Область техники, к которой относится изобретение
Настоящее изобретение относится к векторам, пригодным для экспрессии целевых белков, и к их применению для генной терапии. Настоящее изобретение относится также к векторам и последовательностям нуклеиновых кислот, способствующих лечению мукополисахаридозов (MPS), и в частности, лечению мукополисахаридозов типа III или синдрома Санфилиппо.
Уровень техники
Лизосома представляет собой органеллу, находящуюся в цитоплазме эукариотных клеток, служащую в качестве структуры для хранения многих гидролитических ферментов и в качестве центра деградации и рециклизации компонентов клетки. Эта органелла содержит несколько типов гидролитических ферментов, включая протеазы, нуклеазы, гликозидазы, липазы, фосфолипазы и сульфатазы. Все ферменты представляют собой кислые гидролазы.
Заболевания, связанные с хранением в лизосомах (LSD), обусловлены генетическими дефектами, которые затрагивают один или более лизосомальных ферментов. Эти генетические заболевания возникают обычно в результате недостаточной активности конкретного фермента, присутствующего в лизосоме. В меньшей степени эти заболевания могут быть обусловлены нехваткой белков, вовлеченных в биогенез лизосом.
LSD являются редкими для индивидуумов заболеваниями, хотя в виде группы таких нарушений они относительно обычны в общей популяции. Суммарная распространенность LSD составляет приблизительно 1 на 5000 живых новорожденных. Смотрите Meikle P, et al, JAMA 1999; 281:249-254. Однако некоторые группы в пределах общей популяции особенно страдают от высокой встречаемости LSD. Например, коэффициенты распространенности болезней Гоше и Тея-Сакса у потомков, происходящих от евреев центральной и восточной Европы (ашкенази), составляет 1 на 600 и 1 на 3900 новорожденных, соответственно. Финская популяция также характеризуется необычно высоким коэффициентом распространенности LSD.
Мукополисахаридозы типа III (MPSIII), известные в целом как синдром Санфилиппо, представляют собой LSD, причиной которых является дефицит одного из ферментов, участвующих в разрушении гепаран-сульфата, ведущий к его патологической аккумуляции. MPSIII делятся на четыре подтипа в зависимости от недостаточности определенного фермента. Потеря сульфамидазной активности вызывает подтип IIIA и, как сообщается, он является наиболее тяжелым с наиболее ранним возникновением заболевания и самой короткой выживаемостью. Симптомы MPSIIIA возникают в первые годы жизни и характеризуются тяжелой нейродегенерацией, которая ведет к глубокой задержке умственного развития, агрессивности, гиперактивности и нарушениям сна. Больные постепенно теряют способность говорить, глотать и основную координацию движений. Кроме неврологических симптомов больные MPSIIIA страдают от неневрологических изменений, включая гепато- и спленомегалию, пороки развития скелета и суставов, а также от частой диареи и инфекций дыхательных путей. Прогрессирующее ухудшение симптомов ведет к смерти больного в подростковом возрасте. Смотрите Neufeld E, Muenzer J, «The mucopolysaccharidoses» in Scriver C, et al., Eds., «The metabolic and molecular basis of inherited disease» (McGraw-Hill Publishing Co., New York, NY, US, 2001, pp. 3421-3452).
В настоящее время не существует метода лечения MPSIIIA и, следовательно, существующие типы лечения направлены на контролирование симптомов заболевания, для того, чтобы повысить низкое качество жизни больных. Нарушения MPS может лечить с помощью трансплантации костного мозга или путем заместительной терапии ферментом (ERT). Оба подхода основываются на эндоцитозе лизосомальных ферментов из внеклеточной среды и их направленной доставке к лизосомам через рецептор манноза-6-фосфата (M6PR), присутствующий на клеточной мембране. Тем не менее, трансплантация костного мозга, как показано, является неэффективной для лечения больных MPSIII. Смотрите Sivakamur P, Wraith J, J. Inherit. Metab. Dis. 1999; 22:849-850. ERT, как всесторонне доказано, является эффективной в противодействии неневрологической аккумуляции при других заболеваниях, касающихся хранения в лизосомах, включая MPSI, II и VI. Смотрите Harmatz P, et al., J. Mol. Genet. Metab. 2008; 94:469-475; Muenzer J, et al., Genet. Med. 2006; 8:465-473 и Wraith J, et al., J. Pediatr. 2004; 144:581-588. Кроме высокой стоимости этих типов лечения, показано, что ERT не приводит к коррекции или сохранению функции нейронов из-за неудовлетворительного прохождения предоставляемого экзогенно фермента через гематоэнцефалический барьер (ГЭБ). Смотрите Enns G, Huhn S, Neurosurg. Focus 2008; 24:E12. Позднее было показано, что ERT в высоких дозах приводит к частичному успеху в очистке компартментов хранения в ЦНС при MPS VII, возможно из-за насыщения M6PR и рецепторов маннозы, что ведет к более длительному периоду полужизни белка в циркуляторном русле. Смотрите Vogler C, et al., Proc. Natl. Acad. Sci. USA 2005; 102:14777-14782. В этом исследовании продемонстрировано, что высокие уровни фермента в циркуляторном русле в течение длительных периодов времени коррелируют с лучшей коррекцией патологии. Доставка фермента внутрь мозга или в СМЖ, как было доказано, также является эффективной в отношении снижения патологических изменений ЦНС у мышей с MPS IIIA. Смотрите Hemsley K, et al., Genes Brain Behav. 2008; 53(2):161-8 и Savas P, et al., Mol. Genet. Metab. 2004; 82:273-285. Однако этот подход является высоко инвазивным из-за необходимости множественных повторяющихся инъекций и может повышать риск повреждения и/или инфицирований мозга.
Из-за ограничений имеющихся в настоящее время терапевтических возможностей для лечения MPSIII необходимы альтернативные подходы. С помощью переноса генов можно обеспечить способами достижения постоянной продукции утерянного фермента при одноразовом вмешательстве. Аденоассоциированные векторы (AAV) быстро распространяются в качестве вектора выбора для многих вариантов применения генной терапии из-за высокой эффективности их трансдукции и отсутствия их патогенности. Векторы AAV эффективно трансдуцируют постмитотические клетки, и некоторые доклинические и клинические исследования продемонстрировали возможности переноса генов, опосредованные векторами AAV, для эффективно передаваемой продолжительной экспрессии терапевтических трансгенов при разнообразных заболеваниях. Смотрите Daya S, Berns K, Clin. Microbiol. Rev. 2008; 21:583-593.
Было показано, что введение вектора AAV5, коэкспрессирующего сульфамидазу и активатор сульфатазы SUMF1, в боковые желудочки новорожденных мышей MPSIIIA способно корректировать многие неврологические и поведенческие изменения. Смотрите Fraldi A, et al., Hum. Mol. Genet. 2007; 16:2693-2702. Однако этот предлагаемый образ действия имеет несколько недостатков. Во-первых, используемый промотор CMV, как сообщалось, подавляется. Во-вторых, длительные эффекты коэкспрессии сульфамидазы и SUMF1 не оцениваются вообще. Неясно даже, является ли коэкспрессия SUMF необходимой и предоставляет ли какие-либо дополнительные постоянные преимущества по сравнению с лечением только сульфамидазой. В-третьих, векторы AAV5 характеризуются слабым распределением в пределах паренхимы, и, более важно, что доставка сульфамидазы в мозг с использованием этих векторов не приводит к какой-либо трансдукции ткани мозга, таким образом, при использовании этого подхода не достигается никакой коррекции соматического фенотипа. Наконец Fraldi, 2007, выше, продемонстрировал эффективность переноса генов только у новорожденных мышей MPSIIIA. Не сообщалось ни о каких экспериментах с мышами более старшего возраста. Так как MPSIIIA обычно диагностируется после 3-4-летнего возраста, модель новорожденных животных не является адекватной для прогнозирования эффектов этого лечения у человека.
Принимая во внимание трудности диагностирования MPSIIIA при рождении, предлагается развитие терапевтических вмешательств, начинающихся рано во взрослом состоянии. Сообщалось, что внутривенная доставка лентивирусного вектора, экспрессирующего сульфамидазу, взрослым мышам приводила к небольшому улучшению фенотипа ЦНС вероятно из-за относительно плохого осуществления трансдукции этих векторов in vivo. Смотрите McIntyre C, et al., Mol. Genet. Metab. 2008; 93:411-418. Таким образом, использование вирусных векторов с более высокой эффективностью трансдукции in vivo, таких как векторы AAV, может обеспечить более высокими уровнями сульфамидазы в циркуляторном русле, что потенциально может улучшить или скорректировать неврологическую патологию.
Лечение MPSIIIA с помощью генной терапии требует более эффективных векторов и кодирующих сульфамидазу последовательностей. Следовательно, существует назревшая необходимость эффективного лечения MPSIIIA. Существует также потребность в новых подходах к лечению этого заболевания, которые должны иметь повышенные признаки надежности. MPSIIIA является редким заболеванием и является, следовательно, орфанным заболеванием. Фармакологические агенты, разрабатываемые специально для лечения этого редкого клинического состояния, должны быть орфанными лекарствами.
Краткое изложение сущности изобретения
Настоящее изобретение относится к новой нуклеотидной последовательности для лечения заболеваний, предпочтительно для лечения мукополисахаридозов (MPS). Следовательно, в первом аспекте изобретение относится к нуклеотидной последовательности, которая представляет собой последовательность сульфамидазы человека с оптимизированными кодонами, позволяющую транскрибировать более стабильную мРНК. Эта последовательность транскрибируется с более высокими скоростями и, следовательно, фермент сульфамидаза продуцируется с более высокими выходами. Последовательность характеризуется лучшим профилем экспрессии и является терапевтически более эффективной при сравнении с другими попытками оптимизации кодонов нуклеотидной последовательности сульфамидазы. Следствием этих повышенных уровней экспрессии фермента является повышение сульфамидазной активности в сыворотке, позволяющее снизить аккумуляцию гликозаминогликанов (GAG), которая вызывает заболевание. Указанная последовательность представляет собой SEQ ID NO:1 или последовательность, идентичную последовательности SEQ ID NO:1 по меньшей мере на 85%, которая кодирует белок SEQ ID NO:2.
Во втором аспекте изобретение относится к генетической конструкции, включающей нуклеотидную последовательность согласно первому аспекту изобретения.
Настоящее изобретение также относится к новым векторам AAV серотипа 9, которые способны проходить через гематоэнцефалический барьер (ГЭБ) и проявляют более высокий тропизм в отношении различных структур мозга. Это позволяет сульфамидазной активности специфически повышаться в мозге, снижая аккумуляцию GAG, и следовательно, улучшать неврологические симптомы MPS. AAV серотипа 9 проявляет также неожиданно высокий тропизм в отношении сердца, поджелудочной железы и мышечной ткани, таким образом усиливая суммарные терапевтические преимущества изобретения.
Например, после введения векторов AAV серотипов 8 и 9 (AAV8 и AAV9) взрослым мышам MPSIIIA с помощью внутривенной (в/в) инъекции для направленной доставки в печень или с помощью внутримышечной (в/м) инъекции для направленной доставки в скелетную мышцу, или внутрицистернально (i.c.) для направленной доставки в центральную нервную систему, уровни экспрессии сульфамидазы, достигаемые при внутримышечной доставке вектора не являлись терапевтическими. Внутрицистернальное введение было способно не только повышать уровень сульфамидазы в циркуляторном русле, но также возвращать к исходному состоянию соматический фенотип MPSIIIA в нескольких типах ткани, включая ткань мозга. Подход, направленный на доставку в печень, также обладал способностью создавать высокие уровни сульфамидазной активности в циркуляторном русле, что неожиданно полностью исправляло соматический фенотип MPSIIIA в отношении компартмента хранения и существенно исправляло нейропатологию, связанную с заболеванием. Эти результаты являются доказательством эффективности переноса гена сульфамидазы, опосредованного AAV, у взрослых мышей MPSIIIA, модели заболевания, весьма сходной с клиническими проявлениями у человека. Изобретатели были в состоянии полностью скорректировать как соматические, так и неврологические изменения при MPSIIIA.
Генетические конструкции по настоящему изобретению могут дополнительно включать подходящие промоторы, такие как промоторы CAG или hAAT, для контроля и усиления экспрессии сульфамидазы. Например, промотор CAG является более стабильным, чем промотор CMV, и, следовательно, более приспособлен для индукции экспрессии сульфамидазы в течение более длительных периодов времени. С другой стороны, безопасность и эффективность промотора hAAT делает его идеальным носителем для последующей доставки или поддержания доз сульфамидазы. Контролирование экспрессии SEQ ID NO:1 промоторами CAG или hAAT значительно усиливало ее терапевтические эффекты.
Векторы AAV по настоящему изобретению также повышают сульфамидазную активность, что снижает аккумуляцию GAG и улучшает клинический исход у индивидуумов, страдающих MPS. Может быть достаточным только одно введение, потому что промотор и нуклеотидная последовательность сульфамидазы, локализованные между концевыми обратными повторами (ITR), включаются в геном клеток индивидуума. Следовательно, единственное парентеральное введение достаточно для получения долгосрочного эффекта.
В третьем аспекте изобретение относится к фармацевтической композиции, включающей нуклеотидную последовательность по первому аспекту изобретения, генетическая конструкция или экспрессионный вектор по изобретению.
В четвертом аспекте изобретение относится к нуклеотидной последовательности, генетической конструкции, экспрессионному вектору или фармацевтической композиции по изобретению для применения в качестве лекарственного средства. Лекарственное средство может быть использовано для повышения сульфамидазной активности в организме, для замещающей терапии ферментом, для генной терапии или для лечения MPS.
В пятом аспекте изобретение относится к способу получения экспрессионных векторов по первому и второму аспектам изобретения.
В шестом аспекте изобретение относится к способу получения фармацевтических композиций по третьему аспекту изобретения.
В седьмом аспекте изобретение относится к способу лечения индивидуума, страдающего мукополисахаридозом типа IIIA, по первому, второму и третьему аспектам изобретения.
Настоящее изобретение также относится к применению нуклеотидной последовательности, генетической конструкции, экспрессионного вектора или фармацевтических композиций по изобретению для получения лекарства с целью повышения сульфамидазной активности в организме, для замещающей терапии ферментом, для генной терапии или для лечения мукополисахаридозов или мукополисахаридоза типа IIIA.
