Область техники, к которой относится изобретение
Изобретение относится к области генной терапии связанных с FGF-23 гипофосфатемических заболеваний, в частности, Х-сцепленной гипофосфатемии (XLH). Изобретение касается конструкций из нуклеиновой кислоты для генной терапии связанных с FGF-23 гипофосфатемических заболеваний, в частности генной терапии, направленной на мышечные, печеночные или гемопоэтические ткани, более конкретно на ткани печени. Изобретение также касается векторов, содержащих конструкции из нуклеиновой кислоты, и их применения для лечения связанных с FGF-23 гипофосфатемических заболеваний, в частности XLH, при помощи генной терапии.
Уровень техники
Фактор роста фибробластов 23 (FGF-23 или FGF23) представляет собой фосфатуретический гормон, вырабатываемый в костях, который работает путем связывания с рецепторными комплексами Klotho-FGF. Избыточная активность FGF23 вызывает гипофосфатемические заболевания, включая различные генетические заболевания, такие как Х-сцепленная гипофосфатемия (XLH) и аутосомно-доминантный или рецессивный гипофосфатемический рахит (ADHR, ADHR1, ADHR2), а также приобретенные заболевания, такие как онкогенная остеомаляция (TIO) и хроническая болезнь почек с минеральными и костными нарушениями (CKD-MBD) (см. обзор в Seiji Fukumoto, Calcif. Tissue Int., 2016, 98, 334-340).
Клинические проявления Х-сцепленной гипофосфатемии (XLH, OMIM # 307800) варьируются от изолированной гипофосфатемии до тяжелого искривления нижних конечностей. Заболевание проявляется в первые два года жизни в виде искривления нижних конечностей. У взрослых отмечалась энтезопатия (кальцификация сухожилий), связанная с болью в суставах, спонтанные зубные абсцессы и нейросенсорная тугоухость. XLH возникает из-за мутаций гена фосфат-регулирующей нейтральной эндопептидазы (PHEX), которые вызывают повышение уровня FGF23 в кровотоке. Повышение функции FGF23 ведет к подавлению уровня натрий-фосфатного ко-транспортера в почках. Расположенный в проксимальном канальце почек ко-транспортер обеспечивает реабсорбцию фосфата из мочи. Его подавление приводит к плохой реабсорбции фосфата и к снижению уровня фосфата в крови. Кроме того, повышение FGF23 связано с нарушением синтеза и усилением деградации 1,25(OH)2-витамина D. Снижение уровня фосфата в крови и низкий уровень витамина D ведет к нарушению минерализации костей и переломам.
Классическое лечение XLH заключается в приеме перорального фосфата и высоких доз кальцитриола, активной формы витамина D. На самом деле, реакция на внутривенную фосфатную терапию иногда бывает непредсказуемой, а осложнения включают “чрезмерную” гиперфосфатемию, гипокальциемию и метастатическую кальцификацию, а парентеральные схемы не подходят для хронических заболеваний. Однако пероральная терапия требует высоких доз, что зачастую ведет к диарее или раздражению желудка, а заместительная терапия сама по себе бывает неадекватной при значительном истощении фосфата в почках. Поэтому нужны новые стратегии лечения гипофосфатемии, связанной с FGF23.
Существует животная модель заболевания, происходящая от естественной делеции гена PHEX, модель HypDuk. Эта модель воспроизводит большинство проявлений болезни со снижением уровня фосфата в крови и нарушением роста костей. На мышах HypDuk тестировали различные стратегии терапии. В одном подходе применяются нейтрализующие антитела против FGF23. Недавно было одобрено моноклональное антитело против FGF23 для лечения детской формы XLH (Crysvita®, Ultragenyx). Другая стратегия заключается в использовании усеченной формы FGF23 человека, способной связываться с рецептором FGF23, не вызывая внутриклеточного каскада активации, возникающего при функциональном взаимодействии между рецептором и FGF23. Такой усеченный FGF23 может использоваться в качестве конкурента для снижения повышенной функции FGF23, наблюдаемой при XLH (Goetz R. et al., PNAS, 2009, 107, 407-412).
Пока еще не было сообщений о генной терапии связанных с FGF-23 гипофосфатемических заболеваний типа XLH. Поэтому существует потребность в применении генной терапии для лечения связанных с FGF-23 гипофосфатемических заболеваний, в частности XLH.
Сущность изобретения
Авторы изобретения создали конструкции из нуклеиновой кислоты и производные векторы AAV для генной терапии связанных с FGF-23 гипофосфатемических заболеваний. После однократного введения такого вектора AAV мышам HypDuk у обработанных животных наблюдалась нормализация привеса, размера тела, длины хвоста и циркулирующего фосфата. Коррекция заболевания на биохимическом, макроскопическом и функциональном уровне, наблюдавшаяся после однократной инъекции данного вектора AAV, свидетельствует о высокой эффективности применения генной терапии на основе таких конструкций из нуклеиновой кислоты и производных векторов, в частности векторов AAV, для лечения связанных с FGF-23 гипофосфатемических заболеваний.
Итак, изобретением предусмотрены конструкции из нуклеиновой кислоты для генной терапии связанных с FGF-23 гипофосфатемических заболеваний, которые кодируют слитый белок FGF23, включающий:
(a) сигнальный пептид,
(b) C-концевой пептид FGF23, который связывается с комплексом FGFR/Klotho,
(c) расщепляемый линкер и
(d) молекулу, стабилизирующую белок,
причем сигнальный пептид находится на N-конце слитого белка, а C-концевой пептид FGF23 и молекула, стабилизирующая белок, разделены расщепляемым линкером.
В некоторых воплощениях C-концевой пептид FGF23 включает последовательность из
SEQ ID NO: 1, начиная от какого-либо из положений 175-189 и заканчивая каким-либо из положений 203-251, или последовательность, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична указанной последовательности.
В некоторых воплощениях C-концевой пептид FGF23 включает мотив RXXR в положениях 176-179 SEQ ID NO: 1.
В некоторых предпочтительных воплощениях C-концевой пептид FGF23 включает последовательность SEQ ID NO: 2 или последовательность, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична указанной последовательности.
В некоторых воплощениях сигнальный пептид включает последовательность, выбранную из группы, состоящей из SEQ ID NO: 3-8; предпочтительно это SEQ ID NO: 7.
В некоторых воплощениях молекула, стабилизирующая белок, представляет собой сывороточный альбумин человека, предпочтительно включающий последовательность SEQ ID NO: 9.
В некоторых воплощениях расщепляемый линкер включает последовательность SEQ ID NO: 10.
В некоторых предпочтительных воплощениях конструкция нуклеиновой кислоты кодирует белок FGF23, включающий последовательность SEQ ID NO: 12 или 52 либо последовательность, которая по меньшей мере на 85%, 90%, 91%, 92%, 93%, 94%, 95%. 96%, 97%, 98% или 99% идентична любой из указанных последовательностей.
В некоторых воплощениях конструкция из нуклеиновой кислоты оптимизирована по кодонам для экспрессии в человеке.
В некоторых предпочтительных воплощениях конструкция нуклеиновой кислоты включает последовательность SEQ ID NO: 13, 51 или 57 либо последовательность, которая по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична любой из указанных последовательностей.
В некоторых воплощениях конструкции из нуклеиновой кислоты включают экспрессионную кассету, в которой кодирующая последовательность функционально связана по меньшей мере с одним промотором, который функционален в целевых клетках или тканях индивида, в частности, в мышечных, печеночных или гемопоэтических клетках или тканях. В некоторых предпочтительных воплощениях промотор является специфичным для печени, предпочтительно это промотор α1-антитрипсина человека.
В некоторых воплощениях конструкции из нуклеиновой кислоты также включают один или несколько элементов контроля, выбранных из группы, состоящей из энхансеров, связанных с промотором, предпочтительно участок контроля ApoE человека; интронов, расположенных между промотором и кодирующей последовательностью, предпочтительно модифицированный интрон HBB2 по SEQ ID NO: 17 или модифицированный интрон FIX по SEQ ID NO: 19; и сигналов терминации транскрипции, предпочтительно сигнал полиаденилирования бычьего гормона роста.
В некоторых воплощениях конструкции из нуклеиновой кислоты содержат или состоят из ДНК.
В других воплощениях конструкции из нуклеиновой кислоты содержат или состоят из РНК.
Изобретением также предусмотрены векторы для генной терапии, содержащие конструкции нуклеиновой кислоты по изобретению.
В некоторых воплощениях вектор представляет собой вирусный вектор, в частности вектор AAV или лентивирусный вектор, предпочтительно вектор AAV, содержащий капсид, выбранный из группы, состоящей из капсидов AAV1, AAV2, AAV5, AAV8, AAV2i8, AAV9, AAVrh10, AAVrh39, AAVrh43, AAVrh74, AAV-LK03, AAV2G9, AAV.PHP, AAV-Anc80, AAV3B и их химерных капсидов, в частности капсид AAV8, AAV9 или AAVrh74, как-то капсид AAV8 или AAV9, более предпочтительно капсид AAV8.
В некоторых других воплощениях вектор представляет собой частицу или везикулу, в частности, липидную микро- или нановезикулу либо частицу.
Изобретением предусмотрены клетки, генетически модифицированные конструкцией нуклеиновой кислоты по изобретению или вектором по изобретению, предпочтительно это печеночные, мышечные или гемопоэтические клетки, более предпочтительно клетки печени.
Изобретением также предусмотрена фармацевтическая композиция, содержащая по меньшей мере одно действующее вещество, выбранное из конструкции нуклеиновой кислоты по изобретению, вектора по изобретению или клетки по изобретению, и фармацевтически приемлемый носитель.
Изобретением предусмотрены фармацевтическая композиция по изобретению для применения при лечении связанных с FGF-23 гипофосфатемических заболеваний при помощи генной терапии или клеточной терапии.
В некоторых воплощениях данного применения связанные с FGF-23 гипофосфатемические заболевания представляют собой генетические заболевания, выбранные из группы, включающей: Х-сцепленную гипофосфатемию (XLH), аутосомно-доминантный гипофосфатемический рахит (ADHR), аутосомно-рецессивный гипофосфатемический рахит 1 (ADHR1), аутосомно-рецессивный гипофосфатемический рахит 2 (ADHR2), остеоглофоническую дисплазию, метафизарную хондродисплазию янсеновского типа, гипофосфатемию с зубными аномалиями и эктопической кальцификацией, синдром МакКьюна-Олбрайта/фиброзную дисплазию и гипофосфатемию с поражением кожи и костей, и/или приобретенные заболевания, выбранные из группы, включающей: онкогенную остеомаляцию, гипофосфатемическую остеомаляцию, осложнения при трансплантации почек или парентеральной терапии железом, хроническую болезнь почек и её осложнения типа гиперпаратиреоза; предпочтительно это Х-сцепленный гипофосфатемический рахит.
Раскрытие сущности изобретения
Конструкции из нуклеиновой кислоты
Изобретением предусмотрена конструкция нуклеиновой кислоты для генной терапии связанных с FGF-23 гипофосфатемических заболеваний.
Конструкция нуклеиновой кислоты по изобретению кодирует слитый белок FGF23, включающий по меньшей мере:
(a) сигнальный пептид,
(b) C-концевой пептид FGF23, который связывается с комплексом FGFR/Klotho,
(c) расщепляемый линкер и
(d) молекулу, стабилизирующую белок,
причем сигнальный пептид находится на N-конце слитого белка, а C-концевой пептид FGF23 и стабилизирующая белок молекула разделены расщепляемым линкером.
В настоящем документе термин “фактор роста фибробластов 23” (FGF-23 или FGF23), также известный как фосфатонин или индуцирующий гипофосфатемию фактор опухолевого происхождения, означает белок, кодируемый геном FGF23 в геноме млекопитающих. FGF23 человека имеет последовательность из 251 аминокислоты, номер доступа Q9GZV9.1 в UniProtKB/Swiss-Prot или номер доступа NP_065689 в NCBI (SEQ ID NO: 1). FGF23 экспрессируется в виде предшественника, содержащего N-концевой сигнальный пептид (24 аминокислоты), который отщепляется с образованием зрелого белка (FGF23). Для проявления фосфатуретической активности FGF23 требуется бинарный рецепторный комплекс FGF (FGFR)-Klotho. Кроме того, активность FGF23 регулируется посредством протеолитического расщепления по мотиву 176RXXR179, расположенному на границе между базовым доменом гомологии FGF и C-концевым хвостом FGF23 из 72 остатков. При протеолитическом расщеплении образуются: неактивный N-концевой фрагмент (от Y25 до R179), базовый домен гомологии FGF и C-концевой фрагмент (от S180 до I251). C-концевой фрагмент FGF23 является эндогенным ингибитором или антагонистом FGF23, который конкурирует с полноразмерным лигандом за связывание с комплексом FGFR-Klotho и блокирует передачу сигналов FGF23. Было показано, что C-концевой фрагмент FGF23 (180-251) противодействует фосфатуретической активности FGF23 in vivo. Меньший C-концевой фрагмент FGF (180-205) также проявляет активность антагониста FGF23 (Goetz et al. PNAS, 2010, 107, 407-410). Для активности FGF23 необходимы остатки 189-203 у зрелого FGF23 из 251 остатка, тогда как аминокислоты FGF23 от 3'-конца до остатка 203 не нужны для инициации FGF23-зависимой внутриклеточной сигнализации (Garringer et al., Am. J. Physiol. Endocrinol. Metab., 2008, 295, E929-E937).
В дальнейшем описании остатки обозначаются по стандартному однобуквенному коду аминокислот, а указанные положения определяются по совмещению с SEQ ID NO: 1.
Формы единственного числа включают и значения множественного числа, если из контекста не следует четко иначе. При этом термины “один”, “один или несколько” или “по меньшей мере один” могут применяться здесь взаимозаменяемо, если не указано иначе, а “или” означает “и/или”.
Конструкции из нуклеиновой кислоты могут содержать или состоять из ДНК, РНК либо синтетической или полусинтетической нуклеиновой кислоты, которая может экспрессироваться в целевых клетках или тканях индивида.
C-концевой пептид FGF23, образующийся при расщеплении слитого белка FGF23 in vivo, связывается с комплексом FGFR/klotho. Зрелый слитый белок FGF23 (без сигнального пептида) также может связываться с комплексом FGFR/klotho. Это связывание ингибирует передачу сигналов FGF23 через комплекс FGFR-klotho. Связывание слитого белка FGF23 и производного C-концевого пептида с комплексом FGFR/klotho и ингибирование передачи сигналов FGF23 через комплекс FGFR-klotho можно проверить стандартными методами, которые хорошо известны в данной области и изложены, к примеру, в Goetz et al. PNAS, 2010, 107, 407-410. C-концевой пептид FGF23, образующийся при расщеплении слитого белка FGF23 по изобретению in vivo, является специфическим ингибитором или антагонистом FGFR-klotho-зависимой функции FGF23. Слитый белок FGF23 по изобретению также может быть специфическим ингибитором или антагонистом FGFR-klotho-зависимой функции FGF23. Благодаря способности C-концевого пептида FGF23, а также, возможно, и слитого белка к нейтрализации Klotho-зависимой функции FGF23, слитый белок FGF23 по изобретению может применяться в качестве терапевтического средства для лечения связанных с FGF23 гипофосфатемических заболеваний.
C-концевой пептид FGF23 содержит или состоит из последовательности SEQ ID NO: 1, начиная от какого-либо из положений 175-189 и заканчивая каким-либо из положений 203-251, или последовательности, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична данной последовательности, причем он связывается с комплексом FGFR/klotho. C-концевой пептид FGF23 может включать последовательность от положения 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188 или 189 до положения 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250 или 251 в SEQ ID NO: 1 или последовательность, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична данной последовательности. Предпочтительно C-концевой пептид FGF23 содержит или состоит из последовательности SEQ ID NO: 1, начиная от какого-либо из положений 175-180 и заканчивая каким-либо из положений 205-251, или последовательности, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична данной последовательности, причем она связывается с комплексом FGFR/klotho.
В некоторых конкретных воплощениях C-концевой пептид FGF23 по изобретению содержит мотив 176RXXR179 (положения 176-179 по SEQ ID NO: 1). В некоторых конкретных воплощениях C-концевой пептид FGF23 по изобретению оканчивается в положении 203 SEQ ID NO: 1. В некоторых других конкретных воплощениях C-концевой пептид FGF23 по изобретению оканчивается в положении 204 или в более дальнем положении SEQ ID NO: 1, как-то, к примеру, в положении 232 или 251 SEQ ID NO: 1. В некоторых предпочтительных воплощениях C-концевой пептид FGF23 по изобретению содержит или состоит из последовательности SEQ ID NO: 2 (положения 175-251 SEQ ID NO: 1) или последовательности, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична данной последовательности, причем она связывается с комплексом FGFR/klotho. В некоторых воплощениях C-концевой пептид FGF23 по изобретению содержит мутации, в частности мутации, которые повышают его сродство связывания с комплексом FGFR/klotho.
C-концевой пептид FGF23 по изобретению содержит или состоит из 15-77 аминокислот C-концевого фрагмента FGF23. Поэтому данный C-концевой пептид FGF23 отличается от полноразмерного белка FGF23 и не содержит никакой последовательности из N-концевой области FGF23 (положения 25-174 SEQ ID NO: 1). В некоторых воплощениях C-концевой пептид FGF23 по изобретению содержит или состоит из C-концевого фрагмента FGF23 по меньшей мере из 20, 25, 30 и более аминокислот.
Степень идентичности аминокислотной последовательности или нуклеотидной последовательности определяется как процент аминокислотных остатков или нуклеотидов в сравниваемой последовательности, идентичных эталонной последовательности после совмещения последовательностей и введения пробелов, если нужно, для достижения максимальной идентичности последовательностей и без учета каких-либо консервативных замен в аминокислотных последовательностях в рамках идентичности последовательностей. Идентичность последовательностей рассчитывается по всей длине эталонной последовательности. Совмещение в целях определения степени идентичности аминокислотных последовательностей может осуществляться различными способами, известными специалистам в данной области, к примеру, с помощью общедоступных компьютерных программ типа BLAST (Altschul et al., J. Mol. Biol., 1990, 215, 403). При использовании таких программ предпочтительно используются параметры по умолчанию, например, штраф за пропуск и штраф за его расширение. В программе BLASTP по умолчанию используется длина слова (W), равная 3, и математическое ожидание (E), равное 10.
Слитый белок FGF23 содержит сигнальный пептид на своем N-конце. Сигнальные пептиды (SP) - это короткие пептидные последовательности, которые присутствуют на N-конце секреторных белков и служат для нацеливания белков для секреции. Сигнальные пептиды не состоят из строгой консенсусной последовательности, а имеют трехчастное строение, состоящее из положительно заряженного N-концевого участка (N-участка, 1-5 остатков), гидрофобного центрального участка (H-участка, 7-15 остатков) и нейтрального полярного C-концевого участка (C-участка, 3-5 остатков). В данной области известны и общедоступны многие сигнальные пептиды (в частности, см. сайт Signal Peptide Website и базы данных по последовательностям SPdb; Puzzo et al., Sci. Transl. Med., 2017, 9(418): doi:10.1126). Кроме того, известны способы выбора подходящих последовательностей SP для эффективной секреции белка (в частности, см. Stern et al., BMC Proc., 2011, 5 (suppl. 8): 013).
Сигнальный пептид может быть эндогенным или нативным сигнальным пептидом FGF23 (SEQ ID NO: 3; положения 1-24 по SEQ ID NO: 1) или гетерологичным сигнальным пептидом. В настоящем изобретении гетерологичный сигнальный пептид означает такой сигнальный пептид, который отличается от сигнального пептида FGF23, в частности, от сигнального пептида FGF23 человека. Примеры гетерологичных сигнальных пептидов, которые можно использовать в настоящем изобретении, включают, без ограничения, сигнальные пептиды α1-антитрипсина (SEQ ID NO: 4); синтетический mut1 (SEQ ID NO: 5); синтетический mut3 (SEQ ID NO: 6); химотрипсиногена B2 (CTRB2) (положения 1-18 по номеру доступа Q6GPI1 в Uniprot или номеру доступа NP_001020371 в NCBI или SEQ ID NO: 7) и С1-ингибитора протеаз плазмы (положения 1-22 по номеру доступа P05155 в Uniprot или SEQ ID NO: 8).
В некоторых воплощениях сигнальный пептид представляет собой гетерологичный сигнальный пептид, предпочтительно сигнальный пептид химотрипсиногена В2 (SEQ ID NO: 7).
C-концевой пептид FGF23 соединяется с молекулой, стабилизирующей белок, через расщепляемый линкер. Молекулой, стабилизирующей белок, является любая белковая молекула, которая повышает период полужизни или продолжительность действия терапевтического белка/пептида, присоединенного к ней, и подходит для терапевтического применения. В данной области известны различные стабилизирующие белки молекулы, которые использовались для стабилизации терапевтических белков (к примеру, см. Sven Berger, Peter Lowe & Michael Tesar (2015) Fusion protein technologies for biopharmaceuticals: Applications and challenge. mAbs, 7:3, 456-460, DOI:10.1080/19420862.2015.1019788). Примеры стабилизирующих белок молекул, которые можно использовать в настоящем изобретении, включают, без ограничения: сывороточный альбумин, в частности сывороточный альбумин человека; Fc-фрагмент иммуноглобулина; C-концевой пептид хорионического гонадотропина человека (CTP); рецептор, слитый со своим лигандом (GHR, слитый с GH), и связанный с латентностью пептид TGF-β (соединенный с сайтом расщепления для металлопротеазы).
В некоторых воплощениях стабилизирующая белок молекула отличается от Fc-фрагмента иммуноглобулина.
В некоторых воплощениях стабилизирующая белок молекула происходит из сывороточного транспортного белка. Сывороточные транспортные белки включают, без ограничения: семейство белков альбумина и эволюционно родственные сывороточные транспортные белки, такие, к примеру, как альбумин, альфа-фетопротеин (AFP; Beattie and Dugaiczyk, Gene 1982, 20, 415-422), афамин (AFM; Lichenstein et al., J. Biol. Chem., 1994, 269, 18149-18154) и связывающий витамин D белок (DBP; Cooke and David, J. Clin. Invest., 1985, 76, 2420-2424). Сывороточный транспортный белок может происходить из любых позвоночных, включая млекопитающих, птиц, рыб и других. Изобретение охватывает и функциональные варианты типа природных полиморфных вариантов, а также функциональные фрагменты сывороточных транспортных белков. Функциональный фрагмент или вариант сывороточного транспортного белка означает такой вариант или фрагмент, который способен повысить период полужизни или продолжительность действия терапевтического белка/пептида, присоединенного к нему, и подходит для терапевтического применения.
В некоторых конкретных воплощениях стабилизирующей белок молекулой является альбумин, включая его функциональные фрагменты или варианты, как определено выше. Альбумин может происходить из любых позвоночных, в особенности млекопитающих, к примеру, человека, коровы, овцы или свиньи. Альбумины не млекопитающих включают, без ограничения, курицу и лосося. Альбуминовая часть соединенного с альбумином полипептида может происходить не из того животного, из которого происходит терапевтическая полипептидная часть. В частности, слитые с альбумином белки по изобретению могут включать природные полиморфные варианты альбумина человека (HA) и фрагменты альбумина человека. Альбуминовая часть слитых с альбумином белков может включать полноразмерную последовательность HA (номер доступа NP_000468 в NCBI), предпочтительно содержащую сывороточный альбумин человека без сигнального пептида (положения 25-609 по номеру доступа NP_000468 в NCBI или SEQ ID NO: 9), или же один или несколько его фрагментов, способных стабилизировать или продлевать терапевтическую активность. Такие фрагменты могут состоять из 10 и более аминокислот или же могут включать 15, 20, 25, 30, 50, 70 и более смежных аминокислот из последовательности HA либо некоторые или все домены HA.
В некоторых предпочтительных воплощениях стабилизирующей белок молекулой является сывороточный альбумин человека (номер доступа NP_000468 в NCBI), предпочтительно включающий сывороточный альбумин человека без сигнального пептида (положения 25-609 по номеру доступа NP_000468 в NCBI или SEQ ID NO: 9).
Расщепляемым линкером является любой пептидный линкер, который расщепляется in vivo. В данной области известны различные расщепляемые пептидные линкеры, которые использовались в терапевтических белковых конструкциях. Примеры расщепляемых пептидных линкеров, которые можно использовать в настоящем изобретении, включают, без ограничения: последовательности активации факторов свертывания крови, в частности последовательность активации FIX (а.к. 182-200 или 182-203 по номеру доступа NP_000124 в NCBI): SEQ ID NO: 10 или SEQ ID NO: 11.
В некоторых предпочтительных воплощениях расщепляемый линкер содержит или состоит из последовательности SEQ ID NO: 10.
В некоторых воплощениях сигнальный пептид, C-концевой пептид FGF23, расщепляемый линкер и стабилизирующая белок молекула располагаются от N-конца к C-концу слитого белка FGF23, что означает, что стабилизирующая белок молекула слита с C-концом C-концевого пептида FGF23.
В некоторых воплощениях конструкции из нуклеиновой кислоты содержат или состоят из ДНК.
В некоторых других воплощениях конструкции из нуклеиновой кислоты содержат или состоят из РНК, в частности мРНК.
Примеры предпочтительных конструкций из нуклеиновой кислоты по изобретению включают:
- конструкцию нуклеиновой кислоты, кодирующую белок FGF23, содержащий последовательность SEQ ID NO: 12, которая представлена в примерах настоящей заявки и на фиг. 1A и соответствует конструкции № 12 в таблице 1,
- конструкцию нуклеиновой кислоты, кодирующую белок FGF23, содержащий последовательность SEQ ID NO: 52, которая представлена в примерах настоящей заявки и соответствует конструкции № 10 в таблице 1, и
- конструкцию нуклеиновой кислоты, кодирующую белок FGF23, содержащий последовательность, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 12 или 52; предпочтительно последовательность, которая по меньшей мере на 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична одной из данных последовательностей; более предпочтительно последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична одной из данных последовательностей.
