Заявленная группа технических решений относится к области молекулярной биологии и синтетической биологии, конкретно – к средству и способу синтеза белков в открытых системах вне клеток на основе клеточного метаболизма, скринингу антимикробных веществ, в том числе активных по отношению к трансляционному аппарату Staphylococcus aureus. Заявленные технические решения позволят проводить поиск химических соединений на способность ингибирования синтеза белка, в том числе бактерии Staphylococcus aureus, производить функциональные тесты бактериальных белков, в том числе белков бактерии Staphylococcus aureus, выполнять синтез бактериальных белков для биотехнологии и структурной биологии. Изобретения могут быть использованы, например, на биотехнологических и фармацевтических производствах, в научно-исследовательских организациях, с целью проведения реакций синтеза белка бактерии Staphylococcus aureus в естественном окружении in vitro, тестирования влияния химических веществ (например – антибиотиков) на биосинтез белка Staphylococcus aureus. Синтезируемый белок с использованием заявленной бесклеточной системы может быть использован в биотехнологиях, в том числе – после проведения его выделения, очистки и модификации целевого белка.
Далее в тексте заявителем приведены термины, которые необходимы для облегчения однозначного понимания сущности заявленных материалов и исключения противоречий и/или спорных трактовок при выполнении экспертизы по существу.
АТФ – аденозинтрифосфат.
HEPES – 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота.
ЭДТА –Этилендиаминтетрауксусная кислота.
NTP – нуклеотидтрифосфат.
ЦТФ – цитидинтрифосфат.
ГТФ – гуанозинтрифосфат.
УТФ – уридинтрифосфат.
RNasine – ингибитор РНКаз.
Для решения проблемы антибиотикорезистентности штаммов патогенных микроорганизмов необходима разработка новых лекарственных препаратов на основе комплексного подхода. Одним из наиболее традиционных путей поиска антимикробных препаратов является последовательное тестирование новых природных или синтетических соединений на культуральных жидкостях, после чего производится определение химической и пространственной структуры и поиск механизма действия ингибитора. При этом поиск молекулярной мишени в клетке патогена, на которую воздействует исследуемое вещество с антимикробной активностью, является одним из самых сложных этапов.
На современном этапе развития фармакологии, и биотехнологии на её основе, одной из ключевых задач является получение данных о взаимосвязях и взаимодействии частей, составляющих биологическую систему, а также её кинетических параметров – для этого часто используются in vitro подходы и, в частности, бесклеточные системы.
Программирование бесклеточных систем дает множество преимуществ:
-простоту проведения экспериментов,
-возможность контроля за концентрациями и активностями генов и ферментов,
-простоту в получении количественных измерений,
-возможность управления большим количеством параметров, которые можно оценивать, используя высокопропускные (high-throughput) подходы.
В общем смысле, бесклеточная система синтеза белка — это клетка без мембраны, помещенная в квази-естественную среду, состоящую из буферной системы, метаболитов и ферментов, помогающих метаболизму. В бесклеточной системе синтеза белка воспроизводится набор химических реакций, которые отражают главные метаболические процессы необходимые для синтеза белка. В состав бесклеточной системы синтеза белка входят все компоненты необходимые для транскрипции, трансляции, а также системы синтеза АТФ, обеспечивающие энергию для протекания биохимических реакций.
Бесклеточная система синтеза белка состоит из четырех основных компонент:
-клеточного экстракта как основы метаболизма,
-ДНК/РНК,
-субстратов для транскрипции и трансляции,
-системы регенерации AТФ.
Соединенные воедино компоненты создают условия для протекания основных этапов синтеза белка in vitro. В силу наличия генетических особенностей в строении аппарата синтеза белка у каждого вида организмов необходимо использовать соответствующий клеточный экстракт для конкретного организма и подбирать индивидуальные условия и концентрации компонент бесклеточных систем синтеза белка.
Для скрининга потенциальных ингибиторов исследуемое вещество добавляется в реакционную смесь бесклеточной системы синтеза белка и по количеству сигнального белка, опосредованного через сигнал флюоресценции или люминесценции, оценивается уровень ингибирования синтеза белка. При этом, из-за открытости системы, возможно проводить крупномасштабную проверку и оптимизацию, легко изменяя условия проведения экспериментов, например, концентрацию добавляемого вещества-ингибитора, в рамках одного эксперимента. Также в рамках бесклеточной системы синтеза белка возможно разделять транскрипцию от трансляции и оценивать влияние на отдельные процессы, тем самым уточнять сферу действия вещества-ингибитора. Бесклеточные системы синтеза белка имеют преимущества скорости протекания реакции (от 2 до 10 часов), возможности за один раз автоматизировано проводить большое количество измерений, возможности контролировать реакцию вследствие отсутствия клеточной стенки, возможности добавления любых белков, в том числе токсичных для клетки.
В настоящее время бесклеточные системы синтеза белка активно используются в сфере синтетической биологии для создания модельных минимальных программируемых систем, дизайна белков, поиска биокатализаторов, для масштабной наработки целевых белков и молекул, для исследований процессов ингибирования синтеза белков.
Высокая стоимость бесклеточных систем ранее делала их непривлекательными для высокопроизводительного скрининга. Однако в последнее время бесклеточные системы привлекли внимание исследователей и из других областей, особенно из сферы синтетической биологии. Сейчас технология используется в сфере синтетической биологии для создания модельных минимальных программируемых систем, дизайна белков, поиска биокатализаторов, для масштабной наработки целевых белков и молекул, для поиска ингибиторов. В отличии от тест-систем, основанных на клеточных культурах, в бесклеточной системе синтеза белка возможно проведение точных кинетических экспериментов с добавлением веществ.
Так, известны источники «An Escherichia coli cell-free expression toolbox: application to synthetic gene circuits and artificial cells» (Бесклеточная система экспрессии на основе клеток кишечной палочки E.coli и ее применение в синтетических генных цепочках и искусственных клетках) [Shin J. Et al. An Escherichia coli Cell-Free Expression Toolbox: Application to Synthetic Gene Circuits and Artificial Cells // ACS Synth. Biol, 2012. Vol.1. N. 1. P. 29–41], «A cell-free protein synthesis system for high-throughput proteomics» (Бесклеточная система синтеза белка для высокопроизводительной протеомики) [Sawasaki T. et al. A cell-free protein synthesis system for high-throughput proteomics // Proc Natl Acad Sci USA, 2002. Vol. 99(23). P. 14652-14657], «Microscale Adaptation of In Vitro Transcription/Translation for High-Throughput Screening of Natural Product Extract Libraries» (Микромасштабная адаптация экстракорпоральной транскрипции/трансляции для высокопроизводительного скрининга библиотек экстрактов натуральных продуктов) [Lowell, A. N. et al. Microscale Adaptation of In Vitro Transcription/Translation for High-Throughput Screening of Natural Product Extract Libraries // Chem. Biol. Drug Des. 2015. Vol. 86. P. 1331–1338. Сущностью являются бесклеточные системы для тестирования регуляторных элементов и синтеза белка в грамотрицательной бактерии Escherichia coli.
Недостатком указанных систем является ограниченность применения только для грамотрицательных бактерий Escherichia coli и невозможность синтеза модифицированных белков бактерии Staphylococcus aureus, в силу отсутствия специфических для Staphylococcus aureus компонент транскрипции и трансляции.
Известен источник «Development of a Bacillus subtilis cell-free transcription-translation system for prototyping regulatory elements» (Разработка бесклеточной системы транскрипции-трансляции на основе клеток Bacillus subtilis для прототипирования регуляторных элементов) [Kelwick R. et al. Development of a Bacillus subtilis cell-free transcription-translation system for prototyping regulatory elements // Metab. Eng, 2016. Vol. 38. P. 370-381]. Сущностью является бесклеточная система синтеза белка на основе штамма WB800N бактерии B. subtilis, способная производить 0,8 мкМ флуоресцентного белка GFP. Показана возможность создания библиотеки промоторов B. subtilis, на основе промотора Pgrac (σA) дикого типа, которые демонстрируют ряд сопоставимых транскрипционных активностей in vitro и in vivo.
Недостатком указанной системы является ограниченность применения только для бактерии Bacillus subtilis, что не позволяет проводить синтез модифицированных белков бактерии Staphylococcus aureus, в силу отсутствия специфических для Staphylococcus aureus компонент транскрипции и трансляции.
