ОБЛАСТЬ ТЕХНИКИ
Данное изобретение в целом относится к области молекулярной биологии. Более конкретно, данное изобретение предлагает олигонуклеотиды и способы их применения для обнаружения и/или дифференциации нуклеиновых кислот-мишеней. Олигонуклеотиды и способы находят конкретное применение для одновременной амплификации, обнаружения, различения и/или количественного определения нескольких мишеней одновременно.
УРОВЕНЬ ТЕХНИКИ
Генетический анализ становится повседневной процедурой в клинических условиях для оценки риска заболевания, диагностики заболевания, установления прогноза для пациента или реакции пациента на терапию, а также для наблюдения за ходом заболевания у пациента. Внедрение таких генетических анализов зависит от разработки простых, недорогих и быстрых способов распознавания генетических изменений.
Способы амплификации нуклеиновых кислот in vitro находят широкое применение в генетике и диагностике заболеваний. Такие способы включают полимеразную цепную реакцию (ПЦР), полимеразную цепную реакцию с обратной транскрипцией (ОТ-ПЦР), амплификацию замещения цепи (SDA), реакцию амплификации никирующего фермента (NEAR), геликазозависимую амплификацию (HDA), рекомбиназную полимеразную амплификацию (RPA), петлевую изотермическую амплификацию (LAMP), амплификацию по типу катящегося кольца (RCA), транскрипционно-опосредованную амплификацию (ТМА), самоподдерживающуюся репликацию последовательностей (3SR), амплификацию, основанную на последовательности нуклеиновых кислот (NASBA), лигазную цепную реакцию (LCR) или способ амплификации с разветвлением (RAM). Каждая из этих целевых стратегий амплификации требует применение олигонуклеотидного(ых) праймера(ов). Процесс амплификации приводит к экспоненциальной амплификации ампликонов, которые включают олигонуклеотидные праймеры на своих 5'-концах и которые содержат вновь синтезированные копии последовательностей, расположенных между праймерами.
Обычно применяемые способы мониторинга накопления ампликонов в режиме реального времени или по завершении амплификации включают обнаружение с применением МНКзимов с универсальными субстратными зондами, мишень-специфичных молекулярных маяков, маяков Sloppy, зондов Eclipse, зондов TaqMan или гидролизных зондов, унизондов или бизондов Scorpion, зондов Catcher/Pitcher, зондов двойной гибридизации и/или применением интеркалирующих красителей, таких как SybGreen. Анализ кривой плавления с высоким разрешением может быть выполнен во время или по завершении нескольких из этих протоколов для получения дополнительной информации, поскольку ампликоны с различными последовательностями денатурируют при разных температурах, известных как температура плавления или Tm. Такие протоколы измеряют кривые плавления, которые являются результатом либо а) разделения двух цепей двухцепочечных ампликонов в присутствии интеркалирующего красителя, либо b) разделения одной цепи ампликона и комплементарного зонда, специфичного для мишени, меченного флуорофором и гасителем, или с) разделение дуплексов, не связанных с мишенью, например, дуплексов Catcher, которые образуются только в присутствии мишени. Анализ кривой плавления позволяет получить информацию о кинетике диссоциации двух цепей ДНК при нагревании. Температура плавления (Tm) представляет собой температуру, при которой диссоциирует 50% ДНК. Tm зависит от длины, состава последовательности и содержания G-C в спаренных нуклеотидах. Выяснение информации о ДНК-мишени на основе анализа кривой плавления обычно включает серию измерений флуоресценции, полученных с небольшими интервалами, как правило, в широком диапазоне температур. Температура плавления зависит не только от последовательности оснований. На температуру плавления могут влиять многие факторы, в том числе концентрации олигонуклеотидов, катионов в буфере (как одновалентных (Na+), так и двухвалентных (Mg2+) солей) и/или наличие или отсутствие дестабилизирующих агентов, таких как мочевина или формамид.
В общем, количество доступных флуоресцентных каналов, способных контролировать отдельные длины волн, ограничивает количество мишеней, которые могут быть обнаружены и специфично выявлять в ходе одной реакции на флуоресцентном ридере. Недавно протокол, известный как «Отщепление и удлинение меченого олигонуклеотида» (ТОСЕ), расширяет эту возможность, позволяя анализировать несколько мишеней на одной длине волны. В технологии ТОСЕ используются олигонуклеотиды Pitcher и Catcher. Олигонуклеотиды Pitcher имеют две области: Целевую часть, которая дополняет цель, и Маркирующую часть, которая не является комплементарной и расположена на 5'-конце. Олигонуклеотид Capture имеет двойную метку и имеет область на 3'-конце, комплементарную маркирующей части олигонуклеотида Pitcher. Во время амплификации олигонуклеотид Pitcher связывается с ампликонами, и когда праймеры расширяют экзонуклеазную активность полимеразы, она может отщеплять меченую часть от олигонуклеотида Pitcher. Высвободившийся меченый фрагмент затем связывается с олигонуклеотид ом Catcher и действует как праймер для синтеза комплементарной цепи. Затем температура плавления двухцепочечной молекулы Catcher (Catcher-Tm) действует как суррогатный маркер для исходного темплата. Поскольку можно применять несколько Catcher с разной последовательностью и длиной, плавящихся при разных температурах, можно получить ряд значений Catcher-Tm, указывающих на серию мишеней, при этом продолжая измерение на одной длине волны. Ограничения этого подхода включают присущую ему сложность, поскольку требуется, чтобы высвобождаемый фрагмент инициировал и завершал второе удлинение на искусственной мишени, а анализ после амплификации нескольких мишеней требует сложных алгоритмов для дифференциации или количественного определения доли сигнала, относящегося к каждой конкретной мишени.
Зонды-шпильки или зонды стебель-петля также оказались полезными инструментами для обнаружения нуклеиновых кислот и/или мониторинга амплификации мишени. Один тип зонд-шпилька, дважды меченый парой красителей «флуорофор и гаситель», широко известен в данной области техники как молекулярный маяк. В общем, эти молекулы имеют три особенности; 1) стеблевая структура, образованная путем гибридизации комплементарных 5'- и 3'-концов олигонуклеотида; 2) петлевую область, комплементарную мишени или ампликону-мишени, подлежащим обнаружению; и 3) пару красителей «гаситель/флуорофор», присоединенных к концам молекулярного маяка. Во время ПЦР петлевая область связывается с ампликонами за счет комплементарности, и это приводит к открытию стебля, таким образом разделяя пару красителей «гаситель/флуорофор». Существенной особенностью молекулярных маяков является то, что петлевые области этих молекул остаются интактными во время амплификации и не разрушаются, и не расщепляются в присутствии мишени или целевых ампликонов. Разделение пары красителей, присоединенных к концам открытого молекулярного маяка, вызывает изменение флуоресценции, указывающее на присутствие мишени. Этот способ обычно используется для мультиплексного анализа нескольких мишеней в одном ПЦР-анализе. Как правило, для мультиплексного анализа каждый молекулярный маяк имеет отличную петлевую область, специфичную для мишени, и уникальный флуорофор, так что гибридизацию каждого отдельного молекулярного маяка с каждым видом ампликона можно отслеживать в отдельном канале, т.е. на отдельной длине волны.
Концепция молекулярных маяков была расширена в стратегии, известной как маяки Sloppy. В этом протоколе петлевая область одного маяка является достаточно длинной, чтобы допускать несовпадение основания и, следовательно, связывается с рядом близкородственных мишеней, различающихся по одному или более нуклеотидам. После амплификации выполняется анализ кривой плавления, что позволяет дифференцировать различные виды мишеней в зависимости от температуры, при которой разделяются (плавятся) каждый из дуплексов, образованных путем гибридизации видов мишеней с петлевой областью маяка Sloppy. Таким образом, можно одновременно обнаруживать и различать несколько близкородственных разновидностей при одной длине волны путем определения профиля плавления конкретных мишеней с помощью одного маяка Sloppy. Стандартные молекулярные маяки и маяки Sloppy отличаются от зондов TaqMan и гидролизных зондов тем, что они не предназначены для разрушения или расщепления во время амплификации. Недостаток технологий, основанных на гибридизации ДНК, таких как маяки Sloppy и ТОСЕ, заключается в том, что они могут давать ложноположительные результаты из-за неспецифической гибридизации между зондами и последовательностями нуклеиновых кислот, не являющимися мишенями.
Многие анализы обнаружения нуклеиновых кислот используют анализы кривых плавления либо для определения наличия конкретных последовательностей-мишеней в данном образце, либо для уточнения информации об амплифицированной последовательности. Протоколы анализа кривой плавления включают измерение флуоресценции при различных температурах в постепенно увеличивающемся диапазоне температур. Затем строят график изменения коэффициента угла наклона этой кривой в зависимости от температуры, получая кривую плавления. Этот процесс часто является медленным и обычно занимает от 30 до 60 минут. Кроме того, анализ кривой плавления может требовать интерпретации квалифицированным специалистом и/или применение специального программного обеспечения для интерпретации результатов. Следовательно, существует высокий спрос на более быстрые и/или более простые альтернативы анализу кривых плавления.
Кривые плавления обычно анализируются после ПЦР и поэтому позволяют только качественно определить наличие или отсутствие мишени в образце. Во многих случаях требуется количественное или полуколичественное определение количества геномного материала, присутствующего в образце. Поэтому существует большой спрос на быстрые альтернативы анализу кривой плавления, которые также предоставляют количественную информацию об образце.
Существует потребность в улучшенных композициях и способах одновременного обнаружения, дифференциации и/или количественного определения множественных уникальных ампликонов, полученных с помощью ПЦР или альтернативных протоколов амплификации мишени.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение направлено на устранение одного или более недостатков, существующих в современных анализах, предусматривающих мультиплексное обнаружение.
В настоящем документе предусмотрены способы и композиции, которые расширяют возможности мультиплексирования во время протоколов амплификации. Данные способы сочетают «Стандартные репортеры», которые включают субстраты и зонды, хорошо известные в данной области техники, вместе со структурой (структурами), называемой в настоящем документе LOCS (петли, соединенные со стеблями). Стандартные репортеры включают, но не ограничиваются ими, зонды и субстраты, включая линейные субстраты МНКзимы, зонды TaqMan или гидролизные зонды, молекулярные маяки, маяки Sloppy, зонды Eclipse, унизонды или бизонды Scorpion, олигонуклеотиды Capture/Pitcher и двойные гибридизационные зонды. Комбинация стандартной репортерной системы вместе с одним или более LOCS, причем все виды могут, например, быть помечены одним детектирующим фрагментом (например, одной и той же парой флуорофора и гасителя), позволяет индивидуально различать несколько мишеней в рамках одной реакции. Подход включает измерение сигнала, генерируемого «Стандартным репортером» и одним или более LOCS, при одной или более температурах. Образование сигнала от LOCS может зависеть от нескольких факторов, включая один или более из следующих:
- температура, при которой измеряется сигнал;
- была ли расщеплена или разрушена петлевая область LOCS в ответ на присутствие мишени;
- температура плавления стеблевой области конкретного LOCS в его «интактном» или в расщепленном, или деградированном «разделенном» состоянии.
Температура плавления стеблевой области разделенного LOCS действует как суррогатный маркер для конкретной мишени, которая опосредовала зависимое от мишени расщепление или деградацию петли интактного LOCS. В других способах, включающих структуры стебель-петля, использовалось изменение флуоресцентного сигнала после (а) гибридизации петлевой области с ампликонами-мишенями (например, молекулярные маяки и маяки Sloppy) для увеличения расстояния между парами красителей или (b) путем опосредованного расщепления, позволяющего физическое разделение красителей (например, расщепляемые молекулярные маяки). Расщепляемые молекулярные маяки обычно использовались для генерации положительного или отрицательного сигнала для данной мишени на одной длине волны. Мультиплексное обнаружение мишеней обычно требует обнаружения разных мишеней с помощью сигналов, излучаемых на разных длинах волн. Таким образом, включение различных стеблей в различные расщепляемые молекулярные маяки, помеченные сходными или идентичными обнаруживающими фрагментами и предназначенные для обнаружения разных мишеней, дает возможность различать обнаруживаемые сигналы, указывающие на отдельные мишени, на основе различий в температурах плавления стебля, а не необходимости использования отчетливые обнаруживаемые сигналы между мишенями.
Данное изобретение предлагает улучшения по сравнению с существующими анализами мультиплексного обнаружения, которые возникают по меньшей мере частично за счет управления температурой плавления стеблевой области структур стебель-петля путем изменения длины и/или состава последовательности стебля, так что каждый стебель плавится и генерирует сигнал при другой температуре.
Данное изобретение может включать применение стандартных репортеров вместе с одним репортером LOCS или более репортерами LOCS в одной реакции. Как стандартный, так и репортер LOCS могут быть помечены одинаковым или подобный детектирующим фрагментом(ами), которые могут быть обнаружены по существу одним и тем же способом (например, флуорофоры, излучающие в одной и той же области видимого спектра, наночастицы одинакового размера и/или тип для колориметрического обнаружения или обнаружения ППР, реактивные фрагменты (например, ферменты щелочной фосфатазы или пероксидазы) для хемилюминесцентного обнаружения, электроактивные частицы (например, ферменты ферроцена, метиленового синего или пероксидазы) для электрохимического обнаружения. Когда присутствуют несколько LOCS и помечены, например, одним и тем же детектирующим фрагментом, они могут содержать (а) разные петлевые последовательности, каждая из которых позволяет прямо или косвенно обнаруживать несколько мишеней одновременно, и/или (b) разные стеблевые последовательности, которые плавятся при дискретных температурах и которые можно использовать для идентификации конкретных мишеней, присутствующих в нескольких исследуемых мишенях. В способах согласно настоящему изобретению используют LOCS, которые обеспечивают одно или более преимуществ по сравнению со способами, известными в данной области техники, например, протоколом ТОСЕ, заключающихся в отсутствии необходимости применения отдельных «принимающих» молекул, что уменьшает количество компонентов в реакционной смеси и снижает затраты. Более того, способ ТОСЕ по своей природе более сложен, чем способы согласно настоящему изобретению, поскольку он требует, чтобы высвобожденный фрагмент инициировал и завершал второе удлинение на синтетической мишени.
В некоторых вариантах реализации зонды LOCS могут быть универсальными (независимыми от последовательности-мишени) и/или их можно комбинировать с рядом технологий обнаружения, что обеспечивает широкую применимость в области молекулярной диагностики. Кроме того, температура плавления, используемая в традиционных способах амплификации и обнаружения, основана на гибридизации и плавлении зонда с нуклеиновой кислотой-мишенью. Недостатком этих способов является увеличение количества ложно положительных результатов из-за неспецифичной гибридизации между зондами и неспецифичными нуклеотидными последовательностями. Способы по настоящему изобретению преодолевают это ограничение, поскольку те репортерные зонды LOCS, которые содержат универсальные субстраты, не связываются с последовательностью-мишенью. Наконец, в данной области техники хорошо известно, что внутримолекулярные связи прочнее межмолекулярных связей, и, следовательно, вероятность гибридизации этих нерасщепленных (замкнутых) LOCS с неспецифической мишенью и получения ложноположительного сигнала значительно ниже.
В результате того, что внутримолекулярные связи прочнее межмолекулярных связей, LOCS с двойной меткой плавится при одной температуре в неповрежденном состоянии и при более низкой температурепосле расщепления в зависимости от мишени или деградации петлевой области, которая расщепляет LOCS на два фрагмента. Это свойство нуклеиновых кислот используется в настоящем изобретении для расширения возможностей инструментов по различению нескольких мишеней с применением одного типа детектора, такого как один канал флуоресценции, или определенного режима колориметрии, поверхностного плазмонного резонанса (ППР), хемилюминесцентного или электрохимического обнаружения.
Зависимые от температуры сигналы флуоресценции, продуцируемые репортерами LOCS согласно настоящему изобретению, четко заданы и не зависят от ДНК-мишени. Таким образом, можно получать информацию о ДНК-мишени на основе измерений сигнала флуоресценции, генерируемого при выбранных температурах, а не в диапазоне всего температурного градиента, что дает преимущество в виде снижения времени работы термоциклеров (например, устройств ПЦР). В качестве неограничивающего примера, в системе ПЦР Bio-Rad CFX96 для выполнения традиционного анализа плавления с настройками температуры от 20°С до 90°С с шагом 0,5°С и временем выдержки 5 секунд требуется 141 цикл измерения флуоресценции, что занимает приблизительно 50 минут. При использовании LOCS-зондов информацию о ДНК-мишени можно получить на том же устройстве за 2-6 измерений флуоресценции, что занимает приблизительно 2-5 минут.Без конкретных ограничений, снижение времени работы может быть выгодным при многих вариантах применения, включая, например, диагностику.
В настоящем изобретении зонды LOCS комбинируют со стандартными репортерами или зондами, или субстратами для одновременного обнаружения, дифференциации и/или количественного определения множества мишеней. Отдельные сигналы, указывающие на различные мишени, могут обнаруживаться одними и теми же средствами, такими как, например, посредством сигналов, излучаемых в одном флуоресцентном канале, или обнаруживаемых с помощью определенного режима колориметрического, поверхностного плазмонного резонанса (ППР), хемилюминесцентного или электрохимического обнаружения. При обычной кПЦР количественное определение целевой ДНК выполняют с применением значения цикла количественного определения (Cq) на основе кривой амплификации, полученной путем измерения флуоресценции при одной температуре в каждом цикле амплификации. Значение Cq пропорционально отрицательному логарифмическому значению концентрации ДНК-мишени, и поэтому по экспериментально определенному значению Cq можно определить эту концентрацию. Однако если в одном канале имеется более одного мишень-специфичного зонда, правильное и специфичное количественное определение каждой мишени сопряжено со сложностями, поскольку трудно определить, исходит ли сигнал от конкретного зонда. Решая эту проблему, LOCS позволяет правильно и точно определять количество более чем одной мишени на одном канале при условии, что кривую амплификации получают путем измерения флуоресценции во время амплификации при более чем одной температуре. Это возможно, поскольку репортеры LOCS могут давать существенно различающуюся флуоресценцию при различных температурах. Кроме того, репортеры LOCS можно использовать для обеспечения правильной и специфической количественной оценки первой мишени и одновременного качественного обнаружения второй мишени в одном канале путем регистрации флуоресценции при первой температуре в реальном времени (мишень 1) и при второй температуре до и после амплификации (мишень 2). Преимущество последнего сценария заключается в том, что он не влияет на общее время выполнения протокола амплификации и может не требовать специального программного обеспечения для анализа. Этот подход может быть полезен в сценариях, где количественная оценка или определение Cq требуется только для одной из мишеней.
В некоторых вариантах реализации, где для анализа требуется получение флуоресценции лишь в ограниченное количество моментов времени в ходе ПЦР, например, в начале амплификации или рядом с ней и после амплификации в конечной точке, применение LOCS-структур устраняет необходимость получения данных в каждом цикле. Таким образом, эти варианты реализации хорошо подходят для протоколов с очень быстрой сменой циклов, которые могут сократить время получения результата.
Как отмечалось выше, протоколы анализа кривой плавления подразумевают измерение флуоресценции при различных температурах в постепенно увеличивающемся диапазоне температур (например, между 30°С и 90°С). Затем строят график изменения коэффициента угла наклона этой кривой в зависимости от температуры, получая кривую плавления. Этот процесс часто является медленным и может занимать, например, от 30 до 60 минут. Повышение скорости анализа кривой плавления требует доступа к узкоспециализированным приборам и не достигается с помощью стандартных устройств ПЦР. Таким образом, существует большая потребность в более быстрых альтернативах анализу кривой плавления, которые могут обеспечить одновременное обнаружение нескольких мишеней на одном канале флуоресценции с применением стандартных инструментов. Температура плавления (Tm) LOCS-структур согласно настоящему изобретению является заранее определенной и постоянной для заданных экспериментальных условий (т.е. не зависит от последовательности-мишени или ее концентрации) и, следовательно, не требует постепенного изменения во всем диапазоне градиента температур. Каждая LOCS-структура требует лишь одного измерения флуоресценции при своей специфичной Tm, что исключает необходимость использования полного градиента температур, ускоряет получение результата и, следовательно, позволяет преодолеть вышеуказанные ограничения.
Кроме того, анализ кривой плавления обычно требует интерпретации квалифицированным персоналом или использования специализированного программного обеспечения для интерпретации результатов.
В некоторых вариантах реализации настоящего изобретения измерение флуоресценции при одной температуре после завершения ПЦР устраняет необходимость в субъективной интерпретации кривых плавления и облегчает объективное определение наличия или отсутствия мишеней.
В других вариантах реализации настоящего изобретения для анализа может потребоваться регистрация флуоресценции только при ограниченном количестве моментов времени в пределах ПЦР, например, до ПЦР и после ПЦР, что устраняет необходимость регистрации данных в каждом цикле. Таким образом, эти варианты реализации хорошо подходят для протоколов с очень быстрой сменой циклов, которые могут сократить время получения результата.
Описано несколько способов, включающих регистрацию флуоресценции при нескольких температурах во время ПЦР, включая регистрацию при двух температурах, облегчающую различение полностью совпадающих и несовпадающих зондов. Кроме того, в некоторых протоколах используют несколько температур регистрации после каждого цикла ПЦР для количественного определения концентрации каждой мишени, когда две мишени присутствуют и обнаруживаются на одном канале. Другие способы одновременной количественной оценки двух мишеней достигаются путем построения полной кривой плавления в конце каждого цикла ПЦР.
Данное изобретение использует преимущества объединения LOCS с другими типами репортерных молекул. LOCS-структуры согласно настоящему изобретению могут быть совместимы с большинством, а потенциально со всеми существующими способами анализа ПЦР в реальном времени и конечной точки. Хотя можно проводить анализ, при котором для различения множества мишеней в одной реакции используются только зонды LOCS, может быть выгодно использовать несколько типов зондов в одной реакции. Например, один зонд LOCS можно использовать в сочетании с любой из следующих технологий: линейный субстрат МНКзим, линейный зонд TaqMan, зонды, расщепляемые рестрикционными ферментами, зонд Eclipse, нерасщепляемый молекулярный зонд, нерасщепляемый зонд Sloppy, унизонд Scorpion, бизонд Scorpion, пара зондов двойной гибридизации или зонды, использующие технологию Catcher и Pitcher (например, зонды ТОСЕ).
В различных вариантах реализации настоящего изобретения один зонд LOCS и линейный субстрат МНКзим, линейный зонд TaqMan или нерасщепляемый молекулярный зонд могут быть помечены одинаковым или сходным детектирующим фрагментом. В качестве неограничивающего примера это может включать один и тот же флуорофор для флуорометрического обнаружения, одинаковый размер и/или тип наночастиц (например, золота или серебра) для колориметрического или SPR-детектирования, реактивный фрагмент (например, ферменты щелочной фосфатазы или пероксидазы) для обнаружения хемилюминесценции или электроактивные частицы (например, ферроцен, метиленовый синий или ферменты пероксидазы) для электрохимического обнаружения.
В некоторых вариантах реализации линейный субстрат МНКзим, способный расщепляться первым специфичным для мишени МНКзимом, может быть объединен с одним зондом LOCS, способным расщепляться вторым МНКзимом, специфичным для мишени. Варианты реализации, в которых один линейный субстрат МНКзим и один зонд LOCS используются для обнаружения двух мишеней на одной длине волны видимого спектра или, например, на одной длине волны видимого спектра, могут иметь преимущество перед вариантами реализации, использующими два зонда LOCS, поскольку производство линейных зондов проще и дешевле, чем производство зондов LOCS. Это связано с тем, что линейные субстраты не требуют дополнительной последовательности, необходимой для стеблевой области зонда LOCS, и, следовательно, они короче. Точно так же производство линейных зондов TaqMan может быть дешевле, чем производство зондов LOCS.
Дополнительные преимущества, связанные с применением одного или более LOCS в сочетании с другими типами стандартных репортеров, связаны с присущей линейным зондам разницей в фоновой флуоресценции, в которой временные/пространственные параметры приводят к большему расстоянию между флуорофором и гасителем и, следовательно, более высокая фоновая флуоресценция по сравнению с зондами LOCS, где флуорофор и гаситель удерживаются в непосредственной близости стеблевой областью. Кроме того, разные типы зондов генерируют сигнал, используя разные механизмы, при этом они проявляют разные свойства флуоресценции и гашения при разных температурах. В различных вариантах реализации, приведенных в качестве примеров ниже, эта разница в способности флуоресценции и гашения обеспечивает дополнительный инструмент, с помощью которого исследователь может манипулировать величиной сигнала обнаружения при определенных температурах для обнаружения, различения и/или количественного определения нескольких мишеней на одной длине волны.
В некоторых вариантах реализации настоящее изобретение использует тот факт, что зонды LOCS и зонды Catcher-Pitcher обладают противоположными свойствами флуоресценции/гашения при различных температурах. Например, независимо от наличия или отсутствия мишени, зонды Catcher-Pitcher будут оставаться погашенными при высокой температуре (т.е. выше Tm дуплекса Catcher-Pitcher) из-за денатурации дуплекса и изменения конформации цепи Catcher. И наоборот, зонды LOCS будут оставаться погашенными при низкой температуре (т.е. ниже Tm расщепленного стержня LOCS), независимо от наличия или отсутствия мишени, потому что гибридизированный стебель удерживает флуорофор и гаситель в непосредственной близости. Кроме того, в присутствии мишени зонды Catcher-Pitcher будут генерировать увеличение флуоресценции при низкой температуре (т.е. ниже Tm дуплекса Catcher-Pitcher), тогда как зонды LOCS будут генерировать увеличение флуоресценции при высокой температуре (т.е. выше Tm расщепленного стебля LOCS). Эти противоположные свойства флуоресценции/гашения при высоких и низких температурах позволяют специфически обнаруживать две цели, позволяя обнаруживать одну цель при первой, низкой температуре с помощью зонда Catcher-Pitcher, а другую цель обнаруживать при второй, более высокой температуре, с помощью зонда LOCS.
В различных вариантах реализации настоящего изобретения используются преимущества объединения одного линейного субстрата или зонда, например линейного субстрата МНКзима или зонда TaqMan, с зондом LOCS. Например, преимущество по сравнению с применением пары зондов LOCS с одним стеблем с более низкой и одной с более высокой Tm состоит в том, что как расщепленный линейный субстрат МНКзим, так и деградированный зонд TaqMan производят одинаковые сигналы флуоресценции в широком диапазоне температур. Точно так же нерасщепленный линейный субстрат МНКзим и интактные зонды TaqMan дают аналогичные сигналы флуоресценции в широком диапазоне температур. Таким образом, для обоих типов датчиков отношение сигнал/шум является постоянным в широком диапазоне температур обнаружения. Для сравнения, наблюдаемое отношение сигнал/шум, возникающее при использовании разделенных зондов LOCS с низкой Tm, может уменьшаться при более высоких температурах обнаружения из-за большей фоновой флуоресценции, которая создается денатурацией стеблей интактного LOCS. Это означает, что расщепление линейного субстрата МНКзима или зонда TaqMan можно обнаружить в более широком диапазоне температур обнаружения по сравнению с расщеплением LOCS с низкой Tm, который имеет более ограниченный диапазон температур обнаружения. Это обеспечивает большую гибкость термоциклирования и может быть полезно для более быстрой и упрощенной разработки мультиплексных анализов. Еще одно преимущество связано с возможностью комбинировать один или более зондов LOCS с существующими коммерческими наборами с применением других технологий, таких как зонды TaqMan, и, таким образом, расширять их возможности мультиплексирования.
В других вариантах реализации настоящее изобретение использует преимущества, обеспечиваемые тем фактом, что зонды LOCS и унизонды или бизонды Scorpion также ведут себя по-разному при разных температурах, обеспечивая специфическое обнаружение двух мишеней при двух разных температурах обнаружения. Например, при высокой температуре обнаружения унизонд Scorpion всегда может флуоресцировать (до ПЦР и после ПЦР) независимо от наличия или отсутствия какой-либо мишени, если стебель открыт и флуоресцирует, а петля не может связываться к ампликонам конкретной мишени (мишень 1). Точно так же при высокой температуре детектирования бизонды Scorpion всегда могут быть флуоресцентными (до ПЦР и после ПЦР) независимо от присутствия или отсутствия какой-либо мишени, поскольку комплементарная последовательность гасителя может быть неспособна связываться с зондом, и зонд может быть неспособен связываться с ампликонами конкретной мишени (мишень 1). В обоих случаях (унизонд или бизонд) при одинаковой высокой температуре зонд LOCS будет генерировать флуоресценцию только в присутствии конкретной мишени (мишень 2) из-за расщепления и диссоциации стебля. И наоборот, при низкой температуре детектирования зонд LOCS всегда будет гаситься (до ПЦР и после ПЦР) независимо от присутствия или отсутствия какой-либо мишени, поскольку Tins стеблей как интактного LOCS, так и разделенного LOCS выше этой температуры, тогда как при той же температуре унизонд или бизонд Scorpion будут генерировать флуоресценцию только в присутствии конкретной мишени из-за гибридизации петель или областей зонда соответственно с ампликонами мишени 1. Эти противоположные свойства флуоресценции/гашения при высоких и низких температурах позволяют специфически обнаруживать две мишени, где мишень 1 может быть обнаружена при первой, низкой температуре с помощью либо унизонда Scorpion, либо бизонда Scorpion, а мишень 2 может быть обнаружена при второй, более высокой температуре, с применением зонда LOCS.
Различные типы стандартных репортерных субстратов и зондов будут флуоресцировать либо в широком диапазоне температур, либо только в ограниченном диапазоне. Например, линейные репортерные субстраты или зонды, включая, но не ограничиваясь ими, линейные субстраты МНКзима, зонды Eclipse, зонды TaqMan, гидролизные зонды и другие, как правило, производят флуоресцентный сигнал в широком диапазоне температур. Такие зонды обычно гасят перед ПЦР и флуоресцируют после ПЦР, если присутствует мишень, и эту флуоресценцию можно измерить в широком диапазоне температур. Напротив, зонды LOCS, молекулярные маяки, унизонды Scorpion или бизонды и флуоресцентные системы Pitcher и Catcher (например, зонды ТОСЕ) можно манипулировать таким образом, чтобы они флуоресцировали или гасились в определенных диапазонах температур.
Молекулярные маяки гасятся стеблями, гибридизующимися при температурах ниже той, при которой петля молекулярного маяка связывается с мишенью и флуоресцирует.Напротив, стебель зондов интактного LOCS гибридизуется при температурах, превышающих температуру плавления разделенных LOCS. Кроме того, в то время как многие репортерные системы измеряют увеличение флуоресценции в присутствии мишени, другие технологии, такие как зонды двойной гибридизации, приводят к уменьшению флуоресценции в присутствии мишени. Настоящее изобретение предлагает новые способы комбинирования зондов и настройки параметров, так что увеличение или уменьшение обнаруживаемых сигналов при определенной температуре с конкретными комбинациями зондов позволяет улучшить сценарии мультиплексирования. Таким образом, применение различного поведения различных типов субстратов и зондов стандартного репортера в сочетании с зондами LOCS, помеченными одинаковыми или подобными детектирующими фрагментами и присутствующими в одной реакции, позволяет манипулировать наличием или отсутствием сигнала, например, флуоресценция или гашение при различных температурах, что, в свою очередь, обеспечивает множество преимуществ для анализа мишеней.
Настоящее изобретение относится по меньшей мере частично к следующим вариантам реализации 1-194:
Вариант реализации 1. Способ определения наличия или отсутствия первой и второй мишеней в образце, причем указанный способ включает:
(a) получение реакционной смеси путем приведения образца или его производного, предположительно содержащего первую или вторую мишени с:
- первым олигонуклеотидом для обнаружения первой мишени, и содержащий первый детектирующий фрагмент, способный генерировать первый детектируемый сигнал;
- интактный олигонуклеотид стебель-петля для обнаружения второй мишени, содержащий двуцепочечную стеблевую область, образованную гибридизированными нуклеотидами, противоположные цепи которой связаны неразорванной одноцепочечной петлевой областью, образованной негибридизованными нуклеотидами, при этом стеблевая область содержит второй детектирующий фрагмент, способный генерировать второй детектируемый сигнал,
при этом первый и второй детектирующие фрагменты способны генерировать детектируемые сигналы, которые нельзя различить при одной температуре с применением одного типа детектора; и
- - первый фермент, способный расщеплять один или более негибридизированных нуклеотидов интактного олигонуклеотида стебель-петля только тогда, когда в образце присутствует вторая мишень;
(b) обработку смеси в условиях, подходящих для:
- первой мишени индуцировать модификацию первого олигонуклеотида, тем самым позволяя первому детектирующему фрагменту генерировать первый детектируемый сигнал,
- расщепление одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля первым ферментом только тогда, когда в образце присутствует вторая мишень, с тем чтобы разрушить одноцепочечную петлевую область и получить разделенный олигонуклеотид стебель-петля;
(c) измерение:
- фонового сигнала, обеспечиваемого первой и второй детектирующими фрагментами в смеси или в контрольной смеси;
(d) определение того, в один или более моментов времени во время или после указанной обработки:
- генерируется ли при первой температуре, первый детектируемый сигнал, который отличается от фонового сигнала, возникающий в результате указанной модификации, указывающий на присутствие первой мишени в образце;
- генерируется ли при второй температуре, второй детектируемый сигнал, который отличается от фонового сигнала, указывающий на присутствие второй мишени в образце;
при этом:
при первой температуре второй детектируемый сигнал не отличается от фонового сигнала, и
при второй температуре:
если присутствуют, цепи двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля частично или полностью диссоциированы, позволяя второму детектирующему фрагменту обеспечивать второй детектируемый сигнал; и
- если присутствуют, цепи двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля не могут диссоциировать, тем самым препятствуя тому, чтобы второй детектируемый фрагмент обеспечивал второй детектируемый сигнал.
Вариант реализации 2. Способ по варианту реализации 1, в котором указанное определение в части (d) включает:
применение заданного порогового значения для определения того, отличается ли первый детектируемый сигнал, возникающий в результате упомянутой модификации, от любого упомянутого фонового сигнала при первой температуре; и/или
применение заданного порогового значения для определения того, отличается ли второй детектируемый сигнал от любого упомянутого фонового сигнала при второй температуре.
Вариант реализации 3. Способ по варианту реализации 1 или варианту реализации 2, в котором контрольная смесь не содержит:
- первую мишень; или
- вторую мишень; или
- первую и вторую мишени,
но в остальном эквивалентна указанной смеси.
Вариант реализации 4. Способ по любому из вариантов реализации 1-3, в котором контрольная смесь содержит заданное количество:
- первую мишень; или
- второй мишени; или
- первой и второй мишени,
но в остальном эквивалентна указанной смеси.
Вариант реализации 5. Способ по любому из вариантов реализации 1-4, в котором:
- модификация первого олигонуклеотида позволяет первому детектирующему фрагменту обеспечивать первый детектируемый сигнал при первой температуре или ниже; и
- - образование первого детектируемого сигнала обратимо.
Вариант реализации 6. Способ по варианту реализации 5, в котором:
- часть (с) включает измерение: первого фонового сигнала на уровне или в пределах 1°С, 2°С, 3°С, 4°С или 5°С от первой температуры и второго фонового сигнала на уровне или в пределах 1°С, 2°С, 3°С, 4°С или 5°С от второй температуры;
обеспечиваемого первым и вторым детектирующими фрагментами в смеси или в контрольной смеси; и
- часть (d) включает определение того, в один или более моментов времени во время или после указанной обработки:
- генерируется ли при первой температуре, первый детектируемый сигнал, который отличается от первого фонового сигнала, возникающий в результате указанной модификации, указывающий на присутствие первой мишени в образце;
- генерируется ли при второй температуре, второй детектируемый сигнал, который отличается от второго фонового сигнала, указывающий на присутствие второй мишени в образце.
Вариант реализации 7. Способ по варианту реализации 5 или варианту реализации 6, в котором:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый олигонуклеотид представляет собой олигонуклеотид типа стебель-петля, содержащий двухцепочечную стеблевую область из гибридизированных нуклеотидов на противоположных цепях, которые связаны неразорванной одноцепочечной петлевой областью негибридизированных нуклеотидов, все или часть которых комплементарны первой мишени; и
- - модификация первого олигонуклеотида представляет собой конформационное изменение, возникающее в результате гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида путем комплементарного спаривания оснований.
Вариант реализации 8. Способ по варианту реализации 7, в котором:
- конформационное изменение представляет собой диссоциацию цепей в двухцепочечной стеблевой области первого олигонуклеотида, возникающую в результате указанной гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида путем комплементарного спаривания оснований.
Вариант реализации 9. Способ по варианту реализации 7 или варианту реализации 8, в котором:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая: ниже Tm двухцепочечного дуплекса, образованного в результате указанной гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида, и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- указанный двухцепочечный дуплекс имеет Tm, которая: выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: указанного двухцепочечного дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm: указанного двухцепочечного дуплекса, стеблевой области первого олигонуклеотида и стеблевой области интактного олигонуклеотида стебель-петля, и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- - первая температура ниже второй температуры.
Вариант реализации 10. Способ по варианту реализации 7 или варианту реализации 8, в котором:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая: ниже Tm двухцепочечного дуплекса, образованного в результате указанной гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- указанный двухцепочечный дуплекс имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: указанного двухцепочечного дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm: стеблевой области первого олигонуклеотида и стеблевой области разделенного олигонуклеотида стебель-петля; и ниже Tm: указанного двухцепочечного дуплекса и стеблевой области интактного олигонуклеотида стебель-петля; и
- - первая температура ниже второй температуры.
Вариант реализации 11. Способ по варианту реализации 7 или варианту реализации 8, в котором:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая: ниже Tm двухцепочечного дуплекса, образованного в результате указанной гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля и ниже Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- указанный двухцепочечный дуплекс имеет Tm, которая: ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: указанного двухцепочечного дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm: указанного двухцепочечного дуплекса, стеблевой области первого олигонуклеотида, и стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm: стеблевой области интактного олигонуклеотида стебель-петля; и
- - первая температура ниже второй температуры.
Вариант реализации 12. Способ по варианту реализации 7 или варианту реализации 8, в котором:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая: ниже Tm двухцепочечного дуплекса, образованного в результате указанной гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида, выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- указанный двухцепочечный дуплекс имеет Tm, которая: выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: стеблевой области первого олигонуклеотида и указанного двухцепочечного дуплекса; и выше Tm: стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm: стеблевой области первого олигонуклеотида, указанного двухцепочечного дуплекса и стеблевой области интактного олигонуклеотида стебель-петля; и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- - первая температура выше второй температуры.
Вариант реализации 13. Способ по любому из вариантов реализации 7-12, в котором:
- Tm стеблевой области первого олигонуклеотида находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm указанного двухцепочечного дуплекса; и/или
- - Tm стеблевой области интактного олигонуклеотида стебель-петля находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- - первая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm: стеблевой области первого олигонуклеотида, и/или указанного двухцепочечного дуплекса; и/или
- - вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и/или
- - вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 14. Способ по варианту реализации 5 или варианту реализации 6, в котором:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый олигонуклеотид представляет собой олигонуклеотид стебель-петля, содержащий:
двухцепочечную стеблевую область гибридизированных нуклеотидов, противоположные цепи которого связаны одноцепочечной петлевой областью негибридизированных нуклеотидов, все или часть которых комплементарны первой мишени, и вторую одноцепочечную область, продолжающуюся от одной из указанных противоположных цепей в 3'-направлении и оканчивающейся последовательностью, комплементарной области первой мишени, и
молекула-блокатор, предшествующая указанной последовательности, которая комплементарна области первой мишени;
- смесь дополнительно содержит полимеразу;
- указанная обработка смеси включает:
гибридизацию второй одноцепочечной области с первой мишенью путем комплементарного спаривания оснований;
удлинение второй одноцепочечной области с применением полимеразы и первой мишени в качестве темплатной последовательности с получением двухцепочечной нуклеиновой кислоты, при этом указанная блокирующая молекула предотвращает удлинение полимеразой первой мишени с применением стеблевой области первого олигонуклеотида в качестве темплата; и
- денатурацию двухцепочечной нуклеиновой кислоты и гибридизацию второй одноцепочечной области, удлиненной полимеразой, с одноцепочечной петлевой областью первого олигонуклеотида путем комплементарного спаривания с получением сигнального дуплекса и, таким образом, обеспечение указанной модификации первого олигонуклеотида, активируя первый детектирующий фрагмент для обеспечения первого детектируемого сигнала.
Вариант реализации 15. Способ по варианту реализации 14, в котором:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая: ниже Tm сигнального дуплекса и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая: выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: сигнального дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm: сигнального дуплекса, стеблевой области первого олигонуклеотида и стеблевой области интактного олигонуклеотида стебель-петля; и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- - первая температура является ниже второй температуры.
Вариант реализации 16. Способ по варианту реализации 14, в котором:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая: ниже Tm сигнального дуплекса, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: сигнального дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm: стеблевой области первого олигонуклеотида и стеблевой области разделенного олигонуклеотида стебель-петля; и ниже Tm: сигнального дуплекса и стеблевой области интактного олигонуклеотида стебель-петля; и
- - первая температура является ниже второй температуры.
Вариант реализации 17. Способ по варианту реализации 14, в котором:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая: ниже Tm сигнального дуплекса, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля и ниже Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая: ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: сигнального дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm: сигнального дуплекса, стеблевой области первого олигонуклеотида и стеблевой области разделенного олигонуклеотида стебель-петля; и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и
- - первая температура является ниже второй температуры.
Вариант реализации 18. Способ по варианту реализации 14, в котором:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая: ниже Tm сигнального дуплекса, выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая: выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: стеблевой области первого олигонуклеотида и сигнального дуплекса; и выше Tm: стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm: стеблевой области первого олигонуклеотида, сигнального дуплекса и стеблевой области интактного олигонуклеотида стебель-петля; и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- - первая температура выше второй температуры.
Вариант реализации 19. Способ по любому из вариантов реализации 14-18, в котором:
- Tm стеблевой области первого олигонуклеотида находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm сигнального дуплекса; и/или
- - Tm стеблевой области интактного олигонуклеотида стебель-петля находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- - первая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm: стеблевой области первого олигонуклеотида, и/или сигнального дуплекса; и/или
- - вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и/или
- - вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 20. Способ по любому из вариантов реализации 5-19, в котором:
- первый детектирующий фрагмент представляет собой флуорофор, и модификация увеличивает его расстояние от молекулы гасителя.
Вариант реализации 21. Способ по варианту реализации 20, в котором:
- первый олигонуклеотид содержит молекулу гасителя.
Вариант реализации 22. Способ по варианту реализации 21, в котором:
- флуорофор и молекула гасителя расположены на противоположных цепях двухцепочечной стеблевой области первого олигонуклеотида.
Вариант реализации 23. Способ по варианту реализации 5 или варианту реализации 6, в котором:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый олигонуклеотид содержит:
- первую двухцепочечную область гибридизованных нуклеотидов, первая цепь которой переходит в одноцепочечную область, оканчивающуюся комплементарной последовательностью, способной гибридизоваться с областью первой мишени, при этом первая цепь содержит молекулу-блокатор, предшествующую указанной комплементарной последовательности;
- смесь дополнительно содержит полимеразу;
- указанная обработка смеси включает:
гибридизацию указанной комплементарной последовательности одноцепочечной области с областью первой мишени путем комплементарного спаривания оснований;
удлинение комплементарной последовательности с применением полимеразы и первой мишени в качестве темплатной последовательности для получения второй двухцепочечной области, при этом указанная блокирующая молекула предотвращает удлинение полимеразой первой мишени с применением первой цепи указанной первой двухцепочечной области в качестве темплата;
денатурацию первой и второй двухцепочечных областей; и
- гибридизацию комплементарной последовательности, удлиненной полимеразой, с первой цепью первой двухцепочечной области путем комплементарного спаривания оснований с образованием сигнального дуплекса и, таким образом, обеспечением указанной модификации первого олигонуклеотида, позволяющей первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
Вариант реализации 24. Способ по варианту реализации 23, в котором:
- первая двухцепочечная область имеет температуру плавления (Tm), которая: ниже Tm сигнального дуплекса и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая: выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: сигнального дуплекса, первой двухцепочечной области, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm: сигнального дуплекса, первой двухцепочечной области и стеблевой области интактного олигонуклеотида стебель-петля; и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- - первая температура является ниже второй температуры.
Вариант реализации 25. Способ по варианту реализации 23, в котором:
- первая двухцепочечная область имеет температуру плавления (Tm), которая: ниже Tm сигнального дуплекса, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: сигнального дуплекса, первой двухцепочечной области, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm: первой двухцепочечной области и стеблевой области разделенного олигонуклеотида стебель-петля; и ниже Tm: сигнального дуплекса и стеблевой области интактного олигонуклеотида стебель-петля; и
- - первая температура является ниже второй температуры.
Вариант реализации 26. Способ по варианту реализации 23, в котором:
- первая двухцепочечная область имеет температуру плавления (Tm), которая: ниже Tm сигнального дуплекса, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля и ниже Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая: ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: сигнального дуплекса, первой двухцепочечной области, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm: сигнального дуплекса, первой двухцепочечной области и стеблевой области разделенного олигонуклеотида стебель-петля; и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура является ниже второй температуры.
Вариант реализации 27. Способ по варианту реализации 23, в котором:
- первая двухцепочечная область имеет температуру плавления (Tm), которая: ниже Tm сигнального дуплекса, выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая: выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: первой двухцепочечной области и сигнального дуплекса; и выше Tm: стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm: первой двухцепочечной области, сигнального дуплекса и стеблевой области интактного олигонуклеотида стебель-петля; и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- - первая температура выше второй температуры.
Вариант реализации 28. Способ по любому из вариантов реализации 23-27, в котором:
- Tm первой двухцепочечной области находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm сигнального дуплекса; и/или
- - Tm стеблевой области интактного олигонуклеотида стебель-петля находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- - первая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm: первой двухцепочечной области, и/или сигнального дуплекса; и/или
- - вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и/или
- - вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 29. Способ по любому из вариантов реализации 23-28, в котором:
- первый детектирующий фрагмент представляет собой флуорофор, и модификация увеличивает его расстояние от молекулы гасителя.
Вариант реализации 30. Способ по варианту реализации 29, в котором:
- первый олигонуклеотид содержит молекулу гасителя.
Вариант реализации 31. Способ по варианту реализации 30, в котором:
- флуорофор и молекула гасителя расположены на противоположных цепях первой двухцепочечной области.
Вариант реализации 32. Способ по варианту реализации 5 или варианту реализации 6, в котором:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- смесь дополнительно содержит:
первый праймер, комплементарный первой последовательности в первой мишени,
второй олигонуклеотид, содержащий компонент, комплементарный второй последовательности в первой мишени, который отличается от первой последовательности, и фрагмент метки, не комплементарный первой мишени,
первую полимеразу, обладающую экзонуклеазной активностью, и
необязательно вторую полимеразу, и
- указанная обработка смеси включает: подходящие условия для гибридизации первого праймера и второго олигонуклеотида с первой мишенью,
удлинение первого праймера с применением первой полимеразы и мишени в качестве темплата для отщепления таким образом области метки,
гибридизацию расщепленной области метки с первым олигонуклеотидом путем комплементарного спаривания оснований,
и удлинение области метки с применением первой или второй полимеразы и первого олигонуклеотида в качестве темплата для создания двухцепочечной последовательности, содержащей первый олигонуклеотид, тем самым обеспечивая указанную модификацию первого олигонуклеотида и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
Вариант реализации 33. Способ по варианту реализации 32, в котором:
- двухцепочечная последовательность имеет Tm, которая: выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm двухцепочечной последовательности, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm двухцепочечной последовательности и стеблевой области интактного олигонуклеотида стебель-петля; и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- - первая температура является ниже второй температуры.
Вариант реализации 34. Способ по варианту реализации 32, в котором:
- двухцепочечная последовательность имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm двухцепочечной последовательности, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm: и стеблевой области разделенного олигонуклеотида стебель-петля; и ниже Tm: двухцепочечной последовательности и стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура является ниже второй температуры.
Вариант реализации 35. Способ по варианту реализации 32, в котором:
- двухцепочечная последовательность имеет Tm, которая: ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm двухцепочечной последовательности, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm двухцепочечной последовательности и стеблевой области разделенного олигонуклеотида стебель-петля; и ниже Tm: стеблевой области интактного олигонуклеотида стебель-петля; и
- - первая температура является ниже второй температуры.
Вариант реализации 36. Способ по варианту реализации 32, в котором:
- двухцепочечная последовательность имеет Tm, которая: выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm двухцепочечной последовательности; и выше Tm: стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm двухцепочечной последовательности и стеблевой области интактного олигонуклеотида стебель-петля; и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- первая температура выше второй температуры.
Вариант реализации 37. Способ по любому из вариантов реализации 32-36, в котором:
- Tm стеблевой области интактного олигонуклеотида стебель-петля находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm двухцепочечной последовательности; и/или
- вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и/или
- вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 38. Способ по любому из вариантов реализации 32-37, в котором:
- первый олигонуклеотид содержит флуорофор и молекулу гасителя, и указанное удлинение области метки увеличивает расстояние между флуорофором и молекулой гасителя.
Вариант реализации 39. Способ по варианту реализации 5 или варианту реализации 6, в котором:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый олигонуклеотид комплементарен первому фрагменту мишени;
- смесь дополнительно содержит дополнительный олигонуклеотид, комплементарный второй области первой мишени, при этом первая и вторая области первой мишени фланкируют друг с другом, но не перекрываются;
- указанная обработка смеси включает:
образование дуплексной структуры, включающей:
(i) первый двухцепочечный компонент путем гибридизации первого олигонуклеотида с мишенью путем комплементарного спаривания оснований, и
(ii) второй двухцепочечный компонент путем гибридизации дополнительного олигонуклеотида с мишенью путем комплементарного спаривания оснований,
тем самым сближая первый и последующие олигонуклеотиды и обеспечивая указанную модификацию первого олигонуклеотида, позволяющую первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
Вариант реализации 40. Способ по варианту реализации 39, в котором:
- дуплексная структура имеет Tm, которая: ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm: дуплексной структуры, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm: дуплексной структуры и стеблевой области разделенного олигонуклеотида стебель-петля; и ниже Tm: стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура является ниже второй температуры.
Вариант реализации 41. Способ по варианту реализации 39 варианта реализации 40, в котором:
- Tm стеблевой области интактного олигонуклеотида стебель-петля находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm дуплексной структуры; и/или
- вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и/или
- вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 42. Способ по любому из вариантов реализации 39-41, в котором:
- первый детектируемый фрагмент представляет собой флуорофор, а дополнительный олигонуклеотид содержит гаситель;
- указанное формирование дуплексной структуры дополнительно сближает флуорофор и гаситель; и
- указанный детектируемый сигнал представляет собой уменьшение флуоресценции, обеспечиваемое первым детектирующим фрагментом.
Вариант реализации 43. Способ по варианту реализации 5 или варианту реализации 6, в котором:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый детектирующий фрагмент представляет собой: наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которым связан первый олигонуклеотид;
- указанная обработка смеси включает:
- гибридизацию первой мишени с первым олигонуклеотид ом, чтобы тем самым вызвать модификацию первого олигонуклеотида, позволяющую первому детектирующему фрагменту обеспечить первый детектируемый сигнал, указывающий на присутствие первой мишени в образце;
при этом первый детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета; и/или
(iii) изменение спектра поглощения,
возникающий из первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
Вариант реализации 44. Способ по варианту реализации 5 или варианту реализации 6, в котором:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый детектирующий фрагмент представляет собой электрохимический агент, с которым связан первый олигонуклеотид;
- указанная обработка смеси включает: гибридизацию первой мишени с первым олигонуклеотидом, чтобы тем самым вызвать или облегчить модификацию первого олигонуклеотида, позволяющую первому детектирующему фрагменту обеспечить первый детектируемый сигнал, указывающий на присутствие первой мишени в образце;
при этом первый детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающее из-за первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
Вариант реализации 45. Способ по варианту реализации 44, в котором:
- электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орацетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
Вариант реализации 46. Способ по любому из вариантов реализации 5-19, 23-28, 32-37 и 39-41, в котором:
- первый детектирующий фрагмент представляет собой: наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которым связан первый олигонуклеотид; и
- первый детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета; и/или
(iii) изменение спектра поглощения,
возникающий из первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
Вариант реализации 47. Способ по любому из вариантов реализации 5-19, 23-28, 32-37 и 39-41, в котором:
- первый детектирующий фрагмент представляет собой электрохимический агент, с которым связан первый олигонуклеотид; и
- первый детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающее из-за первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
Вариант реализации 48. Способ по варианту реализации 47, в котором:
- электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орацетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
Вариант реализации 49. Способ по любому из вариантов реализации 43-48, в котором:
- второй детектирующий фрагмент представляет собой: наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которым связан интактный олигонуклеотид стебель-петля; и
- второй детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета; и/или
(iii) изменение спектра поглощения,
возникающие в результате диссоциации указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 50. Способ по любому из вариантов реализации 43-48, в котором:
- второй детектирующий фрагмент представляет собой электрохимический агент, с которым связан интактный олигонуклеотид стебель-петля; и
- второй детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающего в результате диссоциации указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 51. Способ по варианту реализации 50, в котором:
- электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орацетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
Вариант реализации 52. Способ по любому из вариантов реализации 20-22, 29-31, 38 и 42, в котором:
- второй детектирующий фрагмент представляет собой флуорофор, и
- второй детектируемый сигнал, обеспечиваемый указанными цепями двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля, диссоциирует, увеличивая расстояние флуорофора от молекулы гасителя.
Вариант реализации 53. Способ по варианту реализации 52, в котором:
- флуорофор и молекула гасителя расположены на противоположных цепях двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 54. Способ по любому из вариантов реализации 1-4, в котором:
- образование первого детектируемого сигнала необратима;
- модификация первого олигонуклеотида позволяет первому детектирующему фрагменту обеспечивать первый детектируемый сигнал при первой температуре или ниже ее; и
- первый детектируемый сигнал, обеспечиваемый при первой температуре или ниже, остается детектируемым при второй температуре.
Вариант реализации 55. Способ по варианту реализации 54, в котором:
- часть (с) включает измерение:
(i) первого фонового сигнала на уровне или в пределах 1°С, 2°С, 3°С, 4°С или 5°С от первой температуры и второго фонового сигнала на уровне или в пределах 1°С, 2°С, 3°С, 4°С или 5°С от второй температуры; и/или (ii) третьего фонового сигнала при третьей температуре;
обеспечивается первым и вторым детектирующими фрагментами в смеси или в контрольной смеси; и
- часть (d) включает определение того, в один или более моментов времени во время или после указанной обработки:
(i) генерируется ли при первой температуре первый детектируемый сигнал, который отличается от первого или третьего фонового сигнала, возникающий в результате указанной модификации, при этом:
при первой температуре второй детектируемый сигнал не отличается от первого или третьего фонового сигнала, и
обнаружение различия между первым детектируемым сигналом и первым или третьим фоновым сигналом указывает на указанную модификацию первого олигонуклеотида и присутствие первой мишени в образце; и
(ii) генерируется ли при второй температуре второй детектируемый сигнал, который отличается от второго фонового сигнала, указывающий на присутствие второй мишени в образце.
Вариант реализации 56. Способ по варианту реализации 55, в котором:
- когда в образце присутствует первая мишень, упомянутое определение того, генерируется ли второй детектируемый сигнал при второй температуре, включает компенсацию первого детектируемого сигнала, присутствующего при измерении второго детектируемого сигнала.
Вариант реализации 57. Способ по варианту реализации 55 или варианту реализации 56, в котором:
- генерируется первый сигнал, отличающийся от первого фонового сигнала,
- генерируется второй сигнал, отличающийся от второго фонового сигнала, и
- второй детектируемый сигнал отличается от второго фонового сигнала в большей степени, чем первый детектируемый сигнал отличается от первого фонового сигнала,
тем самым указывая, что вторая мишень присутствует в образце.
Вариант реализации 58. Способ по варианту реализации 57, в котором:
- первая температура ниже: второй температуры, Tm двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля и Tm стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 59. Способ по варианту реализации 57, в котором:
- первая температура выше, чем: вторая температура, Tm стеблевой области интактного олигонуклеотида стебель-петля и Tm стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 60. Способ по варианту реализации 55, в котором:
- формируется первый сигнал, отличающийся от третьего фонового сигнала,
- генерируется второй сигнал, отличающийся от третьего фонового сигнала, и
- второй сигнал отличается от третьего фонового сигнала в большей степени, чем первый сигнал отличается от третьего фонового сигнала, тем самым указывая, что вторая мишень присутствует в образце.
Вариант реализации 61. Способ по варианту реализации 55, в котором:
- вторая температура выше первой температуры,
- третья температура ниже Tm двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля,
- генерируется первый детектируемый сигнал, отличающийся от третьего фонового сигнала,
- генерируется второй детектируемый сигнал, отличающийся от третьего фонового сигнала, и
- второй детектируемый сигнал отличается от третьего фонового сигнала в большей степени, чем первый сигнал отличается от третьего фонового сигнала, тем самым указывая, что вторая мишень присутствует в образце.
Вариант реализации 62. Способ по любому из вариантов реализации 55-61, в котором:
- Tm стеблевая область интактного олигонуклеотида стебель-петля выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже второй температуры и ниже Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- вторая температура выше Tm стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля.
Вариант реализации 63. Способ по варианту реализации 62, в котором:
- Tm стеблевой области интактного олигонуклеотида стебель-петля находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже второй температуры; и/или
- первая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm: стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- вторая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже Tm стеблевой области интактного олигонуклеотида стебель-петля.
Вариант реализации 64. Способ по варианту реализации 62 или варианту реализации 63, включающий:
- измерение указанного третьего фонового сигнала, при этом третья температура ниже второй температуры.
Вариант реализации 65. Способ по варианту реализации 64, в котором:
- третья температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С ниже второй температуры.
Вариант реализации 66. Способ по любому из вариантов реализации 55-61, в котором:
- Tm стеблевая область интактного олигонуклеотида стебель-петля выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура выше второй температуры, выше Tm стеблевой области разделенного олигонуклеотида стебель-петля и выше Tm стеблевой области интактного олигонуклеотида стебель-петля; и
- вторая температура выше Tm стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля.
Вариант реализации 67. Способ по варианту реализации 66, в котором:
- Tm стеблевой области интактного олигонуклеотида стебель-петля находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура находится в пределах от 1°С до 10°С, от 1°С до 5°С, от 5°С до 10°С или более чем на 10°С выше второй температуры; и/или
- первая температура находится в пределах от 1°С до 10°С, от 1°С до 5 °С, от 5 °С до 10 °С или более чем на 10 °С выше Tm: стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура находится в пределах от 1 °С до 10 °С, от 1 °С до 5 °С, от 5 °С до 10 °С или более чем на 10 °С выше Tm: стеблевой области интактного олигонуклеотида стебель-петля; и/или
- вторая температура находится в пределах от 1 °С до 10 °С, от 1 °С до 5 °С, от 5°С до 10°С или более чем на 10 °С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- вторая температура находится в пределах от 1 °С до 10 °С, от 1 °С до 5 °С, от 5°С до 10°С или более чем на 10 °С ниже Tm стеблевой области интактного олигонуклеотида стебель-петля.
Вариант реализации 68. Способ по любому из вариантов реализации 54-67, в котором:
- первый олигонуклеотид является субстратом для многокомпонентного фермента нуклеиновой кислоты (МНКзим); смесь дополнительно содержит: МНКзим, способный расщеплять первый олигонуклеотид, когда первая мишень присутствует в образце; и
- указанная обработка смеси дополнительно включает: связывание МНКзима с первой мишенью и гибридизация субстратных плеч МНКзима с первым олигонуклеотидом путем комплементарного спаривания оснований для облегчения расщепления первого олигонуклеотида, тем самым обеспечивая указанную модификацию первого олигонуклеотида и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
Вариант реализации 69. Способ по варианту реализации 68, в котором:
- первая мишень представляет собой последовательность нуклеиновой кислоты; и
- указанная обработка реакционной смеси дополнительно включает: гибридизацию первой мишени с сенсорными плечами МНКзима путем комплементарного спаривания оснований, чтобы тем самым облегчить сборку МНКзима.
Вариант реализации 70. Способ по любому из вариантов реализации 54-67, в котором:
- первый олигонуклеотид представляет собой субстрат для аптазима;
- первая мишень представляет собой аналит, белок, соединение или молекулу;
- смесь дополнительно содержит аптазим, содержащий аптамер, способный связываться с первой мишенью; и
- указанная обработка смеси дополнительно включает: связывание аптазима с первой мишенью и первым олигонуклеотидом для облегчения расщепления первого олигонуклеотида, тем самым обеспечивая указанную модификацию первого олигонуклеотида и позволяя первому детектирующему фрагменту генерировать первый детектируемый сигнал.
Вариант реализации 71. Способ по любому из вариантов реализации 54-67, в котором:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый олигонуклеотид содержит последовательность, комплементарную первой мишени,
- смесь дополнительно содержит:
- праймер, комплементарный области первой мишени, и полимеразу с экзонуклеазной активностью;
указанная обработка смеси включает:
гибридизацию праймера с первой мишенью путем комплементарного спаривания оснований,
гибридизацию первого олигонуклеотида с первой мишенью путем комплементарного спаривания оснований
удлинение праймера с применением полимеразы и первой мишени в качестве темплатной последовательности, чтобы таким образом расщепить первый олигонуклеотид и обеспечить указанную модификацию первого олигонуклеотида, позволяющую первому детектирующему фрагменту генерировать первый детектируемый сигнал.
Вариант реализации 72. Способ по любому из вариантов реализации 54-67, в котором:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- смесь дополнительно содержит:
рестрикционную эндонуклеазу, способную расщеплять двухцепочечный дуплекс, содержащий первую мишень; и
- указанная обработка смеси включает:
гибридизацию первого олигонуклеотида с первой мишенью путем комплементарного спаривания оснований с образованием таким образом двухцепочечного дуплекса,
расщепление дуплекса с применением рестрикционной эндонуклеазы, чтобы тем самым обеспечить указанную модификацию первого олигонуклеотида и сделать возможным получение первым детектируемым фрагментом первого детектируемого сигнала.
Вариант реализации 73. Способ по варианту реализации 72, в котором:
- рестрикционная эндонуклеаза представляет собой разрывающую эндонуклеазу, способную связываться с цепью указанного двухцепочечного дуплекса и расщеплять ее, и указанная цепь содержит весь или часть первого олигонуклеотида.
Вариант реализации 74. Способ по любому из вариантов реализации 54-67, в котором:
- смесь дополнительно содержит ДНКзим или рибозим, требующий кофактора для каталитической активности;
- указанная обработка смеси включает применение условий, подходящих для:
связывания кофактора с ДНКзимом или рибозимом, чтобы сделать его каталитически активным,
гибридизации ДНКзима или рибозима с первым олигонуклеотидом путем комплементарного спаривания оснований, и
каталитической активности ДНКзима или рибозима в расщеплении таким образом первого олигонуклеотида и, таким образом, в обеспечении указанной модификации первого олигонуклеотида, позволяющей первому детектирующему фрагменту обеспечивать первый детектируемый сигнал,
при этом:
- первая цель представляет собой кофактор.
Вариант реализации 75. Способ по варианту реализации 74, в котором кофактор представляет собой ион металла или ион металла, выбранный из: Mg2+, Mn2+, Са2+, Pb2+.
Вариант реализации 76. Способ по любому из вариантов реализации 54-75, в котором:
- первый детектирующий фрагмент представляет собой флуорофор, и модификация первого олигонуклеотида увеличивает расстояние флуорофора от молекулы гасителя.
Вариант реализации 77. Способ по варианту реализации 76, в котором: первый олигонуклеотид содержит молекулу гасителя.
Вариант реализации 78. Способ по варианту реализации 76 или варианту реализации 77, в котором:
- второй детектирующий фрагмент представляет собой флуорофор, и
- второй детектируемый сигнал, обеспечивается диссоциацией указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля, что приводит к увеличению расстояния от флуорофора до молекулы гасителя.
Вариант реализации 79. Способ по варианту реализации 78, в котором:
- флуорофор и молекула гасителя расположены на противоположных цепях двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 80. Способ по любому из вариантов реализации 54-79, в котором:
- первый детектирующий фрагмент представляет собой: наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которым связан первый олигонуклеотид; и
- первый детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета; и/или
(iii) изменение спектра поглощения,
возникающий из первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
Вариант реализации 81. Способ по любому из вариантов реализации 54-79, в котором:
- первый детектирующий фрагмент представляет собой электрохимический агент, с которым связан первый олигонуклеотид; и
- первый детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающее из-за первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
Вариант реализации 82. Способ по варианту реализации 81, в котором:
- электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орацетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
Вариант реализации 83. Способ по любому из вариантов реализации 80-82, в котором:
- второй детектирующий фрагмент представляет собой: наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которым связан интактный олигонуклеотид стебель-петля; и
- второй детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета; и/или
(iii) изменение спектра поглощения,
возникающие в результате диссоциации указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 84. Способ по любому из вариантов реализации 80-82, в котором:
- второй детектирующий фрагмент представляет собой электрохимический агент, с которым связан интактный олигонуклеотид стебель-петля; и
- второй детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающего в результате диссоциации указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 85. Способ по варианту реализации 84, в котором:
- электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орацетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
Вариант реализации 86. Способ по любому из вариантов реализации 1-85, в котором интактный олигонуклеотид стебель-петля не гибридизуется со второй мишенью во время указанного расщепления одного или более негибридизированных нуклеотидов интактного олигонуклеотида стебель-петля первым ферментом.
Вариант реализации 87. Способ по любому из вариантов реализации 1-86, в котором:
- первый фермент представляет собой первый МНКзим, и
- указанная обработка смеси включает:
связывание первого МНКзима со второй мишенью и гибридизация субстратных плеч указанного первого МНКзима с петлевой областью интактного олигонуклеотида стебель-петля с расщеплением, таким образом, одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля и обеспечением разделенного олигонуклеотида стебель-петля.
Вариант реализации 88. Способ по варианту реализации 87, в котором:
- вторая мишень представляет собой последовательность нуклеиновой кислоты; и
- указанная обработка смеси дополнительно включает: гибридизацию второй мишени с сенсорными плечами первого МНКзима путем комплементарного спаривания оснований, чтобы тем самым облегчить сборку первого МНКзима.
Вариант реализации 89. Способ по любому из вариантов реализации 1-86, в котором:
- вторая мишень представляет собой аналит, белок, соединение или молекулу;
- первый фермент представляет собой аптазим, содержащий аптамер, способный связываться со второй мишенью; и
- связывание второй мишени с аптамером способно сделать первый фермент каталитически активным.
Вариант реализации 90. Способ по варианту реализации 89, в котором:
- первый фермент представляет собой любой из следующих: апта-ДНКзим, апта-рибозим, апта-МНКзим.
Вариант реализации 91. Способ по любому из вариантов реализации 1-86, в котором:
- вторая мишень представляет собой аналит, белок, соединение или молекулу;
- первый олигонуклеотид представляет собой субстрат для аптазима;
- первый фермент представляет собой аптазим, содержащий область аптамера, способную связываться со второй мишенью, и область нуклеиновой кислоты с ферментативной активностью, способную расщеплять один или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля
- указанная обработка смеси дополнительно включает:
связывание второй мишени с аптамерной областью аптазима для облегчения активации каталитической активности области нуклеиновой кислоты с ферментативной активностью и гибридизацию интактного олигонуклеотида стебель-петля с активной областью фермента нуклеиновой кислоты для расщепления, таким образом, одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля.
Вариант реализации 92. Способ по любому из вариантов реализации 1-85, в котором:
- вторая мишень представляет собой последовательность нуклеиновой кислоты; и
- первый фермент представляет собой первую рестрикционную эндонуклеазу, и указанная обработка смеси включает:
применение условий, подходящих для гибридизации второй мишени с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований с образованием двухцепочечной последовательности для связывания и расщепления с одним или более негибридизированными нуклеотидами одноцепочечной петлевой области первой рестрикционной эндонуклеазой, тем самым образуя разделенный олигонуклеотид стебель-петля.
Вариант реализации 93. Способ по варианту реализации 92, в котором:
- первая рестрикционная эндонуклеаза представляет собой эндонуклеазу первого разрыва, способную связываться и расщеплять цепь указанной двухцепочечной последовательности для первой рестрикционной эндонуклеазы, и указанная цепь включает весь или часть одноцепочечной петлевой области интактного олигонуклеотида стебель-петля.
Вариант реализации 94. Способ по любому из вариантов реализации 1-85, в котором:
- первый фермент включает полимеразу с экзонуклеазной активностью,
- указанная обработка смеси включает применение условий, подходящих для:
гибридизации второй мишени с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований с образованием первой двухцепочечной последовательности, содержащей область второй мишени,
гибридизации первого праймера олигонуклеотида со второй мишенью с образованием второй двухцепочечной последовательности, расположенной выше относительно первой двухцепочечной последовательности, содержащей область второй мишени,
удлинения праймера с применением полимеразы с экзонуклеазной активностью и применением второй мишени в качестве темплатной последовательности,
при этом первая полимераза, обладающая экзонуклеазной активностью, расщепляет одноцепочечную петлевую область первой двухцепочечной последовательности и, таким образом, образует разделенный олигонуклеотид стебель-петля.
Вариант реализации 95. Способ по любому из вариантов реализации 1-85, в котором:
- первый фермент представляет собой экзонуклеазу, и
- указанная обработка смеси включает применение условий, подходящих для:
гибридизации второй мишени с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований с образованием первой двухцепочечной последовательности, содержащей область второй мишени,
объединения первого фермента, обладающего экзонуклеазной активностью, с двухцепочечной последовательностью, содержащей вторую мишень, и
каталитической активности первого фермента, обладающего экзонуклеазную активность, позволяющей ему расщеплять одноцепочечную петлевую область первой двухцепочечной последовательности, содержащей вторую мишень, и таким образом образовывать разделенный олигонуклеотид стебель-петля.
Вариант реализации 96. Способ по любому из вариантов реализации 1-85, в котором:
- первый фермент представляет собой ДНКзим или рибозим, требующий кофактора для каталитической активности, и указанная обработка смеси включает применение условий, подходящих для:
- связывания кофактора с первым ферментом, чтобы сделать его каталитически активным,
- гибридизации ДНКзима или рибозима с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований,
- каталитической активности ДНКзима или рибозима по расщеплению одного или более негибридизированных нуклеотидов одноцепочечной области петли интактного олигонуклеотида стебель-петля и, таким образом, формированию разделенного олигонуклеотида стебель-петля,
при этом:
- вторую мишень представляет собой кофактор.
Вариант реализации 97. Способ по варианту реализации 96, в котором кофактор представляет собой ион металла или ион металла, выбранный из: Mg2+, Mn2+, Са2+, Pb2+.
Вариант реализации 98. Способ по любому из вариантов реализации 1-97, в котором:
- первая мишень отличается от второй мишени; и/или
- первый олигонуклеотид содержит или состоит из последовательности, которая не находится в пределах области одноцепочечной петли интактного олигонуклеотида стебель-петля.
Вариант реализации 99. Способ по любому из вариантов реализации 1-98, в котором:
- первый фермент не расщепляет вторую мишень.
Вариант реализации 100. Способ по любому из вариантов реализации 1-71, 74-91 или 94-99, в котором:
- ни один из указанных ферментов не расщепляет первую мишень и/или вторую мишень.