SEQ ID NO: 12 содержит от N-конца до C-конца: сигнальный пептид химотрипсиногена B2 (SEQ ID NO: 7), C-концевой пептид FGF23 по SEQ ID NO: 2, расщепляемый линкер по SEQ ID NO: 10 и сывороточный альбумин человека по SEQ ID NO: 9. SEQ ID NO: 52 содержит от N-конца до C-конца: сигнальный пептид химотрипсиногена B2 (SEQ ID NO: 7), C-концевой пептид FGF23, состоящий из последовательности в положениях 180-251 по SEQ ID NO: 1, расщепляемый линкер по SEQ ID NO: 10 и сывороточный альбумин человека по SEQ ID NO: 9.
В некоторых воплощениях конструкция нуклеиновой кислоты содержит последовательность, которая оптимизирована по кодонам для экспрессии у индивида, проходящего лечение при помощи генной терапии, предпочтительно у человека. Соответствующие программы для оптимизации кодонов у требуемого индивида хорошо известны в данной области и общедоступны (например, см. http://www.genscript;com/cgi-bin/rare_codon_analysis; или
https://eu.idtdna.com/site/account/login?returnurl=%2FCodonOpt).
В некоторых предпочтительных воплощениях конструкция из нуклеиновой кислоты содержит нуклеотидную последовательность SEQ ID NO: 13 или SEQ ID NO: 57, которая представляет собой оптимизированную по кодонам последовательность для экспрессии у человека, кодирующую слитый белок FGF23 по SEQ ID NO: 12, нуклеотидную последовательность SEQ ID NO: 51, которая представляет собой оптимизированную по кодонам последовательность для экспрессии у человека, кодирующую слитый белок FGF23 по SEQ ID NO: 52, или последовательность, которая по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична одной из данных последовательностей; предпочтительно последовательность, которая по меньшей мере на 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична одной из данных последовательностей; более предпочтительно последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична одной из данных последовательностей. Данные последовательности предпочтительно представляют собой оптимизированные по кодонам последовательности для экспрессии у человека.
В некоторых воплощениях конструкция из нуклеиновой кислоты содержит экспрессионную кассету, в которой кодирующая последовательность функционально связана с соответствующими регуляторными последовательностями для экспрессии трансгена в целевых клетках или тканях индивида. В некоторых конкретных воплощениях целевая ткань представляет собой мышечные или печеночные клетки или ткани либо гемопоэтические клетки, более конкретно клетки или ткани печени. Такие последовательности, которые хорошо известны в данной области, включают, в частности, промоторы и другие регуляторные последовательности, способные дополнительно контролировать экспрессию трансгенов, как-то, без ограничения, энхансеры, терминаторы, интроны, сайленсеры, в частности тканеспецифичные сайленсеры, и микроРНК.
Промотором может быть тканеспецифичный, универсальный, конститутивный или индуцибельный промотор, который функционален в целевых клетках или тканях индивида, в частности, в мышечных, печеночных либо гемопоэтических клетках или тканях, более конкретно в клетках или тканях печени. Примеры конститутивных промоторов, которые можно использовать в настоящем изобретении, включают, без ограничения: промотор фосфоглицераткиназы (PGK), промотор фактора элонгации-1 альфа (EF-1α), включая укороченную форму данного промотора (EFS), вирусные промоторы, как-то самый ранний энхансер и промотор цитомегаловируса (CMV), энхансер цитомегаловируса/промотор β-актина курицы (CAG), ранний промотор SV40 и ретровирусные 5'- и 3'-промоторы LTR, включая гибридные промоторы LTR. Предпочтительным универсальным промотором является промотор CAG. Примеры индуцибельных промоторов, которые можно использовать в настоящем изобретении, включают регулируемые тетрациклином промоторы. Промоторами предпочтительно являются человеческие промоторы, т.е. промоторы из клеток человека или вирусов человека. Такие промоторы хорошо известны в данной области, а их последовательности имеются в общедоступных базах данных по последовательностям.
В некоторых конкретных воплощениях промотором является специфичный для печени промотор. Неограничительные примеры специфичных для печени промоторов, которые можно использовать в настоящем изобретении, включают промотор α1-антитрипсина человека (hAAT) (SEQ ID NO: 14), промотор транстиретина, промотор альбумина, промотор связывающего тироксин глобулина (TBG), промотор LSP (включающий последовательность промотора связывающего тиреоидный гормон глобулина, две копии последовательности энхансера α1-микроглобулина/бикунина и лидерную последовательность; Charles R. et al., Blood Coag. Fibrinol, 1997, 8: S23-S30) и другие. В данной области известны и другие полезные промоторы, специфичные для печени, например, приведенные в Базе данных по промоторам специфичных для печени генов, составленной в Cold Spring Harbor Laboratory (http://rulai.cshl.edu/LSPD/). Предпочтительным специфичным для печени промотором в настоящего изобретении является промотор hAAT.
В других конкретных воплощениях промотором является специфичный для мышц промотор. Неограничивающие примеры специфичных для мышц промоторов включают промотор мышечной креатинкиназы (MCK). Неограничительные примеры подходящих промоторов мышечной креатинкиназы включают промоторы мышечной креатинкиназы человека и промоторы усеченной мышечной креатинкиназы [(tMCK)] (Wang et al., Gene Therapy, 2008, 15, 1489-99); (репрезентативный номер доступа в GenBank: AF188002). Мышечная креатинкиназа человека имеет идентификатор гена: Gene ID No. 1158 (репрезентативный номер доступа в GenBank: NC_000019.9 по состоянию на 26 декабря 2012 г.). Другие примеры специфичных для мышц промоторов включают: синтетический промотор C5.12 (spC5.12, альтернативно обозначается здесь как “C5.12”) типа промотора spC5.12 (описан в Wang et al., Gene Therapy, 2008, 15, 1489-99); промотор MHCK7 (Salva et al., Mol. Ther., 2007, 15, 320-9); промоторы легкой цепи миозина (MLC), к примеру, MLC2 (Gene ID No. 4624; репрезентативный номер доступа в GenBank: NG_007554.1 по состоянию на 26 декабря 2012 г.); промоторы тяжелой цепи миозина (MHC), например, альфа-MHC (Gene ID No. 4624; репрезентативный номер доступа в GenBank: NG_023444.1 по состоянию на 26 декабря 2012 г.); промоторы десмина (Gene ID No. 1674; репрезентативный номер доступа в GenBank: NG_008043.1 по состоянию на 26 декабря 2012 г.); промоторы сердечного тропонина C (Gene ID No. 7134; репрезентативный номер доступа в GenBank: NG_008963.1 по состоянию на 26 декабря 2012 г.); промоторы тропонина I (Gene ID Nos. 7135, 7136 и 7137; репрезентативные номера доступа в GenBank: NG_016649.1, NG_011621.1 и NG_007866.2 по состоянию на 26 декабря 2012 г.); промоторы семейства генов myoD (Weintraub et al., Science, 251, 761 (1991); Gene ID No. 4654; репрезентативный номер доступа в GenBank: NM_002478 по состоянию на 26 декабря 2012 г.); промоторы α-актина (Gene ID Nos. 58, 59 и 70; репрезентативные номера доступа в GenBank: NG_006672.1, NG_011541.1 и NG_007553.1 по состоянию на 26 декабря 2012 г.); промоторы β-актина (Gene ID No. 60; репрезентативный номер доступа в GenBank: NG_007992.1 по состоянию на 26 декабря 2012 г.); промоторы γ-актина (Gene ID No. 71 и 72; репрезентативные номера доступа в GenBank: NG_011433.1 и NM_001199893 по состоянию на 26 декабря 2012 г.); специфичные для мышц промоторы, расположенные в интроне 1 глазной формы Pitx3 (Gene ID No. 5309) (Coulon et al.; специфичный для мышц промотор соответствует остаткам 11219-11527 по номеру доступа NG_008147 в GenBank, по состоянию на 26 декабря 2012 г.); промоторы, описанные в US Patent Publication US 2003/0157064, и промоторы CK6 (Wang et al., 2008, doi: 10.1038/gt.2008.104). В другом конкретном воплощении специфичным для мышц промотором является промотор E-Syn, описанный в Wang et al. (Gene Therapy, 2008, 15, 1489-99), включающий комбинацию энхансера из MCK и промотора spC5.12. В одном конкретном воплощении изобретения специфичный для мышц промотор выбран из группы, состоящей из промотора spC5.12, промотора MHCK7, промотора E-syn, промотора мышечной креатинкиназы, промотора легкой цепи миозина (MLC), промотора тяжелой цепи миозина (MHC), промотора сердечного тропонина C, промотора тропонина I, промотора семейства генов myoD, промотора α-актина, промотора β-актина, промотора γ-актина, специфичного для мышц промотора, расположенного в интроне 1 глазной формы Pitx3, и промотора CK6. В одном конкретном воплощении специфичный для мышц промотор выбран из группы, состоящей из промоторов spC5.12, десмина и MCK. В другом воплощении специфичный для мышц промотор выбран из группы, состоящей из промоторов spC5.12 и MCK. В одном конкретном воплощении специфичным для мышц промотором является промотор spC5.12.
В некоторых других конкретных воплощениях промотором является универсальный промотор. Типичные универсальные промоторы включают энхансер цитомегаловируса/промотор β-актина курицы (CAG), энхансер/промотор цитомегаловируса (CMV) (необязательно с энхансером CMV) [например, см. Boshart et al., Cell, 41:521-530 (1985)], промотор PGK, ранний промотор SV40, ретровирусный промотор LTR вируса саркомы Рауса (RSV) (необязательно с энхансером RSV), промотор дигидрофолатредуктазы, промотор β-актина, промотор фосфоглицеринкиназы (PGK) и промотор EF1α.
В некоторых других конкретных воплощениях промотором является промотор α-глобина или β-глобина. Промотор β-глобина экспрессируется исключительно в эритроидных клетках.
Еще в других конкретных воплощениях промотором является эндогенный промотор типа промотора альбумина или промотора GDE (фермента, деветвящего гликоген). GDE - это 4-альфа-глюканотрансфераза амило-1,6-глюкозидазы или AGL, которая у человека соответствует Gene ID No. 178 (репрезентативный номер доступа в GenBank: NG_012865 по состоянию на 16 сентября 2018 г.).
В определенных воплощениях промотор связан с последовательностью энхансера типа цис-регуляторного модуля (CRM) или искусственной последовательности энхансера. Применимые в настоящем изобретении CRMs включают описанные в Rincon et al., Mol. Ther., 2015, 23, 43-52; Chuah et al., Mol. Ther. 2014, 22, 1605-13; или Nair et al., Blood, 2014 123, 20, 3195-9. Другие регуляторные элементы, которые, в частности, способны усиливать экспрессию генов, специфичную для мышц, в частности, экспрессию в сердечных мышцах и/или скелетных мышцах, приведены в WO 2015/110449. Конкретные примеры регуляторных элементов из нуклеиновой кислоты, содержащих искусственные последовательности, включают регуляторные элементы, которые получаются при перестройке сайтов связывания факторов транскрипции (TFBS), присутствующих в последовательностях, приведенных в WO 2015/110449. Такие перестройки могут охватывать изменение порядка TFBS и/или изменение положения одного или нескольких TFBS относительно других TFBS и/или изменение количества копий одного или нескольких TFBS. Например, регуляторные элементы нуклеиновой кислоты для усиления специфичной для мышц экспрессии генов, в частности, специфичной для сердечных и скелетных мышц экспрессии генов, могут включать сайты связывания для E2A, HNH1, NF1, C/EBP, LRF, MyoD и SREBP; или для E2A, NF1, p53, C/EBP, LRF и SREBP; или для E2A, HNH1, HNF3a, HNF3b, NF1, C/EBP, LRF, MyoD и SREBP; или для E2A, HNF3a, NF1, C/EBP, LRF, MyoD и SREBP; или для E2A, HNF3a, NF1, CEBP, LRF, MyoD и SREBP; или для HNF4, NF1, RSRFC4, C/EBP, LRF и MyoD; или для NF1, PPAR, p53, C/EBP, LRF и MyoD. Например, регуляторные элементы нуклеиновой кислоты для усиления специфичной для мышц экспрессии генов, в частности, специфичной для скелетных мышц экспрессии генов, также могут включать сайты связывания для E2A, NF1, SRFC, p53, C/EBP, LRF и MyoD; или для E2A, NF1, C/EBP, LRF, MyoD и SREBP; или для E2A, HNF3a, C/EBP, LRF, MyoD, SEREBP и Tal-1b; или для E2A, SRF, p53, C/EBP, LRF, MyoD и SREBP; или для HNF4, NF1, RSRFC4, C/EBP, LRF и SREBP; или для E2A, HNF3a, HNF3b, NF1, SRF, C/EBP, LRF, MyoD и SREBP; или для E2A, CEBP и MyoD. В других примерах эти регуляторные элементы из нуклеиновой кислоты содержат по меньшей мере две, как-то 2, 3, 4 и более копий одного или нескольких TFBS, приведенных ранее. Другие регуляторные элементы, которые, в частности, способны усиливать специфичную для печени экспрессию генов, приведены в WO 2009/130208. Другие примеры энхансеров, которые можно использовать в настоящем изобретении, включают контролирующую область ApoE, в частности, контролирующую область ApoE человека (или локус гена аполипопротеина E/C-I человека, контролирующую область HCR-1 в печени; номер доступа U32510 в Genbank, SEQ ID NO: 15). В некоторых более конкретных воплощениях последовательность энхансера типа контролирующей области ApoE, предпочтительно контролирующей области ApoE человека, связана со специфичным для печени промотором типа приведенных выше, в частности, типа промотора hAAT.
В определенных воплощениях конструкции нуклеиновой кислоты содержат интроны, в частности интроны, расположенные между промотором и кодирующей последовательностью. Интроны вводится для повышения стабильности мРНК и продукции белка. Кроме того, модифицированные интроны, предназначенные для снижения количества или даже полного удаления альтернативных открытых рамок считывания (ARF), которые находятся в таких интронах, могут значительно улучшить экспрессию трансгена. Более того, считается, что при снижении количества ARF в интронах, включенных в конструкции по изобретению, снижается и иммуногенность конструкций. Предпочтительно удаляются такие ARF, длина которых превышает 50 п.н. и содержится стоп-кодон в одной рамке со стартовым кодоном. ARF можно удалять путем замены, вставки или делеции нуклеотидов, предпочтительно путем замены нуклеотидов. Например, в последовательности представляющего интерес интрона можно заменить ATG или GTG на CTG, который не является стартовым кодоном. Примеры интронов, которые можно использовать в настоящем изобретении, включают интрон β-глобина b2 человека (или HBB2; SEQ ID NO: 16), модифицированный интрон HBB2 (SEQ ID NO: 17), интрон фактора свертывания крови IX (FIX), в частности, полученный из первого интрона (SEQ ID NO: 18), и его модифицированный интрон (SEQ ID NO: 19), интрон β-глобина курицы (SEQ ID NO: 20) и его модифицированный интрон (SEQ ID NO: 21) и интрон SV40. Предпочтительными интронами являются модифицированный интрон HBB2 (SEQ ID NO: 17) и модифицированный интрон FIX (SEQ ID NO: 19).
В определенных воплощениях конструкции нуклеиновой кислоты также содержат сигнал терминации транскрипции (сигнал полиаденилирования), функционально связанный с кодирующей последовательностью (т.е. на 3'-конце кодирующей последовательности). Примеры полиА, которые можно использовать в настоящем изобретении, включают полиА бычьего гормона роста (bGH) (SEQ ID NO: 22).
В некоторых предпочтительных воплощениях экспрессионная кассета содержит в ориентации от 5' к 3' специфичный для печени промотор, предпочтительно промотор hAAT; кодирующую последовательность; и сигнал полиаденилирования типа полиА bGH (SEQ ID NO: 22). В некоторых более предпочтительных воплощениях экспрессионная кассета также содержит один или несколько дополнительных регуляторных элементов, выбранных из энхансеров, предпочтительно контролирующей области ApoE человека (SEQ ID NO: 15), и интронов, предпочтительно модифицированного интрона HBB2 (SEQ ID NO: 17). Пример предпочтительной экспрессионной кассеты, приведенной в примерах и на фиг. 1B, включает последовательность SEQ ID NO: 23, содержащую в ориентации от 5' к 3': контролирующую область ApoE (SEQ ID NO: 15), промотор α1-антитрипсина (hAAT) (SEQ ID NO: 14), модифицированный интрон HBB2 (SEQ ID NO: 17), кодирующую последовательность (SEQ ID NO: 13) и полиА bGH (SEQ ID NO: 22). Другой пример предпочтительной экспрессионной кассеты, приведенной в примерах, включает в ориентации от 5' к 3': контролирующую область ApoE (SEQ ID NO: 15), промотор α1-антитрипсина (hAAT) (SEQ ID NO: 14), модифицированный интрон HBB2 (SEQ ID NO: 17), кодирующую последовательность (SEQ ID NO: 51) и полиА bGH (SEQ ID NO: 22). Более предпочтительно экспрессионная кассета включает последовательность SEQ ID NO: 23.
Векторы
Изобретением также предусмотрен вектор, содержащий конструкцию нуклеиновой кислоты, как описано выше.
В изобретении можно использовать любые векторы, подходящие для доставки и экспрессии нуклеиновой кислоты в клетки индивида, в частности, подходящие для генной терапии, более конкретно для адресной генной терапии, направленной на целевую ткань или клетки у индивида. Такие векторы, которые хорошо известны в данной области, включают вирусные и невирусные векторы, причем такие векторы могут быть встраивающимися или не встраивающимися; репликативными или нерепликативными. В некоторых конкретных воплощениях генная терапия направлена на мышечные, печеночные или гемопоэтические клетки или ткани, более конкретно на клетки или ткани печени.
В настоящем изобретении термин “индивид” или “пациент” обозначает млекопитающих. Предпочтительно пациентом или индивидом по изобретению является человек.
Невирусные векторы включают различные (невирусные) средства, которые обычно применяются для введения или сохранения нуклеиновой кислоты в клетках индивида. Средства, которые применяются для введения нуклеиновой кислоты в клетки индивида различными способами, включают, в частности, носители для доставки на основе полимеров, частиц, липидов, пептидов либо их комбинаций, такие, без ограничения, как катионные полимеры, дендримеры, мицеллы, липосомы, экзосомы, микрочастицы и наночастицы, в том числе липидные наночастицы (LNP); и проникающие в клетки пептиды (CPP). В частности, СРР представляют собой катионные пептиды, такие как поли-L-лизин (PLL), олигоаргинин, пептиды Tat, пептиды пенетратина или транспортана и их производные, такие, к примеру, как Pip. Средства, которые применяются для сохранения нуклеиновой кислоты в клетках индивида (встроенной в хромосомы или же во внехромосомной форме), включают, в частности, векторы из “голой” нуклеиновой кислоты типа плазмид, транспозонов и миниколец, а также системы редактирования генов и редактирования РНК. Транспозоны включают, в частности, гиперактивную систему транспозонов Sleeping Beauty (SB100X) (Mates et al., 2009). В системах редактирования генов и редактирования РНК могут применяться любые сайт-специфичные эндонуклеазы типа нуклеазы Cas, TALEN, мегануклеазы, нуклеазы типа цинкового пальца и др. Кроме того, эти подходы можно выгодно комбинировать для введения и сохранения нуклеиновой кислоты по изобретению в клетках индивида.
Вирусные векторы по своей природе способны проникать в клетки и доставлять представляющие интерес нуклеиновые кислоты в клетки в соответствии с процессом, который называется трансдукцией вируса.
В настоящем изобретении термин “вирусный вектор” относится к нереплицирующимся, непатогенным вирусам, созданным для доставки генетического материала в клетки. У вирусных векторов вирусные гены, необходимые для репликации и вирулентности, заменены на экспрессионную кассету для представляющего интерес трансгена. Таким образом, геном вирусного вектора включает экспрессионную кассету для трансгена, фланкированную вирусными последовательностями, необходимыми для продукции вирусного вектора.
В настоящем изобретении термин “рекомбинантный вектор” относится к вирусам, в частности вирусным векторам, полученным стандартными методами технологии рекомбинантной ДНК, известными в данной области.
В настоящем изобретении термин “вирусные частицы” служит для обозначения внеклеточной формы непатогенных вирусов, в частности вирусных векторов, состоящей из генетического материала, состоящего из ДНК или РНК, окруженного белковой оболочкой, называемый капсидом, а в некоторых случаях оболочкой, происходящей из части мембран клеток хозяина и включающей вирусные гликопротеины.
В настоящем изобретении вирусный вектор означает вирусные частицы вектора.
Предпочтительными векторами для доставки нуклеиновой кислоты (конструкции нуклеиновой кислоты) по изобретению являются вирусные векторы, в частности, пригодные для генной терапии, более конкретно для генной терапии, направленной на целевые ткани или клетки у индивида, как-то на мышечные, печеночные или гемопоэтические клетки или ткани, более конкретно на клетки или ткани печени. В частности, вирусный вектор может происходить из непатогенного парвовируса типа аденоассоциированного вируса (AAV), ретровируса типа γ-ретровируса, спумавируса или лентивируса, аденовируса, поксвируса или герпесвируса. Вирусный вектор предпочтительно является встраивающимся вектором типа AAV или лентивирусного вектора, предпочтительно это вектор AAV. Лентивирусный вектор может быть псевдотипирован гликопротеином оболочки из другого вируса для наведения на представляющие интерес клетки/ткани, как-то на мышечные клетки, печеночные клетки или гемопоэтические клетки. В некоторых воплощениях лентивирус псевдотипирован синцитином, как описано в WO 2017/182607.
Вектор содержит вирусные последовательности, необходимые для продукции вирусного вектора, типа последовательностей LTR лентивируса или последовательностей ITR AAV, фланкирующих экспрессионную кассету.
В определенных воплощениях вектор представляет собой частицы или везикулы, в частности микро- или наночастицы на основе липидов либо частицы типа липосом или липидных наночастиц (LNP). В более конкретных воплощениях нуклеиновая кислота представляет собой РНК, а вектор представляет собой частицы или везикулы, как описано выше.
В другом конкретном воплощении вектором является лентивирусный вектор, в частности псевдотипированный лентивирусный вектор, как описано выше.
В другом конкретном воплощении вектором является вектор AAV. Парвовирус человека - аденоассоциированный вирус (AAV) из числа депендовирусов, от природы дефектных по репликации, способен встраиваться в геном инфицированных клеток, порождая латентную инфекцию. Последнее свойство является уникальным среди вирусов млекопитающих, так как встраивание происходит в определенном участке генома человека, называемом AAVS1, расположенном на хромосоме 19 (19q13.3-qter). Поэтому векторы AAV вызвали значительный интерес в качестве векторов для генной терапии человека. Среди благоприятных свойств вируса - отсутствие связи с какими-либо заболеваниями человека, его способность инфицировать как делящиеся, так и неделящиеся клетки, а также широкий спектр клеточных линий, происходящих из различных тканей, которые могут быть инфицированы.
AAV-вирусы можно конструировать стандартными методами молекулярной биологии, что дает возможность оптимизировать эти частицы для специфической доставки в клетки последовательностей нуклеиновых кислот, для минимизации иммуногенности, для настройки стабильности и времени жизни частиц, для эффективной деградации, для точной доставки в ядро.
Как известно в данной области, в конструкции из нуклеиновой кислоты по изобретению можно вводить подходящие дополнительные последовательности для получения функционального вирусного вектора. Подходящие последовательности включают инвертированные концевые повторы (ITR) AAV. Желательные фрагменты AAV для сборки в векторы включают белки cap, в том числе VP1, VP2, VP3, и гипервариабельные области, белки Rep, включая Rep 78, Rep 68, Rep 52 и Rep 40, и последовательности, кодирующие эти белки. Эти фрагменты можно с легкостью использовать в различных векторных системах и клетках хозяина. Рекомбинантные векторы на основе AAV, лишенные белка Rep, встраиваются в геном хозяина с низкой эффективностью и в основном присутствуют в виде стабильных кольцевых эписом, которые могут сохраняться в клетках мишени в течение многих лет.
В настоящем изобретении векторы AAV содержат капсид AAV, способный трансдуцировать представляющие интерес целевые клетки, в частности, мышечные, печеночные или гемопоэтические клетки или ткани, более конкретно клетки или ткани печени. Капсид AAV может быть из одного или нескольких природных или искусственных серотипов AAV.
Среди серотипов AAV, выделенных из человека или других приматов (NHP) и хорошо изученных, серотип 2 человека является первым AAV, который был разработан в качестве вектора для переноса генов. Другие используемые в настоящее время серотипы AAV включают варианты AAV-1, AAV-2 (как-то AAV-2 с оптимизированным капсидом с четырьмя мутациями, содержащий модифицированный капсид с заменами Y44+500+730F+T491V, описанный в Ling et al., 2016 Jul 18, Hum. Gene Ther. Methods), варианты AAV-3 (как-то вариант AAV3-ST, содержащий преобразованный капсид AAV3 с двумя заменами аминокислот, S663V+T492V, описанный в Vercauteren et al., 2016, Mol. Ther. Vol. 24(6), p. 1042), варианты AAV-3B, варианты AAV-4, -5, -6 (как-то вариант AAV6, содержащий капсид AAV6 с тремя мутациями Y731F/Y705F/T492V, описанный в Rosario et al., 2016, Mol. Ther. Methods Clin. Dev. 3, p. 16026), AAV-7, -8, -9, -2G9, -10 типа cy10 и -rh10, -rh39, -rh43, -rh74, -dj, Anc80, LK03, AAV.PHP, AAV2i8, серотипы AAV свиньи типа AAVpo4 и AAVpo6, а также серотипы AAV с мутациями капсида по тирозину, лизину и серину.
