Изобретение в целом относится к медицине, более детально к области применения известных как таковых лекарств и фармацевтических соединений, вызывающих митохондриальную дисфункцию у макрофагов для лечения онкологических заболеваний. Заявленное техническое решение обеспечивает возможность использования известных как таковых лекарств и фармакологических соединений, которые воздействуют на макрофаги, за счет изменения их митохондриальных функций, которые становятся эффективными средствами для лечения онкологических заболеваний. По мнению заявителя, указанное направление является перспективным для лечения и предупреждения развития многих видов онкологических заболеваний.
Далее в тексте заявителем приведены термины, которые необходимы для облегчения однозначного понимания сущности заявленных материалов и исключения противоречий и/или спорных трактовок при выполнении экспертизы по существу.
Ассоциированные с опухолью макрофаги (ТАМ) - представляют собой класс иммунных клеток, присутствующих в большом количестве в микроокружении солидных опухолей. Они активно участвуют в воспалении, связанном с раком. Известно, что макрофаги происходят из моноцитов крови, образующихся в костном мозге (макрофаги, полученные из моноцитов), или предшественников желточного мешка (тканевые макрофаги) [https://en.wikipedia.org/wiki/].
Сóлидные опухоли - это негемопоэтические опухоли, то есть опухоли, развившиеся не из клеток кроветворной системы. Солидные опухоли могут быть доброкачественными и злокачественными. В контексте настоящего описания заявитель подразумевает под указанным термином именно злокачественные опухоли [https://podari-zhizn.ru/ru/get-help/meditsinskii-spravochnik/].
Таргетная терапия - направление в лечении онкологических заболеваний с использованием лекарственных препаратов, блокирующих определенные молекулы опухолевых клеток или клеток опухолевого микроокружения. За счет этого раковые клетки перестают размножаться, ухудшается их кровоснабжение или достигаются другие лечебные эффекты. По сравнению с классическими химиопрепаратами, таргетные препараты действуют более прицельно и почти не затрагивают здоровые клетки [https://www.euroonco.ru/].
Перепрограммирование макрофагов - в контексте настоящего описания заявитель подразумевает под указанным термином процесс смены фенотипа макрофагов.
Митохондриальное дыхание - совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды, а также образование энергии [https://en.wikipedia.org/wiki/].
Chi3L1 - хитиназа-3-подобный белок 1, представляет собой секретируемый гликопротеин, который составляет приблизительно 40k Да по размеру. Экспрессируется и секретируется различными типами клеток, включая макрофаги, хондроциты, фибробластоподобные синовиальные клетки, гладкомышечные клетки сосудов и звездчатые клетки печени [https://ru.abcdef.wiki/wiki/CHI3L1].
Классически активированные макрофаги (М1-поляризация, М1 подтип) - макрофаги, которые приобретают фенотип с более высокой активностью как против патогенов, так и против опухолевых клеток. Они также выделяют воспалительные цитокины [https://ru.wikipedia.org/wiki/].
Альтернативно активированные макрофаги (М2-поляризация, М2 подтип) - макрофаги, которые выделяют минимальное количество провоспалительных цитокинов и обладают более низкой активностью против внутриклеточных патогенов [https://ru.wikipedia.org/wiki/].
Сенсибилизация клеток - приобретение организмом специфической повышенной чувствительности к чужеродным веществам - аллергенам, повышение его чувствительности к воздействию раздражителей [https://ru.wikipedia.org/wiki/]
Кофермент А - фермент, отличающийся своей ролью в синтезе и окислении жирных кислот, а также окислении пирувата в цикле лимонной кислоты [https://en.wikipedia.org/wiki/Coenzyme_A].
Экзосомы - микроскопические внеклеточные везикулы (пузырьки) диаметром 30-100 нанометров, выделяемые в межклеточное пространство клетками различных тканей и органов [https://ru.wikipedia.org/wiki/].
Моноаминоксидаза А - фермент, осуществляющий катаболизм нейромедиаторов и гормонов. Таким образом, этот фермент играет важную роль в поддержании постоянных концентраций эндогенных моноаминов в тканях, что особенно важно для нервной ткани, а также ограничивает их поступление в организм с пищей и участвует в метаболизме опасных биологически активных веществ, структурно сходных с эндогенными моноаминами [https://ru.wikipedia.org/wiki/].
Фармакологическая индукция - абсолютное увеличение количества и активности ферментов метаболизма вследствие воздействия на них определенного химического соединения, в частности лекарственного средства [https://ru.wikipedia.org/wiki/].
Opa1 - мультифункциональный митохондриальный белок, кодирует динамин-подобную ГТФазу. "Активность белка необходима для формирования крист - системы внутренних митохондриальных мембран. Белок управляет процессом слияния и разделения митохондрий, критичным для преодоления стрессовых условий и сохранения энергобаланса клетки. [https://www.genokarta.ru/gene/OPA1]
Трансфекция - процесс введения нуклеиновой кислоты в клетки эукариот невирусным методом [https://web.archive.org/web/20081217035133/].
Малая интерферирующая РНК - это класс двухцепочечных РНК, длиной 20-25 нуклеотидов. Взаимодействие малых интерферирующих РНК с матричной РНК (мРНК) целевого гена приводит к деградации последней (в процессе РНК-интерференции), предотвращая трансляцию мРНК на рибосомах в кодируемый ею белок. В конечном итоге результат действия малых интерферирующих РНК идентичен тому, как если бы просто снижалась экспрессия гена [https://ru.wikipedia.org/wiki\].
ТНР-1 - это моноцитарная клеточная линия человека, полученная от пациента с острым моноцитарным лейкозом. Он используется для тестирования клеточных линий лейкоза в иммуноцитохимическом анализе межбелковых взаимодействий и в иммуногистохимии а также как модельная клеточная линия моноцитов [https/en.wikipedia.org/wiki/THP-1_cell_line].
Кондиционированные среды - питательная среда, содержащая различные физиологически активные продукты жизнедеятельности предварительно культивируемых в ней клеток [http://humbio.ru/humbio/tarantul_sl/00000b02.htm].
Внеклеточные рецепторы - это рецепторы, которые встроены в плазматическую мембрану клеток. Они участвуют в передаче сигналов в клетке, связываясь с внеклеточными молекулами. Это специализированные интегральные мембранные белки, которые обеспечивают связь между клеткой и внеклеточным пространством [https/en.wikipedia.org/wiki/Cell_surface_receptor].
Онкологические заболевания в мире занимают второе место по смертности после сердечно-сосудистой патологии, и несмотря на определенные успехи в диагностике и методах лечения, темпы прироста заболеваемости раком неуклонно растут. Последние научные исследования показали, что эффективный подход к лечению рака требует терапии, а именно:
- терапии направленной на опухолевые клетки (химиотерапия, радиотерапия, хирургическое лечение)
- терапии направленной на микроокружение опухоли (таргетная терапия)
- терапии направленной на иммунные клетки (иммунная терапия).
При этом, следует акцентировать внимание на то, что, опухоль-ассоциированные макрофаги (tumor-associated macrophages = ТАМ) являются основным компонентом этого микроокружения и играют значительную роль в прогрессировании рака.
Во время роста опухоли в организме больного, макрофаги создают благоприятную воспалительную среду, которая способствует более агрессивному росту опухоли, за счёт подавления противоопухолевого иммунного ответа у пациента, что в конечном счете и обеспечивает собственно распространение рака в другие органы (метастазирование) [Mantovani A. et.al. (2022) Macrophages as tools and targets in cancer therapy, Nat Rev Drug Discov., 21(11), 799-820].
Такие солидные раки, как рак легких и рак поджелудочной железы, очень плохо отвечают, как на химиотерапию, так и на иммунную терапию, в основном из-за активного воспалительного процесса внутри опухоли за счёт негативного воздействия раковых клеток на макрофаги больного.
Инфильтрация макрофагов из периферической крови в очаг опухолеобразования является ранним процессом в раке, с изначально благими целями запустить иммунный ответ и уничтожить раковые клетки, однако принимая во внимание известное свойство опухолевых клеток перепрограммировать свойства макрофагов, указанный процесс уничтожения опухолевых клеток макрофагами запускается в противоположном направлении, а именно макрофаги начинают поддерживать рост опухолевых клеток и способствуют прогрессированию опухоли у пациента.
Таким образом, заявителем в вышеизложенном материале выявлена причина и следствие развития онкологических заболеваний, вследствие этого необходимо найти воздействие на макрофаги для остановки поддержания процесса роста опухоли или нахождения метода по прекращению или уничтожению опухолевого процесса в организме больного.
За уничтожение опухолевых клеток отвечают так называемые макрофаги М1 подтипа, за счет того, что они обладают противоопухолевыми свойствами. Однако, опухолевые клетки перепрограммируют макрофаги с М1 в М2 подтип за счёт того, что раковые клетки выделяют в микроокружение вещества: цитокины и хемокины, тем самым обеспечивая рост опухоли и метастазирование.
Таким образом, раковые клетки перепрограммируя макрофаги с М1 в М2 подтип помогают опухоли расти и как следствие выключают противоопухолевый иммунитет организма больного.
Такие перепрограммированные макрофаги называются альтернативно активированные макрофаги подтипа M2, при этом из исследованного уровня техники экспериментально показано что накопление таких макрофагов коррелирует с плохим прогнозом развития заболевания у больных пацтентов рака, особенно с раком легких, аденокарциномы и раком поджелудочной железы [Zhang B. et. al. (2011) M2-polarized tumor-associated macrophages are associated with poor prognoses resulting from accelerated lymphangiogenesis in lung adenocarcinoma, Clin (Sao Paulo), 66, 879-1886].
На дату представления заявленных материалов, для минимизации указанного негативного эффекта развития опухолевого процесса, применяется несколько стратегий, воздействия на раковые макрофаги, а именно:
- элиминация макрофагов из опухоли,
- предотвращение миграции макрофагов (М2) в опухоль,
- подавление функций альтернативно активированных макрофагов М2 подтипа
- перепрограммирование М2 макрофагов в М1подтип макрофагов
Далее заявителем проведён детальный анализ каждого из представленных выше известных стратегий с точки зрения их воздействия на макрофаги.
Недостатком элиминации макрофагов является то, что циркулирующие в крови моноциты непрерывно рекрутируются в опухоли, где они дифференцируются в зрелые макрофаги M2 подтипа и вновь участвуют в прогрессировании опухоли [Zhu Y. et. al. (2017) Tissue-Resident Macrophages in Pancreatic Ductal Adenocarcinoma Originate from Embryonic Hematopoiesis and Promote Tumor Progression, Immunity., 47, 323 - 328].
При этом, главное препятствие в предотвращении миграции макрофагов в опухоль может заключаться в том, что истощение макрофагов совпадает с потерей резидентных в тканях популяций макрофагов, важных для поддержания гомеостаза, т.е. приводит к общему нарушению функционирования организма [Ries C.H. et. al. (2014) Targeting tumor-associated macrophages with anti-CSF-1R antibody reveals a strategy for cancer therapy, Cancer cell., 25(6), P. 846-859].
Другое ограничение стратегий, нацеленных на подавление функций М2 макрофагов, связано с тем, что, если полностью удалить альтернативно активированные макрофаги из организма то, будет очень долго происходить воспалительный процесс в организме, поскольку М2 макрофаги способствуют тканевому восстановлению [Etzerodt A. et. al. (2019) Specific targeting of CD163 mobilizes inflammatory monocytes and promotes T cell-mediated tumor regression, J. Exp. Med., 216(10), 2394-2411].
Исходя из представленного выше анализа можно сделать логичный вывод о том, что перепрограммирование макрофагов из М2 в М1 подтип является наиболее перспективной стратегией комбинированного лечения онкологических заболеваний. Первые доклинические исследования показали, что методы лечения, нацеленные на перепрограммирование макрофагов, не является токсичным для организма пациента, хорошо переносятся, т.к. эта стратегия приводит к клинической активизации иммунной системы пациента в целом, при чём указанная стабилизации заболевания, также наблюдались у пациентов, получавших интенсивное предварительное лечение химиотерапией [[Mantovani A. et.al. (2022) Macrophages as tools and targets in cancer therapy, Nat Rev Drug Discov., 21(11), 799-820].
В процессе перепрограммирования М1 в М2 в организме пациента происходят кардинальные изменения метаболических функций организма пациента, т.е. провоспалительные М1 макрофаги и альтернативно активированные макрофаги М2 обеспечивают массивные метаболические перестройки [Viola A. et. al. (2019) The metabolic signature of macrophage responses, Front. Immunol., 10, 1462], направленные на уничтожение онкологических клеток в организме больного.
Принимая во внимание, что большая часть метаболических путей, необходимых для поляризации макрофагов, связана с митохондриями, представляется перспективным метод воздействие на митохондриальный метаболизм М2 макрофагов.
Известно, что изменения в митохондриальных функциях макрофагов являются отличительным признаком провоспалительных М1 макрофагов [Jha A.K. et.al. (2015) Network integration of parallel metabolic and transcriptional data reveals metabolic modules that regulate macrophage polarization, Immunity., 42(3), 419-30], [Ramond E. et. al. (2019) Pivotal role of mitochondria in macrophage response to bacterial pathogens, Front. Immunol., 10,2461].
Макрофаги M1 не могут использовать митохондриальное дыхание для производства энергии и, следовательно, переключают свой метаболизм на гликолиз [Batista-Gonzalez A. (2020) New Insights on the Role of Lipid Metabolism in the Metabolic Reprogramming of Macrophages, Front. Immunol.].
При этом, макрофаги М2 демонстрируют умеренные уровни гликолиза и используют цикл трикарбоновых кислот, что позволяет им поддерживать высокие уровни окислительного фосфорилирования и рост опухоли.
А при этом провоспалительные М1 макрофаги демонстрируют очень низкие уровни окислительного фосфорилирования из-за нарушения электрон-транспортной цепи (ETC).
Таким образом можно сделать общий вывод о том, что способы перенаправления митохондриального метаболизма в опухолевых макрофагах (М2) из окислительного фосрфорилирования в гликолиз представляется перспективным методом перепрограммирования макрофагов из М2 в М1 при онкологических заболеваниях.
На дату подачи заявленного технического решения заявителем проведены исследования уровня техники и тенденций развития техники в данной области по научной и патентной информации, выявлены существующие проблемы, связанные с отсутствием эффективного подход к лечению рака, направленной как на опухолевые клетки, так и на микроокружение опухоли.
Вследствие изложенного, существует неразрешенная до настоящего времени проблема по выявлению методов воздействия на опухолевые макрофаги в том числе и за счет изменения их митохондриальных функций для лечения рака.
Из исследованного уровня техники выявлено изобретение по патенту № WO 2023287096 A1 (аналог № 1) «Сomposition for inhibiting m2 polarization of macrophages and treating cancer, comprising humanized monoclonal antibody specific to chi3l1» «Композиция для ингибирования м2-поляризации макрофагов и лечения рака, содержащая моноклональное антитело, специфичное к chi3l1».
Сущностью известного технического решения является фармацевтическая композиция, содержащая: моноклональное антитело, специфически связывающееся с Chi3L1 в качестве активного ингредиента, как способ ингибирования поляризации М2 макрофагов, для профилактики, лечения или ингибирования метастазирования рака. Композиция для ингибирования М2 поляризации макрофагов по.1, отличающаяся тем, что моноклональное антитело содержит следующие области:
(a) переменная область тяжелой цепи, состоящая из аминокислотной последовательности SEQ ID NO: 1; и
(b) переменная область легкой цепи, состоящая из аминокислотной последовательности SEQ ID NO: 3.
Композиция для ингибирования М2 поляризации макрофагов по.1, отличающаяся тем, что моноклональное антитело содержит следующие области:
(a) переменная область тяжелой цепи, состоящая из аминокислотной последовательности SEQ ID NO: 2; и
(b) переменная область легкой цепи, состоящая из аминокислотной последовательности SEQ ID NO: 4.
Композиция для ингибирования М2 поляризации макрофагов по.1, при этом композиция ингибирует фосфорилирование STAT6.
Способ ингибирования поляризации макрофагов М2, включающий контакт любой композиции любого из пп.1-4 с клетками, экспрессирующими белок Chi3L1.
Фармацевтическая композиция для профилактики, лечения или ингибирования метастазирования рака, содержащая композицию любого из пп.1-4 в качестве активного ингредиента.
Композиция для диагностики рака, содержащая композицию любого из пп.1-4 в качестве активного ингредиента.
Набор для диагностики рака, содержащий композицию любого из пп.1-4 в качестве активного ингредиента.
Таким образом, кратко сущностью известного технического решения является фармацевтическая композиция, содержащая: моноклональное антитело, специфичное к Chi3L1. Композиция обладает превосходным эффектом ингибирования роста опухоли и метастазирования и, таким образом, может по-разному применяться в качестве композиций для профилактики и лечения рака и ингибирования метастазы рака.
Недостатком известного изобретения по патенту № WO2023287096A1 является то, что моноклональные антитела, направленные против опухолеассоциированных макрофагов, быстро распадаются в организме, и обычно не более 20% введенной дозы взаимодействует с макрофагами [Beckman. et al. (2007) Potential Role for Early Biomarker Testing as Part of a Modern, Multidisciplinary Approach to Sjögren’s Syndrome Diagnosis. Adv Ther, 34, 779-812. - 2017. V 34, 799-812].
Также недостатком является то, что моноклональные антитела могут иметь несколько различных вариантов способа действия на клетки, и фактический механизм действия после введения пациентам не всегда ясен [E. Soderlind. et al. (2001) The Immune Diversity in a Test Tube - Non-Immunised Antibody Libraries and Functional Variability in Defined Protein Scaffolds. Bentham science, 4(5), 409-416]. Таким образом, известное техническое решение является ограниченным по эффективности использования по назначению.
Из исследованного уровня техники выявлено изобретение по патенту № WO 2019143948 A1 (аналог № 2) «Аltering inflammatory states of immune cells in vivo by modulating cellular activation states» «Изменение воспалительных состояний иммунных клеток путем изменения состояний клеточной активации». Сущностью известного технического решения является способ изменения состояния активации макрофагов в опухоли, включающий: введение в участок опухоли наночастиц, содержащих ядро из полиаминоэфира и покрытие из полиглутаминовой кислоты с транскрибируемой мРНК, кодирующей один или несколько регуляторных фактора, инкапсулированные в сердцевине, и при этом состояние активации макрофагов изменяется от классического состояния до активированного состояния. Опухоль представляет собой рак яичника, опухоль глиобластомы или опухоль метастатического рака легкого.
Таким образом, кратко сущностью известного технического решения является использование наночастиц, несущих мРНК, кодирующие M1-поляризующие факторы транскрипции, которые могут перепрограммировать TAM в убивающие опухолевые клетки макрофаги или иным образом вызывать разрушение опухолевых клеток, тем самым лечит рак.
Недостатком известного изобретения по патенту № WO2019143948A1 является то, что нацеливание на IKKβ для блокирования активности M2-подобных макрофагов, не является идеальным подходом при лечении онкологических заболеваний по мнению заявителя, поскольку эти киназы имеют несколько направлений воздействий и могут вызывать в том числе и нежелательные побочные эффекты, а именно ухудшение сердечной функции [Bhullar K.S. et al. (2018) Kinase-targeted cancer therapies: progress, challenges and future directions. Mol Cancer. V. 17 (48)]. Таким образом, известное техническое решение в целом, по мнению заявителя, являются ограниченным по эффективности при их использовании по назначению.
Из исследованного уровня техники выявлено изобретение по патенту № CN 101904837 A (аналог № 3) «Use of rotenone as non-small cell lung cancer cell sensitizer» «Использование ротенона в качестве сенсибилизатора клеток немелкоклеточного рака легкого».
Сущностью известного технического решения является применение ротенона, ингибитора митохондриальной дыхательной цепи, для предотвращения резистентности немелкоклеточных клеток рака легкого к таргетным противоопухолевым препаратам.
Ротенон представляет собой соединение, выделенное из природных растений, и может связываться с комплексом митохондриальной дыхательной цепи I, тем сазаявителем м ингибируя окислительное фосфорилирование митохондрий, уменьшая потенциал митохондриальной мембраны и вызывая повреждение клеток. Данное техническое решение показало, что применение соединения ротенона не оказывает очевидного токсического и побочного эффекта на нормальные клетки (HEK293), и показывает, что ротенон снижает количество опухолевых клеток рака легкого (A549 и NCI-H460) и являются новой лекарственной комбинацией, которая может быть использована для лечения немелкоклеточного рака легкого.
Недостатком известного изобретения по патенту № CN101904837A является то, что ротенон может ингибировать опухолевые клетки, но само соединение может индуцировать апоптоз нервных клеток и вызывать множество токсических и побочных эффектов, а именно рвоту, нарушение координации, мышечный тремор, клонические судороги и дыхательную недостаточность, вследствие чего возможны осложнения или летальный исход.
Заявленное техническое решение решает проблему посредством подбора дозы ротенона и других лекарственных препаратов таким образом, чтобы минимизировать его токсическое воздействие на здоровые клетки и усилит его воздействие на опухолевые клетки.
Из исследованного уровня техники выявлено изобретение по патенту № WO2020006199A2 (аналог № 4) «Methods and agents for modulating inflammation» «Способы и средства для моделирования воспаления».
Сущностью известного технического решения является способ усиления альтернативной активации макрофагов путем введения агента, повышающего внутриклеточные уровни кофермента А. Увеличение количества внутриклеточного кофермента А увеличивает поочередную активацию макрофагов, что приводит к подавлению иммунного ответа, что полезно при лечении воспалительных заболеваний. Снижение внутриклеточного уровня кофермента А снижает альтернативную активацию макрофагов и меняет активность опухолеассоциированных иммунных клеток, что эффективно использовать для лечения рака.
Недостатком известного изобретения по патенту № WO2020006199A2 является то, что он воздействует на один из этапов/процессов митохондриального метаболизма, тогда как в заявленном техническом решении предложен спектр препаратов, которые влияют на различные процессы метаболизма в митохондриях, а именно:
- подавление митохондриального дыхания,
- изменение электронно-транспортной цепи,
- подавление окислительного фосфорилирования,
- подавление карнитина, продукции АТФ.
Из исследованного уровня техники выявлено изобретение по патенту № CA3088009A1 (аналог № 5) «Methods and compositions for macrophage polarization» «Способы и композиции для поляризации макрофагов».
Сущностью известного технического решения является использование внеклеточных везикул, содержащих нуклеиновую кислоту, для изменения поляризации макрофагов, ассоциированных с опухолью.
Внеклеточная везикула, содержащая один или несколько компонентов, которые при контакте с макрофагом избирательно меняют макрофаг из фенотипа М2 в фенотип М1. Внеклеточная везикула представляет собой экзосому и содержит, несколько компонентов, ингибирующих один ген-мишень макрофага. Компонент входящий в состав везикул представляет собой нуклеиновую кислоту - ингибиторную РНК. Анализировали: поглощения внеклеточного везикулы, экспрессию целевого гена, высвобождение цитокинов и поверхность клеток макрофагов. При взаимодействии внеклеточных везикул, содержащих нуклеиновую кислоту, макрофаг M1 подтипа проявлял повышенную секрецию воспалительных цитокинов и хемокинов и пониженную секрецию противовоспалительных цитокинов и хемокинов, по сравнению с макрофагом M2 подтипа.
Таким образом, кратко сущностью известного технического решения является композиция, включающая внеклеточные везикулы, содержащего гены, перепрограммирующие поляризацию макрофагов для лечения рака.
Недостатком известного изобретения по патенту № CA3088009A1 является сложная техническая реализация. Выделение и очистка экзосом с использованием последовательных и воспроизводимых методов остается проблемой. Заявленное техническое решение решает проблему использованием известных фармацевтических препаратов по новому назначению.
Известен патент № WO2022087424A1 (аналог № 6) «Monoamine oxidase blockade therapy for treating cancer through regulating tumor associated macrophages (tams)» «Моноаминоксидазная блокадная терапия для лечения рака путем регуляции опухолеассоциированных макрофагов».
Сущностью известного технического решения является композиция вещества, включающая: химиотерапевтическое средство; ингибитор моноаминоксидазы А; и фармацевтически приемлемый носитель. Композиция по п. 1, отличающаяся тем, что ингибитор моноаминоксидазы А содержит по меньшей мере одно из следующего: фенелзин; моклобемид; хлоргилин; пирлиндол; изокарбоксазид; транилципромид; ипрониазид; кароксазон; бефлоксатон; брофаромин; цимоксатон; эпробемид; эсупрон; метроиндол; или толоксатон. Композиция по п. 1, отличающаяся тем, что композиция содержит липид; и/или композиция содержит ингибитор моноаминоксидазы А, расположенный внутри наночастицы. Композиция по п. 1, отличающаяся тем, что ингибитор моноаминоксидазы А присутствует в композиции в таких количествах, что количество ингибитора моноаминоксидазы А, доступное для ассоциированных с опухолью макрофагов у индивидуума, которому вводят композицию, является достаточным для модулирования фенотипа ассоциированного с опухолью макрофага. Композиция по п. 4, отличающаяся тем, что модуляция фенотипа ассоциированных с опухолью макрофагов включает по меньшей мере одно из следующего: снижение уровней внутриклеточных активных форм кислорода; повышенная иммунореактивность опухоли. Способ модулирования фенотипа опухолеассоциированного макрофага, включающий введение ингибитора моноаминоксидазы А в среду, в которой находится CD8 Т-клетка; при этом количество ингибитора моноаминоксидазы А, вводимого в окружающую среду, выбирают таким образом, чтобы оно было достаточным для модулирования фенотипа ассоциированного с опухолью макрофага. Способ по п. 8, отличающийся тем, что макрофаг, ассоциированный с опухолью, находится у индивидуума, у которого диагностирован рак. Способ по п. 9, отличающийся тем, что индивидуум проходит терапевтическую схему, включающую введение химиотерапевтического агента. Способ по п. 9, отличающийся тем, что рак представляет собой лимфому или рак кожи, молочной железы, яичников, предстательной железы, колоректальный рак или рак легких. Способ по п. 13, отличающийся тем, что ингибитор моноаминоксидазы А расположен внутри наночастицы; необязательно наночастица, содержащая липид. Способ по п. 8 или п. 16, отличающийся тем, что ингибитор моноаминоксидазы А находится в композиции, содержащей сшитую многослойную липосому, имеющую внешнюю поверхность и внутреннюю поверхность, причем внутренняя поверхность образует центральную полость липосомы, при этом многослойная липосома включает по меньшей мере первый липидный бислой и второй липидный бислой, причем первый липидный бислой ковалентно связан со вторым липидным бислоем; и ингибитор моноаминоксидазы А, расположенный внутри липосомы.
Более кратко, сущностью известного технического решения является нацеливание на опухолеассоциированные макрофаги (TAM) как многообещающая стратегия для изменения микроокружения опухоли и улучшения иммунотерапии рака. Моноаминоксидаза А (МАО-А) является ферментом, наиболее известным своей функцией в головном мозге; низкомолекулярные ингибиторы МАО-А клинически используются для лечения неврологических расстройств. Ранее была обнаружена индукция МАО-А в мышиных и человеческих TAM. В данном техническом решении определили, что у мышей с дефицитом МАО-А наблюдалось снижение противовосполительных функций ТАМ, соответствующих усиленному противоопухолевому иммунитету. Данное техническое решение определило, что лечение МАО индуцировало перепрограммирование ТАМ и подавленный рост опухолевых клеток в мышиных и человеческих опухолевых моделях. Также было продемонстрировано, что МАО-А способствует поляризации ТАМ посредством повышения регуляции окислительного стресса. Вместе эти данные идентифицируют МАО-А как регулятора поляризации TAM и возможность применения для улучшения иммунотерапии рака.
Недостатком известного изобретения по патенту № WO2022087424A1 является проблема безопасности при лечении ингибиторами МАО, а именно реакция гипертонического криза и серотониновый синдром. Гипертонические кризы могут возникать в результате фармакодинамического взаимодействия препаратами или при чрезмерном употребления тирамина с пищей. Серотониновый синдром возникает, когда препараты, повышающие доступность серотонина, принимаются при ингибировании активности МАО [Bodkin J.A. et.al. (2019) Moving on With Monoamine Oxidase Inhibitors, Focus (Am Psychiatr Publ), 19(1), 50-52]. Заявленное техническое решение решает проблему, воздействуя на митохондриальный метаболизм, без побочных действий.
Таким образом, на дату представления заявочных материалов заявителем не выявлена какая-либо информация с клиническими и доклиническими исследованиями по использованию фармакологических препаратов, блокирующих процессы окислительного фосфорилирования в митохондриях, для применения при лечении онкологических заболеваний. Таким образом, заявленное техническое решение является дополнительным доказательством эффективности фармакологических методов воздействия на опухолевые макрофаги за счет изменения их митохондриальных функций и, как следствие, является перспективным направлением внедрения эффективных лекарств для лечения рака.
Выявленные аналоги совпадают с заявленным техническим решением по отдельным признакам, поэтому прототип не выявлен и формула изобретения составлена без ограничительной части.
Техническим результатом заявленного технического решения является подбор подходящих лекарств и фармакологических соединений, которые обеспечивают возможность смещения митохондриального метаболизма с окислительного фосфорилирования, свойственного макрофагам М2 типа, в гликолизный тип, характерный для макрофагов М1 типа. Этот подход, по мнению заявителя, является наиболее перспективным способом подавления роста опухолевых клеток у больных онкологическими заболеваниями.
Сущностью заявленного технического решения является способ подбора лекарственных средств для реализации фармакологической индукции митохондриальной дисфункции в макрофагах для противоопухолевой терапии заключающийся в том, что на первом этапе проводят культивирование опухолевых клеток мыши и человека, для чего опухолевые клетки мыши и человека культивируют в питательной среде DМЕМ, содержащей 10% сыворотки крови плодов коровы, L-глутамин и смесь антибиотиков пенициллин-стрептомицин, при температуре 37°C, во влажной атмосфере, содержащей 5% СО2; на втором этапе проводят культивирование и дифференцировка ТНР-1 клеток, для чего моноцитарную клеточную линию человека THP1 в качестве модели макрофагов человека культивируют в среде RPMI-160 содержащей 10% сыворотки, антибиотики и 2 мМ L-глутамина, при этом для дифференциации миелоидных клеток-предшественников до макрофагов добавляют форбол-12-миристат-13-ацетат, PMA в концентрации 2 нг/мл, инкубируют 48 часов при температуре 37°C, во влажной атмосфере, содержащей 5% СО2; далее клетки промывают фосфатно-солевым буфером Дульбекко DPBS и добавляют среду RPMI-160 содержащей 10% сыворотки, антибиотики и 2 мМ L-глутамина и инкубируют при температуре 37°C, во влажной атмосфере, содержащей 5% СО2 в течение двух дней; на третьем этапе проводят получение первичных макрофагов человека из мононуклеарных клеток периферической крови здоровых доноров, для чего мононуклеарные клетки, полученные из периферической крови человека, промывают DPBS 2 раза, далее добавляют питательную среду RPMI-160 с 5% человеческой сывороткой, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ L-глутамина, для дифференцировки до макрофагов добавляют человеческий MCSF в концентрации 20 нг/мл и инкубируют 9 дней при температуре 37°C, во влажной атмосфере, содержащей 5% СО2; на четвертом этапе проводят получение макрофагов из миелоидных клеток-предшественников костного мозга мыши, для чего моноциты, полученные из костного мозга мыши, культивируют в среде RPMI-160 с 10% фетальной бычьей сывороткой FBS, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ L-глутамина при температуре 37°C, во влажной атмосфере, содержащей 5% СО2; для дифференциация миелоидных клеток-предшественников до макрофагов добавляют рекомбинантный макрофагальный колониестимулирующий фактор M-CSF в концентрации 20 нг/мл; на пятом этапе проводят совместное культивирование мышиных макрофагов и опухолевых клеток в сфероидах, для чего для предотвращения клеточной адгезии к пластику, культуральный пластик покрывают тонким слоем 1% агарозы, далее планшет с 1% агарозой инкубируют при температуре 37°C в течение 20 минут для полной полимеризации матрикса, до достижения гелеобразного состояния; далее получают многокомпонентную клеточную ко-культуру путем внесения в лунки заранее подготовленных опухолевых клеток рака легкого мыши и макрофагов, полученных из костного мозга мыши в соотношении 1:3, находящихся в сфероидной среде содержащей, DMEM/F12, 0,4 % бычьего сывороточного альбумина BSA, L-глутамина, смесь антибиотиков пенициллин-стрептомицин, 1 мл добавки B-27 ™ (50X), 20 нг/мл фактора роста фибрабластов FGF2 и 20 нг/мл эпидермального фактор роста EGF; к многокомпонентной клеточной ко-культуре добавляют вещества с исследуемой противоопухолевой активностью в ранее выбранных концентрациях: этомоксир 0,5 мкмоль, циклоспорин А 0,5 мкмоль, диметилфумарат 1 мкмоль, ставудин 1,25 мкмоль, диданозин 2,5 мкмоль, ротенон 0,01 мкмоль, антимицин А 3 наномоль, олигомицин 0,1 мкмоль, карбонилцианид-п-трифторметоксифенилгидразон FCCP 0,15 мкмоль, BPTES 0,5 мкмоль; далее многокомпонентную клеточную ко-культуру инкубируют 5 дней при температуре 37°C, в насыщенной влажной атмосфере, содержащей 5% СО2 до образования клетками сфероподобных структур; морфологию клеток ко-культуры анализируют с использованием фазово-контрастной микроскопии; клетки собирают и центрифугируют для их осаждения; получившийся осадок ресуспендируют и инкубируют 3 мин при температуре 37°C и подсчитывают количество клеток в камере Горяева, для камеры используют формулу: N / 15 * 250 * 10^3, где N - число клеток в пятнадцати квадратах камеры, расположенных по диагонали/объем в мл; на шестом этапе проводят мониторинг роста опухолевых клеток в кондиционированных средах из активированных макрофагов, для чего к макрофагам, полученным из костного мозга мыши, на 5 день дифференцировки добавляют кондиционированную среду от опухолевых клеток рака легкого мыши или рака поджелудочной железы в соотношении 1:3 и препараты в выбранных концентрациях и инкубируют 24 ч при температуре 37°C, во влажной атмосфере, содержащей 5% СО2; на следующий день клетки промывают буфером DPBS и добавляют среду DMEM, содержащей 2% сыворотку крови плодов коровы, L-глутамин и смесь антибиотиков и инкубируют 72 ч; далее кондиционированную среду от активированных макрофагов центрифугируют при 300 g 5 минут и хранят в морозильной камере при температуре минус 20°С; опухолевые клетки рака легкого мыши и поджелудочной железы культивируют в среде DMEM с 10% сывороткой и антибиотиками; на следующий день клетки промывают буфером DPBS и добавляют кондиционированную среду от активированных макрофагов - 1 часть, и среду DMEM, содержащей 0.5 % сыворотки и антибиотики - 2 части; через 9 дней производят подсчет опухолевых клеток; опухолевые клетки рака легкого мыши и поджелудочной железы промывают буфером DPBS, добавляют 200 мкл 0,25% трипсина-ЭДТА, инкубируют 3 мин и подсчитывают количество клеток, после чего определяют рост или подавление опухолевых клеток; на седьмом этапе проводят поляризацию первичных макрофагов, полученных из периферической крови, для чего к дифференцированным макрофагам, полученным из периферической крови доноров, добавляют 20 нг/мл интерлейкина-4, IL-4 и комбинацию 20 нг/мл липополисахарида, LPS + 20 нг/мл интерферона- гамма, IFNγ и препараты в выбранных концентрациях; через 24 часа к клеткам добавляют 1 мл раствора Extra RNA для лизиса и последующего выделения РНК; на восьмом этапе проводят поляризацию мышиных макрофагов, для чего к макрофагам, полученным из костного мозга мыши, на 5 день дифференцировки добавляют 20 нг/мл интерлейкина-4, IL-4 и комбинацию 20 нг/мл липополисахарида, LPS + 20 нг/мл интерферона- гамма, IFNγ и препараты в выбранных концентрациях; на следующий день к клеткам добавляют 1 мл раствора Extra RNA для лизиса и последующего выделения РНК; на девятом этапе проводят поляризацию THP-1 клеток, для чего моноцитарную клеточную линию человека THP1 дифференцируют по стандартному протоколу, к дифференцированным ТНР1 клеткам добавляют 20 нг/мл интерлейкина-4, IL-4 и комбинацию 20 нг/мл липополисахарида, LPS + 20 нг/мл интерферона- гамма, IFNγ и препараты в выбранных концентрациях; на следующий день клетки лизируют в 1 мл Extra RNA раствора для выделения РНК; на десятом этапе проводят выделение РНК, для чего образцы клеток в Extra RNA растворе перемешивают в течение 1 минуты и центрифугируют при 12000 g 5 минут при комнатной температуре, затем перемешивают в течение 1 минуты и центрифугируют при 12000 g 5 мин при комнатной температуре; получившийся супернатант переносят в новую емкость и добавляют к нему 200 мкл хлороформа, перемешивают и инкубируют в течение 5 минут при комнатной температуре, периодически встряхивая образец, после чего центрифугируют при 12000 g 15 мин при
комнатной температуре, затем отбирают верхнюю фазу и переносят ее в новую емкость; добавляют 500 мкл 100% изопропанола, перемешивают и затем инкубируют 10 минут при комнатной температуре; затем центрифугируют при 12000 g 10 минут при комнатной температуре, затем удаляют супернатант и промывают осадок в 1 мл 75% EtOH; затем центрифугируют в течение при 12000 g 5 минут при комнатной температуре, удаляют супернатант и высушивают осадок при комнатной температуре; РНК растворяют в 40 мкл воды стерильной водой свободной от нуклеаз и помещают в морозильную камеру при температуре минус 80°С; на одиннадцатом этапе проводят получение кДНК, для чего предварительно измеряют концентрацию РНК на Nano Drop в концентрации нг/мкл, оценивают чистоту образцов - соотношение пиков поглощения 260 нм/280 нм - соответствуют примесям белков и аминокислот; 260 нм/230 нм - примеси солей и фенола; необходимое количество РНК доводят до 15 мкл стерильной водой свободной от нуклеаз и добавляют 4 мкл 5х iScript Reaction Mix и 1 мкл iScript Reverse Transcriptase; пробирки помещают в амплификатор C1000 Touch (Bio-Rad, США), следуя инструкции производителя; на двенадцатом этапе проводят ПЦР реакцию в реальном времени, для чего готовят Master Mix: к 5 мкл SybrGreen, добавляют 3,6 мкл воды, свободной от нуклеаз и 0,4 мкл праймера для соответствующих генов; готовый Master Mix раскапывают в 96-луночный планшет по 9 мкл; далее раскапывают по 1 мкл образцы ДНК, Standard и Negative Control в трех повторностях; планшет закрывают оптической прозрачной пленкой и помещают в амплификатор; проводят ПЦР, получают данные о пороговых значениях циклов ПЦР, используя которые рассчитывают уровень экспрессии интересующих генов, при этом показывают смещение митохондриального метаболизма с окислительного фосфорилирования, свойственного макрофагам М2 типа, в гликолизный тип, характерный для макрофагов М1 типа.
Заявленное техническое решение иллюстрируется Фиг.1 - Фиг.14.
