ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к антителам, которые связываются с ингибитором пути тканевого фактора (TFPI).
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Гемофилии А и В представляют собой связанные с Х хромосомой генетические расстройства, возникающие в результате функционального дефицита белков плазмы - Фактора VIII (FVIII) или Фактора IX (FIX), соответственно. Клиническая тяжесть гемофилии связана с остаточным уровнем активности фактора свертывания крови. Активность фактора меньше 1% ассоциирована с тяжелым фенотипом, умеренная гемофилия ассоциирована с активностью фактора 2%-5%, а легкая - с активностью фактора 5%-40%.
Стандартом лечения для этих расстройств является замена отсутствующего фактора свертывания крови посредством внутривенных инфузий. Заменяющий фактор обычно представляет собой рекомбинантный белок, такой как Xyntha (Фактор VIII) или BeneFIX (FIX), но продукты разной степени чистоты, полученные из плазмы, все еще используются. Лечение заменяющим фактором может быть либо эпизодическим, лечащим кровотечения по необходимости, по мере того, как они появляются, либо профилактическим, предупреждающим кровотечения посредством поддерживания уровней фактора в защитном интервале. Существуют важные доказательства того, что профилактическое лечение предупреждает кровотечения и ассоциированное повреждение суставов, которые являются главными клиническими проявлениями у пациентов с гемофилией. Эффективное профилактическое лечение требует внутривенной инъекции фактора 3-4 раза в неделю, что приводит к затруднениям в соблюдении схемы и режима лечения и пониженному качеству жизни. Стоимость лечения также является высокой
из-за сложности изготовления факторов свертывания крови. Кроме того, у значительного числа пациентов, вплоть до 32% пациентов с тяжелой гемофилией А, развиваются нейтрализующие антитела к введенным факторам, которые рассматриваются пациентами, которые имеют мутации в данных генах, как чужеродные белки. Для этих пациентов требуются альтернативные способы лечения, такие как обходной фактор - Фактор VIIa (NovoSeven).
Альтернативным подходом к терапии является обход потребности в заменяющих факторах посредством усиления интактного внешнего пути активации свертывания крови. Пациенты с гемофилией имеют некоторую способность к остановке кровотечений посредством их интактного внешнего пути активации свертывания крови; однако, это недостаточно для прекращения больших кровотечений или для предупреждения спонтанных кровотечений. Внешний путь активации свертываемости крови является недостаточным для обеспечения защиты, так как он быстро блокируется ингибитором пути тканевого фактора (TFPI).
Несмотря на то, что в WO 2010/017196 (Bayer), WO 2011/109452 (Bayer), WO 2014/144577 (Bayer), WO 2010/072687 (Novo Nordisk), WO 2012/001087 (Novo Nordisk), WO 2014/140240 (Novo Nordisk) и WO 2015/007880 (Novo Nordisk) раскрыты антитела, которые связываются с человеческим TFPI, в них не предожены антитела по изобретению, которые имеют характеристики, которые делают их новыми потенциальными терапевтическими средствами против гемофилии.
Продукт, который обеспечивал бы профилактическую защиту при уменьшении частоты дозирования факторов свертывания крови, уменьшая количество используемых факторов, обеспечивая альтернативные пути доставки (например, подкожный) и имея пониженный риск образования нейтрализующих антител, удовлетворял бы значительную нереализованную потребность для пациентов с гемофилией.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящем описании раскрыты и проиллюстрированы примерами антитела (и их антигенсвязывающие фрагменты), которые связываются с ингибитором пути тканевого фактора (TFPI).
Специалисты в этой области узнают или смогут установить с использованием не более чем рутинного экспериментирования многие эквиваленты описанных в настоящем описании конкретных воплощений изобретения. Подразумевается то, что такие эквиваленты охватываются следующими воплощениями (В).
В1. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с эпитопом в домене Кунитца 2 (K2) ингибитора пути тканевого фактора (TFPI), где указанный эпитоп содержит остатки Ile105, Arg107 и Leu131 согласно нумерации SEQ ID NO: 2.
В2. Антитело или его антигенсвязывающий фрагмент по В. 1, где указанное антитело или его антигенсвязывающий фрагмент не связывается с доменом Кунитца 1 (K1) TFPI.
В3. Антитело или его антигенсвязывающий фрагмент по В. 1 или 2, где указанный эпитоп дополнительно содержит один или более чем один остаток, выбранный из группы, состоящей из Cys106, Gly108, Cys130, Leu131 и Gly132 согласно нумерации SEQ ID NO: 2.
В4. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-3, где указанный эпитоп дополнительно содержит остатки Cys106, Gly108, Cys130, Leu131 и Gly132 согласно нумерации SEQ ID NO: 2.
В5. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4, где указанный эпитоп дополнительно содержит один или более чем один остаток, выбранный из группы, состоящей из Asp102, Arg112, Tyr127, Gly129, Met134 и Glu138 согласно нумерации SEQ ID NO: 2.
В6. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-5, где указанный эпитоп дополнительно содержит Asp102, Arg112, Tyr127, Gly129, Met134 и Glu138 согласно нумерации SEQ ID NO: 2.
В7. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-6, где указанный эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из E100, E101, P103, Y109, T111, Y113, F114, N116, Q118, Q121, C122, E123, R124, F125, K126 и L 140 согласно нумерации SEQ ID NO: 2.
В8. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-7, где указанный эпитоп не содержит E100, E101, P103, Y109, T111, Y113, F114, N116, Q118, Q121, C122, E123, R124, F125, K126 и L140 согласно нумерации SEQ ID NO: 2.
В9. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-6, где указанный эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из D31, D32, P34, C35, K36, E100, E101, P103, Y109, K126 и G128 согласно нумерации SEQ ID NO: 2.
В10. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-6 и 9, где указанный эпитоп не содержит D31, D32, P34, C35, K36, E100, E101, P103, Y109, K126 и G128 согласно нумерации SEQ ID NO: 2.
В11. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-10, где указанный эпитоп содержит один или более чем один остаток, выбранный из группы, состоящей из Asp102, Gly104, Ile105, Cys106, Arg107, Gly108, Arg112, Tyr127, Gly129, Cys130, Leu131, Gly132, Asn133, Met134 и Glu138 (согласно нумерации SEQ ID NO: 2), где указанный остаток эпитопа имеет ненулевое изменение площади скрытой поверхности (BSA) из-за взаимодействия с указанным антителом или его антигенсвязывающим фрагментом.
В12. Антитело или его антигенсвязывающий фрагмент по В. 11, где указанный эпитоп содержит Asp102, Gly104, Ile105, Cys106, Arg107, Gly108, Arg112, Tyr127, Gly129, Cys130, Leu131, Gly132, Asn133, Met134 и Glu138 (согласно нумерации SEQ ID NO: 2).
В13. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-12, где указанный эпитоп содержит один или более чем один остаток, выбранный из группы, состоящей из Asp102, Arg107, Arg 112, Tyr127 и Leu131 (согласно нумерации SEQ ID NO: 2), где указанный остаток эпитопа участвует в водородной связи с остатком из указанного антитела или его антигенсвязывающего фрагмента.
В14. Антитело или его антигенсвязывающий фрагмент по В. 13, где указанный эпитоп содержит Asp102, Arg107, Arg 112, Tyr127 и Leu131 (согласно нумерации SEQ ID NO: 2).
В15. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-14, где указанный эпитоп содержит один или более чем один контактный остаток, выбранный из группы, состоящей из Asp102, Gly104, Ile105, Cys106, Arg107, Gly108, Arg112, Tyr127, Gly129, Cys130, Leu131, Gly132, Met134 и Glu138 (согласно нумерации SEQ ID NO: 2).
В16. Антитело или его антигенсвязывающий фрагмент по В. 15, где указанный эпитоп содержит Asp102, Gly104, Ile105, Cys106, Arg107, Gly108, Arg112, Tyr127, Gly129, Cys130, Leu131, Gly132, Met134 и Glu138 (согласно нумерации SEQ ID NO: 2).
В17. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-16, содержащее следующие остатки паратопа тяжелой (о) цепи и легкой (L) цепи, которые имеют ненулевой изменение в BSA из-за взаимодействия с TFPI (нумерация согласно Kabat): H33 Ala, H58 Tyr, H95 Leu, H96 Gly, H97 Ala, H98 Thr, H99 Ser, H100 Leu, H100A Ser, L29 Ala, L31 Tyr, L91 Tyr, L95A Ser и L95B Gly.
В18. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-17, содержащее следующие контактные остатки (нумерация согласно Kabat): (a) H47, представляющий собой Trp или Tyr; (б) H58, представляющий собой Tyr; и (в) L91, представляющий собой Tyr или Arg; и возможно содержащее: (г) L96, представляющий собой Gly или Asn.
В19. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-18, содержащее следующие контактные остатки (нумерация согласно Kabat): (a) H33, представляющий собой Ala, Asn, Gly, His, Lys, Met, Phe, Pro, Ser, Thr, Trp или Val; (б) H47, представляющий собой Trp или Tyr; (в) H50, представляющий собой Ala, Arg, Gly, Lys, Met, Phe, Pro, Ser, Thr, Tyr или Val; (г) H51, представляющий собой Ile, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; (д) H52, представляющий собой Ser, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Trp, Tyr или Val; (е) H56, представляющий собой Ser, Arg, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; (ж) H58, представляющий собой Tyr; (з) H95, представляющий собой Leu, Gln, Ile, Phe или Tyr; (и) H96, представляющий собой Gly, Ala, Arg, Asn, Asp, Gln, Ile, Lys, Met, Phe, Pro, Ser, Thr или Val; (к) H97, представляющий собой Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; (л) H98, представляющий собой Thr, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; (м) H99, представляющий собой Ser, Ala, Gly, Phe или Pro; (н) H100, представляющий собой Leu, Arg, His, Ile, Leu, Lys, Phe, Pro, Trp, Tyr или Val; (о) H100A, представляющий собой Ser, Ala, Arg, Asn, Asp, Gln, Glu, His, Leu, Lys, Met, Phe, Pro, Ser, Thr или Trp; (п) L29, представляющий собой Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr or Trp, Tyr, Val; (р) L31, представляющий собой Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; (с) L91, представляющий собой Tyr или Arg; (т) L95A, представляющий собой Ser, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; (у) L95B, представляющий собой Ser, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; и (ф) L95C, представляющий собой Ser, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; и возможно содержащее следующие остатки: (х) L93, представляющий собой Tyr, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; и (ц) L96, представляющий собой Gly или Asn.
В20. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-18, содержащее следующие контактные остатки (нумерация согласно Kabat): (a) H33, представляющий собой Ala или Val; (б) H47, представляющий собой Trp; (в) H50, представляющий собой Ala; (г) H51, представляющий собой Ile; (д) H52, представляющий собой Ser, Arg, Lys, Phe или Tyr; (е) H56, представляющий собой Ser, Arg или Lys; (ж) H58, представляющий собой Tyr; (з) H95, представляющий собой Leu; (и) H96, представляющий собой Gly, Ala, Arg, Asn, Lys, Pro, Ser или Val; (к) H97, представляющий собой Ala; (л) H98, представляющий собой Thr, His, Ile, Leu, Met, Phe или Tyr; (м) H99, представляющий собой Ser; (н) H100, представляющий собой Leu, Phe, Trp или Tyr; (о) H100A, представляющий собой Ser, Arg, Asn, Gln, Glu, His, Leu, Lys, Met, Phe, Pro или Trp; (п) L29, представляющий собой Ala; (р) L31, представляющий собой Tyr; (с) L91, представляющий собой Tyr; (т) L95A, представляющий собой Ser, Phe, Trp или Tyr; (у) L95B, представляющий собой Gly; и (ф) L95C, представляющий собой Ser, Arg, Asn, Gln, Glu, Ile, Leu, Lys, Met, Phe, Trp, Tyr или Val; и возможно содержащее следующие остатки: (х) L93, представляющий собой Ser; и (ц) L96, представляющий собой Gly.
В21. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-18, содержащее следующие контактные остатки (нумерация согласно Kabat): (a) H33, представляющий собой Ala, Val, His или Phe; (б) H47, представляющий собой Trp или Tyr; (в) H50, представляющий собой Ala, Thr, Ser или Phe; (г) H51, представляющий собой Ile, Arg, Lys или Pro; (д) H52, представляющий собой Ser, Phe, Arg или Tyr; (е) H56, представляющий собой Ser, Lys, Tyr или Phe; (ж) H58, представляющий собой Tyr; (з) H95, представляющий собой Leu, Ile, Gln или Phe; (и) H96, представляющий собой Gly, Arg, Asn или Lys; (к) H97, представляющий собой Ala, Leu, Tyr или Ile; (л) H98, представляющий собой Thr, Tyr, Phe или His; (м) H99, представляющий собой Ser, Pro, Ala или Phe; (н) H100, представляющий собой Leu, Tyr, Trp или Phe; (о) H100A, представляющий собой Ser, Arg, Leu или Trp; (п) L29, представляющий собой Ala, Glu, Asp или Gln; (р) L31, представляющий собой Tyr, Glu, Asp или Trp; (с) L91, представляющий собой Tyr или Arg; (т) L95A, представляющий собой Ser, Phe, Tyr или His; (у) L95B, представляющий собой Gly, Glu, Asp или Pro; и (ф) L95C, представляющий собой Ser, Trp, Tyr или Phe; и возможно содержащее следующие остатки: (х) L93, представляющий собой Ser, Glu, Asp или His; и (ц) L96, представляющий собой Gly или Asn.
В22. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-18, содержащее следующие контактные остатки (нумерация согласно Kabat): H33 Ala, H47 Trp, H50 Ala, H51 Ile, H52 Ser, H56 Ser, H58 Tyr, H95 Leu, H96 Gly, H97 Ala, H98 Thr, H99 Ser, H100 Leu, H100A Ser, L29 Ala, L31 Tyr, L91 Tyr, L95A Ser, L95B Gly и L95C Ser; и возможно содержащее следующие остатки: L93 Ser и L96 Gly.
В23. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-22, содержащее вариабельную область тяжелой цепи (VH), которая содержит:
(а) определяющую комплементарность область один VH (CDR-H1), содержащую аминокислотную последовательность SEQ ID NO: 38.
(б) определяющую комплементарность область два VH (CDR-H2), содержащую аминокислотную последовательность SEQ ID NO: 39; и
(в) определяющую комплементарность область три VH (CDR-H3), содержащую аминокислотную последовательность SEQ ID NO: 40.
В24. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-22, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 41.
В25. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-24, содержащее последовательность человеческой каркасной области VH3.
В26. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-24, содержащее последовательность человеческой каркасной области VH1.
В27. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-24, содержащее последовательность человеческой каркасной области VH5.
В28. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-24, содержащее последовательность каркасной области VH человеческой зародышевой линии IGHV3-23 или IGHV1-69.
В29. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-24, содержащее последовательность каркасной области VH человеческой зародышевой линии IGHV3-7.
В30. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-24, содержащее консенсусную последовательность каркасной области VH человеческой зародышевой линии.
В31. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-30, содержащее VH, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 41, 63 и 65.
В32. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-31, содержащее VH, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 41, 63 и 65.
В33. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-32, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 41.
В34. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-32, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 63.
В35. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-32, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 65.
В36. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-35, содержащее вариабельную область легкой цепи (VL), которая содержит:
а) определяющую комплементарность область один VL (CDR-L1), содержащую аминокислотную последовательность SEQ ID NO: 33.
(б) определяющую комплементарность область два VL (CDR-L2), содержащую аминокислотную последовательность SEQ ID NO: 34; и
(в) определяющую комплементарность область три VL (CDR-L3), содержащую аминокислотную последовательность SEQ ID NO: 35.
В37. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-35, содержащее последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 36.
В38. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-37, содержащее последовательность человеческой каркасной области Vκ.
В39. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-37, содержащее последовательность человеческой каркасной области Vλ.
В40. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-37, содержащее последовательность каркасной области VL человеческой зародышевой линии IGKV3-20.
В41. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-37, содержащее последовательность каркасной области VL человеческой зародышевой линии IGKV1-39.
В42. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-37, содержащее консенсусную последовательность каркасной области VL человеческой зародышевой линии.
В43. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-42, содержащее VL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 36.
В44. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-43, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO:36.
В45. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-44, содержащее константную область тяжелой цепи (СН), которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 20.
В46. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-45, содержащее СН, которая содержит аминокислотную последовательность SEQ ID NO: 20.
В47. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-46, содержащее константную область легкой цепи (СL), которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 26.
В48. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-47, содержащее СL, которая содержит аминокислотную последовательность SEQ ID NO: 26.
В49. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-48, содержащее домен Fc.
В50. Антитело или его антигенсвязывающий фрагмент по В. 49, в котором указанный домен Fc представляет собой домен Fc IgA.
B51. Антитело или его антигенсвязывающий фрагмент по В. 50, в котором указанный IgA представляет собой IgA1 или IgA2.
В52. Антитело или его антигенсвязывающий фрагмент по В. 49, в котором указанный домен Fc представляет собой домен Fc IgD.
В53. Антитело или его антигенсвязывающий фрагмент по В. 49, в котором указанный домен Fc представляет собой домен Fc IgЕ.
В54. Антитело или его антигенсвязывающий фрагмент по В. 49, в котором указанный домен Fc представляет собой домен Fc IgМ.
В55. Антитело или его антигенсвязывающий фрагмент по В. 49, в котором указанный домен Fc представляет собой домен Fc IgG.
В56. Антитело или его антигенсвязывающий фрагмент по В. 55, где указанный IgG представляет собой IgG1, IgG2, IgG3 или IgG4.
В57. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-56, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 42.
В58. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-56, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 64.
В59. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-56, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 66.
В60. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-59, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 37.
В61. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-60, содержащее последовательность VH, кодируемую вставкой, присутствующей в плазмиде, депонированной под № АТСС (Американская коллекция типовых культур) РТА-122329.
В62. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-61, содержащее последовательность VL, кодируемую вставкой, присутствующей в плазмиде, депонированной под № АТСС РТА-122328.
В63. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4, где указанный эпитоп дополнительно содержит один или более чем один остаток, выбранный из группы, состоящей из: Glu100, Glu101, Asp102, Gly104 и Tyr109, согласно нумерации SEQ ID NO: 2.
В64. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63, где указанный эпитоп дополнительно содержит Glu100, Glu101, Asp102, Gly104 и Tyr109 согласно нумерации SEQ ID NO: 2.
В65. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-64, где указанный эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из: P103, T111, Y113, F114, N116, Q118, Q121, C122, E123, R124, F125, K126 и L140 (нумерация согласно SEQ ID NO: 2).
В66. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-65, где указанный эпитоп не содержит: P103, T111, Y113, F114, N116, Q118, Q121, C122, E123, R124, F125, K126 и L140 (нумерация согласно SEQ ID NO: 2).
В67. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-64, где указанный эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из: D31, D32, P34, C35, K36, P103, K126, Y127, G128 (нумерация согласно SEQ ID NO: 2).
В68. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4, 63-64 и 67, где указанный эпитоп не содержит: D31, D32, P34, C35, K36, P103, K126, Y127, G128 (нумерация согласно SEQ ID NO: 2).
В69. Антитело или его антигенсвязывающий фрагмент по любому из вв. 1-4 и 63-68, содержащее следующие остатки (нумерация согласно Kabat): H33 Ala, H35 Gln, H52 Ser, H53 Asn, H55 Arg, H56 Ser, H95 Phe, H96 Leu, H97 His, H99 Ser, H101 Asp, L31 Met, L32 Tyr, L34 His, L36 Tyr, L50 Arg, L91 Trp и L96 Tyr.
В70. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-69, содержащее VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 48.
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 49; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 50.
В71. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-69, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 51.
В72. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-71, содержащее человеческую последовательность каркасной области VH3, VH1 или VH5.
В73. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-72, содержащее последовательность каркасной области VH человеческой зародышевой линии IGHV3-23 или IGHV1-69.
В74. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-72, содержащее последовательность каркасной области VH человеческой зародышевой линии IGHV3-7.
В75. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-71, содержащее консенсусную последовательность каркасной области VH человеческой зародышевой линии.
В76. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-75, содержащее VH, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 67, 69, 51 и 79.
В77. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-76, содержащее VH, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 67, 69, 51 и 79.
В78. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-77, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 67.
В79. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-77, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 69.
В80. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-77, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 51.
В81. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-77, содержащее вариабельную область тяжелой цепи (VH), которая содержит аминокислотную последовательность SEQ ID NO: 79.
В82. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-81, содержащее VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 43.
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 44; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 45.
В83. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-81, содержащее последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 46.
В84. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-83, содержащее последовательность человеческой каркасной области Vκ или Vλ.
В85. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-84, содержащее последовательность каркасной области VL человеческой зародышевой линии IGKV3-20 или IGKV1-39.
В86. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-84, содержащее консенсусную последовательность каркасной области VL человеческой зародышевой линии.
В87. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-86, содержащее VL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную последовательности, выбранной из группы, состоящей из SEQ ID NO: 46, 71, 73, 75 и 77.
В88. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-87, содержащее VL, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 46, 71, 73, 75 и 77.
В89. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-88, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 46.
В90. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-88, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 71.
В91. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-88, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 73.
В92. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-88, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 75.
В93. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-88, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 77.
В94. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-93, содержащее СН, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 20.
В95. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-94, содержащее СН, которая содержит аминокислотную последовательность SEQ ID NO: 20.
В96. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-95, содержащее СL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 26.
В97. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-96, содержащее СL, которая содержит аминокислотную последовательность SEQ ID NO: 26.
В98. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-97, содержащее домен Fc.
В99. Антитело или его антигенсвязывающий фрагмент по В. 98, в котором указанный домен Fc представляет собой домен Fc IgA.
В100. Антитело или его антигенсвязывающий фрагмент по В. 99, где указанный IgA представляет собой IgA1 или IgA2.
В101. Антитело или его антигенсвязывающий фрагмент по В. 98, в котором указанный домен Fc представляет собой домен Fc IgD, IgE или IgM.
В102. Антитело или его антигенсвязывающий фрагмент по В. 98, в котором указанный домен Fc представляет собой домен Fc IgG.
В103. Антитело или его антигенсвязывающий фрагмент по ВВ. 102, где указанный IgG представляет собой IgG1, IgG2, IgG3 или IgG4.
В104. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-103, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 52.
В105. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-103, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 68.
В106. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-103, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 70.
В107. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-103, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 80.
В108. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-107, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 47.
В109. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-107, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 72.
В110. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-107, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 74.
В111. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-107, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 76.
В112. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-4 и 63-107, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 78.
В113. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с эпитопом в домене Кунитца 2 (K2) ингибитора пути тканевого фактора (TFPI), где указанный эпитоп содержит остатки Glu101, Pro103, Tyr109, Thr111, Ser119, Gln121, Glu123, Arg124, Lys126 и Leu140 согласно нумерации SEQ ID NO: 2.
В114. Антитело или его антигенсвязывающий фрагмент по В. 113, где указанное антитело или его антигенсвязывающий фрагмент не связывается с доменом Кунитца 1 (K1) TFPI.
В115. Антитело или его антигенсвязывающий фрагмент по В. 113 или 114, где указанный эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из E100, D102, R107, Y113, F114, N116, Q118 и C122 (нумерация согласно SEQ ID NO: 2).
В116. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-115, где указанный эпитоп не содержит: E100, D102, R107, Y113, F114, N116, Q118 и C122 (нумерация согласно SEQ ID NO: 2).
В117. Антитело или его антигенсвязывающий фрагмент по В. 113 или 114, где указанный эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из D31, D32, P34, C35, K36, E100, I105, R107, G108, Y127 и G128 (нумерация согласно SEQ ID NO: 2).
В118. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-114 и 117, где указанный эпитоп не содержит: D31, D32, P34, C35, K36, E100, I105, R107, G108, Y127 и G128 (нумерация согласно SEQ ID NO: 2).
В119. Антитело или его антигенсвязывающий фрагмент по ВВ. 113-118, содержащее следующие остатки (согласно нумерации Kabat): H50 Asp, H57 Thr, H58 Leu, H59 Tyr, H61 Gln, H98 Asp, H99 Tyr, H100 Asp, L30 His, L50 Trp, L92 Tyr, L93 Thr, L94 Thr и L96 Tyr.
В120. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-119, содержащее VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 87;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 88; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 89.
В121. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-119, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 90.
В122. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-121, содержащее последовательность человеческой каркасной области VH3, VH1 или VH5.
В123. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-122, содержащее последовательность каркасной области VH человеческой зародышевой линии IGHV3-23, IGHV1-69 или IGHV3-7.
В124. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-121, содержащее консенсусную последовательность каркасной области VH человеческой зародышевой линии.
В125. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-124, содержащее VH, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 90, 95, 97, 99, 101, 103, 105 и 107.
В126. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-125, содержащее VH, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 90, 95, 97, 99, 101, 103, 105 и 107.
В127. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-126, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 90.
В128. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-126, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 95.
В129. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-126, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 97.
В130. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-126, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 99.
В131. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-126, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 101.
В132. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-126, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 103.
В133. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-126, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 105.
В134. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-126, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 107.
В135. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-134, содержащее VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 81;
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 82; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 83.
В136. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-134, содержащее последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 84.
В137. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-136, содержащее последовательность человеческой каркасной области Vκ или Vλ.
В138. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-137, содержащее последовательность каркасной области VL человеческой зародышевой линии IGKV3-20 или IGKV1-39.
В139. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-136, содержащее консенсусную последовательность каркасной области VL человеческой зародышевой линии.
В140. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-139, содержащее VL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 84, 109 и 111.
В141. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-140, содержащее VL, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 84, 109 и 111.
В142. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-141, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 84.
В143. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-141, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 109.
В144. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-141, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 111.
В145. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-144, содержащее СН, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 20.
В146. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-145, содержащее СН, которая содержит аминокислотную последовательность SEQ ID NO: 20.
В147. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-144, содержащее СН, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 91.
В148. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-144 и 147, содержащее СН, которая содержит аминокислотную последовательность SEQ ID NO: 91.
В149. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-148, содержащее СL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 14.
В150. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-149, содержащее СL, которая содержит аминокислотную последовательность SEQ ID NO: 14.
В151. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-148, содержащее СL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 85.
В152. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-148 и 151, содержащее СL, которая содержит аминокислотную последовательность SEQ ID NO: 85.
В153. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-152, содержащее домен Fc.
В154. Антитело или его антигенсвязывающий фрагмент по В. 153, в котором указанный домен Fc представляет собой домен Fc IgA (например, IgA1 или IgA2).
В155. Антитело или его антигенсвязывающий фрагмент по В. 153, в котором указанный домен Fc представляет собой домен Fc IgD, IgE или IgM.
В156. Антитело или его антигенсвязывающий фрагмент по В. 153, в котором указанный домен Fc представляет собой домен Fc IgG.
В157. Антитело или его антигенсвязывающий фрагмент по В. 156, где указанный IgG представляет собой IgG1, IgG2, IgG3 или IgG4.
В158. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-157, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 92.
В159. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-157, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 94.
В160. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-157, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 96.
В161. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-157, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 98.
В162. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-157, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 100.
В163. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-157, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 102.
В164. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-157, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 104.
В165. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-157, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 106.
В166. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-157, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 108.
В167. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-166, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 86.
В168. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-166, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 93.
В169. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-166, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 110.
В170. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 113-166, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 112.
В171. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом Кунитца 2 (K2) TFPI, содержащее VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 16;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18.
В172. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 19.
В173. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 10.
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 11; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 12.
В174. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 13.
В175. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее:
(1) VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 16;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 17; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 18;
и (2) VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 10;
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 11; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 12.
В176. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 19, и последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 13.
В177. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-176, содержащее человеческие последовательности каркасной области VH3, VH1 или VH5.
В178. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-177, содержащее последовательность каркасной области VH человеческой зародышевой линии IGHV3-23, IGHV1-69 или IGHV3-7.
В179. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-176, содержащее консенсусную последовательность каркасной области VH человеческой зародышевой линии.
В180. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-179, содержащее VH, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 19.
В181. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-180, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 19.
В182. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-181, содержащее последовательность человеческой каркасной области Vκ или Vλ.
В183. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-182, содержащее последовательность каркасной области VL человеческой зародышевой линии IGKV3-20 или IGKV1-39.
В184. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-181, содержащее консенсусную последовательность каркасной области VL человеческой зародышевой линии.
В185. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-184, содержащее VL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 13.
В186. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-185, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 13.
В187. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-186, содержащее СН, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 20.
В188. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-187, содержащее СН, которая содержит аминокислотную последовательность SEQ ID NO: 20.
В189. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-188, содержащее СL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 14.
В190. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-189, содержащее СL, которая содержит аминокислотную последовательность SEQ ID NO: 14.
В191. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-190, содержащее домен Fc.
В192. Антитело или его антигенсвязывающий фрагмент по В. 191, в котором указанный домен Fc представляет собой домен Fc IgA (например, IgA1 или IgA2), IgD, IgE или IgM.
В193. Антитело или его антигенсвязывающий фрагмент по В. 191, в котором указанный домен Fc представляет собой домен Fc IgG.
В194. Антитело или его антигенсвязывающий фрагмент по В. 193, где указанный IgG представляет собой IgG1, IgG2, IgG3 или IgG4.
В195. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-194, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 21.
В196. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 171-195, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 15.
В197. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 28;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 29; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 30.
В198. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 31.
В199. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 22;
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 23; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 24.
В200. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 25.
В201. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее:
(1) VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 28;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 29; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 30;
и (2) VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 22.
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 23; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 24.
В202. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 31, и последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 25.
В203. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-202, содержащее последовательность человеческой каркасной области VH3, VH1 или VH5.
В204. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-203, содержащее последовательность каркасной области VH человеческой зародышевой линии IGHV3-23, IGHV1-69 или IGHV3-7.
В205. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-202, содержащее консенсусную последовательность каркасной области VH человеческой зародышевой линии.
В206. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-205, содержащее VH, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 31.
В207. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-206, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 31.
В208. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-207, содержащее последовательность человеческой каркасной области Vκ или Vλ.
В209. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-208, содержащее последовательность каркасной области VL человеческой зародышевой линии IGKV3-20 или IGKV1-39.
В210. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-207, содержащее консенсусную последовательность каркасной области VL человеческой зародышевой линии.
В211. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-210, содержащее VL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 25.
В212. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-211, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 25.
В213. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-212, содержащее СН, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 20.
В214. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-213, содержащее СН, которая содержит аминокислотную последовательность SEQ ID NO: 20.
В215. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-214, содержащее СL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 26.
В216. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-215, содержащее СL, которая содержит аминокислотную последовательность SEQ ID NO: 26.
В217. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-216, содержащее домен Fc.
В218. Антитело или его антигенсвязывающий фрагмент по В. 217, в котором указанный домен Fc представляет собой домен Fc IgA (например, IgA1 или IgA2), IgD, IgE или IgM.
В219. Антитело или его антигенсвязывающий фрагмент по В. 217, в котором указанный домен Fc представляет собой домен Fc IgG (например, IgG1, IgG2, IgG3 или IgG4).
В220. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-219, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 32.
В221. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 197-220, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 27.
В222. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее вариабельную область тяжелой цепи (VH), которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 58;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 59; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 60.
В223. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 61.
В224. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 53;
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 54; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 55.
В225. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 56.
В226. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее:
(1) VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 58;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 59; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 60;
и (2) VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 53;
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 54; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 55.
В227. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 61, и последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 56.
В228. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-227, содержащее последовательность каркасной области человеческой VH3, VH1 или VH5.
В229. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-228, содержащее последовательность каркасной области VH человеческой зародышевой линии IGHV3-23, IGHV1-69 или IGHV3-7.
В230. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-227, содержащее консенсусную последовательность каркасной области VH человеческой зародышевой линии.
В231. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-230, содержащее VH, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 61.
В232. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-231, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 61.
В233. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-232, содержащее последовательность каркасной области человеческой Vκ или Vλ.
В234. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-233, содержащее последовательность каркасной области VL человеческой зародышевой линии IGKV3-20 или IGKV1-39.
В235. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-232, содержащее консенсусную последовательность каркасной области VL человеческой зародышевой линии.
В236. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-235, содержащее VL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 56.
В237. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-236, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 56.
В238. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-237, содержащее СН, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 20.
В239. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-238, содержащее СН, которая содержит аминокислотную последовательность SEQ ID NO: 20.
В240. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-239, содержащее СL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 26.
В241. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-240, содержащее СL, которая содержит аминокислотную последовательность SEQ ID NO: 26.
В242. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-241, содержащее домен Fc.
В243. Антитело или его антигенсвязывающий фрагмент по В. 242, в котором указанный домен Fc представляет собой домен Fc IgA (например, IgA1 или IgA2), IgD, IgE или IgM.
В244. Антитело или его антигенсвязывающий фрагмент по В. 242, в котором указанный домен Fc представляет собой домен Fc IgG (например, IgG1, IgG2, IgG3 или IgG4).
В245. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-244, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 62.
В246. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 222-245, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 57.
В247. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 118;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 119; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 120.
В248. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 121.
В249. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 113;
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 114; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 115.
В250. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 116.
В251. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее:
(1) VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 118;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 119; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 120;
и (2) VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 113;
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 114; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 115.
В252. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 121, и последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 116.
В253. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-252, содержащее последовательность каркасной области человеческой VH3, VH1 или VH5.
В254. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-253, содержащее последовательность каркасной области VH человеческой зародышевой линии IGHV3-23, IGHV1-69 или IGHV3-7.
В255. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-252, содержащее консенсусную последовательность каркасной области VH человеческой зародышевой линии.
В256. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-255, содержащее VH, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 121.
В257. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-256, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 121.
В258. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-257, содержащее последовательность каркасной области человеческой Vκ или Vλ.
В259. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-258, содержащее последовательность каркасной области VL человеческой зародышевой линии IGKV3-20 или IGKV1-39.
В260. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-257, содержащее консенсусную последовательность каркасной области VL человеческой зародышевой линии.
В261. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-260, содержащее VL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 116.
В262. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-261, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 116.
В263. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-262, содержащее СН, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 91.
В264. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-263, содержащее СН, которая содержит аминокислотную последовательность SEQ ID NO: 91.
В265. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-264, содержащее СL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 85.
В266. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-265, содержащее СL, которая содержит аминокислотную последовательность SEQ ID NO: 85.
В267. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-266, содержащее домен Fc.
В268. Антитело или его антигенсвязывающий фрагмент по В. 267, в котором указанный домен Fc представляет собой домен Fc IgA (например, IgA1 или IgA2), IgD, IgE или IgM.
В269. Антитело или его антигенсвязывающий фрагмент по В. 267, в котором указанный домен Fc представляет собой домен Fc IgG (например, IgG1, IgG2, IgG3 или IgG4).
В270. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-269, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 122.
В271. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 247-270, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 117.
В272. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 128;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 129; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 130.
В273. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 131.
В274. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 123;
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 124; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 125.
В275. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 126.
В276. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее:
(1) VH, которая содержит:
(а) CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 128;
(б) CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 129; и
(в) CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 130;
и (2) VL, которая содержит:
(а) CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 123;
(б) CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 124; и
(в) CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 125.
В277. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, содержащее последовательности CDR-H1, CDR-H2 и CDR-H3 SEQ ID NO: 131, и последовательности CDR-L1, CDR-L2 и CDR-L3 SEQ ID NO: 126.
В278. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-277, содержащее последовательность каркасной области человеческой VH3, VH1 или VH5.
В279. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-278, содержащее последовательность каркасной области VH человеческой зародышевой линии IGHV3-23, IGHV1-69 или IGHV3-7.
В280. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-277, содержащее консенсусную последовательность каркасной области VH человеческой зародышевой линии.
В281. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-280, содержащее VH, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 131.
В282. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-281, содержащее VH, которая содержит аминокислотную последовательность SEQ ID NO: 131.
В283. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-282, содержащее последовательность каркасной области человеческой Vκ или Vλ.
В284. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-283, содержащее последовательность каркасной области VL человеческой зародышевой линии IGKV3-20 или IGKV1-39.
В285. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-282, содержащее консенсусную последовательность каркасной области VL человеческой зародышевой линии.
В286. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-285, содержащее VL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 126.
В287. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-286, содержащее VL, которая содержит аминокислотную последовательность SEQ ID NO: 126.
В288. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-287, содержащее СН, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 91.
В289. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-288, содержащее СН, которая содержит аминокислотную последовательность SEQ ID NO: 91.
В290. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-289, содержащее СL, которая содержит аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 85.
В291. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-290, содержащее СL, которая содержит аминокислотную последовательность SEQ ID NO: 85.
В292. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-291, содержащее домен Fc.
В293. Антитело или его антигенсвязывающий фрагмент по В. 292, в котором указанный домен Fc представляет собой домен Fc IgA (например, IgA1 или IgA2), IgD, IgE или IgM.
В294. Антитело или его антигенсвязывающий фрагмент по В. 292, в котором указанный домен Fc представляет собой домен Fc IgG (например, IgG1, IgG2, IgG3 или IgG4).
В295. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-294, содержащее тяжелую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 132.
В296. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 272-295, содержащее легкую цепь, которая содержит аминокислотную последовательность SEQ ID NO: 127.
В297. Антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, где указанное антитело или его антигенсвязывающий фрагмент конкурирует за связывание с TFPI с антителом или его антигенсвязывающим фрагментом по любому из ВВ. 1-296.
В298. Антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, где указанное антитело или его антигенсвязывающий фрагмент конкурирует за связывание с TFPI с антителом, выбранным из группы, состоящей из TFPI-3, TFPI-21, TFPI-23, TFPI-24, TFPI-26, TFPI-106, TFPI-107, TFPI-108, TFPI-109, TFPI-110, TFPI-111, TFPI-112, TFPI-113, TFPI-114, TFPI-115, TFPI-118, TFPI-119, TFPI-122, TFPI-123, TFPI-126, 4D8.b1, химеры mu-hu 4D8, 4D8-Vk1.0 × VH1.0, 4D8-Vk1.0 × VH1.1, 4D8-Vk1.0 × VH1.2, 4D8-Vk1.0 × VH1.3, 4D8-Vk1.0 × VH1.4, 4D8-Vk1.0 × VH1.5, 4D8-Vk1.0 × VH1.6, 4D8-Vk1.1 × VH1.0, 4D8-Vk1.1 × VH1.1, 4D8-Vk1.1 × VH1.2, 4D8-Vk1.1 × VH1.3, 4D8-Vk1.1 × VH1.4, 4D8-Vk1.1 × VH1.5, 4D8-Vk1.1 × VH1.6, hz4D8, 6B7.c5 и 7A4.D9.
В299. Антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, где указанное антитело или его антигенсвязывающий фрагмент конкурирует за связывание с TFPI с антителом, выбранным из группы, состоящей из TFPI-23, TFPI-24, TFPI-106 и TFPI-118.
В300. Антитело или его антигенсвязывающий фрагмент по В. 299, где указанное антитело или его антигенсвязывающий фрагмент конкурирует за связывание с TFPI с TFPI-23 или TFPI-106.
В301. Антитело или его антигенсвязывающий фрагмент по В. 299, где указанное антитело или его антигенсвязывающий фрагмент конкурирует за связывание с TFPI с TFPI-24 или TFPI-118.
В302. Антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, где указанное антитело или его антигенсвязывающий фрагмент конкурирует за связывание с TFPI с антителом 4D8.
В303. Антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, где указанное антитело или его антигенсвязывающий фрагмент связывается с тем же самым эпитопом TFPI, что и антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-296.
В304. Антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, где указанное антитело или его антигенсвязывающий фрагмент связывается с тем же самым эпитопом TFPI, что и антитело, выбранное из группы, состоящей из TFPI-3, TFPI-21, TFPI-23, TFPI-24, TFPI-26, TFPI-106, TFPI-107, TFPI-108, TFPI-109, TFPI-110, TFPI-111, TFPI-112, TFPI-113, TFPI-114, TFPI-115, TFPI-118, TFPI-119, TFPI-122, TFPI-123, TFPI-126, 4D8.b1, химеры mu-hu 4D8, 4D8-Vk1.0 × VH1.0, 4D8-Vk1.0 × VH1.1, 4D8-Vk1.0 × VH1.2, 4D8-Vk1.0 × VH1.3, 4D8-Vk1.0 × VH1.4, 4D8-Vk1.0 × VH1.5, 4D8-Vk1.0 × VH1.6, 4D8-Vk1.1 × VH1.0, 4D8-Vk1.1 × VH1.1, 4D8-Vk1.1 × VH1.2, 4D8-Vk1.1 × VH1.3, 4D8-Vk1.1 × VH1.4, 4D8-Vk1.1 × VH1.5, 4D8-Vk1.1 × VH1.6, hz4D8, 6B7.c5 и 7A4.D9.
В305. Антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, где указанное антитело или его антигенсвязывающий фрагмент связывается с тем же самым эпитопом TFPI, что и антитело, выбранное из группы, состоящей из TFPI-23, TFPI-24, TFPI-106 и TFPI-118.
В306. Антитело или его антигенсвязывающий фрагмент по В. 305, где указанное антитело или его антигенсвязывающий фрагмент связывается с тем же самым эпитопом TFPI, что и TFPI-23 или TFPI-106.
В307. Антитело или его антигенсвязывающий фрагмент по В. 305, где указанное антитело или его антигенсвязывающий фрагмент связывается с тем же самым эпитопом TFPI, что и TFPI-24 или TFPI-118.
В308. Антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI, где указанное антитело или его антигенсвязывающий фрагмент связывается с тем же самым эпитопом TFPI, что и антитело 4D8.
В309. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 297-308, где указанное антитело или его антигенсвязывающий фрагмент не связывается с доменом K1 TFPI.
В310. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-309, где данное антитело или антигенсвязывающий фрагмент представляет собой слитый белок Fc, монотело, макситело, бифункциональное антитело, scFab, scFv, пептитело или антигенсвязывающий фрагмент любого из вышеупомянтых.
В311. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-310, где указанное антитело или антигенсвязывающий фрагмент связывается с TFPI со значением аффинности связывания (Kd) от примерно 1×10-7 М до примерно 1×10-12 М.
В312. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-311, где указанное антитело или антигенсвязывающий фрагмент связывается с TFPI со значением аффинности связывания (Kd) от примерно 5×10-7 М до примерно 5×10-11 М.
В313. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-312, где указанное антитело или антигенсвязывающий фрагмент связывается с TFPI со значением аффинности связывания (Kd) от примерно 1×10-8 М до примерно 1×10-10 М.
В314. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-313, где указанное антитело или антигенсвязывающий фрагмент: (1) уменьшает время свертывания крови при измерении в анализе протромбинового времени в плазме с разведением (dPT); (2) уменьшает время свертывания в цельной крови при измерении посредством тромбоэластографии или ротационной тромбоэластометрии; (3) увеличивает образование тромбина; (4) увеличивает активность FXa в присутствии TFPI; или (5) приводит к их любой комбинации.
В315. Антитело или его антигенсвязывающий фрагмент по В. 314, где указанное антитело или антигенсвязывающий фрагмент уменьшает время свертывания крови при измерении в анализе протромбинового времени в плазме с разведением.
В316. Антитело или его антигенсвязывающий фрагмент по В. 315, где указанное уменьшение времени свертывания крови при измерении в анализе протромбинового времени в плазме с разведением является дозозависимым.
В317. Антитело или его антигенсвязывающий фрагмент по В. 314, где указанное антитело или антигенсвязывающий фрагмент уменьшает время свертывания в цельной крови при измерении посредством тромбоэластографии или ротационной тромбоэластометрии.
В318. Антитело или его антигенсвязывающий фрагмент по В. 317, где указанное уменьшение времени свертывания крови при измерении посредством тромбоэластографии или ротационной тромбоэластометрии является дозозависимым.
В319. Антитело или его антигенсвязывающий фрагмент по В. 314, где указанное антитело или антигенсвязывающий фрагмент увеличивает образование тромбина.
В320. Антитело или его антигенсвязывающий фрагмент по В. 319, где указанное увеличение образования тромбина является дозозависимым.
В321. Антитело или его антигенсвязывающий фрагмент по В. 314, где указанное антитело или антигенсвязывающий фрагмент увеличивает активность FXa в присутствии TFPI.
В322. Антитело или его антигенсвязывающий фрагмент по В. 321, где указанное увеличение активности FXa в присутствии TFPI является дозозависимым.
В323. Антитело или его антигенсвязывающий фрагмент по В. 322, где указанное антитело усиливает накопление тромбоцитов в присутствии TFPI.
В324. Антитело или его антигенсвязывающий фрагмент по В. 323, где указанное усиление накопления тромбоцитов в присутствии TFPI является дозозависимым.
В325. Антитело или его антигенсвязывающий фрагмент по В. 324, где указанное антитело увеличивает образование фибрина в присутствии TFPI.
В326. Антитело или его антигенсвязывающий фрагмент по В. 325, где указанное увеличение образования фибрина в присутствии TFPI является дозозависимым.
В327. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания в цельной крови определяется с использованием цельной крови, полученной от человеческого пациента, имеющего тяжелую гемофилию А.
В328. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания в цельной крови определяется с использованием цельной крови, полученной от человеческого пациента, имеющего тяжелую гемофилию А и ингибирующие антитела против человеческого Фактора VIII.
В329. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания в цельной крови определяется с использованием цельной крови, полученной от человеческого пациента, имеющего умеренную гемофилию А.
В330. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания в цельной крови определяется с использованием цельной крови, полученной от человеческого пациента, имеющего тяжелую гемофилию В.
В331. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания в цельной крови определяется с использованием цельной крови, полученной от человеческого пациента, имеющего тяжелую гемофилию В и ингибирующие антитела против человеческого Фактора IX.
В332. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания в цельной крови определяется с использованием цельной крови, полученной от человеческого пациента, имеющего умеренную гемофилию В.
В333. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания при измерении в анализе dPT определяется с использованием плазмы, полученной от человеческого пациента, имеющего тяжелую гемофилию А.
В334. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания при измерении в анализе dPT определяется с использованием плазмы, полученной от человеческого пациента, имеющего тяжелую гемофилию А и ингибирующие антитела против человеческого Фактора VIII.
В335. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания при измерении в анализе dPT определяется с использованием плазмы, полученной от человеческого пациента, имеющего умеренную гемофилию А.
В336. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания при измерении в анализе dPT определяется с использованием плазмы, полученной от человеческого пациента, имеющего тяжелую гемофилию В.
В337. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания крови при измерении в анализе dPT определяется с использованием плазмы, полученной от человеческого пациента, имеющего тяжелую гемофилию В и ингибирующие антитела против человеческого Фактора IX.
В338. Антитело или его антигенсвязывающий фрагмент по В. 314, где уменьшение времени свертывания крови при измерении в анализе dPT определяется с использованием плазмы, полученной от человеческого пациента, имеющего умеренную гемофилию В.
В339. Антитело или его антигенсвязывающий фрагмент по В. 314, где увеличение образования тромбина определяется с использованием плазмы, полученной от человеческого пациента, имеющего тяжелую гемофилию А.
В340. Антитело или его антигенсвязывающий фрагмент по В. 314, где увеличение образования тромбина определяется с использованием плазмы, полученной от человеческого пациента, имеющего тяжелую гемофилию А и ингибирующие антитела против человеческого Фактора VIII.
В341. Антитело или его антигенсвязывающий фрагмент по В. 314, где увеличение образования тромбина определяется с использованием плазмы, полученной от человеческого пациента, имеющего умеренную гемофилию А.
В342. Антитело или его антигенсвязывающий фрагмент по В. 314, где увеличение образования тромбина определяется с использованием плазмы, полученной от человеческого пациента, имеющего тяжелую гемофилию В.
В343. Антитело или его антигенсвязывающий фрагмент по В. 314, где увеличение образования тромбина определяется с использованием плазмы, полученной от человеческого пациента, имеющего тяжелую гемофилию В и ингибирующие антитела против человеческого Фактора IX.
В344. Антитело или его антигенсвязывающий фрагмент по В. 314, где увеличение образования тромбина определяется с использованием плазмы, полученной от человеческого пациента, имеющего умеренную гемофилию В.
В345. Антитело или его антигенсвязывающий фрагмент по В. 314, где 100 нМ указанное антитело или его антигенсвязывающий фрагмент является по меньшей мере таким же эффективным в уменьшении времени свертывания цельной крови, полученной от человеческого пациента, имеющего тяжелую гемофилию А, что и количество рекомбинантного человеческого Фактора VIII, которое является достаточным для достижения 5% от нормальной свертывающей активности.
В346. Антитело или его антигенсвязывающий фрагмент по В. 314, где 100 нМ указанное антитело или его антигенсвязывающий фрагмент является по меньшей мере таким же эффективным в увеличении пикового образования тромбина в плазме, обогащенной тромбоцитами, полученной от человеческого пациента, имеющего тяжелую гемофилию А, что и количество рекомбинантного человеческого Фактора VIII, которое является достаточным для достижения 5% от нормальной свертывающей активности.
В347. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-346, где указанный TFPI представляет собой человеческий TFPI.
В348. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-347, где указанный TFPI содержит остатки 91-147 SEQ ID NO: 2.
В349. Выделенная молекула нуклеиновой кислоты или молекулы нуклеиновой кислоты, содержащая одну или более чем одну нуклеотидную последовательность, кодирующую антитело или его антигенсвязывающий фрагмент по любому из вв. 1-348.
В350. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент, которое специфично связывается с TFPI, где указанная нуклеиновая кислота содержит последовательность нуклеиновой кислоты, выбранную из группы, состоящей из следующих: последовательность нуклеиновой кислоты SEQ ID NO:175, последовательность нуклеиновой кислоты SEQ ID NO:176, последовательность нуклеиновой кислоты SEQ ID NO:177, последовательность нуклеиновой кислоты SEQ ID NO:178, последовательность нуклеиновой кислоты вставки вектора, депонированной как mAb-TFPI-106 VL под номером доступа ATCC PTA-122328, и последовательность нуклеиновой кислоты вставки вектора, депонированной как mAb-TFPI-106 VH под номером доступа ATCC PTA-122329.
В351. Вектор, содержащий молекулу нуклеиновой кислоты по ВВ. 349 и 350.
В352. Клетка-хозяин, содержащая молекулу нуклеиновой кислоты по В. 349 или 350, или вектор по В. 351.
В353. Клетка-хозяин по В. 352, где указанная клетка представляет собой клетку млекопитающего.
В354. Клетка-хозяин по В. 353, где указанная клетка представляет собой клетку СНО (яичник китайского хомяка), клетку HEK-293 (клетка эмбриональной почки человека) или клетку Sp2.0.
В355. Способ получения антитела или его антигенсвязывающего фрагмента, включающий культивирование клетки-хозяина по любому из ВВ. 352-354 при условиях, когда указанное антитело или антигенсвязывающий фрагмент экспрессируется указанной клеткой-хозяином.
В356. Способ по В. 355, дополнительно включающий выделение указанного антитела или его антигенсвязывающего фрагмента.
В357. Антитело или его антигенсвязывающий фрагмент, полученное способом по В. 355 или 356.
В358. Фармацевтическая композиция, содержащая антитело или антигенсвязывающий фрагмент по любому из ВВ. 1-347 и 357, и фармацевтически приемлемый носитель или эксципиент.
В359. Способ уменьшения активности ингибитора пути тканевого фактора (TFPI), включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из ВВ. 1-347 и 357 или фармацевтической композиции по В. 358.
В360. Способ сокращения времени кровотечения, включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из ВВ. 1-347 и 357 или фармацевтической композиции по В. 358.
В361. Способ по В. 358 или 360, где указанный субъект представляет собой человека.
В362. Способ по любому из ВВ. 359-361, где указанный субъект страдает от или подвержен недостаточности свертывания крови.
В363. Способ по любому из ВВ. 359-361, где указанный субъект страдает от или подвержен тромбоцитарному расстройству.
В364. Способ по любому из ВВ. 359-361, где указанный субъект страдает от или подвержен гемофилии А, В или С.
В365. Способ по любому из ВВ. 359-361, где указанный субъект страдает от или подвержен гемофилии А или В.
В366. Способ по любому из ВВ. 359-361, где указанный субъект страдает от или подвержен болезни фон Виллебранда (vWD).
В367. Способ по В. 360, дополнительно включающий введение терапевтически эффективного количества FVII.
В368. Способ по В. 367, где указанный способ увеличивает образование тромбина в присутствии TFPI.
В369. Способ по любому из ВВ. 359-368, включающий внутривенное введение указанного антитела или его антигенсвязывающего фрагмента, или фармацевтической композиции.
В370. Способ по любому из ВВ. 359-368, включающий подкожное введение указанного антитела или его антигенсвязывающего фрагмента, или фармацевтической композиции.
