ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Данная заявка заявляет приоритет по предварительной заявке на патент США №62/064375, поданной 15 октября 2014 г., которая включена в данный документ в полном объеме посредством ссылки.
Область изобретения
[0002] Настоящее изобретение относится к области сельского хозяйства и более конкретно к способам и композициям для получения растений томата, которые проявляют устойчивость к болезням с уменьшенным некрозом растений.
Включение перечня последовательностей
[0003] Перечень последовательностей, который содержится в файле с именем «SEMB019WO_ST25», который составляет 11,1 килобайт, как измерено в операционной системе Microsoft Windows, и который был создан 13 октября 2015 года, подается при этом в электронном виде и включен в данный документ посредством ссылки.
[0004] Уровень техники
[0005] Устойчивость к болезням является важной характеристикой для сельского хозяйства, в частности для производства продовольственных культур. Хотя аллели устойчивости к болезням были идентифицированы в неокультуренных линиях томата, попытки внедрения этих аллелей в культурные линии осложнены введением вредных признаков наряду с аллелями устойчивости. Использование маркер-вспомогательной селекции (селекция с помощью маркеров, СПМ) для методов селекции растений предоставило возможность отбора растений на основе генетических маркеров, которые сцеплены с целевыми признаками. Тем не менее, точные маркеры для идентификации или отслеживания желательных признаков у растений часто недоступны, даже если ген, который связан с признаком, охарактеризован. Эти трудности дополнительно осложняются такими факторами, как полигенное или количественное наследование, и часто недостаточным пониманием генетического окружения, которое лежит в основе экспрессии нужного фенотипа. Следовательно, при отсутствии точных и проверенных маркеров для использования в СПМ может не быть возможности производить новые линии растений, которые проявляют определенные фенотипы устойчивости к заболеваниям.
Сущность изобретения
[0006] В одном аспекте данное изобретение относится к растению томата, конкретнее к культивируемому сорту растения томата, которое содержит рекомбинантную интрогрессию из Solanum chilense на хромосоме 6, причем указанная рекомбинантная интрогрессия содержит первый аллель, который придает улучшенную устойчивость к вирусу желтой курчавости листьев томата по сравнению к растением, которое не имеет указанного первого аллеля, и причем указанная рекомбинантная интрогрессия не имеет второго аллеля, который генетически сцеплен с указанным первым аллелем и который вызывает некроз. В некоторых вариантах реализации изобретения рекомбинантная интрогрессия из Solanum chilense находится между приблизительно 30,20 млн п.н. и 30,88 млн п.н. на хромосоме 6, или находится между приблизительно 30,69 млн п.н. и 30,88 млн п.н. на хромосоме 6, или находится между приблизительно 30,71 млн п.н. и 30,88 млн п.н. на хромосоме 6, или находится между приблизительно 30,32 млн п.н. и 30,71 млн п.н. на хромосоме 6, или находится между приблизительно 30,88 млн п.н. и 31,80 млн п.н. на хромосоме 6.
[0007] В других вариантах реализации изобретения растение содержит Solanum chilense аллель в локусе NSLYC008383071 (SEQ ID NO: 1) и не имеет Solanum chilense аллеля в локусе NSLYC005134429 (SEQ ID NO: 3), или растение содержит Solanum chilense аллель в локусе NSLYC008383071 (SEQ ID NO: 1) и не имеет Solanum chilense аллеля в локусе NSLYC009077970 (SEQ ID NO: 2), или растение содержит донорскую ДНК S.chilense в пределах сегмента генома фланкированного NSLYC005134429 (SEQ ID NO: 3) и NSLYC008383071 (SEQ ID NO: 1), или растение содержит донорскую ДНК S.chilense в пределах сегмента генома фланкированного NSLYC009077970 (SEQ ID NO: 2) и NSLYC008383071 (SEQ ID NO: 1), или растение содержит донорскую ДНК S.chilense в пределах сегмента генома фланкированного NSLYC008382908 и NSLYC008383071 (SEQ ID NO: 1), или растение содержит донорскую ДНК S.chilense в пределах сегмента генома фланкированного NSLYC009418370 (SEQ ID NO: 4) и NL231193. В другом варианте реализации данное изобретение относится к части такого растения, в том числе к клетке, семени, корню, стеблю, листу, плоду, цветку или пыльце.
[0008] В другом аспекте данное изобретение относится к способу получения растения томата с улучшенной устойчивостью к вирусу желтой курчавости листьев томата, который включает: а) скрещивание растения томата по п. 1 с самим собой или с другим растением томата с иным генотипом для получения одного или более растений-потомков; и б) отбор растения-потомка, которое содержит указанную рекомбинантную интрогрессию. В одном варианте реализации изобретения отбор растения-потомка включает идентификацию растения-потомка, которое (1) содержит Solanum chilense аллель в локусе, который генетически сцеплен с указанным первым аллелем, и/или не имеет аллеля, который присутствует в соответствующем локусе в растении томата, и (2) не имеет Solanum chilense аллеля в локусе, который генетически сцеплен с указанным вторым аллелем, который вызывает некроз, и/или содержит аллель, который присутствует в соответствующем локусе в растении томата. В другом варианте реализации изобретения отбор указанного растения-потомка включает маркер-вспомогательную селекцию (СПМ). В дополнительном варианте реализации изобретения маркер-вспомогательная селекция (СПМ) включает обнаружение по меньшей мере одного аллеля в локусе, который выбран из группы, состоящей из NSLYC005134429 (SEQ ID № 3), NSLYC009077970 (SEQ ID № 2), NSLYC009418370 (SEQ ID № 4), NSLYC008383071 (SEQ ID № 1), NSLYC008382908 и NL231193. Также в дополнительных вариантах реализации изобретения растение-потомок представляет собой растение-потомок F2-F6, или получение растения-потомка включает возвратное скрещивание, например, после 2-7 поколений возвратного скрещивания.
[0009] В другом аспекте данное изобретение относится к способу получения растения томата, которое проявляет улучшенную устойчивость к вирусу желтой курчавости листьев томата, который включает: а) получение растения томата, гетерозиготного по первому аллелю, который придает устойчивость к вирусу желтой курчавости листьев томата, и который в растении генетически сцеплен со вторым аллелем из Solanum chilense, который вызывает некроз; б) получение потомства растения; и в) отбор по меньшей мере первого растения-потомка, в котором рекомбинация произошла таким образом, что потомство содержит указанный первый аллель, который придает устойчивость к вирусу желтой курчавости листьев томата, но не содержит указанный второй аллель, который вызывает некроз; причем отбор указанного первого растения-потомка включает обнаружение по меньшей мере одного аллеля в локусе, который выбран из группы, состоящей из NSLYC005134429 (SEQ ID № 3), NSLYC009077970 (SEQ ID № 2), NSLYC009418370 (SEQ ID № 4), NSLYC008383071 (SEQ ID № 1), NSLYC008382908 и NL231193. В одном варианте реализации изобретения растение-потомок представляет собой растение-потомок F2-F6, или получение растения-потомка включают возвратное скрещивание, например, после 2-7 поколений возвратного скрещивания. В другом варианте реализации данное изобретение относится к растению, которое получено таким способом, или к части такого растения, которая выбрана из группы, состоящей из клетки, семени, корня, стебля, листа, плода, цветка или пыльцы.
[0010] В другом аспекте данное изобретение относится к растению томата, конкретнее к культивируемому сорту растения томата, который включает TТ аллель в маркере NSLYC008374675 (SEQ ID № 5) и GG аллель в маркере NSLYC008375578 (SEQ ID № 6), причем указанные аллели обеспечивают отсутствие некроза или устойчивость к некрозу. В одном варианте реализации изобретения способ дополнительно включает: г) отбор против аллелей на хромосоме 2, которые связаны с некрозом; причем указанный отбор включает отбор с целью удаления растений, которые включают CC аллель в маркере NSLYC008374675 (SEQ ID № 5) или AA аллель в маркере NSLYC008375578 (SEQ ID № 6), причем наличие указанных аллелей вызывает некроз.
[0011] В другом аспекте данное изобретение относится к способу получения растения томата, которое проявляет устойчивость к некрозу, который включает: а) получение растения томата, гетерозиготного по первому аллелю, который придает устойчивость к некрозу; б) получение потомства растения; и в) отбор по меньшей мере первого растения-потомка, в котором рекомбинация произошла таким образом, что потомство содержит указанный первый аллель, который придает устойчивость к некрозу; причем отбор указанного первого растения-потомка включает обнаружение по меньшей мере одного аллеля в локусе, который выбран из группы, состоящей из маркера NSLYC008374675 (SEQ ID № 5) и маркера NSLYC008375578 (SEQ ID № 6), причем указанные аллели обеспечивают отсутствие некроза или устойчивость к некрозу.
[0012] В другом аспекте данное изобретение относится к способу получения растения томата, которое проявляет улучшенную устойчивость к вирусу желтой курчавости листьев томата, который включает: а) получение растения томата, гетерозиготного по первому аллелю, который придает устойчивость к вирусу желтой курчавости листьев томата и который в растении генетически сцеплен со вторым аллелем из Solanum chilense который вызывает некроз; б) получение потомства растения; и в) отбор по меньшей мере первого растения-потомка, в котором рекомбинация произошла таким образом, что потомство содержит указанный первый аллель, который придает устойчивость к вирусу желтой курчавости листьев томата, но не содержит указанный второй аллель, который вызывает некроз. В соответствии с данным изобретением, отбор растения-потомка может включать отбор растения, которое содержит аллель, который придает устойчивость к вирусу желтой курчавости листьев томата из хромосомы 6 Solanum chilense, и не имеет второго локуса, сцепленного с ним, который вызывает один или более некрозов, уменьшение завязей плодов или уменьшение размера плодов по сравнению с растением, которое не имеет второго локуса. В одном варианте реализации изобретения способ отбора указанного первого растения-потомка включает отбор потомков, у которых рекомбинация произошла между локусом NSLYC009418370 (SEQ ID № 4) и локусом NL231193. В конкретном варианте реализации изобретения способ включает обнаружение по меньшей мере одного аллеля в локусе, который выбран из группы, состоящей из NSLYC008383071 (SEQ ID № 1), NSLYC009418370 (SEQ ID № 4), NSLYC008382908 и NL231193. В другом варианте реализации изобретения способ включает: г) отбор последующего растения-потомка из числа растений, выбранных на стадии в); причем выбор указанного последующего растения-потомка включает обнаружение по меньшей мере одного аллеля в локусе, который выбран из группы, состоящей из NSLYC009418370 (SEQ ID № 4) и NL231193. Еще в одном варианте реализации данное изобретение относится к растению, которое получено любым из вышеуказанных способов.
Краткое описание графических материалов
[0013] ФИГ. 1: проиллюстрировано сравнение рекомбинантов в Mi (нематоды) - Ty (гемини-вирус) области хромосомы 6. Столбцы слева направо: генетические источники, 22 маркерные пробы, включая маркер по нематодному признаку (1) и три Ty-1 маркера по гемини-вирусному признаку (2-4), затем результаты фенотипирования после гемини-вирусной агроинокуляции, полевая устойчивость и оценка некроза листьев, которая представляет собой проявление генетического сопротивления после Ty-1 интрогрессии. Генотипы обозначены оттенением, как проиллюстрировано на легенде к данной фигуре. «нд» указывает на отсутствие данных.
[0014] ФИГ. 2: проиллюстрирован LOD график (количественный показатель сцепления генов), который получен из генотипических и фенотипических данных в популяции F2, которая содержит Ty-1 интрогрессию из S.chilense.