Краткое описание чертежей
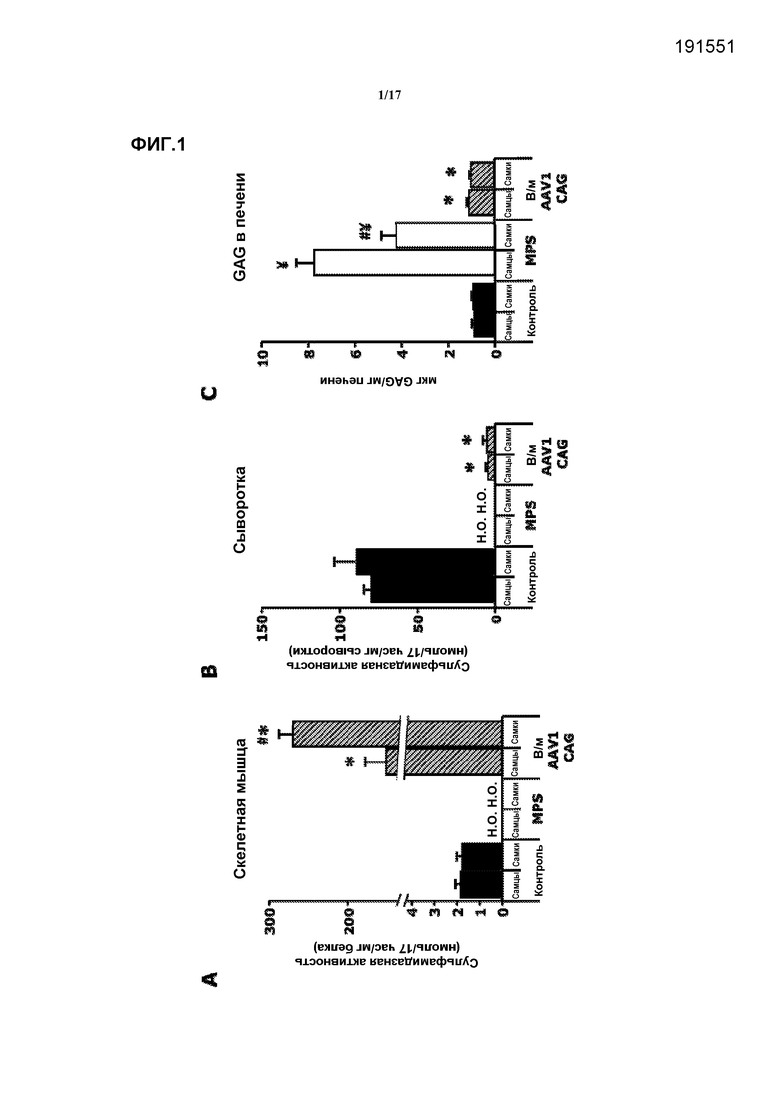
Фиг. 1. Внутримышечная доставка AAV1-CAG-mu-SFMD-WPRE. (A) Сульфамидазная активность в скелетной мышце у мышей в контроле, при MPS и в результате лечения. (B) Сульфамидазная активность в сыворотке у мышей в контроле, при MPS и в результате лечения. (C) Количественное определение гликозаминогликана (GAG) в печени у мышей в контроле, при MPS и в результате лечения. Величины представляют собой среднее ± ст. ош. средн. у от 4 до 8 мышей на группу. ¥ P<0,05 относительно контроля, # P<0,05 относительно самцов, * P<0,05 относительно MPS без лечения. Н.О.: не определяли.
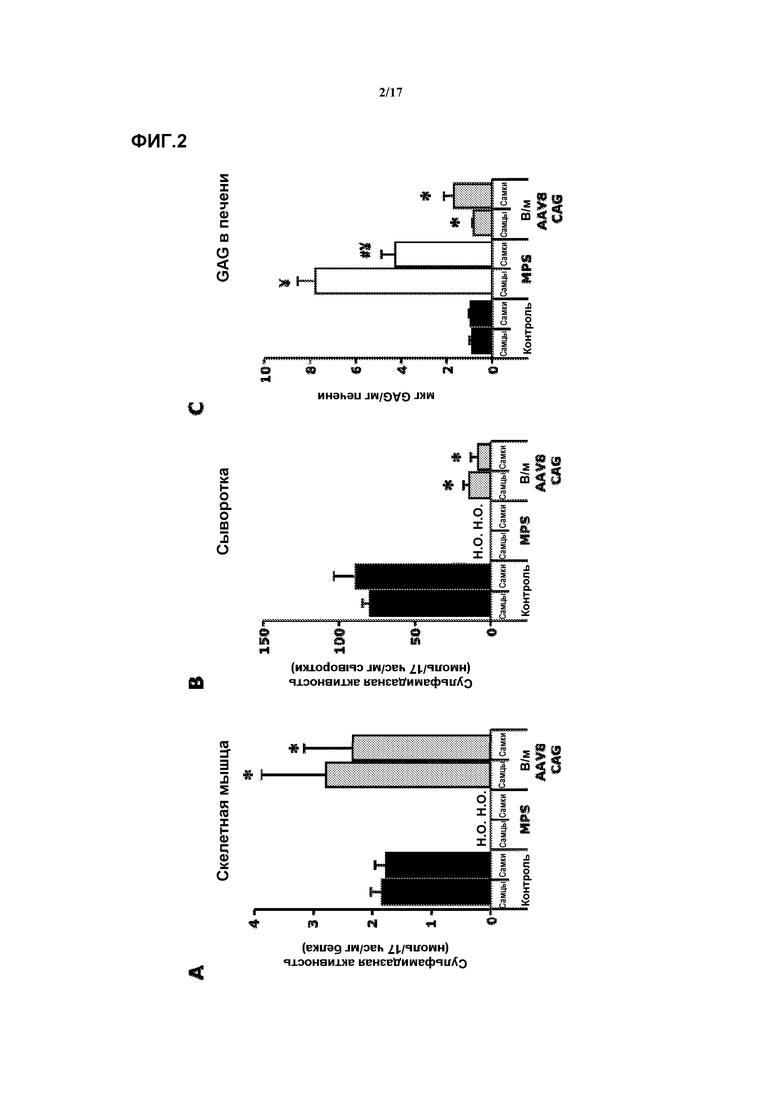
Фиг. 2. Внутримышечная доставка AAV8-CAG-mu-SFMD-WPRE. (A) Сульфамидазная активность в скелетной мышце у мышей в контроле, при MPS и в результате лечения. (B) Сульфамидазная активность в сыворотке у мышей в контроле, при MPS и в результате лечения. (C) Количественное определение гликозаминогликана (GAG) в печени у мышей в контроле, при MPS и в результате лечения. Величины представляют собой среднее ± ст. ош. средн. у от 4 до 8 мышей на группу. ¥ P<0,05 относительно контроля, # P<0,05 относительно самцов, * P<0,05 относительно MPS без лечения. Н.О.: не определяли.
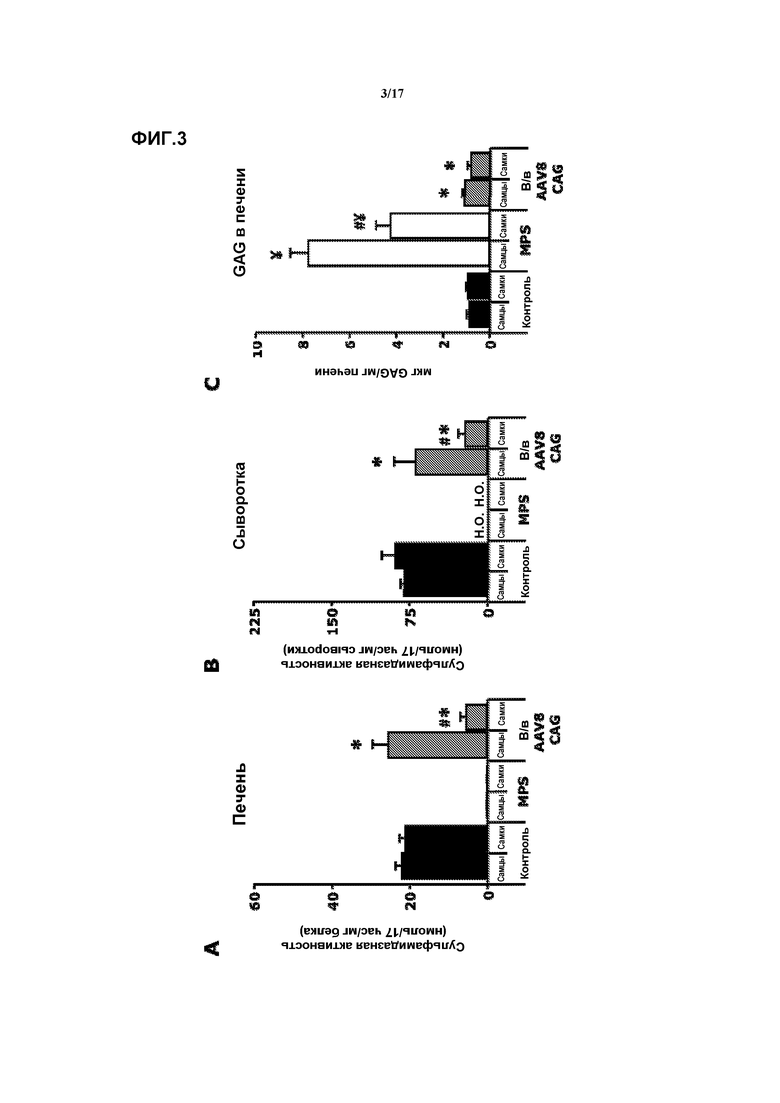
Фиг. 3. Внутривенная доставка AAV8-CAG-mu-SFMD-WPRE. (A) Сульфамидазная активность в печени у мышей в контроле, при MPS и в результате лечения. (B) Сульфамидазная активность в сыворотке у мышей в контроле, при MPS и в результате лечения. (C) Количественное определение гликозаминогликана (GAG) в печени у мышей в контроле, при MPS и в результате лечения. Величины представляют собой среднее ± ст. ош. средн. у от 4 до 8 мышей на группу. ¥ P<0,05 относительно контроля, # P<0,05 относительно самцов, * P<0,05 относительно MPS без лечения. Н.О.: не определяли.
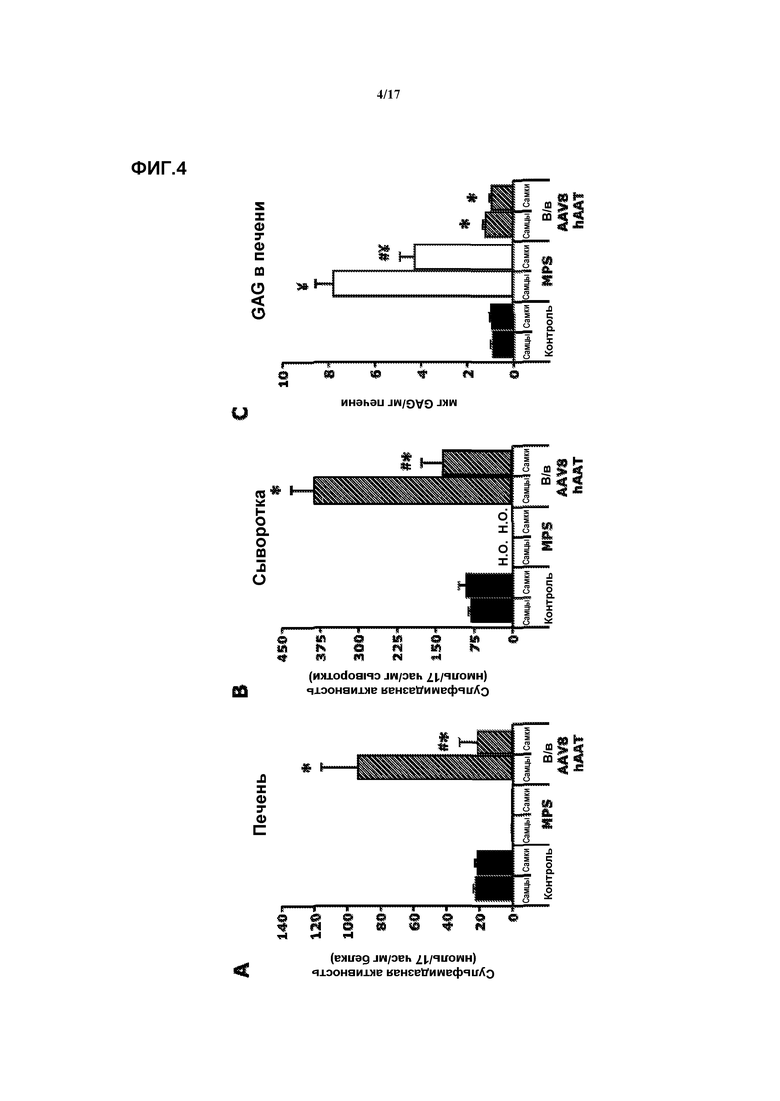
Фиг. 4. Внутривенная доставка AAV8-hAAT-mu-SFMD. (A) Сульфамидазная активность в печени у мышей в контроле, при MPS и в результате лечения. (B) Сульфамидазная активность в сыворотке у мышей в контроле, при MPS и в результате лечения. (C) Количественное определение гликозаминогликана (GAG) в печени у мышей в контроле, при MPS и в результате лечения. Величины представляют собой среднее ± ст. ош. средн. у от 4 до 8 мышей на группу. ¥ P<0,05 относительно контроля, # P<0,05 относительно самцов, * P<0,05 относительно MPS без лечения. Н.О.: не определяли.
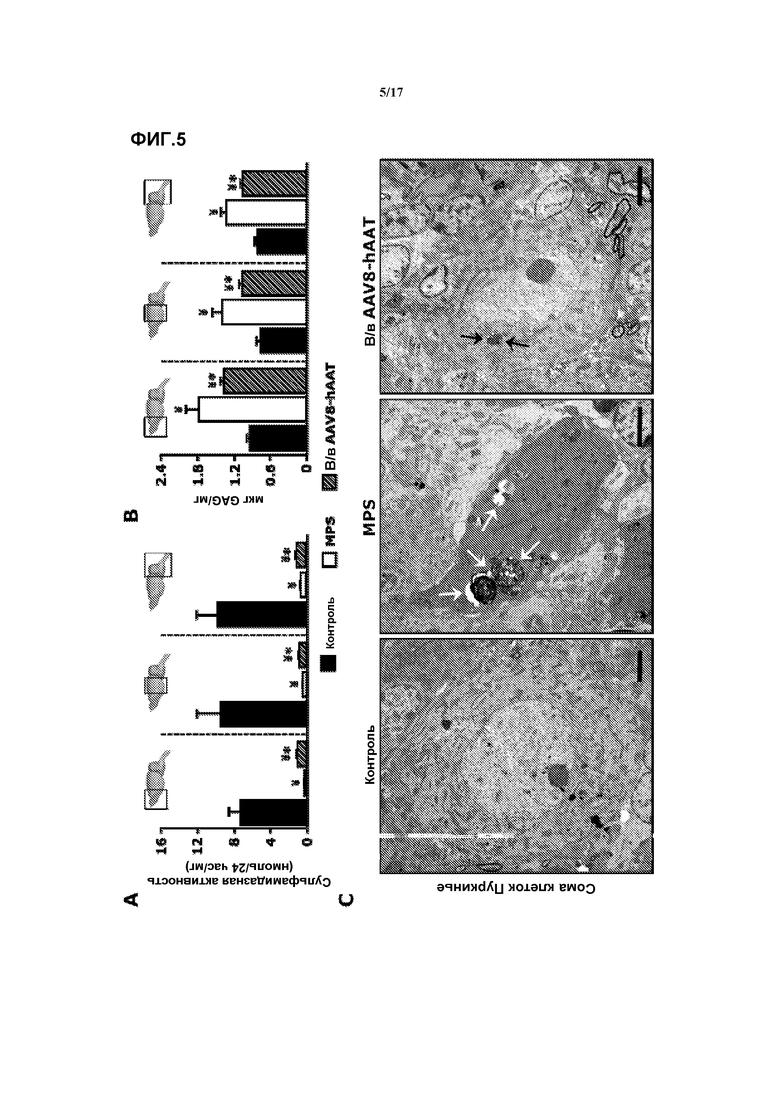
Фиг. 5. Улучшение неврологической патологии у мышей MPSIIIA в результате внутривенной доставки AAV8-hAAT-mu-SFMD. (A) Сульфамидазная активность в различных отделах мозга (изображены на диаграмме) у самцов в контроле, при MPS и в результате лечения. (B) Количественное определение гликозаминогликана (GAG) в тех же отделах мозга. Величины представляют собой среднее ± ст. ош. средн., у от 4 до 8 мышей на группу. ¥ P<0,05 относительно контроля, * P<0,05 относительно MPS без лечения. ND: не определяли. (C) Трансмиссионная электронная микроскопия клеток Пуркинье в мозжечке. Тела нейронов Пуркинье мышей с MPSIIIA без лечения были заполнены множественными электронно-плотными включениями (белые стрелки), тогда как у самцов, которых лечили в/в-AAV8-hAAT, было найдено меньше включений более мелкого размера (черные стрелки).
