ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке испрашивается приоритет Предварительной патентной заявки США No. 62/450917, поданной 26 января 2017 г., полное содержание которой, таким образом, приведено в настоящем описании в качестве ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0002] Настоящее описание относится к области генотерапии, в частности, редактирования генома и направленной доставки кодирующих трансген конструкций в B-клетки для экспрессии обеспечивающих преимущество (терапевтических) белков.
УРОВЕНЬ ТЕХНИКИ
[0003] Генотерапию можно использовать для генетической инженерии клетки, чтобы получить наличие одного или более инактивированных генов и/или чтобы заставить эту клетку экспрессировать продукт, ранее не продуцированный в этой клетке (например, посредством вставки трансгена и/или посредством коррекции эндогенной последовательности). Примеры применений вставки трансгена включают опосредованную нуклеазой модификацию, включающую вставку одного или более генов, кодирующих один или более новых терапевтических белков, включая терапевтические антитела, вставку кодирующей последовательности, кодирующей белок, отсутствующий в клетке или у индивидуума, вставку гена дикого типа в клетку, содержащую мутантную последовательность гена, и/или вставку последовательности, кодирующей структурную нуклеиновую кислоту, такую как микроРНК или миРНК. Эти способы можно также использовать для нокаута экспрессии эндогенного гена и/или для изменения профиля токсичности клетки. См., например, Патенты США No. 9394545; 9150847; 9206404; 9045763; 9005973; 8956828; 8936936; 8945868; 8871905; 8586526; 8563314; 8329986; 8399218; 6534261; 6599692; 6503717; 6689558; 7067317; 7262054; 7888121; 7972854; 7914796; 7951925; 8110379; 8409861; Публикации патентов США 20030232410; 20050208489; 20050026157; 20050064474; 20060063231; 20080159996; 20100218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983 и 20130177960, и 20150056705.
[0004] B-клетки функционируют в рамках гуморального иммунитета посредством секреции антител против множества антигенов. Они являются профессиональными антигенпредставляющими клетками (APC) и подвергаются активации T-клетками-помощниками для дифференцировки в плазматические клетки, продуцирующие большие количества антигенспецифических антител. Развитие B-клеток происходит как в печени, так и в костном мозге плода, и критической стадией в развитии B-клетки является образование B-клеточного рецептора (BCR), комплексной структуры, содержащей уникальные тяжелую и легкую цепи. Процесс образования BCR включает реаранжировку различных фрагментов генов иммуноглобулинов (Ig), как в тяжелой, так и в легкой цепях генов BCR, в ходе про-B-клеточной фазы созревания B-клетки. Про-B-клетки становятся пре-B-клетками после успешного спаривания реаранжированных тяжелой и легкой цепей, где полученный пре-BCR экспрессируется на клеточной поверхности пре-B-клеток. Передача сигналов посредством пре-BCR управляет дальнейшим развитием B-клетки, приводящим к увеличению пре-B-клеток. В конечном счете, большие пре-B-клетки останавливают пролиферацию, и происходит дополнительная реаранжировка легкой цепи, приводящая к экспрессии уникального IgM BCR на клеточной поверхности того, что теперь рассматривают как незрелую B-клетку. Затем эти клетки выходят из костного мозга и циркулируют на периферии как переходные B-клетки.
[0005] Созревание незрелых B-клеток происходит в первую очередь в селезенке, где также происходит отбор, так что B-клетки, продуцирующие антитела с высокой аффинностью к собственным антигенам, разрушаются (см. Naradikian et al. (2014) in Drugs Targeting B-Cells in Autoimmune Diseases, Milestones in Drug Therapy (X. Bosch et al. (eds)) doi 10.1007/978-3-3-0348-0706-7_2, Springer Basel). Циркулирующие B-клетки являются способными входить во вторичные лимфатические узлы и селезенку и получать антиген от фолликулярных дендритных клеток. После входа в лимфатический узел/селезенку и взаимодействия с дендритными клетками, B-клетки интернализуют антиген и процессируют его таким образом, что пептидные фрагменты антигена подвергаются представлению посредством молекул MHC класса II родственным CD4+ T-клеткам. Эти T-клетки являются ранее активированными посредством APC, представляющей тот же самый антиген. Взаимодействие между B- и T-клетками приводит к ряду событий, включая полную активацию T-клетки, приводящую к пролиферации T-клетки. Затем T-клетки продуцируют цитокины, которые действуют непосредственно на B-клетки для индукции пролиферации B-клетки и переключения класса антитела, экспрессированного на поверхности B-клетки. Пролиферирующие B-клетки кластеризуются во временных областях в лимфатических узлах и селезенке, известных как «зародышевые центры». Эти активированные B-клетки дифференцируют либо в специализированные секретирующие антитела клетки (плазмобласт или плазматическая клетка), которые, по-видимому, подвергаются заранее запрограммированному количеству делений (как правило, 5-6) до того, как они завершат конечную стадию своей дифференцировки, чтобы стать непролиферирующими плазматическими клетками. Альтернативно, активированные B-клетки покидают зародышевый центр и дифференцируют в B-клетки памяти (Zhang et al., (2016) Immunol Rev 270(1): 8-19).
[0006] После активации экспрессируется индуцируемая посредством активации фермента геномным мутатором цитидиндезаминаза (AID), которая приводит к соматической гипермутаци в генах антител и рекомбинации с переключением класса (CSR), изменяющим эффекторную функцию антитела. Соматическая гипермутация возникает в гене вариабельной области иммуноглобулина (IgV) с получением репертуара мутантов антитела с различной аффинностью для антигена (Klein and Heise (2015) Curr Opin Hematol 22(4): 379-387). Этот процесс происходит в так называемой «темной зоне» в зародышевом центре. Дифференцированные B-клетки мигрируют в «светлую зону», где B-клетки, продуцирующие антитела с более высокой аффинностью, конкурируют за доступный антиген и/или помощь T-клетки, так что они получают сигналы выживания через B-клеточные рецепторы. B-клетки, продуцирующие антитела с более низкой аффинностью, не получают этих сигналов выживания, поскольку они не могут конкурировать со своими сестринскими B-клетками, продуцирующими антитела с более высокой аффинностью, и таким образом, они подвергаются апоптозу. Затем B-клетки, продуцирующие антитела с более высокой аффинностью, либо могут повторно входить в темную зону для дополнительных циклов пролиферации и соматической гипермутации, могут покидать зародышевый центр и дифференцировать в плазмобласты, либо могут дифференцировать в долгоживущие B-клетки памяти (Recaldin and Fear (2015) Clin and Exp Immunol 183:65-75).
[0007] Таким образом, в очень сложном процессе, B-клетки подвергаются индукции для экспрессии больших количеств антител против антигенов для защиты организма от ряда потенциальных угроз. Интересно, что существует ряд патогенов (например, паразитов, бактерий, вирусов), которые являются способными нарушать ответ антител. Эти патогены включают ряд агентов, ответственных за множество заболеваний человека, включая, но без ограничения, плазмодий, шистосому, микобактерию, HIV, HCV и HBV (Borhis and Richard (2015) BMC Immunology 16:15 doi 10.1186/s12865-015-0079-y). Механизмы, стоящие за опосредованной патогеном супрессией ответа антител, не полностью известны, но по-видимому, определенные патогены индуцируют продукцию необычных подтипов B-клеток, изменяющих клеточное микроокружение, что приводит к супрессии как B-, так и T-клеток. Например, показано, что HBV создает помехи для стимуляции посредством Toll-подобного рецептора 9 (TLR9), так что дендритные клетки продуцируют меньшее количество IFN-α (как известно, индуцирующего B-клетки для пролиферации и секреции IgM). По-видимому, HBV может избирательно ингибировать экспрессию TLR9 в B-лимфоцитах (Vincent et al. (2011) PLoS ONE 6(10):e26315. doi:10.1371/journal.pone.0026315).
[0008] На протяжении последнего десятилетия, исследования предоставили хорошо обоснованное доказательство дискретных подгрупп иммунорегуляторных B-клеток как у мышей, так и у человека. Эти супрессорные B-клетки имеют способность поддерживать иммунную толерантность и супрессировать патологические аутоиммунные и воспалительные иммунные ответы, так же как супрессировать ответы в ходе иммунного надзора злокачественных опухолей, посредством высвобождения противовоспалительных медиаторов, таких как интерлейкин-10 (IL-10), и экспрессии ингибирующих молекул, таких как PD-L1. Эти исследования привели к заключению, что существует пул B-клеток, играющих роль супрессоров в иммунной толерантности, и этот пул в настоящее время обозначен как регуляторные B-клетки или «B-рег». Другие фенотипы, ассоциированные с B-рег человека, включают супрессию аутоиммунного воспаления и роль в толерантности к аллергенам. В более недавних исследованиях показано, что B-клетки могут играть противоречивые роли в прогрессировании злокачественных опухолей. Например, для продуцирующих IL-10 CD1dвысокийCD5+ B-клеток, выделенных от пациентов с CLL, подвергнутых лечению ритуксимабом, выявили, что опосредованное антителами против CD20 истощение B-клеток по большей части обогащало пул B-рег. Предположили, что обогащенные B-рег супрессируют противоопухолевый иммунитет, необходимый для уничтожения связанных с антителами против CD20 клеток опухолей, вызывая у пациентов развитие устойчивости лимфомы против терапии антителами против CD20 и/или, в конечном счете, рецидив, в результате усиленного прогрессирования злокачественной опухоли (Bodogai et al., (2013) Cancer Res 73:2127-2138).
[0009] Наблюдения, что B-клетки человека отрицательно регулируют рост опухолей, отмечены, когда присутствие инфильтрующих CD20+ опухоль лимфоцитов - B-клеток при раке яичника, немелкоклеточной карциноме легкого и раке шейки матки коррелировало с улучшенной выживаемостью и более низкой частотой рецидивов. Эти исследования показали, что инфильтрующие опухоль B-клетки коррелировали с благоприятными исходами. B-рег могут супрессировать разнообразные подтипы клеток, включая T-клетки, посредством секреции противовоспалительных медиаторов, таких как IL-10, и могут способствовать переходу T-клеток в регуляторные T-клетки, таким образом, ослабляя противоопухолевые иммунные ответы. Потенциальные механизмы, лежащие в основе B-клеточного противоопухолевого иммунитета, могут включать секрецию B-клетками эффекторных цитокинов, таких как IFN-γ, которые могут вызывать поляризацию T-клеток в направлении Th1- или Th2-ответа или стимулировать ответы T-клеток благодаря своей роли в качестве антигенпредставляющих клеток (Sarvaria et al. (2017) Cell Mol Immunol 14(8):662-674).
[0010] Однако показано также, что B-клетки человека способствуют прогрессированию опухолей. Можно было показать корреляцию присутствия B-клеток с активированным STAT3 в тканях опухолей человека и тяжести ангиогенеза опухолей. А также, инфильтрация CD19+ B-клеток у пациентов с метастазирующей карциномой яичников; или увеличенная инфильтрация CD20+ и CD138+ B-клеток у пациентов с эпителиальным раком яичника ассоциирована с плохим прогнозом и исходом заболевания. Кроме того, уменьшение опухолевой нагрузки после частичного уменьшения количества B-клеток с помощью ритуксимаба обнаружено у 50% пациентов с колоректальным раком на поздних стадиях. Таким образом, точная роль, которую B-рег играют при злокачественных опухолях все еще неясна, но наиболее вероятно, относится к активности подтипов B-рег (Sarvaria, там же).
[0011] Значительное количество нарушений либо вызваны недостаточностью секретированного продукта гена, либо поддаются лечению посредством секреции терапевтического белка. Нарушения свертывания крови, например, являются весьма распространенными генетическими нарушениями, где факторы в каскаде свертывания крови являются нарушенными некоторым образом, т.е. с отсутствием экспрессии или с продукцией мутантного белка. Большинство нарушений свертывания крови приводят к гемофилиям, таким как гемофилия A (недостаточность фактора VIII), гемофилия B (недостаточность фактора IX) или гемофилия C (недостаточность фактора XI). Лечение этих нарушений часто связано с их тяжестью. Для мягких гемофилий, лечение может включать лекарственные средства, разработанные для увеличения экспрессии фактора с недостаточной экспрессией, в то время как для более тяжелых гемофилий, терапия включает регулярную инфузию отсутствующего фактора свертывания (часто 2-3 раза в неделю посредством заместительной ферментной терапии (ERT)), для предотвращения случаев кровотечения. Пациентов с тяжелой гемофилией часто предостерегают от занятий многими видами спорта, и они должны предпринимать дополнительные предосторожности для избегания повседневных повреждений.
[0012] Недостаточность альфа-1-антитрипсина (A1AT) представляет собой аутосомно-рецессивное заболевание, вызванное недостаточностью продукции альфа-1-антитрипсина, приводящей к неадекватным уровням A1AT в крови и легких. Она может являться ассоциированной с развитием хронического обструктивного заболевания легких (COPD) и нарушений печени. В настоящее время, лечение заболеваний, ассоциированных с этой недостаточностью, может включать инфузию экзогенного A1AT и трансплантацию легкого или печени.
[0013] Заболевания с накоплением лизосом (LSD) представляют собой группу редких метаболических моногенных заболеваний, характеризуемых отсутствием функциональных индивидуальных лизосомных белков, в норме вовлеченных в разрушение ненужных липидов, гликопротеинов и мукополисахаридов. Эти заболевания характеризуются накоплением этих соединений в клетке, поскольку она является неспособной осуществлять их процессинг для рециркуляции из-за неправильного функционирования специфического фермента. Обычные примеры включают болезни Гоше (недостаточность глюкоцереброзидазы - наименование гена: GBA), Фабри (недостаточность α-галактозидазы - GLA), Гунтера (недостаточность идуронат-2-сульфатазы - IDS), Гурлера (недостаточность альфа-L-идуронидазы - IDUA) и Ниманна-Пика (недостаточность сфингомиелинфосфодиэстеразы 1 - SMPD1).
[0014] Диабет I типа представляет собой нарушение, при котором иммуноопосредованное разрушение бета-клеток поджелудочной железы приводит к значительной недостаточности инсулина, первостепенного секретируемого продукта этих клеток. Восстановление фоновых уровней инсулина обеспечивает значительное облегчение множества более серьезных осложнений этого нарушения, которые могут включать «макрососудистые» осложнения, затрагивающие крупные сосуды: ишемическую болезнь сердца (стенокардия и инфаркт миокарда), инсульт и заболевание периферических сосудов, так же как «микрососудистые» осложнения из-за повреждения малых кровеносных сосудов. Микрососудистые осложнения могут включать диабетическую ретинопатию, которая влияет на формирование кровеносных сосудов в сетчатке глаза, и может приводить к зрительным симптомам, нарушению зрения и потенциально, слепоте, и диабетическую нефропатию, которая может включать рубцовые изменения в ткани почки, потерю небольшого или постепенно увеличивающегося количества белка в моче, и в конечном счете, хронической почечной недостаточности, требующей диализа.
[0015] Однако, предоставление терапевтических белков для лечения нарушений у субъекта может быть ограничено собственным иммунным ответом субъекта на терапевтический белок, включая продукцию B-клетками у субъекта антител, которые могут ограничивать эффективность таких способов лечения. Например, у пациентов с гемофилией, подвергаемых ERT фактора (факторов) свертывания, недостаточных или отсутствующих у них (например, фактора VIII, фактора IX и т.д.), могут образовываться антитела против этих необходимых белков (например, антитела против F9). По оценкам, у 15-50% пациентов с гемофилией A развиваются ингибирующие антитела против терапевтического белка фактора 8 (Krudysz-Amblo et al., (2009) Blood 113(11):2587-2594). В некоторых случаях, реакции могут быть тяжелыми (анафилактический шок), приводя к ситуации, когда необходимая ERT вызывает опасные побочные эффекты у пациента (см., например, JM Lusher (2000) Semin Thromb Hemost 26(2):179-188).
[0016] Кроме того, антитела представляют собой секретированные белковые продукты, пластичность связывания которых использовали для разработки широкого разнообразия лекарственных средств. Терапевтические антитела можно использовать для нейтрализации белков-мишеней, непосредственно вызывающих заболевание (например, VEGF при дегенерации желтого пятна), так же как для высокоизбирательного уничтожения клеток, персистенция и воспроизведение которых представляет опасность для хозяина (например, клеток злокачественных опухолей, так же как определенных иммуноцитов при аутоиммунных заболеваниях, включая B-клетки, продуцирующие антитела против собственных антигенов). При таких применениях, терапевтические антитела пользуются преимуществами нормального ответа организма посредством его собственных антител для достижения избирательного уничтожения, нейтрализации или выведения белков-мишеней или клеток, несущих антиген-мишень антитела. Таким образом, терапию с использованием антител широко применяют для множества состояний человека, включая онкологию, ревматологию, трансплантацию и заболевания глаз.
[0017] Таким образом, остается необходимость в дополнительных способах и композициях, которые можно использовать для экспрессии желательного трансгена на терапевтически значимом уровне у субъекта для лечения генетических заболеваний, таких как гемофилии, диабет, заболевания с накоплением лизосом и/или недостаточность A1AT, включая лечение и/или исключение любой ассоциированной токсичности, и которые могут ограничивать экспрессию трансгена или терапевтического белка желательным типом ткани, включая ограничение врожденных ответов B-клеток. Кроме того, остается необходимость в дополнительных способах и композициях для экспрессии желательного трансгена (например, антитела) на терапевтически значимом уровне для лечения других заболеваний, таких как злокачественные опухоли.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0018] Сайт-специфическая модификация B-клеток в одном или более генетических локусах может улучшать функцию B-клеток (включая усиление продукции антител этими клетками и/или нацеливание B-клеток для продукции белков, ограничивающих нежелательные врожденные иммунные ответы), дифференцировку в плазмобласты и способность к приживлению при трансплантации. Контроль B-клеток посредством направленной модификации генов (например, нарушения и/или добавления геномного или эписомного гена) обеспечивает эффективные и менее токсичные виды замещающей белковой терапии и кроме того, обеспечивает коммуникацию внутри зародышевых центров, подлежащих программированию.
[0019] Настоящее изобретение относится к композициям и способам для модуляции экспрессии гена-мишени в B-клетке и/или экспрессии трансгена в B-клетке (включая производные плазмобласты или плазматические клетки). Таким образом, настоящее изобретение относится к генетически модифицированным B-клеткам (включая B-клетки, происходящие из генетически модифицированных гематопоэтических стволовых клеток или других предшественников B-клеток), содержащим одну или более из следующих модификаций: включение одного или более трансгенов в клетку; и/или вставки и/или делеции, модифицирующие (i) гены B-клеточного рецептора и/или (ii) клеточные взаимодействия в зародышевых центрах; и/или (c) модификации, ингибирующие супрессию какой-либо функции B-клетки, ассоциированную с инфекцией патогеном или регуляцией злокачественной опухоли. Трансген(ы) можно экспрессировать вне хромосом (эписомально) и/или можно интегрировать в геном B-клетки (например, посредством опосредованной нуклеазой направленной интеграции, например, в локус области безопасности). В некоторых вариантах осуществления, один или более трансгенов поддерживают эписомально, и один или более трансгенов интегрируют в геном клетки (B-клетки или HSC, подлежащей дифференцировке в B-клетку). В некоторых вариантах осуществления, трансген кодирует белок, вовлеченный в каскад свертывания крови. В других вариантах осуществления, трансген кодирует фермент, дефектный при нарушении с накоплением лизосом, или кодирует терапевтическое антитело. В других вариантах осуществления, трансген кодирует молекулу, нацеленную на B-клетку, продуцирующую нежелательное антитело, например, антитело против терапевтического белка, включая, но без ограничения, антитела против эндогенного белка (например, аутоантитела при аутоиммунных заболеваниях) и/или экзогенного белка (например, белка, поставляемого посредством ERT, такого как фактор свертывания крови). Например, трансген может кодировать антитело, узнающее B-клеточный рецептор на B-клетках, которые являются чувствительными к желательному белку (эндогенному белку при аутоиммунном заболевании и/или белку, поставляемому посредством ERT), для нацеливания на популяцию B-клеток, продуцирующих нежелательные антитела против желательного белка. Неограничивающие примеры таких антител включают антитела, узнающие B-клеточный рецептор, ассоциированный с B-клеткой, продуцирующей антитела против белков, поставляемых посредством ERT, таких как факторы свертывания крови при гемофилии (например, B-клетками, продуцирующими антитела против F9 или против F8); и/или узнающие B-клеточный рецептор на B-клетке, продуцирующей антитела против ауто-/собственных антигенов, включая, но без ограничения, антитела против основного белка миелина (MBP) при MS, противоядерные антитела (ANA) при системной красной волчанке (SLE), антитела против гликопротеинов в сердце, суставах и других тканях при ревматической атаке, антитела против Fc-фрагмента IgG при ревматоидном артрите (RA), так же как на B-клетках, продуцирующих аутоантитела при синдроме Рейтера, синдроме Шегрена, системной склеродермии (склеродермии), воспалительных миопатиях, узелковом полиартериите, болезни Грэйвса, диабете I типа и т.п. Композиции и способы, описанные в настоящем описании, обеспечивают высокие уровни продукции белков как in vitro, так и in vivo, включая уровни, достаточные для проявления клинически значимых (терапевтических) эффектов in vivo.
[0020] В одном аспекте, настоящее изобретение относится к полинуклеотидной экспрессирующей конструкции, содержащей по меньшей мере один специфический для B-клеток промотор, управляющий экспрессией одного или более трансгенов. Промотор B-клетки может быть выбран из любого промотора, который является активным в B-клетках, включая, но без ограничения, промотор цепи каппа иммуноглобулина (Igк, Laurie et al. (2007) Gene Ther 14(23): 1623-31), B29 (Hermanson et al. (1989) Proc Nat'l Acad Sci 86: 7341-7345), BCL6 (Ramachandrareddy et al. (2010) Proc Nat'l Acad Sci 107(26):11930-11935), промотор III CIITA (Deffernnes et al. (2001) J. Immunol 167(1): 98-106), mb-1 (см., например, Malone and Wall (2001) J. Immunol 168(7):3369-3375) и промотор EEK, содержащий промотор легкой цепи (VKp), которому предшествует интронный энхансер (iEκ), MAR и 3′-энхансер (3′Eκ) (см. Патент США 8133727). В некоторых вариантах осуществления, используемый специфический для B-клетки промотор в норме экспрессируется в зародышевом центре, так что трансген экспрессируется, когда клетка находится в зародышевом центре (например, BCL6, Basso et al. (2010) Blood 115(5):975-984). В некоторых вариантах осуществления, трансген вставляют посредством опосредованной нуклеазой направленной интеграции, так что им управляет специфический для B-клетки промотор в геноме клетки. В других вариантах осуществления, конструкция промотор B-клетки-трансген является частью ДНК-вектора, поддерживаемого вне хромосом. В некоторых вариантах осуществления, конструкцию промотор B-клетки-трансген вставляют в транскрипционно молчащий локус и/или локус области безопасности генома B-клетки, например, в ген альбумина или в ген, кодирующий субъединицу T-клеточного рецептора (например, TCRA или TCRB).
