ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Представленное изобретение относится к области рекомбинантных вирусных векторов. В частности, настоящее изобретение касается рекомбинантных вирусных векторов, которые пригодны для доставки терапевтических генов in vivo.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
На сегодняшний день, аденоассоциированный вирус остается одним из наиболее перспективных векторов для доставки терапевтических генов. Значительное количество доклинических и клинических исследований твердо установили, что данный подход пригоден для разработки лекарственных средств на генной основе, которые могут достичь одобрения на рынке.
С самого начала развития AAV2 в качестве вектора для генной терапии в 1980-е годы был достигнут значительный прогресс в оптимизации данной платформы для различных применений и тканей-мишеней. Среди данных разработок, возможно, имеющих наиболее важные последствия, было обнаружено широкий спектр серотипов, из которых десять из двенадцати в настоящее время широко исследуются. Среди наиболее основных характеристик данных различных серотипов выделяют их соответствующий тропизм по отношению к ткани и - в некоторых случаях - способность к нейрональному ретроградному транспорту. Из данных серотипов для доклинических и клинических целей широко используется AAV1-10.
Была разработана более новая платформа, которая включает процессы, которые допускают нацеливание и ненацеливание на конкретные ткани и клеточные подтипы у пациентов. Базовая технология данных подходов основывается на методе проб и ошибок оценки существующих вариантов AAV (серотипов) и in vivo отбора случайным образом введенных мутантов AAV капсида. Вместе, данные два перспективные подходы обеспечивают десятки - если не сотни - потенциальных векторов с различным поведением трансдукции.
Наиболее интригующим аспектом AAV серотипов является их способность эффективно трансдуцировать конкретные ткани в моделях на животных и у человека. На сегодняшний день комплексное молекулярное понимание лежащих в основе механизмов для тропизма ткани до сих пор не выдвигалась, и, таким образом, как правило, предполагается, что общедоступные ткане-специфические рецепторы для каждого серотипа играют центральную роль в эффективной трансдукции различных серотипов.
Соответственно, все еще существует потребность в дополнительных векторах AAV, которые обладают улучшенными свойствами с точки зрения экспрессии трансгена in vivo и тканевой специфичности. В частности, такие векторы имеют эффективность, чтобы обеспечить значительно расширенные преимущества для доставки генов к различным тканям-мишеням в организме человека.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В одном аспекте, представленное изобретение предусматривает рекомбинантный вектор на основе аденоассоциированного вируса (AAV), который содержит: (а) вариантный AAV2 капсидный белок, где вариантный AAV2 капсидный белок содержит, по меньшей мере, четыре аминокислотных замещения по сравнению с немутантному типу AAV2 капсидного белка; где, по меньшей мере, четыре аминокислотных замещения присутствуют в следующих положениях в последовательности AAV2 капсидного белка: 457, 492, 499 и 533; и (b) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт.
В одном варианте осуществления, вариантный AAV капсидный белок содержит последовательность SEQ ID NO: 2, или последовательность, имеющую, по меньшей мере, 95% идентичность последовательности с ней. В другом варианте осуществления, AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 1.
В одном варианте осуществления, вариантный AAV2 капсидный белок содержит один или больше из следующих остатков: М457, А492, D499 и Y533. В предпочтительном варианте осуществления, вариантный AAV2 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с капсидным белком немутантного типа AAV2: Q457M, S492A, E499D и F533Y.
В одном варианте осуществления, вариантный AAV2 капсидный белок дополнительно содержит одно или больше аминокислотных замещений по сравнению с AAV капсидным белком немутантного типа в следующих положениях в последовательности AAV2 капсидного белка: 125, 151, 162 и 205. В предпочтительном варианте осуществления, вариантный AAV2 капсидный белок содержит один или больше из одного или больше из следующих остатков: I125 , А151, S162 и S205. В другом предпочтительном варианте осуществления, вариантный AAV2 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с капсидным белком немутантного типа AAV2: V125I, V151A, A162S и T205S.
В одном варианте осуществления, вариантный AAV2 капсидный белок дополнительно содержит одно или больше аминокислотных замещений по сравнению с AAV капсидным белком немутантного типа в следующих положениях в последовательности AAV2 капсидного белка: 585 и 588. Предпочтительно вариантный AAV2 капсидный белок содержит один или больше из одного или нескольких следующих остатков: S585 и Т588. Более предпочтительно вариантный AAV2 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с немутантным типом AAV2 капсидный белок: R585S и R588T.
В одном варианте осуществления, вариантный AAV2 капсидный белок дополнительно содержит одно или больше аминокислотных замещений по сравнению с AAV капсидным белком немутантного типа в следующих положениях в последовательности AAV2 капсидного белка: 546, 548 и 593. Предпочтительно вариантный AAV2 капсидный белок содержит один или больше из одного или нескольких следующих остатков: D546, G548, и S593. Более предпочтительно вариантный AAV2 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с капсидным белком немутантного типа AAV2: G546D, E548G и A593S.
В одном варианте осуществления, вариантный AAV2 капсидный белок содержит остаток N312, то есть, остаток, который присутствует в немутантном типе AAV2 капсидного белка в положении 312. В данном варианте осуществления, вариантный AAV2 капсидный белок является немутированным в положении 312 по сравнению с немутантным типом последовательности AAV2 капсидного белка.
В другом аспекте, представленное изобретение предусматривает рекомбинантный вектор аденоассоциированного вируса (AAV), содержащий: (а) вариантный AAV8 капсидный белок, в котором вариантный AAV8 капсидный белок содержит аминокислотное замещение по сравнению с AAV8 капсидным белком немутантного типа в положении 315 в последовательности AAV8 капсидного белка; и (b) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт.
В одном варианте осуществления, вариантный AAV капсидный белок содержит последовательность, имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 6. В другом варианте осуществления, AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 6.
В одном варианте осуществления, вариантный AAV8 капсидный белок содержит аминокислотное замещение S315N по сравнению с AAV8 капсидным белком немутантного типа. Предпочтительно последовательность AAV8 капсидного белка содержит одно или больше аминокислотное замещение, присутствующее в одном или больше из следующих положений: 125, 151, 163, 206, 460, 495, 502, 536, 549, 551, 588, 591 и/или 596.
В предпочтительном варианте осуществления, вариантный AAV8 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV8 капсидным белком немутантного типа: (а) V125I, Q151A, K163S, A206S, Т460М, Т495А, N502D, F536Y, N549D, A551G, Q588S и/или G596S; и/или (b) T591R.
В другом аспекте, представленное изобретение предусматривает рекомбинантный вектор аденоассоциированного вируса (AAV), содержащий: (а) вариантный AAV3B капсидный белок, где вариантный AAV3B капсидный белок содержит аминокислотное замещение по сравнению с немутантным типом AAV3B капсидного белка в положении 312 в последовательности AAV3B капсидного белка; и (b) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт.
В одном варианте осуществления, вариантный AAV3B капсидный белок содержит последовательность, имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 11. В другом варианте осуществления, AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 11.
В одном варианте осуществления, вариантный AAV3B капсидный белок содержит аминокислотное замещение S312N по сравнению с немутантным типом AAV3B капсидного белка.
В другом аспекте, представленное изобретение предусматривает рекомбинантный вектор аденоассоциированного вируса (AAV), содержащий (а) вариантный AAV-LK03 капсидный белок, где вариантный AAV-LK03 капсидный белок содержит аминокислотное замещение в положении 312 по сравнению с последовательностью AAV-LK03 капсидного белка, как определено в SEQ ID NO: 12; и (b) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт.
В одном варианте осуществления, вариантный AAV-LK03 капсидный белок содержит последовательность, имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 12.
В другом аспекте, представленное изобретение предусматривает рекомбинантный вектор аденоассоциированного вируса (AAV), содержащий: (а) вариантный AAV капсидный белок, где вариантный AAV капсидный белок содержит, по меньшей мере, одно аминокислотное замещение по сравнению с AAV капсидным белком немутантного типа в положении, отвечающему одному или больше из следующих положений в последовательности AAV2 капсидного белка: 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593; и (b) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт.
В одном варианте осуществления, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV2 капсидного белка: 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593; или в одном или больше соответствующих положениях в альтернативной последовательности AAV капсидного белка.
В одном варианте осуществления, вектор содержит вариантный AAV2 капсидный белок. В другом варианте осуществления, вариантный AAV капсидный белок содержит последовательность SEQ ID NO: 2, или последовательность, имеющую, по меньшей мере, 95% идентичность последовательности с ней. В другом варианте осуществления, AAV капсидный белок немутантного типа является из AAV2. В другом варианте осуществления, AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 1.
В одном варианте осуществления, вариантный AAV2 капсидный белок содержит один или больше из следующих остатков: I125, А151, S162, S205, S312, М457, А492, D499, Y533, D546, G548, S585, Т588 и/или S593. В предпочтительном варианте осуществления, вариантный AAV2 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с капсидным белком немутантного типа AAV2: V125I, V151A, A162S, T205S, N312S, Q457M, S492A, E499D, F533Y, G546D, E548G, R585S, R588T и/или A593S.
В следующих вариантах осуществления, вариантный AAV капсидный белок является из AAV1, AAV5, AAV6, AAV8, AAV9 или AAV10.
В одном варианте осуществления, вектор содержит вариантный AAV1 капсидный белок. В другом варианте осуществления, вариантный AAV капсидный белок содержит последовательность, имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 3. В другом варианте осуществления, AAV капсидный белок немутантного типа является из AAV1. В другом варианте осуществления, AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 3.
В одном варианте осуществления, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV1 капсидного белка: 125, 151, 162, 205, 313, 458, 493, 500, 534, 547, 549, 586, 589 и/или 594. В предпочтительном варианте осуществления, вариантный AAV1 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с капсидным белком немутантного типа AAV1: V125I, Q151A, T162S, N313S, N458M, K493А, N500D, F534Y, S547D, и/или G594S. В альтернативном варианте осуществления, вариантный AAV1 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с капсидным белком немутантного типа AAV1: S205T, G549E, S586R и/или T589R.
В одном варианте осуществления, вектор содержит вариантный AAV5 капсидный белок. В другом варианте осуществления, вариантный AAV капсидный белок содержит последовательность, имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 4. В другом варианте осуществления, AAV капсидный белок немутантного типа является из AAV5. В другом варианте осуществления, AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 4.
В одном варианте осуществления, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV5 капсидного белка: 124, 150, 153, 195, 303, 444, 479, 486, 520, 533, 537, 575, 578 и/или 583. В предпочтительном варианте осуществления, вариантный AAV5 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV5 капсидным белком немутантного типа: V124I, К150А, K153S, A195S, R303S, Т444М, S479A, V486D, T520Y, P533D, и/или G583S. В альтернативном варианте осуществления, вариантный AAV5 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV5 капсидным белком немутантного типа: G537E, S575R и/или T578R.
В одном варианте осуществления, вектор содержит вариантный AAV6 капсидный белок. В другом варианте осуществления, вариантный AAV капсидный белок содержит последовательность, имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 5. В другом варианте осуществления, AAV капсидный белок немутантного типа является из AAV6. В другом варианте осуществления, AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 5.
В одном варианте осуществления, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV6 капсидного белка: 125, 151, 162, 205, 313, 458, 493, 500, 534, 547, 549, 586, 589 и/или 594. В предпочтительном варианте осуществления, вариантный AAV6 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV6 капсидным белком немутантного типа: V125I, Q151A, T162S, N313S, N458M, K493А, N500D, F534Y, S547D, и/или G594S. В альтернативном варианте осуществления, вариантный AAV6 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV6 капсидным белком немутантного типа: S205T, G549E, S586R и/или T589R.
В одном варианте осуществления, вектор содержит вариантный AAV8 капсидный белок. В другом варианте осуществления, вариантный AAV капсидный белок содержит последовательность, имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 6. В другом варианте осуществления, AAV капсидный белок немутантного типа является из AAV8. В другом варианте осуществления, AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 6.
В одном варианте осуществления, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV8 капсидного белка: 125, 151, 163, 206, 315, 460, 495, 502, 536, 549, 551, 588, 591 и/или 596. В предпочтительном варианте осуществления, вариантный AAV8 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV8 капсидным белком немутантного типа: V125I, Q151A, K163S, A206S, Т460М, Т495А, N502D, F536Y, N549D, A551G, Q588S и/или G596S. В альтернативном варианте осуществления, вариантный AAV8 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV8 капсидным белком немутантного типа: S315N и/или T591R.
В одном варианте осуществления, вектор содержит вариантный AAV9 капсидный белок. В другом варианте осуществления, вариантный AAV капсидный белок содержит последовательность, имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 7. В другом варианте осуществления, AAV капсидный белок немутантного типа является из AAV9. В другом варианте осуществления, AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 7.
В одном варианте осуществления, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV9 капсидного белка: 125, 151, 162, 205, 314, 458, 493, 500, 534, 547, 549, 586, 589 и/или 594. В предпочтительном варианте осуществления, вариантный AAV9 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV9 капсидным белком немутантного типа: L125I, Q151A, N314S, Q458M, V493A, E500D, F534Y, G547D, А589Т и/или G594S. В альтернативном варианте осуществления, вариантный AAV9 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV9 капсидным белком немутантного типа: S162A, S205T, G549E и/или S586R.
В одном варианте осуществления, вектор содержит вариантный AAV10 капсидный белок. В другом варианте осуществления, вариантный AAV капсидный белок содержит последовательность, имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 8. В другом варианте осуществления, AAV капсидный белок немутантного типа является из AAV10. В другом варианте осуществления, AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 8.
В одном варианте осуществления, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV10 капсидного белка: 125, 151, 163, 206, 315, 460, 495, 502, 536, 549, 551, 588, 591 и/или 596. В предпочтительном варианте осуществления, вариантный AAV10 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV10 капсидным белком немутантного типа: V125I, Q151A, K163S, A206S, N315S, Т460М, L495A, N502D, F536Y, G549D, Q588S, А591Т и/или G596S. В альтернативном варианте осуществления, вариантный AAV10 капсидный белок содержит следующее аминокислотное замещение по сравнению с AAV10 капсидным белком немутантного типа: G551E.
В одном варианте осуществления, рекомбинантный AAV вектор демонстрирует повышенную трансдукцию нейрональной или ретинальной ткани по сравнению с AAV вектором, содержащим соответствующий AAV капсидный белок немутантного типа.
В другом варианте осуществления, рекомбинантный AAV вектор демонстрирует повышенную трансдукцию ткани печени по сравнению с соответствующим AAV капсидным белком немутантного типа.
В одном варианте осуществления, генный продукт содержит интерферирующую РНК или аптамер. В другом варианте осуществления, генный продукт содержит полипептид. Предпочтительно генный продукт содержит нейропротекторный полипептид, антиангиогенный полипептид, или полипептид который улучшает функцию нейрональной или ретинальной клетки. В предпочтительных вариантах осуществления, генный продукт содержит глиальный нейротрофический фактор, фактор роста фибробластов, фактор роста нервов, нейротрофический фактор головного мозга, родопсин, ретиношизин, RPE65 или периферии.
В другом аспекте, представленное изобретение предусматривает фармацевтическую композищю, содержащую: (а) рекомбинантный AAV вектор, как определено выше; и (b) фармацевтически приемлемый эксципиент.
В другом аспекте, представленное изобретение предусматривает способ доставки генного продукта к ткани у субъекта, где способ включает введение субъекту рекомбинантного AAV вектора или фармацевтической композиции, как определено выше.
В некоторых вариантах осуществления, ткань выбирают из крови, костного мозга, мышечной ткани, нейрональной ткани, ретинальной ткани, ткани поджелудочной железы, ткани печени, почечной ткани, легочной ткани, ткани кишечника или ткани сердца. Предпочтительно ткань представляет собой нейрональную, ретинальную ткань или ткань печени.
В другом аспекте, представленное изобретение предусматривает способ лечения расстройства у субъекта, где способ включает введение субъекту рекомбинантного AAV вектора или фармацевтической композиции, как определено выше. В некоторых вариантах осуществления, расстройство представляет собой неврологическое, глазное расстройство или заболевани печени.
В другом аспекте, представленное изобретение предусматривает рекомбинантный AAV вектор или фармацевтическую композизию, как определено выше, для применения в лечении растройства у субъекта. В некоторых вариантах осуществления, расстройство представляет собой неврологическое, глазное расстройство или заболевани печени. Предпочтительно неврологическое расстройство представляет собой нейродегенеративное заболевание. В альтернативном варианте осуществления, глазное расстройство представляет собой глаукому, пигментную дегенерацию сетчатки, дегенерацию желтого пятна, ретиношизис или диабетическую ретинопатию.
В другом аспекте, представленное изобретение предусматривает выделенный вариантный AAV капсидный белок, где вариантный AAV капсидный белок содержит, по меньшей мере, одно аминокислотное замещение по сравнению с AAV капсидным белком немутантного типа; где, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV2 капсидного белка: 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593; или в одном или больше из соответствующих положения в альтернативной последовательности AAV капсидного белка.
В другом аспекте, представленное изобретение предусматривает выделенную нуклеиновую кислоту, содержащую нуклеотидную последовательность, которая кодирует вариантный AAV капсидный белок, как определено выше.
В другом аспекте, представленное изобретение предусматривает выделенную клетку-хозяина, содержащую нуклеиновую кислоту, как определено выше.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 показывает аминокислотную последовательность капсидного белка VP1 немутантного типа аденоассоциированного вируса 2 (SEQ ID NO: 1; NCBI эталонная последовательность: NC_001401). Остатки V125, V151, А162, Т205, N312, Q457, S492, Е499, F533, G546, Е548, R585, R588 и А593 выделены.
Фигура 2 показывает аминокислотную последовательность капсидного белка VP1 подлинного типа аденоассоциированного вируса 2 (ttAAV2) (SEQ ID NO: 2). Остатки I125 , А151, S162, S205, S312, М457, А492, D499, Y533, D546, G548, S585, Т588, S593 отличаются по сравнению с немутантным типом AAV2 VP1 (SEQ ID NO: 1) и выделены.
Фигура 3 показывает аминокислотную последовательность капсидного белка VP1 немутантного типа аденоассоциированного вируса 1 (SEQ ID NO: 3; NCBI эталонная последовательность: NС_002077). Выделенные остатки: S205 (совпадает с S205 в ttAAV2 (SEQ ID NO: 2)) - G549 (совпадает с G548 в ttAAV2) - S586 (совпадает с S585 в ttAAV2) - Т589 (совпадает с Т588 в ttAAV2).
Фигура 4 показывает аминокислотную последовательность капсидного белка VP1 немутантного типа аденоассоциированного вируса 5 (SEQ ID NO: 4; NCBI эталонная последовательность: AF085716). Выделенные остатки: G537 (совпадает с G548 в ttAAV2) - S575 (совпадает с S585 в ttAAV2) - Т578 (совпадает с Т588 в ttAAV2).
Фигура 5 показывает аминокислотную последовательность капсидного белка VP1 немутантного типа аденоассоциированного вируса 6 (SEQ ID NO: 5; NCBI эталонная последовательность: AF028704). Выделенные остатки: S205 (совпадает с S205 в ttAAV2) - G549 (совпадает с G548 в ttAAV2) - S586 (совпадает с S585 в ttAAV2) -Т589 (совпадает с Т588 в ttAAV2).
Фигура 6 показывает аминокислотную последовательность капсидного белка VP1 немутантного типа аденоассоциированного вируса 8 (SEQ ID NO: 6; NCBI эталонная последовательность: NC_006261). Выделенные остатки: S315 (совпадает с S312 в ttAAV2) - Т591 (совпадает с Т588 в ttAAV2).
Фигура 7 показывает аминокислотную последовательность капсидного белка VP1 немутантного типа аденоассоциированного вируса 9 (SEQ ID NO: 7; NCBI эталонная Последовательность: AY530579). Выделенные остатки: S162 (совпадает с S162 в ttAAV2) - S205 (совпадает с S205 в ttAAV2) - G549 (совпадает с G548 в ttAAV2)- S586 (совпадает с S585 в ttAAV2).
Фигура 8 показывает аминокислотную последовательность капсидного белка VP1 немутантного типа аденоассоциированного вируса 10 (SEQ ID NO: 8). Выделенный остаток: G551 (совпадает с G548 в ttAAV2).
Фигура 9 показывает сравнительный анализ первичной структуры аминокислотных последовательностей AAV капсидного белка VP1.
Фигура 10 Плазмида, которую используют для получения векторов AAV2, представляла собой упаковывающую плазмиду pDG. Выше: pDG с генами немутантного типа AAV2. Ниже: pDG-ttAAV2 с генами подлинного типа AAV2, выделенными являются две ключевые мутации в гепаран-связывающих доменах в положениях 585 и 588. MMTV: промотор, запускающий экспрессию AAV rep, Е2а, E4ORF6 и VA представляют собой гены, экспрессирующий аденовирусные хелперные факторы.
Фигура 11 Количественное определение вирусных титров rAAV2 подлинного типа (ТТ) и немутантного типа (WT) для инъекций in vivo с помощью SDS-PAGE, показывая окрашивание криптоном для разделенных белков, и сканировали с использованием сканера инфракрасной флуоресценции (системы визуализации Odyssey). А: 10 мкл AAV2 вирусных частиц, и 62,5 нг-500 нг BSA разделяли на 12% разделяющем геле, содержащем SDS, и окрашивали криптоновым красителем для белка. Изображение преобразовывали в градации серого. Капсидные белки гена VP1, VP2, VP 3 метили на левой стороне. В: Таблица, показывающая титры из кПЦР (векторный геном [вг/мл]) и SDS-Page (титр капсидов [капсид/мл]).
Фигура 12А. Показаны характерные примеры участков головного мозга крысы, окрашенных GFP-специфическим антителом. Вектор инъекционно вводили в полосатое, тело, как показано стрелкой. В. Показан характерный пример инъекции в черную субстанцию.
Фигура 13 Показана GFP трансдукция глаза, используя ttAAV2 и wtAAV2. А. Показана сетчатка в поперечном сечении после введения вектора ttAAV2 (вверху) и wtAAV2 (внизу). В. Показаны концентрации пунктирных боксов в А.
Фигура 14 Трансдукция мозга мыши после неонатальной инъекции вектора: в.в., внутривенное введение вектора; в.ч., внутричерепная инъекция; AAV-2, wtAAV2; AAV-TT, ttAAV2.
Фигура 15 Трехмерное представление AAV2 капсида. Выделенные остатки соответствуют аминокислотным изменениям между ttAAV2 и частицами немутантного типа, сгруппированным по цвету в зависимости от их положения.
Фигура 16 Представление тройного шиловидного отростка на AAV2 капсиде. остатки соответствуют аминокислотным изменениям между частицами подлинного типа и немутантного типа. Остатки сайтов связывания гепарина выделены зеленым.
Фигура 17 Представление внутренней части AAV2 капсида. Выделенные светло-голубым цветом остатки соответствуют одному аминокислотному изменению в ttAAV2, которое расположено на внутренней стороне капсида.
Фигура 18 Представление тройного шиловидного отростка на AAV2 капсиде. Остатки, выделенными бежевым цветом, соответствуют двум аминокислотным изменениям в векторе подлинного типа, которые пространственно близки и расположены в канавке между двумя трехкратно проксимальными пиками на AAV капсиде.
Фигура 19 Представление тройного шиловидного отростка на AAV2 капсиде. Остаток, выделенный коричневым цветом, соответствует одному выделенному аминокислотному изменению (S593) в векторе подлинного типа, который расположен в канавке между трехкратно проксимальными пиками
Фигура 20 Представление тройного шиповидного отростка на AAV2 капсиде. Четыре аминокислоты, выделенные розовым цветом, участвуют в связывании с рецептором и близко расположены тройных резких скачках.
Фигура 21 Трехмерное представление сравнительного анализа первичной структуры между VP1 капсидным мономером от AAV2 (светло-голубой) и VP1 мономером от AAV1 (оранжевый). Выделенные остатки в средней левой части изображения соответствуют G549 в AAV1 (оранжевые сферы) и Е548 в AAV2 (зелено-голубая сфера). Выделенные остатки в верхнем правом углу изображения соответствуют S586 и Т589 в AAV1 (оранжевые сферы) и R585 и R588b AAV2 (зелено-голубая сфера).
Фигура 22 Трехмерное представление сравнительного анализа первичной структуры между VP1 капсидным мономер от AAV2 (светло-голубой) и VP1 мономером от AAV5 (фиолетовый). Выделенные остатки в средней части изображения соответствуют G537 в AAV5 (фиолетовые сферы) и Е548 в AAV2 (зелено-голубая сфера). Выделенные остатки в верхнем правом углу изображения соответствуют S575 и Т578 в AAV5 (фиолетовые сферы) и R585 и R588 в AAV2 (зелено-голубая сфера).
Фигура 23 Трехмерное представление сравнительного анализа первичной структуры между VP1 капсидным мономером от AAV2 (светло-голубой) и VP1 мономером от AAV6 (желтый). Выделенные остатки в нижней части изображения соответствуют G549 в AAV6 (оранжевые сферы) и Е548 в AAV2 (зелено-голубая сфера). Выделенные остатки в верхнем правом углу изображения соответствуют S586 и Т589 в AAV6 (оранжевые сферы) hR585 и R588b AAV2 (зелено-голубая сфера).
Фигура 24 Трехмерное представление сравнительного анализа первичной структуры между VP1 капсидным мономером от AAV2 (светло-голубой) и VP1 мономером от AAV8 (розовый). Выделенные остатки в верхнем левом углу изображения соответствуют S315 в AAV8 (красные сферы) и N312 в AAV2 (зелено-голубая сфера). Выделенные остатки в нижнем правом углу изображения соответствуют Т591 в AAV8 (красные сферы) и R588 в AAV2 (зелено-голубая сфера).
Фигура 25 Трехмерное представление сравнительного анализа первичной структуры между VP1 капсидным мономером от AAV2 (светло-голубой) и VP1 мономером от AAV9 (зеленый). Выделенные остатки в сере дней части изображения соответствуют G549 в AAV9 (желтые сферы) и Е548 в AAV2 (зелено-голубая сфера). Выделенные остатки в нижнем левом углу изображения соответствуют S586 в AAV9 (желтые сферы) и R585 в AAV2 (зелено-голубая сфера).
