Изобретение относится х молекулярной биологии и биотехнологии, а именно к способам получения полипептидов в бесклеточных системах. Преимущественной областью использования является препаративный синтез биологически активных пептидов и белков.
Известен способ препаративного синтеза полипептидов методом генной инженерии, основанным на внедрении в живую клетку чужеродной ДНК, генетический материал которой экспрессируется аппаратом клетки-хозяина 1-3.
К недостаткам этого известного способа относятся ограничения, накладываемые клеткой на продукты экспрессии гена. Это связано со сложностью выделения продукта экспрессии трансформированными клетками, летальностью некоторых целевых продуктов для клетки-продуцента, элиминированием трансформированных плазмид из клетки, протеолитической деградацией или агрегацией продукта чужеродного гена, В результате только весьма ограниченный набор белков может быть эффективно синтезирован известным спосо- бом.
Известен способ препаративной экспрессии генов, основанный на использовании непрерывной бесклеточной системы сопряженной транскрипции/трансляции 4. Непрерывные бесклеточные системы сопряженной транскрипции/трансляции свободны от ограничений, накладываемых клеткой, и обеспечивают препаративную экспрессию практически любых генов в виде молекулы ДНК, сконструированной нужным образом.
К недостаткам этого известного способа относятся ограничения на использование бесклеточных систем эукариот. Дело в том, что при-экспрессии генов указанным способом используются эндогенные РНК- полимеразы того объекта, из которого приготовлена бесклеточная система сопряженной транскрипции /трансляции. Это приводит к необходимости использования специальных методик выделения клеточного экстракта, обеспечивающих сохранность активности эндогенных РНК- полимераз. Более того, в клетках эукариот процессы транскрипции и трансляции, как правило, разнесены в пространстве и во времени: процесс транскрипции осуществляется в клеточном ядре, а процесс трансляции происходит в цитоплазме клетки после соответствующих модификаций мРНК. В связи с этим получить надежную систему сопряженной системы транскрипции/трансляции на основе экстрактов эукариотических клеток до настоящего времени не удалось. Единственный метод, обеспечивающий надежное получение таких экстрактов, основан на получении экстракта S30 из
бактериальных клеток Esherlchia coli. Однако данный способ также имеет существенный недостаток: всякая плазмида, содержащая интересующий нас клонированный ген, имеет в своем составе ген селекции (ген устойчивости к какому-либо антибиотику), который также находится под промотором РНК-полимеразы Е. coli и экспрессируется столь же эффективно, как и интересующий нас ген. В результате в про5 цессе функционирования дополнительно к нужному продукту в системе экспрессируется побочный продукт.
В качестве прототипа выбран способ препаративного синтеза полипептидов в
0 бесклеточных системах трансляции непрерывного действия с использованием матричных РНК, состоящий в том, что в бесклеточных системах трансляции осуществляют непрерывное удаление из реакци5 онной смеси продуктов реакции, включая синтезированный полипептид, и непрерывное восстановление концентрации низкомолекулярных субстратов 5. Указанный способ позволяет осуществлять препара0 тивный синтез практически любых полипептидов в бесклеточных системах трансляции, полученных из клеток любых организмов.
Так, например, известен способ синтеза кальцитонина в бесклеточной системе
5 трансляции непрерывного действия на основе лизата из зародышей пшеницы. В 1 мл бесклеточной системы содержится 0,5 мл лизатз зародышей пшеницы, 0,1 нмоль мРНК кальцитонина, 64 мкг креатинфосфо0 киназы, 10 мкг ингибитора рибонуклеаз из плаценты человека, по 0,1 мкг ингибиторов протеаз (апротинин, пепстатин, лейпептин) в буфере: 40 мМ HEPES, рН 7,6, 75 мМ Г ацетата, 1,9 мМ Мд2+ ацетата, 0,25 мМ спер5 мидина, 6 мМ дитиотреита, 1,5% глицерина, 2 мМ АТР, 50 мкМ GTP, 8 мМ креатинфосфа- та, 25 мкМ Н лейцина (активность 50 Ки/ммоль) и по 25 мкМ остальных 19 аминокислот.