В качестве альтернативы использованию природных серотипов AAV, в настоящем изобретении можно использовать искусственные серотипы AAV, т.е. с не встречающимся в природе капсидным белком, включая, без ограничения, химерные капсиды AAV, рекомбинантные капсиды AAV или “гуманизованные” капсиды AAV. Такие искусственные капсиды могут быть получены любым подходящим методом, используя выбранные последовательности AAV (например, фрагменты капсидного белка VP1) в сочетании с гетерологичными последовательностями, которые могут быть получены из другого выбранного серотипа AAV, несмежных частей из того же серотипа AAV, из другого вирусного источника, чем AAV, либо из невирусного источника. Модифицированный капсид также может быть получен при модификациях капсида методом подверженной ошибкам ПЦР и/или вставки пептида (например, как описано в Bartel et al., 2011). Кроме того, варианты капсида могут включать изменения отдельных аминокислот типа мутаций тирозина (например, как описано в Zhong et al., 2008). В настоящем изобретении “модифицированным” капсидом может быть химерный капсид или капсид, содержащий один или несколько вариантов капсидных белков VP, происходящих из одного или нескольких капсидных белков VP AAV дикого типа.
В некоторых воплощениях вектор AAV представляет собой химерный вектор, т.е. его капсид содержит капсидные белки VP, происходящие по меньшей мере из двух различных серотипов AAV, или же содержит по меньшей мере один химерный белок VP, объединяющий области или домены белка VP, происходящие по меньшей мере из двух серотипов AAV. Примеры таких химерных векторов AAV, применимых для трансдукции клеток печени, описаны в Shen et al., Molecular Therapy, 2007; и в Tenney et al., Virology, 2014. Например, химерный вектор AAV может происходить из комбинации последовательности капсида AAV8 с последовательностью серотипа AAV, отличного от серотипа AAV8, типа любого из конкретно упомянутых выше. В других воплощениях капсид вектора AAV содержит один или несколько вариантов капсидных белков VP типа описанных в WO 2015/013313, в частности, варианты капсидов RHM4-1, RHM15-1, RHM15-2, RHM15-3/RHM15-5, RHM15-4 и RHM15-6, которые проявляют высокий тропизм к печени.
В других воплощениях вектор AAV представляет собой псевдотипированный вектор, т.е. его геном и капсид происходят из AAV различных серотипов типа приведенных выше серотипов AAV.
Кроме того, геном вектора AAV может быть одноцепочечным или самокомплементарным двухцепочечным (McCarty et al., Gene Therapy, 2003). Самокомплементарные двухцепочечные векторы AAV получают путем удаления концевого сайта расщепления у одного из концевых повторов AAV. Такие модифицированные векторы, у которых реплицирующийся геном составляет половину длины генома AAV дикого типа, имеют тенденцию к упаковке димеров ДНК.
В некоторых воплощениях вектор AAV подходит для генной терапии, направленной на целевые ткани или клетки индивида, в частности, на мышечные, печеночные или гемопоэтические клетки или ткани, более конкретно на клетки или ткани печени. В определенных воплощениях вектор AAV содержит капсид, выбранный из группы, состоящей из капсидов AAV1, AAV2, AAV2i8, AAV5, AAV8, AAV9, AAVrh10, AAVrh39, AAVrh43, AAVrh74, AAV-LK03, AAV2G9, AAV.PHP, AAV-Anc80, AAV3B и их химерных капсидов. В некоторых предпочтительных воплощениях вектор AAV содержит капсид AAV8, AAV9, AAVrh74 или AAV2i8, предпочтительно капсид AAV8, AAV9 или AAVrh74, как-то капсид AAV8 или AAV9, более предпочтительно капсид AAV8. Геном вектора AAV может происходить из другого серотипа (псевдотипированный вектор) и предпочтительно является одноцепочечным.
Изобретением также предусмотрена выделенная клетка, в частности такая клетка индивида, которая генетически модифицирована или трансформирована нуклеиновой кислотой или вектором по изобретению. Индивидом предпочтительно является пациент, подлежащий лечению. В некоторых воплощениях клетка представляет собой печеночную клетку, предпочтительно клетку печени пациента.
В настоящем изобретении “клетка печени” включает первичный гепатоцит, например, из печени взрослого или плода; гепатоцит, созревший in vitro; клеточную линию гепатоцитов; предшественник стволовой клетки печени или плюрипотентную стволовую клетку печени, например, индуцированную плюрипотентную стволовую клетку (iPS-клетку), эмбриональную стволовую клетку, фетальную стволовую клетку и стволовую клетку печени взрослого.
В настоящем изобретении термин “гемопоэтические клетки” относится к клеткам, образующимся при дифференцировке гемопоэтических стволовых клеток (HSC). Гемопоэтические клетки включают HSC, мультипотентные и коммитированные предшественники данной линии, клетки-предшественники и зрелые клетки. Зрелые гемопоэтические клетки включают, без ограничения, лимфоциты (B- и T-лимфоциты), NK-клетки, моноциты, макрофаги, гранулоциты, эритроциты, тромбоциты, плазмацитоидные и миелоидные дендритные клетки и микроглиальные клетки.
В настоящем изобретении термин “гемопоэтические стволовые клетки (HSC)” относится к самовозобновляющимся клеткам, способным восстанавливать краткосрочный или долгосрочный гемопоэз после пересадки.
В настоящем изобретении термин “генетическая модификация” означает вставку, делецию и/или замену одного или нескольких нуклеотидов в геномной последовательности.
В настоящем изобретении “мышечная ткань” включает, в частности, ткани сердечных и скелетных мышц.
В настоящем изобретении термин “мышечные клетки” относится к миоцитам, мышечным трубочкам, миобластам и/или клеткам-сателлитам.
Фармацевтические композиции и терапевтическое применение
Другой аспект изобретения составляют фармацевтические композиции, содержащие по меньшей мере одно действующее средство, выбранное из нуклеиновой кислоты по изобретению, вектора по изобретению или клеток по изобретению, и фармацевтически приемлемый носитель.
Нуклеиновая кислота, вектор и получаемая фармацевтическая композиция по изобретению могут применяться для лечения заболеваний при помощи генной терапии, в частности адресной генной терапии, направленной на мышечные, печеночные или гемопоэтические клетки или ткани, более конкретно на клетки или ткани печени. Клетка и получаемая фармацевтическая композиция по изобретению могут применяться для лечения заболеваний при помощи клеточной терапии, в частности клеточной терапии, направленной на мышечные, печеночные или гемопоэтические клетки или ткани, предпочтительно клеточной терапии, направленной на печень.
В настоящем изобретении “генная терапия” означает такое лечение индивида, которое включает доставку представляющей интерес нуклеиновой кислоты в клетки индивида с целью лечения заболевания. Доставка нуклеиновой кислоты обычно осуществляется с помощью носителя для доставки, также известного как вектор. Для доставки гена в клетки пациента можно использовать вирусные и невирусные векторы.
В настоящем изобретении “клеточная терапия” означает такой процесс, в котором нуждающемуся в этом индивиду вводятся клетки, модифицированные нуклеиновой кислотой или вектором по изобретению, любым подходящим способом, таким, к примеру, как внутривенная инъекция (вливание) или инъекция в представляющую интерес ткань (имплантация или трансплантация). В конкретных воплощениях клеточная терапия включает сбор клеток у индивида, модификацию клеток индивида нуклеиновой кислотой или вектором по изобретению и введение модифицированных клеток обратно пациенту. В настоящем изобретении “клетки” означают выделенные клетки, природные или искусственные агрегаты клеток, биоискусственные клеточные каркасы и биоискусственные органы или ткани.
“Фармацевтически приемлемый носитель” означает такой носитель, в котором вводится лекарственное средство и которое не вызывает побочных, аллергических или других нежелательных реакций при введении млекопитающим, особенно людям, как надлежит. Фармацевтически приемлемый носитель означает нетоксичный твердый или жидкий заполнитель, разбавитель, адъювант, эксципиент, инкапсулирующий материал или рецептурное вспомогательное вещество любого типа. Фармацевтические композиции, составленные по стандартным процедурам, могут иметь вид растворов, суспензий, эмульсий, таблеток, пилюль, капсул, порошков, препаратов с замедленным высвобождением, имплантатов и др.
Фармацевтическая композиция по изобретению содержит терапевтически эффективное количество нуклеиновой кислоты, вектора или лекарственного средства на основе клеток, предпочтительно в очищенном виде, вместе с подходящим количеством носителя с тем, чтобы обеспечить форму для правильного введения субъектам.
В настоящем изобретении терапевтически эффективное количество означает такую дозу, которая достаточна для регрессии, ослабления или торможения развития того заболевания, к которому относится данный термин, либо для регрессии, ослабления или торможения развития одного или нескольких симптомов того заболевания, к которому относится данный термин.
Эффективная доза определяется и корректируется в зависимости от таких факторов, как используемая композиция, способ введения, физические характеристики данного индивида типа пола, возраста и веса, сопутствующее лечение и другие факторы, которые известны специалистам в области медицины. Эффективную дозу можно определить стандартными клиническими методами. Кроме того, можно необязательно использовать анализы in vivo и/или in vitro для предсказания оптимального диапазона доз. При лечении, включающем введение субъектам вирусного вектора типа вектора AAV, типичные дозы вектора составляют по меньшей мере 1×108 векторных геномов на килограмм массы тела (в.г./кг), как-то по меньшей мере 1×109 в.г./кг или 1×1010 в.г./кг или 1×1011 в.г./кг или 1×1012 в.г./кг или 1×1013 в.г./кг или 1×1014 в.г./кг.
В некоторых воплощениях фармацевтические композиции содержат носители, которые фармацевтически приемлемы для препаратов, которые вводятся, в частности, людям. К ним относятся, в частности, стерильные изотонические водные растворы или суспензии типа солевых растворов (мононатриевого или динатриевого фосфата, хлорида натрия, калия, кальция или магния и т.п. либо смеси таких солей) или же сухие, особенно лиофилизированные композиции, которые при добавлении, в зависимости от случая, стерилизованной воды или физиологического раствора позволяют восстановить растворы для инъекций. Растворы или суспензии могут содержать добавки, которые совместимы с нуклеиновыми кислотами и вирусными векторами и не препятствуют проникновению нуклеиновых кислот или частиц вирусного вектора в целевые клетки. Во всех случаях формы должны быть стерильными и должны быть текучими в такой степени, чтобы они легко вводились через шприц. Они должны быть стабильными в условиях производства и хранения и должны быть защищены от загрязняющего действия таких микроорганизмов, как бактерии и грибки. Примером подходящего раствора является буфер типа фосфатно-солевого буфера (PBS) или раствор Рингера с лактатом.
В некоторых конкретных воплощениях нуклеиновые кислоты, векторы или клетки по изобретению составлены в виде композиции, содержащей фосфатно-солевой буфер с добавлением 0,25% сывороточного альбумина человека. В других конкретных воплощениях нуклеиновые кислоты, векторы или клетки по изобретению составлены в виде композиции, содержащей раствор Рингера с лактатом и неионный детергент типа Pluronic F68 в конечной концентрации 0,01-0,0001%, как-то в концентрации 0,001% от общей массы композиции. Препараты также могут содержать сывороточный альбумин, в частности сывороточный альбумин человека, как-то сывороточный альбумин человека в концентрации 0,25%. В данной области известны и другие подходящие составы для хранения или введения, в частности, из WO 2005/118792 или Allay et al., 2011, Hum. Gene Ther., 2011 May, 22(5):595-604.
В некоторых других воплощениях фармацевтическая композиция составлена в виде имплантата. Имплантаты могут быть из пористого, непористого или гелеобразного материала, включая мембраны типа сиаластиковых мембран или волокна. Имплантаты применимы для местного введения фармацевтических композиций по изобретению в область, нуждающуюся в лечении, то есть в печень.
Еще в других воплощениях фармацевтическая композиция составлена в виде системы с контролируемым высвобождением.
Фармацевтические композиции также могут содержать дополнительные терапевтические средства, в частности средства, применимые для лечения связанных с FGF-23 заболеваний, в частности, связанных с FGF-23 гипофосфатемических заболеваний, такие, в частности, как кальцитриол.
В другом аспекте изобретения предусмотрены нуклеиновые кислоты, векторы, клетки, фармацевтические композиции по изобретению для применения в качестве лекарственных средств.
В следующем аспекте изобретения предусмотрены нуклеиновые кислоты, векторы, клетки, фармацевтические композиции по изобретению для применения при лечении связанных с FGF-23 заболеваний, в частности, связанных с FGF-23 гипофосфатемических заболеваний, при помощи генной терапии или клеточной терапии, как описано выше, предпочтительно генной терапии или клеточной терапии, направленной на печень.
В следующем аспекте изобретения предусмотрено применение нуклеиновой кислоты, вектора, клетки, фармацевтической композиции по изобретению при изготовлении лекарственного средства для лечения связанных с FGF-23 заболеваний, в частности, связанных с FGF-23 гипофосфатемических заболеваний, при помощи генной терапии или клеточной терапии, как описано выше, предпочтительно генной терапии или клеточной терапии, направленной на печень.
Конструкция нуклеиновой кислоты, вектор, композицию по изобретению применяются для лечения заболеваний, которые можно лечить путем ингибирования образования рецепторного комплекса FGF23-FGF (FGFR)-Klotho, типа заболеваний, вызванных чрезмерным действием FGF23, в частности, опосредованных посредством взаимодействия FGF23 с комплексом FGFR/klotho.
Заболевания, которые можно лечить, представляют собой, в частности, связанные с FGF-23 гипофосфатемические заболевания. Эти заболевания можно диагностировать по высоким уровням FGF23 на фоне гипофосфатемии, так как у пациентов с гипофосфатемией уровни FGF23 низкие по другим причинам.
Примеры связанных с FGF-23 гипофосфатемических заболеваний, которые можно лечить, включают генетические заболевания, такие как Х-сцепленная гипофосфатемия (XLH), вызванная мутациями гена PHEX; аутосомно-доминантный гипофосфатемический рахит (ADHR), вызванный мутациями гена FGF23; аутосомно-рецессивный гипофосфатемический рахит 1 (ADHR1), вызванный мутациями гена DMP1; аутосомно-рецессивный гипофосфатемический рахит 2 (ADHR2), вызванный мутациями гена ENPP1; остеоглофоническая дисплазия, вызванная мутациями гена FRFR1; метафизарная хондродисплазия янсеновского типа, вызванная мутациями гена PTH1R; гипофосфатемия с зубными аномалиями и эктопической кальцификацией, вызванная мутациями гена FAM20C; синдром МакКьюна-Олбрайта/фиброзная дисплазия, вызванная мутациями гена GNAS1; гипофосфатемия с поражением кожи и костей, вызванная мутациями гена HRAS или NRAS.
Другие примеры связанных с FGF-23 гипофосфатемических заболеваний, которые можно лечить, - приобретенные заболевания, такие как онкогенная остеомаляция (TIO), гипофосфатемическая остеомаляция от сахаратного оксида железа или полимальтозного железа, осложнения при трансплантации почек или парентеральной терапии железом, хроническая болезнь почек и её осложнения типа гиперпаратиреоза.
В некоторых предпочтительных воплощениях связанное с FGF-23 гипофосфатемическое заболевание является генетическим заболеванием, предпочтительно XLH. Такие заболевания предпочтительно лечат при помощи адресной генной терапии, в частности, направленной на мышечные, печеночные или гемопоэтические клетки или ткани, более конкретно на клетки или ткани печени, или же при помощи клеточной терапии, более предпочтительно с помощью AAV- или лентивирусного вектора, в частности, вектора AAV8.
В другом аспекте изобретения предусмотрен способ лечения связанных с FGF-23 гипофосфатемических заболеваний, как описано выше, включающий введение пациентам терапевтически эффективного количества нуклеиновой кислоты, вектора, клеток или фармацевтической композиции, как описано выше.
В настоящем изобретении термин “лечить” или “лечение” означает регрессию, ослабление или торможение развития того заболевания, к которому относится данный термин, либо регрессию, ослабление или торможение развития одного или нескольких симптомов того заболевания, к которому относится данный термин.
Нуклеиновую кислоту, вектор, клетку или фармацевтическую композицию по изобретению обычно вводят в соответствии с известными процедурами, в таких дозах и в течение таких периодов времени, которые эффективно вызывают терапевтические эффекты у пациентов. Нуклеиновые кислоты по изобретению, в составе вектора или без него, можно вводить любым удобным способом, как-то, без ограничения, посредством вливания или болюсной инъекции, всасывания через эпителиальные или кожно-слизистые оболочки (например, слизистую оболочку полости рта, прямой кишки или кишечника). Введение может быть системным или местным; системное включает парентеральное и пероральное, а местное - местное и местно-регионарное. Парентеральное введение преимущественно осуществляется путем инъекции или перфузии типа подкожного (SC), внутримышечного (IM), интраваскулярного типа внутривенного (IV), интраартериального, внутрибрюшинного (IP), интрадермального (ID), интраназального, эпидурального или другого. Кроме того, может потребоваться введение фармацевтических композиций по изобретению в печень субъектов любым подходящим способом.
Нуклеиновая кислота, вектор, клетка или фармацевтическая композиция по изобретению может применяться в комбинации с другими биологически активными средствами, при этом комбинированное применение осуществляется путем одновременного, раздельного или последовательного введения.
В практике настоящего изобретения будут применяться, если не указано иначе, стандартные методы, которые находятся в компетенции специалистов в данной области. Такие методы полностью изложены в литературе.
Далее изобретение будет раскрыто на следующих примерах, которые не являются ограничительными, со ссылкой на прилагаемые чертежи.
Краткое описание фигур
Фиг. 1. Схематическое представление конструкции из нуклеиновой кислоты и экспрессирующего вектора AAV для усеченного слитого белка FGF23 по изобретению.
A. Конструкция нуклеиновой кислоты для усеченного слитого белка FGF23 по изобретению. C-концевые аминокислоты белка FGF человека (а.к. 175-251, cFGF) сливали с сывороточным альбумином человека без сигнального пептида (а.к. 25-609) через расщепляемый линкер, полученный из аминокислот 182-200 фактора свертывания крови IX человека (hFIX). Наконец, с N-концом конструкции сливали сигнальный пептид, полученный из первых 18 аминокислот химотрипсиногена B2 (CTRB2).
B. Экспрессирующий вектор AAV для усеченного слитого белка FGF23 по изобретению. Конструкцию нуклеиновой кислоты вставляли в экспрессирующий вектор AAV, содержащий специфичный для печени промотор (энхансер аполипротеина E (ApoE) - промотор α1-антитрипсина человека (hAAT)), модифицированный интрон HBB2 и сигнал полиаденилирования бычьего гормона роста (bGH).
Фиг. 2. Стабилизация C-концевой части FGF23 при слиянии с альбумином in vitro. Клетки гепатомы печени человека (Huh-7) трансфецировали плазмидами, экспрессирующими различные варианты FGF23 человека, как описано в таблице 1. Через 48 часов после трансфекции собирали среду и анализировали методом вестерн-блоттинга с помощью анантитела против FGF23. (A) Сравнение всех конструкций, показывающее относительную экспрессию различных конструкций и более высокие уровни экспрессии у слитых белков FGF23-C с молекулой альбумина. Слева указаны положения FGF23, FGF23-C или слитых белков FGF23-C с альбумином человека (FGF23-C-альбумин). (B) Сравнение уровней экспрессии, отмеченных после трансфекции в трех экземплярах плазмид, экспрессирующих слитые белки FGF23-C с альбумином в клетках Huh-7.
Фиг. 3. Стабилизация C-концевой части FGF23 при слиянии с альбумином in vivo. Мышам C57BL6/J дикого типа вводили по 1×1012 в.г. на мышь векторов AAV8, экспрессирующих оптимизированный по кодонам вариант C-концевой части FGF23 (coFGF23-C, аминокислоты 175-251), содержащий мотив RRHTR (только конструкции 7, 9 и 12) и сигнальный пептид sp7, слитый (конструкции 9, 11 и 12) или нет (конструкция 7) с молекулой альбумина. Конструкция 12 включала линкер, состоящий из аминокислот 182-200 фактора свертывания крови IX человека, тогда как конструкции 9 и 11 не содержали линкера. Через 1 месяц после инъекции векторов мышей забивали и отбирали плазму и печень для измерения фосфатемии и уровней экспрессии у различных векторов AAV. (A) Фосфатемия при измерении в крови. (B) Число копий векторного генома (VGCN) на 1 диплоидный геном при измерении в печени. (C) Вестерн-блоттинг плазмы с помощью антитела против FGF23. (D) Гистограмма, показывающая относительную интенсивность полос, представленных на панели C. Статистический анализ проводили с помощью t-критерия (*p <0,05 в сравнении с мышами, получавшими ФСБ, n = 4-5 на группу).
Фиг. 4. Опосредованное AAV предотвращение задержки роста у мышей Hypduk. Одномесячным мышам Hypduk вводили либо ФСБ (KO, ФСБ), либо 1×1012 в.г. на мышь вектора AAV, экспрессирующего FGF23-C, слитый с sp7 и альбумином через линкер hFIX и содержащий мотив RRHTR (KO, AAV). В качестве контроля использовали получавших ФСБ мышей дикого типа (WT, ФСБ). A - C. Проводили измерения у мышей через 1, 2 и 3 месяца после введения вектора. Представлен прирост веса (A), длины хвоста (B) и размера тела (C) за три месяца. Статистический анализ проводили методом ANOVA (*p <0,05 в сравнении с KO, ФСБ; # p <0,05 в сравнении с WT, ФСБ; n = 9-11 на группу).
Фиг. 5. Опосредованное AAV предотвращение гипофосфатемии у мышей Hypduk. Одномесячным мышам Hypduk вводили либо ФСБ (KO, ФСБ), либо 1×1012 в.г. на мышь вектора AAV, экспрессирующего FGF23-C, слитый с sp7 и альбумином через линкер hFIX и содержащий мотив RRHTR (KO, AAV). В качестве контроля использовали получавших ФСБ мышей дикого типа (WT, ФСБ). Измеряли уровни фосфата в крови через три месяца после введения вектора. Статистический анализ проводили методом ANOVA (*p <0,05, как указано, NS - не значимо).
Фиг. 6. Опосредованное AAV восстановление функции мышц у мышей Hypduk. Одномесячным мышам Hypduk вводили либо ФСБ (KO, ФСБ), либо 1×1012 в.г. на мышь вектора AAV, экспрессирующего FGF23-C, слитый с sp7 и альбумином через линкер hFIX и содержащий мотив RRHTR (KO, AAV). В качестве контроля использовали получавших ФСБ мышей дикого типа (WT, ФСБ). Измеряли показатели на перевернутой решетке через три месяца после введения вектора. Статистический анализ проводили методом ANOVA (*p <0,05, как указано, NS - не значимо).
Примеры
Материалы и методы
Конструирование нуклеиновых кислот, кодирующих усеченный FGF23
Конструкции нуклеиновой кислоты, кодирующие усеченный FGF23, получали путем слияния аминокислот 175-251 белка FGF23 человека (а.к. 175-251 по NP_065689; SEQ ID NO: 2), содержащих мотив RXXR (аминокислоты 175-179 белка FGF23 человека) с предшествующим R175 (мотив RRHTR; аминокислоты 175-179 белка FGF23 человека), либо аминокислот 180-251 белка FGF человека по SEQ ID NO: 1 (не содержащих данного мотива RXXR), с сывороточным альбумином человека без сигнального пептида (а.к. 25-609 по NP_000468 или SEQ ID NO: 9) через расщепляемый линкер, полученный из фактора свертывания крови IX человека (cFIX; а.к. 182-200 по NP_000124; SEQ ID NO: 10). Наконец, на N-конце конструкций вставляли сигнальный пептид, полученный из первых 18 аминокислот химотрипсиногена B2 (sp7; а.к. 1-18 по NP_001020371 или SEQ ID NO: 7), для обеспечения эффективной секреции (SEQ ID NO: 12 (фиг. 1A); SEQ ID NO: 52). Эти последовательности оптимизировали по кодонам при помощи коммерческого алгоритма. Полученные последовательности представляют собой SEQ ID NO: 57, которая включает CDS по SEQ ID NO: 13, кодирующую слитый белок по SEQ ID NO: 12, фланкированный с 5'-стороны сайтом MluI/Kozak (конструкция № 12 в таблице 1); и SEQ ID NO: 51, которая содержит CDS, кодирующую слитый белок по SEQ ID NO: 52, фланкированный с 5'-стороны сайтом MluI/Kozak (конструкция № 10 в таблице 1). Оптимизированные по кодонам последовательности клонировали в экспрессионную кассету для трансгенов, оптимизированную для экспрессии в печени (фиг. 1B).
Для сравнения получали и другие конструкции (таблица 1). Конструкции нуклеиновой кислоты, кодирующие природный FGF23 человека (SEQ ID NO: 1), получали путем клонирования последовательности FGF23 дикого типа человека или оптимизированной по кодонам последовательности FGF23 в трансгенную экспрессионную кассету, оптимизированную для экспрессии в печени (фиг. 1B).
Конструкции для усеченного C-концевого FGF23 (FGF23-C) получали путем слияния FGF23-C, содержащего или нет мотив RXXR (аминокислоты 175-251 или 180-251 белка FGF23 человека), с сигнальными пептидами нативного FGF23 или sp7. Эти последовательности оптимизировали по кодонам при помощи коммерческого алгоритма и клонировали в экспрессионную кассету для трансгенов, оптимизированную для экспрессии в печени.
Химерные белки получали либо путем слияния FGF23-C, не содержащего мотива RXXR, с сывороточным альбумином человека (без сигнального пептида), без расщепляемого линкера, получаемого из фактора свертывания крови IX человека, либо путем слияния FGF23-C, содержащего мотив RXXR, с сывороточным альбумином человека (без сигнального пептида), без расщепляемого линкера, получаемого из фактора свертывания крови IX человека. На N-конце конструкции вставляли сигнальный пептид, полученный из первых 18 аминокислот химотрипсиногена B2 (sp7), для обеспечения эффективной секреции. Эти последовательности оптимизировали по кодонам при помощи коммерческого алгоритма и клонировали в трансгенную экспрессионную кассету, оптимизированную для экспрессии в печени (фиг. 1B).