[0021] В некоторых аспектах, трансген кодирует фермент, отсутствующий или недостаточный у субъекта. В некоторых вариантах осуществления, трансген кодирует фактор свертывания крови, такой как фактор VII, фактор VIII, фактор IX, фактор X, фактор XI или фактор XII. В других вариантах осуществления, трансген кодирует фермент, недостаточный при заболевании с накоплением лизосом, включая, но без ограничения, глюкоцереброзидазу (GBA), α-галактозидазу (GLA), β-глюкуронидазу (GUSB), идуронат-2-сульфатазу (IDS), альфа-L-идуронидазу (IDUA), сфингомиелинфосфодиэстеразу 1 (SMPD1) или альфа-глюкозидазу (GAA). В некоторых вариантах осуществления, трансген кодирует A1AT. Неограничивающие примеры белков, которые можно экспрессировать, как описано в настоящем описании, включают также фибриноген, протромбин, тканевой фактор, фактор V, фактор фон Виллебранда, прекалликреин, высокомолекулярный кининоген (фактор Фитцджеральда), фибронектин, антитромбин III, кофактор гепарина II, белок C, белок S, белок Z, родственный белку Z ингибитор протеаз, плазминоген, альфа-2-антиплазмин, тканевой активатор плазминогена, урокиназу, ингибитор-1 активатора плазминогена, ингибитор-2 активатора плазминогена, MMAA, MMAB, MMACHC, MMADHC (C2orf25), MTRR, LMBRD1, MTR, пропионил-CoA-карбоксилазу (PCC) (субъединицы PCCA и/или PCCB), белок-транспортер глюкозо-6-фосфата (G6PT) или глюкозо-6-фосфатазу (G6P-азу), рецептор LDL (LDLR), ApoB, LDLRAP-1, PCSK9, митохондриальный белок, такой как NAGS (N-ацетилглутаматсинтетаза), CPS1 (карбамоилфосфатсинтетаза I) и OTC (орнитинтранскарбамилаза), ASS (синтетазу аргининоянтарной кислоты), ASL (лиазу аргининосукцинатной кислоты) и/или ARG1 (аргиназу), и/или белок из семейства переносчиков растворенных веществ 25 (SLC25A13, переносчик аспартата/глутамата), UGT1A1 или полипептид A1 УДФ-глюкуронилтрансферазы, фумарилацетоацетат-гидролиазу (FAH), белок аланин-глиоксилат-аминотрансферазу (AGXT), белок глиоксилатредуктазу/гидроксипируватредуктазу (GRHPR), белок - продукт гена транстиретина (TTR), белок ATP7B, белок фенилаланингидроксилазу (PAH), белок липопротеинлипазу (LPL), сконструированную нуклеазу, сконструированный фактор транскрипции и/или сконструированный одноцепочечный вариабельный фрагмент антитела (диатело, антитело верблюдовых и т.д.). В одном предпочтительном варианте осуществления, трансген кодирует полипептид FVIII. В некоторых вариантах осуществления, полипептид FVIII содержит делецию домена B. В некоторых вариантах осуществления, настоящее изобретение относится к способам и композициям для экспрессии терапевтически значимых уровней одного или более терапевтических белков с одного или более трансгенов. В конкретных вариантах осуществления, экспрессия конструкции трансгена, кодирующей замещающий белок, приводит к 1% от нормальных уровней продуцированного белка, в то время как в других продуцируется 2%, 3%, 4%, 5%, 10%, 15%, 20%, 30%, 50%, 80%, 100%, 150%, 200% или более от нормальных уровней белка. В некоторых вариантах осуществления, трансген кодирует полипептид, преодолевающий ингибирование ответа антител посредством вируса, бактерии или паразита.
[0022] Положительные по CD19 B-клетки подвергаются дифференцировке in vitro до плазмобластов и плазматических клеток, продуцирующих более 10000 нг/мл антител (IgG, IgM, IgA). Таким образом, в некоторых аспектах, трансген кодирует терапевтический белок, такой как одноцепочечное антитело. В некоторых вариантах осуществления, одноцепочечное антитело представляет собой scFv, в то время как в других вариантах осуществления, одноцепочечное антитело представляет собой антитело верблюдовых или наноантитело (см., например, Mejias et al. (2016) Sci Reports 6: srep24913, doi:10:1038). В других аспектах, более одного трансгена экспрессируют в B-клетке. В одном варианте осуществления, более одного трансгена включают последовательности, необходимые для экспрессии полноразмерного антитела или его фрагмента, или другого антигенсвязывающего белка (например, монотела, аптамера, дарпина, аднектинов, аффител, антикалинов, ингибиторов типа Куница и т.д. (Gebauer and Skerra (2009) Curr Opin Chem Biol 13(3):245-55).
[0023] В некоторых вариантах осуществления, популяцию B-клеток, содержащих трансген, кодирующий представляющий интерес терапевтический белок, модифицируют ex vivo и затем повторно вводят нуждающемуся в этом субъекту. Популяцию B-клеток можно модифицировать, как описано в настоящем описании, на любой стадии развития, включая, но без ограничения, форму гематопоэтической стволовой клетки (HSC), лимфоидной клетки-предшественника или зрелой B-клетки. Стволовые клетки или предшественники B-клеток можно модифицировать и затем подвергать дифференцировке in vitro, и вводить в форме предшественников (линейно-коммитированных B-клеток) или зрелых клеток субъекту. Альтернативно, модифицированные стволовые клетки или клетки-предшественники B-клеток могут быть модифицированы in vitro, как описано в настоящем описании, и подвергаться полной дифференцировке в зрелые B-клетки in vivo после введения. Таким образом, для введения ex vivo, популяции B-клеток, как описано в настоящем описании, могут являться гетерогенными в том отношении, что они включают стволовые клетки, клетки-предшественники и/или зрелые B-клетки, находящиеся на различных стадиях развития. Альтернативно, популяции B-клеток могут являться гомогенными и включать только стволовые клетки, клетки-предшественники или зрелые клетки. В других вариантах осуществления, модифицированные B-клетки получают in vivo с использованием вектора для доставки, способного трансдуцировать B-клетки. В следующих вариантах осуществления, вектор для доставки представляет собой вирусный вектор, предпочтительно, аденоассоциированный вирус (AAV). В предпочтительных вариантах осуществления, вектор AAV представляет собой AAV6. В других вариантах осуществления, вектор для доставки является невирусным, например, мРНК, липидная наночастица (LNP) или плазмидный вектор.
[0024] В следующем аспекте, модифицированные B-клетки, как описано в настоящем описании (включая популяции B-клеток), выращивают in vitro для продукции белка, кодированного трансгеном. В предпочтительных вариантах осуществления, белок представляет собой антитело или антигенсвязывающий белок (например, антитело, которое связывается с эндогенными B-клетками, продуцирующими нежелательные антитела у субъекта, включая эндогенные B-клетки, продуцирующие антитела против поставляемых посредством ERT белков, таких как факторы свертывания крови, и/или B-клетки, продуцирующие антитела против собственных белков при аутоиммунных нарушениях), или антитело, нейтрализующее нежелательные антитела. Белок, продуцированный B-клетками, можно выделять и использовать для белковой терапии, такой как заместительная ферментная терапия, и/или в сочетании с заместительной ферментной терапией, для уменьшения и/или исключения врожденной продукции нежелательных антител (например, антител против ERT, образующихся после ERT).
[0025] В некоторых аспектах, изобретение относится к способам и композициям для доставки B-клетки или плазматической клетки, экспрессирующей трансген, пересекающей гематоэнцефалический барьер, которые можно использовать для лечения и/или предотвращения заболевания ЦНС или заболевания, затрагивающего ЦНС. В некоторых вариантах осуществления, трансген кодирует фермент, отсутствующий у субъекта с нарушением с накоплением лизосом. В следующих вариантах осуществления, трансген кодирует глюкоцереброзидазу (GBA), α-галактозидазу (GLA), β-глюкуронидазу (GUSB), идуронат-2-сульфатазу (IDS), альфа-L-идуронидазу (IDUA), сфингомиелинфосфодиэстеразу 1 (SMPD1) или альфа-глюкозидазу (GAA), и его используют для лечения или предотвращения заболевания ЦНС, ассоциированного с болезнью Гоше (Bae et al., (2015) Exp Mol Med 47, e153; doi:10:1038/emm.2014,128), болезнью Фабри, MPS типа VII (Sly et al. (1973) J Pediatr. 1973 Feb; 82(2):249-57), MPS II, MPS I, болезнью Ниманна-Пика или Помпе, соответственно.
[0026] В следующих аспектах, способы и композиции по изобретению включают модифицированную B-клетку или производное B-клетки (плазмобласт, плазматическую клетку), содержащие трансген, а также одну или более дополнительных модификаций. Дополнительная модификация может представлять собой дополнительные эписомные или дополнительные интегрированные последовательности (которые могут быть интегрированы в одну и ту же и/или в различные локализации в геноме). В некоторых вариантах осуществления, содержащие трансген B-клетки содержат дополнительные белковые или пептидные последовательности (или кодирующие их полинуклеотиды), способствующие эффективности пересечения гематоэнцефалического барьера. В некоторых вариантах осуществления, пептид содержит пептид, известный в данной области как облегчающий пересечение барьера к головному мозгу. В следующих вариантах осуществления, пептид представляет собой антитело, экспрессированное на поверхности B-клетки, нацеленное на рецептор трансферрина, в то время как в других вариантах осуществления, пептид представляет собой металлотрансферрин (Karkan et al. (2008) PLoS ONE 3(6):e2469. doi:10.1371/journal.poine.0002469. В некоторых вариантах осуществления, пептид представляет собой рецептор, такой как VLA-4, ICAM-1, IL-8Ra (CXCR1) или IL-8Rb (CXCR2) (Alter et al. (2003) J. Immunol 170:4497-4505). В любом из этих вариантов осуществления, трансген может кодировать фермент, отсутствующий при заболевании с накоплением лизосом, таком как описанные выше, так что фермент доставляют в ЦНС нуждающегося в этом субъекта.
[0027] В некоторых аспектах, модифицированные B-клетки по изобретению содержат дополнительные модификации (например, мутации), способствующие приживлению после трансплантации. В некоторых вариантах осуществления, экспрессию специфических генов ингибируют (например, посредством временной репрессии или постоянного нокаута) для увеличения приживления и/или размера зародышевого центра. Гены, подвергаемые такому ингибированию, включают, но без ограничения, гены инозитол-гексакисфосфат-киназ (Zhang et al. (2014) Basic Res Cardio 109(4): 417), киназы-3β синтазы гликогена (GSK-3β, см. Ko et al. (2011) Stem Cells 29(1):108-18), пептидазы CD26 (DPPIV/дипептидилпептидазы IV), (Tian et al. (2006) Gene Ther 13(7):652-8), RhoA (Ghiaur et al. (2006) Blood 108(6):2087-94), EAF2 (Li et al., (2016) Nat Com 7; doi: 10.1038/ncomms10836), белков аутофагии, таких как Atg5 (Pengo et al., (2013) Nat Immunol 14(3):298-305), и т.п. В следующих вариантах осуществления, модифицированные B-клетки можно дополнительно модифицировать для репрессии (например, нокаута) генов, ассоциированных с индукцией реакции трансплантат против хозяина. В некоторых вариантах осуществления, гены, кодирующие B-клеточный рецептор, подвергают нокауту для предотвращения стимуляции B-клетки у хозяина.
[0028] В других аспектах, модифицированные B-клетки по изобретению содержат дополнительные модификации (мутации, такие как геномные вставки и/или делеции; эписомную экспрессию трансгенов и т.д.), регулирующие клеточные взаимодействия (например, взаимодействия T-клетка-B-клетка) в зародышевых центрах и/или ингибирование супрессии какой-либо функции B-клетки (например, продукции антител, экспрессии цитокинов, передачи сигналов и т.д.), ассоциированных с инфекцией патогеном. В некоторых аспектах, модифицированные B-клетки по изобретению дополнительно содержат белки (или последовательности, кодирующие эти белки) для ингибирования B-клеток, вовлеченных в онкогенную активность. Например, в некоторых вариантах осуществления, модифицированные B-клетки содержат экспрессированное на поверхности антитело против убиквитингидролазы UCH-L1 (включая кодирующий его трансген) для супрессии B-клеток, вовлеченных в некоторые типы крупноклеточной В-клеточной лимфомы (Bedekovics et al. (2016) Blood 127(12):1564-74).
[0029] В другом аспекте, настоящее изобретение относится к фармацевтическим композициям, содержащим одно или более из клеток, экспрессирующих конструкций и/или, необязательно, нуклеаз, описанных в настоящем описании.
[0030] Для опосредованной нуклеазой направленной интеграции экспрессирующих конструкций по настоящему изобретению в подходящей локализации в B-клетке, можно использовать любую нуклеазу, включая, но без ограничения, одну или более нуклеаз с цинковыми пальцами (ZFN), TALEN, нуклеаз CRISPR/Cas и/или нуклеаз TtAgo, так чтобы экспрессирующая конструкция интегрировалась в область (ген), расщепленные нуклеазой (нуклеазами). В конкретных вариантах осуществления, используют одну или более пар нуклеаз. Нуклеазы можно вводить в форме мРНК или можно вводить в клетку с использованием невирусных или вирусных векторов. В некоторых аспектах, полинуклеотиды нуклеазы можно доставлять посредством лентивируса или посредством неинтегрирующего лентивируса. В других аспектах, экспрессирующую кассету можно доставлять посредством AAV и/или ДНК-олигонуклеотидов.
[0031] В любом из описанных композиций и способов, экспрессирующие кассеты и/или нуклеазы может нести вектор AAV, включая, но без ограничения, AAV1, AAV3, AAV4, AAV5, AAV6, AAV8, AAV9 и AAVrh10 или псевдотипированный AAV, такой как AAV2/8, AAV8,2, AAV2/5 и AAV2/6, и т.п. В конкретных вариантах осуществления, полинуклеотиды (экспрессирующих конструкций и/или нуклеаз) доставляют с использованием одинаковых типов вектора AAV. В других вариантах осуществления, полинуклеотиды доставляют с использованием различных типов векторов AAV. Полинуклеотиды можно доставлять с использованием одного или более векторов. В следующих вариантах осуществления, полинуклеотиды доставляют посредством липидной наночастицы (LNP). В конкретных вариантах осуществления, полинуклеотиды доставляют посредством введения в селезенку или лимфатический узел интактного животного. В других вариантах осуществления, полинуклеотиды доставляют посредством внутривенного введения в периферическую вену.
[0032] Способы, описанные в настоящем описании, можно осуществлять на практике in vitro, ex vivo или in vivo. В конкретных вариантах осуществления, композиции вводят живому, интактному млекопитающему. Млекопитающее может находиться на любой стадии развития на время доставки, например, эмбрион, плод, новорожденный, младенец, подросток или взрослый. Кроме того, целевые клетки могут являться здоровыми или пораженными заболеванием. В конкретных вариантах осуществления, одну или более из композиций доставляют в специфическую ткань (например, селезенку или лимфатический узел), внутриартериально, внутрибрюшинно или внутримышечно. Доставку ex vivo можно осуществлять с использованием гомогенных или гетерогенных популяций клеток, включая стволовые клетки, клетки-предшественники B-клеток и/или зрелые B-клетки.
[0033] Настоящее изобретение относится также к набору, содержащему одно или более из экспрессирующих конструкций, векторов AAV, B-клеток и/или фармацевтических композиций, описанных в настоящем описании. Набор может дополнительно содержать нуклеиновые кислоты, кодирующие нуклеазы, (например, молекулы РНК, кодирующие белки ZFN, TALEN или Cas и модифицированную Cas, и направляющие РНК), или аликвоты белков нуклеаз, клеток, инструкции для осуществления способов по изобретению и т.п.
[0034] Эти и другие аспекты будут ясно очевидны специалисту в данной области в свете описания в целом.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0035] Фиг. 1 представляет собой схему, показывающую обзор протокола размораживания и дифференцировки B-клеток in vitro, которому следовали (см. Jourdan et al. (2009) Blood 114:5173-5181).
[0036] Фиг. 2A - 2C представляют собой графики, показывающие способность подвергнутых дифференцировке in vitro B-клеток продуцировать антитела, включая антитела IgM (фиг. 2A), антитела IgG (фиг. 2B) и антитела IgA (фиг. 2C). Образцы обрабатывали с использованием цитокинов («+цитокины») или нет, и затем количество антитела детектировали посредством ELISA. Супернатанты собирали на сутки t4, t7 и t10, и суммарные уровни антител IgM, IgG и IgA оценивали количественно посредством специфического ELISA. Данные представляют собой технические повторы. Планки погрешностей представляют собой стандартное отклонение.
[0037] Фиг. 3A - 3D представляют собой графики, показывающие процент положительных по GFP клеток после электропорации мРНК в B-клетки через 0 суток (фиг. 3A), 1 сутки (фиг. 3B), 2 суток (фиг. 3C) или 3 суток (фиг. 3D) после размораживания. CD19+ B-клетки подвергали электропорации с использованием мРНК на t0, t1, t2 и t3, где t равно количество суток после размораживания, для определения оптимальной временной точки для добавления мРНК. Положительные по CD19+ B-клетки (2,0E+05 клеток) смешивали с мРНК GFP (2 мкг) с последующей электропорацией. Клетки собирали через 24 часа и анализировали посредством проточной цитометрии для оценки уровней GFP. Сутки 2 после размораживания (t2) имели наивысшие уровни GFP и выбраны для дальнейших исследований. Данные представляют собой технические повторы. Планки погрешностей представляют собой стандартное отклонение.
[0038] Фиг. 4A и 4B представляют собой графики, показывающие отбор при проточной цитометрии по экспрессии GFP в трансдуцированных клетках. На фиг. 4A определены области графика, ассоциированные с боковым светорассеянием («SSC») и прямым светорассеянием («FSC»). Фиг. 4B иллюстрирует различия между группой B-клеток после ложной обработки (левая панель) и клеток, подвергнутых электропорации с использованием кодирующей мРНК GFP (правая панель). Как можно видеть на правой панели фигуры 4B, экспрессия мРНК GFP приводит к усилению образованной GFP флуоресценции, которую можно количественно оценивать после отбора.
[0039] Фиг. 5A - 5C представляют собой графики, показывающие редактирование генома в B-клетках. Глубокое секвенирование во множестве локусов показало надежное редактирование генома с использованием нуклеаз с цинковыми пальцами, нацеливающих на AAVS1, CCR5 и TCRA (TRAC). Процент модификации генома рассчитывали посредством деления количества последовательностей, содержащих вставку и делецию (инделы), на общее количество последовательностей. CD19+ B-клетки (2,0E+5 клеток) смешивали с мРНК ZFN (4 мкг) с последующей электропорацией. Клетки собирали в течение времени (t=сутки) после трансфекции. Данные представляют собой технические повторы. Планки погрешностей представляют собой стандартное отклонение. На фигуре 5A изображено редактирование генома в локусе AAVS1 на сутки 4, 7 и 10. На фиг. 5B показан сходный набор данных в локусе CCR5, в то время как на фиг. 5C показаны данные для локуса TCRA (TRAC).
[0040] На Фиг. 6A - 6C показан эффект временного холодового шока на редактирование генома B-клетки. CD19+ B-клетки (2,0E+5 клеток) смешивали с мРНК ZFN (0,75, 1,5, 3 и 6 мкг) с последующей электропорацией. Клетки после электропорации разделяли на две группы. Одну группу помещали в инкубатор при 37°C на 4 суток. Вторую группу помещали при 30°C на ночь, затем переносили в инкубатор при 37°C на 3 суток. Глубокое секвенирование выявило увеличение редактирования генома (% инделов) в клетках, обработанных с использованием временного холодового шока.
[0041] На фиг. 7A и 7B изображена способность к трансдукции нескольких серотипов AAV, тестированных по доставке промотора CMV - донора GFP в CD19+ B-клетки. На фиг. 7A показаны результаты для рекомбинантных серотипов AAV 2, 5, 6, 8 и 9, несущих CMV-GFP, доставляемых в дозах вектора 2,4E+6, 1,2E+6, 6,0E+5, 3,0E+5 геномов вектора (вг)/клетку. Показаны данные из n=2 биологических повторов, где клетки анализировали по экспрессии GFP на сутки 4, 7 и 10 после трансдукции, и они демонстрируют, что AAV6 легко трансдуцирует B-клетки. Планки погрешностей представляют собой стандартное отклонение технических и биологических повторов. Фигура 7B представляет собой схематическое изображение экспрессирующей кассеты, используемой в показанных экспериментах.
[0042] На фиг. 8 изображены использованные иллюстративные экспрессирующие кассеты AAV. Показаны иллюстративные донорные кассеты для вставки в локусы AAVSI, CCR5 и TCRA (TRAC). AAVS1 имеет левое («AAVS1-L») и правое («AAVS1-R») плечи гомологии, имеющие длину 801 и 568 пар оснований, соответственно. CCR5 имеет левое («CCR5-L») и правое («CCR5-R») плечи гомологии, имеющие длину 473 и 1431 пар оснований, соответственно. TCRA («TRAC») имеет левое («TRAC-L») и правое («TRAC-R») плечи гомологии, имеющие длину 925 и 989 пар оснований, соответственно. Доноры содержали либо промотор фосфоглицераткиназы («PGK»), либо специфический для B-клетки промотор («EEK», содержащий промотор легкой цепи (VKp), которому предшествовал интронный энхансер (iEκ), MAR и 3-энхансер (3′Eκ); Патент США 8133727), за которым следовал кодирующий трансген GFP («GFP») и сигнал полиаденилирования бычьего гормона роста («pA»). Инвертированные концевые повторы AAV2 («ITR») использовали для обеспечения упаковки в капсиды AAV.
[0043] Фиг. 9A - 9C представляют собой графики, показывающие, что комбинация мРНК ZFN и векторов rAAV2/6 способствовала высоким уровням добавления трансгена во множестве локусов. Уровни экспрессии GFP измеряли посредством проточной цитометрии. Культуры B-клеток собирали в течение времени (t=сутки) после добавления мРНК ZFN и донорного AAV в B-клетки. Локусы AAVS1 (фиг. 9A), CCR5 (фиг. 9B) и TCRA (TRAC, фиг. 9C) оценивали по добавлению трансгена. Для образцов с ZFN:донором показана длительная экспрессия GFP, в то время как для образцов только с донором показано уменьшение экспрессии GFP с течением времени. Под каждым графиком показан участок-мишень для нуклеаз (например, на фиг. 9A, «ZFN: AAVS1»). Показано также описание трансгена GFP (например, на фиг. 9A, «Донор: PGK-AAVS1» показывает, что трансгеном GFP управлял промотор PGK, и что кодирующая последовательность GFP была фланкирована плечами гомологии с гомологией с участком для нуклеазы в гене AAVS1). На фиг. 9A - 9C, на левой панели показаны результаты после сбора через 4 суток (t4) после трансфекции; на средней панели показаны результаты после сбора через 7 суток (t7) после трансфекции; и на правой панели показаны результаты после сбора через 10 суток после трансфекции.
[0044] Фиг. 10A и 10B представляют собой графики, показывающие процент направленной интеграции донора GFP в локусы AAVS1 (фиг. 10A) и CCR5 (фиг. 10B) в CD19+ B-клетках. Подтверждение направленной интеграции (добавления трансгена) осуществляли посредством глубокого секвенирования. Процент модификации генов рассчитывали посредством деления количества последовательностей, содержащих интегрированную мишень, на общее количество последовательностей. B-клетки собирали в течение времени (t=сутки) после добавления мРНК ZFN и донорного AAV в B-клетки. Данные для AAVSI представляют 3 независимых эксперимента. Данные для CCR5 представляют 2 независимых эксперимента. Планки погрешностей представляют собой стандартное отклонение. Под каждым графиком показан участок-мишень для нуклеаз и конфигурация донора, как описано выше.
[0045] Фиг. 11 представляет собой график, показывающий результаты определения того, гомологичную рекомбинацию (HDR) или связывание концов посредством соединения негомологичных концов (NHEJ) используют B-клетки для направленной интеграции. В таблице под графиком показана специфичность ZFN и конфигурация донора. Во всех донорах использовали промотор PGK, но только в эксперименте #1 присутствовал донорный трансген GFP, фланкированный плечами гомологии, совпадающими с участком разрезания нуклеазой. Для образцов с несовпадающими плечами гомологии ZFN и донора показана экспрессия GFP, сходная с экспрессией только с донором без добавления какой-либо нуклеазы. Наиболее сильная направленная интеграция происходила, когда плечи гомологии совпадали с участком-мишенью нуклеазы, что показывает, что HDR используется для интеграции в B-клетках.
