ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет согласно предварительной заявке на патент США №62/491151, поданной 27 апреля 2017 года, которая включена в данный документ посредством ссылки во всей своей полноте.
ЗАЯВЛЕНИЕ, КАСАЮЩЕЕСЯ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Перечень последовательностей, связанный с настоящей заявкой, представлен в текстовом формате вместо бумажной копии и тем самым включен в настоящее описание посредством ссылки. Текстовый файл, содержащий перечень последовательностей, называется IMCO-006_01WO_ST25.txt. Текстовый файл размером 10 Кбайт был создан 26 апреля 2018 года и подается в электронном виде через EFS-Web.
Предпосылки изобретения
Область техники
Настоящее изобретение относится к применению B-клеток для долговременной доставки in vivo терапевтического средства, такого как антигенспецифическое антитело или белок (например, фермент), и, в частности, к введению однократных и многократных доз B-клеток.
Описание предшествующего уровня техники
Современные способы лечения хронических заболеваний и нарушений включают прямую инфузию терапевтического средства (например, средства заместительной ферментной терапии), средство генной терапии посредством вирусного вектора и адоптивный перенос стволовых клеток (например, перенос гематопоэтических стволовых клеток). Однако каждый из этих способов имеет недостатки. Недостатком инъекции рекомбинантного терапевтического белка является конечный период полураспада белка, а все три способа обеспечивают субоптимальное проникновение в ткань терапевтического средства. Изменение эндогенных тканей с целью получения терапевтического средства, например, путем инъекции векторов на основе рекомбинантного аденоассоциированного вируса (AAV) и лентивируса, обычно приводит к тому, что терапевтическое средство получают из централизованного местоположения. Получение терапевтического средства из одного местоположения увеличивает шансы в отношении локализованной токсичности в продуцирующих тканях. Кроме того, поскольку рекомбинантные вирусы рассматриваются как чужеродные, маловероятно, что вирусные векторы можно вводить множество раз, не вызывая неблагоприятной реакции, что означает, что существует возможность однократной инъекции для достижения правильной дозы терапевтического средства. Учитывая биологические вариации, присущие процедуре, такой как введение нуклеиновых кислот in vivo в клетки с применением вируса, будет очень непросто достичь желаемой дозы в условиях однократной инъекции.
Недавно применение дифференцированных композиций на основе В-клеток для долговременной экспрессии трансгена in vivo было определено как многообещающая стратегия лечения различных заболеваний и нарушений. Однако способы введения модифицированных В-клеток для доставки терапевтических средств с целью достижения терапевтически эффективных уровней средств in vivo еще не были описаны.
Соответственно, в данной области все еще существует потребность в долговременном лечении многих хронических заболеваний и нарушений. В настоящем изобретении представлены способы введения и введения дозы композиций на основе генетически модифицированных В-клеток для лечения хронических заболеваний и нарушений. В настоящем изобретении представлены эти и другие преимущества, как описано в подробном описании.
Краткое описание изобретения
В одном аспекте настоящего изобретения предусмотрен способ введения генетически модифицированных В-клеток субъекту для получения терапевтического средства in vivo, предусматривающий введение двух или более последовательных доз генетически модифицированных В-клеток субъекту.
Один аспект настоящего изобретения относится к способу доставки терапевтического средства во множество тканей in vivo, предусматривающему введение субъекту двух или более доз генетически модифицированных В-клеток.
Один аспект настоящего изобретения относится к способу лечения MPS I, предусматривающему введение субъекту с MPS I двух или более последовательных доз В-клеток, генетически модифицированных с обеспечением продуцирования IDUA.
В одном аспекте настоящего изобретения предусмотрен способ снижения количества гликозаминогликана (GAG) у субъекта с MPS I, предусматривающий введение субъекту двух или более последовательных доз В-клеток, генетически модифицированных с обеспечением продуцирования IDUA.
В одном аспекте настоящего изобретения предусмотрен способ доставки терапевтического средства в одну или более тканей in vivo, предусматривающий введение субъекту одной или более доз генетически модифицированных В-клеток, где генетически модифицированные В-клетки являются мигрирующими.
Один аспект настоящего изобретения относится к способу введения субъекту генетически модифицированных В-клеток для обеспечения синергетического продуцирования in vivo терапевтического средства, предусматривающему: определение оптимальной концентрации однократной дозы модифицированных В-клеток для индуцирования наибольшего продуцирования in vivo терапевтического средства; уменьшение оптимальной концентрации однократной дозы модифицированных B-клеток с получением субоптимальной концентрации однократной дозы модифицированных B-клеток и введение субъекту двух или более доз при субоптимальной концентрации однократной дозы модифицированных В-клеток.
Один аспект настоящего изобретения относится к генетически модифицированной В-клетке, которая была сконструирована для получения терапевтического средства. В некоторых вариантах осуществления терапевтическое средство представляет собой IDUA. В некоторых вариантах осуществления терапевтическое средство представляет собой FIX, LPL или LCAT.
Один аспект настоящего изобретения относится к композиции, содержащей популяцию генетически модифицированных B-клеток, которые были сконструированы для получения терапевтического средства, где генетически модифицированные B-клетки обладают оптимальной миграционной способностью. В некоторых вариантах осуществления терапевтическое средство представляет собой IDUA. В некоторых вариантах осуществления терапевтическое средство представляет собой FIX, LPL или LCAT.
Один аспект настоящего изобретения относится к композиции, содержащей популяцию генетически модифицированных B-клеток, которые были сконструированы для получения терапевтического средства, где генетически модифицированные B-клетки в композиции собирают из культуры в момент времени, когда они не продуцируют значительных количеств воспалительных цитокинов. В некоторых вариантах осуществления терапевтическое средство представляет собой IDUA. В некоторых вариантах осуществления терапевтическое средство представляет собой FIX, LPL или LCAT.
В одном аспекте настоящего изобретения предусмотрен способ введения генетически модифицированных В-клеток субъекту для получения терапевтического средства in vivo, предусматривающий введение субъекту оптимальной однократной дозы генетически модифицированных В-клеток. В некоторых вариантах осуществления терапевтическое средство представляет собой IDUA. В некоторых вариантах осуществления терапевтическое средство представляет собой FIX, LPL или LCAT.
Краткое описание графических материалов
На фигуре 1 представлена диаграмма конструкций/карты транспозона и транспозазы Sleeping Beauty (SB) для обеспечения транспозиции и экспрессии IDUA человека. IDUA регулируется промотором EEK (см. пример 1). Двунаправленный промотор, который включает промотор EF1a выше EEK, регулирует транскрипцию дигидрофолатредуктазы L22Y-F31S (DHFR), обеспечивающей устойчивость к лекарственному средству, в обратном направлении. CMV-регулируемый SB100x обеспечивает транспозазную активность SB. Кэпированная и полиаденилированная mRNA, кодирующая SB100x, была получена путем транскрипции in vitro, предоставленная TriLink. Стрелки: направление транскрипции. Зеленые прямоугольники с темными треугольниками представляют собой последовательности инвертированного повтора/прямого повтора (IR/DR) T2 SB. рА, сигнал полиаденилирования. На фиг. 1А показаны схемы конструкций. На фиг. 1В показана плазмидная карта для конструкции pKT2/EEK-IDUA-DHFR, показанной на фиг. 1А, которая содержит DHFR человека с мутациями L22Y; F31S.
На фигуре 2 показана SB-опосредованная экспрессия IDUA человека в первичных В-клетках человека. Первичные CD19+ В-клетки человека культивировали в течение 2 дней, затем подвергали электропорации с помощью pKT2/EEK-IDUA и pCMV-SB100x как источника транспозазы с применением системы Lonza 4D. Клеточный лизат в день 8 после электропорации анализировали в отношении активности фермента IDUA.
На фигуре 3 представлена серия гистограмм, которые показывают MTX-селективное обогащение IDUA+ клеток во время крупномасштабного размножения B-клеток. В-клетки от двух отдельных доноров (19009 и 2764) подвергали электропорации с помощью транспозона pKT2/EEK-IDUA-DHFR, инкубировали в среде с MTX (нижние две гистограммы) или без него (верхние две гистограммы) от дней 2-4, а затем дополнительно размножали в общей сложности в течение периода 7 дней. Клетки из каждой популяции собирали в день 7 и анализировали в отношении % IDUA-положительных клеток путем внутриклеточного окрашивания в отношении IDUA человека с последующей проточной цитометрией (подсчет клеток/IDUA).
На фигуре 4 показана экспрессия идуронидазы у мышей NSG, которым вводили В-клетки, транспозированные IDUA-DHFR. Мышам NSG IDUA+ вводили i.p. CD4+ Т-клетки в дни -30 и -4, а затем в день 0 вводили посредством либо i.p., либо i.v. инъекции 107 B-клеток pKT2/EEK-IDUA-DHFR, которые прошли отбор с помощью MTX. В качестве контроля некоторым мышам вводили путем i.p. инъекции В-клетки, экспрессирующие GFP. Образцы плазмы анализировали в отношении IDUA в указанные моменты времени. Мыши с 1-8 получали i.p. инфузии В-клеток, экспрессирующих IDUA. Мыши 9, 10, 12 и 42 получали i.v. инфузии В-клеток, экспрессирующих IDUA. Мыши 43 и 48 получали i.p. инфузии В-клеток, экспрессирующих GFP.
На фигуре 5 показано количество идуронидазы (IDUA), присутствующей в плазме крови, с применением мышиной модели MPS I. Мыши получали, сверху вниз обозначений, 3×106 В-клеток, трансдуцированных посредством IDUA (IDUA+ B-клетки) в присутствии CD4+ Т-клеток памяти, 1×107 IDUA+ В-клеток в присутствии CD4+ Т-клеток памяти, 3×107 IDUA+ B-клеток в присутствии CD4+ Т-клеток памяти , только CD4+ Т-клетки памяти или не получали клеток в день 0, а также измеряли уровни активности фермента IDUA в сыворотке крови через 38 дней после введения.
На фигуре 6 показано количество IDUA, присутствующей в плазме, на мышиной модели MPS I с применением многоратных доз B-клеток, трансдуцированных посредством IDUA (IDUA+ B-клетки). В-клетки человека обогащали CD19 из продукта афереза нормального донора и подвергали электропорации с помощью транспозона pKT2/EEK-IDUA плюс mRNA, кодирующей SB100x, во время процесса размножения. CD4+ Т-клетки выделяли у того же донора и вводили животным NSG MPS I внутрибрюшинно (i.p) за одну неделю до инфузии транспозированных IDUA В-клеток. Контрольные группы включали необработанных мышей NSG MPS I («без B-клеток») и мышей NSG MPS I, которым вводили i.v. только IDUA+ B-клетки (т.е. без CD4+ Т-клеток). Мышам NSG MPS I, предварительно обработанным аутологичными CD4+ T-клетками, впоследствии вводили IDUA+ B-клетки либо i.v., либо i.p. в дни 0, 21 и 42 (стрелки). Уровни активности фермента IDUA измеряли в сыворотке крови до дня 56. N=4.
На фигуре 7 показан уровень IgG в плазме крови от тех же мышей NSG MPS I, которые описаны на фигуре 6. N=4.
На фигуре 8 показана активность IDUA в различных тканях от мышей MPS I. Мышам MPS I давали три дозы из 1×107 В-клеток, сконструированных для получения IDUA, (или отсутствие клеток в качестве контроля) в дни 0, 21 и 42 в присутствии CD4+ Т-клеток (или отсутствие клеток в качестве контроля) и измеряли уровни активности фермента IDUA в указанных органах в день 60 после первой инфузии В-клеток. N=4.
На фигуре 9 показано количество гликозаминогликанов (GAG) в различных тканях мышей MPS I. Мышам MPS I давали три дозы из 1×107 В-клеток, сконструированных для получения IDUA, (или отсутствие клеток в качестве контроля) в дни 0, 21 и 42 в присутствии CD4+ Т-клеток (или отсутствие клеток в качестве контроля) в день 0 и измеряли уровни GAG в указанных органах в день 60 после первой инфузии В-клеток. Кроме того, красные горизонтальные столбики показывают среднюю активность фермента IDUA в плазме крови для каждой из групп мышей. N=4.
На фигуре 10 показано, что активность IDUA в плазме крови обнаруживается в течение длительного времени у мышей NSG MPSI, которых обрабатывали с помощью двух доз 2×107 B-клеток, сконструированных с обеспечением продуцирования IDUA. Первую дозу В-клеток давали через одну неделю после введения CD4+ Т-клеток, а вторую дозу В-клеток вводили через 30 дней после первой дозы В-клеток. (IDUA+ B-клетки). Цветные обозначения справа означают группы мышей. На оси X указано время в неделях. На оси Y указана активность фермента IDUA, обнаруженного в образцах плазмы крови мышей.
На фигуре 11 показано количество активности IDUA, присутствующей во множестве тканей у мышей NSG MPSI, обработанных посредством двух доз 2×107 В-клеток, сконструированных для получения IDUA в соответствии с тем же протоколом, что указан на фигуре 10. Цветные обозначения справа означают группы мышей и моменты времени. На оси X показана исследуемая ткань, а на оси Y показаны уровни ферментативной активности IDUA, которые были обнаружены.
На фигуре 12 показано количество гликозаминогликанов (GAG) в различных тканях от мышей NSG MPS I, которым вводили две дозы 2×107 В-клеток, сконструированных для получения IDUA в соответствии с тем же протоколом, что указан на фигурах 10 и 11. Обработка с помощью продукта на основе В-клеток приводит к долговременному снижению уровней GAG во множестве тканей. Цветные обозначения справа означают орган, в котором оценивали GAG. На оси X показана группа мышей и доза клеток. На оси Y показано обнаруженное количество GAG.
На фигуре 13 показана миграция В-клеток, сконструированных с обеспечением экспрессии IDUA, в направлении хемоаттрактанта в двухкамерном анализе Transwell. На фиг. 13А показана миграция сконструированных В-клеток в день 0 в направлении хемоаттрактанта CXCL12. На фиг. 13В показана миграция сконструированных В-клеток в направлении хемоаттрактанта CXCL12 через 4, 5, 6, 7, 8 или 9 дней в культуре после конструирования. На фиг. 13C показана миграция сконструированных В-клеток в направлении хемоаттрактанта CXCL13 через 4, 5, 6, 7, 8 или 9 дней в культуре после конструирования. В случае обеих фиг. 13B и 13C, необходимо обратить внимание, что контроль без хемокинов был реализован только для момента времени день 4. На фиг. 13D показано схематическое изображение анализа Transwell, применяемого для получения данных на фиг. 13A-13C.
На фигуре 14 показаны сводные данные анализа клональности В-клеток, сконструированных с обеспечением экспрессии IDUA, посредством глубокого секвенирования.
На фигуре 15 показан анализ Luminex продуцирования воспалительных цитокинов В-клетками, сконструированными с обеспечением продуцирования IDUA. На фиг. 15A показано продуцирование IL6, IFN-альфа и IFN-гамма в день 2 (D2), день 7 (D7) и день 0 (D0) с IL6 и без него (50 нг/мл). На фиг. 15B показано продуцирование sFAS, TNFRp75, BAFF, HGF и IL5 на базовой среде в D2, D7 и D0.
На фигуре 16 показана экспрессия LCAT (лецитинхолестеринацилтрансферазы) человека, LPL (липопротеинлипазы) человека и FIX (фактора свертывания крови IX) человека в В-клетках человека, сконструированных в соответствии с настоящим изобретением. На фиг. 16А показана активность LCAT в сконструированных плазмабластах/плазматических клетках. На фиг. 16B показана активность LPL в сконструированных плазмабластах/плазматических клетках. На фиг. 16C показана экспрессия белка FIX с помощью ELISA в сконструированных первичных В-клетках.
Подробное описание изобретения
При практическом применении настоящего изобретения применяют, если не указано иное, традиционные способы молекулярной биологии, методики рекомбинантной ДНК, экспрессию белка и химию белка/пептида/углевода в пределах уровня техники, многие из которых описаны ниже с целью иллюстрации. Такие методики подробно описаны в литературе. См., например, Sambrook, et al., Molecular Cloning: A Laboratory Manual (3rd Edition, 2000); DNA Cloning: A Practical Approach, vol. I & II (D. Glover, ed.); Oligonucleotide Synthesis (N. Gait, ed., 1984); Oligonucleotide Synthesis: Methods and Applications (P. Herdewijn, ed., 2004); Nucleic Acid Hybridization (B. Hames & S. Higgins, eds., 1985); Nucleic Acid Hybridization: Modern Applications (Buzdin and Lukyanov, eds., 2009); Transcription and Translation (B. Hames & S. Higgins, eds., 1984); Animal Cell Culture (R. Freshney, ed., 1986); Freshney, R.I. (2005) Culture of Animal Cells, a Manual of Basic Technique, 5th Ed. Hoboken NJ, John Wiley & Sons; B. Perbal, A Practical Guide to Molecular Cloning (3rd Edition 2010); Farrell, R., RNA Methodologies: A Laboratory Guide for Isolation and Characterization (3rd Edition 2005). Публикации, обсуждаемые выше, представлены исключительно для их раскрытия до даты подачи настоящей заявки. Ничто в настоящем документе не должно толковаться как признание того, что настоящее изобретение не имеет права датировать такое раскрытие задним числом ввиду предшествующего изобретения.
ОПРЕДЕЛЕНИЯ И СОКРАЩЕНИЯ
Если не указано иное, все технические и научные выражения, применяемые в данном документе, имеют то же значение, которое обычно подразумевается специалистом в области техники, к которой принадлежит настоящее изобретение. Используемые в описании и прилагаемой формуле изобретения следующие термины, если не указано иное, имеют указанное значение. Что касается данного описания, то всякий раз, когда определение термина, определенное в данном документе, отличается от определения, данного для того же самого термина во включенной ссылке, определение, определенное в данном документе, является верным определением данного термина.
Форма единственного числа обозначает одно или более, если конкретно не указано иное.
Используемый в данном документе термин «приблизительно» относится к количественному параметру, уровню, значению, числу, частоте, проценту, величине, размеру, количеству, массе или длине, которая изменяется на вплоть до 30, 25, 20, 15, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1% от эталонного количественного параметра, уровня, значения, числа, частоты, процента, величины, размера, количества, массы или длины. В любом варианте осуществления, обсуждаемом в контексте числового значения, используемого вместе с термином «приблизительно», конкретно предполагается, что термин «приблизительно» может быть опущен.
«Композиция» может содержать активное средство и носитель, инертный или активный, например, фармацевтически приемлемый носитель, разбавитель или вспомогательное вещество. В конкретных вариантах осуществления композиции являются стерильными, по сути не содержащими эндотоксинов или нетоксичными для реципиентов при применяемой дозе или концентрации.
Если контекст не требует иного, в настоящем описании и формуле настоящего изобретения слово «содержать» и его варианты, такие как «содержит» и «содержащий», следует толковать в открытом и включительном смысле, то есть как «включая без ограничения».
Выражение «состоящий из» подразумевает включение и ограничивается тем, что следует за фразой «состоящий из». Таким образом, фраза «состоящий из» указывает, что перечисленные элементы являются необходимыми или обязательными и что никакие другие элементы не могут присутствовать. Выражение «состоящий по сути из» подразумевает включение любых элементов, перечисленных после фразы, и ограничивается другими элементами, которые не мешают или не способствуют активности или действию, указанным в настоящем изобретении для перечисленных элементов. Таким образом, фраза «состоящий по сути из» указывает, что перечисленные элементы необходимы или обязательны, но что никакие другие элементы не являются необязательными и могут присутствовать или не присутствовать в зависимости от того, влияют ли они на активность или действие перечисленных элементов.
Ссылка в данном описании на «биологическую активность» или «биоактивность» относится к любому ответу, индуцированному в анализе in vitro или в клетке, ткани, органе или организме (например, у животного, или млекопитающего, или человека) как результат введения какого-либо соединения, средства, полипептида, конъюгата, фармацевтической композиции, рассматриваемых в настоящем документе. Биологическая активность может относиться к агонистическим действиям или антагонистическим действиям. Биологическая активность может быть благоприятным эффектом; или биологическая активность может не быть благоприятной, то есть предусматривать токсичность. В некоторых вариантах осуществления биологическая активность будет относиться к положительным или отрицательным эффектам, которые лекарственное средство или фармацевтическая композиция оказывают в отношении живого субъекта, например, млекопитающего, такого как человек. Соответственно, термин «биологически активный» предназначен для описания любого соединения, обладающего биологической активностью, описанной в данном документе. Биологическая активность может быть оценена с помощью любого подходящего способа, известного в настоящее время специалисту в данной области. Такие анализы могут быть качественными или количественными. Специалист в данной области легко поймет необходимость применения различных анализов для оценки активности различных полипептидов; задача, которая является стандартной для среднестатистического исследователя. Такие анализы часто легко реализуются в лабораторных условиях с небольшими требованиями к оптимизации, и чаще всего имеются коммерческие наборы, которые обеспечивают простые, надежные и воспроизводимые показания биологической активности для широкого спектра полипептидов с применением различных технологий, общих для большинства лабораторий. Когда такие наборы недоступны, обычные квалифицированные исследователи могут легко разработать и оптимизировать собственные анализы биологической активности для полипептидов-мишеней без излишних экспериментов, так как это рутинный аспект научного процесса.
Ссылка на термин «например» предназначена для обозначения «например, без ограничения», и, таким образом, следует понимать, что все, что следует, является просто примером конкретного варианта осуществления, но никоим образом не должно рассматриваться как ограничивающий пример. Если не указано иное, использование термина «например» предназначено для явного указания того, что другие варианты осуществления были рассмотрены и охватываются настоящим изобретением.
Ссылка по всему настоящему описанию на «вариант осуществления» или «один вариант осуществления», или «некоторый вариант осуществления», или «некоторые варианты осуществления», или «определенные варианты осуществления» означает, что конкретный признак, структура или характеристика, описанные в связи с вариантом осуществления, включены в по меньшей мере один вариант осуществления настоящего изобретения. Таким образом, появления фраз «в одном варианте осуществления» или «в варианте осуществления» или «в определенных вариантах осуществления» в различных местах по всему данному описанию не обязательно относится к одному и тому же варианту осуществления. Более того, конкретные признаки, структуры или характеристики могут быть объединены любым подходящим способом в одном или более вариантах осуществления.
«Увеличенное» или «повышенное» количество обычно представляет собой «статистически значимое» количество и может включать увеличение, которое составляет в 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2, 2,5, 3, 3,5, 4, 4,5, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40 или 50 или больше раз (например, 100, 500, 1000 раз) (включая все целые числа и десятичные дроби между ними и выше 1, например, 2,1, 2,2, 2,3, 2,4 и т.д.) количества или уровня, описанных в данном документе. Сходным образом, «уменьшенное», или «сниженное», или «меньшее» количество обычно представляет собой «статистически значимое» количество и может включать уменьшение, которое составляет в 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2, 2,5, 3, 3,5, 4, 4,5, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40 или 50 или больше раз (например, 100, 500, 1000 раз) (включая все целые числа и десятичные дроби между ними и выше 1, например, 2,1, 1,5, 1,6, 1,7. 1,8 и т.д.) количества или уровня, описанных в данном документе.
Термины «in vitro», «ex vivo» и «in vivo» представлены в данном документе в своем обыкновенном научном значении. Соответственно, например, «in vitro» предназначен для ссылки на эксперименты или реакции, которые происходят с помощью изолированных клеточных компонентов, такие как, например, ферментативная реакция, проводимая в пробирке с применением подходящего субстрата, фермента, донора и необязательно буферов/кофакторов. «Ex vivo» предназначен для обозначения экспериментов или реакций, проводимых с применением функциональных органов или клеток, которые были удалены из организма или размножены независимо от него. «In vivo» предназначен для обозначения экспериментов или реакций, которые происходят в живом организме в его нормальном интактном состоянии.
«Млекопитающее» включает людей и как домашних животных, таких как лабораторные животные и домашние животные (например, кошки, собаки, свиньи, крупный рогатый скот, овцы, козы, лошади и кролики), так и отличных от домашних животных, таких как дикие животные и тому подобные.
«Необязательный» или «необязательно» означает, что описанное впоследствии событие или обстоятельства могут или не могут произойти, и что описание включает в себя случаи, когда указанное событие или обстоятельство происходит, и случаи, в которых оно не происходит.
«Фармацевтическая композиция» относится к составу из соединения (например, терапевтически применимого полипептида) и среды, общепринятой в данной области техники для доставки соединения животному, например, человеку. Такая среда, таким образом, может включать любые фармацевтически приемлемые носители, разбавители или вспомогательные вещества.
«Фармацевтически эффективные вспомогательные вещества» и «фармацевтически эффективные носители» хорошо известны специалистам в данной области, и способы их получения также очевидны для специалиста в данной области. Такие композиции и способы их получения можно найти, например, в Remington's Pharmaceutical Sciences, 19th Edition (Mack Publishing Company, 1995, включена в настоящий документ).
Термины «полинуклеотид», «нуклеотид», «нуклеотидная последовательность» и «нуклеиновая кислота» используются взаимозаменяемо. Они относятся к полимерной форме нуклеотидов любой длины, либо дезоксирибонуклеотидов, либо рибонуклеотидов, либо их аналогов. Полинуклеотиды могут иметь любую трехмерную структуру и могут выполнять любую функцию, известную или неизвестную. Ниже приведены неограничивающие примеры полинуклеотидов: кодирующие или некодирующие области гена или фрагмента гена, локусы (локус), определенные из анализа сцепления, экзоны, интроны, матричная РНК (mRNA), транспортная РНК, рибосомная РНК, рибозимы, cDNA, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенная ДНК любой последовательности, выделенная РНК любой последовательности, зонды на основе нуклеиновых кислот и праймеры. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды, и нуклеотидные аналоги. В случае присутствия модификации нуклеотидной структуры могут придаваться до или после сборки полимера. Последовательность нуклеотидов может включать отличные от нуклеотидных компоненты. Полинуклеотид может быть дополнительно модифицирован после полимеризации, например, путем конъюгации с метящим компонентом.
«Субъект», как используется в данном документе, включает любое животное, у которого проявляется заболевание или симптом или существует риск возникновения заболевания или симптома, которое можно подвергать лечению с помощью средства по настоящему изобретению. К подходящим субъектам относятся лабораторные животные (такие как мышь, крыса, кролик или морская свинка), сельскохозяйственные животные и домашние животные или питомцы (такие как кошка или собака). Включены отличные от людей приматы и предпочтительно пациенты-люди.
«Существенно» или «по сути» означает достаточное или значительное количество, число, размер; почти полное или целое, например, 95% или больше некоторого заданного количества.
«Терапевтическое средство» относится к любому соединению, которое при введении субъекту (например, предпочтительно млекопитающему, более предпочтительно человеку) в терапевтически эффективном количестве способно воздействовать на лечение заболевания или состояния, как определено ниже.
«Терапевтически эффективное количество» или «терапевтически эффективная доза» относится к количеству соединения по настоящему изобретению, которое при введении субъекту (например, предпочтительно млекопитающему, более предпочтительно человеку) является достаточным для осуществления лечения, как определено ниже, заболевания или состояния у животного. Количество соединения по настоящему изобретению, которое составляет «терапевтически эффективное количество», будет варьироваться в зависимости от соединения, состояния и его тяжести, способа введения и возраста животного, подлежащего лечению, но обычно может быть определено стандартным образом специалистом в данной области техники, принимая во внимание его собственные знания и данное изобретение.
Термин «осуществление лечения» или «лечение», используемый в данном документе, охватывает лечение заболевания или состояния, представляющего интерес, у субъекта, предпочтительно человека, у которого имеется заболевание или состояние, представляющее интерес, и включает в себя: (i) предупреждение или подавление заболевания или состояния у субъекта, в частности, когда такой субъект предрасположен к состоянию, но при этом ему еще не поставлен диагноз наличия заболевания или состояния; (ii) подавление заболевания или состояния, т.е. остановку его развития; (iii) ослабление заболевания или состояния, т.е. обеспечение регресса заболевания или состояния; или (iv) ослабление симптомов, возникающих в результате заболевания или состояния. Используемые в данном документе термины «заболевание», «нарушение» и «состояние» могут использоваться взаимозаменяемо или могут отличаться тем, что конкретная болезнь, травма или состояние могут не иметь известной причины (так что этиология еще не была разработана), и поэтому они еще не признаны травмой или заболеванием, а лишь нежелательным состоянием или синдромом, при котором клиницисты выявили более или менее специфический набор симптомов.
Общий обзор
В настоящем изобретении используют аутологичные и/или аллогенные В-клетки, которые были изменены путем введения нуклеиновых кислот для получения терапевтического средства, и предусмотрены способы введения модифицированных В-клеток. В некоторых вариантах осуществления термины «сконструированная B-клетка», «генетически сконструированная B-клетка», «модифицированная B-клетка» и «генетически модифицированная B-клетка» используются в данном документе взаимозаменяемо для обозначения таких измененных B-клеток, которые содержат одну или более нуклеиновых кислот (например, трансген) для получения терапевтического средства (например, трансгена, который обеспечивает экспрессию полипептида, такого как терапевтический полипептид). В частности, модифицированные В-клетки можно вводить в виде однократной дозы или многократных доз. Неожиданно было обнаружено, что определенные дозы В-клеток способствуют выработке уровней терапевтического средства, превышающих ожидаемые, по сравнению с другими дозами. Кроме того, неожиданно было обнаружено, что применение множества доз B-клеток, доставляемых посредством схемы с многократными дозами, приводит к более высоким уровням терапевтического средства, чем достигается посредством одной дозы, содержащей такое же количество клеток. Кроме того, неожиданно было обнаружено, что модифицированные В-клетки обладают окнами оптимальной миграционной способности по направлению к хемоаттрактантам, и их миграционная способность может снижаться после определенных периодов времени в культуре. Кроме того, было неожиданно обнаружено, что, несмотря на то, что исходная популяция сконструированных В-клеток продуцировала IL6, уровни продуцирования снижались почти до фоновых уровней к концу культивирования, и большинство исследованных воспалительных цитокинов не продуцировалось сконструированными В-клетками. Кроме того, было показано, что конечная популяция сконструированных B-клеток была в значительной степени поликлональной, поскольку в конечной популяции сконструированных B-клеток не было обнаружено наличие какого-либо конкретного клона B-клеток в количестве более чем приблизительно 0,2% от общей популяции B-клеток. Наконец, было обнаружено, что модифицированные В-клетки способны эффективно доставлять лекарственное средство в широкий спектр тканей, таких как легкие, сердце и кишечник, на которые трудно нацеливаться с применением других способов.
Соответственно, способы введения композиций на основе модифицированных В-клеток, описанные в данном документе, являются применимыми для долгосрочной доставки in vivo и экспрессии терапевтических средств. Настоящее изобретение в целом относится к способам достижения достаточного обогащения и количества клеток, продуцирующих терапевтическое средство, и достаточных уровней терапевтического средства in vivo при обеспечении безопасности продукта.
Используемые в данном документе фразы «долгосрочное выживание in vivo» и «долгосрочное выживание» относятся к выживанию модифицированных В-клеток, описанных в данном документе, в течение периода 10 дней или больше после введения субъекту. Долгосрочная выживание можно измерять в днях, неделях или даже годах. В одном варианте осуществления большинство модифицированных В-клеток выживают in vivo в течение периода 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27 , 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50 дней или больше после введения. В одном варианте осуществления большинство модифицированных В-клеток выживают in vivo в течение периода 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 , 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44 , 45, 46, 47, 48, 49, 50, 51, 52 недель или больше после введения. В другом варианте осуществления модифицированные В-клетки выживают in vivo в течение периода 1, 1,5, 2, 2,5, 3, 3,5, 4, 4,5, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30 лет и больше. Кроме того, хотя описанные в данном документе модифицированные В-клетки могут выживать in vivo в течение периода 10 дней или больше, следует понимать, что большинство модифицированных В-клеток выживают in vivo в течение периода 1, 2, 3, 4, 5, 6, 7, 8, 9 дней или больше после введения. Соответственно, предполагают, что модифицированные В-клетки, описанные в данном документе, являются применимыми для способов краткосрочного лечения (например, 4 дня) и долгосрочного лечения (например, 30 или больше дней).
B-клетки
Покинув костный мозг, В-клетка действует как антигенпрезентирующая клетка (АРС) и интернализует антигены. Антиген поглощается В-клеткой посредством рецептор-опосредованного эндоцитоза и подвергается обработке. Антиген перерабатывается в антигенные пептиды, загружается на молекулы MHC II и подвергается презентации на внеклеточной поверхности В-клеток в отношении CD4+ Т-хелперных клеток. Такие Т-клетки связываются с молекулой MHC II/антигена и вызывают активацию B-клетки. После стимуляции Т-клеткой активированная В-клетка начинает дифференцироваться в более специализированные клетки. В-клетки герминального центра могут дифференцироваться в долгоживущие В-клетки памяти или плазматические клетки. Кроме того, вторичная иммунная стимуляция может привести к тому, что В-клетки памяти способствуют образованию дополнительных плазматических клеток. Образованию плазматических клеток из В-клеток памяти или В-клеток, отличных от клеток памяти, предшествует образование плазмобластов-предшественников, которые в конечном итоге дифференцируются в плазматические клетки, которые продуцируют большие объемы антител (см., например, Trends Immunol. 2009 June; 30(6): 277-285; Nature Reviews, 2005, 5: 231-242). Плазмобласты секретируют больше антител, чем В-клетки, но меньше, чем плазматические клетки. Они быстро делятся и продолжают интернализовать антигены и презентировать антигены Т-клеткам. Плазмабласты обладают способностью мигрировать в места продуцирования хемокинов (например, в костном мозге), благодаря чему они могут дифференцироваться в долгоживущие плазматические клетки. В конечном счете, плазмабласт может либо оставаться в виде плазмабласта в течение нескольких дней, а затем погибнуть, или безвозвратно дифференцироваться в зрелую, полностью дифференцированную плазматическую клетку. В частности, плазмабласты, которые способны находиться в тканях, содержащих ниши выживания плазматических клеток (например, в костном мозге), способны вытеснять резидентные плазматические клетки, чтобы стать долгоживущими плазматическими клетками, которые могут продолжать секретировать высокий уровень белков в течение многих лет.