Фигура 26 Анализ экспрессии rAAV2 ТТ и WT в парафасцикулярном ядре после инъекции в полосатое тело головного мозга крыс. А: Приведены типичные изображения срезов головного мозга крысы, показывающие ростральную сторону слева и хвостовую сторону справа. Указывается место инъекции в стриатуме, и площадь проекции в гипоталамусе наблюдается в B и C (парафасцикулярное ядро, pf). B и C: Высокое увеличение изображения экспрессии GFP, обнаруженной в парафасцикулярном ядре (pf) после инъекции в полосатое тело rAAV2 WT (В) или ТТ (С).
Фигура 27 Обзор внутричерепных инъекций rAAV2 ТТ и WT у новорожденных мышей. Показаны характерные примеры срезов неонатального головного мезга, окрашенных GFP- специфическим антителом. 5×1010 вг rAAV2 ТТ (вверху) или rAAV2 WT (в середине) вводили инъекционно в боковой желудочек мозга новорожденных мышей. Неинъекцированный мозг от новорожденных мышей, окрашенный одновременно, представлен в качестве отрицательного контроля (NT, не трансдуцированный).
Фигура 28 Изображения с большим увеличением срезов мозга новорожденных мышей после внутричерепных инъекций rAAV2 ТТ или WT. Показаны срезы неонатального головного мозга, окрашенные GFP-специфическим антителом. 5×1010 вг rAAV2 ТТ (левые панели) или rAAV2 WT (правые панели) вводили инъекционно в боковой желудочек мозга новорожденных мышей. S1BF: поле ствола первичной соматосенсорной коры головного мозга.
Фигура 29 Обзор трансдукции мозга после системной инъекции rAAV2 ТТ и WT у новорожденных мышей. Показаны характерные примеры срезов неонатального головного мезга, окрашенные GFP-специфическим антителом. 2×1011 вг rAAV2 ТТ (вверху) или rAAV2 WT (внизу) вводили инъекционно в яремные вены новорожденных мышей.
Фигура 30 Изображения с большим увеличением срезов мозга новорожденных мышей после системных инъекций rAAV2 ТТ или WT. Показаны срезы неонатального головного мезга, окрашенные GFP-специфическим антителом. 2×1011 вг rAAV2 ТТ (левые панели) или rAAV2 WT (правые панели) вводили инъекционно в яремные вены новорожденных мышей. S1BF: поле ствола первичной соматосенсорной коры головного мозга.
Фигура 31 Изображения с большим увеличением срезов тканей новорожденных мышей после системных инъекций rAAV2 ТТ или WT. 2×1011 вг rAAV2 ТТ или rAAV2 WT вводили инъекционно в яремные вены новорожденных мышей. Неинъекцированные органы мышиные были использованы в качестве отрицательного контроля.
Фигура 32 Изображения с большим увеличением срезов головного мозга взрослых крыс после инъекций в полосатое тело rAAV2 ТТ, WT и HBnull. Показаны характерные примеры срезов головного мозга крыс, окрашенных GFP-специфическим антителом. 3,5×109 вг rAAV2 WT (слева), ТТ (справа) или AAV2-HBnull (в середине) вводили инъекционно в полосатое тело головного мозга взрослых крыс, и характерные изображения были получены для таламуса или черной субстанции (SN).
Фигура 33 Обзор внутричерепных инъекций полного AAV-TT по сравнению с различными ТТ мутантами у новорожденных мышей. Показаны характерные примеры срезов неонатального головного мозга, окрашенных GFP-специфическим антителом. 5×1010 вг rAAV2 ТТ, TT-S312N, TT-S593A или TT-D546G/G548E (TT-DG) вводили инъекционно в боковой желудочек мозга новорожденных мышей. Неинъецированный мозг от новорожденной мыши, окрашенный одновременно, представлен в качестве отрицательного контроля (NT).
Фигура 34 Изображения с большим увеличением срезов мозга новорожденных мышей после внутричерепных инъекций различных ТТ мутантных векторов. Показаны срезы неонатального головного мозга, окрашенные GFP-специфическим антителом. 5×1010 вг векторов вводили инъекционно в боковой желудочек мозга новорожденных мышей. TT-DG: TT-D546G/G548E.
Фигура 35 Обзор внутричерепных инъекций у новорожденных мышей полного AAV-TT по сравнению с TT-S312N мутантом и потенциальным конечным ТТ вектором, содержащим 10 мутаций. Показаны характерные примеры срезов неонатального головного мезга, окрашенные GFP-специфическим антителом. 5×1009 вг rAAV2 ТТ, TT-S312N, ТТ или TT-S312N-D546G/G548E-S593A (TT-S312N-DG-S593A) вводили инъекционно в боковой желудочек мозга новорожденных мышей. Неинъекцированный мозг от новорожденной мыши, окрашенный одновременно, представлен в качестве отрицательного контроля (NT).
Фигура 36 Изображения с большим увеличением срезов мозга новорожденных мышей после внутричерепных инъекций различных ТТ мутантных векторов. Показаны срезы неонатального головного мозга, окрашенные GFP-специфическим антителом. 5×1009 вг векторов вводили инъекционно в боковой желудочек мозга новорожденных мышей.
Фигура 37 ИФА количественное определение белка GFP в головном мозге новорожденных мищей, которым инъекционно вводили полный AAV-TT, TT-S312N мутант или TT-S312N-DG-S593A. 5×1009 вг векторов вводили инъекционно в боковой желудочек мозга новорожденных мишей, и общие белки экстрагировали из цельного собранного головного мозга. GFP-специфическое антитело использовали для детектирования GFP экспрессия GFP в каждом образце головного мозга, и стандартный GFP белок использовали для количественного определения. N=5 животных на каждое условие. Планки погрешностей представляют собой среднее значение ± SEM
Фигура 38 Аминокислотная последовательность VP1 капсидного белка AAV3B. Выделенные остатки представляют собой остатки, которые идентичны остаткам в AAV-tt в соответствующих положениях. Внутренний остаток серина в положении 312 подчеркнут.
Фигура 39 Аминокислотная последовательность VP1 капсидного белка AAV-LK03.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 представляет собой аминокислотную последовательность капсидного белка немутантного типа аденоассоциированного вируса 2 VP1 (смотри фигуру 1).
SEQ ID NO: 2 представляет собой аминокислотную последовательность капсидного белка подлинного типа аденоассоциированного вируса 2 (ttAAV2) (смотри фигуру 2).
SEQ ID NO: 3 представляет собой аминокислотную последовательность капсидного белка немутантного типа аденоассоциированного вируса 1 VP1 (смотри фигуру 3).
SEQ ID NO: 4 представляет собой аминокислотную последовательность капсидного белка немутантного типа аденоассоциированного вируса 5 VP1 (смотри фигуру 4).
SEQ ID NO: 5 представляет собой аминокислотную последовательность капсидного белка немутантного типа аденоассоциированного вируса 6 VP1 (смотри фигуру 5).
SEQ ID NO: 6 представляет собой аминокислотную последовательность капсидного белка немутантного типа аденоассоциированного вируса 8 VP1 (смотри фигуру 6).
SEQ ID NO: 7 представляет собой аминокислотную последовательность капсидного белка немутантного типа аденоассоциированного вируса 9 VP1 (смотри фигуру 7).
SEQ ID NO: 8 представляет собой аминокислотную последовательность Upenn капсидного белка немутантного типа аденоассоциированного вируса 10 VP1 (смотри фигуру 8).
SEQ ID NO: 9 представляет собой аминокислотную последовательность японского капсидного белка немутантного типа аденоассоциированного вируса 10 VP1 (смотри фигуру 9).
SEQ ID NO: 10 представляет собой консенсусную аминокислотную последовательность для аденоассоциированных вирусов, показанных на фигуре 9.
SEQ ID NO: 11 представляет собой аминокислотную последовательность капсидного белка немутантного типа аденоассоциированного вируса 3В VP1 (смотри фигуру 38).
SEQ ID NO: 12 представляет собой аминокислотную последовательность капсидного белка аденоассоциированного вируса LK-03 VP1 (смотри фигуру 39).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В одном аспекте, представленное изобретение касается рекомбинантного вектора аденоассоциированного вируса (AAV). rAAV вектор, как правило, содержит вариантный капсидный белок, который отличается от капсидного белка немутантного типа AAV. Вариантный капсидный белок преимущественно может придавать повышенную инфекционность вектору в головном мозге и/или глазе, что делает вектор особенно подходящим для доставки терапевтических агентов с помощью генной терапии в данные ткани.
РЕКОМБИНАНТНЫЙ AAV ВЕКТОР
Представленное раскрытие предусматривает рекомбинантный вектор аденоассоциированного вируса (rAAV). "AAV" представляет собой сокращение для аденоассоциированного вируса, и может быть использовано для обозначения самого вируса или его производного. Термин охватывает все подтипы и как формы, встречающийся в природе, так и рекомбинантные формы, за исключением тех случаев, когда требуется иное. Сокращение "rAAV" касается рекомбинантного аденоассоциированного вируса, также называемого как рекомбинантный вектор AAV (или "rAAV вектор"). Термин "AAV" включает, например, AAV типа 1 (AAV-1), AAV типа 2 (AAV-2), AAV типа 3 (AAV-3), AAV типа 4 (AAV-4), AAV типа 5 (AAV-5), AAV типа 6 (AAV-6), AAV типа 7 (AAV-7), AAV типа 8 (AAV-8), AAV типа 9 (AAV-9), AAV типа 10 (AAV-10, включая AAVrh10), AAV типа 12 (AAV-12), птичий AAV, бычий AAV, собачий AAV, лошадиный AAV, AAV примат, AAV не примат, и овечий AAV. "Примат AAV" касается AAV, которые инфицируют приматов, "не примат AAV" касается AAV, которые инфицируют млекопитающих не приматов, "бычий AAV" касается AAV, которые инфицируют млекопитающих, крупного рогатого скота, и так далее.
Геномные последовательности различных серотипов AAV, а также последовательности нативных терминальных повторов (TR), Rep белки, и капсидные субединицы известны в данной области с уровня техники. Такие последовательности могут быть найдены в литературе или в общедоступных базах данных, таких как GenBank. Смотри, например, номера доступа GenBank NC-002077 (AAV-1), AF063497 (AAV-1), NC-001401 (AAV-2), AF043303 (AAV-2), NC-001729 (AAV-3), NC-001829 (AAV- 4), U89790 (AAV-4), NC-006152 (AAV-5), AF513851 (AAV-7), AF513852 (AAV-8), и NC-006261 (AAV-8); описания которых включены в данный документ в качестве ссылки. Смотри также, например, Srivistava et al. (1983) J. Virology 45: 555; Chiorini et al. (1998) J. Virology 71: 6823; Chiorini et al. (1999) J. Virology 73: 1309; Bantel-Schaal et al. (1999) J. Virology 73: 939; Xiao et al. (1999) J. Virology 73: 3994; Muramatsu et al. (1996) Virology 221: 208; Shade et al., (1986) J. Virol. 58: 921; Gao et al. (2002) Proc. Nat. Acad. Sci. USA 99: 11854; Moris et al. (2004) Virology 33: 375-383; международные патентные публикации WO 00/28061, WO 99/61601, WO 98/11244; и патент США №6,156,303.
"rAAV вектор", как используется в данном документе, касается AAV вектора, содержащего полинуклеотидную последовательность не AAV происхождения (то есть, полинуклеотидную гетерологичную к AAV), как правило, последовательность, представляющую интерес для генетической трансформации клетки. В некоторых вариантах осуществления, гетерологичный полинуклеотид может быть фланкирован, по меньшей мере, одной, и иногда двумя, AAV последовательностями инвертированного концевого повтора (ITR). Термин rAAV вектор охватывает как векторные частицы rAAV, так и векторные плазмиды rAAV. rAAV вектор может быть либо одноцепочечным (ssAAV) или самокомплементарным (scAAV).
"AAV вирус", или "AAV вирусная частица", или "rAAV векторная частица" касается вирусной частицы, состоящей из, по меньшей мере, одного AAV капсидного белка (как правило, всех капсидных белков немутантного типа AAV) и заключенного в капсид полинуклеотидного rAAV вектора. Если частица содержит гетерологичный полинуклеотид (то есть полинуклеотид, не относящийся к геному AAV немутантного типа, такой как трансген, чтобы быть доставленным в клетку млекопитающего), ее, как правило, называют как "rAAV векторная частица" или просто "rAAV вектор". Таким образом, продуцирование rAAV частицы обязательно включает продуцирование rAAV вектора, поскольку такой вектор содержится внутри rAAV частицы.
"Рекомбинантный," как используется в данном документе, означает, что вектор, полинуклеотид, полипептид или клетка представляет собой продукт различных комбинаций стадий клонирования, рестрикции или лигирования (например, связанных с полинуклеотидом или полипептидом, содержащимся в них), и/или или других процедур, которые в результате приводят к конструкту,, который отличается от продукта, встречающегося в природе. Рекомбинантный вирус или вектор представляет собой вирусную частицу, содержащую рекомбинантный полинуклеотид. Термины соответственно включают репликации исходного полинуклеотидного конструкта и потомство исходного вирусного конструкта.
ВАРИАНТНЫЕ AAV КАПСИДНЫЕ БЕЛКИ
rAAV векторы, описанные в данном документе, содержат вариантный AAV капсидный белок. Под "вариантом" имеется в виду, что AAV капсидный белок отличается от соответствующего AAV капсидного белка немутантного типа того же серотипа. Например, вариантный AAV капсидный белок может содержать одно или больше аминокислотных замещений по сравнению с соответствующим AAV капсидным белком немутантного типа. В данном контексте, "соответствующий" касается капсидного белка того же серотипа, то есть вариантный AAV1 капсидный белок содержит одно или больше аминокислотных замещений по сравнению с соответствующим капсидным белком немутантного типа AAV1, вариантный AAV2 капсидный белок содержит одно или больше аминокислотных замещений по сравнению с соответствующим капсидным белком немутантного типа AAV2, и так далее.
Вариантный AAV капсидный белок может содержать, например, от 1 до 50, от 1 до 30, от 1 до 20 или от 1 до 15 аминокислотных замещений по сравнению с AAV капсидным белком немутантного типа. Предпочтительно вариантный AAV капсидный белок содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14 аминокислотных замещений по сравнению с соответствующим AAV капсидным белком немутантного типа. В предпочтительных вариантах осуществления, вариантный AAV капсидный белок сохраняет, по меньшей мере, 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичность последовательности к капсидному белку немутантного типа.
В вариантах осуществления представленного изобретения, вариантный AAV капсидный белок содержит, по меньшей мере, одно аминокислотное замещение по сравнению с AAV капсидным белком немутантного типа в положении, отвечающем одному или больше из следующих положений в последовательности AAV2 капсидного белка: 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593. В данном контексте, "соответствующий" касается положения в какой-либо последовательности AAV капсидного белка (например, в последовательности AAV2 белка или последовательности не-AAV2 капсидного белка), которое соответствует одному из указанных выше положений в AAV2 капсидном белке. В одном варианте осуществления, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV2 капсидного белка: 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593; или в одном или больше соответствующих положений в альтернативной последовательности AAV капсидного белка.
В общем, AAV капсидные белки включают VP1, VP2 и VP3. В предпочтительном варианте осуществления, капсидный белок содержит AAV капсидный белок VP1.
ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВЫХ КИСЛОТ И АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ, И ИДЕНТИЧНОСТЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Термин "полинуклеотид" касается полимерной формы нуклеотидов какой-либо длины, включая дезоксирибонуклеотиды или рибонуклеотиды, или их аналоги. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и нуклеотидные аналоги, и может прерываться не нуклеотидными компонентами. Если присутствуют, модификации по нуклеотидной структуре могут быть обеспеченными до или после сборки полимера. Термин полинуклеотид, как используется в данном документе, взаимозаменяемо касается двух- и одноцепочечных молекул. Если не указано или не требуется иное, какой-либо вариант осуществления изобретения, описанный в данном документе, который представляет собой полинуклеотид, охватывает как двухцепочечную форму, так и каждую из двух комплементарных одноцепочечных форм, известных или предсказанных для того, чтобы образовывать двухцепочечную форму.
Термины "полипептид," "пептид" и "белок", как используются взаимозаменяемо в данном документе, касаются полимеров аминокислот какой-либо длины. Термины также охватывают аминокислотный полимер, который был модифицирован; например, образованием дисульфидной связи, гликозилированием, липидированием, фосфорилированием или конъюгацией с компонентом маркировки. Полипептиды, такие как антиангиогенные полипептиды, нейропротекторные полипептиды, и тому подобные, при рассмотрении их в контексте доставки генного продукта млекопитающему субъекту, и его композиции, касаются соответствующего интактного полипептида, или какого-либо фрагмента или его производного, полученного методами генной инженерии, который сохраняет требуемую биохимическую интактного белка. Аналогичным образом, ссылки на нуклеиновые кислоты, кодирующие антиангиогенные полипептиды, нуклеиновые кислоты, кодирующие нейропротекторные полипептиды, и другие такие нуклеиновые кислоты для применения в доставке генного продукта млекопитающему субъекту (которые могут быть названы как "трансгены", которые доставляются к клетке-реципиенту), включают полинуклеотиды, кодирующие интактный полипептид, или какой-либо фрагмент, или производное, полученное методами генной инженерии, обладающее желаемой биохимической функцией.
Полинуклеотид или полипептид имеет определенный процент "идентичности последовательности" к другому полинуклеотиду или полипептиду, а это означает, что при выравнивании процент оснований или аминокислот является таким же, как и при сравнении двух последовательностей. Подобность последовательности может быть определена по ряду различных способов. Для того, чтобы определить идентичность последовательности, последовательности могут быть выровнены, используя способы и компьютерные программы, включая BLAST, доступные по всемирной сети на ncbi.nlm.nih.gov/BLAST/. Другим алгоритмом выравнивания является FASTA, доступны в пакете Genetics Computing Group (GCG), от Madison, Wisconsin, USA, стопроцентная дочерняя компания Oxford Molecular Group, Inc. Другие способы выравнивания описаны в Methods in Enzymology, vol. 266: Computer Methods for Macromolecular Sequence Analysis (1996), ed. Doolittle, Academic Press, Inc., подразделение Harcourt Brace & Co., San Diego, California, USA. Особый интерес представляют программы выравнивания, которые позволяют пропуски в последовательности. Алгоритм Смита-Ватермана представляет собой один тип алгоритма, который позволяет пропуски в выравниваниях последовательности. Смотри Meth. Mol. Biol. 70: 173-187 (1997). Кроме того, программа GAP с использованием способа выравнивания Нилдмана-Вунша может быть использована для выравнивания последовательностей. Смотри J. Mol. Biol. 48: 443- 453 (1970).
Представляющей интерес является программа BestFit с использованием алгоритма локальной гомологии Смита-Ватермана (Advances in Applied Mathematics 2: 482-489 (1981) для того, чтобы определить идентичность последовательности. Штраф за генерирование пробела обычно будет находиться в диапазоне от 1 до 5, как правило, от 2 до 4 и во многих вариантах осуществления будет составлять 3. Штраф за продолжение пробела обычно будет находиться в диапазоне от приблизительно 0,01 до 0,20 и во многих случаях будет составлять 0,10. Программа имеет параметры по умолчанию, определяемые последовательностями, введенными для сравнения. Предпочтительно, идентичность последовательности определяется с использованием параметров по умолчанию, определяемых программой. Данная программа доступна также из пакета Genetics Computing Group (GCG), от Madison, Wisconsin, USA.
Другая программа, представляющая интерес, является алгоритмом FastDB. FastDB описывается в Current Methods in Sequence Comparison and Analysis, Macromolecule Sequencing and Synthesis, Selected Methods and Applications, pp. 127-149, 1988, Alan R. Liss, Inc. Процентная идентичность последовательности рассчитывается с помощью FastDB на основании следующих параметров:
Штраф за ошибочное спаривание: 1,00;
Штраф за пропуск в последовательности: 1,00;
Штраф за размер пропуска: 0,33; и
Штраф за слияние: 30,0.
ВАРИАНТНЫЙ AAV2 КАПСИДНЫЙ БЕЛОК
В одном варианте осуществления, вектор содержит вариантный AAV2 капсидный белок. В данном варианте осуществления, вариантный AAV2 капсидный белок содержит, по меньшей мере, одно аминокислотное замещение в одном или больше из следующих положений в последовательности AAV2 капсидного белка: 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593.
Последовательность AAV2 капсидного белка немутантного типа VP1 известна, и показана на фигуре 1 (SEQ ID NO: 1). Последовательности AAV2 капсидного белка немутантного типа также доступны из базы с номерами доступа: NC-001401; UniProt P03 135; NCBI эталонная последовательность: YP_680426.1; GenBank: ААС03780.1.
Предпочтительно, вариантный AAV2 капсидный белок имеет, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность последовательности к SEQ ID NO: 1. В предпочтительном варианте осуществления, вариантный AAV2 капсидный белок содержит последовательность SEQ ID NO: 2, или последовательность, имеющую, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность последовательности с ней.
В одном варианте осуществления, вариантный AAV2 капсидный белок содержит один или больше из следующих остатков: I125, А151, S162, S205, S312, М457, А492, D499, Y533, D546, G548, S585, Т588 и/или S593. В предпочтительном варианте осуществления, вариантный AAV2 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с капсидным белком немутантного типа AAV2: V125I, V151A, A162S, T205S, N312S, Q457M, S492A, E499D, F533Y, G546D, E548G, R585S, R588T и/или A593S.
КОМБИНАЦИИ МУТАЦИЙ В AAV2 КАПСИДНОМ БЕЛКЕ
Вариантный AAV2 капсидный белок может содержать какую-либо комбинацию из указанных выше аминокислотных замещений. Поэтому в конкретных вариантах осуществления, вариантный AAV2 капсидный белок содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14 аминокислотных замещений, выбранных из приведенного выше списка. В одном варианте осуществления, вариантный AAV2 капсидный белок содержит все 14 аминокислотных замещений, раскрытых выше, например, вариантный AAV2 капсидный белок содержит последовательность SEQ ID NO: 2 (то есть ttAAV2 или AAV2-TT как это указано в настоящем документе).
В следующих вариантах осуществления, вариантный AAV2 капсидный белок может содержать подмножество из указанных выше 14 мутаций. Не желая быть связанными теорией, в отдельных вариантах осуществления, вариантный AAV2 капсидный белок может содержать следующие остатки, которые разделяются ниже на функциональные группы:
1) S585 и/или Т588; данные остатки могут быть связаны с уменьшенным гепариновым связыванием и увеличенным распространением вируса в ткани головного мозга богатых на гепаринсульфатпротеогликан;
2) S312; данный внутренний остаток серина может играть определенную роль во взаимодействиях капсид-ДНК;
3) D546 и/или G548; данные остатки могут быть вовлечены во взаимодействиях с нейтрализующими антителами и, таким образом, вносить свой вклад в in vivo характеристики трансдукции;
4) S593; данный остаток расположен в углублении между тройными-прокимальными шиповидными отростками;
5) М457, А492, D499 и/или Y533; данные четыре аминокислоты могут быть вовлечены в связывание с рецептором и близко расположены на тройных шиповидных отростках;
6) I125, А151, S162 и/или S205; данные остатки могут быть связаны с активностью PLA2 и/или направленной миграцией входящего вируса.
Следует принять во внимание, что также рассматриваемыми в настоящем документе являются соответствующие подгруппы, содержащие мутации, отвечающие указанным выше остаткам, когда присутствуют в соответствующих положениях в дальнейших AAV серотипах (смотри ниже).
В предпочтительных вариантах осуществления, вариантный AAV2 капсидный белок содержит четыре или больше мутаций в положениях, которые упоминались выше, которые могут быть связаны со связыванием с рецептором, то есть остатки 457, 492, 499 и 533. Таким образом, особенно предпочтительным является то, что вариантный AAV2 капсидный белок содержит следующие остатки М457, А492, D499 и Y533.
В некоторых предпочтительных вариантах осуществления, вариантный AAV2 капсидный белок является немутированным по сравнению с AAV2 капсидным белком немутантного типа в положении 312, например, вариантный AAV2 капсидный белок содержит остаток N312 (который присутствует в AAV2 капсидном белке немутантного типа). Таким образом в некоторых вариантах осуществления, вариантный AAV2 капсидный белок может содержать от 1 до 13 специфических мутаций, указанных выше, но не мутацию N312S.
ВАРИАНТНЫЕ AAV КАПСИДНЫЕ БЕЛКИ ИЗ ДРУГИХ СЕРОТИПОВ
В следующих вариантах осуществления, вариантный AAV капсидный белок является из альтернативного AAV серотипа, то есть AAV серотипа отличного от AAV2. Например, вариантный AAV капсидный белок может быть получен из AAV1, AAV3B, AAV-LK03, AAV5, AAV6, AAV8, AAV9 или AAV10 (например, AAVrh10) капсидного белка.
В данных вариантах осуществления, вариантный AAV капсидный белок содержит, по меньшей мере, одно аминокислотное замещение в одном или больше положениях, отвечающих тем, что описаны выше по сравнению с AAV2. Другими словами, вариантный AAV капсидный белок содержит, по меньшей мере, одно аминокислотное замещение в положении в последовательности альтернативного (то есть не-AAV2) AAV капсидного белка, которое соответствует положеним 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593в последовательности AAV2 капсидного белка.
Квалифицированным специалистам в данной области известно, на основе сравнения аминокислотных последовательностей капсидных белков различных AAV серотипов, как идентифицировать положения в капсидных белках из альтернативных AAV серотипов, которые соответствуют положениям 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593 в AAV2 капсидном белке. В частности, такие положения могут быть легко идентифицированы с помощью выравниваний последовательностей, как известно в данной области с уровня техники, и описано в данном документе. Например, одно такое выравнивание последовательности приведено на фигуре 9.
