Область техники, к которой относится изобретение
Изобретение относится к молекулярной биологии, генной инженерии и биотехнологии. В частности в изобретении раскрывается способ получения экстракта из растительных соматических зародышей, который может быть использован для синтеза белка, в том числе сопряженного с процессом транскрипции, в бесклеточной системе, которая обеспечивает различные варианты посттрансляционной модификации белкового продукта, характерные для эукариотических клеток. Полученные с использованием изобретения белковые продукты могут быть использованы в медицине, ветеринарии, сельском хозяйстве, пищевой промышленности, например в терапевтических или профилактических целях, для иммунизации, для создания диагностических препаратов (на основе антител, искусственных антигенов, аллергенов), наработки биологически активных соединений белковой природы, получения ферментов, используемых в производстве продуктов питания, а также для научных исследований.
Уровень техники
В последние годы резко возрос интерес к биологически активным полипептидам в связи с открытием совершенно новых их биологических функций. Для примера можно привести такие чрезвычайно интересные и важные объекты, как пептиды-активаторы иммунной системы, пептиды-нейромедиаторы и трансмиттеры, пептиды-регуляторы солевого обмена, пептиды гуморального иммунного ответа, обладающие антибактериальными свойствами и многие другие. Доступность этих пептидов из природных источников практически равна нулю. Ценой колоссальных усилий многих исследователей, используя в основном генетические методы, в лучшем случае, удается установить их структуру и проследить за их генезисом. В большинстве случаев эти пептиды синтезируются в организме в виде достаточно больших белков - предшественников, которые в результате сложного многоступенчатого процессинга превращаются в конечные биологически активные регуляторы тех или иных внутриклеточных процессов. Как следствие этого, резко возрос интерес к созданию новых и совершенствованию существующих методов получения полипептидов. Несмотря на достигнутый в последние годы прогресс в использовании химического синтеза для получения полипептидов, а также использование рекомбинантной техники, ряд важных и интересных биологически активных полипептидов, в первую очередь пептидов длиной в 30-100 аминокислотных остатков, остается до настоящего времени недоступным или малодоступным для широкого использования. Недостатки химического синтеза обусловлены, главным образом, рядом технологических и экономических ограничений. К технологическим ограничениям относятся рацемизация, неполнота удлинения полипептидной цепи на каждой стадии, сложности с выбором защитных групп и их удалением и, что самое важное, сложности с реализацией функционально активной конформации в деблокированном достаточно длинном полипептиде. Экономические ограничения связаны с тем, что стоимость конечного продукта синтеза растет в геометрической прогрессии по мере увеличения длины его полипептидной цепи.
Методы экспрессии чужеродного генетического материала в живых клетках также имеют ряд ограничений, особенно в отношении полипептидов сравнительно небольшой длины (до 50-60 аминокислотных остатков). Ограничения сводятся, в основном, к сложности получения генетического материала, поскольку для многих из таких объектов неизвестен биогенезис, а конечные активные пептиды являются, в большинстве случаев, продуктами сложного процессинга предшественников. В результате, возникают труднопреодолимые сложности в создании генетических конструкций, экспрессия которых в прокариотических или эукариотических клетках приводила бы к активным продуктам. Необходимо отметить, что даже при успешном создании экспрессирующей системы возникает ряд трудностей. Чужеродные гены могут быть нестабильными или плохо экспрессироваться в результате действия регуляторных механизмов клетки хозяина. Во многих случаях продукты чужеродных генов нерастворимы и формируют агрегаты в виде телец включения, что вызывает дополнительные сложности при получении биологически активных молекул. Другие полипептиды являются нестабильными и быстро деградируют под действием внутриклеточных протеаз. Наконец, продукты экспрессии в ряде случаев могут оказывать угнетающее действие на клетку или оказаться токсичными для нее и поэтому не могут быть экспрессированы.
Вместе с тем, успешное развитие методов молекулярной биологии и биохимии открывает принципиально новые возможности и подходы к решению задачи синтеза пептидов, а именно: i) использование бесклеточной системы трансляции для рибосомного синтеза биологически активных пептидов на искусственно синтезированных или естественных матрицах и ii) использование нерибосомных мультиферментных систем синтеза пептидов.
Возможности использования бесклеточных систем трансляции для микропрепаративного синтеза полипептидов обусловлены следующими обстоятельствами:
i) разработаны эффективные методы синтеза заданных дезоксинуклеотидных последовательностей достаточно большой длины (Engels J., Uhlmann E., (1988) in Advances in Biochemical Engineering/Biotechnology, v.37 (A.Fichter eds.) pp.75-127; Erlich H.A. (1989) PCR Technology: Principles and Applications for DNA Amplification, Stockholm, New York). (Здесь и далее все литературные источники включены в описание путем отсылки);
ii) разработаны системы размножения матричных РНК вплоть до препаративных количеств с использованием фаговых полимераз (Campbell J.L., Richardson C.C., Studier F.W. (1978) Proc.Natl.Acad.Sci. USA 75, 2276-2280; Butler E.T., Chamberlin M.J. (1982). J.BioI.Chem. 257, 5772-5778; Melton D.A., Krieg. P.A., Rebagliati M.R., et al. (1984) Nucleic Acids Research 12, 7035-7056);
iii) отлажены прокариотические и эукариотические бесклеточные системы трансляции естественных и искусственных матриц (Pratt J.M. (1984) in Transcription and Translation (Hames B.D., Higgins S.J., eds) pp. 179-210; Clemons M. (1984) in Transcription and Translation (Hames B.D., Higgins S.J. eds) pp.231-270);
iiii) разработаны эффективные системы выделения полипептидов из сложных смесей.
Первые системы для бесклеточного синтеза белков были разработаны в лаборатории Zamechnic (Hoagland M.B., Stephenson M.L., Scott J.F., et al. (1958) J.BioI.Chem, 231, 241-257), где было показано, что пептидный синтез идет на рибосоме и требует АТР, GTP и tRNA. Первый белок с завершенной полипептидной цепью, синтезированный в бесклеточной системе трансляции на основе экстракта из Е.coli, был белком оболочки колифага f2 (Nathans D., Notani G., Schwartz J.H., Zinder N.D. (1962) Proc.Natl. Acad.Sci.USA, 48, 1424-1431).
Несколько позже DeVries и Zubay ((1967) Proc.Natl.Acad.Sci. USA, 57, N 4, 1010-1012) продемонстрировали синтез части полипептидной цепи b-галактозидазы в сопряженной системе транскрипции-трансляции. Со времени первых публикаций (Zubay G. (1973) Ann.Rev.Genet 7, 267-287; DeVries J., Zubay G. (1969) J.Bacterial 97,N 3, 1419-1425; Gold L.M., Schweiger M. (1971) In: Methods in Enzymology, v.20, (PTC), (Moldave K., Grossman eds.) Acad. Press.Inc., New York, 537-542) бесклеточная система трансляции на основе грубых клеточных экстрактов (S-30) из Е.coli или реконструированная система на основе экстракта S-100 претерпела небольшие изменения (Collins J. (1979) Gene 6,N 1, 29-42; Pratt J.M., Boulnois G.J., Darby V., et al. (1981) Nucleic Acid.Res. 9, 4459-4474) и в настоящее время широко используется для исследования механизмов трансляции и ее регуляции, транслокации через или интеграции в мембраны, процессов ко- и посттрансляционного сворачивания белков, модификации и сборки олигомерных белков и многое другое.
Кроме бесклеточных систем на основе бактериальных клеток для трансляции эукариотических матричных РНК используются экстракты многих клеточных культур и перевиваемых клеток асцитных опухолей. Наибольшее распространение получили лизаты ретикулоцитов кролика и экстракты зародышей пшеницы (Clemons M. (1984) in Transcription and Translation (Hames B.D., Higgins S.J. eds) 231-270; Roberts B.E., Paterson B.M. (1973) Proc.Natl.Acad.Sci. USA 70, №4, 2330-2334).