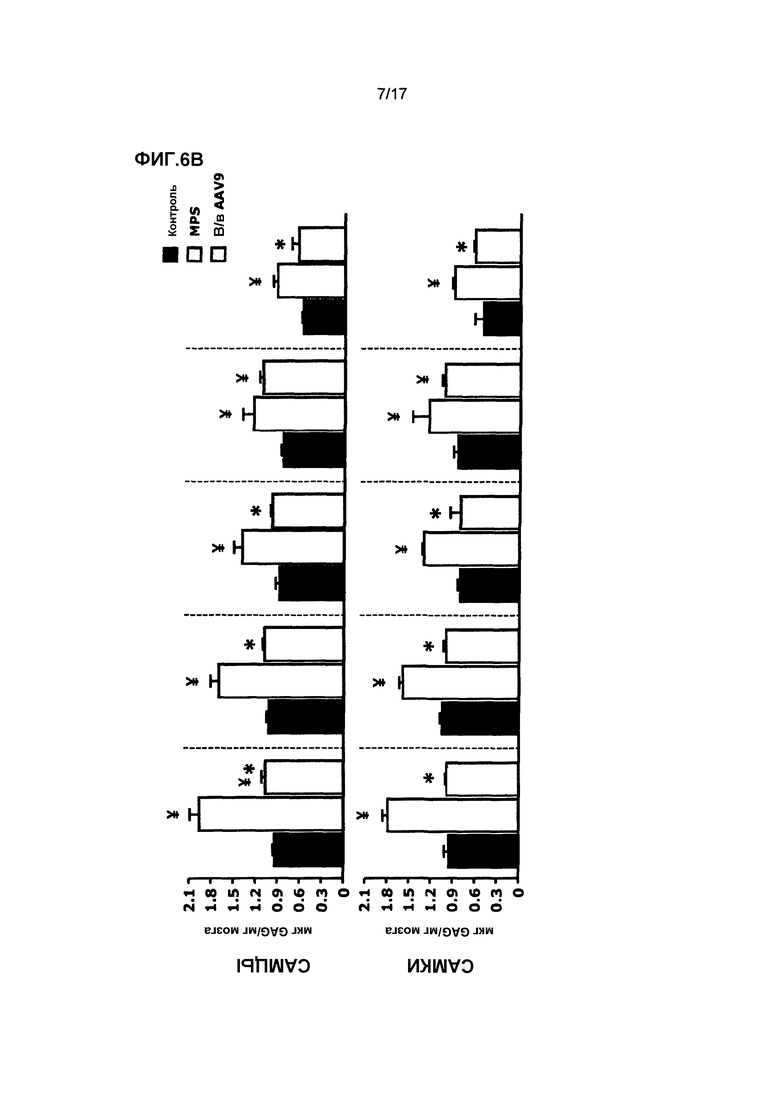
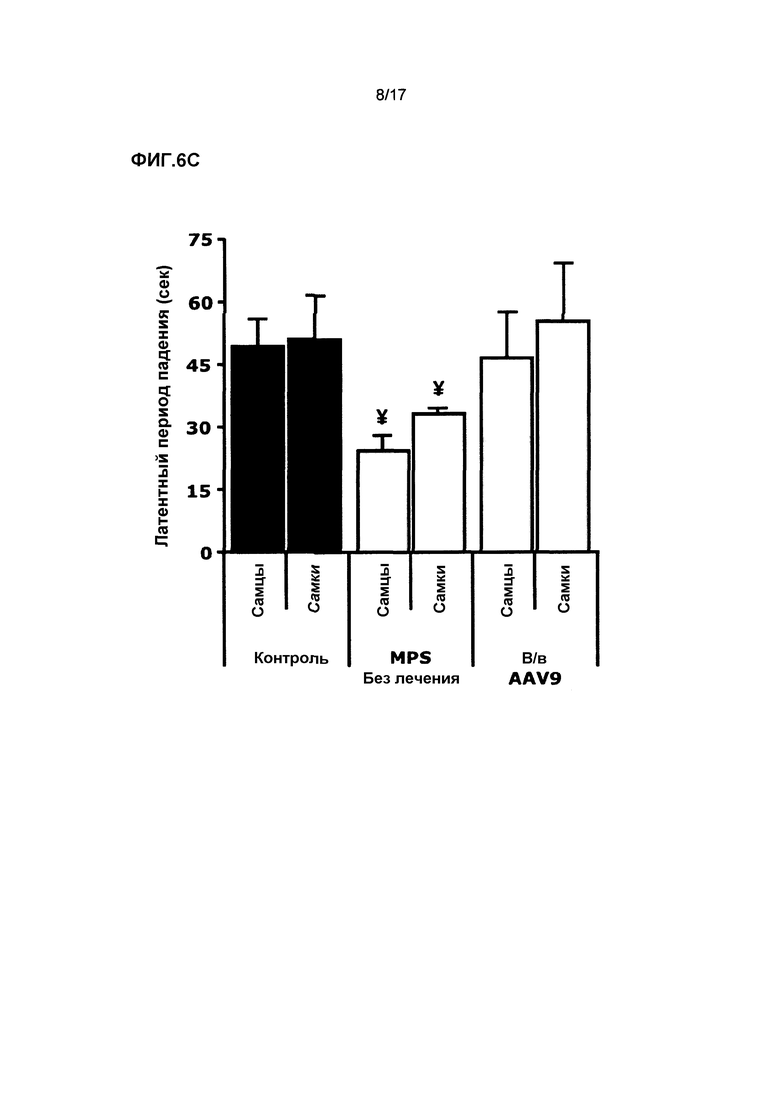
Фиг. 6. Внутривенная доставка AAV9-CAG-mu-SFMD. (A) Сульфамидазная активность в различных отделах мозга (изображены на диаграмме) у мышей в контроле, при MPS и в результате лечения. (B) Количественное определение гликозаминогликана (GAG) в тех же отделах мозга. (C) Оценка моторной функции с помощью теста с вращающимся с ускорением стержня. Величины представляют собой среднее ± ст. ош. средн. у от 4 до 8 мышей на группу. ¥ P<0,05 относительно контроля, * P<0,05 относительно MPS без лечения.
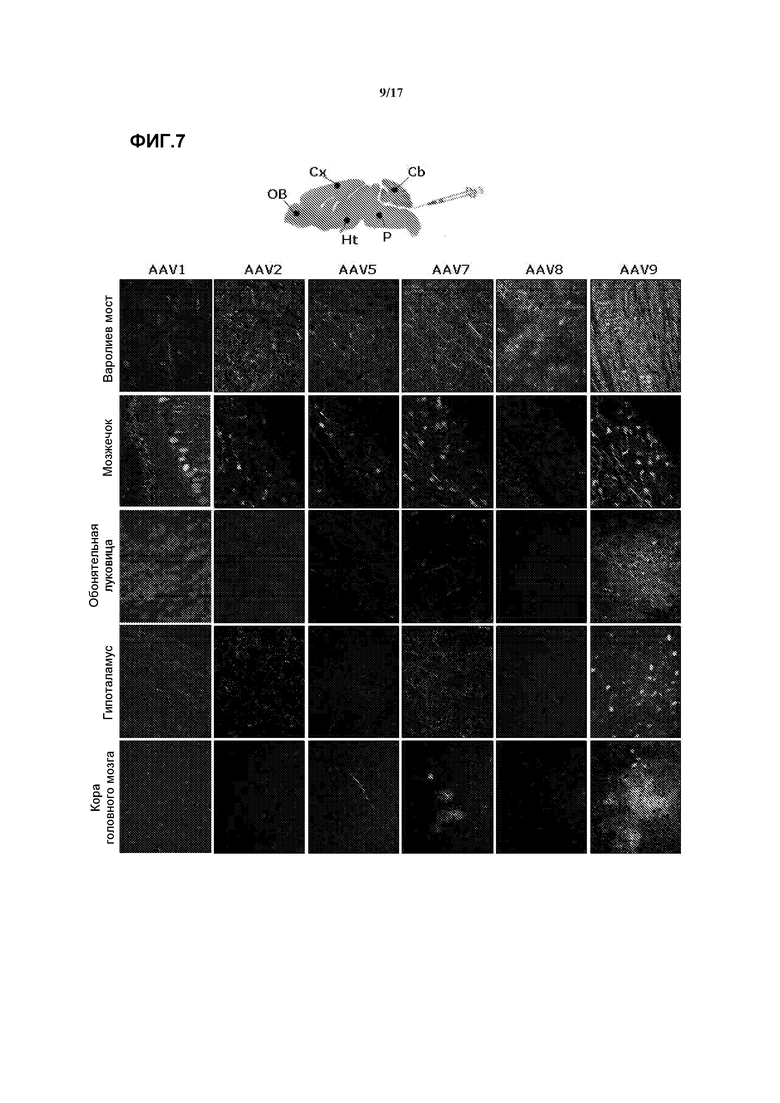
Фиг. 7. Трансдукция в мозг после внутрицистернальной доставки аденоассоциированных вирусных векторов GFP серотипов 1, 2, 5, 7, 8 и 9. Дозу 5×1010 векторных геномов соответствующего вектора вводили внутрицистернально животным 2-месячного возраста, которых через 2 недели забивали и подвергали анализу. Аденоассоциированный вирус серотипа 9 продемонстрировал самую высокую эффективность трансдукции во всех анализируемых областях. Шприц указывает путь доставки вектора, cisterna magna. P: варолиев мост, Cb: мозжечок, OB: обонятельная луковица, Ht: гипоталамус, Cx: кора мозга.
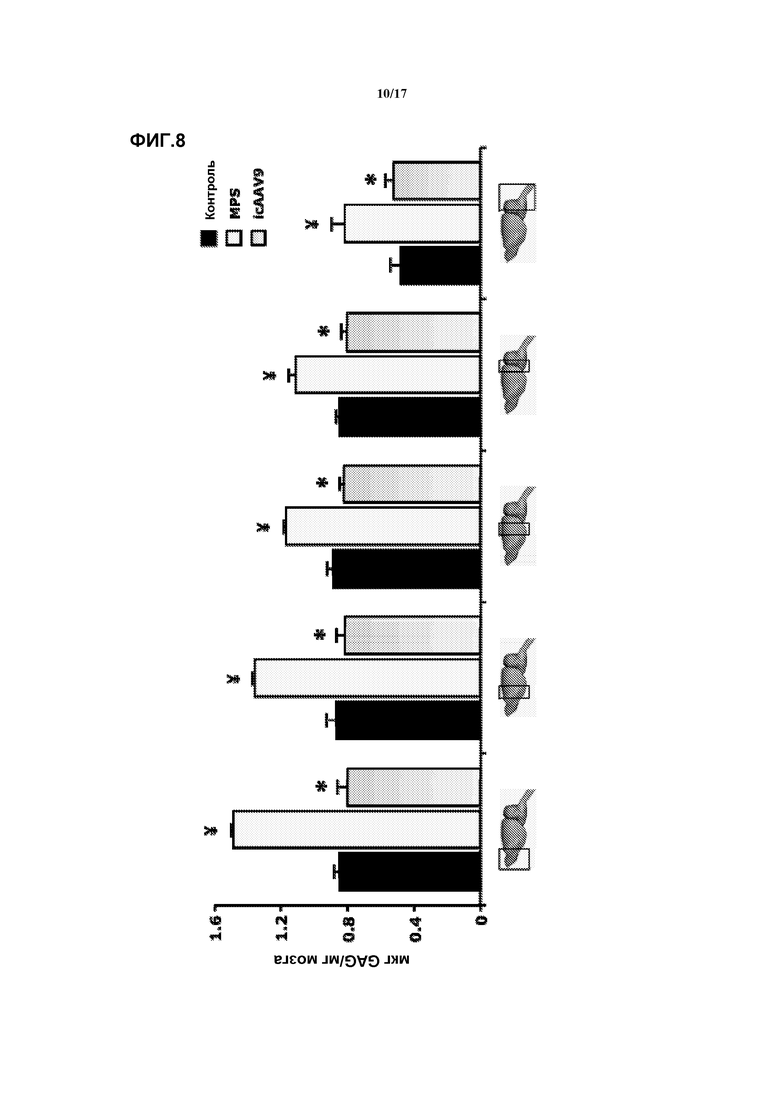
Фиг. 8. Внутрицистернальная (ic) доставка векторов AAV9-CAG-mu-SFMD. Количественное определение гликозаминогликана (GAG) в различных отделах мозга (изображены на диаграмме) у мышей в контроле, при MPS и в результате лечения. Величины представляют собой среднее ± ст. ош. средн. у 3 мышей на группу. ¥ P<0,05 относительно контроля, * P<0,05 относительно MPS без лечения.
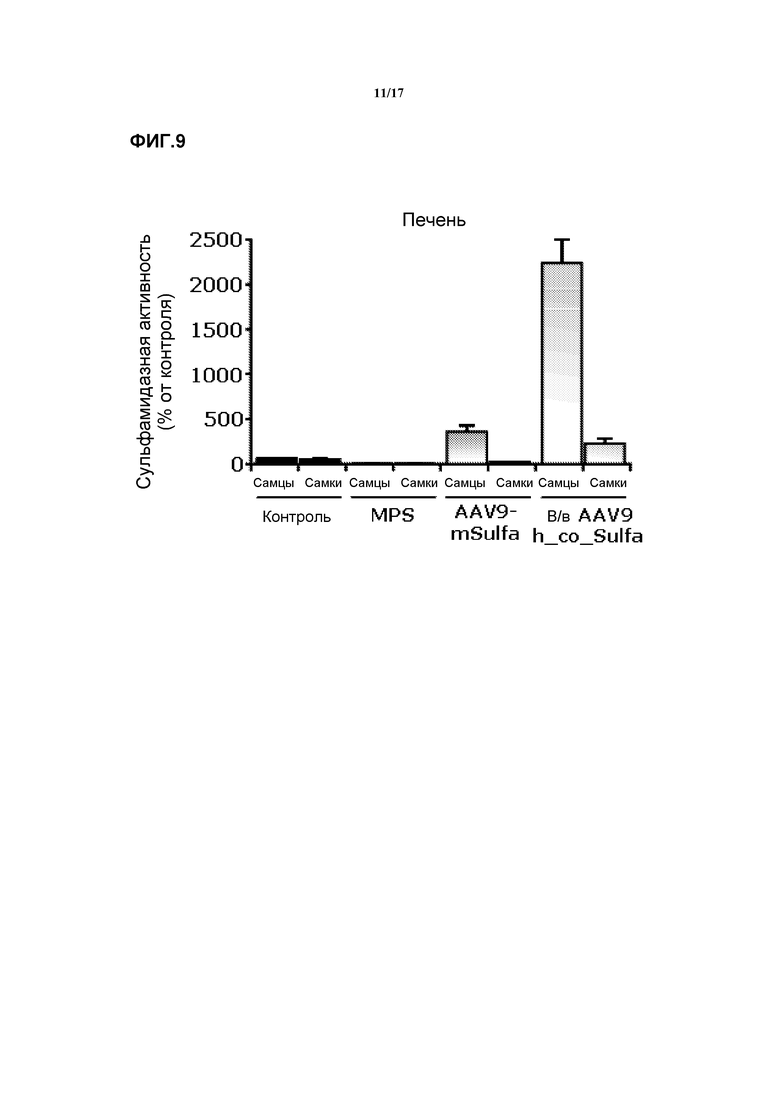
Фиг. 9. Внутривенная доставка AAV9-CAG-hu-co-SFMD. Сульфамидазная активность в печени у контрольных мышей, мышей с MPS и мышей, которых лечили либо AAV9-CAG-mu-SFMD (неоптимизированным геном), либо AAV9-CAG-hu-co-SFMD (оптимизированным геном).