Особое значение в данном контексте представляют собой положения в последовательностях альтернативного AAV капсидного белка, которые соответствуют в трехмерном пространстве положениям 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593 в AAV2 капсидном белке. Способы трехмерного моделирования и выравнивания белковых структур хорошо известны в данной области, и могут быть использованы для идентифицирования таких соответствующих положений в последовательностях не-AAV2 капсидного белка. Иллюстративный 3D сравнительный анализ первичных структур последовательностей AAV2 капсидных белков с последовательностями капсидного белка альтернативных AAV серотипов (например, AAV1, AAV5, AAV6, AAV8 и AAV9) показан на фигурах с 21 по 25 и обсуждаются ниже. Квалифицированный специалист может выполнить аналогичные 3D сравнительные анализы первичной структуры с капсидными белками из последующих серотипов, например, AAV2, AAV3, AAV7, AAV10 и AAV12), и идентифицировать положения в таких последовательностях, которые соответствуют положениям, определенным выше в AAV2.
ВАРИАНТНЫЙ AAV1 КАПСИДНЫЙ БЕЛОК
В одном варианте осуществления, вектор содержит вариантный AAV1 капсидный белок. В данном варианте осуществления, вариантный AAV1 капсидный белок содержит, по меньшей мере, одно аминокислотное замещение в одном или больше из следующих положений в последовательности AAV1 капсидного белка: 125, 151, 162, 205, 313, 458, 493, 500, 534, 547, 549, 586, 589 и/или 594. Данные положения в AAV1 капсидном белке VP1 соответствуют тем, которые раскрыты выше, по отношению к AAV2.
Последовательность AAV1 капсидного белка немутантного типа VP1 известна, и показана на фигуре 3 (SEQ ID NO: 3). Последовательности AAV1 капсидного белка немутантного типа также доступны из базы данных с номером доступа: NC-002077. Предпочтительно, вариантный AAV1 капсидный белок имеет, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность последовательности к SEQ ID NO: 3.
AAV1 капсидный белок немутантного типа VP1 уже содержит следующие остатки в положениях, которые соответствуют аминокислотным остаткам, которые присутствуют в вариантном AAV2 капсидном белке, раскрытых выше (SEQ ID NO: 2, ttAAV2), но не немутантного типа AAV2 (SEQ ID NO: 1): S205 (совпадает с S205 в ttAAV2); G549 (совпадает с G548 в ttAAV2); S586 (совпадает с S585 в ttAAV2); и Т589 (совпадает с Т588 в ttAAV2). Соответственно, в предпочтительном варианте осуществления, вариантный AAV1 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с капсидным белком немутантного типа AAV1: V125I, Q151A, T162S, N313S, N458M, К493А, N500D, F534Y, S547D, и/или G594S. Как правило, такой вариантный AAV1 капсидный белок может разделять одно или больше функциональных свойств с вариантным AAV2 капсидным белком (SEQ ID NO: 2, ttAAV2), например, может обеспечивать повышенную инфекционность и/или трансдукцию нейронов ретинальной ткани по сравнению с капсидным белком немутантного типа AAV1.
В альтернативных вариантах осуществлений, вариантный AAV1 капсидный белок содержит одно или больше аминокислотных замещений, которые соответствуют перестановкам мутаций, присутствующим в ttAAV2 обратно в последовательности немутантного типа AAV2. Например, вариантный AAV1 капсидный белок может содержать одно или больше из следующих замещений: S205T, G549E, S586R и/или T589R. Как правило такой вариантный AAV1 капсидный белок может разделять одно или больше функциональных свойств с AAV2 капсидным белком немутантного типа (SEQ ID NO: 1), например, может обеспечивать сниженную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с капсидным белком немутантного типа AAV1.
ВАРИАНТНЫЙ AAV5 КАПСИДНЫЙ БЕЛОК
В одном варианте осуществления, вектор содержит вариантный AAV5 капсидный белок. В данном варианте осуществления, вариантный AAV5 капсидный белок содержит, по меньшей мере, одно аминокислотное замещение в одном или больше из следующих положений в последовательности AAV5 капсидного белка: 124, 150, 153, 195, 303, 444, 479, 486, 520, 533, 537, 575, 578 и/или 583. Данные положения в AAV5 капсидном белке VP1 соответствуют тем, которые раскрыты выше, по отношению к AAV2.
Последовательность AAV5 капсидного белка немутантного типа VP1 известна и показана на фигуре 4 (SEQ ID NO: 4). Последовательности AAV5 капсидного белка немутантного типа также доступны из базы данных с номером доступа: AF085716. Предпочтительно вариантный AAV5 капсидный белок имеет, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность последовательности к SEQ ID NO: 4.
AAV5 капсидный белок немутантного типа VP1 уже содержит следующие остатки в положениях, которые соответствуют аминокислотным остаткам, которые присутствуют в вариантном AAV2 капсидном белке, раскрытом выше (SEQ ID NO: 2, ttAAV2), но не немутантном типе AAV2 (SEQ ID NO: 1): G537 (совпадает с G548 в ttAAV2); S575 (совпадает с S585 в ttAAV2); Т578 (совпадает с Т588 в ttAAV2). Соответственно, в предпочтительном варианте осуществления, вариантный AAV5 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV5 капсидным белком немутантного типа: V124I, К150А, K153S, A195S, R303S, Т444М, S479A, V486D, T520Y, P533D, и/или G583S. Как правило, такой вариантный AAV5 капсидный белок может разделять одно или больше функциональных свойств с вариантным AAV2 капсидным белком (SEQ ID NO: 2, ttAAV2), например, может обеспечивать повышенную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с AAV5 капсидным белком немутантного типа.
В альтернативных вариантах осуществлений, вариантный AAV5 капсидный белок содержит одно или больше аминокислотных замещений, которые соответствуют перестановкам мутаций, присутствующим в ttAAV2 обратным к AAV2 последовательности немутантного типа. Например, вариантный AAV5 капсидный белок может содержать одно или больше из следующих замещений: G537E, S575R и/или T578R. Как правило, такой вариантный AAV5 капсидный белок может разделять одно или больше функциональных свойств с AAV2 капсидным белком немутантного типа (SEQ ID NO: 1), например, может обеспечивать сниженную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с AAV5 капсидным белком немутантного типа.
ВАРИАНТНЫЙ AAV6 КАПСИДНЫЙ БЕЛОК
В одном варианте осуществления, вектор содержит вариантный AAV6 капсидный белок. В данном варианте осуществления, вариантный AAV6 капсидный белок содержит, по меньшей мере, одно аминокислотное замещение в одном или больше из следующих положений в последовательности AAV6 капсидного белка: 125, 151, 162, 205, 313, 458, 493, 500, 534, 547, 549, 586, 589 и/или 594. Данные положения в AAV6 капсидном белке VP1 соответствуют тем, которые раскрыты выше, по отношению к AAV2.
Последовательность AAV6 капсидного белка немутантного типа VP1 известна и показана на фигуре 5 (SEQ ID NO: 5). Последовательности AAV6 капсидного белка немутантного типа также доступны из базы данных с номером доступа: AF028704. Предпочтительно, вариантный AAV6 капсидный белок имеет, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность последовательности к SEQ ID NO: 5.
AAV6 капсидный белок немутантного типа VP1 уже содержит следующие остатки в положениях, которые соответствуют аминокислотным остаткам, которые присутствуют в вариантном AAV2 капсидном белке, раскрытом выше (SEQ ID NO: 2, ttAAV2), но не немутантном типе AAV2 (SEQ ID NO: 1): S205 (совпадает с S205 в ttAAV2); G549 (совпадает с G548 в ttAAV2); S586 (совпадает с S585 в ttAAV2); Т589 (совпадает с Т588 в ttAAV2). Соответственно, в предпочтительном варианте осуществления, вариантный AAV6 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV6 капсидным белком немутантного типа: V125I, Q151A, T162S, N313S, N458M, K493А, N500D, F534Y, S547D, и/или G594S. Как правило, такой вариантный AAV6 капсидный белок может разделять одно или больше функциональных свойств с вариантным AAV2 капсидным белком (SEQ ID NO: 2, ttAAV2), например, может обеспечивать повышенную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с AAV6 капсидным белком немутантного типа.
В альтернативных вариантах осуществлений, вариантный AAV6 капсидный белок содержит одно или больше аминокислотных замещений, которые соответствуют перестановкам мутаций, присутствующим в ttAAV2 обратным к AAV2 последовательности немутантного типа. Например, вариантный AAV6 капсидный белок может содержать одно или больше из следующих замещений: S205T, G549E, S586R и/или T589R. Как правило, такой вариантный AAV6 капсидный белок может разделять одно или больше функциональных свойств с AAV2 капсидным белком немутантного типа (SEQ ID NO: 1), например, может обеспечивать сниженную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с AAV6 капсидным белком немутантного типа.
ВАРИАНТНЫЙ AAV8 КАПСИДНЫЙ БЕЛОК
В одном варианте осуществления, вектор содержит вариантный AAV8 капсидный белок. В данном варианте осуществления, вариантный AAV8 капсидный белок содержит, по меньшей мере, одно аминокислотное замещение в одном или больше из следующих положений в последовательности AAV8 капсидного белка: 125, 151, 163, 206, 315, 460, 495, 502, 536, 549, 551, 588, 591 и/или 596. Данные положения в AAV8 капсидном белке VP1 соответствуют тем, которые раскрыты выше, по отношению к AAV2.
Последовательность AAV8 капсидного белка немутантного типа VP1 известна и показана на фигуре 6 (SEQ ID NO: 6). Последовательности AAV8 капсидного белка немутантного типа также доступны из базы данных с номером доступа: NC_006261. Предпочтительно, вариантный AAV8 капсидный белок имеет, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность последовательности к SEQ ID NO: 6.
AAV8 капсидный белок немутантного типа VP1 уже содержит следующие остатки в положениях, которые соответствуют аминокислотным остаткам, которые присутствуют в вариантном AAV2 капсидном белке, раскрытом выше (SEQ ID NO: 2, ttAAV2), но не немутантном типе AAV2 (SEQ ID NO: 1): S315 (совпадает с S312 в ttAAV2); Т591 (совпадает с Т588 в ttAAV2). Соответственно, в предпочтительном варианте осуществления, вариантный AAV8 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV8 капсидным белком немутантного типа: V125I, Q151A, K163S, A206S, Т460М, Т495А, N502D, F536Y, N549D, A551G, Q588S и/или G596S. Как правило, такой вариантный AAV8 капсидный белок может разделять одно или больше функциональных свойств с вариантным AAV2 капсидным белком (SEQ ID NO: 2, ttAAV2), например, может обеспечивать повышенную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с AAV8 капсидным белком немутантного типа.
В альтернативных вариантах осуществлений, вариантный AAV8 капсидный белок содержит одно или больше аминокислотных замещений, которые соответствуют перестановкам мутаций, присутствующим в ttAAV2 обратным к AAV2 последовательности немутантного типа. Например, вариантный AAV8 капсидный белок может содержать одно или больше из следующих замещений: S315N и/или T591R. Как правило, такой вариантный AAV8 капсидный белок может разделять одно или больше функциональных свойств с AAV2 капсидным белком немутантного типа (SEQ ID NO: 1), например, может обеспечивать сниженную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с AAV8 капсидным белком немутантного типа.
В одном варианте осуществления, вариантный AAV8 капсидный белок содержит аминокислотное замещение по сравнению с AAV8 капсидным белком немутантного типа в положении 315 в последовательности AAV8 капсидного белка. Например, вариантный AAV8 капсидный белок может содержать остаток N315. Таким образом, в одном варианте осуществления вариантный AAV8 капсидный белок содержит аминокислотное замещение S315N по сравнению с AAV8 капсидным белком немутантного типа.
ВАРИАНТ AAV9 КАПСИДНЫЙ БЕЛОК
В одном варианте осуществления, вектор содержит вариантный AAV9 капсидный белок. В данном варианте осуществления, вариантный AAV9 капсидный белок содержит по меньшей мере одно аминокислотное замещение в одном или больше из следующих положений в последовательности AAV9 капсидного белка: 125, 151, 162, 205, 314, 458, 493, 500, 534, 547, 549, 586, 589 и/или 594. Данные положения в AAV9 капсидном белке VP1 соответствуют тем, которые раскрыты выше, по отношению к AAV2.
Последовательность AAV9 капсидного белка немутантного типа VP1 известна и показана на фигуре 7 (SEQ ID NO: 7). Последовательности AAV9 капсидного белка немутантного типа также доступны из базы данных с номером доступа: AY530579. Предпочтительно вариантный AAV9 капсидный белок имеет, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность последовательности к SEQ ID NO: 7.
AAV9 капсидный белок немутантного типа VP1 уже содержит следующие остатки в положениях, которые соответствуют аминокислотным остаткам, которые присутствуют в вариантном AAV2 капсидном белке, раскрытом выше (SEQ ID NO: 2, ttAAV2), но не в немутантном типе AAV2 (SEQ ID NO: 1): S162 (совпадает с S162 в ttAAV2); S205 (совпадает с S205 в ttAAV2); G549 (совпадает с G548 в ttAAV2); S586 (совпадает с S585 в ttAAV2). Соответственно, в предпочтительном варианте осуществления, вариантный AAV9 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV9 капсидным белком немутантного типа: L125I, Q151A, N314S, Q458M, V493A, E500D, F534Y, G547D, А589Т и/или G594S. Как правило, такой вариантный AAV9 капсидный белок может разделять одно или больше функциональных свойств с вариантным AAV2 капсидным белком (SEQ ID NO: 2, ttAAV2), например, может обеспечивать повышенную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с AAV9 капсидным белком немутантного типа.
В альтернативных вариантах осуществлений, вариантный AAV9 капсидный белок содержит одно или больше аминокислотных замещений, которые соответствуют перестановкам мутаций, присутствующим в ttAAV2, обратным к AAV2 последовательности немутантного типа. Например, вариантный AAV9 капсидный белок может содержать одно или больше из следующих замещений: S162A, S205T, G549E и/или S586R. Как правило, такой вариантный AAV9 капсидный белок может разделять одно или больше функциональных свойств с AAV2 капсидным белком немутантного типа (SEQ ID NO: 1), например может обеспечивать сниженную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с AAV9 капсидным белком немутантного типа.
ВАРИАНТ AAV10 КАПСИДНЫЙ БЕЛОК
В одном варианте осуществления, вектор содержит вариантный AAV10 капсидный белок. Как используется в данном документе, "AAV10" включает AAVrh10. В данном варианте осуществления, вариантный AAV10 (например AAVrh10) капсидный белок содержит, по меньшей мере, одно аминокислотное замещение в одном или больше из следующих положений в последовательности AAV10 капсидного белка: 125, 151, 163, 206, 315, 460, 495, 502, 536, 549, 551, 588, 591 и/или 596. Данные положения в AAV10 капсидном белке VP1 соответствуют тем, которые раскрыты выше, по отношению к AAV2.
Последовательность AAV10 капсидного белка немутантного типа VP1 известна и показана на фигуре 8 (SEQ ID NO: 8). Предпочтительно, вариантный AAV10 капсидный белок имеет, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность последовательности к SEQ ID NO: 8.
AAV10 капсидный белок немутантного типа VP1 уже содержит следующий остаток в положении, которое соответствует аминокислотному остатку, который присутствует в вариантном AAV2 капсидном белке, раскрытом выше (SEQ ID NO: 2, ttAAV2), но не в немутантном типе AAV2 (SEQ ID NO: 1): G551 (совпадает с G548 в ttAAV2). Соответственно, в предпочтительном варианте осуществления, вариантный AAV10 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV10 капсидным белком немутантного типа: V125I, Q151A, K163S, A206S, N315S, Т460М, L495A, N502D, F536Y, G549D, Q588S, А591Т и/или G596S. Как правило, такой вариантный AAV10 капсидный белок может разделять одно или больше функциональных свойств с вариантным AAV2 капсидным белком (SEQ ID NO: 2, ttAAV2), например, может обеспечивать повышенную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с AAV10 капсидным белком немутантного типа.
В альтернативных вариантах осуществлений, вариантный AAV10 капсидный белок содержит аминокислотное замещение, которые соответствует перестановке мутаций, присутствующих в ttAAV2, обратным к AAV2 последовательности немутантного типа. Например, вариантный AAV10 капсидный белок может содержать следующее замещение: G551E. Как правило, такой вариантный AAV10 капсидный белок может разделять одно или больше функциональных свойств с AAV2 капсидным белком немутантного типа (SEQ ID NO: 1), например, может обеспечивать сниженную инфекционность и/или трансдукцию нейрона ретинальной ткани по сравнению с AAV10 капсидным белком немутантного типа.
ВАРИАНТНЫЙ AAV3B КАПСИДНЫЙ БЕЛОК
В одном варианте осуществления, вектор содержит вариантный AAV3B капсидный белок. В данном варианте осуществления, вариантный AAV3B капсидный белок может содержать аминокислотное замещение по сравнению с немутантным типом AAV3B капсидного белка в положении 312. Например, вариантный AAV3B капсидный белок может содержать остаток N312. Таким образом, в одном варианте осуществления вариантный AAV8 капсидный белок содержит аминокислотное замещение S312N по сравнению с AAV8 капсидным белком немутантного типа. В следующих вариантах осуществления, вариантный AAV3B капсидный белок может содержать одну или больше дополнительных мутаций в положениях, которые соответствуют тем, которые раскрыты выше, по отношению к AAV2.
Последовательность AAV3B капсидного белка немутантного типа VP1 известна и показана на фигуре 38 (SEQ ID NO: 11). Последовательность AAV3B капсидного белка немутантного типа также доступна из базы данных NCBI с номером доступа AF028705. Предпочтительно, вариантный AAV3B капсидный белок имеет, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность последовательности к SEQ ID NO: 11.
ВАРИАНТНЫЙ AAV-LK03 КАПСИДНЫЙ БЕЛОК
В одном варианте осуществления, вектор содержит вариантный AAV-LK03 капсидный белок. В данном варианте осуществления, вариантный AAV-LK03 капсидный белок может содержать аминокислотное замещение в положении 312 по сравнению с последовательностью AAV-LK03 капсидного белка, как определено в SEQ ID NO: 12. Например, вариантный AAV-LK03 капсидный белок может содержать остаток N312. Таким образом, в одном варианте осуществления вариантный AAV-LK03 капсидный белок содержит аминокислотное замещение S312N по сравнению с последовательностью AAV-LK03 капсидного белка, как определено в SEQ ID NO: 12. В следующих вариантах осуществления, вариантный ААV-LK03 капсидный белок может содержать одну или больше дополнительных мутаций в положениях, которые соответствуют тем, которые раскрыты выше, по отношению к AAV2.
Последовательность AAV-LK03 капсидного белка немутантного типа VP1 известна и показана на фигуре 39 (SEQ ID NO: 12). Последовательность AAV-LK03 капсидного белка также раскрыта в WO 2013/029030, как последовательность номер 31 в нем. Предпочтительно вариантный AAV-LK03 капсидный белок имеет, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность последовательности к SEQ ID NO: 12.
ГЕННЫЕ ПРОДУКТЫ
В одном варианте осуществления, rAAV дополнительно содержит гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт. "Ген" касается полинуклеотида, содержащего, по меньшей мере, одну открытую рамку считывания, которая способна кодировать специфический белок после того, как транскибируется и транслируется. "Генный продукт" представляет собой молекулу, полученную в результате экспрессии конкретного гена. Генные продукты включают, например, полипептид, аптамер, интерферирующую РНК, мРНК, и тому подобное.
"Гетерологичная" означает полученную из генотипически отличного объекта от того, что остается от объекта, с которым она сравнивается. Например, полинуклеотид, введенный методами генной инженерии в плазмиду или вектор, полученный из различных видов представляет собой гетерологичный полинуклеотид. Связанный промотор, удаленный из его нативной кодирующей последовательности, и функционально связанный с кодирующей последовательностью, с которой он не встречается в природе, представляет собой гетерологичный промотор. Таким образом, например, rAAV, который включает гетерологичную нуклеиновую кислоту, кодирующую гетерологичный генный продукт, представляет собой rAAV, который включает нуклеиновую кислоту, обычно не включенную в AAV немутантного типа, который не встречается в природе, и кодированный гетерологичный генный продукт представляет собой генный продукт, который, как правило, не кодируется немутантным типом AAV, который не встречается в природе.
В некоторых вариантах осуществления, генный продукт представляет собой интерферирующую РНК. В некоторых вариантах осуществления, генный продукт представляет собой аптамер. В некоторых вариантах осуществления, генный продукт представляет собой полипептид.
Интерферирующая РНК
Когда генный продукт представляет собой интерферирующую РНК (РНК-интерференция), приемлемая РНК-интерференция включают РНК-интерференцию, которая понижает уровень апоптического или ангиогенного фактора в клетке. Например, РНК-интерференция может представлять собой кшРНК или миРНК, которая снижает уровень генного продукта, который индуцирует или стимулирует апоптоз в клетке. Гены, чьи генные продукты индуцируют или способствуют апоптозу, упоминаются в данном документа как "про-апоптические гены", и продукты данных генов (мРНК; белок) упоминаются как "про-апоптические генные продукты." Про-апоптические генные продукты включают, например, Вах, Bid, Bak, и Bad генные продукты. Смотри, например, патент США №7,846,730.
Интерферирующие РНК также могут быть против ангиогенного продукта, например, VEGF (например, Cand5; смотри, например, публикацию патента США №2011/0143400; публикацию патента США №2008/0188437; и Reich et al. (2003) Mol. Vis. 9: 210), VEGFR1 (например, миРНК-027; смотри, например, Kaiser et al. (2010) Am. J. Ophthalmol. 150: 33; и Shen et al. (2006) Gene Ther. 13:225), или VEGFR2 (Kou et al. (2005) Biochem. 44: 15064). Смотри также, патенты США №№6,649,596, 6,399,586, 5,661,135, 5,639,872, и 5,639,736; и патенты США№№7,947,659 и 7,919,473.
"Малая интерферирующая", или "короткая интерферирующая РНК", или миРНК представляет собой дуплекс РНК из нуклеотидов, который направляется на интересующий ген ("ген-мишень"). "Дуплекс РНК" касается структуры, образованной комплементарной парой между двумя областями молекулы РНК. миРНК является "мишенной" для гена в том, что нуклеотидная последовательность дуплексной части миРНК является комплементарной к нуклеотидной последовательности гена-мишени. В некоторых вариантах осуществления, длина дуплекса миРНК составляет меньше, чем 30 нуклеотидов. В некоторых вариантах осуществления, дуплекс может составлять 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11 или 10 нуклеотидов в длину. В некоторых вариантах осуществления, длина дуплекса составляет 19-25 нуклеотидов в длину. Дуплексная РНК часть миРНК может представлять собой часть структуры шпильки. В дополнение к дуплексной части, структура шпильки может содержать часть петли, расположенной между двумя последовательностями, которые образуют дуплекс. Петля может варьировать в длину. В некоторых вариантах осуществления петля составляет 5, 6, 7, 8, 9, 10, 11, 12 или 13 нуклеотидов в длину. Структура шпильки может также содержать 3' или 5' «липкие» части. В некоторых вариантах осуществления, «липкий» представляет собой 3' или 5' «липкий» 0, 1, 2, 3, 4 или 5 нуклеотидов в длину.
"Короткая шпилька РНК," или кшРНК, представляет собой полинуклеотидный конструкт, который может быть сделан, чтобы экспрессировать интерферирующую РНК, такую как миРНК.
Аптамеры
Когда генный продукт представляет собой аптамер, иллюстративные аптамеры, представляющие интерес, включают аптамер против фактора роста сосудистого эндотелия (VEGF). Смотри, например, Ng et al. (2006) Nat. Rev. Drug Discovery 5: 123; и Lee et al. (2005) Proc. Natl. Acad. Sci. USA 102: 18902. Также приемлемым для применения является PDGF-специфический аптамер, например, Е10030; смотри, например, Ni and Hui (2009) Ophthalmologica 223: 401; и Akiyama et al. (2006) J. Cell Physiol. 207: 407).
Полипептиды
В одном варианте осуществления, генный продукт представляет собой терапевтический белок. "Терапевтический" пептид или белок представляет собой пептид или белок, который может облегчить или уменьшить симптомы, возникающие в результате отсутствия или дефекта белка в клетке или субъекте. Альтернативно, "терапевтический" пептид или белок представляет собой тот, который в противном случае дает выгоду субъекту, например, анти-дегенеративные эффекты.
Когда генный продукт представляет собой полипептид, полипептид обычно представляет собой полипептид, который улучшает функцию клетки, например, клетки, присутствующей в нейрональной, ретинальной ткани или ткани печени, например, гепатоцита, нейрона, глиальной клетки, палочки или колбочки фоторецепторной клетки, ретинальной ганглиозной клетки, клетки Мюллера, биполярной клетки, амакринной клетки, горизонтальной клетки или ретинальной пигментированной эпителиальной клетки.
Иллюстративные полипептиды включают нейропротекторные полипептиды (например, GDNF, CNTF, NT4, NGF, и NTN); антиангиогенные полипептиды (например, растворимый рецептор фактора роста сосудистого эндотелия (VEGF); VEGF-связывающее антитело; фрагмент VEGF-связывающего антитела (например, одноцепочечное анти-VEGF антитело); эндостатин; тумстатин; ангиостатин; растворимый Fit полипептид (Lai et al. (2005) Mol. Ther. 12: 659); Fc слитый белок, содержащий растворимый Fit полипептид (смотри, например, Pechan et al. (2009) Gene Ther. 16: 10); фактора пигментного эпителя (PEDF); растворимый Tie-2 рецептор; и т.д.); тканевый ингибитор металлопротеиназы-3 (TIMP-3); светочувствительный опсин, например, родопсин; анти-апоптические полипептиды (например, Bcl-2, Вс1-Х1); и тому подобное. Приемлемые полипептиды включают, но не ограничиваются этим, глиальный нейротрофический фактор (GDNF); фактор роста фибробластов 2; нейротурин (NTN); цилиарный нейротрофический фактор (CNTF); фактор роста нервов (NGF); нейротрофин-4 (NT4); нейротрофический фактор головного мозга (BDNF); эпидермальный фактор роста; родопсин; Х-сцепленный ингибитор апоптоза; и звуковой еж. Приемлемые полипептиды раскрыты, например, в публикации WO 2012/145601.