Получение векторов AAV
Векторы AAV получали методом кратковременной трансфекции без аденовируса (Matsushita et al., Gene Therapy, 1998, 5, 938-945) и очищали, как описано ранее (Ayuso et al., Gene Therapy, 2010, 17, 503-510). Титры исходных векторов AAV определяли количественным методом ПЦР в реальном времени (кПЦР) и проверяли по ДНС-ПААГ с последующим окрашиванием гелей на белок SYPRO® Ruby и денситометрией полос.
Исследования in vitro
Клетки Huh-7 при конфлюэнтности 70-80% трансфецировали конструкциями, указанными в таблице 1, с помощью липофектамина. Через два дня после трансфекции собирали среду и определяли уровни FGF23 методом вестерн-блоттинга.
Вестерн-блоттинг
На гель с градиентом полиакриламида 4-15% наносили среду Huh-7 после трансфекции и сыворотку мышей, которым вводились различные векторы AAV, и проводили ДНС-ПААГ. После переноса на нитроцеллюлозу мембрану блокировали и инкубировали с антителом против FGF23. Мембрану инкубировали с соответствующим вторичным антителом и визуализировали на установке для визуализации Odyssey.
Исследования на мышах
Шестинедельным мышам C57BL6/J вводили векторы AAV внутривенно через хвостовую вену. Через 1 месяц после введения вектора определяли уровни FGF23-C в кровотоке методом вестерн-блоттинга. Извлекали печень и определяли число копий векторного генома методом кПЦР.
Одномесячным самцам мышей Hypduk вводили векторы AAV внутривенно через хвостовую вену. В качестве контроля использовали получавших ФСБ однопометных мышей дикого типа и Hypduk. Каждый месяц в течение 3 месяцев после введения вектора мышей взвешивали и измеряли для оценки коррекции заболевания на макроскопическом уровне. Через 3 месяца после введения вектора проводили измерение фосфатемии в крови и функциональную оценку мышечной силы.
Фосфатемия
Для измерения уровня фосфата в кровотоке у мышей брали кровь через три месяца после введения вектора. После центрифугирования при 10000×g в течение 10 мин измеряли содержание неорганического фосфата в сыворотке с помощью стандартного коммерческого набора.
Испытание на перевернутой решетке
Мышей помещали на решетку и давали приспособиться к ней в течение 3-5 секунд, а затем решетку переворачивали и удерживали на расстоянии по меньшей мере 35 см над клеткой для мышей, содержащей 5-7 см мягкой подстилки. Измеряли количество падений на протяжении 3 минут и регистрировали в виде числа падений за 1 минуту.
Результаты
C-концевая часть FGF23 (FGF23-C) человека конкурирует с природным FGF23 и снижает внутриклеточную передачу сигнала после связывания с рецептором.
Одним из важных ограничений применения FGF23-C в качестве средства от XLH является то, что этот пептид очень нестабилен в кровотоке. Для секреции этого пептида в печени и повышения его стабильности был разработан метод генной терапии. Аминокислоты 175-251 белка FGF23 человека (SEQ ID NO: 2), содержащие мотив RXXR с предшествующим R175 (мотив RRHTR; аминокислоты 175-179 белка FGF23 человека), либо аминокислоты 180-251 белка FGF человека по SEQ ID NO: 1 (не содержащие такого мотива RXXR), сливали с сывороточным альбумином человека без сигнального пептида (SEQ ID NO: 9) через расщепляемый линкер, полученный из фактора свертывания крови IX человека (SEQ ID NO: 10). Наконец, на N-конце конструкций вставляли сигнальный пептид, полученный из первых 18 аминокислот химотрипсиногена B2 (SEQ ID NO: 7), для обеспечения эффективной секреции (SEQ ID NO: 12 (фиг. 1A); SEQ ID NO: 52). Эти последовательности оптимизировали по кодонам при помощи коммерческого алгоритма. Полученные последовательности представляют собой SEQ ID NO: 57, которая включает CDS по SEQ ID NO: 13, кодирующую слитый белок по SEQ ID NO: 12, фланкированный с 5'-стороны сайтом MluI/Kozak (конструкция № 12 в таблице 1); и SEQ ID NO: 51, которая содержит CDS, кодирующую слитый белок по SEQ ID NO: 52, фланкированный с 5'-стороны сайтом MluI/Kozak (конструкция № 10 в таблице 1). Оптимизированные по кодонам последовательности клонировали в экспрессионную кассету для трансгенов, оптимизированную для экспрессии в печени (фиг. 1B). Кассета для экспрессии трансгенов, содержащая SEQ ID NO: 13, входящую в SEQ ID NO: 57, имела последовательность SEQ ID NO: 23. А переходный вектор AAV, содержащий экспрессионную кассету по SEQ ID NO: 23, фланкированную ITRs AAV, имел последовательность SEQ ID NO: 24.
Для подтверждения низкой стабильности FGF23-C и проверки подходов к улучшению его секреции и стабильности авторы изобретения трансфецировали клетки гепатомы печени человека различными конструкциями, экспрессирующими как нативный FGF23, так и FGF23-C, под транскрипционным контролем промотора hAAT (таблица 1 и фиг. 1).
Таблица 1. Описание исследуемых конструкций
FGF23, фактор роста фибробластов 23; FGF23-C, C-концевая часть FGF23 (аминокислоты 175-251); spFGF23, нативный сигнальный пептид FGF23; sp7, сигнальный пептид химотрипсиногена B2; RXXRR, аминокислоты 176-179 у FGF23-C; wt, нативная последовательность FGF23; co, оптимизированная по кодонам; cFIX, аминокислоты 182-200 фактора свертывания крови IX человека; альбумин, аминокислоты 25-609 в последовательности сывороточного альбумина человека.
- Конструкция № 1 (нуклеотидная (нт) последовательность SEQ ID NO: 25; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 26, 1 или 27);
- конструкция № 2 (нуклеотидная (нт) последовательность SEQ ID NO: 28; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 29, 1, 26 или 27);
- конструкция № 3 (нуклеотидная (нт) последовательность SEQ ID NO: 30; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 31 или 32);
- конструкция № 4 (нуклеотидная (нт) последовательность SEQ ID NO: 33; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 34 или 35);
- конструкция № 5 (нуклеотидная (нт) последовательность SEQ ID NO: 36; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 37 или 38);
- конструкция № 6 (нуклеотидная (нт) последовательность SEQ ID NO: 39; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 40 или 41);
- конструкция № 7 (нуклеотидная (нт) последовательность SEQ ID NO: 42; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 43 или 44);
- конструкция № 8 (нуклеотидная (нт) последовательность SEQ ID NO: 45; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 46 или 47);
- конструкция № 9 (нуклеотидная (нт) последовательность SEQ ID NO: 48; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 49 или 50);
- конструкция № 10 (нуклеотидная (нт) последовательность SEQ ID NO: 51; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 52, которая идентична SEQ ID NO: 53);
- конструкция № 11 (нуклеотидная (нт) последовательность SEQ ID NO: 54; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 55 или 56);
- конструкция № 12 (нуклеотидная (нт) последовательность SEQ ID NO: 57; соответствующая аминокислотная (а.к.) последовательность SEQ ID NO: 58, которая идентична SEQ ID NO: 12).
Варианты генов FGF-23 и FGF23-C дикого типа и оптимизированные по кодонам тестировали вместе с вариантами, слитыми с нативным сигнальным пептидом FGF23 и гетерологичным сигнальным пептидом химотрипсиногена B2 (SEQ ID NO: 7, sp7). В различных исследуемых последовательностях мотив RRHTR, присутствующий в FGF23 человека и ответственный за его расщепление на N-концевой и C-концевой пептид FGF23, был либо включен, либо нет.
Нативный FGF23 эффективно вырабатывался и секретировался в среде трансфецированных клеток Huh-7, также как и его оптимизированный по кодонам вариант (конструкции № 1-2, фиг. 2A). Ни один из вариантов FGF23-C не вырабатывался в этой среде, за исключением слитого с sp7 FGF23-C без мотива RHTR (конструкция № 8, фиг. 2A). Примечательно, что слияние с молекулой альбумина (SEQ ID NO: 9) усиливало секрецию FGF23-C независимо от состава химерного белка (конструкции № 9-12, фиг. 2A). Слияние FGF23-C с альбумином в отсутствие мотива RHTR и расщепляемого линкера, полученного из фактора свертывания крови IX человека (конструкция № 11), вело к повышению уровня секреции in vitro по сравнению с другими химерными белками, содержащими молекулу альбумина (конструкции № 9, № 10 и № 12, фиг. 2B).
Получали векторы AAV8, экспрессирующие конструкции № 7, № 9, № 11 и № 12: i) FGF23-C, слитый с sp7 и содержащий мотив RRHTR (конструкция № 7), ii) FGF23-C, слитый с sp7 и альбумином и содержащий мотив RRHTR (конструкция № 9), iii) FGF23-C, слитый с sp7 и альбумином без мотива RRHTR (конструкция № 11), и iv) FGF23-C, слитый с sp7 и альбумином через линкер hFIX и содержащий мотив RRHTR (конструкция № 12, таблица 1 и фиг. 1A). Векторы AAV8 получали и вводили 6-недельным мышам C57BL6/J в дозе 1×1012 в.г. на мышь. Через 1 месяц после введения вектора у мышей брали кровь для измерения фосфатемии и уровня FGF23-C методом вестерн-блоттинга. Важно отметить, что повышение уровня фосфора в крови наблюдались только у мышей, получавших вектор AAV8, экспрессирующий конструкцию № 12 (фиг. 3A), несмотря на близкие уровни трансдукции в печени у других конструкций (фиг. 3B) и более высокие уровни экспрессии in vitro у конструкции № 11.
Полосы, соответствующие молекулярной массе химерных белков FGF23-C-альбумин, отмечены только у мышей, получавших векторы AAV8, специфически экспрессирующие конструкции № 9 и № 12 в печени. Напротив, полосы с размером, совместимым с FGF23-C, не отмечались у мышей, получавших векторы AAV8, экспрессирующие конструкцию № 7, то есть FGF23-C, слитый с sp7, что подтверждает нестабильность FGF23-C in vivo (фиг. 3C). Что важно, у мышей, получавших конструкцию № 12, содержащую расщепляемый линкер (hFIX; SEQ ID NO: 10), отмечались значительно более высокие уровни циркулирующего слитого белка FGF-23-C по сравнению с конструкцией № 9 (без линкера; фиг. 3D), несмотря на близкие уровни трансдукции в печени, судя по числу копий векторного генома на клетку (фиг. 3В), и более высокие уровни экспрессии in vitro у конструкции № 11. Эти данные означают, что C-концевая часть FGF23 (FGF23-C) может экспрессироваться в печени только в слитом виде с молекулой альбумина. Они также свидетельствуют, что состав слитого белка влияет на его стабильность и функцию в кровотоке, которые усиливаются при слиянии FGF23-C с молекулой альбумина через расщепляемый линкер, в частности, полученный из фактора свертывания крови IX человека (SEQ ID NO: 10).
Для проверки того, что химерный белок, состоящий из FGF23-C и слитый с sp7 и альбумином через полученный из hFIX линкер по изобретению (конструкция № 12, таблица 1 и фиг. 1A), активен in vivo, одномесячным мышам Hypduk вводили вектор AAV, экспрессирующий эту конструкцию, в дозе 1×1012 в.г. на мышь (KO, AAV). В качестве контроля использовали получавших ФСБ однопометных мышей дикого типа и Hypduk (WT, ФСБ). Оценивали коррекцию фенотипа по ежемесячным измерениям массы тела и длины тела и хвоста, начиная с одного месяца после введения вектора (фиг. 4). В целом мыши Hypduk, получавшие ФСБ, имели меньший вес и длину тела и хвоста (фиг. 4A-C) по сравнению с животными дикого типа. У мышей Hypduk, получавших векторы AAV8, экспрессирующие химерный белок FGF23-C по изобретению, отмечался прирост веса и длины тела и хвоста за 3-месячный период, который был выше, чем у мышей Hypduk, получавших ФСБ, и близкий тому, что наблюдался у животных дикого типа (фиг. 4A-C). Хотя у обработанных AAV животных наблюдалось полное восстановление прироста длины тела, однако прирост веса и длины хвоста восстанавливался лишь частично (фиг. 4A, C). Через три месяца после обработки измеряли уровень фосфата в сыворотке. У получавших AAV мышей Hypduk уровни фосфата значительно отличались от уровней, отмеченных у мышей Hypduk, получавших ФСБ, но не отличались от уровней фосфата у животных дикого типа (фиг. 5). Нарушение роста костей и кальцификация сухожилий может приводить к снижению двигательной функции и мышечной силы. Проводили функциональную оценку фенотипа у мышей Hypduk путем испытания на перевернутой решетке. Наблюдалось повышение числа падений за 1 минуту у мышей Hypduk по сравнению с мышами дикого типа, которое полностью предотвращалось при генной терапии AAV (фиг. 6).
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> GENETHON
INSTITUT NATIONAL DE LA SANTE ET DE LA RECHERCHE MEDICALE
(INSERM)
UNIVERSITE D'EVRY VAL D'ESSONNE
<120> GENE THERAPY OF FIBROBLAST GROWTH FACTOR 23 RELATED
HYPOPHOSPHATEMIC DISEASES
<130> BCT200107 QT
<160> 58
<170> PatentIn version 3.5
<210> 1
<211> 251
<212> PRT
<213> Homo sapiens
<400> 1
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Tyr Pro Asn Ala Ser Pro Leu Leu
20 25 30
Gly Ser Ser Trp Gly Gly Leu Ile His Leu Tyr Thr Ala Thr Ala Arg
35 40 45
Asn Ser Tyr His Leu Gln Ile His Lys Asn Gly His Val Asp Gly Ala
50 55 60
Pro His Gln Thr Ile Tyr Ser Ala Leu Met Ile Arg Ser Glu Asp Ala
65 70 75 80
Gly Phe Val Val Ile Thr Gly Val Met Ser Arg Arg Tyr Leu Cys Met
85 90 95
Asp Phe Arg Gly Asn Ile Phe Gly Ser His Tyr Phe Asp Pro Glu Asn
100 105 110
Cys Arg Phe Gln His Gln Thr Leu Glu Asn Gly Tyr Asp Val Tyr His
115 120 125
Ser Pro Gln Tyr His Phe Leu Val Ser Leu Gly Arg Ala Lys Arg Ala
130 135 140
Phe Leu Pro Gly Met Asn Pro Pro Pro Tyr Ser Gln Phe Leu Ser Arg
145 150 155 160
Arg Asn Glu Ile Pro Leu Ile His Phe Asn Thr Pro Ile Pro Arg Arg
165 170 175
His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val
180 185 190
Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln
195 200 205
Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu
210 215 220
Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly
225 230 235 240
Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
245 250
<210> 2
<211> 77
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide (human FGF23 C-term peptide 175-251)
<400> 2
Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu
1 5 10 15
Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys
20 25 30
Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp
35 40 45
Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly
50 55 60
Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
65 70 75
<210> 3
<211> 24
<212> PRT
<213> artificial sequence
<220>
<223> synthetic peptide (human FGF23 signal peptide)
<400> 3
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala
20
<210> 4
<211> 26
<212> PRT
<213> artificial sequence
<220>
<223> synthetic peptide (alpha-1 anti-trypsin signal peptide)
<400> 4
Met Pro Ser Ser Val Ser Trp Gly Ile Leu Leu Leu Ala Gly Leu Cys
1 5 10 15
Cys Leu Val Pro Val Ser Leu Ala Lys Leu
20 25
<210> 5
<211> 16
<212> PRT
<213> artificial sequence
<220>
<223> synthetic peptide (synthetic mut1 signal peptide)
<400> 5
Met Leu Leu Leu Ser Ala Leu Leu Leu Gly Leu Ala Phe Gly Tyr Ser
1 5 10 15
<210> 6
<211> 16
<212> PRT
<213> artificial sequence
<220>
<223> synthetic peptide (synthetic mut3 signal peptide)
<400> 6
Met Leu Leu Ser Phe Ala Leu Leu Leu Gly Leu Ala Leu Gly Tyr Ser
1 5 10 15
<210> 7
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> synthetic peptide (chymotrypsin B2 (CTRB2) signal peptide)
<400> 7
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly
<210> 8
<211> 22
<212> PRT
<213> artificial sequence
<220>
<223> synthetic peptide (plasma protease C1 signal peptide)
<400> 8
Met Ala Ser Arg Leu Thr Leu Leu Thr Leu Leu Leu Leu Leu Leu Ala
1 5 10 15
Gly Asp Arg Ala Ser Ser
20
<210> 9
<211> 585
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide (human serum albumin 25-609)
<400> 9
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
1 5 10 15
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
20 25 30
Gln Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
35 40 45
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
50 55 60
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
65 70 75 80
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
85 90 95
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
100 105 110
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
115 120 125
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
130 135 140
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
145 150 155 160
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
165 170 175
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
180 185 190
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
195 200 205
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
210 215 220
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
225 230 235 240
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
245 250 255
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
260 265 270
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
275 280 285
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
290 295 300
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
305 310 315 320
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
325 330 335
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
340 345 350
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
355 360 365
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
370 375 380
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
385 390 395 400
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
405 410 415
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
420 425 430
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
435 440 445
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
450 455 460
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
465 470 475 480
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
485 490 495
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
500 505 510
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
515 520 525
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
530 535 540
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
545 550 555 560
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
565 570 575
Ala Ala Ser Gln Ala Ala Leu Gly Leu
580 585
<210> 10
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> synthetic peptide (FIX activation sequence (1))
<400> 10
Ser Val Ser Gln Thr Ser Lys Leu Thr Arg Ala Glu Thr Val Phe Pro
1 5 10 15
Asp Val Asp Gly Ser
20
<210> 11
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> synthetic peptide (FIX activation sequence (2))
<400> 11
Pro Val Ser Gln Thr Ser Lys Leu Thr Arg Ala Glu Thr Val Phe Pro
1 5 10 15
Asp Val
<210> 12
<211> 701
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 12
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp
20 25 30
Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala
35 40 45
Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala
50 55 60
Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala
65 70 75 80
Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Ser
85 90 95
Val Ser Gln Thr Ser Lys Leu Thr Arg Ala Glu Thr Val Phe Pro Asp
100 105 110
Val Asp Gly Ser Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys
115 120 125
Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala
130 135 140
Gln Tyr Leu Gln Gln Cys Pro Phe Glu Asp His Val Lys Leu Val Asn
145 150 155 160
Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu
165 170 175
Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr
180 185 190
Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala
195 200 205
Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp
210 215 220
Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys
225 230 235 240
Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr
245 250 255
Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe
260 265 270
Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala
275 280 285
Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu
290 295 300
Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln
305 310 315 320
Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser
325 330 335
Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr
340 345 350
Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu
355 360 365
Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln
370 375 380
Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu
385 390 395 400
Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala
405 410 415
Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys
420 425 430
Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr
435 440 445
Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg
450 455 460
Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala
465 470 475 480
Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu
485 490 495
Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu
500 505 510
Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr
515 520 525
Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg
530 535 540
Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys
545 550 555 560
Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu
565 570 575
Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys
580 585 590
Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu
595 600 605
Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr
610 615 620
Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys
625 630 635 640
Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr
645 650 655
Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu
660 665 670
Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly
675 680 685
Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<210> 13
<211> 2106
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide codon optimized
<400> 13
atggctttct tgtggcttct cagttgttgg gccttgttgg ggaccacttt cggacgaaga 60
cacacacgct ctgcagagga cgattctgag agggatcctt tgaatgtcct taagccgagg 120
gcccgcatga cgcctgcacc agcatcctgc agtcaagagc tcccttcagc agaagataat 180
agtcctatgg cgtccgaccc cctgggggta gtacgcgggg ggcgagttaa tacgcacgcc 240
gggggaactg gtcctgaagg gtgcaggccc tttgccaagt ttatttcagt gtcacaaacc 300
tctaaactga cacgcgccga gacggtcttt ccggatgttg atggatcaga tgctcataaa 360
tcagaggtag ctcaccgctt caaagacctc ggagaggaga actttaaggc cctcgtcctc 420
atcgcatttg cacagtattt gcaacaatgt ccctttgagg atcatgtcaa attggtaaat 480
gaagttactg agttcgcgaa gacatgcgtg gctgacgagt ccgcagaaaa ctgcgacaag 540
tcccttcata cactgtttgg tgataaactg tgcactgtcg ctactcttag agaaacatat 600
ggagagatgg ctgactgttg tgcgaagcaa gagccagaac ggaatgaatg tttccttcaa 660
cataaagatg ataatccgaa tttgccacgc cttgtacgac cagaggtgga tgttatgtgc 720
accgcgtttc atgacaatga ggaaacgttc ctgaaaaaat atttgtatga gattgctcgg 780
agacatcctt acttttacgc acccgagctc ctgttcttcg ctaagagata caaggcagcc 840
tttacggaat gctgtcaggc cgcggataag gccgcctgct tgctccccaa gctggacgag 900
ctgcgagatg agggcaaggc gtcctctgct aagcaacgac tcaaatgcgc cagcttgcaa 960
aagtttggcg aaagagcgtt taaggcctgg gcagtggcgc gcctgagtca gcggttccct 1020
aaggcggagt ttgccgaggt ctcaaaactg gtgacagact tgacgaaggt acataccgag 1080
tgctgccacg gagatctcct ggagtgcgcg gatgatcgcg ctgacctcgc caaatatata 1140
tgtgagaacc aggattccat ttcctcaaaa cttaaagagt gctgcgaaaa accccttctt 1200
gagaagtcac actgcattgc agaggtcgaa aacgacgaaa tgcctgccga tctgccttcc 1260
cttgcggctg actttgtgga gtctaaggac gtttgcaaaa actatgcaga ggctaaagac 1320
gtattcctgg gcatgtttct ttatgaatac gcgcggcgac atcccgacta ttcagtcgta 1380
ctcctgcttc gactggcgaa aacttacgaa actactctcg agaaatgctg tgctgccgcc 1440
gaccctcatg agtgctatgc taaggtattt gatgaattca aaccgttggt tgaggaaccg 1500
caaaacttga tcaagcagaa ctgtgaactt tttgagcagc tgggggaata taaattccag 1560
aatgcattgc ttgtacgata cacaaagaaa gttccacagg tctccacccc gacacttgtg 1620
gaagttagca ggaacttggg aaaggtcggc agcaagtgtt gtaagcaccc tgaagctaag 1680
cgaatgccat gcgccgaaga ttacctttct gtggtgttga accaactgtg cgttcttcac 1740
gaaaagacgc ccgtatctga tcgggtgacg aaatgctgca cagagtcact tgtgaacaga 1800
cgcccctgct ttagtgcgct cgaggtggac gaaacgtatg tcccgaaaga gtttaacgca 1860
gagacattca cgtttcatgc ggacatctgt accctctctg aaaaggagag acagatcaaa 1920
aagcagacag cgctggttga gcttgttaag cataagccga aggcgacaaa ggagcaactg 1980