Вариант реализации 101. Способ по любому из вариантов реализации 1-100, в котором:
- первая температура отличается от второй температуры более чем на: 1°С, 2°С, 3°С, 4°С, 5°С, 6°С, 7°С, 8°С, 9°С, 10°С, 11°С, 12°С, 13°С, 14°С, 15°С, 16°С, 17°С, 18°С, 19°С, 20°С, 25°С, 30°С, 35°С, 40°С, 45°С, 50°С, 55°С или 60°С.
Вариант реализации 102. Способ по любому из вариантов реализации 1-101, в котором указанное определение включает обнаружение первого детектируемого сигнала и/или любого указанного фонового сигнала (сигналов):
- в один или более моментов времени во время указанной обработки; или
- в один или более моментов времени во время указанной обработки и в один или более моментов времени после указанной обработки.
Вариант реализации 103. Способ по любому из вариантов реализации 1-101, в котором указанное определение включает обнаружение первого детектируемого сигнала и/или любого указанного фонового сигнала (сигналов):
- в один или более моментов времени после указанной обработки.
Вариант реализации 104. Способ по любому из вариантов реализации 1-101, в котором указанное определение включает обнаружение второго детектируемого сигнала и/или любого указанного фонового сигнала (сигналов):
- в один или более моментов времени во время указанной обработки; или
- в один или более моментов времени во время указанной обработки и в один или более моментов времени после указанной обработки.
Вариант реализации 105. Способ по любому из вариантов реализации 1-101, в котором указанное определение включает обнаружение второго детектируемого сигнала и/или любого указанного фонового сигнала (сигналов):
- в один или более моментов времени после указанной обработки.
Вариант реализации 106. Способ по любому из вариантов реализации 1-105, в котором:
- указанное определение наличия или отсутствия первой и второй мишеней включает анализ кривой плавления.
Вариант реализации 107. Способ по варианту реализации 6, в котором:
- указанное определение наличия или отсутствия первой и второй мишеней включает анализ кривой плавления, включающий первый и второй детектируемые сигналы и, необязательно, первый и второй фоновые сигналы.
Вариант реализации 108. Способ по варианту реализации 55, в котором:
- указанное определение наличия или отсутствия первой и второй мишеней включает анализ кривой плавления, включающий первый и второй детектируемые сигналы и, необязательно, первый и второй фоновые сигналы; или
- первый и второй детектируемые сигналы и, необязательно, третий фоновый сигнал.
Вариант реализации 109. Способ по любому из вариантов реализации 1-108, в котором:
- первая мишень и/или вторая мишень представляет собой ампликон нуклеиновой кислоты.
Вариант реализации 110. Способ по любому из вариантов реализации 1-109, в котором:
- первая мишень представляет собой нуклеиновую кислоту и/или вторая мишень представляет собой нуклеиновую кислоту, и
- смесь дополнительно содержит реагенты для амплификации указанной первой и/или второй мишени,
- указанная обработка смеси дополнительно включает условия, подходящие для проведения амплификации первой и/или второй мишени.
Вариант реализации 111. Способ по варианту реализации 110, в котором:
- амплификация представляет собой одну или более из полимеразной цепной реакции (ПЦР), амплификации замещения цепи (SDA), реакции амплификации никирующего фермента (NEAR), геликазозависимой амплификации (HDA), рекомбиназной полимеразной амплификации (RPA), петлевой изотермической амплификации (LAMP), амплификации по типу катящегося кольца (RCA), амплификации, опосредованной транскрипцией (ТМА), самоподдерживающейся репликации последовательности (3SR), амплификации на основе последовательности нуклеиновой кислоты (NASBA), лигазной цепной реакции (LCR) или способа амплификации с разветвлением (RAM) и/или полимеразной цепной реакции с обратной транскрипцией (ОТ-ПЦР).
Вариант реализации 112. Способ по варианту реализации 110 или варианту реализации 111, в котором указанное определение:
- происходит до указанной амплификации или в пределах 1, 2, 3, 4 или 5 циклов от начала указанной амплификации; и/или
- происходит после завершения указанной амплификации.
Вариант реализации 113. Способ по любому из вариантов реализации 110-112, в котором указанное определение:
- происходит до указанной амплификации или в течение 1, 2, 3, 4 или 5 минут после начала указанной амплификации; и/или
- происходит после завершения указанной амплификации.
Вариант реализации 114. Способ по любому из вариантов реализации 110-113, в котором указанное определение происходит:
- в первый момент времени до указанной амплификации; и
- во второй момент времени после завершения указанной амплификации.
Вариант реализации 115. Способ по любому из вариантов реализации 110-114, в котором:
- способ амплификации представляет собой полимеразную цепную реакцию (ПЦР); и
- указанное определение происходит в нескольких циклах, необязательно в каждом цикле.
Вариант реализации 116. Способ по варианту реализации 110 или варианту реализации 111, дополнительно включающий нормализацию:
- первого детектируемого сигнала при первой температуре, измеренный в момент времени во время или после указанной амплификации с применением положительного контрольного сигнала, генерируемого при первой температуре до указанной амплификации и/или до указанной обработки реакции; и/или
- второго детектируемого сигнала при второй температуре, измеренный в момент времени во время или после указанной амплификации с применением положительного контрольного сигнала, генерируемого при второй температуре до указанной амплификации и/или до указанной обработки реакции.
Вариант реализации 117. Способ по варианту реализации 110 или варианту реализации 111, дополнительно включающий нормализацию:
- первый детектируемый сигнал с применением детектируемого сигнала, генерируемого интактным олигонуклеотидом стебель-петля при первой температуре до указанной амплификации и/или до указанной обработки реакции; и/или
- второй детектируемый сигнал с применением детектируемого сигнала, генерируемого интактным олигонуклеотидом стебель-петля при дополнительной температуре до указанной амплификации и/или до указанной обработки реакции;
при этом дополнительная температура превышает Tm интактного олигонуклеотида стебель-петля.
Вариант реализации 118. Способ по любому из вариантов реализации 1-117, дополнительно включающий:
- генерирование сигнала положительного контроля первой мишени с применением известной концентрации первой мишени и/или известной концентрации первого олигонуклеотида после указанной модификации.
Вариант реализации 119. Способ по любому из вариантов реализации 1-118:
- дополнительно включает генерирование сигнала положительного контрольного сигнала для первой мишени путем повторения способа на отдельном контрольном образце, содержащем указанную первую мишень.
Вариант реализации 120. Способ по варианту реализации 119, в котором:
- отдельный контрольный образец, содержащий первую мишень, содержит известную концентрацию первой мишени.
Вариант реализации 121. Способ по варианту реализации 119 или варианту реализации 120, в котором:
- отдельный контрольный образец, содержащий первую мишень, дополнительно содержит вторую мишень.
Вариант реализации 122. Способ по любому из вариантов реализации 1-121, дополнительно включающий:
- генерирование сигнала положительного контроля второй мишени с применением известной концентрации второй мишени и/или известной концентрации олигонуклеотида стебель-петля после указанной модификации.
Вариант реализации 123. Способ по любому из вариантов реализации 1-122, дополнительно включающий:
- генерирование сигнала положительного контрольного второй мишени путем повторения указанного способа на отдельном контрольном образце, содержащем вторую мишень.
Вариант реализации 124. Способ по варианту реализации 123, в котором:
- контрольный образец, содержащий вторую мишень, содержит известную концентрацию второй мишени.
Вариант реализации 125. Способ по варианту реализации 123 или варианту реализации 124, в котором:
- указанный контрольный образец, содержащий вторую мишень, дополнительно содержит указанную первую мишень.
Вариант реализации 126. Способ по любому из вариантов реализации 1-125, дополнительно включающий:
- генерирование комбинированного положительного контрольного сигнала путем повторения указанного способа на отдельном контрольном образце, содержащем первую мишень и вторую мишень.
Вариант реализации 127. Способ по варианту реализации 126, в котором:
- комбинированный контрольный образец содержит известную концентрацию первой мишени и/или известную концентрацию второй мишени.
Вариант реализации 128. Способ по любому из вариантов реализации 116-127, дополнительно включающий:
- нормализацию первого детектируемого сигнала и/или второго детектируемого сигнала с применением любого упомянутого положительного контрольного сигнала.
Вариант реализации 129. Способ по любому из вариантов реализации 116-128, дополнительно включающий:
- оценку уровней отрицательного контрольного сигнала путем повторения способа по любому из вариантов реализации 1-115 на отдельном образце отрицательного контроля, который не содержит:
(i) указанную первую мишень; или
(ii) указанную вторую мишень; или
(iii) указанную первую мишень или указанную вторую мишень.
Вариант реализации 130. Способ по варианту реализации 129, дополнительно включающий:
- нормализацию первого детектируемого сигнала и/или второго детектируемого сигнала с применением указанного отрицательного контрольного сигнала.
Вариант реализации 131. Способ по любому из вариантов реализации 116-130, в котором:
- любой указанный контрольный сигнал является флуоресцентным контрольным сигналом.
Вариант реализации 132. Способ по любому из вариантов реализации 1-131, дополнительно включающий сравнение первого и/или второго обнаружимых сигналов с пороговым значением, при этом:
- пороговое значение генерируется с применением обнаружимых сигналов, полученных из серии образцов или их производных, протестированных в соответствии со способом по любому из вариантов реализации с 1 по 115, и включающих любой один или более из:
(i) нецелевой контроль и первую мишень
(ii) нецелевой контроль и вторую мишень
(iii) нецелевой контроль, первую мишень и вторую мишень,
чтобы таким образом определить указанное присутствие или отсутствие первой и второй мишеней в образце.
Вариант реализации 133. Способ по варианту реализации 132, в котором:
- серии образцов или их производных тестируют, используя известную концентрацию первого олигонуклеотида и/или известную концентрацию интактного олигонуклеотида стебель-петля.
Вариант реализации 134. Способ по любому из вариантов реализации 1-133, в котором:
- образец представляет собой биологический образец, полученный от субъекта.
Вариант реализации 135. Способ по любому из вариантов реализации 1-133:
при этом способ осуществляют in vitro.
Вариант реализации 136. Способ по любому из вариантов реализации 1-133:
при этом способ осуществляют ex vivo.
Вариант реализации 137. Способ по любому из вариантов реализации 1-136, в котором:
- первый и второй обнаружимые фрагменты излучают в одной и той же цветовой области видимого спектра.
Вариант реализации 138. Композиция, содержащая:
- первый олигонуклеотид для обнаружения первой мишени, при этом первая мишень представляет собой нуклеиновую кислоту и комплементарна по меньшей мере части первого олигонуклеотида, и
- первый детектирующий фрагмент, в котором: первый детектирующий фрагмент способен генерировать первый детектируемый сигнал при модификации первого олигонуклеотида, и
модификация индуцируется гибридизацией первой мишени с первым олигонуклеотидом путем комплементарного спаривания оснований;
- интактный олигонуклеотид стебель-петля для обнаружения второй мишени, содержащий двуцепочечную стеблевую область гибридизированных нуклеотидов, противоположные цепи которых связаны неразорванной одноцепочечной петлевой областью негибридизированных нуклеотидов, при этом по меньшей мере одна цепь двухцепочечной стеблевой области содержит второй детектирующий фрагмент; и
- первый фермент, способный расщеплять один или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля только тогда, когда в образце присутствует вторая мишень, с тем чтобы разрушить одноцепочечную петлевую область и получить разделенный олигонуклеотид стебель-петля;
при этом:
- второй детектирующий фрагмент способен генерировать второй детектируемый сигнал при диссоциации двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля, и
- первый и второй детектирующие фрагменты способны генерировать детектируемые сигналы, которые нельзя различить при одной температуре с применением одного типа детектора.
Вариант реализации 139. Композиция по варианту реализации 138, в которой:
- область первого олигонуклеотида, комплементарная первой мишени, имеет другую температуру плавления (Tm) по сравнению с каждой цепью двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля.
Вариант реализации 140. Композиция по варианту реализации 138 или реализации 139, в которой первый олигонуклеотид отличается по последовательности от:
- каждой цепи двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля; и
- одноцепочечной петлевой области интактного олигонуклеотида стебель-петля.
Вариант реализации 141. Композиция по любому из вариантов реализации 138-140, в которой:
- первый олигонуклеотид представляет собой олигонуклеотид стебель-петля, содержащий двухцепочечную стеблевую область из гибридизированных нуклеотидов на противоположных цепях, которые связаны неразорванной одноцепочечной петлевой областью негибридизированных нуклеотидов, все или часть которых комплементарны первой мишени.
Вариант реализации 142. Композиция по варианту реализации 141, в которой:
- первая мишень гибридизуется с первым олигонуклеотидом за счет комплементарного спаривания оснований, вызывающего диссоциацию цепей в двухцепочечной стеблевой области первого олигонуклеотида, что позволяет первой детектирующей молекуле обеспечивать первый детектируемый сигнал.
Вариант реализации 143. Композиция по любому из вариантов реализации 138-140, в которой:
- первый олигонуклеотид представляет собой олигонуклеотид стебель-петля, содержащий:
двухцепочечную стеблевую область гибридизированных нуклеотидов, противоположные цепи которого связаны одноцепочечной петлевой областью негибридизированных нуклеотидов, все или часть которых комплементарны первой мишени, и
вторую одноцепочечную область, отходящую от одной из указанных противоположных цепей в 3'-направлении и оканчивающуюся последовательностью, комплементарной области первой мишени, и
блокирующую молекулу, предшествующую указанной последовательности, которая комплементарна области первой мишени.
Вариант реализации 144. Композиция по варианту реализации 143, в которой:
- первая мишень гибридизуется с ее второй одноцепочечной областью за счет комплементарного спаривания оснований;
композиция дополнительно содержит полимеразу, способную удлинять вторую одноцепочечную часть, используя первую мишень в качестве темплатной последовательности, с получением двухцепочечной нуклеиновой кислоты, причем указанная блокирующая молекула способна предотвращать удлинение полимеразой первой мишени, используя указанную одну противоположную цепь в качестве темплата, и
после денатурации двухцепочечной нуклеиновой кислоты вторая одноцепочечная часть, удлиненная полимеразой, способна гибридизоваться с одноцепочечной петлевой областью первого олигонуклеотида путем комплементарного спаривания оснований с образованием сигнального дуплекса и, таким образом, обеспечивает первый детектирующий фрагмент для обеспечения первого детектируемого сигнала.
Вариант реализации 145. Композиция по любому из вариантов реализации 141-144, в которой:
- первый детектирующий фрагмент представляет собой флуорофор.
Вариант реализации 146. Композиция по варианту реализации 145, в которой:
- первый олигонуклеотид содержит молекулу гасителя, и причем флуорофор и молекула гасителя расположены на противоположных цепях двухцепочечной стеблевой области первого олигонуклеотида.
Вариант реализации 147. Композиция по любому из вариантов реализации 138-140, в которой:
- первый олигонуклеотид содержит: первую двухцепочечную область гибридизованных нуклеотидов, первая цепь которой переходит в одноцепочечную область, оканчивающуюся комплементарной последовательностью, способной гибридизоваться с областью первой мишени, при этом первая цепь содержит молекулу-блокатор, предшествующую указанной комплементарной последовательности.
композиция дополнительно содержит полимеразу;
Вариант реализации 148. Композиция по варианту реализации 147, в которой: область первой мишени гибридизуется с указанной комплементарной
- последовательностью одноцепочечной области путем комплементарного спаривания оснований; и
композиция дополнительно содержит полимеразу, способную удлинять комплементарную последовательность, используя первую мишень в качестве темплатной последовательности, для получения второй двухцепочечной области, при этом указанная блокирующая молекула предотвращает удлинение полимеразой первой мишени с применением одноцепочечной области в качестве темплата; и
- если первая и вторая двухцепочечные области денатурированы, комплементарная последовательность, удлиненная полимеразой, способна гибридизоваться с первой цепью первой двухцепочечной области за счет комплементарного спаривания оснований с образованием сигнального дуплекса и, таким образом, делает возможной первый детектирующий фрагмент для обеспечения первого детектируемого сигнала.
Вариант реализации 149. Композиция по варианту реализации 147 или варианту реализации 148, в которой:
- первый детектирующий фрагмент представляет собой флуорофор, и модификация увеличивает его расстояние от молекулы гасителя.
Вариант реализации 150. Композиция по варианту реализации 149, в которой:
-первый олигонуклеотид содержит молекулу гасителя, и причем флуорофор и молекула гасителя расположены на противоположных цепях первой двухцепочечной области.
Вариант реализации 151. Композиция по любому из вариантов реализации 138-140, в которой:
- первый олигонуклеотид комплементарен первому фрагменту мишени;
- композиция дополнительно содержит дополнительный олигонуклеотид, комплементарный второй области первой мишени, при этом первая и вторая области первой мишени фланкируют друг с другом, но не перекрываются, и каждая способна гибридизоваться с первой мишенью с образованием дуплексной структуры, включающей:
(i) первый двухцепочечный компонент путем гибридизации первого олигонуклеотида с мишенью или путем комплементарного спаривания оснований, и
(ii) второй двухцепочечный компонент путем гибридизации дополнительного олигонуклеотида с мишенью путем комплементарного спаривания оснований,
тем самым сближая первый и дополнительные олигонуклеотиды и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
Вариант реализации 152. Композиция по варианту реализации 151, в которой:
- первый детектируемый фрагмент представляет собой флуорофор, а дополнительный олигонуклеотид содержит гаситель;
- указанное формирование дуплексной структуры дополнительно сближает флуорофор и гаситель; и
- указанный детектируемый сигнал представляет собой уменьшение флуоресценции, обеспечиваемое первым детектирующим фрагментом.
Вариант реализации 153. Способ по любому из вариантов реализации 138-140, в котором:
- первый олигонуклеотид гибридизуется с первой мишенью за счет комплементарного спаривания оснований,
- композиция дополнительно содержит:
праймер, гибрид из ованный с частью первой мишени путем комплементарного спаривания оснований, и
полимераза с экзонуклеазной активностью, способная удлинять праймер с применением первой мишени в качестве темплатной последовательности, тем самым расщепляя первый олигонуклеотид и модифицируя первый олигонуклеотид, позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
Вариант реализации 154. Композиция по любому из вариантов реализации 138-140, в которой:
- первая мишень гибридизуется с первым олигонуклеотидом путем комплементарного спаривания оснований с образованием таким образом двухцепочечного дуплекса,
- композиция дополнительно содержит рестрикционную эндонуклеазу, способную расщеплять двухцепочечный дуплекс, содержащий первую мишень, тем самым модифицируя первый олигонуклеотид и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
Вариант реализации 155. Композиция по варианту реализации 154, в которой:
- рестрикционная эндонуклеаза представляет собой разрывающую эндонуклеазу, способную связываться с цепью указанного двухцепочечного дуплекса и расщеплять ее, и указанная цепь содержит первый олигонуклеотид.
Вариант реализации 156. Композиция по любому из вариантов реализации 153-155, в которой:
- первый детектирующий фрагмент представляет собой флуорофор, и указанная модификация первого олигонуклеотида увеличивает расстояние флуорофора от молекулы гасителя.
Вариант реализации 157. Композиция по варианту реализации 156, в которой:
- первый олигонуклеотид содержит молекулу гасителя.
Вариант реализации 158. Композиция, содержащая:
- первый олигонуклеотид для обнаружения первой мишени, содержащий первый детектирующий фрагмент, причем:
- первый детектирующий фрагмент способен генерировать первый детектируемый сигнал при модификации первого олигонуклеотида, и модификация индуцируется первой мишенью;
- интактный олигонуклеотид стебель-петля для обнаружения второй мишени, содержащий двуцепочечную стеблевую область гибридизированных нуклеотидов, противоположные цепи которых связаны неразорванной одноцепочечной петлевой областью негибридизированных нуклеотидов, при этом по меньшей мере одна цепь двухцепочечной стеблевой области содержит второй детектирующий фрагмент; и
- первый фермент, способный расщеплять один или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля только тогда, когда в образце присутствует вторая мишень, с тем чтобы разрушить одноцепочечную петлевую область и получить разделенный олигонуклеотид стебель-петля;
при этом:
- второй детектирующий фрагмент способен генерировать второй детектируемый сигнал при диссоциации двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля, и
- вторый и второй детектирующие фрагменты способны генерировать детектируемые сигналы, которые нельзя различить при одной температуре с применением одного типа детектора.
Вариант реализации 159. Композиция по варианту реализации 158, в которой первый олигонуклеотид отличается по последовательности от:
каждой цепи двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля; и
- одноцепочечной петлевой области интактного олигонуклеотида стебель-петля.
Вариант реализации 160. Композиция по варианту реализации 158 или варианту реализации 159, в которой:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- композиция дополнительно содержит:
- первый праймер, комплементарный первой последовательности в первой мишени,
второй олигонуклеотид, содержащий компонент, комплементарный второй последовательности в первой мишени, который отличается от первой последовательности, и фрагмент метки, не комплементарный первой мишени,
первую полимеразу, обладающую экзонуклеазной активностью, и
необязательно вторую полимеразу.
Вариант реализации 161. Композиция по варианту реализации 160, в которой:
- каждый из первого праймера и второго олигонуклеотида гибридизуется с первой мишенью путем комплементарного спаривания оснований,
первая полимераза способна удлинять первый праймер, используя мишень в качестве темплата, тем самым отщепляя область метки, позволяя отщепленной области метки гибридизоваться с первым олигонуклеотидом путем комплементарного спаривания оснований, и
первая полимераза или необязательная вторая полимераза способны удлинять область метки с применением первого олигонуклеотида в качестве темплата для создания двухцепочечной последовательности, содержащей первый олигонуклеотид, тем самым модифицируя первый олигонуклеотид и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
Вариант реализации 162. Композиция по варианту реализации 160 или варианту реализации 161, в которой:
- первый олигонуклеотид содержит флуорофор и молекулу гасителя.
Вариант реализации 163. Композиция по варианту реализации 162, в которой:
- первый олигонуклеотид содержит флуорофор и молекулу гасителя, и
- указанное удлинение области метки увеличивает расстояние между флуорофором и молекулой гасителя.
Вариант реализации 164. Композиция по варианту реализации 158 или варианту реализации 159, в которой:
- первая мишень является кофактором каталитической активности ферментов;
- композиция дополнительно содержит ДНКзим или рибозим, которым требуется кофактор для каталитической активности, и
- ДНКазим или рибозим способен связываться с первой мишенью и гибридизоваться с первым олигонуклеотидом за счет комплементарного спаривания оснований, тем самым расщепляя и модифицируя первый олигонуклеотид, позволяя первому детектирующему фрагменту генерировать первый детектируемый сигнал.
Вариант реализации 165. Композиция по варианту реализации 164, в которой кофактор представляет собой ион металла или ион металла, выбранный из: Mg2+, Mn2+, Са2+, Pb2+.
Вариант реализации 166. Способ по варианту реализации 158 или варианту реализации 159, в котором:
- первый олигонуклеотид является субстратом для многокомпонентного фермента нуклеиновой кислоты (МНКзим);
- композиция дополнительно содержит МНКзим, способный расщеплять первый олигонуклеотид, когда первая мишень присутствует в образце; и
- при этом МНКзим способен связываться с первой мишенью и гибридизоваться с первым олигонуклеотидом путем комплементарного спаривания оснований через его субстратные плечи, и указанная гибридизация облегчает расщепление первого олигонуклеотида, тем самым модифицируя его и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
Вариант реализации 167. Композиция по варианту реализации 166, в которой:
- первая мишень представляет собой последовательность нуклеиновой кислоты; и
- первая мишень гибридизуется с сенсорными плечами МНКзима путем комплементарного спаривания оснований, чтобы тем самым облегчить сборку МНКзима.
Вариант реализации 168. Композиция по варианту реализации 158 или варианту реализации 159, в которой:
- первая мишень представляет собой аналит, белок, соединение или молекулу;
- первый олигонуклеотид представляет собой субстрат для аптазима; и композиция дополнительно содержит аптазим, содержащий аптамерную часть, способную связываться с первой мишенью, и ферментативную часть нуклеиновой кислоты, способную расщеплять первый олигонуклеотид и тем самым модифицировать его, позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
Вариант реализации 169. Композиция по варианту реализации 168, в которой:
- первая мишень связывается с аптамерной областью аптазима, и первый олигонуклеотид гибридизуется с активной областью фермента нуклеиновой кислоты за счет комплементарного спаривания оснований, облегчающего расщепление первого олигонуклеотида и, таким образом, модифицирующего его, что позволяет первому детектирующему фрагменту обеспечить первый детектируемый сигнал.
Вариант реализации 170. Композиция по любому из вариантов реализации 166-169, в которой:
- первый детектирующий фрагмент представляет собой флуорофор, и указанная модификация первого олигонуклеотида увеличивает расстояние флуорофора от молекулы гасителя.
Вариант реализации 171. Композиция по варианту реализации 170, в которой:
- первый олигонуклеотид содержит молекулу гасителя.
Вариант реализации 172. Композиция по любому из вариантов реализации 138-144, 147, 148, 151, 153-155, 158-161 и 164-169, в которой:
- первый детектирующий фрагмент представляет собой: наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которым связан первый олигонуклеотид; и
- первый детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета; и/или
(iii) изменение спектра поглощения,
- возникающий из первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
Вариант реализации 173. Композиция по варианту реализации 172, в которой:
- первый детектирующий фрагмент представляет собой электрохимический агент, с которым связан первый олигонуклеотид;
- первый детектируемый сигнал представляет собой изменение электрохимического сигнала.
Вариант реализации 174. Композиция по варианту реализации 173, в которой:
- электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орацетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
Вариант реализации 175. Композиция по любому из вариантов реализации 172-174, в которой:
- второй детектирующий фрагмент представляет собой: наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которым связана по меньшей мере одна цепь двухцепочечного петлевого фрагмента олигонуклеотида, и
- второй детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета; и/или
(iii) изменение спектра поглощения,
возникающая при указанной диссоциации двухцепочечного петлевого фрагмента разделенного олигонуклеотида стебель-петля.
Вариант реализации 176. Композиция по любому из вариантов реализации 172-174, в которой:
- второй детектирующий фрагмент представляет собой электрохимический агент, с которым связан второй олигонуклеотид; и
- второй детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающее при указанной диссоциации двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
Вариант реализации 177. Композиция по варианту реализации 176, в которой:
- электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орацетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
Вариант реализации 178. Композиция по любому из вариантов реализации 145, 146, 149, 150, 152, 156, 157, 162, 163, 170 и 171, в которой:
- второй детектирующий фрагмент представляет собой флуорофор, и второй детектируемый сигнал, обеспечиваемый указанным вторым
- детектирующим фрагментом при диссоциации двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля, увеличивает расстояние флуорофора от молекулы гасителя.
Вариант реализации 179. Композиция по варианту реализации 178, в которой:
- флуорофор и молекула гасителя расположены на противоположных цепях двухцепочечной стеблевой области олигонуклеотида стебель-петля.
Вариант реализации 180. Композиция по любому из вариантов реализации 138-179, в которой:
- первый фермент представляет собой первый МНКзим,
- первый МНКзим связан со второй мишенью,
- субстратные плечи указанного первого МНКзима гибридизуются путем комплементарного спаривания оснований с единичным петлевым фрагментом интактного олигонуклеотида стебель-петля, тем самым облегчая расщепление одного или более негибридизированных нуклеотидов интактного олигонуклеотида стебель-петля и обеспечивая разделенный олигонуклеотид стебель-петля.
Вариант реализации 181. Композиция по варианту реализации 180, в которой:
- вторая мишень представляет собой последовательность нуклеиновой кислоты; и
- вторая мишень гибридизуется с сенсорными плечами первого МНКзима путем комплементарного спаривания оснований, чтобы тем самым облегчить сборку первого МНКзима.
Вариант реализации 182. Композиция по любому из вариантов реализации 138-179, в которой:
- вторая мишень представляет собой аналит, белок, соединение или молекулу;
- первый фермент представляет собой аптазим, содержащий аптамер, способный связываться со второй мишенью; и
- аптамер связывается со второй мишенью, тем самым делая первый фермент каталитически активным.
Вариант реализации 183. Композиция по варианту реализации 182, в которой:
- первый фермент представляет собой любой из следующих: апта-ДНКзим, апта-рибозим, апта-МНКзим.
Вариант реализации 184. Композиция по любому из вариантов реализации 138-179, в которой:
- вторая мишень представляет собой аналит, белок, соединение или молекулу;
- одноцепочечная петлевая область интактного олигонуклеотида стебель-петля является субстратом для аптазима; и
- композиция дополнительно содержит аптазим, содержащий часть аптамера, способную связываться со второй мишенью, и часть фермента нуклеиновой кислоты, способную расщеплять один или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля с образованием, таким образом, разделенного олигонуклеотида стебель-петля.
Вариант реализации 185. Композиция по варианту реализации 184, в которой:
- вторая мишень связана с аптамерной областью аптазима, а одноцепочечную петлевую область интактного олигонуклеотида стебель-петля гибридизуется с активной областью фермента нуклеиновой кислоты за счет комплементарного спаривания оснований, облегчая расщепление одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля с образованием разделенного олигонуклеотида стебель-петля.
Вариант реализации 186. Композиция по любому из вариантов реализации 138-179, в которой:
- вторая мишень представляет собой последовательность нуклеиновой кислоты; и
- первый фермент представляет собой первую рестрикционную эндонуклеазу, и
- вторая мишень гибридизуется с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований с образованием двухцепочечной последовательности для связывания первой рестрикционной эндонуклеазы с одним или более негибридизированными нуклеотидами интактного олигонуклеотид стебель-петля с образованием разделенного олигонуклеотида стебель-петля.
Вариант реализации 187. Композиция по варианту реализации 186, в которой:
- первая рестрикционная эндонуклеаза представляет собой эндонуклеазу первого разрыва, способную связываться и расщеплять цепь указанной двухцепочечной последовательности для первой рестрикционной эндонуклеазы, и указанная цепь содержит интактный олигонуклеотид стебель-петля.
Вариант реализации 188. Композиция по любому из вариантов реализации 138-179, в которой:
- первый фермент включает полимеразу с экзонуклеазной активностью,
- вторую мишень гибридизуют с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований с образованием первой двухцепочечной последовательности, содержащей область второй мишени,
- композиция дополнительно содержит первый праймер-олигонуклеотид, гибрид изо ванный за счет комплементарного спаривания оснований со второй мишенью с образованием второй двухцепочечной последовательности, расположенной выше относительно первой двухцепочечной последовательности, содержащей область второй мишени, и
- праймер можно удлинить, используя полимеразу с экзонуклеазной активностью и вторую мишень в качестве темплатной последовательности, расщепляя одноцепочечную петлевую область первой двухцепочечной последовательности и тем самым образуя разделенный олигонуклеотид стебель-петля.
Вариант реализации 189. Композиция по любому из вариантов реализации 138-179, в которой:
- первый фермент представляет собой экзонуклеазу, и
- вторая мишень гибридизуется путем комплементарного спаривания оснований с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля, образуя первую двухцепочечную последовательность, содержащую область второй мишени, с которой может ассоциироваться первый фермент, обладающий экзонуклеазной активностью, и таким образом расщепляют часть одноцепочечной петлевой области первой двухцепочечной последовательности, содержащей вторую мишень, с образованием разделенного олигонуклеотида стебель-петля.
Вариант реализации 190. Композиция по любому из вариантов реализации 138-179, в которой:
- первый фермент представляет собой ДНКзим или рибозим, требующий кофактор для каталитической активности, и
- вторая мишень является кофактором и связана с ДНКзимом или рибозимом,
- ДНКазим или рибозим гибридизуется с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля за счет комплементарного спаривания оснований, что позволяет ему расщеплять один или более негибридизованных нуклеотидов одноцепочечной петлевой области интактного олигонуклеотида стебель-петля и тем самым образуют разделенный олигонуклеотид стебель-петля.
Вариант реализации 191. Композиция по варианту реализации 190, в которой кофактор представляет собой ион металла или ион металла, выбранный из: Mg2+, Mn2+, Са2+, Pb2+.
Вариант реализации 192. Композиция по любому из вариантов реализации 138-150, 153, 156-158, 166 или 167, в которой:
- первый олигонуклеотид выбран из любого одного или более из: Молекулярный Маяк®, праймера Scorpion®, праймера TaqMan® или субстрата МНКзима.
Вариант реализации 193. Композиция по любому из вариантов реализации 138-192, в которой:
- первая мишень и/или вторая мишень представляет собой ампликон нуклеиновой кислоты.
Вариант реализации 194. Способ определения наличия или отсутствия первой и второй мишеней в образце, причем указанный способ включает:
(а) получение реакционной смеси путем приведения образца или его производного, предположительно содержащего первую или вторую мишени, или их ампликоны, с:
- первым олигонуклеотидом для обнаружения первой мишени или ее ампликона, и содержащий первый детектирующий фрагмент, способный генерировать первый детектируемый сигнал;
- интактным олигонуклеотидом стебель-петля для обнаружения второй мишени или ее ампликона, содержащий двуцепочечную стеблевую область, образованную гибридизированными нуклеотидами, противоположные цепи которого связаны неразорванной одноцепочечной петлевой областью, образованной негибридизованными нуклеотидами, при этом стеблевая область содержит второй детектирующий фрагмент, способный генерировать второй детектируемый сигнал,
при этом первый и второй детектируемые сигналы нельзя различить при одной температуре с применением одного типа детектора; и
- первым ферментом, способным расщеплять один или более негибридизированных нуклеотидов интактного олигонуклеотида стебель-петля только тогда, когда в образце присутствует вторая мишень или ее ампликон;
(b) обработку смеси в условиях, подходящих для:
- первой мишени или ее ампликону индуцировать модификацию первого олигонуклеотида, тем самым позволяя первому детектирующему фрагменту генерировать первый детектируемый сигнал,
- расщепления одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля первым ферментом только тогда, когда в образце присутствует вторая мишень или ее ампликон, с тем чтобы разрушить одноцепочечную петлевую область и получить разделенный олигонуклеотид стебель-петля;
(c) измерение:
- первого фонового сигнала на уровне или в пределах 5°С от первой температуры и второго фонового сигнала на уровне или в пределах 5°С от второй температуры;
- третьего фонового сигнала при третьей температуре; обеспечивается первым и вторым детектирующими фрагментами в смеси или в контрольной смеси;
(d) определение в один или более моментов времени во время или после указанной обработки:
- если первый детектируемый сигнал генерируется при первой температуре, которая отличается от первого или третьего фонового сигнала, при этом:
- второй детектируемый сигнал, генерируемый при первой температуре, не отличается от первого или третьего фонового сигнала, и
обнаружение различия между первым детектируемым сигналом и первым или третьим фоновым сигналом указывает на указанную модификацию первого олигонуклеотида и присутствие в образце первой мишени или ее ампликона; и
- если второй детектируемый сигнал генерируется при второй температуре, которая отличается от второго или третьего фонового сигнала, при этом при второй температуре:
цепи двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля диссоциируют, позволяя второму детектирующему фрагменту обеспечивать второй детектируемый сигнал, указывающий на присутствие второй мишени или ее ампликона в образце; и
цепи двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля не могут диссоциировать, тем самым обеспечивая подавление второго обнаружимого фрагмента и отсутствие второго детектируемого сигнала, указывающего на отсутствие второй мишени в образце.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Далее будут описаны предпочтительные варианты реализации настоящего изобретения будут описаны исключительно в качестве примера со ссылкой на прилагаемые фигуры, представленные ниже.
На Фиг. 1 проиллюстрированы типовой репортер LOCS и его температуры плавления (Tm) в интактной и разделенной конформациях. Репортер LOCS, как показано в качестве примера, можно использовать в сочетании с различными стандартными репортерными зондами и субстратами, хорошо известными в данной области техники для обнаружения нуклеиновых кислот.Типовые интактные репортеры LOCS (Фиг. 1А, LHS; вверху и внизу) имеют петлевую область, которая может быть расщеплена или разрушена, стеблевую область и детектирующий фрагмент, например, пару красителей «флуорофор (F) - гаситель (Q)». Расщепление или разрушение петлевой области в присутствии мишени может привести к образованию разщепленных репортерных структур LOCS (Фиг. 1В, RHS; вверху и внизу). Температуры плавления стеблевых областей интактного LOCS (Tm А) выше, чем Tm стеблевых областей разделенного LOCS (Tm В). Таким образом, стебель интактного LOCS будет плавиться и разделяться при температурах, равных или превышающих Tm А. Напротив, стебель, удерживающий два фрагмента разделенных LOCS, будет плавиться и разделяться при температурах, равных или превышающих Tm В, что приводит к усилению флуоресценции.
На Фиг. 2 показана типичная стратегия обнаружения мишени с применением LOCS-олигонуклеотидов, которые являются универсальными и могут использоваться для обнаружения любой мишени. В этой схеме LOCS-олигонуклеотид содержит стеблевую область, пару красителей «флуорофор - гаситель» и петлевую область. Петлевая область содержит универсальный субстрат для нуклеиновой кислоты, с каталитической активностью, например, МНКзима, также известного в данной области техники как PlexZyme. МНКзимы образуются, когда сенсорные плечи мишени компонента партзимов выравниваются рядом с мишенью. Петлевая область LOCS-олигонуклеотида связывается с ответвлениями связывания субстрата собранного МНКзима, и субстрат внутри петли LOCS расщепляется МНКзимом с образованием структуры разделенного LOCS. Как интактный LOCS, так и разделенный LOCS будут либо гаситься, либо вызывать флуоресценцию, в зависимости от того, выше или ниже температура реакционной среды температуры плавления их стеблей, а именно Tm А и Tm В соответственно. Наличие флуоресценции при температурах между Tm В и Tm А свидетельствует о присутствии мишени, которая облегчает расщепление. Мишень может быть обнаружена напрямую или могут быть обнаружены ампликоны мишеней, полученные с помощью протоколов амплификации мишени.
На Фиг. 3 показан пример стратегии для предпочтительного варианта реализации настоящего изобретения, в котором линейный субстрат МНКзима используется в сочетании с одним зондом LOCS, содержащим субстрат МНКзима в своей петле. Субстрат линейного МНКзима и одиночный зонд LOCS помечены одними и теми же детектирующими фрагментами, например парой специфических красителей «флуорофор (Р)/гаситель (Q)». Линейный субстрат содержит первую последовательность субстрата, расщепляемую первым МНКзимом, который собирается в присутствии первой мишени 1 (Фиг. 3А). В присутствии мишени 1 линейный субстрат расщепляется, что приводит к увеличению флуоресценции, которое можно обнаружить при всех температурах. Зонд LOCS содержит вторую последовательность субстрата в своей петле, которая расщепляется вторым МНКзимом, который собирается в присутствии второй мишени (Фиг. 3В). В присутствии мишени 2 субстрат LOCS расщепляется с образованием разделенного LOCS, который плавится при температуре Tm В, которая ниже температуры плавления интактного LOCS (Tm А). При температурах ниже Tm В стеблевые части разделенного LOCS остаются гибридизованными (закрытыми) и, следовательно, остаются погашенными. При температурах выше Tm В стеблевые части разделенного LOCS диссоциируют (разделяются) и флуоресценция увеличивается. Когда обе мишени присутствуют и флуоресценция измеряется при температурах ниже Tm В, увеличение флуоресценции связано только с мишенью 1; когда флуоресценция измеряется при температурах выше Tm В, но ниже Tm А, увеличение флуоресценции может быть связано с мишенями 1 и/или мишенями 2.
На Фиг. 4 показаны типичные стратегии обнаружения мишени с применением олигонуклеотидов LOCS, специфичных для мишени, которые можно использовать в сочетании с другими типами репортерных зондов или субстратов, такими как стандартные TaqMan, молекулярные маяки, унизонды Scorpion, бизонды Scorpion или линейные субстратные зонды МНКзима. Интактные олигонуклеотиды LOCS могут содержать стеблевую область, пару красителей «флуорофор-гаситель» и петлевую область, которая содержит область, комплементарную ампликону-мишени. В схеме, показанной на Фиг. 4А, петлевая область о лиго нуклеотидов LOCS комплементарна и связывается с ампликонами-мишенями во время амплификации. Во время удлинения праймеров экзонуклеазная активность полимеразы разрушает петлевую область, но оставляет интактной стеблевую область. Стеблевые области комплементарны друг другу, но не своей мишени. Разложение приводит к расщепленному LOCS, при котором стебель остается гибридизированным и погашенным при температуре ниже Tm стебля. Когда температура поднимается выше Tm стебля, цепи разделяются, и может возникать флуоресценция. На схеме, показанной на Фиг. 4В, петлевая область олигонуклеотида LOCS содержит область, комплементарную ампликону-мишени, и дополнительно содержит сайт распознавания для фермента рестрикции, например, никирующего фермента. Петля олигонуклеотида LOCS связывается с мишенью, и фермент расщепляет петлю, оставляя молекулу-мишень интактной. Это разделяет LOCS, и флуоресцентный сигнал излучается при температуре выше температуры плавления стебля. При более низких температурах стеблевые области разделенной структуры LOCS могут отжигать и гасить флуоресценцию. Стратегия может быть использована для непосредственного обнаружения целевых последовательностей или может обнаруживать ампликоны мишеней в сочетании с способом амплификации мишени.
На Фиг. 5 показаны варианты реализации, в которых нерасщепляемый молекулярный маяк можно комбинировать с зондом LOCS, расщепляемым МНКзимом. И молекулярный маяк, и зонд LOCS могут быть помечены одним и тем же флуорофором. Молекулярный маяк может иметь стеблевую область с Tm А и петлевую область, которая может специфически гибридизоваться с первой мишенью 1 с Tm В; где Tm В больше, чем Tm А. Его можно комбинировать с интактным зондом LOCS, который может иметь стеблевую область с Tm С и петлевую область, которая может быть расщеплена МНКзимом в присутствии второй мишени 2, таким образом, генерируя расщепление. LOCS с Tm D; где Tm D меньше, чем Tm С.Присутствие мишени 1 и/или мишени 2 можно различить путем измерения флуоресценции при двух температурах либо в реальном времени; либо с помощью дискретных измерений, полученных при или вблизи, начала усиления и последующего усиления.
На Фиг. 6 проиллюстрированы типовые кривые амплификации ПЦР для количественного анализа первой мишени 1 (CTcry) при первой температуре в присутствии или в отсутствие различных концентраций второй мишени 2 (NGopa). Протокол сочетал один линейный субстрат МНКзим (для мишени 1) с одним репортером LOCS (для мишени 2). Результаты были получены в канале HEX для количественного определения CTcry (мишень 1) при температуре регистрации 52°С для реакций, содержащих 20000 (черная точка), 4000 (черный штрих), 800 (черный квадрат), 160 (серое сплошное пятно) или 32 (серая точка) копии темплата CTry либо отдельно (Фиг. 6А), либо на фоне 20000 (Фиг. 6В), либо 32 (Фиг. 6С) копий NGopa (мишень 2). Данные флуоресценции при 52°С были также собраны для реакций без CTcry, но содержащих либо 20000 (черная линия), либо 32 копии (серая линия) темплата NGopa (Фиг. 6D). Нецелевые контроли (Н2О без нуклеазы) показаны на Фиг. 6А-6С (черная сплошная линия). Кривые амплификации представляют собой средние значения уровня флуоресценции из трех повторных реакций.
На Фиг. 7 показано одновременное качественное обнаружение мишени 1 (CTcry) и/или мишени 2 (NGopa) при двух температурах (D1 и D2) с применением способа анализа конечных точек 1 в канале HEX. Представленные результаты представляют собой средние значения для трех повторных реакций, а планки погрешностей представляют собой стандартное отклонение между этими повторами. В частности, данные показывают изменение флуоресцентного сигнала, когда один линейный субстрат и один зонд LOCS позволяют обнаруживать и различать CTcry (количество копий СТ 20K, 4, 800, 160, 32 или 0) и/или NGopa (количество копий NG 20K, 32 или 0) мишеней соответственно, которые присутствовали на фоне геномной ДНК человека. Кроме того, присутствие гена TFRC человека в геномной ДНК измеряли в канале техасского красного для использования в качестве калибратора. На Фиг. 7А показано изменение сигнала (ΔS; где ΔS=SD-пост-ПЦР-SD-до-ПЦР). ΔS при температуре 1 (ΔSD1; 52°С) показан черно-белым паттерном, a ΔS при температуре 2 (ΔSD2; 70°С) показан серым паттерном. Результаты показывают, что сигнал при температуре 1 (ΔSD1; черные столбцы) пересекает порог 1 (X1), когда в образце присутствует CTcry, но не пересекает этот порог, когда присутствует только NGopa. Таким образом, ΔSD1, превышающее пороговое значение 1 при температуре 1, указывает на наличие первой мишени (CTcry). Результаты на Фиг. 7А также показано, что, когда изменение сигнала при температуре 2 (ΔSD2; серые столбцы) больше, чем при температуре 1 (ΔSD2>ΔSD1), и превышает пороговое значение X1 (ΔSD2>X1), то в образце присутствует NGopa. Результаты на Фиг. 7В и 7С демонстрируют применение сигнала калибратора для калибровки ΔSD1 и ΔSD2. На Фиг. 7В показано изменение сигнала калибратора TFRC (ΔС), измеренного в канале техасского красного, причем значения, превышающие пороговое значение С, указывают на положительный сигнал для ΔС, а значения ниже порогового значения С указывают на отрицательный сигнал для ΔC (NTC). Темплат-калибратор (TFRC) присутствовала в геномной ДНК человека на фоне всех образцов, за исключением нецелевого контроля (NF Н2О). На Фиг. 7С показано изменение сигнала при каждой температуре, откалиброванной по ΔC (ΔS/ΔС). Изменение сигнала при температуре 1 (ΔSD1/ΔC; 52°С) показано черно-белым паттерном, а изменение сигнала при температуре 2 (ΔSD2/ΔC; 70°С) показано серым цветом, где результаты были получены для реакций, положительных для ΔС на Фиг. 7В, но не для реакций, отрицательных для ΔC (обозначенных как «Не применимо» (Н/П)). Кроме того, данные на Фиг. 7С демонстрируют, что калибровка сигналов не изменяет результаты, полученные с применением способа анализа конечных точек 1 (Фиг. 7А), поскольку картина является последовательной.
На Фиг. 8 показано одновременное качественное обнаружение первой мишени (CTcry) и/или второй мишени (NGopa) при двух температурах с применением способа анализа 2. CTcry (количество копий СТ 20K, 4, 800, 160, 32 или 0) и/или NGopa (количество копий NG 20K, 32 или 0) присутствовали на фоне геномной ДНК человека. Данные демонстрируют разницу в изменении флуоресцентного сигнала, полученного в канале HEX при температуре 2 (D2; 70°С) и температуре 1 (D1; 52°С) с применением способа анализа 2 (ΔSD2 минус ΔSD1=AΔSD2ΔSD1). Результаты показывают, что AΔSD2ΔSD1 пересекает порог 2 (Х2), когда в образце присутствует NGopa, но не пересекает этот порог, когда присутствует только CTcry и/или когда NGopa отсутствует (NTC) в образце. Следовательно, разница в изменении флуоресцентного сигнала выше порога 2 (ΔΔSD2ΔSD1>Х2) свидетельствует о наличии NGopa.
На Фиг. 9 показано изменение флуоресцентного сигнала (ΔS), полученного в ходе ПЦР в канале HEX при двух различных температурах (52°С и 70°С) в присутствии первой мишени (CTcry) и/или второй мишени (NGopa) с применением способа анализа конечной точки 3. CTcry (количество копий СТ 20K, 4, 800, 160, 32 или 0) и/или NGopa (количество копий NG 20K, 32 или 0) присутствовали на фоне геномной ДНК человека. На Фиг. 9А показано ΔS при температуре 2 (ΔSD2). Когда ΔSD2 больше порогового значения X1, это указывает на наличие в образце CTcry и/или NGopa. Когда ΔSD2 ниже порога X1, это указывает на отсутствие в реакции как CTcry, так и NGopa. На Фиг. 9В показано отношение ΔSD1:ΔSD2, которое используется для указания того, какие мишени присутствуют в реакции. Когда отношение выше, чем пороговое значение R1, это указывает на наличие CTcry, но не NGopa. Когда отношение ниже порогового значения R2, это указывает на наличие NGopa, но не CTcry. Когда соотношение находится между пороговыми значениями R1 и R2, это указывает на наличие как CTcry, так и NGopa. Когда в реакции не присутствуют ни CTcry, ни NGopa (Фиг. 9А), необходимость расчета соотношения отменяется и обозначается как N/A, как показано на Фиг. 9В.
На Фиг. 10 показаны кривые ПЦР-амплификации, полученные при 52°С в каналах HEX (A-D) и FAM (Е-Н) для различных мишеней. Кривые ПЦР, показанные пунктирной линией, представляют собой присутствие одного гена-мишени на реакцию, тогда как кривые, показанные сплошной линией, представляют присутствие обоих генов-мишеней на реакцию с 20000 копиями (черная линия) и 32 копиями (серая линия) мишени. В канале HEX показаны результаты только для CTcry (А), CTcry и NGopa (В), только NGopa (С) и всех остальных нецелевых контролей, включая 10000 копий TFRC (эндогенный контроль геномной ДНК), TVbtub и MgPa (D). В канале FAM результаты показаны только для TVbtub (Е), TVbtub и MgPa (F), только MgPa (G) и всех остальных нецелевых контролей, включая 10000 копий TFRC (эндогенный контроль геномной ДНК) или 20000 копий и 32 копии CTcry и NGopa (Н). На всех фигурах черная пунктирная линия представляет собой нецелевые контроли (NF Н2О). Кривые амплификации представляют собой средние значения уровня флуоресценции из трех повторных реакций.
На Фиг. 11 показано изменение сигнала (ΔS; где ΔS=Sd-пост-пцр-Sd-до-пцр) для обнаружения (А) CTcry и NGopa в HEX и (В) TVbtub и MgPa в FAM с применением способа анализа конечной точки 1. Результаты представлены черно-белым паттернами для ΔSD1 (52°С) и серым цветом для ΔSD2 (70°С). Значения для каждого образца представляют собой среднее значение трех повторов, а планки погрешностей представляют собой стандартное отклонение между этими повторами.
На Фиг. 12 показано качественное определение конечной точки NGopa в канале HEX (А) и MgPa в канале FAM (В) с применением способа анализа конечных точек 2 (ΔSD2-ΔSD1=AΔSD2ΔSD1). Значения для каждого образца представляют собой средние значения значение трех повторов, а планки погрешностей представляют собой стандартное отклонение между этими повторами.
На Фиг. 13 показаны изменения флуоресцентного сигнала, полученные в каналах HEX (Фиг. 13А и 13В) и FAM (Фиг. 13С и 13D) при двух разных температурах с применением способа анализа конечной точки 3. (Фиг. 13А) Обнаружение CTcry (СТ; 20000 (20 т) или 32 копии) и/или NGopa (NG; 20000 (20 т) или 32 копии) с применением ΔSD2B НЕХ-канале (Фиг. 13В) Дифференциация CTcry и NGopa в HEX с применением соотношения ΔSD1:ΔSD2- Обнаружение только CTcry определяется, когда ΔSD1:ΔSD2>Порог R1. Обнаружение только NGopa определяется, когда ΔSD1:ΔSD2<Порог R2. Обнаружение коинфекции, содержащей обе мишени, определяется, когда ΔSD1:ΔSD2>порога R2 и<порога R1 (Фиг. 13С) Обнаружение TVbtub и/или MgPa с применением ΔSD2B канале FAM (Фиг. 13D). Дифференциация TVbtub и MgPa в FAM с применением соотношения ΔSD1:ΔSD2- Обнаружение только TVbtub определяется, когда ΔSD1:ΔSD2>Порог R1 Обнаружение только MgPa определяется, когда ΔSD1:ΔSD2<Порог R2. Обнаружение коинфекции, содержащей обе мишени, определяется, когда ΔSD1:ΔSD2>порога R2 и<Порог R1 Значения для каждого образца представляют собой средние значения значение трех повторов, а планки погрешностей представляют собой стандартное отклонение между этими повторами.
На Фиг. 14 проиллюстрировано обнаружение TFRC при температуре 1 (D1) (Фиг. 14А) и TPApolA при температуре 2 (D2) (Фиг. 14В) в Су5.5. канале с применением способа анализа конечных точек 2.1 Обнаружение TFRC (Фиг. 14А) достигается путем вычитания флуоресценции до ПЦР при температуре 1 из флуоресценции после ПЦР при температуре 1 (SD1). Обнаружение TPApolA (Фиг. 14В) достигается путем вычитания флуоресценции до ПЦР при температуре 2 из флуоресценции после ПЦР при температуре 2 (ΔSD2) с последующим вычитанием ΔSD1 (AΔSD2ΔSD1). Значения для каждого образца представляют собой средние значения двух повторов для NF Н2О, 10000 копий TPApolA, 40 копий TPApolA, 10000 копий TPApolA и 10000 копий TFRC, 40 копий TPApolA и 10000 копий TFRC. Значения для 10000 копий TFRC являются средними для 48 повторов, поскольку TFRC использовался в качестве эндогенного контроля и присутствует в геномной ДНК при 10000 копий в каждой реакционной лунке, за исключением образцов, содержащих только TPApolA. Столбики погрешностей представляют собой стандартное отклонение между каждой повторностью для каждого образца.
На Фиг. 15 (Фиг. 15А) продемонстрирована сигнатура кривых плавления, полученная расщеплением субстрата 4 в присутствии 10000 копий TFRC (сплошная черная линия). (Фиг. 15В) продемонстрирована сигнатура кривых плавления, полученная расщеплением LOCS-3 в присутствии 10000 копий TPApolA (сплошная черная линия) и 40 копий TPApolA (сплошная серая линия). (С) Сигнатура кривых плавления, полученная расщеплением субстрата 4 и LOCS-3 в присутствии 10000 копий TPApolA плюс 10000 копий TFRC (сплошная черная линия) и 40 копий TPApolA плюс 10000 копий TFRC (сплошная серая линия). Сигнатура кривых плавленияв результате отсутствия обеих мишеней (NF Н2О) представлена на (Фиг. 15А-С) пунктирной линией. Результаты представляют скорость изменения флуоресценции в зависимости от температуры (-d(RFU)/dT).
На Фиг. 16 показаны графики ПЦР-амплификации, полученные для реакций, содержащих 20000 копий CTcry (сплошная черная линия), 20000 копий NGopa (черная пунктирная линия), 20000 копий обеих мишеней (сплошная серая линия) или ни одной мишени (NTC; серая пунктирная линия) при 39°С (А) и 72°С (В). Пороговые значения Порог X и Порог Y указаны для графиков амплификации, полученных при 39°С и 72°С соответственно. Указаны конечные значения флуоресценции, обозначенные Ех1, EX2, Ey1 и EY2.
На Фиг. 17 проиллюстрированы графики ПЦР-амплификации, полученные из реакций, содержащих копии мишени Х/CTcry; а именно либо 0 копий (сплошная черная линия), 32 копии (серая линия) или 20000 копий CTcry (черная пунктирная линия) на фоне целевого объекта Y/NGopa с различными номерами копий, как указано. На графиках A, D и G продемонстрирована флуоресценция при 39°С для реакций, содержащих NGopa в количестве 20000 копий (А), 32 копий (D) или без копий (G). На графиках В, Е и Н продемонстрирована флуоресценция при 74°С для реакции, содержащей NGopa, в количестве 20000 копий (В), 32 копий (Е) или без копий (Н). На графиках С, F и I продемонстрирована флуоресценция при 74°С после нормализации с помощью FΔF для реакций, содержащих NGopa, в количестве 20000 копий (С), 32 копий (F) или без копий (I).
На Фиг. 18 продемонстрированы кривые ПЦР-амплификации для количественного определения GAPDH человека при (D1) 52°С в канале FAM. (Фиг. 18А) Кривые представляют сигнал, полученный только от 10000 копий (серая сплошная линия) и 100 копий (черная пунктирная линия) мишени GAPDH (Фиг. 18В) Результаты получены только для 10000 копий (сплошная серая линия) и 100 копий (черная пунктирная линия) мишени MgPa. (Фиг. 18С) Кривые представляют сигнал, полученный от целевых смесей, содержащих 10000 копий (серая сплошная линия) и 100 копий (черная пунктирная линия) каждого из GAPDH и MgPa. Нецелевой контроль (NF Н2О) представлен на (Фиг. 18А-С) черной сплошной линией. Кривые амплификации представляют собой средние значения уровня флуоресценции из трех повторных реакций.
На Фиг. 19 показано качественное определение GAPDH и MgPa с применением одного зонда TaqMan и одного зонда LOCS соответственно при двух температурах в канале FAM. Результаты были получены с применением способов анализа конечных точек 1-3. Значения для каждого образца представляют собой среднее значение трех повторов, а планки погрешностей представляют собой стандартное отклонение между этими повторами. (Фиг. 19А). Результаты, полученные с помощью способа 1 анализа конечной точки, показывают изменение сигнала (ΔS) между флуоресцентными измерениями после ПЦР и до ПЦР. Результаты представлены черно-белым паттернами для ΔSD1 (52°С) и серым цветом для ΔSD2 (70°С). (Фиг. 19В) Обнаружение только MgPa с результатами, полученными способом анализа конечных точек 2 (AΔSD2ΔSD1). (Фиг. 19С и 19D). Результаты, полученные способом анализа конечных точек 3 (ΔSD1:ΔSD2)- (Фиг. 19С) Обнаружение GAPDH (10000 или 100 копий человека) и/или MgPa (MG 10000 или 100 копий) с применением ΔSD2B канале FAM (Фиг. 19D). Дифференциация GAPDH и MgPa в FAM с применением соотношения ΔSD1:ΔSD2. Обнаружение только GAPDH определяется, когда ΔSD1:ΔSD2>Порог R1 Обнаружение только MgPa определяется, когда ΔSD1:ΔSD2<Порог Pv2. Обнаружение коинфекции, содержащей обе мишени, определяется, когда ΔSD1: ΔSD2>порога R2 и<порога R1 Результаты измеряются в единицах соотношения.
На Фиг. 20 представлены кривые ПЦР-амплификации, полученные в канале FAM для реакций, содержащих либо 25600 копий TVbtub (черная пунктирная линия), 25600 копий MgPa (черная пунктирная линия), либо смесь, содержащую 25600 копий обеих мишеней (серая сплошная линия), либо отсутствие мишени (NF H2O; черная сплошная линия) при 52°С (Фиг. 20А) и 74°С (Фиг. 20В). Увеличение флуоресценции при 52°С (D1) указывает на присутствие TVbtub, обнаруженное с помощью молекулярного маяка, а увеличение флуоресценции при 74°С (D2) указывает на присутствие MgPa, обнаруженное с помощью зонда LOCS. Значения Cq, определенные при D1 и D2, использовались для количественного определения количества TVbtub и MgPa соответственно в образце без необходимости использования специальных способов анализа. Кривые представляют средний уровень флуоресценции из трех повторных реакций.
На Фиг. 21 показана стандартная кривая, полученная при 52°С (D1), которая использовалась для количественного определения TVbtub (Фиг. 20А), и стандартная кривая, полученная при 74°С (D2), которая использовалась для количественного определения MgPa (Фиг. 20В). Для построения стандартных кривых использовали три копии синтетических шаблонов TVbtub и MgPa G-Block в количестве 25600, 6400, 1600, 400 и 100 копий.
На Фиг. 22 показаны варианты реализации, в которых зонд для двойной гибридизации может быть объединен с зондом LOCS, который может расщепляться МНКзимом. И зонд двойной гибридизации, и зонд LOCS могут быть помечены одним и тем же флуорофором. Два зонда двойной гибридизации могут быть способны связываться с мишенью 1 с Tm А и Tm В соответственно. Они могут быть скомбинированы с интактным зондом LOCS, который может иметь стеблевую область с Tm С и петлевую область, которая может быть расщеплена МНКзимом в присутствии второй мишени 2, таким образом, генерируя расщепление. LOCS с Tm D; где Tm D меньше, чем Tm С.Присутствие мишени 1 и/или мишени 2 можно определить путем измерения флуоресценции при двух температурах либо в реальном времени; либо с помощью дискретных измерений, полученных при или вблизи, начала усиления и последующего усиления.
На Фиг. 23 показано одновременное обнаружение по конечной точке двух мишеней (TVbtub и MgPa) в канале FAM с применением одного нерасщепляемого молекулярного маяка и одного зонда LOCS путем измерения изменения флуоресценции во время ПЦР при 52°С (ΔSD1, Фиг. 23А) и 74°С (ΔSD2, Фиг. 23В). Нанесенные на график значения представляют собой среднее значение трех повторных реакций, содержащих различные количества TVbtub и/или MgPa, как указано на графике. На Фиг. 23А, ΔSD1 выше указанного порога указывает на присутствие TVbtub в реакции или на отсутствие, если он ниже. На Фиг. 23В, ΔSD2 Выше указанного порога указывает на присутствие MgPa в реакции или на отсутствие, если он ниже. Ось у представляет собой определенное увеличение флуоресценции при 52°С (ΔSD1, Фиг. 23А) или 74°С (ΔSD2, Фиг. 23В).
На Фиг. 24 показано одновременное обнаружение по конечной точке двух мишеней (TVbtub и MgPa) в канале FAM с применением одного нерасщепляемого молекулярного маяка и одного зонда LOCS путем измерения изменения калиброванного флуоресцентного сигнала во время ПЦР при 52°С (ΔSD1/C, Фиг. 24А) и 74°С (ΔSD2/C, Фиг. 24В). Нанесенные на график значения представляют собой среднее значение повторов, содержащих различные количества шаблонов TVbtub и/или MgPa, как указано на графике, а столбцы погрешностей представляют собой стандартное отклонение между этими повторами. На Фиг. 24А ΔSD1/C выше порога 1 указывает на присутствие TVbtub в реакции или на отсутствие, если он ниже порога. На Фиг. 24В ΔSD2/C выше порога 2 указывает на присутствие MgPa в реакции или на отсутствие, если он ниже порога.
На Фиг. 25 показано одновременное обнаружение по конечной точке двух мишеней (CTcry и NGopa) в канале HEX с применением одного линейного субстрата МНКзима и одного зонда LOCS путем измерения изменения сигнала флуоресценции во время ПЦР при 52°С (ΔSD1, Фиг. 25А) и 70°С (ΔSD2, Фиг. 25В) и калиброванные сигналы при 52°С (ΔSD1/C, Фиг. 25С) и 70°С (ΔSD2/C, Фиг. 25D) на трех протестированных машинах Bio-Rad CFX96 (Машина 1 в черных полосах, Машина 2В сером цвете и Машина 3 в белом). Нанесенные на график значения представляют собой среднее значение повторов, содержащих различные количества шаблонов CTcry и/или NGopa, как указано на графике, а столбцы погрешностей представляют собой стандартное отклонение между этими повторами. Сигнал на Фиг. 25А или откалиброванный сигнал на Фиг. 25С выше порога Ci указывает на присутствие CTry в реакции или отсутствие, если оно ниже. Калиброванный сигнал на Фиг. 25D указывает на отсутствие CTcry и NGopa при значениях ниже порога С2, присутствие как CTcry, так и NGopa при превышении порога Сз и присутствие только одного из CTcry и NGopa при нахождении между порогами С2 и С3. На Фиг. 25В показано, что значение порогового значения С3 варьируется между машинами без калибровки, в отличие от значения, показанного на Фиг. 25D (пороговое значение С2 не показано на Фиг. 25В).
На Фиг. 26 показано одновременное обнаружение конечной точки мишени 1 (CTcry) и/или мишени 2 (NGopa) в канале HEX с применением одного линейного субстрата МНКзима и одного зонда LOCS с применением способа 2 анализа конечной точки. Графики, иллюстрирующие NSm (LHS) и ΔNSD2NSD1 (RHS), были определены для обнаружения CTcry и обнаружения NGopa, соответственно, путем получения сигналов после ПЦР, полученных в D1 (52°С) и D2 (70°С) из экспериментальных образцов, и определения фоновые сигналы в виде измерений флуоресценции до ПЦР (SD3) из той же реакционной лунки при 40°С (Фиг. 26А-В); 52°С (Фиг. 26C-D) и 62°С (Фиг. 26E-F). На Фиг. 26А, 26С и 26Е, где NSD1 выше порогового значения X1, это указывает на присутствие CTry в реакции или отсутствие, если он ниже. На Фиг. 26В, 26D и 26F, где ΔNSD2NSd1 выше порогового значения Х2, это указывает на присутствие NGopa в реакции или отсутствие, если оно ниже.
На Фиг. 27 показано одновременное обнаружение по конечной точке двух мишеней (CTcry и NGopa) в канале HEX с применением одного линейного субстрата МНКзима и одного зонда LOCS с применением способа анализа конечных точек 2. NSD1 (LHS) и ΔNSD2NSD1 (RHS) определяли для обнаружения CTcry и обнаружения NGopa соответственно, принимая пост-ПЦР-сигналы, полученные при D1 (52°С) и D2 (70°С), и определяя фоновые сигналы в качестве измерения флуоресценции пре-ПЦР (SD3) как среднее значение контрольных сигналов отсутствия темплата, измеренных при D1/D3B до ПЦР (Фиг. 27А-В) и при D1 и D2 до ПЦР (Фиг. 27C-D) и после ПЦР (Фиг. 27E-F). На Фиг. 27А, 27С и 27Е, где NSD1 выше порогового значения X1, это указывает на присутствие CTry в реакции или отсутствие, если он ниже. На Фиг. 26В, 26D и 26F, где ΔNSD2NSD1 выше порогового значения Х2, это указывает на присутствие NGopa в реакции или отсутствие, если оно ниже.
Определения
В настоящей заявке формы единственного числа включают упоминание форм множественного числа, если иное явным образом не следует из контекста. Например, фраза «полинуклеотид» также включает множество полинуклеотидов.
В настоящем документе термин «содержащий» означает «включающий». Вариации слова «содержащий», например, «содержать» и «содержит», имеют соответственно различающиеся значения. Таким образом, например, полинуклеотид, «содержащий» последовательность нуклеотидов, может состоять исключительно из этой последовательности нуклеотидов или может содержать один или более из дополнительных нуклеотидов.
В настоящем документе термин «множество» означает более одного. В некоторых конкретных аспектах или вариантах реализации множество может означать 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51 или более, и любое целое число, производное от них, и любой диапазон, производный от них.
В настоящем документе термин «субъект» включает любое животное, имеющее важное экономическое, социальное или исследовательское значение, включая крупный рогатый скот, лошадей, овец, приматов, птиц и грызунов. Следовательно, «субъект» может являться млекопитающим, например, человеком, или млекопитающим, не являющимся человеком. Этот термин также охватывает субъектов-микроорганизмов, включая бактерии, вирусы, грибы/дрожжи, простейших и нематод, но не ограничиваясь ими. «Субъект» в соответствии с настоящим изобретением также включает инфекционные агенты, например, прионы.
В настоящем документе термины «полинуклеотид» и «нуклеиновая кислота» могут использоваться взаимозаменяемо и относятся к одно- или двуцепочечному полимеру дезоксирибонуклеотидных или рибонуклеотидных оснований или их аналогов, производных, вариантов, фрагментов или комбинаций, включая ДНК, метилированную ДНК, алкилированную ДНК, РНК, метилированную РНК, микроРНК, миРНК, кшРНК, мРНК, тРНК, мякРНК, мвРНК, ммРНК, предшественники микроРНК и первичные микроРНК, другие некодирующие РНК, рибосомную РНК, их производные, их ампликоны или любую их комбинацию, но не ограничиваясь ими. В качестве неограничивающего примера, источник нуклеиновой кислоты можно выбрать из группы, состоящей из синтетических соединений, млекопитающих, человека, животных, растений, грибов, бактерий, вирусов, архей или любой их комбинации.
В настоящем документе термин «олигонуклеотид» относится к сегменту ДНК или ДНК-содержащей молекуле нуклеиновой кислоты, или РНК, или РНК-содержащей молекуле, или их комбинации. Примеры олигонуклеотидов включают мишени нуклеиновой кислоты; субстраты, например такие, которые могут быть модифицированы МНКзимом; праймеры, такие как используемые для амплификации мишени in vitro такими способами, как ПЦР; компоненты МНКзимов; и различные другие типы репортерных зондов, включая, но не ограничиваясь ими, TaqMan или гидролизные зонды; молекулярные маяки; маяки Sloppy; зонды Eclipse; унизонд Scorpion, праймеры/зонды бизонд Scorpion, зонды Capture/Pitchers и зонды двойной гибридизации. Термин «олигонуклеотид» включает упоминание любой конкретной последовательности, а также последовательности, комплементарной ей, если не указано иное. Олигонуклеотиды могут содержать по меньшей мере одно добавление или замещение, включая группу, содержащую 4-ацетилцитидин, 5-(карбоксигидроксилметил)уридин, 2'-O-метилцитидин, 5-карбоксиметиламинометилтиоуридин, дигидроуридин, 2'-O-метилпсевдоуридин, бета-D-галактозилкеозин, 2'-O-метилгуанозин, инозин, N6-изопентениладенозин, 1-метиладенозин, 1-метилпсевдоуридин, 1-метил гуанозин, 1-метилинозин, 2,2-диметилгуанозин, 2-метиладенозин, 2-метилгуанозин, 2-метилгуанозин, 3-метилцитидин, 5-метилцитидин, N6-метиладенозин, 7-метил гуанозин, 5-метиламинометилуридин, 5-метоксиаминометил-2-тиоуридин, бета-О-маннозилметилуридин, 5-метоксикарбонилметилуридин, 5-метоксиуридин, 2-метилтио-N6-изопентениладенозин, N-((9-бета-рибофуранозил-2-метилтиопурин-6-ил)карбамоил)треонин, N-((9-бета-рибофуранозилпурин-6-ил)N-метилкарбамоил)треонин, метиловый эфир уридин-5-оксиуксусной кислоты, уридин-5-оксиуксусную кислоту (v), вибутоксозин, псевдоуридин, кеозин, 2-тиоцитидин, 5-метил-2-тиоуридин, 2-тиоуридин, 4-тиоуридин, 5-метилуридин, N-((9-бета-В-рибофуранозилпурин-6-ил)карбамоил)треонин, 2'-O-метил-5-метилуридин, 2-О-метилуридин, вибутозин, 3-(3-амино-3-карбоксипропил)уридин, бета-D-арабинозилуридин, бета-Э-арабинозилтимидин, но не ограничиваясь ими.
Термины «полинуклеотид» и «нуклеиновая кислота», «олигонуклеотид» включают ссылку на любую указанную последовательность, а также на комплементарную ей последовательность, если не указано иное.
В настоящем документе термины «комплементарный», «комплементарность», «совпадение» и «совпадающий» относятся к способности нуклеотидов (например, дезоксирибонуклеотидов, рибонуклеотидов или их комбинаций) гибридизоваться друг с другом посредством спаривания оснований Уотсона-Крика, неканонического спаривание оснований, включая спаривание оснований вобуляции и спаривание оснований Хугстина (например, LNA, PNA или BNA) или неестественное спаривание оснований (UBP). Связи могут образовываться посредством спаривания оснований по Уотсону-Крику между основаниями аденина (А) и основаниями урацила (U), между основаниями аденина (А) и основаниями тимина (Т), между основаниями цитозина (С) и основаниями гуанина (G). Пара оснований вобуляции представляет собой неканоническую пару оснований между двумя нуклеотидами в полинуклеотидном дуплексе (например, гуанин-урацил, инозин-урацил, инозин-аденин и инозин-цитозин). Пары оснований Хугстина представляют собой пары, которые, подобно парам оснований Уотсона-Крика, встречаются между основаниями аденина (А) и основаниями тимина (Т), а также основаниями цитозина (С) и основаниями гуанина (G), но с различной конформацией пурина по отношению к пиримидину по сравнению с парами оснований Уотсона-Крика. Неприродная пара оснований представляет собой искусственную субъединицу, синтезированную в лаборатории и не встречающуюся в природе. Нуклеотиды, обозначаемые как «комплементарные» или являющиеся «комплементарными» друг другу, обладают способностью гибридизоваться друг с другом либо за счет спаривания соответствующих оснований по Уотсону-Крику, либо за счет неоднозначного спаривания оснований, либо путем неканонического спаривания оснований (спаривание оснований колебательного типа, спаривание оснований Хугстина), либо путем неприродного спаривания оснований спаривание оснований (UBP) между их соответствующими основаниями. Последовательность нуклеотидов, которая является «комплементарной» к другой последовательности нуклеотидов в данном документе, может означать, что первая последовательность на 100% идентична комплементарной второй последовательности в области 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100 и более нуклеотидов. Ссылка на последовательность нуклеотидов, которая «по существу комплементарна» другой последовательности нуклеотидов в данном документе, может означать, что первая последовательность составляет по меньшей мере 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98% или 99% идентичных комплементарной второй последовательности в области 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100 или более нуклеотидов.