[0015] ФИГ. 3: проиллюстрирована средняя оценка некроза для нескольких линий F3, которые содержат Ty-1 интрогрессии, наряду с генотипом в нескольких маркерных локусах. «esc» относится к ДНК, которая происходит от S.esculentum у данного маркерного локуса, и «chi» относится к ДНК, которая происходит от S.chilense у данного маркерного локуса.
[0016] ФИГ. 4: проиллюстрирована средняя оценка некрозов для нескольких линий растений, проиллюстрированных на ФИГ. 3, основанная на расположении точки разрыва между донорской ДНК S.chilense и ДНК S.esculentum, определенная с использованием маркеров, проиллюстрированных на ФИГ. 3.
[0017] ФИГ. 5: проиллюстрирована средняя оценка некроза для нескольких линий растений, наряду с генотипами в двух маркерных локусах на хромосоме 2, которые связаны с некрозом, как проиллюстрировано на ФИГ. 2.
[0018] ФИГ. 6: проиллюстрирован генотип ряда рекомбинантных линий F4 в нескольких маркерных локусах в приблизительной области Ty-1 (между 20 и 34 млн п.н. хромосомы 6). Высоко корреляционный анализ TaqMan на основе SNP (single nucleotide polymorphism, единичного нуклеотидного полиморфизма) представляет собой NSLYC008426102. Серый цвет SNP означает идентичный SNP, который присутствует у чувствительных родителей (FIR-16-1078 и FDR-16-0197), а белый цвет SNP означает идентичный SNP, который присутствует у резистентных родителей (FIR-AY06074 и FDR-16-2133). Во второй колонке показаны оценки теста на ВЖКЛТ.
[0019] ФИГ. 7: проиллюстрирован генотип ряда рекомбинантных линий в нескольких маркерных локусах в приблизительной области Ty-1 (между 30,3 млн п.н. и 31,8 млн п.н. хромосомы 6). Серый цвет означает донорный аллель S.chilense, а белый цвет означает реципиентный аллель S.esculentum.
[0020] ФИГ. 8: проиллюстрированы результаты тестирования производительности при завязывании плодов (общее количество зрелых плодов на число растений).
[0021] ФИГ. 9: проиллюстрированы результаты тестирования производительности при завязывании плодов (общий вес урожая в кг на количество плодов).
[0022] ФИГ. 10: проиллюстрировано изображение интрогрессии (около 260 кб).
Подробное описание
[0023] Вирус желтой курчавости листьев томата (ВЖКЛТ) представляет собой растительный патогенный вирус, который обуславливает существенные потери урожая у растений томата. Как известно, некоторые дикие виды томата обладают устойчивостью к ВЖКЛТ, и были предприняты усиленные попытки для интрогрессии аллелей устойчивости к ВЖКЛТ из этих видов в культивируемые линии томатов. Однако эти попытки были затруднены, поскольку аллели устойчивости от диких видов сопровождаются нежелательными агрономическими признаками, такими как некроз. Потеря урожая у растений томата из-за ВЖКЛТ остается серьезной проблемой.
[0024] Впервые, в данном изобретении предложены новые интрогрессии аллелей устойчивости к болезням из Solanum chilense в культивируемые или элитные томатные линии, в результате чего растения томата демонстрируют высокий уровень устойчивости к ВЖКЛТ без вредного некроза, ранее связанного с Ty1 интрогрессиями от диких видов. В конкретном варианте реализации данное изобретение может быть использовано для вида томатов Lycopersicon esculentum (также известный как Solanum lycopersicum). Таким образом, данное изобретение представляет собой значительное достижение в данной области техники. Путем дополнительного обеспечения новыми, точными маркерами для отслеживания интрогрессированных аллелей без ассоциированного некроза во время размножения растений, данное изобретение позволяет интрогрессию устойчивости к болезням в любой желаемый генотип томата.
[0025] Устойчивость к ВЖКЛТ обычно получали через интрогрессии в Ty-1 локус из S.chilense. Однако, Ty-1 интрогрессии, как правило, крупные, и растения, которые содержат эти интрогрессии проявляют агрономически неприемлемые признаки, включая некроз. Попытки снижения частоты возникновения или тяжести некрозов у растений, которые содержат Ty-1 интрогрессии, не увенчались успехом отчасти из-за супрессии рекомбинации в центромерной области хромосомы 6 вблизи Ty-1 локуса. Дополнительно, некоторые генетические области, окружающие Ty-1 локус, инвертируются в линиях диких томатов по сравнению с культивируемыми линиями, дополнительно препятствуя рекомбинации. Интрогрессия Ty-1 аллелей из S.chilense еще дополнительно осложняется отсутствием существующих маркеров и анализов, которые точно коррелируют с генотипом устойчивости и снижением некроза у разнообразных линий томатов.
[0026] Несмотря на многочисленные лимитирующие факторы на пути к успешной интрогрессии Ty-1 аллелей устойчивости из S.chilense в культивируемые линии томатов, авторы настоящего изобретения неожиданно смогли произвести новые интрогрессии из S.chilense, которые придают устойчивость к ВЖКЛТ без вредных признаков, ранее связанных с Ty-1 интрогрессиями. В некоторых вариантах реализации данное изобретение относится к растениям, которые содержат донорскую ДНК S.chilense в маркере NSLYC008383071 (SEQ ID № 1) и реципиентную ДНК в маркере NSLYC009077970 (SEQ ID № 2). В других вариантах реализации данное изобретение относится к растениям, которые содержат донорскую ДНК S.chilense в маркере NSLYC008383071 (SEQ ID № 1) и реципиентную ДНК в маркере NSLYC005134429 (SEQ ID № 3). В других вариантах реализации данное изобретение относится к растениям, которые содержат донорскую ДНК S.chilense в пределах сегмента генома фланкированного NSLYC009077970 (SEQ ID № 2) и NSLYC008383071 (SEQ ID № 1) или в пределах сегмента генома фланкированного NSLYC005134429 (SEQ ID № 3) и NSLYC008383071 (SEQ ID № 1).
[0027] Настоящее изобретение дополнительно относится к новым маркерам, сцепленными с признаком, которые могут быть использованы для получения растений, которые содержат новые рекомбинированные интрогрессии в Ty-1 области хромосомы 6, которая придает устойчивость к ВЖКЛТ без некрозов. В конкретных вариантах реализации в данном изобретении предложены маркеры, показанные в Таблицах 1, 4, 6 и 7. Другие варианты реализации данного изобретения относятся к новым маркерам NSLYC008383071 (SEQ ID № 1), NSLYC009418370 (SEQ ID № 4), NSLYC009077970 (SEQ ID № 2) и NSLYC005134429 (SEQ ID № 3), которые, как было показано, генетически сцеплены с устойчивостью растений к ВЖКЛТ.
[0028] Изобретение дополнительно относится к Ty-1 интрогрессиям, которые могут быть задействованы в гомозиготном состоянии без вредных некрозов или других неприемлемых признаков. Ty-1 аллели обычно задействовали в гетерозиготном состоянии, пытаясь уменьшить тяжесть некротических симптомов, которые проявляют растения, содержащие Ty-1 интрогрессии. Однако, вопреки убеждению в данной области, что гетерозиготное размещение Ty-1 аллелей, которые происходят от S.chilense, в элитной линии, необходимо во избежание негативных признаков, таких как некроз, настоящее изобретение предлагает Ty-1 аллели устойчивости, которые могут быть размещены гомозиготно при отсутствии недопустимых уровней некроза у растений. Следовательно, новые уменьшенные интрогрессии согласно настоящему изобретению обеспечивают значительные преимущества по сравнению с существующими технологиями.
[0029] Дополнительно, данное изобретение идентифицирует новые ЛКП (локусы количественных признаков), которые придают устойчивость к некрозу, на хромосоме 2 томата, а также последовательности нуклеиновых кислот и генетические маркеры, которые связаны с ЛКП. Использование новых маркеров, предложенных в данном документе для отбора растений, которые имеют благоприятные аллели в пределах локуса или генетически сцеплены с этим локусом, дает возможность создания растений с уменьшенными или отсутствующими некротическими симптомами. Следовательно, в некоторых вариантах реализации данное изобретение относится к способам получения растений, у которых наблюдается уменьшение или отсутствие некротических симптомов, путем отбора или размножения растений, которые имеют благоприятные аллели в маркерах на хромосоме 2, которые раскрыты в данном описании. В некоторых вариантах реализации данное изобретение относится к растениям, которые имеют TT аллель в пределах маркера NSLYC008374675 (SEQ ID № 5), и к растениям, которые имеют GG аллель в пределах маркера NSLYC008375578 (SEQ ID № 6), которые демонстрируют уменьшение некрозов по сравнению с растениями, которые не содержат такие аллели.
[0030] Дополнительно, данное изобретение относится к новым маркерам и анализам, которые позволяют точно идентифицировать и отслеживать геномные области, которые предложены в данном документе, в ходе селекции растений. По причине того, что генетически разнообразные линии томатов могут затруднительно скрещиваться из-за частично подавленной рекомбинации в Ty-1 области, введение аллелей устойчивости к ВЖКЛТ без сцепленного некроза из S.chilense в элитные линии томатов или культивируемые линии с использованием традиционных методов селекции потребует чрезмерно больших сегрегационных популяций для скрининга потомства с неопределенным результатом. Следовательно, маркер-вспомогательная селекция (СПМ) представляет собой улучшение для эффективной интрогрессии аллелей диких томатов в элитные сорта. Однако ранее известные маркеры устойчивости к ВЖКЛТ не позволяли отбирать устойчивость к ВЖКЛТ без ассоциированного некроза. В противовес этому, настоящее изобретение позволяет СПМ Ty1 без ассоциированного некроза путем предоставления улучшенных и проверенных маркеров для обнаружения генотипов, которые связаны с устойчивостью к болезням и уменьшенным некрозом, без необходимости выращивать большие популяции растений до стадии зрелости с целью наблюдения фенотипа.
I. Геномные области, аллели и полиморфизмы, связанные с устойчивостью к ВЖКЛТ и уменьшенными некрозами у растений томата
[0031] Изобретение относится к новым интрогрессиям одного или более аллелей, которые сцеплены с устойчивостью к болезни и с уменьшением растительного некроза у растений томата, наряду с полиморфными нуклеиновыми кислотами и сцепленными маркерами для отслеживания интрогрессий во время селекции растений.
[0032] ВЖКЛТ может инфицировать растения томата на любой стадии цикла роста и может привести к существенному снижению урожая и качества культуры томата. Следовательно, были предприняты интенсивные попытки для определения эффективных источников для устойчивости к ВЖКЛТ. Однако ранее известные интрогрессии от диких видов были ассоциированы с некрозом растений. В частности, культивируемые линии томатов, несущие ранее известные интрогрессий генов устойчивости к TYLCV проявляют некроз листьев. Несмотря на многолетний селекционный процесс с целью уменьшения частоты возникновения некрозов, такие эффекты все еще обычно наблюдаются на полях.
[0033] В данной области техники известны дикие виды томатов, которые проявляют устойчивость к ВЖКЛТ, например, S.chilense, и они могут быть использованы в соответствии с некоторыми вариантами реализации данного изобретения. В данной области техники также описаны и известны другие источники для устойчивости к ВЖКЛТ (см., например, Vidavsky et al., Phytopathology, 88(9):910-4, 1998; Hutton et al., HortScience, 47(3):324-327, 2012; Ji et al., HortScience, 44(3):614-618, 2009).