Иллюстративные полипептиды для генной доставки в печень включают, например, PBGD (порфобилиногендезаминазу) IDUA (идуронидазу) Fah (фумарилацетоацетатгидролазу) А1АТ (альфа(1)-антитрипсин), 1A1(hUGT1A1) (уридиндифосфаттлюкуронилтрансферазу), HCCS1 (супрессор гепатоцеллюлярной карциномы 1), CD (цитозиндеаминазу), SOCS3 (супрессор цитокинового сигналинга 3), TNF (фактор некроза опухоли), тимидинкиназу, IL-24 (интерлейкин-24), IL-12 (интерлейкин-12), и TRAIL (родственный фактору некроза опухоли лиганд, индуцирующий апоптоз).
Регуляторные последовательности
В некоторых вариантах осуществления, нуклеотидная последовательность, кодирующая генный продукт, представляющий интерес, функционально связана с конститутивным промотором. В других вариантах осуществления, нуклеотидная последовательность, кодирующая генный продукт, представляющий интерес, функционально связана с индуцируемым промотор. В некоторых случаях, нуклеотидная последовательность, кодирующая генный продукт, представляющий интерес, функционально связана с тканеспецифическим или клеткаспецифическим регуляторным элементом.
Например, в некоторых случаях, нуклеотидная последовательность, кодирующая генный продукт, представляющий интерес, функционально связана с гепатоцит-специфическим, нейрон-специфическим или фоторецептор-специфическим регуляторным элементом (например, фоторецептор-специфическим промотором), например, регуляторным элементом, который обеспечивает селективную экспрессию функционально связанного гена в нейронной или фоторецепторной клетке. Приемлемые фоторецептор-специфические регуляторные элементы включают, например, родопсиновый промотор; родопсинкиназный промотор (Young et al. (2003) Ophthalmol. Vis. Sci. 44: 4076); бета-фосфодиэстеразный генный промотор (Nicoud et al. (2007) J. Gene Med. 9: 1015); промотор гена пигментной дегенерации сетчатки (Nicoud et al. (2007) выше); энхансер гена интерфоторецепторного ретиноид-связывающего белка (IRBP) (Nicoud et al. (2007) supra); промотор гена IRBP (Yokoyama et al. (1992) Exp Eye Res. 55: 225). Приемлемые нейронально-специфические промоторы включают нейрон-специфический энолазный (NSE) промотор, Andersen et al. Cell. Mol. Neurobiol., 13: 503-15 (1993; промотор нейрофиламентного легкоцепочечного гена, Piccioli et al., Proc. Natl. Acad. Sci. USA, 88: 561 1-5 (1991); и промотор нейрон-специфического vgf гена, Piccioli et al., Нейрон, 15: 373-84 (1995)]; среди других. Приемлемые гепатоцит-специфические промоторы включают альбуминовый промотор (Heard et al., Mol Cell Biol 1987; 7: 2425) или альфа 1-антитрипсиновый промотор (Hafenrichter et al. Blood 1994; 84, 3394-404).
"Контрольный элемент" или "контрольная последовательность" представляет собой нуклеотидную последовательность, включенную во взаимодействие молекул, которые способствует функциональной регуляции полинуклеотида, включая репликацию, дупликацию, транскрипцию, сплайсинг, трансляцию или разложение полинуклеотида. Регулирование может влиять на частоту, скорость или специфичность процесса, и может быть повышающим или ингибирующим в природе. Контрольные элементы, известные в данной области с уровня техники включают, например, транскрипционные регуляторные последовательности, такие как промоторы и энхансеры. Промотор представляет собой участок ДНК, способный в определенных условиях связывать РНК полимеразу и инициировать транскрипцию кодирующей области, как правило, расположенной в прямом направлении (в 3' направлении) от промотора.
"Функционально связанный" или "связан функционально" касается смежных генетических элементов, в которых элементы находятся в связи, позволяющей им действовать в ожидаемом порядке. Например, промотор является функционально связанным с кодирующей областью, если промотор помогает инициировать транскрипцию кодирующей последовательности. Там могут существовать промежуточные остатки между промотором и кодирующей областью при условии, что данная функциональная зависимость сохраняется.
Термин "промоторы" или "промотор", как используется в данном документе, может касаться последовательности ДНК, которая расположена рядом с последовательностью ДНК, которая кодирует рекомбинантный продукт. Промотор предпочтительно функционально связан со смежной последовательностью ДНК. Промотор, как правило, повышает количество рекомбинантного продукта, экспрессированного из последовательности ДНК по сравнению с количеством экспрессированного рекомбинантного продукта, когда никакого промотора не существует. Промотор из одного организма может быть использован для повышения экспрессии рекомбинантного продукта из последовательности ДНК, которая походит из другого организма. Например, промотор позвоночного животного может быть использован для экспрессии GFP медузы у позвоночных. Кроме того, один промоторный элемент может увеличивать количество рекомбинантных продуктов, экспрессированных для множественных последовательностей ДНК, прикрепленных в тандеме. Следовательно, один промоторный элемент может усиливать экспрессию одного или нескольких рекомбинантных продуктов. Множественные промоторные элементы хорошо известны квалифицированным специалистам в данной области с уровня техники.
Термин "энхансеры" или "энхансер", как используется в данном документе, может касаться последовательности ДНК, которая расположена рядом с последовательностью ДНК, которая кодирует рекомбинантный продукт. Энхансерные элементы, как правило, расположены в 3'-5' направлении промоторного элемента, или могут быть расположены в 5-3' направлении или в пределах кодирующей последовательности ДНК (например, последовательности ДНК, транскрибированной или транслированной в рекомбинантный продукт или продукты). Следовательно, энхансерный элемент может быть расположен на 100 парах оснований, 200 парах оснований, или 300 или больше пар оснований в 3'-5' направлении или 5'-3' направлении последовательности ДНК, которая кодирует рекомбинантный продукт. Энхансерные элементы могут повышать количество рекомбинантного продукта, экспрессированного из последовательности ДНК, указанной выше, где повышенная экспрессия обеспечивается промоторным элементом. Множественные энхансерные элементы легко доступны квалифицированным специалистам в данной области с уровня техники.
ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ
Представленное раскрытие предусматривает фармацевтическую композицию, содержащую: а) rAAV вектор, как описано выше; и b) фармацевтически приемлемый носитель, разбавитель, эксципиент или буфер. В некоторых вариантах осуществления, фармацевтически приемлемый носитель, разбавитель, эксципиент или буфер пригоден для применения человеком.
Такие эксципиенты, носители, разбавители и буферы включают какой-либо фармацевтический агент, который может быть введен без чрезмерной токсичности.
Фармацевтически приемлемые эксципиенты включают, но не ограничиваются этим, жидкости, такие как вода, солевой раствор, глицерин и этанол.
Фармацевтически приемлемые соли могут быть включены в него, например, соли минеральных кислот, такие как гидрохлориды, гидробромиды, фосфаты, сульфаты и тому подобное; и соли органических кислот, такие как ацетаты, пропионаты, малонаты, бензоаты и тому подобное. Кроме того, вспомогательные вещества, такие как смачивающие или эмульгирующие агенты, pH-буферные вещества и тому подобное, могут присутствовать в таких наполнителях. Широкое разнообразие фармацевтически приемлемых эксципиентов известно в данной области с уровня техники и не нуждается в детальном обсуждении в данном документе. Фармацевтически приемлемые эксципиенты достаточно описаны в различных публикациях, включая, например, A. Gennaro (2000) "Remington: The Science and Practice of Pharmacy," 20th edition, Lippincott, Williams, & Wilkins; Pharmaceutical Dosage Forms and Drug Delivery Systems (1999) H.C. Ansel et al., eds., 7(th) ed., Lippincott, Williams, & Wilkins; and Handbook of Pharmaceutical Excipients (2000) A.H. Kibbe et al., eds., 3 rd ed. Amer. Pharmaceutical Assoc.
В конкретных вариантах осуществления, представленное изобретение предусматривает фармацевтическую композицию, содержащую rAAV вектор, как описано выше, в фармацевтически приемлемом носителе или других лекарственных агентах, фармацевтических агентах, исполнителях, адъювантах, разбавителях, т.д. Для инъекции, носитель, как правило, будет представлять собой жидкость. Для других способов введения, носитель может быть или твердым, или жидким, таким как стерильная, свободная от пирогенов вода, или стерильный свободный от пирогенов фосфатный буферный солевой раствор. Для ингаляционного введения, носитель будет вдыхаемым, и предпочтительно будет находиться в форме твердых или жидких частиц. В качестве инъекционной среды предпочтительным является использовать воду, которая содержит добавки, обычные для инъекционных растворов, такие как стабилизирующие агенты, соли или солевой раствор, и/или буферы.
Под термином "фармацевтически приемлемый" подразумевают материал, который не является биологически или иным образом нежелательными, например, материал, который может быть введен субъекту, не вызывая каких-либо нежелательных биологических эффектов. Таким образом, такая фармацевтическая композиция может быть использована, например, в трансфекции клетки ex vivo или при введении вирусной частицы или клетки непосредственно субъекту.
СПОСОБЫ ДОСТАВКИ ГЕННОГО ПРОДУКТА В ТКАНЬ ИЛИ КЛЕТКУ (НАПРИМЕР, ТКАНЬ ИЛИ КЛЕТКИ ПЕЧЕНИ, НЕЙРОНАЛЬНЫЕ ИЛИ РЕТИНАЛЬНЫЕ ТКАНИ ИЛИ КЛЕТКИ) И СПОСОБЫ ЛЕЧЕНИЯ
Способы согласно представленному изобретению предусматривают способы доставки гетерологичных последовательностей нуклеиновой кислоты в ткань или клетку хозяина, включая как делящиеся, так и не делящиеся клетки. Векторы и другие реагенты, способы и фармацевтические препараты согласно представленному изобретению, кроме того, являются полезными в способе введения белка или пептида субъекту, нуждающемуся в этом, в качестве способа лечения или иным образом. Следовательно, белок или пептид может, таким образом, быть продуцированным in vivo в субъекте. Субъект может нуждаться в белке или пептиде, поскольку субъект имеет дефицит белка или пептида, или поскольку продуцирование белка или пептида у субъекта может обеспечивать некоторый терапевтический эффект, в качестве способа лечения или иным образом, и как описано далее ниже.
Как используется в данном документе, термины "лечение," "лечащий," и тому подобные, касаются получения желаемого фармакологического и/или физиологического эффекта. Эффект может быть профилактическим с точки зрения полной или частичной профилактики его заболевания или симптомов и/или может быть терапевтическим с точки зрения частичного или полного избавления от заболевания и/или неблагоприятного эффекта, относимого к этому заболеванию. "Лечение," как используется в данном документе, охватывает какое-либо лечение заболевания у млекопитающего, в частности у человека, и включает: (а) предупреждение возникновения заболевания у субъекта, который может быть предрасположен к заболеванию или имеет риск заражения болезнью, но у которого еще не диагностировано оно, как имеющееся; (b) сдерживание заболевания, то есть, подавление его развития; и (с) облегчение заболевания, то есть, вызывание ослабления симптомов заболевания.
В общем, представленное изобретение может быть использовано для доставки какой-либо чужеродной нуклеиновой кислоты с биологическим эффектом для лечения или ослабления симптомов, связанных с каким-либо расстройством, связанным с экспрессией гена в каком-либо органе, ткани или клетке, особенно тех, которые связаны с, например, печенью, головным мозгом или глазом. Иллюстративные болезненные состояния включают, но не ограничиваются этим: лизосомную болезнь накопления, острую интермиттирующую порфирию, дефицит орнитинтранскарбамилазы, дефицит альфа(1)-антитрипсина, острую печеночную недостаточность, болезнь Помпа, тирозинемию, синдром Криглера-Наджар, гепатит, цирроз печени, гепатоцеллюлярную карциному, СПИД, болезнь Альцгеймера, болезнь Паркинсона, болезнь Хантингтона, амиотрофический латеральный склероз, эпилепсию, и другие неврологические расстройства, рак (например, рак мозга), дегенеративные заболевания сетчатки и другие заболевания глаз.
Перенос генов имеет значительный потенциал использования в понимании и предоставлении терапии для болезненных состояний. Существует целый ряд наследственных заболеваний, в которых дефектные гены известны и были клонированы. В некоторых случаях функция данных клонированных генов известна. В целом, указанные выше болезненные состояния делятся на два класса: дефицитные состояний, как правило, ферментов, которые обычно наследуются рецессивным образом, и неуравновешенные состояния, которые, по меньшей мере, иногда вовлекают регуляторные или структурные белки, которые наследуются доминирующим образом. Для дефицитных болезненных состояний, перенос генов может быть использован, чтобы принести нормальный ген в пораженные ткани во время заместительной терапии, а также для создания моделей животных для заболевания с использованием антисмысловых мутаций. Для неуравновешенных болезненных состояний, перенос генов может быть использован для создания болезненного состояния в модельной системе, которая затем может быть использована в работе по противодействию болезненному состоянию. Таким образом, способы согласно представленному изобретению позволяют лечение генетических заболеваний. Как используется в данном документе, болезненное состояние лечится частично или полностью, устраняя дефицит или дисбаланс, который вызывает заболевание или делает его более тяжелым. Возможным также является использование сайт-специфической интеграции нуклеиновых последовательностей для того, чтобы вызвать мутации или исправить дефекты.
В одном аспекте представленное изобретение предусматривает способ доставки генного продукта в ткань или клетку (например, ткань или клетку печени, нейрональную или ретинальную ткань или клетку) субъекта, где способ включает введение субъекту rAAV вектора, как описано выше. Генный продукт может представлять собой полипептид или интерферирующую РНК (например, кшРНК, миРНК, и тому подобное), или аптамер, например, как описано выше. Клетка, например, может представлять собой клетку крови, стволовую клетку, клетку костного мозга (например, гематопоэтическую), клетку печени, раковую клетку, сосудистую клетку, клетку поджелудочной железы, нервную клетку, глиальную клетку, глазную или ретинальную клетку, эпителиальную или эндотелиальную клетку, дендритную клетку, фибробласт, легочную клетку, мышечную клетку, клетку сердца, кишечную клетку или почечную клетку. Аналогичным образом ткань, например, может быть выбрана из крови, костного мозга, мышечной ткани (например, скелетной мышцы, сердечной мышцы или гладкой мышцы, включая гладкие мышцы сосудов), ткани центральной или периферической нервной системы (например, такни головного мозга, нейрональной ткани или ретинальной ткани), ткани поджелудочной железы, ткани печени, ткани почек, легочной ткани, ткани кишечника или ткани сердца.
Доставка генного продукта в ретинальную клетку может обеспечивать лечение заболевания сетчатки глаза. Ретинальная клетка может представлять собой фоторецептор, ретинальную ганглиозную клетку, клетку Мюллера, биполярную клетку, амакринную клетку, горизонтальную клетку или ретинальную пигментированную эпителиальную клетку. В некоторых случаях, ретинальная клетка представляет собой фоторецепторную клетку, например, клетки - палочки или колбочки. Аналогичным образом, доставка генного продукта в нейрональную ткань или клетку может обеспечивать лечение неврологического расстройства. Генный продукт может доставляться в клетки различных типов, присутствующие в нейрональной ткани, например, нейроны или глиальные клетки (например, астроциты, олигодендроциты и тому подобные). Доставка генного продукта в печень может обеспечивать лечение заболеваний печени. Генный продукт может доставляться в, например, гепатоциты.
Представленное раскрытие предусматривает способ лечения заболевания (например, заболевания печени, неврологического или глазного заболевания), где способ включает введение субъекту, нуждающемуся в этом, эффективного количества rAAV вектора, как описано выше. Субъекту rAAV вектор может быть введен в виде внутричерепной инъекции, внутримозговой инъекции, внутриглазной инъекции, в виде инъекции в стекловидное тело, ретинальной инъекции, суб-ретинальной инъекции, внутривенной инъекции или каким-либо другим удобным способом или путем введения.
Другие иллюстративные способы введение включают пероральное, ректальное, чресслизистое, местное, чрескожное, ингаляционное, парентеральное (например, внутривенное, подкожное, внутрикожное, внутримышечное и внутрисуставное) введения и тому подобное, а также непосредственное введение в ткань или орган, интратекальные, прямые внутримышечные, внутрижелудочковые, внутривенные, внутрибрюшинные, интраназальные, или внутриглазные инъекции. Инъекции могут быть приготовлены в обычных формах, либо в виде жидких растворов или суспензий, твердых формах, пригодных для приготовления раствора или суспензии в жидкости перед инъекцией, или в виде эмульсий. Альтернативно, может вводиться вирус местным, а не системным способом, например, в виде депо или композиции с замедленным высвобождением.
Рекомбинантные векторы вируса предпочтительно вводят субъекту в количестве, которое достаточно для того, чтобы в результате привести к инфицированию (или трансдукции) и экспрессии гетерологичной последовательности нуклеиновой кислоты в клетках (например, клетках печени, нейрональных или ретинальных клетках) субъекта. Предпочтительно клетки-мишени представляют собой гепатоциты, нервные клетки (включая клетки центральной и периферической нервных систем, в частности, клеток головного мозга) или ретинальные клетки. В некоторых случаях, ретинальная клеткой представляет собой фоторецепторные клетки (например, палочки и/или колючки). В других случаях, ретинальная клеткой представляет собой RGC клетку. В других случаях, ретинальная клеткой представляет собой RPE клетку. В других случаях, ретинальные клетки могут включать амакринные клетки, биполярные клетки, и горизонтальные клетки.
Предпочтительно вектор вводят в терапевтически эффективном количестве. "Терапевтически эффективное" количество, как используется в данном документе, представляет собой количество, которое является достаточным для облегчения (например, смягчения, снижения, уменьшения), по меньшей мере, одного из симптомов, связанных с болезненным состоянием. Альтернативно указано, что "терапевтически эффективное" количество представляет собой количество, которое является достаточным для обеспечения некоторого улучшения состояния субъекта. "Терапевтически эффективное количество" будет попадать в относительно широкий диапазон, который может быть определен экспериментальным путем и/или в ходе клинических испытаний. Например, для инъекций in vivo, терапевтически эффективная доза будет составлять порядка от приблизительно 106 до приблизительно 1015 rAAV вирионов, например, от приблизительно 108 до 1012 rAAV вирионов. Для трансдукции in vitro, эффективное количество rAAV вирионов, которое следует доставить в клетки, будет составлять порядка от приблизительно 108 до приблизительно 1013 rAAV вирионов. Другие эффективные дозировки могут быть легко установлены квалифицированным специалистом в данной области с помощью обычных испытаний, устанавливающих кривые зависимости реакции от дозы.
В некоторых вариантах осуществления, больше чем одно введение (например, два, три, четыре или больше введений) могут быть использованы для достижения желаемого уровня экспрессии гена в течение периода из различных интервалов, например, ежедневно, еженедельно, ежемесячно, ежегодно, и т.д.
Неврологические заболевания, которые можно лечить, включают какое-либо заболевание, связанное с головным мозгом или ЦНС, включая психические заболевания. Заболевания головного мозга делятся на две основные категории: (а) патологические процессы, такие как инфекции, травмы и новообразования; и (b) заболевания, уникальные для нервной системы, которые включают заболевания миелина и дегенерацию нейронов. Заболевания из какой-либо категории могут подвергаться лечению. Например, неврологическое заболевание, может быть выбрано из нейродегенеративных заболеваний, таких как болезнь Альцгеймера, болезнь Паркинсона, амиотрофический латеральный склероз (ALS), спинальная мышечная атрофия и дегенерация мозжечка; шизофрении; эпилепсии; связанных с ишемией заболеваний и инсульта; демиелинизирующих заболеваний, таких как рассеянный склероз, перивенозный энцефалит, лейкодистрофия, такая как метахроматическая лейкодистрофия из-за дефицита арилсульфатазы А, болезнь Краббе из-за дефицита галактоцереброзид-бета-галактозидазы, адренолейкодистрофия и адреномиелонейропатия; после вирусных заболеваний, таких как прогрессивная мультифокальная лейкоэнцефалопатия, острый рассеянный энцефаломиелит, острый некротический геморрагический лейкоэнцефалит; митохондриальных энцефаломиопатий; неврологических видов рака, таких как первичные опухоли головного мозга, включая глиому, менингиому, невриному, аденому гипофиза, медуллобластому, краниофарингиому, гемангиому, эпидермоид, саркому и внутричерепные метастазы из других источников опухолей; неврологических инфекций или неврологических воспалительных состояний.
Глазные заболевания, которые можно лечить у субъекта с использованием способа включают, но не ограничиваются этим, острую макулярную нейроретинопатию; болезнь Бехчета; хороидальную неоваскуляризацию; диабетический увеит; гистоплазмоз; дегенерацию желтого пятна, такую как острая дегенерация желтого пятна, безэкссудативная возрастная дегенерация желтого пятна и экссудативная возрастная дегенерация желтого пятна; отек, такой как отек макулы, кистозный макулярный отек и диабетический макулярный отек; мультифокальный хориоидит; мультифокальный хориоидит; глазную травму, которая влияет на глазной задний участок или местонахождение; глазные опухоли; расстройства сетчатки, такие как центральная ретинальная венозная окклюзия, диабетическая ретинопатия (включая пролиферативную диабетическую ретинопатию), пролиферативная витреоретинопатию (PVR), окклюзионная болезнь артерий сетчатки, отслоение сетчатки, увеальное ретинальное заболевание; симпатическую офтальмию; синдром Фогта-Коянаги-Харада (VKH); увеальную диффузию; заднее глазное состояние, вызванное или находящееся под влиянием лазерного лечения глаз; заднее глазное состояние, вызванное или находящееся под влиянием фотодинамической терапии; фотокоагуляционную, радиационную ретинопатию; эпиретинальные мембранные растройства; разветвленную ретинальную окклюзию вен; переднюю ишемическую невропатию зрительного нерва; нереринопатическую диабетическую дисфункцию сетчатки глаза; ретиношизис; пигментную дегенерацию сетчатки; глаукому; синдром Ушера, колбочко-палочковую дистрофию; болезнь Штаргардта (желтопятнистую абиотрофию сетчатки); наследственную дегенерацию желтого пятна; хориоретинальную дегенерацию; амавроз Лебера; врожденную стационарную ночную слепоту; хороидермию; синдром Барде-Бидля; макулярную телеангиэктазию; наследственную оптическую невропатию Лебера; ретинопатию недоношенных; и нарушения цветового зрения, включая ахроматопсию, протанопию, дейтеранопию и тританопию.
Заболевания печени, которые можно лечить, включают, например, лизосомную болезнь накоплений, например, острую интермиттирующую порфирию, дефицит орнитинтранскарбамилазы, болезнь Вильсона, мукополисахаридозы (например, MPS типа I или MPS типа VI), синдром Слая, болезнь Помпа, тирозинемию, дефицит альфа(1)-антитрипсина, синдром Криглера-Наджар; гепатит A, В или C; цирроз печени; рак печени, например, гепатоцеллюлярную карциному; или острую печеночную недостаточность.
Представленное изобретение находит применение как в ветеринарной, так и медицинской областях. Приемлемые субъекты включают как птиц, так и млекопитающих, где предпочтение отдается млекопитающим. Термин "птицы", как используется в данном документе, включает, но не ограничивается этим, кур, уток, гусей, перепелов, индеек и фазанов. Термин "млекопитающее", как используется в данном документе включает, но не ограничивается этим, людей, коров, овечьих, козьих, лошадиных, кошачих, собачих, зайцеобразных, т.д. Субъекты-люди являются наиболее предпочтительными. Субъекты-люди включают эмбриональные, новорожденные, младенческие, несовершеннолетние и взрослые субъекты.
ТРАНСДУКЦИЯ ТКАНИ (НАПРИМЕР, ТКАНИ ПЕЧЕНИ, НЕЙРОНАЛЬНОЙ ИЛИ РЕТИНАЛЬНОЙ ТКАНИ)
В некоторых вариантах осуществления, rAAV векторы, раскрытые в данном документе, демонстрируют повышенную трансдукцию ткани (например, ткани печени, нейрональной и/или ретинальной тканей), например, по сравнению с соответствующим AAV вектором (из такого же серотипа), содержащим AAV капсидный белок немутантного типа. Например, rAAV вектор может демонстрировать, по меньшей мере, 10%, 50%), 100%, 500% или 1000%) повышенную инфекционность, по сравнению с инфекционностью AAV вирионом, содержащим соответствующий AAV капсидный белок немутантного типа.
В следующих вариантах осуществления, rAAV векторы, раскрытые в данном документе, могут селективно или специфически инфицировать ткань (например, ткань печени, нейрональную или ретинальную ткани), например, демонстрируют повышенную трансдукцию клеток печени, нейрональных или ретинальных клеток по сравнению с другими типами клеток. Например, rAAV вектор может демонстрировать, по меньшей мере, 10%, 50%, 100%), 500% или 1000% повышенную инфекционность конкретного типа клеток (например, клеток печени, нейрональных или ретинальных клеток), по сравнению с другим типом клеток (например, клеток не печени, ненейрональных и/или неретинальных клеток). Например, rAAV вектор может селективно инфицировать гепатоциты, нейронные и/или фоторецепторные клетки по сравнению с клетками за пределами печени, головного мозга и/или глаза.
Когда рекомбинантный AAV вектор демонстрирует повышенную трансдукцию нейрональной или ретинальной ткани, например, когда вектор используется для лечения неврологического или глазного расстройства, вектор предпочтительно содержит вариантный AAV2 капсидный белок.
Когда рекомбинантный AAV вектор демонстрирует повышенную трансдукцию ткани печени, например, когда вектор используется для лечения болезней печени, вектор предпочтительно содержит вариантный AAV3B, AAV-LK03 или AAV8 капсидный белок.
НУКЛЕИНОВЫЕ КИСЛОТЫ И КЛЕТКИ-ХОЗЯЕВА
Представленное раскрытие предусматривает выделенную нуклеиновую кислоту, содержащую нуклеотидную последовательность, которая кодирует вариантный капсидный белок аденоассоциированного вируса (AAV), как описано выше. Выделенная нуклеиновая кислота может содержаться в векторе AAV, например, рекомбинантном векторе AAV.