Так, известно изобретение по патенту РФ №2148649 «Способ получения полипептидов в бесклеточной системе (варианты) и устройство для его осуществления». Сущностью является способ получения полипептидов в бесклеточной системе, предусматривающий приготовление реакционной смеси и питающей смеси, установку внутри реакционного модуля двух пористых барьеров, определяющих границы реакционного объема, введение реакционной смеси в реакционный объем и проведением синтеза при заданных условиях, поддержание синтеза введением питающей смеси, в процессе синтеза выводят из реакционного объема или оставляют внутри реактора синтезируемые целевые полипептиды, отличающийся тем, что перед проведение синтеза подготавливают смесь расходуемых высокомолекулярных компонентов, выбирают соотношение объема питающей смеси и расходуемых высокомолекулярных компонентов к объему фракции, содержащей целевой полипептид, устанавливают режим ввода питающей смеси и расходуемых высокомолекулярных компонентов в реакционный объем, в процессе синтеза продукты синтеза разделяют на фракцию, содержащую низкомолекулярные компоненты, и фракцию, содержащую высокомолекулярные компоненты синтеза, включая целевой полипептид, а основную часть фракции, содержащей низкомолекулярные компоненты, выводят из реакционного объема через одну или обе части второго пористого барьера.
Недостатком известного технического решения является ограничения по времени биосинтеза белков из-за вымывания компонент бесклеточной системы, отсутствующих в питающей смеси.
Известно изобретение по патенту РФ № 2162104 «Способ синтезирования флуоресцентно-меченого белка». Сущностью является способ синтезирования флуоресцентно-меченого белка в прокариотической бесклеточной системе белкового синтеза, предусматривающий следующие стадии: (а) инкубирование пробы рибосом, полученных из бесклеточного экстракта, с плазмидной ДНК, содержащей кодирующую последовательность для представляющего интерес белка, причем пробу инкубируют в среде для сопряженной транскрипции/трансляции вместе с аминоацил-тРНК, имеющей флуоресцентную метку, (б) частичную очистку указанного флуоресцентно-меченого белка отделением вновь синтезированного флуоресцентно-меченого белка от других флуоресцентных компонентов в этой пробе, (в) измерение количества синтезированного белка, (г) определение флуоресценции вновь синтезированного белка, (д) определение биологической активности вновь синтезированного белка.
Недостатком известного технического решения является необходимость дополнительного этапа очистки синтезированного флуоресцентно-меченого белка от других флуоресцентных компонентов в пробе.
Известно изобретение по патенту РФ №2620074 «Рекомбинантная плазмидная ДНК pDUALREP2 и штамм, трансформированный ею для выявления веществ и смесей, ингибирующих биосинтез белка и/или вызывающих SOS-ответ у бактерий». Сущностью является рекомбинантная плазмидная ДНК pDualrep2, предназначенная для идентификации соединений, ингибирующих биосинтез белка и/или вызывающих SOS-ответ у бактерий, включающая: ориджин репликации, способный поддерживать стабильное число копий плазмиды в клетке; селективный маркер для отбора клеток, содержащих плазмиду; гены флюоресцентных белков с различимыми спектрами флюоресценции RFP и Katushka2S; аттенюатор триптофанового оперона, в котором произведены замены триптофановых кодонов UGG на аланиновые кодоны GCG; промотор гена sulA Е. coli, активирующийся при инактивации репрессора SOS-ответа LexA.
Недостатком известного технического решения является ограниченность применения только для грамотрицательных бактерий Escherichia coli и невозможность синтеза модифицированных белков бактерии Staphylococcus aureus, в силу отсутствия специфических для Staphylococcus aureus компонент транскрипции и трансляции.
Известно изобретение (WO 2019035955 A1 опубл. 21.02.2019) «Высокопроизводительная система, использующая бесклеточную систему экспрессии и секвенирование in situ» («High-throughput system using a cell-free expression system and in situ sequencing»). Сущностью является cпособ скрининга множества матричных последовательностей нуклеиновых кислот на продукцию малых молекул, включающий добавление матричной последовательности нуклеиновой кислоты из множества в каждый реакционный объем из множества реакционных объемов, при этом каждый реакционный объем включает бесклеточную систему экспрессии в условиях транскрипции и трансляции; обнаружение образования малой молекулы в целевом реакционном объеме; и секвенирование матричной последовательности нуклеиновой кислоты целевого реакционного объема.
Недостатком известного технического решения является применение клеточных лизатов из актинобактерий Streptomyces lividans, Streptomyces violaceoruber, Streptomyces coelicolor, Streptomyces albovinaceus, грамотрицательных бактерий Vibrio natriegens, Escherichia coli, Pseudomonas aeruginosa и отсутствие описания оптимизированного протокола синтеза белка в грамположительной бактерии Staphylococcus aureus.
Известен источник «Staphylococcus aureus cell extract transcription-translation assay: firefly luciferase reporter system for evaluating protein translation inhibitors» (Анализ транскрипции-трансляции клеточного экстракта Staphylococcus aureus: репортерная система люциферазы светлячка для оценки ингибиторов трансляции белков) [Murray R. W. et al. Staphylococcus aureus cell extract transcription-translation assay: firefly luciferase reporter system for evaluating protein translation inhibitors // Antimicrob Agents Chemother. 2001. Vol. 45(6). P. 1900-1904], описывающий репортерную систему Staphylococcus aureus, состоящую из лизата экстракта клеток Staphylococcus aureus штамма RN4220, выращенных на питательной среде Brain heart, предварительно последовательно промытых в 500 мл холодного буфера А (10 мМ Трис-ацетат, pH 8,0, 14 мМ Mg-ацетат, 1 мМ дитиотреитол), содержащего 1 М KCl, а затем 250 мл буфера A, содержащего 50 мМ KCl, после чего суспензия клеток ресуспендировалась до конечного объема 99 мл буфера B (10 мМ Трис-ацетат, pH 8,0, 20 мМ Mg-ацетат, 50 мМ KCl, 1 мМ дитиотреитол) с последующей инкубацией раствора с коммерческим препаратом фермента лизостафина для разрушения клеточной стенки, далее суспензия инкубировалась при 37 °C в течение 45-60 минут, после чего раствор трижды осаждали центрифугированием при 4°C при 16 000 об/мин (30 000 3 g) в течение 30 мин и отбиралось две трети супернатанта, после чего добавлялось 0,25 объема буфера для преинкубации (670 мМ Трис-ацетат, pH 8,0, 20 мМ Mg-ацетат, 7 мМ Na3-фосфоенолпируват, 7 мМ дитиотреитол, 5,5 мМ АТФ, 70 мМ смеси 20 протеиногенных аминокислот, 75 мг/мл пируваткиназы) и проводилась инкубация смеси при 37°C в течение 30 мин для увеличения активных рибосом после прочтения ими всех имеющихся мРНК в растворе; после чего для дополнительно инкубированного супернатанта проводился диализ в течение 12 часов при 4°C в 2 л диализного буфера (10 мМ Трис-ацетат, pH 8.0, 14 мМ Mg-ацетат, 60 мМ K-ацетат, 1 мМ дитиотреитол) с однократной сменой буфера; для концентрирования до 10 мг/мл (очищенный) лизат подвергали диализу в мешках (Spectra- Por 7; с нижней границей молекулярной массы 3500 Да), содержащих предварительно охлажденный полиэтиленгликоль 8000 (Sigma) при 4 °C; в полученную реакционную смесь добавлялся ПЦР-продукт, содержащий ген люциферазы под контролем промотора гена синтеза капсульного полисахарида Staphylococcus aureus (cap1A), и коммерческого набора Luciferase Assay Reagent (Promega Corporation) для реакции синтеза люминесцентного белка, на конечном этапе регистрировался сигнал люминесценции.
Недостатками известной системы является необходимость проведения этапа инкубации лизата для увеличения активных рибосом после прочтения ими всех имеющихся мРНК в растворе, необходимость применения коммерческого препарата лизостафина для лизирования клеток, необходимость добавления коммерческого набора Luciferase Assay Reagent (Promega Corporation) в реакционную смесь для возможности регистрации сигнала люминесценции, отсутствие возможности синтеза флуоресцентного белка и возможности регистрации флюоресценции.
