ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к пептидам, специфически взаимодействующим с молекулой контроля иммунного ответа В7-2, регулирующим биологическую активность костимулирующего рецептора CD28 путем блокировки молекулы семейства В7 и может быть использовано в области иммунологии, биохимии и молекулярной биологии. Создан пептид, несущий аминокислотную последовательность CLARSLGRC или CH2-LARSLGR-СН2, специфически взаимодействующий с молекулой контроля иммунного ответа В7-2. Пептид может выступать основой для разработки средств, регулирующих иммунный ответ, а также создания эффективных лекарственных форм пептидных препаратов в качестве средств терапии онкологических заболеваний.
УРОВЕНЬ ТЕХНИКИ
Иммунотерапия рака основана на способности иммунной системы воздействовать на опухолеспецифические антигены, чтобы вызвать иммунный ответ. Для активации Т-клеток необходимо распознавание TCR антигенпрезентирующей клетки, несущей антигенный пептид, полученный от инфекционного агента. Вместе с тем, костимулирующие молекулы, экспонированные на поверхности и Т-клетки и АПК должны взаимодействовать. Т-клетки активируются с помощью моноклональных антител CD28. При специфическом связывании с В7-2, CD28 подает сигналы, необходимые для того, чтобы Т-клетки справились с подавлением, опосредованным регуляторными Т-клетками.
Таким образом, В7-2 на активированных клетках может костимулировать Т-клетки через взаимодействие с CD28. Этот начальный ответ требует как связывания пептида МНС/антиген с рецепторным комплексом Т-клетки, так и второго костимулирующего сигнала, где CD86 (В7-2) вместе с CD80 (В7-1) выступают в качестве общих лигандов для первого открытого костимулирующего рецептора CD28 и коингибирующего рецептора CTLA-4. В иммунотерапии онкологии можно использовать механизм взаимодействия CD28/B7 для регулирования иммунного ответа.
Известно, что презентация антигена без Т-клеточной костимуляции приводит к функциональной инактивации Т-клеток. Стимуляция Т-клеток без костимуляции ведет также к индуцированной активации клеточной смерти. Таким образом, костимулирующие пути регулируют процесс вовлечения Т-клеточных рецепторов, что влияет на иммунную активацию или инактивацию. Одним из основных механизмов блокировки индукции является процесс взаимодействия CD28/B7, а вызвать невосприимчивость возможно, если заблокировать молекулы семейства В7 [1].
Применение моноклональных антител безусловно внесло вклад в область исследований и диагностики онкологических заболеваний. Однако, моноклональные антитела не лишены недостатков: стоимостная характеристика; нацеленность на единственный эпитоп, а, следовательно, обеспечение одного типа эффекторной функции, соответствующего его изотопу; специфичность, обладающая антигенной изменчивостью [2].
Прямых аналогов - препаратов на основе пептидов или антител, взаимодействующих с корегуляторной молекулой В7-2 нет. В России одобрены четыре моноклональных антитела, блокирующие другие иммунные контрольные точки, но они не являются полными аналогами препаратов на основе пептидов [3].
Известен аналог, близкий по строению и механизму взаимодействия, являющийся пептидом I1T7, имеющий аминокислотную последовательность INWYKFT и специфически связывающийся с человеческим CTLA-4 и блокирующий взаимодействие с молекулой В7-2. Энергия связывания оптимизированной структуры CTLA-4 с пептидом I1T7 с помощью виртуальный скрининг (МОЕ 2016.0802) с составила -53,2832 [4]. Данный пептид выбран биоинформатическим методом, и поэтому требуется дальнейшая проверка специфичности взаимодействия пептида INWYKFT по отношению к человеческому CTLA-4.
Известны близкие по строению пептидный препарат р2ТА (АВ103) и пептидный препарат, содержащий последовательность LMWPPPWW.
Пептидный препарат, содержащий аминокислотную последовательность LMWPPPWW, относится к низкомолекулярным пептидам-миметикам CD28 лиганда белка CTLA-4.