В-клетки, применяемые в описанных в данном документе способах, включают пан-В-клетки, В-клетки памяти, плазмабласты и/или плазматические клетки. В одном варианте осуществления модифицированные B-клетки представляют собой B-клетки памяти. В одном варианте осуществления модифицированные B-клетки представляют собой плазмабласты. В одном варианте осуществления модифицированные B-клетки представляют собой плазматические клетки.
Окончательно дифференцированные плазматические клетки обычно не экспрессируют общие маркеры пан-В-клеток, такие как CD19 и CD20, и экспрессируют относительно немного поверхностных антигенов. Плазматические клетки экспрессируют CD38, CD78, CD138 и рецептор интерлейкина-6 (IL-6R) и не экспрессируют CD45, и эти маркеры можно применять для идентификации плазматических клеток, например, с помощью проточной цитометрии. CD27 также является показательным маркером для плазматических клеток, также как для наивных B-клеток таковым является CD27-, B-клеток памяти - CD27+, а плазматических клеток - CD27++. Субпопуляции В-клеток памяти также могут экспрессировать поверхностные IgG, IgM и IgD, тогда как плазматические клетки не экспрессируют эти маркеры на клеточной поверхности. CD38 и CD138 экспрессируются при высоких уровнях в плазматических клетках (См. Wikipedia, The Free Encyclopedia., “Plasma cell” ID версии страницы: 404969441; дата последней версии: 30 декабря 2010 года, 09:54 UTC, получена 4 января 2011 года; см. также: Jourdan et al. Blood. 2009 Dec 10; 114(25): 5173-81; Trends Immunol. 2009 June; 30(6): 277-285; Nature Reviews, 2005, 5: 231-242; Nature Med. 2010, 16: 123-129; Neuberger, M. S.; Honjo, T.; Alt, Frederick W. (2004). Molecular biology of B cells. Amsterdam: Elsevier, pp. 189-191 ; Bertil Glader; Greer, John G.; John Foerster; Rodgers, George G.; Paraskevas, Frixos (2008). Wintrobe's Clinical Hematology, 2-Vol. Set. Hagerstwon, MD: Lippincott Williams & Wilkins. pp. 347; Walport, Mark; Murphy, Kenneth; Janeway, Charles; Travers, Paul J. (2008). Janeway's immunobiology. New York: Garland Science, pp. 387-388; Rawstron AC (May 2006). “Immunophenotyping of plasma cells”. Curr Protoc Cytom).
Термин «покоящаяся», как используется в данном документе, относится к состоянию клетки, в котором клетка активно не пролиферирует.
«Активированная», как используется в данном документе, относится к состоянию клетки, в котором клетка активно пролиферирует и/или продуцирует цитокины в ответ на стимул.
Термины «дифференцировать» и «дифференцированный», используемые в данном документе, относятся к изменениям фенотипа клетки от одного типа или состояния клетки к другому типу или состоянию клетки. Например, В-клетка памяти, которая превращается в плазматическую клетку, является дифференцированной.
Термин «субъект» предназначен для включения живых организмов, у которых может быть вызван адаптивный иммунный ответ (например, млекопитающие). Примеры субъектов включают людей, собак, кошек, мышей, крыс и их трансгенных виды. В одном варианте осуществления субъект является человеком. В-клетки могут быть получены из ряда источников, в том числе мононуклеарные клетки периферической крови (РВМС), костный мозг, ткань лимфатического узла, пуповинная кровь, ткань из места инфекции, ткань селезенки и опухоли. В предпочтительном варианте осуществления источником B-клеток являются PBMC. В определенных вариантах осуществления настоящего изобретения может применяться любое количество линий B-клеток, доступных в данной области техники.
В определенных вариантах осуществления способов, описанных в данном документе, B-клетки могут быть получены из единицы крови, собранной у субъекта, с применением любого количества методик, известных специалисту в данной области, таких как разделение посредством FICOLL™ (сополимеры сахарозы и эпихлоргидрина, которые могут быть использованы для получения растворов с высокой плотностью). В одном предпочтительном варианте осуществления клетки из циркулирующей крови индивидуума получают путем афереза или лейкафереза. Продукт афереза обычно содержит лимфоциты, в том числе Т-клетки, моноциты, гранулоциты, В-клетки, другие ядросодержащие белые кровяные клетки, красные кровяные клетки и тромбоциты. В одном варианте осуществления клетки, собранные с помощью афереза, можно промывать для удаления фракции плазмы и помещения клеток в соответствующий буфер или среду для последующих стадий обработки. В одном варианте осуществления способов, описанных в настоящем документе, клетки промывают фосфатно-солевым буфером (PBS). В альтернативном варианте осуществления в растворе для промывания отсутствует кальций, и может отсутствовать магний, и могут отсутствовать многие, если не все, двухвалентные катионы. Специалистам в данной области техники легко понять, что стадию промывки можно выполнять с помощью способов, известных специалистам в данной области, таких как применение полуавтоматической «проточной» центрифуги (например, устройства для обработки клеток Cobe 2991) в соответствии с инструкциями производителя. После промывки клетки можно ресуспендировать в различных биосовместимых буферах, таких как, например, PBS. Альтернативно, нежелательные компоненты образца афереза можно удалять, а клетки непосредственно ресуспендировать в культуральной среде.
В-клетки можно выделять из периферической крови или посредством лейкафереза с применением методик, известных в данной области. Например, РВМС можно выделять с применением FICOLL™ (Sigma-Aldrich, Сент-Луис, Миссури) и CD19+ B-клеток, очищенных с помощью отрицательного или положительного отбора, с применением любого из множества антител, известных в данной области, такого как тетрамерная комплексная система Rosette (StemCell Technologies, Ванкувер, Канада) или MACS™ MicroBead Technology (Miltenyi Biotec, Сан-Диего, Калифорния). В некоторых вариантах осуществления B-клетки памяти выделяют, как описано у Jourdan et al., (Blood. 2009 Dec 10; 114(25): 5173-81). Например, после удаления CD2+ клеток с применением магнитных гранул к CD2 CD19+ CD27+ B-клетки памяти можно отсортировать с помощью FACS. Плазматические клетки костного мозга (BMPC) можно очищать с применением сортировки посредством магнитных гранул к CD138 или других подобных способов и реагентов. В-клетки человека можно выделять, например, с применением микрогранул к CD19 человека (Miltenyi Biotec, Сан-Диего, Калифорния). B-клетку памяти человека можно выделять, например, с применением набора для выделения B-клеток памяти человека (Miltenyi Biotec, Сан-Диего, Калифорния).
Коммерчески доступны другие наборы для выделения, такие как набор для выделения В-клеток человека MagCellect от R&D Systems (Миннеаполис, Миннесота). В некоторых вариантах осуществления покоящиеся В-клетки можно получать путем осаждения на прерывистых градиентах Percoll, как описано в (Defranco et al., (1982) J. Exp. Med. 155: 1523).
В одном варианте осуществления РВМС получают из образца крови с применением очистки на основе градиента (например, FICOLL™). В другом варианте осуществления РВМС получают из коллекции, полученной на основе афереза. В одном варианте осуществления B-клетки выделяют из PBMC путем выделения пан-B-клеток. На стадии выделения можно применять положительный и/или отрицательный отбор. В одном варианте осуществления отрицательный отбор предусматривает истощение Т-клеток с применением конъюгированных микрогранул к CD3 с обеспечением тем самым истощенной фракции Т-клеток. В дополнительном варианте осуществления В-клетки памяти выделяют из пан-В-клеток или истощенной фракции Т-клеток путем положительного отбора в отношении CD27.
В одном конкретном варианте осуществления В-клетки памяти выделяют путем истощения нежелательных клеток и последующего положительного отбора с помощью микрогранул к CD27. Нежелательные клетки, например Т-клетки, NK-клетки, моноциты, дендритные клетки, гранулоциты, тромбоциты и эритроидные клетки можно истощать с применением смеси биотинилированных антител к CD2, CD14, CD16, CD36, CD43 и CD235a (гликофорин A) и микрогранул к биотину.
В одном варианте осуществления получают переключенные B-клетки памяти. Термин «переключенная В-клетка памяти» или «переключенная В-клетка», как используется в данном документе, относится к B-клетке, которая подверглась переключению класса изотипов. В одном варианте осуществления переключенные В-клетки памяти положительно отобраны в отношении IgG. В другом варианте осуществления переключенные В-клетки памяти получают путем истощения экспрессирующих IgD и IgM клеток. Переключенные В-клетки памяти можно выделять, например, с применением набора для переключения В-клеток памяти человека (Miltenyi Biotec, Сан-Диего, Калифорния).
Например, в одном конкретном варианте осуществления нецелевые клетки можно подвергать мечению с помощью смеси биотинилированных CD2, CD14, CD16, CD36, CD43, CD235a (гликофорин A), антител к IgM и к IgD. Эти клетки могут быть впоследствии магнитно помечены с помощью микрогранул к биотину. Переключенные В-клетки памяти с высокой степенью очистки можно получать путем истощения магнитно-меченных клеток.
В дополнительном варианте осуществления последовательность промотора из гена, уникального для В-клеток памяти, такого как, например, ген CD27 (или другой ген, специфический для В-клеток памяти и не экспрессируемый в наивных В-клетках), применяют для управления экспрессией селектируемого маркера, такого как, например, мутированная дигидрофолатредуктаза, с обеспечением возможности положительного отбора В-клеток памяти в присутствии метотрексата. В другом варианте осуществления последовательность промотора из гена пан-В-клетки, такого как, например, ген CD19, применяют для управления экспрессией селектируемого маркера, такого как, например, мутированная дигидрофолатредуктаза, с обеспечением возможности положительного отбора В-клеток памяти в присутствии метотрексата. В другом варианте осуществления Т-клетки истощают с применением CD3 или путем добавления циклоспорина. В другом варианте осуществления CD138+ клетки выделяют из пан-В-клеток путем положительного отбора. В другом варианте осуществления CD138+ клетки выделяют из PBMC путем положительного отбора. В другом варианте осуществления CD38+ клетки выделяют из пан-В-клеток путем положительного отбора. В другом варианте осуществления CD38+ клетки выделяют из PBMC путем положительного отбора. В одном варианте осуществления CD27+ клетки выделяют из PBMC путем положительного отбора. В другом варианте осуществления B-клетки памяти и/или плазматические клетки селективно размножают из PBMC с применением способов культивирования in vitro, доступных в данной области.
Культивирование B-клеток in vitro
В-клетки, такие как В-клетки памяти, можно культивировать с применением способов in vitro для активации и дифференцировки В-клеток в плазматические клетки или плазмабласты или и те и другие. Как будет понятно специалисту, плазматические клетки могут быть идентифицированы по паттернам экспрессии белка на клеточной поверхности с применением стандартных способов проточной цитометрии. Например, окончательно дифференцированные плазматические клетки экспрессируют относительно немного поверхностных антигенов и не экспрессируют общие маркеры пан-В-клеток, такие как CD19 и CD20. Вместо этого плазматические клетки могут быть идентифицированы по экспрессии CD38, CD78, CD138 и IL-6R и отсутствию экспрессии CD45. CD27 также можно применять для идентификации плазматических клеток, поскольку наивные В-клетки являются CD27-, В-клетки памяти являются CD27+, а плазматические клетки являются CD27++. Плазматические клетки экспрессируют высокие уровни CD38 и CD138.
В одном варианте осуществления B-клетки представляют собой CD138- B-клетки памяти. В одном варианте осуществления B-клетки представляют собой CD138+ плазматические клетки. В одном варианте осуществления B-клетки активированы и имеют фенотип клеточной поверхности CD138-, CD27+.
В одном варианте осуществления B-клетки представляют собой CD20-, CD138- B-клетки памяти. В одном варианте осуществления B-клетки представляют собой CD20-, CD138+ плазматические клетки. В одном варианте осуществления B-клетки активированы и имеют фенотип клеточной поверхности CD20-, CD138-, CD27+.
В одном варианте осуществления B-клетки представляют собой CD20-, CD38-, CD138- B-клетки памяти. В одном варианте осуществления B-клетки представляют собой CD20-, CD38+, CD138+ плазматические клетки. В одном варианте осуществления B-клетки активируются и имеют фенотип клеточной поверхности CD20- CD38- CD138- CD27+.
В одном варианте осуществления В-клетки приводят в контакт с одним или более активирующими В-клетки факторами, например, с любым из множества цитокинов, факторов роста или клеточных линий, которые, как известно, активируют В-клетки и/или обеспечивают их дифференцировку (см., например, Fluckiger, et al. Blood 1998 92: 4509-4520; Luo, et al., Blood 2009 1 13: 1422-1431). Такие факторы могут быть выбраны из группы, состоящей без ограничения из IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-1 1 , IL-12, IL-13, IL-14, IL-15, IL-16, IL-17, IL-18, IL-19, IL-20, IL-21, IL-22, IL-23, IL-24, IL-25, IL-26, IL-27, IL-28, IL-29, IL-30, IL-31, IL-32, IL-33, IL-34, и IL-35, IFN-γ, IFN-α, IFN-β, IFN-δ, хемокинов типа C XCL1 и XCL2, хемокинов типа C-C (на сегодняшний день включают CCL1-CCL28) и хемокинов типа CXC (на сегодняшний день включают CXCL1-CXCL17) и представителей суперсемейства TNF (например, TNF-α, лиганд 4-1 BB, фактор активации B-клеток (BLyS), лиганд FAS, sCD40L (в том числе мультимерные версии sCD40L; например, растворимый рекомбинант CD40L с гистидиновой меткой в комбинации с mAb к полигистидину для группировки нескольких молекул sCD40L вместе), лимфотоксина, OX40L, RANKL, TRAIL), CpG и других агонистов toll-подобных рецепторов (например, CpG).
Факторы, активирующие В-клетки, можно добавлять в клеточные культуры in vitro в различных концентрациях для достижения желаемого результата (например, размножения или дифференцировки). В одном варианте осуществления активирующий B-клетки фактор применяют для размножения B-клеток в культуре. В одном варианте осуществления активирующий B-клетки фактор применяют для дифференцировки B-клеток в культуре. В другом варианте осуществления активирующий B-клетки фактор применяют как для размножения, так и для дифференцировки B-клеток в культуре. В одном варианте осуществления активирующий B-клетки фактор предоставляют в одной и той же концентрации для размножения и дифференцировки. В другом варианте осуществления активирующий B-клетки фактор предоставляют в одной концентрации для размножения и в другой концентрации для дифференцировки. Предполагают, что активирующий B-клетки фактор можно 1) использовать для размножения B-клеток, а не для дифференцировки B-клеток, 2) использовать для дифференцировки B-клеток, а не для размножения B-клеток, или 3) использовать для размножения и дифференцировки В-клеток.
Например, B-клетки культивируют с одним или более активирующими B-клетками факторами, выбранными из CD40L, IL-2, IL-4 и IL-10, для размножения B-клеток. В одном варианте осуществления B-клетки культивируют с 0,25-5,0 мкг/мл CD40L. В одном варианте осуществления концентрация CD40L составляет 0,5 мкг/мл. В одном варианте осуществления для создания мультимеров CD40L применяют сшивающее средство (такое как антитело к HIS в комбинации с HIS-меченым CD40L). В одном варианте осуществления молекулы CD40L ковалентно связаны или удерживаются вместе с применением доменов мультимеризации белка (например, Fc-области IgG или домена «лейциновая застежка»). В одном варианте осуществления CD40L конъюгирован с гранулами. В одном варианте осуществления CD40L экспрессируется из питающих клеток. В одном варианте осуществления B-клетки культивируют с 1-10 нг/мл IL-2. В одном варианте осуществления концентрация IL-2 составляет 5 нг/мл. В одном варианте осуществления B-клетки культивируют с 1-10 нг/мл IL-4. В одном варианте осуществления концентрация IL-4 составляет 2 нг/мл. В одном варианте осуществления B-клетки культивируют с 10-100 нг/мл IL-10. В одном варианте осуществления концентрация IL-10 составляет 40 нг/мл.
В одном варианте осуществления B-клетки культивируют с одним или более активирующими B-клетки факторами, выбранными из CD40L, IL-2, IL-4, IL-10, IL-15 и IL-21, для размножения B-клеток. В одном варианте осуществления B-клетки культивируют с 0,25-5,0 мкг/мл CD40L. В одном варианте осуществления концентрация CD40L составляет 0,5 мкг/мл. В одном варианте осуществления для создания мультимеров CD40L применяют сшивающее средство (такое как антитело к HIS в комбинации с HIS-меченым CD40L). В одном варианте осуществления молекулы CD40L ковалентно связаны или удерживаются вместе с применением доменов мультимеризации белка (например, Fc-области IgG или домена «лейциновая застежка»). В одном варианте осуществления CD40L конъюгирован с гранулами. В одном варианте осуществления CD40L экспрессируется из питающих клеток. В одном варианте осуществления B-клетки культивируют с 1-10 нг/мл IL-2. В одном варианте осуществления концентрация IL-2 составляет 5 нг/мл. В одном варианте осуществления B-клетки культивируют с 1-10 нг/мл IL-4. В одном варианте осуществления концентрация IL-4 составляет 2 нг/мл. В одном варианте осуществления B-клетки культивируют с 10-100 нг/мл IL-10. В одном варианте осуществления концентрация IL-10 составляет 40 нг/мл. В одном варианте осуществления B-клетки культивируют с 50-150 нг/мл IL-15. В одном варианте осуществления концентрация IL-15 составляет 100 нг/мл. В одном варианте осуществления B-клетки культивируют с 50-150 нг/мл IL-21. В одном варианте осуществления концентрация IL-21 составляет 100 нг/мл. В конкретном варианте осуществления B-клетки культивируют с CD40L, IL-2, IL-4, IL-10, IL-15 и IL-21 для размножения B-клеток.
Например, в одном варианте осуществления B-клетки культивируют с активирующими B-клетки факторами CD40L, IL-2, IL-4, IL-10, IL-15 и IL-21 для размножения B-клеток, где CD40L является сшитым со сшивающим средством для создания мультимеров CD40L. Такая культуральная система может поддерживаться в течение всего периода культивирования (например, 7-дневного периода культивирования), в котором В-клетки трансфицированы или иным образом сконструированы с обеспечением экспрессии представляющего интерес трансгена (например, экзогенного полипептида, такого как, например, IDUA). Трансген может быть интегрирован в В-клетку (например, с помощью вирусного или невирусного вектора). Трансген может экспрессироваться в В-клетке с помощью транспозона. Трансген может экспрессироваться в В-клетке благодаря нацеленной интеграции трансгена в геном В-клетки. Нацеленная интеграция может осуществляться посредством гомологичной рекомбинации. Гомологичная рекомбинация может происходить в месте двухнитевого разрыва, индуцированного нуклеазой. Нуклеаза может представлять собой, например, нуклеазу с цинковыми пальцами, нуклеазу TALE (TALEN), мегануклеазу (например, хоминг-эндонуклеазу) или CRISPR/CAS9-нуклеазную систему.
В другом примере B-клетки культивируют с одним или более активирующими B-клетки факторами, выбранными из CD40L, IFN-α, IL-2, IL-6, IL-10, IL-15, IL-21 и CpG-олигодезоксинуклеотидов класса P (p-ODN), для дифференцировки В-клеток. В одном варианте осуществления B-клетки культивируют с 25-75 нг/мл CD40L. В одном варианте осуществления концентрация CD40L составляет 50 нг/мл. В одном варианте осуществления B-клетки культивируют с 250-750 ЕД/мл IFN-α. В одном варианте осуществления концентрация IFN-α составляет 500 ЕД/мл. В одном варианте осуществления B-клетки культивируют с 5-50 ЕД/мл IL-2. В одном варианте осуществления концентрация IL-2 составляет 20 ЕД/мл. В одном варианте осуществления B-клетки культивируют с 25-75 нг/мл IL-6. В одном варианте осуществления концентрация IL-6 составляет 50 нг/мл. В одном варианте осуществления B-клетки культивируют с 10-100 нг/мл IL-10. В одном варианте осуществления концентрация IL-10 составляет 50 нг/мл. В одном варианте осуществления B-клетки культивируют с 1-20 нг/мл IL-15. В одном варианте осуществления концентрация IL-15 составляет 10 нг/мл. В одном варианте осуществления B-клетки культивируют с 10-100 нг/мл IL-21. В одном варианте осуществления концентрация IL-21 составляет 50 нг/мл. В одном варианте осуществления B-клетки культивируют с 1-50 мкг/мл p-ODN. В одном варианте осуществления концентрация p-ODN составляет 10 нг/мл.
В одном варианте осуществления В-клетки приводят в контакт с питающими клетками или культивируют на них. В одном варианте осуществления питающие клетки представляют собой линию стромальных клеток, например, линию мышиных стромальных клеток S17 или MS5. В другом варианте осуществления выделенные CD19+ клетки культивируют с одним или более активирующими В-клетки факторами, представляющими собой цитокины, такими как IL-10 и IL-4, в присутствии фибробластов, экспрессирующих лиганд CD40 (CD40L, CD154). В одном варианте осуществления CD40L предусмотрен связанным с поверхностью, такой как планшет для культуры ткани или гранула. В другом варианте осуществления очищенные В-клетки культивируют в присутствии или в отсутствие питающих клеток, с CD40L и одним или более цитокинами или факторами, выбранными из IL-10, IL-4, IL-7, p-ODN, CpG DNA, IL-2, IL-15, IL6 и IFN-α.
В другом варианте осуществления активирующие B-клетки факторы обеспечивают путем трансфекции в B-клетку или другую питающую клетку. В этом контексте можно применять один или более факторов, которые способствуют дифференцировке В-клетки в секретирующую антитело клетку, и/или один или более факторов факторов, которые способствуют продолжительности жизни клетки, продуцирующей антитело. Такие факторы включают, например, Blimp-1, TRF4, антиапоптотические факторы, такие как Bcl-xl или Bcl5, или конститутивно активные мутанты рецептора CD40. Кроме того, факторы, которые способствуют экспрессии расположенных ниже сигнальных молекул, такие как факторы, связанные с рецептором TNF (TRAF), также можно применять для активации/дифференцировки В-клеток. В этом отношении активация клеток, выживание клеток и антиапоптотические функции суперсемейства рецепторов TNF в основном опосредуются TRAF1-6 (см., например, R.H. Arch, et al., Genes Dev. 12 (1998), pp. 2821-2830). Расположенные ниже эффекторы передачи сигналов TRAF включают транскрипционные факторы семейства NF-κB и AP-1, которые могут включать гены, участвующие в различных аспектах клеточных и иммунных функций. Кроме того, было показано, что активация NF-κB и AP-1 обеспечивает защиту клеток от апоптоза посредством транскрипции антиапоптотических генов.
В другом варианте осуществления белки, полученные из вируса Эпштейна-Барра (EBV), применяют для активации и/или дифференцировки В-клеток или для увеличения продолжительности жизни клетки, продуцирующей антитело. Белки, полученные из EBV, включают без ограничения EBNA-1, EBNA-2, EBNA-3, LMP-1, LMP-2, EBER, miRNA, EBV-EA, EBV-MA, EBV-VCA и EBV-AN.
В определенных вариантах осуществления приведение в контакт В-клеток с активирующими В-клетки факторами с применением способов, представленных в настоящем документе, приводит в результате, среди прочего, к пролиферации клеток (т.е. размножению), модуляции фенотипа клеточной поверхности lgM + в сторону такого, который совместим с активированной зрелой В-клеткой, секреции Ig и переключению изотипа. CD19+ B-клетки можно выделять с применением известных и коммерчески доступных наборов для разделения клеток, таких как система разделения клеток MiniMACS™ (Miltenyi Biotech, Бергиш-Гладбах, Германия). В определенных вариантах осуществления фибробласты CD40L облучают перед применением в способах, описанных в настоящем документе. В одном варианте осуществления B-клетки культивируют в присутствии одного или более из IL-3, IL-7, лиганда Flt3, тромбопоэтина, SCF, IL-2, IL-10, G-CSF и CpG. В некоторых вариантах осуществления способы предусматривают культивирование В-клеток в присутствии одного или более из вышеупомянутых факторов в сочетании с трансформированными стромальными клетками (например, MS5) с обеспечением низкого уровня заякоренных CD40L и/или CD40L, связанных с планшетом или гранулой.
Как обсуждалось выше, активирующие В-клетки факторы индуцируют размножение, пролиферацию или дифференцировку В-клеток. Соответственно, В-клетки приводят в контакт с одним или более активирующими В-клетки факторами, перечисленными выше, с целью размножения популяции клеток. Популяцию клеток можно размножать до трансфекции. Альтернативно или дополнительно, клеточную популяцию можно размножать после трансфекции. В одном варианте осуществления размножение популяции В-клеток предусматривает культивирование клеток с IL-2, IL-4, IL-10 и CD40L (см., например, Neron et al. PLoS ONE, 2012 7(12): e51946). В одном варианте осуществления размножение популяции B-клеток предусматривает культивирование клеток с IL-2, IL-10, CpG и CD40L. В одном варианте осуществления размножение популяции B-клеток предусматривает культивирование клеток с IL-2, IL-4, IL-10, IL-15, IL-21 и CD40L. В одном варианте осуществления размножение популяции B-клеток предусматривает культивирование клеток с IL-2, IL-4, IL-10, IL-15, IL-21 и мультимеризованным CD40L.
В другом варианте осуществления размножение популяции B-клеток индуцируют и/или усиливают с помощью трансгена, введенного в B-клетки. Например, В-клетка, которая содержит рекомбинантный рецептор или сконструированный рецептор, который индуцирует сигнальный путь клетки (например, передачу сигнала ниже CD40) при связывании своего лиганда (например, растворимого лиганда или лиганда, экспрессируемого на клеточной поверхности). В одном варианте осуществления B-клетка сверхэкспрессирует CD40 вследствие экспрессии трансгена CD40. В другом варианте осуществления B-клетка экспрессирует сконструированный рецептор, в том числе, например, рекомбинантно сконструированное антитело. В одном варианте осуществления сконструированный рецептор подобен химерному антигенному рецептору (CAR) и содержит слитый белок scFv и внутриклеточную сигнальную часть рецептора B-клетки (например, CD40).
В одном варианте осуществления размножение популяции B-клеток индуцируется и/или усиливается посредством низкомолекулярного соединения, добавленного в клеточную культуру. Например, соединение, которое связывается с и димеризует CD40, можно применять для запуска сигнального пути CD40.
В настоящих способах можно применять любую из множества питательных сред, известных специалисту в данной области (см., например, Current Protocols in Cell Culture, 2000-2009, John Wiley & Sons, Inc.). В одном варианте осуществления среда для применения в способах, описанных в настоящем документе, включает без ограничения среду Дульбекко в модификации Искова (с фетальной бычьей сывороткой или без нее или с другой подходящей сывороткой). Иллюстративные среды также включают в себя без ограничения IMDM, RPMI 1640, AIM-V, DMEM, MEM, a-MEM, F-12, X-Vivo 15 и X-Vivo 20. В других вариантах осуществления среда может содержать поверхностно-активное вещество, антитело, плазманат или восстановитель (например, N-ацетилцистеин, 2-меркаптоэтанол), один или более антибиотиков и/или добавки, такие как инсулин, трансферрин, селенит натрия и циклоспорин. В некоторых вариантах осуществления также можно применять IL-6, растворимый CD40L и усилитель сшивки.
В-клетки культивируют в условиях и в течение достаточных периодов времени для достижения желаемой дифференцировки и/или активации. В некоторых вариантах осуществления B-клетки культивируют в условиях и в течение достаточных периодов времени для обеспечения, например, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60% 65%, 70%, 75%, 80%, 85%, 90%, 95% или даже 100% дифференцировки и/или активации В-клеток по необходимости. В одном варианте осуществления B-клетки активируются и дифференцируются с получением смешанной популяции плазмабластов и плазматических клеток. Как будет понятно специалисту в данной области, плазмабласты и плазматические клетки могут быть идентифицированы по паттернам экспрессии белка на клеточной поверхности с применением стандартных способов проточной цитометрии, как описано в другом месте данного документа, таких как экспрессия одного или более из CD38, CD78, IL-6R, CD27high и CD138 и/или отсутствие или снижение экспрессии одного или более из CD19, CD20 и CD45. Как должно быть понятно специалисту, В-клетки памяти, как правило, представляют собой CD20+ CD19+ CD27+ CD38- клетки, тогда как ранние плазмабласты представляют собой CD20- CD19+ CD27++ CD38++ клетки. В одном варианте осуществления клетки, культивируемые с применением способов, описанных в настоящем документе, являются CD20-, CD38+, CD138-. В другом варианте осуществления клетки имеют фенотип CD20-, CD38+, CD138+. В определенных вариантах осуществления клетки культивируют в течение периода 1-7 дней. В дополнительных вариантах осуществления клетки культивируют в течение периода 7, 14, 21 дня или дольше. Таким образом, клетки можно культивировать в соответствующих условиях в течение периода 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 дней или больше. Клетки повторно высевают, а среды и добавки можно добавлять или изменять по мере необходимости с применением методик, известных в данной области.
В некоторых вариантах осуществления B-клетки культивируют в условиях и в течение достаточных периодов времени для обеспечения, например, по меньшей мере 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% дифференцировки и активации клеток для получения Ig и/или с обеспечением экспрессии трансгена.
Индукцию активации B-клеток можно измерять с помощью методик, таких как включение 3H-уридина в РНК (при дифференциации B-клеток увеличение синтеза РНК) или включение 3H-тимидина для измерения синтеза ДНК, связанного с пролиферацией клеток. В одном варианте осуществления для усиления пролиферации B-клеток в культуральную среду можно добавлять интерлейкин-4 (IL-4) в подходящей концентрации (например, приблизительно 10 нг/мл).
Альтернативно, активацию В-клеток измеряют в зависимости от секреции иммуноглобулина. Например, CD40L добавляют к покоящимся B-клеткам вместе с IL-4 (например, 10 нг/мл) и IL-5 (например, 5 нг/мл) или другими цитокинами, которые активируют B-клетки. Для измерения маркеров клеточной поверхности, типичных для активированных В-клеток, можно также применять проточную цитометрию. См., например, Civin CI, Loken MR, Int'l J. Cell Cloning 987; 5: 1-16; Loken, MR, et al, Flow Cytometry Characterization of Erythroid, Lymphoid and Monomyeloid Lineages in Normal Human Bone Marrow, in Flow Cytometry in Hematology, Laerum OD, Bjerksnes R. eds., Academic Press, New York 1992; pp. 31-42; and LeBein TW, et ai, Leukemia 1990; 4: 354-358.
После культивирования в течение соответствующего периода времени, такого как 2, 3, 4, 5, 6, 7, 8, 9 дней или больше, как правило, около 3 дней, можно добавлять дополнительный объем культуральной среды. Супернатант из отдельных культур можно собирать в разное время в ходе культивирования и количественно определять в отношении IgM и IgG1, как описано в Noelle et al., (1991) J. Immunol. 146: 1118-1124. В одном варианте осуществления культуру собирают и оценивают в отношении экспрессии представляющего интерес трансгена с применением проточной цитометрии, ферментно-связанного иммуносорбентного анализа (ELISA), ELISPOT или другого анализа, известного в данной области.
В другом варианте осуществления для оценки продуцирования изотипа антител, например, IgM, или продукта представляющего интерес трансгена применяют ELISA. В некоторых вариантах осуществления определения IgG проводят с применением коммерчески доступных антител, таких как антитело козы к IgG человека, в качестве антитела захвата с последующим обнаружением с применением любого из множества подходящих реагентов для обнаружения, таких как биотинилированное козье антитело к Ig человека, стрептавидин-щелочная фосфатаза и субстрат.
В некоторых вариантах осуществления B-клетки культивируют в условиях и в течение достаточных периодов времени, в результате чего количество клеток в 1, 10, 25, 50, 75, 100, 125, 150, 175, 200, 250, 300, 350, 400 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, в 1000 раз или больше превышает количество В-клеток в начале культивирования. В одном варианте осуществления количество клеток в 10-1000 раз превышает, включая последовательные целые числа, количество В-клеток в начале культивирования. Например, размноженная популяция В-клеток в по меньшей мере 10 раз превышает исходную выделенную популяцию В-клеток. В другом варианте осуществления размноженная популяция В-клеток в по меньшей мере 100 раз превышает исходную выделенную популяцию В-клеток. В одном варианте осуществления размноженная популяция В-клеток в по меньшей мере 500 раз превышает исходную выделенную популяцию В-клеток.
Конструирование В-клеток
В одном варианте осуществления генетически модифицированные В-клетки трансфицированы посредством трансгена. Иллюстративные способы трансфекции В-клеток представлены в WO 2014/152832 и WO 2016/100932, оба из которых включены в настоящий документ посредством ссылки во всей своей полноте. Трансфекцию В-клеток можно осуществлять с применением любого из множества способов, доступных в данной области, для введения ДНК или РНК в В-клетку. Подходящие методики могут включать трансфекцию с фосфатом кальция, DEAE-декстран, электропорацию, опосредованную давлением трансфекцию или «сжатие клеток» (например, микрожидкостная система CellSqueeze, SQZ Biotechnologies), трансфекцию, опосредованную наночастицами или липосомами, трансдукцию с применением ретровируса или других вирусов, например, коровьей оспы. См., например, Graham et al., 1973, Virology 52: 456; Sambrook et al., 2001, Molecular Cloning, a Laboratory Manual, Cold Spring Harbor Laboratories; Davis et al., 1986, Basic Methods in Molecular Biology, Elsevier; Chu et al., 1981, Gene 13: 197; US 5,124,259; US 5,297,983; патенты США №5283185; 5661018; 6878548; 7799555; 8551780; и 8633029. Одним из примеров коммерчески доступной методики электропорации, подходящей для В-клеток, является технология трансфекции Nucleofector™.
Трансфекция может иметь место до или во время культивирования in vitro выделенных В-клеток в присутствии одного или более активирующих и/или дифференцирующих факторов, описанных выше. Например, клетки подвергают трансфекции в день 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38 или 39 культуры in vitro. В одном варианте осуществления клетки подвергают трансфекции в день 1, 2 или 3 культивирования in vitro. В конкретном варианте осуществления клетки подвергают трансфекции в день 2. Например, клетки подвергают электропорации в день 2 культивирования in vitro для доставки, например, плазмиды, транспозона, миникольца или самореплицирующейся РНК. В другом варианте осуществления клетки подвергают трансфекции в день 4, 5, 6 или 7 культивирования in vitro. В конкретном варианте осуществления клетки подвергают трансфекции в день 6 культивирования in vitro. В другом варианте осуществления клетки подвергают трансфекции в день 5 культивирования in vitro.
В одном варианте осуществления клетки подвергают трансфекции или иным образом конструируют (например, посредством нацеленной интеграции трансгена) перед активацией. В другом варианте осуществления клетки подвергают трансфекции или иным образом конструируют (например, посредством нацеленной интеграции трансгена) во время активации. В одном варианте осуществления клетки подвергают трансфекции или иным образом конструируют (например, посредством нацеленной интеграции трансгена) после активации. В одном варианте осуществления клетки подвергают трансфекции или иным образом конструируют (например, посредством нацеленной интеграции трансгена) перед дифференцировкой. В другом варианте осуществления клетки подвергают трансфекции или иным образом конструируют (например, посредством нацеленной интеграции трансгена) во время дифференцировки. В одном варианте осуществления клетки подвергают трансфекции или иным образом конструируют (например, посредством нацеленной интеграции трансгена) после дифференцировки.
В одном варианте осуществления для доставки ДНК или РНК в В-клетки памяти и/или плазматические клетки применяют невирусный вектор. Например, системы, которые могут облегчать трансфекцию В-клеток памяти и/или плазматических клеток без необходимости системы вирусной интеграции, включают без ограничения транспозоны (например, систему транспозонов Sleeping Beauty), нуклеазы с «цинковыми пальцами» (ZFN), подобные активаторам транскрипции эффекторные нуклеазы (TALEN), короткие палиндромные повторы, регулярно расположенные группами (CRISPR), мегануклеазы, миникольца, репликоны, искусственные хромосомы (например, искусственные бактериальные хромосомы, искусственные хромосомы млекопитающих и дрожжевые искусственные хромосомы), плазмиды, космиды и бактериофаг.
В некоторых вариантах осуществления такие независимые от вирусов векторные системы также можно доставлять с помощью вирусного вектора, известного в данной области техники или описанного ниже. Например, в некоторых вариантах осуществления вирусный вектор (например, ретровирус, лентивирус, аденовирус, аденоассоциированный вирус) применяют для доставки одного или более невирусных векторов (таких как, например, один или более факторов из указанных выше нуклеаз с «цинковыми пальцами» (ZFN), подобных активаторам транскрипции эффекторных нуклеаз (TALEN), коротких палиндромных повторов, регулярно расположенных группами (CRISPR), или любых других ферментных/комплементарных векторов, полинуклеотидов и/или полипептидов, способных облегчать нацеленную интеграцию). Соответственно, в некоторых вариантах осуществления клетка (например, B-клетки, такие как B-клетки памяти и/или плазматические клетки) может быть сконструирована с обеспечением экспрессии экзогенной последовательности (например, последовательности, кодирующей терапевтический полипептид, такой как IDUA) посредством способа нацеленной интеграции. Такие способы известны в данной области техники и могут предусматривать расщепление эндогенного локуса в клетке с применением одной или более нуклеаз (например, ZFN, TALEN, CRISPR/Cas, мегануклеазы) и введение трансгена в клетку таким образом, чтобы он интегрировался в эндогенный локус и экспрессировался в клетке. Трансген может состоять из донорной последовательности, которая интегрируется в ДНК клетки-хозяина в точке расщепления нуклеазой или вблизи нее.
Интеграция экзогенной последовательности (например, последовательности, кодирующей терапевтический полипептид, такой как IDUA) может происходить посредством рекомбинации. Как будет понятно специалисту в данной области, «рекомбинация» относится к процессу обмена генетической информацией между двумя полинуклеотидами, в том числе без ограничения захвату донора негомологичным присоединением конца (NHEJ) и гомологичной рекомбинации. Рекомбинация может представлять собой гомологичную рекомбинацию. Для целей настоящего изобретения термин «гомологичная рекомбинация (HR)» относится к специализированной форме такого обмена, который имеет место, например, во время восстановления двухнитевых разрывов в клетках с помощью направляемых гомологией механизмов репарации. В данном процессе используется гомология нуклеотидной последовательности, в результате чего «донорная» молекула (например, донорная полинуклеотидная последовательность или донорный вектор, содержащий такую последовательность) используется механизмом репарации ДНК клетки как матрица для репарации молекулы-«мишени» (т.е. та, которая претерпела двухнитевый разрыв), и тем самым вызывает передачу генетической информации от донора к мишени. В некоторых вариантах осуществления HR-направляемой интеграции донорная молекула может содержать по меньшей мере 2 области гомологии с геномом («плечи гомологии»). В некоторых вариантах осуществления плечи гомологии могут иметь длину, например, по меньшей мере 50-100 пар оснований. Плечи гомологии могут иметь существенную гомологию ДНК с областью геномной ДНК, фланкирующей сайт расщепления, где должна происходить нацеленная интеграция. Плечи гомологии молекулы донора могут фланкировать ДНК, которая должна быть интегрирована в геном-мишень или локус-мишень ДНК. Разрушение хромосомы с последующим восстановлением с применением гомологичной области плазмидной ДНК в качестве матрицы может привести к переносу промежуточного трансгена, фланкированного плечами гомологии, в геном. См., например, Koller et al. (1989) Proc. Nat'l. Acad. Sci. USA 86(22): 8927-8931; Thomas et al. (1986) Cell 44(3): 419-428. Частота этого типа направляемой гомологией нацеленной интеграции может быть увеличена на величину до 105 раз путем преднамеренного создания двухнитевого разрыва вблизи области-мишени (Hockemeyer et al. (2009) Nature Biotech. 27(9): 851-857; Lombardo et al. (2007) Nature Biotech. 25(11): 1298-1306; Moehle et al. (2007) Proc. Nat'l Acad. Sci. USA 104(9): 3055-3060; Rouet et al. (1994) Proc. Nat'l Acad. Sci. USA 91(13): 6064-6068.
Для конструирования клетки (например, В-клетки памяти или плазмобласта) согласно настоящему изобретению можно применять любую нуклеазу, способную опосредовать нацеленное расщепление геномного локуса, так что трансген может быть интегрирован в геном клетки-мишени (например, путем рекомбинации, такой как HR).
Двухнитевый разрыв (DSB) или ник может быть создан с помощью сайт-специфической нуклеазы, такой как нуклеаза с «цинковыми пальцами» (ZFN), TAL-эффекторная нуклеаза (TALEN), мегануклеаза, или с применением системы CRISPR/Cas9 со сконструированной crRNA/tract RNA (одиночная направляющая РНК) для направления специфического расщепления. См., например, Burgess (2013) Nature Reviews Genetics 14: 80-81, Urnov et al. (2010) Nature 435(7042): 646-51; патентные публикации США 20030232410; 20050208489; 20050026157; 20050064474; 20060188987; 20090263900; 20090117617; 20100047805; 20110207221; 20110301073 и международная публикация WO 2007/014275, раскрытие которых включено в данный документ посредством ссылки во всей своей полноте для всех целей.
В некоторых вариантах осуществления клетка (например, В-клетка памяти или плазмабласт) сконструирована с помощью опосредованной нуклеазой с «цинковыми пальцами» нацеленной интеграции донорной конструкции. Нуклеаза с «цинковыми пальцами» (ZFN) представляет собой фермент, который способен распознавать и расщеплять нуклеотидную последовательность-мишень со специфичностью благодаря связыванию «ДНК-связывающего белка с цинковыми пальцами» (ZFP) (или связывающего домена), который связывает ДНК специфическим в отношении последовательности образом с помощью одного или более цинковых пальцев и нуклеазный фермент. ZFN могут содержать любые подходящие домены расщепления (например, нуклеазный фермент), функционально связанные с ДНК-связывающим доменом ZFP, чтобы образовать сконструированный ZFN, который может способствовать сайт-специфическому расщеплению последовательности ДНК-мишени (см., например, Kim et al. (1996) Proc Natl Acad Sci USA 93(3): 1156-1160). Например, ZFN могут содержать специфичный в отношении мишени ZFP, связанный с ферментом FOK1 или частью фермента FOK1. В некоторых вариантах осуществления ZFN, используемая в ZFN-опосредованном подходе нацеленной интеграции, использует две отдельные молекулы, каждая из которых содержит субъединицу фермента FOK1, каждая из которых связана с ZFP, каждый ZFP со специфичностью в отношении последовательности ДНК, фланкирующей сайт-мишень расщепления, и, когда два ZFP связываются со своими соответствующими сайтами-мишенями ДНК, субъединицы фермента FOK1 сближаются друг с другом, и они связываются вместе, активируя нуклеазную активность, посредством которой происходит расщепление сайта-мишени расщепления. ZFN применяли для модификации генома в ряде организмов (например, патентные публикации США 20030232410; 20050208489; 20050026157; 20050064474; 20060188987; 20060063231; и международная публикация WO 07/014275, включенные в настоящий документ посредством ссылки во всей своей полноте). Специально изготовленные ZFP и ZFN коммерчески доступны, например, от Sigma Aldrich (Сент-Луис, Миссури), и с применением таких ZFN любое местоположение ДНК можно подвергать регулярному нацеливанию и расщеплению.
В некоторых вариантах осуществления клетка (например, В-клетка памяти или плазмабласт) сконструирована с помощью опосредованной нуклеазой интеграции донорной конструкции CRISPR/Cas (например, CRISPR Cas9). Нуклеазная система CRISPR (короткие палиндромные повторы, регулярно расположенные группами)/Cas (ассоциированный с CRISPR) представляет собой сконструированную нуклеазную систему, основанную на бактериальной системе, которую можно применять для конструирования генома. Она основывается на части адаптивного иммунного ответа многих бактерий и архей. Когда вирус или плазмида проникают в бактерию, сегменты ДНК-захватчика преобразуются в РНК CRISPR (crRNA) посредством «иммунного» ответа. Затем эта crRNA связывается посредством области частичной комплементарности с другим типом РНК, называемой tracrRNA, для направления нуклеазы Cas9 в область, гомологичную crRNA, в ДНК-мишени, называемой «протоспейсер». Cas9 расщепляет ДНК с получением тупых концов в DSB в сайтах, указанных с помощью 20-нуклеотидной направляющей последовательности, содержащейся в транскрипте crRNA. Cas9 требует как crRNA, так и tracrRNA для сайт-специфического распознавания и расщепления ДНК. В настоящее время эта система разработана таким образом, что crRNA и tracrRNA могут быть объединены в одну молекулу («одиночная направляющая РНК»), а эквивалентная часть crRNA одиночной направляющей РНК может быть сконструирована так, чтобы направлять нуклеазу Cas9 для нацеливания на любую желаемую последовательность (см. Jinek et al (2012) Science 337, p. 816-821, Jinek et al, (2013), eLife 2: e00471, и David Segal, (2013) eLife 2: e00563). Таким образом, система CRISPR/Cas может быть сконструирована для создания DSB с желаемой целью в геноме, а на репарацию DSB может влиять применение ингибиторов репарации с целью вызвать повышение подверженности ошибкам репарации.
В некоторых вариантах осуществления в интеграции, опосредованной нуклеазой CRISPR/Cas, используется CRISPR типа II. CRISPR типа II является одной из наиболее хорошо охарактеризованных систем и выполняет нацеленный двухнитевый разрыв ДНК в четыре последовательные стадии. Во-первых, две некодирующие РНК, матрица pre-crRNA и tracrRNA, транскрибируются из локуса CRISPR. Во-вторых, tracrRNA гибридизуется с повторяющимися участками pre-crRNA и опосредует процессинг pre-crRNA в зрелые crRNA, содержащие отдельные спейсерные последовательности. В-третьих, зрелый комплекс crRNA:tracrRNA направляет Cas9 к ДНК-мишени посредством спаривания оснований Уотсона-Крика между спейсером на crRNA и протоспейсером на ДНК-мишени рядом с мотивом, смежным с протоспейсером (PAM), что является дополнительным требованием для распознавания цели. В-четвертых, Cas9 опосредует расщепление ДНК-мишени с получением двухнитевого разрыва в пределах протоспейсера.
Связанная с Cas9 система CRISPR/Cas содержит два некодирующих РНК-компонента: tracrRNA и матрицу pre-crRNA, содержащую направляющие последовательности (спейсеры) нуклеазы, разделенные одинаковыми прямыми повторами (DR). Чтобы применять систему CRISPR/Cas для выполнения конструирования генома, должны присутствовать обе функции этих РНК (см. Cong et al, (2013) Sciencexpress 1/10.1126/science 1231143). В некоторых вариантах осуществления tracrRNA и pre-crRNAs обеспечиваются через отдельные экспрессирующие конструкции или в виде отдельных РНК. В других вариантах осуществления конструируют химерную РНК, где сконструированную зрелую crRNA (придающую специфичность к цели) сливают с tracrRNA (обеспечивающей взаимодействие с Cas9) с созданием гибрида химерной cr-РНК-tracrRNA (также называемого одиночной направляющей РНК). (См. Jinek ibid and Cong, ibid).
В некоторых вариантах осуществления можно конструировать одиночную направляющую РНК, содержащую как crRNA, так и tracrRNA, для направления нуклеазы Cas9 для нацеливания на любую желаемую последовательность (например, Jinek et al (2012) Science 337, p. 816-821, Jinek et al, (2013), eLife 2: e00471, David Segal, (2013) eLife 2: e00563). Таким образом, система CRISPR/Cas может быть сконструирована для создания DSB в желаемой цели в геноме.
Специально созданные системы CRISPR/Cas коммерчески доступны, например, от Dharmacon (Lafayette, CO), и любое местоположение ДНК можно подвергать регулярному нацеливанию и расщеплению с применением таких специально созданных последовательностей одиночной направляющей РНК. Однонитевые ДНК-матрицы для рекомбинации могут быть синтезированы (например, с помощью способов синтеза олигонуклеотидов, известных в данной области техники и коммерчески доступных) или предоставлены в векторе, например, вирусном векторе, таком как AAV.
В некоторых вариантах осуществления клетка (например, B-клетка памяти или плазмабласт) сконструирована посредством опосредованной нуклеазой TALE (TALEN) нацеленной интеграции донорной конструкции. «ДНК-связывающий домен TALE» или «TALE» представляет собой полипептид, содержащий один или более факторов повторяющихся доменов/единиц TALE. Повторяющиеся домены участвуют в связывании TALE с его родственной последовательностью-мишенью ДНК. Одна «повторяющаяся единица» (также называемая «повтор») обычно имеет длину 33-35 аминокислот и проявляет по меньшей мере некоторую степень гомологии последовательности с другими повторяющимися последовательностями TALE в пределах встречающегося в природе белка TALE. TAL-эффекторы могут содержать последовательность ядерной локализации, кислый домен активации транскрипции и централизованный домен тандемных повторов, где каждый повтор содержит примерно 34 аминокислоты, которые являются ключевыми для специфичности связывания ДНК этих белков. (например, Schornack S, et al (2006) J Plant Physiol 163(3): 256-272). TAL-эффекторы зависят от последовательностей, обнаруживаемых в тандемных повторах, которые содержат приблизительно 102 п.о., и эти повторы обычно на 91-100% гомологичны друг другу (например, Bonas et al. (1989) MoI Gen Genet 218: 127-136). Такие ДНК-связывающие повторы могут быть сконструированы в белки с новыми комбинациями и количеством повторов для создания искусственных факторов транскрипции, которые способны взаимодействовать с новыми последовательностями и активировать экспрессию неэндогенного репортерного гена (например, Bonas et al. (1989) MoI Gen Genet 218: 127-136). Сконструированные белки TAL могут быть связаны с половинным доменом расщепления FokI с получением слитого белка нуклеазы с эффекторным доменом TAL (TALEN) для расщепления специфической последовательности-мишени ДНК (например, Christian et al. (2010) Genetics epub 10.1534/genetics.110.120717).
Специально созданные TALEN коммерчески доступны, например, от Thermo Fisher Scientific (Уолтем, Массачусетс), и любое местоположение ДНК можно подвергать регулярному нацеливанию и расщеплению.
В некоторых вариантах осуществления клетка (например, В-клетка памяти или плазмабласт) сконструирована посредством опосредованной мегануклеазой нацеленной интеграции донорной конструкции. Мегануклеаза (или «хоминг-эндонуклеаза») представляет собой эндонуклеазу, которая связывает и расщепляет двухнитевую ДНК с последовательностью узнавания, которая превышает 12 пар оснований. Встречающиеся в природе мегануклеазы могут быть мономерными (например, I-SceI) или димерными (например, I-CreI). Встречающиеся в природе мегануклеазы распознают сайты расщепления с 15-40 парами оснований и обычно сгруппированы в четыре семейства: семейство LAGLIDADG, семейство GIY-YIG, семейство His-Cyst box и семейство HNH. Иллюстративные хоминг-эндонуклеазы включают I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I- TevI, I-TevII и I-TevIII. Их последовательности распознавания являются известными. См. также патент США №5420032; патент США №6833252; Belfort et al. (1997) Nucleic Acids Res. 25: 3379-3388; Dujon et al. (1989) Gene 82: 115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12: 224-228; Gimble et al. (1996) J. Mol. Biol. 263: 163-180; Argast et al. (1998) J. Mol. Biol. 280: 345-353 и каталог New England Biolabs. Термин «мегануклеаза» включает мономерные мегануклеазы, димерные мегануклеазы и мономеры, которые связываются с образованием димерных мегануклеаз.
В определенных вариантах осуществления в способах и композициях, описанных в настоящем документе, применяют нуклеазу, которая содержит сконструированную (не встречающуюся в природе) хоминг-эндонуклеазу (мегануклеазу). Последовательности распознавания хоминг-эндонуклеаз и мегануклеаз, таких как I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII, известны. См. также патент США №5420032; патент США №6833252; Belfort et al. (1997) Nucleic Acids Res. 25: 3379-3388; Dujon et al. (1989) Gene 82: 115-118; Perler et al. (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12: 224-228; Gimble et al. (1996) J. Mol. Biol. 263: 163-180; Argast et al. (1998) J. Mol. Biol. 280: 345-353 и каталог New England Biolabs. Кроме того, для связывания неприродных сайтов-мишеней можно конструировать ДНК-связывающую специфичность хоминг-эндонуклеаз и мегануклеаз. См., например, Chevalier et al. (2002) Molec. Cell 10: 895-905; Epinat et al. (2003) Nucleic Acids Res. 31: 2952-2962; Ashworth et al. (2006) Nature 441: 656-659; Paques et al. (2007) Current Gene Therapy 7: 49-66; публикацию патента США №20070117128. ДНК-связывающие домены хоминг-эндонуклеаз и мегануклеаз могут быть изменены в контексте нуклеазы в целом (т.е. таким образом, что нуклеаза включает в себя родственный домен расщепления) или могут быть слиты с гетерологичным доменом расщепления. Специально созданная мегануклеаза коммерчески доступна, например, от New England Biolabs (Ипсвич, Массачусетс), и любое местоположение ДНК можно подвергать регулярному нацеливанию и расщеплению.
Конструирование В-клетки может включать введение одной или более нуклеаз (например, ZFN, TALEN, CRISPR/Cas, мегануклеазы) в В-клетку, например, посредством одного или более векторов, кодирующих нуклеазы, таким образом, что векторы, содержащие кодируемые нуклеазы, попадают в B-клетки. Векторы могут представлять собой вирусные векторы.
В некоторых вариантах осуществления нуклеазы расщепляют специфический эндогенный локус (например, ген safe harbor или локус, представляющий интерес) в клетке (например, В-клетке памяти или плазматической клетке), и вводят одну или более экзогенных (донорных) последовательностей (например, трансгенов) (например, один или более векторов, содержащих эти экзогенные последовательности). Нуклеаза может индуцировать двухнитевый (DSB) или однонитевый разрыв (ник) в ДНК-мишени. В некоторых вариантах осуществления нацеленная вставка донорного трансгена может осуществляться посредством направляемой гомологией репарации (HDR), механизмов негомологической репарации (например, NHEJ-опосредованного захвата конца) или вставок и/или делеции нуклеотидов (например, эндогенной последовательности) в сайт интеграции трансгена в геном клетки.
В одном варианте осуществления способ трансфекции B-клетки включает электропорацию B-клетки перед приведением в контакт B-клетки с вектором. В одном варианте осуществления клетки подвергают электропорации в день 1, 2, 3, 4, 5, 6, 7, 8 или 9 культивирования in vitro. В одном варианте осуществления для доставки плазмиды клетки подвергают электропорации в день 2 культивирования in vitro. В одном варианте осуществления клетки подвергают трансфекции с применением транспозонов в день 1, 2, 3, 4, 5, 6, 7, 8 или 9 культивирования in vitro. В другом варианте осуществления клетки подвергают трансфекции с применением миникольца в день 1, 2, 3, 4, 5, 6, 7, 8 или 9 культивирования in vitro. В одном варианте осуществления электропорацию транспозона Sleeping Beauty осуществляют в день 2 культивирования in vitro.
В одном варианте осуществления B-клетки приводят в контакт с вектором, содержащим нуклеиновую кислоту, представляющую интерес, функционально связанную с промотором, в условиях, достаточных для трансфекции по меньшей мере части B-клеток. В одном варианте осуществления B-клетки приводят в контакт с вектором, содержащим нуклеиновую кислоту, представляющую интерес, функционально связанную с промотором, в условиях, достаточных для трансфекции по меньшей мере 5% B-клеток. В следующем варианте осуществления В-клетки приводят в контакт с вектором в условиях, достаточных для трансфекции по меньшей мере 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или даже 100% В-клеток. В одном конкретном варианте осуществления B-клетки, культивируемые in vitro, как описано в данном документе, подвергают трансфекции, и в этом случае культивируемые B-клетки приводят в контакт с вектором, как описано в данном документе, в условиях, достаточных для трансфекции по меньшей мере 5%, 10%,15%, 20. %, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или даже 100% В-клеток.
Вирусные векторы можно применять для трансдукции В-клеток памяти и/или плазматических клеток. Примеры вирусных векторов включают без ограничения векторы на основе аденовирусов, векторы на основе аденоассоциированных вирусов (AAV), ретровирусные векторы, ретровирусно-аденовирусные векторы и векторы, полученные из вирусов простого герпеса (HSV), в том числе векторы-ампликоны, векторы на основе дефектного по репликации HSV и ослабленного HSV (см., например, Krisky, Gene Ther. 5: 1517-30, 1998; Pfeifer, Annu. Rev. Genomics Hum. Genet. 2: 177-211, 2001, каждая из которых включена посредством ссылки во всей своей полноте).
В одном варианте осуществления клетки подвергают трансдукции вирусным вектором (например, лентивирусным вектором) в день 1, 2, 3, 4, 5, 6, 7, 8 или 9 культивирования in vitro. В конкретном варианте осуществления клетки подвергают трансдукции вирусным вектором в день 5 культивирования in vitro. В одном варианте осуществления вирусный вектор представляет собой лентивирус. В одном варианте осуществления клетки подвергают трансдукции лентивирусом, псевдотипированным вирусом кори, в день 1 культивирования in vitro.
В одном варианте осуществления В-клетки подвергают трансдукции ретровирусными векторами с применением любого из множества известных в данной области способов (см., например, Science 12 April 1996 272: 263-267; Blood 2007, 99: 2342-2350; Blood 2009, 1 13: 1422-1431 ; Blood 2009 Oct 8; 1 14(15): 3173-80; Blood. 2003; 101 (6): 2167-2174; Current Protocols in Molecular Biology or Current Protocols in Immunology, John Wiley & Sons, New York, N.Y.(2009)). Дополнительное описание вирусной трансдукции В-клеток можно найти в WO 2011/085247 и WO 2014/152832, каждая из которых включена в настоящий документ посредством ссылки во всей своей полноте.
Например, PBMC, B- или T-лимфоциты от доноров и другие B-клеточные раковые клетки, такие как B-CLL, можно выделять и культивировать в среде IMDM или RPMI 1640 (GibcoBRL Invitrogen, Окленд, Новая Зеландия) или другой подходящей среде, такой как описанные в данном документе, либо без сыворотки, либо с добавлением сыворотки (например, 5-10% FCS, сыворотки AB человека и заменителей сыворотки) и с добавлением пенициллина/стрептомицина и/или других подходящих добавок, таких как трансферрин и/или инсулин. В одном варианте осуществления клетки высевают в количестве 1×105 клеток в 48-луночные планшеты и добавляют концентрированный вектор в различных дозах, которые могут быть обычно оптимизированы специалистом с применением стандартных методологий. В одном варианте осуществления B-клетки переносят в монослой клеток MS5 в RPMI, дополненной 10% AB сывороткой, 5% FCS, 50 нг/мл rhSCF, 10 нг/мл rhlL-15 и 5 нг/мл rhlL-2, и периодически обновляют среду по мере необходимости. Как будет понятно специалисту в данной области, по желанию можно применять другие подходящие носители и добавки.
Некоторые варианты осуществления относятся к применению ретровирусных векторов или векторов, полученных из ретровирусов. «Ретровирусы» представляют собой РНК-вирусы с оболочкой, которые способны инфицировать клетки животных и которые используют фермент обратную транскриптазу на ранних стадиях инфекции для получения копии ДНК из своего РНК-генома, которая затем обычно интегрируется в геном хозяина. Примерами ретровирусных векторов являются полученные из вируса мышиного лейкоза Молони (MLV) векторы, ретровирусные векторы на основе вируса стволовых клеток мыши, который обеспечивает долговременную стабильную экспрессию в клетках-мишенях, таких как гематопоэтические клетки-предшественники и их дифференцированное потомство (см., например, Hawley et al., PNAS USA 93: 10297-10302, 1996; Keller et al., Blood 92: 877-887, 1998), гибридные векторы (см., например, Choi, et al., Stem Cells 19: 236-246, 2001) и сложные векторы, полученные из ретровирусов, такие как лентивирусные векторы.
В одном варианте осуществления B-клетки приводят в контакт с ретровирусным вектором, содержащим нуклеиновую кислоту, представляющую интерес, функционально связанную с промотором, в условиях, достаточных для трансдукции по меньшей мере части B-клеток. В одном варианте осуществления B-клетки приводят в контакт с ретровирусным вектором, содержащим нуклеиновую кислоту, представляющую интерес, функционально связанную с промотором, в условиях, достаточных для трансдукции по меньшей мере 2% B-клеток. В следующем варианте осуществления В-клетки приводят в контакт с вектором в условиях, достаточных для трансдукции по меньшей мере 2%, 3%, 4%, 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или даже 100% полученных в результате В-клеток. В одном конкретном варианте осуществления дифференцированные и активированные B-клетки, культивируемые in vitro, как описано в настоящем документе, подвергают трансдукции, и в этом случае культивируемые дифференцированные/активированные B-клетки приводят в контакт с вектором, как описано в настоящем документе, в условиях, достаточных для трансдукции по меньшей мере 2%, 3%, 4%, 5%, 10% 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или даже 100% дифференцированных и активированных В-клеток.
В некоторых вариантах осуществления перед трансдукцией клетки предварительно стимулируют Staphylococcus Aureus Cowan (SAC; Calbiochem, Сан-Диего, Калифорния) и/или IL-2 в соответствующих концентрациях, известных специалисту и стандартно оптимизируемых. Можно применять другие активирующие В-клетки факторы (например, РМА), которые известны специалисту в данной области техники и описаны в данном документе.
Как отмечено выше, в некоторых вариантах осуществления применяют лентивирусные векторы. Термин «лентивирус» относится к роду сложных ретровирусов, которые способны инфицировать как делящиеся, так и неделящиеся клетки. Примеры лентивирусов включают HIV (вирус иммунодефицита человека; в том числе HIV типа 1 и HIV типа 2), висна-маеди, вирус артрита-энцефалита коз, вирус инфекционной анемии лошадей, вирус иммунодефицита кошек (FIV), вирус иммунодефицита крупного рогатого скота (BIV) и вирус иммунодефицита обезьян (SIV). Лентивирусные векторы могут быть получены из любого одного или более этих лентивирусов (см., например, Evans et al., Hum Gene Ther. 10: 1479-1489, 1999; Case et al., PNAS USA 96: 2988-2993, 1999; Uchida et al., PNAS USA 95: 1 1939-1 1944, 1998; Miyoshi et al., Science 283: 682-686, 1999; Sutton et al., J Virol 72: 5781-5788, 1998; и Frecha et al., Blood. 1 12: 4843-52, 2008, каждая из которых включена посредством ссылки во всей своей полноте).
Было задокументировано, что покоящиеся Т- и В-клетки можно подвергать трансдукции посредством покрытого VSVG LV, несущего большинство вспомогательных белков HIV (vif, vpr, vpu и nef) (см., например, Frecha et al., 2010 Mol. Therapy 18: 1748). В некоторых вариантах осуществления ретровирусный вектор содержит определенные минимальные последовательности из генома лентивируса, такого как геном HIV или геном SIV. Геном лентивируса обычно организован на область длинного концевого 5'-повтора (LTR), ген gag, ген pol, ген env, вспомогательные гены (например, nef, vif, vpr, vpu, tat, rev) и 3'-LTR-область. Вирусный LTR разделен на три области, называемые U3, R (повтор) и U5. U3-область содержит энхансерные и промоторные элементы, U5-область содержит сигналы полиаденилирования, а R-область разделяет области U3 и U5. Транскрибированные последовательности R-области находятся как на 5'-, так и на 3'-концах вирусной РНК (см., например, «РНК-вирусы: A Practical Approach» (Alan J. Cann, Ed., Oxford University Press, 2000); O Narayan, J. Gen. Virology. 70: 1617-1639, 1989; Fields et al., Fundamental Virology Raven Press., 1990; Miyoshi et al., J Virol. 72: 8150-7,1998; и патент США № 6013516, каждый изкоторых включен посредством ссылки во всей своей полноте). Лентивирусные векторы могут содержать любой один или более из этих элементов лентивирусного генома для регулирования активности вектора по необходимости, или они могут содержать делеции, вставки, замены или мутации в одном или более из этих элементов, например, с целью уменьшения патологических эффектов репликации лентивируса или ограничения лентивирусного вектора до одного раунда инфекции.
Как правило, минимальный ретровирусный вектор содержит определенные последовательности 5'-LTR и 3'-LTR, один или более представляющих интерес генов (с обеспечением экспрессии в клетке-мишени), один или более промоторов и цис-действующую последовательность для упаковки РНК. Могут быть включены другие регуляторные последовательности, описанные в данном документе и известные в данной области. Вирусный вектор обычно клонируют в плазмиду, которая может быть трансфицирована в пакующую клеточную линию, такую как эукариотическая клетка (например, 293-HEK), и также обычно содержит последовательности, применимые для репликации плазмиды в бактериях.
В некоторых вариантах осуществления вирусный вектор содержит последовательности из 5'- и/или 3'-LTR ретровируса, такого как лентивирус. Последовательности LTR могут представлять собой последовательности LTR из любого лентивируса из любого вида. Например, они могут представлять собой последовательности LTR из HIV, SIV, FIV или BIV. Предпочтительно последовательности LTR представляют собой последовательности LTR HIV.