Рекомбинантный AAV вектор, содержащий такую кодирующую последовательность вариантного AAV капсидного белка может быть использован для получения рекомбинантного вириона AAV (то есть рекомбинантной векторной частицы AAV). Таким образом, представленное раскрытие предусматривает рекомбинантный AAV вектор который, когда введен в приемлемую клетку, может обеспечивать продуцирование рекомбинантного AAV вириона.
Представленное изобретение, кроме того, предусматривает клетки-хозяива, например, выделенные (генетически модифицированные) клетки-хозяива, содержащие нуклеиновую кислоту субъекта. Клетка-хозяин субъекта может представлять собой выделенную клетку, например, клетку в культуре in vitro. Клетка-хозяин субъекта является полезной для продуцирования rAAV вириона субъекта, как описано ниже. Когда клетка-хозяин субъекта используется для получения rAAV вириона субъекта, она называется, как "упаковывающая клетка." В некоторых вариантах осуществления, клетка-хозяин субъекта является стабильно генетически модифицированной с нуклеиновой кислотой субъекта. В других вариантах осуществления, клетка-хозяин субъекта является временно генетически модифицированной с нуклеиновой кислотой субъекта.
Нуклеиновая кислота субъекта вводится стабильно или временно в клетку-хозяина, с использованием установленных методов, включая, но не ограничиваясь этим, электропорацию, осаждение фосфатом кальция, опосредованную липосомами трансфекцию, и тому подобное. Для стабильной трансформации, нуклеиновая кислота субъекта будет, как правило, дополнительно включать селективный маркер, например, какой-либо из нескольких хорошо известных селективных маркеров, таких как резистентность к неомицину, и тому подобное.
Клетка-хозяин субъекта генерируется путем введения нуклеиновой кислоты субъекта в какие-либо из множества клеток, например, из множества клеток, включая, например, мышиные клетки, и клетки приматов (например, клетки человека). Приемлемые клетки млекопитающих включают, но не ограничиваются этим, первичные клетки и клеточные линии, где приемлемые клеточные линии включают, но не ограничиваются этим, 293 клетки, COS клетки, HeLa клетки, Vero клетки, 3Т3 мышиные фибробласты, С3Н10Т1/2 фибробласты, СНО клетки, и тому подобное. Неограничивающие примеры приемлемых клеток-хозяев включают, например, HeLa клетки (например, American Type Culture Collection (АТСС) No. CCL-2), СНО клетки (например, АТСС №№CRL9618, CCL61, CRL9096), 293 клетки (например, АТСС №CRL-1573), Vero клетки, NIH 3Т3 клетки (например, АТСС №CRL-1658), Huh-7 клетки, ВНК клетки (например, АТСС №CCL10), РС12 клетки (АТСС №CRL1721), COS клетки, COS-7 клетки (АТСС №CRL1651), RATI клетки, мышиные L клетки (АТСС №CCLI.3), человеческие эмбриональные клетки почек (НЕК) (АТСС №CRL1573), HLHepG2 клетки, и тому подобное. Клетка-хозяин субъекта также может быть создана с использованием бакуловируса для того, чтобы инфицировать клетки насекомых, такие как Sf9 клетки, которые продуцируют AAV (смотри, например, патент США №7,271,002; заявка на патент США 12/297,958).
В некоторых вариантах осуществления, генетически модифицированная клетка-хозяин субъекта включает, в дополнение к нуклеиновой кислоте, содержащей нуклеотидную последовательность, кодирующую вариантный AAV капсидный белок, как описано выше, где нуклеиновая кислота содержит нуклеотидную последовательность, кодирующую один или больше AAV rep белков. В других вариантах осуществления, клетка-хозяин субъекта дополнительно содержит anrAAV вектор. AnrAAVвирион может быть получен с использованием клетки-хозяина субъекта. Способы получения anrAAV вириона описаны в, например, публикации патента США №2005/0053922 и публикации патента США №2009/0202490.
Как используется в данном документе, "упаковывающая" касается серии внутриклеточных событий, которые в результате приводят к сборке и заключению в капсид AAV частицы. Гены AAV "rep" и "cap" касаются полинуклеотидных последовательностей, кодирующих репликацию и заключение в капсид белков аденоассоциированного вируса. AAV rep и cap упоминаются в данном документе как AAV "упаковывающие гены." Сборка ассоциированного белка (ААР) представляет собой продукт открытой рамки считывания в пределах cap гена, и также может нуждаться в упаковке.
"Хелперный вирус" для AAV относится к вирусу, который позволяет AAV (например, немутантный тип AAV) реплицироваться и упаковываться с помощью клетки млекопитающего. Разнообразие таких хелперных вирусов для AAV известны в данной области, включая аденовирусы, герпесвирусы и вирусы оспы, такие как вирус коровьей оспы. Аденовирусы охватывают ряд различных подгрупп, хотя, аденовирус типа 5 подгруппы C наиболее часто используется. Многочисленные аденовирусы человека, млекопитающего нечеловека и птичьего происхождения известны и доступны из депозитариев, таких как АТСС. Вирусы семейства герпеса включают, например, вирусы простого герпеса (HSV) и вирусы Эпштейна-Барра (EBV), а также цитомегаловирусы (CMV) и вирус ложного бешенства (PRV); которые также доступны из депозитариев, таких как АТСС.
"Функция(и) хелперного вируса" касается функции(ий), закодированной(ых) в геноме хелперного вируса, которая позволяет AAV репликацию и упаковку (в сочетании с другими требованиями к репликации и упаковке, описанными в данном документе). Как описано в данном документе, "функция хелперного вируса" может обеспечиваться несколькими способами, включая предоставление хелперного вируса или предоставление, например, полинуклеотидных последовательностей, кодирующих требуемую(ые) функцию(ии), клетке-производителю в транс. Например, плазмида или другой вектор экспрессии, содержащие нуклеотидные последовательности, кодирующие один или больше аденовирусных белков, трансфицируются в клетку-производитель вместе с anrAAV вектором.
"Инфекционный" вирус или вирусная частица представляет собой ту, которая содержит компетентно собранный вирусный капсид и способен доставлять полинуклеотидный компонент в клетку, для которой вид вируса является тропным. Термин не обязательно подразумевает какую-либо способность к репликации вируса. Анализы для подсчета инфекционных вирусных частиц описаны в другой части в данном раскрытии и в уровне техники. Инфекционность вируса может быть выражена, как соотношение инфекционных вирусных частиц к общему количеству вирусных частиц. Способы определения соотношения инфекционной вирусной частицы к общей вирусной частице известны в данной области с уровня техники. Смотри, например, Grainger et al. (2005) Mol. Ther. 11: S337 (описывающий анализ TCID50 инфекционного титра); и Zolotukhinet al. (1999) Gene Ther. 6: 973.
"Репликационно-компетентный" вирус (например, репликационно-компетентный AAV) касается фенотипического вируса немутантного типа, который является инфекционным, и также способен реплицироваться в инфицированной клетке (то есть в присутствии хелперного вируса или функций хелперного вируса). В случае AAV, компетенция репликации, как правило, требует присутствие функциональных генов ААВ упаковки. В общем, rAAV векторы, как описано в данном документе, являются репликационно-некомпетентный в клетках млекопитающих (особенно в клетках человека) в силу отсутствия одного или нескольких генов упаковки AAV. Как правило, у таких rAAV векторов недостает каких-либо последовательностей генов упаковки AAV для того, чтобы минимизировать возможность того, что репликационно-компетентный AAV генерируются путем рекомбинации между генами упаковки AAV и входящим rAAV вектором. Во многих вариантах осуществления, препараты rAAV вектора, как описано в данном документе, представляю собой такие, которые содержат мало, если вообще содержит какой-либо репликационно-компетентный AAV (rcAAV, также упоминается как RCA) (например, меньше, чем приблизительно 1 rcAAV на 102 rAAV частиц, меньше, чем приблизительно 1 rcAAV на 104 rAAV частиц, меньше, чем приблизительно 1 rcAAV на 108 rAAV частиц, меньше, чем приблизительно 1 rcAAV на 1012 rAAV частиц, или никакого rcAAV).
"Выделенная" нуклеиновая кислота, вектор, вирус, вирион, клетка-хозяина, или другое вещество касается препарата вещества, лишенного, по меньшей мере, некоторые других компонентов, которые могут также присутствовать, когда вещество или подобное вещество встречается в природе или изначально получено из такового. Таким образом, например, выделенное вещество может быть получено с использованием технологии очистки для обогащения его из исходной смеси. Обогащение может быть измерено на абсолютной основе, такой как масса, на объем раствора, или оно может быть измерено по отношению ко второму, потенциально интерферирующему веществу, присутствующему в исходной смеси. Возрастающие обогащения согласно вариантам осуществления данного раскрытия являются все в большей степени выделенными. Выделенная нуклеиновая кислота, вектор, вирус, клетка-хозяина, или другое вещество в некоторых вариантах осуществления являются очищенными, например, от приблизительно 80% до приблизительно 90% чистоты, по меньшей мере, приблизительно 90% чистоты, по меньшей мере, приблизительно 95% чистоты, по меньшей мере, приблизительно 98% чистоты, или, по меньшей мере, приблизительно 99%, или больше, чистоты.
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют такое же значение, которое обычно понимается квалифицированным специалистом в данной области, к которой принадлежит данное изобретение. Хотя какие-либо способы и материалы, подобные или эквивалентные тем, которые описаны в данном документе, также могут быть использованы на практике или при тестировании представленного изобретения, предпочтительные способы и материалы описаны ниже. Все публикации, упомянутые в данном документе, включены в настоящее описание в качестве ссылки для раскрытия и описания способов и/или материалов в связи, с которыми эти публикации цитируются.
Следует понимать, что определенные признаки изобретения, которые для ясности описаны в контексте отдельных вариантов осуществления, также могут быть представлены в комбинации в одном варианте осуществления. Наоборот, различные признаки изобретения, которые для краткости описаны в контексте одного варианта осуществления, также могут быть представлены отдельно или в какой-либо подходящей подкомбинации. Все комбинации вариантов осуществления, относящихся к настоящему изобретению, в частности, охватываются представленным изобретением и раскрыты в данном документе так, если бы каждая комбинация была индивидуально и явным образом раскрыта. Кроме того, все суб-комбинации различных вариантов осуществления и их элементов, также конкретно охватываются представленным изобретением и раскрыты в данном документе так, если бы каждая такая субкомбинация была индивидуально и в явном виде раскрыта в данном документе.
Следующий пример представлен для иллюстрации некоторых вариантов осуществления настоящего изобретения. Он ни в коем случае не предназначен для ограничения изобретения.
ПРИМЕРЫ
Пример 1
В данном примере, авторы изобретения разработали и сконструировали новый AAV2 вектор, обозначенный ttAAV2 (как в истинном типе). Кроме того, новый вектор исследовали в ряде моделей на животных (крысах, мышах и новорожденных мышах) для того, чтобы оценить насколько ttAAV2 вели себя отлично от культуры ткани, адаптированного (немутантного типа) AAV2. Авторы изобретения продемонстрировали, что ttAAV2 имеет преимущества для генов при доставке AAV2, и являются особенно полезными для in vivo трансдукции тканей головного мозга или глаза с гетерологичными последовательностями.
Способы
1. Клонирование: Капсидный ген wtAAV2 был взят от нашей плазмиды-производителя pDG (фигура 10). Данная плазмида содержит wtAAV2 rep и cap гены. Субфрагменты капсидного гена (pDG нуклеотидов А: 3257-3759, В: 4025-4555, С: 4797-5287 и D: 5149-5425, соответственно) субклонировали в pBS для последующего мутагенеза. Четыре мутации были введены в фрагмент A, используя сайт-направленный мутагенез, который в результате приводит к конструкту, который кодирует аминокислотные (АА) изменения V125I, V151A, A162S и T205S. Фрагмент B был мутирован для кодирования одного АА изменения, N312S. Фрагмент C был мутирован для кодирования АА изменений Q457M, S492A, E499D, F533Y, G546D, E548G, R585S, R588T, и A593S. После подтверждения успешных фрагментов мутагенеза A-C были повторно клонированы в pDG, что в результате приводит к плазмиде-производителю (pDG-ttAAV2), которая будет поддерживать продуцирование рекомбинантного вируса, который заключен в капсид с помощью HAAV2 капсида.
2. Очистка и титрование продуцирования вирусного вектора ttAAV2-GFP.
Продуцирование вектора основывалось на следующих стандартных протоколах, использующих котрансфекции rAAV плазмид с pDG, который обеспечивает как Ad хелперные функции, а также AAV гер и cap гены. Разнообразие rAAV плазмид использовалось для получения рекомбинантных плазмид.
pTR-UF11 (CAG-GFP) использовали как rAAV плазмиду. 8×108 293 клеток высевали на клеточную фабрику (CF10). Через 14-18 часов клетки трансфицировали с pDG или pDG-rrAAV2 и prAAV (например, pTR-UF11), используя способ соосаждения CaPO4. Через 72 часа клетки собирали и снова суспендировали в буфере для лизиса (20 мМ Tris-HCl, рН8, 150 мМ NaCl, 0,5% дезоксихолат). Клеточные пеллеты лизировали, используя четыре цикла замораживания и оттаивания для высвобождения вируса, где каждый цикл состоит из 30 минут при -80°C с последующими 30 минутами при 37°C. После последнего оттаивания лизат обрабатывали бензоназой при концентрации 50 ед./мл и инкубировали в течение 30 минут при 37°C. Рекомбинантный вирус чистили с использованием колонки самотеком.
Очистка. В качестве первой стадии, неочищенный лизат осветляли центрифугированием на 4000 g в течение 15 минут и наносили на предварительно образованный йодиксанольный ступенчатый градиент. Вирусную фракцию затем собирали и повторно буферили в лактированном растворе Рингера, а также концентрировали с использованием фильтров центрифугирования Amicon.
Затем, оценивали чистоту вирусных препаратов с помощью SDS полиакриламидного электрофореза и титровали, используя методы ПЦР в реальном времени. Неочищенный экстракт содержал 4,5×1012 частиц; собранные вирусные фракции содержали 1,5×1012 частиц. В этот момент способ очистки восстанавливал приблизительно 33% вируса, присутствующего в неочищенном экстракте.
3. Продуцирование и очистка rAAV вектора (альтернативный способ)
В альтернативном варианте осуществления том, который указан в пункте 2 выше, rAAV2 вектор получали следующим образом. Для того, чтобы получить rAAV2 вирионы, 5×108 293Т клеток высевали на клеточную фабрику (CF10). Через 14-18 часов, клетки в два раза трансфектировали с GFP-coдержащим вектором PD10-pST2-CMV-GFP, и или pDG, или pDG капсидным мутантом (pDG-ttAAV2), чтобы продуцировать AAV2-CMV-GFP векторы немутантного типа или подлинного типа, соответственно. Двойные трансфекции были реализованы с использованием PEI-max от Polysciences в соотношении 3,5 мл PEI на мг ДНК. Клетки собирали после 72 часов инкубирования при 37°C путем центрифугирования сркд и клеток на 2200 оборотах в минуту в течение 10 минут при 4°C. Супернатант удаляли и сохраняли для дальнейшей обработки, и клеточные пеллеты снова суспендировали в буфере для лизиса (0,15 М NaCl, 50 мМ Tris-HCl [pH 8,8]).
Клеточные пеллеты лизировали, используя четыре цикла замораживания и оттаивания для высвобождения вируса, где каждый цикл состоит из 30 минут при -80°C с последующими 30 минутами при 37°C. После последнего оттаивания лизат обрабатывали бензоназой при концентрации 150 ед./мл и инкубировали в течение 30 минут при 37°C. Лизат затем откручивали на 2000 об./мин. в течение 20 минут для осветления лизата. Супернатант фильтровали, используя 0,22 мкм целлюлозноацетатный фильтр, и препараты рекомбинантного AAV2 вируса чистили с помощью FPLC, используя  хроматографическую систему (GE Healthcare) и колонку с AVB-сефарозной аффинностью (буфер A: PBS, pH 8; буфер В: 0,5 М глицин, pH 2,7). Собранные фракции диализовали против PBS в течение ночи, и затем вирусные препараты титровали, используя SDS полиакриламидный электрофорез и методы. ПНР в реальном времени.
хроматографическую систему (GE Healthcare) и колонку с AVB-сефарозной аффинностью (буфер A: PBS, pH 8; буфер В: 0,5 М глицин, pH 2,7). Собранные фракции диализовали против PBS в течение ночи, и затем вирусные препараты титровали, используя SDS полиакриламидный электрофорез и методы. ПНР в реальном времени.
Результаты
In vivo трансдукция и распространение ttAAV2
Вектор ttAAV2 исследовали in vivo для того, чтобы оценить биологическую активность модифицированного вируса в таком контексте. Образцы вирусов AAV2-CMV-GFP WT и ТТ были подготовлены для инъекций на ряд моделей in vivo. Для этой цели мы концентрировали вирусы, поскольку только ограниченные объемы векторов могут быть инъекционно введены in vivo. Затем мы провели количественную ПЦР (кПЦР) и SDS-PAGE для того, чтобы оценить новые титре концентрированных векторов (фигура 11).
После кПЦР и анализа белок белкового геля мы получили следующие новые титры: AAV2-CMV-GFP ТТ - 1,33×1012 вирусных геномов/мл и AAV2-CMV-GFP WT - 1,25×1012 вирусных геномов/мл. Титры капсидов были следующими: AAV2-CMV-GFP ТТ - 8,89×1012 капсидов/мл - AAV2-CMV-GFP WT at 7,83×1012 капсидов/мл. Титры между геномными копиями и капсидными копиями различаются, поскольку SDS-PAGE также показывает пустые капсиды, которые, как правило, генерируются во время продуцирования рекомбинантных AAV векторов, следовательно, титр капсида является выше, чем титр вирусного генома, полученный по кПЦР.
Трансдукция в головном мозге крысы.
Вирусы rAAV2 ТТ и WT вводили инъекционно в черную субстанцию или в полосатое тело крыс немутантного типа, при чем инъекционно вводили 3 крысам на одно состояние, в дозе 2×109 вг или 3,5×109 вг на инъекцию. Через 28 дней головной мозг препарировали и срезы ткани подготовили для иммунофлюоресцентного анализа. Первичные данные показаны на фигуре 12 и фигуре 26.
Оба вируса rAAV2 ТТ и WT были способны трансдуцировать нейрональные и глиальные клетки с каждого места инъекции, хотя и с разными эффективностями. При сравнении, мы наблюдали, что ТТ вектор трансдуцировал ткани головного мозга более эффективно и больше распространялся от места инъекции, чем WT вектор. Кроме того, мы наблюдали присутствие трансдуцированных нейронов в парафасцикулярном ядре, области гипоталамуса, после инъекции в полосатое тело rAAV2 ТТ. Это указывает на то, что ТТ вектор был способен мигрировать из трансдуцированных клеточных тел в месте инъекции в гипоталамус путем активного транспорта по нейронным проекциям, подчеркивая сильную способность к ретроградному транспорту. Данная способность к ретроградному транспорту имеет была утрачена в тканевой культуре адаптированного WT rAAV2 вектора, поскольку никакие трансдуцированные клетки не могли наблюдаться в той же области (смотри фигуру 26).
Взятые вместе, данные результаты по головному мозгу крыс указывают на значительно более высокое распространение и эффективность трансдукции от ttAAV2 по сравнению с титр-совпадающим wtAAV2. Кроме того, AAV2 ТТ демонстрирует доказательство очень хорошей ретроградной транспортной способности, которая имеет была потеряна у ААV2 WT вируса.
Трансдукция в модели мышиного глаза.
ttAAV2 и wt-AAV2 из той же серии, как и те, которые использовали для наших исследований головного мозга крыс, инъекционно вводили в глаза взрослых мышей в дозе 2×109 вг на глаз. Для того, чтобы избежать вариабельности от животного к животному, каждая мышь получала инъекцию rAAV2 ТТ в один глаз и инъекцию rAAV2 WT в глаз, расположенный на противоположной стороне. Были проанализированы три различных пути внутриглазных инъекций: интракамеральный, интравитреальный и субретинальный. Животных собирали и оценивали GFP экспрессию с помощью иммунофлуоресценции через 6 недель. Результаты показаны на фигуре 13. В совокупности эти данные указывают на заметное увеличение трансдукции фоторецепторных клеток после субретинальной инъекции ttAAV2, с точки зрения, как уровня, так и количества трансдуцированных фоторецепторных клеток, по сравнению с wtAAV2 (который был использован в успешном клиническом исследовании RPE65).
Трансдукция в модели новорожденных мышей.
Таким образом, оба вектора как ttAAV2, так и wtAAV2 GFP вводили инъекционно новорожденным мышам. Были протестированы два способа введения инъекции, внутривенные инъекции и внутричерепные инъекции. Через 4 недели животных умерщвляли и собирали все ткани со всех мышей. Мы проанализировали мозг, который после сбора визуализировали путем прямой флюоресценции органа на флуоресцентном микроскопе. Результаты показаны на фигуре 14. Результаты внутричерепных и системных инъекций обсуждаются более подробно ниже.
Внутричерепные инъекции
5×1010 вг какого-либо вектора вводили инъекционно в боковой желудочек Р1 новорожденных. Животных умерщвляли через 4 недели после инъекции, и головной мозг иссекали, разрезали и окрашивали, используя анти-GFP антитело. Результаты показаны на фигуре 27.
Как наблюдалось в головном мозге взрослых крыс, эти данные показывают, что AAV2 ТТ демонстрирует повышенную трансдукцию тканей головного мозга мышей и более высокое распространение после внутричерепной инъекции по сравнению с AAV2 WT вектором. При наблюдении окрашенных срезов тканей при более высоком увеличении, были дополнительно выделены различия в эффективности трансдукции между двумя векторами: ТТ вектор демонстрировал себя лучше, как сточки зрения уровня экспрессии, так и количества трансдуцированных клеток. AAV2 ТТ и WT, как оказалось, имеют одну и ту же аффинность клеточного типа, каждый из которых показывает трансдукцию нейрональных, а также глиальных клеток, предполагая, что наблюдаемые различия представляют собой различия в эффективности, а не в специфичности клеточного типа (фигура 28). Взятые в совокупности эти данные указывают на то, что ttAAV2 показывает намного большую повышенную трансдукцию тканей головного мозга мышей после внутрикожной (в.к.) инъекции по сравнению с векторами на основе wtAAV2. Кроме того, некоторые доказательства наводит на мысль касательно трансдукции эпендимального клеточного слоя, выстилающего желудочки, когда используются векторы ttAAV2. Данный феномен не был обнаружен с вектором wtAAV2.
Системные инъекции
Внутрияремные инъекции 2×1011 вг какого-либо вектора были сделаны в Р1 новорожденных. Животных умерщвляли через 4 недели после инъекции, и различные органы собирали и оценивали касательно GFP трансдукции с помощью иммуногистохимического анализа с использованием анти-GFP антитела (головной мозг, печень, сердце, мышцы, легкие, селезенка и почки). Результаты окрашивания головного мозга показаны на фигуре 29 и изображения с большим увеличением представлены на фигуре 30.
Мы наблюдали хорошую экспрессию трансгена в ЦНС после системной инъекции AAV2 ТТ. Вектор AAV2 WT проявил себя хуже по сравнению с наблюдаемыми лишь несколькими трансдуцированными нейронами.
Для того чтобы оценить общее биораспределение вектора AAV2 ТТ мы оценили уровень трансдукции, полученный в различных тканях после системной инъекции. Собранные органы фиксировали, парафин разрезали и окрашивали касательно экспрессии GFP (фигура 31).
Эти данные указывают на то, что вектор AAV-TT, как оказывается, не имеет сильной аффинности к другим органам, но вместо этого показывает специфичность в основном для нейрональных тканей. Данное наблюдение может оказаться полезным для лечения нейрональных генетических нарушений путем внутривенной инъекции AAV, поскольку это гарантирует то, что вектор не будет трансдуцировать нецелевые периферические органы, а, главным образом, только мозг при данном способе инъекции.
В совокупности, наши in vivo данные предполагает, что ttAAV2 имеет необыкновенные характеристики трансдукции в тканях глаза и головного мозга, демонстрируя почти исключительную специфичность в отношении нейрональных тканей.
Пример 2
Дополнительные рассмотрения
Не желая быть связанными теорией, полагают, что мутации, присутствующие в ttAAV2 по сравнению с wtAAV2 содержат следующее функциональные группы:
1) связывающее гепарин остатки, расположенные на тройных шиловидных отростках капсида AAV2 (S585 и Т588); полагают, что данные остатки являются ответственными за гепариновое связывание wtAAV2 капсида. В ttAAV2 это заменяется, и мы предполагаем, что данная замена поддерживает распространение вируса в ткани головного мозга, богатой на гепарансульфатпротеогликан.
2) одно аминокислотное изменение в ttAAV2, которое располагается на внутренней стороне капсида (S312); этот внутренний остаток серина может играть определенную роль во взаимодействиях капсид-DNA, тем самым потенциально способствуя, либо стабильности вируса, упаковке генома или высвобождению генома во время инфекции.
3) две пространственно близких аминокислоты (D546 и G548), расположенных в углублении между тройными проксимальными шиловидными отростками на структуре AAV капсида; данные остатки могут быть вовлечены во взаимодействия с нейтрализующими антителами и, таким образом, вносят свой вклад в характеристики in vivo трансдукции.
4) одно выделенное аминокислотное изменение (S593), расположенное в углублении между тройными проксимальными шиловидными отростками;
5) четыре аминокислоты, как полагают, участвуют в связывании с рецептором и тесно расположены на тройных шиловидных отростках (М457, А492, D499 и Y533);
6) четыре оставшиеся аминокислотные изменения, расположенные в VP1/VP2 первичной последовательности (I125 , А151, S162 и S205); данные остатки находятся в пределах области капсида так, что не являются частью VP3. Известно, что VP1/VP2 специфические области в пределах вирусного капсида имеют активность PLA2, которая может быть вовлечена в направленную миграцию входящего вируса. Изменения в этой области, как было показано влияют на инфекционность вируса.