Аминокислотная последовательность LMWPPPWW имитирует последовательность MYPPPY, фрагмента белка CD28 участок связывания на CTLA-4. Этот пептид способен ингибировать взаимодействие CD28/CTLA-4 с CD80 (В7-1) и CD86 (В7-2) [5]. Пептидный препарат р2ТА (АВ103) является пептидом-миметиком мембранного белка CD28, экспрессированного на Т-лимфоцитах, участвующим в костимуляции, необходимой для активации Т-клеток. Обладая иммуномодулирующей активностью, пептид р2ТА (АВ103) ослабляет сигнальный путь CD28 в Т-клетках, но не блокирует его полностью, оставляя без воздействия нормальный гуморальный иммунный ответ. Однократное введение пептида р2ТА (АВ103), после тотальной обработки гамма-лучами в дозе 8 Гр мышей, показало значительное снижение на 7 сутки повышенного из-за облучения интерлекина 6 и фибриногена [6].
Недостатком пептидного препарата р2ТА (АВ103) и пептидного препарата, содержащий последовательность LMWPPPWW является отсутствие высокой специфичности в ингибировании взаимодействия лиганда CD28 на поверхности белка CTLA-4 с рецепторами CD80 и CD86.
Известен химерный полипептид, близкий по механизму действия, содержащий в направлении от N-конца к С-концу домены PD-L2, CTLA-4 и константную область иммуноглобулина человека. Этот слитый полипептид способен одновременно воздействовать на иммунные пути CD80/86-CD28 и PD-1/PDL1-PDL2 на Т-клетках, тем самым сильно ингибируя активацию Т-клеток. Слитый полипептид показал эффективность против предотвращения отторжения трансплантатов и значительно более высокую выживаемость (р<0,05) у группы крыс, получавших индукционную монотерапию иммуносупрессией слитым в дозе 500 мкг (НВ-500) внутрибрюшинно за 2 часа до трансплантации с последующим введением той же суточной дозы в течение следующих последовательных 6 дней подряд по сравнению с контрольной группой крыс, не получавшей лечения. Недостатком данного химерного полипептида является отсутствие высокой специфичности в ингибировании активации Т-клеток [7].
Наиболее близким аналогом (прототипом близким по специфичности взаимодействия) является APVO210 - биспецифический слитый белок, состоящий из антитела к CD86, слитого с мономерным рецептором интерлейкина 10 (ADAPTIR™ от Aptevo Therapeutics). APVO210 специфически индуцирует передачу сигналов рецептора интерлейкина 10 в CD86+антигенпрезентирующих клетках, но не в Т- и В-клетках. APVO210 индуцирует дифференцировку толерогенных дендритных клеток (DC-A210), которые продуцировали высокие уровни интерлейкина 10, экспрессировали CD86, HLA-G и промежуточные уровни CD14 и CD16. Основным недостатком препарата является сложность получения векторной конструкции, позволяющей экспрессировать слитый белок в эукариотических системах с достаточно большим входом [8].
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Техническим результатом заявляемого изобретения является расширение спектра пептидов, специфически взаимодействующих с молекулой контроля иммунного ответа В7-2 и создание альтернативных средств моноклональным антителам, устраняющих их недостатки.
Указанный технический результат достигается созданием пептида, специфически взаимодействующем с молекулой контроля иммунного ответа В7-2 (CD86) человека, регулирующим взаимодействие с костимулирующей молекулой CD28 и несущим аминокислотную последовательность CLARSLGRC или CH2-LARSLGR-CH2.
Изобретение представляет собой пептид, способный специфически связываться с корегуляторной молекулой В7-2 (CD86) и, таким образом, регулировать взаимодействие с ее лигандами CD28 и антигеном CD152.
Пептид выявлен из синтезированных продуктов с помощью технологии фагового дисплея. Метод фагового дисплея широко используется для быстрой характеристики белок-белковых взаимодействии среди миллиардов кандидатов (11). Библиотеки сконструированы так, что чужеродный пептид, находясь на N-конце рекомбинантного белка, располагается на поверхности вириона и доступен для взаимодействия с комплексом белком-лигандом, который используется для аффинной селекции [9]. Популярность применения технологии фагового дисплея определяется наличием физической связи между фенотипом (отображаемый пептид) и генотипом (последовательность ДНК, кодирующая отображаемый пептид) [10]. Связывание пептида с В7-2 была выявлена с помощью аффинной селекции. Пептид может служить материалом, на основе которого возможен расчет и конструирование искусственных иммуногенов, используемых для создания иммунотерапевтических средств против раковых заболеваний.
