Заявленное техническое решение относится к области синтетической биологии, конкретно – к синтезу белков в открытых системах вне клеток на основе клеточного метаболизма. Заявленное техническое решение позволяет проводить поиск эукариотических регуляторных элементов генома, производить функциональные тесты эукариотических белков, выполнять синтез эукариотических белков для биотехнологии и структурной биологии. Может быть использовано, например, на биотехнологических и фармацевтических производствах, в научно-исследовательских организациях, с целью тестирования регуляторных элементов для изучения регуляции метаболизма эукариот и аппарата синтеза белка у птиц, проведение реакций синтеза в естественном окружении in vitro, тестирование влияния химических веществ, используемых в биотехнологиях и животноводстве (например – антибиотиков) и проведение исследований птиц (например, вирусов птиц) и разработку биотехнологий на их основе. Синтезируемый белок с использованием заявленной бесклеточной системы, может быть использован в биотехнологиях, в том числе – после проведения его выделения, очистки и модификации целевого белка.
Далее в тексте заявителем приведены термины, которые необходимы для облегчения однозначного понимания сущности заявленных материалов и исключения противоречий и/или спорных трактовок при выполнении экспертизы по существу.
АТФ – аденозинтрифосфат.
HEPES – 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота.
NTP – нуклеотидтрифосфат.
ЦТФ – цитидинтрифосфат.
ГТФ – гуанозинтрифосфат.
УТФ – уридинтрифосфат.
RNasine – ингибитор РНКаз.
На современном этапе развития синтетической биологии и биотехнологий на её основе одной из ключевых задач является получение данных о взаимосвязях и взаимодействии частей, составляющих биологическую систему, а также её кинетических параметров – для этого в научных исследованиях часто используются in vitro подходы и, в частности, бесклеточные системы.
Программирование бесклеточных систем дает множество преимуществ:
– простоту проведения экспериментов,
– возможность контроля за концентрациями и активностями генов и ферментов,
– простоту в получении количественных измерений,
– возможность управления большим количеством параметров, которые можно оценивать, используя высокопропускные (high-throughput) подходы.
В общем смысле, бесклеточная система синтеза белка — это клетка без мембраны помещенная в квази-естественную среду состоящую из буферной системы, метаболитов и ферментов, помогающих метаболизму.
Эта псевдо-клеточная структура обладает всеми метаболическими функциями, кроме репликации ДНК, биогенеза рибосом и функций связанных с мембраной, включая деление. В бесклеточной системе синтеза белка воспроизводится набор химических реакций, которые отражают главные метаболические процессы необходимые для синтеза белка. В состав бесклеточной системы синтеза белка входят все компоненты необходимые для транскрипции, трансляции, а также системы синтеза АТФ, обеспечивающей энергию для протекания биохимических реакций.
Бесклеточная система синтеза белка состоит из четырех основных компонент:
– клеточного экстракта как основы метаболизма,
– ДНК/РНК,
– субстратов для транскрипции и трансляции
– и системы регенерации AТФ.
Соединенные воедино компоненты создают условия для протекания основных этапов синтеза белка in vitro. В силу наличия генетических особенностей в строении аппарата синтеза белка у каждого вида организмов необходимо использовать соответствующий клеточный экстракт для конкретного организма и подбирать индивидуальные условия и концентрации компонент бесклеточных систем синтеза белка.
Клеточный экстракт – это главная часть бесклеточной системы синтеза белка. Он содержит все ферменты необходимые для метаболизма, включая аминоацил-тРНК-синтетазу и трансляционные факторы, а также функциональные рибосомы и большую часть метаболитов. Для анализа влияния мутаций в строении рибосомы в бесклеточные системы синтеза белка используется клеточный экстракт, очищенный от рибосом, экзогенные мутантные рибосомы добавляются в смесь отдельно вместе с другими компонентами. Приготовление клеточного экстракта — это самый времязатратный этап при проведении экспериментов с бесклеточными системами синтеза белка.
Этапы приготовления клеточных экстрактов в последнее время были оптимизированы за счет удаления дорогостоящих реагентов и пропускания некоторых этапов выделения, отсутствие которых не оказывает значительного влияния на синтез белка в целом.
Для получения клеточного экстракта клетки собираются в наиболее активной фазе жизнедеятельности организма – например, в середине логарифмической фазы роста для микроорганизмов; на 5-10 день образования эмбриона (например, для домашней курицы Gallus gallus). Клетки далее переносят в первичный буфер, который является базовой среды для клеточного экстракта, после чего производят лизис (разрушение) клеточной стенки. Клетки высших эукариот лизируются с помощью размораживания, пропускания через шприцевые насадки, лизирующих растворов, воздействием ультразвука, а также посредством растирания клеток в пробирках для гомогенизации со стеклянными шариками. Далее, после этапа лизиса раствор очищают от остатков разрушенной клеточной стенки и крупных агрегатов центрифугированием и отбирают надосадочную жидкость, в которой содержатся все необходимые биомолекулы для реакции синтеза белка. Для увеличения активности клеточного экстракта обычно проводят этап дотранслирования – т.е. увеличение количества вакантных рибосом за счет дополнительной инкубации в буфере с добавлением всех реагентов для синтеза белка и ферментов, чтобы все рибосомы закончили транслировать мРНК, содержащуюся в клеточном экстракте, и освободились от нее. За счет оптимизации условий получения компонент бесклеточных систем синтеза белка существует возможность снизить затраты на дорогостоящие компоненты, необходимые для дотранслирования, и в результате либо пропустить данный этап, либо осуществить его, не используя ферменты (“пустое дотранслирование”). В итоге полученный клеточный экстракт очищают от растворенных низкомолекулярных соединений при помощи диализа в конечном буферном растворе и замораживают в жидком азоте для длительного хранения до проведения экспериментов.
Другая важная часть бесклеточной системы синтеза белка — это ДНК, которая кодирует репортерную систему (белок-репортер) для детекции параметров, которая необходима для синтеза целевого белка. Наиболее использующиеся репортеры – различные варианты флуоресцентных белков, таких как зеленый флуоресцентный белок (Green fluorescent protein или GFP) флуоресцирующий на длине волны 395 нм или люминесцентные белки, такие как люцифераза. За счет регистрации уровня флюоресценции или люминесценции количественно определяют уровень синтеза белка в бесклеточной системе. Поскольку для бесклеточных систем синтеза белка необходимо использовать большое количество молекул ДНК, для каждого вида организма необходимо разрабатывать индивидуальный протокол ее получения. В заявленном техническом решении использовалась ДНК содержащая следующие элементы: промотор T7-полимеразы, Ω-сайт вируса табачной мозаики, ген sfGFP и поли(А)-хвост из 50 аденинов (поли(А)-хвост - обязательная часть мРНК эукариот). Тип элементов не ограничивается T7 регуляторными областями - можно тестировать любые другие наборы, включая естественные для исследованного организма области, а также другие вирусные последовательности, в том числе вирусов специфичных для птиц.
