони-Биккеля; липогранулематоз Фарбера; фиброматоз; гелеофизическую дисплазию; болезнь накопления гликогена I; болезнь накопления гликогена lb; болезнь накопления гликогена 1 с; болезнь накопления гликогена III; болезнь накопления гликогена IV; болезнь накопления гликогена V; болезнь накопления гликогена VI; болезнь накопления гликогена VII; болезнь накопления гликогена 0; иммуно-остеоидную дисплазию, тип Шимке; липидоз; дефицит липазы Ь; муколипидоз II; муколипидоз II, включая вариантную форму; муколипидоз IV; дефицит нейраминидазы с дефицитом бета-галактозидазы; муколипидоз I; болезнь Ниманна-Пика (дефицит сфингомиелиназы); болезнь Ниманна-Пика без дефицита сфингомиелиназы (дефицит гена npcl, кодирующего фермента, метаболизирующий холестерин); болезнь Рефсума; болезнь голубых гистиоцитов; болезнь накопления сиаловой кислоты; сиалурию; множественную сульфатазную недостаточность; болезнь накопления триглицеридов с нарушенным окислением длинноцепочечных жирных кислот; синдром Винчестера; болезнь Вольмана (дефицит кислой гидролазы эфиров холестерина); нарушение, связанное дезоксирибонуклеазой I (Deoxyribonuclease I-like 1); нарушение, связанное с арилсульфатазой Е; нарушение, связанное с субъединицей 1 лисзосомной Н+-АТФазы; болезнь накопления гликогена lib; Ras-пссоциированное нарушение белка гаЬ9; точечную эпифизарную дисплазию 1, рецессивное расстройство, связанной с X-хромосомой; болезнь накопления гликогена VIII; связанные с лизосомой нарушение мембранного белка 2; синдром Менкеса; врожденное нарушение гликозилирования, тип Ic; и сиалурию. Изменение, составляющее менее чем 20%, например, менее чем 10% или около 1%-5% от уровней лизосомного фермента, связанного с болезнью лизосомного накопления, у здоровых млекопитающих, может предупреждать, ингибировать или лечить неврологические симптомы, такие как неврологическая дегенерация у млекопитающих.
В одном варианте осуществления способы, описанные здесь, включают доставку в CNS иммунокомпетентного человека, нуждающегося в лечении, композиции, содержащей эффективное количество вектора rAAV9, содержащего открытую рамку считывания, кодирующую IDUA. Пути введения в CNS/головной мозг включают, но без ограничения, интратекальное введение, интракраниальное введение, например,
интрацеребровентрикулярное введение или боковое церебровентрикулярное введение, интраназальное введение, эндоваскулярное введение и интрапаренхимальное введение.
Другие вирусные векторы могут быть использованы в способах согласно изобретению, например, вирусные векторы, такие как векторы на основе ретровируса, лентивируса, аденовируса, вируса леса Семлики или вируса простого герпеса.
Краткое описание чертежей
Фигура 1. Дизайн эксперимента для идуронидаза-дефицитных мышей, которым интрацеребровентрикулярно (ICV) или интратекально вводили AAV9-IDUA. Для предотвращения иммунного ответа животных либо подвергали иммуносупрессии циклофосфамидом (CP), индукции иммунологической толерантности при рождении путем внутривенного введения человеческого белка идуронидазы (альдуразим), или инъекции выполняли у иммунодефицитных мышей NOD-SCID, которые также были дефицитными по идуронидазе. Животных умерщвляли в указанное время после лечения, головной мозг подвергали микродессекции и экстракты анализировали на активность идуронидазы.
Фигура 2. Активность IDUA у иммунодефицитных, IDUA-дефицитных животных.
Фигура 3. Активность IDUA у иммуносупрессированных животных, которым вводили вектор AAV путем ICV-введения.
Фигура 4. Активность IDUA у иммуносупрессированных животных, которым вводили вектор AAV путем ГГ-введения.
Фигура 5. Активность IDUA у животных, подвергнутых индукции иммунологической толерантности, которым вводили вектор AAV путем ICV-введения.
Фигура 6. Компиляция всех средних уровней активности IDUA для параллельного сравнения.
Фигура 7. Данные сгруппированы в соответствии с областью головного мозга.
Фигура 8. Анализ на материал накопления GAG в различных отделах головного мозга для всех четырех тестируемых групп.
Фигура 9. Схема дизайна эксперимента.
Фигура 10. Интракраниальная инфузия AAV9-IDUA иммунодефицитным мышам с MPS I. Взрослым животным инъецировали 1011 геномов вектора и оценивали на экспрессию идуронидазы в головном мозге через 10 недель. Уровни активности фермента в головном мозге были значительно более высокими, чем в головном мозге животных дикого типа, то есть в 30-300 раз выше по сравнению с диким типом.
Фигура 11. Интракраниальное введение AAV9-IDUA иммунокомпетентным, IDUA-дефицитным мышам. Взрослым животным инъецировали 1011 геномов вектора и подвергали иммунодепрессии путем еженедельной инъекции циклофосфамида (CP). Инъекции CP прекращали через 6 недель после инъекции вектора из-за слабого здоровья, и животных умерщвляли через 8 недель после инъекции. Головной мозг подвергали микродессекции и анализировали на активность фермента IDUA.
Фигура 12. Интракраниальная инфузия AAV9-IDUA подвергнутым индукции иммунологической толерантности мышам с MPS I. У мышей с MPS I индуцировали иммунологическую толерантность с помощью однократной дозы Альдуразима при рождении или многократных доз, вводимых еженедельно, начиная с рождения. Мышам инфузировали вектор в возрасте 4-х месяцев и умерщвляли через 11 недель после введения. Головной мозг подвергали микродессекции и анализировали на экспрессию идуронидазы. Активность фермента в среднем от 10 до 1000 раз была более высокой по сравнению с уровнями у мышей дикого типа.
Фигура 13. Интратекальное введение AAV9-IDUA иммунокомпетентным, IDUA-дефицитным животным. Взрослым мышам с MPS I интратекально инъецировали AAV9-IDUA с последующим еженедельным режимом иммунодепрессии циклофосфамидом. Животных умерщвляли через 11 недель после инъекции, а затем головной мозг и спинной мозг анализировали на активность фермента IDUA.
Фигура 14. Интратекальная инфузия AAV9-IDUA мышам с MPS I, подвергнутым индукции иммунологической толерантности. У IDUA-дефицитных животных индуцировали иммунологическую толерантность при рождении с помощью однократной дозы Альдуразима или многократных доз, вводимых еженедельно, начиная с рождения. В возрасте 4 -х месяцев животным инфузировали интратекально вектор AAV9-IDUA, и через 10 недель после введения животных умерщвляли, головной мозг подвергали микродиссекции и анализировали на активность идуронидазы. Наблюдалось восстановление активности фермента во всех отделах головного мозга, при этом активности в мозжечке были от 200 до 1500 раз более высокими по сравнению с уровнями активности у животных дикого типа. Уровни активности фермента в обонятельной луковице и мозжечке (справа от пунктирной линии) относятся к правой оси Y.
Фигура 15. Интратекальная инфузия AAV9-IDUA иммунокомпетентным животным с MPS I. Контрольным животным с MPS I инъецировали вектор AAV9-IDUA, но не подвергали иммуносупрессии или индукции иммунологической толерантности. Животных умерщвляли через 11 недель после инъекции вектора, а затем их головной мозг анализировали на активность идуронидазы. Уровни фермента были восстановлены до уровней у животных дикого типа во всех отделах головного мозга, но были значительно ниже, чем у животных, которые были подвергнуты иммуносупрессии или индукции иммунологической толерантности.
Фигура 16. Нормализация уровней гликозаминогликанов (GAG) после интракраниальной или интратекальной инфузии AAV9. AAV9 - IDUA инъецировали интракраниально или интратекально иммунодефицитным, подвергнутым иммуносупрессии или индукции иммунологической толерантности мышам с MPS I, как указано. Животных умерщвляли через 8-11 недель после инъекции, затем головной мозг подвергали микродиссекции и анализировали на уровни GAG. Накопление GAG восстановилось до уровней у животных дикого типа или уровней, близких к уровням у животных дикого типа, во всех подвергнутых анализу группах.
Фигура 17. Копии вектора IDUA в головном мозге. Подвергнутый микродиссекции головной мозг анализировали на последовательности вектора IDUA с помощью QPCR. Число копий у интракраниально и интратекально инъецированных мышей коррелирует с уровнями активности фермента, показанными на фигурах 11 и 13.
Фигура 18. ICV-инфузия AAV8-MCI взрослым животным.
Фигура 19. Интраназальное введение AAV9-IDUA иммунокомпетентным, IDUA-дефицитным животным. Взрослым мышам с MPS I интраназально инфузировали AAV9-IDUA с последующим еженедельным режимом иммуносупрессии циклофосфамидом. Животных умерщвляли через 12 недель после введения и головной мозг анализировали на активность фермента IDUA.
Фигура 20. Копии вектора IDUA в головном мозге. Подвергнутый микродиссекции головной мозг анализировали на последовательности вектора IDUA с помощью QPCR. Число копий у интраназально инъецированных мышей коррелирует с уровнями фермента на фигуре 19.
Фигура 21. Протокол с иммунодепрессантом или индукцией иммунологической толерантности с использованием IN-доставки AAV9-IDUA.
Фигура 22. Восстановление активности IDUA после IN-доставки AAV9-IDUA.
Фигура 23. Активность GAG после IN-доставки AAV9-IDUA.
Фигура 24. Иммунофлуоресценция IDUA в головном мозге после IN-доставки AAV9-IDUA.
Фигура 25. Иммунофлуоресценция GFP в головном мозге после IN-доставки AAV9-GFP.
Фигуры 26A-D. А) Окрашивание толуидиновым синим. В) Сводные данные по патологии ткани у контрольных гетерозиготных и гомозиготных мышей с MPS I и мышей, подвергнутых лечению путем IN-доставки IDUA AAV9-MCI. С) Лабиринт Барнеса. D) Данные по лабиринту Барнеса.
Фигуры 27А-В. А) Схематическое изображение векторов AAV9. В) Краткое описание in vivo тестируемых групп для IT- и IV-доставки векторов AAV9.hIDS.
Фигура 28. Активность IDS в плазме мышей, которым вводили векторы AAV9.hIDS путем IT- и IV-введения.
Фигура 29. Активность IDS в CNS после ГГ-инъекции AAV9-hIDS. Для каждой группы мышей данные для следующих тканей представлены слева направо: спинной мозг, таламус/ствол головного мозга, мозжечок, кора головного мозга, гиппокамп и стриатум.
Фигуры 30A-D. А) Активность ffiS в CNS после ICV-инъекции AAV9-hIDS. Для каждой группы мышей данные для следующих тканей представлены слева направо: спинной мозг; таламус/ствол головного мозга, мозжечок, кора головного мозга, гиппокамп, стриатум, обонятельная луковица левой стороны и обонятельная луковица, стриатум, гиппокамп, кора головного мозг, мозжечок и таламус/ствол головного мозга правой стороны. В) Активность IDS в плазме после ICV-инъекции AAV9-hIDS. С) Активность IDS в периферических органах после ICV-инъекции AAV9-hIDS. Для каждой группы мышей данные для следующих органов представлены слева направо: печень, сердце, легкое, селезенка и почки. D) Содержание GAG после ICV-инъекции. Для каждой группы мышей данные для следующих тканей представлены слева направо: спинной мозг, остальные отделы, мозжечок, кора головного мозга, гиппокамп, стриатум и обонятельная луковица.
Фигуры 31А-В. А) Лабиринт Барнеса. В) Выполнение на 1-й день и 4-й день. Фигура 32. Сравнение неврологической функции у мышей дикого типа и мышей с MPS П.
Фигура 33. Дизайн эксперимента для IV-доставки генов с использованием AAV9 или AAVrhlO.
Фигуры 34А-С. Восстановление активности IDUA в плазме (А), периферических тканях (В) и CNS (С) и после IV-введения.
Фигуры 35А-С. Активность GAG в моче (А), периферических тканях (В) и CNS
(С).
Фигуры 36A-F. Иммунофлуоресценция IDUA в срезах тканей. А) Печень. В) Сердце. С) Легкое. D) Таламус.Е) Гиппокамп.F) Мозжечок.
Фигура 37. Уровни активности фермента IDUA после IN-инфузии AAV9-IDUA. Высокие уровни активности фермента IDUA наблюдался в обонятельной луковице (в 10-100 раз выше, чем у нормального животного) и нормализованные (масс.) уровни в других отделах головного мозга.
Фигура 38. Сокращение материала накопления у подвергнутых лечению мышей.
Фигуры 39A-D. IDUA (A), GFP (В); и иммунофлуоресценция обонятельной луковицы (С). (D) Совместное окрашивание с обонятельным маркерным белком (Olfactory Marker Protein) в обонятельной луковице.
Фигура 40. Улучшенная нейрокогнитивная функция у мышей, подвергнутых лечению путем IN-введения.
Фигуры 41А-В. Нейрохимические профили в мозжечке (А) и гиппокампе (В). Контрольные мыши (CTR) представлены левым столбцом, мыши с MPS I (не подвергнутые лечению) представлены столбцом в середине, и подвергнутые лечению мыши с MPS I представлены правым столбцом для каждого нейрохимического профиля.
Фигура 42. Окрашивание хороидного сплетения после IN-доставки AAVrh10-GFP.
Подробное описание изобретения
Определения
Используемый здесь термин «индивидуум» (как в субъекте лечения) означает млекопитающее. Млекопитающие включают, например, человека; не относящихся к человеку приматов, например, человекообразных обезьян и мартышек; и не относящихся к приматам млекопитающих, например, собак, кошек, крыс, мышей, крупный рогатый скот, лошадей, овец и коз. Не млекопитающие включают, например, рыб и птиц.
Термины «заболевание» и «нарушение» используются взаимозаменяемо и означают заболевания или состояния, в которых отсутствие или пониженное содержание специфического генного продукта, например, лизосомного фермента, связанного с лизосомной болезнью накопления, обуславливает заболевание таким образом, что терапевтически благоприятный эффект может быть достигнут путем восполнения, например, по меньшей мере до 1% от нормальных уровней.
Используемый здесь термин «по существу» означает полностью или почти полностью; например, композиция является «по существу свободной» от компонента, когда не содержит этот компонент или содержит такое следовое количество, присутствие которого не нарушает любое важное функциональное свойство композиции, или соединение является «по существу чистым», когда присутствуют лишь незначительные (следовые) количества примесей.
Термин «подвергание лечению» или «лечение» в том смысле, в котором он используется здесь, относится к ослаблению симптомов, связанных с нарушением или заболеванием, «ингибирование» означает замедление дальнейшего прогрессирования или ухудшения симптомов, связанных с нарушением или заболеванием, и «предупреждение» относится к предупреждению возникновения симптомов, связанных с нарушением или заболеванием.
Используемое здесь «эффективное количество» или «терапевтически эффективное количество» агента согласно изобретению, например, рекомбинантного AAV, кодирующего генный продукт, означает количество агента, которое полностью или частично ослабляет симптомы, связанные с нарушением или состоянием; или останавливает или замедляет дальнейшее прогрессирование или ухудшение этих симптомов; или предупреждает или обеспечивает профилактику нарушения или состояния; например, количество, которое является эффективным для предупреждения, ингибирования или лечения одного или нескольких неврологических симптомов у индивидуума.
В частности, «терапевтически эффективное количество» относится к такому количеству, которое в назначенной дозировке и в течение определенного промежутка времени позволяет достичь желательного терапевтического результата. Под терапевтически эффективным количеством подразумевается также такое количество, при котором любые токсические или вредные эффекты соединений согласно изобретению перевешиваются их полезными терапевтическими эффектами.
Используемый здесь термин «вектор» относится к макромолекуле или ассоциации макромолекул, которая содержит или ассоциирует с полинуклеотидом, и может быть использована с целью опосредования доставки полинуклеотида в клетку, in vitro или in vivo. Иллюстративные векторы включают, например, плазмиды, вирусные векторы, липосомы и другие носители для доставки генов. Полинуклеотид, подлежащий доставке, иногда называемый как «целевой полинуклеотид» или «трансген», может содержать кодирующую последовательность, представляющую интерес для генной терапии (например, кодирующий белок ген, представляющий терапевтический интерес), и/или селектируемый или детектируемый маркер.
Аббревиатура «AAV» обозначает аденоассоциированный вирус и может быть использована для обозначения самого вируса или его производных. Термин охватывает все подтипы, серотипы и псевдотипы, и как природные, так и рекомбинантные формы, за исключением случаев, когда указано иное. Используемый здесь термин «серотип» относится к AAV, который идентифицирован и отличается от других AAV исходя из его свойств связывания, например, существует одиннадцать серотипов AAV1-AAV11, включая AAV2, AAV5, AAV8, AAV9 и AAVrhlO, и термин охватывает псевдотипы с такими же свойствами связывания. Так, например, серотипы AAV9 включают AAV со свойствами связывания AAV9, например, псевдотипированный AAV, содержащий капсид AAV9 и геном rAAV, который не происходит или не получен из AAV9, или геном которого является химерным. Аббревиатура «rAAV» относится к рекомбинантному аденоассоциированному вирусу, также называемому рекомбинантным вектором AAV (или «вектор rAAV»).
«Вирус AAV» относится к частице вируса, состоящей из по меньшей мере одного капсидного белка AAV и инкапсидированного полинуклеотида. Если частица содержит гетерологичный полинуклеотид (т.е. полинуклеотид, отличный от полинуклеотида генома AAV дикого типа, такой как трансген, подлежащий доставке в клетку млекопитающего), то указанная частица обычно называется как «rAAV». «Капсидный белок» AAV включает капсидный белок AAV дикого типа, а также модифицированные формы капсидного белка AAV, которые структурно и/или функционально способны упаковывать геном rAAV и связываются по меньшей мере с одним специфическим клеточным рецептором, который может отличаться от рецептора, используемого AAV дикого типа. Модифицированный капсидный белок AAV включает химерный капсидный белок AAV, например, содержащий аминокислотные последовательности от двух или более серотипов AAV; например, капсидный белок, образованный из части капсидного белка от AAV9, слитой или связанной с частью капсидного белка от AAV-2; и капсидный белок AAV, имеющий метку или другой детектируемый не-AAV капсидный пептид или белок, слитый или связанный с капсидным белком AAV, например, часть молекулы антитела, которая связывается с рецептором, отличными от рецептора для AAV9, таким как рецептор трансферрина, может быть рекомбинантно слита с капсидным белком AAV9.
«Псевдотипированный» rAAV представляет собой инфекционный вирус, содержащий любую комбинацию капсидного белка AAV и генома AAV. Капсидные белки из любого серотипа AAV могут быть использованы с геномом rAAV, который происходит или может быть получен из генома AAV дикого типа другого серотипа, или который является химерным геномом, то есть образованным из ДНК AAV от двух или более различных серотипов, например, химерным геномом, содержащим 2 инвертированных концевых повтора (ITR), при этом каждый ITR происходит от различных серотипов или химерных ITR. Использование химерных геномов, таких как химерные геномы, содержащие ITR от двух серотипов AAV или химерные ITR, может привести к направленной рекомбинации, которая может дополнительно повысить продукцию транскрипционно активных межмолекулярных конкатамеров. Таким образом, 5' и 3' инвертированные концевые повторы (ITR) в пределах вектора rAAV согласно изобретению могут быть гомологичными, то есть от одного и того же серотипа, гетерологичными, т.е. от различных серотипов, или химерными, то есть инвертированный концевой повтор (ITR), который имеет последовательности ITR от более чем одного серотипа AAV.
Векторы rAAV
Аденоассоциированные вирусы любого серотипа являются подходящими для получения rAAV, так как различные серотипы являются функционально и структурно родственными, даже на генетическом уровне. Все серотипы AAV, по-видимому, проявляют сходные свойства репликации, опосредованной гомологичными генами rep; и все, как правило, несут три родственных капсидных белка, таких, которые экспрессируются в AAV2. Степень родства дополнительно подтверждается гетеродуплексным анализом, который показал широкую перекрестную гибридизацию между серотипами по длине генома; и присутствием схожих самоотжигаемых сегментов на концах, которые соответствуют инвертированным концевым повторам (ITR). Аналогичные паттерны инфекционности также позволяют предположить, что функции репликации в каждом серотипе находятся под аналогичным регуляторным контролем. Среди различных серотипов AAV чаще всего используют AAV2.
Вектор AAV согласно изобретению обычно содержит полинуклеотид, который является гетерологичным AAV. Полинуклеотид, как правило, представляет интерес из-за способности обеспечить клетку-мишень функцией в контексте генной терапии, такой как ап- или даунрегулирование экспрессии определенного фенотипа. Такой гетерологичный полинуклеотид или «трансген», как правило, имеет достаточную длину для обеспечения желательной функции или кодирующей последовательности.
Когда транскрипция гетерологичного полинуклеотида является желательной в предполагаемой клетке-мишени, он может быть функционально связан со своим собственным или гетерологичным промотором, в зависимости, например, от желаемого уровня и/или специфичности транскрипции в клетках-мишенях, как известно в данной области. Различные типы промоторов и энхансеров являются подходящими для использования в этом контексте. Конститутивные промоторы обеспечивают постоянный уровень транскрипции гена, и может быть предпочтительным, когда это желательно, чтобы экспрессия терапевтического или профилактического полинуклеотида происходила на постоянной основе. Как правило, индуцируемые промоторы обладают низкой активностью в отсутствие индуктора, и апрегулированы в присутствии индуктора. Они могут быть предпочтительными, когда экспрессия желательна только в определенные моменты времени или в определенных локализациях, или когда желательно регулировать уровень экспрессии с использованием индуктора. Промоторы и энхансеры могут быть также тканеспецифическими: то есть они проявляют свою активность только в определенных типах клеток, что предположительно обусловлено регуляторными элементами гена, обнаруживаемыми исключительно в этих клетках.
Примерами промоторов являются поздний промотор SV40 вируса обезьян 40, полиэдронный энхансерный/промоторный элемент бакуловируса, тимидинкиназа вируса простого герпеса (HSV tk), сверхранний промотор цитомегаловируса (CMV) и различные ретровирусные промоторы, включая элементы LTR. Индуцируемые промоторы включают промоторы, индуцируемые ионами тяжелых металлов (такие как промотор мышиного вируса рака молочной железы (mMTV) или различные промоторы гормона роста), а также промоторы фага Т7, активные в присутствии РНК-полимеразы фага Т7. В качестве иллюстрации, примеры тканеспецифических промоторов включают различные промоторы сурфактина (для экспрессии в легком), промоторы миозина (для экспрессии в мышцах), а также промоторы альбумина (для экспрессии в печени). Множество других различных промоторов известно и, как правило, доступно в данной области, и последовательности многих таких промоторов содержатся в базах данных последовательностей, таких как база данных GenBank.
В случае, когда трансляция также желательна в предполагаемой клетке-мишени, гетерологичный полинуклеотид будет также предпочтительно содержать контрольные элементы, которые содействуют трансляции (такие как сайт связывания с рибосомой или «RBS» и сигнал полиаденилирования). Соответственно, гетерологичный полинуклеотид обычно содержит по меньшей мере один кодирующий участок, функционально связанный с подходящим промотором, и может также содержать, например, функционально связанный энхансер, сайт связывания с рибосомой и сигнал полиаденилирования. Гетерологичный полинуклеотид может содержать один кодирующий участок или несколько кодирующих участков, находящихся под контролем одного и того же или различных промоторов. Вся структурная единица, содержащая комбинацию контрольных элементов и кодирующего участка, часто называется как экспрессионная кассета.
Гетерологичный полинуклеотид интегрируют с помощью рекомбинантных технологий внутрь или вместо кодирующего участка генома AAV (т.е., вместо генов rep и cap AAV), но, как правило, является фланкированным с каждой стороны инвертированными концевыми повторами (ITR) AAV. Это означает, что инвертированные концевые повторы (ITR) расположены на 5' и 3' концах кодирующих последовательностей, либо в непосредственной близости, например (хотя и не обязательно) без какой-либо встраиваемой последовательности AAV-происхождения для уменьшения вероятности рекомбинации, которая может регенерировать компетентный по репликации геном AAV. Тем не менее, может быть достаточно одного инвертированного концевого повтора (ITR) для осуществления функций, которые обычно связаны с конфигурациями, содержащими два ITR (см., например, WO 94/13788), и, таким образом, векторные конструкции, содержащие только один ITR, могут быть использованы в сочетании со способами упаковки и получения согласно настоящему изобретению.
Нативные промоторы для rep являются саморегулируемыми и могут ограничивать количество продуцируемых частиц AAV. Ген rep может быть также функционально связан с гетерологичным промотором, независимо от того, является ли rep частью векторной конструкции, или расположен отдельно. Любой гетерологичный промотор, который сильно не даунрегулирован экспрессией гена rep, является подходящим; но индуцируемые промоторы могут быть предпочтительными, поскольку непрерывная экспрессия гена rep может оказаться гибельной для клетки-хозяина. Большое разнообразие индуцируемых промоторов известно в данной области; включая, в качестве иллюстрации, промоторы, индуцируемые ионами тяжелых металлов (например, промоторы металлотионеина); промоторы, индуцируемые стероидным гормоном (такие как промотор MMTV или промоторы гормона роста); и промоторы, такие как промоторы фага Т7, которые являются активными в присутствии РНК-полимеразы Т7. Один подкласс индуцируемых промоторов включает промоторы, которые индуцируются вирусом-помощником, который используют для дополнения репликации и упаковки вектора rAAV. Описан целый ряд индуцируемых вирусом-помощником промоторов, включая промотор ранних генов аденовируса, индуцируемый белком Е1А аденовируса; главный поздний промотор аденовируса; промотор вируса герпеса, индуцируемый белками вируса герпеса, такими как VP 16 или 1СР4; а также промоторы, индуцируемые осповакциной или поксвирусом.
Описаны способы идентификации и тестирования промоторов, индуцируемых вирусом-помощником (см., например, WO 96/17947). Таким образом, в данной области известны способы определения, являются или нет промоторы-кандидаты индуцируемыми вирусом-помощником, и будут ли они полезными для генерации клеток с высокой эффективностью упаковки. Вкратце, один такой способ включает замену промотора р5 гена rep AAV на предполагаемый промотор, индуцируемый вирусом-помощником (известным в данной области или идентифицированным с использованием хорошо известных методов, таких как соединение с «репортерными» генами, не несущими промотор). Гены rep-cap AAV (с заменой р5), например, соединенные с положительным селектируемым маркером, таким как ген устойчивости к антибиотикам, затем стабильно интегрировали в подходящую клетку-хозяина (такую как клетки HeLa или А549, проиллюстрированные ниже). Клетки, обладающие способностью расти относительно хорошо в условиях отбора (например, в присутствии антибиотика), затем тестировали на их способность экспрессировать гены rep и cap после добавления вируса-помощника. В качестве начального тестирования на экспрессию rep и/или cap, клетки могут быть легко подвергнуты скринингу с использованием иммунофлуоресценции для детекции белков Rep и/или Сар. Подтверждение способностей к упаковке и эффективности может быть затем получено с помощью функциональных тестов на репликацию и упаковку экзогенных векторов rAAV. С использованием этой методологии индуцируемый вирусом-помощником промотор гена мышиного металлотионеина был идентифицирован в качестве подходящей замены для промотора р5, и использован для продукции частиц rAAV в высоких титрах (как описано в WO 96/17947).
Удаление одного или нескольких генов AAV в любом случае желательно для уменьшения вероятности образования компетентного по репликации AAV («RCA»). Соответственно, кодирующие или промоторные последовательности для rep, cap или обоих могут быть удалены, поскольку функции, обеспечиваемые этими генами, могут быть обеспечены in trans, например, в стабильной линии или посредством котрансфекции.