ОПИСАНИЕ ФИГУР
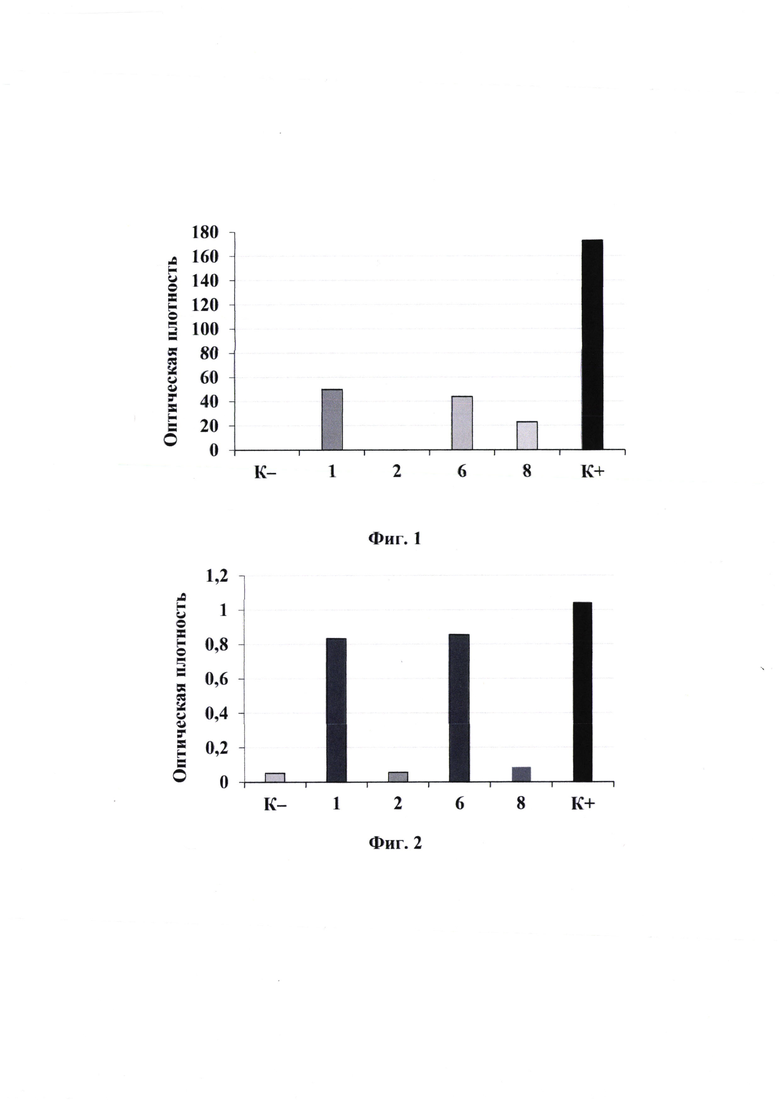
На Фиг. 1 приведена гистограмма, показывающая взаимодействие фаговых клонов №1, 2, 6 и 8 с рекомбинантным белком В7-2 в дот-блоте. Фаговый клон №1 несет аминокислотную последовательность CLARSLGRC. Дот-блот взаимодействия фаговых клонов с B7-2Fc. B7-2Fc нанесен на нитроцеллюлозную бумагу. Далее были добавлены фаговые клоны в концентрации 108 БОЕ/мкл. Взаимодействие В7-2 с пептидами на бактериофагах было определено с помощью антител против р8 бактериофага М13. Графическое представление оптической плотности образцов выполнено в приложении ImageJ (https://imagej.nih.gov/ij/). На фиг.2 представлена гистограмма, показывающая взаимодействия фаговых клонов №1, 2, 6 и 8 с рекомбинантным белком В7-2 в ИФА. Фаговый клон №1 несет аминокислотную последовательность CLARSLGRC. Фаговые клоны были иммобилизованы на 96-луночный планшет в концентрации 108 БОЕ/мкл. Связывание B7-2Fc определено с помощью вторичных антител против IgG человека.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Терминология. Под пептидом, описываемом в данном тексте, подразумевается короткий полимер, образованный вследствие межмолекулярного взаимодействия α - аминокислот в определенном порядке. Все аминокислоты, входящие в состав пептида, относятся к L-ряду.