В настоящем документе термины «некомплементарный», «не комплементарный», «несоответствие» и «несоответствующий» относятся к нуклеотидам (например, дезоксирибонуклеотидам, рибо нуклеотидам и их комбинациям), не обладающим способностью гибридизоваться друг с другом за счет спаривания соответствующих оснований по Уотсону-Крику либо за счет неоднозначного спаривания оснований. Последовательность нуклеотидов, которая является «некомплементарной» другой последовательности нуклеотидов в данном документе, может означать, что первая последовательность на 0% идентична комплементарной второй последовательности в области 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100 и более нуклеотидов.
Ссылка на последовательность нуклеотидов, которая «по существу не комплементарна» другой последовательности нуклеотидов в данном документе, может означать, что первая последовательность менее чем на 1%, 2%, 3%, 4%, 5%, 10%, 15%, 20%, 25%, 30%, 35% или 40% идентична комплементарной второй последовательности в области 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100 или более нуклеотидов.
В настоящем документе термин «мишень» относится к любой молекуле или аналиту, присутствующему в образце, для обнаружения которого можно использовать способы по настоящему изобретению. Следует понимать, что термин «мишень» включает мишени из нуклеиновых кислот и мишени из ненуклеиновых кислот, такие как, например, белки, пептиды, аналиты, лиганды и ионы (например, ионы металлов).
В настоящем документе термин «фермент» относится к любой молекуле, способной катализировать химическую реакцию (например, амплификацию полинуклеотид а, расщепление полинуклеотид а и т.д.). Неограничивающие примеры ферментов, подходящих для использования в настоящем изобретении, включают ферменты нуклеиновых кислот и белковые ферменты. Неограничивающие примеры подходящих ферментов нуклеиновых кислот включают МНКзимы, ДНКзимы и аптазимы. Неограничивающие примеры подходящих белковых ферментов включают экзонуклеазы и эндонуклеазы. Ферменты обычно обеспечивают каталитическую активность, способствующую осуществлению одного или более способов, описанных в настоящем документе. В качестве неограничивающего примера, экзонуклеазная активность может представлять собой каталитическую активность, присущую, например, полимеразе. В качестве неограничивающего примера, эндонуклеазная активность может представлять собой каталитическую активность, присущую, например, ферменту рестрикции, включая никующую эндонуклеазу, рибоэндонуклеазу или нуклеазу, специфичную к двуцепочечной структуре (DSN).
В настоящем документе термин «ампликон» относится к нуклеиновой кислоте (например, ДНК или РНК или их комбинации), являющейся продуктом естественной или искусственной амплификации или репликации нуклеиновых кислот, включая, помимо прочего, ПЦР, ОТ-ПЦР, SDA, NEAR, HDA, RPA, LAMP, RCA, ТМА, LCR, RAM, 3SR, NASBA и любую их комбинацию.
В настоящем документе следует понимать, что термин «олигонуклеотид типа стебель-петля означает ДНК или молекулу, содержащую ДНК, или РНК, или молекулу, содержащую РНК, или их комбинацию (т.е. гибридную молекулу или комплекс ДНК-РНК), содержащую или состоящую из двуцепочечного стеблевого компонента, соединенного с одноцепочечным петлевым компонентом. Двуцепочечный стеблевой компонент содержит прямую цепь, гибридизованную за счет спаривания комплементарных оснований с комплементарной обратной цепью, причем 3'-нуклеотид прямой цепи соединен с 5'-нуклеотидом компонента одноцепочечной петли, а 3'-нуклеотид обратной цепи соединен с 3'-нуклеотидом компонента одноцепочечной петли. Двуцепочечный стеблевой компонент может дополнительно содержать один или более фрагментов обнаружения, включая, помимо прочего, флуорофор на одной цепи (например, прямой цепи) и один или более гасителей на противоположной цепи (например, обратной цепи).Другие неограничивающие примеры включают наночастицы золота или серебра на обеих цепях для колориметрического обнаружения, иммобилизацию одной цепи на поверхности золота (например, прямой цепи) и наночастицы золота на противоположной цепи (например, обратной цепи) для ППР-детекции, и иммобилизацию одной цепи на поверхности электрода (например, прямой цепи) и молекулы метиленового синего на противоположной цепи (например, обратной цепи) для электрохимического обнаружения.
В настоящем документе термины «олигонуклеотид стебель-петля» следует понимать как включающий «LOCS», также упоминаемый в настоящем документе как «LOCS-олигонуклеотид», «LOCS-структура», «LOCS-репортер», «интактный LOCS», «замкнутый LOCS» и «LOCS-зонды». Одноцепочечный петлевой компонент LOCS может содержать область, способную служить субстратом для нуклеиновой кислоты, с каталитической активностью, такой как, например, МНКзима, ДНКзима, рибозима, апта-МНКзима или аптазима. Дополнительно или альтернативно, одноцепочечный петлевой компонент может содержать область, комплементарную нуклеиновой кислоте-мишени (например, мишени для обнаружения, количественной оценки и т.п.) и/или полученным из нее ампликонам, которая может дополнительно служить в качестве субстрата для фермента-экзонуклеазы. В качестве неограничивающего примера, экзонуклеазная активность может быть присуща ферменту-полимеразе. В качестве дополнения или альтернативы, область одноцепочечного петлевого компонента может содержать область, которая может: (i) быть комплементарной мишени, подлежащей обнаружению, (ii) содержать одну цепь двуцепочечного сайта, распознаваемого ферментом рестрикции; и (iii) служить субстратом для фермента рестрикции, например, эндонуклеазы разрыва. В настоящем документе термины «раскрытый олигонуклеотид типа стебель-петля, «раскрытый LOCS», «раскрытый LOCS-олигонуклеотид», «раскрытая LOCS-структура», «раскрытые LOCS-репортеры», «раскрытые LOCS-зонды», «раскрытый LOCS», «расщепленный LOCS» и «разрушенный LOCS» используются взаимозаменяемо и относятся к «LOCS», в котором одноцепочечный петлевой компонент расщеплен и/или разрушен (например, ферментом, описанным в настоящем документе), так что по меньшей мере одна связь между соседними нуклеотидами в составе петли удалена, что приводит к образованию раскрытой структуры в петлевой области. В разделенном LOCS прямая и обратная цепи двуцепочечного стеблевой области могут сохранять способность гибридизироваться друг с другом с образованием стебля в зависимости от температуры.
LOCS предназначены для включения расщепляемой петлевой области, обеспечивающей зависимое от мишени расщепление петлевой области ферментом, генерирующим расщепленную LOCS. Это, в свою очередь, может облегчить обнаружение мишени по детектируемому сигналу, генерируемому при определенной температуре (температурах) после ассоциации (гибридизации) или диссоциации стеблевой части интактных или расщепленных LOCS. Напротив, молекулярный маяк, используемый в настоящем документе, относится к олигонуклеотиду стебель-петля, предназначенному для включения петлевой области, которая не расщепляется во время описанных в данном документе способов. Молекулярные маяки могут способствовать обнаружению цели, генерируя обнаруживаемый сигнал при определенных температурах после ассоциации (гибридизации) или диссоциации (разделения) петлевой части зонда с целью, которую необходимо обнаружить. Таким образом, основное различие между этими двумя типами петлевых стеблевых структур в контексте настоящего изобретения заключается в том, что LOCS отслеживают путем измерения изменений в сигналах из-за гибридизации или диссоциации стеблевой области интактных или расщепленных LOCS, тогда как молекулярные маяки являются отслеживают путем измерения изменений сигнала из-за гибридизации или диссоциации петлевой области и мишени.
В настоящем документе термин «универсальный стебель» относится к двуцепочечной последовательности, которую можно включить в любую LOCS-структуру. Один и тот же «универсальный стебель» можно использовать в LOCS, содержащих петли, содержащие субстраты каталитических нуклеиновых кислот либо последовательность, комплементарную рассматриваемой мишени. Один универсальный стебель можно использовать в качестве суррогатного маркера для любой мишени, способной облегчить раскрытие конкретного LOCS. Ряд универсальных стеблей можно внедрить в ряд LOCS, предназначенных для анализа любого набора мишеней.
В настоящем документе термин «универсальный LOCS» относится к LOCS-структуре, содержащей «универсальный стебель» и «универсальную петлю», содержащую универсальный субстрат нуклеиновой кислоты, с каталитической активностью, который может расщепить любой МНКзим с дополнительными субстрат-связывающими цепями независимо от последовательностей цепей-датчиков мишеней в составе МНКзима. Один универсальный LOCS можно использовать в качестве суррогатного маркера для любой мишени, способной облегчить раскрытие конкретного LOCS. Ряд универсальных LOCS можно внедрить в любой мультиплексный анализ, предназначенный для анализа любого набора мишеней.
Используемые в настоящем документе термины «фермент нуклеиновой кислоты», «каталитическая нуклеиновая кислота», «нуклеиновая кислота с каталитической активностью» и «фермент нуклеиновой кислоты, с каталитической активностью» используются в данном документе взаимозаменяемо и означают ДНК или ДНК-содержащую молекулу или комплекс, или РНК или РНК-содержащую молекулу или комплекс, или их комбинацию (т.е. гибридная молекула или комплекс ДНК-РНК), которые могут распознавать по меньшей мере один субстрат и катализировать модификацию (такую как расщепление) по меньшей мере одного субстрата. Нуклеотидные остатки в каталитических нуклеиновых кислотах могут содержать основания А, С, G, Т и U, а также их производные и аналоги. Вышеприведенные термины включают одномолекулярные нуклеиновые кислоты с ферментативной активностью, которые могут содержать одну ДНК или ДНК-содержащую молекулу (также известную в данной области техники как «ДНК-фермент», «дезоксирибозим» или «ДНКзим») или РНК, или молекулу, содержащую РНК (также известную в данной области техники как «рибозим») или их комбинацию, представляющую собой ДНК-РНК-гибридную молекулу, способную распознавать по меньшей мере один субстрат и катализировать модификацию (например, расщепление) по меньшей мере одного субстрата. Вышеприведенные термины включают нуклеиновые кислоты с ферментативной активностью, содержащие ДНК или ДНК-содержащий комплекс, или РНК, или РНК-содержащий комплекс, или их комбинацию, являющуюся ДНК-РНК-гибридным комплексом, способным распознавать по меньшей мере один субстрат и катализировать модификацию (например, расщепление) по меньшей мере одного субстрата. Термины «нуклеиновая кислота с ферментативной активностью», «каталитическая нуклеиновая кислота», «нуклеиновая кислота с каталитической активностью» и «каталитическая нуклеиновая кислота с ферментативной активностью» включают в своем значении МНКзимы.
В настоящем документе термины «МНКзим» и «многокомпонентный нуклеозим» имеют одинаковое значение и относятся к двум или более олигонуклеотидным последовательностям (например, партзимам), которые лишь в присутствии соединения, облегчающего сборку МНКзима (например, мишени), образуют активную нуклеиновую кислоту с ферментативной активностью, способную каталитически модифицировать субстрат.«МНКзим» также известен в данной области техники как «PlexZyme». МНКзимы могут катализировать ряд реакций, включая расщепление субстрата и другие ферментативные модификации субстрата или субстратов. МНКзимы с эндонуклеазной или расщепляющей активностью также известны как «расщепляющие МНКзимы». Каждый из компонентов-партзимов, партзимы А и В, связывается с соединением, облегчающим сборку (например, с последовательностью-мишенью ДНК или РНК) посредством спаривания оснований. МНКзим образуется только тогда, когда сенсорные плечи партзимов А и В гибридизуются рядом друг с другом на фасилитаторе сборки.
Субстратные плечи МНКзима взаимодействуют с субстратом, модификацию (например, расщепление) которого катализирует каталитическое ядро МНКзима, образованное за счет взаимодействия каталитических доменов партзимов А и В. МНКзимы могут расщеплять субстраты ДНК/РНК-химерного репортера. Расщепление МНКзимом субстрата между парой красителей «флуорофор и гаситель» может приводить к появлению флуоресцентного сигнала. Термины «многокомпонентная нуклеиновая кислота с ферментативной активностью» и «МНКзим» включают двухкомпонентные структуры, состоящие из двух молекул, или трехкомпонентные структуры, состоящие из трех молекул нуклеиновой кислоты, или другие многокомпонентные структуры, например, образованные четырьмя или более молекулами нуклеиновой кислоты.
Следует понимать, что термины «МНКзим» и «многокомпонентная нуклеиновая кислота с ферментативной активностью», используемые в настоящем документе, охватывают все известные МНКзимы и модифицированные МНКзимы, включая соединения, описанные в одной или более публикациях патентов РСТ №WO/2007/041774, WO/2008/040095, WO2008/122084 и родственных публикациях патентов США №2007-0231810, 2010-0136536 и 2011-0143338 (содержание каждого из этих документов полностью включено в настоящий документ посредством ссылки). Неограничивающие примеры МНКзимов и модифицированных МНКзимов, охватываемых терминами «МНКзим» и «многокомпонентная нуклеиновая кислота с ферментативной активностью», включают МНКзимы с расщепляющей каталитической активностью (представленные в настоящем документе), разобранные или частично собранные МНКзимы, содержащие один или более ингибиторов сборки, МНКзимы, содержащие один или более аптамеров («апта-МНКзимы»), МНКзимы, содержащие одну или более укороченную сенсорную цепь и, необязательно, один или более из стабилизирующих ол иго нуклеотидов, МНКзимы, содержащие один или более из ингибиторов активности, многокомпонентные неактивные пронуклеиновые кислоты с ферментативной активностью (MHKi), каждый из которых подробно описан в одном или более из патентов WO/2007/041774, WO/2008/040095, US 2007-0231810, US 2010-0136536 и/или US 2011-0143338.
В настоящем документе термины «партзим», «компонент-партзим» и «партзим-компонент» относятся к ДНК-содержащим или РНК-содержащим, или ДНК-РНК-содержащим олигонуклеотидам, два или более из которых только в присутствии соединения, облегчающего сборку МНКзима, описанного в настоящем документе, могут совместно образовывать «МНКзим». В некоторых предпочтительных вариантах реализации один или более, предпочтительно по меньшей мере два компонента-партзима, могут содержать три области или домена: «каталитический» домен, образующий часть каталитического ядра, катализирующего модификацию; домен «сенсорной цепи», способный ассоциировать с соединением, облегчающим сборку и/или связываться с ним; и домен «субстратной цепи», способный ассоциировать с субстратом и/или связываться с ним. Термины «сенсорное плечо», «плечо-датчик мишени» или «плечо, чувствительное к мишени» или «плечо-мишень» могут взаимозаменяемо использоваться для описания домена партзимов, который связывается с соединением, облегчающим сборку, например, с мишенью. Партзимы могут содержать по меньшей мере один дополнительный компонент, включая аптамер, называемый в настоящем документе «апта-партзимом», но не ограничиваясь им. Партзим может содержать несколько компонентов, включая партзим-компонент с укороченным сенсорным плечом и компонент стабилизирующего плеча, стабилизирующий структуру МНКзима посредством взаимодействия либо с соединением, облегчающим сборку, либо с субстратом, но не ограничиваясь ими.
Термины «молекула, облегчающая сборку», «соединение, облегчающее сборку», «молекула, облегчающая сборку МНКзима» и «соединение, облегчающее сборку МНКзима», используемые в настоящем документе, относятся к объектам, которые могут облегчать самосборку компонентов-партзимов с образованием каталитически активного МНКзима посредством взаимодействия с сенсорными плечами МНКзима. В настоящем документе соединения, облегчающие сборку, могут облегчать сборку МНКзимов, обладающих расщепляющей или другой ферментативной активностью. В предпочтительных вариантах реализации для самосборки МНКзима требуется соединение, облегчающее сборку. Соединение, облегчающее сборку, может состоять из одной молекулы или может состоять из двух или более «компонентов, облегчающих сборку», которые могут спариваться или связываться с сенсорными плечами одного или более олигонуклеотидных «партзимов». Соединение, облегчающее сборку, может содержать один или более из нуклеотидных компонентов, не обладающий комплементарностью последовательности с сенсорным плечом/ами МНКзима. Соединение, облегчающее сборку, может являться мишенью. Мишень может представлять собой нуклеиновую кислоту, выбранную из группы, состоящей из ДНК, метилированной ДНК, алкилированной ДНК, РНК, метилированной РНК, микроРНК, миРНК, кшРНК, тРНК, мРНК, мякРНК, мвРНК, ммРНК, предшественников микроРНК и первичных микроРНК, других некодирующих РНК, рибосомной РНК, их производных, ампликонов или любых их комбинаций. Нуклеиновую кислоту можно амплифицировать. Амплификация может включать один или более из следующих способов: ПЦР, ОТ-ПЦР, SDA, NEAR, HDA, RPA, LAMP, RCA, ТМА, RAM, LCR, 3SR или NASBA.
МНКзимы способны расщеплять линейные субстраты и/или субстраты, которые присутствуют в петлевой области структур репортерного зонда LOCS типа стебель-петля. Расщепление линейного субстрата может разделить флуорофор и гаситель, что позволит обнаружить мишень. Расщепление петлевой области LOCS под действием МНКзима может привести к образованию разделенной структуры LOCS, состоящей из двух фрагментов, которые могут оставаться гибридизованными и связанными при температурах ниже температуры плавления стебля и которые могут разделяться и диссоциировать при температуре выше температуры плавления стебля разделенного LOCS.
Термины «обнаружимый эффект» и «детектируемый сигнал» используются в данном документе взаимозаменяемо, и следует понимать, что они имеют одно и то же значение. Термины относятся к сигналу или эффекту, генерируемому детектирующим фрагментом, который присоединен или иным образом связан с олигонуклеотидом по настоящему изобретению (например, зондом, репортером или субстратом), обычно при модификации олигонуклеотида для изменения его конформации, структуры, ориентации, положения относительно других объектов и т.п.Модификация может быть, например, вызвана присутствием мишени, для обнаружения которой предназначен олигонуклеотид. Неограничивающие примеры таких модификаций (например, вызванных присутствием мишени) включают открытие части стержня-петли молекулярного маяка, открытие двухцепочечной области унизондов и бизондов Scorpion, связывание двойной гибридизации зондов к последовательности-мишени, производство Catcher дуплекс и расщепление/деструкция линейного субстрата МНКзима или зонда TaqMan и т.п. Обнаружимый эффект можно обнаруживать различными способами, включая флуоресцентную спектроскопию, поверхностный плазмонный резонанс (ППР), масс-спектроскопию, ЯМР, электронный спиновый резонанс, поляризационную флуоресцентную спектроскопию, круговой дихроизм, иммуноанализ, хроматографию, радиометрию, фотометрию, сцинтиграфию, электронные способы, электрохимические способы, УФ-спектроскопию, спектроскопию в видимой части спектра или инфракрасную спектроскопию, ферментативные способы или любую их комбинацию. Обнаруживаемый сигнал/эффект может быть обнаружен или определен количественно, а его величина может свидетельствовать о наличии и/или количестве входных данных, таких как количество молекулы-мишени, присутствующей в образце. Кроме того, величину детектируемого сигнала/эффекта, обеспечиваемого детектирующим фрагментом, можно модулировать путем изменения условий реакции, в которой используется олигонуклеотид, содержащий детектирующий фрагмент, включая, но не ограничиваясь этим, температуру реакции. Таким образом, можно модулировать способность обнаружимых фрагментов, присоединенных к олигонуклеотидам или иным образом связанных с ними, генерировать зависимый от мишени сигнал и/или независимый от мишени фоновый сигнал.
В данном документе термины «фоновый сигнал» и «базовый сигнал» используются взаимозаменяемо, и следует понимать, что они имеют одно и то же значение. Термины относятся к сигналу, генерируемому обнаруживаемым фрагментом, присоединенным к олигонуклеотиду по настоящему изобретению или иным образом связанным с ним, который не зависит от присутствия или отсутствия конкретной мишени, для измерения или обнаружения которой олигонуклеотид предназначен в конкретных условиях измерения.
В настоящем документе термины «полинуклеотидный субстрат», «олигонуклеотидный субстрат» и «субстрат» включают любой одноцепочечный или двуцепочечный полимер из дезоксирибонуклеотидных или рибонуклеотидных оснований или их аналогов, производных, вариантов, фрагментов или комбинаций, которые может распознавать, модифицировать или на которые может воздействовать фермент, в том числе каталитическая нуклеиновая кислота с ферментативной активностью. «Полинуклеотидный субстрат» или «олигонуклеотидный субстрат», или «субстрат» можно модифицировать за счет различных ферментативных активностей, включая расщепление, но не ограничиваясь им. Расщепление или разрушение «полинуклеотидного субстрата» или «олигонуклеотидного субстрата», или «субстрата» может обеспечить «обнаружимый эффект» для мониторинга каталитической активности фермента. «Полинуклеотидный субстрат» или «субстрат» может расщепляться или разрушаться одним или более ферментами, в том числе каталитическими нуклеиновыми кислотами с ферментативной активностью, например, МНКзимами, апта-МНКзимами, ДНКзимами, аптазимами, рибозимами и/или белковыми ферментами, например, экзонуклеазами или эндонуклеазами, но не ограничиваясь ими.
В настоящем документе «репортерный субстрат» представляет собой субстрат, в особенности адаптированный для облегчения измерения расщепления субстрата или появления расщепленного продукта катализируемой реакции. Репортерные субстраты могут находиться в растворе в свободном или связанном (или «прикрепленном») состоянии, например, к поверхности или к другой молекуле. Репортерный субстрат можно метить любым из множества способов, в том числе, например, флуорофорами (с одним или более дополнительными компонентами, например, гасителями, или без них), радиоактивными метками, биотином (например, путем биотинилирования) или хемилюминесцентными метками.
В настоящем документе термин «линейный субстрат МНКзим» представляет собой субстрат, например, репортерный субстрат, который распознается множеством МНКзимов и действует на него каталитически. «Линейный субстрат МНКзим» не содержит последовательности на своих 5'- или 3'-концах, которые способны гибридизоваться с образованием стебля. Альтернативно, субстраты МНКзима могут присутствовать в петлевой области зонда LOCS.
В настоящем документе «универсальный субстрат» представляет собой субстрат, например, репортерный субстрат, распознаваемый и подверженный каталитическому действию множества МНКзимов, каждый из которых может распознавать свое соединение, облегчающее сборку. Применение таких субстратов облегчает разработку отдельных способов анализа для обнаружения, идентификации или количественной оценки широкого спектра соединений, облегчающих сборку, с применением структурно родственных МНКзимов, каждый из которых распознает универсальный субстрат.Каждый из этих универсальных субстратов можно независимо метить одной или более метками. В предпочтительных вариантах реализации независимо обнаруживаемые метки применяют для мечения одного или более универсальных субстратов, что позволяет создать удобную систему для независимого или одновременного обнаружения множества соединений, облегчающих сборку, с применением МНКзимов. В некоторых вариантах реализации субстраты могут быть подвержены каталитической модификации ДНКзимами, каталитически активными в присутствии кофактора, например, кофактора-иона металла, например, свинца или ртути. В некоторых вариантах реализации субстраты могут подвергаться каталитической модификации аптазами, которые могут становиться каталитически активными в присутствии аналита, белка, соединения или молекулы, способных связываться с аптамерной областью аптазима, тем самым активируя каталитический потенциал части нуклеиновой кислоты с ферментативной активностью.
Термины «зонд» и «репортер», используемые в настоящем документе, относятся к олигонуклеотиду, который используется для обнаружения молекулы-мишени (например, нуклеиновой кислоты или аналита). Неограничивающие примеры стандартных зондов или репортеров, которые хорошо известны в данной области техники, включают, но не ограничиваются ими, линейные субстраты МНКзим, зонды TaqMan или гидролизные зонды, молекулярные маяки, маяки Sloppy, зонды Eclipse, унизонд Scorpion, праймеры/зонды бизонд Scorpion, олигонуклеотиды capture/pitcher и зонды двойной гибридизации. Варианты реализации настоящего изобретения объединяют стандартные зонды с зондами LOCS. Некоторые зонды LOCS содержат субстраты ферментов нуклеиновых кислот в петлевых областях, которые могут быть универсальными и которые способны к каталитическому расщеплению ферментами нуклеиновых кислот, такими как МНКзимы, ДНКзимы и аптазимы. Другие зонды LOCS содержат специфические последовательности-мишени в петлевой области, которые способны к каталитическому расщеплению белковыми ферментами, включая эндонуклеазы и экзонуклеазы.
Термин «продукт» относится к новой молекуле или молекулам, которые образуются в результате ферментативной модификации субстрата. В настоящем документе термин «продукт расщепления» относится к новой молекуле, полученной в результате расщепления или эндонуклеазной активности фермента. В некоторых вариантах реализации продукты ферментативного расщепления или разрушения интактной замкнутой LOCS-структуры содержат два олигонуклеотидных фрагмента, вместе называемых расщепленными LOCS, где два олигонуклеотидных фрагмента могут быть способны либо к гибридизации, либо к диссоциации/разделению в зависимости от температуры реакции.
В настоящем документе термины «температура плавления» и «Tm» в контексте полинуклеотид а следует понимать как упоминание температуры плавления (Tm), рассчитанной с применением правила Уоллеса, согласно которому Tm=2°С (А+Т)+4°С (G+C) (см. Wallace et al., (1979) Nucleic Acids Res. 6, 3543), если специально не указано иное. Влияние состава последовательности на температуру плавления можно понять, используя способ ближайшего соседа, который определяется следующей формулой: Tm (°С)=ΔН° / (ΔS°+R lnfoligo]) - 273,15. Известно, что на температуру плавления, помимо длины стебля и состава последовательности, влияют и другие факторы, включая ионную силу и концентрацию олигонуклеотидов. Более высокая концентрация олигонуклеотида и/или иона увеличивает вероятность образования двуцепочечной структуры, что приводит к повышению температуры плавления. И наоборот, более низкая концентрация олигонуклеотида и/или иона способствует диссоциации стебля, что приводит к снижению температуры плавления.
В настоящем документе термин «гаситель» включает любую молекулу, которая, находясь в непосредственной близости от флуорофора, принимает энергию излучения, генерируемую флуорофором, и либо рассеивает энергию в виде тепла, либо излучает свет с большей длиной волны, чем длина волны излучения флуорофора. Неограничивающие примеры гасителей включают Dabcyl, TAMRA, графен, FRET-флуорофоры, гасители ZEN, гасители АТТО, гасители «черные дыры» (BHQ) и гасители типа Black Berry (BBQ).
Следует понимать, что в настоящем документе термин «основание» в контексте нуклеиновой кислоты имеет то же значение, что и термин «нуклеотид».
В настоящем документе термин «блокатор» или «блокирующая молекула» относится к любой молекуле или функциональной группе, которые могут быть включены в олигонуклеотид для предотвращения использования полимеразой части олигонуклеотида в качестве темплата для синтеза комплементарной цепи. В качестве неограничивающего примера, блокатор гексатиленгликоля может быть включен, например, в зонд Scorpion, чтобы связать его 5'-зондирующую последовательность с его 3'-праймирующей последовательностью, при этом блокатор действует для предотвращения полимеразы с применением зондирующей последовательности как темплат.
В настоящем документе термины «нормализовать», «нормализующий» и «нормализованный» относятся к преобразованию измеренного сигнала (например, детектируемого сигнала, генерируемого детектирующим фрагментом) в шкалу относительно известного и воспроизводимого значения или в контрольное значение.
В настоящем документе термин «набор» относится к любой системе доставки материалов. Такие системы доставки включают системы, которые позволяют хранить, транспортировать или доставлять реагенты (например, метки, эталонные образцы, материал подложки и т.д. в соответствующих контейнерах) и/или вспомогательные материалы (например, буферы, письменные инструкции по выполнению анализа и т.д.) из одного места в другое. Например, наборы могут содержать один или более контейнеров, например, коробок, содержащих соответствующие реагенты и/или вспомогательные материалы. Термин «набор» включает как фрагментированные, так и комбинированные наборы.
В настоящем документе термин «фрагментированный набор» относится к системе доставки, включающей два или более отдельных контейнера, каждый из которых содержит часть всех компонентов набора. Контейнеры можно доставлять предполагаемому получателю вместе или по отдельности. Любая система доставки, включающая два или более отдельных контейнера, каждый из которых содержит часть всех компонентов набора, включена в значение термина «фрагментированный набор».
В настоящем документе термин «комбинированный набор» относится к системе доставки, содержащей все компоненты для анализа реакции в одном контейнере (например, в одной коробке, содержащей каждый из нужных компонентов).
Следует понимать, что применение в настоящем документе термина «около» по отношению к указанному численному значению включает указанное численное значение и численные значения в пределах плюс-минус десять процентов от указанного значения.
Следует понимать, что применение в настоящем документе термина «между» при упоминании диапазона численных значений охватывает численные значения в каждой конечной точке диапазона. Например, полипептид длиной от 10 до 20 остатков включает полипептид длиной 10 остатков и полипептид длиной 20 остатков.
В настоящем документе любое описание документов, описывающих существующий уровень техники, или утверждений, полученных из этих документов или основанных на них, не является признанием того, что документы или полученных из них утверждения являются частью общеизвестных знаний в соответствующей области.
В целях описания все документы, упомянутые в настоящем документе, полностью включены в настоящий документ посредством ссылки, если не указано иное.
Сокращения
В настоящем документе и во всем описании заявки используются следующие сокращения:
LOCS: петля, соединенная со стеблями
МНКзим: многокомпонентная нуклеиновая кислота с ферментативной активностью;
Партзим: частичный фермент, содержащий олигонуклеотид;
ПЦР: полимеразная цепная реакция;
гДНК: геномная ДНК
NTC: Нецелевой контроль
кПЦР: количественная ПЦР в реальном времени
Ct; Пороговый цикл
Cq; Цикл количественного определения
R2; Коэффициент корреляции
нМ; Наномолярный
мМ; Миллимолярный
мкл; Микролитр
мкМ; Микромолярный
dNTP; Дезоксирибонуклеотидтрифосфат
NF-H2O: вода, не содержащая нуклеаз;
LNA: закрытая нуклеиновая кислота;
F: флуорофор;
Q: гаситель;
N=А, С, Т, G или любой их аналог;
N'=любой нуклеотид, комплементарный N или способный образовывать пару оснований с N;
(N)x: любое количество N;
(N')x: любое количество N;
W: А или Т;
R: A, G или АА;
rN: любое рибонуклеотидное основание;
(rN)x: любое количество rN;
rR: А или G;
rY: С или U;
М: А или С;
Н: А, С или Т;
D: G, А или Т;
JOE или 6-JOE: 6-карбокси-4',5'-дихлор-2',7'-диметоксифлуоресцеин;
FAM или 6-FAM: 6-карбоксифлуоресцеин.
BHQ1: Гаситель - «черная дыра» 1
BHQ2: Гаситель - «черная дыра» 2
ОТ-ПЦР: полимеразная цепная реакция с обратной транскрипцией
SDA: амплификация с замещением цепи
NEAR: Реакция никинг-ферментной амплификации
HDA: амплификация, зависимая от хеликазы
RPA: рекомбиназно-полимеразная амплификация
LAMP: петлевая изотермическая амплификация
RCA: амплификация по типу катящегося кольца
ТМА: амплификация, опосредованная транскрипцией
3SR: самоподдерживающаяся репликация последовательности
NASBA: амплификация на основе нуклеотидной последовательности
LCR: Лигазная цепная реакция
RAM: амплификации в варианте рамификации
IB: Iowa Black® FQ
IBR: Iowa Black® RQ
кшРНК: короткая шпилечная РНК
миРНК: малая интерферирующая РНК
мРНК: матричная РНК
тРНК: транспортная РНК
мякРНК: малая ядрышковая РНК
мвРНК: малая временная РНК
ммРНК: малая модулирующая РНК
пре-микроРНК: предшественник микроРНК
перв. микроРНК: первичная микроРНК
LHS: Левая сторона
RHS: Правая сторона
ДЦО: двуцепочечный олигонуклеотид
Tm: Температура плавления
RFU: Относительные единицы флуоресценции
СТ: Хламидия трахоматис
NG: Гонококковая нейссерия
ППР: поверхностный плазмонный резонанс
НЧЗ: наночастицы золота
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В следующем подробном описании приведена информация о типичных вариантах реализации настоящего изобретения в достаточных подробностях, чтобы дать возможность специалистам в данной области техники реализовать настоящее изобретение на практике. Особенности или ограничения различных описанных вариантов реализации не обязательно ограничивают другие варианты реализации настоящего изобретения или настоящее изобретение в целом. Следовательно, последующее подробное описание не ограничивает сущность настоящего изобретения, которая определяется исключительно формулой изобретения.
Настоящее изобретение относится к способам и композициям для улучшенного мультиплексного обнаружения мишеней (например, нуклеиновых кислот, белков, анализируемых веществ, соединений, молекул и т.п.). В каждом из способов и композиций используются комбинации олигонуклеотида LOCS вместе с другими олигонуклеотидными репортерами, зондами или субстратами, которые можно использовать в комбинации с различными другими агентами.
Репортеры, зонды и субстраты Согласно настоящему изобретению мультиплексное обнаружение молекул-мишеней облегчается с применением LOCS в сочетании с другим олигонуклеотидом, подходящим для использования в качестве зонда в анализе мультиплексного обнаружения.
Многие олигонуклеотиды для обнаружения нуклеиновых кислот-мишеней были описаны и хорошо известны в данной области техники. Подходящие олигонуклеотиды, которые можно использовать в сочетании с LOCS, включают, но не ограничиваются ими, субстраты МНКзимы, зонды TaqMan или гидролизные зонды, молекулярные маяки, маяки Sloppy, зонды Eclipse, унизонды Scorpion, бизонды Scorpion, двойные гибридизационные зонды и зонды Capture/Pitcher.
В некоторых вариантах реализации эти олигонуклеотиды связываются непосредственно с мишенью или целевым ампликоном для облегчения их обнаружения, однако субстраты МНКзима и олигонуклеотиды Capture/Pitcher представляют собой исключение, поскольку они могут быть универсальными и подходящими для обнаружения любой мишени.
В некоторых вариантах реализации олигонуклеотиды генерируют флуоресценцию в присутствии мишени вследствие ферментативно опосредованного расщепления или деградации, например, субстраты МНКзима и зонды TaqMan, или гидролиза.
В других вариантах реализации олигонуклеотиды обеспечивают различные уровни флуоресцентного сигнала в результате изменения конформации, вызванного связыванием с мишенью или целевым ампликоном (например, молекулярные маяки, маяки Sloppy, зонды Eclipse, унизонды Scorpion, бизонды Scorpion и двойные гибридизационные зонды).
В системе ТОСЕ Catcher изменяет флуоресценцию в результате конформационных изменений, вызванных связыванием и расширением Pitcher, который активируется и высвобождается только в присутствии мишени.
Любой или все из этих типов репортерных олигонуклеотидов подходят для использования в сочетании с зондами LOCS для обнаружения множественных мишеней путем измерения изменений, связанных с одним детектирующим фрагментом, включая, помимо прочего, изменение флуоресценции, измеренное при одной длине волны.
Олигонуклеотиды для комбинации с LOCS можно синтезировать в соответствии со стандартными протоколами. Например, они могут быть синтезированы фосфорамидитной химией с применением нуклеозидных и ненуклеозидных фосфорамидитов в последовательных синтетических циклах, которые включают удаление защитной группы, связывание фосфорамидитов, блокирование и окисление либо в твердой фазе, либо в фазе раствора и, необязательно, в автоматическом синтезаторе. Альтернативно, они могут быть приобретены из коммерческих источников. Неограничивающие примеры коммерческих источников, из которых можно приобрести субстраты МНКзима, зонды TaqMan или гидролиза, молекулярные маяки, маяки Sloppy, зонды Eclipse, унизонды Scorpion, бизонды Scorpion, двойные гибридизационные зонды и зонды Capture/Pitcher или полученное иным образом, включают: Субстраты МНКзима можно приобрести у SpeeDx (plexpcr.com); TaqMan и зонды для гидролиза можно приобрести у компаний Thermo Fisher Scientific (www.thermofisher.com), Sigma Aldrich (www.sigmaaldrich.com), Promega (www.promega.com), Generi Biotech (www.generi-biotech.com); Молекулярные маяки и маяки Sloppy можно приобрести в компаниях Integrated DNA Technologies (www.idtdna.com), Eurofins (www.eurofinsgenomics.com), Sigma Aldrich (www.sigmaaldrich.com) и TriLink BioTechnologies (www.trilinkbiotech.com); зонды Eclipse можно приобрести в компании Integrated DNA Technologies (www.idtdna.com); унизонд Scorpion можно приобрести у компаний Sigma Aldrich (www.sigmaaldrich.com) и Bio-Synthesis (https://www.biosyn.com); бизонды Scorpion можно приобрести в компании Bio-Synthesis (https://www.biosyn.com); зонды для двойной гибридизации можно приобрести у компаний Bio-Synthesis (https://www.biosyn.com), Sigma Aldrich (www.sigmaaldrich.com) и Eurofins (www.eurofinsgenomics.com), а анализы Catcher Pitcher можно приобрести в компании Seegene (www.seegene.com).
LOCS-олигонуклеотиды
Типовые LOCS-олигонуклеотиды согласно настоящему изобретению проиллюстрированы на Фиг. 1. Типовые интактные LOCS-олигонуклеотиды (Фиг. 1А, LHS) содержат петлевую область, стеблевую область и пару красителей «флуорофор (Е)/гаситель (Q)». Хотя в качестве примера можно привести пару флуорофор/гаситель, специалисту в данной области техники будет понятно, что для той же цели можно использовать любые другие подходящие детектирующие фрагменты. Петлевая область содержит область субстрата, которая поддается ферментативному расщеплению или деградации в присутствии мишени или ампликонов-мишеней. Расщепление или деградация петли в интактном LOCS генерирует дуплекс Split LOCS (Фиг. 1B, RHS).
В некоторых вариантах реализации температура плавления («Tm») интактного олигонуклеотида LOCS выше, чем Tm разделенной структуры LOCS.
Поскольку внутримолекулярные связи прочнее межмолекулярных связей, стеблевые области интактных LOCS-структур обычно плавятся при более высоких температурах, чем стебли раскрытых, расщепленных или разрушенных структур LOCS-олигонуклеотидов. Например, стебель интактного LOCS А плавится при Tm А, превышающей Tm В, которая является температурой, при которой плавится стебель разделенного LOCS (Фиг. 1В). Наличие флуоресценции при температуре, при которой возможно плавление разделенных LOCS, но не интактных LOCS, указывает на присутствие мишени или целевых ампликонов. В отношении типового LOCS, изображенного на Фиг. 2, последовательность петлевой области LOCS-олигонуклеотида может, например, являться субстратом МНКзима или другой нуклеиновой кислоты, с каталитической активностью/кислот.
Типовой LOCS, пригодный для использования в изобретении, может содержать петлевую область, содержащую субстрат для нуклеиновой кислоты, с каталитической активностью, проиллюстрированную на Фиг. 2. В этих вариантах реализации олигонуклеотиды LOCS содержат универсальные субстраты, которые можно использовать для обнаружения любой мишени. Олигонуклеотид LOCS содержит стеблевую область, пару флуорофорный гаситель/краситель (могут быть использованы альтернативные фрагменты детекции, как описано в настоящем документе) и промежуточную петлевую область, которая содержит универсальный субстрат для нуклеиновой кислоты, с каталитической активностью, такой как МНКзим. МНКзим может обнаруживать мишень напрямую или может использоваться для обнаружения ампликонов, генерируемых во время амплификации мишени. МНКзим образуется, когда каждое сенсорное плечо мишени партзимов гибридизуется с мишенью или ампликонами-мишенями путем комплементарного спаривания оснований с образованием активного каталитического ядра МНКзима. Петлевая область олигонуклеотида LOCS гибридизуется с ответвлениями МНКзима, связывающими субстрат, за счет комплементарного спаривания оснований, и субстрат внутри петли расщепляется МНКзимом. Это создает структуру разделенного LOCS, которая имеет стебель с Tm В, которая ниже, чем Tm А интактного LOCS. Измерение флуоресцентного сигнала при температурах выше Tm В, но ниже Tm А свидетельствует о присутствии мишени в реакции. Специалистам в данной области техники понятно, что мишени можно обнаружить в реальном времени или в конце реакции.
Что касается примерного варианта реализации, показанного на Фиг. 3, показан стандартный линейный субстрат МНКзим, который используется вместе с одним зондом LOCS, содержащим субстрат МНКзим в своей петле. Субстрат линейного МНКзима и одиночный зонд LOCS могут быть помечены одинаковыми (или подобными) детектирующими фрагментами, например определенной парой красителей флуорофор (Р)/гаситель (Q). Могут быть использованы альтернативные фрагменты детекции, описанные в настоящем документе. Линейный субстрат содержит первую последовательность субстрата, расщепляемую первым МНКзимом, который собирается в присутствии первой мишени (Фиг. 3А). В присутствии первой мишени линейный субстрат расщепляется первым МНКзимом, что приводит к увеличению флуоресценции, которую можно обнаружить в широком диапазоне температур. LOCS содержит вторую последовательность субстрата в своей петле, которая расщепляется вторым МНКзимом, который собирается в присутствии второй мишени (Фиг. 3В). В присутствии второй мишени LOCS расщепляется с образованием разделенного LOCS, который плавится при температуре Tm В, которая ниже температуры плавления интактного LOCS (Tm А). При температурах ниже Tm В стеблевая область разделенного LOCS остаются гибридизованными, и, следовательно, флуорофор гасится из-за близости к молекуле гасителя. При температурах выше Tm В стеблевая область разделенного LOCS диссоциируют и отделяют флуорофор от молекулы гасителя, что приводит к увеличению флуоресценции. Когда присутствуют обе мишени, и флуоресценция измеряется при первой температуре ниже Tm В, увеличение флуоресценции связано только с первой мишенью 1. Когда флуоресценцию измеряют при второй температуре выше Tm В, но ниже TmA, увеличение флуоресценции связано с первой мишенью и/или второй мишенью. Когда присутствуют и мишень 1, и мишень 2, наблюдаемое изменение флуоресценции во время амплификации при второй температуре больше, чем изменение при первой температуре, что позволяет определить, присутствует ли в реакции мишень 1, или мишень 2, или мишени 1 и 2, или ни одна из мишеней.
В других вариантах реализации настоящего изобретения можно использовать альтернативные структуры LOCS, подходящие для комбинации со стандартными репортерными зондами. Как показано на Фиг. 4А и Фиг. 4В, петлевая область олигонуклеотида LOCS может содержать специфичную для мишени последовательность, которая полностью или частично комплементарна мишени, подлежащей обнаружению, и которая, будучи двухцепочечной, может служить субстратом для деградации экзонуклеазой, например, экзонуклеазной активностью, присущей полимеразе (Фиг. 3А). В еще одном варианте реализации, показанном на Фиг. 3В, специфическая для мишени последовательность в петле может дополнительно содержать одну цепь сайта узнавания двухцепочечного фермента рестрикции. Гибридизация петлевой последовательности с последовательностью-мишенью может привести к функциональному расщепляемому сайту рестрикции. В предпочтительных вариантах реализации фермент рестрикции представляет собой фермент, способный расщеплять петлевую цепь олигонуклеотида LOCS, оставляя мишень интактной.
В различных вариантах реализации настоящего изобретения различные комбинации хорошо известных репортерных зондов или субстратов могут быть объединены с зондом LOCS в одной реакции. В качестве неограничивающего примера, реакция обнаружения двух мишеней может включать любую комбинацию первого зонда, выбранного из группы 1, включая, но не ограничиваясь ими, линейные субстраты МНКзимы, зонды TaqMan или гидролизные зонды, молекулярные маяки, маяки Sloppy, маяки Eclipse, унизонд Scorpion, бизонд Scorpion, олигонуклеотиды Capture/Pitcher и зонды двойной гибридизации вместе со вторым зондом, выбранным из группы 2, включая, но не ограничиваясь этим, зонд LOCS, содержащий универсальный субстрат МНКзима, LOCS зонд, содержащий специфичный для мишени субстрат для экзонуклеазы, и зонд LOCS, содержащий специфичный для мишени субстрат для эндонуклеазы, такой как никирующий фермент.
Любая комбинация зонда группы 1 с зондом группы 2 может быть использована для измерения нескольких мишеней в одной реакции в соответствии со способами по настоящему изобретению.
Вариант реализации, показанный на Фиг. 3, иллюстрирует примерную комбинацию субстрата линейного МНКзима, расщепляемого первым МНКзимом, в сочетании с зондом LOCS, который расщепляется вторым МНКзимом. Другие неограничивающие варианты реализации настоящего изобретения проиллюстрированы на Фиг. 5, на которой нерасщепляемый молекулярный маяк может быть объединен с зондом LOCS, расщепляемым МНКзимом. И молекулярный маяк, и зонд LOCS могут быть помечены одним и тем же (или сходным) детектирующим компонентом, например, одним и тем же флуорофором или флуорофорами, излучающими на одинаковых длинах волн. Молекулярный маяк может иметь стеблевую область с Tm А и петлевую область, которая может специфически гибридизоваться с первой мишенью 1 с Tm В; где Tm В больше, чем Tm А. Его можно комбинировать с интактным зондом LOCS, который может иметь стеблевую область с Tm С и петлевую область, которая может быть расщеплена МНКзимом в присутствии второй мишени 2, таким образом, генерируя расщепление. LOCS с Tm D, где Tm D меньше, чем Tm С.Присутствие мишени 1 и/или мишени 2 можно различить путем измерения флуоресценции при двух температурах либо в реальном времени, либо с помощью дискретных измерений, полученных при или вблизи, начала усиления и последующего усиления.
Комбинации LOCS, включающие обратимые зонды
В некоторых вариантах реализации композиции и способы по настоящему изобретению включают комбинацию LOCS и олигонуклеотидного зонда, способного генерировать детектируемые сигналы, зависящие от мишени, которые могут быть обратимо модулированы температурой. Кроме того, LOCS и олигонуклеотидный зонд могут поддаваться модуляции независимой от мишени генерации сигнала температурой, что позволяет манипулировать фоновым шумом или базовыми уровнями.
Например, олигонуклеотидный зонд может принимать первую конформацию или расположение в отсутствие мишени, при котором испускание детектируемого сигнала подавляется, и вторую конформацию или расположение в присутствии мишени, которое облегчает испускание детектируемого сигнала свидетельствует о присутствии мишени. Неограничивающие примеры олигонуклеотидных зондов в этой категории включают молекулярные маяки, зонды Sloppy, унизонды Scorpion, бизонд Scorpion и олигонуклеотиды Capture/Pitcher.
В качестве альтернативы, в случае зондов с двойной гибридизацией, два дополнительных олигонуклеотида в дополнение к LOCS могут принимать расположение, при котором детектируемый сигнал подавляется в присутствии мишени и при котором детектируемый сигнал генерируется, когда мишень отсутствует.
В вариантах реализации, приведенных выше, интактный LOCS подвергается расщеплению, зависящему от мишени, с получением разделенного LOCS. Двухцепочечная часть стебля разделенного LOCS может быть спроектирована так, чтобы диссоциировать при температуре, которая отличается от температуры, при которой генерируется зависимое от мишени изменение конформации или расположения первого(их) олигонуклеотида(ов) и связанный с ним обнаруживаемый сигнал.
В некоторых вариантах реализации олигонуклеотид представляет собой молекулярный маяк. Следующие сценарии представляют собой неограничивающие примеры анализов мультиплексного обнаружения в соответствии с вариантами реализации настоящего изобретения. В этих сценариях молекулярный маяк может использоваться в сочетании с LOCS для обнаружения мишеней 1 и 2 соответственно. Молекулярный маяк может содержать Tm А, представляющую собой температуру плавления его двухцепочечной области стебля, и Tm В, представляющую собой температуру плавления дуплекса, образованного между его одноцепочечным петлевым дуплексом и мишенью 1. LOCS может включать Tm С, представляющую собой температуру плавления его двухцепочечной области стебля, когда он неповрежден, и Tm D, являющуюся температурой плавления его двухцепочечной области стебля, когда он расщеплен. Противоположные цепи двухцепочечной области стебля молекулярного маяка могут быть помечены флуорофором и гасителем, как и LOCS. Флуорофор молекулярного маяка может быть таким же или излучать в той же области видимого спектра, что и флуорофор LOCS. В альтернативных вариантах реализации можно использовать различные детектирующие фрагменты, включая, например, наночастицы одинакового или сходного размера и/или типа для колориметрического или ППР-детектирования, реактивные фрагменты (например, ферменты щелочной фосфатазы или пероксидазы) для хемилюминесцентного детектирования, электроактивные частицы (например, ферменты ферроцен, метиленовый синий или пероксидаза) для электрохимического обнаружения. Специалист в данной области техники легко поймет, что молекулярный маяк в этих случаях можно заменить на зонд Sloppy, унизонд Scorpion или бизонд Scorpion.
Между температурами, при которых измеряют реакцию, и температурами плавления различных областей репортерных зондов может существовать различное соотношение. Четыре сценария подробно описаны ниже в контексте реакций, включающих один молекулярный маяк и один зонд LOCS, а не ограничивающие примерные температуры для таких сценариев приведены в Таблице 1 ниже.
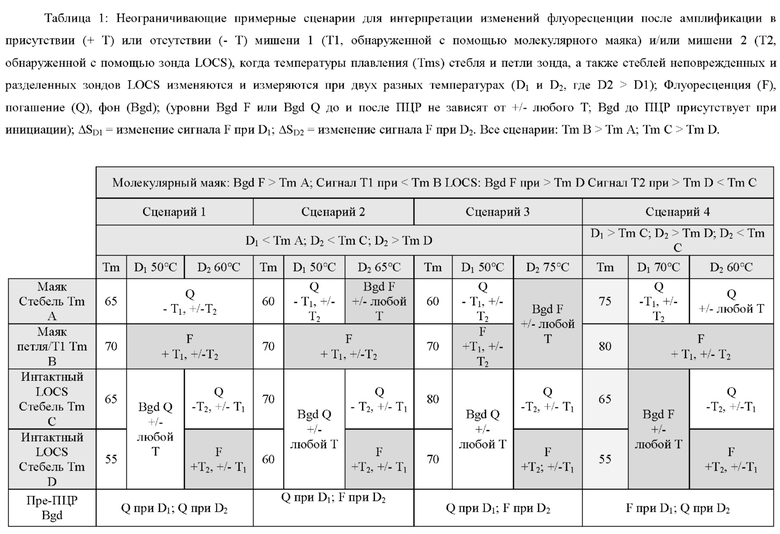

Сценарий 1
В первом неограничивающем примере Tm А может быть больше, чем Tm D, а Tm В может быть больше, чем Tm D. Наличие мишени 1 и/или мишени 2 можно отличить путем измерения флуоресценции при двух температурах, полученных либо в или около начала амплификации и последующей амплификации. В присутствии мишени 1 и/или мишени 2 измерение флуоресценции при первой температуре 1, которая может быть меньше Tm А, Tm В и Tm D, может генерировать сигнал, указывающий только на присутствие мишени 1. При этой температуре молекулярный маяк гибридизуется с мишенью 1 (если она присутствует) и флуоресцирует; но в отсутствие мишени 1 его стебель останется внутренне гибрид изо ванным и, следовательно, погашенным. При температуре 1 как целые, так и/или разделенные виды LOCS будут гаситься из-за гибридизации их соответствующих стеблей при этой температуре. Кроме того, в присутствии мишени 1 и/или мишени 2 измерение флуоресценции при второй температуре 2, которая выше как температуры 1, так и Tm D, но меньше как Tm В, так и Tm С, может свидетельствовать о присутствии мишени 1 и/или мишени 2. При этой второй температуре молекулярный маяк гибридизуется с мишенью 1 (если она присутствует) и флуоресцирует, но в отсутствие мишени 1 его стержень останется внутренне гибрид изо ванным и, следовательно, гасится. Кроме того, при этой второй температуре, если мишень 2 отсутствует, зонд LOCS останется интактным и гасится, но будет расщеплен МНКзимом, специфичным для мишени 2 (если он присутствует), и его стебель будет диссоциирован для генерации флуоресценции. В этом сценарии, если флуоресценция при температуре 1 увеличивается во время амплификации, это указывает на присутствие мишени 1. Кроме того, если увеличение флуоресценции, наблюдаемое при температуре 2В ходе амплификации, больше, чем наблюдаемое при температуре 1, это указывает на присутствие мишени 2.
Сценарий 2
Во втором неограничивающем примере Tm А может быть подобна Tm D, Tm В может быть подобна Tm С, а Tm В может быть больше, чем Tm D. Наличие мишени 1 и/или мишени 2 можно отличить путем измерения флуоресценции при двух температурах либо в режиме реального времени; либо с помощью дискретных измерений, полученных при или вблизи, начала усиления и последующего усиления. В присутствии мишени 1 и/или мишени 2 измерение флуоресценции при первой температуре 1, которая может быть меньше чем Tm А, и меньше чем Tm В, и меньше чем Tm D, может генерировать сигнал, указывающий только на присутствие мишени 1. При этой температуре молекулярный маяк гибридизуется с мишенью 1 (если она присутствует) и флуоресцирует, но его стебель останется внутренне гибридизованным в отсутствие мишени 1 и будет погашенным. При этой первой температуре 1 как интактные, так и/или расщепленные виды LOCS будут гаситься из-за гибридизации области стебля при этой температуре. Кроме того, в присутствии мишени 1 и/или мишени 2 измерение флуоресценции при второй температуре 2, которая выше как температуры 1 и Tm А, так и Tm D, но меньше как Tm В, так и Tm С, может свидетельствовать о присутствии мишени 2. При этой второй температуре молекулярный маяк гибридизуется с мишенью 1 (если она присутствует) и флуоресцирует; или будет флуоресцировать независимо от мишени из-за диссоциации и открытия его стебля при этой температуре. Таким образом, молекулярный маяк будет флуоресцировать независимо от наличия или отсутствия какой-либо мишени, обеспечивая уровень фоновой флуоресценции при этой температуре, который может оставаться неизменным во время амплификации. Кроме того, при этой второй температуре, если мишень 2 отсутствует, зонд LOCS останется интактным и гасится, но будет расщеплен МНКзимом, специфичным для мишени 2 (если он присутствует), и его стебель будет диссоциирован и, таким образом, генерировать флуоресценцию. В этом сценарии, если флуоресценция при температуре 1 увеличивается во время амплификации, это указывает на то, что мишень 1 присутствует и обнаружена молекулярным маяком, в то время как увеличение флуоресценции при температуре 2Во время амплификации указывает на то, что мишень 2 присутствует и обнаружена зондом LOCS. Качественные данные могут быть получены с применением дискретных измерений температуры в/в начале и в конце ПЦР; и/или количественные данные могут быть считаны непосредственно с двух кривых амплификации, полученных при каждой температуре во время ПЦР. В качестве альтернативы, если Tm А и Tm D не одинаковы, то же соотношение между уровнем флуоресценции при двух температурах и наличием или отсутствием мишени (мишеней) сохраняется, если и Tm А, и Tm D больше, чем температура 1, и меньше, чем температура 2.
Вышеописанный подход может обеспечить основные преимущества по сравнению с другими способами, известными в данной области техники, которые используют измерения при нескольких температурах для различения нескольких мишеней на одной длине волны, таких как, например, ТОСЕ. ТОСЕ измеряет флуоресценцию первой мишени при первой температуре и измеряет флуоресценцию двух мишеней при второй температуре (первая мишень плюс вторая мишень). Эти данные анализируют, чтобы математически вычесть величину флуоресценции, связанную с первой мишенью при второй температуре, для количественной оценки второй мишени в комплексном анализе, который дополнительно требует корректировки для учета внутренней разницы флуоресценции, которая связана с температурой как таковой. В описанных вариантах реализации настоящего изобретения используют молекулярный маяк и зонд LOCS в способе, который устраняет необходимость сложного пост-ПЦР-анализа, поскольку он позволяет проводить прямое количественное определение первой мишени по первой кривой амплификации, полученной при первой температуре, и прямое количественное определение второй мишени по второй кривой амплификации, полученной при второй температуре. В данных вариантах реализации измеряют каждую мишень по отдельности, и, кроме того, нет необходимости в корректировке для учета разницы в выходе флуоресценции одних и тех же молекул, поскольку каждая мишень будет генерировать сигнал, который можно обнаружить только выше фона при одной из двух температур, выбранных для сбора данных.
Сценарий 3
В третьем неограничивающем примере Tm А может быть меньше чем Tm D, Tm В может быть подобна Tm D, а Tm С может быть больше, чем Tm В. Наличие мишени 1 и/или мишени 2 можно отличить путем измерения флуоресценции при двух температурах либо в режиме реального времени; либо с помощью дискретных измерений, полученных при или вблизи, начала усиления и последующего усиления. В присутствии мишени 1 и/или мишени 2 измерение флуоресценции при первой температуре 1, которая может быть меньше чем Tm А, и меньше чем Tm В, и меньше чем Tm D, может генерировать сигнал, указывающий только на присутствие мишени 1. При этой температуре молекулярный маяк гибридизуется с мишенью 1 (если она присутствует) и флуоресцирует, но его стебель останется внутренне гибридизованным в отсутствие мишени 1 и будет погашенным. При данной температуре 1 как целые, так и/или разделенные виды LOCS будут гаситься из-за гибридизации их стеблей при этой температуре. Кроме того, в присутствии мишени 1 и/или мишени 2 измерение флуоресценции при второй температуре 2, которая выше как температуры 1 и Tm D, и Tm А, так и Tm В, но меньше Tm С, может свидетельствовать о присутствии мишени 2. При этой второй температуре молекулярный маяк не может гибридизоваться с мишенью 1 и всегда будет иметь открытую диссоциированный стебель и, следовательно, будет флуоресцировать независимо от присутствия или отсутствия любой мишени, обеспечивая уровень фоновой флуоресценции при этой температуре, который может оставаться неизменным во время амплификации. Кроме того, при этой второй температуре, если мишень 2 отсутствует, зонд LOCS останется интактным и гасится, но будет расщеплен МНКзимами, специфичным для мишени 2 (если он присутствует), и его стебель будет диссоциирован и, таким образом, генерировать флуоресценцию. В этом сценарии, если флуоресценция при температуре 1 увеличивается во время амплификации, это указывает на то, что мишень 1 присутствует и обнаружена молекулярным маяком, в то время как увеличение флуоресценции при температуре 2 указывает на то, что мишень 2 присутствует и обнаружена зондом LOCS. Качественные данные могут быть получены с применением дискретных измерений температуры в/в начале и в конце ПЦР; и/или количественные данные могут быть считаны непосредственно с двух кривых амплификации, полученных при каждой температуре во время ПЦР. В качестве альтернативы, когда Tm А и Tm D одинаковы или Tm А больше, чем Tm D, такое же соотношение между уровнем флуоресценции при двух температурах и наличием или отсутствием мишени (мишеней) сохраняется, если и Tm А, и Tm D больше, чем температура 1, и меньше, чем температура 2.
Сценарий 4
В четвертом неограничивающем примере как Tm А, так и Tm В может быть больше, чем Tm С и Tm D. Наличие мишени 1 и/или мишени 2 можно отличить путем измерения флуоресценции при двух температурах, полученных либо в или около начала амплификации и последующей амплификации. В присутствии мишени 1 и/или мишени 2 измерение флуоресценции при первой температуре 1, которая может быть меньше чем Tm А и Tm В, но больше, чем Tm С и Tm D, может генерировать сигнал, указывающий на присутствие мишени 1. При этой температуре молекулярный маяк гибридизуется с мишенью 1 (если она присутствует) и флуоресцирует, но его стебель останется внутренне гибридизованным в отсутствие мишени 1 и будет погашен. При этой температуре LOCS будет флуоресцировать независимо от присутствия или отсутствия мишени 2 и, следовательно, будет вносить вклад только в фон, который останется неизменным во время амплификации. В отсутствие мишени 2 стебель интактного LOCS будет диссоциировать и флуоресцировать, и аналогичным образом в присутствии мишени 2 стебель разделенного LOCS будет диссоциировать и флуоресцировать. Кроме того, увеличение флуоресценции в ходе амплификации при температуре 2, которая меньше температуры 1 и меньше, чем Tm С, Tm А и Tm В, но больше, чем Tm D, может свидетельствовать о присутствии мишени 1 и/или мишени 2. При этой второй температуре молекулярный маяк будет гибридизован с мишенью 1 (если она присутствует) и, следовательно, будет флуоресцировать, или останется погашенным с гибридизированным стеблем в отсутствие мишени 1. Кроме того, при этой второй температуре, если мишень 2 отсутствует, стебельь интактного зонда LOCS останется гибридизированным и погашенным, или, если мишень 2 присутствует, LOCS будет расщеплен МНКзимом, специфичным для мишени 2, а стебель разделенного LOCS будет диссоциировать и флуоресцировать. В этом сценарии, если флуоресценция при температуре 2 увеличивается во время амплификации, это указывает на присутствие мишени 1 и/или мишени 2. Кроме того, если увеличение флуоресценции, наблюдаемое в ходе амплификации при температуре 2, больше, чем наблюдаемое при температуре 1, это указывает на присутствие мишени 2.
Комбинации LOCS, содержащие зонды Catcher-Pitcher В некоторых вариантах реализации композиции и способы по настоящему изобретению могут включать комбинацию LOCS и первого олигонуклеотида, который действует как компонент Catcher анализа ТОСЕ. Эта комбинация может, например, обеспечить одновременное обнаружение и количественную оценку двух мишеней в одном флуоресцентном канале путем получения показаний флуоресценции при двух температурах в режиме реального времени во время ПЦР. В качестве альтернативы, при отсутствии мониторинга в реальном времени, этот подход может применяться к флуоресцентным данным, собранным в дискретные моменты времени, например, вблизи или в начале амплификации и после амплификации.
В одном варианте реализации первый олигонуклеотид, содержащий Catcher, можно комбинировать с зондом LOCS, оба из которых могут быть помечены одними и теми же фрагментами флуорофора и гасителя для одновременной детекции в одном и том же флуоресцентном канале. Реакция может также содержать Pitcher, содержащий одноцепочечный олигонуклеотид, который включает 5'-меточную область, комплементарную Catcher, и 3'-сенсорную область, комплементарную первой мишени 1. Catcher может содержать одноцепочечный олигонуклеотид, помеченный гасителем на 5'-конце и флуорофором, расположенным ниже гасителя, и 3'-областью, комплементарной метке части Pitcher. Когда Catcher находится в одноцепочечной конформации, флуорофор оказывается в непосредственной близости от гасителя, и сигнал гасится.
В качестве неограничивающего примера реакции амплификации, такой как ПЦР, праймеры и 3'-сенсорная область Pitcher могут гибридизоваться с мишенью 1. Во время удлинения праймера с применением мишени 1 или ее ампликонов в качестве темплата Pitcher может быть расщеплен экзонуклеазной активностью ДНК-полимеразы, что приводит к высвобождению части метки. Затем высвобожденная метка может гибридизоваться с комплементарной 3'-частью Catcher и удлиняться с помощью ДНК-полимеразы, образуя таким образом двухцепочечный дуплекс Catcher с Tm А, в котором флуорофор и гаситель разделены, что приводит к усилению флуоресценции, указывающему на присутствие мишени 1. Кроме того, реакция может содержать интактный зонд LOCS со стеблевой областью с Tm С и петлевой областью, которая может быть расщеплена МНКзимом в присутствии второй мишени 2 с образованием разделенного LOCS с Tm D ниже, чем Tm А. Могут существовать различные соотношения между температурами, при которых измеряются реакции, и температурами плавления дуплекса Catcher и репортеров LOCS. Три сценария подробно описаны ниже в контексте реакций, включающих один зонд Catcher и один зонд LOCS, а не ограничивающие типовые температуры для таких сценариев приведены в Таблице 2 ниже.
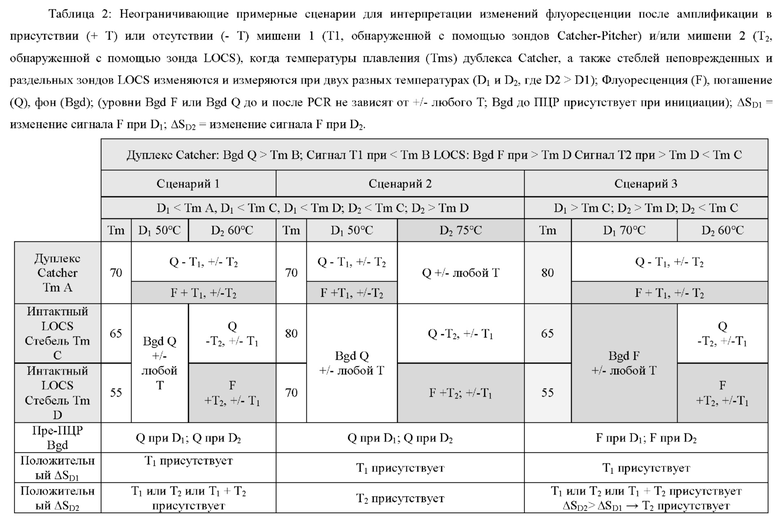

В первом неограничивающем сценарии (см. Таблицу 2), когда сигнал измеряется при первой температуре обнаружения, которая ниже, чем Tm А, Tm С и Tm D, тогда в присутствии мишени 1 может образоваться дуплекс Catcher и флуоресцировать, в то время как в отсутствие мишени 1 одноцепочечный Catcher останется погашенным. При этой температуре как стебли интактного, так и разделенного LOCS будут гибридизоваться и, следовательно, будут гаситься независимо от присутствия или отсутствия мишени 2. Таким образом, увеличение флуоресценции во время амплификации указывает на мишень 1. Кроме того, измерение флуоресценции при второй температуре обнаружения, которая выше, чем Tm D, но ниже, чем Tm А и Tm С, может свидетельствовать о присутствии мишени 1 и/или мишени 2. При этой второй температуре Catcher остается одноцепочечным и гасится в отсутствие мишени и образует дуплекс и флуоресцирует в присутствии мишени. Кроме того, при этой второй температуре, если мишень 2 отсутствует, зонд LOCS останется интактным и гасится, но будет расщеплен в присутствии мишени 2, и его стебель диссоциирует, вызывая флуоресценцию. В этом сценарии, если флуоресценция при температуре 1 увеличивается во время амплификации, это указывает на присутствие мишени 1. Кроме того, если увеличение флуоресценции, наблюдаемое при температуре 2В ходе амплификации, больше, чем наблюдаемое при температуре 1, это указывает на присутствие мишени 2. Увеличение флуоресценции при второй температуре, но не при первой, указывает на присутствие только мишени 2.
Во втором неограничивающем сценарии (см. Таблицу 2) специфическое обнаружение первой мишени при первой температуре обнаружения может быть достигнуто в соответствии со сценарием 1 выше. Кроме того, при второй температуре детектирования, которая выше, чем Tm А и Tm D, но ниже, чем Tm С, дуплекс Catcher диссоциирует (т.е. становится одноцепочечным) и гасится независимо от присутствия или отсутствия мишени 1 и/или мишени 2, так как температура выше той, при которой может образоваться дуплекс Catcher. При отсутствии мишени 2Все LOCS будут интактными и погашенными; однако, когда присутствует мишень 2, будут генерироваться расщепленные LOCS, их стебли будут диссоциировать, и можно будет наблюдать увеличение флуоресценции. Следовательно, увеличение флуоресценции во время ПЦР при первой температуре указывает на присутствие мишени 1 независимо от наличия или отсутствия мишени 2; и, наоборот, увеличение флуоресценции во время ПЦР при второй температуре свидетельствует о присутствии мишени 2 независимо от наличия или отсутствия мишени 1. Таким образом, комбинация LOCS и зондов Catcher-Pitcher может позволить обнаруживать мишень 1 только с применением зондов Catcher-Pitcher, о чем свидетельствует увеличение флуоресценции над фоном при первой температуре; и обнаружение мишени 2 только с применением зондов LOCS, о чем свидетельствует увеличение флуоресценции над фоном при второй температуре.
В третьем неограничивающем сценарии (см. Таблицу 2), когда сигнал измеряется при первой температуре обнаружения, которая ниже, чем Tm А, но выше, чем Tm С и Tm D, тогда в присутствии только мишени 1 может образоваться дуплекс Catcher и флуоресцировать, в то время как в отсутствие мишени 1 одноцепочечный Catcher останется погашенным. При этой температуре как стебли интактного, так и разделенного LOCS будут диссоциированы и будут генерировать высокий уровень фоновой флуоресценции, независимо от наличия или отсутствия мишени 2. Таким образом, увеличение флуоресценции выше фоновой флуоресценции во время амплификации указывает только на мишень 1. Кроме того, измерение флуоресценции при второй температуре обнаружения, которая ниже, чем Tm А и Tm С, но выше, чем Tm D, может свидетельствовать только о присутствии мишени 2. При этой второй температуре Catcher остается одноцепочечным и гасится в отсутствие мишени и образует дуплекс и флуоресцирует в присутствии мишени. Кроме того, при этой второй температуре, если мишень 2 отсутствует, зонд LOCS останется интактным и гасится, но будет расщеплен в присутствии мишени 2, и его стебель диссоциирует, вызывая флуоресценцию. В этом сценарии, если флуоресценция при температуре 1 увеличивается во время амплификации, это указывает на присутствие только мишени 1, а если флуоресценция при температуре 2 увеличивается во время амплификации, это указывает на присутствие только мишени 2.
В другом неограничивающем формате, например, при обнаружении с помощью ППР, Catcher может быть прикреплен к наночастице золота (НЧЗ) и свободно находиться в растворе, в то время как Pitcher может быть прикреплен к поверхности золота. В присутствии мишени 1 и при температурах ниже Tm А дуплекс Catcher может образоваться и приблизить НЧЗ к поверхности золота, что приведет к измеримому сдвигу сигнала ППР. Однако в отсутствие мишени 1 Catcher был бы одноцепочечным и свободным в растворе (т.е. не находился бы в непосредственной близости от поверхности золота) и, следовательно, не вызывал бы какого-либо измеримого сдвига сигнала ППР по сравнению с исходным сигналом ППР. Следовательно, любой измеримый сдвиг сигнала ППР при этой температуре будет свидетельствовать о присутствии мишени 1 в образце. Кроме того, LOCS может быть прикреплен одним концом к НЧЗ, а другим концом к поверхности золота. В отсутствие мишени 2 НЧЗ всегда будет присоединен, тогда как при наличии мишени 2 будет генерироваться расщепленный LOCS, и поэтому в этом случае НЧЗ будет находиться вблизи поверхности золота только при температуре обнаружения ниже, чем у расщепленных LOCS. Как и прежде, в сценарии, аналогичном сценарию 2 выше, если первая температура обнаружения ниже Tm В, Tm С и Tm D, то изменение сигнала укажет на присутствие мишени 1, поскольку НЧЗ на Catcher будет близок к золотой поверхности. Если вторая температура обнаружения ниже Tm С, но выше Tm А и Tm D, то изменение сигнала укажет на присутствие мишени 2, так как НЧЗ на расщепленном LOCS будет удаляться от поверхности золота.
В еще одном формате, например, колориметрическом детектировании, как Catcher, так и зонд LOCS могут быть помечены на обоих концах НЧЗ. При первой температуре ниже Tm А, Tm С и Tm D присутствие мишени 1 приведет к измеримому изменению цвета с фиолетового (агрегированный НЧЗ) на красную (рассеянный НЧЗ), независимо от присутствия или отсутствия мишени 2. При второй температуре выше Tm А и Tm D, но ниже Tm С присутствие мишени 2 приведет к измеримому изменению цвета с фиолетового (агрегированный НЧЗ) на красный (рассеянный НЧЗ), независимо от присутствия или отсутствия мишени 1.
В еще одном формате catcher может быть помечен электроактивным компонентом, таким как метиленовый синий или ферроцен, и pitcher может быть прикреплен к поверхности электрода. При первой температуре (ниже Tm А) на поверхности электрода (при наличии мишени 1) будет образовываться дуплекс Catcher, в результате чего электроактивный фрагмент окажется в непосредственной близости от поверхности электрода, что вызовет измеримый сдвиг электрохимического сигнала (т.е. ток окисления или восстановления). Однако в отсутствие мишени 1 catcher будет находиться в свободном состоянии в растворе, а не в непосредственной близости от поверхности электрода, и, следовательно, не будет производить какого-либо измеримого сдвига электрохимического сигнала (т.е. тока окисления или восстановления) выше базового сигнала. Следовательно, любой измеримый сдвиг электрохимического сигнала при этой температуре будет свидетельствовать о присутствии мишени 1 в образце.
LOCS в сочетании с двойными зондами гибридизации
В некоторых вариантах реализации композиции и способы по настоящему изобретению включают комбинацию LOCS и олигонуклеотидного зонда, который содержит два компонента, специфичных для мишени.
Двойные гибридизационные зонды могут содержать первый олигонуклеотид с Tm А и второй олигонуклеотид с Tm В, где Tm А и Tm В могут быть одинаковыми, или Tm А и Tm В могут быть разными. Первый олигонуклеотид может быть помечен на его 3'-конце флуорофором, а второй олигонуклеотид может быть помечен на его 5'-конце гасителем. (В качестве альтернативы один олигонуклеотид может быть помечен на его 3'-конце гасителем, а другой может быть помечен на его 5'-конце флуорофором.) Два олигонуклеотидных зонда гибридизуются рядом или практически рядом (например, менее чем в 2, 3, 4 или 5 нуклеотидов) на мишени 1 и образуют дуплексную структуру с Tm, равной наименьшему из Tm А и Tm В. В этом сценарии, подходящая первая температура для сбора данных может быть ниже Tm А и Tm В, и ниже Tm С и Tm D; а именно Tins стеблей интактного и разделенного LOCS соответственно. При этой первой температуре первый и второй олигонуклеотиды находятся в свободном состоянии в растворе и флуоресцируют до амплификации или в отсутствие мишени 1, но гибридизация с мишенью 1, если она присутствует, сближает флуорофор и гаситель, вызывая гашение. Далее при первой температуре будет происходить гашение как стебля интактного, так и разделенного LOCS. Таким образом, наблюдаемое снижение флуоресценции при первой температуре указывает на присутствие мишени 1, независимо от наличия или отсутствия мишени 2.
Зонд LOCS может расщепляться МНКзимом в присутствии второй мишени 2. Сбор данных можно проводить при второй температуре, которая выше Tm А и/или Tm В, выше Tm D, но ниже Tm С. В этом сценарии интактный LOCS остается погашенным до амплификации и в отсутствие мишени, но в присутствии мишени 2 расщепленный стебель LOCS будет диссоциировать, что приведет к увеличению флуоресценции. При этой температуре первый и второй олигонуклеотид будут флуоресцировать независимо от присутствия или отсутствия мишени 1, поскольку они не могут гибридизоваться с мишенью 1 при этой температуре. Эта флуоресценция вносит свой вклад в фоновый сигнал при этой температуре. Таким образом, увеличение флуоресценции выше фона после амплификации указывает на присутствие мишени 2 независимо от присутствия или отсутствия мишени 1.
Таким образом, комбинация позволяет обнаруживать мишень 1 с применением зондов двойной гибридизации, что определяется как уменьшение флуоресценции при первой температуре; и обнаружение мишени 2 только с применением зондов LOCS, определяемое по увеличению флуоресценции при второй температуре.
В другом формате, например, при обнаружении с помощью ППР, первый олигонуклеотид может быть присоединен к наночастице золота (НЧЗ), а второй олигонуклеотид может быть присоединен к поверхности золота. При первой температуре два олигонуклеотидных зонда могут гибридизоваться рядом на мишени 1 (если она присутствует), образуя дуплексную структуру и приближая НЧЗ к поверхности золота, что может вызывать измеримый сдвиг сигнала ППР. Однако в отсутствие мишени 1 первый олигонуклеотид может быть свободным в растворе и не находиться в непосредственной близости от поверхности золота и, следовательно, не вызывать какого-либо измеримого сдвига сигнала ППР по сравнению с исходным сигналом ППР. Следовательно, любой измеримый сдвиг сигнала ППР при этой температуре может свидетельствовать о присутствии мишени 1 в образце.
В еще одном формате, например, колориметрическом обнаружении, как первый, так и второй олигонуклеотиды могут быть помечены наночастицами золота. При первой температуре первый и второй олигонуклеотиды могут быть свободными в растворе и проявлять красный цвет в отсутствие мишени 1. Однако в присутствии мишени 1 два олигонуклеотидных зонда могут гибридизоваться рядом с мишенью, образуя дуплексную структуру и приближая НЧЗ друг к другу, вызывая измеримое изменение цвета с красного на фиолетовый. Следовательно, измеримое изменение цвета с красного на фиолетовый может свидетельствовать о присутствии мишени 1 в образце.
В еще одном формате первый олигонуклеотид может быть помечен электроактивным компонентом, таким как метиленовый синий или ферроцен, а второй олигонуклеотид может быть прикреплен к поверхности электрода. При первой температуре два олигонуклеотидных зонда могут гибридизоваться рядом на мишени 1 (если она присутствует), образуя дуплексную структуру на поверхности электрода и приближая электроактивный фрагмент к поверхности электрода, что может вызвать измеримый сдвиг электрохимического сигнала (то есть ток окисления или восстановления). Однако в отсутствие мишени 1 первый олигонуклеотид может быть в свободном состоянии в растворе, а не в непосредственной близости от поверхности электрода, и, следовательно, не будет производить какого-либо измеримого сдвига электрохимического сигнала (т.е. тока окисления или восстановления) выше базового сигнала. Следовательно, любой измеримый сдвиг электрохимического сигнала при этой температуре может свидетельствовать о присутствии мишени 1 в образце.
Комбинации LOCS, содержащие расщепляемые зонды В некоторых вариантах реализации композиции и способы по настоящему изобретению включают комбинацию LOCS и олигонуклеотидного зонда, который генерирует детектируемые сигналы, зависящие от мишени, которые не могут быть обратимо модулированы температурой. Тем не менее, LOCS по-прежнему допускает модуляцию генерации сигнала температурой независимым от цели способом, что позволяет манипулировать фоновым шумом или базовыми уровнями.
Например, первый олигонуклеотидный зонд для первой мишени 1 может подвергаться мишень-зависимой модификации, обеспечивающей детектируемый сигнал, который нельзя подавить при изменении температуры детектирования. Зависимой от мишени модификацией может быть расщепление или расщепление первого олигонуклеотида, что приводит к запуску детектируемого сигнала, указывающего на присутствие мишени. Неограничивающие примеры олигонуклеотидных зондов в этой категории включают зонды TaqMan, субстраты МНКзима и зонды, которые расщепляются ферментами рестрикции в зависимости от мишени. После модификации сигнал, генерируемый этими зондами, можно измерять в широком диапазоне температур.
В этих вариантах реализации неповрежденные LOCS, предназначенные для обнаружения второй мишени 2, могут подвергаться расщеплению с образованием расщепленных LOCS только в присутствии этой мишени. Измерение мишени 2 требует детектирования при температуре ниже Tm стебля интактного LOCS, но выше Tm стебля интактного LOCS. Напротив, температура, при которой обнаруживается первая мишень, может быть либо ниже Tms стеблей как интактного, так и разделенного LOCS, или выше Tins стеблей как интактного, так и разделенного LOCS. Когда первая температура обнаружения ниже Tms стеблей, фоновый сигнал будет подавляться, так как и интактный, и/или разделенный стебель останутся связанными и погашенными, тогда как, когда первая температура обнаружения выше Tms стеблей, фоновый сигнал будет выше из-за диссоциации как интактных, так и/или расщепленных стеблей. Таким образом, обнаружение сигнала при первой температуре может быть специфичным для мишени 1, независимо от наличия или отсутствия мишени 2. Обнаружение сигнала при второй температуре указывает на присутствие мишени 1 и/или мишени 2; однако, если увеличение сигнала, наблюдаемое при второй температуре, больше, чем наблюдаемое при первой температуре, то это указывает на присутствие мишени 2. Если изменение сигнала наблюдается только при второй температуре, а не при первой, это будет свидетельствовать о наличии только второй мишени.
Следующие сценарии представляют собой неограничивающие примеры анализов мультиплексного обнаружения в соответствии с вариантами реализации настоящего изобретения и суммированы в Таблице 3 ниже. В этих сценариях LOCS может включать Tm А, представляющую собой температуру плавления его двухцепочечной области стебля, когда он неповрежден, и Tm В, являющуюся температурой плавления его двухцепочечной области стебля, когда он разделен. Олигонуклеотидный зонд (например, субстрат МНКзима) может быть помечен флуорофором и гасителем, как и противоположные цепи двухцепочечной стеблевой части LOCS. Флуорофор олигонуклеотидного зонда может быть таким же или излучать в той же области видимого спектра, что и флуорофор LOCS. В альтернативных вариантах реализации можно использовать различные детектирующие фрагменты, включая, например, наночастицы одинакового или сходного размера и/или типа для колориметрического или SPR-детектирования, реактивные фрагменты (например, ферменты щелочной фосфатазы или пероксидазы) для хемилюминесцентного детектирования или электроактивные частицы (например, ферменты ферроцен, метиленовый синий или пероксидаза) для электрохимического обнаружения. Специалисту в данной области техники легко понять, что олигонуклеотидный зонд может, например, представлять собой субстрат МНКзим, зонд TaqMan, зонд гидролиза или зонд, расщепляемый рестрикционным ферментом в зависимости от мишени (зонд RE).
Изменение сигнала из-за присутствия или отсутствия первой мишени может быть получено путем измерения флуоресценции после амплификации (после ПЦР) при первой температуре, нормализованной по отношению к фоновой флуоресценции, полученной либо в начале амплификации до ПЦР, либо вблизи нее (пре-ПЦР) при той же первой температуре; в то время как изменение сигналов из-за присутствия или отсутствия второй мишени может быть получено путем получения полной флуоресценции после ПЦР при второй температуре, нормализованной по отношению к фоновой флуоресценции, измеренной до ПЦР при этой второй температуре. В качестве альтернативы, изменение сигнала из-за присутствия или отсутствия первой и второй мишени соответственно может быть получено путем измерения общей флуоресценции после амплификации при первой и второй температуре, обе из которых могут быть нормализованы относительно фоновой флуоресценции при третьей температуре, полученной либо пре-ПЦР в той же реакции или в любое время в контрольной реакции, которая эквивалентна, но в которой отсутствует мишень. Измерение фона можно проводить при третьей температуре при условии, что первая температура меньше второй температуры, а третья температура меньше Tm интактного стебля.
Во всех сценариях присутствие или отсутствие сигнала, указывающего на первую мишень, измеряют при первой температуре с применением первого зонда, который может быть, например, субстратом МНКзимом, зондом TaqMan или зондом RE. Перед усилением этот зонд будет погашен; однако, если присутствует мишень 1, зонд будет расщеплен во время амплификации, что приведет к увеличению флуоресценции, обнаруживаемой как при первой, так и при второй температуре. Наличие или отсутствие второй мишени определяют при второй температуре с помощью зонда LOCS, где вторая температура всегда выше Tm разделенного LOCS (Tm В), но ниже Tm интактного LOCS (Tm А).
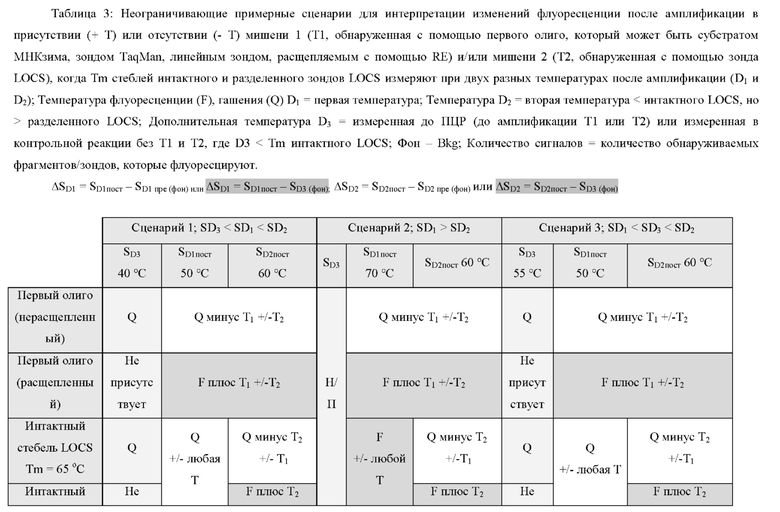
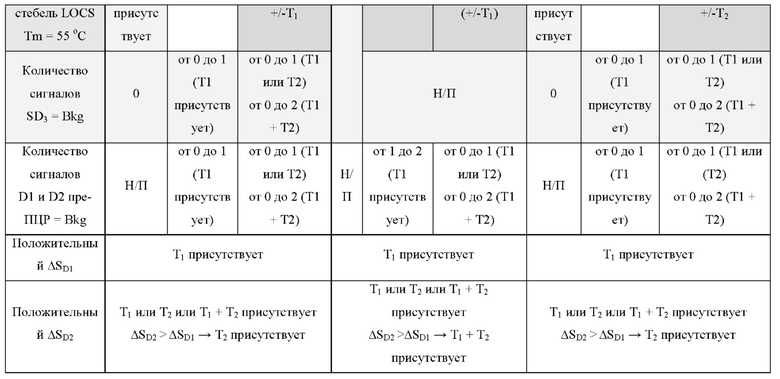
Сценарий 1.
В первом неограничивающем примере, как описано в Таблице 3, первая температура ниже, чем вторая температура, и ниже, чем Tm А и Tm В. Увеличение флуоресценции при первой температуре будет указывать на присутствие мишени 1 и фоновый сигнал будет отражать наличие погашенных LOCS, где как интактные, так и/или разделенные стебли остаются связанными. Фон при первой температуре и второй температуре можно измерить перед ПЦР при каждой температуре соответственно. В качестве альтернативы фоновое измерение может быть измерено при третьей температуре, где третья температура ниже как первой, так и второй температуры.
Сценарий 2
Во втором неограничивающем примере, как описано в Таблице 3, первая температура выше, чем вторая температура, а также выше, чем Tm А и Tm В. Увеличение флуоресценции при первой температуре будет указывать на присутствие мишени 1 и фоновый сигнал будет отражать флуоресценцию от интактных и/или разделенных LOCS с диссоциированными стеблями. Фон при первой температуре и второй температуре можно измерить перед ПЦР при каждой температуре соответственно.
Сценарий 3
В третьем неограничивающем примере, как описано в Таблице 3, первая температура ниже, чем вторая температура, и ниже, чем Tm А и Tm В. Увеличение флуоресценции при первой температуре будет указывать на присутствие мишени 1 и фоновый сигнал будет отражать наличие погашенных LOCS, где как интактные, так и/или разделенные стебли остаются связанными. Фон при первой температуре и второй температуре можно измерить перед ПЦР при каждой температуре соответственно. В качестве альтернативы фоновое измерение может быть измерено при третьей температуре, где третья температура ниже второй температуры, но выше первой температуры.
В других форматах, например, при обнаружении с помощью ППР, первый олигонуклеотид и зонд LOCS могут быть присоединены к наночастице золота (НЧЗ) одним концом и прикреплены к поверхности золота другим концом. В отсутствие мишени первый олигонуклеотид может оставаться интактным, а НЧЗ будет находиться в непосредственной близости от поверхности золота, создавая базовый уровень сигнала ППР. В присутствии мишени 1 первый олигонуклеотид может расщепляться, отделяя НЧЗ от поверхности золота и вызывая измеримый сдвиг сигнала ППР, указывающий на присутствие первой мишени в образце. Это может быть измерено при первой температуре ниже Tm А и Tm В, так что НЧЗ, прикрепленная или связанная либо с интактными, либо с расщепленными LOCS, может оставаться на поверхности. Измерение изменения сигнала ППР при второй температуре выше Tm разделенного LOCS может указывать на присутствие первой и/или второй мишени (мишеней). Кроме того, если изменение сигнала наблюдается при первой температуре, а большее изменение наблюдается при второй температуре, это указывает на присутствие мишени 2. В качестве альтернативы, если сигнал обнаруживается при второй температуре, но не при первой, это также будет указывать только на вторую мишень.
В другом формате, например при колориметрическом обнаружении, первый олигонуклеотидный зонд и LOCS могут быть помечены на обоих концах наночастицами золота. При первой температуре ниже Tm А и Tm В первый олигонуклеотид может оставаться интактным в отсутствие мишени 1, где НЧЗ могут находиться в агрегированном состоянии в непосредственной близости друг от друга, проявляя фиолетовый цвет.Однако первый олигонуклеотид может расщепляться в присутствии мишени 1, отделяя НЧЗ и демонстрируя заметное изменение цвета с фиолетового на красный. Следовательно, измеримое изменение цвета с фиолетового на красный может свидетельствовать о присутствии мишени 1 в образце. При второй температуре, которая выше первой температуры и выше Tm В, но ниже Tm А, в присутствии только второй мишени возможно расщепление LOCS и НЧЗ будут находиться в дисперсном состоянии, а цвет изменится с фиолетового до красного. Если присутствуют обе мишени, изменение цвета может быть более интенсивным.
В еще одном формате первый олигонуклеотид может быть помечен электроактивным фрагментом, таким как метиленовый синий или ферроцен, на одном конце, присоединенным к поверхности электрода на другом конце. При первой температуре и в отсутствие мишени 1 первый олигонуклеотид может оставаться интактным, а электроактивные частицы могут находиться в непосредственной близости от поверхности электрода, создавая базовый уровень электрохимического сигнала (т.е. ток окисления или восстановления). Однако в присутствии мишени 1 первый олигонуклеотид может быть расщеплен, и электроактивная часть может быть высвобождена в раствор, вызывая измеримый сдвиг электрохимического сигнала (т.е. тока окисления или восстановления) выше исходного сигнала. Следовательно, любой измеримый сдвиг электрохимического сигнала при этой температуре может свидетельствовать о присутствии мишени 1 в образце.
Дополнительные типовые варианты реализации
В некоторых вариантах реализации репортерные олигонуклеотиды, включая LOCS-олигонуклеотиды по настоящему изобретению, можно использовать для непосредственного обнаружения мишени. В других иллюстративных вариантах реализации репортерные зонды или субстраты могут быть использованы для обнаружения ампликонов мишеней, полученных с помощью технологий амплификации мишени, включая, помимо прочего, ПЦР, ОТ-ПЦР, SDA, NEAR, HDA, RPA, LAMP, RCA, ТМА, 3SR, LCR, RAM или NASBA. Расщепление или деградация, приводящая к расщеплению LOCS, могут происходить в реальном времени во время амплификации мишени или после амплификации, в конечной точке реакции. Петлевую область можно раскрыть в результате мишень-зависимого расщепления или разрушения, опосредованного ферментативной активностью нуклеиновой кислоты, с каталитической активностью, включая МНКзим, ДНКзим, рибозим или ферментативной активностью белкового фермента, включая экзонуклеазу или эндонуклеазу, но не ограничиваясь ими. В качестве неограничивающего примера, экзонуклеазная активность может представлять собой каталитическую активность, присущую, например, полимеразе. В качестве неограничивающего примера, эндонуклеазная активность может представлять собой каталитическую активность, присущую, например, ферменту рестрикции, включая никующую эндонуклеазу, рибоэндонуклеазу или нуклеазу, специфичную к двуцепочечной структуре (DSN).
Реакционные смеси по настоящему изобретению предназначены для одновременного обнаружения нескольких мишеней с применением одного олигонуклеотида LOCS в сочетании с другими типами репортерных зондов. Как будет очевидно специалистам в данной области техники, стандартные репортерные зонды могут дополнительно комбинироваться с дополнительными LOCS, например, при этом каждый LOCS может содержать другой универсальный субстрат в своей петле и другую стеблевую область, способную плавиться при другой температуре после расщепления субстрата/петли различными МНКзимами.
Реакционная смесь может дополнительно содержать дополнительные репортерные зонды или субстраты в сочетании с LOCS, меченными различными парами флуорофора и гасителя. В качестве неограничивающего примера, репортерный олигонуклеотид 1 и интактный олигонуклеотид LOCS 2 могут быть помечены флуорофором А, а репортерный олигонуклеотид 3 и интактный олигонуклеотид LOCS 4 могут быть помечены флуорофором В. В варианте реализации, репортерные олигонуклеотиды 1 и 3 представляют собой линейные субстраты МНКзим, причем МНКзим 1 может образовываться в присутствии мишени 1 и расщеплять репортерный олигонуклеотид 1, вызывая флуоресценцию при всех температурах; и МНКзим 2 может образовываться в присутствии мишени 2 и расщеплять субстрат 2Внутри LOCS-олигонуклеотида 2, что приводит к расщепленной, расщепленной структуре LOCS 2, содержащей стебель 2, который плавится при температуре X. МНКзим 3 может образовываться в присутствии мишени 3 и расщеплять репортерный олигонуклеотид 3 для генерации флуоресценции при всех температурах. МНКзим 4 может образовываться в присутствии мишени 4 и расщеплять субстрат 4 внутри LOCS-олигонуклеотида 4, что приводит к расщепленной структуре LOCS 4, содержащей стебель 4, который плавится при температуре Y.
При анализе флуоресценции при температуре ниже как X, так и Y, возбуждение при длине волны флуорофора А указывает на присутствие мишени 1, а возбуждение при длине волны флуорофора В указывает на присутствие мишени 3. При анализе флуоресценции при температуре выше как X, так и Y, но ниже температуры плавления обоих интактных олигонуклеотидов LOCS 2 и 4, тогда возбуждение при длине волны флуорофора А указывает на присутствие мишени 1 и/или мишени 2, а возбуждение при длине волны Флуорофора В указывает на присутствие мишени 3 и/или мишени 4.
При анализе профиля плавления реакционной смеси при длине волны возбуждения флуорофора А, наличие пика при первой температуре X и отсутствие пика при второй температуре 2, которая выше температуры X, указывает на то, что репортер LOCS 2 был разделен/расщеплен МНКзимом в присутствии мишени 2. Альтернативно, отсутствие пика при температуре X и наличие пика при температуре 2 указывает на то, что репортер LOCS 2 остается интактным и не расщепляется МНКзимом из-за отсутствия мишени 2В образце. При анализе профиля плавления реакции при длине волны возбуждения флуорофора В наличие пика при третьей температуре Y и отсутствие пика при четвертой температуре 4, которая выше температуры Y, указывает на расщепление репортера LOCS 4 с помощью МНКзима в присутствии мишени 4, тогда как отсутствие пика при температуре Y и наличие пика при температуре 4 указывает на то, что репортер 4 LOCS остается интактным и не расщепляется МНКзимом из-за отсутствия мишени 4 в образце. Поскольку такой анализ при двух длинах волн, считываемый по двум каналам на приборе, может использоваться в качестве подтверждающего инструмента для обнаружения и дифференциации мишеней при условии, что присутствие оставшихся двух мишеней определяется с применением других средств обнаружения, таких как обнаружение в реальном времени. Специалисту в данной области техники будет понятно, что эта стратегия может быть расширена для мониторинга расщепления более чем двух мишеней на одной конкретной длине волны, и, кроме того, количество анализируемых флуорофоров может быть увеличено до количества, определяемого максимальной способностью доступного прибора различать отдельные длины волн.
В другом примерном варианте реализации можно комбинировать линейный субстрат МНКзим и олигонуклеотид LOCS, при этом оба содержат одну и ту же пару флуорофор/гаситель, а участки субстрата специфичны для ДНКзима или рибозима, например, ДНКзима или рибозима, которые могут только быть каталитически активным в присутствии определенного иона металла. В данной области техники известно, что специфические ДНКзимы и рибозимы требуют наличия ко фактора-катиона металла для обеспечения каталитической активности. Например, некоторые ДНКзимы и рибозимы могут быть каталитически активными только в присутствии, например, свинца или ртути. Такие металлы могут присутствовать, например, в образце из окружающей среды. Реакция может включать один линейный субстрат МНКзим для ДНКзима, который, например, зависит от ртути, при этом присутствие ртути в образце может привести к расщеплению линейного субстрата МНКзима и генерации флуоресцентного сигнала. Та же реакция может также включать репортер LOCS, который содержит петлю, содержащую субстрат для ДНКзима, который, например, зависит от свинца, при этом присутствие свинца в образце может привести к расщеплению LOCS и генерации флуоресцентного сигнала при температуре выше Tm разделенного LOCS. Увеличение флуоресценции при первой температуре, которая ниже Tm разделенного LOCS, указывало бы на присутствие ртути. Увеличение флуоресценции при второй температуре, которая выше Tm разделенного LOCS, но ниже Tm интактного LOCS, указывает на присутствие ртути и/или свинца. Если бы ртуть была обнаружена при первой температуре, то дальнейшее усиление флуоресценции при второй температуре указывало бы на присутствие свинца. Это можно подтвердить, дав реакционной смеси охладиться, обеспечивая возможность повторного отжига интактного стебля разделенной LOCS-структуры, а затем можно выполнить анализ кривой плавления, чтобы определить наличие или отсутствие пиков, указывающих на присутствие структур разделенного LOCS. Специалист в данной области легко поймет, что несколько зондов, расщепляемых в присутствии определенных кофакторов металлов, могут быть объединены в одну реакцию и обнаружены либо в реальном времени, либо по окончании реакции.
Неограничивающие примеры нуклеиновых кислот-мишеней (т.е. полинуклеотидов), которые могут быть обнаружены с применением олигонуклеотидов LOCS в сочетании с другими хорошо известными типами зондов, могут включать ДНК, метилированную ДНК, алкилированную ДНК, комплементарную ДНК (кДНК), РНК, метилированную РНК, микроРНК, миРНК, кшРНК, мРНК, тРНК, мякРНК, мвРНК, ммРНК, предшественники микроРНК и первичные микроРНК, другие некодирующие РНК, рибосомные РНК, их производные, их ампликоны или любые их комбинации (включая смешанные полимеры дезоксирибонуклеотидных и рибонуклеотидных оснований).
Образование детектируемых сигналов
В способах и композициях по настоящему изобретению используются детектирующие фрагменты для обеспечения обнаружимых сигналов. Природа детектируемого сигнала, который способны продуцировать фрагменты, будет зависеть от типа детектирующего фрагмента и/или конформации олигонуклеотида, с которым он связан.
Можно использовать любую подходящую обнаружимую часть, которая способна обеспечить детектируемый сигнал при модификации олигонуклеотида, с которым она связана. Неограничивающие примеры подходящих обнаружимых молекул включают флуорофоры для генерации флуоресцентного сигнала, наночастицы для генерации колориметрического сигнала или сигнала ППР, реакционноспособные фрагменты (например, ферменты щелочной фосфатазы или пероксидазы) для генерации хемилюминесцентного сигнала, электроактивные частицы для генерации электрохимического сигнала и любую их комбинацию. В качестве неограничивающего примера подходящие электроактивные соединения включают метиленовый синий, толуоловый синий, орасетовый синий, ферроцен, Hoechst 33258, [Ru(phen)3]2+или дауномицин, а наиболее распространенные электродные материалы включают золото, стеклоуглерод, карандашный графит или углеродную ионную жидкость. Способы обнаружения и измерения флуоресцентных, хемилюминесцентных, колориметрических сигналов, сигналов поверхностного плазмонного резонанса (ППР) и электрохимических сигналов хорошо известны специалистам в данной области техники.
В качестве неограничивающего примера олигонуклеотиды по настоящему изобретению, включая LOCS, могут иметь присоединенный один или более флуорофоров. Обнаруживаемый сигнал, изначально генерируемый флуорофором, может подавляться из-за близости к одной или более молекулам гасителя. Например, но не ограничиваясь этим, флуорофор(ы) может быть присоединен к одной цепи двухцепочечной области стебля (например, на 5'- или 3'-конце) молекулярного маяка или LOCS, а гаситель(и) может быть присоединен к противоположной цепи двухцепочечной области стебля (например, на 5'-или 3'-конце). В качестве альтернативы, гаситель(и) может быть присоединен к другому объекту (например, к поверхности или другому олигонуклеотиду), с которым связан олигонуклеотид, таким образом, чтобы обнаруживаемый сигнал, изначально генерируемый флуорофором, мог гаситься. В присутствии мишени олигонуклеотид может подвергаться модификации, которая отдаляет флуорофор(ы) от молекулы(молекул) гасителя, тем самым генерируя детектируемый сигнал.
Дополнительно или альтернативно олигонуклеотиды (включая LOCS) могут быть присоединены к НЧЗ для колориметрического обнаружения. Когда наночастицы золота собраны в непосредственной близости друг от друга, они приобретают фиолетовый цвет (т.е. поглощают при большей длине волны), а когда наночастицы золота разделены, они приобретают красный цвет (т.е. поглощение на более короткой длине волны), при этом измеримое изменение цвета с фиолетового на красный (например, LOCS, линейные субстраты МНКзима, зонды Catcher-Pitcher, зонды TaqMan и зонды рестрикционных ферментов) или, альтернативно, от красного до фиолетового (например, зонды двойной гибридизации) указывают на присутствие в образце конкретной мишени.
Дополнительно или альтернативно, олигонуклеотиды (включая LOCS) и/или олигонуклеотидные компоненты могут быть присоединены к НЧЗ и/или поверхности золота для обнаружения мишени в образце с помощью ППР. Когда НЧЗ перемещаются в непосредственной близости или, альтернативно, когда они удаляются от поверхности золота, они могут генерировать изменение измеримого сигнала ППР, когда уменьшение сигнала ППР с применением некоторых подходов (например, LOCS, линейных субстратов МНКзима, зондов TaqMan и зондов рестрикционных ферментов) может свидетельствовать о присутствии конкретной мишени в образце или, альтернативно, увеличение сигнала ППР при использовании других подходов (например, зондов Catcher-Pitcher и зондов двойной гибридизации) может свидетельствовать о присутствии конкретной мишени в образце.
Дополнительно или альтернативно олигонуклеотидный репортер и зонды (включая LOCS) и/или олигонуклеотидные компоненты могут быть присоединены к электроактивным веществам и/или к поверхности электрода для электрохимического обнаружения. Когда олигонуклеотиды, присоединенные к электроактивным частицам, перемещаются в непосредственной близости от поверхности электрода или, альтернативно, когда они удаляются от нее, они могут генерировать измеримое изменение тока окисления или восстановления. В некоторых вариантах реализации (например, LOCS, линейные субстраты МНКзима, зонды TaqMan и зонды рестрикционных ферментов) результирующий измеримый сигнал, возникающий при удалении электроактивных частиц от поверхности электрода, указывает на присутствие конкретной мишени в образце. Альтернативно, в других вариантах реализации (например, зонды Catcher-Pitcher и зонды двойной гибридизации) результирующий измеримый сигнал, возникающий в результате перемещения электроактивных частиц в непосредственной близости от поверхности электрода, указывает на присутствие конкретной мишени в образце.
В некоторых вариантах реализации в композициях и способах по настоящему изобретению используются LOCS, присоединенные к определенному детектирующему фрагменту, в сочетании с другим олигонуклеотидным зондом, который присоединен к тому же детектирующему фрагменту, или аналогичный детектирующий фрагмент, который генерирует обнаруживаемый сигнал, способный обнаруживаться одновременно с сигналом, генерируемым обнаруживаемой частью LOCS (например, с применением одного типа детектора, такого как один флуоресцентный канал, или определенного режима колориметрического, поверхностного плазмонного резонанса (ППР), хемилюминесцентного или электрохимического обнаружения).
Анализ флуоресцентных сигналов
Без ограничения и только в качестве примера обнаружимые фрагменты, используемые в соответствии с настоящим изобретением, включают флуоресцентные сигналы, генерируемые этими детектируемыми фрагментами при модификации, расщеплении или разрушении олигонуклеотидных зондов, к которым они присоединены, объединены или связаны иным образом, включая диссоциацию расщепленные структуры LOCS, можно анализировать любым подходящим способом для обнаружения, дифференциации и/или количественного определения молекул-мишеней в соответствии со способами по настоящему изобретению.
Несмотря на то, что можно использовать стандартные анализы кривой плавления, в настоящем документе описаны и проиллюстрированы различные другие подходы (см. «Примеры»), которые можно легко применять для анализа в различных форматах.
В качестве неограничивающего примера, в различные моменты времени в ходе реакции, подходящей для обнаружения расщепления или разрушения петлевых областей LOCS-олигонуклеотидов, можно выполнить измерения сигнала флуоресценции при одной температуре или при нескольких температурах. В качестве неограничивающих примеров, эти моменты времени могут включать (i) момент времени в начале реакции или близкий к нему, и/или (i) один момент времени или несколько моментов времени в ходе реакции; и/или (iii) момент завершения или конечной точки реакции.
В некоторых вариантах реализации измерение флуоресцентного сигнала можно выполнять при двух или более температурах в каждом цикле во время реакции амплификации, такой как ПЦР-амплификация. Анализ можно выполнять путем сравнения уровней флуоресценции, полученных при первой и/или второй температуре и/или при другой температуре.
В других вариантах реализации измерение флуоресцентного сигнала можно выполнять при двух температурах во время реакций, адаптированных для измерения двух мишеней на одной и той же длине волны.
В некоторых вариантах осуществления первая мишень может быть обнаружена с применением первого олигонуклеотидного репортерного зонда или субстрата, где флуоресцентные сигналы могут быть измерены при нескольких моментах времени или в нескольких циклах, например, в каждом цикле во время ПЦР. В той же реакции вторая мишень может быть обнаружена с применением зондов LOCS путем сравнения уровней флуоресценции до и после ПЦР. В таких вариантах реализации количественные данные могут быть определены для первой мишени, в то время как качественные данные могут быть получены для второй мишени. В качестве неограничивающего примера первый олигонуклеотидный зонд может представлять собой субстрат МНКзима, расщепляемый первым МНКзимом в присутствии первой мишени и контролируемый в режиме реального времени; тогда как зонд LOCS может расщепляться вторым МНКзимом в присутствии второго и контролироваться с помощью анализа обнаружения конечной точки. Примеры сочетания количественного анализа в реальном времени и качественного анализа конечной точки включают Пример 1, в котором используется один линейный субстрат МНКзим и один зонд LOCS, и Пример 6, в котором используется один зонд TaqMan и один зонд LOCS.
В некоторых вариантах реализации увеличение флуоресценции при первой температуре свидетельствует о присутствии первой мишени, а увеличение флуоресценции при второй температуре свидетельствует о присутствии первой и/или второй мишени. В других вариантах реализации увеличение флуоресценции при первой температуре свидетельствует о присутствии первой мишени, а увеличение флуоресценции при второй температуре свидетельствует о присутствии второй мишени. В еще других вариантах реализации уменьшение флуоресценции при первой температуре свидетельствует о присутствии первой мишени, а увеличение флуоресценции при второй температуре свидетельствует о присутствии второй мишени. В других вариантах реализации измерение флуоресцентного сигнала может быть получено при двух или более температурах в каждом цикле во время ПЦР, и кривые амплификации могут быть построены для каждой серии измерений, полученных при каждой температуре. Пороговые значения флуоресценции могут быть присвоены каждому графику амплификации для каждой конкретной температуры, а значения Cq могут быть измерены как номер цикла, в котором графики амплификации пересекают пороговые значения. В вариантах реализации, в которых первый зонд представляет собой стандартный линейный субстрат МНКзим для обнаружения мишени 1, а второй зонд представляет собой репортер LOCS для обнаружения мишени 2; и в которых измерение флуоресцентного сигнала получают при двух температурах в каждом цикле во время ПЦР, Cq, измеренное с применением флуоресцентного сигнала от линейного субстрата МНКзима при более низкой температуре, которая ниже, чем Tm разделенного LOCS, может позволить провести прямую количественную оценку исходной концентрации первой цели; и Cq, измеренное с применением общего флуоресцентного сигнала как от линейного субстрата МНКзима, так и от репортера LOCS при более высокой температуре, которая выше Tm разделенного LOCS, но ниже Tm интактного LOCS, может быть проанализировано, как показано в Примере 4, что позволяет количественно определить начальную концентрацию второй мишени.
В других вариантах реализации измерение флуоресцентного сигнала может быть получено при двух или более температурах в каждом цикле во время ПЦР, и кривые амплификации могут быть построены для каждой серии измерений, полученных при каждой температуре. Пороговые значения флуоресценции могут быть присвоены каждому графику амплификации для каждой конкретной температуры, а значения Cq могут быть измерены как номер цикла, в котором графики амплификации пересекают пороговые значения. В вариантах реализации, в которых первый зонд для обнаружения первой мишени, являющийся либо стандартным молекулярным маяком, либо зондом Catcher/Pitcher, либо унизонд Scorpion, либо бизонд Scorpion, объединен со вторым зондом для обнаружения второй мишени, которая может быть репортером LOCS; и в которых измерение флуоресцентного сигнала получают при двух температурах в каждом цикле во время ПЦР, Cq, измеренное с применением флуоресцентного сигнала от первого зонда при более низкой температуре, которая ниже Tm разделенного LOCS, может позволить прямое количественное определение исходной концентрации первой мишени; и Cq, измеренная с применением флуоресцентного сигнала от репортера LOCS при более высокой температуре, которая выше Tm разделенного LOCS, но ниже Tm интактного LOCS, может обеспечить прямое количественное определение исходной концентрации второй мишени. Конкретные комбинации первого стандартного зонда со вторым зондом LOCS могут иметь дополнительные требования к Tm конкретных областей первого зонда и к двум температурам, при которых собираются данные, как указано выше и как показано в качестве примера для молекулярных маяков в Примерах 5 и 7; для зондов Scorpion в Примере 9 и для олигонуклеотид о в Catcher Pitcher в Примере 11.
В качестве неограничивающего примера базовый сигнал флуоресценции может быть получен путем измерения флуоресценции при выбранных температурах, например, при первой и второй температуре, в момент времени, который находится либо в момент начала реакции, либо вблизи него, например, до ПЦР. Перед ПЦР и при первой температуре стандартный репортерный зонд, например, линейный субстрат МНКзим, или зонд TaqMan, или молекулярный маяк, и интактный LOCS должны быть погашены и не давать значительного флуоресцентного сигнала, при условии, что эта температура ниже Tm стебля интактного LOCS (и молекулярного маяка при его присутствии). Анализ можно проводить путем сравнения уровней флуоресценции, полученных при первой и второй температуре в момент времени в момент инициации реакции (например, до ПЦР), и уровней флуоресценции, полученных при первой и второй температуре в момент времени во время и/или или после реакции (например, во время ПЦР или после ПЦР).
Как показано в Примере 1, расщепление линейного субстрата МНКзима, измеренное при первой температуре после ПЦР, дает значительный сигнал флуоресценции по сравнению с сигналом интактного линейного субстрата МНКзима, измеренным при первой температуре перед ПЦР. Этот относительный сигнал (ΔSD1) пересекает первый заранее определенный порог, который указывает на присутствие первой мишени в образце. При этой первой температуре интактный LOCS и/или расщепленный, разделенный LOCS не способствует выработке значительного флуоресцентного сигнала по сравнению с сигналом, полученным до ПЦР, из-за Tm стебля обеих структур LOCS. При второй температуре, которая выше первой температуры, и в момент времени после ПЦР расщепление LOCS дает значительный сигнал флуоресценции по сравнению с сигналом, полученным при второй температуре в момент времени до ПЦР. Этот относительный сигнал (ΔSD2) пересекает второй заранее определенный порог и больше, чем относительный сигнал при первой температуре (ΔSD1), как показано в Примере 1 (способ анализа конечной точки 1), независимо от того, расщепляется ли линейный субстрат МНКзимом или нет и, таким образом, указывает на обнаружение разделенного LOCS и, следовательно, на присутствие второй мишени. При этой второй температуре и в момент времени после ПЦР интактный LOCS не способствует выработке значительного сигнала флуоресценции по сравнению с моментом времени до ПЦР при той же второй температуре и не превышает заранее установленный порог.Альтернативно, разницу между относительным сигналом, полученным до и после ПЦР при второй температуре, и относительным сигналом, полученным до и после ПЦР при первой температуре (ΔSD2 - ΔSD1), можно сравнить с заранее определенным порогом для определения наличие разделенных, расщепленных LOCS. Как показано в Примере 1 (способ анализа конечных точек 2), наличие расщепленного, разделенного LOCS, указывающего на присутствие второй мишени в образце, может быть определено, когда разница превышает предварительно определенный порог (ΔSD2 - ΔSD1>порог). Отсутствие второй цели в образце можно определить, когда значение разности ниже заранее установленного порога (ΔSD2 - ΔSD1<порог). Кроме того, когда разница между относительным сигналом, полученным до и после ПЦР при второй температуре, и относительным сигналом, полученным до и после ПЦР при первой температуре (ΔSD1 -ΔSD1), пересекает предварительно определенный порог, уникальное соотношение изменения флуоресценции от температуры 1 до температуры 2 (ΔSD1:ΔSD2) или обратное соотношение (ΔSD2:ΔSD1) можно использовать для определения того, присутствует ли мишень 1, мишень 2 или обе мишени 1 и 2Внутри образца. Как показано в Примере 1 (способ анализа конечных точек 3), если ΔSD1:ΔSD2 больше порогового значения R1, это означает, что присутствует только мишень 1; если ΔSD1:ΔSD2 меньше порогового значения R2, это означает, что присутствует только мишень 2; и если ΔSD1:ΔSD2 меньше порогового значения R1, но больше порогового значения R2, это указывает на то, что присутствуют как мишень 1, так и мишень 2.
Примеры применения олигонуклеотидов LOCS в сочетании с другими стандартными репортерами/зондами
Детектирование мишеней во время или после амплификации Мишени
Олигонуклеотиды LOCS по данному изобретению можно применять для определения присутствия амплифицированных последовательностей нуклеиновых кислот мишеней. Не существует конкретных ограничений в отношении способов амплификации, к которым могут применяться репортеры LOCS. Ампликоны, образующиеся в результате различных реакций, могут быть обнаружены репортерами LOCS при условии, что присутствие ампликонов мишеней может способствовать расщеплению или деградации репортера LOCS с образованием разделенный структур LOCS. Неограничивающие примеры способов, пригодных для расщепления или деградации петлевых областей, содержащихся в структурах LOCS, включают расщепление МНКзимами, ДНКзимами, рибозимами, рестрикционными ферментами, эндонуклеазами или расщепление экзонуклеазами, включая, но не ограничиваясь экзонуклеазной активностью полимеразы.
Как правило, в способах амплификации нуклеиновых кислот используются ферменты (например, полимеразы) для создания копий нуклеиновой кислоты мишени, которая специфически связывается одним или более олигонуклеотидными праймерами.
Неограничивающие примеры способов амплификации, в которых можно применять олигонуклеотиды LOCS, включают одну или более из полимеразной цепной реакции (ПЦР), полимеразной цепной реакции с обратной транскрипцией (ОТ-ПЦР), амплификации со смещением цепи (SDA), зависимой от геликазы амплификации (HDA), рекомбиназно-полимеразной амплификации (RPA), петлевой изотермической амплификации (LAMP), амплификации по типу катящегося кольца (RCA), транскрипционно-опосредованной амплификации (ТМА), самоподдерживающейся репликации последовательности (3SR), амплификации на основе последовательности нуклеиновой кислоты (NASBA), лигазной цепной реакции (LCR) или способа разветвленной амплификации (RAM).
Квалифицированный специалист легко поймет, что применение олигонуклеотидов LOCS, описанное выше, предоставлено только с целью неограничивающей иллюстрации. Описанные олигонуклеотиды LOCS можно применять в любом способе амплификации нуклеиновых кислот на основе праймеров, и изобретение не ограничивается этими конкретно описанными вариантами реализации.
Детектирование ампликонов, полученных с помощью репортеров LOCS
Как обсуждалось выше, репортеры LOCS по данному изобретению можно применять в любом способе амплификации полинуклеотидов, не ограничивающие примеры которой включают ПЦР, ОТ-ПЦР, SDA, HDA, RPA, LAMP, RCA, ТМА, RAM, LCR, 3SR или NASBA.
Ампликоны, полученные с помощью этих способов, могут быть обнаружены с использованием репортеров LOCS, которые могут быть расщеплены или деградированы с использованием любого подходящего способа, известного в данной области. Неограничивающие примеры включают использование каталитических нуклеиновых кислот, экзонуклеаз, эндонуклеаз и т.п.
МНКзимы можно применять для создания разделенных LOCS репортеров путем детектирования ампликонов, полученных с помощью таких способов, как ПЦР, ОТ-ПЦР, SDA, HDA, RPA, ТМА, LAMP, RCA, LCR, RAM, 3SR и NASBA. МНКзим может содержать один или более партзимов. МНКзимы представляют собой многокомпонентные нуклеиновые кислоты с ферментативной активностью, которые собираются и проявляют каталитическую активность только в присутствии облегчающего сборку фактора, которым может быть, например, детектируемая мишень, такая как ампликон, полученный из полинуклеотидной последовательности с использованием праймеров. МНКзимы состоят из нескольких частичных ферментов или партзимов, которые самособираются в присутствии одного или более посредников сборки и образуют активные МНКзимы, которые каталитически модифицируют субстраты. Субстрат и субстрат облегчающий сборку (мишень) представляют собой отдельные молекулы нуклеиновых кислот.Партзимы имеют несколько доменов, включая (i) сенсорные плечи, которые связываются с субстратом облегчающим сборку (таким как нуклеиновая кислота мишень); (ii) плечи субстрата, которые связывают субстрат, и (iii) частичные последовательности каталитического ядра, которые при сборке объединяются для получения полного каталитического ядра. МНКзимы можно сконструировать так, чтобы они распознавали широкий спектр субстратов облегчающих сборку, включая, например, различные последовательности нуклеиновых кислот мишеней. В ответ на присутствие посредника сборки МНКзимы модифицируют свои субстраты. Эта модификация субстрата может быть связана с генерированием сигнала, и, таким образом, МНКзимы могут генерировать ферментативно амплифицированный исходящий сигнал. Субстрат облегчающий сборку может представлять собой нуклеиновую кислоту мишень, присутствующую в биологическом образце или образце из окружающей среды (например, ампликон, полученный из полинуклеотид а мишени с использованием праймеров). В таких случаях детектирование модификации субстрата посредством активности МНКзима свидетельствует о присутствии мишени. В данной области известно несколько МНКзимов, способных расщеплять субстраты нуклеиновые кислоты. МНКзимы и их модифицированные формы известны в данной области и описаны в публикациях патентов РСТ WO/2007/041774, WO/2008/040095, WO2008/122084 и соответствующих публикациях патентов США 2007-0231810, 2010-0136536 и 2011- 0143338 (содержание каждого из этих документов полностью включено в данное описание посредством ссылки).
Использование LOCS репортеров в качестве внутреннего калибратора при вариаций от инструмента к инструменту или от лунки к лунке
Способ калибратора, продемонстрированный в Примере 12, имеет несколько преимуществ, в том числе то, что он не требует добавления в реакцию дополнительных реагентов и не требует использования данных, полученных из других лунок. Этот способ служит для калибровки и корректировки возможных вариаций от лунки к лунке. Кроме того, калибровка выполняется с использованием данных, полученных в одном и том же канале, и поэтому на нее не влияют какие-либо различия между каналами, которые могут присутствовать между приборами. Если для мультиплексной реакции используется несколько каналов, каждый канал можно калибровать независимо по калибровочному сигналу LOCS в каждом канале. Это благоприятно для сценария, в котором сигналы калибруются по сигналам в другом канале, например, от внутреннего контроля или эндогенного контроля, поскольку калибровка поддается отрицательному влиянию, если отношение ожидаемой интенсивности сигнала между каналами значительно отличается между инструментами, вызывая вариации от канала к каналу.
Диагностические применения
Способы с использованием стандартных репортерных зондов в сочетании с олигонуклеотидами LOCS можно применять для диагностических и/или прогностических целей в соответствии с описанными в данном документе способами. Диагностические и/или прогностические способы могут быть выполнены ex vivo или in vitro. Однако способы по данному изобретению необязательно должны применяться в диагностических и/или прогностических целях, и, следовательно, также рассматриваются применения, которые не являются диагностическими или прогностическими.
В некоторых вариантах реализации описанные в данном документе способы можно применять для диагностики инфекции у субъекта. Например, способы могут быть использованы для диагностики инфицирования субъекта бактериями, вирусами, грибками/дрожжами, протестами и/или нематодами. В одном варианте реализации вирус может представлять собой энтеровирус.Субъектом может быть крупный рогатый скот, лошадь, овца, примат, птица или грызун. Например, субъект может быть млекопитающим, таким как человек, собака, кошка, лошадь, овца, коза или корова. Субъект может быть поражен заболеванием, вызванным инфекцией. Например, у субъекта может быть менингит, возникающий в результате энтеровирусной инфекции. Соответственно, способы по данному изобретению в некоторых вариантах реализации могут быть использованы для диагностики менингита.
Способы по данному изобретению могут быть выполнены на образце. Образец может быть получен из любого источника. Например, образец может быть получен из источника окружающей среды, промышленного источника или путем химического синтеза.
Следует понимать, что рассматриваемый в данном документе термин «образец» включает образец, модифицированный по сравнению с исходным состоянием, например, путем очистки, разбавления или добавления любого другого компонента или компонентов.
Способы по данному изобретению, включая, но не ограничиваясь ими, диагностические и/или прогностические способы, можно применять на биологическом образце. Биологический образец может быть взят у субъекта. Можно также применять хранящиеся биологические образцы. Неограничивающие примеры подходящих биологических образцов включают цельную кровь или ее компонент (например, клетки крови, плазму, сыворотку), мочу, стул, слюну, лимфу, желчную жидкость, мокроту, слезы, спинномозговую жидкость, жидкость бронхиоальвеолярного лаважа, синовиальную жидкость, сперму, асцитическую опухолевую жидкость, грудное молоко и гной.
Наборы
Данное изобретение предлагает наборы, включающие одно или более средств для выполнения способов по данному изобретению. Обычно наборы для выполнения способов по данному изобретению содержат все необходимые реагенты для реализации способа.
В некоторых вариантах реализации наборы могут включать олигонуклеотидные компоненты, способные образовывать МНКзим в присутствии соответствующего субстрата облегчающего сборку (например, ампликона, как описано в настоящем документе). Например, набор может включать по меньшей мере первый и второй олигонуклеотидный компонент, содержащий первый и второй партзимы, и второй контейнер, содержащий субстрат, при этом для самосборки первого и второго партзимов и субстрата в МНКзима требуется ассоциация субстрата облегчающего сборку (например, ампликона), присутствующего в тестируемом образце. Соответственно, в таком варианте реализации первый и второй партзимы и олигонуклеотид LOCS, содержащий субстрат в области петли, могут быть применены к тестируемому образцу для определения присутствия одного или более ампликонов мишеней. Как правило, наборы содержат по меньшей мере один представленный в данном документе олигонуклеотид LOCS.
Как правило, наборы по данному изобретению также будут включать другие реагенты, реагенты для промывки, ферменты и/или другие реагенты, необходимые для выполнения способов по данному изобретению, таких как ПЦР или другие способы амплификации нуклеиновых кислот.
Наборы могут быть фрагментированными наборами или комбинированными наборами, как определено в данном документе.
Фрагментированные наборы состоят из реагентов, помещенных в отдельные контейнеры, и могут включать небольшие стеклянные контейнеры, пластиковые контейнеры или полоски из пластика или бумаги. Такие контейнеры могут обеспечивать эффективную передачу реагентов из одного отсека в другой, избегая при этом перекрестного загрязнения образцов и реагентов, а также количественное добавление агентов или растворов каждого контейнера из одного отсека в другой.
Такие наборы могут также включать контейнер, в который помещается тестируемый образец, контейнер, содержащий реагенты, используемые в анализе, контейнеры, содержащие реагенты для промывки, и контейнеры, содержащие реагент для детектирования.
Комбинированные наборы содержат все компоненты реакционного анализа в одном контейнере (например, в одной коробке, содержащей каждый из желаемых компонентов).
Набор по данному изобретению может также включать инструкции по использованию компонентов набора для применения соответствующих способов. Наборы и способы по изобретению можно применять в сочетании с оборудованием и системами автоматизированного анализа, например, включая, но не ограничиваясь ими, инструментовы для ПЦР в реальном времени.
Для применения для амплификации, детекции, идентификации или количественного определения различных мишеней может быть применим один набор по изобретению или, альтернативно, могут потребоваться разные наборы, например, содержащие реагенты, специфичные для каждой мишени. Способы и наборы по данному изобретению находят применение в любых обстоятельствах, при которых желательно обнаружить, идентифицировать или количественно определить любой объект.
Специалистам в данной области техники будет понятно, что в данное изобретение могут быть внесены многочисленные изменения и/или модификации, как раскрыто в конкретных вариантах реализации, без отклонения от сущности или объема данного изобретения, как описано в общих чертах. Таким образом, настоящие варианты реализации во всех отношениях следует рассматривать как иллюстративные, а не ограничивающие.
Примеры
Далее данное изобретение будет описано более подробно со ссылкой на следующие конкретные примеры, которые не следует рассматривать как ограничивающие каким-либо образом объем изобретения.
Пример 1: Способ анализа нескольких мишеней на одной длине волны с использованием одного линейного субстрата МНКзима и одного зонда LOCS в формате, позволяющем либо одновременную количественную оценку одной мишени в реальном времени в сочетании с качественным определением конечной точки для второй мишени, либо одновременный качественный анализ двух мишеней на каждом канале.
В следующем примере демонстрируется подход, в котором один линейный субстрат и один репортер LOCS используются в комбинации для детектирования и дифференциации двух мишеней (CTcry и NGopa) и для количественного определения одной мишени (CTcry) на одном флуоресцентном канале без необходимости анализа кривой плавления. Анализ разработан таким образом, что CTcry может быть обнаружен и количественно определен с использованием линейного субстрата МНКзима, a NGopa может быть обнаружен и дифференцирован в конечной точке с использованием репортера LOCS, который содержит другой субстрат МНКзима в своей петле.
Во время ПЦР МНКзим 1 может расщеплять линейный субстрат 1 в присутствии CTcry, чтобы разделить флуорофор и гаситель, и вызвать усиление сигнала, которое можно обнаружить в широком диапазоне температур. В этом примере детектирование и количественная оценка CTcry в реальном времени достигается путем получения флуоресценции в каждом цикле во время ПЦР при температуре 1 (D1) (52°С). Стебель LOCS-1 как в интактной, так и в разделенной конфигурации имеет температуру плавления (Tm), которая выше температуры 1 (52°С), поэтому LOCS-1 не дает сигнала при температуре 1 независимо от наличия или отсутствия NGopa в пробе.
В присутствии NGopa МНКзим 2 может расщеплять LOCS-1 во время ПЦР. Детектирование конечной точки NGopa может быть достигнуто путем сравнения сигнала флуоресценции при более высокой температуре 2 (70°С) до и после амплификации (ΔSD2)- Поскольку Tm интактного стебля LOCS-1 выше температуры 2 (Tm>70°С), и Tm разделенного стебля LOCS-1 ниже температуры 2 (Tm<70°С), то увеличение флуоресценции во время ПЦР выше той, которая связана с расщепленным линейным субстратом 1 при этой температуре (если она присутствует), связано с разделенным LOCS-1 с диссоциированными стеблями и указывает на присутствие NGopa.
В настоящем примере демонстрируется количественная оценка мишени 1 (CTcry) в реальном времени в сочетании с детектированием конечной точки Мишени 2 (NGopa). Кроме того, в этом примере демонстрируется детектирование конечной точки как мишени 1 (CTcry), так и мишени 2 (NGopa) наряду с несколькими альтернативными способами анализа этих данных.
Следующие способы анализа конечной точки (от 1 до 3) требуют измерения флуоресценции только в дискретные моменты времени, а именно; в момент инициации реакции амплификации (флуоресценция пре-ПЦР) и после амплификации (конечная точка или флуоресценция пост-ПЦР). В эти моменты времени снимаются показания при различных температурах (D1 и D2), и анализ позволяет выяснить наличие мишени 1, или мишени 2, или мишеней 1 и 2, или отсутствие мишени 1 и мишени 2.
Способ Анализа Конечной Точки 1
При температуре 1 (D1) расщепление субстрата 1, опосредованное мишенью, вызывает значительное увеличение сигнала флуоресценции во время ПЦР (ΔSD1), которое измеряется как разница между флуоресценцией пост-ПЦР в D1 (SD1-пост-ПЦР) и флуоресценцией пре-ПЦР в D1 (SD1-пре-ПЦР), и которая превышает первый порог (X1), такой, что SD1-пост-ПЦР - SD1-пре-ПЦР=ΔSD1>X1. Напротив, при этой температуре, если произошло расщепление LOCS-1, это не приводит к значительному увеличению сигнала флуоресценции и, таким образом, не превышает порог (X1). Это происходит потому, что Tm разделенного стебля выше D1 и структура остается погашеной. Таким образом, сравнение измерений флуоресценции пре-ПЦР и пост-ПЦР при температуре 1 позволяет специфически обнаруживать расщепленный субстрат 1 и, следовательно, мишень 1.
При температуре 2 (D2), которая выше температуры 1 (D1), опосредованное мишенью расщепление LOCS-1 приводит к значительному увеличению флуоресцентного сигнала во время амплификации. При наличии разделенного LOCS-1 увеличение сигнала на D2 (ΔSD2) превышает порог Х1. Кроме того, увеличение сигнала в D2 (ΔSD2) больше, чем в D1 (ΔSD1), независимо от того, расщеплен ли субстрат 1 или нет. Поэтому, когда величина увеличения флуоресценции в D2 (ΔSD2), измеренная как разница между флуоресценцией пост-ПЦР в D2 (SD2-пост-ПЦР) и флуоресценцией пре-ПЦР в D2 (SD2-пре-ПЦР), такая, что SD2-пост-ПЦР - SD2-пре-ПЦР, превышает как при D1 (ΔSD1), так и порог X1, так что ΔSD2>ΔSD1 и ΔSD2>порог X1, это указывает на детектирование разделенного LOCS-1 и, следовательно, на присутствие мишени 2.
Способ Анализа Конечной Точки 2
При температуре 1 (D1) расщепление субстрата 1 вызывает значительное увеличение сигнала флуоресценции (ΔSD1) во время ПЦР, который превышает первый порог (X1), такой как ΔSD1>X1; однако расщепление LOCS-1 не приводит к значительному увеличению сигнала флуоресценции при D1 и не превышает порог (X1) из-за высокой Tm стрежня, которая превышает D1 как в интактном, так и в разделенном состоянии. Поэтому сравнение измерений флуоресценции пре-ПЦР и пост-ПЦР в D1 (ΔSD1) позволяет специфически обнаруживать расщепленный субстрат 1 и, следовательно, мишень 1.
При температуре 2 (D2), которая выше температуры 1 (D1), расщепление LOCS-1 в ходе ПЦР приводит к изменению сигнала флуоресценции (ΔSD2), которое больше, чем изменение, наблюдаемое при температуре 1 (ΔSD1), где разница между ΔSD2 и ΔSD1 (ΔSD2 -ΔSD1) превышает второй порог (Х2); такое, что ΔSD2 - ΔSD1=ΔΔSD2 ΔSD1>Х2. Напротив, расщепление одного только субстрата 1 приводит к сходным значениям ΔSD2 и ΔSD1, причем разница между этими двумя значениями (ΔΔSD2ΔSD1) не имеет значения и не превышает второго порога (Х2). Следовательно, анализ флуоресценции при температуре 2 таким образом позволяет специфически обнаруживать расщепленный, разделенный LOCS-1, что указывает на присутствие мишени 2.
Способ Анализа Конечной Точки 3
Когда в образце присутствует либо мишень 1 и/или мишень 2, усиление сигнала во время амплификации при температуре 2 (ΔSD2) является значительным и превышает порог Хь Поэтому, когда ΔSD2>X1, это свидетельствует о присутствии в образце либо CTcry, и/или NGopa. Кроме того, когда ΔSD2<X1, это свидетельствует об отсутствии в образце CTcry, и NGopa.
Кроме того, когда ΔSD2>X1, можно применять уникальное соотношение изменений флуоресценции при температуре 1 и температуре 2 (ΔSD1:ΔSD2) или обратное соотношение (ΔSD2:ΔSD1), чтобы определить, присутствуют ли в образце мишень 1, мишень 2 или обе мишени 1 и 2.
В этом случае, если ΔSD1:ΔSD2 больше, чем порог R1, это указывает, что присутствует только мишень 1; если ΔSD1:ΔSD2 меньше, чем порог R2, это указывает, что присутствует только мишень 2; и если ΔSD1:ΔSD2 меньше, чем порог R1, но больше, чем порог R2, это указывает, что присутствуют как мишень 1, так и мишень 2.
Способы Анализа Конечной точки 1, 2 и 3
Кроме того, в приведенных выше способах анализа конечной точки 1-3 изменения сигналов при температурах 1 и 2 (ΔSD1 и ΔSD2) также можно откалибровать по сигналу калибратора (С) или по разнице между результатами пост-ПЦР и сигналу пре-ПЦР калибратора (ΔС). В качестве неограничивающего примера калибратор может включать эндогенный контроль, внутренний контроль или специальные олигонуклеотиды-калибраторы, которые можно измерять в том же канале или в другом канале при заданной температуре или температурах. Изменения сигналов при температурах 1 и 2 (ΔSD1 и ΔSD2) могут быть выражены как отношение к изменению сигнала от калибратора (например, ΔSD2/ΔC, ΔSD1/ΔC) или, альтернативно, как обратное отношение (например, ΔC/ΔSD2, ΔС/ΔSD1), где ΔС определяется как положительный сигнал, такой, что ΔС больше порога С (ΔС>порога С).
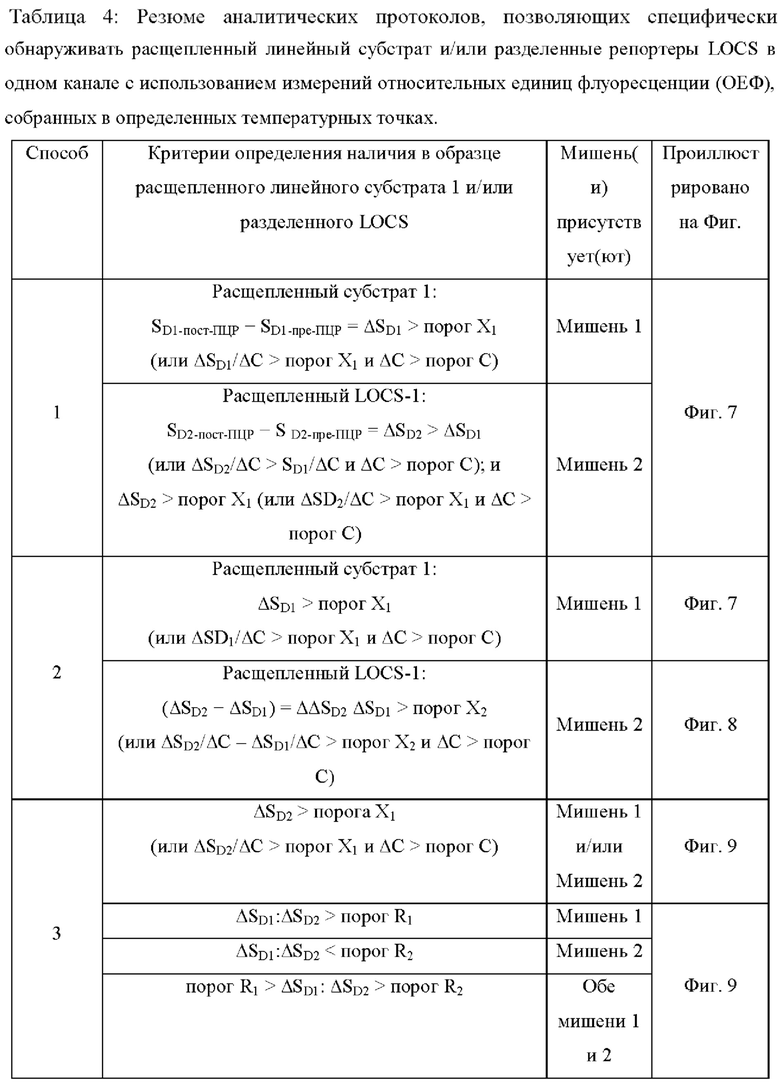
Олигонуклеотиды
Олигонуклеотиды, специфичные для этого эксперимента, включают; прямой праймер 1 (SEQ ID NO: 1), обратный праймер 1 (SEQ ID NO: 2), партзим A1 (SEQ ID NO: 3), партзим B1 (SEQ ID NO: 4), прямой праймер 2 (SEQ ID NO: 5), обратный праймер 2 (SEQ ID NO: 6), партзим A2 (SEQ ID NO: 7), партзим B2 (SEQ ID NO: 8), прямой праймер 3 (SEQ ID NO: 9), обратный праймер 3 (SEQ ID NO: 10), партзим A3 (SEQ ID NO: 11), партзим B3 (SEQ ID NO: 12), линейный субстрат МНКзима 1 (SEQ ID NO: 13), LOCS-1 (SEQ ID NO: 14) и линейный субстрат МНКзима 2 (SEQ ID NO: 15). Последовательности перечислены в списке последовательностей.
Олигонуклеотиды, специфичные для амплификации и детектирования мишени 1 (CTcry), представляют собой субстрат 1, партзим А1, партзим В1 (МНКзим 1), прямой праймер 1 и обратный праймер 1. Олигонуклеотиды, специфичные для амплификации и детектирования мишени 2 (NGopa), представляют собой LOCS-1, партзим А2, партзим В2 (МНКзим 2), прямой праймер 2 и обратный праймер 2. Олигонуклеотиды, специфичные для амплификации и детектирования гена калибратора (TFRC), представляют собой субстрат 2, партзим A3, партзим ВЗ (МНКзим 3), прямой праймер 3 и обратный праймер 3.
Условия реакции
Амплификацию и детектирование в реальном времени проводили в общем реакционном объеме 20 мкл с использованием термоциклера BioRad® CFX96. Параметры циклирования и точки сбора флуоресцентных данных (DA) были следующими: 1 цикл 95°С в течение 2 минут, 52°С в течение 15 секунд (DA), 70°С в течение 15 секунд (DA); 10 циклов: 95°С в течение 5 секунд и 61°С в течение 30 секунд (уменьшение на 0,5°С за цикл); 40 циклов 95°С в течение 5 секунд и 52°С в течение 40 секунд (DA в каждом цикле); и 1 цикл 70°С в течение 15 секунд (DA). Все реакции проводили в трех повторностях и содержали 40 нМ каждого прямого праймера, 200 нМ каждого обратного праймера, 200 нМ каждого партзима А, 200 нМ каждого партзима В, 200 нМ каждого субстрата, 200 нМ LOCS-1 и lx SensiFast буфера (Bioline). Реакции либо не содержали мишени (NF H2O), либо содержали синтетический G-блок CTcry (20000, 4000, 800, 160 или 32 копии); или ген NGopa (20000 или 32 копии); или различные концентрации гена CTry (20000, 4000, 800, 160 или 32 копии) на фоне гена NGopa (20000 и 32 копии). Все реакции, кроме контроля без мишени (NF H2O), дополнительно содержали фон из 34,5 нг (10000 копий) геномной ДНК человека. Наконец, дополнительный контроль содержал только геномную ДНК без какого-либо блока CTcry или NGopa G.
Полученные результаты
В этом примере три МНКзима (МНКзимы 1-3) использовали в одной ПЦР для одновременного детектирования и дифференциации трех целевых нуклеиновых кислот (CTcry, NGopa и TFRC человека соответственно) с использованием только двух флуоресцентных каналов (HEX и Техаский красный). Наличие или отсутствие CTcry и/или NGopa определяли и дифференцировали в канале HEX, а присутствие гена TFRC определяли в канале Техаский красный. Присутствие генов CTcry и TFRC определяли в режиме реального времени по увеличению сигнала флуоресценции при 52°С (D1) в каналах HEX и Техаский красный соответственно, а также определяли в конечной точке при 52°С. Присутствие NGopa было обнаружено с помощью дискретных измерений при 70°С (D2) в канале HEX, который отслеживает расщепление LOCS-1.
Результаты, представленные на Фиг. 6, иллюстрируют кривые ПЦР-амплификации, полученные в канале HEX при температуре сбора данных 52°С в реакциях, содержащих 20000 (черная точка), 4000 (черный штрих), 800 (черный квадрат), 160 (серая сплошная) или 32 (серая точка) копии темплата CTcry либо отдельно (Фиг. 6А), либо на фоне 20000 (Фиг. 6В) или 32 (Фиг. 6С) копий темплата NGopa. Флуоресцентные данные при 52°С были также собраны для реакций без CTcry, но содержащих либо 20000 (черная линия), либо 32 копии (серая линия) матрицы NGopa (Фиг. 6D). Нецелевые контроли (NF H2O) показаны на Фиг. 6А-6С (черная сплошная линия). Кривые амплификации представляют собой средние значения уровня флуоресценции из трех повторных реакций. Рассчитанные числа копий CTcry для каждой из приведенных выше реакций показаны в Таблице 5, где Не Применимо (Н/П) относится к случаям, когда значение Cq, определенное при 52°С, не соответствует отсутствию CTcry в этих реакциях. Значения Cq определяли с использованием режима регрессии в программном обеспечении BioRad(Baseline SubtractedCurve Fit).
Результаты в Таблице 5 показывают, что стандартные кривые CTry имели значения R выше 0,99 и что эффективность ПЦР была высокой (100-106%). Присутствие 20000 или 32 копий NGopa в реакции существенно не изменило рассчитанное число копий гена CTcry (парный критерий Стьюдента, р-значение 0,144 и 0,315 соответственно), что свидетельствует о том, что детектирование в реальном времени при 52°С может применяться для прямого количественного анализа CTry в образце.