В371. Способ по любому из ВВ. 359-368, в котором указанное антитело или его антигенсвязывающий фрагмент, или фармацевтическая композиция вводится один раз каждые 3 суток, один раз каждые 4 суток, один раз каждые 5 суток, один раз каждые 6 суток, один раз в неделю или дважды в неделю.
В372. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-347 и 357, или фармацевтическая композиция по В. 358 для применения в качестве лекарственного средства.
В373. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-347 и 357, или фармацевтическая композиция по В. 358 для применения в уменьшении активности TFPI у субъекта.
В374. Антитело или его антигенсвязывающий фрагмент по любому из ВВ. 1-347 и 357, или фармацевтическая композиция по В. 357 для применения в сокращении времени кровотечения у субъекта.
В375. Антитело или антигенсвязывающий фрагмент, или фармацевтическая композиция по любому из ВВ. 372-374, где указанный субъект представляет собой человека.
В376. Антитело или антигенсвязывающий фрагмент, или фармацевтическая композиция по любому из ВВ. 372-375, где указанный субъект страдает от или подвержен недостаточности свертывания крови.
В377. Антитело или антигенсвязывающий фрагмент, или фармацевтическая композиция по любому из ВВ. 372-375, где указанный субъект страдает от или подвержен гемофилии А, В или С.
В378. Антитело или антигенсвязывающий фрагмент, или фармацевтическая композиция по любому из ВВ. 372-375, где указанный субъект страдает от или подвержен гемофилии А или В.
В379. Антитело или антигенсвязывающий фрагмент, или фармацевтическая композиция по любому из ВВ. 372-375, где указанный субъект страдает от или подвержен болезни фон Виллебранда (vWD).
В380. Антитело или антигенсвязывающий фрагмент, или фармацевтическая композиция по любому из ВВ. 372-375, где указанный субъект страдает от или подвержен тромбоцитарному расстройству.
В381. Применение антитела или его антигенсвязывающего фрагмента по любому из ВВ. 1-348 и 357, или фармацевтической композиции по В. 358 для уменьшения активности TFPI у субъекта.
В382. Применение антитела или его антигенсвязывающего фрагмента по любому из ВВ. 1-348 и 357, или фармацевтической композиции по В. 358 в изготовлении лекарственного средства для уменьшения активности TFPI у субъекта.
В383. Применение антитела или его антигенсвязывающего фрагмента по любому из ВВ. 1-348 и 357, или фармацевтической композиции по В. 358 для сокращения времени кровотечения у субъекта.
В384. Применение антитела или его антигенсвязывающего фрагмента по любому из ВВ. 1-348 и 357, или фармацевтической композиции по В. 358 в изготовлении лекарственного средства для сокращения времени кровотечения у субъекта.
В385. Применение по любому из ВВ. 381-384, где указанный субъект представляет собой человека.
В386. Применение по любому из ВВ. 381-384, где указанный субъект страдает от или подвержен недостаточности свертывания крови.
В387. Применение по любому из ВВ. 381-384, где указанный субъект страдает от или подвержен гемофилии А, В или С.
В388. Применение по любому из ВВ. 381-384, где указанный субъект страдает от или подвержен гемофилии А или В.
В389. Применение по любому из ВВ. 381-384, где указанный субъект страдает от или подвержен болезни фон Виллебранда (vWD).
В390. Применение по любому из вв. 381-384, где указанный субъект страдает от или подвержен тромбоцитарному расстройству.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
ФИГ. 1А-1Е представляют собой графические материалы, показывающие структуры сокристаллов разных антител против TFPI и домена K2 TFPI. В частности, как показано на ФИГ. 1Е, все раскрытые в настоящем описании примеры антител - TFPI-23, TFPI-24 и 4D8 - связываются с неперекрывающимися эпитопами домена K2 по сравнению с другими контрольными антителами. TFPI-106 связывается с тем же самым сайтом, что и TFPI-23, а TFPI-118 связывается с тем же самым сайтом, что и TFPI-24. Обозначения «R&D» или «R&D Fab» относятся к антителу Mab 2974 от R&D Systems. Антитело Novo2021 также обозначено «hz4F36». Обозначение «Клон 23» относится к TFPI-23; обозначение «клон 24» относится к TFPI-24.
ФИГ. 2А-2Д представляют собой диаграммы, показывающие взаимодействия между остатками эпитопов в пределах домена K2 TFPI и остатками паратопов из разных антител против TFPI. Обозначения «R&D» или «R&D Fab» относятся к антителу Mab 2974 от R&D Systems. Обозначение «Клон 23» относится к TFPI-23; обозначение «клон 24» относится к TFPI-24.
На ФИГ. 3А-3Д показаны эффективности in vivo разных антител против TFPI в модели мышиной раны. На ФИГ. 3А и 3Б показана продолжительность уменьшающегося кровотечения у мышей с гемофилией А с недостаточностью Фактора VIII (FVIII -/-) при введении антител 2А8-200 и 2А8 (используемых в этом исследовании в качестве контрольных антител). На ФИГ. 3В показана продолжительность эффекта у мышей с гемофилией А при введении антител TFPI 4D8 (контроль), TFPI-21, TFPI-23 и TFPI-24. На ФИГ. 3Г и 3Д показана продолжительность эффекта у мышей с гемофилией А при введении антител TFPI-106 и TFPI-118. Антитела вводили мышам с гемофилией А
(FVIII -/-) посредством внутривенной инъекции в дозе 6 мг/кг в указанные моменты времени (часы (ч)) до ранения. Затем измеряли общий объем кровопотери (мкл) после перерезания хвоста. Мыши с гемофилией А, обработанные носителем (физиологическим раствором), служили в качестве контроля. Все измерения представлены в виде среднего значения плюс/минус SEM (стандартная ошибка среднего). * - Р меньше 0,05. FVIII +/+ мыши (дикий тип) получали физиологический раствор. n равно 5 на группу.
На ФИГ. 4 показана продолжительность кровотечения у мышей с гемофилией В после перерезания хвоста при введении антитела TFPI-106. Антитела вводили мышам с гемофилией В посредством внутривенной инъекции в дозе 6 мг/кг в указанные моменты времени (часы (ч)) до ранения. Затем измеряли общий объем кровопотери (мкл) после перерезания хвоста. Мыши с гемофилией А, обработанные носителем (физиологическим раствором), служили в качестве контроля. Все измерения представлены в виде среднего значения плюс/минус SEM. * - Р меньше 0,05. n равно 4-5 на группу.
На ФИГ. 5, содержащей панели А и Б, каждая из которых содержит шесть панелей (ФИГ. 5А, 1-6 и ФИГ. 5Б, 1-6), показаны микрофотографии прижизненной микроскопии (IVM), демонстрирующие то, что TFPI выявляется в тромбоцитарном тромбе и вдоль эндотелия in vivo в месте ранениясосуда у мыши дикого типа. На Фиг. 5А показано увеличение тромбоцитарного тромба в месте раненияпри выявлении с использованием CD42c, меченного Dylight 649, который связывается с GP1bβ на тромбоцитах. Присутствие или отсутствие тромбоцитов демонстрируется по флуоресцентному сигналу, выявляемому на панели 1 (0 с); панели 2 (15 с); панели 3 (30 с); панели 4 (60 с); панели 5 (90 с) и панели 6 (120 с). Также вводили IgG негативного контроля, меченный Alexa 488, и не выявляли флуоресценции. На Фиг. 5Б, содержащей панели 1-6, показаны микрофотографии IVM, демонстрирующие то, что TFPI присутствует в тромбоцитарном тромбе и вдоль эндотелия после индуцированного лазером ранения сосуда у мышей дикого типа. Меченный Alexa 488 TFPI (зеленый сигнал, показанный серым) не выявляется в 0 секунд (ФИГ. 5Б, панель 1), и бледный сигнал можно увидеть в 15 секунд (ФИГ. 5Б, панель 2). В 30 секунд (ФИГ. 5Б, панель 3) зеленый флуоресцентный сигнал усиливался, и можно увидеть бледный красный сигнал (CD42c, меченный Dylight 649), указывающий выявление накопления тромбоцитов приблизительно в том же самом месте, где выявляется TFPI. На ФИГ. 5Б, панель 4 (60 секунд) показаны сильный зеленый и красный флуоресцентные сигналы (оба светло-серые, где красную флуоресценцию можно увидеть слева от места ранения сосуда, и зеленый сигнал, главным образом, выявляется в направлении правой стороны места ранения), демонстрируя то, что и накопление тромбоцитов, и TFPI выявляется в месте ранения. На ФИГ. 5Б, панель 5, показаны и красный (тромбоциты), и зеленый (TFPI) флуоресцентный сигналы в месте ранения спустя 60 секунд, когда оба сигнала сильнее, чем спустя 30 секунд. На ФИГ. 5Б, панель 6, показан пониженный красный сигнал (тромбоциты), и пониженный зеленый сигнал (TFPI), оба все еще выявляемые в месте ранения спустя 120 секунд.
ФИГ. 6А представляет собой график, показывающий гемостатический эффект TFPI-106 у мышей с гемофилией А после индуцированного лазером ранения сосудов при оценке с использованием IVM, где количество тромбоцитарного тромба выражается как площадь под кривой (AUC) (* показывает Р меньше 0,005). На ФИГ. 6А показано накопление тромбоцитов в месте ранения мышей дикого типа (WT) спустя 0,5 часа после ранения, где мыши получали только контроль в виде физиологического раствора, по сравнению с отсутствием накопления тромбоцитов у мыши с гемофилией А в 0,5 часов, где вводился физиологический раствор. Накопление тромбоцитов выявляли у мышей с гемофилией А в 0,5 часов, где вводился рекомбинантный фактор VIII (rFVIII) или TFPI-106. Эффект накопления тромба все еще выявлялся в 168 часов у мышей с гемофилией А, которым вводился TFPI-106.
ФИГ. 6Б представляет собой график, показывающий гемостатический эффект TFPI-106 у мышей с гемофилией А после индуцированного лазером ранения сосуда при оценке с использованием IVM, где количество образовавшегося фибрина выражается как площадь под кривой (AUC) (* показывает Р меньше 0,005). На ФИГ. 6Б показано образование фибрина в месте ранения у мышей дикого типа (WT) спустя 0,5 часа после ранения, где мыши получали только контроль в виде физиологического раствора, по сравнению с отсутствием выявляемого образования фибрина у мыши с гемофилией А спустя 0,5 часа, где вводился физиологический раствор. Образование фибрина выявляли у мышей с гемофилией А спустя 0,5 часов, где вводился рекомбинантный фактор VIII (rFVIII) или TFPI-106. Эффект образования фибрина все еще выявлялся спустя 168 часов у мышей с гемофилией А, которым вводился TFPI-106.
На ФИГ. 7 изображен график, показывающий эффект введения TFPI-106 и рекомбинантного фактора VIIa (rFVIIa) на анализ образования тромбина (TGA) в плазме при тяжелой гемофилии А в присутствии 1 пМ тканевого фактора и 4 мкМ фосфолипидов. На этом графике показаны тромбограммы плазмы при гемофилии А с введением одного TFPI-106 (16 мкг/мл) и в комбинации с rFVIIa (20 мкг/мл; 2 мкг/мл или 0,2 мкг/мл). Также показаны тромбограммы плазмы при гемофилии А и негемофилической плазмы при отсутствии введения и TFPI-106, и rFVIIa.
На ФИГ. 8А изображены тромбограммы, показывающие эффект на образование тромбина в присутствии 1 пМ тканевого фактора и 4 мкМ фосфолипидов в плазме при гемофилии А в присутствии TFPI-106 с или без rFVIIa, или в присутствии только rFVIIa. Негемофилическая плазма включена в качестве контроля.
На ФИГ. 8Б изображены тромбограммы, показывающие эффект на образование тромбина в присутствии трех единиц Бетезда (3 BU) ингибитора в цитратной обедненной тромбоцитами плазме при гемофилии А в присутствии TFPI-106 с или без rFVIIa, или в присутствии только rFVIIa. Негемофилическая плазма включена в качестве контроля. Также включен контроль негемофилической плазмы, в которую был добавлен TFPI-106 (16 мкг/мл).
На ФИГ. 8В изображены тромбограммы, показывающие эффект на образование тромбина в присутствии трех единиц Бетезда (3 BU) ингибитора в цитратной обедненной тромбоцитами плазме при гемофилии В в присутствии TFPI-106 с или без rFVIIa, или в присутствии только rFVIIa. Негемофилическая плазма включена в качестве контроля. Также включен контроль негемофилической плазмы, в которую был добавлен TFPI-106 (16 мкг/мл).
На ФИГ. 9А показан эффект на кровопотерю по сравнению с контролем введения антитела TFPI-106 (6 мг/кг) и отдельно рекомбинантного Фактора VIII (200 единиц/кг) мышам с гемофилией А сразу после перерезания хвоста. На ФИГ. 9Б показан эффект на кровопотерю по сравнению с контролем введения трех разных доз антитела TFPI-106 (6 мг/кг) и отдельно рекомбинантного Фактора VIII (200 единиц/кг) мышам с гемфилией А через 2 минуты после перерезания хвоста.
На ФИГ. 10А показан эффект разных концентраций TFPI 106 по сравнению с рекомбинантным Фактором VIII на время свертывания цельной крови от человеческого пациента с тяжелой гемофилией А. На ФИГ. 10Б показан эффект разных концентраций TFPI 106 по сравнению с рекомбинантным Фактором VIII на пиковое образование тромбина в плазме, обогащенной тромбоцитами, от человеческого пациента с тяжелой гемофилией А.
На ФИГ. 11А показан эффект разных концентраций TFPI 106 и ингибиторов FVIII на время свертывания цельной крови от человеческого пациента с тяжелой гемофилией А. На ФИГ. 11Б показан эффект разных концентраций TFPI 106 и ингибиторов FVIII на пиковое образование тромбина в плазме, обогащенной тромбоцитами, от человеческого пациента с тяжелой гемофилией А. На ФИГ. 11В показан эффект разных концентраций TFPI 106 и ингибиторов FVIII на время свертывания плазмы, обедненной тромбоцитами, от человеческого пациента с тяжелой гемофилией А.
На ФИГ. 12А показан эффект разных концентраций TFPI 106 по сравнению с рекомбинантным фактором VIII на время свертывания цельной крови от человеческого пациента с умеренной гемофилией А. На ФИГ. 12Б показан эффект разных концентраций TFPI 106 по сравнению с рекомбинантным фактором VIII на пиковое образование тромбина в плазме, обогащенной тромбоцитами, от человеческого пациента с умеренной гемофилией А. На ФИГ. 12В показан эффект разных концентраций TFPI 106 на время свертывания плазмы, обедненной тромбоцитами, от человеческого пациента с умеренной гемофилией А.
На ФИГ. 13А показан эффект разных концентраций TFPI 106 по сравнению с рекомбинантным фактором IX на время свертывания цельной крови от человеческого пациента с умеренной гемофилией В. На ФИГ. 13Б показан эффект разных концентраций TFPI 106 по сравнению с рекомбинантным фактором IX на пиковое образование тромбина в плазме, обогащенной тромбоцитами, от человеческого пациента с умеренной гемофилией В. На ФИГ. 13В показан эффект разных концентраций TFPI 106 на время свертывания плазмы, обедненной тромбоцитами, от человеческого пациента с умеренной гемофилией В.
На ФИГ. 14А показан эффект разных концентраций TFPI 106 по сравнению с рекомбинантным Фактором VIII на время свертывания цельной крови от многих человеческих пациентов с гемофилией А. На ФИГ. 14Б показан эффект разных концентраций TFPI 106 по сравнению с рекомбинантным Фактором VIII на пиковое образование тромбина в плазме, обогащенной тромбоцитами, от многих человеческих пациентов с гемофилией А. На ФИГ. 14В показан эффект разных концентраций TFPI 106 на время свертывания плазмы, обедненной тромбоцитами, от многих человеческих пациентов с гемофилией В.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
1. Обзор
Как отмечалось выше, пациенты с гемофилией имеют некоторую способность останавливать кровотечения посредством их интактного внешнего пути; однако, внешний путь является недостаточным для обеспечения защиты, так как он быстро останавливается ингибитором пути тканевого фактора (TFPI). Блокирование/нейтрализация ингибирования TFPI у данных пациентов может компенсировать неадекватное образование FXa и нормализовать предрасположенность к кровотечению. Соответственно, в настоящем описании раскрыты и проиллюстрированы примерами антитела и их антигенсвязывающие фрагменты, которые специфично связываются с TFPI и ингибируют его активность.
2. Определения
Под «уменьшением активности TFPI» подразумевается то, что антитело или его антигенсвязывающий фрагмент может: (1) уменьшать время свертывания по сравнению со временем свертывания в отсутствии антитела при измерении, например, посредством анализа протромбинового времени в плазме с разведением; (2) уменьшать время свертывания в цельной крови по сравнению со временем свертывания в отсутствии антитела при измерении, например, посредством тромбоэластографии или ротационной тромбоэластометрии; (3) увеличивать образование тромбина; (4) увеличивать активность FXa в присутствии TFPI; (5) увеличивать накопление тромбоцитов в присутствии TFPI; (6) увеличивать образование фибрина в присутствии TFPI; или (7) приводить к любой их комбинации. Ингибирующие активности антитела или антигенсвязывающего фрагмента могут быть, но не обязательно являются дозозависимыми (например, вызывая дозозависимое уменьшение времени свертывания при измерении в анализе протромбинового времени в плазме с разведением).
Кроме того, как раскрыто и проиллюстрировано в настоящем описании примерами, были получены структуры сокристаллов антител против TFPI и домена Кунитца 2 (домен K2) TFPI. Структурный анализ показывает то, что типичные антитела по изобретению распознают уникальные эпитопы TFPI посравнению с другими публично раскрытыми антителами против TFPI (которые были использованы в качестве контрольных антител в Примерах). Например, как показано на Фиг. 1А-1А-1Е и Фиг. 2А-2Д, по сравнению с несколькими контрольными антителами против TFPI (R&D (Mab2479) Fab, Novo2021 (также именуемое hz4F36) Fab, 2A8 Fab), антитела TFPI-23, TFPI-24 и 4D8 связываются с неперекрывающимися сайтами в домене K2 TFPI.
Соответственно, в некоторых воплощениях раскрытые в настоящем описании антитела (и антигенсвязывающие фрагменты) распознают уникальный эпитоп TFPI, расположенный в домене K2 TFPI. На основе структуры сокристалла и расчетного аланинового сканирования данный эпитоп содержит три остатка, которые являются важными для взаимодействия антитело-антиген: Ile105, Arg107 и Leu131 (согласно нумерации человеческого TFPI, как показано в SEQ ID NO: 2). Мутирование данных трех остатков на аланин приводит к потере связывания антитела. Например, все антитела TFPI-23 и его варианты (например, TFPI-106 и TFPI-107) распознают данный эпитоп.
В некоторых воплощениях распознавание ключевых раскрытых в настоящем описании остатков эпитопа позволяет антителам (и их антигенсвязывающим фрагментам) уменьшать активность TFPI. В частности, кристаллическая структура показывает то, что домен K2 TFPI принимает конусообразную структуру с вершиной конуса (особенно Arg107), связывающейся с FXa. И TFPI-23, и TFPI-24 распознают вершину этой конусообразной области и блокируют связывание TFPI с FXa. Антитело 4D8 распознает другой эпитоп в домене K2. 4D8, хотя и не взаимодействует непосредственно с остатками на вершине конуса, тем не менее, блокирует связывание TFPI с FXa. В Таблице 15 обобщены остатки неперекрывающихся эпитопов, распознаваемых типичными раскрытыми в настоящем описании антителами, по сравнению с другими общеизвестными антителами против TFPI.
Кроме того, в некоторых воплощениях раскрытые в настоящем описании антитела и их антигенсвязывающие фрагменты продемонстрировали требуемые фармакологические активности и фармакокинетические свойства для лечения дефицитов свертывания крови (таких как гемофилия) и для уменьшения времени кровотечения.
Антитело, которое «преимущественно связывается» или «специфично связывается» (используются в настоящем описании взаимозаменяемо) с эпитопом, представляет собой термин, хорошо понятный в этой области, и способы определения такого специфичного или предпочтительного связывания также хорошо известны в этой области. Говорят, что молекула демонстрирует «специфичное связывание» или «преимущественное связывание», если она взаимодействует или ассоциирует чаще, быстрее, с большей продолжительностью и/или большей аффинностью с конкретной клеткой или веществом, чем с альтернативными клетками или веществами. Антитело «специфично связывается» или «преимущественно связывается» с мишенью, если оно связывается с большей аффинностью, авидностью, легче и/или с большей продолжительностью, чем с другими веществами. Также антитело «специфично связывается» или «предпочтительно связывается» с мишенью, если оно связывается с большей аффинностью, авидностью, легче и/или с большей продолжительностью с этой мишенью в образце, чем с другими веществами в этом образце. Например, антитело, которое специфично или предпочтительно связывается с эпитопом TFPI, представляет собой антитело, которое связывается с этим эпитопом с большей аффинностью, авидностью, легче и/или с большей продолжительностью, чем с другими эпитопами TFPI или эпитопами, не являющимися эпитопами TFPI. При прочтении данного определения также понятно то, что, например, антитело (или группировка, или эпитоп), которое специфично или преимущественно связывается с первой мишенью, может или не может специфично или преимущественно связываться со второй мишенью. «Специфичное связывание» или «преимущественное связывание», как таковое, не обязательно требует (хотя оно и может включать) исключительного связывания. В общем, но не обязательно, ссылка на связывание означает преимущественное связывание. «Специфичное связывание» или «преимущественное связывание» включает соединение, например, белок, нуклеиновую кислоту, антитело или тому подобное, которое распознает и связывается с конкретной молекулой, но оно по существу не распознает или не связывается с другими молекулами в образце. Например, антитело или рецептор пептида, которое распознает и связывается с когнатным лигандом или партнером связывания (например, антитело против TFPI, которое связывается с TFPI) в образце, но по существу не распознает и не связывается с другими молекулами в образце, специфично связывается с данным когнатным лигандом или партнером связывания. Таким образом, при назначенных условиях анализа специфичная связывающая группировка (например, антитело или его антигенсвязывающая часть, или рецептор, или его лигандсвязывающая часть) предпочтительно связывается с конкретной молекулой-мишенью и не связывается в значительном количестве с другими компонентами, присутствующими в тестируемом образце.
Для отбора антитела или пептида, который специфично связывается с интересующей молекулой, можно использовать множество форматов анализа. Например, твердофазный иммуноанализ ELISA, иммунопреципитация, Biacore™ (GE Healthcare, Piscataway, NJ), KinExA, флуоресцентная сортировка клеток (FACS), Octet™ (FortéBio, Inc., Menlo Park, CA) и вестерн-блоттинг находятся среди многих анализов, которые можно использовать для идентификации антитела, которое специфично реагирует с антигеном, или рецептора, или его лигандсвязывающей части, который специфично связывается с когнатным лигандом или партнером связывания. Типично специфичная или селективная реакция будет по меньшей мере вдвое превышать фоновый сигнал или шум, более типично - более чем в 10 раз превышать фон, еще более типично - более чем в 50 раз превышать фон, более типично - более чем в 100 раз превышать фон, еще более типично - более чем в 500 раз превышать фон, еще более типично - более чем в 1000 раз превышать фон, и еще более типично - более чем в 10000 раз превышать фон. Также говорят, что антитело «специфично связывается» с антигеном, когда равновесная константа диссоциации (KD) меньше или равна 7 нМ.
Термин «аффинность связывания» используется в настоящем описании как мера силы нековалентного взаимодействия между двумя молекулами, например, антителом или его фрагментом и антигеном. Термин «аффинность связывания» используется для описания одновалентных взаимодействий (собственная активность).
Аффинность связывания между двумя молекулами, например, антителом или его фрагментом и антигеном через одновалентное взаимодействие, может количественно определяться посредством определения константы диссоциации (KD). В свою очередь, KD может определяться измерением кинетики образования и диссоциации комплекса с использованием, например, способа поверхностного плазмонного резонанса (SPR) (Biacore). Константы скорости, соответствующие ассоциации и диссоциации одновалентного комплекса, именуются константой скорости ассоциации ka (или kon) и константой скорости диссоциации kd (или koff) соответственно. KD соотносится с ka и kd через уравнение KD = kd/ka. Значение константы диссоциации можно определять непосредственно хорошо известными способами и можно рассчитывать даже для сложных смесей способами, такими как, например, способы, изложенные в Caceci et al. (1984, Byte 9: 340-362). Например, KD может быть установлена с использованием анализа связывания на фильтре на основе двойного нитроцеллюлозного фильтра, как, например, раскрыто Wong & Lohman (1993, Proc. Natl. Acad. Sci. USA 90: 5428-5432). В этой области известны другие стандартные анализы для оценки способности к связыванию лигандов, таких как антитела, в отношении антигенов-мишеней, включающие, например, ELISA, вестерн-блоттинг, RIA (радиоиммуноанализ) и анализ проточной цитометрией, и другие анализы, проиллюстрированные в настоящем описании примерами в других местах. Кинетику связывания и аффинность связывания антитела также можно оценивать стандартными анализами, известными в этой области, такими как поверхностный плазмонный резонанс (SPR), например, с использованием системы Biacore™, или KinExA.
Можно проводить конкурентный анализ связывания, в котором связывание антитела с антигеном сравнивается со связыванием мишени другим лигандом этой мишени, таким как другое антитело или растворимый рецептор, который в иных условиях связывается с мишенью. Концентрация, при которой происходит 50%-ное ингибирование, известна как Ki. При идеальных условиях Ki эквивалентна KD. Значение Ki никогда не будет меньше, чем KD, таким образом, измерение Ki может быть с удобством заменено с получением верхней границы KD.
Следуя приведенному выше определению, аффинности связывания, ассоциированные с разными молекулярными взаимодействиями, например, сравнение аффинности связывания разных антител в отношении данного антигена, могут сравниваться посредством сравнения значений KD для индивидуальных комплексов антитело/антиген. Значения KD для антител или других партнеров связывания могут быть определены с использованием хорошо установленных в этой области способов. Одним способом определения KD является применение поверхностного плазмонного резонанса, типично с использованием системы биосенсора, такой как система Biacore®.
Аналогичным образом, специфичность взаимодействия может оцениваться определением и сравнением значения KD для интересующего взаимодействия, например, специфичного взаимодействия между антителом и антигеном, со значением KD неинтересующего взаимодействия, например, контрольного антитела, для которого известно, что оно не связывается с TFPI.
Антитело, которое специфично связывается с его мишенью, может связываться с его мишенью с высокой аффинностью, то есть, демонстрируя низкую KD, как обсуждалось выше, и может связываться с другими молекулами, не являющимися мишенями, с меньшей аффинностью. Например, антитело может связываться с молекулами, не являющимися мишенями, с KD 1×10-6 М или более, более предпочтительно - 1×10-5 М или более, более предпочтительно - 1×10-4 М или более, более предпочтительно - 1×10-3 М или более, даже более предпочтительно - 1×10-2 М или более. Антитело по изобретению предпочтительно способно связываться с его мишенью с аффинностью, которая по меньшей мере вдвое, 10-кратно, 50-кратно, 100-кратно, 200-кратно, 500-кратно, 1000-кратно или 10000-кратно или более превышает его аффинность в отношении связывания с другой молекулой, не являющейся TFPI.
В общем, антителу против TFPI необходимо связываться с TFPI с высокой аффинностью для того, чтобы эффективно уменьшать активности TFPI. Однако когда аффинность связывания антитела является слишком высокой, антитело может подвергаться быстрой интернализации и деградации клеткой-хозяином. Это потенциально может приводить к короткому периоду полужизни и повторным инъекциям. Например, антитело TFPI-23 демонстрирует меньшую аффинность связывания (Kd) по сравнению с TFPI-24 и, при определенных обстоятельствах, оказывается более желательным для клинических применений, так как оно имеет меньшую скорость интернализации и более продолжительный период полужизни. Соответственно, обычно желательными являются аффинности связывания (Kd) от 5×10-7 М до примерно 5×10-11 М, в частности, от примерно 1×10-8 М до примерно 1×10-10 М, особенно для лечения хронического состояния (например, гемофилии), для которого требуются повторные инъекции. Не желая быть связанными какой-либо конкретной теорией, считается то, что данный интервал аффинности устанавливает равновесие между (1) аффинностями связывания, которые необходимы для эффективного ингибирования активностей TFPI, и (2) более длительным периодом полужизни и пониженной интернализацией антитела.
Положения конкретных аминокислотных остатков в TFPI нумеруются согласно SEQ ID NO: 2 (человеческий TFPIα K1K2K3). Однако настоящее изобретение не ограничивается SEQ ID NO: 2. Соответствующие остатки из других гомологов TFPI, изоформ, вариантов или фрагментов могут быть идентифицированы в соответствии с выравниванием последовательностей или выравниванием структур, которое известно в этой области. Например, выравнивания можно делать вручную или с использованием хорошо известных программ выравнивания последовательностей, таких как ClustalW2 или “BLAST 2 Sequences”, с использованием параметров по умолчанию. Например, Arg107 SEQ ID NO: 2 соответствует Arg104 мышиного TFPI K1K2 (SEQ ID NO: 4).
«Антигенсвязывающий фрагмент» антитела относится к фрагменту полноразмерного антитела, который сохраняет способность специфично связываться с антигеном (предпочтительно с по существу такой же аффинностью связывания). Примеры антигенсвязывающего фрагмента включают (1) фрагмент Fab - одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (2) фрагмент F(ab')2 - двухвалентный фрагмент, содержащий два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области; (3) фрагмент Fd, состоящий из доменов VH и СН1; (4) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела; (5) фрагмент dAb (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH; и (6) выделенную определяющую комплементарность область (CDR), Fv, связанные дисульфидной связью фрагменты Fv (dsFv), и антиидиотипические (анти-Id) антитела и интратела. Кроме того, хотя два домена фрагмента Fv, представляющие собой VL и VH, и кодируются отдельными генами, они могут быть соединены с использованием способов генной инженерии посредством синтетического линкера, который позволяет им сделать одну белковую цепь, в которой области VL и VH образуют пару с формированием одновалентных молекул (известных как одноцепоченый Fv (scFv)); см., например, Bird et al. Science 242:423-426 (1988) и Huston et al. Proc. Natl. Acad. Sci. USA 85:5879-5883 (1988)). Также охватываются другие формы одноцепочечных антител, такие как диатела. Диатела представляют собой двухвалентные биспецифичные антитела, в которых домены VH и VL экспрессируются на одной полипептидной цепи, но с использованием линкера, который является слишком коротким для обеспечения образования пары между двумя доменами на той же самой цепи, принуждая данные домены, посредством этого, образовать пару с комплементарными доменами другой цепи и создавая два антигенсвязывающих сайта (см., например, Holliger et al. Proc. Natl. Acad. Sci. USA 90:6444-6448 (1993); Poljak et al., 1994, Structure 2:1121-1123).
Термин «вариабельный домен» антитела относится к вариабельной области легкой цепи антитела (VL) или вариабельной области тяжелой цепи антитела (VН) - либо к одной, либо в комбинации. Как известно в этой области, каждая из вариабельных областей тяжелой и легкой цепи состоит из четырех каркасных областей (FR), соединенных тремя определяющими комплементарность областями (CDR), и способствует образованию антигенсвязывающего сайта антител.
Остатки в вариабельном домене пронумерованы согласно Kabat, что представляет собой систему нумерации, используемую для вариабельных доменов тяжелой цепи или вариабельных доменов легкой цепи компиляции антител. См., Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). С использованием этой системы нумерации реальная линейная аминокислотная последовательность может содержать меньшее число или дополнительные аминокислоты, соответствующие укорачиванию или вставке в FR или CDR вариабельного домена. Например, вариабельный домен тяжелой цепи может включать вставку одной аминокислоты (остаток 52а согласно Kabat) после остатка 52 Н2 и вставленные остатки (например, остатки 82а, 82b и 82с согласно Kabat) после остатка 82 FR тяжелой цепи. Нумерация остатков по Kabat может быть определена для данного антитела посредством выравнивания в областях гомологии последовательности антитела со «стандартной» последовательностью, пронумерованной по Kabat. Доступны разные алгоритмы для присвоения нумерации по Kabat. В настоящей заявке для присвоения нумерации по Kabat вариабельным областям, если не отмечено иное, используется алгоритм, применяемый в Abysis выпуска 2012 года (www.abysis.org).
Положения конкретных аминокислотных остатков в антителе (такие как раскрытые в настоящем описании остатки паратопа) также пронумерованы согласно Kabat.
«Определяющие комплементарность области» (CDR) могут быть идентифицированы согласно определениям Kabat, Chothia, сочетанию и Kabat, и Chothia, AbM, контактному и/или конформационному определениям или любому способу определения CDR, хорошо известному в этой области. См., например, Kabat et al., 1991, Sequences of Proteins of Immunological Interest, 5th ed. (определяющие комплементарность области); Chothia et al., 1989, Nature 342:877-883 (структуры структурных петель). Определение CDR AbM является компромиссом между Kabat и Chothia, и в нем используется программа для моделирования антител AbM от Oxford Molecular (Accelrys®). «Контактное» определение CDR основывается на наблюдаемых контактах с антигенами, изложенных в MacCallum et al., 1996, J. Mol. Biol., 262:732-745. «Конформационное» определение CDR основывается на остатках, которые делают энтальпический вклад в связывание антигена (см., например, Makabe et al., 2008, Journal of Biological Chemistry, 283:1156-1166). Другие определения границ CDR все же могут строго не следовать одному из приведенных выше подходов, но, тем не менее, перекрываются по меньшей мере с частью CDR по Kabat, хотя они могут быть сокращены или удлинены в свете прогнозирования или экспериментальных данных о том, что конкретные остатки или группы остатков или даже целые CDR значимо не влияют на связывание антигена. Термин «CDR» в том виде, в котором он в настоящем описании используется, может относиться к CDR, определенным любым подходом, известным в этой области, включая комбинации подходов.
В Примерах (см. Таблицы 3 и 4) CDR определяются следующим образом (нумерация согласно Kabat; H: тяжелая цепь; L: легкая цепь):
CDR-H1: H26-H35B; CDR-H2: H50-H65; CDR-H3: H95-H102
CDR-L1: L24-L34; CDR-L2: L50-L56; CDR-L3: L89-L97
Остатки «каркасной области» (FR) представляют собой остатки вариабельного домена антитела, отличные от остатков CDR. Каркасная область домена VH или VL содержит четыре каркасные подобласти: FR1, FR2, FR3 и FR4, перемежающиеся с CDR в следующей структуре: FR1 - CDR1 - FR2 - CDR2 - FR3 - CDR3 - FR4. В Примерах (см. Таблицы 3 и 4) остатки FR включают следующие (нумерация согласно Kabat; H: тяжелая цепь; L: легкая цепь):
Термин «эпитоп» относится к поверхности или области антигена (Ag), с которой специфично связывается антитело, например, к поверхности или области, содержащей остатки, которые взаимодействуют с антителом (Ab). Эпитопы могут быть линейными или конформационными. В линейном эпитопе все точки взаимодействия между белком и взаимодействующей молекулой (такой как антитело) находятся линейно вдоль первичной аминокислотной последовательности белка. «Нелинейный эпитоп» или «конформационный эпитоп» содержит несмежные полипептиды (или аминокислоты) в пределах антигенного белка, с которым связывается антитело, специфичное к данному эпитопу. Термин «эпитоп» в том виде, в котором он в настоящем описании используется, определяется как часть антигена, с которой может специфично связываться антитело, при определении любым способом, хорошо известным в этой области, например, традиционными иммуноанализами. В качестве альтернативы, получение и характеристика антител во время процесса открытия могут прояснять информацию о желательных эпитопах. Из этой информации затем можно подвергать антитела конкурентному скринингу на связывание с тем же самым эпитопом. Подходом для достижения этого является проведение конкурентных и перекрестноконкурентных исследований для нахождения антител, которые конкурируют или перекрестно конкурируют друг с другом за связывание с TFPI. То есть, антитела конкурируют за связывание с антигеном таким образом, что данные антитела конкурируют за связывание с антигенсвязывающим сайтом антитела против TFPI по настоящему изобретению.
Термин «паратоп» происходит из приведенного выше определения «эпитоп» посредством обращения ракурса и относится к поверхности или области молекулы антитела, которая участвует в связывании антигена, например, к площади или области, содержащей остатки, которые взаимодействуют с антигеном. Паратоп может быть линейным или конформационным (таким как прерывающиеся остатки в CDR).
Эпитоп/паратоп для этой пары связывания антитело/антиген может быть определен и охарактеризован с разными уровнями детализации с использованием целого ряда экспериментальных и расчетных методов картирования эпитопов. Экспериментальные методы включают мутагенез, рентгеноструктурный анализ, спектроскопию ядерного магнитного резонанса (ЯМР), масс-спектрометрию с обменом водорода/дейтерия (ВД-МС) и разные способы кокурентного связывания. Поскольку каждый способ полагается на уникальный принцип, описание эпитопа тесно связано со способом, посредством которого он был определен. Таким образом, эпитоп/паратоп для этой пары антитело/антиген будет определен по-разному, в зависимости от использованного способа картирования.
На самом детализированном уровне эпитоп/паратоп для взаимодействия между Ag (антиген) и Ab (антитело) может быть определен посредством пространственных координат, определяющих контакты атомов, присутствующие во взаимодействии Ag-Ab, а также информации об их относительных вкладах в термодинамику связывания. На одном уровне остаток эпитопа/паратопа может быть охарактеризован пространственными координатами, определяющими контакты атомов между Ag и Ab. В одном аспекте остаток эпитопа/паратопа может быть определен по конкретному критерию, например, расстоянию между атомами в Ab и Ag (например, расстояние, равное или меньшее, чем 4Å от тяжелого атома когнатного антитела до тяжелого атома антигена («контактные» остатки)). В другом аспекте остаток эпитопа/паратопа может быть охарактеризован как участвующий во взаимодействии на основе водородной связи с когнатным антителом/антигеном или с молекулой воды, которая также связывается водородной связью с когнатным антителом/антигеном (опосредованное водой связывание водородными связями). В другом аспекте остаток эпитопа/паратопа может быть охарактеризован как образующий солевой мостик с остатком когнатного антитела/антигена. В еще одном другом аспекте остаток эпитопа/паратопа может быть охарактеризован как остаток, имеющий ненулевое изменение в скрытой области поверхности (BSA), из-за взаимодействия с когнатным антителом/антигеном. На другом менее подробном уровне эпитоп/паратоп может быть охарактеризован через функцию, например, посредством конкурентного связывания с другими Ab. Эпитоп/паратоп также может быть определен более общим способом, как содержащий аминокислотные остатки, для которых замена другой аминокислотой будет изменять характеристики взаимодействия между Ab и Ag (например, аланиновое сканирование).
В контексте полученной рентгеноструктурным анализом кристаллической структуры, определенной пространственными координатами комплекса между антителом, например, фрагментом Fab или двумя фрагментами Fab, и его антигеном, если не определено иначе, остаток эпитопа относится к остатку TFPI, (1) имеющему тяжелый атом (т.е. атом, не являющийся атомом водорода), который находится в пределах расстояния 4 Å от тяжелого атома когнатного антитела (также именуемые «контактные» остатки); (2) участвующему в водородной связи с остатком когнатного антитела или с молекулой воды, которая также связывается водородной связью с когнатным антителом (опосредованное водой связывание водородными связями), (3) участвующему в солевом мостике с остатком когнатного антитела, и/или (4) имеющему ненулевое изменение в скрытой области поверхности (BSA) из-за взаимодействия с когнатным антителом. В общем, для BSA налагается пороговое значение для того, чтобы избегать включения остатков, которые имеют минимальные взаимодействия. Следовательно, если не определено иначе, остатки эпитопа в категории (4) отбираются, если они имеют BSA 20 Å2 или больше, или участвуют в элетростатических взаимодействиях при связывании антитела с TFPI. Аналогичным образом, в контексте кристаллической структуры, полученной рентгеноструктурным анализом, если не определено иначе или не противоречит контексту, остаток паратопа относится к остатку антитела, (1) имеющему тяжелый атом (т.е. атом, не являющийся атомом водорода), который находится в пределах расстояния 4 Å от тяжелого атома TFPI (также именуемые «контактные» остатки); (2) участвующему в водородной связи с остатком TFPI или с молекулой воды, которая также связывается водородной связью с TFPI (опосредованное водой связывание водородными связями), (3) участвующему в солевом мостике с остатком TFPI, и/или (4) имеющему ненулевое изменение в скрытой области поверхности, из-за взаимодействия с TFPI. Опять-таки, если не определено иначе, остатки паратопа в категории (4) отбираются, если они имеют BSA 20 Å2 или больше, или участвуют в электростатических взаимодействиях при связывании антитела с TFPI.
Из того факта, что описания и определения эпитопов зависят от использованного способа картирования эпитопов и получаются с разными уровнями детализации следует то, что сравнения эпитопов для разных Ab на одном и том же Ag, аналогичным образом, могут проводиться с разными уровнями детализации. Например, говорят, что эпитопы, описанные на уровне аминокислот, например, определенные из рентгеновской структуры, являются идентичными, если они содержат тот же самый набор аминокислотных остатков. Говорят, что эпитопы являются отдельными (уникальными), если между эпитопами нет общего аминокислотного остатка. Говорят, что эпитопы, характеризуемые конкурентным связыванием, являются перекрывающимися, если связывание соответствующих антител является взаимоисключающим, т.е. связывание одного антитела исключает одновременное или последовательное связывание другого антитела; и говорят, что эпитопы являются отдельными (уникальными), если антиген способен обеспечивать одновременное связывание обоих соответствующих антител.
Эпитоп и паратоп для этой пары антитело/антиген могут быть идентифицированы традиционными методами. Например, общее расположение эпитопа может быть определено посредством оценки способности антитела к связыванию с разными фрагментами или вариантами полипептидов TFPI, как в настоящем описании было ранее более подробно описано в других местах. Конкретные остатки в пределах TFPI, которые образуют контакт с конкретными остатками в пределах антитела, также могут быть определены с использованием традиционных методов, таких как методы, описанные в примерах. Например, может быть закристаллизован коплекс антитело/антиген. Кристаллическая структура может быть определена и использована для идентификации специфических сайтов взаимодействия между антителом и антигеном.
Термин «конкурировать» в том виде, в котором он используется в настоящем описании в отношении антитела, означает то, что связывание первого антитела или его антигенсвязывающей части с антигеном уменьшает последующее связывание того же самого антигена вторым антителом или его антигенсвязывающей частью. В общем, связывание первого антитела создает стерическое препятствие, конформационное измерение или связывание с общим эпитопом (или его частью) таким образом, что связывание второго антитела с тем же самым антигеном уменьшаеся. Для определения того, конкурируют ли друг с другом два антитела, можно использовать стандартные конкурентные анализы. Один подходящий анализ для конкуренции антител включает применение технологии Biacore, в которой может измеряться степень взаимодействий с использованием технологии поверхностного плазмонного резонанса (SPR), как правило, с использованием системы биосенсора (такой как система BIACORE®). Например, SPR можно использовать в анализах конкурентного ингибирования связывания in vitro для определения способности одного антитела ингибировать связывание второго антитела. В другом анализе для измерения конкуренции антител используется подход на основе ELISA. Кроме того, высокопроизводительный способ «сортировки» антител на основе их конкуренции описан в международной патентной заявке № WO2003/48731. Конкуренция присутстует, если одно антитело (или фрагмент) уменьшает связывание другого антитела (или фрагмента) с TFRI. Например, можно использовать конкурентный анализ последовательного связывания, с последовательным добавлением разных антител. Первое антитело может добавляться с достижением связывания, которое близко к насыщению. Затем добавлется второе антитело. Если связывание второго антитела с TFPI не выявляется или значительно снижается (например, снижение по меньшей мере примерно на 10%, по меньшей мере примерно на 20%, по меньшей мере примерно на 30%, по меньшей мере примерно на 40%, по меньшей мере примерно на 50%, по меньшей мере примерно на 60%, по меньшей мере примерно на 70%, по меньшей мере примерно на 80% или по меньшей мере примерно на 90%) по сравнению с параллельным анализом в отсутствие первого антитела (значение для которого может быть принято за 100%), два антитела считаются конкурирующими друг с другом. Типичный конкурентный анализ антител (и анализ перекрывающихся эпитопов) посредством SPR приводится в Примере 6.
Антитело против TFPI по изобретению может иметь способность конкурировать или перекрестно конкурировать с другим антителом по изобретению за связывание с TFPI, как описано в настоящем описании. Например, антитело по изобретению может конкурировать или перекрестно конкурировать с описанными в настоящем описании антителами за связывание с TFPI или с подходящим фрагментом или вариантом TFPI, который связывается раскрытыми в настоящем описании антителами.
То есть, если первое антитело против TFPI конкурирует со вторым антителом за связывание с TFPI, но оно не конкурирует, когда с TFPI сначала связывается второе антитело, оно считается «конкурирующим» со вторым антителом (что также называется односторонней конкуренцией). Когда антитело конкурирует с другим антителом, независимо от того, какое антитело сначала связывается с TFPI, тогда антитело «перекрестно конкурирует» за связывание с TFPI с другим антителом. Такие конкурирующие или перекрестноконкурирующие антитела могут быть идентифицированы на основе их способности конкурировать/перекрестно конкурировать с известным антителом по изобретению в стандартных анализах связывания. Для демонстрации конкуренции/перекрестной конкуренции можно использовать, например, SPR, например, с использованием системы Biacore™, анализы ELISA или проточную цитометрию. Такая конкуренция/перекрестная конкуренция может свидетельствовать о том, что два антитела связываются с идентичными, перекрывающимися или аналогичными эпитопами.
Антитело против TFPI по настоящему изобретению, следовательно, может быть идентифицировано способом, который включает анализ связывания, в котором оценивается то, способно ли или нет тестируемое антитело конкурировать/перекрестно конкурировать с контрольным антителом, раскрытым в описании (например, TFPI-3, TFPI-21, TFPI-23, TFPI-24, TFPI-26, TFPI-106, TFPI-107, TFPI-108, TFPI-109, TFPI-110, TFPI-111, TFPI-112, TFPI-113, TFPI-114, 4D8, 6B7.c5, 7A4.D9), за сайт связывания на молекуле-мишени.
Белок «слияние с Fc» представляет собой белок, в котором один или более чем один полипептид связан функциональным образом с полипептидом Fc. Слияние с Fc объединяет область Fc иммуноглобулина с партнером слияния.
Аффинность связывания антитела может быть выражена в виде значения Kd, которое относится к скорости диссоциации конкретного взаимодействия антиген-антитело. Kd представляет собой отношение скорости диссоциации, также именуемой koff, к скорости ассоциации или kon. Таким образом, Kd равна koff / kon и выражается в виде молярной концентрации (н), и чем меньше Kd, тем сильнее аффинность связывания. Значения Kd для антител можно определять с использованием способов, хорошо отработанных в этой области. Одним типичным способом измерения Kd является поверхностный плазмонный резонанс (SPR), типично с использованием системы биосенсора, такой как система BIACORE®. Кинетический анализ BIAcore включает анализирование связывания и диссоциации антигена от чипов с иммобилизованными молекулами (например, молекулами, содержащими эпитопсвязывающие домены) на их поверхности. Другим способом определения Kd антитела является использование интерферометрии Bio-Layer, как правило, с использованием технологии OCTET® (система Octet QKe, ForteBio). Альтернативно или дополнительно, также можно использовать анализ KinExA® (анализ кинетического исключения), доступный от Sapidyne Instruments (Boise, Id.).
Термин «терапевтически эффективное количество» означает количество антитела против TFPI или его фрагмента, или комбинации, содержащей такое антитело или его фрагмент, которое является достаточным количеством для достижения намеченной цели, такой как увеличение свертывания, или, в случае гемофилии, уменьшение времени свертывания, или принесение заметной пользы in vivo субъекту, нуждающемуся в этом, иным способом. Точное количество будет зависеть от многочисленных факторов, включающих компоненты и физические характеристики терапевтической композиции, намеченную популяцию пациентов, особенности индивидуального пациента и тому подобное, но не ограничивающихся ими, и может быть определено специалистом в этой области.
Под термином «синергическое терапевтически эффективное количество» подразумевается количество антитела против TFPI или его антигенсвязывающего фрагмента, которое, при предоставлении со вторым терапевтическим агентом, например, фактором VIIa (FVIIa), обеспечивает заметную пользу (например, уменьшенное время свертывания, уменьшенное время кровотечения, усиленное образование фибрина, повышенное накопление тромбоцитов и тому подобное), которая больше, чем аддитивный измеримый эффект каждого терапевтического агента или антитела, вводимых поодиночке.
Термин «лечение» включает профилактическое и/или терапевтическое лечения. Если введение осуществляетсяч до клинического проявления состояния, то лечение считается профилактическим. Терапевтическое лечение включает, например, уменьшение интенсивности или ослабление тяжести заболевания, или сокращение продолжительности заболевания.
Термин «примерно» в том виде, в котором он в настоящем описании используется, относится к плюс/минус 10% от значения.
3. Антитела против TFPI
В настоящем описании раскрыты и проиллюстрированы примерами антитела (и их антигенсвязывающие фрагменты), которые связываются с ингибитором пути тканевого фактора (TFPI). Данные антитела и фрагменты антител связываются с уникальными эпитопами TFPI. В некоторых воплощениях распознавание определенных остатков эпитопов в TFPI позволяет антителам (и их антигенсвязывающим фрагментам) уменьшать активность TFPI. Кроме того, в некоторых воплощениях раскрытые в настоящем описании антитела (и их антигенсвязывающие фрагменты) продемонстрировали желательные фармакологические активности и фармакокинетические свойства для лечения недостаточностей свертывания и уменьшения времени кровотечения.
А. Ингибитор пути тканевого фактора (TFPI)
TFPI представляет собой ингибитор протеаз, содержащий многовалентный домен Кунитца. Типичные последовательности TFPI человека, мыши, яванского макака, кролика и крысы приведены в Таблице 2.
Человеческий TFPI представляет собой внеклеточный гликопротеин с двумя преобладающими формами - TFPI-альфа и TFPI-бета. TFPI-альфа, который представляет собой гликозилированный белок из 276 аминокислот (MW (молекулярная масса) 43 кДа), является наибольшей формой TFPI и состоит из трех доменов, подобных домену Кунитца, и основной карбоксиконцевой области. Альтернативный сплайсинг дает TFPI-бета, который содержит домен Кунитца 1 (K1) и домен Кунитца 2 (K2), но содержит и альтернативную С-концевую часть, не имеющую домена Кунитца 3 (K3) и основной области. TFPI-бета заякоривается в клеточных мембранах посредством посттрансляционной модификации гликозилфосфатидилинозитольным (GPI) якорем.
Первичными мишенями TFPI являются протеазы Фактор Xa (FXa) и Фактор VIIa (FVIIa), которые являются ключевыми факторами на стадии инициации каскада свертывания крови. Биохимический анализ выявил то, что K2 является ингибитором FXa, тогда как K1 ингибирует комплекс FVIIa-тканевой фактор. Роль K3 не ясна, так как он, по-видимому, не имеет прямой активности протеазного ингибитора, но может служить в качестве сайта распознавания для кофактора белка S. С-концевой домен, уникальный для TFPI-альфа, может участвовать в распознавании протромбиназы на поверхности тромбоцитов.
Домен Кунитца 1 (K1) соответстует аминокислотным остаткам 26-76 SEQ ID NO: 2, и домен Кунитца 2 (K2) соответстует остаткам 91-147 SEQ ID NO: 2. Домены K1 и K2 из других гомологов, изоформ, вариантов или фрагментов TFPI могут быть идентифицированы посредством выравнивания последовательностей или структурного выравнивания относительно SEQ ID NO: 2.
TFPI согласно настоящему описанию включает любую встречающуюся в природе форму TFPI, которая может происходить из любого подходящего организма. Например, TFPI может представлять собой TFPI млекопитающего, как, например, TFPI человека, мыши, крысы, примата, не являющегося человеком, коровы, овцы, собаки, кошки или свиньи. В некоторых воплощениях TFPI представляет собой человеческий TFPI. TFPI может представлять собой зрелую форму TFPI (т.е. белок TFPI, который подвергся посттрансляционному процессингу в подходящей клетке). Такой зрелый белок TFPI, например, может быть гликозилирован.
TFPI согласно настоящему описанию включает любые функциональные фрагменты или варианты, происходящие из TFPI, встречающегося в природе. Функциональный фрагмент TFPI может представлять собой любую часть или фрагмент TFPI, который сохраняет активность TFPI, такую как способность ингибировать Фактор Xa (FXa), ингибировать активность комплекса FVIIa-тканевой фактор и/или функцию, в качестве отрицательного регулятора свертывания или гемостаза. Например, функциональный фрагмент может содержать домен Кунитца, такой как домен K1, домен K2 или оба домена K1 и K2 TFPI.