Специфичность, упоминаемая в данном документе, обозначает антигенную способность пептида избирательно связываться с молекулой В7-2. Применение пептидов данного изобретения
Пептид обладает сродством к молекуле В7-2 и способен регулировать иммунную активность ко-регуляторной молекулы В7-2.
Примеры, подтверждающие заявляемый технический результат.
ПРИМЕР 1. Отбор специфических пептидов, способных связываться с В7-2.
Аффинную селекцию фаговой библиотеки против рекомбинантного белка В7-2, включающую три цикла, проводили с помощью магнитных частиц (МЧ) DynabeadsTM Protein G (Invitrogen, Thermo Fisher Scientific, США). Фаговая пептидная библиотека GerLab на основе нитчатого бактериофага fd была получена из лаборатории профессора Дж. М. Гершони, и состоит из смеси восьми фаговых пептидных библиотек, изготовленных на основе фагмидного вектора типа р88 и экспонирующих в составе главного поверхностного белка pVIII чужеродные рандомизированые пептиды длиной 6, 8, 10, 12 а.о., а также пептиды, замкнутые в петлю за счет фланкирующих вставку цистеинов (С).
Селекцию начинали с промывки МЧ промывочным раствором (0,1%-ный полисорбат 20 в Трис-буферном растворе). Затем добавляли 1 мл блокирующего раствора (5%-ный бычий сывороточный альбумин (БСА) в Трис-буферном растворе) и оставляли на 1 ч при 4°С. В это время смешивали фаговую библиотеку и рекомбинантный белок В7-2 в промывочном растворе и инкубировали 20 мин при комнатной температуре. Полученную смесь добавляли к промытым МЧ, инкубировали в течение 15 мин при комнатной температуре. Образованный комплекс МЧ-фаг-мишень отмывали от несвязавшихся бактериофагов, связавшиеся же фаги элюировали. Далее удаляли МЧ и добавляли нейтрализующий раствор (1 М Трис-HCl). Полученный элюат (50 мкл) инокулировали в 5 мл культуры, измеряли оптическую плотность на спектрофотометре NanoPhotometer N50 (Implen, Германия) при длине волны 600 нм (она составляла 0,6) и инкубировали в течение ночи при 37°С.
Для отслеживания обогащения популяции фагов после каждого цикла определяли биологический титр бактериофагов, элюированных с магнитных частиц, и после их амплификации в бактериальной культуре по методу Грациа (4). После 2-го цикла наблюдалось увеличение титра бактериофагов в элюате. После 3-го цикла увеличения титра не происходило, что, возможно, свидетельствует о достижении предела насыщения популяции фагов. С чашек Петри, содержащих единичные бляшки фагового элюата 3-го цикла, отобрали девять клонов для выделения ДНК и последующего секвенирования участка ДНК, кодирующего чужеродный пептид.
Одноцепочечную ДНК (оцДНК) выделяли осаждением с помощью йодида натрия. В 500 мкл суспензии бактериофага добавили 100 мкл иодидного буфера и 250 мкл 96%-ного этилового спирта и ресуспендировали осадок, и инкубировали 10 мин при комнатной температуре. Далее осаждали оцДНК центрифугированием в течение 10 минут при 14000 об/мин при 4°С. После удаляли супернатант и промывали осадок 500 мкл 70%-ного этилового спирта. Снова осаждали оцДНК центрифугированием в течение 10 минут при 14000 об/мин при 4°С. Далее сушили осадок оцДНК при вакууме и растворяли в 30 мкл воды. Секвенирование проводилось по методу Сэнгера. Чужеродная нуклеотидная последовательность, кодирующая отображаемый пептид, ограничена прямым праймером 5'-GCCGCTGGGGCCGACCCTG-3' и обратным праймером 5'-GGCGGCCAACGTGGC-3'. Биоинформатический анализ последовательностей после секвенирования проводили с помощью программного обеспечения BioEdit. Затем провели иммуноферментный анализ и дот-блот взаимодействия фаговых клонов с В7-2.