[0046] Фиг. 12 представляет собой график, показывающий сравнение промотора PGK, управляющего экспрессией GFP, со специфическим для B-клетки промотором EEK, управляющим экспрессией GFP. B-клетки смешивали с мРНК ZFN (4 мкг), нацеливающей на локус TCRA (TRAC), с последующей электропорацией. После электропорации, CD19+ B-клетки затем трансдуцировали с использованием AAV6, содержащего плечи гомологии TRAC, фланкирующего экспрессирующую трансген кассету и либо PGK, либо специфический для B-клетки промотор (EEK), управляющий экспрессией трансгена GFP. Их доставляли в дозе вектора 2,4E+06 вг/клетку. Данные показывают, что при использовании специфического для B-клетки промотора (EEK) показано небольшое увеличение экспрессии GFP по сравнению с промотором PGK.
[0047] Фиг. 13A - 13D представляют собой графики, показывающие уровень экспрессии трансгена GFP в CD19+ B-клетках в диапазоне доз донорного AAV. Во всех случаях, трансген GFP был интегрирован в локус TCRA (TRAC) с использованием специфических для TCRA нуклеаз. На каждом графике показаны также результаты для экспрессии GFP, когда трансдукцию осуществляли в отсутствие нуклеаз. Донорные конструкции содержали специфические для TCRA плечи гомологии и либо промотор EEK, как описано выше, либо промотор PGK. Использованный диапазон AAV включал 3,0E+05вг/клетку (фиг. 13D), 6,0E+05вг/клетку (фиг. 13C), 1,2E+06вг/клетку (фиг. 13B) и 2,4E+06вг/клетку (фиг. 13A). Положительные по CD19+ B-клетки смешивали с кодирующей ZFN мРНК (4 мкг), нацеливающий на локус TCRA, с последующей электропорацией. После электропорации, CD19+ B-клетки трансдуцировали с использованием AAV, содержащего плечи гомологии TCRA, либо с PGK, либо со специфическим для B-клеток промотором (EEK), управляющим экспрессией GFP. Их доставляли при дозах вектора 2,4E+06, 1,2E+06, 6,0E+05 и 3,0E+05 вг/клетку. Процент экспрессии GFP, управляемой промотором PGK, уменьшался с уменьшением дозы, в то время как специфический для B-клетки промотор сохранял экспрессию GFP на протяжении 8-кратного разведения. Присутствовало почти 5-кратное различие при 3,0E+05 вг/клетку между двумя промоторами.
[0048] Фиг. 14A - 14C представляют собой графики, показывающие влияние на продукцию антител после манипуляций редактирования генома, демонстрирующие отсутствие большой потери продукции IgG in vitro, как измерено посредством ELISA, в результате манипуляций. Общие уровни секретированных IgG являлись сходным образом независимыми в ходе эксперимента (представляющие объединенные уровни секретированных IgG на сутки 4, 7 и 10), что показывает, что электропорация и трансдукция не оказывают отрицательного влияния на продукцию IgG. Добавление цитокинов является необходимым для продукции IgG. CD19+ B-клетки обрабатывали с использованием специфической для AAVS1 ZFN (фиг. 14A), специфической для CCR5 ZFN (фиг. 14B) или специфической для TCRA (TRAC) ZFN (фиг. 14C). Различные условия, использованные для каждого набора данных, включали специфическую ZFN, образующую пару с трансгеном GFP с использованием совпадающих плеч гомологии («ZFN:донор»), трансген GFP и плечи гомологии («Донор»), только специфическую ZFN («ZFN»), CD19+ B-клетки, обработанные в устройстве BTX с использованием только буфера («BTX клетки»), необработанные CD19+ B-клетки плюс цитокины («B-клетки») и необработанные CD19+ B-клетки в отсутствие цитокинов («B-клетки - цитокины»). ZFN:донор, Донор, ZFN, BTX клетки и B-клетки все обрабатывали с использованием цитокинов.
[0049] Фиг. 15A - 15C представляют собой графики, показывающие влияние на продукцию антител после манипуляций редактирования генома, демонстрирующие отсутствие большой потери продукции IgM in vitro, как измерено посредством ELISA. CD19+ B-клетки обрабатывали с использованием специфической для AAVS1 ZFN (фиг. 15A), специфической для CCR5 ZFN (фиг. 15B) или специфической для TCRA (TRAC) ZFN (фиг. 15C). Образцы являются такими, как описано выше на фиг. 14.
[0050] Фиг. 16 представляет собой график, показывающий продукцию IgM в дифференцированных CD19+ B-клетках, обработанных с использованием цитокинов от одного донора-человека. CD19+ B-клетки подвергали обработке с использованием вируса AAV2, 5, 6, 8 или 9. В присутствии добавленных цитокинов, продукция IgM, как измерено посредством ELISA, подвергалась «бустер-эффекту» при обработке только AAV2. Преобладание антител по сравнению с AAV дикого типа в человеческой популяции является сильным и не ассоциировано с заболеванием. В настоящем описании показан потенциальный бустер-эффект на продукцию антител из-за того, что можно рассматривать как повторную инфекцию AAV.
[0051] Фиг. 17A и 17B представляют собой иллюстрации, изображающие потенциальный механизм увеличения экспрессии IgM в результате «бустер-эффекта» AAV2. На левой панели (фиг. 17A) изображен упрощенный сценарий продукции антител в B-клетке после инфекции AAV. Панель, показанная справа (фиг. 17B), представляет собой пример, обозначающий, каким образом можно задействовать AAV для функционирования в качестве бустера для увеличения экспрессии вставленного трансгена, управляемого промотором антитела в модифицированной B-клетке.
ПОДРОБНОЕ ОПИСАНИЕ
[0052] Настоящее изобретение относится к способам и композициям для генной инженерии B-клетки, включая нокаут эндогенных генов и вставку (стабильно или эписомально) экспрессирующих кассет для экспрессии трансгена. Способы можно осуществлять in vitro, ex vivo или in vivo и можно использовать для экспрессии любого трансгена(трансгенов) для лечения и/или предотвращения любого заболевания или нарушения, которые можно облегчать посредством предоставления одного или более из трансгенов.
Общее описание
[0053] В практическом осуществлении способов, так же как в получении и применении композиций, описанных в настоящем описании, используют, если не указано иное, общепринятые способы молекулярной биологии, биохимии, структуры и анализа хроматина, компьютерной химии, культуры клеток, рекомбинантной ДНК и родственных областей, находящиеся в компетенции специалистов в данной области. Эти способы полностью объяснены в литературе. См., например, Sambrook et al. MOLECULAR CLONING: A LABORATORY MANUAL, Second edition, Cold Spring Harbor Laboratory Press, 1989 и Third edition, 2001; Ausubel et al., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John Wiley & Sons, New York, 1987 и периодические обновления; серии METHODS IN ENZYMOLOGY, Academic Press, San Diego; Wolffe, CHROMATIN STRUCTURE AND FUNCTION, Third edition, Academic Press, San Diego, 1998; СПОСОБЫ IN ENZYMOLOGY, Vol. 304, «Chromatin» (P.M. Wassarman и A. P. Wolffe, eds.), Academic Press, San Diego, 1999; и METHODS IN MOLECULAR BIOLOGY, Vol. 119, «Chromatin Protocols» (P.B. Becker, ed.) Humana Press, Totowa, 1999.
Определения
[0054] Термины «нуклеиновая кислота», «полинуклеотид» и «олигонуклеотид» используют взаимозаменяемо, и они относятся к дезоксирибонуклеотидному или рибонуклеотидному полимеру, в линейной или кольцевой конформации, и либо в одноцепочечной, либо в двухцепочечной форме. Для целей по настоящему описанию, эти термины не предназначены в качестве ограничивающих по отношению к длине полимера. Термины могут включать известные аналоги природных нуклеотидов, так же как нуклеотиды, которые являются модифицированными в группах основания, сахара и/или фосфата (например, фосфоротиоатных остовов). Как правило, аналог конкретного нуклеотида обладает такой же специфичностью спаривания оснований; т.е. аналог основания A спаривается с основанием T.
[0055] Термины «полипептид», «пептид» и «белок» используют взаимозаменяемо для обозначения полимера из аминокислотных остатков. Термин относится также к аминокислотным полимерам, в которых одна или более аминокислот представляют собой химические аналоги или модифицированные производные соответствующих природных аминокислот.
[0056] «Рекомбинация» относится к процессу обмена генетической информацией между двумя полинуклеотидами, включая, но без ограничения, связывание посредством соединения негомологичных концов (NHEJ) и гомологичную рекомбинацию. Для целей по этому описанию, «гомологичная рекомбинация (HR)» относится к специализированной форме такого обмена, который имеет место, например, в ходе репарации двухцепочечных разрывов в клетках посредством механизмов направляемой гомологией репарации.
[0057] В конкретных способах по описанию, одна или более направленных нуклеаз, как описано в настоящем описании, образует двухцепочечный разрыв (DSB) в последовательности-мишени (например, клеточном хроматине) в предопределенном участке (например, гене альбумина). DSB опосредует интеграцию конструкции, как описано в настоящем описании. Необязательно, конструкция имеет гомологию с нуклеотидной последовательностью в области разрыва. Экспрессирующая конструкция может быть физически интегрирована или, альтернативно, экспрессирующую кассету используют в качестве матрицы для репарации разрыва посредством гомологичной рекомбинации, приводящей к введению всей или части нуклеотидной последовательности в качестве экспрессирующей кассеты в клеточный хроматин. Таким образом, первая последовательность в клеточном хроматине может быть изменена и, в конкретных вариантах осуществления, может быть конвертирована в последовательность, присутствующую в экспрессирующей кассете. Таким образом, использование терминов «заменить» или «замена» можно понимать как представляющее замену одной нуклеотидной последовательности на другую, (т.е., замену последовательности в информационном смысле), и оно не обязательно требует физической или химической замены одного полинуклеотида другим.
[0058] В любом из способов, описанных в настоящем описании, экзогенная нуклеотидная последовательность («экспрессирующая конструкция» или «экспрессирующая кассета», или «вектор») может содержать последовательности, которые являются гомологичными, но не идентичными, геномным последовательностям в представляющей интерес области, таким образом, стимулируя гомологичную рекомбинацию для вставки не идентичной последовательности в представляющую интерес область. Таким образом, в конкретных вариантах осуществления, части последовательности экспрессирующей кассеты, которые являются гомологичными последовательностям в представляющей интерес области, имеют между приблизительно 80 и 99% (или любое целое число между ними) идентичности последовательности с геномной последовательностью, которую она заменяет. В других вариантах осуществления, гомология между экспрессирующей кассетой и геномной последовательностью выше 99%, например, если только 1 нуклеотид отличается между областями гомологии экспрессирующей кассеты и геномных последовательностей из более 100 непрерывных пар оснований. В конкретных случаях, негомологичная часть экспрессирующей кассеты может содержать последовательности, не присутствующие в представляющей интерес области, так что новые последовательности вводят в представляющую интерес область. В этих случаях, негомологичная последовательность, как правило, фланкирована последовательностями из 50-1000 пар оснований (или любого целого числа между ними) или любого количества пар оснований более 1000, которые являются гомологичными или идентичными последовательностям в представляющей интерес области.
[0059] Термин «последовательность» относится к нуклеотидной последовательности любой длины, которая может представлять собой ДНК или РНК; может являться линейной, кольцевой или разветвленной и может являться либо одноцепочечной, либо двухцепочечной. Термин «трансген» относится к нуклеотидной последовательности, которую вставляют в геном. Трансген может иметь любую длину, например, длину между 2 и 100000000 нуклеотидами (или любое целое значение между ними или выше них), предпочтительно, длиной между приблизительно 100 и 100000 нуклеотидами (или любое целое значение между ними), более предпочтительно, длиной между приблизительно 2000 и 20000 нуклеотидами (или любое целое значение между ними) и даже более предпочтительно, между приблизительно 5 и 15 т.п.о. (или любое значение между ними).
[0060] «Хромосома» представляет собой комплекс хроматина, содержащий весь или часть генома клетки. Геном клетки часто характеризуют по его кариотипу, который представляет собой набор всех хромосом, содержащих геном клетки. Геном клетки может содержать одну или более хромосом.
[0061] «Эписома» представляет собой реплицирующуюся нуклеиновую кислоту, нуклеопротеиновый комплекс или другую структуру, содержащую нуклеиновую кислоту, которая не является частью хромосомного кариотипа клетки. Примеры эписом включают плазмиды и определенные вирусные геномы. Специфические для печени конструкции, описанные в настоящем описании, можно поддерживать эписомально или, альтернативно, можно стабильно интегрировать в клетку.
[0062] «Экзогенная» молекула представляет собой молекулу, которая в норме не присутствует в клетке, но которую можно вводить в клетку посредством одного или более генетических, биохимических или других способов. «Нормальное присутствие в клетке» определяют применительно к конкретной стадии развития и условиям окружения клетки. Таким образом, например, молекула, присутствующая только в ходе эмбрионального развития мышцы, является экзогенной молекулой применительно к клетке взрослой мышцы. Подобным образом, молекула, индуцированная тепловым шоком, является экзогенной молекулой применительно к не подвергнутой тепловому шоку клетке. Экзогенная молекула может содержать, например, функциональный вариант неправильно функционирующей эндогенной молекулы или неправильно функционирующий вариант функционирующей в норме эндогенной молекулы.
[0063] Экзогенная молекула может представлять собой, среди других молекул, малую молекулу, такую как получают способом комбинаторной химии, или макромолекулу, такую как белок, нуклеиновая кислота, углевод, липид, гликопротеин, липопротеин, полисахарид, любое модифицированное производное вышеуказанных молекул, или любой комплекс, содержащий одну или более вышеуказанных молекул. Нуклеиновые кислоты включают ДНК и РНК, могут являться одно- или двухцепочечными; могут являться линейными, разветвленными или кольцевыми; и могут иметь любую длину. Нуклеиновые кислоты включают нуклеиновые кислоты, способные формировать дуплексы, также как формирующие триплекс нуклеиновые кислоты. См., например, Патенты США No. 5176996 и 5422251. Белки включают, но без ограничения, ДНК-связывающие белки, факторы транскрипции, факторы ремоделирования хроматина, связывающие метилированную ДНК белки, полимеразы, метилазы, деметилазы, ацетилазы, деацетилазы, киназы, фосфатазы, лигазы, деубиквитиназы, интегразы, рекомбиназы, лигазы, топоизомеразы, гиразы и хеликазы.
[0064] Экзогенная молекула может относиться к такому же типу молекул, как эндогенная молекула, например, экзогенный белок или нуклеиновая кислота. Например, экзогенная нуклеиновая кислота может содержать инфекционный вирусный геном, плазмиду или эписому, введенные в клетку, или хромосому, которая в норме не присутствует в клетке. Способы введения экзогенных молекул в клетки известны специалисту в данной области и включают, но без ограничения, опосредованный липидами перенос (т.е. липосомы, содержащие нейтральные и катионные липиды), электропорацию, прямую инъекцию, слияние клеток, бомбардировку частицами, совместную преципитацию с фосфатом кальция, опосредованный DEAE-декстраном перенос и опосредованный вирусным вектором перенос. Экзогенная молекула может также относиться к такому же типу молекул, как эндогенная молекула, но являться выделенной из вида, отличного от того, из которого происходит клетка. Например, последовательность нуклеиновой кислоты человека можно вводить в линию клеток, исходно происходящую из мыши или хомяка.
[0065] В отличие от этого, «эндогенная» молекула представляет собой молекулу, которая в норме присутствует в конкретной клетке на конкретной стадии развития в конкретных условиях внешней среды. Например, эндогенная нуклеиновая кислота может содержать хромосому, геном митохондрии, хлоропласта или другой органеллы, или природную эписомную нуклеиновую кислоту. Дополнительные эндогенные молекулы могут включать белки, например, факторы транскрипции и ферменты.
[0066] В рамках изобретения, термин «продукт экзогенной нуклеиновой кислоты» включает как полинуклеотидные, так и полипептидные продукты, например, продукты транскрипции (полинуклеотиды, такие как РНК) и продукты трансляции (полипептиды).
[0067] «Слитая» молекула представляет собой молекулу, в которой две или более молекул субъединиц связаны, предпочтительно, ковалентно. Молекулы субъединиц могут представлять собой один и тот же химический тип молекул, или могут представлять собой различные химические типы молекул. Примеры слитых молекул включают, но без ограничения, слитые белки (например, слитый белок между ДНК-связывающим доменом и расщепляющим доменом белка), слитые молекулы между ДНК-связывающим доменом полинуклеотида (например, сгРНК), функционально связанным с расщепляющим доменом, и слитые нуклеиновые кислоты (например, нуклеиновую кислоту, кодирующую слитый белок).
[0068] Экспрессия слитого белка в клетке может происходить в результате доставки слитого белка в клетку или в результате доставки полинуклеотида, кодирующего слитый белок, в клетку, где полинуклеотид транскрибируется и транскрипт транслируется, для получения слитого белка. Транс-сплайсинг, расщепление полипептида и лигирование полипептида могут быть также вовлечены в экспрессию белка в клетке. Способы доставки полинуклеотида и полипептида в клетки представлены в другом месте в этом описании.
[0069] «Ген», для целей по настоящему описанию, включает область ДНК, кодирующую продукт гена (см. ниже), так же как все области ДНК, регулирующие получение продукта гена, вне зависимости от того, являются или нет такие регуляторные последовательности соседними с кодирующими и/или транскрибируемыми последовательностями. Соответственно, ген включает, но без ограничения, промоторные последовательности, терминаторы, регуляторные последовательности для трансляции, такие как участки связывания рибосом и внутренние участки связывания рибосом, энхансеры, сайленсеры, инсуляторы, пограничные элементы, точки начала репликации, участки присоединения матрикса и контрольные области локусов.
[0070] «Экспрессия гена» относится к переводу информации, содержащейся в гене, в продукт гена. Продукт гена может представлять собой непосредственно продукт транскрипции гена (например, мРНК, тРНК, рРНК, антисмысловую РНК, рибозим, структурную РНК или любой другой тип РНК) или белок, полученный посредством трансляции мРНК. Продукты генов включают также РНК, модифицированные посредством таких процессов, как кэпирование, полиаденилирование, метилирование и редактирование, и белки, модифицированные, например, посредством метилирования, ацетилирования, фосфорилирования, убиквитинилирования, АДФ-рибозилирования, миристилирования и гликозилирования.
[0071] «Модуляция» экспрессии гена относится к изменению активности гена. Модуляция экспрессии может включать, но без ограничения, активацию гена и репрессию гена. Редактирование генома (например, расщепление, изменение, инактивацию, случайную мутацию) можно использовать для модуляции экспрессии. Инактивация гена относится к любому уменьшению экспрессии гена по сравнению с клеткой, не содержащей ZFP, TALE или системы CRISPR/Cas, как описано в настоящем описании. Таким образом, инактивация гена может являться частичной или полной. «Генетически модифицированная» клетка включает клетки с любым изменением генетического материала в клетке, включая, но без ограничения, эписомальные и/или геномные модификации. Неограничивающие примеры генетических модификаций включают вставки и/или делеции (например, эписомальная и/или направленная интеграция одного или более трансгенов, РНК или некодирующих последовательностей) и/или мутации (например, точечные мутации, замены и т.д.), изменяющие экспрессию белка в клетке).
[0072] «Представляющая интерес область» представляет собой любую область клеточного хроматина, например, такую как ген или некодирующая последовательность внутри гена или по соседству с геном, в которой желательно связывание экзогенной молекулы. Связывание может происходить с целью направленного расщепления ДНК и/или направленной рекомбинации. Представляющая интерес область может присутствовать, например, на хромосоме, эписоме, в геноме органеллы (например, митохондрии, хлоропласта) или в инфицирующем вирусном геноме. Представляющая интерес область может находиться внутри кодирующей области гена, внутри транскрибируемых некодирующих областей, например, таких как лидерные последовательности, трейлерные последовательности или интроны, или внутри нетранскрибируемых областей, либо выше, либо ниже кодирующей области. Представляющая интерес область может составлять настолько мало, как отдельная пара нуклеотидов, или вплоть до 2000 пар нуклеотидов в длину, или любое целое число пар нуклеотидов.
[0073] «Эукариотические» клетки включают, но без ограничения, клетки грибов (таких как дрожжи), клетки растений, клетки животных, клетки млекопитающих и клетки человека (например, B-клетки), включая стволовые клетки (плюрипотентные и мультипотентые).
[0074] Термины «функциональная связь» и «функционально связанный» (или «оперативно связанный») используют взаимозаменяемо по отношению к смежному положению двух или более компонентов (таких как элементы последовательности), в котором компоненты организованы таким образом, что оба компонента функционируют нормально и обеспечивают возможность того, что по меньшей мере один из компонентов может опосредовать функцию, оказывающую воздействие по меньшей мере на один из других компонентов. В качестве иллюстрации, последовательность регуляции транскрипции, такая как промотор, является функционально связанной с кодирующей последовательностью, если последовательность регуляции транскрипции контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или более факторов регуляции транскрипции. Последовательность регуляции транскрипции, как правило, является функционально связанной в цис- с кодирующей последовательностью, но не обязательно должна непосредственно соседствовать с ней. Например, энхансер представляет собой последовательность регуляции транскрипции, функционально связанную с кодирующей последовательностью, даже если они не являются непрерывными.
[0075] «Функциональный фрагмент» белка, полипептида или нуклеиновой кислоты представляет собой белок, полипептид или нуклеиновую кислоту, последовательность которого не является идентичной полноразмерному белку, полипептиду или нуклеиновой кислоте, еще сохраняющий такую же функцию, как полноразмерный белок, полипептид или нуклеиновая кислота. Функциональный фрагмент может обладать большим, меньшим или одинаковым количеством остатков, как соответствующая нативная молекула, и/или может содержать одну или более замен аминокислот или нуклеотидов. Способы определения функции нуклеиновой кислоты, (например, кодирующей функции, способности гибридизоваться с другой нуклеиновой кислотой) хорошо известны в данной области. Подобным образом, способы определения функции белка хорошо известны. Например, фактор VIII человека с делетированным B-доменом представляет собой функциональный фрагмент полноразмерного белка фактора VIII.
[0076] Полинуклеотидные «вектор» или «конструкция» являются способными переносить последовательности генов к клеткам-мишеням. Как правило, «векторная конструкция», «экспрессирующий вектор», «экспрессирующая конструкция», «экспрессирующая кассета» и «вектор для переноса генов» обозначает любую конструкцию нуклеиновой кислоты, которая способна управлять экспрессией представляющего интерес гена и которая может переносить последовательности генов к клеткам-мишеням. Таким образом, термин включает клонирующие и экспрессирующие векторы, так же как интегрирующие векторы.
[0077] Термины «субъект» и «пациент» используют взаимозаменяемо, и они относятся к млекопитающим, таким как пациенты-люди и нечеловекообразные приматы, так же как экспериментальные животные, такие как кролики, собаки, кошки, крысы, мыши и другие животные. Соответственно, термин «субъект» или «пациент», в рамках изобретения, обозначает любого пациента или субъекта из числа млекопитающих, которому можно вводить экспрессирующие кассеты по изобретению. Субъекты по настоящему изобретению включают субъектов с нарушением.
Экспрессирующие конструкции для B-клеток
[0078] Настоящее изобретение относится к экспрессирующим кассетам (конструкциям) для использования для управления экспрессией трансгена в B-клетке (включая плазмобласты и плазматические клетки), включая in vivo, после введения экспрессирующей кассеты (кассет) субъекту (например, внутривенной доставки). Экспрессирующие конструкции можно поддерживать эписомально, и они могут управлять экспрессией трансгена вне хромосом или, альтернативно, экспрессирующую конструкцию можно интегрировать в геном B-клетки, например, посредством опосредованной нуклеазой направленной интеграции.