Полученный вектор называется как «дефектный» по этим функциям. Для репликации и упаковки вектора отсутствующие функции дополняются пакующим геном или множеством пакующих генов, которые вместе кодируют необходимые функции для различных отсутствующих продуктов гена rep и/или cap.Пакующие гены или генные кассеты в одном варианте осуществления не фланкированы инвертированными концевыми повторами (ITR) AAV и в одном варианте осуществления не проявляют какой-либо существенной гомологии с геномом rAAV. Таким образом, для того, чтобы свести к минимуму гомологичную рекомбинацию, происходящую в ходе процесса репликации, между последовательностью вектора и отдельно обеспеченными пакующими генами, желательно избежать перекрытия двух полинуклеотидных последовательностей. Уровень гомологии и соответствующая частота рекомбинации увеличиваются с увеличением длины гомологичных последовательностей и их уровня общей идентичности. Уровень гомологии, который будет представлять проблему в данной системе, может быть определен теоретически и подтвержден экспериментально, как известно в данной области. Однако обычно рекомбинации может быть значительно уменьшена или устранена, если перекрывающаяся последовательность представляет собой последовательность из менее чем около 25 нуклеотидов, если она по меньшей мере на 80% идентична по всей свой длине, или последовательность из менее чем около 50 нуклеотидов, если она по меньшей мере на 70% идентична по всей своей длине. Конечно, даже более низкие уровни гомологии являются предпочтительными, так как они будут дополнительно уменьшать вероятность рекомбинации. Оказалось, что, даже при отсутствии какой-либо гомологии перекрывающихся последовательностей, имеется некоторая остаточная частота генерации RCA. Даже дополнительные сокращения частоты генерации RCA (например, путем негомологичной рекомбинации) могут быть получены путем «расщепления» функций репликации и инкапсидирования AAV, как описано в WO 98/27204 автора Allen et al.
Векторная конструкция на основе rAAV и комплементарные упаковывающие генетические конструкции могут быть реализованы в данном изобретении в ряде различных форм. Вирусные частицы, плазмиды и стабильно трансформированные клетки-хозяева могут быть использованы для введения таких конструкций в упаковывающую клетку, временно или стабильно.
В некоторых вариантах осуществления настоящего изобретения вектор AAV и комплементарный упаковывающий ген(ы), если таковые имеются, представлены в форме бактериальных плазмид, частиц AAV или любой их комбинации. В других вариантах осуществления последовательность вектора AAV, упаковывающий ген(ы), или и то и другое, представлены в форме генетически измененных (предпочтительно наследственно измененных) эукариотических клеток. Разработка клеток-хозяев, наследственно измененных для экспрессии последовательности вектора AAV, упаковывающих AAV генов, или того и другого, обеспечивает получение источника материала, который экспрессируется на надежном уровне.
Таким образом, множество различных генетически измененных клеток может быть использовано в контексте настоящего изобретения. В качестве иллюстрации, клетка-хозяин млекопитающего может быть использована по меньшей мере с одной интактной копией стабильно интегрированного вектора rAAV. Упаковывающая AAV плазмида, содержащая, по меньшей мере ген rep AAV, функционально связанный с промотором, может быть использована для обеспечения функций репликации (как описано в патенте США 5658776). Альтернативно, стабильная линия клеток млекопитающих с геном rep AAV, функционально связанным с промотором, может быть использована для обеспечения функций репликации (см., например, Trempe et al., WO 95/13392); Burstein et al. (WO 98/23018); и Johnson et al. (патент США 5656785). Ген cap AAV, обеспечивающий капсидирование белков, как описаны выше, может быть обеспечен вместе с геном rep AAV или отдельно (см., например, вышеуказанные заявки и патенты, а также Allen et al. (WO 98/27204). Другие комбинации возможны и включены в объем настоящего изобретения.
Пути доставки
Несмотря на огромную мозговую сосудистую сеть, системная доставка терапевтических средств к центральной нервной системе (CNS) является неэффективной для более 98% малых молекул и почти 100% крупных молекул (Partridge, 2005). Отсутствие эффективности обусловлено присутствием гематоэнцефалического барьера (ВВВ), который препятствует поступлению в головной мозг большинства чужеродных веществ, даже многих полезных терапевтических средств из циркулирующей крови. Несмотря на то, что некоторые низкомолекулярные, пептидные и белковые терапевтические средства, вводимые системно, достигают паренхимы головного мозга путем пересечения ВВВ (Banks, 2008), как правило, высокие системные дозы требуются для достижения терапевтических уровней, что может привести к возникновению неблагоприятных эффектов в организме. Терапевтические средства могут быть введены непосредственно в CNS путем интрацеребровентрикулярных или интрапаренхимальных инъекций. Интраназальная доставка обходит ВВВ и нацеливает терапевтические средства непосредственно на CNS, используя пути вдоль обонятельного и тройничного нервов, иннервирующих носовые проходы (Frey II, 2002; Thorne et al., 2004; Dhanda et al., 2005).
Любой путь введения rAAV может быть использован при условии, что этот путь и введенное количество являются профилактически или терапевтически полезными. В одном примере пути введения в CNS включают интратекальное и интракраниальное введение. Интракраниальное введение может представлять собой введение в мозжечково-мозговую цистерну или желудочек. Термин «мозжечково-мозговая цистерна» включает доступ к пространству вокруг и под мозжечком через отверстие между черепом и верхней частью позвоночника. Термин «желудочек мозга» включает полости в головном мозге, которые являются продолжением центрального канала спинного мозга. Интракраниальное введение осуществляют путем инъекции или инфузии, и подходящие диапазоны доз для интракраниального введения, как правило, составляют около 103-1015
инфекционных единиц вирусного вектора на микролитр, доставляемых в 1-3000 микролитрах объема единичной инъекции. Например, геномы вируса или инфекционные единицы вектора на микролитр, как правило, будут содержать около 104, 105, ΙΟ6, ΙΟ7, 108, 109, 1010, 1011, ΙΟ12, 1013, 10 м, 1015, 1016, или 1017 геномов вируса или инфекционных единиц вирусного вектора, доставляемых в около 10, 50, 100, 200, 500, 1000 или 2000 микролитрах. Следует понимать, что вышеуказанная дозировка является лишь иллюстративной и специалистам в данной области будет понятно, что эта дозировка может варьировать. Эффективные дозы могут быть экстраполированы из кривых доза-ответ, полученных в тестовых системах in vitro или in vivo.
AAV, доставляемый в интратекальных способах лечения согласно настоящему изобретению, можно вводить любым общепринятым путем, обычно используемым для интратекального введения. Например, интратекальное введение может быть осуществлено посредством медленной инфузии препарата в течение приблизительно одного часа. Интратекальное введение осуществляют посредством инъекции или инфузии, и подходящие диапазоны доз для интратекального введения, как правило, составляют около 103-1015 инфекционных единиц вирусного вектора на микролитр, доставляемых, например, в 1, 2, 5, 10, 25, 50, 75 или 100 миллилитрах или более, например, 1-10000 миллилитрах или 0,5-15 миллилитрах объема единичной инъекции. Например, геномы вируса или инфекционные единицы вектора на микролитр, как правило, будут содержать около 104, 105, 106, 107, 108, 109, 1010, 1011, 1012, 1013 или 1014 геномов вируса или инфекционных единиц вирусного вектора.
AAV, доставляемый в интраназальных способах лечения согласно настоящему изобретению, может быть введен в дозах, находящихся в подходящих диапазонах, как правило, около 103-1015 инфекционных единиц вирусного вектора на микролитр, доставляемых, например, в 1, 2, 5, 10, 25, 50, 75 или 100 миллилитрах или более, например, 1-10000 миллилитрах или 0,5-15 миллилитрах. Например, геномы вируса или инфекционные единицы вектора на микролитр, как правило, будут содержать около 104, 105, 106, 107, 108, 109, 1010, 1011, 1012, 1013, 10 м, 1015, 1016 или 1017 геномов вируса или инфекционных единиц вирусного вектора, например, по меньшей мере 1,2×101 Геномов или инфекционных единиц, например, по меньшей мере от 2×1011 до около 2×1012 геномов или инфекционных единиц, или от около 1×1013 до около 5×1016 геномов или инфекционных единиц. В одном варианте осуществления AAV, используемый для интраназальной доставки, представляет собой такой AAV, который связывается с гликанами, содержащими концевые остатки галактозы, и в одном варианте осуществления доза в 2-8 раз превышает дозу, составляющую от 9×1010 до менее чем 1 χ 1011 геномов AAV8 или инфекционных единиц вирусного вектора.
Терапия, в случае экспрессии лизосомного фермента, связанного с лизосомной болезнью накопления, такого как IDUA, приводит к нормализации лизосомных гранул накопления в нейрональной и/или менингеальной ткани субъектов, как обсуждалась выше. Предполагается, что осаждение гранул накопления из нейрональной и глиальной ткани уменьшается, тем самым ослабляя задержку и регрессию развития, наблюдаемую у индивидуумов, страдающих лизосомной болезнью накопления. Другие эффекты терапии могут включать нормализацию лизосомных гранул накопления в оболочках головного мозга вблизи арахноидальной грануляции, присутствие которой в лизосомной болезни накопления приводит к гидроцефалии с высоким давлением. Способы согласно изобретению можно также применять для лечения компрессии спинного мозга позвоночника, возникшей вследствие присутствия лизосомных гранул накопления в шейных сегментах спинного мозга С1-С5 или где-либо в других местах в спинном мозге. Способы согласно изобретению также направлены на лечение кист, которые вызваны периваскулярным накоплением лизосомных гранул накопления вокруг сосудов головного мозга. В других вариантах осуществления терапия может также успешно приводить к нормализации объема печени и экскреции гликозаминогликанов с мочой, уменьшению размера селезенки и событий апноэ/гипопноэ, увеличению длины тела и скорости роста у субъектов в препубертатный период, увеличению сгибания рук в плечевом суставе и увеличению разгибания в локтевом и коленном суставах, и уменьшению трикуспидальной регургитации или легочной регургитации.
Интратекальное введение согласно настоящему изобретению может включать введение композиции в поясничную область. Любое такое введение может быть осуществлено посредством болюсной инъекции. В зависимости от тяжести симптомов и отвечаемости субъекта на терапию, болюсную инъекцию можно вводить раз в неделю, раз в месяц, раз в 6 месяцев или ежегодно. В других вариантах осуществления интратекальное введение достигается за счет использования инфузионного насоса. Специалистам в данной области техники известны устройства, которые могут быть использованы для осуществления интратекального введения композиции. Композицию можно вводить интратекально, например, путем единичной инъекции или непрерывной инфузии. Следует понимать, что режим дозирования может представлять собой схему на основе однократной дозы или многократных доз.
Используемый здесь термин «интратекальное введение» включает доставку фармацевтической композиции непосредственно в спинномозговую жидкость субъекта способами, включающими латеральную церебровентрикулярную инъекцию через трепанационное отверстие или посредством цистернальной или люмбальной пункции, или т.п.Термин «поясничная область» предусматривает включение области между третьим и четвертым поясничными позвонками (позвонками нижнего отдела спины) и, в частности, L2-S1 область позвоночника.
Введение композиции в соответствии с настоящим изобретением в любой из вышеуказанных участков может быть достигнуто путем прямой инъекции композиции или путем использования инфузионных насосов. Для инъекции композиция может быть составлена в виде жидких растворов, например, в физиологически совместимых буферах, таких как раствор Хэнкса, раствор Рингера или фосфатный буфер. Кроме того, фермент может быть составлен в твердой форме и повторно растворен или суспендирован непосредственно перед применением. Лиофилизированные формы также включены. Инъекция может быть осуществлена, например, в форме болюсной инъекции или непрерывной инфузии (например, с использованием инфузионных насосов) фермента.
В одном варианте осуществления изобретения rAAV вводят путем латеральной церебровентрикулярной инъекции в головной мозг субъекта. Инъекция может быть проведена, например, через трепанационное отверстие, сделанное в черепе субъекта. В другом варианте осуществления фермент и/или другой фармацевтический препарат вводят через хирургический введенный шунт в желудочке головного мозга субъекта. Например, инъекция может быть сделана в латеральные желудочки, которые больше по размеру, хотя может быть выполнена также инъекция путем введения в третий и четвертый, меньшие по размеру, желудочки. В еще одном варианте осуществления композиции, используемые в настоящем изобретении, вводят путем инъекции в мозжечково-мозговую цистерну или поясничную область субъекта.
Несмотря на отсутствие полного понимания конкретных механизмов, лежащих в основе интраназальной доставки лекарственного средства в CNS, накопленный объем данных показывает, что пути, в которые вовлечены нервы, соединяющие носовые проходы с головным мозгом и спинным мозгом, являются важными. Кроме того, пути, в которые вовлечена васкулатура, спинномозговая жидкость и лимфатическая система, задействованы в транспорте молекул из носовой полости в CNS. Вероятно, что сочетание этих путей, хотя в зависимости от свойств терапевтического средства один путь может преобладать, обуславливает характеристики используемого препарата и устройства для доставки.
Терапевтические средства могут быстро получать доступ к CNS после интраназального введения вдоль путей обонятельных нервов, ведущих из носовой полости непосредственно в CNS. Пути обонятельных нервов являются основным компонентом интраназальной доставки, что подтверждается тем, что флуоресцентные метки связываются с обонятельными нервами, так только они пересекают решетчатую пластину (Jansson и др., 2002), при этом концентрации лекарственного средства в обонятельных луковицах обычно являются одними из самых высоких концентраций, наблюдаемых в CNS (Thorne et al., 2004; Banks et al., 2004; Graff et al., 2005a); Nonaka et al., 2008; Ross et al., 2004; Ross et al., 2008; Thorne et al., 2008), и существует сильная положительная корреляция между концентрациями в обонятельном эпителии и обонятельных луковицах (Dhuria et al., 2009а).
Обонятельные пути начинаются в верхней части носовых проходов, в обонятельной области, где обонятельные рецепторные нейроны (ORN) располагаются между вспомогательными клетками (поддерживающими клетками), микровиллярными клетками и базальными клетками. Обонятельные рецепторные нейроны (ORN) опосредуют обоняние путем передачи чувствительной информации с периферии в CNS (Clerico et al., 2003). Под эпителием собственная пластинка содержит выделяющие секрет железы Боумена, аксоны, кровеносные сосуды, лимфатические сосуды и соединительную ткань. Дендриты ORN погружены в слой слизи, покрывающий обонятельный эпителий, при этом аксоны этих биполярных нейронов проходят в полость черепа через собственную пластинку и через отверстия в продырявленной пластине решетчатой кости, которая отделяет носовую полость от полости черепа. Аксоны ORN проходят через субарахноидальное пространство, содержащее CSF, и оканчиваются на митральных клетках в обонятельных луковицах. Оттуда нейронные проекции распространяются на множество областей головного мозга, включая обонятельный тракт, переднее обонятельное ядро, грушевидную кору, миндалину и гипоталамус (Buck, 2000). В дополнении к ORN, хемосенсорные нейроны, расположенные на переднем конце носовой полости в ганглионе Грюнеберга, ведут в обонятельные луковицы (Fuss et al., 2005; Koos et al., 2005).
Уникальные характеристики обонятельных рецепторных нейронов (ORN) способствуют динамическому клеточному окружению, важному для интраназальной доставки в CNS. Из-за непосредственного контакта с токсинами во внешней среде, обонятельные рецепторные нейроны (ORN) регенерируют каждые 3-4 недели из базальных клеток, находящихся в обонятельном эпителии (Mackay-Sim, 2003). Специальные клетки, подобные шванновской клетке, называемые обкладочными нейроэпителиальными клетками (ОЕС), обволакивают аксоны обонятельных рецепторных нейронов (ORN) и играют важную роль в регенерации, возобновлении роста и ремиелинизации аксонов (Field et al., 2003; Li et al., 2005a; Li et al., 2005b). Обкладочные нейроэпителиальные клетки ОЕС создают непрерывные, заполненные жидкостью периневральные каналы, которые, что примечательно, остаются открытыми, несмотря на дегенерацию и регенерацию обонятельных рецепторных нейронов (ORN) (Williams et al., 2004).
Благодаря уникальному окружению обонятельного эпителия, интраназально вводимые терапевтические средства могут достигать CNS посредством внеклеточных или внутриклеточных механизмов транспорта по обонятельным нервам. Механизмы внеклеточного транспорта включают быстрое движение молекул между клетками в назальном эпителии, требующее лишь от нескольких минут до 30 минут для достижения лекарственным средством обонятельных луковиц и других областей CNS после интраназального введения (Frey II, 2002; Balin et al., 1986). Транспорт, по всей вероятности, включает механизмы объемного потока (Thorne et al., 2004; Thorne et al., 2001) в каналах, образованных обкладочными нейроэпителиальными клетками (ОЕС). Лекарственные средства могут также перемещаться по этим каналам за счет структурных изменений, которые происходят во время деполяризации и аксонального распространения потенциала действия в соседних аксонах (Luzzati et al., 2004). Механизмы внутриклеточного транспорта включают поглощение молекул обонятельными рецепторными нейронами (ORN) путем пассивной диффузии, рецептор-опосредованного эндоцитоза или адсорбционного эндоцитоза с последующим более медленным аксональным транспортом, при этом требуется от нескольких часов до нескольких дней для появления лекарственного средства в обонятельных луковицах и других областях головного мозга (Baker et al., 1986; Broadwell et al., 1985; Kristensson et al., 1971). Внутриклеточный транспорт в ORN был продемонстрирован для малых липофильных молекул, таких как частицы золота (de Lorenzo, 1970; Gopinath et al., 1978), соли алюминия (Perl et al., 1987), а также для веществ с рецепторами на ORN, таких как WGA-HRP (Thorne et al., 1995; Baker et al., 1986; Itaya et al., 1986; Shipley, 1985). Несмотря на то, что внутриклеточные механизмы являются важными для определенных терапевтических средств, маловероятно, что они будут являться преобладающим механизмом транспорта в CNS. В то время как некоторые крупные молекулы, такие как галанин-подобный пептид (GALP), проявляют пути насыщаемого транспорта в CNS (Nonaka et al., 2008), для других крупных молекул, таких как фактор роста нервов (NGF) и инсулиноподобный фактор роста-I (IGF-I), интраназальная доставка в головной мозг является ненасыщаемой и не опосредованной рецептором (Thorne et al., 2004; Chen et al., 1998; Zhao et al., 2004).
Часто игнорируемый, но важный путь, соединяющий носовые проходы с CNS, включает тройничный нерв, который обеспечивает иннервацию дыхательного и обонятельного эпителия носовых проходов и входит в CNS на уровне варолиева моста (Clerico et al., 2003; Graff et al., 2003). Примечательно, что небольшая часть тройничного нерва также оканчивается в обонятельных луковицах (Schaefer et al., 2002). Клеточный состав дыхательной области носовых проходов отличается от клеточного состава обонятельной области тем, что клетки ресничного эпителия распределены среди продуцирующих слизь бокаловидных клеток. Эти клетки способствуют механизмам мукоцилиарного клиренса, обеспечивающим удаление слизи, а также чужеродных субстанций из носовой полости в носоглотку. Тройничный нерв передает сенсорную информацию из носовой полости, ротовой полости, век и роговицы в CNS через глазную ветвь (VI), верхнечелюстную ветвь (V2) или нижнечелюстную ветвь (V3) тройничного нерва (Clerico et al., 2003; Gray, 1978). Ветви глазного нерва, являющегося ветвью тройничного нерва, обеспечивают иннервацию слизистой оболочки дорсального носового хода и передней части носа, при этом верхнечелюстная ветвь обеспечивают иннервацию боковых стенок слизистой оболочки носа. Нижнечелюстная ветвь тройничного нерва проходит к нижней челюсти и зубам без прямых входов нейронов в носовую полость. Три ветви тройничного нерва сходятся в ганглии тройничного нерва, который входит в головной мозг в области варолиева моста, заканчиваясь в спинномозговом ядре тройничного нерва в стволе головного мозга. Уникальная особенность тройничного нерва состоит в том, что он входит в головной мозг из респираторного эпителия носовых проходов в двух местах: (1) через переднее рваное отверстие вблизи варолиева моста и (2) через решетчатую пластинку вблизи обонятельных луковиц, создавая точки входа в хвостовые и ростральные области головного мозга после интраназального введения. Кроме того, вероятно, что другие нервы, которые иннервируют лицо и голову, такие как лицевой нерв, или другие сенсорные структуры в полости носа, такие как ганглии Грюнеберга, могут обеспечить точки входа для интраназально применяемых терапевтических средств в CNS.
Обычно интраназальный путь введения используют для доставки лекарственных средств в большой круг кровообращения посредством абсорбции в капиллярных кровеносных сосудах, выстилающих слизистую оболочку полости носа. Слизистая оболочка полости носа является высоковаскулярной, получающей кровоснабжение из ветвей верхнечелюстной, глазной и лицевой артерий, которые выходят из сонной артерии (Clerico et al., 2003; Cauna, 1982). Обонятельная слизистая оболочка получает кровь из малых ветвей глазной артерии, тогда как дыхательная слизистая оболочка получает кровь из артериальной ветви крупного калибра верхнечелюстной артерии (DeSesso, 1993). Относительная плотность кровеносных сосудов является более высокой в респираторной слизистой оболочке по сравнению с обонятельной слизистой оболочкой, что делает первую область идеальным участком абсорбции в крови (DeSesso, 1993). Васкулатура в дыхательной области содержит смесь непрерывного и фенестрированного эндотелия (Grevers et al., 1987; Van Diest et al., 1979), что позволяет как малым, так и крупным молекулам входить в большой круг кровообращения после назального введения.
Доставка в CNS после абсорбции в большом круге кровообращения и последующего транспорта через ВВВ являются возможной, особенно для низкомолекулярных липофильных лекарственных средств, которые легче входят в кровоток и пересекают ВВВ по сравнению с высокомолекулярными гидрофильными терапевтическими средствами, такими как пептиды и белки.
Появляется все больше данных, позволяющих предположить, что механизмы, в которые вовлечены каналы, связанные с кровеносными сосудами, или периваскулярные каналы, задействованы в интраназальной доставке лекарственных средств в CNS. Периваскулярные пространства расположены между наружным слоем кровеносных сосудов и базальной мембраной окружающей ткани (Pollock et al., 1997). Эти периваскулярные пространства действуют в качестве лимфатической системы для головного мозга, при этом субстанции, происходящие из нейронов, выводятся из интерстициальной жидкости головного мозга путем вхождения в периваскулярные каналы, связанные с кровеносными сосудами головного мозга. Периваскулярный транспорт обусловлен механизмами объемного потока, в отличие от взятой в отдельности диффузии (Cserr et al., 1981; Groothuis et al., 2007), и пульсации артерий также являются движущей силой для периваскулярного транспорта (Rennels et al., 1985; Rennels et al., 1985). Интраназально применяемые лекарственные средства могут перемещаться в периваскулярные пространства в носовых проходах или после достижения головного мозга, и широкое распределение, наблюдаемое в CNS, может быть обусловлено механизмами периваскулярного транспорта (Thorne et al., 2004).
Пути, соединяющие субарахноидальное пространство, содержащее CSF, периневральные пространства, охватывающие обонятельные нервы, и лимфатические сосуды носовой полости, являются важными для дренажа CSF, и эти же самые пути обеспечивают доступ интраназально применяемых терапевтических средств к CSF и другим областям CNS. Несколько исследований подтверждают, что метки, введенные в CSF в желудочках мозга или субарахноидальном пространстве, дренируют в нижнюю часть обонятельных луковиц в каналы, связанные с обонятельными нервами, пересекающими решетчатую пластинку, и достигают лимфатической системы носовой полости и шейных лимфатических узлов (Bradbury et al., 1983; Hatterer et al., 2006; Johnston et al., 2004a); Kida et al., 1993; Walter et al., 2006a; Walter et al., 2006b). Лекарственные средства могут получить доступ к CNS посредством этих же путей после интраназального введения, двигаясь от носовых проходов к CSF в интерстициальные пространства головного мозга и периваскулярные пространства для распределения по всему головному мозгу. Эти дренажные пути являются важными у животных некоторых видов (овцы, кролики и крысы), составляя приблизительно 50% клиренса CSF (Bradbury et al., 1981; Boulton et al., 1999; Boulton et al., 1996; Cserr et al., 1992). Пути между носовыми проходами и CSF все еще являются важными и функциональными у человека, о чем свидетельствует тот факт, что терапевтические средства непосредственно доставляются в CSF после интраназальной доставки без поступления в значительной степени в кровь (Born et al., 2002). Ряд исследований интраназальной доставки показывает, что лекарственные средства получают прямой доступ к CSF из носовой полости с последующим распределением в головном и спинном мозге. Многие интраназально введенные молекулы быстро входят в CSF, и этот транспорт зависит от липофильности, молекулярной массы и степени ионизации молекул (Dhanda et al., 2005; Born et al., 2002; Kumar et al., 1974; Sakane et al., 1995; Sakane et al., 1994; Wang et al., 2007). Оценка распределения в CSF может дать информацию о механизме интраназальной доставки.
Оптимальная доставка в CNS вдоль нервных путей связана с доставкой агента в верхнюю треть полости носа (Hanson et al., 2008). Хотя положение «лежа на спине» может быть использовано, другим положением для нацеливания на обонятельную область является положение «praying to Месса», когда голова направлена вниз и вперед. Положение «лежа на спине» с головой, наклоненной под углом 70° или 90°, может быть подходящим для эффективной доставки в CSF с использованием трубки, вставленной в ноздри для доставки лекарственного средства посредством интраназального введения (van den Berg et al., (2002)).
Для интраназального введения лекарственного средства назальные капли можно вводить в течение 10-20 минут попеременно в каждую ноздрю каждые 1-2 минуты для обеспечения абсорбции раствора в назальном эпителии (Thorne et al., 2004; Capsoni et al., 2002; Ross et al., 2004; Ross et al., 2008; Dhuria et al., 2009a; Dhuria et al., 2009b; Francis et al., 2008; Martinez et al., 2008). Этот неинвазивный способ не включает помещение устройства в ноздри. Вместо этого капли помещают у открытия ноздри, позволяя индивидууму вдыхать каплю в носовую полость. Другие способы введения у подвергнутого анестезии индивидуума включают блокирование пищевода и вставку дыхательной трубки в трахею, чтобы предотвратить проглатывание назального препарата и устранить проблемы, связанные с дыхательной недостаточностью (Chow et al., 1999; Chow et al., 2001; Fliedner et al., 2006; Dahlin et al., 2001). Гибкая трубка может быть вставлена в ноздри для локализованной доставки небольшого объема раствора лекарственного средства в дыхательный или обонятельный эпителий, в зависимости от длины трубки (Chow et al., 1999; Van den Berg et al., 2003; van den Berg et al., 2004a; Banks et al., 2004; van den Berg et al., 2002; Vyas et al., 2006a; Charlton et al., 2007a; Gao et al., 2007a).
Устройства для назальной доставки, такие как спреи, капли для носа или безыгольные шприцы, могут быть использованы для нацеливания агента в различные области носовой полости. OptiMist представляет собой активируемое вдохом устройство, которое нацеливает жидкие или порошкообразные назальные препараты в носовую полость, включая обонятельную область, без осаждения в легких или пищеводе (Djupesland et al., 2006). Устройство ViaNase™ также может быть использовано для нацеливания назального спрея в обонятельный и дыхательный эпителий носовой полости. Назальные капли имеют тенденцию к осаждению на носовой перегородке и подвергаются быстрому мукоцилиарному клиренсу, тогда как назальные спреи распределяются в среднюю часть носового хода слизистой оболочки носа (Scheibe et al., 2008).