Однако различные варианты бесклеточных систем трансляции, описанные к настоящему времени, обладают общим недостатком - крайне низкой эффективностью синтеза полипептида. Обычно с одной молекулы мРНК синтезируется не более 2-3 молекул полипептида. Это обстоятельство в значительной степени снижает потенциальные возможности бесклеточной экспрессирующей системы для выполнения тех исследований, для осуществления которых необходимы значительные количества изучаемого полипептида.
Для расширения возможностей использования бесклеточной экспрессирующей системы возможно реализовать два подхода. Первый заключается в разработке таких методических приемов, которые позволили бы получать информацию, например, о структуре вновь синтезированного полипептида, используя для этого нанограммовые его количества (Fedorov A.N., Dolgikh D.A., Chemeris et al. (1992) J.Mol.Biol.225, 927-931). Второй подход заключается в модернизации самой бесклеточной системы трансляции с целью резкого повышения ее эффективности.
После того как была показана возможность работы бесклеточной системы трансляции в непрерывном режиме, стало возможным говорить о ее биотехнологическом применении и прежде всего для получения токсичных для клеток, но важных с точки зрения практического использования, биологически активных пептидов. В последнее время стало возможным использование бесклеточной системы трансляции также для препаративной наработки белков с целью их структурных исследований (ЯМР, рентгеноструктурный анализ и т.д.).
Среди используемых в настоящий момент систем бесклеточная система из зародышей пшеницы является более доступной и дешевой по сравнению с системой из ретикулоцитов кролика и более предпочтительной по сравнению с S30-системой Е.coli, так как позволяет в целом ряде случаев успешно решать проблему правильного сворачивания эукариотических белков. При помощи этой системы, главным образом, решаются научные задачи, связанные с изучением биосинтеза белка.
Одной из основных причин, обусловливающих ограниченность использования бесклеточных систем на основе растительных зиготических зародышей для синтеза белка в течение длительного времени, является наличие примесных нуклеазных и протеазных активностей. В нативных семенах эти активности обычно приурочены к запасающим тканям или специализированным органам зиготических зардышей, таким как семядоли. Достижения последних лет в области культуры клеток растений открывают новые возможности для получения соматических зародышей без запасающих тканей семян и индукции прорастания из состояния покоя. В связи с этим для получения экстрактов для длительного бесклеточного синтеза белков обезвоженные соматические зародыши представляются наиболее привлекательным и перспективным исходным материалом. Другим дополнительным преимуществом такого подхода является возможность стабильного, не зависящего от времени года и погодных условий получения свежего гомогенного материала для приготовления экстрактов.
Успех использования соматических зародышей для получения экстрактов для бесклеточного синтеза белка зависит от двух обстоятельств. Во-первых, необходимо, чтобы полученные соматические зародыши морфологически были высокого качества. Во-вторых, должны быть решены проблемы индукции состояния покоя, неразрывно связанные с синхронизацией развития зародышей.
Для получения in vitro большого количества соматических зародышей наиболее обещающим представляется подход, основанный на использовании каллусных и суспензионных культур. Этот подход активно разрабатывается в последнее время в приложении к таким двудольным растениям, как морковь, люцерна, табак, картофель (Gray DJ., Purohit A. 1991 Critical Reviews in Plant Sci 10 (1): 33-61; Mc Kersie B.D., Senarata Т., Bowley S.R et al. (1989) In vitro Cell. Dev. Biol. 25: 1183-1188; Janic J., Kitto S.L., Yong-Hwan. (1989) In vitro Cell. Dev. Biol. 25: 1167-1172; Carman JG. (1990) In vitro Cell. Dev. Biol. 26: 746-753).
В случае однодольных растений, таких как пшеница, получение суспензионных культур, способных к постоянному воспроизводству соматических зародышей, является крайне трудоемким и занимает много времени. Лишь в ограниченном числе лабораторий во всем мире были разработаны такие протоколы для пшеницы (Redway F.A., V. Vasil, D. Lu & I.K. Vasil, 1990 Plant Cell Rep. 8:714-717; Wang W.C. and Nguyen H.T. 1990 Plant Cell Rep. 8: 639-642; Yang Y.M., He DG, Scott KJJ. 1994 Plant Cell Reports 13: 176-179), однако во многих случаях они оказались невоспроизводимыми и обеспечивали крайне низкий выход обособленных хорошо сформированных биполярных соматических зародышей. Во всех этих протоколах исходным материалом для получения суспензионной культуры служил длительно культивируемый высоко эмбриогенный каллус, который сам по себе может служить источником для соматических зародышей. Такой каллус может быть получен из различных тканей, например незрелых зиготических зародышей, зачатков соцветия и зрелых семян. Поддержание эмбриогенной способности в каллусной культуре пшеницы длительное время зависит от многих факторов, в числе которых генотип, тип питательной среды, гормональный состав, условия и длительность культивирования.
Незрелые зиготические зародыши являются излюбленным исходным материалом для получения непосредственно соматических зародышей in vitro, а также для получения длительно поддерживаемого эмбриогенного каллуса (Maddock, S.E., V.A. Lancaster, R. Risiott & J. Franklin, 1983. J Exp Bot 34: 915-926; Borelli, G.M., Lupotto, E., Locatelli, F. and Wittmer, G. (1991) Plant Cell Rep. 10: 296-299; Vasit V, Redway FA & Vasil IK (1990) Bio/TechnoIogy 8: 429-434; Mohamand A.S., Nabors M.W. 1991. Plant Cell Tissue Organ Cult 26: 185-187; Felfoldi К., and Purnhauser 1. 1992. Cereal Res Comm 20: 273-277). Однако экспланты такого рода доступны лишь в течение ограниченного периода в течение года, или же донорное растение должно выращиваться в тепличных условиях, что увеличивает трудоемкость. Обычно такое выращивание занимает 3-4 месяца. В том случае, когда используются зачатки соцветий, продолжительность тепличного выращивания сокращается всего на 2,5-3 недели. В случае зрелых семян, которые представляются весьма перспективным исходным материалом в связи с их неограниченной доступностью независимо от времени года, их использование ограничивается пока весьма низкой эмбриогенной способностью (Fennell, S., N. Bohorova, M. van Ginkel, J. et al. 1996. Theor Appl Genet 92: 163-169; Bartok Т., and Sagi F. 1990. Plant Cell Tissue Organ Cult 22:37-41;  Sancak C. (1998) Plant Cell Reports 18:331-335).
Sancak C. (1998) Plant Cell Reports 18:331-335).
Сущность изобретения
Настоящее изобретение обеспечивает экстракты из соматических растительных зародышей, полученных в условиях искусственного культивирования in vitro, для бесклеточных систем трансляции и/или сопряженной транскрипции-трансляции. Получение соматических зародышей в условиях in vitro основано на способности тканей растений к регенерации органов/целых растений из соматических клеток. Этот принцип является универсальным для большого числа видов растений, независимо от их принадлежности к двудольным или однодольным. В частности, изобретением предусмотрены экстракты из соматических зародышей пшеницы, ячменя и кукурузы.
В соответствии с другим аспектом настоящего изобретения обеспечивается способ получения экстрактов из соматических растительных зародышей для бесклеточных систем трансляции и/или сопряженной транскрипции-трансляции, включающий:
a). получение первичного материала из соматических тканей;
b). индукцию соматического эмбриогенеза;
c). приготовление экстракта из соматических зародышей.
Полученный из соматических тканей растения первичный материал (эксплант) помещают на питательную среду для индукции соматического эмбриогенеза в условиях in vitro. При культивировании эксплантов образуется эмбриогенный каллус, из которого по мере культивирования формируются соматические зародыши. При развитии зародышей до стадии формирования первого листа их отделяют от каллуса и используют для приготовления экстрактов.
Еще один аспект относится к экстракту из соматических растительных зародышей, полученному способом по изобретению в соответствии с предыдущим аспектом.
Следующий аспект изобретения предусматривает способ получения экстрактов из соматических растительных зародышей для бесклеточных систем трансляции и/или сопряженной транскрипции-трансляции, включающий:
a). получение первичного материала из соматических тканей;
b). получение высокоэмбриогенного каллуса;
c). получение эмбриогенной суспензионной культуры;
d). индукцию вторичного соматического эмбриогенеза;
e). приготовление экстракта из соматических зародышей.
Соответственно еще один аспект предусматривает экстракт из соматических растительных зародышей, полученный способом настоящего изобретения в соответствии с предыдущим аспектом.
В следующем аспекте изобретение предусматривает применение экстракта из растительных соматических зародышей, полученных в условиях искусственного культивирования in vitro, для бесклеточной системы трансляции и/или сопряженной транскрипции-трансляции. В частности, предусмотрено применение экстракта по изобретению для бесклеточного синтеза олигопептидов, полипептидов, белков.
Следующий аспект изобретения предусматривает применение экстракта из растительных соматических зародышей, полученных в условиях искусственного культивирования in vitro, для производства диагностической системы для обнаружения биологически активных соединений, обладающих модулирующей активностью на процессы транскрипции и/или трансляции, и/или для скрининга на такие соединения.
Еще один аспект изобретения относится к диагностической системе на основе экстракта из растительных соматических зародышей для обнаружения биологически активных соединений, обладающих модулирующей активностью на процессы транскрипции и/или трансляции, и/или для скрининга на такие соединения.
В следующем аспекте изобретения предусмотрено применение такой диагностической системы на основе экстракта из растительных соматических зародышей для обнаружения биологически активных соединений, обладающих модулирующей активностью на процессы транскрипции и/или трансляции, и/или для скрининга на такие соединения.
По сравнению со стандартным экстрактом, под которым понимается экстракт, приготовленный из зиготических зародышей зрелых семян пшеницы, являющийся в настоящее время наиболее широко применяемой эукариотической системой трансляции растительного происхождения, настоящее изобретение обеспечивает следующие преимущества:
1. Получение экстрактов из растительных соматических зародышей, выращенных в условиях in vitro, позволяет преодолеть видовые ограничения при получении подобных экстрактов из зиготических зародышей различных культур.
2. Получение соматических зародышей может осуществляться круглогодично, что позволяет преодолеть ярко выраженную сезонность при получении экстрактов из зиготических зародышей.
3. Способ получения растительных соматических зародышей настоящего изобретения обеспечивает получение зародышей морфологически высокого качества. Настоящим изобретением решается проблема получения экстракта гарантированно высокого качества путем индукции состояния покоя сформировавшихся зародышей, обеспечивающей синхронизацию развития зародышей.
4. Использование эмбриогенной суспензионной культуры позволяет преодолеть большие материальные и трудовые затраты на выращивание и подготовку большого количества высококачественного семенного материала. Эмбриогенная суспензионная культура растений позволяет получать большое количество соматических зародышей.
5. Экстракт изобретения обеспечивает более высокую эффективность трансляции и сопряженной транскрипции-трансляции в бесклеточной системе. Экстракт из соматических зародышей позволяет увеличить выход синтезируемого в бесклеточной системе продукта трансляции по меньшей мере на 30%.
6. Экстракт изобретения меняет кинетику бесклеточного синтеза белков, делая ее особенно эффективной на начальных этапах биосинтеза, что позволяет сократить время реакции.
7. Активность экстракта, полученного из соматических зародышей, одинаково высока независимо от сорта, что позволяет преодолеть ярко выраженную сортовую специфику экстракта, приготовленного из зиготических зародышей.
8. Получаемый из соматических зародышей экстракт одинаково успешно работает с различными матрицами; синтезируемые белки являются полноразмерными и обладают функциональной биологической активностью.
Перечень фигур
Фиг.1. Трансляция мРНК обелина (80 пмоль/мл) в различных экстрактах.
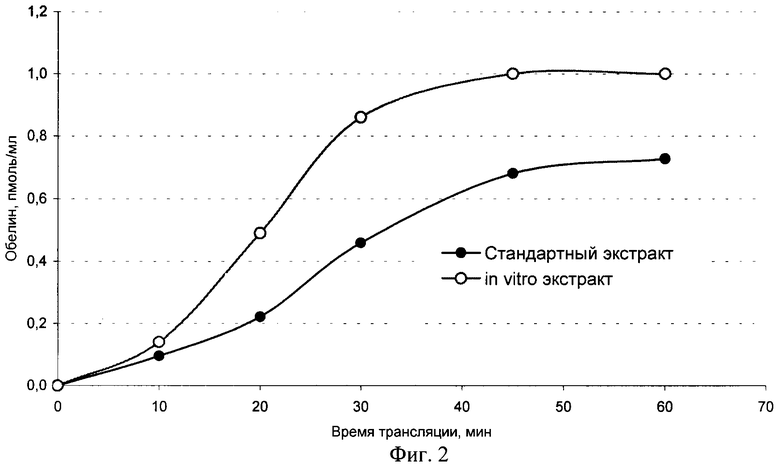
Фиг.2. Кинетика трансляции мРНК обелина (80 пмоль/мл).
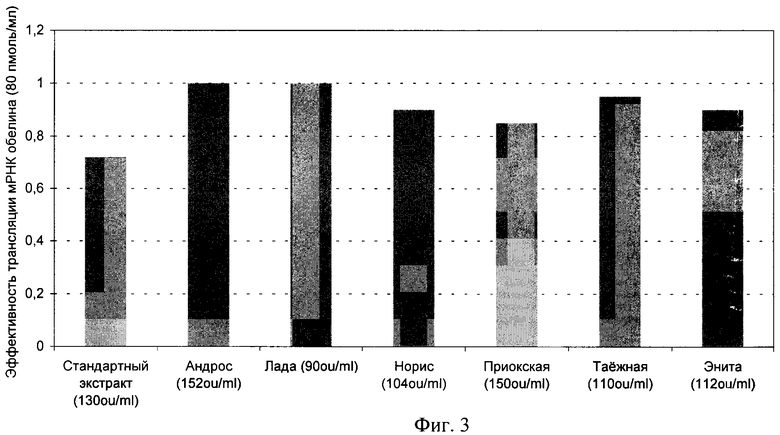
Фиг.3. Синтез обелина в экстрактах, полученных из различных сортов пшеницы (продолжительность трансляции 60 мин).
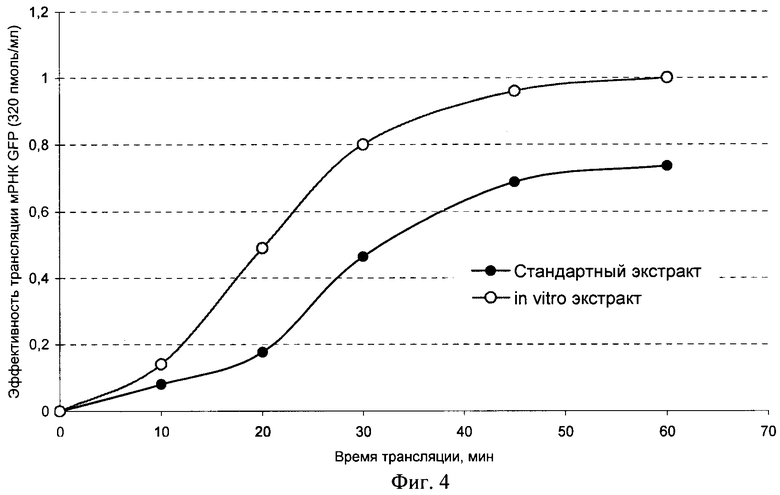
Фиг.4. Кинетика синтеза зеленого флуоресцентного белка GFP.
Фиг.5. Кинетика синтеза белка GFP в сопряженной системе транскрипции-трансляции.
Сведения, подтверждающие возможность осуществления изобретения
В настоящем описании используются общепринятые в данной области термины и понятия. Однако, с целью обеспечить их толкование в том смысле и объеме, как это предполагается в контексте настоящего описания, ниже приводятся определения используемых терминов.
Под «соматическими тканями» в настоящем описании понимается ткани вегетативных органов растений (листья, корни, стебли и др.).
«Зиготические зародыши (эмбрионы)» - это зародыши, образовавшиеся в результате слияния половых клеток при оплодотворении.
«Соматические зародыши (эмбрионы)» - зародыши растений, сформировавшиеся в результате простого деления клеток (митоз) соматических тканей без слияния половых клеток.
«Зрелые зародыши (эмбрионы)» - под этим термином подразумевается полностью сформировавшиеся зародыши, способные к дальнейшему самостоятельному прорастанию в естественных условиях без создания дополнительных условий.
«Незрелые зародыши (эмбрионы)» - под этим термином подразумеваются несформировавшиеся зародыши, не способные к самостоятельному прорастанию без создания дополнительных условий.
Под «каллусом» в настоящем описании понимается недифференцированная растительная ткань, образующаяся в результате простого деления клеток.
Под «первичным соматическим эмбриогенезом» подразумевается формирование соматических зародышей (эмбрионов) из первичных тканей экспланта в достаточно короткий срок (до 2,5 мес.)
Под «вторичным соматическим эмбриогенезом» подразумевается формирование соматических зародышей (эмбрионов) из длительно поддерживаемого каллуса (3-4 мес и более), первоначально образовавшегося из тканей первичного экспланта.
Термин «индукция» понимается в том смысле, что производится изменение условий культивирования растительных тканей с целью искусственного стимулирования процессов соматического эмбриогенеза.
Способы получения экстракта из растительных зародышей настоящего изобретения предусматривают получение первичного материала (экспланта) из соматической растительной ткани. Предварительно растительную ткань поверхностно обрабатывают стерилизующими агентами, известными в данной области. Примерами стерилизующих агентов, но не ограничиваясь только ими, могут служить 70% этанол и/или 0,5% гипохлорит натрия. Выделение первичного материала из растительной ткани проводят под бинокулярным микроскопом.
В качестве первичного материала (экспланта) для получения каллуса in vitro могут служить различные соматические ткани - незрелые зиготические зародыши, зародыши зрелых семян, зачатки соцветия, основания молодых листьев, пыльники и др. ткани, в зависимости от используемого вида растения. В предпочтительном воплощении настоящего изобретения в качестве экспланта используются незрелые зиготические зародыши, получаемые из незрелых семян.
Первичный материал может быть получен из разнообразных двудольных и однодольных растений. В предпочтительном воплощении изобретения такими растениями являются пшеница, ячмень и кукуруза.
Эксплант помещается на питательную среду для индукции соматического эмбриогенеза в условиях in vitro. При культивировании эксплантов образуется первичный эмбриогенный каллус, из которого по мере культивирования формируются соматические зародыши. При развитии зародышей до стадии формирования первого листа они отделяются от каллуса и используются для приготовления экстрактов. Этот процесс может осуществляться круглогодично, что позволяет преодолеть ярко выраженную сезонность при получении экстрактов из зиготических зародышей.
Индукция соматического эмбриогенеза (как первичного, так и вторичного) происходит в контролируемых условиях на искусственной питательной среде независимо от внешних условий. Это позволяет преодолеть климатические, географические и сезонные факторы, приводящие к непредсказуемой вариабельности в качестве экстракта, полученного из зиготических зародышей.
Искусственные питательные среды, используемые на разных стадиях получения соматических эмбрионов, могут быть различными или одинаковыми, в зависимости от применяемого способа получения экстрактов из растительных соматических зародышей, вида растения и других факторов. Питательная среда может быть жидкой, полужидкой или твердой и включает в себя несколько компонентов: минеральные соли, включая микроэлементы, органические добавки различного состава и концентрации (сахара, аминокислоты, витамины, гидролизаты белков, осморегуляторы и др), фитогормоны, а также при необходимости агар и его природные или искусственные аналоги.
В качестве минеральной составляющей питательной среды для получения соматических зародышей могут быть использованы различные композиции солей, используемые для культуры клеток и тканей растений (Chu C.C., Wang C.C., Sun C.S., et al. 1975. Sci. Sin. 18:659-668; Gamborg O.L., Miller R.A., Ojima K. 1968. Expt. Cell. Res. 50: 151-158). В предпочтительном воплощении настоящего изобретения используются композиция солей по Murashige и Scoog (Murashige Т., Skoog F. 1962 Physiol. Plantarum. 15: 473-497), однако и другие известные из уровня техники композиции могут быть с равным успехом использованы в настоящем изобретении.
Кроме стандартных композиций солей в искусственные среды в зависимости от вида растения могут добавляться различные микроэлементы. Например, при индукции первичного каллуса в случае ячменя в питательную среду предпочтительно добавляют CuSO4.
В качестве витаминов можно использовать различные композиции. Предпочтительной композицией витаминов по настоящему изобретения является пропись, предложенная в (Murashige Т., Skoog F. 1962 Physiol. Plantarum. 15: 473-497), однако и другие известные из уровня техники прописи могут быть с равным успехом использованы в настоящем изобретении.
В качестве сахаров, одного из основных компонентов питательной среды, могут использоваться различные типы моно- и дисахаридов, а также их сочетания. В предпочтительном воплощении настоящего изобретения используются глюкоза в концентрации от около 1 до около 3%, мальтоза в концентрации от около 2 до около 5%, или сахароза в концентрации от около 2 до около 4%. Наиболее предпочтительным по настоящему изобретению является использование глюкозы или сахарозы в концентрации от около 2 до около 3%, или мальтозы в концентрации от около 2 до около 5%.
В качестве аминокислот могут использоваться различные природные L-аминокислоты, например, но не ограничиваясь только ими, пролин, аспарагин.
В качестве гидролизатов белков могут использоваться различные ферментативные или кислотные гидролизаты белков, например, известные под торговыми наименованиями Триптон, Пептон, казаминовые кислоты. Предпочтительным по настоящему изобретению является использование гидролизата казеина.
Фитогормоны необходимы для направленной индукции соматического эмбриогенеза. В качестве основных индукторов эмбриогенеза in vitro используют ауксины, например: 2,4-дихлорфеноксиуксусную кислоту (2,4-D), 2,4,5-трихлорфеноксиуксусную кислоту (2,4,5-Т), 3,6-дихлор-О-анисовую кислоту (Dicamba) and р-хлорфеноксиуксусную кислоту (СРА), а также цитокинины кинетин и 6-бензиламинопурин. В предпочтительном варианте настоящего изобретения используют 2,4-D в концентрации около 2 мг/л питательной среды.
Для увеличения эффективности соматического эмбриогенеза в среду также добавляют другие фитогормоны, такие как цитокинины и гиббереллины и др. В предпочтительном варианте настоящего изобретения используют Daminozide в концентрации около 25 мг/л.
Эффективность эмбриогенеза может также зависеть от осмотического шока, который может создаваться введением в состав культуральной среды таких компонентов, как полиэтиленгликоль, или увеличением концентрации сахаров. В предпочтительном воплощении настоящего изобретения используется полиэтиленгликоль 6000 (PEG 6000) в концентрации от около 0,5 до около 1 г/л.
Для придания консистентных свойств питательной среде могут использоваться агар бактериологический, агароза и искусственные заменители агара (gelrite, plant agar).
Культивирование тканей in vitro позволяет направленно контролировать качество получаемого экстракта путем искусственной индукции состояния покоя у соматических зародышей. Перевод активно растущих соматических зародышей в состояние покоя может осуществляться путем изменения условий культивирования с целью имитации условий, сопутствующих образованию семян в природе, в частности, путем увеличения температуры культивирования, изменения осмотического баланса питательной среды, принудительного подсушивания соматических зародышей и др. Альтернативным способом является использование фитогормонов. В предпочтительном воплощении настоящего изобретения применяют абсцизовую кислоту (АВА) и ее аналоги. Абсцизовую кислоту или ее аналоги вводят в питательную среду через определенный промежуток времени, в частности после того, как уже произошла закладка соматических зародышей в эмбриогенном каллусе. В предпочтительном воплощении настоящего изобретения абсцизовая кислота (АВА) в концентрации около 1 мг/л вводится в питательную искусственную среду приблизительно за 15 дней до отделения соматических эмбрионов от каллуса.
Предусмотренное одним из способов настоящего изобретения использование высокотехнологичных методов культуры клеток и тканей растений in vitro позволяет преодолеть большие материальные и трудовые затраты на выращивание и подготовку большого количества высококачественного семенного материала. Таким предпочтительным методом настоящего изобретения является эмбриогенная суспензионная культура растений, позволяющая получать большое количество соматических зародышей, которые могут формироваться как непосредственно при культивировании клеток в жидкой питательной среде, так и через стадию переноса каллуса, образованного в суспензионной культуре, на твердую среду и индукции вторичного эмбриогенеза.
Процедура приготовления экстракта из соматических зародышей в своих основных этапах полностью повторяет классическую (Roberts B.E., Paterson В.М. (1973) Proc. Natl. Acad. Sci. USA 70,N 4, 2330-2334; Erickson, A.H., BIobel, G. (1983) Meth. EnzymoL 96, 38-50) с использованием зиготических зрелых зародышей пшеницы и включает:
1 этап - гомогенизация зародышей, замороженных в жидком азоте,
2 этап - экстракция буферным раствором,
3 этап - центрифугирование,
4 этап - хроматография.
Активность получаемых из соматических эмбрионов экстрактов проверяют в стандартной бесклеточной системе трансляции мРНК. Для сравнения используют экстракт из зиготических зрелых зародышей пшеницы. Оптическая плотность экстрактов была примерно одинакова (А260=100-150 ОЕ). В качестве тестовых матриц контроля хода трансляции могут быть использованы любые подходящие искусственные РНК или природные мРНК. Для облегчения контроля за ходом трансляции могут быть использованы, например, мРНК Са2+ - активируемого фотобелка обелина или зеленого флуоресцентного белка (GFP). Синтез данных белков в бесклеточной системе трансляции позволяет контролировать ход трансляции не только по включению радиоактивной метки, но и по люминесцентной реакции (в случае обелина) или по флуоресценции GFP. Все сравнения экстрактов проводят при равных концентрациях мРНК в системе трансляции.
Экстракт из соматических зародышей в соответствии с настоящим изобретением позволяет увеличить выход синтезируемого в бесклеточной системе продукта трансляции по меньшей мере на 30%. Получаемый экстракт одинаково успешно работает при синтезе различных белков (GFP, обелин), все продукты трансляции полноразмерны и обладают биологической активностью. Наличие функциональной биологической активности у продукта бесклеточной трансляции служит подтверждением правильной пространственной укладки полипептидной цепи (фолдинга) и сборки фермента в активной конформации. Таким образом, конформация продукта трансляции и/или сопряженной транскрипции-трансляции, получаемого в бесклеточной системе настоящего изобретения, является идентичной или по существу идентичной его нативной конформации.
Экстракт настоящего изобретения, полученный из растительных соматических зародышей, полученных в условиях культивирования in vitro, изменяет кинетику синтеза данных белков, делая ее более эффективной по сравнению со стандартным экстрактом. Наиболее заметна разница в эффективности трансляции мРНК на начальных этапах биосинтеза (до 20 минут).
Активность экстракта по настоящему изобретению, полученного из соматических зародышей, одинаково высока независимо от сорта, что позволяет преодолеть ярко выраженную сортовую специфику экстракта, приготовленного из зиготических зародышей.
Полученные в соответствии с настоящим изобретением экстракты из растительных соматических зародышей могут быть, в частности, применены для препаративной наработки белков, полипептидов, олигопептидов. Данная система позволяет преодолеть технологические и экономические ограничения, присущие химическому синтезу и рекомбинантной технике, использующимся в настоящее время. В частности становится возможным получение для широкого использования пептидов длиной в 30-100 аминокислотных остатков. При этом становится возможным преодолеть неполноту удлинения полипептидной цепи, избежать сложностей с выбором защитных групп и их удалением и, что самое важное, сложностей с реализацией функционально-активной конформации в длинном полипептиде. Это в свою очередь позволит получать относительно недорогой конечный продукт. Представленная система также позволяет преодолеть и ряд трудностей, возникающих при создании генетических конструкций для экспрессии в прокариотических или эукариотических клетках для получения биологически активных молекул, связанных с нестабильностью или плохой экспрессией в результате действия регуляторных механизмов клетки хозяина, нестабильности и деградации формирующихся агрегатов или их токсичности для клеток.
В качестве информационной РНК для бесклеточного синтеза белкового продукта могут использоваться природные мРНК, синтетические РНК, тотальная РНК, а также РНК-транскрипты, наработанные в условиях in vitro. Для получения РНК-транскриптов in vitro могут быть использованы общеизвестные подходы, основанные на применении высокоактивных и высокоспецифичных РНК-полимераз фагов Т3, Т7 или SP6. Для этого в последовательность ДНК, кодирующую белковый продукт, в upstream-положение (против направления транскрипции) вводят последовательности соответствующих промоторов, узнаваемые фаговыми РНК-полимеразами.
Бесклеточный экстракт настоящего изобретения может быть с равным успехом использован для осуществления сопряженной транскрипции-трансляции. В этом случае в систему в качестве матрицы вводят экзогенную ДНК, кодирующую белковый продукт, которая может быть представлена, например, фрагментом ДНК, продуктом полимеразной цепной реакции (ПЦР), лигированными взаимно перекрывающимися олигонуклеотидами. Такая кодирующая белковый продукт последовательность ДНК должна содержать сигналы транскрипции и трансляции. Непременным условием является присутствие эффективного транскрипционного промотора. Например, ДНК может содержать последовательность 35S-промотора РНК вируса мозаики цветной капусты, клонированного в upstream-положении по отношению к кодирующей последовательности. Альтернативно, промотор в составе ДНК-матрицы может быть представлен промотором РНК-полимераз фагов Т3, Т7 или SP6, и кодирующая белковый продукт последовательность ДНК располагается по ходу транскрипции. В последнем случае для обеспечения эффективной транскрипции ДНК-матрицы в систему сопряженной транскрипции-трансляции на основе бесклеточного экстракта настоящего изобретения добавляют экзогенный фермент - РНК-полимеразу фага Т3, Т7 или SP6, который доступен из коммерческих источников или может быть легко получен в лабораторных условиях. При необходимости ДНК-матрица также может содержать сигнал полиаденилирования.
Весьма перспективным представляется другое направление практического использования систем бесклеточного синтеза белка - разработка и производство на основе бесклеточной системы трансляции и/или сопряженной транскрипции-трансляции новых диагностических тест-систем, предназначенных для обнаружения различных биологически активных веществ, способных модулировать процессы транскрипции и/или трансляции, и/или для скрининга на такие соединения. Возможность производства таких тест-систем основывается на том, что различные факторы биологической и химической природы, модулирующие, т.е. ингибирующие или активирующие, процесс биосинтеза белка на различных его этапах (транскрипция, трансляция), могут быть выявлены при помощи бесклеточной системы биосинтеза по уровню выхода продукта трансляции или сопряженной транскрипции/трансляции. К таким соединениям относятся, например, различные яды, токсины, антибиотики, тяжелые металлы и т.п. Например, токсическое действие целого ряда грибных токсинов (наиболее ярким представителем которых является α-аманитин) обусловлено тем, что они полностью блокируют транскрипцию под действием эукариотических РНК-полимераз II и III. При скрининге, направленном на соединения, способные модулировать процессы транскрипции и/или трансляции, могут использоваться, например, но не ограничиваясь только перечисленным, природные смеси, вытяжки, экстракты, среды после культивирования различных микроорганизмов, искусственно созданные библиотеки индивидуальных соединений, в том числе комбинаторные библиотеки. Очевидно, что такого рода тест-системы на основе экстракта настоящего изобретения могут найти широкое применение в области биологии, медицины, фармакологии, токсикологии, экологии и т.п.
В качестве примера такой диагностической системы, основанной на использовании системы бесклеточной трансляции, может служить описанная Gorokhovatsky и соавторами (Biotechnol. Appl. Biochem. (1998) 27, 259-263) лиофилизированная система, полученная из зародышей пшеницы, которая была успешно использована для анализа различных соединений, ингибирующих процесс биосинтеза белка на различных его стадиях: дифтерийного токсина и антибиотиков - тетрациклина, канамицина и пуромицина. Специалисту очевидно, что аналогичным образом могут анализироваться и вещества, активирующие процесс биосинтеза белка на различных его стадиях. Среди преимуществ таких диагностических систем на основе систем бесклеточной трансляции авторы публикации отмечают, что при использовании биолюминесцентного способа детекции образующегося продукта трансляции, т.е. когда продуктом реакции является природный люминесцентный белок, как, например, активируемый Са2+ фотобелок обелин, система обеспечивает чрезвычайно высокую чувствительность, превосходящую на три порядка чувствительность при использовании радиоизотопного способа детекции продукта. Дополнительно можно отметить, что использование люминесцирующих белков (как, например, обелина, зеленого флуоресцентного белка GFP и его вариантов и т.п.) в качестве детектируемых продуктов трансляции также обеспечивает еще одно преимущество, а именно экологическую безопасность диагностической системы.
Примеры.
Нижеследующие примеры даются с целью продемонстрировать возможность осуществления изобретения, однако любому специалисту в данной области ясно, что они приведены в качестве иллюстрации и не могут быть использованы для ограничения объема притязаний заявителя.
Пример 1. Получение первичного материала из незрелых зиготических зародышей.
Пример 1.1. Получение первичного материала из незрелых зиготических зародышей пшеницы.
Зиготические зародыши выделяют из незрелых семян пшеницы (стадия полупрозрачности). Для этого собирают формирующиеся семена с молодых колосьев через 12-15 дней после выброса пыльников. Незрелые семена поверхностно обрабатывают стерилизующими агентами (этанол 70%, гипохлорит натрия 0,5%), после чего незрелые зародыши отделяют от семян под бинокулярным микроскопом и помещают на искусственную питательную среду для индукции эмбриогенного каллуса.
Пример 1.3. Получение первичного материала из незрелых семян кукурузы.
Незрелые зиготические зародыши кукурузы инбредной линии №91 размером 1,5-2,5 мм выделяют из незрелых семян, собранных с молодых початков кукурузы через 9-12 дней после выброса пыльников. Незрелые семена поверхностно обрабатывают стерилизующими агентами (этанол 70%, гипохлорит натрия 0,5%), после чего незрелые зародыши отделяют от семян под бинокулярным микроскопом и помещают на искусственную питательную среду для индукции каллуса.
Пример 2. Индукция соматического эмбриогенеза.
Пример 2.1. Индукция соматического эмбриогенеза у пшеницы
Пример 2.1.1. Искусственная питательная среда для получения соматических эмбрионов пшеницы включает в себя следующие компоненты: минеральные соли по Murashige and Scoog (Табл.1), 3% сахароза, 150 мг/л аспарагин, витамины (Табл.2), фитогормоны: ауксин 2,4-D в концентрации 2 мг/л среды, абсцизовая кислота (АВА) в концентрации 1 мг/л среды (добавляют для стимулирования состояния покоя у сформировавшихся соматических зародышей через 15 дней после начала индукции эмбриогенеза), 0,7% агар бактериологический. Культивирование материала in vitro проводят в течение 25-30 дней при температуре 26°С в темноте. При достижении зародышами стадии начала формирования первого листа их отделяют под бинокулярным микроскопом от неэмбриогенного каллуса и хранят в замороженном виде при температуре - 70°С.
Пример 2.1.2. Индукция соматического эмбриогенеза происходит при тех же условиях, за одним исключением: в состав питательной среды включают глюкозу вместо сахарозы.
Пример 2.1.3. Индукция соматического эмбриогенеза происходит при тех же условиях, за одним исключением: в состав питательной среды входит мальтоза вместо сахарозы.
Пример 2.1.4. Индукция соматического эмбриогенеза происходит при тех же условиях, за одним исключением: в состав питательной среды включают ауксин Dicamba вместо 2,4-D.
Пример 2.1.5. Индукция соматического эмбриогенеза происходит при тех же условиях, за одним исключением: в состав питательной среды дополнительно включают 1% PEG 6000.
Пример 2.1.6. Индукция соматического эмбриогенеза происходит при тех же условиях, за одним исключением: в состав питательной среды дополнительно включают Daminozide (succinic acid mono(2,2-dimethylhydrazide)) в концентрации 25 мг/л.
Пример 2.1.7. Индукция соматического эмбриогенеза происходит при тех же условиях, за одним исключением: в состав питательной среды включают Daminozide в концентрации 200 мг/л вместо АВА.
Пример 2.2. Индукция соматического эмбриогенеза у ячменя
Пример 2.2.1 Индукция соматического эмбриогенеза у ячменя состоит из двух последовательных культивирований:
А. Индукция первичного каллуса
Б. Индукция эмбриогенеза из первичного каллуса.
Для индукции первичного каллуса незрелые зиготические зародыши ячменя помещают на питательную среду следующего состава: минеральные соли по Murashige and Scoog и (Табл. 1), CuSO4 в концентрации 0,5 мМ, витамины (Табл.2), 3% мальтоза, ауксин 2,4-D в концентрации 2 мг/л среды, 0,7% агар бактериологический. Незрелые зиготические зародыши культивируют 3 недели при температуре 26°С в темноте.
Для индукции соматического эмбриогенеза плотные очаги каллуса, образовавшегося в результате культивирования незрелых зиготических зародышей, перемещают на питательную среду следующего состава: минеральные соли по Murashige and Scoog (Табл.1), 3% сахароза, витамины (Табл.2), цитокинин кинетин в концентрации 1 мг/л среды, 0,7% агар бактериологический. Культивирование материала in vitro проводят в течение 25-30 дней при температуре 26°С в темноте. При достижении зародышами стадии начала формирования первого листа их отделяют под бинокулярным микроскопом от неэмбриогенного каллуса и хранят в замороженном виде при температуре - 70°С.
Пример 2.2.2 Индукция соматического эмбриогенеза происходит при тех же условиях, за одним исключением: в состав питательной среды для эмбриогенеза дополнительно включают ауксин NAA.
Пример 2.2.3 Индукция соматического эмбриогенеза происходит при тех же условиях, за одним исключением: в состав питательной среды для эмбриогенеза включают цитокинин ВА вместо кинетина.
Пример 2.3. Индукция соматического эмбриогенеза у кукурузы.
Для индукции эмбриогенного каллуса незрелые зиготические зародыши кукурузы помещают на питательную среду следующего состава: минеральные соли по Murashige and Scoog (Табл.1), витамины (Табл.2), 2% сахароза, гидролизат казеина в концентрации 100 мг/л среды, L-пролин в концентрации 3 г/л среды, ауксин 2,4-D в концентрации 2 мг/л среды, 0,7% агар бактериологический. Незрелые зиготические зародыши культивируют 5 недель при температуре 26°С в темноте. Полученный каллус культивируют каждые 4 недели на свежей питательной среде того же состава. При достижении зародышами стадии начала формирования первого листа их отделяют под бинокулярным микроскопом от каллуса и хранят в замороженном виде при температуре - 70°С.
Все приведенные в примере 2 варианты воплощения изобретения свидетельствуют о том, что изобретение с одинаковым успехом осуществимо во всем объеме притязаний, как он сформулирован в формуле изобретения.
Пример 3. Получение эмбриогенной суспензионной культуры
Пример 3.1 Получение эмбриогенной суспензионной культуры пшеницы
Пример 3.1.1. Для получения эмбриогенной суспензии используют высокоэмбриогенный каллус, полученный из незрелых зиготических зародышей. Каллус культивируют на среде индукции эмбриогенеза (см. Пример 2) не менее 1,5-2 мес. Свежий эмбриогенный каллус, раздробленный на мелкие кусочки размером 0,2-0,5 см, переносят в колбы Эрленмеера на 250 мл. В каждую колбу наливают 50 мл жидкой питательной среды, которая включает в себя минеральные соли по Murashige and Scoog (Табл.1), 3% сахарозу, витамины (Табл.2) и ауксин 2,4-D в концентрации 2 мг/л среды и добавляют около 1 г каллуса. Первичную суспензию культивируют в течение 4 недель. После этого верхнюю фазу среды (25 мл) с молодыми делящимися клетками переносят в новую колбу, содержащую 25 мл свежей питательной среды. Через каждые 10-14 дней проводят обновление суспензионной культуры путем разбавления свежей средой в соотношении 1:1. Колбы культивируют в темноте при 26°С на вращающейся качалке (120 об/мин). После установления суспензионной культуры (2,5-3 мес.) образующиеся в культуральной среде клеточные агрегаты размером более 1 мм переносят на чашки Петри для индукции вторичного соматического эмбриогенеза. Для этого используют питательную среду следующего состава: минеральные соли по Murashige and Scoog и (Табл.1), 3% сахароза, витамины (Табл.2), ауксин 2,4-D в концентрации 1 мг/л среды), 0,7% агар бактериологический. Агрегаты культивируют в течение 30-45 дней при температуре 26°С в темноте. По мере формирования соматических зародышей на каллусах, образованных из суспензионных агрегатов, их отделяют под бинокулярным микроскопом от неэмбриогенного каллуса и хранят в замороженном виде при температуре - 80°С.
Пример 3.1.2. Получение эмбриогенной культуры происходит при тех же условиях, за одним исключением: в состав жидкой питательной среды включают мальтозу вместо сахарозы.
Пример 3.1.3. Получение эмбриогенной культуры происходит при тех же условиях, за одним исключением: в состав жидкой питательной среды включают ауксин 2,4,5-Т вместо ауксина 2,4-D.
Пример 3.1.4. Получение вторичных соматических эмбрионов из каллуса, сформированного из суспензионных агрегатов, происходит при тех же условиях, за одним исключением: для индукции вторичного эмбриогенеза в состав питательной среды включают ауксин Dicamba вместо ауксина 2,4-D.
Пример 3.1.5. Получение вторичных соматических эмбрионов из каллуса, сформированного из суспензионных агрегатов, происходит при тех же условиях, за одним исключением: для индукции вторичного эмбриогенеза в состав питательной среды включают ауксин 2,4,5-Т вместо ауксина 2,4-D.
Пример 3.2. Получение эмбриогенной суспензионной культуры кукурузы
Для получения эмбриогенной суспензии используют высокоэмбриогенный каллус кукурузы, культивирующийся на среде индукции эмбриогенеза (см. Пример 2.3.) не менее 3 мес. Свежий эмбриогенный каллус, раздробленный на мелкие кусочки размером 0,2-0,5 см, переносят в колбы Эрленмеера на 250 мл. В каждую колбу наливают 50 мл жидкой питательной среды, которая включает в себя минеральные соли по Murashige and Scoog (Табл.1), 3% сахарозу, витамины (Табл.2), гидролизат казеина в концентрации 200 мг/л, инозитол в концентрации 100 мг/л, L-пролин в концентрации 3 г/л и ауксин 2,4-D в концентрации 2 мг/л среды, и добавляют около 1 г каллуса. Первичную суспензию культивируют в течение 7 недель, проводя обновление культуры через каждые 7 дней. После этого верхнюю фазу среды (25 мл) с молодыми делящимися клетками переносят в новую колбу, содержащую 25 мл свежей питательной среды. Через каждые 7-10 дней проводят обновление суспензионной культуры путем разбавления свежей средой в соотношении 1:1. Колбы культивируют в темноте при 26°С на вращающейся качалке (120 об/мин). После установления суспензионной культуры (2,5-3 мес.) образующиеся в культуральной среде клеточные агрегаты размером более 1 мм переносят на чашки Петри для индукции вторичного соматического эмбриогенеза. Для этого используют питательную среду, описанную в примере 2.3.1. Агрегаты культивируют в течение 30-45 дней при температуре 26°С в темноте. По мере формирования соматических зародышей на каллусах, образованных из суспензионных агрегатов, их отделяют под бинокулярным микроскопом от неэмбриогенного каллуса и хранят в замороженном виде при температуре - 70°С.
Все приведенные в примере 3 варианты воплощения изобретения свидетельствуют о том, что изобретение с одинаковым успехом осуществимо во всем объеме притязаний, как они сформулированы в формуле изобретения.
Пример 4. Приготовление экстрактов из соматических зародышей.
5 г зародышей, замороженных в жидком азоте, растирают фарфоровым пестиком в ступке 2 минуты. Помещают ступку в лед, через 10 минут (после окончательного оттаивания) добавляют 1 мл буферного раствора следующего состава: 160 мМ HEPES, рН 7,6, 480 мМ ацетат калия, 20 мМ ацетат магния, 24 мМ дитиотрейтол (DTT). Экстракцию проводят 15 минут.
После двухкратного центрифугирования (20 минут, 30000 g, осадки отбрасывают) супернатант (2,5 мл) наносят на колонку с Sephadex G-25 Medium (NAP 25 Column), уравновешенную буферным раствором, содержащим 40 мМ HEPES, рН 7,6, 120 мМ ацетат калия, 5 мМ ацетат магния, 6 мМ DTT. Фракцию, соответствующую свободному объему колонки (2,5 мл), собирают и центрифугируют при тех же условиях. Оптическая плотность экстрактов составляет 60-150 ОЕ при 260 нм. Экстракты замораживают в жидком азоте аликвотами и хранят при температуре - 70°С.
Пример 5. Сравнение ативности экстрактов, полученных из тканей пшеницы.
Активность экстрактов, полученных из соматических эмбрионов, выращенных в условиях искусственного культивирования in vitro как в отсутствии, так и в присутствии абсцизовой кислоты, проверяют в стандартной бесклеточной системе трансляции мРНК. Для сравнения используют экстракт из зиготических зрелых зародышей пшеницы. Оптическая плотность экстрактов примерно одинакова (А260=100-150 ОЕ). В качестве тестовой матрицы контроля хода трансляции выбирают мРНК Са2+ - активируемого фотобелка обелина. Синтез данного белка в бесклеточной системе трансляции позволяет контролировать ход трансляции не только по включению радиоактивной метки, но и по люминесцентной реакции. Все сравнения экстрактов проводят при равных концентрациях мРНК в системе трансляции.
Как видно на фиг.1, экстракт настоящего изобретения из соматических зародышей, полученных в условиях искусственного культивирования in vitro в присутствии абсцизовой кислоты, обладает наибольшей активностью в системе бесклеточной трансляции мРНК.
Пример 6. Изучение кинетики синтеза белка с использованием экстракта, полученного из соматических зародышей.
Активность получаемых из соматических эмбрионов экстрактов проверяют в стандартной бесклеточной системе трансляции мРНК. Для эксперимента используют соматические зародыши, полученные в присутствии абсцизовой кислоты. Для сравнения используют экстракт из зиготических зрелых зародышей пшеницы. Оптическая плотность экстрактов составляет А260=130 ОЕ для стандартного экстракта и А260=152 ОЕ для экстракта из соматических зародышей. В качестве тестовой матрицы контроля хода трансляции выбирают мРНК Са2+ - активируемого фотобелка обелина. Все сравнения экстрактов проводят при равных концентрациях мРНК в системе трансляции.
Как видно из фиг.2, экстракт из соматических эмбрионов, полученный в соответствии с настоящим изобретением, обеспечивает более высокий уровень синтеза конечного белкового продукта - обелина, причем трансляция идет с большей скоростью, что наиболее отчетливо видно на начальных этапах (до 20-30 мин).
Пример 7. Сравнение экстрактов, полученных из соматических зародышей шести сортов пшеницы.
Активность получаемых из соматических эмбрионов экстрактов проверяют в стандартной бесклеточной системе трансляции мРНК. Для эксперимента используют соматические зародыши шести сортов пшеницы, полученные в присутствии абсцизовой кислоты. Для сравнения используют экстракт из зиготических зрелых зародышей пшеницы. Оптическая плотность экстрактов примерно одинакова (А260=90-152 ОЕ). В качестве тестовой матрицы контроля хода трансляции выбирают мРНК Са2+-активируемого фотобелка обелина. Все сравнения экстрактов проводят при равных концентрациях мРНК (80 пмоль/мл) в системе трансляции через 60 мин после начала трансляции.
Как видно из фиг.3, независимо от сорта экстракты в соответствии с настоящим изобретением обеспечивают стабильно более высокую эффективность трансляции, чем стандартный экстракт.