[0079] Любую подходящую промоторную последовательность можно использовать в экспрессирующих кассетах по изобретению. В конкретных вариантах осуществления, промотор представляет собой конститутивный промотор. В других вариантах осуществления, промотор является индуцируемым и/или является специфическим для B-клетки промотором. Беспромоторные конструкции, в которых трансген управляется эндогенным промотором B-клетки, также предусмотрены для генетической модификации клеток, как описано в настоящем описании.
[0080] Очевидно, что любой трансген можно использовать в конструкциях, описанных в настоящем описании. Кроме того, индивидуальные компоненты экспрессирующей конструкции (промотор, энхансер, инсулятор, интрон, трансген и т.д.) из конструкций, описанных в настоящем описании, могут присутствовать или нет, и их можно смешивать и сочетать в любой комбинации.
[0081] Конструкции, описанные в настоящем описании, могут содержаться в любом вирусном или невирусном векторе. Конструкции можно поддерживать эписомально или можно интегрировать в геном клетки (например, посредством опосредованной нуклеазой направленной интеграции).
[0082] Невирусные векторы включают ДНК- или РНК-плазмиды, мкДНК, голую нуклеиновую кислоту и нуклеиновую кислоту в комплексе с носителем для доставки, таким как липосома, липидная наночастица, наночастица или полоксамер. Вирусные векторы, которые можно использовать для переноса экспрессирующих кассет, описанных в настоящем описании, включают, но без ограничения, ретровирусный, лентивирусный, аденовирусный, аденоассоциированный вирусный векторы, векторы на основе вируса осповакцины и вируса простого герпеса. Интеграция в геном хозяина является возможной с использованием способов переноса генов посредством ретровируса, лентивируса и аденоассоциированного вируса, и как описано в настоящем описании, может быть облегчена посредством опосредованной нуклеазой интеграции.
[0083] В конкретных предпочтительных вариантах осуществления, конструкции включены в аденоассоциированный вирусный («AAV») вектор или векторную систему, которые можно поддерживать эписомально или интегрировать в геном B-клетки (например, посредством опосредованной нуклеазой направленной интеграции). Конструирование рекомбинантных векторов AAV приведено в ряде публикаций, включая Патент США No. 5173414; Tratschin et al., Mol. Cell. Biol. 5:3251-3260 (1985); Tratschin, et al., Mol. Cell. Biol. 4:2072-2081 (1984); Hermonat & Muzyczka, PNAS 81:6466-6470 (1984); и Samulski et al., J. Virol. 63:03822-3828 (1989).
[0084] Таким образом, в конкретных вариантах осуществления, экспрессирующая конструкция переносится в конструкции AAV и дополнительно содержит 5'- и 3'-ITR, фланкирующие элементы экспрессирующих конструкций (например, энхансер, промотор, необязательно, интрон, трансген и т.д.), как описано в настоящем описании. Необязательно, молекулы спейсера также включены между одним или более из компонентов экспрессирующей конструкции, например, между 5'-ITR и энхансером и/или между сигналом полиаденилирования и 3'-ITR. Спейсеры могут функционировать как плечи гомологии для облегчения рекомбинации в локус области безопасности (например, альбумина). В конкретных вариантах осуществления, конструкция представляет собой конструкцию, как показано на фигуре 8.
[0085] В конкретных вариантах осуществления, векторы AAV, как описано в настоящем описании, могут происходить из любого AAV. В конкретных вариантах осуществления, вектор AAV происходит из дефектного и непатогенного парвовирусного аденоассоциированного вируса типа 2. Все такие векторы происходят из плазмиды, сохраняющей только инвертированные концевые повторы AAV 145 п.о., фланкирующие экспрессирующую кассету для трансгена. Эффективный перенос генов и стабильная доставка трансгенов благодаря интеграции в геномы трансдуцированных клеток являются ключевыми признаками этой векторной системы. (Wagner et al., Lancet 351:9117 1702-3 (1998), Kearns et al., Gene Ther. 9:748-55 (1996)). Другие серотипы AAV, включая AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9 и AAVrh.10, и любой новый серотип AAV можно также использовать в соответствии с настоящим изобретением. Особенно предпочтительными являются серотипы AAV6. В некоторых вариантах осуществления, используют химерный AAV, где вирусные точки начала последовательностей ITR из вирусной нуклеиновой кислоты являются гетерологичными вирусной точке начала капсидных последовательностей. Неограничивающие примеры включают химерный вирус с ITR, происходящими из AAV2, и капсидами, происходящими из AAV5, AAV6, AAV8 или AAV9 (т.е. AAV2/5, AAV2/6, AAV2/8 и AAV2/9, соответственно).
[0086] Ретровирусные векторы включают векторы на основе вируса лейкоза мышей (MuLV), вируса лейкоза обезьяны гиббона (GaLV), вируса иммунодефицита обезьян (SIV), вируса иммунодефицита человека (HIV) и их комбинаций (см., например, Buchscher et al., J. Virol. 66:2731-2739 (1992); Johann et al., J. Virol. 66:1635-1640 (1992); Sommerfelt et al., Virol. 176:58-59 (1990); Wilson et al., J. Virol. 63:2374-2378 (1989); Miller et al., J. Virol. 65:2220-2224 (1991); PCT/US94/05700).
[0087] Конструкции, описанные в настоящем описании, можно также включать в аденовирусную векторную систему. Векторы на основе аденовирусов являются способными к очень высокой эффективности трансдукции во множестве типов клеток и не требуют деления клеток. С такими векторами получены высокий титр и высокие уровни экспрессии. Этот вектор можно получать в больших количествах в относительно простой системе.
[0088] pLASN и MFG-S являются примерами ретровирусных векторов, которые использовали в клинических исследованиях (Dunbar et al., Blood 85:3048-305 (1995); Kohn et al., Nat. Med. 1:1017-102 (1995); Malech et al., PNAS 94:22 12133-12138 (1997)). PA317/pLASN был первым терапевтическим вектором, использованным в генотерапевтическом исследовании. (Blaese et al., Science 270:475-480 (1995)). Эффективность трансдукции 50% или более наблюдали для упакованных векторов MFG-S. (Ellem et al., Immunol Immunother. 44(1):10-20 (1997); Dranoff et al., Hum. Gene Ther. 1:111-2 (1997).
[0089] Дефектные по репликации рекомбинантные аденовирусные векторы (Ad) можно также использовать с полинуклеотидами, описанными в настоящем описании. Большинство аденовирусных векторов конструируют таким образом, что трансген заменяет гены Ad E1a, E1b и/или E3; затем дефектный по репликации вектор размножают в клетках 293 человека, обеспечивающих функцию делетированного гена в транс-. Векторами Ad можно трансдуцировать множество типов тканей in vivo, включая неделящиеся, дифференцированные клетки, такие как клетки, обнаруженные в печени, почке и мышце. Общепринятые векторы Ad обладают большой емкостью переноса. Пример использования вектора Ad в клинических исследованиях включает терапию полинуклеотидом для противоопухолевой иммунизации с помощью внутримышечной инъекции (Sterman et al., Hum. Gene Ther. 7:1083-9 (1998)). Дополнительные примеры использования аденовирусных векторов для переноса генов в клинических исследованиях включают Rosenecker et al., Infection 24:1 5-10 (1996); Sterman et al., Hum. Gene Ther. 9:7 1083-1089 (1998); Welsh et al., Hum. Gene Ther. 2:205-18 (1995); Alvarez et al., Hum. Gene Ther. 5:597-613 (1997); Topf et al., Gene Ther. 5:507-513 (1998); Sterman et al., Hum. Gene Ther. 7:1083-1089 (1998).
[0090] Упаковывающие клетки используют для формирования вирусных частиц, способных инфицировать клетку-хозяина. Такие клетки включают клетки HEK293 и Sf9, которые можно использовать для упаковки AAV и аденовируса, и клетки ψ2 или клетки PA317, упаковывающие ретровирус. Вирусные векторы, используемые в генотерапии, обычно получают в линии клеток-продуцентов, упаковывающих нуклеиновую кислоту вектора в вирусную частицу. Векторы, как правило, содержат минимальные вирусные последовательности, необходимые для упаковки и последующей интеграции в хозяина (если это применимо), где другие вирусные последовательности заменены экспрессирующей кассетой, кодирующей белок, подлежащий экспрессии. Отсутствующие функции вируса обеспечивает упаковывающая линия клеток в транс-. Например, векторы AAV, используемые в генотерапии, как правило, обладают только последовательностями инвертированных концевых повторов (ITR) из генома AAV, необходимыми для упаковки и интеграции в геном хозяина. Вирусную ДНК упаковывают в линии клеток, содержащей плазмиду-помощника, кодирующую другие гены AAV, а именно, rep и cap, но лишенную последовательностей ITR. Линию клеток инфицируют также аденовирусом в качестве помощника. Вирус-помощник стимулирует репликацию вектора AAV и экспрессию генов AAV с плазмиды-помощника. Плазмида-помощник не упаковывается в значительных количествах из-за отсутствия последовательностей ITR. Контаминацию аденовирусом можно снижать посредством, например, термической обработки, к которой аденовирус является более чувствительным, чем AAV. В некоторых вариантах осуществления, AAV получают с использованием бакуловирусной системы экспрессии (см., например, Патенты США 6723551 и 7271002).
[0091] Очистка частиц AAV из 293 или бакуловирусной системы, как правило, включает выращивание клеток, продуцирующих вирус, с последующим сбором вирусных частиц из супернатанта клеток, или лизис клеток и сбор вируса из неочищенного лизата. Затем AAV очищают способами, известными в данной области, включая ионообменную хроматографию (например, см. Патенты США 7419817 и 6989264), ионообменную хроматографию и центрифугирование в градиенте плотности CsCl (например, публикацию PCT WO2011094198A10), иммуноаффинную хроматографию (например, WO2016128408) или очистку с использованием сефарозы AVB (например, GE Healthcare Life Sciences).
[0092] Для многих генотерапевтических применений является желательным, чтобы вектор для генотерапии был доставлен с высокой степенью специфичности в конкретный тип ткани. Соответственно, можно модифицировать вирусный вектор для обладания специфичностью к данному типу клеток посредством экспрессии лиганда в форме белка, слитого с белком вирусной оболочки на внешней поверхности вируса. Выбирают лиганд, обладающий аффинностью для рецептора, как известно, присутствующего на клетках представляющего интерес типа. Например, в Han et al., Proc. Natl. Acad. Sci. USA 92:9747-9751 (1995), опубликовано, что вирус лейкоза мышей Молони можно модифицировать для экспрессии херегулина человека, слитого с gp70, и рекомбинантный вирус инфицирует конкретные клетки рака молочной железы, экспрессирующие рецептор эпидермального фактора роста человека. Этот принцип можно распространять на другие пары вирус - клетка-мишень, в которых клетка-мишень экспрессирует рецептор, и вирус экспрессирует слитый белок, содержащий лиганд для рецептора поверхности клеток. Например, можно сконструировать нитевидный фаг для экспонирования фрагментов антитела (например, FAB или Fv), имеющих специфическую аффинность связывания фактически для любого выбранного клеточного рецептора. Хотя приведенное выше описание относится к в первую очередь к вирусным векторам, те же принципы можно применять к невирусным векторам. Такие векторы можно конструировать с содержанием конкретных последовательностей для поглощения, благоприятствующих поглощению специфическими клетками-мишенями.
[0093] Полинуклеотиды, описанные в настоящем описании, могут включать одно или более неприродных оснований и/или остовов. В частности, экспрессирующая кассета, как описано в настоящем описании, может включать метилированные остатки цитозина для достижения состояния транскрипционного молчания в представляющей интерес области.
[0094] Кроме того, экспрессирующие конструкции, как описано в настоящем описании, могут также включать дополнительные регуляторные последовательности транскрипции или трансляции, или другие последовательности, например, последовательности Козак, дополнительные промоторы, энхансеры, инсуляторы, интроны, внутренние участки связывания рибосом, последовательности, кодирующие пептиды 2A, участки расщепления фурином и/или сигналы полиаденилирования. Кроме того, контрольные элементы представляющих интерес генов можно функционально связывать с репортерными генами для получения химерных генов (например, экспрессирующих репортер кассет).
Модификации
[0095] Настоящее изобретение относится к генетически модифицированным B-клеткам, содержащим одну или более из следующих модификаций: (a) предоставление в клетке одного или более трансгенов (эписомных и/или интегрированных в любых комбинациях); (b) вставки и/или делеции в одном или более генах, модифицирующие (i) гены B-клеточного рецептора и/или (ii) клеточные взаимодействия в зародышевых центрах; и/или (c) модификации (мутации), ингибирующие супрессию какой-либо функции B-клетки, ассоциированную с инфекцией патогеном или регуляцией злокачественной опухоли. Генетически модифицированные B-клетки, как описано в настоящем описании, могут также происходить из HSC, содержащих одну или более из этих генетических модификаций.
[0096] В конкретных вариантах осуществления, конструкции, описанные в настоящем описании, можно использовать для экспрессии в B-клетке любого трансгена(трансгенов). Один или более трансгенов можно экспрессировать эписомально в модифицированных B-клетках и/или после опосредованной нуклеазой направленной интеграции одного или более трансгенов. Иллюстративные трансгены (также обозначенные как представляющие интерес гены и/или экзогенные последовательности) включают, но без ограничения, любую кодирующую полипептид последовательность (например, кДНК), промоторные последовательности, энхансерные последовательности, эпитопные метки, маркерные гены, участки узнавания расщепляющих ферментов и/или различные типы экспрессирующих конструкций. Маркерные гены включают, но без ограничения, последовательности, кодирующие белки, придающие устойчивость к антибиотику (например, устойчивость к ампициллину, устойчивость к неомицину, устойчивость к G418, устойчивость к пуромицину), последовательности, кодирующие окрашенные или флуоресцентные, или люминесцентные белки (например, зеленый флуоресцентный белок, улучшенный зеленый флуоресцентный белок, красный флуоресцентный белок, люциферазу), и белки, опосредующие усиленный рост клеток и/или амплификацию генов (например, дигидрофолатредуктазу). Эпитопные метки включают, например, одну или более копий FLAG, His, myc, Tap, HA или любой поддающейся детекции аминокислотной последовательности.
[0097] В предпочтительном варианте осуществления, трансген содержит полинуклеотид, кодирующий любой полипептид, экспрессия которого в клетке является желательной, включая, но без ограничения, антитела, антигены, ферменты, рецепторы (клеточной поверхности или ядерные), гормоны, лимфокины, цитокины, репортерные полипептиды, факторы роста и функциональные фрагменты любых из вышеуказанных. Кодирующие последовательности могут представлять собой, например, кДНК.
[0098] В конкретных вариантах осуществления, трансген(ы) кодирует(кодируют) функциональные варианты белков, отсутствующих или недостаточных при каком-либо генетическом заболевании, включая, но без ограничения, нарушения с накоплением лизосом (например, Гоше, Фабри, Гунтера, Гурлера, Ниманна-Пика и т.д.), метаболические нарушения и/или нарушения крови, такие как гемофилии и гемоглобинопатии, и т.д. См., например, Патентные публикации США No. 20140017212 и 20140093913; Патенты США No. 9255250 и 9175280.
[0099] Например, трансген может содержать последовательность, кодирующую полипептид, отсутствующий или нефункциональный у субъекта, страдающего генетическим заболеванием, включая, но без ограничения, любое из следующих генетических заболеваний: ахондроплазия, ахроматопсия, недостаточность кислой мальтазы, недостаточность аденозиндезаминазы (OMIM No.102700), адренолейкодистрофия, синдром Экарди, недостаточность альфа-1-антитрипсина, альфа-талассемия, синдром нечувствительности к андрогенам, синдром Аперта, аритмогенная дисплазия правого желудочка, телеангиоэктатическая атаксия, синдром Барта, бета-талассемия, синдром голубых эластичных пузырчатых невусов, болезнь Канавана, хронические гранулематозные заболевания (CGD), синдром кошачьего крика, кистозный фиброз, болезнь Деркума, эктодермальная дисплазия, анемия Фанкони, прогрессирующая оссифицирующая фибродисплазия, синдром ломкой Х-хромосомы, галактоземия, болезнь Гоше, генерализованный ганглиозидоз (например, GM1), гемохроматоз, мутация гемоглобина С в 6-м кодоне бета-глобина (HbC), гемофилия, болезнь Гентингтона, синдром Гурлера, гипофосфатазия, синдром Клайнфельтера, болезнь Краббе, синдром Лангера-Гиедиона, нарушение адгезии лейкоцитов (LAD, OMIM No. 116920), лейкодистрофия, синдром удлинения интервала QT, синдром Марфана, синдром Мебиуса, мукополисахаридоз (MPS), синдром ногтей-надколенника, несахарный почечный диабет, нейрофиброматоз, болезнь Ниманна-Пика, несовершенный остеогенез, порфирия, синдром Прадера-Вилли, прогерия, синдром Протея, ретинобластома, синдром Ретта, синдром Рубинштейна-Тэйби, синдром Санфилиппо, тяжелый комбинированный иммунодефицит (SCID), синдром Швачмана, заболевание серповидного изменения эритроцитов (серповидно-клеточная анемия), синдром Смита-Магениса, синдром Стиклера, болезнь Тея-Сакса, синдром тромбоцитопении с отсутствием лучевой кости (TAR), синдром Тричера-Коллинза, трисомия, туберозный склероз, синдром Тернера, нарушение цикла мочевины, болезнь фон Гиппеля-Линдау, синдром Ваарденбурга, синдром Вильямса, болезнь Вильсона, синдром Вискотта-Олдрича, X-сцепленный лимфопролиферативный синдром (XLP, OMIM No. 308240), приобретенные иммунодефициты, заболевания с накоплением лизосом (например, болезнь Гоше, GM1, болезнь Фабри и болезнь Тея-Сакса), мукополисахаридоз (например, болезнь Гунтера, болезнь Гурлера), гемоглобинопатии (например, заболевания серповидного изменения эритроцитов, HbC, α-талассемия, β-талассемия) и гемофилии.
[0100] Неограничивающий примеры белков (включая их функциональный фрагменты, такие как укороченные варианты), которые можно экспрессировать, как описано в настоящем описании, включают фибриноген, протромбин, тканевой фактор, фактор V, фактор VII, фактор VIII, фактор IX, фактор X, фактор XI, фактор XII (фактор Хагемана), фактор XIII (фибрин-стабилизирующий фактор), фактор фон Виллебранда, прекалликреин, высокомолекулярный кининоген (фактор Фитцджеральда), фибронектин, антитромбин III, кофактор гепарина II, белок C, белок S, белок Z, родственный белку Z ингибитор протеаз, плазминоген, альфа-2-антиплазмин, тканевой активатор плазминогена, урокиназу, ингибитор-1 активатора плазминогена, ингибитор-2 активатора плазминогена, глюкоцереброзидазу (GBA), α-галактозидазу A (GLA), идуронат-сульфатазу (IDS), идуронидазу (IDUA), кислую сфингомиелиназу (SMPD1), MMAA, MMAB, MMACHC, MMADHC (C2orf25), MTRR, LMBRD1, MTR, пропионил-CoA-карбоксилазу (PCC) (субъединицы PCCA и/или PCCB), белок-транспортер глюкозо-6-фосфата (G6PT) или глюкозо-6-фосфатазу (G6P-азу), рецептор LDL (LDLR), ApoB, LDLRAP-1, PCSK9, митохондриальный белок, такой как NAGS (N-ацетилглутаматсинтетаза), CPS1 (карбамоилфосфатсинтетаза I) и OTC (орнитинтранскарбамилаза), ASS (синтетазу аргининоянтарной кислоты), ASL (лиазу аргининосукцинатной кислоты) и/или ARG1 (аргиназу), и/или белок из семейства переносчиков растворенных веществ 25 (SLC25A13, переносчик аспартата/глутамата) белок, UGT1A1 или полипептид A1 УДФ-глюкуронилтрансферазы, фумарилацетоацетат-гидролиазу (FAH), белок аланин-глиоксилат-аминотрансферазу (AGXT), белок глиоксилатредуктазу/гидроксипируватредуктазу (GRHPR), белок - продукт гена транстиретина (TTR), белок ATP7B, белок фенилаланингидроксилазу (PAH), белок липопротеинлипазу (LPL), сконструированную нуклеазу, сконструированный фактор транскрипции и/или терапевтическое одноцепочечное антитело.
[0101] В других вариантах осуществления, модифицированные B-клетки, описанные в настоящем описании, содержат один или более трансгенов, кодирующих одно или более антител, представляющих собой сконструированные молекулы, разработанные для нацеливания на иммуноциты посредством специфических молекулярных мишеней, экспрессированных на клеточных поверхностях. В некоторых вариантах осуществления, модифицированные B-клетки экспрессируют антитела, разработанные для нацеливания на эндогенные B-клетки. Эти антитела могут индуцировать опосредованное антителами уничтожение (например, посредством ADCC или опосредованного комплементом уничтожения) B-клеток или других иммуноцитов, вовлеченных в ослабление иммунного ответа.
[0102] B-клетки, как описано в настоящем описании, можно генетически модифицировать для продукции одного или более антител, специфических для B-клеток, продуцирующих нежелательные антитела. Неограничивающие примеры B-клеток, продуцирующих нежелательные антитела, включают B-клетки, продуцирующие антитела против белков, введенных при ERT (факторов свертывания крови, таких как F8, F9 и т.д., у пациентов с гемофилией, и/или белков, отсутствующих или недостаточных при нарушениях с накоплением лизосом). Кодирующие антитело конструкции вводят в предшественника B-клетки или в B-клетку ex vivo, так что когда клетку повторно вводят пациенту, продуцирующие антитело B-клетки специфически нацеливают на клетки (B-клетки), продуцирующие белок (например, антитело), связываемый сконструированным антителом. В конкретных вариантах осуществления, сконструированное антитело является специфическим для антитела, направленного против терапевтического белка, поставляемого экзогенно (посредством ERT и/или генотерапии), так что антитела против терапевтических белков нейтрализуют. Таким образом, композиции и способы, описанные в настоящем описании, включают сконструированные B-клетки, которые продуцируют антитела, специфически нацеленные на антитела (например, антитела против F9), продуцированные у пациента. Модифицированные B-клетки из этих композиций и способов можно вводить субъекту в форме зрелых B-клеток или в форме клеток-предшественников (таких как HSC или лимфоидные клетки-предшественники), которые подвергаются дифференцировке у субъекта после введения или, альтернативно, могут подвергаться генетической модификации in vivo. В дополнительных вариантах осуществления, белки, продуцированные с трансгенов (например, антитела против ERT), из генетически модифицированных B-клеток выделяют для введения нуждающемуся в этом субъекту, например, пациенту, нуждающемуся в антителах к антителам против ERT, продуцируемых его организмом.
[0103] В дополнительных вариантах осуществления, трансген может представлять собой антитело, специфическое для B-клетки, чувствительной к белку, вовлеченному в аутоиммунное заболевание. Термин «аутоиммунное заболевание» относится к любому заболеванию или нарушению, при котором у субъекта возникает деструктивный иммунный ответ против его собственных тканей. Аутоиммунные нарушения могут поражать почти каждую систему органов у субъекта (например, человека), включая, но без ограничения, заболевания нервной, желудочно-кишечной и эндокринной систем, так же как кожи и других соединительных тканей, глаз, крови и кровеносных сосудов. Примеры аутоиммунных заболеваний включают, но без ограничения, тиреоидит Хашимото, системную красную волчанку, синдром Шегрена, болезнь Грэйвса, склеродермию, ревматоидный артрит, рассеянный склероз, миастению и диабет. Таким образом, B-клетки, как описано в настоящем описании, могут содержать молекулу (например, сконструированное антитело), нацеленное на популяцию B-клеток у субъекта, которая является чувствительной к (и продуцирует антитела к) аутоантигену, вовлеченному в аутоиммунное заболевание, включая, но без ограничения, основной белок миелина (MBP), инсулин, ANA, белки суставов или мышц, белки щитовидной железы и т.п.
[0104] В конкретных вариантах осуществления, трансген может содержать маркерный ген (описанный выше), позволяющий отбор клеток, которые подверглись направленной интеграции, и связанную последовательность, кодирующую дополнительную функциональность. Неограничивающие примеры маркерных генов включают GFP, маркер(ы) для отбора с лекарственным средством и т.п.
[0105] Конструкции, описанные в настоящем описании, можно также использовать для доставки некодирующих трансгенов. Последовательности, кодирующие антисмысловые РНК, РНКи, кшРНК и микро-РНК (мкРНК) также можно использовать для направленных вставок.
[0106] В конкретных вариантах осуществления, трансген включает последовательности (например, кодирующие последовательности, также обозначенные как трансгены) длиной более 1 т.п.о., например, между 2 и 200 т.п.о., между 2 и 10 т.п.о. (или любое значение между ними). Трансген может также включать один или более участков-мишеней для нуклеазы. Трансген может также содержать одно или более плеч гомологии. Плечи гомологии содержат последовательности с высокой степенью гомологии с последовательностями, фланкирующими участок-мишень для расщепления нуклеазой. Плечо гомологии может содержать 50, 100, 200, 500, 1000, 2000 или более нуклеотидов, или любое значение между ними.
[0107] При интеграции (например, посредством опосредованной нуклеазой интеграции), трансген может быть вставлен в эндогенный ген таким образом, что экспрессируется весь эндогенный ген, некоторая его часть, или эндогенный ген не экспрессируется.
Нуклеазы
[0108] Как отмечено выше, экспрессирующие кассеты можно поддерживать эписомально или можно интегрировать в геном клетки. Интеграция может являться случайной. В конкретных вариантах осуществления, интеграцию конструкции(конструкций) трансгенов нацеливают на указанный ген после расщепления гена-мишени одной или более нуклеазами (например, нуклеазами с цинковыми пальцами («ZFN»), TALEN, TtAgo, системами нуклеаз CRISPR/Cas и эндонуклеазами хоминга), и конструкцию интегрируют способами, управляемыми либо посредством зависимой от гомологии репарации (HDR), либо посредством связывания концов соединением негомологичных концов (NHEJ). См., например, Патенты США No. 9394545; 9150847; 9206404; 9045763; 9005973; 8956828; 8936936; 8945868; 8871905; 8586526; 8563314; 8329986; 8399218; 6534261; 6599692; 6503717; 6689558; 7067317; 7262054; 7888121; 7972854; 7914796; 7951925; 8110379; 8409861; Публикации патентов США 20030232410; 20050208489; 20050026157; 20050064474; 20060063231; 20080159996; 20100218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983 и 20130177960, и 20150056705, полное содержание описаний которых приведено в качестве ссылки для всех целей.
[0109] Любую нуклеазу можно использовать для направленной интеграции экспрессирующей трансген конструкции.
[0110] В конкретных вариантах осуществления, нуклеаза содержит нуклеазу с цинковыми пальцами (ZFN), содержащую ДНК-связывающий домен с цинковыми пальцами и расщепляющий (нуклеазный) домен. См., например, Патенты США No. 9255250; 9200266; 9045763; 9005973; 9150847; 8956828; 8945868; 8703489; 8586526; 6534261; 6599692; 6503717; 6689558; 7067317; 7262054; 7888121; 7972854; 7914796; 7951925; 8110379; 8409861.
[0111] В других вариантах осуществления, нуклеаза содержит TALEN, содержащую TAL-эффекторный ДНК-связывающий домен и расщепляющий (нуклеазный) домен. См., например, Патент США No. 8586526 и Патентную публикацию США No. 20130196373.
[0112] В дополнительных вариантах осуществления, нуклеаза содержит систему нуклеазы CRISPR/Cas, включающую одиночную направляющую РНК для узнавания участка-мишени и один или более расщепляющих доменов. См., например, Публикацию патента США No. 20150056705. В некоторых вариантах осуществления, используют систему CRISPR-Cpf1 (см. Fagerlund et al., (2015) Genom Bio 16:251). Понятно, что термин система «CRISPR/Cas» относится к обеим системам CRISPR/Cas и CRISPR/Cfp1.
[0113] Расщепляющие домены нуклеаз могут представлять собой домены дикого типа или мутантные домены, включая неприродные (сконструированные) расщепляющие домены, формирующие облигатные гетеродимеры. См., например, Патенты США No. 8623618; 7888121; 7914796; и 8034598 и U.S. Publication No. 20110201055.
[0114] Нуклеаза(нуклеазы) могут осуществлять один или более двухцепочечных и/или одноцепочечных разрезов в участке. В конкретных вариантах осуществления, нуклеаза содержит каталитически неактивный расщепляющий домен (например, белок FokI и/или Cas). См., например, Патент США No. 9200266; 8703489 и Guillinger et al. (2014) Nature Biotech. 32(6):577-582. Каталитически неактивный расщепляющий домен может, в комбинации с каталитически активным доменом, действовать в качестве никазы для осуществления одноцепочечного разреза. Таким образом, две никазы можно использовать в комбинации для получения двухцепочечного разреза в специфической области. Дополнительные никазы также известны в данной области, например, McCaffery et al. (2016) Nucleic Acids Res. 44(2):e11. doi: 10.1093/nar/gkv878. Epub 2015 Oct 19.
[0115] В конкретных вариантах осуществления, нуклеаза расщепляет ген области безопасности (например, CCR5, Rosa, альбумина, AAVS1, TCRA, TCRB и т.д. См., например, Патенты США No. 7888121; 7972854; 7914796; 7951925; 8110379; 8409861; 8586526; Публикации патентов США 20030232410; 20050208489; 20050026157; 20060063231; 20080159996; 201000218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983 и 20130177960. В предпочтительных вариантах осуществления, нуклеаза расщепляет эндогенный ген альбумина таким образом, что экспрессирующая кассета интегрирует в эндогенный локус альбумина клетки печени. Специфические для альбумина нуклеазы описаны, например, в Патенте США No. 9150847; и Патентных публикациях США No. 20130177983 и 20150056705.
Доставка
[0116] Конструкции, описанные в настоящем описании (и/или нуклеазы) можно доставлять in vivo посредством любых пригодных способов в любой тип клеток, предпочтительно, в селезенку или вторичные лимфатические узлы. Подобным образом, при использовании в комбинации с нуклеазами для направленной интеграции, нуклеазы можно доставлять в форме полинуклеотида и/или белка, например, с использованием невирусного вектора(векторов), вирусного вектора(векторов), и/или в форме РНК, например, в форме мРНК.
[0117] Способы невирусной доставки нуклеиновых кислот включают электропорацию, липофекцию, микроинъекцию, биолистику, виросомы, липосомы, липидные наночастицы, иммунолипосомы, другую наночастицу, конъюгаты поликатион или липид: нуклеиновая кислота, голую ДНК, искусственные вирионы и усиленное агентами поглощение ДНК. Сонопорацию с использованием, например, системы Sonitron 2000 (Rich-Mar) также можно использовать для доставки нуклеиновых кислот. Дополнительные иллюстративные системы доставки нуклеиновой кислоты включают системы, представленные Amaxa Biosystems (Cologne, Germany), Maxcyte, Inc. (Rockville, Maryland), BTX Molecular Delivery Systems (Holliston, MA) и Copernicus Therapeutics Inc. (см., например, US6008336).
[0118] В предпочтительных вариантах осуществления, экспрессирующие конструкции представляют собой векторы AAV. Необязательно, нуклеазы можно вводить в форме мРНК или с использованием одного или более вирусных векторов (AAV, Ad и т.д.). Введение можно проводить любыми способами, в которых полинуклеотиды доставляют в желательные клетки-мишени. Предусмотрены способы как in vivo, так и ex vivo. Внутривенная инъекция в периферический кровеносный сосуд является предпочтительным способом введения. Другие способы введения in vivo включают, например, прямую инъекцию в ткани, содержащие B-клетки, включая лимфатические узлы, костный мозг, плазму, лимфатическую систему и селезенку.
[0119] В системах, включающих доставку более одного полинуклеотида (например, конструкции, как описано в настоящем описании, и нуклеазы в форме полинуклеотида), два или более полинуклеотида(полинуклеотидов) доставляют с использованием одного или более одинаковых и/или различных векторов. Например, нуклеазу в форме полинуклеотида можно доставлять в форме мРНК, и специфические для B-клеток конструкции, как описано в настоящем описании, можно доставлять с помощью других средств, таких как вирусные векторы (например, AAV), миникольцевая ДНК, плазмидная ДНК, линейная ДНК, липосомы, липидные наночастицы, наночастицы и т.п.
[0120] Фармацевтически приемлемые носители частично определяются конкретной вводимой композицией, так же как конкретным способом, используемым для введения композиции. Соответственно, существует широкое множество доступных пригодных составов фармацевтических композиций, как описано ниже (см., например, Remington's Pharmaceutical Sciences, 17th ed., 1989).
[0121] Эффективное количество экспрессирующей кассеты (и, необязательно, нуклеазы (нуклеаз) и/или модифицированных клеток), подлежащее введению, может меняться от пациента к пациенту. Соответственно, эффективные количества лучше всего определять терапевту, вводящему композиции (например, клетки), и соответствующие дозы может легко определить специалист в данной области. Посредством анализа в сыворотке, плазме или другой ткани уровней терапевтического полипептида и сравнения с исходным уровнем до введения можно определять, является ли вводимое количество слишком низким, находящимся в правильном диапазоне или слишком высоким. Подходящие режимы начального и последующих введений также можно менять, однако типичным примером является исходное введение с последующими введениями затем, если необходимо. Последующие введения можно осуществлять с различными интервалами, в диапазоне от ежесуточного до ежегодного, до раза в каждые несколько лет. Специалисту в данной области понятно, что пригодные способы иммуносупрессии могут быть рекомендованы для избегания ингибирования или блокирования трансдукции посредством иммуносупрессии векторов для доставки, см., например, Vilquin et al., (1995) Human Gene Ther., 6:1391-1401.
[0122] Составы для введений как ex vivo, так и in vivo, включают суспензии (например, генетически модифицированных клеток, липосом, липидных наночастиц или наночастиц) в жидкости или эмульгированных жидкостях. Активные ингредиенты часто смешивают с наполнителями, которые являются фармацевтически приемлемыми и совместимыми с активным ингредиентом. Пригодные наполнители включают, например, воду, солевой раствор, декстрозу, глицерин, этанол или т.п., и их комбинации. Кроме того, композиция может содержать незначительные количества вспомогательных веществ, таких как увлажняющие или эмульгирующие средства, забуферивающие pH средства, стабилизаторы или другие реагенты, усиливающие эффективность фармацевтической композиции.
Применения
[0123] Способы и композиции, описанные в настоящем описании, предназначены для обеспечения терапии какого-либо заболевания посредством предоставления трансгена, который экспрессирует продукт, отсутствующий или недостаточный при этом заболевании, или иным образом осуществляет лечение или предотвращение заболевания. Клетку можно модифицировать in vivo или можно модифицировать ex vivo и затем вводить субъекту. Таким образом, способы и композиции обеспечивают лечение и/или предотвращение таких генетических заболеваний. Кроме того, способы и композиции, описанные в настоящем описании, позволяют модификацию B-клеток таким образом, что эти клетки обладают модифицированной токсичностью, продукцией антител и/или производственными характеристиками.
[0124] Следующие примеры включают иллюстративные варианты осуществления по настоящему описанию, в которых необязательно используемая нуклеаза содержит нуклеазу с цинковыми пальцами (ZFN). Следует понимать, что это представлено только с целью примера, и что можно использовать другие нуклеазы, например, TALEN, системы CRISPR/Cas, эндонуклеазы хоминга (мегануклеазы) со сконструированными ДНК-связывающими доменами и/или белки, слитые с ДНК-связывающими доменами природных или сконструированных эндонуклеаз хоминга (мегануклеаз) и гетерологичными расщепляющими доменами, и/или белки, слитые с мегануклеазами и белками TALE. Кроме того, следует понимать, что экспрессирующие конструкции, как описано в настоящем описании, можно переносить в других векторах (отличных от AAV) для получения таких же результатов в лечении и/или предотвращении нарушений, вызванных недостаточностью продукции белков.
ПРИМЕРЫ
Пример 1: Способы
Культивирование клеток
[0125] Замороженные CD19+ B-клетки из периферической крови человека закупали из STEMCELL Technologies (Vancouver, Canada). Система для дифференцировки культуры B-клеток in vitro (см. фигуру 1) описана ранее (Jourdan et al., там же). Все культивирование проводили в среде Дульбекко в модификации Искова (Corning, Corning, NY) и 10% эмбриональной бычьей сыворотке (VWR, Radnor, PA).
[0126] Клетки культивировали в 24-луночном планшете при плотности 2,0E+5 клеток на лунку в 0,5 мл культуральной среды. Клетки размораживали и культивировали в течение 4 суток в среде для активации B-клеток, содержащей Ab против His (5 мкг/мл), ODN (10 мкг/мл), sCD40L (50 нг/мл), IL-2 (10 нг/мл), IL-10 (50 нг/мл) и IL-15 (10 нг/мл). На сутки 4 культивирования, клетки собирали, собирали супернатанты, клетки промывали с использованием DPBS и затем переносили в среду для получения плазмобластов (PB), содержащую IL-2 (10 нг/мл), IL-6 (40-50 нг/мл), IL-10 (50 нг/мл) и IL-15 (10 нг/мл). На сутки 7 культивирования, клетки собирали, супернатанты собирали, клетки промывали DPBS и затем переносили в среду для получения плазматических клеток (PC), содержащую IL-6 (40-50 нг/мл), IL-15 (10 нг/мл), IFN-α (500 ед./мл). На сутки 10 культивирования, клетки собирали, и супернатанты собирали.
Генетическая модификация B-клеток
Реагенты ZFN:
[0127] ZFN разрабатывали для нацеливания на TCRA (TRAC, SBS53909 и SBS53885, см. Публикацию патента США No. US-2017-0211075-A1), CCR5 (SBS8266 и SBS8196, см. Патент США No. 7925921) и AAVS1 (SBS30035 и SBS30054, см. Патент США No. 8110379). Кодирующие последовательности ZFN для CCR5 и AAVS1 клонировали в модифицированный вариант плазмиды pGEM4Z (Promega, Madison, WI), содержащий последовательность из 64 остатков аденина на 3' от вставленной последовательности гена (Boczkowski et al. (2000) Canc Res 60:1028-1034), который линеаризовали посредством расщепления SpeI для получения матриц для синтеза мРНК. мРНК ZFN для TRAC получали с линейных ДНК-матриц (одной для каждой ZFN) посредством амплификации ПЦР кодирующей ZFN последовательности с использованием набора с ДНК-полимеразой Accuprime PFX (Invitrogen, Carlsbad, CA). Продукты ПЦР использовали в качестве матриц для синтеза мРНК. мРНК получали с использованием набора mMESSAGE mMACHINE T7 ULTRA (Life Technologies, Carlsbad, CA), по протоколу производителя.
[0128] Кратко, 1,0 мкг ДНК, кодирующей ZFN, используемой в качестве матрицы для синтеза мРНК, инкубировали при 37°C в течение часов поставленного буфера, с последующим расщеплением ДНКазой, поставленной с набором. Реакцию присоединения поли-A-хвоста in vitro не проводили, поскольку поли-T-хвост включали в ДНК-матрицу в ходе получения матрицы TRAC посредством ПЦР. Матрицы AAVS1 и CCR5 содержат поли-T-матрицу в векторе. Затем мРНК очищали с использованием набора RNeasy Mini (Qiagen, Carlsbad, CA), по протоколу производителя, и количественно оценивали в Nanodrop 8000 (ThermoScientific, Waltham, MA). Праймеры, использованные для матриц мРНК, представляли собой прямой праймер: 5' GCAGAGCTCTCTGGCTAACTAGAG (SEQ ID NO:1) и обратный праймер: 5'TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTCTGGCAACTAGAAGGCACAG (SEQ ID NO:2).
Векторы AAV:
[0129] Все векторы AAV получали в Sangamo Therapeutics, как описано ниже. Донорные матрицы AAV для AAVS1, CCR5 и TRAC содержали плечи, гомологичные их локусам-мишеням. AAVS1 имела левое и правое плечи гомологии длиной 801 и 568 пар оснований, соответственно. CCR5 имела левое и правое плечи гомологии длиной 473 и 1431 пар оснований, соответственно. TRAC имела левое и правое плечи гомологии длиной 925 и 989 пар оснований, соответственно. Экспрессирующую кассету GFP, содержащую промотор, последовательность GFP и сигнал полиаденилирования гормона роста человека (hGHpA), клонировали между правым и левым плечами гомологиии. Промотор представлял собой либо промотор фосфоглицераткиназы (PGK), либо специфический для B-клеток промотор (EEK). Специфический для B-клеток промотор (EEK) состоял из 3'-энхансера, MAR, и интронного энхансера выше промотора легкой цепи κ человека (Luo et al. (2009) Blood 113:1422-1431). Донорную матрицу TRAC клонировали в вектор pAAV. Донорные матрицы AAVS1 и CCR5 клонировали в сделанную на заказ плазмиду pRS165 (Lombardo et al. (2011) Nat Methods 8:861-869; Wang et al. (2012) Genome Res 22:1316-1326), полученную из pAAV-MCS (Agilent Technologies, Santa Clara, CA). Инвертированные концевые повторы (ITR) AAV2 использовали для обеспечения упаковки, как для векторов AAV, с использованием способа тройной трансфекции (Xiao and Samulski (1998) J. Virol 72:2224-2232). Кратко, клетки HEK 293 рассевали в 10-слойные камеры CellSTACK (Corning, Acton, MA), выращивали 3 суток до плотности 80%, затем трансфицировали, с использованием способа с фосфатом кальция, плазмидой-помощником AAV, экспрессирующей гены AAV2 Rep и специфический для серотипа Cap, аденовирусной плазмидой-помощником, и содержащим ITR донорным плазмидным вектором. Через 3 суток клетки лизировали посредством трех циклов замораживания/размораживания, и клеточный дебрис удаляли центрифугированием. Векторы AAV преципитировали из лизатов с использованием полиэтиленгликоля и очищали посредством ультрацентрифугирования в течение ночи в градиенте хлорида цезия. Составы векторов получали посредством диализа и стерилизовали фильтрацией.
ELISA IgM, IgG, IgA:
[0130] Супернатанты собирали в конце стадий культивирования активации B-клеток, получения плазмобластов и получения плазматических клеток. IgM, IgG, IgA анализировали с использованием коммерческих наборов для твердофазного иммуноферментного анализа (ELISA) (Bethyl Laboratories; Montgomery, TX), в соответствии с протоколом производителя. Кратко, супернатант добавляли в планшет, инкубировали с качанием при комнатной температуре в течение одного часа, с последующей промывкой четыре раза с использованием буфера, поставленного в наборе. Добавляли детектирующее антитело, поставленное с набором, и инкубировали в течение 1 часа при комнатной температуре, с последующей промывкой четыре раза с использованием буфера для промывки, поставленного в наборе. Пероксидазу хрена (HRP), поставленную с набором, добавляли и инкубировали в течение 30 минут при комнатной температуре, с последующей промывкой четыре раза с использованием буфера, поставленного в наборе. Добавляли субстрат тетраметилбензидин (TMB), поставленный с набором, и позволяли проявление в течение 30 минут. Реакцию останавливали с использованием раствора для остановки, поставленного с набором, и считывали оптическую плотность при 450 нМ с использованием считывателя для планшетов.
Пример 2: Продукция антител в культивируемых in vitro B-клетках
[0131] CD19+ B-клетки размораживали и культивировали, как описано выше и проиллюстрировано на фиг. 1. Культуральные супернатанты собирали на сутки t4, t7 и t10, и тотальные IgM, IgG и IgA детектировали посредством ELISA, как описано выше.
[0132] Результаты (фиг. 2A-2C) показали, что клетки являются способными отвечать на стимуляцию цитокинами продукции антител и продуцируют антитела, как можно было ожидать.
Пример 3: Электропорация мРНК
[0133] Культивированые B-клетки обрабатывали с использованием мРНК, кодирующей трансген (GFP), для определения наилучших временных рамок для введения мРНК. CD19+ B-клетки (2,0E+05 клеток) подвергали электропорации с использованием 2 мкг мРНК GFP на сутки t0, t1, t2 или t3, где t0 представляет собой сутки размораживания клеток (фиг. 1). Подвергнутые электропорации клетки анализировали посредством анализа FACs, где отбор проводили, как показано на фиг. 4. Результаты (фигуры 3A-3D) показали, что электропорация на сутки t2 приводила к наивысшей экспрессии GFP, так что эти временные рамки выбрали для последующих исследований.
Пример 4: Расщепление нуклеазой в культивированных B-клетках
[0134] ZFN, специфические для трех локусов, AAVS1, CCR5 и TCRA, использовали для расщепления их мишеней в культивированных B-клетках. CD19+ B-клетки размораживали и культивировали в течение 2 суток в среде для активации B-клеток. Клетки промывали 2 раза с использованием DPBS, затем ресуспендировали в растворе для высокоэффективной электропорации BTXpress (Harvard Apparatus, Holliston, MA) до конечной плотности 2,0E+6 клеток/мл. Клетки (100 мкл) и раствор для электропорации смешивали с мРНК ZFN (4 мкг) с последующей электропорацией в электропораторе с прямоугольными импульсами BTX ECM830 (Harvard Apparatus) в 96-луночном многолуночном планшете для электропорации MOS 2 мм (Harvard Apparatus). После электропорации клетки переносили в среду для активации B-клеток в 24-луночном планшете на двое суток. Через 2 суток, клетки собирали, супернатанты собирали, и клетки промывали с использованием DPBS, затем переносили в среду для получения плазмобластов. Через 3 суток, клетки собирали, супернатанты собирали, и клетки промывали с использованием DPBS, затем переносили в среду для получения плазматических клеток. Клетки собирали для выделения геномной ДНК (гДНК) на сутки t4, t7 и t10, для анализа активности ZFN посредством глубокого секвенирования. Кратко, CD19+ B-клетки (2,0E+5 клеток) смешивали с мРНК ZFN (4 мкг), с последующей электропорацией.
[0135] Для измерения активности ZFN в локусах TCRA (TRAC), CCR5 и AAVS1, ДНК выделяли с использованием набора QIAamp DNA mini (Qiagen, Carlsbad, CA) по инструкциям производителя. Использовали сто нанограмм геномной ДНК (гДНК). Затем проводили двухстадийную ПЦР для локусов AAVS1 и TRAC с использованием полимеразы Phusion®Hot Start Flex (New England Biolabs, Ipswich, MA). Трехстадийную ПЦР использовали для локусов CCR5. С использованием глубокого секвенирования Illumina измеряли инделы в каждом локусе. Праймеры, использованные для каждого локуса, показаны ниже:
Праймеры для AAVS1:
[0136] AAVS1 Прямой: GACGTGTGCTCTTCCGATCTNNNNCCGGTTAATGTGGCTCTGGT (SEQ ID NO:3)
[0137] AAVS1 Обратный: ACACGACGCTCTTCCGATCTNNNNGACTAGGAAGGAGGAGGCCT (SEQ ID NO:4).
[0138] Ампликон AAVS1 представлял собой: 5'NNNNGACTAGGAAGGAGGAGGCCTAAGGATGGGGCTTTTCTGTCACCAATCCTGTCCCTAGTGGCCCCACTGTGGGGTGGAGGGGACAGATAAAAGTACCCAGAACCAGAGCCACATTAACCGGNNNN (SEQ ID NO:5).
Праймеры для CCR5:
[0139] CCR5 Прямой 1: CTGTGCTTCAAGGTCCTTGTCTGC (SEQ ID NO:6),
[0140] CCR5 Обратный 1: CTCTGTCTCCTTCTACAGCCAAGC (SEQ ID NO:7),
[0141] CCR5 Прямой 2: CTGCCTCATAAGGTTGCCCTAAG (SEQ ID NO:8),
[0142] CCR5 Обратный 2: CCAGCAATAGATGATCCAACTCAAATTCC (SEQ ID NO:9),
[0143] CCR5 Прямой 3: ACACGACGCTCTTCCGATCTNNNNNGCCAGGTTGAGCAGGTAGATG (SEQ ID NO:10),
[0144] CCR5 Обратный 3: AGACGTGTGCTCTTCCGATCTGCTCTACTCACTGGTGTTCATCTTT (SEQ ID NO:11).
[0145] Ампликон CCR5 представлял собой:
5'NNNNNGCCAGGTTGAGCAGGTAGATGTCAGTCATGCTCTTCAGCCTTTTGCAGTTTATCAGGATGAGGATGACCAGCATGTTGCCCACAAAACCAAAGATGAACACCAGTGAGTAGAGC (SEQ ID NO:12).
Праймеры для TCRA (TRAC):
[0146] TCRA Прямой: 5'ACACGACGCTCTTCCGATCTNNNNCCTCTTGGTTTTACAGATACGAAC (SEQ ID NO:13)
[0147] TCRA Обратный: 5'GACGTGTGCTCTTCCGATCTCTCACCTCAGCTGGACCAC (SEQ ID NO:14)
[0148] Ампликон TCRA представлял собой:
5'NNNNCCTCTTGGTTTTACAGATACGAACCTAAACTTTCAAAACCTGTCAGTGATTGGGTTCCGAATCCTCCTCCTGAAAGTGGCCGGGTTTAATCTGCTCATGACGCTGCGGCTGTGGTCCAGCTGAGGTGAG (SEQ ID NO:15).
[0149] Результаты этих исследований показаны на фиг. 5A-5C и демонстрируют, что нуклеазы являлись активными в культивированных B-клетках с использованием этих способов, и что более чем 80% модификация достигнута в множестве локусов.
[0150] Проводили также эксперименты, тестирующие влияние временного (в течение ночи) холодового шока (см. Патент США 8772008) на расщепляющую активность нуклеаз. В этих исследованиях, использовали диапазон вводимых количеств мРНК от 0,75 до 6 мкг. После электропорации, культуры разделяли, и одну часть помещали в инкубатор при 37°C на 4 суток. Вторую группу помещали при 30°C в течение ночи, затем переносили в инкубатор при 37°C на 3 суток. Глубокое секвенирование проводили для измерения % инделов, детектированных в результате расщепления нуклеазой.
[0151] Результаты (как показано на фигурах 6A-6C) показали, что способ холодового шока увеличивал общую активность расщепления.
Пример 5: Сравнение серотипов AAV при трансдукции B-клеток
[0152] Вирус AAV, содержащий экспрессирующую кассету трансгена (GFP), использовали для сравнения способности различных серотипов AAV для трансдукции культивированных B-клеток. Кратко, клетки размораживали и культивировали в течение 2 суток в среде для активации B-клеток в 24-луночном планшете при плотности 2,0E+5 клеток/лунку. Клетки собирали, подсчитывали и затем рассевали в 24-луночный планшет при плотности 2,0E+5 клеток/лунку. B-клетки трансдуцировали с использованием серотипов AAV 2, 5, 6, 8 и 9 при дозах вектора 2,4E+6, 1,2E+6, 6,0E+5, 3,0E+5 геномов вектора (вг)/клетку. Геномы вектора AAV содержали управляемую промотором CMV экспрессирующую кассету eGFP и инвертированные концевые повторы (ITR), см. фигуру 7B. Векторы AAV получали в Sangamo Therapeutics. Культуру клеток (25 мкл из 500 мкл в одной лунке 24-луночного планшета) собирали и смешивали с DPBS (175 мкл) на сутки t4, t7 и t10, и анализировали по экспрессии GFP с использованием Guava EasyCyte 5HT (EMD Millipore, Billerica, MA, USA). Данные анализировали с использованием InCyte версии 2.5 (EMD Millipore).
[0153] Результаты (фигура 7A) показали, что AAV6 являлся наиболее эффективным серотипом AAV при трансдукции культивированных B-клеток в ходе процесса дифференцировки в плазмобласты и плазматические клетки.
Пример 6: Управляемая нуклеазой направленная интеграция
[0154] Нуклеазы, описанные выше, затем использовали в комбинации с донором трансгена (GFP, белков, отсутствующих или недостаточных у субъекта, и/или представляющих интерес терапевтических антител) для тестирования способности системы поддерживать направленную интеграции донора в геном. Получено несколько иллюстративных доноров с GFP (фиг. 8), содержащих трансген GFP, фланкированный плечами гомологии, где плечи имеют гомологию с областями, окружающими мишень расщепления любого из AAVS1, CCR5 или TCRA. Кроме того, тестировали два различных промотора, либо PGK, либо EEK.
[0155] CD19+ B-клетки размораживали и культивировали в течение 2 суток в среде для активации B-клеток. Использовали комбинацию мРНК ZFN и донорного AAV или донора мРНК, нацеленного на такие же локусы (AAVS1, TCRA, CCR5, альбумин, HPRT и т.д.). Клетки промывали 2 раза с использованием DPBS, затем ресуспендировали в растворе для высокоэффективной электропорации BTXpress (Harvard Apparatus, Holliston, MA) до конечной плотности 2,0E+6 клеток/мл. Клетки (100 мкл) и раствор для электропорации смешивали с мРНК ZFN (4 мкг) с последующей электропорацией в электропораторе с прямоугольными импульсами BTX ECM830 (Harvard Apparatus) в 96-луночном многолуночном планшете для электропорации MOS 2 мм (Harvard Apparatus). После электропорации клетки переносили в 0,5 мл среды для активации B-клеток в 24-луночном планшете. AAV, содержащий гомологичные донорные матрицы для локусов-мишеней, затем добавляли при 2,4×106 вг/клетку. Планшеты осторожно встряхивали в течение 2 минут. Через 2 суток, культуры клеток собирали, 25 мкл культуры клеток отбирали, смешивали с DPBS (175 мкл) для анализа проточной цитометрии, остальную культуру клеток осаждали центрифугированием в настольной центрифуге, супернатанты собирали, и клетки промывали с использованием DPBS перед переносом в среду для получения плазмобластов. Через 3 суток, культуру клеток собирали, 25 мкл культуры клеток отбирали, смешивали с DPBS (175 мкл) для анализа проточной цитометрии, остальную культуру клеток осаждали центрифугированием в настольной центрифуге, супернатанты собирали, и клетки промывали с использованием DPBS перед переносом в среду для получения плазматических клеток. Через 3 суток проведения эксперимента, культуру клеток (25 мкл из 500 мкл в одной лунке 24-луночного планшета) собирали и смешивали с PBS (175 мкл) для анализа проточной цитометрии, остальную культуру клеток осаждали центрифугированием в настольной центрифуге, супернатанты собирали, клетки промывали с использованием DPBS и собирали для гДНК.
[0156] Для проточной цитометрии, культуру клеток (25 мкл из 500 мкл в одной лунке 24-луночного планшета) собирали и смешивали с PBS (175 мкл) на сутки 2, 5 и 8 после введения донора мРНК и AAV. Экспрессию GFP анализировали с использованием Guava EasyCyte 5HT (EMD Millipore, Billerica, MA, USA). Данные анализировали с использованием InCyte версии 2.5 (EMD Millipore). Результаты (фиг. 9A-9C) показывают, что присутствовала интеграция трансгена GFP во всех случаях, и что использование специфических нуклеаз приводило к наивысшему проценту положительных по GFP клеток.
[0157] Для измерения направленной интеграции доноров CCR5 и AAVS1, ДНК выделяли посредством набора QIAamp DNA mini (Qiagen, Carlsbad CA), по инструкциям производителя. Использовали сто нанограмм гДНК, и затем проводили трехстадийную ПЦР с использованием полимеразы Phusion®Hot Start Flex (New England Biolabs, Ipswich, MA) и набора готовых реакционных смесей HotStartTaq (Qiagen, Carlsbad, CA). С использованием глубокого секвенирования Illumina измеряли направленную интеграцию в каждом локусе. Праймеры для каждой стадии показаны ниже:
Праймеры для AASV1:
Праймеры для стадии 1 ПЦР:
[0158] AAVS1 Прямой 1: 5'CGGAACTCTGCCCTCTAACG (SEQ ID NO:16).
[0159] AAVS1 Обратный 1: 5'GTGTGTCACCAGATAAGGAATCTG (SEQ ID NO:17).
Праймеры для стадии 2 ПЦР:
[0160] AAVS1 Прямой 2: 5'CGTCTCTCTCCTGAGTCCG (SEQ ID NO:18).
[0161] AAVS1 Обратный 2: 5'GTGTGTCACCAGATAAGGAATCTG (SEQ ID NO:17).
Праймеры для стадии 3 ПЦР:
[0162] AAVS1 Прямой 3: 5'CTCTTTCCCTACACGACGCTCTTCCGATCTNNNNCTCTGGTTCTGGGTACTTTTATCTG (SEQ ID NO:19).
[0163] AAVS1 Обратный 3: 5'AGACGTGTGCTCTTCCGATCTGTGTGTCACCAGATAAGGAATCTG (SEQ ID NO:20).
[0164] Последовательность ампликона AAVS1 дикого типа:
5'NNNNCTCTGGTTCTGGGTACTTTTATCTGTCCCCTCCACCCCACAGTGGGGCCACTAGGGACAGGATTGGTGACAGAAAAGCCCCATCCTTAGGCCTCCTCCTTCCTAGTCTCCTGATATTGGGTCTAACCCCCACCTCCTGTTAGGCAGATTCCTTATCTGGTGACACAC (SEQ ID NO:21).
[0165] Последовательность AAVS1 GFP-TI (SEQ ID NO:22):
5'NNNNCTCTGGTTCTGGGTACTTTTATCTGTCCCCTCCACCCCACAGTGGGGCAAGCTTCGAGCCATCAGGGCCTGGTTCTTTCCGCCTCAGAAGGCCTTTTGCAGTTTATCAGGATGAGGATGACCAGCATGTTGCCCACAAAACCAAAGATGAACACCAGATTCCTTATCTGGTGACACAC
Праймеры для CCR5
Праймеры для стадии 1 ПЦР:
[0166] CCR5 Прямой 1: 5'GCTCTACTCACTGGTGTTCATCTTT (SEQ ID NO:12).
[0167] CCR5 Обратный 1: 5'CTCTGTCTCCTTCTACAGCCAAGC (SEQ ID NO:7).
Праймеры для стадии 2 ПЦР:
[0168] CCR5 Прямой 2: 5'GCTCTACTCACTGGTGTTCATCTTT (SEQ ID NO:12).
[0169] CCR5 Обратный 2: 5'CCAGCAATAGATGATCCAACTCAAATTCC (SEQ ID NO:9).
Праймеры для стадии 3 ПЦР:
[0170] CCR5 Прямой 3: 5'ACACTCTTTCCCTACACGACGCTCTTCCGATCTNNNNNGCCAGGTTGAGCAGGTAGATG (SEQ ID NO:23).
[0171] CCR5 Обратный 3: 5'AGACGTGTGCTCTTCCGATCTGCTCTACTCACTGGTGTTCATCTTT (SEQ ID NO:11).
[0172] Последовательность ампликона CCR5 дикого типа:
5'NNNNNGCCAGGTTGAGCAGGTAGATGTCAGTCATGCTCTTCAGCCTTTTGCAGTTTATCAGGATGAGGATGACCAGCATGTTGCCCACAAAACCAAAGATGAACACCAGTGAGTAGAGC (SEQ ID NO:12)
[0173] Последовательность CCR5 TI-GFP:
5'NNNNNGCCAGGTTGAGCAGGTAGATGTCAGTCATGCTCTTCAGCCTTTTGCAGTTTCTCGAGCCATCAGGGCCTGGTTCTTTCCGCCTCAGAAGTAGAAAGATGAACACCAGTGAGTAGAGC (SEQ ID NO:24)
[0174] Результаты (как показано на фиг. 10A и 10B) показали направленную интеграцию между 38% и 50% для этих локусов.
[0175] Проводили также эксперименты с использованием доноров несовпадающих трансгенов. Плечи гомологии в этих донорах несовпадающих трансгенов не имели гомологии с последовательностями, фланкирующими участок-мишень нуклеазы для вводимой совместно нуклеазы. Например, специфические для TCRA (TRAC) ZFN использовали в комбинации с донором трансгена GFP, содержащим плечи гомологии CCR5. Этот донор использовали также в комбинации со специфической для AAVS1 ZFN. Увеличение интеграции, обнаруженное с использованием совпадающих участка-мишени ZFN и плеч гомологии доноров, может указывать на то, что, по меньшей мере частично, интеграция трансгена основана на реакции гомологичной рекомбинации. Результаты (фиг. 11) показали, что увеличение интеграции донора происходит, когда донор фланкирован плечами гомологии, совпадающими с областью, окружающей участок расщепления нуклеазой, и таким образом, показывают, что культивированые B-клетки преимущественно используют гомологичную рекомбинацию для направленной интеграции. Наблюдали интеграцию на низком уровне для несовпадающих доноров, показывающую, что интеграция может также происходить посредством связывания концов с использованием NHEJ.
[0176] Сравнивали также клетки, трансдуцированные с использованием донорных конструкций, содержащих два альтернативных промотора. Экспрессию GFP анализировали посредством проточной цитометрии, как описано выше (см. фигуру 12), и результаты показали, что промотор EEK вызывал более высокую экспрессию GFP в B-клетках, чем промотор PGK.
[0177] Титрование, сравнивающее различные количества конструкции AAV-донора, проводили с использованием постоянной дозы мРНК ZFN. Культуру B-клеток обрабатывали с использованием 4 мкл специфической для TCRA (TRAC) ZFN посредством электропорации, и затем трансдуцировали с использованием диапазона донорных AAV, от 3,0E+05 до 2,4E+06 вг/клетку. Кроме того, два промотора также сравнивали в этих условиях. Результаты (фиг. 13A-13D) показали, что при более низких концентрациях донора, промотор EEK сохранял экспрессию GFP на протяжении 8-кратного разведения в ходе прогрессирования B-клетки до плазмобласта и плазматической клетки. Эксперимент подтвердил также, что использование нуклеаз для управления направленной интеграцией приводило к более высокой экспрессии GFP в B-клетках.
Пример 7: Экспрессия антител в ходе редактирования генома
[0178] Уровни IgG и IgM анализировали посредством ELISA, как описано выше, для культивированных B-клеток, которые подвергали электропорации для доставки пар ZFN, а также донора GFP. Результаты (фигуры 14 и 15) показывают, что уровни антител, продуцированных B-клетками, не подвергались большому влиянию электропорации или электропорации с последующей трансдукцией AAV-донора.
Пример 8: Потенциальная бустерная функция AAV в культивированных B-клетках
[0179] Возможно, что в культивированных B-клетках, экспрессирующих антитело против AAV, повторное подвергание B-клетки воздействию серотипа AAV, против которого B-клетка является реактивной, может вызывать «бустерный» эффект и индуцировать увеличение продукции антител против AAV. Для одной популяции CD19+ B-клеток от одного донора-человека показано увеличение секреции IgM после обработки с использованием AAV2. Известно, что антитела против AAV2 имеют большое преобладание в человеческой популяции из-за повсеместного присутствия AAV2. Таким образом, в этом исследовании показано, что в популяции CD19+ B-клеток от одного донора-человека, потенциально экспрессирующего антитела против AAV2, индуцировано резкое увеличение продукции IgM после подвергания воздействию AAV2, но не других серотипов AAV (фиг. 16).
[0180] Потенциальный механизм резкого увеличения экспрессии антител показан на фигуре 17 и демонстрирует использование этой системы для экспрессии представляющего интерес трансгена.
Пример 9: Бустерный эффект на экспрессию трансгена после подвергания воздействию AAV2 in vivo
[0181] Кассеты доноров трансгенов конструировали для вставки трансгена ниже B-клеточного промотора. B-клетки обрабатывали ex vivo с использованием специфической нуклеазы и донорной конструкции, содержащей специфический промотор антитела, связанный с представляющим интерес трансгеном. Ранее подтвердили, что B-клетки, выбранные для этой работы, продуцируют антитела против AAV2. Клетки повторно вводили субъекту, и после короткого периода времени для приживления, субъекта подвергали воздействию пептидов AAV2 или AAV2. Бустер AAV осуществляет повышающую регуляцию промотора антитела, вызывающую резкое увеличение экспрессии трансгена.
Пример 10: Модификация B-клеток посредством направленной интеграции специфического для B-клеток антитела
[0182] Кассеты доноров трансгенов (AAV, мРНК, плазмиду и т.д.) конструировали для вставки кодирующих антитело трансгенов, в которых антитело является(являются) специфическим для B-клетки, продуцирующей нежелательные антитела (например, ингибиторы) против белка, доставляемого посредством ERT (или аутоантигена), например, B-клетки, продуцирующей антитела против фактора свертывания крови, такого как F9 (антитела против F9). Донорные кассеты могут включать плечи гомологии с локусами-мишенями нуклеазы (например, альбумина, TCRA, CCR5, AAVS1 и т.д.), и их вводят in vivo в комбинации с подходящей нуклеазой и/или ex vivo в популяции B-клеток (популяции зрелых клеток, стволовых клеток и/или клеток-предшественников B-клеток) нуждающемуся в этом субъекту (пациенту с гемофилией с антителами против F9).
[0183] После модификации ex vivo или in vivo, продуцирующие антитело B-клетки секретируют нацеленные антитела, которые связываются с B-клетками, продуцирующими нежелательные антитела. Эти нацеленные антитела затем опосредуют лизис посредством мобилизации и активации антителозависимых цитотоксических клеток или посредством опосредованного комплементом лизиса. Таким образом, у пациента эти введенные B-клетки вызывают уменьшение количества эндогенных B-клеток, продуцирующих нежелательные антитела, например, против белков, доставляемых посредством ERT, или аутоантигена.
[0184] Полное содержание всех патентов, патентных заявок и публикаций, упомянутых в настоящем описании, таким образом, приведено в качестве ссылки.
[0185] Несмотря на то, что описание представлено в некоторых подробностях посредством иллюстрации и примера с целью ясности понимания, специалисту в данной области очевидно, что различные изменения и модификации можно осуществлять на практике без отклонения от содержания или объема описания. Соответственно, приведенные выше описание и примеры не следует рассматривать как ограничивающие.