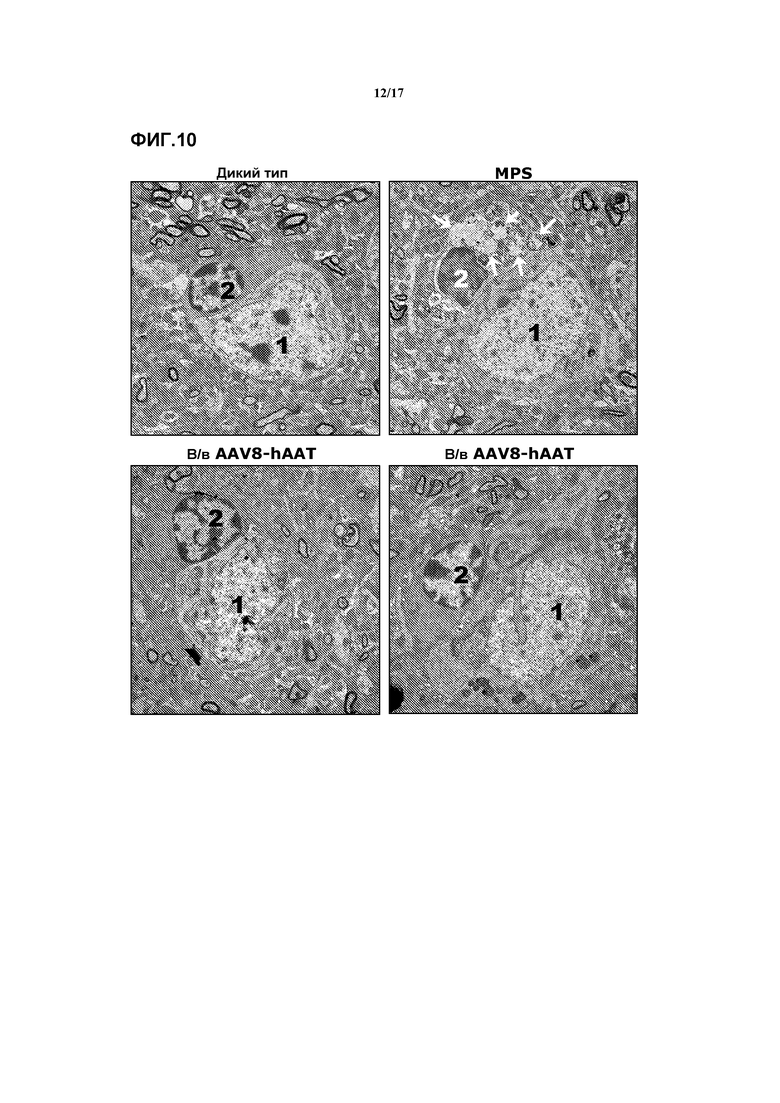
Фиг. 10. Снижение патологии лизосом у окружающих нейроны глиальных клеток затылочной коры. Трансмиссионная электронная микроскопия, изображающая кортикальные нейроны затылочной коры и ассоциированные с ними глиальные клетки. Лизосомная патология была намного сильнее выражена в окружающих нейроны глиальных клетках по сравнению с нейронами. Показано присутствие больших электронно-прозрачных вакуолей в образцах глиальных клеток от мышей MPSIIIA, не подвергавшихся лечению (белые стрелки, правая верхняя панель), и их отсутствие в образцах дикого типа (левая верхняя панель). Это увеличение лизосомного компартмента было существенно снижено у мышей, которых лечили в/в-AAV8hAAT, и большинство окружающих нейроны глиальных клеток в этих образцах имели вид, сходный с видом у дикого типа (нижние панели). (1) нейрон, (2) окружающая нейрон глиальная клетка.
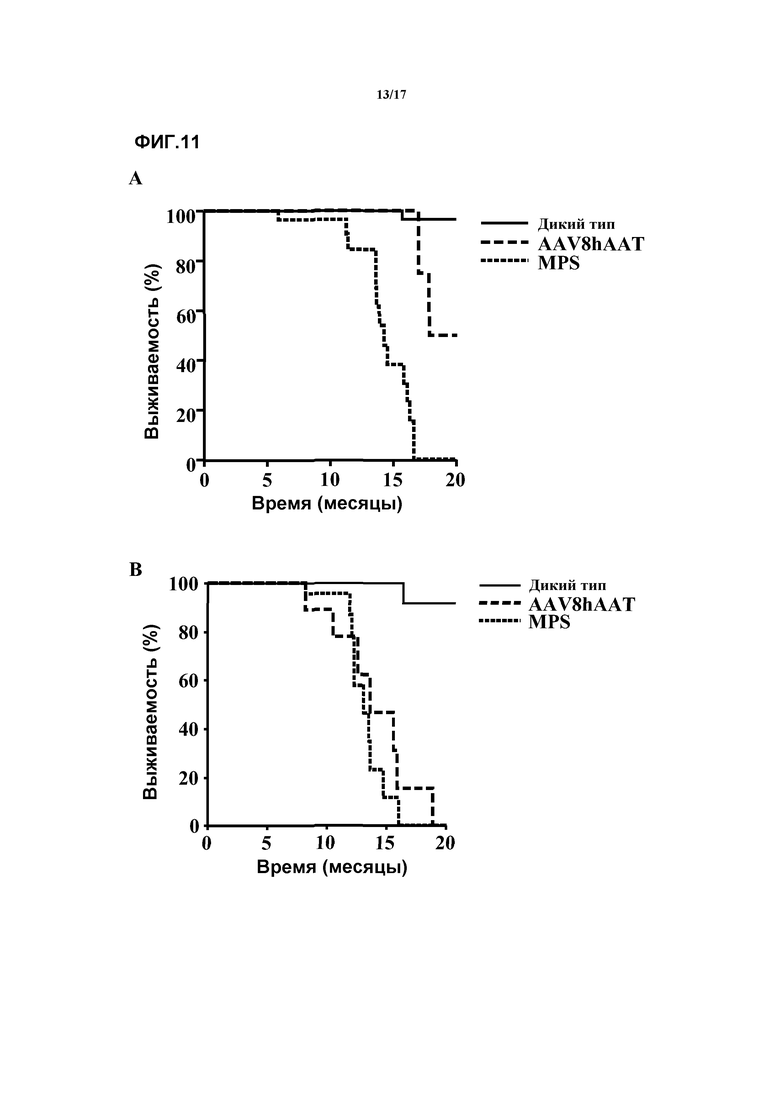
Фиг. 11. Выживаемость у самцов и самок, которых лечили внутривенной доставкой AAV8-hAAT-SFMD. (A) Анализ с помощью кривых выживаемости Каплана-Мейера для самцов дикого типа, самцов с MPSIIIA и самцов, которых лечили с помощью внутривенной доставки AAV8-hAAT-SFMD. Лечение с помощью направленной на печень генной терапии, опосредуемой AAV, значительно увеличивало продолжительность жизни животных с MPSIIIA (p<0,001). (B) Анализ с помощью кривых выживаемости Каплана-Мейера для самок дикого типа, самок с MPSIIIA и самок, которых лечили в/в-AAV8-hAAT-SFMD. Лечение с помощью направленной на печень генной терапии, опосредуемой AAV, не увеличивало продолжительность жизни животных с MPSIIIA (p=0,467).
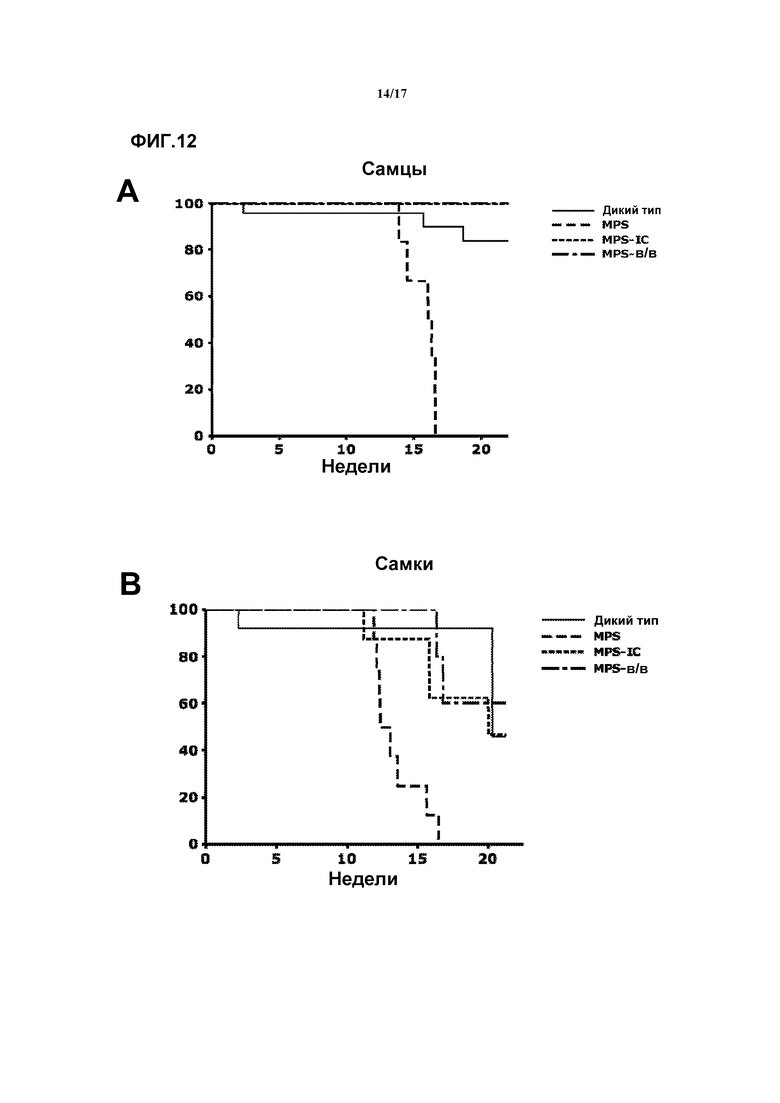
Фиг. 12. Выживаемость у самцов и самок, которых лечили внутрицистернальной (ic) и внутривенной доставкой AAV9-CAG-mu-SFMD. (A) Анализ с помощью кривых выживаемости Каплана-Мейера самцов (A) и самок (B) дикого типа, с MPSIIIA и получавших лечение AAV9. Как внутрицистернальное, так и внутривенное лечение с помощью опосредуемой AAV генной терапии увеличивало продолжительность жизни животных с MPSIIIA.
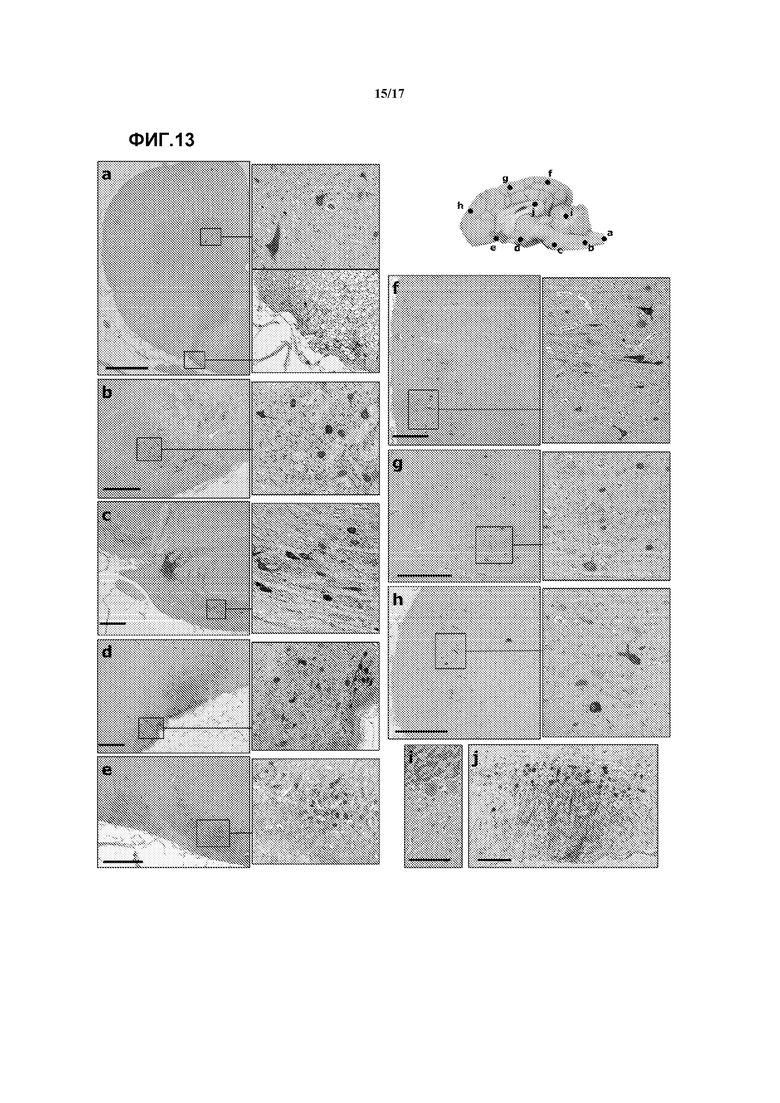
Фиг. 13. Внутрицистернальное введение векторов AAV9 собакам приводит к трансдукции широко распространенных областей ЦНС и печени. Иммуногистохимическое определение GFP на срезах ЦНС и печени собаки, которой вводили AAV9-GFP через cisterna magna. Изображения соответствуют: (a) спинному мозгу, (b) оливе продолговатого мозга, (c) ядрам шва варолиева моста, (d) гипоталамическим ядрам, (e) обонятельному мозгу, (f) затылочной коре, (i) мозжечку, (j) зубчатой извилине гиппокампа. Отрезок масштаба: 1 мм для (a), 500 мкм для (b)-(h), 100 мкм для (i)-(j).
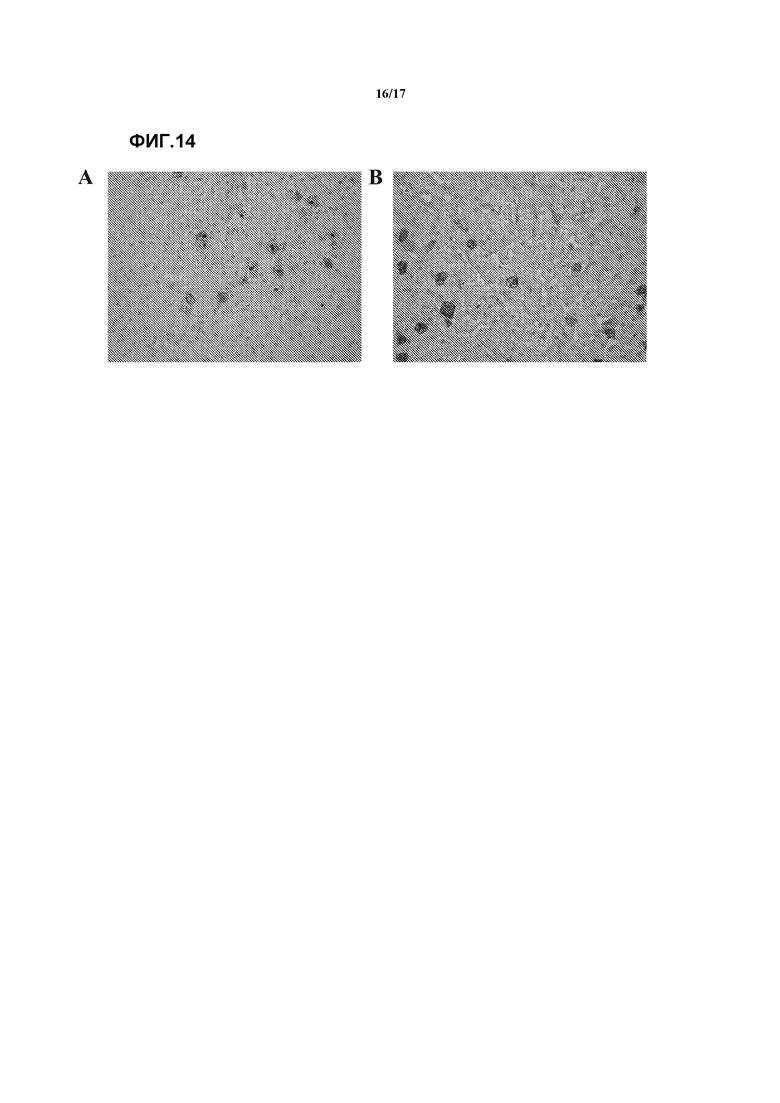
Фиг. 14. Трансдукция в печень после внутрицистернальной доставки вектора AAV9-GFP здоровым собакам породы Бигль. Иммуногистохимическое определение GFP на срезах печени, подкрашенных гематоксилином. Показаны репрезентативные изображения у собаки 1 (A) и собаки 2 (B). Исходное увеличение 200Х.