В определенных вариантах осуществления вирусный вектор содержит последовательности R и U5 из 5'-LTR лентивируса и инактивированный или «самоинактивирующийся»" 3'-LTR из лентивируса. «Самоинактивирующийся 3'-LTR» представляет собой длинный концевой 3'-повтор (LTR), который содержит мутацию, замену или делецию, которая не позволяет последовательностям LTR управлять экспрессией гена, расположенного ниже. Копия области U3 из 3'-LTR действует как матрица для получения обоих LTR в интегрированном провирусе. Таким образом, когда 3'-LTR с инактивирующей делецией или мутацией интегрируется как 5'-LTR провируса, транскрипция из 5'-LTR невозможна. Это устраняет конкуренцию между вирусным энхансером/промотором и любым внутренним энхансером/промотором. Самоинактивирующиеся 3'-LTR описаны, например, в Zufferey et al., J. Virol. 72: 9873-9880, 1998; Miyoshi et al., J Virol. 72: 8150-8157, 1998; и Iwakuma et al., J Virology 261: 120-132, 1999, каждая из которых включена посредством ссылки во всей своей полноте. Самоинактивирующиеся 3'-LTR можно получать посредством любого способа, известного в данной области техники. В некоторых вариантах осуществления U3-элемент 3'-LTR содержит делецию своей энхансерной последовательности, предпочтительно TATA-бокса, сайтов Spl и/или NF-каппа B. В результате самоинактивирующейся 3'-LTR провирус, который интегрирован в геном клетки-хозяина, будет содержать инактивированную 5'-LTR.
Представленные в данном документе векторы обычно содержат ген, который кодирует белок (или другую молекулу, такую как siRNA), который желательно экспрессировать в одной или более клетках-мишенях. В вирусном векторе ген, представляющий интерес, предпочтительно расположен между последовательностями 5'-LTR и 3'-LTR. Кроме того, ген, представляющий интерес, предпочтительно находится в функциональной взаимосвязи с другими генетическими элементами, например, регуляторными последовательностями транскрипции, такими как промоторы и/или энхансеры, с целью регуляции экспрессии гена, представляющего интерес, определенным образом после того, как ген включен в клетку-мишень. В некоторых вариантах осуществления применимыми транскрипционными регуляторными последовательностями являются те, которые в большой степени регулируются в отношении активности как во времени, так и в пространстве.
В определенных вариантах осуществления один или более дополнительных генов могут быть включены в качестве меры безопасности, главным образом для обеспечения возможности избирательного уничтожения трансфицированных клеток-мишеней в гетерогенной популяции, такой как у пациента-человека. В одном иллюстративном варианте осуществления выбранный ген представляет собой ген тимидинкиназы (TK), экспрессия которого делает клетку-мишень восприимчивой к действию лекарственного средства ганцикловира. В дополнительном варианте осуществления суицидальный ген представляет собой суицидальный ген каспазы 9, активируемый димеризующим лекарственным средством (см., например., Tey et al., Biology of Blood and Marrow Transplantation 13: 913-924, 2007).
В определенных вариантах осуществления ген, кодирующий маркерный белок, может быть размещен до или после первичного гена в вирусном или невирусном векторе для обеспечения идентификации и/или отбора клеток, которые экспрессируют желаемый белок. Определенные варианты осуществления предусматривают включение флуоресцентного маркерного белка, такого как зеленый флуоресцентный белок (GFP) или красный флуоресцентный белок (RFP), наряду с первичным геном, представляющим интерес. В случае когда включены один или более дополнительных репортерных генов, также могут быть включены последовательности IRES или элементы 2А, отделяющие первичный ген, представляющий интерес, от репортерного гена и/или любого другого гена, представляющего интерес.
В некоторых вариантах осуществления можно применять гены, которые кодируют один или более селектируемых маркеров. Примеры включают селектируемые маркеры, которые эффективны в эукариотической клетке или прокариотической клетке, такие как ген лекарственной устойчивости, который кодирует фактор, необходимый для выживания или роста трансформированных клеток-хозяев, выращенных в селективной культуральной среде. Иллюстративные гены для отбора кодируют белки, которые придают устойчивость к антибиотикам или другим токсинам, например, G418, гигромицину B, пуромицину, зеоцину, уабаину, бластицидину, ампициллину, неомицину, метотрексату или тетрациклину, компенсируют ауксотрофный дефицит или пополняют, могут присутствовать в отдельных плазмидах, и их вводят путем совместной трансфекции с вирусным вектором. В одном варианте осуществления ген кодирует мутантную дигидрофолатредуктазу (DHFR), которая придает устойчивость к метотрексату. В некоторых других вариантах осуществления можно применять гены, которые кодируют один или более рецепторов клеточной поверхности, которые можно применять для мечения и обнаружения или очистки трансфицированных клеток (например, низкоаффинного рецептора фактора роста нервов (LNGFR) или других таких рецепторов, применимых в качестве систем для мечения трансдукции). См., например, Lauer et al., Cancer Gene Ther. 2000 Mar; 7(3): 430-7.
В определенных вирусных векторах, таких как ретровирусные векторы, применяют один или более гетерологичных промоторов, энхансеров или и то и другое. В некоторых вариантах осуществления последовательность U3 из ретровирусного или лентивирусного 5'-LTR может быть заменена промоторной или энхансерной последовательностью в вирусной конструкции. В некоторых вариантах осуществления применяют «внутренний» промотор/энхансер, который расположен между последовательностями 5'-LTR и 3'-LTR вирусного вектора и функционально связан с геном, представляющим интерес.
«Функциональная связь» и «функционально связанный» означают без ограничения, что ген находится в правильном положении и ориентации по отношению к промотору и/или энхансеру, за счет чего приведение в контакт промотора и/или энхансера с соответствующими регуляторными молекулами будет оказывать влияние на экспрессию гена. Можно применять любую комбинацию энхансер/промотор, посредством которой происходит либо регуляция (например, увеличение, уменьшение) экспрессии генома вирусной РНК в пакующей линии клеток, либо регуляция экспрессии выбранного гена, представляющего интерес, в инфицированной клетке-мишени, или и то и другое.
Промотор представляет собой элемент контроля экспрессии, образованный последовательностью ДНК, который обеспечивает связывание с полимеразой и осуществление транскрипции. Промоторы представляют собой нетранслируемые последовательности, которые расположены выше (5') от стартового кодона выбранного гена, представляющего интерес (обычно в пределах от приблизительно 100 до 1000 п.о.), и контролируют транскрипцию и трансляцию кодирующей полинуклеотидной последовательности, с которой они функционально связаны. Промоторы могут быть индуцируемыми или конститутивными. Индуцибельные промоторы инициируют повышенные уровни транскрипции ДНК под их контролем в ответ на некоторые изменения условий культивирования, такие как изменение температуры. Промоторы могут быть однонаправленными или двунаправленными. Двунаправленные промоторы можно применять для совместной экспрессии двух генов, например, гена, представляющего интерес, и селективного маркера. Альтернативно, можно применять конфигурацию двунаправленного промотора, предусматривающую два промотора, каждый из которых контролирует экспрессию другого гена, в противоположной ориентации в одном и том же векторе.
В данной области известны различные промоторы, а также способы функционального связывания промотора с полинуклеотидной кодирующей последовательностью. Для направления экспрессии выбранного гена, представляющего интерес, можно применять как нативные промоторные последовательности, так и многие гетерологичные промоторы. В определенных вариантах осуществления применяют гетерологичные промоторы, поскольку они, как правило, обеспечивают большую транскрипцию и более высокие выходы желаемого белка по сравнению с нативным промотором.
В некоторых вариантах осуществления можно применять гетерологичные вирусные промоторы. Примеры таких промоторов включают промоторы, полученные из геномов вирусов, таких как вирус полиомы, вирус оспы птиц, аденовирус, вирус папилломы крупного рогатого скота, вирус птичьей саркомы, цитомегаловирус, ретровирус, вирус гепатита В и вирус обезьян 40 (SV40). В некоторых вариантах осуществления можно применять гетерологичный промотор млекопитающего, такой как промотор актина, промотор иммуноглобулина, промотор теплового шока или промотор, который связан с нативной последовательностью гена, представляющего интерес. Как правило, промотор совместим с клеткой-мишенью, такой как активированный B-лимфоцит, плазматическая B-клетка, B-клетка памяти или другая клетка-мишень, представляющая собой лимфоцит.
В некоторых вариантах осуществления можно применять один или более промоторов РНК-полимеразы II и III. Подходящий выбор промоторов РНК-полимеразы III можно найти, например, в Paule and White. Nucleic Acids Research., Vol. 28, pp 1283-1298, 2000, которая включена в данный документ посредством ссылки во всей своей полноте. Промоторы РНК-полимеразы II и III также включают любые синтетические или сконструированные фрагменты ДНК, которые могут направлять РНК-полимеразу II или III соответственно для транскрибирования ее последовательностей, кодирующих расположенные ниже РНК. Кроме того, промотор или промоторы РНК-полимеразы II или III (Pol II или III), используемые как часть вирусного вектора, могут быть индуцибельными. Любой подходящий индуцибельный промотор Pol II или III можно применять в описанных в данном документе способах. Иллюстративные промоторы Pol II или III включают чувствительные к тетрациклину промоторы, представленные в Ohkawa and Taira, Human Gene Therapy, Vol. 11, pp 577-585, 2000; и Meissner et al., Nucleic Acids Research, Vol. 29, pp 1672-1682, 2001, каждый из которых включен в данный документ посредством ссылки во всей полноте.
Неограничивающие примеры конститутивных промоторов, которые можно применять, включают промотор для убиквитина, промотор CMV (см., например, Karasuyama et al., J. Exp. Med. 169: 13, 1989), β-актин (см., например, Gunning et al., PNAS USA 84: 4831-4835, 1987), промотор фактора элонгации-1 альфа (EF-1 альфа), промотор CAG и промотор pgk (см., например, Adra et al., Gene 60: 65-74, 1987); Singer-Sam et al., Gene 32: 409-417, 1984; и Dobson et al., Nucleic Acids Res. 10: 2635-2637, 1982, каждый из которых включен в данный документ посредством ссылки). Неограничивающие примеры тканеспецифических промоторов включают промотор lck (см., например, Garvin et al., Mol. Cell Biol. 8: 3058-3064, 1988; и Takadera et al., Mol. Cell Biol. 9: 2173-2180, 1989), промотор миогенина (Yee et al., Genes and Development 7: 1277-1289. 1993) и тиловый промотор (см., например, Gundersen et al., Gene 1 13: 207-214, 1992).
Дополнительные примеры промоторов включают промотор убиквитина С, промотор тяжелой цепи μ человека или промотор тяжелой цепи Ig (например, МН) и промотор легкой цепи κ человека или промотор легкой цепи Ig (например, EEK), которые являются функциональными в В-лимфоцитах. Промотор MH содержит промотор тяжелой цепи μ человека, которому предшествует энхансер iEμ, фланкированный областями прикрепления к матриксу, а промотор EEK содержит промотор легкой цепи κ, предшествующий интронному энхансеру (iEκ), области прикрепления к матриксу и 3'-энхансеру (3Eκ) (см., например, Luo et al., Blood. 1 13: 1422-1431, 2009, публикацию заявки на патент США № 2010/0203630). Соответственно, в определенных вариантах осуществления можно использовать один или более из этих промоторных или энхансерных элементов.
В одном варианте осуществления один промотор управляет экспрессией селектируемого маркера, а второй промотор управляет экспрессией гена, представляющего интерес. Например, в одном варианте осуществления промотор EF-1 альфа управляет продуцированием селективного маркера (например, DHFR), а миниатюрный промотор CAG (см., например, Fan et al. Human Gene Therapy 10: 2273-2285, 1999) управляет экспрессией гена, представляющего интерес (например, IDUA).
Как отмечено выше, в некоторых вариантах осуществления применяют энхансерные элементы, такие как внутренний энхансер, для увеличения экспрессии гена, представляющего интерес. Энхансеры представляют собой цис-действующие элементы ДНК, обычно длиной от 10 до 300 п.о., которые воздействуют на промотор с увеличением его транскрипции. Последовательности энхансеров могут быть получены из генов млекопитающих (например, глобина, эластазы, альбумина, α-фетопротеина, инсулина), такие как энхансер ιεμ, интронный энхансер ιεκ и энхансер 3' εκ. Также включены энхансеры из вируса эукариот, в том числе энхансер SV40 на поздней части точки начала репликации (100-270 п.о.), энхансер раннего промотора цитомегаловируса, энхансер полиомы на поздней части точки начала репликации и энхансеры аденовируса. Энхансеры могут встраиваться в вектор в положении 5' или 3' по отношению к антигенспецифической полинуклеотидной последовательности, но предпочтительно расположены в направлении 5' от промотора. Специалисты в данной области могут выбрать соответствующий энхансер на основе желаемого паттерна экспрессии.
В некоторых вариантах осуществления промоторы выбирают с целью обеспечения индуцибельной экспрессии гена. В данной области техники известен ряд систем для индуцибельной экспрессии, в том числе система, чувствительная к тетрациклину, и система lac оператор-репрессор. Также предполагают, что для получения желаемой экспрессии гена, представляющего интерес, можно применять комбинацию промоторов. Специалист в данной области сможет выбрать промотор на основе желаемого паттерна экспрессии гена в организме и/или клетке-мишени, представляющей интерес.
Некоторые вирусные векторы содержат цис-действующие пакующие последовательности, способствующие включению геномной вирусной РНК в вирусную частицу. Примеры включают в себя пси-последовательности. Такие цис-действующие последовательности известны в данной области. В определенных вариантах осуществления описанные в данном документе вирусные векторы могут экспрессировать два или более генов, что может быть достигнуто, например, путем включения внутреннего промотора, который функционально связан с каждым отдельным геном за пределами первого гена, путем включения элемента, который способствует совместной экспрессии, такого как элемент последовательности внутренней посадки рибосомы (IRES) (патент США №4937190, включенный с помощью ссылки), или элемент 2A, или и тот и другой. В качестве иллюстрации можно использовать элементы IRES или 2А, когда один вектор содержит последовательности, кодирующие каждую цепь молекулы иммуноглобулина с желаемой специфичностью. Например, первая кодирующая область (кодирующая либо тяжелую, либо легкую цепь) может быть расположена непосредственно ниже промотора, а вторая кодирующая область (кодирующая другую цепь) может быть расположена ниже первой кодирующей области с элементом IRES или 2А, расположенным между первой и второй кодирующими областями, предпочтительно непосредственно перед второй кодирующей областью. В других вариантах осуществления элемент IRES или 2А используется для совместной экспрессии неродственного гена, такого как репортерный ген, селектируемый маркер или ген, который усиливает иммунную функцию. Примеры последовательностей IRES, которые можно применять, включают без ограничения элементы IRES вируса энцефаломиелита (EMCV), вируса ящура (FMDV), вируса мышиного энцефаломиелита Тейлера (TMEV), риновирус человека (HRV), вируса Коксаки (CSV), полиовируса (POLIO), вируса гепатита A (HAV), вируса гепатита C (HCV) и пестивирусов (например, вируса свиной холеры (HOCV) и вируса вирусной диареи крупного рогатого скота (BVDV)) (см., например, Le et al. . Virus Genes 12: 135-147, 1996; Le et al., Nuc. Acids Res. 25: 362-369, 1997, каждая из которых включена посредством ссылки во всей своей полноте). Один пример элемента 2А включает последовательность F2A из вируса ящура.
В определенных вариантах осуществления векторы, представленные в настоящем документе, также содержат дополнительные генетические элементы для достижения желаемого результата. Например, некоторые вирусные векторы могут включать сигнал, который способствует ядерному проникновению вирусного генома в клетку-мишень, такой как флэп-сигнал HIV-1. Как дополнительный пример, определенные вирусные векторы могут включать элементы, которые облегчают характеристику сайта интеграции провируса в клетке-мишени, такие как амбер-супрессорная последовательность tRNA. Некоторые вирусные векторы могут содержать один или более генетических элементов, предназначенных для усиления экспрессии гена, представляющего интерес. Например, в конструкцию можно включать элемент, чувствительный к вирусу гепатита сурков (WRE), (см., например, Zufferey et al., J. Virol. 74: 3668-3681, 1999; и Deglon et al., Hum. Gene Ther. 11: 179-190, 2000, каждая из которых включена посредством ссылки во всей своей полноте). В качестве другого примера, в конструкцию также может быть включен инсулятор β-глобина курицы. Было показано, что этот элемент снижает вероятность сайленсинга встроенной ДНК в клетке-мишени вследствие эффектов метилирования и гетерохроматизации. Кроме того, инсулятор может защищать внутренний энхансер, промотор и экзогенный ген от положительного или отрицательного позиционного воздействия окружающей ДНК в сайте интеграции на хромосоме. В определенных вариантах осуществления применяют каждый из этих генетических элементов. В другом варианте осуществления вирусные векторы, представленные в настоящем документе, также могут содержать универсальный хроматинраскрывающий элемент (UCOE) для увеличения экспрессии (см., например, Zhang F, et al., Molecular Therapy: The journal of the American Society of Gene Therapy 2010 Sep;18(9): 1640-9.)
В некоторых вариантах осуществления вирусные векторы (например, ретровирусный, лентивирусный), представленные в настоящем документе, «псевдотипированы» с одним или более выбранными вирусными гликопротеинами или белками оболочки, главным образом для нацеливания на выбранные типы клеток. Псевдотипирование относится, как правило, к включению одного или более гетерологичных вирусных гликопротеинов в вирусную частицу клеточной поверхности, часто с обеспечением вирусной частице инфицирования выбранной клетки, которая отличается от ее нормальных клеток-мишеней. «Гетерологичный» элемент происходит от вируса, отличного от вируса, из которого происходит геном РНК вирусного вектора. Как правило, гликопротеин-кодирующие области вирусного вектора были генетически изменены, например, путем делеции с целью предотвратить экспрессию собственного гликопротеина. Как иллюстрация, гликопротеины оболочки gp41 и/или gp120 из лентивирусного вектора, полученного из HIV, обычно удаляют перед псевдотипированием гетерологичным вирусным гликопротеином.
В некоторых вариантах осуществления вирусный вектор псевдотипируют с гетерологичным вирусным гликопротеином, который нацеливается на В-лимфоциты. В некоторых вариантах осуществления вирусный гликопротеин обеспечивает избирательную инфекцию или трансдукцию покоящихся или молчащих В-лимфоцитов. В некоторых вариантах осуществления вирусный гликопротеин обеспечивает избирательную инфекцию плазматических клеток B-лимфоцитов, плазмабластов и активированных B-клеток. В определенных вариантах осуществления вирусный гликопротеин обеспечивает инфекцию или трансдукцию молчащих В-лимфоцитов, плазмабластов, плазматических клеток и активированных В-клеток. В некоторых вариантах осуществления вирусный гликопротеин обеспечивает инфекцию клеток В-клеточного хронического лимфоцитарного лейкоза. В одном варианте осуществления вирусный вектор псевдотипирован с помощью VSV-G. В другом варианте осуществления гетерологичный вирусный гликопротеин получен из гликопротеина вируса кори, такого как вирус кори Эдмонтона. В некоторых вариантах осуществления псевдотипируют с помощью гликопротеинов вируса кори гемагглютинина (H), белка слияния (F) или и того и другого (см., например, Frecha et al., Blood). 1 12: 4843-52, 2008; и Frecha et al., Blood. 1 14: 3173-80, 2009, каждая из которых включена посредством ссылки во всей своей полноте). В одном варианте осуществления вирусный вектор псевдотипирован вирусом лейкемии гиббоновых обезьян (GALV). В одном варианте осуществления вирусный вектор псевдотипирован эндогенным ретровирусом кошек (RD114). В одном варианте осуществления вирусный вектор псевдотипирован эндогенным ретровирусом бабуинов (BaEV). В одном варианте осуществления вирусный вектор псевдотипирован вирусом мышиного лейкоза (MLV). В одном варианте осуществления вирусный вектор псевдотипирован вирусом лейкемии гиббоновых обезьян (GALV). В других вариантах осуществления вирусный вектор содержит встроенный связывающий домен антитела, например, одну или более вариабельных областей (например, вариабельных областей тяжелой и легкой цепи), который служит для нацеливания вектора на конкретный тип клеток.
Получение вирусных векторов может быть осуществлено с применением любых подходящих методик генной инженерии, известных в данной области, в том числе без ограничения стандартных методик расщепления рестрикционной эндонуклеазой, лигирования, трансформации, очистки плазмиды, ПЦР-амплификации и секвенирования ДНК, например, как описано в Sambrook et al. (Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, N.Y. (1989)), Coffin et al. (Retroviruses. Cold Spring Harbor Laboratory Press, N.Y. (1997)) and “RNA Viruses: A Practical Approach” (Alan J. Cann, Ed., Oxford University Press, (2000)).
Для получения подходящих ретровирусных частиц, геном которых содержит РНК-копию вирусного вектора, можно применять любой ряд способов, известных в данной области. В качестве одного способа вирусный вектор может быть введен в линию пакующих клеток, которая пакует вирусную геномную РНК на основе вирусного вектора в вирусные частицы с желаемой специфичностью к клеткам-мишеням. Линия пакующих клеток обычно обеспечивает транс-вирусные белки, которые необходимы для упаковки вирусной геномной РНК в вирусные частицы и инфицирования клетки-мишен, включая структурные белки gag, ферментативные белки pol и гликопротеины оболочки.
В некоторых вариантах осуществления линия пакующих клеток стабильно экспрессирует определенные необходимые или желаемые вирусные белки (например, gag, pol) (см., например, патент США №6218181, включенный в данный документ посредством ссылки. В некоторых вариантах осуществления линию пакующих клеток транзиентно подвергают трансфекции плазмидами, которые кодируют определенные необходимые или желаемые вирусные белки (например, gag, pol, гликопротеин), включая последовательности гликопротеина вируса кори, описанные в данном документе. В одном примерном варианте осуществления линия пакующих клеток стабильно экспрессирует последовательности gag и pol, и клеточную линию затем подвергают трансфекции плазмидой, кодирующей вирусный вектор, и плазмидой, кодирующей гликопротеин. После введения желаемых плазмид вирусные частицы собирают и обрабатывают соответствующим образом, например, посредством ультрацентрифугирования, с целью получения концентрированного запаса вирусных частиц. Иллюстративные линии пакующих клеток включают клеточные линии 293 (ATCC CCL X), HeLa (ATCC CCL 2), D17 (ATCC CCL 183), MDCK (ATCC CCL 34), BHK (ATCC CCL-10) и Cf2Th (ATCC CRL 1430).
Терапевтическое средство
Используемый в данном документе термин «ген, представляющий интерес», или «ген», или «нуклеиновая кислота, представляющая интерес» относится к трансгену, который подлежит экспрессии в трансфицированной клетке-мишени. Несмотря на то, что можно использовать термин «ген», это не означает, что это ген, который обнаруживается в геномной ДНК и который используется взаимозаменяемо с термином «нуклеиновая кислота». Как правило, нуклеиновая кислота, представляющая интерес, предоставляет подходящую нуклеиновую кислоту для кодирования терапевтического средства и может содержать cDNA или ДНК и может включать или не включать интроны, но, как правило, не включает интроны. Как отмечено в другом месте в данном документе, нуклеиновая кислота, представляющая интерес, функционально связана с последовательностями контроля экспрессии для эффективной экспрессии представляющего интерес белка в клетке-мишени. В определенных вариантах осуществления векторы, описанные в данном документе, могут содержать один или более генов, представляющих интерес, и могут включать 2, 3, 4 или 5 или более генов, представляющих интерес, таких как, например, тяжелые и легкие цепи иммуноглобулина, которые могут быть упорядочены с применением внутреннего промотора, описанного в данном документе.
Используемое в данном документе выражение «полинуклеотид» или «нуклеиновая кислота» обозначает mRNA, РНК, cRNA, cDNA или ДНК. Термин, как правило, относится к полимерной форме нуклеотидов длиной по меньшей мере 10 оснований, либо рибонуклеотидам, либо дезоксинуклеотидам, либо модифицированной форме нуклеотидов любого типа. Данный термин охватывает одно- и двухнитевые формы ДНК и РНК. Нуклеиновая кислота или ген, представляющие интерес, могут представлять собой любую нуклеиновую кислоту, кодирующую белок, представляющий интерес.
Терапевтическое средство, которое должно быть доставлено с помощью генетически модифицированной В-клетки, как описано в данном документе, может представлять собой белок. Белок, представляющий интерес для применения, описанного в данном документе, предусматривает любой белок, обеспечивающий желаемую активность. В этом отношении белок, представляющий интерес, включает без ограничения антитело или его антигенсвязывающий фрагмент, рецептор клеточной поверхности, секретируемый белок, такой как цитокин (лимфокины, интерлейкины, интерфероны или хемокины), другие секретируемые сигнальные молекулы, такие как TGF-бета и фактор роста фибробластов, антигенный фрагмент белка, кодируемая ДНК малая молекула (см., например, Nature Chemical Biology 5, 647-654 (2009)), фермент, фактор свертывания крови и адгезивная молекула. В одном варианте осуществления нуклеиновая кислота кодирует антитело или его антигенсвязывающий фрагмент. Иллюстративные антигенсвязывающие фрагменты включают доменные антитела, sFv, scFv, Fab, Fab', F(ab')2 и Fv. В одном варианте осуществления нуклеиновая кислота кодирует белок, представляющий интерес, в виде слитого белка, содержащего расщепляемый линкер. Например, тяжелая цепь антитела и легкая цепь могут быть экспрессированы с помощью саморасщепляемого линкерного пептида, например, F2A.
В одном варианте осуществления антитело, кодируемое нуклеиновой кислотой, содержит по меньшей мере антигенсвязывающий домен HIV-нейтрализующего антитела, b12 (см., например, J Virol 2003, 77: 5863-5876; J Virol. 1994 Aug; 68(8): 4821-8; Proc Natl Acad Sci US A. 1992, 89: 9339-9343; иллюстративные последовательности представлены под номерами доступа в GenBank для легкой цепи b12 (AAB26306.1 Gl 299737) и тяжелой цепи (AAB26315.1 Gl 299746)). В дополнительном варианте осуществления антитело, кодируемое нуклеиновой кислотой, представляющей интерес, содержит Fuzeon(TM) (T-20/энфувиртид/пентафузид/DP-178). DP-178 представляет собой аминокислотную последовательность gp41 на HIV и влияет на способность HIV сливаться со своей клеткой-мишенью. Фузеон можно получать синтетическим путем с применением способов, известных специалисту (см., например, 2001 J. Virol. 75: 3038-3042; следует отметить, что весьма маловероятно, что способы, описанные в данной статье, привели в результате к выделению терапевтической дозы пептида DP-178).
В одном конкретном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует иммунологически активный белок. В некоторых вариантах осуществления нуклеиновая кислота, представляющая интерес, кодирует белок или его биологически активный фрагмент (например, антигенный фрагмент), который индуцирует иммунную вакциноподобную реакцию посредством презентации белка на поверхности B-клетки, T-клетки или другой иммунной клетки. В определенных вариантах осуществления белок, представляющий интерес, влияет на регуляцию В-клеток, например, без ограничения посредством стимуляции деления клеток, дифференцировки в различные В-линии, инактивации или уничтожения клеток, или регулирует продуцирование или активность других введенных элементов ДНК. Интерлейкины известны специалисту и на сегодняшний день включают IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-14, IL-15, IL-16, IL-17, IL-18, IL-19, IL-20, IL-21 , IL-22, IL-23, IL-24, IL- 25, IL-26, IL-27, секретируемую форму субъединицы p28 IL27, IL-28, IL-29, IL-30, IL-31, IL-32, IL-33, IL-34 и IL-35. Интерфероны включают IFN-γ, IFN-α, IFN-β и IFN-ο. Хемокины, рассматриваемые для применения в настоящем документе, включают хемокины типа C XCL1 и XCL2, хемокины типа C-C (на сегодняшний день включают CCL1-CCL28) и хемокины типа CXC (на сегодняшний день включают CXCL1-CXCL17). В качестве гена, представляющего интерес, также рассматривают представителей суперсемейства TNF (например, TNF-α, лиганд 4-1 BB, фактор активации B-клеток, лиганд FAS, лимфотоксин, OX40L RANKL и TRAIL).
В определенных вариантах осуществления белок, представляющий интерес, индуцирует иммунологическую толерантность. В этом отношении белок, представляющий интерес, может предусматривать слитый белок IgG-антиген (см., например, Cellular Immunology 235(1), 2005, 12-20). В некоторых вариантах осуществления экспрессия белка, представляющего интерес, может сопровождаться стимуляцией клеток такими факторами, как TGF-β, IL-10 и LPS. В некоторых вариантах осуществления такие факторы, как IL-10 или факторы транскрипции, которые индуцируют толерантность, экспрессируются культивируемыми В-клетками.
В дополнительном варианте осуществления ген(ы), представляющий(-ие) интерес, кодирует(-ют) один или более факторов, которые способствуют дифференцировке В-клетки в клетку, секретирующую антитело, и/или один или более факторов, которые способствуют продолжительности жизни клетки, продуцирующей антитело. Такие факторы включают, например, Blimp-1, Xbp1, IRF4, Zbtb20, TRF4, антиапоптотические факторы, такие как Bcl-xl, Bcl-2, Mcl-1 или Bcl5, и конститутивно активные мутанты рецептора CD40. Другие гены, представляющие интерес, кодируют факторы, которые способствуют экспрессии расположенных ниже сигнальных молекул, такие как факторы, связанные с рецептором TNF (TRAF). В этом отношении активация клеток, выживание клеток и антиапоптотические функции суперсемейства рецепторов TNF в основном опосредуются TRAF 1-6 (см., например, R.H. Arch, et al., Genes Dev. 12 (1998), pp. 2821-2830). Расположенные ниже эффекторы передачи сигналов TRAF включают транскрипционные факторы семейства NF-KB и AP-1, которые могут включать гены, участвующие в различных аспектах клеточных и иммунных функций. Кроме того, было показано, что активация NF-κβ и AP-1 обеспечивает защиту клеток от апоптоза посредством транскрипции антиапоптотических генов. В дополнительном варианте осуществления кодируемый фактор, такой как IL-10, IL-35, TGF-бета или Fc-слитый белок, связан с индукцией иммунной толерантности.
В дополнительном варианте осуществления нуклеиновая(-ые) кислота(-ы), представляющая(-ие) интерес, кодирует(-ют) один или более белков, полученных из вируса Эпштейна-Барра (EBV). Белки, полученные из EBV, включают без ограничения EBNA-1, EBNA-2, EBNA-3, LMP-1, LMP-2, EBER, EBV-EA, EBV-MA, EBV-VCA и EBV-AN. В одном конкретном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует антитело или его антигенсвязывающий фрагмент. В этом отношении антитело может представлять собой природное антитело или специально созданное рекомбинантно сконструированное антитело. Предполагается, что слитые белки, содержащие антитело или его часть, кодируются векторами, описанными в данном документе.
В одном варианте осуществления антитело или его фрагмент согласно настоящему изобретению характеризуются аминокислотной последовательностью антитела к HIV, такого как антитело к HIV m36 (см., например, Proc Natl Acad Sci US A. 2008 Nov 4; 105 (44): 17121 - 6), или аминокислотной последовательностью, характеризующейся по меньшей мере 60%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью антитела к HIV, такого как m36. В частности, специально рассматриваются слитые белки, содержащие m36 или его производные, такие как m36L2CD4Fc (см., Antiviral Research volume 88, Issue 1, October 2010, Pages 107-1 15). В одном варианте осуществления антитело к HIV представляет собой нейтрализующее моноклональное антитело VRC01 широкого спектра действия (см., например, Wu et al., Science, 2010, 329(5993): 856861, и Li et al., J Virol, 2011, 85(17): 8954-8967).
В дополнительном варианте осуществления антитело, кодируемое трансгеном по настоящему изобретению, связывается с аутоантигеном. В определенных вариантах осуществления аутоантиген в этом отношении связан с развитием рассеянного склероза или диабета 1 типа, в том числе без ограничения MBP, альфа-B-кристаллин, S100-бета, протеолипидный белок (PLP), HSP105, антиген 1 (BPAG1-е) эпителиальной изоформы буллезного пемфигоида (BP), липиды и изоформы гликопротеина (MOG)-альфа и MOG-бета миелинового олигодендроцита или любой из множества аутоантигенов островковых клеток (например, сиалогликолипид, глутаматдекарбоксилаза, инсулин, рецептор инсулина, 38 кДа, бычий сывороточный альбумин, переносчик глюкозы, hsp 65, карбоксипептидаза H, 52 кДа, ICA 12/ICA512, 150 кДа и RIN полярный). Антитела к этим аутоантигенам известны в данной области и могут быть секвенированы и получены рекомбинантным путем с применением обычных методик (см., например, J. Clin. Invest. 107(5): 555-564 (2001)).
В следующем варианте осуществления антитело связывается с антигеном, связанным с раком. Связанные с раком антигены могут быть получены из ряда опухолевых белков. Иллюстративные опухолевые белки, применяемые в настоящем изобретении, включают без ограничения один или более из p53, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A6, MAGE-A10, MAGE-A12, BAGE, DAM-6, -10, GAGE-1, -2, -8, GAGE-3, -4, -5, -6, -7B, NA88-A, NY-ESO-1, MART-1 , MC1 R, Gp100, PSA, PSM, тирозиназы, TRP-1, TRP-2, ART-4, CAMEL, CEA, Cyp-B, Her2/neu (например, антитело может быть получено из Her2-специфического mAb, герцептина (R)), hTERT, hTRT, iCE, MUC1, MUC2, PRAME, P15, RU1, RU2, SART-1, SART-3, WT1, -катенин/m, каспаза-8/m, CEA, CDK-4/m, ELF2M, GnT-V, G250, HSP70-2M, HST-2, KIAA0205, MUM-1, MUM-2, MUM- 3, миозин/m, RAGE, SART-2, TRP-2/INT2, 707-AP, аннексин II, CDC27/m, TPI/mbcr-abl, ETV6/AML, LDLR/FUT, Pml/RARa и TEL/AML1. Эти и другие опухолевые белки известны специалисту в данной области.
В других вариантах осуществления нуклеиновая кислота, представляющая интерес, кодирует пептид или другой связывающий домен с определенным функциональным свойством, таким как без ограничения ингибирующая активность, способность вызывать клеточную гибель у раковых клеток или способность замедлять или подавлять пролиферацию раковых клеток. В этом отношении в одном варианте осуществления пептид или связывающий домен, кодируемый нуклеиновой кислотой, представляющей интерес, может связывать любой из белков-мишеней, описанных в данном документе, таких как связанный с раком антиген, описанный выше, CD4, gp120 HIV или другой вирусный белок, ICAM-3, DC-SIGN (см., например, патент США №7301010). В некоторых вариантах осуществления пептиды можно получать из патогенных и непатогенных бактерий и зеленых растений. Иллюстративные пептиды раскрыты в патентах США №№7084105, 7301010, 7338766, 7381701, 7491394, 7511117, 7556810. В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует азурин-p28 (NSC745104), пептидный ингибитор убиквитинирования p53 (см., например, Cancer Chemother Pharmacol 2010, DOI 10.1007/S00280-010-1518-3; патент США №7084105). В дополнительном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует фактор, известный как грелин, который вызывает аппетит и может применяться для лечения раковых пациентов (см., например, Obes Facts. 2010 3: 285-92; FASEB J. 18 (3): 439-56). В другом варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует связывающий пептид, который связывается с ангиопоэтином 1 и 2 и ингибирует его (см., например, AMG386, Fc-фрагмент антитела (пептититела), применяемый для лечения рака; в некоторых вариантах осуществления опухолевые антигены можно идентифицировать непосредственно от индивидуума, у которого имеется рак. В связи с этим скрининги можно выполнять с применением множества известных технологий. Например, в одном варианте осуществления у пациента берут биопсию опухоли, выделяют РНК из опухолевых клеток и подвергают скринингу с применением генного чипа (например, от Affymetrix, Санта-Клара, Калифорния) и идентифицируют опухолевый антиген. Как только опухолевый антиген-мишень идентифицирован, его можно затем клонировать, экспрессировать и очищать с применением методик, известных в данной области.
В одном конкретном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует фермент. В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует фермент для лечения нарушения лизосомального накопления. В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует идуронидазу (IDUA) для лечения или предупреждения мукополисахаридоза типа I (MPS I). В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует идурсульфазу для лечения или предупреждения мукополисахаридоза типа II (MPS II). В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует галсульфазу для лечения или предупреждения мукополисахаридоза типа VI (MPS VI). В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует альфа-элосульфазу для лечения или предупреждения мукополисахаридоза типа IVA (MPS IVA). В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует бета-агальсидазу для лечения или предупреждения болезни Фабри. В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует альфа-агальсидазу для лечения или предупреждения болезни Фабри. В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует альфа-1-антитрипсин для лечения или предупреждения дефицита альфа-1-антитрипсина. В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует альфа-N-ацетилглюкозаминидазу для лечения или предупреждения мукополисахаридоза типа IIIB (MPS IIIB). В другом варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует фактор VII для лечения или предупреждения гемофилии. В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует лецитинхолестеринацилтрансферазу (LCAT), применимую для лечения или предупреждения, например, дефицита LCAT и атеросклероза. В другом варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует аполипопротеин A-1 Milano (ApoA-1 Milano) для лечения или предупреждения сердечно-сосудистых заболеваний и нарушений, таких как, например, атеросклероз. В одном варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует липопротеинлипазу (LPL) для лечения или предупреждения дефицита LPL. В другом варианте осуществления нуклеиновая кислота, представляющая интерес, кодирует нейтрализующее антитело (bNAb) широкого спектра действия или его слитый белок, который связывает и нейтрализует множество штаммов HIV-1 (например, b12). В еще одном варианте нуклеиновая кислота, представляющая интерес, кодирует фенилаланингидроксилазу для лечения или предупреждения феникетонурии (PKU).
Используемый в данном документе термин «антитело» включает как поликлональные, так и моноклональные антитела; приматизированные (например, гуманизированные); мышиные; мышь-человек; мышь-примат и химерные; и может представлять собой интактную молекулу, ее фрагмент (такой как фрагменты scFv, Fv, Fd, Fab, Fab'и F(ab)'2) или мультимеры или агрегаты интактных молекул и/или фрагментов; и могут быть встречающимися в природе или полученными, например, путем иммунизации, синтеза или генной инженерии; «фрагмент антитела», как используется в данном документе, относится к фрагментам, полученным из антитела или родственным ему, которые связывают антиген и которые, в некоторых вариантах осуществления, могут быть дериватизированы с проявлением структурных особенностей, которые облегчают клиренс и поглощение, например, путем введения галактозных остатков. Они включают, например, F(ab), F(ab)'2, scFv, вариабельную область легкой цепи (VL), вариабельную область тяжелой цепи (VH) и их комбинации. Источники включают последовательности генов антител из различных видов (которые могут быть отформатированы как антитела, sFv, scFv или Fab, например, в фаговой библиотеке), в том числе человеческие, верблюжьи (из верблюдов, дромадеров или лам; Hamers-Casterman et al. (1993) Nature, 363: 446, и Nguyen et al. (1998) J. Mol. Biol., 275: 413), акульи (Roux et al. (1998) Proc. Nat'l. Acad. Sci. (USA) 95: 1 1804), рыбьи (Nguyen et al. (2002) Immunogenetics, 54: 39), крысиные, птичьи, овечьи последовательности, которые кодируют случайные пептидные библиотеки или последовательности, которые кодируют полученное посредством конструирования разнообразие аминокислот в петлевых областях альтернативных каркасов, не относящихся к антителам, таких как домены фибриногена (см., например, Weisel et al. (1985) Science 230: 1388), домены Кунитца (см., например, патент США № 6423498), липокалиновые домены (см., например, WO 2006/095164), V-подобные домены (см., например, публикацию заявки на патент США № 2007/0065431), лектиновые домены С-типа (Zelensky and Gready (2005) FEBS J. 272: 6179) и т.д. (см., например, публикацию патентной заявки РСТ № WO 2007/098934; WO 2006/072620) или тому подобное.
Термины, понимаемые специалистами в данной области как относящиеся к технологии антител, имеют значение, приобретенное в данной области техники, если оно явно не определено в данном документе. Например, термины «VL» и «VH» относятся к вариабельной связывающей области, полученной из легкой и тяжелой цепи антитела соответственно. Вариабельные связывающие области состоят из дискретных, четко определенных подобластей, известных как «определяющие комплементарность области» (CDR) и «каркасные области» (FR). Термины «CL» и «CH» относятся к «константной области иммуноглобулина», то есть константной области, полученной из легкой или тяжелой цепи антитела соответственно, причем последняя область, как считается, дополнительно делится на домены константной области Cm, CH2, CH3 и СН4 в зависимости от изотипа антитела (IgA, IgD, IgE, IgG, IgM), из которого получена область. Часть доменов константной области составляет Fc-область (область «кристаллизуемого фрагмента»), которая содержит домены, ответственные за эффекторные функции иммуноглобулина, такие как ADCC (антителозависимая клеточноопосредованная цитотоксичность), CDC (комплементзависимая цитотоксичность) и фиксация комплемента, связывание с Fc-рецепторами, увеличенное время полужизни in vivo по сравнению с полипептидом, лишенным Fc-области, связывание с белком A и, возможно, даже плацентарный перенос (см. Capon et al. (1989) Nature, 337: 525). Кроме того, полипептид, содержащий Fc-область, обеспечивает димеризацию или мультимеризацию полипептида.
Доменная структура иммуноглобулинов поддается конструированию, так как антигенсвязывающие домены и домены, обеспечивающие эффекторные функции, могут подвергаться обмену между классами и подклассами иммуноглобулинов. Например, аминокислотные замены (например, делеции, вставки, замены) могут обеспечивать изменение посттрансляционных процессов иммуноглобулина, таких как изменение количества или положения сайтов гликозилирования и/или фукозилирования. Способы усиления ADCC посредством гликозилирования известны в данной области и предполагаются для применения в данном документе. Например, ферменты, которые усиливают гликозилирование, могут экспрессироваться совместно с антителом. В одном варианте осуществления MGAT3 сверхэкспрессируется в клетках, продуцирующих антитело, для усиления гликозилирования антитела и его функции ADCC. В одном варианте осуществления подавление Fut8 посредством, например, siRNA, обеспечивает усиление гликозилирования антитела и ADCC.
Структура и функция иммуноглобулинов рассматриваются, например, в Harlow et al., Eds., Antibodies: A Laboratory Manual, Chapter 14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, 1988). Обширное введение, а также подробную информацию обо всех аспектах технологии рекомбинантных антител можно найти в учебнике Recombinant Antibodies (John Wiley & Sons, NY, 1999). Всестороннее собрание подробных протоколов лабораторного конструирования антител можно найти в R. Kontermann и S. Dubel, Eds., The Antibody Engineering Lab Manual (Springer Verlag, Heidelberg/New York, 2000). Другие соответствующие протоколы также доступны в Current Protocols in Immunology (August 2009), опубликованном John Wiley & Sons, Inc., Boston, MA. Способы получения ферментов и конструирования белков (например, IDUA) также известны в данной области и предназначены для применения в настоящем документе.
Таким образом, в данном изобретении предусмотрены полинуклеотиды (выделенные, или очищенные, или чистые полинуклеотиды), кодирующие терапевтические средства (например, белки, представляющие интерес) по настоящему изобретению для генетического модифицирования В-клеток, векторы (в том числе векторы клонирования и векторы экспрессии), содержащие такие полинуклеотиды, и клетки (например, клетки-хозяева), трансформированные или трансфицированные полинуклеотидом или вектором согласно настоящему изобретению. В определенных вариантах осуществления рассмотрен полинуклеотид (ДНК или РНК), кодирующий белок, представляющий интерес, по данному изобретению. Также в настоящем документе рассматрены кассеты экспрессии, кодирующие белки, представляющие интерес.
Настоящее изобретение также относится к векторам, которые содержат полинуклеотид по данному изобретению, и, в частности, к рекомбинантным экспрессионным конструкциям. В одном варианте осуществления в данном изобретении рассматрен вектор, содержащий полинуклеотид, кодирующий белок по данному изобретению, наряду с другими полинуклеотидными последовательностями, которые вызывают или облегчают транскрипцию, трансляцию и процессинг таких кодирующих белок последовательностей. Подходящие векторы клонирования и экспрессии для применения с прокариотическими и эукариотическими хозяевами описаны, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor, NY, (1989). Иллюстративные векторы клонирования/экспрессии включают векторы клонирования, челночные векторы и экспрессионные конструкции, которые могут быть основаны на плазмидах, фагмидах, фазмидах, космидах, вирусах, искусственных хромосомах или любом носителе нуклеиновой кислоты, известном в данной области техники, подходящем для амплификации, переноса и/или экспрессии содержащегося в нем полинуклеотида.
При использовании в данном документе, если не указано иное, в отношении вирусных векторов, «вектор» означает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой ее связали. Иллюстративные векторы включают плазмиды, миникольца, транспозоны (например, транспозон Sleeping Beauty), дрожжевые искусственные хромосомы, самореплицирующиеся РНК и вирусные геномы. Определенные векторы могут автономно реплицироваться в клетке-хозяине, тогда как другие векторы могут быть интегрированы в геном клетки-хозяина и, таким образом, реплицироваться с геномом хозяина. Кроме того, некоторые векторы в данном документе называют «рекомбинантными векторами экспрессии» (или просто «векторами экспрессии»), которые содержат последовательности нуклеиновых кислот, которые функционально связаны с последовательностью контроля экспрессии и, следовательно, способны направлять экспрессию этих последовательностей. В некоторых вариантах осуществления экспрессионные конструкции получены из плазмидных векторов. Иллюстративные конструкции включают модифицированный вектор pNASS (Clontech, Пало-Альто, Калифорния), который содержит последовательности нуклеиновой кислоты, кодирующие ген устойчивости к ампициллину, сигнал полиаденилирования и сайт промотора T7; pDEF38 и pNEF38 (CMC ICOS Biologies, Inc.), которые содержат промотор CHEF1; и pD18 (Lonza), который содержит промотор CMV. Другие подходящие векторы экспрессии млекопитающих хорошо известны (см., например, Ausubel et al., 1995; Sambrook et al., выше; см. также, например, каталоги от Invitrogen, Сан-Диего, Калифорния; Novagen, Мадисон, Висконсин; Pharmacia, Пискатауэй, Нью-Джерси).
Можно получать применимые конструкции, которые включают в себя последовательность, кодирующую дигидрофолатредуктазу (DHFR), при подходящем регуляторном контроле для стимулирования повышенных уровней продуцирования слитых белков, уровни которых являются результатом амплификации генов после применения подходящего средства для отбора (например, метотрексата). В одном варианте осуществления применение бифункционального транспозона, кодирующего терапевтический ген (например, IDUA), наряду с устойчивым к лекарственным средствам DHFR в комбинации с инкубацией в метотрексате (MTX) для обогащения успешно транспозированных В-клеток дает более эффективный продукт.
Как правило, рекомбинантные векторы экспрессии будут включать в себя точки начала репликации и селектируемые маркеры, позволяющие трансформировать клетку-хозяина, и промотор, полученный из высокоэкспрессированного гена, для прямой транскрипции структурной последовательности, расположенной ниже, как описано выше. Вектор в функциональной связи с полинуклеотидом согласно данному изобретению обеспечивает конструкцию для клонирования или экспрессии. Иллюстративные конструкции для клонирования/экспрессии содержат по меньшей мере один элемент контроля экспрессии, например, промотор, функционально связанный с полинуклеотидом по настоящему изобретению. Дополнительные элементы контроля экспрессии, такие как энхансеры, специфические в отношении фактора сайты связывания, терминаторы и сайты связывания рибосом, также рассматриваются в векторах и конструкциях для клонирования/экспрессии в соответствии с настоящим изобретением. Гетерологичную структурную последовательность полинуклеотида в соответствии с настоящим изобретением собирают в соответствующей фазе с последовательностями инициации и терминации трансляции. Таким образом, например, кодирующие нуклеиновые кислоты, представленные в настоящем документе, могут быть включены в любую из множества конструкций вектора экспрессии (например, миникольца) в качестве рекомбинантной экспрессионной конструкции с обеспечением экспрессии такого белка в клетке-хозяине.
Соответствующая(-ие) последовательность(-и) ДНК может(могут) быть вставлена(-ы) в вектор, например, посредством ряда процедур. Как правило, последовательность ДНК встраивают в соответствующий(-ие) сайт(ы) расщепления рестрикционной эндонуклеазой с помощью процедур, известных в данной области. Рассмотрены стандартные способы клонирования, выделения, амплификации и очистки ДНК для ферментативных реакций с участием ДНК-лигазы, ДНК-полимеразы, рестрикционных эндонуклеаз и тому подобного, а также различные методики разделения. Ряд стандартных методик описан, например, в Ausubel et al. (Current Protocols in Molecular Biology, Greene Publ. Assoc. Inc. & John Wiley & Sons, Inc., Boston, MA, 1993); Sambrook et al. (Molecular Cloning, Second Ed., Cold Spring Harbor Laboratory, Plainview, NY, 1989); Maniatis et al. (Molecular Cloning, Cold Spring Harbor Laboratory, Plainview, NY, 1982); Glover (Ed.) (DNA Cloning Vol. I and II, IRL Press, Oxford, UK, 1985); Hames and Higgins (Eds.) (Nucleic Acid Hybridization, IRL Press, Oxford, UK, 1985); и в других источниках.
Последовательность ДНК в векторе экспрессии функционально связана по меньшей мере с одной подходящей последовательностью контроля экспрессии (например, конститутивным промотором или регулируемым промотором) для направления синтеза mRNA. Репрезентативные примеры таких последовательностей контроля экспрессии включают промоторы эукариотических клеток или их вирусов, как описано выше. Промоторные области могут быть выбраны из любого необходимого гена с применением векторов CAT (хлорамфениколтрансфераза), векторов канамицина или других векторов с селективными маркерами. Эукариотические промоторы включают немедленно-ранний промотор CMV, промотор тимидинкиназы HSV, ранний и поздний промотор SV40, LTR из ретровируса и мышиный металлотионеин-1. Выбор подходящего вектора и промотора находится в пределах уровня специалиста в данной области, и получение определенных особенно предпочтительных рекомбинантных экспрессирующих конструкций, содержащих по меньшей мере один промотор или регулируемый промотор, функционально связанный с нуклеиновой кислотой, кодирующей белок или полипептид, в соответствии с настоящим изобретением описано в данном документе.
Например, в одном варианте осуществления вектор может представлять собой плазмиду, имеющую структуру, показанную на фиг. 1b. В одном варианте осуществления плазмида может содержать последовательность под SEQ ID NO: 1. В одном варианте осуществления плазмида может состоять из последовательности под SEQ ID NO: 1. В одном варианте осуществления плазмида может содержать последовательность, которая на по меньшей мере приблизительно 60% идентична последовательности под SEQ ID NO: 1, или состоять из нее. В одном варианте осуществления плазмида может содержать последовательность, которая на по меньшей мере приблизительно 85% идентична последовательности под SEQ ID NO: 1 или на по меньшей мере приблизительно 90%, 95%, 96%, 97%, 98%, 99% или более чем 99% идентична SEQ ID NO: 1, или состоять из нее.
Варианты полинуклеотидов по данному изобретению также рассматриваются. Вариантные полинуклеотиды на по меньшей мере 60%, 65%, 70%, 75%, 80%, 85%, 90% и предпочтительно на 95%, 96%, 97%, 98%, 99% или 99,9% идентичны одному из полинуклеотидов определенной последовательности, описанной в данном документе, или которые гибридизуются с одним из этих полинуклеотидов определенной последовательности в жестких условиях гибридизации с 0,015 М хлорида натрия, 0,0015 М цитрата натрия при приблизительно 65-68°С или 0,015 М хлорида натрия, 0,0015 М цитрата натрия и 50% формамида при приблизительно 42°С. Варианты полинуклеотидов сохраняют способность кодировать связывающий домен или его слитый белок, характеризующийся функциональностью, описанной в данном документе.
Термин «жесткие» используется для обозначения условий, которые обычно понимаются в данной области как жесткие. Жесткость гибридизации в основном определяется температурой, ионной силой и концентрацией денатурирующих средств, таких как формамид. Примерами жестких условий для гибридизации и промывки являются 0,015 М хлорида натрия, 0,0015 М цитрата натрия при приблизительно 65-68°С или 0,015 М хлорида натрия, 0,0015 М цитрата натрия и 50% формамида при приблизительно 42°С (см. Sambrook et ai, Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., 1989). Также можно применять более жесткие условия (такие как более высокая температура, более низкая ионная сила, более высокая концентрация формамида или другого денатурирующего средства); однако будет оказываться влияние на скорость гибридизации. В случаях, когда речь идет о гибридизации дезоксиолигонуклеотидов, дополнительные иллюстративные жесткие условия гибридизации включают промывку в 6x SSC, 0,05% пирофосфате натрия при 37°C (для 14-основных олигонуклеотидов), 48°C (для 17-основных олигонуклеотидов), 55°C (для 20-основных олигонуклеотидов) и 60°C (для 23-основных олигонуклеотидов).
Дополнительный аспект данного изобретения относится к клетке-хозяину, трансформированной или трансфицированной или иным образом содержащей любые из полинуклеотидов или векторных/экспрессионных конструкций по данному изобретению. Полинуклеотиды или конструкции для клонирования/экспрессии по настоящему изобретению вводят в подходящие клетки с применением любого способа, известного в данной области, в том числе трансформации, трансфекции и трансдукции. Клетки-хозяева включают клетки субъекта, подвергающиеся клеточной терапии ex vivo, в том числе, например, генной терапии ex vivo. Эукариотические клетки-хозяева рассматриваются как аспект данного изобретения, когда они несут полинуклеотид, вектор или белок в соответствии с данным изобретением в дополнение к собственным клеткам субъекта (например, собственным клеткам пациента), клеткам VERO, клеткам HeLa, клеточным линиям яичников китайского хомячка (СНО) (в том числе модифицированным клеткам СНО, способным модифицировать профиль гликозилирования экспрессируемых мультивалентных связывающих молекул, см. публикацию патентной заявки США № 2003/01 15614), клеткам COS (такие как COS-7), W138, BHK, HepG2, 3T3, RIN, MDCK, A549, PC12, K562, клеткам HEK293, клеткам HepG2, N-клеткам, клеткам 3T3, клеткам Spodoptera frugiperda (например, клетки Sf9), клеткам Saccharomyces cerevisiae и любым другим эукариотическим клеткам, известным в данной области техники в качестве применимых для обеспечения экспрессии и необязательно выделения белка или пептида в соответствии с настоящим изобретением. Также рассмотрены прокариотические клетки, в том числе Escherichia coli, Bacillus subtilis, Salmonella typhimurium, Streptomycete или любая прокариотическая клетка, известная в данной области, которая подходит для обеспечения экспрессии и необязательно выделения белка или пептида в соответствии с настоящим изобретением. В частности, при выделении белка или пептида из прокариотических клеток предполагается, что можно применять известные в данной области способы выделения белка из телец включения. Выбор подходящего хозяина находится в пределах компетенции специалистов в данной области из приведенных в данном документе идей. Рассмотрены клетки-хозяева, которые гликозилируют слитые белки по данному изобретению.
Термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин») относится к клетке, содержащей рекомбинантный вектор экспрессии. Следует понимать, что такие термины предназначены для обозначения не только конкретной клетки-субъекта, но и потомства такой клетки. Поскольку в последующих поколениях могут возникать определенные модификации вследствие мутации или влияния окружающей среды, такое потомство может фактически не быть идентичным родительской клетке, но все же включено в объем термина «клетка-хозяин», используемого в данном документе. Рекомбинантные клетки-хозяева можно культивировать в обычной питательной среде, модифицированной в зависимости от необходимости для активации промоторов, отбора трансформантов или амплификации определенных генов. Условия культивирования для конкретных клеток-хозяев, выбранных для обеспечения экспрессии, такие как температура, рН и тому подобное, будут очевидны для специалиста в данной области. Для экспрессии рекомбинантного белка также можно применять различные системы культивирования клеток млекопитающих. Примеры систем экспрессии млекопитающих включают линии COS-7 фибробластов почек обезьян, описанные Gluzman (1981) Cell 23: 175, и другие линии клеток, способные экспрессировать совместимый вектор, например, C127, 3T3, CHO, HeLa и клеточные линии BHK. Векторы экспрессии млекопитающих будут содержать точку начала репликации, подходящий промотор и необязательно энхансер, а также любые необходимые сайты связывания рибосом, сайт полиаденилирования, сайты донора и акцептора сплайсинга, последовательности терминации транскрипции и 5'-фланкирующие нетранскрибируемые последовательности, например, как описано в настоящем документе в отношении получения поливалентных связывающих белковых экспрессионных конструкций. Последовательности ДНК, полученные в результате сплайсинга SV40, и сайты полиаденилирования можно применять для обеспечения необходимых нетранскрибируемых генетических элементов. Введение конструкции в клетку-хозяина может быть осуществлено посредством различных способов, с которыми знакомы специалисты в данной области, в том числе трансфекции фосфатом кальция, DEAE-декстран-опосредованной трансфекции или электропорации (Davis et al. (1986) Basic Methods in Molecular Biology).
Клетки и композиции
В одном варианте осуществления модифицированные В-клетки, описанные в данном документе, были активированы/дифференцированы in vitro и трансфицированы с обеспечением экспрессии терапевтического средства, описанного в данном документе. В одном варианте осуществления модифицированные В-клетки, описанные в данном документе, были активированы/дифференцированы in vitro и сконструированы (например, с применением подхода нацеленной интеграции трансгена, такого как опосредованная нуклеазой с «цинковыми пальцами», TALEN, мегануклеазой или CRISPR/CAS9 интеграция трансгена) с обеспечением экспрессии терапевтического средства, описанного в данном документе. В одном варианте осуществления композиции содержат В-клетки, которые дифференцировались в плазматические В-клетки, подверглись трансфекции или иным образом сконструированы и экспрессируют один или более белков, представляющих интерес. Популяции клеток-мишеней, такие как трансфицированные или иным образом сконструированные и активированные В-клеточные популяции по настоящему изобретению, можно вводить либо отдельно, либо в виде фармацевтической композиции в комбинации с разбавителями и/или другими компонентами, такими как цитокины или клеточные популяции.
В одном варианте осуществления модифицированные B-клетки, которые были сконструированы с обеспечением экспрессии одного или более белков, представляющих интерес, собирают из культуры после активации/дифференцировки in vitro в момент времени, когда модифицированные B-клетки обладают оптимальной миграционной способностью в отношении конкретного хемоатрактанта. В некоторых вариантах осуществления оптимальная миграционная способность может быть в день 7, 8 или 9 культивирования В-клеток. В некоторых вариантах осуществления оптимальная миграционная способность может быть в день 5, 6 или 7 культивирования В-клеток после трансфекции или конструирования. В некоторых вариантах осуществления оптимальная миграционная способность может быть в день 8 культивирования В-клеток после трансфекции или конструирования или позднее дня 8 культивирования после трансфекции или конструирования (например, день 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или позднее дня 20). В некоторых вариантах осуществления оптимальная миграционная способность может быть до дня 10 культивирования B-клеток. В некоторых вариантах осуществления оптимальная миграционная способность может быть до дня 8 культивирования B-клеток после трансфекции или конструирования. В некоторых вариантах осуществления оптимальная миграционная способность может быть в день 6 или 7 культивирования В-клеток. В некоторых вариантах осуществления оптимальная миграционная способность может быть в день 4 или 5 культивирования В-клеток после трансфекции или культивирования. В некоторых вариантах осуществления оптимальная миграционная способность может быть до дня 9 культивирования B-клеток. В некоторых вариантах осуществления оптимальная миграционная способность может быть до дня 7 культивирования B-клеток после трансфекции или конструирования. В некоторых вариантах осуществления оптимальная миграционная способность является оптимальной для хоуминга модифицированных B-клеток к CXCL12. В некоторых вариантах осуществления оптимальная миграционная способность является оптимальной для хоуминга модифицированных В-клеток в костный мозг субъекта, получающего одно или более введений модифицированных В-клеток. В некоторых вариантах осуществления В-клетки собирают для введения субъекту при оптимальной миграционной способности к CXCL12 и/или к костному мозгу субъекта в период с приблизительно дня 7 до приблизительно дня 9 культивирования. В некоторых вариантах осуществления В-клетки собирают для введения субъекту при оптимальной миграционной способности к CXCL12 и/или к костному мозгу субъекта в период с приблизительно дня 5 до приблизительно дня 7 культивирования после трансфекции или конструирования. В некоторых вариантах осуществления В-клетки собирают для введения субъекту при оптимальной миграционной способности к CXCL12 и/или к костному мозгу субъекта до приблизительно дня 10 культивирования. В некоторых вариантах осуществления В-клетки собирают для введения субъекту при оптимальной миграционной способности к CXCL12 и/или к костному мозгу субъекта до приблизительно дня 8 культивирования после трансфекции или конструирования. В некоторых вариантах осуществления оптимальная миграционная способность является оптимальной для хоуминга модифицированных B-клеток к CXCL13. В некоторых вариантах осуществления оптимальная миграционная способность является оптимальной для хоуминга модифицированных В-клеток в участок воспаления у субъекта, получающего одно или более введений модифицированных В-клеток. В некоторых вариантах осуществления В-клетки собирают для введения субъекту при оптимальной миграционной способности к CXCL13 и/или участку воспаления у субъекта в приблизительно день 6 или приблизительно день 7 культивирования. В некоторых вариантах осуществления В-клетки собирают для введения субъекту при оптимальной миграционной способности к CXCL13 и/или участку воспаления у субъекта в приблизительно день 4 или приблизительно день 5 культивирования после трансфекции или конструирования. В некоторых вариантах осуществления В-клетки собирают для введения субъекту при оптимальной миграционной способности к CXCL13 и/или участку воспаления у субъекта до приблизительно дня 10 культивирования. В некоторых вариантах осуществления В-клетки собирают для введения субъекту при оптимальной миграционной способности к CXCL13 и/или участку воспаления у субъекта до приблизительно дня 8 культивирования после трансфекции или конструирования.
В некоторых вариантах осуществления оптимальная миграционная способность является оптимальной для хоуминга модифицированных B-клеток и к CXCL12, и к CXCL13. В некоторых вариантах осуществления B-клетки собирают при оптимальной миграционной способности для хоуминга к CXCL12 и CXCL13 в день 7 культивирования B-клеток. В некоторых вариантах осуществления B-клетки собирают при оптимальной миграционной способности для хоуминга к CXCL12 и CXCL13 в день 5 культивирования B-клеток после трансфекции и конструирования.
В некоторых вариантах осуществления сконструированные В-клетки собирают, когда по меньшей мере приблизительно 20% В-клеток мигрируют в анализе хемотаксиса к конкретному хемоаттрактанту. Например, без ограничения сконструированные B-клетки (например, которые продуцируют IDUA) можно собирать, когда по меньшей мере приблизительно 20% B-клеток мигрируют в анализе хемотаксиса к CXCL12. Или в другом не ограничивающем примере сконструированные B-клетки (например, которые продуцируют IDUA) можно собирать, когда по меньшей мере приблизительно 20% B-клеток мигрируют в анализе хемотаксиса к CXCL13. Кроме того, сконструированные В-клетки (например, которые продуцируют IDUA) можно собирать, когда по меньшей мере приблизительно 30% В-клеток мигрируют в анализе хемотаксиса к конкретному хемоаттрактанту (например, CXCL12 или CXCL13) или когда по меньшей мере приблизительно 40% 45%, 50%, 55%, 60%, 65% или по меньшей мере приблизительно 70% В-клеток мигрируют в анализе хемотаксиса к конкретному хемоаттрактанту (например, CXCL12 или CXCL13). Кроме того, сконструированные B-клетки (например, которые продуцируют IDUA) можно собирать, когда более 70% B-клеток мигрируют в анализе хемотаксиса. Такие анализы хемотаксиса известны в данной области и описаны в данном документе (см., например, пример 6 в настоящем документе).
Вкратце, композиции на основе клеток по настоящему изобретению могут содержать дифференцированную и активированную популяцию В-клеток, которую подвергли трансфекции и которая экспрессирует терапевтическое средство, описанное в данном документе, в комбинации с одним или более фармацевтически или физиологически приемлемыми носителями, разбавителями или вспомогательными веществами. Такие композиции могут содержать буферы, такие как физиологический раствор с нейтральным буфером, физиологический раствор с фосфатным буфером, раствор Рингера с лактатом и тому подобное; углеводы, такие как глюкоза, манноза, сахароза или декстран, маннит; белки; полипептиды или аминокислоты, такие как глицин; антиоксиданты; хелатирующие средства, такие как EDTA или глутатион; адъюванты (например, гидроксид алюминия) и консерванты. Композиции по настоящему изобретению предпочтительно составлены для внутривенного или подкожного введения.
В одном варианте осуществления клеточную композицию оценивают в отношении чистоты перед введением. В другом варианте осуществления клеточную композицию исследуют в отношении устойчивости продуцирования терапевтического средства. В одном варианте осуществления клеточную композицию исследуют в отношении стерильности. В другом варианте осуществления клеточную композицию подвергают проверке для подтверждения ее соответствия субъекту-реципиенту.
В одном варианте осуществления сконструированную популяцию В-клеток оценивают в отношении поликлональности перед введением субъекту. Обеспечение поликлональности конечного клеточного продукта является важным параметром безопасности. В частности, появление доминантного клона рассматривается как потенциально способствующее in vivo опухолеобразованию или аутоиммунному заболеванию. Поликлональность можно оценивать с помощью любого способа, известного в данной области техники или описанного в данном документе. Например, в некоторых вариантах осуществления поликлональность оценивают путем секвенирования (например, путем глубокого секвенирования) рецепторов В-клеток, экспрессируемых в сконструированной популяции В-клеток. Поскольку рецептор В-клеток претерпевает изменения во время развития В-клеток, что делает его уникальным среди В-клеток, этот способ позволяет количественно определить, какое количество клеток имеют одну и ту же последовательность В-клеточного рецептора (то есть они являются клональными). Таким образом, в некоторых вариантах осуществления, чем больше В-клеток в сконструированной популяции В-клеток, которые экспрессируют одну и ту же последовательность рецептора В-клеток, тем более клональной является популяция и, следовательно, тем менее безопасна популяция для введения субъекту. Наоборот, в некоторых вариантах осуществления, чем меньше В-клеток в сконструированной популяции В-клеток, которые экспрессируют одну и ту же последовательность рецептора В-клеток, тем менее клональной является популяция (т.е. она является более поликлональной) и, следовательно, тем более безопасна популяция для введения субъекту.
В некоторых вариантах осуществления сконструированные В-клетки вводят субъекту после того, как было определено, что они являются достаточно поликлональными. Например, сконструированные В-клетки можно вводить субъекту после того, как будет определено, что ни один конкретный клон В-клеток в конечной популяции не составляет более приблизительно 0,2% от общей популяции В-клеток. Сконструированные В-клетки можно вводить субъекту после того, как будет определено, что ни один конкретный клон В-клеток в конечной популяции не составляет более приблизительно 0,1% от общей популяции В-клеток, или более приблизительно 0,09%, 0,08%, 0,07%, 0,06%, 0,05%, или приблизительно 0,04% от общей популяции В-клеток. В конкретных вариантах осуществления сконструированные B-клетки (например, которые продуцируют IDUA) вводят субъекту после того, как было определено, что ни один конкретный клон B-клеток в конечной популяции не составляет более приблизительно 0,03% от общей популяции B-клеток.
В одном варианте осуществления клеточную композицию хранят и/или транспортируют при 4°С. В другом варианте осуществления клеточную композицию замораживают для хранения и/или транспортировки. Композицию клеток можно замораживать, например, при -20°С или -80°С. В одном варианте осуществления стадия замораживания клеточной композиции предусматривает применение жидкого азота. В одном варианте осуществления клеточную композицию замораживают с применением программируемого криозамораживателя. Соответственно, способы, описанные в данном документе, могут дополнительно предусматривать стадию размораживания.
Способы применения
Один аспект настоящего изобретения направлен на долговременную доставку терапевтического средства in vivo. В конкретных вариантах осуществления модифицированные В-клетки применяют в способах лечения и/или предупреждения хронических заболеваний и нарушений.
Модифицированные В-клетки, описанные в настоящем документе, можно вводить с помощью способа, подходящего для заболевания или нарушения, подлежащего лечению или предупреждению. Количество и частота введения будут определяться такими факторами, как состояние пациента, а также тип и тяжесть заболевания пациента, хотя соответствующие дозы могут быть определены посредством соответствующих клинических испытаний.
В одном варианте осуществления субъекту вводят однократную дозу модифицированных В-клеток. В одном варианте осуществления субъекту последовательно вводят две или более доз модифицированных В-клеток. В одном варианте осуществления субъекту последовательно вводят три дозы модифицированных В-клеток. В одном варианте осуществления дозу модифицированных В-клеток вводят субъекту еженедельно, раз в две недели, ежемесячно, раз в два месяца, ежеквартально, раз в полгода, ежегодно или раз в два года. В одном варианте осуществления вторую или последующую дозу модифицированных B-клеток вводят субъекту, когда количество терапевтического средства, продуцируемого модифицированными B-клетками, уменьшается.
В одном варианте осуществления дозу модифицированных В-клеток вводят субъекту с определенной частотой (например, еженедельно, раз в две недели, ежемесячно, раз в два месяца или ежеквартально) до тех пор, пока у субъекта не будет обнаружено желаемое количество (например, эффективное количество) терапевтического средства. В одном варианте осуществления у субъекта контролируют количество терапевтического средства. В одном варианте осуществления следующую дозу модифицированных B-клеток вводят субъекту, когда количество терапевтического средства, продуцируемого модифицированными B-клетками, меньше желаемого количества. В одном варианте осуществления желаемое количество представляет собой диапазон, который обеспечивает получение желаемого эффекта. Например, в способе уменьшения количества гликозаминогликанов (GAG) у индивидуума с MPS I желаемое количество IDUA представляет собой количество, которое обеспечивает снижение уровня GAG в определенной ткани по сравнению с уровнем GAG в отсутствие IDUA.
Когда указано «эффективное количество», «противоопухолевое эффективное количество», «подавляющее опухоль эффективное количество» или «терапевтическое количество», точное количество композиций по настоящему изобретению для введения может быть определено врачом с учетом индивидуальных различий в возрасте, весе, размере опухоли, степени инфекции или метастазирования и состоянии пациента (субъекта). Композиции на основе В-клеток также можно вводить многократно в подходящей(-их) дозе(-ах). Клетки можно вводить с помощью инфузионных методик, которые обычно известны в иммунотерапии (см., например, Rosenberg et al., New Eng. J. of Med. 319: 1676, 1988).
Специалист в области медицины может определить оптимальную дозировку и режим лечения для конкретного пациента путем мониторинга пациента на наличие признаков заболевания и соответствующей корректировки лечения. Лечение также можно корректировать после измерения уровней терапевтического средства (например, гена или белка, представляющего интерес) в биологическом образце (например, жидкости организма или образце ткани), это также можно применять для оценки эффективности лечения, а лечение может быть скорректировано соответственно в сторону увеличения или уменьшения. Как правило, в связанных исследованиях адоптивной иммунотерапии антигенспецифические Т-клетки вводят пациенту в количестве от примерно 2×109 до 2×1011 клеток. См., например, патент США №5057423).
В некоторых аспектах настоящего изобретения оптимальная доза модифицированных B-клеток в случае схемы с многократными дозами может быть определена сначала путем определения оптимальной концентрации однократной дозы B-клеток для субъекта с уменьшением количества B-клеток, присутствующих при оптимальной концентрации однократной дозы, с обеспечением субоптимальной концентрации однократной дозы модифицированных В-клеток и введения субъекту двух или более доз при субоптимальной концентрации однократной дозы модифицированных В-клеток. В некоторых аспектах субъекту вводят 2, 3 или более доз при субоптимальной концентрации однократной дозы модифицированных В-клеток. В некоторых аспектах введение субъекту 2, 3 или более доз при субоптимальной концентрации однократной дозы модифицированных В-клеток приводит к синергетическому продуцированию in vivo терапевтического полипептида, с обеспечением экспрессии которого сконструированы модифицированные В-клетки. В некоторых аспектах субоптимальная концентрация однократной дозы составляет на 1/2 или в 3, 4, 5, 6, 7, 8, 9, 10 раз меньше, чем оптимальная концентрация однократной дозы. В некоторых аспектах терапевтический полипептид представляет собой IDUA. В некоторых аспектах терапевтический полипептид представляет собой фактор свертывания крови крови человека X (FIX). В некоторых аспектах терапевтический полипептид представляет собой лецитинхолестеринацилтрансферазу человека (LCAT). В некоторых аспектах терапевтический полипептид представляет собой липопротеинлипазу человека (LCAT).
В некоторых аспектах настоящего изобретения можно вводить меньшие количества трансфицированных В-клеток по настоящему изобретению, в диапазоне 106/килограмм (106-1011 на пациента). В определенных вариантах осуществления В-клетки вводят субъекту в концентрации 1×104, 5×104, 1×105, 5×105, 1×106, 5×106, 1×107, 5×107, 1×108, 5×108, 5×109, 1×1010, 5×1010, 1×1011, 5×1011 или 1×1012. Композиции на основе В-клеток можно вводить многократно в дозах в пределах таких диапазонов. Клетки могут быть аутологичными или гетерологичными (например, аллогенными) для пациента, подвергающегося терапии. При необходимости лечение может также включать введение митогенов (например, PHA) или лимфокинов, цитокинов и/или хемокинов (например, GM-CSF, IL-4, IL-6, IL-13, IL-21, Flt3-L, RANTES, MIP1α, BAFF и т.д.), как описано в данном документе, для усиления индуцирования иммунного ответа и приживления инфузированных В-клеток.
Введение рассматриваемых композиций можно осуществлять любым удобным способом, в том числе посредством вдыхания аэрозоля, инъекции, приема внутрь, трансфузии, имплантации или трансплантации. Композиции, описанные в данном документе, можно вводить пациенту подкожно, внутрикожно, внутриопухолево, интранодально, интрамедуллярно, интратекально, внутримышечно, внутривенно (i.v.) или интраперитонеально. Композиции, описанные в данном документе, можно вводить пациенту непосредственно в нервную систему. В одном варианте осуществления композиции на основе В-клеток по настоящему изобретению вводят пациенту путем внутрикожной или подкожной инъекции. В другом варианте осуществления композиции на основе В-клеток, описанные в данном документе, предпочтительно вводят посредством i.v. инъекции. Композиции на основе В-клеток можно вводить непосредственно в опухоль, лимфатический узел, костный мозг или место инфекции.
В еще одном варианте осуществления фармацевтическую композицию можно доставлять с помощью системы с контролируемым высвобождением. В одном варианте осуществления можно применять насос (см. Langer, 1990, Science 249: 1527-1533; Sefton 1987, CRC Crit. Ref. Biomed. Eng. 14: 201; Buchwald et al., 1980; Surgery 88: 507; Saudek et al., 1989, N. Engl. J. Med. 321: 574). В другом варианте осуществления можно применять полимерные материалы (см. Medical Applications of Controlled Release, 1974, Langer and Wise (eds.), CRC Pres., Boca Raton, Fla.; Controlled Drug Bioavailability, Drug Product Design and Performance, 1984, Smolen and Ball (eds.), Wiley, New York; Ranger and Peppas, 1983; J. Macromol. Sci. Rev. Macromol. Chem. 23: 61 ; см. также Levy et al., 1985, Science 228: 190; During et al., 1989, Ann. Neurol. 25: 351 ; Howard et al., 1989, J. Neurosurg. 71: 105). В еще одном варианте осуществления система с контролируемым высвобождением может быть размещена в непосредственной близости от терапевтической мишени, таким образом, требуется только часть системной дозы (см, например, Medical Applications of Controlled Release, 1984, Langer and Wise (eds.), CRC Pres., Boca Raton, Fla., vol. 2, pp. 1 15-138).
Композиции на основе В-клеток по настоящему изобретению также можно вводить с применением любого количества матриц. Матрицы применялись в течение ряда лет в контексте конструирования тканей (см., например, Principles of Tissue Engineering (Lanza, Langer, and Chick (eds.)), 1997. В настоящем изобретении применяют такие матрицы в новом контексте действия как искусственный лимфоидный орган для поддержки и сохранения В-клеток. Соответственно, в настоящем изобретении можно использовать композиции и составы на основе матрицы, которые продемонстрировали применимость в конструировании тканей. Соответственно, тип матрицы, который можно использовать в композициях, устройствах и способах по настоящему изобретению, практически не ограничен и может включать как биологические, так и синтетические матрицы. В одном конкретном примере применяют композиции и устройства, изложенные в патентах США №№5980889; 5913998; 5902745; 5843069; 5787900 или 5626561. Матрицы предусматривают признаки, обычно связанные с биосовместимостью при введении хозяину-млекопитающему. Матрицы можно получать из природных и/или синтетических материалов. Матрицы могут быть небиоразлагаемыми в случаях, когда желательным является введение постоянных структур или съемных структур в организм животного, таких как имплантат; или могут быть биоразлагаемыми. Матрицы могут быть в форме губок, имплантатов, трубок, прокладок из тельфы, волокон, полых волокон, лиофилизированных компонентов, гелей, порошков, пористых композиций или наночастиц. Кроме того, матрицы могут быть сконструированы c обеспечением возможности замедленного высвобождения высеянных клеток или продуцированного цитокина или другого активного средства. В определенных вариантах осуществления матрица по настоящему изобретению является гибкой и эластичной и может быть описана как полутвердый каркас, который является проницаемым для таких веществ, как неорганические соли, водные жидкости и растворенные газообразные средства, в том числе кислород.
Марица, применяемая в данном документе, является примером биоразлагаемого вещества. Однако настоящее изобретение не ограничено матрицами и, таким образом, везде, где появляется термин «матрица» или «матрицы», эти термины должны считаться как включающие устройства и другие вещества, которые обеспечивают удержание клеток или перенос клеток, являются биосовместимыми и обеспечивающими перенос макромолекул либо непосредственно через вещество, в результате чего вещество само по себе представляет собой полупроницаемую мембрану, либо используется совместно с конкретным полупроницаемым веществом.
В определенных вариантах осуществления настоящего изобретения B-клетки, трансфицированные и активированные с применением способов, описанных в настоящем документе, или других способов, известных в данной области техники, вводят пациенту в сочетании с любым количеством соответствующих методов лечения (например, до, одновременно или после), в том числе без ограничения лечения с помощью таких средств, как противовирусные средства, средство химиотерапии, средство лучевой терапии, иммунодепрессанты, такие как циклоспорин, бисульфин, бортезомиб, азатиоприн, метотрексат, микофенолат и FK506, антитела или другие иммуноабляционные средства, такие как САМРАТ, антитела к CD3 или другие средства терапии на основе антител, цитоксин, флударибин, циклоспорин, FK506, рапамицин, микофеноловая кислота, стероиды, FR901228, цитокины и облучение. Такие лекарственные средства подавляют либо кальцийзависимую фосфатазу кальциневрин (циклоспорин и FK506), протеасому (бортезомиб), либо подавляют киназу p70S6, которая важна для передачи сигналов, индуцируемой фактором роста (рапамицин). (Liu et al., Cell 66: 807-815, 1991; Henderson et al., Immun. 73: 316-321, 1991; Bierer et al., Curr. Opin. Immun. 5: 763-773, 1993; Isoniemi (выше)). В дополнительном варианте осуществления композиции на основе клеток по настоящему изобретению вводят пациенту в сочетании с (например, до, одновременно или после) трансплантацией костного мозга, абляционным средством терапии на основе Т-клеток с применением любых химиотерапевтических средств, таких как флударабин, наружная дистанционная лучевая терапия (XRT), циклофосфамид или антитела, такие как OKT3 или CAMPATH. В одном варианте осуществления композиции на основе клеток по настоящему изобретению вводят после средства абляционной терапии на основе B-клеток, такого как средства, которые реагируют с CD20, например, Rituxan®. В одном варианте осуществления композиции на основе клеток по настоящему изобретению вводят после средства абляционной терапии на основе B-клеток с применением средства, такого как бортезомиб. Например, в одном варианте осуществления субъекты могут проходить стандартное лечение посредством химиотерапии с высокой дозой с последующей трансплантацией стволовых клеток периферической крови. В определенных вариантах осуществления после трансплантации субъекты получают инфузию размноженных иммунных клеток по настоящему изобретению. В дополнительном варианте осуществления размноженные клетки вводят до или после хирургического вмешательства.
Дозы вышеупомянутых средств лечения, которые подлежат введению пациенту, будут варьироваться в зависимости от точного характера состояния, подвергаемого лечению, и реципиента, получающего лечение. Масштабирование доз для введения человеку может быть выполнено в соответствии с принятыми в данной области практиками.
Модифицированные В-клетки можно применять для лечения или предупреждения различных инфекционных заболеваний, разновидностей рака, дегенеративных заболеваний и иммунологических нарушений.
Композиции, содержащие модифицированные В-клетки, описанные в настоящем документе, можно применять для лечения любого из множества инфекционных заболеваний, вызываемых инфекционными организмами, такими как вирусы, бактерии, паразиты и грибки. Инфекционные организмы могут включать вирусы (например, РНК-вирусы, ДНК-вирусы, вирус иммунодефицита человека (HIV), вирус гепатита A, B и C, вирус простого герпеса (HSV), цитомегаловирус (CMV), вирус Эпштейна-Барр (EBV), вирус папилломы человека (HPV), паразиты (например, простейшие и метазойные патогены, такие как виды Plasmodia, виды Leishmania, виды Schistosoma, виды Trypanosoma), бактерии (например, микобактерии, в частности, M. tuberculosis, Salmonella, Streptococci, E. coli, Staphylococci), грибы (например, виды Candida, виды Aspergillus), Pneumocystis carinii и прионы (известные прионы заражают животных, вызывая скрепи, трансмиссивное, дегенеративное заболевание нервной системы овец и коз, а также губчатую энцефалопатию крупного рогатого скота (BSE), или «коровье бешенство», и кошачью губчатую энцефалопатию у кошек). Четыре прионных заболевания, которые, как известно, поражают человека, представляют собой (1) куру, (2) болезнь Крейтцфельда-Якоба (CJD), (3) болезнь Герстмана-Штраусслера-Шейнкера (GSS) и (4) фатальная семейная бессонница (FFI)). Используемый в данном документе термин «прион» включает все формы прионов, вызывающие все или некоторые из этих заболеваний или другие заболевания у любых используемых животных и, в частности, у людей и домашних сельскохозяйственных животных. Иллюстративные инфекционные заболевания включают без ограничения токсоплазмоз, гистоплазмоз, CMV, EBV, кокцидиомикоз, туберкулез, HIV и тому подобное.
В определенных вариантах осуществления композиции на основе модифицированных В-клеток, описанные в настоящем документе, также можно применять для предупреждения или лечения различных видов рака. В связи с этим в некоторых вариантах осуществления композиции, содержащие трансфицированные В-клетки, применимы для предупреждения или лечения меланомы, неходжкинской лимфомы, болезни Ходжкина, лейкемии, плазмоцитомы, саркомы, глиомы, тимомы, рака молочной железы, рака простаты, колоректального рака, рака почки, почечно-клеточного рака, рака матки, рака поджелудочной железы, рака пищевода, рака головного мозга, рака легкого, рака яичника, рака шейки матки, рака яичка, рака желудка, рака пищевода, множественной миеломы, гепатомы, острого лимфобластного лейкоза (ALL), острого миелогенного лейкоза (AML), хронического миелогенного лейкоза (CML) и хронического лимфоцитарного лейкоза (CLL) или других видов рака.
В одном варианте осуществления модифицированные В-клетки также можно применять для лечения иммунологических нарушений, таких как синдром приобретенного иммунодефицита (AIDS), агаммаглобулинемия, гипогаммаглобулинемия, других иммунодефицитов, иммуносупрессии и тяжелого комбинированного иммунодефицита (SCID).
В одном варианте осуществления модифицированные В-клетки, описанные в настоящем документе, также можно применять для лечения аутоиммунных заболеваний, таких как без ограничения ревматоидный артрит, рассеянный склероз, инсулинозависимый диабет, болезнь Аддисона, целиакия, синдром хронической усталости, воспалительные заболевания кишечника, язвенный колит, болезнь Крона, фибромиалгия, системная красная волчанка, псориаз, синдром Шегрена, гипертиреоз/болезнь Грейвса, гипотиреоз/болезнь Хашимото, инсулинозависимый диабет (тип 1), миастения гравис, эндометриоз, склеродермия, пернициозная анемия, синдром Гудпасчера, болезнь Вегенера, гломерулонефрит, апластическая анемия, пароксизмальная ночная гемоглобинурия, миелодиспластический синдром, идиопатическая тромбоцитопеническая пурпура, аутоиммунная гемолитическая анемия, синдром Эванса, синдром ингибитора фактора VIII, системный васкулит, дерматомиозит, полимиозит и ревматизм. Таким образом, в одном варианте осуществления способы в настоящем документе включают способы лечения заболевания, предусматривающие введение субъекту или пациенту, нуждающемуся в этом, терапевтически эффективного количества композиций, содержащих модифицированные В-клетки, описанные в настоящем документе, с осуществлением тем самым лечения заболевания.
В одном варианте осуществления модифицированные В-клетки, описанные в настоящем документе, также можно применять для лечения заболеваний и нарушений, связанных с дефицитом ферментов, таких как без ограничения MPS I, MPS II, MPS III, MP IV, MPS V, MPS VI, MPS VII, лизосомные нарушения накопления, болезнь Нимана-Пика (типы A, B и C), болезнь Гуахера (типы I, II и III), болезнь Тея-Сакса и болезнь Помпе.
В одном варианте осуществления предусмотрен способ лечения MPS I у индивидуума, предусматривающий введение В-клетки, генетически модифицированной с обеспечением экспрессии IDUA (IDUA+ B-клетки), субъекту, у которого имеется или предположительно имеется MPS I. В одном варианте осуществления субъекту вводят однократную максимально эффективную дозу IDUA+ B-клеток. В другом варианте осуществления субъекту вводят две или более доз IDUA+ B-клеток, тем самым обеспечивая увеличение до максимума количества приживленных IDUA+ B-клеток. В некоторых вариантах осуществления две или более доз IDUA+ B-клеток, которые вводят субъекту, содержат меньше IDUA+ B-клеток, чем однократная максимально эффективная доза IDUA+ B-клеток. В некоторых вариантах осуществления, когда две или более доз IDUA+ B-клеток вводят субъекту при дозировке IDUA+ В-клеток, которая ниже максимально эффективной однократной дозы IDUA+ B-клеток, в результате происходит синергетическое увеличение продуцирования IDUA. В одном варианте осуществления введение IDUA+ В-клеток субъекту приводит к нормальным уровням IDUA, наблюдаемым у здорового контрольного субъекта. В одном варианте осуществления введение IDUA+ В-клеток субъекту приводит к превышению нормальных уровней IDUA у субъекта. В одном варианте осуществления введение IDUA+ В-клеток субъекту приводит к снижению уровней GAG у субъекта до нормального уровня. В одном варианте осуществления введение IDUA+ В-клеток субъекту приводит к снижению уровней GAG у субъекта до уровня, который ниже нормального уровня GAG у субъекта.
ПРИМЕРЫ
ПРИМЕР 1
Получение B-клеток, экспрессирующих IDUA
Получали конструкции транспозона и транспозазы Sleeping Beauty для транспозиции и экспрессии IDUA человека. Транспозоны, собранные для достижения интеграции и экспрессии гена IDUA в В-клетках, показаны на фигуре 1. Авторы настоящего изобретения применяли промотор EEK, состоящий из промоторных и энхансерных элементов из гена иммуноглобулина человека, а также других регуляторных элементов, описанных ранее, для достижения высокого уровня экспрессии в В-клетках. Для исследования транспозиции и экспрессии IDUA В-клетки человека выделяли от двух отдельных доноров и размножали в культуре с осуществлением электропорации в день 2 с помощью pKT2/EEK-IDUA плюс pCMV-SB100x. Клеточные лизаты, полученные через 8 дней после электропорации, характеризовались активностью IDUA приблизительно 60 нмоль/ч./мг, что приблизительно в 50 раз превышает уровень IDUA, обнаруживаемый в клетках дикого типа, что демонстрирует эффективность системы транспозонов SB для достижения высокого уровня экспрессии IDUA в размноженных B-клетках человека (фигура 2).
Для обогащения клеток, экспрессирующих IDUA, авторы также создали бифункциональный транспозон (pKT2/EEK-IDUA-DHFR, фигура 1), кодирующий IDUA человека наряду с дигидрофолатредуктазой человека, синтезированной для кодирования нового варианта фермента (L22Y, F31S), который является устойчивым к антагонисту фолата метотрексату (MTX: (McIvor RS. 1996. Bone Marrow Transplantation 18:S50-54.)). Плазмидная карта для этой конструкции показана на фиг. 1B, и ее последовательность представлена под SEQ ID NO: 1.
Авторы настоящего изобретения также разработали методику с применением коммерчески доступного антитела к IDUA человека для идентификации клеток, экспрессирующих высокие уровни IDUA, посредством пермебиализации и внутриклеточного окрашивания с последующей проточной цитометрией. С применением транспозона GFP-DHFR, сходного с pKT2/EEK-IDUA-DHFR, авторы сначала установили условия для селективного роста транспозированных B-клеток путем инкубации при нескольких различных концентрациях MTX в дни 2-4 клеточного размножения. Авторы настоящего изобретения обнаружили, что концентрация 200 нМ обеспечивала наиболее эффективные условия для селективного роста В-клеток, экспрессирующих репортерный трансген GFP (таблица 1). Затем авторы настоящего изобретения применили эти условия в отношении размножения подвергнутых электропорации клеток pKT2/EEK-IDUA-DHFR + pCMV-SB100x с оценкой % IDUA+ клеток путем внутриклеточного окрашивания в день 7. В то время как частота популяции IDUA+ клеток, составляющая 10-12%, наблюдалась в популяции транспозированных клеток, эта частота увеличивалась до 25% или больше путем применения отбора посредством MTX (фигура 3).
Таблица 1. Условия отбора с применением метотрексата*

*% GFP-положительных клеток в день 7 после отбора с помощью MTX в дни 2-4
Исходные исследования in vivo начинали с использованием мышиной линии MPS I, которую подвергали обратному скрещиванию с NOD-SCID, но не обнаружили никаких доказательств сохранения B-клеток через неделю после их инфузии этой линии. Эта проблема была в конечном итоге решена путем скрещивания мышей NSG-MPS I с нокаутным аллелем гамма-C-рецептора IL-2, как описано ниже в примере 2, но в то же время авторы настоящего изобретения исследовали адоптивный перенос отобранных с помощью MTX IDUA+ В-клеток (как показано на фигуре 3) у IDUA+ мышей NSG. Аутологичные клетки периферической крови обогащали CD4+ и вводили инфузией интраперитонеально (i.p.) в дни -30 и -4 для обеспечения поддержания трансплантированных В-клеток pKT2/EEK-IDUA-DHFR, введенных либо i.p., либо внутривенно в день 0. Авторы наблюдали у животных, которым вводили как i.p., так и i.v. В-клетки, экспрессирующие IDUA, IDUA в плазме крови в диапазоне от уровня дикого типа до уровня, в 200 раз превышающего уровень дикого типа. (Фигура 4). Авторы настоящего изобретения также наблюдали чрезвычайно высокие уровни иммуноглобулина человека (в среднем от 1 до 4 мг/мл) в плазме крови - убедительное доказательство успешного адоптивного переноса B-клеток человека. Несмотря на отсутствие таких экспериментов с животными, которые характеризуются дефицитом IDUA, результаты этого эксперимента, тем не менее, дают пример уровней IDUA человека, которые могут быть достигнуты после введения высокоэффективной популяции IDUA+ В-клеток мышам NSG.
ПРИМЕР 2
Получение IDUA in vivo у мышей MPS I
С целью определить, является ли линейной взаимосвязь между количеством введенных модифицированных В-клеток и количеством полученного терапевтического средства, в мышиной модели мукополисахаридоза типа I (MPS I) применяли аллогенные В-клетки, генетически модифицированные с обеспечением экспрессии идуронидазы (IDUA).
Мышей NSG (NOD-SCID с дефицитом гамма-C) скрещивали с мышами NOD-SCID с дефицитом IDUA с целью собрать аллели с дефицитом гамма-C и IDUA в одной и той же линии и получить мышей NSG MPS I, также называемых в настоящем документе «мыши MPS I». После получения достаточного количества мышей NSG MPS I этим животным вводили i.p. 3×106 CD4+ Т-клеток в день -7, а затем 3×106, 1×107 или 3×107 транспозированных pKT2/EEK-IDUA В-клеток (примерно 10% IDUA+ согласно внутриклеточному окрашиванию) i.v. в день 0. Мышам MPS I давали однократную дозу В-клеток, сконструированных с обеспечением продуцирования IDUA, (или отсутствие клеток в качестве контроля) в присутствии CD4+ Т-клеток памяти (или отсутствие клеток в качестве контроля) и уровней активности фермента IDUA, измеренных в сыворотке крови до дня 38 (фигура 5) с применением протокола анализа фермента IDUA, ранее описанного Hopwood JJ, et al., Clin Chim Acta. 1979 Mar 1; 92(2): 257-65, содержание которой включено в данный документ посредством ссылки во всей своей полноте. Поскольку у мышей отсутствуют факторы, необходимые для долговременного выживания В-клеток, CD4+ Т-клетки памяти вводили мышам интраперитонеально также для того, чтобы способствовать выживанию В-клеток.
Неожиданно эти результаты продемонстрировали, что связь между количеством клеток и уровнями IDUA в сыворотке крови не была линейной при определенных дозах клеток. Без ограничения какой-либо теорией, это может быть связано с тем, что В-клетки взаимодействуют друг с другом in vivo, что приводит к различным результатам выживаемости при разных концентрациях клеток. Таким образом, эти результаты указывают на то, что существует идеальная доза, которая обеспечивает достаточное продуцирование терапевтического средства при введении наименьшего количества необходимых клеток. Эта концепция проиллюстрирована на фигуре 5, где показано, что доза клеток 3×107 привела к более чем 3-кратному увеличению уровней активности IDUA в плазме крови по сравнению с дозой 1×107 клеток.
ПРИМЕР 3
Многократные дозы B-клеток, продуцирующих IDUA, у мышей MPS I
С целью определить, влияет ли доза B-клеток на количество терапевтического средства, продуцируемого in vivo, мышам MPS I вводили серию из 3 доз B-клеток, продуцирующих IDUA.
Мышам MPS I давали серию из 3 доз 1×107 В-клеток, сконструированных с обеспечением продуцирования IDUA, (или отсутствие клеток в качестве контроля) в присутствии CD4+ Т-клеток памяти (или отсутствие клеток в качестве контроля) в день 0 и измеряли уровни активности фермента IDUA в сыворотке крови до дня 56 (фигура 6). В частности, мышам MPS I вводили i.p. 3×106 CD4+ T-клеток в день -7, а затем 107 транспозированных pKT2/EEK-IDUA B-клеток (примерно 10% IDUA+ согласно внутриклеточному окрашиванию) либо i.p., либо i.v. в день 0. Животным вводили дополнительные инфузии 107 транспозированных pKT2/EEK-IDUA В-клеток тем же путем введения в дни 21 и 42 после первой инъекции.
С применением такой процедуры авторы настоящего изобретения обнаружили, что уровни IDUA дикого типа в плазме крови (приблизительно 1 нмоль/ч./мл) достигались у большинства животных, обработанных В-клетками, и что для этого требовалось предварительное введение CD4+ Т-клеток (фигура 6). Авторы настоящего изобретения обнаружили, что уровни IgG человека в плазме крови варьировались от 200 мкг/мл до 1 мг/мл, что служит доказательством адоптивного переноса В-клеток (фигура 7).
Эти результаты демонстрируют, что введение одинакового количества модифицированных В-клеток в виде нескольких доз приводит к более высоким конечным уровням терапевтического средства по сравнению с результатами, которые достигаются путем введения всех или модифицированных В-клеток в однократной дозе. Как видно из фигуры 6, начальные уровни IDUA после первой дозы 1×107 B-клеток были низкими по сравнению с уровнями, достигнутыми после введения третьей дозы, что неожиданно привело к тому, что уровни IDUA в сыворотке крови значительно превышали 3-кратные уровни IDUA в сыворотке крови, наблюдаемые после введения первой дозы. Это явление также можно наблюдать при сравнении данных при 1×107 клеток/мышь на фигуре 5 с данными на фигуре 6, поскольку каждая доза составляет 1×107. Хотя на фигуре 5 показано, что однократная доза 1×107 клеток, введенных внутривенно, приводила к снижению уровней IDUA с помощью D38, на фигуре 6 ясно, что уровни IDUA, являющиеся результатом 3 доз 1×107 клеток, введенных внутривенно, приводили к уровням экспрессии, которые продолжали как значительно увеличиваться, так и значительно превышать 3-кратный уровень однократной дозы 1×107 и уровень однократной дозы 3×107 в один и тот же момент времени. Неожиданно этот синергизм среди многократных доз наблюдался только у этих групп мышей, которым внутривенно вводили сконструированные В-клетки, но не у мышей, которые получали сконструированные В-клетки посредством интраперитонеальной инъекции.
В дополнение к демонстрации того, что многократные дозы приводили к тому, что уровни терапевтического средства были выше, чем ожидалось от однократной дозы одного и того же количества клеток, простое представление о том, что многократные дозы приводили к более высоким уровням терапевтического средства, является преимущественным и неожиданным. Что касается механизма действия, считается, что дифференцированные В-клетки занимают предельный уровень ниш выживания, и когда возникают новые дифференцированные клетки, они вытесняют некоторые из старых клеток. Следовательно, можно ожидать, что последующие дозы B-клеток не приведут к сопутствующему увеличению уровней терапевтического средства в сыворотке крови. Однако авторы показали, что это не тот случай и что дополнительные инфузии В-клеток приводят к более высоким равновесным уровням терапевтического средства в плазме крови, которые они обеспечивают. Это явление применимо в случае правильной дозировки терапевтического лекарственного средства in vivo при требуемой минимизации дозы клеток.
В частности, пациентам можно вводить дозу, а затем измерять уровни лекарственного средства в плазме крови. В случае, если уровни ниже желаемых, можно вводить дополнительную дозу клеток. Учитывая средний период полужизни большинства инъекционных биопрепаратов, эти результаты, безусловно, не применимы к прямой инфузии биопрепарата. Кроме того, эти результаты вряд ли будут достижимы с применением других способов, таких как вирусная доставка лекарственных средств, которая также может вызывать иммунный ответ на вектор, тем самым снижая эффективность/препятствуя будущим попыткам введения дополнительной дозы.
Соответственно, способы доставки терапевтического средства, раскрытые в данном документе, обеспечивают преимущества по сравнению со способами предшествующего уровня техники, включая возможность введения более чем одной дозы модифицированных В-клеток и суммирование доз, что происходит в результате этого.
ПРИМЕР 4
Доставка во множество тканей с использованием B-клеток
Все из прямой инфузии терапевтического средства, доставки посредством вирусных векторов и переноса гематопоэтических стволовых клеток не могут обеспечить успешную доставку терапевтического средства во множество тканей. С целью определить, можно ли применять модифицированные B-клетки для доставки терапевтического средства in vivo во множество тканей, мышам MPS I вводили серии из 3 доз B-клеток, продуцирующих IDUA, как описано в предыдущем примере.
В соответствии с протоколом, описанным в предыдущем примере, мышам MPS I вводили дозы согласно схеме, включающей 3 дозы 1×107 B-клеток, сконструированных с обеспечением продуцирования IDUA, (или отсутствие клеток в качестве контроля) в присутствии CD4+ T-клеток (или отсутствие клеток в качестве контроля). Животных подвергали эвтаназии и собирали ткани через 60 дней после первой инфузии B-клеток, а уровни активности фермента IDUA измеряли в печени, легком, селезенке, почке, кишечнике, мышце, головном мозге, сердце, перитонеальном смыве и костном мозге (фигура 8).
Проточная цитометрия показала от 20% до 35% CD 45+ клеток человека и от 2% до 10% CD19+ клеток в селезенке и лимфатических узлах животных, которым вводили инфузией Т-клетки и В-клетки человека, экспрессирующие и IDUA. Наблюдали существенную метаболическую перекрестную коррекцию по восстановленным показателей активности IDUA в периферических тканях и даже в головном мозге фигура 8).
IDUA разрушает GAG, а количество присутствующих GAG увеличивается в отсутствие IDUA, например, в тканях мышей MPS I. Соответственно, уровни гликозаминогликанов (GAG) также измеряли в головном мозге, легком, печени, сердце, почке, мышце, селезенке и кишечнике в день 60 с целью определить, разрушает ли IDUA GAG в этих тканях (фигура 9). В результате метаболической перекрестной коррекции, вызванной IDUA, уровни гликозаминогликанов в тканях были значительно снижены у животных, обработанных В-клетками, которым также вводили инфузией CD4+ Т-клетки (фигура 9).
Эти результаты продемонстрировали, что доставка IDUA посредством модифицированных В-клеток обеспечивает как усиленную доставку терапевтических средств в ткани, так и активность терапевтических средств в этих тканях. На фигуре 8 показаны уровни активности фермента IDUA в тканях в результате инфузии В-клеток, продуцирующих IDUA. На фигуре 9 показаны уровни GAG у тех же мышей. GAG являются токсичными клеточными продуктами, которые накапливаются в тканях мышей MPS I и которые разрушаются с помощью ферментативной активности IDUA. Как можно видеть на обеих фигурах, продуцирование IDUA и снижение уровня GAG эффективно происходили во множестве тканей, включая легкое, селезенку, печень, сердце и кишечник. Следует отметить, что инфузия IDUA, как считается, не позволяет адекватно бороться с проявлениями заболевания в таких тканях, как сердце, селезенка и печень. Эти результаты впервые продемонстрировали эффективность человеческих B-клеток, экспрессирующих IDUA, для метаболической коррекции MPS I. Таким образом, полученные данные подтверждают, что доставка фермента in vivo с помощью генетически модифицированных В-клеток характеризуется тенденцией к усилению лечения различных органов в организме.
ПРИМЕР 5
Долгосрочная эффективность доставки терапевтического средства на основе B-клеток
С целью определить, приводит ли доставка терапевтических средств на основе B-клеток к долгосрочному продуцированию белка, авторы настоящего изобретения проанализировали показатели долгосрочной активности IDUA в плазме крови и тканях мышей NSG MPSI, которые получили B-клетки, продуцирующие IDUA.
В-клетки, сверхэкспрессирующие IDUA, получали, как описано ранее. Животным NSG MPS I интраперитонеально вводили инфузией 3e6 CD4+ T-клеток памяти за одну неделю перед инфузиями B-клеток. Затем животным вводили инфузией 2е7 В-клеток, экспрессирующих IDUA, в дни 0 и 30.
Сначала авторы настоящего изобретения анализировали активность фермента IDUA в плазме крови у мышей NSG MPSI, которые получили В-клетки, продуцирующие IDUA, как описано в предыдущих примерах. Необработанных мышей NSG MPSI также анализировали в течение этого периода в качестве контроля. У этих животных собирали кровь каждые 2 недели в течение 6,5 месяца и анализировали в отношении ферментативной активности IDUA, как описано ранее.
Как показано на фигуре 10, активность IDUA в плазме крови сильно индуцировалась у мышей, получающих сконструированные B-клетки, при этом максимальные ответы в плазме крови имели место через около 5 недель после инфузии, и активность IDUA у этих животных оставалась более высокой до момента умерщвления через 6,5 месяца после инфузии.
Более того, долговременная обработка с помощью продукта на основе В-клеток также приводила к в результате к длительному повышению уровней IDUA во множестве тканей, анализированных через 3, 6 и 6,5 месяца после инфузии с помощью сконструированных В-клеток (фигура 11), что демонстрирует то, что усиленная доставка в ткани IDUA, которую авторы наблюдали в вышеприведенном примере 4, не просто временное явление, а сохраняется в течение длительного времени.
Кроме того, через 3 месяца, 6 месяцев и 6,5 месяца после первой инфузии В-клеток животных подвергали эвтаназии и анализировали экстракты из тканей в отношении уровней GAG, как описано в примере 4, и наблюдали показатели долговременного снижения уровней GAG в множестве тканей (фиг. 12). В качестве положительного контроля снижения уровня GAG анализировали мышей NSG-IDUA+/- (которые гетерозиготны по IDUA и являются фенотипически нормальными). В качестве отрицательного контроля снижения уровня GAG группа мышей NSG MPSI не получала клеток.
Таким образом, эти данные показывают, что доставка терапевтических средств посредством модифицированных В-клеток обеспечивает длительное повышение уровней активности фермента in vivo не только в плазме крови, но и в тканях, таких как сердце, селезенка и печень, которые обычно не поддаются лечению с помощью инфузии IDUA.
ПРИМЕР 6
Оптимизация миграционной способности сконструированных B-клеток
Общепринято считается, что для того, чтобы плазматические клетки выживали в течение длительного времени, клетки-предшественники (то есть плазмабласты) должны обладать способностью мигрировать в ниши долгосрочного выживания плазматических клеток, обнаруживаемые в таких областях, как костный мозг. Напротив, общепринято считается, что, как только плазматические клетки завершают дифференцировку, у них снижается их способность мигрировать. Следовательно, при намерении создания популяции плазматических клеток из инфузированной популяции плазмабластов считается важным, чтобы клетки обладали сильной миграционной способностью. В частности, миграция в направлении CXCL12 важна для миграции предшественников плазматических клеток в костный мозг. Кроме того, хемокины, такие как CXCL13, могут быть важны для миграции в места воспаления и ткани, такие как селезенка.
С целью определить, способствуют ли условия культивирования получению мигрирующих В-клеток, и определить, зависит ли миграционная способность В-клеток, сконструированных с обеспечением экспрессии терапевтического белка, от количества времени, в течение которого сконструированные В-клетки остаются в культуре после конструирования, но до сбора для введения субъекту, авторы настоящего изобретения получали В-клетки, сверхэкспрессирующие IDUA, как описано ранее, и авторы поддерживали сконструированные В-клетки в культуре в течение периода четырех-девяти дней перед анализированием их миграционной способности.
Анализ проводили с применением двухкамерных культуральных сосудов, в которых камеры соединены (например, планшет Transwell®). Сконструированные B-клетки высевали в одну камеру, а в другую камеру загружали 100 нг/мл CXCL12, который представляет собой хемоаттрактант, который привлекает B-клетки в костный мозг, или CXCL13, который представляет собой хемоаттрактант, который привлекает B-клетки к участкам воспаления, а также к тканям, таким как селезенка. После обеспечения мигрирования В-клеток в течение 3 часов, В-клетки собирали из второй лунки и подсчитывали. В каждом анализе применяли отрицательный контроль, для которого во вторую камеру не добавляли хемоатрактанта (CXCL12 или CXCL13). Схема анализа в отношении CXCL12 представлена на фигуре 13D. Тот же самый анализ применяли для CXCL13.
Исследуемые группы представляли собой В-клетки, которые подвергали воздействию системы культивирования, которая, как авторы настоящего изобретения показали ранее, обеспечивает значительное повышение миграционной способности сконструированных В-клеток (см. PCT/US2015/066908, полностью включенную в настоящий документ посредством ссылки). Такая система культивирования, которую применяли для всех описанных в данном документе экспериментов (если не указано иное), включает CD40L (HIS-меченый для мультимеризации), сшивающее средство CD40L (антитело к HIS, которое индуцирует мультимеризацию CD40L), IL-2, IL-4, IL-10, IL-15 и IL-21. В-клетки культивировали в течение соответствующего количества дней после конструирования до анализа Transwell.
Неожиданно авторы настоящего изобретения обнаружили, что В-клетки, сконструированные в предложенных ими условиях культивирования, обладают оптимальным миграционным потенциалом в направлении CXCL12, который достигает максимума через около 7-9 дней (в культуре или через 5-7 дней после конструирования), и оптимальным миграционным потенциалом в направлении CXCL13, который достигает максимума через около 6-7 дней (в культуре или через около 4-5 дней после конструирования) (фигуры 13A-C). Эти данные дают основание предполагают, что в некоторых случаях может быть преимущественным собирать и вводить сконструированные В-клетки субъекту в пределах этого окна, чтобы обеспечить оптимальный миграционный потенциал и индуцировать оптимальную миграцию в ткани-мишени.
ПРИМЕР 7
Оценка клональности конечных B-клеток, сконструированных с обеспечением экспрессии IDUA
Обеспечение поликлональности конечного продукта на основе клеток является важным параметром безопасности. В частности, появление доминантного клона рассматривается как потенциально способствующее in vivo опухолеобразованию или аутоиммунному заболеванию. Для оценки конечного продукта на основе В-клеток клетки выращивали в культуре, как описано ранее (см. условия культивирования, представленные в примере 6), конструировали с обеспечением экспрессии IDUA с применением способа, описанного в примере 1, собирали в день 7 культивирования и подвергали криоконсервации. Выделяли ДНК и подвергали высокопроизводительному глубокому секвенированию В-клеточного рецептора. Поскольку рецептор В-клеток претерпевает изменения во время развития В-клеток, что делает его уникальным среди В-клеток, этот способ позволяет количественно определить, какое количество клеток имеют одну и ту же последовательность В-клеточного рецептора (то есть они являются клональными).
Результаты глубокого секвенирования показаны на фигуре 14 и в таблице 2 ниже. Проанализировали более 248000 молекул-матриц и обнаружили около 241000 уникальных перестроек, представляющих максимальную частоту клональности (т.е. распространенность какого-либо конкретного клона В-клеток), составляющую 0,03%.
Таблица 2 Сводная информация в отношении секвенирования при секвенировании клональности В-клеток

Этот эксперимент демонстрирует, что конечный продукт на основе В-клеток является высокополиклональным, и что ни один клон не представляет значительной части клеточной популяции. В частности, результаты секвенирования демонстрируют, что ни один клон не представляет более 0,03% конечной популяции. Таким образом, ожидается, что конечные сконструированные В-клетки будут достаточно поликлональными для терапевтических целей и, по-видимому, не содержать какого-либо преобладающего конкретного клона. Поскольку вероятно, что условия культивирования, модификации генома и присутствие посторонних факторов (например, эмбриональная бычья сыворотка, присутствующая в питательных средах) могут вызывать появление клональности, авторы настоящего изобретения рассматривают этот результат как неожиданный по причине того, что ни один клон не достиг значительной встречаемости.
ПРИМЕР 8
Воспалительный потенциал сконструированных В-клеток.
Как и большинство терапевтических средств, терапевтическое средство по настоящему изобретению характеризуется потенциалом стимуляции иммунной системы, что может потенциально привести к неблагоприятным побочным эффектам и/или нейтрализации антител, что может снизить или устранить эффективность. Одним из применимых прогностических параметров того, может ли быть вызвана такая неблагоприятная иммунная реакция, является то, продуцируют ли конечные продукты на основе сконструированных В-клеток воспалительные цитокины. Таким образом, авторы настоящего изобретения измеряли, продуцирует ли конечный продукт культивирования на основе В-клеток (В-клеток, сконструированных с обеспечением продуцирования IDUA в соответствии с приведенными выше примерами) какой-либо из различных воспалительных цитокинов.
В-клетки культивировали в течение 7 дней в культуре, как описано ранее (см. условия культивирования, представленные в примере 6), конструировали с обеспечением экспрессии IDUA с применением способа, описанного в примере 1, собирали в день 7 и подвергали криоконсервации. В течение множества моментов времени во время культивирования отбирали образцы сред и анализировали в отношении панели цитокинов с применением устройства Luminex. В частности, анализировали в отношении наличия следующих факторов: IL6, IFN-альфа, IFN-гамма, sFAS, TNFRp75, BAFF, HGF, IL5, IL2R-альфа, TNF-альфа, IL1ra, TNFRp55, VEGF, IL-1-альфа, sIL6R. Поскольку состав среды включает в себя FBS, который может содержать эти факторы, среду без В-клеток применяли как отрицательный контроль. В случае IL6 рекомбинантный белок (в некоторых случаях) добавляли в среду без В-клеток, чтобы выступал в качестве положительных контролей.
Удивительно, что в большинстве случаев к концу культивирования В-клеток исследуемый фактор либо не определялся, либо его уровень не превышал таковой в контрольных средах (фигуры 15А и В). Этот результат был неожиданным, учитывая, что сконструированные клетки являются иммунными клетками, подвергающимися значительной стимуляции в процессе культивирования и конструирования. Исключениями этого были IL6 и sFAS (фигура 15A, первая группа слева, и фигура 15B, первая группа слева). Единственный фактор, который, как обнаружено, увеличивался во время культивирования, представлял собой sFAS. В случае IL6, хотя повышенные уровни были обнаружены в день 2 культивирования, к дню 7 уровни IL6 были очень близки к уровням, обнаруженным в средах отдельно.
В целом, эти результаты иллюстрируют, что конечный продукт культивирования на основе В-клеток не продуцирует значительных уровней исследуемых воспалительных цитокинов. Учитывая, что авторы настоящего изобретения обеспечивали ряд стимулирующих цитокинов В-клеткам, было неожиданным, что конечный продукт не продуцирует значительных уровней этих цитокинов. Совершенно неожиданно авторы настоящего изобретения обнаружили, что со временем B-клетки снижали свое продуцирование IL6 до уровней, близких к фоновым. Это очень важно для клинической реализации, поскольку известно, что IL-6 является мощным иммуностимулирующим сигналом. В целом, авторы настоящего изобретения полагают, что эти результаты отражают, что конечный продукт на основе B-клеток, как ожидается, будет безопасным in vivo с точки зрения потенциала, позволяющего избежать значительной стимуляции иммунной системы в целом.
ПРИМЕР 9
Получение B-клеток, экспрессирующих LCAT или FIX человека
Конструкции транспозона и транспозазы Sleeping Beauty для транспозиции и экспрессии LCAT человека, LPL человека и FIX человека получали, как описано в примере 1, и трансфицировали первичные В-клетки памяти посредством электропорации в день 2 в культуре. Среду собирали через 2 дня после электропорации (день 4 в культуре, «D4»), а также через 6 дней после электропорации в случае LCAT и LPL (день 8 в культуре, «D8») и анализировали в отношении экспрессии.
Экспрессию LCAT в трансфицированных В-клетках подтверждали с применением анализа активности фермента LCAT на основе флуоресценции (см. способ ниже). Среда, собранная от трансфицированных клеток, характеризовалась сильной активностью LCAT как в D4, так и в D8, тогда как не наблюдалась значительной активности только в контрольной среде (фигура 16A).
Экспрессию LPL в трансфицированных В-клетках подтверждали с применением анализа активности фермента LPL на основе флуоресценции (см. способ ниже). Среда, собранная от трансфицированных клеток, характеризовалась сильной активностью LPL как в D4, так и в D8, тогда как не наблюдалась значительной активности только в контрольной среде (фигура 16B).
Для выявления лецитинхолестеринацилтрансферазы (LCAT) и липопротеинлипазы (LPL) B-клетки культивировали (см. условия культивирования, представленные в примере 6) и подвергали электропорации в день 2 культивирования (как в примере 1) с помощью транспозона, кодирующего либо LCAT, либо LPL, а также конструкции, кодирующей источник транспозазы (SB100x, см. пример 1 и фигуру 1). Образцы среды отбирали в день 4 и 8 культивирования и анализировали в отношении присутствия LCAT и LPL с применением флуорометрического ферментного анализа. В частности, применяли субстрат 4-метилумбеллиферилпальмитат (4-MUP), а расщепление субстрата с помощью LCAT и LPL определяли путем измерения увеличения флуоресценции при длинах волн 340, 390 и 460 нм. Получали реакционный буфер, содержащий 100 мМ фосфатно-натриевого буфера (рН 7,4), и помещали при 37°С. 4-MUP разбавляли посредством смешивания с 2 мг 4-MUP с 16 мг Triton X-100. 4-MUP и реакционный буфер объединяли и общий объем 150 мкл добавляли в лунку в 96-луночном планшете вместе с 2 мкл активаторного соединения (например, п-нитрофенилбутирата). Получали серийные разведения сред для культивирования В-клеток и затем к смеси добавляли 50 мкл разведений и инкубировали при 37°С. Измерения флуоресценции начинали сразу после добавления среды для культивирования и собирали каждую минуту в течение общего периода 30 минут.
Экспрессию FIX в трансфицированных В-клетках подтверждали с помощью ELISA. Более конкретно, клеточные лизаты В-клеток получали и выявляли с помощью ELISA с применением коммерчески доступного набора ELISA (FIX-EIA, Enzyme Research Laboratories, Саут-Бенд, Индиана) в соответствии с рекомендованным протоколом производителя. Среду собирали через 2 дня после электропорации, и среда содержала белок FIX в концентрации примерно 15 нг/мл, тогда как белок FIX не обнаруживался в клетках отрицательного контроля (В-клетки, трансфицированные GPF) (фигура 16C).
Таким образом, эти данные демонстрируют, что В-клетки можно применять в способах, раскрытых в данном документе, с обеспечением экспрессии и доставки субъекту широкого спектра полипептидов.
Различные варианты осуществления, описанные выше, могут быть объединены с обеспечением дополнительных вариантов осуществления. Все патенты США, публикации заявок на патент США, заявки на патент США, иностранные патенты, заявки на иностранные патенты и непатентные публикации, упомянутые в настоящем описании и/или перечисленные в информационном листке заявки, включены в данный документ посредством ссылки во всей своей полноте. Аспекты вариантов осуществления могут быть модифицированы, если необходимо использовать концепции различных патентов, заявок и публикаций для обеспечения еще дополнительных вариантов осуществления.
Эти и другие изменения можно осуществлять в вариантах осуществления в свете вышеизложенного подробного описания. В целом, в следующей формуле изобретения используемые условия не должны толковаться как ограничивающие формулу изобретения конкретными вариантами осуществления, раскрытыми в описании и формуле изобретения, но должны толковаться как включающие все возможные варианты осуществления вместе с полным объемом эквивалентов, на который распространяется эта формула изобретения. Соответственно, данная формула изобретения не ограничена настоящим раскрытием.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Иммусофт Корпорейшн
ШОЛЬЦ, Мэттью, Рейн
ХЕРБИГ, Эрик, Дж.
МАКИВОР, Р., Скотт
ДЕ ЛАТ, Риан
ОЛСОН, Эрик
<120> В-КЛЕТКИ ДЛЯ ДОСТАВКИ ТЕРАПЕВТИЧЕСКИХ СРЕДСТВ IN VIVO И ИХ ДОЗЫ
<130> IMCO-006/01WO 312423-2025
<150> US 62/491151
<151> 27-04-2017
<160> 1
<170> PatentIn, версия 3.5
<210> 1
<211> 7322
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Полученная в лаборатории плазмидная конструкция pKT2/EEK-IDUA-DHFR
<400> 1
cctggatcca gatccctata cagttgaagt cggaagttta catacactta agttggagtc 60
attaaaactc gtttttcaac tactccacaa atttcttgtt aacaaacaat agttttggca 120
agtcagttag gacatctact ttgtgcatga cacaagtcat ttttccaaca attgtttaca 180
gacagattat ttcacttata attcactgta tcacaattcc agtgggtcag aagtttacat 240
acactaagtt gactgtgcct ttaaacagct tggaagctgc gcactaggca agttaactaa 300
ctcctctgaa tgtcagtatt tccatctgta agatgaacac agtggggctc caattccata 360
ccacatttgt agaggtttta cttgctttaa aaaacctccc acacctcccc ctgaacctga 420
aacataaaat gaatgcaatt gttgttgtta acttgtttat tgcagcttat aatggttaca 480
aataaagcaa tagcatcaca aatttcacaa ataaagcatt tttttcactg cattctagtt 540
gtggtttgtc caaactcatc aatgtatctt atcatgtctg gccagctaga gcggccgctt 600
aatcattctt ctcatatact tcaaatttgt acttaatgcc tttctcctcc tggacatcag 660
agagaacacc tgggtattct ggcagaagtt tatatttctc caaatcaatt tctggaaaaa 720
acgtgtcact ttcaaagtct tgcatgatcc ttgtcacaaa tagtttaaga tggcctgggt 780
gattcatggc ttccttataa acagaactgc caccaactat ccagaccatg tctactttat 840
ttgctaattc tggttgttca gtaagtttta aggcatcatc tagacttctg gaaagaaaat 900
gagctccttg tggaggttcc ttgagttctc tgctgagaac taaattaatt ctacccttta 960
aaggtcgatt cttctcagga atggagaacc aggtcttctt acccataatc accagattct 1020
gtttaccttc tactgaagag gttgtggtca ttctctggaa atatctggat tcattcctga 1080
gcggtggcca gggatagtcc ccgttcttgc cgatgcccat gttctgggac acagcgacga 1140
tgcagtttag cgaaccaacc atgatggaag ctactgtaca ccaacctgtc aggagaggaa 1200
agagaagaag gttagtacaa ttgtctaggg ctgcagggtt catagtgcca cttttcctgc 1260
actgccccat ctcctgccca ccctttccca ggcatagaca gtcagtgact taccaaactc 1320
acaggaggga gaaggcagaa gcttgaatgt tcacagagac tactgcactt atatatggtt 1380
ctcccccacc ctggggaaaa aggtggagcc agtacaccac atcactttcc cagtttaccc 1440
aagccccacc ttctctaggc accagttcaa ttgcccaccc ctccccccaa cttctcaggg 1500
actgtgggcc atgtgctctc tgcccactga ggggcactca gccctcaagc atgctcttct 1560
ccactagtca cccctattga ccttatgtat gtgccaataa tgggaaaaac ccattgactc 1620
accccctatt gaccttttgt actgggcaaa acccaatgga aagtccctat tgactcagtg 1680
tacttggctc caatgggact ttcctgttga ttggcgcgcc cgggggatcc agtttggtta 1740
attaaaccgg tgagtttcat ggttacttgc ctgagaagat taaaaaaagt aatgctacct 1800
tatgagggag agtcccaggg accaagatag caactgtcat agcaaccgtc acactgcttt 1860
ggtcaaggag aagacccttt ggggaactga aaacagaacc ttgagcacat ctgttgcttt 1920
cgctcccatc ctcctccaac agggctgggt ggagcactcc acaccctttc accggtcgta 1980
cggctcagcc agagtaaaaa tcacacccat gacctggcca ctgagggctt gatcaattca 2040
ctttgaattt ggcattaaat accattaagg tatattaact gattttaaaa taagatatat 2100
tcgtgaccat gtttttaact ttcaaaaatg tagctgccag tgtgtgattt tatttcagtt 2160
gtacaaaata tctaaaccta tagcaatgtg attaataaaa acttaaacat attttccagt 2220
accttaattc tgtgatagga aaattttaat ctgagtattt taatttcata atctctaaaa 2280
tagtttaatg atttgtcatt gtgttgctgt cgtttacccc agctgatctc aaaagtgata 2340
tttaaggaga ttattttggt ctgcaacaac ttgatagggc tcagcctctc ccacccaacg 2400
ggtggaatcc cccagagggg gatttccaag aggccacctg gcagttgctg agggtcagaa 2460
gtgaagctag ccacttcctc ttaggcaggt ggccaagatt acagttgacc cgtacgtgca 2520
gctgtgccca gcctgcccca tcccctgctc atttgcatgt tcccagagca caacctcctg 2580
ccctgaagcc ttattaatag gctggtcaca ctttgtgcag gagtcagact cagtcaggac 2640
acagctctag agtcgagaat tcggccatgc gtcccctgcg cccccgcgcc gcgctgctgg 2700
cgctcctggc ctcgctcctg gccgcgcccc cggtggcccc ggccgaggcc ccgcacctgg 2760
tgcaggtgga cgcggcccgc gcgctgtggc ccctgcggcg cttctggagg agcacaggct 2820
tctgcccccc gctgccacac agccaggctg accagtacgt cctcagctgg gaccagcagc 2880
tcaacctcgc ctatgtgggc gccgtccctc accgcggcat caagcaggtc cggacccact 2940
ggctgctgga gcttgtcacc accagggggt ccactggacg gggcctgagc tacaacttca 3000
cccacctgga cgggtacctg gaccttctca gggagaacca gctcctccca gggtttgagc 3060
tgatgggcag cgcctcgggc cacttcactg actttgagga caagcagcag gtgtttgagt 3120
ggaaggactt ggtctccagc ctggccagga gatacatcgg taggtacgga ctggcgcatg 3180
tttccaagtg gaacttcgag acgtggaatg agccagacca ccacgacttt gacaacgtct 3240
ccatgaccat gcaaggcttc ctgaactact acgatgcctg ctcggagggt ctgcgcgccg 3300
ccagccccgc cctgcggctg ggaggccccg gcgactcctt ccacacccca ccgcgatccc 3360
cgctgagctg gggcctcctg cgccactgcc acgacggtac caacttcttc actggggagg 3420
cgggcgtgcg gctggactac atctccctcc acaggaaggg tgcgcgcagc tccatctcca 3480
tcctggagca ggagaaggtc gtcgcgcagc agatccggca gctcttcccc aagttcgcgg 3540
acacccccat ttacaacgac gaggcggacc cgctggtggg ctggtccctg ccacagccgt 3600
ggagggcgga cgtgacctac gcggccatgg tggtgaaggt catcgcgcag catcagaacc 3660
tgctactggc caacaccacc tccgccttcc cctacgcgct cctgagcaac gacaatgcct 3720
tcctgagcta ccacccgcac cccttcgcgc agcgcacgct caccgcgcgc ttccaggtca 3780
acaacacccg cccgccgcac gtgcagctgt tgcgcaagcc ggtgctcacg gccatggggc 3840
tgctggcgct gctggatgag gagcagctct gggccgaagt gtcgcaggcc gggaccgtcc 3900
tggacagcaa ccacacggtg ggcgtcctgg ccagcgccca ccgcccccag ggcccggccg 3960
acgcctggcg cgccgcggtg ctgatctacg cgagcgacga cacccgcgcc caccccaacc 4020
gcagcgtcgc ggtgaccctg cggctgcgcg gggtgccccc cggcccgggc ctggtctacg 4080
tcacgcgcta cctggacaac gggctctgca gccccgacgg cgagtggcgg cgcctgggcc 4140
ggcccgtctt ccccacggca gagcagttcc ggcgcatgcg cgcggctgag gacccggtgg 4200
ccgcggcgcc ccgcccctta cccgccggcg gccgcctgac cctgcgcccc gcgctgcggc 4260
tgccgtcgct tttgctggtg cacgtgtgtg cgcgccccga gaagccgccc gggcaggtca 4320
cgcggctccg cgccctgccc ctgacccaag ggcagctggt tctggtctgg tcggatgaac 4380
acgtgggctc caagtgcctg tggacatacg agatccagtt ctctcaggac ggtaaggcgt 4440
acaccccggt cagcaggaag ccatcgacct tcaacctctt tgtgttcagc ccagacacag 4500
gtgctgtctc tggctcctac cgagttcgag ccctggacta ctgggcccga ccaggcccct 4560
tctcggaccc tgtgccgtac ctggaggtcc ctgtgccaag agggccccca tccccgggca 4620
atccatgagc ctgtgctgag ccccagtggg atcctctaga gtcgagaatt cactcctcag 4680
gtgcaggctg cctatcagaa ggtggtggct ggtgtggcca atgccctggc tcacaaatac 4740
cactgagatc tttttccctc tgccaaaaat tatggggaca tcatgaagcc ccttgagcat 4800
ctgacttctg gctaataaag gaaatttatt ttcattgcaa tagtgtgttg gaattttttg 4860
tgtctctcac tcggaaggac atatgggagg gcaaatcatt taaaacatca gaatgagtat 4920
ttggtttaga gtttggcaac atatgccata tgctggctgc catgaacaaa ggtggctata 4980
aagaggtcat cagtatatga aacagccccc tgctgtccat tccttattcc atagaaaagc 5040
cttgacttga ggttagattt tttttatatt ttgttttgtg ttattttttt ctttaacatc 5100
cctaaaattt tccttacatg ttttactagc cagatttttc ctcctctcct gactactccc 5160
agtcatagct gtccctcttc tcttatgaag atccctcgac ctgcataccg gtcaagctag 5220
cgatatcaat taaccctcac taaagggaga ccaagttaaa caatttaaag gcaatgctac 5280
caaatactaa ttgagtgtat gtaaacttct gacccactgg gaatgtgatg aaagaaataa 5340
aagctgaaat gaatcattct ctctactatt attctgatat ttcacattct taaaataaag 5400
tggtgatcct aactgaccta agacagggaa tttttactag gattaaatgt caggaattgt 5460
gaaaaagtga gtttaaatgt atttggctaa ggtgtatgta aacttccgac ttcaactgta 5520
tagggatctg gtaccattta aatctgttcc gcttcctcgc tcactgactc gctgcgctcg 5580
gtcgttcggc tgcggcgagc ggtatcagct cactcaaagg cggtaatacg gttatccaca 5640
gaatcagggg ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccaggaac 5700
cgtaaaaagg ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac 5760
aaaaatcgac gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg 5820
tttccccctg gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac 5880
ctgtccgcct ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat 5940
ctcagttcgg tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag 6000
cccgaccgct gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac 6060
ttatcgccac tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt 6120
gctacagagt tcttgaagtg gtggcctaac tacggctaca ctagaaggac agtatttggt 6180
atctgcgctc tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc 6240
aaacaaacca ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga 6300
aaaaaaggat ctcaagaaga tcctttgatc ttttctacgg ggtctgacgc tcagtggaac 6360
gaaaactcac gttaagggat tttggtcatg agattatcaa aaaggatctt cacctagatc 6420
ctttttgcca gtgttacaac caattaacca attctgatta gaaaaactca tcgagcatca 6480
aatgaaactg caatttattc atatcaggat tatcaatacc atatttttga aaaagccgtt 6540
tctgtaatga aggagaaaac tcaccgaggc agttccatag gatggcaaga tcctggtatc 6600
ggtctgcgat tccgactcgt ccaacatcaa tacaacctat taatttcccc tcgtcaaaaa 6660
taaggttatc aagtgagaaa tcaccatgag tgacgactga atccggtgag aatggcaaaa 6720
gtttatgcat ttctttccag acttgttcaa caggccagcc attacgctcg tcatcaaaat 6780
cactcgcatc aaccaaaccg ttattcattc gtgattgcgc ctgagcgaga cgaaatacgc 6840
gatcgctgtt aaaaggacaa ttacaaacag gaatcgaatg caaccggcgc aggaacactg 6900
ccagcgcatc aacaatattt tcacctgaat caggatattc ttctaatacc tggaatgctg 6960
tttttccggg gatcgcagtg gtgagtaacc atgcatcatc aggagtacgg ataaaatgct 7020
tgatggtcgg aagaggcata aattccgtca gccagtttag tctgaccatc tcatctgtaa 7080
catcattggc aacgctacct ttgccatgtt tcagaaacaa ctctggcgca tcgggcttcc 7140
catacaagcg atagattgtc gcacctgatt gcccgacatt atcgcgagcc catttatacc 7200
catataaatc agcatccatg ttggaattta atcgcggcct cgacgtttcc cgttgaatat 7260
ggctcataac accccttgta ttactgttta tgtaagcaga cagttttatt gttcatgatg 7320
ca 7322
<---

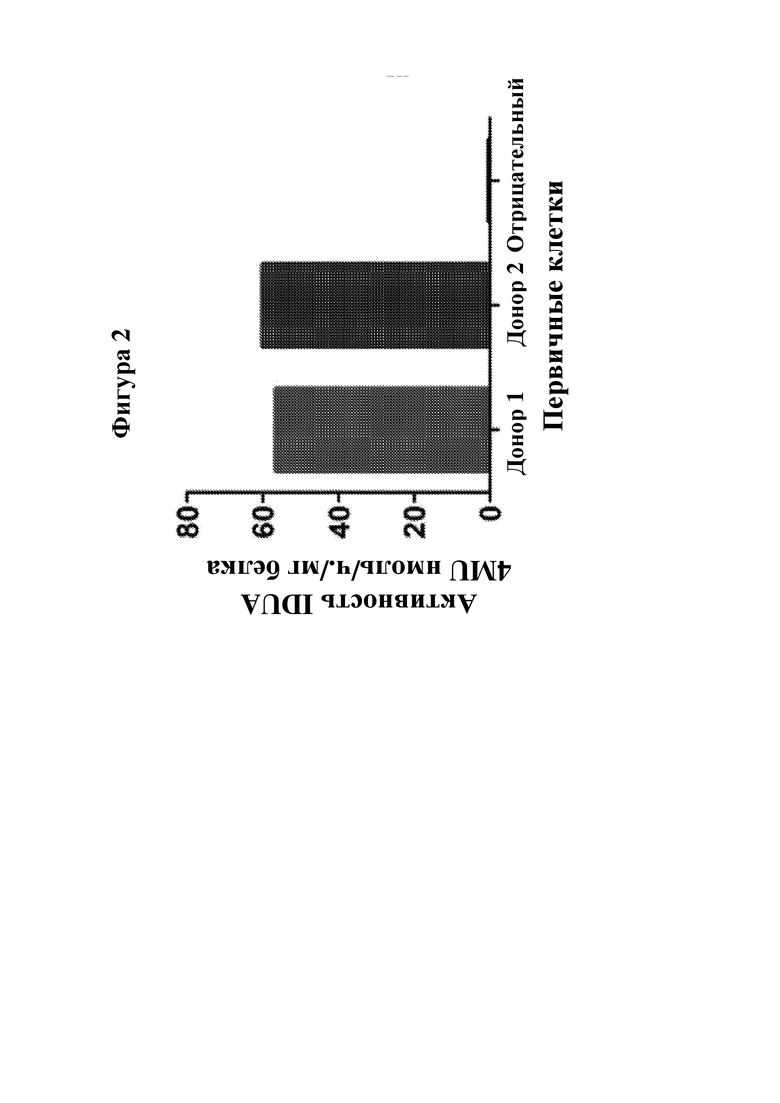

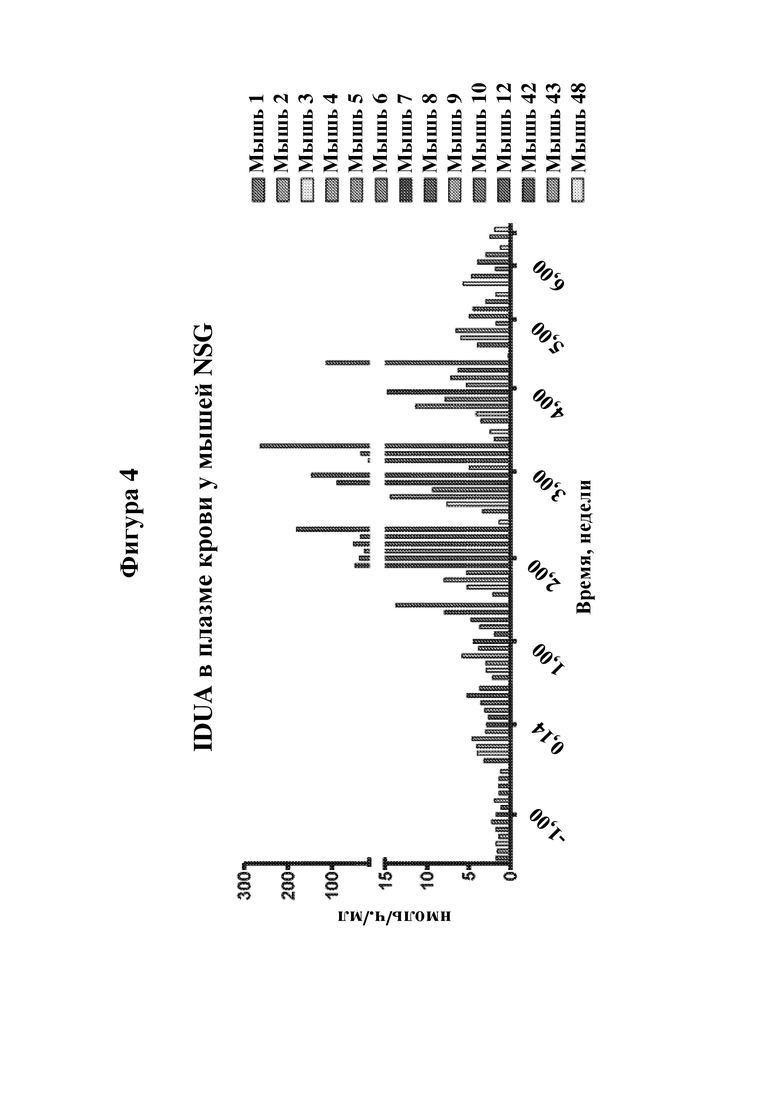


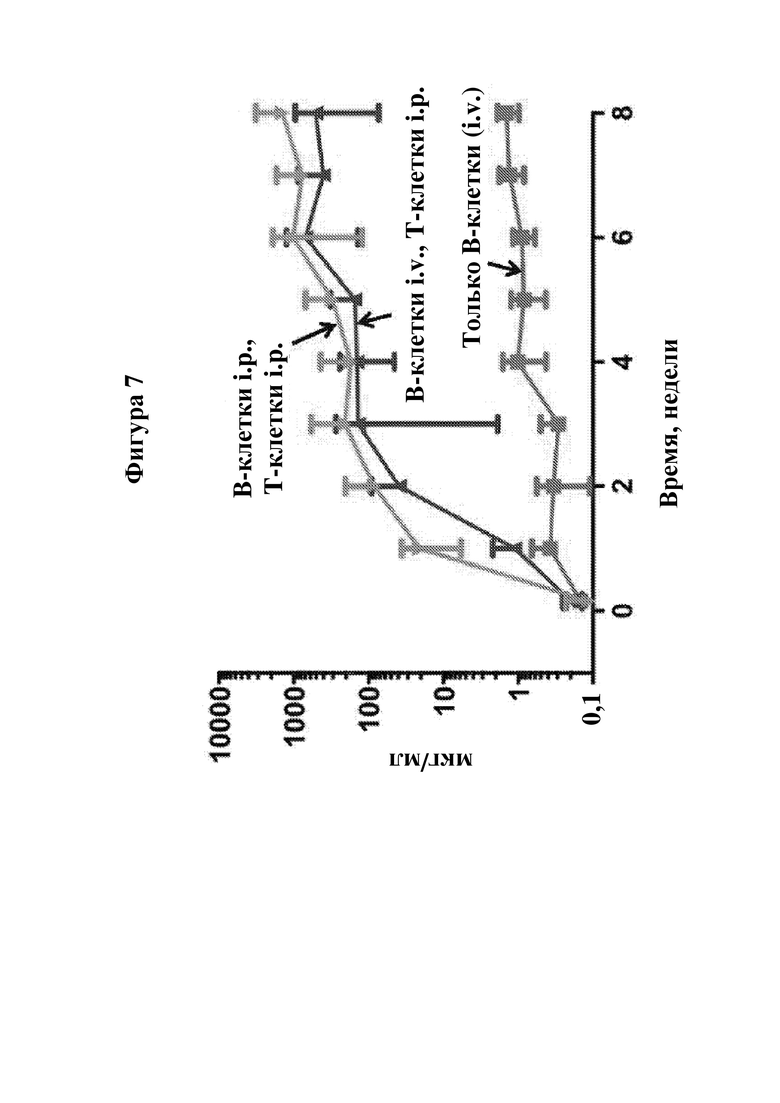

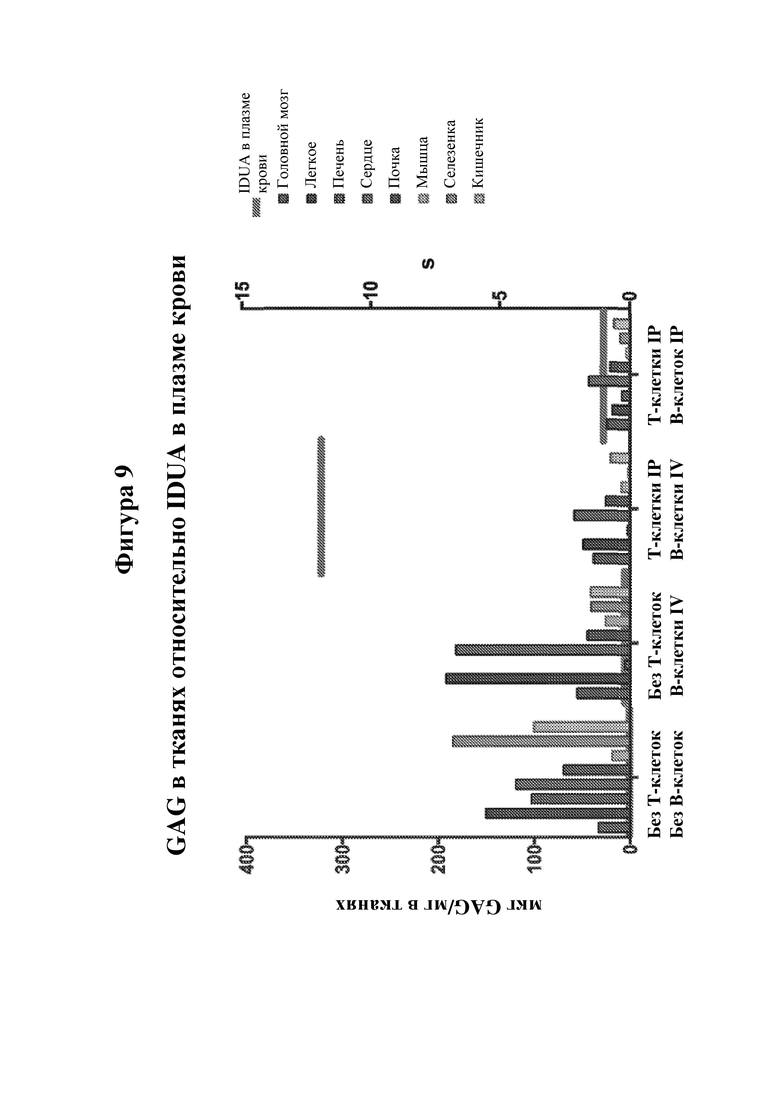
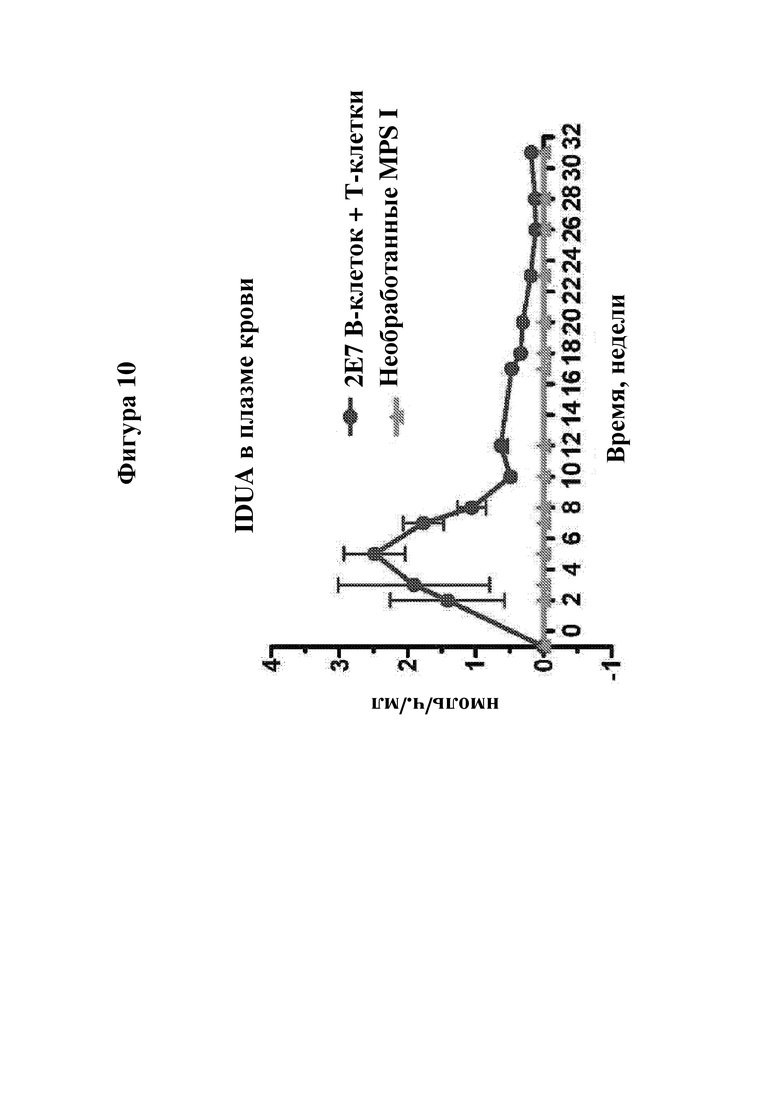



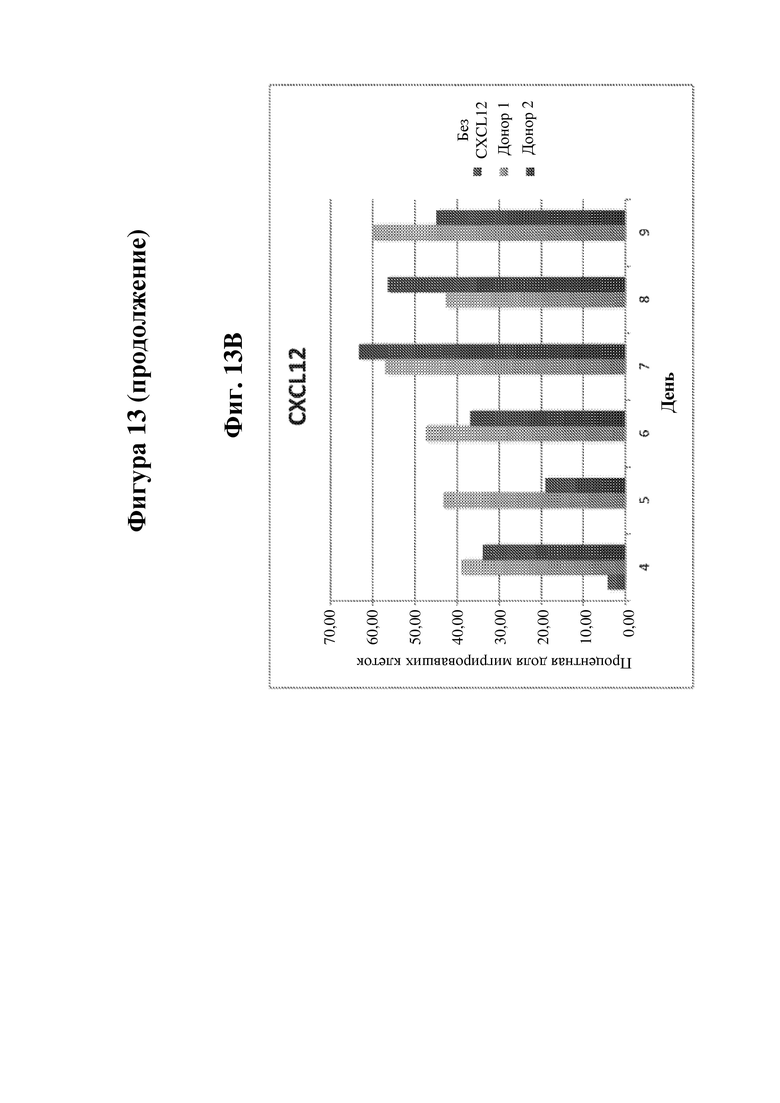

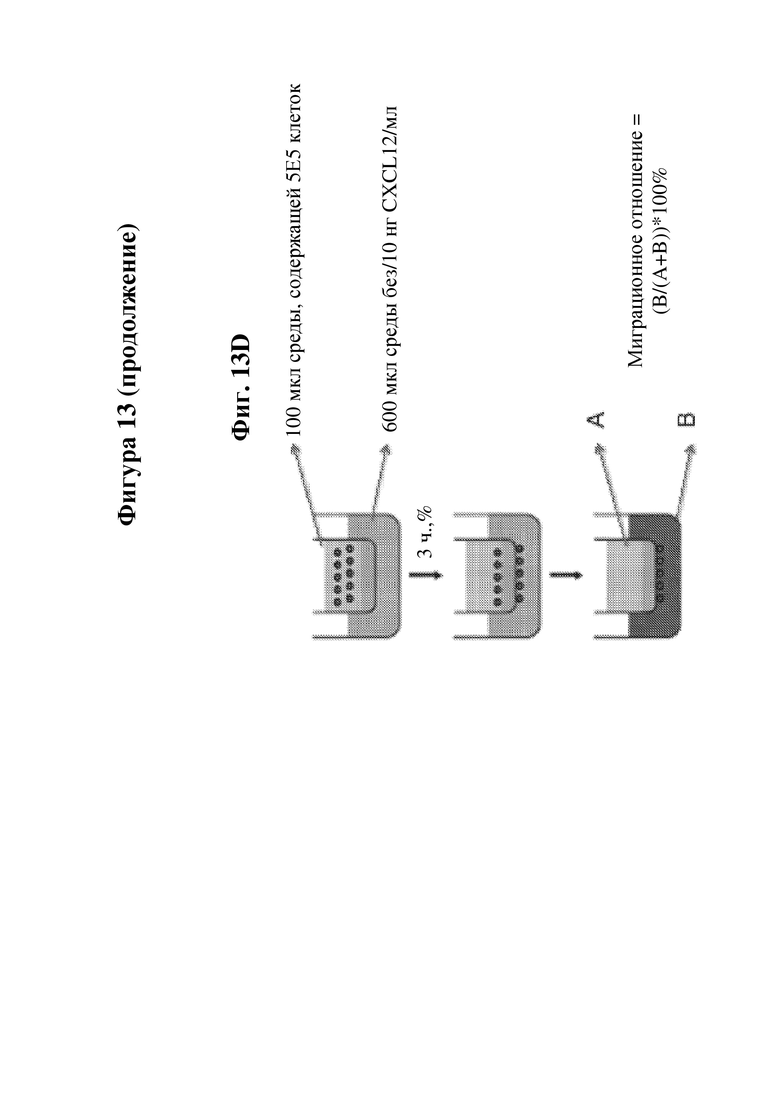
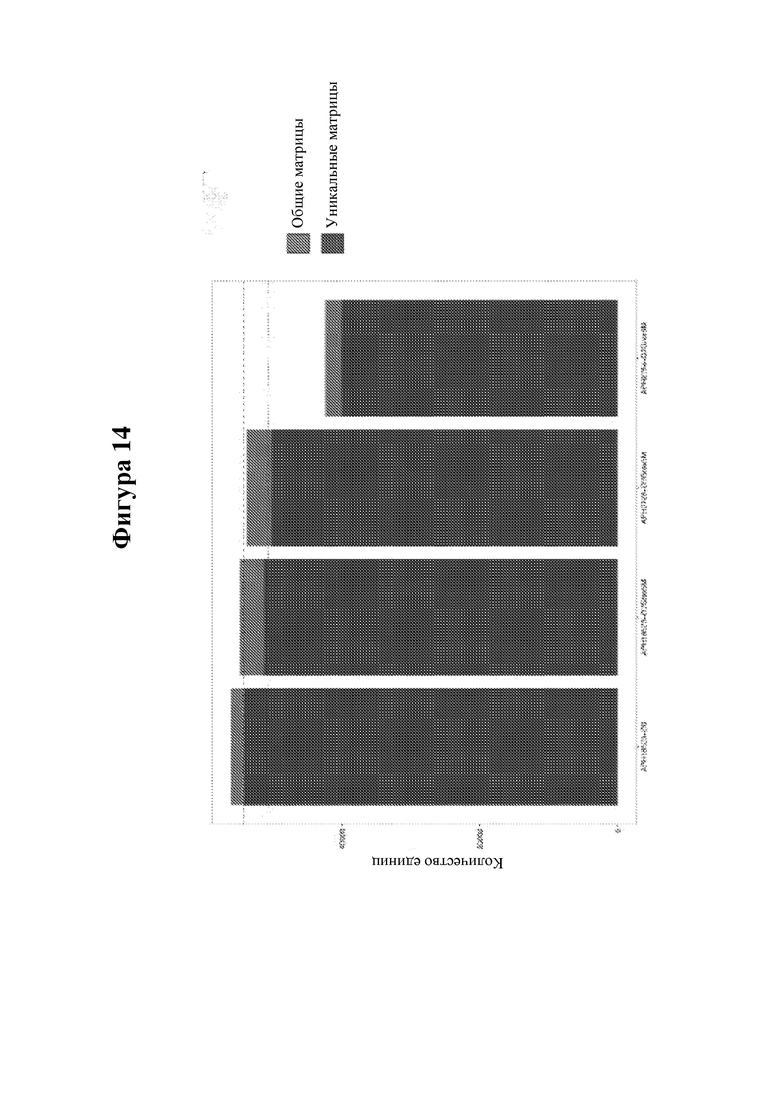
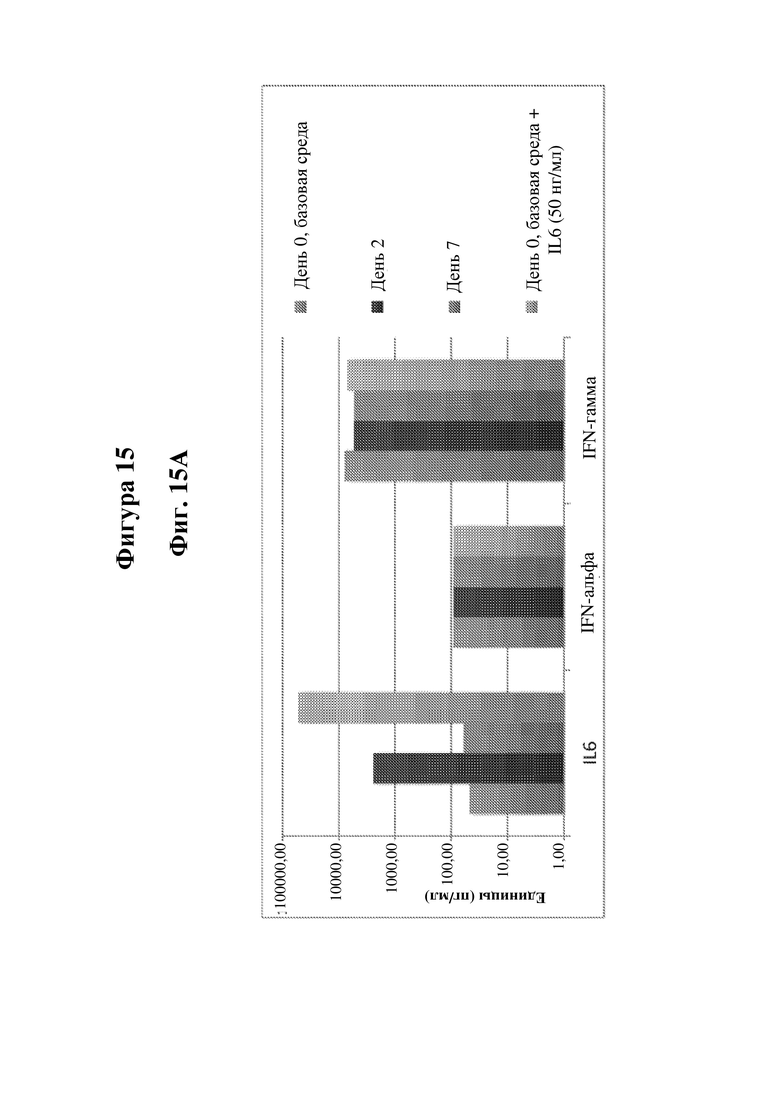

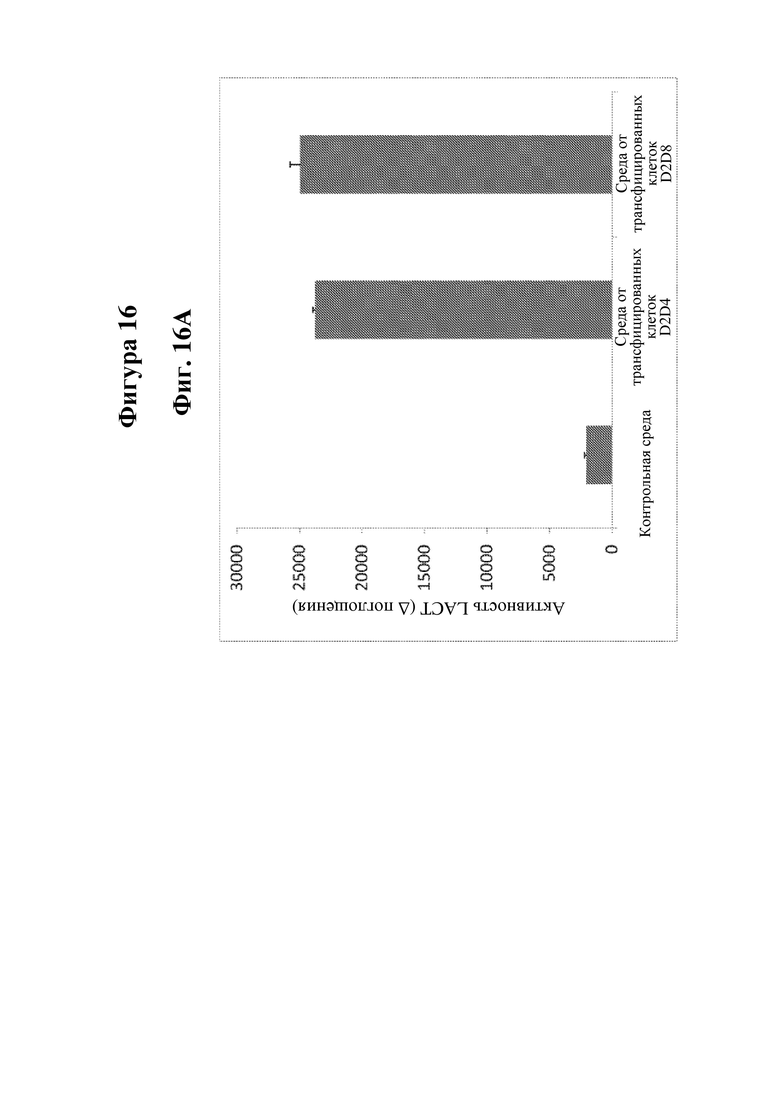
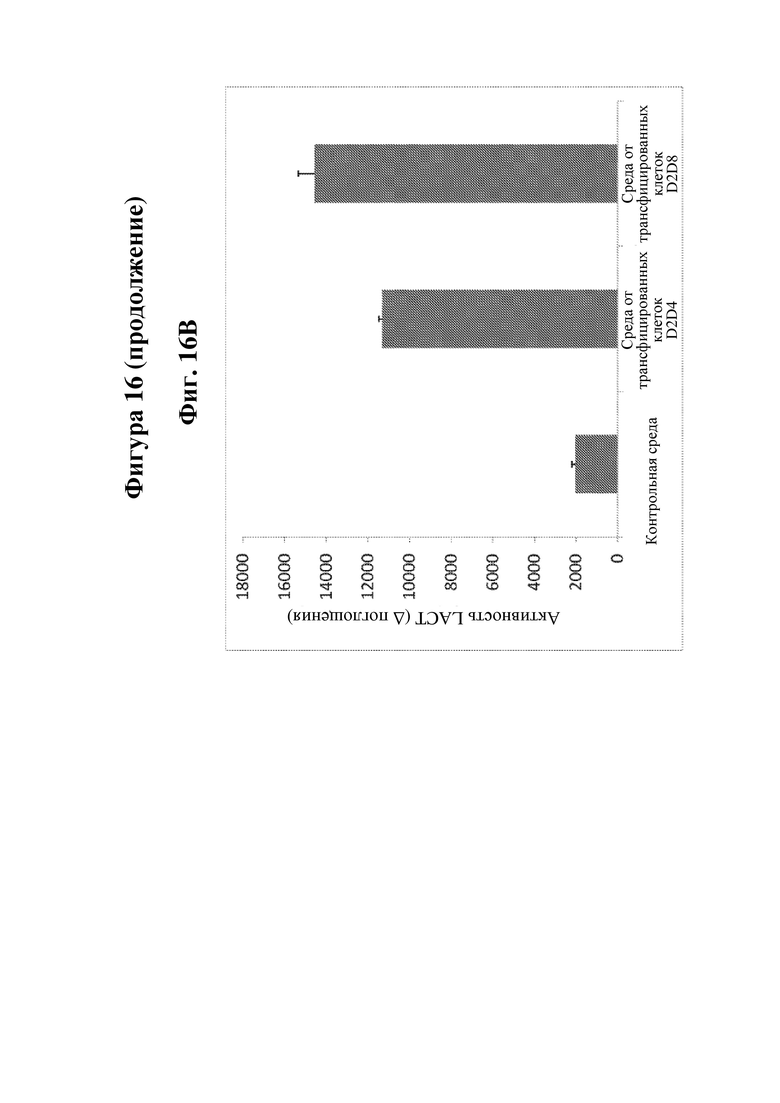

Изобретение относится к биотехнологии, в частности к способам введения генетически модифицированных В-клеток с обеспечением продуцирования терапевтического средства, такого как терапевтический белок, путем введения однократной максимально эффективной дозы генетически модифицированных В-клеток. Изобретение применимо для долговременной доставки терапевтического средства in vivo. 8 з.п. ф-лы, 2 табл., 16 ил., 9 пр.
1. Способ введения генетически модифицированных В-клеток субъекту с обеспечением продуцирования in vivo терапевтического средства для лечения MPS I у субъекта, предусматривающий:
определение оптимальной концентрации однократной дозы генетически модифицированных В-клеток, где указанная оптимальная однократная доза генетически модифицированных В-клеток представляет собой дозу, которая приводит к наибольшим уровням in vivo указанного терапевтического средства в пересчете на клетку в контексте однократной инфузии B-клеток указанных генетически модифицированных В-клеток, где указанное терапевтическое средство представляет собой идуронидазу (IDUA) человека и указанные генетически модифицированные В-клетки содержат полинуклеотидную последовательность, которая кодирует указанную IDUA человека, и сконструированы с обеспечением экспрессии и секреции IDUA человека;
уменьшение оптимальной концентрации однократной дозы генетически модифицированных B-клеток с получением субоптимальной концентрации однократной дозы генетически модифицированных B-клеток, где указанная субоптимальная концентрация однократной дозы составляет на 1/2 или меньше, чем оптимальная концентрация однократной дозы; и
введение субъекту двух или более внутривенных доз при субоптимальной концентрации однократной дозы генетически модифицированных В-клеток.
2. Способ по п. 1, где проводят исследование с использованием множества однократных доз генетически модифицированных B-клеток, в результате чего определяют оптимальную концентрацию однократной дозы модифицированных B-клеток.
3. Способ по п. 2, где увеличение дозы генетически модифицированных В-клеток, присутствующих в некоторой концентрации однократной дозы модифицированных В-клеток, приводит к линейному повышению продуцирования терапевтического средства.
4. Способ по п. 1, где проводят исследование с использованием множества субоптимальных концентраций однократной дозы генетически модифицированных В-клеток, в результате чего определяют оптимальную дозу, при этом полученная в результате доза приводит к большему, чем линейное, повышению по сравнению с более низкими дозами.
5. Способ по п. 1, где субоптимальная концентрация однократной дозы составляет половину или одну треть дозы от оптимальной концентрации однократной дозы.
6. Способ по п. 1, где субоптимальная концентрация однократной дозы составляет менее одной трети дозы от оптимальной концентрации однократной дозы.
7. Способ по любому из пп. 1-6, где указанная полинуклеотидная последовательность, которая кодирует указанную IDUA человека, имеет последовательность, которая идентична SEQ ID NO: 1.
8. Способ по любому из пп. 1-6, где указанная полинуклеотидная последовательность, которая кодирует указанную IDUA человека, имеет последовательность, которая на по меньшей мере приблизительно 85% идентична последовательности под SEQ ID NO: 1 или на по меньшей мере приблизительно 90%, 95%, 96%, 97%, 98%, 99% или более чем 99% идентична SEQ ID NO: 1.
9. Способ по любому из пп. 1-6, где указанная полинуклеотидная последовательность, которая кодирует указанную IDUA человека, представляет собой бифункциональный трансген или транспозон.
| US 20160046908 A1, 18.02.2016 | |||
| US 20130143267 A1, 06.06.2013 | |||
| КОМПОЗИЦИЯ ДЛЯ СТИМУЛЯЦИИ ИММУННОГО ОТВЕТА (ВАРИАНТЫ), СПОСОБЫ ЕЕ ПОЛУЧЕНИЯ, ПРИМЕНЕНИЯ И СПОСОБЫ СТИМУЛЯЦИИ ИММУННОГО ОТВЕТА С ИСПОЛЬЗОВАНИЕМ ЭТИХ КОМПОЗИЦИЙ | 2002 |
|
RU2316347C2 |