| название | год | авторы | номер документа |
|---|---|---|---|
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2017 |
|
RU2788133C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ОПОСРЕДОВАННОЙ НУКЛЕАЗОЙ НАПРАВЛЕННОЙ ИНТЕГРАЦИИ ТРАНСГЕНОВ | 2013 |
|
RU2650819C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2020 |
|
RU2822369C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2015 |
|
RU2691102C2 |
| РЕДАКТИРОВАНИЕ ГЕНОВ С ИСПОЛЬЗОВАНИЕМ МОДИФИЦИРОВАННОЙ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (зкДНК) | 2018 |
|
RU2811724C2 |
| МОДУЛЯТОРЫ ТАУ И СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ИХ ДОСТАВКИ | 2017 |
|
RU2789459C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2014 |
|
RU2693891C1 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ГЕМОФИЛИИ В | 2011 |
|
RU2608643C2 |
| ПЕЧЕНЬ-СПЕЦИФИЧНЫЕ КОНСТРУКЦИИ, ЭКСПРЕССИОННЫЕ КАССЕТЫ ФАКТОРА VIII И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2774874C2 |
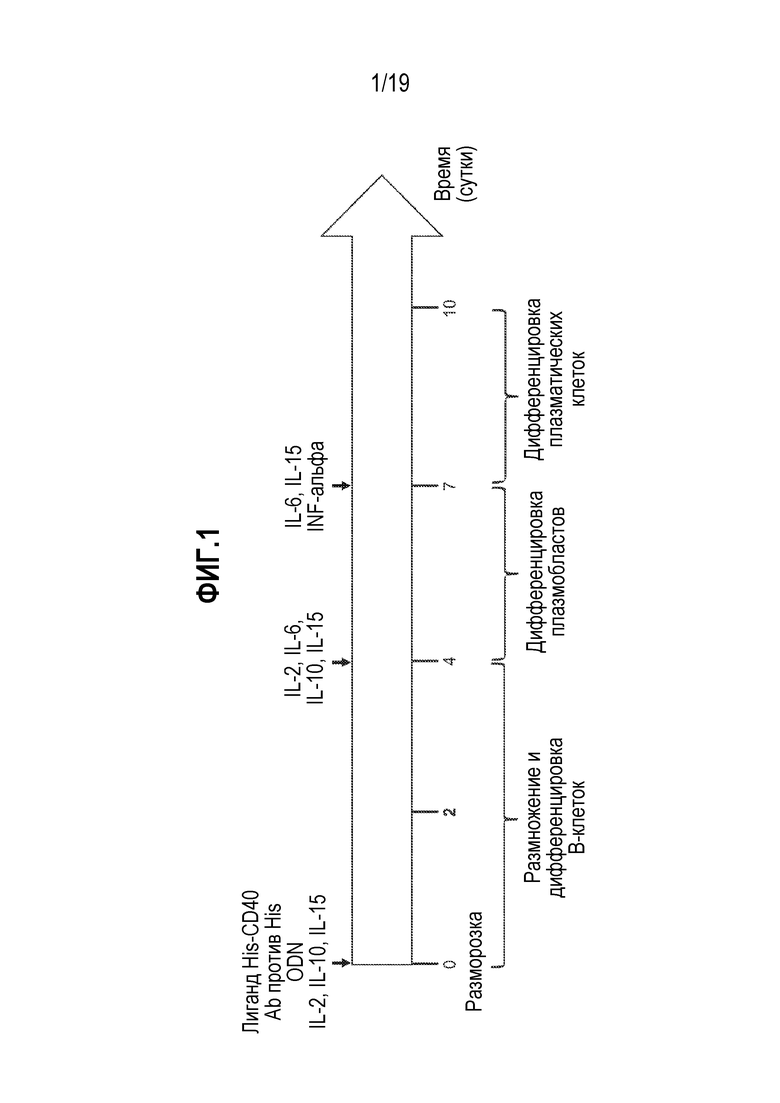
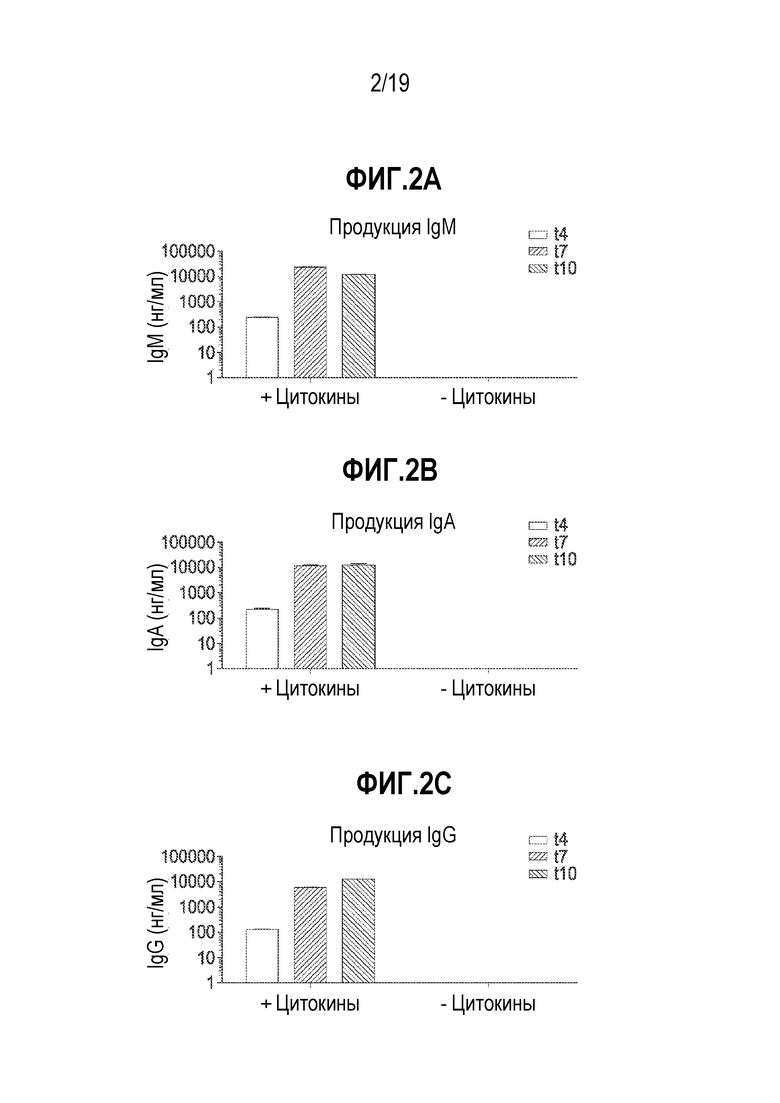
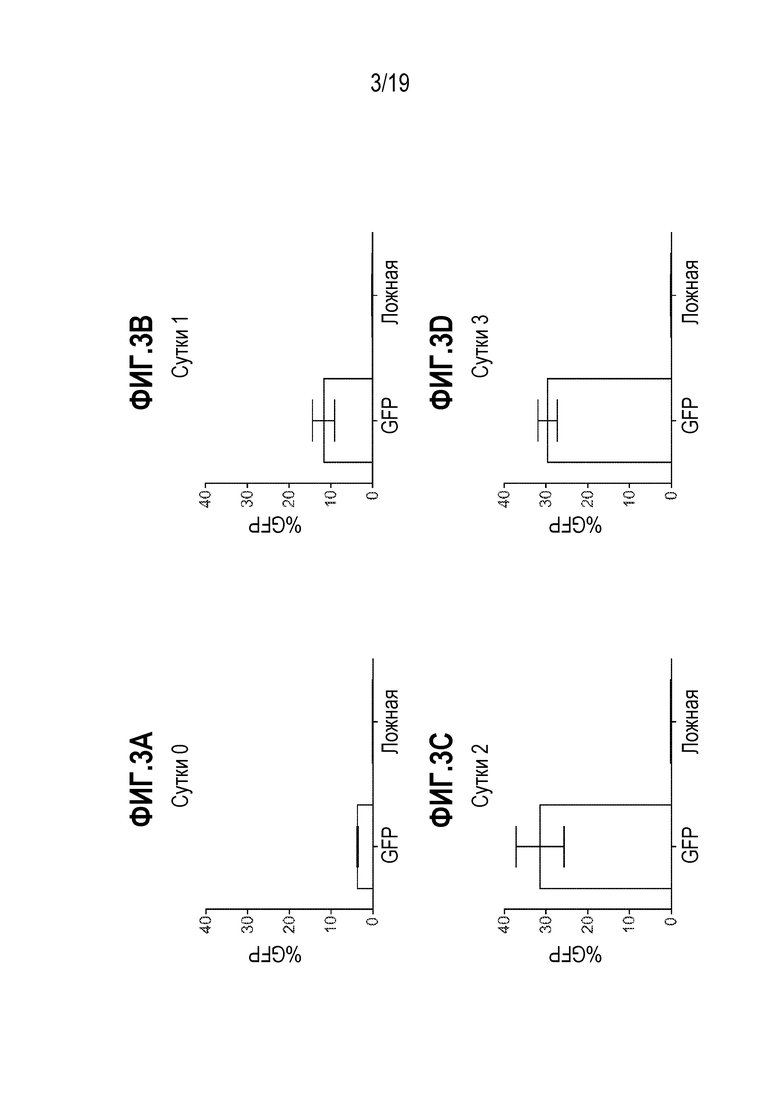
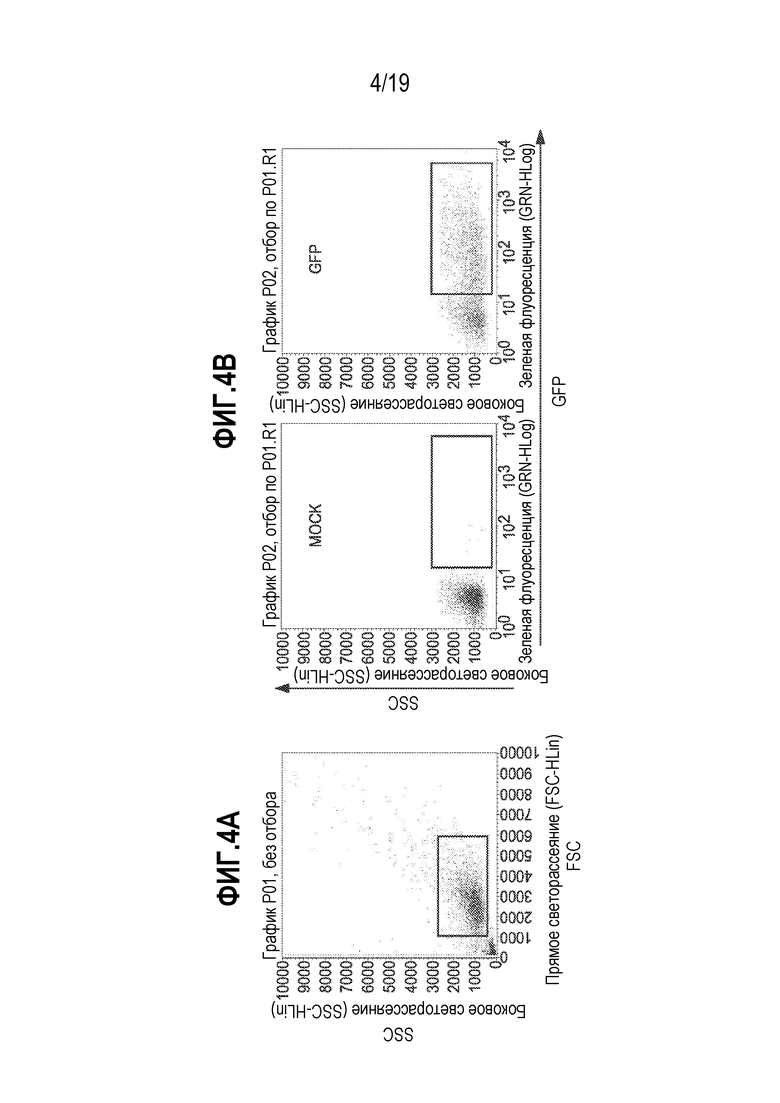

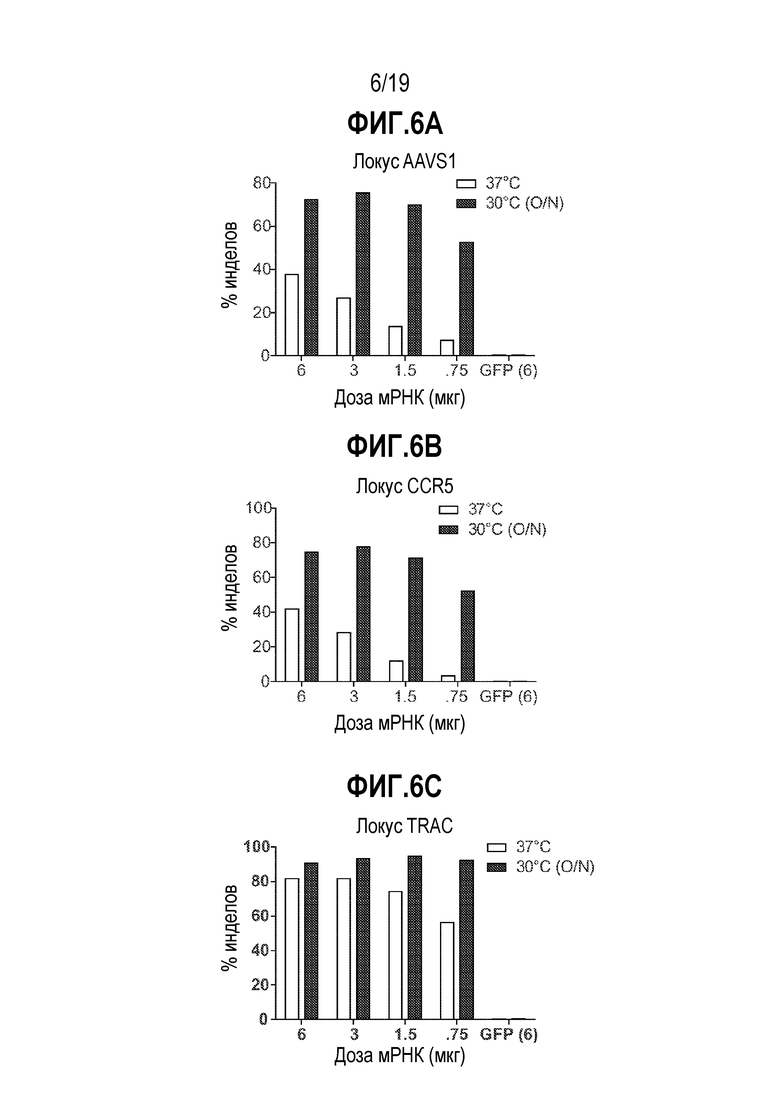
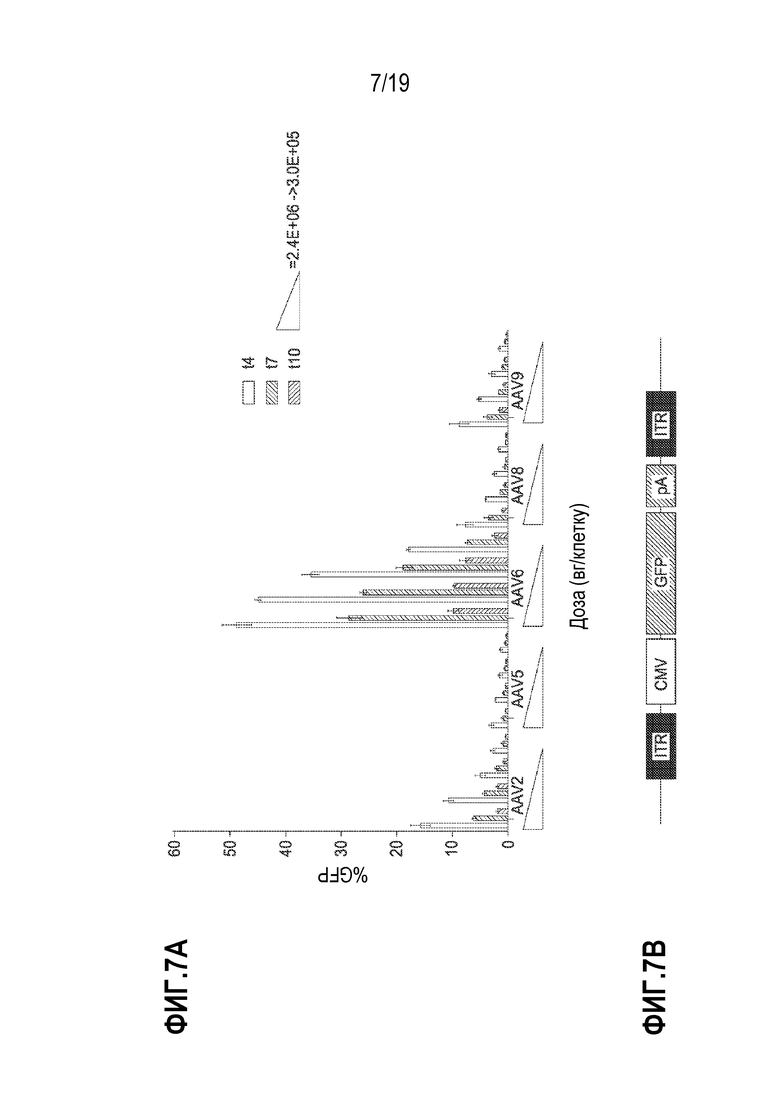
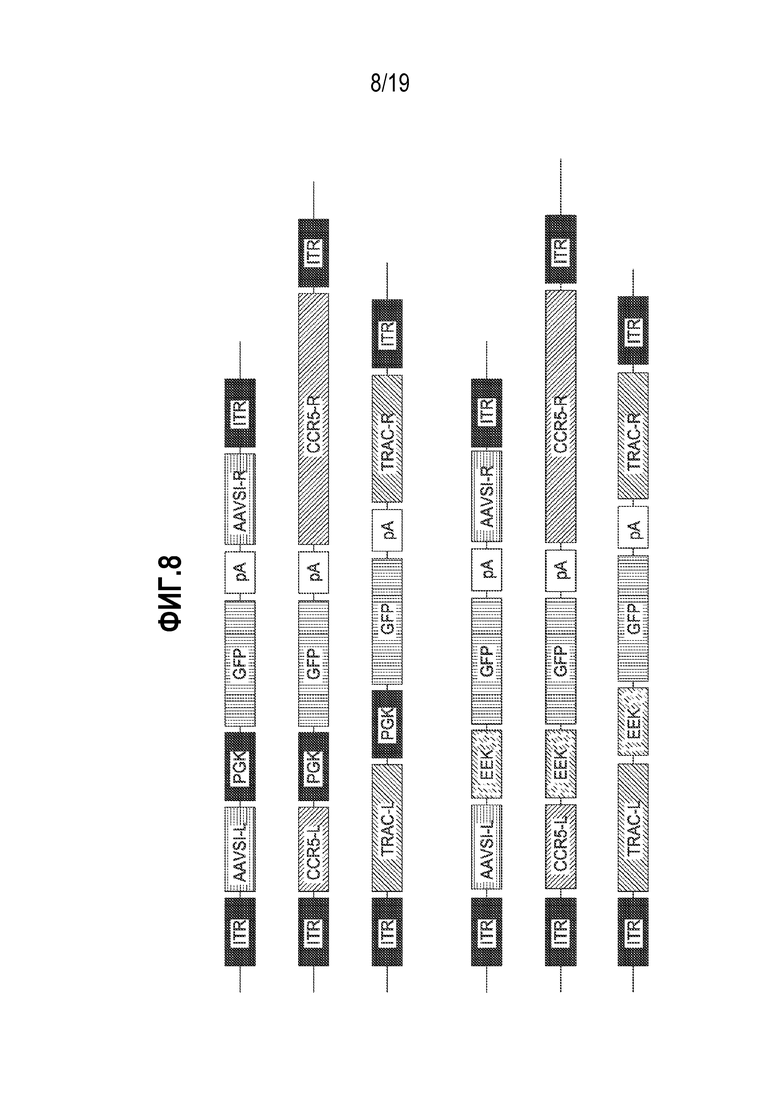
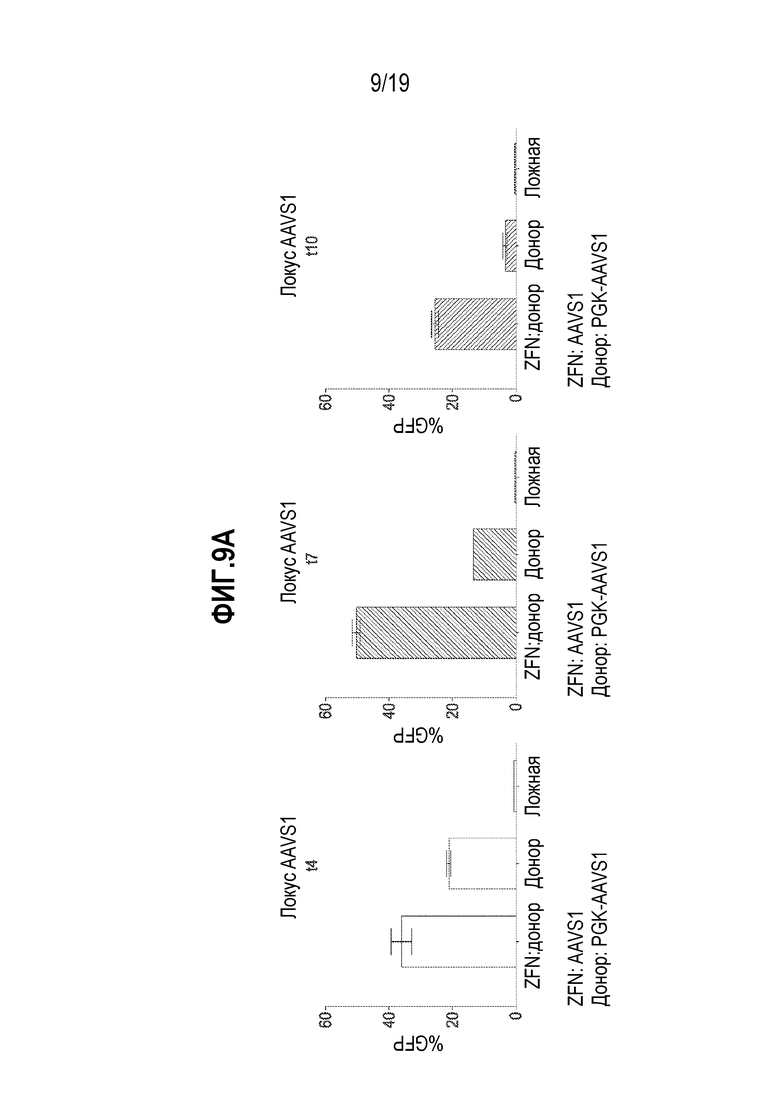
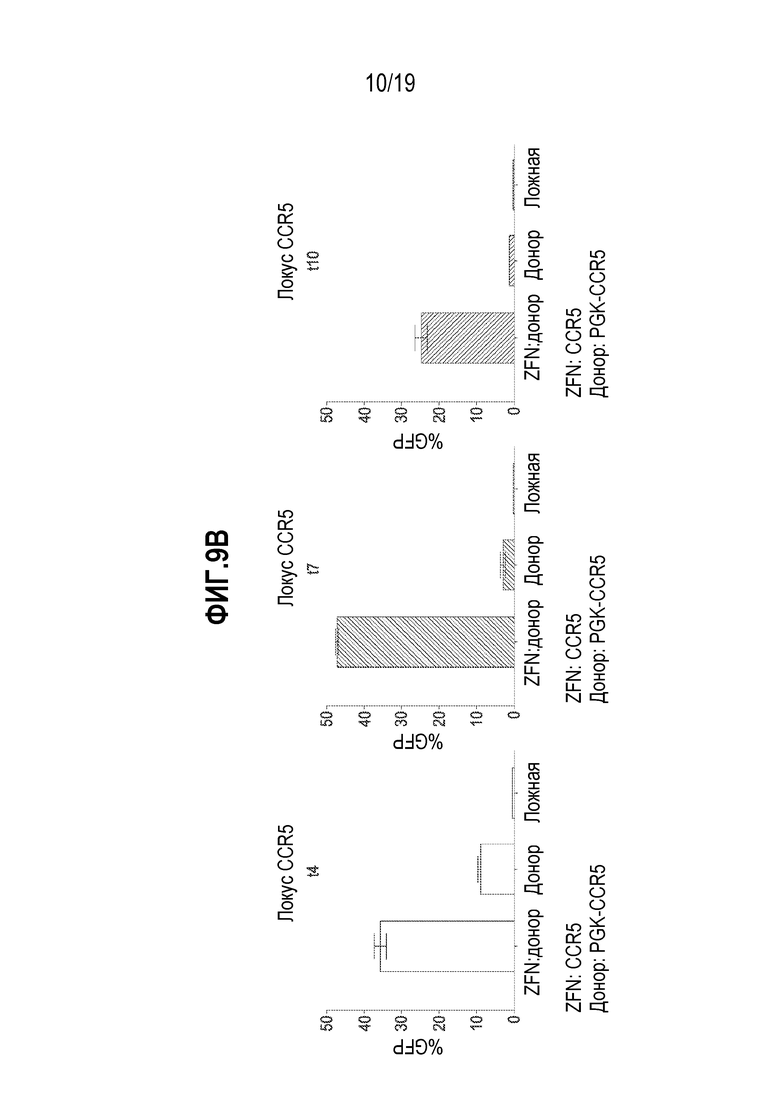

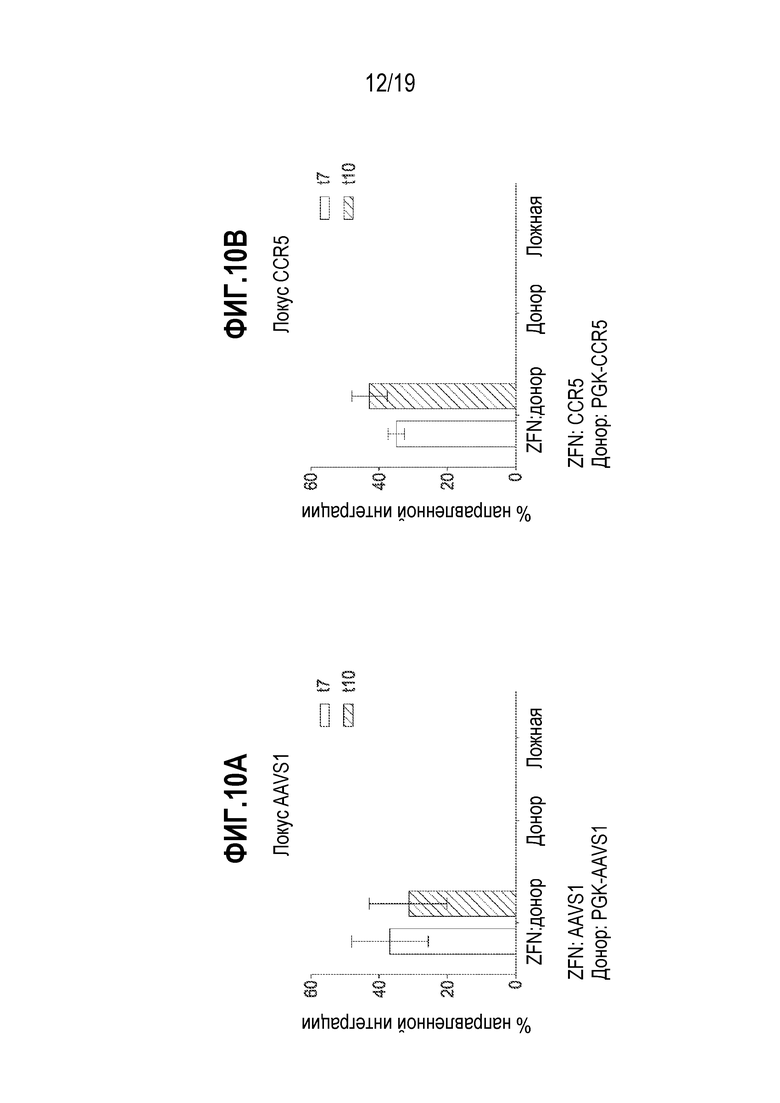
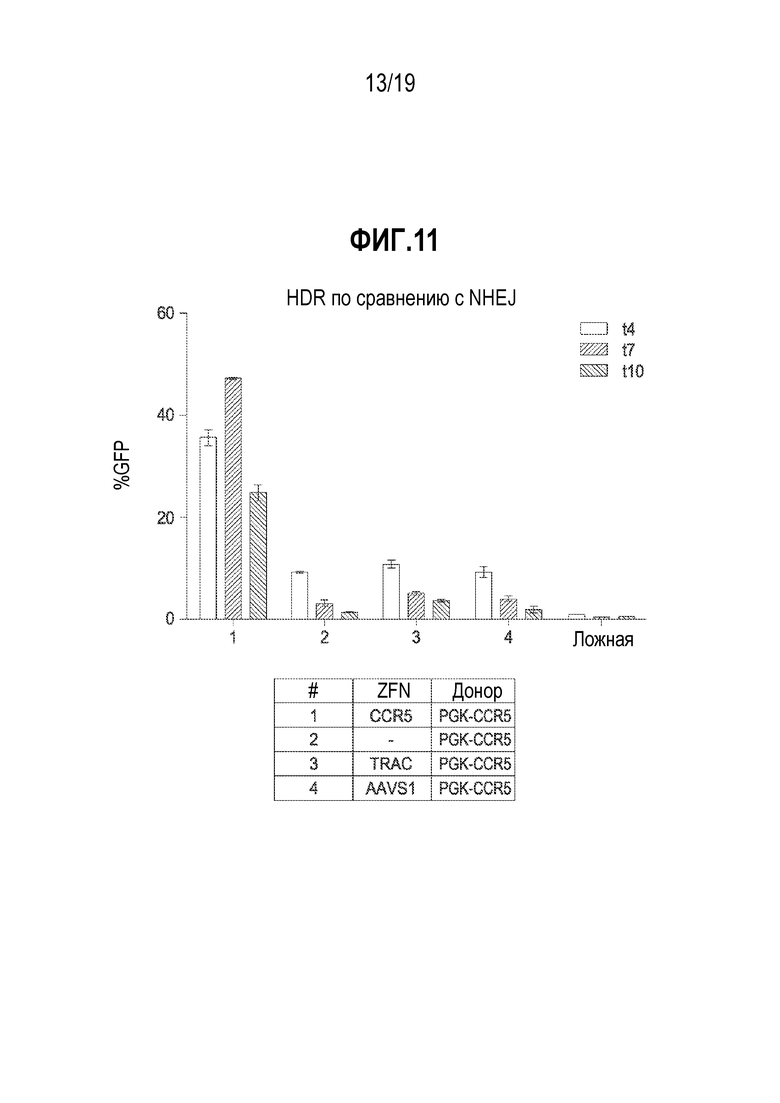
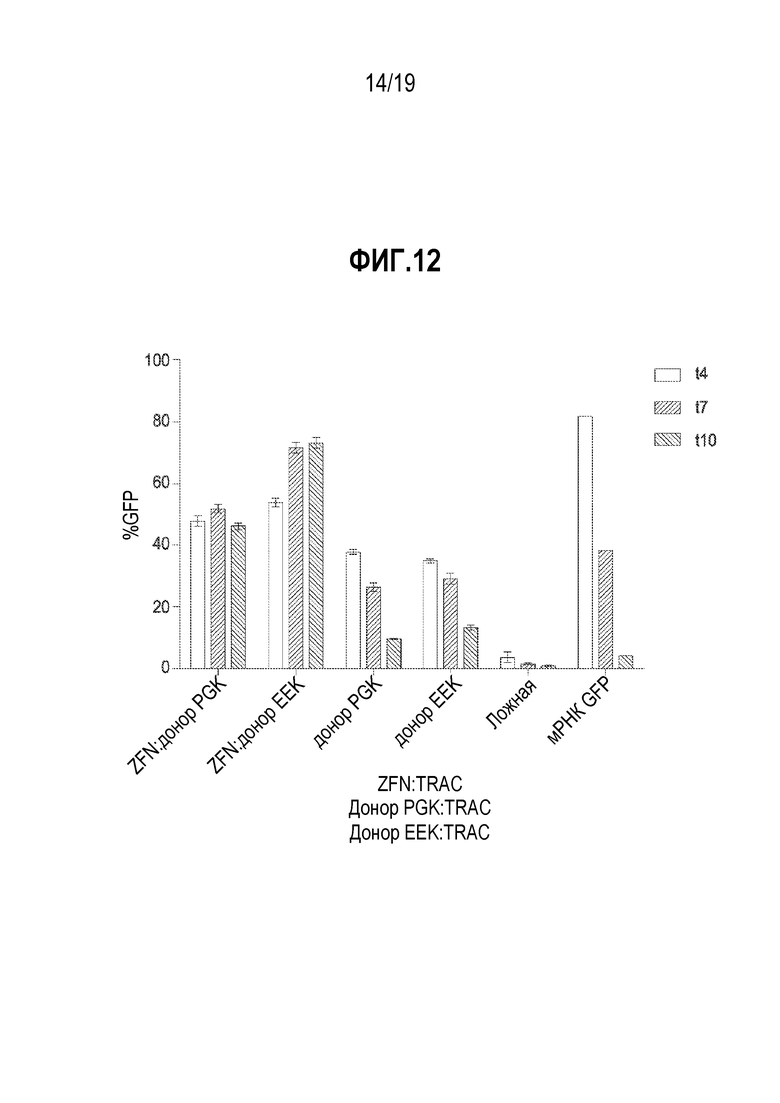
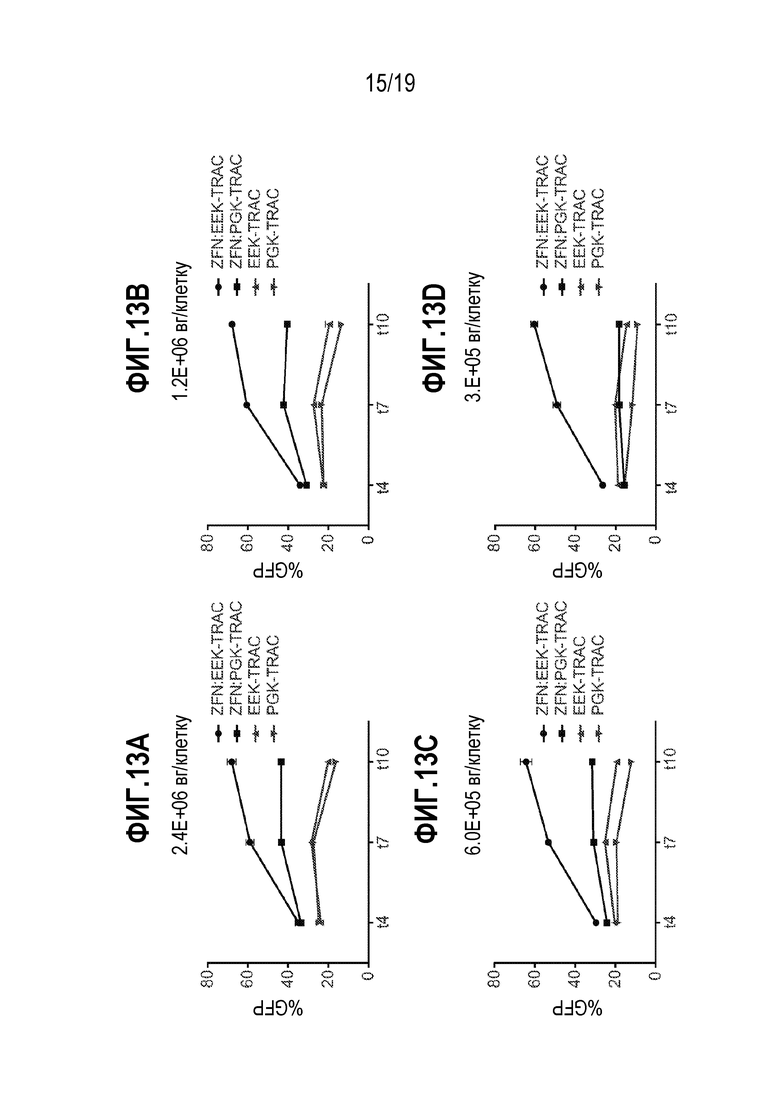
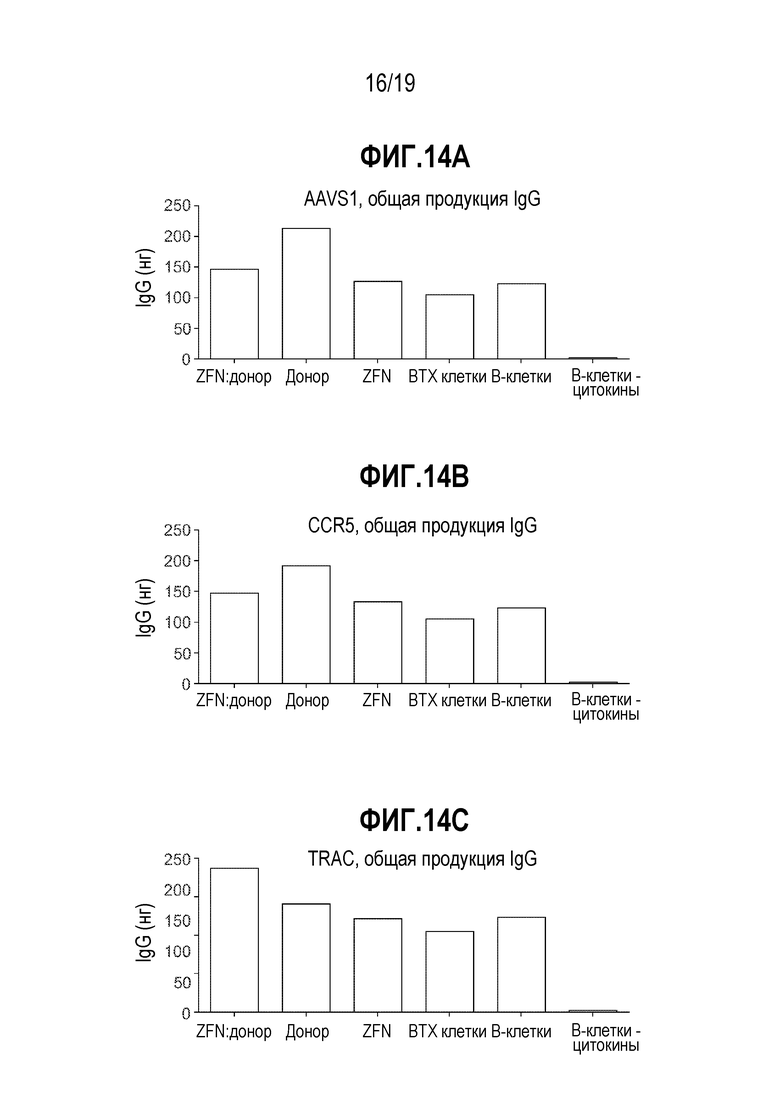
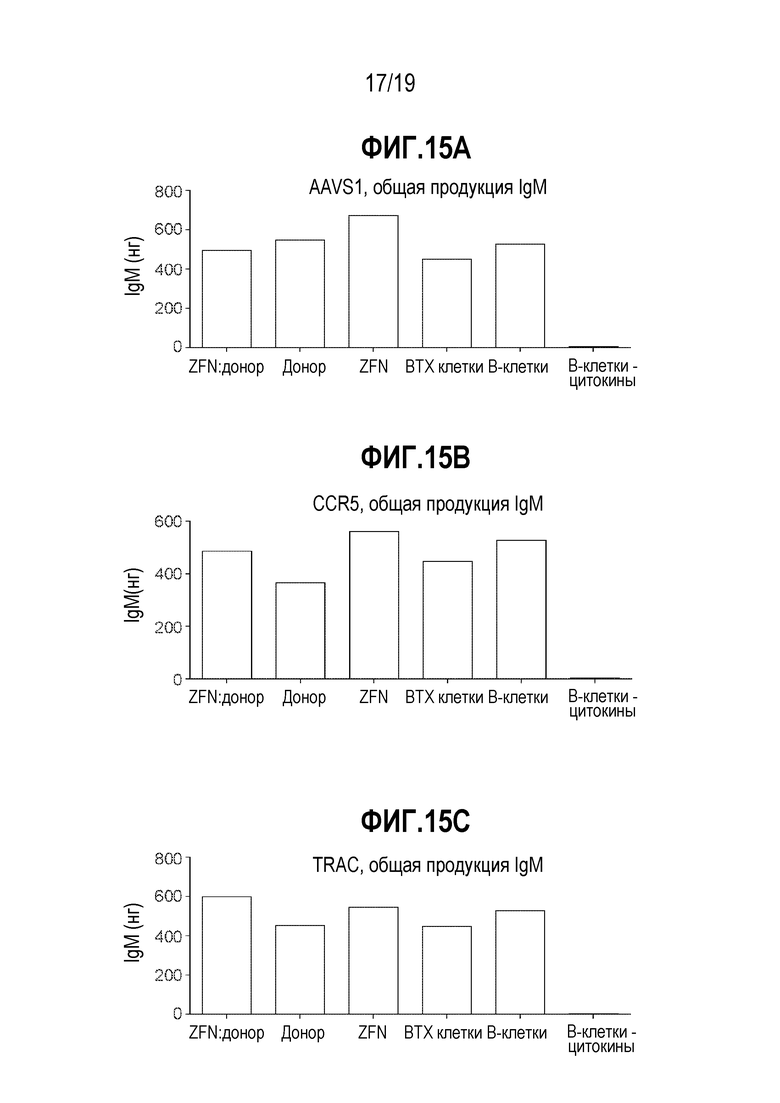
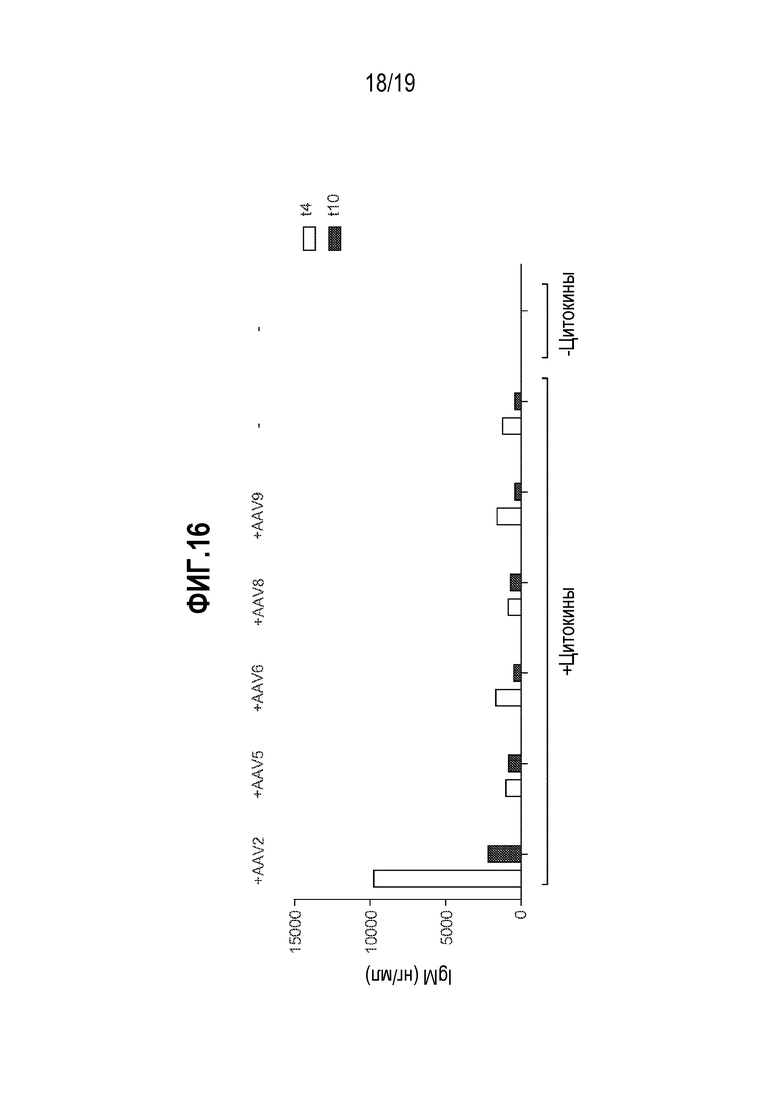
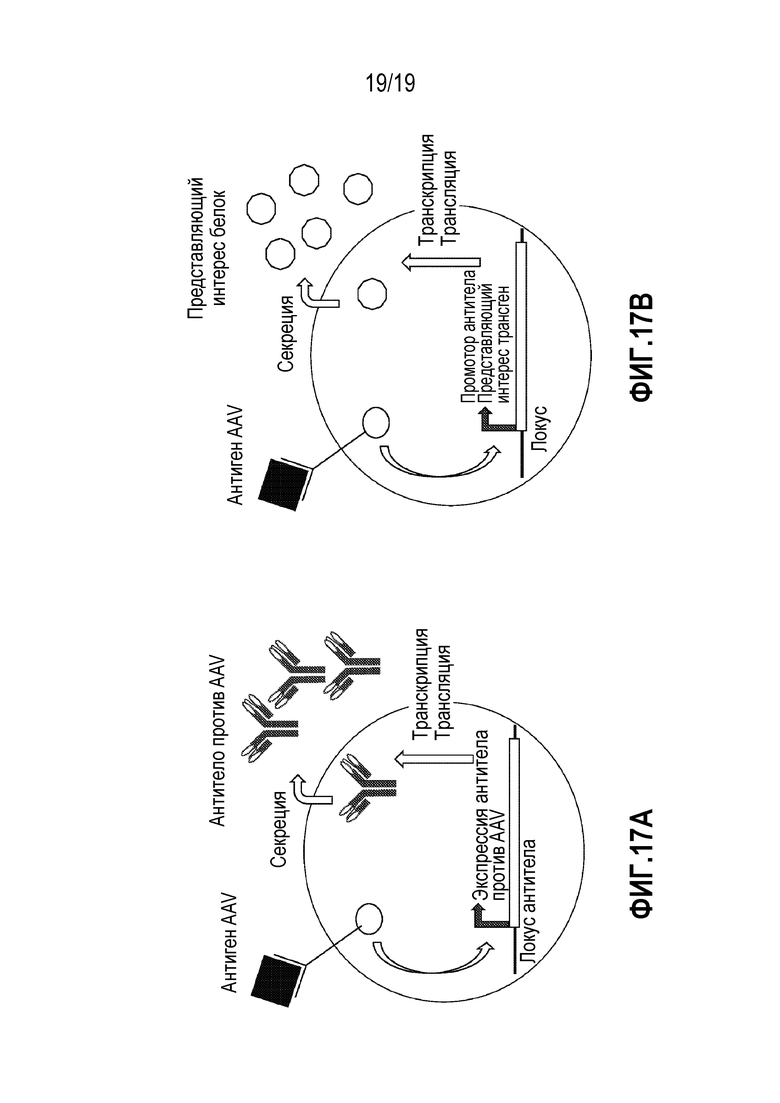
Настоящее изобретение относится к биотехнологии и клеточной биологии, в частности к генетически модифицированной В-клетке, композиции для её доставки, способу продукции белка и применения указанной клетки в способе продукции белка. Указанная генетически модифицированная B-клетка продуцирует ингибиторные антитела против фактора свертывания крови или белка, который принимает участие в аутоиммунном заболевании и способна экспрессировать трансген (антитело, специфичное в отношении B-клетки). Она содержит один или более трансгенов и одну или более дополнительных модификаций, которые осуществляют нокаут генов B-клеточного рецептора, и/или модифицируют клеточные взаимодействия в зародышевых центрах, и модификаций, ингибирующих супрессию какой-либо функции B-клетки, ассоциированную с инфекцией патогеном или регуляцией злокачественной опухоли. При этом указанный трансген экспрессируется эписомально в указанных модифицированных B-клетках или интегрирован в локус области безопасности, выбранный из группы, состоящей из CCR5, Rosa, альбумина, AAVS1, TCRA, TCRB. Настоящее изобретение позволяет расширить арсенал средств для экспрессии желательного трансгена на терапевтически значимом уровне у субъекта для лечения генетических заболеваний, таких как гемофилия, диабет, заболевания с накоплением лизосом, недостаточность A1AT, и злокачественные опухоли. 4 н. и 14 з.п. ф-лы, 10 пр., 39 ил.
1. Генетически модифицированная B-клетка, способная экспрессировать трансген, содержащая один или более трансгенов и одну или более дополнительных модификаций, выбранных из:
(a) вставок и/или делеций, которые (i) осуществляют нокаут генов B-клеточного рецептора, и/или (ii) модифицируют клеточные взаимодействия в зародышевых центрах, и
(b) модификаций, ингибирующих супрессию какой-либо функции B-клетки, ассоциированную с инфекцией патогеном или регуляцией злокачественной опухоли,
где указанный трансген экспрессируется эписомально в указанных модифицированных B-клетках или интегрирован в локус области безопасности, выбранный из группы, состоящей из CCR5, Rosa, альбумина, AAVS1, TCRA, TCRB, и