Иммуносупрессант или агент, индуцирующий иммунологическую толерантность, может быть введен любым путем, включая парентеральный. В одном варианте осуществления иммуносупрессант или агент, индуцирующий иммунологическую толерантность, может быть введен путем подкожной, внутримышечной или внутривенной инъекции, перорально, интратекально, интракраниально или интраназально, или путем замедленного высвобождения, например, путем использования подкожного имплантата. Иммуносупрессант или агент, индуцирующий иммунологическую толерантность, может быть растворен или диспергирован в жидком носителе. Для парентерального введения активное вещество может быть подходящим образом смешано с приемлемым носителем, например, на основе различных растительных масел, таких как арахисовое масло, хлопковое масло и т.п. Могут быть также использованы другие носители для парентерального введения, такие как органические композиции с использованием солкеталя, глицерина, формаля, и водные препараты для парентерального введения. Для парентерального введения путем инъекции композиции могут содержать водный раствор водорастворимой фармацевтически приемлемой соли активных кислот в соответствии с изобретением, предпочтительно при концентрации 0,01-10%, и, необязательно, также стабилизирующий агент и/или буферные вещества в водном растворе. Единицы дозирования раствора могут быть удобным образом заключены в ампулы.
Композиция, например, rAAV-содержащая композиция, содержащая иммуносупрессант композиция или композиция, индуцирующая иммунологическую толерантность, может быть представлена в форме единичной инъекционной дозы. Примеры носителей или разбавителей, подходящих для изготовления таких инъекционных доз, включают разбавители, такие как вода, этиловый спирт, макрогол, пропиленгликоль, этоксилированный изостеариловый спирт, полиоксилированный изостеариловый спирт и эфиры полиоксиэтиленсорбитана и жирных кислот, агенты или буферы, регулирующие рН, такие как цитрат натрия, ацетат натрия и фосфат натрия, стабилизаторы, такие как пиросульфит натрия, EDTA, тиогликолевая кислота и тиомолочная кислота, изотонические агенты, такие как хлорид натрия и глюкоза, местные анестетики, такие как прокаина гидрохлорид и лидокаина гидрохлорид. Кроме того, могут быть добавлены обычные солюбилизирующие агенты и анальгетики. Инъекционные лекарственные формы могут быть изготовлены путем добавления таких носителей к ферменту или другому активному веществу в соответствии с процедурами, хорошо известными специалистам в данной области. Подробное описание фармацевтически приемлемых вспомогательных веществ содержится в REMINGTON'S PHARMACEUTICAL SCIENCES (Mack Pub. Co., N.J. 1991). Фармацевтически приемлемые препараты могут быть легко суспендированы в водных носителях и введены с помощью обычных игл для подкожных инъекций или с использованием инфузионных насосов. Перед введением препараты могут быть простерилизованы предпочтительно гамма-облучением или стерилизацией пучком электронов.
Когда иммуносупрессант или агент, индуцирующий иммунологическую толерантность, вводят в форме подкожного имплантата, соединение суспендируют или растворяют в медленно диспергируемом материале, известном специалистам в данной области, или вводят в устройство, которое медленно высвобождает активное вещество посредством использования постоянной движущей силы, такой как осмотический насос.В таких случаях возможно введение в течение продолжительного периода времени.
Дозы, при которых вводят композицию, содержащую иммуносупрессант или агент, индуцирующий иммунологическую толерантность, могут изменяться в широком диапазоне и зависят от различных факторов, таких как тяжесть заболевания, возраст пациента и т.д., и могут быть индивидуально скорректированы. Возможный диапазон для количества, которое может быть введено в день, составляет примерно от 0,1 мг до примерно 2000 мг или примерно от 1 мг до примерно 2000 мг.Композиции, содержащие иммуносупрессант или агент, индуцирующий иммунологическую толерантность, могут быть составлены таким образом, чтобы обеспечивать дозы в пределах этих диапазонов в виде единичных дозированной формы или множества дозированных форм. Дополнительно к содержанию иммуносупрессанта, рассматриваемые препараты могут содержать один или несколько rAAV, кодирующих терапевтический генный продукт.
Композиции, описанные здесь, можно применять в комбинации с другим лекарственным средством. Композиции могут быть представлены в стандартных формах, например, в виде аэрозолей, растворов, суспензий, или композиций для местного применения, или в лиофилизированной форме.
Типичные композиции включают rAAV, иммуносупрессант, усилитель проницаемости или их комбинацию и фармацевтически приемлемое вспомогательное вещество, которое может представлять собой носитель или разбавитель. Например, активный агент(ы) может быть смешан с носителем или разбавлен носителем, или заключен в носитель. Когда активный агент смешан с носителем, или когда носитель служит в качестве разбавителя, он может быть твердым, полутвердым или жидким материалом, который действует в качестве носителя, вспомогательного вещества или среды для активного агента. Некоторые примеры подходящих носителей включают воду, растворы солей, спирты, полиэтиленгликоли, полигидроксиэтоксилированное касторовое масло, арахисовое масло, оливковое масло, желатин, лактозу, каолин, сахарозу, декстрин, карбонат магния, сахар, циклодекстрин, амилозу, стеарат магния, тальк, желатин, агар, пектин, аравийскую камедь, стеариновую кислоту или низшие алкиловые эфиры целлюлозы, кремниевую кислоту, жирные кислоты, амины жирных кислот, моноглицериды и диглицериды жирных кислот, сложные эфиры пентаэритритола и жирных кислот, полиоксиэтилен, гидроксиметилцеллюлозу и поливинилпирролидон. Аналогично, носитель или разбавитель может включать любое вещество с замедленным высвобождением, известное в данной области, такое как глицерилмоностеарат или глицерилдистеарат, взятый в отдельности или в смеси с воском.
Препараты могут быть смешаны со вспомогательными веществами, которые не взаимодействуют пагубным образом с активным агентом(ами). Такие добавки могут включать смачивающие агенты, эмульгаторы и суспендирующие агенты, соли для влияния на осмотическое давление, буферы и/или окрашивающие вещества, консервирующие агенты, подслащивающие агенты или ароматизирующие агенты. Композиции также могут быть стерилизованы, по необходимости.
При использовании жидкого носителя препарат может быть представлен в форме жидкости, такой как водная жидкая суспензия или раствор. Приемлемые растворители или носители включают стерилизованную воду, раствор Рингера или изотонический водный солевой раствор.
Агент(ы) может быть представлен в форме порошка, подходящего для восстановления с помощью соответствующего раствора, описанного выше. Примеры включают, но без ограничения, порошки, полученные путем сушки замораживанием, ротационной сушки, сушки распылением, аморфные порошки, гранулы, осадки или твердые частицы. Композиция может необязательно содержать стабилизаторы, модификаторы рН, поверхностно-активные вещества, модификаторы биологической доступности и их комбинации. Стандартная лекарственная форма может содержаться в индивидуальных контейнерах или в контейнерах, рассчитанных на множество доз.
Композиции, рассматриваемые в настоящем изобретении, могут включать, например, мицеллы или липосомы, или какую-либо другую инкапсулированную форму, или могут быть введены в виде лекарственной формы с пролонгированным высвобождением для обеспечения длительного хранения и/или длительного эффекта доставки, например, с использованием биоразлагаемых полимеров, например, полилактид-полигликолида. Примеры других биоразлагаемых полимеров включают сложные поли(ортоэфиры) и поли(ангидриды).
Полимерные наночастицы, например, состоящие из гидрофобного ядра полимолочной кислоты (PLA) и гидрофильной оболочки метокси-поли(этиленгликоля) (MPEG), могут обладать улучшенной растворимостью и нацеливать на CNS. Региональные различия в нацеливании между композициями в виде микроэмульсий и наночастиц могут быть обусловлены различиями в размерах частиц.
Липосомы представляют собой очень простые структуры, состоящие из одного или более липидных бислоев амфифильных липидов, т.е. фосфолипидов или холестерина. Липофильные фрагменты бислоев направлены по отношению друг к другу и образуют внутреннюю гидрофобную среду в мембране. Липосомы являются подходящими носителями для некоторых липофильных лекарственных средств, которые могут быть связаны с неполярными частями липидных бислоев, если они подходят по размеру и геометрии. Размер липосом варьирует от 20 нм до нескольких мкм.
Смешанные мицеллы являются эффективными структурами детергента, которые состоят из солей желчных кислот, фосфолипидов, три-, ди- и моноглицеридов, жирных кислот, свободного холестерина и жирорастворимых питательных микроэлементов. Так как длинноцепочечные фосфолипиды, как известны, образуют бислой при диспергировании в воде, предпочтительной фазой короткоцепочечных аналогов является сферическая мицеллярная фаза. Мицеллярный раствор представляет собой термодинамически стабильную систему, образованную спонтанно в воде и органических растворителях. Взаимодействие между мицеллами и гидрофобными/липофильными лекарственными средствами приводит к образованию смешанных мицелл (ММ), часто называемых также «набухаемыми» мицеллами. В человеческом организме они включают гидрофобные соединения с низкой растворимостью в воде и действуют в качестве резервуара для продуктов пищеварения, например, моноглицеридов.
Липидные микрочастицы включают липидные нано- и микросферы. Микросферы, как правило, определяются как малые сферические частицы, изготовленные из любого материала, имеющие размер примерно 0,2-100 мкм. Сферы с меньшим размером, составляющим менее 200 нм, как правило, называются наносферами. Липидные микросферы представляют собой гомогенные микроэмульсии масло в воде, подобные коммерчески доступным жировым эмульсиям, и получены с помощью интенсивной ультразвуковой обработки или эмульгированием при высоком давлении (методы измельчения). Лецитин, природное поверхностно-активное вещество, снижает поверхностное натяжение жидкости, таким образом, действуя в качестве эмульгатора для образования стабильной эмульсии. Структура и состав липидных наносфер схожи с липидными микросферами, но имеют меньший диаметр.
Полимерные наночастицы служат в качестве носителей различных ингредиентов. Активные компоненты могут быть растворены в полимерной матрице или же захвачены или адсорбированы на поверхности частицы. Полимеры, подходящие для изготовления органических наночастиц, включают производные целлюлозы и сложные полиэфиры, такие как поли(молочная кислота), поли(гликолевая кислота) и их сополимеры. Благодаря их малому размеру, высокому соотношению площади поверхности к объему и возможности функционализации поверхности раздела полимерные наночастицы являются идеальными носителями и системами высвобождения. Если размер частиц составляет менее 50 нм, они не распознаются как частицы многими биологическими, а также синтетическими барьерными слоями, но действуют аналогично молекулярным дисперсным системам.
Таким образом, композиция согласно настоящему изобретению может быть составлена для обеспечения быстрого, замедленного, контролируемого или отсроченного высвобождения, или любой их комбинации, активного агента после введения индивидууму с использованием процедур, хорошо известных в данной области. В одном варианте осуществления фермент содержится в изотоническом или гипотоническом растворе. В одном варианте осуществления для ферментов, которые не растворимы в воде, могут быть использованы носители для доставки на основе липидов, например, микроэмульсии, описанной в WO 2008/049588, раскрытие которого включено в данное описание путем ссылки, или липосомы.
В одном варианте осуществления препарат может содержать агент, растворенный или суспендированный в жидком носителе, таком как водный носитель, для применения в виде аэрозоля. Носитель может содержать добавки, такие как солюбилизирующие агенты, например пропиленгликоль, поверхностно-активные вещества, усилители абсорбции, такие как лецитин (фосфатидилхолин) или циклодекстрин, или консерванты, такие как парабены. Например, дополнительно к растворимости, эффективная доставка в CNS после интраназального введения может зависеть от проницаемости мембраны. Для ферментов, парацеллюлярный транспорт которых затруднен из-за размера и полярности, улучшение проницаемости мембран может улучшить механизмы внеклеточного транспорта в CNS вдоль обонятельного и тройничного нервов. Один из подходов к модификации проницаемости мембран внутри назального эпителия состоит в использовании усилителей проницаемости, такие как поверхностно-активные вещества, например, лауроилкарнитин (LC), соли желчных кислот, липиды, циклодекстрины, полимеры или модификаторы плотного сочленения
Как правило, активные агенты вносят в виде стандартной лекарственной формы, включающей активный ингредиент вместе с фармацевтически приемлемым носителем на стандартную дозу. Обычно лекарственные формы, пригодные для назального введения, включают примерно от 125 мкг до 125 мг, например, примерно от 250 мкг до 50 мг, или примерно от 2,5 мг до 25 мг соединений, смешанных с фармацевтически приемлемым носителем или разбавитедем.
Лекарственные формы можно вводить ежедневно или несколько раз в день, например, два или три раза в день. Кроме того, лекарственные формы можно вводить реже, чем ежедневно, например, через день или раз в неделю, если это является целесообразным по мнению лечащего врача.
Изобретение будет описано с помощью следующих не ограничивающих примеров.
Пример I
Опосредованная вектором AAV доставка гена идуронидазы в мышиной модели мукополисахаридоза I типа: сравнение различных путей доставки в CNS
Мукополисахаридоз I типа (MPS I) представляет собой наследственное заболевание обмена веществ, обусловленное дефицитом лизосомного фермента альфа-Ь-идуронидазы (IDUA). Системное и аномальное накопление гликозаминогликанов связано с задержкой роста, органомегалией, дисплазией скелета и заболеванием сердца и легких. Индивидуумы с наиболее тяжелой формой заболевания (синдром Гурлер) страдают нейродегенерацией, задержкой умственного развития и преждевременно умирают.Разработанные на сегодняшний день два метода лечения MPS I (трансплантация гемопоэтических стволовых клеток и ферментная заместительная терапия) не могут эффективно лечить все проявления заболевания со стороны центральной нервной системы (CNS)
Что касается генной терапии, ранее было продемонстрировано, что интраваскулярная доставка AAV9 у взрослых мышей не достигает широкого прямого нацеливания на нейроны (см. Foust et al, 2009). В предыдущих работах также было показано, что прямая инъекция AAV8-IDUA в CNS взрослых IDUA-дефицитных мышей приводит к низкой частоте или низкому уровню экспрессии трансгена (см. фигуру 18).
Следующие примеры, в которых использованы доклинические модели для лечения MPS1, неожиданно показали, что прямая инъекция AAV9-IDUA в CNS иммунокомпетентных взрослых ШиА-дефицитных мышей приводит к экспрессии и активности фермента IDUA, которые являются такими же или более высокими, чем экспрессия и активность фермента IDUA у взрослых мышей дикого типа (см. фигуру 15 ниже). Способы
Получение AAV9-IDUA. Векторная конструкция AAV-IDUA (MCI) описана ранее (Wolf et al., 2011) (промотор mCags). Плазмидная ДНК AAV-IDUA упакована в вирионы AAV9 в лаборатории University of Florida Vector Core, с получением титра 3×ΙΟ13 геномов вектора на миллилитр.
ICV-инфузии. Взрослых Idua-/- мышей анестезировали смесью кетамина и ксилазина (100 мг кетамина+10 мг ксилазина на кг) и помещали на стереотаксическую рамку. Десять микролитров AAV9-IDUA вводили путем инфузии в правый боковой желудочек (стереотаксические координаты АР 0.4, ML 0.8, DV 2.4 мм от брегмы), используя шприц Гамильтона. Животных возвращали в их клетки на грелки-подушки для восстановления.
Интратекальные инфузии. Инфузии молодым взрослым мышам проводили путем инъекции 10 мкл раствора, содержащего вектор AAV, между позвонками L5 и L6 через 20 минут после внутривенной инъекции 0,2 мл 25% маннита.
Индукция иммунологической толерантности. Новорожденным IDUA-дефицитным мышам через лицевую височную вену инъецировали 5 мкл, содержащих 5,8 мкг рекомбинантного белка идуронидазы (Альдуразим), а затем животных возвращали в их клетки.
Иммуносупрессия циклофосфамидом. Для индукции иммуносупрессии животным вводили циклофосфамид раз в неделю в дозе 120 мг/кг, начиная со следующего дня после инфузии вектора AAV9-IDUA.
Животные. Животных анестезировали смесью кетамина/ксилазина (100 мг кетамина+10 мг ксилазина на кг) и транскардиально перфузировали 70 мл PBS перед умерщвлением. Головной мозг извлекали и подвергали микродиссекции на льду на мозжечок, гиппокамп, стриатум, кору головного мозга и ствол головного мозга/таламус («остальные отделы»). Образцы замораживали на сухом льду и затем хранили при температуре -80°С. Образцы оттаивали и гомогенизировали в присутствии 1 мл буфера PBS с помощью автоматического пестика и пермеабилизировали в 0,1%-ном растворе Triton Х-100. Активность IDUA определяли с помощью флуориметрического анализа с использованием в качестве субстрата 4Ми-идуронида. Активность выражали в единицах (процент превращения субстрата в продукт в минуту) на мг белка, как определено с помощью анализа методом Бредфорда (BioRad).
Ткани. Гомогенаты тканей осветляли путем центрифугирования в течение 3 мин при 13000 об/мин на настольной центрифуге Eppendorf 5415D (Eppendorf) и инкубировали в течение ночи с протеиназой К, ДНКазой 1 и РНКазой. Концентрацию GAG определяли методом Blyscan Sulfated Glycosaminoglycan Assay (Accurate Chemical) в соответствии с инструкциями изготовителя.
Результаты
На фигуре 1 показан дизайн эксперимента для идуронидаза-дефицитных мышей, которым вводили AAV интрацеребровентрикулярно (ICV) или интратекально (IT). Для предотвращения иммунного ответа животных подвергали иммуносупрессии циклофосфамидом (CP), индуцировали иммунологическую толерантность при рождении путем внутривенного введения человеческого белка идуронидазы (Альдуразим) или инъекции проводили у иммунодефицитных мышей NOD-SCID, которые были также дефицитными по идуронидазе. Животных умерщвляли в указанное время после лечения, головной мозг подвергали микродиссекции и экстракты анализировали на активность идуронидазы.
На фигуре 2 представлены данные для иммунодефицитных, IDUA-дефицитных животных, которым инъецировали ICV вектор AAV-IDUA. Эти животные показали высокие уровни экспрессии IDUA (в 10-100 раз выше по сравнению с диким типом) во всех областях головного мозга, при этом самый высокий уровень наблюдался в стволе головного мозга и таламуса («остальные отделы»).
Иммуносупрессированные животные, которым вводили вектор AAV ICV-путем, имели относительно более низкий уровень фермента в головном мозге по сравнению с иммунодефицитными животными (фигура 3). Следует отметить, что иммуносупрессия может быть ослаблена у этих животных из-за отмены циклофосфамида (CP) за 2 недели до умерщвления по причине слабого здоровья.
На фигуре 4 представлены данные для иммуносупрессированных животных, которым вводили вектор AAV IT-путем. Животные, подвергнутые индукции иммунологической толерантности, которым вводили вектор AAV ICV-путем, проявили широкую активность IDUA во всех отделах головного мозга (фигура 5), аналогичную активности, наблюдаемой у иммунодефицитных животных, что указывает на эффективность процедуры индукции иммунологической толерантности.
Фигура 6 представляет собой компиляцию всех средних уровней активности IDUA для параллельного сравнения, и на фигуре 7 показаны данные, сгруппированные в соответствии с областью головного мозга.
Материал накопления GAG анализировали в различных срезах головного мозга у всех четырех тестируемых групп.Для каждой группы среднее значение для каждого отдела головного мозга показано слева, значения для каждого индивидуального животного показаны справа (фигура 8). IDUA-дефицитные животные (крайние слева) содержали высокие уровни GAG по сравнению с животными дикого типа (пурпурный столбец). Уровни GAG были на уровне дикого типа или ниже дикого типа для всех отделов головного мозга во всех группах AAV-обработанных животных. Уровни GAG были немного, хотя и не значительно выше, чем у дикого типа в коре головного мозга и стволе головного мозга животных, которым вводили AAV9-IDUA интратекально.
Выводы
Результаты показали высокое и широкое распределение IDUA в головном мозге независимо от пути доставки (ICV или IT), хотя экспрессия IDUA в стриатуме и гиппокампе была более низкой у животных, инъецированных IT, по сравнению с ICV. По-видимому, присутствует иммунный ответ, поскольку иммунодефицитные мыши имеют более высокие уровни экспрессии по сравнению с иммунокомпетентными мышами. Что касается ICV инъекции, в случае преждевременной отмены CP экспрессия IDUA является более низкой. Кроме того, индукция иммунологической толерантности была эффективной в отношении восстановления высоких уровней активности фермента. Кроме того, уровни GAG были восстановлены до нормальных уровней во всех подвергнутых лечению экспериментальных группах мышей.
Пример II
Способы
Получение AAV9 - IDUA. Плазмиду AAV-IDUA упаковывали в вирионы AAV9 в лаборатории University of Florida vector core или в лаборатории University of Pennsylvania vector core, получая титр 1-3×1013 геномов вектора на миллилитр.
ICV-инфузии. См. пример I.
Интратекальные инфузии. См. пример I.
Индукция иммунологической толерантности. Как в примере I, за исключением следующего: для многократной индукции иммунологической толерантности новорожденным IDUA-дефицитным мышам в лицевую височную вену инъецировали первую дозу Альдуразима с последующими 6 еженедельными инъекциями, вводимыми интраперитонеально.
Иммуносупрессия циклофосфамидом. См. пример I.
Животные. Животные анестезировали смесью кетамина/ксилазина (100 мг кетамина+10 мг ксилазина на кг) и транскардиально перфузировали 70 мл PBS перед умерщвлением. Головной мозг извлекали и подвергали микродиссекции на льду на мозжечок, гиппокамп, стриатум, кору и ствол головного мозга/таламус («остальные отделы»). Образцы замораживали на сухом льду и хранили при температуре -80°С.
Активность IDUA в тканях. Образцы тканей оттаивали и гомогенизировали в водном растворе хлорида натрия в гомогенизаторе тканей. Гомогенаты тканей осветляли путем центрифугирования при 15000 об/мин на настольной центрифуге Эппендорфа при температуре 4°С в течение 15 минут.Лизаты тканей (супернатант) собирали и анализировали на уровни активности IDUA и накопления GAG.
Уровни GAG в тканях. Лизаты тканей инкубировали в течение ночи с протеиназой К, РНКазой и ДНКазой. Уровни GAG определяли методом Blyscan Sulfated Glycosaminoglycan Assay в соответствии с инструкциями изготовителя.
Копии вектора IDUA. Гомогенаты тканей использовали для выделения ДНК и последующей QPCR, как описано в работе Wolf et al. (2011).
Результаты
На фигуре 9 показан дизайн эксперимента и группы. Животным вводили вектор AAV9-IDUA путем интрацеребровентрикулярной (ICV) или интратекальной (IT) инфузии. Введение вектора осуществляли у иммунодефицитных (ID) мышей NOD-SCID, которые были также дефицитными по IDUA, или у Ши А-дефицитных мышей, которые были подвергнуты иммуносупрессии циклофосфамидом (CP) или индукции иммунологической толерантности при рождении путем однократной инъекции или многократных инъекций человеческого белка идуронидазы (Альдуразим). На фигуре 9 показаны моменты времени, в которые проводили лечение вектором и умерщвление. Все введения вектора проводили у взрослых животных в возрасте 3-4,5 месяцев. Животным инъецировали 10 мкл вектора в дозе 3 χ 1011 геномов вектора на 10 микролитров.
На фигуре 10 показаны активности фермента IDUA у интракраниально инфузированных, иммунодефицитных, Ши А-дефицитных мышей. Во всех областях головного мозга наблюдались высокие уровни активности фермента, которые в 30-300 раз превышали уровни активности, наблюдаемые у мышей дикого типа. Наиболее высокие уровни экспрессии фермента наблюдались в таламусе и стволе мозга, а также в гиппокампе.
Животные, которые были инъецированы интракраниально и подвергнуты иммуносупрессии циклофосфамидом (CP), показали значительно более низкие уровни активности фермента по сравнению с другими группами (фигура 11). Однако введение CP в этом случае вынуждены были отменить за 2 недели до умерщвления из-за слабого здоровья животных.
Уровни фермента IDUA у животных, подвергнутых индукции толерантности при рождении с помощью белка IDUA (Альдуразим), и которым интракраниально вводили вектор, показаны на фигуре 12. Все животные показали во всех отделах головного мозга высокие уровни фермента, которые в 10-1000 раз превышали уровни, наблюдаемые у мышей дикого типа, аналогично уровням, достигнутым у иммунодефицитных животных, что указывает на эффективность процедуры индукции иммунологической толерантности.
На фигуре 13 показаны уровни фермента IDUA у мышей, которые были интратекально инъецированы и которым вводили CP на еженедельной основе. Повышенные уровни IDUA наблюдались во всех частях головного мозга, особенно в мозжечке и спинном мозге. Уровни фермента были самыми низкими в стриатуме и гиппокампе, при этом значения активности находились на уровнях, наблюдаемых у животных дикого типа.
IDUA-дефицитных мышей подвергали индукции иммунологической толерантности Альдуразимом, как описано, и инъецировали интратекально вектором (фигура 14). Наблюдалась широкая активность фермента IDUA во всех отделах головного мозга, при этом самые высокие уровни активности наблюдались в стволе мозга и таламусе, обонятельной луковице, спинном мозге и мозжечке. Аналогично данным, представленным на фигуре 13, самые низкие уровни активности фермента наблюдались в стриатуме, коре головного мозга и гиппокампе.
Контрольным иммунокомпетентным IDUA-дефицитным животным инфузировали интратекально вектор, без иммуносупрессии или индукции иммунологической толерантности (фигура 15). Полученные результаты свидетельствуют о том, что хотя показатели активности фермента соответствовали или немного превышали уровни активности у мышей дикого типа, они были значительно ниже показателей активности, наблюдаемых у животных, подвергнутых иммуномодулированию. Снижения уровней фермента были особенно значительными в мозжечке, обонятельной луковице и таламусе, а также стволе мозга, то есть областях, которые экспрессировали самые высокие уровни фермента у иммуномодулированных животных.
Животных анализировали на материал накопления GAG, как показано на фигуре 16. Все группы продемонстрировали клиренс накопления GAG, при этом уровни GAG были сходными с уровнями, наблюдаемыми у животных дикого типа. Животные, подвергнутые иммуносупрессии и интратекально инъецированные вектором AAV9-IDUA, показали уровни GAG в коре головного мозга, которые были немного более высокими по сравнению с диким типом, но все еще гораздо более низкими по сравнению с IDUA-дефицитными мышами, не подвергнутыми лечению.
Присутствие вектора AAV9-IDUA у животных, подвергнутых индукции иммунологической толерантности и инъецированных интракраниально или интратекально вектором, оценивали с помощью QPCR, как показано на фигуре 16. Число копий IDUA на клетку было выше у животных, инфузированных интракраниально, по сравнению с животными, инфузированными интратекально, что согласуется с более высоким уровнем активности фермента, наблюдаемым у животных, инъецированных интракраниально.
Выводы
Высокие, широко распространенные и терапевтические уровни IDUA наблюдались во всех областях головного мозга после введения интрацеребровентрикулярным и интратекальным путем AAV9-IDUA взрослым мышам. Показатели активности фермента восстановились до уровней, наблюдаемых у животных дикого типа, или немного более высокого уровня у иммунокомпетентных IDUA-дефицитных животных, интратекально инфузированных AAV-IDUA. Наблюдались значительно более высокие уровни фермента IDUA для обоих путей введения вектора у животных, подвергнутых индукции иммунологической толерантности при рождении путем введения белка IDUA.