Технической проблемой, на решение которой направлена заявляемая группа изобретений, является расширение арсенала средств и способов для синтеза белка, а также для скрининга веществ на способность ингибировать синтез белка бактерий, в том числе Staphylococcus aureus.
Технический результат заявленной группы изобретений заключается в
– возможности синтеза белков грамположительной бактерии Staphylococcus aureus в естественном генетическом и метаболическом окружении, что позволяет получать функциональные белки Staphylococcus aureus с посттрансляционными модификациями за счет наличия в экстракте всех необходимых ферментов Staphylococcus aureus;
- оптимальном составе реакционной смеси, что обеспечивает возможность получения экстракта без этапа инкубации смеси для увеличения активных рибосом после прочтения ими всех имеющихся мРНК в растворе;
- возможности регистрации ингибирующей активности химических соединений как с помощью регистрации сигнала флуоресценции репортерного белка, так и люминесценции.
Указанная техническая проблема решается, и технический результат достигается заявляемой бесклеточной системой синтеза белка на основе клеток Staphylococcus aureus, полученной путем смешения компонентов реакционной смеси в следующем порядке и в следующем объемном соотношении (об.%):
вода – 7,85,
100 мМ HEPES-KOH pH 8,0 – 5,00,
2% полиэтиленгликоль 8000 – 4,00,
0,1 мМ ЭДТА pH 8,0 – 0,5,
2 мМ дитиотреитол – 0,20,
9,1 мМ магний ацетат – 0,91,
228,5 мМ калий ацетат – 5,71,
0,5 мМ раствор из 20 протеиногенных аминокислот – 12,50,
16,7 мМ раствор из аминокислот в эквимольном соотношении: аргинин, цистеин, триптофан, метионин, аспарагиновая кислота, глутаминовая кислота – 5,99,
0,05% азид натрия – 1,67,
1% раствор ингибитора протеаз Complete – 1,00,
ингибитор РНКаз RNasine – 0,75,
тотальная тРНК из Escherichia coli – 1,25,
2,11 мМ фолиновая кислота – 1,00,
20 мМ ацетил фосфат литиево-калиевая соль – 4,00,
20 мМ фосфоенолпируват дикалиевая соль – 4,0,
NTP микс: 90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ, pH 7,0 – 1,33,
T7 полимераза – 2,50;
S30 экстракт клеток Staphylococcus aureus RN4220 – 35;
плазмидная ДНК – 4,44.
Состав заявляемой бесклеточной системы синтеза белка является оптимальным, его изменение или иной, чем заявлен, порядок добавления компонентов приводит к ухудшению работы системы и снижению уровня синтеза белка. Заявляемый состав также обеспечивает возможность получения экстракта без этапа инкубации смеси для увеличения активных рибосом после прочтения ими всех имеющихся мРНК в растворе.
Также техническая проблема решается, и технический результат достигается заявляемым способом синтеза белка в бесклеточной системе синтеза белка на основе клеток Staphylococcus aureus, включающий инкубацию при температуре 37°С вышеописанной бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus.
Кроме того, техническая проблема решается, и технический результат достигается заявляемым способом скрининга ингибиторов синтеза белка, включающим инкубацию при 37°С потенциальных ингибиторов синтеза белка совместно с заявляемой бесклеточной системой синтеза белка на основе клеток Staphylococcus aureus, количественное измерение синтезированного белка по сигналу люминесцентного или флуоресцентного излучения и определение способности к ингибированию синтеза белка по падению светового сигнала.
В случае использования люминесценции после инкубирования потенциальных ингибиторов синтеза белка совместно с заявляемой бесклеточной системой в реакционную смесь добавляют люциферин.
Предложенная группа изобретений может стать платформой для поиска ингибиторов синтеза белка, в том числе селективных в отношении бактерии Staphylococcus aureus.
Заявленные изобретения имеют следующие преимущества:
- уменьшается количество этапов получения бесклеточной системы синтеза белка, что снижает расходы и трудозатраты;
- возможность применения не только люминесценции, но и флуоресценции при количественном определении белка, что позволяет оценить синтез белка в динамике;
- отсутствие необходимости использования коммерческого препарата люциферина для люминесценции за счет собственного синтеза флуоресцентного белка.
Также расходы на получение заявленной бесклеточной системы могут быть снижены за счет самостоятельного синтеза рекомбинантного белка лизостафина без использования коммерческого препарата лизостафина для лизиса клеток.
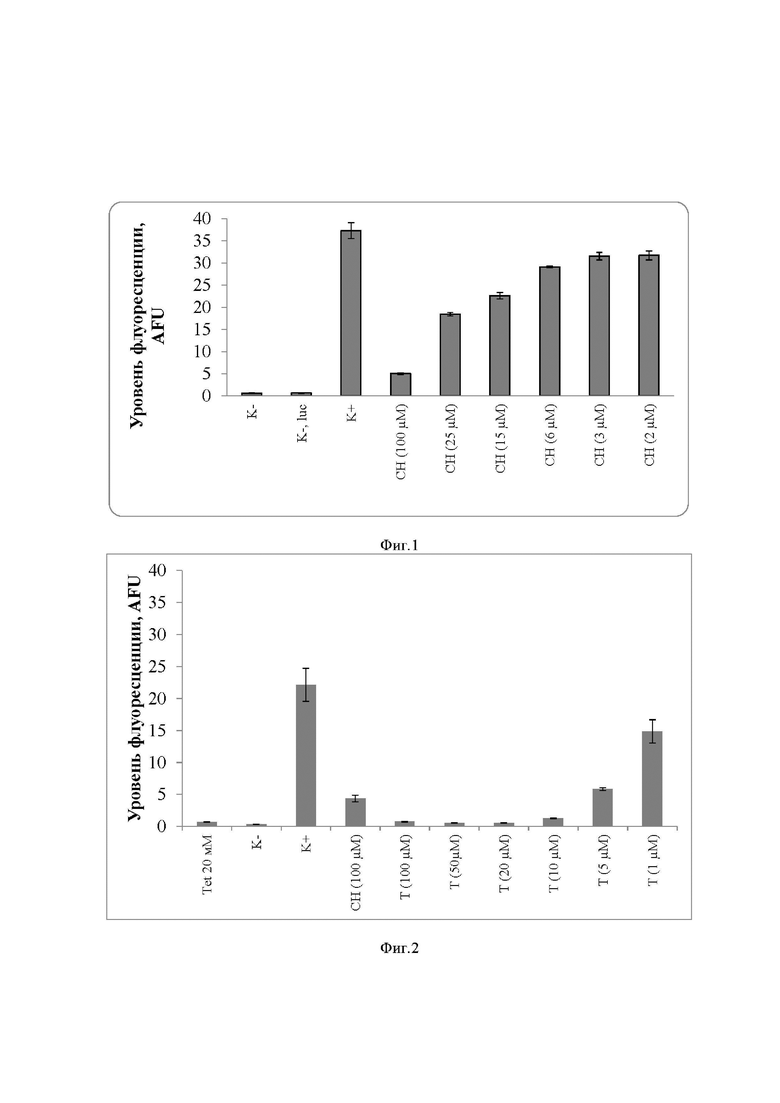
Заявленное техническое решение иллюстрируется фигурами 1 и 2, где представлены результаты тестирования заявляемой бесклеточной системы синтеза белка Staphylococcus aureus в отсутствии и присутствии известных трансляционных ингибиторов:
на фиг. 1 – хлорамфеникола, где: K- – реакция с H2O вместо репортерной плазмиды (негативный контроль); K-, luc – реакция с добавлением нерепортерной плазмиды (используется, чтобы исключить влияние добавления нерепортеной плазмиды на уровень флюоресценции); K+ – реакция с репортерной плазмидой sfGFP (сигнал принимается за 100% уровень синтеза белка); 2-100 мкМ – образцы с добавлением хлорамфеникола в различных концентрациях. При увеличении концентрации хлорамфеникола подавляется синтез белка GFP и снижается уровень флуоресценции. Измерение проводили по финальной точке через 8 часов после начала реакции. AFU – относительные единицы флюоресценции (Arbitrary Fluorescence Units).
на фиг. 2 – тетрациклина, где Tet 20 mM – контроль флюоресценции раствора тетрациклина с концентрацией 20 мМ; K- – реакция с H2O вместо репортерной плазмидой (негативный контроль); K+ – реакция с репортерной плазмидой sfGFP (сигнал принимается за 100% уровень синтеза белка); СН – хлорамфеникол, Т – тетрациклин, в скобках указана концентрация в мкМ. При увеличении концентрации тетрациклина подавляется синтез белка GFP и снижается уровень флуоресценции. AFU – относительные единицы флюоресценции (Arbitrary Fluorescence Units).