0 Бесклеточную систему помещают в
ячейку для ультрафильтрации фирмы
Amicon и проводят синтез полипептидов
при температуре 25°С. Отбор продуктов
трансляции, включающих целевой продукт
5 и продукты распада, (AMP, GDP, пирофос- фат и неорганический фосфат) осуществляют через полупроницаемую мембрану с одновременной подачей субстратов в виде АТР, GTP и аминокислот в течение 20 ч. В результате за время работы бесклеточной
истемы трансляции получают 10 нмоль альцитонина, что составляет 100 пмоль олипептида на 1 пмоль мРНК,
К недостаткам этого способа относится евозможность экспрессии генетического атериала в виде ДНК. Основанный на бес- леточных системах трансляции, данный пособ использует матричные РНК. Это оз- ачает, что для реализации способа необхо- има дополнительная работа по синтезу атричных РНК тем или иным способом. В о же время основная масса полипептидов белков в живых клетках закодирована в иде молекул ДНК. Различными генно-ин- енерными манипуляциями в настоящее ремя выделено огромное число генов в ви- е молекул ДНК. Кроме того, синтез и раз- ножение искусственных генов в виде олекул ДНК представляет собой несрав- енно более простую задачу, чем наработка атричных РНК.
Целью изобретения является унифика- ия способа препаративного получения олипептидов в бесклеточных системах и
В бесклеточных системах трансляции прерывного действия, в процессе работы орых осуществляется непрерывное вы- дение продуктов трансляции, в том числе ипептидов, AMP, ADP, GDP, CDP, UDP, фосфатов и неорганических фосфатов, дновременным введением в систему суб- эатов в виде аминокислот, АТР, GTP, СТР, Р для поддержания их исходной концен-
к;
вышение выхода и чистоты целевого про-
п Д
KI BI
п п с
С
U
т ации, используются гены белков в виде м элекул ДНК, имеющих промоторныеучаст- Ki, специфичные к чужеродным экзогенным Р К-полимеразам, а в систему трансляции добавляются экзогенные РНК-полимеразы, ответствующие выбранным промоторам. Сущность предложенного способа за- очается в следующем. Экстракты прока- отических или эукариотических клеток, держащие рибосомы и все компоненты парата трансляции, но свободные от эн- енных мРНК и ДНК, готовят известными тодами. К экстракту добавляют низкомоД м
лекулярные компоненты транскрипции и трансляции (аминокислоты, АТР, GTP, СТР, U Р), ген в виде молекулы ДНК, имеющие лидерный участок, являющийся промоторов для экзогенной РНК-полимеразы, причем такая конструкция может быть использована как в виде фрагмента ДНК, та : и в виде плазмиды, экзогенную РНК- шмеразу, специфичную для выбранного мотора, а также в случае необходимости
ибиторыэндогенных РНК-полимераэор- изма, из которого получен экстракт. Ре5
5
0
0 5
акционную смесь вносят в резервуар, ограниченный от окружающего пространства полупроницаемой перегородкой, где и протекает процесс сопряженной транскрипции/трансляции. Полупроницаемая перегородка может быть как органического, так и неорганического происхождения. В качестве полупроницаемой перегородки могут быть использованы ультрафильтраци- 0 онные мембраны, полые волокна, микрс ап- сулы или пленки, оболочка которых представляет собой полиэлектролитные комплексы. Для предотвращения остановки процесса из резервуара через полупрони- 5 цаемую перегородку непрерывно отводят продукты транскрипции (неорганические фосфаты, ADP, GDP, CDP, UDP) и трансляции (синтезированный полипептид, AMP, GDP, неорганические фосфаты и пирофос- фаты).