Функциональный вариант может содержать одну или более чем одну мутацию по сравнению с встречающимся в природе TFPI и все еще сохранять активность встречающегося в природе TFPI, такую как способность ингибировать Фактор Xa (FXa) или способность ингибировать активность комплекса FVIIa-тканевой фактор. Например, вариант может иметь разные степени идентичности последовательности с SEQ ID NO: 1, 2, 3, 4, 5, 6 или 7, например быть по меньшей мере на 60%, 70%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичным последовательности, приведенной в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 или SEQ ID NO: 7.
Фрагменты, варианты, изоформы и гомологи TPFI по изобретению должны сохранять важные остатки эпитопа (такие как Ile105, Arg107 и Leu131, если используются антитела TFPI-23 и TFPI-24), как описано в настоящем описании. Кроме того, TFPI может содержать пять или более чем пять, восемь или более чем восемь, десять или более чем десять, двенадцать или более чем двенадцать, или пятнадцать или более чем пятнадцать доступных с поверхности остатков домена K2 TFPI. Доступный с поверхности остаток представляет собой остаток, имеющий больше чем 40%-ную относительную доступность.
Например, для домена K2 TFPI (см., например, SEQ ID NO: 2) следующие аминокислотные остатки имеют больше чем 40%-ную относительную доступность: 94-95, 98, 100-110, 118-121, 123-124, 131, 134, 138-142 и 144-145. TFPI может содержать пять или более чем пять, восемь или более чем восемь, десять или более чем десять, двенадцать или более чем двенадцать, или пятнадцать или более чем пятнадцать данных остатков, как, например, фрагмент TFPI, который включает пять или более чем пять, восемь или более чем восемь, десять или более чем десять, двенадцать или более чем двенадцать, или пятнадцать или более чем пятнадцать данных остатков
Б. Антитела против TFPI
Антитело или его антигенсвязывающий фрагмент по изобретению специфично связывается с доменом K2 TFPI и может ингибировать его взаимодействие с FXa и/или уменьшать активности TFPI.
TFPI-23 и варианты
В одном аспекте данное изобретение включает антитело TFPI-23 и варианты
TFPI-23, которые были созданы для увеличения содержания остатков каркасной области человеческой зародышевой линии («модифицированные на уровне зародышевой линии»). Например, TFPI-106 содержит мутации H1Q на Е и H3V на L (нумерация по Kabat), и TFPI-107 содержит мутации H1Q на Е, H5V на L и H94I на K (нумерация по Kabat). Для целей данного изобретения родительское антитело TFPI-23 и вариант зародышевой линии TFPI-106 являются взаимозаменимыми в взаимодействиях остатка эпитопа и остатка паратопа.
В одном аспекте согласно изобретению предложено выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с эпитопом в домене Кунитца 2 (K2) ингибитора пути тканевого фактора (TFPI), где указанный эпитоп содержит остатки Ile105, Arg107 и Leu131 согласно нумерации SEQ ID NO: 2. В некоторых воплощениях антитело или его антигенсвязывающий фрагмент не связывается с доменом Кунитца 1 (K1) TFPI.
Как раскрыто и проиллюстрировано примерами в настоящем описании, на основе структуры сокристалла и расчетного аланинового сканирования, были обнаружены уникальные эпитопы в домене K2 TFPI. В частности, кристаллическая структура показывает то, что домен K2 TFPI принимает конусообразную структуру с вершиной конуса (в особенности, Arg107), связывающейся с FXa. Все TFPI-23, TFPI-24 и их варианты распознают остатки около вершины этой конусообразной области и блокируют связывание TFPI с FXa. Следовательно, антитела, распознающие остатки эпитопа, расположенные около вершины конуса, являются особенно полезными для ингибирования активностей TFPI.
В некоторых воплощениях согласно изобретению раскрыты эпитопы TFPI, которые содержат три остатка, которые являются важными для взаимодействия антитело-антиген: Ile105, Arg107 и Leu131 (согласно нумерации человеческого TFPI, как показано в SEQ ID NO: 2). Мутирование данных трех остатков на аланин приводит к потере связывания TFPI-23, TFPI-24 и их вариантов. См. Таблицу 28, в которой обобщены результаты аланинового сканирования.
Также были идентифицированы дополнительные остатки TPFI, участвующие в связывании с антителом, но эти остатки могут быть мутированы на аланин без значительного дестабилизирующего эффекта. См. Таблицу 28, в которой обобщены результаты аланинового сканирования.
Соответственно, в некоторых воплощениях эпитопы TFPI дополнительно содержат один или более чем один остаток, выбранный из группы, состоящей из Cys106, Gly108, Cys130, Gly132 (согласно нумерации SEQ ID NO: 2), и их любой комбинации. Данные остатки эпитопа распознаются TFPI-23, TFPI-24 и их вариантами. См. Таблицу 27, в которой показаны общие остатки эпитопов для TFPI-23 и TFPI-24.
В некоторых воплощениях эпитоп дополнительно содержит один или более чем один остаток, выбранный из группы, состоящей из: Asp102, Arg112, Tyr127, Gly129, Met134, Glu138 (согласно нумерации SEQ ID NO: 2) и любой их комбинации. Эти остатки эпитопов распознаются TFPI-23 и его вариантами (например, TFPI-106, TFPI-107), но не TFPI-24 (и его вариантами). См. Таблицу 27.
В некоторых воплощениях эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из: E100, E101, P103, Y109, T111, Y113, F114, N116, Q118, Q121, C122, E123, R124, F125, K126, L140 (нумерация согласно SEQ ID NO: 2) и их любой комбинации. См. Таблицу 27. Согласно WO201007269 (Novo Nordisk), контрольное антитело 4F36 распознает эпитоп, содержащий E100, E101, P103, Y109, T111, Y113, F114, N116, Q118, Q121, C122, E123, R124, F125, K126 и L140.
В некоторых воплощениях эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из: D31, D32, P34, C35, K36, E100, E101, P103, Y109, K126, G128 (нумерация согласно SEQ ID NO: 2) и их любой комбинации. См. Таблицу 27. Согласно Таблице 27 контрольные антитела 2А8 и 2А8-200 распознают эпитоп, содержащий D31, D32, P34, C35, K36, E100, E101, P103, Y109, K126 и G128.
В некоторых воплощениях эпитоп может относиться к одному или более чем одному «контактному» остатку TFPI (имеющему тяжелый атом (т.е. атом, не являющийся атомом водорода), который находится в пределах расстояния 4 Å от тяжелого атома когнатного антитела) и содержит один или более чем один остаток, выбранный из группы, состоящей из Asp102, Gly104, Ile105, Cys106, Arg107, Gly108, Arg112, Tyr127, Gly129, Cys130, Leu131, Gly132, Met134, Glu138 (согласно нумерации SEQ ID NO: 2) и любой их комбинации. См. Таблицу 29Б.
В некоторых воплощениях эпитоп может относиться к одному или более чем одному остатку TFPI, участвующему в водородной связи с остатком антитела или с молекулой воды, которая также связана водородной связью с когнатным антителом (опосредованное водой связывание водородными связями), и он содержит один или более чем один остаток, выбранный из группы, состоящей из: Asp102, Arg107, Arg112, Tyr127 и Leu131 (согласно нумерации SEQ ID NO: 2) и любой их комбинации. Данные остатки эпитопа участвуют в водородной связи с когнатным антителом. См., Таблицу 29Б.
В некоторых воплощениях эпитоп может относиться к остаткам, имеющим ненулевое изменение в скрытой области поверхности (BSA), из-за взаимодействия с когнатным антителом, и он содержит один или более чем один остаток, выбранный из группы, состоящей из: Asp102, Gly104, Ile105, Cys106, Arg107, Gly108, Arg112, Tyr127, Gly129, Cys130, Leu131, Gly132, Asn133, Met134, Glu138 (согласно нумерации SEQ ID NO: 2) и любой их комбинации. См., Таблицу 29Б.
Данным изобретением также охватывается любая комбинация этих разных категорий остатков эпитопа.
В некоторых воплощениях эпитоп содержит по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13 или все описанные выше остатки эпитопа, или любую комбинацию разных категорий описанных выше остатков эпитопа.
Остатки паратопа TFPI-23 (и вариантов) могут относиться к следующим контактным остаткам (в пределах 4 Å от остатка эпитопа TFPI): H33 Ala, H47 Trp, H50 Ala, H51 Ile, H52 Ser, H56 Ser, H58 Tyr, H95 Leu, H96 Gly, H97 Ala, H98 Thr, H99 Ser, H100 Leu, H100A Ser, L29 Ala, L31 Tyr, L91 Tyr, L95A Ser, L95B Gly, и L95C Ser; и возможно L93 Ser и L96 Gly (нумерация согласно Kabat). L93 Ser (4,07 Å) и L96 Gly (4,03 Å) являются опцией из-за того, что расстояния незначительно превышают 4 Å, но являются достаточно близкими для округления до 4 Å.
Заметьте то, что приведенные выше контактные остатки представляют собой исходные остатки из антитела TFPI-23. Однако на основе структурного анализа и аланинового сканирования, считается, что целый ряд контактных остатков в TFPI-23 может быть заменен на другой остаток без значительного влияния на связывание антигена. Например, в Таблице 29А показано то, что целый ряд контактных остатков в TFPI-23 может быть заменен на другие остатки и приводит к эффекту на связывание или стабильность, составляющему лишь меньше, чем 0,02 кДж/моль («меньше, чем 0,02 кДж/моль» означает то, что замена имеет нейтральный эффект на связывание). В частности, как показано в Таблице 29А, столбец 4, три положения CDR и 1 положение каркасной области: H47, H58, L91 и L96 (нумерация согласно Kabat) допускают только один или два остатка: (a) H47 представляет собой Trp или Tyr; (б) H58 представляет собой Tyr; (в) L91 представляет собой Tyr или Arg; и (г) L96 представляет собой Gly или Asn. Другие положения CDR могут допускать больше замен, как обобщено в Таблице 29А, столбец 4.
Соответственно, в некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие остатки (нумерация согласно Kabat):
(a) H33 представляет собой Ala, Asn, Gly, His, Lys, Met, Phe, Pro, Ser, Thr, Trp или Val;
(б) H47 представляет собой Trp или Tyr;
(в) H50 представляет собой Ala, Arg, Gly, Lys, Met, Phe, Pro, Ser, Thr, Tyr или Val;
(г) H51 представляет собой Ile, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val;
(д) H52 представляет собой Ser, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Trp, Tyr или Val;
(е) H56 представляет собой Ser, Arg, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val;
(ж) H58 представляет собой Tyr;
(з) H95 представляет собой Leu, Gln, Ile, Phe или Tyr;
(и) H96 представляет собой Gly, Ala, Arg, Asn, Asp, Gln, Ile, Lys, Met, Phe, Pro, Ser, Thr или Val;
(к) H97 представляет собой Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val;
(л) H98 представляет собой Thr, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val;
(м) H99 представляет собой Ser, Ala, Gly, Phe или Pro;
(н) H100 представляет собой Leu, Arg, His, Ile, Leu, Lys, Phe, Pro, Trp, Tyr или Val;
(о) H100A представляет собой Ser, Ala, Arg, Asn, Asp, Gln, Glu, His, Leu, Lys, Met, Phe, Pro, Ser, Thr или Trp;
(п) L29 представляет собой Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe Pro, Ser, Thr, Trp, Tyr или Val;
(р) L31 представляет собой Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val;
(с) L91 представляет собой Tyr или Arg;
(т) L95A представляет собой Ser, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val;
(у) L95B представляет собой Ser, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; и
(ф) L95C представляет собой Ser, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val;
и возможно содержит (х) L93, представляющий собой Tyr, Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val; и
(х) L96, представляющий собой Gly или Asn.
Среди данных остатков Н47 представляет собой остаток каркасной области; а все другие представляют собой остатки CDR.
При наложении более строгого критерия замены - замена должна приводить к аффинности меньше -0,02 кДж/моль, означая то, что данная замена должна иметь позитивный (нейтральный/стабилизирущий) эффект - контактные остатки являются следующими (нумерация согласно Kabat) (Таблица 29А, столбец 5):
(a) H33 представляет собой Ala или Val;
(б) H47 представляет собой Trp;
(в) H50 представляет собой Ala;
(г) H51 представляет собой Ile;
(д) H52 представляет собой Ser, Arg, Lys, Phe или Tyr;
(е) H56 представляет собой Ser, Arg или Lys;
(ж) H58 представляет собой Tyr;
(з) H95 представляет собой Leu;
(и) H96 представляет собой Gly, Ala, Arg, Asn, Lys, Pro, Ser или Val;
(к) H97 представляет собой Ala;
(л) H98 представляет собой Thr, His, Ile, Leu, Met, Phe или Tyr;
(м) H99 представляет собой Ser;
(н) H100 представляет собой Leu, Phe, Trp или Tyr;
(о) H100A представляет собой Ser, Arg, Asn, Gln, Glu, His, Leu, Lys, Met, Phe, Pro или Trp;
(п) L29 представляет собой Ala;
(р) L31 представляет собой Tyr;
(с) L91 представляет собой Tyr;
(т) L95A представляет собой Ser, Phe, Trp или Tyr;
(у) L95B представляет собой Gly; и
(ф) L95C представляет собой Ser, Arg, Asn, Gln, Glu, Ile, Leu, Lys, Met, Phe, Trp, Tyr или Val;
и возможно: (х) L93 представляет собой Ser; и
(ц) L96 представляет собой Gly.
В качестве альтернативы или дополнительно, выбор подходящих замен может быть основан на столбце 6 Таблицы 29А, где верхние 3 остатка отбираются на основе их влияния на аффинность (т.е. 3 лидирующих прогнозируемых сайта с самым стабилизирующим эффектом на аффинность). При этих критериях контактными остатками являются следующие (нумерация согласно Kabat):
(a) H33 представляет собой Ala, Val, His или Phe;
(б) H47 представляет собой Trp или Tyr;
(в) H50 представляет собой Ala, Thr, Ser или Phe;
(г) H51 представляет собой Ile, Arg, Lys или Pro;
(д) H52 представляет собой Ser, Phe, Arg или Tyr;
(е) H56 представляет собой Ser, Lys, Tyr или Phe;
(ж) H58 представляет собой Tyr;
(з) H95 представляет собой Leu, Ile, Gln или Phe;
(и) H96 представляет собой Gly, Arg, Asn или Lys;
(к) H97 представляет собой Ala, Leu, Tyr или Ile;
(л) H98 представляет собой Thr, Tyr, Phe или His;
(м) H99 представляет собой Ser, Pro, Ala или Phe;
(н) H100 представляет собой Leu, Tyr, Trp или Phe;
(о) H100A представляет собой Ser, Arg, Leu или Trp;
(п) L29 представляет собой Ala, Glu, Asp или Gln;
(р) L31 представляет собой Tyr, Glu, Asp или Trp;
(с) L91 представляет собой Tyr или Arg;
(т) L95A представляет собой Ser, Phe, Tyr или His;
(у) L95B представляет собой Gly, Glu, Asp или Pro; и
(ф) L95C представляет собой Ser, Trp, Tyr или Phe;
и, возможно: (х) L93 представляет собой Ser, Glu, Asp или His;
(ц) L96 представляет собой Gly или Asn.
Остатки паратопа TFPI-23 (и вариантов) также могут относиться к остаткам, участвующим в водородной связи с остатком TFPI или с молекулой воды, которая также связана водородной связью с TFPI, и включают следующие: H58 Tyr, H96 Gly, H97 Ala, H98 Thr, H99 Ser, H100 Leu, L29 Ala, L31 Tyr и L95B Gl (нумерация согласно Kabat). См., Таблицу 29Б.
Остатки паратопа TFPI-23 (и вариантов) также могут относиться к остаткам, имеющим ненулевое изменение в BSA из-за взаимодействия с TFPI, и включают следующие (нумерация согласно Kabat): H33 Ala, H58 Tyr, H95 Leu, H96 Gly, H97 Ala, H98 Thr, H99 Ser, H100 Leu, H100A Ser, L29 Ala, L31 Tyr, L91 Tyr, L93 Ser, L95A Ser и L95B Gly. Применяется пороговое значение (BSA 20 Å2 или больше, или участие в электростатическом взаимодействии) для того, чтобы избежать включения остатков, которые имеют минимальные взаимодействия. См. Таблицу 29Б.
Если не применяется пороговое значение BSA, остатки паратопа включают следующие: H33 Ala, H34 Met, H47 Trp, H50 Ala, H51 Ile, H52 Ser, H56 Ser, H58 Tyr, H95 Leu, H96 Gly, H97 Ala, H98 Thr, H99 Ser, H100 Leu, H100A Ser, L28 Gly, L29 Ala, L31 Tyr, L91 Tyr, L93 Ser, L94 Ser, L95A Ser, L95B Gly, L95C Ser и L96 Gly. См. Таблицу 29В.
Антитело или его антигенсвязывающий фрагмент по изобретению может связываться с тем же самым эпитопом или доменом TFPI, что и антитела, которые конкретно проиллюстрированы в настоящем описании примерами. Например, антитело или его антигенсвязывающий фрагмент может быть идентифицировано сравнением его связывания с TFPI со связыванием TFPI-23 или гуманизированных вариантов (например, TFPI-106 и TFPI-107); или сравнением функции данных антител с TFPI-23 и его вариантами. Анализы и тесты, которые можно использовать с целью такой идентификации, включают анализы, оценивающие конкуренцию за связывание с TFPI, и они иллюстрируются в Примерах.
В одном воплощении антитело или его антигенсвязывающий фрагмент по изобретению может связываться с тем же самым эпитопом или областью, что и описанные в настоящем описании антитела, такие как TFPI-23 и его варианты. Это может включать нахождение в контакте с определенным остатком TFPI, как описано выше. Например, антитело или антигенсвязывающий фрагмент по изобретению может связываться с TFPI таким образом, что оно находится в контакте (в пределах 4 Å) с остатком, выбранным из группы, состоящей из: Asp102, Gly104, Ile105, Cys106, Arg107, Gly108, Arg112, Tyr127, Gly129, Cys130, Leu131, Gly132, Met134, Glu138 (согласно нумерации SEQ ID NO: 2) и любой их комбинации. Антитело или антигенсвязывающий фрагмент по изобретению может быть способно к связыванию с эпитопом, содержащим один или более чем один остаток, выбранный из группы, состоящей из Asp102, Gly104, Ile105, Cys106, Arg107, Gly108, Arg112, Tyr127, Gly129, Cys130, Leu131, Gly132, Met134, Glu138 (согласно нумерации SEQ ID NO: 2) и любой их комбинации.
Антитело и его антигенсвязывающий фрагмент может содержать по меньшей мере один остаток паратопа (нумерация согласно Kabat), который находится в пределах 4,0 Å от по меньшей мере одного остатка эпитопа TFPI (нумерация согласно SEQ ID NO: 2) следующим образом: остаток эпитопа 102 Asp находится в пределах 4,0 Å от остатка паратопа H58 Tyr; остаток эпитопа 104 Gly находится в пределах 4,0 Å от остатка паратопа H58 Tyr; остаток эпитопа 105 Ile находится в пределах 4,0 Å от остатков паратопа H33 Ala, H50 Ala, H51 Ile, H52 Ser, H56 Ser, H58 Tyr, H95 Leu; остаток эпитопа 106 Cys находится в пределах 4,0 Å от остатков паратопа H100 Leu, H100A Ser; остаток эпитопа 107 Arg находится в пределах 4,0 Å от остатков паратопа H96 Gly, H97 Ala, H98 Thr, H99 Ser, H100 Leu; остаток эпитопа 108 Gly находится в пределах 4,0 Å от остатка паратопа H100 Leu; остаток эпитопа 112 Arg находится в пределах 4,0 Å от остатка паратопа L29 Ala, L31 Tyr; остаток эпитопа 127 Tyr находится в пределах 4,0 Å от остатка паратопа L31 Tyr; остаток эпитопа 129 Gly находится в пределах 4,0 Å от остатка паратопа L31 Tyr; остаток эпитопа 130 Cys находится в пределах 4,0 Å от остатка паратопа L91 Tyr, L95B Gly; остаток эпитопа 131 Leu находится в пределах 4,0 Å от остатка паратопа H47 Trp, H50 Ala, H58 Tyr, L95A Ser, L95B Gly, L95C Ser; остаток эпитопа 132 Gly находится в пределах 4,0 Å от остатка паратопа H58 Tyr, L95A Ser; H58 Tyr, L95A Ser; остаток эпитопа 134 Met находится в пределах 4,0 Å от остатка паратопа L95A Ser; и остаток эпитопа 138 Glu находится в пределах 4,0 Å от остатка паратопа L29 Ala. См. Таблицы 29А и 29Б.
Антитело или его антигенсвязывающий фрагмент также может содержать по меньшей мере один остаток паратопа (нумерация согласно Kabat), который может образовать водородную связь с остатком эпитопа TFPI (нумерация согласно SEQ ID NO: 2) следующим образом: остаток эпитопа 102 Asp может образовать водородную связь с остатком паратопа H58 Tyr; остаток эпитопа 107 Arg может образовать водородную связь по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из H96 Gly, H97 Ala, H98 Thr, H99 Ser и H100 Leu; остаток эпитопа 112 Arg может образовать водородную связь с остатком паратопа L29 Ala; остаток эпитопа 127 Tyr может образовать водородную связь с остатком паратопа L31 Tyr; и остаток эпитопа 131 Leu может образовать водородную связь с остатком паратопа L95B Gly. См. Таблицу 29Б.
Антитело или его антигенсвязывающий фрагмент также может содержать по меньшей мере один остаток паратопа (нумерация согласно Kabat), имеющий ненулевое изменение в BSA из-за взаимодействия с остатком эпитопа (нумерация согласно SEQ ID NO: 2), следующим образом: остаток эпитопа 102 Asp взаимодействует с остатком паратопа Н58 Tyr; остаток эпитопа 104 Gly взаимодействует с остатком паратопа H58 Tyr; остаток эпитопа 105 Ile взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из H33 Ala, H34 Met, H50 Ala, H51 Ile, H52 Ser, H56 Ser, H58 Tyr и H95 Leu; остаток эпитопа 106 Cys взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из H95 Leu, H100 Leu, H100A Ser и L91 Tyr; остаток эпитопа 107 Arg взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из H96 Gly, H97 Ala, H98 Thr, H99 Ser и H100 Leu; остаток эпитопа 108 Gly взаимодействует с остатком паратопа H100 Leu; остаток эпитопа 112 Arg взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из L29 Ala, L31 Tyr и L93 Ser; остаток эпитопа 127 Tyr взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из L31 Tyr и L95B Gly; остаток эпитопа 129 Gly взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из H100A Ser, L31 Tyr и L91 Tyr; остаток эпитопа 130 Cys взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из H95 Leu, H100A Ser, L31 Tyr, L91 Tyr и L95B Gly; остаток эпитопа 131 Leu взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из H47 Trp, H50 Ala, H58 Tyr, H95 Leu, L31 Tyr, L91 Tyr, L95A Ser, L95B Gly, L95C Ser и L96 Gly; остаток эпитопа 132 Gly взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из H58 Tyr и L95A Ser; остаток эпитопа 133 Asn взаимодействует с остатком паратопа L95A Ser; остаток эпитопа 134 Met взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из L93 Ser, L94 Ser и L95A Ser; и остаток эпитопа 138 Glu взаимодействует по меньшей мере с одним остатком паратопа, выбранным из группы, состоящей из L28 Gly, L29 Ala и L93 Ser.
Антитело или его антигенсвязывающий фрагмент по изобретению может представлять собой любое антитело или антигенсвязывающий фрагмент, которое содержит любой из описанных выше остатков паратопа, который взаимодействует по меньшей мере с одним остатком эпитопа, перечисленным выше.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13 или все описанные выше остатки паратопа, или любую комбинацию разных категорий описанных выше остатков паратопа. Кроме того, в данные остатки паратопа могут быть введены консервативные замены. Например, антитело или его антигенсвязывающий фрагмент может содержать 1, 2, 3, 4, 5, 6, 7 или 8 консервативных замен согласно Таблице 34.
Антитело по изобретению может иметь способность конкурировать с другим антителом за связывание с TFPI, как описано в настоящем описании. Например, антитело по изобретению может перекрестно конкурировать с TFPI-23 и его описанными в настоящем описании вариантами за связывание с TFPI или с подходящим фрагментом или вариантом TFPI, который связывается антителами TFPI-23. Такие перекрестно конкурирующие антитела могут быть идентифицированы на основе их способности перекрестно конкурировать с проиллюстрированным примером антителом по изобретению в стандартном анализе связывания. Например, для демонстрации перекрестной конкуренции можно использовать SPR (например, с использованием системы Biacore™), анализы ELISA или проточную цитометрию. Такая перекрестная конкуренция может свидетельствовать о том, что два антитела связываются с идентичными, перекрывающимися или аналогичными эпитопами.
Антитело по изобретению может, следовательно, быть идентифицировано способом, который включает анализ связывания, в котором оценивается то, способно ли или нет тестируемое антитело конкурировать с проиллюстрированным примером антителом по изобретению (таким как TFPI-23 или любым его вариантом, или фрагментом, как описано в настоящем описании) за сайт связывания на молекуле-мишени.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, содержащую SEQ ID NO: 38, CDR-H2, содержащую SEQ ID NO: 39 и CDR-H3, содержащую SEQ ID NO: 40; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, содержащую SEQ ID NO: 33, CDR-L2, содержащую SEQ ID NO: 34 и CDR-L3, содержащую SEQ ID NO: 35. В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 38, CDR-H2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 39 и CDR-H3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 40; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 33, CDR-L2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 34 и CDR-L3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 35. В некоторых воплощениях в каждом CDR делается не более чем 10, не более чем 9, не более чем 8, не более чем 7, не более чем 6, не более чем 5, не более чем 4, не более чем 3, не более чем 2 или не более чем одна замена по отношению к SEQ ID NO: 38, 39, 40, 33, 34 и 35 соответственно. В некоторых воплощениях данная замена представляет собой консервативную замену согласно Таблице 34. В некоторых воплощениях замена делается согласно Таблице 29А, столбец 4, столбец 5 или столбец 6.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит последовательность человеческой каркасной области. Например, последовательность каркасной области тяжелой цепи может происходить из человеческой VH3 зародышевой линии, VH1 зародышевой линии, VH5 зародышевой линии или VH4 зародышевой линии. Предпочтительные каркасные области тяжелой цепи человеческой зародышевой линии представляют собой каркасные области, происходящие из зародышевых линий VH1, VH3 или VH5. Например, можно использовать каркасные области VH из следующих зародышевых линий: IGHV3-23, IGHV3-7 или IGHV1-69 (названия зародышевых линий основаны на определении зародышевой линии IMGT). Предпочтительные каркасные области легкой цепи человеческой зародышевой линии представляют собой каркасные области, происходящие из зародышевых линий VΚ или Vλ. Например, можно использовать каркасные области VL из следующих зародышевых линий: IGKV1-39 или IGKV3-20 (названия зародышевых линий основаны на определении зародышевой линии IMGT). Альтернативно или дополнительно, последовательность каркасной области может представлять собой консенсусную последовательность каркасной области человеческой зародышевой линии, такую как каркасная область человеческой консенсусной последовательности Vλ1, консенсусной последовательности VΚ1, консенсусной последовательности VΚ2, консенсусной последовательности VΚ3, консенсусная последовательность VH3 зародышевой линии, консенсусная последовательность VH1 зародышевой линии, консенсусная последовательность VH5 зародышевой линии или консенсусная последовательность VH4 зародышевой линии.
Последовательности каркасных областей человеческой зародышевой линии доступны из разных общедоступных баз данных, таких как V-base, IMGT, NCBI или Abysis.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит: (1) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 41, 63 и 65; и/или (2) VL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 36. Любая комбинация данных последовательностей VL и VH также охватывается данным изобретением.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит: (1) СH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 20; и/или (2) СL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 26. Любая комбинация данных последовательностей СH и СL также охватывается данным изобретением.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит домен Fc. Домен Fc может происходить из IgA (например, IgA1 или IgA2), IgG, IgE или IgG (например, IgG1, IgG2, IgG3 или IgG4).
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит: (1) тяжелую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 42, SEQ ID NO: 64 или SEQ ID NO: 66; и/или (2) легкую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 37. Любая комбинация данных последовательностей тяжелой цепи и легкой цепи также охватывается данным изобретением.
TFPI-24 и варианты
Структуры сокристаллов показывают то, что TFPI-24 (и его варианты) имеют целый ряд общих остатков эпитопа с TFPI-23. Среди них Ile105, Arg107 и Leu131 (согласно нумерации человеческого TFPI, как показано SEQ ID NO: 2) считаются важными для взаимодействия антитело-антиген. Другие общие остатки эпитопа включают: Cys106, Gly108, C130, L131 и G132 (согласно нумерации SEQ ID NO: 2).
Остатки эпитопа, которые являются специфичными в отношении TFPI-24 (и его вариантов) включают: Glu100, Glu101, Asp102, Gly104 и Tyr109. TFPI-23 и его варианты не связываются с данными остатками. См. Таблицу 27. Соответственно, согласно данному изобретению предложено выделенное антитело или его антигенсвязывающий фрагмент, которое специфично связывается с эпитопом в K2 TFPI, где указанный эпитоп (1) содержит остатки Ile105, Arg107 и Leu131; (2) возможно содержит один или более чем один остаток, выбранный из группы, состоящей из: Cys106, Gly108, Cys130, Leu131 и Gly132; и (3) дополнительно возможно содержит один или более чем один остаток, выбранный из группы, состоящей из: Glu100, Glu101, Asp102, Gly104 и Tyr109 (согласно нумерации SEQ ID NO: 2).
В некоторых воплощениях эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из: P103, T111, Y113, F114, N116, Q118, Q121, C122, E123, R124, F125, K126, L140 (нумерация согласно SEQ ID NO: 2) и любой их комбинации. См., Таблицу 27. Согласно WO201007269 (Novo Nordisk), контрольное антитело 4F36 распознает эпитоп, содежащий P103, T111, Y113, F114, N116, Q118, Q121, C122, E123, R124, F125, K126 и L140.
В некоторых воплощениях эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из: D31, D32, P34, C35, K36, P103, K126, Y127, G128 (нумерация согласно SEQ ID NO: 2) и любой их комбинации. См., Таблицу 27. Согласно Таблице 27 контрольные антитела 2А8 и 2А8-200 распознают эпитоп, содержащий D31, D32, P34, C35, K36, P103, K126, Y127 и G128.
Остатки паратопа из TFPI-24 (на основе BSA) также были охарактеризованы (см. Таблицу 24) и включают следующие: H33 Ala, H35 Gln, H52 Ser, H53 Asn, H55 Arg, H56 Ser, H95 Phe, H96 Leu, H97 His, H99 Ser, H101 Asp, L31 Met, L32 Tyr, L34 His, L36 Tyr, L50 Arg, L91 Trp и L96 Tyr. В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17 или все из этих остатков паратопа. Кроме того, в данные остатки паратопа могут быть введены консервативные замены. Например, антитело или его антигенсвязывающий фрагмент может содержать 1, 2, 3, 4, 5, 6, 7 или 8 консервативных замен согласно Таблице 34.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, содержащую SEQ ID NO: 48, CDR-H2, содержащую SEQ ID NO: 49, и CDR-H3, содержащую SEQ ID NO: 50; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, содержащую SEQ ID NO: 43, CDR-L2, содержащую SEQ ID NO: 44, и CDR-L3, содержащую SEQ ID NO: 45. В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 48, CDR-H2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 49, и CDR-H3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 50; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 43, CDR-L2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 44, и CDR-L3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 45. В некоторых воплощениях в каждой CDR по отношению к SEQ ID NO: 48, 49, 50, 43, 44 и 45 соответственно делается не больше, чем 10, не больше, чем 9, не больше, чем 8, не больше, чем 7, не больше, чем 6, не больше, чем 5, не больше, чем 4, не больше, чем 3, не больше, чем 2 или не больше, чем одна замена. В некоторых воплощениях данная замена представляет собой консервативную замену согласно Таблице 34.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит последовательность человеческой каркасной области. Например, последовательность каркасной области тяжелой цепи может происходить из человеческой VH3 зародышевой линии, VH1 зародышевой линии, VH5 зародышевой линии или VH4 зародышевой линии, как описано выше. Предпочтительные каркасные области легкой цепи человеческой зародышевой линии представляют собой каркасные области, происходящие из зародышевых линий VΚ и Vλ, как описано выше. Также можно использовать консенсусные последовательности каркасной области человеческой зародышевой линии, как описано выше.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 67, 69, 51 и 79; и/или (2) VL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 46, 71, 73, 75 и 77. Данным изобретением также охватывается любая комбинация из данных последовательностей VL и VH.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит: (1) СH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 20; и/или (2) СL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 26. Данным изобретением также охватывается любая комбинация из данных последовательностей СН и CL.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит домен Fc. Данный домен Fc может происходить из IgA (например, IgA1 или IgA2), IgG, IgE или IgG (например, IgG1, IgG2, IgG3 или IgG4).
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент сдержит: (1) тяжелую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 52, SEQ ID NO: 68, SEQ ID NO: 70 или SEQ ID NO: 80; и/или (2) легкую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 47, SEQ ID NO: 72, SEQ ID NO: 74, SEQ ID NO: 76 или SEQ ID NO: 78. Данным изобретением также охватывается любая комбинация данных последовательностей тяжелой цепи и легкой цепи.
4D8 и варианты
Структуры сокристалла также выявляют информацию об эпитопе и паратопе для антитела 4D8 и вариантов. Остатки эпитопа для 4D8 и его вариантов включают: Glu101, Pro103, Tyr109, Thr111, Ser119, Gln121, Glu123, Arg124, Lys126 и Leu140 согласно нумерации SEQ ID NO: 2.
В некоторых воплощениях эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из: E100, D102, R107, Y113, F114, N116, Q118, C122 (нумерация согласно SEQ ID NO: 2) и любой их комбинации. См. Таблицу 27. Согласно WO201007269 (Novo Nordisk), контрольное антитело 4F36 распознает эпитоп, содержащий E100, D102, R107, Y113, F114, N116, Q118 и C122.
В некоторых воплощениях эпитоп не содержит один или более чем один остаток, выбранный из группы, состоящей из: D31, D32, P34, C35, K36, E100, I105, R107, G108, Y127, G128 (нумерация согласно SEQ ID NO: 2) и любой их комбинации. См. Таблицу 27. Согласно Таблице 27, контрольные антитела 2A8 и 2А8-200 распознают эпитоп, содержащий D31, D32, P34, C35, K36, E100, I105, R107, G108, Y127 и G128.
Также были охарактеризованы остатки паратопа из 4D8 (на основе BSA) (см. Таблицу 20), и они включают следующие: H50 Asp, H57 Thr, H58 Leu, H59 Tyr, H61 Gln, H98 Asp, H99 Tyr, H100 Asp, L30 His, L50 Trp, L92 Tyr, L93 Thr, L94 Thr и L96 Tyr. В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13 или все из этих остатков паратопа. Кроме того, в данные остатки паратопа могут быть введены консервативные замены. Например, антитело или его антигенсвязывающий фрагмент может содержать 1, 2, 3, 4, 5, 6, 7 или 8 консервативных замен согласно Таблице 34.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, содержащую SEQ ID NO: 87, CDR-H2, содержащую SEQ ID NO: 88 и CDR-H3, содержащую SEQ ID NO: 89; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, содержащую SEQ ID NO: 81, CDR-L2, содержащую SEQ ID NO: 82 и CDR-L3, содержащую SEQ ID NO: 83. В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 87, CDR-H2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 88 и CDR-H3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 89; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 81, CDR-L2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 82 и CDR-L3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 83. В некоторых воплощениях в каждой CDR делается не более чем 10, не более чем 9, не более чем 8, не более чем 7, не более чем 6, не более чем 5, не более чем 4, не более чем 3, не более чем 2 или не более чем одна замена по отношению к SEQ ID NO: 87, 88, 89, 81, 82 и 83 соответственно. В некоторых воплощениях данная замена представляет собой консервативную замену согласно Таблице 34.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит последовательность человеческой каркасной области. Например, последовательность каркасной области тяжелой цепи может происходить из человеческой VH3 зародышевой линии, VH1 зародышевой линии, VH5 зародышевой линии или VH4 зародышевой линии, как описано выше. Предпочтительные каркасные области легкой цепи человеческой зародышевой линии представляют собой каркасные области, происходящие из зародышевых линий VΚ или Vλ, как описано выше. Также могут быть использованы консенсусные последовательности каркасной области человеческой зародышевой линии, как описано выше.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 90, 95, 97, 99, 101, 103, 105 и 107; и/или (2) VL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 84, 109 и 111. Данным изобретением также охватывается любая комбинация из данных последовательностей VL и VH.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) СH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 20; и/или (2) СL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 91 или SEQ ID NO: 85. Данным изобретением также охватывается любая комбинация из данных последовательностей СН и CL.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит домен Fc. Домен Fc может происходить из IgA (например, IgA1 или IgA2), IgG, IgE или IgG (например, IgG1, IgG2, IgG3 или IgG4).
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) тяжелую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 92, SEQ ID NO: 94, SEQ ID NO: 96, SEQ ID NO: 98, SEQ ID NO: 100, SEQ ID NO: 102, SEQ ID NO: 104, SEQ ID NO: 106, SEQ ID NO: 108; и/или (2) легкую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 86, SEQ ID NO: 93, SEQ ID NO: 110 или SEQ ID NO: 112. Данным изобретением также охватывается любая комбинация из данных последовательностей тяжелой цепи и легкой цепи.
TFPI-3 и варианты
В настоящем описании также предложены TFPI-3 и его варианты. Соответственно, антитело или его антигенсвязывающий фрагмент на основе TFPI-3 содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, содержащую SEQ ID NO: 16, CDR-H2, содержащую SEQ ID NO: 17 и CDR-H3, содержащую SEQ ID NO: 18; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, содержащую SEQ ID NO: 10, CDR-L2, содержащую SEQ ID NO: 11 и CDR-L3, содержащую SEQ ID NO: 12. В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 16, CDR-H2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 17 и CDR-H3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 18; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 10, CDR-L2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 11 и CDR-L3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 12. В некоторых воплощениях в каждой CDR делается не более чем 10, не более чем 9, не более чем 8, не более чем 7, не более чем 6, не более чем 5, не более чем 4, не более чем 3, не более чем 2 или не более чем одна замена по отношению к SEQ ID NO: 16, 17, 18, 10, 11 и 12 соответственно. В некоторых воплощениях замена представляет собой консервативную замену согласно Таблице 34.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит последовательность человеческой каркасной области. Например, последовательность каркасной области тяжелой цепи может происходить из человеческой VH3 зародышевой линии, VH1 зародышевой линии, VH5 зародышевой линии или VH4 зародышевой линии, как описано выше. Предпочтительные каркасные области легкой цепи человеческой зародышевой линии представляют собой каркасные области, происходящие из зародышевых линий VΚ или Vλ, как описано выше. Также могут быть использованы консенсусные последовательности каркасной области человеческой зародышевой линии, как описано выше.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 19; и/или (2) VL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 13. Данным изобретением также охватывается любая комбинация из данных последовательностей VL и VH.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) СH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 20; и/или (2) СL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 91 или SEQ ID NO: 14. Данным изобретением также охватывается любая комбинация из данных последовательностей СН и CL.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит домен Fc. Домен Fc может происходить из IgA (например, IgA1 или IgA2), IgG, IgE или IgG (например, IgG1, IgG2, IgG3 или IgG4).
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) тяжелую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 21; и/или (2) легкую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 15. Данным изобретением также охватывается любая комбинация из данных последовательностей тяжелой цепи и легкой цепи.
TFPI-21 и варианты
В настоящем описании также предложены TFPI-21 и его варианты. Соответственно, антитело или его антигенсвязывающий фрагмент на основе TFPI-21 содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, содержащую SEQ ID NO: 28, CDR-H2, содержащую SEQ ID NO: 29, и CDR-H3, содержащую SEQ ID NO: 30; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, содержащую SEQ ID NO: 22, CDR-L2, содержащую SEQ ID NO: 23, и CDR-L3, содержащую SEQ ID NO: 24. В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 28, CDR-H2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 29, и CDR-H3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 30; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 22, CDR-L2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 23, и CDR-L3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 24. В некоторых воплощениях в каждой CDR делается не более чем 10, не более чем 9, не более чем 8, не более чем 7, не более чем 6, не более чем 5, не более чем 4, не более чем 3, не более чем 2 или не более чем одна замена по отношению к SEQ ID NO: 28, 29, 30, 22, 23 и 24 соответственно. В некоторых воплощениях данная замена представляет собой консервативную замену согласно Таблице 34.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит последовательность человеческой каркасной области. Например, последовательность каркасной области тяжелой цепи может происходить из человеческой VH3 зародышевой линии, VH1 зародышевой линии, VH5 зародышевой линии или VH4 зародышевой линии, как описано выше. Предпочтительные каркасные области легкой цепи человеческой зародышевой линии представляют собой каркасные области, происходящие из зародышевых линий VΚ или Vλ, как описано выше. Также могут быть использованы консенсусные последовательности каркасной области человеческой зародышевой линии, как описано выше.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 31; и/или (2) VL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 25. Данным изобретением также охватывается любая комбинация из данных последовательностей VL и VH.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) СH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 20; и/или (2) СL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 91 или SEQ ID NO: 26. Данным изобретением также охватывается любая комбинация из данных последовательностей СН и CL.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит домен Fc. Домен Fc может происходить из IgA (например, IgA1 или IgA2), IgG, IgE или IgG (например, IgG1, IgG2, IgG3 или IgG4).
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) тяжелую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 32; и/или (2) легкую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 27. Данным изобретением также охватывается любая комбинация из данных последовательностей тяжелой цепи и легкой цепи.
TFPI-26 и варианты
В настоящем описании также предложены TFPI-26 и его варианты. Соответственно, антитело или его антигенсвязывающий фрагмент на основе TFPI-26 содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, содержащую SEQ ID NO: 58, CDR-H2, содержащую SEQ ID NO: 59, и CDR-H3, содержащую SEQ ID NO: 60; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, содержащую SEQ ID NO: 53, CDR-L2, содержащую SEQ ID NO: 54, и CDR-L3, содержащую SEQ ID NO: 55. В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 58, CDR-H2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 59, и CDR-H3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 60; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 53, CDR-L2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 54, и CDR-L3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 55. В некоторых воплощениях в каждой CDR делается не более чем 10, не более чем 9, не более чем 8, не более чем 7, не более чем 6, не более чем 5, не более чем 4, не более чем 3, не более чем 2 или не более чем одна замена по отношению к SEQ ID NO: 58, 59, 60, 53, 54 и 55 соответственно. В некоторых воплощениях данная замена представляет собой консервативную замену согласно Таблице 34.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит последовательность человеческой каркасной области. Например, последовательность каркасной области тяжелой цепи может происходить из человеческой VH3 зародышевой линии, VH1 зародышевой линии, VH5 зародышевой линии или VH4 зародышевой линии, как описано выше. Предпочтительные каркасные области легкой цепи человеческой зародышевой линии представляют собой каркасные области, происходящие из зародышевых линий VΚ или Vλ, как описано выше. Также могут быть использованы консенсусные последовательности каркасной области человеческой зародышевой линии, как описано выше.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 61; и/или (2) VL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 56. Данным изобретением также охватывается любая комбинация из данных последовательностей VL и VH.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) СH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 20; и/или (2) СL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 26. Данным изобретением также охватывается любая комбинация из данных последовательностей СН и CL.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит домен Fc. Домен Fc может происходить из IgA (например, IgA1 или IgA2), IgG, IgE или IgG (например, IgG1, IgG2, IgG3 или IgG4).
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) тяжелую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 62; и/или (2) легкую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 57. Данным изобретением также охватывается любая комбинация из данных последовательностей тяжелой цепи и легкой цепи.
6В7.с5 и варианты
В настоящем описании также предложены 6В7.с5 и его варианты. Соответственно, антитело или его антигенсвязывающий фрагмент на основе 6В7.с5 содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, содержащую SEQ ID NO: 118, CDR-H2, содержащую SEQ ID NO: 119, и CDR-H3, содержащую SEQ ID NO: 120; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, содержащую SEQ ID NO: 113, CDR-L2, содержащую SEQ ID NO: 114, и CDR-L3, содержащую SEQ ID NO: 115. В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 118, CDR-H2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 119, и CDR-H3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 120; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 113, CDR-L2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 114, и CDR-L3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 115. В некоторых воплощениях в каждой CDR делается не более чем 10, не более чем 9, не более чем 8, не более чем 7, не более чем 6, не более чем 5, не более чем 4, не более чем 3, не более чем 2 или не более чем одна замена по отношению к SEQ ID NO: 118, 119, 120, 113, 114 и 115 соответственно. В некоторых воплощениях замена представляет собой консервативную замену согласно Таблице 34.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит последовательность человеческой каркасной области. Например, последовательность каркасной области тяжелой цепи может происходить из человеческой VH3 зародышевой линии, VH1 зародышевой линии, VH5 зародышевой линии или VH4 зародышевой линии, как описано выше. Предпочтительные каркасные области легкой цепи человеческой зародышевой линии представляют собой каркасные области, происходящие из зародышевых линий VΚ или Vλ, как описано выше. Также могут быть использованы консенсусные последовательности каркасной области человеческой зародышевой линии, как описано выше.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 121; и/или (2) VL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 116. Данным изобретением также охватывается любая комбинация из данных последовательностей VL и VH.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) СH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 91; и/или (2) СL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 91 или SEQ ID NO: 85. Данным изобретением также охватывается любая комбинация из данных последовательностей СН и CL.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит домен Fc. Домен Fc может происходить из IgA (например, IgA1 или IgA2), IgG, IgE или IgG (например, IgG1, IgG2, IgG3 или IgG4).
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) тяжелую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 122; и/или (2) легкую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 117. Данным изобретением также охватывается любая комбинация из данных последовательностей тяжелой цепи и легкой цепи.
7A4.D9 и варианты
В настоящем описании также предложены 7А4.D9 и его варианты. Соответственно, антитело или его антигенсвязывающий фрагмент на основе 7А4.D9 содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, содержащую SEQ ID NO: 128, CDR-H2, содержащую SEQ ID NO: 129, и CDR-H3, содержащую SEQ ID NO: 130; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, содержащую SEQ ID NO: 123, CDR-L2, содержащую SEQ ID NO: 124, и CDR-L3, содержащую SEQ ID NO: 125. В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (1) CDR-H1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 128, CDR-H2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 129, и CDR-H3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 130; и/или (2) следующие последовательности CDR легкой цепи: CDR-L1, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 123, CDR-L2, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 124, и CDR-L3, по меньшей мере на 85%, по меньшей мере на 90% или по меньшей мере на 95% идентичную SEQ ID NO: 125. В некоторых воплощениях в каждой CDR делается не более чем 10, не более чем 9, не более чем 8, не более чем 7, не более чем 6, не более чем 5, не более чем 4, не более чем 3, не более чем 2 или не более чем одна замена по отношению к SEQ ID NO: 128, 129, 130, 123, 124 и 125 соответственно. В некоторых воплощениях замена представляет собой консервативную замену согласно Таблице 34.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит последовательность человеческой каркасной области. Например, последовательность каркасной области тяжелой цепи может происходить из человеческой VH3 зародышевой линии, VH1 зародышевой линии, VH5 зародышевой линии или VH4 зародышевой линии, как описано выше. Предпочтительные каркасные области легкой цепи человеческой зародышевой линии представляют собой каркасные области, происходящие из зародышевых линий VΚ или Vλ, как описано выше. Также могут быть использованы консенсусные последовательности каркасной области человеческой зародышевой линии, как описано выше.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) VH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 131; и/или (2) VL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 126. Данным изобретением также охватывается любая комбинация из данных последовательностей VL и VH.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) СH, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 91; и/или (2) СL, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 91 или SEQ ID NO: 85. Данным изобретением также охватывается любая комбинация из данных последовательностей СН и CL.
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит домен Fc. Домен Fc может происходить из IgA (например, IgA1 или IgA2), IgG, IgE или IgG (например, IgG1, IgG2, IgG3 или IgG4).
В некоторых воплощениях описанное в настоящем описании антитело или его антигенсвязывающий фрагмент содержит (1) тяжелую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 132; и/или (2) легкую цепь, содержащую аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична SEQ ID NO: 127. Данным изобретением также охватывается любая комбинация из данных последовательностей тяжелой цепи и легкой цепи.
Также раскрыто антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI и конкурирует за связывание с TFPI с любым описанным в настоящем описании антителом или его антигенсвязывающим фрагментом, таким как любое из антител, перечисленных в Таблице 3 (или их антигенсвязывающий фрагмент). Например, если связывание антитела или его антигенсвязывающей части с TFPI мешает последующему связыванию TFPI-23 или TFPI-106 с TFPI, данное антитело или его антигенсвязывающая часть конкурирует с TFPI-23 или TFPI-106 за связывание с TFPI.
Также раскрыто антитело или его антигенсвязывающий фрагмент, которое специфично связывается с доменом K2 TFPI и связывается с тем же самым эпитопом TFPI, что и любое описанное в настоящем описании антитело или его антигенсвязывающий фрагмент, такое как любое из антител, перечисленных в Таблице 3, или его антигенсвязывающий фрагмент.
Типичный конкурентный анализ антител (и анализ перекрывающихся эпитопов) посредством SPR приводится в Примере 6.
Раскрытые в настоящем описании антитела и антигенсвязывающие фрагменты включают моноклональные антитела, поликлональные антитела, фрагменты антител (например, Fab, Fab', F(ab')2, Fv, Fc и т.д.), химерные антитела, биспецифичные антитела, гетероконъюгатные антитела, одноцепочечные (scFv), их мутанты, слитые белки, содержащие часть антитела, доменные антитела (dAb), гуманизированные антитела и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит сайт распознавания антигена требующейся специфичности, включая гликозилированные варианты антител, варианты антител по аминокислотной последовательности и ковалентно модифицированные антитела. Антитела и антигенсвязывающие фагменты могут быть мышиными, крысиными, человеческими или любого другого происхождения (включая химерные или гуманизированные антитела). В некоторых воплощениях антитело представляет собой моноклональное антитело. В некоторых воплощениях антитело представляет собой химерное, гуманизированное или человеческое антитело. В некоторых воплощениях антитело представляет собой человеческое антитело. В некоторых воплощениях антитело представляет собой гуманизированное антитело.
В некоторых воплощениях раскрытое в настоящем описании антитело или его антигенсвязывающий фрагмент имеет значение аффинности (Kd) не больше, чем примерно 1×10-3 М, как, например, не больше, чем примерно 5×10-4 М, не больше, чем примерно 4×10-4 М, не больше, чем примерно 3×10-4 М, не больше, чем примерно 2×10-4 М, не больше, чем примерно 1×10-4 М, не больше, чем примерно 9×10-5 М, не больше, чем примерно 8×10-5 М, не больше, чем примерно 7×10-5 М, не больше, чем примерно 6×10-5 М, не больше, чем примерно 5×10-5 М, не больше, чем примерно 4×10-5 М, не больше, чем примерно 3×10-5 М, не больше, чем примерно 2×10-5 М, не больше, чем примерно 1×10-5 М, не больше, чем примерно 9×10-6 М, не больше, чем примерно 8×10-6 М, не больше, чем примерно 7×10-6 М, не больше, чем примерно 6×10-6 М, не больше, чем примерно 5×10-6 М, не больше, чем примерно 4×10-6 М, не больше, чем примерно 3×10-6 М, не больше, чем примерно 2×10-6 М, не больше, чем примерно 1×10-6 М, не больше, чем примерно 9×10-7 М, не больше, чем примерно 8×10-7 М, не больше, чем примерно 7×10-7 М, не больше, чем примерно 6×10-7 М, не больше, чем примерно 5×10-7 М, не больше, чем примерно 4×10-7 М, не больше, чем примерно 3×10-7 М, не больше, чем примерно 2×10-7 М, не больше, чем примерно 1×10-7 М, не больше, чем примерно 9×10-8 М, не больше, чем примерно 8×10-8 М, не больше, чем примерно 7×10-8 М, не больше, чем примерно 6×10-8 М, не больше, чем примерно 5×10-8 М, не больше, чем примерно 4×10-8 М, не больше, чем примерно 3×10-8 М, не больше, чем примерно 2×10-8 М, не больше, чем примерно 1×10-8 М, не больше, чем примерно 9×10-9 М, не больше, чем примерно 8×10-9 М, не больше, чем примерно 7×10-9 М, не больше, чем примерно 6×10-9 М, не больше, чем примерно 5×10-9 М, не больше, чем примерно 4×10-9 М, не больше, чем примерно 3×10-9 М, не больше, чем примерно 2×10-9 М, не больше, чем примерно 1×10-9 М, от примерно 1×10-3 М до примерно 1×10-13 М, от 1×10-4 М до примерно 1×10-13 М, от 1×10-5 М до примерно 1×10-13 М, от примерно 1×10-6 М до примерно 1×10-13 М, от примерно 1×10-7 М до примерно 1×10-13 М, от примерно 1×10-8 М до примерно 1×10-13 М, от примерно 1×10-9 М до примерно 1×10-13 М, от 1×10-3 М до примерно 1×10-12 М, от 1×10-4 М до примерно 1×10-12 М, от 1×10-5 М до примерно 1×10-12 М, от примерно 1×10-6 М до примерно 1×10-12 М, от примерно 1×10-7 М до примерно 1×10-12 М, от примерно 1×10-8 М до примерно 1×10-12 М, от примерно 1×10-9 М до примерно 1×10-12 М, от 1×10-3 М до примерно 1×10-11 М, от 1×10-4 М до примерно 1×10-11 М, от примерно 1×10-5 М до примерно 1×10-11 М, от примерно 1×10-6 М до примерно 1×10-11 М, от примерно 1×10-7 М до примерно 1×10-11 М, от примерно 1×10-8 М до примерно 1×10-11 М, от примерно 1×10-9 М до примерно 1×10-11 М, от 1×10-3 М до примерно 1×10-10 М, от 1×10-4 М до примерно 1×10-10 М, от примерно 1×10-5 М до примерно 1×10-10 М, от примерно 1×10-6 М до примерно 1×10-10 М, от примерно 1×10-7 М до примерно 1×10-10 М, от примерно 1×10-8 М до примерно 1×10-10 М или от примерно 1×10-9 М до примерно 1×10-10 М.
В некоторых воплощениях константа диссоциации измеряется с использованием способа поверхностного плазмонного резонанса (SPR) (Biacore). Термин «поверхностный плазмонный резонанс» относится к оптическому явлению, которое обеспечивает анализ в реальном времени биоспецифических взаимодейсвий посредством выявления изменений концентрации белка в матрице биосенсора, например, с использованием системы BIACORE™. В некоторых воплощениях измерение SPR проводится с использованием прбора Biacore T100 или Т200.
Например, стандартные условия анализа для поверхностного плазмонного резонанса могут быть основаны на иммобилизации лиганда - приблизительно 100 резонансных единиц (RU) IgG на чипе SPR. Очищенные белки-мишени разводятся в буфере до интервала конечных концентраций и инъецируются с требующейся скоростью тока (например, 10-100 мкл/мин) для обеспечения расчета Ka. Давали идти диссоциации для установления скорости диссоциации (Kd), с последующим 5 с импульсом 3 М MgCl2 (или 20 мМ NaOH) для регенерации поверхности чипа. Сенсограммы затем анализируются с использованием программного пакета для оценки кинетики.
В типичном воплощении анализ SPR осуществляется согласно условиям, как изложено в Примере 1 под подзаголовком «Поверхностный плазмонный резонанс (SPR)».
В некоторых воплощениях константа диссоциации измеряется с использованием анализа кинетического исключения на основе раствора (KinExA™). В конкретном воплощении измерение KinExA проводится с использованием прибора KinExA™ 3200 (Sapidyne). Анализ кинетического исключения (KinExA™) представляет собой многоцелевую платформу для иммуноанализа (по существу - проточный спектрофлуориметр), который способен к измерению равновесных констант диссоциации и констант скорости ассоциации и диссоциации для взаимодействий антиген/антитело. Поскольку KinExA™ проводится после получения равновесия, он является преимущественной методикой для применения при измерении Kd высокоаффинных взаимодействий, где скорость диссоциации взаимодействия может быть очень медленной. Методологию KinExA™ обычно можно осуществлять, как описано в Drake et al (2004) Analytical Biochemistry 328, 35-43.
В общем, антителу против TFPI необходимо связываться с TFPI с высокой аффинностью для того, чтобы эффективно блокировать активности TFPI. Однако, поскольку TFPI также экспрессируется на поверхности клетки, при слишком высокой аффинности связывания антитела данное антитело может быстро интернализоваться и деградировать клеткой-хозяином. Это потенциально могло бы приводить к короткому периоду полувыведения и повторным инъекциям. Например, антитело TFPI-23 демонстрирует меньшую аффинность связывания (Kd) по сравнению с TFPI-24 и, при определенных обстоятельствах, является более желательным, так как оно имеет меньшую скорость интернализации и более продолжительный период полувыведения. Соответственно, обычно желательными являются аффинности связывания (Kd) от 5×10-7 М до примерно 5×10-11 М, в частности, от примерно 1×10-8 М до примерно 1×10-10 М (от 0,1 нМ до 10 нМ), если желателен более длительный период полувыведения. Считается, что в этом интервале достигается баланс между (1) аффинностями связывания, которые необходимы для эффективного ингибирования активностей TFPI, и (2) более длительным периодом полувыведения и пониженной интернализацией антитела.
В частности, считается, что для поддержания еженедельного подкожного дозирования 3 мг/кг желательным является значение Kd от примерно 1×10-8 М до примерно 1×10-10 М (от 0,1 нМ до 10 нМ).
То, уменьшает ли антитело или его антигенсвязывающий фрагмент активность TFPI, или уменьшает связывание TFPI с физиологическим субстратом (например, FXa), может быть определено посредством измерения уменьшения аффинности связывания TFPI с указанным физиологическим субстратом, например, посредством сравнения (1) аффинности связывания TFPI с его субстратом в присутствии антитела против TFPI (или его антигенсвязывающего фрагмента) с (2) аффинностью связывания TFPI с тем же самым субстратом в отсутствие антитела против TFPI. Уменьшение в связывании TFPI с физиологическим субстратом (например, с FXa) может составлять по меньшей мере примерно 10%, по меньшей мере примерно 20%, по меньшей мере примерно 30%, по меньшей мере примерно 40%, по меньшей мере примерно 50%, по меньшей мере примерно 60%, по меньшей мере примерно 70%, по меньшей мере примерно 80%, по меньшей мере примерно 90%, по меньшей мере примерно 95%, по меньшей мере примерно 96%, по меньшей мере примерно 97%, по меньшей мере примерно 98% или по меньшей мере примерно 99% в присутствии антитела против TFPI (или его антигенсвязывающего фрагмента). Ожидаемое связывание TFPI с его физиологическим субстратом в отсутствие антитела (или фрагмента) может быть принято за 100%.
Активности, ингибирующие TFPI, также именуемые в настоящем описании «уменьшение активности TFPI» антитела против TFPI или его антигенсвязывающего фрагмента также можно оценивать в модели in vivo и/или in vitro c использованием, например, плазматических систем. Например, ингибирующие активности (или уровень уменьшения активности TFPI) антитела можно оценивать посредством: (1) уменьшения времени свертывания крови при измерении в анализе протромбинового времени в плазме с разведением; (2) уменьшения времени свертывания в цельной крови при измерении посредством тромбоэластографии; (3) увеличения образование тромбина; (4) увеличения активности FXa в присутствии TFPI; (5) усиленного накопления тромбоцитов в присутствии TFPI; (6) увеличенного образования фибрина в присутствии TFPI; или (7) их любой комбинации. Ингибирующие активности антитела или антигенсвязывающего фрагмента могут быть дозозависимыми (например, вызывая дозозависимое уменьшение времени свертывания крови при измерении в анализе протромбинового времени в плазме с разведением).
Несколько типичных анализов для оценки ингибирующей активности антитела в отношении TFPI подробно описаны в Примерах. Например, анализ протромбинового времени (РТ) в плазме с разведением представляет собой модифицированный анализ РТ с использованием разведенного тромбопластина или тканевого фактора для увеличения времени свертывания крови и динамического интервала анализа. Ингибирующее/нейтрализующее антитело против TFPI должно уменьшать протромбиновое время при разведении.
Другой типичной модельной системой для определения активности, ингибирующей TFPI, является внешний теназный анализ, в котором тестируется способность антитела или его антигенсвязывающего фрагмента восстанавливать опосредованную внешним комплексом активацию FX в присутствии TFPI. Другой модельной системой для характеристики активности, ингибирующей TFPI, является анализ ингибирования FXa, в котором активность FXa измеряется в присутствии TFPI (см. Sprecher et al., Proc. Nat. Acad. Sci. USA 91:3353-3357 (1994)).
Ингибирующие активности антитела или его антигенсвязывающего фрагмента также можно оценивать в анализе в плазме. Образование тромбина может запускаться в плазме, по существу не имеющей активности FVIII или FIX (например, остаточная активность фактора свертывания крови меньше, чем 1%), в присутствии антитела против TFPI или его антигенсвязывающего фрагмента. Образование тромбина можно выявлять с использованием флуорогенного или хромогенного субстрата. Превращение протромбина можно измерять с использованием, например, Thrombograph™ (Thermo Scientific, Waltham, Mass.), и полученные в результате данные можно объединять в калиброванную автоматическую тромбограмму, генерированную программой Thrombinoscope™, доступной от Thrombinoscope BV.
Например, антитело или антигенсвязывающий фрагмент может улучшать регулируемое TFPI образование тромбина в отстуствие FVIII (например, в плазме, обедненной FVIII) до по меньшей мере 1% уровня TFPI-зависимого образования тромбина в нормальной плазме. В общем, нормальная (непораженная заболеванием) плазма содержит примерно от 0,5 U/мл до примерно 2 U/мл фактора VIII. Соответственно, в некоторых случаях антитело или антигенсвязывающий фрагмент по изобретению будет усиливать образование тромбина в отсутствие FVIII по меньшей мере до примерно 1% образования тромбина, наблюдающегося в присутствии от 0,5 U/мл до 2 U/мл FVIII. В других воплощениях антитело (или его антигенсвязывающий фрагмент) усиливает образование тромбина в отсутствие FVIII по меньшей мере до примерно 2%, по меньшей мере примерно 3%, по меньшей мере примерно 5%, по меньшей мере примерно 7% или по меньшей мере примерно 10% уровня образования тромбина в нормальной плазме, т.е. в присутствии физиологических уровней Фактора VIII.
Антитело или антигенсвязывающий фрагмент также можно вводить в животную модель тромбиновой недостаточности или гемофилии для характеристики активностей, ингибирующих TFPI, in vivo. Такие модели in vivo известны в этой области и вкючают, например, мышей, которым вводятся антитела против FVIII для индукции гемофилии А (Tranholm et al., Blood, 102, 3615-3620 (2003)); модели, нокаутированные по фактору свертывания крови, такие как мыши, нокаутированные по FVIII (Bi et al., Nat. Genet., 10(1), 119-121 (1995)), и мыши, нокаутированные по FIX (Wang et al., Proc. Nat. Acad. Sci. USA 94(21):11563-11566 (1997)), но не ограничивающиеся ими; индуцированную гемофилию-A у кроликов (Shen et al., Blood, 42(4):509-521 (1973)); и собак Chapel Hill HA (Lozier et al., Proc. Nat. Acad. Sci. USA 99:12991-12996 (2002)).
В некоторых воплощениях раскрытые в настоящем описании антитела (или антигенсвязывающие фрагменты) усиливают активность FXa в присутствии TFPI с полумаксимальной эффективной концентрацией (EC50) не больше чем 1×10-4 М, не больше чем 1×10-5 М, не больше чем 1×10-6 М, не больше чем 1×10-7 М, не больше чем 1×10-8 М, не больше чем 1×10-9 М, не больше чем 1×10-10 М, не больше чем 1×10-11 М или не больше чем 1×10-12 М. Предпочтительно EC50 составляет от примерно 5×10-7 М до 1×10-11 М, как, например, от примерно 1×10-7 М до 5×10-10 М, от примерно 1×10-7 М до 1×10-10 М, от 1×10-7 М до 5×10-9 М, от 5×10-7 М до 5×10-10 М, от примерно 5×10-7 М до 1×10-10 М или от примерно 5×10-7 М до 5×10-9 М.
В некоторых воплощениях раскрытые в настоящем описании антитела (или антигенсвязывающие фрагменты) нейтрализуют ингибирование TFPI FVIIa/TF-опосредованной активации FX с полумаксимальной эффективной концентрацией (EC50) не более чем 1×10-4 М, не более чем 1×10-5 М, не более чем 1×10-6 М, не более чем 1×10-7 М, не более чем 1×10-8 М, не более чем 1×10-9 М, не более чем 1×10-10 М, не более чем 1×10-11 М или не более чем 1×10-12 М. Предпочтительно EC50 составляет от примерно 5×10-7 М до 1×10-11 М, как, например, от примерно 1×10-7 М до 5×10-10 М, от примерно 1×10-7 М до 1×10-10 М, от 1×10-7 М до 5×10-9 М, от 5×10-7 М до 5×10-10 М, от примерно 5×10-7 М до 1×10-10 М или от примерно 5×10-7 М до 5×10-9 М.
В некоторых воплощениях раскрытые в настоящем описании антитела (или антигенсвязывающие фрагменты) уменьшают время свертывания крови при измерении в анализе протромбинового времени в плазме с разведением с полумаксимальной эффективной концентрацией (ЕС50) не более чем 1×10-4 М, не более чем 1×10-5 М, не более чем 1×10-6 М, не более чем 1×10-7 М, не более чем 1×10-8 М, не более чем 1×10-9 М, не более чем 1×10-10 М, не более чем 1×10-11 М или не более чем 1×10-12 М. Предпочтительно EC50 составляет от примерно 5×10-7 М до 1×10-11 М, как, например, от примерно 1×10-7 М до 5×10-10 М, от примерно 1×10-7 М до 1×10-10 М, от 1×10-7 М до 5×10-9 М, от 5×10-7 М до 5×10-10 М, от примерно 5×10-7 М до 1×10-10 М или от примерно 5×10-7 М до 5×10-9 М.
В некоторых воплощениях раскрытые в настоящем описании антитела (или антигенсвязывающие фрагменты) увеличивают индекс скорости образования тромбина с полумаксимальной эффективной концентрацией (ЕС50) не более чем 1×10-4 М, не более чем 1×10-5 М, не более чем 1×10-6 М, не более чем 1×10-7 М, не более чем 1×10-8 М, не более чем 1×10-9 М, не более чем 1×10-10 М, не более чем 1×10-11 М или не более чем 1×10-12 М. Предпочтительно EC50 составляет от примерно 5×10-7 М до 1×10-11 М, как, например, от примерно 1×10-7 М до 5×10-10 М, от примерно 1×10-7 М до 1×10-10 М, от 1×10-7 М до 5×10-9 М, от 5×10-7 М до 5×10-10 М, от примерно 5×10-7 М до 1×10-10 М или от примерно 5×10-7 М до 5×10-9 М.
В некоторых воплощениях раскрытые в настоящем описании антитело и фрагменты антитела также можно дополнительно оценивать другими анализами биологической активности, например, для того, чтобы оценить его эффективность, фармакологическую активность и потенциальную эффективность в качестве терапевтического агента. Такие анализы известны в этой области и зависят от антигена-мишени и намеченного применения антитела. Примеры включают, например, анализы ингибирования роста опухолевых клеток; анализы антителозависимой клеточной цитотоксичности (ADCC) и комплементзависимой цитотоксичности (CDC); анализы агонистической активности или антагонистической активности.
В. Полинуклеотиды, векторы и клетки-хозяева
Согласно изобретению также предложены полинуклеотиды, кодирующие любое из антител, связывающихся с TFPI по настоящему изобретению включая описанные в настоящем описании фрагменты антител и модифицированные антитела, такие как, например, антитела, имеющие ослабленную эффекторную функцию Fc. В другом аспекте согласно изобретению предложен способ получения любого из описанных в настоящем описании полинуклеотидов. Полинуклеотиды можно получать и экспрессировать методиками, известными в этой области. Соответственно, согласно изобретению предложены полинуклеотиды или композиции, включающие фармацевтические композиции, содержащие полинуклеотиды, кодирующие любое из антител против TFPI и их антигенсвязывающих фрагментов по изобретению.
В одном воплощении домены VH и VL или их антигенсвязывающие фрагменты, или полноразмерные НС или LC кодируются отдельными полинуклеотидами. В качестве альтернативы, и VH, и VL или их антигенсвязывающий фрагмент, или НС и LC кодируются одним полинуклеотидом.
Согласно изобретению предложены полинуклеотиды или композиции, содержащие полинуклеотиды, кодирующие любое из антител против TFPI и их антигенсвязывающих фрагментов по изобретению, включая TFPI-23, TFPI-24, TFPI-106, TFPI-107 и 4D8, но не ограничиваясь ими, где последовательность данного полинуклеотида охватывает последовательность SEQ ID NO:175 (кодирующую область VH TFPI-106), SEQ ID NO:176 (кодирующую область VL TFPI-106), SEQ ID NO:177 (кодирующую тяжелую цепь TFPI-106) и SEQ ID NO:178 (кодирующую легкую цепь TFPI-106).
В другом аспекте согласно изобретению предложена выделенная нуклеиновая кислота, кодирующая область VH антитела или его антигенсвязывающую часть, которая специфично связывается с TFPI, содержащая последовательность нуклеиновой кислоты вставки, присутствующей в плазмиде, депонированной под № АТСС (Американская коллекция типовых клеточных культур) РТА-122329.
В другом аспекте согласно изобретению предложена выделенная нуклеиновая кислота, кодирующая область VL антитела или его антигенсвязывающую часть, которая специфично связывается с TFPI, содержащая последовательность нуклеиновой кислоты вставки, присутствующей в плазмиде, депонированной под № АТСС РТА-122328.
В другом аспекте согласно изобретению предложены полинуклеотиды и их варианты, кодирующие антитело против TFPI или его часть, где такие варианты полинуклеотидов содержат последовательность нуклеиновой кислоты, имеющую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 87%, по меньшей мере 89%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности с любой из раскрытых в настоящем описании специфических последовательностей нуклеиновой кислоты. Степень идентичности последовательности по любой длине нуклеотидной последовательности может быть рассчитана с использованием способов, известных обычным специалистам в этой области. В одном неограничивающем воплощении процент идентичности последовательностей между двумя или более чем двумя родственными нуклеотидными пследовательностями может быть определен с использованием нуклеотидного сервера BLAST, доступного от Национальной библиотеки медицины (http://blast.ncbi.nlm.nih.gov/). В этой программе предоставлены разные установки, которые обычный специалист в этой области может использовать для оптимизации сравнений последовательностей в зависимости от таких факторов, как длина, сложность и другие факторы.
Согласно изобретению предложена молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, кодирующую аминокислотную последовательность любого антитела против TFPI и его антигенсвязывающего фрагмента по изобретению, включая аминокислотную последовательность антитела или его антигенсвязывающего фрагмента, приведенную в Таблице 33 (например, аминокислотную последовательность SEQ ID NO:21-174), и антитела, которое связывается с тем же самым эпитопом, что и и/или конкурирует за связывание с TFPI с антителом по изобретению, но не ограничиваясь ими.
В другом аспекте согласно изобретению предложены полинуклеотиды и их варианты, кодирующие антитело против TFPI, где такие варианты полинуклеотидов кодируют аминокислотную последовательность, имеющую по меньшей мере 70%-ную, по меньшей мере 75%-ную, по меньшей мере 80%-ную, по меньшей мере 85%-ную, по меньшей мере 87%-ную, по меньшей мере 89%-ную, по меньшей мере 90%-ную, по меньшей мере 91%-ную, по меньшей мере 92%-ную, по меньшей мере 93%-ную, по меньшей мере 94%-ную, по меньшей мере 95%-ную, по меньшей мере 96%-ную, по меньшей мере 97%-ную, по меньшей мере 98%-ную или по меньшей мере 99%-ную идентичность последовательности с любой раскрытой в настоящем описании аминокислотной последовательностью антитела против TFPI.
В других воплощениях степень родства между нуклеиновыми кислотами, содержащими вариант полинуклеотидной последовательности, кодирующий антитело против TFPI или его часть, и любой из конкретных раскрытых в настоящем описании полинуклеотидных последовательностей может быть определена посредством тестирования того, гибридизуется ли вариант последовательности (или комплементарная ей последовательность) со специфической нуклеотидной последовательностью (или комплементарной ей последовательностью) при умеренных или сильно жестких условиях в формате анализа норзерн-блоттинг или саузерн-блоттинг. Типичные и неограничивающие «умеренно жесткие условия» включают предварительную промывку в растворе 5 × SSC (хлорид натрия и цитрат натрия), 0,5% SDS (додецилсульфат натрия), 1,0 мM EDTA (этилендиаминтетрауксусная кислота) (pH 8,0); гибридизацию при 50°C-65°C, 5 × SSC, в течение ночи; с последующей двухкратной промывкой при 65°C в течение 20 минут, каждый раз 2×, 0,5× и 0,2× SSC, содержащим 0,1 % SDS.
Типичные и неограничивающие «сильно жесткие условия» или «условия сильной жесткости» представляют собой условия, при которых: (1) для промывки используется низкая ионная сила и высокая температура, например, 0,015 М хлорид натрия/0,0015 М цитрат натрия/0,1% додецилсульфат натрия при 50°C; (2) во время гибридизации используется денатурирующий агент, такой как формамид, например 50%-ный (об./об.) формамид с 0,1%-ным бычьим сывороточным альбумином/0,1%-ным фиколлом/0,1%-ным поливинилпиролидоном/50 мМ натрий-фосфатным буфером при рН 6,5 с 750 мМ хлоридом натрия, 75 мМ цитратом натрия при 42°С; или (3) используется 50%-ный формамид, 5 × SSC (0,75 M NaCl, 0,075 M цитрат натрия), 50 мМ фосфат натрия (рН 6,8), 0,1%-ный пирофосфат натрия, 5× раствор Денхардта, обработанная ультразвуком ДНК спермы лосося (50 мкг/мл), 0,1%-ный SDS и 10%-ный декстрана сульфат при 42°С, с промывками при 42°С в 0,2 × SSC (хлорид натрия/цитрат натрия) и 50%-ном формамиде при 55°С, с последующей помывкой в сильно жестких условиях, состоящей из 0,1 × SSC, содержащего EDTA, при 55°С. Специалисту будет понятно как откорректировать температуру, ионную силу и т.д., как необходимо для приспособления к таким факторам, как длина зонда и тому подобное. Обычный специалист в этой области также будет знаком со стандартными методиками для проведения анализа норзерн-блоттинг или саузерн-блоттинг с целью выявления степени родства между вариантом нуклеотидной последовательности и специфической нуклеотидной последовательностью по настоящему изобретению.
Настоящим изобретением также охватываются полинуклеотиды, комплементарные любым таким последовательностям. Полинуклеотиды могут быть одноцепочечными (кодирующими или антисмысловыми) или двухцепочечными и могут представлять собой молекулы ДНК (геномной, кДНК или синтетической) или РНК. Молекулы РНК включают молекулы гяРНК (гетероядерная РНК), которые содержат интроны и соответствуют молекуле ДНК один к одному, и молекулы мРНК, которые не содержат интроны. Дополнительные кодирующие или некодирующие последовательности могут, но не обязательно присутствуют в пределах полинуклеотида по настоящему изобретению.
Варианты также или альтернативно могут быть по существу гомологичными природному гену или его части, или комплементарной ему последовательности. Такие варианты полинуклеотида способны гибридизоваться с встречающейся в природе последовательностью ДНК, кодирующей природное антитело, (или с комплементарной ей последовательностью) при умеренно жестких условиях.
Обычным специалистам в этой области будет понятно то, что в результате вырожденности генетического кода существует много нуклеотидных последовательностей, способных кодировать любое из раскрытых в настоящем описании антител против TFPI или их частей. Некоторые из этих полинуклеотидов могут иметь относительно низкую степень идентичности последовательности с любой из конкретных приведенных в настоящем описании нуклеотидных последовательностей антител против TFPI при кодировании той же самой аминокислотной последовательности. Тем не менее, в настоящем изобретении конкретно рассматриваются полинуклеотиды, которые отличаются из-за различий в использовании кодонов.
Полинуклеотиды по настоящему изобретению могут быть получены с использованием химического синтеза, способов генной инженерии или ПЦР (полимеразной цепной реакцией). Способы химического синтеза полинуклеотидов хорошо известны в этой области, и их не нужно в настоящем описании подробно описывать. Специалист в этой области может использовать предложенные в настоящем описании последовательности и имеющийся в продаже синтезатор ДНК для получения требуемой последовательности ДНК.
Для получения полинуклеотидов с использованием способов генной инженерии полинуклеотид, содержащий желательную последовательность, может быть вставлен в подходящий вектор, и данный вектор, в свою очередь, может быть введен в подходящую клетку-хозяина для репликации и амплификации, как обсуждается в настоящем описании далее. Полинуклеотиды могут быть вставлены в клетки-хозяева посредством любых способов, известных в этой области. Клетки могут быть трансформированы введением экзогенного полинуклеотида посредством прямого поглощения, эндоцитоза, трансфекции, F-спаривания или электропорации. Сразу после введения экзогенный полинуклеотид может поддерживаться в клетке в виде неинтегрированного вектора (такого как плазмида) или интегрированного в геном клетки-хозяина. Полинуклеотид, амплифицированный таким образом, может быть выделен из клетки-хозяина способами, хорошо известными в этой области. См., например, Sambrook et al., 1989.
В качестве альтернативы, репродукцию последовательностей ДНК обеспечивает ПЦР. Технология ПЦР хорошо известна в этой области и описывается в патентах США № 4683195, 4800159, 4754065 и 4683202, а также в PCR: The Polymerase Chain Reaction, Mullis et al. eds., Birkauswer Press, Boston, 1994.
РНК может быть получена с использованием выделенной ДНК в подходящем векторе и вставки его в подходящую клетку-хозяина. При репликации клетки и транскрипции ДНК в РНК данная РНК может быть затем выделена с использованием способов, хорошо известных специалистам в этой области, как изложено, например, в Sambrook et al., 1989, выше.
Подходящие векторы для клонирования могут быть сконструированы согласно стандартным методикам или могут быть выбраны из большого числа векторов для клонирования, доступных в этой области. В то время как выбранный вектор для клонирования может варьировать согласно клетке-хозяину, намеченной для применения, полезные векторы для клонирования обычно будут иметь способность к саморепликации, могут обладать одной мишенью для конкретной эндонуклеазы рестрикции и/или могут нести гены маркера, который можно использовать при отборе клонов, содержащих вектор. Подходящие примеры включают плазмиды и бактериальные вирусы, например, pUC18, pUC19, Bluescript (например, pBS SK+) и его производные, mp18, mp19, pBR322, pMB9, ColE1, pCR1, RP4, фаговые ДНК и челночные векторы, такие как pSA3 и pAT28. Эти и многие другие векторы для клонирования доступны у коммерческих поставщиков, таких как BioRad, Stratagene и Invitrogen.
Кроме того, предложены экспрессионные векторы. Экспрессионные векторы обычно представляют собой реплицируемые полинуклеотидные конструкции, которые содержат полинуклеотид согласно изобретению. Подразумевается то, что экспрессионный вектор должен быть реплицируемым в клетках-хозяевах либо в виде эписом, либо в виде интегральной части хромосомной ДНК. Подходящие экспрессионные векторы включают плазмиды, вирусные векторы, включающие аденовирусы, аденосателлитные вирусы, ретровирусы, космиды и экспрессионный(ные) вектор(ры), раскрытый(тые) в публикации РСТ № WO 87/04462, но не ограничиваются ими. Компоненты вектора обычно могут включать одно или более чем одно из следующего: сигнальную последовательность; репликатор; один или более чем один маркерный ген; подходящие транскрипционные контролирующие элементы (такие как промоторы, энхансеры и терминатор), но не ограничиваются ими. Для экспрессии (т.е. трансляции) обычно также требуется один или более чем один трансляционный контролирующий элемент, такой как сайты связывания рибосомы, сайты инициации трансляции и терминирующие кодоны.
Векторы, содержащие интересующие полинуклеотиды и/или сами полинуклеотиды могут быть введены в клетку-хозяина любым из целого ряда подходящих способов, включающих электропорацию, трансфекцию с использованием хлорида кальция, хлорида рубидия, фосфата кальция, DEAE-декстрана или других веществ; бомбардировку микрочастицами; липофекцию и инфекцию (например, когда вектор представляет собой инфекционный агент, такой как вирус осповакцины). Выбор вводимых векторов или полинуклеотидов часто будет зависеть от характеристик клетки-хозяина.
Согласно изобретению также предложены клетки-хозяева, содержащие любые из описанных в настоящем описании полинуклеотидов. С целью выделения генов, кодирующих интересующие антитело, полипептид или белок можно использовать любые клетки-хозяева, способные сверхэкспрессировать гетерологичные ДНК. Неограничивающие примеры клеток-хозяев млекопитающих включают клетки обезьяны COS (клетки почки обезьяны), человеческие HeLa, клетки человеческой эмбриональной почки (HEK) 293, Sp2.0 и клетки яичника китайского хомяка (СНО), но не ограничиваются ими. См. также публикацию РСТ № WO 87/04462. Подходящие клетки-хозяева, не являющиеся клетками млекопитающих, включают прокариот (таких как E. coli или B. subtillis) и дрожжей (таких как S. cerevisae, S. pombe или K. lactis). Скрининг клеток-хозяев, экспрессирующих антитело против TFPI или его антигенсвязывающую часть, можно проводить с использованием анализа иммунного связывания, такого как ELISA (твердофазный иммуноферментный анализ), FACS (флуоресцентная сортировка клеток) или другой анализ, знакомый обычным специалистам в этой области.
Таким образом, антитело (или его антигенсвязывающий фрагмент) по изобретению может быть продуцировано рекомбинантно с использованием подходящей клетки-хозяина. Нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент, может быть клонирована в экспрессионный вектор, который может быть затем введен в клетку-хозяина, такую как клетка E. coli, дрожжевая клетка, клетка насекомого, клетка COS, клетка СНО или миеломная клетка, которая иным образом не продуцирует иммуноглобулиновый белок, с получением синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Типичные клетки-хозяева включают клетку СНО, клетки HEK 293 и Sp2.0.
Экспрессионный вектор можно использовать для управления экспрессией антитела против TFPI. Специалист в этой области знаком с введением экспрессионных векторов для получения экспрессии экзогенного белка in vivo. См., например, патенты США № 6436908; 6413942 и 6376471. Введение экспрессионных векторов включает местное или системное введение, включающее инъекцию, пероральное введение, генную пушку, введение через катетер и местное введение. Согласно некоторым неограничивающим воплощениям экспрессионный вектор вводится непосредственно в печень, скелетные мышцы, костный мозг или в другие ткани.
Также можно использовать таргетную доставку терапевтических композиций, содержащих экспрессионный вектор или субгеномные полинуклеотиды. Методики опосредованной рецептором доставки ДНК описываются, например, в Findeis et al., Trends Biotechnol., 1993, 11:202; Chiou et al., Gene Therapeutics: Methods And Applications Of Direct Gene Transfer, J.A. Wolff, ed., 1994; Wu et al., J. Biol. Chem., 1988, 263:621; Wu et al., J. Biol. Chem., 1994, 269:542; Zenke et al., Proc. Natl. Acad. Sci. USA, 1990, 87:3655; Wu et al., J. Biol. Chem., 1991, 266:338. Терапевтические композиции, содержащие полинуклеотид, вводятся в интервале от примерно 100 нг до примерно 200 мг ДНК для местного введения в протоколе генотерапии. На протяжении протокола генотерапии также можно использовать интервалы концентрации от примерно 500 нг до примерно 50 мг, от примерно 1 мкг до примерно 2 мг, от примерно 5 мкг до примерно 500 мкг и от примерно 20 мкг до примерно 100 мкг ДНК. Терапевтические полинуклеотиды и полипептиды можно доставлять с использованием носителей для доставки генов. Носитель для доставки гена может иметь вирусное или невирусное происхождение (см., в общем, Jolly, Cancer Gene Therapy, 1994, 1:51; Kimura, Human Gene Therapy, 1994, 5:845; Connelly, Human Gene Therapy, 1995, 1:185 и Kaplitt, Nature Genetics, 1994, 6:148). Экспрессию таких кодирующих последовательностей можно индуцировать с использованием эндогенных промоторов млекопитающих или гетерологичных промоторов. Экспрессия кодирующей последовательности может быть либо конститутивной, либо регулируемой.
Векторы на основе вирусов для доставки желательного полинуклеотида и экспрессии в желательной клетке хорошо известны в этой области. Типичные носители на основе вируса включают рекомбинантные ретровирусы (см., например, публикации РСТ № WO 90/07936; WO 94/03622; WO 93/25698; WO 93/25234; WO 93/11230; WO 93/10218; WO 91/02805; патенты США № 5219740 и 4777127; патент Великобритании № 2200651 и европейский патент № 0345242), векторы на основе альфавируса (например, векторы на основе вируса Синдбис, вируса леса Семлики (ATCC VR-67; ATCC VR-1247), вируса реки Росс (ATCC VR-373; ATCC VR-1246) и вируса венесуэльского энцефалита лошадей (ATCC VR-923; ATCC VR-1250; ATCC VR 1249; ATCC VR-532)), и векторы на основе аденосателлитного вируса (AAV) (см., например, публикации PCT № WO 94/12649, WO 93/03769; WO 93/19191; WO 94/28938; WO 95/11984 и WO 95/00655), но не ограничиваются ими. Также можно использовать ведение ДНК, связанной с умерщвленным аденовирусом, как описано в Curiel, Hum. Gene Ther., 1992, 3:147.
Также можно использовать носители и способы для невирусной доставки, включающие поликатионную конденсированную ДНК, связанную или не связанную с одним умерщвленным аденовирусом (см., например, Curiel, Hum. Gene Ther., 1992, 3:147); ДНК, связанную с лигандом (см., например, Wu, J. Biol. Chem., 1989, 264:16985); эукариотические клетки-носители для доставки (см., например, патент США № 5814482; публикации РСТ № WO 95/07994; WO 96/17072; WO 95/30763 и WO 97/42338) и нейтрализацию заряда нуклеиновых кислот или слияние с клеточными мембранами, но не ограничивающиеся ими. Также можно использовать голую ДНК. Типичные способы введения на основе голой ДНК описываются в публикации РСТ № WO 90/11092 и патенте США № 5580859. Липосомы, которые могут действовать в качестве носителей для доставки генов, описываются в патенте США № 5422120; публикациях PCT № WO 95/13796; WO 94/23697; WO 91/14445 и EP 0524968. Дополнительные подходы описываются в Philip, Mol. Cell Biol., 1994, 14:2411 и в Woffendin, Proc. Natl. Acad. Sci., 1994, 91:1581.
Последовательность желательного антитела (или его антигенсвязывающего фрагмента) и нуклеиновой кислоты, кодирующей такое антитело (или его антигенсвязывающий фрагмент), может быть определена с использованием стандартных методик секвенирования. Последовательность нуклеиновой кислоты, кодирующей желательное антитело (или фрагменты), может быть вставлена в другие векторы (такие как векторы для клонирования и экспрессионные векторы) для рекомбинантной продукции и характеристики. Тяжелая цепь (или фрагмент тяжелой цепи) и легкая цепь (или фрагмент легкой цепи) могут быть клонированы в тот же самый вектор или в разные векторы.
Подходящие векторы для клонирования и экспрессионные векторы могут включать целый ряд компонентов, таких как промотор, энхансер и другие транскрипционные регуляторные последовательности. Также может быть сконструирован вектор для обеспечения перемещения вариабельного домена антитела между разными веторами.
Фрагменты антитела могут быть получены протеолитической или другой деградацией антител, рекомбинантными способами или химическим синтезом. Полипептиды антител, особенно более короткие полипептиды из вплоть до примерно 50 аминокислот, с удобством получаются посредством химического синтеза. Способы химического синтеза известны в этой области и коммерчески доступны.
Раскрытое в настоящем описании антитело или его антигенсвязывающий фрагмент может иметь созревшую аффинность. Например, антитела с созревшей аффинностью могут быть получены методиками, известными в этой области (Marks et al., 1992, Bio/Technology, 10:779-783; Barbas et al., 1994, Proc. Nat. Acad. Sci. USA 91:3809-3813; Schier et al., 1995, Gene, 169:147-155; Yelton et al., 1995, J. Immunol., 155:1994-2004; Jackson et al., 1995, J. Immunol., 154(7):3310-3319; Hawkins et al., 1992, J. Mol. Biol., 226:889-896 и WO2004/058184).
4. Композиции и применения
Описанные в настоящем описании антитела или антигенсвязывающие фрагменты могут быть приготовлены в виде фармацевтических композиций. Фармацевтическая композиция может дополнительно содержать фармацевтически приемлемые носители, эксципиенты или стабилизаторы (Remington: The Science and practice of Pharmacy 20th Ed., 2000, Lippincott Williams and Wilkins, Ed. K. E. Hoover) в виде лиофилизированных препаратов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципентов при используемых дозировках и концентрациях и могут содержать такие буферы, как буферы на основе фосфата, цитрата и других органических кислот; антиоксиданты, включающие аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид, бензэтония хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (меньше, чем примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстраны; хелатирующие агенты, такие как EDTA (этилендиаминтетрауксусная кислота); сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (ПЭГ). В настоящем описании дополнительно описаны фармацевтически приемлемые эксципиенты.
Описанные в настоящем описании антитела или антигенсвязывающие фрагменты можно использовать для разных терапевтических или диагностических целей. Например, антитело или его антигенсвязывающий фрагмент можно использовать в качестве агентов для аффинной очистки (например, для очистки TFPI in vitro), в качестве диагностического агента (например, для выявления экспрессии TFPI в конкретных клетках, тканях или сыворотке).
Типичные терапевтические применения антитела и фрагментов антитела по изобретению включают лечение тромбоцитопении, тромбоцитарных расстройств (нарушений функции или числа тромбоцитов) и расстройств, связанных с кровотечением (например, гемофилии А, гемофилии В и гемофилии С). Данные антитела и фрагменты антител также можно использовать для лечения неконтролируемого кровотечения при таких показаниях, как травма и геморрагический инсульт. Антитела и фрагменты антител также можно использовать в профилактическом лечении (например, перед хирургическими вмешательствами).
В частности, описанные в настоящем описании антитела или антигенсвязывающие фрагменты можно использовать для лечения недостаточностей или дефектов свертывания крови. Например, описанные в настоящем описании антитела или антигенсвязывающие фрагменты можно использовать для уменьшения или ингибирования взаимодействия TFPI с FXa, или для уменьшения TFPI-зависимого ингибирования активности TF/FVIIa/FXa.
Для терапевтических применений описанные в настоящем описании антитела или антигенсвязывающие фрагменты можно вводить млекопитающему, особенно человеку, традиционными методиками, как, например, внутривенно (в виде болюса или посредством непрерывной инфузии на протяжении определенного периода времени), внутримышечно, внутрибрюшинно, в спинной мозг, подкожно, внутрь сустава, интрасиновиально, подоболочечно, перорально, местно или посредством ингаляции. Антитела или антигенсвязывающие фрагменты также подходящим образом вводятся внутриопухолевыми, околоопухолевыми, внутриочаговыми или околоочаговыми путями.
Соответственно, в одном аспекте согласно изобретению предложен способ уменьшения активности ингибитора пути тканевого фактора (TFPI), включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества описанных в настоящем описании антител или антигенсвязывающих фрагментов. В другом аспекте согласно изобретению предложен способ сокращения времени кровотечения, включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества описанных в настоящем описании антител или антигенсвязывающих фрагментов.
В некоторых воплощениях субъект представляет собой человека.
В некоторых воплощениях данный субъект страдает от или является подверженным недостаточности свертывания крови. Недостаточность в свертывании крови включает, например, болезнь фон Виллебранда (vWD), гемофилии А, В или С и другие тромбоцитарные расстройства (такие как врожденные тромбоцитарные дефекты, врожденная и приобретенная недостаточность пула тромбоцитов, удлиненное время кровотечения).
В некоторых воплощениях описанные в настоящем описании антитела или антигенсвязывающие фрагменты вводятся подкожно. В некоторых воплощениях описанные в настоящем описании антитела или антигенсвязывающие фрагменты вводятся внутривенно.
Фармацевтические композиции могут вводиться субъекту, нуждающемуся в этом, с частотой, которая может варьировать в зависимости от тяжести эпизода кровотечения или, в случае профилактической терапии, может варьировать в зависимости от тяжести недостаточности свертывания крови пациента.
Композиции могут вводиться нуждающимся пациентам в виде болюса или посредством непрерывной инфузии. Например, болюсное введение антитела, присутствующего в виде фрагмента Fab, может осуществляться в количестве от 0,0025 до 100 мг/кг массы тела, от 0,025 до 0,25 мг/кг, от 0,010 до 0,10 мг/кг или 0,10-0,50 мг/кг. Для непрерывной инфузии антитело, присутствующее в виде фрагмента Fab, может вводиться дозировке от 0,001 до 100 мг/кг массы тела/минуту, от 0,0125 до 1,25 мг/кг/мин, от 0,010 до 0,75 мг/кг/мин, от 0,010 до 1,0 мг/кг/мин или 0,10-0,50 мг/кг/мин в течение периода 1-24 часов, 1-12 часов, 2-12 часов, 6-12 часов, 2-8 часов или 1-2 часов.
Для введения антитела, присутствующего в виде полноразмерного антитела (с полными константными областями), количества дозировки могут составлять от примерно 1 мг/кг до примерно 10 мг/кг, от примерно 2 мг/кг до примерно 10 мг/кг, от примерно 3 мг/кг до примерно 10 мг/кг, от примерно 4 мг/кг до примерно 10 мг/кг, от примерно 5 мг/кг до примерно 10 мг/кг, от примерно 1 мг/кг до примерно 20 мг/кг, от примерно 2 мг/кг до примерно 20 мг/кг, от примерно 3 мг/кг до примерно 20 мг/кг, от примерно 4 мг/кг до примерно 20 мг/кг, от примерно 5 мг/кг до примерно 20 мг/кг, примерно 1 мг/кг или больше, примерно 2 мг/кг или больше, примерно 3 мг/кг или больше, примерно 4 мг/кг или больше, примерно 5 мг/кг или больше, примерно 6 мг/кг или больше, примерно 7 мг/кг или больше, примерно 8 мг/кг или больше, примерно 9 мг/кг или больше, примерно 10 мг/кг или больше, примерно 11 мг/кг или больше, примерно 12 мг/кг или больше, примерно 13 мг/кг или больше, примерно 14 мг/кг или больше, примерно 15 мг/кг или больше, примерно 16 мг/кг или больше, примерно 17 мг/кг или больше, примерно 19 мг/кг или больше, или примерно 20 мг/кг или больше. Частота введения зависела бы от тяжести состояния. Частота могла бы варьировать от трех раз в неделю до одного раза каждые две или три недели.
Дополнительно композиции могут вводиться пациентам посредством подкожной инъекции. Например, доза от 1 до 100 мг антитела против TFPI может вводиться пациентам посредством подкожной инъекции один раз в сутки, один раз в 2 суток, один раз в 3 суток, один раз в 4 суток, один раз в 5 суток, один раз в 6 суток, дважды в неделю, еженедельно, раз в две недели или ежемесячно.
В некоторых воплощениях фармацевтическая композиция вводится подкожно посредством еженедельной схемы с дозой от примерно 0,1 мг/кг до примерно 10 мг/кг, от примерно 0,5 мг/кг до примерно 10 мг/кг, от примерно 1 мг/кг до примерно 10 мг/кг, от примерно 1,5 мг/кг до примерно 10 мг/кг, от примерно 2 мг/кг до примерно 10 мг/кг, от примерно 0,1 мг/кг до примерно 8 мг/кг, от примерно 0,5 мг/кг до примерно 8 мг/кг, от примерно 1 мг/кг до примерно 8 мг/кг, от примерно 1,5 мг/кг до примерно 8 мг/кг, от примерно 2 мг/кг до примерно 8 мг/кг, от примерно 0,1 мг/кг до примерно 5 мг/кг, от примерно 0,5 мг/кг до примерно 5 мг/кг, от примерно 1 мг/кг до примерно 5 мг/кг, от примерно 1,5 мг/кг до примерно 5 мг/кг, от примерно 2 мг/кг до примерно 5 мг/кг, примерно 0,5 мг/кг, примерно 1,0 мг/кг, примерно 1,5 мг/кг, примерно 2,0 мг/кг, примерно 2,5 мг/кг, примерно 3,0 мг/кг, примерно 3,5 мг/кг, примерно 4,0 мг/кг, примерно 4,5 мг/кг, примерно 5,0 мг/кг, примерно 5,5 мг/кг, примерно 6,0 мг/кг, примерно 6,5 мг/кг, примерно 7,0 мг/кг, примерно 7,5 мг/кг, примерно 8,0 мг/кг, примерно 8,5 мг/кг, примерно 9,0 мг/кг, примерно 9,5 мг/кг или примерно 10,0 мг/кг.
В некоторых воплощениях фармацевтическая композиция вводится подкожно посредством еженедельной схемы с дозой примерно 2,0 мг/кг. В некоторых воплощениях фармацевтическая композиция вводится подкожно посредством еженедельной схемы с дозой примерно 3,0 мг/кг.
Описанные в настоящем описании антитела и фрагменты антител можно использовать в качестве монотерапии или в комбинации с другими терапиями для лечения гемостатического расстройства. Например, совместное введение одного или более чем одного антитела (или фрагмента антитела) по изобретению со свертывающим агентом, таким как фактор VIIa, фактор VIII, фактор IX или транексамовая кислота, может быть полезным для лечения гемофилии.
В одном воплощении предложен способ лечения недостаточности в свертывании или сокращении времени кровотечения, включающий введение (а) первого количества антитела или антигенсвязывающего фрагмента по изобретению и (б) второго количества фактора VIII или фактора IX. Возможно фактор VII не вводится совместно. В другом воплощении предложен способ лечения недостаточности в свертывании или сокращении времени кровотечения, включающий введение (а) первого количества антитела или антигенсвязывающего фрагмента по изобретению и (б) второго количества фактора VIII или фактора IX. Возможно фактор VII не вводится совместно. Специалист в этой области понял бы, что при лечении недостаточности в свертывании сокращение времени кровотечения также может называться сокращением времени свертывания.
Данное изобретение также включает фармацевтическую композицию, содержащую терапевтически эффективное количество комбинации антитела (или фрагмента антитела) по изобретению и фактора VIII или фактора IX, где данная композиция не содержит фактор VII. «Фактор VII» включает фактор VII и фактор VIIa.
БИОЛОГИЧЕСКОЕ ДЕПОНИРОВАНИЕ
Репрезентативные материалы по настоящему изобретению были депонированы в Американскую коллекцию типовых культур, 10801 University Boulevard, Manassas, Va. 20110-2209, США, 22 июля 2015 г. Плазмидный вектор mAb-TFPI-106 VH, имеющий № АТСС РТА-122329, содержит вставку ДНК, кодирующую вариабельную область тяжелой цепи антитела TFPI-106, и плазмидный вектор mAb-TFPI-106 VL, имеющий № АТСС РТА-122328, содержит вставку ДНК, кодирующую вариабельную область легкой цепи антитела TFPI-106. Депонирования осуществляли согласно положениям Будапештского соглашения по международному признанию депонирования микроорганизмов для цели патентной процедуры и инструкции на его основе (Будапештское соглашение). Это обеспечивает поддержание жизнеспособной культуры депонирования в течение 30 лет с даты депонирования. Депонирование будет сделано доступным АТСС согласно условиям Будапештского соглашения и с соблюдением соглашения между Pfizer Inc. и АТСС, что обеспечивает постоянную и неограниченную доступность потомства культуры депонирования для публики при выдаче подходящего патента США или при выкладывании в открытый доступ для публики любой патентной заявки США или иностранной патентной заявки, вне зависимости от того, что происходит сначала, и гарантирует доступность потомства для того, кто определен комиссаром США по патентам и торговым маркам как имеющий право на это согласно разделу 122 капитула 35 U.S.C. (свод законов США) и правам комиссара согласно им (включая раздел 1.14 капитула 37 C.F.R. (свод федеральных правил США) с особой ссылкой на 886 OG 638).
Правоприобретатель настоящей заявки согласен с тем, что, если культура депонированных материалов погибнет или будет потеряна, или повреждена при культивировании при подходящих условиях, данные материалы будут быстро заменены по уведомлению другими материалами такого же типа. Доступность депонированного материала не следует истолковывать как лицензию на воплощение данного изобретения на практике в нарушение прав, предоставляемых властью любого правительства согласно его патентному праву.
ПРИМЕРЫ
Данное изобретение далее подробно описано посредством ссылки на следующие экспериментальные примеры. Данные примеры приведены лишь для целей иллюстрации и не предназначены для того, чтобы быть ограничивающими, если не определено иначе. Таким образом, данное изобретение ни в коей мере не следует истолковывать как ограниченное следующими примерами, но скорее следует истолковывать как охватывающее все и любые вариации, которые становятся очевидными как результат предложенных здесь идей.
Пример 1. Экспериментальные материалы и методы
1. Белковые реагенты TFPI
Белковые реагенты, используемые для иммунизации, селекции на основе фагового дисплея и характеризации антител против TFPI, перечислены в Таблице 1, и ID их последовательностей перечислены в Таблице 2.
Конструкции TFPI (вектор pSMED2) временно экспрессировались в клетках HEK293F, и кондиционированные среды отбирали через 120 часов после трансфекции. Интересующий белок захватывали из кондиционированных сред с использованием никель сефарозы НР и далее очищали посредством гель-фильтрации. Фактор Xa и фактор Х были приобретены у Haematologic Technologies, Inc. Хромогенный субстрат для амидолитического анализа Фактора Ха, представляющий собой Spectrozyme®ТFXa, был приобретен у Sekisui Diagnostics.
Белковые реагенты, использованные для иммунизации, селекции на основе фагового дисплея и идентификации антител против TFPI
Идентификационные номера последовательностей реагентов TFPI, описания и последовательности
остатки 29-177)
2. Реагенты - антитела
Моноклональные антитела, используемые для целей сравнения (эталонные антитела 2A8, 2A8-200, 3F18, hz4F36), перечислены в Таблице 3. Описания, источники и последовательности антител перечислены в Таблице 4 (Фиг. 5). Последовательности легкой цепи и тяжелой цепи клонировали в подходящие векторы, временно экспрессировали в клетках человеческой эмбриональной почки-293 (HEK-293) и очищали посредством белок А-сефарозы и гель-фильтрации. Mab 2974 приобретали у R&D Systems (каталожный № MAB2974).
Идентификационные номера последовательности (SeqID) антител для CDR1,2,3 легкой цепи (LC), вариабельной области легкой цепи (VL), константной области легкой цепи (СL), легкой цепи (LC), CDR1,2,3 тяжелой цепи (НC), вариабельной области тяжелой цепи (VН), константной области тяжелой цепи (СН) и тяжелой цепи (НС). Состав последовательностей для номеров SeqID приводится в Таблице 4.
CDR1
CDR3
CDR1
CDR2
CDR3
3. ELISA связывания TFPI
Рекомбинантные humTFPI K1K2, murTFPI K1K2, cynTFPI K1K2, ratTFPI K1K2, rabTFPI K1K2 или TFPI2 биотинилировали с использованием системы AviTag™ и захватывали на покрытых стрептавидином 96-луночных планшетах Greiner в концентрации 1 × 10-8 М в буфере для анализа ELISA. Очищенные антитела против TFPI разводили до 1 мкг/мл в буфере для анализа ELISA, затем серийно трехкратно разводили с получением восьмиточечных серий разведения. Разведенные антитела добавляли в объеме 100 мкл на лунку. Планшеты инкубировали при комнатной температуре в течение 2 часов. После промывки планшетов PBS (фосфатно-солевой буферный раствор)/0,05% Tween 20 планшеты инкубировали с поликлональным антителом козы против мышиного IgG-Fc, конъюгированным с пероксидазой хрена (Pierce) в разведении 1:10000. После 1 часа инкубации связавшееся антитело выявляли добавлением раствора суб страта TMB (тетраметилбензидин). Поглощения считывали при 450 нм, и данные анализировали посредством программы GraphPad Prism.
4. Поверхностный плазмонный резонанс (SPR)
Сенсорный чип против человеческого Fc получали посредством аминного связывания антитела против человеческого IgG (каталожный номер BR-1008-39, GE Healthcare) со всеми четырьмя проточными ячейками сенсорного чипа, покрытого карбоксиметилированным декстраном (СМ5) (каталожный номер BR100530, GE Healthcare). Проточные ячейки активировали инъецированием смеси 1:1 400 мМ 1-этил-3-(3-диметиламинопропил)карбодиимида гидрохлорида (EDC) и N-гидроксисукцинимида (NHS) в течение 7 минут при скорости тока 10 мкл/минуту. Антитело против человеческого IgG разводили до 25 мкг/мл в 10 мМ ацетате натрия, рН 5,0, и инъецировали над всеми проточными ячейками в течение 7 минут со скоростью 10 мкл/минуту. Все проточные ячейки блокировали 1 М этаноламин-HCl (ETH) в течение 7 минут при скорости тока 10 мкл/минуту. Конечный уровень иммобилизации захватывающего антитела составлял приблизительно 10000 резонансных единиц (RU). Проточным буфером для иммобилизации и определения кинетики был 10 мM HEPES, pH 7,4, 150 мM NaCl, 3 мM EDTA, 0,05% (об./об.) Tween-20 (HBS-EP+). Для характеризации связывания антител против TFPI с человеческим TFPI антитела разводили до 0,5 мкг/мл в HBS-EP+ и захватывали посредством антитела против человеческого IgG, иммобилизованного на проточных ячейках 2, 3 и 4 в течение 30 секунд - 1 минуты при скорости тока 5 мкл/минуту с достижением уровня захвата от 70 до 300 RU. Проточную ячейку 1 использовали в качестве контрольной поверхности. После захвата антитела скорость тока увеличивали до 50 мкл/минуту, и инъецировали буфер или человеческий TFPI в концентрации, варьирующей от 0,2 нМ до 200 нМ, в HBS-EP+ через все проточные ячейки на протяжении 1,0 минуты ассоциации, и обеспечивали диссоциацию в течение
10-15 минут. Циклы с буфером для каждого захваченного антитела использовали для двойного контроля (Myszka,D.G. J.Mol.Recognit. 12, 279-284 (1999)). В конце каждого цикла всю поверхность против IgG регенерировали посредством 60-секундного импульса 3 М MgCl2. Кинетические анализы проводили при 25°С при скорости получения сигнала 10 Гц на установке BIAcore T200 (GE Healthcare). Константы скорости и аффинности определяли посредством аппроксимации данных к модели 1:1 в программе BIAcore T200 Evaluation, версия 1.0 (GE).
5. Анализ обращения ингибирования Фактора Ха посредством TFPI
Способность очищенных антител против TFPI восстанавливать активность Фактора Ха в присутствии ингибирующих концентраций TFPI оценивали in vitro. Антитела против TFPI, разведенные в PBS в концентрациях, варьирующих от 1 нМ до 500 нМ, предварительно инкубировали со следующими белками: 10 нМ рекомбинантный человеческий TFPI K1K2 или 10 нМ кроличий TFPI K1K2 в буфере для определения активности (20 мM HEPES, pH 8,0, 150 мM NaCl, 5 мM CaCl2, 0,5 мг/мл BSA) в течение 30 минут при 37°С. Добавляли 2 нМ Фактор Ха, полученный из человеческой плазмы, и реакционные смеси инкубировали при 37°С в течение 30 минут. Добавляли хромогенный субстрат спектрозим Ха до конечной концентрации 500 мкМ для конечного объема реакции 100 мкл. Контрольные реакции включали реакции без Фактора Ха для контроля фона анализа, без TFPI, обеспечивая максимальное образование FXa (100%-ная активность), или без антитела против TFPI (один PBS). Поглощения реакций немедленно считывали на многорежимном планшет-ридере SpectraMax M5e при длине волны 405 нм с 2-минутными интервалами на протяжении периода 60 минут. ЕС50 рассчитывали с использованием программы Prism Graph Pad.
6. Двухэтапный анализ обращения ингибирования TF-FVIIa-FX
Способность очищенных антител против TFPI восстанавливать активность Фактора VIIa-тканевого фактора в присутствии ингибирующих концентраций TFPI оценивали in vitro. Антитела против TFPI в концентрациях, варьирующих от 1 нМ до 500 нМ, предварительно инкубировали с 10 нМ рекомбинантными белками TFPI в буфере для определения активности в течение 30 минут при 37°С. В реакции добавляли приблизительно 1 пМ липидированный тканевой фактор и 1 нМ рекомбинантный Фактор VIIa (NovoSeven) и инкубировали при 37°С в течение 5 минут. В реакции вводили 150 нМ человеческий Фактор Х. Хромогенный субстрат спектрозим Ха добавляли до конечной концентрации 500 мкМ в каждую лунку для конечного объема реакции 100 мкл. Контрольные реакции включали реакции без Фактора VIIa, без тканевого фактора, без Фактора Х, без TFPI или без антитела против TFPI (один PBS). Поглощения реакций немедленно считывали на многорежимном планшет-ридере SpectraMax M5e при длине волны 405 нм с 2-минутными интервалами на протяжении периода 60 минут. ЕС50 рассчитывали с использованием программы Prism Graph Pad.
7. Анализ образования тромбина (TGA)
Способность очищенных антител против TFPI восстанавливать образование тромбина в плазме с ослабленной активностью Фактора VIII оценивали в анализах образования тромбина с использованием системы калиброванной автоматической тромбограммы (САТ). Антитела против TFPI в концентрациях 1 нМ - 500 нМ, разведенные в PBS, вводили в реакционные смеси, содержащие человеческую плазму, дефицитную по Фактору VIII, и реагент РРР-Low (обедненная тромбоцитами плазма), содержащий 4 мкМ фосфолипид и 1 пМ тканевой фактор. В контрольных реакциях использовался PBS без антитела. Реакции запускали добавлением буфера Fluca, содержащего флуорогенный тромбиновый субстрат и CaCl2. Флуоресценцию каждой реакции немедленно считывали с использованием планшет-ридера Fluoroskan Ascent с использованием программы Thrombinoscope с 20 секундным интервалом в течение 60 минут. Каждую реакцию сравнивали с калибраторной контрольной лункой, содержащей PBS, тромбин - калибровочный стандарт, дефицитную по FVIII плазму и буфер FLUCA. Кривые образования тромбина, полученные с использованием Thrombinoscope (нМ тромбина относительно времени), анализировали для извлечения лаг-перода, высоты пика, времени до пика и площади под кривой или эндогенного тромбинового потенциала (ЕТР) с использованием программы Thrombinoscope (Thrombinoscope, версия BV). Данные использовали для расчета индекса скорости (пиковая концентрация тромбина/ время до пика - лаг-период).
Также оценивали способность очищенных антител против TFPI восстанавливать образование тромбина в плазме кролика с ослабленной активностью FVIIIa. Нормальную плазму белого новозеландского кролика обрабатывали антителом против FVIII (GM-8015) в конечной концентрации 100 мкг/мл или контрольным мышиным антителом против человеческого IgG2a в концентрации 100 мкг/мл в течение 60 минут при 37°С. Непосредственно перед добавлением в реакционные лунки плазму кролика разводили 1:3 в буфере (20 мМ HEPES, 140 мМ NaCl). Анализы образования тромбина с кроличьей плазмой с нейтрализованным FVIII проводили, как описано выше.
8. Получение домена K2 TFPI яванского макака для структурных исследований
TFPI K1K2 яванского макака (cyno) (Таблица 1) экспрессировали в клетках HEK293, и кондиционированные среды отбирали через 120 часов после трансфекции. Очищенный TFPI K1K2 cyno инкубировали с человеческой эластазой нейтрофилов (HNE) в соотношении 1:70 (мольное отношение HNE:TFPI) в течение 120 мин при RT (комнатная температура) для того, чтобы происходило расщепление. Отделение K1 cyno от K2 cyno проводили с использованием анионообменной хроматографии на HQ50 (Poros). В качестве конечной стадии очистки проводили гель-фильтрацию с использованием Superdex 75. Эндопротеиназу AspN использовали для отрезания остаточной AviTag от С-конца домена K2 cyno.
9. Образование комплексов Fab антитела/K2 TFPI cyno для структурных исследований
Антитела против TFPI, представляющие собой 4D8.b1, TFPI-23, TFPI-24, 2A8-200 (Таблица 3) и Mab 2974 (R&D systems), расщепляли иммобилизованным папаином согласно протоколу изготовителя (Thermo/Pierce). Использовали MabSelect SuRe для очистки из продуктов расщеления Fab, которые затем использовали для образования комплекса с K2 TFPI cyno. Затем комплексы Fab/K2 TFPI cyno концентрировали приблизительно до 16 мг/мл. Концентрированные комплексы затем использовали для скрининга на условия кристаллизации белка.
Пример 2. Образование мышиных антител против TFPI
1. Иммунизация мышей и получение гибридомы
Каждую из групп из пяти мышей BALB/c подкожно иммунизировали смесью 5 мкг humTFPI K1K2 и 5 мкг murTFPI K1K2, эмульгированный в полном адъюванте Фрейнда. Затем мышей иммунизировали дважды в неделю смесью белка, поочередно эмульгированной в неполном адъюванте Фрейнда или разведенной в PBS. Образцы крови отбирали на сутки 17 и на сутки 27 (после пятых и седьмых иммунизаций соответственно), и сыворотки тестировали на присутствие циркулирующих антител против TFPI посредством ELISA. К суткам 27 каждая мышь получала конечную внутрибрюшинную бустерную иммунизацию смесью белка (10 мкг). Через четверо суток отбирали дренирующие лимфатические узлы (подмышечные, паховые и подколенные), и объединенные клетки лимфатических узлов смешивали в соотношении 1:1 с клетками P3X63.Ag8.653 и подвергали электрическому слиянию клеток. Слитые клетки высаживали на чашки в среды RPMI1640, дополненные FBS (фетальная телячья сыворотка) (25%), NCTC-109 (12,5%), Glutamax (1%), пенициллином-стрептомицином (1%), добавкой для клонирования гибридомы (5%) и HAT (1 × 10-4 М гипоксантина, 4 × 10-7 М аминоптерина и 1,6 × 10-5 М тимидина). Через четырнадцать суток после слияния супернатанты культуры гибридомы тестировали на связывание с humTFPI K1K2 посредством ELISA. На основе их активности связывания и их эффективности в функциональных анализах для дальнейшей характеристики были отобраны антитела из трех гибридом: 4D8, 6B7 и 7А4.
2. Клонирование и секвенирование антител против TFPI, полученных из гибридомы
Получали РНК из гибридом 4D8, 6B7 и 7А4, и последовательности ДНК вариабельной области из экспрессированных антител получали посредством клонирования ПЦР-ОТ (полимеразная цепная реакция с обратной транскриптазой).
ПЦР-продукты клонировали в вектор для клонирования ТОРО-ТА, затем секвенировали традиционными способами. Из гибридом 6B7 и 7А4 выявляли одну пару кДНК тяжелой цепи и легкой цепи. Из 4D8 выявляли две кДНК тяжелой цепи и легкой цепи.
Пример 3. Характеристика антител против TFPI мышиной гибридомы
Родительские гибридомы 4D8, 6B7 и 7А4 субклонировали посредством ограниченного разведения с получением моноклональных линий клеток гибридомы. Позитивные субклоны идентифицировали посредством скрининга ELISA на реактивность в отношении humTFPI K1K2 и размножали. Далее характеризовали очищенные антитела из одного субклона каждой гибридомы.
1. Связывание с TFPI
Очищенные антитела против TFPI 4D8.B1, 6B7.C5 и 7A4.D9 тестировали на связывание с рекомбинантными человеческим и кроличьим белками TFPI посредством ELISA связывания белка. Значения EC50 каждого антитела в отношении как humTFPI K1K2-aviHis10, так и rabTFPI K1K2 показаны в Таблице 5.
Значения ЕС50 (нМ) мышиных MAb против TFPI в отношении человеческого и кроличьего TFPI
Проводили эксперименты поверхностного плазмонного резонанса для оценки аффинности очищенных мышиных антител против TFPI в отношении человеческого и кроличьего белков TFPI K1K2. Значения ka, kd и KD для связывания каждого антитела с человеческим и кроличьим TFPI K1K2 приводятся в Таблице 6.
Кинетические измерения в отношении связывания мышиных гибридомных клонов против TFPI с человеческим и кроличьим TFPI
2. Анализы активности in vitro
Мышиные моноклональные антитела против TFPI тестировали на активность в анализах обращения ингибирования FXa и TF-FXa-FVIIa и в анализе образования тромбина (TGA). Было выбрано самое мощное антитело, представляющее собой 4D8.B1, для продвижения в дальнейшие исследования.
Активность мышиных моноклональных антител против TFPI в анализах обращения ингибирования FXa и TF-FXa-FVIIa и в анализе образования тромбина (TGA)
Пример 4. Получение химерных и гуманизированных антител из клона 4D8
1. Получение мышинно-человеческого химерного антитела 4D8
кДНК вариабельной области, полученные из гибридомы 4D8, субклонировали в экспрессионные векторы млекопитающего с получением химерных антител, в которых вариабельная область тяжелой цепи мыши была слита в рамке считывания с человеческим IgG1 3M (SEQ 20, Таблица 4), и вариабельная область легкой цепи мыши была слита в рамке считывания с константной областью каппа человеческого Ig (SEQ 62, Таблица 4). Химерные конструкции временно трансфицировали в клетки HEK293. Для идентификации правильной пары тяжелой и легкой цепи из гибридомы 4D8 провели всего четыре временные трансфекции всеми возможными комбинациями тяжелой и легкой цепи. Антитело, полученное из одной из трансфекций, было названо химера hu-mu 4D8 (Таблицы 3 и 4).
2. Характеристика мышинно-человеческого химерного антитела 4D8 (mu-hu 4D8)
Химеру mu-hu 4D8 тестировали на ее способность связываться и с человеческим, и с кроличьим белками TFPI K1K2 посредством ELISA связывания белка (Таблица 8) и SPR (Таблица 9). Значения KD и ЕС50 были хорошо сопоставимыми со значениями, измеренными с очищенным мышиным MAb 4D8.B1, демонстрируя то, что пересадка мышиных вариабельных областей, пересаженных на каркасную область человеческого IgG1, сохраняла активность связывания.
Значения ЕС50 (нМ) химеры mu-hu 4D8 для человеческого и кроличьего TFPI
Кинетические измерения для связывания химеры mu-hu 4D8 с человеческим и кроличьим TFPI
mu-hu 4D8
mu-hu 4D8
3. Гуманизация химеры hu-mu 4D8
Последовательность химеры hu-mu 4D8 гуманизировали посредством пересадки CDR на человеческие акцепторные последовательности каркасной области. Выбрали каркасную область DP54 и каркасную область DPK9. Затем экспрессировали комбинации конструкций тяжелой и легкой цепи (см. Таблицу 3). Антитела тестировали на связывание с человеческим и кроличьим TFPI в анализе связывания посредством ELISA (Таблица 10) и на связывание с человеческим TFPI в анализе связывания SPR (Таблица 11).
Значения ЕС50 (нМ) связывания гуманизированных антител 4D8 c человеческим и кроличьим белками TFPI K1K2
Анализ SPR связывания гуманизированных антител 4D8 c белком humTFPI K1K2
На основе этих данных 4D8 Vk 1.1 × VH 1.4 отбирали для дальнейшей характеристики и обозначали hz4D8 (Таблица 3). Гуманизированное антитело против TFPI (hz4D8) сравнивали с мышиным 4D8.B1 в отношении активности в анализе обращения ингибирования FXa, двухэтапном анализе ингибирования TF-FVIIa-FX и анализе образования тромбина. Данные в Таблице 12 показывают то, что данное гуманизированное антитело имело улучшенную активность во всех трех анализах по сравнению с мышиным антителом, указывая на то, что у гуманизированного антитела полностью сохранялась активность связывания с TFPI.
Сравнение антител 4D8.B1 и hz4D8 в анализах обращения ингибирования FXa и Fxa-FVIIa и в анализе образования тромбина (TGA)
при 20 нM
Пример 5. Получение дополнительных антител против TFPI посредством фагового дисплея
1. Отбор антител против TFPI посредством фагового дисплея
Идентифицировали антитела на основе одноцепочечного вариабельного фрагмента (scFv), которые связываются с рекомбинантным человеческим и мышиным TFPI K1K2, после четырех циклов селекции с использованием библиотеки фагового дисплея фрагментов антитела scFv, полученных от неиммунизированных человеческих доноров. Отборы фагов поводили в растворе с использованием стрептавидиновых шариков. Связавшийся фаг элюировали посредством инкубации со 140 мМ триэтаноламином (ТЕА), рН 11,5, или 50 мМ MES (морфолино-этансульфоновая кислота), рН 5,5, в течение 10 мин при комнатной температуре на ротационном шейкере и нейтрализовали 1 М Tris-HCl, pH 7,5.
Пул элюированного фага использовали для инфекции 10 мл культуры E. coli ER2738, которая была выращена до средней логарифмической фазы (соответствующей OD600 приблизительно 0,5). Бактерии инфицировали фагом в течение 30 минут при 37°С без встряхивания, концентрировали центрифугированием и высаживали на чашки, с последующим ростом в течение ночи при 30°С. Для следующего цикла отбора фаги спасали посредством инокуляции 25 мл 2× TYAG/тетрациклина до OD600 ~0,1, выращивали при 37°С до OD600 0,3-0,5. Клетки сверхинфицировали хелперным фагом MK13K07 в соотношении клетка/хелперный фаг 1:20 и инкубировали при 37°С без встряхивания в течение 30 минут, затем встряхивая при 150 об./мин в течение 60 минут. Клетки затем центрифугировали, и осадок ресуспендировали в среде, содержащей канамицин и не содержащей глюкозу. Данную культуру выращивали в течение ночи при 25°С. Фаг отбирали в супернатанте после центрифугирования и использовали в следующем цикле селекции.
2. Получение неочищенного периплазматического материала для применения в анализе ELISA
Фрагменты антитела scFv могут экспрессироваться либо на поверхности фаговой частицы, либо в растворе в бактериальном периплазматическом пространстве, в зависимости от используемых условий роста. Для индукции высвобож дения фрагментов антитела scFv в периплазму, 96-глубоколуночные планшеты, содержащие 2× среды TY с 0,1% глюкозы/100 мкг/мл ампициллина, инокулировали из оттаявших глицериновых маточных растворов и выращивали при 37°С в течение приблизительно 4 часов. Содержимое бактериальной периплазмы (перипрепараты) высвобождали осмотическим шоком. Планшеты центрифугировали и отбирали супернатант, содержащий scFv.
3. ELISA для измерения связывания экспрессированных в периплазме scFv с человеческим и мышиным TFPI K1K2.
Всего из циклов 2, 3 и 4 всех ветвей селекции случайным образом отбирали 1984 клона. Агенты на основе scFv, связывающие TFPI, идентифицировали посредством ELISA связывания периплазматического препарата (перипрепарат). Биотинилированным человеческим и мышиным TFPI K1K2 покрывали 384-луночные стрептавидиновые планшеты Nunc Maxisorp в концентрации 1 мкг/мл в PBS. Растворы TFPI K1K2 удаляли, и планшеты блокировали в течение 1 часа при комнатной температуре в 0,05% Tween 20/1% BSA/PBS. Получали перипрепараты и блокировали в течение 1 часа при комнатной температуре в равном объеме 6% молока/1% BSA. 20 мкл/лунку блокированных периплазматических scFv и контрольных антител переносили в подходящие планшеты и инкубировали в течение 1 часа при комнатной температуре. Добавляли 1:2000 разведение антитела против myc, конъюгированного с пероксидазой хрена (HRP) или разведение 1:10000 вторичного антитела козы против человеческого антитела-HRP для выявления связавшихся scFv или контрольных антител против TFPI. Сигнал проявляли с использованием 3,3',5,5'-тетраметилбензидина со считыванием поглощения при 450 нм на планшет-ридере Envision (Perkin Elmer). Всего 883 клона scFv было идентифицировано в качестве агентов, связывающихся с TFPI. Данные 883 scFv, связывающиеся с TFPI, секвенировали для идентификации уникальных клонов. Было выбрано 288 уникальных клонов для тестирования конкурентного связывания TFPI/FXa.
4. ELISA для идентификации scFv, которые конкурируют с FXa за связывание с человеческим и мышиным TFPI K1K2
Всего 288 уникальных клонов тестировали в ELISA конкурентного связывания FXa/TFPI. Человеческим FXa в течение ночи покрывали 384-луночные планшеты Nunc Maxisorp в концентрации 1 мкг/мл в PBS. Раствор FXa удаляли, и поверхность планшета блокировали в течение 1 часа при комнатной температуре в 0,05% Tween 20/1% BSA/PBS. Получали перипрепараты и блокировали в течение 1 часа при комнатной температуре в равном объеме 6% молока/1% BSA. 20 мкл/лунку блокированных периплазматических scFv и контрольных антител смешивали с биотинилированным humTFPI K1K2 и давали инкубироваться в течение 1 часа при комнатной температуре. Данную смесь переносили в планшеты, покрытые FXa, и инкубировали в течение 1 часа при комнатной температуре. Для выявления связавшегося TFPI добавляли разведение 1:2000 стрептавидин-пероксидаза хрена. Сигнал проявляли с использованием 3,3',5,5'-тетраметилбензидина со считыванием поглощения при 450 нм на планшет-ридере Envision. Всего 48 антител scFv было классифицировано в качестве конкурентных ингибиторов связывания TFPI/FXa.
5. Превращение scFv до человеческого IgG
Для субклонирования в человеческие векторы для клонирования IgG-3M отбирали всего 48 антител scFv с уникальными последовательностями, которые демонстрировали связывание с TFPI и ингибирование в конкурентном ELISA с TFPI/FXa. Вкратце, фрагменты амплифицировали стандартной ПЦР. Фрагменты VH или VL очищали на геле и лигировали в экспрессионный вектор млекопитающих, содержащий либо человеческий IgG1-3M (VH), либо константную область каппа или лямбда (VK/VL). Затем использовали векторы для парной экспрессии VH и VK/VL для кратковременной экспрессии у млекопитающих и очистки в клетках HEK 293.
6. Характеристика человеческих антител на основе IgG-3M против TFPI
48 антител против TFPI ранжировали в разлиных анализах, включая анализы обращения ингибирования FXa и TF/FVIIa/FXa. TFPI-3, TFPI-21, TFPI-23, TFPI-24 и TFPI-26 имели желательные свойства, такие как связывание с TFPI из разных видов, слабое или отсутствие связывания с humTFPI2 K1K2K3 (Таблица 13). Данные по анализу обращения ингибирования FXa и TF/FVIIa/FXa для этих тех же самых 5 антител показаны в Таблице 14, а данные по связыванию SPR - в Таблице 15.
Антитела против TFPI, которые демонстрировали связывание с TFPI многих видов (человека, обезьяны, мышиным, кроличьим и крысиным) и отсутствие или слабое связывание с человеческим TFPI2
K1K2
Активность антитела против TFPI в анализах обращения ингибирования TF/FVIIa/FXa и образования тромбина
SPR кинетики связывания антитела против TFPI с человеческим и мышиным TFPI
Пример 6. Картирование эпитопа антител против TFPI посредством SPR
Сэндвич-анализ посредством SPR использовали для картирования эпитопов антител против TFPI, открытых и изложенных в этом документе (TFPI-21, TFPI-23, TFPI-24, 4D8, 6B7.c5 и 7A4.D9). Другие известные контрольные антитела (hz4F36, 2A8-200 и Mab2974) также были включены в эксперимент по картированию эпитопов. Антитело 1 иммобилизовали на чипе Biacore CM5 с использование химии NHS
(N-гидрокси-сукцинимид). Человеческий TFPI (humTFPI K1K2) исходно инъецировали на чип, пока связывание не было около кажущегося равновесия. Немедленно после остановки инъекции человеческого TFPI над чипом инъецировали антитело 2. Если антитело 2 связывается с комплексом антитела 1 и человеческого TFPI на поверхности чипа СМ5, тогда антитело 2 имеет отличный и неперекрывающийся эпитоп связывания TFPI относительно антитела 1 (оценивается как +). Если антитело 2 не демонстрирует связывания, тогда он оценивается как значимо перекрывающийся эпитоп (негативный (-)) относительно антитела 1. Если антитело 2 демонстрирует слабое связывание, тогда считается, что антитела 1 и 2 имеют некоторое частичное перекрывание в эпитопах TFPI (оценивается как +/-). Как показано в Таблице 16, TFPI-21 и TFPI-23 имеют аналогичные эпитопы, и эти данные также показывают то, что TFPI-21 и TFPI-23 имеют эпитопы, которые полностью отличаются от mab2974 и hz4F36.
Картирование эпитопов антител против TFPI с использование сэндвич-анализа посредством SPR
Антитело 1 иммобилизовали на поверхности чипа CM5. Затем инъецировали на поверхность человеческий TFPI (humTFPI K1K2), пока измерение не было близким к кажущемуся равновесию. Немедленно после остановки инъекции TFPI инъецировали антитело 2 для измерения связывания с комплексом антитело 1/TFPI. Оценка «+» дается парам антитела 1 и 2, которые имеют полностью отличные эпитопы. Оценка «-» дается парам антител, которые имеют сильно перекрывающиеся эпитопы. Если антитело 2 демонстрирует слабое связывание, тогда считается, что антитела 1 и 2 имеют некоторое частичное перекрывание в эпитопах TFPI (оценивается как +/-).
Пример 7. Модифицирование человеческих каркасных областей антитела TFPI-23 до последовательности зародышевой линии
Получали два варианта TFPI-23 для увеличения содержания остатков каркасной области человеческой зародышевой линии. TFPI-106 содержит мутации от H1Q до E и от H5V до L (нумерация по Kabat), и TFPI-107 (Таблицы 3 и 4) содержало мутации от H1Q до E, от H5V до L и от H94I до K (нумерация по Kabat). TFPI-106, TFPI-107 и TFPI-23 экспрессировали, очищали и тестировали на связывание с humTFPI K1K2 посредством SPR. Данные в Таблице 17 показывают то, что, по сравнению с родительским антителом TFPI-23, вариант зародышевой линии TFPI-106 сохранял полную аффинность связывания.
Определенная посредством SPR кинетика связывания с человеческим TFPI вариантов TFPI-23 с каркасными областями человеческими зародышевой линии
TFPI-106 продемонстрировало умеренное улучшение в связывании по сравнению с родительским антителом TFPI-23.
Пример 8. Модифицирование человеческих каркасных областей антитела TFPI-24 до последовательности зародышевой линии
Получали четыре варианта VL TFPI-24 (TFPI-110, TFPI-111, TFPI-112, TFPI-113) и формировали пару с последовательностью VH TFPI-24. Получали три варианта VН
TFPI-24 (TFPI-108, TFPI-109, TFPI-114) и формировали пару с последовательностью VL TFPI-24. На основе этих данных образовали пару наилучшего варианта VL, представляющего собой TFPI-113, и наилучшего варианта VН, представляющего собой TFPI-108, с получением антитела TFPI-118. TFPI-118 и TFPI-24 тестировали на связывание с человеческим TFPI посредством SPR, и результаты в Таблице 18 демонстрируют сопоставимую кинетику связывания.
Кинетику связывания TFPI-24 и варианта с человеческой каркасной областью, представляющего собой FPI-118, с человеческим TFPI сравнивали с использованием SPR
TFPI-118 демонстрировало сопоставимую кинетику связывания с родительским антителом TFPI-23.
Пример 9. Определенная посредством SPR кинетика связывания антител против TFPI с TFPI из разных видов
Антитела против TFPI (TFPI-106, TFPI-118 и hz4F36) анализировали посредством SPR для определения кинетики связывания с TFPI из разных видов животных (человека (huTFPI K1K2), яванского макака (cynTFPI K1K2), кролика (rabTFPI K1K2), мыши (murTFPI K1K2) и крысы (ratTFPI K1K2); Таблица 1). В данный эксперимент также включали три антитела для сравнения (hz4F36, 2A8 и 2A8-200).
Определенная посредством SPR кинетика связывания антитела против TFPI с TFPI человека, яванского макака, кролика и мыши (значения Kd представляют собой средние из 2 экспериментов)
Пример 10. Структуры комплекса антитела против TFPI/TFPI
1. Структура комплекса Fab 4D8.b1 / K2 TFPI яванского макака
Fab 4D8.b1 / K2 TFPI яванского макака смешивали в мольном отношении 1:1 с образованием комплекса. Конечную очистку проводили с использованием колонки Superdex 200. Данный комплекс концентрировали до 12,6 мг/мл для структурных исследований. Кристаллы K2 TFPI+Fab 4D8 получали в 100 мМ Tris-HCl, pH 8,5, 20% ПЭГ 10000 (полиэтиленгликоль 10000). Это давало стержневидные кристаллы, которые дифрагировали до 2,9 Å. Кристаллы временно подвергали криозащите, и отбор данных синхротрона осуществляли удаленно в Advanced Photon Source. Кадры изображений обрабатывали с использованием программы AutoPROC (Global Phasing Ltd). Данные принадлежат к пространственной группе P212121 со следующей элементарной ячейкой:
a = 62,102 Å, b = 82,284 Å, c = 103,628 Å, α = β = γ = 90°, с одним комплексом на асимметричную ячейку. Поиски молекулярного замещения с использованием моделей гомологии Fab 4D8, а также общедоступных структур (банк данных белков RSCB; коды PDB 1TFX и 4DTG) доменов K2 TFPI дали убедительные решения для каждого компонента. Уточнение осуществляли с использованием программы autoBUSTER (Global Phasing Ltd), и конечные множители R/Rсвободный при 2,9 Å составляют 0,1707 и 0,2424 соответственно с RMSD (среднеквадратичное отклонение) связи 0,010 Å, RMSD углов 1,26°. На основе скрытой области поверхности (BSA) и процента BSA (% BSA) для остатков на поверхности контакта Fab / K2 TFPI были определены эпитоп и паратоп Fab 4D8. Следующие остатки в домене K2 TFPI участвуют в непосредственном контакте с Fab 4D8 (эпитоп согласно BSA): E101, P103, Y109, I110, T111, Y113, F114, S119, Q121, C122, E123, R124, F125, K126 и L140. Следующие остатки в тяжелой цепи Fab 4D8 составляют паратоп тяжелой цепи: D50, T57, L58, Y59, Q61, K64, D98, Y99 и D100. Следующие остатки в легкой цепи Fab 4D8 составляют паратоп легкой цепи: H30, W50, H91, Y92, T93, T94, P95 и Y96. Значения BSA и % BSA для остатков эпитопа и паратопа показаны в Таблице 20.
Остатки эпитопа и паратопа антитела 4D8.b1 против TFPI, определенные по скрытой области поверхности (BSA) и проценту BSA (% BSA) остатков поверхности контакта в структуре комплекса Fab 4D8.b1 / K2 TFPI яванского макака (остатки легкой цепи (LC) и тяжелой цепи (НС) антитела пронумерованы с использованием определений Kabat). В анализе BSA применяется пороговое значение (BSA 20 Å2 или больше, или участие в электростатическом взаимодействии).
2. Структуры комплекса Fab 2A8 и 2A8-200 / K1K2 яванского макака
Fab 2A8 и K1K2 TFPI яванского макака смешивали в мольном соотношении 1:1 с образованием комплекса. Конечную очистку проводили с использованием колонки Superdex 200. Для структурных исследований данный комплекс концентрировали до 10,8 мг/мл. Кристаллы комплекса, содержащего Fab 2A8 и K1K2 TFPI, получали при следующих двух условиях: (1) 100 мМ HEPES, pH 7,5, 12,5% ПЭГ 8000, что давало игловидные кристаллы, которые дифрагировали до 3,0 Å; (2) 100 мМ HEPES, pH 7,5, 1600 мМ сульфат аммония, 2% ПЭГ 1000, что давало глыбовидные кристаллы, которые дифрагировали до 3,3 Å. Кристаллы временно подвергали криозащите, и отбор данных синхротрона осуществляли удаленно в Advanced Photon Source. Кадры изображений обрабатывали с использованием программы AutoPROC (Global Phasing Ltd). Данные принадлежат к пространственной группе P3221 со следующими элементарными ячейками: a = b = 196,146 Å, c = 41,262 Å, α = β= 90°, γ = 120°, с одним комплексом на асимметричную ячейку. Поиски молекулярного замещения с использованием моделей гомологии Fab 2А8, а также общедоступных структур (банк данных белков RSCB; коды PDB 1TFX и 4DTG) доменов K2 TFPI дали убедительные решения для каждого компонента. Уточнение осуществляли с использованием программы PHENIX, и конечные множители R/Rсвободный при 3,0 Å составляют 0,1667 и 0,2088, соответственно, с RMSD связи 0,011 Å, RMSD углов 1,474°. На основе скрытой области поверхности (BSA) и процента BSA (% BSA) для остатков на поверхности контакта Fab K1K2 TFPI были определены эпитоп и паратоп данного комплекса. Следующие остатки в доменах K1K2 TFPI участвуют в непосредственном контакте с Fab 2А8 (эпитоп согласно BSA): D31, D32, G33, P34, C35, K36, E100, E101, P103, G104, I105, C106, R107, G108, Y109, E123, K126, Y127 и G128. Следующие остатки в тяжелой цепи Fab 4D8 составляют паратоп тяжелой цепи: G26, T28, S31, Y32, Y96, R97, Y98, W99 и D101 (нумерация по Kabat). Следующие остатки в легкой цепи Fab 2A8 составляют паратоп легкой цепи: L28, R29, N30, Y31, Y32, Y49, Y50, D51 и N66 (нумерация по Kabat). Значения BSA и % BSA для остатков эпитопа и паратопа показаны в Таблице 21. Структура очень близкородственного антитела, представляющего собой 2А8-200, в комплексе с K1K2 TFPI также была определена с использованием по существу идентичных способов. Эпитоп и паратоп данного антитела были идентичными эпитопу и паратопу 2А8.
Остатки эпитопа и паратопа антитела 2А8 против TFPI, определенные по скрытой области поверхности (BSA) и проценту BSA (% BSA) остатков поверхности контакта в структуре комплекса Fab 2А8 / K2 TFPI яванского макака (остатки легкой цепи (LC) и тяжелой цепи (НС) антитела пронумерованы с использованием определений Kabat). В анализе BSA применяется пороговое значение (BSA 20 Å2 или больше, или участие в электростатическом взаимодействии).
3. Структура комплекса Fab Mab 2974 / K2 TFPI
Fab Mab 2974 (R&D Systems) и K2 TFPI яванского макака смешивали в мольном соотношении 1:1,2 с образованием комплекса. Конечную очистку проводили с использованием колонки Superdex 200. Для структурных исследований данный комплекс концентрировали до 17,5 мг/мл. Кристаллы комплекса, содержащего Fab Mab 2974 и K2 TFPI, получали в 100 мМ цитрате натрия, pH 5,6, 20% изопропанола, 20% ПЭГ 4000, что давало глыбовидные кристаллы, которые дифрагировали до 2,15 Å. Кристаллы временно подвергали криозащите, и отбор данных синхротрона осуществляли удаленно в Advanced Photon Source. Кадры изображений обрабатывали с использованием программы AutoPROC (Global Phasing Ltd). Данные комплекса принадлежат к пространственной группе P212121 со следующими элементарными ячейками: a = 82,075 Å, b = 117,829 Å, c = 170,945 Å, α = β= γ = 90°, с тремя комплексами на асимметричную ячейку. Поскольку последовательность Fab Mab 2947 не была доступна, для расшифровки последовательности белка был отобран набор данных высокого разрешения для одного Fab (1,63 Å), наряду с биоинформационным анализом. Поиски молекулярного замещения с использованием структуры Fab Mab 2947, а также общедоступных структур (банк данных белков RSCB; коды PDB 1TFX и 4DTG) доменов K2 TFPI дали убедительные решения для каждого компонента. Уточнение осуществляли с использованием программы autoBUSTER, и конечные множители R/Rсвободный при 2,15 Å составляют 0,1702 и 0,2161, соответственно, с RMSD связи 0,010 Å, RMSD углов 1,13°. На основе скрытой области поверхности (BSA) и процента BSA (% BSA) для остатков на поверхности контакта Fab K2 TFPI был определен эпитоп данного комплекса. Следующие остатки в домене K2 TFPI участвуют в непосредственном контакте с Fab Mab 2974 (эпитоп согласно BSA): E100, E101, P103, R107, Y109, T111, N116, Q118, S119, Q121, E123, R124, F125 и K126. Значения BSA и % BSA для остатков эпитопа показаны в Таблице 22.
Остатки эпитопа антитела Mab 2974 против TFPI, определенные по скрытой области поверхности (BSA) и проценту BSA (% BSA) остатков поверхности контакта в структуре комплекса Fab Mab 2974 / K2 TFPI яванского макака. В анализе BSA применяется пороговое значение (BSA 20 Å2 или больше, или участие в электростатическом взаимодействии)
4. Структура комплекса Fab TFPI-23 / K2 TFPI яванского макака
Fab TFPI-23 и K2 TFPI яванского макака смешивали в мольном соотношении 1:2 с образованием комплекса. Конечную очистку проводили с использованием колонки Superdex 200. Для структурных исследований данный комплекс концентрировали до 12,4 мг/мл. Кристаллы комплекса K2 TFPI+Fab 4D8 получали в 100 мМ Bis-Tris, pH 6,5, 20% PEGMME5000. Это давало волокнообразные кристаллы, которые дифрагировали до 2,9 Å. Кристаллы временно подвергали криозащите, и отбор данных синхротрона осуществляли удаленно в Advanced Photon Source. Кадры изображений обрабатывали с использованием программы AutoPROC (Global Phasing Ltd). Данные принадлежат к пространственной группе P1 со следующими элементарными ячейками: a = 74,669 Å, b = 101,372 Å, c = 119,275 Å, α = 101,83, β = 92,27°, γ = 96,78°, с шестью копиями комплексами на асимметричную ячейку. Поиски молекулярного замещения с использованием моделей гомологии Fab TFPI-23, а также общедоступных структур (банк данных белков RSCB; коды PDB 1TFX и 4DTG) доменов K2 TFPI дали убедительные решения для каждого компонента. Уточнение осуществляли с использованием программы autoBUSTER, и конечные множители R/Rсвободный при 2,9 Å составляют 0,1961 и 0,2344, соответственно, с RMSD связи 0,010 Å, RMSD углов 1,22°. На основе BSA и процента BSA (% BSA) для остатков на поверхности контакта Fab K2 TFPI был определен эпитоп и паратоп данного комплекса. Следующие остатки в домене K2 TFPI участвуют в непосредственном контакте с Fab TFPI-23 (эпитоп согласно BSA): D102, I105, C106, R107, G108, R112, Y127, G129, C130, L131, G132, M134 и E138. Следующие остатки в тяжелой цепи Fab 4D8 составляют паратоп тяжелой цепи: A33, W47, A50, I51, S52, S56, Y58, L95, G96, A97, T98, S99, L100 и S100A. Следующие остатки в легкой цепи Fab 4D8 составляют паратоп легкой цепи: A29, Y31, Y91, S95A, G95B и S95C. Значения BSA и % BSA для остатков эпитопа и паратопа показаны в Таблице 23.
Остатки эпитопа и паратопа антитела против TFPI TFPI-23, определенные по скрытой области поверхности (BSA) и проценту BSA (% BSA) остатков поверхности контакта в структуре комплекса Fab TFPI-23 / K2 TFPI яванского макака (остатки легкой цепи (LC) и тяжелой цепи (НС) антитела пронумерованы с использованием определений Kabat). В анализе BSA применяется пороговое значение (BSA 20 Å2 или больше, или участие в электростатическом взаимодействии).
5. Структура комплекса Fab TFPI-24 / K2 TFPI яванского макака
Fab TFPI-24 и K2 TFPI яванского макака смешивали в мольном соотношении 1:2 с образованием комплекса. Конечную очистку проводили с использованием колонки Superdex 200. Для структурных исследований данный комплекс концентрировали до 12,2 мг/мл. Кристаллы комплекса K2 TFPI / Fab TFPI-24 получали в 20% PEG3350, 200 мМ нитрате аммония. Это давало кристаллы, которые дифрагировали до 1,75 Å. Кристаллы временно криозащищали, и отбор данных синхротрона осуществляли удаленно в Advanced Photon Source. Кадры изображений обрабатывали с использованием программы AutoPROC. Данные принадлежат к пространственной группе P212121 со следующими элементарными ячейками: a = 42,817 Å, b = 71,362 Å, c = 148,729 Å, α = β = γ = 90°, с одним комплексом на асимметричную ячейку. Поиски молекулярного замещения с использованием моделей гомологии Fab TFPI-24, а также общедоступных структур (банк данных белков RSCB; коды PDB 1TFX и 4DTG) доменов K2 TFPI дали убедительные решения для каждого компонента. Уточнение осуществляли с использованием программы autoBUSTER, и конечные множители R/Rсвободный при 1,75 Å составляют 0,1900 и 0,2269, соответственно, с RMSD связи 0,010 Å, RMSD углов 1,18°. На основе скрытой области поверхности (BSA) и процента BSA (% BSA) для остатков на поверхности контакта Fab / K2 TFPI были определены эпитоп и паратоп данного комплекса. Следующие остатки в домене K2 TFPI участвуют в непосредственном контакте с Fab TFPI-24 (эпитоп согласно BSA): E100, E101, D102, G104, I105, C106, R107, G108, Y109, I110, G129, C130, L131 и G132. Следующие остатки в тяжелой цепи Fab TFPI-24 составляют паратоп тяжелой цепи: A33, Q35, W47, G50, I51, S52, N53, R55, S56, I57, G58, F95, L96, H97, S99 и D101. Следующие остатки в легкой цепи Fab TFPI-24 составляют паратоп легкой цепи: M31, Y32, H34, Y36, L46, R50, W91 и Y96. Значения BSA и % BSA для остатков эпитопа и паратопа показаны в Таблице 24.
Остатки эпитопа и паратопа антитела против TFPI TFPI-24, определенные по скрытой области поверхности (BSA) и проценту BSA (% BSA) остатков поверхности контакта в структуре комплекса Fab TFPI-24 / K2 TFPI яванского макака (остатки легкой цепи (LC) и тяжелой цепи (НС) антитела пронумерованы с использованием определений Kabat). В анализе BSA применяется пороговое значение (BSA 20 Å2 или больше, или участие в электростатическом взаимодействии).
6. Анализ эпитопа hz4F36
Структура Fab hz4F36 в комплексе с доменом K2 человеческого TFPI доступна в банке данных белков (код доступа PDB - 4DTG). На основе BSA и процента BSA (%BSA) остатков поверхности контакта в структуре комплекса hz4F36 / K2 TFPI были определены остатки эпитопа, как показано в Таблице 25.
Таблица 25
Остатки эпитопа антитела против TFPI hz4F36, определенные по скрытой области поверхности (BSA), и проценту BSA (% BSA) остатков поверхности контакта в структуре комплекса Fab hz4F36 / K2 TFPI яванского макака (код доступа PDB - 4DTG). В анализе BSA применяется пороговое значение (BSA 20 Å2 или больше, или участие в электростатическом взаимодействии).
7. Сравнение эпитопов антител против TFPI
Эпитопы антител против TFPI, показанные в Таблицах 20-25, сравниваются в Таблицах 26 и 27. В Таблице 26 показаны эпитопы антител, которые являются специфичными в отношении домена K2 TFPI. В Таблицу 27 включены 2 дополнительных антитела (2А8 и 2А8-200), которые связываются и с K1, и с K2 доменами.
Остатки эпитопов антител против TFPI на основе данных в Таблицах 20, 22-25
ческого TFPI
Х обозначает аминокислотные остатки TFPI, которые являются частью эпитопа. Х (жирный шрифт) обозначает новые остатки эпитопа для антител, раскрытых в этом изобретении. Следует отметить, что TFPI-23 не конкурирует за связывание с TFPI с hz4F36, 4D8 или mab2974, но действительно конкурирует с TFPI-24 за связывание с TFPI. Антитела 2А8 и 2А8-200 не включены в эту таблицу, так как они требуют для связывания обоих доменов K1 и K2 и не связываются с одним доменом K2.
Сравнение остатков эпитопов антител против TFPI на основе данных, показанных в Таблицах 20-25
ческого TFPI
Х обозначает аминокислотные остатки TFPI, которые являются частью эпитопа. Х (жирный шрифт) обозначает новые остатки эпитопа для антител, раскрытых в этом изобретении. Антитела 2А8 и 2А8-200 требуют для связывания обоих доменов K1 и K2 и не связываются с одним доменом K2 или K1.
Прогнозирование ключевых остатков эпитопа TFPI для TFPI-23 посредством аланинового сканирования с использованием расчетных способов (Accelrys Discovery Studio 4.1)
На основе произвольного порогового значения больше 1 ккал/моль (4,184 кДж/моль) прогнозируется то, что 3 остатка TFPI (Ile105, Arg107 и Leu131), при мутировании до аланина, значимо способствуют связыванию TFPI-23 с TFPI.
CDR TFPI-23 и остатки каркасной области в пределах 4 ангстрем от эпитопа K2 TFPI
(4.07 Å)
(4,03 Å)
С использованием расчетных способов прогнозирования (Accelrys Discovery Studio 4.1) также демонстрируются возможные аминокислотные замены для каждого остатка CDR/паратопа, которые, как прогнозируется, не оказывают негативного влияния на стабильность TFPI-23 или аффинность в отношении TFPI (меньше 0,5 ккал/моль, столбцы 4-6). Они классифицируются на три группы: (1) замены по меньшей мере с нейтральным эффектом на связывание (аффинность меньше 0,5 ккал/моль, столбец 4), (2) замены, которые имеют нейтральный/стабилизирующий эффект на связывание (аффинность меньше -0,5 ккал/моль, столбец 5), и (3) верхние 3 прогнозируемых сайта с самым стабилизирующим эффектом на аффинность. Для всех остатков используется нумерация по Kabat (столбец 6). 138Glu/L93Ser и 131Leu/L96Gly включаются как возможные контактные остатки, так как расстояние незначительно превышает 4 Å (4,07 Å и 4,03 Å соответственно), но является достаточно близким к округленным 4 Å.
Остатки эпитопа TFPI и соответствующие остатки паратопа TFPI-23
Применяется пороговое значение (BSA 20 Å2 или больше, или участие в электростатическом взаимодействии)
Остатки эпитопа TFPI и соответствующие остатки паратопа TFPI-23, определенные посредством BSA (не применяется пороговое значение или применяется минимальная BSA)
Пример 11. Протромбиновое время с разведением (dPT)
Способность антител против TFPI ингибировать эндогенный TFPI в человеческой плазме, дефицитной по FVIII (гемофилия А), исследовали с использованием анализа протромбинового времени (РТ) с разведением. РТ с разведением представляет собой модифицированный анализ РТ с использованием разведенного тканевого фактора (инновина) для продления времени свертывания крови.
Для анализа dPT в человеческой плазме, дефицитной по FVIII (George King Biomedical), реактив Innovin® разводили 1:3000 в буфере для разведения (50 мМ имидазол, 0,1 М хлорид натрия, 1 мг/мл BSA (бычий сывороточный альбумин), 8,34 мМ хлорида кальция, рН 7,4) и предварительно инкубировали при 37°С. Плазму размораживали в водной бане при 37°С в течение 5 минут немедленно перед анализом. Разведения антител против TFPI готовили в PBS, добавляли в плазму, и плазму инкубировали в течение 20 минут при комнатной температуре. После этой инкубации 50 мкл плазмы инкубировали в течение 1 минуты при 37°С, и немедленно инициировали реакцию свертывания посредством добавления 50 мкл разведения 1:3000 реактива Innovin®, нагретого до 37°С. Время до свертывания определяли при 37°С с использованием анализатора свертывания STart® 4. Точки данных получали в двойной повторности, вводили в Microsoft Excel, и 50%-ную эффективную концентрацию (ЕС50) оценивали с использованием GraphPad Prism®. Результаты показаны в Таблице 30.
TFPI осуществляет понижающую регуляцию внешнего пути свертывания крови FVIIa/TF/FXa, уменьшая образование FXa и, в конечном итоге, тромбина. dPT измеряет эффекты на внешний путь свертывания крови. Данные показывают то, что добавление антител против TFPI к плазме страдающего от гемофилии А дозозависимо сокращало время свертывания. Контрольный IgG в концентрации 300 нМ не имел влияния на время свертывания.
TFPI-23
TFPI-106
TFPI-24
TFPI-118
против
TFPI 4F36
против
TFPI h4D8
Пример 12. Тромбоэластография (TEG)
Тромбоэластография (TEG) представляет собой глобальный гемостатический анализ, в котором измеряется кинетика образования сгустков крови в цельной крови. Цельную кровь выделяли у здоровых человеческих доноров, отбирая в пластмассовые пробирки для сбора крови, содержащие 3,2% цитрата натрия, и первую пробирку для отбора крови отбрасывали для минимизации введения активаторов свертывания, таких как тканевой фактор. Цельную кровь с цитратом обрабатывали в течение одного часа контрольным мышиным антителом против человеческого IgG2 (100 мкг/мл) или ингибирующим антителом против FVIII (GM1805 (Green Mountain), 100 мкг/мл) для ингибирования эндогенного FVIII, индуцируя гемофилия А-подобный фенотип. Цельную кровь (320 мкл) с дозированными антителами против TFPI или контрольным антителом IgG1 добавляли в реакционную чашку TEG®, содержащую 20 мкл 0,2 М хлорида кальция и 20 мкл липидированного тканевого фактора (Innovin®), разведенного в 20 мМ HEPES, 150 мМ хлориде натрия, рН 7,4, что приводило к конечному разведению липидированного тканевого фактора 1:200000 в каждой реакции. Реакции проводили в двойной повторности и немедленно начинали при добавлении цельной крови в чашку TEG. Анализ проводили на анализаторах TEG® 5000 Hemostasis с использованием программы TEG® согласно инструкциям изготовителя после калибровки с использованием контролей Level I и Level II (Haemonetics). Реакции проводили при 37°С в течение 60 минут. См. Таблицу 31.
Параметры TEG в крови с гемофилией А (НА), индуцированной антителом, обработанной антителами против TFPI
мин
градусы
мм
В Таблице 31 показано то, что обработка цельной крови антителом против FVIII значительно пролонгировала значение TEG-R до 41,5 минут. В присутствии цельной крови антитело TFPI 106 демонстрировало благоприятный профиль относительно 2А8-200 и 4F36. Добавление TFPI-106, 2А8-200 или 4F36 (300 нМ) приводило к сокращению значения TEG-R до 17,85 минут, 20,4 минут и 24,05 минут соответственно. Антитело TFPI 106 стимулировало свертывание в гемофиличной крови, что демонстрируется уменьшением значения TEG-R и наблюдаемым увеличением угла TEG-альфа.
Пример 13. Нейтрализация TFPI и образование тромбина
Нейтрализацию TFPI антителами против TFPI измеряли с использованием двух хромогенных анализов: прямого анализа активности Фактора Xa и двухэтапного анализа FVIIa/TF/FXa, основанного на ингибировании TFPI образования FXa посредством TF-FVIIa. В первом анализе TFPI-106, TFPI-118, hz4D8 и два контрольных антитела - 4F36 или 2А8-200 - предварительно инкубировали при разных концентрациях (0-500 нМ) с фиксированной концентрацией человеческого рекомбинантного TFPI K1K2 и FXa для обеспечения образования комплекса. Активность FXa оценивали с использованием хромогенного субстрата FXa. Добавление антител против TFPI по изобретению вызывало дозозависимое увеличение активности FXa в этом анализе (см. Таблицу 32, анализ ингибирования Ха, значения ЕС50).
Двумя преобладающими формами TFPI in vivo являются TFPI-альфа (K1K2K3) и TFPI-бета (K1K2). Способность антител против TFPI ингибировать рекомбинантный TFPI K1K2 или TFPI K1K2K3 оценивали в двухэтапном анализе FVIIa/TF/FXa. В этом анализе измеряются объединенные эффекты нейтрализации ингибирования TFPI и на FXa, и на FVIIa/TF/FXa. Антитела в возрастающих концентрациях (0-500 нМ) инкубировали с TFPI, добавляли в среду для анализа с FVIIa/TF/FX, и активность FXa измеряли с использованием хромогенного субстрата FXa. Антитела против TFPI по изобретению нейтрализовали ингибирование TFPI K1K2 опосредованной FVIIa/TF активации FX (см. Таблицу 32, значения ЕС50 FVIIa/TF/FXa). Типичные антитела по изобретению также были эффективными в ингибировании TFPI K1K2K3 (ЕС50 для TFPI-106 составляет 8,47 нМ для нейтрализации ингибирования FVIIa/TF/FXa посредством TFPI K1K2K3). Эти данные демонстрируют ингибирование полноразмерного и усеченного TFPI.
Клиническая тяжесть гемофилии относится к остаточному уровню активности фактора свертывания. Активность фактора меньше 1% ассоциирована с тяжелым фенотипом, умеренная гемофилия ассоциирована с активностью фактора 2-5%, и легкая - с активностью фактора больше 5% - меньше 40%. Дефекты во внутреннем пути свертывания крови при гемофилии приводят к неадекватному образованию тромбина. Анализ образования тромбина (TGA) использовали для проверки ингибирования эндогенного TFPI в гемофилической плазме, обедненной тромбоцитами. В анализе TGA измеряется исходная фаза, фаза активации и фаза инактивации образования тромбина. Способности антител против TFPI восстанавливать образование тромбина в гемофилической плазме, обедненной тромбоцитами, тестировали с использованием калиброванного автоматического анализа образования тромбина (САТ). TFPI-106,
TFPI-118, hz4D8 и два контрольных антитела, представляющих собой 4F36 или 2A8-200 (0-500 нМ), инкубировали в гемофилической плазме для нейтрализации TFPI до добавления в среду для анализа. По сравнению с нормальной человеческой объединенной плазмой, образование тромбина заметно уменьшается в человеческой гемофилической плазме. При введении в плазму гемофилии А нормального контроля FACT, который представляет собой стандартизированный в концентрации 1U/мл FVIII, наблюдали дозозависимый ответ. Аналогичным образом, добавление FVIII с делецией В-домена в концентрации 200 нг/мл (1U/мл) восстанавливало образование тромбина до 100 нМ. На протяжении 60-минутного хода анализа в гемофилической плазме наблюдали минимальное образование тромбина. Инкубация плазмы с антителами по изобретению приводила к дозозависимому увеличению пика тромбина, эндогенного тромбинового потенциала и индекса скорости. Контрольные антитела, представляющие собой 2А8-200 и 4F36, также анализировали для сравнения (Таблица 32, значения ЕС50 скорости TGA).
Пример 14. Эффективность in vivo в мышиных моделях гемофилии
Эффективность некоторых антител против TFPI в качестве прокоагулянтов тестировали с использованием анализа острого перерезания хвоста у гемофилических мышей. В этом анализе ампутировали дистальную часть хвоста, что приводит к существенной потере крови, которая может быть уменьшена при введении гемостатического агента до или вскоре после осуществления перерезания. Гемофилические мыши получали одну внутривенную (в.в.) дозу антител против TFPI (6 мг/кг) через хвостовую вену в объеме 4 мл/кг, неспецифичный контрольный IgG (6 мг/кг) или носитель - физиологический раствор. В разное время после дозирования эффект антител на кровотечение оценивали следующим образом.
Мышей внутрибрюшинно анестезировали смесью кетамина/ксилазина. Хвосты погружали в 50 мл предварительно нагретого фосфатно-солевого буферного раствора (PBS) при 37°С на 2 минуты. Осуществляли отрезание 3 мм хвоста, и кровь собирали в PBS в течение 10-минутного периода. Затем количественно определяли объем потери крови посредством измерения содержания гемоглобина в PBS с использованием следующей методики. Пробирки центрифугировали для сбора эритроцитов, ресуспендировали в 5 мл лизирующего буфера (8,3 г/л хлорида аммония, 1,0 г/л бикарбоната калия и 0,037 г/л EDTA), и измеряли поглощение образцов при 575 нМ посредством спектрофотометра. Значения поглощения пересчитывали в общую кровопотерю (мкл) с использованием стандартной кривой. Статистическую значимость различия между средними оценивали посредством дисперсионного анализа (ANOVA) с последующим критерием множественных сравнений Даннета с использованием программы GraphPad®Prism. Результаты выражены как среднее плюс/минус стандартная ошибка среднего (SEM). В графических материалах, описанных ниже, статистическая значимость определяется как значение Р меньше 0,05 и указывается звездочкой над данными.
На Фиг. 3А показан эффект на кровопотерю у мышей с гемофилией А (обозначены FVIII -/-) после перерезания хвоста дозирования антитела 2А8-200 в разное время перед перерезанием хвоста по сравнению с контролем в виде носителя (физиологичекий раствор). На Фиг. 3Б показан эффект на кровопотерю у мышей с гемофилией А (обозначены FVIII -/-) после перерезания хвоста дозирования антитела 2А8 в разное время перед перерезанием хвоста по сравнению с контролем в виде носителя (физиологичекий раствор) и неспецифического человеческого IgG1 (обозначен hIgG1). На Фиг. 3Б также показан эффект на кровопотерю у нормальных мышей (обозначены FVIII +/+) после перерезания хвоста дозирования антитела 2А8 в два разных момента времени перед перерезанием хвоста. На Фиг. 3В показан эффект на кровопотерю у мышей с гемофилией А после перерезания хвоста дозирования антител 4D8, 21, 23 и 24 в разное время перед перерезанием хвоста по сравнению с контролем в виде носителя (физиологичекий раствор). На Фиг. 3Г показан эффект на кровопотерю у мышей с гемофилией А после перерезания хвоста дозирования антитела 106 в разное время перед перерезанием хвоста по сравнению с контролем в виде носителя (физиологичекий раствор). На Фиг. 3Д показан эффект на кровопотерю у мышей с гемофилией А после перерезания хвоста дозирования антитела 118 в разное время перед перерезанием хвоста по сравнению с контролем в виде носителя (физиологичекий раствор). На Фиг. 4 показан эффект на кровопотерю у мышей с гемофилией В после перерезания хвоста дозирования антитела 106 в разное время перед перерезанием хвоста по сравнению с контролем в виде носителя (физиологичекий раствор).
Как показано на Фиг. 3Г и Фиг. 4, введение 6 мг/кг антитела 106 против TFPI мышам с гемофилией А или гемофилией В уменьшало кровопотерю в модели острого травматического повреждения при введении антител в разное время до повреждения хвоста в виде перерезания. У мышей с гемофилией А гемостатический эффект сохранялся в течение по меньшей мере 189 часов после введения, хотя к 240 часам после введения данный эффект возвращался к контрольным уровням. У мышей с гемофилией В данный эффект сохранялся в течение по меньшей мере 72 часов после введения и возвращался к исходному уровню к 192 часам после введения.
Описанные выше данные и результаты свидетельствуют о том, что протестированные антитела, включая антитело TFPI 106, могут профилактически вводиться субъектам с гемофилией А или гемофилией В для уменьшения кровотечения перед травматическим повреждением или другим типом эпизода кровотечения.
Эффект антитела TFPI 106 в качестве гемостатического средства у мышей с гемофилией А также был протестирован для определения того, могло ли оно уменьшать кровотечение при введении вскоре после перерезания хвоста. Данные эксперименты проводили с использованием аналогичной методологии, что и эксперименты, тестирующие эффект антител при введении до перерезания хвоста, за исключением того, что немедленно после перерезания хвоста инфундировали в.в. дозу TFPI 106 (6 мг/кг) или рекомбинантного Фактора VIII (200 единиц/кг) через канюлю, вставленную в яремную вену, после чего отбирали кровь в течение 10 минут до количественного измерения кровопотери. Во второй серии экспериментов разные дозы TFPI 106 и Фактора VIII (200 Ед/кг) раздельно вводили мышам с гемофилией А через 2 минуты после обрезания хвоста, и затем отбирали кровь в течение 10 минут до количественного измерения кровопотери.
На Фиг. 9А показано то, что 6 мг/мл антитела TFPI 106, введенного немедленно после перерезания хвоста, было эффективным в уменьшении кровопотери у мышей с гемофилией А по сравнению с контролем в виде носителя, хотя и не в такой же степени, что 200 Ед/кг Фактора VIII, введенного таким же способом. На Фиг. 9Б показано то, что антитело TFPI 106 дозозависимо уменьшает кровотечение у мышей с гемофилией А при введении через 2 минуты после повреждения посредством перерезания хвоста, и что наивысшая протестированная доза (6 мг/кг) TFPI 106 была приблизительно такой же эффективной, что и рекомбинантный Фактор VIII в дозе 200 Ед/кг, когда оба были введены через 2 мин после перерезания хвоста. Данные и результаты, описанные на этих Фигурах, свидетельствуют о том, что антитело TFPI 106 может быть эффективным в качестве лечения по требованию против кровотечения, которое уже началось у субъектов из-за травмы или некоторой другой причины.
Пример 15. Фармакокинетика и метаболизм продукта
Фармакокинетику (PK) и/или токсикокинетику (TK) TFPI-106 охарактеризовывали после внутривенного (в.в.) и/или подкожного (п.к.) дозирования у крыс Wistar Han, новозеландских белых кроликов и яванских макаков.
Антитело против TFPI в плазме кролика с цитратом натрия количественно определяли с использованием сэндвич-иммуноанализа на Gyrolab. Единицы ответа считывали посредством прибора Gyrolab при установке фотоумножителя (PMT) на 1%. Концентрации образца определяли посредством интерполяции из стандартной кривой, которая была аппроксимирована с использованием 5-параметрической аппроксимации логистической кривой с весом ответа 1/у2. Стандартные точки в буфере для анализа содержали 5% объединенной кроличьей плазмы с цитратом натрия, с содержанием, варьирующим от 0,78 нг/мл до 891 нг/мл, и интервал количественного измерения в 100% матриксе кроличьей плазмы составлял от 90 нг/мл до 5500 нг/мл. Пять образцов антитела против TFPI в концентрации 0,45; 8,0; 47,15; 153 и 275 нг/мл в буфере для анализа, содержащем 5% объединенной кроличьей плазмы с цитратом натрия, служили в качестве контроля качества.
Антитело против TFPI в крысиной плазме с цитратом натрия также количественно определяли с использованием сэндвич-иммуноанализа на Gyrolab, как описано выше. Стандартные точки в буфере для анализа, содержащем 5% объединенной крысиной плазмы с цитратом натрия, имели содержание, варьирующее от 0,78 нг/мл до 891 нг/мл, и интервал количественного измерения в 100%-ном матриксе крысиной плазмы составлял от 90 нг/мл до 5500 нг/мл. Пять образцов антитела против TFPI в концентрации 0,45; 8,1; 47,15; 153 и 275 нг/мл в буфере для анализа, содержащем 5% объединенной крысиной плазмы с цитратом натрия, служили в качестве контроля качества.
Для количественного определения антитела против TFPI в плазме яванского макака использовали количественный анализ связывания лиганда общих человеческих Ig с использованием платформы для анализа Meso-Scale Discovery (MSD). Связавшееся антитело против TFPI выявляли с использованием рутениилированного мышиного антитела против Fc человеческого IgG с получением электрохемилюминисцентного сигнала в приборе MSD. Концентрации образца определяли посредством интерполяции из стандартной кривой, которая представляет собой аппроксимацию с использованием
5-параметрического логистического уровнения, формулой взвешивания для стандартной кривой является 1/у^2. Стандартные точки в 5%-ной плазме обезьяны варьировали от 0,999 нг/мл до 1156 нг/мл антитела против TFPI, и интервал количественного измерения в 100%-ной плазме составлял от 64,8 нг/мл до 7136 нг/мл. Пять образцов антитела против TFPI в концентрации 64,8, 117, 680, 3964 и 7136 нг/мл в 100%-ной плазме, разведенной до MRD 1:20 (3,24; 5,83; 34,0; 198 и 357 нг/мл в 5%-ной плазме соответственно), служили в качестве контроля качества.
У новозеландских белых кроликов TFPI-106 демонстрировало более быстрый клиренс (CL) по сравнению с изотипическим контрольным моноклональным антителом (mAb). У яванских макаков TFPI-106 демонстрировало нелинейную кинетику PK при низких дозах, согласующуюся с опосредованным мишенью распределением лекарственного средства (TMDD), наблюдаемым для антител против TFPI. См. Таблицу 32.
В Таблице 32 обобщены определенные фармакокинетические свойства и фармакологические активности типичных антител против TFPI по изобретению (hum4D8, TFPI-106 и TFPI-108), а также двух контрольных антител (4F36 и 2A8-200).
Фармакокинетические свойства и фармакологические активности типичных антител против TFPI
96 ч
196 ч
39%
~ 6%
39%
19%
32%°
Эффективность в 0,5 ч
Продолжитель-ность по aPTT/ PT с разведением
>48 часов
>48 часов
> 48 часов
> 48 часов
>48 часов
Пример 16. Гемостатический эффект ингибирования антителом TFPI усиливает накопление тромбоцитов и образование фибрина in vivo в модели индуцированного лазером повреждения у гемофилических мышей
Как утверждалось в настоящем описании ранее, ингибитор пути тканевого фактора (TFPI) представляет собой ингибитор сериновых протеаз плазмы, который непосредственно связывается с комплексом тканевой фактор (TF)/Фактор VIIa/Фактор Xa и ингибирует его, а также модулирует инициацию свертывания крови, индуцированную TF. Блокирование TFPI может потенциально облегчать гемостаз, инициированный TF/FVIIa, компенсируя потерю фактора VIII (FVIII) или фактора IX при гемофилии А или В. Антитела по изобретению ингибируют TFPI с широкой видовой перекрестной реактивностью. Здесь оценивали гемостатический эффект TFPI-106 на образование тромбоцитарного сгустка и отложение фибрина in vivo с использованием прижизненной микроскопии (IVM) у мышей с гемофилией А и В.
Материалы и методы. Антитело TFPI-106, рекомбинантный Фактор VIII и носитель (фосфатно-солевой буферный раствор) получали, как описано в настоящем описании выше. Антитело против CD42c, конъюгированное с Dylight-649 приобретали у Emfret Analytics (Германия). Антитело против фибрина клона 59D8 (Hui KY et al. (1983) Science 222(4628):1129-1132) метили Alexa Fluor 488 с использованием набора для мечения белка согласно инструкциям изготовителя (Life Technologies, Carlsbad, CA). Самцов мышей с гемофилией А (F8 KO), весящих в среднем от 30 до 35 граммов, приобретали из частной линии, поддерживаемой в Charles River laboratories (Wilmington, MA). Мышей акклиматизировали в течение по меньшей мере 3 суток до проведения экспериментальных процедур.
Самцам мышей с гемофилией А, гемофилией В или дикому типу C57BL/6J (WT) дозировали одну внутривенную дозу TFPI-106 (6 мг/кг), носитель, рекомбинантный человеческий FVIII (rFVIII, 200 IU (международная единица)/кг) или TFPI-106, меченное Alexa-488 (0,7 мг/кг). Микроциркуляцию в кремастере у анестезированных мышей наблюдали с использованием IVM. Накопление тромбоцитов и образование фибрина количественно определяли после повреждения стенки сосуда артерии кремастера посредством нагревания лазером. Тромбоциты визуализировали с использованием антитела против CD42c, конъюгированного с Dylight-649 (GP1bβ), и фибрин выявляли посредством антитела против фибрина клона 59D8, конъюгированного с Alexa-488.
Более конкретно для подготовки мышей с гемофилией А для визуализации прижизненной микроскопией животных анестезировали смесью кетамина/ксилазина, доставленной внутрибрюшинно. Вводили канюлю в яремную вену, и в качестве поддерживающей анестезии использовали пентобарбитал натрия (5 мг/кг, внутривенно). Вводили канюлю в трахею для поддержания открытых дыхательных путей. Затем для визуализации микрососудов экспонировали мышцу кремастер. Животных поддерживали на теплом согревающем коврике, используя теплый буферизованный раствор, в котором инкубировалась экспонированная ткань кремастера на протяжении всего периода визуализации. Через канюлю, вставленную в яремную вену, осуществляли инфузию меченых антител против CD42c тромбоцитов, конъюгированных с Dylight 649 (GP1bβ), и антитела против фибрина, которое не имеет перекрестной реактивности с фибриногеном (антитело против фибрина клона 59D8, конъюгированное с Alexa Fluor 488). Сфокусированный пучок лазерного излучения (532 нанометра) инициировал повреждение микрососудов кремастера мыши. Каждая мышь получала 2 повреждения, индуцированных лазером. Первое повреждение делали у необработанной мыши для того, чтобы оно служило в качестве контроля. Второе повреждение делали у той же самой мыши, и немедленно вводили TFPI-106 (в дозе 6 мг/кг) внутривенно через канюлю, вставленную в яремную вену. При обоих повреждениях образование сгустка отслеживали из интенсивностей флуоресценции накопления тромбоцитов и отложения фибрина.
Точки данных (интенсивности флуоресценции) отбирали и анализировали с использованием программы SlideBook (версия 6.0). Медианные интенсивности флуоресценции откладывали на графике как функцию времени для каждого индивидуального сгустка, и соответствующую площадь под кривой измеряли с использованием программы GraphPad®Prism (версия номер 6.03). Статистическую значимость определяли с использованием критерия Манна-Уитни с использованием программы GraphPad®Prism (версия номер 6.03).
Результаты: TFPI выявляли у мышей WT в месте накопления тромбоцитов в пределах сгустка тромбоцитов и выстилки эндотелия с использованием TFPI-106, меченного Alexa-488 (Фиг. 5). Фиг. 5А содержит шесть верхних панелей (пронумерованных от 1 до 6), демонстрирующих выявление тромбоцитов с использованием антитела против CD42c, конъюгированного с Dylight 649, у мышей WT, которым был введен контрольный IgG, меченный с использованием Alexa 488, в 0 (панель 5А-1), 15 (Фиг. 5А-2), 30 (Фиг. 5А-3), 60 (Фиг. 5А-4), 90 (Фиг. 5А-5) и 120 (Фиг. 5А-6) секунд после повреждения, индуцированного лазером. Метка Alexa 488 не была выявлена в тромбоцитарном тромбе (выявлен с использованием антитела против CD42c, конъюгированного с Dylight 649) в месте повреждения. На Фиг. 5Б, содержащей шесть панелей (пронумерованных 1-6) вдоль нижней части этой Фиг., показано то, что TFPI-106, меченное Alexa-488, выявлялось, начиная примерно в 15 секунд (Фиг. 5Б-2) и увеличивая интенсивность флуоресценции в примерно 30 (Фиг. 5Б-3), 60 (Фиг. 5Б-4) и 90 (Фиг. 5Б-5) секунд, и затем поддерживая интенсивность примерно в 120 секунд (Фиг. 5Б-6) в тромбоцитарном тромбе вдоль эндотелия после повреждения, индуцированного лазером, у мышей WT, где тромбоциты были выявлены с использованием антитела против CD42c, меченного Dylight 649.
TFPI-106 усиливало накопление тромбоцитов и образование фибрина у мышей с гемофилией А (F8 KO) по сравнению с мышами с гемофилией А, обработанными носителем, в 0,5 часов, и данный эффект сохранялся примерно в 168 часов. Более конкретно, у мышей F8 KO (с гемофилией А), обработанных TFPI-106, IVM показала повышенную зеленую флуоресценцию (антитело против фибрина клона 59D8, конъюгированное с Alexa 488) и красную флуоресценцию (антитело против CD42c, меченное Dylight 649 - GP1bβ, выявлющее тромбоциты) в месте повреждения примерно в 0,5 часов, которая сохранялась примерно в 168 часов, и картина флуоресценции была аналогичной картине флуоресценции, наблюдаемой у мышей WT, обработанных носителем. Однако такая флуоресценция не была выявлена у мышей с гемофилией А, обработанных носителем, где не наблюдали ни накопления тромбоцитов, ни образования фибрина.
Мыши с гемофилией А демонстрировали увеличение накопления тромбоцитов (Фиг. 6А) и отложения фибрина (Фиг. 6Б) примерно в 0,5 часов после дозирования TFPI-106, аналогичное уровням, достигаемым посредством rFVIII (* - P меньше 0,005 указана на каждом графике). Гемостатический эффект сохранялся с TFPI-106 у мышей с гемофилией А в течение вплоть до 168 часов. Аналогичным образом, мыши с гемофилией В, обработанные TFPI-106, аналогично демонстрировали улучшенный гемостаз примерно в 30 минут после дозирования с увеличением накопления тромбоцитов и образования фибрина по сравнению с контролями, обработанными носителем. Ни TFPI-106, ни rFVIII, которые дозировали гемофилическим мышам, не достигали максимального уровня отложения фибрина, наблюдаемого у негемофилических мышей WT. Таким образом, эти данные показывают то, что после повреждения эндотелия лазером TFPI выявлялся в месте повреждения. Более важно то, что эти данные демонстрируют то, что введение TFPI-106 гемофилическим мышам приводит к значительному и стойкому улучшению гемостаза в модели повреждения лазером.
Пример 17. Эффект на образование тромбина ингибирования против TFPI в комбинации с FVIIa при гемофилии А и В в анализе TGA
Способы: анализы образования тромбина (TGA)
Оценивали влияние антитела против TFPI, представляющего собой TFPI-106, на образование тромбина самого по себе и в комбинации с рекомбинантным FVIIa (rFVIIa) в гемофилических плазмах с помощью анализа образования тромбина (TGA). Цитратная, обедненная тромбоцитами при тяжелой гемофилии А (дефицитная по FVIII) плазма, плазма с ингибитором при тяжелой гемофилии А или при гемофилии В (дефицитная по FIX) от доноров с врожденной недостаточностью были приобретены у George King Biomedical (Overland Park, KS) и HRF (Raleigh, NC). Нормальная объединенная плазма (негемофилическая) была приобретена у George King Biomedical. Реактивы для образования тромбина; PPP-Reagent LOW (4 мкл фосфолипида и 1 пМ тканевой фактор - конечная концентрация в объеме реакционной среды); буфер FluCa (содержащий хлорид кальция и флуорогенный субстрат) и калибратор тромбина были приобретены у Diagnostica Stago (Parsippany, NJ). Анализы образования тромбина осуществляли с использованием калиброванной автоматической тромбограммы (CAT), включающей флуоресцентный планшет-ридер Fluoroskan Ascent (Thrombinoscope BV, Маастрихт, Нидерланды).
Рекомбинантный FVIIa (rFVIIa в 20 мМ HEPES, 150 мМ хлориде натрия, 1% бычьем сывороточном альбумине (BSA), pH 7,4) добавляли в 70 мкл плазмы при гемофилии А в концентрациях вплоть до 20 мкг/мл. Двадцать мкл PPP-Reagent LOW и 10 мкл антитела TFPI-106 добавляли в реакционные лунки в конечной концентрации 16 мкг/мл. Контрольные реакционные смеси, откалиброванные по эталону, включали 20 мкл калибратора, представляющего собой тромбин, и 70 мкл плазмы. Для тестирования концентрации 0 мкг/мл использовали носитель (фосфатно-солевой буферный раствор (PBS)). Десять мкл носителя добавляли в контрольные лунки с калибратором. Во втором наборе реакций rFVIIa (5 мкл) в буфере HEPES или буфер HEPES (5 мкл) добавляли в 70 мкл образцов гемофилической плазмы (гемофилия А, гемофилия А с ингибитором или гемофилия В) для конечной концентрации rFVIIa в плазме 2 мкг/мл. Двадцать мкл PPP-Reagent LOW и 5 мкл антитела TFPI-106 (разведенного в PBS) или PBS (5 мкл) добавляли в образцы гемофилической плазмы с дозированием rFVIIa (75 мкл) и в гемофилическую плазму, обработанную буфером HEPES (75 мкл), до конечных концентраций 16 мкг/мл. Кроме того, антитело TFPI-106 (в концентрации 16 мкг/мл) анализировали в негемофилической плазме, обработанной буфером HEPES (75 мкл). В анализ была включена контрольная необработанная негемофилическая плазма (80 мкл). Параллельно осуществляли реакции, откалиброванные по эталону (20 мкл калибратора, представляющего собой тромбин, с 80 мкл носителя с дозированной гемофилической или негемофилической плазмой или 80 мкл необработанной негемофилической плазмы). Все реакции проводили в двойной повторности. Образцы инкубировали при 37°С в течение 5 минут, и затем инициировали реакции добавлением 20 мкл FluCa в общем объеме реакции 120 мкл. Флуоресценцию реакций в плазме считывали при 37°С с 20-секундными интервалами на флуориметре Fluoroscan Ascent и сравнивали с реакциями эталонного калибратора, представляющего собой тромбина, для определения концентраций тромбина. Интенсивность флуоресцентного сигнала непрерывно отслеживали при 37°С с использованием CAT (компьютерная томография).
Результаты: недостаточности фактороы свертывания крови FVIII (гемофилия А) и FIX (гемофилия В) препятствуют достаточному образованию тромбина для превращения фибриногена в фибрин для развития стабильного сгустка. Антитело TFPI-106, новое моноклональное антитело, которое специфично связывается с и ингибирует и/или нейтрализует ингибирующую активность TFPI, имеет в качестве мишени путь свертывания крови внешнего тканевого фактора/FVIIa. Пациенты с ингибиторами, получающие TFPI-106, также могут получать rFVIIa для лечения кровотечения в период между инфузиями фактора свертывания. Таким образом, эффект TFPI-106 на образование тромбина в присутствии и в отсутствие rFVIIa в гемофилической плазме проверяли in vitro c использованием признанного в этой области анализа TGA (анализ образования тромбина).
Исследования in vitro c использованием анализа образования тромбина в человеческой плазме с цитратом, обедненной тромоцитами, дефицитной по фактору VIII проводили для исследования активности TFPI-106 в присутствии rFVIIa. В одном исследовании антитело TFPI-106 добавляли в человеческую плазму, дефицитную по фактору VIII, в фиксированной концентрации (16 мкг/мл), представляющей собой концентрацию rFVIIa, которая, как известно, увеличивает образование тромбина в плазме при гемофилии А. rFVIIa добавляли в среду для анализа в возрастающих концентрациях вплоть до 20 мкг/мл. Неожиданно, комбинация TFPI-106 и rFVIIa восстанавливала образование тромбина до уровней, наблюдаемых в нормальной плазме. Пиковые уровни тромбина, наблюдаемые с комбинацией TFPI-106 и ряда концентраций rFVIIa (эптаког альфа) - 0,2, 2 и 20 мкг/мл - были аналогичными. Данные, как показано на Фиг. 7, демонстрируют то, что TFPI-106 улучшало ответ в виде образования тромбина в плазме при тяжелой гемофилии А, в которую дозировали rFVIIa (Фиг. 7, сплошная черная, темно-серая и светло-серая линии).
Эффект TFPI-106 (16 мкг/мл; Фиг. 7, темно-серая пунктирная линия) на образование тромбина также измеряли в негемофилической плазме, которая имела полный набор факторов свертывания крови. В соответствии с ингибирующей активностью TFPI-106 на внешний путь TFPI, добавление антитела TFPI-106 приводило к увеличению образования тромбина с уменьшением lag-периода и увеличением пикового тромбина. Пиковый тромбин ~200 нМ при добавлении TFPI-106 и rFVIIa находится в пределах интервала, описанного для нормальной негемофилической плазмы.
TFPI-ингибирующая активность TFPI-106 на образование тромбина в присутствии и в отсутствие rFVIIa была дополнительно исследована в дополнительных образцах плазмы от гемофилии А (Фиг. 8А; 1 пМ тканевой фактор и 4 мкМ фосфолипиды), плазмы от гемофилии В (Фиг. 8В; ингибитор 3 BU) и в плазме от гемофилии А (Фиг. 8Б; ингибитор 3 BU) с ингибитором, и данные результаты графически показаны на Фиг. 8А, 8В и 8Б соответственно. TFPI-106 добавляли в образцы плазмы в конечной концентрации 16 мкг/мл. Конечная концентрация rFVIIa в данных исследованиях составляла 2 мкг/мл. Добавление 2 мкг/мл rFVIIa в гемофилическую плазму приводило к умеренному увеличению образования тромбина по сравнению с контролем в виде носителя. Добавление 16 мкг/мл TFPI-106 одного или в комбинации с rFVIIa приводило к увеличению образования тромбина, включающему более высокую пиковую концентрацию тромбина и сокращение лаг-периода по сравнению с добавлением одного rFVIIa. Минимальный аддитивный эффект в образовании тромбина наблюдали после совместной обработки rFVIIa и TFPI-106. Пиковые уровни тромбина, достигаемые при дозировании 16 мкг/мл TFPI-106 самого по себе или в комбиации с rFVIIa, были сравнимыми с уровнями, наблюдаемыми в негемофилической плазме, и не превышали уровень, наблюдаемый в негемофилической плазме, в которую дозировали TFPI-106. Эти данные демонстрируют то, что TFPI-106 было эффективным в обходе недостаточностей образования тромбина при гемофилии А, гемофилии В и в плазме при гемофилии А с ингибиторами.
Пример 18. Прокоагулирующая активность в человеческой гемофилической крови и плазме
В этом примере описана гемостатическая активность антитела TFPI-106 при тестировании с использованием цельной крови и плазмы, полученных от человеческих субъектов, имеющих гемофилию А и В, по сравнению с рекомбинантными факторами свертывания - Фактором VIII и Фактором IX.
Материалы и методы. Цельная кровь и плазма (обогащенная тромбоцитами и обедненная тромбоцитами) были получены от волонтеров - пациентов с гемофилией в возрасте по меньшей мере 18 лет согласно протоколу, одобренному комиссией по биомедицинской этике. Эти субъекты имели умеренную или тяжелую недостаточность Фактора VIII (FVIII) или Фактора IX (FIX), с ингибирующими антителами или без них, но в иных отношениях были здоровыми в состоянии без кровотечения. Волонтеров исключали, если они использовали какую-либо заместительную терапию в отношении факторов в пределах предыдущих 48 часов перед поступлением в исследование, имели активное кровотечение или имели медицинскую или семейную историю тромбоза. Из 11 волонтеров 5 имели тяжелую недостаточность FVIII, 2 имели умеренную недостаточность FVIII, 1 имел умеренную недостаточность FIX, и 3 имели тяжелую недостаточность FVIII с ингибитором FVIII. Для завершения всех целей данного исследования 46 мл крови (10 пробирок для крови) отбирали у каждого волонтера посредством асептической венопункции в опорожняемые пробирки, содержащие 3,2% цитрата натрия.
Исследуемые препараты включали TFPI 106, совпадающее по изотипу антитело против человеческого IgG1 негативного контроля, рекомбинантный человеческий Фактор VIII и рекомбинантный человеческий Фактор IX, которые добавляли в цельную кровь или плазму, в зависимости от эксперимента. В зависимости от анализа, TFPI 106 тестировали в концентрации 1, 5, 20, 50 или 100 нМ, контрольное антитело IgG1 в концентрации 100 нМ, и рекомбинантный FVIII или FIX в концентрациях , при которых достигалось бы 5%, 10% или 40% нормальной активности фактора на основе анализа активированного частичного тромбопластинового времени (aPTT). Для достижения требуемых коцентраций исследуемые препараты разводили буфером для разведения с рН 7,4, содержащим 20 мМ HEPES, 150 мМ NaCl и 0,5% бычьего сывороточного альбумина.
Для определения прокоагулирующего эффекта исследуемых препаратов использовали три типа анализов, включая ротационную тромбоэластографию (ROTEM), анализ образования тромбина (TGA) и анализ протромбинового времени с разведением (dPT).
ROTEM измеряет вязкоэластические свойства образца цельной крови по мере его свертывания при условиях низкого срезывающего усилия. По ходу свертывания вязкость образца возрастает, что может быть проанализировано графически. ROTEM проводили с использованием анализатора ROTEM (Pentapharm GmbH, Мюнхен, Германия), используя программу Pentapharm 1.0.04 для оценки свертывания в цельной крови. Свертывание инициировали добавлением 0,020 мл разведения 1:2333 липидированного тканевого фактора (Innovin, Siemens Healthcare) для конечного разведения реакционной среды 1:42000 и 0,020 мл CaCl2 до 0,300 мл цельной крови с цитратом. Все реакции проводили в двойной повторности. Отслеживали параметры ROTEM, и анализировали время свертывания (СТ) ROTEM. Данные, отобранные программой устройства, экспортировали в Microsoft Excel 2010 и/или в GraphPad Prism (версия 6) для анализа. Для каждого волонтера рассчитывали процент изменения времени свертывания ROTEM обработанных образцов по отношению к необработанному образцу от того же самого волонтера.
С использованием анализа образования тромбина (TGA) оценивали кинетику образования тромбина в плазме, обогащенной тромбоцитами (PRP), и плазме, обедненной тромбоцитами (РРР), полученной из крови волонтера согласно способам Hemker, et al., Calibrated automated thrombin generation measurement in clotting plasma. Pathophysiolol Haemost Thromb, 33:4-15 (2003). Вкратце, образцы цельной крови, в которые были дозированы исследуемые препараты, центрифугировали при 150 g в течение 10 минут при комнатной температуре с получением РRР. Неиспользованную плазму замораживали и хранили при -80°С. В PRP плазме число тромбоцитов доводили до 150000 тромбоцитов на микролитр с использованием аутологичной РРР. Немедленно измеряли образование тромбина в свежей PRP. Для образцов PRP в лунки 96-луночного планшета для микротитрования в тройной повторности добавляли 20 мкл 1 пМ тканевого фактора (реактив PRP, Diagnostica Stago, Inc., Parsippany, NJ) и 80 мкл PRP. Образование тромбина инициировали добавлением 20 мкл буфера FLUCa (конечная концентрация 16,7 ммоль/л CaCl2 и 417 ммоль/л флуорогенного тромбинового субстрата Z-Gly-Gly-Arg-AMC). Интенсивность флуоресценции выявляли с использованием флуориметра Fluoroskan Ascent. Для образцов РРР вместо реактива PRP использовали реактив РРР LOW (конечная концентрация 1 пМ тканевой фактор и 4 микромолярные прокоагулирующие фосфолипиды) и прогоняли, как описано для образцов PRP. Образование тромбина рассчитывали с использованием программы Thrombinoscope в версии 5.0.0.742 (Thrombinoscope BV, Маастрихт, Нидерланды). Данные, полученные в программе Thrombinoscope, экспортировали в Microsoft Excel 2010 и/или GraphPad Prism (версия 6) для анализа. Для каждого волонтера рассчитывали процентное изменение пиковой концентрации тромбина TGA обработанных образцов по отношению к необработанному обрацу от того же самого волонтера.
Анализы протромбинового времени с разведением (dPT) проводили на анализаторе свертывания STart 4 (Diagnostica Stago, Parsipanny, NJ). Замороженную РРР размораживали, и дозировали в нее антитело TFPI 106 в концентрациях 1, 5, 20 и 100 нМ или изотипическое контрольное антитело в концентрации 100 нМ. Дозированные образцы плазмы инкубировали при 37°С в течение 30 минут до проведения анализа dPT. Пятьдесят микролитров плазмы добавляли в кювету STart 4 и инкубировали в течение 60 секунд при 37°С. Реакцию активировали добавлением разведения 1:6000 реактива на основе тканевого фактора (Innovin), приготовленного в буфере для разведения (50 мМ имидазол, 0,1 М хдорид натрия, 1 мг/мл бычий сывороточный альбумин и 8,34 мМ CaCl2, pH 7,4), и предварительно инкубировали при 37°С. Время до свертывания измеряли при 37°С с реакциями, идущими в двойной повторности. Время свертывания экспортировали в Microsoft Excel 2010 или GraphPadPrism (версия 6) для анализа. Процентное изменение в dPT свертывания для образцов, обработанных исследуемым препаратом, рассчитывали относительно времени свертывания dPT необработанного образца для каждого волонтера.
Как описано в графических материалах, антитело TFPI-106 вызывало дозозависимое увеличение свертывания цельной крови, полученной от волонтеров с гемофилией А или В, при измерении с использованием способа ROTEM. В частности, антитело уменьшало время свертывания и увеличивало максимальную твердость сгустка по сравнению с негативными контролями. Кроме того, TFPI-106 приводило к дозозависимым увеличениям образования тромбина при добавлении как к обогащенной тромбоцитами, так и к обедненной тромбоцитами плазме, полученной от гемофилических пациентов, что доказывается уменьшениями лаг-периода и повышенной пиковой концентрацией образованного тромбина по сравнению с негативными контролями.
На Фиг. 10А и 10Б проиллюстрирован прокоагулирующий эффект TFPI-106 в цельной крови и плазме, полученных от волонтеров с тяжелой гемофилией А. На Фиг. 10А показан эффект на свертывание крови в анализе ROTEM двух концентраций TFPI 106 по сравнению с IgG негативного контроля и концентраций рекомбинантного FVIII, достаточных для достижения 5%, 10% и 40% от нормальной активности. На Фиг. 10Б показано пиковое образование тромбина в обогащенной тромбоцитами плазме при трех концентрациях TFPI-106 по сравнению с негативными контролями и FVIII. Эти анализы демонстрируют дозозависимый эффект пониженного времени свертывания и повышенного пикового образования тромбина из-за TFPI 106.
На Фиг. 11А, 11Б и 11В проиллюстрирован прокоагулирующий эффект TFPI-106 в цельной крови и плазме от волонтера с тяжелой гемофилией А и ингибирующими антителами (ингибиторами) FVIII. На Фиг. 11А показан эффект на свертывание крови в анализе ROTEM двух концентраций TFPI 106 по сравнению с IgG негативного контроля. На Фиг. 11Б показано пиковое образование тромбина в обогащенной тромбоцитами плазме трех концентраций TFPI-106 по сравнению с IgG негативного контроля. На Фиг. 11В показан эффект четырех концентраций TFPI-106 на образование сгустка из обедненной тромбоцитами плазмы в анализе РТ с разведением по сравнению с IgG негативного контроля. Эти анализы показывают дозозависимый эффект пониженного времени свертывания и повышенного пикового образования тромбина из-за TFPI 106.
На Фиг. 12А, 12Б и 12В проиллюстрирован прокоагулирующий эффект TFPI-106 в цельной крови и плазме от волонтера с умеренной гемофилией А. На Фиг. 12А показан эффект на свертывание крови в анализе ROTEM двух концентраций TFPI 106 по сравнению с IgG негативного контроля и концентрациями рекомбинантного FVIII, достаточными для достижения 5%, 10% и 40% нормальной активности. На Фиг. 12Б показано пиковое образование тромбина в обогащенной тромбоцитами плазме трех концентраций TFPI-106 по сравнению с негативными контролями и FVIII. На Фиг. 12В показан эффект четырех концентраций TFPI-106 на образование сгустка из обедненной тромбоцитами плазмы в анализе РТ с разведением по сравнению с IgG негативного контроля. Эти анализы показывают дозозависимый эффект пониженного времени свертывания и повышенного пикового образования тромбина из-за TFPI 106.
На Фиг. 13А, 13Б и 13В проиллюстрирован прокоагулирующий эффект TFPI-106 в цельной крови и плазме от волонтера с умеренной гемофилией В. На Фиг. 13А показан эффект на свертывание крови в анализе ROTEM двух концентраций TFPI-106 по сравнению с IgG негативного контроля и концентрациями рекомбинантного Фактора IX (FIX), достаточными для достижения 5%, 10% и 40% нормальной активности. На Фиг. 13Б показано пиковое образование тромбина в обогащенной тромбоцитами плазме трех концентраций TFPI-106 по сравнению с негативными контролями и FIX. На Фиг. 13В показан эффект четырех концентраций TFPI 106 на образование сгустка из обедненной тромбоцитами плазмы в анализе РТ с разведением по сравнению с IgG негативного контроля. Эти анализы показывают дозозависимый эффект пониженного времени свертывания и повышенного пикового образования тромбина из-за TFPI-106.
На Фиг. 14А, 14Б и 14В проиллюстрирован прокоагулирующий эффект TFPI-106 в цельной крови и плазме от волонтеров с умеренной гемофилией А. На Фиг. 14А показан эффект на свертывание крови в анализе ROTEM трех концентраций TFPI-106 по сравнению с концентрациями рекомбинантного FVIII, достаточными для достижения 5%, 10% и 40% нормальной активности. Каждая точка данных представляет собой процентное уменьшение времени свертывания образца крови одного волонтера, обработанного TFPI-106 или FVIII, относительно времени свертывания необработанного образца крови от того же самого волонтера. Самая широкая горизонтальная планка среди точек данных для каждой обработки представляет среднее процентное уменьшение времени свертывания всех протестированных образцов волонтеров. На Фиг. 14Б показан эффект на пиковое образование тромбина в обогащенной тромбоцитами плазме трех концентраций TFPI 106 по сравнению с FVIII. Каждая точка данных представляет собой процентное увеличение пикового образования тромбина образца плазмы одного волонтера, обработанного TFPI-106 или FVIII, относительно пикового образования тромбина необработанного образца плазмы от того же самого волонтера. Самая широкая горизонтальная планка среди точек данных для каждой обработки представляет среднее процентное увеличение пикового образования тромбина всех протестированных образцов волонтеров. На Фиг. 14В показан эффект четырех концентраций TFPI 106 на образование сгустка из обедненной тромбоцитами плазмы в анализе РТ с разведением. Каждая точка данных представляет собой процентное уменьшение времени свертывания образца плазмы одного волонтера, обработанного TFPI-106, относительно времени свертывания необработанного образца плазмы от того же самого волонтера. Самая широкая горизонтальная планка среди точек данных для каждой обработки представляет среднее процентное уменьшение времени свертывания всех протестированных образцов волонтеров. Анализ РТ с разведением продемонстрировал дозозависимое уменьшение времени свертывания, вызванное TFPI-106, и анализ образования тромбина продемонстрировал дозозависимое увеличение пикового образования тромбина, вызванное TFPI-106.
ПОСЛЕДОВАТЕЛЬНОСТИ
CDR1, CDR2, CDR3 подчеркнуты
CDR1, 2, 3
подчеркнуты Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркуты
Мутации эффекторной функции: L117A, L118A, G120A подчеркнуты C-концевой лизин удален
CDR1, CDR2 и CDR3
подчеркнуты Последовательность вариабельной области выделена курсивом Мутации эффекторной функции выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, 2, 3 подчеркнуты Последовательность вариабельной области выделена курсивом
CDR1, CDR2 и CDR3 подчеркнуты Последовательность вариабельной области выделена курсивом Мутации эффекторной функции выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, 2, 3 подчеркнуты Последовательность вариабельной области выделена курсивом
CDR1, CDR2 и CDR3 подчеркнуты Последовательность вариабельной области выделена курсивом Мутации эффекторной области выделены жирным шрифтом
CDR1, 2, 3 подчеркнуты Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2 и CDR3 подчеркнуты Последовательность вариабельной области выделена курсивом Мутации эффекторной области выделены жирным шрифтом
SLSPG
CDR1, CDR2, CDR3 подчеркнуты
CDR1, 2, 3 подчеркнуты Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2 и CDR3 подчеркнуты Последовательность вариабельной области выделена курсивом Мутации эффекторной области выделены жирным шрифтом
SEQ 26 с мутацями Q1E, V5L, выделенными жирным шрифтом
CDR1, CDR2 и CDR3 подчеркнуты Последовательность вариабельной области выделена курсивом Мутации Q1E,V5L выделены жирным шрифтом
SEQ 26 с мутациями Q1E, V5L, I94K, выделенными жирным шрифтом
CDR1, CDR2 и CDR3 подчеркнуты Последовательность вариабельной области выделена курсивом Мутации Q1E, V5L, I94K выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
SEQ 38 с мутациями S11L, R40A, L67F, I82M, D82aN, D85E, выделенными жирным шрифтом
CDR1, CDR2 и CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом Мутации S11L, R40A, L67F, I82M, D82aN, D85E выделены жирным шрифтом
SLSPG
CDR1, CDR2, CDR3 подчеркнуты
SEQ 38 с мутациями S11L, V24A, R40A, I82M, D82aN, D85E, выделенными жирным шрифтом
CDR1, CDR2 и CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом Мутации S11L, V24A, R40A, I82M, D82aN, D85E выделены жирным шрифтом
SLSPG
CDR1, CDR2, CDR3 подчеркнуты
SEQ 32 с мутациями V10A, V39L, G71A, выделенными жирным шрифтом
CDR1, 2, 3 подчеркнуты
Последовательность вариабельной области выделена курсивом Мутации V10A, V39L, G71A выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
SEQ 32 с мутациями V10A, H38Q, V39L, выделенными жирным шрифтом
CDR1, 2, 3 подчеркнуты Последовательность вариабельной области выделена курсивом Мутации V10A, H38Q, V39L выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
SEQ 32 с мутациями V10A, V39L, выделенными жирным шрифтом
CDR1, 2, 3 подчеркнуты
Последовательность вариабельной области выделена курсивом Мутации V10A, V39L выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
SEQ 32 с мутациями V10A, H38Q, V39L, G71A, выделенными жирным шрифтом
CDR1, 2, 3 подчеркнуты
Последовательность вариабельной области выделена курсивом Мутации V11A, H38Q, V39L, G71A выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
SEQ 38 с мутациями S11L, R40A, I82M, D82AN, D85E, выделенными жирным шрифтом
CDR1, CDR2 и CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом Мутации S11L, R40A, I82M, D82AN, D85E выделены жирным шрифтом
SLSPG
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
PGK
CDR1, 2, 3 подчеркнуты Последовательность вариабельной области выделена курсивом
CDR1, 2, 3 подчеркнуты Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
Обратные мутации A49G, F67A, R71V, N73Q, L78A выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом Обратные мутации A49G, F67A, R71V, N73Q, L78A выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
Обратная мутация A49G выделена жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом Обратная мутация A49G выделена жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
Обратная мутация F67A выделена жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом Обратная мутация F67A выделена жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты Обратная мутация N73Q выделена жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты Последовательность вариабельной области выделена курсивом Обратная мутация N73Q выделена жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты Обратная мутация L78A выделена жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом Обратная мутация L78A выделена жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
Обратные мутации A49G, F67A, R71V, N73Q, L78A, A93S, R94I выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом Обратные мутации A49G, F67A, R71V, N73Q, L78A, A93S, R94I выделены жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
VL выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты Обратная мутация F71Y выделена жирным шрифтом
CDR1, CDR2, CDR3 подчеркнуты
VL выделена курсивом
CDR1, CDR2, CDR3
подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3
подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3
подчеркнуты
CDR1, CDR2, CDR3
подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
CDR1, CDR2, CDR3 подчеркнуты
Константная область тяжелой цепи человеческого IgG4
CDR1, CDR2, CDR3 подчеркнуты
Последовательность вариабельной области выделена курсивом
нуклеиновые кислоты, кодирующие CDR1, CDR2, CDR3, подчеркнуты
нуклеиновые кислоты, кодирующие CDR1, CDR2, CDR3, подчеркнуты
нуклеиновые кислоты, кодирующие CDR1, CDR2, CDR3, подчеркнуты;
Последовательность вариабельной области выделена курсивом
нуклеиновые кислоты, кодирующие CDR1, CDR2, CDR3, подчеркнуты;
Последовательность вариабельной области выделена курсивом
Описание и состав последовательности для номеров SeqID (идентификационный номер последовательности) антител. (При названии конкретных остатков используется нумерация по Kabat. Точки начала и конца CDR VH и VL определяются с использованием определений Kabat).
Разные характеристики и воплощения настоящего изобретения, относящиеся к приведенным выше индивидуальным разделам, применяются, сообразно обстоятельствам, к другим разделам, с учетом необходимых изменений. Следовательно, характеристики, определенные в одном разделе, могут быть объединены с характеристиками, определенными в других разделах, сообразно обстоятельствам. Все процитированные в настоящем описании ссылки, включая патенты, патентные заявки, статьи, учебники и процитированные номера доступа последовательностей, и ссылки, процитированные в них, являются тем самым включенными посредством ссылки во всей их полноте. В случае, когда одна или более чем одна включенная литературная ссылка и аналогичные материалы отличаются от или противоречат этой заявке, включая определенные термины, применение терминов, описанные методики или тому подобное, но не ограничиваясь ими, данная заявка будет иметь преимущество.
--->
Перечень последовательностей
SEQUENCE LISTING
<110> Pfizer Inc.
Pittman, Debra
Apgar, James R.
Juo, Zong Sean
Jin, Macy
Stahl, Mark
Carven, Gregory J.
Holsti, Matthew
Benard, Susan
Hett, Sunita R.
Jasuja, Reema
<120> TISSUE FACTOR PATHWAY INHIBITOR ANTIBODIES AND USES THEREOF
<130> PC72096A
<150> US 62/207,229
<151> 2015-08-19
<150> US 62/360,205
<151> 2016-07-08
<160> 178
<170> PatentIn version 3.5
<210> 1
<211> 149
<212> PRT
<213> Homo sapiens
<400> 1
Asp Ser Glu Glu Asp Glu Glu His Thr Ile Ile Thr Asp Thr Glu Leu
1 5 10 15
Pro Pro Leu Lys Leu Met His Ser Phe Cys Ala Phe Lys Ala Asp Asp
20 25 30
Gly Pro Cys Lys Ala Ile Met Lys Arg Phe Phe Phe Asn Ile Phe Thr
35 40 45
Arg Gln Cys Glu Glu Phe Ile Tyr Gly Gly Cys Glu Gly Asn Gln Asn
50 55 60
Arg Phe Glu Ser Leu Glu Glu Cys Lys Lys Met Cys Thr Arg Asp Asn
65 70 75 80
Ala Asn Arg Ile Ile Lys Thr Thr Leu Gln Gln Glu Lys Pro Asp Phe
85 90 95
Cys Phe Leu Glu Glu Asp Pro Gly Ile Cys Arg Gly Tyr Ile Thr Arg
100 105 110
Tyr Phe Tyr Asn Asn Gln Thr Lys Gln Cys Glu Arg Phe Lys Tyr Gly
115 120 125
Gly Cys Leu Gly Asn Met Asn Asn Phe Glu Thr Leu Glu Glu Cys Lys
130 135 140
Asn Ile Cys Glu Asp
145
<210> 2
<211> 254
<212> PRT
<213> Homo sapiens
<400> 2
Asp Ser Glu Glu Asp Glu Glu His Thr Ile Ile Thr Asp Thr Glu Leu
1 5 10 15
Pro Pro Leu Lys Leu Met His Ser Phe Cys Ala Phe Lys Ala Asp Asp
20 25 30
Gly Pro Cys Lys Ala Ile Met Lys Arg Phe Phe Phe Asn Ile Phe Thr
35 40 45
Arg Gln Cys Glu Glu Phe Ile Tyr Gly Gly Cys Glu Gly Asn Gln Asn
50 55 60
Arg Phe Glu Ser Leu Glu Glu Cys Lys Lys Met Cys Thr Arg Asp Asn
65 70 75 80
Ala Asn Arg Ile Ile Lys Thr Thr Leu Gln Gln Glu Lys Pro Asp Phe
85 90 95
Cys Phe Leu Glu Glu Asp Pro Gly Ile Cys Arg Gly Tyr Ile Thr Arg
100 105 110
Tyr Phe Tyr Asn Asn Gln Thr Lys Gln Cys Glu Arg Phe Lys Tyr Gly
115 120 125
Gly Cys Leu Gly Asn Met Asn Asn Phe Glu Thr Leu Glu Glu Cys Lys
130 135 140
Asn Ile Cys Glu Asp Gly Pro Asn Gly Phe Gln Val Asp Asn Tyr Gly
145 150 155 160
Thr Gln Leu Asn Ala Val Asn Asn Ser Leu Thr Pro Gln Ser Thr Lys
165 170 175
Val Pro Ser Leu Phe Glu Phe His Gly Pro Ser Trp Cys Leu Thr Pro
180 185 190
Ala Asp Arg Gly Leu Cys Arg Ala Asn Glu Asn Arg Phe Tyr Tyr Asn
195 200 205
Ser Val Ile Gly Lys Cys Arg Pro Phe Lys Tyr Ser Gly Cys Gly Gly
210 215 220
Asn Glu Asn Asn Phe Thr Ser Lys Gln Glu Cys Leu Arg Ala Cys Lys
225 230 235 240
Lys Gly Phe Ile Gln Arg Ile Ser Lys Gly Gly Leu Ile Lys
245 250
<210> 3
<211> 189
<212> PRT
<213> Homo sapiens
<400> 3
Asp Ala Ala Gln Glu Pro Thr Gly Asn Asn Ala Glu Ile Cys Leu Leu
1 5 10 15
Pro Leu Asp Tyr Gly Pro Cys Arg Ala Leu Leu Leu Arg Tyr Tyr Tyr
20 25 30
Asp Arg Tyr Thr Gln Ser Cys Arg Gln Phe Leu Tyr Gly Gly Cys Glu
35 40 45
Gly Asn Ala Asn Asn Phe Tyr Thr Trp Glu Ala Cys Asp Asp Ala Cys
50 55 60
Trp Arg Ile Glu Lys Val Pro Lys Val Cys Arg Leu Gln Val Ser Val
65 70 75 80
Asp Asp Gln Cys Glu Gly Ser Thr Glu Lys Tyr Phe Phe Asn Leu Ser
85 90 95
Ser Met Thr Cys Glu Lys Phe Phe Ser Gly Gly Cys His Arg Asn Arg
100 105 110
Ile Glu Asn Arg Phe Pro Asp Glu Ala Thr Cys Met Gly Phe Cys Ala
115 120 125
Pro Lys Lys Ile Pro Ser Phe Cys Tyr Ser Pro Lys Asp Glu Gly Leu
130 135 140
Cys Ser Ala Asn Val Thr Arg Tyr Tyr Phe Asn Pro Arg Tyr Arg Thr
145 150 155 160
Cys Asp Ala Phe Thr Tyr Thr Gly Cys Gly Gly Asn Asp Asn Asn Phe
165 170 175
Val Ser Arg Glu Asp Cys Lys Arg Ala Cys Ala Lys Ala
180 185
<210> 4
<211> 146
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 4
Leu Ser Glu Glu Ala Asp Asp Thr Asp Ser Glu Leu Gly Ser Met Lys
1 5 10 15
Pro Leu His Thr Phe Cys Ala Met Lys Ala Asp Asp Gly Pro Cys Lys
20 25 30
Ala Met Ile Arg Ser Tyr Phe Phe Asn Met Tyr Thr His Gln Cys Glu
35 40 45
Glu Phe Ile Tyr Gly Gly Cys Glu Gly Asn Glu Asn Arg Phe Asp Thr
50 55 60
Leu Glu Glu Cys Lys Lys Thr Cys Ile Pro Gly Tyr Glu Lys Thr Ala
65 70 75 80
Val Lys Ala Ala Ser Gly Ala Glu Arg Pro Asp Phe Cys Phe Leu Glu
85 90 95
Glu Asp Pro Gly Leu Cys Arg Gly Tyr Met Lys Arg Tyr Leu Tyr Asn
100 105 110
Asn Gln Thr Lys Gln Cys Glu Arg Phe Val Tyr Gly Gly Cys Leu Gly
115 120 125
Asn Arg Asn Asn Phe Glu Thr Leu Asp Glu Cys Lys Lys Ile Cys Glu
130 135 140
Asn Pro
145
<210> 5
<211> 149
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 5
Asp Ser Glu Glu Asp Glu Glu Tyr Thr Ile Ile Thr Asp Thr Glu Leu
1 5 10 15
Pro Pro Leu Lys Leu Met His Ser Phe Cys Ala Phe Lys Pro Asp Asp
20 25 30
Gly Pro Cys Lys Ala Ile Met Lys Arg Phe Phe Phe Asn Ile Phe Thr
35 40 45
Arg Gln Cys Glu Glu Phe Ile Tyr Gly Gly Cys Gly Gly Asn Gln Asn
50 55 60
Arg Phe Glu Ser Met Glu Glu Cys Lys Lys Val Cys Thr Arg Asp Asn
65 70 75 80
Val Asn Arg Ile Ile Gln Thr Ala Leu Gln Lys Glu Lys Pro Asp Phe
85 90 95
Cys Phe Leu Glu Glu Asp Pro Gly Ile Cys Arg Gly Tyr Ile Thr Arg
100 105 110
Tyr Phe Tyr Asn Asn Gln Ser Lys Gln Cys Glu Arg Phe Lys Tyr Gly
115 120 125
Gly Cys Leu Gly Asn Met Asn Asn Phe Glu Thr Leu Glu Glu Cys Lys
130 135 140
Asn Thr Cys Glu Asp
145
<210> 6
<211> 149
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 6
Ala Ala Glu Glu Asp Glu Glu Phe Thr Asn Ile Thr Asp Ile Lys Pro
1 5 10 15
Pro Leu Gln Lys Pro Thr His Ser Phe Cys Ala Met Lys Val Asp Asp
20 25 30
Gly Pro Cys Arg Ala Tyr Ile Lys Arg Phe Phe Phe Asn Ile Leu Thr
35 40 45
His Gln Cys Glu Glu Phe Ile Tyr Gly Gly Cys Glu Gly Asn Glu Asn
50 55 60
Arg Phe Glu Ser Leu Glu Glu Cys Lys Glu Lys Cys Ala Arg Asp Tyr
65 70 75 80
Pro Lys Met Thr Thr Lys Leu Thr Phe Gln Lys Gly Lys Pro Asp Phe
85 90 95
Cys Phe Leu Glu Glu Asp Pro Gly Ile Cys Arg Gly Tyr Ile Thr Arg
100 105 110
Tyr Phe Tyr Asn Asn Gln Ser Lys Gln Cys Glu Arg Phe Lys Tyr Gly
115 120 125
Gly Cys Leu Gly Asn Leu Asn Asn Phe Glu Ser Leu Glu Glu Cys Lys
130 135 140
Asn Thr Cys Glu Asn
145
<210> 7
<211> 148
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 7
Leu Pro Glu Glu Asp Asp Asp Thr Ile Asn Thr Asp Ser Glu Leu Arg
1 5 10 15
Pro Met Lys Pro Leu His Thr Phe Cys Ala Met Lys Ala Glu Asp Gly
20 25 30
Pro Cys Lys Ala Met Ile Arg Ser Tyr Tyr Phe Asn Met Asn Ser His
35 40 45
Gln Cys Glu Glu Phe Ile Tyr Gly Gly Cys Arg Gly Asn Lys Asn Arg
50 55 60
Phe Asp Thr Leu Glu Glu Cys Arg Lys Thr Cys Ile Pro Gly Tyr Lys
65 70 75 80
Lys Thr Thr Ile Lys Thr Thr Ser Gly Ala Glu Lys Pro Asp Phe Cys
85 90 95
Phe Leu Glu Glu Asp Pro Gly Ile Cys Arg Gly Phe Met Thr Arg Tyr
100 105 110
Phe Tyr Asn Asn Gln Ser Lys Gln Cys Glu Gln Phe Lys Tyr Gly Gly
115 120 125
Cys Leu Gly Asn Ser Asn Asn Phe Glu Thr Leu Glu Glu Cys Arg Asn
130 135 140
Thr Cys Glu Asp
145
<210> 8
<211> 35
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 8
Gly Gly Gly Ser Gly Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys
1 5 10 15
Ile Glu Trp His Glu Gly Gly Pro Pro His His His His His His His
20 25 30
His His His
35
<210> 9
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 9
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly
20
<210> 10
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 10
Arg Ala Ser Gln Gly Ile Ser Ser Ser Leu Ala
1 5 10
<210> 11
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 11
Ala Ala Ser Thr Leu Gln Ser
1 5
<210> 12
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 12
Gln Gln Leu Asp Ser Tyr Pro Leu Ser
1 5
<210> 13
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 13
Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asp Ser Tyr Pro Leu
85 90 95
Ser Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 14
<211> 107
<212> PRT
<213> Homo sapiens
<400> 14
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 15
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 15
Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asp Ser Tyr Pro Leu
85 90 95
Ser Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 16
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 16
Gly Tyr Thr Phe Thr Gly Tyr Tyr Met His
1 5 10
<210> 17
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 17
Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 18
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 18
Gly Ile Ala Arg Leu Gln Trp Leu Pro Thr Glu Ala Asp Phe Asp Tyr
1 5 10 15
<210> 19
<211> 125
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 19
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ile Ala Arg Leu Gln Trp Leu Pro Thr Glu Ala Asp Phe
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 20
<211> 329
<212> PRT
<213> Homo sapiens
<400> 20
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 21
<211> 454
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 21
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ile Ala Arg Leu Gln Trp Leu Pro Thr Glu Ala Asp Phe
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly
450
<210> 22
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 22
Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10
<210> 23
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 23
Gly Asn Ser Asn Arg Pro Ser
1 5
<210> 24
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 24
Gln Ser Tyr Asp Ser Ser Leu Ser Gly Ser Val Val
1 5 10
<210> 25
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 25
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Phe Tyr Cys Gln Ser Tyr Asp Ser Ser
85 90 95
Leu Ser Gly Ser Val Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu
100 105 110
Gly
<210> 26
<211> 105
<212> PRT
<213> Homo sapiens
<400> 26
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
35 40 45
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
50 55 60
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
65 70 75 80
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
85 90 95
Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 27
<211> 218
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 27
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Phe Tyr Cys Gln Ser Tyr Asp Ser Ser
85 90 95
Leu Ser Gly Ser Val Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 28
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 28
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1 5 10
<210> 29
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 29
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 30
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 30
Leu Gly Ala Thr Ser Leu Ser Ala Phe Asp Ile
1 5 10
<210> 31
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 31
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Leu Gly Ala Thr Ser Leu Ser Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 32
<211> 449
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 32
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Leu Gly Ala Thr Ser Leu Ser Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 33
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 33
Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10
<210> 34
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 34
Gly Asn Ser Asn Arg Pro Ser
1 5
<210> 35
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 35
Gln Ser Tyr Asp Ser Ser Leu Ser Gly Ser Gly Val
1 5 10
<210> 36
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 36
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser
85 90 95
Leu Ser Gly Ser Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly
<210> 37
<211> 218
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 37
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser
85 90 95
Leu Ser Gly Ser Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 38
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 38
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1 5 10
<210> 39
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 39
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 40
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 40
Leu Gly Ala Thr Ser Leu Ser Ala Phe Asp Ile
1 5 10
<210> 41
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 41
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Leu Gly Ala Thr Ser Leu Ser Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 42
<211> 449
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 42
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Leu Gly Ala Thr Ser Leu Ser Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 43
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 43
Ser Gly Ser Thr Ser Asn Ile Gly Thr Met Tyr Val His
1 5 10
<210> 44
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 44
Arg Asn Asn His Arg Pro Ser
1 5
<210> 45
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 45
Leu Ala Trp Asp Asp Thr Leu Arg Ala Tyr Val
1 5 10
<210> 46
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 46
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Thr Met
20 25 30
Tyr Val His Trp Tyr Gln His Val Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Gly Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Asp Asp Thr Leu
85 90 95
Arg Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly
100 105 110
<210> 47
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 47
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Thr Met
20 25 30
Tyr Val His Trp Tyr Gln His Val Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Gly Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Asp Asp Thr Leu
85 90 95
Arg Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 48
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 48
Gly Leu Thr Ile Asp Asn Tyr Ala Met Gln
1 5 10
<210> 49
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 49
Gly Ile Ser Gly Asn Ser Arg Ser Ile Gly Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 50
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 50
Phe Leu His Glu Ser Asp Tyr
1 5
<210> 51
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 51
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Leu Thr Ile Asp Asn Tyr
20 25 30
Ala Met Gln Trp Val Arg Gln Arg Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Asn Ser Arg Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Leu Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Ile Asp Ser Leu Arg Ala Asp Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ile Phe Leu His Glu Ser Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 52
<211> 445
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 52
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Leu Thr Ile Asp Asn Tyr
20 25 30
Ala Met Gln Trp Val Arg Gln Arg Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Asn Ser Arg Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Leu Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Ile Asp Ser Leu Arg Ala Asp Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ile Phe Leu His Glu Ser Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 53
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 53
Thr Gly Ser Ser Ser Asn Leu Gly Ala Asp Tyr Asp Val Gln
1 5 10
<210> 54
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 54
Gly Asn Asn Asn Arg Pro Ser
1 5
<210> 55
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 55
Gln Ser Tyr Asp Arg Ser Leu Ser Gly Ser Met Val
1 5 10
<210> 56
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 56
Gln Ser Val Leu Thr Gln Pro Pro Ser Leu Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Leu Gly Ala Asp
20 25 30
Tyr Asp Val Gln Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Phe Gly Asn Asn Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Arg Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asn Tyr Tyr Cys Gln Ser Tyr Asp Arg Ser
85 90 95
Leu Ser Gly Ser Met Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly
<210> 57
<211> 218
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 57
Gln Ser Val Leu Thr Gln Pro Pro Ser Leu Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Leu Gly Ala Asp
20 25 30
Tyr Asp Val Gln Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Phe Gly Asn Asn Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Arg Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asn Tyr Tyr Cys Gln Ser Tyr Asp Arg Ser
85 90 95
Leu Ser Gly Ser Met Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 58
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 58
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1 5 10
<210> 59
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 59
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 60
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 60
Asn Gly Ala Ala Ala Ala Trp Asp Tyr
1 5
<210> 61
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 61
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Asn Asn Gly Ala Ala Ala Ala Trp Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 62
<211> 447
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 62
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Asn Asn Gly Ala Ala Ala Ala Trp Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 63
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 63
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Leu Gly Ala Thr Ser Leu Ser Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 64
<211> 449
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 64
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Leu Gly Ala Thr Ser Leu Ser Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 65
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 65
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Leu Gly Ala Thr Ser Leu Ser Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 66
<211> 449
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 66
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Leu Gly Ala Thr Ser Leu Ser Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 67
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 67
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Leu Thr Ile Asp Asn Tyr
20 25 30
Ala Met Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Asn Ser Arg Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ile Phe Leu His Glu Ser Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 68
<211> 445
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 68
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Leu Thr Ile Asp Asn Tyr
20 25 30
Ala Met Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Asn Ser Arg Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ile Phe Leu His Glu Ser Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 69
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 69
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Ile Asp Asn Tyr
20 25 30
Ala Met Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Asn Ser Arg Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Leu Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ile Phe Leu His Glu Ser Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 70
<211> 445
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 70
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Ile Asp Asn Tyr
20 25 30
Ala Met Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Asn Ser Arg Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Leu Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ile Phe Leu His Glu Ser Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 71
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 71
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Thr Met
20 25 30
Tyr Val His Trp Tyr Gln His Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Asp Asp Thr Leu
85 90 95
Arg Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly
100 105 110
<210> 72
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 72
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Thr Met
20 25 30
Tyr Val His Trp Tyr Gln His Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Asp Asp Thr Leu
85 90 95
Arg Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 73
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 73
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Thr Met
20 25 30
Tyr Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Gly Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Asp Asp Thr Leu
85 90 95
Arg Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly
100 105 110
<210> 74
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 74
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Thr Met
20 25 30
Tyr Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Gly Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Asp Asp Thr Leu
85 90 95
Arg Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 75
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 75
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Thr Met
20 25 30
Tyr Val His Trp Tyr Gln His Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Gly Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Asp Asp Thr Leu
85 90 95
Arg Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly
100 105 110
<210> 76
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 76
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Thr Met
20 25 30
Tyr Val His Trp Tyr Gln His Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Gly Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Asp Asp Thr Leu
85 90 95
Arg Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 77
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 77
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Thr Met
20 25 30
Tyr Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Asp Asp Thr Leu
85 90 95
Arg Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly
100 105 110
<210> 78
<211> 216
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 78
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Thr Met
20 25 30
Tyr Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn His Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Leu Ala Trp Asp Asp Thr Leu
85 90 95
Arg Ala Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 79
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 79
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Leu Thr Ile Asp Asn Tyr
20 25 30
Ala Met Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Asn Ser Arg Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Leu Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ile Phe Leu His Glu Ser Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 80
<211> 445
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 80
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Leu Thr Ile Asp Asn Tyr
20 25 30
Ala Met Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Asn Ser Arg Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Leu Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ile Phe Leu His Glu Ser Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 81
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 81
Lys Ala Ser Gln Asp Val His Thr Ala Val Ala
1 5 10
<210> 82
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 82
Trp Ala Ser Thr Arg His Thr
1 5
<210> 83
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 83
Gln Gln His Tyr Thr Thr Pro Tyr Thr
1 5
<210> 84
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 84
Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val His Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Cys Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Leu Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Met Lys
100 105
<210> 85
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 85
Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln
1 5 10 15
Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr
20 25 30
Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln
35 40 45
Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg
65 70 75 80
His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro
85 90 95
Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<210> 86
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 86
Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val His Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Cys Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Leu Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Met Lys Ala Asp Ala Ala Pro
100 105 110
Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly Gly
115 120 125
Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile Asn
130 135 140
Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu Asn
145 150 155 160
Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser Ser
165 170 175
Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr Thr
180 185 190
Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser Phe
195 200 205
Asn Arg Asn Glu Cys
210
<210> 87
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 87
Gly Tyr Thr Phe Thr Asp Tyr Asn Leu Asp
1 5 10
<210> 88
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 88
Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 89
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 89
Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr
1 5 10
<210> 90
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 90
Glu Val Leu Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Gln Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Ile Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Ser Val Thr Val Ser Ser
115
<210> 91
<211> 324
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 91
Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala
1 5 10 15
Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr Val
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro
100 105 110
Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu
115 120 125
Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser
130 135 140
Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu
145 150 155 160
Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr
165 170 175
Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn
180 185 190
Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro
195 200 205
Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln
210 215 220
Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val
225 230 235 240
Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val
245 250 255
Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln
260 265 270
Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn
275 280 285
Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val
290 295 300
Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His
305 310 315 320
Ser Pro Gly Lys
<210> 92
<211> 443
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 92
Glu Val Leu Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Gln Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Ile Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Ser Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser
180 185 190
Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys
210 215 220
Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr
245 250 255
Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser
260 265 270
Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile
290 295 300
Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu
340 345 350
Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe
355 360 365
Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala
370 375 380
Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr
385 390 395 400
Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly
405 410 415
Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His
420 425 430
Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 93
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 93
Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val His Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Cys Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Leu Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Met Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 94
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 94
Glu Val Leu Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Gln Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Ile Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 95
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 95
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 96
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 96
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 97
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 97
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Ile Ser Val Asp Gln Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 98
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 98
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Ile Ser Val Asp Gln Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 99
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 99
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 100
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 100
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 101
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 101
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 102
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 102
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 103
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 103
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Gln Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 104
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 104
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Gln Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 105
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 105
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 106
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 106
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 107
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 107
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Ile Ser Val Asp Gln Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Ile Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 108
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 108
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Leu Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Asn Pro Ile Asn Gly Ala Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Ile Ser Val Asp Gln Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Ile Tyr Tyr Gly Asp Tyr Asp Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 109
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 109
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val His Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 110
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 110
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val His Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 111
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 111
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val His Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 112
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 112
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val His Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 113
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 113
Lys Ala Ser Gln Asp Val Ile Thr Ala Val Ala
1 5 10
<210> 114
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 114
Trp Ala Ser Thr Arg His Thr
1 5
<210> 115
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 115
Gln Gln His Tyr Ser Thr Pro Tyr Thr
1 5
<210> 116
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 116
Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val Ile Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Val Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ile Ser Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Leu Tyr Tyr Cys Gln Gln His Tyr Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 117
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 117
Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val Ile Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Val Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ile Ser Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Leu Tyr Tyr Cys Gln Gln His Tyr Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Ala Asp Ala Ala Pro
100 105 110
Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly Gly
115 120 125
Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile Asn
130 135 140
Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu Asn
145 150 155 160
Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser Ser
165 170 175
Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr Thr
180 185 190
Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser Phe
195 200 205
Asn Arg Asn Glu Cys
210
<210> 118
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 118
Gly Tyr Thr Phe Thr Asp Tyr Thr Met Asp
1 5 10
<210> 119
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 119
Asp Ile Asn Pro Ser Asn Gly Gly Ser Ile Tyr Asn Arg Lys Phe Lys
1 5 10 15
Gly
<210> 120
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 120
Met His Tyr Asn Tyr Asp Gly Phe Pro Tyr
1 5 10
<210> 121
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 121
Glu Val Leu Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Ser Asn Gly Gly Ser Ile Tyr Asn Arg Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met His Tyr Asn Tyr Asp Gly Phe Pro Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala
115
<210> 122
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 122
Glu Val Leu Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Ser Asn Gly Gly Ser Ile Tyr Asn Arg Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met His Tyr Asn Tyr Asp Gly Phe Pro Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 123
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 123
Arg Ala Ser Lys Ser Val Ser Thr Ser Gly Tyr Thr Tyr Met His
1 5 10 15
<210> 124
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 124
Leu Ala Ser Asn Leu Glu Ser
1 5
<210> 125
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 125
Gln His Ile Arg Glu Leu Pro Phe Thr
1 5
<210> 126
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 126
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Lys Ser Val Ser Thr Ser
20 25 30
Gly Tyr Thr Tyr Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Ala Ala Ala Tyr Tyr Cys Gln His Ile Arg
85 90 95
Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 127
<211> 217
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 127
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Lys Ser Val Ser Thr Ser
20 25 30
Gly Tyr Thr Tyr Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Ala Ala Ala Tyr Tyr Cys Gln His Ile Arg
85 90 95
Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Ala
100 105 110
Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu
115 120 125
Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro
130 135 140
Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn
145 150 155 160
Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His
180 185 190
Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile
195 200 205
Val Lys Ser Phe Asn Arg Asn Glu Cys
210 215
<210> 128
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 128
Gly Tyr Thr Phe Thr Ser Tyr Val Met His
1 5 10
<210> 129
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 129
Tyr Leu Asn Pro Tyr Asn Asp Gly Thr Lys Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly
<210> 130
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 130
Thr Leu Leu Tyr Ala Met Asp Tyr
1 5
<210> 131
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 131
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Val Met His Trp Val Lys Gln Lys Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Leu Asn Pro Tyr Asn Asp Gly Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Ser Leu Ile Ser Asp Lys Ser Ser Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Thr Leu Leu Tyr Ala Met Asp Tyr Trp Gly Gln Gly Ser Ser
100 105 110
Val Thr Val Ser Ser
115
<210> 132
<211> 358
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 132
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Val Met His Trp Val Lys Gln Lys Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Leu Asn Pro Tyr Asn Asp Gly Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Ser Leu Ile Ser Asp Lys Ser Ser Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Thr Leu Leu Tyr Ala Met Asp Tyr Trp Gly Gln Gly Ser Ser
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met
355
<210> 133
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 133
Ser Gly Asp Asn Leu Arg Asn Tyr Tyr Ala His
1 5 10
<210> 134
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 134
Tyr Asp Asn Asn Arg Pro Ser
1 5
<210> 135
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 135
Gln Ser Trp Asp Asp Gly Val Pro Val
1 5
<210> 136
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 136
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Leu Arg Asn Tyr Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Val Val Ile Tyr
35 40 45
Tyr Asp Asn Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp Asp Gly Val Pro Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 137
<211> 212
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 137
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Leu Arg Asn Tyr Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Val Val Ile Tyr
35 40 45
Tyr Asp Asn Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp Asp Gly Val Pro Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 138
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 138
Gly Phe Thr Phe Arg Ser Tyr Gly Met Ser
1 5 10
<210> 139
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 139
Ser Ile Arg Gly Ser Ser Ser Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 140
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 140
Lys Tyr Arg Tyr Trp Phe Asp Tyr
1 5
<210> 141
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 141
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Arg Gly Ser Ser Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Lys Tyr Arg Tyr Trp Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 142
<211> 446
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 142
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Arg Gly Ser Ser Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Lys Tyr Arg Tyr Trp Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 143
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 143
Ser Gly Asp Asn Leu Arg Asn Tyr Tyr Ala His
1 5 10
<210> 144
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 144
Tyr Asp Val Asn Arg Pro Ser
1 5
<210> 145
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 145
Gln Ser Trp Trp Asp Gly Val Pro Val
1 5
<210> 146
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 146
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Leu Arg Asn Tyr Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Val Val Ile Phe
35 40 45
Tyr Asp Val Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Trp Asp Gly Val Pro Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 147
<211> 212
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 147
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Leu Arg Asn Tyr Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Val Val Ile Phe
35 40 45
Tyr Asp Val Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Trp Asp Gly Val Pro Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 148
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 148
Gly Phe Thr Phe Arg Ser Tyr Gly Met Asp
1 5 10
<210> 149
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 149
Ser Ile Arg Gly Ser Arg Ser Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 150
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 150
Leu Tyr Arg Tyr Trp Phe Asp Tyr
1 5
<210> 151
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 151
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Arg Gly Ser Arg Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Tyr Arg Tyr Trp Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 152
<211> 446
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 152
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Arg Gly Ser Arg Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Tyr Arg Tyr Trp Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Ala Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 153
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 153
Ser Gly Asp Asn Leu Arg Asn Tyr Tyr Ala His
1 5 10
<210> 154
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 154
Tyr Asp Asn Asn Arg Pro Ser
1 5
<210> 155
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 155
Gln Ser Trp Asp Asp Gly Val Pro Val
1 5
<210> 156
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 156
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Leu Arg Asn Tyr Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Val Val Ile Tyr
35 40 45
Tyr Asp Asn Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp Asp Gly Val Pro Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 157
<211> 105
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 157
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
35 40 45
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
50 55 60
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
65 70 75 80
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
85 90 95
Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 158
<211> 212
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 158
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Leu Arg Asn Tyr Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Val Val Ile Tyr
35 40 45
Tyr Asp Asn Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Trp Asp Asp Gly Val Pro Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 159
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 159
Gly Phe Thr Phe Ser Asn Tyr Ala Leu Ser
1 5 10
<210> 160
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 160
Ser Ile Ser Ser Gly Gly Ala Thr Tyr Tyr Pro Asp Ser Val Glu Gly
1 5 10 15
<210> 161
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 161
Gly Ala Tyr Gly Ser Asp Tyr Phe Asp Tyr
1 5 10
<210> 162
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 162
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Leu Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Ala Thr Tyr Tyr Pro Asp Ser Val Glu
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Arg Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Ser Ser Leu Gln Ser Glu Asp Thr Ala Met Tyr Tyr Cys Thr
85 90 95
Arg Gly Ala Tyr Gly Ser Asp Tyr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 163
<211> 441
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 163
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Leu Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Ala Thr Tyr Tyr Pro Asp Ser Val Glu
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Arg Asn Ile Leu Tyr Leu
65 70 75 80
Gln Met Ser Ser Leu Gln Ser Glu Asp Thr Ala Met Tyr Tyr Cys Thr
85 90 95
Arg Gly Ala Tyr Gly Ser Asp Tyr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro
115 120 125
Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly
130 135 140
Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn
145 150 155 160
Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr
180 185 190
Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser
195 200 205
Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro
210 215 220
Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys
245 250 255
Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp
260 265 270
Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu
275 280 285
Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser
305 310 315 320
Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
325 330 335
Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln
340 345 350
Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe
355 360 365
Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu
370 375 380
Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe
385 390 395 400
Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn
405 410 415
Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His Thr
420 425 430
Glu Lys Ser Leu Ser His Ser Pro Gly
435 440
<210> 164
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 164
Lys Ser Ser Gln Ser Leu Leu Glu Ser Asp Gly Lys Thr Tyr Leu Asn
1 5 10 15
<210> 165
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 165
Leu Val Ser Ile Leu Asp Ser
1 5
<210> 166
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 166
Leu Gln Ala Thr His Phe Pro Gln Thr
1 5
<210> 167
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 167
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Glu Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Val Ser Ile Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Leu Gln Ala
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 168
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 168
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Glu Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Val Ser Ile Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Leu Gln Ala
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 169
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 169
Gly Phe Thr Phe Ser Asn Tyr Ala Met Ser
1 5 10
<210> 170
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 170
Thr Ile Ser Arg Ser Gly Ser Tyr Ser Tyr Phe Pro Asp Ser Val Gln
1 5 10 15
Gly
<210> 171
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 171
Leu Gly Gly Tyr Asp Glu Gly Asp Ala Met Asp Ser
1 5 10
<210> 172
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 172
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Arg Ser Gly Ser Tyr Ser Tyr Phe Pro Asp Ser Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Gly Tyr Asp Glu Gly Asp Ala Met Asp Ser Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 173
<211> 327
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 173
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 174
<211> 448
<212> PRT
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 174
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Arg Ser Gly Ser Tyr Ser Tyr Phe Pro Asp Ser Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Gly Tyr Asp Glu Gly Asp Ala Met Asp Ser Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 175
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 175
gaggtgcagc tgctggagtc tggcggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtat attactgtgc gattctggga 300
gctacttcgt tatcggcttt tgatatctgg ggccaaggga caatggtcac cgtctcgagc 360
<210> 176
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 176
cagtctgtgc tgacgcagcc gccctcagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gcaggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gcaatcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagtcctatg acagcagcct gagtggttca 300
ggggtattcg gcggagggac caagctgacc gtccta 336
<210> 177
<211> 1347
<212> DNA
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 177
gaggtgcagc tgctggagtc tggcggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtat attactgtgc gattctggga 300
gctacttcgt tatcggcttt tgatatctgg ggccaaggga caatggtcac cgtctcgagc 360
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaagc cgctggggca 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc cccgggt 1347
<210> 178
<211> 654
<212> DNA
<213> Artificial Sequence
<220>
<223> Синтетическая конструкция
<400> 178
cagtctgtgc tgacgcagcc gccctcagtg tctggggccc cagggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gcaggttatg atgtacactg gtaccagcag 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gcaatcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgagg atgaggctga ttattactgc cagtcctatg acagcagcct gagtggttca 300
ggggtattcg gcggagggac caagctgacc gtcctaggtc agcccaaggc tgccccctcg 360
gtcactctgt tcccgccctc ctctgaggag cttcaagcca acaaggccac actggtgtgt 420
ctcataagtg acttctaccc gggagccgtg acagtggcct ggaaggcaga tagcagcccc 480
gtcaaggcgg gagtggagac caccacaccc tccaaacaaa gcaacaacaa gtacgcggcc 540
agcagctatc tgagcctgac gcctgagcag tggaagtccc acagaagcta cagctgccag 600
gtcacgcatg aagggagcac cgtggagaag acagtggccc ctacagaatg ttca 654
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕЖИМ ДОЗИРОВАНИЯ ДЛЯ АНТАГОНИСТОВ TFPI | 2019 |
|
RU2780590C1 |
| АНТИТЕЛА К ТАУ И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2787779C2 |
| АНТИ-GDF15 АНТИТЕЛА, КОМПОЗИЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2019 |
|
RU2795457C2 |
| ТРИСПЕЦИФИЧЕСКИЕ И/ИЛИ ТРИВАЛЕНТНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С ПРИМЕНЕНИЕМ КРОССОВЕРНОГО ФОРМАТА С ДВОЙНЫМ ВАРИАБЕЛЬНЫМ ДОМЕНОМ (CODV) ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИИ, ВЫЗВАННОЙ HIV | 2020 |
|
RU2820164C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2018 |
|
RU2810094C2 |
| CD47-АНТИГЕНСВЯЗЫВАЮЩАЯ ЕДИНИЦА И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2780859C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЙ ПОЛИПЕПТИД, СВЯЗЫВАЮЩИЙСЯ С CD47, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2833294C2 |
| ТЕРАПЕВТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЛИГАНДА CD40 | 2018 |
|
RU2770209C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
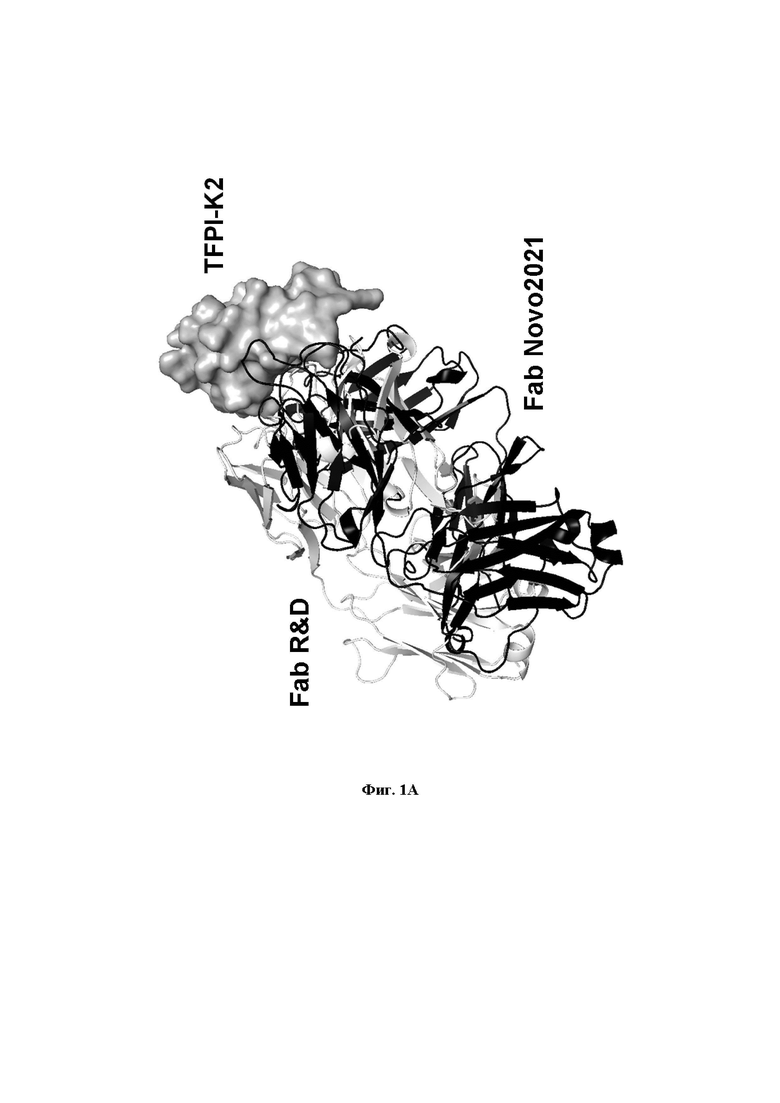
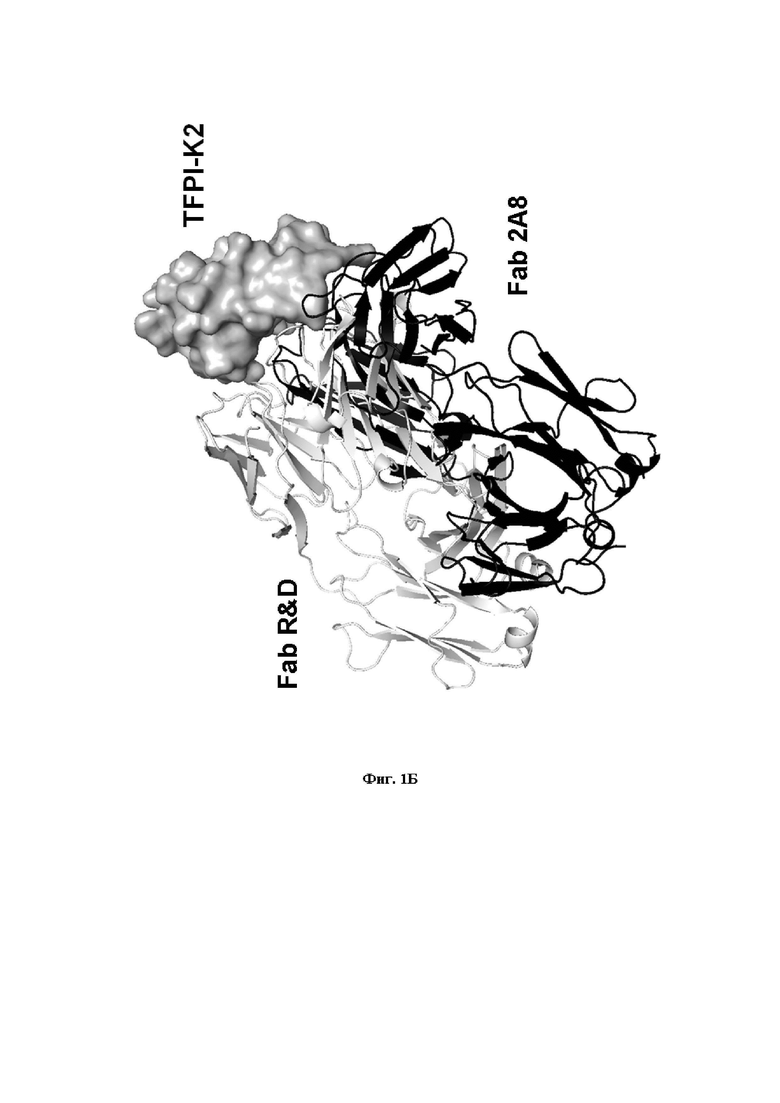
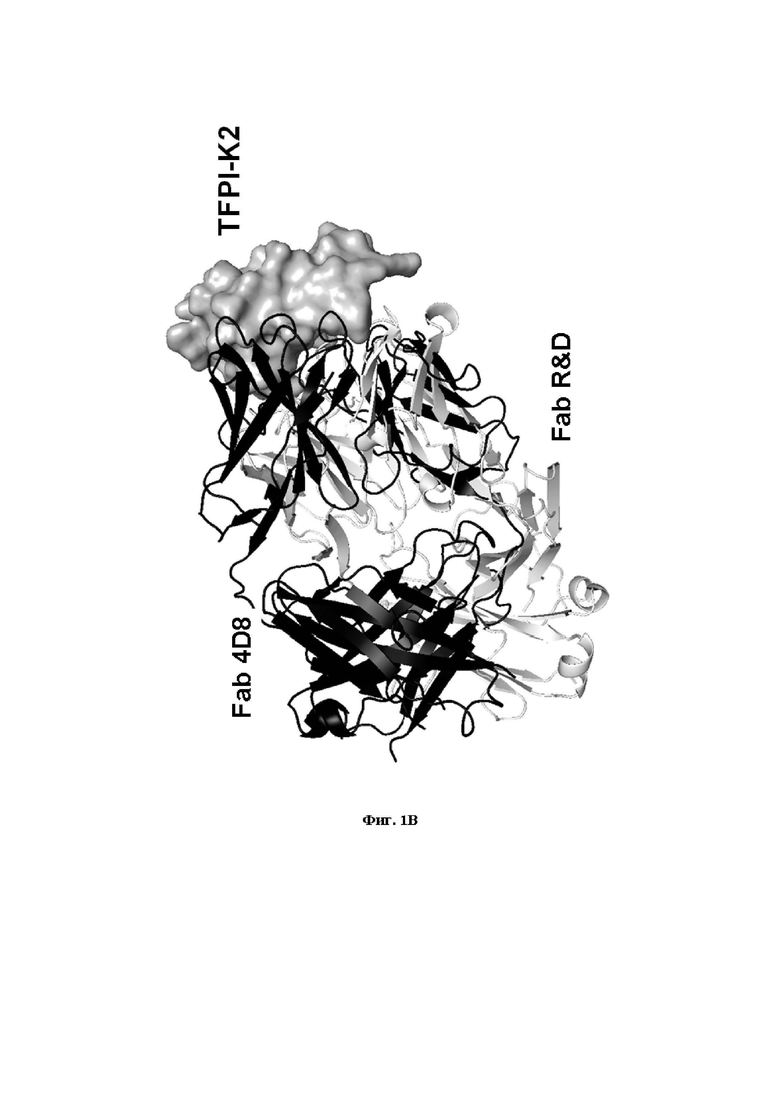
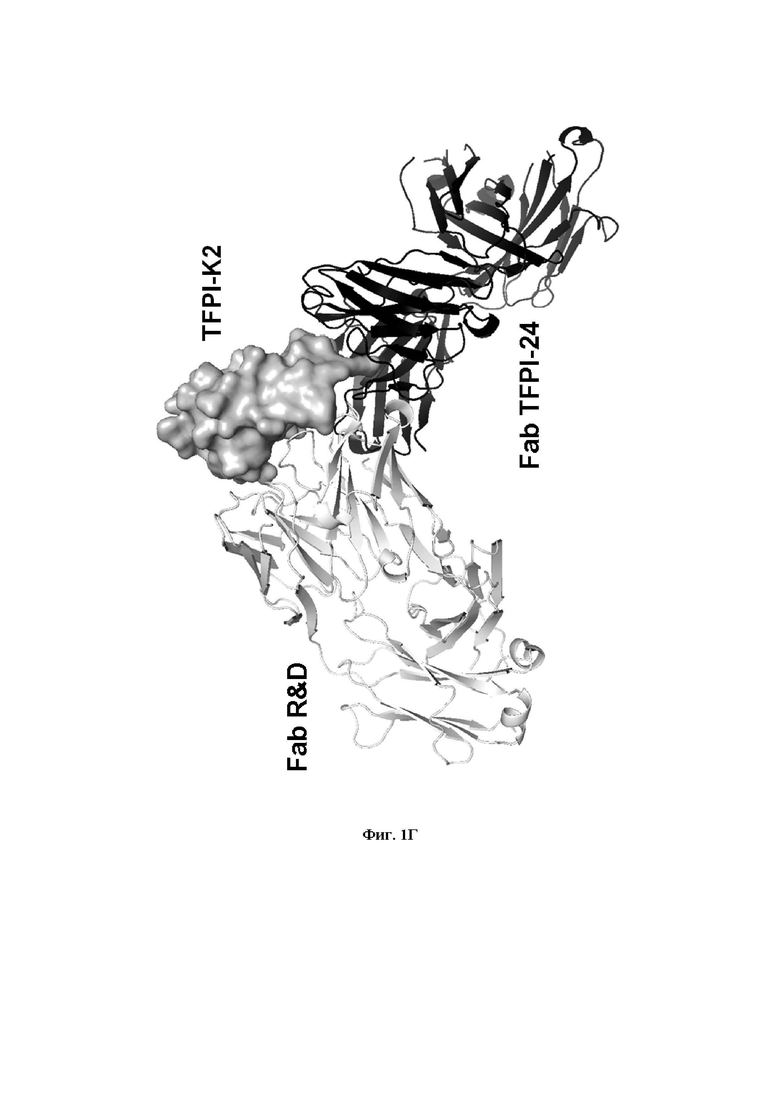
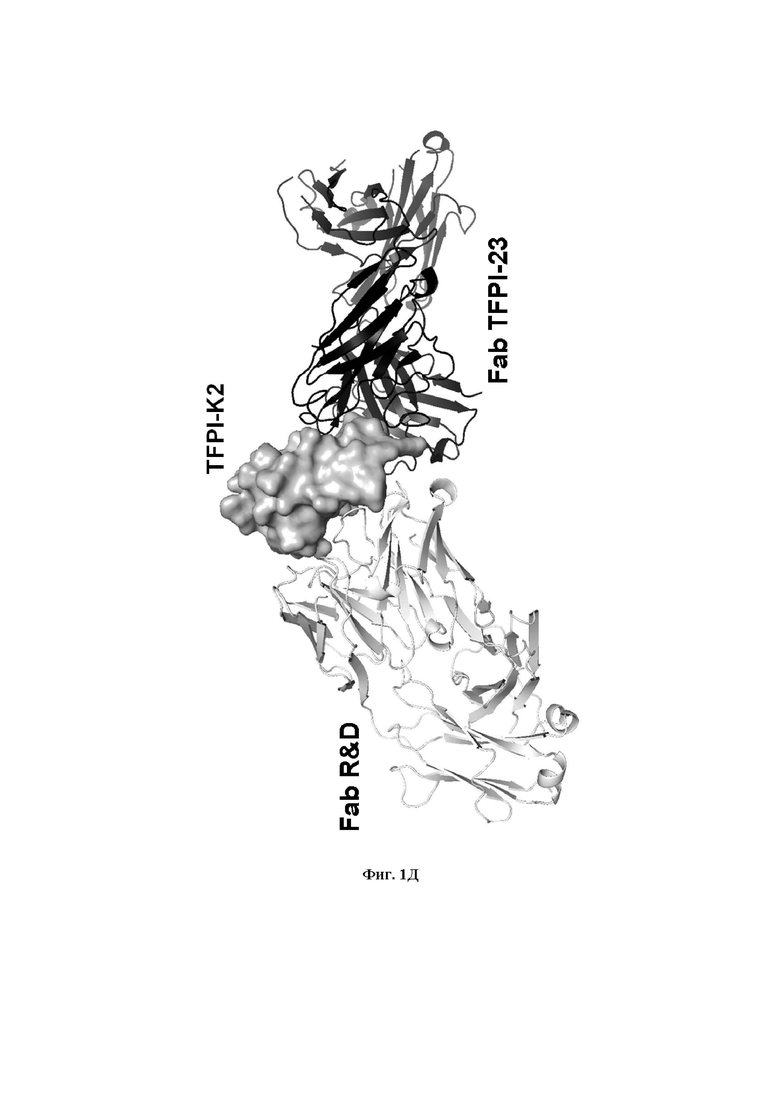
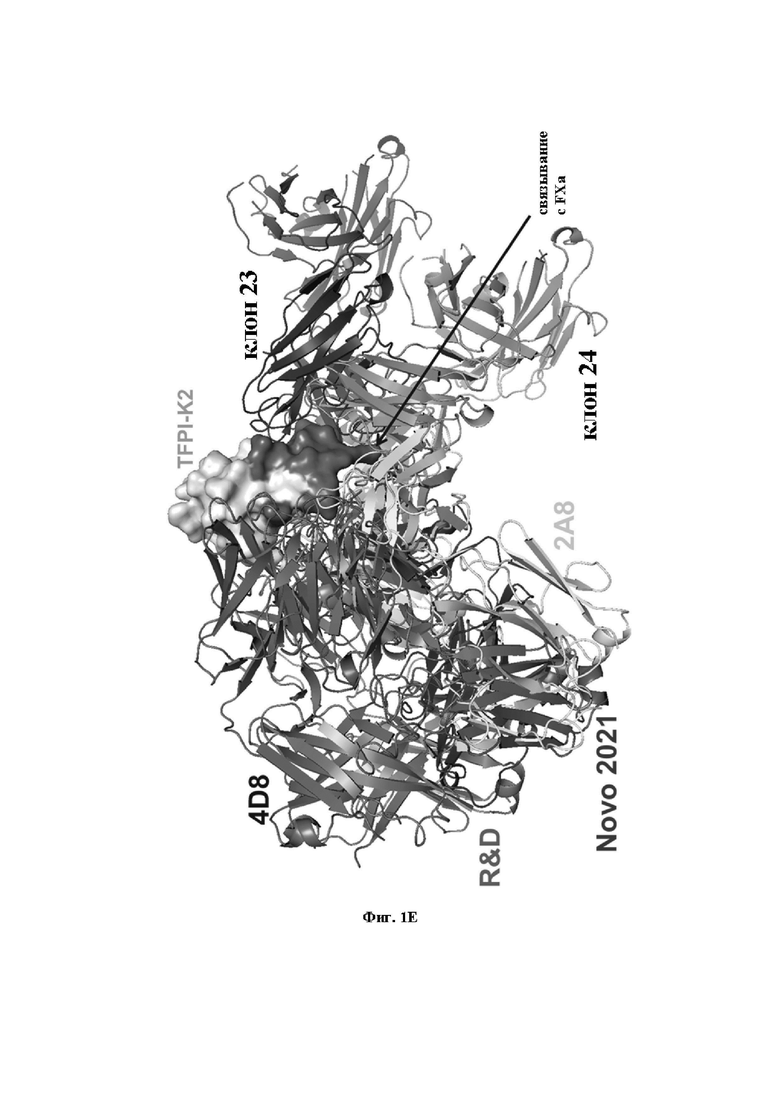
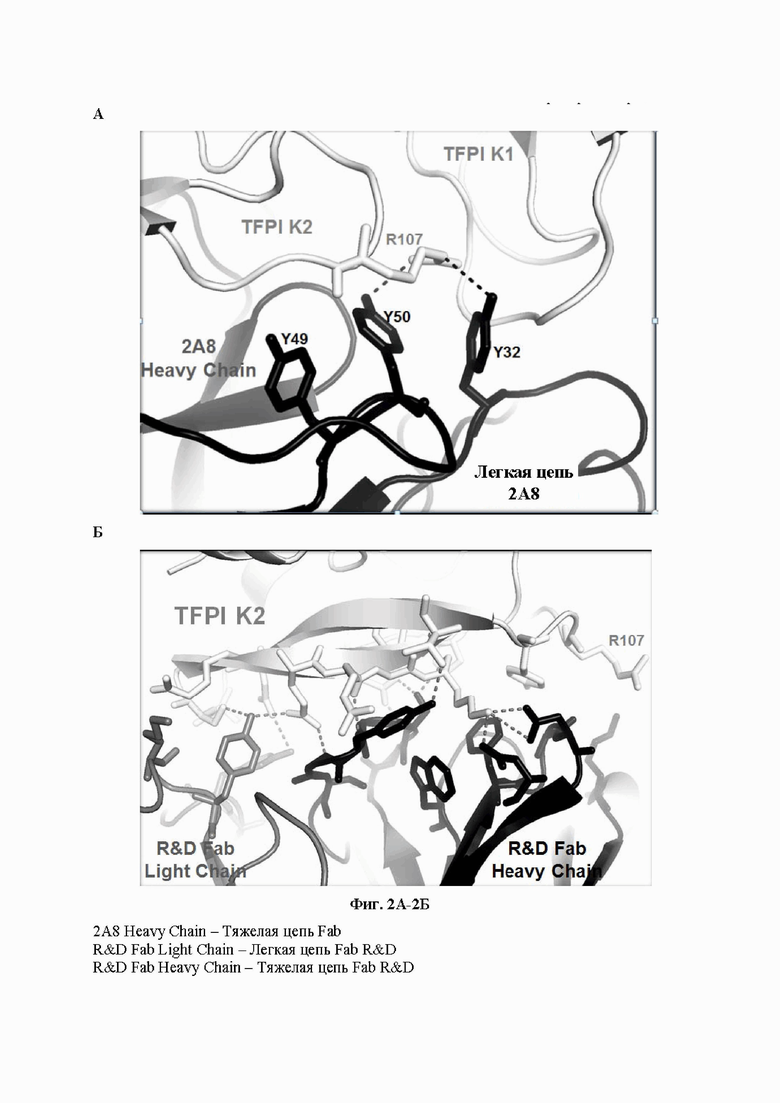
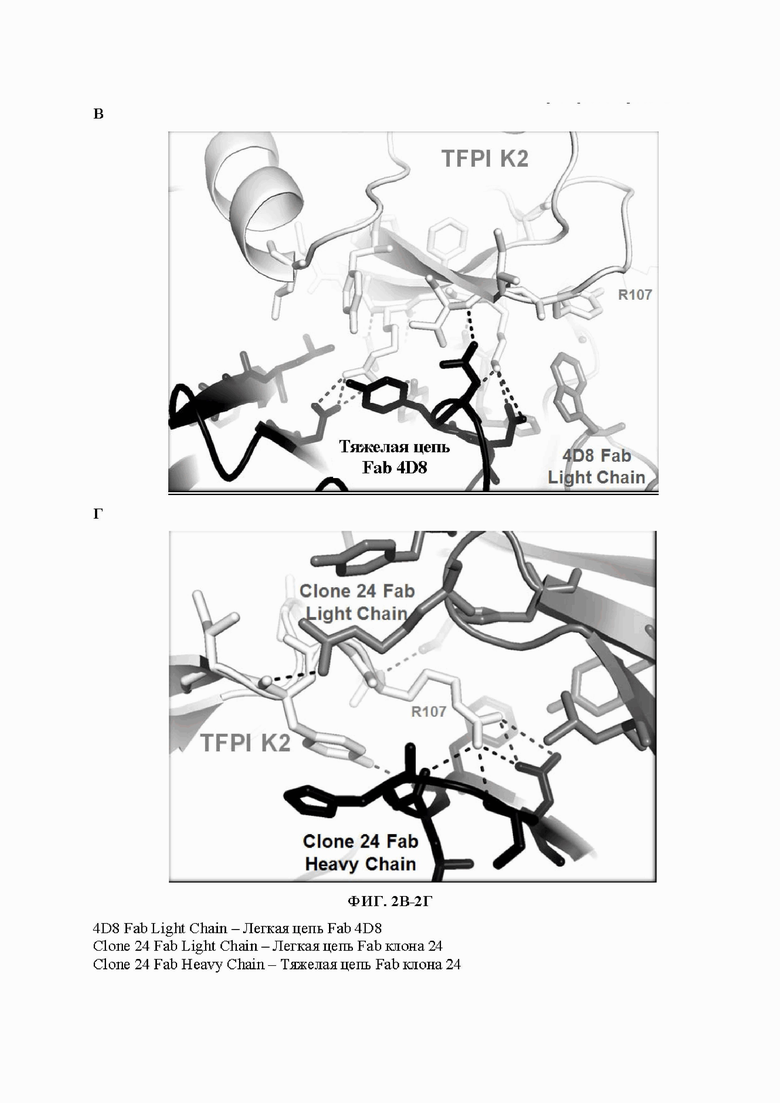
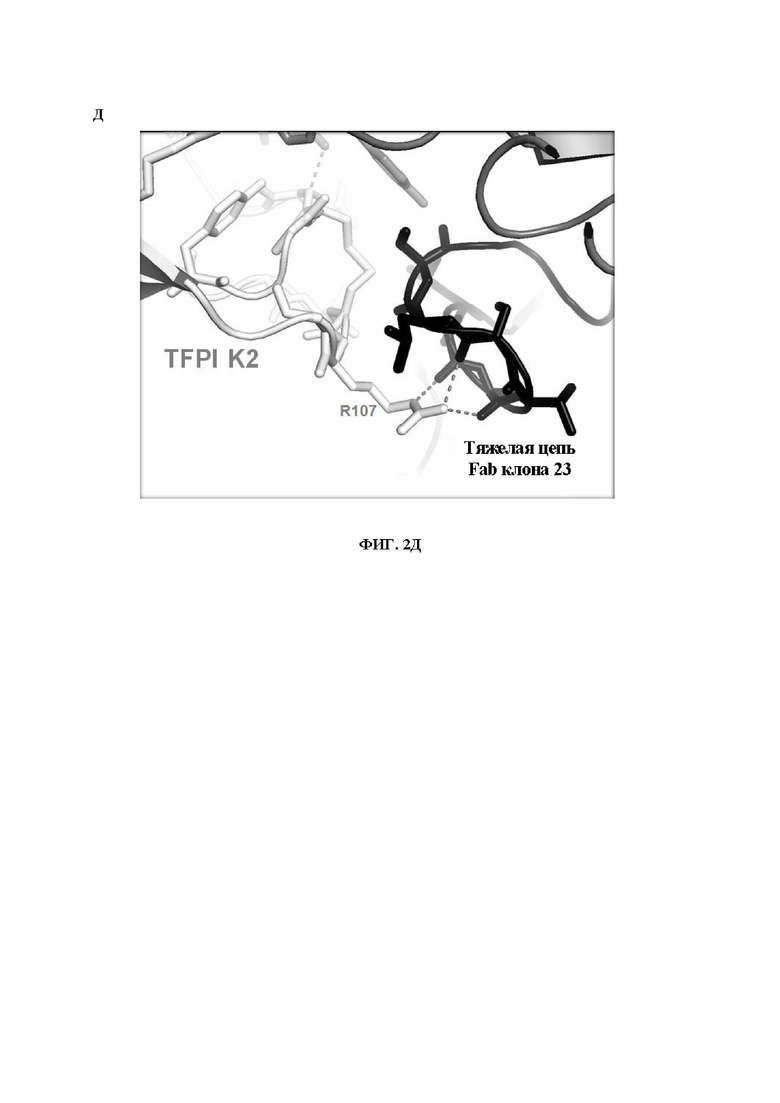
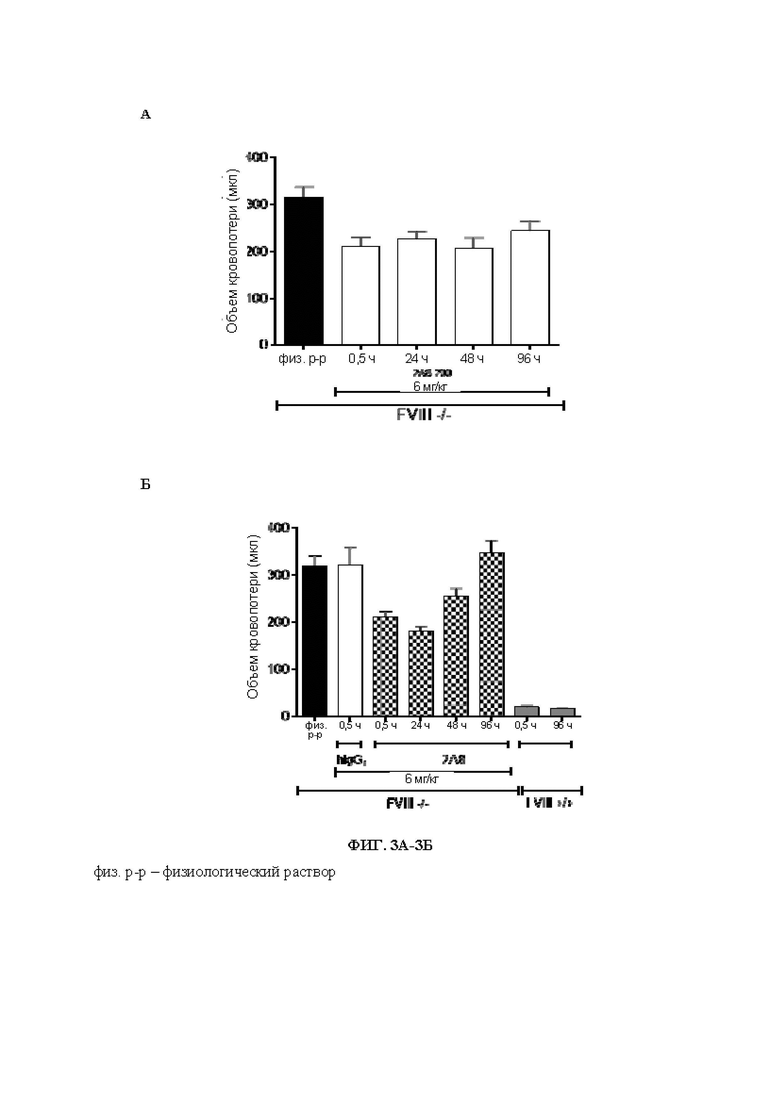
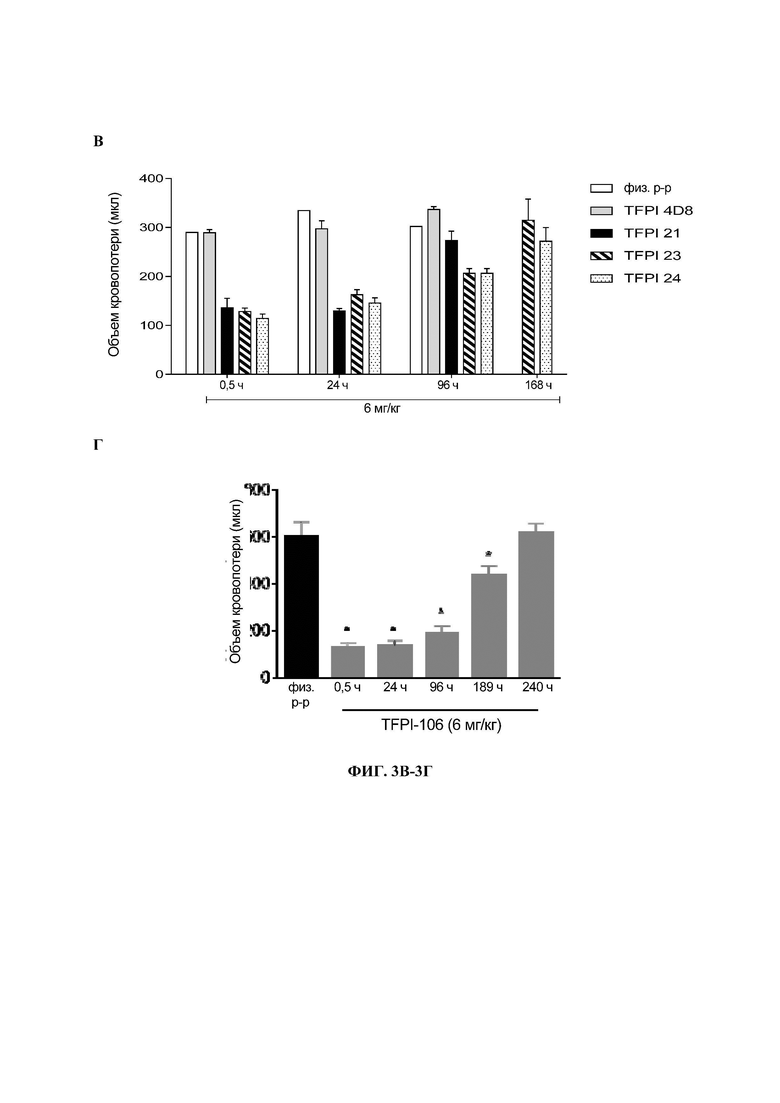
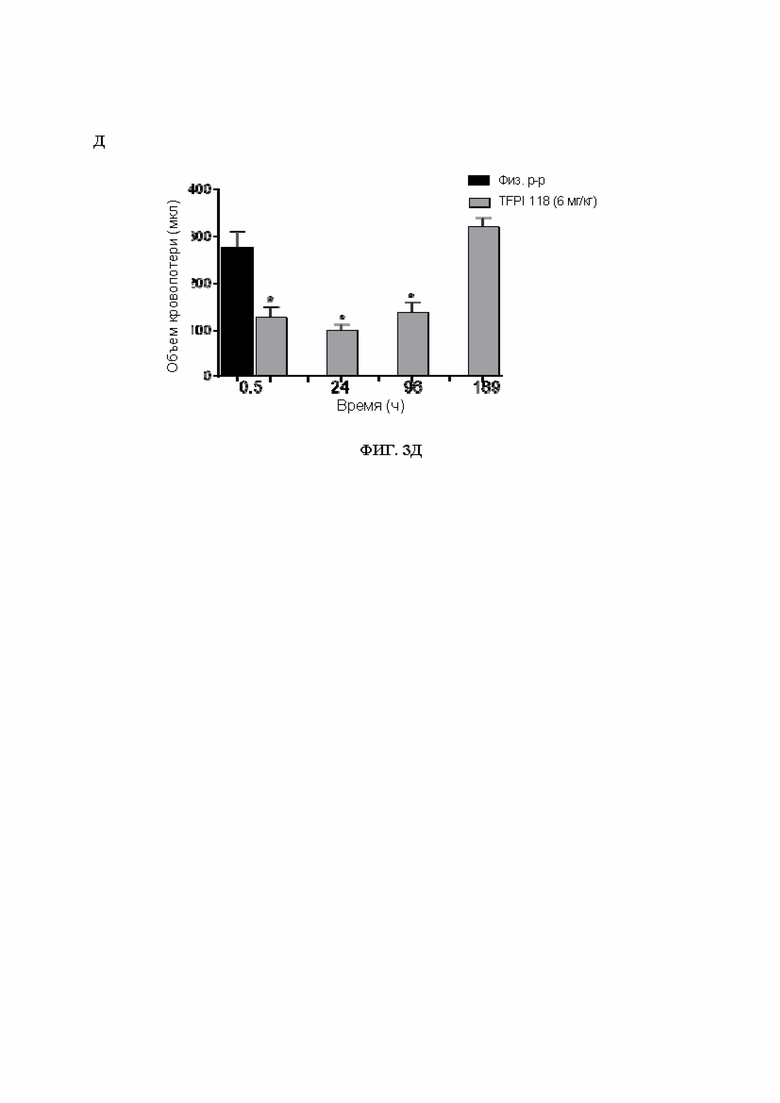
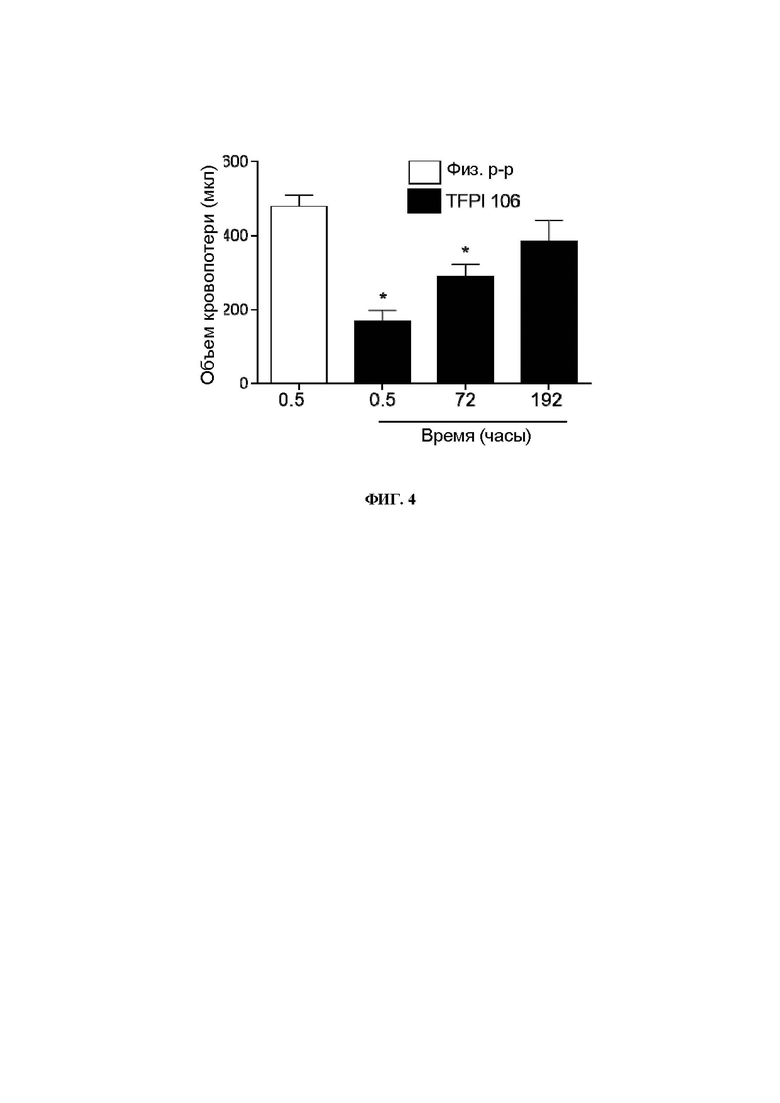


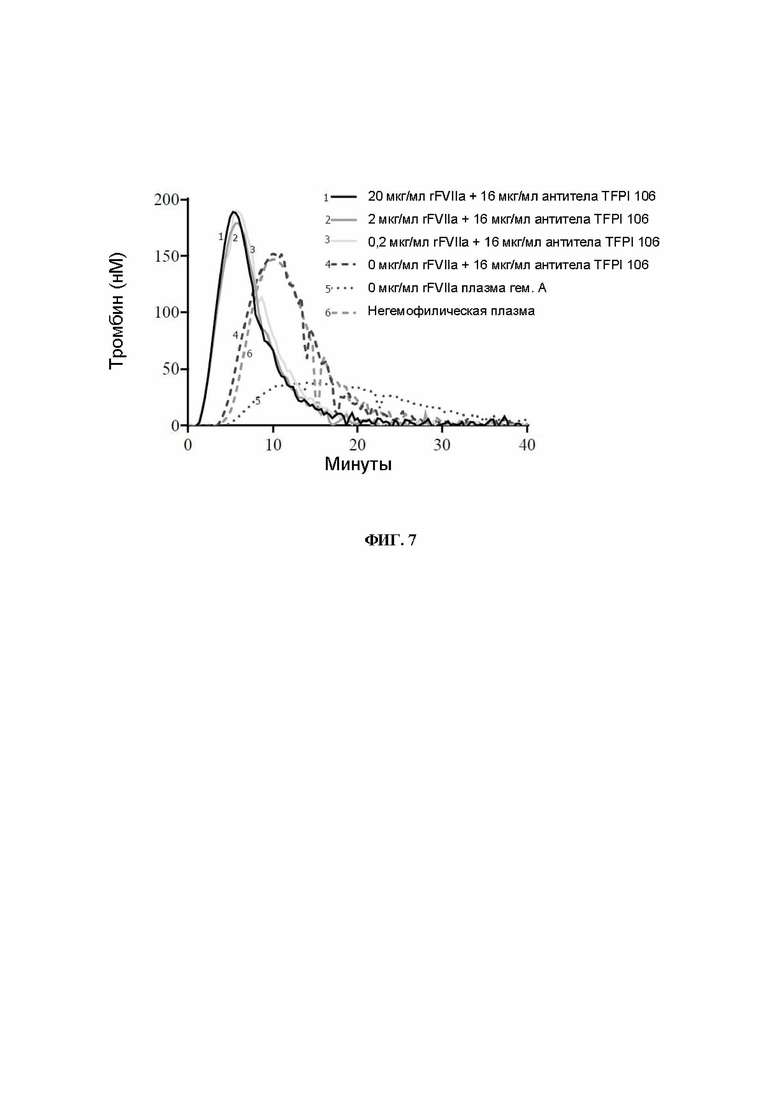
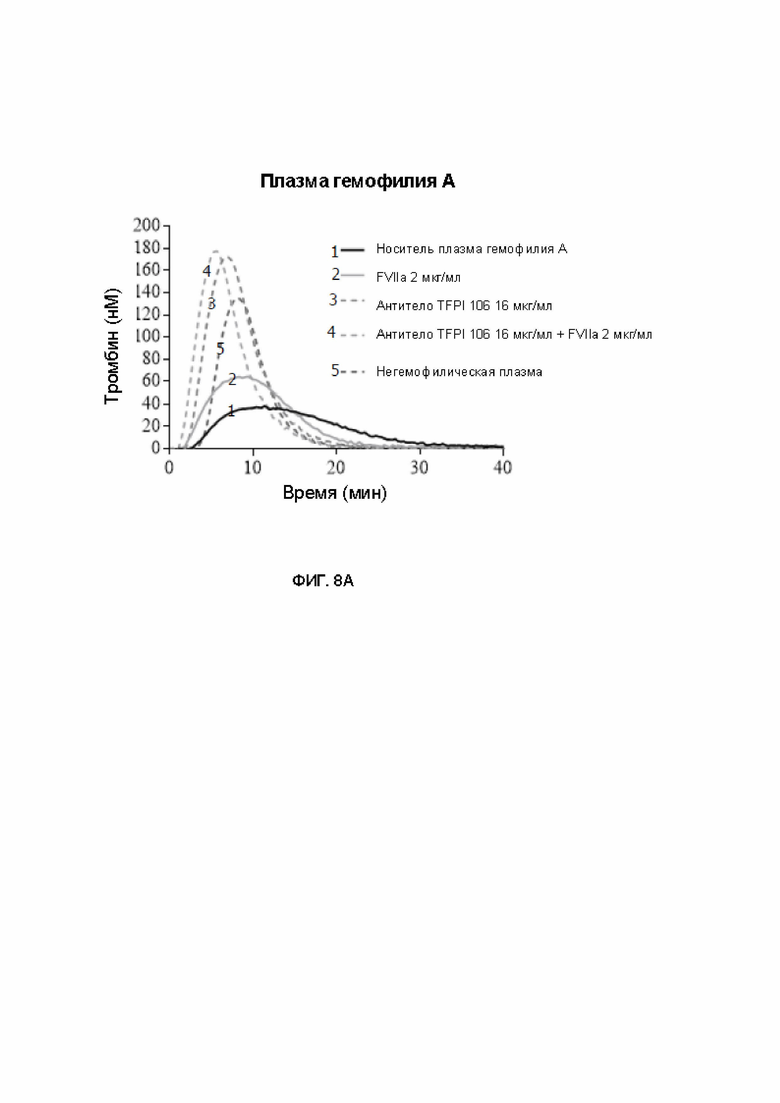
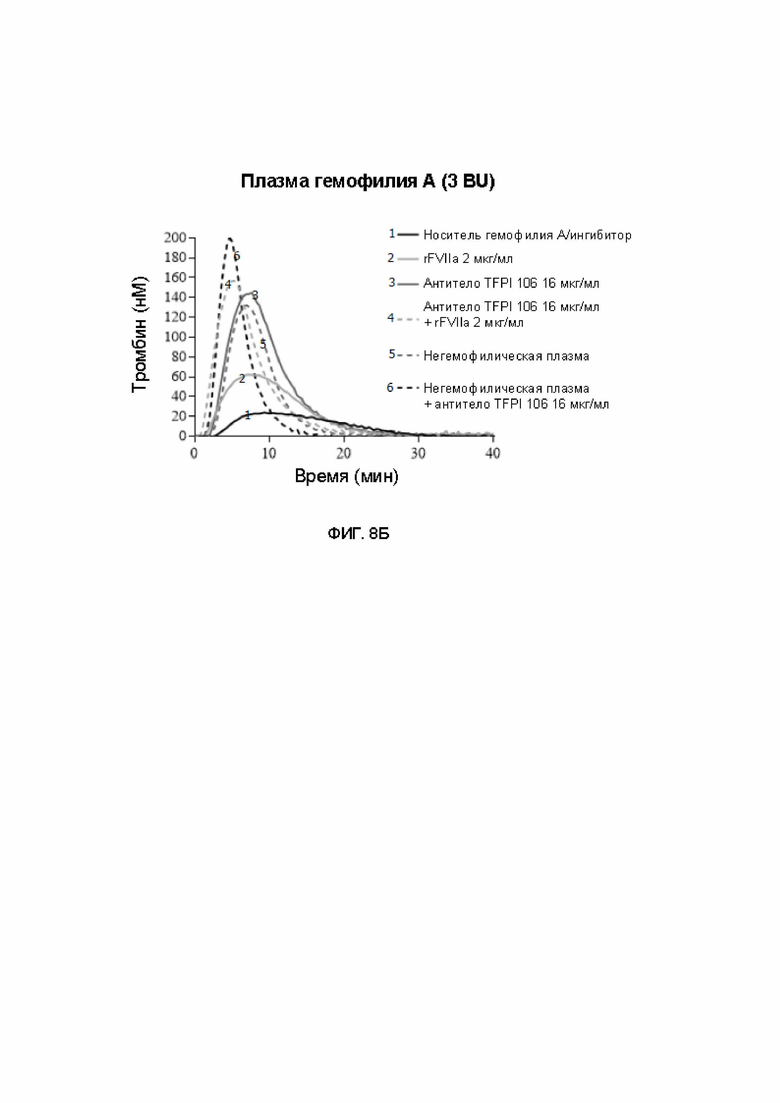
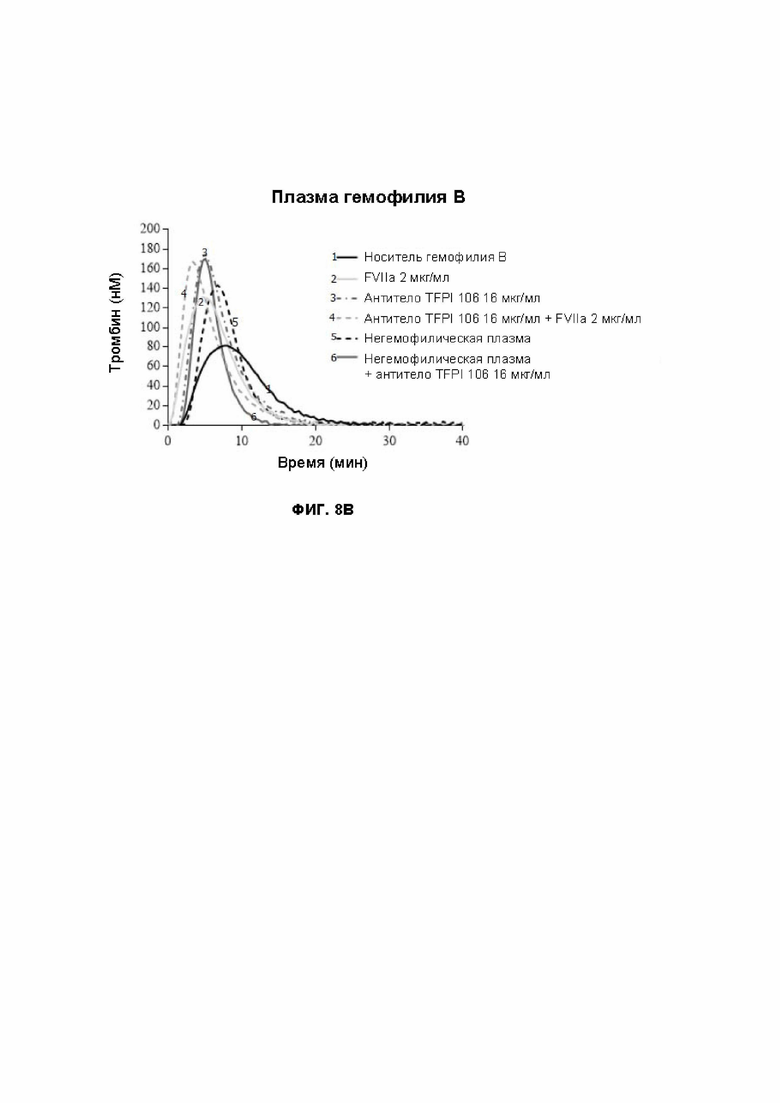
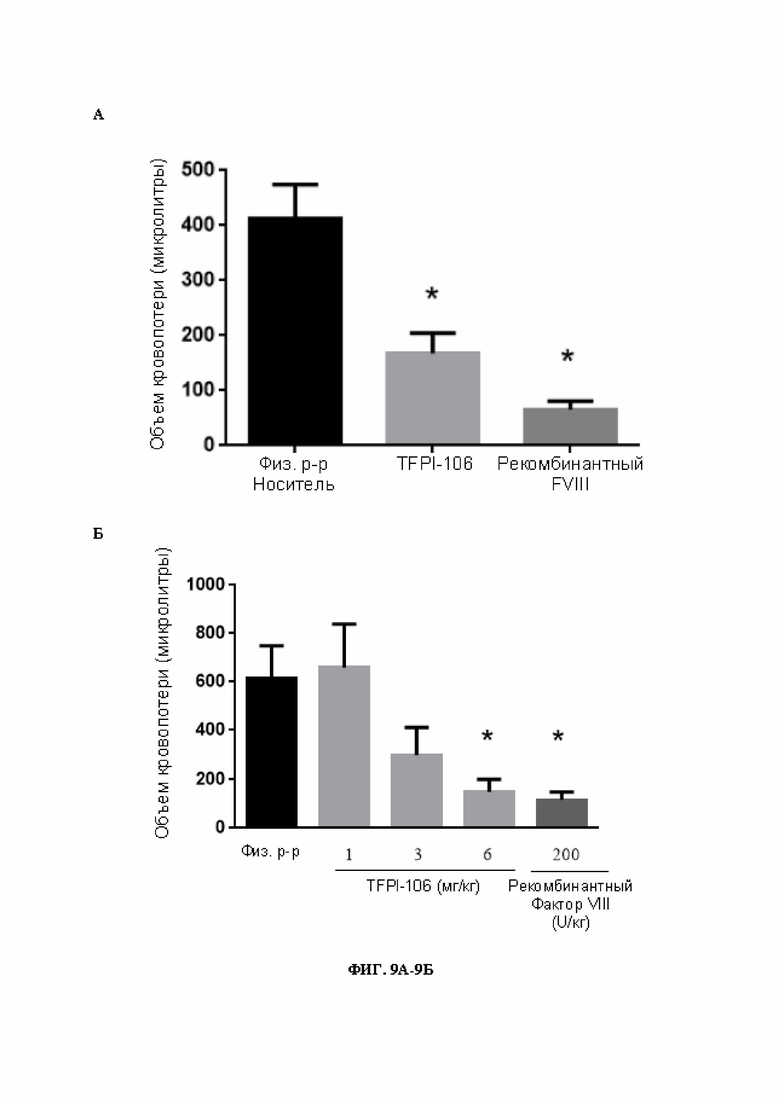
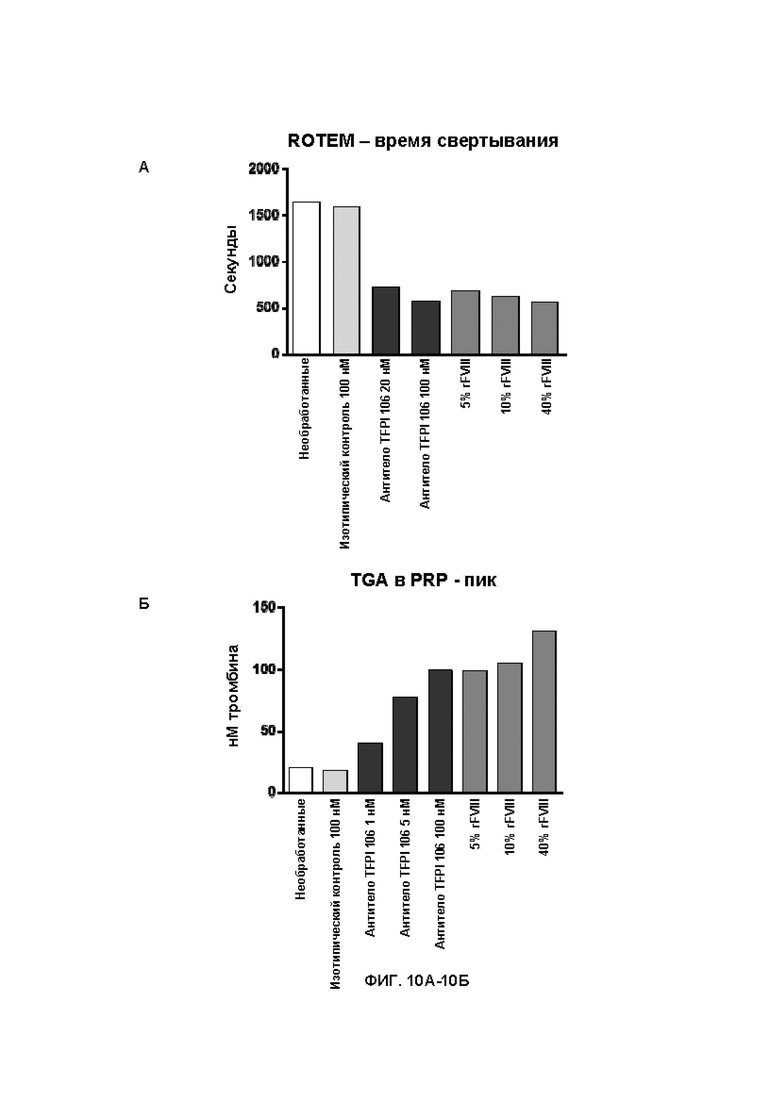
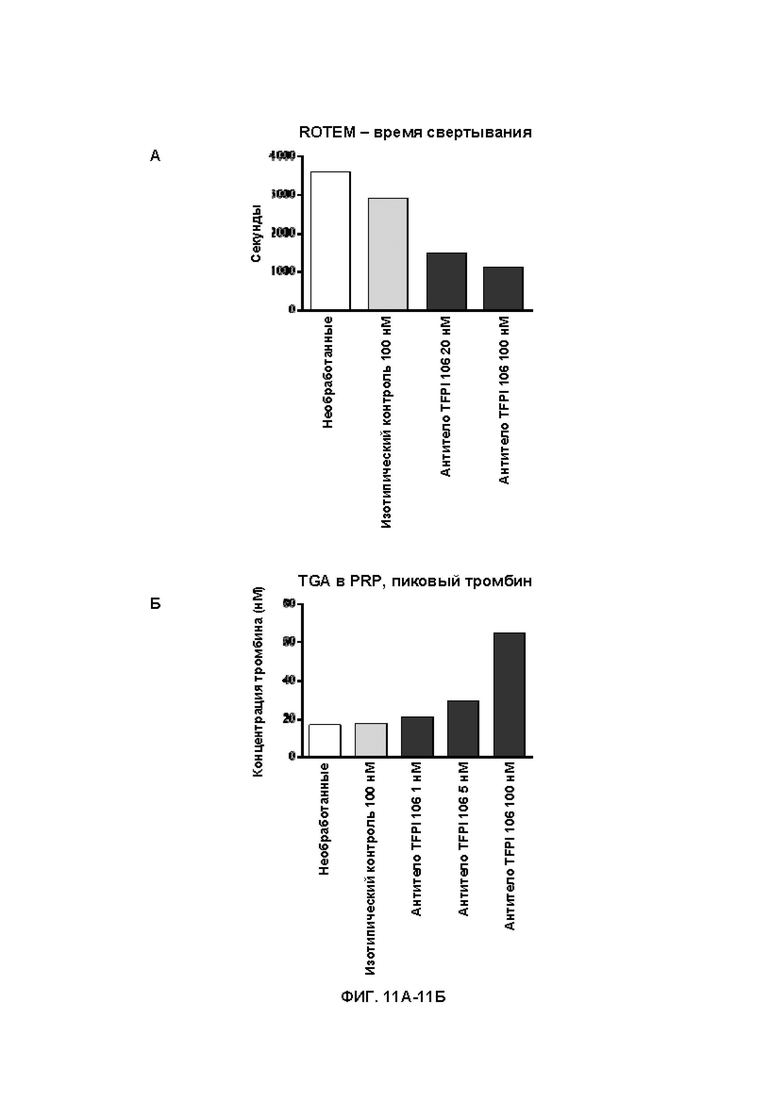
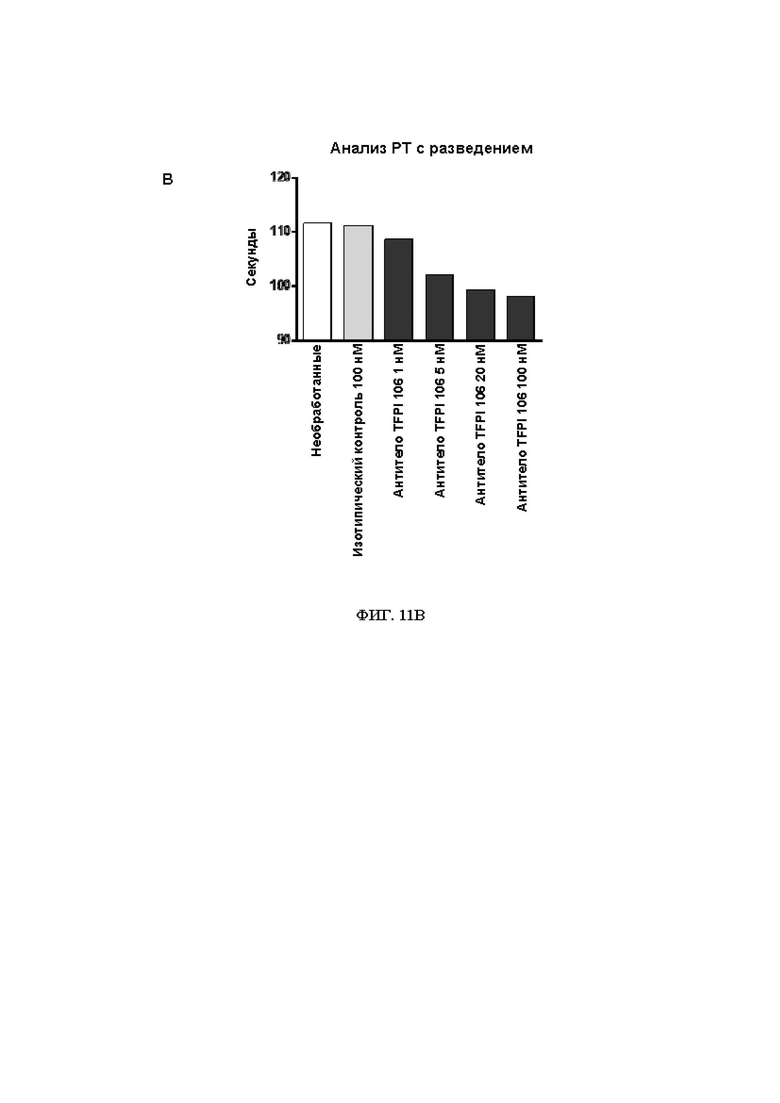
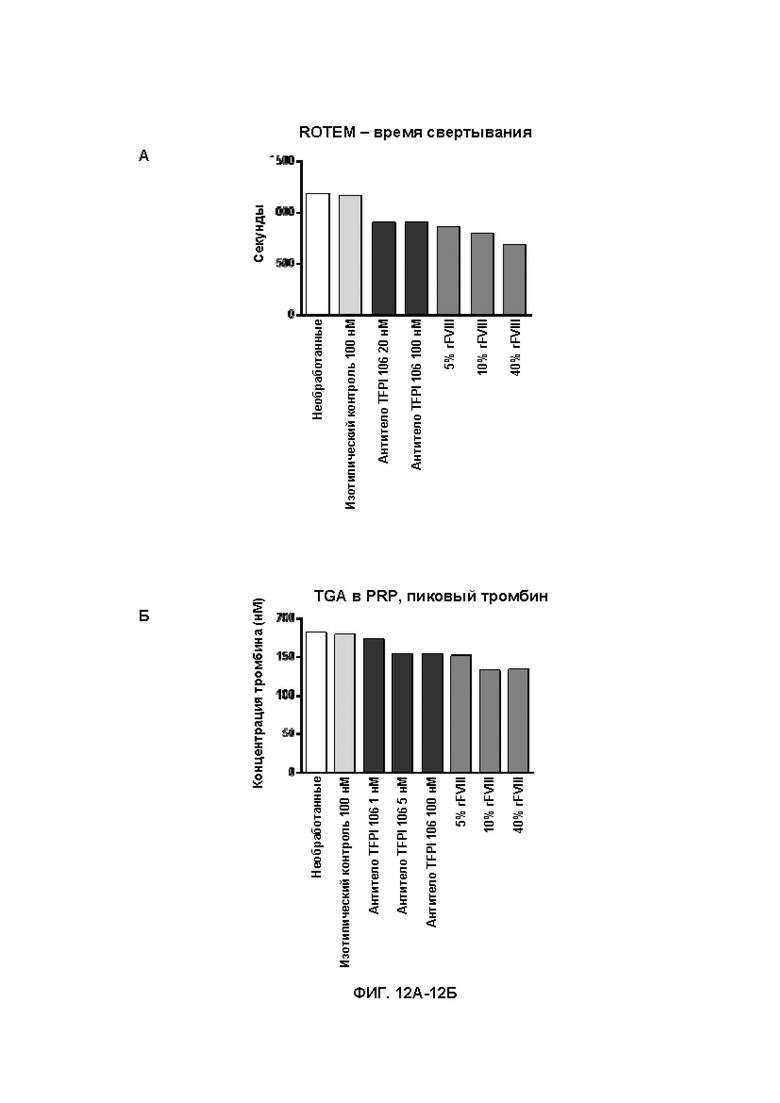
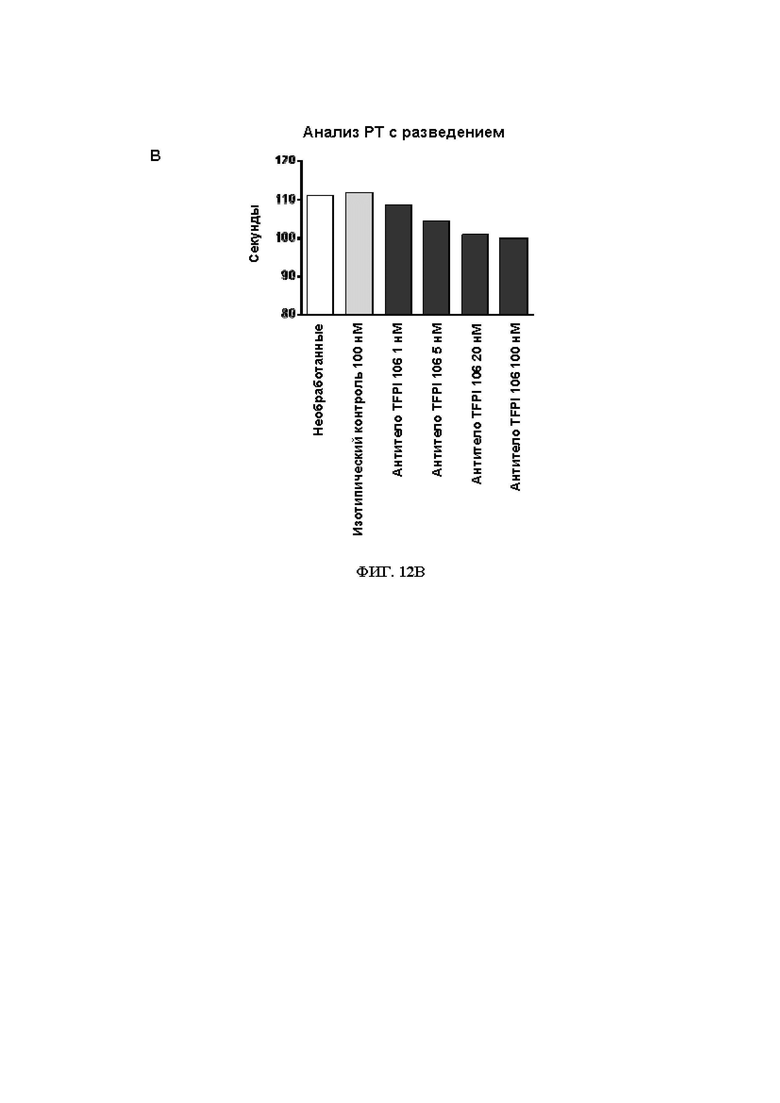
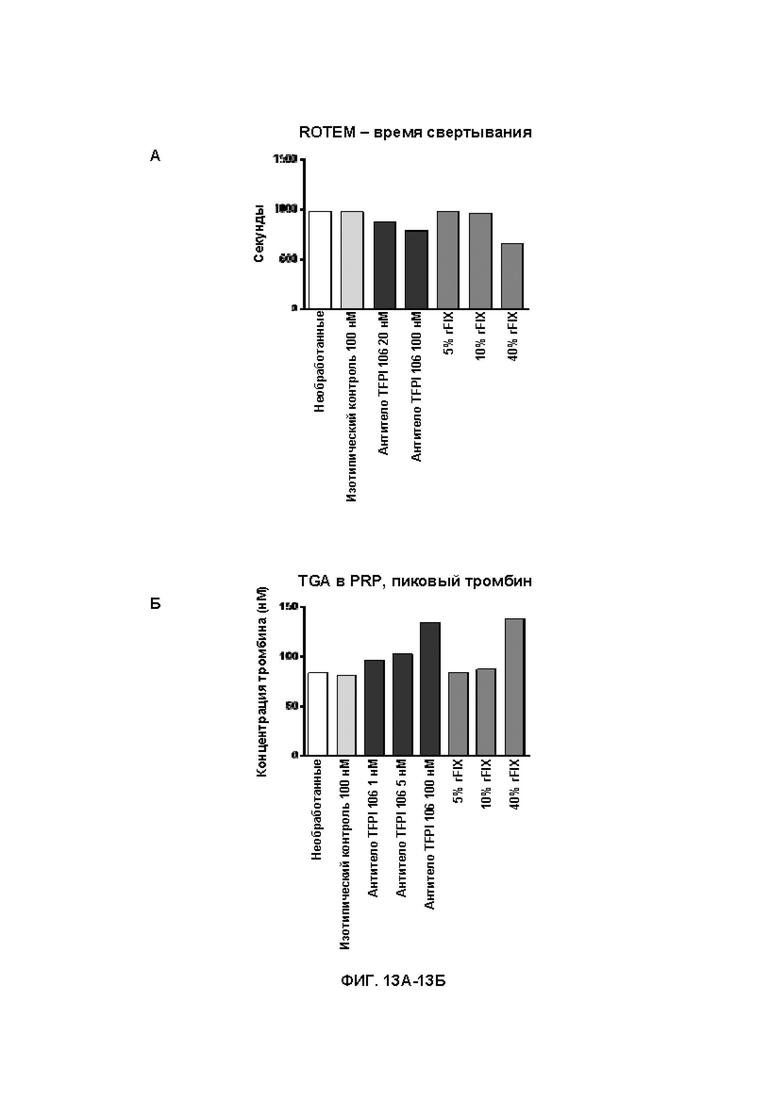
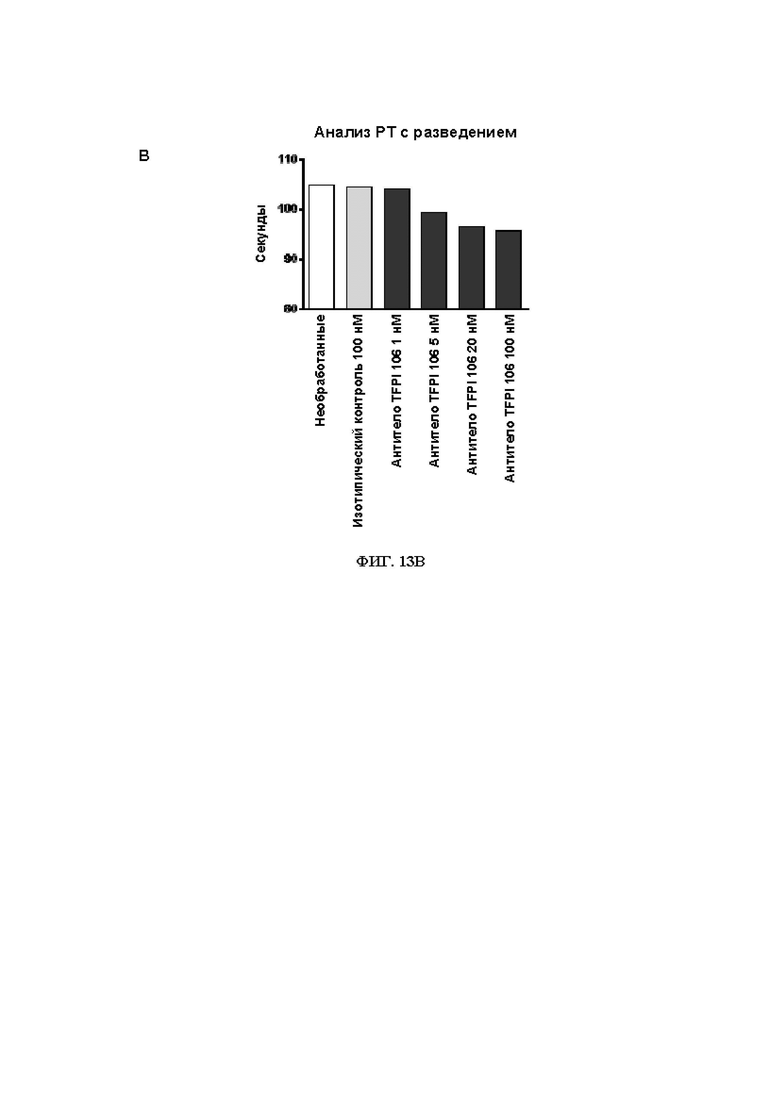
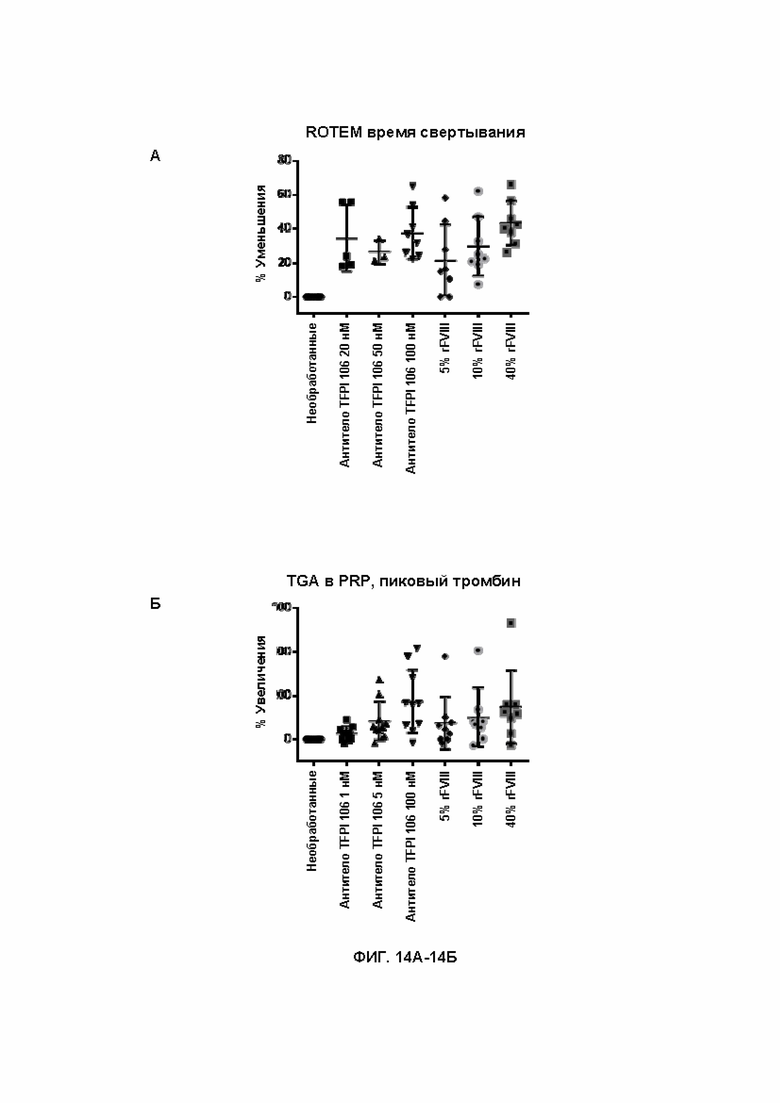
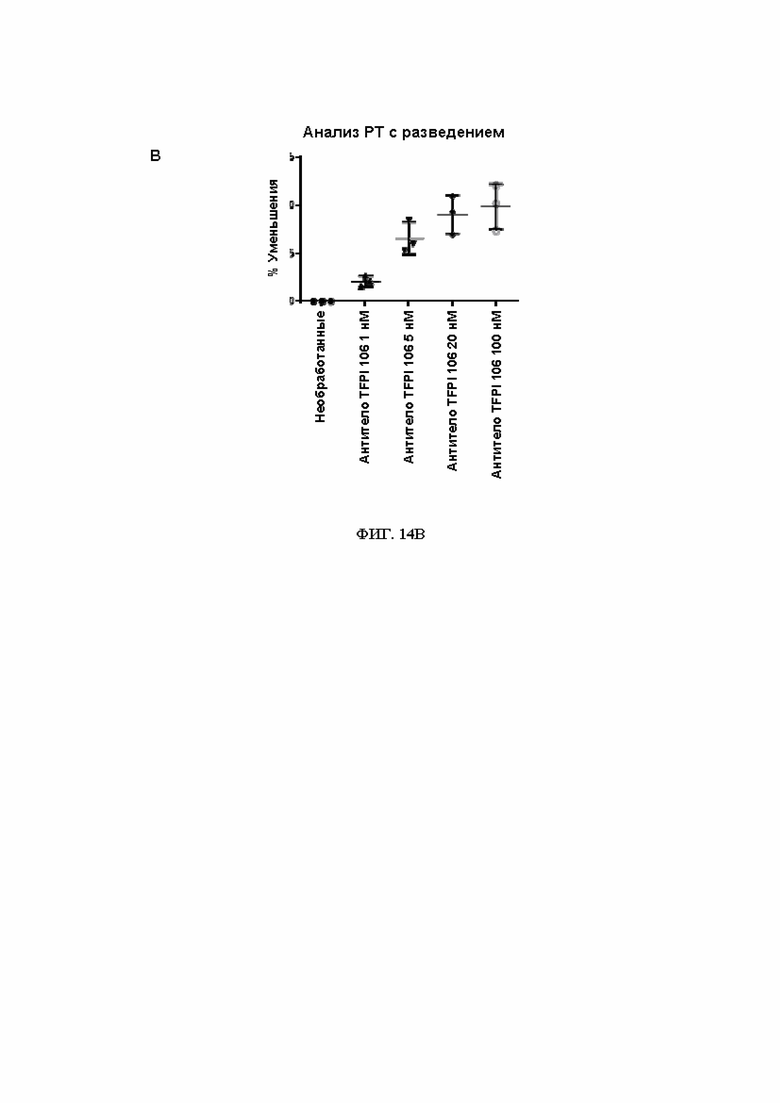
Группа изобретений относится к биотехнологии. Представлены: экспрессионные вектора, содержащие молекулу нуклеиновой кислоты или молекулы нуклеиновых кислот, содержащую(ие) одну или более чем одну нуклеотидную последовательность, экспрессирующая клетка-хозяин, содержащая данные вектора и способ получения антитела или его антигенсвязывающего фрагмента, включающий культивирование клетки-хозяина. Изобретения используются для лечения расстройств кровотечения и для сокращения времени свертывания крови. 4 н. и 3 з.п. ф-лы, 33 табл., 29 ил., 18 пр.
1. Экспрессионный вектор, содержащий молекулу нуклеиновой кислоты или молекулы нуклеиновых кислот, содержащую(ие) одну или более чем одну нуклеотидную последовательность:
(1) кодирующую VH, содержащую CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 38, CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 39, и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 40, и VL, содержащую CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 33, CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 34, и CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 35;
(2) кодирующую VH, содержащую аминокислотную последовательность SEQ ID NO: 63, и VL, содержащую аминокислотную последовательность SEQ ID NO: 36;
(3) кодирующую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:64, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 37; или
(4) содержащую последовательность SEQ ID NO: 175, последовательность SEQ ID NO: 176, последовательность SEQ ID NO: 177, или последовательность SEQ ID NO: 178.
2. Экспрессионный вектор, содержащий молекулу нуклеиновой кислоты или молекулы нуклеиновых кислот, содержащую(ие) одну или более чем одну нуклеотидную последовательность, кодирующую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 64, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 37.
3. Экспрессирующая клетка-хозяин, содержащая вектор по любому из пп. 1-2.
4. Клетка-хозяин п. 3, где указанная клетка представляет собой клетку млекопитающих.
5. Клетка-хозяин по п. 4, где указанная клетка-хозяин представляет собой клетку СНО, клетку HEK-293 или клетку Sp2.0.
6. Способ получения антитела или его антигенсвязывающего фрагмента, включающий культивирование клетки-хозяина по любому из пп. 3-5 при условиях, когда указанное антитело или антигенсвязывающий фрагмент экспрессируется указанной клеткой-хозяином.
7. Способ по п. 6, дополнительно включающий выделение указанного антитела или его антигенсвязывающего фрагмента.
| WO 2012135671 A2, 04.10.2012 | |||
| WO 2012001087 A1, 05.01.2012 | |||
| ABUMIYA T | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| RU 2011129068 A, 27.01.2013. | |||