Далее заявителем приведено описание заявленного технического решения.
Заявленные изобретения предназначены для синтеза белков в бесклеточной системе Staphylococcus aureus, выявления ингибиторов синтеза белка на основе бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus.
Заявленный технический результат в целом достигается тем, что на основе клеток Staphylococcus aureus штамма RN4220 подготавливают экстракт, смешивают компоненты реакционной смеси, проводят реакцию синтеза белка и используют синтезированный белок, например, для детекции сигнала с целью оценки эффективности синтеза.
Все используемые реагенты являются коммерчески доступными.
Заявленная бесклеточная система синтеза белка без учета порядка добавления компонентов включает, об.%:
- S30 экстракт клеток Staphylococcus aureus RN4220 – 35;
- оптимизированный набор компонентов для проведения реакции синтеза – 56,16, в состав которого входит:
вода – 7,85,
100 мМ HEPES-KOH pH 8,0 – 5,00,
2% полиэтиленгликоль 8000 – 4,00,
0,1 мМ ЭДТА pH 8,0 – 0,5,
2 мМ дитиотреитол – 0,20,
9,1 мМ магний ацетат – 0,91,
228,5 мМ калий ацетат – 5,71,
0,5 мМ раствор из 20 протеиногенных аминокислот – 12,50,
16,7 мМ раствор из аминокислот в эквимольном соотношении: аргинин, цистеин, триптофан, метионин, аспарагиновая кислота, глутаминовая кислота – 5,99,
0,05% азид натрия – 1,67,
1% раствор ингибитора протеаз Complete – 1,00,
ингибитор РНКаз RNasine – 0,75,
тотальная тРНК из Escherichia coli – 1,25,
2,11 мМ фолиновая кислота – 1,00,
20 мМ ацетил фосфат литиево-калиевая соль – 4,00,
NTP микс: 90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ, pH 7,0 – 1,33,
T7 полимераза – 2,50;
плазмидная ДНК – 4,44;
система регенерации АТФ – 4,4, включающая:
20 мМ фосфоенолпируват дикалиевую соль – 4,0,
0,18 мМ пируваткиназу – 0,4.
При реализации изобретения были использованы следующие реактивы и оборудование:
вода (>99,9999%, ОСТ 11.029.003-80), HEPES-KOH (≥99.5%, арт. H0527, Sigma-Aldrich), полиэтиленгликоль 8000 (≥99.0%, арт. HR2-535, Hampton Research), ЭДТА (>99.0%, арт. H-E5134-0.1, Хеликон), дитиотреитол (≥ 99.0 %, арт. R0861, Thermo Scientific), магний ацетат (>99.0%, арт. Am-O131-0.5, Хеликон), калий ацетат (>99.0%, арт. P1190-100G, Sigma-Aldrich), раствор из 20 протеиногенных аминокислот (>98%, арт. L4461, Promega), аминокислоты: аргинин (≥99.0%, арт. 1015420100, Sigma-Aldrich), цистеин (≥99.0%, арт. 1028380025, Sigma-Aldrich), триптофан (≥99.0%, арт. 1083740010, Sigma-Aldrich), метионин (≥99.0%, арт. 1057070025, Sigma-Aldrich), аспарагиновая кислота (≥99.0%, арт. 1001260100, Sigma-Aldrich), глутаминовая кислота (≥99.0%, арт. 1002910250, Sigma-Aldrich), азид натрия (≥99.5%, арт. 71289-5G, Sigma-Aldrich), ингибитор протеаз Complete (>95%, арт. 11697498001, Roche), ингибитор РНКаз RNasine (>95%, арт. N2111, Promega), тотальная тРНК из Escherichia coli (арт. rnamrero, Roche), фолиновая кислота (≥99.0%, арт. 47612-250MG, Sigma-Aldrich), ацетил фосфат литиево-калиевая соль (≥85% арт. A0262-5G, Sigma-Aldrich), NTP микс: АТФ (≥99.0%, арт. R0441, Thermo Scientific), ЦТФ(≥99.0%, арт. R0451, Thermo Scientific), ГТФ (≥99.0%, арт. R0461, Thermo Scientific), УТФ (≥99.0%, арт. R0471, Thermo Scientific), T7 полимераза (>95%, арт. EP0113, Thermo Scientific), Плазмидная ДНК (#102634, Addgene), 20 мМ фосфоенолпируват дикалиевая соль (99%, арт. 860077-1G, Sigma-Aldrich), Пируваткиназа (>95%, арт. P1506-1KU, Sigma-Aldrich), планшетный спектрофотометр-люминометр Varioskan LUX (Thermo Scientific).
Представленные выше исходные компоненты заявленной системы получают следующим образом:
- экстракт на основе клеток Staphylococcus aureus штамма RN4220 подготавливают с помощью ферментативного лизиса лизостафином, ультрацентрифугирования и диализа, без проведения этапа дотранслирования;
- плазмидную ДНК-репортер получают из трансформированных клеток Escherichia coli штамма DH5a (#102634, Addgene), с помощью метода щелочного лизиса и переосаждения в этиловом спирте и ПЭГ/СaCl2;
- оптимизированный набор компонентов для проведения реакции синтеза получают путем растворения компонентов в деионизованной воде.
Световой сигнал детектируют с помощью измерения флюоресценции или люминесценции в планшетном спектрофотометре Varioskan LUX (Thermo Scientific).
Таким образом, заявленное техническое решение включает этапы подготовки экстракта клеток Staphylococcus aureus штамма RN4220, проведения реакции синтеза и детекции сигнала. Экстракт для проведения реакций изготавливают из клеток Staphylococcus aureus штамма RN4220 путем ферментативного лизиса лизостафином, полученным рекомбинантным способом. Полученный экстракт выступает основой для проведения программируемой реакции по синтезу белка. Для программирования системы используется плазмидная ДНК, кодирующая репортерный белок (например, флуоресцентный белок sfGFP). Описание изобретения содержит протокол проведения реакции синтеза с оптимизированным набором реагентов, добавляемых в реакцию синтеза, условия проведения реакции и условия измерения сигнала эквивалентному количеству репортерного белка по уровню флюоресценции на планшетном спектрофотометре.
Далее заявителем приведен способ получения заявленной бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus, заявленный способ синтеза белка на основе бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus и заявленный способ выявления ингибиторов синтеза белка на основе бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus.
Штамм Staphylococcus aureus RN4220 высевают в агаризированную твердую среду и инкубируют в течение 12 часов при 37 °C. Одиночную колонию клеток Staphylococcus aureus высевают в среду LB и выращивают клеточную массу в течение 12 ч. Далее культуру инокулируют в свежую среду и выращивают клеточную массу до достижения середины логарифмической фазы роста. Затем клеточную массу осаждают центрифугированием и удаляют супернатант. Полученный преципитат ресуспендируют в буфере А (10мМ Tрис-Aцетат pH 8.2, 14 мМ Mg(CH3COO)2, 60 мM KСl, 1 мМ DTT) и проводят отмывку клеток. Полученный осадок хранят при температуре -80 °С.
Для препаративного выделения плазмидной ДНК-репортера используют комбинированный протокол, состоящий из стандартного протокола выделения плазмидной ДНК и протокола выделения плазмидной ДНК с помощью CaCl2 и ПЭГ 8000. Данный протокол позволяет получить плазмидную ДНК без примесей РНК, без использования РНКаз и в достаточной концентрации (1 мкг/мкл) для бесклеточной системы в короткий промежуток времени (48 часов). Для in vivo экспрессии репортерного белка sfGFP (superfolded GFP) под контролем Т7 промотора используется плазмидная ДНК pJL1-sfGFP от компании AddGene (Plasmid #102634), кодирующая последовательность белка sfGFP, который применятся в качестве стандартного репортерного высокостабильного и быстро фолдирующегося белка. Предпочтительно использовать плазмидные ДНК, кодирующие флуоресцентные белки — GFP, YFP, CFP, mCherry и их аналоги. При использовании других плазмидных ДНК необходимо использовать векторы высокой копийности, с целью увеличения выхода плазмидной ДНК при выделении.