Аминокислотная последовательность CLARCLGRC (клон №1, циклический пептид) представлена в 66.7% анализированных фаговых клонов, в то время как последовательности CPSASSGLTC (клон №2, циклический пептид), QMPALMQQ (клон №6, линейный пептид) и AHIGVVSP (клон №8, линейный пептид) - в 11.1% клонов.
ПРИМЕР 2. Оценка и подтверждение специфичности заявляемого пептида с помощью дот-блот анализа.
На нитроцеллюлозную мембрану однократно наносили фаговые клоны (1 мкл, 108 вирионов) и высушивали на воздухе. В качестве отрицательного контроля (К-) - использовали бактериофаг fd (1 мкл, 108 вирионов), не содержащий чужеродной вставки; в качестве положительного контроля (К+) - рекомбинантный белок В7-2 (Sino Biological, КНР; 1 мкл, 2.5 мкг/мл).
Целлюлозу инкубировали с блокирующим Трис-буферным раствором (15 мл), содержащим 1% БСА и 0.1% полисорбата 20, для предотвращения неспецифического взаимодействия. После трехкратной промывки Трис-буферным раствором с 0.1% полисорбата 20 наносили 1.5 мл рекомбинантного белка В7-2 (2.5 мкг/мл) в блокирующем растворе и инкубировали при комнатной температуре в течение 15 мин. После трехкратной промывки промывочным раствором наносили 15 мл раствора антител козы против Fc-участка IgG человека, конъюгированных с щелочной фосфатазой (Invitrogen, США), в рабочем разведении 1:5000 в блокирующем растворе и инкубировали при комнатной температуре в течение 15 мин. После трехкратной промывки Трис-буферным раствором с 0.1% полисорбата 20 наносили смесь 5-бром-4-хлор-3-индолил-фосфата (BCIP, 0.21 мг/мл) и нитросинего тетразолия (NBT, 0.42 мг/мл в воде) объемом 15 мл, инкубировали при комнатной температуре в течение 15 мин в темноте. После трехкратной промывки водой нитроцеллюлозную мембрану высушивали на воздухе в недоступном для света месте. Сигналы на мембране переводили в компьютерное изображение с помощью приложения ImageJ (https://imagej.nih.gov/ij/).
Наибольшее значение оптической плотности (фиг.1), равное 50, наблюдалось у клона №1 (пептид CLARCLGRC). У клонов №6 (пептид AHIEVVSP) и №8 (пептид QMPALMQQ) оптическая плотность была ниже и составила 44 и 23 соответственно, что свидетельствует о меньшей аффинности этих пептидов к рекомбинантному белку В7-2. У клона №2 (пептид CPSASSQLTC) оптическая плотность составила - 10, следовательно, данный пептид не связывается с рекомбинантным белком В7-2. Значения оптической плотности у положительного и отрицательного контролей составили 173 и 0 соответственно.
ПРИМЕР 3. Оценка и подтверждение специфичности заявляемого пептида с помощью иммуноферментного анализа.
Для ИФА использовали среднесорбционные 96-луночные планшеты (Jet Biofil, КНР). В лунки вносили по 100 мкл раствора рекомбинантного белка В7-2 (Sino Biological, КНР) в концентрации 2.5 мкг/мл в Трис-буферном растворе (рН 8.6) и сорбировали при 4°С в течение ночи. Затем блокировали сайты неспецифического связывания добавлением 200 мкл Трис-буферного раствора (рН 7.0), содержащего 5% БСА, инкубировали в термошейкере для планшетов PST-60HL (BioSan, Латвия) при 37°С и 200 об/мин в течение 2 ч. После удаления блокирующего раствора в лунки вносили бактериофаги (100 мкл в блокирующем растворе, 108 вирионов) и инкубировали в течение 1 ч при 37°С. В качестве отрицательного контроля (К-) - использовали бактериофаг fd (100 мкл в блокирующем растворе, 108 вирионов), не содержащий чужеродной вставки; в качестве положительного контроля (К+) - рекомбинантный белок В7-2 (Sino Biological, КНР; 100 мкл, 2.5 мкг/мл). После трехкратной промывки промывочным раствором, содержащим Трис-буферный раствор с 0.5% полисорбата 20, добавляли 100 мкл конъюгата моноклонального антитела против нитчатого бактериофага М13 (GE Healthcare Life Sciences, США), меченого пероксидазой хрена, в выбранном рабочем разведении 1:5000 и инкубировали в течение 1 ч при 37°С. После шестикратной промывки промывочным раствором в лунки добавляли субстрат на основе 3,3',5,5'-тетраметилбензидина (ТМВ; Sigma-Aldrich, США). Реакцию останавливали добавлением 50 мкл 1 М серной кислоты в каждую лунку. Детекцию результатов проводили на планшетном фотометре iMark (Bio-Rad, США) при длине волны 450 нм. Вычисляли средние значения оптической плотности и ошибки средних значений (М±S.E.), по полученным данным строили диаграммы с указанием величин стандартного отклонения.
Результаты ИФА представлены на фиг.2. Наибольшие значения оптической плотности наблюдались у клонов №1 (пептид CLARCLGRC) и №6 (пептид AHIEVVSP): 0.83±0.02 и 0.86±0.03 соответственно, что подтверждает высокое сродство данных пептидов к рекомбинантному белку В7-2. ИФА с клонами №2 и 4 показал низкое специфическое взаимодействие с мишенью В7-2 (0.06±0.00 и 0.09±0.01 соответственно).
ПРИМЕР 4. Оценка и подтверждение специфичности различных вариантов пептида с помощью иммуноферментного анализа.
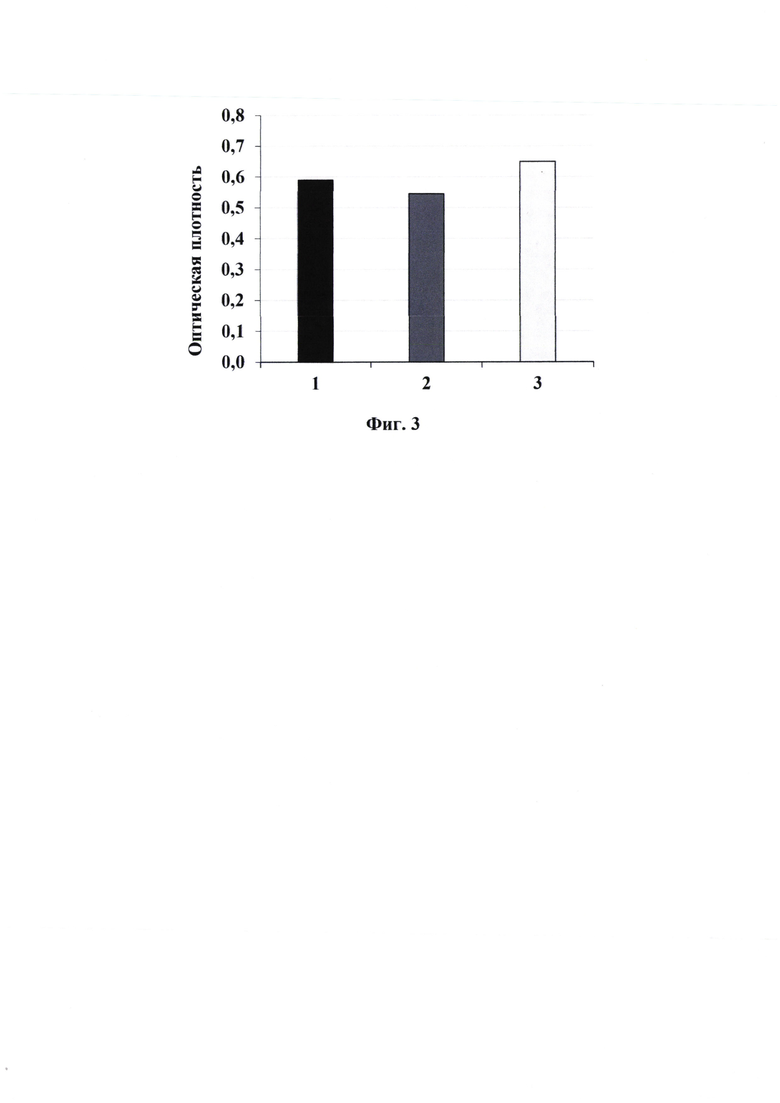
Для ИФА использовали среднесорбционные 96-луночные планшеты (Jet Biofil, КНР). В лунки вносили по 100 мкл раствора рекомбинантного белка В7-2 (Sino Biological, КНР) в концентрации 2.5 мкг/мл в Трис-буферном растворе (рН 8.6) и сорбировали при 4°С в течение ночи. Затем блокировали сайты неспецифического связывания добавлением 200 мкл Трис-буферного раствора (рН 7.0), содержащего 5% БСА, инкубировали в термошейкере для планшетов PST-60HL (BioSan, Латвия) при 37°С и 200 об/мин в течение 2 ч. После удаления блокирующего раствора в лунки вносили синтезированные пептиды (№1 - пептид CLARCLGRC; №2 - пептид CLARSLGRC; №3 - пептид СН2-LARSLGRC-CH2), содержащие биотин (100 мкл в блокирующем растворе, 2.5 мкг/мл) и инкубировали в течение 1 ч при 37°С. После трехкратной промывки промывочным раствором, содержащим Трис-буферный раствор с 0.5% полисорбата 20, добавляли 100 мкл конъюгата стрептавидина (Invitrogen, США), меченого пероксидазой хрена, в выбранном рабочем разведении 1:5000 и инкубировали в течение 1 ч при 37°С. После шестикратной промывки промывочным раствором в лунки добавляли субстрат на основе 3,3',5,5'-тетраметилбензидина (ТМВ; Sigma-Aldrich, США). Реакцию останавливали добавлением 50 мкл 1 М серной кислоты в каждую лунку. Детекцию результатов проводили на планшетном фотометре iMark (Bio-Rad, США) при длине волны 450 нм. Вычисляли средние значения оптической плотности и ошибки средних значений (М±S.E.), по полученным данным строили диаграммы с указанием величин стандартного отклонения.
Результаты ИФА представлены на фиг.3. Средние значения оптических плотностей составили 0.59±0.03, 0.55±0.06, 0.65±0.03 соответственно у пептидов №1, №2 и №3. ИФА показал, что изменение фланкирующих цистеинов (С), на любые аминокислотные остатки, обеспечивающие циклизацию пептида и изменение центрального цистеина (С) на серии (S) для упрощения синтеза пептидов в аминокислотной последовательности CLARCLGRC или CLARSLGRC или СН2-LARSLGRC-CH2 незначительно отражаются на оптической плотности.
Таким образом заявляемый технический результат подтверждается примерами 1-4 описания изобретения.
Источники научно-технической и патентной информации.
1. Linsley P.S., Ledbetter J.A. The Role of the CD28 Receptor During T Cell Responses to Antigen // Annual Review of Immunology, 1993. - No11(1). - P. 191-212.
2. Nagarajan Т., Marissen W., Rupprecht C. Monoclonal antibodies for the prevention of rabies: theory and clinical practice // Antibody Technology Journal. 2014. - No1.
3. Karateev, D., Belokoneva, N. Evaluation of Physicians' Knowledge and Attitudes Towards Biosimilars in Russia and Issues Associated with Their Prescribing // Biomolecules. 2019. - No9(2). - P. 57.
4. Патент CN №112646004A, 13.04.2021. Yan Zhongyl, Guo Xiangqian, Han Yali, Zhang Lu, Xie Longxiang, Li Huimin. 7 peptide for blocking CTLA4 in immunotherapy.
5. Патент AU №743939B2, 30.12.1998. Steven Blechner, Nabil E1 Tayar, Brad Jameson, Mark Tepper. CD28/CTLA-4 inhibiting peptidomimetics, pharmaceutical compositions thereof, and method of using same.
6. Mirzoeva S. et al. Single administration of p 2TA (AB103), a CD28 antagonist peptide, prevents inflammatory and thrombotic reactions and protects against gastrointestinal injury in total-body irradiated mice //PLoS One. - 2014. - T. 9. - №. 7. - C. e101161.
7. Патент WO №2020144332A1, 16.07.2020. Josep Maria 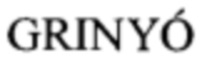 BOIRA, Oriol BESTARD MATAMOROS, Juan Torras Ambros. Fusion protein with immunosuppressive activity.
BOIRA, Oriol BESTARD MATAMOROS, Juan Torras Ambros. Fusion protein with immunosuppressive activity.
8. Pellerin L. et al. APVO210: a bispecific anti-CD86-IL-10 fusion protein (ADAPTIR™) to induce antigen-specific T regulatory type 1 cells //Frontiers in immunology. - 2018. - T. 9. -C. 881 (прототип).
9. Garaizar J., Brena S., Bikandi J., et al. Use of DNA microarray technology and gene expression profiles to investigate the pathogenesis, cell biology, antifungal susceptibility and diagnosis of Candida albicans // FEMS Yeast Research, 2006. - No6(7). - P. 987-998.
10. Aghebati-Maleki L., Bakhshinejad В., Baradaran В., et al. Phage display as a promising approach for vaccine development // Journal of Biomedical Science, 2016. - No23(1). - P. 1-18.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3"
fileName="st26-annex-iii-sequence-listing-specimen.xml"
softwareName="WIPO Sequence" softwareVersion="1.2.0"
productionDate="2022-11-07" originalFreeTextLanguageCode="ru"
nonEnglishFreeTextLanguageCode="ru">
<ApplicantFileReference>none</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>IB</IPOfficeCode>
<ApplicationNumberText></ApplicationNumberText>
<FilingDate>2022-11-07</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Алтайский государственный
университет</ApplicantName>
<ApplicantNameLatin>Altai state university</ApplicantNameLatin>
<InventorName languageCode="ru">Евгения Колосова</InventorName>
<InventorNameLatin>Evgeniia Kolosova</InventorNameLatin>
<InventionTitle languageCode="ru">Пептид специфически
взаимодействующий с молекулой контроля иммунного ответа B7-2 (CD86)
человека</InventionTitle>
<InventionTitle languageCode="en">Peptide that specifically
interacts with the human immune response control molecule B7-2
(CD86)Mus musculus abcd-1 gene for efg protein</InventionTitle>
<SequenceTotalQuantity>1</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>9</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..9</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q1">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>synthetic peptide that specifically
interacts with the human immune response control molecule B7-2
(CD86)</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>UNSURE</INSDFeature_key>
<INSDFeature_location>1...9</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>S or C</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>XLARXLGRX</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
Пептид, специфически взаимодействующий с молекулой
контроля иммунного ответа B7-2 (CD86) человека
Приложение
Перечень последовательностей
<110> Федеральное государственное бюджетное образовательное учреждение высшего образования «Алтайский государственный университет» (ФГБОУ ВО «Алтайский государственный университет»)
<120> Пептид, специфически взаимодействующий с молекулой контроля иммунного ответа B7-2 (CD86).
<160> SEQ ID NO 1
<210> SEQ ID NO: 1
<211> 12
<212> Peptide
<213> Synthetic construct
<223> Аминокислотная последовательность пептида, специфически взаимодействующая с молекулой контроля иммунного ответа B7-2 (CD86).
<400> 1
XLAR(S/С)LGRX
Изобретение относится к области биотехнологии, конкретно к пептидным антагонистам В7-2 (CD 28). Предложен пептид, специфически взаимодействующий с молекулой контроля иммунного ответа В7-2 (CD86) человека, регулирующий взаимодействие с костимулирующей молекулой CD28, выбранный из циклического пептида с аминокислотной последовательностью CLARSLGRC или CH2-LARSLGR-CH2. Пептид может выступать основой для разработки средств, регулирующих иммунный ответ, а также создания эффективных лекарственных форм пептидных препаратов в качестве средств терапии онкологических заболеваний. 3 ил., 4 пр.
Пептид, специфически взаимодействующий с молекулой контроля иммунного ответа В7-2 (CD86) человека, регулирующий взаимодействие с костимулирующей молекулой CD28, выбранный из циклического пептида с аминокислотной последовательностью CLARSLGRC или CH2-LARSLGR-CH2.
| Колосова Е.А | |||
| и др | |||
| "Поиск пептидов, специфически связывающихся с корегуляторной мишенью B7-2", Биоорганическая химия, 2021, т | |||
| Способ очищения сернокислого глинозема от железа | 1920 |
|
SU47A1 |
| Останов в машинах для обработки пряжи | 1924 |
|
SU762A1 |
| Шаповал А | |||
| И | |||
| и др | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |