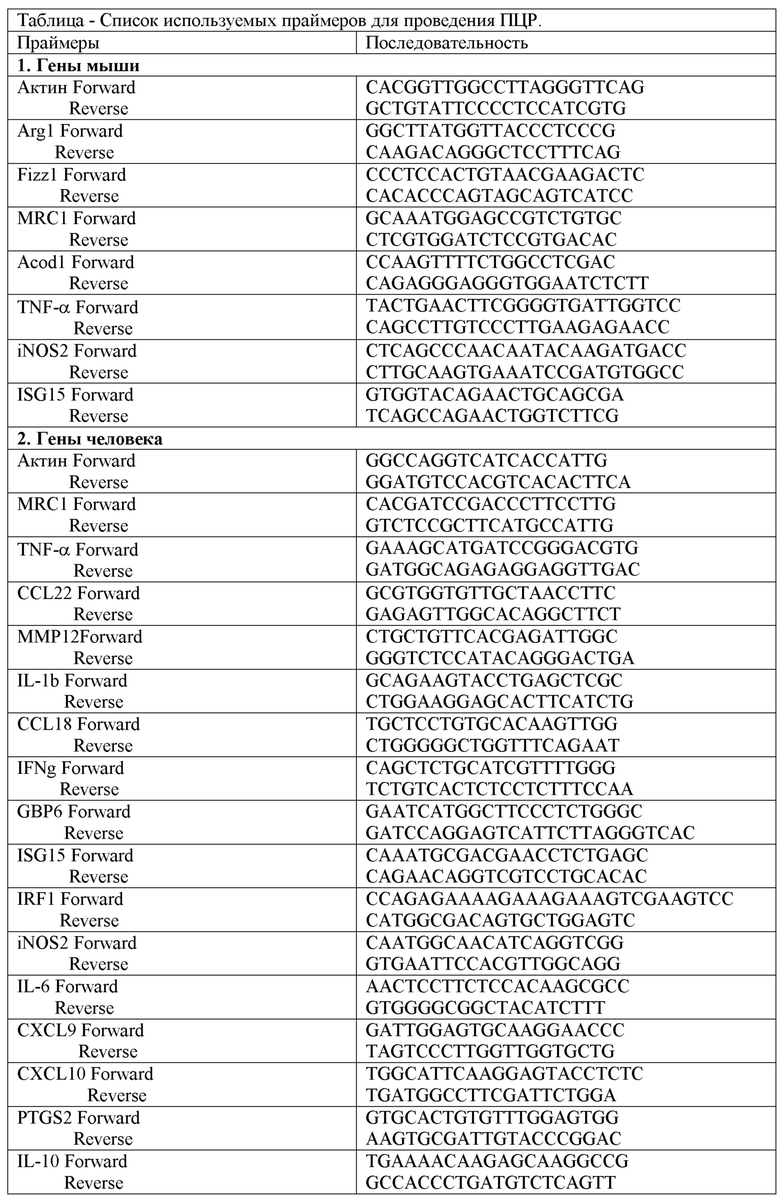
На Фиг. 1 представлен анализ профиля поляризации макрофагов дикого типа и OPA1ΔM, где:
1A - qPCR-анализ экспрессии генов, являющихся маркерами M2 макрофагов мыши, стимулированных IL-4 в течение 24 часов. Необработанные M0 макрофаги мыши указаны, как контроль (co).
1B - qPCR-анализ экспрессии генов, являющихся маркерами M1 макрофагов мыши, стимулированных рекомбинантным IFNγ и LPS в течение 24 часов.
1C - qPCR-анализ экспрессии генов, являющихся маркерами M2 и M1 макрофагов мыши, стимулированных 24 часа кондиционированной средой, собранной с опухолевых клеток линии KPC.
Значения являются средними значениями ± SD. n.s. = несущественный; *p<0.05; **p<0.01; ***p<0.001; n= 3.
Condition medium (CM) - кондиционная среда; relative expression (fold) - относительная экспрессия (сумма); WT - макрофаги, полученные от мышей Opa1flox / flox; KO - макрофаги, полученные от мышей Opa1flox / flox Lyz2Cre / Cre; М1-like - М1 подтип макрофагов; М2-like - М2 подтип макрофагов; со - контроль; Arg1, Ym1, Fizz1 - праймеры к М2- подтип макрофагам; TNF-α, iNOS, Acod1, CXCL9, ISG15, IRF1, GBP6, IL6; CXCL1 - праймеры к М1 - подтип макрофагам; IL4 - макрофаги, поляризованные IL-4; IFN - макрофаги, поляризованные IFNγ и LPS; СМ - кондиционная среда.
На Фиг. 2 представлен анализ профиля поляризации контрольных и трансфицированных малой интерферирующей РНК-Opa1 (миРНК-Opa1) макрофагов человека, где:
2A - Уровень мРНК OPA1 в макрофагах человека после трансфекции миРНК-OPA1.
2B - qPCR-анализ генов, экспрессия которых является маркером M2 макрофагов человека, стимулированных IL-4 в течение 24 часов.
2C - qPCR-анализ генов, экспрессия которых является маркером M1 макрофагов человека, стимулированных IFNγ и LPS в течение 24 часов. Необработанные M0 макрофаги мыши указаны, как контроль (co). Значения являются средними значениями ± SD. n.s. = несущественный; *p<0.05; **p<0.01; ***p<0.001; **** p<0.0001 n = 3.
Relative expression (fold) - относительная экспрессия (сумма), M2- markers - М2- маркеры, M1- markers- М1- маркеры, si-scr - малая интерферирующая РНК, si-OPA1 - малая интерферирующая РНК-Opa1; ОРА1 - праймеры к белку ОРА1; МRС1, СD163, CCL18 - праймеры к М2-подтип макрофагам; TNF-α, CXCL10, IRF1,- праймеры к М1 - подтип макрофагам; со - контроль; IL4 - макрофаги, поляризованные IL-4; IFN - макрофаги, поляризованные IFNγ и LPS.
На Фиг. 3 представлен анализ, подтверждающий неспособность макрофагов OPA1ΔM поддерживать рост опухолевых клеток при совместном со-культивировании, где:
3A - Схема эксперимента по совместному культивированию.
3B - Микроскопическая оценка сфероидов на 5-й день после посева клеток.
3C - Количество опухолевых клеток на 5-й день после посева. Культура опухолевых клеток взята в качестве контроля (control); серые столбики - опухолевые клетки, выращенные с макрофагами дикого типа (wtM); красные столбики - опухолевые клетки, выращенные с макрофагами OPA1ΔM (koM). *p<0.05; **p<0.01; ***p<0.001; n = 3.
Tumor cells - опухолевые клетки; mouse macrophages - мышиные макрофаги; Co- culture with tumor cells 1:3 (in spheroid medium) - ко-культивирование с опухолевыми клетками 1:3 (в сфероидной среде); 5 days - 5 дней; spheroids - сфероиды; count cells - подсчет клеток; breast cancer E0771 - рак молочной железы, E0771; direct co-culture with macrophages - прямое ко-культивирование с макрофагами; pancreatic cancer, КРС - рак поджелудочной железы; lung cancer, (KrasG12D/p53-/-) - рак легкого, (KrasG12D/p53-/-); macrophages - макрофаги; control - контроль; + WT M0 - совместное культивирование опухолевых клеток с макрофагами дикого типа; +KO M0 - совместное культивирование опухолевых клеток с макрофагами OPA1ΔM; cell number x103 - количество клеток x103; wtM - опухолевые клетки, выращенные с макрофагами дикого типа; koM - опухолевые клетки, выращенные с макрофагами OPA1ΔM.
На Фиг. 4 представлен анализ фагоцитоза макрофагов методом проточной цитометрии, где:
4A - Схема, изображающая анализ процесса фагоцитоза макрофагами раковых клеток методом проточной цитофлуориметрии.
4B - Фагоцитоз опухолевых клеток линий KPC и E0771 диким типом и OPA1ΔM макрофагами.
4C - Анализ фагоцитоза макрофагами человека. Макрофаги метили красным красителем; опухолевые клетки - зеленым красителем. Фагоцитарные макрофаги являются дважды положительными клетками. Значения являются средними значениями ±SD. n.s. = несущественный; n = 3.
CellTraker™Green - зеленый клеточный краситель; CellTraker™Deep Red - красный клеточный краситель; tumor cells - опухолевые клетки; mouse macrophages - мышиные макрофаги; 8 h - 8 часов; phagocytosis - фагоцитоз; cancer cells - раковые клетки; B525 FITC-A - флуоресцентный краситель; R660 APC-A - флуоресцентный краситель; phagocytic E0771 - фагоцитоз Е0771; phagocytic КРС - фагоцитоз КРС macrophages - макрофаги; human macrophages - человеческие макрофаги; % of phagocytic macrophages - % фагоцитирующих макрофагов; si-scr - малая интерферирующая РНК, si-OPA1 - малая интерферирующая РНК-Opa1; WT - макрофаги, полученные от мышей Opa1flox / flox; KO - макрофаги, полученные от мышей Opa1flox / flox Lyz2Cre / Cre; ОРА-1 - белок ОРА-1.
- На Фиг. 5 представлена схема эксперимента: культивирование опухолевых клеток мыши в кондиционированной среде, собранной с активированных макрофагов (полученные из костного мозга мыши), предварительно проинкубированные с кондиционированной средой из опухолевых клеток, где:
5A - Схема, изображающая эксперимент по совместному культивированию.
5B - Микроскопическая оценка сфероидов на 5-й день, культивированных в кондиционированной среде от макрофагов дикого типа (WT) или OPA1ΔM макрофагов (KO). Культура опухолевых клеток в питательной среде помечена как контроль (control).
5C - Количество опухолевых клеток на 5-й день после посева, культивируемых в кондиционированных средах от макрофагов. Серые столбцы - опухолевые клетки, выращенные с макрофагами дикого типа (wtM); красные столбцы - опухолевые клетки, выращенные с макрофагами OPA1ΔM (KO). *p<0.05; **p<0.01; n=3.
Tumor cells - опухолевые клетки; CM from activated macrophages - кондиционная среда от опухолевых клеток; 3 days - 3 дня; mouse macrophages - мышиные макрофаги; 24 hrs - 24 часа; CM from tumor cells - кондиционная среда от опухолевых клеток; growth for 5 day - рост в течении 5 дней; spheroids 1:1 CM + media - сфероиды 1:1 кондиционная среда+ среда; lung cancer KrasG12D;p53-/- - рак легкого KrasG12D;p53-/-; with CM from macrophages - с кондиционной средой от макрофагов; control - контроль; + WT М0 - кондиционированная среда от макрофагов дикого типа; + КО М0 - кондиционированная среда от OPA1ΔM макрофагов; culture in conditioned medium from macrophages - культура в кондиционной среде от макрофагов; pancreatic cancer, КРС - рак поджелудочной железы, КРС, СМ from macrophages; cell number x103 - количество клеток x103; wtM - опухолевые клетки, выращенные с макрофагами дикого типа; koM - опухолевые клетки, выращенные с макрофагами OPA1ΔM.
На Фиг. 6 представлен список фармацевтических соединений, воздействующих на митохондриальные функции, и оптимизированные нетоксичные концентрации для исследований in vitro.
На Фиг. 7 представлена оптимизация концентраций фармацевтических соединений, вызывающих митохондриальную дисфункцию в макрофагах и опухолевых клетках.
Mouse macrophages - мышиные макрофаги; human macrophages - человеческие макрофаги; THP-1 - линия клеток ТНР-1; А549 - опухолевые клетки рака легкого человека; КРС - опухолевые клетки поджелудочной железы мыши; lung cancer - рак легкого; number of tumor cells - количество опухолевых клеток; control - контроль; etomoxir - этомоксир; dimethyl fumarate - диметил фумарат; stavidine - ставудин; didanosine - диданозин; cyclosporine A (CyA) - циклоспорин А, rotenone - ротенон; antimycin A - антимицин А; oligomycin - олигомицин; FCCP - карбонилцианид-п-трифторметоксифенилгидразон; DMSO - диметилсульфоксид; number of tumor cells, x10 - количество опухолевых клеток.
На Фиг. 8 представлен анализ активации макрофагов, дифференцированных из костного мозга мыши и поляризованных в макрофаги M2 с помощью IL4 и в M1 с помощью IFNg+LPS в присутствии препаратов, вызывающих митохондриальную дисфункцию, где:
8A - RT-qPCR анализ экспрессии mRNA для M2-маркерных генов в макрофагах мыши дифференцированных с M-CSF в течение 5 дней и обработанных 20 нг/мл рекомбинантного IL4 в течение 24 часов в присутствии ДМСО (контроль) или выбранных препаратов.
8B - RT-qPCR анализ экспрессии mRNA для M1- маркерных генов в макрофагах мыши дифференцированных с M-CSF в течение 5 дней и обработанных 20 нг/мл IFNg и 100 нг/мл LPS в течение 24 часов в присутствии ДМСО (контроль) или выбранных препаратов. ns - несущественный, *p<0.05; **p<0.01; ***p<0.001; n=3.
Relative expression (fold) - относительная экспрессия (сумма), M2- genes - М2- гены, IL-4 - интерлейкин 4; M1- genes- М1- гены; IFNg+LPS - интерферон гамма+ липополисахарид; didanosine - диданозин; cyclosporine A (CyA) - циклоспорин А, rotenone - ротенон; oligomycin - олигомицин; FCCP - карбонилцианид-п-трифторметоксифенилгидразон, DMSO - диметилсульфоксид; ng/ml - нанограмм на милилитр; Arg1, Fizz1, Acod1, МRС1 - праймеры к М2- подтип макрофагам; TNF-α, iNOS, ISG15 - праймеры к М1 - подтип макрофагам.
На Фиг. 9 представлен анализ влияния препаратов, вызывающих митохондриальную дисфункцию, на поляризацию макрофагов, дифференцированных из моноцитов периферической крови человека. Влияние препаратов, вызывающих митохондриальную дисфункцию, на поляризацию макрофагов, дифференцированных из моноцитов переферической крови человека, где:
9А - RT-qPCR анализ экспрессии mRNA в донорских моноцитах, дифференцированных в течение 8 дней и обработанных 20 нг/мл рекомбинантного IL4 (M2-маркерные гены)
9В - или 20 нг/мл IFNg и 100 нг/мл LPS в течение 24 часов (M1-маркерные гены) в присутствии ДМСО (контроль) или выбранных препаратов. ns - несущественный, *p<0.05; **p<0.01; ***p<0.001; n=3.
Relative expression (fold) - относительная экспрессия (сумма); M2- genes - М2- гены; M1- genes- М1- гены; IL-4 - интерлейкин 4; M1- genes- М1- гены; didanosine - диданозин; cyclosporine A (CyA) - циклоспорин А; rotenone - ротенон; oligomycin - олигомицин; FCCP - карбонилцианид-п-трифторметоксифенилгидразон; DMSO - диметилсульфоксид; MYLS22 - ингибитор ОРА-1; Alox15, CCL18, CD206, MMP12, CD163 - праймеры к М2- подтип макрофагам; TNF-α, IRG1, IL12b, IL1b, GBP6 - праймеры к М1 - подтип макрофагам.
На Фиг. 10 представлено тестирование фармацевтических соединений, вызывающих митохондриальную дисфункцию, в условиях со-культивировании клеток рака легкого с макрофагами. Клетки первичного рака легкого мыши культивировали в сфероидах либо поодиночке (только wt/ko) или с макрофагами (+macro) в присутствии ДМСО (-) или ингибиторов (+) в указанных концентрациях. Представлено количество раковых клеток на 5-й день после посева. ns - не значительно; *p<0.05; **p<0.01; ***p<0.001; n=4.
10А - Тестирование расширенной группы препаратов.
10В - Представлено нормализованное число клеток из трех независимых экспериментов. ns- несущественный, *p<0.05; **p<0.01; ***p<0.001; n=3.
Group - группа; number of tumor cells - количество опухолевых клеток; control - контроль; etomoxir - этомоксир; dimethyl fumarate - диметил фумарат; stavidine - ставудин; didanosine - диданозин; cyclosporine A (CyA) - циклоспорин А; rotenone - ротенон; antimycin A - антимицин А; oligomycin - олигомицин; FCCP - карбонилцианид-п-трифторметоксифенилгидразон; DMSO - диметилсульфоксид; only wt/ko - только клетки рака легкого; wt/ko + macro - клетки рака легкого и макрофаги; drug addition - добавление препарата; μM - микромоль; nM - наномоль; only tumor cells - только опухолевые клетки; tumor cells + macro - опухолевые клетки и макрофаги; number of tumor cells; x50 - количество опухолевых клеток; number of tumor cells (fold) - количество опухолевых клеток (сумма).
На Фиг. 11 представлено тестирование фармацевтических соединений, вызывающих митохондриальную дисфункцию, в условиях культивировании мышиных раковых клеток в кондиционной среде от макрофагов. Клетки первичного рака легкого мыши (А) и поджелудочной железы (В) культивировали в кондиционной среде, собранной с активированных макрофагов, обработанных митохондриальными препаратами. Представлено количество раковых клеток на 7-й день после обработки кондиционной средой. ns - не значительно; *p<0.05; **p<0.01; ***p<0.001; n=3.
Cell number, x104 - количество клеток, x104; control - контроль; didanosine - диданозин; rotenone - ротенон; oligomycin - олигомицин; FCCP - карбонилцианид-п-трифторметоксифенилгидразон; lung cancer - рак легкого; КРС - опухолевые клетки рака поджелудочной железы мыши; CM from mouse macro - кондиционная среда от макрофагов.
На Фиг. 12 представлено влияние факторов, продуцируемых макрофагами THP1, на рост опухолевых клеток рака легкого A549. Макрофаги THP1 дифференцировали с помощью РМА в концентрации 2 нг/мл и обрабатывали кондиционированной средой от клеток рака легкого A549 в течение 24 часов. После этого среду меняли на 0,5% сыворотку. Через три дня кондиционированную среду (СМ) из макрофагов THP1 собирали и добавляли к клеткам A549, культивируемых на среде с 0.5% сыворотки в среде (соотношение 1 нормальная среда: 2 СМ). Клетки культивировали еще 8 дней. Количество контрольных необработанных клеток A549 (control) и клеток, выращенных в CM из THP1, было подсчитано и представлено на графике. ****p<0.0001;
Cell number, x104 - количество клеток; control - контроль; CM - кондиционная среда; A549 - опухолевые клетки рака легкого человека.
На Фиг. 13 представлены препараты, влияющие на митохондриальный метаболизм, и способность ТНР1 макрофагов индуцировать пролиферацию раковых клеток. Клетки рака легкого человека A549 (А) и NCI-H69 (В) культивировали в условиях дефицита сыворотки (0,5% сыворотки) либо в контрольной среде (control), либо в кондиционированной среде из активированных макрофагов THP1 (CM из THP1), предварительно обработанных указанными лекарственными препаратами. Количество клеток, выросших за 8 дней, представлено на графике. **p<0.01; ***p<0.001; n=2-4.
Number of tumor cells, x103 - количество опухолевых клеток, x103; number of tumor cells, fold change - количество опухолевых клеток, кратное изменение; co - контроль; didanosine - диданозин; rotenone - ротенон; oligomycin - олигомицин; FCCP - карбонилцианид-п-трифторметоксифенилгидразон; ; Conditioned medium - кондиционная среда; A549 - опухолевые клетки рака легкого человека; NCI-H69- опухолевые клетки рака легкого человека.
На Фиг. 14 представлены препараты, влияющие на митохондриальный метаболизм, и способность первичных макрофагов индуцировать пролиферацию раковых клеток. Клетки рака легкого человека A549 (А) и рака поджелудочной железы Panc1 (В) культивировали в условиях дефицита сыворотки (0,5% сыворотки) либо в контрольной среде (control), либо в кондиционированной среде из активированных первичных макрофагов, дифференцированных из моноцитов переферической крови здоровых доноров (conditioned media from primary macrophages), предварительно обработанных указанными лекарственными препаратами. Количество клеток, выросших за 8 дней, представлено на графике. *p<0.05; **p<0.01; ***p<0.001; n=3.
Number of cells, x104 - количество клеток, x104; relative cell density (fold) - относительная плотность клеток (сумма); control - контроль; didanosine - диданозин; cyclosporine A (CyA) - циклоспорин А, rotenone - ротенон; oligomycin - олигомицин; FCCP - карбонилцианид-п-трифторметоксифенилгидразон; DMSO - диметилсульфоксид; MYLS22 - ингибитор ОРА-1; conditioned medium from prime macrophages - кондиционная среда от активированных макрофагов; А549 - опухолевые клетки рака легкого человека; Panc1 - опухолевые клетки рака поджелудочной железы человека.
Далее заявителем приведено описание заявленного технического решения.
В заявленном техническом решении идентифицированы фармацевтические соединения, воздействующие на митохондриальные функции в макрофагах. Заявителем осуществлены исследования по подбору лекарств и фармакологических соединений, известных из доступной информации на дату представления заявочных материалов и литературных данных, а именно таких как этомоксир, циклоспорин А, диметилфумарат, ставудин, диданозин, ротенон, антимицин А, олигомицин, карбонилцианид-п-трифторметоксифенилгидразон (FCCP), BPTES, MYLS22, которые обладают свойствами по изменению митохондриальных функций макрофагов.
При этом следует акцентировать внимание на том, что некоторые из этих препаратов уже используется для лечения таких заболеваний как:
- сахарный диабет (этомоксир),
- ВИЧ (ставудин, диданозин),
- аутоиммунные заболевания (циклоспорин А, диметилфумарат).
Таким образом, заявителем проведены исследования по применению известных препаратов и соединений по новому назначению, а именно - для лечения онкологических заболеваний.
Представленные на основе заявленного технического решения лекарственные препараты и фармацевтические соединения после проведения доклинических и клинических испытаний могут стать перспективными препаратами для лечения онкологических заболеваний. При этом имеется возможность использовать эти лекарства и соединения при различных комбинациях лечения, в том числе и при химиотерапии и/или иммунной терапией для усиления их терапевтического воздействия на опухоль. В исследованиях заявителя были протестированы два типа рака - рак легкого и рак поджелудочной железы, являющимися наиболее агрессивными типами рака. При этом имеется высокая вероятность того, что эти лекарства могут быть также эффективны и для лечения других типов рака в силу того, что все виды онкологических заболеваний также зависят от воздействия макрофагов на опухолевые и здоровые клетки больного, например, рак молочной железы или рак кишечника и иные виды онкологических заболеваний, с явно выраженным вкладом макрофагов.
Далее заявителем представлены систематизированные данные о производителе и каталожных номерах препаратов, использованных для проведения экспериментов.
фенилгидразон
(нет перевода на русский язык)
(нет перевода на русский язык)
Далее заявителем приведено подробное описание заявленного технического решения, которое проводят в двенадцать этапов.
Первый этап (культивирование опухолевых клеток мыши и человека).
В экспериментах используют опухолевые культуры клеток E0771, А549, NCI-H69, Panc1, Patu8988T которые получают из Американской коллекции клеточных культур (от англ. American Type Culture Collection, ATCC). Клеточные линии рака легкого (KrasG12D и делетированный р53) и рака поджелудочный железы (линия KPC c мутациями KrasG12D и p53 R172H) могут быть выделены из опухолей мышей линии C57Black/6.
Опухолевые клетки мыши и человека культивируют в питательной среде DМЕМ (ПанЭко, Россия), содержащей 10% сыворотки крови плодов коровы (Biosera США), L-глутамин (ПанЭко, Россия) и смесь антибиотиков пенициллин-стрептомицин (ПанЭко, Россия) при температуре 37°C, во влажной атмосфере, содержащей 5% СО2.
Второй этап (культивирование и дифференцировка ТНР-1 клеток).
В ходе исследования также используют моноцитарную клеточную линию человека THP1 в качестве модели макрофагов человека (American Type Culture Collection (ATCC). Клетки воспроизводят основные морфологические, фенотипические и функциональные характеристики моноцитов, а именно способность к про- и противовоспалительной активации, продукции цитокинов, антиген-презентирующей активности. Клетки линии THP-1 представляют собой суспензионную культуру, требующую пересева 1 раз в 2-3 дня.
Моноцитарную клеточную линию человека THP1 в качестве модели макрофагов человека культивируют в среде RPMI-160 (ПанЭко, Россия) содержащей 10% сыворотки (Biosera США), антибиотики и 2 мМ L-глутамина (ПанЭко, Россия). При этом для дифференциации миелоидных клеток-предшественников до макрофагов добавляют форбол-12-миристат-13-ацетат, PMA (Сигма, Германия) в концентрации 2 нг/мл, инкубируют 48 часов при температуре 37°C, во влажной атмосфере, содержащей 5% СО2. Далее клетки промывают фосфатно-солевым буфером Дульбекко DPBS (ПанЭко, Россия) и добавляют среду RPMI-160 (ПанЭко, Россия) содержащей 10% сыворотки, антибиотики и 2 мМ L-глутамина (ПанЭко, Россия) и инкубируют при температуре 37°C, во влажной атмосфере, содержащей 5% СО2 в течение двух дней. Через два дня клетки готовы к экспериментам.
Третий этап (получение первичных макрофагов человека из мононуклеарных клеток периферической крови здоровых доноров).
Мононуклеарные клетки из крови человека получают методом осаждения в градиенте плотности фиколла. Для выделения мононуклеарных клеток используют периферическую кровь здорового донора. Все манипуляции по непосредственной работе с кровью с целью нивелирования риска контаминации проводят в асептических условиях в ламинарном боксе. Выделение ядросодержащих клеток проводят в 50 мл центрифужных пробирках (ПанЭко, Россия). В каждую пробирку вносят по 25 мл фиколла плотностью 1,077 г/мл (ПанЭко, Россия) и аккуратно, при помощи автоматического дозатора, наслаивают равный объём крови. Далее пробирку центрифугируют при 1080 g 30 мин, при комнатной температуре (без торможения). После центрифугирования получают чёткое разделение крови на 3 фракции: эритроциты, лейкоциты (мононуклеарные клетки) и плазму. Мононуклеарные клетки отбирают в отдельную 50 мл центрифужную пробирку (ПанЭко, Россия), добавляют 40 мл буфера DPBS (ПанЭко, Россия), полученную смесь ресуспендируют пипетированием и центрифугируют при 300 g в течение 5 минут, при комнатной температуре. Супернатант удаляют. Полученный клеточный осадок ресуспендируют в 40 мл DPBS (ПанЭко, Россия) и повторно центрифугируют при 10 g в течение 10 минут, при комнатной температуре. Супернатант удаляют, а осадок ресуспендируют в 10 мл DPBS (ПанЭко, Россия). Клетки считают в камере Горяева и 2-3*107 клеток переносят в стерильную 10 см культуральную чашку Петри и культивируют 30 минут в инкубаторе при 37°С во влажной атмосфере с 5 % содержанием СО2.
Мононуклеарные клетки, полученные из периферической крови человека, промывают DPBS (ПанЭко, Россия) 2 раза. Далее добавляют питательную среду RPMI-160 (ПанЭко, Россия) с 5% человеческой сывороткой (SCI store, Россия), 100 ед/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ L-глутамина (ПанЭко, Россия), для дифференцировки до макрофагов добавляют человеческий MCSF в концентрации 20 нг/мл (SCI store, Россия) и инкубируют 9 дней при температуре 37°C, во влажной атмосфере, содержащей 5% СО2.
Четвертый этап (получение макрофагов из миелоидных клеток-предшественников костного мозга мыши).
Моноциты получают из аллогенного материала - костного мозга мыши. Для забора костного мозга мыши C57Black/6 6-8 недель (СТЕЗАР, Россия) выполняют антисептическую обработку в брюшной полости и задних ногах. Хирургические манипуляции на мышах проводят после их эвтаназии СО2. В асептических условиях извлекают большеберцовые и бедренные кости, удаляют окружающие ткани и собирают в стерильный контейнер с буфером DPBS (ПанЭко, Россия) и с 100 ед/мл пенициллином, 100 мкг/мл стрептомицином (ПанЭко, Россия). Костный мозг вымывали из концов костей с помощью иглы 26 калибра и шприцом на 1 мл, наполненный средой RPMI-1640 (ПанЭко, Россия) с антибиотиками 100 ед/мл пенициллина, 100 мкг/мл стрептомицина (ПанЭко, Россия) и пропускают через 70-мкм клеточный фильтр (SPL, Южная Корея). Суспензию клеток центрифугируют при 300 g в течение 5 мин при комнатной температуре для осаждения клеток. Супернатант выливают и осадок клеток ресуспендируют в 3 мл буфере для лизиса эритроцитов (50 мл дистиллированной воды, 0.449 г NH4Cl, 0.05 г KHCO3, 1.85 мг EDTA, pH=7.3) и инкубируют 3 мин, а затем центрифугируют при 300 g в течение 5 мин при комнатной температуре.
Моноциты, полученные из костного мозга мыши, сажают на чашку Петри, например, на 10-ти сантиметровую, или аналогичный, 6-ти или 12-ти луночный планшет в расчете моноциты из одной мыши на одну чашку/планшет и культивируют в среде RPMI-160 (ПанЭко, Россия) с 10% фетальной бычьей сывороткой (FBS), 100 ед/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ L-глутамина (ПанЭко, Россия) при температуре 37°C, во влажной атмосфере, содержащей 5% СО2. Для дифференциация миелоидных клеток-предшественников до макрофагов добавляют рекомбинантный макрофагальный колониестимулирующий фактор M-CSF в концентрации 20 нг/мл (SCI store, Россия). Оставшиеся клетки замораживают в 90% сыворотки крови плодов коровы (Biosera США) и 10% ДМСО (ПанЭко, Россия) в расчете 2 мл на моноциты из одной мыши (2 ампулы) и хранят при минус 80°C в течение ночи, а затем в хранилище с жидким азотом.
Пятый этап (совместное культивирование мышиных макрофагов и опухолевых клеток в сфероидах).
Для предотвращения клеточной адгезии к пластику, культуральный пластик покрывают тонким слоем 1% агарозы (Carl Roth, Германия). Для этого 1% агарозу вносят в лунки 6-луночного планшета в объеме 1 мл/лунка и сразу же отбирают для предотвращения преждевременной полимеризации. Далее планшет с 1% агарозой инкубируют при температуре 37°C в течение 20 минут для полной полимеризации матрикса, до достижения гелеобразного состояния. Далее получают многокомпонентную клеточную ко-культуру путем внесения в лунки заранее подготовленных опухолевых клеток рака легкого мыши и макрофагов, полученных из костного мозга мыши в соотношении 1:3 (для 6-ти луночного планшета по 2*104 опухолевых клеток и 6*104 макрофагов), находящихся в сфероидной среде содержащей, DMEM/F12 (ПанЭко, Россия), 0,4 % бычьего сывороточного альбумина BSA (Диаэм, Россия), L-глутамина (ПанЭко, Россия), смесь антибиотиков пенициллин-стрептомицин (ПанЭко, Россия), 1 мл добавки B-27 ™ (50X), 20 нг/мл фактора роста фибрабластов FGF2 (SCI store, Россия) и 20 нг/мл эпидермального фактор роста EGF (SCI store, Россия). К многокомпонентной клеточной ко-культуре добавляют вещества с исследуемой противоопухолевой активностью в ранее выбранных концентрациях: этомоксир (0,5 мкмоль), циклоспорин А (0,5 мкмоль), диметилфумарат (1 мкмоль), ставудин (1,25 мкмоль), диданозин (2,5 мкмоль), ротенон (0,01 мкмоль), антимицин А (3 наномоль), олигомицин (0,1 мкмоль), карбонилцианид-п-трифторметоксифенилгидразон (FCCP) (0,15 мкмоль), BPTES (0,5 мкмоль). Далее многокомпонентную клеточную ко-культуру инкубируют 5 дней при температуре 37°C, в насыщенной влажной атмосфере, содержащей 5% СО2 до образования клетками сфероподобных структур. Морфологию клеток ко-культуры анализируют с использованием фазово-контрастной микроскопии. Клетки собирают в микроцентрифужную пробирку на 2 мл (Химмед, Россия) и центрифугируют при 300 g в течение 5 мин при комнатной температуре, для осаждения клеток. Получившийся осадок ресуспендируют в 200 мкл 0,25% трипсина-ЭДТА (ПанЭко, Россия) и инкубируют 3 мин при температуре 37°C и подсчитывают количество клеток в камере Горяева или автоматического счетчика клеток CytoSmart (Corning, США). Для камеры используют формулу: N / 15 * 250 * 10^3, где N - число клеток в пятнадцати квадратах камеры, расположенных по диагонали/объем в мл.
При этом показывают, что классически активированные макрофаги не способны способствовать росту опухолевых клеток.
Шестой этап (мониторинг роста опухолевых клеток в кондиционированных средах из активированных макрофагов).
К макрофагам, полученным из костного мозга мыши, на 5 день дифференцировки добавляют кондиционированную среду от опухолевых клеток рака легкого мыши или рака поджелудочной железы в соотношении 1:3 и препараты в выбранных концентрациях и инкубируют 24 ч при температуре 37°C, во влажной атмосфере, содержащей 5% СО2. На следующий день клетки промывают буфером DPBS (ПанЭко, Россия) и добавляют среду DMEM (ПанЭко, Россия), содержащей 2% сыворотку крови плодов коровы (Biosera США), L-глутамин (ПанЭко, Россия) и смесь антибиотиков и инкубируют 72 ч. Далее кондиционированную среду от активированных макрофагов собирают в микроцентрифужные пробирки на 1,5 мл (Химмед, Россия), центрифугируют при 300 g 5 минут и хранят в морозильной камере при температуре минус 20°С. Опухолевые клетки рака легкого мыши и поджелудочной железы сажают в 24-луночных планшет плотностью 500 клеток на лунку в среде культивируют в среде DMEM (ПанЭко, Россия) с 10% сывороткой и антибиотиками. На следующий день клетки промывают буфером DPBS (ПанЭко, Россия) и добавляют кондиционированную среду от активированных макрофагов (1 часть) и среду DMEM (ПанЭко, Россия), содержащей 0.5 % сыворотки и антибиотики (2 части). Через 9 дней производят подсчет опухолевых клеток с использованием стандартных/известных методов клеточной биологии, например, подсчетом клеток на камере Горяева. Опухолевые клетки рака легкого мыши и поджелудочной железы промывают буфером DPBS, добавляют 200 мкл 0,25% трипсина-ЭДТА (ПанЭко, Россия), инкубируют 3 мин и подсчитывают количество клеток, после чего определяют рост или подавление опухолевых клеток.
Количество клеток возможно определить, например, стандартным методом с помощью камеры Горяева.).
Кроме этого, количество клеток можно определить с помощью автоматического счетчика клеток CytoSmart (Corning, США).
Кроме этого, количество клеток можно определить с помощью метода генциан фиолетовый (crystal violet). Для этого, опухолевые клетки рака легкого мыши и клетки поджелудочной железы фиксируют 350 мкл 10% формалина (Biovitrum, Россия) и инкубируют 15 мин при комнатной температуре. Затем клетки промывают буфером DPBS и добавляют 250 мкл 0,1% генциан фиолетовый (Panreac, Испания) и инкубируют 3 мин при комнатной температуре. Далее клетки дважды промывают MQ, насухо отбирают и сушат планшеты с клетками 30 мин при комнатной температуре. Затем добавляют 500 мкл уксусной кислоты и перемешивают на орбитальном шейкере (Biosan, Латвия) в течении 30 мин, пока цвет не станет однородным без участков плотной окраски на дне лунок. Далее на микропланшетном ридере Infinite M200 pro (Tecan, Швейария) измеряют абсорбцию каждой лунки при длине волны 570 нм. Результаты интерпретируют согласно абсорбции.
При этом показывают, что классически активированные макрофаги не способны способствовать росту опухолевых клеток.
Седьмой этап (поляризация первичных макрофагов, полученных из периферической крови).
К дифференцированным макрофагам, полученным из периферической крови доноров, добавляют 20 нг/мл интерлейкина-4, IL-4 (SCI store, Россия) и комбинацию 20 нг/мл липополисахарида, LPS (SCI store, Россия) + 20 нг/мл интерферона- гамма, IFNγ (SCI store, Россия) и препараты в выбранных концентрациях. Через 24 часа к клеткам добавляют 1 мл раствора Extra RNA для лизиса и последующего выделения РНК. Образцы замораживают при минус 80°С.
Восьмой этап (поляризация мышиных макрофагов).
К макрофагам, полученным из костного мозга мыши, на 5 день дифференцировки добавляют 20 нг/мл интерлейкина-4, IL-4 (SCI store, Россия) и комбинацию 20 нг/мл липополисахарида, LPS (SCI store, Россия) + 20 нг/мл интерферона- гамма, IFNγ (SCI store, Россия) и препараты в выбранных концентрациях. На следующий день к клеткам добавляют 1 мл раствора Extra RNA для лизиса и последующего выделения РНК. Образцы замораживают при минус 80°С.
Девятый этап (поляризация THP-1 клеток).
Моноцитарную клеточную линию человека THP1 сажают на 6-луночную плату в концентрации 2 млн на лунку и дифференцируют по стандартному протоколу, описанному выше. К дифференцированным ТНР1 клеткам добавляют 20 нг/мл интерлейкина-4, IL-4 (SCI store, Россия) и комбинацию 20 нг/мл липополисахарида, LPS (SCI store, Россия) + 20 нг/мл интерферона- гамма, IFNγ (SCI store, Россия) и препараты в выбранных концентрациях. На следующий день клетки лизируют в 1 мл Extra RNA раствора (Евроген, Россия) для выделения РНК и замораживают при 80°C.
Десятый этап (выделение РНК).
Образцы клеток в Extra RNA растворе перемешивают на вортексе в течение 1 минуты и центрифугируют при 12000 g, 5 мин, при комнатной температуре. После перемешивают содержимое пробирок на вортексе в течение 1 минуты и центрифугируют при 12000 g, 5 мин, при комнатной температуре. Получившийся супернатант переносят в новую пробирку и добавляют к нему 200 мкл хлороформа, перемешивают и инкубируют в течение 5 минут при комнатной температуре, периодически встряхивая образец. После этого центрифугируют при 12000 g, 15 мин, при комнатной температуре, затем аккуратно отбирают верхнюю фазу и переносят ее в новую пробирку. Добавляют 500 мкл 100% изопропанола, перемешивают на вортексе и затем инкубируют 10 минут при комнатной температуре. Пробирки центрифугируют при 12000 g, 10 минут, при комнатной температуре, затем осторожно удаляют супернатант и промывают осадок в 1 мл 75% EtOH. Снова центрифугируют в течение при 12000 g, 5 минут, при комнатной температуре, удаляют супернатант и высушивают осадок при комнатной температуре. РНК растворяют в 40 мкл воды стерильной водой свободной от нуклеаз (Евроген, Россия) и помещают в морозильную камеру при температуре минус 80°С.
Одиннадцатый этап (получение кДНК).
Для синтеза кДНК предварительно измеряют концентрацию РНК на Nano Drop (Thermo scientific, США) в концентрации нг/мкл, также оценивают чистоту образцов - соотношение пиков поглощения 260 нм/280 нм - соответствуют примесям белков и аминокислот; 260 нм/230 нм - примеси солей и фенола. Необходимое количество РНК доводят до 15 мкл стерильной водой свободной от нуклеаз (BIO-RAD, США) и добавляют 4 мкл 5х iScript Reaction Mix (BIO-RAD, США) и 1 мкл iScript Reverse Transcriptase (BIO-RAD, США). Пробирки помещают в амплификатор C1000 Touch (Bio-Rad, США), следуя инструкции производителя. После окончания образцы оставляют в морозильной камере при температуре минус 20°С.
Двенадцатый этап (ПЦР реакция в реальном времени).
Для проведения ПЦР в реальном времени готовят Master Mix: к 5 мкл SybrGreen (Евроген, Россия) добавляют 3,6 мкл воды, свободной от нуклеаз (Евроген, Россия) и 0,4 мкл праймера (Евроген, Россия) для соответствующих генов (таблица). Готовый Master Mix раскапывают в 96-луночный планшет по 9 мкл. Далее раскапывают по 1 мкл образцы ДНК, Standard и Negative Control в трех повторностях. Планшет закрывают оптической прозрачной пленкой и помещают в амплификатор.
