Уровень техники
Настоящее изобретение относится к растениеводству и к молекулярной биологии. Более конкретно, настоящее изобретение относится к способу детекции локуса количественного признака (QTL), ассоциированного с устойчивостью к Botrytis cinerea в томатах, к способу получения с помощью этого устойчивого к Botrytis растения томата и к устойчивым к Botrytis растениям томата, полученным таким образом, и их частям.
Предпосылки изобретения
Botrytis cinerea представляет собой некротрофный патогенный гриб с исключительно широким диапазоном хозяев, включающим по меньшей мере 235 возможных хозяев. Вследствие его широкого диапазона хозяев и вследствие того, что он поражает экономически значимые части растения, B. cinerea является большой проблемой для многих коммерчески выращиваемых сельскохозяйственных культур. Среди овощеводов гриб обычно называют Botrytis. Культивируемый томат (Solanum lycopersicum; ранее Lycopersicon esculentum) также является восприимчивым к инфекции Botrytis, и гриб поражает, главным образом, стебель, листья и плоды растения томата. В обогреваемых теплицах встречаемость инфекций Botrytis на стеблях является особенно частой.
Botrytis активно уничтожает инфицированные клетки, вызывая мягкую гниль, увядание, пятнистость листьев, черную ножку и злокачественные опухоли стебля. Пораженные листья покрываются конидиофорами и конидиями, а затем поникают и увядают. Гриб будет прорастать из пораженных листьев в стебель и образовывать сухие светло-коричневые очаги повреждения длиной от нескольких миллиметров до нескольких сантиметров. Очаги повреждения также могут образовываться на рубцах от переломов на стебле. Очаги повреждения на стебле также могут быть покрыты серой гнилью. В тяжелых случаях инфекция опоясывает стебель и уничтожает растение. Старая, увядающая ткань растения томата обычно является более восприимчивой к атаке Botrytis, чем молодая ткань.
В целях предотвращения развития Botrytis в выращиваемых в теплицах томатах температуру и относительную влажность необходимо тщательно регулировать. Кроме того, важно предоставлять воду без смачивания листьев. Для выращиваемых в поле растений следует применять хорошую дренажную систему и борьбу с сорняками. Более того, необходимо поддерживать высокий уровень питательных веществ растений. Однако эти профилактические меры не могут полностью предотвратить возникновение значительной потери урожая в случае инфекции.
Для борьбы с Botrytis в томатах, выращиваемых как в теплицах, так и в поле, доступны фунгициды. Примеры некоторых фунгицидов включают Dowicide A® и хлорталонил, которые можно наносить на плоды томата после сбора. Однако известно, что Botrytis обладает приобретенной устойчивостью против некоторых обычно используемых фунгицидов. Кроме того, применение фунгицидов является нежелательным как вследствие экономической, так и вследствие экологической перспективы. В настоящее время существует необходимость в коммерческих сортах томата, которые проявляют устойчивость к Botrytis.
Частичная устойчивость к Botrytis была выявлена в некоторых диких видах томата (Egashira et al. 2000; Nicot et al. 2002; Urbasch 1986). Однако эти растения не продуцируют коммерческие культуры томата.
В WO 02/085105 описана генетическая область на 10 хромосоме генома S. habrochaites, которая, как полагают, вовлечена в частичную устойчивость к Botrytis. Было показано, что интрогрессия этого генетического материала в культивируемые сорта томата приводит к культивируемым растениям томата, которые являются частично устойчивыми к Botrytis.
Однако до настоящего времени программы разведения, нацеленные на обеспечение устойчивости Botrytis в томате, имели ограниченный успех. Причина этих плохих результатов в настоящее время не ясна. Частично это может быть следствием недостаточного знания о генетической основе и наследовании устойчивости к Botrytis. С другой стороны это может быть следствием отсутствия надлежащих биологических анализов для оценки уровней устойчивости к Botrytis в растениях томата, полученных в программах разведения. Недостаточность знания и способов также осложняет селекцию растений как среди диких изолятов, так и среди растений-потомков, которые содержат гены, вовлеченные в устойчивость к Botrytis.
В более раннем исследовании авторы настоящего изобретения выявили, что устойчивость к Botrytis в томатах наследуется полигенно и что это частично может объяснить плохие результаты разведения устойчивых растений.
Целью настоящего изобретения является повышение успеха программ разведения, нацеленных на получение коммерческих сортов томата, которые являются устойчивыми к Botrytis. Кроме того, целью настоящего изобретения является обеспечение дополнительной и/или повышенной устойчивости к Botrytis в коммерческих сортах томата. Другой целью настоящего изобретения является предоставление дополнительного генетического материала в геноме диких изолятов томата, который вовлечен в устойчивость к Botrytis в таких растениях. Такой дополнительный генетический материал можно использовать для расширения базиса для продукции устойчивых к Botrytis сортов культивируемых томатов.
Сущность изобретения
Авторы настоящего изобретения выявили, что в геноме S. habrochaites имеется генетический материал на ряде хромосом, которые ранее не были идентифицированы как вовлеченные в устойчивость к Botrytis. В действительности, авторы настоящего изобретения успешно идентифицировали локусы количественных признаков (QTL) в геноме линии дикого родственника томата, т.е. в Solanum habrochaites LYC 4/78. Эти дополнительные QTL были открыты посредством применения интрогрессивных линий.
После этого авторы изобретения получили возможность вывести устойчивые к Botrytis растения томата скрещиванием растений этих устойчивых к Botrytis диких (донорных) линий томата с неустойчивыми реципиентными растениями томата. Эти растения проявляли более высокий уровень устойчивости, чем любое культивируемое растение томата, полученное к настоящему времени.
Усовершенствование относительно предшествующего уровня техники состоит в доступности дополнительных критериев скрининга, посредством которых можно подвергать мониторингу процесс разведения и управлять им. Растения-потомки, полученные скрещиванием между устойчивым диким Solanum habrochaites и восприимчивым культивируемым томатом, можно отбирать по наличию одной или нескольких, или даже всех из геномных областей, вовлеченных в устойчивость к Botrytis в диком изоляте. Результатом этого является разработка способа получения растения томата, при котором требуемый генетический состав можно лучше контролировать. Преимущество способа по изобретению состоит в том, с генетической точки зрения, что можно получить потомство, более сходное с изолятом дикого типа по требуемому признаку. Следовательно, признак устойчивости в культивируемом растении томата может быть более стабильно внесен в него, т.е. в настоящее время предоставлены возможности для оценки того, какие геномные области можно легко подвергать интрогрессии и которые трудно переносить в растения-потомки, какие из геномных областей являются необходимыми, какие из них являются совместно действующими и какие можно использовать для дальнейшего повышения уровней устойчивости в частично устойчивых линиях культивируемых томатов.
Посредством оценки уровня устойчивости к Botrytis в различных интрогрессивных линиях, в каждой из которых произошла конкретная геномная интрогрессия от S. habrochaites LYC 4/78, в зависимости от наличия молекулярных маркеров в донорном растении, авторы настоящего изобретения получили способность идентифицировать дополнительные QTL, связанные с устойчивостью к Botrytis, в устойчивых диких линиях томата и, таким образом, установить расположение ряда придающих устойчивость последовательностей ДНК в геноме. В приведенном ниже описании локус количественного признака (QTL), ассоциированный с устойчивостью к Botrytis в томате, будет кратко обозначен как QTL для устойчивости к Botrytis или QTL, ассоциированный с устойчивостью к Botrytis.
В дикой линии томата S. habrochaites было выявлено всего 5 новых QTL для устойчивости к Botrytis. Один QTL был расположен на 4 хромосоме, которая ранее была идентифицирована, как имеющая QTL, ассоциированный с устойчивостью к Botrytis. Кроме того, было выявлено, что область на этой хромосоме, которую ранее ассоциировали с устойчивостью, в действительности содержит два отдельных QTL. Другие QTL были идентифицированы на 6, 9, 11 и 12 хромосомах. Все новые QTL могли быть связаны с количественным параметром, который отражал способность растения снижать исходное установление инфекции, в дальнейшем в настоящем описании называемым параметром заболеваемости (DI), а также с другим количественным параметром, который отражал способность растения замедлять прогрессирование инфекции, в дальнейшем в настоящем описании называемым параметром скорости роста очага повреждения (LG). Также наличие QTL на хромосоме 10, как описано на предшествующем уровне техники, нельзя было подтвердить используемыми способами. Все QTL, протестированные к настоящему времени, можно подтвердить оценкой устойчивости к заболеванию в потомках BC5S1 или BC5S2 (возвратное скрещивание 5, однократное или двукратное самоопыление), сегрегирующих по исследуемым QTL.
В первом аспекте настоящее изобретение относится к способу получения устойчивого к Botrytis растения томата, при этом указанный способ включает стадии:
a) получения устойчивого к Botrytis донорного растения томата, предпочтительно устойчивого к Botrytis растения вида S. habrochaites, более предпочтительно устойчивого к Botrytis растения линии S. habrochaites LYC 4/78;
b) переноса нуклеиновой кислоты из указанного донорного растения в одно или несколько восприимчивых к Botrytis реципиентных растений томата, предпочтительно в растение вида S. lycopersicum, где указанный перенос приводит к внесению геномного материала из донорного растения в соответствующую область генома указанных одного или нескольких восприимчивых реципиентных растений;
c) селекции среди указанных реципиентных растений томата (или из дополнительно самоопыленного полученного растения, подвергнутого возвратному скрещиванию с указанным реципиентным растением томата; т.е. из рекомбинантных растений, полученных после указанного переноса) растения, которое содержит в его геноме по меньшей мере один QTL для устойчивости к Botrytis из указанного устойчивого к Botrytis донорного растения томата, где указанная селекция включает детекцию на 4, 6, 9, 11 и/или 12 хромосоме указанного реципиентного растения томата по меньшей мере одного генетического маркера, связанного с указанным по меньшей мере одним QTL для устойчивости к Botrytis;
- где на расположение указанного QTL на 4 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры CT50, C2At1g74970, P14M49-283e, P14M48-74e, P14M50-67e, CT1739 и P14M50-85h на 4 хромосоме S. habrochaites или на 4 хромосоме S. lycopersicum, более предпочтительно на 4 хромосоме S. habrochaites LYC 4/78 или на 4 хромосоме S. lycopersicum cv. Moneymaker.
В предпочтительных вариантах осуществления на расположение указанного QTL на 6 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P22M50-188h, P14M48-521e, P15M48-386h, P18M51-199h, P18M51-103h, P22M50-103e, P18M51-388e, Pl5M48-395e, P22M50-124e, P14M48-160e и P22M50-513h на 6 хромосоме S. habrochaites или на 6 хромосоме S. lycopersicum, более предпочтительно на 6 хромосоме S. habrochaites LYC 4/78 или на 6 хромосоме S. lycopersicum cv. Moneymaker.
В других предпочтительных вариантах осуществления на расположение указанного QTL на 9 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P18M50-141, P14M49-240, TG254, TG223, TG10, P18M50-134h, P14M49-243h, P18M50-599, P14M60-222h, P22M51-417h, P14M50-174h, P14M60-157h, P14M60-107h, P15M48-138h, P14M48-113h, Tm2a, P18M51-146h, Pl4M48-282h и P14M50-276h на 9 хромосоме S. habrochaites или на 9 хромосоме S. lycopersicum, более предпочтительно на 9 хромосоме S. habrochaites LYC 4/78 или на 9 хромосоме S. lycopersicum cv. Moneymaker.
В других предпочтительных вариантах осуществления на расположение указанного QTL на 11 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P14M60-215e, P14M61-173h, P14M50-307h, TG47, P14M50-29xCD, P18M51-358h, P18M50-27xCD, P18M51-136h, P22M50-488h, TG393, P14M61-396h, P22M51-235h и P22M51-174e на 11 хромосоме S. habrochaites или на 11 хромосоме S. lycopersicum, более предпочтительно на 11 хромосоме S. habrochaites LYC 4/78 или на 11 хромосоме S. lycopersicum cv. Moneymaker.
В других предпочтительных вариантах осуществления на расположение указанного QTL на 12 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры CT19, TG68, P14M48-411e, P18M50-244h, P18M50-273h, P14M61-420h, P14M61-406h, Pl4M61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296, P22M50-131h и P22M51-135h, предпочтительно геномная область, содержащая генетические маркеры P14M61-420h, P14M61-406h, P14M61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296 и P22M50-131h на 12 хромосоме S. habrochaites или на 12 хромосоме S. lycopersicum, более предпочтительно на 12 хромосоме S. habrochaites LYC 4/78 или на 12 хромосоме S. lycopersicum cv. Moneymaker.
Перенос нуклеиновой кислоты, содержащей по меньшей мере один QTL для устойчивости к Botrytis, или его придающую устойчивость к Botrytis часть, можно пригодным образом проводить скрещиванием указанного устойчивого к Botrytis донорного растения томата с восприимчивым к Botrytis реципиентным растением томата с получением растений-потомков.
Предпочтительный способ селекции, таким образом, включает селекцию на основе маркера (MAS) (см., например, Tanksley et al. 1998) указанной интрогрессивной ДНК, где в растениях-потомках проводят детекцию одного или нескольких маркеров, ассоциированных с указанным QTL. Например, MAS можно проводить выделением генетического материала из указанных растений-потомков и определением наличия в нем молекулярными технологиями одного или нескольких маркеров донорного растения. Альтернативно способы детекции молекулярных маркеров можно применять без предшествующего выделения генетического материала. Необязательно, в дополнение к детекции маркера в целях селекции пригодных растений можно проводить фенотипический тест устойчивости к Botrytis. Высокопригодным тестом, таким образом, является количественный биологический анализ, как описано в настоящем описании, при котором определяют такие параметры, как заболеваемость и/или скорость роста очагов повреждения. Подтверждение наличия по меньшей мере одного маркера из QTL для устойчивости к Botrytis в сочетании с установлением наличия устойчивого фенотипа обеспечивает доказательство успешного переноса нуклеиновой кислоты, содержащей по меньшей мере один QTL, или его устойчивой к Botrytis части из донорного растения в реципиентное растение.
В альтернативном варианте осуществления способа получения устойчивого к Botrytis растения томата указанный перенос нуклеиновой кислоты можно пригодным образом проводить трансгенными способами (например, трансформацией), слиянием протопластов, способами двойных гаплоидов или спасением зародышей.
В предпочтительном варианте осуществления способа получения устойчивого к Botrytis растения томата донорные растения представляют собой Solanum habrochaites LYC 4/78, и нуклеиновая кислота, перенесенная из этих донорных растений в реципиентные растения, предпочтительно содержит по меньшей мере один QTL для устойчивости к Botrytis, выбранный из группы, состоящей из QTL на 4, 6, 9, 11 и/или 12 хромосомах Solanum habrochaites LYC 4/78, ассоциированных с устойчивостью к Botrytis, или их придающей устойчивость к Botrytis части.
В другом предпочтительном варианте осуществления способа получения устойчивого к Botrytis растения томата способ включает скрещивание указанного устойчивого к Botrytis донорного растения томата с восприимчивым к Botrytis реципиентным растением томата с получением растений-потомков первого поколения; селекцию из растений-потоков первого поколения растения, которое содержит в его геноме нуклеиновую кислоту, подвергшуюся интрогрессии из указанного донорного растения томата, где указанная подвергшаяся интрогрессии нуклеиновая кислота содержит по меньшей мере один QTL, предпочтительно два, более предпочтительно более двух QTL для устойчивости к Botrytis в соответствии с этим изобретением, или их придающую устойчивость к Botrytis часть; скрещивание указанного выбранного растения-потомка с пригодной коммерческой линией томата с получением растений-потомков второго поколения; селекцию из растений-потомков второго поколения растения, которое содержит в его геноме нуклеиновую кислоту, подвергшуюся интрогрессии из указанного растения томата-потомка первого поколения, где указанная подвергшаяся интрогрессии нуклеиновая кислота содержит по меньшей мере один QTL, предпочтительно два, более предпочтительно более двух QTL для устойчивости к Botrytis в соответствии с этим изобретением, или их придающую устойчивость к Botrytis часть, и необязательно получение последующих поколений растений-потомков. Упомянутые два, более предпочтительно более двух QTL для устойчивости к Botrytis, которые подвергаются интрогрессии в растениях-потомках, могут представлять собой QTL для заболеваемости, QTL для скорости роста очагов повреждения или сочетание этих типов.
В наиболее предпочтительном варианте осуществления стадия c) включает селекцию растения, которое содержит в своем геноме по меньшей мере 4 QTL для устойчивости к Botrytis, выбранных из группы, состоящей QTL на 1, 2, 4, 6, 9, 11 и 12 хромосоме Solanum habrochaites, предпочтительно линии LYC 4/78, ассоциированных с устойчивостью к Botrytis.
В другом аспекте настоящее изобретение относится к устойчивому к Botrytis растению томата, или его части, получаемому способом по настоящему изобретению.
Кроме того, настоящее изобретение относится к QTL для устойчивости к Botrytis в томате, где указанный QTL выбран из группы, состоящей из QTL на 4, 6, 9, 11 и 12 хромосомах Solanum habrochaites, предпочтительно линии LYC 4/78, ассоциированных с устойчивостью к Botrytis. Эти QTL находятся в положениях генома, которые ранее не ассоциировали с устойчивостью к Botrytis. Характеристики этих QTL более подробно описаны в настоящем описании ниже.
Аллели, находящиеся в положениях генома, определяемых этими QTL, являются аспектом настоящего изобретения.
QTL по настоящему изобретению может находиться в форме выделенной, предпочтительно двухцепочечной последовательности нуклеиновой кислоты, содержащей указанный QTL или его придающую устойчивость часть. Является в значительной степени пригодным, чтобы размер последовательности нуклеиновой кислоты, которая, например, может быть выделена из хромосомы пригодного донорного растения, соответствовал генетическому расстоянию 1-100 сМ, предпочтительно, 10-50 сМ на указанной хромосоме. Указанная нуклеиновая кислота может содержать по меньшей мере 50, более предпочтительно по меньшей мере 500, более предпочтительно по меньшей мере 1000, более предпочтительно по меньшей мере 5000 пар оснований. Одна или несколько последовательностей нуклеиновых кислот, содержащих QTL или его придающую устойчивость часть в соответствии с этим изобретением, может, в свою очередь, находиться в конструкции нуклеиновой кислоты, при этом указанная конструкция может далее содержать участки, которые фланкируют указанную одну или несколько последовательностей нуклеиновых кислот, и эти участки могут встраиваться в пригодный вектор для переноса указанной одной или нескольких последовательностей нуклеиновых кислот в пригодное восприимчивое к Botrytis реципиентное растение томата. Кроме того, вектор может содержать пригодные промоторные области или другие регуляторные последовательности. Также QTL могут находиться в форме, представленной в геноме растения томата. QTL по настоящему изобретению предпочтительно содержат по меньшей мере один маркер, предпочтительно, два, более предпочтительно, три, более предпочтительно, четыре, более предпочтительно, более четырех маркеров, ассоциированных с устойчивостью к Botrytis, выбранных из группы, состоящей из маркеров, представленных в таблицах 1-5, которые сцеплены с указанным QTL.
В другом аспекте настоящее изобретение относится к способу детекции QTL для устойчивости к Botrytis, включающему детекцию по меньшей мере одного маркера, сцепленного с QTL для устойчивости к Botrytis на 4, 6, 9, 11 и/или 12 хромосоме предполагаемого устойчивого к Botrytis растения томата, где на расположение указанного QTL на 4 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры CT50, C2At1g74970, P14M49-283e, P14M48-74e, P14M50-67e, CT1739 и P14M50-85h на 4 хромосоме S. habrochaites или на 4 хромосоме S. lycopersicum, более предпочтительно, на 4 хромосоме S. habrochaites LYC 4/78 или на 4 хромосоме S. lycopersicum cv. Moneymaker.
В предпочтительном варианте осуществления способа детекции QTL по настоящему изобретению на расположение указанного QTL на 6 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P22M50-188h, P14M48-521e, P15M48-386h, P18M51-199h, P18M51-103h, P22M50-103e, P18M51-388e, P15M48-395e, P22M50-124e, P14M48-160e и P22M50-513h на 6 хромосоме S. habrochaites или на 6 хромосоме S. lycopersicum, более предпочтительно на 6 хромосоме S. habrochaites LYC 4/78 или на 6 хромосоме S. lycopersicum cv. Moneymaker.
В другом предпочтительном варианте осуществления способа детекции QTL по настоящему изобретению на расположение указанного QTL на 9 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P18M50-141, P14M49-240, TG254, TG223, TG10, P18M50-134h, P14M49-243h, P18M50-599, P14M60-222h, P22M51-417h, P14M50-174h, P14M60-157h, P14M60-107h, P15M48-138h, P14M48-113h, Tm2a, P18M51-146h, P14M48-282h и P14M50-276h на 9 хромосоме S. habrochaites или на 9 хромосоме S. lycopersicum, более предпочтительно, на 9 хромосоме S. habrochaites LYC 4/78 или на 9 хромосоме S. lycopersicum cv. Moneymaker.
В другом предпочтительном варианте осуществления способа детекции QTL по настоящему изобретению на расположение указанного QTL на 11 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P14M60-215e, P14M61-173h, P14M50-307h, TG47, P14M50-29xCD, P18M51-358h, P18M50-27xCD, P18M51-136h, P22M50-488h, TG393, P14M61-396h, P22M51-235h и P22M51-174e на 11 хромосоме S. habrochaites или на 11 хромосоме S. lycopersicum, более предпочтительно, на 11 хромосоме S. habrochaites LYC 4/78 или на 11 хромосоме S. lycopersicum cv. Moneymaker.
В другом предпочтительном варианте осуществления способа детекции QTL по настоящему изобретению на расположение указанного QTL на 12 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры CT19, TG68, P14M48-411e, P18M50-244h, P18M50-273h, P14M61-420h, P14M61-406h, P14M61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296, P22M50-131h и P22M51-135h, предпочтительно, геномная область, содержащая генетические маркеры P14M61-420h, P14M61-406h, P14M61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296 и P22M50-131h на 12 хромосоме S. habrochaites или на 12 хромосоме S. lycopersicum, более предпочтительно, на 12 хромосоме S. habrochaites LYC 4/78 или на 12 хромосоме S. lycopersicum cv. Moneymaker.
В другом аспекте настоящее изобретение относится к устойчивому к Botrytis растению вида S. lycopersicum, или его части, содержащему в своем геноме по меньшей мере один QTL, или его придающую устойчивость к Botrytis часть, где указанный QTL выбран из группы, состоящей из QTL на 4, 6, 9, 11 и 12 хромосоме Solanum habrochaites, предпочтительно, линии LYC 4/78, ассоциированных с устойчивостью к Botrytis, где на расположение указанного QTL на 4 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры CT50, C2At1g74970, P14M49-283e, P14M48-74e, P14M50-67e, CT1739 и P14M50-85h на 4 хромосоме S. habrochaites или на 4 хромосоме S. lycopersicum, более предпочтительно, на 4 хромосоме S. habrochaites LYC 4/78 или на 4 хромосоме S. lycopersicum cv. Moneymaker, где указанный QTL или его придающая устойчивость к Botrytis часть находятся не в их природном генетическом окружении и где указанное растение дополнительно необязательно содержит один или несколько дополнительных QTL, или их придающую устойчивость к Botrytis часть, ассоциированных с устойчивостью к Botrytis, выбранных из QTL на 1, 2 и/или 4 хромосоме Solanum habrochaites, предпочтительно, Solanum habrochaites линии LYC 4/78, где на расположение указанного дополнительного QTL на 4 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P18M51-169.5e, P18M51-305.4h, P14M60-262.9e, P14M61-292.7h, TG609, P14M48-345e, P14M48-177e и P18M50-147e на 4 хромосоме S. habrochaites или на 4 хромосоме S. lycopersicum, более предпочтительно, на 4 хромосоме S. habrochaites LYC 4/78 или на 4 хромосоме S. lycopersicum cv. Moneymaker.
В другом аспекте настоящее изобретение относится к способу получения устойчивого к Botrytis инбредного растения томата. Способ включает стадии получения устойчивого к Botrytis растения томата в соответствии со способом по этому изобретению, как описано выше, самоопыление указанного растения, выращивание полученного из указанного самоопыленного растения новых растений; идентификацию растений, которые проявляют устойчивость к Botrytis и обладают коммерчески желательными характеристиками в указанных новых растениях, и повторение стадий самоопыления и селекции с получением инбредного растения томата, которое проявляет устойчивость к Botrytis и обладает коммерчески желательными характеристиками.
Способ получения устойчивого к Botrytis инбредного растения томата, кроме того, может включать дополнительную стадию селекции гомозиготных инбредных растений томата, которые проявляют устойчивость к Botrytis и обладают коммерчески желательными характеристиками.
В следующем аспекте настоящее изобретение относится к устойчивому к Botrytis инбредному растению томата, или к его частям, получаемым способом по этому изобретению.
В следующем аспекте настоящее изобретение относится к гибридному растению томата, или к его частям, которое проявляет устойчивость к Botrytis, где указанное растение томата получают скрещиванием устойчивого к Botrytis инбредного растения томата, получаемого способом по этому изобретению, с инбредным растением томата, которое проявляет коммерчески желательные характеристики.
Кроме того, это изобретение относится к тканевой культуре регенерируемых клеток растений томата по настоящему изобретению. В предпочтительном варианте осуществления такой тканевой культуры клетки или протопласты указанных клеток выделены из ткани, выбранной из группы, состоящей из листьев, пыльцы, зародышей, корней, кончиков корней, пыльников, цветков, плодов и стеблей и семян.
Кроме того, это изобретение относится к применению маркера, выбранного из группы, состоящей из маркеров, приведенных в таблицах 1-5, для детекции QTL для устойчивости к Botrytis в соответствии с этим изобретением и/или для детекции устойчивых к Botrytis растений томата.