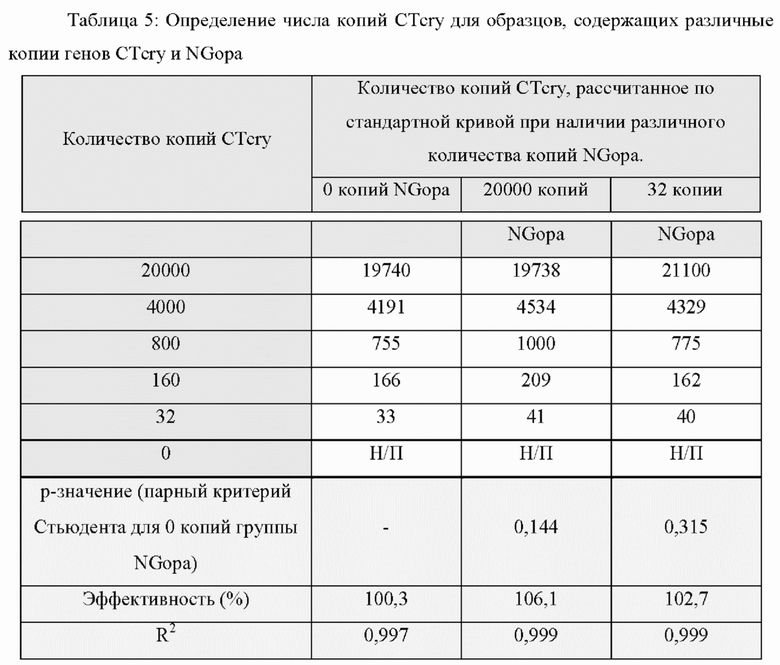
Результаты на Фиг. 7 иллюстрируют изменение флуоресцентного сигнала во время ПЦР, измеренное в канале HEX при двух разных температурах с использованием Способа Анализа Конечных Точек 1. На Фиг. 7А изменение сигнала при температуре 1 (ΔSD1; 52°С) показано черно-белым паттерном, а изменение сигнала при температуре 2 (ΔSD2; 70°С) показано серым цветом. Результаты показывают, что изменение сигнала при температуре 1 (ΔSD1; черные и белые полосы) превышает порог 1 (X1), когда в образце присутствует CTcry, независимо от того, присутствует или отсутствует NGopa, но не превышает этот порог только при наличии NGopa и/или при отсутствии CTcry в образце. Следовательно, изменение сигнала в конечной точке больше, чем порог 1 при температуре 1, указывает на наличие CTcry. Результаты на Фиг. 7А также показывают, что когда изменение сигнала при температуре 2 (ΔSD2; серые столбцы) больше, чем изменение сигнала при температуре 1 (ΔSD2>ΔSD1), а также больше, чем порог X1 (ΔSD2>X1), то в образце присутствует NGopa.
В качестве альтернативы изменение сигнала при температурах 1 и 2 можно откалибровать по изменению сигнала калибратора (ΔС). На Фиг. 7В показано изменение сигнала калибратора TFRC (ΔС), измеренного в канале Техаский красный, где значения, превышающие порогС, указывают на положительный сигнал для ΔС, тогда как значения ниже порога С указывают на отрицательный сигнал для ΔС. На Фиг. 7С изменение сигнала при температуре 1, откалиброванное по ΔC (ΔSD1/ΔC; 52°С), показано черно-белым паттерном, а изменение сигнала при температуре 2 откалибровано по ΔC (ΔSD2/ΔC; 70°С) показано серым цветом для реакций, признанных положительными для ΔС на Фиг. 7В, где неприменимо (Н/П) относится к реакциям, признанным отрицательными для ΔС на Фиг. 7В. Результаты на Фиг. 7С можно анализировать так же, как и на Фиг. 7А, который поясняет тот же вывод об обнаружении мишеней 1 и 2.
Результаты на Фиг. 8 иллюстрируют разницу в изменении флуоресцентного сигнала, полученного в канале HEX при температуре 2 (ΔSD2) и температуре 1 (ΔSD1) с использованием Способа Анализа Конечных Точек 2 (AΔSD2ΔSD1). Результаты показывают, что ΔΔSD2ASD1 превышает порог 2 (Х2), когда в образце присутствует NGopa, но не превышает этот порог, когда в образце присутствует только CTcry и/или когда в образце отсутствует NGopa (только NTC и геномная ДНК). Таким образом, разница в флуоресцентном сигнале конечной точки выше порога 2 (AΔSD2ΔSD1>X2) указывает на присутствие NGopa.
Результаты на Фиг. 9 иллюстрируют изменение флуоресцентного сигнала, полученного в канале HEX при двух разных температурах с использованием Способа Анализа Конечных Точек 3. На Фиг. 9А, где ΔSD2 больше, чем порог X1, это указывает на присутствие в образце CTcry и/или NGopa. Если ΔSD2 ниже порога X1, это указывает на отсутствие в реакции CTcry, и NGopa (только NTC и геномная ДНК). На Фиг. 9В показано отношение ΔSD1:ΔSD2, которое можно применять для указания того, какие мишени присутствуют в реакции. Когда отношение выше порога R1, это указывает на наличие CTcry, но не NGopa. Если отношение ниже порога R2, это указывает на наличие NGopa, но не CTcry. Когда отношение находится между порогами R1 и R2, это указывает на наличие как CTcry, так и NGopa. Когда в реакции не присутствуют ни CTry, ни NGopa (Фиг. 9А), необходимость вычисления соотношения отменяется и указывается как Н/П, как показано на Фиг. 9В.
В целом этот пример демонстрирует, что две мишени могут быть обнаружены в одном флуоресцентном канале при двух разных температурах путем мониторинга флуоресценции в реальном времени и в дискретные моменты времени (до ПЦР и после ПЦР). С помощью этого способа одна мишень может быть определена количественно с использованием линейного субстрата МНКзима, а другая мишень обнаружена с использованием зонда LOCS. Простой способ не требует анализа кривой плавления после ПЦР. Пример также демонстрирует, что качественные данные могут быть получены для нескольких мишеней на одной длине волны путем сравнения значений флуоресценции до и после ПЦР при различных температурах. Кроме того, для анализа этих данных можно применять несколько способов анализа.
Пример 2: Способ одновременного количественного определения двух мишеней в реальном времени и качественного детектирования двух мишеней в каждом из двух флуоресцентных каналов с использованием двух линейных субстратов МНКзима и двух репортеров LOCS.
Следующий пример демонстрирует использование двух флуоресцентных каналов, HEX и FAM, для одновременного детектирования и дифференциации четырех мишеней, при этом определение Cq может быть выполнено для двух из этих мишеней в одной реакции. В канале HEX детектирование в реальном времени и определение Cq одной мишени (CTcry), а также детектирование конечной точки второй мишени (NGopa) были достигнуты с использованием одного линейного субстрата МНКзима и одного репортера LOCS соответственно. Точно так же в канале FAM детектирование в реальном времени и определение Cq третьей мишени (TVK), а также детектирование конечной точки четвертой мишени (MgPa) были достигнуты с использованием второго линейного субстрата МНКзима и второго репортера LOCS соответственно. Наконец, пятая мишень, ген TFRC человека, была обнаружена в фоновой геномной ДНК человека с использованием третьего линейного субстрата МНКзима, считанного в канале Техаский Красный.
Опосредованное мишенью расщепление линейных субстратов соответствующими МНКзимами вызывает разделение флуорофора и гасителя, что приводит к увеличению сигнала в широком диапазоне температур. В этом примере сигнал от линейных субстратов может быть обнаружен в режиме реального времени путем регистрации флуоресценции в каждом цикле во время ПЦР при Температуре 1 (D1=52°С) или в конечной точке путем измерения увеличения флуоресценции, которое происходит во время ПЦР, когда мишень присутствует.При этой температуре как разделенные, так и интактные репортеры LOCS для NGopa и MgPa не дают какого-либо детектируемого сигнал, поскольку Tm их стеблевых областей выше, чем Температура 1. Следовательно, значения Cq, полученные при температуре 1 на каждой длине волны во время ПЦР, отражают начальное количество мишеней Ctcry, TVK и TFRC (независимо от присутствия или отсутствия других мишеней) и, таким образом, могут применяться для количественного определения.
Опосредованное мишенью расщепление репортеров LOCS их соответствующими МНКзимами вызывает усиление сигнала в определенном диапазоне температур. В этом примере этот сигнал флуоресценции измеряется при Температуре 2 (D2=70°С), которая выше, чем Tm обоих разделенных репортеров LOCS, но ниже, чем Tm обоих интактных репортеров LOCS (Tm интактных репортеров LOCS>Температура 2>Tm разделенного репортера LOCS), и, следовательно, интактные репортеры LOCS не дают значимого сигнала при Температуре 2. Следовательно, увеличение сигнала при Температуре 2 отражает присутствие разделенного репортера LOCS, что подтверждает присутствие специфических мишеней NGopa и MgPa в каналах HEX и FAM соответственно.
Пример демонстрирует одновременное получение смешанных количественных и качественных данных в одной реакции с мониторингом в реальном времени и определением Cq для трех мишеней в сочетании с простым детектированием двух других мишеней, определенных путем сравнения показаний флуоресценции до и после ПЦР. Кроме того, пример демонстрирует качественное детектирование четырех мишеней в двух флуоресцентных каналах в сочетании со способами анализа конечной точки 1-3 (как указано в Примере 1).
Ол иго нукл е отиды
Олигонуклеотиды, специфичные для этого эксперимента, включают; прямой праймер 1 (SEQ ID NO: 1), обратный праймер 1 (SEQ ID NO: 2), партзим A1 (SEQ ID NO: 3), партзим B1 (SEQ ID NO: 4), прямой праймер 2 (SEQ ID NO: 5), обратный праймер 2 (SEQ ID NO: 6), партзим A2 (SEQ ID NO: 7), партзим B2 (SEQ ID NO: 8), прямой праймер 3 (SEQ ID NO: 9), обратный праймер 3 (SEQ ID NO: 10), партзим A3 (SEQ ID NO: 11), партзим B3 (SEQ ID NO: 12), линейный субстрат МНКзима 1 (SEQ ID NO: 13), LOCS-1 (SEQ ID NO: 14), линейный субстрат МНКзима 2 (SEQ ID NO: 15), прямой праймер 4 (SEQ ID NO: 16), обратный праймер 4 (SEQ ID NO: 17), партзим A4 (SEQ ID NO: 18), партзим B4 (SEQ ID NO: 19), прямой праймер 5 (SEQ ID NO: 20), обратный праймер 5 (SEQ ID NO: 21), партзим A5 (SEQ ID NO: 22), партзим B5 (SEQ ID NO: 23), линейный субстрат МНКзима 3 (SEQ ID NO: 24), LOCS-2 (SEQ ID NO: 25). Последовательности перечислены в перечне последовательностей.
Олигонуклеотиды, специфичные для амплификации и детектирования CTcry, представляют собой субстрат 1, партзим А1, партзим В1 (МНКзим 1), прямой праймер 1 и обратный праймер 1. Олигонуклеотиды, специфичные для амплификации и детектирования NGopa, представляют собой LOCS-1, партзим А2, партзим В2 (МНКзим 2), прямой праймер 2 и обратный праймер 2. Олигонуклеотиды, специфичные для амплификации и детектирования TFRC, представляют собой субстрат 2, партзим A3, партзим ВЗ (МНКзим 3), прямой праймер 3 и обратный праймер 3. Олигонуклеотиды, специфичные для амплификации и детектирования TVK, представляют собой субстрат 3, партзим А4, партзим В4 (МНКзим 4), прямой праймер 4 и обратный праймер 4. Олигонуклеотиды, специфичные для амплификации и детектирования MgPa, представляют собой LOCS-2, партзим А5, партзим В5 (МНКзима 5), прямой праймер 5 и обратный праймер 5.
Условия реакции
Амплификацию и детектирование в реальном времени проводили в общем реакционном объеме 20 мкл с использованием термоциклера BioRad® CFX96. Параметры циклирования и точки сбора флуоресцентных данных (DA) были следующими: 1 цикл 95°С в течение 2 минут, 52°С в течение 15 секунд (DA), 70°С в течение 15 секунд (DA), 10 циклов 95°С в течение 5 секунд и 61°С в течение 30 секунд (уменьшение на 0,5°С за цикл), 40 циклов 95°С в течение 5 секунд и 52°С в течение 40 секунд (DA в каждом цикле), и 70°С в течение 15 секунд (DA). Все реакции проводили в двух повторностях и содержали 40 нМ каждого прямого праймера, 200 нМ каждого обратного праймера, 200 нМ каждого партзима А, 200 нМ каждого партзима В, 200 нМ каждого субстрата, 200 нМ LOCS-1, 300 нМ LOCS-2 и lx SensiFast буфер (Bioline). Реакции не содержали мишени (NF H2O), либо содержали синтетический G-блок CTcry (20000 или 32 копии), ген NGopa (20000 или 32 копии), как CTcry, так и NGopa (20000 или 32 копии каждого), TVK (20000 копий или 32 копии), ген MgPa (20000 или 32 копии) или как TVK, так и MgPa (по 20000 или 32 копии каждого). Все реакции, кроме контроля без мишени (NF Н2О), содержали фон из 10000 копий геномной ДНК человека. Дополнительный контроль содержал только геномную ДНК.
Полученные результаты
В этом примере пять МНКзимов (МНКзимы 1-5) использовали в одной реакции ПЦР для одновременного детектирования и дифференциации пяти нуклеиновых кислот мишеней (CTcry, NGopa, TFRC, TVK и MgPa соответственно) с использованием только трех флуоресцентных каналов (HEX, Техаский Красный и FAM). Наличие или отсутствие CTcry и/или NGopa выявляли и дифференцировали в канале HEX, присутствие гена TFRC в геномной ДНК выявляли в канале Техаский Красный, и присутствие или отсутствие TVK и/или MgPa выявляли и дифференцировали в канале FAM. Наличие генов CTcry, TFRC и TVK определяли в режиме реального времени по увеличению сигнала флуоресценции при 52°С в каналах HEX, Техаский Красный и FAM соответственно, и также определяли в конечной точке при 52°С.Присутствие NGopa и MgPa определяли в конечной точке при 70°С в каналах HEX и FAM соответственно, которые контролируют расщепление LOCS-1 и LOCS-2 соответственно.
Результаты, показанные на Фиг. 10, иллюстрируют кривые ПЦР-амплификации, полученные в каналах HEX (Фиг. 10A-D) и FAM (Фиг. 10Е-Н) при температуре сбора данных 52°С.Полученные кривые амплификации представляют собой 20000 копий и 32 копии только темплата CTry, полученного в канале HEX (Фиг. 10А), или только TVK (Фиг. ЮЕ), полученного в канале FAM. Кривые амплификации также были получены для матричных смесей, содержащих либо 20000 копий и 32 копии каждого из темплатов CTcry и NGopa (Фиг. 10В), полученных в канале HEX, либо 20000 копий и 32 копии каждого из темплатов TVK и MgPa (Фиг. 10F), полученный в канале FAM. Не наблюдалось амплификации в канале HEX для образцов, содержащих 20000 копий и 32 копии только NGopa (Фиг. ЮС), и не было зарегистрировано амплификации для любой концентрации остальных мишеней, включая TVK, MgPa и TFRC (эндогенный контроль) (Фиг. 10D). Аналогичным образом, в канале FAM не было зарегистрировано амплификации для образцов, содержащих 20000 копий и 32 копии только матрицы MgPa (Фиг. 10G), и не было зарегистрировано амплификации для любой концентрации остальных мишеней, включая CTcry, NGopa и TFRC (Фиг. 10Н). Не было увеличения сигнала для всех реакций без контроля мишени (NF Н2О), и результаты показаны в виде черной пунктирной линии (Фиг. 10А-Н). Кривые амплификации представляют собой средние значения для трех повторных реакций и построены с использованием Microsoft Excel (версия 14). Значения Cq определяли с использованием способа одного порога, установленного на уровне 100 ОЕФ (аппроксимация базовой вычтенной кривой).
Значения Cq для каждой из вышеуказанных реакций показаны в Таблице 6, где неприменимо (Н/П) относится к случаям, когда значение Cq не определено при 52°С, что согласуется с отсутствием CTry и TVK в этих реакциях. Результаты показывают, что на значения Cq для CTcry и TVK не влияет присутствие темплатов NGopa в канале FAM или темплата MgPa в канале HEX соответственно. Кроме того, каждая из нецелевых реакций не давала поддающихся обнаружению кривых амплификации или значений Cq. Таким образом, значения Cq, полученные по каналам HEX и FAM, можно применять для прямого количественного анализа CTry и TVK в образце соответственно.
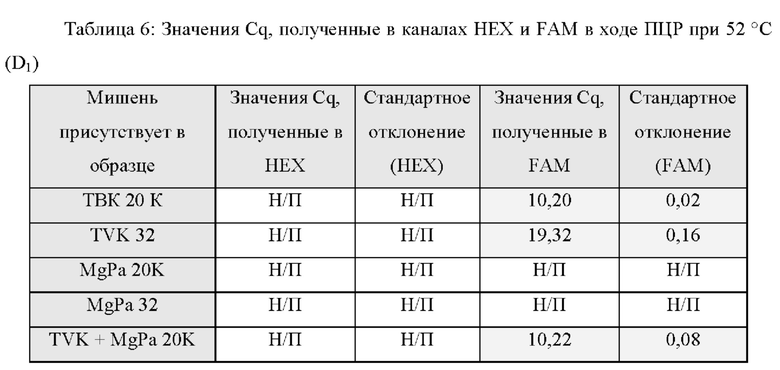
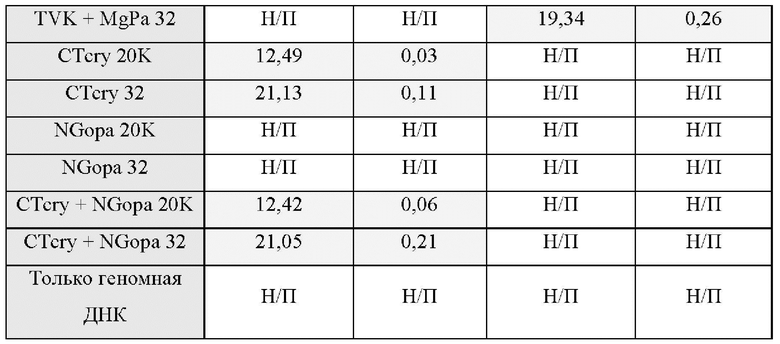
Результаты на Фиг. 11 иллюстрируют изменения флуоресцентного сигнала во время амплификации (пост-ПЦР минус пре-ПЦР), полученные в каналах HEX (Фиг. ПА) и FAM (Фиг. 11В) при двух разных температурах с использованием Способа Анализа Конечных Точек 1. Изменение сигнала при Температуре 1 (ΔSD1; 52°С) показано черно-белым паттерном, а изменение сигнала при Температуре 2 (ΔSD2; 70°С) показано серым цветом. На Фиг. 11А показано, что ΔSD1 превышает Порог X1, когда в образце присутствует CTcry, но не превышает этот порог, когда в образце отсутствует CTcry. Следовательно, ΔSD1>X1 свидетельствует о наличии CTcry. Результаты на Фиг. 11А также показывают, что, когда изменение сигнала при Температуре 2 (ΔSD2) больше, чем изменение при Температуре 1 (ΔSD2>ΔSD1), а также превышает Порог X1 (ΔSD2>X1), то в образце присутствует NGopa. Когда ΔSD2 превышает Порог X1, a ΔSD1 нет, это указывает на присутствие только NGopa. Аналогичным образом, на Фиг. 11В показано, что изменение сигнала при Температуре 1 (ΔSD1) превышает Порог X1, когда в образце присутствует TVK, но не превышает этого порога, когда в образце отсутствует TVK. Следовательно, ΔSD1>X1 свидетельствует о наличии TVK. Результаты на Фиг. 11В также показывают, что когда изменение сигнала при Температуре 2 (ΔSD2) значительно превышает изменение сигнала при Температуре 1 (ΔSD2>ΔSD1), а также превышает Порог X1 (ΔSD2>XI), то в образце присутствует MgPa. Когда SD2 превышает Порог Х1, и SD1 нет, это указывает на присутствие только MgPa.
Результаты на Фиг. 12 иллюстрируют различия в увеличении флуоресцентного сигнала в конечной точке, полученные в каналах HEX (Фиг. 12А) и FAM (Фиг. 12В) при Температуре 2 и Температуре 1 с использованием Способа Анализа Конечных Точек 2 (AΔSD2 ΔSD1). Результаты на Фиг. 12А показывают, что AΔSD2 ΔSD1 превышает Порог Х2, когда в образце присутствует NGopa, но не превышает этот порог, когда в образце отсутствует NGopa. Аналогично, результаты на Фиг. 12В показывают, что AΔSD2 ΔSD1 превышает Порог Х2, когда в образце присутствует MgPa, но не превышает этот порог, когда в образце отсутствует MgPa. Следовательно, когда ΔΔSD2 ΔSD1>Х2В каналах HEX и FAM, это указывает на присутствие NGopa и MgPa соответственно.
Результаты на Фиг. 13 иллюстрируют изменение флуоресцентного сигнала, полученного в каналах HEX (Фиг. 13А, В) и FAM (Фиг. 13С, D) при двух разных температурах с использованием Способа Анализа Конечных Точек 3. На Фиг. 13А, где ΔSD2B канале HEX больше, чем Порог X1, это указывает на присутствие в образце CTcry и/или NGopa. Если ΔSD2B канале HEX ниже, чем Порог X1, это указывает на отсутствие в реакции CTcry и NGopa. На Фиг. 13В показано отношение ΔSD2: ΔSD1 в канале HEX, которое указывает, какие мишени присутствуют в реакции. Если отношение выше, чем Порог R1, это указывает на наличие CTcry, но не NGopa. Если отношение ниже Порога R2, это указывает на наличие NGopa, но не CTcry. Если соотношение находится между Порогами R1 и R2, это указывает на наличие как CTcry, так и NGopa. На Фиг. 13А показано, когда в реакции не присутствуют ни CTcry, ни NGopa, поэтому необходимость вычисления соотношения для Фиг. 13В отменяется и обозначается как Н/П. На Фиг. 13С ΔSD2B канале FAM больше Порога X1, что указывает на присутствие TVK и/или MgPa в образце. Если ΔSD2B канале FAM ниже Порога X1, это указывает на отсутствие в реакции в TVK, и MgPa. На Фиг. 13D показано отношение ΔSD2: ΔSD1 в канале FAM, которое указывает, какие мишени присутствуют в реакции. Если отношение выше, чем Порог R1, это указывает на наличие TVK, но не MgPa. Если отношение ниже Порога R2, это указывает на наличие MgPa, но не TVK. Если соотношение находится между Порогами R1 и R2, это указывает на присутствие как MgPa, так и TVK. На Фиг. 13С показано, когда в реакции не присутствуют и TVK, и MgPa, поэтому необходимость расчета соотношения для Фиг. 13D отменяется и обозначается как Н/П.
Пример 3 - Детектирование конечной точки и дифференциация двух мишеней в одном канале с использованием одного линейного субстрата МНКзима и одного репортера LOCS с дополнительным подтверждением с помощью анализа кривой плавления.
В следующем примере демонстрируется подход, в котором один линейный субстрат МНКзима и один репортер LOCS используются в комбинации для детектирования и дифференциации двух мишеней (TPPolA и TFRC) в одном флуоресцентном канале с использованием анализа конечных точек и анализа кривой плавления. Анализ разработан таким образом, что TFRC может быть обнаружен и дифференцирован в конечной точке с использованием линейного субстрата МНКзима, и TPApolA может быть обнаружен и дифференцирован в конечной точке с использованием репортера LOCS. Подтверждающее детектирование TPApolA также может быть достигнуто с помощью тандемного анализа кривой плавления.
В этом примере TFRC включен в качестве эндогенного контроля в разовой концентрации 10000 копий на реакцию. TPApolA тестировали в двух разных концентрациях: 10000 копий на реакцию и 40 копий на реакцию. Одновременное детектирование как TPApolA, так и TFRC тестировали при следующих концентрациях: 10000 копий TFRC вместе с 10000 копий TPApolA на реакцию и 10000 копий TFRC вместе с 40 копиями TPApolA на реакцию. Этот анализ представляет собой мультиплекс с 10 мишенями, который также содержит праймеры, партзимы и LOCS для амплификации и детектирования восьми других мишеней, включая: CTry, LGV, NGopa, NGporA, MgPa, TVbtub, HSV-1 hHSV-2.
Во время ПЦР МНКзим 6 может расщеплять линейный Субстрат 4 в присутствии TFRC, чтобы разделить флуорофор и гаситель, чтобы вызвать усиление сигнала, которое можно обнаружить в широком диапазоне температур. В этом примере детектирование конечной точки TFRC достигается путем получения флуоресценции до и после цикла ПЦР при Температуре 1 (48°С).
Во время ПЦР МНКзим 7 может расщеплять LOCS-3 в присутствии TPApolA. Анализ разработан таким образом, что стебель LOCS-3 как для интактной, так и для разделенной конфигурации имеет температуру плавления (Tm), которая выше, чем температура 1 (48°С), и, следовательно, не дает сигнала при Температуре 1 независимо от наличия или отсутствия TPApolA в образце. Кроме того, анализ разработан таким образом, чтобы интактный LOCS-3 имел Tm выше температуры 2 (Tm>68°С), и разделенная конфигурация имела Tm равную или ниже температуры 2 (Tm<68°С). Таким образом, детектирование конечной точки и дифференциация TPApolA достигается путем измерения флуоресценции до и после цикла ПЦР при температуре 2 (68°С).
В этом примере детектирование и дифференциация TFRC и TPApolA достигается путем измерения флуоресценции до и после цикла ПЦР при Температуре 1 (48°С) и Температуре 2 (68°С) соответственно. Значения флуоресценции, полученные до цикла ПЦР, вычитаются из значений флуоресценции, полученных после цикла ПЦР при каждой температуре, чтобы удалить фоновую флуоресценцию, не связанную с инициируемым мишенью расщеплением МНКзимом 6 или МНКзимом 7. Затем проводится дальнейший анализ:
Как описано ранее в способе анализа 2 (Пример 1), при температуре 1 (D1) расщепление Субстрата 4 приводит к значительному увеличению сигнала флуоресценции (ΔSD1) во время ПЦР, которое превышает первый порог (X1), так что ΔSD1>X1; однако расщепление LOCS-3 не приводит к значительному увеличению флуоресцентного сигнала при D1 и не превышает порога (X1) из-за высокой Tm стрежня, которая превышает D1 как в неповрежденном, так и в разделенном состоянии. Таким образом, сравнение измерений флуоресценции до ПЦР и после ПЦР в D1 (ΔSD1) позволяет специфически обнаруживать расщепленный субстрат 4 и, следовательно, мишень TFRC.
При температуре 2 (D2), которая выше температуры 1 (D1), расщепление LOCS-3 в ходе ПЦР приводит к большему изменению сигнала флуоресценции (ΔSD2), чем изменение, наблюдаемое при температуре 1 (ΔSD1), при этом разница между ΔSD2 и ΔSD1 (ΔSD2 - ΔSD1) пересекает второй порог (Х2); так, что ΔSD2 - ΔSD1=AΔSD2 ΔSD1>Х2. Напротив, расщепление одного только субстрата 4 дает сходные значения ΔSD2 и ΔSD1, причем разница между этими двумя значениями (AΔSD2 ΔSD1) не имеет значения и не превышает второго порога (Х2). Таким образом, анализ флуоресценции в D2 позволяет специфически обнаруживать расщепленный, разделенный LOCS-3, что указывает на присутствие TPApolA.
Детектирование и дифференциация TPApolA также может быть подтверждена с помощью анализа кривой плавления. Уникальная кривая плавления получается, когда в образце присутствует TPApolA, что отличается от реакций, в которых TPApolA отсутствует в образце. Это позволяет визуально подтвердить, присутствует ли TPApolA в образце или нет.
Ол иго нукл е отиды
Олигонуклеотиды, специфичные для этого эксперимента, включают: Прямой праймер 3 (SEQ ID NO: 9), обратный праймер 3 (SEQ ID NO: 10), партзим А6 (SEQ ID NO: 26), партзим B6 (SEQ ID NO: 27), субстрат 4 (SEQ ID NO: 28), прямой праймер 6 (SEQ ID NO: 29), обратный праймер 6 (SEQ ID NO: 30), партзим A7 (SEQ ID NO: 31), партзим B7 (SEQ ID NO: 32), LOCS-3 (SEQ ID NO: NO: 33), прямой праймер 1 (SEQ ID NO: 1), прямой праймер 2 (SEQ ID NO: 2), партзим A1 (SEQ ID NO: 3), партзим B1 (SEQ ID NO: 4), LOCS-4 (SEQ ID NO: 34), прямой праймер 2 (SEQ ID NO: 5), прямой праймер 3 (SEQ ID NO: 6), партзим A8 (SEQ ID NO: 35), партзим B8 (SEQ ID NO: 36), LOCS-5 (SEQ ID NO: 37), прямой праймер 7 (SEQ ID NO: 38), обратный праймер 7 (SEQ ID NO: 39), партзим A9 (SEQ ID NO: 40), партзим B9 (SEQ ID NO: 41), LOCS-6 (SEQ ID NO: 42), прямой праймер 8 (SEQ ID NO: 43), обратный праймер 8 (SEQ ID NO: 44), партзим A10 (SEQ ID NO: 45), партзим BIO (SEQ ID NO: IDNO: 46), LOCS-7 (SEQ ID NO: 47), прямой праймер 9 (SEQ ID NO: 48), обратный праймер 9 (SEQ ID NO: 49), партзим All (SEQ ID NO: 50), партзим Bll (SEQ ID NO: 51), LOCS-8 (SEQ ID NO: 52), прямой праймер 10 (SEQ ID NO: 53), обратный праймер 10 (SEQ ID NO: 54), партзим A12 (SEQ ID NO: 55), партзим B12 (SEQ ID NO: 56), LOCS-9 (SEQ ID NO: 57), прямой праймер 11 (SEQ ID NO: 58), обратный праймер 11 (SEQ ID NO: 59), партзим A13 (SEQ ID NO: IDNO: 60), партзим B13 (SEQ ID NO: 61), LOCS-10 (SEQ ID NO: 62), прямой праймер 12 (SEQ ID NO: 63), обратный праймер 12 (SEQ ID NO: 56), партзим A14 (SEQ ID NO: 65), партзим В14 (SEQ ID NO: 66), LOCS-11 (SEQ ID NO: 67). Последовательности перечислены в списке последовательностей.
Олигонуклеотиды, специфичные для амплификации и детектирования TFRC, представляют собой субстрат 4, партзим А6, партзим В6 (МНКзим 6), прямой праймер 3 и обратный праймер 3. Олигонуклеотиды, специфичные для амплификации и детектирования TPApolA, представляют собой LOCS-3, партзим А7, партзим В7 (МНКзим 7), прямой праймер 6 и обратный праймер 6. Олигонуклеотиды, специфичные для амплификации и детектирования CTcry, представляют собой LOCS-4, прямой праймер 1, обратный праймер 1, партзим А1 и партзим В1 (МНКзим 1). Олигонуклеотиды, специфичные для амплификации и детектирования LGV, представляют собой LOCS-6, прямой праймер 7, обратный праймер 7, партзим А9 и партзим В9 (МНКзим 9). Олигонуклеотиды, специфичные для амплификации и детектирования NGopa, представляют собой LOCS-5, прямой праймер 2, обратный праймер 2, партзим А8 и партзим В8 (МНКзим 8). Олигонуклеотиды, специфичные для амплификации и детектирования NGporA, представляют собой LOCS-7, прямой праймер 8, обратный праймер 8, партзим А10 и партзим В10 (МНКзим 10). Олигонуклеотиды, специфичные для амплификации и детектирования MgPa, представляют собой LOCS-10, прямой праймер 11, обратный праймер 11, партзим А13 и партзим В13 (МНКзим 13). Олигонуклеотиды, специфичные для амплификации и детектирования TVbtub, представляют собой LOCS-11, прямой праймер 12, обратный праймер 12, партзим А14 и партзим В14 (МНКзим 14). Олигонуклеотиды, специфичные для амплификации и детектирования HSV-1, представляют собой LOCS-8, прямой праймер 9, обратный праймер 9, партзим All и партзим В11 (МНКзим 11). Олигонуклеотиды, специфичные для амплификации и детектирования HSV-2, представляют собой LOCS-9, прямой праймер 10, обратный праймер 10, партзим А12 и партзим В12 (МНКзим 12).
Условия реакции
Амплификацию и детектирование в реальном времени проводили в общем реакционном объеме 20 мкл с использованием термоциклера BioRad® CFX96. Параметры циклирования и точки сбора флуоресцентных данных (DA) были следующими: I цикл каждые 42°С в течение 20 секунд (DA), 48°С в течение 20 секунд (DA), 65°С в течение 20 секунд (DA), 68°С в течение 20 секунд (DA) и 95°С в течение 2 минут; 10 циклов 95°С в течение 5 секунд и 61°С в течение 30 секунд (уменьшение на 0,5°С за цикл); 40 циклов: 95°С в течение 5 секунд, 52°С в течение 40 секунд и 65°С в течение 5 секунд (DA в каждом цикле); и 1 цикл каждые: 30°С в течение 20 секунд (DA), 42°С в течение 20 секунд (DA), 48°С в течение 20 секунд (DA), 65°С в течение 20 секунд (DA), 68°С в течение 20 секунд (DA). Параметры кривой плавления представляли собой шаг 0,5°С от 20°С до 95°С с выдержкой 5 секунд (DA в выдержке). Все реакции проводили в двух повторностях и содержали 40 нМ каждого прямого праймера, 200 нМ каждого обратного праймера, 200 нМ каждого партзима А, 200 нМ каждого партзима В, 200 нМ субстрата 4, 300 нМ каждого LOCS-3, LOCS-4, LOCS-7, LOCS-8, 200 нМ LOCS-5, LOCS-6, LOCS-9, 240 нМ LOCS-10, 120 нМ LOCS-11, 8 мМ MgC12 (Bioline), 0,2 мМ dNTP (Bioline), 2 единицы полимеразы MyTaq (Bioline) и 1х буфер NH4 (Bioline). Реакции содержали либо синтетическую матрицу G-Блок (10000 или 40 копий), гомологичную генам TPApolA, NGopa и/или рогА, gpdH/или gpD3, TV-Btub и/или MgPa, CTcry и/или LGV, либо не содержали мишени (NF Н2О). Все реакции, кроме реакций только с TPApolA, содержали фон из 10000 копий геномной ДНК человека, которая содержит ген-мишень TFRC, который служит эндогенным контролем. Детектирование гена TFRC контролировали в каждой реакции (за исключением реакций только с TPApolA) в качестве внутреннего контроля по увеличению флуоресценции в канале Су5.5.
Полученные результаты
В этом примере два МНКзима (МНКзимы 6 и 7) использовали в одной реакции ПЦР для одновременного детектирования и дифференциации двух целевых нуклеиновых кислот (TFRC и TPApolA соответственно) с использованием одного флуоресцентного канала (Су5.5). Присутствие или отсутствие TPApolA и/или TFRC определяли и дифференцировали в канале Су5.5 с помощью анализа конечных точек. TPApolA обнаруживали при 68°С (D2) по увеличению флуоресценции, вызванной расщеплением и плавлением LOCS-3, и TFRC обнаруживали при 48°С (D1) по расщеплению субстрата 4. Присутствие TPApolA в образце было подтверждено при анализе кривой плавления по наличию пика плавления при 68°С (D2), отражающего флуоресценцию, вызванную расщепленным и разделенным LOCS-3. Отсутствие TPApolA в образце было подтверждено на кривых плавления наличием пика плавления при 85°С, отражающего флуоресценцию нерасщепленного LOCS-3.
В этом примере восемь дополнительных мишеней (по две мишени на канал) были успешно амплифицированы, обнаружены и дифференцированы с использованием анализа кривой плавления (данные не показаны). В то время как TFRC и TPApolA были обнаружены и дифференцированы в канале Су5.5, CTcry и LGV были обнаружены и дифференцированы в канале Су5, NGopa и NGporA были обнаружены и дифференцированы в канале FAM, MgPa и TVbtub были обнаружены и дифференцированы в канале Техаский Красный, и HSV-1 и HSV-2 были обнаружены и дифференцированы в канале JOE (данные не показаны).
Результаты на Фиг. 14 иллюстрируют разницу в флуоресцентном сигнале конечной точки, полученном в канале Су5.5 при Температуре 1 (D1) и Температуре 2 (D2) с использованием анализа конечной точки (ΔSD1 и AΔSD2 ΔSD1 соответственно). Результаты представляют собой средние значения повторных реакций и нанесены на график с использованием Microsoft Excel (версия 14). Результаты показывают, что ΔSD1 превышает Порог 1 (X1), когда в образце присутствует TFRC, но не превышает порог, когда присутствует только TPApolA и/или когда TFRC (и TPApolA) отсутствуют (Фиг. 14А). Аналогично, AΔSD2 ΔSD1 превышает Порог 2 (Х2), когда в образце присутствует TPApolA, но не превышает этот порог, когда в образце присутствует только TFRC и/или когда TPApolA (и TFRC) отсутствуют (Фиг. 14В). Таким образом, сигнал флуоресценции в конечной точке выше Порога 1 (ΔSD1>X1) указывает на присутствие TFRC, а разница в сигнале флуоресценции в конечной точке между ΔSD2 и ΔSD1 превышающее Порог 2 (AΔSD2 ΔSD1>Х2) указывает на присутствие TPApolA независимо от того, присутствует ли в реакции TPApolA отдельно или вместе с TFRC.
В этом примере после ПЦР-амплификации и анализа конечной точки следовал цикл плавления после ПЦР, в ходе которого присутствие или отсутствие TPApolA подтверждалось на основании пиков плавления LOCS либо при 68°С (разделенный LOCS), либо при 85°С (интактный LOCS), соответственно. Результаты, показанные на Фиг. 15, иллюстрируют соответствующие сигнатуры кривых плавления, полученные после амплификации для реакций, содержащих 10000 копий TFRC (Фиг. 15А), 10000 копий и 40 копий TPApolA (Фиг. 15В), и коинфекций, содержащих 10000 копий TPApolA с 10000 копий TFRC, и 40 копий TPApolA с 10000 копий TFRC (Фиг. 15С). Результаты представляют собой средние значения из повторных реакций для всех образцов, содержащих TPApolA, и для реакций, не содержащих мишень (NF H2O), а результаты только для TFRC представляют собой средние значения из 48 повторов, где 10000 копий TFRC использовались в качестве эндогенного контроля. Кривые были построены с использованием Microsoft Excel (версия 14). Результаты демонстрируют, что сигнатура плавления, продуцируемая Субстратом 4 в присутствии мишени гена TFRC или без мишени (пик при Tm 85°С), отличается от сигнатуры плавления, продуцируемой LOCS-3 в присутствии TPApolA или в присутствии TPApolA или присутствии как TFRC, так и TPApolA (пик при Tm 68°С). Результаты обобщены в Таблице 7.
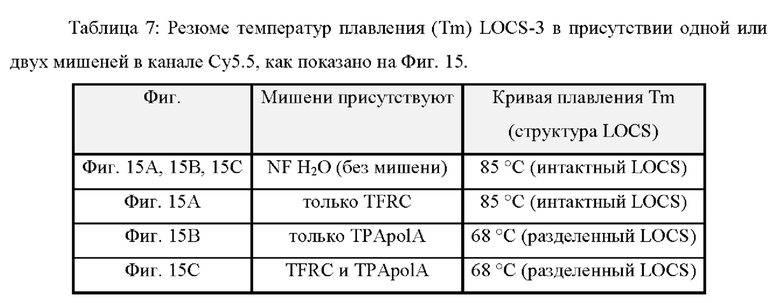
Хотя для этого примера кривая плавления была построена в широком диапазоне температур от 20°С до 90°С, а измерения проводились через каждые полградуса, в этом нет необходимости при использовании репортеров LOCS. Поскольку Tm нерасщепленного и расщепленного, разделенного LOCS не изменяется при различных целевых концентрациях, для получения подтверждающих кривых плавления можно применять меньшие диапазоны температур. Tm расщепленного, разделенного LOCS-3 и нерасщепленного Интактного LOCS-3 составляют 68°С и 85°С соответственно, поэтому анализ кривой плавления можно проводить в диапазоне от ~50°С до 90°С, чтобы зафиксировать оба пика и уменьшить время до результата. Точно так же точки сбора данных могут быть ограничены сбором данных, например, каждые 1°С, а не каждые 0,5°С, чтобы упростить анализ кривой плавления и теоретически вдвое сократить время, необходимое для получения кривой плавления. Это выгодно, когда анализ кривой плавления используется в качестве подтверждающего инструмента для поддержки анализа конечной точки.
Этот пример демонстрирует, что две мишени могут быть обнаружены в одном флуоресцентном канале при двух разных температурах с использованием анализа конечной точки с необязательным анализом кривой плавления, где одна мишень обнаруживается с использованием линейного субстрата МНКзима, а другая обнаруживается с использованием репортера LOCS. Все целевые сценарии можно было обнаружить с помощью анализа конечной точки, а наличие или отсутствие TPApolA можно было дополнительно подтвердить сигнатурами пиков плавления. В этом примере представлены два способа детектирования нескольких мишеней в одном флуоресцентном канале, при этом способ измерения конечной точки флуоресценции можно применять независимо или в тандеме с анализом кривой плавления для подтверждения результатов.
Пример 4: Способ одновременного детектирования и количественного определения двух мишеней в одном флуоресцентном канале с использованием одного линейного субстрата МНКзима и одного репортера LOCS.
В следующем примере демонстрируется подход, в котором комбинация одного линейного субстрата МНКзима и одного репортера LOCS позволяет одновременно обнаруживать и количественно определять две мишени в одном флуоресцентном канале путем получения показаний флуоресценции при двух температурах в режиме реального времени во время ПЦР. Кроме того, в этом примере описывается способ количественного определения количества одной и/или обоих мишеней, если они присутствуют в образце.
В этом примере один линейный субстрат МНКзима и один репортер LOCS использовались для одновременного детектирования, дифференциации и количественного определения двух мишеней (мишень X и мишень Y) в канале JOE путем измерения флуоресценции в реальном времени при двух разных температурах (D1 и D2) в каждом цикле ПЦР. Анализ разработан таким образом, что мишень X отслеживается с использованием линейного субстрата X (расщепляемый МНКзимом X только в присутствии мишени X), а мишень Y отслеживается с использованием LOCS-Y (расщепляемый МНКзимом Y только в присутствии мишени Y). Кроме того, анализ разработан таким образом, чтобы разделенный LOCS-Y имел Tm, которая выше, чем более низкая температура детектирования (D1).
Более низкую температуру детектирования (D1) выбирают таким образом, чтобы расщепленный линейный субстрат X флуоресцировал; тогда как нерасщепленный линейный Субстрат X, разделенный LOCS-Y и интактный LOCS-Y останутся погашенными. Более высокую температуру (D2) выбирают таким образом, чтобы стебель разделенного LOCS-Y плавился (диссоциировал), что приводило бы к усилению флуоресценции, в то время как стебель интактного LOCS-Y оставался связанным и гасился.
Можно построить две кривые ПЦР-амплификации, используя измерения флуоресценции, полученные при температурах D1 и D2. Пороговые значения для определения наличия мишеней X и/или Y устанавливаются как для графика D1 (Порог X), так и для графика D2 (Порог Y). Пороги (Порог X и Порог Y) и различные конечные точки, в которых известно, что реакции выходят на плато, могут быть заранее определены на основе предыдущих экспериментов, в которых реакции, содержащие только целевое плато X в конечной точке X1 (EX1) в D1 или в конечной точке Х2 (EX2) в точке D2; реакции, содержащие только целевое плато Y в конечной точке Y1 (EY1) в D2, и реакции, содержащие как целевое X, так и целевое плато Y в конечной точке Y2 (EY2) в D2. Необязательно конечные точки EX1, EX2, EY1 и EY2 могут быть получены из положительных контролей, запущенных параллельно с экспериментальными образцами.
Что касается графика амплификации D1, если ПЦР дает кривую амплификации, которая превышает Порог X и выходит на плато в конечной точке EX1 которая превышает Порог X, этот результат будет указывать на присутствие расщепленного линейного субстрата X, связанного с мишенью X. Если мишень Y также присутствует, кривая амплификации не должна быть затронута, поскольку расщепленный LOCS-Y не вызывает флуоресценции в D1 Таким образом, значения Cq, полученные из кривой амплификации, пересекающей Порог X, позволяют количественно определить мишень X в образце.
Что касается графика амплификации D2, если ПЦР дает кривую амплификации, которая превышает порог Y и выходит на плато либо при EY1, либо при EY2, это указывает на присутствие мишени Y. Порог Y устанавливается выше значения EX2, так что кривая амплификации реакции, содержащей только мишень X, не превышает этот порог.Когда обе мишени X и Y присутствуют, расщепление линейного субстрата X и LOCS-Y будет давать кривую амплификации, которая превышает порог Y и выходит на плато в конечной точке EY2, которая больше, чем конечная точка EY1, которая связана с расщеплением LOCS-Y в наличие только мишени Y. Поскольку EX2<Порог Y<EY1<EY2 и Е X2 ≠ Порог Y ≠ EY1 ≠ EY2, конечные точки, в которых кривая усиления выходит на плато, могут указывать на присутствие мишени X, Y или обоих. Однако на значения Cq, полученные из кривых амплификации D2, пересекающих Порог Y, будет влиять количество как мишени X, так и Y, и поэтому без дальнейшей обработки данных результаты будут только полуколичественными для мишени Y. Аналитический способ расчета количества мишени Y, присутствующей в образце, где также присутствует мишень X, описан ниже.
Способ количественного определения мишени Y
Две кривые амплификации, полученные только для расщепленного линейного субстрата МНКзима X в D1 (39°С) и D2 (72°С), имеют одинаковую эффективность и Cq, но плато в разных конечных точках, EX1 и EX2 соответственно. Следовательно, сигнал флуоресценции от расщепленного субстрата X (Sx) при 72°С ((SXD2) можно экстраполировать, используя сигнал, возникающий от расщепленного субстрата X при 39°С (SxDi), применяя фактор корректировки флуоресценции (ФКФ). ФКФ представляет собой отношение конечных точек EX1 и EX2 (ФКФ=EX2/EX1). Общий сигнал флуоресценции на кривой амплификации при 72°С (SXYD2) включает сигнал, возникающий как от расщепленного линейного субстрата X (SXD2), если он присутствует, так и от разделенного LOCS-Y (SYD2), если он присутствует. Исходя из этого, сигнал при 72°С, возникающий только от LOCS-Y (SYD2), можно экстраполировать следующим образом:
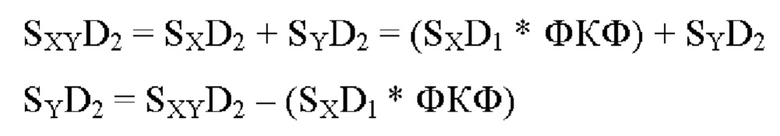
Таким образом, значения Cq, полученные из кривой амплификации, построенной из рассчитанных значений SYD2. В каждом цикле, могут предоставить количественные данные для мишени Y. Формулу можно применять независимо от того, содержит ли образец мишень X, Y, и X, и Y, или не содержит мишени, правильно определить количество мишени Y в образце.
Олигонуклеотиды
Олигонуклеотиды, специфичные для этого эксперимента, включают; линейный субстрат МНКзима 1 (SEQ ID NO: 13), LOCS-1 (SEQ ID NO: 14), партзим A1 (SEQ ID NO: 3), партзим B1 (SEQ ID NO: 4), партзим A2 (SEQ ID NO: 7), партзим B2 (SEQ ID NO: 8), прямой праймер 1 (SEQ ID NO: 1), обратный праймер 1 (SEQ ID NO: 2), прямой праймер 2 (SEQ ID NO: 5) и обратный праймер 2 (SEQ ID NO: 6). Последовательности перечислены в списке последовательностей. Олигонуклеотиды, специфичные для амплификации и количественного определения мишени Х(CTcry), представляют собой субстрат 1, партзимы А1 и В1 (МНКзим 1; МНКзим X), прямой праймер 1 и обратный праймер 1. Олигонуклеотиды, специфичные для амплификации и количественного определения мишени Y (NGopa), представляют собой LOCS-1, партзимы А2 и В2 (МНКзим 2; МНКзим Y), прямой праймер 2 и обратный праймер 2.
Условия реакции
Детекцию целевой последовательности в режиме реального времени проводили в общем реакционном объеме 20 мкл с использованием термоциклера BioRad® CFX96. Параметры цикла: 95°С в течение 2 минут, затем 10 циклов: 95°С в течение 5 секунд и 61°С в течение 30 секунд (уменьшение на 0,5°С за цикл) и 40 циклов 95°С в течение 5 секунд, 52°С в течение 40 секунд, 39°С в течение 5 секунд и 72°С в течение 5 секунд (данные собраны на стадиях как 39°С, так и 72°С). Все реакции проводили в двух повторностях и содержали 40 нМ каждого прямого праймера, 200 нМ каждого обратного праймера, 200 нМ каждого партзима, 100 нМ линейного субстрата МНКзима 1, 200 нМ LOCS-1 и lx PlexMastermix (Bioline). Реакции либо не содержали мишени (NF H2O), либо содержали синтетический G-блок CTcry (20000, 4 000, 800, 160 или 32 копии), синтетический G-блок гена NGopa (20000, 4 000, 800, 160 или 32 копии), различные концентрации синтетического G-блока гена CTcry (20000, 4 000, 800, 160 или 32 копии) на фоне синтетического G-блока гена NGopa (20000, 4 000, 800, 160 или 32 копии) или содержали различные концентрации синтетического G-блока гена NGopa (20000, 4 000, 800, 160 или 32 копии) на фоне синтетического G-блока гена CTcry (20000, 4 000, 800, 160 или 32 копии).
Полученные результаты
Во время ПЦР два МНКзима (МНКзима 1 и МНКзима 12) используются для мониторинга амплификации целевых нуклеиновых кислот в режиме реального времени посредством расщепления линейного субстрата МНКзима 1 и LOCS 1 соответственно. МНКзим 1 был разработан для детектирования последовательностей, гомологичных CTcry (мишень X), для детектирования Chlamydia trachomatis и для расщепления субстрата 1. МНКзим 2 был разработан для детектирования последовательностей, гомологичных NGopa (мишень Y), для детектирования Neisseria gonorrhoeae, а также для отщепления и разделения LOCS-1.
Результаты, показанные на Фиг. 16, иллюстрируют сравнительные кривые амплификации, полученные в канале JOE в реакциях, содержащих либо 20000 копий CTcry или NGopa, либо 20000 копий обоих генов-мишеней, измеренных при 39°С (D1) (Фиг. 16А) и 72°С (D2) (Фиг. 16В), соответственно. Кривые амплификации представляют собой средние значения для повторных реакций и построены с использованием Microsoft Excel (версия 14). Значения Cq определяли с использованием способа одного порога, установленного на 400 ОЕФ и 500 ОЕФ для D1 (39°С) и D2 (68°С) соответственно (подгонка кривой с вычитанием исходного уровня).При 39°С образцы, содержащие 20000 копий матрицы CTcry (сплошная черная линия), показывают кривые амплификации с уровнями флуоресценции, которые превышают Порог X (черная горизонтальная линия) и достигают конечной точки (Е xi) (черная горизонтальная линия обозначена как EX1). При 39°С образцы, содержащие 20000 копий матриц CTcry и NGopa (сплошная серая линия), также показывают кривые амплификации с уровнями флуоресценции, которые превышают Порог X и достигают EX1 При 39°С образцы, которые содержат только матрицу NGopa (черная пунктирная линия) или не содержат матрицу CTcry (NTC; серая пунктирная линия), не превышают Порог X и не достигают EX1. Таким образом, кривая амплификации при 39°С, выходящая на плато при EX1 подтверждает наличие CTcry в образце. На графике показаны образцы, содержащие только 20000 копий CTcry (сплошная черная линия) и 20000 копий как CTcry, так и NGopa (сплошная серая линия), имеют сопоставимые значения Cq. Реакции, содержащие только NGopa (штриховая черная линия) или не содержащие мишени (штриховая серая линия), не показали увеличения флуоресценции во время ПЦР.
Поскольку на значения Cq при 39°С не влияет присутствие NGopa, его можно применять для количественного определения количества CTcry в образце. Результаты количественного определения CTcry в образцах всех комбинаций 0, 32, 160, 800, 4000 и 20000 копий CTry вместе с 0, 32, 160, 800, 4000 и 20000 копий генов-мишеней NGopa суммированы в Таблице 8, где Н/П означает отсутствие значения Cq, определенного при 39°С, и, следовательно, отсутствие CTcry в образце.
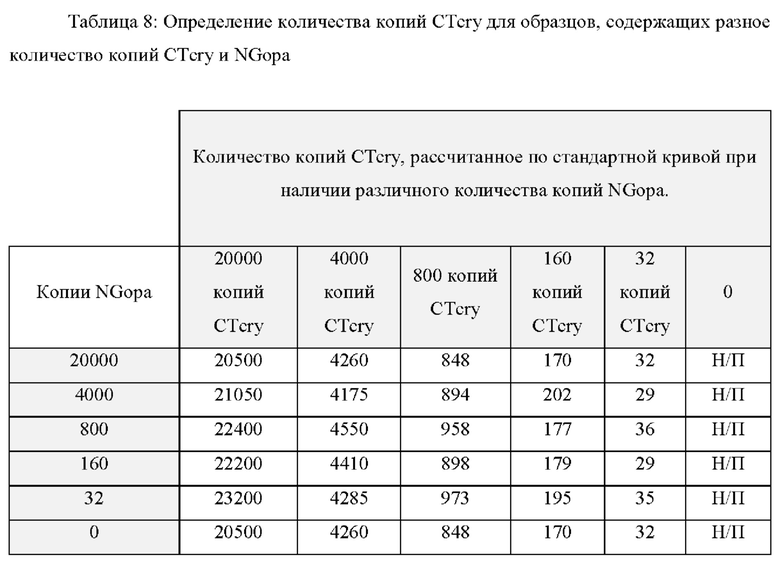
На Фиг. 16В показаны кривые амплификации при 72°С для образцов, содержащих 20000 копий матрицы NGopa (черная пунктирная линия) с уровнями флуоресценции, превышающими Порог Y (черная горизонтальная линия), и плато в первой конечной точке Y (EY1) (черная горизонтальная линия обозначена как EY1). При 72°С образцы, содержащие 20000 копий CTcry и NGopa (сплошная серая линия), демонстрируют кривые амплификации с уровнями флуоресценции, превышающими Порог Y, и плато во второй конечной точке Y (EY2) (черная горизонтальная линия обозначена как EY2). При 72°С образцы, которые не содержат матрицу NGopa (сплошная черная линия), но содержат CTcry, не превышают Порог Y и не достигают EY1 и EY2, а вместо этого выходят на плато в конечной точке EX2 (черная горизонтальная линия обозначена как конечная точка EX1)-При 72°С образцы, не содержащие ни NGopa, ни CTcry-матриц (NTC; серая пунктирная линия), не показывают никакой кривой амплификации и, следовательно, не превышают Порог Y и не достигают EY1 и EY2. Таким образом, кривая амплификации при 72°С, выходящая на плато при EX2, подтверждает наличие в образце мишени CTcry, но не NGopa; плато при EY1 подтверждает наличие в образце NGopa, но не CTcry; плато при EY2 подтверждает присутствие в образце как CTcry, так и NGopa.
При 72°С значение Cq для образцов, содержащих 20000 копий как CTcry, так и NGopa (сплошная серая линия), не совпадает со значением Cq для образцов, содержащих только 20000 копий темплата NGopa (штриховая черная линия). Следовательно, значения Cq при 72°С без нормализации нельзя применять для точного определения концентрации NGopa в образце, когда также присутствует CTcry.
Результаты, представленные на Фиг. 17, иллюстрируют сравнительные кривые амплификации, полученные в канале JOE из реакций, содержащих все возможные комбинации 0, 32 и 20000 копий CTry и 0, 32 и 20000 копий мишеней гена NGopa, измеренных при 39°С (Фиг. 17А, 17D и 17G) и 72°С (Фиг. 17В, 17Е и 17Н) соответственно. Результаты нанесены на график в виде средних значений для повторных реакций и нанесены на график с использованием Microsoft Excel (версия 14). Значения Cq определяли с использованием способа одного порога, установленного на 400 ОЕФ и 500 ОЕФ для D1 (39°С) и D2 (72°С) соответственно (базовая кривая вычитания). Образцы, содержащие 20000 копий CTcry, представлены пунктирной черной линией, образцы, содержащие 32 копии CTcry, представлены сплошной серой линией, а образцы, содержащие 0 копий CTcry, представлены сплошной черной линией.
На Фиг. 17А, 17D и 17G показана флуоресценция при 39°С (D1) для реакций, содержащих NGopa (сплошная черная линия), при 20000 копий (Фиг. 17А), 32 копиях (Фиг. 17D) или без копий (Фиг. 17G). Результаты показывают, что нет увеличения сигнала при 39°С в присутствии NGopa (Фиг. 17А и 17D), и сигнал сравним с реакциями, которые не содержат NGopa или CTry (NTC; Фиг. 17G), таким образом демонстрируя, что детектирование в реальном времени при 39°С (D1) можно применять для определения присутствия CTcry в образце.
Фиг. 17В, 17Е и 17Н показывают флуоресценцию при 74°С (D2) для реакций, содержащих NGopa (сплошная черная линия) при 20000 копий (Фиг. 17В), 32 копиях (Фиг. 17Е) или без копий (Фиг. 17Н). Результаты показывают, что ненормализованные кривые амплификации NGopa (Фиг. 17В и 17Е) смещаются, когда в образце присутствует 20,00 копий (штриховая черная линия) или 32 копии (серая линия) CTcry.
Фиг. 17С, 17F и 171 показывают флуоресценцию при 74°С (D2) после нормализации с помощью ФКФ для реакций, содержащих NGopa (черная сплошная линия), при 20000 копий (Фиг. 17С), 32 копиях (Фиг. 17F) или без копий (Фиг. 171). Результаты показывают, что нормализованные кривые амплификации при 72°С (D2) демонстрируют сходные значения Cq, где присутствует одинаковое количество матриц NGopa, независимо от количества CTcry в образце (Фиг. 17С и 17F). На Фиг. 171 показано, что экстраполированные кривые амплификации при 72°С не показывают какой-либо значительной амплификации, если образцы не содержат каких-либо темплатов NGopa. Влияние вышеупомянутого способа нормализации ФКФ дополнительно продемонстрировано в Таблице 9 (значения SYD2 Cq, где SYD2=SXYD2 - 8×Э2ФКФ) и Таблице 10 (количественное определение NGopa после SYD2=SXYD2-SXD2ФКФ нормализации), которые представляют собой анализ амплификации при 72°С (D2) образцов всех комбинаций 0, 32, 160, 800, 4000 и 20000 копий CTcry и 0, 32, 160, 800, 4000 и 20000 копий гена-мишени NGopa после нормализации путем вычитания SYD2=SXYD2-SXD2ФКФ. Неприменимо (Н/П) означает, что значение Cq не определено при 72°С и, следовательно, в образце отсутствует NGopa.
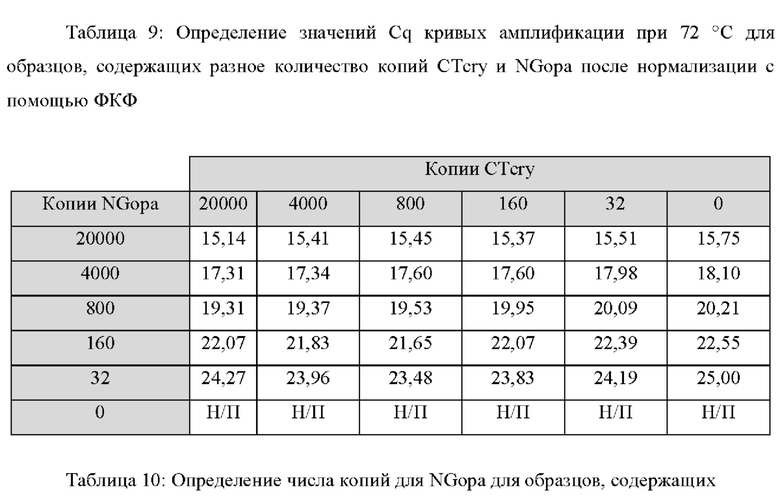
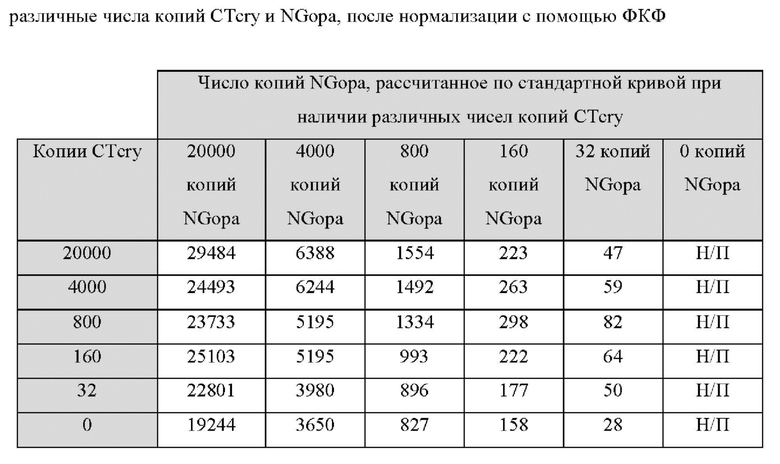
Пример возможного применения 5: Детектирование и различение нескольких мишеней на одной длине волны с использованием комбинации молекулярных маяков и LOCS.
Нерасщепляемый молекулярный маяк можно комбинировать с зондом LOCS, расщепляемым МНКзимом, для детектирования и различения нескольких мишеней на одной длине волны. Как показано на Фиг. 5, и молекулярный маяк, и зонд LOCS могут быть помечены одним и тем же детектирующим фрагментом, например одним и тем же флуорофором. Молекулярный маяк может иметь стеблевую область с Tm А и петлевую область, которая может специфически гибридизоваться с первой мишенью 1 с Tm В; где Tm В больше, чем Tm А (Tm В>Tm А). Это может быть объединено с интактным зондом LOCS, который может иметь стеблевую область с Tm С и петлевую область, которая может быть расщеплена МНКзимом в присутствии второй мишени 2, таким образом генерируя разделенный LOCS с Tm D; где Tm D меньше Tm С (Tm D<Tm С). Присутствие мишени 1 и/или мишени 2 можно определить путем измерения флуоресценции при двух температурах (D1 и D2) либо в режиме реального времени; или с использованием одиночных измерений, полученных либо в начале амплификации, либо почти в начале амплификации и после амплификации. Варианты конечной точки или измерения флуоресцентного выхода в реальном времени, а также стратегия интерпретации результатов зависят от относительных температур Tm А, Tm В, TmC, Tm D по отношению друг к другу и по отношению к D1 и D2.
Следующие сценарии обобщены в Таблице 1, в которой представлены примерные Tm для молекулярных маяков и LOCS, а также температуры сбора данных для Сценариев 1^, а также ожидаемые результаты.
Сценарий 1
В первом протоколе Tm А может быть 65°С, Tm В может быть 70°С, Tm С может быть 65°С и Tm D может быть 55°С (Tm А>Tm D и Tm В>TmC, Tm В>Tm D). ПЦР-амплификацию можно проводить для амплификации мишени 1 и/или мишени 2, если они присутствуют, и измерение флуоресценции можно проводить при двух температурах в начале амплификации или почти в начале амплификации и снова после амплификации. В присутствии мишени 1 и/или мишени 2 увеличение флуоресценции при первой температуре (D1 50°С), которая меньше Tm А, Tm В и Tm D (D1<Tm A, D1<Tm В, D1<Tm D), будет указывать на присутствие только мишени 1. При этой температуре Молекулярный Маяк будет флуоресцировать в ответ на гибридизацию с мишенью 1 или будет гаситься в ее отсутствие и поддерживать внутренне гибридизованный стебель. При одной и той же температуре D1 как интактные, так и разделенные виды LOCS будут гаситься из-за гибридизации их соответствующих стеблей при этой температуре, независимо от присутствия или отсутствия мишени 1 или мишени 2В реакции. Далее, увеличение флуоресценции при второй температуре (D2 60°С), которая выше как первой температуры D1; так и Tm D, но ниже как Tm В, так и Tm С (D2>D1, D2>Tm D, D2<Tm В, D2<Tm С), будет указывать на присутствие мишени 1 и/или мишени 2. При этой второй температуре Молекулярный Маяк будет флуоресцировать в ответ на гибридизацию с мишенью 1 или будет гаситься в отсутствие мишени 1 и поддерживать внутренне гибридизованный стебель. На Молекулярный Маяк не будет влиять наличие или отсутствие мишени 2. Кроме того, в D2, если мишень 2 отсутствует, зонд LOCS остается интактным и гасится. Если бы мишень 2 присутствовала, флуоресценция увеличилась бы, поскольку расщепление МНКзимами, специфичными для мишени 2, привело бы к образованию разделенных LOCS со стеблевыми областями, которые диссоциируют при этой температуре. Таким образом, увеличение флуоресценции при температуре 2Во время ПЦР, превышающее увеличение флуоресценции, наблюдаемое при температуре 1, будет указывать на присутствие мишени 2 (ΔF D2>ΔF D1).
Сценарий 2
Во втором протоколе Tm А может быть 60°С, Tm В может быть 70°С, Tm С может быть 70°С и Tm D может быть 60°С (Tm А ≈ Tm D, Tm В ≈ TmC, и Tm В>Tm D). ПЦР-амплификацию можно проводить для одновременной амплификации мишени 1 и/или мишени 2, если они присутствуют, и измерение флуоресценции можно проводить при двух температурах либо в режиме реального времени, либо в начале/почти в начале амплификации и снова после амплификации. В присутствии мишени 1 и/или мишени 2 увеличение флуоресценции при первой температуре (D1 50°С), которая меньше Tm А, Tm В и Tm D (D1<Tm A, D1<Tm В, D1>Tm D), будет указывать на присутствие только мишени 1. При этой температуре Молекулярный Маяк будет флуоресцировать в ответ на гибридизацию с мишенью 1 или будет гаситься в ее отсутствие и поддерживать внутренне гибридизованный стебель. При одной и той же температуре D1 как интактные, так и разделенные виды LOCS будут гаситься из-за гибридизации их соответствующих стеблей при этой температуре, независимо от присутствия или отсутствия мишени 1 или мишени 2В реакции. Далее усиление флуоресценции при второй температуре (D2 65°С), которая больше как первой температуры D1 и Tm А и Tm D, но меньше Tm В и Tm С (D2>D1, D2>Tm A, D2>Tm В, D2>TmC, D2<Tm D), будет указывать на наличие мишени 1 и/или мишени 2. При этой второй температуре Молекулярный Маяк будет флуоресцировать в ответ на гибридизацию с мишенью 1 или будет флуоресцировать из-за диссоциации его стебля при этой температуре независимо от присутствия или отсутствия мишени 2В реакции. Таким образом, флуоресценция Молекулярного Маяка будет обеспечивать фоновый уровень флуоресценции при этой температуре до, во время и после амплификации. Кроме того, в D2, если мишень 2 отсутствует, зонд LOCS остается интактным и гасится. Если бы мишень 2 присутствовала, флуоресценция превышала бы фоновые уровни во время ПЦР, поскольку расщепление МНКзимами, специфичными для мишени 2, приводило бы к образованию разделенных LOCS со стрежневыми областями, диссоциированными при этой температуре. Таким образом, увеличение флуоресценции при температуре 2Во время ПЦР будет указывать на присутствие мишени 2. В целом увеличение флуоресценции в D1 во время ПЦР указывает на присутствие мишени 1, обнаруженной с помощью Молекулярного Маяка, а увеличение флуоресценции во время ПЦР в D2 указывает на присутствие мишени 2, обнаруженной зондом LOCS.
Сценарий 3
В третьем протоколе Tm А может быть 60°С, Tm В может быть 70°С, Tm С может быть 80°С и Tm D может быть 70°С (Tm А<Tm D, Tm А<Tm С и Tm В ≈ Tm D). ПЦР-амплификацию можно проводить для амплификации мишени 1 и/или мишени 2, если они присутствуют, и измерение флуоресценции можно проводить при двух температурах либо в режиме реального времени; либо в начале/почти в начале амплификации и снова после амплификации. В присутствии мишени 1 и/или мишени 2 увеличение флуоресценции при первой температуре (D1 50°С), которая меньше Tm А и Tm В и Tm С и Tm D (D1<Tm А, D1<Tm В, D1<TmC, D1<Tm D), будет указывать на присутствие только мишени 1. При этой температуре Молекулярный Маяк будет флуоресцировать в ответ на гибридизацию с мишенью 1 или будет гаситься в ее отсутствие и поддерживать внутренне гибридизованный стебель. При одной и той же температуре D1 как интактные, так и разделенные виды LOCS будут гаситься из-за гибридизации их соответствующих стеблей при этой температуре, независимо от присутствия или отсутствия мишени 1 или мишени 2В реакции. Далее усиление флуоресценции при второй температуре (D2 75°С), которая больше как первой температуры D1 и Tm А, так и Tm В и Tm D, но меньше Tm С (D2>D1, D2>Tm A, D2>Tm В, D2>Tm D, D2<Tm С), будет указывать на присутствие мишени 2. При этой второй температуре Молекулярный Маяк не сможет гибридизоваться с мишенью 1, но будет флуоресцировать из-за диссоциации его стебля при этой температуре независимо от присутствия или отсутствия мишени 1 или мишени 2В реакции. Таким образом, флуоресценция Молекулярного Маяка будет обеспечивать фоновый уровень флуоресценции при этой температуре до, во время и после амплификации. Кроме того, в D2, если мишень 2 отсутствует, зонд LOCS оставутся интактным и гасится. Если бы мишень 2 присутствовала, флуоресценция превышала бы фоновые уровни во время ПЦР, поскольку расщепление МНКзимами, специфичными для мишени 2, приводило бы к образованию разделенных LOCS со стрежневыми областями, диссоциированными при этой температуре. Таким образом, увеличение флуоресценции при температуре 2. Во время ПЦР будет указывать на присутствие мишени 2. В целом увеличение флуоресценции при D1 во время ПЦР указывает на присутствие мишени 1, обнаруженной с помощью Молекулярного Маяка, а увеличение флуоресценции во время ПЦР при D2 указывает на присутствие мишени 2, обнаруженной зондом LOCS.
Подобно сценарию 2, этот вариант реализации обеспечивает большое преимущество по сравнению с другими известными в данной области способами, в которых используются измерения при различных температурах. Этот конкретный вариант реализации сочетает в себе Молекулярные Маяки и зонд LOCS в способе, который устраняет необходимость сложного пост-ПЦР-анализа. Формат позволяет проводить прямое количественное определение первой мишени по первой кривой амплификации, полученной при первой температуре, и прямое количественное определение второй мишени по второй кривой амплификации, полученной при второй температуре.
Сценарий 4
В четвертом протоколе Tm А может быть 75°С, Tm В может быть 80°С, Tm С может быть 65°С и Tm D может быть 55°С (Tm А>TmC, Tm А>Tm D и Tm В>Tm С). ПЦР-амплификацию можно проводить для амплификации мишени 1 и/или мишени 2, если они присутствуют, и измерение флуоресценции можно проводить при двух температурах в начале/почти в начале амплификации и снова после амплификации. В присутствии мишени 1 и/или мишени 2 увеличение флуоресценции при первой температуре (D1 70°С), которая меньше, чем Tm А и Tm В, но больше, чем Tm С и Tm D(D1<Tm A, D1<Tm В, D1>TmC, D1>Tm D), будет указывать на присутствие мишени 1. При этой температуре Молекулярный Маяк будет флуоресцировать в ответ на гибридизацию мишени 1 или будет гаситься в отсутствие мишени 1 и поддерживать внутренне гибридизованный стебель. При одной и той же температуре D1 как интактный, так и/или разделенный LOCS будут флуоресцировать из-за диссоциации их соответствующих стеблевых областей при этой температуре. Далее, увеличение флуоресценции при второй температуре (D2 60°С), которая больше Tm D, но меньше первой температуры, Tm А, Tm В и Tm С (D2<D1, D2<Tm AD2<Tm В, D2<TmC, D2>Tm D) будет указывать на присутствие мишени 1 и/или мишени 2. При этой второй температуре интактные виды LOCS будут гаситься из-за гибридизации их стебля при этой температуре. Если бы мишень 2 присутствовала, флуоресценция увеличилась бы во время ПЦР, поскольку расщепление МНКзимами, специфичными для мишени 2, привело бы к образованию разделенных LOCS со стеблевыми областями, диссоциированными при этой температуре. Молекулярный Маяк будет флуоресцировать при этой температуре в ответ на гибридизацию мишени 1 в присутствии мишени 1 или будет гаситься в отсутствие мишени 1 и поддерживать внутренне гибридизованный стебель. На Молекулярный Маяк не будет влиять наличие или отсутствие мишени 2. Таким образом, наблюдаемое увеличение флуоресценции при температуре 2Во время ПЦР, превышающее наблюдаемое увеличение флуоресценции при температуре 1 (ΔSD2>ΔSD1), будет указывать на присутствие мишени 2.