Работа бесклеточной системы синтеза белка требует больших количеств АТФ. И хотя бесклеточная система синтеза белка в какой-то мере, и сама может воспроизводить АТФ, с помощью присутствующих в ней собственных ферментных систем, для повышения эффективности синтеза белка необходимо добавление дополнительных компонент для регенерации АТФ. Самый распространенный подход – добавление экзогенных субстратов и соответствующих им ферментов: например, фосфоенол пируват и пируват киназа, ацетил фосфат и креатин фосфокиназа.
Для запуска реакции синтеза белка в смеси необходим подбор соотношений и концентраций индивидуальных компонент для каждого организма. Для транскрипции добавляют рибонуклеозидтрифосфаты или нуклеозидмонофосфат, а для трансляции дополнительный набор из протеиногенных аминокислот. Также можно добавлять: фолиновую кислоту, как донор формильных групп для фМет-тРНК; экзогенную T7 полимеразу для транскрипции информации с T7 промоторов; полиэтилен гликоль, чтобы имитировать внутриклеточную высоконцетрированную среду (концентрация белков в цитоплазме около 100-200 мг/мл); буферную систему для поддержания pH (HEPES, TRIS); соли калия и магния для контроля и регуляции их концентрации в системе.
В течение длительного периода времени технологии бесклеточных систем использовались как инструмент для решения узких задач в сфере фундаментальных исследований синтеза белка. В настоящее время бесклеточные системы синтеза белка активно используются в сфере синтетической биологии для создания модельных минимальных программируемых систем, дизайна белков, поиска биокатализаторов, для масштабной наработки целевых белков и молекул, для исследований процессов ингибирования синтеза белков.
Так, известны источники «An E. coli cell-free expression toolbox: application to synthetic gene circuits and artificial cells» (Бесклеточная система экспрессии на основе клеток кишечной палочки E.coli и ее применение в синтетических генных цепочках и искусственных клетках) [Shin J. Et al. An E. coli Cell-Free Expression Toolbox: Application to Synthetic Gene Circuits and Artificial Cells // ACS Synth. Biol, 2012. Vol.1. N. 1. P. 29–41] и «Development of a Bacillus subtilis cell-free transcription-translation system for prototyping regulatory elements» (Разработка бесклеточной системы транскрипции-трансляции на основе клеток Bacillus subtilis для прототипирования регуляторных элементов) [Kelwick R. et al. Development of a Bacillus subtilis cell-free transcription-translation system for prototyping regulatory elements // Metab. Eng, 2016. Vol. 38. P. 370-381] для тестирования элементов созданных на основе прокариот. Сущностью являются бесклеточные системы для тестирования регуляторных элементов и создания генетических конструкций, логических схем, осцилляторов бактерий.
Недостатком указанных систем является возможность применения только для бактерий и невозможность применения для эукариот в силу отличий в процессах транскрипции и трансляции, и компонент белоксинтезирующего аппарата.
Дальнейшее развитие синтетической биологии и создание биотехнологий на её основе требуют создания новых методов для тестирования регуляторных элементов генома и схем регуляции.
В настоящее время существует насущная необходимость в области технологий бесклеточного синтеза – несмотря на наличие множества бесклеточных систем синтеза белка для различных организмов, включая млекопитающих, все еще не существует таковой системы для птиц, которые являются важнейшей группой животных как с практической, так и с научной точки зрения. Птицы являются важнейший частью животноводства и чрезвычайно широко применяются в биотехнологиях – например, для производства вакцин. До сих пор не существует бесклеточной системы, в которой можно проводить исследования генетической регуляции в естественном генетическом и метаболическом окружении для клеток птиц. Кроме того, известные бесклеточные системы синтеза белка для млекопитающих устроены на основе культур клеток, что делают их недоступными для множества лабораторий, ввиду необходимости в дорогом лабораторном оборудовании. Данная проблема задерживает прогресс в исследовании общности регуляции, регуляции метаболизма эукариот и аппарата синтеза белка, создания in silico моделей метаболизма.
Известны источники «Wheat germ systems for cell-free protein expression» (Бесклеточная система экспрессии белка на основе клеток зародышей пшеницы) [Harbers M. Wheat germ systems for cell-free protein expression // FEBS Lett., 2014, Vol.25, 588(17), P. 2762-2773], «A versatile coupled cell-free transcription-translation system based on tobacco BY-2 cell lysates» (Сопряженная бесклеточная система транскрипции-трансляции на основе клеточных лизатов растений табака BY-2) [Buntru M. et al. A versatile coupled cell-free transcription-translation system based on tobacco BY-2 cell lysates // Biotechnol Bioeng, 2015. Vol. 112(5). P.867-878] и «Arabidopsis cell-free extract, ACE, a new in vitro translation system derived from Arabidopsis callus cultures» (Бесклеточный экстракт арабидопсиса, новая система in vitro трансляции на основе растительной культуры Arabidopsis callus) [Murota K. et al. Arabidopsis cell-free extract, ACE, a new in vitro translation system derived from Arabidopsis callus cultures // Plant Cell Physiol , 2011. Vol. 52(8). P. 1443-1453]. Сущностью данных технических решений в целом, являются бесклеточные системы, созданные на основе клеток растений, которые являются кеп-независимыми системами и уровень синтеза белка в которых регистрируется по сигналу желтого флуоресцентного белка (eYFP), зеленого флуоресцентного белка (GFP) и люминесценции люциферазы светлячка.
Недостатком указанных систем является то, что на их основе нельзя проводить исследования регуляторных элементов животных в естественном окружении, в частности птиц в силу отличий в процессах транскрипции и трансляции, и компонент белоксинтезирующего аппарата птиц по сравнению с растениями.
Известны источники «Leishmania cell-free protein expression system» (Бесклеточная система экспресии белка на основе клеток Leishmania tarentolae) [Kovtun O. et al. Leishmania cell-free protein expression system // Methods, 2011. Vol. 55(1). P. 58-64] и «Cell-free protein synthesis system prepared from insect cells by freeze-thawing» (Бесклеточная система синтеза белка из клеток насекомых путем заморозки-оттаивания) [Ezure T. Et al. Cell-free protein synthesis system prepared from insect cells by freeze-thawing // Biotechnol Prog, 2006. Vol. 22(6). P. 1570-1577]. Сущностью данных изобретений являются бесклеточные системы на основе лизатов клеток протистов (эукариотические организмы, не входящие в состав животных, грибов и растений) и насекомых с применением комбинации синтеза на основе ПЦР и инженерии трансляционных матриц в комбинированной системе транскрипции-трансляции.
Недостатком указанных систем является то, что на их основе нельзя проводить исследования регуляторных элементов животных в естественном генетическом и метаболическом окружении, в частности птиц, в силу отличий в процессах транскрипции и трансляции, и компонент белоксинтезирующего аппарата птиц по сравнению с протистами и насекомыми.
Известен источник «Cell-free protein synthesis enhancement from real-time NMR metabolite kinetics» (Бесклеточная система усиления синтеза белка на основе кинетики метаболитов в реальном времени по данным ядерного магнитного резонанса) [Panthu B. et al. Cell-free protein synthesis enhancement from real-time NMR metabolite kinetics: redirecting energy fluxes in hybrid RRL systems // ACS Synth. Biol, 2018. Vol. 7., N.1., P. 218-226]. Сущностью известного технического решения является оптимизированный метод in vitro синтеза белка на основе лизата ретикулоцитов кролика, анализируемый методом спектроскопии ядерного магнитного резонанса. Показано, что уменьшение доли рибосом в лизате ретикулоцитов кролика (RRL), приводит к созданию гибридных систем, состоящих из супернатанта RRL без рибосом, дополненного рибосомами из различных типов клеток млекопитающих. Гибридные системы RRL сохраняют трансляционные свойства исходных типов клеток с рибосомами и обеспечивают уровни экспрессии белков, аналогичные RRL. Показано, что способность к синтезу белка имеет верхний предел при концентрации нативной рибосомы и что меньшее количество рибосомальной фракции оптимизирует потоки энергии на трансляцию белка, следовательно, увеличивая выход бесклеточной системы синтеза белка из лизата ретикулоцитов кролика.
Недостатком известного технического решения является необходимость использования устаревших методов производства экстракта эритроцитов и невозможность проведения исследований регуляторных элементов птиц, в силу отличий в процессах транскрипции и трансляции, и компонент белоксинтезирующего аппарата птиц по сравнению с кроликами.
Известны источники «An efficient mammalian cell-free translation system supplemented with translation factors» (Эффективная бесклеточная система трансляции на основе клеток млекопитающих с добавлением трансляционных факторов) на основе культур человеческих раковых клеток HeLa [Mikami S. et al. An efficient mammalian cell-free translation system supplemented with translation factors // Protein Expr. Purif, 2006. Vol. 46(2). P. 348-357] и культур клеток яичника китайского хомячка «Cell-free protein expression based on extracts from CHO cells» (Бесклеточная экспрессия белка на основе экстрактов клеток яичников китайского хомячка) [Brödel A. K. et al. Cell-free protein expression based on extracts from CHO cells // Biotechnol. Bioeng, 2014. Vol. 111(1). P. 25-36]. Сущностью данных технических решений являются бесклеточные системы, созданные с использованием экстракта клеток HeLa, дополненным факторами трансляции eIF2, eIF2B, eIF4E и p97 (гомолог С-концового участка белка eIF4G) с усиленной трансляцией некэппированных мРНК, а также экстракта клеток яичников китайского хомячка, с применением специфического экспрессионного вектора in vitro, включающий участок внутренней посадки рибосомы (IRES) из межгенной области вируса паралича сверчка (CrPV), непосредственно связывающиеся с эукариотической 40S рибосомальной субъединицей, минуя процесс инициации трансляции, который часто является основным узким местом в бесклеточных системах.
Недостатком указанных технических решений является то, что для их подготовки необходим доступ к оборудованию для клеточных культур, а также то, что на их основе нельзя проводить исследования регуляторных элементов птиц в естественном генетическом и метаболическом окружении, в силу отличий в процессах транскрипции и трансляции, и компонент белоксинтезирующего аппарата птиц по сравнению с раковыми клетками человека и клетками хомяков.
Известен источник «A combined cell-free transcription-translation system from Saccharomyces cerevisiae for rapid and robust protein synthesis» (Комбинированная бесклеточная система транскрипции-трансляции из клеток дрожжей S.cerevisiae для быстрого и эффективного синтеза белка) [Gan R. et al. A combined cell-free transcription-translation system from Saccharomyces cerevisiae for rapid and robust protein synthesis // Biotechnol J, 2014. Vol. 9(5). P. 641-645]. Сущностью является бесклеточная система с возможностью работать с эукариотическими организмами с помощью бактериальных методов, что позволяет легко проводить генетические манипуляции и выращивать клетки в ферментерах и колбах.
Недостатком указанной системы является то, что на ее основе нельзя проводить исследования для животных в естественном генетическом и метаболическом окружении, в частности птиц, в силу отличий в процессах транскрипции и трансляции, и компонент белоксинтезирующего аппарата птиц по сравнению с клетками дрожжей.
Известно изобретение по патенту РФ №2148649 «Способ получения полипептидов в бесклеточной системе (варианты) и устройство для его осуществления». Сущностью является способ получения полипептидов в бесклеточной системе, предусматривающий приготовление реакционной смеси и питающей смеси, установку внутри реакционного модуля двух пористых барьеров, определяющих границы реакционного объема, введение реакционной смеси в реакционный объем и проведением синтеза при заданных условиях, поддержание синтеза введением питающей смеси, в процессе синтеза выводят из реакционного объема или оставляют внутри реактора синтезируемые целевые полипептиды, отличающийся тем, что перед проведение синтеза подготавливают смесь расходуемых высокомолекулярных компонентов, выбирают соотношение объема питающей смеси и расходуемых высокомолекулярных компонентов к объему фракции, содержащей целевой полипептид, устанавливают режим ввода питающей смеси и расходуемых высокомолекулярных компонентов в реакционный объем, в процессе синтеза продукты синтеза разделяют на фракцию, содержащую низкомолекулярные компоненты, и фракцию, содержащую высокомолекулярные компоненты синтеза, включая целевой полипептид, а основную часть фракции, содержащей низкомолекулярные компоненты, выводят из реакционного объема через одну или обе части второго пористого барьера.
Недостатком известного технического решения является ограничения по времени биосинтеза белков из-за вымывания компонент бесклеточной системы, отсутствующих в питающей смеси.
Известно изобретение по патенту РФ № 2162104 «Способ синтезирования флуоресцентно-меченого белка». Сущностью является способ синтезирования флуоресцентно-меченого белка в прокариотической бесклеточной системе белкового синтеза, предусматривающий следующие стадии: (а) инкубирование пробы рибосом, полученных из бесклеточного экстракта, с плазмидной ДНК, содержащей кодирующую последовательность для представляющего интерес белка, причем пробу инкубируют в среде для сопряженной транскрипции/трансляции вместе с аминоацил-тРНК, имеющей флуоресцентную метку, (б) частичную очистку указанного флуоресцентно-меченого белка отделением вновь синтезированного флуоресцентно-меченого белка от других флуоресцентных компонентов в этой пробе, (в) измерение количества синтезированного белка, (г) определение флуоресценции вновь синтезированного белка, (д) определение биологической активности вновь синтезированного белка.
Недостатком известного технического решения является возможность применения только для прокариот и невозможность применения для эукариот в силу отличий в процессах транскрипции и трансляции, и компонент белоксинтезирующего аппарата, необходимость дополнительного этапа очистки синтезированного флуоресцентно-меченого белка от других флуоресцентных компонентов в пробе.
Известно изобретение по патенту РФ № 2620074 «Рекомбинантная плазмидная ДНК pDUALREP2 и штамм, трансформированный ею для выявления веществ и смесей, ингибирующих биосинтез белка и/или вызывающих SOS-ответ у бактерий». Сущностью является рекомбинантная плазмидная ДНК pDualrep2, предназначенная для идентификации соединений, ингибирующих биосинтез белка и/или вызывающих SOS-ответ у бактерий, включающая: ориджин репликации, способный поддерживать стабильное число копий плазмиды в клетке; селективный маркер для отбора клеток, содержащих плазмиду; гены флюоресцентных белков с различимыми спектрами флюоресценции RFP и Katushka2S; аттенюатор триптофанового оперона, в котором произведены замены триптофановых кодонов UGG на аланиновые кодоны GCG; промотор гена sulA Е. coli, активирующийся при инактивации репрессора SOS-ответа LexA.
Недостатком известного технического решения является возможность применения только для бактерий и невозможность применения для эукариот в силу отличий в процессах транскрипции и трансляции, и компонент белоксинтезирующего аппарата.
Известно изобретение по патенту WO 2019035955 A1 опубл. 21.02.2019 «Высокопроизводительная система, использующая бесклеточную систему экспрессии и секвенирование in situ» («High-throughput system using a cell-free expression system and in situ sequencing»). Сущностью является метод скрининга множества матричных последовательностей нуклеиновых кислот для производства малых молекул, включающий в себя: синтез малых молекул, включающий добавление последовательности матричной нуклеиновой кислоты из набора в каждый реакционный объем в пределах множества реакционных объемов, где каждый реакционный объем включает бесклеточную экспрессионную систему в условиях транскрипции и трансляции; детектирование синтеза малой молекулы в реакционном объеме; секвенирование последовательности матричной нуклеиновой кислоты из реакционного объема.
Недостатком известного технического решения является применение клеточных лизатов из бактериальных клеток, дрожжевых клеток, клеток растений, клеток насекомых или клеток млекопитающих и невозможность применения для клеток птиц в силу отличий в процессах транскрипции и трансляции, и компонент белоксинтезирующего аппарата.
Известно изобретение по патенту US8012712B2 «Cell-free system for synthesis of proteins derived from cultured mammalian cells» (Бесклеточная система синтеза белков на основе культур клеток млекопитающих). Сущностью является Способ получения белка в системе бесклеточного синтеза белка, включающий добавление матричной мРНК и эукариотического фактора инициации трансляции к экстракту, полученному из культивируемых клеток млекопитающих, и инкубация их смеси, Способ по п. 1, отличающийся тем, что указанный эукариотический фактор инициации трансляции представляет собой один или несколько факторов, выбранных из группы, состоящей из эукариотических факторов инициации трансляции 4E (eIF4E), 2 (eIF2) и 2B (eIF2B), Способ по п. 1, отличающийся тем, что указанная матричная мРНК имеет кэп-структуру на своем 5'-конце, Способ по п. 1, отличающийся тем, что указанная матричная мРНК содержит внутренний сайт посадки рибосомы (IRES) вируса энцефаломиокардита (EMCV), а указанный эукариотический фактор инициации трансляции представляет собой один или оба эукариотических фактора инициации трансляции 2 (eIF2) и 2B (eIF2B), Способ получения белка в системе бесклеточного синтеза белка, включающий добавление некэпированной мРНК и одного или нескольких, выбранных из группы, состоящей из эукариотических факторов инициации трансляции 2 (eIF2) и 2B (eIF2B), и эукариотического регулятора трансляции p97, в экстракт, полученный из культивируемых клеток млекопитающих, и инкубирование их смеси, Способ по п. 1, отличающийся тем, что указанные культивируемые клетки млекопитающих представляют собой культивируемые клетки человека, Способ по п. 5, отличающийся тем, что указанные культивируемые клетки млекопитающих представляют собой культивируемые клетки человека, Способ по п. 6, отличающийся тем, что указанные культивируемые клетки человека представляют собой культивируемые клетки HeLa S3, Способ по п. 7, отличающийся тем, что указанные культивируемые клетки человека представляют собой культивируемые клетки HeLa S3, Композиция для получения белка с использованием бесклеточной системы, содержащая экстракт культивируемых клеток млекопитающих и один или несколько эукариотических факторов инициации трансляции, выбранных из группы, состоящей из факторов инициации трансляции 4E (eIF4E), 2 (eIF2) и 2B (eIF2B), Композиция по п. 10, дополнительно содержащая эукариотический регулятор трансляции р97, указанная композиция используется для бесклеточного синтеза белка с использованием некэпированной мРНК, Композиция по п. 11, отличающаяся тем, что содержащиеся в ней эукариотические факторы инициации трансляции представляют собой как eIF2, так и eIF2B, Композиция по п. 10, отличающаяся тем, что указанные культивируемые клетки млекопитающих представляют собой культивируемые клетки человека, Композиция по п. 13, отличающаяся тем, что указанные культивируемые клетки человека представляют собой культивируемые клетки HeLa S3.
Недостатками известного технического решения в отношении композиции являются: необходимость добавления дополнительных белков (eIF4E, eIF2, eIF2B, p97); невозможность применения для клеток птиц, так как известное изобретение предназначено для работы с клетками человека.
Основываясь на изложенном выше возможно сделать вывод о том, что известные технические решения совпадают с заявленным техническим решением по отдельным признакам, однако не совпадают по назначению, поэтому формула изобретения составлена без ограничительной части.
Техническим результатом заявленного технического решения является расширение арсенала средств указанного назначения путем создания бесклеточной системы синтеза белка на основе эмбриональных клеток домашней курицы Gallus gallus и проведение синтеза белка на её основе, при этом достигается:
-возможность проведения исследования регуляторных элементов птиц в естественном генетическом и метаболическом окружении.
-отсутствие необходимости добавления дополнительных белковых факторов (eIF4E, eIF2, eIF2B, p97);
-отсутствие необходимости производства экстракта эритроцитов.
Кроме того, предложенная группа изобретений может стать платформой для тестирования влияния антибиотиков, используемых в животноводстве. Система потенциально способна показать, насколько новый антибиотик может быть токсичен для сельскохозяйственных птиц, например, Gallus gallus. Изобретение также может стать основой для проведения исследований вирусов птиц.
Сущностью заявленного технического решения является бесклеточная система синтеза белка на основе эмбриональных клеток Gallus gallus содержащая, об.%: экстракт на основе эмбриональных клеток Gallus gallus из инкубированных яиц – 35; оптимизированный набор компонентов для проведения реакции синтеза – 44,93, включающий: HEPES-KOH pH7.5 - 2,50, дитиотреитол - 2,00, NTP микс: 90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ, pH~7 - 1,67, магний глутамат - 5,30, калий глутамат - 4,03, раствор из 20 протеиногенных аминокислот - 2,50, тРНК из Saccharomyces cerevisiae - 0,23, полиэтиленгликоль 8000 - 4,00, спермидин - 0,75, RNasine - 0,75, T7 полимераза в глицерине 50% - 2,50, вода - 18,46; – ПЦР-продукт, кодирующий флуоресцентный белок – 12,37; – система регенерации АТФ – 7,7, включающая: креатин фосфат – 5,0, креатин фосфокиназу – 2,7. Способ синтеза белка с помощью бесклеточной системы синтеза белка на основе эмбриональных клеток Gallus gallus по п.1, заключающийся в том, что исходные компоненты смешиваются в следующей последовательности и об.%: вода - 18,46; HEPES-KOH pH7.5 - 2,50; дитиотреитол - 2,00; магний глутамат - 5,30; калий глутамат - 4,03; раствор из 20 протеиногенных аминокислот - 2,50; тРНК из Saccharomyces cerevisiae - 0,23; полиэтиленгликоль 8000 - 4,00; спермидин - 0,75; креатин фосфат – 5,0; NTP микс: 90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ, pH~7 - 1,67; RNasine - 0,75; креатин фосфокиназа – 2,7; T7 полимераза в глицерине 50% - 2,50; экстракт на основе эмбриональных клеток Gallus gallus из инкубированных яиц – 35; ПЦР-продукт, кодирующий флуоресцентный белок – 12,37; выдерживают при температуре 37 °С в течение 6 часов, измеряют флюоресценцию, получают значение количества синтезированного белка.
Заявленное техническое решение иллюстрируется Фиг.1 – Фиг.3.
На Фиг 1. представлена Таблица, в которой приведен протокол приготовления 100 мкл реакционной смеси.
На Фиг. 2 представлена схема линейного участка ДНК, состоящего из T7-промотора, Ω-сайт TMV, гена sfGFP и поли(А)-хвоста длинной 50 нуклеотидов. Данный фрагмент получается после двух последовательных ПЦР реакций, где: 5’ UTR –5′-нетранслируемая область, ORF - открытая рамка считывания, 3’ UTR –3′-нетранслируемая область, PolyA – полиадениновый хвост. Стрелками отмечено начало и конец диапазона.
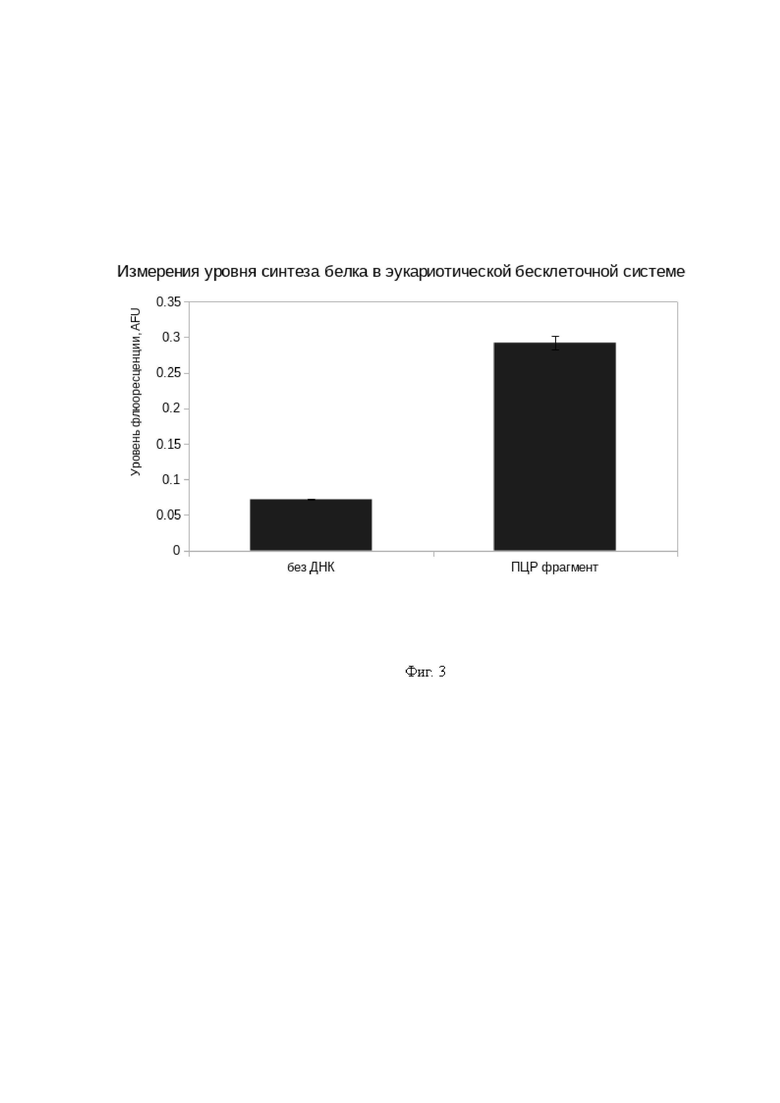
На Фиг. 3 представлены результаты тестирования бесклеточной системы синтеза белка по конечной точке после 6 часов инкубации. Без ДНК - негативный контроль без добавления ДНК; ПЦР фрагмент – тестируемый образец с добавлением матрицы для синтеза белка-репортера sfGFP. AFU – Arbituary Fluorescense Units (Относительные единицы измерения флуоресценции)
Далее заявителем приведено описание заявленного технического решения.
Заявленное изобретение предназначено для синтеза белков в бесклеточной системе на основе эмбриональных клеток домашней курицы Gallus gallus – например, для тестирования регуляторных элементов для изучения регуляции метаболизма эукариот и аппарата синтеза белка у птиц.
Заявленный технический результат в целом достигается тем, что на основе эмбриональных клеток Gallus gallus подготавливают экстракт, подготавливают и (при необходимости) оптимизируют систему синтеза белка на основе полученного экстракта, проводят реакцию синтеза белка и используют синтезированный белок, например – для детекции сигнала с целью оценки эффективности синтеза.
Все используемые реагенты являются коммерчески доступными.
Заявленная бесклеточная система синтеза белка содержит:
– экстракт на основе эмбриональных клеток Gallus gallus из инкубированных яиц – 35;
– оптимизированный набор компонентов для проведения реакции синтеза – 44,93, включающий:
HEPES-KOH pH7.5 - 2,50,
дитиотреитол (DTT) - 2,00,
NTP микс (90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ), pH~7 - 1,67,
магний глутамат - 5,30,
калий глутамат - 4,03,
раствор из 20 протеиногенных аминокислот - 2,50,
тРНК из Saccharomyces cerevisiae - 0,23,
полиэтиленгликоль (ПЭГ) 8000 - 4,00,
спермидин - 0,75,
RNasine - 0,75,
T7 полимераза в глицерине 50% - 2,50,
вода - 18,46;
– ПЦР-продукт с возможностью изменения регуляторных элементов – 12,37;
– система регенерации АТФ – 7,7, включающая:
креатин фосфат – 5,0,
креатин фосфокиназу – 2,7.
При этом представленные выше исходные компоненты заявленной системы получают следующим образом:
-экстракт на основе эмбриональных клеток Gallus gallus из инкубированных яиц подготавливают с помощью ультрацентрифугирования и диализа, без проведения этапа дотранслирования;
-клетки лизируют с помощью пробирки Поттера;
-ПЦР-продукт с возможностью изменения регуляторных элементов получают выделением ДНК из агарозного геля и реакционных смесей;
-оптимизированный набор компонентов для проведения реакции синтеза получают путем растворения компонентов в деионизованной воде;
– сигнал детектируется с помощью измерения флюоресценции или люминесценции в планшетном спектрофотометре.
Таким образом, заявленное техническое решение включает этапы подготовки экстракта эукариотических клеток домашней курицы Gallus gallus, проведения реакции синтеза и детекции сигнала. Экстракт для проведения реакций изготавливается из клеток эмбриона куриц, полученных из инкубированных яиц. Полученный экстракт выступает основой для проведения программируемой реакции по синтезу белка. Для программирования системы используется линейная ДНК, кодирующая репортерный белок (например, флуоресцентный белок sfGFP) и регуляторные элементы – промоторы, терминаторы, сайленсеры, энхансеры - которые можно заменять с помощью молекулярного клонирования для их тестирования. Описание изобретения содержит протокол проведения реакции синтеза с оптимизированным набором реагентов, добавляемых в реакцию синтеза, условия проведения реакции и условия измерения сигнала эквивалентному количеству репортерного белка по уровню флюоресценции на планшетном спектрофотометре.
Далее заявителем приведен способ получения заявленной бесклеточной системы синтеза белка на основе эмбриональных клеток Gallus gallus и заявленный способ синтеза белка на основе бесклеточной системы синтеза белка эмбриональных клеток.
Куриные оплодотворенные яйца инкубируют при 37 °С и относительной влажности 60% в течение 5 дней. Яйца отмывают и вскрывают, при присутствии эмбриона ножницами эксцизируют все лишнее и переносят в пробирку, содержащую буфер А (20 мM HEPES–KOH pH 7.5, 100 мМ калий ацетат, 2 мM магний ацетат, 2 мM DTT, без ингибиторов протеаз). В среднем из 10 инкубированных оплодотворенных яиц только 5-7 яиц содержат эмбрионы. Далее отобранные эмбрионы отмывают в буфере А несколько раз. Эмбрионы лизируются в гомогенизаторе Поттера с добавлением буфера А с ингибиторами протеаз. Отобранный лизат далее центрифугируют на скорости 9000 g, при 4 °С в течение 15 мин – далее работают с супернатантом. Отобраный супернатант (экстракт) диализуется 12 часов против буфера А без добавления ингибиторов протеаз. Далее экстракт центрифугируют при 20000 g, при 4 °C в течение 10 мин, измеряют концентрацию, экстракт аликвотируют и замораживают в жидком азоте для дальнейшего использования.
Для приготовления линейного продукта ДНК для программирования бесклеточной системы проводят две полимеразных цепных реакции (ПЦР) с праймерами, которые добавляют необходимые регуляторные элементы в два этапа (Фиг. 2). В качестве матрицы для проведения ПЦР используют плазмиду (например, pJL1-sfGFP) или участок ДНК, содержащие необходимый белок-репортер (например, белок sfGFP). После проведения первой полимеразной реакции проводят очистку необходимого фрагмента с помощью электрофореза и очистки из геля с помощью набора для выделения ДНК из агарозного геля и реакционных смесей. После второй реакции ПЦР также проводят очистку реакционной смеси с помощью набора.
В заявленном техническом решении используются ген репортера sfGFP, полученный из плазмиды pJL1-sfGFP (Addgene, #102634). sfGFP это часто используемый белок-репортер для бесклеточных систем синтеза белка. Его отличительные характеристики – способность давать сигнал флюоресценции, который можно легко детектировать почти всеми планшетными спектрофотометрами, а также быстрое время созревания, что позволяет отслеживать синтез белка в динамике. В системе можно использовать и другие репортеры.
В заявленном техническом решении используются специализированные вирусные сайты (IRES), которые выступают сайтами связывания рибосомы бактериального типа (RBS). При такой схеме, кепирование мРНК не требуется, что удешевляет систему. При этом остается возможность использовать и кепированную мРНК.
Реакцию синтеза в разрабатываемой бесклеточной системе проводят, например, в планшетах с объемом реакционной смеси в лунке около 10-20 мкл. Реакционные реагенты добавляют согласно оптимизированному набору компонентов (Таблица на Фиг. 1), а именно: вода, HEPES-KOH, дитиотреитол, магний глутамат, калий глутамат, раствор из 20 протеиногенных аминокислот, тРНК из Saccharomyces cerevisiae, полиэтиленгликоль 8000, спермидин, креатин фосфат, NTP микс, Rnasine, креатин фосфокиназа, T7 полимераза в глицерине 50%, экстракт на основе эмбриональных клеток Gallus gallus из инкубированных яиц, ПЦР-продукт с возможностью изменения регуляторных элементов. Реакционную смесь выдерживают при температуре 37 °С в течение 6 часов, например, в луночном планшете. Измеряют уровнень флуоресценции, например, в спектрофотометре. Таким образом, получают измерение флуоресценции по конечной точке. Получают значение количества синтезированного белка.
Далее приведены примеры конкретного осуществления изобретения и реализации назначения, при этом заявитель подразумевает, что приведенные примеры не ограничивают предполагаемый объем заявленного технического решения.
Пример 1. Получение заявленной бесклеточной системы синтеза белка на основе эмбрионального белка домашней курицы Gallus gallus.
1. Получают экстракт из эмбриональных клеток Gallus gallus – компонент заявленной системы.
Для этого куриные оплодотворенные яйца, например - породы "араукан", полученные из ИП«Серебряное копытце» (Республике Татарстан, г. Казани) закладывают в инкубатор при 37,5 °С и относительной влажности 60%, с автоматическим пятикратным переворачиванием яиц. Заложенные оплодотворенные яйца породы кур "араукан", инкубируют в течение 5 дней, контролируя эмбриональное развитие с помощью овоскопа качества яиц «Алиса». Оплодотворенные яйца отбирают из инкубатора и доставляют лабораторию в инсулированном боксе без поддержки температуры. Яйца отмывают теплой водой с мылом, промывают водой и дистиллятом, протирают спиртом и осушают на комнатной температуре. Сухие яйца вскрывают с заостренной стороны с помощью стерильных ножниц и содержимое аккуратно выливают в чашку Петри. При присутствии эмбриона ножницами отрезают все лишнее и с помощью 1 мл купированного носика для пипетки содержимое переносят в пробирку-фалькон на 50 мл, содержащий Буфер А (20 мM HEPES–KOH pH 7.5, 100 мМ калий ацетат, 2 мM магний ацетат, 2 мM DTT, без ингибиторов протеаз). В среднем из 10 инкубированных оплодотворенных яиц эмбрионы содержат около 5-7 яиц. После сбора всех эмбрионов в одну пробирку-фалькон, эмбрионы отмывают в буфере: заполняют буфером А до максимального объема и аккуратно декантируют – повторяют 3-5 раз. Далее эмбрионы переносят в пробирку-гомогенизатор Поттера для лизиса растиранием по 2-3 эмбриона, добавляли 1 объём буфера А с добавлением Protase Inhibitor Cocktail (PIC, Roshe) и протирают шпателем около 20 раз до полной гомогенизации. Лизат собирают в отдельную пробирку и промывают лизирующую пробирку буфером А для добора материала со стенок. Весь лизат далее ресуспендируют и аликвотируют по пробиркам на 2 мл, центрифугируют на скорости 9000 g, 4 °С в течение 15 мин и отбирают супернатант. Отобранный супернатант (экстракт) переносят в новую пробирку и далее в мешок для диализа. Диализ проводят в диализных мешочках c мембраной RC 12-14 кДа (например - Spectrum Labs Spectra/Por), мембрану промывают дважды дистилированной H2O, кипятят 5 минут в 2% NaHCO3 с 1 мкМ EDTA и снова промывают дважды дистилированной H2O и деионизованной водой (milliQ). Проводят диализ против буфера А без PIC (20 мM HEPES–KOH pH 7.5, 100 мМ калий ацетат, 2 мM магний ацетат, 2 мM DTT). Объём образца к буферу - 1:100. Для этого меняют буфер в конце первого и второго часа, далее диализируют в течение 12 часов. Полученный экстракт переносят в 1.5 мл пробирки типа эппендорф и центрифугируют при 20000 g, 4 °C в течение 10 мин, измеряют концентрацию (А260, А280, и методом Бредфорда), аликвотируют и замораживают в жидком азоте.
2. Получают ПЦР продукт – компонент заявленной системы.
Для этого проводят первую ПЦР с праймерами QEGFP-f (TTACTATTTACAATTACATATGAGCAAAGGTGAAGAAC) и QEGFP-r (AGCAGCCGGATCTCAGTTTATTTTTCGAACTGCGGA). Состав реакционной смеси для проведения ПЦР реакции: mQ вода, праймер 1, праймер 2, смесь нуклеотидов dNTPs (например – «Евроген», Москва), плазмида (например – pJL1 – sfGFP, Addgene , #102634), полимераза, например – смесь Phusion Green HS Polymerase High-Fidelity PCR mix («Thermo Scientific», #00696696, США). ПЦР проводят на амплификаторе, например – T100 производства компании «BioRad» (США). Температурный режим ПЦР: 1x – 30 сек, 98 °C; 35х - 10 сек, 95 °C + 15 сек, 44 °C + 30 сек, 72 °C; 1x – 240 сек, 72 °C. Продукт ПЦР: 759 п.н., концентрация 146,6 нг/мкл.
Для проведения второй ПЦР используют праймеры QET7Ome-f: AC-GCTGCCCGAGATCTCGATCCCGCGAAATTAATACGACTCACTATAGGGAG-TATTTTTACAACAATTAC-CAACAACAACAAACAACAAACAACATTACAATTACTATTTACAATTACAT и QEGFP-r: TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTGTTAG-CAGCCGGATCTCAGT. Состав реакционной смеси для проведения ПЦР реакции: mQ вода, праймер 1, праймер 2, смесь нуклеотидов dNTPs (например – «Евроген», Москва), плазмида (например – pJL1 – sfGFP, Addgene , #102634), полимераза, например – смесь Phusion Green HS Polymerase High-Fidelity PCR mix («Thermo Scientific», #00696696, США). Температурный режим ПЦР: 1x – 30 сек, 98 °C; 5х - 10сек, 95 °C + 15 сек, 35 °C + 30 сек, 72 °C; 30х - 10 сек, 95 °C + 15 сек, 61 °C + 30 сек, 72 °C; 1x – 240 сек, 72 °C. Продукт ПЦР: 913 п.н., концентрация 173,1 нг/мкл.
Далее проводят очистку полученного ПЦР продукта. Для этого после горизонтального электрофореза нуклеиновых кислот в 1,5% агарозном геле, например – на системе для электрофореза «BioRad». ПЦР продукт далее выделяют из геля, например – с помощью коммерческого набора для выделения ДНК из агарозного геля и реакционных смесей («Евроген», Москва).
3. Берут оптимизированный набор компонентов для проведения реакции синтеза, включающий систему регенерации АТФ из креатин фосфата и креатин фосфокиназы (Таблица на Фиг.1).
4. Смешивают указанные выше компоненты при комнатной температуре согласно последовательности, указанной в Таблице на Фиг.1, а именно: вода, HEPES-KOH, дитиотреитол, магний глутамат, калий глутамат, раствор из 20 протеиногенных аминокислот, тРНК из Saccharomyces cerevisiae, полиэтиленгликоль 8000, спермидин, креатин фосфат, NTP микс, Rnasine, креатин фосфокиназа, T7 полимераза в глицерине 50%, экстракт на основе эмбриональных клеток Gallus gallus из инкубированных яиц, ПЦР-продукт с возможностью изменения регуляторных элементов – до получения однородного раствора. Получают заявленную систему при следующем соотношении компонентов, об.% (Таблица на Фиг.1):
– экстракт на основе эмбриональных клеток Gallus gallus из инкубированных яиц – 35;
– оптимизированный набор компонентов для проведения реакции синтеза – 44,93, включающий:
HEPES-KOH pH7.5 - 2,50,
дитиотреитол (DTT) - 2,00,
NTP микс (90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ), pH~7 - 1,67,
магний глутамат - 5,30,
калий глутамат - 4,03,
раствор из 20 протеиногенных аминокислот - 2,50,
тРНК из Saccharomyces cerevisiae - 0,23,
полиэтиленгликоль (ПЭГ) 8000 - 4,00,
спермидин - 0,75,
RNasine - 0,75,
T7 полимераза в глицерине 50% - 2,50,
вода - 18,46;
– ПЦР-продукт с возможностью изменения регуляторных элементов – 12,37;
– система регенерации АТФ – 7,7, включающая:
креатин фосфат – 5,0,
креатин фосфокиназу – 2,7.
Пример 2. Проведение синтеза белка на основе бесклеточной системы синтеза белка эмбриональных клеток.
Проводят реакцию по синтезу белка в разрабатываемой бесклеточной системе. Для этого готовят реакционную смесь в планшетах на 384 лунки (например - Grenier), объем лунок 20мкл, объем реакционной смеси в лунке 15 мкл. Список реагентов и последовательность смешения реагентов приведены в Таблице на Фиг.1 и в Примере 1. Измерение проводят по конечной точке: реакцию проводят в пробирках, например, на 1,5 мл (Eppendorf) и инкубируют на температуре 37 °С в течение 6 часов, далее реакционную смесь раскапывают, например, в луночный планшет и ставят в спектрофотометр, например - Varioskan LUX, Thermo для измерения флюоресценции с параметрами Ex = 485 нм, Em = 510 нм.
Наличие сигнала флуоресценции при внесении ДНК в виде ПЦР-фрагмента, превышающего сигнал без внесения ДНК (Фиг. 3) свидетельствует о наличии флуоресцентного белка sfGFP и, тем самым, об успешном синтезе белка с использованием бесклеточной системы на основе эмбриональных клеток домашней курицы Gallus gallus, полученной в соответствии с описанием выше.
Таким образом, проведение манипуляций в соответствии с описанием выше, позволяет достичь заявленной цели – получить бесклеточную системы синтеза белка на основе эмбриональных клеток Gallus gallus и провести синтез белка на её основе.
Приведенные примеры осуществления заявленного изобретения показывают успешное создание бесклеточной системы синтеза белка на основе эмбриональных клеток Gallus gallus и проведение синтеза белка с на её основе, с использованием стандартного оборудования и материалов, полезность для применения в биотехнологиях, например – для получения репортерных белков.
На основании изложенного выше можно сделать вывод, что заявителем достигнут заявленный технический результат, а именно - расширен арсенал средств указанного назначения путем создания бесклеточной системы синтеза белка на основе эмбриональных клеток домашней курицы Gallus gallus и проведен синтез белка на её основе, при этом достигнуто:
-возможность проведения исследования регуляторных элементов птиц в естественном генетическом и метаболическом окружении;
-отсутствие необходимости добавления дополнительных белковых факторов (eIF4E, eIF2, eIF2B, p97);
-отсутствие необходимости производства экстракта эритроцитов.
Кроме того, предложенная группа изобретений может стать платформой для тестирования влияния антибиотиков, используемых в животноводстве. Система потенциально способна показать, насколько новый антибиотик может быть токсичен для сельскохозяйственных птиц, например, Gallus gallus. Изобретение также может стать основой для проведения исследований вирусов птиц.
Заявленное техническое решение удовлетворяет критерию «новизна», предъявляемому к изобретениям, так как при определении уровня техники не выявлены средства, которым присущи признаки, идентичные (то есть совпадающие по исполняемой ими функции и форме выполнения этих признаков) всем признакам, перечисленным в формуле предлагаемого изобретения, включая характеристику назначения.
Заявленное техническое решение удовлетворяет критерию «изобретательский уровень», т.к. не является очевидным для специалиста в данной области техники и из исследованного уровня техники заявителем не выявлены технические решения, совпадающие по технической сущности с предлагаемым решением, и не установлена известность влияния отличительных признаков на полученный технический результат.
Заявленное техническое решение соответствует условию патентоспособности «промышленная применимость», предъявляемому к изобретениям, так как может быть изготовлено с использованием известных материалов и оборудования.


Изобретение относится к области биотехнологии. Описана бесклеточная система синтеза белка на основе эмбриональных клеток Gallus gallus, содержащая экстракт на основе эмбриональных клеток Gallus gallus из инкубированных яиц, а также оптимизированный набор компонентов для проведения реакции синтеза. Также описан способ синтеза белка с помощью бесклеточной системы синтеза белка на основе эмбриональных клеток Gallus gallus. Техническим результатом заявленного технического решения является расширение арсенала средств указанного назначения путем создания бесклеточной системы синтеза белка на основе эмбриональных клеток домашней курицы Gallus gallus и проведения синтеза белка на её основе. 2 н.п. ф-лы, 3 ил., 2 пр.
1. Бесклеточная система синтеза белка на основе эмбриональных клеток Gallus gallus, содержащая, об.%:
– экстракт на основе эмбриональных клеток Gallus gallus из инкубированных яиц - 35;
– оптимизированный набор компонентов для проведения реакции синтеза - 44,93, включающий:
HEPES-KOH pH 7.5 - 2,50,
дитиотреитол - 2,00,
NTP микс: 90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ, pH~7 - 1,67,
магний глутамат - 5,30,
калий глутамат - 4,03,
раствор из 20 протеиногенных аминокислот - 2,50,
тРНК из Saccharomyces cerevisiae - 0,23,
полиэтиленгликоль 8000 - 4,00,
спермидин - 0,75,
RNasine - 0,75,
T7 полимераза в глицерине 50% - 2,50,
вода - 18,46;
– ПЦР-продукт, кодирующий флуоресцентный белок - 12,37;
– система регенерации АТФ - 7,7, включающая:
креатин фосфат - 5,0,
креатин фосфокиназу - 2,7.
2. Способ синтеза белка с помощью бесклеточной системы синтеза белка на основе эмбриональных клеток Gallus gallus по п.1, заключающийся в том, что исходные компоненты смешиваются в следующей последовательности и об.%: вода - 18,46; HEPES-KOH pH 7.5 - 2,50; дитиотреитол - 2,00; магний глутамат - 5,30; калий глутамат - 4,03; раствор из 20 протеиногенных аминокислот - 2,50; тРНК из Saccharomyces cerevisiae - 0,23; полиэтиленгликоль 8000 - 4,00; спермидин - 0,75; креатин фосфат - 5,0; NTP микс: 90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ, pH~7 - 1,67; RNasine - 0,75; креатин фосфокиназа - 2,7; T7 полимераза в глицерине 50% - 2,50; экстракт на основе эмбриональных клеток Gallus gallus из инкубированных яиц - 35; ПЦР-продукт, кодирующий флуоресцентный белок - 12,37; выдерживают при температуре 37°С в течение 6 часов, измеряют флюоресценцию, получают значение количества синтезированного белка.
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДОВ В БЕСКЛЕТОЧНОЙ СИСТЕМЕ | 1999 |
|
RU2169154C2 |
| US 8012712 B2 (HIROAKI IMATAKA YOKOHAMA JP JP OMITTED SATOSHI MIKAMI YOKOHAMA JP JP OMITTED SHIGEYUKI YOKOYAMA YOKOHAMA JP JP OMITTED ), 06.09.2011 | |||
| WO 2014144583 A3 (NORTHWESTERN UNIVERSITY 633 CLARK STREET, EVANSTON, IL 60208 US 60208 US US ), 18.09.2014 | |||
| Gan R | |||
| et al | |||
| A combined cell-free | |||