Пример III
Взрослых иммунокомпетентных Ши А-дефицитных мышей (в возрасте 12 недель) анестезировали смесью кетамина/ксилазина с последующей интраназальной инфузией вектора AAV9-IDUA. Вектор вводили путем внесения восьми капель по 3 мкл с помощью микропипетки в носовую полость, попеременно в каждую ноздрю, с 2-минутными интервалами между каждым внесением. В общей сложности каждому взрослому животному вводили 2,4-7×1011 геномов вектора, в зависимости от источника вектора. Для подавления иммунного ответа у мышей на человеческий IDUA, продуцируемый вектором AAV9-IDUA, животных подвергали иммуносупрессии путем еженедельного введения 120 мг/кг циклофосфамида, начиная со следующего дня после введения вектора. Однако иммуносупрессия у субъектов-людей необязательна и специалист в данной области будет знать, в каких случаях ее необходимо применять в соответствии с надлежащей/стандартной медицинской практикой. Мышей умерщвляли через 12 недель после инфузии вектора, животных анализировали на экспрессию фермента IDUA и копии вектора в головном мозге (фигуры 19 и 20).
Пример IV
Идуронидаза-дефицитным мышам, модели человеческого мукополисахаридоза I типа (MPS I), интраназально вводили приблизительно 1011 копий вектора AAV9-IDUA. Через четыре недели животных умерщвляли, и головной мозг подвергали микродиссекции и экстрагировали для анализа на фермент идуронидазу. Как показано на фигуре 22 (среднее значение+Z-SD показано слева, индивидуальные животные показаны справа), высокий уровень активности фермента IDUA (почти в 100 раз более высокий по сравнению с диким типом) наблюдался в обонятельной луковице, при этом во всех других областях головного мозга наблюдались уровни фермента, соответствующие животным дикого типа. На фигуре 23 показана активность GAG.
Аналогично обработанных животных умерщвляли, и срезы тканей окрашивали на присутствие человеческого белка IDUA, используя антитело к IDUA. Как показано на фигуре 24 слева, сильное окрашивание белка IDUA наблюдается в назальном эпителии и обонятельной луковице у четырех животных, подвергнутых интраназальному лечению вектором, при этом справа отсутствует окрашивание у контрольных нормальных или IDUA-дефицитных животных, которым не вводили вектор. Окрашивание не наблюдалось в других отделах головного мозга (гиппокамп, мозжечок, кора головного мозга, стриатум, таламус и ствол мозга) животных, подвергнутых лечению AAV-MCI, демонстрируя, что трансдукции была ограничена обонятельной луковицей и назальным эпителием.
Животных обрабатывали интраназально приблизительно 1011 геномами вектора AAV9-GFP в качестве репортерной системы для идентификации локализации трансдуцированной клетки. Через две недели животных умерщвляли, собирали ткани, и срезы окрашивали на экспрессию GFP, используя антитело к GFP (зеленый), а также окрашивали DAPI (синий) для идентификации клеточных ядер (фигура 25). Наблюдалось сильное окрашивание белка GFP в обонятельном эпителии (левые две панели) и обонятельной луковице (верхние две панели) у подвергнутых лечению животных по сравнению с не подвергнутыми лечению контрольными животными (правые панели для обонятельного эпителия и нижняя панель для обонятельной луковицы). Окрашивания GFP не наблюдалось в других отделах головного мозга. Эти результаты согласуются с результатами окрашивания на IDUA, демонстрируя, что опосредованный AAV перенос гена и экспрессия ограничивается назальным эпителием и обонятельной луковицей после интраназального введения вектора AAV9-MCI. Эти результаты позволяют рассматривать диффузию IDUA, экспрессирующегося на высоких уровнях в переднем мозге, в качестве механизма, посредством которого уровни IDUA, соответствующие уровням IDUA у животных дикого типа, достигаются во всех областях головного мозга после интраназального введения вектора AAV9-MCI. Этот подход к достижению высокого уровня экспрессии терапевтического белка в переднем мозге, то есть неинвазивное интраназальное введение вектора AAV с последующим его распространением в головном мозге, применим не только для лечения MPS и связанных метаболических заболеваний, но и других более распространенных неврологических расстройств, таких как болезнь Паркинсона и болезнь Альцгеймера.
Пример V.
Мукополисахаридоз II типа (MPS II; синдром Хантера) представляет собой лизосомную болезнь накопления с Х-сцепленным рецессивным типом наследования, обусловленную дефицитом идуронат-2-сульфатазы (IDS), приводящим к накоплению гликозаминогликанов (GAG) дерматан- и гепарансульфата. Пораженные индивидуумы проявляют манифестации различной степени тяжести физически, неврологически, и укороченную продолжительность жизни. Например, пораженные индивидуумы проявляют манифестации различной степени тяжести, такие как органомегалия, дисплазия скелета, сердечно-легочная обструкция, нейрокогнитивный дефицит и укороченная продолжительность жизни. На сегодняшний день лечение для MPS II отсутствует.Существующим на сегодняшний день стандартом лечения является ферментная заместительная терапия (ELAPSRASE; идурсульфаза), которую применяют для контроля прогрессирования заболевания. Тем не менее, ферментная заместительная терапия (ERT) не приводит к неврологическому улучшению. Так как трансплантация гемопоэтических стволовых клеток (HSCT) не показала неврологического эффекта для MPS II, в настоящее время не существует клинической возможности для пациентов, проявляющих неврологические манифестации данного заболевания, и крайне необходимы новые методы лечения.
AAV9 векторы разработаны для доставки последовательности, кодирующей IDS человека (AAV9-hIDS), в центральную нервную систему мышей с MPS II для восстановления уровней IDS в головном мозге и предупреждения возникновения нейрокогнитивных дефицитов у подвергнутых лечению животных (фигура 27А). В частности, создавали серии векторов, кодирующих IDS человека с модифицирующим сульфатазу фактором-1 (SUMF-1) человека или без него, требуемым для активации активного центра сульфатазы. В этих экспериментах использовали три пути введения: интратекальное (IT) (фигуры 28-29), интрацеребровентрикулярное (ICV) (фигуры 30A-D) и внутривенное (IV) (фигуры 28-29). Существенных различий в уровне фермента не было обнаружено между мышами, которым вводили вектор AAV9, трансдуцирующий только hIDS, и мышами, которым вводили вектор AAV9, кодирующий IDS и SUMF-1 человека, независимо от пути введения. Мыши NOD.SCID (IDS Y+) и C57BL/6 (IDS Y+), подвергнутые IT-введению, не показали повышенной активности IDS в головном мозге и спинном мозге по сравнению с животными, не подвергнутыми лечению, при этом уровни в плазме в десять раз (NOD.SCID) и 150 раз (C57BL/6) превышали уровни, наблюдаемые у животных, не подвергнутых лечению. IDS-дефицитные мыши, которым внутривенно вводили AAV9-hIDS, показали активности IDS во всех органах, сопоставимые с активностями, наблюдаемыми у животных дикого типа. Кроме того, активность фермента в плазме IV-инъецированных животных в 100 раз превышала активность, наблюдаемую у животных дикого типа. IDS-дефицитные мыши, которым ICV-путем вводили AAV9-hlDUA, показали сопоставимые с диким типом активности IDS в большинстве областей головного мозга и периферических тканях, при этом некоторые отделы головного мозга показали активность, которая в два-четыре раза превышала активность, наблюдаемую у животных дикого типа. Кроме того, активность IDS в плазме была в 200 раз выше, чем у животных дикого типа. Удивительно, что активность фермента IDS в плазме у всех подвергнутых лечению животных показала устойчивость в течение по меньшей мере 12 недель после инъекции; следовательно, фермент IDS не был иммуногенными по меньшей мере на мышах C57BL/6. Дополнительное тестирование нейроповеденческих функций проводили с использованием лабиринта Барнеса, чтобы дифференцировать нейрокогнитивные дефициты не подвергнутых лечению животных с MPS II от нейрокогнитивных дефицитов однопометных животных дикого типа. Было обнаружено, что способность к обучению пораженных животных заметно медленнее, чем способность к обучению, наблюдаемая у однопометных животных. Таким образом, лабиринт Барнеса использовали для определения благоприятного эффекта этих методов лечения в мышиной модели MPS II (фигуры 31А-В и 32). Эти результаты указывают на возможный терапевтический эффект ААУ9-опосредованного переноса гена IDS человека в CNS для предотвращения неврологического дефицита в MPS П.
В целом, интрацеребровентрикулярная (ICV) инъекция AAV9-hIDS привела к системной коррекции дефицита фермента IDS, включая уровни IDS в головном мозге, соответствующие уровням IDS у животных дикого типа. Совместная доставка hIDS с hSUMF-Ι не повысила активность IDS в тканях. Экспрессия hIDS была неиммуногенной у мышей C57BL/6 дикого типа (WT) и имеющих MPS П.
Пример VI
Мукополисахаридоз типа I (MPS I) представляет собой метаболическое заболевание с аутосомно-рецессивным типом наследования, обусловленное дефицитом α -L-идуронидазы (IDUA), что приводит к накоплению гликозаминогликанов (GAG) гепарина и дерматансульфата. Индивидуумы с наиболее тяжелой формой заболевания (синдром Гурлер) страдают нейродегенерацией, задержкой умственного развития и смерть наступает к 10 годам. Современные методы лечения этого заболевания включают в аллогенную трансплантацию гемопоэтических стволовых клеток (HSCT) и ферментную заместительную терапию (ERT). Однако, эти методы лечения недостаточно эффективны в отношении лечения манифестаций заболевания в CNS.
Целью является улучшение терапии для тяжелого MPS I путем дополнения существующих на сегодняшний день ERT и HSCT переносом гена IDUA в CNS, тем самым предотвращая неврологические манифестации заболевания. В этом исследовании тестировали способность внутривенно вводимых векторов AAV серотипов 9 и rhlO (AAV9 и AAVrhlO) пересекать гематоэнцефалический барьер для доставки и экспрессии гена IDUA в CNS (фигура 33). Животным в возрасте 4-5 месяцев, имеющим MPS I, внутривенно инфузировали через хвостовую вену вектор AAV9 или вектор AAVrhlO, кодирующий ген IDUA человека. Образцы крови и мочи собирали на еженедельной основе до момента умерщвления животных на 10 неделе после инъекции. Активности IDUA в плазме у подвергнутых лечению животных были почти в 1 ООО раз более высокими по сравнению с гетерозиготными контролями через 3 недели после инъекции (фигура 34). Головной мозг, спинной мозг и периферические органы анализировали на активность IDUA, клиренс накопления GAG и иммунофлуоресценцию IDUA в срезах тканей (фигуры 34-36). Подвергнутые лечению животные продемонстрировали широкое восстановление активности фермента IDUA во всех органах, включая CNS. Эти данные демонстрируют эффективность системной инфузии вектора AAV9 и AAVrhlO в отношении противодействия манифестациям MPS I в CNS.
Пример VII
Мукополисахаридоз I типа (MPS I) представляет собой прогрессирующее, мультисистемное, наследственное метаболическое заболевание, обусловленное дефицитом α -L-идуронидазы (IDUA). Наиболее тяжелая форма этого заболевания (синдром Гурлер) приводит к смерти в возрасте 10. Существующие на сегодняшний день методы лечения этого заболевания являются неэффективными в отношении лечения заболевания CNS из-за неспособности лизосомных ферментов пересекать гематоэнцефалический барьер. Целью является дополнение существующей на сегодняшний день терапии и лечение манифестаций заболевания в CNS путем AAV-опосредованной доставки и экспрессии гена IDUA.
Неинвазивный и эффективный подход к лечению заболевания CNS состоял в интраназальном введении IDUA-кодирующего вектора AAV9. Взрослых IDUA-дефицитных мышей подвергали индукции иммунологической толерантности при рождении с помощью человеческой идуронидазы для предотвращения анти-IDUA иммунного ответа, и в возрасте 3-х месяцев интраназально инфузировали вектором AAV9-IDUA, регулируемым С AGS (энхансер CMV/промотор бета-актина/интрон глобина). Животные, умерщвленные через 3 месяца после инфузии, показали уровни активности фермента IDUA, которые в 100 раз превышали уровни активности, наблюдаемые у животных дикого типа, в обонятельной луковице, при этом уровни фермента у животных дикого типа восстанавливались во всех других отделах головного мозга (фигура 37). Интраназальное лечение AAV9-IDUA также привело к сокращению материалов накопления GAG в тканях во всех отделах головного мозга (фигура 38). Тестирование нейрокогнитивной функции с использованием лабиринта Барнса показало, что подвергнутые лечению IDUA-дефицитные мыши не отличались от нормальных контрольных животных, тогда как не подвергнутые лечению IDUA-дефицитные мыши показали значительный дефицит научения (фигура 40). Непораженные гетерозиготные животные показали улучшенное выполнение в этом тесте, тогда как мыши с MPS I проявили дефицит в поиске отверстия. Примечательно, что мыши с MPS I, подвергнутые интраназальному введению AAV9-IDUA, показали поведение, сходное с гетерозиготными контролями, демонстрируя предотвращение нейрокогнитивного дефицита, наблюдаемого у не подвергнутых лечению животных с MPS I (фигура 40D).
Наблюдалось сильное иммунофлуоресцентное окрашивание IDUA в срезах тканей назального эпителия и обонятельной луковицы, при этом окрашивание не наблюдалось в других отделах головного мозга (фигуры 39А-В). Это указывает на то, что широкое распределение фермента IDUA, скорее всего, является результатом диффузии фермента из участков трансдукции и экспрессии IDUA в обонятельной луковице и назальном эпителий в более глубоких областях головного мозга. Для увеличения доступа, доставки и распределения вектора по всему головному мозгу, IDUA-дефицитных животных предварительно подвергали интраназальным инфузиям усилителя абсорбции. В разные моменты времени после предварительной обработки животным интраназально инфузировали вектор AAV9 или AAVrhlO, кодирующий IDUA. Животных умерщвляли через 2 месяца после инфузии, головной мозг подвергали микродиссекции и анализировали на активность фермента IDUA, клиренс гликозаминогликанов и иммунофлюоресцентное окрашивание на IDUA и GFP. Эту новую неинвазивную стратегию интраназального введения AAV9-IDUA потенциально можно применять для лечения манифестаций MPS I в CNS и других лизосомных болезней.
Пример VIII
Мукополисахаридоз I типа (MPS I) представляет собой аутосомно-рецессивную лизосомную болезнь накопления, обусловленную дефицитом альфа-Ь-идуронидазы (IDUA), что приводит к накоплению гликозаминогликанов. Манифестации заболевания включают мультисистемные расстройства и в случае тяжелой формы заболевания (синдром Гурлер) приводят к смерти в возрасте до 10 лет.Существующие на сегодняшний день методы лечения, такие как ферментная заместительная терапия и аллогенная трансплантация гемопоэтических стволовых клеток, оказались неэффективными для лечения CNS. В данном исследовании использовали интратекальную доставку вектора на основе аденоассоциированного вируса серотипа 9, трансдуцирующего ген IDUA (AAV9-IDUA), в CNS в мышиной нокаутной модели MPS I. Целью данного исследования является оценка способности AAV-опосредованной генной терапии предотвращать патологические нейрохимические изменения, связанные с болезнью MPS I.
Способы
В качестве хорошо изученной модели синдрома Гурлер использовали нокаутных мышей C57BL/6, дефицитных по IDUA. Вектор AAV9 -IDUA доставляли интратекально мышам с MPS I в возрасте 12 недель. Перед введением AAV мышам инъецировали маннит для открытия гематоэнцефалического барьера и подвергали индукции иммунологической толерантности с помощью ларонидазы для предотвращения анти-IDUA иммунного ответа. !Н MP-спектры получали in vivo в гиппокампе и мозжечке мышей с MPS I, подвергнутых лечению геном AAV9-IDUA (подвергнутые лечению, имеющие MPS I, N=11), мышей с MPS I, не подвергнутых лечению (MPS I, N=12) и гетерозиготных однопометных мышей (контроль, N=12) в возрасте 9 месяцев. Все протонные спектры !Н МРС получали при 9.4Т с помощью методики FastMap и с помощью пространственно локализованной одновоксельной спектроскопии методом STEAM (stimulated echo acquisition mode) с параметрами импульсной последовательности ТЕ=2 мс в сочетании с подавлением сигнала воды в спектрах. Для определения количественного состава метаболитов использовался программный пакет LCModel со спектром макромолекул с быстрой релаксацией, включенных в основной набор. Дышащих самостоятельно животных анестезировали 1,0 - 1,5% изофлураном.
Результаты
Спектральный состав, полученный в данном исследовании, позволил произвести количественную оценку пятнадцати метаболитов головного мозга. Небольшие, но значимые увеличения концентраций аскорбата (Asc,+0,6 мкмоль/г, ρ=0,003) и Ν-ацетиласпартилглутамата (NAAG,+0,3 моль/г, ρ=0,015) наблюдались в гиппокампе не подвергнутых лечению мышей с MPS I относительно контролей. Кроме того, наблюдалась тенденция к увеличению уровня глутатиона (GSH,+0,2 мкмоль/г, ρ=0,054). Различия между нейрохимическими профилями мозжечка у не подвергнутых лечению мышей с MPS I и контролей включали увеличение NAAG (0,25 мкмоль/г, ρ=0,026) и уменьшение фосфоэтаноламина (РЕ, -0,44 мкмоль/г, ρ=0,04). Нейрохимические профили мышей с MPS I, подвергнутых лечению AAV9-IDUA, показали значительное сходство с нейрохимическими профилями контрольных мышей (фигуры 42А-В). В гиппокампе подвергнутых лечению мышей с MPS I уровни Asc, NAAG и GSH нормализовались; только лактат (Lac) показал небольшое различие относительно контроля. В мозжечке подвергнутых лечению мышей с MPS I уровень РЕ, но не NAAG, нормализовался. Небольшие, но значимые различия между подвергнутыми лечению и контрольными мышами наблюдались для Asc, Lac, таурина (Таи) и общего креатина (Cr+РСг). За исключением Asc, изменения в концентрациях метаболитов у подвергнутых лечению мышей с MPS I были всегда противоположны изменениям, наблюдаемым у не подвергнутой лечению группы. Кроме того, для ряда метаболитов, которые не показали существенных изменений между не подвергнутыми лечению мышами с MPS I и контролями (например, глюкоза, глутамат, NAA), оказалось, что уровни метаболитов, обнаруженные у подвергнутых лечению мышей с MPS I, были более близкими контролям, чем не подвергнутым лечению мышам с MPS I. Обсуждение
Значительно увеличенные концентрации Asc и тенденция к увеличению GSH в гиппокампе у не подвергнутых лечению мышей с MPS I указывают на защитную реакцию против окислительного стресса, наблюдаемого в лизосомных болезнях. В свою очередь, пониженный уровень РЕ в мозжечке и повышенный уровень NAAG в обеих областях головного мозга у не подвергнутых лечению мышей с MPS I может указывать на демиелинизацию. Аналогичная картина пониженного уровня РЕ и повышенного уровня NAAG наблюдалась в модели с дефицитом железа, в которой была подтверждена измененная миелинизация. Сравнение нейрохимических профилей гиппокампа и мозжечка у подвергнутых лечению мышей с MPS I против не подвергнутых лечению мышей с MPS I и контрольных мышей ясно демонстрирует, что прямой перенос отсутствующего гена IDUA в CNS с использованием интратекальной доставки AAV9 (в возрасте 12 недель) предотвращает нейрохимические изменения (в возрасте 9 месяцев), связанные с нейродегенеративными процессами, в этой мышиной модели MPS I. Эти нейрохимические результаты согласуются с аналогичными подходами генной терапии, апробированными на мышиной модели MPS I.
Генная терапия, основанная на прямой доставке AAV9-IDUA в CNS, показала, что окислительный стресс и демиелинизация, связанные с этой мышиной моделью MPS I, могут быть предотвращены.
Все публикации, патенты и патентные заявки включены в настоящий документ посредством ссылки. Хотя в вышеизложенном описании данное изобретение описано в связи с конкретными предпочтительными вариантами его осуществления, и многие детали представлены в целях иллюстрации, специалистам в данной области будет очевидно, что могут быть предложены дополнительные варианты осуществления, и что определенные детали, описанные здесь, могут значительно варьировать без отступления от основных принципов изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВЕКТОР НА ОСНОВЕ АДЕНОАССОЦИИРОВАННОГО ВИРУСА ДЛЯ ТЕРАПЕВТИЧЕСКОЙ ДОСТАВКИ В ЦЕНТРАЛЬНУЮ НЕРВНУЮ СИСТЕМУ | 2016 |
|
RU2751952C2 |
| ОПОСРЕДОВАННЫЙ АДЕНОАССОЦИИРОВАННЫМ ВИРУСОМ ПЕРЕНОС ГЕНОВ В ЦЕНТРАЛЬНУЮ НЕРВНУЮ СИСТЕМУ | 2014 |
|
RU2692251C2 |
| ОПОСРЕДОВАННЫЙ АДЕНОАССОЦИИРОВАННЫМ ВИРУСОМ ПЕРЕНОС ГЕНОВ В ЦЕНТРАЛЬНУЮ НЕРВНУЮ СИСТЕМУ | 2014 |
|
RU2801511C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ MPS1 | 2014 |
|
RU2708318C2 |
| КОМПОЗИЦИИ, СПОСОБЫ И ПРИМЕНЕНИЯ ПЕРЕНОСА ГЕНОВ ДЛЯ ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНЫХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2805606C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНЕЙ МОЗГА | 2014 |
|
RU2664471C2 |
| ВЕКТОР АДЕНОАССОЦИИРОВАННОГО ВИРУСА | 2015 |
|
RU2743382C2 |
| ИНТРАТЕКАЛЬНОЕ ВВЕДЕНИЕ ВЕКТОРОВ НА ОСНОВЕ АДЕНОАССОЦИИРОВАННЫХ ВИРУСОВ ДЛЯ ГЕННОЙ ТЕРАПИИ | 2016 |
|
RU2775138C2 |
| СРЕДСТВО ЛЕЧЕНИЯ БОЛЕЗНИ ГЕНТИНГТОНА НА ОСНОВЕ AAV | 2017 |
|
RU2749971C2 |
| ВЕКТОРЫ И ПОСЛЕДОВАТЕЛЬНОСТИ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 2011 |
|
RU2588667C2 |
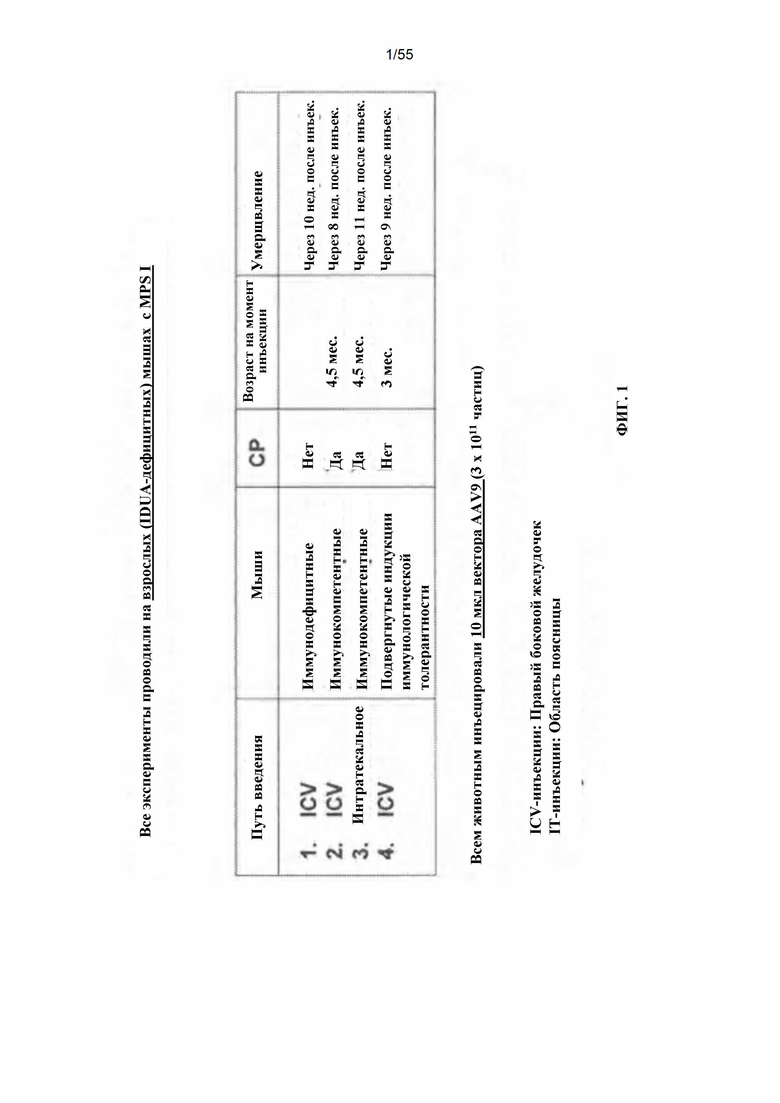
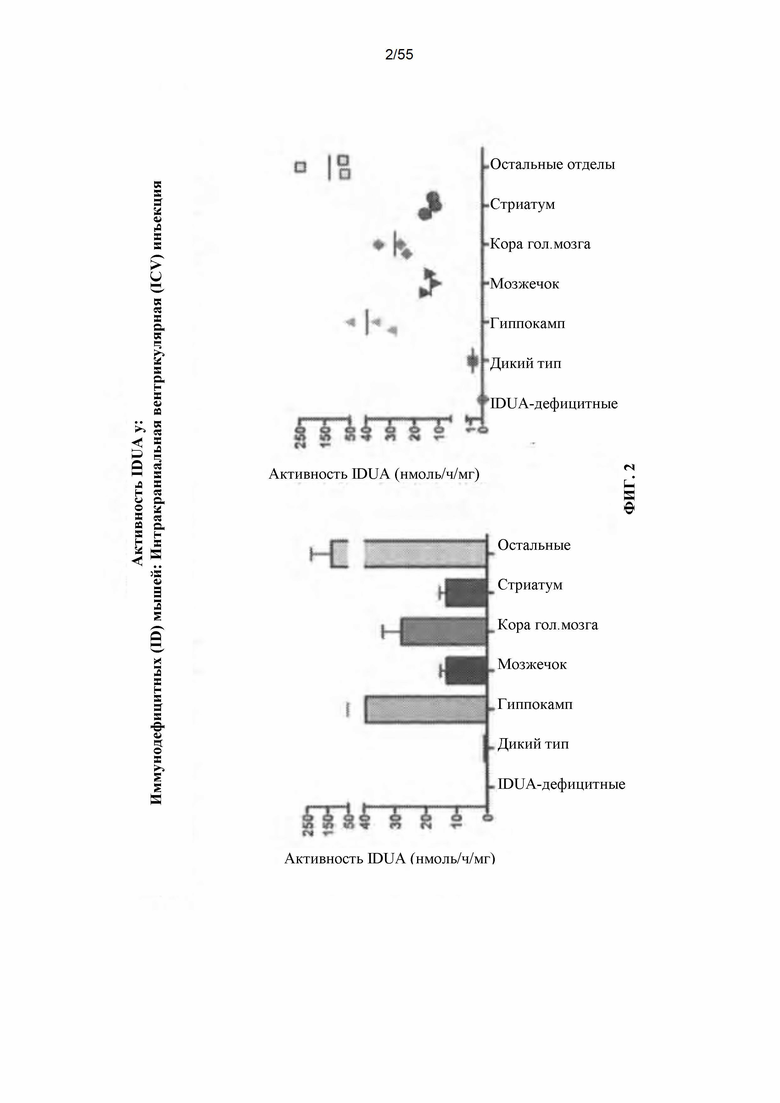
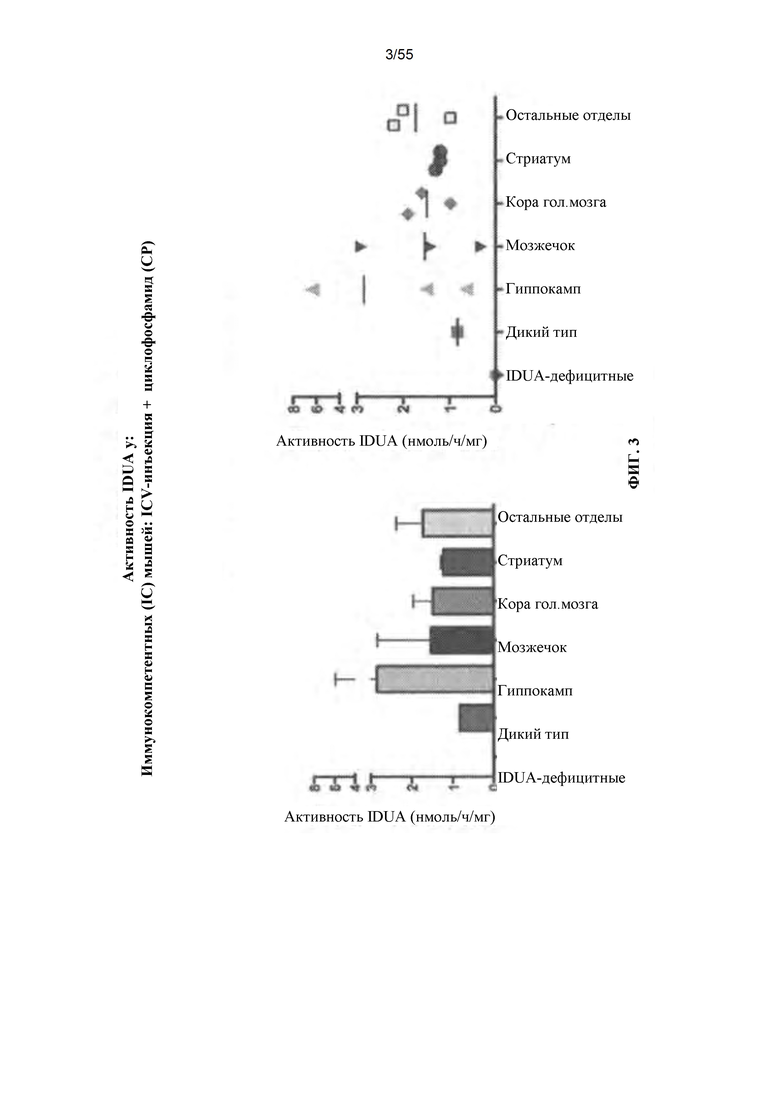
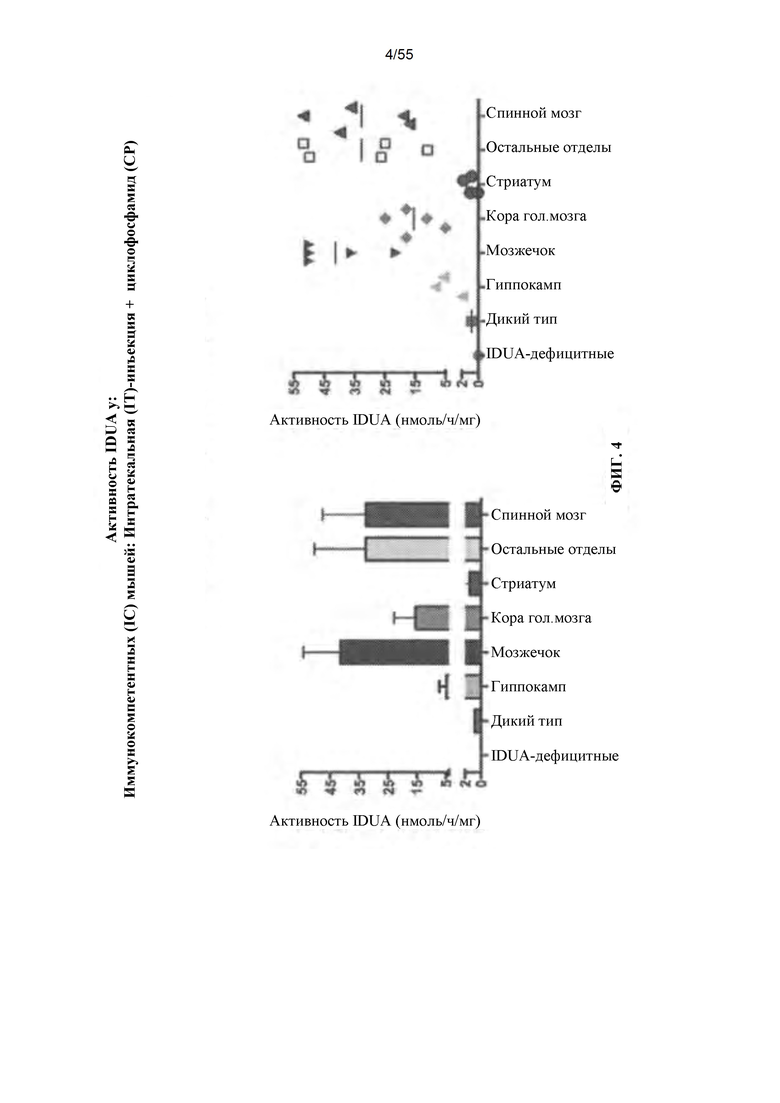
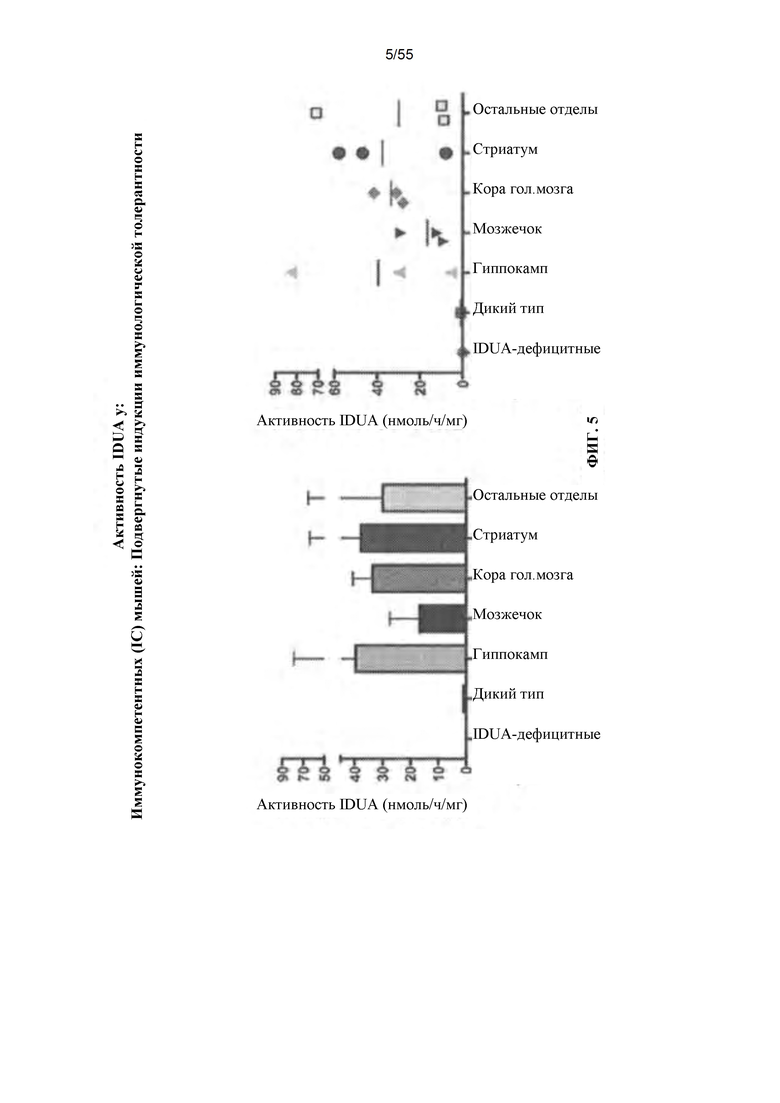
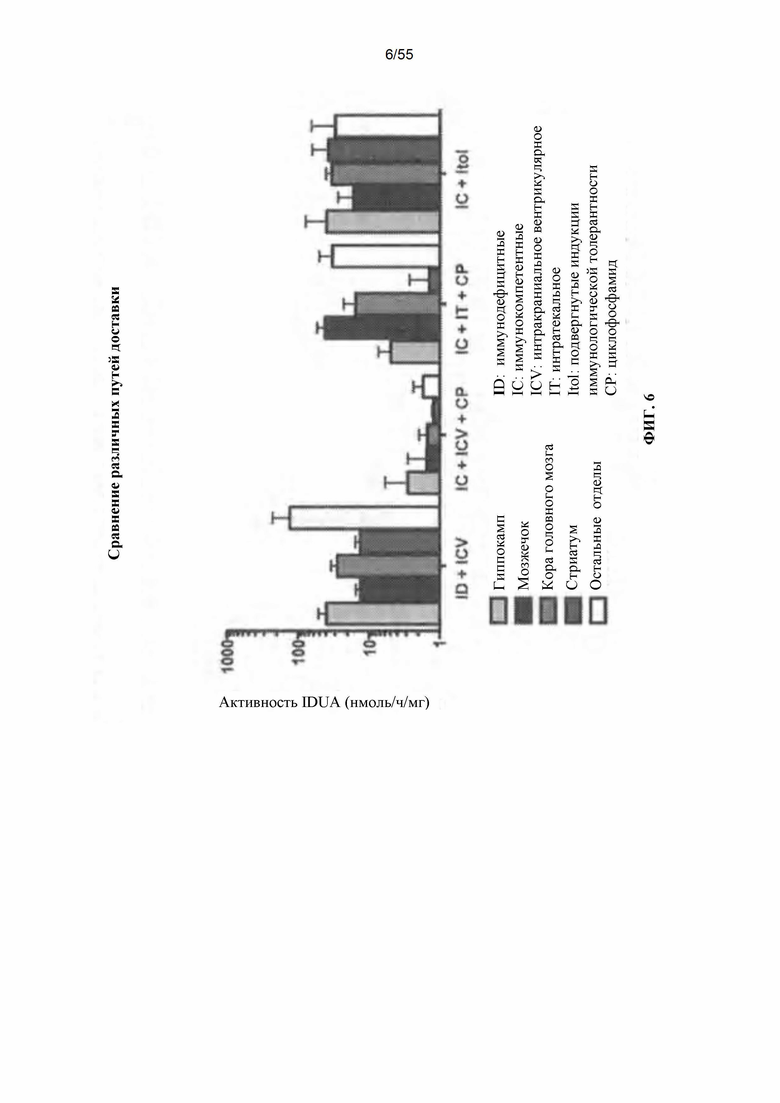