aaagcggtga tggatgactt cgccgcgttc gttgaaaaat gctgcaaagc cgacgataag 2040
gaaacctgtt ttgctgaaga aggtaaaaaa ctcgttgctg cgagtcaggc cgccctgggg 2100
ctgtga 2106
<210> 14
<211> 397
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide (hAAT promoter)
<400> 14
gatcttgcta ccagtggaac agccactaag gattctgcag tgagagcaga gggccagcta 60
agtggtactc tcccagagac tgtctgactc acgccacccc ctccaccttg gacacaggac 120
gctgtggttt ctgagccagg tacaatgact cctttcggta agtgcagtgg aagctgtaca 180
ctgcccaggc aaagcgtccg ggcagcgtag gcgggcgact cagatcccag ccagtggact 240
tagcccctgt ttgctcctcc gataactggg gtgaccttgg ttaatattca ccagcagcct 300
cccccgttgc ccctctggat ccactgctta aatacggacg aggacagggc cctgtctcct 360
cagcttcagg caccaccact gacctgggac agtgaat 397
<210> 15
<211> 321
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide (ApoE enhancer)
<400> 15
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg g 321
<210> 16
<211> 441
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide (HBB2 intron)
<400> 16
gtacacatat tgaccaaatc agggtaattt tgcatttgta attttaaaaa atgctttctt 60
cttttaatat acttttttgt ttatcttatt tctaatactt tccctaatct ctttctttca 120
gggcaataat gatacaatgt atcatgcctc tttgcaccat tctaaagaat aacagtgata 180
atttctgggt taaggcaata gcaatatttc tgcatataaa tatttctgca tataaattgt 240
aactgatgta agaggtttca tattgctaat agcagctaca atccagctac cattctgctt 300
ttattttatg gttgggataa ggctggatta ttctgagtcc aagctaggcc cttttgctaa 360
tcatgttcat acctcttatc ttcctcccac agctcctggg caacgtgctg gtctgtgtgc 420
tggcccatca ctttggcaaa g 441
<210> 17
<211> 441
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide (modified HBB2 intron)
<400> 17
gtacacatat tgaccaaatc agggtaattt tgcatttgta attttaaaaa atgctttctt 60
cttttaatat acttttttgt ttatcttatt tctaatactt tccctaatct ctttctttca 120
gggcaataat gatacaatgt atcatgcctc tttgcaccat tctaaagaat aacagtgata 180
atttctgggt taaggcaata gcaatatttc tgcatataaa tatttctgca tataaattgt 240
aactgatgta agaggtttca tattgctaat agcagctaca atccagctac cattctgctt 300
ttattttctg gttgggataa ggctggatta ttctgagtcc aagctaggcc cttttgctaa 360
tcttgttcat acctcttatc ttcctcccac agctcctggg caacctgctg gtctctctgc 420
tggcccatca ctttggcaaa g 441
<210> 18
<211> 1438
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide (FIX intron)
<400> 18
ggtttgtttc cttttttaaa atacattgag tatgcttgcc ttttagatat agaaatatct 60
gatgctgtct tcttcactaa attttgatta catgatttga cagcaatatt gaagagtcta 120
acagccagca cgcaggttgg taagtactgg ttctttgtta gctaggtttt cttcttcttc 180
atttttaaaa ctaaatagat cgacaatgct tatgatgcat ttatgtttaa taaacactgt 240
tcagttcatg atttggtcat gtaattcctg ttagaaaaca ttcatctcct tggtttaaaa 300
aaattaaaag tgggaaaaca aagaaatagc agaatatagt gaaaaaaaat aaccacatta 360
tttttgtttg gacttaccac tttgaaatca aaatgggaaa caaaagcaca aacaatggcc 420
ttatttacac aaaaagtctg attttaagat atatgacatt tcaaggtttc agaagtatgt 480
aatgaggtgt gtctctaatt ttttaaatta tatatcttca atttaaagtt ttagttaaaa 540
cataaagatt aacctttcat tagcaagctg ttagttatca ccaacgcttt tcatggatta 600
ggaaaaaatc attttgtctc tatgtcaaac atcttggagt tgatatttgg ggaaacacaa 660
tactcagttg agttccctag gggagaaaag cacgcttaag aattgacata aagagtagga 720
agttagctaa tgcaacatat atcactttgt tttttcacaa ctacagtgac tttatgtatt 780
tcccagagga aggcatacag ggaagaaatt atcccatttg gacaaacagc atgttctcac 840
aggaagcatt tatcacactt acttgtcaac tttctagaat caaatctagt agctgacagt 900
accaggatca ggggtgccaa ccctaagcac ccccagaaag ctgactggcc ctgtggttcc 960
cactccagac atgatgtcag ctgtgaaatc gacgtcgctg gaccataatt aggcttctgt 1020
tcttcaggag acatttgttc aaagtcattt gggcaaccat attctgaaaa cagcccagcc 1080
agggtgatgg atcactttgc aaagatcctc aatgagctat tttcaagtga tgacaaagtg 1140
tgaagttaac cgctcatttg agaactttct ttttcatcca aagtaaattc aaatatgatt 1200
agaaatctga ccttttatta ctggaattct cttgactaaa agtaaaattg aattttaatt 1260
cctaaatctc catgtgtata cagtactgtg ggaacatcac agattttggc tccatgccct 1320
aaagagaaat tggctttcag attatttgga ttaaaaacaa agactttctt aagagatgta 1380
aaattttcat gatgttttct tttttgctaa aactaaagaa ttattctttt acatttca 1438
<210> 19
<211> 1438
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide (modified FIX intron)
<400> 19
ggtttgtttc cttttttaaa atacattgag tatgcttgcc ttttagatat agaaatatct 60
gatgctgtct tcttcactaa attttgatta catgatttga cagcaatatt gaagagtcta 120
acagccagca cgcaggttgg taagtactgg ttctttgtta gctaggtttt cttcttcttc 180
atttttaaaa ctaaatagat cgacattgct tttgttgcat ttatgtttaa taaacactgt 240
tcagttcatg atttggtcat gtaattcctg ttagaaaaca ttcatctcct tggtttaaaa 300
aaattaaaag tgggaaaaca aagaaatagc agaatatagt gaaaaaaaat aaccacatta 360
tttttgtttg gacttaccac tttgaaatca aattgggaaa caaaagcaca aacaatggcc 420
ttatttacac aaaaagtctg attttaagat atatgacatt tcaaggtttc agaagtatgt 480
aatgaggtgt gtctctaatt ttttaaatta tatatcttca atttaaagtt ttagttaaaa 540
cataaagatt aacctttcat tagcaagctg ttagttatca ccaacgcttt tcatggatta 600
ggaaaaaatc attttgtctc tttgtcaaac atcttggagt tgatatttgg ggaaacacaa 660
tactcagttg agttccctag gggagaaaag cacgcttaag aattgacata aagagtagga 720
agttagctat tgcaacatat atcactttgt tttttcacaa ctacagtgac tttttgtatt 780
tcccagagga aggcatacag ggaagaaatt atcccatttg gacaaacagc ttgttctcac 840
aggaagcatt tatcacactt acttgtcaac tttctagaat caaatctagt agctgacagt 900
accaggatca ggggtgccaa ccctaagcac ccccagaaag ctgactggcc ctgtggttcc 960
cactccagac atgatgtcag ctgtgaaatc gacgtcgctg gaccataatt aggcttctgt 1020
tcttcaggag acatttgttc aaagtcattt gggcaaccat attctgaaaa cagcccagcc 1080
agggtgttgg atcactttgc aaagatcctc attgagctat tttcaagtgt tgacaaagtg 1140
tgaagttaac cgctcatttg agaactttct ttttcatcca aagtaaattc aaatatgatt 1200
agaaatctga ccttttatta ctggaattct cttgactaaa agtaaaattg aattttaatt 1260
cctaaatctc catgtgtata cagtactgtg ggaacatcac agattttggc tccatgccct 1320
aaagagaaat tggctttcag attatttgga ttaaaaacaa agactttctt aagagatgta 1380
aaattttctt gttgttttct tttttgctaa aactaaagaa ttattctttt acatttca 1438
<210> 20
<211> 881
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide (chicken beta globin intron)
<400> 20
gcgggagtcg ctgcgttgcc ttcgccccgt gccccgctcc gccgccgcct cgcgccgccc 60
gccccggctc tgactgaccg cgttactccc acaggtgagc gggcgggacg gcccttctcc 120
tccgggctgt aattagcgct tggtttaatg acggcttgtt tcttttctgt ggctgcgtga 180
aagccttgag gggctccggg agggcccttt gtgcgggggg agcggctcgg ggggtgcgtg 240
cgtgtgtgtg tgcgtgggga gcgccgcgtg cggctccgcg ctgcccggcg gctgtgagcg 300
ctgcgggcgc ggcgcggggc tttgtgcgct ccgcagtgtg cgcgagggga gcgcggccgg 360
gggcggtgcc ccgcggtgcg gggggggctg cgaggggaac aaaggctgcg tgcggggtgt 420
gtgcgtgggg gggtgagcag ggggtgtggg cgcgtcggtc gggctgcaac cccccctgca 480
cccccctccc cgagttgctg agcacggccc ggcttcgggt gcggggctcc gtacggggcg 540
tggcgcgggg ctcgccgtgc cgggcggggg gtggcggcag gtgggggtgc cgggcggggc 600
ggggccgcct cgggccgggg agggctcggg ggaggggcgc ggcggccccc ggagcgccgg 660
cggctgtcga ggcgcggcga gccgcagcca ttgcctttta tggtaatcgt gcgagagggc 720
gcagggactt cctttgtccc aaatctgtgc ggagccgaaa tctgggaggc gccgccgcac 780
cccctctagc gggcgcgggg cgaagcggtg cggcgccggc aggaaggaaa tgggcgggga 840
gggccttcgt gcgtcgccgc gccgccgtcc ccttctccct c 881
<210> 21
<211> 881
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide (Modified chicken beta-globin intron)
<400> 21
gcgggagtcg ctgcgttgcc ttcgccccgt gccccgctcc gccgccgcct cgcgccgccc 60
gccccggctc tgactgaccg cgttactccc acaggtgagc gggcgggacg gcccttctcc 120
tccgggctgt aattagcgct tggtttaatg acggcttgtt tcttttctgt ggctgcgtga 180
aagccttgag gggctccggg agggcccttt gtgcgggggg agcggctcgg ggggtgcgtg 240
cgtgtgtgtg tgcgtgggga gcgccgcgtg cggctccgcg ctgcccggcg gctgtgagcg 300
ctgcgggcgc ggcgcggggc tttgtgcgct ccgcagtgtg cgcgagggga gcgcggccgg 360
gggcggtgcc ccgcggtgcg gggggggctg cgaggggaac aaaggctgcg tgcggggtgt 420
gtgcgtgggg gggtgagcag ggggtgtggg cgcgtcggtc gggctgcaac cccccctgca 480
cccccctccc cgagttgctg agcacggccc ggcttcgggt gcggggctcc gtacggggcg 540
tggcgcgggg ctcgccgtgc cgggcggggg gtggcggcag gtgggggtgc cgggcggggc 600
ggggccgcct cgggccgggg agggctcggg ggaggggcgc ggcggccccc ggagcgccgg 660
cggctgtcga ggcgcggcga gccgcagcca ttgccttttt tggtaatcgt gcgagagggc 720
gcagggactt cctttgtccc aaatctgtgc ggagccgaaa tctgggaggc gccgccgcac 780
cccctctagc gggcgcgggg cgaagcggtg cggcgccggc aggaaggaat tgggcgggga 840
gggccttcgt gcgtcgccgc gccgccgtcc ccttctccct c 881
<210> 22
<211> 169
<212> DNA
<213> artificial seqeunce
<220>
<223> synthetic polynucleotide (bGH polyA)
<400> 22
tgtgccttct agttgccagc catctgttgt ttgcccctcc cccgtgcctt ccttgaccct 60
ggaaggtgcc actcccactg tcctttccta ataaaatgag gaaattgcat cgcattgtct 120
gagtaggtgt cattctattc tggggggtgg ggtgggggct agctctaga 169
<210> 23
<211> 3598
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide (expression cassette)
<400> 23
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg ggtacccggg gatcttgcta ccagtggaac agccactaag 360
gattctgcag tgagagcaga gggccagcta agtggtactc tcccagagac tgtctgactc 420
acgccacccc ctccaccttg gacacaggac gctgtggttt ctgagccagg tacaatgact 480
cctttcggta agtgcagtgg aagctgtaca ctgcccaggc aaagcgtccg ggcagcgtag 540
gcgggcgact cagatcccag ccagtggact tagcccctgt ttgctcctcc gataactggg 600
gtgaccttgg ttaatattca ccagcagcct cccccgttgc ccctctggat ccactgctta 660
aatacggacg aggacagggc cctgtctcct cagcttcagg caccaccact gacctgggac 720
agtgaataga tcctgagaac ttcagggtga gtctatggga cccttgatgt tttctttccc 780
cttcttttct atggttaagt tcatgtcata ggaaggggag aagtaacagg gtacacatat 840
tgaccaaatc agggtaattt tgcatttgta attttaaaaa atgctttctt cttttaatat 900
acttttttgt ttatcttatt tctaatactt tccctaatct ctttctttca gggcaataat 960
gatacaatgt atcatgcctc tttgcaccat tctaaagaat aacagtgata atttctgggt 1020
taaggcaata gcaatatttc tgcatataaa tatttctgca tataaattgt aactgatgta 1080
agaggtttca tattgctaat agcagctaca atccagctac cattctgctt ttattttctg 1140
gttgggataa ggctggatta ttctgagtcc aagctaggcc cttttgctaa tcttgttcat 1200
acctcttatc ttcctcccac agctcctggg caacctgctg gtctctctgc tggcccatca 1260
ctttggcaaa gcacgcgtgc caccatggct ttcttgtggc ttctcagttg ttgggccttg 1320
ttggggacca ctttcggacg aagacacaca cgctctgcag aggacgattc tgagagggat 1380
cctttgaatg tccttaagcc gagggcccgc atgacgcctg caccagcatc ctgcagtcaa 1440
gagctccctt cagcagaaga taatagtcct atggcgtccg accccctggg ggtagtacgc 1500
ggggggcgag ttaatacgca cgccggggga actggtcctg aagggtgcag gccctttgcc 1560
aagtttattt cagtgtcaca aacctctaaa ctgacacgcg ccgagacggt ctttccggat 1620
gttgatggat cagatgctca taaatcagag gtagctcacc gcttcaaaga cctcggagag 1680
gagaacttta aggccctcgt cctcatcgca tttgcacagt atttgcaaca atgtcccttt 1740
gaggatcatg tcaaattggt aaatgaagtt actgagttcg cgaagacatg cgtggctgac 1800
gagtccgcag aaaactgcga caagtccctt catacactgt ttggtgataa actgtgcact 1860
gtcgctactc ttagagaaac atatggagag atggctgact gttgtgcgaa gcaagagcca 1920
gaacggaatg aatgtttcct tcaacataaa gatgataatc cgaatttgcc acgccttgta 1980
cgaccagagg tggatgttat gtgcaccgcg tttcatgaca atgaggaaac gttcctgaaa 2040
aaatatttgt atgagattgc tcggagacat ccttactttt acgcacccga gctcctgttc 2100
ttcgctaaga gatacaaggc agcctttacg gaatgctgtc aggccgcgga taaggccgcc 2160
tgcttgctcc ccaagctgga cgagctgcga gatgagggca aggcgtcctc tgctaagcaa 2220
cgactcaaat gcgccagctt gcaaaagttt ggcgaaagag cgtttaaggc ctgggcagtg 2280
gcgcgcctga gtcagcggtt ccctaaggcg gagtttgccg aggtctcaaa actggtgaca 2340
gacttgacga aggtacatac cgagtgctgc cacggagatc tcctggagtg cgcggatgat 2400
cgcgctgacc tcgccaaata tatatgtgag aaccaggatt ccatttcctc aaaacttaaa 2460
gagtgctgcg aaaaacccct tcttgagaag tcacactgca ttgcagaggt cgaaaacgac 2520
gaaatgcctg ccgatctgcc ttcccttgcg gctgactttg tggagtctaa ggacgtttgc 2580
aaaaactatg cagaggctaa agacgtattc ctgggcatgt ttctttatga atacgcgcgg 2640
cgacatcccg actattcagt cgtactcctg cttcgactgg cgaaaactta cgaaactact 2700
ctcgagaaat gctgtgctgc cgccgaccct catgagtgct atgctaaggt atttgatgaa 2760
ttcaaaccgt tggttgagga accgcaaaac ttgatcaagc agaactgtga actttttgag 2820
cagctggggg aatataaatt ccagaatgca ttgcttgtac gatacacaaa gaaagttcca 2880
caggtctcca ccccgacact tgtggaagtt agcaggaact tgggaaaggt cggcagcaag 2940
tgttgtaagc accctgaagc taagcgaatg ccatgcgccg aagattacct ttctgtggtg 3000
ttgaaccaac tgtgcgttct tcacgaaaag acgcccgtat ctgatcgggt gacgaaatgc 3060
tgcacagagt cacttgtgaa cagacgcccc tgctttagtg cgctcgaggt ggacgaaacg 3120
tatgtcccga aagagtttaa cgcagagaca ttcacgtttc atgcggacat ctgtaccctc 3180
tctgaaaagg agagacagat caaaaagcag acagcgctgg ttgagcttgt taagcataag 3240
ccgaaggcga caaaggagca actgaaagcg gtgatggatg acttcgccgc gttcgttgaa 3300
aaatgctgca aagccgacga taaggaaacc tgttttgctg aagaaggtaa aaaactcgtt 3360
gctgcgagtc aggccgccct ggggctgtga ctcgagagat ctaccggtga attcaccgcg 3420
ggtttaaact gtgccttcta gttgccagcc atctgttgtt tgcccctccc ccgtgccttc 3480
cttgaccctg gaaggtgcca ctcccactgt cctttcctaa taaaatgagg aaattgcatc 3540
gcattgtctg agtaggtgtc attctattct ggggggtggg gtgggggcta gctctaga 3598
<210> 24
<211> 7559
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide (AAV gene transfer vector
pAAV hAAT sp7 FGF23-DNco)
<400> 24
cagcagctgc gcgctcgctc gctcactgag gccgcccggg caaagcccgg gcgtcgggcg 60
acctttggtc gcccggcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc 120
atcactaggg gttccttgta gttaatgatt aacccgccat gctacttatc tacgtagcca 180
tgctctagag gatcaaggct cagaggcaca caggagtttc tgggctcacc ctgccccctt 240
ccaacccctc agttcccatc ctccagcagc tgtttgtgtg ctgcctctga agtccacact 300
gaacaaactt cagcctactc atgtccctaa aatgggcaaa cattgcaagc agcaaacagc 360
aaacacacag ccctccctgc ctgctgacct tggagctggg gcagaggtca gagacctctc 420
tgggcccatg ccacctccaa catccactcg accccttgga atttcggtgg agaggagcag 480
aggttgtcct ggcgtggttt aggtagtgtg agaggggtac ccggggatct tgctaccagt 540
ggaacagcca ctaaggattc tgcagtgaga gcagagggcc agctaagtgg tactctccca 600
gagactgtct gactcacgcc accccctcca ccttggacac aggacgctgt ggtttctgag 660
ccaggtacaa tgactccttt cggtaagtgc agtggaagct gtacactgcc caggcaaagc 720
gtccgggcag cgtaggcggg cgactcagat cccagccagt ggacttagcc cctgtttgct 780
cctccgataa ctggggtgac cttggttaat attcaccagc agcctccccc gttgcccctc 840
tggatccact gcttaaatac ggacgaggac agggccctgt ctcctcagct tcaggcacca 900
ccactgacct gggacagtga atagatcctg agaacttcag ggtgagtcta tgggaccctt 960
gatgttttct ttccccttct tttctatggt taagttcatg tcataggaag gggagaagta 1020
acagggtaca catattgacc aaatcagggt aattttgcat ttgtaatttt aaaaaatgct 1080
ttcttctttt aatatacttt tttgtttatc ttatttctaa tactttccct aatctctttc 1140
tttcagggca ataatgatac aatgtatcat gcctctttgc accattctaa agaataacag 1200
tgataatttc tgggttaagg caatagcaat atttctgcat ataaatattt ctgcatataa 1260
attgtaactg atgtaagagg tttcatattg ctaatagcag ctacaatcca gctaccattc 1320
tgcttttatt ttctggttgg gataaggctg gattattctg agtccaagct aggccctttt 1380
gctaatcttg ttcatacctc ttatcttcct cccacagctc ctgggcaacc tgctggtctc 1440
tctgctggcc catcactttg gcaaagcacg cgtgccacca tggctttctt gtggcttctc 1500
agttgttggg ccttgttggg gaccactttc ggacgaagac acacacgctc tgcagaggac 1560
gattctgaga gggatccttt gaatgtcctt aagccgaggg cccgcatgac gcctgcacca 1620
gcatcctgca gtcaagagct cccttcagca gaagataata gtcctatggc gtccgacccc 1680
ctgggggtag tacgcggggg gcgagttaat acgcacgccg ggggaactgg tcctgaaggg 1740
tgcaggccct ttgccaagtt tatttcagtg tcacaaacct ctaaactgac acgcgccgag 1800
acggtctttc cggatgttga tggatcagat gctcataaat cagaggtagc tcaccgcttc 1860
aaagacctcg gagaggagaa ctttaaggcc ctcgtcctca tcgcatttgc acagtatttg 1920
caacaatgtc cctttgagga tcatgtcaaa ttggtaaatg aagttactga gttcgcgaag 1980
acatgcgtgg ctgacgagtc cgcagaaaac tgcgacaagt cccttcatac actgtttggt 2040
gataaactgt gcactgtcgc tactcttaga gaaacatatg gagagatggc tgactgttgt 2100
gcgaagcaag agccagaacg gaatgaatgt ttccttcaac ataaagatga taatccgaat 2160
ttgccacgcc ttgtacgacc agaggtggat gttatgtgca ccgcgtttca tgacaatgag 2220
gaaacgttcc tgaaaaaata tttgtatgag attgctcgga gacatcctta cttttacgca 2280
cccgagctcc tgttcttcgc taagagatac aaggcagcct ttacggaatg ctgtcaggcc 2340
gcggataagg ccgcctgctt gctccccaag ctggacgagc tgcgagatga gggcaaggcg 2400
tcctctgcta agcaacgact caaatgcgcc agcttgcaaa agtttggcga aagagcgttt 2460
aaggcctggg cagtggcgcg cctgagtcag cggttcccta aggcggagtt tgccgaggtc 2520
tcaaaactgg tgacagactt gacgaaggta cataccgagt gctgccacgg agatctcctg 2580
gagtgcgcgg atgatcgcgc tgacctcgcc aaatatatat gtgagaacca ggattccatt 2640
tcctcaaaac ttaaagagtg ctgcgaaaaa ccccttcttg agaagtcaca ctgcattgca 2700
gaggtcgaaa acgacgaaat gcctgccgat ctgccttccc ttgcggctga ctttgtggag 2760
tctaaggacg tttgcaaaaa ctatgcagag gctaaagacg tattcctggg catgtttctt 2820
tatgaatacg cgcggcgaca tcccgactat tcagtcgtac tcctgcttcg actggcgaaa 2880
acttacgaaa ctactctcga gaaatgctgt gctgccgccg accctcatga gtgctatgct 2940
aaggtatttg atgaattcaa accgttggtt gaggaaccgc aaaacttgat caagcagaac 3000
tgtgaacttt ttgagcagct gggggaatat aaattccaga atgcattgct tgtacgatac 3060
acaaagaaag ttccacaggt ctccaccccg acacttgtgg aagttagcag gaacttggga 3120
aaggtcggca gcaagtgttg taagcaccct gaagctaagc gaatgccatg cgccgaagat 3180
tacctttctg tggtgttgaa ccaactgtgc gttcttcacg aaaagacgcc cgtatctgat 3240
cgggtgacga aatgctgcac agagtcactt gtgaacagac gcccctgctt tagtgcgctc 3300
gaggtggacg aaacgtatgt cccgaaagag tttaacgcag agacattcac gtttcatgcg 3360
gacatctgta ccctctctga aaaggagaga cagatcaaaa agcagacagc gctggttgag 3420
cttgttaagc ataagccgaa ggcgacaaag gagcaactga aagcggtgat ggatgacttc 3480
gccgcgttcg ttgaaaaatg ctgcaaagcc gacgataagg aaacctgttt tgctgaagaa 3540
ggtaaaaaac tcgttgctgc gagtcaggcc gccctggggc tgtgactcga gagatctacc 3600
ggtgaattca ccgcgggttt aaactgtgcc ttctagttgc cagccatctg ttgtttgccc 3660
ctcccccgtg ccttccttga ccctggaagg tgccactccc actgtccttt cctaataaaa 3720
tgaggaaatt gcatcgcatt gtctgagtag gtgtcattct attctggggg gtggggtggg 3780
ggctagctct agagcatggc tacgtagata agtagcatgg cgggttaatc attaactaca 3840
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 3900
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 3960
gagcgcgcag ctggcgtaat agcgaagagg cccgcaccga tcgcccttcc caacagttgc 4020
gcagcctgaa tggcgaatgg cgattccgtt gcaatggctg gcggtaatat tgttctggat 4080
attaccagca aggccgatag tttgagttct tctactcagg caagtgatgt tattactaat 4140
caaagaagta ttgcgacaac ggttaatttg cgtgatggac agactctttt actcggtggc 4200
ctcactgatt ataaaaacac ttctcaggat tctggcgtac cgttcctgtc taaaatccct 4260
ttaatcggcc tcctgtttag ctcccgctct gattctaacg aggaaagcac gttatacgtg 4320
ctcgtcaaag caaccatagt acgcgccctg tagcggcgca ttaagcgcgg cgggtgtggt 4380
ggttacgcgc agcgtgaccg ctacacttgc cagcgcccta gcgcccgctc ctttcgcttt 4440
cttcccttcc tttctcgcca cgttcgccgg ctttccccgt caagctctaa atcgggggct 4500
ccctttaggg ttccgattta gtgctttacg gcacctcgac cccaaaaaac ttgattaggg 4560
tgatggttca cgtagtgggc catcgccctg atagacggtt tttcgccctt tgacgttgga 4620
gtccacgttc tttaatagtg gactcttgtt ccaaactgga acaacactca accctatctc 4680
ggtctattct tttgatttat aagggatttt gccgatttcg gcctattggt taaaaaatga 4740
gctgatttaa caaaaattta acgcgaattt taacaaaata ttaacgctta caatttaaat 4800
atttgcttat acaatcttcc tgtttttggg gcttttctga ttatcaaccg gggtacatat 4860
gattgacatg ctagttttac gattaccgtt catcgattct cttgtttgct ccagactctc 4920
aggcaatgac ctgatagcct ttgtagagac ctctcaaaaa tagctaccct ctccggcatg 4980
aatttatcag ctagaacggt tgaatatcat attgatggtg atttgactgt ctccggcctt 5040
tctcacccgt ttgaatcttt acctacacat tactcaggca ttgcatttaa aatatatgag 5100
ggttctaaaa atttttatcc ttgcgttgaa ataaaggctt ctcccgcaaa agtattacag 5160
ggtcataatg tttttggtac aaccgattta gctttatgct ctgaggcttt attgcttaat 5220
tttgctaatt ctttgccttg cctgtatgat ttattggatg ttggaatcgt cctgatgcgg 5280
tattttctcc ttacgcatct gtgcggtatt tcacaccgca tatggtgcac tctcagtaca 5340
atctgctctg atgccgcata gttaagccag ccccgacacc cgccaacacc cgctgacgcg 5400
ccctgacggg cttgtctgct cccggcatcc gcttacagac aagctgtgac cgtctccggg 5460
agctgcatgt gtcagaggtt ttcaccgtca tcaccgaaac gcgcgagacg aaagggcctc 5520
gtgatacgcc tatttttata ggttaatgtc atgataataa tggtttctta gacgtcaggt 5580
ggcacttttc ggggaaatgt gcgcggaacc cctatttgtt tatttttcta aatacattca 5640
aatatgtatc cgctcatgag acaataaccc tgataaatgc ttcaataata ttgaaaaagg 5700
aagagtatga gtattcaaca tttccgtgtc gcccttattc ccttttttgc ggcattttgc 5760
cttcctgttt ttgctcaccc agaaacgctg gtgaaagtaa aagatgctga agatcagttg 5820
ggtgcacgag tgggttacat cgaactggat ctcaacagcg gtaagatcct tgagagtttt 5880
cgccccgaag aacgttttcc aatgatgagc acttttaaag ttctgctatg tggcgcggta 5940
ttatcccgta ttgacgccgg gcaagagcaa ctcggtcgcc gcatacacta ttctcagaat 6000
gacttggttg agtactcacc agtcacagaa aagcatctta cggatggcat gacagtaaga 6060
gaattatgca gtgctgccat aaccatgagt gataacactg cggccaactt acttctgaca 6120
acgatcggag gaccgaagga gctaaccgct tttttgcaca acatggggga tcatgtaact 6180
cgccttgatc gttgggaacc ggagctgaat gaagccatac caaacgacga gcgtgacacc 6240
acgatgcctg tagcaatggc aacaacgttg cgcaaactat taactggcga actacttact 6300
ctagcttccc ggcaacaatt aatagactgg atggaggcgg ataaagttgc aggaccactt 6360
ctgcgctcgg cccttccggc tggctggttt attgctgata aatctggagc cggtgagcgt 6420
gggtctcgcg gtatcattgc agcactgggg ccagatggta agccctcccg tatcgtagtt 6480
atctacacga cggggagtca ggcaactatg gatgaacgaa atagacagat cgctgagata 6540
ggtgcctcac tgattaagca ttggtaactg tcagaccaag tttactcata tatactttag 6600
attgatttaa aacttcattt ttaatttaaa aggatctagg tgaagatcct ttttgataat 6660
ctcatgacca aaatccctta acgtgagttt tcgttccact gagcgtcaga ccccgtagaa 6720
aagatcaaag gatcttcttg agatcctttt tttctgcgcg taatctgctg cttgcaaaca 6780
aaaaaaccac cgctaccagc ggtggtttgt ttgccggatc aagagctacc aactcttttt 6840
ccgaaggtaa ctggcttcag cagagcgcag ataccaaata ctgtccttct agtgtagccg 6900
tagttaggcc accacttcaa gaactctgta gcaccgccta catacctcgc tctgctaatc 6960
ctgttaccag tggctgctgc cagtggcgat aagtcgtgtc ttaccgggtt ggactcaaga 7020
cgatagttac cggataaggc gcagcggtcg ggctgaacgg ggggttcgtg cacacagccc 7080
agcttggagc gaacgaccta caccgaactg agatacctac agcgtgagct atgagaaagc 7140
gccacgcttc ccgaagggag aaaggcggac aggtatccgg taagcggcag ggtcggaaca 7200
ggagagcgca cgagggagct tccaggggga aacgcctggt atctttatag tcctgtcggg 7260
tttcgccacc tctgacttga gcgtcgattt ttgtgatgct cgtcaggggg gcggagccta 7320
tggaaaaacg ccagcaacgc ggccttttta cggttcctgg ccttttgctg gccttttgct 7380
cacatgttct ttcctgcgtt atcccctgat tctgtggata accgtattac cgcctttgag 7440
tgagctgata ccgctcgccg cagccgaacg accgagcgca gcgagtcagt gagcgaggaa 7500
gcggaagagc gcccaatacg caaaccgcct ctccccgcgc gttggccgat tcattaatg 7559
<210> 25
<211> 768
<212> DNA
<213> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(768)
<400> 25
acgcgtgcca cc atg ttg ggg gcc cgc ctc agg ctc tgg gtc tgt gcc ttg 51
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu
1 5 10
tgc agc gtc tgc agc atg agc gtc ctc aga gcc tat ccc aat gcc tcc 99
Cys Ser Val Cys Ser Met Ser Val Leu Arg Ala Tyr Pro Asn Ala Ser
15 20 25
cca ctg ctc ggc tcc agc tgg ggt ggc ctg atc cac ctg tac aca gcc 147
Pro Leu Leu Gly Ser Ser Trp Gly Gly Leu Ile His Leu Tyr Thr Ala
30 35 40 45
aca gcc agg aac agc tac cac ctg cag atc cac aag aat ggc cat gtg 195
Thr Ala Arg Asn Ser Tyr His Leu Gln Ile His Lys Asn Gly His Val
50 55 60
gat ggc gca ccc cat cag acc atc tac agt gcc ctg atg atc aga tca 243
Asp Gly Ala Pro His Gln Thr Ile Tyr Ser Ala Leu Met Ile Arg Ser
65 70 75
gag gat gct ggc ttt gtg gtg att aca ggt gtg atg agc aga aga tac 291
Glu Asp Ala Gly Phe Val Val Ile Thr Gly Val Met Ser Arg Arg Tyr
80 85 90
ctc tgc atg gat ttc aga ggc aac att ttt gga tca cac tat ttc gac 339
Leu Cys Met Asp Phe Arg Gly Asn Ile Phe Gly Ser His Tyr Phe Asp
95 100 105
ccg gag aac tgc agg ttc caa cac cag acg ctg gaa aac ggg tac gac 387
Pro Glu Asn Cys Arg Phe Gln His Gln Thr Leu Glu Asn Gly Tyr Asp
110 115 120 125
gtc tac cac tct cct cag tat cac ttc ctg gtc agt ctg ggc cgg gcg 435
Val Tyr His Ser Pro Gln Tyr His Phe Leu Val Ser Leu Gly Arg Ala
130 135 140
aag aga gcc ttc ctg cca ggc atg aac cca ccc ccg tac tcc cag ttc 483
Lys Arg Ala Phe Leu Pro Gly Met Asn Pro Pro Pro Tyr Ser Gln Phe
145 150 155
ctg tcc cgg agg aac gag atc ccc cta att cac ttc aac acc ccc ata 531
Leu Ser Arg Arg Asn Glu Ile Pro Leu Ile His Phe Asn Thr Pro Ile
160 165 170
cca cgg cgg cac acc cgg agc gcc gag gac gac tcg gag cgg gac ccc 579
Pro Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro
175 180 185
ctg aac gtg ctg aag ccc cgg gcc cgg atg acc ccg gcc ccg gcc tcc 627
Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser
190 195 200 205
tgt tca cag gag ctc ccg agc gcc gag gac aac agc ccg atg gcc agt 675
Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser
210 215 220