Пример 6: Способ анализа нескольких мишеней на одной длине волны с использованием одного зонда TaqMan и одного зонда LOCS в формате, позволяющем либо одновременную количественную оценку в реальном времени одной мишени и качественное определение конечной точки второй мишени, либо одновременный качественный анализ конечной точки двух мишеней на канал.
В следующем примере демонстрируется подход, при котором один зонд TaqMan и один репортер LOCS используются вместе для детектирования и дифференциации двух генов-мишеней (GAPDH и MgPa) с определением Cq одной мишени (GAPDH) в одном флуоресцентном канале без необходимости анализа кривой плавления. Анализ разработан таким образом, что ген GAPDH можно обнаружить в режиме реального времени с помощью зонда TaqMan, а ген MgPa можно обнаружить и дифференцировать в конечной точке с помощью репортера LOCS.
Во время ПЦР зонд TaqMan расщепляется в присутствии GAPDH, чтобы разделить флуорофор и гаситель, чтобы получить усиление сигнала, которое можно обнаружить в широком диапазоне температур. В этом примере детектирование в реальном времени и определение Cq GAPDH достигается путем получения флуоресценции в каждом цикле во время ПЦР при температуре 1 (52°С). Стебель зонда LOCS как в исходной, так и в разделенной конфигурации имеет температуру плавления (Tm), которая выше температуры 1 (52°С), поэтому репортер LOCS не дает сигнала при температуре 1 независимо от наличия или отсутствия MgPa в образце.
В присутствии MgPa МНКзим 5 может расщеплять зонд LOCS во время ПЦР. Качественное определение MgPa достигается путем измерения флуоресценции при более высокой температуре 2 (70°С) как до, так и после амплификации. Поскольку Tm стебля интактного LOCS выше температуры 2 (Tm>70°С), а Tm стебля разделенного LOCS ниже температуры 2 (Tm<70°С), то увеличение флуоресценции связано с разделенным и диссоциированным LOCS и указывает на присутствие MgPa. Это увеличение флуоресценции должно быть выше любой фоновой флуоресценции, вызванной деградировавшим зондом TaqMan при этой температуре, чтобы считаться показателем присутствия мишени MgPa.
Этот пример демонстрирует смешанное количественное измерение в реальном времени первой мишени с качественным дискретным определением температуры второй мишени в одном флуоресцентном канале (FAM). Кроме того, пример демонстрирует качественный анализ тех же двух мишеней путем измерения дискретных измерений флуоресценции с использованием Способов Анализа Конечных Точек 1-3, как описано в Примере 1.
Ол иго нукл е отиды
Олигонуклеотиды, специфичные для этого эксперимента, включают прямой праймер 5 (SEQ ID NO: 20), обратный праймер 5 (SEQ ID NO: 21), партзим А5 (SEQ ID NO: 22), партзим B5 (SEQ ID NO: 23), LOCS-2 (SEQ ID NO: 25), которые специфичны для амплификации и детектирования MgPa. Олигонуклеотиды, специфичные для амплификации и детектирования GAPDH, включены в запатентованный анализ экспрессии генов TaqMan™ (FAM) Human GAPDH (AppliedBiosystems).
Условия реакции
Амплификацию и детектирование в реальном времени проводили в общем реакционном объеме 20 мкл с использованием термоциклера BioRad® CFX96. Параметры циклирования и точки сбора флуоресцентных данных (DA) были следующими: 1 цикл 95°С в течение 2 минут, 52°С в течение 15 секунд (DA), 70°С в течение 15 секунд (DA); 10 циклов: 95°С в течение 5 секунд и 61°С в течение 30 секунд (уменьшение на 0,5°С за цикл); 40 циклов 95°С в течение 5 секунд и 52°С в течение 40 секунд (DA в каждом цикле при 52°С); и 1 цикл 70°С в течение 15 секунд (DA). Все реакции проводили в трех повторностях и содержали 40 нМ прямого праймера 5, 200 нМ обратного праймера 5, 200 нМ партзима А5, 200 нМ партзима В5, 300 нМ LOCS-2, lx TaqMan™ Анализ экспрессии генов (FAM) Человека GAPDH (AppliedBiosystems) и lx буфер SensiFast (Bioline). Реакции либо не содержали мишени (NF Н2О), либо содержали геномную ДНК человека (10000 или 100 копий), либо синтетический G-блок, содержащий участок гена MgPa (10000 или 100 копий), либо как геномную ДНК человека, так и синтетический MgPa G-Блок (10000 или 100 копий каждого).
Полученные результаты
В этом примере один зонд TaqMan и один зонд LOCS, расщепляемый МНКзимом 5, объединяли в одной реакции ПЦР для одновременного детектирования и дифференциации двух мишеней GAPDH и MgPa соответственно с использованием только одного флуоресцентного канала (FAM). Наличие гена GAPDH определяли в режиме реального времени по увеличению сигнала флуоресценции при 52°С в канале FAM, а также определяли анализом флуоресценции в дискретные моменты времени при 52°С до и после циклирования ПЦР. Присутствие MgPa определяли путем мониторинга расщепления LOCS-2 посредством анализа флуоресценции в дискретные моменты времени при 70°С до и после цикла ПЦР в одном и том же канале.
Результаты, показанные на Фиг. 18А, иллюстрируют кривые ПЦР-амплификации, полученные в канале FAM при температуре получения 52°С из реакционных смесей, содержащих 10000 (серая линия) и 100 (пунктирная линия) копий только матриц GAPDH человека (Фиг. 18А); 10000 (серая линия) и 100 (пунктирная линия) копий только MgPa (Фиг. 18В); и 10000 (серая линия) и 100 (пунктирная линия) копий каждого из темплатов GAPDH и MgPa человека (Фиг. 18С). Нецелевые контроли (NF H2O) показаны на Фиг. 18А-С (черная сплошная линия).
Значения Cq, полученные в результате вышеуказанных реакций, показаны в Таблице 11, где Неприменимо (Н/П) относится к случаю, когда значение Cq не определено при 52°С, что согласуется с отсутствием GAPDH человека. Значения Cq определяли с использованием одного порога в 200 ОЕФ. Результаты показывают, что значения Cq для GAPDH человека сопоставимы независимо от наличия или отсутствия MgPa. Поэтому значения Cq, полученные в канале FAM, можно применять для прямого количественного анализа GAPDH человека в образце.
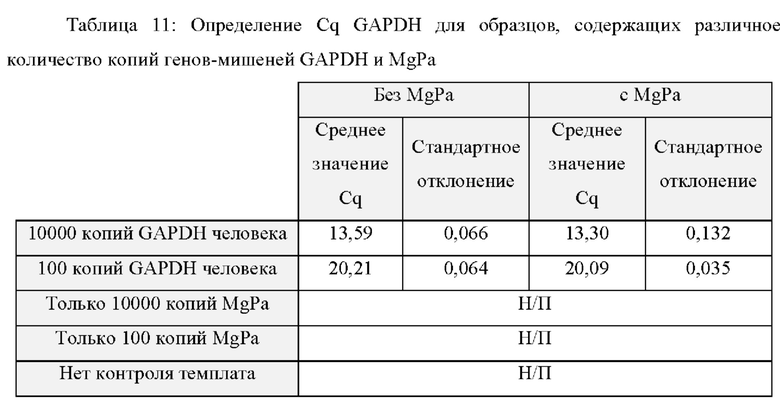
Результаты на Фиг. 19 иллюстрируют результаты качественного детектирования генов GAPDH и MgPa в канале FAM. Результаты для Способа Анализа Конечной Точки 1 представлены на Фиг. 19А, которая иллюстрирует изменение флуоресцентного сигнала, полученного при двух разных температурах. ΔSD1 (измерено при 52°С) показано черно-белым паттерном, a ΔSD2 (измерено при 70°С) показано серым цветом. Результаты показывают, что ΔSD1 превышает Порог X1, когда в реакции присутствует GAPDH, но не превышает порог, когда GAPDH отсутствует.Таким образом, ΔSD1 выше Порога X1 указывает на наличие GAPDH. Результаты также показывают, что в присутствии MgPa ΔSD2 превышает Порог X1 и превышает ΔSD1 в реакции (ΔSD2>ΔSD1).
Результаты на Фиг. 19В иллюстрируют разницу во флуоресцентном сигнале, полученном в канале FAM при Температуре 2 и Температуре 1 с использованием Способа Анализа Конечных Точек 2 (AΔSD2 ΔSD1) только для детектирования MgPa. Результаты показывают, что AΔSD2 ΔSD1 превышает Порог Х2, когда в реакции присутствует MgPa, но не превышает этого порога, когда MgPa отсутствует в образце. Следовательно, AΔSD2 ΔSD1 выше Порога Х2 указывает на присутствие MgPa.
Результаты на Фиг. 19С и 19D иллюстрируют изменение флуоресцентного сигнала, полученного в канале FAM при двух разных температурах с использованием Способа Анализа Конечных Точек 3. На Фиг. 19С, где ΔSD2 больше Порога Хь это указывает на то, что в реакции присутствуют GAPDH и/или MgPa. Если ΔSD2 ниже Порога X1, это указывает на отсутствие в реакции ни GAPDH, ни MgPa. На Фиг. 19D показано отношение ΔSD1: ΔSD2, которое можно применять для указания того, какие мишени присутствуют в реакции. Если отношение выше, чем Порог R1, это указывает на присутствие GAPDH, но не на MgPa. Если отношение ниже Порога R2, это указывает на присутствие MgPa, но не на GAPDH. Если соотношение находится между Порогами R1 и R2, это указывает на присутствие как GAPDH, так и MgPa. Когда в реакции не присутствует ни GAPDH, ни MgPa (определено из Фиг. 19С), необходимость расчета соотношения отменяется и указывается как Н/П, как показано на Фиг. 19D.
В целом этот пример демонстрирует, что две мишени могут быть обнаружены в одном флуоресцентном канале при двух разных температурах, при этом одна мишень может быть определена количественно, а другая мишень обнаружена в конечной точке с использованием одного зонда TaqMan и одного зонда LOCS соответственно. Этот простой способ не требует анализа кривой плавления. Пример также демонстрирует, что качественные данные могут быть получены для нескольких мишеней на одной длине волны путем сравнения значений флуоресценции до и после ПЦР при нескольких дискретных температурах, и что для анализа этих данных можно применять несколько альтернативных способов. Пример демонстрирует еще одно преимущество, а именно возможность комбинировать один или более зондов LOCS с существующими коммерческими наборами с использованием других технологий, таких как зонды TaqMan, и, таким образом, расширять их возможности мультиплексирования.
Пример 7: Детектирование и количественная оценка двух мишеней в режиме реального времени на одной длине волны с использованием комбинации Молекулярных Маяков и LOCS.
В следующем примере демонстрируется подход, в котором комбинация одного нерасщепляемого молекулярного маяка и одного репортера LOCS позволяет одновременно обнаруживать и количественно определять две мишени в одном флуоресцентном канале путем получения показаний флуоресценции при двух температурах в режиме реального времени во время ПЦР. Эта стратегия устраняет потребность в специализированных способах анализа после амплификации, как показано в Примере 4. Как показано на Фиг. 5, и Молекулярный Маяк, и зонд LOCS помечены одними и теми же фрагментами флуорофора и гасителя для одновременного детектирования в одном и том же флуоресцентном канале. Молекулярный Маяк содержит стеблевую область с Tm А и петлевую область, которая может специфически гибридизоваться с мишенью 1 (TVbtub) с Tm В; где Tm В больше, чем Tm А (Tm В>Tm А). В этом примере интактный зонд LOCS имеет стеблевую область с Tm С (82°С) и петлевую область, которая при расщеплении МНКзимом в присутствии второй мишени 2 (MgPa) генерирует разделенный LOCS с Tm D(62°С); где Tm D меньше Tm С (Tm D<Tm С). Наличие мишени 1 (TVbtub) и/или мишени 2 (MgPa) можно различить путем измерения флуоресценции при двух температурах (D1 и D2; 52°С и 74°С) либо в реальном времени после каждого цикла ПЦР; или с использованием одиночных измерений, полученных либо в начале амплификации, либо почти в начале и после амплификации. В следующем примере Tm А составляет -60°С, Tm В составляет -68°С, Tm С составляет -82°С и Tm D составляет -62°С, что соответствует Сценарию 3, описанному в Примере 5, где D1<Tm А<Tm В<D2 и D1<Tm D<D2<Tm C.
Олигонуклеотиды
Олигонуклеотиды, специфичные для этого эксперимента, включают: Прямой праймер 12 (SEQ ID NO: 63), обратный праймер 12 (SEQ ID NO: 64) и молекулярный маяк 1 (SEQ ID NO: 68) для амплификации и количественного определения Мишени 1 (TVbtub). Прямой праймер 5 (SEQ ID NO: 20) и обратный праймер 5 (SEQ ID NO: 21), партзим А5 (SEQ ID NO: 22), партзим B5 (SEQ ID NO: 23) и LOCS-2 (SEQ ID NO: 25) для амплификации и количественного определения Мишени 2 (MgPa). Последовательности перечислены в списке последовательностей.
Условия реакции
Детекцию целевой последовательности в режиме реального времени проводили в общем реакционном объеме 20 мкл с использованием термоциклера BioRad® CFX96. Параметры циклов: 95°С в течение 2 минут, 52°С в течение 15 секунд, 74°С в течение 15 секунд (данные собраны с шагом 52°С и 74°С), после чего следует 10 циклов при 95°С в течение 5 секунд и 61°С в течение 30 секунд (уменьшение на 0,5°С за цикл) и 40 циклов 95°С в течение 5 секунд, 52°С в течение 40 секунд и 74°С в течение 5 секунд (данные собраны как при 52°С, так и при с шагом 74°С). Каждая реакция содержала 20 нМ прямого праймера 12, 40 нМ прямого праймера 5, 200 нМ каждого обратного праймера, 200 нМ каждого партзима, 200 нМ молекулярного маяка 1, 200 нМ репортера LOCS-1 и 1х PlexMastermix (Bioline).
Реакции либо не содержали мишень (NF H2O), либо содержали синтетический G-блок, содержащий участок гена TVbtub (25600, 6400, 1600, 400 или 100 копий), синтетический G-блок, содержащий участок гена MgPa (25600, 6400, 1600, 400 или 100 копий), различные концентрации синтетического TVbtub G-блока (25600, 6400, 1600, 400 или 100 копий) на фоне синтетического MgPa G-блока (25600 копий) или различные концентрации синтетического TVbtub G-блока (25600, 6400, 1600, 400 или 100 копий) на фоне синтетического MgPa G-Блока (25600 копий). Четыре реакции содержали обе мишени в различных концентрациях: 12800 копий TVbtub с 200 копиями MgPa, 200 копий TVbtub с 12800 копиями MgPa, 3200 копий TVbtub с 800 копиями MgPa и 800 копий TVbtub с 3200 копиями MgPa.
Полученные результаты
Во время ПЦР-амплификации измеряли флуоресценцию при двух температурах в режиме реального времени для детектирования и количественной оценки присутствия мишени 1 (TVbtub) и/или мишени 2 (MgPa). Молекулярный Маяк был разработан для детектирования последовательностей, гомологичных TVbtub, для детектирования Trichomonas vaginalis (TV). МНКзим был разработан для расщепления LOCS-2В присутствии последовательностей, гомологичных MgPa, для детектирования Mycoplasma genitalium. Наличие или отсутствие специфического сигнала во время ПЦР обобщено в Таблице 12, и общая схема аналогична схеме, описанной в Сценарии 3 (Таблица 1) Примера 5.
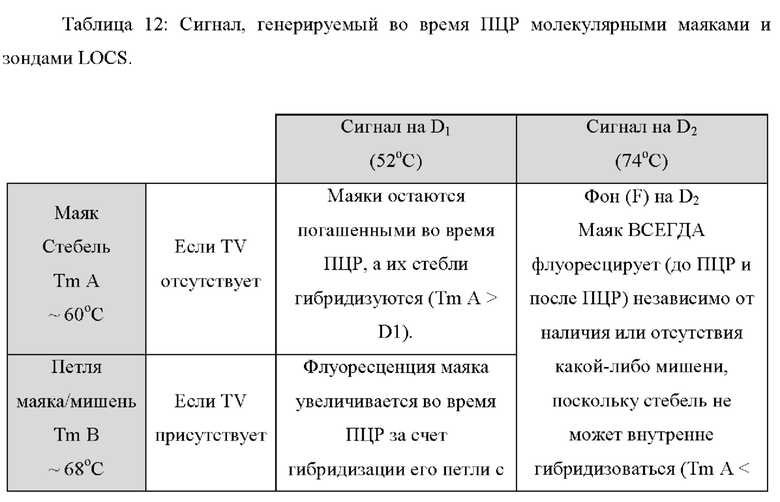
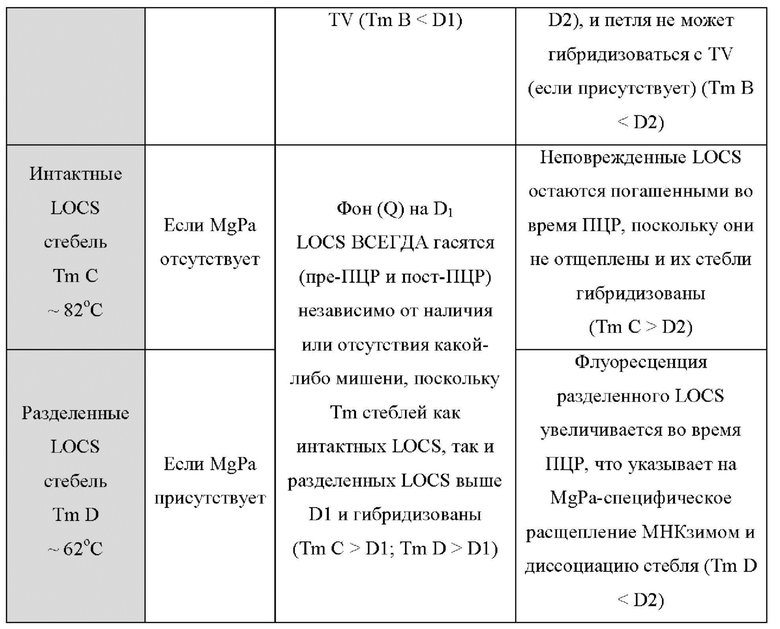
Результаты, показанные на Фиг. 20, иллюстрируют сравнительные кривые усиления флуоресценции при 52°С (D1, Фиг. 20А) и 74°С (D2, Фиг. 20В), измеренные в канале FAM для реакций, содержащих либо 25600 копий TVbtub (черная пунктирная линия) или MgPa (черная пунктирная линия), смесь 25600 копий обоих генов-мишеней (серая сплошная линия) или отсутствие генов-мишеней (NF H2O; черная сплошная линия). Результаты нанесены на график как средние из трех повторов реакций.
На Фиг. 20А показано, что при 52°С (D1) наблюдается усиление флуоресценции в присутствии TVbtub (черная пунктирная линия) или обоих TVbtub и MgPa (серая сплошная линия) в реакции, в то время как увеличения флуоресценции в отсутствие TVbtub не происходит, включая реакции, содержащие только MgPa (черная пунктирная линия). При этой температуре Молекулярный Маяк флуоресцирует в ответ на гибридизацию с геном-мишенью TVbtub, когда он присутствует, или гасится в отсутствие своей мишени и поддерживает внутренне гибридизованный стебель. При одной и той же температуре D1 как интактные, так и/или разделенные виды LOCS гасятся из-за гибридизации их соответствующих стеблей при этой температуре, независимо от присутствия или отсутствия TVbtub или MgPa в реакции. Число циклов, при котором флуоресценция начинает экспоненциально увеличиваться, сравнимо в реакциях, содержащих только TVbtub и TVbtub плюс MgPa, что указывает на то, что присутствие MgPa не влияет на количественное определение TVbtub в D1. Следовательно, увеличение флуоресценции при 52 0 С (D1) указывает на присутствие TVbtub, а значение Cq, полученное при D1, можно применять для прямого количественного определения TVbtub независимо от присутствия или отсутствия MgPa в реакции.
На Фиг. 20В показано, что при 74°С (D2) наблюдается усиление флуоресценции в присутствии MgPa (черная пунктирная линия) или как TVbtub, так и MgPa (серая сплошная линия) в реакции, в то время как увеличения флуоресценции в отсутствии MgPa не происходит, включая реакции, содержащие только TVbtub (черный пунктир). При D2 Молекулярный Маяк не способен гибридизоваться с TVbtub, даже если он присутствует, и всегда флуоресцирует из-за диссоциации его стебля при этой температуре, независимо от присутствия или отсутствия какой-либо мишени в реакции. Таким образом, флуоресценция Молекулярного Маяка обеспечивает уровень фоновой флуоресценции при этой температуре до, во время и после амплификации. Кроме того, при D2, когда MgPa отсутствует, зонд LOCS остается интактным и гасится. Когда в реакции присутствует MgPa, флуоресценция увеличивается выше фоновых уровней во время ПЦР, поскольку расщепление MgPa-специфическими МНКзимами генерирует разделенные LOCS со стеблевыми областями, диссоциированными при этой температуре. Таким образом, увеличение флуоресценции на D2 во время ПЦР указывает на присутствие MgPa. Число циклов, при котором флуоресценция начинает увеличиваться экспоненциально, сопоставимо в присутствии только MgPa и в присутствии коинфекции, содержащей как MgPa, так и TVbtub, что указывает на то, что присутствие TVbtub не влияет на количественный анализ MgPa в D2. Следовательно, увеличение флуоресценции при 72°С (D2) указывает на присутствие MgPa, а значение Cq, полученное при D2, можно применять для прямого количественного определения MgPa независимо от присутствия в реакции TVbtub.
Стандартные реакции, содержащие 25600, 6400, 1600, 400 и 100 копий TVbtub, использовали для построения стандартной кривой (Фиг. 21 А), используемой для количественного определения исходных концентраций TVbtub при 52°С (D1) в образцах, содержащих TVbtub в наличие или отсутствие MgPa. Таблица 13 показывает, что присутствие 25600 копий MgPa не оказывало существенного влияния на количественную оценку TVbtub, обнаруженную при 52°С (парный критерий Стьюдента, значение р=0,418). Аналогичным образом, стандарты, содержащие 25600, 6400, 1600, 400 и 100 копий MgPa, использовали для построения стандартной кривой (Фиг. 21В), используемой для количественного определения исходных концентраций MgPa при 74°С (D2) в образцах, содержащих MgPa в наличие или отсутствие TVbtub. В Таблице 13 показано, что присутствие 25600 копий TVbtub существенно не влияло на количественную оценку MgPa, обнаруженную при 74°С (парный критерий Стьюдента, значение р=0,150). Образцы, содержащие случайные концентрации генов-мишеней TVbtub и MgPa, также количественно анализировали, и полученные в результате оценочные числа копий были сравнимы с известной концентрацией TVbtub, добавленной к образцу (Таблица 14). Это указывает на то, что TVbtub может быть точно определен количественно при D1 (52°С) независимо от присутствия MgPa, и аналогичным образом MgPa может быть точно определен количественно при D2 (74°С) независимо от присутствия TVbtub.
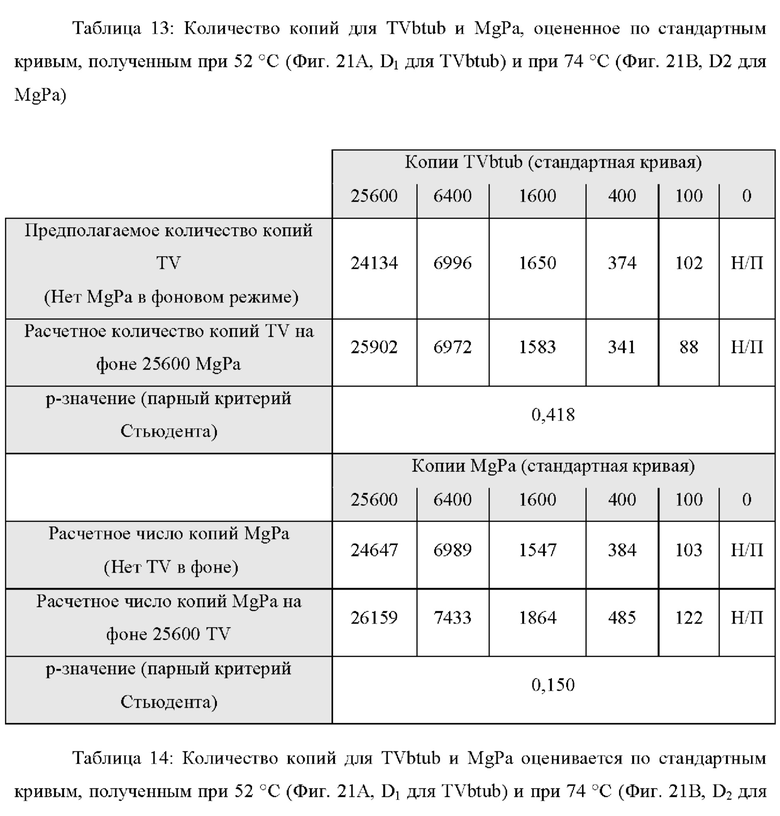
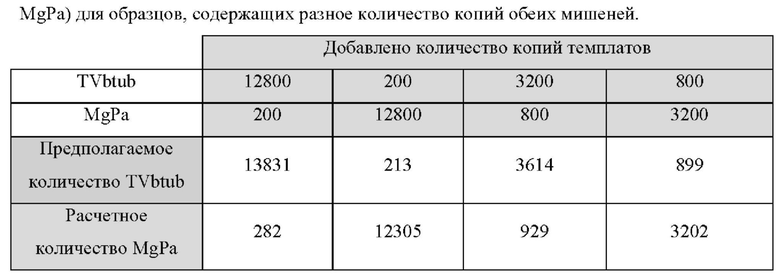
Этот способ обеспечивает большое преимущество по сравнению с другими известными в данной области способами, в которых используются измерения при различных температурах с последующим анализом с использованием Поправочного Коэффициента Флуоресценции (ФКФ) (например, ТОСЕ) для различения нескольких мишеней на одной длине волны.
Вариант реализации в этом примере не требует корректировки для учета связанных с температурой различий в выходе флуоресценции одних и тех же молекул. В отличие от других способов, которые обнаруживают одну мишень при первой температуре и две мишени при второй температуре, показанный в данном документе пример, объединяющий LOCS и Молекулярный Маяк, позволяет обнаруживать и количественно определять одну мишень при одной температуре, а также обнаруживать и количественно определять вторую мишень при второй температуре без вмешательства со стороны первой мишени (и наоборот). Точно так же в отсутствие мониторинга в реальном времени наблюдаемое увеличение флуоресценции, измеренное в дискретные моменты времени (пост-ПЦР минус пре-ПЦР), указывает на присутствие мишени 1 только при первой температуре и мишени 2 только при второй температуре.
Пример возможного применения 8: Детектирование и количественная оценка двух мишеней в режиме реального времени на одной длине волны с использованием комбинации зондов двойной гибридизации и LOCS.
Следующий пример иллюстрирует подход, при котором комбинация одной пары зондов двойной гибридизации и одного репортера LOCS может обеспечить одновременное детектирование и количественную оценку двух мишеней в одном флуоресцентном канале путем получения показаний флуоресценции при двух температурах в режиме реального времени во время ПЦР. В качестве альтернативы, при отсутствии мониторинга в реальном времени этот подход может применяться к флуоресцентным данным, собранным в дискретные моменты времени, например, почти в начале или в начале амплификации и после амплификации. Эта стратегия устранит потребность в специализированных способах анализа после амплификации, как показано в Примере 4.
Как показано на Фиг. 22, как зонды двойной гибридизации, так и зонды LOCS могут быть помечены одними и теми же фрагментами флуорофора и гасителя для одновременного детектирования в одном и том же флуоресцентном канале. Двойной гибридизационный зонд может содержать первый зонд с Tm А и второй зонд с Tm В, где Tm А и Tm В могут быть равными, или Tm А и Tm В могут быть разными. Один зонд может быть помечен на его 3'-конце флуорофором, а другой зонд может быть помечен на его 5'-конце гасителем. (В качестве альтернативы один зонд может быть помечен на его 3'-конце гасителем, а другой зонд может быть помечен на его 5'-конце флуорофором). Двойные гибридизационные зонды могут быть разработаны для гибридизации рядом на мишени 1 с Tm А, например, 60°С и Tm В, например, 60°С. В этом сценарии зонды двойной гибридизации будут флуоресцировать до амплификации, но будут гаситься после амплификации, если присутствует мишень 1 и температура детектирования ниже Tm А и Tm В. Таким образом, наблюдаемое снижение флуоресценции при первой температуре (D1), например, при 50°С будет свидетельствовать о присутствии мишени 1.
Кроме того, реакция может содержать интактный зонд LOCS, который имеет стеблевую область с TmC, например, 80°С, и петлевую область, которая при расщеплении МНКзимом в присутствии второй мишени 2 может генерировать разделенный LOCS с Tm D, например, 60°С (Tm D<Tm С). В этом сценарии интактные LOCS будут гаситься до амплификации, но флуоресценция будет увеличиваться после амплификации, только если присутствует мишень 2 и температура детектирования выше Tm D, но ниже Tm С и выше Tm А и Tm В. Таким образом, наблюдаемое увеличение флуоресценции при второй температуре (D2), например, при 70°С, будет свидетельствовать о присутствии мишени 2.
Как таковая, комбинация позволила бы обнаруживать мишень 1 только с использованием зондов двойной гибридизации, определяемой как уменьшение флуоресценции при первой температуре (D1); и детектирование мишени 2 только с использованием зондов LOCS, определяемое по увеличению флуоресценции при второй температуре (D2).
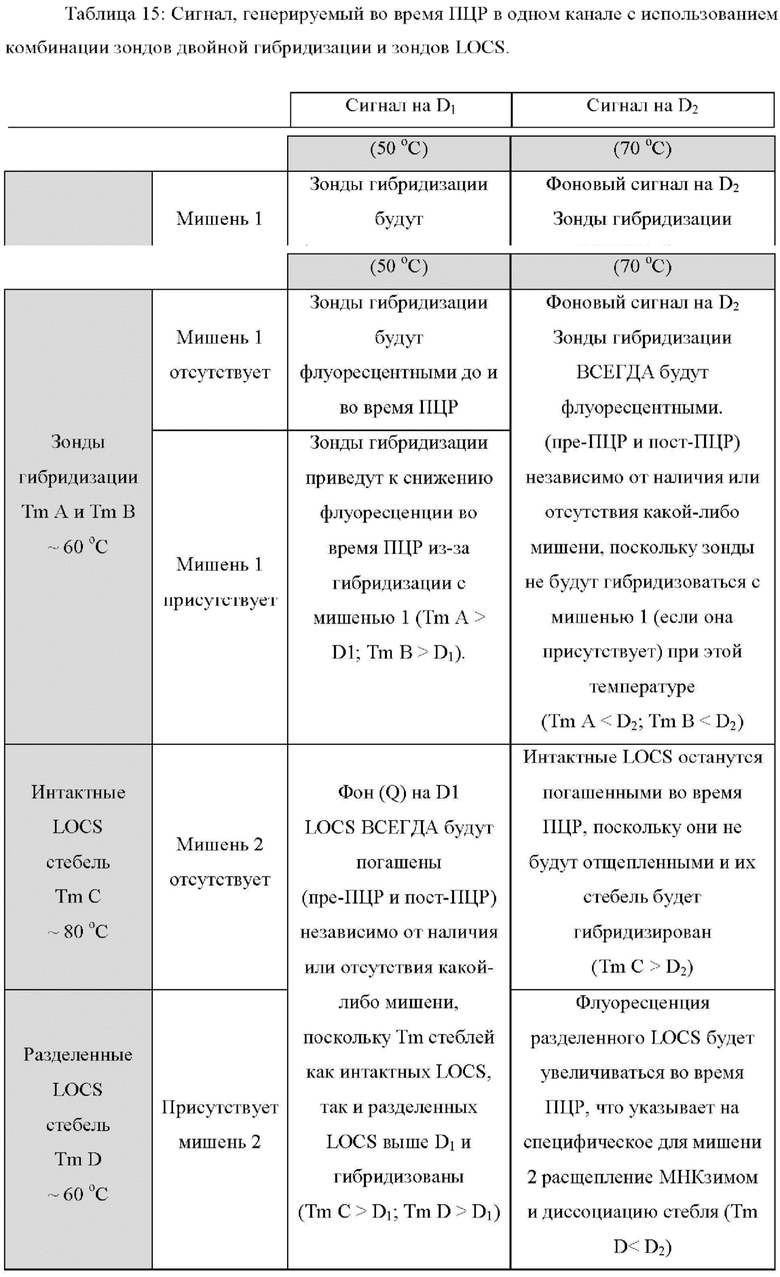
Пример возможного применения 9: Детектирование и количественная оценка двух мишеней в режиме реального времени на одной длине волны с использованием комбинации зонда Scorpion и зонда LOCS.
Следующий пример иллюстрирует подход, при котором комбинация одного зонда Scorpion и одного репортера LOCS может обеспечить одновременное детектирование и количественную оценку двух мишеней в одном флуоресцентном канале путем получения показаний флуоресценции при двух температурах в режиме реального времени во время ПЦР. В качестве альтернативы, при отсутствии мониторинга в реальном времени этот подход может применяться к флуоресцентным данным, собранным в дискретные моменты времени, например, вблизи или в начале амплификации и после амплификации. Эта стратегия устранит потребность в специализированных способах анализа после амплификации, как описано в Примере 4.
В этом варианте реализации как зонд Scorpion, так и зонд LOCS могут быть помечены одними и теми же фрагментами флуорофора и гасителя для одновременного детектирования в одном и том же флуоресцентном канале. В этой стратегии можно применять два типа зондов Scorpion, а именно унизонды Scorpion и бизонды Scorpion. Унизонд Scorpion может состоять из одноцепочечной последовательности флуоресцентного зонда с двойной меткой, удерживаемой в конформации стебель-петля с 5'-концом репортера и внутреннего гасителя, непосредственно связанного с 5'-концом праймера для ПЦР через блокатор. Во время ПЦР часть праймера может быть сконструирована так, чтобы гибридизироваться с мишенью 1 и расширять ее, при этом стеблевая область стеблевой петли может иметь Tm А, например, 55°С, а петлевая область стеблевой петли может связываться с ампликонами мишени 1 с Tm В, например, 60°С. Во время ПЦР петля-шпилька могла разворачиваться, и петлевая область унизонда могла внутримолекулярно гибридизоваться с вновь синтезированной последовательностью-мишенью 1, разделяя, таким образом, флуорофор и гаситель. Таким образом, наблюдаемое во время ПЦР увеличение флуоресценции при первой температуре (D1), например, при 50°С, будет свидетельствовать о присутствии мишени 1.
Кроме того, реакция может содержать интактный зонд LOCS, который имеет стеблевую область с Tm С, например, 80°С, и петлевую область, которая при расщеплении МНКзимом в присутствии второй мишени 2 может генерировать разделенный LOCS с Tm D, например, 60°С (Tm D<Tm С). В этом сценарии интактный LOCS будет гаситься до амплификации, но флуоресценция будет увеличиваться после амплификации, только если присутствует мишень 2 и температура детектирования выше Tm D, но ниже Tm С и выше Tm А и Tm В. Таким образом, наблюдаемое увеличение флуоресценции при второй температуре (D2), например, при 65°С, будет свидетельствовать о присутствии мишени 2.
Как таковая, комбинация позволила бы обнаруживать мишень 1 только с использованием унизондов Scorpion, о чем свидетельствует увеличение флуоресценции при первой температуре (D1); и детектирование мишени 2 только с использованием зондов LOCS, что контролируется увеличением флуоресценции при второй температуре (D2).
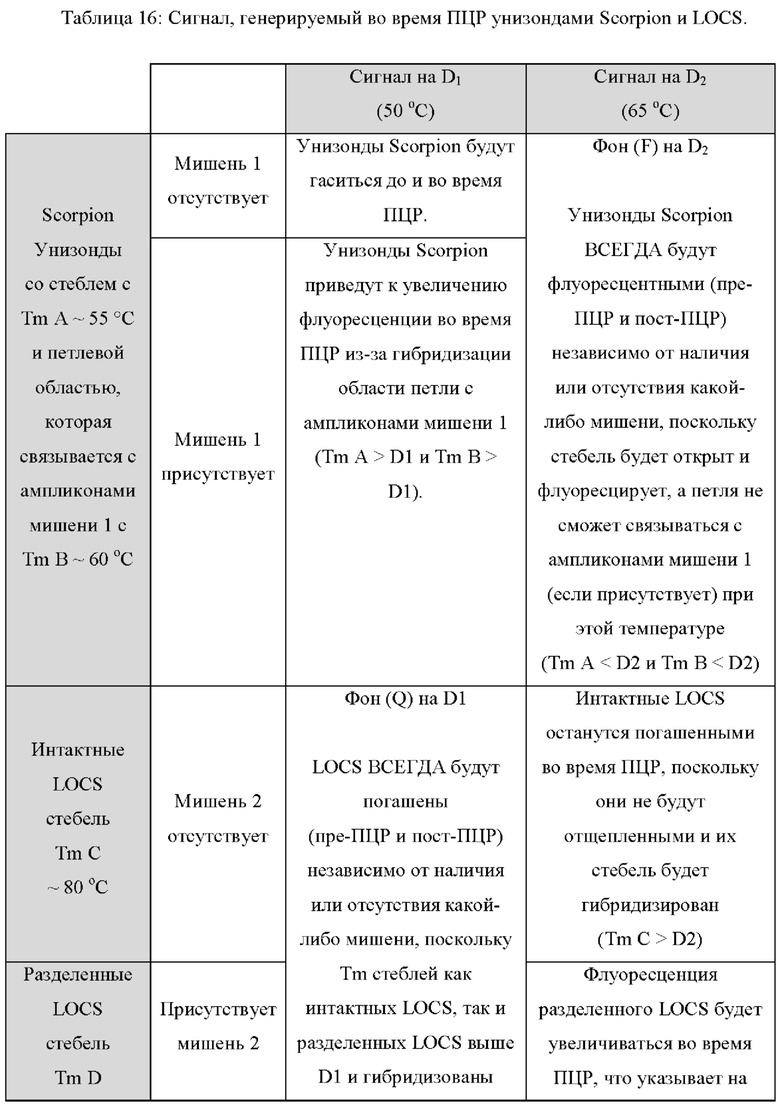
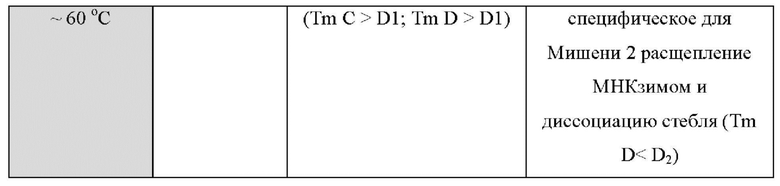
Точно так же бизонды Scorpion можно комбинировать с зондами LOCS. Бизонд Scorpion может состоять из последовательности одноцепочечного флуоресцентного зонда, непосредственно связанного с 5'-концом праймера для ПЦР через блокатор. Кроме того, последовательность, комплементарная зонду и меченная гасителем, может связываться с молекулой праймера/зонда с Tm А, образуя дуплекс, который гасится до ПЦР или в отсутствие мишени. Во время ПЦР часть праймера может быть сконструирована таким образом, чтобы гибридизироваться с мишенью 1 и расширять ее, при этом область зонда и комплементарная последовательность гасителя могут иметь Tm А, например, 55°С, и область зонда может связываться с ампликонами мишени 1 с Tm В, например, 60°С. Во время ПЦР комплементарная последовательность гасителя может диссоциировать, и бизонд может внутримолекулярно гибридизоваться с вновь синтезированной последовательностью-мишенью 1, тем самым блокируя связывание комплементарной последовательности гасителя и таким образом генерируя флуоресценцию. Таким образом, наблюдаемое во время ПЦР увеличение флуоресценции при первой температуре (D1), например, при 50°С, будет свидетельствовать о присутствии Мишени 1.
Кроме того, реакция может содержать интактный зонд LOCS, который имеет стеблевую область с Tm С, например, 80°С, и петлевую область, которая при расщеплении МНКзимом в присутствии второй мишени 2 может генерировать разделенный LOCS с Tm D, например, 60°С (Tm D<Tm С). В этом сценарии интактный LOCS будет гаситься до амплификации, но флуоресценция будет увеличиваться после амплификации, только если присутствует мишень 2 и температура детектирования выше Tm D, но ниже Tm С и выше Tm А и Tm В. Таким образом, наблюдаемое увеличение флуоресценции при второй температуре (D2), например, при 65°С, будет свидетельствовать о присутствии Мишени 2.
Как таковая, комбинация позволила бы обнаруживать мишень 1 только с использованием бизондов Scorpion, о чем свидетельствует увеличение флуоресценции при первой температуре (D1); и детектирование мишени 2 только с использованием зондов LOCS, что контролируется увеличением флуоресценции при второй температуре (D2).
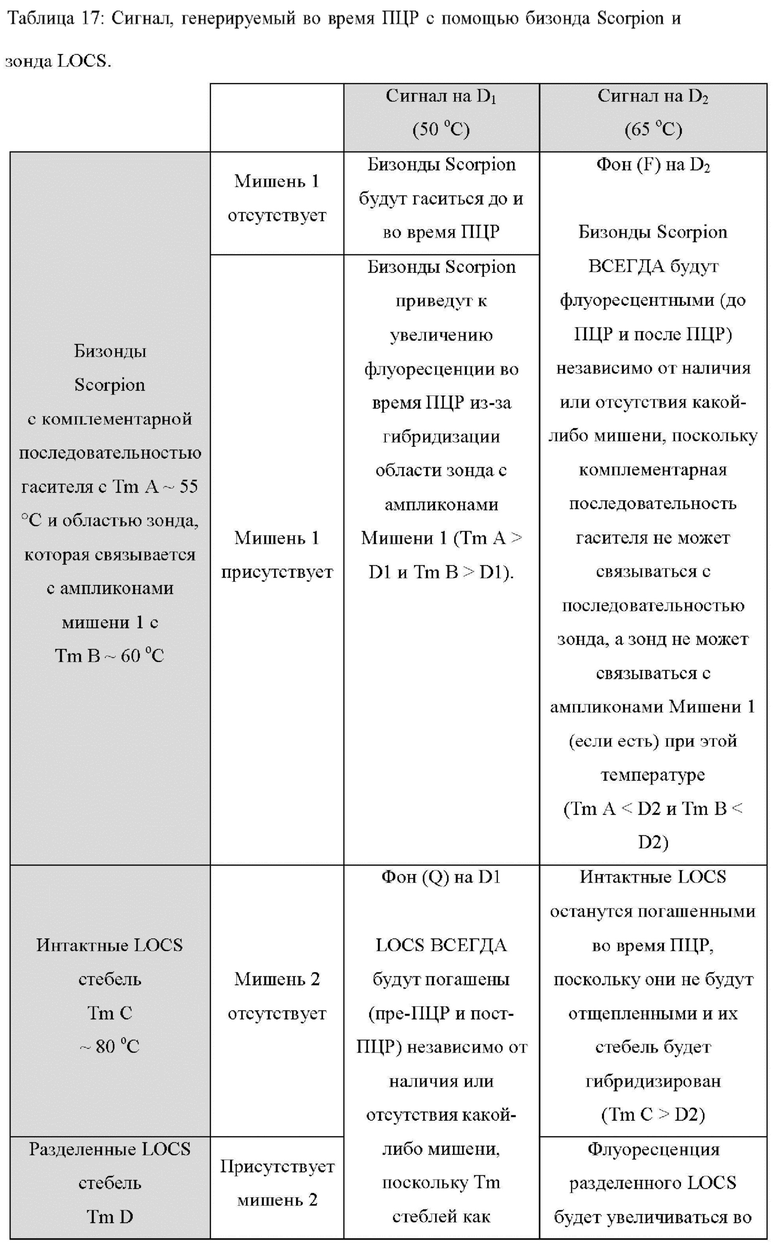

Пример 10: Способы анализа нескольких мишеней на одной длине волны с использованием одного молекулярного маяка и одного репортера LOCS с внутренней калибровкой флуоресценции и без нее.
В следующем примере демонстрируется способ анализа нескольких мишеней на одной длине волны с использованием одного нерасщепляемого молекулярного маяка и одного репортера LOCS путем получения показаний флуоресценции до и после усиления при двух температурах. Эта стратегия устраняет потребность в специализированных способах анализа конечных точек, продемонстрированных в Примере 1. Кроме того, эта стратегия устраняет необходимость сбора данных в каждом цикле, необходимом для детектирования в реальном времени, и, следовательно, помогает сократить общее время получения результата. Как показано на Фиг. 5, как Молекулярный Маяк, так и зонд LOCS помечены одними и теми же фрагментами флуорофора и гасителя для одновременного детектирования в одном и том же флуоресцентном канале. Молекулярный Маяк содержит стеблевую область с Tm А и петлевую область, которая может специфически гибридизоваться с мишенью 1 (TVbtub) с Tm В; где Tm В больше, чем Tm А (Tm В>Tm А). В этом примере интактный зонд LOCS имеет стеблевую область с Tm С (82°С) и петлевую область, которая при расщеплении МНКзимом в присутствии мишени 2 (MgPa) генерирует разделенный LOCS с Tm D(62°С); где Tm D меньше Tm С (Tm D<Tm С). Наличие мишени 1 (TVbtub) и/или мишени 2 (MgPa) можно различить путем измерения увеличение флуоресценции при двух температурах (D1 и D2; 52°С и 74°С) с использованием одиночных измерений, полученных либо в начале амплификации, либо почти в начале и после амплификации. В следующем примере Tm А составляет -60°С, Tm В составляет -68°С, Tm С составляет -82°С и Tm D составляет -62°С, что соответствует Сценарию 3, описанному в Примере 5, где D1<Tm А<Tm В<D2 и D1<Tm D<D2<Tm C.
Олигонуклеотиды
Олигонуклеотиды, специфичные для этого эксперимента, включают: прямой праймер 12 (SEQ ID NO: 63), обратный праймер 12 (SEQ ID NO: 64) и молекулярный маяк 1 (SEQ ID NO: 68) для амплификации и количественного определения мишени 1 (TVbtub); прямой праймер 5 (SEQ ID NO: 20), обратный праймер 5 (SEQ ID NO: 21), партзим А5 (SEQ ID NO: 22), партзим B5 (SEQ ID NO: 23), субстрат 3 (SEQ ID NO: 24).), LOCS-2 (SEQ ID NO: 25) для амплификации и количественного определения мишени 2 (MgPa). Последовательности перечислены в списке последовательностей.
Условия реакции
Детекцию целевой последовательности в режиме реального времени проводили в общем реакционном объеме 20 мкл с использованием термоциклера BioRad® CFX96. Параметры циклов и точки сбора флуоресцентных данных (DA) были следующими: 1 цикл 95°С в течение 2 минут, 52°С в течение 15 секунд (DA) и 74°С в течение 15 секунд (DA); 50 циклов 95°С в течение 1 секунды и 60°С в течение 20 секунд; и 1 цикл 52°С в течение 5 минут (DA) и 74°С в течение 15 секунд (DA). Все реакции проводили в трех повторностях, за исключением реакций, содержащих 10 копий матриц, для которых использовали 6 повторов. Каждая реакция содержала 40 нМ каждого прямого праймера, 200 нМ каждого обратного праймера, 200 нМ каждого партзима, 200 нМ молекулярного маяка 1, 300 нМ репортера LOCS-2 и lx PlexMastermix (Bioline). Реакции на первом планшете либо не содержали мишени (NF H2O), либо содержали синтетический G-блок TVbtub (10000 или 200 копий), синтетический G-блок гена MgPa (10000 или 200 копий), либо различные комбинации синтетического G-блока гена TVbtub (10000 или 200 копий) на фоне синтетического G-блока гена MgPa (10000 или 200 копий). Реакции на втором планшете либо не содержали мишени (NF H2O), либо содержали синтетический G-блок TVbtub (200, 100, 50, 25 или 10 копий), синтетический G-блок гена MgPa (200, 100, 50, 25 или 10 копий) или оба синтетических G-блока генов TVbtub и MgPa (200, 100, 50, 25 или 10 копий каждого).
Полученные результаты
Увеличение флуоресценции определяли путем расчета разницы между измерениями пре-ПЦР и пост-ПЦР (ΔS) при двух температурах (D1 и D2; 52°С и 74°С), чтобы определить присутствие мишени 1 (TVbtub) и мишени 2 (MgPa), соответственно. Молекулярный Маяк был разработан для детектирования последовательностей, гомологичных TVbtub, для детектирования Trichomonas vaginalis (TV). МНКзим был разработан для расщепления LOCS-1 в присутствии MgPa для детектирования Mycoplasma genitalium. Наличие или отсутствие специфического сигнала во время ПЦР аналогично описанному в Сценарии 3 Примера 5.
Фиг. 23 иллюстрирует сводку по конечной точке детектирования TVbtub и MgPa путем определения увеличения флуоресценции при 52°С (ΔSD1, Фиг. 23А) и 74°С (ΔSD2, Фиг. 23В), соответственно, как средние значения реакции в трех повторностях, измеренные с использованием первого 96-луночного планшета. Когда ΔSD1 превышает указанный порог (Порог 1) на Фиг. 23А, это указывает на присутствие в реакции TVbtub. Все реакции, содержащие 10000 или 200 копий TVbtub, были правильно определены как положительные для TVbtub, независимо от присутствия или отсутствия MgPa в реакции. Во всех реакциях, где TVbtub отсутствует, ΔSD1 остается ниже указанного порога и аналогичен контролю без матрицы (NTC). Точно так же, когда ΔSD2 выше определенного порога (Порог 2) на Фиг. 23В, это указывает на присутствие MgPa в реакции. Все реакции, содержащие 10000 или 200 копий MgPa, были правильно определены как положительные для MgPa, независимо от присутствия или отсутствия TVbtub в реакции. Во всех реакциях, где MgPa отсутствует, ΔSD2 остается ниже указанного порога и аналогичен контролю без матрицы (NTC).
На Фиг. 24 показана сводка по конечной точке детектирования TVbtub и MgPa, измеренных на втором 96-луночном планшете. Результаты представляют собой среднее значение повторов и показывают увеличение калиброванного сигнала при 52°С (ΔSD1/C, Фиг. 24А) и 74°С (ΔSD2/C, Фиг. 24В). Калибровочный коэффициент определяли как разницу во флуоресценции до ПЦР между 52°С и 74°С (С=S D2-пре-ПЦР - S D1-пре-ПЦР), что указывает на флуоресценцию, возникающую в результате диссоциации стебля маяка в закрытой и открытой конформациях. Этот сигнал, возникающий от молекулярного маяка в точке D2, присутствует во всех реакциях в одинаковом количестве и не зависит от присутствия любой из мишеней. Таким образом, вариации сигналов пре-ПЦР в D1 и D2 (С) между лунками отражают истинные вариации от лунки к лунке в используемом канале. Измеренные сигналы ΔSD1 и ΔSD2 калибровали путем деления каждого на калибровочный коэффициент (С) (ΔSD1/C и ΔSD2/C). На Фиг. 24А показано, что калиброванный сигнал ΔSD1/C выше заданного порога (Порог 1) во всех реакциях, где присутствует TVbtub, или ниже, если TVbtub отсутствует.Во всех реакциях, где отсутствует TVbtub, ΔSD1/C остается ниже указанного порога и аналогичен контролю без темплата (NTC). На Фиг. 24В показано, что калиброванный сигнал ΔSD2/C выше заданного порога (порог 2) во всех реакциях, где присутствует MgPa, или ниже, если MgPa отсутствует.Во всех реакциях, где MgPa отсутствует, ΔSD2/C остается ниже указанного порога и аналогичен контролю без темплата (NTC). Фиг. 24 демонстрирует высокую чувствительность анализа к обеим мишеням, поскольку их можно было обнаружить с 10 копиями на реакцию.
Этот пример демонстрирует использование одного молекулярного маяка и одного репортера LOCS для быстрого качественного детектирования конечной точки двух мишеней в одном флуоресцентном канале путем определения ΔSD1 (с указанием только мишени 1) и ΔSD2 (с указанием только мишени 2). Такой подход устраняет необходимость в специализированных способах анализа конечных точек. В этом примере было также продемонстрировано, что сигналы ΔSD1 и ΔSD2 можно калибровать в одном и том же канале, не требуя дополнительного реагента-калибратора, используя формулы ΔSD1/C и ΔSD2/C. Этот подход выгоден тем, что вариации от анализа к анализу и от инструмента к инструменту могут быть нормализованы для более согласованных результатов флуоресценции. Кроме того, в этом примере не требовался сбор данных в реальном времени, что сокращает время выполнения, которое в этом примере составило в общей сложности 54 минуты.
Пример возможного применения 11: Детектирование и количественная оценка двух мишеней в режиме реального времени на одной длине волны с использованием комбинации зондов Catcher и Pitcher и LOCS.
Следующий пример иллюстрирует подход, при котором комбинация одной пары Catcher и Pitcher, и одного репортера LOCS может обеспечить одновременное детектирование и количественную оценку двух мишеней в одном флуоресцентном канале путем получения показаний флуоресценции при двух температурах в режиме реального времени во время ПЦР. В качестве альтернативы, при отсутствии мониторинга в реальном времени этот подход может применяться к флуоресцентным данным, собранным в дискретные моменты времени, например, почти в начале или в начале амплификации и после амплификации. Эта стратегия устранит потребность в специализированных способах анализа после амплификации, таких как те, что продемонстрированы в Примере 4.
В этом варианте реализации как Catcher, так и зонд LOCS могут быть помечены одними и теми же фрагментами флуорофора и гасителя для одновременного детектирования в одном и том же флуоресцентном канале. Pitcher может состоять из одноцепочечного олигонуклеотида, который включает 5"-маркирующую область, комплементарную Catcher, и 3''-сенсорную область, комплементарную первой мишени 1. Catcher может состоять из одноцепочечного олигонуклеотида, помеченного гасителем на 5'-конце, и флуорофора ниже гасителя и может включать 3'-маркирующую область, комплементарную pitcher. Когда Catcher находится в одноцепочечной конформации, флуорофор будет находиться в непосредственной близости от гасителя и останется погашенным. Во время ПЦР праймеры и 3'-сенсорная область Pitcher могут гибридизоваться с мишенью. Во время удлинения праймера Pitcher может быть расщеплен экзонуклеазной активностью ДНК-полимер азы и может высвободить маркирующую область Pitcher. Высвобожденная маркирующая область может затем гибридизоваться с комплементарной 3'-маркирующей областью Catcher. Удлинение меченой области ДНК-полимеразой во время ПЦР может генерировать двухцепочечный Catcher дуплекс, в котором флуорофор и гаситель разделены. Разделение флуорофора и гасителя может привести к увеличению флуоресценции, что может указывать на присутствие мишени 1. При первой температуре детектирования (D1), например, 50°С, которая ниже, чем Tm двухцепочечного Catcher дуплекса (Tm А, 60°С), может наблюдаться увеличение флуоресценции, поскольку флуорофор и гасители разделяются, когда они находятся в дуплексной конформации Catcher. При второй температуре детектирования (D2), например, 70°С, которая выше, чем Tm двухцепочечного Catcher дуплекса (Tm А, 60°С), Catcher и Pitcher могут диссоциировать так, что они оба находятся в одноцепочечной конформации. Это может привести к уменьшению флуоресценции, поскольку флуорофор и гаситель больше не разделены. Следовательно, увеличение флуоресценции во время ПЦР в D1 можно применять для указания на присутствие двухцепочечного Catcher дуплекса и, следовательно, на присутствие мишени 1. В отсутствие мишени 1 Catcher может оставаться одноцепочечным и погашеным и, следовательно, не будет способствовать увеличению флуоресценции в D1. На флуоресценцию в D2 не будет влиять присутствие или отсутствие мишени 1, потому что Catcher всегда будет оставаться одноцепочечным и гасится при температуре выше Tm двухцепочечного Catcher дуплекса (Tm А, 60°С).
Кроме того, реакция может содержать интактный зонд LOCS, который имеет стеблевую область с Tm В, например, 80°С, и петлевую область, которая при расщеплении МНКзимом в присутствии второй мишени 2 может генерировать разделенныйй LOCS с Tm С, например, 60°С (Tm С<Tm В). В этом сценарии Intact LOCS будет гаситься до амплификации, но флуоресценция будет увеличиваться после амплификации, если присутствует мишень 2 и температура детектирования выше Tm С, Tm А, но ниже Tm В. Таким образом, наблюдаемое увеличение флуоресценции при второй температуре (D2), например, при 70°С, будет свидетельствовать о присутствии Мишени 2. Как интакный, так и разделенный LOCS останутся погашенными при первой температуре детектирования (D1), поскольку их Tm выше (D1<Tm В и D1<Tm С), и, следовательно, присутствие Мишени 2 не будет способствовать изменению в сигнале при первой температуре.
Таким образом, комбинация зондов LOCS и Catcher-Pitcher может позволить обнаружить мишень 1 только с использованием зондов Catcher-Pitcher, о чем свидетельствует увеличение флуоресценции при первой температуре; и детектирование мишени 2 только с использованием зондов LOCS, о чем свидетельствует увеличение флуоресценции при второй температуре.
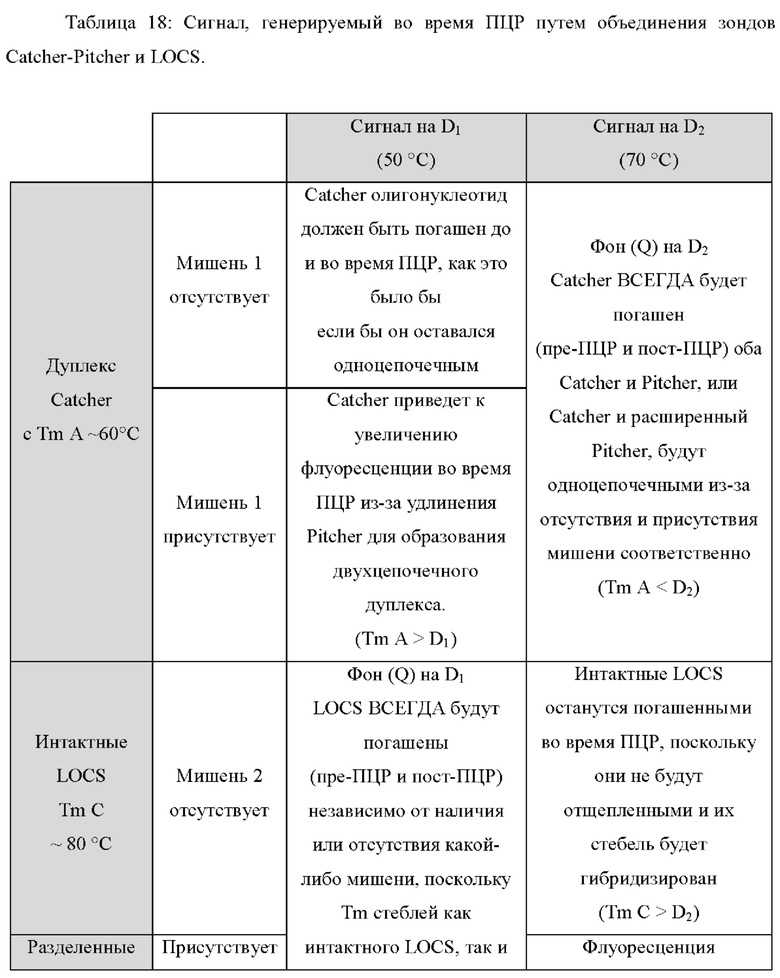

Пример 12: Способ анализа конечных точек нескольких мишеней на одной длине волны с использованием одного линейного субстрата МНКзима и одного зонда LOCS с использованием одного и того же зонда LOCS в качестве калибратора для сведения к минимуму межинструментной изменчивости.
В следующем примере демонстрируется подход, в котором один линейный субстрат МНКзима и один репортер LOCS используются для детектирования и дифференциации двух генов-мишеней (CTcry и NGopa) в одном флуоресцентном канале без необходимости анализа кривой плавления. В данном документе калибровочный коэффициент определяется из сигнала предварительного усиления от репортера LOCS при двух разных температурах и используется для нормализации данных, чтобы свести к минимуму вариации от анализа к анализу и от инструмента к инструменту.
Во время ПЦР линейный субстрат МНКзима расщепляется в присутствии CTcry для разделения флуорофора и гасителя, что приводит к усилению сигнала, которое можно обнаружить в широком диапазоне температур. В этом примере детектирование в реальном времени и определение Cq CTcry достигается за счет получения флуоресценции в каждом цикле во время ПЦР при температуре 1 (52°С, D1). Качественное детектирование CTcry также достигается путем измерения флуоресценции в D1 как до, так и после амплификации. Зонд LOCS как в интактной, так и в разделенной конфигурациях имеет температуру плавления (Tm), которая выше, чем D1 (52°С), поэтому репортер LOCS не дает сигнала в D1 независимо от присутствия или отсутствия NGopa в образец.
В присутствии NGopa МНКзим 2 может расщеплять зонд LOCS во время ПЦР. Качественное детектирование NGopa достигается путем измерения флуоресценции при более высокой температуре 2 (70°С, D2) как до, так и после амплификации. Поскольку Tm интактного LOCS стебля выше D2 (Tm>70°С), а Tm разделенного LOCS стебля ниже D2 (Tm<70°С), то увеличение флуоресценции связано с разделенным и диссоциированным LOCS и указывает на присутствие NGopa. Это увеличение флуоресценции должно быть выше любой фоновой флуоресценции, вызванной расщеплением линейного субстрата МНКзимом при этой температуре, чтобы считаться показателем присутствия мишени NGopa.
В этом примере сигнал флуоресценции (ΔS) откалиброван по сигналу флуоресценции до амплификации при двух разных температурах (S D1-пре-пцр и S эз-пре-пцр), что объясняет различия в сигнале, возникающие из-за двух разных конформационных состояний интактного LOCS (т.е. интактный гибридизированный LOCS в D1 и интактный диссоциированный LOCS в D3). В этом примере диссоциация интактных LOCS происходит при температуре 3, (D3; 85°С), которая выше Tm интактных LOCS (Tm<85°С). Калибровочный коэффициент (С) рассчитывается как разница между сигналом до амплификации при D3 (SD3-пре-ПЦР), когда стебель интактного LOCS диссоциирован и флуоресцирует, и сигналом при D1 (SD1-пре-ПЦР), где стебель интактного LOCS гибридизуется и гасится (С=SD3-пре-ПЦР-SD1-пре-ПЦР). Калибровочный коэффициент (С) представляет собой относительное соотношение между отрицательным и положительным сигналом (т.е. динамический диапазон) для каждой реакции, на которую не должно влиять присутствие какой-либо цели. Любое изменение, наблюдаемое в калибровочном коэффициенте (С), может затем отражать любое изменение от лунки к лунке в каждом конкретном канале. Таким образом, этот калибровочный коэффициент (С) можно применять для минимизации вариаций от анализа к анализу и от инструмента к инструменту. В этом примере сигнал флуоресценции, полученный в результате каждой реакции, калибруется путем деления ΔS при каждой температуре (ΔSD1 и ΔSD2) на калибровочный коэффициент (ΔSD1/C и ΔSD2/C).
Этот пример демонстрирует дополнительную функцию LOCS, которая служит калибратором для минимизации анализ к анализу и инструмент к инструменту изменений. При использовании измерений до ПЦР, полученных с помощью репортера LOCS, в качестве калибратора, этот пример демонстрирует одновременный качественный анализ двух мишеней в одном канале путем измерения флуоресценции при дискретных температурах, взятых до и после ПЦР и определения Cq одной цели.
Олигонуклеотиды
Олигонуклеотиды, специфичные для этого эксперимента, включают; прямой праймер 1 (SEQ ID NO: 1), обратный праймер 1 (SEQ ID NO: 2), партзим A1 (SEQ ID NO: 3), партзим B1 (SEQ ID NO: 4), прямой праймер 2 (SEQ ID NO: 5), обратный праймер 2 (SEQ ID NO: 6), партзим A2 (SEQ ID NO: 7), партзим B2 (SEQ ID NO: 8), субстрат 1 (SEQ ID NO: 13) и LOCS-1 (SEQ ID NO: №: 14). Последовательности перечислены в списке последовательностей.
Олигонуклеотиды, специфичные для амплификации и детектирования мишени 1 (CTcry), представляют собой субстрат 1, партзим А1, партзим В1 (МНКзим 1), прямой праймер 1 и обратный праймер 1. Олигонуклеотиды, специфичные для амплификации и детектирования мишени 2 (NGopa), представляют собой LOCS-1, партзим А2, партзим В2 (МНКзим 2), прямой праймер 2 и обратный праймер 2.
Условия реакции
Амплификацию и детекцию способом ПЦР в реальном времени проводили при общем реакционном объеме 20 мкл с использованием трех разных термоциклеров BioRad® CFX96 на трех планшетах (Инструменты 1-3). Параметры циклирования и точки сбора флуоресцентных данных (DA): 1 цикл 95°С в течение 2 минут, 52°С в течение 15 секунд (DA), 70°С в течение 15 секунд (DA), 85°С в течение 15 секунд (DA); 10 циклов: 95°С в течение 5 секунд и 61°С в течение 30 секунд (уменьшение на 0,5°С за цикл); 40 циклов: 95°С в течение 5 секунд и 52°С в течение 40 секунд (DA в каждом цикле); и 1 цикл 70°С в течение 15 секунд (DA). Все реакции на каждом планшете проводились в трех повторностях и содержали 40 нМ каждого прямого праймера, 200 нМ каждого обратного праймера, 200 нМ каждого партзима А, 200 нМ каждого партзима В, 200 нМ каждого субстрата, 200 нМ LOCS-1 и lx SensiFast буфера (Bioline). Реакции либо не содержали мишени (NF H2O), либо содержали синтетический G-блок CTcry (10000 или 40 копий); или CTcry (10000 или 40 копий); или оба гена CTry и NGopa (по 10000 или 40 копий каждого). Все реакции, кроме контроля без мишени (NF H2O), дополнительно содержали фон 34,5 нг (10000 копий гена) геномной ДНК человека.
Полученные результаты
В этом примере один линейный субстрат МНКзима, расщепляемый МНКзимом 1, и один зонд LOCS, расщепляемый МНКзимом 2, объединяли в одной реакции ПЦР для одновременного детектирования и дифференциации двух мишеней CTcry и NGopa соответственно с использованием только одного флуоресцентного канала (HEX). Присутствие гена CTcry было обнаружено путем мониторинга расщепления линейного Субстрата-1 посредством анализа флуоресценции в дискретные моменты времени при 52°С до и после циклов ПЦР (ΔSD1) в НЕХ-канале. Кроме того, значения Cq, полученные в результате сбора данных в режиме реального времени при 52°С, указывают на количество CTry в реакции (данные не показаны). Присутствие NGopa определяли путем мониторинга расщепления LOCS-1 посредством анализа флуоресценции в дискретные моменты времени при 70°С до и после цикла ПЦР в одном и том же канале (ΔSD2)-Калибровочный коэффициент (С) определяли как разницу между сигналами флуоресценции до ПЦР при 52°С (D1) и 85°С (D3).
Средние значения ΔSD1 и ΔSD2 для трех повторных реакций, содержащих различные количества темплатов CTcry и NGopa на трех инструментах Bio-RadCFX96, показаны в Таблице 19 и Таблице 20 соответственно. Кроме того, эти значения были нормализованы с использованием калибровочного коэффициента, как показано в Таблице 21 (ΔSD1/C) и Таблице 22 (ΔSD2/C), где результаты представляют собой среднее значение для трех повторных реакций. В Таблице 19 и Таблице 20 показаны большие различия в значениях ΔSD1 и ΔSD2 для реакций, содержащих одну и ту же матрицу, на трех протестированных инструментах, о чем свидетельствует высокое значение коэффициента вариации (CV) (CV>23,05%). Эти вариации от анализа к анализу и/или от инструмента к инструменту затрудняют размещение единого порога для определения наличия или отсутствия мишени для каждой из трех инструментов. Однако после нормализации с использованием калибровочного коэффициента различия в результатах, полученных на трех разных инструментах, значительно меньше, о чем свидетельствует уменьшение значений CV (CV<4,13%), как показано в Таблице 21 и Таблице 22. Результаты показывают, что нормализация с использованием С является эффективным способом уменьшения вариаций от анализа к анализу или от инструмента к инструменту.