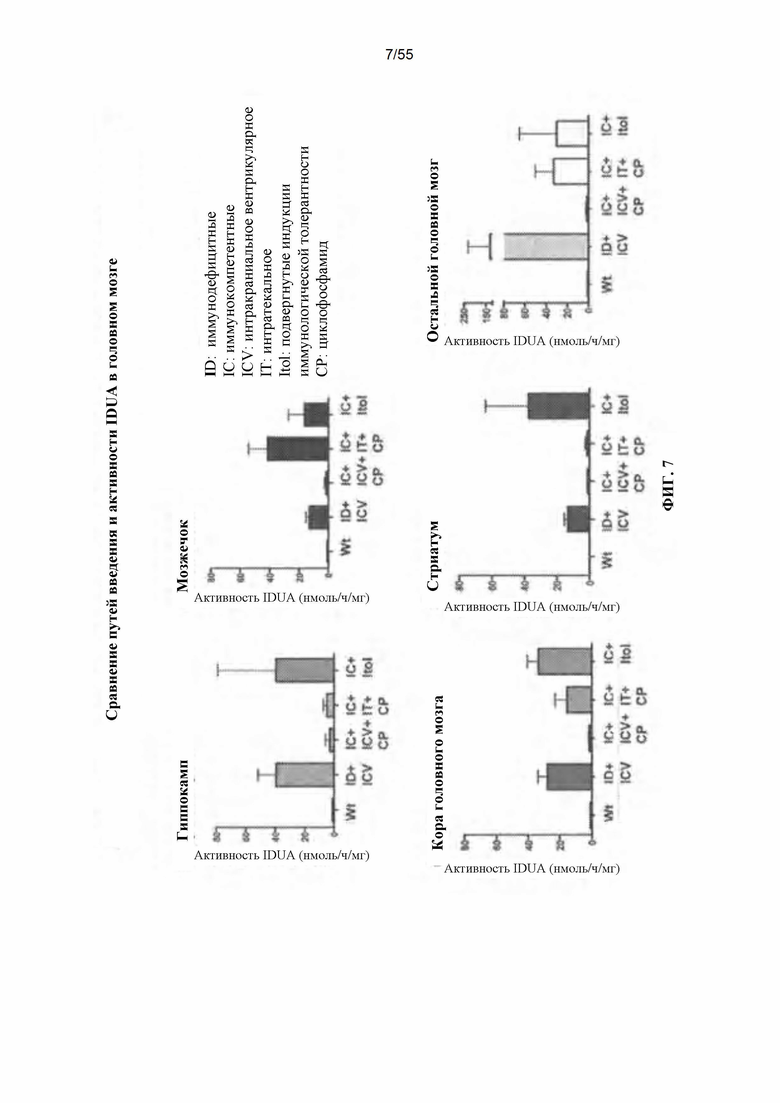
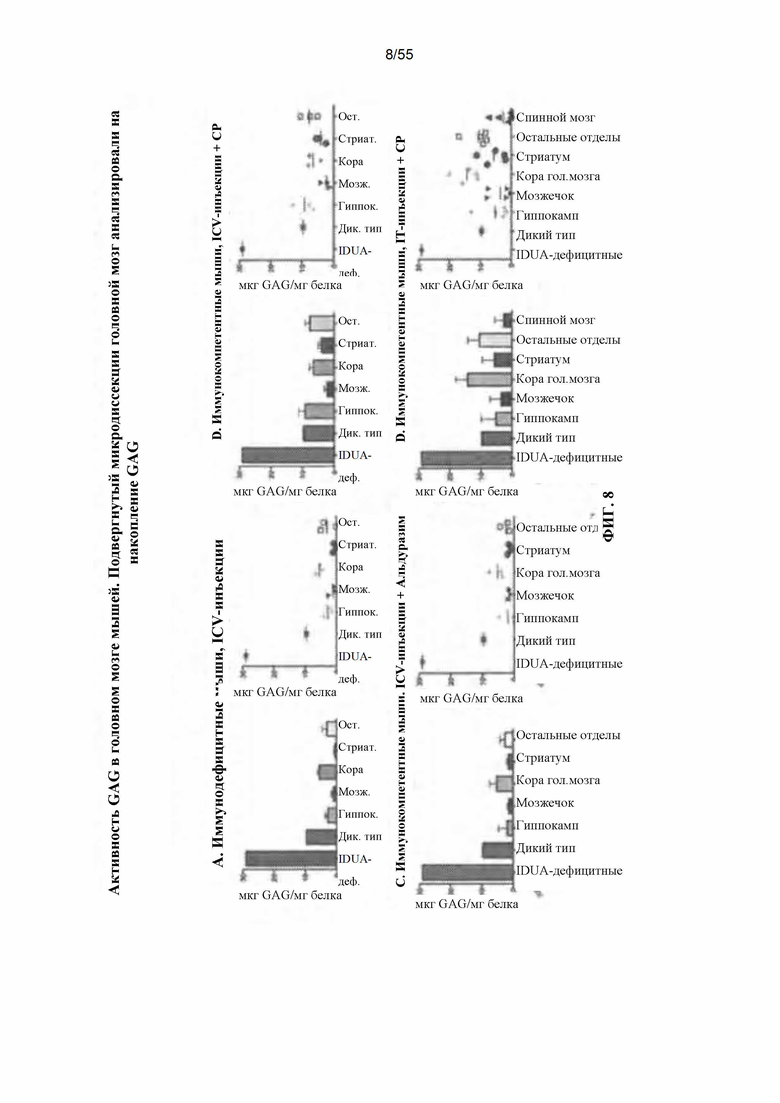
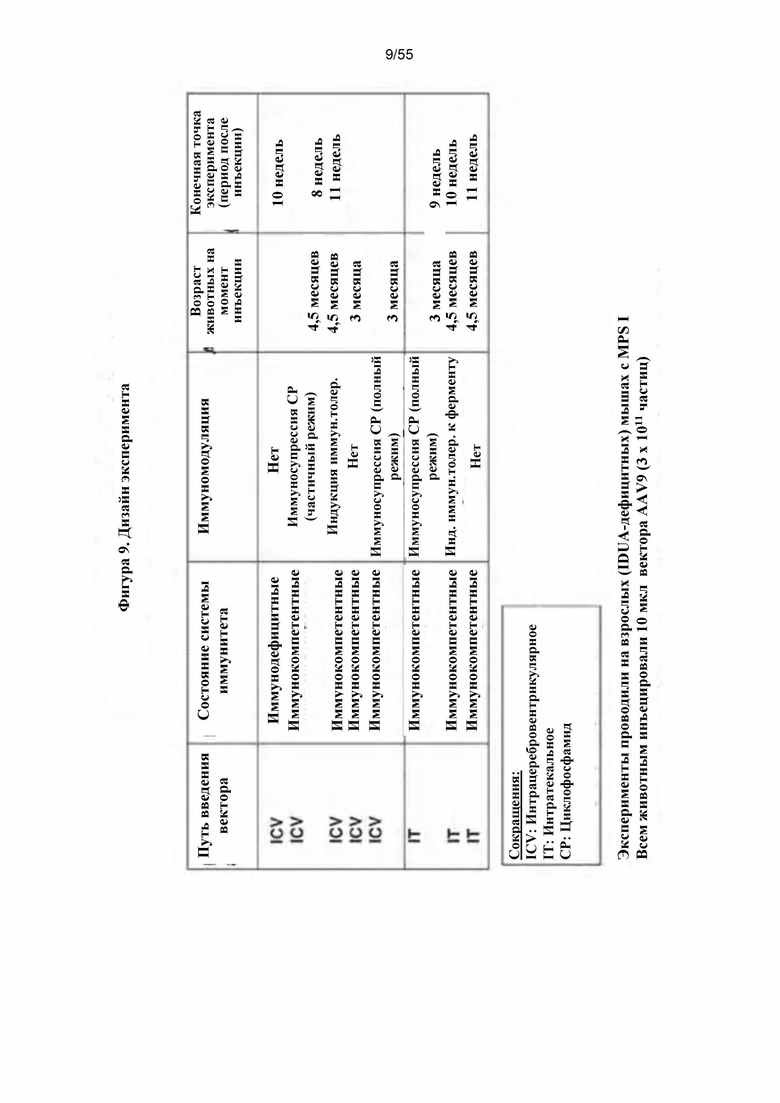
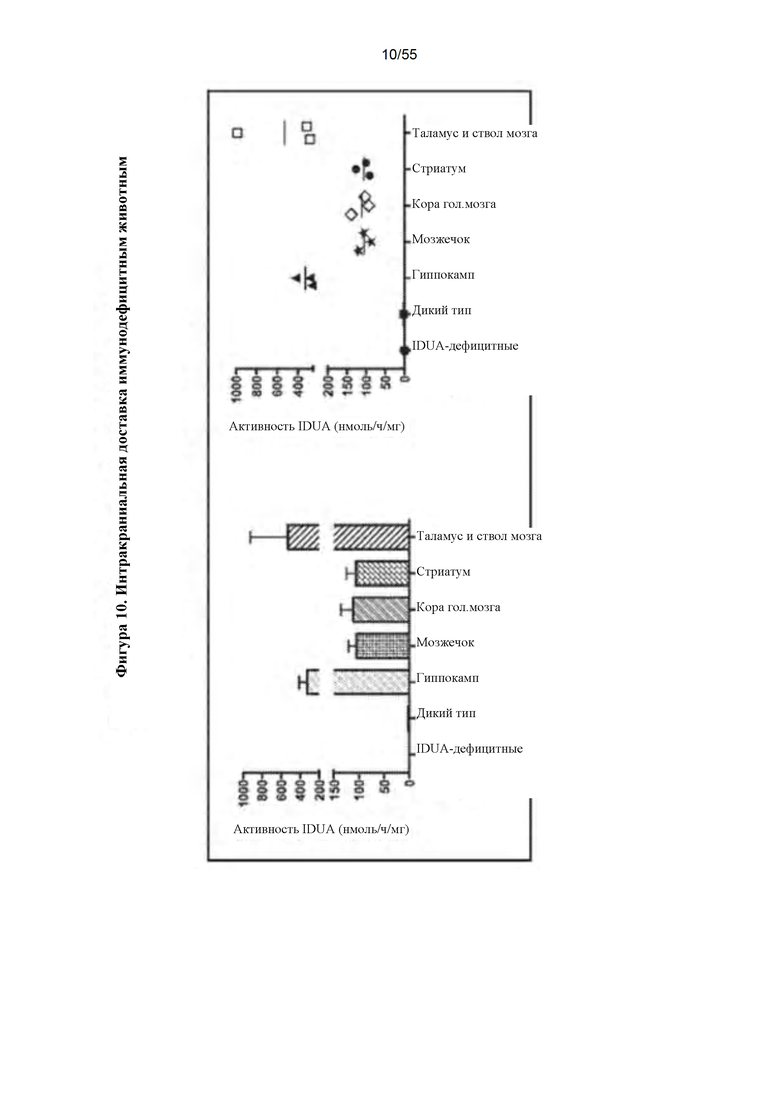
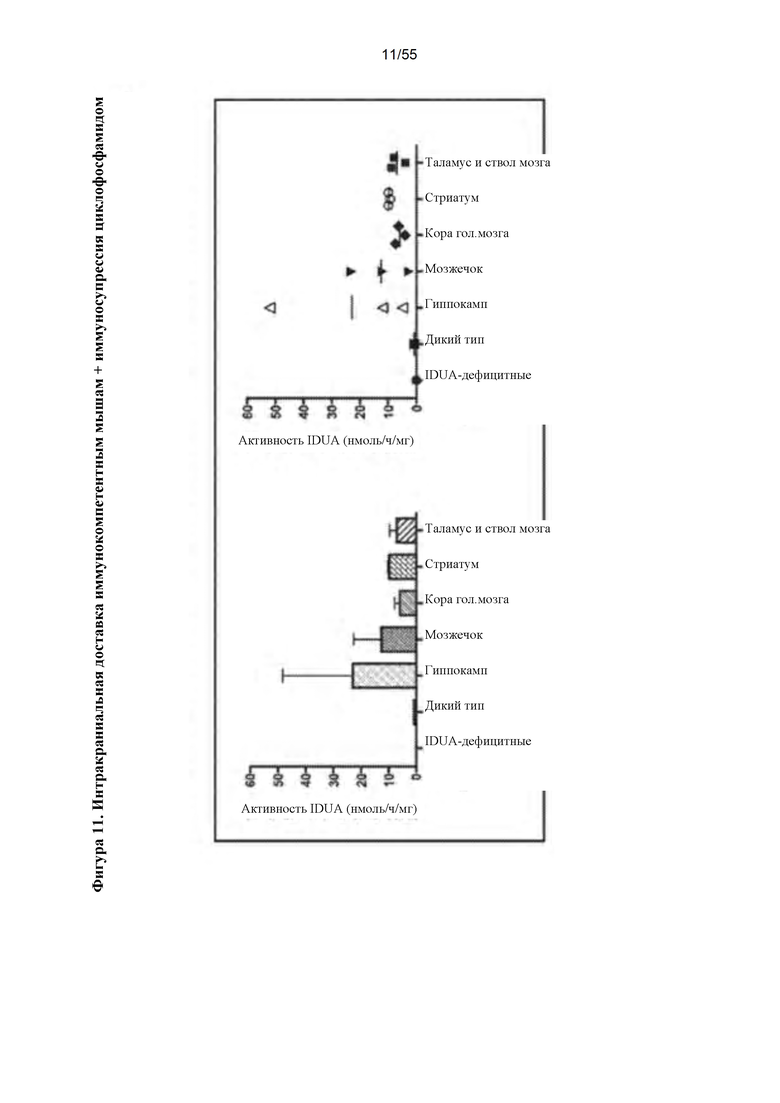
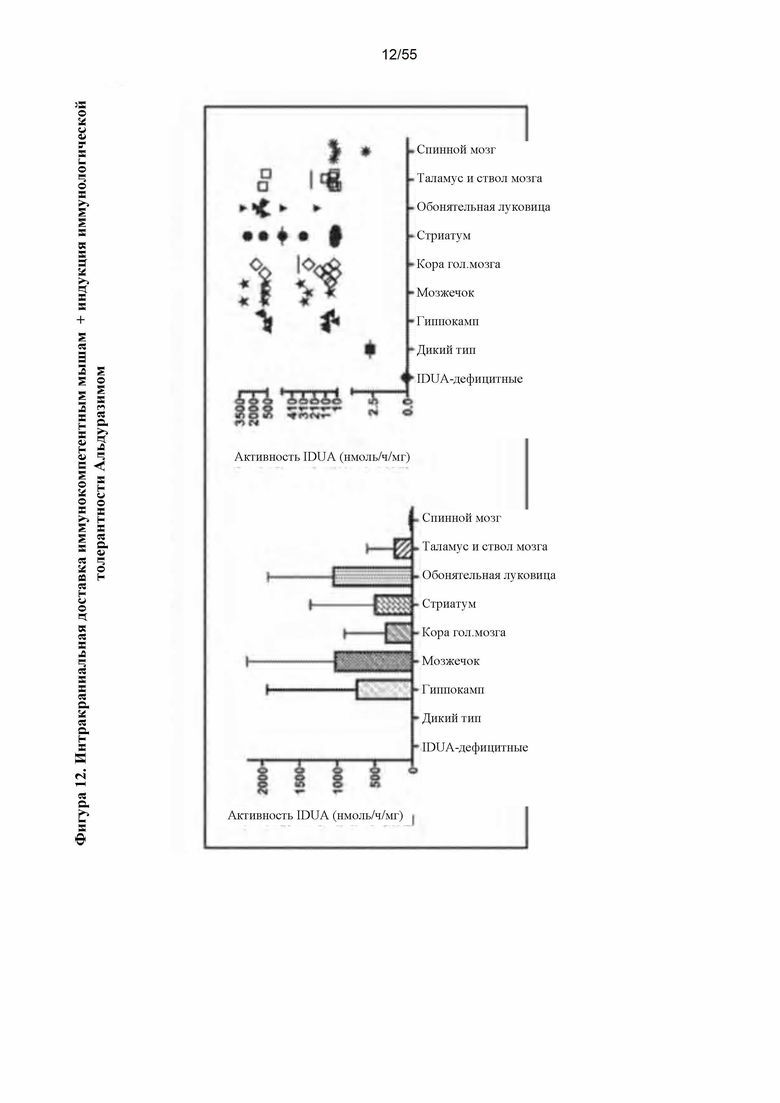
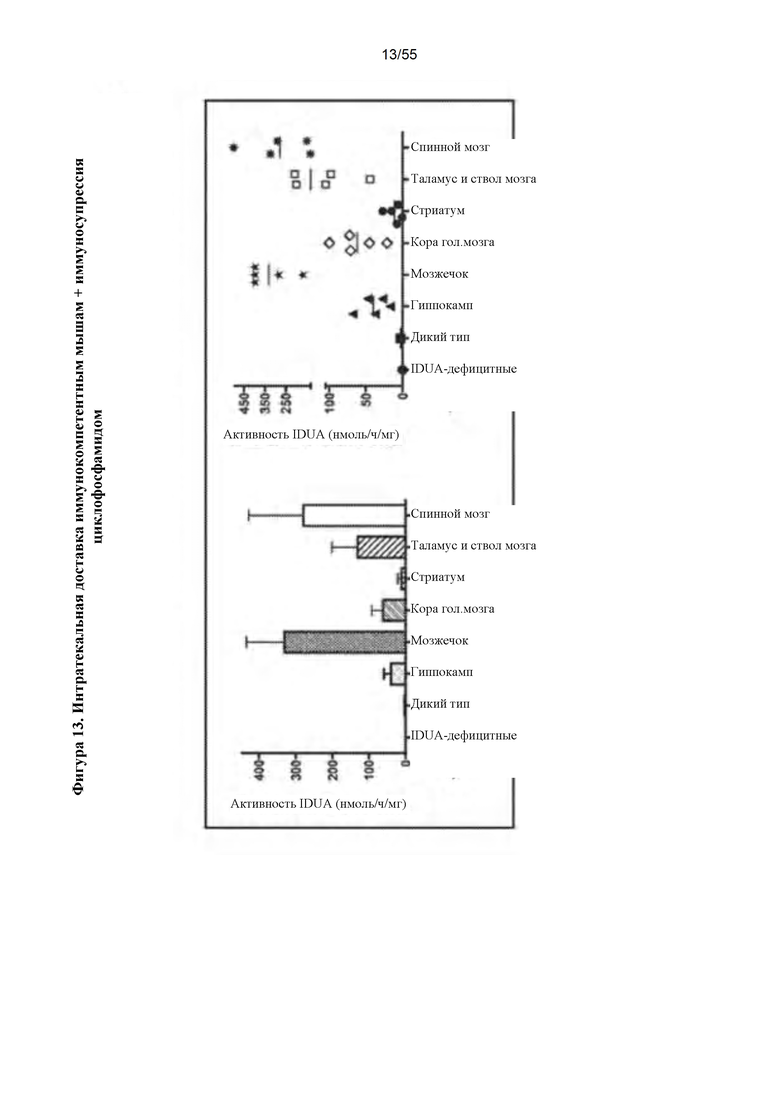
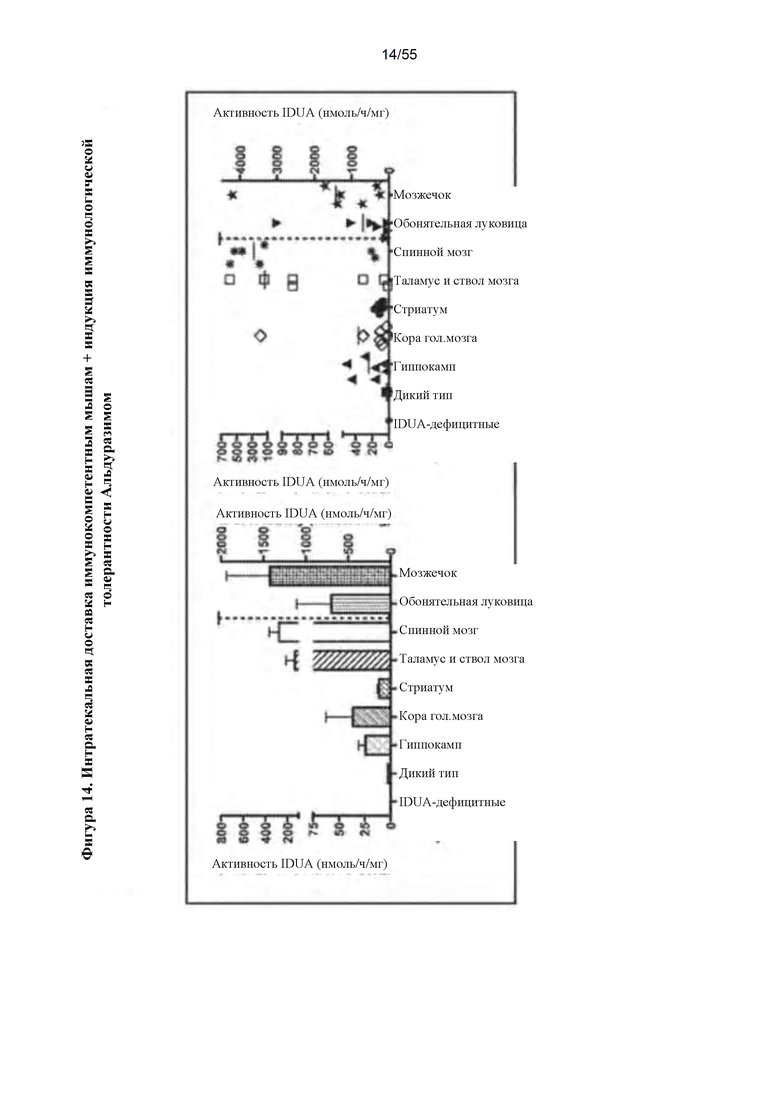
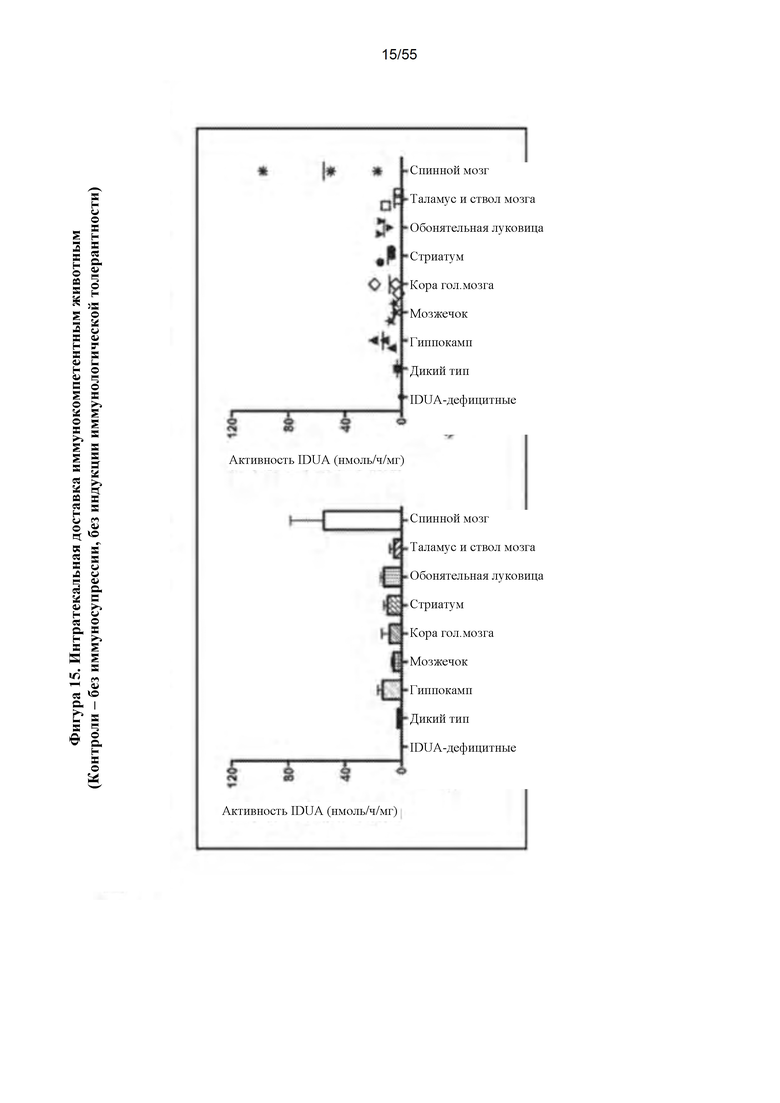
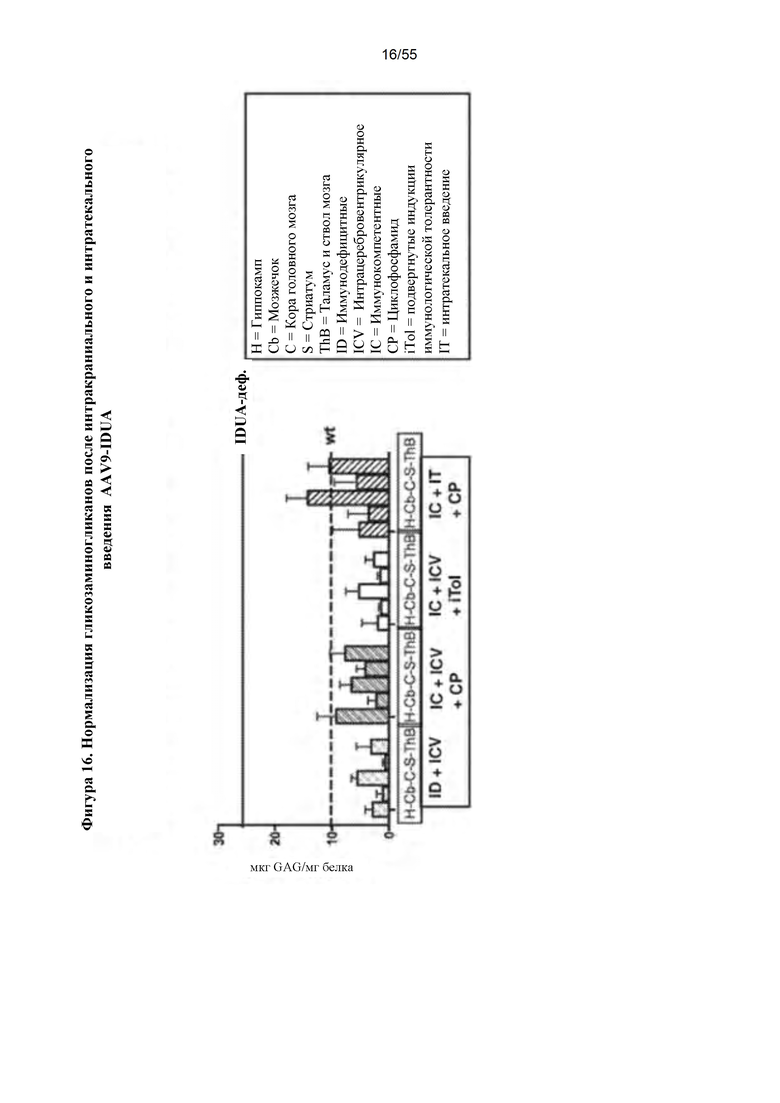
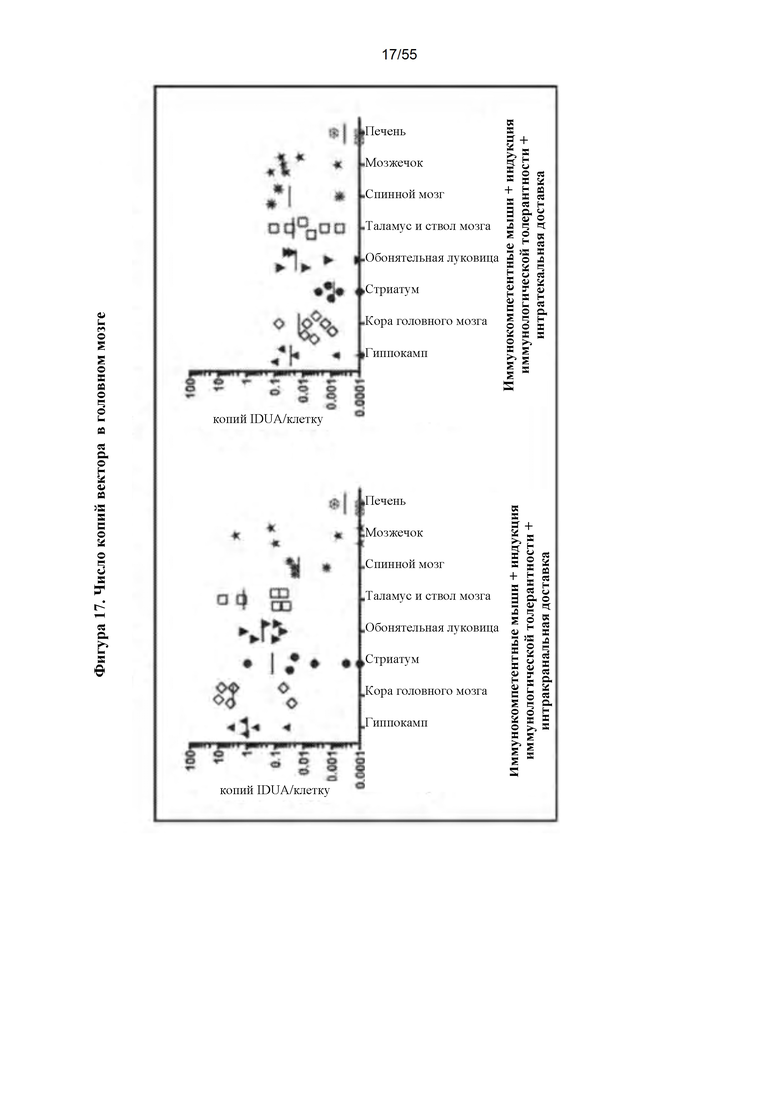

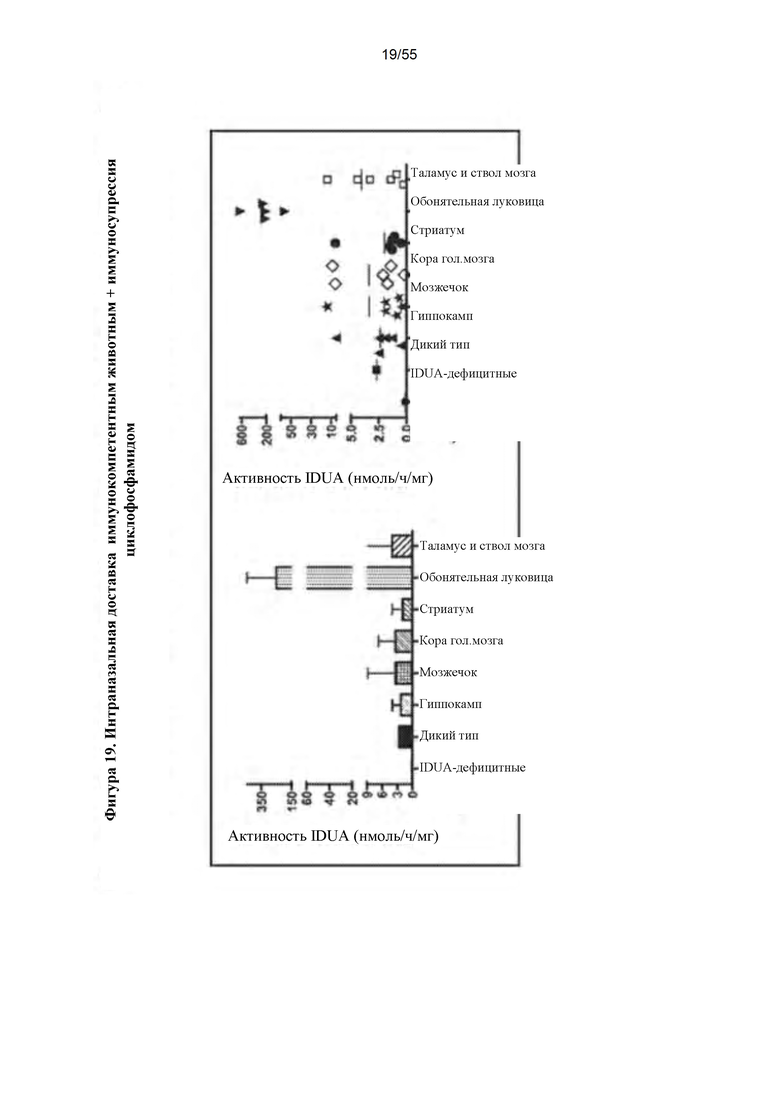


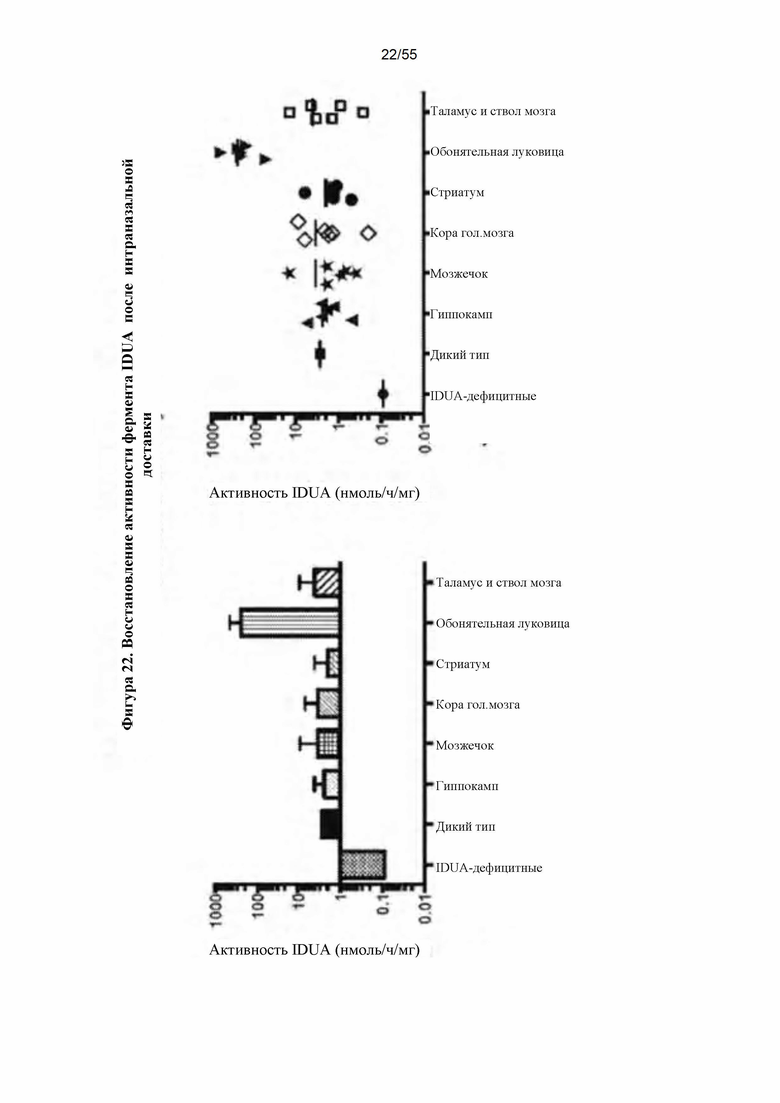
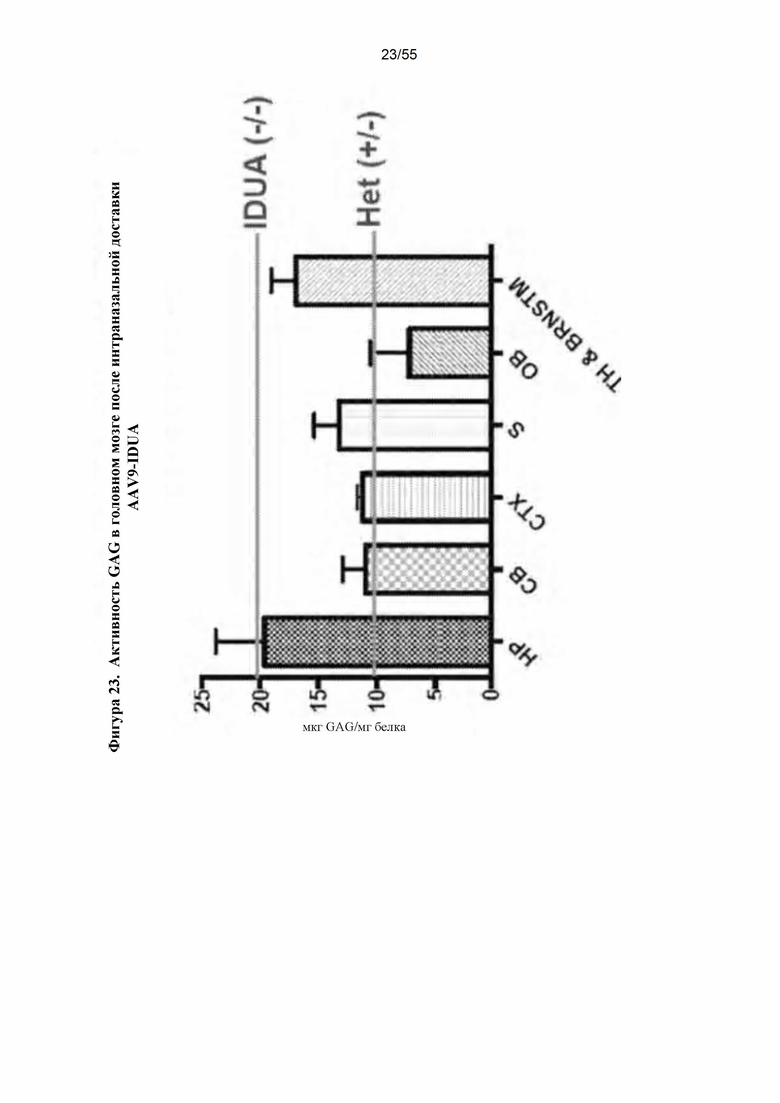

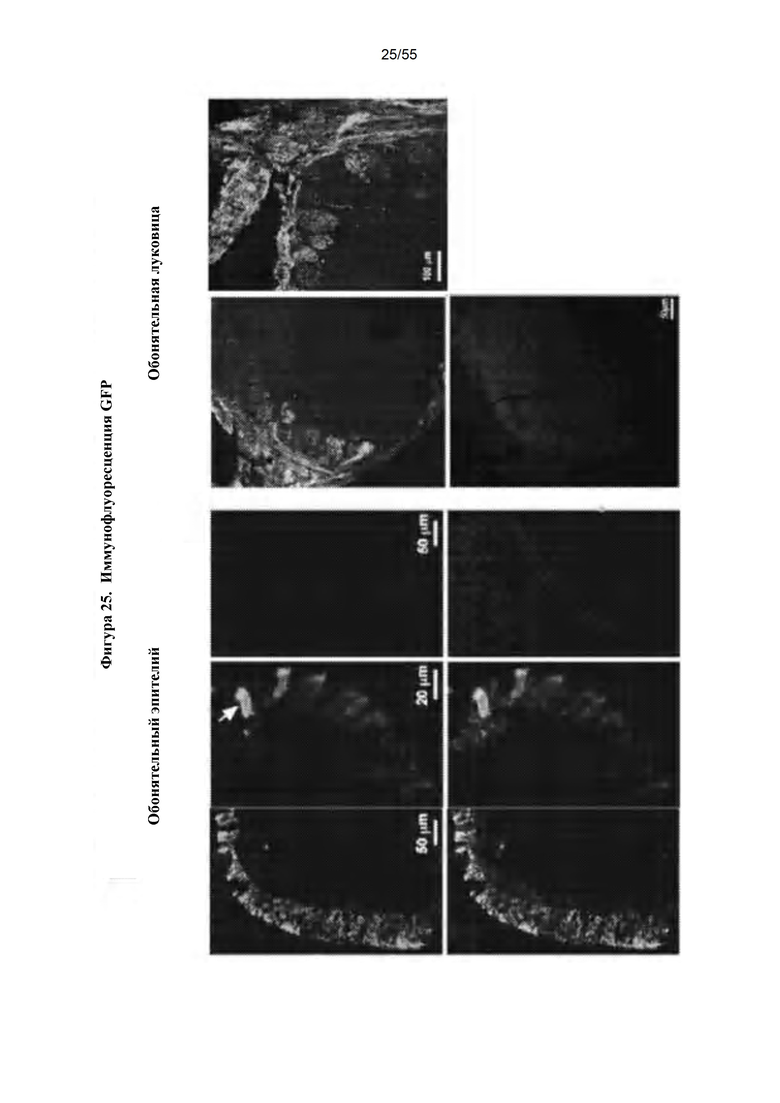
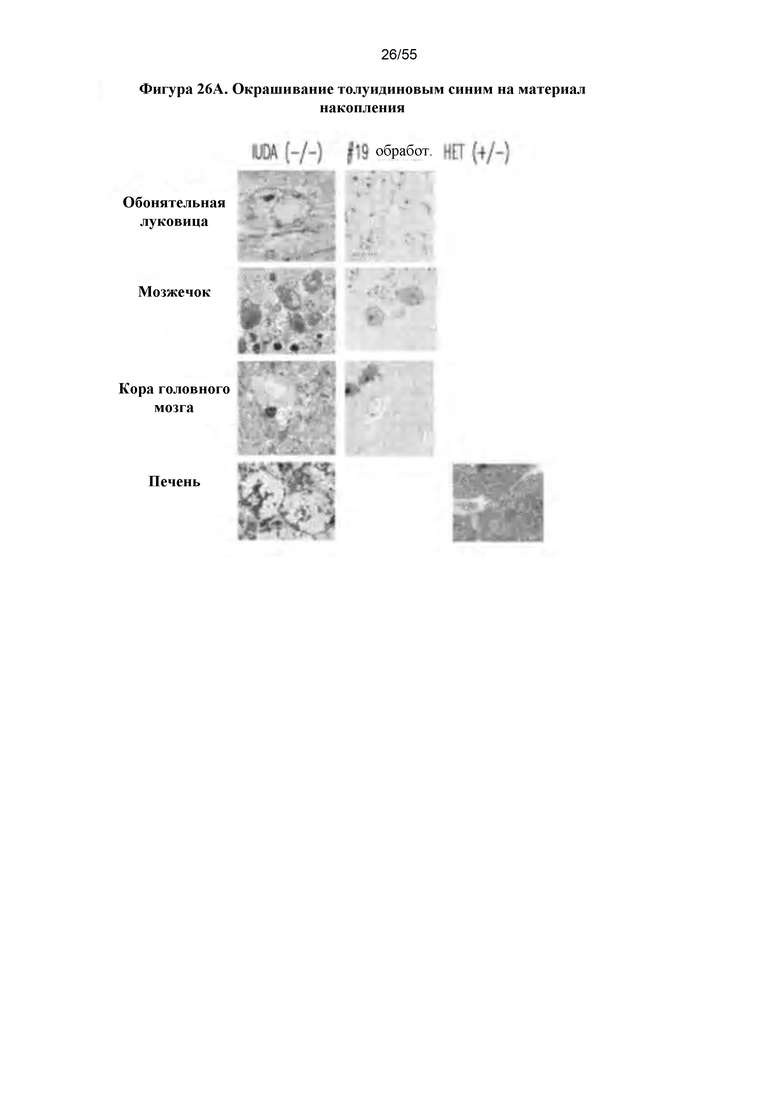

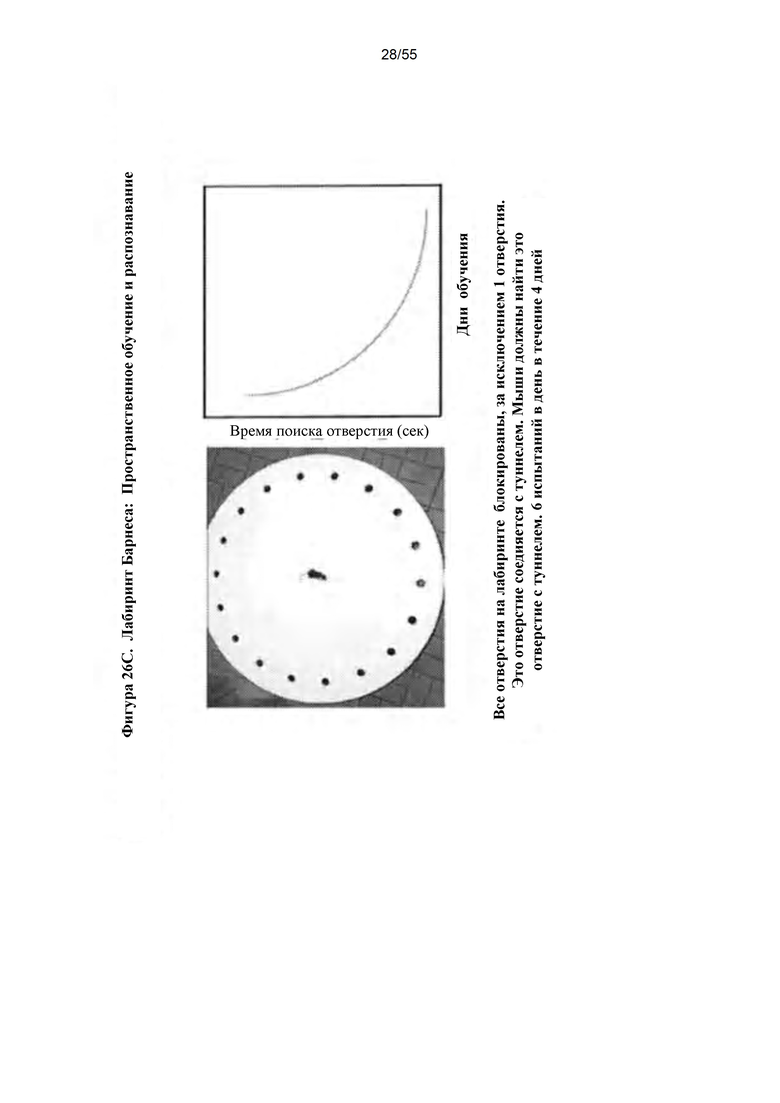
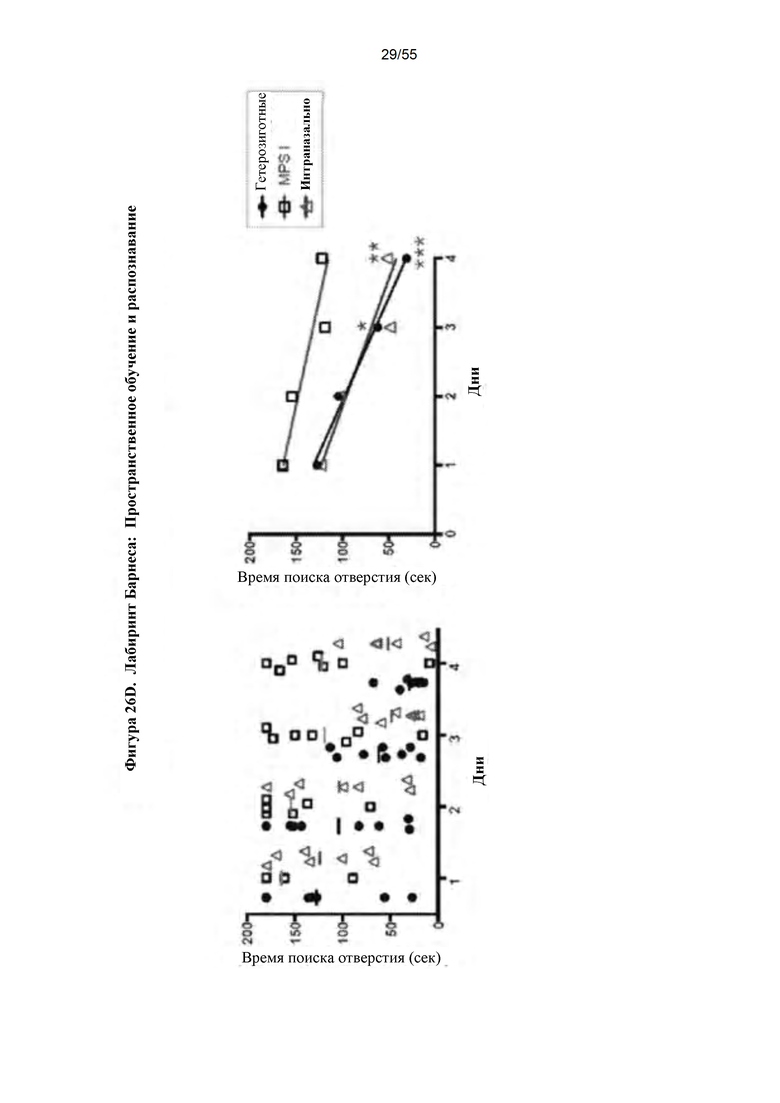
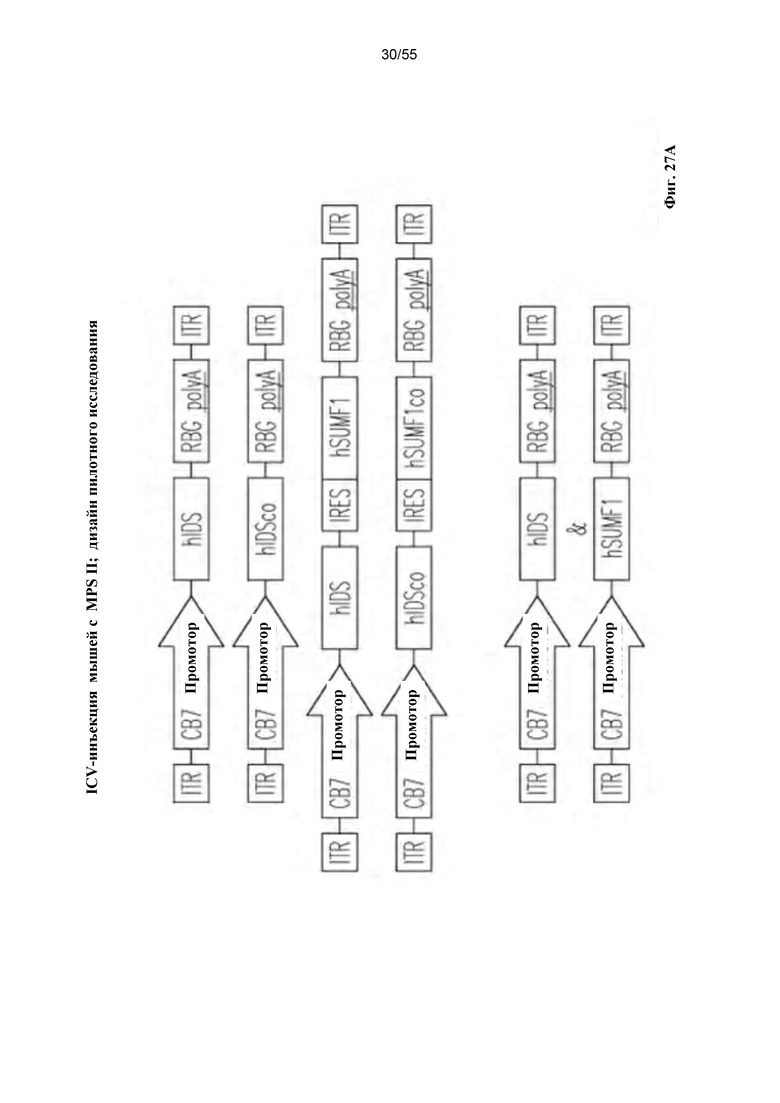
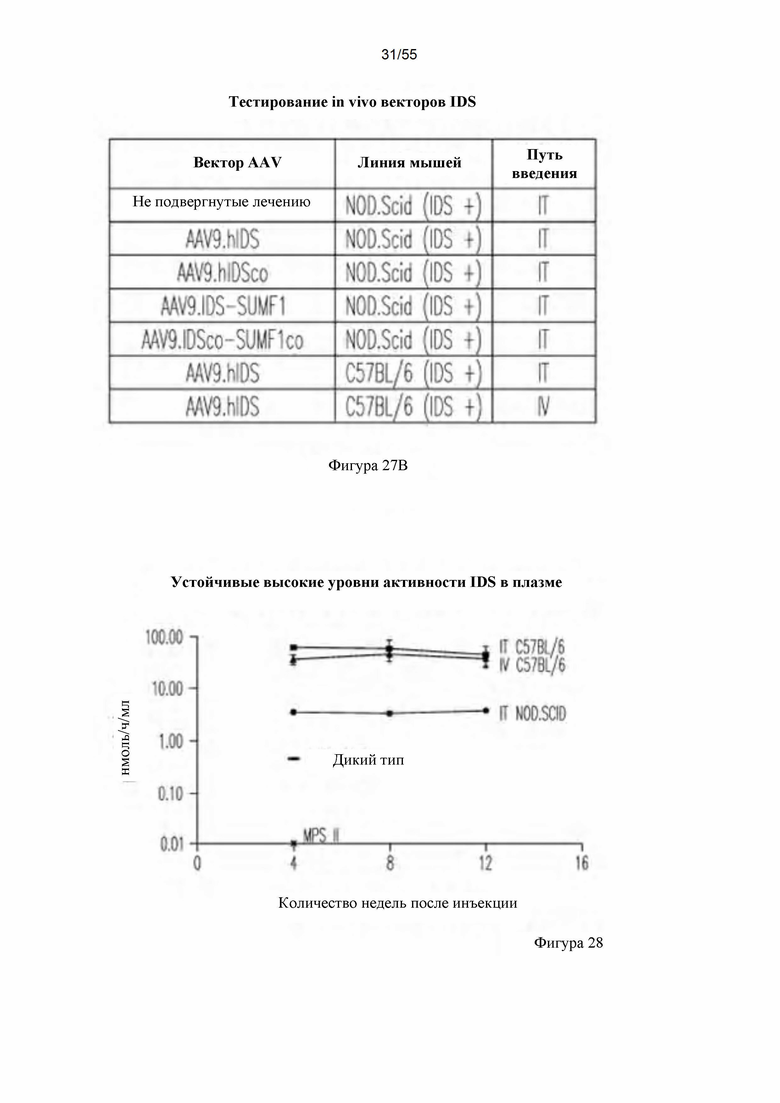
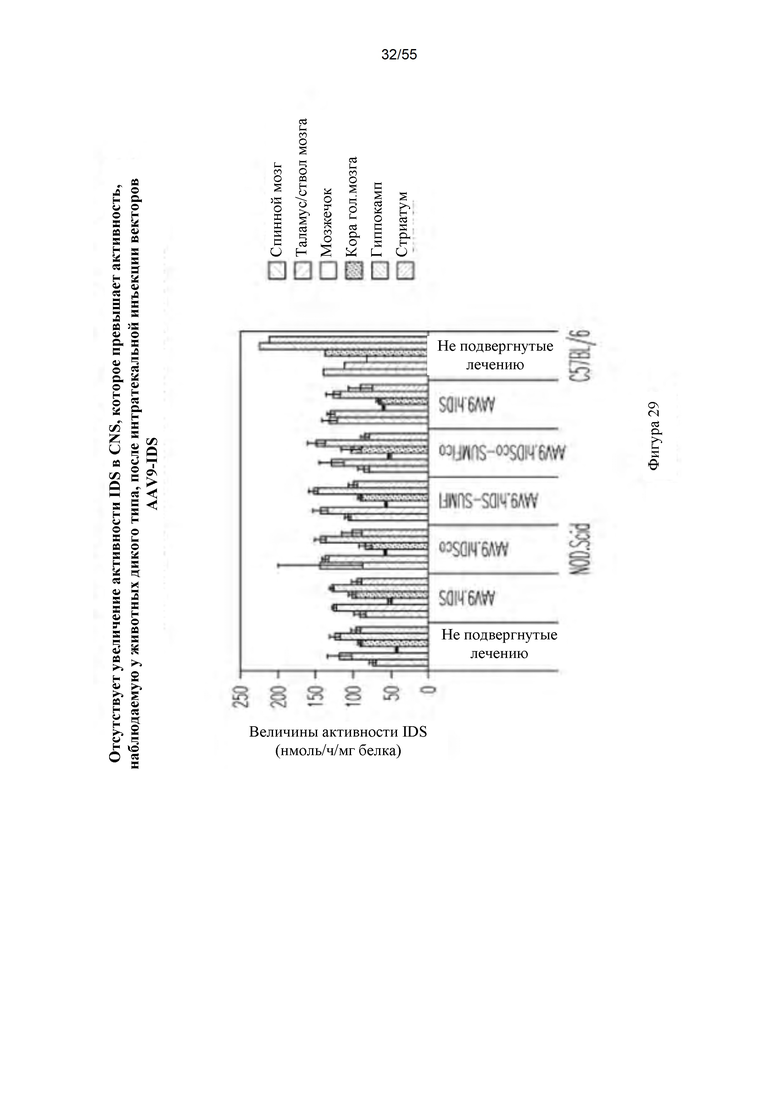
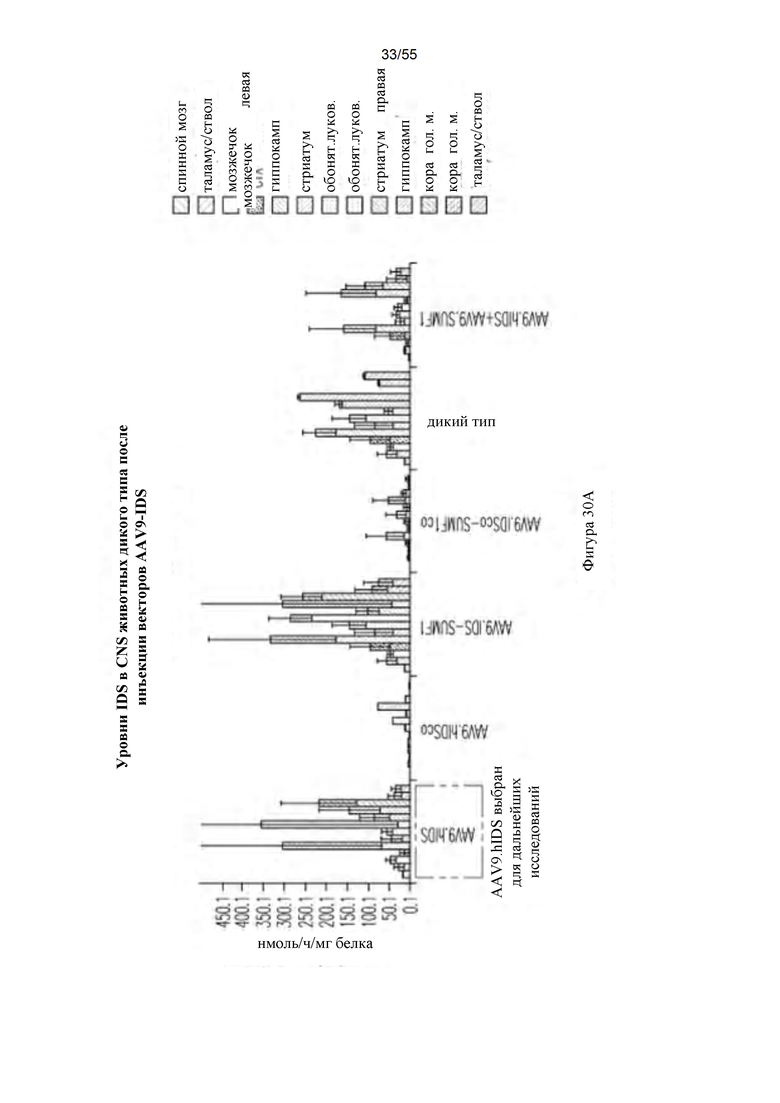
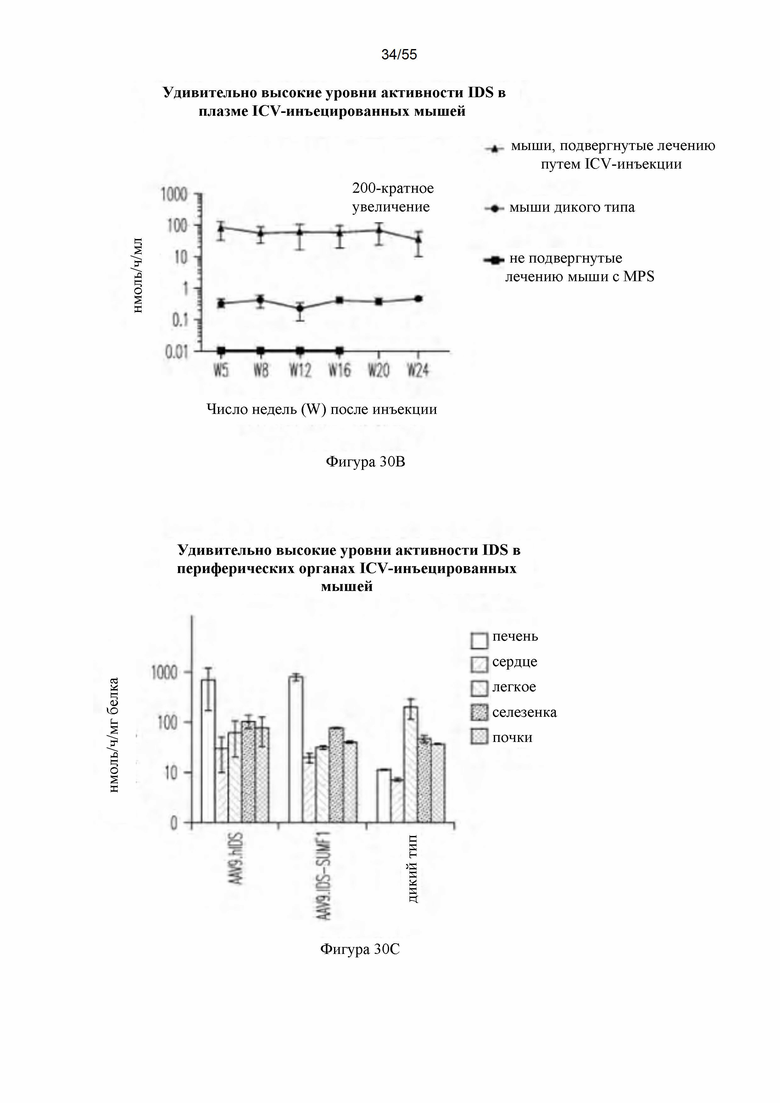
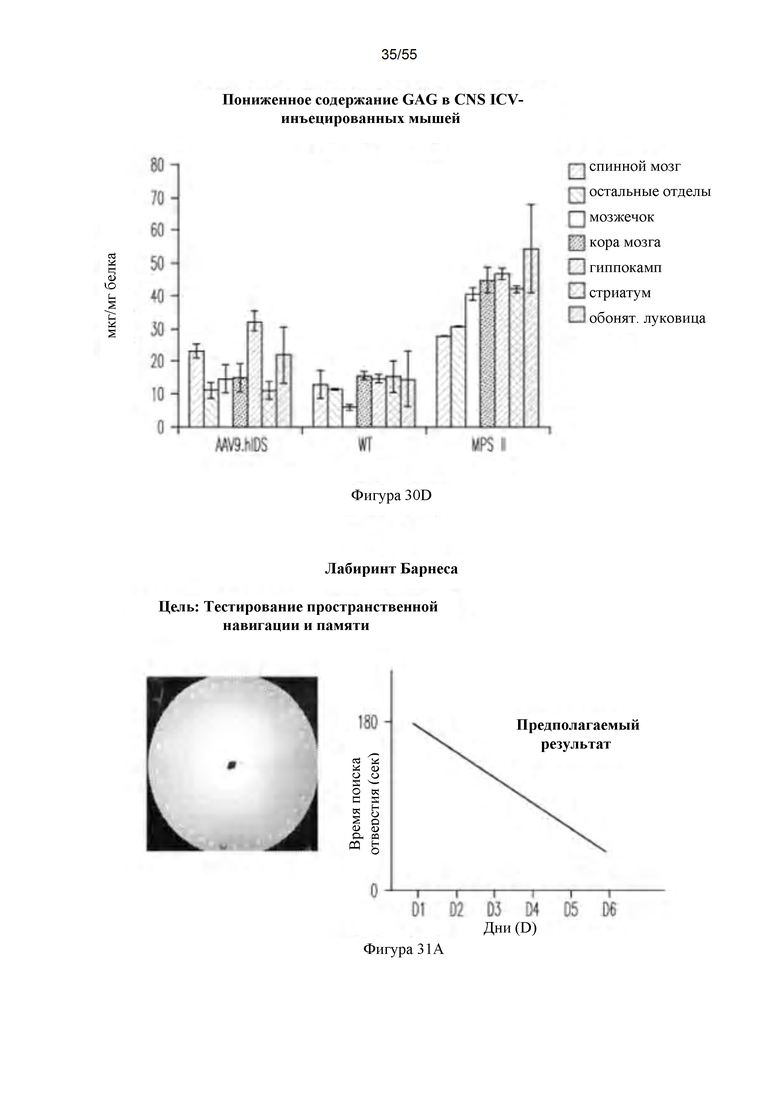
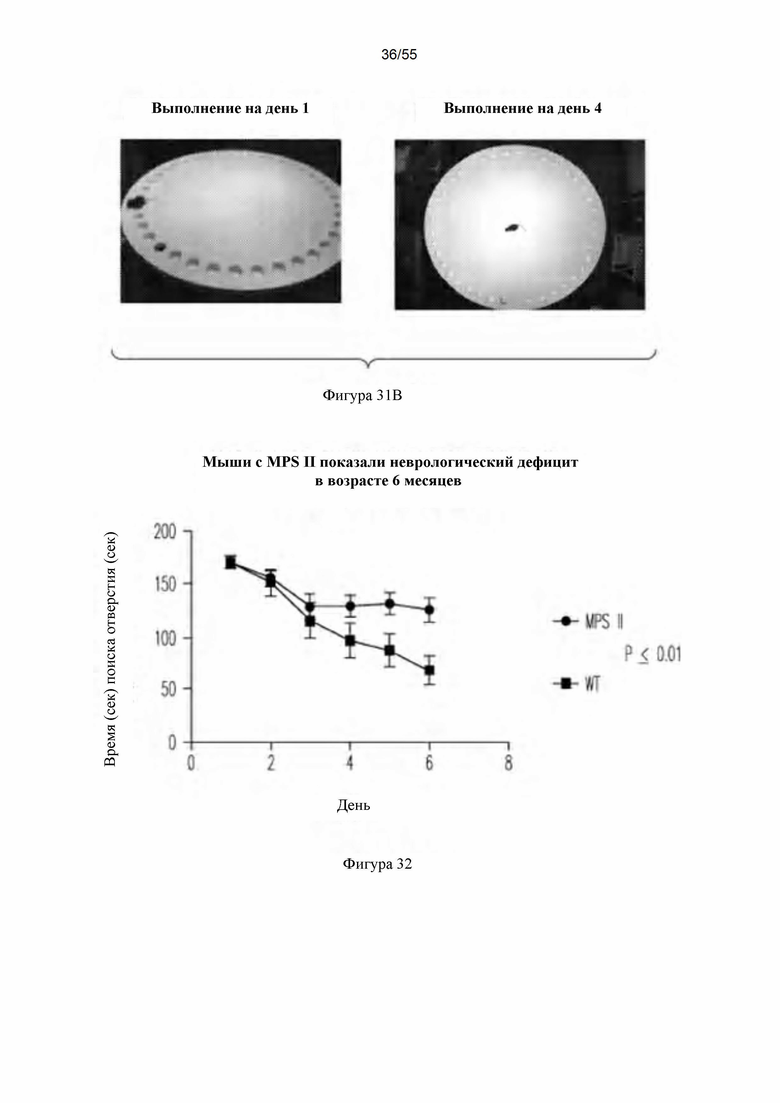
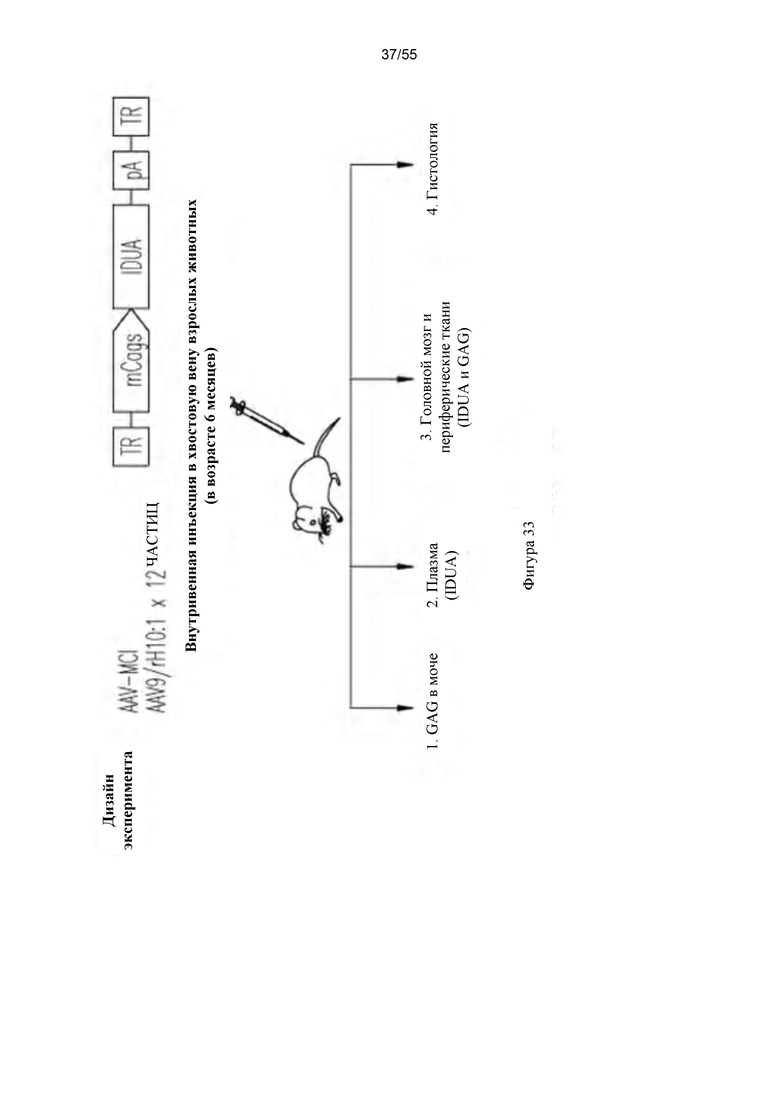
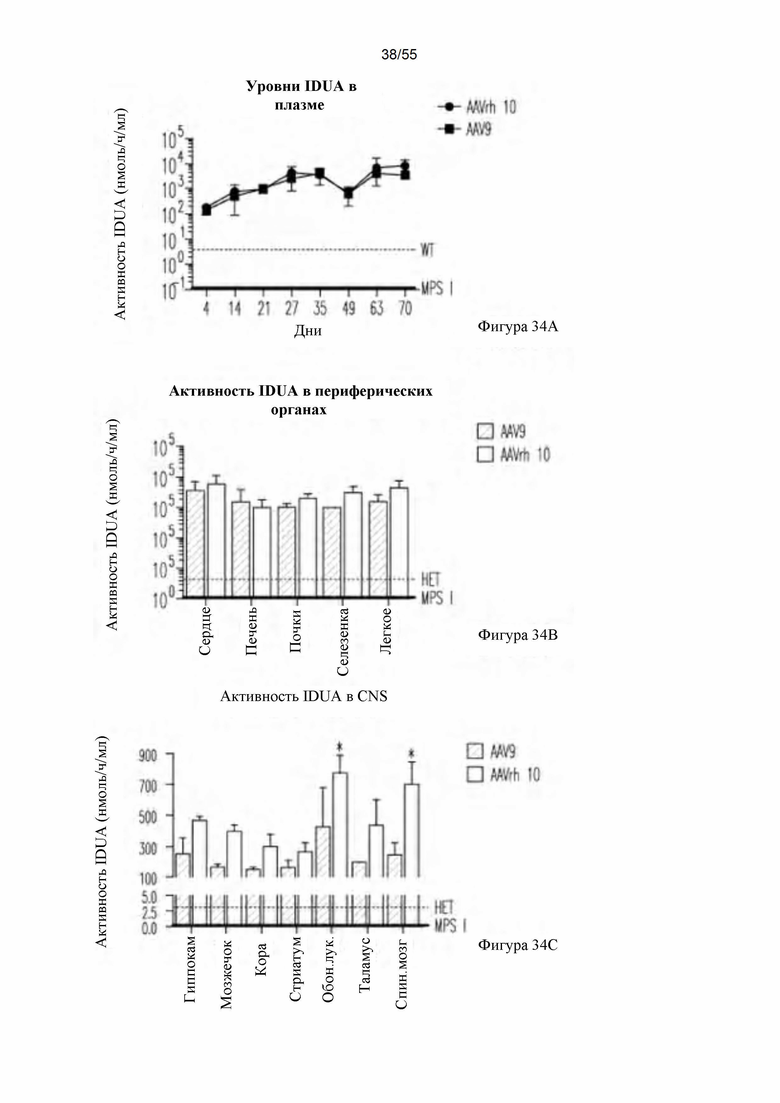
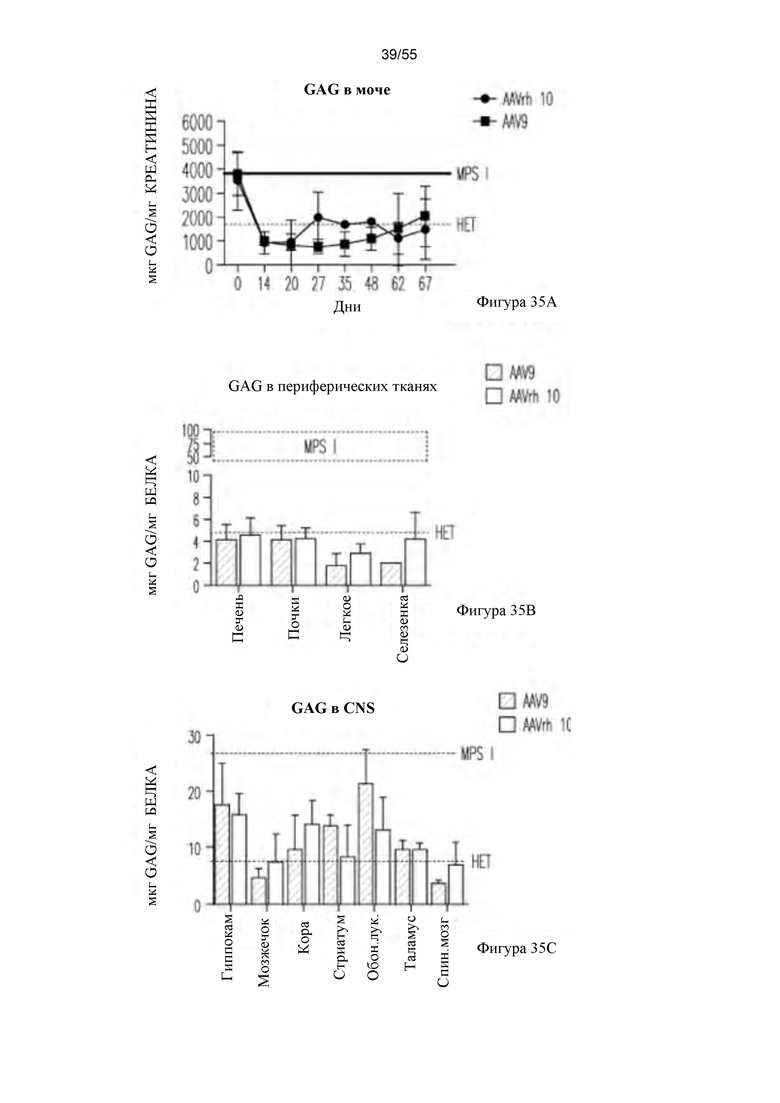






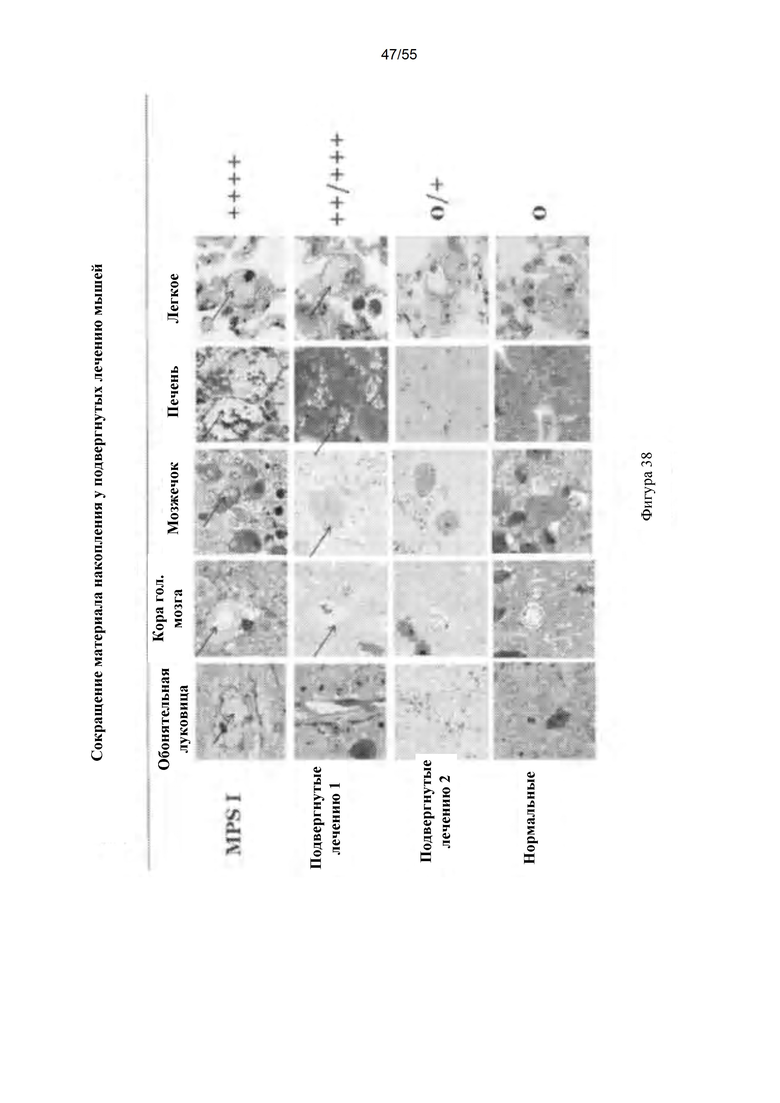

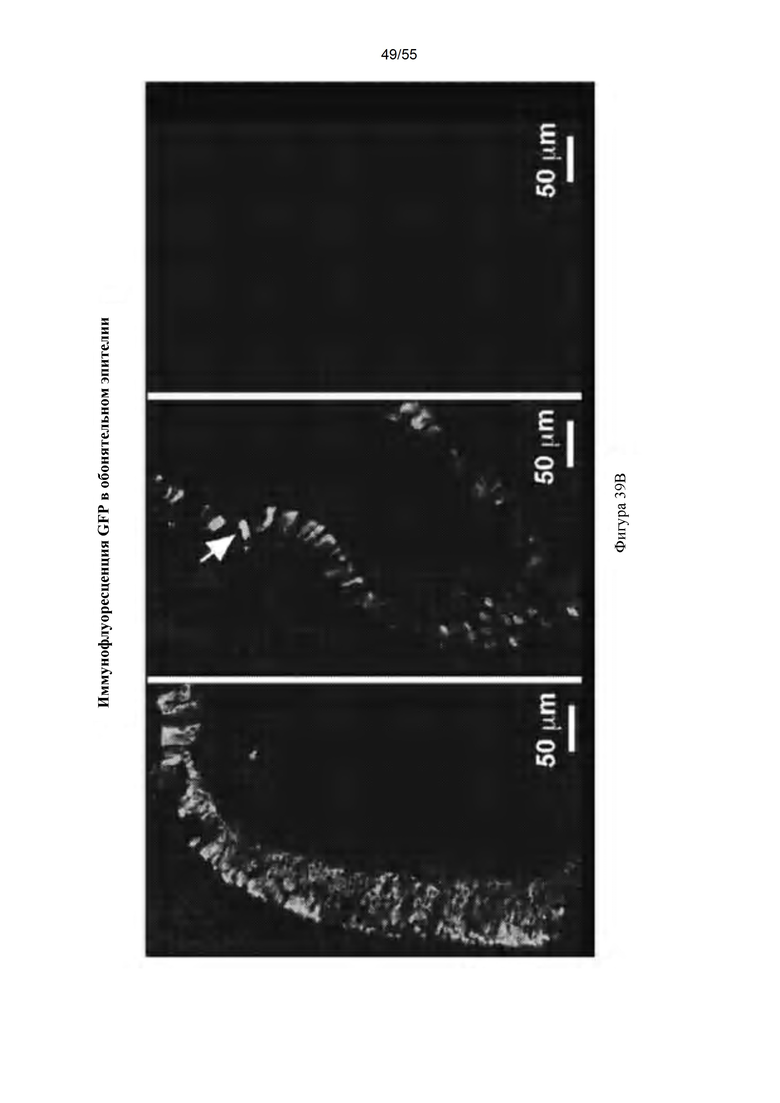
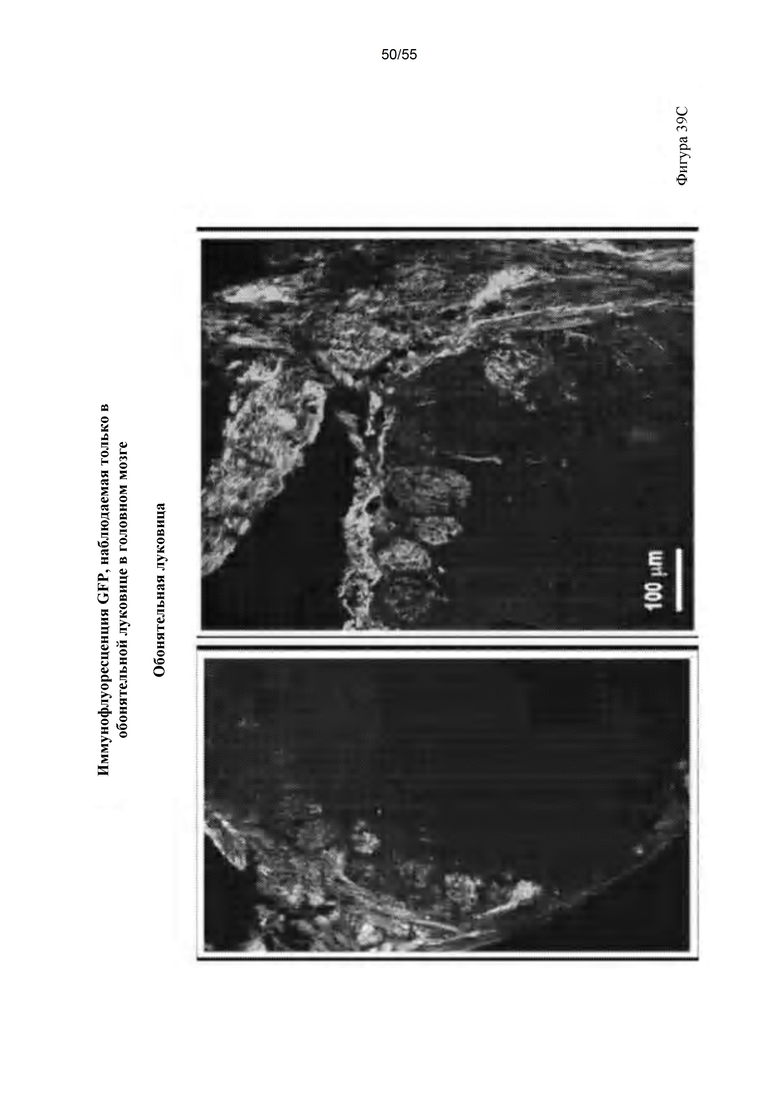
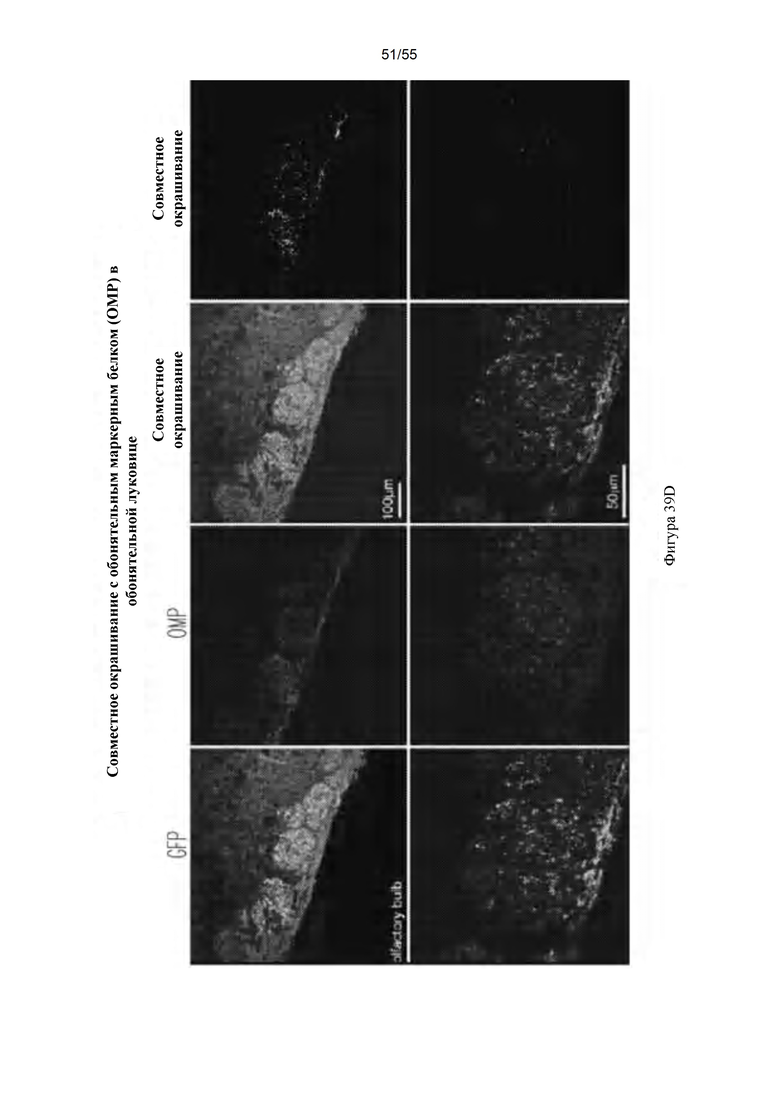
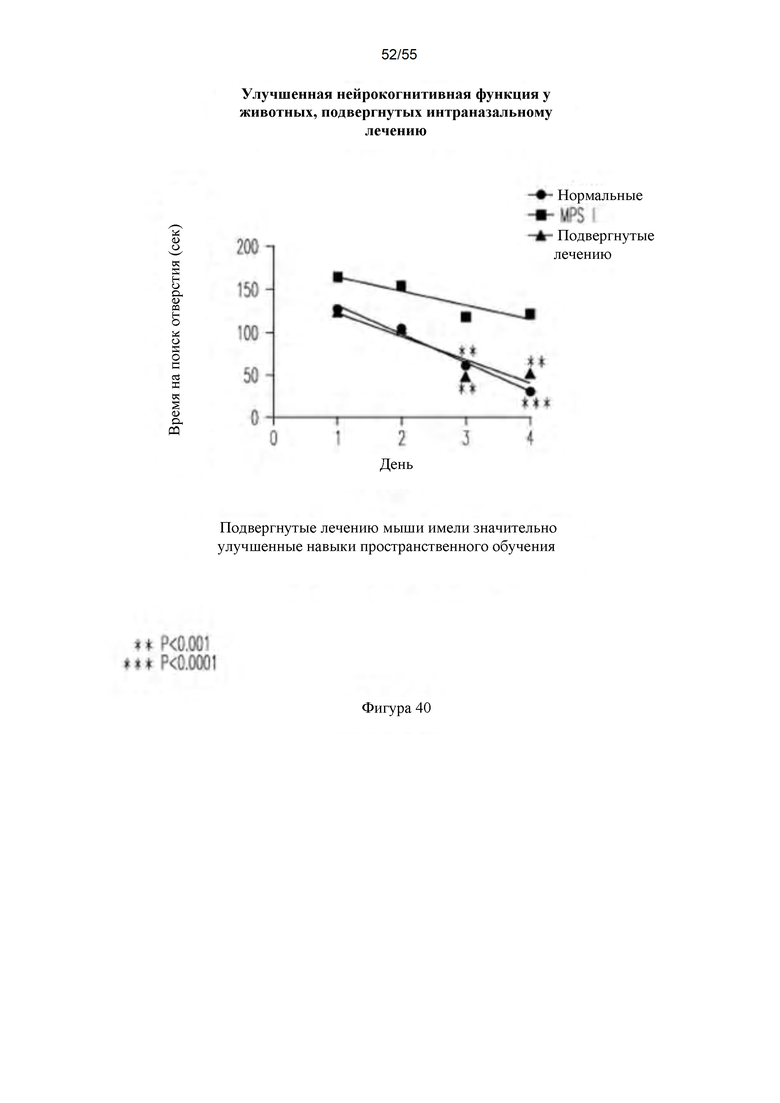
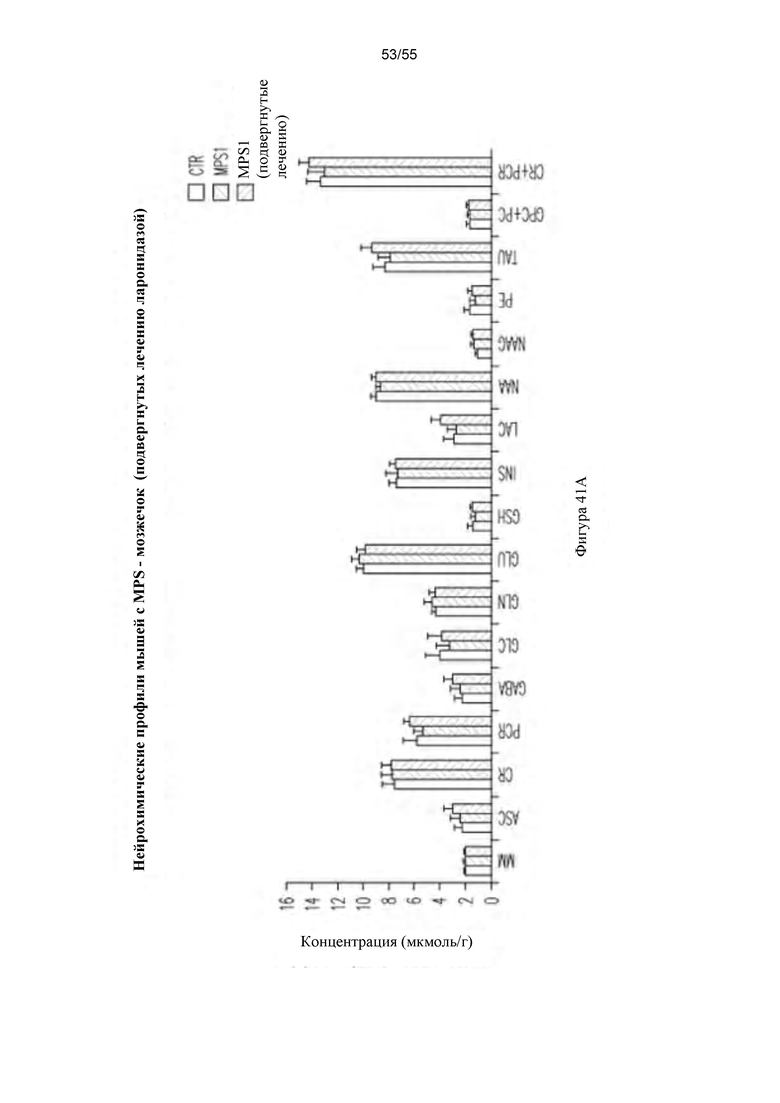
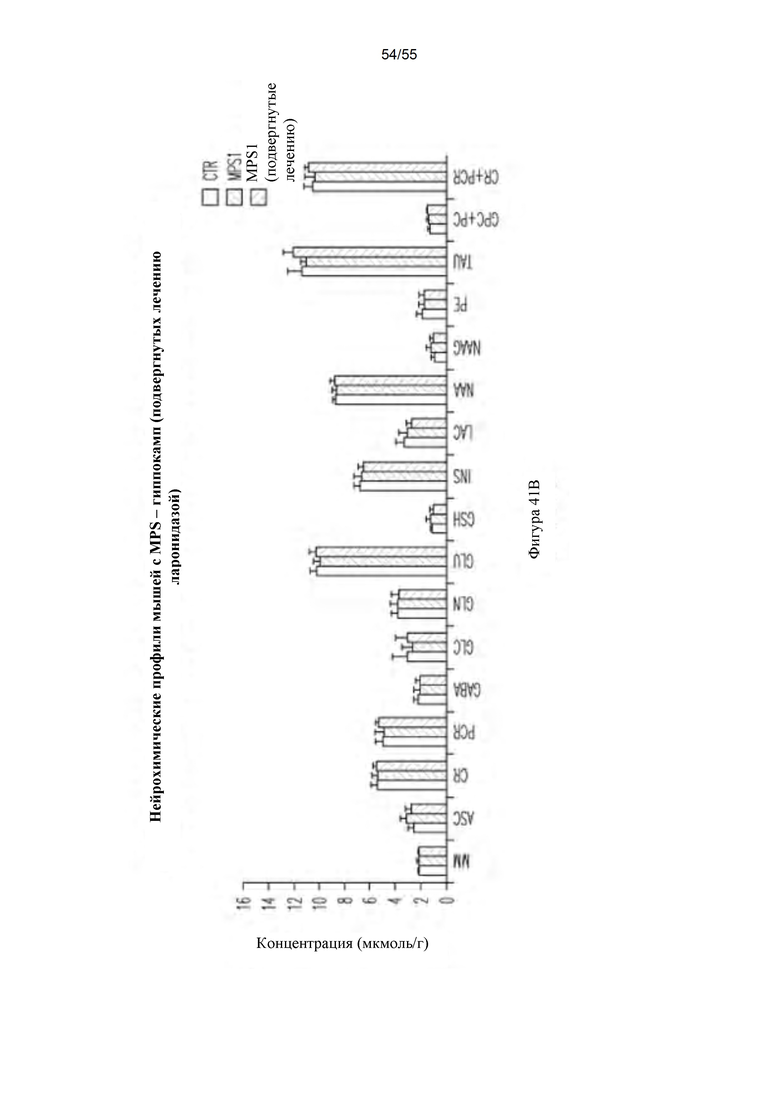

Изобретение относится к биотехнологии. Описан способ усиления нейрокогнитивной функции у млекопитающего, имеющего лизосомную болезнь накопления, включающий: интратекальное введение млекопитающему композиции, содержащей количество рекомбинантного аденоассоциированного вируса (rAAV), содержащего открытую рамку считывания, кодирующую альфа-L-идуронидазу или идуронат-2-сульфатазу, эффективное для усиления нейрокогнитивной функции у млекопитающего по сравнению с млекопитающим, имеющим лизосомную болезнь накопления, которому не вводили rAAV, где rAAV имеет капсид rAAV9 или rAAVrh10. Изобретение расширяет арсенал средств усиления нейрокогнитивной функции у млекопитающего. 20 з.п. ф-лы, 42 ил., 8 пр.
1. Способ усиления нейрокогнитивной функции у млекопитающего, имеющего лизосомную болезнь накопления, включающий: интратекальное введение млекопитающему композиции, содержащей количество рекомбинантного аденоассоциированного вируса (rAAV), содержащего открытую рамку считывания, кодирующую альфа-L-идуронидазу или идуронат-2-сульфатазу, эффективное для усиления нейрокогнитивной функции у млекопитающего по сравнению с млекопитающим, имеющим лизосомную болезнь накопления, которому не вводили rAAV, где rAAV имеет капсид rAAV9 или rAAVrh10.
2. Способ по п. 1, в котором млекопитающее не подвергают лечению иммуносупрессантом.
3. Способ по п. 1, в котором млекопитающее подвергают лечению иммуносупрессантом.
4. Способ по п. 3, в котором иммуносупрессант представляет собой циклофосфамид.
5. Способ по п. 3, в котором иммуносупрессант представляет собой глюкокортикоид, цитостатические агенты, включая алкилирующий агент, антиметаболит, цитотоксический антибиотик, антитело или агент, связывающийся с иммунофилином.
6. Способ по п. 3, в котором иммуносупрессант представляет собой азотистый иприт, нитрозомочевину, соединение платины, метотрексат, азатиоприн, меркаптопурин, фторурацил, дактиномицин, антрациклин, митомицин С, блеомицин, митрамицин, направленные на рецептор IL-2 (CD25-) или CD3-антитела, антитела к IL-2, циклоспорин, такролимус, сиролимус, IFN-β, IFN-γ, опиоид или связывающее TNF-α (фактор некроза опухоли-альфа) средство.
7. Способ по любому из пп. 4-6, в котором rAAV и иммуносупрессант вводят совместно или иммуносупрессант вводят после rAAV.
8. Способ по любому из пп. 1-7, в котором млекопитающее не подвергают индукции иммунологической толерантности до введения rAAV.
9. Способ по любому из пп. 1-8, в котором млекопитающее подвергают индукции иммунологической толерантности до введения rAAV.
10. Способ по п. 1, в котором млекопитающее представляет собой иммунокомпетентное взрослое млекопитающее.
11. Способ по любому из пп. 1-10, в котором rAAV имеет капсид rAAV9.
12. Способ по любому из пп. 1-11, в котором открытая рамка считывания, кодирует идуронат-2-сульфатазу.
13. Способ по любому из пп. 1-12, в котором млекопитающее представляет собой человека.
14. Способ по любому из пп. 1-13, в котором млекопитающее является дефицитным по идуронат-2-сульфатазе.
15. Способ по любому из пп. 1-13, в котором млекопитающее имеет нарушение микополисахаридоз I типа.
16. Способ по любому из пп. 1–11, в котором открытая рамка считывания, кодирует альфа-L-идуронидазу.
17. Способ по любому из пп. 1–11, в котором млекопитающее имеет нарушение микополисахаридоз II типа.
18. Способ по любому из пп. 1-17, в котором вводят многократные дозы.
19. Способ по любому из пп. 1-17, в котором композицию вводят раз в неделю.
20. Способ по любому из пп. 1-19, в котором определенное количество ингибирует задержку роста, ингибирует гепатоспленомегалию, ингибирует заболевание сердца и легких или ингибирует дисплазию скелета, или любую их комбинацию.
21. Способ по любому из пп. 1-20, в котором rAAV имеет капсид rAAVrh10.
| US 20140088179 A1, 27.03.2014 | |||
| WO 2015013148 A2, 29.01.2015 | |||
| Супотницкий М | |||
| В., Генотерапевтические векторные системы на основе вирусов, БИОпрепараты | |||
| Профилактика, диагностика, лечение., 2011, номер 3 (43), стр | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| WO 2014186579 A1, 20.11.2014. | |||