Одновременно в резервуар подают аминокислоты, АТР, GTP, СТР, UTP для восстановления их исходной концентрации. Синтезированный полипептид собирают на колонке с сорбентом, а продукты транскрипции и трансляции собирают в специальном резервуаре для их последующей регенерации. Соотношения компонентов в реакционной смеси, ионные и температурные условия синтеза определяются свойствами организмов, из которых приготовлены бесклеточные системы и экзогенные РНК- полимеразы. Диапазон этих условий достаточно широк. Способ позволяет осуществлять сопряженную транскрипцию/трансляцию в экстрактах из таких организмов, для которых сопряжение процессов трансляции и транскрипции in vivo невозможно, Предлагаемый способ обеспечивает препаративный синтез полипептидов с постоянно высокой скоростью в течение десятков часов с выходом функционально активного продукта (полипептида) 1-10 нмоль с 1 мл реакционной смеси.
На фиг.1,2,3,4,5 изображены графики зависимости количества синтезируемого полипептида (нмоль) от времени синтеза (ч).
Для катализа реакции из бесклеточной системы транскрипции/трансляции из ре- 0 тикулоцитов кролика взято 10 мл реакционной смеси до и через 0,5 ч после начала работы.
Для катализа реакции из непрерывной бесклеточной системы транскрипции/трансляции из ретикулоцитов кролика взято также 10 мкл раствора А, содержащего продукты экспрессии, через 5.7,9 и 12 ч после начала работы непрерывной системы.
Пример 1. Сопряженная транскрипция/трансляция фермента дигидрофолат
редуктазы с плазмиды, содержащей ген ди- гидрофолат редуктазы под промотором SP6 полимеразы, в бесклеточной прокариотиче- ской системе трансляции из Е. coll.
Получают плазмиду, содержащую ген дигидрофолат редуктазы под промотором SP6 полимеразы.
Сопряженную систему транскрипции/трансляции готовят следующим образом.
1 мл реакционной смеси содержит 350 мкл S30 экстракта из Е. coll, 0,2 мг тРНК, 0,1 мг плазмиды, 20000 U SP6 полимеразы, 0,1 мг пируват киназы, 50 U рибонуклеазного ингибитора из плаценты человека, по 5 мкг ингибиторов протеаз (леупептина, химости- на) и с:2-макроглобулина в буфере А: 50 мМ , рН 7,5, 14 мМ MgCte, 100 мМ КАс, 2 мМ СаАс2, 1 мМ АТР, 0,4 мМ GTP, 0,4 мМ СТР, 0,4 мМ UTP, 10 мМ фосфоенолпирува- та, 4,0 мМ дитиотреита, 50 мкМ спермидина, 10 мкг лейковорина, 40 мкМ рифампицина, ингибитора РНК-полимеразы Е. coll., 30 мкМ 35S Met с удельной радиоактивностью 800 МКи/ммоль и по 30 мкМ каждой из остальных 19 аминокислот.
Синтез проводят при температуре 37°С в реакционной ячейке. Питательную смесь пропускают со скоростью 1.5 мл в 1 ч. Отбор целевого продукта и продуктов распада осуществляют путем отбора питательной смеси,прошедшей через ультрафильтрационную мембрану, с одновременной подачей в ячейку буфера А в течение 24 ч.
В течение всего времени синтез продукта проходит с постоянной скоростью (фиг. 1). В результате синтезируют 680 пмоль фермента дигидрофолат редуктазы за 24 ч рабо- ты. Синтезированный фермент функционально активен. Удельная активность полученного фермента составляет 0, ед. активности на 1 пмоль синтезированного фермента.
В данном случае плазмида содержит ген дигидрофолат редуктазы под промотором SP6 полимеразы и ген/3-лактамазы под промотором РНК-полимераэы Е, coll. Поскольку в систему вводят ингибитор РНК- полимеразы Е. coli рифампицин, в системе синтезировался только целевой продукт дигидрофолат редуктаза высокой чистоты, не содержащий каких-либо примесей / -лакта- мазы.
Пример 2. Сопряженная транскрипция/трансляция /3 -лактамазы с фрагмента ДНК, содержащего ген / -лактамазы и короткий участок промотора Т7 полимеразы, в
бесклеточной прокариотической системе трансляции из Е. coll.
Получают плазмиду, содержащую ген предшественника/3-лактамазы и промотор
для Т7 полимеразы.
Сопряженную систему транскрипции/трансляции готовят следующим образом.
1 мл реакционной смеси содержит 350
мкл S30 экстракта из Е. coli, 0,1 мг тРНК, 0,04 мг фрагмента ДНК, 30000 U T7 полимеразы, 0,1 мг пируват киназы, 50 U рибонуклеазного ингибитора из плаценты человека, по 5 мкг ингибиторов протеаз {леупептина,
химостина)ио:2-макроглобулина в буфере А: 50 мМ , рН 7,5,14 мМ MgCfc, 100 мМ КАс, 2 мМ СаАс2, 1 мМ АТР, 0,4 мМ GTP, 0,4 мМ СТР, 0,4 мМ UTP, 10 мМ фосфоенолпи- рувата, 4,0 мМ дитиотреита, 50 мкМ спермидина, 10 мкг лейковорина, 40 мкМ рифампицина, 30 мкМ 3Н Leu с удельной радиоактивностью 1,7 Ки/ммоль и по 30 мкм каждой из остальных 19 аминокислот.
Синтез проводят при температуре 37°С
в реакционной ячейке. Питательную смесь пропускают со скоростью 2 мл в 1 ч. Отбор целевого продукта и продуктов распада осуществляют путем отбора питательной смеси,прошедшей через
ультрафильтрационную мембрану, с одновременной подачей в ячейку буфера А в течение 20 ч.
В течение всего времени синтез продукта проходит с постоянной скоростью (фиг.2).
В результате синтезируют 250 пмоль/3 -лактамазы за 20 ч работы системы.
Пример 3. Сопряженная транскрипция/трансляция дигидрофолат редуктазы с плазмиды, содержащей ген дигидрофолат
редуктазы под промотором SP6 полимеразы в бесклеточной эукариотической системе трансляции из зародышей пшеницы.
Получают плазмиду, содержащую ген дигидрофолат редуктазы под промотором
SP6 полимеразы.
Сопряженную систему транскрипции/трансляции готовят следующим образом.
1 мл инкубационной смеси содержит
320 мкл экстракта из зародышей пшеницы, 0.1 мг плазмиды, содержащей ген дигидрофолат редуктазы, 20000 U SP6 полимеразы, 0,1 мг пируват киназы, 50 U рибонуклеазного ингибитора из плаценты человека, по 5
мкг ингибиторов протеаз (леупептина, химо- статина) и а 2-макроглобулина в буфере А: 40 мМ HEPES, рН 7,6, 2,5 мМ МдАса, 70 мМ КАс. 1 мМ АТР. 0,4 мМ GTP, 0,4 мМ СТР, 0,4 мМ UTP, 0,25 мМ спермидина, 4,0 мМ дитиотреита, 6 мМ креатинфосфата, 20 мкМ 14С Leu с удельной радиоактивностью 21 мКи/ммоль, 20 мкМ каждой из 19 остальных аминокислот.