где указанный трансген кодирует терапевтическое антитело, специфичное в отношении B-клетки, которая продуцирует ингибиторные антитела против фактора свертывания крови или белка, который принимает участие в аутоиммунном заболевании.
2. Генетически модифицированная B-клетка по п.1, где один или более трансгенов интегрированы в локус области безопасности B-клетки.
3. Генетически модифицированная B-клетка по п.1 или 2, где трансген кодирует белок, отсутствующий или недостаточный у субъекта с гемофилией, с заболеванием с накоплением лизосом, терапевтическое антитело и/или пептид, облегчающий пересечение гематоэнцефалического барьера при слиянии с терапевтическим белком.
4. Генетически модифицированная B-клетка по п.3, где терапевтическое антитело является специфическим для B-клетки, которая образует ингибирующие антитела к фактору свертывания крови или белку, который принимает участие в аутоиммунном заболевании.
5. Генетически модифицированная B-клетка по п.3, где терапевтическое антитело является специфическим для регуляторной B-клетки (B-рег), способной ослаблять противоопухолевый ответ.
6. Генетически модифицированная B-клетка по п.2, где локус области безопасности представляет собой TCRA.
7. Генетически модифицированная B-клетка по п.6, где фактор свертывания крови представляет собой фактор IX (F9).
8. Генетически модифицированная B-клетка по любому из пп.1-7, где трансген дополнительно содержит промотор, управляющий экспрессией трансгена.
9. Генетически модифицированная B-клетка по п.8, где промотор представляет собой конститутивный промотор или специфический для линии B-клеток промотор.
10. Генетически модифицированная B-клетка по п.8, где специфический для линии B-клеток промотор представляет собой промотор цепи каппа иммуноглобулина, B29, BCL6, промотор III CIITA, mb-1 или промотор EEK.
11. Композиция для доставки B-клетки, экспрессирующей трансген, содержащая генетически модифицированную B-клетку по любому из пп.1-10.
12. Композиция по п.11, где генетически модифицированная B-клетка происходит из генетически модифицированной гематопоэтической стволовой клетки.
13. Способ продукции белка у нуждающегося в этом субъекта, включающий введение субъекту B-клетки по любому из пп.1-10 или композиции по п.11 или 12.
14. Способ по п.13, где белок модулирует ответ антител у субъекта.
15. Применение генетически модифицированной B-клетки по любому из пп.1-12 в способе продукции белка у субъекта, включающем: введение субъекту B-клеток или их клеток-предшественников в таких условиях, что B-клетка продуцирует белок у субъекта.
16. Применение по п.15, где белок представляет собой белок, отсутствующий или недостаточный при заболевании или нарушении, выбранном из гемофилии или заболевания с накоплением лизосом, или аутоиммунного заболевания, или антитело, специфическое для B-клетки, продуцирующей антитела против терапевтического белка, поставляемого посредством заместительной ферментной терапии (ERT).
17. Применение по п.16, где терапевтический белок, поставляемый посредством ERT, представляет собой фактор свертывания крови, и антитело является специфическим для B-клеток, продуцирующих антитела против фактора свертывания крови.
18. Применение по п.17, где фактор свертывания крови представляет собой фактор IX (F9).
| US 20170016027 A1, 19.01.2017 | |||
| US 20110287532 A1, 24.11.2011 | |||
| US 9290786 B2, 22.03.2016 | |||
| РУМЯНЦЕВ А.Г | |||
| и др., Гемофилия в практике врачей различных специальностей, Москва, издательская группа "ГЭОТАР-Медиа", 2012, 132 с., с.69-75 | |||
| SHAMMAH S., et al., Generation of human monoclonal antibodies by transformation of lymphoblastoid B cells with ras |