Устойчивое к Botrytis донорное растение томата, используемое в способах по настоящему изобретению, предпочтительно выбрано из группы, состоящей из Lycopersicon cerasiforme, Lycopersicon cheesmanii, Lycopersicon chilense, Lycopersicon chmielewskii, Lycopersicon Lycopersicum, Lycopersicon habrochaites, Lycopersicon parviflorum, Lycopersicon pennellii, Lycopersicon peruvianum, Lycopersicon pimpinellifolium и Solanum lycopersicoides, более предпочтительно, в качестве донорного растения используют изолят дикого томата. Высокопредпочтительными донорными растениями являются Solanum habrochaites, в частности Solanum habrochaites LYC 4/78.
Восприимчивое к Botrytis реципиентное растение томата, используемое в способах по настоящему изобретению, предпочтительно представляет собой растение вида Solanum lycopersicum, более предпочтительно культивар S. lycopersicum, который обладает коммерчески желательными характеристиками, или другую коммерческую линию томата.
Краткое описание чертежей
На фиг.1 представлено положение локусов количественных признаков (QTL) для устойчивости к B. cinerea, источником которых является S. habrochaites LYC 4/78, с картами сцепления, соответствующими 4 хромосоме (предполагаемое положение QTL представлено в качестве темных сегментов). Положения на карте приведены в cM. Эти данные дают возможность предположительной идентификации двух отдельных QTL, один из которых сосредоточен в области маркера TG62 (см. пример 1) и идентифицирован в IL 4-1 в примере 4, и другой, отдельный от них, не включающий маркер TG62, но сосредоточенный в области маркера P14M49-283e и идентифицированный в IL 4-3 в примере 4. QTL, выявленные на 4 хромосоме, снижают как скорость роста очага повреждения, так и заболеваемость. Коды для маркеров AFLP более подробно описаны в таблице 1. Все маркеры, указанные в качестве ассоциированных с QTL на фигурах, иллюстрирующих настоящее изобретение, можно как по отдельности, так и в сочетании использовать в качестве маркеров в его аспектах.
На фиг.2 представлено положение локуса количественного признака (QTL) для устойчивости к B. cinerea, источником которых является S. habrochaites LYC 4/78, с картами сцепления, соответствующими 6 хромосоме (предполагаемое положение QTL представлено в качестве темного сегмента). QTL на 6 хромосоме снижает как скорость роста очага повреждения, так и заболеваемость.
На фиг.3 представлено положение локуса количественного признака (QTL) для устойчивости к B. cinerea, источником которого является S. habrochaites LYC 4/78, с картами сцепления, соответствующими 9 хромосоме (предполагаемое положение QTL представлено в качестве темного сегмента). QTL на 9 хромосоме снижает как скорость роста очага повреждения, так и заболеваемость. Кроме того, на фигуре представлена интрогрессия на 9 хромосоме линий IL 11-2 и 12-3.
На фиг.4 представлено положение локуса количественного признака (QTL) для устойчивости к B. cinerea, источником которого является S. habrochaites LYC 4/78, с картами сцепления, соответствующими 11 хромосоме (предполагаемое положение QTL представлено в качестве темного сегмента). QTL на 11 хромосоме снижает как скорость роста очага повреждения, так и заболеваемость. Карта сцепления соответствует линии IL 11-2 примера 4.
На фиг.5 представлено положение локусов количественных признаков (QTL) для устойчивости к B. cinerea, источником которых является S. habrochaites LYC 4/78, с картами сцепления, соответствующими 12 хромосоме (предполагаемое положение QTL представлено в качестве темного сегмента). QTL на 12 хромосоме снижают как скорость роста очага повреждения, так и заболеваемость.
На фиг.6 представлена стратегия возвратного скрещивания и селекции, используемая для получения популяции IL S. habrochaites LYC 4/78, подвергнутой интрогрессии в генетическом фоне S. lycopersicum cv. Moneymaker, описанная в примере 4.
На фиг.7 графически представлен генотип популяции интрогрессивной линии S. lycopersicum cv. Moneymaker x S. habrochaites LYC 4/78, используемой в примере 4. Все хромосомы изображены в масштабе сегментов размером 20 сМ в соответствии с генетической картой сцепления F2. К концам 3, 4, 5 и 9 хромосом были добавлены некоторые участки (маркеры CAPS). Гомозиготные интрогрессии из S. habrochaites показаны черным цветом, в то время как гетерозиготные интрогрессии обозначены с использованием диагонального паттерна (серый).
На фиг.8 представлено положение локусов количественных признаков (QTL) для устойчивости к B. cinerea, источником которых является S. habrochaites LYC 4/78, с картами сцепления, соответствующими 1, 2 и 4 хромосомам, и указаны QTL, представленные в этой заявке как QTL-1h, QTL-2h и QTL-4hA.
Подробное описание изобретения
Определения
В рамках изобретения термин "Botrytis" означает Botrytiscinerea, также известный как серая гниль или серая пятнистость, заболевание, часто встречающееся в стебле, листьях и плодах томатов. Главным образом считается, что патогенный гриб растения Sclerotinia sclerotiorum обладает механизмом инфицирования, сходным с механизмом B. cinerea (Prins et al., 2000). Несмотря на то что инфекция S. sclerotiorum в томатах экономически значительно менее значима, чем инфекция B. cinerea, оба гриба секретируют спектр протеаз, ферменты деградации клеточной стенки, токсины, а также щавелевую кислоту. Известно, что некоторые из этих факторов участвуют в стратегии инфицирования обоих грибов. В результате считается, что механизмы и гены, которые придают устойчивость к Botrytis, являются равно эффективными в отношении обеспечения устойчивости к инфекции S. sclerotiorum. Таким образом, когда в настоящем описании упоминается "устойчивость к Botrytis", следует понимать, что такая устойчивость включает устойчивость к любому грибу семейства Sclerotiniaceae, предпочтительно устойчивость к S. sclerotiorum и B. cinerea, более предпочтительно устойчивость к B. cinerea.
В рамках изобретения термин "аллель(и)" означает любую одну или несколько альтернативных форм гена, при этом все из аллелей связаны по меньшей мере с одним признаком или характеристикой. В диплоидной клетке или организме два аллеля данного гена занимают соответствующие локусы на паре гомологичных хромосом. Поскольку настоящее изобретение относится к QTL, т.е. к геномным областям, которые могут содержать один или несколько генов, а также регуляторные последовательности, их в некоторых случаях более точно относить к "гаплотипу" (т.е. аллелю сегмента хромосомы) вместо "аллеля", однако следует понимать, что в этих случаях термин "аллель" включает термин "гаплотип".
"Ген" определяют в настоящем описании как единицу наследования, состоящую из последовательности ДНК, которая занимает конкретное положение на хромосоме и которая содержит генетическую инструкцию для конкретных характеристик или признака в организме.
"Локус" определяют в настоящем описании как положение, которое занимает данный ген на хромосоме данного вида.
В рамках изобретения термин "гетерозиготный" означает генетическое состояние, существующее, когда на соответствующих локусах гомологичных хромосом расположены различные аллели.
В рамках изобретения термин "гомозиготный" означает генетическое состояние, существующее, когда в соответствующих локусах на гомологичных хромосомах расположены идентичные аллели.
В рамках изобретения термин "гибрид" означает потомка скрещивания между двумя генетически отличающимися индивидами, включая, но не ограничиваясь ими, скрещивание между двумя инбредными линиями.
В рамках изобретения термин "инбредный" означает по существу гомозиготный индивид или линию.
В этой заявке "процесс рекомбинации" понимают как мейотический кроссинговер.
В рамках изобретения термины "интрогрессия", "подвергшийся интрогрессии" и "подвергающийся интрогрессии" относятся к как природному, так и к искусственному процессу, при котором гены одного вида, сорта или культивара перемещаются в геном другого вида, сорта или культивара посредством скрещивания этих видов. Процесс необязательно может быть осуществлен посредством возвратного скрещивания с рекуррентной родительской формой.
Термины "генетическая инженерия", "трансформация" и "генетическая модификация" используют в настоящем описании в качестве синонимов для переноса выделенных и клонированных генов в ДНК, как правило, в хромосомную ДНК или геном другого организма.
В рамках изобретения термин "молекулярный маркер" относится к индикатору, который используют в способах визуализации различий характеристик последовательностей нуклеиновых кислот. Примерами таких индикаторов являются маркеры полиморфизма длин рестрикционных фрагментов (RFLP), маркеры полиморфизма длин амплифицированных фрагментов (AFLP), однонуклеотидные полиморфизмы (SNP), микросателлитные маркеры (например, SSR), маркеры амплифицированного участка с охарактеризованной последовательностью (SCAR), маркеры расщепленной амплифицированной полиморфной последовательности (CAPS) или маркеры изозимов, или сочетания маркеров, описанных в настоящем описании, которые определяют конкретное генетическое и хромосомное положение.
Термины "устойчивый" и "устойчивость" охватывают как частичную, так и полную устойчивость к инфекции. Восприимчивое к Botrytis растение томата либо может быть неустойчивым, либо оно может иметь низкие уровни устойчивости к инфекции Botrytis.
В рамках изобретения термин "часть растения" указывает на часть растения томата, включая отдельные клетки и ткани из клеток, такие как клетки растений, которые являются интактными в растениях, клеточные массы и культуры тканей, из которых можно регенерировать растения томата. Примеры частей растений включают, но не ограничиваются ими, отдельные клетки и ткани из пыльцы, семяпочки, листьев, зародышей, корней, концов корней, пыльников, цветков, плодов, побегов стеблей и семян; а также пыльцу, семяпочку, листья, зародыши, корни, концы корней, пыльники, цветки, плоды, стебли, побеги, прививочные черенки, корневище, семена, протопласты, каллюсы и т.п.
В рамках изобретения термин "популяция" означает генетически гетерогенную коллекцию растений, обладающих общим генетическим происхождением.
В рамках изобретения термин "томат" означает любое растение, линию или популяцию, ранее известную под родовым названием Lycopersicon, включая, но не ограничиваясь ими, Lycopersicon cerasiforme, Lycopersicon cheesmanii, Lycopersicon chilense, Lycopersicon chmielewskii, Lycopersicon esculentum (в настоящее время Solanum lycopersicum), Lycopersicon hirsutum, Lycopersicon parviflorum, Lycopersicon pennellii, Lycopersicon peruvianum, Lycopersicon pimpinellifolium или Solanum lycopersicoides. Новым предложенным научным названием для Lycopersicon esculentum является Solanum lycopersicum. Аналогично могут измениться названия диких видов. L. pennellii стал Solanum pennellii, L. hirsutum может стать S. habrochaites, L. peruvianum может быть разделен на S. "N peruvianum" и S. "Callejon de Huayles", S. peruvianum и S. corneliomuelleri, L. parviflorum может стать S.neorickii, L. chmielewskii может стать S. chmielewskii, L. chilense может стать S. chilense, L. cheesmaniae может стать S. cheesmaniae или S. Galapagense и L. pimpinellifolium может стать S. pimpinellifolium (Solanacea Genome Network (2005) Spooner and Knapp; http://www.sgn.cornell.edu/help/about/solanum_nomenclature.html).
В особенности, отмечается, что S. habrochaites можно определить как вид томата, который обладает волосистыми плодами, в то время как S. lycopersicum представляет собой вид томата, имеющий плоды без волосков.
В рамках изобретения термин "сорт" или "культивар" означает группу сходных растений, которые по структурным или генетическим признакам и/или внешнему виду можно отличить от других сортов того же вида.
Термин "QTL" используют в настоящем описании в его известном в данной области значении. Термин "QTL, ассоциированный с устойчивостью к B. cinerea в томате", а также более короткий термин "QTL для устойчивости к Botrytis" относятся к области, расположенной на конкретной хромосоме томата, которая ассоциирована по меньшей мере с одним геном, который кодирует устойчивость к Botrytis, или по меньшей мере с регуляторной областью, т.е. с областью хромосомы, которая контролирует экспрессию одного или нескольких генов, вовлеченных в устойчивость к Botrytis. Фенотипическую экспрессию этого гена, например, можно наблюдать в качестве сниженной скорости роста очага повреждения и/или в качестве сниженной заболеваемости. Например, QTL может содержать один или несколько генов, продукты которых придают генетическую устойчивость. Альтернативно QTL может содержать, например, регуляторные гены или последовательности, продукты которых влияют на экспрессию генов на других локусах в геноме растения, придавая посредством этого устойчивость к Botrytis. QTL по настоящему изобретению могут быть определены указанием их генетического положения в геноме соответствующего изолята томата дикого типа с использованием одного или нескольких молекулярных геномных маркеров. Один или несколько маркеров, в свою очередь, указывают на конкретный локус. Расстояния между локусами, как правило, определяют по частоте кроссинговера между локусами на одной хромосоме. Чем дальше друг от друга расположены локусы, тем более вероятно, что между ними произойдет кроссинговер. Напротив, если два локуса расположены близко друг к другу, вероятность того, что между ними произойдет кроссинговер, является меньшей. Как правило, один сантиморган (сМ) равен 1% рекомбинации между локусами (маркерами). Когда QTL может быть указан посредством множества маркеров, генетическое расстояние между концевыми маркерами является показателем размера QTL.
Термин "восприимчивое к Botrytis реципиентное растение томата" используют в настоящем описании для указания на растение томата, которое подлежит получению ДНК из донорного растения томата, которая содержит QTL для устойчивости к Botrytis. Указанное "восприимчивое к Botrytis реципиентное растение томата" может уже содержать один или несколько QTL для устойчивости к Botrytis или может не содержать их, и в этом случае термин указывает на растение, которое подлежит получению дополнительного QTL.
Термин "природный генетический фон" используют в настоящем описании для указания на исходный генетический фон для QTL. Такой фон может представлять собой, например, геном дикого изолята томата с устойчивостью к Botrytis. Например, QTL по настоящему изобретению были выявлены в конкретных областях на 4, 6, 9, 11 и 12 хромосоме Solanum habrochaites LYC 4/78. В качестве примера Solanum habrochaites LYC 4/78 соответствует природному генетическому фону QTL на 4, 6, 9, 11 и 12 хромосомах Solanum habrochaites LYC 4/78. Также Solanum habrochaites LYC 4/78 соответствует природному генетическому фону указанных QTL. Напротив, способ, который вовлекает перенос ДНК, содержащей QTL, или его придающую устойчивость часть, из 4 хромосомы Solanum habrochaites LYC 4/78 в то же положение на 4 хромосоме другого вида томата, особенно S. lycopersicum, приведет к QTL, или указанной его придающей устойчивость части, находящемуся не в его природном генетическом фоне.
Термин "заболеваемость" определяют в настоящем описании как параметр, который отражает способность растения снижать развитие инфекции, и она может быть установлена, например, определением успеха достижения инфицирования растения при контакте с инфекционным агентом.
Термин "скорость роста очага повреждения" или "скорость роста для очага повреждения" определяют в настоящем описании как параметр, который отражает способность растения замедлять или снижать прогрессирование инфекции, и она может быть установлена, например, определением скорости роста растущих очагов повреждения.
Термин "количественное определение" определяют в настоящем описании как установление или оценку способом, вовлекающим измерение, в частности, измерение аспектов, поддающихся измерению с точки зрения количеств и числа. Определения в степенях тяжести и указания на превышающий, больший, меньший или равный или на повышение или снижение величины не относятся к термину "количественное определение", который в конечном итоге подразумевает наличие объективного механизма подсчета для определения абсолютных величин. Таким образом, "количественное определение заболеваемости и/или скорости роста очага повреждения" предпочтительно включает определение процентного количества всех потенциально инфекционных контактов между растением и инфекционным агентом, которые приводят к поддающимся измерению очагам повреждения (в целях оценки заболеваемости), и/или определение увеличения диаметра, окружности, площади поверхности или объема одного или нескольких из указанных очагов повреждения с течением времени в благоприятных условиях для роста грибов (в целях оценки скорости роста очага повреждения).
Термины "стандартные условия практики", "стандартные условия теплицы" и "стандартные условия" относятся к условиям света, влажности, температуры и т.д., в которых растения выращивают или инкубируют, например, для целей фенотипической охарактеризации устойчивости к заболеванию, в качестве стандартных. Например, для теплиц они относятся к 16-часовой длительности дня, 15°C-25°C. В более общем смысле термины относятся к стандартным и контрольным условиям выращивания со световым периодом от 8 до 24 ч (фотосинтетический поток фотонов (PPF) от 50 до 1000 мкмоль м-2 с-1), предпочтительно со световым режимом 16 часов на свету и 8 часов в темноте, с температурой воздуха приблизительно 19°C днем и 15°C ночью, с дефицитом давления водяного пара приблизительно 4,4 г м-3, соответствующим относительной влажности (RH) приблизительно 60%-85%, при 600-700 м.д. CO2 и атмосферной концентрации O2 и при атмосферном давлении воздуха (как правило, 1008 гПа). Воду и питательные вещества можно давать капельно вблизи стебля или в форме аэрозоля или орошения. Стандартные экспериментальные условия биологического анализа, такого как анализ длины повреждения стебля, определение заболеваемости и скорости роста очага повреждения, далее указаны в разделе "Примеры" ниже. Более подробно анализ средней длины повреждения стебля проводят, как описано в примерах 3.10 и 3.11.
Идентификация QTL, ассоциированных с устойчивостью к Botrytis в томате
Известно, что дикие виды томата обеспечивают пригодные источники признаков устойчивости к заболеванию и паразитам, и наличие частичной устойчивости к B. cinerea в листьях дикого томата было документировано (Urbasch, 1986). Два фактора препятствовали выведению устойчивости к B. cinerea в томатах в прошлом. Во-первых, скрещивание для частичной устойчивости в коммерческих выращиваемых линиях имело ограниченный успех. Во-вторых, отсутствовали надежные и воспроизводимые способы анализа заболевания, которые могли обеспечить идентификацию и определение локализации генетического материала, ответственного за придание устойчивости.
Например, Urbasch (Urbasch, 1986) инфицировал листья мицелием с использованием пробки из агара, предоставляющей гриб с избытком пищевых добавок, которые строго влияют на процесс инъекции. Другие исследователи использовали субъективные признаки заболевания растений, которые не пригодны для количественного анализа, требуемого для идентификации локусов количественных признаков (QTL).
Инфекция Botrytis cinerea в Solanum lycopersicum в лабораторных условиях относительно хорошо изучена (например, Benito et al., 1998). Капельная инокуляция листьев и последующая инкубация при умеренных температурах (15-20°С) приводит к быстрому (через 16-24 часов после инфекции (hpi)) развитию некротических пятен в участке инокуляции. Инфекция ограничивается по времени в этой области в течение приблизительно 48 ч. Начиная с этого момента и далее доля очагов повреждения (как правило, 5-10%) начинает возрастать. Разрастание этих так называемых "растущих очагов повреждения" сопровождается повышением биомассы гриба и приводит к колонизации всего листа в последующие 48 ч.
Авторы настоящего изобретения ранее выявили, что конкретные QTL, ассоциированные с устойчивостью к Botrytis в томате, можно идентифицировать с использованием биологического анализа для определения устойчивости, где количественно измеряют скорость прогрессирования инфекции и/или успех достижения инфицирования при контакте с инфекционным агентом на частях растения томата, предпочтительно на отделенных частях, более предпочтительно на сегментах стебля.
Кроме того, можно использовать тест, как описано в примере 4, где исследуют устойчивость целых растений и где стебель механически повреждали и на рану наносили инокулят Botrytis.
С использованием интрогрессивных линий, каждая из которых содержит определенный интрогрессивный сегмент генома S. habrochaites LYC 4/78 в фоне S. lycopersicum cv. Moneymaker, авторы изобретения неожиданно обнаружили дополнительные QTL, ассоциированные с устойчивостью к Botrytis в S. habrochaites LYC 4/78. Таким образом, настоящее изобретение далее относится к способу получения культивируемого растения томата, которое является устойчивым к Botrytis, включающему:
- получение серии интрогрессивных линий культивируемого томата, где каждая интрогрессивная линия обладает конкретной геномной интрогрессией (хромосомной областью) из устойчивого к Botrytis дикого изолята томата, предпочтительно S. habrochaites LYC 4/78, и при этом серии совместно охватывают геном указанного устойчивого изолята томата дикого типа;
- идентификацию QTL, ассоциированных с устойчивостью к Botrytis в растениях каждой из указанных интрогрессивных линий посредством проведения количественного биологического анализа для определения устойчивости к Botrytis в растениях каждой из указанных отдельных интрогрессивных линий, предпочтительно посредством определения заболеваемости и/или скорости роста очагов повреждения, и установление генетической карты сцепления, которая связывает наблюдаемую устойчивость к Botrytis с наличием хромосомных маркеров в указанных интрогрессивных линиях и отнесение сопряженных маркеров на указанной карте, которые сцеплены с повышенной устойчивостью (например, со сниженной заболеваемостью и/или сниженной скоростью роста очага повреждения) к локусу количественного признака;
- и применение интрогрессивной линии, которая содержит QTL, ассоциированный с устойчивостью к Botrytis, для получения культивируемого растения томата, которое является устойчивым к Botrytis, или применение информации маркеров для QTL, идентифицированных таким образом, в селекции на основе маркера предполагаемых устойчивых к Botrytis растений томата.
Было неожиданно выявлено, что в геномах устойчивых к Botrytis растениях томата представлено множество QTL для устойчивости к Botrytis, в то время как способы предшествующего уровня техники привели к ориентировочной идентификации QTL на 1, 2, 4 и 10 хромосомах. Более того, QTL, которые были выявлены с использованием настоящих способов, были расположены на хромосомах, которые ранее не ассоциировали с устойчивостью растений томата к Botrytis даже с использованием сходного генетического фона (WO 02/085105). Таким образом, способы по настоящему изобретению обеспечили дополнительное понимание того, что применение IL может привести к более детальному изучению генетического происхождения устойчивости к Botrytis в томатах и может привести к идентификации хромосомного материала, вовлеченного в такую устойчивость, ранее не идентифицированного в них.
Способ детекции локуса количественного признака (QTL), ассоциированного с устойчивостью к Botrytis в томатах в соответствии с настоящим изобретением, иначе называемый способом идентификации или определения локализации локуса количественного признака (QTL), требует доступности (частично) устойчивого к Botrytis растения томата. Такое растение можно получить способами, известными в данной области, и с использованием любого способа определения наличия указанной (частичной) устойчивости в указанном растении. Получение (частично) устойчивого к Botrytis растения томата (которое далее будет служить в качестве донорного растения в способе по настоящему изобретению) обеспечивает установление или предоставление хромосомных маркеров, предпочтительно маркеров AFLP, CAPS и/или SCAR, наиболее предпочтительно маркеров CAPS и/или SCAR, по меньшей мере для одной, но предпочтительно для всех хромосом указанного растения. Посредством установления совокупности хромосомных маркеров на протяжении всей длины указанных хромосом различные участки указанных хромосом могут быть эффективно маркированы. Такие способы хорошо известны в данной области, и иллюстративные способы будут более подробно описаны в настоящем описании ниже.
Способ детекции локуса количественного признака (QTL), ассоциированного с устойчивостью к Botrytis в томатах в соответствии с настоящим изобретением, может включать стадию определения устойчивости к Botrytis в растении. Для этого растение контактируют с инфекционным количеством Botrytis. Такое количество может варьировать между растениями и между тестируемыми видами грибов. Как правило, достаточным является количество приблизительно от 1-10 до количества приблизительно 500-5000 конидий указанного гриба.
Определение устойчивости к Botrytis может включать количественное определение заболеваемости и/или скорости роста очага повреждения в растении.
Стадию контактирования растения с инфекционным количеством Botrytis и количественного определения устойчивости предпочтительно проводят в качестве части биологического анализа устойчивости на сегментах стебля или листьях, как описано в настоящем описании, предпочтительно, биологического анализа устойчивости на сегментах стебля, более предпочтительно, на целых растениях, как описано в примере 4. Квалифицированный специалист поймет, что возможны варианты этих анализов, как описано в настоящем описании ниже.
Биологический анализ устойчивости на сегментах стебля главным образом можно проводить следующим образом: сначала семена растений-потомков высевают и выращивают до проростков/растений ростом соответственно приблизительно 50 см. Верхние 5-10 см и нижние 5-10 см стебля растений можно удалять и оставшиеся 30 см можно нарезать на равные сегменты по 5-6 см. Сегменты стебля предпочтительно помещают вертикально на сетке с основанием стебля на влажной фильтровальной бумаге. Перед инокуляцией сегменты стебля пригодным образом опрыскивают водой в целях обеспечения равномерного распределения инокулята на поверхности раны. Затем на каждый сегмент стебля можно инокулировать суспензию конидий B. cinerea. Пригодное количество инокулята, например, одну каплю, приблизительно 5 мкл, содержащую приблизительно 106·конидий·мл-1, можно наносить к тому же на верхушку каждого сегмента. Затем сегменты стебля инкубируют при пригодной температуре, составляющей приблизительно 16°С, предпочтительно в темноте и предпочтительно при высокой влажности (например, 100% RH). Прогрессирование инфицирования можно определять количественно измерением максимального развития симптома гниения через различные промежутки времени после инокуляции с помощью штангенциркуля с нониусом. Затем через ряд подходящих промежутков времени, например, через 96, 120 и 144 часов после инфицирования (hpi), стебли исследуют в отношении образования очагов повреждения (заболеваемости) и роста очага повреждения количественным образом. Наиболее пригодные параметры включают определение размера очага повреждения, например, с использованием штангенциркуля. В целях коррекции отклонений, являющихся следствием времени года или культивирования растений, количественные показатели биологических анализов можно сопоставлять со сравнимыми показателями в восприимчивых контрольных или референсных линиях. Заболеваемость можно пригодным образом определять делением общего количества растущих очагов на общее количество инокуляционных капель. Затем долю растущих очагов повреждения на конкретном генотипе можно делить на долю растущих очагов повреждения, выявленных в контрольном или референсном генотипе, и выражать в процентах. Альтернативно или дополнительно можно определять скорости роста очагов посредством вычисления увеличения размера очага повреждения (например, в мм) в течение пригодного периода, например в течение 24 ч. Данные для нерастущих очагов повреждения можно удалять из количественного анализа. Затем полученную скорость роста очага повреждения можно необязательно делить на скорость роста очагов повреждения, выявленную в контрольном или референсном генотипе, и выражать в процентах или в качестве абсолютного значения, например в миллиметрах.