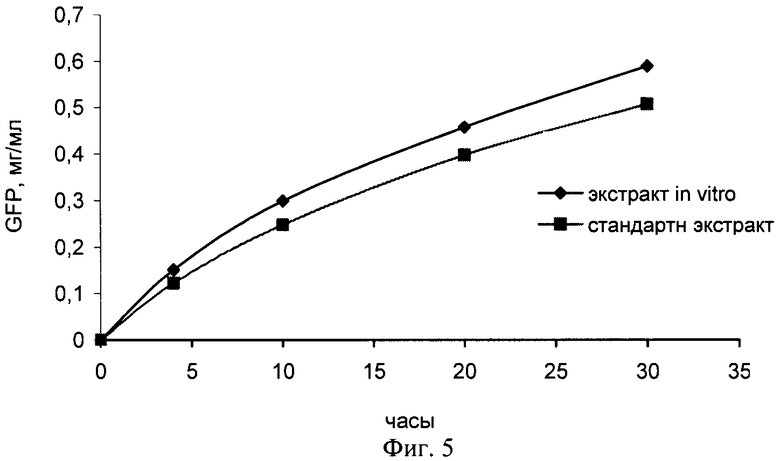
Пример 8. Изучение кинетики синтеза белка GFP с использованием экстракта. полученного из соматических зародышей
Для подтверждения возможности синтеза различных белков с использованием экстрактов, полученных их соматических эмбрионов, проверяют активность в стандартной бесклеточной системе трансляции мРНК. Для эксперимента использовали соматические зародыши, полученные в присутствии абсцизовой кислоты. Для сравнения используют экстракт из зиготических зрелых зародышей пшеницы. Оптическая плотность экстрактов составляет А260=130 ОЕ для стандартного экстракта и А260=152 ОЕ для экстракта из соматических зародышей. В качестве тестовой матрицы контроля хода трансляции выбирают мРНК зеленого флуоресцентного белка (GFP). Все сравнения экстрактов проводят при равных концентрациях мРНК (320 пмоль/мл) в системе трансляции.
Как видно из фиг.4, экстракт настоящего изобретения обеспечивает более эффективную трансляцию мРНК GFP, чем стандартный экстракт. Вместе с результатами, приведенными на фиг.1 и 2, представленные результаты свидетельствуют о том, что экстракт настоящего изобретения обеспечивает стабильно более эффективную трансляцию в бесклеточной системе по сравнению со стандартным экстрактом из зрелых зиготических зародышей и, таким образом, подтверждают, что изобретение осуществимо во всем объеме притязаний, сформулированных в формуле изобретения.
Пример 9. Синтез белка GFP в сопряженной системе транскрипции-трансляции в режиме CECF (continuous-exchange cell-free).
Для подтверждения возможности синтеза белков в сопряженной системе транскрипции-трансляции с использованием экстрактов настоящего изобретения, полученных из соматических зародышей, синтез белка GFP в CECF-системе из соматических зародышей пшеницы проводили в реакторе с фиксированным реакционным объемом 50 мкл с использованием диализной мембраны с размером пор 12000-14000 кДа. Реакционная смесь содержала экстракт из зародышей пшеницы (30% вес/объем), 7000 Ед/мл Т7 РНК полимеразы, 100 мкг/мл линеаризованной плазмиды рТ7ОВЕ-1, 500 Ед/мл ингибитора РНКазы, 100 мкг/мл креатинфосфокиназы, 50 мкг/мл дрожжевой суммарной тРНК, 0,2 мМ каждой аминокислоты, 1 мМ АТФ, 0,6 мМ ГТФ, 0,4 мМ ЦТФ и УТФ каждой, 16 мМ креатинфосфат, 40 мМ HEPES-KOH рН 7,6, 4 мМ Mg(OAc)2, 100 мМ КОАс, 3 мМ NaN3, 2,5 мМ DTT, 0,25 мМ спермидин, 2% глицерин. Питательная смесь (0,5 мл) содержала те же компоненты, за исключением экстракта из зародышей пшеницы, РНК полимеразы, плазмиды, креатинфосфокиназы, ингибитора РНКазы; концентрация Mg(OAc)2 снижена до 2,5 мМ. Реакцию синтеза проводили при 25°С с перемешиванием. Питательную смесь заменяли на свежую каждые 10 часов. Концентрацию белка GFP в реакционной смеси измеряли по флюоресценции при 510 нм.
Как видно из Фиг.5, экстракт настоящего изобретения обеспечивает сопряженную транскрипцию-трансляцию ДНК GFP даже с большей эффективностью, чем стандартный экстракт. Полученные результаты свидетельствуют о том, что экстракт настоящего изобретения способен обеспечить синтез различных белков в бесклеточной системе аналогично стандартному экстракту из зрелых зиготических зародышей и, таким образом, подтверждают, что изобретение осуществимо во всем объеме притязаний, сформулированных в формуле изобретения.
Должно быть понятно, что аналогичным образом, с учетом раскрытия изобретения в описании специалистом в данной области могут быть получены другие варианты изобретения, которые охватываются формулой, приводимой ниже.
Настоящее изобретение относится к биотехнологии, в частности к экстрактам из растительных соматических зародышей для бесклеточных систем трансляции и/или сопряженной транскрипции-трансляции. Из первичного материала получают эмбриогенный каллус, далее эмбриогенную суспензионную культуру. После индукции вторичного соматического эмбриогенеза приготовляют экстракт из соматических зародышей. На основе экстракта создают диагностическую систему для обнаружения биологически активных соединений. Изобретение позволяет преодолеть видовые ограничения и сортовую специфику, а также достичь высокой эффективности бесклеточной системы трансляции и сопряженной транскрипции-трансляции. 9 н. и 40 з.п. ф-лы, 5 ил., 2 табл.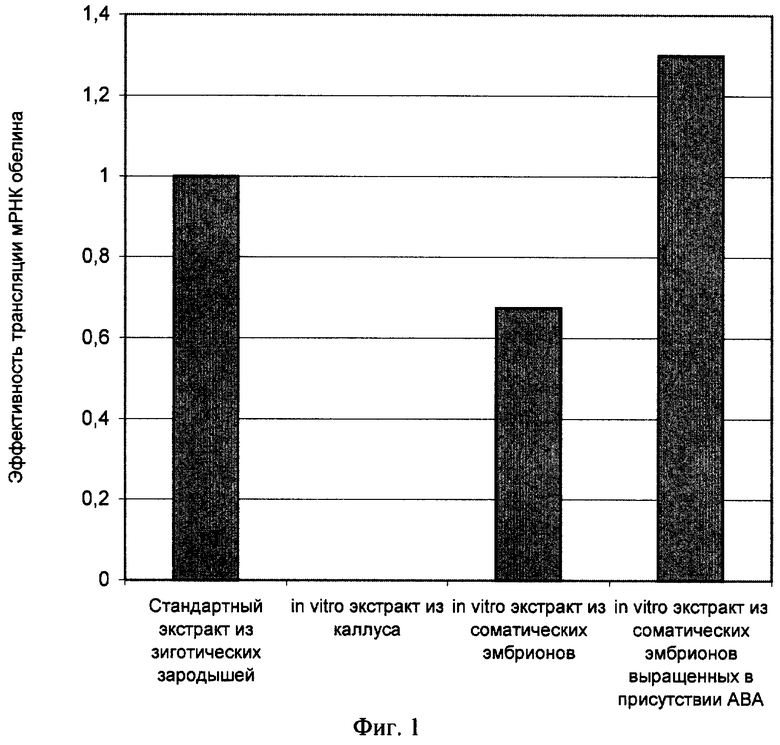
а) получение первичного материала из соматических тканей;
b) индукцию соматического эмбриогенеза;
с) приготовление экстракта из соматических зародышей.
а) получение первичного материала из соматических тканей;
b) получение высокоэмбриогенного каллуса;
с) получение эмбриогенной суспензионной культуры;
d) индукцию вторичного соматического эмбриогенеза;
е) приготовление экстракта из соматических зародышей.
| Способ получения полипептидов в бесклеточной системе трансляции | 1990 |
|
SU1839191A1 |
| DAVIES, PAUL KAESBERG "Translation of virus mRNA: Synthesis of bacteriophage QB proteins in a cell-free extract from wheat embryo", Journal of Virology, Dec.1973, p.1434-1441. | |||