[0034] Используя улучшенные генетические маркеры и анализы по данному изобретению, заявители смогли успешно идентифицировать новую область устойчивости к ВЖКЛТ из С. chilense, которая связана с меньшим количеством вредных признаков при интрогрессии в культурную линию. В конкретных вариантах реализации данное изобретение относится к растениям, которые содержат донорскую ДНК S.chilense для маркера NSLYC008383071 (SEQ ID № 1) и реципиентную ДНК для маркера NSLYC009077970 (SEQ ID № 2). В других вариантах реализации данное изобретение относится к растениям, которые содержат донорскую ДНК S.chilense в маркере NSLYC008383071 (SEQ ID № 1) и реципиентную ДНК в маркере NSLYC005134429 (SEQ ID № 3). В других вариантах реализации данное изобретение относится к растениям, которые содержат донорскую ДНК S.chilense в пределах сегмента генома фланкированного NSLYC009077970 (SEQ ID № 2) и NSLYC008383071 (SEQ ID № 1) или в пределах сегмента генома фланкированного NSLYC005134429 (SEQ ID № 3) и NSLYC008383071 (SEQ ID № 1).
[0035] В изобретении дополнительно идентифицированы и предложены геномные сегменты приблизительно от 30,20 млн п.н. (NSLYC005134429, SEQ ID № 3) до 30,88 млн п.н. (NSLYC009418370, SEQ ID № 4), приблизительно от 30,69 млн п.н. (NSLYC009077970, SEQ ID № 2) до 30,88 млн п.н. (NSLYC009418370, SEQ ID № 4) или приблизительно от 30,71 млн п.н. (NSLYC008383071, SEQ ID № 1) до 30,88 млн п.н. (NSLYC009418370, SEQ ID № 4) на хромосоме 6, которые связаны с устойчивостью к ВЖКЛТ, но не связаны с некрозом, после интрогрессии в культивируемые линии томатов. В конкретных вариантах реализации данное изобретение относится к растению, которое содержит реципиентную ДНК по одному или более маркерам NL0232061, NL0216350, NL0244835, NSLYC005134413, NSLYC005134429 (SEQ ID № 3) и NSLYC008382908, и которое содержит донорскую ДНК S.chilense по одному или более маркерам NSLYC008373232, NSLYC009078368, NSLYC009077969, NSLYC009077970 (SEQ ID № 2), NSLYC008383071 (SEQ ID № 1), NSLYC009418370 (SEQ ID № 4), NSLYC008426177 и NL231193, которое проявляет устойчивость к ВЖКЛТ и не проявляет некрозы.
[0036] В другом варианте реализации данное изобретение относится к новым маркерам, которые могут быть использованы для идентификации локуса, как описано в данном документе, таких как маркеры, которые предложены в таблицах 1, 4, 6 и 7. Другие варианты реализации данного изобретения относятся к новым маркерам NSLYC008383071 (SEQ ID № 1), NSLYC009418370 (SEQ ID № 4), NSLYC009077970 (SEQ ID № 2) и NSLYC005134429 (SEQ ID № 3), которые, как было показано, генетически сцеплены с устойчивостью растений к ВЖКЛТ.
II. Интрогрессия геномных областей, связанных с устойчивостью к болезни
[0037] Маркер-вспомогательная интрогрессия включает перенос хромосомной области, которая определена одним или более маркерами, из первого генетического окружения во второе. Потомство от скрещивания, которое содержит интрогрессированную геномную область, может быть идентифицировано с помощью комбинации маркеров, характерных для желаемой интрогрессированной геномной области из первого генетического окружения, и как сцепленными, так и несцепленными маркерами, характерными для второго генетического окружения.
[0038] Данное изобретение относится к новым точным маркерам для идентификации и отслеживания интрогрессии одной или более геномных областей S.chilense, раскрытых в данном документе, в культивируемых линиях. Дополнительно, данное изобретение относится к маркерам для идентификации и отслеживания новых интрогрессий, раскрытых в данном документе, в ходе селекции растений, в том числе маркеров, которые предложены в Таблицах 1, 4, 6 и 7. Другие варианты реализации данного изобретения относятся к новым маркерам NSLYC008383071 (SEQ ID № 1), NSLYC009418370 (SEQ ID № 4), NSLYC009077970 (SEQ ID № 2) и NSLYC005134429 (SEQ ID № 3), которые, как было показано, генетически сцеплены с устойчивостью растений к ВЖКЛТ.
[0039] Маркеры в пределах геномных интервалов или сцепленные с любым из геномных интервалов по настоящему изобретению могут быть полезны для различных селекционных действий, которые включают интрогрессию геномных областей, которыеассоциированы с устойчивостью к болезни, в желаемое генетическое окружение. Например, маркер в пределах 40 cM (сантиморган), 20 cM, 15 cM, 10 cM, 5 cM, 2 cM или 1 cM от маркера, который связан с устойчивостью к болезням, описанного в данном документе, может быть использован для маркер-вспомогательной интрогрессии геномных областей, которые связаны с фенотипом, устойчивым к болезни.
[0040] Также предлагаются растения томатов, которые содержат одну или более интрогрессированных областей, которые ассоциированы с желаемым фенотипом, причем по меньшей мере 10%, 25%, 50%, 75%, 90% или 99% оставшихся геномных последовательностей несут маркеры, характерные для зародышевой плазмы. Также предлагаются растения томатов, которые содержат интрогрессированную область, которая содержит области, тесно сцепленные или смежные с геномными областями, и маркеры, предложенные в данном документе и связанные с фенотипом устойчивости к болезни.
III. Создание устойчивых к болезни сортов томатов
[0041] Для большинства селективных целей, коммерческие селекционеры работают с зародышевой плазмой, которая представляет собой «культивируемую», «культивируемый тип» или «элитную». «Элитными» сортами называются сорта, которые получили в результате селекции и отбора для получения наилучших агрономических характеристик, включая урожайность и другие выбранные признаки. Аналогичным образом, «культивируемые» сорта или «культивары» представляют собой растения или группы растений, которые отобраны по желательным характеристикам для выращивания в сельском хозяйстве и которые могут поддерживаться путем размножения сорта. Элитные сорта или культивары легче размножать, потому что они, как правило, имеют хорошие качества при оценке плодоовощной продуктивности. Создан ряд культивируемых и элитных типов томатов (S. lycopersicum или L. esculentum), которые являются улучшенными с агрономической точки зрения и пригодными для коммерческого культивирования. Однако преимущество продуктивности, которое обеспечивается культивируемой зародышевой плазмой, может быть сведено на нет из-за отсутствия аллельного разнообразия. Селекционеры обычно принимают этот компромисс, поскольку прогресс идет быстрее при работе с культивируемым материалом, чем при селекции с генетически разнообразными источниками. Таким образом, в некоторых вариантах реализации настоящее изобретение относится к новым интрогрессиям аллелей устойчивости к болезням из Solanum chilense в элитную линию томата или в культурную линию томата, что в результате приводит к тому, что растения томатов демонстрируют высокий уровень устойчивости к ВЖКЛТ при отсутствии вредного некроза, ранее связанного с интрогрессиями Ty1 из диких видов. Такие интрогрессии могут обеспечить улучшенные растения томата, которые могут обладать рядом агрономических элитных признаков, в дополнение к новой интрогрессии по настоящему изобретению.
[0042] В отличие от этого, когда культурную зародышевую плазму скрещивают с некультивируемой зародышевой плазмой, селекционер может получить доступ к новым аллелям из некультивируемого типа. Однако этот подход представляет значительные трудности из-за проблем фертильности, которые сцеплены со скрещиваниями между различными линиями, и отрицательного привнесенного сцепления от некультивируемого родителя. Растения томатов некультивируемых типов, таких как S.chilense, могут предоставлять аллели, которые связаны с устойчивостью к болезням. Однако, эти некультивируемые типы могут иметь плохие плодоовощные качества, такие как восприимчивость к некрозу или дефектам плодов.
[0043] Процесс интрогрессии желаемых генов устойчивости от некультивируемых линий в элитные культивируемые линии, с избеганием проблем привнесенного сцепления или низкой наследуемости, представляет собой длительный и зачастую трудоемкий процесс. Следовательно, успех применения аллелей, которые получены от диких родственников, сильно зависит от минимальных или укороченных интрогрессий, которые не имеют вредных эффектов, и от надежных маркерных анализов, которые заменяют фенотипические скрининги. Дальнейший успех определяется путем упрощения генетики для ключевых характеристик для того, чтобы позволить сосредоточиться на генетической выгоде для количественных признаков, таких как устойчивость к болезням. Более того, процесс интрогрессии геномных областей из некультивируемых линий может быть значительно облегчен благодаря наличию точных маркеров для СПМ.
[0044] Таким образом, специалисту в данной области техники будет очевидно, что аллели, полиморфизмы и маркеры, предложенные в данном изобретением, позволяют отслеживать и интродуцировать любую из геномных областей, которые определены в данном документе, в любое генетическое окружение. Кроме того, раскрытые в данном документе геномные области, которые связаны с устойчивость к болезни, могут быть интрогрессированы из одного генотипа в другой и прослежены с использованием СПМ. Таким образом, обнаружение заявителями точных маркеров, которые связаны с устойчивостью к болезням, будет способствовать созданию растений томатов с полезными фенотипами. Например, семена могут быть генотипированы с использованием маркеров по настоящему изобретению с целью отбора растений, которые содержат желаемые геномные области, которые связаны с устойчивостью к болезни. Кроме того, СПМ позволяет идентифицировать растения, гомозиготные или гетерозиготные по желаемой интрогрессии.
[0045] Межвидовые скрещивания также могут приводить к супрессии рекомбинации и получению растений с низкой фертильностью или плодовитостью. Например, супрессия рекомбинации наблюдалась для гена устойчивости к томатной нематоде Mi, генов ячменя Mla и Mlg, генов пшеницы Yr17 и Lr20, гена виноградной лозы Run1 и гена арахиса Rma. Мейотическая рекомбинация имеет важное значение для классической селекции, поскольку она позволяет переносить благоприятные аллели в другие генетические окружения, удалять вредные геномные фрагменты и пирамидировать генетически сцепленные признаки. Поэтому при отсутствии точных маркеров супрессированная рекомбинация вынуждает селекционеров увеличивать сегрегацию популяций при скрининге потомства.
[0046] Фенотипическая оценка больших популяций требует много времени, ресурсов и не воспроизводится в любой среде. Маркер-вспомогательная селекция предлагает приемлемую альтернативу. Молекулярные анализы, которые разработаны для обнаружения уникальных полиморфизмов, таких как SNP, являются универсальными. Однако, они могут не различать аллели внутри и среди видов томатов в одном анализе. Структурные перестройки хромосом, такие как делеции, ухудшают гибридизацию и расширение синтетически меченых олигонуклеотидов. В случае событий дубликации многочисленные копии амплифицируются в одной реакции без разграничения. Поэтому разработка и проверка точных и высокопрогнозируемых маркеров представляются важными для успешных селекционных программ СПМ.
IV. Молекулярно-вспомогательные методы селекции
[0047] Генетические маркеры, которые могут быть использованы в практике настоящего изобретения, включают, но не ограничиваются ими, полиморфизмы длин рестрикционных фрагментов (RFLP), полиморфизмы длин амплифицированных фрагментов (AFLP), простые повторяющиеся последовательности (SSR), полиморфизмы длины простой последовательности (SSLP), одиночные нуклеотидные полиморфизмы (SNP), полиморфизмы вставки/делеции (Indel), тандемные повторы с вариабельным числом (VNTR) и случайно амплифицированную полиморфную ДНК (RAPD), изозимы и другие маркеры, известные специалистам в данной области техники. Открытие и создание маркеров в сельскохозяйственных растениях обеспечивает начальную основу для применения маркер-вспомогательной селекционной деятельности (публикации патентов США №№2005/0204780, 2005/0216545, 2005/0218305 и 2006/00504538). Полученная «генетическая карта» является представлением положения характеризуемых локусов (полиморфных маркеров нуклеиновой кислоты или любого другого локуса, для которого могут быть идентифицированы аллели) относительно друг к другу.
[0048] Полиморфизмы, которые содержат всего лишь одну нуклеотидную замену, можно анализировать несколькими способами. Например, детектирование может быть выполнено электрофоретическими методами, включая конформационный полиморфизм однонитевой ДНК (Orita et al. (1989) Genomics, 8(2), 271-278), денатурирующий градиентный гель-электрофорез (Myers (1985) EPO 0273085) или полиморфизмы длин рестрикционных фрагментов (Life Technologies, Inc., Gathersberg, MD), но с повсеместной доступностью секвенирования ДНК часто легче сделать непосредственно простой сиквенс амплифицированных продуктов. После того, как известно различие полиморфных последовательностей, могут быть разработаны быстрые анализы для тестирования потомства, как правило, с использованием какой-либо версии ПЦР амплификации специфических аллелей (PASA; Sommer, et al. (1992) Biotechniques 12(1), 82-87) или ПЦР амплификации множественных специфических аллелей (PAMSA; Dutton and Sommer (1991) Biotechniques, 11(6), 700-7002).
[0049] Полиморфные маркеры служат полезным инструментом для анализа растений с целью определения степени идентичности линий или сортов (патент США № 6207367). Эти маркеры являются основой для определения ассоциаций с фенотипами и могут быть использованы для стимулирования генетической выгоды. В некоторых вариантах реализации способов по данному изобретению полиморфные нуклеиновые кислоты могут быть использованы для обнаружения в растении томата генотипа, ассоциированного с устойчивостью к болезням, идентификации растения томата с генотипом, ассоциированным с устойчивостью к болезням, и для отбора растения томата с генотипом, ассоциированным с устойчивостью к болезням. В некоторых вариантах реализации способов по данному изобретению полиморфные нуклеиновые кислоты могут быть использованы для получения растения томата, которое содержит в своем геноме интрогрессированный локус, связанный с устойчивостью к болезням. В некоторых вариантах реализации данного изобретения полиморфные нуклеиновые кислоты могут быть использованы для размножения потомства растений томата, которое содержит локус, связанный с устойчивостью к болезням.
[0050] Генетические маркеры могут включать «доминантные» или «кодоминантные» маркеры. «Кодоминантные» маркеры обнаруживают наличие двух или более аллелей (два на диплоидный индивидуум). «Доминантные» маркеры обнаруживают наличие только одного аллеля. Маркеры предпочтительно наследуются кодоминантным способом, так что присутствие обоих аллелей в диплоидном локусе, или множественных аллелей в триплоидных или тетраплоидных локусах, легко обнаруживается, и они не ограничены изменением окружающей среды, то есть, их наследуемость равна 1. Маркерный генотип обычно содержит два маркерных аллеля в каждом локусе диплоидного организма. Маркерная аллельная композиция каждого локуса может быть либо гомозиготной, либо гетерозиготной. Гомозиготность является условием, при котором оба аллеля в локусе характеризуются одной и той же нуклеотидной последовательностью. Гетерозиготностью называют различные состояния аллеля в локусе.
[0051] Анализы на основе нуклеиновых кислот, для определения наличия или отсутствия генетического полиморфизма (т. е. генотипирования), могут быть использованы в селекционных программах для идентификации, отбора, интрогрессии и т. п. Широкий спектр генетических маркеров для анализа генетических полиморфизмов доступен и известен специалистам в данной области техники. Анализ может быть использован для отбора генов, позиций генов, ЛКП, аллелей или геномных областей, которые содержат или сцеплены с генетическим маркером, который сцеплен или связан с устойчивостью к болезням у растений томата.
[0052] Используемые в данном документе способы анализа нуклеиновых кислот включают, но не ограничиваются ими, методы детектирования на основе ПЦР (например, анализы TaqMan), методы микрочипов, методы на основе масс-спектрометрии и/или методы секвенирования нуклеиновых кислот, включая полное секвенирование генома. В некоторых вариантах реализации изобретения обнаружение полиморфных сайтов в образце ДНК, РНК или кДНК может быть облегчено с помощью способов амплификации нуклеиновых кислот. Такие способы специально увеличивают концентрацию полинуклеотидов, которые охватывают полиморфный сайт или включают этот сайт и последовательности, которые расположены либо дистально, либо проксимально по отношению к нему. Такие амплифицированные молекулы могут быть легко обнаружены с помощью гель-электрофореза, методами детектирования флуоресценции или другими способами.
[0053] Один из способов достижения такой амплификации использует полимеразную цепную реакцию (ПЦР) (Mullis et al.1986 Cold Spring Harbor Symp. Quant. Biol. 51: 263-273; Европейский патент 50424; Европейский патент 84796; Европейский патент 258017; Европейский патент 237362; Европейский патент 201184; патент США 4683202; патент США 4582788; и патент США 4683194), с использованием пары праймеров, которые способны гибридизоваться с проксимальными последовательностями, которые определяют полиморфизм в его двухцепочечной форме. Могут также использоваться способы типирования ДНК на основе масс-спектрометрии. Такие способы раскрыты в патентах США 6613509 и 6503710 и в содержащихся в них ссылках.
[0054] Полиморфизмы в последовательностях ДНК могут быть обнаружены или типированы с помощью множества эффективных способов, хорошо известных в данной области техники, включая, но не ограничиваясь ими, способы, раскрытые в патентах США №№ 5468613, 5217863; 5210015; 5876930; 6030787; 6004744; 6013431; 5595890; 5762876; 5945283; 5468613; 6090558; 5800944; 5616464; 7312039; 7238476; 7297485; 7282355; 7270981 и 7250252, все из которых включены в данный документ в полном объеме посредством ссылки. Однако композиции и способы по настоящему изобретению могут быть использованы в сочетании с любым методом типирования полиморфизма для типирования полиморфизмов в образцах геномной ДНК. Эти использованные образцы геномной ДНК включают, но не ограничиваются этими, геномную ДНК, изолированную непосредственно из растения, клонированную геномную ДНК или амплифицированную геномную ДНК.
[0055] Например, полиморфизмы в последовательностях ДНК могут быть обнаружены путем гибридизации с аллель-специфическими олигонуклеотидными (ASO) зондами, как описано в патентах США №№ 5468613 и 5217863. Патент США №5468613 раскрывает аллель-специфические олигонуклеотидные гибридизации, в которых единичные или множественные нуклеотидные вариации в последовательности нуклеиновой кислоты могут быть обнаружены в нуклеиновых кислотах с помощью процесса, при котором последовательность, которая содержит нуклеотидную вариацию, амплифицируют, наносят на мембрану и обрабатывают меченым олигонуклеотидным зондом, специфическим к данной последовательности.
[0056] Целевая последовательность нуклеиновой кислоты также может быть обнаружена методами лигирования зондов, например, как описано в патенте США № 5800944, где целевая последовательность амплифицируется и гибридизируется с зондами с последующим лигированием для обнаружения меченой части зонда.
[0057] Также для обнаружения полиморфизма могут использоваться микрочипы, в которых наборы олигонуклеотидных зондов собраны перекрывающимся способом для того, чтобы представить единичную последовательность таким образом, что различие в целевой последовательности в одной точке приведет к частичной гибридизации зонда (Borevitz et al., Genome Res. 13:513-523 (2003); Cui et al., Bioinformatics 21:3852-3858 (2005). Ожидается, что на любом одном микрочипе будет множество целевых последовательностей, которые могут представлять гены и/или некодирующие области, причем каждая целевая последовательность представлена серией перекрывающихся олигонуклеотидов, а не одним зондом. Эта платформа обеспечивает высокопроизводительный скрининг множества полиморфизмов. Типирование целевых последовательностей способами на основе микрочипов описано в патентах США 6799122, 6913879 и 6996476.
[0058] Другие способы для обнаружения SNP и Indel включают способы достройки по одному основанию (single base extension, SBE). Примеры SBE способов включают, но не ограничиваются ими, примеры, которые раскрыты в патентах США №№ 6004744; 6013431; 5595890; 5762876 и 5945283.
[0059] В другом способе детектирования полиморфизмов SNP и Indel могут быть обнаружены способами, которые описаны в патентах США №№ 5210015, 5876930 и 6030787, в которых олигонуклеотидный зонд имеет 5'- флуоресцентный репортерный краситель и 3'- гасящий краситель, ковалентно сцепленные с 5'- и 3'- концами зонда. Когда зонд остается неповрежденным, близость репортерного красителя к гасящему красителю приводит к супрессии флуоресценции репортерного красителя, например, путем переноса энергии по Ферстеровскому типу. Во время ПЦР прямой и обратный праймеры гибридизуются со специфической последовательностью целевой ДНК, которая фланкирует полиморфизм, тогда как гибридизационный зонд гибридизуется с полиморфизмосодержащей последовательностью в амплифицированном продукте ПЦР. В последующем цикле ПЦР ДНК-полимераза с 5' → 3' экзонуклеазной активностью гидролизирует зонд и отделяет репортерный краситель от гасящего красителя, что в результате приводит к повышенной флуоресценции репортера.
[0060] В другом варианте реализации изобретения целевой локус или локусы могут быть непосредственно секвенированы с использованием технологий секвенирования нуклеиновых кислот. Способы секвенирования нуклеиновой кислоты, известные в данной области техники, включают технологии, представленные 454 Life Sciences (Branford, CT), Agencourt Bioscience (Beverly, MA), Applied Biosystems (Foster City, CA), LI-COR Biosciences (Lincoln, NE), NimbleGen Systems (Madison, WI), Illumina (San Diego, CA) и VisiGen Biotechnologies (Houston, TX). Такие технологии секвенирования нуклеиновых кислот включают такие форматы, как параллельные массивы бусин (parallel bead arrays), секвенирование путем лигирования, капиллярный электрофорез, электронные микрочипы, «биочипы», микрочипы, параллельные микрочипы и одномолекулярные массивы (single-molecule arrays).
Определения
[0061] Нижеследующие определения приведены для лучшего описания настоящего изобретения и в качестве руководства для специалистов в данной области техники при применении настоящего изобретения. Если не указано иное, термины следует понимать в соответствии с обычным использованием специалистами в данной области техники.
[0062] Используемый в данном документе термин «растение» включает растительные клетки, протопласты растений, растительные клетки культуры ткани, из которых могут быть регенерированы растения томата, каллусы растений, группы растений и растительные клетки, которые являются интактными в растениях или частях растений, таких как пыльца, цветки, семена, листья, стебли и тому подобное.
[0063] Используемый в данном документе термин «популяция» означает генетически гетерогенную совокупность растений, которые имеют общее родительское происхождение.
[0064] Используемые в данном документе термины «сорт» и «культивар» означают группу сходных растений, которые по своим генетическим источникам происхождения и характеристикам могут быть отличены от других сортов внутри одного и того же вида.
[0065] Используемый в данном документе термин «аллель» относится к одной из двух или более альтернативных форм геномной последовательности в данном локусе на хромосоме.
[0066] «Локусы количественных признаков (ЛКП)» представляет собой хромосомное расположение, которое кодирует по меньшей мере первую аллель, которая влияет на экспрессивность фенотипа.
[0067] Используемый в данном документе термин «маркер» означает детектируемую характеристику, которая может быть использована для различения организмов. Примеры таких характеристик включают, но не ограничиваются ими, генетические маркеры, биохимические маркеры, метаболиты, морфологические характеристики и агрономические характеристики.
[0068] Используемый в данном документе термин «фенотип» означает детектируемые характеристики клетки или организма, на которые может повлиять экспрессия гена.
[0069] Используемый в данном документе термин «генотип» означает конкретные аллельные составляющие растения.
[0070] Используемый в данном документе термин «элитный» сорт означает любой сорт, который получили в результате селекции и отбора по улучшенным агрономическим характеристикам, включая урожайность и другие выбранные признаки. Термин «элитное растение» относится к растению, которое принадлежит к элитному сорту и которое обладает различными желаемыми признаки. Многочисленные элитные сорта доступны и известны специалистам в области селекции томатов. «Элитная популяция» представляет собой множество элитных особей или сортов, которые могут быть использованы для представления состояния в области техники с точки зрения агрономически улучшенных генотипов данного вида сельскохозяйственной культуры, такой как томат. Аналогичным образом «элитная зародышевая плазма» или элитный штамм зародышевой плазмы представляет собой агрономически улучшенную зародышевую плазму.
[0071] «Культивированный» сорт или «культивар» представляет собой растение или группу растений, которые отобраны по желаемым характеристикам при выращивании в сельском хозяйстве, и которые могут поддерживаться путем размножения сорта. Культивируемый сорт не встречается в естественных условиях в природе, а является результатом вмешательства человека для получения желаемых характеристик для культивирования.
[0072] Используемый в данном документе термин «интрогрессированный», когда он используется в контексте генетического локуса, относится к генетическому локусу, который был введен в новое генетическое окружение, например, путем возвратного скрещивания. Интрогрессия генетического локуса может быть достигнута посредством методов селекции растений и/или молекулярно-генетическими методами. Такие молекулярно-генетические методы включают, но не ограничиваются ими, различные способы трансформации растений и/или способы, которые обеспечивают гомологичную рекомбинацию, негомологичную рекомбинацию, сайт-специфическую рекомбинацию и/или геномные модификации, которые обеспечивают замену локуса или преобразование локуса.
[0073] Используемый в данном документе термин «сцепленный», когда он используется в контексте маркеров нуклеиновой кислоты и/или геномных областей, означает, что маркеры и/или геномные области расположены в одной и той же группе сцепления или на одной и той же хромосоме, так что они проявляют тенденцию к совместной сегрегации при мейозе.
[0074] Используемый в данном документе термин «некроз», когда речь идет о растении или растительной ткани, относится к ткани, которая становится бурой и отмирает. Используемый в данном документе термин «некроз» также может включать возникновение хлорозов, при которых хлоротическая ткань отмечается как пожелтение или белое обесцвечивание зеленой в нормальном состоянии ткани.
[0075] Используемый в данном документе термин «локус устойчивости» означает локус, который связан с устойчивостью или толерантностью к болезням. Например, локус устойчивости, согласно настоящему изобретению, в одном варианте реализации изобретения может контролировать резистентность или чувствительность к вирусу желтой курчавости листьев томата (ВЖКЛТ).
[0076] Используемый в данном документе термин «аллель устойчивости» означает последовательность нуклеиновой кислоты, которая связана с устойчивостью или толерантностью к болезням.
[0077] Используемый в данном документе термин «устойчивость» или «улучшенная устойчивость» растения к болезненному состоянию является показателем того, что растение в большей мере способно снижать нагрузку от заболевания, чем неустойчивое или менее устойчивое растение. Устойчивость представляет собой относительный термин, который указывает на то, что «устойчивое» растение в большей мере способно снижать нагрузку от заболевания по сравнению с другим (менее устойчивым) растением (например, различные сорта растений), которое выращено в сходных болезненных состояниях. Специалисту в данной области будет понятно, что устойчивость растений к болезням сильно варьирует и может представлять собой спектр более устойчивых или менее устойчивых фенотипов. Однако, путем простого наблюдения специалист, как правило, может определить относительную устойчивость различных растений, сортов растений или семейств растений при болезненных состояниях и, кроме того, также распознает фенотипические градации «устойчивости».
[0078] Специалисту в данной области будет понятно, что устойчивость растений к болезням сильно варьирует и может представлять собой спектр более устойчивых или менее устойчивых фенотипов. Однако, путем простого наблюдения специалист, как правило, может определить относительную устойчивость или чувствительность различных растений, линий растений или семейств растений при болезненных состояниях и, кроме того, также распознает фенотипические градации «устойчивости».
[0079] Термин «около» используется для указания того, что значение включает стандартное отклонение ошибки для устройства или способа, который используется для определения значения. Использование термина «или» в формуле изобретения используется в значении «и/или», если он точно не указывается только для ссылок на альтернативы или альтернативы являются взаимоисключающими, хотя раскрытие изобретения поддерживает определение, которое относится как к только альтернативным вариантам, так и к «и/или». В формуле изобретения определения в единственном числе в сочетании со словом «содержащий» или другим открытым текстом означают «один или более», если конкретно не указано другое. Термины «содержать», «иметь» и «включать» являются открытыми связующими глаголами. Любые формы или времена одного или более из этих глаголов, таких как «содержит», «содержащий», «имеет», «имеющий», «включает» и «включающий» также являются открытыми. Например, любой способ, который «содержит», «имеет» или «включает» один или более этапов, не ограничен тем, что обладает только этими одним или более этапами, а также охватывает другие, не включенные в список, этапы. Аналогичным образом, любое растение, которое «содержит», «имеет» или «включает» один или более признаков, не ограничено тем, что обладает только этими одним или более признаками, а охватывает другие, не включенные в список, признаки.
ПРИМЕРЫ
Пример 1
Идентификация рекомбинантных линий томата, которые содержат Ty-1
[0080] Ty-1 локус устойчивости к ВЖКЛТ из S.chilense LA1969 интрогрессировали в элитные растения S. lycopersicon, что привело к появлению нескольких селекционных линий с высоким уровнем устойчивости к ВЖКЛТ. Однако, эти линии обнаруживали нежелательные агрономические признаки, которые связаны с Ty-1 интрогрессией из S.chilense, включая некроз. Молекулярные маркеры, которые приведены в Таблице 1, были разработаны для идентификации рекомбинантов, которые поддерживают устойчивость к ВЖКЛТ при сниженной частоте возникновения некроза.
Таблица 1. Маркеры на хромосоме 6 для идентификации Ty-1 рекомбинантов.
[0081] Использование этих маркеров, сцепленных с Ty-1, привело к идентификации линий томатов 11039-1, 09GA0361-11 и 08TK01-26-2. Как проиллюстрировано на ФИГ. 1, линия 11039-1 содержит рекомбинацию между маркерами Q_NL5113515 и Q_NL5113488. Эта линия чувствительна к ВЖКЛТ, но все еще обнаруживает некроз листьев, указывая на то, что Ty-1 локус устойчивости расположен справа (южнее на хромосоме) от маркера Q_NL5113515, хотя, по меньшей мере, частично локус некроза из S.chilense интрогрессии расположен слева (севернее на хромосоме) от маркера Q_NL5113488. Линия 09GA0361-11 содержит рекомбинацию справа от CD67 и устойчива к ВЖКЛТ, и при этом проявляет уменьшенный некроз листьев. Еще меньшая интрогрессия была идентифицирована в линии 08TK01-26-2, которая проявляет устойчивость к ВЖКЛТ в полевых условиях и в более контролируемых условиях (агроинокуляция ВЖКЛТ).
Пример 2
Картирование популяций
[0082] Родительские сорта Petit (FIR-191-PETIT) и Screen (FIR-191-SCREEN) скрестили для создания F2 популяции для дальнейшего анализа некрозов и других признаков у растений, которые содержат Ty-1 интрогрессии. FIR-191-PETIT представляет собой родителя, который содержит chilense интрогрессию. Девяносто две сотни семян из F2 популяции высевали и анализировали с помощью маркеров NL0232070 и NL0231193, в результате чего было получено 209 проверенных рекомбинантов. Из этих рекомбинантов 42 были выращены в теплице для получения самоопыленных (F3) семян, и F3 растения фенотипировали по некрозам. Двадцать пять F3 линий, с установленным событием рекомбинации между NL0232070 и NL0231193, были выращены в тепличных условиях для анализа Ty-1 интрогрессии.
Пример 3
Оценка устойчивости к ВЖКЛТ и некрозу в картированных популяциях
[0083] Растения оценивали по устойчивости к ВЖКЛТ, используя нижеследующий протокол инокуляции с помощью белокрылки. Растения подвергались воздействию белокрылок (Bemisia tabaci (Q биотип)), которые переносили ВЖКЛТ в течение 48 часов на стадии первого настоящего листа (приблизительно через 3 недели после посева). Растения оценивали по устойчивости к ВЖКЛТ согласно шкале, приведенной в Таблице 2, и оценивали некрозы, с использованием шкалы, приведенной в Таблице 3. Оценку выполняли через 15 и 30 дней после инокуляции, когда, по крайней мере, 90% чувствительных контрольных растений были инфицированы ВЖКЛТ (по оценке 9).
Таблица 2. Шкала оценки ВЖКЛТ
3=несколько желтых пятен или полос по краям верхних листьев.
5=очевидное пожелтение верхних листьев, отсутствие курчавости.
7=очевидное пожелтение и курчавость верхних листьев. Может наблюдаться некоторое замедление роста растения.
9=значительное замедление роста растения, значительная курчавость и пожелтение верхних листьев.
Таблица 3. Шкала оценки некрозов. Все листья оставляют на растении для оценки.
2=край листа имеет некрозы; оставшаяся часть листа все еще остается зеленой.
3=незначительные некрозы; несколько явных некротических пятен на верхних листьях. Отсутствие хлорозов. Зеленый лист.
4=незначительные некрозы; светлые пятна на листе и явные некротические пятна на листе в нижней части растения.
5=умеренные некрозы; явные пятна на листе и явные некротические пятна на листе в нижней части растения.
6=некрозы и незначительные хлорозы листа в нижней части растения.
7=много некротических пятен и хлорозов на листе, поражено около 75% растения.
8=значительные некрозы и хлорозы. Только верхняя часть растения имеет несколько оставшихся зеленых листьев.
9=все растение с некрозами и хлорозами.
Пример 4
Идентификация некротических ЛКП
[0084] Семена из F2 популяции, которые описаны в примере 2, были высеяны, и растения выращивали в тепличных условиях зимой без искусственного освещения. Световые дни были < 12 ч, ночная температура составляла > 15 °C, а максимальная дневная температура составляла < 25 °C. Некроз оценивали согласно протоколу, который описан в примере 3. F2 растения отбирали индивидуально и генотипировали с помощью набора TaqMan анализных маркеров, распределенных по геному. Условия анализа были следующими: 20 секунд при 95 °C, далее 35 циклов по 3 секунды при 95°C и 20 секунд при 60°C и хранение при 20 °C.
[0085] Объединяя генотипические и фенотипические данные, можно было обнаружить два ЛКП со значением логарифмом шансов (LOD) > 3,0 с помощью R/ лкп (qtl), используя стандартные настройки. Основной ЛКП располагался на хромосоме 6, как проиллюстрировано на ФИГ. 2. В Таблице 4 показаны значения LOD для маркеров, которые фланкируют идентифицированные ЛКП на хромосомах 2 и 6.
Таблица 4. Данные LOD для маркеров, которые значимые для некрозов.
Пример 5
Картирование локусов некрозов в пределах Ty-1 интрогрессии на хромосоме 6
[0086] F3 популяции, которые описаны в примере 2, содержащие Ty-1 интрогрессии из S.chilense, были дополнительно проанализированы для выявления локусов, которые связаны с некрозом, происходящим от Ty-1 интрогрессий. В Таблице 5 представлены результаты оценки некрозов для F3 популяций, а также средние оценки некрозов для каждой линии в апреле и мае. Был разработан ряд маркеров для генотипирования этих F3 популяций в области, которая окружает Ty-1, и эти маркеры приведены в Таблице 6.
[0087] При анализе Ty-1 области этими маркерами было отмечено, что наблюдали почти полное отсутствие рекомбинации в центромерной области хромосомы 6, где расположена Ty-1. Разработка дополнительных маркеров в области 5-25 сМ хромосомы 6 увеличило возможность обнаружения событий рекомбинации. Эти новые молекулярные маркеры также позволяют скринировать большие популяции для отбора растений, которые содержат уменьшенные Ty-1 интрогрессии, которые проявляют высокие уровни устойчивости к ВЖКЛТ, которую придает Ty-1 ген устойчивости, при этом также уменьшается или исключается появление нежелательных агрономических признаков, ранее связанных с Ty-1 интрогрессиями, таких как некроз.
[0088] На ФИГ. 3 проиллюстрированы данные генотипирования для контрольных линий и F3 линий, которые содержат рекомбинационные события между маркерами NL0232061 и NL231193. Также показаны средние оценки некрозов в апреле и мае. На ФИГ. 4 проиллюстрирована средняя оценка некрозов для нескольких линий растений, проиллюстрированных на ФИГ. 3, основанная на расположении точки разрыва между донорской ДНК S.chilense и ДНК S.esculentum, определенная с использованием маркеров, проиллюстрированных на ФИГ. 3. Эти данные четко показывают, что растения, которые включают Ty-1 интрогрессию, в которой S.chilense Ty-1 донорская ДНК заменяется на ДНК S.esculentum из верхней части хромосомы до маркера NSLYC008382908 не обнаруживают повышенных уровней некрозов, ранее наблюдавшихся для растений с Ty-1 интрогрессиями.
Таблица 5. Результаты оценки некрозов для F3 растений.
2ESC контроль
3одно растение гетерозиготное
4chi контроль
Таблица 6. Последовательности, которые связаны с маркерами в Ty-1 области на хромосоме 6 томатов.
позиция
позиция
Пример 6
Идентификация ЛКП, связанного с некрозом на хромосоме 2
[0089] Как проиллюстрировано на ФИГ. 2 и в Таблице 4, ЛКП, который связан с некрозом, был идентифицирован на хромосоме 2. Гаплотип TT_GG (аллель TT в маркере NSLYC008374675 (SEQ ID № 5) и аллель GG в маркере NSLYC008375578 (SEQ ID № 6) ассоциируется с уменьшенным или отсутствующим некрозом, как показано в Таблице 7 и на ФИГ. 5. Поэтому некроз у растений, которые имеют гаплотип TT_GG, связан с другим некротическим ЛКП. Гаплотип CC_AA (аллель CC в маркере NSLYC008374675 (SEQ ID № 5) и аллель AA в маркере NSLYC008375578 (SEQ ID № 6 ) ассоциируется с некрозом. Линии томатов, которые имеют этот аллель, могут обнаруживать некрозы, хотя, как правило, на промежуточных уровнях (ФИГ. 5). Последовательности зонда VIC, последовательности зонда FAM и прямые и обратные праймеры для маркера NSLYC008374675 и маркера NSLYC008375578 представлены в SEQ ID № 7-8, 9-10, 11-12 и 13-14, соответственно.
Таблица 7. Маркеры, связанные с некрозом на хромосоме 2 томата.
позиция
Пример 7
TaqMan анализ
[0090] Анализ TaqMan был разработан для обнаружения SNP на хромосоме 6 (положение основания 32987053) с использованием маркера NSLYC008426102. Результаты генотипирования ряда рекомбинантных F4 линий с помощью анализов TaqMan, которые расположены в Ty-1 области между 20 и 34 млн п.н. хромосомы 6. Анализ TaqMan показал 100% корреляцию с существующей рекомбинантной популяцией (ФИГ. 6).
Пример 8
Интрогрессия признака устойчивости к ВЖКЛТ без некроза
[0091] Линию томата, которая включает обычную Ty-1 интрогрессию и проявляет некрозы, скрещивают с линией томата, которая содержит уменьшенную Ty-1 интрогрессию, описанную в данном документе, и проявляет незначительные некрозы или их отсутствие. Для растений потомства от этого скрещивания проводят возвратное скрещивание для образования BC1 поколения. BC1 потомство растений отобрано по S.esculentum аллелю в маркере NSLYC009077970 (SEQ ID № 2), что указывает на наличие уменьшенной Ty-1 интрогрессии по данному изобретению. BC1 потомство растений или последующее потомство растений отобрано по наличию S.chilense аллеля в маркере NSLYC009418370 (SEQ ID № 4), чтобы иметь Ty-1 интрогрессию. Дополнительно, частота возникновения некрозов может быть дополнительно модулирована путем отбора аллелей, которые связаны с отсутствием некрозов в описанном интервале ЛКП хромосомы 2, с использованием маркеров NSLYC008374675 (SEQ ID № 5) и NSLYC008375578 (SEQ ID № 6).
Пример 9
Рекомбинантные растения томата с уменьшенным размером Ty-1 интрогрессии
[0092] Для создания растений томата с Ty-1 интрогрессией, которая имеет укороченную донорскую ДНК S.chilense, начальный отбор для рекомбинации между маркерами NSLYC008382908 (8,51 cM) и NSLYC008383071 (9,31 cM, SEQ ID № 1) был выполнен с целью разрыва сцепления между Ty-1 и нежелательным привнесенным некрозом (см. ФИГ. 7). Затем выбирают рекомбинацию между маркером NSLYC009418370 (9,64 сМ, SEQ ID № 4) и NL231193 (11,58 сМ), чтобы минимизировать количество S.chilense ДНК и тем самым уменьшить размер интрогрессии. Последовательности для маркеров NSLYC008382908 и NL231193 приведены в Таблице 9. Отобранные рекомбинанты тестировали на устойчивость к ВЖКЛТ путем инокуляции с помощью белокрылки в 4 повторах по 10 растений в каждом. Дополнительно, подсчет урожая зрелых плодов со всего растения и общий вес урожая и количество плодов выполняли в 4 повторах по 6 растений в каждом. Результаты тестирования продуктивности показаны в Таблице 8 и на ФИГ. 8 и 9.
Таблица 8. Средние значения однофакторного дисперсионного анализа эффективности (one-way ANOVA).
[0093] Улавливание последовательности выполняли для характеристики интрогрессии на хромосоме 6 томата (ФИГ. 10). Было определено, что интрогрессия составляет приблизительно 260 кб (килобаз), а растения, в которые была введена эта интрогрессия, сохраняют устойчивость к ВЖКЛТ и при этом демонстрируют отсутствие некрозов, и отсутствие уменьшения размера плода и завязи плода.
Таблица 9. Последовательности маркера.
* * * * * * * * * * * * *
[0094] Все композиции и/или способы, которые раскрыты и заявлены в данном документе, могут быть изготовлены и выполнены без излишнего экспериментирования в свете данного описания. Хотя композиции и способы данного изобретения были описаны в отношении предпочтительных вариантов реализации изобретения, специалистам в данной области техники будет очевидно, что вариации могут быть применены для композиций и/или способов, и для этапов или последовательности этапов способа, описанного в данном изобретении, без отклонения от концепции, сущности и объема изобретения. Более конкретно, будет очевидно, что некоторые агенты, которые как химически, так и физиологически родственные, могут быть заменены на агенты, которые описаны в данном изобретении, при этом будут достигнуты такие же или сходные результаты. Все подобные аналогичные замены и модификации, очевидные для специалистов в данной области техники, считаются находящимися в пределах сущности, объема и концепции изобретения, как определено прилагаемой формулой изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Seminis Vegetable Seeds, Inc.
<120> РАСТЕНИЯ ТОМАТА С УЛУЧШЕННОЙ УСТОЙЧИВОСТЬЮ К БОЛЕЗНЯМ
<130> SEMB:019WO
<150> 62/064375
<151> 2014-10-15
<160> 22
<170> PatentIn версия 3.5
<210> 1
<211> 764
<212> ДНК
<213> Solanum tomato
<220>
<221> misc_feature
<222> (46)..(46)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (48)..(48)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (76)..(76)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (83)..(83)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (97)..(99)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (102)..(104)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (106)..(110)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (192)..(192)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (204)..(204)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (293)..(293)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (347)..(347)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (461)..(461)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (520)..(520)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (534)..(534)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (543)..(543)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (545)..(545)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (595)..(595)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (643)..(643)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (655)..(655)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (686)..(686)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (698)..(698)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (709)..(709)
<223> n представляет собой a, c, g, или t
<400> 1
cataaccatg acacctcacc ttgaatttgt cgtgtcaatg catccntnac caaggctaat 60
aaaaaaggct aagggntgat tcntggtgca accccannna annngnnnnn tgctcagagt 120
ctcctcccag tgtccttaca tgagtattgg cttcatcata tataccctta ttttcctagt 180
gtaagccaca tntatacctc tagnctccaa tcaactccat agaaccttcc ttgggacttc 240
atggtaagcc ttctctaggt ggatgaatat catcggtaag tccttcgctt ccntatattg 300
ctccaccagt ctttttacga gatgaatgac ttctgtcatt caggccntga catgaatcta 360
aacaggtttt caaaaatagc cacgcccctc ctccctctaa tctccaccac cctctcccaa 420
accttcatat tgtagcttag cagcttgata ccctttggtt nttgcaattt tagatgtcac 480
ccttgttcat gaacaacaaa atcattgtac ttcacctccn ttcttcaggc atcnttgtgt 540
ccnanaaacg acattaaaca actctgtcaa gcactccaca cctgccctgt ctgcnacctt 600
ccaaaattca actgggatct cgtctggccc agtcactcta ccnttatgcc agggncgaac 660
ctacatagtg tcaagagggt ccatangaac ccacttcnta aaaaaattna tattgtatat 720
ataggtagat ttggtttgtc ttctggacaa tacatgtata gata 764
<210> 2
<211> 121
<212> ДНК
<213> Solanum tomato
<400> 2
aaaccaattc acaacaaaga tgatgatatc ctttaactac tgtaaggcta gggagtacct 60
gtatttcaag ttatacggaa aacaatgcat cactccagtt gcaatcatga caagtgcagc 120
a 121
<210> 3
<211> 747
<212> ДНК
<213> Solanum tomato
<220>
<221> misc_feature
<222> (59)..(59)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (69)..(70)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (86)..(86)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (91)..(91)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (126)..(126)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (159)..(159)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (162)..(162)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (187)..(187)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (206)..(206)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (249)..(255)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (277)..(277)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (306)..(306)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (351)..(351)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (368)..(368)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (381)..(381)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (384)..(384)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (414)..(415)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (417)..(424)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (528)..(528)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (530)..(530)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (685)..(685)
<223> n представляет собой a, c, g, или t
<400> 3
caatttctct ttgttctcct tgcttattta taatcaaaat catcagtaaa caaaaaaant 60
aattaaacnn aaatgtaacc taaatnaact naccttctcc aaatcttctt gtaaatatct 120
aacaanggag gcaaccattt atattttcca ttttggtcnt anattcccac atttttctta 180
aaatacnttt taggaattaa tattancttc tttatatgtg caacatcttt cacatatttt 240
gttatatann nnnnnatata tgagatttgt ctacaantca aagtaacaaa atgttataaa 300
aaaaanctca taagataatt gaattttcct atactccatc atcaatccat ngctaaatgg 360
gattaacntg acatttttat natntagcta tgaaatttta tctgtcgcta attnngnnnn 420
nnnnagaaag agatgatgat atatacatac cttaatagat ggattggatg aagaagcaga 480
aagaagtcca gttattgttt tattatcact tccttttgta ttttcaanan gatcaacctt 540
gttggtagtc aacgtaatta ttgttctagc tccttcgtaa caagctgcac atattgtatt 600
tcttggtggc cttaaaataa atggcatcgc gctacatata gaacaatcca tatttatttg 660
tgcgcttcga ttaatttttt tctgntcaat attcgaagaa gaaattactg aactcaaatg 720
atcaatgaag aagagacgat gatcgat 747
<210> 4
<211> 338
<212> ДНК
<213> Solanum tomato
<220>
<221> misc_feature
<222> (4)..(4)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (23)..(23)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (29)..(31)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (94)..(94)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (119)..(119)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (139)..(139)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (172)..(172)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (179)..(179)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (191)..(191)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (202)..(203)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (213)..(213)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (230)..(230)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (234)..(234)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (244)..(244)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (253)..(253)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (257)..(257)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (262)..(262)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (271)..(271)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (277)..(277)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (280)..(280)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (284)..(285)
<223> n представляет собой a, c, g, или t
<220>
<221> misc_feature
<222> (313)..(314)
<223> n представляет собой a, c, g, или t
<400> 4
aagnattata aagatgcata canaataann nataccagag aatcaaaaca cttaaaaaga 60
cagatatgtt tttctatcat tctgttcctg cacnaaaagt aataaatttt gtaagcatnt 120
gttcttcaca aatgatatnc tccctccgtt gcaatttgtt tgtttgatat tnacttgana 180
cggagtttaa naaagtgaag anngcttttg aanattgtgg tcttgaaagn aatntgttga 240
attnaccaaa atntccntca ancttatggt nttaaanagn ctanntggaa agttaaaatt 300
aaatagttgt cannaaaagg aaagaggtct taaacatg 338
<210> 5
<211> 101
<212> ДНК
<213> Solanum tomato
<400> 5
agtcttcctg ctcctggccc ttccgaaaga agtcaacaca gtacttctct tcctcctccc 60
catcagaaag ctcacccctc accagcttat catataactt a 101
<210> 6
<211> 101
<212> ДНК
<213> Solanum tomato
<400> 6
gaagatttgt gggtattgat gcaaaagaag aatgttgatg ctgacttggg aagttacacc 60
attagattac aaggattggt tgcgaataac caggttaacg a 101
<210> 7
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Probe
<400> 7
cagtacttct cttcctcctc 20
<210> 8
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Probe
<400> 8
tctaatggtg taacttccca ag 22
<210> 9
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Probe
<400> 9
agtacttctc tccctcctc 19
<210> 10
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Probe
<400> 10
ctaatggtgt aactccccaa g 21
<210> 11
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 11
cctggccctt ccgaaagaa 19
<210> 12
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 12
gtgggtattg atgcaaaaga agaatgt 27
<210> 13
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 13
gggtgagctt tctgatggg 19
<210> 14
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 14
tcgttaacct ggttattcgc aacc 24
<210> 15
<211> 15
<212> ДНК
<213> Lycopersicon esculentum
<400> 15
ctcaatggca aacaa 15
<210> 16
<211> 16
<212> ДНК
<213> Lycopersicon esculentum
<400> 16
cctcaatgac aaacaa 16
<210> 17
<211> 24
<212> ДНК
<213> Lycopersicon esculentum
<400> 17
agaaaaatgt ggccatgggt aact 24
<210> 18
<211> 25
<212> ДНК
<213> Lycopersicon esculentum
<400> 18
ggtagcctta gatgcaatag tgtga 25
<210> 19
<211> 17
<212> ДНК
<213> Lycopersicon esculentum
<400> 19
tctacacaaa agaatgc 17
<210> 20
<211> 17
<212> ДНК
<213> Lycopersicon esculentum
<400> 20
tctacacaaa cgaatgc 17
<210> 21
<211> 24
<212> ДНК
<213> Lycopersicon esculentum
<400> 21
cttgggagat actctctgtt gctt 24
<210> 22
<211> 26
<212> ДНК
<213> Lycopersicon esculentum
<400> 22
gcccaacaga tgatctttaa gaatgg 26
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЯ ТОМАТОВ С УЛУЧШЕННЫМИ ПРИЗНАКАМИ | 2019 |
|
RU2821992C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ ГЕНА RLM2 РЕЗИСТЕНТНОСТИ К ЧЕРНОЙ НОЖКЕ BRASSICA NAPUS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2717017C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПОЛУЧЕНИЯ УКОРОЧЕННЫХ РАСТЕНИЙ КУКУРУЗЫ | 2016 |
|
RU2745987C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ ГЕНА RLM4 РЕЗИСТЕНТНОСТИ К ЧЕРНОЙ НОЖКЕ BRASSICA NAPUS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2718584C2 |
| Участки генов и гены, ассоциированные с повышенной урожайностью у растений | 2016 |
|
RU2758718C2 |
| SNP-МАРКЕРЫ И ОТБОР В ОТНОШЕНИИ НИЗКОГО СОДЕРЖАНИЯ КЛЕТЧАТКИ В ПРЕДСТАВИТЕЛЯХ РОДА BRASSICA | 2019 |
|
RU2834673C2 |
| РАСТЕНИЕ ТОМАТА, ОБРАЗУЮЩЕЕ ПЛОДЫ С МОДИФИЦИРОВАННЫМ СОДЕРЖАНИЕМ САХАРОВ | 2019 |
|
RU2817600C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ, АССОЦИИРОВАННЫЕ С УСТОЙЧИВОСТЬЮ ПОДСОЛНЕЧНИКА К OROBANCHE | 2018 |
|
RU2776361C2 |
| ИНТРОГРЕССИЯ QTL УРОЖАЯ В РАСТЕНИЯ CUCUMIS SATIVUS | 2016 |
|
RU2741964C2 |
| РАСТЕНИЯ ОГУРЦА, УСТОЙЧИВЫЕ К ЗАБОЛЕВАНИЯМ | 2006 |
|
RU2418405C2 |


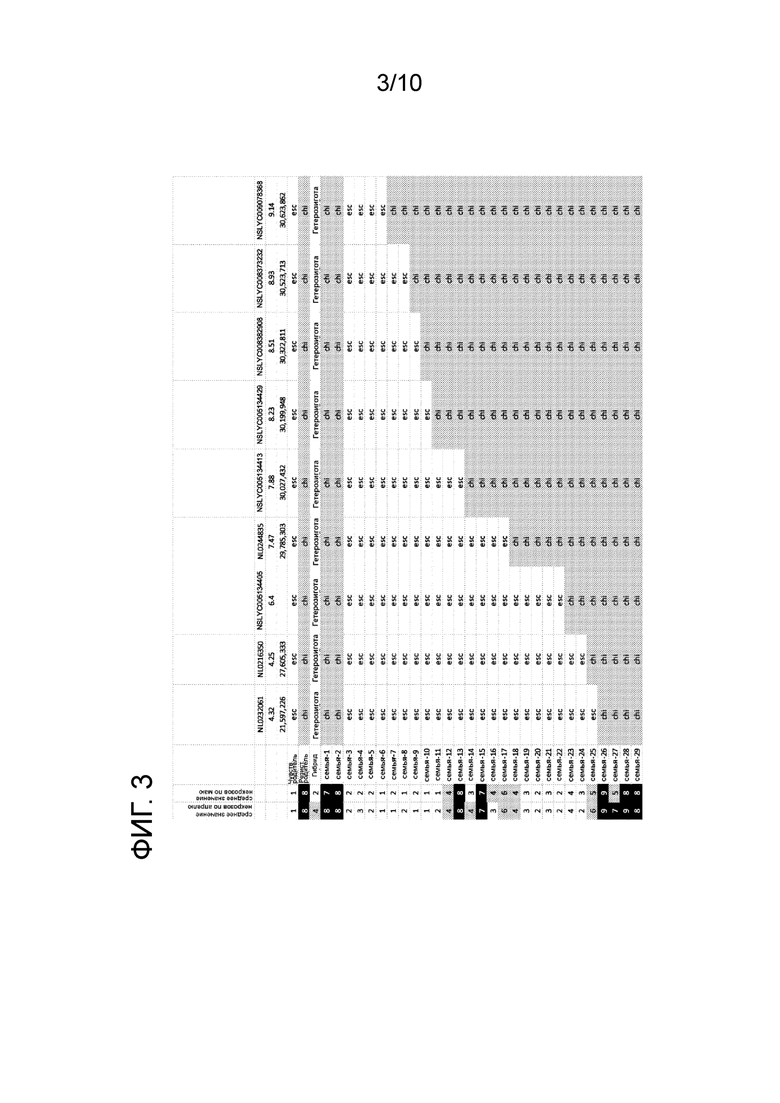
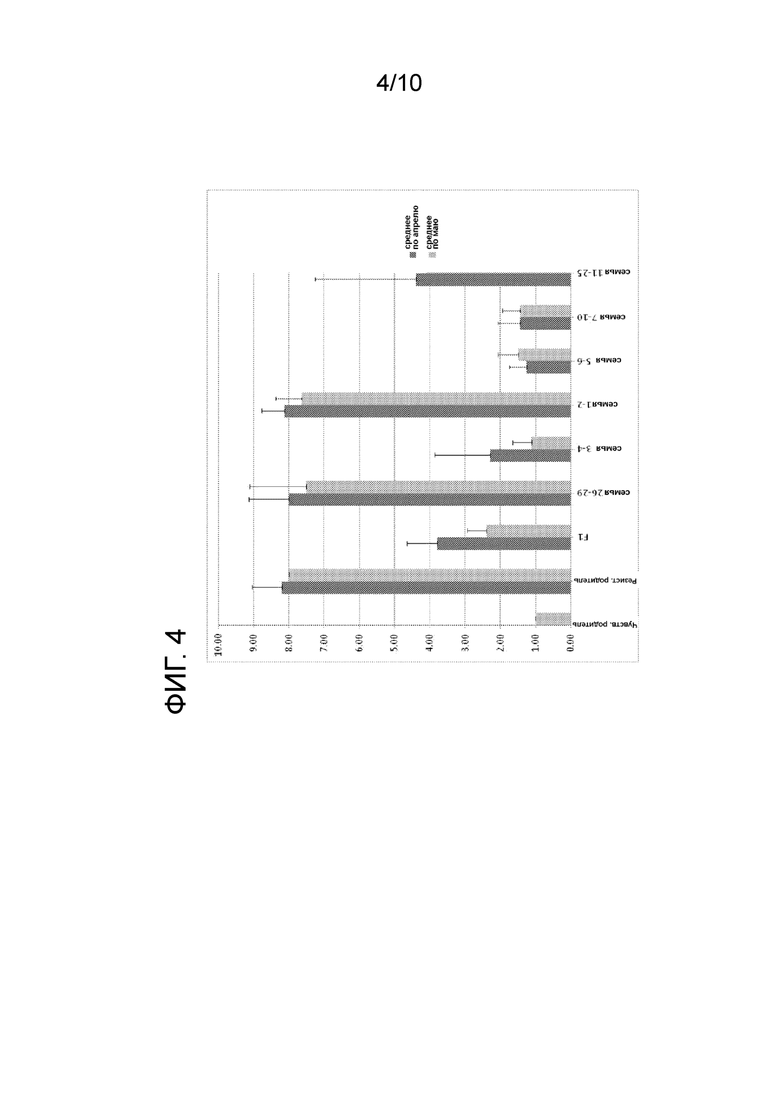

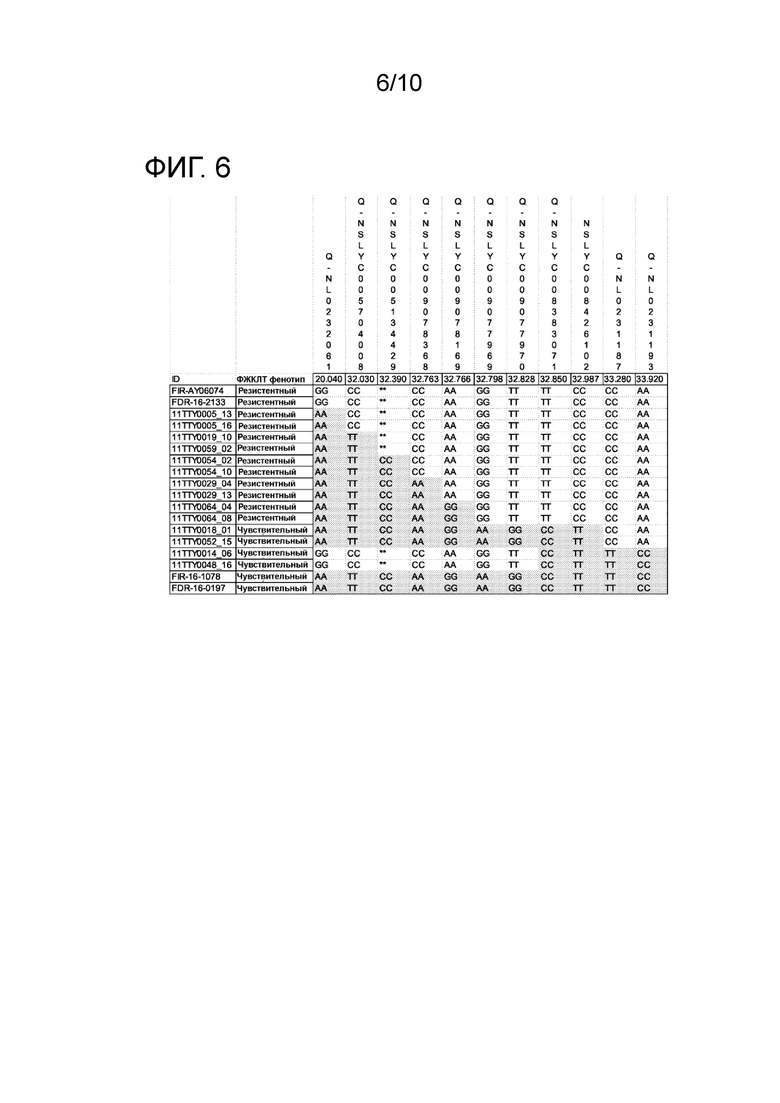




Изобретение относится к области биохимии, в частности к растению томата, имеющему улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеющему некроз, а также к его части. Также раскрыт способ получения растения томата с улучшенной устойчивостью к вирусу желтой курчавости листьев томата и не имеющее некроз. Изобретение позволяет эффективно получать растение томата, имеющее улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеющее некроз. 9 н. и 22 з.п. ф-лы, 10 ил., 9 табл., 9 пр.
1. Растение томата, имеющее улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеющее некроз, где указанное растение томата содержит донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и не содержит донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 3, причем указанное растение томата имеет улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеет некроз относительно растения, не содержащего указанную донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и содержащего указанную донорскую ДНК, содержащую маркерную SEQ ID NO: 3.
2. Растение томата по п. 1, отличающееся тем, что рекомбинантная интрогрессия из Solanum chilense находится между приблизительно 30,20 млн п.н. и 30,88 млн п.н. на хромосоме 6.
3. Растение томата по п. 2, отличающееся тем, что рекомбинантная интрогрессия из Solanum chilense находится между приблизительно 30,69 млн п.н. и 30,88 млн п.н. на хромосоме 6.
4. Растение томата по п. 3, отличающееся тем, что рекомбинантная интрогрессия из Solanum chilense находится между приблизительно 30,71 млн п.н. и 30,88 млн п.н. на хромосоме 6.
5. Растение томата по п. 1, отличающееся тем, что рекомбинантная интрогрессия из Solanum chilense находится между приблизительно 30,32 млн п.н. и 30,71 млн п.н. на хромосоме 6.
6. Растение томата по п. 1, отличающееся тем, что рекомбинантная интрогрессия из Solanum chilense находится между приблизительно 30,88 млн п.н. и 31,80 млн п.н. на хромосоме 6.
7. Растение томата по п. 1, отличающееся тем, что растение содержит Solanum chilense аллель в маркерной SEQ ID NO: 1 и не имеет Solanum chilense аллеля в маркерной SEQ ID NO: 2.
8. Растение томата по п. 1, отличающееся тем, что растение содержит донорскую ДНК S.chilense в пределах сегмента генома, фланкированного маркерной SEQ ID NO: 3 и маркерной SEQ ID NO: 1.
9. Растение томата по п. 1, отличающееся тем, что растение содержит донорскую ДНК S.chilense в пределах сегмента генома фланкированного маркерной SEQ ID NO: 2 и маркерной SEQ ID NO: 1.
10. Растение томата по п. 1, отличающееся тем, что растение содержит донорскую ДНК S.chilense в пределах сегмента генома, фланкированного маркером, выявляемым праймерами SEQ ID NO: 17 и SEQ ID NO: 18, и маркерной SEQ ID NO: 1.
11. Растение томата по п. 1, отличающееся тем, что растение содержит донорскую ДНК S.chilense в пределах сегмента генома, фланкированного маркерной SEQ ID NO: 4 и маркером, выявляемым праймерами SEQ ID NO: 21 и SEQ ID NO: 22.
12. Часть растения, имеющая улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеющая некроз, где указанная часть растения является частью растения по п. 1, где указанная часть растения имеет улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеет некроз относительно части растения, не содержащей донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и содержащей указанную донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 3, где указанная часть растения содержит донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и не содержит донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 3.
13. Часть растения по п. 12, отличающаяся тем, что часть растения представляет собой клетку, семя, корень, стебель, лист, плод, цветок или пыльцу.
14. Способ получения растения томата с улучшенной устойчивостью к вирусу желтой курчавости листьев томата и не имеющее некроз, который включает:
а) скрещивание растения томата по п. 1 с самим собой или с другим растением томата с иным генотипом для получения одного или более растений-потомков; и
б) отбор растения-потомка, которое содержит донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1 и не содержащую донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 3, с помощью маркер-вспомогательной селекции (СПМ),
где растение-потомок проявляет улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеющее некроз относительно растения томата, не содержащего донорскую ДНК Solanum chilense, содержащей маркерную SEQ ID NO: 1, и содержащего донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 3.
15. Способ по п. 14, отличающийся тем, что маркер-вспомогательная селекция (СПМ) включает обнаружение по меньшей мере одного аллеля в локусе, который выбран из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 1, маркера, выявляемого праймерами SEQ ID NO: 17 и SEQ ID NO: 18, и маркера, выявляемого праймерами SEQ ID NO: 21 и SEQ ID NO: 22.
16. Способ по п. 14, отличающийся тем, что растение-потомок представляет собой растение-потомок F2-F6.
17. Способ по п. 14, отличающийся тем, что получение растения-потомка включает возвратное скрещивание.
18. Способ по п. 17, отличающийся тем, что возвратное скрещивание включает 2-7 поколений возвратного скрещивания.
19. Способ получения растения томата, которое проявляет улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеющее некроз, который включает:
а) получение растения томата, гетерозиготного по донорской ДНК Solanum chilense в пределах сегмента генома, фланкированного маркерной SEQ ID NO: 3 и маркерной SEQ ID NO: 1;
б) получение потомства растения; и
в) отбор по меньшей мере первого растения-потомка, содержащего донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и не содержащего донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 3;
причем отбор указанного первого растения-потомка включает обнаружение по меньшей мере одного аллеля в локусе, который выбран из группы, состоящей SEQ ID NO: 3, SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 1, маркера, выявляемого праймерами SEQ ID NO: 17 и SEQ ID NO: 18, и маркера, выявляемого праймерами SEQ ID NO: 21 и SEQ ID NO: 22 с помощью маркер-вспомогательной селекции (СПМ).
20. Способ по п. 19, отличающийся тем, что растение-потомок представляет собой растение-потомок F2-F6.
21. Способ по п. 19, отличающийся тем, что получение растения-потомка включает возвратное скрещивание.
22. Способ по п. 21, отличающийся тем, что возвратное скрещивание включает 2-7 поколений возвратного скрещивания.
23. Растение томата, которое получено способом по п. 19, где указанное растение томата содержит донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и не содержит донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 3, причем указанное растение томата проявляет улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеет некроз относительно растения, не содержащего указанную донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и содержащего указанную донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 3.
24. Часть растения по п. 23, которая выбрана из группы, состоящей из клетки, семени, корня, стебля, листа, плода, цветка или пыльцы, где указанная часть растения содержит донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и не содержит донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 3, и причем указанная часть растения проявляет улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеет некроз относительно растения, не содержащего указанную донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и содержащего указанную донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 3.
25. Растение томата, имеющее улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеющее некроз, где растение содержит TT аллель в маркерной SEQ ID NO: 5 и GG аллель в маркерной SEQ ID NO: 6, причем указанное растение томата имеет улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеет некроз относительно растения, не содержащего указанные аллели.
26. Способ по п. 19, который дополнительно включает:
г) отбор против аллелей на хромосоме 2, которые связаны с некрозом;
причем указанный отбор включает отбор с целью удаления растений, которые включают CC аллель в маркерной SEQ ID NO: 5 или AA аллель в маркерной SEQ ID NO: 6, причем наличие указанных аллелей вызывает некроз.
27. Способ получения растения томата, которое проявляет улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеющее некроз, который включает:
а) получение растения томата, гетерозиготного по донорской ДНК Solanum chilense в пределах сегмента генома, фланкированного маркерной SEQ ID NO: 1 и маркером, выявляемым праймерами SEQ ID NO: 21 и SEQ ID NO: 22;
б) получение потомства растения; и
в) отбор по меньшей мере первого растения-потомка, содержащего донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и не содержащего донорскую ДНК Solanum chilense, содержащую маркер, выявляемый праймерами SEQ ID NO: 21 и SEQ ID NO: 22.
28. Способ по п. 27, в котором указанный отбор включает обнаружение по меньшей мере одного аллеля в локусе, который выбран из группы, состоящей из SEQ ID NO: 1, маркера, выявляемого праймерами SEQ ID NO: 17 и SEQ ID NO: 18, SEQ ID NO: 4, маркера, выявляемого праймерами SEQ ID NO: 21 и SEQ ID NO: 22.
29. Способ по п. 27, который дополнительно включает:
г) отбор последующего растения-потомка из числа растений, выбранных на стадии в), или его последующего потомства, содержащего донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 4, и не содержащего донорскую ДНК Solanum chilense, содержащую маркер, выявляемый праймерами SEQ ID NO: 21 и SEQ ID NO: 22.
30. Способ по п. 27, отличающийся тем, что отбор включает идентификацию растения-потомка, у которого отсутствует один или более признак, выбранный из таких, как некроз, пониженная завязь плодов и уменьшенный размер плодов, по сравнению с растением, не содержащим указанную донорскую ДНК, содержащую маркерную SEQ ID NO: 1, и содержащим указанную донорскую ДНК, содержащую маркер, выявляемый праймерами SEQ ID NO: 21 и SEQ ID NO: 22.
31. Растение томата, которое получено способом по п. 27, где указанное растение содержит донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и не содержит донорскую ДНК Solanum chilense, содержащую маркер, выявляемый праймерами SEQ ID NO: 21 и SEQ ID NO: 22, причем указанное растение проявляет улучшенную устойчивость к вирусу желтой курчавости листьев томата и не имеет некроз относительно растения, не содержащего указанную донорскую ДНК Solanum chilense, содержащую маркерную SEQ ID NO: 1, и содержащего указанную донорскую ДНК Solanum chilense, содержащую маркер, выявляемый праймерами SEQ ID NO: 21 и SEQ ID NO: 22.
| MAARTEN G | |||
| VERLAAN et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| TAO LIN et al | |||
| Genomic analyses provide insights into the history of tomato breeding, Nature Genetics, 12 October 2014, Vol.46, N.11, | |||