Трансформация клеток Escherichia coli штамма DH5a производится целевой плазмидной ДНК. Далее культуру клеток выращивают в течение 8 ч и инокулируют в свежую среду в колбах большего объема и выращивают клеточную массу до OD600 равной 0,5. Клетки осаждают центрифугированием и ресуспендируют осадок первоначально в растворе для ресуспендирования, затем в лизирующем растворе NaOH и SDS, после чего в образец добавляют ацетат аммония для нейтрализации. Образец осаждают центрифугированием и добавляют изопропиловый спирт, инкубируют и центрифугируют. Преципитат ресуспендируют в TЕ буфере и добавляют ацетат аммония, после чего смесь инкубируют и центрифугируют. К полученному супернатанту добавляют изопропиловый спирт, инкубируют и далее осаждают образец центрифугированием. Образец отмывают этиловым спиртом и высушивают. Полученный осадок ресуспендируют в высокоочищенной деионнизированной воде (mQ). Для удаления высокомолекулярной РНК обрабатывают образец с помощью CaCl2, после чего центрифугируют, отбирают супернатант, и проводят осаждение с помощью изопропилового спирта и ацетата калия, осадок промывают этиловым спиртом и сушат. Для удаления низкомолекулярной РНК осадок растворяют в буфере TE с ПЭГ/NaCl, перемешивают и центрифугируют. Осадок промывают этиловым спиртом и высушивают. Плазмидную ДНК растворяют в малом объёме высокоочищенной деионнизированной воды (mQ). Для бесклеточных систем также можно использовать плазмидные ДНК, очищенные при помощи наборов со спин-колонками – в этом случае необходимо провести дополнительный этап очистки фенол-хлороформом, для удаления РНКазы.
Экстракт для бесклеточной системы получают путем разрушения клеточной стенки Staphylococcus aureus с помощью фермента лизостафина. Лизостафин получают с помощью рекомбинантного синтеза в клетках Escherichia coli, что снижает затраты на приобретение коммерческих препаратов. Гены, кодирующие аминокислотные последовательности лизостафина без сигнального пептида и Т7 полимеразы, клонируются в вектор pET28A для экспрессии в Escherichia coli Bl21. Одиночная колония клеток Escherichia coli штамма BL21, трансформированного плазмидной ДНК, с чашки Петри высевается в 25 мл среды LB с соответствующими маркеру устойчивости антибиотиками. Клеточная масса выращивается в течение 12 ч при интенсивном перемешивании 180 об/мин и 37°С. Далее 20 мл «ночной» культуры пересевается в 1 литр среды LВ и растят клеточную массу до OD600 0,5 при 180 об/мин и 37 °С. Экспрессию индуцируют добавлением 0,5 мМ IPTG и растят при перемешивании в течение 4 ч при 180 об/мин и 37 °С. Осаждение клеточной массы производят центрифугированием в течение 15 мин при 5 400 об/мин. Клеточную массу Escherichia coli, содержащую рекомбинантный белок лизостафин, ресуспендируют в 50 мл буфера CRB (20 мМ Tris-HCl (pH 7,6), 0,5 M NH4Cl) и осаждают центрифугированием 15 мин при 3220 об/мин. Клетки Escherichia coli, содержащие рекомбинантный белок лизостафин, размораживают в бане со льдом и ресуспендируют 5 мл холодного буфера CRB (20 мМ Tris-HCl (pH 7,6), 0,5 M NH4Cl), в который добавляется смесь ингибиторов протеиназ Complete (1 таблетка на 1 мл) и PMSF (0,5 мМ). Клетки лизируют при помощи ультразвукового гомогенизатора (амплитуда 40%, 3 раза по 8 минут, чередовали 3 минуты ультразвук включен, 5 минут пауза). Лизат осветляют центрифугированием при 25000 об/мин в течение 30 мин. Надосадочную жидкость переносят в новые пробирки и повторно очищают центрифугированием при 45000 об/мин в течение 45 мин. Далее проводят очистку рекомбинантных белков методом металл хелатной аффинной хроматографии. В колонку, содержащую Ni-NTA- агарозу, вносится лизат клеток Escherichia coli, содержащих рекомбинантный белок лизостафин, и промывается 15 мл буфера “wash А” (20 мМ Tris-HCl (pH 7,6), 1 M NH4Cl), затем 15 мл буфера “wash B” (20 мМ Tris-HCl (pH 7,6), 0,5 M NH4Cl, 5 мМ имидазол). Белок смывают с сорбента 5 мл элюирующего буфера (20 мМ Tris-HCl (pH 7,8), 250 мM NH4Cl, 300 мМ имидазол). Дополнительную очистку лизостафина проводят с помощью эксклюзионной хроматографии на FPLC хроматографе NGC Discover (BioRad) и колонке Superdex 75 10/300 (производитель GE, Великобритания). Эксклюзионную хроматографию проводят в потоке фосфатного буфера (25 мМ NaH2PO4, 25 мМ Na2HPO4, 250 мM NH4Cl) при 4 °C со скоростью нанесения 0,8 мл/мин, и собирают фракции объемом 1 мл. За разделением белков следят по профилю поглощения А280. Фракции концентрируют с помощью концентраторов Amicon-Ultra (Merck Millipore, Германия).
Замороженные клетки Staphylococcus aureus размораживают и добавляют буфер с лизостафином, объем доводят до 15 мл для достижения финальной концентрация экстракта 30 мг/мл на конечной стадии. На данном этапе для снижения вязкости раствора не применяются ДНКазы, поскольку это приведет к гидролизу кодирующей плазмидной ДНК в получаемой бесклеточной системе. Для лизиса клеток полученную смесь инкубируют на водяной бане. Для уменьшения вязкости и расщепления остаточной ДНК образец гомогенизируют ультразвуком при температуре 4 °C, после чего производят центрифугирование. Полученный клеточный экстракт инкубируют при 37 °C в течение 60 мин в инкубаторе. Для определения концентрации экстракта применяют метод Бредфорда.
Для проведения реакции в бесклеточной системе синтеза белка Staphylococcus aureus производится последовательное добавление реагентов согласно оптимизированному набору компонентов, а именно: вода, HEPES-KOH pH 8,0, полиэтиленгликоль 8000, ЭДТА pH 8,0, дитиотреитол, магний ацетат, калий ацетат, раствор из 20 протеиногенных аминокислот, смесь аминокислот в эквимольном соотношении: аргинин, цистеин, триптофан, метионин, аспарагиновая кислота, глутаминовая кислота, азид натрия, ингибитор протеаз Complete, ингибитор РНКаз RNasine, тотальная тРНК из Escherichia coli, фолиновая кислота, ацетил фосфат литиево-калиевая соль, фосфоенолпируват дикалиевая соль, NTP микс, пируваткиназа, T7 полимераза, S30 экстракт клеток Staphylococcus aureus RN4220, плазмидная ДНК, разделяют реакционную смесь по 15 мкл в 384 луночный планшет, выдерживают при температуре 37 °С и измеряют флюоресценцию или люминесценцию, получают значение количества синтезированного белка. Способность к ингибированию синтеза белка химическими соединениями, потенциально являющимися ингибиторами синтеза белка, определяют по падению сигнала флуоресценции.
Далее приведены примеры конкретного осуществления изобретения и реализации назначения, при этом заявитель подразумевает, что приведенные примеры не ограничивают предполагаемый объем заявленного технического решения.
Пример 1. Получение заявленной бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus .
1. Получают экстракт из клеток Staphylococcus aureus – компонент заявленной системы.
Клетки Staphylococcus aureus штамма RN4220, в котором имеются мутации по генам sau1 hsdR, что позволяет трансформировать данный штамм плазмидными ДНК из Escherichia coli, высевают в агаризированную твердую среду BHI (Difco) на чашки Петри и инкубируют в течение 12 часов при 37 °C. Одиночную колонию клеток Staphylococcus aureus высевают в 20 мл среды Lysogeny broth (LB) и растят клеточную массу в течение 12 ч при интенсивном перемешивании 180 об/мин и 37 °С. Далее культуру инокулируют в свежую среду в колбы большего объема в соотношении 1:100 к свежей среде LB и растят клеточную массу до OD600 = 1,0 (до достижения середины логарифмической фазы роста) при 180 об/мин и 37 °С. Для остановки роста клеток колбы переносят на 15 мин в ледяную баню. Затем клеточную массу осаждают центрифугированием при 4 °C в течение 15 мин при 2500 g и удаляют супернатант. Полученный преципитат ресуспендируют в 50 мл буфера А (10мМ Tрис-Aцетат pH 8.2, 14 мМ Mg(CH3COO)2, 60 мM KСl, 1 мМ DTT), центрифугируют при 4 °C в течение 15 мин при 2500 g, и повторяют данную процедуру трижды для удаления из раствора остатков компонент питательной среды. Полученный осадок взвешивают и хранят при температуре -80 °С.
В заявленном техническом решении лизис клеток Staphylococcus aureus получают с помощью обработки рекомбинантно синтезированным в клетках Escherichia coli ферментом лизостафином, что снижает затраты на приобретение коммерческого препарата (например, L7386-1MG, Sigma).
Гены, кодирующие аминокислотные последовательности лизостафина без сигнального пептида и Т7 полимеразы, клонируются в вектор pET28A для экспрессии в Escherichia coli Bl21. Одиночная колония клеток Escherichia coli штамма BL21, трансформированного плазмидной ДНК, с чашки Петри высевается в 25 мл среды LB с соответствующими маркеру устойчивости антибиотиками. Клеточная масса выращивается в течение 12 ч при интенсивном перемешивании 180 об./мин и 37°С. Далее 20 мл «ночной» культуры пересевается в 1 литр среды LВ и растят клеточную массу до OD600 0,5 при 180 об/мин и 37°С. Экспрессию индуцируют добавлением 0,5 мМ IPTG и растят при перемешивании в течение 4 ч при 180 об/мин и 37 °С. Осаждение клеточной массы производят центрифугированием в течение 15 мин при 5400 об/мин. Клеточную массу Escherichia coli, содержащую рекомбинантный белок лизостафин, ресуспендируют в 50 мл буфера CRB (20 мМ Tris-HCl (pH 7,6), 0,5 M NH4Cl) и осаждают центрифугированием 15 мин при 3220 об/мин. Клетки Escherichia coli, содержащие рекомбинантный белок лизостафин, размораживают в бане со льдом и ресуспендируют 5 мл холодного буфера CRB (20 мМ Tris-HCl (pH 7,6), 0,5 M NH4Cl), в который добавляется смесь ингибиторов протеиназ Complete (1 таблетка на 1 мл) и PMSF (0,5 мМ). Клетки лизируют при помощи ультразвукового гомогенизатора (амплитуда 40%, 3 раза по 8 минут, чередовали 3 минуты ультразвук включен, 5 минут пауза). Лизат осветляют центрифугированием при 25000 об/мин в течение 30 мин. Надосадочную жидкость переносят в новые пробирки и повторно очищают центрифугированием при 45000 об/мин в течение 45 мин. Далее проводят очистку рекомбинантных белков методом металл хелатной аффинной хроматографии. В колонку, содержащую Ni-NTA- агарозу, вносится лизат клеток Escherichia coli, содержащих рекомбинантный белок лизостафин, и промывается 15 мл буфера “wash А” (20 мМ Tris-HCl (pH 7,6), 1 M NH4Cl), затем 15 мл буфера “wash B” (20 мМ Tris-HCl (pH 7,6), 0,5 M NH4Cl, 5 мМ имидазол). Белок смывают с сорбента 5 мл элюирующего буфера (20 мМ Tris-HCl (pH 7,8), 250 мM NH4Cl, 300 мМ имидазол). Дополнительную очистку лизостафина проводят с помощью эксклюзионной хроматографии на FPLC хроматографе NGC Discover (BioRad) и колонке Superdex 75 10/300 (производитель GE, Великобритания). Эксклюзионную хроматографию проводят в потоке фосфатного буфера (25 мМ NaH2PO4, 25 мМ Na2HPO4, 250 мM NH4Cl) при 4°C со скоростью нанесения 0,8 мл/мин, и собирают фракции объемом 1 мл. За разделением белков следят по профилю поглощения А280. Фракции концентрируют с помощью концентраторов Amicon-Ultra (Merck Millipore, Германия).
Для приготовления экстракта используют подготовленные клетки Staphylococcus aureus штамма RN4220. Замороженные клетки размораживают и добавляют буфер с лизостафином, объем доводят до 5-15 мл для достижения финальной концентрация экстракта 10-30 мг/мл на конечной стадии. На данном этапе для снижения вязкости раствора не применяются ДНКазы, поскольку это приведет к гидролизу кодирующей плазмидной ДНК в получаемой бесклеточной системе. Для лизиса клеток полученную смесь инкубируют на водяной бане. Для уменьшения вязкости и расщепления остаточной ДНК образец гомогенизируют ультразвуком при температуре 4 °C, после чего производят центрифугирование. Полученный клеточный экстракт инкубируют при 37 °C в течение 60 мин в инкубаторе. На данном этапе вместо дополнительного проведения этапа инкубации лизата для увеличения активных рибосом после прочтения ими всех имеющихся мРНК в растворе и применения многокомпонентных растворов с ферментами и системой регенерации АТФ, проводят диализ экстракта в финальном буфере в диализных мешках c мембраной 14 кДа, центрифугируют и замораживают в жидком азоте. Для определения концентрации экстракта применяют метод Бредфорда. Поскольку индивидуальные аминокислоты плохо растворяются, используют суспензии, которые добавляются после тщательного перемешивания в финальный объем реакционной смеси.
2. Получают плазмидную ДНК – компонент заявленной системы.
В заявленном техническом решении для in vivo экспрессии репортерного белка sfGFP (superfolded GFP) под контролем Т7 промотора используется плазмидная ДНК pJL1-sfGFP от компании AddGene (Plasmid #102634), кодирующая последовательность белка sfGFP, который применятся в качестве стандартного репортерного высокостабильного и быстро фолдирующегося белка. Для приготовления плазмидной ДНК-репортера используется комбинированный протокол, состоящий из стандартного протокола выделения плазмидной ДНК и протокола выделения плазмидной ДНК с помощью CaCl2 и ПЭГ 8000. Данный протокол позволяет получить плазмидную ДНК без примесей РНК, без использования РНКаз и концентрации 1 мкг/мкл для бесклеточной системы в 48 часов.
Одиночную колонию клеток Escherichia coli штамма DH5a, трансформированного плазмидой pJL1-sfGFP, с чашки Петри с твердой агаризированной средой LB высевают в 25 мл среды LB с соответствующими маркеру устойчивости антибиотиками. Клеточную массу выращивают в течение 8 ч при интенсивном перемешивании 180 об/мин и 37 °С. Далее культуру инокулируют в свежую среду в колбах большего объема в соотношении 1:100 к свежей среде LB и наращивают клеточную массу до OD600 0,5 при 180 об/мин и 37 °С. Для осаждения клеточной массы применяют центрифугирование при 4 °C в течение 15 мин при 4000 g, удаляют супернатант и работают с преципитатом. Затем клеточную массу Escherichia coli ресуспендируют в 40 мл буфера Раствора I (5 мМ Tris-HCl pH 8,0, 10 мМ ЭДТА, 50 мМ глюкозы) и инкубируют на льду в течение 5 мин. Далее добавляют 80 мл Раствора II (0,2 М NaOH, 1% SDS), перемешают раствор и повторно проводят инкубацию на льду в течение 5 мин. Затем в образец добавляют 80 мл охлажденного (0 °С) 10 М ацетата аммония и перемешивают раствор до гомогенного состояния. Затем образец инкубируют на льду в течение 5 мин и осаждают центрифугированием при 4 °C в течение 30 мин при 15000 об/мин. Супернатант фильтруют через стерильный медицинский бинт и добавляют к 0,6 объема изопропилового спирта, после чего инкубируют на комнатной температуре в течение 10 мин. Далее производят центрифугирование при 4 °C в течение 30 мин при 15000 об/мин, супернатант удаляют и преципитат ресуспендируют в 0,8 мл среды TЕ. Затем добавляют 0,2 мл 10 М ацетата аммония и инкубируют смесь на льду в течение 5 мин, после чего производят центрифугирование при 4 °C в течение 5 мин при 15000 об/мин и отбирают супернатант. К полученному образцу добавляют 0,6 объема изопропилового спирта для преципитации плазмидной ДНК и инкубируют при комнатной температуре в течение 10 мин. Далее осаждают центрифугированием при 15000 об/мин, после чего пробирки с образцом переворачивают над фильтровальной бумагой и остатки жидкости удаляют с помощью пипетки. Затем образец отмывают 70% этиловым спиртом, и высушивают при 50 °C в пробирках с открытой крышкой. Полученный сухой осадок ресуспендируют в высокоочищенной деионнизированной воде и инкубируют в течение 15 мин при 50 °C до полного растворения осадка. Для удаления остаточной РНК применяют протокол для выделения плазмид с помощью CaCl2 и ПЭГ 8000. Для этого сначала преципитируют высокомолекулярную РНК с помощью добавления CaCl2 до 1,4 M и перемешивают раствор, после производится центрифугирование при комнатной температуре при 15000 об/мин в течение 10 мин и отбирают супернатант. Затем в супернатант добавляют 0,6 объема изопропилового спирта и 0,1 объем 3 M ацетата калия, образец перемешивают и снова осаждают центрифугированием при комнатной температуре в течение 10 мин при 15000 об/мин. Полученный осадок промывают 70% этиловым спиртом и высушивают на воздухе в инкубаторе при 37 °C. Далее растворяют осадок в среде TE и добавляют 1 объем раствора ПЭГ/NaCl (20% ПЭГ 8000, 500 мМ NaCl), перемешивают и центрифугируют при комнатной температуре течение 10 мин при 14000 об/мин. Затем осадок промывают холодным (0 °С) 70% этиловым спиртом, высушивают на воздухе в инкубаторе при 37 °C и повторяют данную процедуру дважды. Полученную таким образом плазмидную ДНК растворяют в объёме 100 мкл высокоочищенной деионнизированной воды и инкубируют при 37 °C в течение 15 минут до полного растворения. Концентрацию плазмидной ДНК определяют на спектрофотометре для нано объёмов по поглощению на длине волны 260 нм. Чистоту образца и соответствие молекулярной массе определяют по картине электрофореза в агарозном геле.