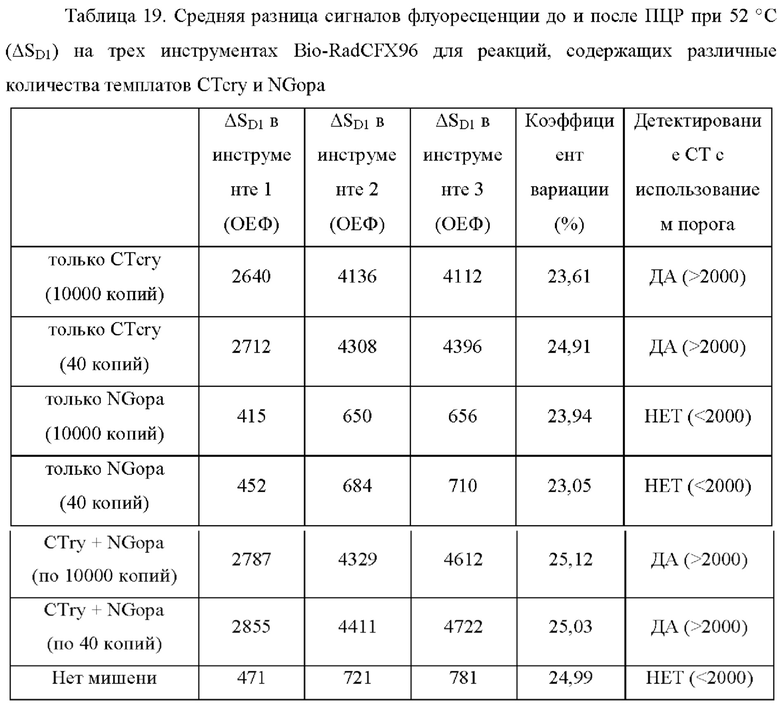
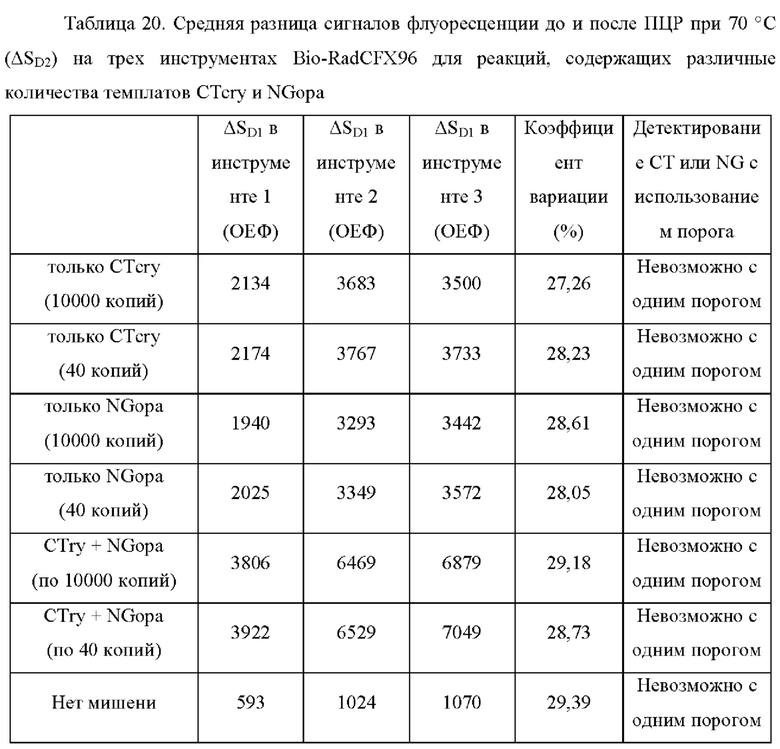
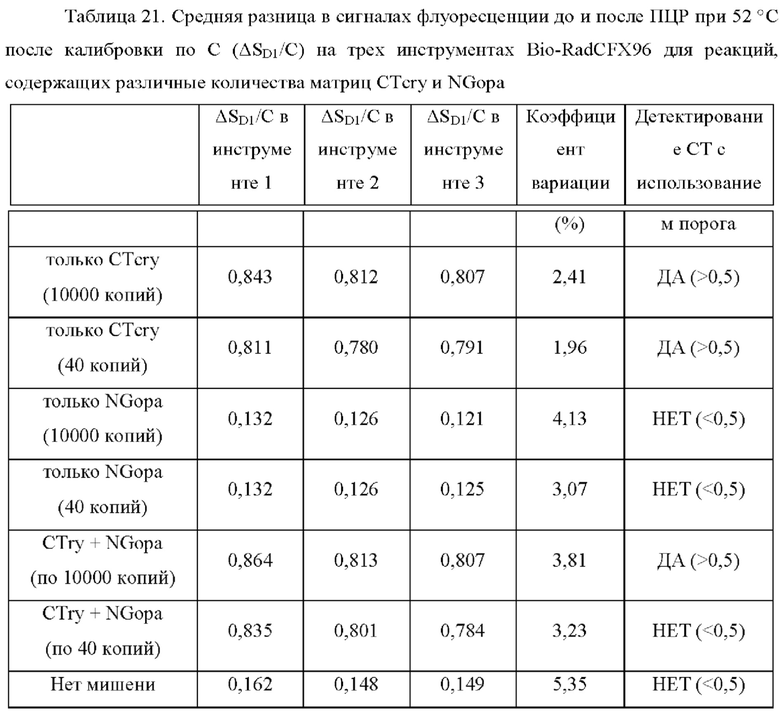
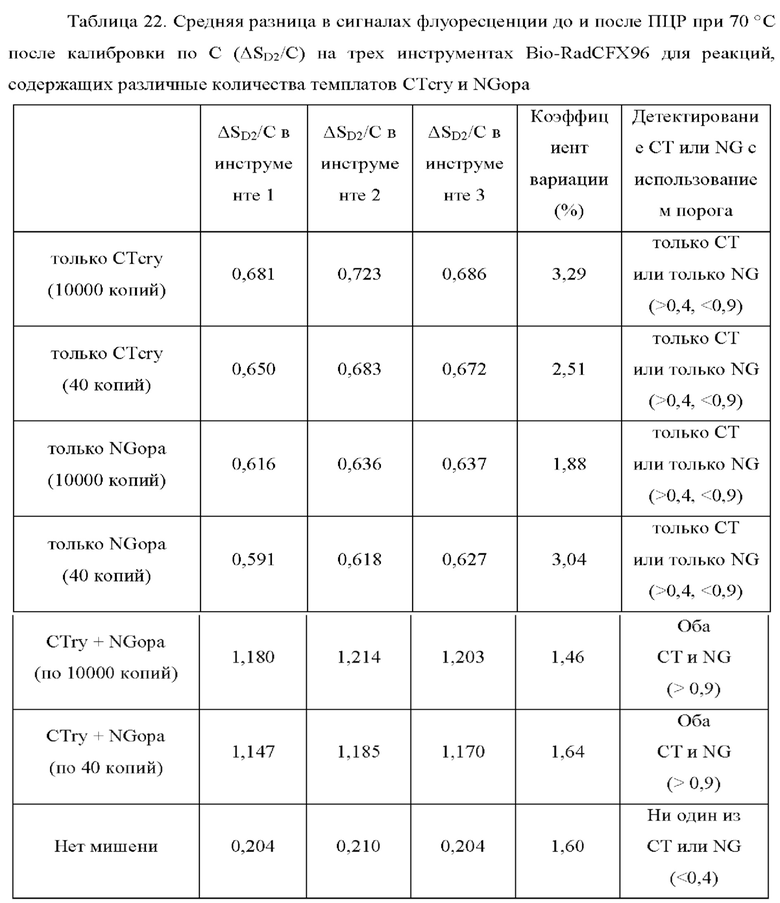
Нормализация с использованием С позволяет применять альтернативный способ для точного определения наличия или отсутствия CTcry, NGopa или обоих CTcry и NGopa в образце с фиксированными порогами, используемыми на всех инструментах. На Фиг. 25 показаны ΔSD1 (Фиг. 21А), ΔSD2 (Фиг. 25В), ΔSD1/C (Фиг. 25С) и ΔSD2/C (Фиг. 25D) для трех протестированных инструментов Bio-RadCFX96 (среднее значение из трех повторностей; инструмент 1 с черными полосами, инструмент 2 с серым цветом и инструмент 3 с белым цветом). На Фиг. 25С показано, что ΔSD1/C больше 0,4 (порог С1) указывает на присутствие CTry в реакции во всех трех инструментах. На Фиг. 25D показаны значения ΔSD2/C от 0,4 (порог С2) до 0,9 (порог С3), что указывает на присутствие только одного из CTry или NGopa, присутствующих в реакции во всех трех инструментах. Если ΔSD2/C выше 0,9 (порог С3), это указывает на то, что в реакции присутствуют как CTcry, так и NGopa во всех трех инструментах. Если ΔSD2/C ниже 0,4, это означает, что ни один из CTcry или NGopa не присутствует в реакции во всех трех инструментах. Эти результаты суммированы в Таблице 21 и Таблице 22. Объединенная информация от ΔSD1/C и ΔSD2/C может затем применяться для правильного определения присутствия CTry и/или NGopa в реакции. Для значений ΔSD1 без нормализации на Фиг. 25А показано, что можно применять один и тот же порог С1 (2000 ОЕФ) для определения наличия CTcry, несмотря на различия между инструментами, как показано в Таблице 19. Однако из-за наблюдаемых изменений ΔSD2 от анализа к анализу или от инструмента к инструменту на Фиг. 25В показано, что для разных инструментов требуются разные пороговые значения С3, и дальнейший анализ невозможен с использованием фиксированного порога. Таким образом, нормализация с использованием С позволяет применять фиксированные пороговые значения для точного определения наличия или отсутствия целей, независимо от инструмент к инструменту или анализ к анализу вариаций.
Этот пример демонстрирует дополнительную функцию LOCS, которая служит калибратором для минимизации изменчивости от анализа к анализу и от инструмента к инструменту, что, в свою очередь, позволяет применять альтернативный способ анализа, который может точно определить наличие или отсутствие двух мишеней в одном канале путем измерения дискретной флуоресценции до и после ПЦР с использованием фиксированных пороговых значений и определением Cq одной мишени.
Способ калибратора, продемонстрированный в этом примере, имеет несколько преимуществ, включая то, что он не требует добавления в реакцию дополнительных реагентов и не требует использования данных, полученных из других лунок. Этот способ служит для калибровки и корректировки возможных вариаций от лунки к лунке. Кроме того, калибровка выполняется с использованием данных, полученных в одном и том же канале, и поэтому на нее не влияют какие-либо различия между каналами, которые могут присутствовать между приборами. Если для мультиплексной реакции используется несколько каналов, каждый канал можно калибровать независимо по калибровочному сигналу LOCS в каждом канале. Это благоприятно для сценария, в котором сигналы калибруются по сигналам в другом канале, например, от внутреннего контроля или эндогенного контроля, поскольку калибровка поддается отрицательному влиянию, где отношение ожидаемой интенсивности сигнала между каналами значительно отличается между инструментами, вызывая вариации от канала к каналу.
Пример Возможного Применения 13: Способ одновременного колориметрического детектирования двух мишеней с использованием одного линейного субстрата МНКзима и одного зонда LOCS.
Следующий пример демонстрирует использование наночастиц золота для одновременного колориметрического детектирования и дифференциации двух мишеней в одной реакции, что может быть достигнуто с использованием одного линейного субстрата МНКзима, детектируемого при одной температуре (D1), и одного репортера LOCS, детектируемого при второй температуре (D2).
Как линейные субстраты МНКзима, так и репортеры LOCS могут быть помечены на каждом конце наночастицами золота (НЧЗ), при этом НЧЗ будут находиться в агрегированном состоянии, когда субстрат и репортеры LOCS остаются интактными и нерасщепленными. В агрегированном состоянии НЧЗ будут демонстрировать поглощение на более длинных волнах из-за связи их отдельных локализованных плазмонов и будут визуализированы как фиолетовый цвет.Этот фиолетовый цвет можно наблюдать как невооруженным глазом, так и с помощью спектроскопии в области поглощения УФ/видимой области. Расщепление линейных субстратов их соответствующим МНКзимом в присутствии конкретной Мишени 1 вызовет разделение НЧЗ, что вызовет изменение цвета с фиолетового на красный, которое можно будет наблюдать в широком диапазоне температур. В этом примере изменение цвета линейного субстрата МНКзима можно обнаружить путем измерения спектроскопического сдвига оптической плотности в УФ/видимой области спектра при Температуре 1 (D1=52°С), до и после амплификации (ΔSD1), где абсорбция на более короткой длине волны указывает на присутствие Мишени 1. При Температуре 1 (D1) как разделенный, так и интактный репортер LOCS для второй мишени (Мишень 2) не будут вызывать каких-либо изменений цвета или детектируемого спектроскопического сдвига поглощения, поскольку Tm его стеблевой области выше, чем Температура 1 (D1). Следовательно, любое изменение цвета или спектроскопический сдвиг поглощения, полученный при температуре 1 (D1) после ПЦР, будет отражать присутствие Мишени 1 в реакции, независимо от присутствия или отсутствия Мишени 2.
Опосредованное мишенью расщепление репортера LOCS соответствующим МНКзимом также будет вызывать изменение цвета (от фиолетового к красному) и/или спектроскопический сдвиг поглощения, однако это будет происходить только в определенном диапазоне температур, которые выше, чем Tm стежневой области LOCS. В этом примере изменение цвета и/или спектроскопический сдвиг поглощения можно измерить при Температуре 2 (D2=70°С) до и после амплификации (ΔSD2), которая выше, чем Tm разделенного репортера LOCS, но ниже, чем Tm интактных репортеров LOCS (Tm интактного репортера LOCS>Температура 2>Tm разделенного репортера LOCS). Таким образом, интактный репортер LOCS не будет вызывать какого-либо изменения цвета и/или спектроскопического сдвига поглощения при температуре 2 (D2) и, следовательно, повышенного сдвига поглощения во время ПЦР по сравнению со сдвигом, связанным с отщепленным линейным субстратом 1 при этой температуре (если присутствует), будет связано с разделенным LOCS-1 и будет свидетельствовать о присутствии Мишени 2.
Пример Возможного Применения 14: Способ одновременного детектирования двух мишеней способом поверхностного плазмонного резонанса (SPR) с использованием одного линейного субстрата МНКзима и одного зонда LOCS.
В следующем примере демонстрируется использование наночастиц золота для одновременного детектирования поверхностным плазмонным резонансом (SPR) и дифференциации двух мишеней в одной реакции, что может быть достигнуто с использованием одного линейного субстрата МНКзима, детектируемого при одной температуре (D1), и одного репортера LOCS, детектируемого при второй температуре (D2).
Как линейные субстраты МНКзима, так и репортеры LOCS могут быть прикреплены к поверхности золота на одном конце и помечены на другом конце наночастицами золота (НЧЗ). Когда линейный субстрат МНКзима и LOCS не расщеплены и интактны, НЧЗ останутся в непосредственной близости от поверхности золота и будут демонстрировать измеримый базовый сигнал ППР. Расщепление линейных субстратов их соответствующим МНКзимом в присутствии конкретной Мишени 1 вызовет отделение НЧЗ от поверхности золота, что приведет к измеримому сдвигу сигнала SPR. В этом примере сдвиг сигнала SPR от линейного субстрата МНКзима может быть обнаружен при первой Температуре 1 (D1=52°С) до и после амплификации (ΔSD1), при этом измеримый сдвиг сигнала SPR указывает на наличие Мишени 1. При Температуре 1 (D1) как разделенный, так и интактный репортер LOCS для второй мишени (Мишень 2) не будут давать какого-либо измеримого сдвига в сигнале SPR, поскольку Tm его стеблевой области выше, чем Температура 1 (D1), а НЧЗ останется гибридизированной с поверхностью золота. Таким образом, любой измеримый сдвиг SPR, полученный при Температуре 1 (D1) после ПЦР, будет отражать присутствие Мишени 1 в реакции, независимо от присутствия или отсутствия Мишени 2.
Опосредованное мишенью расщепление репортера LOCS, например, МНКзимом, также может вызывать измеримый сдвиг в сигнале SPR, однако это может происходить только в определенном диапазоне температур, которые выше, чем Tm стеблевой области LOCS. В этом примере дальнейшее измеримое смещение сигнала SPR может быть измерено при Температуре 2 (например, при D2=70°С) до и после амплификации (ΔSD2), которая выше, чем Tm разделенного репортера LOCS, но ниже, чем Tm интактных репортеров LOCS (Tm интактного репортера LOCS>Температура 2>Tm разделенного репортера LOCS). Таким образом, интактный репортер LOCS не будет производить какого-либо измеримого сдвига сигнала SPR при Температуре 2 (D2) и, следовательно, повышенного измеримого сдвига сигнала SPR во время ПЦР по сравнению со сдвигом, связанным с расщепленным линейным субстратом 1 при этой температуре (если он присутствует), будет связано с разделенным LOCS-1 и будет свидетельствовать о наличии Мишени 2.
Пример Возможного Применения 15: Способ одновременного электрохимического детектирования двух мишеней с использованием одного линейного субстрата МНКзима и одного зонда LOCS.
Следующий пример демонстрирует использование окислительно-восстановительных активных частиц, таких как метиленовый синий, для одновременного электрохимического детектирования и дифференциации двух мишеней в одной реакции, что может быть достигнуто с использованием одного линейного субстрата МНКзима, детектируемого при одной температуре (D1), и одного репортера LOCS, детектируемого при второй температуре (D2).
Как линейные субстраты МНКзима, так и репортеры LOCS могут быть иммобилизованы на поверхности электрода, такого как золотой электрод, с помощью связей Au-S на одном конце и помечены на другом конце метиленовым синим, который представляет собой электрохимически активную молекулу. Когда линейный субстрат МНКзима и LOCS не расщеплены и интактны, молекула метиленового синего будет находится в непосредственной близости от поверхности электрода и будет генерировать большой ток, который может быть обнаружен электрохимическим считывающим устройством.
Расщепление линейных субстратов соответствующим МНКзимом в присутствии конкретной Мишени 1 может привести к отделению молекул метиленового синего от поверхности электрода, что приведет к значительному снижению тока. В этом примере снижение тока от расщепленного линейного субстрата МНКзима может быть обнаружено при первой Температуре 1 (D1=52°С) до и после амплификации (ΔSD1), при этом это снижение тока может быть использовано, чтобы указать на присутствие Мишени 1. При Температуре 1 (D1) как разделенный, так и интактный репортер LOCS для второй мишени (Мишень 2) не будут давать какого-либо измеримого снижения тока, поскольку Tm его стеблевой области выше, чем Температура 1 (D1) и молекула метиленового синего останется гибридизированной и будет находиться в непосредственной близости от поверхности электрода. Следовательно, любое измеримое снижение тока, полученное при Температуре 1 (D1) после ПЦР, будет отражать присутствие Мишени 1 в реакции, независимо от присутствия или отсутствия Мишени 2.
Опосредованное мишенью расщепление репортера LOCS соответствующим МНКзимом также может вызывать измеримое снижение тока, однако это может происходить только в определенном диапазоне температур, которые выше, чем Tm стеблевой области LOCS. В этом примере дальнейшее измеримое снижение тока может быть измерено при Температуре 2 (D2=70°С) до и после амплификации (ΔSD2), которая выше, чем Tm разделенного репортера LOCS, но ниже, чем Tm интактных репортеров LOCS (Tm интактного репортера LOCS>Температура 2>Tm разделенного репортера LOCS). Таким образом, интактный репортер LOCS не будет вызывать какого-либо измеримого снижения тока при Температуре 2 (D2) и, таким образом, дальнейшее снижение тока во время ПЦР, превышающее любое, связанное с расщепленным линейным субстратом 1 (если он присутствует) при этой температуре, будет связанным с разделенным LOCS-1 и будет указывать на наличие Мишени 2.
Пример 16: Способы одновременного детектирования и количественного определения нескольких мишеней на одной длине волны с использованием одного молекулярного маяка и одного репортера LOCS, а также полимеразы, замещающей цепи, лишенной с 5' на 3' экзонуклеазной активности.
В следующем примере демонстрируется способ одновременного детектирования и количественного определения двух мишеней в одном флуоресцентном канале путем получения измерений флуоресценции при двух температурах в режиме реального времени во время ПЦР с использованием одного нерасщепляемого молекулярного маяка и одного репортера LOCS. Этот эксперимент демонстрирует использование полимеразы, замещающей цепь, лишенной 5'-экзонуклеазной активности, для устранения деградации молекулярного маяка в присутствии мишени. Эта стратегия не требует использования специализированных способов анализа, продемонстрированных в Примере 4. Как показано на Фиг. 5, как молекулярный маяк, так и зонд LOCS помечены одними и теми же фрагментами флуорофора и гасителя для одновременного детектирования в одном и том же флуоресцентном канале. Молекулярный Маяк содержит стеблевую область с Tm А и петлевую область, которая может специфически гибридизоваться с мишенью 1 (TVbtub) с Tm В; где Tm В больше, чем Tm А (Tm В>Tm А). В этом примере интактный зонд LOCS имеет стеблевую область с Tm С (82°С) и петлевую область, которая при расщеплении МНКзимом в присутствии мишени 2 (MgPa) генерирует разделенный LOCS с Tm D(62°С); где Tm D меньше Tm С (Tm D<Tm С). Наличие мишени 1 (TVbtub) и/или мишени 2 (MgPa) можно определить путем измерения флуоресценции при двух температурах (D1 и D2; 50°С и 72°С) либо в реальном времени после каждого цикла ПЦР. В следующем примере Tm А составляет -60°С, Tm В составляет -68°С, Tm С составляет -82°С и Tm D составляет -62°С, что соответствует Сценарию 3, описанному в Примере 5, где D1<Tm А<Tm В<D2 и D1<Tm D<D2<Tm C.
Олигонуклеотиды
Олигонуклеотиды, специфичные для этого эксперимента, включают: Прямой праймер 12 (SEQ ID NO: 63), обратный праймер 12 (SEQ ID NO: 64) и молекулярный маяк 1 (SEQ ID NO: 68) для амплификации и количественного определения мишени 1 (TVbtub); прямой праймер 5 (SEQ ID NO: 20), обратный праймер 5 (SEQ ID NO: 21), партзим А5 (SEQ ID NO: 22), партзим B5 (SEQ ID NO: 23), LOCS-2 (SEQ ID NO: 25) для амплификации и количественного определения мишени 2 (MgPa). Последовательности перечислены в списке последовательностей.
Условия реакции
Детекцию целевой последовательности в режиме реального времени проводили в общем реакционном объеме 20 мкл с использованием термоциклера BioRad® CFX96. Параметры циклов и точки сбора флуоресцентных данных (DA) были следующими: 1 цикл 92°С в течение 2 минут, 50°С в течение 15 секунд (DA) и 72°С в течение 15 секунд (DA); 50 циклов: 92°С в течение 5 секунд, 50°С в течение 40 секунд (DA) и 72°С в течение 5 секунд (DA). Все реакции проводили в трех повторностях. Каждая реакция содержала 40 нМ каждого прямого праймера, 200 нМ каждого обратного праймера, 200 нМ каждого партзима, 200 нМ молекулярного маяка 1, 200 нМ репортера LOCS-2, 2 единицы SD-полимеразы Hotstart (Bioron), 800 мкМ смеси dNTP (Bioline), 8 мМ MgCb (Bioron) и lx буфера NH4 (Bioline). Реакции на первом планшете либо не содержали мишени (NF Н2О), либо содержали различные концентрации синтетического G-блока TVbtub (25600, 6400, 1600, 400 или 100 копий) на фоне 0 или 25600 копий гена MgPa, либо синтетического G-блока гена MgPa (25600, 6400, 1600, 400 или 100 копий) на фоне 0 или 25600 копий гена TVbtub.
Полученные результаты
Во время ПЦР-амплификации измеряли флуоресценцию при двух температурах в режиме реального времени для детектирования и количественной оценки присутствия мишени 1 (TVbtub) и/или мишени 2 (MgPa). Молекулярный Маяк был разработан для детектирования последовательностей, гомологичных TVbtub, для детектирования Trichomonas vaginalis (TV). МНКзим был разработан для расщепления LOCS-2В присутствии MgPa для детектирования Mycoplasma genitalium. Номер цикла (Cq), при котором флуоресценция пересекает динамический порог (установленный на уровне 20% от максимальной флуоресценции), определяли по регистрации флуоресценции в реальном времени при 50°С и 72°С для детектирования и количественного определения TVbtub и MgPa соответственно. Наличие или отсутствие специфического сигнала во время ПЦР аналогично описанному в Сценарии 3 Примера 5.
Стандартные реакции, содержащие 25600, 6400, 1600, 400 и 100 копий TVbtub, использовали для построения стандартной кривой (R=0,989; Е=139,14%) для количественного определения исходных концентраций TVbtub при 50°С (D1) в образцах, которые содержали TVbtub в присутствии или в отсутствие MgPa. В Таблице 23 суммировано число копий TVbtub, определенное по стандартной кривой, полученной из данных в реальном времени, полученных при 50°С.На рассчитанное количество копий TVbtub не влияет присутствие в реакции 25600 копий MgPa, поскольку значение р, равное 0,297, подтверждает статистическую незначимость (парный t-критерий Стьюдента). Аналогично, стандарты, содержащие 25600, 6400, 1600, 400 и 100 копий MgPa, использовали для построения стандартной кривой (R2=0,990; Е=116,49%) для количественного определения исходных концентраций MgPa при 72°С (D2) в образцах, содержащих MgPa в присутствии или в отсутствие TVbtub. В Таблице 24 суммировано число копий MgPa, определенное по стандартной кривой, полученной из данных в реальном времени, полученных при 72°С.На рассчитанные числа копий MgPa не влияет присутствие в реакции 25600 копий TVbtub, поскольку значение р, равное 0,319, подтверждает статистическую незначимость (парный t-критерий Стьюдента).
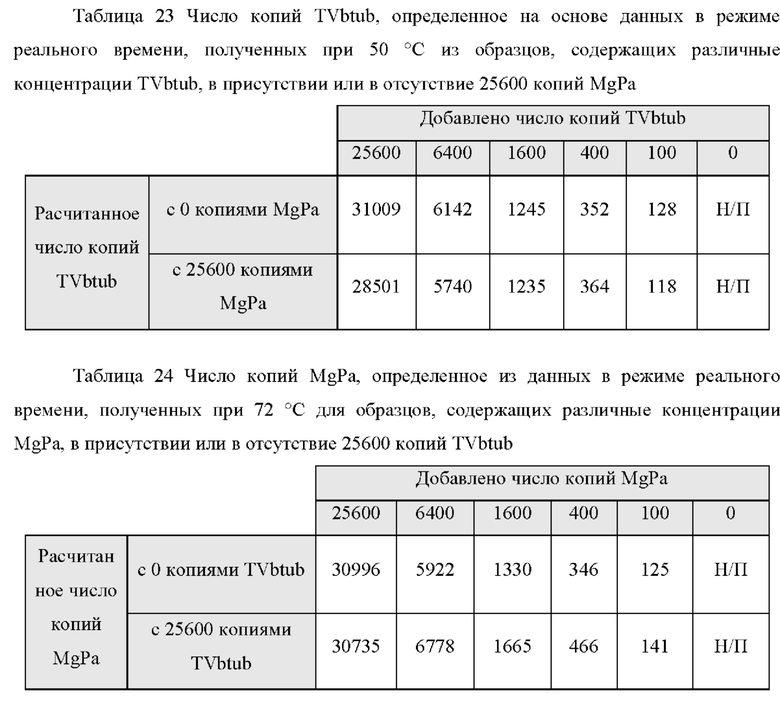
Пример 17. способы определения фонового сигнала с использованием одного или более измерений, полученных до или после усиления; где фоновый сигнал измеряется либо в экспериментальной реакции, либо в эквивалентной контрольной реакции без мишени.
В следующем примере демонстрируются различные стратегии, позволяющие проводить качественный анализ нескольких мишеней на одной длине волны. Способ анализа, показанный в этом примере, аналогичен Способу Анализа Конечной Точки 2, показанному в Примере 1, но этот пример демонстрирует, что фоновый сигнал может быть определен другими способами, а не только с использованием показаний предварительного усиления в D1 и D2B той же реакционной лунке.
Анализ разработан таким образом, что мишень 1 (CTcry) может быть обнаружена и дифференцирована с использованием линейного субстрата МНКзима, а мишень 2 (NGopa) может быть обнаружена и дифференцирована с использованием репортера LOCS, который содержит другой субстрат МНКзима в своей петле. Во время ПЦР МНКзим 1 может расщеплять линейный субстрат 1 в присутствии CTcry, чтобы разделить флуорофор и гаситель, и вызвать усиление сигнала, которое можно обнаружить в широком диапазоне температур. В этом примере определение конечной точки CTcry может быть достигнуто путем определения нормализованного сигнала флуоресценции (NSD1), который выполняет функцию, аналогичную ΔSD1 в Примере 1, и определяется как разница между сигналом после ПЦР при температуре 1 (D1; 52°С) и фоновым сигналом, который определяли с использованием нескольких различных подходов, как показано в Таблице 25 ниже. Стебель LOCS-1 как в интактной, так и в разделенной конфигурации имеет Tm выше D1, и, следовательно, NSD1 остается незатронутым расщеплением LOCS-1 и, следовательно, не зависит от присутствия или отсутствия NGopa в образце. Таким образом, когда NSD1 превышает порог (X1), это указывает на присутствие расщепленного линейного субстрата 1 МНКзима и, следовательно, на мишень 1, CTcry.
В присутствии NGopa МНКзим 2 может расщеплять LOCS-1 во время ПЦР. Нормированный сигнал флуоресценции (NSD2) при температуре 2 (D2, 70°С) можно рассчитать как разницу между сигналом пост-ПЦР при температуре 2 и фоновым сигналом, снова измеренным в соответствии с Таблицей 25. NSD2 имеет функцию, аналогичную ΔSD2B Примере 1. Поскольку расщепление LOCS-1 в ходе ПЦР способствует увеличению нормализованного сигнала флуоресценции при температуре 2 (NSD2), но не влияет на NSD1, где разница между NSD2 и NSD1 (NSD2 - NSD1) пересекает второй порог (Х2); так, что NSD2 - NSD1=ΔNSD2NSD1>Х2, указывает на наличие разделенного LOCS-1. Напротив, расщепление только субстрата 1 способствует увеличению значений NSD2 и NSD1, причем разница между этими двумя значениями (ΔNSD2NSD1) меньше второго порога (Х2). Поэтому, когда разница между NSD2 и NSD1 (ΔNSD2NSD1=NSD2 - NSD1) больше второго порога (Х2), это указывает на специфическое детектирование разделенного LOCS-1 и, следовательно, мишени 2, NGopa.
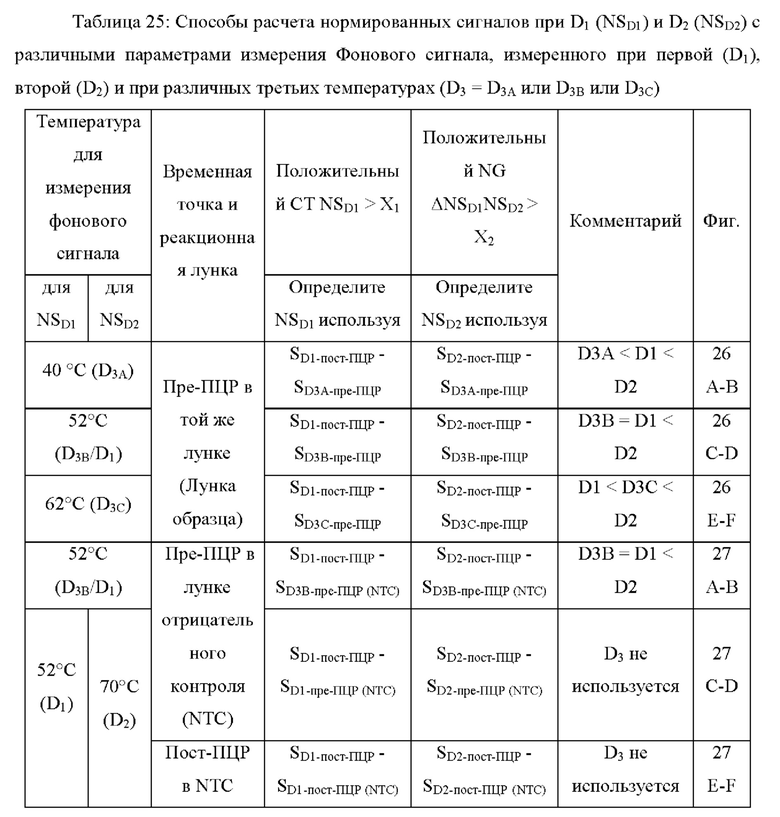
Следующий пример демонстрирует, что фоновые сигналы (SD3), измеренные перед ПЦР при третьей температуре (D3) в той же лунке, можно применять для расчета NSD1 и NSD2. Были испытаны три различные температуры пре-ПЦР, а именно: D3A=40°С; D3B/D1=52°С и D3C=60°С (Таблица 25). Эта стратегия устраняет необходимость в нескольких точках сбора данных до ПЦР (т.е. один раз в D3 вместо двух в D1 и D2) и сокращает время и сложность экспериментов. Кроме того, следующий пример демонстрирует, что фоновые сигналы могут быть измерены в отдельных реакциях отрицательного контроля без матрицы. В одном случае одно пре-ПЦР фоновое измерение выполненое при D3B=52°С=D1 использовали для расчета как NSD1, так и NSD2; в то время как в других случаях были выполнены два фоновых измерения при D1 (52°С) и D2 (70°С) с получением данных либо до, либо после ПЦР из отдельных реакций отрицательного контроля.
Ол иго нукл е отиды
Олигонуклеотиды, специфичные для этого эксперимента, включают: Прямой праймер 1 (SEQ ID NO: 1), обратный праймер 1 (SEQ ID NO: 2), партзим A1 (SEQ ID NO: 3), партзим B1 (SEQ ID NO: 4) и линейный субстрат МНКзима 1 (SEDID: 13) для усиления и детектирования Мишени 1 (CTcry); прямой праймер 2 (SEQ ID NO: 5), обратный праймер 2 (SEQ ID NO: 6), партзим А5 (SEQ ID NO: 7), партзим B5 (SEQ ID NO: 8) и LOCS-1 (SEQ ID NO: 14) для усиления и детектирования Мишени 2 (NGopa). Последовательности перечислены в списке последовательностей.
Условия реакции
Детекцию целевой последовательности в режиме реального времени проводили в общем реакционном объеме 20 мкл с использованием термоциклера BioRad® CFX96. Параметры циклирования и точки сбора флуоресцентных данных (DA): 1 цикл 95°С в течение 2 минут, 40°С в течение 15 секунд (DA), 52°С в течение 15 секунд (DA), 62°С в течение 15 секунд (DA) и 70°С в течение 15 секунд (DA); 10 циклов: 95°С в течение 5 секунд и 61°С в течение 30 секунд (уменьшение на 0,5°С за цикл); 40 циклов 95°С в течение 5 секунд и 52°С в течение 40 секунд; и 1 цикл 52°С в течение 15 секунд (DA) и 70°С в течение 15 секунд (DA). Все реакции проводили в трех повторностях, и каждая реакция содержала 40 нМ каждого прямого праймера, 200 нМ каждого обратного праймера, 200 нМ каждого партзима, 200 нМ линейного субстрата МНКзима 1, 200 нМ репортера LOCS-1 и lx PlexMastermix (Bioline). Реакции либо не содержали мишени (NF H2O), либо содержали синтетический G-блок CTcry (10000 или 40 копий); или ген NGopa (10000 или 40 копий); или различные концентрации гена CTry (10000 или 40 копий) на фоне гена NGopa (10000 и 40 копий). Все реакции, кроме отрицательного (без мишени) контроля (NF H2O), дополнительно содержали фон из 34,5 нг (10000 копий) геномной ДНК человека.
Полученные результаты
Результаты на Фиг. 26 иллюстрируют расчетные значения для NSD1 (левый) и ΔNSD2 NSD1 (правый) для детектирования CTcry и NGopa, соответственно, с использованием фонового сигнала, определенного с использованием измерений флуоресценции до ПЦР (SD3) из той же реакционной лунки при 40°С (Фиг. 26А-В); 52°С (Фиг. 26C-D) и 62°С (Фиг. 26E-F). Результаты на Фиг. 27 иллюстрируют расчетные значения для NSD1 (левый) и ΔNSD2NSD1 (правый) для детектирования CTcry и NGopa соответственно с использованием значений фонового сигнала, определенных из отдельных реакций отрицательного контроля. Фоновые сигналы определяли как среднее значение контрольных сигналов при отсутствии темплата, измеренных в D3B/D1 до ПЦР (Фиг. 27А-В) и в D1 и D2 до ПЦР (Фиг. 27C-D) и после ПЦР (Фиг. 27E-F).
Результаты на Фиг. 26А, Фиг. 26С, Фиг. 26Е, Фиг. 27А, Фиг. 27С и Фиг. 27Е показывают, что для всех сценариев нормированный сигнал при температуре 1 (NSD1) больше, чем порог 1 (X1) когда CTcry присутствует в образце независимо от того, присутствует или отсутствует NGopa, но не превышает этот порог только при наличии NGopa и/или когда CTcry отсутствует в образце. Следовательно, нормализованный сигнал конечной точки, превышающий порог 1 при температуре 1, указывает на наличие CTcry. Результаты на Фиг. 26В, Фиг. 26D, Фиг. 26F, Фиг. 27В, Фиг. 27D и Фиг. 27F иллюстрируют для всех шести сценариев ΔNSD2NSD1 больше порога 2 (Х2), когда NGopa присутствует в образце, но не превышает этого порога, когда в образце присутствует только CTcry и/или когда в образце отсутствует NGopa (NTC). Следовательно, разница в нормированных флуоресцентных сигналах конечной точки, NSD2 и NSD1, превышающая порог 2 (ΔNSD2 NSD1>Х2), свидетельствует о наличии NGopa.
Этот пример демонстрирует использование одного линейного субстрата МНКзима и одного репортера LOCS для определения конечной точки и дифференциации двух мишеней в одном флуоресцентном канале с гибкостью определения фонового сигнала, подлежащего измерению при третьей температуре и/или по данным отдельного отрицательного контроля реакции.
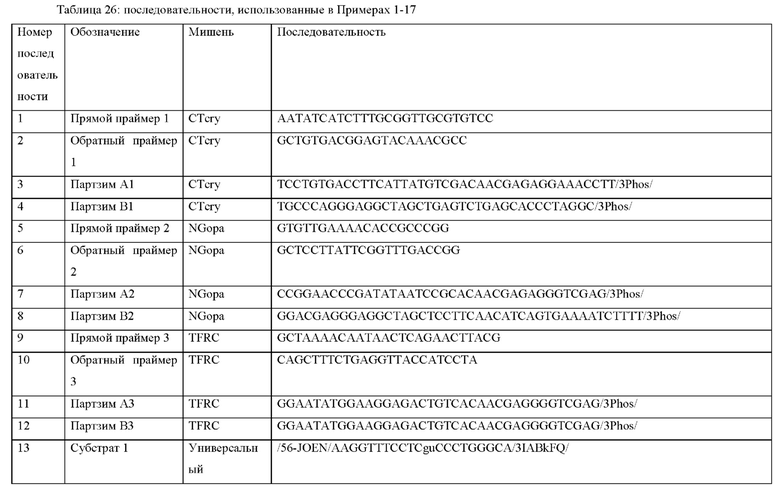
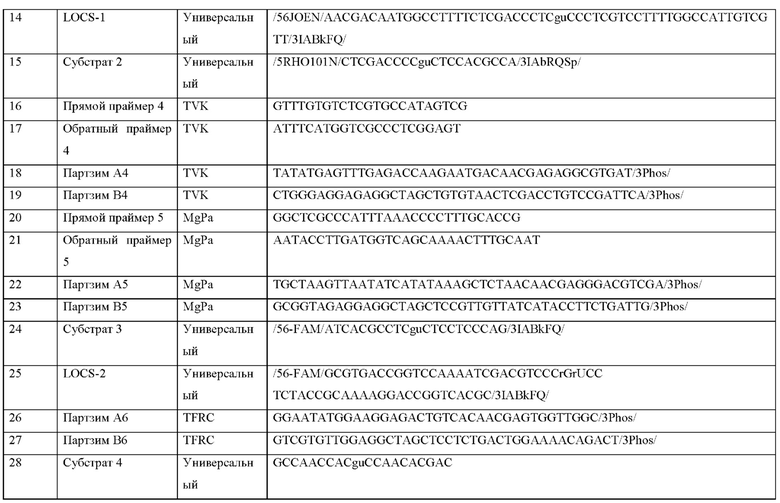
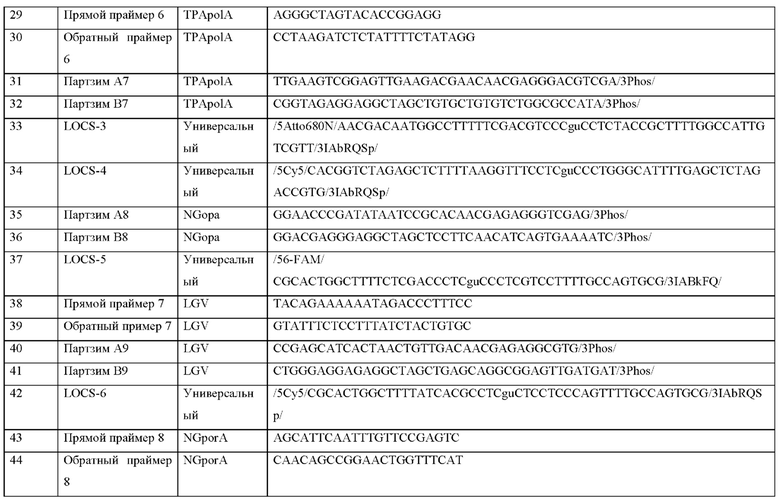

Описание олигопоследовательностей в Таблице 26
Последовательности олигонуклеотид о в перечислены от 5' до 3'. ЗАГЛАВНЫЕ ОСНОВАНИЯ обозначают ДНК, а строчные - РНК. /56-FAM/ указывает местоположение флуорофора FAM, /56-JOEN7 указывает местоположение флуорофора JOE, /5RHO101N/ указывает местоположение флуорофора АТТО RholOl, /5Atto680N/ указывает местоположение флуорофора АТТО 680 и /5Су5/ указывает местоположение флуорофора су5.5. /3IABkFQ/ указывает местоположение гасителя Iowa Black FQ, способного поглощать флуоресценцию в диапазоне 420-620 нм, и /3IAbRQSp/ указывает местоположение гасителя Iowa Black RQ, используемого для поглощения флуоресценции в диапазоне 500-700 нм. /3Phos/ указывает на 3'-фосфатную группу.
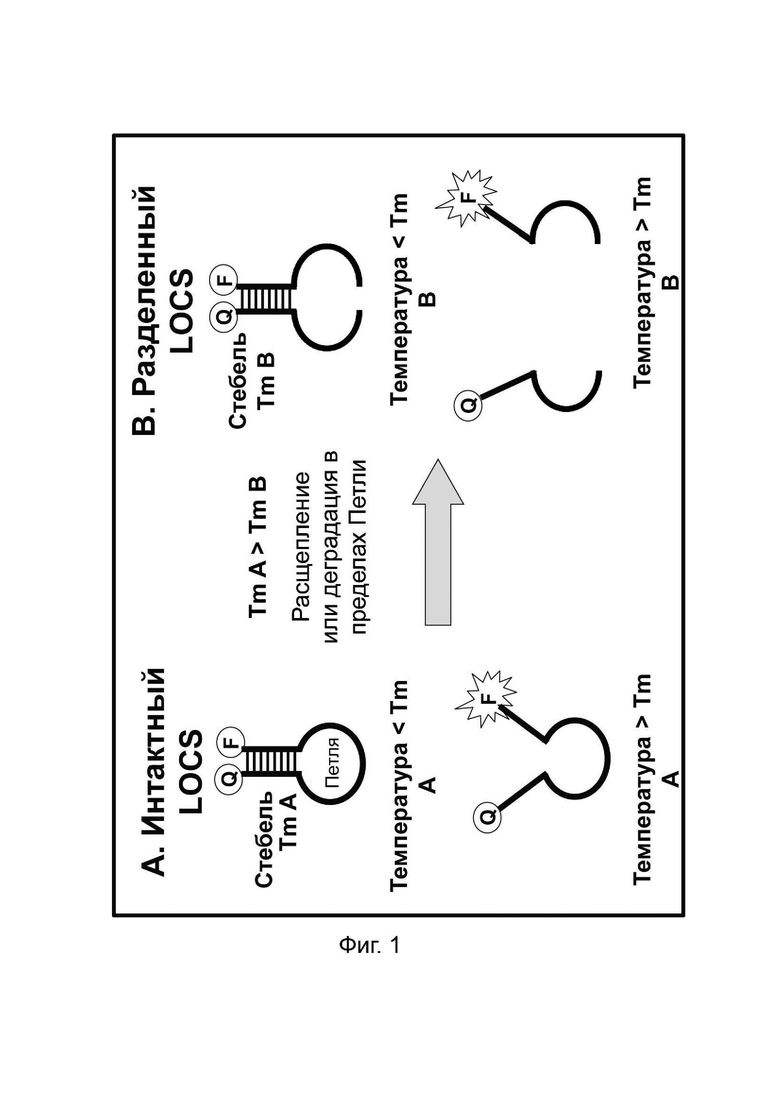
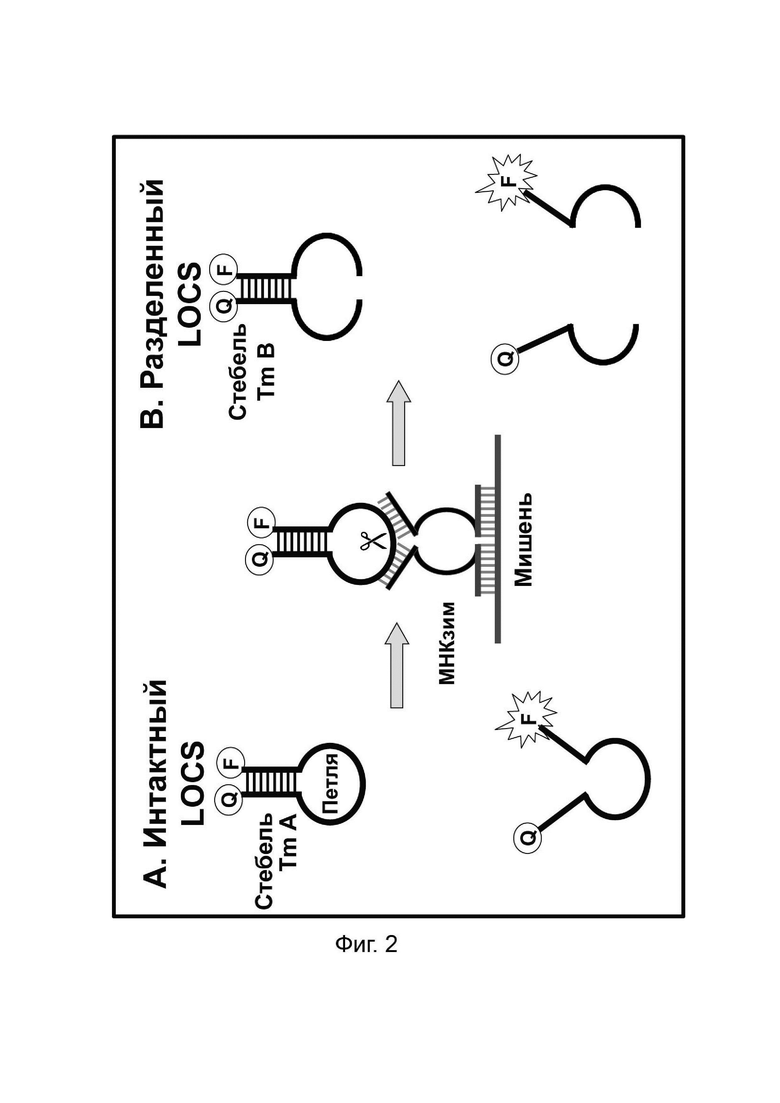
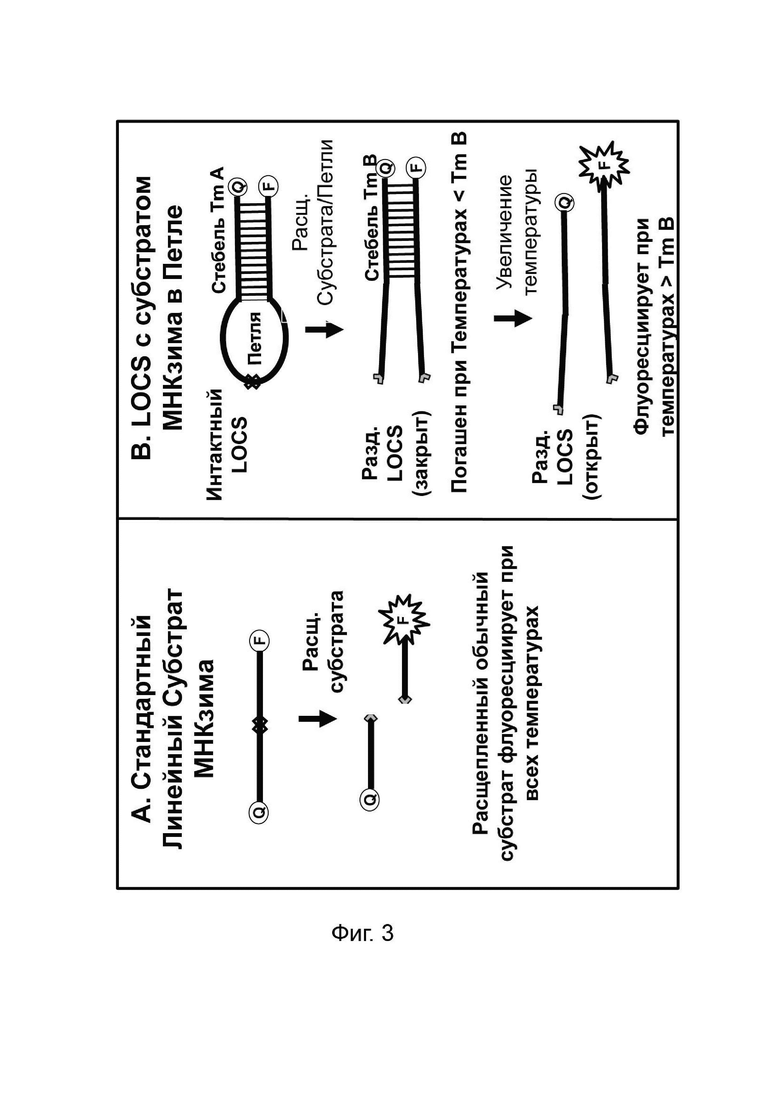
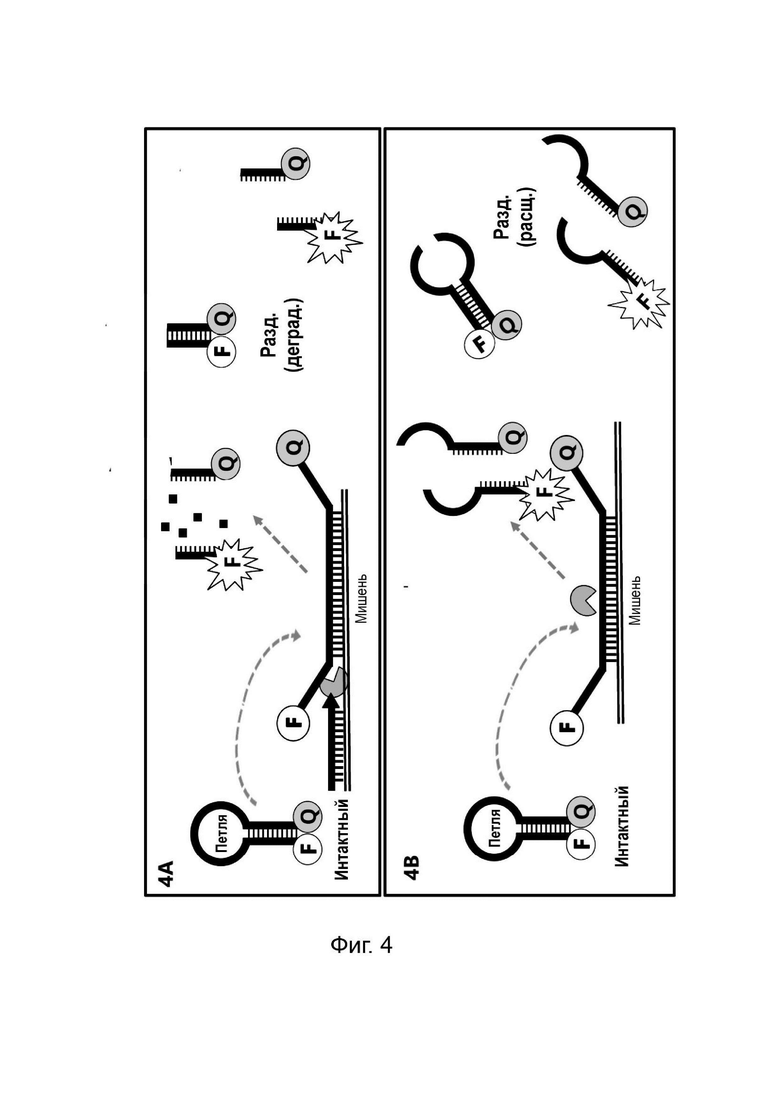
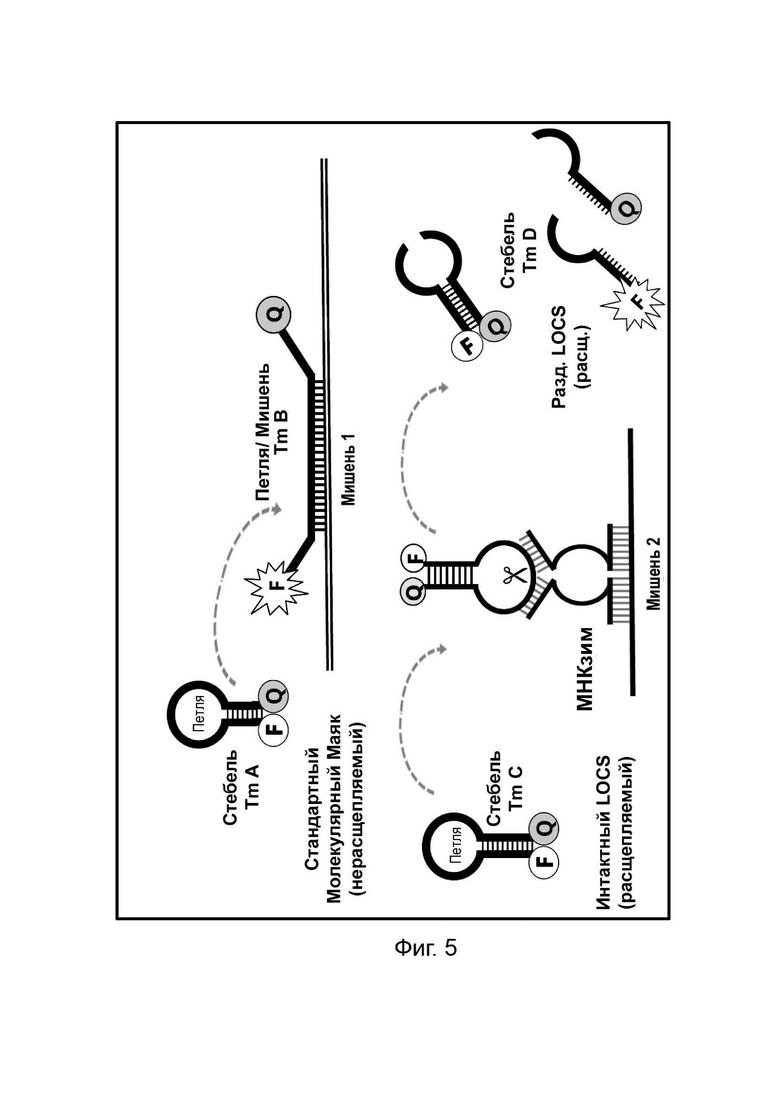
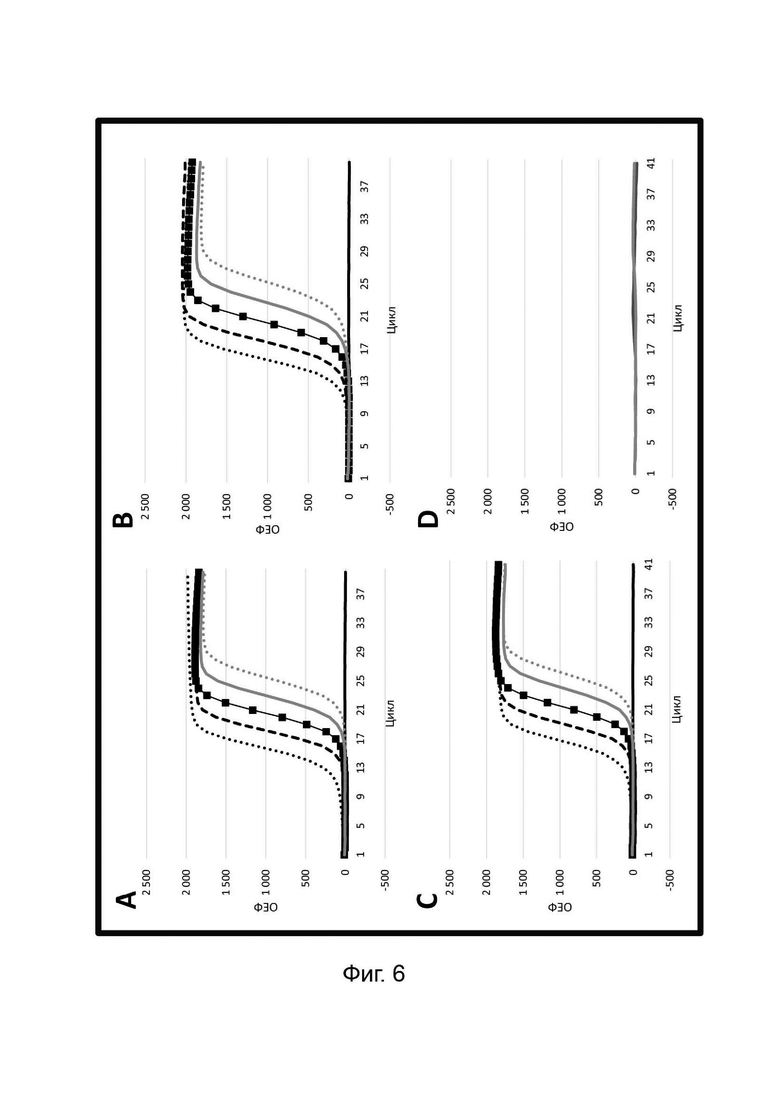
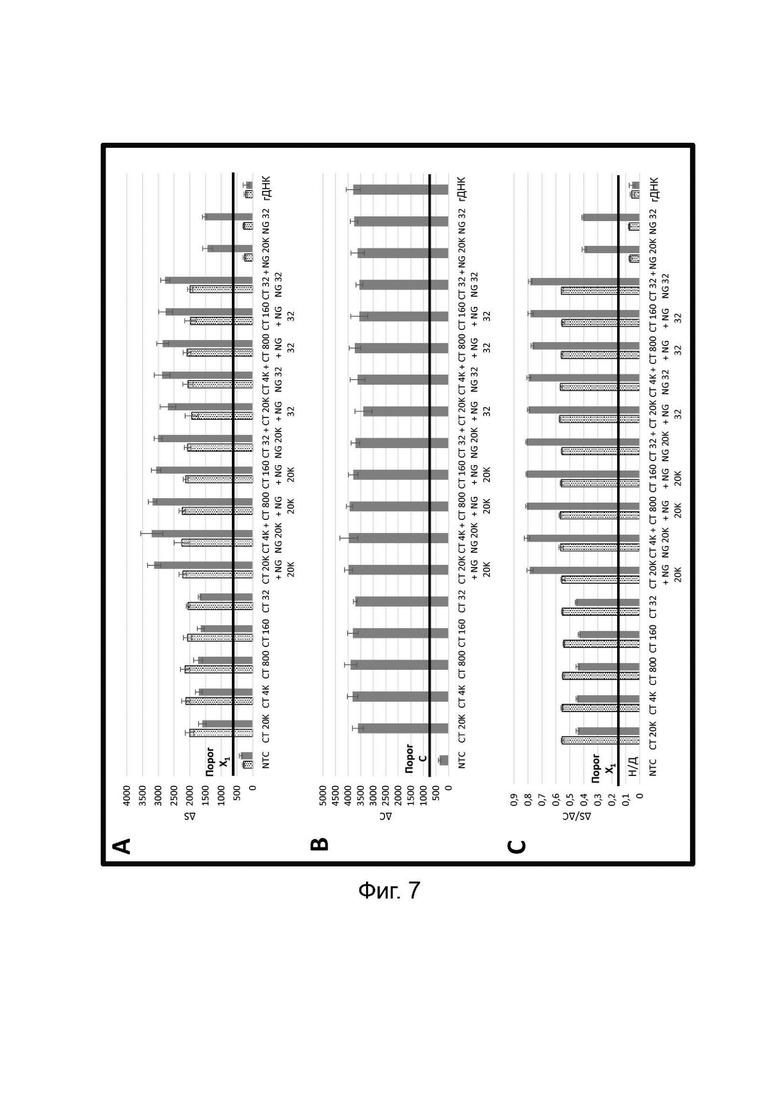
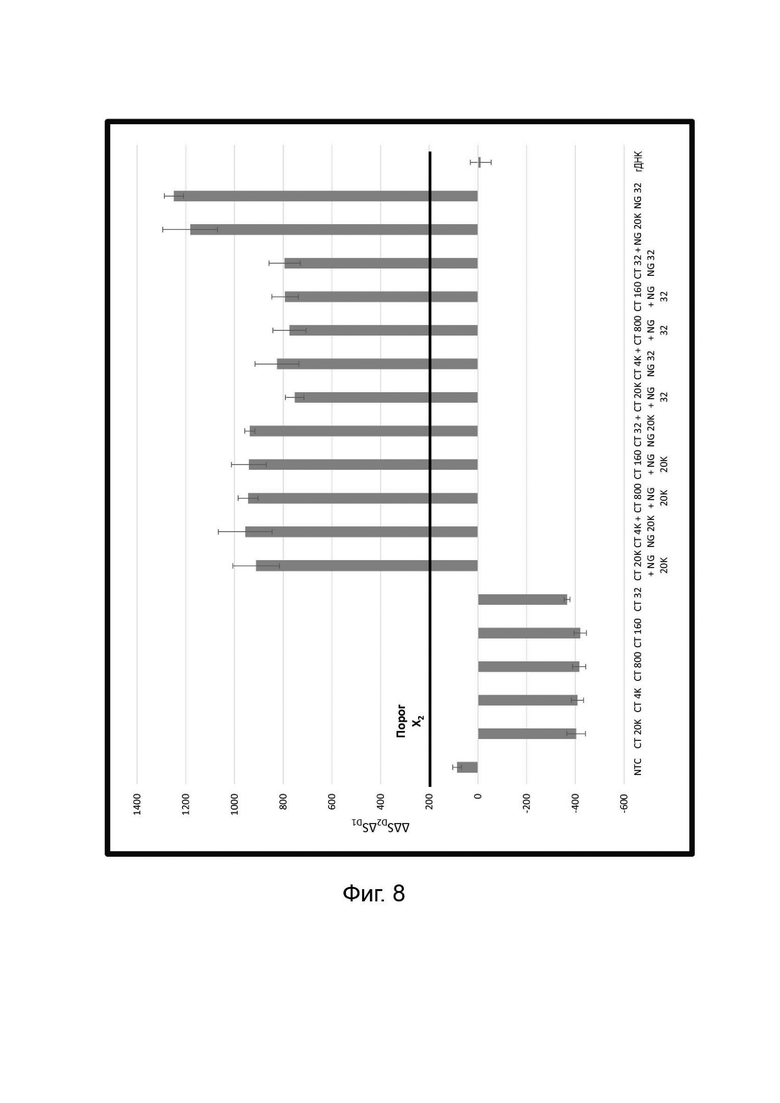

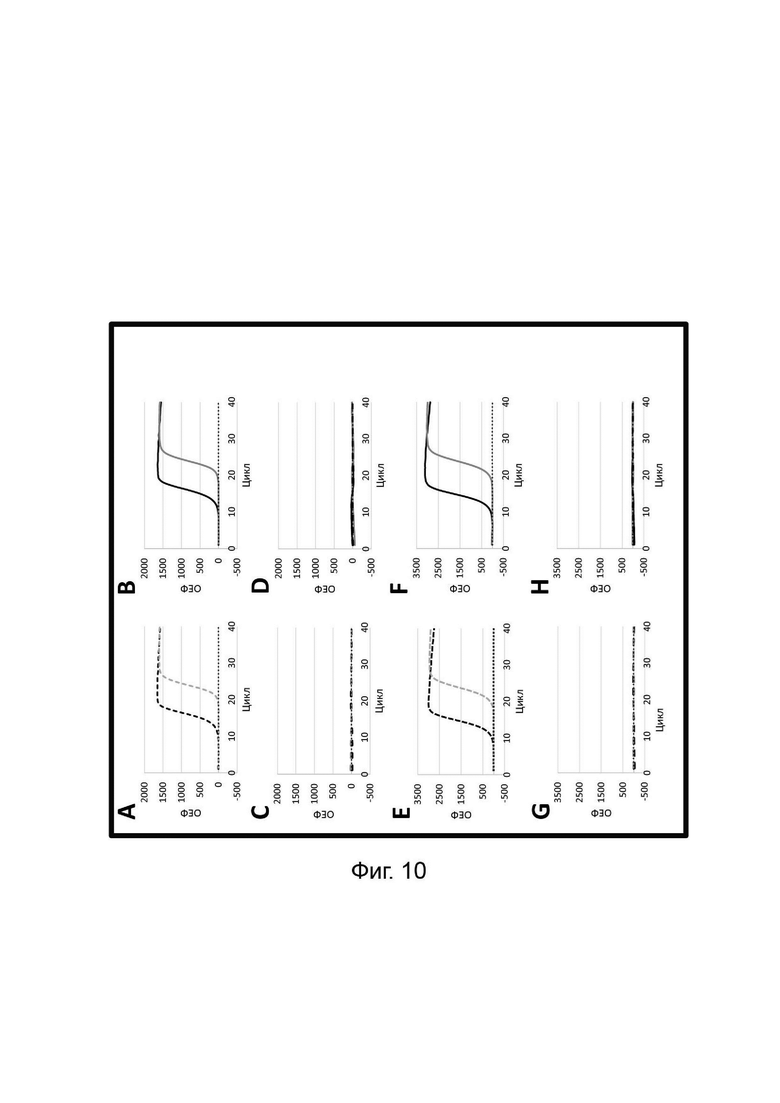
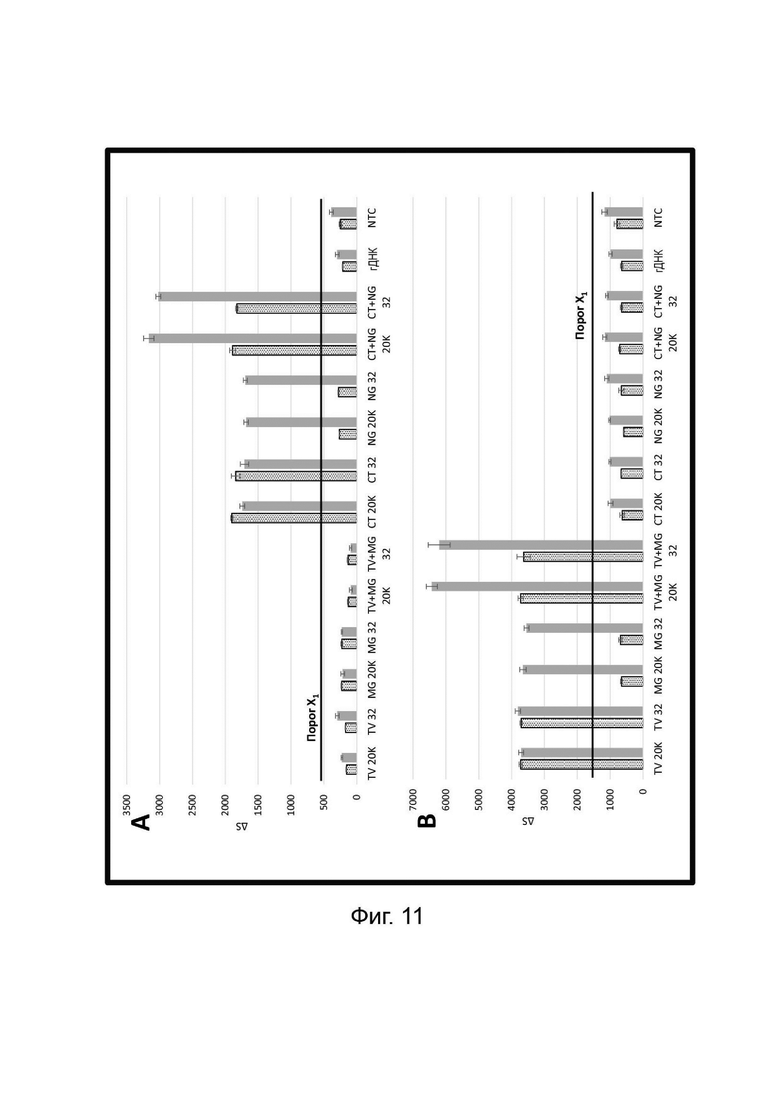

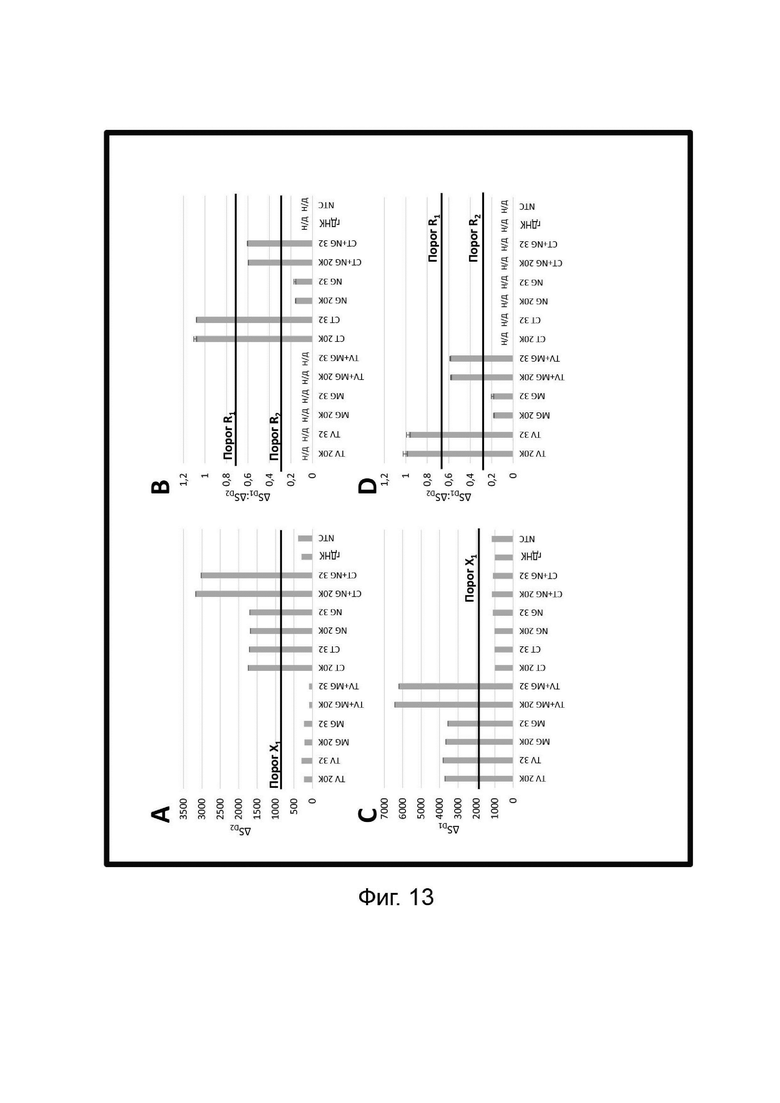

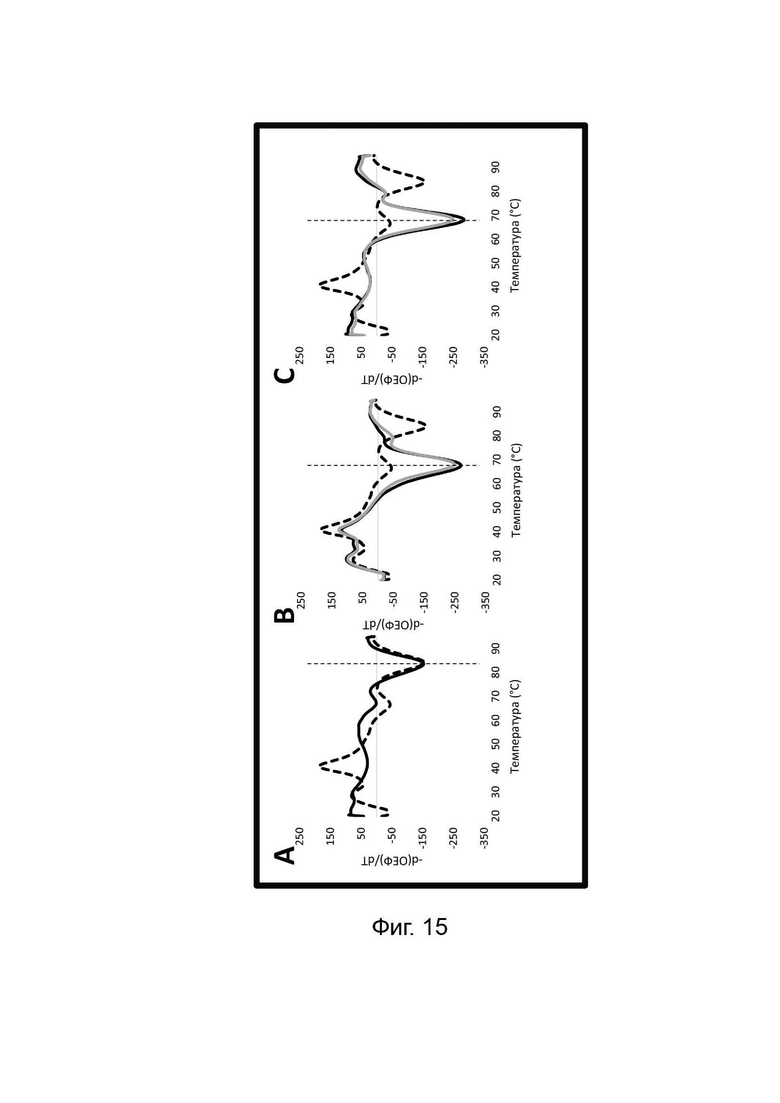
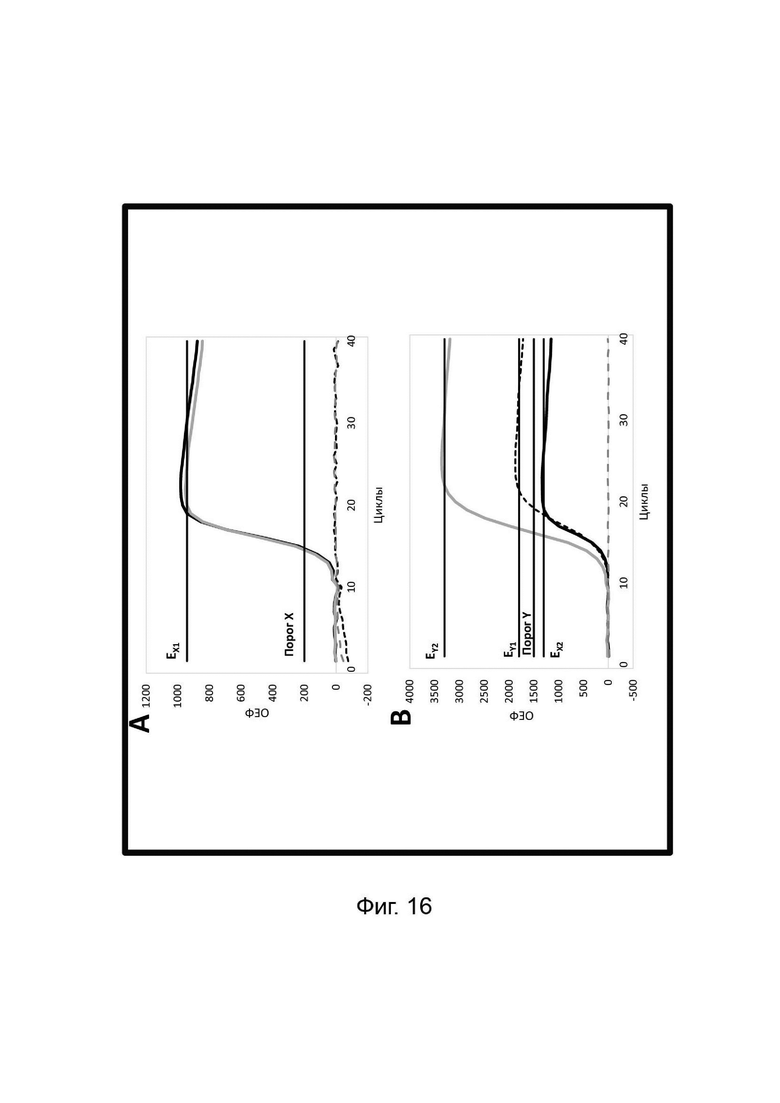
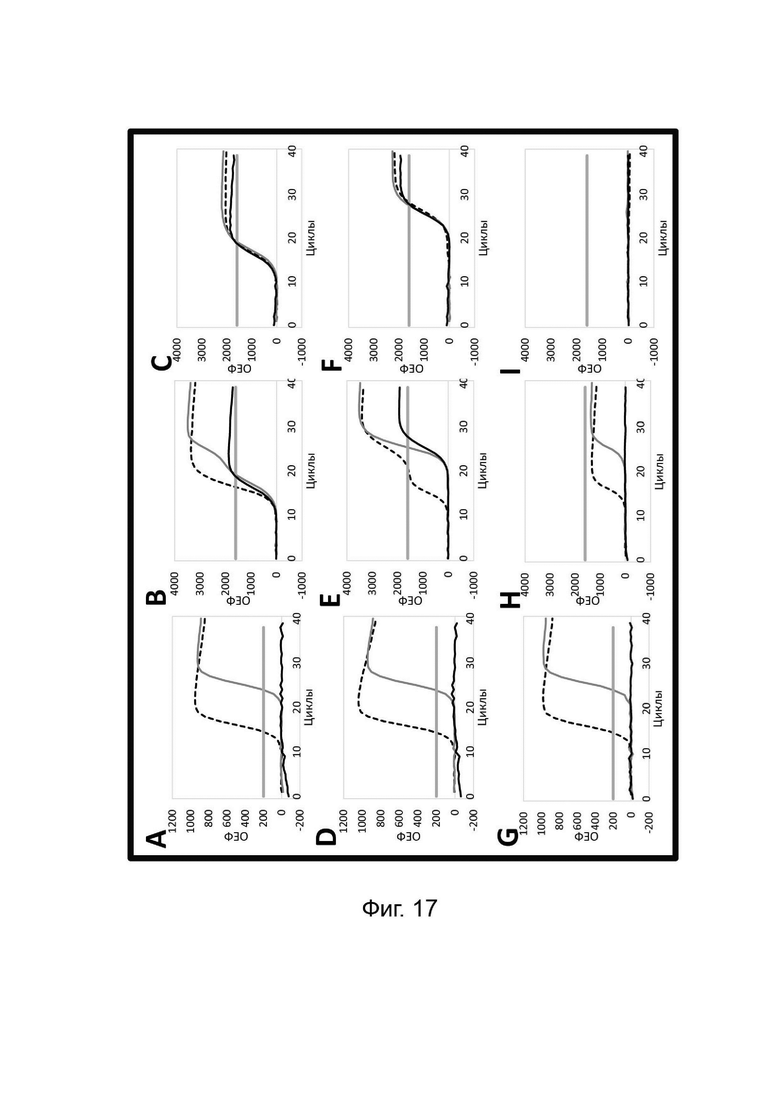
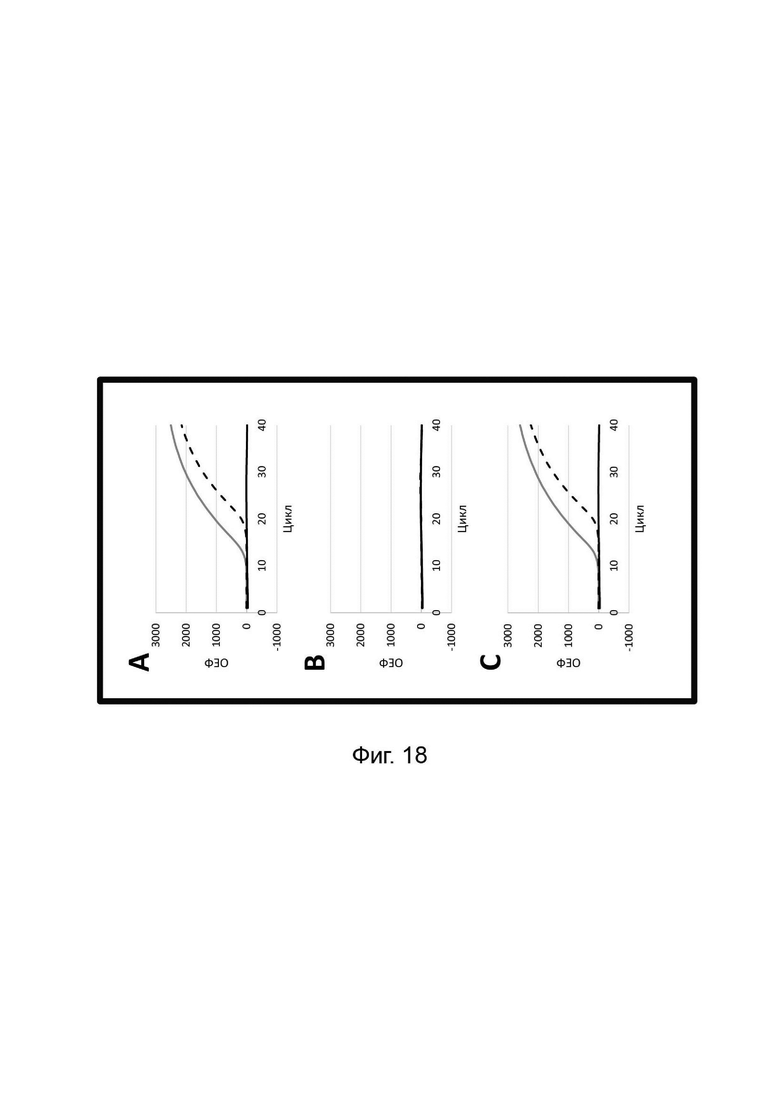
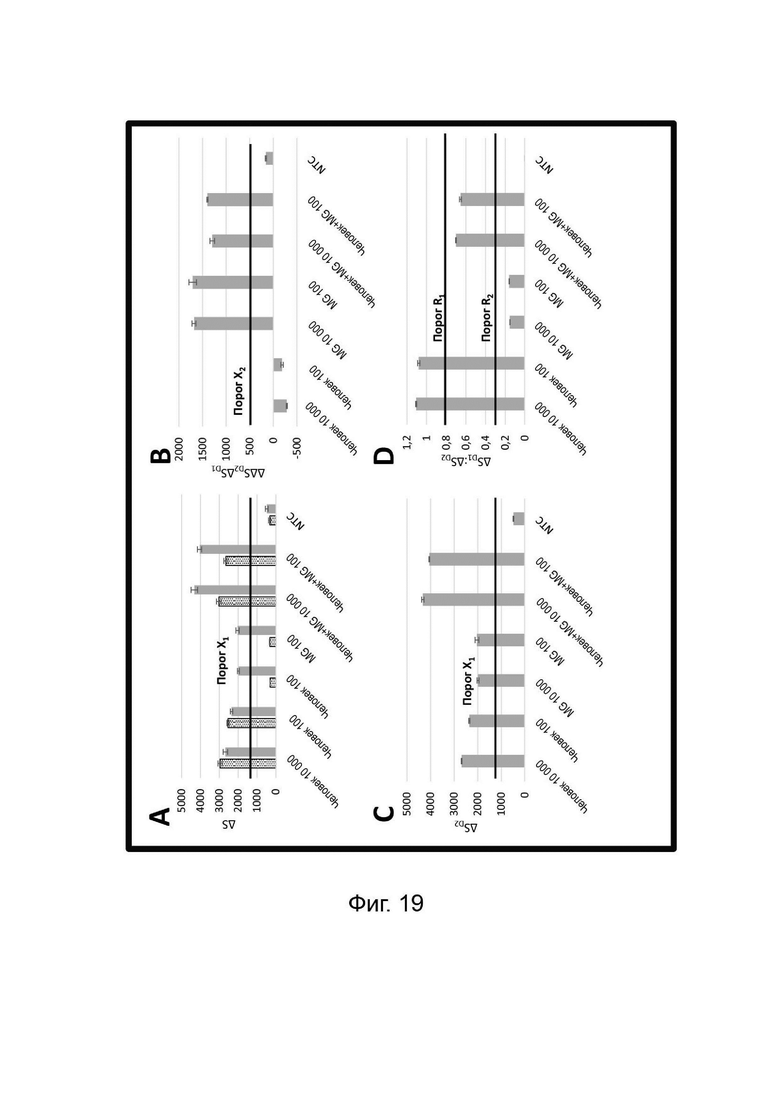
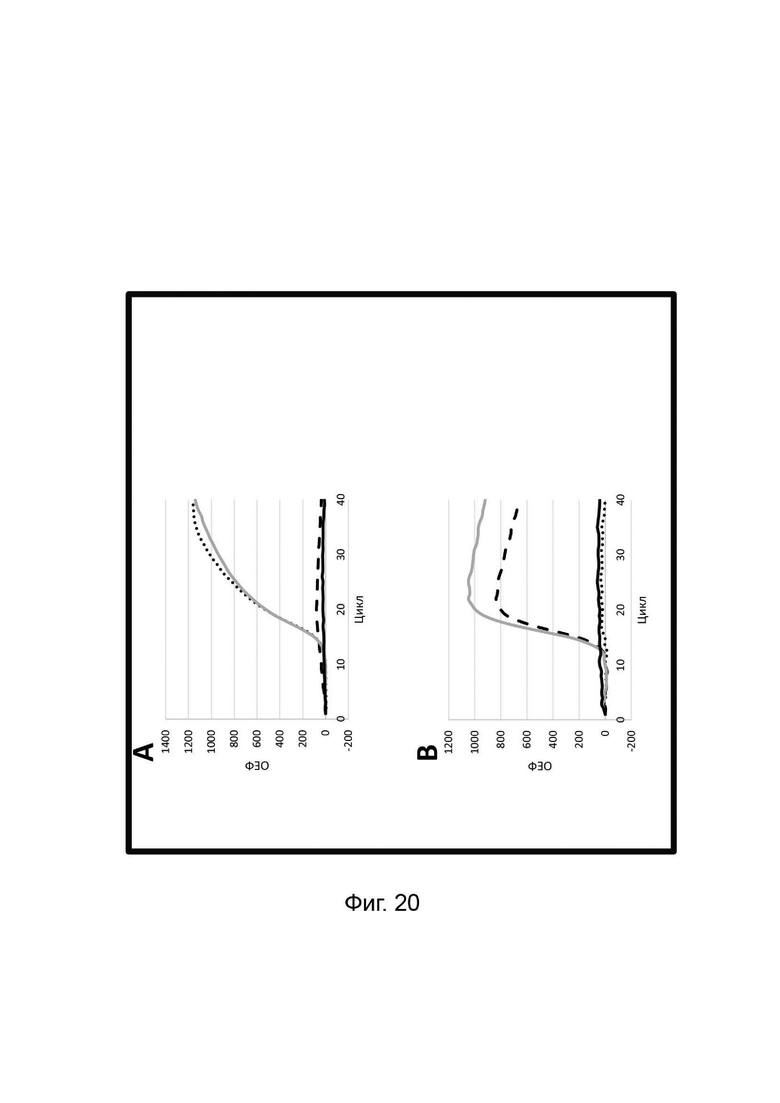
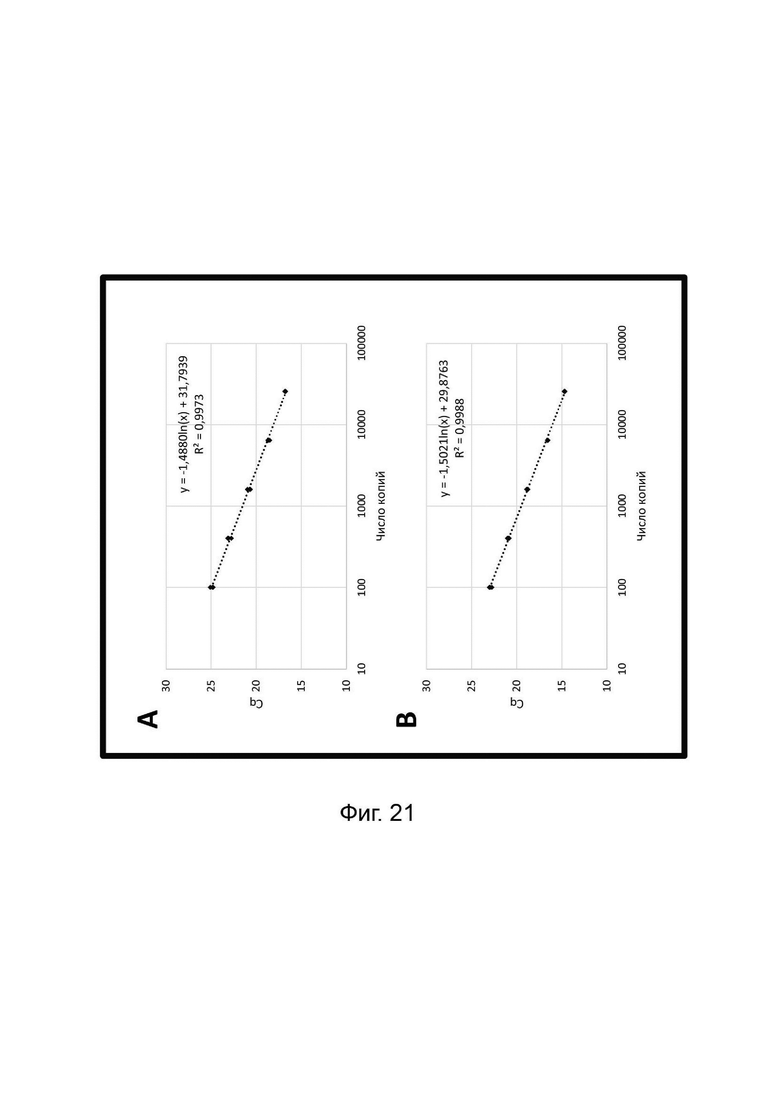
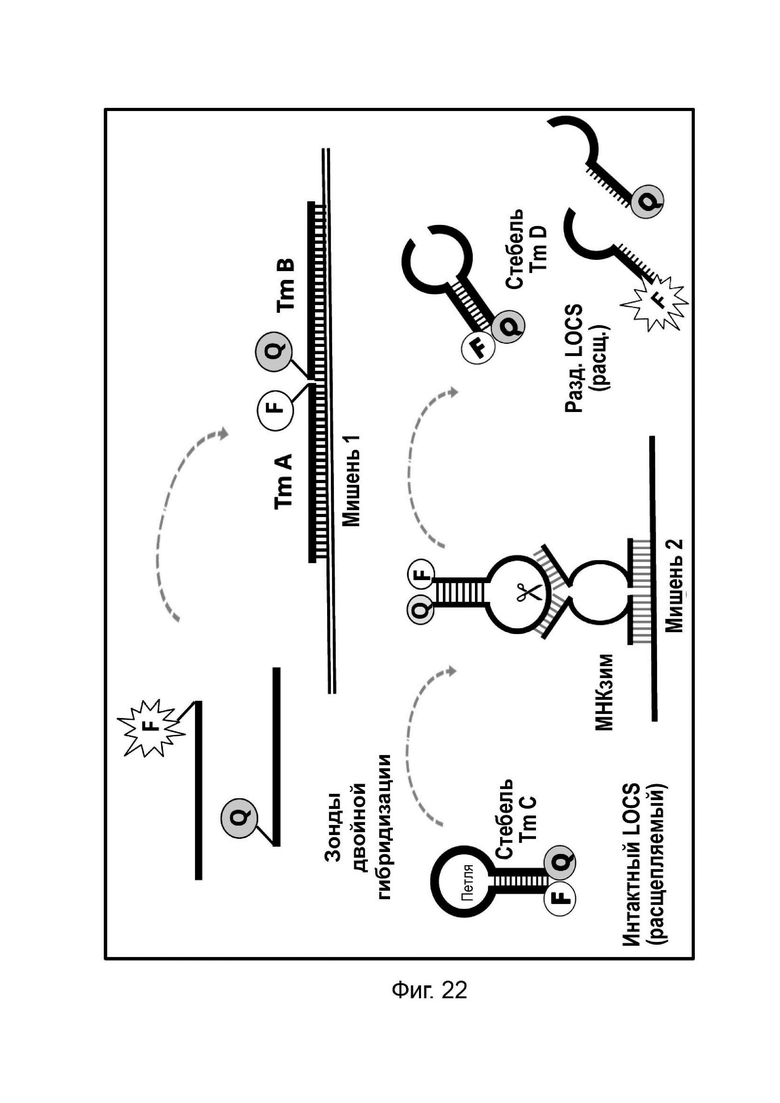
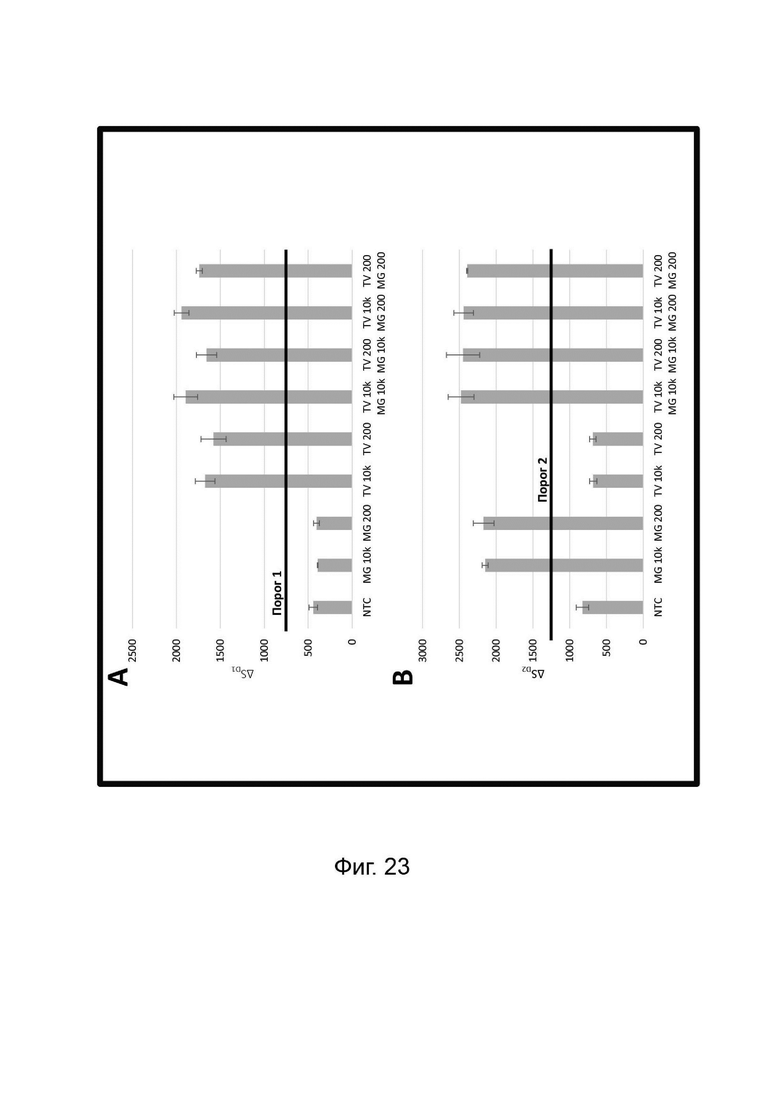

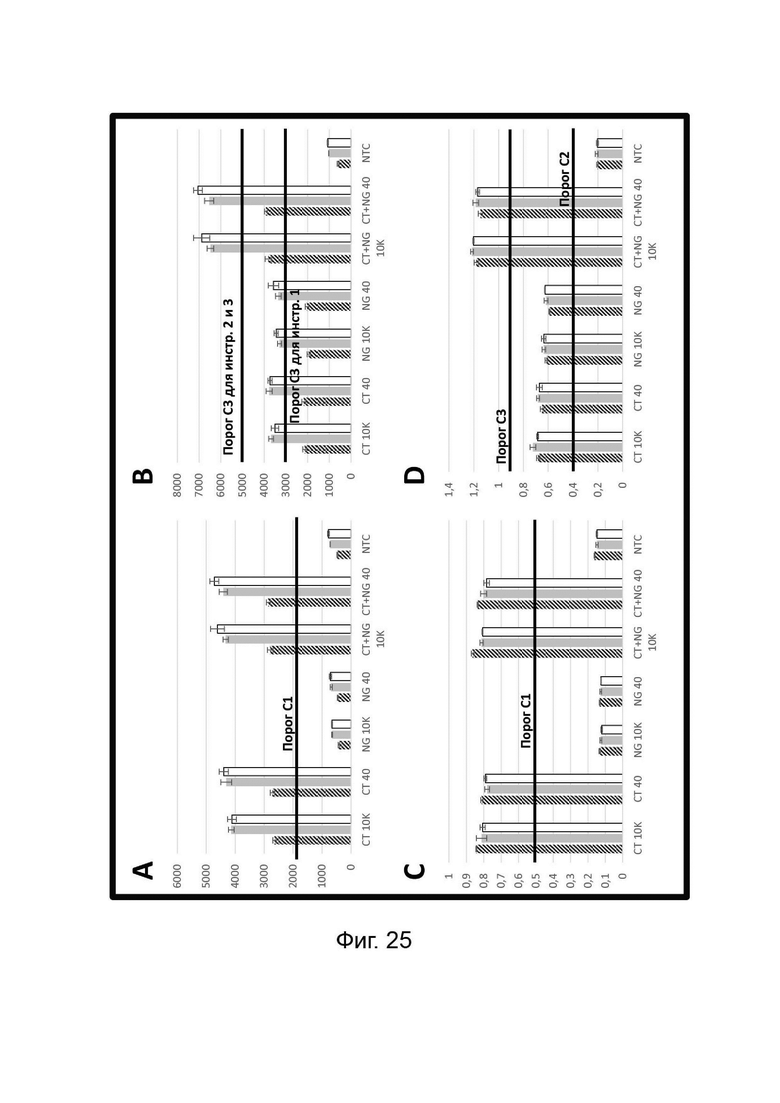

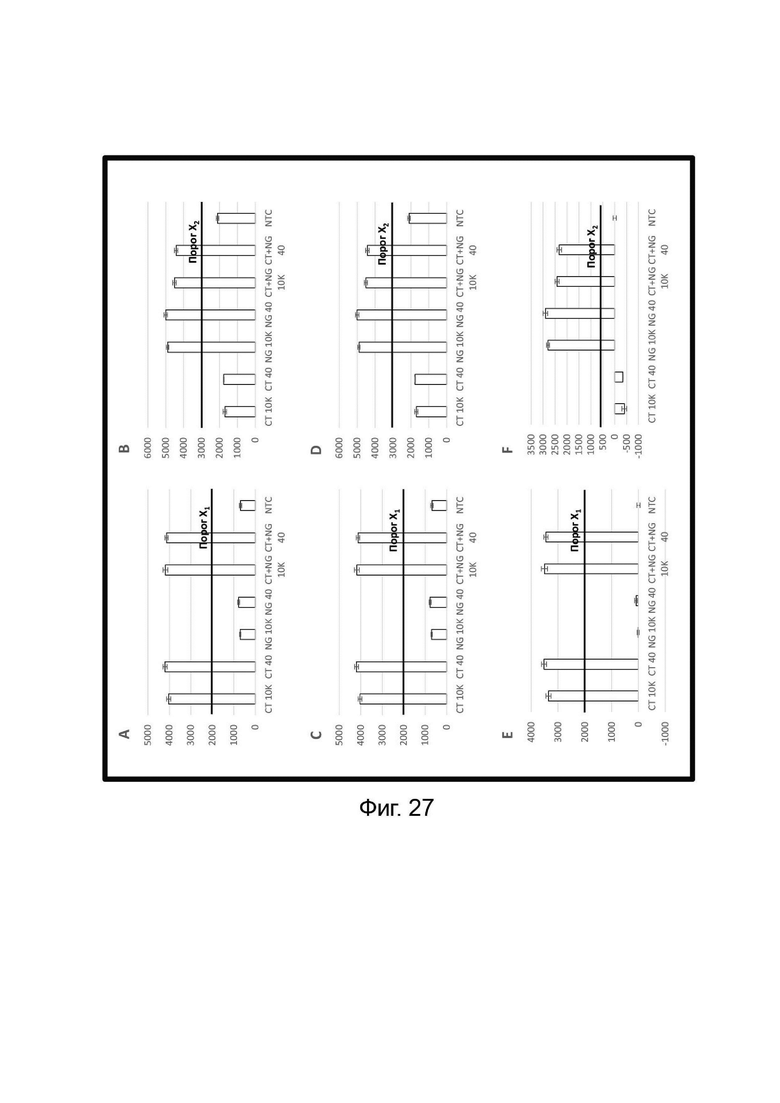
Настоящее изобретение относится к области молекулярной биологии. Описан способ определения наличия или отсутствия первой и второй мишеней в образце, включающий: приготовление смеси для реакции путем приведения в контакт образца, предположительно содержащего первую и вторую мишени с олигонуклеотидами и ферментом; обработку смеси; измерение фонового сигнала, обеспечиваемого первым и вторым детектирующими фрагментами в смеси или в контрольной смеси; определение генерации первого и второго сигнала в определённых условиях. Также описаны композиции для реализации способа. Технический результат заключается в расширении арсенала средств мультиплексного анализа нуклеиновых кислот. 3 н. и 190 з.п. ф-лы, 27 ил., 26 табл., 17 пр.
1. Способ определения наличия или отсутствия первой и второй мишеней в образце, включающий:
(a) приготовление смеси для реакции путем приведения в контакт образца, предположительно содержащего первую и вторую мишени, с:
- первым олигонуклеотидом для детектирования первой мишени, содержащим первый детектирующий фрагмент, способный генерировать первый детектируемый сигнал;
- интактным олигонуклеотидом стебель-петля для детектирования второй мишени, содержащим двухцепочечную стеблевую область из гибридизированных нуклеотидов, противоположные цепи которого связаны неразорванной одноцепочечной петлевой областью из негибридизированных нуклеотидов, при этом стеблевая область содержит второй детектирующий фрагмент, способный генерировать второй детектируемый сигнал,
причем первый и второй детектирующие фрагменты способны генерировать детектируемые сигналы, которые нельзя различить при одной температуре с использованием одного типа детектора; и
- первым ферментом, способным расщеплять один или более негибридизированных нуклеотидов интактного олигонуклеотида стебель-петля только тогда, когда в образце присутствует вторая мишень;
(b) обработку смеси в условиях, подходящих для:
- модификации первого олигонуклеотида вызванной первой мишенью, тем самым позволяя первому детектирующему фрагменту генерировать первый детектируемый сигнал,
- расщепления одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля первым ферментом только тогда, когда в образце присутствует вторая мишень, с тем чтобы разрушить одноцепочечную петлевую область и получить разделенный олигонуклеотид стебель-петля;
(c) измерение
фонового сигнала, обеспечиваемого первым и вторым детектирующими фрагментами в смеси или в контрольной смеси;
(d) определение того, в один или более моментов времени во время или после указанной обработки:
- генерируется ли при первой температуре первый детектируемый сигнал, который отличается от фонового сигнала, возникающий в результате указанной модификации, указывающий на присутствие первой мишени в образце;
- генерируется ли при второй температуре второй детектируемый сигнал, который отличается от фонового сигнала, указывающий на присутствие второй мишени в образце;
при этом:
при первой температуре второй детектируемый сигнал не отличается от фонового сигнала и
при второй температуре:
если они присутствуют, цепи двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля частично или полностью диссоциированы, позволяя второму детектирующему фрагменту обеспечивать второй детектируемый сигнал; и
если они присутствуют, цепи двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля не могут диссоциировать, тем самым препятствуя второму детектируемому фрагменту обеспечивать второй детектируемый сигнал.
2. Способ по п. 1, отличающийся тем, что указанное определение в части (d) включает:
- применение заданного порога для определения того, отличается ли первый детектируемый сигнал, возникающий в результате упомянутой модификации, от любого упомянутого фонового сигнала при первой температуре; и/или
- применение заданного порога для определения того, отличается ли второй детектируемый сигнал от любого упомянутого фонового сигнала при второй температуре.
3. Способ по п. 1 или 2, отличающийся тем, что контрольная смесь не содержит:
- первую мишень; или
- вторую мишень; или
- первую и вторую мишени,
но в остальном она эквивалентна смеси.
4. Способ по п. 1 или 2, отличающийся тем, что контрольная смесь содержит заданное количество:
- первой мишени; или
- второй мишени; или
- первой и второй мишеней,
но в остальном она эквивалентна смеси.
5. Способ по любому из пп. 1-4, отличающийся тем, что:
- модификация первого олигонуклеотида позволяет первому детектирующему фрагменту обеспечивать первый детектируемый сигнал при первой температуре или ниже ее; и
- образование первого детектируемого сигнала обратимо.
6. Способ по п. 5, отличающийся тем, что:
- часть (c) включает измерение
первого фонового сигнала на уровне или в пределах 1 °C, 2 °C, 3 °C, 4 °C или 5 °C от первой температуры и второго фонового сигнала на уровне или в пределах 1 °C, 2 °C, 3 °C, 4 °C или 5 °C от второй температуры,
обеспечиваемого первым и вторым детектирующими фрагментами в смеси или в контрольной смеси; и
- часть (d) включает определение того, в один или более моментов времени во время или после указанной обработки:
генерируется ли при первой температуре первый детектируемый сигнал, который отличается от первого фонового сигнала, возникающий в результате указанной модификации, указывающий на присутствие первой мишени в образце;
генерируется ли при второй температуре второй детектируемый сигнал, который отличается от второго фонового сигнала, указывающий на присутствие второй мишени в образце.
7. Способ по п. 5 или 6, отличающийся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый олигонуклеотид представляет собой олигонуклеотид типа стебель-петля, содержащий двухцепочечную стеблевую область из гибридизированных нуклеотидов на противоположных цепях, которые связаны неразорванной одноцепочечной петлевой областью негибридизированных нуклеотидов, все или часть которых комплементарны первой мишени; и
- модификация первого олигонуклеотида представляет собой конформационное изменение, возникающее в результате гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида путем комплементарного спаривания оснований.
8. Способ по п. 7, отличающийся тем, что
конформационное изменение представляет собой диссоциацию цепей в двухцепочечной стеблевой области первого олигонуклеотида, возникающую в результате указанной гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида путем комплементарного спаривания оснований.
9. Способ по п. 7 или 8, отличающийся тем, что:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая ниже Tm двухцепочечного дуплекса, образованного в результате указанной гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида, и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- указанный двухцепочечный дуплекс имеет Tm, которая выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm указанного двухцепочечного дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm указанного двухцепочечного дуплекса, стеблевой области первого олигонуклеотида и стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
10. Способ по п. 7 или 8, отличающийся тем, что:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая ниже Tm двухцепочечного дуплекса, образованного в результате указанной гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- указанный двухцепочечный дуплекс имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm указанного двухцепочечного дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm стеблевой области первого олигонуклеотида и стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm указанного двухцепочечного дуплекса и стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
11. Способ по п. 7 или 8, отличающийся тем, что:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая ниже Tm двухцепочечного дуплекса, образованного в результате указанной гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля и ниже Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- указанный двухцепочечный дуплекс имеет Tm ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm указанного двухцепочечного дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm указанного двухцепочечного дуплекса, стеблевой области первого олигонуклеотида и стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
12. Способ по п. 7 или 8, отличающийся тем, что:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая ниже Tm двухцепочечного дуплекса, образованного в результате указанной гибридизации мишени с одноцепочечной петлевой областью первого олигонуклеотида, выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- указанный двухцепочечный дуплекс имеет Tm, которая выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm стеблевой области первого олигонуклеотида и указанного двухцепочечного дуплекса и выше Tm стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm стеблевой области первого олигонуклеотида, указанного двухцепочечного дуплекса и стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- первая температура выше второй температуры.
13. Способ по любому из пп. 7-12, отличающийся тем, что:
- Tm стеблевой области первого олигонуклеотида на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm указанного двухцепочечного дуплекса; и/или
- Tm стеблевой области интактного олигонуклеотида стебель-петля на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm стеблевой области первого олигонуклеотида и/или указанного двухцепочечного дуплекса; и/или
- вторая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и/или
- вторая температура на 1-10 °С, 1-5 °С, 5-10 °С или более чем на 10 °С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля.
14. Способ по п. 5 или 6, отличающийся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый олигонуклеотид представляет собой олигонуклеотид стебель-петля, содержащий:
двухцепочечную стеблевую область из гибридизированных нуклеотидов, противоположные цепи которой связаны одноцепочечной петлевой областью негибридизированных нуклеотидов, все или часть которых комплементарны первой мишени, и вторую одноцепочечную область, продолжающуюся от одной из указанных противоположных цепей в 3'-направлении и оканчивающейся последовательностью, комплементарной области первой мишени, и
молекулу-блокатор, предшествующую указанной последовательности, которая комплементарна области первой мишени;
- смесь дополнительно содержит полимеразу;
- указанная обработка смеси включает:
гибридизацию второй одноцепочечной области с первой мишенью путем комплементарного спаривания оснований;
удлинение второй одноцепочечной области с использованием полимеразы и первой мишени в качестве темплатной последовательности с получением двухцепочечной нуклеиновой кислоты, при этом указанная блокирующая молекула предотвращает удлинение полимеразой первой мишени с использованием стеблевой области первого олигонуклеотида в качестве темплата; и
денатурацию двухцепочечной нуклеиновой кислоты и гибридизацию второй одноцепочечной области, удлиненной полимеразой, с одноцепочечной петлевой областью первого олигонуклеотида путем комплементарного спаривания с получением сигнального дуплекса и, таким образом, обеспечение указанной модификации первого олигонуклеотида, активируя первый детектирующий фрагмент для обеспечения первого детектируемого сигнала.
15. Способ по п. 14, отличающийся тем, что:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая ниже Tm сигнального дуплекса и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm сигнального дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm сигнального дуплекса, стеблевой области первого олигонуклеотида и стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
16. Способ по п. 14, отличающийся тем, что:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая ниже Tm сигнального дуплекса, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm сигнального дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm стеблевой области первого олигонуклеотида и стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm сигнального дуплекса и стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
17. Способ по п. 14, отличающийся тем, что:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая ниже Tm сигнального дуплекса, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля и ниже Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая ниже Tm стеблевой области интактного олигонуклеотида стебель-петля,
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm сигнального дуплекса, стеблевой области первого олигонуклеотида, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm сигнального дуплекса, стеблевой области первого олигонуклеотида и стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