gac cca tta ggg gtg gtc agg ggc ggt cga gtg aac acg cac gct ggg 723
Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly
225 230 235
gga acg ggc ccg gaa ggc tgc cgc ccc ttc gcc aag ttc atc tag 768
Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
240 245 250
<210> 26
<211> 251
<212> PRT
<213> synthetic polynucleotide
<400> 26
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Tyr Pro Asn Ala Ser Pro Leu Leu
20 25 30
Gly Ser Ser Trp Gly Gly Leu Ile His Leu Tyr Thr Ala Thr Ala Arg
35 40 45
Asn Ser Tyr His Leu Gln Ile His Lys Asn Gly His Val Asp Gly Ala
50 55 60
Pro His Gln Thr Ile Tyr Ser Ala Leu Met Ile Arg Ser Glu Asp Ala
65 70 75 80
Gly Phe Val Val Ile Thr Gly Val Met Ser Arg Arg Tyr Leu Cys Met
85 90 95
Asp Phe Arg Gly Asn Ile Phe Gly Ser His Tyr Phe Asp Pro Glu Asn
100 105 110
Cys Arg Phe Gln His Gln Thr Leu Glu Asn Gly Tyr Asp Val Tyr His
115 120 125
Ser Pro Gln Tyr His Phe Leu Val Ser Leu Gly Arg Ala Lys Arg Ala
130 135 140
Phe Leu Pro Gly Met Asn Pro Pro Pro Tyr Ser Gln Phe Leu Ser Arg
145 150 155 160
Arg Asn Glu Ile Pro Leu Ile His Phe Asn Thr Pro Ile Pro Arg Arg
165 170 175
His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val
180 185 190
Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln
195 200 205
Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu
210 215 220
Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly
225 230 235 240
Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
245 250
<210> 27
<211> 251
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 27
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Tyr Pro Asn Ala Ser Pro Leu Leu
20 25 30
Gly Ser Ser Trp Gly Gly Leu Ile His Leu Tyr Thr Ala Thr Ala Arg
35 40 45
Asn Ser Tyr His Leu Gln Ile His Lys Asn Gly His Val Asp Gly Ala
50 55 60
Pro His Gln Thr Ile Tyr Ser Ala Leu Met Ile Arg Ser Glu Asp Ala
65 70 75 80
Gly Phe Val Val Ile Thr Gly Val Met Ser Arg Arg Tyr Leu Cys Met
85 90 95
Asp Phe Arg Gly Asn Ile Phe Gly Ser His Tyr Phe Asp Pro Glu Asn
100 105 110
Cys Arg Phe Gln His Gln Thr Leu Glu Asn Gly Tyr Asp Val Tyr His
115 120 125
Ser Pro Gln Tyr His Phe Leu Val Ser Leu Gly Arg Ala Lys Arg Ala
130 135 140
Phe Leu Pro Gly Met Asn Pro Pro Pro Tyr Ser Gln Phe Leu Ser Arg
145 150 155 160
Arg Asn Glu Ile Pro Leu Ile His Phe Asn Thr Pro Ile Pro Arg Arg
165 170 175
His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val
180 185 190
Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln
195 200 205
Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu
210 215 220
Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly
225 230 235 240
Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
245 250
<210> 28
<211> 768
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(768)
<400> 28
acgcgtgcca cc atg ttg ggc gcc agg ctt cgg ctc tgg gtc tgt gca ctg 51
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu
1 5 10
tgc agt gtc tgt tcc atg agt gtg ttg cgg gct tac cct aac gca tca 99
Cys Ser Val Cys Ser Met Ser Val Leu Arg Ala Tyr Pro Asn Ala Ser
15 20 25
ccg ctg ttg ggc tct tcc tgg ggt gga ctc ata cac ctt tat acg gcg 147
Pro Leu Leu Gly Ser Ser Trp Gly Gly Leu Ile His Leu Tyr Thr Ala
30 35 40 45
act gcg cgc aat agc tac cat ttg caa ata cac aag aat ggg cat gtt 195
Thr Ala Arg Asn Ser Tyr His Leu Gln Ile His Lys Asn Gly His Val
50 55 60
gat ggt gct ccc cac cag act atc tac tct gct ctt atg att agg tca 243
Asp Gly Ala Pro His Gln Thr Ile Tyr Ser Ala Leu Met Ile Arg Ser
65 70 75
gaa gac gcg ggg ttc gta gtt att act ggt gtt atg tcc cgc aga tat 291
Glu Asp Ala Gly Phe Val Val Ile Thr Gly Val Met Ser Arg Arg Tyr
80 85 90
ctc tgc atg gac ttc cgc ggt aac atc ttt ggg tca cac tat ttc gac 339
Leu Cys Met Asp Phe Arg Gly Asn Ile Phe Gly Ser His Tyr Phe Asp
95 100 105
cca gag aac tgt cgc ttc cag cac cag acg ttg gag aat ggt tat gac 387
Pro Glu Asn Cys Arg Phe Gln His Gln Thr Leu Glu Asn Gly Tyr Asp
110 115 120 125
gtg tat cac tcc ccg cag tac cat ttc ttg gtc tct ctg ggc cgg gcg 435
Val Tyr His Ser Pro Gln Tyr His Phe Leu Val Ser Leu Gly Arg Ala
130 135 140
aaa agg gcc ttt ctg cca ggt atg aat cct ccg cct tat agt cag ttc 483
Lys Arg Ala Phe Leu Pro Gly Met Asn Pro Pro Pro Tyr Ser Gln Phe
145 150 155
ctt agc agg cgg aac gaa att cct ctg att cac ttc aac acg ccg atc 531
Leu Ser Arg Arg Asn Glu Ile Pro Leu Ile His Phe Asn Thr Pro Ile
160 165 170
cct cgc agg cat acg cga tcc gca gaa gac gat tct gag cga gat cct 579
Pro Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro
175 180 185
ctc aat gtt ctg aaa ccc cgc gcc cga atg acg cca gct ccg gcc agt 627
Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser
190 195 200 205
tgt agt cag gag ttg ccg tct gcc gaa gat aat agt ccc atg gcc tcc 675
Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser
210 215 220
gat cca ctt ggc gtg gta agg gga ggt aga gtt aac acc cac gcc ggt 723
Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly
225 230 235
gga act ggc cct gaa ggg tgt cgg cct ttt gcc aaa ttc ata tag 768
Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
240 245 250
<210> 29
<211> 251
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 29
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Tyr Pro Asn Ala Ser Pro Leu Leu
20 25 30
Gly Ser Ser Trp Gly Gly Leu Ile His Leu Tyr Thr Ala Thr Ala Arg
35 40 45
Asn Ser Tyr His Leu Gln Ile His Lys Asn Gly His Val Asp Gly Ala
50 55 60
Pro His Gln Thr Ile Tyr Ser Ala Leu Met Ile Arg Ser Glu Asp Ala
65 70 75 80
Gly Phe Val Val Ile Thr Gly Val Met Ser Arg Arg Tyr Leu Cys Met
85 90 95
Asp Phe Arg Gly Asn Ile Phe Gly Ser His Tyr Phe Asp Pro Glu Asn
100 105 110
Cys Arg Phe Gln His Gln Thr Leu Glu Asn Gly Tyr Asp Val Tyr His
115 120 125
Ser Pro Gln Tyr His Phe Leu Val Ser Leu Gly Arg Ala Lys Arg Ala
130 135 140
Phe Leu Pro Gly Met Asn Pro Pro Pro Tyr Ser Gln Phe Leu Ser Arg
145 150 155 160
Arg Asn Glu Ile Pro Leu Ile His Phe Asn Thr Pro Ile Pro Arg Arg
165 170 175
His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val
180 185 190
Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln
195 200 205
Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu
210 215 220
Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly
225 230 235 240
Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
245 250
<210> 30
<211> 315
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(315)
<400> 30
acgcgtgcca cc atg ttg ggg gcc cgc ctc agg ctc tgg gtc tgt gcc ttg 51
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu
1 5 10
tgc agc gtc tgc agc atg agc gtc ctc aga gcc cga aga cac aca cgc 99
Cys Ser Val Cys Ser Met Ser Val Leu Arg Ala Arg Arg His Thr Arg
15 20 25
gcc gag gac gac tcg gag cgg gac ccc ctg aac gtg ctg aag ccc cgg 147
Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val Leu Lys Pro Arg
30 35 40 45
gcc cgg atg acc ccg gcc ccg gcc tcc tgt tca cag gag ctc ccg agc 195
Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro Ser
50 55 60
gcc gag gac aac agc ccg atg gcc agt gac cca tta ggg gtg gtc agg 243
Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu Gly Val Val Arg
65 70 75
ggc ggt cga gtg aac acg cac gct ggg gga acg ggc ccg gaa ggc tgc 291
Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly Pro Glu Gly Cys
80 85 90
cgc ccc ttc gcc aag ttc atc tag 315
Arg Pro Phe Ala Lys Phe Ile
95 100
<210> 31
<211> 100
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 31
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Arg Arg His Thr Arg Ala Glu Asp
20 25 30
Asp Ser Glu Arg Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met
35 40 45
Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp
50 55 60
Asn Ser Pro Met Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg
65 70 75 80
Val Asn Thr His Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe
85 90 95
Ala Lys Phe Ile
100
<210> 32
<211> 100
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 32
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Arg Arg His Thr Arg Ala Glu Asp
20 25 30
Asp Ser Glu Arg Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met
35 40 45
Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp
50 55 60
Asn Ser Pro Met Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg
65 70 75 80
Val Asn Thr His Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe
85 90 95
Ala Lys Phe Ile
100
<210> 33
<211> 300
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(300)
<400> 33
acgcgtgcca cc atg ttg ggg gcc cgc ctc agg ctc tgg gtc tgt gcc ttg 51
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu
1 5 10
tgc agc gtc tgc agc atg agc gtc ctc aga gcc gcc gag gac gac tcg 99
Cys Ser Val Cys Ser Met Ser Val Leu Arg Ala Ala Glu Asp Asp Ser
15 20 25
gag cgg gac ccc ctg aac gtg ctg aag ccc cgg gcc cgg atg acc ccg 147
Glu Arg Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro
30 35 40 45
gcc ccg gcc tcc tgt tca cag gag ctc ccg agc gcc gag gac aac agc 195
Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser
50 55 60
ccg atg gcc agt gac cca tta ggg gtg gtc agg ggc ggt cga gtg aac 243
Pro Met Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn
65 70 75
acg cac gct ggg gga acg ggc ccg gaa ggc tgc cgc ccc ttc gcc aag 291
Thr His Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys
80 85 90
ttc atc tag 300
Phe Ile
95
<210> 34
<211> 95
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 34
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Ala Glu Asp Asp Ser Glu Arg Asp
20 25 30
Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala
35 40 45
Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala
50 55 60
Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala
65 70 75 80
Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
85 90 95
<210> 35
<211> 95
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 35
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Ala Glu Asp Asp Ser Glu Arg Asp
20 25 30
Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala
35 40 45
Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala
50 55 60
Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala
65 70 75 80
Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
85 90 95
<210> 36
<211> 321
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(321)
<400> 36
acgcgtgcca cc atg ttg ggc gcc agg ctt cgg ctc tgg gtc tgt gca ctg 51
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu
1 5 10
tgc agt gtc tgt tcc atg agt gtg ttg cgg gct cga aga cac aca cgc 99
Cys Ser Val Cys Ser Met Ser Val Leu Arg Ala Arg Arg His Thr Arg
15 20 25
tct gca gag gac gat tct gag agg gat cct ttg aat gtc ctt aag ccg 147
Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val Leu Lys Pro
30 35 40 45
agg gcc cgc atg acg cct gca cca gca tcc tgc agt caa gag ctc cct 195
Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro
50 55 60
tca gca gaa gat aat agt cct atg gcg tcc gac ccc ctg ggg gta gta 243
Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu Gly Val Val
65 70 75
cgc ggg ggg cga gtt aat acg cac gcc ggg gga act ggt cct gaa ggg 291
Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly Pro Glu Gly
80 85 90
tgc agg ccc ttt gcc aag ttt att gtt taa 321
Cys Arg Pro Phe Ala Lys Phe Ile Val
95 100
<210> 37
<211> 102
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 37
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Arg Arg His Thr Arg Ser Ala Glu
20 25 30
Asp Asp Ser Glu Arg Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg
35 40 45
Met Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu
50 55 60
Asp Asn Ser Pro Met Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly
65 70 75 80
Arg Val Asn Thr His Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro
85 90 95
Phe Ala Lys Phe Ile Val
100
<210> 38
<211> 102
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 38
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Arg Arg His Thr Arg Ser Ala Glu
20 25 30
Asp Asp Ser Glu Arg Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg
35 40 45
Met Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu
50 55 60
Asp Asn Ser Pro Met Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly
65 70 75 80
Arg Val Asn Thr His Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro
85 90 95
Phe Ala Lys Phe Ile Val
100
<210> 39
<211> 306
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(306)
<400> 39
acgcgtgcca cc atg ttg ggc gcc agg ctt cgg ctc tgg gtc tgt gca ctg 51
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu
1 5 10
tgc agt gtc tgt tcc atg agt gtg ttg cgg gct tct gca gag gac gat 99
Cys Ser Val Cys Ser Met Ser Val Leu Arg Ala Ser Ala Glu Asp Asp
15 20 25
tct gag agg gat cct ttg aat gtc ctt aag ccg agg gcc cgc atg acg 147
Ser Glu Arg Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr
30 35 40 45
cct gca cca gca tcc tgc agt caa gag ctc cct tca gca gaa gat aat 195
Pro Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn
50 55 60
agt cct atg gcg tcc gac ccc ctg ggg gta gta cgc ggg ggg cga gtt 243
Ser Pro Met Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val
65 70 75
aat acg cac gcc ggg gga act ggt cct gaa ggg tgc agg ccc ttt gcc 291
Asn Thr His Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala
80 85 90
aag ttt att gtt taa 306
Lys Phe Ile Val
95
<210> 40
<211> 97
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 40
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Ser Ala Glu Asp Asp Ser Glu Arg
20 25 30
Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro
35 40 45
Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met
50 55 60
Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His
65 70 75 80
Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
85 90 95
Val
<210> 41
<211> 97
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 41
Met Leu Gly Ala Arg Leu Arg Leu Trp Val Cys Ala Leu Cys Ser Val
1 5 10 15
Cys Ser Met Ser Val Leu Arg Ala Ser Ala Glu Asp Asp Ser Glu Arg
20 25 30
Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro
35 40 45
Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met
50 55 60
Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His
65 70 75 80
Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile
85 90 95
Val
<210> 42
<211> 303
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(303)
<400> 42
acgcgtgcca cc atg gct ttc ttg tgg ctt ctc agt tgt tgg gcc ttg ttg 51
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu
1 5 10
ggg acc act ttc gga cga aga cac aca cgc tct gca gag gac gat tct 99
Gly Thr Thr Phe Gly Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser
15 20 25
gag agg gat cct ttg aat gtc ctt aag ccg agg gcc cgc atg acg cct 147
Glu Arg Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro
30 35 40 45
gca cca gca tcc tgc agt caa gag ctc cct tca gca gaa gat aat agt 195
Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser
50 55 60
cct atg gcg tcc gac ccc ctg ggg gta gta cgc ggg ggg cga gtt aat 243
Pro Met Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn
65 70 75
acg cac gcc ggg gga act ggt cct gaa ggg tgc agg ccc ttt gcc aag 291
Thr His Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys
80 85 90
ttt att gtt taa 303
Phe Ile Val
95
<210> 43
<211> 96
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 43
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp
20 25 30
Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala
35 40 45
Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala
50 55 60
Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala
65 70 75 80
Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Val
85 90 95
<210> 44
<211> 96
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 44
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp
20 25 30
Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala
35 40 45
Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala
50 55 60
Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala
65 70 75 80
Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Val
85 90 95
<210> 45
<211> 288
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(288)
<400> 45
acgcgtgcca cc atg gct ttc ttg tgg ctt ctc agt tgt tgg gcc ttg ttg 51
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu
1 5 10
ggg acc act ttc gga tct gca gag gac gat tct gag agg gat cct ttg 99
Gly Thr Thr Phe Gly Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu
15 20 25
aat gtc ctt aag ccg agg gcc cgc atg acg cct gca cca gca tcc tgc 147
Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys
30 35 40 45
agt caa gag ctc cct tca gca gaa gat aat agt cct atg gcg tcc gac 195
Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp
50 55 60
ccc ctg ggg gta gta cgc ggg ggg cga gtt aat acg cac gcc ggg gga 243
Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly
65 70 75
act ggt cct gaa ggg tgc agg ccc ttt gcc aag ttt att gtt taa 288
Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Val
80 85 90
<210> 46
<211> 91
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 46
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val Leu
20 25 30
Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu
35 40 45
Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu Gly
50 55 60
Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly Pro
65 70 75 80
Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Val
85 90
<210> 47
<211> 91
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 47
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val Leu
20 25 30
Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu
35 40 45
Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu Gly
50 55 60
Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly Pro
65 70 75 80
Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Val
85 90
<210> 48
<211> 2055
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(2055)
<400> 48
acgcgtgcca cc atg gct ttc ttg tgg ctt ctc agt tgt tgg gcc ttg ttg 51
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu
1 5 10
ggg acc act ttc gga cga aga cac aca cgc tct gca gag gac gat tct 99
Gly Thr Thr Phe Gly Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser
15 20 25
gag agg gat cct ttg aat gtc ctt aag ccg agg gcc cgc atg acg cct 147
Glu Arg Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro
30 35 40 45
gca cca gca tcc tgc agt caa gag ctc cct tca gca gaa gat aat agt 195
Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser
50 55 60
cct atg gcg tcc gac ccc ctg ggg gta gta cgc ggg ggg cga gtt aat 243
Pro Met Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn
65 70 75
acg cac gcc ggg gga act ggt cct gaa ggg tgc agg ccc ttt gcc aag 291
Thr His Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys
80 85 90
ttt att gat gct cat aaa tca gag gta gct cac cgc ttc aaa gac ctc 339
Phe Ile Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu
95 100 105
gga gag gag aac ttt aag gcc ctc gtc ctc atc gca ttt gca cag tat 387
Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr
110 115 120 125
ttg caa caa tgt ccc ttt gag gat cat gtc aaa ttg gta aat gaa gtt 435
Leu Gln Gln Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val
130 135 140
act gag ttc gcg aag aca tgc gtg gct gac gag tcc gca gaa aac tgc 483
Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys
145 150 155
gac aag tcc ctt cat aca ctg ttt ggt gat aaa ctg tgc act gtc gct 531
Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala
160 165 170
act ctt aga gaa aca tat gga gag atg gct gac tgt tgt gcg aag caa 579
Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln
175 180 185
gag cca gaa cgg aat gaa tgt ttc ctt caa cat aaa gat gat aat ccg 627
Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro
190 195 200 205
aat ttg cca cgc ctt gta cga cca gag gtg gat gtt atg tgc acc gcg 675
Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala
210 215 220
ttt cat gac aat gag gaa acg ttc ctg aaa aaa tat ttg tat gag att 723
Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile
225 230 235
gct cgg aga cat cct tac ttt tac gca ccc gag ctc ctg ttc ttc gct 771
Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala
240 245 250
aag aga tac aag gca gcc ttt acg gaa tgc tgt cag gcc gcg gat aag 819
Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys
255 260 265
gcc gcc tgc ttg ctc ccc aag ctg gac gag ctg cga gat gag ggc aag 867
Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys
270 275 280 285
gcg tcc tct gct aag caa cga ctc aaa tgc gcc agc ttg caa aag ttt 915
Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe
290 295 300
ggc gaa aga gcg ttt aag gcc tgg gca gtg gcg cgc ctg agt cag cgg 963
Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg
305 310 315
ttc cct aag gcg gag ttt gcc gag gtc tca aaa ctg gtg aca gac ttg 1011
Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu
320 325 330
acg aag gta cat acc gag tgc tgc cac gga gat ctc ctg gag tgc gcg 1059
Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala
335 340 345
gat gat cgc gct gac ctc gcc aaa tat ata tgt gag aac cag gat tcc 1107
Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser
350 355 360 365
att tcc tca aaa ctt aaa gag tgc tgc gaa aaa ccc ctt ctt gag aag 1155
Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys
370 375 380
tca cac tgc att gca gag gtc gaa aac gac gaa atg cct gcc gat ctg 1203
Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu
385 390 395
cct tcc ctt gcg gct gac ttt gtg gag tct aag gac gtt tgc aaa aac 1251
Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn
400 405 410
tat gca gag gct aaa gac gta ttc ctg ggc atg ttt ctt tat gaa tac 1299
Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr
415 420 425
gcg cgg cga cat ccc gac tat tca gtc gta ctc ctg ctt cga ctg gcg 1347
Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala
430 435 440 445
aaa act tac gaa act act ctc gag aaa tgc tgt gct gcc gcc gac cct 1395
Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro
450 455 460
cat gag tgc tat gct aag gta ttt gat gaa ttc aaa ccg ttg gtt gag 1443
His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu
465 470 475
gaa ccg caa aac ttg atc aag cag aac tgt gaa ctt ttt gag cag ctg 1491
Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu
480 485 490
ggg gaa tat aaa ttc cag aat gca ttg ctt gta cga tac aca aag aaa 1539
Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys
495 500 505
gtt cca cag gtc tcc acc ccg aca ctt gtg gaa gtt agc agg aac ttg 1587
Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu
510 515 520 525
gga aag gtc ggc agc aag tgt tgt aag cac cct gaa gct aag cga atg 1635
Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met
530 535 540
cca tgc gcc gaa gat tac ctt tct gtg gtg ttg aac caa ctg tgc gtt 1683
Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val
545 550 555
ctt cac gaa aag acg ccc gta tct gat cgg gtg acg aaa tgc tgc aca 1731
Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr
560 565 570
gag tca ctt gtg aac aga cgc ccc tgc ttt agt gcg ctc gag gtg gac 1779
Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp
575 580 585
gaa acg tat gtc ccg aaa gag ttt aac gca gag aca ttc acg ttt cat 1827
Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His
590 595 600 605
gcg gac atc tgt acc ctc tct gaa aag gag aga cag atc aaa aag cag 1875
Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln
610 615 620
aca gcg ctg gtt gag ctt gtt aag cat aag ccg aag gcg aca aag gag 1923
Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu
625 630 635
caa ctg aaa gcg gtg atg gat gac ttc gcc gcg ttc gtt gaa aaa tgc 1971
Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys
640 645 650
tgc aaa gcc gac gat aag gaa acc tgt ttt gct gaa gaa ggt aaa aaa 2019
Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys
655 660 665
ctc gtt gct gcg agt cag gcc gcc ctg ggg ctg tga 2055
Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
670 675 680
<210> 49
<211> 680
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 49