Альтернативно растения можно подвергать скринингу с использованием анализа инфицирования листьев следующим образом: сначала семена высевают и выращивают до проростков/растений. Для каждого отдельного растения один или два сложных листа можно отрезать от основного стебля и перенести в предварительно увлажненную пену для цветов. Затем пену для цветов помещают в чашку Петри, содержащую водопроводную воду, а затем помещают в увлажненный распылением контейнер, содержащий влажную фильтровальную бумагу. Пригодный инокулят, содержащий конидии B. cinerea, можно получать способами, известными в данной области, например, как описано Benito et al., 1998. Затем на сложные листья инокулируют суспензией конидий B. cinerea нанесением некоторого количества капель, например, пригодным является количество от 6 до 10 капель по 2 мкл каждая, на верхнюю поверхность листьев. Затем контейнер закрывают и листья инкубируют при температуре, являющейся пригодной между 15°C-20°C, предпочтительно в темноте и предпочтительно при высокой влажности. Затем через ряд пригодных промежутков времени, например, через 96, 120 и 144 hpi, листья можно исследовать в отношении заболеваемости и роста очага повреждения количественным образом, как описано выше для биологического анализа стебля.
Способ детекции локуса количественного признака (QTL), ассоциированного с устойчивостью к Botrytis в томате в соответствии с настоящим изобретением, включает стадии установления генетической карты сцепления, которая связывает наблюдаемую устойчивость с наличием хромосомных маркеров донорного растения томата в реципиентных растениях IL, и отнесения соседних маркеров на указанной карте, которые сцеплены с усиленной устойчивостью, к локусу количественного признака.
Генетическая карта сцепления, которая связывает наблюдаемую повышенную устойчивость с наличием хромосомных маркеров донорного растения томата в указанных растениях IL, может быть установлена любым способом, известным в данной области. Квалифицированному специалисту известны способы идентификации молекулярных маркеров, сцепленных с локусами количественных признаков (QTL) для устойчивости, и картирования этих маркеров на генетической карте сцепления (см., например, Bai et al., 2003; Foolad et al., 2002; van Heusden et al., 1999). Выявление ассоциации между устойчивым к Botrytis фенотипом и генотипом маркеров можно пригодным образом проводить с использованием таких пакетов программ, как JoinMap® и MapQTL® (см. примеры), или любого стандартного статистического пакета, который может проводить дисперсионный анализ. Молекулярные маркеры можно использовать для конструирования генетических карт сцепления и для идентификации локусов количественных признаков (QTL) для устойчивости к Botrytis. Пригодные типы молекулярных маркеров и способы их получения описаны более подробно в настоящем описании ниже.
Молекулярные маркеры и QTL
Молекулярные маркеры используют для визуализации отличий в последовательностях нуклеиновых кислот. Эта визуализация является возможной благодаря способам гибридизации ДНК-ДНК (RFLP) и/или благодаря способам с использованием полимеразной цепной реакции (например, STS, анализ микросателлитов, AFLP). Все отличия между двумя родительскими генотипами будут сегрегировать в подвергаемой картированию популяции (например, BC1, F2; см. фиг.2) на основе скрещивания этих родительских генотипов. Сегрегацию различных маркеров можно сравнивать и можно вычислять частоты рекомбинации. Частоты рекомбинации молекулярных маркеров на различных хромосомах, как правило, составляют 50%. Между молекулярными маркерами, расположенными на одной хромосоме, частота рекомбинации зависит от расстояния между маркерами. Низкая частота рекомбинации соответствует небольшому расстоянию между маркерами на хромосоме. Сравнение всех частот рекомбинации приведет к наиболее логичному порядку молекулярных маркеров на хромосомах. Этот наиболее логичный порядок можно изображать на карте сцепления (Paterson, 1996). Группа соседних или смежных маркеров на карте сцепления, которая ассоциирована со сниженной заболеваемостью и/или со сниженной скоростью роста очага, указывает на положение QTL.
При идентификации QTL эффект QTL (устойчивость) можно подтверждать, например, посредством оценки устойчивости к Botrytis в потомках BC2S1, сегрегирующих по исследуемым QTL. Оценку устойчивости к Botrytis можно пригодным образом проводить с использованием биологического анализа стебля или листьев, как описано в настоящем описании.
QTL для устойчивости к Botrytis в томатах, получаемых с использованием способа по этому изобретению, являются аспектом настоящего изобретения. Характеристикой таких QTL является то, что, когда они присутствуют в растениях, они являются показателем наличия сниженной заболеваемости и/или сниженной скорости роста очага повреждения при контакте указанного растения с инфекционным количеством материала Botrytis, при этом материал может находиться в любой форме, например, в форме конидий или мицелия.
Настоящее изобретение также относится к QTL для устойчивости к Botrytis в томатах, где указанный QTL выбран из группы, состоящей из QTL на 4, 6, 9, 11 и 12 хромосомах Solanum habrochaites LYC 4/78, ассоциированных с устойчивостью к Botrytis. Эти QTL могут быть более четко определены или показаны посредством маркеров, приведенных в таблицах 1-5. В этих таблицах геномная область, где расположены QTL, указана посредством приведенных маркеров AFLP. QTL по настоящему изобретению содержат генетическую информацию в форме ДНК, отвечающей за обеспечение (частичное) заболеваемости Botrytis и/или сниженной скорости роста очагов повреждения Botrytis в растении томата. Генетическая информация может содержать, например, ген или регуляторный элемент.
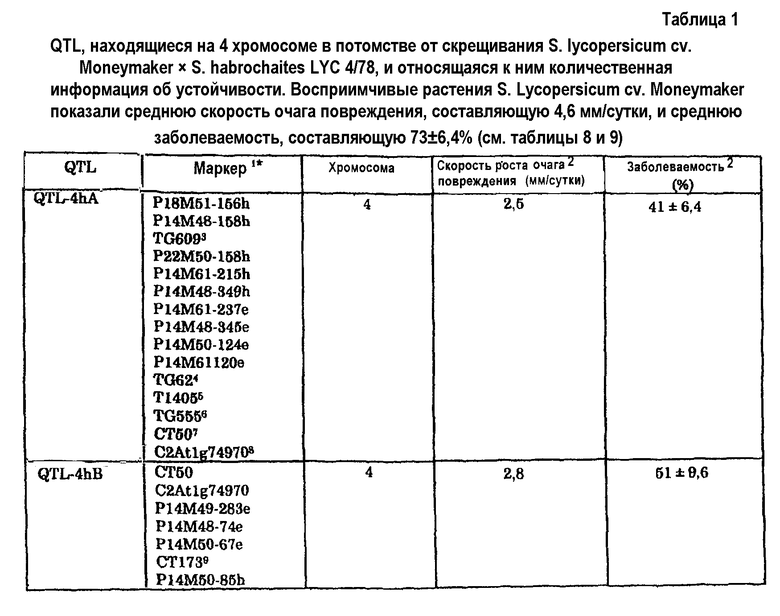
1номенклатура маркеров: коды, посредством которых обычно показывают сочетание праймеров AFLP, например P18M51-156h, где P и M представляют собой общие последовательности праймеров PstI и MseI или универсальных праймеров (Vos et al., 1995; Bai et al., 2003), за которыми следуют 2 или 3 дополнительных селективных оснований, как показывают кодами удлинения из двух цифр. Коды удлинения из двух цифр являются такими, как следующие коды: 14: AT; 15: CA; 18: CT; 22: GT; 48: CAC; 49: CAG; 50: CAT; 51: CCA; 60: CTC; 61: CTG. 156h представляет собой приблизительный размер в количестве пар оснований полученного полиморфного фрагмента (приведенный размер ± 2 пары оснований). Размер обычно округляют, однако также он может быть приведен в виде десятичной дроби. Этот фрагмент амплифицирован либо в S. lycopersicum cv Moneymaker (e), либо в S. habrochaites LYC 4/78 (h). Наличие маркера указывает на то, что имеется по меньшей мере один аллель из указанного источника. Праймеры и соединительные последовательности подробно описаны Bai et al. 2003. 2Заболеваемость и рост очага повреждения определяют с использованием способов, подробно разъясненных в разделе "Примеры". 3TG609: см. таблицу 10. 4TG62: см. таблицу 11. 5T1405: см. таблицу 20. 6TG555: см. таблицу 12. 7CT50: см. таблицу 13. 8C2_At1g74970: см. таблицу 14. 9CT173: см. таблицу 21.

1TG254: см. таблицу 22. 2TG223: см. таблицу 23. 3TG10: см. таблицу 17. 4Tm2a: см. таблицу 18 или Sorbir, O.T. et al. (2000).

1TG47: см. таблицу 24. 2TG393: см. таблицу 25.
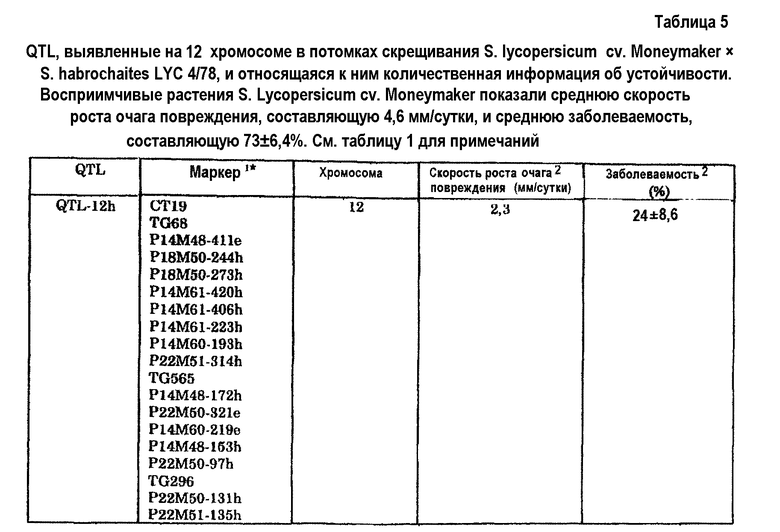
1CT19: см. таблицу 26. 2TG68: см. таблицу 27. TG565: см. таблицу 28. TG296: см. таблицу 29.
Наиболее вероятно, геномная область, где расположен QTL-4hA, находится между маркерами P18M51-156h и C2_At1g74970 (таблица 14), как показано на фиг.1. Таким образом, для оценки наличия QTL в геноме растения можно использовать любой маркер, расположенный в этой области, а также любой маркер, о котором известно, что он расположен в этом участке, исходя из общедоступной информации, например, из консенсусных карт Tomato-EXPEN 1992 (Tanksley et al., 1992), Tomato-EXHIR 1997 (Bernacchi и Tanksley, 1997), Tomato-EXPEN 2000 (Fulton et al., 2002) или Tomato-EXPIMP 2001 (Grandillo и Tanksley, 1996; Tanksley et al. 1996, Doganlar et al. 2002).
Наиболее вероятно, геномная область, где расположен QTL-4hB, расположена между маркерами CT50 (таблица 13) и P14M50-85h, как показано на фиг.1. Таким образом, для оценки наличия QTL в геноме растения можно использовать любой маркер, расположенный в этой области, а также любой маркер, о котором известно, что он расположен в этом участке, исходя из общедоступной информации.
Наиболее вероятно, геномная область, где расположен QTL-6h, расположена между маркерами P22M50-188h и P22M50-513h, как показано на фиг.2. Таким образом, для оценки наличия QTL в геноме растения можно использовать любой маркер, расположенный в этой области, а также любой маркер, о котором известно, что он расположен в этом участке, исходя из общедоступной информации.
Наиболее вероятно, геномная область, где расположен QTL-9h, расположена между маркерами P18M50-141 и P14M50-276h, как показано на фиг.3. Таким образом, для оценки наличия QTL в геноме растения можно использовать любой маркер, расположенный в этой области, а также любой маркер, о котором известно, что он расположен в этом участке, исходя из общедоступной информации.
Наиболее вероятно, геномная область, где расположен QTL-11h, расположена между маркерами P14M60-215e и P22M51-174e, как показано на фиг.4. Таким образом, для оценки наличия QTL в геноме растения можно использовать любой маркер, расположенный в этой области, а также любой маркер, о котором известно, что он расположен в этом участке, исходя из общедоступной информации.
Наиболее вероятно, геномная область, где расположен QTL-12h, расположена между маркерами P14M61-420h и P22M50-131h, как показано на фиг.5. Таким образом, для оценки наличия QTL в геноме растения можно использовать любой маркер, расположенный в этой области, а также любой маркер, о котором известно, что он расположен в этом участке, исходя из общедоступной информации.
Предпочтительно QTL по настоящему изобретению содержит по меньшей мере один маркер, приведенный в таблицах 1-5, ассоциированный с указанным QTL. Вследствие того, что последовательность нуклеиновой кислоты QTL, которая отвечает за придание устойчивости к Botrytis, может представлять собой только часть целого указанного в настоящем описании QTL, маркеры указывают только на сцепленное наследование генетических областей или на отсутствие наблюдаемой рекомбинации в таких генетических областях. Таким образом, следует отметить, что маркеры, приведенные в таблицах 1-5, указывают на хромосомную область, где QTL по этому изобретению расположен в геноме указанных линий Solanum, и что эти маркеры не обязательно определяют границы или структуру этого QTL. Таким образом, часть QTL, которая содержит основную придающую устойчивость последовательность(и) нуклеиновой кислоты, может быть значительно меньшей, чем указано посредством соседних маркеров, приведенных для конкретного QTL. Такую часть называют в настоящем описании "придающей устойчивость частью" QTL. В результате придающая устойчивость часть QTL не обязательно должна содержать любой из указанных приведенных маркеров. Также для указания на различные QTL можно использовать другие маркеры при условии, что такие маркеры являются генетически сцепленными с QTL, и квалифицированный специалист может выявить или использовать QTL, которые являются аналогичными QTL по настоящему изобретению, но где один или несколько маркеров, приведенных в таблицах 1-5 и указанных в качестве сцепленных с указанным QTL, отсутствуют.
Придающую устойчивость к Botrytis часть QTL для устойчивости против Botrytis в томате можно идентифицировать с использованием способа молекулярных маркеров, например, с одним или несколькими из маркеров для QTL, представленных в таблицах 1-5, в качестве связанных с указанным QTL, предпочтительно в сочетании с биологическим анализом устойчивости. Растения томата, которые не содержат придающую устойчивость к Botrytis часть QTL по настоящему изобретению, являются относительно восприимчивыми к инфицированию Botrytis.
Маркеры, обеспечиваемые настоящим изобретением, можно использовать для детекции наличия одного или нескольких QTL по этому изобретению в предполагаемом устойчивом к Botrytis растении томата, и, таким образом, их можно использовать в способах, вовлекающих разведение на основе маркеров и селекцию устойчивых к Botrytis растений томата. Предпочтительно детекцию наличия QTL по этому изобретению проводят с помощью по меньшей мере одного из маркеров для QTL, показанных в таблицах 1-5, в качестве сцепленных с указанным QTL. Настоящее изобретение, таким образом, относится в другом аспекте к способу детекции наличия QTL для устойчивости к Botrytis, включающему детекцию наличия последовательности нуклеиновой кислоты указанного QTL в предполагаемом устойчивом к Botrytis растении томата, при этом наличие можно выявлять посредством применения указанных маркеров.
Последовательность нуклеиновой кислоты QTL по настоящему изобретению можно определять способами, известными специалисту. Например, последовательность нуклеиновой кислоты, содержащую указанный QTL или его придающую устойчивость часть, можно выделять из устойчивого к Botrytis донорного растения посредством фрагментации генома указанного растения и селекции тех фрагментов, которые обладают одним или несколькими маркерами, указывающими на конкретный QTL. Впоследствии, или альтернативно, маркерные последовательности (или их части), указывающие на конкретный QTL, можно использовать в качестве праймеров для амплификации (ПЦР), в целях амплификации последовательности нуклеиновой кислоты, содержащей указанный QTL из образца геномной нуклеиновой кислоты или фрагмента генома, полученного из указанного растения. Затем амплифицированную последовательность можно очищать в целях получения выделенного QTL. Затем нуклеотидную последовательность QTL, и/или любых дополнительных маркеров, содержащихся в ней, можно получать стандартными способами секвенирования.
Таким образом, настоящее изобретение также относится к выделенной последовательности нуклеиновой кислоты (предпочтительно ДНК), которая содержит QTL по настоящему изобретению, или его придающую устойчивость к Botrytis часть. Таким образом, маркеры, которые указывают на различные QTL, описанные в настоящем описании, можно использовать для идентификации, выделения и очистки одного или нескольких генов из томата, которые кодируют устойчивость к Botrytis.
Нуклеотидную последовательность QTL по настоящему изобретению, например, также можно анализировать посредством определения нуклеотидной последовательности одного или нескольких маркеров, ассоциированных с указанным QTL, и разработки внутренних праймеров для указанных маркерных последовательностей, которые впоследствии можно использовать для определения последовательности QTL, снаружи указанных маркерных последовательностей. Например, нуклеотидную последовательность маркеров AFLP из таблиц 1-5 можно получать выделением указанных маркеров из электрофоретического геля, используемого для определения наличия указанных маркеров в геноме представляющего интерес растения, и определением нуклеотидной последовательности указанных маркеров, например, посредством способов с дидезокситерминацией цепи, хорошо известных в данной области.
В вариантах осуществления таких способов детекции наличия QTL в предполагаемом устойчивом к Botrytis растении томата способ также может включать стадии получения олигонуклеотида или полинуклеотида, способного гибридизоваться в строгих условиях гибридизации с последовательностью нуклеиновой кислоты маркера, сцепленного с указанным QTL, предпочтительно выбранного из маркеров, приведенных в таблицах 1-5 в качестве сцепленных с указанным QTL, контактирования указанного олигонуклеотида или полинуклеотида с геномной нуклеиновой кислотой предполагаемого устойчивого к Botrytis растения томата и определения наличия специфичной гибридизации указанного олигонуклеотида или полинуклеотида с указанной геномной нуклеиновой кислотой. Предпочтительно указанный способ проводят на образце нуклеиновой кислоты, полученном из указанного предполагаемого устойчивого к Botrytis растения томата, хотя также можно использовать способы гибридизации in situ. Альтернативно и в более предпочтительном варианте осуществления специалист может, после определения нуклеотидной последовательности QTL, сконструировать специфичные зонды для гибридизации или олигонуклеотиды, способные гибридизоваться в строгих условиях гибридизации с последовательностью нуклеиновой кислоты указанного QTL, и может применять такие зонды для гибридизации в способах детекции наличия QTL по этому изобретению в предполагаемом устойчивом к Botrytis растении томата.
Выражение "строгие условия гибридизации" относится к условиям, при которых зонд или полинуклеотид будет гибридизоваться с его подпоследовательностью-мишенью, как правило, в комплексной смеси нуклеиновых кислот, но по существу не с другими последовательностями. Строгие условия являются зависимыми от последовательности, и они будут отличаться в различных обстоятельствах. Более длинные последовательности специфично гибридизуются при более высоких температурах. Подробное руководство по гибридизации нуклеиновых кислот можно найти в Tijssen (Thijssen, 1993). Как правило, строгие условия выбирают, чтобы они были приблизительно на 5-10°С ниже, чем температура плавления (Tm) для конкретной последовательности при определенной ионной силе и pH. Tm представляет собой температуру (при определенной ионной силе, pH и концентрации нуклеиновых кислот), при которой 50% зондов, комплементарных мишени, равновесно гибридизуются с последовательностью-мишенью (поскольку последовательности-мишени представлены в избытке, при Tm, в равновесном состоянии 50% зондов занято). Строгие условия будут представлять собой условия, в которых концентрация соли составляет менее чем приблизительно 1,0 М концентрацию ионов натрия, как правило, приблизительно от 0,01 до 1,0 M ионов натрия (или других солей) при pH от 7,0 до 8,3, и температура составляет по меньшей мере приблизительно 30°С для коротких зондов (например, от 10 до 50 нуклеотидов) и по меньшей мере приблизительно 60°С для длинных зондов (например, более 50 нуклеотидов). Строгие условия также могут быть достигнуты посредством добавления дестабилизирующих веществ, таких как формамид. Для селективной или специфичной гибридизации положительный сигнал по меньшей мере в два раза превышает фон, предпочтительно в 10 превышает фоновую гибридизацию. Иллюстративные строгие условия гибридизации часто представляют собой: 50% формамид, 5xSSC и 1% SDS, инкубация при 42°C, или 5xSSC, 1% SDS, инкубация при 65°C, с промыванием в 0,2xSSC, и 0,1% SDS при 65°C. Для ПЦР температура приблизительно 36°C является типичной для амплификации с низкой строгостью, хотя температура отжига может варьировать между приблизительно 32°С и 48°С в зависимости от длины праймеров. Дополнительные руководства для определения параметров гибридизации представлены во множестве источников литературы, например, Current Protocols in Molecular Biology, eds. Ausubel, et al. 1995.
"Нуклеиновая кислота" или "олигонуклеотид", или "полинуклеотид" или грамматические эквиваленты, используемые в настоящем описании, означают по меньшей мере два нуклеотида, ковалентно связанных друг с другом. Олигонуклеотиды, как правило, обладают длиной от приблизительно 7, 8, 9, 10, 12, 15, 18, 20 25, 30, 40, 50 или вплоть до приблизительно 100 нуклеотидов. Нуклеиновые кислоты и полинуклеотиды представляют собой полимеры любой длины, включая большие длины, например, 200, 300, 500, 1000, 2000, 3000, 5000, 7000, 10000 и т.д. Нуклеиновая кислота по настоящему изобретению главным образом будет содержать фосфодиэфирные связи, хотя в некоторых случаях включаются аналоги нуклеиновых кислот, которые могут обладать альтернативными остовами, содержащими, например, фосфорамидатные, фосфоротиоатные, фосфородитиоатные или O-метилфосфорамидитные связи (см. Eckstein, 1991), и остовы и связи пептидных нуклеиновых кислот. Можно использовать смеси встречающихся в природе нуклеиновых кислот и аналогов. Особенно предпочтительные аналоги для олигонуклеотидов представляют собой пептидные нуклеиновые кислоты (PNA).
Получение устойчивых к Botrytis растений томата трансгенными способами
В соответствии с другим аспектом настоящего изобретения последовательность нуклеиновой кислоты (предпочтительно ДНК), содержащую по меньшей мере один QTL по настоящему изобретению или его придающую устойчивость к Botrytis часть, можно использовать для получения устойчивого к Botrytis растения томата. В этом аспекте это изобретение относится к применению QTL по настоящему изобретению или его придающих устойчивость к Botrytis частей для получения устойчивого к Botrytis растения томата, при этом применение включает введение последовательности нуклеиновой кислоты, содержащей указанный QTL, в восприимчивое к Botrytis реципиентное растение томата. Как упомянуто выше, источником указанной последовательности нуклеиновой кислоты может быть устойчивое к Botrytis донорное растение томата. Два пригодных устойчивых к Botrytis донорных растений томата, способных предоставить последовательность нуклеиновой кислоты, содержащую по меньшей мере один из описанных в настоящем описании выше QTL, или его придающие устойчивость к Botrytis части, представляют собой S. habrochaites LYC 4/78. Другие сходные растения томата, которые обладают устойчивостью к Botrytis и содержат один или несколько генов, которые кодируют устойчивость к Botrytis, также можно использовать в качестве устойчивых к Botrytis донорных растений, поскольку для настоящего изобретения описано, каким образом этот материал можно идентифицировать. Другие изоляты видов томата можно исследовать в отношении устойчивости к Botrytis, включая, но не ограничиваясь ими, Lycopersicon cerasiforme, Lycopersicon cheesmanii, Lycopersicon chilense, Lycopersicon chmielewskii, Solanum lycopersicum, Lycopersicon hirsutum, Lycopersicon parviflorum, Lycopersicon pennellii, Lycopersicon peruvianum, Lycopersicon pimpinellifolium и Solanum lycopersicoides.
После идентификации в пригодном донорном растении томата последовательность нуклеиновой кислоты, которая содержит QTL для устойчивости к Botrytis в соответствии с настоящим изобретением, или его придающую устойчивость к Botrytis часть, можно переносить в пригодное реципиентное растение любым доступным способом. Например, указанную последовательность нуклеиновой кислоты можно переносить скрещиванием устойчивого к Botrytis донорного растения томата с восприимчивым реципиентным растением томата (т.е. посредством интрогрессии), посредством трансформации, посредством слияния протопластов, посредством технологии двойных гаплоидов или посредством спасения зародышей или с помощью любой другой системы переноса нуклеиновой кислоты, необязательно с последующей селекцией растений-потомков, содержащих QTL и проявляющих устойчивость к Botrytis. Для трансгенных способов переноса последовательность нуклеиновой кислоты, содержащую QTL для устойчивости к Botrytis в соответствии с настоящим изобретением, или его придающую устойчивость к Botrytis часть, можно выделять из указанного донорного растения с использованием способов, известных в данной области, и выделенную таким образом последовательность нуклеиновой кислоты можно переносить в реципиентное растение трансгенными способами, например, посредством вектора, в гамете, или в любом другом пригодном элементе для переноса, таком как баллистическая частица, на которую нанесена указанная последовательность нуклеиновой кислоты.
Трансформация растений, как правило, вовлекает конструирование экспрессирующего вектора, который будет функционировать в клетках растений. В настоящем изобретении такой вектор содержит последовательность нуклеиновой кислоты, которая содержит QTL для устойчивости к Botrytis по настоящему изобретению, или его придающую устойчивость к Botrytis часть, при этом вектор может содержать придающий устойчивость к Botrytis ген, который находится под контролем регуляторного элемента, такого как промотор, или функционально связан с ним. Экспрессирующий вектор может содержать одно или несколько таких функционально связанных сочетаний гена/регуляторного элемента при условии, что по меньшей мере один из генов, содержащихся в сочетании, кодирует устойчивость к Botrytis. Вектор(ы) может находиться в форме плазмиды, и его можно использовать, отдельно или в сочетании с другими плазмидами, для получения трансгенных растений, которые являются устойчивыми к Botrytis, с использованием способов трансформации, известных в данной области, таких как система для транформации Agrobacterium.
Экспрессирующие векторы могут включать по меньшей мере один маркерный ген, функционально связанный с регуляторным элементом (таким как промотор), который дает возможность выделения трансформированных клеток либо посредством отрицательной селекции (посредством ингибирования роста клеток, которые не содержат ген селективного маркера), либо посредством положительной селекции (посредством скрининга продукта, кодируемого маркерным геном). Множество обычно используемых генов селективных маркеров для трансформации растений известны в данной области и включают, например, гены, которые кодируют ферменты, которые метаболически детоксифицируют селективное химическое вещество, которое может представлять собой антибиотик или гербицид, или гены, которые кодируют измененную мишень, которая является нечувствительной к ингибитору. В данной области известно несколько способов положительной селекции, такие как селекция с маннозой. Альтернативно можно использовать трансформацию без маркера для получения растений без упомянутых выше маркерных генов, технологии которой хорошо известны в данной области.
Один способ внесения экспрессирующего вектора в растение основан на природной системе трансформации Agrobacterium (см., например, Horsch et al., 1985). A. tumefaciens и A. rhizogenes представляют собой патогенные для растений бактерии почвы, которые генетически трансформируют клетки растений. Плазмиды Ti и Ri A. tumefaciens и A. rhizogenes, соответственно, обладают генами, ответственными за генетическую трансформацию растения (см., например, Kado, 1991). Способы внесения экспрессирующих векторов в ткань растения включают прямое инфицирование или кокультивирование клеток растений с Agrobacterium tumefaciens (Horsch et al., 1985). Описание векторных систем Agrobacterium и способов опосредуемого Agrobacterium переноса генов предоставлено Gruber and Crosby, 1993 и Moloney et al., 1989. Также см. патент США № 5591616. Основное описание экспрессирующих векторов растений и репортерных генов и протоколов трансформации и описания векторных систем Agrobacterium и способов опосредуемого Agrobacterium переноса генов могут быть найдены в Gruber and Crosby, 1993. Основные способы культивирования тканей растений предоставлены, например Miki et al., 1993 и Phillips, et al., 1988. Пригодным справочным руководством по способам молекулярного клонирования и пригодным экспрессирующим векторам является Sambrook and Russell (2001).
Другой способ внесения экспрессирующего вектора в растения основан на опосредуемой микроснарядами трансформации, где ДНК находится на поверхности микроснарядов. Экспрессирующий вектор вводят в ткани растения посредством биолистического устройства, которое ускоряет микроснаряды до скоростей от 300 до 600 м/с, которые являются достаточными для проникновения через стенки и мембраны клеток растений (см. Sanford et al., 1987, 1993; Sanford, 1988, 1990; Klein et al., 1988, 1992). Другой способ введения ДНК в растения представляет собой облучение ультразвуком клеток-мишеней (см. Zhang et al., 1991). Альтернативно для введения экспрессирующих векторов в растения использовали слияние с липосомами или сферопластами (см., например, Deshayes et al., 1985 и Christou et al., 1987). Также описаны прямой захват ДНК в протопластах с использованием осаждения посредством CaCl2, поливинилового спирта или поли-L-орнитина (см., например, Hain et al., 1985 и Draper et al., 1982). Также описана электропорация протопластов и цельных клеток и тканей (D'Halluin et al., 1992 и Laursen et al, 1994).
После трансформации тканей-мишеней томата экспрессия описанных выше генов селективных маркеров обеспечивает предпочтительную селекцию трансформированных клеток, тканей и/или растений с использованием способов регенерации и селекции, в настоящее время хорошо известных в данной области. Маркеры, приведенные в таблицах 1-5, также можно использовать для этой цели.
Получение устойчивых к Botrytis растений томата нетрансгенными способами
В альтернативном варианте осуществления для получения устойчивого к Botrytis растения томата, в целях переноса нуклеиновых кислот из донорного растения в реципиентное растение можно использовать слияние протопластов. Слияние протопластов представляет собой индуцированное или спонтанное объединение, такое как соматическая гибридизация, между двумя или более протопластами (клетками, клеточные стенки которых удалены ферментативной обработкой), с получением одной двухъядерной или многоядерной клетки. Слитая клетка, которую можно получать даже в случае видов растений, которые не могут скрещиваться между собой в природе, представляет собой ткань, культивируемую в гибридное растение, проявляющее требуемое сочетание признаков. Более конкретно, первый протопласт можно получать из растения томата или другой линии растений, которая обладает устойчивостью к инфицированию посредством Botrytis. Например, можно использовать протопласт из S. habrochaites LYC 4/78. Второй протопласт можно получить из второго томата или другого сорта томата, предпочтительно линии томата, которая содержит коммерчески желательные характеристики, такие как, но не ограничиваясь ими, устойчивость к заболеванию, устойчивость к насекомым, ценные характеристики плодов и т.д. Затем протопласты подвергают слиянию с использованием традиционных способов слияния протопластов, которые известны в данной области.
Альтернативно можно использовать спасение зародышей при переносе нуклеиновой кислоты, содержащей один или несколько QTL по настоящему изобретению, из донорного растения в реципиентное растение. Спасение зародышей можно использовать в качестве способа выделения зародышей от скрещиваний, где растения не продуцируют жизнеспособные семена. В этом процессе оплодотворенная завязь или незрелое семя растения представляют собой ткань, культивируемую для создания новых растений (Pierik, 1999).
Настоящее изобретение также относится к способу получения устойчивого к Botrytis растения томата, включающему стадии проведения способа детекции наличия локуса количественного признака (QTL), ассоциированного с устойчивостью к B. cinerea в донорном растении томата в соответствии с этим изобретением, как описано выше, и перенос последовательности нуклеиновой кислоты, содержащей по меньшей мере один QTL, выявленный таким образом, или его придающую устойчивость к Botrytis часть, из указанного донорного растения в восприимчивое к Botrytis реципиентное растение томата. Перенос указанной последовательности нуклеиновой кислоты можно проводить любым из способов, описанных ранее в настоящем описании.
Предпочтительный вариант осуществления такого способа включает перенос посредством интрогрессии указанной последовательности нуклеиновой кислоты из устойчивого к Botrytis донорного растения томата в восприимчивое к Botrytis реципиентное растение томата скрещиванием указанных растений. Этот перенос, таким образом, можно пригодным образом проводить с использованием традиционных способов разведения. QTL предпочтительно подвергают интрогрессии в коммерческие сорта томата с использованием разведения на основе маркера (MAS). Разведение на основе маркера или селекция на основе маркера вовлекает применение одного или нескольких молекулярных маркеров для идентификации и селекции тех растений-потомков, которые содержат один или несколько из генов, которые кодируют требуемый признак. В данном случае такая идентификация и селекция основаны на селекции QTL по настоящему изобретению или маркеров, ассоциированных с ними. MAS также можно использовать для разработки практически изогенных линий (NIL), обладающих представляющим интерес QTL, обеспечивая более подробное изучение эффекта каждого QTL, и также он представляет собой эффективный способ для создания популяций инбредных линий, полученных возвратным скрещиванием (BIL) (см., например, Nesbitt et al., 2001; van Berloo et al., 2001). Растения томата, созданные в соответствии с этим предпочтительным вариантом осуществления, могут преимущественно получить большинство из их признаков из реципиентного растения, а устойчивость к Botrytis получают из донорного растения.
Поскольку устойчивость к B. cinerea наследуется полигенно, предпочтительно, чтобы посредством пригодного способа переноса в отдельное реципиентное растение встраивали по меньшей мере два, предпочтительно три QTL или их придающие устойчивость к Botrytis части, т.е. чтобы в геноме реципиентного растения объединялось множество QTL. Полагают, что объединение двух или более QTL по этому изобретению может приводить к повышенной устойчивости к Botrytis. Как легко поймет квалифицированный специалист, объединение можно проводить любым способом, например, трансформацией растения конструкцией нуклеиновой кислоты, содержащей множество QTL по этому изобретению. Альтернативно по меньшей мере один QTL может быть представлен в каждом родительском растении при скрещивании, так чтобы в полученном гибриде находилось по меньшей мере два QTL. Посредством объединения этих признаков устойчивости можно получить устойчивые растения. Такие растения являются высокопредпочтительными вариантами осуществления настоящего изобретения.
Как кратко рассмотрено выше, для интрогрессии последовательности нуклеиновой кислоты, кодирующей устойчивость к Botrytis, в восприимчивое к Botrytis реципиентное растение томата можно использовать традиционные способы разведения. В одном способе, который называется чистопородным разведением, донорное растение томата, которое обладает устойчивостью к Botrytis и содержит последовательность нуклеиновой кислоты, кодирующую устойчивость к Botrytis, скрещивают с восприимчивым к Botrytis реципиентным растением томата, которое предпочтительно обладает коммерчески желательными характеристиками, такими как, но не ограничиваясь ими, устойчивость к заболеваниям, устойчивость к насекомым, ценные характеристики плодов и т.д. Затем полученную популяцию растений (соответствующую гибридам F1) подвергают самоопылению и семена сажают (семена F2). Затем растения F2, выращенные из семян F2, подвергают скринингу в отношении устойчивости к Botrytis. Популяцию можно подвергать скринингу множеством различных способов.
Во-первых, популяцию можно подвергать скринингу с использованием традиционных способов скрининга заболевания. Такие способы скрининга заболевания известны в данной области. Предпочтительно используют количественный биологический анализ стебля или листьев, предпочтительно применяют биологический анализ, используемый в способах по настоящему изобретению, как более подробно описано в настоящем описании выше и в примерах. Во-вторых, селекцию на основе маркера можно проводить с использованием одного или нескольких из описанных в настоящем описании ранее молекулярных маркеров для идентификации тех потомков, которые содержат последовательность нуклеиновой кислоты, кодирующую устойчивость к Botrytis. Можно использовать другие способы, названные выше в настоящем описании способами детекции наличия QTL. Также для подтверждения результатов, полученных из количественных биологических анализов, можно использовать селекцию на основе маркера и, таким образом, также можно использовать несколько способов в сочетании.
Устойчивые к Botrytis растения и семена томата
Устойчивое к Botrytis растение томата по настоящему изобретению характеризуется наличием высокого уровня устойчивости. Его определяют как уровень устойчивости, который превышает устойчивость, наблюдаемую в восприимчивых контрольных растениях. В действительности, растения по этому изобретению имеют уровень устойчивости, который превышает уровень в любом коммерческом сорте томата, т.е. в сорте, обладающем желательными характеристиками, известными к настоящему времени. Растение по этому изобретению обладает восприимчивостью к Botrytis cinerea, которая по меньшей мере в 3 раза ниже, чем в восприимчивом контрольном растении при измерении посредством биологического анализа, например, при измерении посредством биологического анализа, где среднюю длину очага повреждения стебля, возникшего вследствие инфекции Botrytis cinerea во взрослых растениях, измеряют в ходе трехнедельного периода в условиях стандартной практики, как более подробно описано ниже в примерах 3.10 и 3.11. Как правило, растение по этому изобретению имеет уровень устойчивости, который приводит к средней длине очага повреждения стебля Botrytis cinerea во взрослых растениях менее 3,2 см через три недели после инокуляции с использованием условий стандартной практики в биологическом анализе устойчивости, разработанном для определения устойчивости, исходя из таких характеристик. Более типично, растение по этому изобретению показывает среднюю длину очага повреждения стебля менее 2,9 см. Некоторые растения по этому изобретению показывают даже среднюю длину очага повреждения стебля, составляющую 2,0 см. Учитывая, что указанные числа выражают длину очага повреждения, включающую исходную рану при инокуляции размером 2 см, можно сделать вывод, что в растениях по этому изобретению наблюдается высокий уровень устойчивости, и даже полная устойчивость в случаях некоторых QTL. Для сравнения, восприимчивые контрольные растения показывают среднюю длину очага повреждения стебля в тех же условиях, составляющую от приблизительно 3,6 см до приблизительно 6,0 см, со средним значением 4,85 см (см. таблицу 7). Также для сравнения, S. habrochaites LA 1777, содержащий QTL-10, частично устойчивый к Botrytis источник из WO02/085105, показывает среднюю длину очага повреждения стебля в тех же условиях, составляющую приблизительно 4,3 см. В общем растения по этому изобретению показывают суммарные очаги повреждения стебля в указанном выше биологическом анализе устойчивости, которые главным образом составляют менее 30% (0,9/2,85×100%) от суммарной длины восприимчивых контрольных растений и главным образом менее приблизительно 40% (0,9/2,3×100%) от суммарной длины в частично устойчивых S. habrochaites LA 1777.
Таким образом, растение по настоящему изобретению обладает восприимчивостью к Botrytis cinerea при измерении посредством биологического анализа, которая в 3 раза меньше, чем в восприимчивом контрольном растении, или которая составляет менее 1/3 от уровня в нем. Аналогично растение по этому изобретению является более чем в 3 раза более устойчивым, чем восприимчивое контрольное растение, как определено в настоящем описании и как измеряют посредством описанного биологического анализа. При QTL-1h выявлена полная устойчивость. Восприимчивое контрольное растение определяют как растение, обладающее нормальной восприимчивостью, или не обладающее устойчивостью, к инфекции посредством Botrytis cinerea. Примером восприимчивого контрольного растения является растение гибрида Solanum lycopersicum cv. "Moneyberg" (De Ruiter Seeds R&D BV, Bergschenhoek, The Netherlands).
Устойчивое к Botrytis растение томата, или его часть, получаемое способом по этому изобретению, также является аспектом настоящего изобретения.
Другой аспект настоящего изобретения относится к устойчивому к Botrytis растению томата, или к его части, содержащему в его геноме по меньшей мере один QTL, или его придающую устойчивость к Botrytis часть, выбранный из группы, состоящей из QTL на 4, 6, 9, 11 и 12 хромосомах Solanum habrochaites LYC 4/78, ассоциированных с устойчивостью к Botrytis, где указанный QTL или указанная его придающая устойчивость к Botrytis часть находятся не в их природном генетическом фоне. Устойчивые к Botrytis растения томата по настоящему изобретению могут представлять собой растения любого типа, такие как инбредное, гибридное, гаплоидное, дигаплоидное, партенокарпное или трансгенное растения. Кроме того, растения по настоящему изобретению могут быть гетерозиготными или гомозиготными по признаку устойчивости, предпочтительно они являются гомозиготными. Несмотря на то что QTL по настоящему изобретению, а также QTL, получаемые способом по этому изобретению, а также их придающие устойчивость к Botrytis части можно переносить в любое растение в целях получения устойчивого к Botrytis растения, способы и растения по этому изобретению предпочтительно относятся к растениям семейства Solanaceae, более предпочтительно к томату.
В дополнение к QTL на 4 (QTL-4hB), 6, 9, 11 и 12 хромосомах Solanum habrochaites LYC 4/78, ассоциированным с устойчивостью к Botrytis, как идентифицировано в настоящем описании, растение по настоящему изобретению необязательно может дополнительно содержать один или несколько QTL из S. habrochaites, идентифицированных в настоящем описании в качестве QTL-1h, QTL-2h и QTL-4hA и описанных более подробно в находящихся на рассмотрении заявках PCT/NL2005/000762 и EP 1652930A (европейская патентная заявка 04077931.6), на которые в настоящем описании приведена прямая ссылка.
Предполагается, что сочетание QTL, либо QTL, как описано в настоящем описании, либо QTL, как описано в PCT/NL2005/000762, повышает устойчивость к заболеванию в растениях, обладающих таким сочетанием. Однако каждый из QTL также обладает генетической информацией из фонового предка S. habrochaites, что означает, что в потомстве со слишком большим количеством генетической информацией этого фона будет невозможно получить растения с желательными оптимальными агрономическими свойствами.
Инбредные устойчивые к Botrytis линии растений томата можно получить с использованием способов рекуррентной селекции и возвратного скрещивания, самоопыления и/или дигаплоидов или любого другого способа, используемого для получения родительских линий. В способе селекции и возвратного скрещивания устойчивость к Botrytis можно вносить интрогрессией в реципиентное растение-мишень (которое называется рекуррентным родителем) посредством скрещивания рекуррентного родительского растения с первым донорным растением (которое отличается от рекуррентного родительского растения и обозначается в настоящем описании как "нерекуррентное родительское растение"). Рекуррентное родительское растение представляет собой растение, которое является неустойчивым или проявляет низкий уровень устойчивости к Botrytis и обладает коммерчески желательными характеристиками, такими как, но не ограничиваясь ими, устойчивость к заболеванию, устойчивость к насекомым, ценные характеристики плодов и т.д. Нерекуррентное родительское растение обладает устойчивостью к Botrytis и содержит последовательность нуклеиновой кислоты, которая кодирует устойчивость к Botrytis. Нерекуррентное родительское растение может представлять собой любой сорт растений или инбредную линию, которая является фертильной в отношении скрещивания с рекуррентным родителем. Потомство, возникающее в результате скрещивания между рекуррентным родителем и нерекуррентным родителем, подвергают возвратному скрещиванию с рекуррентным родителем. Затем полученную популяцию растений подвергают скринингу. Скрининг популяции можно проводить рядом различных способов. Например, скрининг популяции можно проводить с использованием количественных биологических анализов стебля, как описано в настоящем описании выше. Затем гибридные растения F1, которые обладают устойчивым к Botrytis фенотипом, содержат требуемую последовательность нуклеиновой кислоты, кодирующую устойчивость к Botrytis, и обладают коммерчески желательными характеристиками, подвергают селекции и самоопылению в течение ряда поколений в целях обеспечения возрастающей инбредности растения томата. Этот процесс продолжающегося самоопыления и селекции можно проводить в течение от двух до пяти или более поколений. Результат такого разведения и селекции представляет собой получение линий, которые являются генетически гомогенными по генам, ассоциированным с устойчивостью к Botrytis, а также по другим генам, ассоциированным с признаками, представляющими коммерческий интерес. Вместо применения фенотипического скрининга патологии в биологических анализах MAS можно проводить с использованием одного или нескольких из описанных в настоящем описании выше молекулярных маркеров, зондов для гибридизации или полинуклеотидов в целях идентификации тех потомков, которые содержат последовательность нуклеиновой кислоты, кодирующую устойчивость к Botrytis. Альтернативно MAS можно использовать для подтверждения результатов, полученных из количественных биологических анализов. После проведения соответствующей селекции процесс повторяют. Процесс возвратного скрещивания с рекуррентным родителем и селекции по устойчивости к Botrytis повторяют в течение приблизительно пяти или более поколений. Потомство, появляющееся в результате этого процесса, является гетерозиготным по одному или нескольким генам, которые кодируют устойчивость к Botrytis. Затем последнее поколение от возвратного скрещивания подвергают самоопылению в целях обеспечения чистого разведения потомков, гомозиготных по устойчивости к Botrytis.
Устойчивые к Botrytis инбредные линии томата, описанные в настоящем описании, можно использовать в дополнительных скрещиваниях для создания устойчивых к Botrytis гибридных растений. Например, первое устойчивое к Botrytis инбредное растение томата по этому изобретению можно скрещивать со вторым инбредным растением томата, обладающим коммерчески желательными признаками, такими как, но не ограничиваясь ими, устойчивость к заболеваниям, устойчивость к насекомым, желательные характеристики плодов и т.д. Эта вторая инбредная линия томата может быть устойчивой к Botrytis или не устойчивой к нему.
Другой аспект настоящего изобретения относится к способу получения семян, которые можно выращивать в устойчивые к Botrytis растения томата. В одном варианте осуществления способ включает стадии получения устойчивого к Botrytis растения томата по этому изобретению, скрещивания указанного устойчивого к Botrytis растения с растением Solanum lycopersicum и создания коллекции семян, полученных от указанного скрещивания, которые при посеве приводят к устойчивым к Botrytis растениям томата.
В другом варианте осуществления способ включает стадии представления устойчивого к Botrytis растения томата по этому изобретению, скрещивания указанного устойчивого к Botrytis растения с растением Solanum lycopersicum, создания коллекции семян, полученных в результате указанного скрещивания, регенерации указанных семян в растения, селекции устойчивых к Botrytis растений любыми способами, описанными в настоящем описании, самоопыления выбранных растений в течение достаточного количества поколений с получением растений, в которых фиксирован аллель, который придает устойчивость к Botrytis в растениях, возвратного скрещивания растений, полученных таким образом, с растениями S. lycopersicum, имеющими желательные фенотипические признаки, в течение достаточного количества поколений с получением растений S. lycopersicum, которые являются устойчивыми к Botrytis и обладают желательными фенотипическими признаками, и коллекционирования семян, продуцированных растениями, полученными в результате последнего скрещивания, которые при высевании приводят к растениям томата, которые являются устойчивыми к Botrytis.
В качестве примера, а не для ограничения, далее приведены примеры по настоящему изобретению.
ПРИМЕРЫ
Пример 1
Способ идентификации QTL, ассоциированных с устойчивостью к Botrytis cinerea
1.1. Введение
В этом примере представлена разработка количественного биологического анализа для оценки устойчивости к Botrytis cinerea коллекции диких генотипов томата.
Частичная устойчивость к Botrytis cinerea описана для диких видов томата, однако эти данные в большей степени были описательными и качественными. Идентификация частично устойчивых генотипов может обеспечить перспективы для интрогрессии устойчивости в коммерчески выращиваемые линии с получением линий с управляемыми уровнями устойчивости. Доступность воспроизводимого, объективного и количественного анализа, а также идентификация генотипов с генетически определенной (частичной) устойчивостью к серой гнили обеспечивает способ для выведения устойчивости в культивируемых сортах томата.
В этом примере описан количественный анализ заболевания. Анализ проводят на листьях (анализ инокуляции листьев) и на сегментах стебля (анализ инокуляции стебля). Исследовали два параметра восприимчивости к заболеванию. Первым измеряемым параметром была заболеваемость (DI), т.е. доля инокуляций, которые приводили к увеличению очага повреждения. Если (частичная) неспособность первичного очага повреждения B. cinerea к распространению на конкретном генотипе хозяина является генетическим признаком растения, такой признак является важным, поскольку он непосредственно ограничивает количество очагов заболевания в культуре. Вторым тестируемым параметром была скорость роста очага повреждения в течение 24 ч (рост очага повреждения, LG). Оказалось, что очаги повреждения, которые разрастались из первичного инокуляционного пятна, распространялись с равной скоростью (в мм/сутки) в течение периода времени, пока очаг не достигал края листа или нижнего конца сегмента стебля. Данные способы анализа обеспечивают количественное определение как заболеваемости (встречаемости), так и развития (роста очага повреждения) инфекции B. cinerea, что приводит к двум наборам данных количественных признаков. Анализ использовали для скрининга коллекции видов томата (в дальнейшем в настоящем описании также называемых "изолятами") в отношении наличия в них устойчивости.
1.2. Растения
Генотипы тестированных растений приведены в таблице 6.
1DRS: De Ruiter Seeds, Bergschenhoek, The Netherlands; WU PPW: Plantkundig Proefcentrum Wageningen, Wageningen University, Wageningen, The Netherlands; LoPB: Laboratory of Plant Breeding, Wageningen University, Wageningen, The Netherlands; MPIZK: Max Planck Institut für Züchtungsforschung an Kulturpflanze, Köln, Germany; TGRC: Tomato Genetics Resource Center, University of California at Davis, Davis CA, USA; IPK: Institut für Pflanzengenetik und Kulturpflanzenforschung, Gatersleben, Germany.
2Y указывает на то, что генотип тестировали в конкретном анализе, S указывает на то, что генотип служил в качестве восприимчивого референсного контроля.
(3) Ранее опубликован в качестве устойчивого к B. cinerea.
Растения выращивали в почве для горшечных культур в 12-см горшках в теплице с минимальной температурой 15°C. Применяли искусственное освещение натриевой лампой (16 ч/сутки) с октября по март. Через 5-7 суток после проращивания добавляли 10 мл раствора FeNaEDTA (3,5 г/л), а через 3 суток 10 мл раствора микронутриентов (0,286 г/л H3BO3; 0,1558 г/л MnSO4·H2O; 0,008 г/л CuO4·H2O; 0,022 г/л ZnSO4; 0,00196 (NH4)6Mo7O24·4H2O). Через две недели после начала прорастания добавляли 5 мл раствора Hoagland (5 мМ Ca(NO3)2; 5 мМ KNO3; 2 мМ MgSO4; 1 мМ KH2PO4) еженедельно.
1.3. Анализ листьев
Инокулят штамма B. cinerea B05.10 получали в соответствии с Benito (1998). Для каждого отдельного растения один или два сложных листа, которые были полностью вытянутыми, отделяли от основного стебля острым лезвием бритвы и переносили на предварительно смоченную пену для цветов. Пену для цветов помещали в чашку Петри, содержащую водопроводную воду, а затем помещали в увлажненный распылением контейнер, содержащий влажную фильтровальную бумагу. Затем сложные листья инокулировали суспензией конидий B. cinerea осторожным пипетированием всего от 6 до 10 капель инокулята (2 мкл) на верхней поверхности листьев. Затем контейнеры закрывали смоченной опрыскиванием крышкой и листья инкубировали при 15°C в темноте при 100% RH, по существу, как описано Benito et al., 1998. Было приемлемым разделение одного сложного листа на четыре листика, и где на каждый листик инокулировали 10 капель по 2 мкл каждая, содержащие 2000 конидий. Мониторинг как стремительно распространяющихся очагов повреждения (заболеваемость), так и скорости роста очага повреждения проводили в течение нескольких дней.
Для коррекции по отклонениям, связанным со временем года или культивированием растений, заболеваемость конкретного генотипа в каждом эксперименте соотносили с заболеваемостью в Moneymaker, тестируемом в том же эксперименте.
Размеры очагов повреждения измеряли через 96, 120 и 144 hpi с использованием штангенциркуля. Заболеваемость определяли делением общего количества разрастающихся очагов повреждения на общее количество инокуляционных капель. Скорости роста очага повреждения определяли вычислением увеличения размера очага повреждения (в мм) в течение 24 ч. Данные для нерастущих очагов повреждения не включали в количественный анализ.
1.4. Анализ стебля (стандартизованный способ)
Анализ стебля проводили следующим образом: Верхние 5-10 см и нижние 5-10 см стебля растений высотой приблизительно 50 см удаляли и оставшиеся 30 см нарезали на равные сегменты по 5-6 см. Каждый сегмент стебля помещали вертикально на сетке с основанием стебля на влажной фильтровальной бумаге. Перед инокуляцией сегменты стебля опрыскивали водой в целях обеспечения равномерного распределения инокулята на поверхности раны. Инокулят получали, как описано для анализа листьев. Одну каплю инокулята объемом приблизительно 5 мкл, содержащую приблизительно 106 конидий·мл-1, наносили на вершину каждого сегмента стебля. Инкубации проводили при 15±2°С со 100% относительной влажностью. Прогрессирование инфицирования определяли посредством измерения максимального развития симптома гниения через различные промежутки времени после инокуляции посредством штангенциркуля с нониусом.
Для каждого генотипа вычисляли процентное количество инфицированных сегментов стебля. Заболеваемость определяли делением общего количества сегментов стебля с разрастающимися очагами повреждения на общее количество инокулированных сегментов. Скорости роста очагов повреждения определяли вычислением увеличения размера очага повреждения в течение 24 ч, при этом данные для нерастущих очагов повреждения были исключены из анализа.
1.5. Результаты
Заболеваемость и рост очага повреждения в экспериментах с инфицированием отделенных листьев определяли для каждого генотипа в течение нескольких суток, как правило, в течение 2-4 суток после инфицирования. Заболеваемость в S. lycopersicum cv. Moneymaker, которая служит в качестве контроля, варьировала между 15 и 78% в этих экспериментах. За исключением генотипов 82/2577 и 83/2896 (оба вида S. lycopersicum) тестированные генотипы показали во всех экспериментах более низкую заболеваемость, чем Moneymaker. Генотипы G1.1556, G1.1560 и G1.1601 показали низкую заболеваемость в трех независимых экспериментах, варьирующую от 0 до 21%. Статистический анализ показал, что заболеваемость в генотипах 78/1604, 91/4311, 96/4326, G1.1556, GI 1558, G1.1560, G1.1601, LA716 и LYC 4/78 была значимо ниже, чем в контрольной линии S. lycopersicum cv. Moneymaker (p<0,05). Однако существовало значительное колебание между неделями, и некоторые из отличий, выявленных в анализах открепленных листьев, в действительности могут не быть высоконадежными вследствие колебаний в заболеваемости между экспериментами/неделями (15-78%).
В этих устойчивых генотипах (с заболеваемостью, значительно меньшей, чем заболеваемость в контроле Moneymaker) очаги повреждения, которые часто успешно разрастались, разрастались со скоростью, сходной со скоростью Moneymaker (например, 96/4326, G1.1560, LA716). Обратная ситуация не наблюдалась: ни один из генотипов не проявлял заболеваемость, сходную с заболеваемостью Moneymaker, но скорость роста, меньшую, чем для Moneymaker.
Скорость роста очага повреждения в течение 24 ч (между 48 и 72 hpi) в большинстве генотипов находилась в том же диапазоне, что и в Moneymaker. Пять изолятов (91/4311, 160/79, G1.1556, G1.1601 и LYC 4/78) показали более медленную скорость роста очага повреждения, которая со статистической значимостью отличалась от скорости роста очага повреждения S. lycopersicum cv. Moneymaker.
Анализ инфекции сегмента стебля оказался более надежным, чем анализ листьев с точки зрения воспроизводимости между экспериментами, проводимыми в различные времена года. Даже несмотря на то, что количество результатов для сегментов стебля (5-8 сегментов на растение) значительно меньше, чем для анализа листьев (40 инокуляционных капель на сложный лист, можно тестировать один или два листа на растение), вариабельность между экспериментами была значительно ниже в анализе сегмента стебля. Заболеваемость в анализе сегмента стебля для контрольного генотипа S. lycopersicum cv. Moneymaker варьировала от 52-95%. Заболеваемость в 17 генотипах сравнивали с заболеваемостью контрольной линии S. lycopersicum cv. Moneymaker, определенной в том же эксперименте/неделе. Большинство генотипов показали заболеваемость в сходном диапазоне с контрольной линией Moneymaker. Генотипы G1.1556 (29% и 41%) и G1.1560 (28% и 7%) показали сниженную заболеваемость. Только G1.1560 отличался со статистической значимостью (p<0,05) от контроля.
Скорости роста очагов повреждения в анализе стебля для контрольного генотипа S. lycopersicum cv. Moneymaker варьировали от 5,4 до 9,2 мм/сутки. Скорости роста очагов повреждения многих генотипов находились в сходном диапазоне с контролем. Однако в образцах 89/3793, G1.1601, LYC 4/78, T566-81 скорость роста очага повреждения статистически отличалась (p<0,01) от контрольного cv. Moneymaker.
Для ряда генотипов, которые были определены как устойчивые в анализе сегмента стебля, проводили количественные анализы на целых растениях, выращенных в теплице на Rockwool®. Целью была оценка того, действительно ли генотипы, которые оказались устойчивыми в сегментах стебля в лабораторных условиях, были более устойчивыми, чем контрольные линии, в полукоммерческой системе для выращивания. Растения выращивали в случайном порядке в рядах Rockwool®, помещение теплицы наполняли плодами цитрусов, инфицированными B. cinerea в момент споруляции. В помещении теплицы поддерживали высокую влажность, орошая пол два раза в сутки водопроводной водой и оставляя двери и окна закрытыми. Через регулярные промежутки времени создавали раны рассечением на всех растениях и проводили мониторинг встречаемости серой гнили с течением времени.
Идентифицировали количество диких изолятов томата, которые проявляли выраженное снижение обоих параметров, таким образом, обеспечивая потенциальные источники для интрогрессии двух потенциально независимых механизмов частичной устойчивости в S. lycopersicum.
Пример 3
Картирование частичной устойчивости к Botrytis cinerea в межвидовой популяции томатов (S. lycopersicum cv Moneymaker × изолят S. habrochaites LYC 4/78)
В этом примере представлены два локуса QTL, придающих частичную устойчивость к B. cinerea, источником которых является S. habrochaites LYC 4/78. Подтверждение результатов получали посредством оценки уровня устойчивости к B. cinerea в двух популяциях BC2S1, сегрегирующих по одному из двух локусов QTL соответственно.
3.1. Растительный материал
Семена Solanum habrochaites LYC 4/78 (в дальнейшем в настоящем описании обозначаемого как LYC 4/78) получали из банка генов, находящегося в Institute for Plant Genetics and Crop Plant Research, Gatersleben, Germany.
Семена Solanum lycopersicum cv. Moneymaker (в дальнейшем в настоящем описании обозначаемого как Moneymaker) получали из банка семян De Ruiter Seeds R&D BV, Bergschenhoek, The Netherlands.
Проводили межвидовое скрещивание между Moneymaker и LYC 4/78 с получением семян F1. Семена F1 выращивали в растения F1. Семена F2, образованные посредством самоопыления одного растения F1, высевали с получением популяции F2 из 174 отдельных растений. Популяцию BC2 (возвратное скрещивание 2) из 59 отдельных растений получали посредством двух раундов возвратного скрещивания с Moneymaker в качестве рекуррентного и женского родительского растения. С использованием MAS отбирали генотипы BC2, BC3 и BC4, содержащие один из двух идентифицированных QTL, и некоторые BC2 подвергали самоопылению с получением семян BC2S1 (см. фиг.2). Выращивали две популяции BC2S1: одну из 60 отдельных растений BC2S1, которые сегрегировали по QTL для заболеваемости, и другую из 47 отдельных растений BC2S1, которые сегрегировали по QTL для роста очага повреждения.
3.2. Анализ стебля
Инокулят из штамма B. cinerea B05.10 получали в соответствии с Benito (1998). Анализ стебля проводили, как описано в примере 1.
3.3. Выделение ДНК и анализ маркеров
Геномную ДНК выделяли из двух молодых (свернутых) листов с использованием протокола на основе бромида цетилтриметиламмония (CTAB) в соответствии со Steward and Via (1993), скорректированного для высокопроизводительного выделения ДНК с использованием микронных пробирок объемом один мл (Micronic B.V., Lelystad, The Netherlands), и измельченных с использованием 300-мм устройства для встряхивания Retsch при максимальной скорости (Retsch B.V., Ochten, The Netherlands). Проводили анализ AFLP (Vos et al., 1995) популяций F2, BC2, BC3, BC4 и BC2S1, и фрагменты AFLP разделяли на ДНК-секвенаторе LI-COR 4200, по существу в соответствии со способом, опубликованным Myburg (Myburg et al. 2001). Селективный праймер Pst метили флуоресцентной меткой IRD 700 или IRD 800. Изображения геля AFLP оценивали с использованием пакета программ AFLP-Quantar Pro (Keygene B.V., Wageningen, The Netherlands). Для генотипирования использовали следующие сочетания десяти праймеров и соединительные последовательности: P14M48, P14M49, P14M50, P14M60, P14M61, P15M48, P18M50, P18M51, P22M50 и P22M51, как описано Bai et al. (2003).
3.4. Фенотипический анализ популяции F 2
Наблюдали вариабельность заболеваемости между различными анализами Botrytis (см. пример 1, выше). Таким образом, проводили семь независимых последовательных анализов заболевания стебля на 172 из 174 отдельных растениях популяции F2, полученной скрещиванием между Moneymaker x LYC 4/78. Это привело по меньшей мере к пяти независимым оценкам биологического анализа заболевания практически для каждого генотипа F2. В каждом отдельном биологическом анализе заболевания в вычисление роста очага повреждения включали шесть сегментов стеблей. Средние значения для заболеваемости и роста очага повреждения в популяции F2 показали нормальное распределение (данные не представлены). Средняя заболеваемость для Moneymaker составляет 59% с ростом очага повреждения, составляющего 9,2 мм/сутки. Средняя заболеваемость в популяции F2 варьировала между 10% и 97% со средним значением для популяции, составляющим 48%. Рост очага повреждения варьировал между 3,3 мм и 11,5 мм/сутки со средним значением 7,8 мм/сутки.
Средняя заболеваемость в каждом отдельном эксперименте варьировала от 31% до 73%, в то время как средний рост очага повреждения варьировал от 6,2 до 7,9 мм/сутки (данные не представлены). Рост очага повреждения можно вычислить, только если инфекция существует по меньшей мере в одном из шести сегментах стеблей. Следовательно, увеличение количества информативных генотипов для роста очага повреждения можно наблюдать при более высокой заболеваемости. Например, при низкой средней заболеваемости (31%) только 52% генотипов были информативными в отношении роста очага повреждения.
3.5. Молекулярные маркеры и генетическая карта сцепления
Генетическую карту сцепления вычисляли для популяции F2 (n=174), полученной скрещиванием Moneymaker × LYC 4/78. Использовали сочетания десяти праймеров с получением 218 маркеров полиморфизма длин амплифицированных фрагментов (AFLP) в популяции F2 (n=174). Всего 69 маркеров (31,7%) можно было легко оценить кодоминантно, таким образом, обеспечивая вычисление объединенной генетической карты сцепления F2. Анализ маркеров, проводимый на генотипах BC2, BC3 и BC2S1, обеспечил добавление дополнительных 145 маркеров AFLP. Всего 102 из этих 145 дополнительных маркеров AFLP ранее не оценивали вследствие комплексности гелей F2. Общая генетическая карта сцепления состояла из 315 маркеров AFLP из 14 групп сцепления и обладала общей длиной 958 сМ. Поскольку совместно мигрирующие маркеры AFLP вида, как правило, являются аллель-специфическими, для отнесения групп сцепления к хромосомам использовали совместную линейность с другими картами сцепления AFLP. Некоторые специфичные для Moneymaker маркеры AFLP были общими с генетическими картами сцепления, как опубликовано (Haanstra et al. 1999; Bai et al. 2003), и, таким образом, некоторые группы сцепления можно приписать к хромосомам, включая группы сцепления, обладающие идентифицированными QTL. Для усовершенствования карты сцепления в интервалах QTL в этих участках добавляли диагностические маркеры CAPS, исходя из опубликованной карты S. lycopersicum × L. pennellii (Tanksley et al. 1992; Haanstra et al. 1999).
3.6. Анализ сцепления и картирование QTL
Данные о маркерах анализировали и вычисляли генетическую карту сцепления, как описано в разделе 3.5.
Общая длина карты сцепления F2 составляла 958 сМ, которая является меньшей, чем другие опубликованные межвидовые карты Lycopersicon с генетической длиной, варьирующей от 1200-1400 сМ (Foolad et al. 2002; Haanstra et al. 1999; Tanksley et al. 1992). Дополнительные маркеры AFLP оценивали с использованием данных о маркерах AFLP, полученных от популяций возвратного скрещивания и BC2S1. Несмотря на то что на карту сцепления было нанесено на 46% больше маркеров, длина генетической карты сцепления не увеличилась. Причина этого состоит в том, что используемые данные были получены от нескольких небольших подсемейств и, таким образом, не являлись информативными для вычисления генетических расстояний, однако оценка положения является возможной посредством визуального исследования представленных в виде графика генотипов (Van Berloo, 1999).
3.7. Картирование QTL в популяции F 2
Фенотипические данные и данные о маркерах использовали для идентификации QTL посредством картирования расстояний (IM, см. раздел 3.5). IM применяли как к данным, полученным для отдельных повторений, так и к средним значениям для повторений.
Заболеваемость
Картирование расстояний для заболеваемости в популяции F2 проводили для отдельных тестов заболевания со средней заболеваемостью менее 50% и для средних данных, полученных во всех тестах заболевания. Средние данные для всех тестов привели в способе картирования расстояний к одному значимому QTL для заболеваемости (показатель для шансов (LOD) должен превышать 3,4 для полногеномного уровня достоверности P<0,05). Этот QTL обладал показателем LOD, составляющим 4,5, и объяснял 13% от общей фенотипической вариабельности. Источником аллеля, приводящего к устойчивости, было устойчивое родительское растение LYC 4/78. Картирование QTL в каждом отдельном эксперименте привело во всех четырех случаях к одному участку QTL. В каждом независимом эксперименте иногда наблюдали другие "минорные QTL".
Рост очага повреждения
Рост очага повреждения можно наилучшим образом измерить в тестах заболевания с высокой заболеваемостью. Для картирования QTL использовали среднее значение для всех 7 тестов заболевания, и был идентифицирован один QTL для роста очага повреждения B. cinerea выше порогового значения (LOD 3,4 для полногеномного уровня достоверности P<0,05). Этот QTL имел показатель LOD, составляющий 4,2, и объяснял 12% общей фенотипической вариабельности. Положительный эффект происходил от устойчивого родительского растения LYC 4/78. Проиллюстрирована необходимость проведения многократных тестов заболевания, поскольку только в одном отдельном повторе был выявлен профиль LOD выше порогового значения.
QTL для роста очага повреждения был выявлен на 1 хромосоме (QTL-1h), и QTL для заболеваемости был выявлен на 2 хромосоме (QTL-2h). Эти QTL, а также QTL, обозначенный QTL4hA, являются объектом находящейся на рассмотрении заявки PCT/NL2005/000762.
3.8. Подтверждение QTL в биологическом анализе
Растение F1 от скрещивания Moneymaker × LYC 4/78 два раза подвергали возвратному скрещиванию с Moneymaker, и 59 растений-потомков подвергали скринингу в отношении наличия двух идентифицированных областей QTL (QTL-1h и QTL-2h) с использованием маркеров APLP. Растения, гетерозиготные по одному из двух идентифицированных QTL, подвергали селекции и самоопылению с получением двух популяций BC2S1. Для каждого генотипа BC2S1 проводили всего по четыре биологических анализа заболевания. Данные для обеих субпопуляций BC2S1, проанализированные посредством SPSS, показали нормальное распределение для роста очага повреждения, но не для заболеваемости, поскольку было выявлено несколько подклассов.
Все растения BC2S1 подвергали генотипированию AFLP с теми же 10 сочетаниями праймеров, которые описаны для популяции F2 в разделе 3.3 выше. Средний рост очага повреждения в популяции, сегрегирующей по локусу роста очага повреждения, составлял 5,3 мм/сутки, в то время как в другой популяции наблюдали средний рост очага повреждения 6,3 мм/сутки. Ни одно из растений не обладало настолько низким ростом очага повреждения, как устойчивое родительское растение LYC 4/78. Однако для заболеваемости были выявлены растения с более низкой заболеваемостью, чем в устойчивом родительском растении LYC 4/78. Средняя заболеваемость для обеих популяций BC2S1 была равной (57-59%).
Положительный эффект каждого QTL подтверждали в популяциях BC2S1. QTL для заболеваемости снижал вероятность инфицирования на 17% (46% от родительской вариабельности), и QTL для роста очага повреждения снижал рост грибов на 1,3 мм/сутки (33% от родительской вариабельности).
Только часть вариабельности можно объяснить эффектом обоих QTL. Были идентифицированы некоторые дополнительные ("минорные") локусы QTL.
В ходе анализа данных тестов заболевания полученных генотипов от как F2, так и BC2S1, был идентифицирован один основной QTL для заболеваемости (QTL-2h). Помимо этого QTL, были идентифицированы другие "предполагаемые" локусы QTL для заболеваемости. С использованием этой информации были отобраны кофакторы для проведения ограниченного способа "множественного картирования QTL" (MQM) на наборе данных F2. В этом анализе был идентифицирован один дополнительный "минорный" локус QTL для заболеваемости (QTL-4hA). QTL обозначают как "минорный", когда его показатель является ниже порога значимости LOD 3,4. Однако полагают, что эффекты являются истинными эффектами QTL.
QTL-4hA расположен на 4 хромосоме и снижает заболеваемость.
3.9 Заключения для анализа заболевания и картирования QTL
Было показано, что биологический анализ для определения устойчивости к B. cinerea является полезным инструментом. Однако оказалось, что, тем не менее, большая и неизвестная вариабельность влияет на инфекционный процесс. Эту крупную негенетическую вариабельность можно минимизировать с использованием стандартизованных способов и посредством проведения множества независимых повторений. Вариабельность может быть вызвана изменением тепличных условий из недели в неделю (длина дня, количество часов солнечного света и температура), приводящим к отличиям в физиологическом состоянии стебля. Также небольшая вариабельность при получении инокулята грибов может играть роль в вариабельности инфекционного процесса. Другое наблюдение состоит в том, что на развитие заболевания также может влиять микроклимат в лотках, в которые помещены сегменты стебля. Для биологических анализов BC2S1 использовали десять различных экспериментальных лотков. Статистический анализ использовали для компенсации вариабельности между экспериментами и в экспериментах. Эксперименты с наиболее высокой средней заболеваемостью были наиболее информативными для измерения роста очага повреждения, в то время как эксперименты с более умеренной заболеваемостью были более информативными. Заболеваемость и рост очага повреждения являются независимыми признаками, поскольку нельзя выявить линейную корреляцию между двумя признаками.
Локусы количественных признаков для устойчивости к B. cinerea в томате были идентифицированы в F2. Эти идентифицированные QTL были подтверждены в популяциях BC2S1 и объясняли 46% и 33% родительской вариабельности заболеваемости и роста очага повреждения соответственно. Эти результаты подразумевают, что не все QTL, придающие устойчивость к B. cinerea, были выявлены в исходном картировании популяции F2. В обеих популяциях BC2S1 были выявлены растения с более высокими уровнями устойчивости, чем в случае устойчивого родительского растения LYC 4/78. Это указывает на наличие дополнительных локусов устойчивости, сегрегирующих в популяции BC2S1. Дополнительная сегрегация устойчивости была удивительной, поскольку можно было ожидать, что крупные части генома двух популяций BC2S1 уже были гомозиготными по Moneymaker.
3.10 Подтверждение эффекта отдельных QTL в тепличных условиях
Растения, содержащие любой из QTL, описанных выше, помещали в фон S. lycopersicum с использованием способа, описанного на фиг.2. Линии BC2S2 помещали в теплицу в почву и выращивали в общепринятых в Нидерландах условиях. Через 3 месяца растения инокулировали, помещая диск агара, содержащий Botrytis, в рану основного стебля. После этого рану закрывали с использованием Parafilm®. Через три недели после инокуляции измеряли длину очага повреждения стебля (в см) (для более подробного описания см. ниже). Результаты приведены в таблице 7. Очевидно, линии, содержащие QTL для роста очага повреждения, показывают значительное снижение размера очага повреждения.
***) a, b, c и d представляют собой повторы, при этом каждый повтор соответствует 5 растениям; e и f представляют собой повторы, при этом каждый повтор соответствует 3 растениям; GT представляет собой Moneyberg с устойчивостью к TMV; Durintha представляет собой гибрид с частичной устойчивостью, по утверждению овощеводов; Tradiro представляет собой гибрид, восприимчивый к Botrytis, по утверждению овощеводов; BChirs указывает на линии от возвратного скрещивания, полученные посредством интрогрессий S. habrochaites LYC 4/78; LA 1777 представляет собой изолят дикого типа S. habrochaites LA 1777; BC chrs 10 указывает на линии от возвратного скрещивания с интрогрессией в 10 хромосоме от S. habrochaites LA 1777; parv указывает на линии, полученные вследствие интрогрессий S. neorickii.
3.11. Уровень устойчивости к Botrytis, обеспечиваемый QTL S. habrochaites LYC 4/78, превышает уровень устойчивости, обеспечиваемый QTL S. habrochaites LA 1777 в 10 хромосоме.
Уровень устойчивости в растениях, содержащих QTL S. habrochaites LYC 4/78, описанные в настоящем описании, сравнивали с уровнем S. habrochaites LA1777, источника из WO02/085105, который содержит QTL для частичной устойчивости к Botrytis на 10 хромосоме, и с интрогрессивными линиями, образованными из него, с интрогрессиями на 10 хромосоме.
Линии помещали в теплицу в почву и выращивали в стандартных в Нидерландах условиях. Через 3 месяца растения инокулировали помещением агарового диска размером 0,5 см × 0,5 см, содержащего Botrytis, на вертикальную рану стебля длиной 2 см в основном стебле. Затем рану закрывали с использованием Parafilm®. Через три недели после инокуляции определяли длину очага повреждения стебля (длину обесцвеченной ткани с пятнами роста грибов) (в см) от верхней части очага повреждения до нижней части очага повреждения. Результаты представлены в таблице 7. Было выявлено, что линии, содержащие QTL из S. habrochaites LYC 4/78, показали более высокий уровень устойчивости к Botrytis, чем источник LA 1777 и IL-линии. Первые линии показали меньший рост очага повреждения на стебле, и, таким образом, они проявляют более высокий уровень устойчивости к Botrytis, чем линии, образованные из LA 1777 (см. таблицу 7). В случае, где зарегистрирована длина очага повреждения, составляющая 2,0 см, можно было измерить только исходную рану и не было выявлено роста грибов, что указывает на высокий уровень устойчивости. Таким образом, длина очага повреждения стебля, составляющая 2 см, указывает на отсутствие фактического роста.
Пример 4
Картирование частичной устойчивости к Botrytis cinerea в межвидовой популяции томата (S. lycopersicum cv Moneymaker x изолят S. habrochaites LYC 4/78)
Введение
В целях обеспечения более эффективного процесса выращивания, вовлекающего селекцию родительских растений-кандидатов, имеющих надлежащую генетическую конституцию, необходимо иметь в наличии один или несколько генетических маркеров, которые указывают на наличие этой генетической конституции по меньшей мере в одном из родительских растений-кандидатов. Этот процесс, который включает скрещивание отобранных растений и называется селекцией на основе маркеров (MAS), эффективно переносит благоприятные родительские аллели от донорной в реципиентную популяцию и обеспечивает, чтобы выведение более не зависело от совпадений и являлось более экономически эффективным с точки зрения затрат на разработку.
Устойчивость к B. cinerea была идентифицирована в диком изоляте Solanum habrochaites LYC 4/78 (Urbasch, 1986; пример 1). Для исследования генетики, определяющей эту устойчивость, было проведено картирование популяции F2 (n=174) от скрещивания между S. lycopersicum cv. Moneymaker и S. habrochaites LYC 4/78 (см. пример 3). Исходно в исследовании по картированию F2 было идентифицировано два QTL для устойчивости к B. cinerea (QTL-1h и QTL-2h, как описано выше). Позднее был выявлен третий QTL (QTL-4hA) в сегрегирующих потомках BC2S1, QTL, эффект которого можно было наблюдать в отсутствие QTL-2h. С использованием 2-стороннего анализа ANOVA было идентифицировано значимое эпистатическое взаимодействие между обоими QTL в наборе данных F2. Некоторые генотипические классы представлены с низкой частотой, и, таким образом, требуются крупные популяции F2 для детекции взаимодействий QTL (Tanksley, 1993). В популяции F2, полученной авторами настоящего изобретения, три растения представляли собой гомозиготные по QTL-1h и QTL-2h S. lycopersicum, в то время как 12 растений представляли собой гомозиготные по обоим из них S. habroсhaites. С использованием 2-стороннего ANOVA было выявлено значимое взаимодействие между обоими локусами. Затем анализ средних значений каждого из классов показал, что в случае гомозиготности по QTL2 в S. habroсhaites отсутствует дополнительный эффект QTL-4hA.
Одним недостатком QTL-картирования в межвидовых сегрегиующих популяциях F2 является широкая вариабельность фенотипов, которая легко имитирует QTL с незначительными эффектами. Другим недостатком является невозможность проведения повторяющихся тестов, поскольку каждое растение F2 обладает уникальным генотипом. Альтернативно для картирования можно использовать генетическую библиотеку, состоящую из ряда интрогрессивных линий (IL). Каждая IL в идеальном случае обладает одним определенным сегментом хромосомы, источником которого является донорный вид в единообразном в ином случае генетическом фоне (Zamir, 2001). Такие линии обладают повышенной способностью к идентификации QTL, поскольку: I) фенотипическая вариабельность между линией и контрольным культиваром ассоциирована с интрогрессивным сегментом; II) каждая линия содержит главным образом более 95% рекуррентного культивируемого родительского генома, и можно легко идентифицировать незначительные количественные эффекты посредством сравнения с рекуррентным родителем; III) эпистатические эффекты, вызываемые другими областями дикого генома, не представлены. Несвязанные отрицательные эпистатические взаимодействия, таким образом, могут привести к идентификации новых QTL (Eshed et al., 1995); IV) каждая линия является гомозиготной и бессмертной и, таким образом, позволяет многократное тестирование (во множестве условий окружающей среды), и V) проблемы стерильности практически отсутствуют вследствие того факта, что генетическая конституция каждой линии в большой степени идентична культивируемому сорту.
Первая полученная популяция IL относится уже к 1965 (Wehrhahn et al.), большинство популяций IL были получены в ходе последнего десятилетия. Помимо томата были получены популяции IL для ячменя (von Korff et al., 2004), капусты (Ramsay et al., 1996), салата (Jeuken et al., 2004), дыни (Eduardo et al., 2005), риса (Lin et al., 1998) и пшеницы (Pestsova et al., 2001). Все эти популяции IL были разработаны с использованием селекции на основе маркера (MAS), однако использовали различные стратегии, определяемые различным количеством возвратных скрещиваний и поколений близкородственного скрещивания для получения IL. Вторым отличием в стратегии был выбор используемой маркерной системы (т.е. типа маркеров) для получения популяции IL. Четыре из популяций, упомянутых выше, были получены с использованием маркеров SSR.
В Solanum, IL была получена для Solanum pennellii LA716 (Eshed et al., 1994), S. habrochaites LA1777 (Monforte et al., 2000a) и Solanum lycopersicoides LA2951 (Canady et al., 2005). Было показано, что такие популяции являются высокополезными для идентификации количественных признаков (Eshed et al., 1995; Rousseaux et al., 2005), точного картирования QTL (Fridman et al., 2004; Monforte et al., 2001; Monforte et al., 2000b) и клонирования QTL (Frary et al., 2000; Fridman et al., 2000; Ku et al., 2001).
В настоящее время одна популяция IL S. habrochaites LA1777 существует в детерминатном растущем S. lycopersicum E6203 (Monforte et al., 2000a).
В этом примере авторы настоящего изобретения описывают получение второй популяции IL S. habrochaites, в данном случае, основанной на интрогрессиях от S. habrochaites LYC 4/78 в фоне недетерминатно растущего культивируемого томата S. lycopersicum cv. Moneymaker, и применение линий для идентификации QTL для устойчивости к B. cinerea.
Материалы и способы
Растительный материал и получение IL
Межвидовое скрещивание между S. lycopersicum cv. Moneymaker (в дальнейшем в настоящем описании обозначаемым как SL) и S. habrochaites LYC 4/78 (в дальнейшем в настоящем описании обозначаемым как SH; партия семян 1978 года) проводили для получения семян F1. Семена SH получали из банка генов, находящегося в Institute for Plant Genetics and Crop plant research, Gatersleben, Germany. Одно растение F1 подвергали самоопылению с получением семян F2 и возвратному скрещиванию с SL с получением семян BC1. Семена F2 первоначально использовали для создания генетической карты сцепления. Семена BC1 использовали для получения IL (фиг.6).
Анализ маркеров
Геномную ДНК выделяли из двух молодых (свернутых) листов с использованием протокола на основе CTAB в соответствии со Steward et al. (1993), скорректированным для высокопроизводительного выделения ДНК с использованием микронных пробирок объемом один мл (Micronic BV, Lelystad, The Netherlands), и измельченных с использованием 300-мм устройства для встряхивания Retsch при максимальной скорости (Retsch BV, Ochten, The Netherlands).
Проводили анализ AFLPTM (Vos et al., 1995) каждого возвратного скрещивания и IL, и фрагменты AFLP разделяли на ДНК-секвенаторе LI-COR 4200, по существу в соответствии со способом, опубликованным Myburg (2001). Селективный праймер Pst метили посредством флуоресцентной метки IRD700 или IRD800. Изображения на геле для AFLP оценивали с использованием пакета программ AFLP-QuantarTM Pro (http://www.keygene-products.com). Последовательности праймеров и соединительные последовательности описаны Bai et al. (2003).
Наборы праймеров CAPS были получены посредством "Solanaceae Genomics Website" (http://sgn.cornell.edu) или сконструированы на основе последовательностей клонов геномной ДНК или кДНК, доступных из того же источника. Полиморфизмы между S. habrochaites и S. lycopersicum определяли с использованием подхода с расщеплением CAPS, описанного Brugmans et al. (2003). Последовательности маркеров, условия ПЦР и специфичные эндонуклеазы рестрикции, используемые для генотипирования, представлены в таблице 30. Продукты ПЦР в большинстве случаев разделяли с использованием 2,5% агарозного геля. В таблице 31 различные продукты расщепления, которые определяют отличия между S. lycopersicum и S. habrochaites, указаны для каждого из маркеров, приведенных в таблице 30, выявленных в представляющих интерес QTL.
Графическое изображение генотипов
Графическое изображение генотипов для каждого возвратного скрещивания и IL получали с использованием программного обеспечения GGT (van Berloo, 1999). Для вычисления размера интрогрессии и геномных процентов полуинтервалы, фланкирующие маркерный локус, считали одной и той же интрогрессией, как выполняется на программном обеспечении GGT. Пропущенные данные о маркерах оценивали из фланкирующих маркеров; считалось, что они обладают одним и тем же генотипом с двумя фланкирующими маркерами, если они обладали идентичными генотипами. Если два фланкирующих маркера обладали противоположными генотипами, тогда данные записывали как потерянные.
Оценка заболевания
Для оценки устойчивости 16 неполных случайных блоков использовали всего в 11 повторениях для каждой IL. Каждый блок содержал по меньшей мере два растения SE и одно растение S. lycopersicum cv. Durinta. Durinta представляет собой коммерческий культивар, продуцирующий гроздья томатов с длительным сроком хранения, и он проявляет определенный (но не высокий) уровень устойчивости. Через шесть недель после высевания растения переносили в полные отделы для почвы и выращивали в режиме 15 градусов ночью/19 градусов в течение дня и со световым периодом, составляющим 16 часов. Через 11 недель с использованием кухонного ножа в стебле каждого растения вырезали два очага повреждения приблизительно по 15 мм. На каждую рану инокулировали 1-см тампон с B. cinerea B05.10, содержащий агар (Benito et al. (1998)), и закрывали пленкой. Вторую инокуляцию проводили через две недели. В ходе теста растения поливали в начале вечера для поддержания увлажненного климата в течение ночи. Прогрессирование заболевания определяли через 9, 12 и 22 суток после инокуляции с использованием штангенциркуля. Прогрессирование заболевания описывали в соответствии со следующими параметрами: скорректированный размер очага повреждения (LS) через 12 суток (т.е. общая длина очага повреждения минус 15 мм раны), процентное количество увеличенных очагов повреждения (DI) и скорость роста очага повреждения, выраженная в качестве отличий в скорректированном размере очага повреждения, измеренного на 9 и 12 сутки и выраженного в мм/сутки (LG).
Статистический анализ
Статистический анализ проводили с использованием пакета программ SPSS 12.0 (SPSS Inc, Chicago, U.S.A.). С использованием способа общей линеаризованной модели (GLM) оценивали средние значения для каждой IL/признак. Измеренные средние значения признаков сравнивали с контрольным генотипом SL с использованием теста Даннета (Dunnett, 1955), и значения вероятности, меньшие 0,05, считали значимыми. В целях анализа LG и LS применяли преобразование квадратного корня для нормализации данных обоих признаков. Корреляции между признаками вычисляли с использованием коэффициента корреляции Пирсона.
Результаты
Популяция IL
Получали популяцию интрогрессивной линии (IL) S. habrochaites LYC 4/78 (SH) в генетическом фоне S. lycopersicum cv. Moneymaker (SL). Одно растение F1, полученное от скрещивания между SL и SH, подвергали возвратному скрещиванию с SL (фиг.6). Затем случайный набор из 14 растений BC1 подвергали возвратному скрещиванию с SL с получением потомства BC2 (n=59). Для всех растений BC2 проводили генотипирование, и отобранный набор подвергали возвратному скрещиванию с SL. Этот набор был выбран таким образом, чтобы комбинированные интрогрессии охватывали геном SH настолько, насколько это возможно, при селекции рекомбинантных растений таким образом, чтобы каждая чужеродная хромосома была представлена тремя IL. Этот процесс селекции и возвратного скрещивания повторяли до BC5. 31 выбранное растение BC5, главным образом обладающее одной или двумя интрогрессиями, подвергали самоопылению. Вплоть до 12 растений из каждого из 31 семейств BC5S1 подвергали самоопылению и скринингу с маркерами AFLP с получением потомства BC5S2 (n=44), гомозиготного по интрогрессии. Маркеры 44 IL подвергали скринингу еще один раз, и был выбран основной набор из 30 IL. Этот основной набор соответствует максимальному охвату генома SH в минимальном возможном количестве IL (фиг.7). Основной набор состоит из 15 IL, обладающих одной интрогресией, 10 IL, обладающих двумя интрогрессиями, 4 IL, обладающих тремя интрогрессиями, в то время как еще одна IL обладала четырьмя гомозиготными интрогрессиями. В среднем каждая IL содержала 60 сМ (= 5,2%) генома SH, и длина интрогрессий варьировала между 20 (1,7%) и 122 сМ (10,6%). Популяция IL, полученная авторами настоящего изобретения, охватывает 95% от длины исходной карты сцепления F2. Однако авторы настоящего изобретения понимают, что эта карта сцепления F2 не полностью охватывает геном. Это иллюстрируется дополнительным анализом CAPS на 3 (верхняя часть короткого плеча), 4 (верхняя часть короткого плеча), 5 (длинное плечо) и 9 (верхняя часть короткого плеча) хромосомах, где маркеры CAPS показали интрогрессии без маркеров в AFLP на основе карты сцепления F2. Размер этих интрогрессий оценивали исходя из плотности карты RFLP (Tanksley et al., 1992; http://www.sgn.cornell.edu). Поскольку ранее не проводили скрининга для верхней части 3 хромосомы, IL по этой области является гетерозиготной. Растения, отобранные, чтобы они представляли собой гомозиготные SH для IL5-1 и 5-2, не образовывали семена, таким образом, эти линии поддерживали в их гетерозиготном состоянии. Не было представлено IL, содержащих верхнюю часть короткого плеча 8 хромосомы и нижнюю часть длинного плеча 2 хромосомы. Интрогрессии на верхней части короткого плеча 7 и 9 хромосом представлены во множестве IL. Селекция по верхней части 9 хромосомы была возможна только после получения маркеров CAPS, специфичных для этой области.
Оценка заболевания
Популяцию из 30 IL выращивали в одиннадцати повторениях в неполном случайном расположении делянок, инокулировали и оценивали в отношении симптомов заболевания. Набор явно устойчивых и восприимчивых контролей был включен в тест. На 9, 12 и 22 сутки после инокуляции прогрессирование заболевания определяли и оценивали сравнением следующих трех параметров: вероятность роста инфекции, или заболеваемость (DI), скорректированный размер увеличивающего очага повреждения (LS) и скорость роста очага повреждения (LG). Устойчивое родительское растение SH не показало каких-либо симптомов (таблица 8, таблица 9), в то время как 73% очагов повреждения SL были растущими.
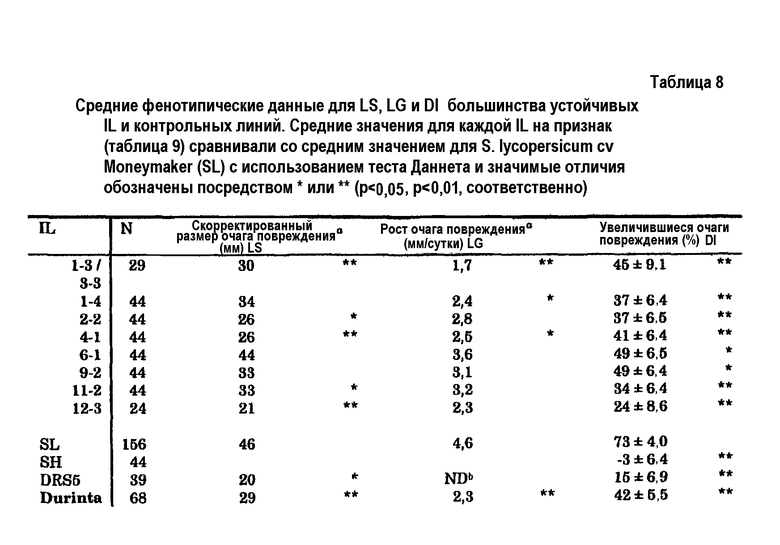
aПервое исследование поддающегося измерению очагу повреждения было проведено через 12 суток. bНет данных.
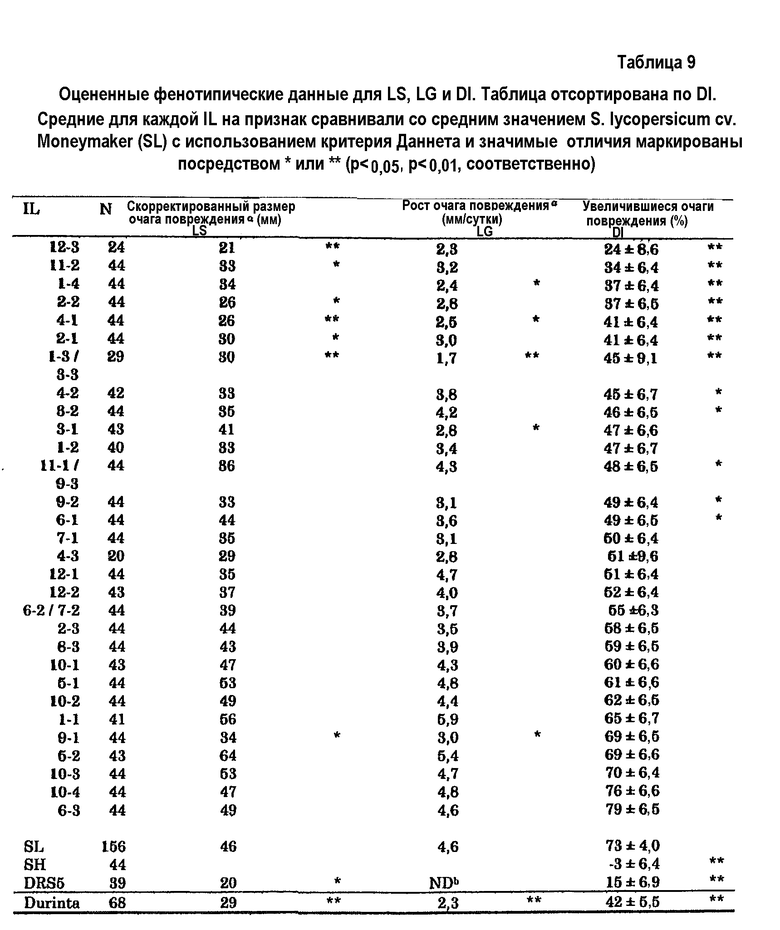
В популяции IL 14 IL были идентифицированы с уменьшенными симптомами заболевания (таблица 9). Всего 12 IL показали значительно меньшие DI. Семь IL имели значительно сниженный LS, и пять имели значительно меньший LG. IL4-1 и IL 1-3/3-3 показали значительное снижение всех трех параметров (DI, LG и LS). В IL со значительно сниженным DI диапазон в процентных количествах растущих очагов составлял 24-49%. LS и LG варьировали для значительно варьирующих IL между 21-34 мм и 1,7-3,0 мм/сутки соответственно. Контрольный S. lycopersicum cv. Durinta с определенным уровнем устойчивости также обладал значимо меньшими всеми тремя параметрами, и устойчивость в каждом из семи идентифицированных IL является более или менее сравнимой с этим уровнем. В предшествующих экспериментах был отобран высокоустойчивый генотип BC2S2 (DRS 5, см. таблицы 8 и 9). Эта линия содержит три гомозиготных интрогрессии, соответствующих суммарно 18% от генома SH (фиг.7). Эта линия представляла собой наиболее устойчивую линию в тесте, описанном выше. Она имела значительно более низкий DI (15%), и LS также был значительно снижен. По сравнению с S. lycopersicum cv. Durinta, сниженные в 2,8 раза DI этой линии, в значительно меньшей степени показывают потенциал в отношении выстраивания пирамиды из множества аллелей, придающих устойчивость к B. cinerea.
Эффект объединенных интрогрессий
При анализе различных интрогрессий, представленных в отдельных линиях IL (фиг.7), и их сравнении с их индивидуальными эффектами на устойчивость, как показано в таблице 9, можно сделать заключение об эффектах объединения. Как можно видеть в линиях 9-1, 9-2, 11-2 и 12-3, объединение интрогрессий может оказывать эффект.
Из примера следует, что интрогрессия в 6 хромосоме в линии 9-2 не производит эффект, поскольку идентичная интрогрессия в линии IL6-3 не придает устойчивость.
Аналогично интрогрессия в 7 хромосоме в линии 11-2 не производила эффект (сравнить с линиями 7-1 и 8-2).
Два паттерна устойчивости (DI и LS), представленные в 11-2, не являются результатом интрогрессии только на 11 хромосоме. Снижение размера очага повреждения (LS) в этой линии может быть следствием интрогрессии с 9 хромосомы (сравнить со сходными интрогрессиями в 9-1), в то время как снижение заболеваемости ни разу не было выявлено в ассоциации с интрогрессией 9-1 отдельно. Таким образом, снижение заболеваемости должно быть следствием интрогрессии с 11 хромосомы. Таким образом, в линии 11-2 общая устойчивость является результатом различных интрогрессий.
Аналогично снижение % увеличивающихся очагов повреждения в линии 12-3 по сравнению с линией 9-1 может быть следствием интрогрессии в 12 хромосоме, в то время как сниженный скорректированный размер очага повреждения может быть следствием интрогрессии с 9 хромосомы. Таким образом, здесь также комбинированное наличие множественных интрогрессий приводит к повышенной устойчивости.
Сцепление между IL и данными о заболевании
С использованием биологического анализа в теплице авторы настоящего изобретения идентифицировали набор IL, обладающих интрогрессиями, ответственными за повышенную устойчивость к B. cinerea. Эти участки, расположенные на 2, 4 и 6 хромосомах, однозначно содержат ген(ы) для повышенной устойчивости. Другие IL обладают множественными интрогрессиями, что делает трудным определение генов устойчивости. IL9-2 обладает интрогрессиями на 6 и 9 хромосомах. Интрогрессия на 6 хромосоме в IL9-2 является сходной с интрогрессией хромосомы 6 в IL6-3, IL, настолько же восприимчивой, как и SL. Таким образом, авторы настоящего изобретения полагают, что эффект IL9-2 вызван интрогрессией на 9 хромосоме, а не на 6 хромосоме. IL11-2 обладает интрогрессией на 9 хромосоме, меньшей, чем представлено в IL9-1. Таким образом, полагают, что DI вызван локусом на 11 хромосоме. Две IL 1-4 и 12-3 обладают интрогрессиями, перекрывающими интрогрессию на 9 хромосоме IL9-2. Только IL12-3 является значительно более устойчивой, чем IL9-2, предполагая сочетанный эффект интрогрессий на 9 и 12 хромосомах. Поскольку IL 1-4 и 11-2 не являются значительно более устойчивыми, чем IL9-2, авторы настоящего изобретения не могут исключить, что устойчивость в этих двух линиях является результатом интрогрессии 9 хромосомы.
Обобщая, авторы настоящего изобретения идентифицировали интрогрессии, расположенные на 1, 2, 4 (2x), 6, 9, 11 и 12 хромосомах, которые отвечают за повышенную устойчивость к B. cinerea. Эффекты на 2 хромосоме и на 4 хромосоме ранее были выявлены в ходе анализа сегрегирующих популяций F2 и BC2S1 от этого скрещивания (см. примеры 1-3).
Сегрегация идентифицированных локусов в F 2
Для всех участков, в которых выявлена ассоциация между интрогрессией и повышенной устойчивостью к B. cinerea, исходный набор данных F2 проверяли для того, чтобы найти возможное объяснение того, почему QTL исходно был пропущен. Проверяли как смещение данных о маркерах, так и результаты анализа QTL. Интрогрессии на 1 хромосоме (1:6:6), 2 хромосоме (1:3:2) и 9 хромосоме (1:6) значительно отклонялись от ожидаемых соотношений 1:2:1 или 1:3. Для всех трех участков выявлено отсутствие гомозиготных аллелей S. lycopersicum. Данные анализа QTL, как для картирования расстояний, так и для анализа отдельных маркеров с использованием теста Крускала-Валлиса, проверяли, но не было выявлено данных о существовании значимых QTL в наборе данных популяции F2 на 6, 9, 11 и 12 хромосомах.
Последовательности маркеров, используемые в настоящем описании.
В следующих таблицах представлена подробная информация о различных маркерах RFLP и COS-II, как указано в различных картах сцепления и как указано для ассоциации с QTL по настоящему изобретению. Информацию копировали непосредственно из базы данных SOL Genomic Network (SGN), находящейся в Cornell University, версия 7 октября 2005 года.




















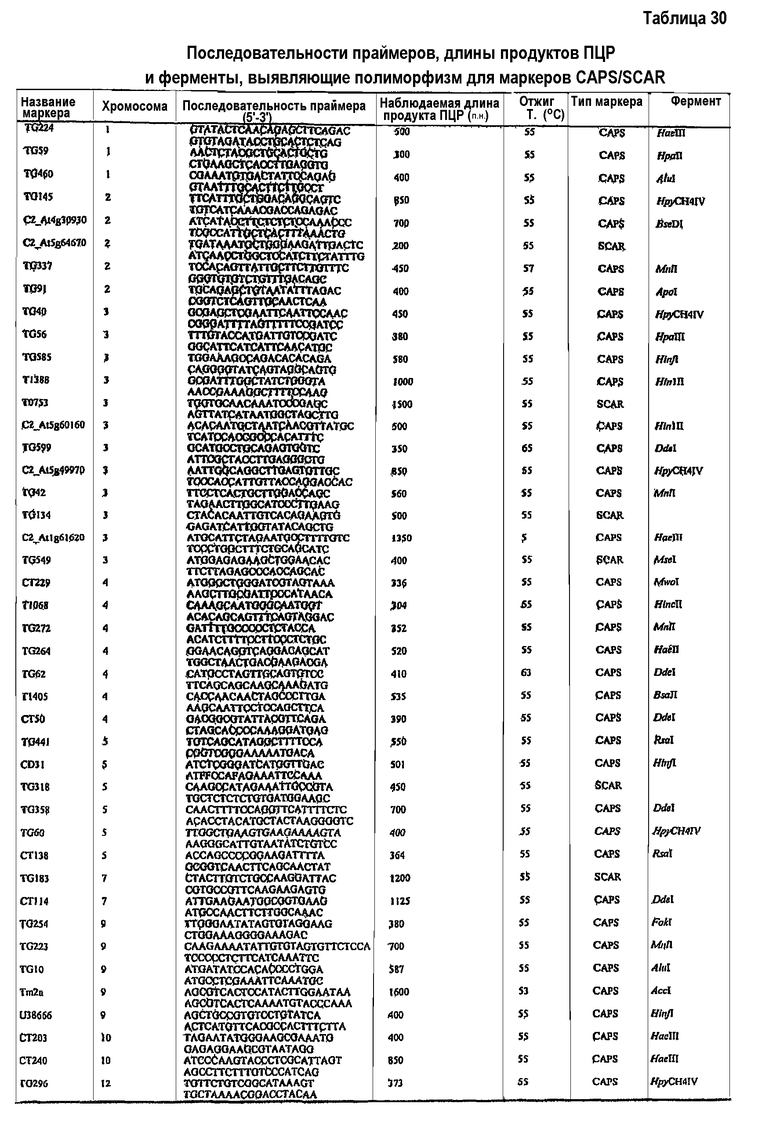
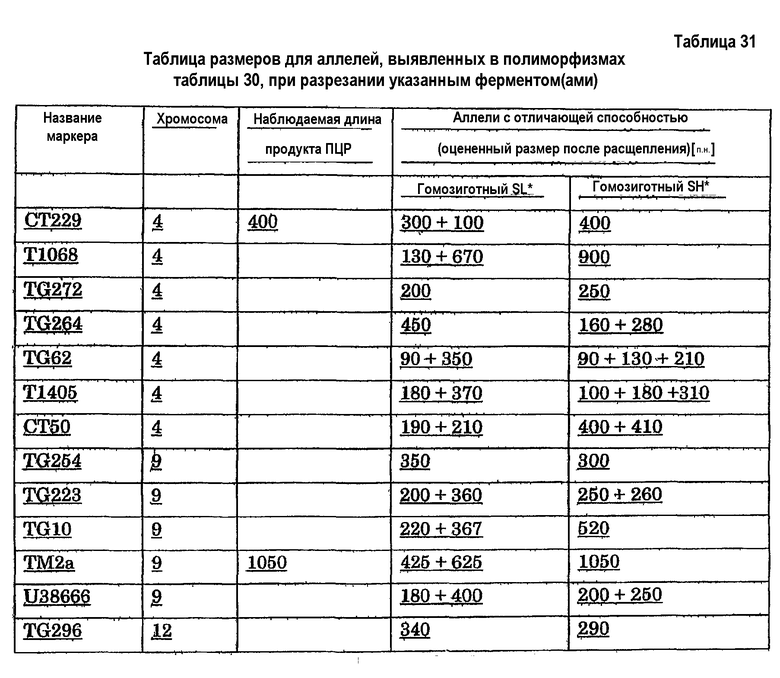
*SL = Solanum lycopersicum. SH = Solanum habrochaites. В гетерозиготных растениях выявлены расщепленные продукты как SL, так и SH. Как в таблице 30, так и в таблице 31 выявленная длина продукта ПЦР оценена из полос на агарозном геле
Список литературы
Ausubel F.M, Brent R, Kingston R.E, Moore D.D, Seidman J.G, Smith J.A, Struhl K. (1995). "Current Protocols in Molecular Biology", 4th edition, John Wiley and Sons Inc., Mew York, N.Y.
Bai Y.L, Huang C.C, van der Hulst R, Meijer Dekens F, Bonnema G, Lindhout P (2003) QTLs for tomato powdery mildew resistance (Oidium lycopersiti) in Lycopersicon parviflorum G1.1601 co-localize with two qualitative powdery mildew resistance genes. Mol. plant microbe interactions 16:169-176.
Benito E.P, ten Have A, van 't Klooster J.W, van Kan JAL (1998) Fungal and plant gene expression during synchronized infection of tomato leaves by Botrytis cinerea, Eur. J. Plant Pathol. 104:207-220.
Bernacchi D, Tanksley S.D. (1997) An interspecific backcross of Licopersicon lycopersicum × L. hirsutum: Linkage analysis and a QTL study of sexual compatibility factors and floral traits. Genetics 147:861-877.
Brugmans B, van der Hulst RGM, Visser RGF, Lindhout P, van Eck H.J. (2003) A new and versatile method for the successful conversion of AFLP (TM) markers into simple single locus markers. Nucleic acids research 31: Nil_9-Nil_17.
Canady M.A, Meglic V, Chetelat R.T (2005) A library of Solanum lycopersicoides introgression lines in cultivated tomato. Genome 48:685-697.
Christon P, Murphy J.E, and Swain W.F (1987) Stable transformation of soybean by electroporation and root formation from transformed callus. Proc. Natl. Acad. Sci. USA 84:3962-3966.
Churchill G.A., Doerge R.W. (1994) Empirical threshold values for Quantitative trait mapping. Genetics 188:963-971.
Deshayes A, Herrera-Estrella L, Caboche M. (1985) Liposome-mediated transformation of tobacco mesophyll protoplasts by an Escherichia coli plasmid. EMBO J. 4:2731-2737.
D'Halluin K, Bonne E, Bossut M, De Beuckeleer M, Leemans J. (1992) Plant. Cell 4:1495-1505.
Dik A.J., Koning G, Kohl J. (1999) Evaluation of microbial antagonists for biological control of Botrytis cinerea stem infection in cucumber and tomato. Eur. J. Plant Pathol. 105:115-122.
Doganlar S, Frary A, Ku H.M. and Tanksley S.D. (2002) Mapping Quantitative Trait Loci in Inbred Backcross Lines of Lycopersicon pimpinellifolium (LA1589). Genome 45:1189-1202.
Draper J, Davey M.R., Freeman J.P., Cocking E.C. and Cox B.J. (1982) Ti plasmid homologous sequences present in tissues from Agrobacterium plasmid-transformed Petunia protoplasts. Plant and Cell Physiol. 23:451-458.
Dunnett C.W. (1955) A multiple comparison procedure for comparing several treatments with a control. Journal of the American Statistical Association 50:1096-1121.
Eckstein F (ed) (1901) Oligonucleotides and Analogues, A Practical Approach. Oxford Univ. Press, NY 1991.
Eduardo I, Arus P, Monforte A.J. (2005) Development of a genomic library of near isogenic lines (NILs) in melon (Cucumis melo L.) from the exotic accession PI161375. Theor Appl. Genet 112:139-148.
Egashira H, Kuwashima A, Ishiguro H, Fukushima K, Kaya T, Imanishi S. (2000) Screening of wild accessions resistant to gray mold (Botrytis cinerea Pers.) in Lycopersicon. Acta physiologiae plantarum 22:324-326.
Eshed V, Zamir D. (1994) A genomic library of Lycopersicon pennellii in S. lycopersicum: a tool for fine mapping of genes. Euphytica, Dordrecht: Kluwer Academic Publishers. 1994 79:175-179.
Eshed Y, Zamir D. (1995) An introgression line population of Lycopersicon pennellii in the cultivated tomato enables the identification and fine mapping of yield-associated QTL. Genetics. Bethesda, Md.: Genetics Society of America. Nov 1995 141:1147-1162.
Foolad M.R, Zhang L.P, Khan A.A, Nino Liu D, Liln G.Y (2002) Identification of QTLs for early blight (Alternaria solani) resistance in tomato using backcross populations of a Lycopersicon lycopersicum × L. hirsutum cross. Theor, Appl. Genetics 104:945-958.
Frary A, Doganlar S, Frampton A, Fulton T, Uhlig J, Yates H, Tanksley S. (2003) Fine mapping of quantitative trait loci for improved fruit characteristics from Lycopersicon chmielewskii chromosome 1. Genome 46:235-243.
Frary A, Nesbitt T.C, Grandillo S, Knaap Evd, Cong B, Liu J, Meller J, Elber R, Alpert K.B, Tanksley S.D (2000) fw2/2: a quantitative trait locus key to the evolution of tomato fruit size. Science Washington. 2000; 289:85-88.
Fridman E, Carrari F, Liu Y.S, Fernie A.R, Zamir D. (2004) Zooming in on a quantitative trait for tomato yield using interspecific introgressions. Science 305:1786-1789.
Fridman E, Pleban T, Zamir D (2000) A recombination hotspot delimits a wild-species quantitative trait locus for tomato sugar content to 484 bp within an invfectase gene. Proc. Natl. Acad Sci USA. Washington, D.C.: National Academy of Sciences. Apr 25, 2000 97:4718-4723.
Fulton T, van der Hoeven R, Eannetta N, Tanksley S (2002). Identification, Analysis and Utilization of a Conserved Ortholog Set (COS) Markers for Comparative Genomics in Higher Plants. The Plant Cell 14(7):1457-1467.
Godoy G, Steadman J.R, Dickman M.B, Dam R. (1990) Use of mutants to demonstrate the role of oxalic acid in pathogenicity of Sclerotinia sclerotiorum on Phaseolus vulgaris. Physiological Molecular Plant Pathology 87, 179-191.
Grandillo S, Tanksley S.D. (1996) QTL analysis of horticultural traits differentiating the cultivated tomato from the closely related species Lycopersicon pimpinellifolium. Theor Appl Genet 92:935-951.
Gruber M.Y, Crosby W.L, (1993) Vectors for Plant Transformation. In: Glick B.R, and Thompson J.E, (Eds.) Methods in Plant Molecular Biology & Biotechnology, CRC Press, pp.89-119.
Haanstra JPW, Wye C, Verbakel H, Meijer Dekens F, van den Berg P, Odinot P, van Heusden A.W, Tanksley S, Lindhout P, Peleman J. (1999) An integrated high density RFLP-AFLP map of tomato based on two Lycopersicon lycopersicum × L. pennellii F2 populations. Theor. Appl. Genetics 99:254-271.
Hain R, Stabel P, Czernilofsky A.P, Steinbliss H.H, Herrera-Estrella L, Schell J. (1985) Uptake, integration, expression and genetic transmission of a selectable chimaeric gene to plant protoplasts. Mol. Gen. Genet. 199:161-168.
Horsph R.B, Fry J.E, Hoffman N.L, Eichholts D, Rogers S.G, Fraley R.T, (1985) A simple method for transferring genes into plants. Science 227:1229-1231.
Jansen R.C. (1993) Interval Mapping of Multiple Quantitative Trait Loci. Genetics 135:205-211.
Jansen R.C. (1994) Controlling the Type I and Type II Errors in Mapping Quantitative Trait Loci. Genetics 138:871-881.
Jeuken MJW, Lindhout P. (2004) The development of lettuce backcross inbred lines (BILs) for exploitation of the Lactuca saligna (wild lettuce) germplasm. Theor Appl Genet 109:394-401.
Kado C.I, (1991) Molecular mechanisms of crown gall tumorigenesis. Crit. Rev. Plant Sci. 10:1-32.
Klein T.M, Gradziel T, Fromm M.E, Sanford J.C, (1988). Factors influencing gene delivery into zea mays cells by high velocity microprojectiles. Biotechnology 6:559-563.
Klein T.M, Arentzen R, Lewis P.A, and Fitzpatrick- McElligott S. (1992) Transformation of microbes, plants and animals by particle bombardment, Bio/Technology 10:286-291.
Kosambi D.D. (1944) The estimation of map distances from recombination values. Ann. Eugen. 12:172-175.
Ku H.M, Liu J, Doganlar S, Tanksley S.D. (2001) Exploitation of Arabidopsis-tomato synteny to construct a high-resolution map of the ovate-containing region in tomato chromosome 2. Gzenome. Ottawa, Ontario, Canada: National Research Council of Canada. June 2001, 44:470-475.
Laursen C.M, Krzyzek R.A, Flick C.E, Anderson P.C, Spencer T.M. (1994) Production of fertile transgenic maize by electroporation of suspension culture cells. Plant Mol. Biol. 24(1):51-61.
Lin S.Y, Sasaki T, Yano M. (1998) Mapping quantitative trait loci controlling seed dormancy and heading date in rice, Oryza sativa L., using backcross inbred lines. Theor Appl. Genet 96:997-1003.
Miki B.L, Fobert P.F, Charest P.J, Iyer V.N. (1993) Procedures for Introducing Foreign DNA into Plants. In: Glick B.R. and Thompson J.E. (Eds.) Methods in Plant Molecular Biology & Biotechnology, CRC Press, pp.67-88.
Moloney M.M, Walker J.M, Sharma K.K. (1989) High efficiency transformation of Brassica napus using Agrobacterium vectors. Plant Cell Reports 8:238-242.
Monforte A.J, Friedman E, Zamir D, Tanksley S.D. (2001) Comparison of a set of allelic QTL-NILs for chromosome 4 of tomato: deductions about natural variation and implications for germplasm utilization. Theor appl genet. Berlin; Springer Verlag. Mar 2001? 102:572-590.
Monforte A.J, Tanksley S.D. (2000a) Development of a set of near isogenic and backcross recombinant inbred lines containing most of the Lycopersicon hirsutum genome in a L. esculentum genetic background: A tool for gene mapping and gene discovery. Genome 43:803-813.
Monforte A.J, Tanksley S.D. (2000b) Fine mapping of a quantitative trait locus (QTL) from Lycopersicon hitsutum chromosome 1 affecting fruit characteristics and agronomic traits: breaking linkage among QTLs affecting different traits and dissection of heterosis for yield. Theor appl genet. Berlin; Springer Verlag. Feb 2000, 100:471-479.
Myburg A.A, Remington D.L, O'Malley D.M., Sederoff R.R, Whetten R.W. (2001) High-throughput AFLP analysis using infrared dye-labeled primers and an automated DNA sequencer. Biotechniques 30:348-357.
Nesbitt T.C, Tanksley S.D. (2001) fw2.2 directly affects the size of developing tomato fruit, with secondary effects on fruit number and photosynthate distribution. Plant Physiol. 127:575-583.
Nicot P.C, Moretti A, Romiti C, Bardin M, Caranta C, Ferriere H. (2002) Differences in susceptibility of pruning wounds and leaves to infection by Botrytis cinerea among wild tomato accessions. TGC Report 52:24-26.
Paterson A.H. (ed.) (1996) Genome Mapping in Plants, Academic Press Inc San Diego, CA, USA.
Pestsova E.G, Borner A, Roder M.S. (2001) Development of a set of Triticum aestivum-Aegilops tauschii introgression lines. Hereditas 135:139-143.
Phillips R.L, Somers D.A, Hibberd K.A. 1988. Cell/tissue culture and in vitro manipulation. In: G.F. Sprague & J.W. Dudley, eds. Corn and corn improvement, 3rd ed., p.p.345-387. Madison, WI, USA, American Society of Agronomy.
Pierik RLM (1999) In vitro Culture of Higher Plants, 4th edition, 360 pages, ISBN:0-7923-5267-X.
Prins T.W, Tudzynski P, von Tiedemann A, Tudzynski B, ten Have A, Hansen M.E, Tenberge K, van Kan JAL (2000) Infection strategies of Botrytis cinerea and related necrotrophic pathogens. In "Fungal Pathology" (J. Kronstad, editor). Kluwer Academic Publishers, pp.33-64.
Ramsay L.D, Jennings D.E, Bohuon EJR, Arthur A.E, Lydiate D.J, Kearsey M.J, Marshall D.F. (1996) The construction of a substitution library of recombinant backcross lines in Brassica oleracea for the precision mapping of quantitative trait loci. Genome 39:558-667.
Roupe van der Voort JNAM, van Zandvoort P, van Eck H.J, Folkertsma R.T, Hutten RCB, Draaistra J, Gommers F.J, Jacobsen E, Helder J, Bakker J. (1997) Use of allele specificity of comigrating AFLP markers to align genetic maps from different potato genotypes. Mol. Gen Genetics 256:438-447.
Rousseaux M.C, Jones C.M, Adams D, Chetelat R, Bennett A, Powell A. (2005) QTL analysis of fruit antioxidants in tomato using Lycopersicon pennellii introgression lines. Theor Appl Genet 111:1396-1408.
Sambrook J, and Russell D.W. (2001). Molecular Cloning: A Laboratory Manual, New York, NY, USA., Cold Spring Harbor Laboratory Press.
Sanford J.C, Klein T.M, Wolf E.D, Allen N. (1987), Delivery of substances into cells and tissues using a particle bombardment process. J. Particulate Sci. Technol. 5:27-37.
Sanford J.C. (1988) The biolistic process. Trends in Biotechnology 6:299-302.
Sanford J.C. (1990) Biolistic plant transformation. Physiologica Plantarum 79:206-209.
Sanford J.C, Smith F.D, and Russell J.A. (1993) Optimizing the biolistic process for different biological applications. Methods in Enzymology 217:483-509.
Sobir O.T, Murata M, and Motoyoshi F. (2000) Molecular characterization of the SCAR markers tightly linked to the TM-2 locus of the genus Lycopersicon. Theor. Appl. Genet. 101:64-69.
Steward C.N, Via L.E. (1993) A rapid CTAB DNA isolation technique useful for RAPD fingerprinting and other PCR applications. Biotechniques 14:748-750.
Tanksley S.D. (1993) Mapping polygenes. Annu Rev Genet 27:205-233.
Tanksley S.D, Ganal M.W, Prince J.P, de Vicente M.C, Bonierbale M.W, Broun P, Fulton T.M, Giovannoni J.J, Grandillo S, Martin G.B. (1992) High density molecular linkage maps of the tomato and potato genomes. Genetics 132:1141-1160.
Tanksley S.D, Grandillo S, Fulton T.M, Zamir D, Eshed Y, Petiard V, Lopez J. and Beck-Bunn T. (1996) Advanced backcross QTL analysis in a cross between an elite processing line of tomato and its wild relative L. pimpinellifolium. Theor Appl Genet 92:213-224.
Tanksley S.D, Young N.D, Paterson A.H, Bonierbale M.W. (1998) RFLP mapping in plant breeding: New tools for an old science. Bio/technology 7:257-263.
Tijssen P. (1993) Hybridisation With Nucleic Acid Probes. Part I. Theory and Nucleic Acid Preparation. In: Laboratory Techniques in Biochemistry and Molecular Biology. Elsevier.
Urbashn I. (1986) Resistenz verschiedener Kultur- und Wildtomatenpflanzen (Licopersicon spp.) gegenüber Botrytis cinerea Pers. J. Phytopathol 116:344-351.
Utkhede R, Bogdanoff C, McNevin J. (2001) Effects of biological and chemical treatments on Botrytis stem canker and fruit yield of tomato under greenhouse conditions. Can. J. Plant Pathol 23:253-259.
Utkhede R.S, Mathur S. (2002) Biological control of stem canker of greenhouse tomatoes caused by Botrytis cinerea. Can. J. Microbiol. 48:550-554.
Van Berloo R. (1999) GGT: Software for the display of graphical genotypes. J. Heredity 90:328-329.
Van Berloo R, Aalbers H, Werkman A, Niks R.E. (2001) Resistance QTL confirmed through development of QTL-NILs for barley leaf rust resistance. Mol. Breeding 8:187-195.
Van Heusden A.W, Koornneef M, Voorrips R.E, Bruggemann W, Pet G, Vrielink van Ginkel R, Chen X, Lindhout P. (1999) Three QTLs from Lycopersicon peruvianum confer a high level of resistance to Clavibacter michiganensis ssp michiganensis. Theor. Appl. Genetics 99:1068-1074.
Von Korff M, Wang H, Leon J, Pillen K. (2004) Development of candidate introgression lines using an exotic barley accession (Hordeum vulgare ssp spontaneum) as donor. Theor Appl Genet 109:1736-1745.
Voorrips R.E. (2002) MapChart: software for the graphical presentation of linkage maps and QTLs. J. Heredity 93:77-78.
Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Hornes M, Frijters A, Pot J, Peleman J, Kuiper M/ (1995) AFLP: a new technique for DNA fingerprinting. Nucl. Acids Res, 23:4407-4414.
Wehrhahn C, Allard R.W. (1965) The detection and measurement of the effects of individual genes involved in inheritance of a quantitative character in wheat. Genetics 51:109-119.
Zamir D. (2001) Improving plant breeding with exotic genetic libraries. Nature reviews genetics 2:983-989.
Zhang L, Cheng L, Xu N, Zhao M, Li C, Yuan J, and Jia S. (1991) Efficient transformation of tobacco by ultrasonication. Biotechnology 9:996-997.
| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЯ ТОМАТОВ, ИМЕЮЩИЕ БОЛЕЕ ВЫСОКИЕ УРОВНИ УСТОЙЧИВОСТИ К BOTRYTIS | 2005 |
|
RU2403709C2 |
| РАСТЕНИЕ ТОМАТА, ОБРАЗУЮЩЕЕ ПЛОДЫ С МОДИФИЦИРОВАННЫМ СОДЕРЖАНИЕМ САХАРОВ | 2019 |
|
RU2817600C2 |
| РАСТЕНИЯ ТОМАТОВ С УЛУЧШЕННЫМИ ПРИЗНАКАМИ | 2019 |
|
RU2821992C2 |
| РАСТЕНИЯ ТОМАТА С УЛУЧШЕННОЙ УСТОЙЧИВОСТЬЮ К БОЛЕЗНЯМ | 2015 |
|
RU2721952C2 |
| СПОСОБЫ ОТПУГИВАНИЯ БЕЛОКРЫЛОК | 2008 |
|
RU2484628C2 |
| ИНТРОГРЕССИЯ ДВУХ QTL УРОЖАЯ В РАСТЕНИЯ CUCUMIS SATIVUS | 2017 |
|
RU2746936C2 |
| РАСТЕНИЯ ОГУРЦА, УСТОЙЧИВЫЕ К ЗАБОЛЕВАНИЯМ | 2006 |
|
RU2418405C2 |
| ГЕН, ПРИДАЮЩИЙ УСТОЙЧИВОСТЬ К PHYTOPHTHORA INFESTANS (ФИТОФТОРОЗУ) В СЕМЕЙСТВЕ SOLANACEAE | 2003 |
|
RU2361920C2 |
| ГЕНЫ И СПОСОБЫ ОБЕСПЕЧЕНИЯ УСТОЙЧИВОСТИ К ФИТОФТОРОЗУ ПАСЛЕНОВЫХ | 2008 |
|
RU2511423C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ ГЕНА RLM2 РЕЗИСТЕНТНОСТИ К ЧЕРНОЙ НОЖКЕ BRASSICA NAPUS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2717017C2 |
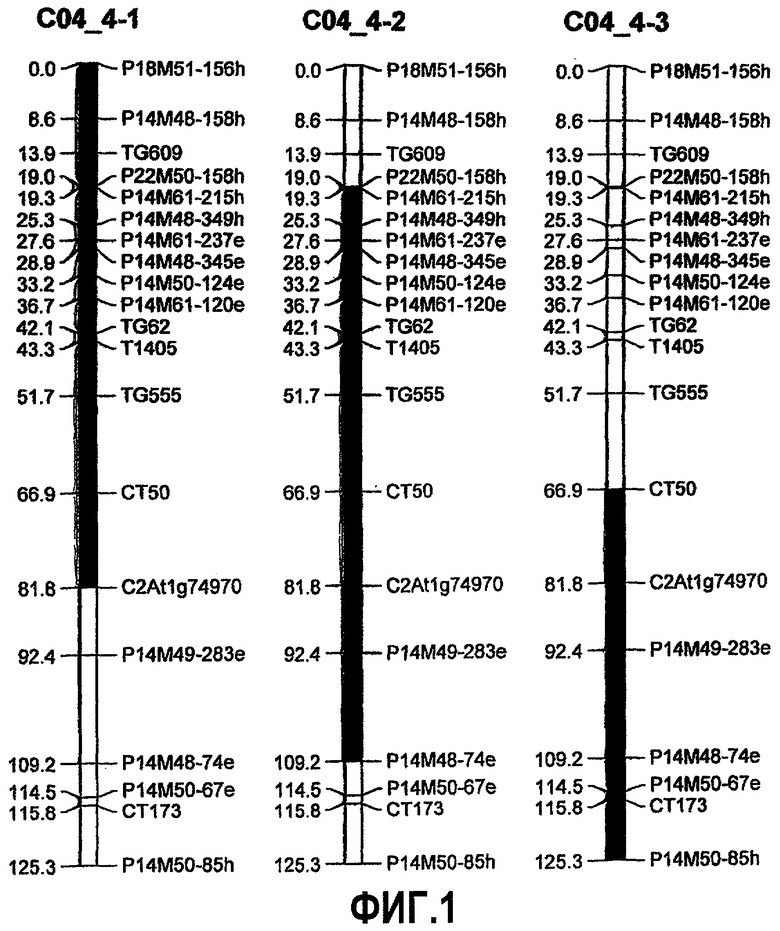
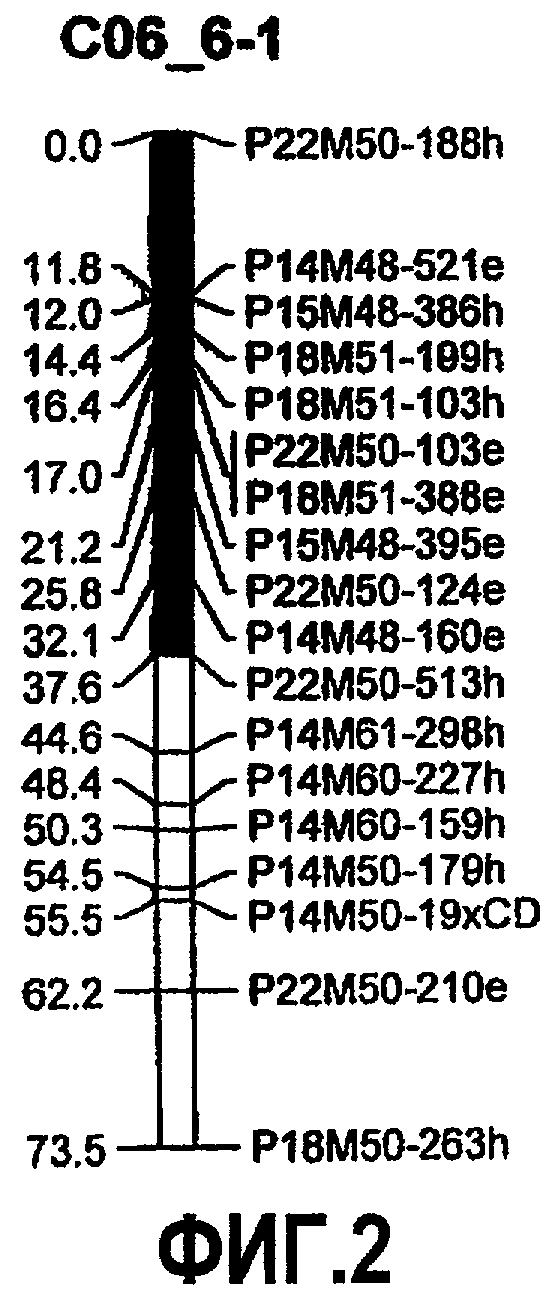
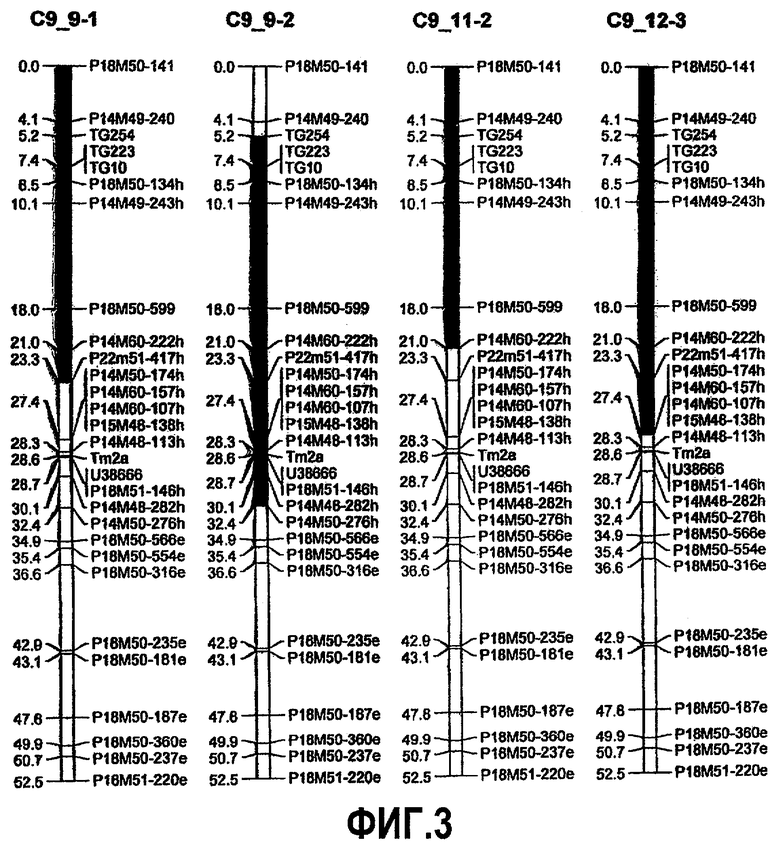
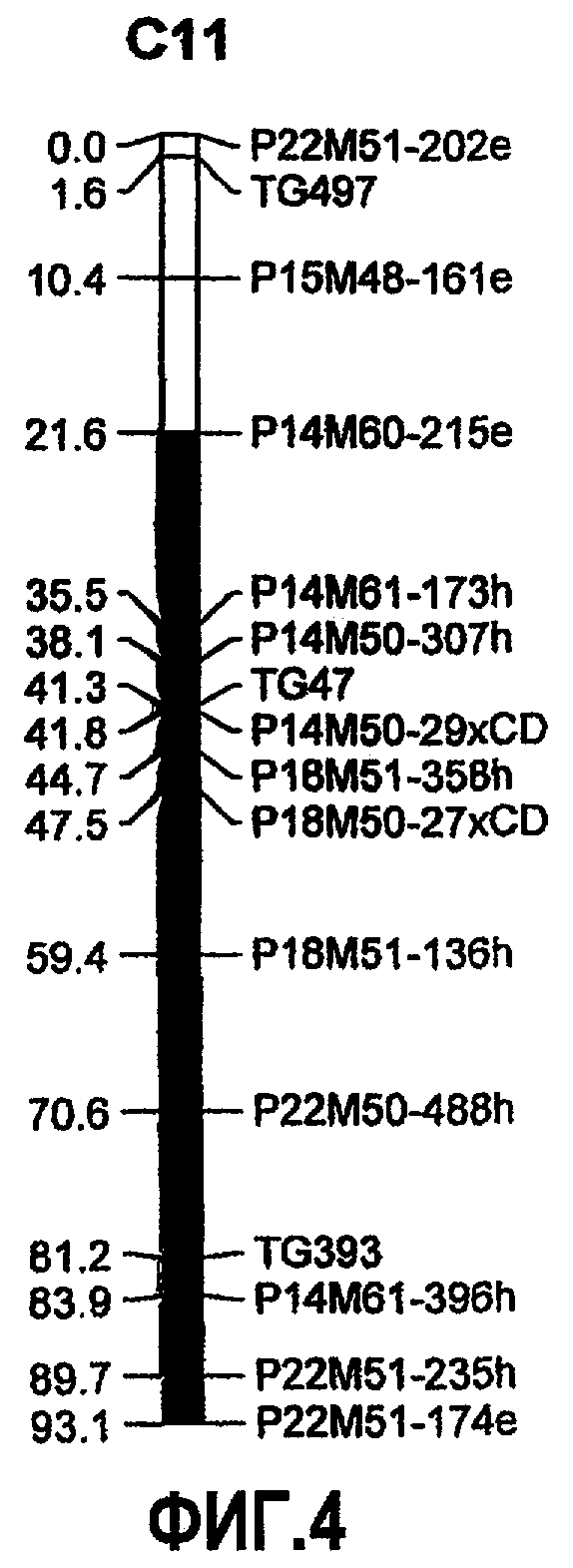

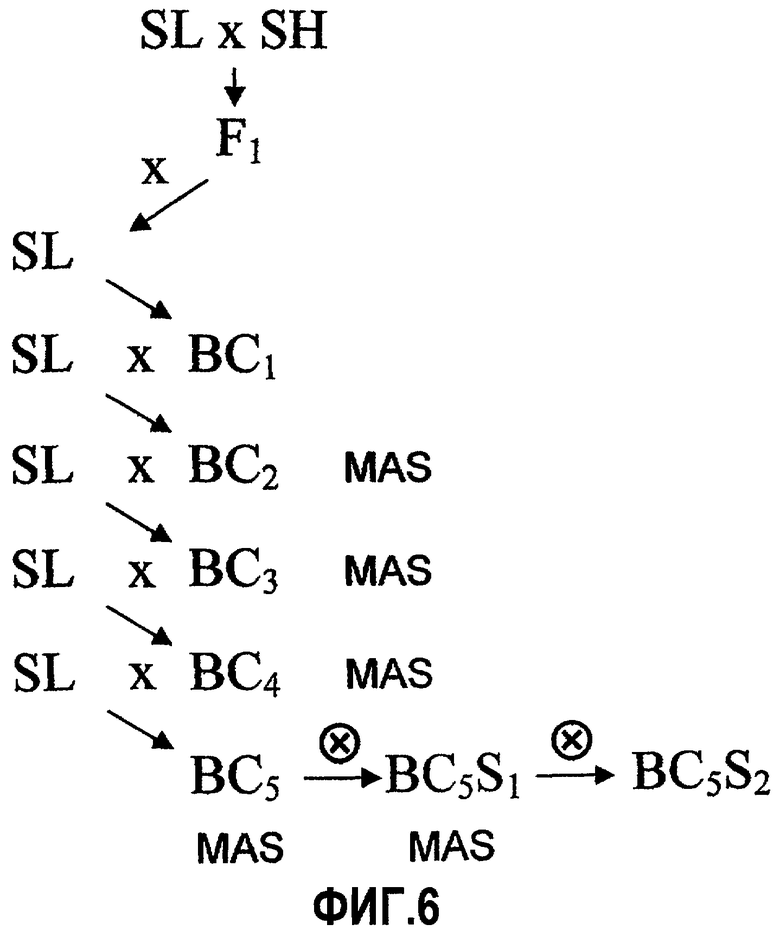

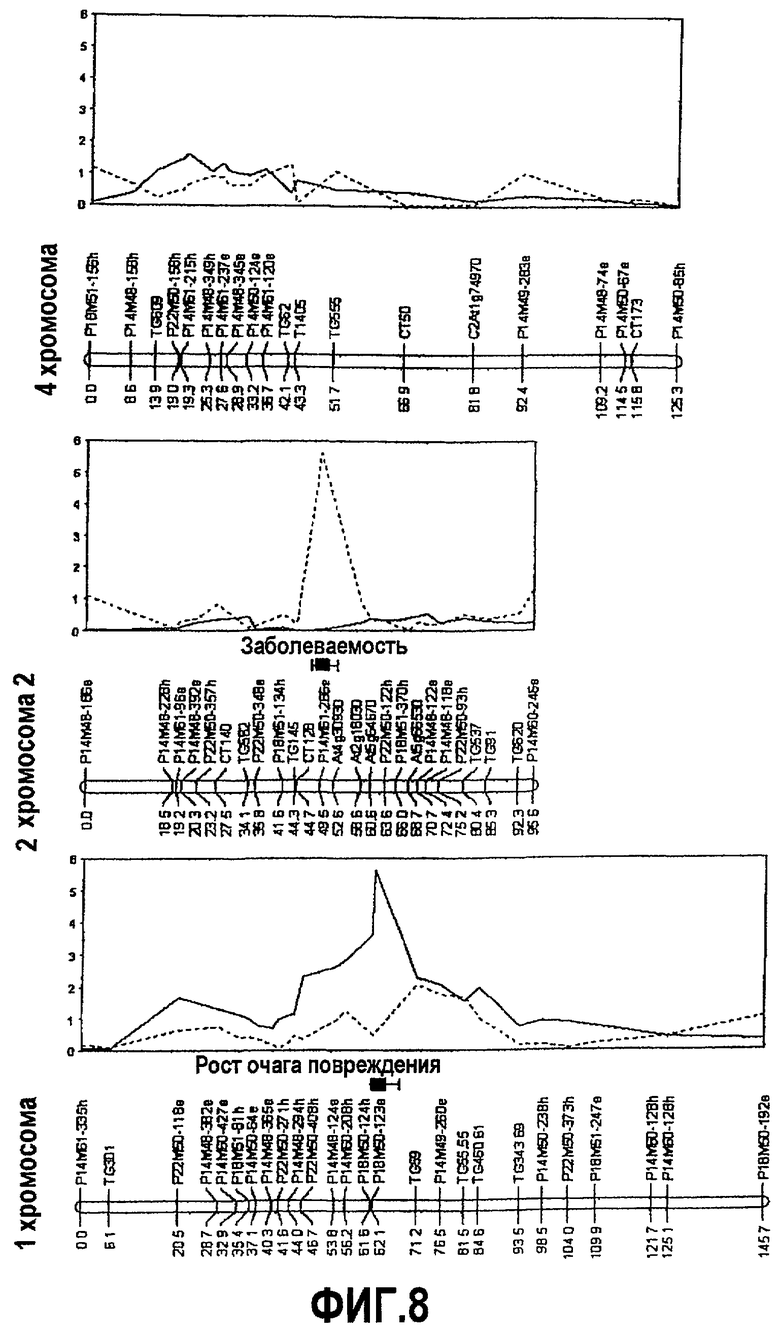
Изобретение относится к области биотехнологии. Описан способ получения томата, устойчивого к Botrytis, включающий а) получения устойчивого к Botrytis донорного растения томата предпочтительно вида S. Habrochaites или линии S. habrochaites LYC 4/78, b) переноса нуклеиновой кислоты из указанного донорного растения в одно или несколько восприимчивых к Botrytis реципиентных растений томата, с) отбора среди реципиентных растений томата растения, которое содержит в геноме по меньшей мере один локус количественного признака (QTL) для устойчивости к Botrytis из указанного устойчивого к Botrytis донорного растения, где указанный отбор включает детекцию на 4, 6, 9, 11 и/или 12 хромосоме указанного реципиентного растения томата по меньшей мере одного генетического маркера, связанного с указанным по меньшей мере одним QTL для устойчивости к Botrytis. Предложен способ получения инбредного растения томата, включающий скрещивание устойчивого к Botrytis растения томата с этим же растением или с другим растением томата. Кроме того, описана ДНК, полученная из растения томата вида S. lycopersicum, содержащая QTL, выбранный из группы, состоящей из QTL на хромосомах 4, 6, 9, 11 и 12 Solanum habrochaites LYC 4/78; устойчивое к Botrytis растение томата и устойчивая к Botrytis часть растения томата. Также представлен способ детекции QTL для устойчивости к Botrytis, включающий детекцию по меньшей мере одного генетического маркера, сцепленного с QTL для устойчивости к Botrytis, образованного из S. habrochaites LYC 4/78 на 4, 6, 9, 11 и/или 12 хромосоме. Предложено применение генетического маркера, выбранного из группы, состоящей из генетических маркеров, представленных в таблицах 1-5, имеющихся в описании, для детекции QTL для устойчивости к Botrytis. Изобретение позволяет получить устойчивое к Botrytis растение томата. 11 н. и 9 з.п. ф-лы, 31 табл., 8 ил., 4 пр.
1. Способ получения устойчивого к Botrytis растения томата, включающий стадии:
a) получения устойчивого к Botrytis донорного растения томата, предпочтительно устойчивого к Botrytis растения вида S. habrochaites, более предпочтительно устойчивого к Botrytis растения линии S. habrochaites LYC 4/78;
b) переноса нуклеиновой кислоты из указанного донорного растения в одно или несколько восприимчивых к Botrytis реципиентных растений томата, предпочтительно в растение вида S. lycopersicum, где указанный перенос приводит к внесению геномного материала из донорного растения в соответствующую область генома указанных одного или нескольких восприимчивых реципиентных растений, причем указанный перенос нуклеиновой кислоты проводят трансгенными способами (например, посредством трансформации), посредством слияния протопластов, способом двойных гаплоидов или посредством спасения зародышей;
c) отбора среди указанных реципиентных растений томата растения, которое содержит в его геноме по меньшей мере один локус количественного признака (QTL) для устойчивости к Botrytis из указанного устойчивого к Botrytis донорного растения томата, где указанный отбор включает детекцию на 4, 6, 9, 11 и/или 12 хромосоме указанного реципиентного растения томата по меньшей мере одного генетического маркера, связанного с указанным по меньшей мере одним QTL для устойчивости к Botrytis;
- где на расположение указанного QTL на 4 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры СТ50, C2Atlg74970, Р14М49-283е, Р14М48-74е, Р14М50-67е, СТ1739 и P14M50-85h на 4 хромосоме S. habrochaites или на 4 хромосоме S. Lycopersicum,
- где на расположение указанного QTL на 6 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P22M50-188h, Р14М48-521е, P15M48-386h, P18M51-199h, P18M51-103h, Р22М50-103е, Р18М51-388е, Р15М48-395е, Р22М50-124е, Р14М48-160е и P22M50-513h на 6 хромосоме S. habrochaites или на 6 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 9 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры Р18М50-141, Р14М49-240, TG254, TG223, TG10, P18M50-134h, Р14М49-243h, Р18М50-599, P14M60-222h, P22M51-417h, P14M50-174h, P14M60-157h, P14M60-107h, P15M48-138h, P14M48-113h, Tm2a, P18M51-146h, P14M48-282h и P14M50-276h на 9 хромосоме S. habrochaites или на 9 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 11 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры Р14М60-215е, P14M61-173h, P14M50-307h, TG47, P14M50-29xCD, Р18М51-358h, P18M50-27xCD, P18M51-I36h, P22M50-488h, TG393, P14M61-396h, P22M51-235h и P22M51-174e на 11 хромосоме S. habrochaites или на 11 хромосоме S. lycopersicum, и
- где на расположение указанного QTL на 12 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры СТ19, TG68, Р14М48-411е, P18M50-244h, P18M50-273h, P14M61-420h, P14M61-406h, P14M61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296, P22M50-131h и P22M51-135h, предпочтительно геномная область, содержащая генетические маркеры P14M61-420h, P14M61-406h, Р14М61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296 и P22M50-131h на 12 хромосоме S. habrochaites или на 12 хромосоме S. lycopersicum.
2. Способ по п.1, где стадию с) проводят посредством детекции указанного генетического маркера в ДНК, выделенной из указанных реципиентных растений томата.
3. Способ по п.1 или 2, где указанная стадия с) дополнительно включает проведение биологического анализа указанных растений для определения устойчивости к Botrytis в указанных растениях.
4. Способ по любому из пп.1-3, где стадия с) дополнительно включает селекцию растения, которое содержит в его геноме по меньшей мере один QTL для устойчивости к Botrytis, выбранный из QTL, расположенных на 1, 2 и 4 хромосоме S. habrochaites LYC 4/78,
- где на расположение указанного QTL на 1 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P22M50-412h, P14M50-349h, P14M60-69h, P14M49-192h, P14M49-232h, Р14М49-260е, P14M50-503h, P18M50-124h и P14M49-114h на 1 хромосоме S. habrochaites или на 1 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 2 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P14M60-537h, Р15М48-257е, P14M49-327h, P14M49-325h, Р14М61-286е, P14M61-125h, P18M51-134h и СТ128 на 2 хромосоме S. habrochaites или на 2 хромосоме S. lycopersicum, и
- где на расположение указанного QTL на 4 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P18M51-169.5e, P18M51-305.4h, Р14М60-262.9е и P14M61-292.7h на 2 хромосоме S. habrochaites или на 2 хромосоме S. lycopersicum.
5. Способ по любому из пп.1-4, где стадия с) включает отбор
- растения, которое содержит в своем геноме по меньшей мере 4 QTL для устойчивости к Botrytis, выбранных из группы, состоящей из QTL, определенных в пп.1, 2 и 7, или
- растения, которое содержит в своем геноме по меньшей мере 2 QTL для устойчивости к Botrytis, выбранных из группы, состоящей из QTL, определенных в пп.1 и 2.
6. Способ по любому из пп.1-5, где указанный способ дополнительно включает стадию d) скрещивания указанного растения, отобранного на стадии с), с растением культивируемой линии томата с получением растений-потомков.
7. Устойчивое к Botrytis растение томата, которое может быть получено способом по любому из пп.1-6.
8. Выделенный образец ДНК, полученный из растения томата вида S. lycopersicum, содержащий QTL, выбранный из группы, состоящей из QTL на хромосомах 4, 6, 9, 11 и 12 Solanum habrochaites LYC 4/78, ассоциированных с устойчивостью к Botrytis, причем положение QTL является таким, как определено в п.1, для идентификации одного или более генов, которые кодируют устойчивость к Botrytis, или для получения трансгенного растения.
9. Способ детекции QTL для устойчивости к Botrytis, включающий детекцию по меньшей мере одного генетического маркера, сцепленного с QTL для устойчивости к Botrytis, образованного из S. habrochaites LYC 4/78 на 4, 6, 9, 11 и/или 12 хромосоме предполагаемого устойчивого к Botrytis растения томата,
- где на расположение указанного QTL на 4 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры СТ50, C2Atlg74970, Р14М49-283е, Р14М48-74е, Р14М50-67е, СТ1739 и P14M50-85h на 4 хромосоме S. habrochaites или на 4 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 6 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P22M50-188h, Р14М48-521е, P15M48-386h, P18M51-199h, P18M51-103h, Р22М50-103е, Р18М51-388е, Р15М48-395е, Р22М50-124е, Р14М48-160е и P22M50-513h на 6 хромосоме S. habrochaites или на 6 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 9 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры Р18М50-141, Р14М49-240, TG254, TG223, TG10, P18M50-134h, Р14М49-243h, Р18М50-599, P14M60-222h, P22M51-417h, P14M50-174h, P14M60-157h, P14M60-107h, P15M48-138h, P14M48-113h, Tm2a, P18M51-146h, P14M48-282h и P14M50-276h на 9 хромосоме S. habrochaites или на 9 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 11 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры Р14М60-215е, P14M61-173h, P14M50-307h, TG47, P14M50-29xCD, Р18М51-358h, P18M50-27xCD, P18M51-136h, P22M50-488h, TG393, P14M61-396h, P22M51-235h и P22M51-174e на 11 хромосоме S. habrochaites или на 11 хромосоме S. lycopersicum, и
- где на расположение указанного QTL на 12 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры СТ19, TG68, Р14М48-411е, P18M50-244h, P18M50-273h, P14M61-420h, P14M61-406h, P14M61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296, P22M50-131h и P22M51-135h, предпочтительно геномная область, содержащая генетические маркеры P14M61-420h, P14M61-406h, Р14М61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296 и P22M50-131h на 12 хромосоме S. habrochaites или на 12 хромосоме S. lycopersicum.
10. Устойчивое к Botrytis растение вида S. lycopersicum, содержащее в геноме по меньшей мере один QTL, где указанный QTL выбран из группы, состоящей из QTL на 4, 6, 9, 11 и 12 хромосоме Solanum habrochaites, ассоциированных с устойчивостью к Botrytis,
- где на расположение указанного QTL на 4 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры СТ50, C2Atlg74970, Р14М49-283е, Р14М48-74е, Р14М50-67е, СТ1739 и P14M50-85h на 4 хромосоме S. habrochaites или на 4 хромосоме S. lycopersicum, где указанный QTL или указанная его придающая устойчивость к Botrytis часть находятся не в их природном генетическом фоне,
- где на расположение указанного QTL на 6 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P22M50-188h, Р14М48-521е, P15M48-386h, Р18М5l-199h, Р18М5l-103h, Р22М50-103е, Р18М51-388е, Р15М48-395е, Р22М50-124е, Р14М48-160е и P22M50-513h на 6 хромосоме S. habrochaites или на 6 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 9 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры Р18М50-141, Р14М49-240, TG254, TG223, TG10, P18M50-134h, Р14М49-243h, Р18М50-599, P14M60-222h, Р22М51-417h, P14M50-174h, P14M60-157h, P14M60-107h, P15M48-138h, P14M48-113h, Tm2a, P18M51-146h, P14M48-282h и P14M50-276h на 9 хромосоме S. habrochaites или на 9 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 11 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры Р14М60-215е, P14M61-173h, P14M50-307h, TG47, P14M50-29xCD, Р18М51-358h, P18M50-27xCD, P18M51-136h, P22M50-488h, TG393, P14M61-396h, P22M51-235h и P22M51-174e на 11 хромосоме S. habrochaites или на 11 хромосоме S. lycopersicum, и
- где на расположение указанного QTL на 12 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры СТ19, TG68, Р14М48-411е, P18M50-244h, P18M50-273h, P14M61-420h, P14M61-406h, P14M61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296, P22M50-131h и P22M51-135h, предпочтительно геномная область, содержащая генетические маркеры P14M61-420h, P14M61-406h, Р14М61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296 и P22M50-131h на 12 хромосоме S. habrochaites или на 12 хромосоме S. lycopersicum, и
- где указанное растение необязательно дополнительно содержит один или несколько дополнительных QTL, или их придающую устойчивость к Botrytis часть, ассоциированных с устойчивостью к Botrytis, выбранных из QTL на 1, 2 и/или 4 хромосоме Solanum habrochaites, предпочтительно Solanum habrochaites линии LYC 4/78, где на расположение указанного дополнительного QTL на 4 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры Р18М51-169.5е, P18M51-305.4h, Р14М60-262.9е, P14M61-292.7h, TG609, Р14М48-345е, Р14М48-177е и Р18М50-147е на 4 хромосоме S. habrochaites или на 4 хромосоме S. lycopersicum, более предпочтительно на 4 хромосоме S. habrochaites LYC 4/78 или на 4 хромосоме S. lycopersicum cν. Moneymaker.
11. Растение по п.10, где расположение указанных QTL на 1, 2, 6, 9, 11 и/или 12 хромосомах является таким, как определено в п.4.
12. Способ получения устойчивого к Botrytis инбредного растения томата, включающий
а) получение устойчивого к Botrytis растения томата по любому из пп.1-6;
b) скрещивание указанного устойчивого к Botrytis растения томата с этим же растением или с другим растением томата с получением семени потомка томата;
c) выращивание указанного семени потомка томата стадии b) с получением устойчивых к Botrytis растений томата;
d) повторение стадий скрещивания и выращивания от 0 до 7 раз с получением устойчивого к Botrytis инбредного растения томата.
13. Способ по п.12, где стадия с) дополнительно включает стадию идентификации растений, которые проявляют устойчивость к Botrytis и обладают коммерчески желательными характеристиками.
14. Способ по п.12 или 13, где указанный способ дополнительно включает стадию селекции гомозиготных инбредных растений томата.
15. Устойчивое к Botrytis инбредное растение томата, которое может быть получено способом по любому из пп.12-14.
16. Гибридное растение томата, которое проявляет устойчивость к Botrytis, где указанное гибридное растение томата получают скрещиванием устойчивого к Botrytis инбредного растения томата, получаемого способом по любому из пп.12-14, с инбредным растением томата, которое обладает коммерчески желательными характеристиками.
17. Применение генетического маркера, выбранного из группы, состоящей из генетических маркеров, представленных в таблицах 1-5, для детекции QTL для устойчивости к Botrytis, и/или для детекции устойчивых к Botrytis растений томата.
18. Способ получения устойчивого к Botrytis растения томата, включающий стадии:
- высевания семян растения томата и выращивание указанных семян до проростков/растений, причем указанное растение томата получают путем переноса нуклеиновой кислоты из указанного устойчивого к Botrytis донорного растения томата, предпочтительно устойчивого к Botrytis растения вида S. habrochaites, более предпочтительно устойчивого к Botrytis растения линии S. habrochaites LYC 4/78, в восприимчивое к Botrytis реципиентное растение томата, предпочтительно в растение вида S. lycopersicum, где указанный перенос приводит к внесению геномного материала из донорного растения в соответствующую область генома указанного реципиентного растения, и
- отбора растения томата, которое содержит в своем геноме по меньшей мере один QTL для устойчивости к Botrytis из указанного устойчивого к Botrytis донорного растения томата, где указанный отбор включает детекцию на 4, 6, 9, 11 и/или 12 хромосоме указанного реципиентного растения томата по меньшей мере одного генетического маркера, связанного с указанным по меньшей мере одним QTL для устойчивости к Botrytis,
- где на расположение указанного QTL на 4 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры СТ50, C2Atlg74970, Р14М49-283е, Р14М48-74е, Р14М50-67е, СТ1739 и P14M50-85h на 4 хромосоме S. habrochaites или на 4 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 6 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры P22M50-188h, Р14М48-521е, P15M48-386h, P18M51-199h, P18M51-103h, Р22М50-103е, Р18М51-388е, Р15М48-395е, Р22М50-124е, Р14М48-160е и P22M50-513h на 6 хромосоме S. habrochaites или на 6 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 9 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры Р18М50-141, Р14М49-240, TG254, TG223, TG10, P18M50-134h, Р14М49-243h, Р18М50-599, P14M60-222h, Р22М51-417h, P14M50-174h, P14M60-157h, P14M60-107h, P15M48-138h, P14M48-113h, Tm2a, P18M51-146h, P14M48-282h и P14M50-276h на 9 хромосоме S. habrochaites или на 9 хромосоме S. lycopersicum,
- где на расположение указанного QTL на 11 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры Р14М60-215е, P14M61-173h, P14M50-307h, TG47, P14M50-29xCD, Р18М51-358h, P18M50-27xCD, P18M51-136h, P22M50-488h, TG393, P14M61-396h, P22M51-235h и P22M51-174e на 11 хромосоме S. habrochaites или на 11 хромосоме S. lycopersicum, и
- где на расположение указанного QTL на 12 хромосоме указанного растения указывает геномная область, содержащая генетические маркеры СТ19, TG68, Р14М48-411е, P18M50-244h, P18M50-273h, P14M61-420h, P14M61-406h, P14M61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296, P22M50-131h и P22M51-135h, предпочтительно геномная область, содержащая генетические маркеры P14M61-420h, Р14М61-406h, Р14М61-223h, P14M60-193h, P22M51-314h, TG565, P14M48-172h, P22M50-321e, P14M60-219e, P14M48-153h, P22M50-97h, TG296 и P22M50-131h на 12 хромосоме S. habrochaites или на 12 хромосоме S. lycopersicum.
19. Устойчивая к Botrytis часть растения томата по пп.7, 10, 15 или 16.
20. Часть растения томата по п.19, представляющая собой семя.
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ С ПОВЫШЕННОЙ УСТОЙЧИВОСТЬЮ К ФИТОПАТОГЕНАМ | 2002 |
|
RU2261275C2 |