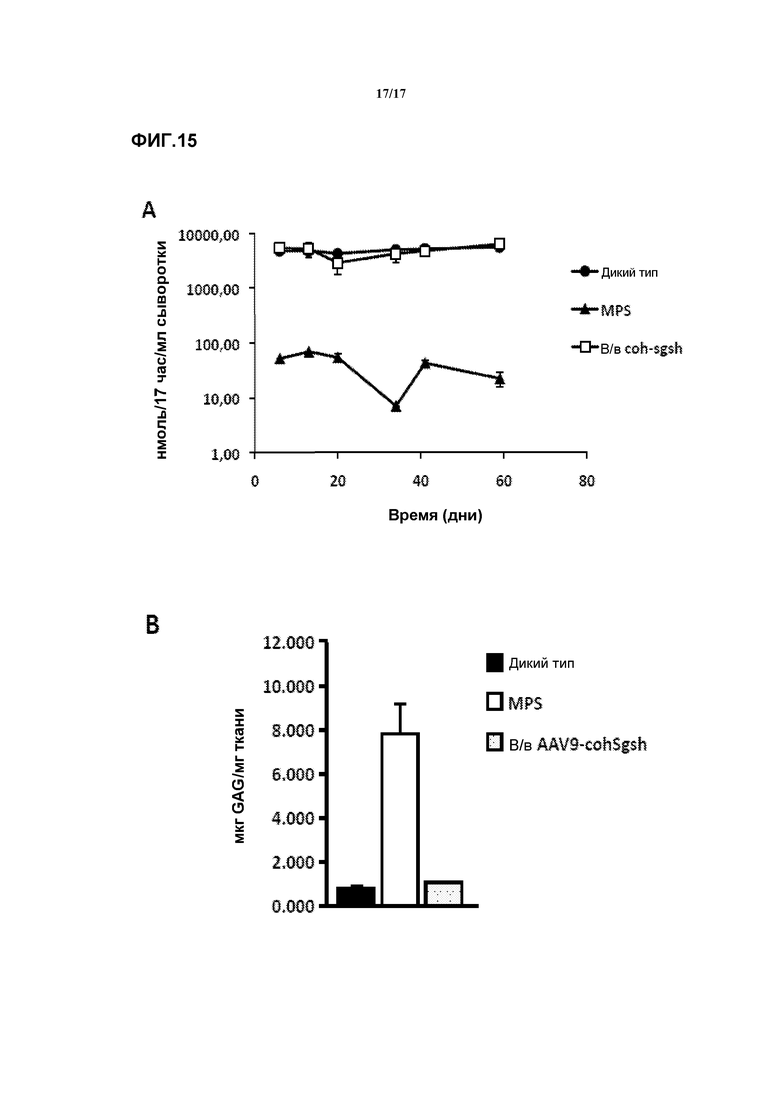
Фиг. 15. Сульфамидазная активность сыворотки и содержание GAG в печени у животных с внутривенным введением AAV9-co-hu-SFMD. (A) Сульфамидазная активность в сыворотке, измеренная с флуорогенным субстратом. (B) Запасание GAG в печени через 2 месяца после введения вектора.
Депонирование микроорганизмов
Плазмида pAAV-CAG-co-hu-SFMD была депонирована 16 мая 2011 года под номером доступа DSM 24817 в DSMZ - Deutsche Sammlung von Mikroorganismen und Zellkulturen, Inhoffenstβe 7 B, D-38124 Braunschweig, Федеративная республика Германия.
Плазмида pAAV-CAG-mu-SFMD была депонирована 16 мая 2011 года под номером доступа DSM 24818 в DSMZ - Deutsche Sammlung von Mikroorganismen und Zellkulturen, Inhoffenstβe 7 B, D-38124 Braunschweig, Федеративная республика Германия.
Плазмида pGG2-hAAT-mu-SFMD была депонирована 16 мая 2011 года под номером доступа DSM 24819 в DSMZ - Deutsche Sammlung von Mikroorganismen und Zellkulturen, Inhoffenstβe 7 B, D-38124 Braunschweig, Федеративная республика Германия.
Определения
Термин «нуклеотидная последовательность» относится к молекуле нуклеиновой кислоты, либо ДНК, либо РНК, содержащей дезоксирибонуклеотиды или рибонуклеотиды. Нуклеиновая кислота может быть двухцепочечной, одноцепочечной или содержать части как двухцепочечной, так и одноцепочечной последовательности.
Термин «% идентичности последовательностей» относится к процентному содержанию нуклеотидов кандидатной последовательности, которые идентичны нуклеотидам в SEQ ID NO:1 после выравнивания последовательностей с достижением максимального % идентичности последовательностей. % идентичности последовательностей может быть определен с помощью любых методов или алгоритмов, установленных в данной области техники, таких как алгоритмы ALIGN, BLAST и BLAST 2.0. Смотрите Altschul S, et al., Nuc. Acids Res. 1977; 25:3389-3402 и Altschul S, et al., J. Mol. Biol. 1990; 215:403-410.
В настоящем описании % идентичности последовательностей рассчитывается путем деления количества нуклеотидов, которые после выравнивания идентичны SEQ ID NO:1 и кандидатной последовательности, на общее количество нуклеотидов в SEQ ID NO:1 и умножения результата на 100.
Термин «кодировать» относится к генетическому коду, который определяет как транслируется нуклеотидная последовательность в полипептид или белок. Порядок нуклеотидов в последовательности определяет порядок аминокислот в полипептиде или белке.
Термин «белок» относится к линейной цепи аминокислот или полипептиду, которые укладываются в глобулярную структуру. Белки могут претерпевать посттрансляционные модификации, такие как превращение цистеинового остатка в 3-оксоаланин, гликозилирование или связывание металла. Гликозилирование белка представляет собой добавление различных углеводов, которые ковалентно связываются с аминокислотной цепью.
Термин «эффективное количество» относится к количеству вещества, достаточному для достижения предназначенной цели. Например, эффективное количество экспрессионного вектора для повышения сульфамидазной активности представляет собой количество, достаточное для снижения аккумуляции гликозаминогликана. «Терапевтически эффективное количество» экспрессионного вектора для лечения заболевания или нарушения представляет собой количество экспрессионного вектора, достаточное для снижения или устранения симптомов заболевания или нарушения. Эффективное количество данного вещества должно варьироваться в связи с такими факторами, как природа вещества, путь введения, размер и вид животного, получающего вещество, и цель назначения вещества. Эффективное количество в каждом индивидуальном случае может быть определено эмпирически специалистом в данной области техники в соответствии с методами, принятыми в данной области техники.
Термин «индивидуум» относится к произвольному животному, предпочтительно к человеку или млекопитающему, не являющемуся человеком, более предпочтительно к мыши, крысе, другим грызунам, кролику, собаке, кошке, свинье, корове, лошади или примату, еще более предпочтительно к человеку.
Термин «функционально связанный» относится к функциональному отношению и локализации промоторной последовательности по отношению к целевому гену (например, промотор или энхансер является функционально связанным с кодирующей последовательностью, если он воздействует на транскрипцию последовательности). Обычно функционально связанный промотор примыкает к интересующей последовательности. Однако энхансер может быть не примыкающим к интересующей последовательности для контроля ее экспрессии.
Термин «тропизм» относится к пути, в котором различные вирусы эволюционировали до предпочтительной направленности на конкретные виды хозяев или конкретные типы клеток в пределах этих видов.
Термин «генная терапия» относится к переносу интересующего генетического материала (например, ДНК или РНК) в организм хозяина для лечения или предотвращения генетического или приобретенного заболевания или состояния. Интересующий генетический материал кодирует продукт (например, белок, полипептид, пептид или функционирующую РНК), продукция которого in vivo желательна. Например, интересующий генетический материал может кодировать фермент, гормон, рецептор или полипептид, имеющий терапевтическую ценность.
Термин «промотор CAG» относится к сочетанию, образованному ранним энхансерным элементом цитомегаловируса и промотором β-актина цыпленка (т.е. SEQ ID NO:3). Смотрите Alexopoulou A, et al., BMC Cell Biology 2008; 9(2): 1-11.
Термин «промотор hATT» относится к промотору альфа1-антитрипсина человека (т.е. SEQ ID NO:4). Смотрите Hafenrichter H, et al., Blood 1994; 84: 3394-3404.
Термин «вирусная векторная частица» относится к генетически модифицированному вирусу, используемому для доставки генов в организм. Вирусные векторные частицы несут вирусный геном. Вирусный геном включает нуклеотидную последовательность, которая локализована между ITR в экспрессионном векторе, используемом для продукции вирусных векторных частиц. Аденоассоциированные вирусные векторные частицы называются AAV.
Термин «вектор AAV» относится к аденоассоциированным вирусным векторным частицам.
Подробное описание изобретения
В предпочтительном варианте осуществления последовательность по первому аспекту изобретения идентична по меньшей мере на 85% последовательности SEQ ID NO:1, которая кодирует белок SEQ ID NO:2. Предпочтительно идентичность последовательностей составляет по меньшей мере 87%. Более предпочтительно идентичность последовательностей составляет по меньшей мере 90%. Еще более предпочтительно идентичность последовательностей составляет по меньшей мере 95%. Еще более предпочтительно идентичность последовательностей составляет по меньшей мере 98%. Наиболее предпочтительно идентичность последовательностей составляет по меньшей мере 99%. В более предпочтительном варианте осуществления последовательность по первому аспекту изобретения представляет собой нуклеотидную последовательность SEQ ID NO:1. В другом варианте осуществления изобретение относится к нуклеотидной последовательности SEQ ID NO:1 или биологически активному варианту этой последовательности. Биологически активный вариант включает молекулу, обладающую той же самой биологической активностью, что и SEQ ID NO:1 и по меньшей мере 85% идентичностью последовательностей. Биологическая активность обозначает тот факт, что нуклеотидная последовательность SEQ ID NO:1 может транскрибироваться в матричную РНК, которая обладает повышенной стабильностью и, следовательно, дает высокие скорости трансляции, в результате чего делает возможным экспрессию высоких уровней активной сульфамидазы человека.
В предпочтительном варианте осуществления второго аспекта генетическая конструкция включает нуклеотидную последовательность, идентичную последовательности SEQ ID NO:1 по меньшей мере на 85%, предпочтительно на 87%, 90%, 95%, 98%, 99%. В более предпочтительном варианте осуществления генетическая конструкция включает нуклеотидную последовательность SEQ ID NO:1. Генетическая конструкция представляет собой молекулу нуклеиновой кислоты, в которой различные элементы сконструированы конкретным и желаемым образом. Эти элементы могут представлять собой, среди прочего, реплицирующиеся последовательности, контролирующие последовательности, кодирующие последовательности, мультиклонирующие последовательности или рекомбинантные последовательности. В предпочтительном варианте осуществления генетическая конструкция представляет собой вектор. Вектор представляет собой молекулу нуклеиновой кислоты, используемую для переноса генетического материала в клетку. Кроме указанного генетического материала вектор может также содержать различные функциональные элементы, которые включают, элементы, контролирующие транскрипцию, такие как промоторы или операторы, области связывания транскрипционных факторов или энхансеры, и контролирующие элементы для инициации или терминации трансляции. Векторы включают, но не ограничиваются этим: плазмиды, космиды, вирусы, фаги, рекомбинантные экспрессионные кассеты и транспозоны. Аденоассоциированные векторы (AAV) представляют собой вирусные векторные частицы, следовательно, они не являются молекулой нуклеиновой кислоты, но представляют собой генетически модифицированный вирус, используемый для доставки генов в организм.
В предпочтительном варианте осуществления второго аспекта изобретения генетическая конструкция представляет собой вектор, который используется для трансляции и транскрипции интересующего гена, обычно контролируемого промотором. Промотор представляет собой нуклеотидную последовательность, которая контролирует трансляцию интересующего гена. Промотор функционально связан с интересующим геном.
Другим предпочтительным вектором является аденоассоциированный вектор. В предпочтительном варианте осуществления аденоассоциированный вектор используется для продукции аденоассоциированных частиц на основе серотипов 1, 2, 5, 7, 8 или 9. В более предпочтительном варианте осуществления серотип представляет собой серотип 9. Аденоассоциированный вектор представляет собой вектор, происходящий из генома аденоассоциированного вируса (AAV) семейства Parvoviridae. Геном AAV построен из одноцепочечной дезоксирибонуклеиновой кислоты (ssDNA). AAV инфицирует людей, но является непатогенным (т.е. не вызывает заболевания). Эти вирусы могут инфицировать делящиеся и неделящиеся клетки, и их тропизм изменяется в зависимости от серотипа. Под серотипом понимается подразделение вирусов на группы в зависимости от их капсидных антигенов. Серотип AAV, определенный по его капсидным белкам, ограничивает тропизм вируса и позволяет ему входить в конкретный тип клеток. Продукция аденоассоциированных вирусных частиц описана ниже.
В первом предпочтительном варианте осуществления второго аспекта экспрессионный вектор включает промотор CAG, функционально связанный с SEQ ID NO:1.
Предпочтительный вектор представляет собой экспрессионный вектор, включающий промотор CAG, причем последовательность промотора представляет собой SEQ ID NO:3. Следовательно, в одном варианте осуществления второго аспекта изобретения предлагается экспрессионный вектор, включающий промотор CAG, причем последовательность промотора представляет собой SEQ ID NO:3, что подходит для лечения MPS.
Во втором предпочтительном варианте осуществления второго аспекта экспрессионный вектор включает специфичный для печени промотор hAAT, функционально связанный с SEQ ID NO:1.
Предпочтительный вектор представляет собой экспрессионный вектор, включающий специфичный для печени промотор hAAT, причем последовательность промотора представляет собой SEQ ID NO:4. Следовательно, в одном варианте осуществления второго аспекта изобретения предлагается экспрессионный вектор, включающий специфичный для печени промотор hAAT, причем последовательность промотора представляет собой SEQ ID NO:4, что подходит для лечения MPS.
Другой аспект настоящего изобретения относится к вирусной векторной частице, также называемой экспрессионным вектором, которая несет нуклеотидные последовательности по первому аспекту изобретения или генетическую конструкцию, или экспрессионный вектор по второму аспекту изобретения.
Предпочтительный экспрессионный вектор имеет серотип 1, 2, 5, 7, 8 или 9. Более предпочтительная вирусная векторная частица имеет серотип 9.
Предпочтительный экспрессионный вектор имеет серотип 9 и включает вирусный геном, включающий промотор CAG, функционально связанный с SEQ ID NO:1.
Предпочтительный экспрессионный вектор имеет серотип 8 или 9 и включает вирусный геном, включающий промотор hAAT, функционально связанный с SEQ ID NO:1.
Предпочтительный экспрессионный вектор имеет серотип 9 и включает вирусный геном, включающий промотор hAAT, функционально связанный с SEQ ID NO:1.
В предпочтительном варианте осуществления экспрессионный вектор представляет собой AAV-CAG-co-hu-SFMD и более предпочтительно AAV9-CAG-co-hu-SFMD.
В еще одном предпочтительном варианте осуществления экспрессионный вектор представляет собой AAV-hAAT-co-hu-SFMD и более предпочтительно AAV8-hAAT-co-hu-SFMD или pAAV9-hAAT-co-hu-SFMD. Наиболее предпочтительный вектор, используемый, когда применяется промотор hAAT, представляет собой AAV9-hAAT-co-hu-SFMD.