Трехмерное положения данных мутаций в AAV2 капсидном белке известны и показаны на фигурах 15-20. Соответствующие положения могут быть идентифицированы в AAV капсидных белках из других серотипов (смотри ниже). Для дальнейшей характеристики ttAAV2 и роли индивидуальных аминокислотных изменений, которые отвечают за улучшенный фенотип ttAAV2, ttAAV2 капсид может быть мутирован для того, чтобы полностью изменить отдельные выбранные аминокислоты (или группы аминокислот, например, на основе групп 1-6, которые обсуждались выше) до их соответствующей последовательности в капсиде немутантного типа AAV2. Каждый мутантный вектор затем может быть проанализирован с использованием методов, как описано в примерах выше.
Например, различные мутантные векторы, экспрессирующие GFP, могут быть представлены для первого скрининга в модели на животных в виде внутричерепной (IС) инъекции у CD1 новорожденных мышей. Сигнал GFP, полученный в головном мозге, в который была сделана инъекция, от каждого мутантного вектора затем наблюдался с помощью флуоресцентной микроскопии и сравнивался с тем, который получен от исходных GFP-экспрессирующих wtAAV2 и ttAAV2 векторов. После идентификации нового фенотипа (то есть отмена ttAAV2-специфической сильной GFP экспрессии, когда мутирует конкретная аминокислотная группа), затем дополнительно делали срезы и исследовали головной мозг, в который была сделана инъекция, используя иммуногистохимические методы.
Параллельно, отобранные мутантные векторы предоставляются на второй скрининг для внутричерепных инъекций взрослым крысам. Кроме того, соответствующие мутантные комбинации анализируют, используя внутривенные (IV) инъекции новорожденным мышам для того, чтобы оценить биораспределение векторов. Отдельные органы (сердце, легкие, печень, селезенку, почки, мышцы) затем обрабатываются для иммуногистохимии и оценки GFP экспрессии.
Анализ вклада каждого ТТ-специфического остатка в эффективность и биораспределение вектора
Для дальнейшей характеристики AAV2 ТТ и определения аминокислотных изменений, которые отвечают за улучшенный ТТ фенотип, AAV2 ТТ капсид был мутирован шаг за шагом для того, чтобы полностью изменить выбранные аминокислоты до их соответствующих последовательностей в капсиде немутантного типа AAV2. Эта стратегия позволила нам более конкретно различать вклад каждого из 14 аминокислотных мутаций в фенотип AAV2 ТТ и определить минимальный вектор истинного типа, содержащий только необходимые мутации.
Как обсуждалось выше, мы сгруппировали 14 ТТ-специфических остатков в группы на основе их положения на AAV капсиде и ассоциированных с ними потенциальных вкладов в профиль трансдукции. Различные AAV-TT мутанты подвергали скринингу с использованием внутричерепных инъекций в головной мозг новорожденных мышей или в головной мозг взрослых крыс для того, чтобы пронаблюдать была ли перестановка некоторых ТТ-специфических остатков на WT эквиваленты связана с потерей фенотипа, тем самым, идентифицируя важные аминокислотные изменения среди 14 ТТ остатков.
Сайт связывания гепарин (HBS)
Было показано, что остатки 585 и 588 являются ответственными за связывание гепарина AAV2 WT капсидом. В AAV-TT они заменяются, и мы предположили, что эта замена поддерживает распространение вируса в ткани головного мозга, обогащенной гепаринсульфатпротеогликаном. Эти два остатка расположены на тройных прокимальных шиповидных отростках AAV2 капсидной структуры (смотри фигуру 16).
Мы упразднили способность связывания гепарина AAV2 WT путем инженерии изменений R585S и R588T на WT капсиде (AAV2-HBnull), то есть мутируя остатки до эквивалентов истинного типа. Этот первый анализ был проведен для того, чтобы убедиться, что AAV-TT не был просто AAV2 без сайта связывания гепарина, способный распространяться больше, но не так, как некоторые из других 12 аминокислотных замен также играют определенную роль в улучшенном AAV-TT профиле трансдукции.
Внутричерепные инъекции взрослой крысе
С соответствующим титром векторы AAV2-TT, AAV2-WT и AAV2-HBnull вводили инъекционно в черную субстанцию или в полосатое тело крыс немутантного типа в дозе 3,5×109 вг на инъекцию. Через 28 дней головной мозг препарировали и срезы ткани подготовили для иммуногистохимического анализа касательно GFP экспрессии (фигура 32).
Мы наблюдали сильную GFP экспрессию в таламусе и в черной субстанции после инъекции в полосатое тело AAV-TT вируса, выделяя сильную ретроградную транспортную способность вектора. Для сравнения, AAV2-HBnull и AAV2 WT демонстрировали намного меньший - если вообще был какой-либо - ретроградный транспорт, чем AAV-TT, поскольку очень немногие клеточные тела трансдуцировались в данные области после инъекций в полосатое тело. Данное наблюдение показало, что AAV2-HBnull отличается от AAV-TT, и что отсутствие способности связывать гепарин на AAV2 капсиде способствует хорошему распространению вектора истинного типа в головном мозге. Однако, этого недостаточно, чтобы объяснить его улучшенный профиль трансдукции.
Остатки S312, D546-G548 и S593
S312 остаток представляет собой только ТТ-специфическое изменение, которое расположено на внутренней стороне AAV2 капсида. Наше предположение было таким, что данный внутренний остаток может играть определенную роль во взаимодействиях капсид-ДНК, тем самым потенциально способствуя либо стабильности вируса, упаковке генома или высвобождению генома во время инфекции.
D и G остатки в положениях 546 и 548, соответственно, расположены в углублении между прокимальными 3-ными пиками.
Один изолированный серии, S593, находится в углублении между тройными прокимальными шиловидными отростками.
Положения данных ТТ-специфических остатков на трехмерной структуре AAV2 капсида представлены на фигурах 17-19. Принимая во внимание их менее видное положение в структуре, мы предположили, что данные остатки не могли бы внести свой вклад в трансдукцию фенотипа ttAAV2.
Внутричерепные инъекции новорожденным мышам
Мутация S312N была сконструирована на AAV2-TT капсиде для того, чтобы создать TT-S312N мутант. Мутации D546G и G548E были сконструированы на AAV2-TT капсиде для того, чтобы создать AAV-TT-DG мутант. Мутация S593A была сконструирована на AAV2-TT капсиде для того, чтобы создать AAV-TT-S593А мутант.
При возвращении данных выбранных ТТ аминокислот до их соответствующей последовательности в капсиде немутантного типа AAV2 мы стремились определить вклад данных остатков в улучшенный AAV-TT фенотип.
5×1010 вг каждого мутантного вектора вводили инъекционно в боковой желудочек Р1 новорожденных. Животных умерщвляли через 4 недели после инъекции, и головной мозг иссекали, разрезали и окрашивали, используя анти-GFP антитело. Результаты показаны на фигуре 33.
Интересно отметить, что эти данные свидетельствуют о том, что AAV TT-S312N демонстрирует повышенную трансдукцию тканей головного мозга мышей по сравнению с полным AAV2 ТТ вектором. С другой стороны, аминокислотные изменения S593A и D546E/G548D, как оказалось, не влияют на ТТ фенотип, поскольку аналогичные профили трансдукции можно было наблюдать по всему мозгу. При наблюдении окрашенных участков при большем увеличении, различия в эффективности трансдукция были дополнительно выделены (фигура 34).
На изображении с большим увеличением, мы могли наблюдать, что, как оказалось, AAV TT-S312 трансдуцирует нейрональные ткани с более высокой эффективностью, чем исходный AAV-TT с 14 аминокислотными изменениями. В частности, мы могли видеть более сильную экспрессию трансгена в ростральной части мозга (кора головного мозга, полосатое тело, гиппокампус) после инъекции TT-S312N вектора, как с точки зрения уровня, так и количества трансдуцированных клеток. Несмотря на высокую вариабельность в головном мозге новорожденного, в который была сделана инъекция, из-за трудностей, связанных с нацеливанием в место инъекции, данное наблюдение подтверждалось у всех проанализированных животных. С другой стороны, перестановки S593A или D546E/G548D, как оказалось, не имеют большого влияния на фенотип трансдукции ТТ вектора.
Пример 3
Направленные аминокислотные мутации на AAV2 капсиде подлинного типа, выбранном из результатов, полученных с комбинациями мутантов в примере 2
На основании результатов мутаций аминокислотной группы на полном AAV ТТ капсиде, мы могли определить то, что мутация S312N, как представляется, является полезной для ТТ фенотипа, дальнейшего повышения эффективности его трансдукции в головном мозге. Кроме того, мы наблюдали, что перестановки S593A и D546G/G548E, как оказалось, не имеют влияния на нейрональный фенотип AAV-TT. Мы, поэтому, предположили, что ТТ-специфические остатки S593, D546 и G548 могли быть исключены из последовательности капсида истинного типа, оставляя вместо AAV2 остатки WT в данных положениях, чтобы получить конечный ТТ вектор только с 10 аминокислотными изменениями.
Для проверки этих гипотез, мы сконструировали TT-S312N-D546G-G548D-S593A вектор и исследовали эффективность его трансдукции, используя инъекции в головной мозг новорожденным мышам. Поскольку последние внутричерепные инъекции новорожденным, как оказалось, приводят к насыщенному сигналу в детектируемой GFP экспрессии, мы решили, кроме того, вводить ТТ и TT-S312N векторы вместе с данным "до-окончательным" ТТ, используя дозу в 10 раз более низкую, чем использовали ранее. При использовании данной более низкой дозы мы стремились избежать достижения уровней насыщения GFP окрашивания в трансдуцированном головном мозге и избежать трудностей при сравнении эффективности трансдукции между различными мутантами.
5×1009 вг каждого мутантного вектора вводили инъекционно в боковой желудочек Р1 новорожденных. Животных умерщвляли через 4 недели после инъекции, и головной мозг иссекали, разрезали и окрашивали, используя анти-GFP антитело. Результаты показаны на фигуре 35.
Как наблюдалось ранее, эти данные свидетельствуют о том, что AAV TT-S312N демонстрирует повышенную трансдукцию мышиных тканей головного мозга по сравнению с полным AAV2 ТТ вектором. С другой стороны, минимальный TT-S312N-D546G/G548E-S593A вектор, как оказалось, не достигал этих более высоких уровней трансдукции, даже если он также содержал внутреннюю мутацию S312N. Это свидетельствует о том, что одна или больше аминокислотных замен играет определенную роль в ТТ фенотипе. При полном изменении данных остатков обратно к эквивалентам AAV2 WT, мы потеряли часть способности к повышенной трансдукции. При наблюдении окрашенных участков при более высоком увеличении, данные наблюдения были подтверждены (фигура 36).
Мы решили исследовать далее эффективность трансдукции, полученную каждым из данных векторов, используя количественный анализ общей GFP экспрессии, полученной в головном мозге, в который была сделана инъекция, с помощью твердофазного иммуноферментного анализа (ИФА) на полных белковых экстрактах головного мозга. Если коротко, то через 4 недели после инъекции 5×1009 вг векторов, животных умерщвляли, целый головной мозг собирали и лизировали, и экстрагировали общий белок мозга. Используя GFP белковый стандарт, затем мы могли количественно определить количества GFP белка, экспрессированные в каждом головном мозге, в который была сделана инъекция (фигура 37).
Мы могли подтвердить, что внутренний мутант TT-S312N трансдуцирует головной мозг мышей мышиные с большей эффективностью, чем полный ТТ вектор, поскольку это приводит к большей GFP экспрессии в целом во всем головном мозге, в который была сделана инъекция. С другой стороны, минимальный ТТ вектор, TT-S312N-D546G/G548E-S593A, как оказалось, приводит к более низким уровням трансдукции: хотя среднее количество экспрессированного GFP на головной мозг кажется на этом графике выше, это было связано с экстремальными значениями GFP, измеренными в одном мозге, как проиллюстрировано высокой планкой погрешности для данного состояния. С помощью этого минимального вектора ТТ, вариабельность между животными была очень высокой, и только у одного животного из пяти работает лучше, чем у животных, трансдуцированных TT-S312N. С помощью этого минимального вектора ТТ, изменчивость между животными Данная высокая вариабельность привела нас к рассмотрению данного временного минимального ТТ вектора с осторожностью, особенно потому, что иммуногистохимические анализы также показали, что TT-S312N вариант выполнялся лучше, чем TT-S312N-DG-S593A.
Поэтому мы выбрали TT-S312N вариант как наш наиболее предпочтительный AAV ТТ вектор, который состоит из следующих 13 аминокислотных мутаций по сравнению с немутантным типом AAV2: V125I, V151A, A162S, T205S, Q457M, S492A, E499D, F533Y, G546D, E548G, R585S, R588T и A593S.
Хотя приведенные выше исследования предполагают TT-S312N в качестве наиболее предпочтительного AAV-TT вектора, данные исследования иллюстрируют отдельные функции и вклад ряда ТТ-специфических остатков. В частности, четыре аминокислоты тесно расположены на тройном шиловидном отростке капсида, вероятно, участвуют в связывании рецептора. В некоторых вариантах осуществления, данные остатки возвращают AAV-TT обратно к AAV2 WT соответствующим остаткам. Например, мутации M457Q, A492S, D499E и Y533F могут быть сконструированы на AAV-TT капсиде, этот мутантный вектор анализировали, как описано ранее, для того, чтобы проиллюстрировать роль данных остатков. В следующих вариантах осуществления, четыре аминокислоты, расположенные в VP1/VP2 первичной последовательности, которые, вероятно, могут быть вовлечены в направленную миграцию вируса, могут быть возвращены в AAV-TT обратно к AAV2 WT соответствующим остаткам (I125V, A151V, S162A, S205T) и проанализированы аналогичным образом.
Пример 4
Конструирование вариантов AAV вектора в других серотипах
Функция ttААV2-специфических аминокислотных изменений в контексте других, не-AAV2 серотипов также может быть определена.
Капсидные аминокислотные последовательности основных аденоассоциированных вирусов (AAV), а именно AAV1, 5, 6, 8, 9 и rh10, могут быть приведены в соответствие с одним из ttAAV2, например, как показано на фигуре 9. Данная возможность идентификации, из которых ttAAV2-специфические аминокислоты уже присутствуют в тех же положениях в других серотипах. Соответствующие остатки в различных серотипах затем мутировали в соответствующие остатки в wtAAV2. Данные изменения демонстрируют важность ttAAV2-специфических остатков для эффективности и биораспределения каждого серотипа.
Как обсуждалось выше, различные мутантные векторы, экспрессирующие GFP, затем подают на первый скрининг по внутричерепной (1С) инъекции CD1 новорожденным мышам. GFP сигнал, полученный в головном мозге, в который была сделана инъекция, затем сравнивали с тем, который получен от контролей соответствующего GFP-экспрессирующего серотипа. Уменьшение или увеличение GFP экспрессии при мутировании идентифицированных аминокислот в их соответствующие AAV2 остатки демонстрирует важность данных конкретных остатков в данные специфические положения. Там, где это применимо, головной мозг, в который была сделана инъекция, затем дополнительно делали срезы и исследовали с помощью иммуногистохимических анализов. Кроме того, выбранные мутантные серотипы анализировали с помощью внутривенных (IV) инъекций новорожденным мышам для того, чтобы оценить биораспределение векторов и сравнить его с исходными, немутировавшими дубликатами.
В следующих вариантах осуществления соответствующие аминокислоты, идентефицированные в ttAAV2 вставлены в качестве ключевых мутаций в другие известные серотипы в релевантных положениях. Вставка ttAAV2-специфических остатков в другие AAV подтипы позволяет нам улучшить трансдукцию и профили биораспределение каждого серотипа.
Пример 5
Модификация ttAAV2-специфических остатков, сохраняющихся в других AAV серотипах в соответствующих остатках AAV2.
Сравнительный анализ аминокислотных последовательностей капсида существующих аденоассоциированных серотипов с одним из ttAAV2 впервые позволило нам идентифицировать ttAAV2-специфические остатки, которые являются консервативными в других серотипах (смотри фигуру 9). Данные остатки состоят из S162, S205, S312, G548, S585 и Т588. Каждый не-AAV2 серотип содержит один или комбинацию нескольких из данных остатков в соответствующем аминокислотном положении в своей последовательности.
В конкретных вариантах осуществления, каждый из данных остатков в различных серотипах преобразуется в соответствующую(ие) аминокислоту(ы) немутантного типа AAV2, и исследуется эффективность трансдукции данных новых мутантов.
1) Модификация AAV1 серотипа
AAV1 содержит остатки S205, G549, S586 и Т589, которые соответствуют следующим остаткам в ttAAV2: S205, G548, S585 и Т588. Когда VP1 мономер из AAV1 выравнивали трехмерно с VP1 от AAV2, мы могли контролировать, что соответствующие остатки в wtAAV2, а именно Т205, Е548, R585 и R588, находятся преимущественно в соответствующих положениях на 3D структуре (фигура 21). Таким образом, мы пришли к выводу, что было бы существенным преобразовать каждый из ttAAV2-специфических остатков в AAV1 в соответствующие эквиваленты немутантного типа AAV2 без влияния трехмерной структуры белка. В конкретных вариантах осуществления следующее мутации были сделаны в последовательности AAV1 капсида: S205T, G549E, S586R, T589R.
2) Модификация AAV5 серотипа
AAV5 содержит остатки G537, S575 и Т578, которые соответствуют следующим остаткам в ttAAV2: G548, S585 и Т588. R585 и R588 остатки в AAV2 находятся в соответствующих положениях с S575 и Т578 в AAV5 на 3D структуре. Хотя остаток Е548 в AAV2 преимущественно не соответствовал остатку G537 в AAV5 в соответствии с трехмерной структурой (фигура 22), мы все же решили включить его в исследование, поскольку оба остатка находятся пространственно относительно близко. Поэтому в конкретных вариантах осуществления следующие мутации были сделаны в последовательности AAV5 капсида: G537E, S575R, T578R.
3) Модификация AAV6 серотипа
AAV6 содержит остатки S205, G549, S586 и Т589, которые соответствуют следующим остаткам в ttAAV2: S205, G548, S585 и Т588. Соответствующие остатки в wtAAV2, а именно Т205, Е548, R585 и R588 (фигура 23), находятся преимущественно в соответствующих положениях на VP1 3D структурах. Поэтому в конкретных вариантах осуществления следующие мутации были сделаны в последовательности AAV6 капсида: S205T, G549E, S586R, T589R.
4) Модификация AAV8 серотипа
AAV8 содержит остатки S315 и Т591, которые соответствуют следующим остаткам в ttAAV2: S312 и Т588. Соответствующие остатки в wtAAV2, а именно N312 и R588, находятся преимущественно в соответствующих положениях на VP1 3D структурах (Фигура 24). Поэтому в конкретных вариантах осуществления следующие мутации были сделаны в последовательности AAV8 капсида: S315N, T591R.
В одном варианте осуществления, улучшенная эффективность трансдукции, сообщаемая S312N мутацией в ТТ AAV2, может быть передана AAV8 серотипу путем применения аминокислотного изменения S315N.
Мы мутировали последовательность AAV8 капсида с помощью сайт-направленного мутагенеза, и тем самым сконструировали AAV8-S315N векторную плазмиду. Данную плазмида использовали для продуцирования рекомбинантных AAV8-S315N векторов, экспрессирующий ITR-содержащий CMV-GFP трансген путем двойной трансфекции 293Т клеток. Вектор потом чистили из клеточного лизата и из собранного культурального супернатанта с помощью FPLC аффинной хроматографии, с использованием AVB сефарозной смолы. Титр капсида и титр векторного генома оценивали, используя SDS-PAGE и кПЦР, соответственно.
Мутантный AAV8-S315N вектор, экспрессирующий GFP, подвергали скринингу с помощью системных инъекций CD1 новорожденным мышам. Титр соответствующего AAV8 вектора используется в качестве контроля. Затем анализируют GFP экспрессию, полученную в различных органах после инъекции внутрь яремной вены 2×1011 вг, сосредоточиваясь в первую очередь на печени, где AAV8 ранее продемонстрировал некоторую сильную эффективность трансдукции.
5) Модификация AAV9 серотипа
AAV9 содержит остатки S162, S205, G549 и S586, которые соответствуют следующим остаткам в ttAAV2: S162, S205, G548 и S585. Соответствующие остатки в AAV2, а именно А162, Т205, Е548 и R585, находятся преимущественно в соответствующих положениях на VP1 3D структурах (фигура 25). Поэтому в конкретных вариантах осуществления следующие мутации были сделаны в последовательности AAV8 капсида: S162A, S205T, G549E и S586R.
6) Модификация AAVrh10 серотипа
AAVrh10 содержит остаток G551, который соответствует G548 с ttAAV2. Учитывая то, как сохранившийся данный остаток и положение появляется среди различных серотипов, мы предполагаем, что G551 в AAVrh10 будет выравниваться с Е548 в wtAAV2 трехмерно. Поэтому в одном варианте осуществления следующая мутация сделана в последовательности AAVrh10 капсида: G551E.
7) Модификация AAV3B и AAV-LK03 серотипов
Подобно серотипу AAV8, мы наблюдали, что AAV3B серотип также содержит внутренний серии в положении 312 после выравнивания последовательности капсидного белка VP1 с одной из AAV-TT (смотри последовательность AAV3B капсида на фигуре 38).
Недавно описанный LK03 AAV вектор, химерный капсид, состоящий из пяти различных родительская AAV капсидов, сконструированных М.А. Кау с помощью ДНК-перетасовки, также содержит остаток S312 во внутренней части капсида (смотри Lisowski et al., Selection and evaluation of clically relevant AAV variants in a xenograft liver model, Nature 506, 382-386 (2014)). Капсидная последовательность AAV-LK03 раскрыта в WO 2013/029030 и показана на фигуре 39.
Поэтому в следующих вариантах осуществления, AAV3B и AAV-LK03 векторы мутируют путем применения аминокислотного изменения S312N в обоих векторах. Данные новые мутанты, и соответствующие им AAV контрольные серотипы, также исследуют с помощью внутри яремной инъекции новорожденным мышам. GFP экспрессию, полученную в различных собранных органах, затем анализируют.
Пример 6
Идентификация ttAAV2-специфических аминокислот, которые являются передаваемыми между AAV серотипами
В следующих вариантах осуществления, ключевые аминокислоты, идентефицированные в ходе определения характеристик ttAAV2 вставляют в другие известные серотипы в соответствующие положения. Вновь сконструированные векторы затем исследуют с использованием соответствующих немутированных серотипов в качестве контролен. Это подтверждает важность отдельных аминокислот в конкретных положениях на AAV капсидах, независимо от серотипа.
1) Остатки S585, Т588, S312, D546, G548 и S593
AAV1, AAV5 и AAV6, естественно, содержат один и тот же аминокислотный остаток в положениях в последовательностях их капсидного белка, отвечающих G548, S585 и Т588 в ttAAV2. Поэтому в следующих вариантах осуществления, капсидные белки в данных серотипах мутируют в соответствующих положениях для того, чтобы включить другие остатки S312, D546 и S593, присутствующие в ttAAV2. Аналогичным образом AAV8, который уже содержит такой же аминокислотный остаток как в ttAAV2 в положениях, отвечающих S312 и Т588 в ttAAV2, в дальнейшем мутируют для того, чтобы содержать остатки, отвечающие S585, D546, G548 и S593 в ttAAV2. AAV9, который уже содержит соответствующие остатки G548 и S585 в ttAAV2, мутируют для того, чтобы включить остатки, отвечающие Т588, S312, D546 и S593 в ttAAV2. Наконец, AAV10, который уже содержит остаток, отвечающий G548, является дополнительно модифицированным также для того, чтобы содержать остатки, отвечающие S585, Т588, S312, G548 и S593.
2) Остатки I125, А151, S162, S205, М457, А492, D499 и Y533
AAV1 и AAV6 уже, естественно, содержат остаток S205. Поэтому в следующих вариантах осуществления данные серотипы мутируют в положениях, отвечающих остаткам I125, А151, S162, М457, А492, D499 и Y533 в ttAAV2. Аналогичный AAV9, который уже содержит остатки S162 и S205, в дальнейшем мутируют для того, чтобы содержать остатки, отвечающие I125, А151, М457, А492, D499 и Y533 в ttAAV2. Наконец, AAV5, 8 и 10 являются модифицированными для того, чтобы продемонстрировать остатки, отвечающие I125 , А151, S162, S205, М457, А492, D499 и Y533 в ttAAV2.
Положения мутаций, присутствующих в ttAAV2, и соответствующие остатки, присутствующие в последовательностях капсидного белка немутантного типа VP1 других AAV серотипов, показаны в таблице 1 ниже. В общем, вариант не-AAV2 векторов может быть сконструирован путем мутирования какого-либо из остатков, показанных в таблице 1 для данных серотипов. Остатки, показанные курсивом, представляют собой остатки, которые уже присутствуют в HAAV2 в соответствующем положении. В предпочтительном варианте осуществления, He-AAV2 серотипы являются мутированными в одном или больше остатках, показанных некурсивным шрифтом. В этом смысле, предпочтительные свойства, показанные ttAAV2 могут быть перенесены в альтернативные AAV серотипы.