Проводят ПЦР, например, при следующих параметрах: 1) предварительная денатурация в течение 3 минут при 95°С, 2) денатурация в течение 10 секунд при температуре 95°С, 3) отжиг в течение 30 секунд, температура которого варьируется в зависимости от праймера, 4) плавление при температуре 72°С, длительностью 30 секунд на амплификаторе Real-Time CFX96 Touch (Bio-Rad, США). Такой цикл повторяется 40 раз, после чего получают данные о пороговых значениях циклов ПЦР, используя которые рассчитывают уровень экспрессии интересующих генов.
При этом показывают смещение митохондриального метаболизма с окислительного фосфорилирования, свойственного макрофагам М2 типа, в гликолизный тип, характерный для макрофагов М1 типа.
Далее заявителем приведены Примеры осуществления заявленного технического решения.
Заявителем проведены следующие эксперименты в условиях лаборатории, которые доказывают эффективность заявленного способа для лечения онкологических заболеваний в целом:
Пример 1. Выполнен анализ митохондриальной дисфункции в мышиных макрофагах и макрофагов, полученных из мононуклеарных клеток периферической крови донора, вызванных отсутствием гена Oра1, и было показано нарушение поляризацию макрофагов, в М2 подтип.
Пример 2. Выполнен эксперимент, подтверждающий указанное предположение, а именно, неспособность макрофагов М1 (OPA1ΔM) стимулировать рост опухолевых клеток при совместном культивировании макрофагов с опухолевыми клетками.
Пример 3. Выполнен поиск наиболее эффективных фармакологических препаратов, вызывающие митохондриальную дисфункцию.
Пример 4. Выполнены исследования, подтверждающие факт того, что отобранные на предыдущем этапе фармацевтические препараты, действительно подавляют поляризацию мышиных макрофагов и макрофагов человека, и как следствие указанного приводят к изменению метаболизма митохондрий.
Пример 5. Выполнены исследования, подтверждающие факт того, что отобранные заявителем фармацевтические препараты, изменяющие метаболизм митохондрий, действительно подавляют макрофагальную функцию мышиных макрофагов М2 подтипа, которые поддерживают рост опухолевых клеток.
Пример 6. Выполнены исследования, подтверждающие факт того, что отобранные фармацевтические средства, влияющие на митохондриальный метаболизм, блокируют онкогенные свойства макрофаг подобной клеточной линии человека ТНР1.
Пример 7. Выполнены исследования, подтверждающие факт того, что отобранные фармацевтические средства, индуцирующие митохондриальную дисфункцию в первичных макрофагах человека, снижают пролиферацию опухолевых клеток.
Далее заявителем более детально приведены описания экспериментов, подтверждающие представленные выше последовательность выполненных экспериментов.
Пример 1. Выполнение анализа митохондриальной дисфункции в мышиных макрофагах и макрофагов, полученных из мононуклеарных клеток периферической крови донора, вызванных отсутствием гена Oра1.
Для того чтобы проверить, могут ли митохондриальные перестройки вызывать метаболическое перепрограммирование в опухолеассоциированных макрофагах (ТАМ) проводят четвертый этап - получение макрофагов из миелоидных клеток-предшественников костного мозга мыши по алгоритму, описанному выше. Заявителем использована мышиная модель, в которой макрофаги имеют митохондриальную дисфункцию. Для этого в мышах был специфически инактивирован ген Optic Atrophy 1 (Opa1), причем эта делеция проявляется только в макрофагах. В результате скрещивания, заявителем были получены мыши, несущих аллель Opa1 flox у которых, из-за делеции экзонов 2 и 3, происходит аберрантная транскрипция exon1-exon4. Полученные мыши Opa1flox / flox Lyz2Cre / Cre были обозначены как мыши Opa1ΔM (или Ора1-KO), а макрофаги, полученные из этих мышей, как макрофаги OPA1ΔM, соответственно. Во всех проведенных экспериментах макрофаги OPA1ΔM (упоминаются в тексте как Ора1-KO) сравнивали с макрофагами, полученными от мышей Opa1flox / flox (далее в тексте упоминаются как контрольные или макрофаги дикого типа WT). Заявителем предположено, что макрофаги, дефицитные по OPA1, могут демонстрировать митохондриальные аберрации, которые радикально перестраивают их метаболизм, воспроизводят метаболические программы макрофагов M1, которые, в свою очередь, способны эффективно бороться с раковыми клетками.
Первоначально, чтобы оценить функциональное состояние поляризации в макрофагах OPA1ΔM, заявителем решено оценить экспрессию известных маркеров поляризации. Экспрессия этих маркеров часто коррелирует с функциональной способностью макрофагов к поляризации. Для этого заявителем выполнены восьмой этап - поляризация мышиных марофагов; десятый этап - выделение РНК; одиннадцатый этап - получение кДНК и двенадцатый этап - проведение полимеразной цепной реакции в режиме реального времени по алгоритму, описанному выше.
Заявитель проверил методом qPCR экспрессию маркеров M1, индуцированных IFNg и LPS, и маркеров M2, индуцированных IL-4. Заявителем обнаружено, что как маркеры М2 (Arg1, Ym1, Fizz1), так и многочисленные воспалительные маркеры М1 не индуцировались в OPA1-дефицитных макрофагах в той же степени, что и в клетках дикого типа (Фиг. 1A и Фиг. 1B). Интересно, что отсутствие OPA1 в макрофагах также препятствовало экспрессии генов M2 и M1 подтипа в ответ на кондиционированную среду из опухолевых клеток (Фиг. 1). В целом, эти данные продемонстрировали, что митохондриальная дисфункция, вызванная делецией гена Opa1 в макрофагах, значительно влияет на программирование макрофагов в проопухолевый М2 подтип. Ранее сообщалось, что признаки М2-макрофагов коррелируют с худшим прогнозом у онкологических больных. Поэтому значительное подавление признаков М2 подтипа в опухолеассоциированных макрофагах может быть преимуществом для пациентов для осуществления эффективного противоопухолевого ответа.
В результате достигнут заявленный технический результат - митохондриальная дисфункция, вызванная делецией гена Opa1 в макрофагах, значительно влияет на программирование макрофагов в проопухолевый М2 подтип.
Далее, заявителем проверена роль Opa1 в макрофагах человека. Для этого был проведен третий этап - получение первичных макрофагов человека их мононуклеарных клеток периферической крови здоровых доноров. Мононуклеарные клетки периферической крови высевали на пластик и культивировали в течение 9 дней в 5% человеческой сыворотке. Для нокдауна макрофаги были трансфицированы siRNA, нацеленной на OPA1, что обеспечило эффективный нокдаун (Фиг. 2 А).
Для анализа статуса поляризации макрофагов человека проводили седьмой этап. Для этого первичные макрофаги человека стимулировали IL-4 и IFN-γ и LPS. Далее проводили десятый этап - выделение РНК; одиннадцатый этап - получение кДНК и двенадцатый этап - проведение полимеразной цепной реакции в режиме реального времени по алгоритму, описанному выше. Подобно мышиным макрофагам при активации IL-4, М2-гены также демонстрировали ослабленную экспрессию в макрофагах человека с нокаутом OPA1 (Фиг. 2В). Однако гены M1, в свою очередь, сильно повышались под действием IFN-γ и LPS в макрофагах с дефицитом OPA1 (Фиг. 2С). Эти данные также подтверждают данные о том, что OPA1 необходим для проопухолевой активации макрофагов.
В результате достигнут заявленный технический результат - показано нарушение поляризации макрофагов в М2 подтип.
Пример 2. Выполнен эксперимент, подтверждающий указанное предположение, а именно не способность макрофагов М1 (OPA1ΔM) стимулировать рост опухолевых клеток при совместном культивировании макрофагов с опухолевыми клетками.
Далее заявителем проанализировано, как макрофаги с OPA1ΔM могут влиять на рост опухолевых клеток. Для этого заявителем были провидены первый этап - культивирование опухолевых клеток мыши; четвертый этап - получение макрофагов из миелоидных клеток-предшественников костного мозга мыши и пятый этап - совместное культивирование мышиных макрофагов и опухолевых клеток в сфероидах по алгоритму, описанному выше (Фиг. 3А). Культивирование опухолевых клеток вместе с дифференцированными макрофагами в условиях сфероидного роста позволяет оценить эффект прямого контакта между опухолевыми клетками и макрофагами и проанализировать влияние секретома макрофагов на пролиферацию опухолевых клеток. Примечательно, что в условиях прямого совместного культивирования макрофаги OPA1ΔM не могли стимулировать пролиферацию трех линий опухолевых клеток мыши. Этот эффект привел к образованию сфероидов меньшего размера (Фиг. 3B) и значительному снижению числа клеток (Фиг. 3C). Эти данные четко подтверждают, что OPA1ΔM макрофаги не способны способствовать росту опухолевых клеток. Это означает, что OPA1 и нормальные функции митохондрий имеют решающее значение для проопухолевого перепрограммирования макрофагов. Эти результаты также коррелируют с данными qPCR активированных макрофагов. Поэтому препараты, вызывающие дисфункцию митохондрий в макрофагах, могут стать перспективными кандидатами для противораковой терапии. Заявитель считает, что, скорее всего, макрофаги OPA1ΔM не производят достаточного количества факторов/метаболитов, стимулирующих пролиферацию опухолевых клеток, или выделяют некоторые цитотоксические молекулы.
Существует несколько возможных причин, объясняющих, почему макрофаги OPA1ΔM меньше поддерживают рост опухолевых клеток: либо их метаболическая дисфункция приводит к изменению производства секретируемых факторов и метаболитов, необходимых для стимулирования пролиферации опухолевых клеток; либо макрофаги OPA1ΔM более цитотоксичны в плане уничтожения опухолевых клеток. Сначала заявитель решил проанализировать фагоцитарную способность макрофагов дикого типа и OPA1ΔM. Для этого опухолевые клетки были помечены зеленым красителем и высеяны поверх макрофагов, помеченных красным красителем. Процент дважды положительных клеток, представляющих макрофаги с опухолевыми клетками, измеряли методом проточной цитометрии через 8 часов после посева (Фиг. 4A). В результате, заявителем не обнаружено существенной разницы между макрофагами дикого типа и OPA1-дефицитными макрофагами в их способности фагоцитировать опухолевые клетки, среди всех трех протестированных клеточных линий (Фиг. 4B).
В результате достигнут заявленный технический результат - OPA1ΔM макрофаги не способствуют росту опухолевых клеток.
Далее заявителем проанализировано, как макрофаги, полученные из периферической крови здоровых доноров с OPA1ΔM могут влиять на рост опухолевых клеток человека. Для этого заявителем были провидены первый этап - культивирование опухолевых клеток человека; третий этап - получение первичных макрофагов человека из мононуклеарных клеток периферической крови здоровых доноров и пятый этап - совместное культивирование макрофагов и опухолевых клеток в сфероидах по алгоритму, описанному выше. Заявителем получены аналогичные результаты для макрофагов человека с siRNA-нокаутом OPA1 (Фиг. 4C).
В результате достигнут заявленный технический результат - макрофаги OPA1ΔM не производят достаточного количества факторов/метаболитов, стимулирующих пролиферацию опухолевых клеток.
Чтобы проверить, регулируют ли митохондриальная дисфункция и OPA1 продукцию секретируемых макрофагами факторов, необходимых для опухолевого роста, заявителем были использованы четвертый этап - получение макрофагов из миелоидных клеток-предшественников костного мозга мыши; первый этап - культивирование опухолевых клеток мыши; пятый этап - совместное культивирование мышиных макрофагов и опухолевых клеток в сфероидах; шестой этап - мониторинг роста опухолевых клеток в кондиционированных средах из активированных макрофагов по алгоритму, описанному выше. Заявителем были получены мышиные макрофаги и далее были обработаны кондиционированной средой, собранной от опухолевых клеток. После этого макрофаги промывали и выращивали в течение трех дней в сфероидной среде для обогащения ее секретом макрофагов. Затем опухолевые клетки помещали либо в среду сфероидов, либо в среду, собранную с макрофагов (Фиг. 5 А). Через 5 дней заявителем были сделаны снимки сфероидов и подсчитаны клетки сфероидов. Примечательно, что заявителем обнаружено, что кондиционированная среда из макрофагов значительно стимулировала пролиферацию опухолевых клеток, но факторы, продуцируемые активированными макрофагами OPA1ΔM, были недостаточны для поддержания роста опухолевых клеток (Фиг. 5B и Фиг. 5C).
В результате достигнут заявленный технический результат - митохондрильный метаболизм регулирует выработку макрофагами секретируемых факторов, необходимых для опухолевого роста.
Пример 3. Выполнен поиск наиболее эффективных фармакологических препаратов, вызывающие митохондриальную дисфункцию.
Далее, заявителем составлен список фармацевтических препаратов ( Фиг. 6), которые могли бы воздействовать на функции митохондрий в макрофагах с физиологическим эффектом, подобным нокауту гена Opa1. Заявителем выбрано несколько фармакологических соединений на основе наших предыдущих наблюдений и литературного поиска: этомоксир, циклоспорин А, диметилфумарат, ставудин, диданозин, ротенон, антимицин А, олигомицин, карбонилцианид-п-трифторметоксифенилгидразон, BPTES и новый ингибитор Ора1 MYLS22. Заявителем были выполнены первый этап - культивирование опухолевых клеток мыши и человека; второй этап - культивирование и дифференцировка ТНР-1 клеток; третий этап - получение первичных макрофагов человека из мононуклеарных клеток периферической крови здоровых доноров и четвертый этап - получение макрофагов из миелоидных клеток-предшественников костного мозга мыши. Заявителем были протестированы эти соединения на мышиных макрофагах, макрофагах, дифференцированных из периферической крови здоровых доноров, марофагальной клеточной линии ТНР1 и нескольких мышиных раковых клетках (первичный KrasG12D/p53-/- рак легких, рак поджелудочной железы KPC - KrasG12D и мутантный р53R172H, и клеточная линия рака молочной железы E0771; человеческие опухолевых клеточные линии (A549, PANC1, NCI-Н69).
В результате достигнут заявленный технический результат - выявлена концентрация препарата, которая не является токсичной для клеток (Фиг. 7).
Пример 4. Выполнены исследования, подтверждающие факт того, что отобранные на предыдущем этапе фармацевтические препараты, действительно подавляют поляризацию мышиных макрофагов и макрофагов человека, и как следствие указанного приводят к изменению метаболизма митохондрий.
Чтобы объяснить, как именно препараты, нацеленные на метаболизм митохондрий, влияют на функции ТАМs, заявителем выполнены третий этап - получение первичных макрофагов; четвертый этап - получение макрофагов из миелоидных клеток-предшественников костного мозга мыши; протестированы их действие на поляризацию макрофагов, полученных из костного мозга мыши и макрофагов человека, дифференцированных из моноцитов периферической крови; седьмой этап - поляризация первичных макрофагов, полученных из периферической крови; восьмой этап - поляризация мышиных макрофагов; десятый этап - выделение РНК; одиннадцатый этап - получение кДНК; двенадцатый этап - ПЦР реакция в реальном времени по алгоритму, описанному выше. Обработка цитокином IL4 была использована для поляризации макрофагов в подтип M2 (про-туморогенные макрофаги), а комбинация интерферона гамма и липополисахаридов (IFNg- γ и LPS) - для поляризации в противоопухолевый подтип M1. Таким образом, заявителем показано, что все выбранные препараты, вызывающие митохондриальную дисфункцию, предотвращали активацию нескольких M2-подобных маркеров как в макрофагах мыши (Фиг. 8А), так и в макрофагах человека (Фиг. 9А). Важно отметить, что экспрессия маркеров поляризации M1 в макрофагах мыши и человека либо повышалась (TNFa, IL12, GBP6), либо не изменялась (Isg15, IRF1) в ответ на фармакологические соединения, вызывающие митохондриальную дисфункцию (Фиг. 8B и 9B). Однако, были отмечены некоторые различия между препаратами. Тем не менее, олигомицин, ротенон и диданозин, по-видимому, являются наиболее многообещающими препаратами ингибирующие функции опухоль-ассоциированных макрофагов. Наши данные указывают на важность митохондриальных функций для активации проонкогенных ТАМs и доказывают противоопухолевое действия выбранных митохондриальных препаратов. Так, олигомицин, ротенон, диданозин, FCCP и новый синтетический ингибитор Opa1 MYLS22 ингибируют проонкогенную М2-поляризацию ТАМs и не затрагивают противоопухолевую активацию М1 макрофагов, а некоторые вещества даже усиливают экспрессию воспалительных генов TNFa и iNOS2, участвующих в механизмах по подавлению опухоли.
В результате достигнут заявленный технический результат - выбранные препараты, вызывающие митохондриальную дисфункцию, предотвращали активацию нескольких M2-подобных маркеров как в макрофагах мыши, так и в макрофагах человека.
Пример 5. Выполнены исследования, подтверждающие факт того, что отобранные заявителем фармацевтические препараты, изменяющие метаболизм митохондрий, действительно подавляют макрофагальную функцию мышиных макрофагов М2 подтипа, которые поддерживают рост опухолевых клеток.
Далее, заявителем протестированы препараты на предмет блокирования функции макрофагов, стимулировать пролиферацию опухолевых клеток in vitro. Заявителем использованы первый этап - метод культивирования клеток рака легкого мыши с мышиными макрофагами, дифференцированными из клеток костного мозга; четвертый этап получение макрофагов из миелоидных клеток-предшественников костного мозга мыши; пятый этап - совместное культивирование мышиных макрофагов и опухолевых клеток в сфероидах по алгоритму, описанному выше. Результаты показали, что циклоспорин А, ротенон и олигомицин, диданозин, антимицин А и FCCP достаточно эффективно ингибировали рост опухолевых клеток в условиях культивирования с макрофагами (Фиг. 10А), но особо не влияли на рост опухолевых клеток, культивируемых без макрофагов. Четыре препарата (этомоксир, диметилфумарат, ставудин и BPTES) не продемонстрировали такого эффекта и были исключены из дальнейших исследований. Концентрации препаратов, использованных в экспериментах по совместному культивированию, не были токсичными для макрофагов, полученных из костного мозга мыши, за исключением циклоспорина А. Поэтому заявитель продолжил тестировать следующие фармацевтические соединения: ротенон, олигомицин; диданозин, антимицин А и FCCP и повторил не менее трех раз эксперимент по совместному культивированию опухолевых клеток с макрофагами в присутствии выбранных препаратов (Фиг.10В).
В результате достигнут заявленный технический результат - выбранные препараты, вызывающие митохондриальную дисфункцию, ингибировали рост опухолевых клеток в условиях культивирования с макрофагами.
Помимо непосредственного культивирования макрофагов с опухолевыми клетками, заявителем протестирован рост опухолевых клеток в присутствии кондиционной среды, собранной с активированных мышиных макрофагов, предварительно обработанных препаратами, вызывающими митохондриальную дисфункцию. Для этого заявителем были выполнены первый этап - культивирование опухолевых клеток мыши; четвертый этап - получение макрофагов из миелоидных клеток-предшественников костного мозга мыши; шестой этап - мониторинг роста опухолевых клеток в кондиционированных средах из активированных макрофагов по алгоритму, описанному выше. Заявителем включены в анализ две мышиные клеточные линии: рак легкого и рак поджелудочной железы. Результаты оказались сходными с экспериментами при прямом культивировании макрофагов и раковых клеток, за исключением вещества FCCP, которое не показало подавление роста опухолевых клеток (Фиг. 11). Так, препараты диданозин, ротенон и олигомицин являются наиболее перспективными потенциальными противоопухолевыми лекарствами, влияющие на функции макрофагов.
В результате достигнут заявленный технический результат - выбранные препараты, вызывающие митохондриальную дисфункцию, ингибировали рост опухолевых клеток в присутствии кондиционной среды, собранной с активированных мышиных макрофагов, предварительно обработанных препаратами.
Пример 6. Выполнены исследования, подтверждающие факт того, что отобранные фармацевтические средства, влияющие на митохондриальный метаболизм, блокируют онкогенные свойства макрофаг подобной клеточной линии человека ТНР1.
Чтобы транслировать наши результаты на клетки человека, заявитель использовал моноцитарную клеточную линию THP1 в качестве модели человеческих макрофагов. Заявитель выполнил первый этап - культивирование опухолевых клеток человека; второй этап - культивирование и дифференцировка ТНР-1 клеток; шестой этап - мониторинг роста опухолевых клеток в кондиционированных средах от активированных макрофагов по алгоритму, описанному выше. Шестой этап - совместное культивирование макрофагов и опухолевых клеток в сфероидах не подходит для наших целей из-за высокого процента фагоцитоза у THP1, что будет мешать точной интерпретации данных.
В результате достигнут заявленный технический результат - факторы, продуцируемые дифференцированными после обработки PMA макрофагами THP1, усиливали пролиферацию клеток A549 в низко сывороточной среде (Фиг. 12).
Заявителем был использованы первый этап - культивирование опухолевых клеток человека; второй этап - культивирование и дифференцировка ТНР-1 клеток; шестой этап - мониторинг роста опухолевых клеток в кондиционированных средах от активированных макрофагов; девятый этап - поляризация ТНР-1 клеток по алгоритму, описанному выше, для мониторинга того, как факторы, секретируемые макрофагами, стимулируют пролиферацию раковых клеток человека. Заявителем протестирован набор митохондриальных препаратов, которые заявитель уже опробовали на мышиной системе. Важно отметить, что все четыре препарата - ротенон, диданозин, олигомицин и FCCP, которые ингибировали пролиферацию раковых клеток мыши, также были эффективны в блокировании проонкогенной функции макрофагов THP1 человека (Фиг. 13) по отношению к росту клеток рака легкого человека линии А549 (Фиг. 13А) и линии NCI-H69 (Фиг. 13В).
В результате достигнут заявленный технический результат - препараты, вызывающих митохондриальную дисфункцию в макрофагах, ингибировали пролиферацию раковых клеток.
Пример 7. Выполнены исследования, подтверждающие факт того, что отобранные фармацевтические средства, индуцирующие митохондриальную дисфункцию в первичных макрофагах человека, снижают пролиферацию опухолевых клеток.
Получив убедительные данные по клеточной линии ТНР1, заявитель перешел на первичные макрофаги человека, дифференцированные из моноцитов периферической крови здоровых доноров. Заявитель выполнил первый этап - культивировал опухолевые клетки человека; третий этап - получение первичных макрофагов из мононуклеарных клеток периферической крови здоровых доноров; шестой этап - мониторинг роста опухолевых клеток в кондиционированных средах от активированных макрофагов по алгоритму, описанному выше. Макрофаги дифференцировались в течение девяти дней, после чего, они были активированы кондиционной средой от соответствующих раковых клеток (рак легкого А549 и рак поджелудочной железы Panc1) в присутствии растворителя ДМСО как контроль, или же препаратов, вызывающих митохондриальную дисфункцию. Как и в экспериментах с макрофагами мыши и линией ТНР1, препараты диданозин, олигомицин, ротенон, FCCP, циклоспорин А, а также ингибитор Ора1 MYLS22 значительно подавляли функцию первичных макрофагов человека в поддержании роста опухолевых клеток, как в клетках А549, так и Panc1 (Фиг. 14).
В результате достигнут заявленный технический результат - препараты, вызывающих митохондриальную дисфункцию в макрофагах, ингибировали пролиферацию раковых клеток.
На дату подачи настоящей заявки заявитель не выявил источники, которые бы демонстрировали сходное действие вышеперечисленных препаратов на функции макрофагов, ассоциированных с опухолями. Роль Ора1 в опухоле-ассоциированных макрофагах также не выявлена. При этом выявлено in vitro, что препараты, вызывающие митохондриальную дисфункцию, непосредственно в опухолевых клетках, при использовании в высоких дозах препятствуют их росту [Zamberlan M. et. al. (2022) Inhibition of the mitochondrial protein Opa1 curtails breast cancer growth, Exp Clin Cancer Res., 41(1), 95], [Herkenne S. et. al. (2020) Developmental and Tumor Angiogenesis Requires the Mitochondria-Shaping Protein Opa1, Cell Metab., 31(5), 987-1003]. Эти данные позволяют предположить, что при сочетании действия препаратов в условиях in vivo может наблюдаться усиление в их действии на макрофаги (исследования заявителя) и на опухолевые клетки (публикации).
Таким образом, базируясь на приведенных экспериментальных данных, возможно сделать выводы о том, заявителем достигнут заявленный технический результат, а именно: произведен подбор подходящих лекарств и фармакологических соединений, которые обеспечивают возможность смещения митохондриального метаболизма с окислительного фосфорилирования, свойственного макрофагам М2 типа, в гликолизный тип, характерный для макрофагов М1 типа. Этот подход, по мнению заявителя, является наиболее перспективным способом подавления роста опухолевых клеток у больных онкологическими заболеваниями.
Показано, что метаболизм митохондрий в макрофагах регулирует их опухолевые свойства, связанные с изменениями внеклеточных рецепторов (контактов) и с продукцией секретируемых факторов, необходимых для усиленного роста опухолевых клеток. Фармацевтические соединения, вызывающие митохондриальную дисфункцию ТАМs, могут быть использованы в качестве потенциальных противораковых препаратов. Заявителем выявлены и протестированы лекарства и фармакологические вещества диданозин, олигомицин, ротенон, FCCP, циклоспорин А, а также ингибитор Ора1 MYLS22, которые блокируют работу транспортной цепи электронов в макрофагах и значительно снижают их способность индуцировать пролиферацию опухолевых клеток. Заявитель считает, что данные лекарственные препараты можно использовать в доклинических испытаниях как противораковые средства и вещества, влияющие на опухолево-ассоциированные макрофаги. Возможно, эти лекарства и вещества могут быть использованы в комбинации с химиотерапией или иммунной терапией для усиления их терапевтического воздействия на опухоль. В заявленном техническом решении протестированы два типа рака - рак легкого и рак поджелудочной железы. Однако по мнению заявителя эти лекарства и вещества могут быть также эффективны для других типов рака, которые также зависят от функций макрофагов, например, рака молочной железы или рака кишечника.
Заявленное техническое решение соответствует условию патентоспособности «новизна», предъявляемому к изобретениям, так как на дату предоставления заявочных материалов заявителем из исследованного уровня техники не выявлены источники, обладающие совокупностью признаков, идентичными совокупности признаков заявленного технического решения.
Заявляемое техническое решение соответствует условию патентоспособности «изобретательский уровень», предъявляемому к изобретениям, так как не является очевидным для специалиста в анализируемой области техники.
Заявленное техническое решение соответствует условию патентоспособности «промышленная применимость», предъявляемому к изобретениям, т.к. может быть осуществлено с использованием стандартного оборудования и известных приемов.
| название | год | авторы | номер документа |
|---|---|---|---|
| Применение мембранных везикул мультипотентных стромальных клеток, индуцированных цитохалазином В, для восстановления и повышения митохондриальной функции | 2019 |
|
RU2727540C1 |
| Способ получения линий первичных макрофагов с провоспалительными свойствами | 2022 |
|
RU2792602C1 |
| СПОСОБ ТОРМОЖЕНИЯ РОСТА ПОДКОЖНОГО ТРАНСПЛАНТАТА ЭКСПЕРИМЕНТАЛЬНОЙ ГЛИОБЛАСТОМЫ ЧЕЛОВЕКА U-87, ПЕРЕВИТОГО ИММУНОДЕФИЦИТНЫМ МЫШАМ NU/J | 2019 |
|
RU2717218C1 |
| Способ идентификации функционального М1 и М2 фенотипа макрофагов человека, генерированных in vitro из моноцитов крови | 2019 |
|
RU2717024C1 |
| СПОСОБ ПОДАВЛЕНИЯ РОСТА ОПУХОЛИ В ЭКСПЕРИМЕНТЕ | 2015 |
|
RU2599438C1 |
| ИСПОЛЬЗОВАНИЕ ПРОСТАГЛАНДИНА Е1 (АЛПРОСТАДИЛА) ДЛЯ АКТИВАЦИИ ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТИ МАКРОФАГОВ | 2024 |
|
RU2832882C1 |
| СПОСОБ ПОДАВЛЕНИЯ РОСТА ОПУХОЛИ ОСНОВЕ IN VITRO РЕПРОГРАММИРОВАНИЯ МАКРОФАГОВ НА М1 ФЕНОТИП С ПОВЫШЕННОЙ СПОСОБНОСТЬЮ К ПРОДУКЦИИ ОКСИДА АЗОТА В ЭКСПЕРИМЕНТЕ | 2015 |
|
RU2599545C1 |
| ИСПОЛЬЗОВАНИЕ ОКСИДА ГРАФЕНА ДЛЯ АКТИВАЦИИ ФАГОЦИТАРНОЙ ФУНКЦИИ МАКРОФАГОВ | 2022 |
|
RU2809372C1 |
| КОМБИНАЦИЯ СИРОСИНГОПИНА И МИТОХОНДРИАЛЬНЫХ ИНГИБИТОРОВ ДЛЯ ЛЕЧЕНИЯ РАКА И ДЛЯ ИММУНОСУПРЕССИИ | 2012 |
|
RU2602937C2 |
| Способ подавления воспалительной реакции в тканях пародонта в эксперименте | 2017 |
|
RU2677642C1 |
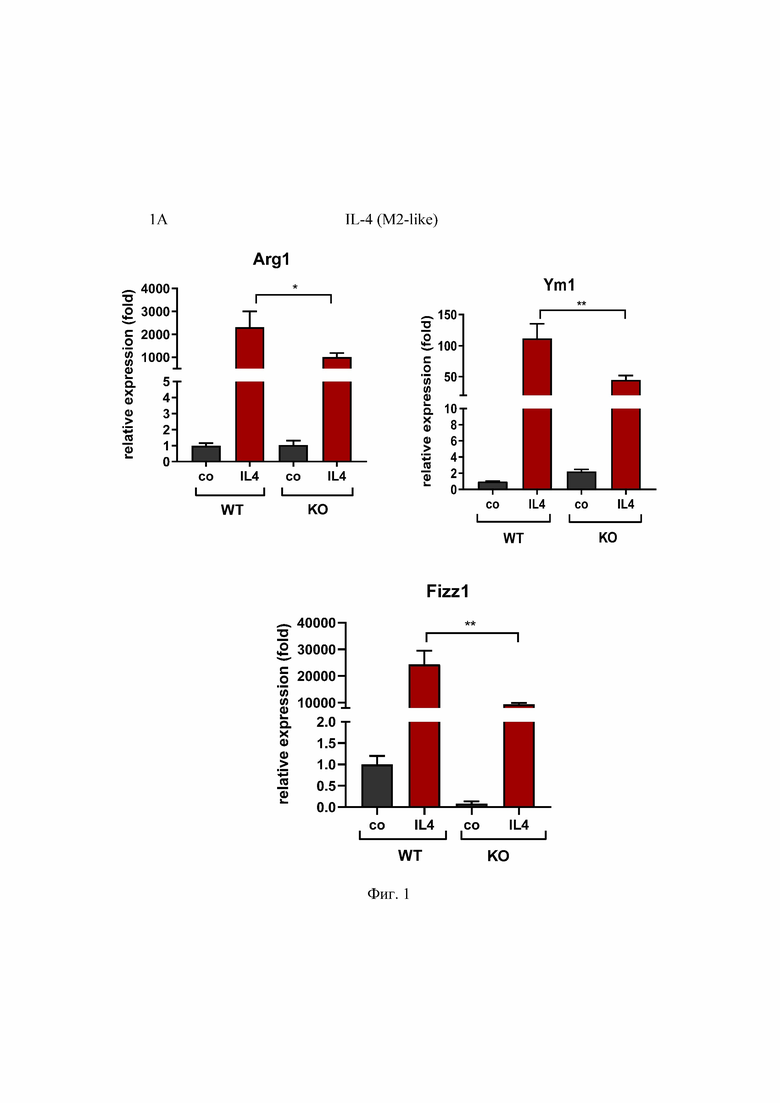
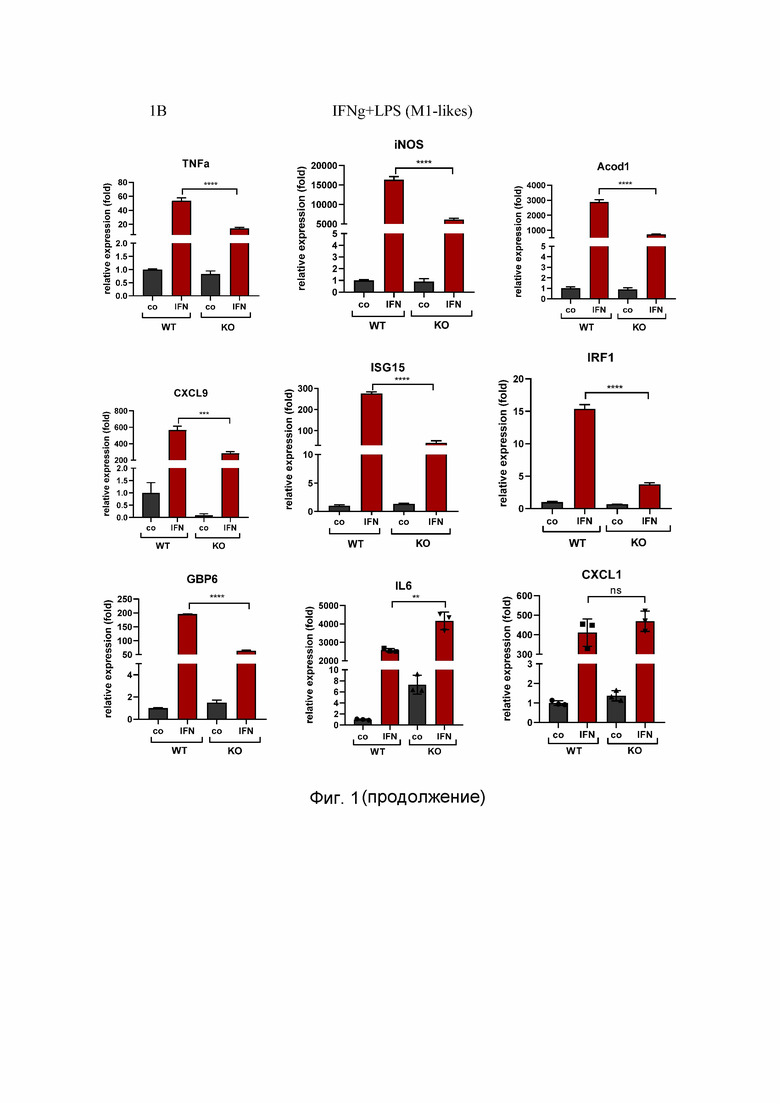
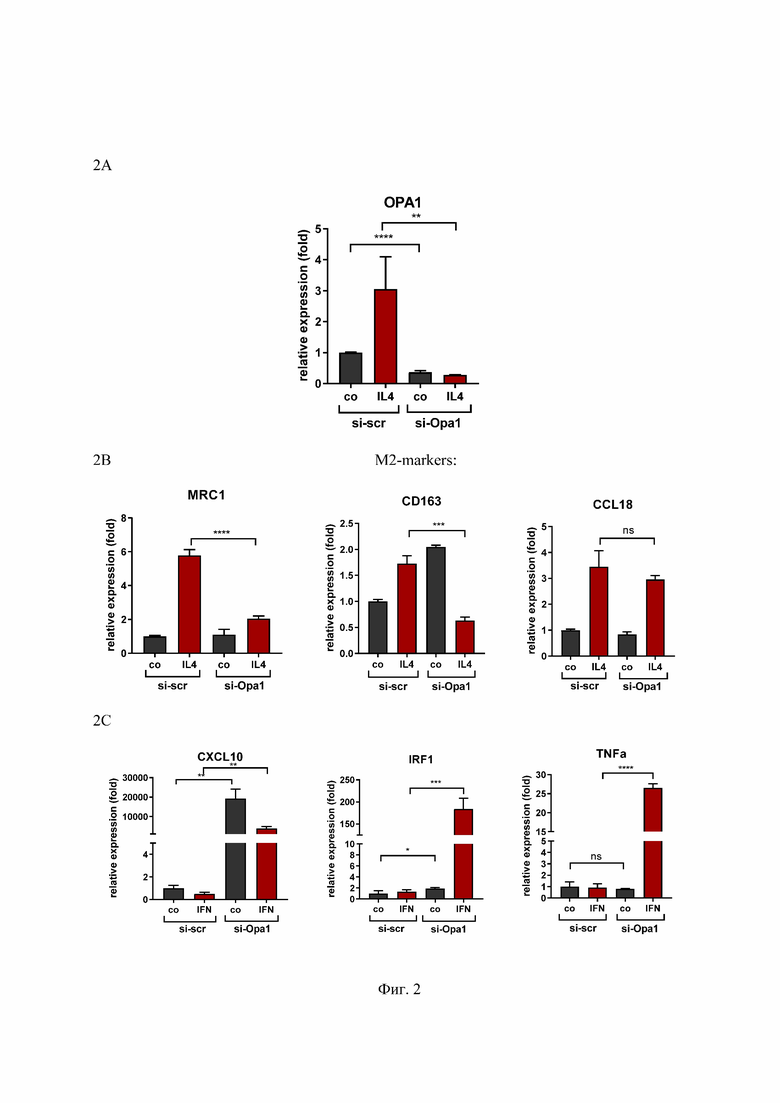
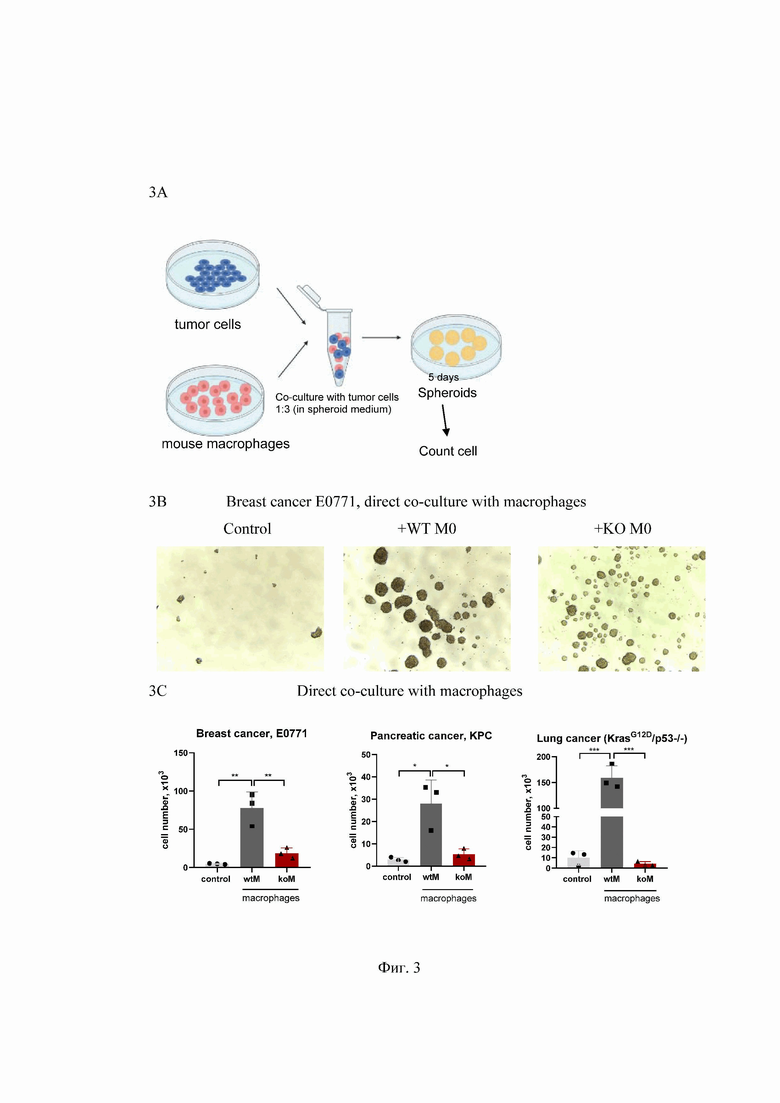
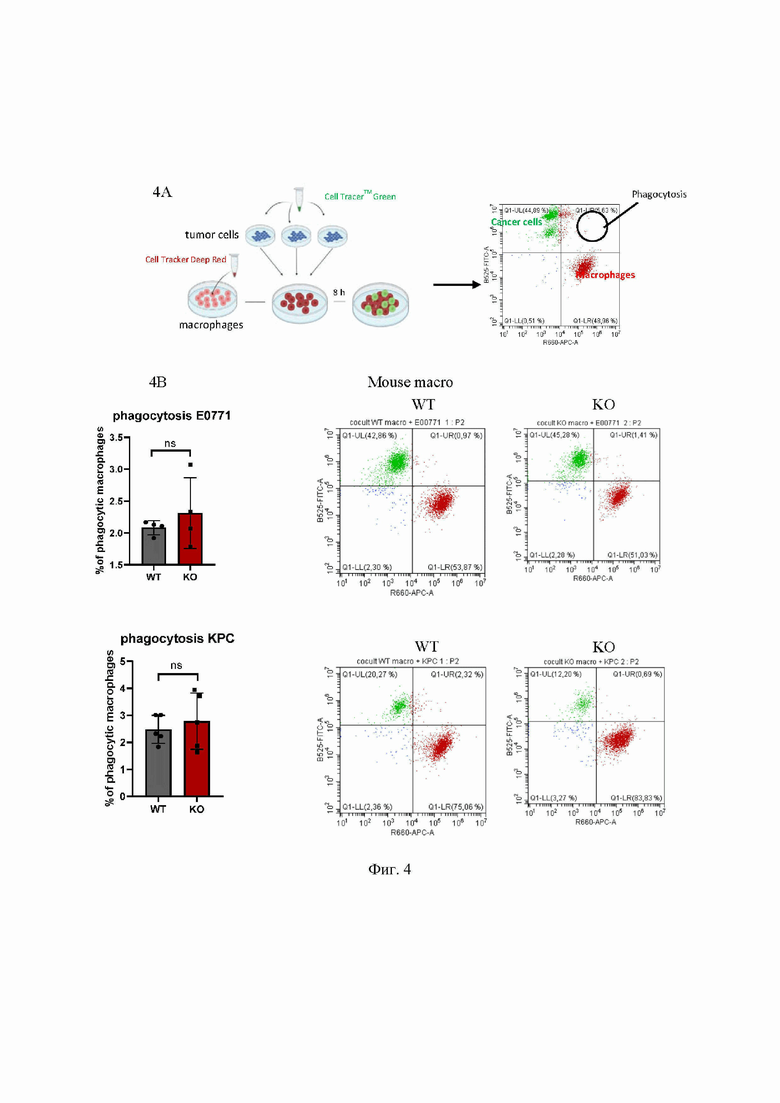
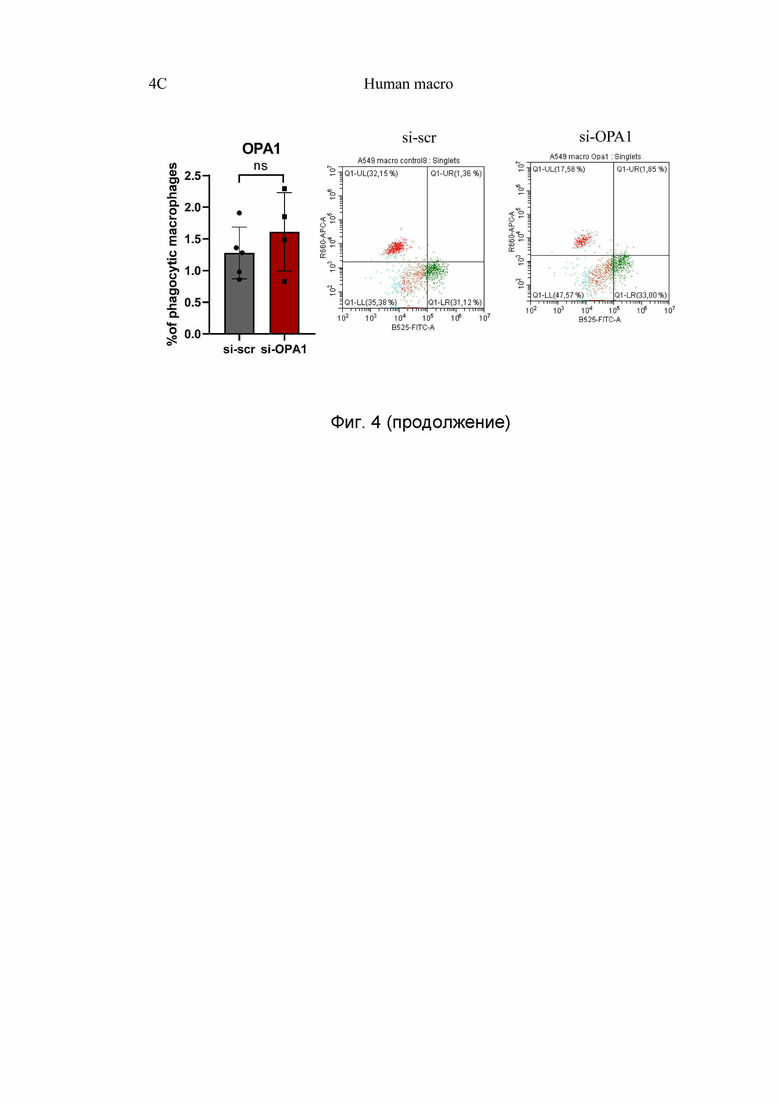


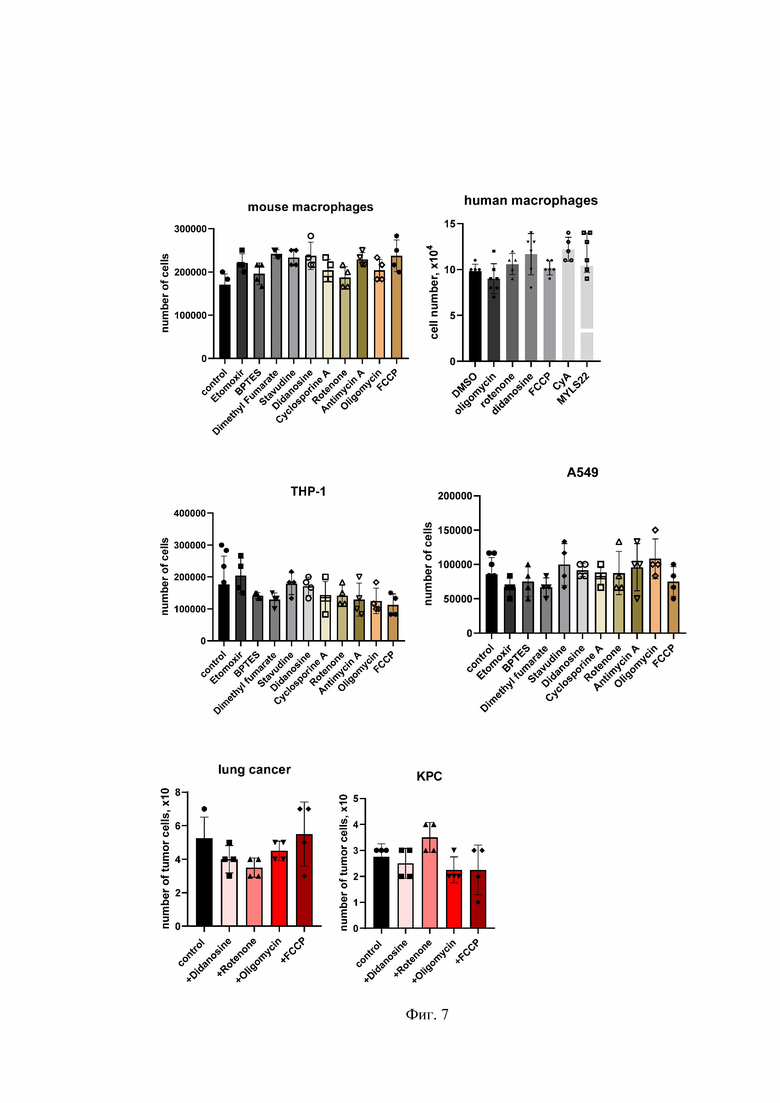
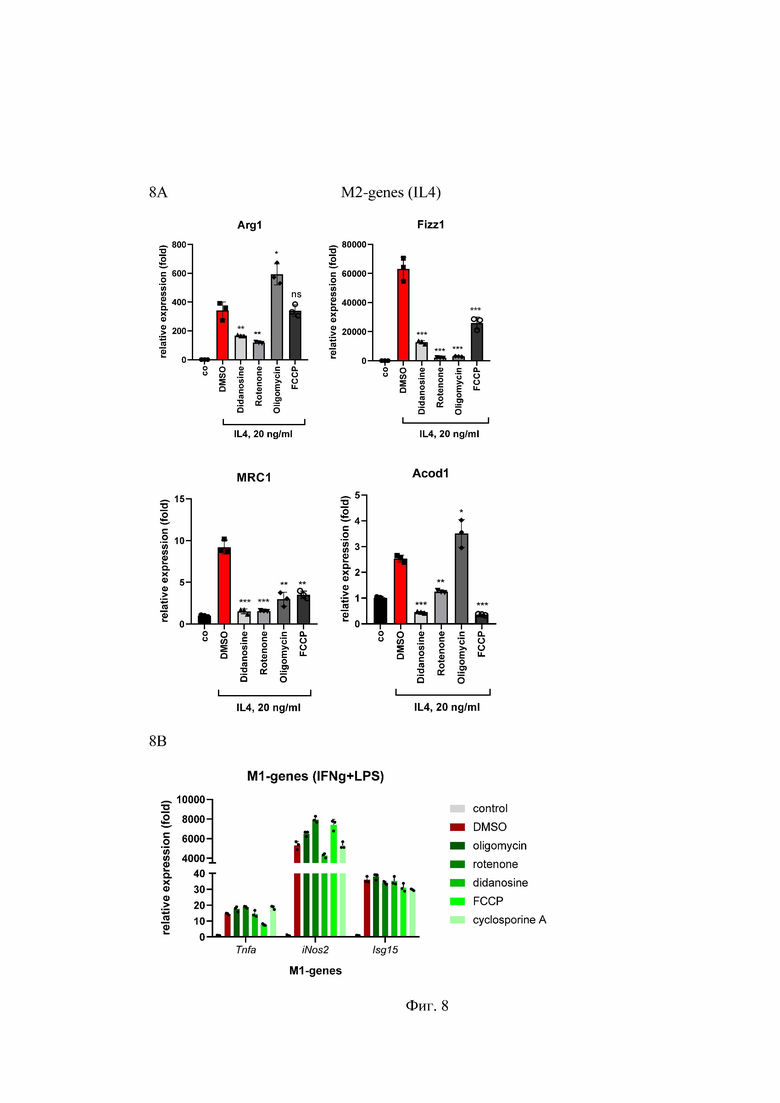
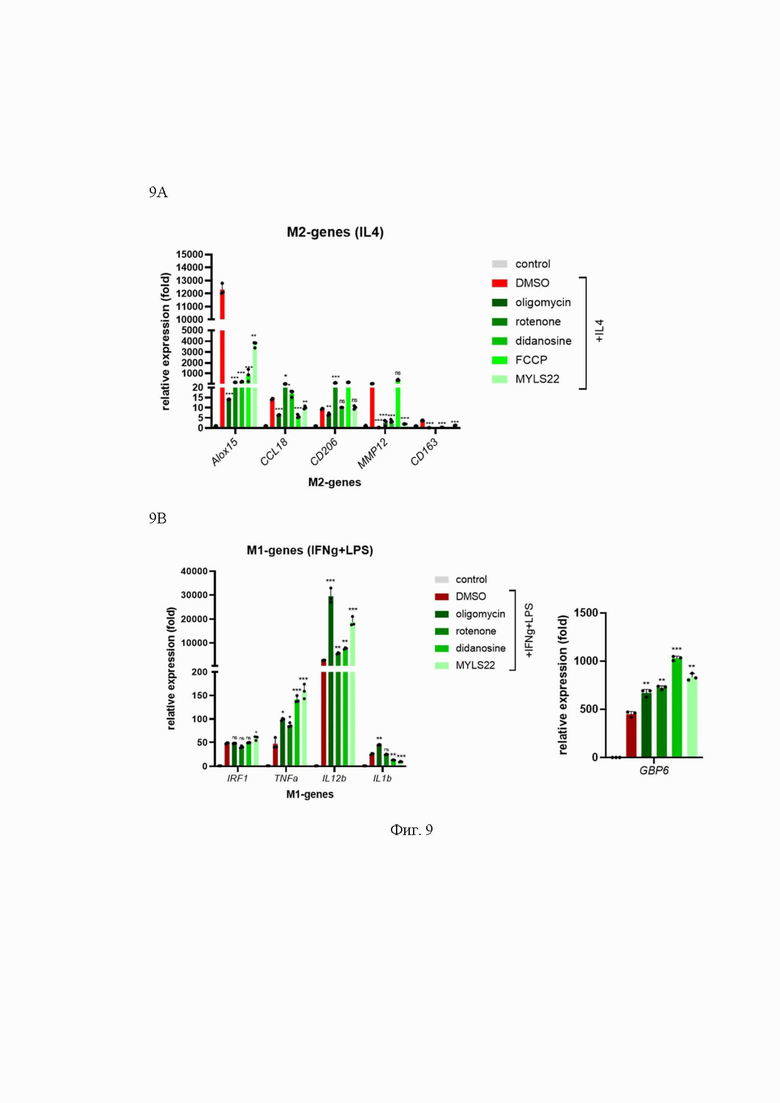
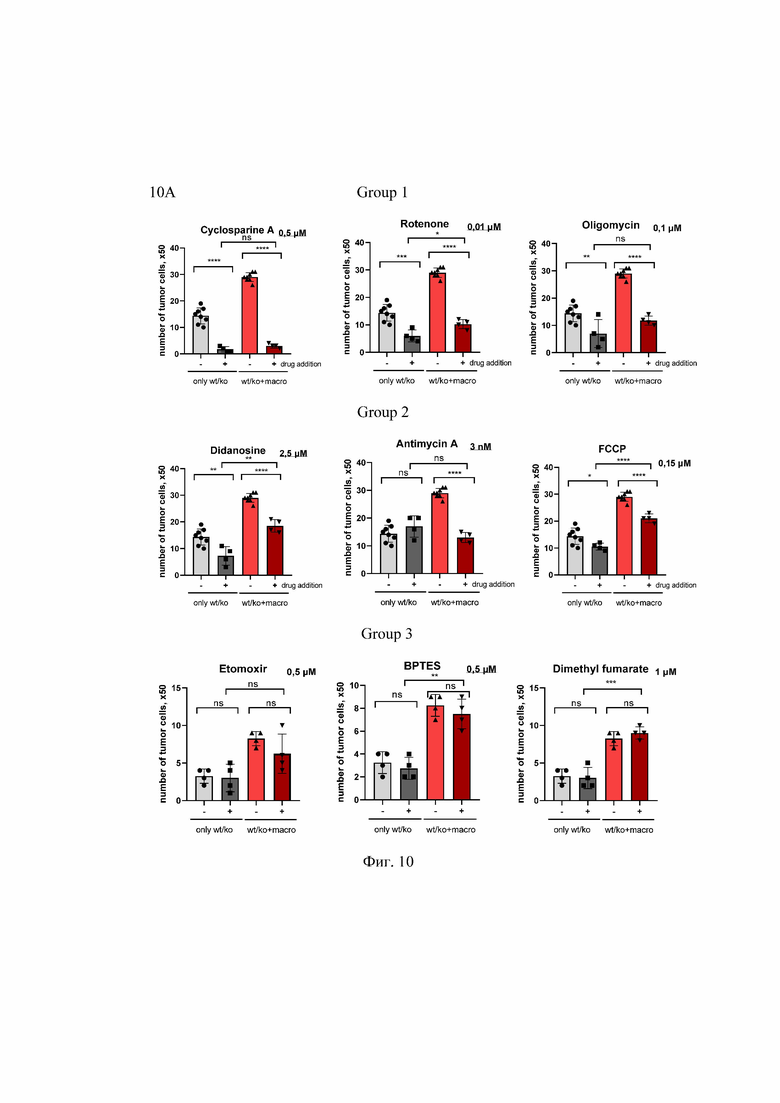
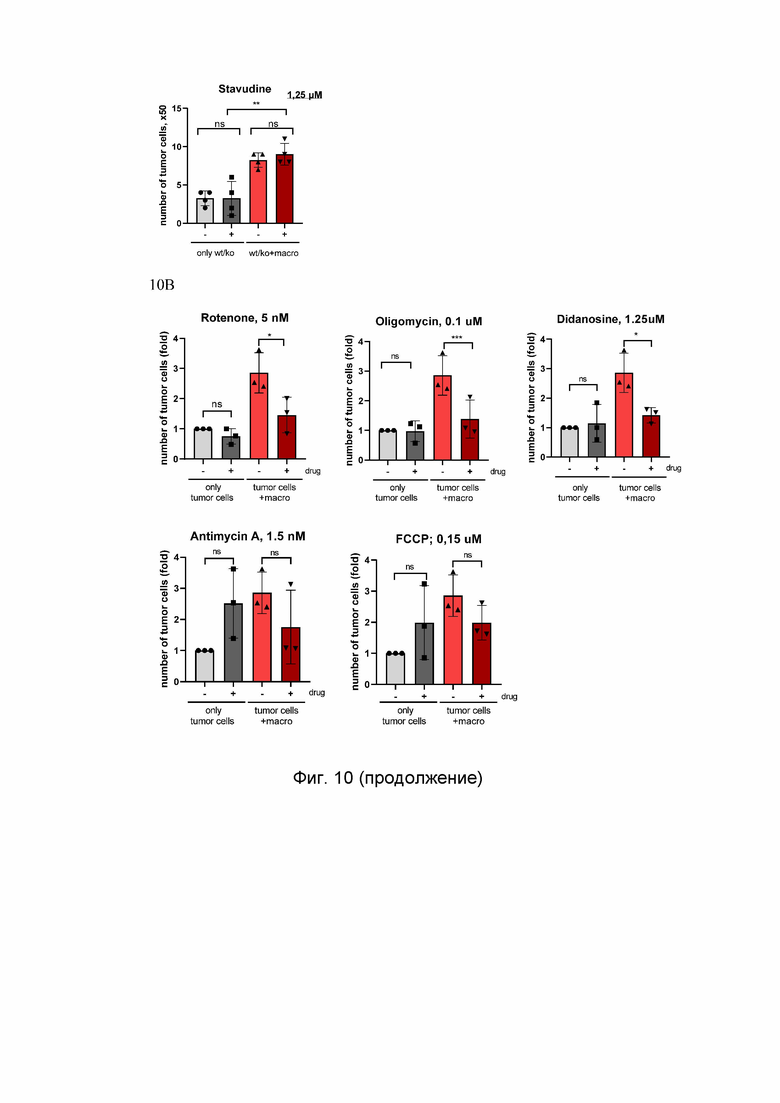
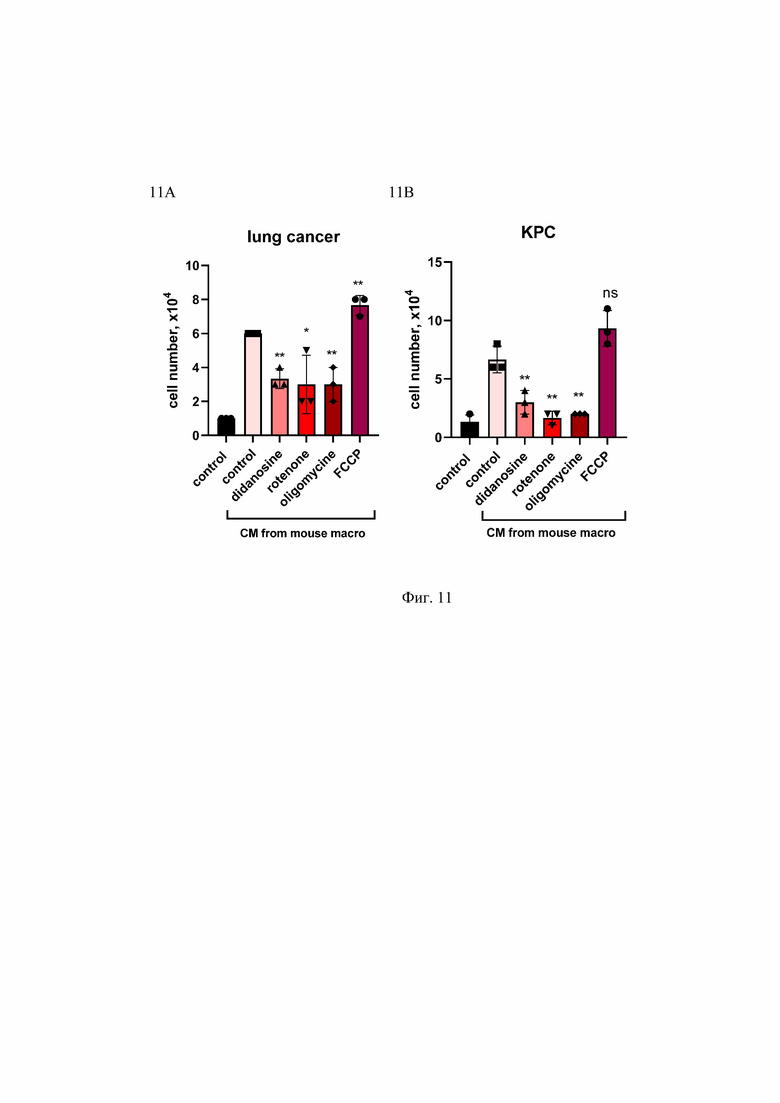
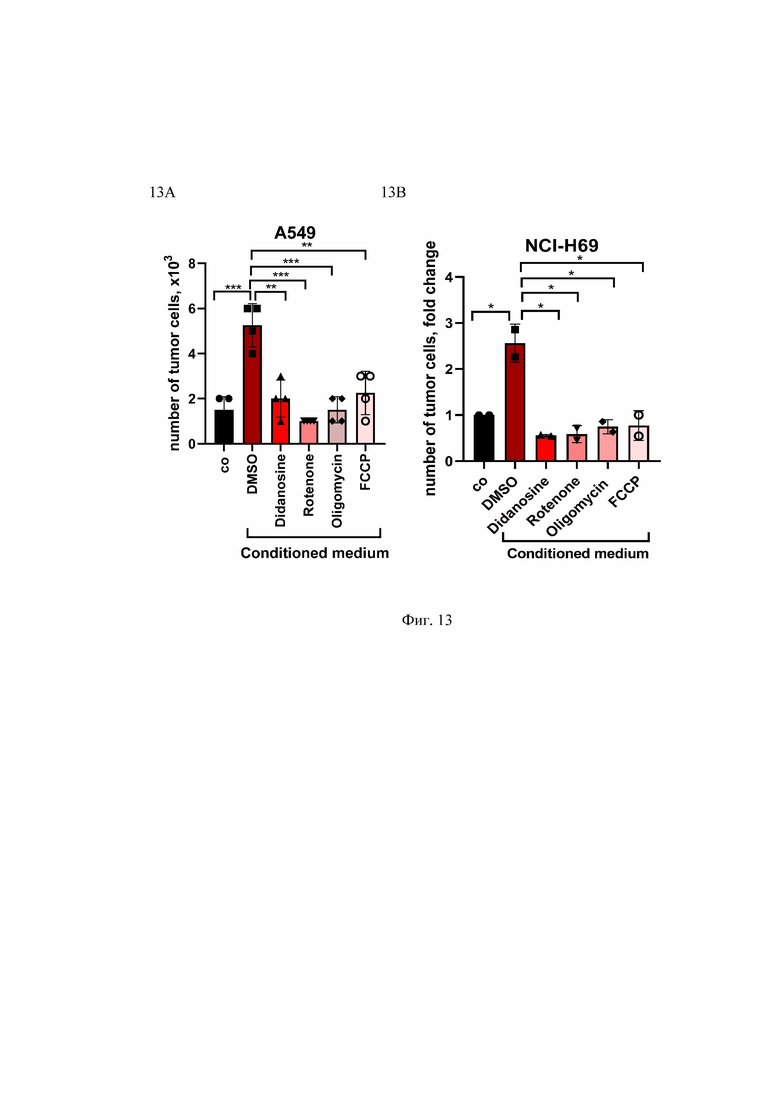
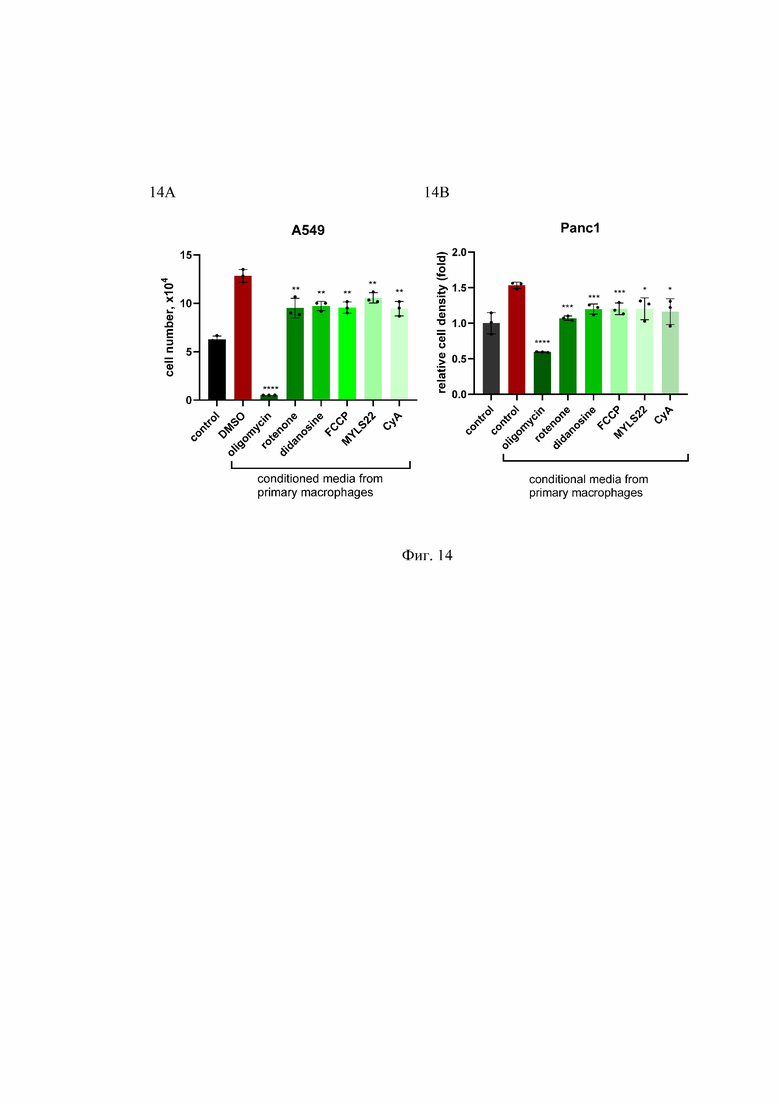
Изобретение относится к биотехнологии, к способам подбора и применения известных лекарств и фармацевтических соединений, вызывающих митохондриальную дисфункцию у макрофагов для противоопухолевой терапии. Заявленное изобретение обеспечивает возможность использования известных как таковых лекарств и фармакологических соединений, которые воздействуют на макрофаги, за счет изменения их митохондриальных функций, а именно смещения митохондриального метаболизма с окислительного фосфорилирования, свойственного макрофагам М2 типа, в гликолизный тип, характерный для макрофагов М1 типа, для лечения онкологических заболеваний. Изобретение является перспективным для лечения рака, а также может быть использовано в перспективе и для предупреждения возникновения онкологических заболеваний. 14 ил., 2 табл., 7 пр.
Способ подбора лекарственных средств для реализации фармакологической индукции митохондриальной дисфункции в макрофагах для противоопухолевой терапии, заключающийся в том, что
на первом этапе проводят культивирование опухолевых клеток мыши и человека, для чего опухолевые клетки мыши и человека культивируют в питательной среде DМЕМ, содержащей 10 % сыворотки крови плодов коровы, L-глутамин и смесь антибиотиков пенициллин-стрептомицин, при температуре 37 °C, во влажной атмосфере, содержащей 5 % СО2;
на втором этапе проводят культивирование и дифференцировка ТНР-1 клеток, для чего моноцитарную клеточную линию человека THP1 в качестве модели макрофагов человека культивируют в среде RPMI-160 содержащей 10 % сыворотки, антибиотики и 2 мМ L-глутамина, при этом для дифференциации миелоидных клеток-предшественников до макрофагов добавляют форбол-12-миристат-13-ацетат, PMA в концентрации 2 нг/мл, инкубируют 48 ч при температуре 37 °C, во влажной атмосфере, содержащей 5 % СО2; далее клетки промывают фосфатно-солевым буфером Дульбекко DPBS и добавляют среду RPMI-160, содержащей 10 % сыворотки, антибиотики и 2 мМ L-глутамина и инкубируют при температуре 37 °C, во влажной атмосфере, содержащей 5 % СО2 в течение двух дней;
на третьем этапе проводят получение первичных макрофагов человека из мононуклеарных клеток периферической крови здоровых доноров, для чего мононуклеарные клетки, полученные из периферической крови человека, промывают DPBS 2 раза, далее добавляют питательную среду RPMI-160 с 5 % человеческой сывороткой, 100 ед./мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ L-глутамина, для дифференцировки до макрофагов добавляют человеческий MCSF в концентрации 20 нг/мл и инкубируют 9 дней при температуре 37 °C, во влажной атмосфере, содержащей 5 % СО2;
на четвертом этапе проводят получение макрофагов из миелоидных клеток-предшественников костного мозга мыши, для чего моноциты, полученные из костного мозга мыши, культивируют в среде RPMI-160 с 10 % фетальной бычьей сывороткой FBS, 100 ед./мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ L-глутамина при температуре 37 °C, во влажной атмосфере, содержащей 5 % СО2; для дифференциации миелоидных клеток-предшественников до макрофагов добавляют рекомбинантный макрофагальный колониестимулирующий фактор M-CSF в концентрации 20 нг/мл;
на пятом этапе проводят совместное культивирование мышиных макрофагов и опухолевых клеток в сфероидах, для чего для предотвращения клеточной адгезии к пластику культуральный пластик покрывают тонким слоем 1 % агарозы, далее планшет с 1 % агарозой инкубируют при температуре 37 °C в течение 20 мин для полной полимеризации матрикса, до достижения гелеобразного состояния; далее получают многокомпонентную клеточную ко-культуру путем внесения в лунки заранее подготовленных опухолевых клеток рака легкого мыши и макрофагов, полученных из костного мозга мыши в соотношении 1:3, находящихся в сфероидной среде, содержащей DMEM/F12, 0,4 % бычьего сывороточного альбумина BSA, L-глутамина, смесь антибиотиков пенициллин-стрептомицин, 1 мл добавки B-27 ™ (50X), 20 нг/мл фактора роста фибрабластов FGF2 и 20 нг/мл эпидермального фактор роста EGF; к многокомпонентной клеточной ко-культуре добавляют вещества с исследуемой противоопухолевой активностью в ранее выбранных концентрациях: этомоксир 0,5 мкмоль, циклоспорин А 0,5 мкмоль, диметилфумарат 1 мкмоль, ставудин 1,25 мкмоль, диданозин 2,5 мкмоль, ротенон 0,01 мкмоль, антимицин А 3 наномоль, олигомицин 0,1 мкмоль, карбонилцианид-п-трифторметоксифенилгидразон FCCP 0,15 мкмоль, BPTES 0,5 мкмоль; далее многокомпонентную клеточную ко-культуру инкубируют 5 дней при температуре 37 °C, в насыщенной влажной атмосфере, содержащей 5 % СО2, до образования клетками сфероподобных структур; морфологию клеток ко-культуры анализируют с использованием фазово-контрастной микроскопии; клетки собирают и центрифугируют для их осаждения; получившийся осадок ресуспендируют и инкубируют 3 мин при температуре 37 °C и подсчитывают количество клеток в камере Горяева, для камеры используют формулу: N / 15 ⋅ 250 ⋅ 10^3, где N – число клеток в пятнадцати квадратах камеры, расположенных по диагонали/объем, мл;
на шестом этапе проводят мониторинг роста опухолевых клеток в кондиционированных средах из активированных макрофагов, для чего к макрофагам, полученным из костного мозга мыши, на 5 день дифференцировки добавляют кондиционированную среду от опухолевых клеток рака легкого мыши или рака поджелудочной железы в соотношении 1:3 и препараты в выбранных концентрациях и инкубируют 24 ч при температуре 37 °C, во влажной атмосфере, содержащей 5 % СО2; на следующий день клетки промывают буфером DPBS и добавляют среду DMEM, содержащей 2 % сыворотку крови плодов коровы, L-глутамин и смесь антибиотиков и инкубируют 72 ч; далее кондиционированную среду от активированных макрофагов центрифугируют при 300 g 5 мин и хранят в морозильной камере при температуре минус 20 °С; опухолевые клетки рака легкого мыши и поджелудочной железы культивируют в среде DMEM с 10 % сывороткой и антибиотиками; на следующий день клетки промывают буфером DPBS и добавляют кондиционированную среду от активированных макрофагов – 1 часть, и среду DMEM, содержащей 0.5 % сыворотки и антибиотики – 2 части; через 9 дней производят подсчет опухолевых клеток; опухолевые клетки рака легкого мыши и поджелудочной железы промывают буфером DPBS, добавляют 200 мкл 0,25 % трипсина-ЭДТА, инкубируют 3 мин и подсчитывают количество клеток, после чего определяют рост или подавление опухолевых клеток;
на седьмом этапе проводят поляризацию первичных макрофагов, полученных из периферической крови, для чего к дифференцированным макрофагам, полученным из периферической крови доноров, добавляют 20 нг/мл интерлейкина-4, IL-4 и комбинацию 20 нг/мл липополисахарида, LPS + 20 нг/мл интерферона-гамма, IFNγ и препараты в выбранных концентрациях; через 24 ч к клеткам добавляют 1 мл раствора Extra RNA для лизиса и последующего выделения РНК;
на восьмом этапе проводят поляризацию мышиных макрофагов, для чего к макрофагам, полученным из костного мозга мыши, на 5 день дифференцировки добавляют 20 нг/мл интерлейкина-4, IL-4 и комбинацию 20 нг/мл липополисахарида, LPS + 20 нг/мл интерферона-гамма, IFNγ и препараты в выбранных концентрациях; на следующий день к клеткам добавляют 1 мл раствора Extra RNA для лизиса и последующего выделения РНК;
на девятом этапе проводят поляризацию THP-1 клеток, для чего моноцитарную клеточную линию человека THP1 дифференцируют по стандартному протоколу, к дифференцированным ТНР1 клеткам добавляют 20 нг/мл интерлейкина-4, IL-4 и комбинацию 20 нг/мл липополисахарида, LPS + 20 нг/мл интерферона-гамма, IFNγ и препараты в выбранных концентрациях; на следующий день клетки лизируют в 1 мл Extra RNA раствора для выделения РНК;
на десятом этапе проводят выделение РНК, для чего образцы клеток в Extra RNA растворе перемешивают в течение 1 мин и центрифугируют при 12 000 g 5 мин при комнатной температуре, затем перемешивают в течение 1 мин и центрифугируют при 12000 g 5 мин при комнатной температуре; получившийся супернатант переносят в новую емкость и добавляют к нему 200 мкл хлороформа, перемешивают и инкубируют в течение 5 мин при комнатной температуре, периодически встряхивая образец, после чего центрифугируют при 12000 g 15 мин при комнатной температуре, затем отбирают верхнюю фазу и переносят ее в новую емкость; добавляют 500 мкл 100 % изопропанола, перемешивают и затем инкубируют 10 мин при комнатной температуре; затем центрифугируют при 12000 g 10 мин при комнатной температуре, затем удаляют супернатант и промывают осадок в 1 мл 75 % EtOH; затем центрифугируют в течение при 12000 g 5 мин при комнатной температуре, удаляют супернатант и высушивают осадок при комнатной температуре; РНК растворяют в 40 мкл воды стерильной водой свободной от нуклеаз и помещают в морозильную камеру при температуре минус 80 °С;
на одиннадцатом этапе проводят получение кДНК, для чего предварительно измеряют концентрацию РНК на Nano Drop в концентрации нг/мкл, оценивают чистоту образцов – соотношение пиков поглощения 260 нм/280 нм – соответствуют примесям белков и аминокислот; 260 нм/230 нм – примеси солей и фенола; необходимое количество РНК доводят до 15 мкл стерильной водой свободной от нуклеаз и добавляют 4 мкл 5х iScript Reaction Mix и 1 мкл iScript Reverse Transcriptase; пробирки помещают в амплификатор C1000 Touch (Bio-Rad, США), следуя инструкции производителя;
на двенадцатом этапе проводят ПЦР-реакцию в реальном времени, для чего готовят Master Mix: к 5 мкл SybrGreen добавляют 3,6 мкл воды, свободной от нуклеаз, и 0,4 мкл праймера для соответствующих генов; готовый Master Mix раскапывают в 96-луночный планшет по 9 мкл; далее раскапывают по 1 мкл образцы ДНК, Standard и Negative Control в трех повторностях; планшет закрывают оптической прозрачной пленкой и помещают в амплификатор; проводят ПЦР, получают данные о пороговых значениях циклов ПЦР, используя которые рассчитывают уровень экспрессии интересующих генов: маркеры макрофагов мыши М1 типа TNFa, iNOS, Acod1, CXCL9, ISG15, IRF1, GBP6, IL6, CXCL1 и макрофагов мыши М2 типа Arg1, Fizz1, MRC1, Ym1, Acod1; маркеры макрофагов человека М1 типа IRF1, GBP6, TNFa, IL12, IL1b и М2 типа ALOX15, CCL18, CD206, MMP12, CD169; полученные данные показывают подавление поляризации макрофагов в М2 тип и усиление поляризации в М1 тип, что подтверждает смещение митохондриального метаболизма с окислительного фосфорилирования, свойственного макрофагам М2 типа, в гликолизный тип, характерный для макрофагов М1 типа, а различия между экспрессией этих генов при добавлении препаратов позволяют в дополнение к данным о пролиферации опухолевых клеток с кондиционной средой от макрофагов определить, какое лекарственное средство является подходящим для реализации фармакологической индукции митохондриальной дисфункции в макрофагах для противоопухолевой терапии.
| Способ идентификации функционального М1 и М2 фенотипа макрофагов человека, генерированных in vitro из моноцитов крови | 2019 |
|
RU2717024C1 |
| RU 2739572 C1, 25.12.2020 | |||
| US 20220087950 A1, 24.03.2022 | |||
| WO 2018149978 A1, 23.08.2018 | |||
| ФЕДОРОВ А.А., ЕРМАК Н.А., ГЕРАЩЕНКО Т.С., ТОПОЛЬНИЦКИЙ Е.Б., ШЕФЕР Н.А., РОДИОНОВ Е.О., СТАХЕЕВА М.Н | |||
| Поляризация макрофагов: механизмы, маркеры и факторы индукции | |||
| Сибирский онкологический журнал | |||
| Способ получения продуктов конденсации фенолов с формальдегидом | 1924 |
|
SU2022A1 |
| - doi: | |||