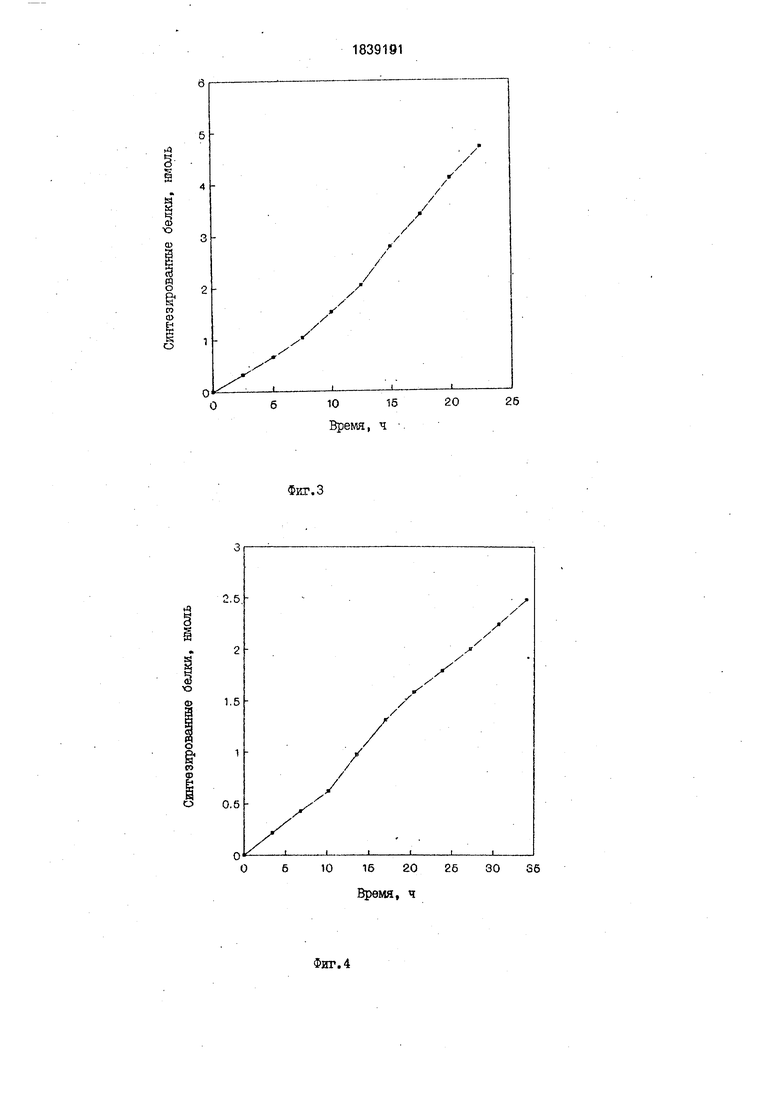
Синтез проводят при температуре 24°С в реакционной ячейке. Питательную смесь пропускают со скоростью 2 мл в 1 ч. Отбор целевого продукта и продуктов распада осуществляют путем отбора питательной смеси,прошедшей через ультрафильтрационную мембрану, с одновременной подачей в ячейку буфера А в течение 24 ч.
В течение всего времени синтез продукта проходит с постоянной скоростью (фиг.З). В результате синтезируют 5 нмоль фермента дигидрофолат редуктазы за 24 ч работы. Синтезированный фермент функционально активен. Удельная активность полученного фермента составляет 0, ед. активно- сти на 1 пикомоль синтезированного фермента.
Пример 4. Сопряженная транскрипция/трансляция хлорамфеникол ацетилт- рансферазы с плазмиды, содержащей ген хлорамфеникол ацетилтрансферазы под промотором SP6 полимеразы, в бесклеточной эукариотмческой системе трансляции из ретикулоцитов кролика.
Получают плазмиду, содержащую ген хлорамфеникол ацетилтрансферазы под пр мотором SP6 полимеразы.
Сопряженную систему транскрипции/трансляции готовят следующим образом.
1 мл инкубационной смеси содержит 600 мкл лизата из ретикулоцитов кролика, 0,1 мг плазмиды, содержащей ген хлорамфеникол ацетилтрансферазы под промотором SP6 РНК-полимеразы, 30000 U SP6 полимеразы, 0,1 мг пируват киназы, 50 U рибонуклеазного ингибитора из плаценты человека, по 5 мкг ингибиторов протеаз (ле- упептина, химостатина) и «2-макроглобули- на в буфере А: 25 мМ HEPES, рН 7,6, 1,5мМ МдАс2, 100 мМ КАс, 1 мМ АТР, 0,4 мМ GTP, 0,4 мМ СТР, 0,4 мМ UTP, 0,25 мМ спермиди- на, 4,0 мМ дитиотреита, 6 мМ креатинфосфата, 20 мкМ 35S Met с удельной радиоактивностью 800 мКи/ммоль, 20 мкМ каждой из 19 остальных аминокислот.
Синтез проводят при температуре 34°С в реакционной ячейке. Питательную смесь пропускают со скоростью 1,5 мл в 1 ч. Отбор целевого продукта и продуктов распада осу- ществляют путем отбора питательной смеси,прошедшей через ультрафильтрационную мембрану, с одновременной подачей в ячейку буфера А в течение 34 ч.