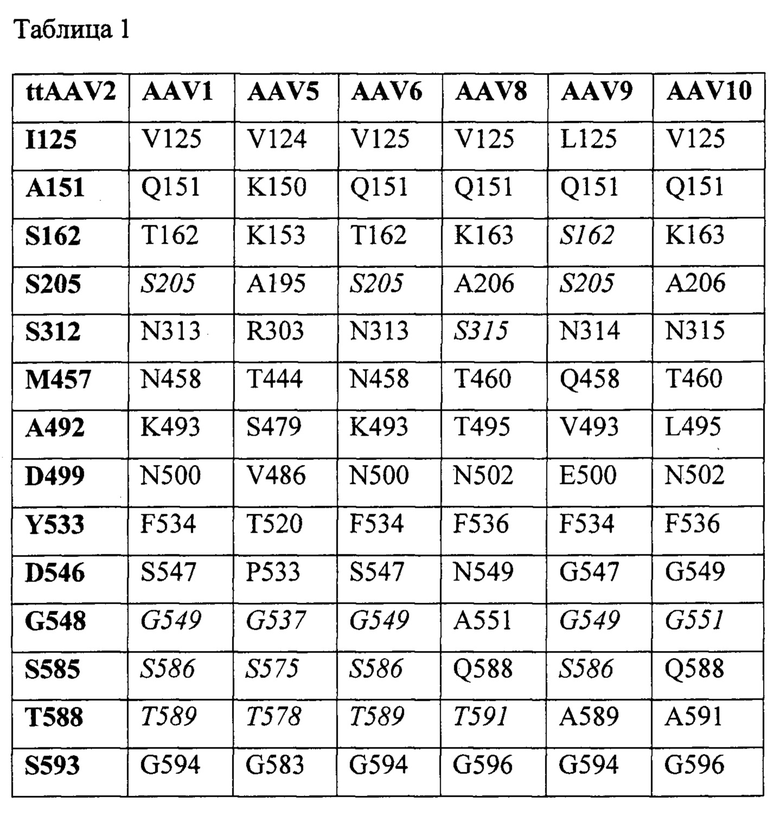
Дополнительные варианты осуществления изобретения
Изобретение, кроме того, касается дополнительных аспектов, как определено в следующих кратких пунктах:
1. Рекомбинантный вектор аденоассоциированного вируса (AAV), содержащий:
(а) вариантный AAV капсидный белок, где вариантный AAV капсидный белок содержит, по меньшей мере, одно аминокислотное замещение по сравнению с AAV капсидным белком немутантного типа; где, по меньшей мере, одно аминокислотное замещение присутствует в положении, отвечающем одному или больше из следующих положений в последовательности AAV2 капсидного белка: 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593; и
(b) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт.
2. Рекомбинантный AAV вектор в соответствии с пунктом 1, где (i) вектор содержит вариантный AAV2 капсидный белок; (ii) вариантный AAV капсидный белок содержит последовательность SEQ ID NO: 2, или последовательность, имеющую, по меньшей мере 95% идентичность последовательности с ней; (iii) AAV капсидный белок немутантного типа является из AAV2; и/или (iv) AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 1.
3. Рекомбинантный AAV вектор в соответствии с пунктом 2, где вариантный AAV2 капсидный белок содержит один или больше из следующих остатков: I125, А151, S162, S205, S312, М457, А492, D499, Y533, D546, G548, S585, Т588 и/или S593.
4. Рекомбинантный AAV вектор в соответствии с пунктом 2 или пунктом 3, где вариантный AAV2 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с капсидным белком немутантного типа AAV2: V125I, V151A, A162S, T205S, N312S, Q457M, S492A, E499D, F533Y, G546D, E548G, R585S, R588T и/или A593S.
5. Рекомбинантный AAV вектор в соответствии с каким-либо из пунктов 1-3, где вариантный AAV капсидный белок является из AAV1, AAV5, AAV6, AAV8, AAV9 или AAV10.
6. Рекомбинантный AAV вектор в соответствии с пунктом 5, где (i) вектор содержит вариантный AAV1 капсидный белок, (ii) вариантный AAV капсидный белок содержит последовательность, имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO:3; (iii) AAV капсидный белок немутантного типа является из AAV1; и/или (iv) AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 3;
и где, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV1 капсидного белка: 125, 151, 162, 205, 313, 458, 493, 500, 534, 547, 549, 586, 589 и/или 594.
7. Рекомбинантный AAV вектор в соответствии с пунктом 6, где вариантный AAV1 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с капсидным белком немутантного типа AAV1:
(a) V125I, Q151A, T162S, N313S, N458M, К493А, N500D, F534Y, S547D, и/или G594S; и/или
(b) S205T, G549E, S586R и/или T589R.
8. Рекомбинантный AAV вектор в соответствии с пунктом 5, где (i) вектор содержит вариантный AAV5 капсидный белок, (ii) вариантный AAV капсидный белок содержит последовательность имеющую по меньшей мере 95% идентичность последовательности к SEQ ID NO: 4; (iii) AAV капсидный белок немутантного типа является из AAV5; и/или (iv) AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 4;
и где, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV5 капсидного белка: 124, 150, 153, 195, 303, 444, 479, 486, 520, 533, 537, 575, 578 и/или 583.
9. Рекомбинантный AAV вектор в соответствии с пунктом 8, где вариантный AAV5 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV5 капсидным белком немутантного типа:
(a) V124I, K150А, K153S, A195S, R303S, Т444М, S479A, V486D, T520Y, P533D, и/или G583S; и/или
(b) G537E, S575R и/или T578R.
10. Рекомбинантный AAV вектор в соответствии с пунктом 5, где (i) вектор содержит вариантный AAV6 капсидный белок, (ii) вариантный AAV капсидный белок содержит последовательность имеющую, по меньшей мере, 95%» идентичность последовательности к SEQ ID NO: 5; (iii) AAV капсидный белок немутантного типа является из AAV6; и/или (iv) AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 5;
и где, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV6 капсидного белка: 125, 151, 162, 205, 313, 458, 493, 500, 534, 547, 549, 586, 589 и/или 594.
11. Рекомбинантный AAV вектор в соответствии с пунктом 10, где вариантный AAV6 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV6 капсидным белком немутантного типа:
(a) V125I, Q151A, T162S, N313S, N458M, K493А, N500D, F534Y, S547D, и/или G594S; и/или
(b) S205T, G549E, S586R и/или T589R.
12. Рекомбинантный AAV вектор в соответствии с пунктом 5, где (i) вектор содержит вариантный AAV8 капсидный белок, (ii) вариантный AAV капсидный белок содержит последовательность имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 6; (iii) AAV капсидный белок немутантного типа является из AAV8; и/или (iv) AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 6;
и где, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV8 капсидного белка: 125, 151, 163, 206, 315, 460, 495, 502, 536, 549, 551, 588, 591 и/или 596.
13. Рекомбинантный AAV вектор в соответствии с пунктом 12, где вариантный AAV8 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV8 капсидным белком немутантного типа:
(a) V125I, Q151A, K163S, A206S, Т460М, Т495А, N502D, F536Y, N549D, A551G, Q588S и/или G596S; и/или
(b) S315N и/или T591R.
14. Рекомбинантный AAV вектор в соответствии с пунктом 5, где (i) вектор содержит вариантный AAV9 капсидный белок, (ii) вариантный AAV капсидный белок содержит последовательность имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 7; (iii) AAV капсидный белок немутантного типа является из AAV9; и/или (iv) AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 7;
и где, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV9 капсидного белка: 125, 151,162, 205, 314,458,493, 500, 534, 547, 549, 586, 589 и/или 594.
15. Рекомбинантный AAV вектор в соответствии с пунктом 14, где вариантный AAV9 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAV9 капсидным белком немутантного типа:
(a) L125I, Q151A, N314S, Q458M, V493A, E500D, F534Y, G547D, А589Т и/или G594S; и/или
(b) S162A, S205T, G549E и/или S586R.
16. Рекомбинантный AAV вектор в соответствии с пунктом 5, где (i) вектор содержит вариантный AAVrh10 капсидный белок, (ii) вариантный AAV капсидный белок содержит последовательность имеющую, по меньшей мере, 95% идентичность последовательности к SEQ ID NO: 8; (iii) AAV капсидный белок немутантного типа является из AAVrh10; и/или (iv) AAV капсидньш белок немутантного типа содержит последовательность SEQ ID NO: 8;
и где, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV10 капсидного белка: 125, 151, 163, 206, 315, 460,495, 502, 536, 549, 551, 588, 591 и/или 596.
17. Рекомбинантный AAV вектор в соответствии с пунктом 16, где вариантный AAVrh10 капсидный белок содержит одно или больше из следующих аминокислотных замещений по сравнению с AAVrhl0 капсидным белком немутантного типа:
(a) V125I, Q151A, K163S, A206S, N315S, Т460М, L495A, N502D, F536Y, G549D, Q588S, А591Т и/или G596S; и/или
(b) G551E.
18. Рекомбинантный AAV вектор в соответствии с каким-либо из предыдущих пунктов, где рекомбинантный AAV вектор демонстрирует повышенную трансдукцию нейрональной или ретинальной ткани по сравнению с AAV вектором, содержащим соответствующий AAV капсидный белок немутантного типа.
19. Рекомбинантный AAV вектор в соответствии с каким-либо из предыдущих пунктов, где генный продукт содержит интерферирующую РНК или аптамер.
20. Рекомбинантный AAV вектор в соответствии с каким-либо из пунктов 1-18, где генный продукт содержит полипептид.
21. Рекомбинантный AAV вектор в соответствии с пунктом 20, где генный продукт содержит нейропротекторный полипептид, антиангиогенный полипептид или полипептид, который улучшает функцию нейрональной или ретинальной клетки.
22. Рекомбинантный AAV вектор в соответствии с пунктом 21, где генный продукт содержит глиальный нейротрофический фактор, фактор роста фибробластов, фактор роста нервов, нейротрофический фактор головного мозга, родопсин, ретиношизин, RPE65 или периферии.
23. Фармацевтическая композиция, содержащая:
(a) рекомбинантный AAV вектор в соответствии с каким-либо из предыдущих пунктов; и
(b) фармацевтически приемлемый эксципиент.
24. Способ доставки генного продукта в нейрональную или ретинальную ткань у субъекта, где способ включает введение субъекту рекомбинантного AAV вектора или фармацевтической композиции в соответствии с каким-либо из предыдущих пунктов.
25. Способ лечения неврологического или глазного расстройства, где способ включает введение субъекту рекомбинантного AAV вектора или фармацевтической композиции в соответствии с каким-либо из предыдущих пунктов.
26. Рекомбинантный AAV вектор или фармацевтическая композиция в соответствии с каким-либо из пунктов 1-23, для применения в лечении неврологического или глазного расстройства.
27. Способ, рекомбинантный AAV вектор или фармацевтическая композиция для применения в соответствии с каким-либо из пунктов 24-26, где неврологическое расстройство представляет собой нейродегенеративное заболевание.
28. Способ, рекомбинантный AAV вектор или фармацевтическая композиция для применения в соответствии с каким-либо из пунктов 24-26, где глазное расстройство представляет собой глаукому, пигментную дегенерацию сетчатки, дегенерацию желтого пятна, ретиношизис или диабетическую ретинопатию.
29. Выделенный вариантный AAV капсидный белок, где вариантный AAV капсидный белок содержит, по меньшей мере, одно аминокислотное замещение по сравнению с AAV капсидным белком немутантного типа; где, по меньшей мере, одно аминокислотное замещение присутствует в одном или больше из следующих положений в последовательности AAV2 капсидного белка: 125, 151, 162, 205, 312, 457, 492, 499, 533, 546, 548, 585, 588 и/или 593; или в одном или больше соответствующих положениях в альтернативной последовательности AAV капсидного белка.
30. Выделенная нуклеиновая кислота, содержащая нуклеотидную последовательность, которая кодирует вариантный AAV капсидный белок как определено в параграфе 29.
31. Выделенная клетка-хозяин, содержащая нуклеиновую кислоту как определено в параграфе 30.



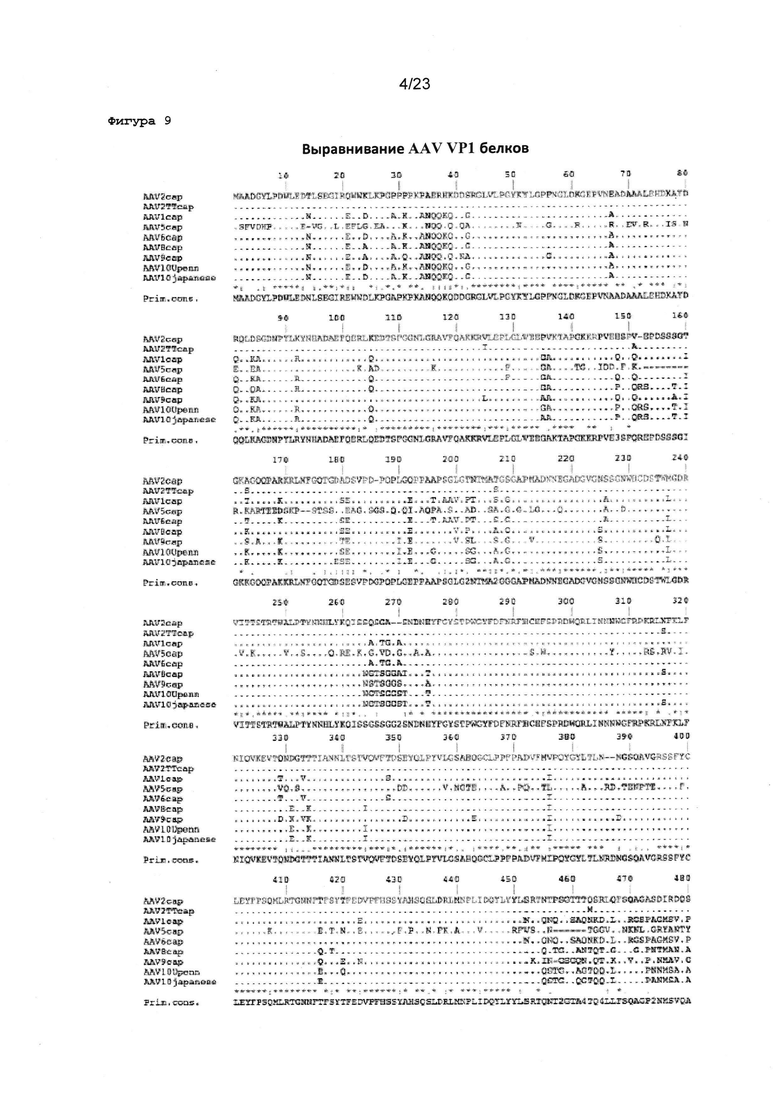
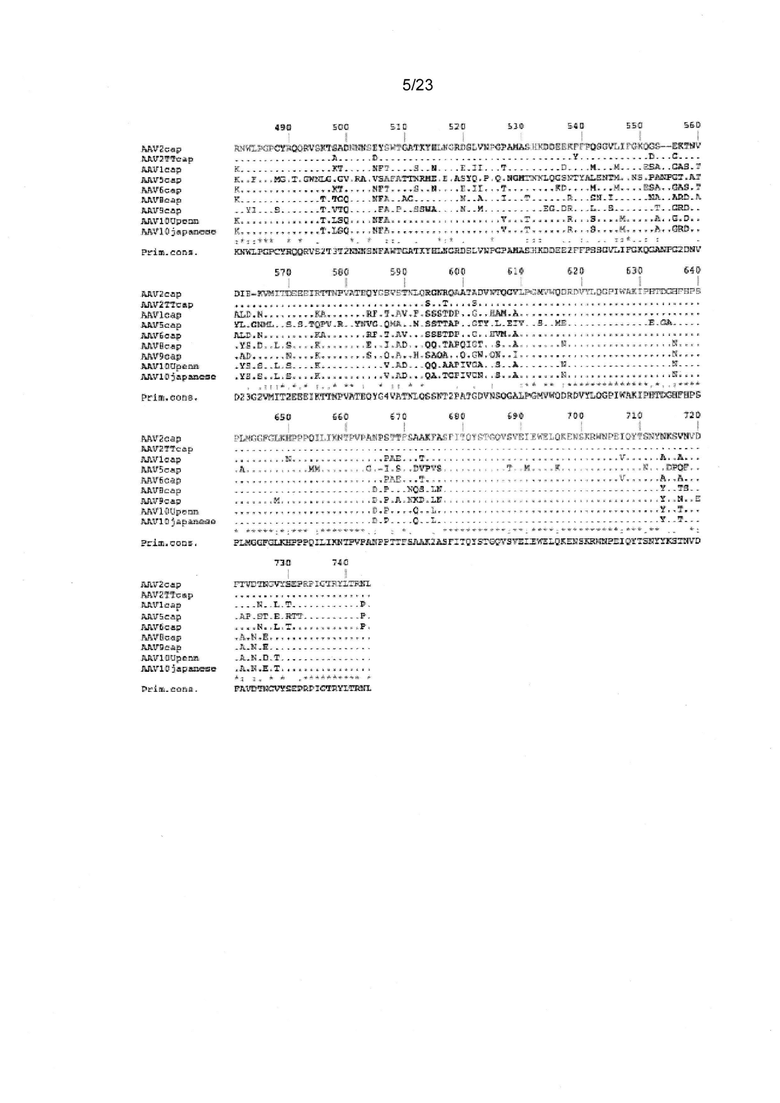
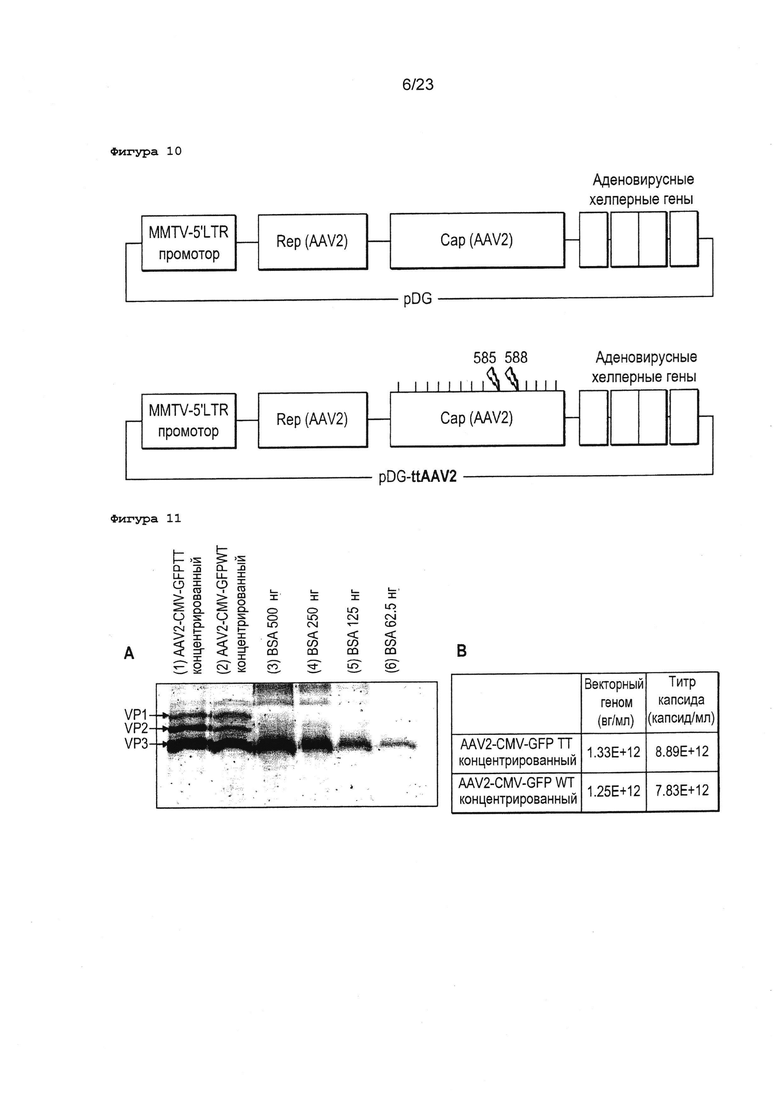
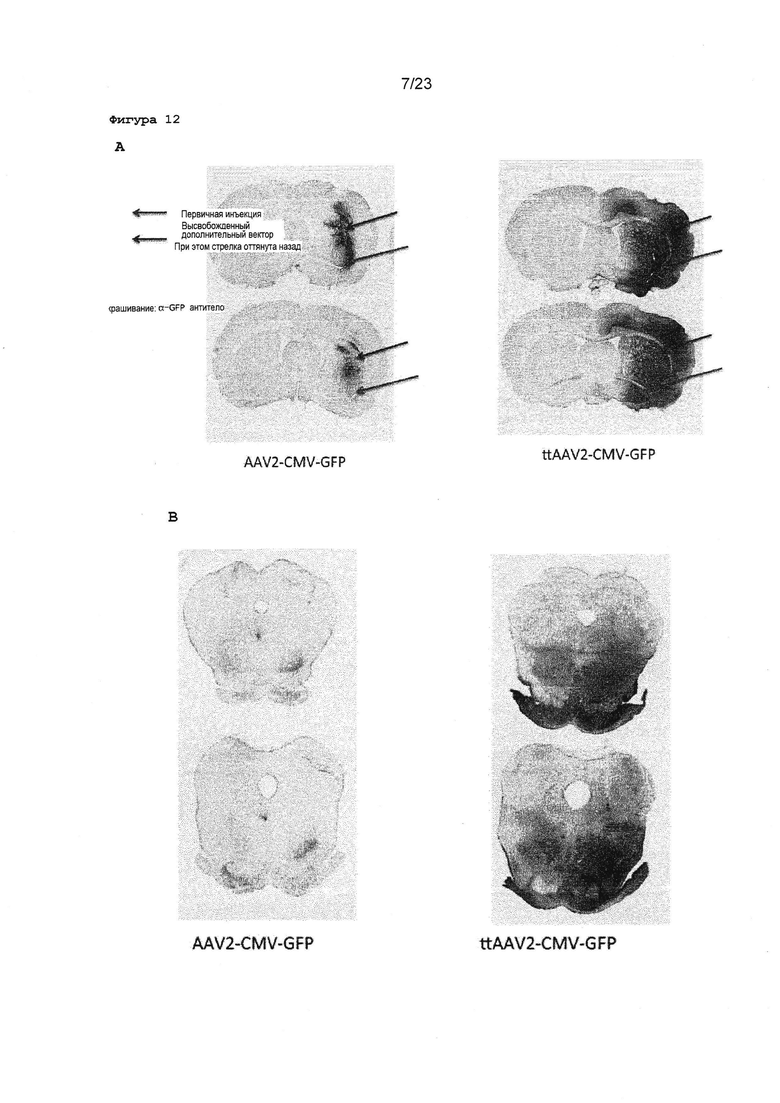
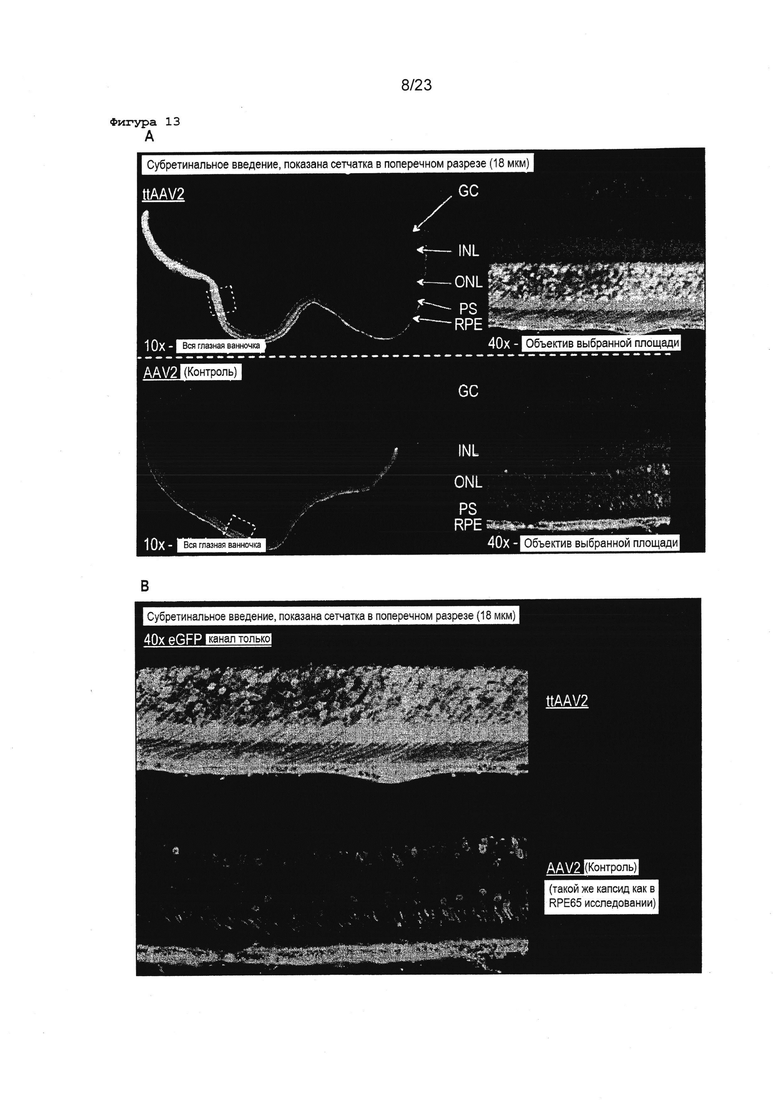
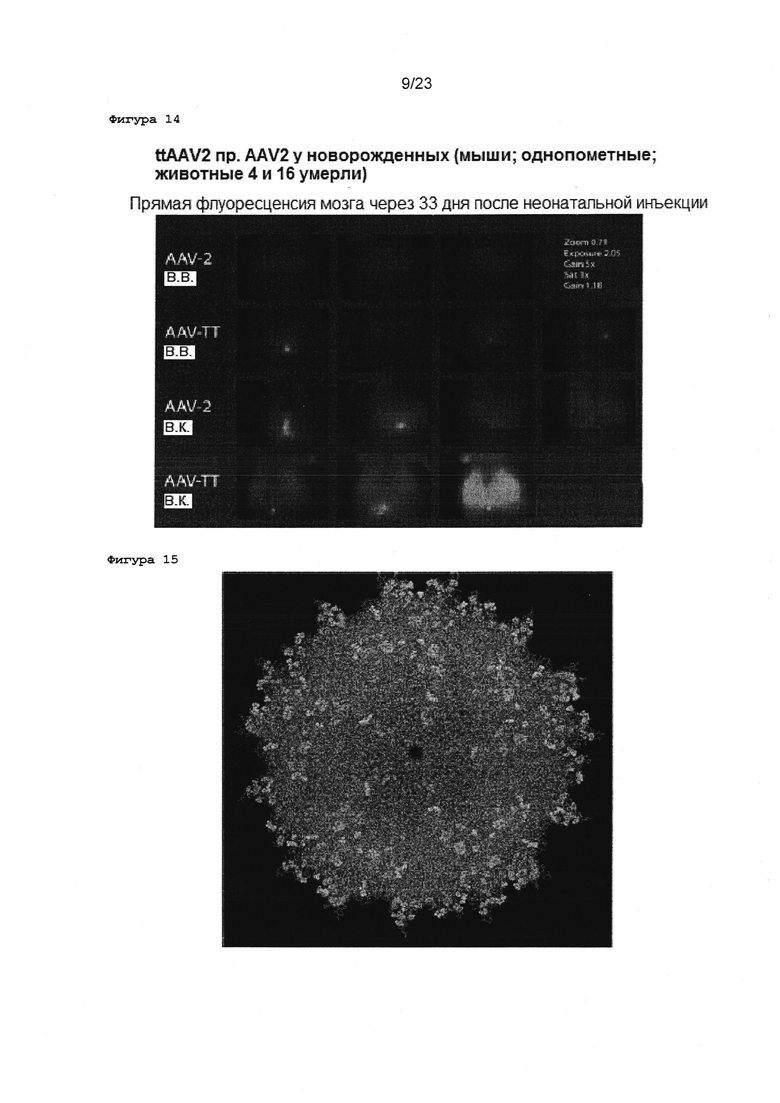
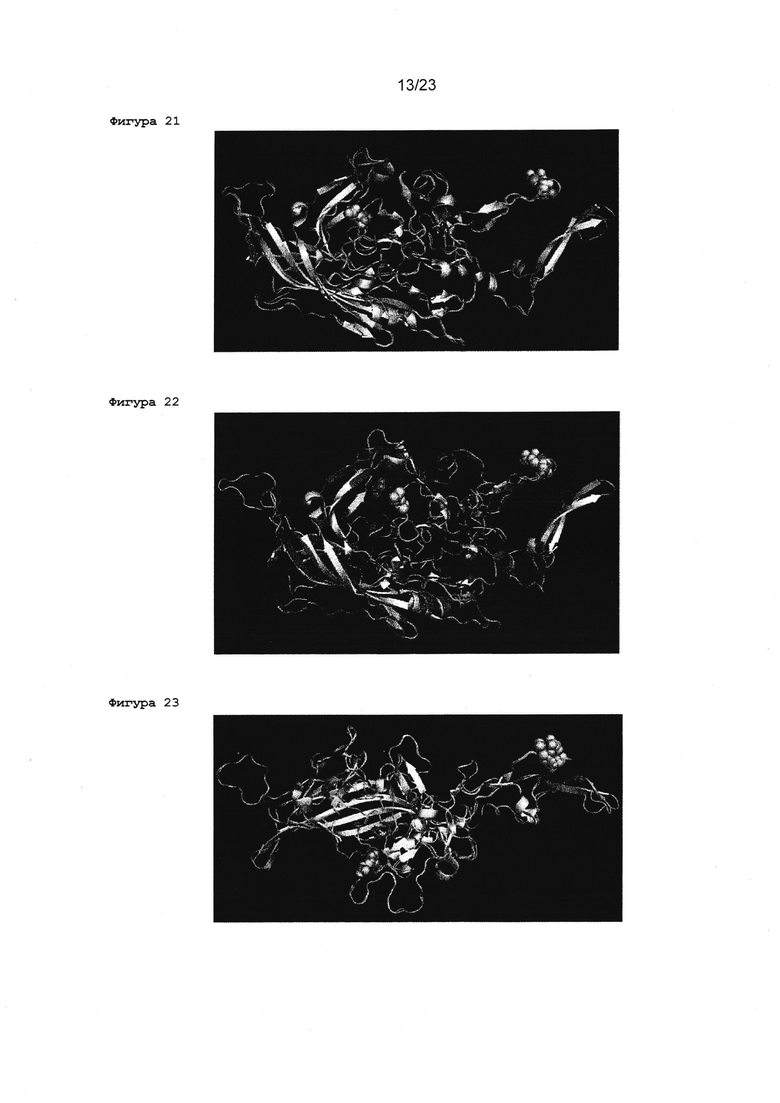

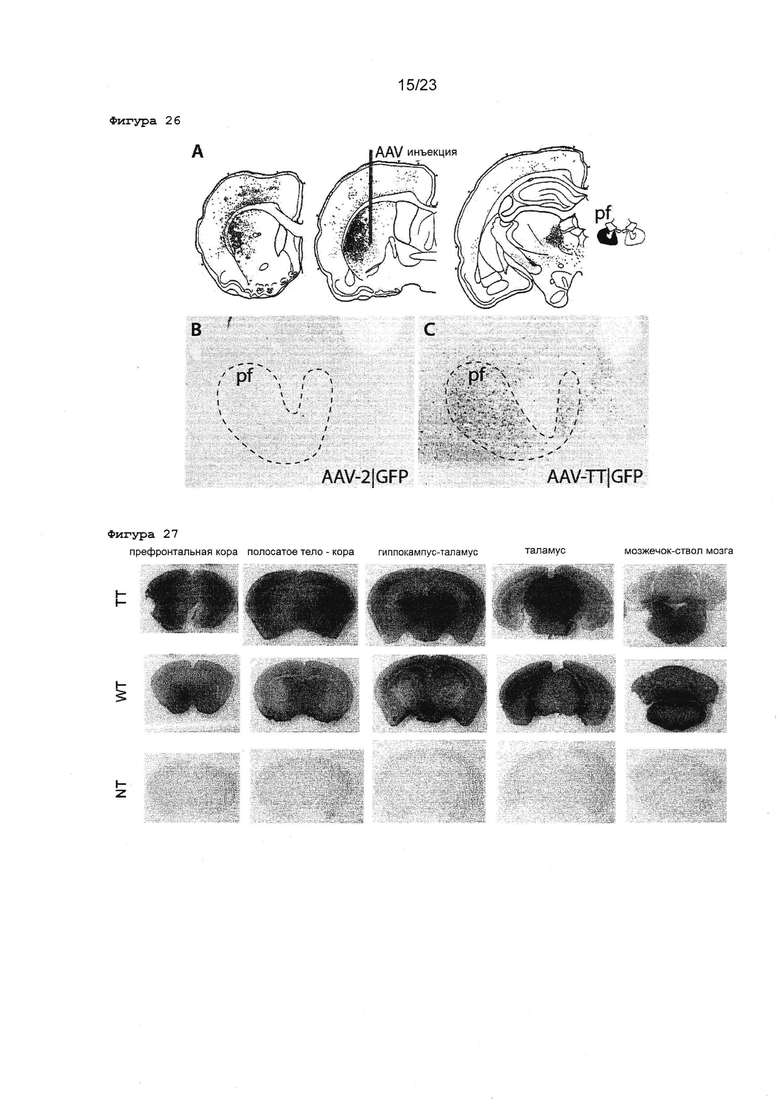
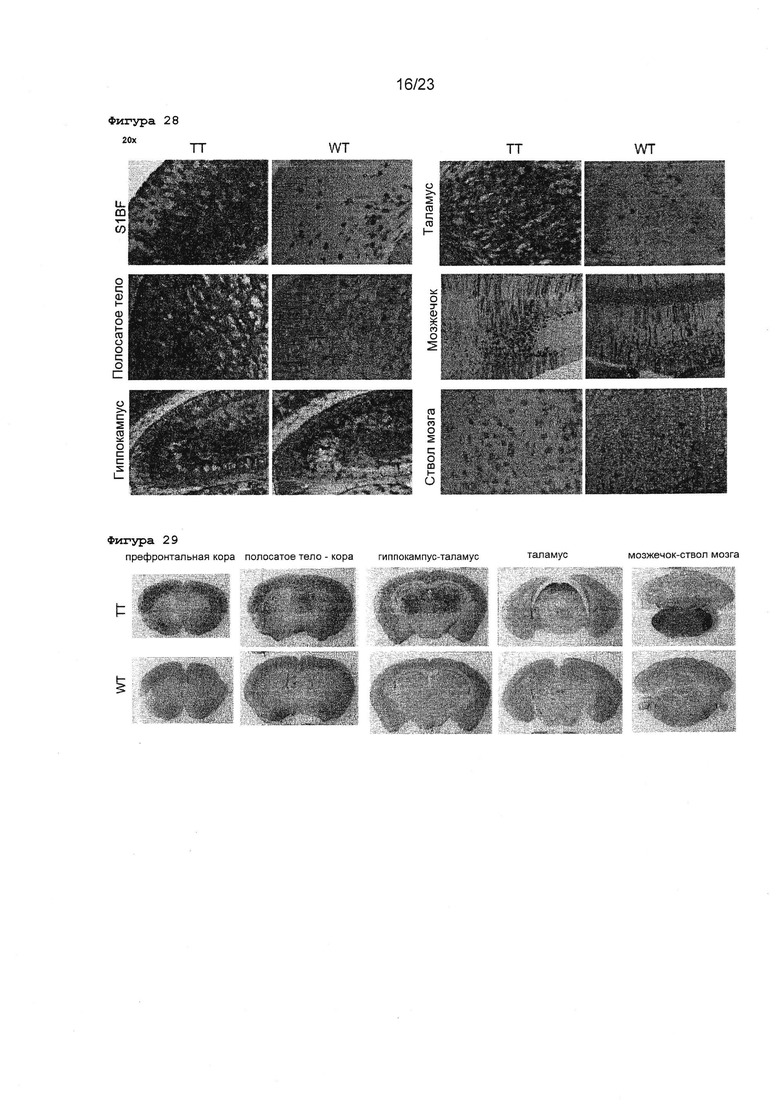

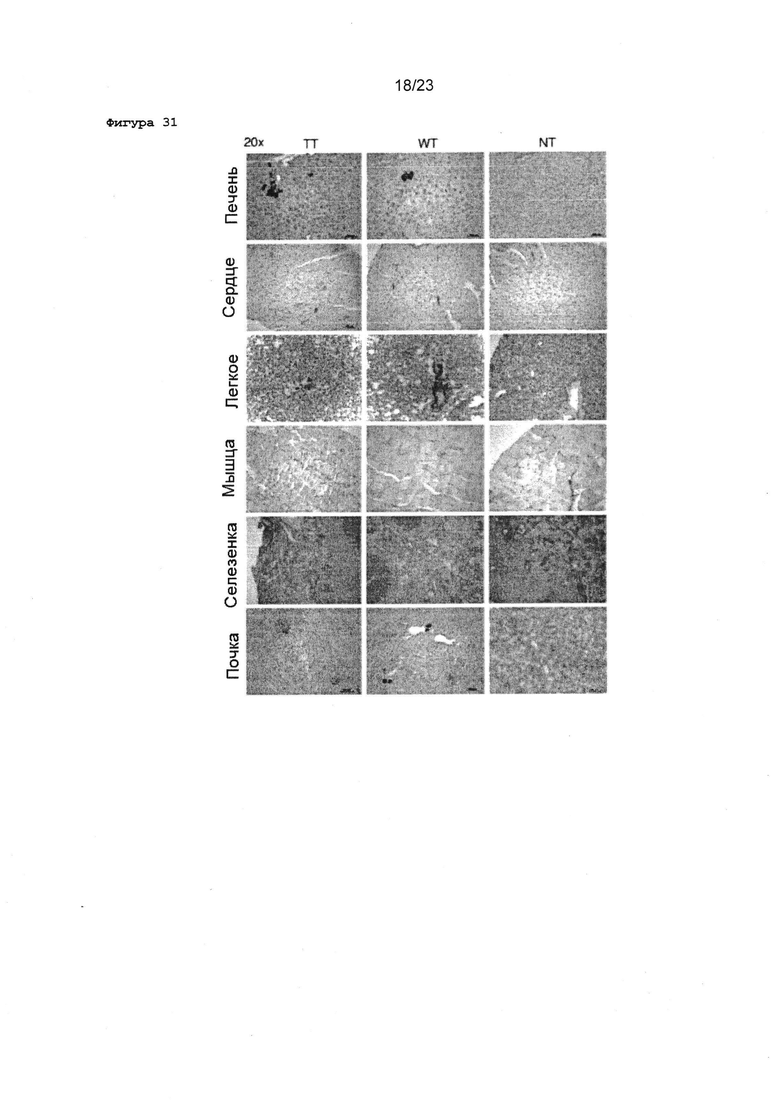
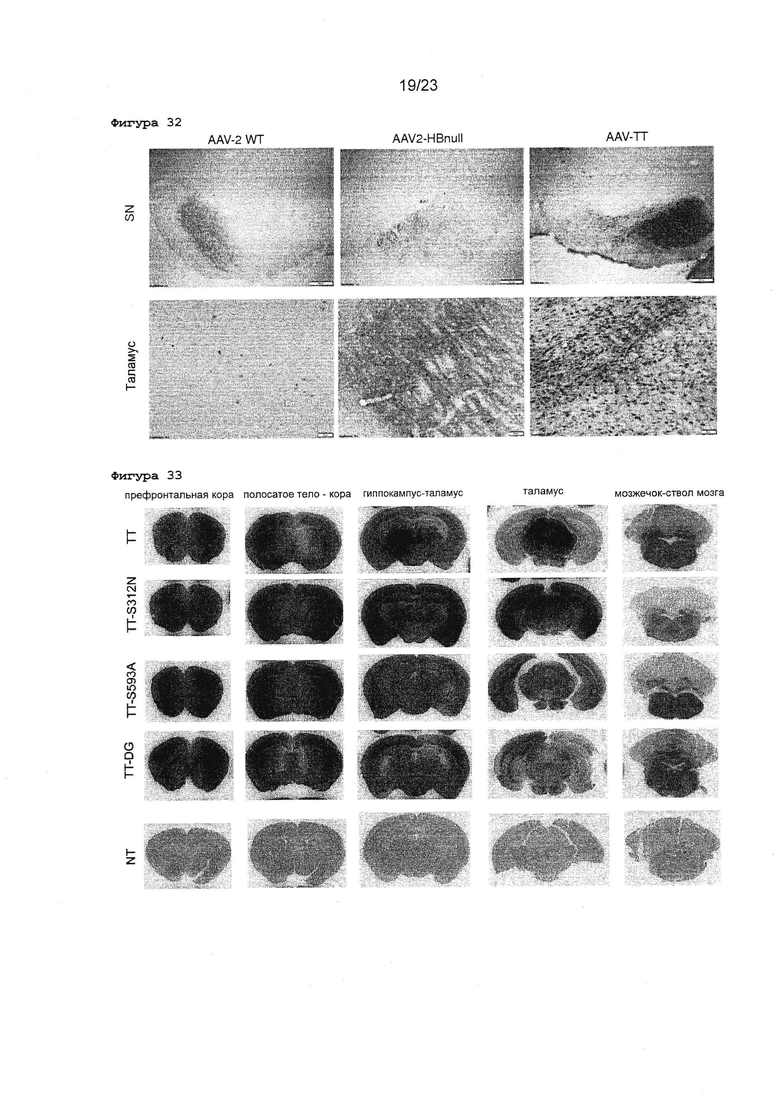
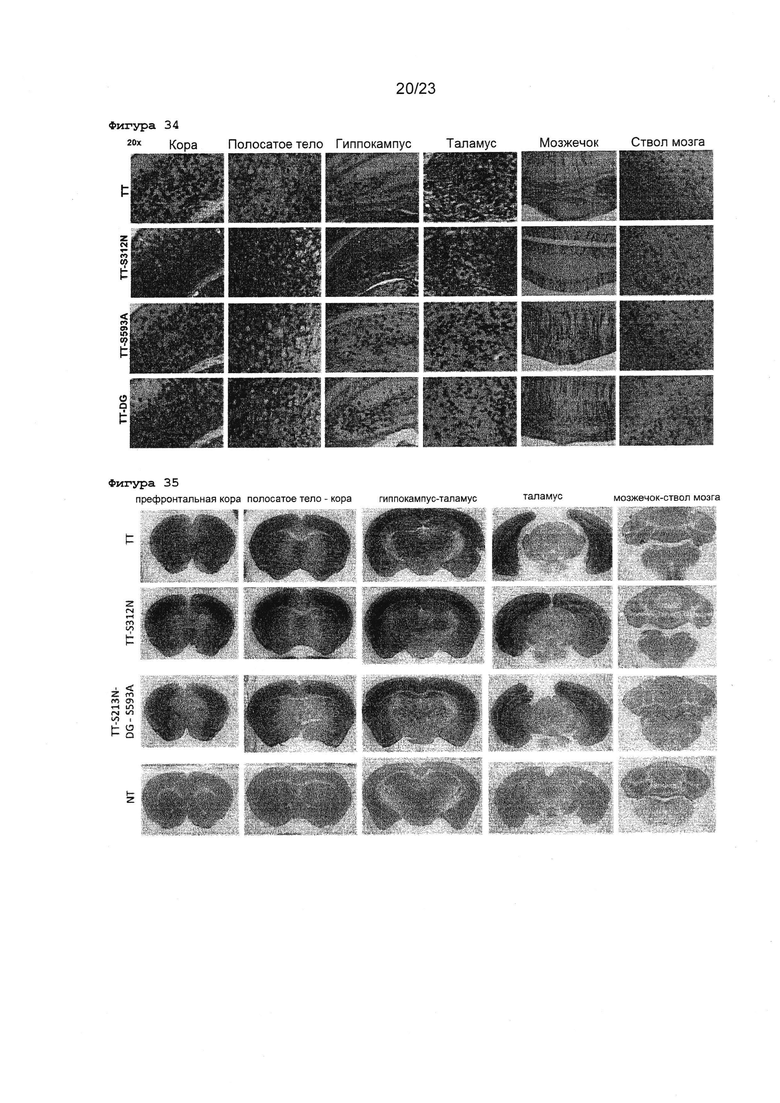
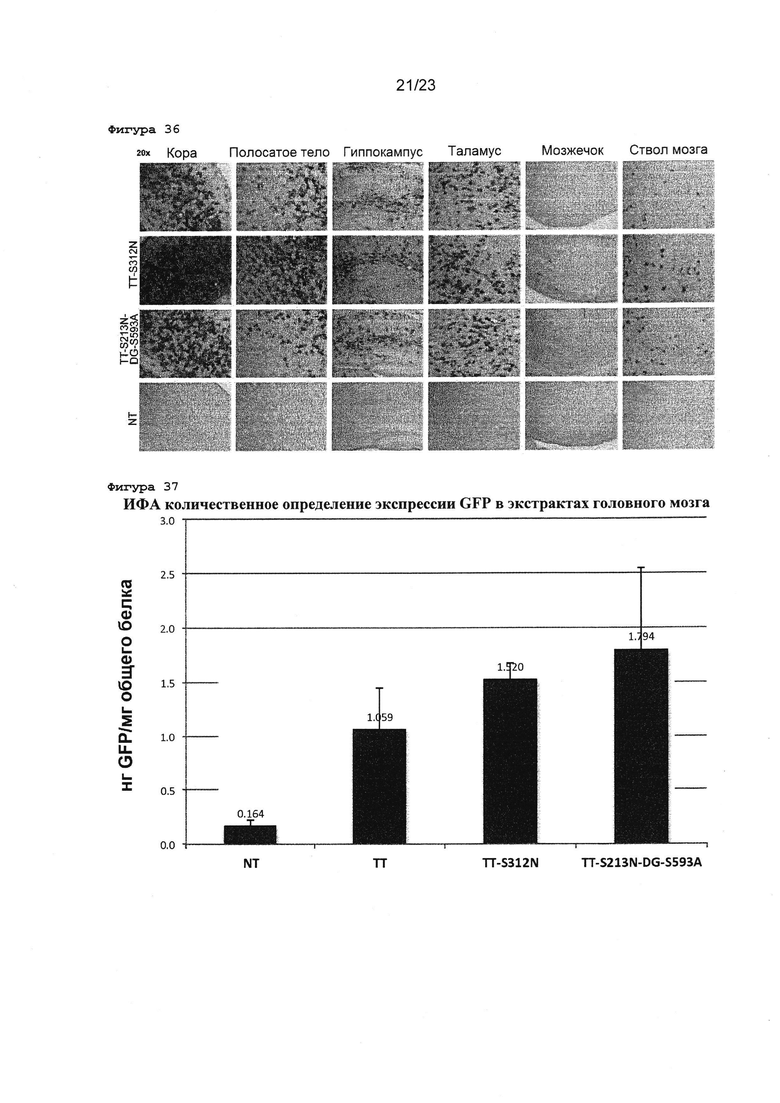


Изобретение относится к биотехнологии. Описан рекомбинантный вектор аденоассоциированного вируса (AAV), содержащий (а) вариантный AAV2 капсидный белок, где вариантный AAV2 капсидный белок содержит, по меньшей мере, четыре аминокислотных замещения по сравнению с капсидным белком немутантного типа AAV2; где, по меньшей мере, четыре аминокислотных замещения присутствуют в следующих положениях в последовательности AAV2 капсидного белка: 457, 492, 499 и 533; и (b) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую генный продукт. Изобретение может быть использовано для лечения неврологического и глазного заболевания, а также печени. 5 н. и 12 з.п. ф-лы, 39 ил., 1 табл., 6 пр.
1. Рекомбинантный вектор аденоассоциированного вируса (AAV) для доставки терапевтических генов in vivo, содержащий:
(a) вариантный AAV2 капсидный белок, где вариантный AAV2 капсидный белок содержит аминокислотные замещения по сравнению с капсидным белком немутантного типа AAV2 SEQ ID NO:1; где аминокислотные замещения присутствуют в следующих положениях в последовательности капсидного белка AAV2: Q457M, S492A, E499D и F533Y; и
(b) гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность для кодирования генного продукта.
2. Рекомбинантный AAV2 вектор по п. 1, где (i) вариантный AAV2 капсидный белок содержит последовательность SEQ ID NO:2, или последовательность, имеющую, по меньшей мере, 95% идентичность последовательности с ней; и/или (ii) AAV капсидный белок немутантного типа содержит последовательность SEQ ID NO: 1.
3. Рекомбинантный AAV2 вектор по какому-либо из пп. 1 или 2, где вариантный AAV2 капсидный белок содержит аминокислотные замещения по сравнению с AAV капсидным белком немутантного типа SEQ ID NO:1 в следующих положениях в последовательности AAV2 капсидного белка: Q457M, S492A, E499D, F533Y и V125I, V151A, A162S, T205S.
4. Рекомбинантный AAV вектор по какому-либо из пп. 1-3, где вариантный AAV2 капсидный белок содержит аминокислотные замещения по сравнению с AAV капсидным белком немутантного типа SEQ ID NO:1 в следующих положениях в последовательности AAV2 капсидного белка: (a) Q457M, S492A, E499D, F533Y и R585S, R588T или (b) Q457M, S492A, E499D, F533Y и V125I, V151A, A162S, T205S и R585S, R588T.
5. Рекомбинантный AAV2 вектор по какому-либо из пп. 1-4, где вариантный AAV2 капсидный белок содержит аминокислотные замещения по сравнению с AAV капсидным белком немутантного типа SEQ ID NO:1 в следующих положениях в последовательности AAV2 капсидного белка: (a) Q457M, S492A, E499D, F533Y и G546D, E548G, A593S, (b) Q457M, S492A, E499D, F533Y и V125I, V151A, A162S, T205S и G546D, E548G, A593S, (с) Q457M, S492A, E499D, F533Y и R585S, R588T и G546D, E548G, A593S, (d) Q457M, S492A, E499D, F533Y и V125I, V151A, A162S, T205S и R585S, R588T и G546D, E548G, A593S.
6. Рекомбинантный AAV вектор по какому-либо из пп. 1-5, где вариантный AAV2 капсидный белок содержит также остаток N312.
7. Рекомбинантный AAV2 вектор по какому-либо из пп. 1-6, где рекомбинантный AAV2 вектор демонстрирует повышенную трансдукцию нейрональной или ретинальной ткани по сравнению с AAV2 вектором, содержащим соответствующий AAV2 капсидный белок немутантного типа SEQ ID NO:1.
8. Рекомбинантный AAV2 вектор по какому-либо из предыдущих пунктов, где генный продукт содержит интерферирующую РНК или аптамер.
9. Рекомбинантный AAV2 вектор по какому-либо из пп. 1-8, где генный продукт содержит полипептид.
10. Рекомбинантный AAV2 вектор по п. 9, где генный продукт содержит нейропротекторный полипептид, антиангиогенный полипептид или полипептид, который улучшает функцию нейрональной или ретинальной клетки.
11. Рекомбинантный AAV2 вектор по п. 10, где генный продукт содержит глиальный нейротрофический фактор, фактор роста фибробластов, фактор роста нервов, нейротрофический фактор головного мозга, родопсин, ретиношизин, RPE65 или периферии.
12. Способ доставки генного продукта к ткани у субъекта, где способ включает введение субъекту рекомбинантного AAV2 вектора по какому-либо из предыдущих пунктов.
13. Способ по п. 12, в котором ткань выбирают из крови, костного мозга, мышечной ткани, нейрональной ткани, ретинальной ткани, ткани поджелудочной железы, ткани печени, почечной ткани, легочной ткани, ткани кишечника или ткани сердца.
14. Способ по п. 13, в котором ткань представляет собой нейрональную, ретинальную ткань или ткань печени.
15. Применение рекомбинантного AAV2 вектора по какому-либо одному из пп. 1-11 в лечении неврологического расстройства у субъекта, где неврологическое расстройство представляет собой нейродегенеративное заболевание.
16. Применение рекомбинантного AVV2 вектора по какому-либо одному из пп. 1-11 в лечении глазного расстройства, где глазное расстройство представляет собой глаукому, пигментную дегенерацию сетчатки, дегенерацию желтого пятна, ретиношизис или диабетическую ретинопатию.
17. Применение рекомбинантного AVV2 вектора по какому-либо одному из пп. 1-11 в лечении заболевания печени.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Супотницкий М.В | |||
| Генотерапевтические векторные системы на основе вирусов// Биопрепараты | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| - с | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |