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp
20 25 30
Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala
35 40 45
Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala
50 55 60
Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala
65 70 75 80
Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Asp
85 90 95
Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu
100 105 110
Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln
115 120 125
Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe
130 135 140
Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser
145 150 155 160
Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg
165 170 175
Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu
180 185 190
Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro
195 200 205
Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp
210 215 220
Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg
225 230 235 240
His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr
245 250 255
Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys
260 265 270
Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser
275 280 285
Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg
290 295 300
Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys
305 310 315 320
Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val
325 330 335
His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg
340 345 350
Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser
355 360 365
Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys
370 375 380
Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu
385 390 395 400
Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu
405 410 415
Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg
420 425 430
His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr
435 440 445
Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys
450 455 460
Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln
465 470 475 480
Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr
485 490 495
Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln
500 505 510
Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val
515 520 525
Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala
530 535 540
Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu
545 550 555 560
Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu
565 570 575
Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr
580 585 590
Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile
595 600 605
Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu
610 615 620
Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys
625 630 635 640
Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala
645 650 655
Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala
660 665 670
Ala Ser Gln Ala Ala Leu Gly Leu
675 680
<210> 50
<211> 680
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 50
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp
20 25 30
Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala
35 40 45
Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala
50 55 60
Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala
65 70 75 80
Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Asp
85 90 95
Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu
100 105 110
Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln
115 120 125
Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe
130 135 140
Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser
145 150 155 160
Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg
165 170 175
Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu
180 185 190
Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro
195 200 205
Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp
210 215 220
Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg
225 230 235 240
His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr
245 250 255
Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys
260 265 270
Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser
275 280 285
Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg
290 295 300
Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys
305 310 315 320
Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val
325 330 335
His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg
340 345 350
Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser
355 360 365
Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys
370 375 380
Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu
385 390 395 400
Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu
405 410 415
Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg
420 425 430
His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr
435 440 445
Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys
450 455 460
Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln
465 470 475 480
Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr
485 490 495
Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln
500 505 510
Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val
515 520 525
Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala
530 535 540
Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu
545 550 555 560
Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu
565 570 575
Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr
580 585 590
Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile
595 600 605
Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu
610 615 620
Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys
625 630 635 640
Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala
645 650 655
Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala
660 665 670
Ala Ser Gln Ala Ala Leu Gly Leu
675 680
<210> 51
<211> 2103
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(2103)
<400> 51
acgcgtgcca cc atg gct ttc ttg tgg ctt ctc agt tgt tgg gcc ttg ttg 51
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu
1 5 10
ggg acc act ttc gga tct gca gag gac gat tct gag agg gat cct ttg 99
Gly Thr Thr Phe Gly Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu
15 20 25
aat gtc ctt aag ccg agg gcc cgc atg acg cct gca cca gca tcc tgc 147
Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys
30 35 40 45
agt caa gag ctc cct tca gca gaa gat aat agt cct atg gcg tcc gac 195
Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp
50 55 60
ccc ctg ggg gta gta cgc ggg ggg cga gtt aat acg cac gcc ggg gga 243
Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly
65 70 75
act ggt cct gaa ggg tgc agg ccc ttt gcc aag ttt att tca gtg tca 291
Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Ser Val Ser
80 85 90
caa acc tct aaa ctg aca cgc gcc gag acg gtc ttt ccg gat gtt gat 339
Gln Thr Ser Lys Leu Thr Arg Ala Glu Thr Val Phe Pro Asp Val Asp
95 100 105
gga tca gat gct cat aaa tca gag gta gct cac cgc ttc aaa gac ctc 387
Gly Ser Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu
110 115 120 125
gga gag gag aac ttt aag gcc ctc gtc ctc atc gca ttt gca cag tat 435
Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr
130 135 140
ttg caa caa tgt ccc ttt gag gat cat gtc aaa ttg gta aat gaa gtt 483
Leu Gln Gln Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val
145 150 155
act gag ttc gcg aag aca tgc gtg gct gac gag tcc gca gaa aac tgc 531
Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys
160 165 170
gac aag tcc ctt cat aca ctg ttt ggt gat aaa ctg tgc act gtc gct 579
Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala
175 180 185
act ctt aga gaa aca tat gga gag atg gct gac tgt tgt gcg aag caa 627
Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln
190 195 200 205
gag cca gaa cgg aat gaa tgt ttc ctt caa cat aaa gat gat aat ccg 675
Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro
210 215 220
aat ttg cca cgc ctt gta cga cca gag gtg gat gtt atg tgc acc gcg 723
Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala
225 230 235
ttt cat gac aat gag gaa acg ttc ctg aaa aaa tat ttg tat gag att 771
Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile
240 245 250
gct cgg aga cat cct tac ttt tac gca ccc gag ctc ctg ttc ttc gct 819
Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala
255 260 265
aag aga tac aag gca gcc ttt acg gaa tgc tgt cag gcc gcg gat aag 867
Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys
270 275 280 285
gcc gcc tgc ttg ctc ccc aag ctg gac gag ctg cga gat gag ggc aag 915
Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys
290 295 300
gcg tcc tct gct aag caa cga ctc aaa tgc gcc agc ttg caa aag ttt 963
Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe
305 310 315
ggc gaa aga gcg ttt aag gcc tgg gca gtg gcg cgc ctg agt cag cgg 1011
Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg
320 325 330
ttc cct aag gcg gag ttt gcc gag gtc tca aaa ctg gtg aca gac ttg 1059
Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu
335 340 345
acg aag gta cat acc gag tgc tgc cac gga gat ctc ctg gag tgc gcg 1107
Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala
350 355 360 365
gat gat cgc gct gac ctc gcc aaa tat ata tgt gag aac cag gat tcc 1155
Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser
370 375 380
att tcc tca aaa ctt aaa gag tgc tgc gaa aaa ccc ctt ctt gag aag 1203
Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys
385 390 395
tca cac tgc att gca gag gtc gaa aac gac gaa atg cct gcc gat ctg 1251
Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu
400 405 410
cct tcc ctt gcg gct gac ttt gtg gag tct aag gac gtt tgc aaa aac 1299
Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn
415 420 425
tat gca gag gct aaa gac gta ttc ctg ggc atg ttt ctt tat gaa tac 1347
Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr
430 435 440 445
gcg cgg cga cat ccc gac tat tca gtc gta ctc ctg ctt cga ctg gcg 1395
Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala
450 455 460
aaa act tac gaa act act ctc gag aaa tgc tgt gct gcc gcc gac cct 1443
Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro
465 470 475
cat gag tgc tat gct aag gta ttt gat gaa ttc aaa ccg ttg gtt gag 1491
His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu
480 485 490
gaa ccg caa aac ttg atc aag cag aac tgt gaa ctt ttt gag cag ctg 1539
Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu
495 500 505
ggg gaa tat aaa ttc cag aat gca ttg ctt gta cga tac aca aag aaa 1587
Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys
510 515 520 525
gtt cca cag gtc tcc acc ccg aca ctt gtg gaa gtt agc agg aac ttg 1635
Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu
530 535 540
gga aag gtc ggc agc aag tgt tgt aag cac cct gaa gct aag cga atg 1683
Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met
545 550 555
cca tgc gcc gaa gat tac ctt tct gtg gtg ttg aac caa ctg tgc gtt 1731
Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val
560 565 570
ctt cac gaa aag acg ccc gta tct gat cgg gtg acg aaa tgc tgc aca 1779
Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr
575 580 585
gag tca ctt gtg aac aga cgc ccc tgc ttt agt gcg ctc gag gtg gac 1827
Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp
590 595 600 605
gaa acg tat gtc ccg aaa gag ttt aac gca gag aca ttc acg ttt cat 1875
Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His
610 615 620
gcg gac atc tgt acc ctc tct gaa aag gag aga cag atc aaa aag cag 1923
Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln
625 630 635
aca gcg ctg gtt gag ctt gtt aag cat aag ccg aag gcg aca aag gag 1971
Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu
640 645 650
caa ctg aaa gcg gtg atg gat gac ttc gcc gcg ttc gtt gaa aaa tgc 2019
Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys
655 660 665
tgc aaa gcc gac gat aag gaa acc tgt ttt gct gaa gaa ggt aaa aaa 2067
Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys
670 675 680 685
ctc gtt gct gcg agt cag gcc gcc ctg ggg ctg tga 2103
Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695
<210> 52
<211> 696
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 52
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val Leu
20 25 30
Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu
35 40 45
Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu Gly
50 55 60
Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly Pro
65 70 75 80
Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Ser Val Ser Gln Thr Ser
85 90 95
Lys Leu Thr Arg Ala Glu Thr Val Phe Pro Asp Val Asp Gly Ser Asp
100 105 110
Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu
115 120 125
Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln
130 135 140
Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe
145 150 155 160
Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser
165 170 175
Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg
180 185 190
Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu
195 200 205
Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro
210 215 220
Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp
225 230 235 240
Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg
245 250 255
His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr
260 265 270
Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys
275 280 285
Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser
290 295 300
Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg
305 310 315 320
Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys
325 330 335
Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val
340 345 350
His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg
355 360 365
Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser
370 375 380
Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys
385 390 395 400
Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu
405 410 415
Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu
420 425 430
Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg
435 440 445
His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr
450 455 460
Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys
465 470 475 480
Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln
485 490 495
Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr
500 505 510
Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln
515 520 525
Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val
530 535 540
Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala
545 550 555 560
Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu
565 570 575
Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu
580 585 590
Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr
595 600 605
Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile
610 615 620
Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu
625 630 635 640
Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys
645 650 655
Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala
660 665 670
Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala
675 680 685
Ala Ser Gln Ala Ala Leu Gly Leu
690 695
<210> 53
<211> 696
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 53
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val Leu
20 25 30
Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu
35 40 45
Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu Gly
50 55 60
Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly Pro
65 70 75 80
Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Ser Val Ser Gln Thr Ser
85 90 95
Lys Leu Thr Arg Ala Glu Thr Val Phe Pro Asp Val Asp Gly Ser Asp
100 105 110
Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu
115 120 125
Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln
130 135 140
Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe
145 150 155 160
Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser
165 170 175
Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg
180 185 190
Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu
195 200 205
Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro
210 215 220
Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp
225 230 235 240
Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg
245 250 255
His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr
260 265 270
Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys
275 280 285
Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser
290 295 300
Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg
305 310 315 320
Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys
325 330 335
Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val
340 345 350
His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg
355 360 365
Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser
370 375 380
Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys
385 390 395 400
Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu
405 410 415
Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu
420 425 430
Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg
435 440 445
His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr
450 455 460
Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys
465 470 475 480
Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln
485 490 495
Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr
500 505 510
Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln
515 520 525
Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val
530 535 540
Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala
545 550 555 560
Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu
565 570 575
Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu
580 585 590
Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr
595 600 605
Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile
610 615 620
Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu
625 630 635 640
Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys
645 650 655
Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala
660 665 670
Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala
675 680 685
Ala Ser Gln Ala Ala Leu Gly Leu
690 695
<210> 54
<211> 2040
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(2040)
<400> 54
acgcgtgcca cc atg gct ttc ttg tgg ctt ctc agt tgt tgg gcc ttg ttg 51
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu
1 5 10
ggg acc act ttc gga tct gca gag gac gat tct gag agg gat cct ttg 99
Gly Thr Thr Phe Gly Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu
15 20 25
aat gtc ctt aag ccg agg gcc cgc atg acg cct gca cca gca tcc tgc 147
Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys
30 35 40 45
agt caa gag ctc cct tca gca gaa gat aat agt cct atg gcg tcc gac 195
Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp
50 55 60
ccc ctg ggg gta gta cgc ggg ggg cga gtt aat acg cac gcc ggg gga 243
Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly
65 70 75
act ggt cct gaa ggg tgc agg ccc ttt gcc aag ttt att gat gct cat 291
Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Asp Ala His
80 85 90
aaa tca gag gta gct cac cgc ttc aaa gac ctc gga gag gag aac ttt 339
Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe
95 100 105
aag gcc ctc gtc ctc atc gca ttt gca cag tat ttg caa caa tgt ccc 387
Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Cys Pro
110 115 120 125
ttt gag gat cat gtc aaa ttg gta aat gaa gtt act gag ttc gcg aag 435
Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys
130 135 140
aca tgc gtg gct gac gag tcc gca gaa aac tgc gac aag tcc ctt cat 483
Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His
145 150 155
aca ctg ttt ggt gat aaa ctg tgc act gtc gct act ctt aga gaa aca 531
Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr
160 165 170
tat gga gag atg gct gac tgt tgt gcg aag caa gag cca gaa cgg aat 579
Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn
175 180 185
gaa tgt ttc ctt caa cat aaa gat gat aat ccg aat ttg cca cgc ctt 627
Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu
190 195 200 205
gta cga cca gag gtg gat gtt atg tgc acc gcg ttt cat gac aat gag 675
Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn Glu
210 215 220
gaa acg ttc ctg aaa aaa tat ttg tat gag att gct cgg aga cat cct 723
Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro
225 230 235
tac ttt tac gca ccc gag ctc ctg ttc ttc gct aag aga tac aag gca 771
Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala
240 245 250
gcc ttt acg gaa tgc tgt cag gcc gcg gat aag gcc gcc tgc ttg ctc 819
Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu
255 260 265
ccc aag ctg gac gag ctg cga gat gag ggc aag gcg tcc tct gct aag 867
Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys
270 275 280 285
caa cga ctc aaa tgc gcc agc ttg caa aag ttt ggc gaa aga gcg ttt 915
Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe
290 295 300
aag gcc tgg gca gtg gcg cgc ctg agt cag cgg ttc cct aag gcg gag 963
Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu
305 310 315
ttt gcc gag gtc tca aaa ctg gtg aca gac ttg acg aag gta cat acc 1011
Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His Thr
320 325 330
gag tgc tgc cac gga gat ctc ctg gag tgc gcg gat gat cgc gct gac 1059
Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp
335 340 345
ctc gcc aaa tat ata tgt gag aac cag gat tcc att tcc tca aaa ctt 1107
Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu
350 355 360 365
aaa gag tgc tgc gaa aaa ccc ctt ctt gag aag tca cac tgc att gca 1155
Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala
370 375 380
gag gtc gaa aac gac gaa atg cct gcc gat ctg cct tcc ctt gcg gct 1203
Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala
385 390 395
gac ttt gtg gag tct aag gac gtt tgc aaa aac tat gca gag gct aaa 1251
Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys
400 405 410
gac gta ttc ctg ggc atg ttt ctt tat gaa tac gcg cgg cga cat ccc 1299
Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro
415 420 425
gac tat tca gtc gta ctc ctg ctt cga ctg gcg aaa act tac gaa act 1347
Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr
430 435 440 445
act ctc gag aaa tgc tgt gct gcc gcc gac cct cat gag tgc tat gct 1395
Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala
450 455 460
aag gta ttt gat gaa ttc aaa ccg ttg gtt gag gaa ccg caa aac ttg 1443
Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu
465 470 475
atc aag cag aac tgt gaa ctt ttt gag cag ctg ggg gaa tat aaa ttc 1491
Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe
480 485 490
cag aat gca ttg ctt gta cga tac aca aag aaa gtt cca cag gtc tcc 1539
Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser
495 500 505
acc ccg aca ctt gtg gaa gtt agc agg aac ttg gga aag gtc ggc agc 1587
Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser
510 515 520 525
aag tgt tgt aag cac cct gaa gct aag cga atg cca tgc gcc gaa gat 1635
Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp
530 535 540
tac ctt tct gtg gtg ttg aac caa ctg tgc gtt ctt cac gaa aag acg 1683
Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr
545 550 555
ccc gta tct gat cgg gtg acg aaa tgc tgc aca gag tca ctt gtg aac 1731
Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn
560 565 570
aga cgc ccc tgc ttt agt gcg ctc gag gtg gac gaa acg tat gtc ccg 1779
Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro
575 580 585
aaa gag ttt aac gca gag aca ttc acg ttt cat gcg gac atc tgt acc 1827
Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr
590 595 600 605
ctc tct gaa aag gag aga cag atc aaa aag cag aca gcg ctg gtt gag 1875
Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu
610 615 620
ctt gtt aag cat aag ccg aag gcg aca aag gag caa ctg aaa gcg gtg 1923
Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val
