[0001] Перечень последовательностей в текстовом формате ASCII, предоставленный в соответствии с 37 C.F.R. § 1.821, под названием "81547_ST25.txt", размером 305 килобайт, созданный 14 марта 2018 г. и поданный с помощью EFS-Web, представлен вместо бумажной копии. Данный перечень последовательностей тем самым включен посредством ссылки в описание данного документа для его раскрытия.
ОБЛАСТЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
[0002] Настоящее изобретение относится к областям белковой инженерии, молекулярной биологии растений и контроля вредителей. Более конкретно, настоящее изобретение относится к новому белку и его вариантам, характеризующимся инсектицидной активностью, нуклеиновым кислотам, экспрессия которых приводит к образованию инсектицидных белков, а также способам получения и способам применения инсектицидных белков и соответствующих нуклеиновых кислот для контроля насекомых.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0003] Насекомые-вредители являются главной причиной потерь урожая. Только в США ежегодные потери, вызванные заражением различными видами насекомых, составляют миллиарды долларов. В дополнение к потерям урожая полевых культур, насекомые-вредители также являются отрицательным фактором для плантаторов, выращивающих овощи и фрукты, специалистов по выращиванию декоративных цветов, и они являются неприятностью для садоводов и домовладельцев.
[0004] Наиболее пагубными вредителями кукурузы считаются разные виды кукурузного корневого жука. В Соединенных Штатах тремя важными видами являются Diabrotica virgifera virgifera, западный кукурузный корневой жук, D. longicornis barberi, северный кукурузный корневой жук и D. undecimpunctata howardi, южный кукурузный корневой жук. Основными вредителями кукурузы в кукурузном поясе США считаются только западный и северный кукурузные корневые жуки. Дополнительно, важным коревым жуком-вредителем кукурузы в южных штатах США является мексиканский кукурузный корневой жук Diabrotica virgifera zeae. Наиболее существенный вред растению наносят личинки кукурузного корневого жука, поскольку они питаются фактически исключительно корнями кукурузы. Это поражение выражается в увеличении полегания растений, снижении урожая зерна и растительной массы, а также в изменении содержания питательных веществ в зерне. Поедание личинками также оказывает косвенные воздействия на кукурузу тем, что в его результате в корнях открываются ходы для заражений бактериями и грибами, что приводит к заболеваниям, выражающимся в гниении корня и стебля. Взрослые особи кукурузного корневого жука активны на кукурузных полях поздним летом, когда они питаются початками, кистями нитей рыльца и пыльцой, что препятствует нормальному опылению.
[0005] Кукурузных корневых жуков контролируют, главным образом, путем усиленного применения химических пестицидов, которые проявляют активность путем подавления роста насекомых, препятствования поеданию насекомыми или их размножению, или вызывая гибель. Таким путем может быть достигнут надлежащий контроль кукурузного корневого жука, однако иногда эти химические вещества также могут воздействовать и на другие, полезные организмы. Другой проблемой, возникающей в результате широкого применения химических пестицидов, является возникновение устойчивых видов насекомых. Еще одна проблема вызвана тем фактом, что личинки кукурузного корневого жука питаются под землей, что затрудняет нанесение "спасательных" обработок инсектицидами. Поэтому в большинстве случаев нанесения инсектицидов осуществляются профилактически, во время посева. Результатом такой практики является большая нагрузка на окружающую среду. Частично эту нагрузку ослабляют за счет применения различных способов ведения фермерского хозяйства, однако существует растущая потребность в альтернативных механизмах контроля вредителей.
[0006] Биологические средства для контроля вредителей, такие как штаммы Bacillus thuringiensis (Bt), экспрессирующие пестицидные токсины, как, например, δ-эндотоксины (дельта-эндотоксины; также называемые кристаллическими токсинами или белками Cry), были применены к сельскохозяйственным культурам с удовлетворительными результатами в отношении насекомых-вредителей. δ-эндотоксины представляют собой белки, содержащиеся в кристаллической матрице, которые, как известно, обладают инсектицидной активностью при поглощении некоторыми насекомыми. Несколько нативных белков Cry из Bacillus thuringiensis или сконструированных белков Cry были экспрессированы в трансгенных сельскохозяйственных культурах и использовались в коммерческих целях для контроля некоторых чешуекрылых и жесткокрылых насекомых-вредителей. Например, начиная с 2003 года в США коммерчески доступны трансгенные гибриды кукурузы, которые осуществляют контроль кукурузного корневого жука посредством экспрессии белка Cry3Bb1, Cry34Ab1/Cry35Ab1 или модифицированного белка Cry3A (mCry3A) или Cry3Ab (eCry3.1Ab).
[0007] Хотя было показано, что использование трансгенных растений, экспрессирующих белки Cry, является чрезвычайно эффективным, известны насекомые-вредители, которые теперь характеризуются устойчивостью к белкам Cry, экспрессируемым в некоторых трансгенных растениях. Следовательно, остается необходимость в идентификации новых и эффективных средств для контроля вредителей, которые обеспечивают экономическую выгоду фермерам и которые являются экологически приемлемыми. В особенности, необходимыми являются белки, которые токсичны по отношению к видам Diabrotica, главному вредителю кукурузы, способ действия которых отличается от существующих продуктов для контроля насекомых в качестве средства для ослабления развития устойчивости. Кроме того, необходима доставка средств для контроля насекомых с помощью продуктов, которые сводят к минимуму нагрузку на окружающую среду, как и с помощью трансгенных растений.
КРАТКОЕ ОПИСАНИЕ
[0008] Ввиду этих потребностей настоящее изобретение предусматривает новые инсектицидные белки, а именно NitromobCRW, а также белки, которые в значительной степени идентичны NitromobCRW и его вариантам. Белки по настоящему изобретению характеризуются токсичностью по отношению к кукурузному корневому жуку (Diabrotica spp). Белки по настоящему изобретению также могут характеризоваться токсичностью по отношению к другим жесткокрылым и/или чешуекрылым. Настоящее изобретение дополнительно относится к молекулам нуклеиновой кислоты, которые кодируют NitromobCRW или его варианты, комплементарным им
последовательностям, или которые в значительной степени идентичны NitromobCRW и его вариантам.
[0009] Настоящее изобретение также предусматривает векторы, содержащие такие рекомбинантные (или комплементарные им) нуклеиновые кислоты; растение или микроорганизм, которые содержат и обеспечивают экспрессию таких нуклеиновых кислот; растения, трансформированные такими нуклеиновыми кислотами, например трансгенные растения кукурузы; потомство таких растений, которое содержит нуклеиновые кислоты, стабильно встроенные и наследуемые по менделевскому принципу, и/или семена таких растений и такого потомства. Настоящее изобретение также предусматривает способы селекции для введения трансгена, содержащего молекулу нуклеиновой кислоты по настоящему изобретению, в потомство растения и в различные зародышевые плазмы.
[0010] Настоящее изобретение также предусматривает композиции и составы, содержащие NitromobCRW или его варианты, которые способны к ингибированию способности насекомых-вредителей к выживанию, росту и/или размножению, или к ограничению связанного с насекомыми вреда или потери урожая сельскохозяйственных культур, например, при применении NitromobCRW или его вариантов в качестве части композиций или составов в отношении зараженных насекомыми зон или растений, или для профилактической обработки чувствительных к насекомым зон или растений для обеспечения защиты от насекомых-вредителей.
[0011] Настоящее изобретение дополнительно относится к способу получения NitromobCRW или его вариантов и к способам применения нуклеиновых кислот, например, в микроорганизмах для контроля насекомых или в трансгенных растениях для обеспечения защиты от вреда, наносимого насекомыми.
[0012] Описанные в данном документе новые белки являются активными в отношении насекомых. Например, в вариантах осуществления белки по настоящему изобретению могут применяться для контроля экономически важных насекомых-вредителей, в том числе жесткокрылых насекомых, таких как западный кукурузный корневой жук (WCR), северный кукурузный корневой жук (NCR), южный кукурузный корневой жук (SCR) и/или мексиканский кукурузный корневой жук (D. virgifera zeae). Инсектицидные белки по настоящему изобретению могут применяться по отдельности или в комбинации с другими стратегиями для контроля насекомых, чтобы обеспечить повышенную эффективность контроля вредителей против тех же насекомых-вредителей и/или расширить спектр целевых насекомых с минимальным воздействием на окружающую среду.
[0013] Другие аспекты и преимущества по настоящему изобретению станут очевидными для специалистов в данной области в результате рассмотрения нижеследующего описания изобретения и неограничивающих примеров.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ В ПЕРЕЧНЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0014] Последовательности нуклеиновых кислот, перечисленные в сопутствующем перечне последовательностей, показаны с применением стандартных буквенных сокращений для нуклеотидных оснований, как определено в 37 C.F.R.§1.822. Перечисленные последовательности нуклеиновых кислот и аминокислотные последовательности определяют молекулы (т.е. полинуклеотиды и полипептиды, соответственно), содержащие нуклеотидные и аминокислотные мономеры, расположенные описанным образом. Каждая из перечисленных последовательностей нуклеиновых кислот и аминокислотных последовательностей также определяет тип полинуклеотидов или полипептидов, которые содержат нуклеотидные и аминокислотные мономеры, расположенные описанным образом. Ввиду избыточности генетического кода следует понимать, что нуклеотидная последовательность, содержащая кодирующую последовательность, также описывает тип полинуклеотидов, кодирующих тот же полипептид, как полинуклеотид, состоящий из эталонной последовательности. Дополнительно следует понимать, что аминокислотная последовательность описывает тип ORF полинуклеотида, кодирующего этот полипептид.
[0015] Показана только одна нить каждой последовательности нуклеиновой кислоты, но комплементарная нить подразумевается как включенная при любом упоминании показанной нити. Поскольку последовательности, комплементарные и обратно-комплементарные первичной последовательности нуклеиновой кислоты, обязательно раскрываются первичной последовательностью, комплементарная последовательность и обратно-комплементарная последовательность относятся к последовательность нуклеиновой кислоты, если явно не указано иное (или если иное ясно из контекста, в котором находится последовательность). Кроме того, как известно из уровня техники, нуклеотидная последовательность нити РНК определяется последовательностью ДНК, с которой она транскрибирована (кроме замены урациловых (U) нуклеиновых оснований на тимин (Т)), последовательность РНК включена при любом упоминании кодирующей ее последовательности ДНК. В сопутствующем перечне последовательностей:
SEQ ID NO: 1 представляет собой оптимизированную нуклеотидную последовательность NitromobCRW Е. coli.
SEQ ID NO: 2 представляет собой нуклеотидную последовательность варианта NitromobCRW I98L.
SEQ ID NO: 3 представляет собой нуклеотидную последовательность варианта NitromobCRW V99L.
SEQ ID NO: 4 представляет собой нуклеотидную последовательность варианта NitromobCRW I175L.
SEQ ID NO: 5 представляет собой нуклеотидную последовательность варианта NitromobCRW I208L.
SEQ ID NO: 6 представляет собой нуклеотидную последовательность варианта NitromobCRW 1215L.
SEQ ID NO: 7 представляет собой нуклеотидную последовательность варианта NitromobCRW 1215F.
SEQ ID NO: 8 представляет собой нуклеотидную последовательность варианта NitromobCRW I215 Y.
SEQ ID NO: 9 представляет собой нуклеотидную последовательность варианта NitromobCRW Y213L/I215L.
SEQ ID NO: 10 представляет собой нуклеотидную последовательность варианта NitromobCRW I245L.
SEQ ID NO: 11 представляет собой нуклеотидную последовательность варианта NitromobCRW I255L.
SEQ ID NO: 12 представляет собой нуклеотидную последовательность варианта NitromobCRW I265L.
SEQ ID NO: 13 представляет собой нуклеотидную последовательность варианта NitromobCRW I257L.
SEQ ID NO: 14 представляет собой нуклеотидную последовательность варианта NitromobCRW G216A.
SEQ ID NO: 15 представляет собой нуклеотидную последовательность варианта NitromobCRW G216L.
SEQ ID NO: 16 представляет собой нуклеотидную последовательность варианта NitromobCRW V122L.
SEQ ID NO: 17 представляет собой нуклеотидную последовательность варианта NitromobCRW V167L.
SEQ ID NO: 18 представляет собой нуклеотидную последовательность варианта NitromobCRW V220L.
SEQ ID NO: 19 представляет собой нуклеотидную последовательность варианта вставки NitromobCRW L214-Leu-I215.
SEQ ID NO: 20 представляет собой нуклеотидную последовательность варианта вставки NitromobCRW I215-Leu-G216.
SEQ ID NO: 21 представляет собой нуклеотидную последовательность варианта NitromobCRW Y213F/I215L.
SEQ ID NO: 22 представляет собой нуклеотидную последовательность варианта NitromobCRW I175L/I215L.
SEQ ID NO: 23 представляет собой нуклеотидную последовательность варианта NitromobCRW I208L/I215L.
SEQ ID NO: 24 представляет собой нуклеотидную последовательность варианта NitromobCRW I215L/I255L.
SEQ ID NO: 25 представляет собой нуклеотидную последовательность варианта NitromobCRW I255L/I257L.
SEQ ID NO: 26 представляет собой нуклеотидную последовательность варианта NitromobCRW L214S/I215L.
SEQ ID NO: 27 представляет собой нуклеотидную последовательность варианта NitromobCRW V203S/M204L.
SEQ ID NO: 28 представляет собой нуклеотидную последовательность варианта NitromobCRW T218L.
SEQ ID NO: 29 представляет собой нуклеотидную последовательность варианта NitromobCRW T218F.
SEQ ID NO: 30 представляет собой нуклеотидную последовательность варианта NitromobCRW V185L.
SEQ ID NO: 31 представляет собой нуклеотидную последовательность варианта NitromobCRW V193L/I215L.
SEQ ID NO: 32 представляет собой нуклеотидную последовательность варианта NitromobCRW E196L/I215L.
SEQ ID NO: 33 представляет собой нуклеотидную последовательность варианта NitromobCRW E186L/I215L.
SEQ ID NO: 34 представляет собой нуклеотидную последовательность варианта NitromobCRW V177L/I215L.
SEQ ID NO: 35 представляет собой нуклеотидную последовательность варианта NitromobCRW Y213L.
SEQ ID NO: 36 представляет собой нуклеотидную последовательность варианта NitromobCRW V203S/M204L/I215L.
SEQ ID NO: 37 представляет собой нативную нуклеотидную последовательность NitromobCRW.
SEQ ID NO: 38 представляет собой кодон-оптимизированную нуклеотидную последовательность варианта NitromobCRW Y213L/I215L маиса.
SEQ ID NO: 39 представляет собой нативную аминокислотную последовательность NitromobCRW.
SEQ ID NO: 40 представляет собой аминокислотную последовательность варианта NitromobCRW I98L.
SEQ ID NO: 41 представляет собой аминокислотную последовательность варианта NitromobCRW V99L.
SEQ ID NO: 42 представляет собой аминокислотную последовательность варианта NitromobCRW I175L.
SEQ ID NO: 43 представляет собой аминокислотную последовательность варианта NitromobCRW I208L.
SEQ ID NO: 44 представляет собой аминокислотную последовательность варианта NitromobCRW I215L.
SEQ ID NO: 45 представляет собой аминокислотную последовательность варианта NitromobCRW I215F.
SEQ ID NO: 46 представляет собой аминокислотную последовательность варианта NitromobCRW I215Y.
SEQ ID NO: 47 представляет собой аминокислотную последовательность варианта NitromobCRW Y213L/I215L.
SEQ ID NO: 48 представляет собой аминокислотную последовательность варианта NitromobCRW I245L.
SEQ ID NO: 49 представляет собой аминокислотную последовательность варианта NitromobCRW I255L.
SEQ ID NO: 50 представляет собой аминокислотную последовательность варианта NitromobCRW I265L.
SEQ ID NO: 51 представляет собой аминокислотную последовательность варианта NitromobCRW I257L.
SEQ ID NO: 52 представляет собой аминокислотную последовательность варианта NitromobCRW G216A.
SEQ ID NO: 53 представляет собой аминокислотную последовательность варианта NitromobCRW G216L.
SEQ ID NO: 54 представляет собой аминокислотную последовательность варианта NitromobCRW V122L.
SEQ ID NO: 55 представляет собой аминокислотную последовательность варианта NitromobCRW V167L.
SEQ ID NO: 56 представляет собой аминокислотную последовательность варианта NitromobCRW V220L.
SEQ ID NO: 57 представляет собой аминокислотную последовательность варианта вставки NitromobCRW L214-Leu-I215.
SEQ ID NO: 58 представляет собой аминокислотную последовательность варианта вставки NitromobCRW I215-Leu-G216.
SEQ ID NO: 59 представляет собой аминокислотную последовательность варианта NitromobCRW Y213F/I215L.
SEQ ID NO: 60 представляет собой аминокислотную последовательность варианта NitromobCRW I175L/I215L.
SEQ ID NO: 61 представляет собой аминокислотную последовательность варианта NitromobCRW I208L/I215L.
SEQ ID NO: 62 представляет собой аминокислотную последовательность варианта NitromobCRW I215L/I255L.
SEQ ID NO: 63 представляет собой аминокислотную последовательность варианта NitromobCRW I255L/I257L.
SEQ ID NO: 64 представляет собой аминокислотную последовательность варианта NitromobCRW L214S/I215L.
SEQ ID NO: 65 представляет собой аминокислотную последовательность варианта NitromobCRW V203S/M204L.
SEQ ID NO: 66 представляет собой аминокислотную последовательность варианта NitromobCRW T218L.
SEQ ID NO: 67 представляет собой аминокислотную последовательность варианта NitromobCRW T218F.
SEQ ID NO: 68 представляет собой аминокислотную последовательность варианта NitromobCRW V185L.
SEQ ID NO: 69 представляет собой аминокислотную последовательность варианта NitromobCRW V193L/I215L.
SEQ ID NO: 70 представляет собой аминокислотную последовательность варианта NitromobCRW E196L/I215L.
SEQ ID NO: 71 представляет собой аминокислотную последовательность варианта NitromobCRW E186L/I215L.
SEQ ID NO: 72 представляет собой аминокислотную последовательность варианта NitromobCRW V177L/I215L.
SEQ ID NO: 73 представляет собой аминокислотную последовательность варианта NitromobCRW Y213L.
SEQ ID NO: 74 представляет собой аминокислотную последовательность варианта NitromobCRW V203S/M204L/I215L.
SEQ ID NO: 75 представляет собой нуклеотидную последовательность NitromobCRW-Cterm-SUMO.
SEQ ID NO: 76 представляет собой аминокислотную последовательность удлиненного пептида NitromobCRW-Cterm-SUMO.
SEQ ID NO: 77 представляет собой аминокислотную последовательность NitromobCRW Y213L/I215L-Cterm-SUMO.
ОПРЕДЕЛЕНИЯ
[0016] Для ясности определенные термины, используемые в данном описании, определены и представлены как указано далее.
[0017] "Активность" инсектицидных белков по настоящему изобретению означает, что инсектицидные белки функционируют как активные при пероральном поглощении средства для контроля насекомых, характеризуются токсическим эффектом и/или способны нарушать или сдерживать поедание насекомыми, что может вызывать или не вызывать смерть насекомого. В случае если инсектицидный белок по настоящему изобретению доставляется в организм насекомого, то результатом обычно является гибель насекомого или же насекомое не поедает источник, который делает инсектицидный белок доступным для насекомого. "Пестицидной" называется токсическая биологическая активность, способная осуществлять контроль вредителя, такого как насекомое, нематода, гриб, бактерии или вирус, предпочтительно путем их уничтожения или разрушения. "Инсектицидной" называется токсическая биологическая активность, способная осуществлять контроль насекомых, предпочтительно путем их уничтожения. "Пестицидное средство" представляет собой средство, которое характеризуется пестицидной активностью. "Инсектицидное средство" представляет собой средство, которое характеризуется инсектицидной активностью.
[0018] Термин "ассоциированная с/функционально связанная" относится к двум нуклеиновым кислотам, которые связаны физически или функционально. Например, говорят, что промотор или регуляторная последовательность ДНК "ассоциированы с" последовательностью ДНК, которая кодирует РНК или белок, если эти две последовательности функционально связаны или расположены так, что регуляторная последовательность ДНК будет влиять на уровень экспрессии кодирующей или структурной последовательности ДНК.
[0019] "Кодирующая последовательность" представляет собой последовательность нуклеиновой кислоты, которая транскрибируется в РНК, такую как mRNA, rRNA, tRNA, snRNA, смысловая РНК или антисмысловая РНК. Предпочтительно, РНК затем транслируется в организме с продуцированием белка.
[0020] "Контролировать" насекомых означает подавлять посредством токсического эффекта способность насекомых-вредителей к выживанию, росту, поеданию и/или размножению или ограничивать связанные с насекомыми вред или потерю урожая сельскохозяйственных культур. "Контроль" насекомых может означать или может не означать уничтожение насекомых, хотя предпочтительно означает уничтожение насекомых.
[0021] "Доставлять" инсектицидный белок означает, что инсектицидный белок вступает в контакт с насекомым, что приводит к токсическому эффекту и контролю насекомого. Инсектицидный белок может быть доставлен многими известными путями, например, с помощью трансгенного растения, экспрессирующего инсектицидный белок, составленной(-ых) композиции(-й) с белком, распыляемой(-ых) композиции(-й) с белком, матрицы с приманкой или с помощью любой другой известной из уровня техники системы для доставки токсина.
[0022] "Эффективное для контроля насекомых количество" означает концентрацию инсектицидного белка, которая подавляет посредством токсического эффекта способность насекомых выживать, расти, поедать и/или размножаться или ограничивает связанные с насекомыми вред или потерю урожая сельскохозяйственных культур. "Эффективное для контроля насекомых количество" может означать или может не означать уничтожение насекомых, хотя предпочтительно оно означает уничтожение насекомых.
[0023] "Кассета экспрессии", используемая в данном документе, означает последовательность нуклеиновой кислоты, способную направлять экспрессию конкретной нуклеотидной последовательности в соответствующую клетку-хозяина, содержащую промотор, функционально связанный с представляющей интерес нуклеотидной последовательностью, которая функционально связана с сигналами терминации. Она также обычно содержит последовательности, необходимые для правильной трансляции нуклеотидной последовательности. По меньшей мере один из компонентов кассеты экспрессии, содержащей представляющую интерес нуклеотидную последовательность, может быть гетерологичным по отношению к по меньшей мере одному из других ее компонентов. Кассета экспрессии может также представлять собой последовательность, которая встречается в природе, но была получена в рекомбинантной форме, применимой для гетерологичной экспрессии. Однако, как правило, кассета экспрессии является гетерологичной по отношению к хозяину, т.е. конкретная последовательность нуклеиновой кислоты в кассете экспрессии не встречается в клетке-хозяине в природе, и ее необходимо было ввести в клетку-хозяина или предка клетки-хозяина с помощью события трансформации. Экспрессия нуклеотидной последовательности в кассете экспрессии может находиться под контролем конститутивного промотора или индуцируемого промотора, который инициирует транскрипцию только тогда, когда клетка-хозяин подвергается воздействию некоторого определенного внешнего стимула. В случае многоклеточного организма, такого как растение, промотор также может быть специфичным по отношению к конкретной ткани, или органу, или стадии развития.
[0024] Кассета экспрессии, содержащая представляющую интерес нуклеотидную последовательность, может быть химерной, что означает, что по меньшей мере один из ее компонентов является гетерологичным по отношению к по меньшей мере одному из ее остальных компонентов. Кассета экспрессии также может представлять собой таковую, которая содержит нативный промотор, управляющий ее нативным геном, однако она была получена в рекомбинантной форме, применимой для гетерологичной экспрессии. Такое использование кассеты экспрессии обеспечивает то, что она является не встречающейся в природе в клетке, в которую была введена.
[0025] Кассета экспрессии также необязательно может содержать участок терминации транскрипции и/или трансляции (т.е. участок терминации), функционирующий в растениях. Разнообразные терминаторы транскрипции доступны для применения в кассетах экспрессии, и они отвечают за терминацию транскрипции за пределами представляющей интерес гетерологичной нуклеотидной последовательности и правильное полиаденилирование mRNA. Участок терминации может быть нативным по отношению к участку инициации транскрипции, может быть нативным по отношению к функционально связанной представляющей интерес нуклеотидной последовательности, может быть нативным по отношению к растению-хозяину или может быть получен из другого источника (т.е. быть чужеродным или гетерологичным по отношению к промотору, представляющей интерес нуклеотидной последовательности, растению-хозяину или любой их комбинации). Соответствующие терминаторы транскрипции включают без ограничения терминатор 35S CaMV, терминатор гена tm1, терминатор гена нопалинсинтазы и/или терминатор гена rbcs-E9 гороха. Они могут применяться как у однодольных, так и у двудольных растений. В дополнение, может применяться нативный терминатор транскрипции кодирующей последовательности. В контексте настоящего изобретения может применяться любой доступный терминатор, о котором известно, что он функционирует в растениях.
[0026] Термин "экспрессия" в случае использования со ссылкой на полинуклеотид, такой как ген, ORF или ее часть или трансген в растениях, относится к процессу преобразования генетической информации, кодируемой геном, в РНК (например, mRNA, rRNA, tRNA или snRNA) посредством "транскрипции" гена (т.е. за счет ферментативного действия РНК-полимеразы), и в белок, если это применимо (например, если ген кодирует белок), посредством "трансляции" mRNA. Экспрессия гена может регулироваться на многих стадиях в ходе этого процесса. Например, в случае антисмысловых конструкций или конструкций dsRNA соответственно, экспрессия может относиться к транскрипции только антисмысловой РНК или только dsRNA. В вариантах осуществления "экспрессия" относится к транскрипции и стабильному накоплению смысловой (mRNA) или функциональной РНК. "Экспрессия" может также относиться к продуцированию белка.
[0027] "Ген" представляет собой определенный участок, который расположен внутри генома и содержит кодирующую последовательность нуклеиновой кислоты и, как правило, также содержит другие, преимущественно регуляторные нуклеиновые кислоты, ответственные за контроль экспрессии, иными словами транскрипции и трансляции кодирующей части. Ген может также содержать другие 5'- и 3'-нетранслируемые последовательности и последовательности терминации. Дополнительными элементами, которые могут присутствовать, являются, например, интроны. Регуляторная последовательность нуклеиновой кислоты гена в норме может не быть функционально связанной с ассоциированной последовательностью нуклеиновой кислоты, как это встречается в природе, и в таком случае ген будет химерным геном.
[0028] "Представляющий интерес ген" относится к любой молекуле нуклеиновой кислоты, которая, в случае переноса в растение, придает растению требуемый признак, такой как устойчивость к антибиотикам, устойчивость к вирусам, устойчивость к насекомым, устойчивость к заболеваниям или устойчивость к другим вредителям, переносимость гербицидов, переносимость абиотического стресса, мужская стерильность, модифицированный метаболизм жирных кислот, модифицированный метаболизм углеводов, улучшенная пищевая ценность, улучшенные характеристики при промышленном способе или измененная репродуктивная способность. "Представляющий интерес ген" также может являться таким, который переносят в растения для получения коммерчески ценных ферментов или метаболитов в растении.
[0029] "Гетерологичная" последовательность нуклеиновой кислоты или молекула нуклеиновой кислоты представляет собой последовательность нуклеиновой кислоты или молекулу нуклеиновой кислоты, которая в природе не ассоциирована с клеткой-хозяином, в которую ее вводят, в том числе не встречающиеся в природе множественные копии встречающейся в природе последовательности нуклеиновой кислоты. Гетерологичная последовательность нуклеиновой кислоты или молекула нуклеиновой кислоты может содержать химерную последовательность, такую как химерная кассета экспрессии, где промотор и кодирующих участок получены из нескольких исходных организмов. Последовательность промотора может представлять собой последовательность конститутивного промотора, последовательность тканеспецифичного промотора, последовательность химически индуцируемого промотора, последовательность индуцируемого повреждением промотора, последовательность индуцируемого стрессом промотора или последовательность специфичного для стадии развития промотора.
[0030] "Гомологичная" последовательность нуклеиновой кислоты представляет собой последовательность нуклеиновой кислоты, в природных условиях связанную с клеткой-хозяином, в которую ее вводят.
[0031] "Гомологичная рекомбинация" представляет собой взаимный обмен фрагментами нуклеиновой кислоты между гомологичными молекулами нуклеиновой кислоты.
[0032] "Идентичность" или "процентная идентичность" относится к степени сходства между двумя последовательностями нуклеиновых кислот или последовательностями белка. При сравнении последовательностей одна последовательность обычно выступает в качестве эталонной последовательности, с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей тестируемую и эталонную последовательности вводят в компьютер, при необходимости задают координаты подпоследовательности, и задают программные параметры алгоритма сравнения последовательностей. Затем с помощью алгоритма сравнения последовательностей на основе заданных программных параметров вычисляют процентную идентичность последовательностей для тестируемой(-ых) последовательности(-ей) относительно эталонной последовательности. Фраза "в значительной степени идентичные" в контексте двух последовательностей нуклеиновых кислот или двух аминокислотных последовательностей относится к двум или более последовательностям или подпоследовательностям, которые характеризуются по меньшей мере приблизительно 50% идентичностью по нуклеотидам или аминокислотным остаткам при сравнении и выравнивании для максимального соответствия, что определяют с применением одного из следующих алгоритмов сравнения последовательностей или путем визуального осмотра. В определенных вариантах осуществления в значительной степени идентичные последовательности характеризуются по меньшей приблизительно 60%, или по меньшей мере приблизительно 70%, или по меньшей мере приблизительно 80%, или по меньшей мере приблизительно 85% или даже по меньшей мере приблизительно 90% или 95% идентичностью по нуклеотидам или аминокислотным остаткам. В определенных вариантах осуществления значительная степень идентичности существует на протяжении участка последовательностей, длина которого составляет по меньшей мере приблизительно 50 остатков, или на протяжении участка, состоящего из по меньшей мере приблизительно 100 остатков, или последовательности в значительной степени идентичны на протяжении по меньшей мере приблизительно 150 остатков. В дополнительных вариантах осуществления последовательности являются в значительной степени идентичными, когда они идентичны на протяжении всей длины кодирующих участков.
[0033] Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с помощью алгоритма поиска локальной гомологии из Smith & Waterman, Adv. Appl. Math. 2: 482 (1981), с помощью алгоритма выравнивания областей гомологии из Needleman & Wunsch, J. Mol. Biol. 48: 443 (1970), с помощью способа поиска сходства из Pearson & Lipman, Proc. Nat'l. Acad. Sci. USA 85: 2444 (1988), с помощью программной реализации данных алгоритмов (GAP, BESTFIT, FASTA и TFASTA из Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Мадисон, Висконсин) или с помощью визуального осмотра (см. в общем Ausubel et al., ниже).
[0034] Одним примером алгоритма, который подходит для определения процента идентичности последовательностей и сходства последовательностей, является алгоритм BLAST, который описан в Altschul et al., J. Mol. Biol. 215. 403-410 (1990). Программное обеспечение для осуществления анализов BLAST находится в открытом доступе благодаря Национальному центру биотехнологической информации (http://www.ncbi.nlm.nih.gov/). Данный алгоритм предусматривает вначале идентификацию пар последовательностей с высоким показателем сходства (HSP) путем идентификации коротких "слов" длиной W в запрашиваемой последовательности, которые либо совпадают, либо удовлетворяют некоторому положительному пороговому показателю Т при выравнивании со "словом" такой же длины в последовательности из базы данных. Т называется пороговым значением показателя соседнего "слова" (Altschul et al., 1990). Эти первоначальные совпадения соседних "слов" выступают в качестве затравки, чтобы инициировать поиски для обнаружения более длинных HSP, содержащих их. Совпадения "слов" затем продлеваются в обоих направлениях вдоль каждой последовательности до тех пор, пока может увеличиваться совокупный показатель выравнивания. В случае нуклеотидных последовательностей совокупные показатели рассчитывают с применением параметров М (балл-вознаграждение, начисляемый за пару совпадающих остатков; всегда >0) и N (штрафной балл, начисляемый за несовпадающие остатки; всегда <0). В случае аминокислотных последовательностей для расчета совокупного показателя применяют матрицу замен. Продление совпадений "слов" в каждом направлении прекращается, когда совокупный показатель выравнивания снижается на величину X от своего максимального достигнутого значения, при этом совокупный показатель падает до нуля или ниже вследствие накопления одного или нескольких выравниваний остатков с отрицательными показателями, или при достижении конца одной из последовательностей. Параметры W, Т и X алгоритма BLAST определяют чувствительность и скорость выравнивания. В программе BLASTN (для нуклеотидных последовательностей) по умолчанию используется длина "слова" (W), составляющая 11, ожидаемое значение (Е), составляющее 10, пороговое значение, составляющее 100, М=5, N= -4 и сравнение по обеим нитям. В случае аминокислотных последовательностей программа BLASTP использует по умолчанию длину "слова" (W), составляющую 3, ожидаемое значение (Е), составляющее 10, а также матрицу замен BLOSUM62 (см. Henikoff & Henikoff, Proc. Natl. Acad. Sci USA 89: 10915 (1989)).
[0035] В дополнение к расчету процента идентичности последовательностей алгоритм BLAST также выполняет статистический анализ сходства между двумя последовательностями (см., например, Karlin & Altschul, Proc. Nat'l. Acad. Sci. USA 90: 5873-5787 (1993)). Одной мерой сходства, предоставляемой алгоритмом BLAST, является наименьшая суммарная вероятность (P(N)), которая является показателем того, что совпадения между двумя нуклеотидными или аминокислотными последовательностями будут возникать случайным образом. Например, считается, что тестируемая последовательность нуклеиновой кислоты сходна с эталонной последовательностью, если наименьшая суммарная вероятность при сравнении тестируемой последовательности нуклеиновой кислоты с эталонной последовательностью нуклеиновой кислоты составляет менее приблизительно 0,1, более предпочтительно менее приблизительно 0,01 и наиболее предпочтительно менее приблизительно 0,001.
[0036] Другой широко применяемой и признанной компьютерной программой для осуществления выравниваний последовательностей является CLUSTALW v1.6 (Thompson, et al. Nuc. Acids Res., 22: 4673-4680, 1994). Число совпадающих оснований или аминокислот делят на общее число оснований или аминокислот и умножают на 100 с получением процента идентичности. Например, если бы две последовательности из 580 пар оснований имели 145 совпавших оснований, то они были бы идентичны на 25 процентов. Если две сравниваемые последовательности имеют разную длину, то число совпадений делят на более короткую из двух длин. Например, если в белках из 200 и 400 аминокислот было 100 совпадавших аминокислот, то они идентичны на 50 процентов с учетом более короткой последовательности. Если длина более короткой последовательность составляет менее 150 оснований или 50 аминокислот, то число совпадений делят на 150 (в случае нуклеиновых оснований) или 50 (в случае аминокислот), и умножают на 100 с получением процента идентичности.
[0037] Другим показателем того, что две нуклеиновые кислоты являются в значительной степени идентичными, служит то, что эти две молекулы гибридизируются друг с другом в жестких условиях. Фраза "гибридизируется специфически с" относится к связыванию, образованию дуплекса или гибридизации молекулы только с определенной нуклеотидной последовательностью в жестких условиях, когда такая последовательность присутствует в сложной смеси (например, общих клеточных) ДНК или РНК. "В значительной степени связывается" относится к комплементарной гибридизации между нуклеиновой кислотой-зондом и целевой нуклеиновой кислотой и охватывает незначительные несовпадения, которые могут быть исправлены за счет снижения жесткости среды для гибридизации, чтобы добиться необходимого выявления целевой последовательности нуклеиновой кислоты.
[0038] "Жесткие условия гибридизации" и "жесткие условия отмывки при гибридизации" в контексте экспериментов с гибридизацией нуклеиновых кислот, таких как саузерн- и нозерн-гибридизации, зависят от последовательности и отличаются при разных параметрах окружающей среды. Более длинные последовательности специфично гибридизируются при более высоких температурах. Подробное руководство по гибридизации нуклеиновых кислот можно найти в Tijssen (1993) Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes part I chapter 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays" Elsevier, New York. Как правило, условия гибридизации и отмывки высокой жесткости выбирают так, чтобы температура была приблизительно на 5°С ниже точки плавления (Tm) для конкретной последовательности при определенных ионной силе и значении рН. Как правило, при "жестких условиях" зонд будет гибридизироваться со своей целевой последовательностью, но ни с какими другими последовательностями.
[0039] Значение Tm представляет собой температуру (при определенных ионной силе и значении рН), при которой 50% целевой последовательности гибридизируется с точно совпадающим зондом. Для очень жестких условий выбирают температуру, равную Tm для конкретного зонда. Примером жестких условий гибридизации для гибридизации комплементарных нуклеиновых кислот, которые имеют более чем 100 комплементарных остатков на фильтре в саузерн- или нозерн-блоте, является 50% формамид с 1 мг гепарина при 42°С, причем гибридизация проводится в течение ночи. Примером условий отмывки высокой жесткости является 0,15 М NaCl при 72°С в течение приблизительно 15 минут. Примером жестких условий отмывки является отмывка с помощью 0,2х SSC при 65°С в течение 15 минут (см. Sambrook, ниже, в отношении описания буфера SSC). Часто с целью избавления от фонового сигнала зонда отмывке в условиях высокой жесткости предшествует отмывка в условиях низкой жесткости. Примерными условиями отмывки средней жесткости для дуплекса, например с более чем 100 нуклеотидами, являются 1x SSC при 45°С в течение 15 минут. Примерными условиями отмывки низкой жесткости для дуплекса, например с более чем 100 нуклеотидами, являются 4-6х SSC при 40°С в течение 15 минут. Для коротких зондов (например, длиной от приблизительно 10 до 50 нуклеотидов) жесткие условия обычно предусматривают концентрации солей, составляющие менее чем приблизительно 1,0 М ионов Na, как правило концентрацию, составляющую приблизительно 0,01-1,0 М ионов Na (или других солей), при рН 7,0-8,3, а также температуру, как правило, составляющую по меньшей мере приблизительно 30°С. Жесткие условия также могут быть достигнуты с добавлением дестабилизирующих средств, таких как формамид. В целом, соотношение сигнал-шум, в 2 (или более) раза превышающее наблюдаемое для несвязанного зонда при конкретном гибридизационном анализе, указывает на выявление специфической гибридизации. Нуклеиновые кислоты, которые не гибридизируются друг с другом в жестких условиях, все еще являются в значительной степени идентичными, если белки, которые они кодируют, в значительной степени идентичны. Например, это происходит в том случае, когда копию нуклеиновой кислоты создают с применением максимальной вырожденности кодонов, допускаемой генетическим кодом.
[0040] Примеры наборов условий гибридизации/отмывки, которые можно применять для клонирования гомологичных нуклеотидных последовательностей, которые в значительной степени идентичны эталонным нуклеотидным последовательностям по настоящему изобретению, являются следующими: эталонная нуклеотидная последовательность предпочтительно гибридизируется с эталонной нуклеотидной последовательностью в 7% додецилсульфате натрия (SDS), 0,5 М NaPO4, 1 мМ EDTA при 50°С с отмывкой в 2Х SSC, 0,1% SDS при 50°С, более желательно в 7% додецилсульфате натрия (SDS), 0,5 М NaPO4, 1 мМ EDTA при 50°С с отмывкой в 1X SSC, 0,1% SDS при 50°С, еще более желательно в 7% додецилсульфате натрия (SDS), 0,5 М NaPO4, 1 мМ EDTA при 50°С с отмывкой в 0,5Х SSC, 0,1% SDS при 50°С, предпочтительно в 7% додецилсульфате натрия (SDS), 0,5 М NaPO4, 1 мМ EDTA при 50°С с отмывкой в 0,1X SSC, 0,1% SDS при 50°С, более предпочтительно в 7% додецилсульфате натрия (SDS), 0,5 М NaPO4, 1 мМ EDTA при 50°С с отмывкой в 0,1X SSC, 0,1% SDS при 65°С.
[0041] Дополнительным свидетельством того, что две нуклеиновые кислоты или два белка являются в значительной степени идентичными, является то, что белок, кодируемый первой нуклеиновой кислотой, является иммунологически перекрестно реактивным с белком, закодированным второй нуклеиновой кислотой, или специфически связывается с ним. Таким образом, как правило, белок является в значительной степени идентичным второму белку, например, если два белка отличаются только консервативными заменами.
[0042] Последовательность нуклеиновой кислоты является "изокодонной с" эталонной последовательностью нуклеиновой кислоты, когда последовательность нуклеиновой кислоты кодирует полипептид, имеющий такую же аминокислотную последовательность, что и полипептид, кодируемый эталонной последовательностью нуклеиновой кислоты.
[0043] "Выделенная" молекула нуклеиновой кислоты или выделенный токсин, рукой человека, представляют собой молекулу нуклеиновой кислоты или токсин, которые существуют независимо от своей природной среды, а следовательно не является природным продуктом. Выделенные молекула нуклеиновой кислоты или токсин могут существовать в очищенной форме или могут существовать в неприродной среде, например, без ограничения, такой как рекомбинантная микробная клетка, растительная клетка, растительная ткань или растение.
[0044] "Молекула нуклеиновой кислоты" или "последовательность нуклеиновой кислоты" представляет собой сегмент одно- или двухнитевой ДНК или РНК, который может быть выделен из любого источника. В контексте настоящего изобретения молекула нуклеиновой кислоты обычно представляет собой сегмент ДНК. В некоторых вариантах осуществления молекулы нуклеиновой кислоты по настоящему изобретению представляют собой выделенные молекулы нуклеиновой кислоты.
[0045] Термины "белок", "пептид" и "полипептид" используются в данном документе взаимозаменяемо.
[0046] Используемое в данном документе выражение "кодон-оптимизированная" последовательность означает нуклеотидную последовательность рекомбинантного, трансгенного или синтетического полинуклеотида, в котором кодоны выбраны так, чтобы отражать склонность к определенным кодонам, которая может наблюдаться в клетке-хозяине. Это выполняется таким образом, чтобы сохранить аминокислотную последовательность полипептида, кодируемого кодон-оптимизированным полинуклеотидом. В некоторых вариантах осуществления нуклеотидная последовательность рекомбинантной ДНК-конструкции содержит последовательность, которая была кодон-оптимизирована для клетки (например, клетки животного, растения или гриба), в которой конструкция будет экспрессироваться. Например, в конструкции, которая будет экспрессироваться в растительной клетке, могут быть кодон-оптимизированы для экспрессии в растении вся последовательность или ее части (например, первый элемент для супрессии гена или элемент для экспрессии гена). См., например, патент США №6121014, включенный в данный документ посредством ссылки.
[0047] "Растение" представляет собой любое растение на любой стадии развития, в частности семенное растение.
[0048] "Растительная клетка" представляет собой структурную и физиологическую единицу растения, содержащую протопласт и клеточную стенку. Растительная клетка может быть в виде выделенной одиночной клетки или культивируемой клетки или в качестве части более высокоорганизованной единицы, такой как, например, растительная ткань, орган растения или целое растение.
[0049] "Культура растительных клеток" означает культуры единиц растения, таких как, например, протопласты, клетки в клеточной культуре, клетки в растительных тканях, пыльца, пыльцевые трубки, семязачатки, зародышевые мешки, зиготы и зародыши на различных стадиях развития.
[0050] "Растительный материал" относится к листьям, стеблям, корням, цветкам или частям цветков, плодам, пыльце, яйцеклеткам, зиготам, семенам, черенкам, клеточным или тканевым культурам, или к любой другой части или продукту растения.
[0051] "Орган растения" представляет собой отдельную и визуально структурированную и дифференцированную часть растения, такую как корень, стебель, лист, цветочная почка или зародыш.
[0052] "Растительная ткань", применяемая в данном документе, означает группу растительных клеток, организованных в структурную и функциональную единицу. Включена любая ткань растения in planta или в культуре. Данный термин включает без ограничения целые растения, органы растений, семена растений, тканевую культуру и любые группы растительных клеток, организованных в структурные и/или функциональные единицы. Использование данного термина в сочетании с любым специфическим типом растительной ткани, приведенным выше или иным образом охваченным данным определением, или без такового, не предназначено для исключения любого другого типа растительной ткани.
[0053] "Промотор" представляет собой нетранслируемую последовательность ДНК выше кодирующей области, которая содержит сайт связывания для РНК-полимеразы и инициирует транскрипцию ДНК. Промоторный участок может также содержать другие элементы, которые выступают в качестве регуляторов экспрессии генов.
[0054] "Регуляторные элементы" относятся к последовательностям, вовлеченным в контроль экспрессии нуклеотидной последовательности. Регуляторные элементы предусматривают промотор, функционально связанный с представляющей интерес нуклеотидной последовательностью, и сигналы терминации. Как правило, они охватывают также последовательности, необходимые для надлежащей трансляции нуклеотидной последовательности.
[0055] "Трансформация" представляет собой процесс введения гетерологичной нуклеиновой кислоты в клетку- или организм-хозяин. В конкретных вариантах осуществления "трансформация" означает стабильную интеграцию молекулы ДНК в геном (ядерный или пластидный) представляющего интерес организма.
[0056] "Трансформированный/трансгенный/рекомбинантный" относится к организму-хозяину, такому как бактерия или растение, в который была введена гетерологичная молекула нуклеиновой кислоты. Молекулу нуклеиновой кислоты можно стабильно интегрировать в геном хозяина, или же молекула нуклеиновой кислоты также может присутствовать в виде внехромосомной молекулы. Такая внехромосомная молекула может быть автореплицирующейся. Подразумевается, что трансформированные клетки, ткани или растения охватывают не только конечный продукт процесса трансформации, но также и их трансгенное потомство. "Нетрансформированный", "нетрансгенный" или "нерекомбинантный" хозяин относится к организму дикого типа, например бактерии или растению, которые не содержат гетерологичную молекулу нуклеиновой кислоты.
[0057] Нуклеотиды обозначены по их основаниям с помощью следующих стандартных сокращений: аденин (А), цитозин (С), тимин (Т) и гуанин (G). Аналогичным образом, аминокислоты обозначены с помощью следующих стандартных сокращений: аланин (Ala; А), аргинин (Arg; R), аспарагин (Asn; N), аспарагиновая кислота (Asp; D), цистеин (Cys; С), глутамин (Gin; Q), глутаминовая кислота (Glu; Е), глицин (Gly; G), гистидин (His; Н), изолейцин (Не; 1), лейцин (Leu; L), лизин (Lys; K), метионин (Met; М), фенилаланин (Phe; F), пролин (Pro; Р), серии (Ser; S), треонин (Thr; Т), триптофан (Тгр; W), тирозин (Туг; Y) и валин (Val; V).
ПОДРОБНОЕ ОПИСАНИЕ
[0058] Настоящее изобретение относится к новым инсектицидным белкам, которые характеризуются активностью против жесткокрылых, например, Diabrotica virgifera virgifera (западного кукурузного корневого жука; WCR), Diabrotica barberi (северного кукурузного корневого жука; NCR) и/или Diabrotica undecimpunctata howardi (южного кукурузного корневого жука; SCR), и/или других видов Diabrotica, в том числе Diabrotica virgifera zeae (мексиканского кукурузного корневого жука) и/или других жесткокрылых насекомых-вредителей, таких как колорадский жук. В вариантах осуществления новый инсектицидный белок по настоящему изобретению может характеризоваться активностью против видов чешуекрылых. Настоящее изобретение также относится к нуклеиновым кислотам, экспрессия которых приводит к образованию инсектицидных белков по настоящему изобретению, а также к получению и применению инсектицидных белков для контроля насекомых-вредителей. В вариантах осуществления экспрессия нуклеиновых кислот приводит к образованию инсектицидных белков, которые можно применять для контроля жесткокрылых насекомых, таких как западный, северный и/или южный кукурузный корневой жук, в частности при экспрессии в трансгенном растении, таком как трансгенное растение кукурузы.
[0059] Настоящее изобретение дополнительно охватывает молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая кодирует инсектицидный белок по настоящему изобретению. Нуклеотидная последовательность может быть оптимизирована для экспрессии в бактериях, таких как Escherichia coli, или для экспрессии в растении, таком как Zea mays. Нуклеотидная последовательность, оптимизированная для экспрессии в гетерологичном организме, таком как виды бактерий, отличные от тех, из которых она получена, или растение, не встречается в природе. В одном аспекте данного варианта осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность любой из SEQ ID NO: 1-38 или комплементарную ей последовательность. В частности, иллюстративные рекомендации в отношении способов получения молекул нуклеиновой кислоты, которые кодируют инсектицидные белки по настоящему изобретению, могут быть найдены в примерах данной заявки. Специалистам в данной области будет понятно, что в иллюстративных способах получения инсектицидных белков, охватываемых настоящим изобретением, могут быть сделаны модификации.
[0060] Специалисту в данной области техники будет понятно, что трансген для коммерческого применения, такой как молекула нуклеиновой кислоты, которая содержит любую из SEQ ID NO: 1-38 или комплементарную ей последовательность, может характеризоваться относительно незначительными модификациями по отношению к последовательности нуклеиновой кислоты для соответствия требованиям государственных регулятивных норм. Такие модификации не будут влиять на функцию полученной молекулы, которая будет в значительной степени идентичной SEQ ID NO: 1-38. Специалисту в данной области будет понятно, что модифицированная молекула нуклеиновой кислоты будет фактически такой же, как и исходная молекула, и охвачена настоящим изобретением.
[0061] Настоящее изобретение также охватывает молекулу нуклеиновой кислоты, которая содержит (а) нуклеотидную последовательность под любым из SEQ ID NO: 1-38; (b) нуклеотидную последовательность, которая представляет собой последовательность на по меньшей мере 45% идентичную, по меньшей мере 50% идентичную, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или на 100% идентичную любой из нуклеотидных последовательностей под SEQ ID NO: 1-38; (с) нуклеотидную последовательность, которая кодирует полипептид, где аминокислотная последовательность полипептида содержит SEQ ID NO: 39-74 и характеризуется активностью в отношении контроля насекомых; (d) нуклеотидную последовательность, которая кодирует полипептид, где аминокислотная последовательность полипептида на по меньшей мере 45% идентична, по меньшей мере 50% идентична, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или на 100% идентична любой из аминокислотных последовательностей под SEQ ID NO: 39-74; или (е) нуклеотидную последовательность, которая комплементарна нуклеотидной последовательности любой из (a)-(d), приведенных выше.
[0062] Настоящее изобретение дополнительно охватывает кассету экспрессии, содержащую промотор, функционально связанный с гетерологичной нуклеотидной последовательностью, которая содержит: (а) нуклеотидную последовательность под любым из SEQ ID NO: 1-38; (b) нуклеотидную последовательность, которая на по меньшей мере 45% идентична, по меньшей мере 50% идентична, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или на 100% идентична нуклеотидной последовательности под любым из SEQ ID NO: 1-38; (с) нуклеотидную последовательность, которая кодирует полипептид, где аминокислотная последовательность полипептида содержит SEQ ID NO: 39-74 и характеризуется активностью в отношении контроля насекомых; (d) нуклеотидную последовательность, которая кодирует полипептид, где аминокислотная последовательность полипептида на по меньшей мере 45% идентична, по меньшей мере 50% идентична, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или на 100% идентична аминокислотной последовательности под любым из SEQ ID NO: 39-74; или (е) нуклеотидную последовательность, которая комплементарна нуклеотидной последовательности любой из (a)-(d), приведенных выше. В некоторых вариантах осуществления настоящее изобретение охватывает кассету экспрессии, содержащую гетерологичную молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая кодирует полипептид, где аминокислотная последовательность полипептида на по меньшей мере 93% идентична аминокислотной последовательности под SEQ ID NO: 39-74. Кассета экспрессии содержит промотор, функционально связанный с гетерологичной нуклеотидной последовательностью и не встречающийся в природе.
[0063] Настоящее изобретение также охватывает рекомбинантные векторы или конструкции, которые также могут называться векторами или конструкциями, содержащими кассеты экспрессии и/или молекулы нуклеиновой кислоты по настоящему изобретению. В таких векторах нуклеиновые кислоты предпочтительно находятся в кассетах экспрессии, содержащих регуляторные элементы для экспрессии нуклеотидных молекул в клетке-хозяине, способной экспрессировать нуклеотидные молекулы. Такие регуляторные элементы обычно содержат промотор и сигналы терминации и, предпочтительно, также содержат элементы, дающие возможность выполнять эффективную трансляцию полипептидов, кодируемых нуклеиновыми кислотами по настоящему изобретению. Векторы, содержащие нуклеиновые кислоты, могут быть способны к репликации в конкретных клетках-хозяевах, предпочтительно как внехромосомные молекулы, и таким образом используются для амплификации нуклеиновых кислот по настоящему изобретению в клетках-хозяевах. Настоящее изобретение также охватывает клетку-хозяина, которая содержит кассету экспрессии или молекулу нуклеиновой кислоты по настоящему изобретению. В одном варианте осуществления клетками-хозяева для таких векторов являются микроорганизмы, такие как бактерии, в частности Bacillus thuringiensis или Е. coli, или такие как грибы, как, например, дрожжи. В другом варианте осуществления клетки-хозяева для таких рекомбинантных векторов представляют собой эндофиты или эпифиты. В еще одном варианте осуществления такие векторы являются вирусными векторами и применяются для репликации нуклеотидных последовательностей в конкретных клетках-хозяевах, например в клетках насекомых или растительных клетках. Рекомбинантные векторы также используются для трансформации нуклеотидных молекул по настоящему изобретению в клетки-хозяева, вследствие чего нуклеотидные молекулы стабильно интегрируются в ДНК трансгенного хозяина. В одном варианте осуществления трансгенным хозяином является растение, например однодольное растение, такое как растение кукурузы или растение пшеницы. В вариантах осуществления трансгенное растение-хозяин представляет собой двудольное растение, такое как растение сои или растение хлопка.
[0064] В другом варианте осуществления по меньшей мере одна из нуклеиновых кислот по настоящему изобретению вводится в соответствующую кассету экспрессии, содержащую промотор и сигнал терминации. Экспрессия нуклеиновой кислоты может быть конститутивной, или может применяться индуцибельный промотор, отвечающий на различные типы стимулов для инициации транскрипции. В другом варианте осуществления клетка, в которой экспрессируется инсектицидный белок по настоящему изобретению, представляет собой микроорганизм, такой как вирус, бактерии или гриб. Еще в одном варианте осуществления вирус, такой как бакуловирус, содержит в своем геноме нуклеиновую кислоту по настоящему изобретению и экспрессирует большие количества соответствующего инсектицидного белка после заражения соответствующих эукариотических клеток, которые пригодны для репликации вируса и экспрессии нуклеиновой кислоты. Полученный таким образом инсектицидный белок применяется в качестве инсектицидного средства. В качестве альтернативы, бакуловирусы, сконструированные с возможностью содержания нуклеиновой кислоты, применяются для инфицирования насекомых in vivo, и они уничтожают их либо посредством экспрессии инсектицидного токсина, либо посредством комбинации вирусной инфекции и экспрессии инсектицидного токсина. В дополнительном варианте осуществления настоящее изобретение также охватывает способ получения полипептид с инсектицидной активностью, включающий культивирование клетки-хозяина в условиях, при которых экспрессируется молекула нуклеиновой кислоты, кодирующая полипептид.
[0065] Бактериальные клетки также являются клетками-хозяевами для экспрессии нуклеиновых кислот по настоящему изобретению. В одном варианте осуществления применяются непатогенные симбиотические бактерии, способные жить и размножаться в растительных тканях, так называемые эндофиты, или непатогенные симбиотические бактерии, способные колонизировать филлосферу или ризосферу, так называемые эпифиты. Такие бактерии включают бактерий родов Agrobacterium, Alcaligenes, Azospirillum, Azotobacter, Bacillus, Clavibacter, Enterobacter, Erwinia, Flavobacter, Klebsiella, Pseudomonas, Rhizobium, Serratia, Streptomyces и Xanthomonas. Симбиотические грибы, такие как Trichoderma и Gliocladium, также являются возможными хозяевами для экспрессии нуклеиновых кислот по настоящему изобретению для той же цели.
[0066] Методики для этих генетических манипуляций являются специальными для различных доступных хозяев и известны из уровня техники. Например, векторы экспрессии pKK223-3 и pKK223-2 могут использоваться для экспрессии гетерологичных генов в Е. coli либо в транскрипционном, либо трансляционном слиянии под действием промотора tac или trc. Для экспрессии оперонов, кодирующих несколько ORF, самой простой процедурой является введение оперона в вектор, такой как pKK223-3, в виде транскрипционного слияния, что дает возможность применять когнатный сайт связывания рибосом гетерологичных генов. Методики для сверхэкспрессии у грамположительных видов, таких как Bacillus, также известны из уровня техники и могут использоваться в контексте настоящего изобретения (Quax et al. в: Industrial Microorganisms: Basic and Applied Molecular Genetics, Eds. Baltz et al., American Society for Microbiology, Washington (1993)). Альтернативные системы сверхэкспрессии основаны, например, на дрожжевых векторах и включают использование Pichia, Saccharomyces и Kluyveromyces (Sreekrishna, в: Industrial microorganisms: basic and applied molecular genetics, Baltz, Hegeman, and Skatrud eds., American Society for Microbiology, Washington (1993); Dequin & Barre, Biotechnology L2:173-177(1994); van den Bergef a/, Biotechnology 8:135-139 (1990)).
[0067] Некоторые инсектицидные белки были экспрессированы в растениях, и семена таких растений ежегодно продаются фермерам для применения для контроля различных насекомых-вредителей. Такие самозащищенные инсектицидные продукты рассматриваются и регистрируются различными регулирующими органами, включая, например, Агентство по охране окружающей среды США (ЕРА).
[0068] Воздействие при поступлении с пищей является основным путем, за счет которого люди могут подвергаться воздействию инсектицидных белков, экспрессируемых трансгенными растениями. Острая пероральная токсичность для млекопитающих и перевариваемость белка являются конечными точками оценки риска для здоровья человека, определяемыми ЕРА. Дополнительным научным доказательством безопасности инсектицидных белков является то, что они, как было показано, быстро расщепляются in vitro под действием искусственного желудочного сока. Например, результаты семи анализов in vitro, проведенных с иллюстративными белками Cry1, Cry2 и Cry3, показали, что белки быстро расщепляются, как правило, в течение 30 секунд. Эти результаты подтверждают более широкий вывод о том, что представители этих групп белков Cry (которые обладают значительной идентичностью аминокислотных последовательностей), вероятно, быстро расщепляются после поедания человеком. Подобные тесты проводят для каждого трансгенного белка, экспрессируемого в растениях. Другая область рассмотрения охватывает определение того, способны ли инсектицидные белки вызывать аллергическую реакцию. Продемонстрированное быстрое расщепление трансгенного инсектицидного белка in vitro должно сводить к минимуму возможность такого явления. Для сравнения, пищевые аллергены обычно персистируют в in vitro модели желудочно-кишечного тракта, тогда как обычные белки пищи, для которых не показана аллергенность, быстро расщепляются в искусственном желудочном соке (Metcalfe et al., 1996).
[0069] С помощью анализа с применением искусственного желудочного сока (SGF) измеряют in vitro перевариваемость тестируемого белка в строго контролируемых условиях, характерных для верхних отделов пищеварительного тракта млекопитающих. Например, полученный с помощью бактерий тестируемый белок Cry (в концентрации 0,5-5 мг/мл) подвергали воздействию фермента пепсина (из слизистой оболочки желудка свиньи, растворенного в 2 мг/мл NaCl, рН 1,2) в соотношении 10 единиц активности пепсина/мкг тестируемого белка в течение периода времени, составляющего один час, при 37°С. Образцы отбирали в моменты времени 1, 2, 5, 10, 30 и 60 минут и немедленно блокировали посредством добавления предварительно нагретого (95°С - 2 минуты) стоп-буфера (65% 0,5 М бикарбоната натрия, рН 11, 35% трициновый загрузочный буфер) для немедленной инактивации пепсина, и снова нагревали в течение дополнительных 5 минут. После завершения анализа образцы, взятые в определенные моменты времени, и контроли (только тестируемый белок, только пепсин) исследовали с помощью SDS-PAGE в 10-20% трис-трициновом геле (при этом пептиды были визуально различимы вплоть до 1 кДа) для отслеживания кинетики и уровня переваривания, осуществляемого пепсином. Если тестируемый белок или значительный полипептидный фрагмент тестируемого белка является визуально различимым, например, в моменты времени 5 и/или 10 минут, то он является неперевариваемым или не полностью перевариваемым в анализе с применением SGF и может быть качественно оценен как "нет" или "неперевариваемый". Если тестируемый белок и любой значительный полипептидный фрагмент не является визуально различимым, например, в момент времени 5 минут, то он является перевариваемым в анализе с применением SGF и может быть качественно оценен как "да" или "перевариваемый".
[0070] Настоящее изобретение также охватывает полипептид, содержащий аминокислотную последовательность, на по меньшей мере 45% идентичную, по меньшей мере 50% идентичную, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99% идентичную или на 100% идентичную любой из SEQ ID NO: 39-74 и дополнительно содержащую введенный сайт расщепления протеазой. Введенный сайт расщепления протеазой не встречается в природе и вводится в полипептидную последовательность в результате мутации по типу замены или в результате мутации по типу вставки или делеции. Введенный сайт расщепления протеазой может быть введен посредством вставки по меньшей мере одного остатка лейцина в полипептидную последовательность, содержащую любую из SEQ ID NO: 39-74. Введенная мутация может дестабилизировать полипептид таким образом, что протеаза может получать доступ к сайту расщепления, к которому она ранее не имела доступ вследствие плотной и/или стабильной укладки белка или стерического несоответствия. Введенный сайт расщепления протеазой может представлять собой введенную мутацию в полипептидной последовательности, которая распознается протеазой, такой как химотрипсин, трипсин или пепсин, в качестве сайта протеолитического расщепления. В некоторых вариантах осуществления введенный сайт расщепления протеазой может изменять существующий сайт расщепления протеазой таким образом, что он распознается другой протеазой. Сайты расщепления протеазой для химотрипсина, трипсина и пепсина широко известны из уровня техники. Химотрипсин преимущественно расщепляет пептидные амидные связи, в которых с карбоксильной стороны от амидной связи (в положении Р1) расположена крупная гидрофобная аминокислота (тирозин, триптофан и фенилаланин). Трипсин расщепляет пептидные цепи главным образом с карбоксильной стороны аминокислот лизина или аргинина, за исключением случаев, когда после какой-либо из них расположен пролин. Пепсин является наиболее эффективным при расщеплении пептидных связей между гидрофобными и предпочтительно ароматическими аминокислотами, такими как фенилаланин, триптофан, тирозин и лейцин. Эти сайты расщепления являются преимущественными сайтами расщепления и не включают все сайты расщепления, распознаваемые химотрипсином, трипсином или пепсином и, кроме того, не включают все сайты расщепления для всех протеаз.
[0071] Примером полипептида, сконструированного таким образом, что он содержит введенный сайт расщепления протеазой, является вариант NitromobCRW Y213L/I215L (SEQ ID NO: 47). Эта мутация по типу замены изменяет мотив с "YNAYLIG" на "YNALLL". Этот введенный сайт расщепления протеазой может распознаваться пепсином и/или химотрипсином и отсутствует в последовательности белка NitromobCRW дикого типа. В некоторых вариантах осуществления введенный сайт расщепления протеазой может находиться в сайте мутации или рядом с ним, например, в остатках 190-230 полипептида. Вариант NitromobCRW Y213L/I215L может характеризоваться измененной или менее стабильной третичной структурой по сравнению с NitromobCRW дикого типа. В некоторых вариантах осуществления введенный сайт расщепления протеазой может быть расположен дистально относительно введенной мутации. Например, введенная мутация Y213 и/или I215 может "ослаблять" трехмерную укладку полипептида NitromobCRW, тем самым делая сайт расщепления протеазой, который ранее был недоступным (и, следовательно, не расщеплялся), доступным для протеазы. Это приводит к тому, что в результате введения мутации вводится сайт расщепления протеазой, который не существовал в неизмененном полипептиде. В некоторых вариантах осуществления введенная мутация и/или введенный сайт расщепления протеазой расположены в участке, соответствующем аминокислотным остаткам 1-300 любой из SEQ ID NO: 39-74. В некоторых вариантах осуществления введенная мутация и/или введенный сайт расщепления протеазой расположены в участке, соответствующем аминокислотным остаткам 97-300 любой из SEQ ID NO: 39-74. В дополнительных вариантах осуществления введенная мутация и/или введенный сайт расщепления протеазой расположены в участке, соответствующем аминокислотным остаткам 97-266 любой из SEQ ID NO: 39-74. В дополнительных вариантах осуществления введенная мутация и/или введенный сайт расщепления протеазой расположены в участке, соответствующем аминокислотным остаткам 175-266 любой из SEQ ID NO: 39-74. В дополнительных вариантах осуществления введенная мутация и/или введенный сайт расщепления протеазой расположены в участке, соответствующем аминокислотным остаткам 185-250 любой из SEQ ID NO: 40-74. В дополнительных вариантах осуществления введенная мутация и/или введенный сайт расщепления протеазой расположены в участке, соответствующем аминокислотным остаткам 200-230 любой из SEQIDNO: 40-74.
[0072] Настоящее изобретение также охватывает полипептид, содержащий аминокислотную последовательность, на по меньшей мере 45% идентичную, по меньшей мере 50% идентичную, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99% идентичную или на 100% идентичную любой из SEQ ID NO: 39-74 и дополнительно содержащую введенную мутацию, которая улучшает перевариваемость в анализе с применением SGF по сравнению с полипептидом, содержащим аминокислотную последовательность под SEQ ID NO: 39. Мутация может представлять собой мутацию по типу замены, вставку или делецию. Мутация может представлять собой вставку по меньшей мере одного остатка лейцина.
[0073] Настоящее изобретение также включает способ улучшения перевариваемости полипептида, на по меньшей мере 45% идентичного, по меньшей мере 50% идентичного, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99% идентичного или на 100% идентичного любой из SEQ ID NO: 39-74, включающий введение по меньшей мере одной мутации в аминокислотную последовательность полипептида. В вариантах осуществления эта введенная мутация улучшает перевариваемость полипептида в анализе с применением SGF. Мутация может улучшать перевариваемость за счет введения сайта расщепления протеазой. В других вариантах осуществления мутация может улучшать перевариваемость за счет изменения специфичности протеазы в этом сайте. Например, таким образом то, что могло быть сайтом для химотрипсина или трипсина, подвергается мутации с образованием сайта для пепсина. В других вариантах осуществления мутация может дестабилизировать белок таким образом, что сайт становится доступным для расщепления протеазой. Сайт, становящийся доступным для протеазы, может располагаться дистально относительно введенной мутации. В предпочтительных вариантах осуществления мутация не изменяет или в значительной степени не изменяет активность или инсектицидную активность полипептида. В некоторых вариантах осуществления полипептид с введенной мутацией обладает по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% от инсектицидной активности NitromobCRW. Данный способ пояснен в примерах в настоящем описании, где, например, было обнаружено, что вариант NitromobCRW Y213L/I215L характеризуется улучшенной перевариваемостью в анализе с применением SGF. Он также сохранял очень высокую инсектицидную активность.
[0074] В некоторых вариантах осуществления способа, описанного выше, введенная(введенные) мутация(мутации) может(могут) быть
расположена(расположены) в участке, соответствующем аминокислотным остаткам 1-300 любой из SEQ ID NO: 39-74. В дополнительных вариантах осуществления введенная(введенные) мутация(мутации) может(могут) быть
расположенафасположены) в участке, соответствующем аминокислотным остаткам 97-300 любой из SEQ ID NO: 39-74. В дополнительных вариантах осуществления введенная(введенные) мутация(мутации) может(могут) быть
расположенафасположены) в участке, соответствующем аминокислотным остаткам 97-266 любой из SEQ ID NO: 39-74. В дополнительных вариантах осуществления введенная(введенные) мутация(мутации) может(могут) быть
расположенафасположены) в участке, соответствующем аминокислотным остаткам 175-266 любой из SEQ ID NO: 39-74. В дополнительных вариантах осуществления введенная(введенные) мутация(мутации) может(могут) быть
расположенафасположены) в участке, соответствующем аминокислотным остаткам 185-250 любой из SEQ ID NO: 39-74. В дополнительных вариантах осуществления введенная(введенные) мутация(мутации) может(могут) быть
расположенафасположены) в участке, соответствующем аминокислотным остаткам 200-230 любой из SEQ ID NO: 39-74.
[0075] В других вариантах осуществления мутация может быть введена в Y213 и/или I215 SEQ ID NO: 39 или проксимально относительно них. В дополнительных вариантах осуществления мутация может представлять собой Y213L/I215L. В других вариантах осуществления мутация может представлять собой вставку или делецию аминокислотного остатка, такую как, например, вставка по меньшей мере одного остатка лейцина. Данный(данные) остаток(остатки) может(могут) быть прилегающим(прилегающими) к Y213 и/или I215 или соседним(соседними) по отношению к ним в SEQ ID NO: 39, например, как в вариантах NitromobCRW L214-Leu-I215 (SEQ ID NO: 57) или I215-Leu-G216 (SEQ ID NO: 58). Остатки лейцина также могут быть вставлены проксимально относительно Y213 и/или I215, где "проксимально" может означать на расстоянии по меньшей мере 1, по меньшей мере 2, по меньшей мере 4, по меньшей мере 6, по меньшей мере 8, по меньшей мере 10 или по меньшей мере 20 аминокислот от Y213 и/или I215.
[0076] Инсектицидные белки по настоящему изобретению характеризуются активностью в отношении контроля насекомых при тестировании против насекомых-вредителей в биоанализах. В одном варианте осуществления инсектицидные белки по настоящему изобретению активны против жесткокрылых и/или чешуекрылых насекомых. Специалисту в данной области техники будет понятно, что белок по настоящему изобретению может характеризоваться другим диапазоном инсектицидной активности по сравнению с другими белками по настоящему изобретению. В некоторых вариантах осуществления мутантный вариант NitromobCRW может характеризоваться инсектицидной активностью в отношении более широкого диапазона насекомых-вредителей, как, например, в отношении большего количества видов жесткокрылых или чешуекрылых, по сравнению с другими вариантами NitromobCRW. В других вариантах осуществления вариант NitromobCRW может характеризоваться инсектицидной активностью в отношении видов чешуекрылых, но не в отношении видов жесткокрылых. В некоторых вариантах осуществления вариант NitromobCRW может характеризоваться более широким диапазоном инсектицидной активности в отношении видов жесткокрылых или чешуекрылых по сравнению с немодифицированным NitromobCRW (SEQ ID NO: 39).
[0077] Насекомые в отряде Lepidoptera включают без ограничения любое известное в настоящее время или идентифицированное позже насекомое, которое классифицируется как чешуекрылое, в том числе те виды насекомых, которые относятся к подотрядам Zeugloptera, Glossata и Heterobathmiina, а также любую их комбинацию. Иллюстративные чешуекрылые насекомые включают без ограничения Ostrinia spp., такой как О. nubilalis (огневка кукурузная); Plutella spp., такой как Р. xylostella (капустная моль); Spodoptera spp., такой как S. frugiperda (совка травяная), S. ornithogalli (желтополосая совка), S. praefica (западная желтополосая совка), S. eridania (южная совка) и S. exigua (совка малая); Agrotis spp., такой как A. ipsilon (совка-ипсилон), A. segetum (озимая совка обыкновенная), A. gladiaria (озимая совка рода Agrotis) и A. orthogonia (совка прямоугольная); Striacosta spp., такой как S. albicosta (западная бобовая совка); Helicoverpa spp., такой как//, zea (совка кукурузная), Н. punctigera (совка хлопковая австралийская), S. littoralis (совка хлопчатника египетского) иН. armigera (совка хлопковая); Heliothis spp., такой как Н. virescem (табачная листовертка); Diatraea spp., такой как, grandiosella (огневка кукурузная юго-западная) и D. saccharalis (точильщик стеблей сахарного тростника); Trichoplusia spp., такой как Т. ni (совка капустная); Sesamia spp., такой как S. nonagroides (мотылек кукурузный средиземноморский); Pectinophora spp., такой как P. gossypiella (розовый коробочный червь); Cochylis spp., такой как С. hospes (полосатая подсолнечная моль); Manduca spp., такой как М. sexta (табачный бражник) и М quinquemaculata (бражник пятиточечный); Elasmopalpus spp., такой как E. lignosellus (точильщик стеблей кукурузы малый); Pseudoplusia spp., такой как P. includens (соевая совка); Anticarsia spp., такой как A. gemmatalis (совка бархатных бобов); Plathypena spp., такой как Р. scabra (совка клеверная); Pieris spp., такой как P. brassicae (белянка капустная), Papaipema spp., такой как P. nebris (точильщик Papaipema nebris); Pseudaletia spp., такой как P. unipuncta (совка луговая обыкновенная); Peridroma spp., такой как P. saucia (совка маргаритковая); Keiferia spp., такой как K. lycopersicella (томатная острица); Artogeia spp., такой как A rapae (белянка репная); Phthorimaea spp., такой как P. operculella (картофельная моль); Crymodes spp., такой как С. devastator (стеклянница); Feltia spp., такой как F. ducens (гусеница совки Feltia subgothica); и любую комбинацию вышеуказанных. В одном аспекте данного варианта осуществления инсектицидные белки по настоящему изобретению активны против совки-ипсилон, огневки сахарного тростника и/или огневки кукурузной юго-западной.
[0078] Насекомые в отряде Coleoptera включают без ограничения любое жесткокрылое насекомое, известное в настоящее время или идентифицированное позже, в том числе насекомых из подотрядов Archostemata, Myxophaga, Adephaga и Polyphaga, а также любую их комбинацию.
[0079] В одном аспекте данного варианта осуществления инсектицидные белки по настоящему изобретению активны против Diabrotica spp. Diabrotica представляет собой род жуков отряда Coleoptera, обычно называемый "кукурузными корневыми жуками" или "огуречными жуками". Типичные виды Diabrotica включают без ограничения Diabrotica barberi (северного кукурузного корневого жука), D. virgifera virgifera (западного кукурузного корневого жука), D. undecimpunctata howardii (южного кукурузного корневого жука), D. balteata (жука-блошку окаймленного), D. undecimpunctata undecimpunctata (жука-блошку одиннадцатиточечную), D. significata (3-точечного листоеда), D. speciosa (диабротику особую), D. virgifera zeae (мексиканского кукурузного корневого жука), D. beniensis, D. cristata, D. curviplustalata, D. dissimilis, D. elegantula, D. emorsitans, D. graminea, D. hispanloe, D. lemniscata, D. linsleyi, D. milleri, D. nummularis, D. occlusal, D. porrecea, D. scutellata, D. tibialis, D. trifasciata и D. viridula и любую их комбинацию.
[0080] Другие неограничивающие примеры жесткокрылых насекомых-вредителей в соответствии с настоящим изобретением включают Leptinotarsa spp., такой как Р. decemlineata (колорадский жук); Chrysomela spp., такой как С. scripta (листоед тополевый); Hypothenemus spp., такой как Н. hampei (жук кофейный); Sitophilus spp., такой как S. zeamais (маисовый долгоносик); Epitrix spp., такой как P. hirtipennis (жук-блошка шершавая) и P. cucumeris (блошка картофельная); Phyllotreta spp., такой как Р. cruciferae (блошка крестоцветная) и P. pusilla (западная черная блошка); Anthonomus spp., такой как A. eugenii (перечный долгоносик); Hemicrepidus spp., такой как H. memnonius (проволочники); Melanotus spp., такой как М communis (проволочник); Ceutorhychus spp., такой как С. assimilis (рапсовый семенной скрытнохоботник); Phyllotreta spp., такой как P. cruciferae (блошка крестоцветная); Aeolus spp., такой как А. mellillus (проволочник); Aeolus spp., такой как A. mancus (пшеничный проволочник); Horistonotus spp., такой как H. uhlerii (песчаный проволочник); Sphenophorus spp., такой как S. maidis (долгоносик маисовый), S. zeae (долгоносик тимофеечный), S. parvulus (мятликовый долгоносик) и S. callosus (долгоносик клубеньковый эспарцетовый); Phyllophaga spp. (личинки хрущей); Chaetocnema spp., такой как С.pulicaria (блошка стеблевая хлебная); Popillia spp., такой как P. japonica (японский жук); Epilachna spp., такой как P. varivestis (мексиканский бобовый жук); Cerotoma spp., такой как С.trifurcate (жук-листоед); Epicauta spp., такой как P. pestifera и Е. lemniscata (шпанские мушки), и любую комбинацию вышеуказанных.
[0081] Инсектицидные белки по настоящему изобретению также могут быть активными против полужесткокрылых, двукрылых, Lygus spp. и/или других колющих и сосущих насекомых, например из отряда Orthoptera или Thysanoptera. Насекомые в отряде Diptera включают без ограничения любое двукрылое насекомое, известное в настоящее время или идентифицированное позже, в том числе без ограничения Liriomyza spp., такой как P. trifolii (листовой минер) и P. sativae (листовой минер овощей); Scrobipalpula spp., такой как S. absoluta (листовой минер томатов); Delia spp., такой как D. platura (личинка мухи ростковой), D. brassicae (личинка мухи капустной весенней) и P. radicum (муха капустная весенняя); Psilia spp., такой как P. rosae (муха морковная); Tetanops spp., такой как Т. myopaeformis (личинка тли свекловичной), и любую комбинацию вышеуказанных.
[0082] Насекомые в отряде Orthoptera включают без ограничения любое известное в настоящее время или идентифицированное позже прямокрылое насекомое, в том числе без ограничения Melanoplus spp., такой как М. differentialis (кобылка отличительная), М. femurrubrum (краснобедрая кобылка), М. bivittatus (кобылка двухполосая) и любую их комбинацию.
[0083] Насекомые в отряде Thysanoptera включают без ограничения любое известное в настоящее время или идентифицированное позже бахромчатокрылое насекомое, в том числе без ограничения Frankliniella spp., такой как P. occidentalis (западный цветочный трипе) и P. fusca (табачный трипе); и Thrips spp., такой как Т. tabaci (трипе луковый), Т. palmi (пальмовый трипе), и любую комбинацию вышеуказанных.
[0084] Инсектицидные белки по настоящему изобретению также могут быть активными против нематод. Используемый в данном документе термин "нематода" охватывает любой известный в настоящее время или идентифицированный позже организм, который классифицирован в царстве животных, тип нематоды, в том числе без ограничения нематоды в пределах класса Adenophorea (в том числе, например, отряды Enoplida, Isolaimida, Mononchida, Dorylaimida, Trichocephalida, Mermithida, Muspiceida, Araeolaimida, Chromadorida, Desmoscolecida, Desmodorida и Monhysterida) и/или класса Secementea (в том числе, например, отряды Rhabdita, Strongylida, Ascaridida, Spirurida, Camallanida, Diplogasterida, Tylenchida и Aphelenchida).
[0085] Нематоды включают без ограничения паразитические нематоды, такие как клубеньковые нематоды, цистообразующие нематоды и/или ранящие нематоды. Иллюстративный роды нематод в соответствии с настоящим изобретением включают без ограничения Meloidogyne (клубеньковые нематоды), Heterodera (цистообразующие нематоды), Globodera (цистообразующие нематоды), Radopholus (роющие нематоды), Rotylenchulus (почковидные нематоды), Pratylenchus (ранящие нематоды), Aphelenchoides (нематоды листовые), Helicotylenchus (нематоды спиральные), Hoplolaimus (ланцетообразные нематоды), Paratrichodorus (нематоды, вызывающие тупоконечность корней), Longidorus, Nacobbus (ложно-клубеньковые нематоды), Subanguina, Belonlaimus (жалящие нематоды), Criconemella, Criconemoides (кольцевые нематоды), Ditylenchus, Dolichodorus, Hemicriconemoides, Hemicycliophora, Hirschmaniella, Hypsoperine, Macroposthonia, Melinius, Punctodera, Quinisulcius, Scutellonema, Xiphinema (кинжальные нематоды), Tylenchorhynchus (карликовые нематоды), Tylenchulus, Bursaphelenchus (круглые кишечные черви) и любую их комбинацию.
[0086] Типичные паразитические нематоды растений в соответствии с настоящим изобретением включают без ограничения Belonolaimus gracilis, Belonolaimus longicaudatus, Bursaphelenchus xylophilus (нематоду древесины хвойных пород), Criconemoides ornata, Ditylenchus destructor (нематоду корней картофеля), Ditylenchus dipsaci (стеблевую и луковичную нематоду), Globodera pallida (картофельную цистообразующую нематоду), Globodera rostochiensis (золотистую картофельную нематоду), Heterodera glycines (соевую цистообразующую нематоду), Heterodera schachtii (цистообразующую нематоду сахарной свеклы); Heterodera zeae (кукурузную цистообразующую нематоду), Heterodera avenae (цистообразующую нематоду злаковых культур), Heterodera carotae, Heterodera trifolii, Hoplolaimus columbus, Hoplolaimus galeatus, Hoplolaimus magnistylus, Longidorus breviannulatus, Meloidogyne arenaria, Meloidogyne chitwoodi, Meloidogyne hapla, Meloidogyne incognita, Meloidogyne javanica, Mesocriconema xenoplax, Nacobbus aberrans, Naccobus dorsalis, Paratrichodorus christiei, Paratrichodorus minor, Pratylenchus brachyurus, Pratylenchus crenatus, Pratylenchus hexincisus, Pratylenchus neglectus, Pratylenchus penetrans, Pratylenchus projectus, Pratylenchus scribneri, Pratylenchus tenuicaudatus, Pratylenchus thornei, Pratylenchus zeae, Punctodera chaccoensis, Quinisulcius acutus, Radopholus similis, Rotylenchulus reniformis, Tylenchorhynchus dubius, Tylenchulus semipenetrans (цитрусовую нематоду), Siphinema americanum, X. Mediterraneum и любую комбинацию вышеуказанных.
[0087] В другом варианте осуществления настоящее изобретение охватывает способ получения инсектицидного белка, который является активным против насекомых, включающий: (а) получение клетки-хозяина, содержащей ген, который сам содержит кассету экспрессии и/или молекулу нуклеиновой кислоты по настоящему изобретению; и (b) выращивание трансгенной клетки-хозяина таким образом, чтобы экспрессировать инсектицидный белок, который является активным против насекомых.
[0088] В еще одном дополнительном варианте осуществления изобретение охватывает способ осуществления контроля насекомых, включающий доставку насекомым эффективного для контроля насекомых количества инсектицидного белка по настоящему изобретению.
[0089] В одном варианте осуществления по меньшей мере один из инсектицидных белков по настоящему изобретению экспрессируется в высшем организме, таком как растение. В данном случае трансгенные растения экспрессируют эффективные для контроля насекомых количества инсектицидного белка для самозащиты от насекомых-вредителей. В случае если насекомое начинает поедать такое трансгенное растение, оно также поглощает экспрессированный инсектицидный белок. Это будет удерживать насекомое от дальнейшего вгрызания в растительную ткань и/или даже может причинять вред насекомому или уничтожать его. Нуклеиновая кислота по настоящему изобретению вводится в кассету экспрессии, которая затем может быть устойчиво интегрирована в геном растения. В другом варианте осуществления нуклеиновую кислоту включают в непатогенный самореплицирующийся вирус.Растения, трансформированные в соответствии с настоящим изобретением, могут быть однодольными или двудольными растениями и включают без ограничения кукурузу, пшеницу, овес, газонные травы, пастбищные травы, лен, ячмень, рожь, сладкий картофель, фасоль, горох, цикорий, салат-латук, капусту, цветную капусту, брокколи, репу, редьку, шпинат, спаржу, лук, чеснок, перец, сельдерей, тыкву крупноплодную, тыкву, коноплю, цукини, яблоню, грушевое дерево, айву, дыню, сливовое дерево, вишню, персиковое дерево, нектарин, абрикосовое дерево, клубнику, виноград, малину, ежевику, ананас, авокадо, папайю, манго, банановое дерево, сою, томат, сорго, сахарный тростник, сахарную свеклу, подсолнечник, рапс, клевер, табак, морковь, хлопчатник, люцерну, рис, картофель, баклажан, огурец, арабидопсис и древесные растения, такие как хвойные и лиственные деревья.
[0090] В другом варианте осуществления настоящее изобретение охватывает способ получения растения или части растения с повышенной устойчивостью к насекомым по сравнению с контрольными растением или частью растения, включающий: (а) введение молекулы нуклеиновой кислоты, содержащей кассету экспрессии по настоящему изобретению; и (b) выращивание части растения с получением растения, которое экспрессирует гетерологичную молекулу нуклеиновой кислоты кассеты экспрессии и которое характеризуется повышенной устойчивостью к насекомым по сравнению с контрольными растением или частью растения, которые не были трансформированы молекулой нуклеиновой кислоты, содержащей кассету экспрессии. В предпочтительном варианте осуществления кассета экспрессии может кодировать полипептид, содержащий аминокислотную последовательность, которая на по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или на 100% идентична или сходна с SEQ ID NO: 39-74. В предпочтительном варианте осуществления кассета экспрессии может кодировать полипептид, содержащий аминокислотную последовательность, которая на по меньшей мере 60% идентична SEQ ID NO: 47. "Повышенная" устойчивость к насекомым может быть измерена как повышение инсектицидной активности. Повышенная устойчивость к насекомым может быть больше 0%, на по меньшей мере 1%, на по меньшей мере 2%, на по меньшей мере 3%, на по меньшей мере 4%, на по меньшей мере 5%, на по меньшей мере 10%, на по меньшей мере 15%, на по меньшей мере 20%, на по меньшей мере 25%, на по меньшей мере 30%, на по меньшей мере 40%, на по меньшей мере 50%, на по меньшей мере 60%, на по меньшей мере 70%, на по меньшей мере 80%, на по меньшей мере 90%, на по меньшей мере 100%, на по меньшей мере 125%, на по меньшей мере 150%, на по меньшей мере 200%, на по меньшей мере 300%, на по меньшей мере 400%, на по меньшей мере 500%, на по меньшей мере 600%, на по меньшей мере 700%, на по меньшей мере 800%, на по меньшей мере 900% или на по меньшей мере 1000% больше инсектицидной активности по сравнению с контрольным растением. Растение или часть растения, характеризующиеся повышенной устойчивостью к насекомым по сравнению с контрольными растением или частью растения, могут быть получены с помощью способов трансформации растения, культивирования растительной ткани или селекции. Растение или часть растения могут быть получены с помощью способов полового или бесполого размножения. Могут использоваться любые подходящие контрольные растение или часть растения, например растение с таким же или аналогичным генетическим фоном, выращенное в той же среде. В вариантах осуществления контрольные растение или часть растения имеют тот же генетический фон и растут в той же среде, что и описанное растение, однако они не содержат молекулу по настоящему изобретению, в то время как описанное растение содержит молекулу по настоящему изобретению.
[0091] В другом варианте осуществления настоящее изобретение охватывает способ повышения устойчивости к насекомым у растения или части растения по сравнению с контрольным растением или частью растения, включающий обеспечение экспрессии в растении или части растения молекулы нуклеиновой кислоты или кассеты экспрессии по настоящему изобретению, где экспрессия гетерологичной нуклеиновой кислоты кассеты экспрессии приводит к повышенной устойчивости к насекомым у растения или части растения по сравнению с контрольным растением или частью растения. В вариантах осуществления кассета экспрессии или молекула нуклеиновой кислоты содержит промотор, функционально связанный с гетерологичной молекулой нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая содержит: (а) нуклеотидную последовательность под любым из SEQ ID NO: 1-38; (b) нуклеотидную последовательность, которая на по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или на 100% идентична нуклеотидной последовательности под любым из SEQ ID NO: 1-38; (с) нуклеотидную последовательность, которая кодирует полипептид, где аминокислотная последовательность полипептида содержит SEQ ID NO: 39-74 и характеризуется активностью в отношении контроля насекомых; (d) нуклеотидную последовательность, которая кодирует полипептид, где аминокислотная последовательность полипептида на по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или на 100% идентична аминокислотной последовательности под SEQ ID NO: 39-74; или (е) нуклеотидную последовательность, которая комплементарна нуклеотидной последовательности любой из (a)-(d), приведенных выше. Молекула нуклеиновой кислоты или кассета экспрессии может быть введена в растение. В некоторых вариантах осуществления молекула нуклеиновой кислоты или кассета экспрессии может быть введена в часть растения, и при этом растение, содержащее молекулу нуклеиновой кислоты или кассету экспрессии, может быть получено из части растения.
[0092] В другом варианте осуществления настоящее изобретение охватывает способ получения растения с повышенной устойчивостью к насекомым по сравнению с контрольным растением, включающий выявление в части растения гетерологичной нуклеиновой кислоты, предусматривающей молекулу нуклеиновой кислоты или кассету экспрессии по настоящему изобретению, и получение растения из части растения, за счет чего обеспечивается получение растения с повышенной устойчивостью к насекомым по сравнению с контрольным растением. В дополнительном варианте осуществления настоящее изобретение охватывает способ идентификации растения или части растения, характеризующегося повышенной устойчивостью к насекомым по сравнению с контрольным растением или частью растения, включающий выявление в растении или части растения молекулы нуклеиновой кислоты или кассеты экспрессии по настоящему изобретению, за счет чего обеспечивается идентификация растения или части растения с повышенной устойчивостью к насекомым. В дополнительном варианте осуществления кассету экспрессии или ее диагностический фрагмент выявляют в продукте амплификации из образца нуклеиновой кислоты из растения или части растения. Диагностический фрагмент может представлять собой молекулу нуклеиновой кислоты длиной по меньшей мере 10 смежных нуклеотидов, которая является уникальной для кассеты экспрессии по настоящему изобретению.
[0093] В еще одном варианте осуществления настоящее изобретение охватывает способ получения растения, характеризующегося повышенной устойчивостью к насекомым по сравнению с контрольным растением или частью растения, включающий скрещивание первого родительского растения со вторым родительским растением, где по меньшей мере первое родительское растение содержит в своем геноме гетерологичную нуклеиновую кислоту, которая предусматривает молекулу нуклеиновой кислоты или кассету экспрессии по настоящему изобретению, и получение поколения потомства, где поколение потомства содержит по меньшей мере одно растение, которое содержит гетерологичную нуклеиновую кислоту в своем геноме и которое проявляет повышенную устойчивость к насекомым по сравнению с контрольным растением.
[0094] В предпочтительных вариантах осуществления способы по настоящему изобретению придают повышенную устойчивость к насекомым растению или части растения против жесткокрылого и/или чешуекрылого насекомого-вредителя. Контроль насекомых, представляющих собой жесткокрылых насекомых-вредителей, продемонстрирован в примерах. В дополнительных вариантах осуществления способы по настоящему изобретению придают повышенную устойчивость к насекомым растению или части растения против видов Diabrotica, в том числе Diabrotica virgifera virgifera, Diabrotica barberi, Diabrotica undecimpunctata howardi, Diabrotica virgifera zeae и/или Diabrotica speciosa, и/или родственных видов.
[0095] В предпочтительных вариантах осуществления способы по настоящему изобретению придают повышенную устойчивость к насекомым однодольному растению.
[0096] Настоящее изобретение дополнительно охватывает трансгенное растение, содержащее гетерологичную молекулу нуклеиновой кислоты или кассету экспрессии по настоящему изобретению, которая при транскрипции и трансляции придает повышенную устойчивость к насекомым. В предпочтительных вариантах осуществления гетерологичная молекула нуклеиновой кислоты содержит последовательность, на по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичную любой из SEQ ID NO: 1-38 или комплементарную ей последовательность. В дополнительном варианте осуществления трансгенное растение содержит гетерологичную молекулу нуклеиновой кислоты, содержащую последовательность, на по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичную SEQ ID NO: 1-38 или комплементарную ей последовательность. В вариантах осуществления трансгенное растение представляет собой двудольное растение. В предпочтительных вариантах осуществления трансгенное растение представляет собой однодольное растение. В дополнительных вариантах осуществления трансгенное растение представляет собой люцерну, укроп, яблоню, абрикосовое дерево, артишок, рукколу, спаржу, авокадо, банановое дерево, бобовые, свеклу, ежевику, чернику, брокколи, брюссельскую капусту, кочанную капусту, канолу, мускусную дыню, морковь, кассаву, цветную капусту, сельдерей, вишню, кинзу, цитрусовое растение, клементин, кофейное дерево, кукурузу, хлопчатник, огурец, Дугласову пихту, баклажан, эндивий, салат эскариоль, эвкалипт, фенхель, разновидности фигового дерева, тыкву бутылочную, виноград, грейпфрут, дыню мускатную белую, хикаму, киви, салат-латук, разновидности лука порея, лимон, лайм, сосну ладанную, манговое дерево, дыню, гриб, орех, окру, лук, апельсин, декоративное растение, папайю, петрушку, горох, персиковое дерево, земляной орех, грушевое дерево, перец, хурму, сосну, ананас, плантайн, сливовое дерево, гранатовое дерево, тополь, картофель, тыкву, айву, сосну лучистую, радиккио, редис, малину, рис, рожь, сорго, южную сосну, сою, шпинат, тыкву большую столовую, клубнику, сахарную свеклу, подсолнечник, сладкий картофель, ликвидамбар, танжерин, чай, табак, томат, газонную траву, виноград культурный, арбуз, ямс или цуккини. В предпочтительных вариантах осуществления трансгенное растение представляет собой просо, просо прутьевидное, маис, сорго, пшеницу, овес, газонную траву, пастбищную траву, лен, рис, сахарный тростник, масличный рапс или ячмень.
[0097] В еще одном варианте осуществления трансгенное растение по настоящему изобретению содержит гетерологичную молекулу нуклеиновой кислоты, содержащую последовательность промотора. В еще одном варианте осуществления трансгенное растение по настоящему изобретению может содержать гетерологичную молекулу нуклеиновой кислоты, которая кодирует по меньшей мере один дополнительный требуемый признак. Дополнительный признак может быть закодирован в той же гетерологичной молекуле нуклеиновой кислоты, что и молекула по настоящему изобретению, или он может быть закодирован во второй гетерологичной молекуле нуклеиновой кислоты. Дополнительный требуемый признак может придавать устойчивость к насекомым ко второму насекомому-вредителю, устойчивость к насекомым к тому же насекомому-вредителю, переносимость абиотического стресса, мужскую стерильность, устойчивость к гербицидам, устойчивость к бактериальным заболеваниям, устойчивость к грибковым заболеваниям, устойчивость к вирусным заболеваниям, устойчивость к нематодам, модифицированный метаболизм жирных кислот, модифицированный метаболизм углеводов, улучшенную пищевую ценность, улучшенные характеристики при промышленном способе или измененную репродуктивную способность. Дополнительный требуемый признак также может стимулировать продуцирование в растении ценного с коммерческой точки зрения фермента или метаболита.
[0098] В вариантах осуществления требуемый дополнительный признак представляет собой второе пестицидное средство. Второе пестицидное средство может быть активным по отношению к любому вредителю растения, включающему насекомых, нематод, грибы, вирусы или бактерии. Примеры насекомых-вредителей растений включают, и не ограничиваются ими, Nilaparvata spp. (например, N. lugens (бурую рисовую цикадку)); Laodelphax spp. (например, L. striatellus (малую бурую рисовую цикадку)); Nephotettix spp. (например, N. virescens или N. cincticeps (зеленую цикадку) или N. nigropictus (рисовую цикадку)); Sogatella spp. (например, S. furcifera (цикадку белоспинную)); Blissus spp. (например,. В. leucopterus leucopterus (клопа постельного)); Scotinophora spp. (например, S. vermidulate (рисового долгоносика)); Acrosternum spp. (например,. A. hilare (зеленого жука-вонючку)); Parnara spp. (например,. P. guttata (рисовую толстоголовку)); Chilo spp. (например, С. suppressalis (рисового полосатого стеблееда), С. auricilius (золотистого стеблееда) или С. polychrysus (черноголового стеблееда)); Chilotraea spp. (например, С. polychrysa (рисового стеблееда)); Sesamia spp. (например, S. inferens (розового рисового точильщика)); Tryporyza spp. (например,. Т. innotata (белого рисового точильщика) или Т. incertulas (желтого рисового точильщика)); Cnaphalocrocis spp. (например, С. medinalis (листовертку рисовую)); Agromyza spp. (например, A. oryzae (листового минера), или A. parvicornis (кукурузного точкового листового минера)); Diatraea spp. (например, D. saccharalis (точильщика стеблей сахарного тростника) или D. grandiosella ((огневку кукурузную юго-западную)); Narnaga spp. (например, N. aenescens (зеленую рисовую гусеницу)); Xanthodes spp. (например, X transversa (зеленую гусеницу)); Spodoptera spp. (например, S. frugiperda (совку травяную), S. exigua (совку малую), S. littoralis (египетскую хлопковую совку) или S. praefwa (западную желтополосую совку)); Mythimna spp. (например, Mythmna (Pseudaletia) seperata (гусеницу)); Helicoverpa spp. (например, H. zea (совку кукурузную)); Colaspis spp. (например, С. brunnea (листоеда виноградного)); Lissorhoptrus spp. (например,. L. oryzophilus (долгоносика рисового водяного)); Echinocnemus spp. (например, Е. squamos (долгоносика растений риса)); Diclodispa spp. (например, D. armigera (рисовую шипоноску)); Oulema spp. (например, О. oryzae (листогрыза); Sitophilus spp. (например, S. oryzae (рисового долгоносика)); Pachydiplosis spp. (например,. P. oryzae (рисовую галлицу)); Hydrellia spp. (например, Н. griseola (малого рисового листового минера) или Н. sasakii (личинку, поедающую рисовые стебли)); Chlorops spp. (например,. С.o ryzae (личинку, поедающую стебли)); Diabrotica spp. (например, D. virgifera virgifera (западного кукурузного корневого жука), D. barberi (северного кукурузного корневого жука), D. undecimpunctata howardi (южного кукурузного корневого жука), D. virgifera zeae (мексиканского кукурузного корневого жука); D. balteata (жука-блошку окаймленную)); Ostrinia spp. (например,. О. nubilalis (огневку кукурузную)); Agrotis spp. (например,. A. ipsilon (совку-ипсилон)); Elasmopalpus spp. (например, Е. lignosellus (точильщика стеблей кукурузы малого)); Melanotus spp. (проволочников); Cyclocephala spp. (например, С.borealis (северного майского жука) или С. immaculata (южного майского жука)); Popillia spp. (например, Р. japonica (хрущика японского)); Chaetocnema spp. (например, С.pulicaria (кукурузного жука-блошку)); Sphenophorus spp. (например, S. maidis (долгоносика маисового)); Rhopalosiphum spp. (например,. R. maidis (тлю кукурузную листовую)); Anuraphis spp. (например, A maidiradicis (тлю кукурузную корневую)); Melanoplus spp. (например, М. femurrubrum (краснобедрую кобылку) М. differentialis (кобылку отличительную) или М. sanguinipes (мигрирующую кобылку)); Hylemya spp. (например,. Н. platura (личинку мухи ростковой)); Anaphothrips spp. (например, A. obscrurus (трипса злакового)); Solenopsis spp. (например, S. milesta (муравья-вора)); или spp. (например,. Т. urticae (обыкновенного паутинного клеща), Т. cinnabarinus (красного паутинного клеща); Helicoverpa spp. (например, Н. zea (совку кукурузную) или Н. armigera (совку хлопковую)); Pectinophora spp. (например,. P. gossypiella (розового коробочного червя)); Earias spp. (например, E. vittella (точечного коробочного червя хлопка)); Heliothis spp. (например, Н. virescens (табачную листовертку)); Anthonomus spp. (например, A grandis (долгоносика хлопкового)); Pseudatomoscelis spp. (например, Р. seriatus (марокканскую кобылку)); Trialeurodes spp. (например, Т. abutiloneus (белокрылку с полосами на крыльях) Т. vaporariorum (белокрылку тепличную)); Bemisia spp. (например,. В. argentifolii (белокрылку серебрянокрылую)); Aphis spp. (например, A gossypii (тлю хлопковую)); Lygus spp. (например, L. lineolaris (клопа полевого) или L. hesperus (западного клопа лугового)); Euschistus spp. (например, Е. conspersus (маскирующегося жука-вонючку)); Chlorochroa spp. (например, С. sayi (жука-вонючку Say)); Nezara spp. (например, N. viridula (южного зеленого жука-вонючку)); Thrips spp. (например, Т. tabaci (трипса лукового)); Frankliniella spp. (например,. F. fusca (трипса табачного) или F. occidentalis (западного трипса обыкновенного)); Leptinotarsa spp. (например, L. decemlineata (колорадского жука), L. juncta (ложного картофельного жука) или L. texana (техасского ложного картофельного жука)); Lema spp. (например, L. trilineata (картофельного жука с тремя полосками)); Epitrix spp. (например, Е. cucumeris (картофельного жука-блошку), Е. hirtipennis (жука-блошку) или E. tuberis (жука-блошку клубневого)); Epicauta spp. (например, Е. vittata (шпанку полосатую)); Phaedon spp. (например, P. cochleariae (горчичного листогрыза)); Epilachna spp. (например, Е. varivetis (мексиканского жука бобового)); Acheta spp. (например, A. domesticus (сверчка домового)); Empoasca spp. (например, Е. fabae (цикадку картофельную)); Myzus spp. (например, М. persicae (тлю персиковую зеленую)); Paratrioza spp. (например, P. cockerelli (листоблошку)); Conoderus spp. (например, С. falli (южного картофельного проволочника) или С. vespertinus (табачного проволочника)); Phthorimaea spp. (например, P. operculella (картофельную моль)); Macrosiphum spp. (например, М. euphorbiae (тлю картофельную обыкновенную)); Thyanta spp. (например, Т. pallidovirens (красноплечего жука-вонючку)); Phthorimaea spp. (например, P. operculella (картофельную моль)); Helicoverpa spp. (например, Н. zea (совку томатную); Keiferia spp. (например, K. lycopersicella (томатную острицу)); Limonius spp. (проволочников); Manduca spp. (например, М. sexta (табачного бражника) или М. quinquemaculata (бражника пятиточечного)); Liriomyza spp. (например, L. sativae, L. trifolli или L. huidobrensis (листового минера)); Drosophilla spp. (например, D. melanogaster, D. yakuba, D. pseudoobscura или D. simulans); Carabus spp. (например, С. granulatus); Chironomus spp. (например,. С. tentanus); Ctenocephalides spp. (например, С. felis (блоху кошачью)); Diaprepes spp. (например, D. abbreviatus (скосаря люцернового)); Ips spp. (например, I. pini (короеда соснового)); Tribolium spp. (например,. Т. castaneum (хрущака малого булавоусого)); Glossina spp. (например, G. morsitans (муху цеце)); Anopheles spp. (например, A. gambiae (комара малярийного)); Helicoverpa spp. (например, H. armigera (африканскую совку хлопковую)); Acyrthosiphon spp. (например, A. pisum (тлю гороховую)); Apis spp. (например,. А. melifera (медоносную пчелу)); Homalodisca spp. (например, Н. coagulate ((цикадку с зеркальными крыльями)); Aedes spp. (например, Ае. aegypti (желтолихорадочного комара)); Bombyx spp. (например, В. mori (шелковичного червя)); Locusta spp. (например, L. migratoria (мигрирующую саранчу)); Boophilus spp. (например, В. microplus (клеща кольчатого)); Acanthoscurria spp. (например, A gomesiana (рыжего шоколадного птицееда)); Diploptera spp. (например, D. punctata (тихоокеанского жука-таракана)); Heliconius spp. (например, Н. erato (красную бабочку-почтальона) или Н. melpomene (бабочку-почтальона)); Curculio spp. (например, С. glandium (плодожила дубового)); Plutella spp. (например, P. xylostella (капустную моль)); Amblyomma spp. (например, A. variegatum (клеща кольчатого)); Anteraea spp. (например, A. yamamai (шелкопряда)); nArmigeres spp. (например, A subalbatus).
[0099] Инсектицидные белки по настоящему изобретению могут использоваться в комбинации с другими пестицидными средствами (например, белками Cry Bt) с расширением целевого диапазона вредителей. Кроме того, применение инсектицидных белков по настоящему изобретению в комбинации с инсектицидным средством, которое характеризуется превосходным механизмом действия или целенаправленно воздействует на другой рецептор в кишечнике насекомого, характеризуется особой полезностью в отношении предупреждения появления устойчивости у насекомых и/или управления ею.
[00100] Второе пестицидное средство может представлять собой инсектицидный белок, полученный из Bacillus thuringiensis. Инсектицидный белок В. thuringiensis может представлять собой любой из нескольких инсектицидных белков, в том числе без ограничения белок Cry1, белок Cry3, белок Cry 6, белок Cry7, белок Cry8, белок Cry9, белок Cry11, белок Cry22, белок Cry 23, белок Cry 36, белок Cry37, белок Cry34 вместе с белком Cry35, бинарный инсектицидный белок СгуЕТ33 и CryET34, бинарный инсектицидный белок TIC100 и TIC101, бинарный инсектицидный белок PS149B1, VIP (вегетативный инсектицидный белок, раскрытый в патентах США 5849870 и 5877012, включенных в данный документе посредством ссылки), TIC900 или родственный белок, TIC901, TIC1201, TIC407, TIC417, модифицированный белок Cry3A или гибридные белки или химеры, полученные из любых вышеуказанных инсектицидных белков. В других вариантах осуществления инсектицидный белок В. thuringiensis выбран из группы, состоящей из Cry3Bb1, Cry34Ab1 вместе с Cry35Ab1, mCry3A (патент США №7276583, включенный в данный документ посредством ссылки), eCry3.1Ab (патент США №8309516, включенный в данный документ посредством ссылки) и белки Vip3A, в том числе Vip3Aa (патент США №6137033, включенный в данный документ посредством ссылки).
[00101] В других вариантах осуществления трансгенное растение по настоящему изобретению может содержать второе пестицидное средство, которое может быть получено из источников, отличных от В. thuringiensis. Второе инсектицидное средство может представлять собой средство, выбранное из группы, содержащей α-амилазу, пероксидазу, холестериноксидазу, пататин, протеазу, ингибитор протеазы, уреазу, ингибитор альфа-амилазы, порообразующий белок, хитиназу, лектин, сконструированное антитело или фрагмент антитела, инсектицидный белок Bacillus cereus, инсектицидный белок Xenorhabdus spp. (такого как Х. nematophila или Х. bovienii), инсектицидный белок Photorhabdus spp. (такого как P. luminescens или Р. asymobiotica), инсектицидный белок Brevibacillus spp. (такого как В. laterosporous), инсектицидный белок Lysinibacillus spp. (такого как L. sphearicus), инсектицидный белок Chromobacterium spp. (такого как С. subtsugae или С. piscinae), инсектицидный белок Yersinia spp. (такого как Y. entomophaga), инсектицидный белок Paenibacillus spp. (такого как P. propylaea), инсектицидный белок Clostridium spp. (такого как С. bifermentans), Pseudomonas spp. (такого как Р. fluorescens) и лигнин. В других вариантах осуществления второе средство может представлять собой по меньшей мере один инсектицидный белок, полученный из инсектицидного токсинового комплекса (Тс) из Photorhabdus, Xenorhabus, Serratia или Yersinia. В других вариантах осуществления инсектицидный белок может представлять собой ADP-рибозилтрансферазу, полученную из инсектицидных бактерий, таких как Photorhabdus ssp. В еще других вариантах осуществления инсектицидный белок может представлять собой Axmi205 или полученный из Axmi205 (патент США №8575425 и №9394345, каждый включен в данный документ посредством ссылки). В других вариантах осуществления инсектицидный белок может представлять собой VIP-белок, такой как VIP1 и/или VIP2, из В. cereus. В еще одних вариантах осуществления инсектицидный белок может представлять собой бинарный токсин, полученный из инсектицидных бактерий, такой как ISP1A и ISP2A из В. laterosporous или BinA и BinB из L. sphaericus. В других вариантах осуществления инсектицидный белок может представлять собой белок или вариант белка LachbCRW (заявка согласно РСТ № PCT/US 2017/045256), HmassCRW (заявка согласно РСТ № PCT/US 2017/058179) или WoodsCRW (заявка согласно РСТ № PCT/US 2018/012730). В еще одних вариантах осуществления инсектицидный белок может быть сконструированным, или может быть гибридным или химерой из любых предыдущих инсектицидных белков.
[00102] В некоторых вариантах осуществления трансгенное растение по настоящему изобретению может содержать и/или экспрессировать по меньшей мере второе пестицидное средство, которое отличается от молекул белковой природы. В некоторых вариантах осуществления второе пестицидное средство может присутствовать на поверхности растения, например, в качестве средства для местного применения. В предпочтительных вариантах осуществления второе пестицидное средство представляет собой молекулу интерферирующей РНК. Как правило, интерферирующая РНК содержит по меньшей мере фрагмент РНК, соответствующий целевому гену, спейсерную последовательность и второй фрагмент РНК, который комплементарен первому, за счет чего может образовываться двухнитевая структура РНК. РНК-интерференция (RNAi) происходит, когда организм распознает молекулы двухнитевой РНК (dsRNA) и гидролизует их. Полученные продукты гидролиза представляют собой небольшие фрагменты РНК длиной приблизительно 19-24 нуклеотидов, называемые малыми интерферирующими РНК (siRNA). Затем siRNA распространяются или разносятся по всему организму, в том числе через клеточные мембраны, где они гибридизируются с mRNA (или другими РНК) и вызывают гидролиз РНК. Интерферирующие РНК распознаются комплексом сайленсинга РНК-интерференции (RISC), в который вводится эффекторная нить (или "направляющая нить") РНК. Данная направляющая нить действует как матрица для распознавания и разрушения дуплексных последовательностей. Данный процесс повторяется каждый раз, когда siRNA гибридизируется с комплементарной ей целевой РНК, эффективно предотвращая трансляцию таких mRNA, и таким образом "сайленсингу" подвергается экспрессия специфических генов, с которых mRNA были транскрибированы. Интерферирующие РНК известны из уровня техники как применяемые для контроля насекомых (см., например, публикацию WO 2013/192256, включенную в данный документ посредством ссылки). Интерферирующая РНК, предназначенная для применения в контроле насекомых, обеспечивает получение не встречающейся в природе двухнитевой РНК, которая пользуется нативными путями RNAi у насекомого для запуска понижающей регуляции целевых генов, что может приводить к прекращению питания и/или роста и может приводить к гибели насекомого-вредителя. Молекула интерферирующей РНК может придавать устойчивость к насекомым против того же целевого вредителя, что и белок по настоящему изобретению, или может целенаправленно воздействовать на другого вредителя. Целевое насекомое-вредитель растения может питаться путем жевания, сосания или прокалывания. Интерферирующие РНК известны из уровня техники как применимые для контроля насекомых. В вариантах осуществления dsRNA, пригодная для контроля насекомых, описана в публикациях WO №№ WO2018/026770, WO 2018/026773 и WO 2018/026774, включенных в данный документ посредством ссылки. В вариантах осуществления dsRNA, пригодная для контроля насекомых, описана в патентах США №№92388223, 9340797 или 8946510, включенных в данный документ посредством ссылки. В вариантах осуществления dsRNA, пригодная для контроля насекомых, описана в заявках на патент США №№12/868994, 13/831230, 14/207313 или 14/207318, включенных в данный документ посредством ссылки. В других вариантах осуществления интерферирующая РНК может придавать устойчивость против вредителя растений, отличного от насекомого, такого как нематода-вредитель или вирус-вредитель.
[00103] Совместная экспрессия более чем одного пестицидного средства в одном трансгенном растении может быть достигнута путем получения одиночного рекомбинантного вектора, содержащего кодирующие последовательности более чем одного пестицидного средства в так называемом молекулярном стеке, и генетического конструирования растения, которое бы содержало и экспрессировало все эти пестицидные средства в трансгенном растении. Такие молекулярные стеки также могут быть получены с применением мини-хромосом, которые описаны например в патенте США 7235716. Как альтернатива, трансгенное растение, содержащее одну нуклеиновую кислоту, кодирующую первое пестицидное средство, может быть повторно трансформировано с помощью другой нуклеиновой кислоты, кодирующей второе пестицидное средство и т.д. Как альтернатива, растение, родитель 1, может быть получено с помощью способов генной инженерии для экспрессии генов по настоящему изобретению. Второе растение, родитель 2, может быть получено с помощью способов генной инженерии для экспрессии второго пестицидного средства. Путем скрещивания родителя 1 с родителем 2 получают растения-потомки, которые экспрессируют все гены, введенные в родителей 1 и 2.
[00104] Трансгенные растения или семена, содержащие и/или экспрессирующие инсектицидный белок по настоящему изобретению, также могут быть обработаны инсектицидом или инсектицидным покрытием для семян, которые описаны в патентах США №№5849320 и 5876739, включенных в данный документ посредством ссылки. В вариантах осуществления, в которых как инсектицид или инсектицидное покрытие для семян, так и трансгенное растение или семя по настоящему изобретению являются активными против одного и того же целевого насекомого, например, жесткокрылого вредителя или целевого вредителя Diabrotica, данная комбинация является применимой (i) в способе дополнительного усиления активности композиции по настоящему изобретению против целевого насекомого и/или (ii) в способе предупреждения развития устойчивости к композиции по настоящему изобретению посредством обеспечения еще одного механизма действия против целевого насекомого. Таким образом, в вариантах осуществления настоящее изобретение предусматривает способ усиления контроля популяции насекомых Diabrotica, включающий обеспечение трансгенного растения или семени по настоящему изобретению и применение по отношению к растению или семени инсектицида или инсектицидного покрытия для семян, предназначенного для трансгенного растения или семени по настоящему изобретению.
[00105] Даже в случае, если инсектицид или инсектицидное покрытие для семян являются активными против другого насекомого, инсектицид или инсектицидное покрытие для семян являются пригодными для расширения диапазона контроля насекомых, например, посредством добавления инсектицида или инсектицидного покрытия для семян, которые характеризуются активностью против чешуекрылых насекомых, к трансгенному семени по настоящему изобретению, которое в некоторых вариантах осуществления характеризуется активностью против жесткокрылых и некоторых чешуекрылых насекомых, при этом полученное покрытое трансгенное семя обеспечивает контроль как чешуекрылых, так и жесткокрылых насекомых-вредителей.
[00106] Примеры таких инсектицидов и/или инсектицидных покрытий для семян включают без ограничения карбамат, пиретроид, фосфорорганическое соединение, фрипрол, неоникотиноид, хлорорганическое соединение, нереистоксин или их комбинацию. В другом варианте осуществления инсектицид или инсектицидное покрытие для семян выбраны из группы, состоящей из карбофурана, карбарила, метомила, бифентрина, тефлутрина, перметрина, цифлутрина, лямбда-цигалотрина, циперметрина, дельтаметрина, хлорпирифоса, хлорэтоксифоса, диметоата, этопрофоса, малатиона, метил-паратиона, фората, тербуфоса, тебупиримифоса, фипронила, ацетамиприда, имидаклоприда, тиаклоприда, тиаметоксама, эндосульфана и их комбинации. Коммерческие продукты, содержащие такие инсектициды и инсектицидные покрытия для семян, включают без ограничения Furadan® (карбофуран), Lanate® (мезомил, метомил, месомил), Sevin® (карбарил), Talstar® (бифентрин), Force® (тефлутрин), Ammo® (киперметрин), Cymbush® (циперметрин), Delta Gold® (дельтаметрин), Karate® (лямбда-цигалотрин), Ambush® (перметрин), Pounce® (перметрин), Brigade® (бифентрин), Capture® (бифентрин), ProShield® (тефлутрин), Warrior® (лямбда-цигалотрин), Dursban® (хлорпирифос), Fortress® (хорэтоксифос), Мосар® (этопроп), Thimet® (форат), AAstar® (форат, флуцитинат), Rampart® (форат), Counter® (тербуфос), Cygon® (диметоат), Dicapthon, Regent® (фипронил), Cruiser® (тиаметоксам), Gaucho® (имидаклоприд), Prescribe® (имидаклоприд), Poncho® (клотианидин) и Aztec® (цифлутрин, тебупиримфос).
[00107] Настоящее изобретение также охватывает композицию, содержащую эффективное для контроля насекомых количество инсектицидного белка в соответствии с настоящим изобретением. В дополнительных вариантах осуществления композиция содержит подходящий сельскохозяйственный носитель и полипептид по настоящему изобретению с инсектицидной активностью. Сельскохозяйственный носитель может предусматривать вспомогательные вещества, вещества, облегающие смешивание, усиливающие средства и т.д., целесообразные для применения активного ингредиента, такого как полипептид по настоящему изобретению, в том числе полипептид, содержащий аминокислотную последовательность, которая на по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или 100% идентична любой из SEQ ID NO: 39-74. Подходящие носители не должны быть фитотоксичными по отношению к ценным культурам, в частности при концентрациях, используемых для применения композиций в присутствии культур, и не должны вступать в химическую реакцию с соединениями активного ингредиента, описанного в данном документе, а именно, полипептидом по настоящему изобретению или другими ингредиентами композиции. Такие смеси могут быть разработаны для непосредственного применения по отношению к культурам или могут представлять собой концентраты или составы, которые обычно разбавляют дополнительными носителями и вспомогательными средствами перед применением. Они могут включать инертные или активные компоненты и могут представлять собой твердые вещества, такие как, например, дусты, порошки, гранулы, диспергируемые в воде гранулы или смачиваемые порошки, или жидкостями, такими как, например, эмульгируемые концентраты, растворы, эмульсии или суспензии. Подходящие сельскохозяйственные носители могут включать жидкие носители, например, воду, толуол, ксилол, лигроин, растительное масло, ацетон, метилэтилкетон, циклогексанон, трихлорэтилен, перхлорэтилен, этилацетат, амилацетат, бутилацетат, монометиловый эфир пропиленгликоля и монометиловый эфир диэтиленгликоля, метанол, этанол, изопропанол, амиловый спирт, этиленгликоль, пропиленгликоль, глицерин и т.п. Как правило, лучшим выбором в отношении носителя для разбавления концентратов является вода. Подходящие твердые носители могут включать тальк, пирофиллитовую глину, кремнезем, аттапульгитовую глину, кизельгур, мел, диатомовую землю, известь, карбонат кальция, бентонитовую глину, фуллерову землю, шелуху семян хлопчатника, пшеничную муку, соевую муку, пемзу, древесную муку, муку из скорлупы грецкого ореха, лигнин и т.п.В другом варианте осуществления полипептид по настоящему изобретению может быть инкапсулирован в синтетической матрице, такой как полимер, и нанесен на поверхность хозяина, такого как растение. Поглощение насекомым клеток-хозяев обеспечивает доставку в насекомое средств для контроля насекомых и приводит к токсическому эффекту у насекомого-вредителя.
[00108] В дополнительных вариантах осуществления композиция по настоящему изобретению может представлять собой порошок, дуст, пеллету, гранулу, жидкость для распыления, эмульсию, коллоидное вещество или раствор. Композицию по настоящему изобретению можно получать посредством обезвоживания, лиофилизации, гомогенизации, экстракции, фильтрации, центрифугирования, осаждения или концентрирования культуры бактериальных клеток. Композиция по настоящему изобретению может содержать по меньшей мере 1%, по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97% или по меньшей мере 99% по весу полипептида по настоящему изобретению.
[00109] В вариантах осуществления композиция по настоящему изобретению может содержать по меньшей мере второе пестицидное средство (например, которое может быть экспрессировано трансгенным путем в растении и/или включено в композицию), которое может являться инсектицидным, нематоцидным, фунгицидным или бактерицидным. По меньшей мере второе пестицидное средство может быть инсектицидным по отношению к тому же насекомому, что и полипептид по настоящему изобретению, или к другому насекомому. Второе пестицидное средство может представлять собой полипептид. Пестицидное средство может представлять собой интерферирующую РНК (например, dsRNA). Второе пестицидное средство может представлять собой микроорганизм, такой как бактерии, который содержит молекулу нуклеиновой кислоты, кодирующую пестицидное средство, и/или содержит пестицидное средство, такое как полипептид или интерферирующая РНК. Микроорганизм может быть аттенуированным, инактивированным нагреванием или лиофилизированным. Микроорганизм может быть мертвым или не способным к воспроизведению. Второе пестицидное средство может представлять собой инсектицид, например, карбофуран, карбарил, метомил, бифентрин, тефлутрин, перметрин, цифлутрин, лямбда-цигалотрин, циперметрин, дельтаметрин, хлорпирифос, хлорэтоксифос, клотианидин, диметоат, этопрофос, малатион, метилпаратион, форат, тербуфос, тебупиримифос, фипронил, ацетамиприд, имидаклоприд, тиаклоприд, тиаметоксам, эндосульфан, бенсултап или их комбинацию или коммерческий продукт, содержащий такие инсектициды и инсектицидные покрытия для семян, описанные выше.
[00110] Композиция по настоящему изобретению, например композиция, содержащая полипептид по настоящему изобретению и приемлемый с точки зрения сельского хозяйства носитель, может применяться в традиционных агротехнических способах. Приемлемый с точки зрения сельского хозяйства носитель представляет собой состав, применимый для нанесения на растение или семя композиции, содержащей полипептид по настоящему изобретению. Например, композиции по настоящему изобретению можно смешивать с водой и/или удобрениями и можно применять в отношении требуемого места произрастания/обитания до появления всходов и/или после появления всходов любым способом, как например, с помощью распылительных баков самолетов, оросительного оборудования, распылительного оборудования непосредственного впрыскивания, ранцевых распылительных баки, бочек для купания скота, фермерского оборудования, применяемого в наземном распылении (например, штанговые распылители, ручные распылители) и т.п. Требуемое место произрастания/обитания может представлять собой почву, растения и т.п.
[00111] Композицию по настоящему изобретению можно применять в отношении семени или части растения для вегетативного размножения в любом физиологическом состоянии, в любое время между сбором семян и посевом семян; во время или после посева и/или после прорастания. Предпочтительно, чтобы семя или часть растения для вегетативного размножения находились в достаточно устойчивом состоянии, чтобы в процессе обработки они не подвергались повреждению или подвергались минимальному повреждению, включая физическое повреждение или биологическое повреждение. Состав может применяться по отношению к семенам или частям растения для вегетативного размножения с применением традиционных методик и устройств для нанесения покрытия, таких как методики псевдоожиженного слоя, способ вальцовой мельницы, ротостатические протравливатели семян и барабаны для нанесения покрытий.
[00112] Настоящее изобретение также включает способ контроля популяции чешуекрылых и/или жесткокрылых вредителей, включающий приведение указанной популяции в контакт с эффективным для контроля насекомых количеством полипептида по настоящему изобретению с инсектицидной активностью, где полипептид на по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или на 100% идентичен SEQ ID NO: 39-74. Приведение в контакт предусматривает представителей популяции вредителей, питающихся или поглощающих полипептид. Полипептид может быть введен в рацион насекомого или может экспрессироваться в растительной ткани, которую насекомое затем поглощает, или присутствовать на ней. В дополнительных вариантах осуществления осуществление контроля популяций чешуекрылых и/или жесткокрылых вредителей включает уничтожение насекомых путем приведения насекомых в контакт с эффективным для контроля насекомых количеством полипептида по настоящему изобретению.
[00113] Настоящее изобретение также предусматривает способ защиты растения от насекомого-вредителя, включающий экспрессирование в растении или растительной клетке нуклеотидной последовательности или кассеты экспрессии, которая кодирует инсектицидный полипептид по настоящему изобретению. В вариантах осуществления нуклеотидная последовательность на по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или на 100% идентична нуклеотидной последовательности под SEQ ID NO: 2-36 или кодирует полипептид, содержащий аминокислотную последовательность, которая на по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или на 100% идентична SEQ ID NO: 39-74. В дополнительных вариантах осуществления растение или растительная клетка продуцируют инсектицидный полипептид с инсектицидной активностью против чешуекрылого и/или жесткокрылого вредителя.
[00114] Настоящее изобретение также предусматривает способ повышения урожайности у растения, включающий выращивание в поле растения или его семени, содержащих стабильно встроенную в его геном молекулу нуклеиновой кислоты кассеты экспрессии по настоящему изобретению, и где указанное поле инфицировано вредителем, против которого указанный полипептид характеризуется инсектицидной активностью.
[00115] После того как требуемая нуклеиновая кислота трансформирована в конкретный вид растения, она может распространяться в этом виде или переходить в другие сорта этого же вида с помощью традиционных технологий разведения, в особенности включая коммерчески важные сорта.
[00116] В вариантах осуществления нуклеиновая кислота по настоящему изобретению экспрессируется в трансгенных растениях, обуславливая таким образом биосинтез соответствующего инсектицидного белка в трансгенных растениях. Таким путем создаются трансгенные растения с повышенной устойчивостью к насекомым, в частности кукурузному корневому жуку. Для их экспрессии в трансгенных растениях нуклеиновые кислоты по настоящему изобретению необязательно могут быть модифицированы и оптимизированы. Хотя во многих случаях гены из микробных организмов могут экспрессироваться в растениях на высоких уровнях без модификации, в трансгенных растениях может происходить низкая экспрессия микробных нуклеиновых кислот, содержащих кодоны, которые не являются предпочтительными в растениях. Из уровня техники известно, что все организмы имеют специфические предпочтения в отношении использования кодонов, и при этом кодоны нуклеиновых кислот, описанных в настоящем изобретении, могут быть изменены в соответствии с характерными для растений предпочтениями, с сохранением в то же время кодируемых ими аминокислот. Кроме того, высокий уровень экспрессии в растениях лучше всего достигается в случае кодирующих последовательностей с содержанием GC, составляющим по меньшей мере приблизительно 35%, предпочтительно более чем приблизительно 45%, более предпочтительно более чем приблизительно 50% и наиболее предпочтительно более чем приблизительно 60%. Микробные нуклеиновые кислоты, которые характеризуются низкими содержаниями GC, могут экспрессироваться в растениях на недостаточном уровне вследствие наличия мотивов АТТТА, которые могут дестабилизировать транскрипты, и мотивов ААТААА, которые могут вызывать ненадлежащее полиаденилирование. В вариантах осуществления последовательности могут быть модифицированы, принимая во внимание конкретные предпочтения в отношении кодонов и предпочтения в отношении содержания GC однодольных или двудольных, поскольку было показано, что эти предпочтения отличаются (Murray et al. Nucl. Acids Res. 17:477-498 (1989)). Кроме того, осуществляется скрининг нуклеиновых кислот на наличие нестандартных сайтов сплайсинга, которые могут вызвать усечение транскрипта. Все изменения, которые требуется выполнить внутри нуклеиновых кислот, такие как описанные выше, могут быть выполнены с применением широко известных методик сайт-направленного мутагенеза, ПЦР и конструирования синтетических генов, например, с применением способов, описанных в опубликованных патентных заявках ЕР 0385962, ЕР 0359472 и WO 93/07278.
[00117] В одном варианте осуществления настоящего изобретения кодирующая последовательность инсектицидного белка по настоящему изобретению получена в соответствии с процедурой, раскрытой в патенте США 5625136, включенном в данный документ посредством ссылки. В этой процедуре применяют предпочтительные для маиса кодоны, т.е. один кодон, который чаще всего кодирует данную аминокислоту в маисе. Предпочтительный для маиса кодон для конкретной аминокислоты может быть получен, например, из известной генной последовательности маиса. Частота использования кодонов для маиса для 28 генов растений маиса встречается в Murray et al., Nucleic Acids Research 17:477-498 (1989), раскрытие которого включено в данный документ посредством ссылки.
[00118] Подобным образом нуклеотидные последовательности могут быть оптимизированы для экспрессии в любом растении. Понятно, что вся последовательность гена или любая ее часть могут быть оптимизированными или синтетическими. То есть также могут применяться синтетические или частично оптимизированные последовательности.
[00119] Для более эффективной инициации трансляции можно модифицировать последовательности, прилегающие к инициирующему метионину. Например, они могут быть модифицированы путем включения последовательностей, которые, как известно, являются эффективными в растениях. Joshi предложил подходящую консенсусную последовательность для растений (NAR 15:6643-6653 (1987)), a Clontech предлагает дополнительную консенсусную последовательность, являющуюся инициатором трансляции (каталог 1993/1994, стр. 210). Эти консенсусные последовательности пригодны для использования с нуклеиновыми кислотами по настоящему изобретению. В вариантах осуществления последовательности встроены в конструкции, содержащие нуклеиновые кислоты, вплоть до и включая ATG (оставляя при этом вторую аминокислоту немодифицированной) или в альтернативном варианте вплоть до и включая GTC, следующую за ATG (с возможностью модифицирования второй аминокислоты данного трансгена).
[00120] Экспрессия нуклеиновых кислот в трансгенных растениях управляется промоторами, которые функционируют в растениях. Выбор промотора будет изменяться в зависимости от временных и пространственных требований для экспрессии, а также в зависимости от целевого вида. Таким образом, предпочтительной является экспрессия нуклеиновых кислот по настоящему изобретению в листьях, стеблях или стволах, в початках, в соцветиях (например, в шипах, метелках, косточках и т.п.), в корнях и/или саженцах. Однако, во многих случаях требуется защита от более чем одного типа насекомого-вредителя, а следовательно желательной является экспрессия в нескольких тканях. Хотя, как было показано, многие промоторы из двудольных растений функционируют у однодольных, и наоборот, в идеальном случае для экспрессии в двудольных растениях выбирают промоторы двудольных, а для экспрессии в однодольных растениях выбирают промоторы однодольных. Однако не существует каких-либо ограничений относительно происхождения выбранных промоторов; достаточно, чтобы они эффективно управляли экспрессией нуклеиновых кислот в требуемой клетке.
[00121] В одном варианте осуществления применяются промоторы с конститутивной экспрессией, в том числе промоторы генов актина или убиквитина, или промоторы СМР, или промоторы CaMV 35S и 19S. Нуклеиновые кислоты по настоящему изобретению также могут экспрессироваться под контролем промоторов, которые регулируются химическим путем. Предпочтительная технология для химической индукции экспрессии генов подробно изложена в опубликованной заявке ЕР 0332104 (Ciba-Geigy) и патенте США №5614395. Предпочтительным промотором для химической индукции является промотор табака PR-1а.
[00122] В другом варианте осуществления может применяться категория промоторов, индуцируемых повреждением. Были описаны многочисленные промоторы, которые экспрессируются в участках ранения, а также в участках инфицирования фитопатогеном. Оптимально, такой промотор должен быть активным только локально в местах инфекции, и таким образом инсектицидные белки по настоящему изобретению накапливаются исключительно в клетках, в которых необходимо синтезировать белки для уничтожения инвазивных насекомых-вредителей. Предпочтительные промоторы этого типа включают описанные Stanford et al. Mol. Gen. Genet. 215:200-208 (1989), Xu et al. Plant Molec. Biol. 22:573-588 (1993), Logemann et al. Plant Cell 1:151-158 (1989), Rohrmeier & Lehle, Plant Molec. Biol. 22:783-792 (1993), Firek etal. Plant Molec. Biol. 22:129-142 (1993) и Warner et al. Plant J. 3:191-201 (1993).
[00123] Тканеспецифичными или предпочтительными для ткани промоторами, применимыми для экспрессии генов, кодирующих инсектицидные белки по настоящему изобретению в растениях, в частности кукурузе, являются промоторы, которые управляют экспрессией в корне, сердцевине, листе или пыльце, в частности в корне. Такие промоторы, например выделенные из РЕРС или trpA, раскрыты в патенте США №5625136, или MTL, раскрытого в патенте США №5466785. Оба патента США включены в данный документ посредством ссылки в полном своем объеме.
[00124] Кроме того, могут применяться промоторы, функционирующие в пластидах. Неограничивающие примеры таких промоторов включают промотор 5'-UTR гена 9 бактериофага Т3 и другие промоторы, раскрытые в патенте США №7579516. Другие промоторы, пригодные в соответствии с настоящим изобретением, включают без ограничения промотор малой субъединицы S-E9 RuBP-карбоксилазы и промотор гена ингибитора трипсина Кунитца (Kti3).
[00125] В дополнительных аспектах нуклеотидные последовательности по настоящему изобретению могут быть функционально связанны с промотором, индуцируемым нанесением повреждения или индуцируемым инфицированием вредителями или патогенами (например, насекомым- или нематодой-вредителем растения). Были описаны многочисленные промоторы, которые обеспечивают экспрессию в местах повреждения и/или в участках нападения вредителей (например, в местах питания насекомых/нематод) или фитопатогенной инфекции. Оптимально, такой промотор должен быть активным только локально в участках нападения или рядом с ними, и таким образом экспрессия нуклеотидных последовательностей по настоящему изобретению будет сосредоточена на клетках, которые подвергаются внедрению в них или поеданию их. Такие промоторы включают без ограничения таковые, описанные в Stanford et al., Mol. Gen. Genet. 215:200-208 (1989), Xu et al. Plant Molec. Biol. 22:573-588 (1993), Logemann et al. Plant Cell 1:151-158 (1989), Rohrmeier and Lehle, Plant Molec. Biol. 22:783-792 (1993), Firek et al. Plant Molec. Biol. 22:129-142 (1993), Warner et al. Plant J. 3:191-201 (1993), патенте США №5750386, патенте США №5955646, патенте США№6262344, патенте США №6395963, патенте США №6703541, патенте США№7078589, патенте США №7196247, патенте США №7223901 и публикации заявки на выдачу патента США 2010043102.
[00126] В некоторых вариантах осуществления настоящего изобретения применяют "минимальный промотор" или "основной промотор". Минимальный промотор может привлекать и связывать комплекс РНК-полимеразы II и ее вспомогательных белков для обеспечения инициации транскрипции и элонгации. В некоторых вариантах осуществления минимальный промотор конструируют таким образом, что он содержит только нуклеотиды/нуклеотидные последовательности из выбранного промотора, которые требуются для связывания транскрипционных факторов и транскрипции представляющей интерес нуклеотидной последовательности, которая функционально связана с минимальным промотором, включая без ограничения последовательности ТАТА-бокса. В других вариантах осуществления в минимальном промоторе отсутствуют действующие в цис-положении последовательности, которые привлекают и связывают транскрипционные факторы, модулирующие транскрипцию (например, усиливают, подавляют, придают тканеспецифичность, придают способность к индуцированию или способность к подавлению). Как правило, минимальный промотор помещают выше (т.е. в направлении 5') нуклеотидной последовательности, подлежащей экспрессии. Следовательно, для применения в качестве минимального промотора могут быть выбраны нуклеотиды/нуклеотидные последовательности из любого промотора, применимого в соответствии с настоящим изобретением.
[00127] В описанные в настоящем изобретении кассеты экспрессии могут быть встроены многие другие последовательности. Они предусматривают последовательности, которые, как показано, усиливают экспрессию, как например, интронные последовательности (например, из Adhl и bronzel) и вирусные лидерные последовательности (например, из TMV, MCMV и AMV).
[00128] Может быть предпочтительным направлять экспрессию нуклеиновых кислот по настоящему изобретению в различные клеточные локализации в растении. В некоторых случаях может быть желательна локализация в цитозоле, тогда как в других случаях предпочтительной может быть локализация в определенной субклеточной органелле. Субклеточная локализация ферментов, кодируемых трансгеном, выполняется с помощью методик, хорошо известных из уровня техники. Как правило, ДНК, кодирующую целевой пептид из известного продукта гена, направленного на органеллу, обрабатывают и подвергают слиянию выше нуклеиновой кислоты. Большинство таких целевых последовательностей известны для хлоропласта, и было показано их функционирование в гетерологичных конструкциях. Экспрессия нуклеиновых кислот по настоящему изобретению также направлена в эндоплазматический ретикулум или в вакуоли клеток-хозяев. Методики для достижения этого хорошо известны из уровня техники.
[00129] Векторы, подходящие для трансформации растений, широко известны из уровня техники. Для опосредованной Agrobacterium трансформации пригодны бинарные векторы или векторы, несущие по меньшей мере одну Т-ДНК граничную последовательность, тогда как для прямого переноса генов пригоден любой вектор, и предпочтительной может быть линейная ДНК, содержащая только представляющую интерес конструкцию. В случае прямого переноса генов может применяться трансформация с помощью одного вида ДНК или совместная трансформация (Schocher et al. Biotechnology 4:1093-1096 (1986)). Как в случае прямого переноса генов, так и в случае опосредованного Agrobacterium переноса, трансформацию обычно (но не обязательно) осуществляют с селектируемым маркером, который может обеспечивать устойчивость к антибиотику (канамицину, гигромицину или метотрексату) или гербициду (баста). Векторы трансформации растений, содержащие молекулы нуклеиновой кислоты по настоящему изобретению, могут также содержать гены (например, изомеразу фосфоманнозы; PMI), обеспечивающие положительную селекцию трансгенных растений, как описано в патентах США 5767378 и 5994629, включенных в данный документ посредством ссылки. Однако, выбор селективного маркера не является определяющим для настоящего изобретения.
[00130] В вариантах осуществления нуклеиновая кислота может быть трансформирована в ядерный геном. В другом варианте осуществления нуклеиновая кислота по настоящему изобретению непосредственно трансформируется в геном пластид. Большим преимуществом трансформации пластид является то, что пластиды обычно способны к экспрессии бактериальных генов без существенной оптимизации кодонов, и при этом пластиды способны к экспрессии множественных открытых рамок считывания под контролем одного промотора. Технология трансформации пластид подробно описана в патентах США №№5451513, 5545817 и 5545818, в заявке согласно РСТ №WO 95/16783 и в McBride et al. (1994) Proc. Nati. Acad. Sci. USA 91, 7301-7305. Основная методика трансформации хлоропластов предусматривает введение участков клонированной пластидной ДНК, фланкирующих селектируемый маркер, вместе с представляющим интерес геном в подходящую целевую ткань, например, с применением биолистики или трансформации протопласта (например, трансформации, опосредованной хлоридом кальция или PEG). Фланкирующие участки размером 1-1,5 т.о., называемые нацеливающими последовательностями, содействуют гомологичной рекомбинации с геномом пластид и, таким образом, обеспечивают возможность замещения или модификации специфических участков пластома. Изначально в качестве селектируемых маркеров для трансформации используются точечные мутации в 16S rRNA и генах rps12 хлоропластов, придающие устойчивость к спектиномицину и/или стрептомицину (Svab, Z., Hajdukiewicz, P., and Maliga, P. (1990) Proc. Nati. Acad. Sci. USA 87, 8526-8530; Staub, J. M., и Maliga, P. (1992) Plant Cell 4, 39-45). Это приводило к получению стабильных гомоплазмических трансформантов с частотой, составляющей приблизительно одна трансформация на 100 бомбардировок целевых листьев. Присутствие сайтов клонирования между данными маркерами обеспечило возможность создания нацеливающегося на пластиды вектора для введения чужеродных генов (Staub, J. М., and Maliga, P. (1993) EMBO J. 12, 601-606). Существенного повышения частоты трансформации достигали путем замещения рецессивных генов rRNA или r-белка, обеспечивающих устойчивость к антибиотикам, на доминантный селектируемый маркер, бактериальный ген aadA, кодирующий фермент аминогликозид-3'-аденилтрансферазу, обезвреживающий спектиномицин (Svab, Z., and Maliga, P. (1993) Proc. Natl. Acad. Sci. USA 90, 913-917). Ранее данный маркер успешно применялся для трансформации пластидного генома зеленой водоросли Chlamydomonas reinhardtii с высокой частотой (Goldschmidt-Clermont, М. (1991) Nucl. Acids Res. 19:4083-4089). Другие селектируемые маркеры, применимые для трансформации пластид, известны из уровня техники и включены в объем настоящего изобретения. Как правило, для достижения гомопластидного состояния требуется около 15-20 циклов клеточного деления после трансформации. Экспрессия в пластидах, при которой за счет гомологической рекомбинации гены встроены во все несколько тысяч копий кольцевого генома пластид, присутствующих в каждой растительной клетке, использует преимущество огромного числа копий в сравнении с генами, экспрессируемыми в ядре, с обеспечением уровней экспрессии, которые легко могут превышать 10% от общего количества растворимого растительного белка. В предпочтительном варианте осуществления нуклеиновую кислоту по настоящему изобретению вводят в направленный на пластиды вектор, и при этом она трансформируется в геном пластид требуемого растения-хозяина. Получают растения, гомопластические в отношении геномов пластид, содержащие нуклеиновую кислоту по настоящему изобретению, и при этом они предпочтительно способны к высокой экспрессии нуклеиновой кислоты.
ПРИМЕРЫ
[00131] Настоящее изобретение далее будет описано посредством ссылки на следующие подробные примеры. Эти примеры предоставлены только для целей иллюстрации и не предназначены для ограничения, если не указано иное. Стандартные методики с использованием рекомбинантной ДНК и молекулярного клонирования, применяемые в данном документе, хорошо известны из уровня техники и описаны J. Sambrook, et al., Molecular Cloning: A Laboratory Manual, 3d Ed., Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press (2001); by T.J. Silhavy, M.L. Berman, and L.W. Enquist, Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1984) и Ausubel, F.M. et al., Current Protocols in Molecular Biology, New York, John Wiley and Sons Inc., (1988), Reiter, et al., Methods in Arabidopsis Research, World Scientific Press (1992), и Schultz et al., Plant Molecular Biology Manual, Kluwer Academic Publishers (1998).
Пример 1. Идентификация белка с инсектицидной активностью против западного кукурузного корневого жука
[00132] Инсектицидный белок (SEQ ID NO: 39) идентифицировали из Nitrococcus mobilis. Синтезировали оптимизированный вариант этого гена Е. coli (SEQ ID NO: 1), и ген клонировали в вектор рЕТ29а с образованием конструкции p(Nitromob). Конструкцией p(Nitromob) трансформировали Е. coli BL21* (DE3), и экспрессию белка осуществляли в бульоне Лурия-Бертани с индукцией с помощью IPTG при 18°С в течение ночи. Растворимую фракцию лизатов получали из этих культур с применением пресса Френча с последующим центрифугированием лизатов целых клеток при 20000 х g в течение тридцати минут. Супернатант (растворимую фракцию) затем тестировали на биоактивность по отношению к западному кукурузному корневому жуку (WCR; Diabrotica virgifera).
[00133] Анализы биоактивности осуществляли с применением способа включения в рацион. Вкратце, лизаты Е. coli BL21*(DE3) смешивали с равным объемом нагретого искусственного рациона для насекомых (Bioserv, Inc., Френчтаун, Нью-Джерси) в центрифужных пробирках объемом 1,5 мл, а затем вносили в небольшие чашки Петри. После того как смесь образца и питательной среды охлаждали и обеспечивали возможность отвердевания, в каждый планшет добавляли 12 личинок WCR. Планшеты запечатывали и выдерживали в окружающих условиях лаборатории с учетом температуры, освещения и относительной влажности. Лизаты культур Е. coli BL21* (DE3), несущих пустой вектор рЕТ29а, применяли в качестве отрицательных контролей. Смертность оценивали на 4-й день и 7-й день или необязательно на 3-й день и 6-й день. В этой и всех последующих таблицах, в которых показана инсектицидная активность в отношении CRW, аббревиатуры в столбце "Примечания" расшифровываются следующим образом: s = небольшие личинки, sm = небольшие/средние личинки, m = средние личинки, mb = средние/большие личинки, b = большие личинки, vb = очень большие личинки. В этой и всех последующих таблицах показана инсектицидная активность NitromobCRW или его варианта, "SEQ ID NO." относится к аминокислотной последовательности белка. Как показано в таблице 1, лизат культуры, экспрессирующей p(Nitromob), демонстрировал сильную биоактивность против WCR. Белок N. mobilis был переименован в NitromobCRW.

Пример 2. Варианты NitromobCRW обладают инсектицидной активностью против WCR
[00134] В NitromobCRW вводили мутации и анализировали стабильность белка и инсектицидную активность бактериальных лизатов, содержащих мутантный вариант NitromobCRW. Мутации включают изменения различных аминокислотных остатков, а также вставку остатков лейцина. Эти мутации вводили для определения того, можно ли разработать мутантный вариант NitromobCRW, который сохранял бы инсектицидную активность, но был бы перевариваемым в анализе с применением искусственного желудочного сока (SGF). Такой вариант NitromobCRW может иметь коммерческое значение, например, для придания инсектицидных свойств растению посредством трансгенной экспрессии в растении.
[00135] Инсектицидную активность определяли с помощью анализов с включением в рацион, осуществляемых фактически как описано в примере 1, с применением 12 личинок WCR в каждом экспериментальном анализе. Результаты показаны в таблицах 2-6. SEQ ID NO соответствуют аминокислотной последовательности варианта. Обработки, показанные в таблицах 5 и 6, также указывают на разбавление применяемого бактериального лизата. Все мутантные варианты NitromobCRW демонстрируют инсектицидную активность к 6-му или 7-му дню.

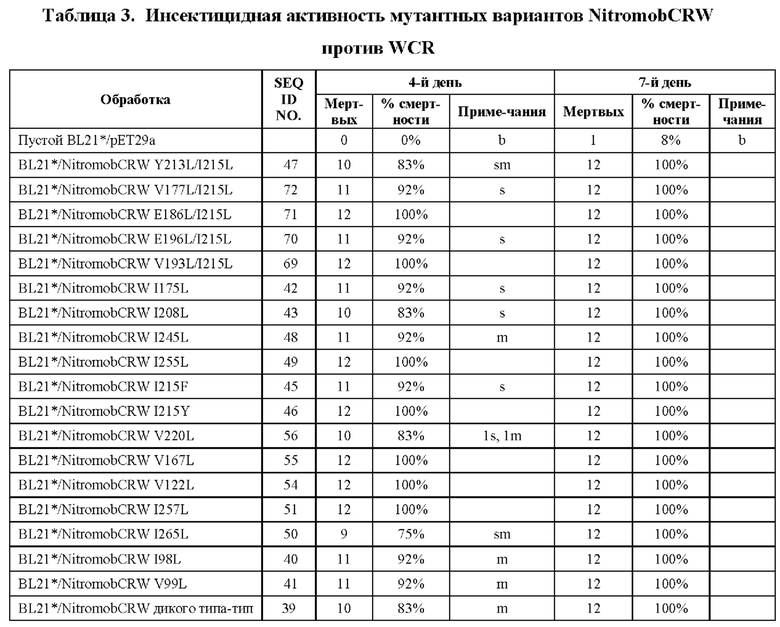
Пример 3. Тестирование воздействия искусственного желудочного сока на препараты на основе лизатов Е. coli
[00136] В данном примере описывается анализ, осуществляемый для определения перевариваемости под действием SGF. Каждый вариант белка NitromobCRW получали в штамме BL21* (DE3) Е. coli. Уровень экспрессии варианта в бактериальном штамме и растворимость варианта указаны в таблице 7. Бактериальные лизаты в 50 мМ фосфата калия, рН 7,0, 50 мМ хлорида натрия разбавляли до 3 мг/мл (концентрации общего белка) для анализа перевариваемости. Реакцию переваривания инициировали посредством добавления 15 мкл лизата к 285 мкл искусственного желудочного сока [10 единиц пепсина/мкг белка или примерно 1579 единиц пепсина/мл в растворе G-Con (2 мг/мл хлорида натрия, рН 1,2)] при 37°С. Через 5 минут отбирали 100 мкл реакционной смеси лизат-SGF и посредством добавления ее к 100 мкл предварительно нагретого (95°С) останавливающего раствора, содержащего 65% трицинового загрузочного буфера (Bio-rad, 2х трициновый загрузочный буфер с 10% β-меркаптоэтанолом) и 35% 500 мМ бикарбоната натрия, рН 11,0, реакцию останавливали. Образец в нулевой момент времени (Т0) получали посредством добавления 5 мкл тестируемого лизата к 100 мкл предварительно нагретого (95°С) останавливающего раствора и 95 мкл искусственного желудочного сока. Все образцы нагревали при 95°С в течение 5 минут, а затем хранили на льду до проведения анализа с помощью SDS-PAGE. Тридцать микролитров каждой реакционной смеси загружали в 10-20% трис-трициновый гель для пептидов перед проведением стандартного гель-электрофореза белков. Трис-трициновый гель фиксировали в течение 20 минут с помощью смеси 40% метанола и 10% уксусной кислоты непосредственно после электрофореза. Гель затем окрашивали с помощью красителя для белков GelCode Blue в течение 1 часа при комнатной температуре. Через 1 час полиакриламидный гель обесцвечивали с помощью дистиллированной воды в течение по меньшей мере 12 часов. Результаты качественной оценки показаны в таблице 7. "Неудовлетворительный" в случае теста Т5 означает, что интактный или частично переваренный вариант белка NitromobCRW поддавался выявлению с помощью красителя для белков GelCode Blue после гель-электрофореза, что указывает на то, что белок не был полностью перевариваемым в анализе с применением SGF. "Удовлетворительный" в случае теста Т5 означает, что интактный вариант белка NitromobCRW не поддавался выявлению, что указывает на то, что вариант белка NitromobCRW был перевариваемым в анализе с применением SGF. Также указана инсектицидная активность ("активный") варианта белка NitromobCRW, причем "да" указывает на инсектицидную активность, как показано в предыдущих примерах.
[00137] Их всех полученных вариантов NitromobCRW только NitromobCRW Y213L/I215L (SEQ ID NO: 47) убедительно прошел тест Т5 с анализом с применением SGF. NitromobCRW I215L (SEQ ID NO: 44) продемонстрировал лучшую перевариваемость, чем белок дикого типа, но данный вариант не прошел тест Т5. Варианты NitromobCRW 214-Leu-215, 215-Leu-216, G216A, G216L и L214S/I215L были нерастворимыми, а вариант NitrobCRW I265L характеризовался низкой растворимостью. Эти данные позволяют предположить, что домен, мотив, или укладка в данном участке белка критичны для функционирования белка и/или стабильности белка.
Пример 4. Очищенный вариант NitromobCRW Y213L/I215L обладает инсектицидной активностью против WCR
[00138] Данный вариант дополнительно характеризовали в отношении его инсектицидных свойств. Два литра клеток Е. coli BL21* (DE3), несущих рЕТ-NitromobCRW Y213L/I215L, выращивали при 37°С в среде LB. IPTG (1 мМ) добавляли к культурам при достижении O.D., составляющей 0,8-1,0, и затем культуры перемещали на 18°С в течение 18 часов. Осадок клеток собирали и ресуспендировали в 20 мМ Трис, рН 8,5, с 10% глицерина. Клетки лизировали с применением пресса Френча; лизат затем центрифугировали при 100k х g в ультрацентрифуге. Супернатант собирали и затем фильтровали перед загрузкой в колонку для анионообменной хроматографии HiPrepQ, которую предварительно уравновешивали с помощью 20 мМ Трис, рН 8,5, с 10% глицерина. Колонка HiPrepQ связывала NitromobCRW Y213L/I215L эффективно; белок элюировали с колонки с применением линейного градиента NaCl. Буфер с высокой концентрацией соли состоял из 20 мМ Трис, рН 8,5, 0,5 М NaCl с 10% глицерина. Наиболее чистые фракции объединяли и затем концентрировали до примерно 2 мл. Белок загружали в колонку для гель-фильтрации Sephadex 200, которую предварительно уравновешивали с помощью IX PBS. Фракции с колонки Sephadex 200 анализировали в отношении чистоты с помощью SDS-PAGE (NitromobCRW Y213L/I215L (SEQ ID NO: 47) характеризуется прогнозируемым молекулярным весом 32,1 кДа). Наиболее чистые фракции объединяли и затем концентрировали до 7,2 мг/мл перед хранением при -80°С. Чистый белок затем тестировали против 12 личинок WCR в диапазоне концентраций в способе включения в рацион, фактически как описано в примере 1. Как показано в таблице 8, NitromobCRW Y213L/I215L является эффективным против WCR; NitromobCRW Y213L/I215L при 50 мкг/мл вызывал по меньшей мере 75% смертность на 6-й день.
Пример 5. NitromobCRWCRW Y213L/I215L обладает инсектицидной активностью против устойчивых к Cry разновидностей западного кукурузного корневого жука
[00139] Для определения того, проявляется ли токсичность NitromobCRW Y213L/I215L (SEQ ID NO: 47) посредством механизма действия, отличающегося от белков, родственных Cry3, лизат NitromobCRW Y213L/I215L очищали как в примере 4 и тестировали в отношении эффективности против разновидности WCR, которая устойчива к токсину eCry3.1Ab (eCry3.1 Ab-R; см. таблицу 9), разновидности WCR, которая устойчива к модифицированному токсину Cry3A (mCry3A) (mCry3A-R; см. таблицу 10), и разновидности WCR, которая устойчива к токсину Cry3Bb (Cry3Bb-R; см. таблицу 11). Анализы с включением в рацион осуществляли в диапазоне белка NitromobCRW Y213L/I215L фактически как описано в примере 4 и смертность оценивали либо на 3-й день и 6-й день (таблица 9), либо на 2-й день и 7-й день (таблица 10). NitrobmobCRW Y213L/I215L тестировали дважды при ряде концентраций (мкг/мл), как указано в таблицах 9, 10 и 11. Отрицательный контроль содержал только IX PBS. Каждый анализ осуществляли с 12 личинками WCR. Как показано в таблицах 9, 10 и 11, NitromobCRW Y213L/I215L демонстрирует инсектицидную активность против устойчивых к Cry разновидностей WCR.

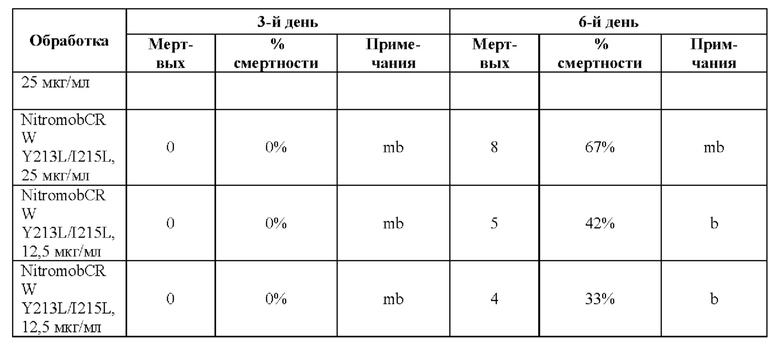
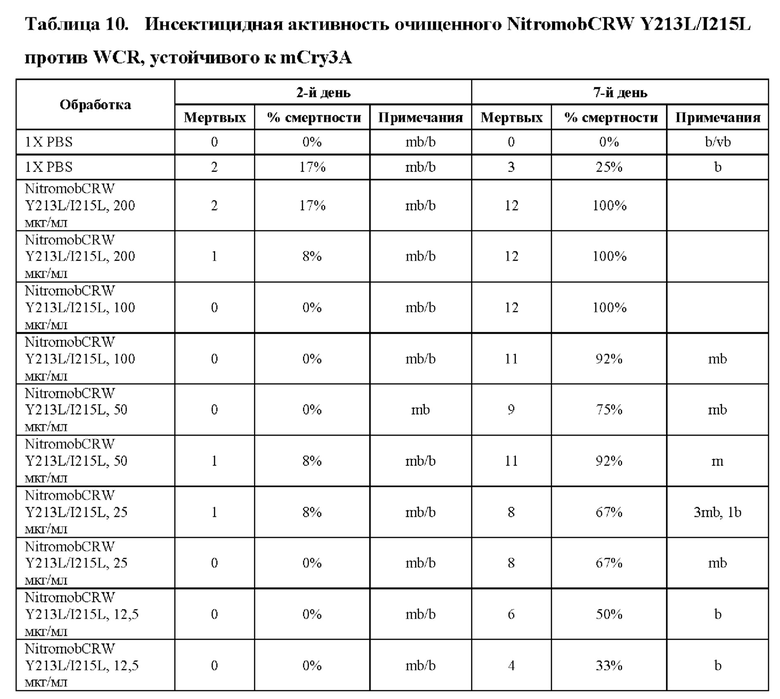

Пример 6. NitromobCRW Y213L/I215L не обладает инсектицидной активностью против чешуекрылых
[00140] Лизаты бактериальных культур, экспрессирующих NitromobCRW Y213L/I215L (SEQ ID NO: 47), тестировали на биоактивность на панели чешуекрылых насекомых-вредителей с применением биоанализов с перекрыванием рационов. Каждого из огневки кукурузной (ЕСВ), совки-ипсилон (BCW), а также совки хлопковой (CEW) и совки травяной (FAW) тестировали в отношении инсектицидной активности NitromobCRW с помощью анализа с включением в рацион, аналогичного таковому из примера 1. В каждом эксперименте тестировали 12 личинок L1 с применением лизатов бактериальных культур В121* (DE3), несущих ген, кодирующий NitromobCRW Y213L/I215L (SEQ ID NO: 9). Положительный контрольный образец BCW, CEW и FAW состоял из личинок, подвергнутых воздействию лизатов Е. coli BL21* (DE3), экспрессирующих белок Vip3. Только IX PBS и лизаты бактериальных культур BL21* (DE3), несущих пустой вектор рЕТ29, применяли в качестве отрицательных контролей. Смертность оценивали на 7-й день. В отношении личинок, которые достигали стадии L3, обработка не оказывала значительного воздействия. Если личинки достигали только стадии L2, то возможно, что обработка вызывала подавление роста. Если личинки оставались на стадии L1 на протяжении всей обработки, то происходило подавление роста. Это также может считаться "эффективной смертностью", так как личинки не будут развиваться после стадии L1, даже если они остаются живыми. В таблицах 11-14 L1=1-я личиночная стадия, L2=2-я личиночная стадия, L3=3-я личиночная стадия. NitromobCRW не являлся активным против тестируемых чешуекрылых насекомых-вредителей в данных экспериментальных условиях (таблицы 12-15).
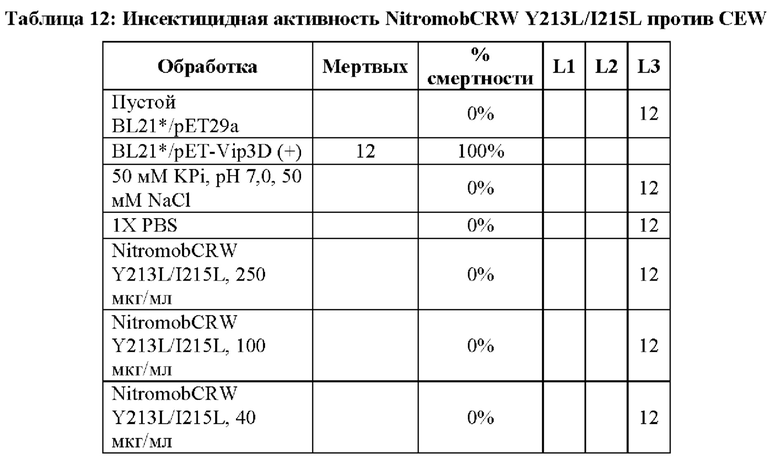
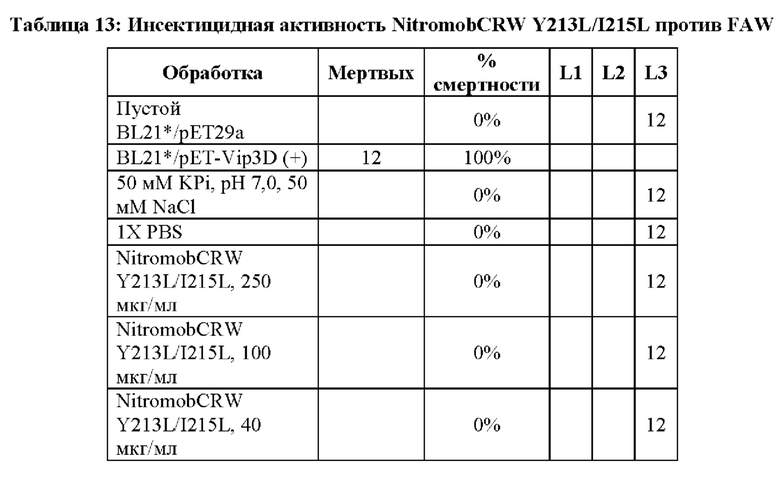
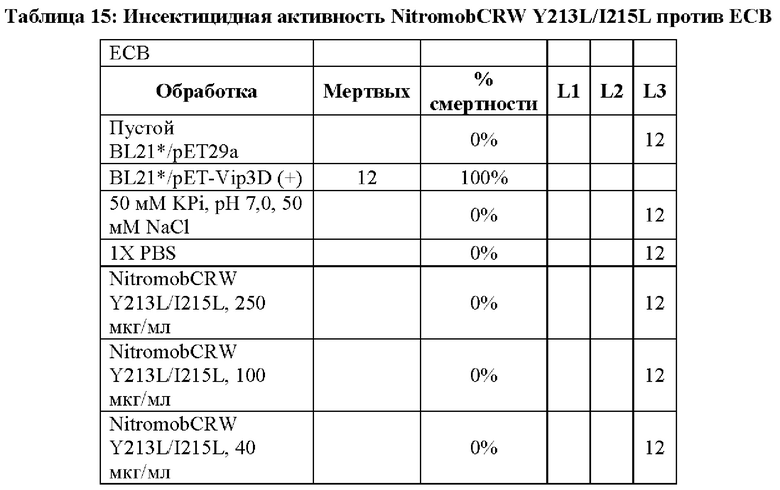
Пример 7. NitromobCRW Y213L/I215L обладает инсектгщпдной активностью против северного кукурузного корневого жука
[00141] NitromobCRW Y213L/I215L очищали как в примере 1 и тестировали в отношении эффективности против 12 личинок северного кукурузного корневого жука (NCR) для каждой концентрации в анализе с включением в рацион, осуществляемом фактически как описано в примере 1. Отрицательный контроль содержал только 1X PBS.
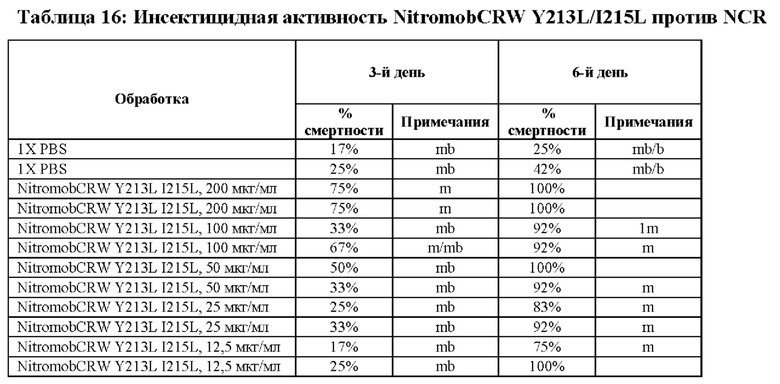
Пример 8. NitromobCRW Y213L/I215L обладает инсектицидной активностью против южного кукурузного корневого жука
[00142] NitromobCRW Y213L/I215L очищали как в примере 1 и тестировали в отношении эффективности против 12 личинок южного кукурузного корневого жука (SCR) в анализе с включением в рацион, осуществляемом фактически как описано в примере 1. NitromobCRW Y213L/I215L (SEQ ID NO: 47) тестировали в диапазоне концентраций 100-400 мкг/мл. Отрицательный контроль содержал только IX PBS. Как показано в таблице 16, NitromobCRW Y213L/I215L демонстрирует инсектицидную активность против SCR.
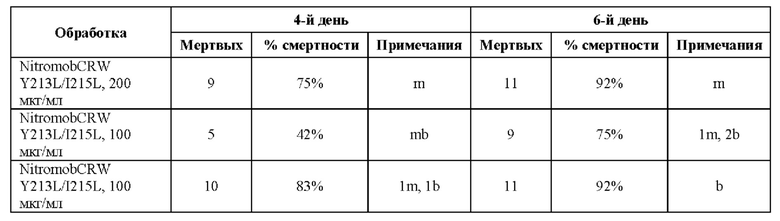
Пример 9. Трансформация маиса с помощью NitromobCRW Y213L/I215L
[00143] Получали бинарную векторную конструкцию, подходящую для опосредованной Agrobacterium трансформации с помощью NitromobCRW Y213L/I215L. Бинарный вектор содержит оптимизированную для маиса кодирующую последовательность NitromobCRW Y213L/I215L (SEQ ID NO: 38), функционально связанную на 5'-конце с промотором, подходящим для управления экспрессией в растениях, и функционально связанную на 3'-конце с терминаторной последовательностью. Оптимизацию кодонов для маиса осуществляют, например, с применением способов, описанных в патенте США №6320100 (включенном в данный документ посредством ссылки). Конструкцией трансформировали Agrobacterium tumefaciem с применением стандартных методик молекулярной биологии, известных специалистам в данной области техники. Для получения агробактерий для трансформации клетки культивировали в жидкой среде YPC при 28°С и 220 об/мин в течение ночи. Опосредованную Agrobacterium трансформацию незрелых зародышей маиса осуществляют фактически как описано в Negrotto et al., 2000, (Plant Cell Reports 19: 798-803). В случае данного примера все компоненты среды представляли собой компоненты, описываемые главным образом в Negrotto et al. выше. Однако они могут быть заменены различными компонентами среды, известными из уровня техники.
[00144] После трансформации, отбора и регенерации растения анализируют в отношении присутствия гена, кодирующего селектируемый маркер и кодон-оптимизированную для маиса кодирующую последовательность NitromobCRW Y213L/I215L, с применением анализа TaqMan®. Растения также тестировали в отношении присутствия остова вектора. Растения, отрицательные в отношении остова вектора и содержащие одну копию трансгена, переносили в теплицу и анализировали в отношении устойчивости к повреждениям, наносимым WCR.
Пример 10. Растения маиса, экспрессирующие NitromobCRW Y213L/I215L, характеризуются инсектицидной активностью против WCR
Присутствие NitromobCRW Y213L/I215L выявляли посредством ELISA в нг/мг общего растворимого белка (TSP) в ткани листа или корня от каждого трансформанта. Инсектицидную активность определяли с помощью биоанализа сегментов корней. Вкратце, образцы ткани корня маиса отбирали у каждого трансформанта, когда трансформанты маиса, экспрессирующие вариант NitromobCRW, достигали стадии V3-V4. Ткань корня маиса помещали в чашку Петри, а затем инфицировали с помощью 12 личинок WCR. Два образца ткани корня (повторность 1 и повторность 2) оценивали в отношении отверстий, образующихся при поедании (FH), и повреждений в виде рубцов на 3-й день. Ткань корня из нетрансформированного (нулевого) маиса служила в качестве отрицательного контроля. Оценку повреждений, наносимых насекомыми, проводят с применением следующего: ND = не выявлено; FH = отверстия, образующиеся при поедании; L = слабое рубцевание; М = среднее рубцевание; Н = сильное рубцевание; ++ = объект с превосходной эффективностью; + = объект с хорошей эффективностью; - = объект с низкой эффективностью

Пример 11: NitromobCRW Y213L/I215L в комбинации с интерферирующей РНК, характеризующейся инсектицидной активностью против WCR
[00145] NitromobCRW и/или вариант NitromobCRW очищают как бактериальные лизаты в примере 1 или очищают как белки аналогично примеру 4. Получали dsRNA против важной мишени и, как известно, характеризующуюся инсектицидной активностью. В неограничивающих примерах dsRNA может нацеливаться на ген, кодирующий вакуолярную АТФ-синтазу, бета-тубулин, белок р28, представляющий собой субъединицу протеасомы 26S, EF1α 48D, тропонин I, тетраспанин, гамма-коатомер, бета-коатомер и/или эпоксидгидролазу ювенильного гормона (публикации WO №№ WO 2018/026770, WO 2018/026773 и WO 2018/026774; патент США №7812219; каждый из которых включен в данный документ посредством ссылки). dsRNA и очищенный белок NitromobCRW тестируют в отношении инсектицидной эффективности против WCR в анализе с включением в рацион, осуществляемом фактически как описано в примере 1, но с добавлением dsRNA в искусственный рацион.
[00146] Asdadasdas
Пример 12. NiromobCRW с С-концевым удлинением активен против CRW.
[00147] В данном примере описываются влияния присоединения С-концевого пептида к белку NitromobCRW. Конструкцию pET-NitromobCRW-Y213L/I215L с С-концевым удлинением, кодирующую SEQ ID NO: 77, клонировали в Е. coli. С-концевой удлиненный пептид содержит SEQ ID NO: 76 и представляет собой комбинацию линкерной последовательности (аминокислоты 1-35 SEQ ID NO: 76) и SUMO-метки (аминокислоты 36-133 SEQ ID NO: 76). SUMO представляет собой небольшой убиквитин-подобный модифицирующий белок, который при слиянии с представляющим интерес белком усиливает образование функционального белка в прокариотических и эукариотических системах экспрессии на основе в значительной степени улучшенной стабильности и растворимости белка. После экспрессии и очистки слитого белка SUMO-метку как правило отщепляют специфическими (SUMO) протеазами посредством их эндопептидазной активности in vitro для получения требуемого высвобожденного белка-партнера. В случае данного примера С-концевой удлиненный пептид (SEQ ID NO: 76) не отщепляли от белка NitromobCRW и интактный удлиненный белок (NitromobCRW-Cterm-SUMO; SEQ ID NO: 77) тестировали в отношении перевариваемости под действием SGF и инсектицидной активности против WCR.
[00148] Анализ NitromobCRW-Y213L-I215L-Cterm-SUMO с применением SGF был таким, как описано выше. Перевариваемость NitromobCRW-Y213L-I215L-Cterm-SUMO сравнивали с NitromobCRW-Y213L-I215L без С-концевого удлинения в качестве контроля. Белок NitromobCRW-Y213L-I215L-Cterm-SUMO также тестировали в отношении активности против WCR, как описано выше.
[00149] Результаты анализа перевариваемости с применением SGF продемонстрировали, что NitromobCRW-Y213L-1215L-Cterm-SUMO переваривается до 5-минутного момента времени. Следовательно, добавление метки Cterm SUMO не оказало влияния на перевариваемость белка NitromobCRW. SGF-гели в случае меченого и немеченого белка были почти идентичными (данные не показаны). Результаты биоанализа, показанные в таблице 18, демонстрируют, что белок NitromobCRW с С-концевым удлиненным пептидом является таким же активным, как и белок NitromobCRW без С-концевого удлиненного пептида.
[00150] Следует понимать, что примеры и варианты осуществления, описанные в данном документе, предназначены только для иллюстративных целей, и что различные модификации или изменения с учетом данного описания будут предполагаться специалистами в данной области техники и подлежат включению в сущность и содержание данной заявки, а также объем прилагаемой формулы изобретения.
[00151] Все публикации и патентные заявки, упомянутые в данном описании, показывают уровень компетентности специалистов в области техники, к которой относится настоящее изобретение. Все публикации и патентные заявки включены в данный документ посредством ссылки в той же степени, как если бы каждая отдельная публикация или патентная заявка была конкретно и отдельно включена посредством ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЗИНГЕНТА ПАРТИСИПЕЙШНС АГ
<120> ИНСЕКТИЦИДНЫЕ БЕЛКИ
<130> 81547-US-L-ORG-NAT-1
<160> 7
<170> PatentIn версия 3.5
<210> 1
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 1
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 2
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 2
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gttagttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 3
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 3
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattttaaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 4
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 4
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccctgacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 5
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 5
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tctgcggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 6
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 6
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 7
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 7
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc tttttggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 8
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 8
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc tttatggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 9
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 9
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgctttac ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 10
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 10
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggctgactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 11
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 11
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atctggaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 12
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 12
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aattagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 13
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 13
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaatt aggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 14
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 14
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttatagctaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 15
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 15
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttatattaaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 16
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 16
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcgttaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 17
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 17
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttgggggtt agggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 18
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 18
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgcttta 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 19
<211> 906
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 19
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttctgatagg taacaccgct 660
gtgaattaca acccaaccta taaggaccat catttctgga gccttggggt tgccggagtc 720
atggctaagg gtgggatcac taactccgta cagtcaactg aggatatcga aatcggctat 780
tactctaatt caaaaataga attgaaagac aaggctacag gcgcattaaa ggcggcgtat 840
aatatggccg acgccccagg gcagtcggca gctgagtcgc gtcaacctgc gctggacgaa 900
gcataa 906
<210> 20
<211> 906
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 20
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttatattagg taacaccgct 660
gtgaattaca acccaaccta taaggaccat catttctgga gccttggggt tgccggagtc 720
atggctaagg gtgggatcac taactccgta cagtcaactg aggatatcga aatcggctat 780
tactctaatt caaaaataga attgaaagac aaggctacag gcgcattaaa ggcggcgtat 840
aatatggccg acgccccagg gcagtcggca gctgagtcgc gtcaacctgc gctggacgaa 900
gcataa 906
<210> 21
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 21
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttttc ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 22
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 22
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat cccttacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 23
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 23
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tctgcggtat aacgcttacc ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 24
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 24
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atctggaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 25
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 25
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atctggaact gggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 26
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 26
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttaca gtctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 27
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 27
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agagggagtc ttaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 28
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 28
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa cttagctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 29
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 29
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa ctttgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 30
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 30
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cgctggaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 31
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 31
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcttgc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 32
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 32
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccctgct tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 33
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 33
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtactttt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 34
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 34
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacact tggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgcttacc ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 35
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 35
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agaggggtta tgaaagttcg tatacggtat aacgctttac ttataggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 36
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 36
atgggtatta gcataagtat agttgcaggc cacgataaat cggcgtctag tgtgaacgct 60
acagggactg ttcaacacgt tatcaccgat caggagcgca ctaccttcca cttgggtgat 120
aaacagttga aggacgccgt taaggcttat tttggaaagt ctcccaacga cgtgtatttg 180
cactctccga cgccctgggg tgacttgtac aaaaagtatt catggcccca agtccagatg 240
atactggtcg ttcagtcggc agaaatcctt ggtatcacta gcgagccggt gattgttaag 300
acccaggaat ttgtcaacaa ttcaagacaa aaggggacgt tcaatgtggc gataacagag 360
tcggtaaata atacgacgtc gtctaattgg agtacgggag ggacccttac gatcggccaa 420
aaattctctt acggggttaa gttcctggga gccggagcgg aaggagaaac ctcgttatcc 480
tacagccaaa gttggggggt ggggggacag gaatctaaat ccatcacagt cggctcatcc 540
agcggcgttt cggtagaatt agaccctggt gaaagcgttc ttgccgaact tagtgcctcg 600
agagggagtc ttaaagttcg tatacggtat aacgcttacc ttctgggtaa caccgctgtg 660
aattacaacc caacctataa ggaccatcat ttctggagcc ttggggttgc cggagtcatg 720
gctaagggtg ggatcactaa ctccgtacag tcaactgagg atatcgaaat cggctattac 780
tctaattcaa aaatagaatt gaaagacaag gctacaggcg cattaaaggc ggcgtataat 840
atggccgacg ccccagggca gtcggcagct gagtcgcgtc aacctgcgct ggacgaagca 900
taa 903
<210> 37
<211> 903
<212> ДНК
<213> Nitrococcus mobilis
<400> 37
atgggcatca gtatcagcat cgtcgccggt cacgacaaat ccgcatcaag cgtaaacgcc 60
accggaaccg tgcagcacgt cattacggat caggaacgga cgactttcca tcttggcgat 120
aagcaactca aggacgccgt caaggcgtat ttcgggaaaa gcccgaacga cgtgtacttg 180
catagcccga cgccgtgggg tgatctctac aagaagtaca gttggccgca ggtccaaatg 240
atcctggtcg tccagagcgc ggagattctc ggcatcacct cggagccggt gatcgtcaag 300
actcaggagt tcgtgaataa tagccgccag aaaggcacct ttaacgttgc catcacggag 360
tcggtgaaca acaccacgtc ctccaactgg agtaccggcg gcacgcttac gatcgggcag 420
aagttctcct acggtgtcaa gtttctcggt gccggggccg agggggagac ttcgctgtcg 480
tacagccagt cgtggggagt cggggggcag gagtcgaaat cgatcactgt aggttcatcg 540
tccggggtga gtgtagagct tgatcccggc gagtccgttc tcgccgagct ctctgccagt 600
cggggcgtga tgaaagtacg aattcgctat aacgcctacc tcatcggcaa tacagccgtg 660
aattacaacc ccacctataa ggaccatcat ttctggagct tgggtgtcgc gggtgtcatg 720
gcgaaaggcg gcattaccaa ttcggtgcaa tcgaccgaag acatcgagat cggttattac 780
tccaattcca agatcgagct caaagataag gcgacgggtg ccttgaaggc cgcctacaat 840
atggccgacg cgcccgggca atcggccgcg gagtctcgcc agcctgctct cgatgaggcc 900
tag 903
<210> 38
<211> 903
<212> ДНК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 38
atgggcatca gtatcagcat cgtcgccggt cacgacaaat ccgcatcaag cgtaaacgcc 60
accggaaccg tgcagcacgt cattacggat caggaacgga cgactttcca tcttggcgat 120
aagcaactca aggacgccgt caaggcgtat ttcgggaaaa gcccgaacga cgtgtacttg 180
catagcccga cgccgtgggg tgatctctac aagaagtaca gttggccgca ggtccaaatg 240
atcctggtcg tccagagcgc ggagattctc ggcatcacct cggagccggt gatcgtcaag 300
actcaggagt tcgtgaataa tagccgccag aaaggcacct ttaacgttgc catcacggag 360
tcggtgaaca acaccacgtc ctccaactgg agtaccggcg gcacgcttac gatcgggcag 420
aagttctcct acggtgtcaa gtttctcggt gccggggccg agggggagac ttcgctgtcg 480
tacagccagt cgtggggagt cggggggcag gagtcgaaat cgatcactgt aggttcatcg 540
tccggggtga gtgtagagct tgatcccggc gagtccgttc tcgccgagct ctctgccagt 600
cggggcgtga tgaaagtacg aattcgctat aacgcctacc tcatcggcaa tacagccgtg 660
aattacaacc ccacctataa ggaccatcat ttctggagct tgggtgtcgc gggtgtcatg 720
gcgaaaggcg gcattaccaa ttcggtgcaa tcgaccgaag acatcgagat cggttattac 780
tccaattcca agatcgagct caaagataag gcgacgggtg ccttgaaggc cgcctacaat 840
atggccgacg cgcccgggca atcggccgcg gagtctcgcc agcctgctct cgatgaggcc 900
tag 903
<210> 39
<211> 300
<212> БЕЛОК
<213> Nitrococcus mobilis
<400> 39
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 40
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 40
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Leu Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 41
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 41
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Leu Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 42
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 42
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Leu Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 43
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 43
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Leu
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 44
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 44
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 45
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 45
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Phe Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 46
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 46
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Tyr Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 47
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 47
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Leu Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 48
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 48
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Leu Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 49
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 49
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Leu Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 50
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 50
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Leu Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 51
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 51
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Leu Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 52
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 52
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Ala Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 53
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 53
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Leu Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 54
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 54
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Leu Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 55
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 55
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Leu Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 56
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 56
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Leu Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 57
<211> 301
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 57
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn
210 215 220
Pro Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val
225 230 235 240
Met Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile
245 250 255
Glu Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala
260 265 270
Thr Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln
275 280 285
Ser Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 58
<211> 301
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 58
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Leu Gly Asn Thr Ala Val Asn Tyr Asn
210 215 220
Pro Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val
225 230 235 240
Met Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile
245 250 255
Glu Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala
260 265 270
Thr Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln
275 280 285
Ser Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 59
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 59
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Phe Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 60
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 60
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Leu Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 61
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 61
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Leu
195 200 205
Arg Tyr Asn Ala Tyr Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 62
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 62
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Leu Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 63
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 63
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Leu Glu
245 250 255
Leu Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 64
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 64
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Ser Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 65
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 65
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Ser Leu Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 66
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 66
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Leu Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 67
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 67
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Phe Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 68
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 68
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Leu Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 69
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 69
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Leu Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 70
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 70
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Leu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 71
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 71
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Leu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 72
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 72
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Leu Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 73
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 73
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Val Met Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Leu Leu Ile Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<210> 74
<211> 300
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> получен из Nitrococcus mobilis
<400> 74
Met Gly Ile Ser Ile Ser Ile Val Ala Gly His Asp Lys Ser Ala Ser
1 5 10 15
Ser Val Asn Ala Thr Gly Thr Val Gln His Val Ile Thr Asp Gln Glu
20 25 30
Arg Thr Thr Phe His Leu Gly Asp Lys Gln Leu Lys Asp Ala Val Lys
35 40 45
Ala Tyr Phe Gly Lys Ser Pro Asn Asp Val Tyr Leu His Ser Pro Thr
50 55 60
Pro Trp Gly Asp Leu Tyr Lys Lys Tyr Ser Trp Pro Gln Val Gln Met
65 70 75 80
Ile Leu Val Val Gln Ser Ala Glu Ile Leu Gly Ile Thr Ser Glu Pro
85 90 95
Val Ile Val Lys Thr Gln Glu Phe Val Asn Asn Ser Arg Gln Lys Gly
100 105 110
Thr Phe Asn Val Ala Ile Thr Glu Ser Val Asn Asn Thr Thr Ser Ser
115 120 125
Asn Trp Ser Thr Gly Gly Thr Leu Thr Ile Gly Gln Lys Phe Ser Tyr
130 135 140
Gly Val Lys Phe Leu Gly Ala Gly Ala Glu Gly Glu Thr Ser Leu Ser
145 150 155 160
Tyr Ser Gln Ser Trp Gly Val Gly Gly Gln Glu Ser Lys Ser Ile Thr
165 170 175
Val Gly Ser Ser Ser Gly Val Ser Val Glu Leu Asp Pro Gly Glu Ser
180 185 190
Val Leu Ala Glu Leu Ser Ala Ser Arg Gly Ser Leu Lys Val Arg Ile
195 200 205
Arg Tyr Asn Ala Tyr Leu Leu Gly Asn Thr Ala Val Asn Tyr Asn Pro
210 215 220
Thr Tyr Lys Asp His His Phe Trp Ser Leu Gly Val Ala Gly Val Met
225 230 235 240
Ala Lys Gly Gly Ile Thr Asn Ser Val Gln Ser Thr Glu Asp Ile Glu
245 250 255
Ile Gly Tyr Tyr Ser Asn Ser Lys Ile Glu Leu Lys Asp Lys Ala Thr
260 265 270
Gly Ala Leu Lys Ala Ala Tyr Asn Met Ala Asp Ala Pro Gly Gln Ser
275 280 285
Ala Ala Glu Ser Arg Gln Pro Ala Leu Asp Glu Ala
290 295 300
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2018 |
|
RU2761666C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2817591C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2816526C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2017 |
|
RU2765304C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2740312C2 |
| НОВЫЕ БЕЛКИ, ОБЛАДАЮЩИЕ ИНГИБИРУЮЩЕЙ АКТИВНОСТЬЮ В ОТНОШЕНИИ НАСЕКОМЫХ | 2015 |
|
RU2740313C2 |
| КОМПОЗИЦИИ И СООТВЕТСТВУЮЩИЕ СПОСОБЫ ДЛЯ СЕЛЬСКОГО ХОЗЯЙСТВА | 2018 |
|
RU2805081C2 |
| ГЕНЫ ТОКСИНОВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2825305C2 |
| КОНТРОЛЬ ЖЕСТКОКРЫЛЫХ ВРЕДИТЕЛЕЙ С ПРИМЕНЕНИЕМ МОЛЕКУЛ РНК | 2017 |
|
RU2783144C2 |
| ГЕНЫ ТОКСИНОВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2723717C2 |
Настоящее изобретение относится к области биотехнологии и может быть использовано в сельском хозяйстве. Изобретение раскрывает новые инсектицидные белки, характеризующиеся токсичностью в отношении представителей отряда жесткокрылых, конкретно в отношении западного, северного и южного кукурузных корневых жуков. Изобретение может быть использовано, например, для получения трансгенных растений с целью обеспечения их защиты от вреда, наносимого указанными насекомыми. 12 н. и 3 з.п. ф-лы, 18 табл., 12 пр.
1. Кассета экспрессии, содержащая промотор, функционально связанный с гетерологичной молекулой нуклеиновой кислоты, содержащей:
(a) нуклеотидную последовательность, которая кодирует полипептид, где аминокислотная последовательность полипептида содержит SEQ ID NO: 39-74;
(b) нуклеотидную последовательность, которая кодирует полипептид, где аминокислотная последовательность полипептида по меньшей мере на 99% идентична аминокислотной последовательности SEQ ID NO: 39-74;
(c) нуклеотидную последовательность, которая комплементарна нуклеотидной последовательности любого из (а) или (b), приведенных выше.
2. Полипептид для обеспечения устойчивости к западному кукурузному корневому жуку, северному кукурузному корневому жуку или южному кукурузному корневому жуку, содержащий аминокислотную последовательность под любым из SEQ ID NO: 40-74.
3. Экспрессионный вектор, содержащий кассету экспрессии по п. 1.
4. Клетка-хозяин для получения полипептида по п. 2, которая содержит кассету экспрессии по п. 1.
5. Способ получения растения или части растения, характеризующихся повышенной устойчивостью к насекомым: западному кукурузному корневому жуку, северному кукурузному корневому жуку или южному кукурузному корневому жуку по сравнению с контрольными растением или частью растения без устойчивости к насекомым, включающий:
(a) введение в часть растения молекулы нуклеиновой кислоты, содержащую кассету экспрессии по п. 1; и
(b) выращивание части растения с получением растения, которое экспрессирует молекулу нуклеиновой кислоты и которое характеризуется повышенной устойчивостью к насекомым по сравнению с контрольными растением или частью растения, которые не содержат молекулу нуклеиновой кислоты, содержащей кассету экспрессии по п. 1.
6. Способ идентификации растения или части растения, характеризующихся повышенной устойчивостью к насекомым: западному кукурузному корневому жуку, северному кукурузному корневому жуку или южному кукурузному корневому жуку по сравнению с контрольными растением или частью растения без устойчивости к насекомым, включающий выявление в растении или части растения нуклеиновой кислоты, кодирующей полипептид по п. 2, за счет чего обеспечивается идентификация растения или части растения с повышенной устойчивостью к насекомым: западному кукурузному корневому жуку, северному кукурузному корневому жуку или южному кукурузному корневому жуку.
7. Способ по п. 6, где указанную нуклеиновую кислоту или ее диагностический фрагмент выявляют в продукте амплификации образца нуклеиновой кислоты из растения или части растения.
8. Способ получения растения с повышенной устойчивостью к насекомым: западному кукурузному корневому жуку, северному кукурузному корневому жуку или южному кукурузному корневому жуку по сравнению с контрольным растением или частью растения без устойчивости к насекомым-вредителям, включающий скрещивание первого родительского растения со вторым родительским растением, где по меньшей мере первое родительское растение содержит в своем геноме молекулу нуклеиновой кислоты, содержащую кассету экспрессии по п. 1 и дает потомство, при этом потомство содержит по меньшей мере одно растение, которое содержит нуклеиновые кислоты в своем геноме и которое проявляет повышенную устойчивость к насекомым: западному кукурузному корневому жуку, северному кукурузному корневому жуку или южному кукурузному корневому жуку по сравнению с контрольным растением.
9. Трансгенное растение, с повышенной устойчивостью к насекомым: западному кукурузному корневому жуку, северному кукурузному корневому жуку или южному кукурузному корневому жуку, содержащее молекулу нуклеиновой кислоты, которая содержит кассету экспрессии по п. 1.
10. Композиция для обеспечения устойчивости к западному кукурузному корневому жуку, северному кукурузному корневому жуку или южному кукурузному корневому жуку, содержащая подходящий сельскохозяйственный носитель и полипептид с инсектицидной активностью, где полипептид выбран из группы, состоящей из:
a) полипептида, содержащего аминокислотную последовательность SEQ ID NO: 39-74; а также
b) полипептида, содержащего аминокислотную последовательность, по меньшей мере на 99% идентичную аминокислотной последовательности SEQ ID NO: 39-74, где указанная аминокислотная последовательность обладает инсектицидной активностью.
11. Способ борьбы с популяцией западного кукурузного корневого жука, северного кукурузного корневого жука или южного кукурузного корневого жука, включающий приведение указанной популяции в контакт с эффективным для борьбы с насекомыми количеством полипептида с инсектицидной активностью, где полипептид выбран из группы, состоящей из:
a) полипептида, содержащего аминокислотную последовательность SEQ ID NO: 39-74; и
b) полипептида, содержащего аминокислотную последовательность, по меньшей мере на 99% идентичную аминокислотной последовательности SEQ ID NO: 39-74.
12. Полипептид для обеспечения устойчивости к западному кукурузному корневому жуку, северному кукурузному корневому жуку или южному кукурузному корневому жуку, содержащий аминокислотную последовательность, по меньшей мере на 99% идентичную любой из SEQ ID NO: 39-74, и дополнительно содержащий введенный сайт расщепления протеазой.
13. Полипептид по п. 12, в котором введенный сайт расщепления протеазой расположен между аминокислотными остатками с 97 по 266 любой из SEQ ID NO: 39-74.
14. Полипептид по п. 12, в котором введенный сайт расщепления протеазой введен путем вставки остатка лейцина.
15. Полипептид для обеспечения устойчивости к западному кукурузному корневому жуку, северному кукурузному корневому жуку или южному кукурузному корневому жуку, содержащий аминокислотную последовательность, по меньшей мере на 99% идентичную любой из SEQ ID NO: 40-74, содержащий мутацию, которая улучшает усвояемость в анализе SGF по сравнению с полипептидом, содержащим аминокислотную последовательность SEQ ID NO: 39.
| US 20160031949 A1, 04.02.2016 | |||
| Приспособление для графления на пишущей машине | 1929 |
|
SU14836A1 |
| НАВЕС ДЛЯ СУШКИ СЫРЦА | 1926 |
|
SU3346A1 |
| RU 2015142582 A1, 10.04.2017. | |||