В предпочтительном варианте осуществления третьего аспекта фармацевтическую композицию вводят парентеральным путем введения. Парентеральное введение относится к пути введения фармацевтической композиции в виде инъекции или инфузии. Примерами парентерального введения являются внутривенное, подкожное, внутрицистернальное и внутримышечное введение. Предпочтительно фармацевтическую композицию вводят с помощью внутривенного или внутрицистернального пути введения.
В другом предпочтительном варианте осуществления фармацевтическая композиция включает терапевтически эффективное количество нуклеотидной последовательности, генетической конструкции, вирусной векторной частицы или экспрессионного вектора по изобретению.
В предпочтительном варианте осуществления четвертого аспекта нуклеотидную последовательность, генетическую конструкцию, экспрессионный вектор, вирусную векторную частицу или фармацевтическую композицию по изобретению используют в качестве лекарства. В предпочтительном варианте осуществления они используются для повышения сульфамидазной активности в организме.
В другом предпочтительном варианте осуществления нуклеотидную последовательность, генетическую конструкцию, экспрессионный вектор, вирусную векторную частицу или фармацевтическую композицию по изобретению используют в качестве лекарства для замещения фермента или в качестве генной терапии, предпочтительно в качестве генной терапии. Изобретатели предлагают новый подход к генной терапии для терапевтического лечения MPSIIIA, который является более эффективным, чем другие, известные в данной области техники. Этот подход основывается на векторах AAV, экспрессирующих сульфамидазу. Замещающая терапия ферментом (ERT) представляет собой терапевтическое лечение, которое состоит в замещении фермента больным, у которых конкретного фермента недостаточно или он отсутствует. Фермент обычно продуцируется в виде рекомбинантного белка и вводится больному.
В дополнительном варианте осуществления нуклеотидную последовательность, генетическую конструкцию, экспрессионный вектор, вирусные векторные частицы или фармацевтическую композицию по изобретению предпочтительно используют для лечения мукополисахаридозов, более предпочтительно мукополисахаридоза типа III или синдрома Санфилиппо, предпочтительно с помощью генной терапии. В пределах синдрома мукополисахаридоза типа III подтип A особенно поддается реакции на лечение по настоящему изобретению.
В предпочтительном варианте осуществления пятого аспекта заявлен способ получения экспрессионных векторов по изобретению. Способ включает стадии:
i) получения первого вектора, включающего SEQ ID NO:1, вставленную между первым концевым повтором AAV и вторым концевым повтором AAV, промотора CAG или hAAT, функционально связанного с SEQ ID NO:1; второго вектора, включающего AAV rep ген и AAV cap ген; и третьего вектора, включающего ген со вспомогательной аденовирусной функцией;
ii) совместного трансфицирования компетентных клеток векторами стадии i);
iii) культивирования трансфицированных клеток стадии ii); и
iv) очистки экспрессионных векторов из культуры стадии iii).
В предпочтительном варианте осуществления первый и второй концевые повторы AAV первого вектора представляют собой ITR от AAV серотипа 2. В другом предпочтительном варианте осуществления AAV rep гены второго вектора взяты от AAV серотипа 2. В еще одном предпочтительном варианте осуществления AAV cap гены второго вектора взяты от AAV серотипов 1, 2, 5, 7, 8 и 9. Более предпочтительно AAV cap гены второго вектора взяты от AAV серотипа 9. В другом предпочтительном варианте осуществления компетентные клетки представляют собой клетки HEK293.
Вирусные векторы вводят в достаточных количествах для трансфекции клеток и для получения достаточных уровней переноса и экспрессии гена, достаточной для предоставления терапевтического преимущества без чрезмерных неблагоприятных эффектов или с терапевтически приемлемыми физиологическими эффектами, которые могут быть определены специалистами в данной области медицины.
В предпочтительном варианте осуществления шестого аспекта заявлен способ получения фармацевтических композиций по изобретению. Этот способ включает объединение любых нуклеотидных последовательностей, генетических конструкций, вирусных векторных частиц или экспрессионных векторов по изобретению и фармацевтически приемлемого наполнителя или носителя для облегчения введения с получением фармацевтических композиций по изобретению. Носитель представляет собой, например, воду или забуференный физиологический раствор с консервантом или без него. Фармацевтические композиции могут быть лиофилизованными для ресуспендирования в момент введения или в виде раствора.
В предпочтительном варианте осуществления седьмого аспекта заявлен способ лечения индивидуума, страдающего мукополисахаридозом типа IIIA, нуклеотидными последовательностями, генетическими конструкциями, вирусными векторными частицами или экспрессионными векторами, или фармацевтическими композициями по изобретению. Схемы и дозы вводимых нуклеотидных последовательностей, генетических конструкций, векторов, экспрессионных векторов или фармацевтических композиций по настоящему изобретению могут быть определены в соответствии с протоколами дозировок, известными в данной области техники. В предпочтительном варианте осуществления нуклеотидные последовательности, генетические конструкции, вирусные векторные частицы, экспрессионные векторы или фармацевтические композиции по настоящему изобретению вводят однократно.
В одном дополнительном варианте осуществления терапевтическое лечение путем генной терапии MPS с помощью фармацевтической композиции состоит в парентеральном введении экспрессионного вектора, включающего нуклеотидную последовательность, идентичную на 90% последовательности SEQ ID NO:1.
В другом дополнительном варианте осуществления вирусный вектор, включающий промотор CAG и нуклеотидную последовательность, идентичную на 95% последовательности SEQ ID NO:1, используют для генной терапии при лечении заболевания, касающегося хранения в лизосомах (LSD), при введении путем внутримышечной инъекции.
В другом дополнительном варианте осуществления вектор AAV серотипа 1, включающий промотор CAG и нуклеотидную последовательность, идентичную на 87% последовательности SEQ ID NO:1, используют в качестве лекарства для лечения MPS и вводят внутривенно.
В другом дополнительном варианте осуществления фармацевтическую композицию, включающую нуклеотидную последовательность, идентичную на 98% последовательности SEQ ID NO:1, и распространенный промотор, вводят парентерально для лечения заболевания.
После описания изобретения в общих чертах оно должно быть легче понято при ссылке на последующие примеры, которые представлены в качестве иллюстрации и не предназначены для ограничения настоящего изобретения.
Общие процедуры
1. Рекомбинантные векторы AAV
Описанные в настоящем описании векторы AAV были сконструированы посредством тройной трансфекции. Материалы, необходимые для получения векторов, включали: клетки HEK293 (экспрессирующие гены E1), вспомогательную плазмиду с функцией аденовируса, вспомогательную плазмиду, содержащую гены AAV rep от серотипа 2 и гены cap от желаемого серотипа (т.е. AAV1, AAV2, AAV5, AAV7, AAV8, AAV9), и, наконец, основную плазмиду с ITR и интересующий конструкт.
Для создания экспрессирующих сульфамидазу векторов AAV кДНК сульфамидазы мыши клонировали в основную плазмиду AAV под контролем универсального гибридного промотора CAG или специфичного для печени промотора hAAT.
Векторы (частицы вирусного вектора) получали путем вспомогательной безвирусной трансфекции клеток HEK293 с использованием трех плазмид, с модификациями. Смотрите Matsushita T, et al., Gene Ther. 1998; 5:938-945 и Wright J, et al., Mol. Ther. 2005; 12:171-178. Клетки культивировали до 70% конфлуентности во вращающихся сосудах (RB) (Corning, Corning, NY, США) в DMEM (модифицированной по Дульбекко среде Игла) с добавкой 10% FBS (сыворотки плодов телят) и затем совместно трансфицировали: 1) плазмидой, несущей экспрессионную кассету с расположенными по обе стороны вирусными ITR (описанными выше); 2) вспомогательной плазмидой, несущей AAV rep2 и соответствующий cap (гены cap1 и cap9); и 3) плазмидой, обеспечивающей вспомогательные функции аденовируса. Плазмиды очищали посредством двух последовательных градиентов хлорида цезия с использованием либо стандартного протокола, либо оптимизированного протокола, как описано ранее. Смотрите Ayuso E, et al., Gene Ther. 2010; 17:503-510. Векторы диализовали против PBS, фильтровали, титровали с помощью qPCR (количественной полимеразной цепной реакции) и хранили при -80°С до использования.
a. Конструирование pAAV-CAG-mu-SFMD-WPRE
В качестве исходного материала использовали кДНК сульфамидазы мыши (Clone ID: D330015N16; Riken, Saitama, JP). кДНК находилась внутри плазмиды pFLCI-Sgsh. Для амплификации кодирующей области сульфамидазы проводили высокоточную ПЦР с праймерами, которые включали сайты рестрикции MluI на обоих концах. Использовали смысловой и антисмысловой праймеры со следующими последовательностями, соответственно: SEQ ID NO:5 (Fw) CTTACTTATGACGCGTATGCACTGCCCGGGACTG и SEQ ID NO:6 (Rv) TATCCTATCGACGCGTTCAGAGTTCATTGTGAAGCGGTC.
Основная плазмида pAAV-CAG-WPRE была создана ранее и содержала обе ITR из генома AAV2, промотор CAG, элемент WPRE и сигнал polyA β-глобина кролика. Промотор CAG представляет собой гибридный промотор, состоящий из раннего/немедленного энхансера CMV и промотора β-актина кур. Этот промотор способен стимулировать высокую экспрессию повсеместно. Посттранскрипционный регуляторный элемент вируса гепатита североамериканского лесного сурка (WPRE) представляет собой гепаднавирусную последовательность, которая широко используется в качестве действующего внутри гена регуляторного модуля в разнообразных плазмидных или вирусных векторах генов. При помещении WPRE в 3'-нетранслируемую область кассет из переносимых генов эта последовательность повышает продукцию трансгена путем повышения уровней мРНК в ядре и цитоплазме. Смотрите Zanta-Boussif М, et al., Gene Ther. 2009; 16: 605-619.
Амплифицированную с помощью ПЦР кодирующую область сульфамидазы клонировали в сайт рестрикции MluI основной плазмиды AAV pAAV-CAG-WPRE, и полученная плазмида была названа pAAV-CAG-mu-SFMD-WPRE. Смотрите SEQ ID NO: 7.
b. Конструирование pAAV-CAG-mu-SFMD
Элемент WPRE в плазмиде pAAV-CAG-mu-SFMD-WPRE окружен двумя сайтами рестрикции EcoRI. Для создания плазмиды pAAV-CAG-mu-SFMD (номер регистрации DSM 24818) плазмиду pAAV-CAG-mu-SFMD-WPRE гидролизовали EcoRI для удаления последовательности WPRE и затем вновь лигировали. Смотрите SEQ ID NO: 8.
c. Конструирование pAAV-CAG-co-hu-SFMD
Плазмиду pAAV-CAG-mu-SFMD гидролизовали MluI и EcoRI для удаления области, кодирующей сульфамидазу мыши. После этого гидролизовали кДНК сульфамидазы человека с оптимизированными кодонами (co-hu-SFMD) и клонировали в те же самые сайты рестрикции с получением плазмиды pAAV-CAG-co-hu-SFMD (номер регистрации DSM 24817). Смотрите SEQ ID NO: 9.
d. Конструирование AAV9-CAG-co-hu-SFMD
Вектор AAV9-CAG-co-hu-SFMD получали путем совместной трансфекции клеток 293НЕК плазмидой pAAV-CAG-co-hu-SFMD, плазмидой, кодирующей аденовирусную вспомогательную функцию, и плазмидой, кодирующей гены AAV2 rep и AAV9 cap.
e. Конструирование pGG2-hAAT-mu-SFMD
Кодирующую область сульфамидазы мыши вырезали из плазмиды pAAV-CAG-mu-SFMD-WPRE путем гидролиза MluI. Эту область затем клонировали в сайт MluI основной плазмиды AAV pGG2-hAAT сполучением плазмиды pGG2-hAAT-mu-SFMD (номер регистрации DSM 24819). Смотрите SEQ ID NO:10.
f. Конструирование pGG2-hAAT-co-hu-SFMD
Область, кодирующую сульфамидазу человека с оптимизированными кодонами, вырезали из плазмиды pAAV-CAG-co-hu-SFMD (номер регистрации DSM 24817) путем гидролиза MluI-EcoRI. Плазмиду pGG2-hAAT-mu-SFMD (номер регистрации DSM 24819) гидролизовали с помощью MluI для удаления гена mu-SFMD, и затем область, кодирующую сульфамидазу человека с оптимизированными кодонами, клонировали в этот сайт посредством лигирования тупых концов. Полученная плазмида была названа pGG2-hAAT-co-hu-SFMD. Смотрите SEQ ID NO:11.
Плазмида pGG2-hAAT-co-hu-SFMD содержала обе AAV2-ITRs, промотор hAAT и сигнал полиаденилирования из SV40.
g. Конструирование pAAV9-hAAT-co-hu-SFMD и pAAV8-hAAT-co-hu- SFMD
Векторы получали с помощью вспомогательной безвирусной трансфекции клеток HEK293 с использованием трех плазмид с модификациями. Смотрите Matsushita, 1998, выше и Wright, 2005, выше. Клетки культивировали до 70% конфлуентности во вращающихся сосудах (RB) (Corning, Corning, NY, США) в DMEM с добавкой 10% FBS и затем совместно трансфицировали: 1) плазмидой, несущей экспрессионную кассету с расположенными по обе стороны вирусными ITR (pGG2-hAAT-co-hu-SFMD); 2) вспомогательной плазмидой, несущей AAV rep2 и соответствующие гены cap (cap8 или 9); и 3) плазмидой, обеспечивающей вспомогательные функции аденовируса. Векторы очищали посредством двух последовательных градиентов хлорида цезия с использованием либо стандартного протокола, либо оптимизированного протокола, как описано ранее. Смотрите Ayuso, 2010, выше. Векторы диализовали против PBS, фильтровали, титровали с помощью qPCR и хранили при -80°С до использования.
Плазмида pGG2-hAAT-mu-SFMD содержала обе AAV2-ITR, промотор hAAT и сигнал полиаденилирования из SV40. Промотор hAAT представляет собой гибридный промотор, состоящий из 4 тандемных повторов энхансера контролируемой в гепатоцитах области (HCR) из промотора аполипопротеина E и α-антитрипсина человека. Его экспрессия ограничена гепатоцитами. Смотрите Mingozzi F, et al., J. Clin. Invest. 2003; 111:1347-1356.
Векторы по настоящему изобретению конструировали в соответствии с методами молекулярной биологии, хорошо известными в данной области техники. Смотрите Brown T, "Gene Cloning" (Chapman & Hall, London, GB, 1995); Watson R, et al., "Recombinant DNA", 2nd Ed. (Scientific American Books, New York, NY, US, 1992); Alberts B, et al., "Molecular Biology of the Cell" (Garland Publishing Inc., New York, NY, US, 2008); Innis M, et al., Eds., "PCR Protocols. A Guide to Methods and Applications" (Academic Press Inc., San Diego, CA, US, 1990); Erlich H, Ed., "PCR Technology. Principles and Applications for DNA Amplification" (Stockton Press, New York, NY, US, 1989); Sambrook J, et al., "Molecular Cloning. A Laboratory Manual" (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, US, 1989); Bishop T, et al., "Nucleic Acid and Protein Sequence. A Practical Approach" (IRL Press, Oxford, GB, 1987); Reznikoff W, Ed., "Maximizing Gene Expression" (Butterworths Publishers, Stoneham, MA, US, 1987); Davis L, et al., "Basic Methods in Molecular Biology" (Elsevier Science Publishing Co., New York, NY, US, 1986), Schleef M, Ed., "Plasmid for Therapy and Vaccination" (Wiley-VCH Verlag GmbH, Weinheim, DE, 2001).
2. Животные
Использовали колонию (MPSIIIA) родственных мышей C57BI/6 с недостаточностью сульфамидазы. Смотрите Crawley A, et al., Brain Res. 2006; 1104:1-17. Мышей с повреждением MPSIIIA и контрольных здоровых мышей получали близкородственным скрещиванием потомков гетерозиготных основателей рода. Генотип определяли с помощью анализа ПЦР на геномной ДНК из образцов, полученных при надрезе хвоста, с амплификацией последовательности, включающей мутацию, и ее последующим гидролизом ферментом рестрикции MspAI, как описано ранее. Смотрите Bhattacharyya R, et al., Glycobiology 2001; 11:99-103. Мышей кормили без ограничения стандартным кормом (Panlab, Barcelona, ES) и содержали в условиях 12-час светового цикла освещение-темнота (освещение с 9:00 утра).
3. Введение векторов и сбор образцов
Для внутривенной доставки векторов AAV 2-месячным животным MPSIIIA через хвостовую вену вводили соответствующий вектор AAV в суммарной дозе 1012 геномов вектора. Для внутримышечной инъекции 2-месячных животных MPSIIIA анестезировали смесью кетамина (100 мг/кг) и ксилацина (10 мг/кг), и соответствующий вектор AAV инъецировали в 6 мышц задних конечностей (четырехглавую, икроножную и переднюю большеберцовую обеих лап). В возрасте 10 месяцев животных анестезировали и затем через сердце перфузировали 10 мл PBS для полного отмывания тканей от крови. Выделяли мозг целиком и множество соматических тканей (включая печень, селезенку, поджелудочную железу, почку, легкое, сердце, скелетную мышцу и семенники) и либо замораживали в жидком азоте и хранили при -80°С, либо погружали в формалин для последующих гистологических анализов.
4. Анализ РНК
Суммарную мРНК получали из образцов скелетной мышцы и печени с помощью реагента для выделения TriPure (Roche Diagnostics, Barcelona, ES) и анализировали с помощью нозерн-блотинга. Отпечатки гибридизовали с зондом сульфамидазы мыши, меченным 32P-dCTP методом случайной затравки с использованием готовых к употреблению шариков для промечивания ДНК (Amersham Biosciences, Piscataway, NJ, US).
5. Сульфамидазная активность и количественное определение гликозаминогликанов
Образцы печени, скелетной мышцы и мозга обрабатывали ультразвуком в воде, и в супернатантах определяли сульфамидазную активность с помощью флуорогенного субстрата, производного 4-метилумбеллиферона (Moscerdam Substrates, Oegstgeest, NL), как описано ранее. Смотрите Karpova E, et al., J. Inherit. Metab. Dis. 1996; 19:278-285. Уровни активности сульфамидазы нормализовали по суммарному белку, измеряемому с помощью анализа белка по Брэдфорду (Bio-Rad, Hercules, CA, US).
Для количественного определения гликозаминогликанов (GAG) образцы тканей взвешивали и гидролизовали протеиназой K, и экстракты осветляли центрифугированием и фильтрованием. Уровни GAG в экстрактах тканей и моче определяли с помощью набора для сульфированных гликозаминогликанов Blyscan (Biocolor, Carrickfergus, County Antrim, GB) с использованием в качестве стандарта хондроинин-4-сульфата. Уровни GAG в тканях нормализовали по сырой массе ткани, а в моче по концентрации креатинина, измеряемой с помощью специального набора (Horiba ABX, Irvine, CA, US).
6. Гистологические анализы
Ткани фиксировали в течение 12-24 час в формалине, заключали в парафин и получали срезы с последующим индуцируемым нагреванием восстановлением эпитопов (цитратный буфер, pH 6). Для иммуногистохимического определения Lamp1 парафиновые срезы инкубировали в течение ночи при 4°С с антителом крысы против Lamp1 (1D4B; Santa Cruz Biotechnology, Santa Cruz, CA, US), разбавленным 1:100, и затем инкубировали с биотинилированным антителом кролика против мыши (Dako, Glostrup, DK), разведенном 1:300. Сигнал Lamp1 усиливали путем инкубации срезов с набором для окрашивания ABC-пероксидаза (Thermo Scientific, Waltham, MA, US), разведенным 1:100, и визуализировали с использованием 3,3-диаминобензидина (Sigma-Aldrich, St. Louis, MO, US) в качестве хромогена. Изображения в светлом поле получали на оптическом микроскопе (Eclipse E800; Nikon, Tokyo, JP). Для иммуноокрашивания парвальбумина и кальбиндина парафиновые срезы инкубировали в течение ночи при 4°С с антителом кролика против кальбиндина D28k (Swant, Marly, CH), разбавленным 1:2000, или с антителом кролика против парвальбумина (Swant, Marly, CH), разбавленным 1:100. После этого образцы инкубировали с биотинилированными IgG козы против кролика (Vector Labs., Burlingame, CA, US) и затем со стрептавидин-Alexa 488 (1:100, Molecular Probes, Invitrogen, Carlsbad, CA, US), а ядра окрашивали TOPRO-3. Изображения получали с помощью конфокального микроскопа (Leica Microsystems, Heidelberg, DE).
Для двойного иммуноокрашивания LAMP1 и Mac2 срезы сначала инкубировали в течение ночи при 4°С с антителом крысы против Lamp1 в разведении 1:100, затем с биотинилированным антителом кролика против мыши в разведении 1:300 с последующей инкубацией со стрептавидин-Alexa 488 (1:300). После этого срезы инкубировали с антителом кролика против Mac2 в разведении 1:50, затем с биотинилированными антителами козы против кролика в разведении 1:300 с последующей инкубацией со стрептавидин-Alexa 488 (1:300; Molecular Probes, Invitrogen, Carlsbad, CA, US). Наконец, ядра окрашивали с помощью Hoechst (1:100; Sigma- Aldrich, St. Louis, MO, US).
7. Анализ иммуноблотингом
Половинки мозжечка гомогенизировали в буфере для лизиса белка. Десять микрограмм белка разгоняли в 10% (масс./об.) SDS-PAGE, переносили на мембраны поливинилидендифторида и зондировали в течение ночи при 4°С первыми антителами против кальбиндина (Swant, Marly, CH) и α-тубулина (Abeam, Cambridge, MA, US). Выявление осуществляли с помощью меченного пероксидазой хрена антитела свиньи против кролика (Dako, Glostrup, DK) и реагента для проявления иммуноблотов ECL Plus (Amersham Biosciences, Piscataway, NJ, US).
8. Анализ трансмиссионной электронной микроскопией
Мышей забивали передозировкой изофлуорана (Isofluo, Labs. Esteve, Barcelona, ES) и перфузировали через нижнюю полую вену 1 мл 2,5% глутаральдегида и 2% параформальдегида. Из небольшой части (приблизительно 1 мм3) боковой доли печени и вершины червя мозжечка готовили срезы и инкубировали в течение 2 часов при 4°С в том же фиксаторе. После промывки в холодном какодилатном буфере образцы дополнительно фиксировали в 1% тетроксиде осмия, окрашивали в водном уранилацетате, затем дегидратировали проводкой через последовательные разведения этанола и заключали в эпоксидную смолу. Ультратонкие срезы (600-800 Å) блоков со смолой окрашивали цитратом свинца и анализировали с помощью трансмиссионного электронного микроскопа (H-7000; Hitachi, Tokyo, JP).
9. Статистический анализ
Все результаты представляли в виде среднее±SEM. Статистические сравнения проводили с использованием либо t-теста, либо одностороннего ANOVA. Статистически значимым считали различие при P<0,05.
Примеры
Пример 1
Внутримышечная доставка AAV1-CAG-mu-SFMD-WPRE
В 6 мышц задних конечностей (четырехглавую, икроножную и переднюю большеберцовую обеих лап) 2-месячных самцов и самок мышей MPSIIIA инъецировали вектор AAV1-CAG-mu-SFMD-WPRE в суммарной дозе 1012 геномов вектора.
Через восемь месяцев после введения в инъецированных мышцах выявлялись высокие уровни направляемой вектором экспрессии и активности сульфамидазы, но в сыворотке уровни активности сульфамидазы были очень низкими (6-7% от контрольных мышей), что указывает на низкую эффективность секреции из скелетной мышцы. Смотрите фиг.1A и 1B. Кроме того, очень низкая, но значимая направляемая вектором экспрессия сульфамидазы наблюдалась в печени этих мышей, что указывает на то, что вектор вытекал из скелетной мышцы в кровоток и трансдуцировал печень. Даже при низких достигнутых уровнях сульфамидазной активности в кровотоке наблюдались коррекция аккумуляции GAG в печени и значительное снижение в некоторых других соматических тканях (селезенке, сердце, поджелудочной железе), но не в других (почке, легком). Смотрите фиг.1C. В мозгу не происходило снижение запасания GAG.
Пример 2
Внутримышечная доставка AAV8-CAG-mu-SFMD-WPRE
В 6 мышц задних конечностей (четырехглавую, икроножную и переднюю большеберцовую обеих лап) 2-месячных самцов и самок мышей MPSIIIA инъецировали вектор AAV8-CAG-mu-SFMD-WPRE в суммарной дозе 1012 геномов вектора.
Через восемь месяцев после введения в инъецированных мышцах сульфамидазная активность выявлялась на уровне, сходном с активностью у здоровых контрольных животных. Смотрите фиг.2A. Низкие уровни сульфамидазной активности наблюдались в сыворотке (10-15% от контрольных мышей). Смотрите фиг.2B. Также наблюдалась утечка вектора в печень, поскольку в печени обнаруживалась направляемая вектором экспрессия и активность сульфамидазы, и даже на более высоком уровне, чем у мышей, получавших внутримышечную инъекцию AAV1. Смотрите пример 1. Коррекция аккумуляции GAG наблюдалась в печени и селезенке, и более значительное снижение наблюдалось в других соматических тканях (сердце, поджелудочной железе, мочевом пузыре), но почка и легкие оставались практически без коррекции. Смотрите фиг.2C. Не достигалось снижение запасов GAG в мозгу.
Пример 3
Внутривенная доставка AAV8-CAG-mu-SFMD-WPRE
2-месячным мышам MPSIIIA через хвостовую вену инъецировали вектор AAV8-CAG-mu-SFMD-WPRE в суммарной дозе 1012 геномов вектора.
Через восемь месяцев после введения у получавших лечение самцов в печени проявлялась сульфамидазная активность на уровне, сходном с уровнем у контрольных мышей, но у самок активность была в 4 раза ниже. Смотрите фиг.3A. Соответствующим образом, сульфамидазная активность в кровотоке была высокой у самцов (сходно с уровнями у контрольных мышей) и более низкой у самок (25% от контрольных мышей). Смотрите фиг.3B. Эти высокие уровни сульфамидазы в кровотоке были способны корректировать аккумуляцию GAG в печени, сердце, селезенке, поджелудочной железе и мочевом пузыре и значительно снижать в легких, но не в почке. Смотрите фиг.3C по поводу содержания GAG в печени. В мозгу снижения запасов GAG не наблюдалось.
Пример 4
Внутривенная доставка AAV8-hAAT-mu-SFMD
2-месячным мышам MPSIIIA через хвостовую вену инъецировали вектор AAV8-hAAT-mu-SFMD в суммарной дозе 1012 геномов вектора.
Через восемь месяцев после введения у подвергнутых лечению самцов активность сульфамидазы в печени обнаруживалась на уровне, на 500% превышающем уровень у контрольных животных. У женских особей уровень активности сульфамидазы в печени достигал уровня у контрольных особей. Смотрите фиг.4A. Сульфамидазная активность в кровотоке была соответственно выше у самцов по сравнению с самками (500% у самцов против 160% у самок). Смотрите фиг.4B. Эти сверхфизиологические уровни сульфамидазы в кровотоке обеспечивали коррекцию аккумуляции GAG во всех соматических органах, включая почку. Смотрите фиг.4C по поводу количественного определения GAG.
У подвергнутых лечению самцов наблюдались низкие уровни сульфамидазной активности и сниженная аккумуляция GAG в мозгу. Смотрите фиг.5A и B. В клетках Пуркинье мозжечка подвергнутых лечению самцов наблюдалось меньше электронно-плотных включений при оценке с помощью электронной микроскопии. Смотрите фиг.5C. Лечение внутривенным введением вектора AAV8-hAAT-mu-SFMD (“в/в-AAV8-hAAT-mu-SFMD”) обеспечивало достижение коррекции соматической патологии, но лишь облегчало нейродегенерацию, свойственную мышам MPSIIIA.
Ультраструктуру коры анализировали с помощью трансмиссионной электронной микроскопии. Не было отмечено выявляемых различий в ультраструктуре нейронов затылочной коры особей MPSIIIA, подвергнутых и не подвергнутых лечению. В перинейрональных глиальных клетках мышей MPSIIIA без лечения наблюдалось явное увеличение лизосомального компартмента, которое практически отсутствовало у животных, подвергнутых лечению. Смотрите фиг.10. Эти результаты позволяют предполагать, что продолжительная высокая активность сульфамидазы в кровотоке предотвращает дегенерацию нейронов у особей MPSIIIA.
К 17-месячному возрасту все самцы MPSIIIA без лечения погибли, в то время как 100% самцов, подвергнутых лечению в/в-AAV8-hAAT-mu-SFMD, оставались живы (медиана выживания=14,2±0,5 против 18,8±0,9 месяцев для самцов MPSIIIA без лечения и подвергнутых лечению, соответственно, p=0,001). Улучшение не было очевидным в группе самок, где и подвергнутые лечению особи, и особи без лечения продемонстрировали сходные показатели выживания (медиана выживания=13,1±0,5 против 13,9±1,2 месяцев для самок MPSIIIA без лечения и подвергнутых лечению, соответственно, p=0,467). Этот результат находится в соответствии с более низкими уровнями сульфамидазной активности, измеренными в сыворотке и мозгу и меньшей степенью снижения GAG, наблюдаемой у женских особей. Смотрите фиг.11.
Большая выживаемость повергнутых лечению в/в-AAV8-hAAT-mu-SFMD самцов MPSIIIA дополнительно продемонстрировала терапевтический потенциал продолжительно действующих сверхфизиологических уровней сульфамидазы в кровотоке, полученных посредством направленного на печень переноса гена. Лечение в/в-AAV8-hAAT-mu-SFMD увеличивало продолжительность жизни мужских особей MPSIIIA. Смотрите фиг.11.
Пример 5
Внутривенная доставка AAV9-CAG-mu-SFMD
2-месячным мышам MPSIIIA через хвостовую вену инъецировали вектор AAV9-CAG-mu-SFMD в суммарной дозе 1012 геномов вектора.
И у самцов, и у самок, подвергнутых лечению, были обнаружены высокие уровни сульфамидазы в кровотоке (500% от контрольного уровня у самцов и 150% у самок), которые вызывали эффективную коррекцию всех соматических тканей у животных обоего пола. Кроме того, благодаря высокой эффективности переноса в мозг AAV серотипа 9 значительная сульфамидазная активность наблюдалась в мозгу животных обоего пола, которая эффективно корректировала аккумуляцию GAG во всех областях мозга. Смотрите фиг.6A и 6B. Нейровоспаление (астроглиоз и микроглиоз), характерное для MPSIIIA, было полностью нормализовано у лечившихся AAV9 мышей. Кроме того, лечившиеся AAV9 мыши лучше выполняли тест с вращающимся стержнем по сравнению с животными без лечения. Смотрите фиг.6C.
Лечение внутривенным введением вектора AAV9-CAG-mu-SFMD (“в/в-AAV9-CAG-mu-SFMD”) увеличивало продолжительность жизни животных MPSIIIA. Смотрите фиг.12. К 17-месячному возрасту все самцы MPSIIIA без лечения погибли, в то время как 100% самцов, подвергнутых лечению в/в-AAV9-CAG-mu-SFMD, оставались живы в 20-месячном возрасте (p<0,001 и p=0,037 для подвергнутых лечению самцов MPSIIIA против самцов без лечения, соответственно). Смотрите фиг.12. В группе самок было продемонстрировано сходное, но менее впечатляющее улучшение (p=0,063 и p=0,057 для подвергнутых лечению самок MPSIIIA против самок без лечения, соответственно). Этот результат находится в соответствии с более низкими уровнями сульфамидазной активности, измеренными в сыворотке самок животных после лечения в/в-AAV9-CAG-mu-SFMD.
Пример 6
Внутрицистернальная доставка AAV9-CAG-mu-SFMD
2-месячным анестезированным мышам MPSIIIA в cisterna magna инъецировали вектор AAV9-CAG-mu-SFMD в суммарной дозе 5×1010 геномов вектора в суммарном объеме 5 мкл.
Через три месяца после введения достигалась полная коррекция аккумуляции GAG во всем мозгу подвергнутых лечению животных. Смотрите фиг.8. Определяемая вектором экспрессия сульфамидазы была обнаружена также в печени подвергнутых лечению животных, что позволяет предполагать, что после внутрицистернальной доставки некоторые векторы попадают в кровоток и захватываются печенью. В соответствии с этим результатом аккумуляция GAG нормализовалась также и в печени.
Внутрицистернальная (ic) доставка вектора AAV9-CAG-mu-SFMD (“ic-AAV9-CAG-mu-SFMD”) увеличивала продолжительность жизни животных MPSIIIA. Смотрите фиг.12. К 17-месячному возрасту все самцы MPSIIIA без лечения погибли, в то время как 100% самцов, подвергнутых лечению ic-AAV9-CAG-mu-SFMD, оставались живы в 20-месячном возрасте (p<0,001 и p=0,037 для подвергнутых лечению самцов MPSIIIA против самцов без лечения, соответственно). Смотрите фиг.12. В группе самок было продемонстрировано сходное, но менее впечатляющее улучшение (p=0,063 и p=0,057 для подвергнутых лечению самок MPSIIIA против самок без лечения, соответственно). Этот результат находится в соответствии с более низкими уровнями сульфамидазной активности, измеренными в сыворотке самок животных после лечения ic-AAV9-CAG-mu-SFMD.
Пример 7
Внутривенная доставка AAV9-CAG-co-hu-SFMD (сульфамидазы человека с оптимизированными кодонами)
Используемые кодоны сульфамидазы человека были оптимизированы для снижения дозы вводимого вектора. Целями данного подхода являлись стабилизация мРНК сульфамидазы и повышение ее трансляции, что должно было способствовать более высокой продукции сульфамидазы при той же дозе вектора.
2-месячным мышам MPSIIIA через хвостовую вену инъецировали внутривенно вектор AAV9-CAG-co-hu-SFMD в количестве 1012 вирусных геномов (vg). По сравнению с неоптимизированным геном было получено по меньшей мере 3-кратное увеличение уровня сульфамидазы в печени. Смотрите фиг.9.
Пример 8
Внутривенная доставка AAV9-hAAT-co-hu-SFMD
В соответствии с тем же протоколом, что и описанный в примере 7, двухмесячных мышей MPSIIIA лечат 1012 vg вектора AAV9-hAAT-co-hu-SFMD путем внутривенного введения в хвостовую вену. Уровень сульфамидазы измеряют таким же образом, как и в примере 7. Результаты показывают существенное повышение по сравнению с неоптимизированным геном.
Пример 9
Внутрицистернальная доставка различных серотипов AAV-CAG-GFP-WPRE
Для оценки тропизма в мозгу разных серотипов AAV при введении в спинномозговую жидкость 5×1010 векторных геномов векторов AAV1, AAV2, AAV5, AAV7, AAV8 и AAV9, несущих репортерный ген GFP (конструкт CAG-GFP-WPRE), вводили мышам MPSIIIA внутрицистернально.
Значительная трансдукция клеток была достигнута в варолиевом мосту при использовании всех серотипов при наибольшей и наименьшей эффективности, достигнутой при использовании AAV9 и AAV1, соответственно. В мозжечке сигнал репортера был приурочен преимущественно к аксонам, идентифицированным морфологически как мшистые волокна, особенно при использовании AAV1 и AAV9, а также при использовании других серотипов за исключением AAV8 в нейронах Пуркинье. Более значительные различия между серотипами в эффективности переноса гена наблюдались в отдаленных областях мозга. Многие клетки в коре головного мозга, обонятельной луковице и гиппокампе были трансдуцированы в группе с инъекцией AAV9, в меньшей степени - в группе AAV7, тогда как в случаях серотипов AAV1, AAV2, AAV5 или AAV8 позитивные по GFP тела клеток в этих областях не обнаруживались. В гипоталамусе серотип AAV9 эффективно трансдуцировал нейроны, а серотип AAV1 давал немногочисленные разбросанные клетки GFP+. Во всех группах можно было наблюдать по всему мозгу случайные GFP-позитивные аксоны, которые, вероятно, проецировались от инфицированных нейронов, расположенных рядом с cisterna magna. Среди различных серотипов векторы AAV9 проявляли наибольшую эффективность трансдукции. Смотрите фиг.7.
Пример 10
Масштабируемость внутрицистернальной доставки AAV9-CAG-co-hu-SFMD для клинического применения
В качестве первого шага в направлении потенциального клинического применения внутрицистернальной доставки AAV9 была произведена оценка того, сохраняется ли паттерн трансдукции, наблюдавшийся у мышей, у животного с более подходящим размером мозга. С этой целью 1,5×1012 vg/кг AAV9-CAG-GFP-WPRE вводили в cisterna magna здоровых гончих собак. Инъекции были сделаны в общей сложности 4 собакам; у двух животных (собаки № 1 и 4) использовали насос для инфузии раствора вирусного вектора со скоростью, сходной со скоростью образования CFS (1 мл/10 минут), а двум другим (собаки № 2 и 3) инфузия вектора была произведена за несколько секунд. На фиг.13 показано иммунологическое выявление GFP в образцах от собаки 1. Сильное окрашивание наблюдалось в областях, расположенных рядом с cisterna magna, таких как продолговатый мозг, варолиев мост и гипоталамус. Смотрите фиг.13b, c и d. В мозжечке, несмотря на близость к месту введения, трансдуцировались лишь немногие отдельные клетки Пуркинье, тогда как в гиппокампе, области, отдаленной от цистерны, имела место эффективная трансдукция зубчатой извилины. Смотрите фиг.13i и j. Распространение вируса по CSF обеспечивало трансдукцию областей, отдаленных от места инъекции, таких как обонятельный мозг и лобная, теменная и затылочная кора, где более поверхностно расположенные области проявляли большую трансдукцию. Смотрите фиг.13e, h и f. Наконец, вектор достигал также спинного мозга, и сигнал GFP выявлялся в вентральных мотонейронах и астроцитах близлежащих ганглиев. Смотрите фиг.13a. Полуколичественное сравнение локализации GFP у четырех собак позволило предположить, что скорость инфузии раствора вируса не оказывает существенного влияния на эффективность распространения вектора AAV9. Смотрите таблицу 1.
Наконец, сходно с наблюдениями, сделанными на мышах после внутрицистернальной доставки AAV9, GFP также выявлялся в печени гончих собак, где было трансдуцировано в среднем 3,7% гепатоцитов. Смотрите фиг.14. Эти результаты позволили предположить системное распространение вектора AAV9 после внутрицистернальной доставки.
Полуколичественный анализ трансдукции мозга после ic доставки векторов AAV9-GFP здоровым гончим собакам. Несколько изображений каждой области мозга оценивались тремя независимыми наблюдателями, и представлены средние значения. Использовали следующие полуколичественные критерии: (+) менее 10 GFP-позитивных клеток/10× поле зрения микроскопа; (++) 10-30 GFP-позитивных клеток/10× поле зрения микроскопа; (+++) 30-60 GFP-позитивных клеток/10× поле зрения микроскопа и (++++) более 60 GFP-позитивных клеток/10× поле зрения микроскопа; Н.О. - не определяли.
Пример 11
Функциональная эффективность сульфамидазы человека с оптимизированными кодонами (co-hu-SFMD)
Были сконструированы и получены экспрессионные кассеты, включающие версию последовательности кДНК сульфамидазы человека с оптимизированными кодонами (co-hu-SFMD). Оптимизацию кодонов производили для повышения эффективности продукции белка SFMD у человека путем использования наиболее представленных тРНК для типов кодонов и с учетом конкретного профиля трансляции. Для целей эксперимента использовали мышей благодаря их сходству с человеком и предсказательному потенциалу мышиной животной модели.
Для гарантии того, что данная последовательность ведет к продукции активной сульфамидазы, самцам мышей MPSIIIA внутривенно инъецировали 1×1012 vg вектора AAV9, в котором co-hu-SFMD экспрессировалась под контролем универсального промотора CAG. Активность сульфамидазы в сыворотке этих мышей достигала уровня, сходного с уровнем здоровых животных дикого типа, и сохранялась на протяжении всего исследования (2 месяца). Смотрите фиг.15a. Эта продолжительная сульфамидазная активность обеспечивала нормализацию содержания GAG в печени этих животных, подобно тому, как это наблюдалось при доставке мышиного трансгена с помощью AAV9. Смотрите фиг.15b.
Все упомянутые выше в настоящем описании публикации включены таким образом в качестве ссылки в полном объеме.
Хотя изложенное выше изобретение было описано несколько подробно для целей ясности и понятности, из прочтения данного раскрытия специалисту в данной области техники должно быть ясно, что могут быть осуществлены различные изменения в форме и деталях без выхода за пределы истинного объема изобретения и прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| АДЕНОАССОЦИИРОВАННЫЕ ВИРУСНЫЕ ВЕКТОРЫ ДЛЯ ЛЕЧЕНИЯ МУКОПОЛИСАХАРИДОЗОВ ТИПА IV A | 2019 |
|
RU2794960C2 |
| АДЕНОАССОЦИИРОВАННЫЕ ВИРУСНЫЕ ВЕКТОРЫ ДЛЯ ЛЕЧЕНИЯ ЛИЗОСОМНЫХ БОЛЕЗНЕЙ НАКОПЛЕНИЯ | 2015 |
|
RU2718248C2 |
| ВИРУСНАЯ ЭКСПРЕССИОННАЯ КОНСТРУКЦИЯ, СОДЕРЖАЩАЯ КОДИРУЮЩУЮ ПОСЛЕДОВАТЕЛЬНОСТЬ ФАКТОРА РОСТА ФИБРОБЛАСТОВ 21 (FGF21) | 2018 |
|
RU2839576C2 |
| ВЕКТОРЫ НА ОСНОВЕ АДЕНОАССОЦИИРОВАННЫХ ВИРУСОВ ДЛЯ ЛЕЧЕНИЯ МУКОПОЛИСАХАРИДОЗОВ | 2016 |
|
RU2744593C2 |
| Кодон-оптимизированная последовательность нуклеотидов, кодирующая hAIPL1, и её содержащий экспрессионный вектор | 2021 |
|
RU2785621C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ MPS1 | 2014 |
|
RU2708318C2 |
| ВЕКТОР АДЕНОАССОЦИИРОВАННОГО ВИРУСА | 2015 |
|
RU2743382C2 |
| ВАРИАНТ СРЕДСТВА ДЛЯ RNAi | 2018 |
|
RU2789647C2 |
| ЛЕЧЕНИЕ ЗАБОЛЕВАНИЙ ПОСРЕДСТВОМ ЭКСПРЕССИИ ФЕРМЕНТА, ОБЛАДАЮЩЕГО ДЕЗОКСИРИБОНУКЛЕАЗНОЙ (ДНКазной) АКТИВНОСТЬЮ, В ПЕЧЕНИ | 2019 |
|
RU2773691C2 |
| ВЕКТОР | 2017 |
|
RU2749479C1 |





























Изобретение относится к биохимии. Описана молекула нуклеиновой кислоты для генетической терапии мукополисахаридозов. Также описаны векторы и фармацевтические композиции для лечения заболеваний мукополисахаридозов. Изобретение расширяет арсенал средств для лечения мукополисахаридозов. 7 н. и 8 з.п. ф-лы, 15 ил., 1 табл., 11 пр.
1. Выделенная нуклеиновая кислота, имеющая нуклеотидную последовательность согласно SEQ ID NO: 1, для генной терапии мукополисахаридозов, кодирующая белок SEQ ID NO: 2.
2. Аденоассоциированный вектор для генной терапии мукополисахаридозов, содержащий нуклеиновую кислоту по п. 1.
3. Аденоассоциированный вектор по п. 2, где серотип представлен серотипом 1, 2, 5, 7, 8 или 9.
4. Аденоассоциированный вектор по п. 3, где серотип представлен серотипом 9.
5. Аденоассоциированный вектор по п. 2, включающий промотор CAG, функционально связанный с SEQ ID NO: 1.
6. Аденоассоциированный вектор по п. 5, представляющий собой аденоассоциированный вирус серотипа 9, AAV9, содержащий ген с оптимизированными кодонами, co-hu-SFMD, SEQ ID NO: 1, функционально связанный с ранним энхансерным элементом цитомегаловируса и промотором β-актина цыпленка, CAG, идентифицируемый как AAV9-CAG-co-hu-SFMD.
7. Плазмида pAAV-CAG-co-hu-SFMD для генной терапии мукополисахаридозов с номером регистрации DSM 24817, содержащая ген с оптимизированными кодонами, co-hu-SFMD, SEQ ID NO: 1, по п. 1, функционально связанный с ранним энхансерным элементом цитомегаловируса и промотором β-актина цыпленка, CAG.
8. Аденоассоциированный вектор по п. 2, включающий промотор hAAT, функционально связанный с SEQ ID NO: 1.
9. Плазмида pAAV-hAAT-co-hu-SFMD для генной терапии мукополисахаридозов, содержащая ген с оптимизированными кодонами, co-hu-SFMD, SEQ ID NO: 1, по п. 1, функционально связанный с промотором альфа1-антитрипсина человека, hATT.
10. Фармацевтическая композиция для генной терапии мукополисахаридозов, включающая терапевтически эффективное количество аденоассоциированного вектора по любому из пп. 2-6 или 8 и фармацевтически приемлемый носитель или основу.
11. Фармацевтическая композиция по п. 10 для парентерального введения, предпочтительно для внутривенного или внутрицистернального введения.
12. Аденоассоциированный вектор по любому из пп. 2-6 или 8 или фармацевтическая композиция по любому из пп. 10 или 11 для повышения сульфамидазной активности в организме.
13. Аденоассоциированный вектор по любому из пп. 2-6 или 8 или фармацевтическая композиция по любому из пп. 10-11 для лечения мукополисахаридозов, предпочтительно мукополисахаридоза типа IIIA или синдрома Санфилиппо.
14. Способ получения аденоассоциированных векторов по п. 2, включающий стадии:
i) получения первого вектора, включающего SEQ ID NO: 1, вставленную между первым концевым повтором AAV и вторым концевым повтором AAV, промотор CAG или hAAT, функционально связанный с SEQ ID NO: 1; второго вектора, включающего ген AAV rep и ген AAV cap; и третьего вектора, включающего аденовирусную вспомогательную функцию;
ii) совместной трансфекции компетентных клеток векторами стадии i);
iii) культивирования трансфицированных клеток стадии ii); и
iv) очистки экспрессионных векторов из культуры стадии iii).
15. Выделенная клетка для генной терапии мукополисахаридозов, содержащая аденоассоциированный вектор по любому из пп. 2-6 или 8.
| FRALDI A | |||
| ET AL., Functional correction of CNS lesions in an MPS-IIIA mouse model by intracerebral AAV-mediated delivery of sulfamidase and SUMF1 genes, HUMAN MOLECULAR GENETICS, 2007, v | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| Способ сульфирования жиров | 1923 |
|
SU2693A1 |
| FOSTER H | |||
| ET AL., Codon and mRNA sequence optimization of microdystrophin transgenes improves expression and physiological outcome in | |||