3. Берут оптимизированный набор компонентов для проведения реакции синтеза, включающий систему регенерации АТФ из фосфоенолпируват дикалиевой соли и пируваткиназы.
4. Смешивают указанные выше компоненты при комнатной температуре согласно последовательности: вода, HEPES-KOH pH 8,0, полиэтиленгликоль 8000, ЭДТА pH 8,0, дитиотреитол, магний ацетат, калий ацетат, раствор из 20 протеиногенных аминокислот, смесь аминокислот в эквимольном соотношении: аргинин, цистеин, триптофан, метионин, аспарагиновая кислота, глутаминовая кислота, азид натрия, ингибитор протеаз Complete, ингибитор РНКаз RNasine, тотальная тРНК из Escherichia coli, фолиновая кислота, ацетил фосфат литиево-калиевая соль, фосфоенолпируват дикалиевая соль, NTP микс, пируваткиназа, T7 полимераза, S30 экстракт клеток Staphylococcus aureus RN4220, плазмидная ДНК pJL1-sfGFP – до получения однородного раствора. Получают заявленную бесклеточную систему синтеза белка на основе клеток Staphylococcus aureus заявленного состава.
Пример 2. Проведение синтеза белка на основе бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus .
Проводят реакцию по синтезу белка в разрабатываемой бесклеточной системе. Для этого готовят реакционную смесь в черных непрозрачных планшетах на 384 лунки (например, Grenier), объем лунок 20 мкл, объем реакционной смеси в лунке 15 мкл. Список реагентов и последовательность смешения реагентов приведены в Примере 1. Количественное определение синтезируемого белка осуществляют по флуоресцентному сигналу, количество белка зависит от времени синтеза, в данном случае 8 часов. Флуоресценцию можно измерять как в режиме кинетики (в течение 2-10 часов с измерением каждые 30 минут), так и по финальной точке. Измерение флюоресценции производится с помощью спектрофотометра-люминометра, например, Varioskan LUX (Thermo Scientific), на длинах волн 485 нм (возбуждение), 510 нм (испускание) в случае с репортером sfGFP. Помимо регистрации уровня синтеза по флюоресценции также возможно проводить изменение уровня синтеза по сигналу люминесценции. В этом случае используется репортерная плазмидная ДНК, кодирующая последовательность люциферазы, а также добавляется в реакционную смесь люциферин до концентрации 0,1 мМ.
Наличие сигнала флуоресценции при внесении плазмидной ДНК, превышающего сигнал без внесения плазмидной ДНК, свидетельствует о наличии флуоресцентного белка sfGFP и, тем самым, об успешном синтезе белка с использованием бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus.
Пример 3. Определение способности хлорамфеникола и тетрациклина ингибировать синтез белка в бесклеточной системе синтеза белка на основе клеток Staphylococcus aureus .
Проводят синтез флуоресцентного белка GFP в разрабатываемой бесклеточной системе в черных непрозрачных планшетах на 384 лунки (Grenier), объемом лунок 20 мкл и объемом реакционной смеси в лунке 15 мкл. Последовательность действий по приготовлению планшетов в приведена в Примере 2. В разные планшеты в трех повторах, в реакционную смесь добавляли известные ингибиторы синтеза белка – хлорамфеникол (растворенный в ДМСО) и тетрациклин (растворенный в mQ) – в различных концентрациях. Для образца, растворенного в ДМСО, с целью исключения влияния растворителя на процесс трансляции в контрольный образец (К+) также добавляли ДМСО до концентрации 1,66%. Объем реакционной смеси для реакции в 384 луночном планшете составлял 15 мкл на лунку (объем лунки 20 мкл). Флуоресценцию измеряли по финальной точке при температуре 30 °C после 8 часов. Для измерения флюоресценции использовали черные непрозрачные планшеты на 384 лунки (Grenier, США). Измерение флюоресценции производили на длинах волн 485 нм (возбуждение), 510 нм (испускание) (Varioskan LUX, Thermo).
Результаты представлены на фигурах 1 и 2, где приводятся уровни синтеза белка в заявляемой бесклеточной системе трансляции Staphylococcus aureus в отсутствии и присутствии указанных известных трансляционных ингибиторов в различных концентрациях. Для контроля одновременно проводят реакцию с H2O вместо репортерной плазмидной ДНК (негативный контроль) и реакцию с добавлением нерепортерной плазмидной ДНК, чтобы исключить влияние добавления нерепортерной плазмидной ДНК на уровень флюоресценции. Реакция с репортерной плазмидной ДНК sfGFP без добавления ингибиторов, принимается за 100% уровень синтеза. Принимая интенсивность сигнала флуоресценции за 100% уровень синтеза белка для лунки с репортерной плазмидой sfGFP видно, что при инкубировании реакционной смеси с хлорамфениколом с концентрацией 2 мкМ, 3 мкМ, 6 мкМ, 15 мкМ, 25 мкМ, 100 мкМ уровень синтеза белка снижается на 15%, 15%, 22%, 39%, 51% и 87% соответственно. А при инкубировании реакционной смеси с тетрациклином с концентрацией 1 мкМ, 5 мкМ, 10 мкМ, 20 мкМ, 50 мкМ, 100 мкМ уровень синтеза белка снижается на 33%, 74%, 94%, 97%, 97% 97%.
Таким образом, проведение манипуляций в соответствии с описанием выше позволяет достичь заявленной цели – получить бесклеточную систему синтеза белка на основе клеток Staphylococcus aureus, провести синтез белка, выявить ингибиторы синтеза белка на её основе. Приведенные примеры осуществления заявленного изобретения показывают успешное создание бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus, проведение синтеза белка и выявление ингибиторов синтеза белка с её применением, что демонстрирует полезность применения заявленных изобретений в фармакологии, например, для поиска ингибиторов синтеза белка, при этом с использованием стандартного оборудования и материалов.
На основании изложенного выше можно сделать вывод, что заявителем достигнут заявленный технический результат, а именно расширен арсенал средств указанного назначения путем создания бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus, проведен синтез белка и выявление ингибиторов синтеза белка на её основе, при этом достигнуто:
- возможность синтеза белков грамположительной бактерии Staphylococcus aureus в естественном генетическом и метаболическом окружении;
- возможность получения экстракта без этапа инкубации смеси для увеличения активных рибосом после прочтения ими всех имеющихся мРНК в растворе;
- возможность синтеза рекомбинантного белка лизостафина без использования коммерческого препарата лизостафина для лизиса клеток;
- возможность регистрации ингибирующей активности химических соединений как с помощью регистрации сигнала флуоресценции репортерного белка, так и люминесценции.
Кроме того, предложенная группа изобретений может стать платформой для поиска ингибиторов синтеза белка у бактерии Staphylococcus aureus.
Заявленное техническое решение удовлетворяет критерию «новизна», предъявляемому к изобретениям, так как при определении уровня техники не выявлены средства, которым присущи признаки, идентичные (то есть совпадающие по исполняемой ими функции и форме выполнения этих признаков) всем признакам, перечисленным в формуле предполагаемого изобретения, включая характеристику назначения.
Заявленное техническое решение удовлетворяет критерию «изобретательский уровень», поскольку не является очевидным для специалиста в данной области техники – из исследованного уровня техники заявителем не выявлены технические решения, совпадающие по технической сущности с предлагаемым решением, и не установлена известность влияния отличительных признаков на полученный технический результат.
Заявленное техническое решение соответствует условию патентоспособности «промышленная применимость», предъявляемому к изобретениям, так как может быть изготовлено с использованием известных материалов и оборудования.
Группа изобретений относится к биотехнологии. Предложены бесклеточная система синтеза белка на основе клеток Staphylococcus aureus, полученная путем смешения компонентов реакционной смеси в заданном порядке и объемном соотношении (об.%); способ синтеза белка в бесклеточной системе на основе клеток Staphylococcus aureus, включающий инкубацию заявленной бесклеточной системы при 37 °С; способ выявления ингибиторов синтеза белка на основе клеток Staphylococcus aureus, включающий инкубацию при 37 °С потенциальных ингибиторов синтеза белка совместно с бесклеточной системой синтеза белка на основе клеток Staphylococcus aureus, количественное измерение синтезированного белка по сигналу люминесцентного или флуоресцентного излучения и определение способности к ингибированию синтеза белка по падению светового сигнала. Изобретения обеспечивают расширение арсенала средств для синтеза белков в открытых системах вне клеток на основе клеточного метаболизма, для скрининга антимикробных веществ на биотехнологических и фармацевтических производствах. 3 н. и 3 з.п. ф-лы, 2 ил., 3 пр.
1. Бесклеточная система синтеза белка на основе клеток Staphylococcus aureus, полученная путем смешения компонентов реакционной смеси в следующем порядке и в следующем объемном соотношении, об.%:
вода – 7,85,
100 мМ HEPES-KOH pH 8,0 – 5,00,
2% полиэтиленгликоль 8000 – 4,00,
0,1 мМ ЭДТА pH 8,0 – 0,5,
2 мМ дитиотреитол – 0,20,
9,1 мМ магний ацетат – 0,91,
228,5 мМ калий ацетат – 5,71,
0,5 мМ раствор из 20 протеиногенных аминокислот – 12,50,
16,7 мМ раствор из аминокислот в эквимольном соотношении: аргинин, цистеин, триптофан, метионин, аспарагиновая кислота, глутаминовая кислота – 5,99,
0,05% азид натрия – 1,67,
1% раствор ингибитора протеаз Complete – 1,00,
ингибитор РНКаз RNasine – 0,75,
тотальная тРНК из Escherichia coli – 1,25,
2,11 мМ фолиновая кислота – 1,00,
20 мМ ацетил фосфат литиево-калиевая соль – 4,00,
20 мМ фосфоенолпируват дикалиевая соль – 4,0,
0,18 мМ пируваткиназа – 0,4,
NTP микс: 90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ, pH 7,0 – 1,33,
T7 полимераза – 2,50;
S30 экстракт клеток Staphylococcus aureus RN4220 – 35;
плазмидная ДНК – 4,44.
2. Бесклеточная система синтеза белка на основе клеток Staphylococcus aureus по п.1, отличающаяся тем, что в случае количественного определения синтезированного белка методом флуоресценции в качестве плазмидной ДНК используют pJL1-sfGFP.
3. Бесклеточная система синтеза белка на основе клеток Staphylococcus aureus по п.1, отличающаяся тем, что в случае количественного определения синтезированного белка методом люминисценции в качестве плазмидной ДНК используют pT7luc.
4. Способ синтеза белка в бесклеточной системе синтеза белка на основе клеток Staphylococcus aureus, включающий инкубацию при температуре 37 °С бесклеточной системы синтеза белка на основе клеток Staphylococcus aureus, полученной путем смешения компонентов реакционной смеси в следующем порядке и в следующем объемном соотношении, об.%:
воды – 7,85,
100 мМ HEPES-KOH pH 8,0 – 5,00,
2% полиэтиленгликоль 8000 – 4,00,
0,1 мМ ЭДТА pH 8,0 – 0,5,
2 мМ дитиотреитол – 0,20,
9,1 мМ магний ацетат – 0,91,
228,5 мМ калий ацетат – 5,71,
0,5 мМ раствор из 20 протеиногенных аминокислот – 12,50,
16,7 мМ раствор из аминокислот в эквимольном соотношении: аргинин, цистеин, триптофан, метионин, аспарагиновая кислота, глутаминовая кислота – 5,99,
0,05% азид натрия – 1,67,
1% раствор ингибитора протеаз Complete – 1,00,
ингибитор РНКаз RNasine – 0,75,
тотальная тРНК из Escherichia coli – 1,25,
2,11 мМ фолиновая кислота – 1,00,
20 мМ ацетил фосфат литиево-калиевая соль – 4,00,
20 мМ фосфоенолпируват дикалиевая соль – 4,0,
0,18 мМ пируваткиназа – 0,4,
NTP микс: 90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ, pH 7,0 – 1,33,
T7 полимераза – 2,50;
S30 экстракт клеток Staphylococcus aureus RN4220 – 35;
плазмидная ДНК – 4,44.
5. Способ выявления ингибиторов синтеза белка, включающий инкубацию при 37 °С потенциальных ингибиторов синтеза белка совместно с бесклеточной системой синтеза белка на основе клеток Staphylococcus aureus, полученной путем смешения компонентов реакционной смеси в следующем порядке и в следующем объемном соотношении, об.%:
воды – 7,85,
100 мМ HEPES-KOH pH 8,0 – 5,00,
2% полиэтиленгликоль 8000 – 4,00,
0,1 мМ ЭДТА pH 8,0 – 0,5,
2 мМ дитиотреитол – 0,20,
9,1 мМ магний ацетат – 0,91,
228,5 мМ калий ацетат – 5,71,
0,5 мМ раствор из 20 протеиногенных аминокислот – 12,50,
16,7 мМ раствор из аминокислот в эквимольном соотношении: аргинин, цистеин, триптофан, метионин, аспарагиновая кислота, глутаминовая кислота – 5,99,
0,05% азид натрия – 1,67,
1% раствор ингибитора протеаз Complete – 1,00,
ингибитор РНКаз RNasine – 0,75,
тотальная тРНК из Escherichia coli – 1,25,
2,11 мМ фолиновая кислота – 1,00,
20 мМ ацетил фосфат литиево-калиевая соль – 4,00,
20 мМ фосфоенолпируват дикалиевая соль – 4,0,
0,18 мМ пируваткиназа – 0,4,
NTP микс: 90 мМ АТФ, 60 мМ ЦТФ, 60 мМ ГТФ, 60 мМ УТФ, pH 7,0 – 1,33,
T7 полимераза – 2,50;
S30 экстракт клеток Staphylococcus aureus RN4220 – 35;
плазмидная ДНК – 4,44,
количественное измерение синтезированного белка по сигналу люминесцентного или флуоресцентного излучения и определение способности к ингибированию синтеза белка по падению светового сигнала.
6. Способ выявления ингибиторов синтеза белка по п.5, отличающийся тем, что в случае количественного измерения синтезированного белка по сигналу люминесцентного излучения после инкубирования потенциальных ингибиторов синтеза белка совместно с бесклеточной системой синтеза белка на основе клеток Staphylococcus aureus в реакционную смесь добавляют люциферин.
| MURRAY R.W | |||
| et al | |||
| "Staphylococcus aureus cell extract transcription-translation assay: firefly luciferase reporter system for evaluating protein translation inhibitors"; Antimicrobial agents and chemotherapy, 2001, v.45, N 6, p.1900-1904 | |||
| ОСТЕРМАН И.А | |||
| "Поиск и изучение новых антибиотиков ингибиторов синтеза белка"; автореферат диссертации на |