625 630 635
atg gat gac ttc gcc gcg ttc gtt gaa aaa tgc tgc aaa gcc gac gat 1971
Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp
640 645 650
aag gaa acc tgt ttt gct gaa gaa ggt aaa aaa ctc gtt gct gcg agt 2019
Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Ala Ser
655 660 665
cag gcc gcc ctg ggg ctg tga 2040
Gln Ala Ala Leu Gly Leu
670 675
<210> 55
<211> 675
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 55
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val Leu
20 25 30
Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu
35 40 45
Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu Gly
50 55 60
Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly Pro
65 70 75 80
Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Asp Ala His Lys Ser Glu
85 90 95
Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu
100 105 110
Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Cys Pro Phe Glu Asp
115 120 125
His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val
130 135 140
Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe
145 150 155 160
Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu
165 170 175
Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe
180 185 190
Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro
195 200 205
Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe
210 215 220
Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr
225 230 235 240
Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr
245 250 255
Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu
260 265 270
Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu
275 280 285
Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp
290 295 300
Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu
305 310 315 320
Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys
325 330 335
His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys
340 345 350
Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys
355 360 365
Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu
370 375 380
Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val
385 390 395 400
Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe
405 410 415
Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser
420 425 430
Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu
435 440 445
Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe
450 455 460
Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln
465 470 475 480
Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala
485 490 495
Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr
500 505 510
Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys
515 520 525
Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser
530 535 540
Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser
545 550 555 560
Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro
565 570 575
Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe
580 585 590
Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu
595 600 605
Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys
610 615 620
His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp
625 630 635 640
Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr
645 650 655
Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala
660 665 670
Leu Gly Leu
675
<210> 56
<211> 675
<212> PRT
<213> artificial sequence
<220>
<223> synthetic polypeptide
<400> 56
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Ser Ala Glu Asp Asp Ser Glu Arg Asp Pro Leu Asn Val Leu
20 25 30
Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala Ser Cys Ser Gln Glu
35 40 45
Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala Ser Asp Pro Leu Gly
50 55 60
Val Val Arg Gly Gly Arg Val Asn Thr His Ala Gly Gly Thr Gly Pro
65 70 75 80
Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Asp Ala His Lys Ser Glu
85 90 95
Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu
100 105 110
Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Cys Pro Phe Glu Asp
115 120 125
His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val
130 135 140
Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe
145 150 155 160
Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu
165 170 175
Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe
180 185 190
Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro
195 200 205
Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe
210 215 220
Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr
225 230 235 240
Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr
245 250 255
Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu
260 265 270
Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu
275 280 285
Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp
290 295 300
Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu
305 310 315 320
Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys
325 330 335
His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys
340 345 350
Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys
355 360 365
Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu
370 375 380
Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val
385 390 395 400
Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe
405 410 415
Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser
420 425 430
Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu
435 440 445
Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe
450 455 460
Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln
465 470 475 480
Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala
485 490 495
Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr
500 505 510
Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys
515 520 525
Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser
530 535 540
Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser
545 550 555 560
Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro
565 570 575
Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe
580 585 590
Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu
595 600 605
Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys
610 615 620
His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp
625 630 635 640
Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr
645 650 655
Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala
660 665 670
Leu Gly Leu
675
<210> 57
<211> 2118
<212> DNA
<213> artificial sequence
<220>
<223> synthetic polynucleotide
<220>
<221> CDS
<222> (13)..(2118)
<400> 57
acgcgtgcca cc atg gct ttc ttg tgg ctt ctc agt tgt tgg gcc ttg ttg 51
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu
1 5 10
ggg acc act ttc gga cga aga cac aca cgc tct gca gag gac gat tct 99
Gly Thr Thr Phe Gly Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser
15 20 25
gag agg gat cct ttg aat gtc ctt aag ccg agg gcc cgc atg acg cct 147
Glu Arg Asp Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro
30 35 40 45
gca cca gca tcc tgc agt caa gag ctc cct tca gca gaa gat aat agt 195
Ala Pro Ala Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser
50 55 60
cct atg gcg tcc gac ccc ctg ggg gta gta cgc ggg ggg cga gtt aat 243
Pro Met Ala Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn
65 70 75
acg cac gcc ggg gga act ggt cct gaa ggg tgc agg ccc ttt gcc aag 291
Thr His Ala Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys
80 85 90
ttt att tca gtg tca caa acc tct aaa ctg aca cgc gcc gag acg gtc 339
Phe Ile Ser Val Ser Gln Thr Ser Lys Leu Thr Arg Ala Glu Thr Val
95 100 105
ttt ccg gat gtt gat gga tca gat gct cat aaa tca gag gta gct cac 387
Phe Pro Asp Val Asp Gly Ser Asp Ala His Lys Ser Glu Val Ala His
110 115 120 125
cgc ttc aaa gac ctc gga gag gag aac ttt aag gcc ctc gtc ctc atc 435
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
130 135 140
gca ttt gca cag tat ttg caa caa tgt ccc ttt gag gat cat gtc aaa 483
Ala Phe Ala Gln Tyr Leu Gln Gln Cys Pro Phe Glu Asp His Val Lys
145 150 155
ttg gta aat gaa gtt act gag ttc gcg aag aca tgc gtg gct gac gag 531
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
160 165 170
tcc gca gaa aac tgc gac aag tcc ctt cat aca ctg ttt ggt gat aaa 579
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
175 180 185
ctg tgc act gtc gct act ctt aga gaa aca tat gga gag atg gct gac 627
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
190 195 200 205
tgt tgt gcg aag caa gag cca gaa cgg aat gaa tgt ttc ctt caa cat 675
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
210 215 220
aaa gat gat aat ccg aat ttg cca cgc ctt gta cga cca gag gtg gat 723
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
225 230 235
gtt atg tgc acc gcg ttt cat gac aat gag gaa acg ttc ctg aaa aaa 771
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
240 245 250
tat ttg tat gag att gct cgg aga cat cct tac ttt tac gca ccc gag 819
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
255 260 265
ctc ctg ttc ttc gct aag aga tac aag gca gcc ttt acg gaa tgc tgt 867
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
270 275 280 285
cag gcc gcg gat aag gcc gcc tgc ttg ctc ccc aag ctg gac gag ctg 915
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
290 295 300
cga gat gag ggc aag gcg tcc tct gct aag caa cga ctc aaa tgc gcc 963
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
305 310 315
agc ttg caa aag ttt ggc gaa aga gcg ttt aag gcc tgg gca gtg gcg 1011
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
320 325 330
cgc ctg agt cag cgg ttc cct aag gcg gag ttt gcc gag gtc tca aaa 1059
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
335 340 345
ctg gtg aca gac ttg acg aag gta cat acc gag tgc tgc cac gga gat 1107
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
350 355 360 365
ctc ctg gag tgc gcg gat gat cgc gct gac ctc gcc aaa tat ata tgt 1155
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
370 375 380
gag aac cag gat tcc att tcc tca aaa ctt aaa gag tgc tgc gaa aaa 1203
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
385 390 395
ccc ctt ctt gag aag tca cac tgc att gca gag gtc gaa aac gac gaa 1251
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
400 405 410
atg cct gcc gat ctg cct tcc ctt gcg gct gac ttt gtg gag tct aag 1299
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
415 420 425
gac gtt tgc aaa aac tat gca gag gct aaa gac gta ttc ctg ggc atg 1347
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
430 435 440 445
ttt ctt tat gaa tac gcg cgg cga cat ccc gac tat tca gtc gta ctc 1395
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
450 455 460
ctg ctt cga ctg gcg aaa act tac gaa act act ctc gag aaa tgc tgt 1443
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
465 470 475
gct gcc gcc gac cct cat gag tgc tat gct aag gta ttt gat gaa ttc 1491
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
480 485 490
aaa ccg ttg gtt gag gaa ccg caa aac ttg atc aag cag aac tgt gaa 1539
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
495 500 505
ctt ttt gag cag ctg ggg gaa tat aaa ttc cag aat gca ttg ctt gta 1587
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
510 515 520 525
cga tac aca aag aaa gtt cca cag gtc tcc acc ccg aca ctt gtg gaa 1635
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
530 535 540
gtt agc agg aac ttg gga aag gtc ggc agc aag tgt tgt aag cac cct 1683
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
545 550 555
gaa gct aag cga atg cca tgc gcc gaa gat tac ctt tct gtg gtg ttg 1731
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
560 565 570
aac caa ctg tgc gtt ctt cac gaa aag acg ccc gta tct gat cgg gtg 1779
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
575 580 585
acg aaa tgc tgc aca gag tca ctt gtg aac aga cgc ccc tgc ttt agt 1827
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
590 595 600 605
gcg ctc gag gtg gac gaa acg tat gtc ccg aaa gag ttt aac gca gag 1875
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
610 615 620
aca ttc acg ttt cat gcg gac atc tgt acc ctc tct gaa aag gag aga 1923
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
625 630 635
cag atc aaa aag cag aca gcg ctg gtt gag ctt gtt aag cat aag ccg 1971
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
640 645 650
aag gcg aca aag gag caa ctg aaa gcg gtg atg gat gac ttc gcc gcg 2019
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
655 660 665
ttc gtt gaa aaa tgc tgc aaa gcc gac gat aag gaa acc tgt ttt gct 2067
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
670 675 680 685
gaa gaa ggt aaa aaa ctc gtt gct gcg agt cag gcc gcc ctg ggg ctg 2115
Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
tga 2118
<210> 58
<211> 701
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 58
Met Ala Phe Leu Trp Leu Leu Ser Cys Trp Ala Leu Leu Gly Thr Thr
1 5 10 15
Phe Gly Arg Arg His Thr Arg Ser Ala Glu Asp Asp Ser Glu Arg Asp
20 25 30
Pro Leu Asn Val Leu Lys Pro Arg Ala Arg Met Thr Pro Ala Pro Ala
35 40 45
Ser Cys Ser Gln Glu Leu Pro Ser Ala Glu Asp Asn Ser Pro Met Ala
50 55 60
Ser Asp Pro Leu Gly Val Val Arg Gly Gly Arg Val Asn Thr His Ala
65 70 75 80
Gly Gly Thr Gly Pro Glu Gly Cys Arg Pro Phe Ala Lys Phe Ile Ser
85 90 95
Val Ser Gln Thr Ser Lys Leu Thr Arg Ala Glu Thr Val Phe Pro Asp
100 105 110
Val Asp Gly Ser Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys
115 120 125
Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala
130 135 140
Gln Tyr Leu Gln Gln Cys Pro Phe Glu Asp His Val Lys Leu Val Asn
145 150 155 160
Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu
165 170 175
Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr
180 185 190
Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala
195 200 205
Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp
210 215 220
Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys
225 230 235 240
Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr
245 250 255
Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe
260 265 270
Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala
275 280 285
Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu
290 295 300
Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln
305 310 315 320
Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser
325 330 335
Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr
340 345 350
Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu
355 360 365
Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln
370 375 380
Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu
385 390 395 400
Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala
405 410 415
Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys
420 425 430
Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr
435 440 445
Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg
450 455 460
Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala
465 470 475 480
Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu
485 490 495
Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu
500 505 510
Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr
515 520 525
Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg
530 535 540
Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys
545 550 555 560
Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu
565 570 575
Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys
580 585 590
Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu
595 600 605
Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr
610 615 620
Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys
625 630 635 640
Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr
645 650 655
Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu
660 665 670
Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly
675 680 685
Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
690 695 700
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2020 |
|
RU2811752C2 |
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И ВЕКТОРЫ ДЛЯ ГЕНОТЕРАПИИ ДЛЯ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ВИЛЬСОНА И ДРУГИХ СОСТОЯНИЙ | 2015 |
|
RU2745567C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ ПОЧЕЧНЫХ КЛЕТОК-ПРЕДШЕСТВЕННИКОВ | 2016 |
|
RU2730861C2 |
| Вакцина против герпеса | 2019 |
|
RU2731073C1 |
| СПОСОБ УПРАВЛЕНИЯ РИТМОМ СЕРДЦА И СОКРАЩЕНИЕМ ОТДЕЛЬНЫХ КАРДИОМИОЦИТОВ ПРИ ПОМОЩИ ТЕРМОГЕНЕТИКИ | 2022 |
|
RU2802995C1 |
| ИММУНОИНДУЦИРУЮЩЕЕ СРЕДСТВО | 2016 |
|
RU2744843C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДОТВРАЩЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2714205C2 |
| ПРОДУКТЫ И СПОСОБЫ ДЛЯ УХОДА ЗА ПОЛОСТЬЮ РТА, СОДЕРЖАЩИЕ HLP | 2016 |
|
RU2759134C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БИОЛОГИЧЕСКОГО ПОЛУЧЕНИЯ ЛАКТАТА ИЗ С1-СОЕДИНЕНИЙ С ПРИМЕНЕНИЕМ ТРАНСФОРМАНТОВ ЛАКТАТ ДЕГИДРОГЕНАЗЫ | 2014 |
|
RU2710714C2 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2802222C2 |

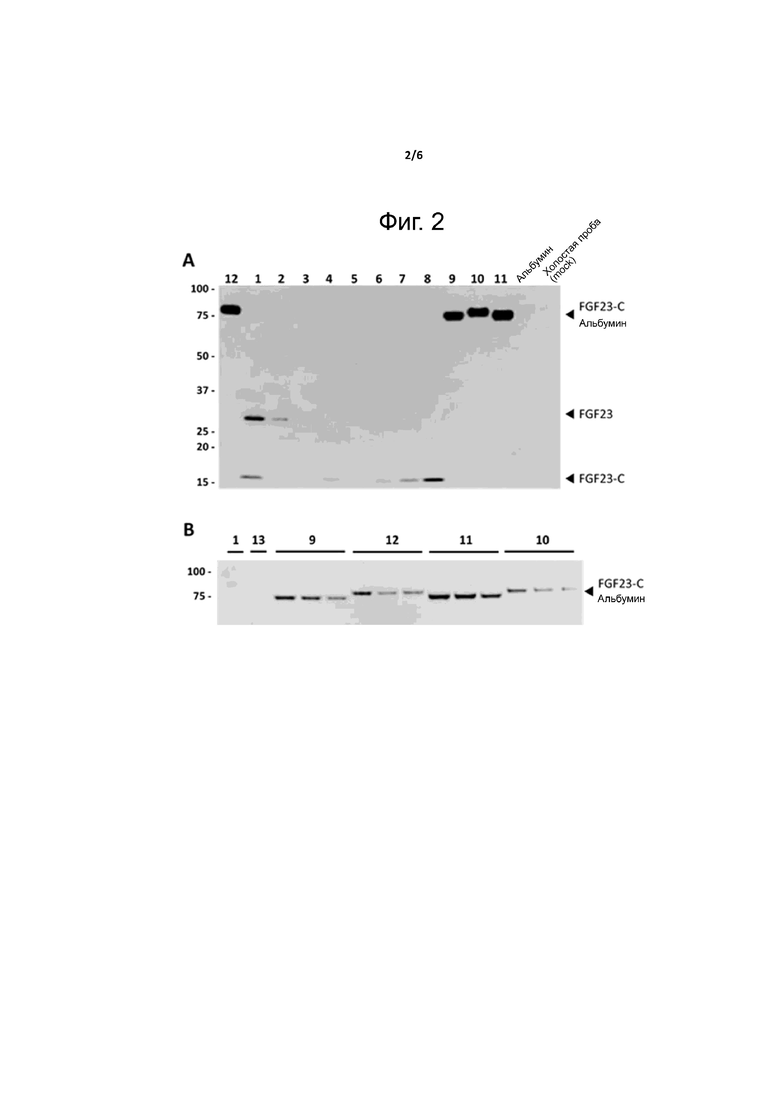
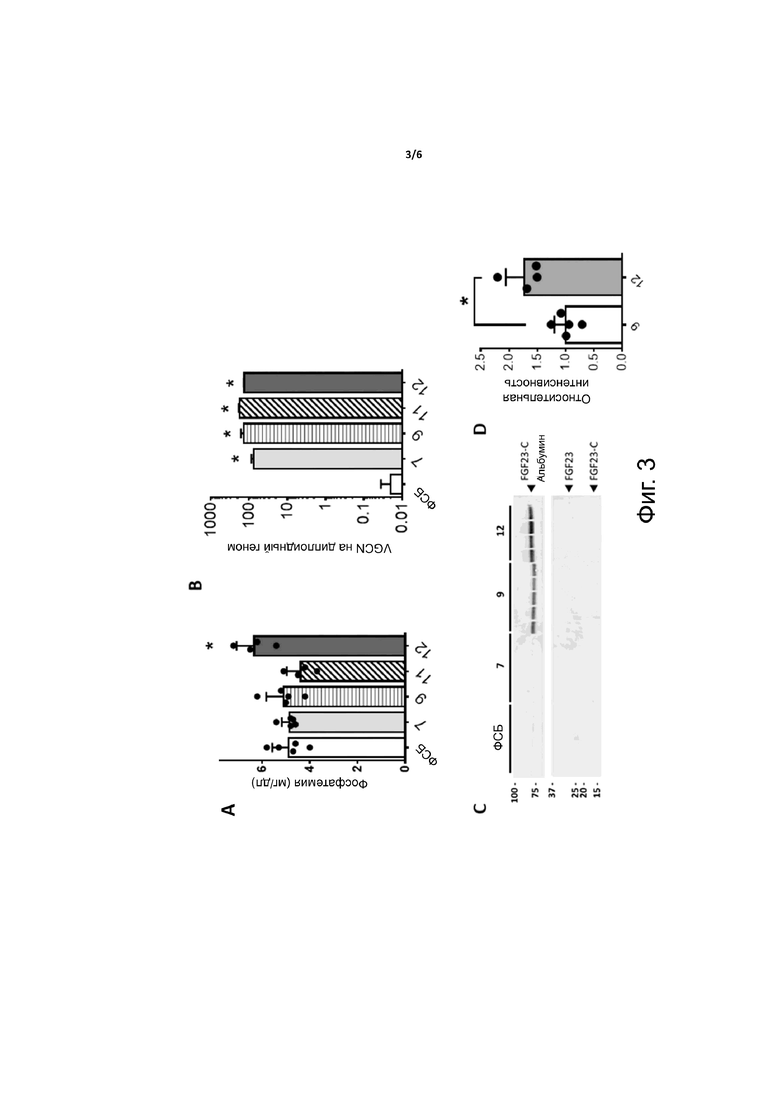
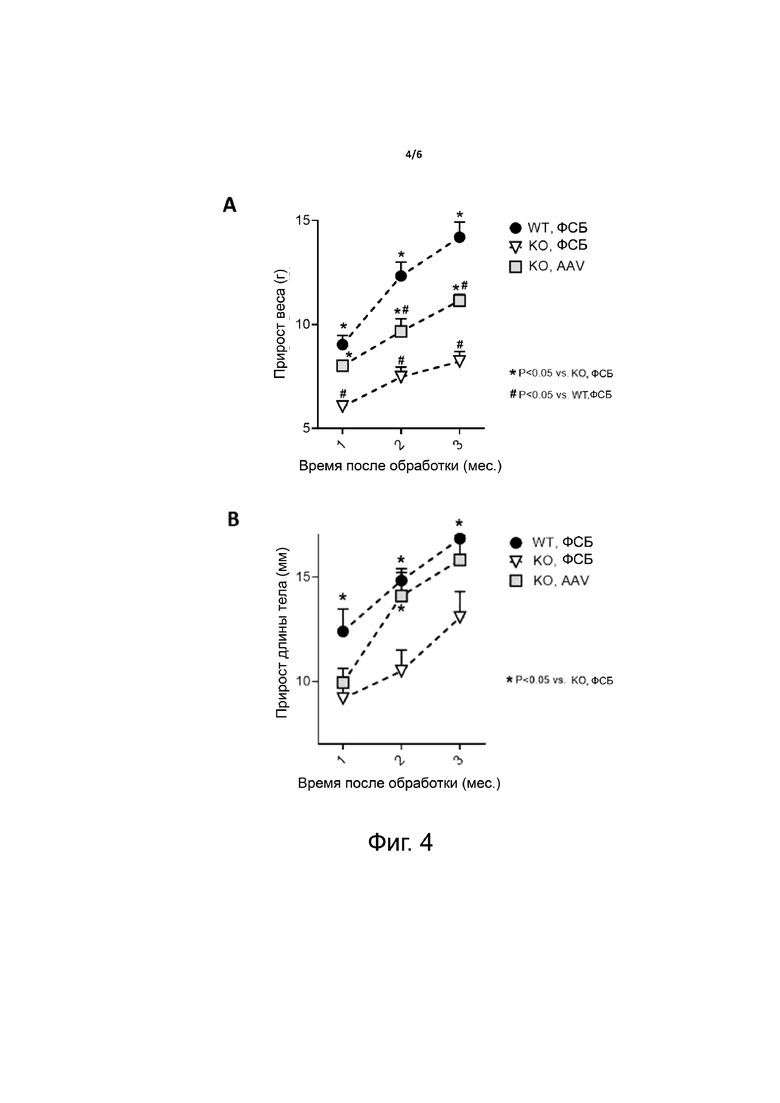
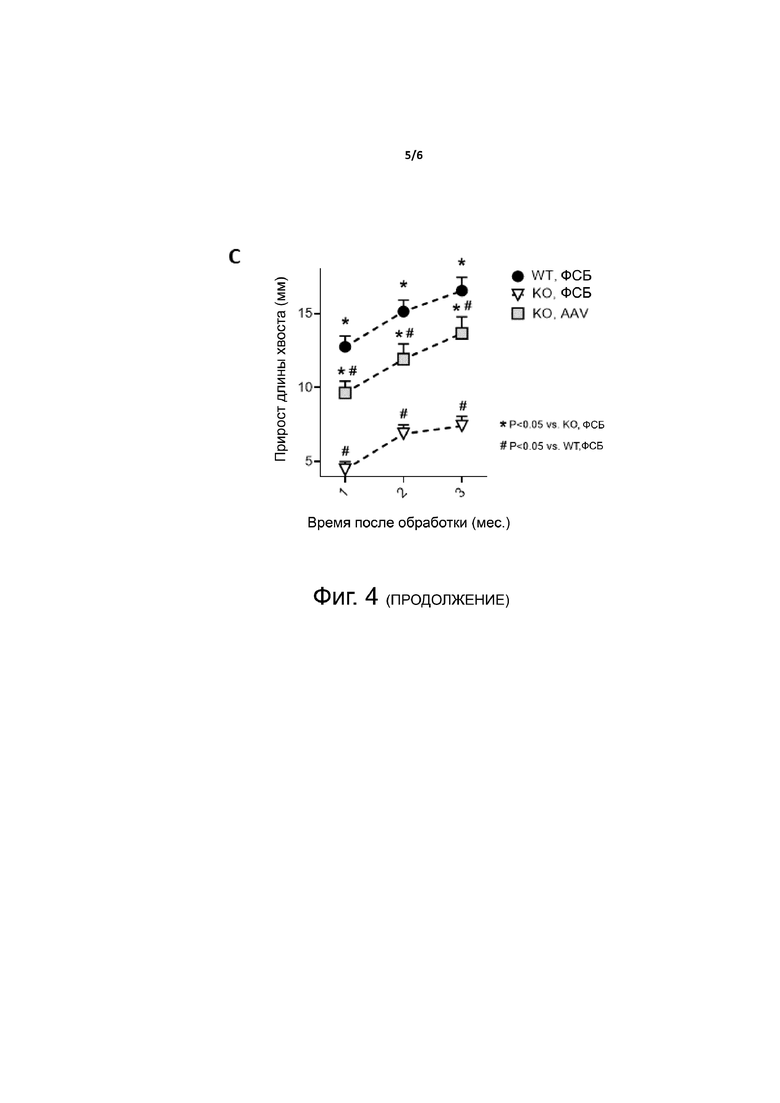
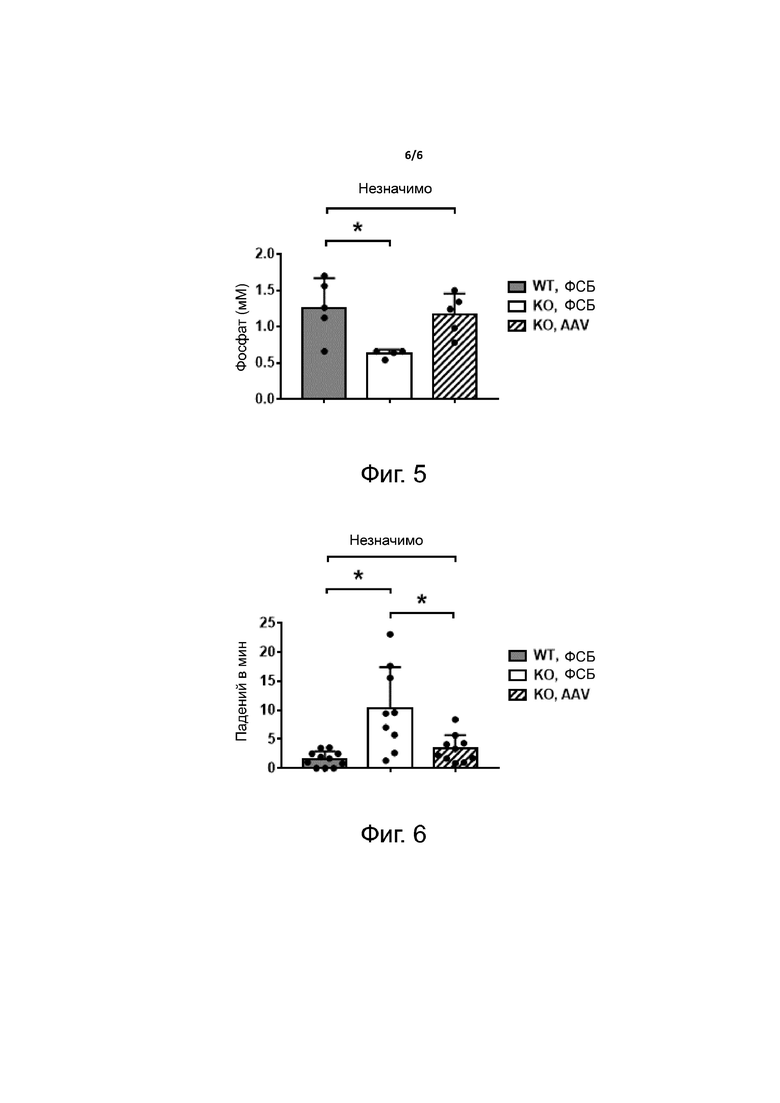
Группа изобретений относится к биотехнологии. Представлены: вектор, содержащий конструкцию нуклеиновой кислоты, клетка и фармацевтическая композиция для лечения связанных с FGF-23 гипофосфатемических заболеваний, в частности XLH, при помощи генной терапии. Изобретение может эффективно применяться для генной терапии связанных с FGF-23 гипофосфатемических заболеваний, в частности генной терапии, направленной на мышечную, печеночную или гемопоэтическую ткань, более конкретно, на ткань печени. 3 н. и 14 з.п. ф-лы, 1 табл., 6 ил., 1 пр.
1. Вектор для генной терапии связанных с FGF-23 гипофосфатемических заболеваний, включающий конструкцию нуклеиновой кислоты, экспрессируемую в целевых клетках или тканях индивидуума, где указанная конструкция нуклеиновой кислоты имеет нуклеотидную последовательность, кодирующую слитый белок FGF23, включающий:
(a) сигнальный пептид,
(b) C-концевой пептид FGF23, который связывается с комплексом FGFR/klotho,
(c) расщепляемый линкер, который расщепляется in vivo, и
(d) молекулу, стабилизирующую белок,
причем сигнальный пептид находится на N-конце слитого белка, а C-концевой пептид FGF23 и стабилизирующая белок молекула разделены расщепляемым линкером.
2. Вектор по п. 1, при этом C-концевой пептид FGF23 включает последовательность от какого-либо из положений 175-189 до какого-либо из положений 203-251 в SEQ ID NO: 1 или последовательность, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична указанной последовательности.
3. Вектор по п. 1 или 2, при этом C-концевой пептид FGF23 включает мотив RXXR в положениях 176-179 в SEQ ID NO: 1.
4. Вектор по любому из пп. 1-3, при этом C-концевой пептид FGF23 включает последовательность SEQ ID NO: 2 или последовательность, которая по меньшей мере на 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична указанной последовательности.
5. Вектор по любому из пп. 1-4, при этом сигнальный пептид включает последовательность, выбранную из группы, состоящей из SEQ ID NO: 3-8, предпочтительно SEQ ID NO: 7; и/или расщепляемый линкер включает последовательность SEQ ID NO: 10.
6. Вектор по любому из пп. 1-5, при этом стабилизирующая белок молекула представляет собой сывороточный альбумин человека, предпочтительно включающий последовательность SEQ ID NO: 9.
7. Вектор по любому из пп. 1-6, где конструкция нуклеиновой кислоты кодирует белок FGF23, включающий последовательность SEQ ID NO: 12 или 52 либо последовательность, которая по меньшей мере на 85%, 90%, 91%, 92%, 93%, 94%, 95%. 96%, 97%, 98% или 99% идентична какой-либо из указанных последовательностей.
8. Вектор по любому из пп. 1-7, где конструкция нуклеиновой кислоты оптимизирована по кодонам для экспрессии у человека.
9. Вектор по п. 7 или 8, где конструкция нуклеиновой кислоты включает последовательность SEQ ID NO: 13, 51 или 57 либо последовательность, которая по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична какой-либо из указанных последовательностей.
10. Вектор по любому из пп. 1-9, где конструкция нуклеиновой кислоты включает экспрессионную кассету, в которой кодирующая последовательность функционально связана по меньшей мере с одним промотором, который функционален в мышечных, печеночных или гемопоэтических клетках или тканях, в частности, специфичным для печени промотором, предпочтительно промотором α1-антитрипсина человека.
11. Вектор по п. 10, где конструкция нуклеиновой кислоты дополнительно включает один или несколько элементов контроля, выбранных из группы, состоящей из энхансеров, связанных с промотором, предпочтительно участок контроля ApoE человека; интронов, расположенных между промотором и кодирующей последовательностью, предпочтительно модифицированный интрон HBB2 SEQ ID NO: 17 или модифицированный интрон FIX из SEQ ID NO: 19; и сигналов терминации транскрипции, предпочтительно сигнал полиаденилирования бычьего гормона роста.
12. Вектор по любому из пп. 1-11, где конструкция нуклеиновой кислоты представляет собой ДНК или РНК.
13. Вектор по любому из пп. 1-12, который представляет собой вирусный вектор, в частности AAV или лентивирусный вектор, предпочтительно вектор AAV, содержащий капсид, выбранный из группы, состоящей из капсидов AAV1, AAV2, AAV5, AAV8, AAV2i8, AAV9, AAVrh10, AAVrh39, AAVrh43, AAVrh74, AAV-LK03, AAV2G9, AAV.PHP, AAV-Anc80, AAV3B и их химерных капсидов, более предпочтительно капсид AAV8.
14. Вектор по любому из пп. 1-12, который представляет собой частицу или везикулу, в частности, основанную на липидах микро- или нановезикулу либо частицу, предпочтительно содержащие конструкцию из РНК по п. 12.
15. Клетка для генной терапии связанных с FGF-23 гипофосфатемических заболеваний, которая генетически модифицирована вектором по любому из пп. 1-14, в частности мышечная, печеночная или гемопоэтическая клетка, предпочтительно клетка печени.
16. Фармацевтическая композиция для генной терапии связанных с FGF-23 гипофосфатемических заболеваний, содержащая по меньшей мере одно действующее вещество, выбранное из вектора по любому из пп. 1-14 или клетки по п. 15, и фармацевтически приемлемый носитель.
17. Фармацевтическая композиция по п. 16 для применения при лечении связанных с FGF-23 гипофосфатемических заболеваний при помощи генной терапии или клеточной терапии, причем связанное с FGF-23 гипофосфатемическое заболевание предпочтительно представляет собой генетическое заболевание, выбранное из группы, включающей: Х-сцепленную гипофосфатемию, аутосомно-доминантный гипофосфатемический рахит, аутосомно-рецессивный гипофосфатемический рахит 1, аутосомно-рецессивный гипофосфатемический рахит 2, остеоглофоническую дисплазию, метафизарную хондродисплазию янсеновского типа, гипофосфатемию с зубными аномалиями и эктопической кальцификацией, синдром МакКьюна-Олбрайта/фиброзную дисплазию и гипофосфатемию с поражением кожи и костей, или же приобретенное заболевание, выбранное из группы, включающей: онкогенную остеомаляцию, гипофосфатемическую остеомаляцию, осложнения при трансплантации почек или парентеральной терапии железом, хроническую болезнь почек и её осложнения типа гиперпаратиреоза; предпочтительно указанное заболевание представляет собой Х-сцепленную гипофосфатемию.
| WO 2015149069 A1, 01.10.2015 | |||
| FUKUMOTO S., Rickets/Osteomalacia | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| Пружинный двигатель с сдвоенными пружинными барабанами | 1924 |
|
SU1313A1 |
| QUARLES L | |||
| Darryl, FGF-23 and α-Klotho Co-Dependent and Independent Functions, Curr Opin Nephrol Hypertens, Jan 2019, 28(1), pp | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| ЕСАЯН А.М | |||
| и др., Фактор роста фибробластов | |||