В течение всего времени синтез продукта проходил с постоянной скоростью (фиг.4). В результате синтезируют 2,5 нмоль фермента хлорамфеникол ацетилтрансферазы за 34 ч работы. Синтезированный фермент функционально активен.
Пример 5. Сопряженная транскрипция/трансляция фермента дигидрофолат редуктазы с плазмиды, содержащей ген дигидрофолат редуктазы под промотором SP6 полимеразы, в бесклеточной эукариотиче- ской системе трансляции из ретикулоцитов кролика.
Получают плазмиду, содержащую ген дигидрофолат редуктазы под промотором SP6 полимеразы.
Сопряженную систему транскрипции/трансляции готовят следующим образом.
1 мл инкубационной смеси содержит 600 мкл лизата из ретикулоцитов кролика, 0,1 мг плазмиды, содержащей ген дигидрофолат редуктазы, 30000 U SP6 полимеразы, 0,1 мг пируват киназы, 50 U рибонуклеазного ингибитора из плаценты человека, по 5 мкг ингибиторов протеаз (леупептина, химостатина) и о2-макроглобулина в буфере А: 25мМНЕРЕ5,рН7,6, 1,5мММдАс2, 100 мМ КАс, 1 мМ АТР, 0,4 мМ GTP, 0,4 мМ СТР, 0,4 мМ UTP, 0,25 мМ спермидина, 4,0 мМ дитиотреита, 6 мМ креатинфосфата, 20 мкМ 14С Leu с удельной радиоактивностью 21 мКи/ммоль, 20 мкМ каждой из 19 остальных аминокислот.
Синтез проводят при температуре 34°С в реакционной ячейке. Питательную смесь пропускают со скоростью 2 мл в 1 ч. Отбор целевого продукта и продуктов распада осуществляют путем отбора питательной смеси,прошедшей через ультрафильтрационную мембрану, с одновременной подачей в ячейку буфера А в течение 20 ч.
В течение всего времени синтез продукта проходит с постоянной скоростью (фиг.5). В результате синтезируют 7,0 нмоль фермента дигидрофолат редуктазы за 20 ч работы. Синтезированный фермент функционально активен. Удельная активность полученного фермента составляет 0, ед. активности на 1 пмоль синтезированного фермента.
Использование изобретения дает возможность препаративной экспрессии практически любых генов в виде молекул ДНК в любых бесклеточных системах трансляции. Способ может служить важным дополнением, а в некоторых случаях и альтернативой генно-инженерным методам и методу препаративной экспрессии генов в бесклеточной системе. Способ не требует специальной наработки матричных РНК, что значительно упрощает и удешевляет его в сравнении с известным способом препара- тивного синтеза полипептидов и белков в бесклеточных системах трансляции.
(56) 1. Graham F.L., van der Eb A.J. - Virology, 1973, v.52, p.456.
2. Grammar W.J. - In: Genetic Engineering, 1982, v.3, p.53. (ed. Wllllamson R.), Academic Press Inc., London.
3. Frazer A.C., Curtlss R. - Curr. Top. Microbiol. Immunol., 1975, v.69, p.1.
4. Baranov V.I,, Morozov I.Yu., Ortlepp S.A., Spirln A.S. - Gene, 1989, v.84, p.463.
5. Spirln A.S., Baranov V.I., Ryabova LA,, Ovodov S.Yu., Alakhov Yu.B. - Science, 1988, v.242, p.1162.







Формула изобретения
е
9
to
р го
р со
р .ь.
О ел
р Ь)
р
CD
ел
I
СО
со со
а
СП
го о
w
О
Синтезированные белки, ймоль
Л -,
i
к
§
5 о
о
I,
0
510 16
Время, ч
Редактор
Составитель Л. Рябова
Техред М.МоргенталКорректор С. Юско
Заказ 3404Тираж Подписное
НПО Поиск Роспатента 113035, Москва, Ж-35, Раушская наб., 4/5
/
/
20
26