18. Способ по п. 14, отличающийся тем, что:
- стеблевая область первого олигонуклеотида имеет температуру плавления (Tm), которая ниже Tm сигнального дуплекса, выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm стеблевой области первого олигонуклеотида и сигнального дуплекса и выше Tm стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm стеблевой области первого олигонуклеотида, сигнального дуплекса и стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- первая температура выше второй температуры.
19. Способ по любому из пп. 14-18, отличающийся тем, что:
- Tm стеблевой области первого олигонуклеотида на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm сигнального дуплекса; и/или
- Tm стеблевой области интактного олигонуклеотида стебель-петля на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm: стеблевой области первого олигонуклеотида, и/или сигнального дуплекса; и/или
- вторая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm стеблевой области неповрежденного олигонуклеотида стебель-петля; и/или
- вторая температура на 1-10 °С, 1-5 °С, 5-10 °С или более чем на 10 °С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля.
20. Способ по любому из пп. 5-19, отличающийся тем, что
первый детектирующий фрагмент представляет собой флуорофор и модификация увеличивает расстояние от флуорофора до молекулы гасителя.
21. Способ по п. 20, отличающийся тем, что
первый олигонуклеотид содержит молекулу гасителя.
22. Способ по п. 21, отличающийся тем, что
флуорофор и молекула гасителя расположены на противоположных цепях двухцепочечной стеблевой области первого олигонуклеотида.
23. Способ по п. 5 или 6, отличающийся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый олигонуклеотид содержит
первую двухцепочечную область гибридизированных нуклеотидов, первая цепь которой переходит в одноцепочечную область, оканчивающуюся комплементарной последовательностью, способной гибридизоваться с областью первой мишени, при этом первая цепь содержит молекулу-блокатор, предшествующую указанной комплементарной последовательности;
- смесь дополнительно содержит полимеразу;
- указанная обработка смеси включает:
гибридизацию указанной комплементарной последовательности одноцепочечной области с областью первой мишени путем комплементарного спаривания оснований;
удлинение комплементарной последовательности с использованием полимеразы и первой мишени в качестве целевой последовательности для получения второй двухцепочечной области, при этом указанная блокирующая молекула предотвращает удлинение полимеразой первой мишени с использованием первой цепи указанной первой двухцепочечной области в качестве темплата;
денатурацию первой и второй двухцепочечных областей; и
гибридизацию комплементарной последовательности, удлиненной полимеразой, с первой цепью первой двухцепочечной области путем комплементарного спаривания оснований с образованием сигнального дуплекса и, таким образом, обеспечением указанной модификации первого олигонуклеотида, позволяющей первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
24. Способ по п. 23, отличающийся тем, что:
- первая двухцепочечная область имеет температуру плавления (Tm), которая ниже Tm сигнального дуплекса и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm сигнального дуплекса, первой двухцепочечной области, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm сигнального дуплекса, первой двухцепочечной области и стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
25. Способ по п. 23, отличающийся тем, что:
- первая двухцепочечная область имеет температуру плавления (Tm), которая ниже Tm сигнального дуплекса, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm сигнального дуплекса, первой двухцепочечной области, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm первой двухцепочечной области и стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm сигнального дуплекса и стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
26. Способ по п. 23, отличающийся тем, что:
- первая двухцепочечная область имеет температуру плавления (Tm), которая ниже Tm сигнального дуплекса, ниже Tm стеблевой области интактного олигонуклеотида стебель-петля и ниже Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm сигнального дуплекса, первой двухцепочечной области, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm сигнального дуплекса, первой двухцепочечной области и стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
27. Способ по п. 23, отличающийся тем, что:
- первая двухцепочечная область имеет температуру плавления (Tm), которая ниже Tm сигнального дуплекса, выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- сигнальный дуплекс имеет Tm, которая выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm первой двухцепочечной области и сигнального дуплекса и выше Tm стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm первой двухцепочечной области, сигнального дуплекса и стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- первая температура выше второй температуры.
28. Способ по любому из пп. 23-27, отличающийся тем, что:
- Tm первой двухцепочечной области на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm сигнального дуплекса; и/или
- Tm стеблевой области интактного олигонуклеотида стебель-петля на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm первой двухцепочечной области и/или сигнального дуплекса; и/или
- вторая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm стеблевой области неповрежденного олигонуклеотида стебель-петля; и/или
- вторая температура на 1-10 °С, 1-5 °С, 5-10 °С или более чем на 10 °С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля.
29. Способ по любому из пп. 23-28, отличающийся тем, что
первый детектирующий фрагмент представляет собой флуорофор и модификация увеличивает расстояние от флуорофора до молекулы гасителя.
30. Способ по п. 29, отличающийся тем, что
первый олигонуклеотид содержит молекулу гасителя.
31. Способ по п. 30, отличающийся тем, что
флуорофор и молекула гасителя расположены на противоположных цепях первой двухцепочечной области.
32. Способ по п. 5 или 6, отличающийся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- смесь дополнительно содержит:
первый праймер, комплементарный первой последовательности в первой мишени,
второй олигонуклеотид, содержащий компонент, комплементарный второй последовательности в первой мишени, которая отличается от первой последовательности, и область метки, не комплементарную первой мишени,
первую полимеразу, обладающую экзонуклеазной активностью, и
необязательно вторую полимеразу; и
- указанная обработка смеси включает:
подходящие условия для гибридизации первого праймера и второго олигонуклеотида с первой мишенью,
удлинение первого праймера с использованием первой полимеразы и мишени в качестве темплата для отщепления таким образом области метки,
гибридизацию расщепленной области метки с первым олигонуклеотидом путем комплементарного спаривания оснований и
удлинение области метки с использованием первой или второй полимеразы и первого олигонуклеотида в качестве темплата для создания двухцепочечной последовательности, содержащей первый олигонуклеотид, тем самым обеспечивая указанную модификацию первого олигонуклеотида и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
33. Способ по п. 32, отличающийся тем, что:
- двухцепочечная последовательность имеет Tm, которая выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm двухцепочечной последовательности, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm двухцепочечной последовательности и стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
34. Способ по п. 32, отличающийся тем, что:
- двухцепочечная последовательность имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm двухцепочечной последовательности, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm двухцепочечной последовательности и стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
35. Способ по п. 32, отличающийся тем, что:
- двухцепочечная последовательность имеет Tm, которая ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm двухцепочечной последовательности, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm двухцепочечной последовательности и стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
36. Способ по п. 32, отличающийся тем, что:
- двухцепочечная последовательность имеет Tm, которая выше Tm стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm двухцепочечной последовательности и выше Tm стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура ниже Tm двухцепочечной последовательности и стеблевой области интактного олигонуклеотида стебель-петля и выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- первая температура выше второй температуры.
37. Способ по любому из пп. 32-36, отличающийся тем, что:
- Tm стеблевой области интактного олигонуклеотида стебель-петля на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm двухцепочечной последовательности; и/или
- вторая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm стеблевой области неповрежденного олигонуклеотида стебель-петля; и/или
- вторая температура на 1-10 °С, 1-5 °С, 5-10 °С или более чем на 10 °С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля.
38. Способ по любому из пп. 32-37, отличающийся тем, что:
- первый олигонуклеотид содержит флуорофор и молекулу гасителя и
- указанное удлинение области метки увеличивает расстояние между флуорофором и молекулой гасителя.
39. Способ по п. 5 или 6, отличающийся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый олигонуклеотид комплементарен первой области мишени;
- смесь дополнительно содержит дополнительный олигонуклеотид, комплементарный второй области первой мишени, при этом первая и вторая области первой мишени фланкируют друг с другом, но не перекрываются;
- указанная обработка смеси включает:
образование дуплексной структуры, включающей:
(i) первый двухцепочечный компонент путем гибридизации первого олигонуклеотида с мишенью путем комплементарного спаривания оснований и
(ii) второй двухцепочечный компонент путем гибридизации дополнительного олигонуклеотида с мишенью путем комплементарного спаривания оснований,
тем самым сближая первый и последующие олигонуклеотиды и обеспечивая указанную модификацию первого олигонуклеотида, позволяющую первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
40. Способ по п. 39, отличающийся тем, что:
- дуплексная структура имеет Tm, которая ниже Tm стеблевой области интактного олигонуклеотида стебель-петля;
- стеблевая область интактного олигонуклеотида стебель-петля имеет Tm, превышающую Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже Tm дуплексной структуры, стеблевой области интактного олигонуклеотида стебель-петля и стеблевой области разделенного олигонуклеотида стебель-петля;
- вторая температура выше Tm дуплексной структуры и стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля; и
- первая температура ниже второй температуры.
41. Способ по пп. 39 и 40, отличающийся тем, что:
- Tm стеблевой области интактного олигонуклеотида стебель-петля на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm дуплексной структуры; и/или
- вторая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm стеблевой области неповрежденного олигонуклеотида стебель-петля; и/или
- вторая температура на 1-10 °С, 1-5 °С, 5-10 °С или более чем на 10 °С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля.
42. Способ по любому из пп. 39-41, отличающийся тем, что:
- первый детектирующий фрагмент представляет собой флуорофор и дополнительный олигонуклеотид содержит гаситель;
- указанное формирование дуплексной структуры дополнительно сближает флуорофор и гаситель; и
- указанный детектируемый сигнал представляет собой уменьшение флуоресценции, обеспечиваемое первым детектируемым фрагментом.
43. Способ по п. 5 или 6, отличающийся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый обнаруживающий фрагмент представляет собой: наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которой связан первый олигонуклеотид;
- указанная обработка смеси включает
гибридизацию первой мишени с первым олигонуклеотидом, чтобы тем самым вызвать модификацию первого олигонуклеотида, позволяющую первому детектирующему фрагменту обеспечить первый детектируемый сигнал, указывающий на присутствие первой мишени в образце;
при этом первый детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета и/или
(iii) изменение спектра поглощения,
возникающий из первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
44. Способ по п. 5 или 6, отличающийся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый детектирующий фрагмент представляет собой электрохимический агент, с которым связан первый олигонуклеотид;
- указанная обработка смеси включает:
гибридизацию первой мишени с первым олигонуклеотидом, чтобы тем самым вызвать или облегчить модификацию первого олигонуклеотида, позволяющую первому детектирующему фрагменту обеспечить первый детектируемый сигнал, указывающий на присутствие первой мишени в образце;
при этом первый детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающее из-за первого детектируемого фрагмента после указанной модификации первого олигонуклеотида.
45. Способ по п. 44, отличающийся тем, что
электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орасетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
46. Способ по любому из пп. 5-19, 23-28, 32-37 и 39-41, отличающийся тем, что:
- первый детектирующий фрагмент представляет собой наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которой связан первый олигонуклеотид; и
- первый детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета и/или
(iii) изменение спектра поглощения,
возникающий из первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
47. Способ по любому из пп. 5-19, 23-28, 32-37 и 39-41, отличающийся тем, что:
- первый детектирующий фрагмент представляет собой электрохимический агент, с которым связан первый олигонуклеотид; и
- первый детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающее из-за первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
48. Способ по п. 47, отличающийся тем, что
электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орасетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
49. Способ по любому из пп. 43-48, отличающийся тем, что:
- второй детектирующий фрагмент представляет собой наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которой связан интактный олигонуклеотид стебель-петля; и
- второй детектируемый сигнал:
(i) изменение показателя преломления,
(ii) изменение цвета и/или
(iii) изменение спектра поглощения,
возникающие в результате диссоциации указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
50. Способ по любому из пп. 43-48, отличающийся тем, что:
- второй детектирующий фрагмент представляет собой электрохимический агент, с которым связан интактный олигонуклеотид стебель-петля; и
- второй детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающее из-за диссоциации указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
51. Способ по п. 50, отличающийся тем, что
электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орасетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
52. Способ по любому из пп. 20-22, 29-31, 38 и 42, отличающийся тем, что:
- второй детектирующий фрагмент представляет собой флуорофор и
- второй детектируемый сигнал, обеспечивается диссоциацией указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля, что приводит к увеличению расстояния от флуорофора до молекулы гасителя.
53. Способ по п. 52, отличающийся тем, что
флуорофор и молекула гасителя расположены на противоположных цепях двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
54. Способ по любому из пп. 1-4, отличающийся тем, что:
- образование первого детектируемого сигнала необратимо;
- модификация первого олигонуклеотида позволяет первому детектирующему фрагменту обеспечивать первый детектируемый сигнал при первой температуре или ниже ее; и
- первый детектируемый сигнал, обеспечиваемый при первой температуре или ниже, остается детектируемым при второй температуре.
55. Способ по п. 54, отличающийся тем, что:
- часть (c) включает измерение:
(i) первого фонового сигнала на уровне или в пределах 1 °C, 2 °C, 3 °C, 4 °C или 5 °C от первой температуры и второго фонового сигнала на уровне или в пределах 1 °C, 2 °C, 3 °C, 4 °C или 5 C от второй температуры; и/или
(ii) третьего фонового сигнала при третьей температуре;
обеспечиваемых первым и вторым детектирующими фрагментами в смеси или в контрольной смеси; и
- часть (d) включает определение того, в один или более моментов времени во время или после указанной обработки:
(i) генерируется ли при первой температуре первый детектируемый сигнал, который отличается от первого или третьего фонового сигнала, возникающий в результате указанной модификации, при этом
при первой температуре второй детектируемый сигнал не отличается от первого или третьего фонового сигнала и
детектирование разницы между первым детектируемым сигналом и первым или третьим фоновым сигналом указывает на указанную модификацию первого олигонуклеотида и присутствие первой мишени в образце; и
(ii) генерируется ли при второй температуре второй детектируемый сигнал, который отличается от второго фонового сигнала, указывающий на присутствие второй мишени в образце.
56. Способ по п. 55, отличающийся тем, что,
когда в образце присутствует первая мишень, упомянутое определение того, генерируется ли второй детектируемый сигнал при второй температуре, включает компенсацию первого детектируемого сигнал, присутствующего при измерении второго детектируемого сигнала.
57. Способ по п. 55 или 56, отличающийся тем, что:
- генерируется первый сигнал, отличающийся от первого фонового сигнала,
- генерируется второй сигнал, отличающийся от второго фонового сигнала, и
- второй детектируемый сигнал отличается от второго фонового сигнала в большей степени, чем первый детектируемый сигнал отличается от первого фонового сигнала,
тем самым указывая, что вторая мишень присутствует в образце.
58. Способ по п. 57, отличающийся тем, что
первая температура ниже второй температуры, Tm двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля и Tm стеблевой области разделенного олигонуклеотида стебель-петля.
59. Способ по п. 57, отличающийся тем, что
первая температура выше второй температуры, Tm стеблевой области интактного олигонуклеотида стебель-петля и Tm стеблевой области разделенного олигонуклеотида стебель-петля.
60. Способ по п. 55, отличающийся тем, что:
- генерируется первый сигнал, отличающийся от третьего фонового сигнала,
- генерируется второй сигнал, отличающийся от третьего фонового сигнала, и
- второй сигнал отличается от третьего фонового сигнала в большей степени, чем первый сигнал отличается от третьего фонового сигнала,
тем самым указывая, что вторая мишень присутствует в образце.
61. Способ по п. 55, отличающийся тем, что:
- вторая температура выше первой температуры,
- третья температура ниже Tm двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля,
- генерируется первый детектируемый сигнал, отличающийся от третьего фонового сигнала,
- генерируется второй детектируемый сигнал, отличающийся от третьего фонового сигнала, и
- второй детектируемый сигнал отличается от третьего фонового сигнала в большей степени, чем первый сигнал отличается от третьего фонового сигнала,
тем самым указывая, что вторая мишень присутствует в образце.
62. Способ по любому из пп. 55-61, отличающийся тем, что:
- Tm стеблевой области интактного олигонуклеотида стебель-петля выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура ниже второй температуры и ниже Tm стеблевой области разделенного олигонуклеотида стебель-петля; и
- вторая температура выше Tm стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля.
63. Способ по п. 62, отличающийся тем, что:
- Tm стеблевой области интактного олигонуклеотида стебель-петля на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже второй температуры; и/или
- первая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- вторая температура на 1-10 °С, 1-5 °С, 5-10 °С или более чем на 10 °С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- вторая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm стеблевой области неповрежденного олигонуклеотида стебель-петля.
64. Способ по п. 62 или 63, включающий
измерение указанного третьего фонового сигнала, при этом третья температура ниже второй температуры.
65. Способ по п. 64, отличающийся тем, что
третья температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже второй температуры.
66. Способ по любому из пп. 55-61, отличающийся тем, что:
- Tm стеблевой области интактного олигонуклеотида стебель-петля выше Tm стеблевой области разделенного олигонуклеотида стебель-петля;
- первая температура выше второй температуры, выше Tm стеблевой области разделенного олигонуклеотида стебель-петля и выше Tm стеблевой области интактного олигонуклеотида стебель-петля; и
- вторая температура выше Tm стеблевой области разделенного олигонуклеотида стебель-петля и ниже Tm стеблевой области интактного олигонуклеотида стебель-петля.
67. Способ по п. 66, отличающийся тем, что:
- Tm стеблевой области интактного олигонуклеотида стебель-петля на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C выше второй температуры; и/или
- первая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- первая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C выше Tm стеблевой области интактного олигонуклеотида стебель-петля; и/или
- вторая температура на 1-10 °С, 1-5 °С, 5-10 °С или более чем на 10 °С выше Tm стеблевой области разделенного олигонуклеотида стебель-петля; и/или
- вторая температура на 1-10 °C, 1-5 °C, 5-10 °C или более чем на 10 °C ниже Tm стеблевой области неповрежденного олигонуклеотида стебель-петля.
68. Способ по любому из пп. 54-67, отличающийся тем, что:
- первый олигонуклеотид является субстратом для многокомпонентного фермента нуклеиновой кислоты (МНКзима);
- смесь дополнительно содержит
МНКзим, способный расщеплять первый олигонуклеотид, когда первая мишень присутствует в образце; и
- указанная обработка смеси дополнительно включает:
связывание МНКзима с первой мишенью и гибридизацию субстратных плеч МНКзима с первым олигонуклеотидом путем комплементарного спаривания оснований для облегчения расщепления первого олигонуклеотида, тем самым обеспечивая указанную модификацию первого олигонуклеотида и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
69. Способ по п. 68, отличающийся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты; и
- указанная обработка реакционной смеси дополнительно включает
гибридизацию первой мишени с сенсорными плечами МНКзима путем комплементарного спаривания оснований, чтобы тем самым облегчить сборку МНКзима.
70. Способ по любому из пп. 54-67, отличающийся тем, что:
- первый олигонуклеотид является субстратом для аптазима;
- первая мишень является аналитом, белком, соединением или молекулой;
- смесь дополнительно содержит аптазим, содержащий аптамер, способный связываться с первой мишенью; и
- указанная обработка смеси дополнительно включает
связывание аптазима с первой мишенью и первым олигонуклеотидом для облегчения расщепления первого олигонуклеотида, тем самым обеспечивая указанную модификацию первого олигонуклеотида и позволяя первому детектирующему фрагменту генерировать первый детектируемый сигнал.
71. Способ по любому из пп. 54-67, отличающийся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- первый олигонуклеотид содержит последовательность, комплементарную первой мишени,
- смесь дополнительно содержит
праймер, комплементарный области первой мишени и
полимеразу с экзонуклеазной активностью;
- указанная обработка смеси включает:
гибридизацию праймера с первой мишенью путем комплементарного спаривания оснований,
гибридизацию первого олигонуклеотида с первой мишенью путем комплементарного спаривания оснований,
удлинение праймера с использованием полимеразы и первой мишени в качестве темплатной последовательности, чтобы, таким образом, расщепить первый олигонуклеотид и обеспечить указанную модификацию первого олигонуклеотида, позволяющую первому детектирующему фрагменту генерировать первый детектируемый сигнал.
72. Способ по любому из пп. 54-67, отличающийся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- смесь дополнительно содержит
рестрикционную эндонуклеазу, способную расщеплять двухцепочечный дуплекс, содержащий первую мишень; и
- указанная обработка смеси включает:
гибридизацию первого олигонуклеотида с первой мишенью путем комплементарного спаривания оснований с образованием таким образом двухцепочечного дуплекса,
расщепление дуплекса с использованием рестрикционной эндонуклеазы, чтобы тем самым обеспечить указанную модификацию первого олигонуклеотида и сделать возможным генерирование первым детектирующим фрагментом первого детектируемого сигнала.
73. Способ по п. 72, отличающийся тем, что
рестрикционная эндонуклеаза представляет собой разрывающую эндонуклеазу, способную связываться с цепью указанного двухцепочечного дуплекса и расщеплять ее, и указанная цепь содержит всю или часть первого олигонуклеотида.
74. Способ по любому из пп. 54-67, отличающийся тем, что:
- смесь дополнительно содержит ДНКзим или рибозим, требующий кофактор для каталитической активности;
- указанная обработка смеси включает применение условий, подходящих для:
связывания кофактора с ДНКзимом или рибозимом, чтобы сделать его каталитически активным,
гибридизации ДНКзима или рибозима с первым олигонуклеотидом путем комплементарного спаривания оснований, и
каталитической активности ДНКзима или рибозима в расщеплении таким образом первого олигонуклеотида и, таким образом, в обеспечении указанной модификации первого олигонуклеотида, позволяющей первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
при этом первая мишень представляет собой кофактор.
75. Способ по п. 74, отличающийся тем, что кофактор представляет собой ион металла или ион металла, выбранный из Mg2+, Mn2+, Ca2+, Pb2+.
76. Способ по любому из пп. 54-75, отличающийся тем, что
первый детектирующий фрагмент представляет собой флуорофор и модификация первого олигонуклеотида увеличивает расстояние от флуорофора до молекулы гасителя.
77. Способ по п. 76, отличающийся тем, что
первый олигонуклеотид содержит молекулу гасителя.
78. Способ по п. 76 или 77, отличающийся тем, что:
- второй детектирующий фрагмент представляет собой флуорофор и
- второй детектируемый сигнал обеспечивается диссоциацией указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля, что приводит к увеличению расстояния от флуорофора до молекулы гасителя.
79. Способ по п. 78, отличающийся тем, что
флуорофор и молекула гасителя расположены на противоположных цепях двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
80. Способ по любому из пп. 54-79, отличающийся тем, что:
- первый обнаруживающий фрагмент представляет собой наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которой связан первый олигонуклеотид; и
- первый детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета и/или
(iii) изменение спектра поглощения,
возникающий из первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
81. Способ по любому из пп. 54-79, отличающийся тем, что:
- первый детектирующий фрагмент представляет собой электрохимический агент, с которым связан первый олигонуклеотид; и
- первый детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающее из-за первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
82. Способ по п. 81, отличающийся тем, что
электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орасетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
83. Способ по любому из пп. 80-82, отличающийся тем, что:
- второй детектирующий фрагмент представляет собой наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которой связан интактный олигонуклеотид стебель-петля; и
- второй детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета и/или
(iii) изменение спектра поглощения,
возникающие в результате диссоциации указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
84. Способ по любому из пп. 80-82, отличающийся тем, что:
- второй детектирующий фрагмент представляет собой электрохимический агент, с которым связан интактный олигонуклеотид стебель-петля; и
- второй детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающее из-за диссоциации указанных цепей двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
85. Способ по п. 84, отличающийся тем, что
электрохимический агент выбран из любой одной или более наночастиц, метиленового синего, толуолового синего, орасетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
86. Способ по любому из пп. 1-85, отличающийся тем, что интактный олигонуклеотид стебель-петля не гибридизуется со второй мишенью во время указанного расщепления одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля первым ферментом.
87. Способ по любому из пп. 1-86, отличающийся тем, что:
- первый фермент представляет собой первый МНКзим и
- указанная обработка смеси включает
связывание первого МНКзима со второй мишенью и гибридизацию субстратных плеч указанного первого МНКзима с петлевой областью интактного олигонуклеотида стебель-петля с расщеплением, таким образом, одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля и обеспечением разделенного олигонуклеотида стебель-петля.
88. Способ по п. 87, отличающийся тем, что:
- вторая мишень представляет собой последовательность нуклеиновой кислоты и
- указанная обработка смеси дополнительно включает
гибридизацию второй мишени с сенсорными плечами первого МНКзима путем комплементарного спаривания оснований, чтобы тем самым облегчить сборку первого МНКзима.
89. Способ по любому из пп. 1-86, отличающийся тем, что:
- вторая мишень является аналитом, белком, соединением или молекулой;
- первый фермент представляет собой аптазим, содержащий аптамер, способный связываться со второй мишенью; и
- связывание второй мишени с аптамером способно сделать первый фермент каталитически активным.
90. Способ по п. 89, отличающийся тем, что
первый фермент является любым из следующих: апта-ДНКзим, апта-рибозим, апта-МНКзим.
91. Способ по любому из пп. 1-86, отличающийся тем, что:
- вторая мишень является аналитом, белком, соединением или молекулой;
- первый олигонуклеотид является субстратом для аптазима;
- первый фермент представляет собой аптазим, содержащий область аптамера, способную связываться со второй мишенью, и область нуклеиновой кислоты с ферментативной активностью, способную расщеплять один или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля;
- указанная обработка смеси дополнительно включает
связывание второй мишени с аптамерной областью аптазима для облегчения активации каталитической активности области нуклеиновой кислоты с ферментативной активностью и гибридизацию интактного олигонуклеотида стебель-петля с активной областью фермента нуклеиновой кислоты для расщепления, таким образом, одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля.
92. Способ по любому из пп. 1-85, отличающийся тем, что:
- вторая мишень представляет собой последовательность нуклеиновой кислоты;
- первый фермент представляет собой первую рестрикционную эндонуклеазу; и
- указанная обработка смеси включает
применение условий, подходящих для гибридизации второй мишени с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований с образованием двухцепочечной последовательности для связывания и расщепления с одним или более негибридизированными нуклеотидами одноцепочечной петлевой области первой рестрикционной эндонуклеазой, тем самым образуя разделенный олигонуклеотид стебель-петля.
93. Способ по п. 92, отличающийся тем, что
первая рестрикционная эндонуклеаза представляет собой эндонуклеазу первого разрыва, способную связываться и расщеплять цепь указанной двухцепочечной последовательности для первой рестрикционной эндонуклеазы и указанная цепь включает всю или часть одноцепочечной петлевой области интактного олигонуклеотида стебель-петля.
94. Способ по любому из пп. 1-85, отличающийся тем, что:
- первый фермент включает полимеразу с экзонуклеазной активностью,
- указанная обработка смеси включает применение условий, подходящих для:
гибридизации второй мишени с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований с образованием первой двухцепочечной последовательности, содержащей область второй мишени,
гибридизации первого праймера олигонуклеотида со второй мишенью с образованием второй двухцепочечной последовательности, расположенной выше относительно первой двухцепочечной последовательности, содержащей область второй мишени,
удлинения праймера с использованием полимеразы с экзонуклеазной активностью и использованием второй мишени в качестве темплатной последовательности,
при этом первая полимераза, обладающая экзонуклеазной активностью, расщепляет одноцепочечную петлевую область первой двухцепочечной последовательности и, таким образом, образует разделенный олигонуклеотид стебель-петля.
95. Способ по любому из пп. 1-85, отличающийся тем, что:
- первый фермент представляет собой экзонуклеазу и
- указанная обработка смеси включает применение условий, подходящих для:
гибридизации второй мишени с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований с образованием первой двухцепочечной последовательности, содержащей область второй мишени,
ассоциации первого фермента, обладающего экзонуклеазной активностью, с двухцепочечной последовательностью, содержащей вторую мишень, и
каталитической активности первого фермента, включающей экзонуклеазную активность, позволяющую ему расщеплять одноцепочечную петлевую область первой двухцепочечной последовательности, содержащей вторую мишень, и, таким образом, образовывать разделенный олигонуклеотид стебель-петля.
96. Способ по любому из пп. 1-85, отличающийся тем, что:
- первый фермент представляет собой ДНКзим или рибозим, требующий кофактора для каталитической активности, и указанная обработка смеси включает применение условий, подходящих для:
- связывания кофактора с первым ферментом, чтобы сделать его каталитически активным,
- гибридизации ДНКзима или рибозима с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований,
- каталитической активности ДНКзима или рибозима по расщеплению одного или более негибридизированных нуклеотидов одноцепочечной области петли интактного олигонуклеотида стебель-петля и, таким образом, образованию разделенного олигонуклеотида стебель-петля,
при этом второй мишенью является кофактор.
97. Способ по п. 96, отличающийся тем, что кофактор представляет собой ион металла или ион металла, выбранный из Mg2+, Mn2+, Ca2+, Pb2+.
98. Способ по любому из пп. 1-97, отличающийся тем, что:
- первая мишень отличается от второй мишени; и/или
- первый олигонуклеотид содержит или состоит из последовательности, которая не находится в пределах одноцепочечной петли интактного олигонуклеотида стебель-петля.
99. Способ по любому из пп. 1-98, отличающийся тем, что
первый фермент не расщепляет вторую мишень.
100. Способ по любому из пп. 1-71, 74-91 или 94-99, отличающийся тем, что
ни один из указанных ферментов не расщепляет первую мишень и/или вторую мишень.
101. Способ по любому из пп. 1-100, отличающийся тем, что
первая температура отличается от второй температуры более чем на: 1 °C, 2 °C, 3 °C, 4 °C, 5 °C, 6 °C, 7 °C, 8 °C, 9 °C, 10 °C, 11 °C, 12 °C, 13 °C, 14 °C, 15 °C, 16 °C, 17 °C, 18 °C, 19 °C, 20 °C, 25 °C, 30 °C, 35 °C, 40 °C, 45 °C, 50 °C, 55 °C или 60 °C.
102. Способ по любому из пп. 1-101, отличающийся тем, что указанное определение включает детектирование первого детектируемого сигнала и/или любого указанного фонового сигнала (сигналов):
- в один или более моментов времени во время указанной обработки; или
- в один или более моментов времени во время указанной обработки и в один или более моментов времени после указанной обработки.
103. Способ по любому из пп. 1-101, отличающийся тем, что указанное определение включает детектирование первого детектируемого сигнала и/или любого указанного фонового сигнала (сигналов):
- в один или более моментов времени после указанной обработки.
104. Способ по любому из пп. 1-101, отличающийся тем, что указанное определение включает детектирование второго детектируемого сигнала и/или любого указанного фонового сигнала (сигналов):
- в один или более моментов времени во время указанной обработки; или
- в один или более моментов времени во время указанной обработки и в один или более моментов времени после указанной обработки.
105. Способ по любому из пп. 1-101, отличающийся тем, что указанное определение включает детектирование второго детектируемого сигнала и/или любого указанного фонового сигнала (сигналов):
- в один или более моментов времени после указанной обработки.
106. Способ по любому из пп. 1-105, отличающийся тем, что
указанное определение наличия или отсутствия первой и второй мишеней включает анализ кривой плавления.
107. Способ по п. 6, отличающийся тем, что
указанное определение наличия или отсутствия первой и второй мишеней включает анализ кривой плавления, включающей первый и второй детектируемые сигналы и, необязательно, первый и второй фоновые сигналы.
108. Способ по п. 55, отличающийся тем, что:
- указанное определение наличия или отсутствия первой и второй мишеней включает анализ кривой плавления, включающей первый и второй детектируемые сигналы и, необязательно, первый и второй фоновые сигналы; или
- первый и второй детектируемые сигналы и, необязательно, третий фоновый сигнал.
109. Способ по любому из пп. 1-108, отличающийся тем, что
первая мишень и/или вторая мишень представляет собой ампликон нуклеиновой кислоты.
110. Способ по любому из пп. 1-109, отличающийся тем, что:
- первая мишень представляет собой нуклеиновую кислоту и/или вторая мишень представляет собой нуклеиновую кислоту, и
- смесь дополнительно содержит реагенты для амплификации указанной первой и/или второй мишени,
- указанная обработка смеси дополнительно включает условия, подходящие для проведения амплификации первой и/или второй мишени.
111. Способ по п. 110, отличающийся тем, что
амплификация представляет собой одну или более из полимеразной цепной реакции (ПЦР), амплификации замещения цепи (SDA), реакции амплификации никирующего фермента (NEAR), геликазозависимой амплификации (HDA), рекомбиназной полимеразной амплификации (RPA), петлевой изотермической амплификации (LAMP), амплификации по типу катящегося кольца (RCA), амплификации, опосредованной транскрипцией (TMA), самоподдерживающейся репликации последовательности (3SR), амплификации на основе последовательности нуклеиновой кислоты (NASBA), лигазной цепной реакции (LCR) или способа амплификации с разветвлением (RAM) и/или полимеразной цепной реакции с обратной транскрипцией (ОТ-ПЦР).
112. Способ по п. 110 или 111, отличающийся тем, что указанное определение:
- происходит до указанной амплификации или в пределах 1, 2, 3, 4 или 5 циклов от начала указанной амплификации; и/или
- происходит после завершения указанной амплификации.
113. Способ по любому из пп. 110-112, отличающийся тем, что указанное определение:
- происходит до указанной амплификации или в пределах 1, 2, 3, 4 или 5 минут от начала указанной амплификации; и/или
- происходит после завершения указанной амплификации.
114. Способ по любому из пп. 110-113, отличающийся тем, что указанное определение происходит:
- в первый момент времени до указанной амплификации; и
- во второй момент времени после завершения указанной амплификации.
115. Способ по любому из пп. 110-114, отличающийся тем, что:
- способ амплификации представляет собой полимеразную цепную реакцию (ПЦР); и
- указанное определение происходит в нескольких циклах, необязательно в каждом цикле.
116. Способ по п. 110 или 111, дополнительно включающий нормализацию:
- первого детектируемого сигнала при первой температуре, измеренного в момент времени во время или после указанной амплификации с использованием положительного контрольного сигнала, генерируемого при первой температуре до указанной амплификации и/или до указанной обработки реакции; и/или
- второго детектируемого сигнала при второй температуре, измеренного в момент времени во время или после указанной амплификации с использованием положительного контрольного сигнала, генерируемого при второй температуре до указанной амплификации и/или до указанной обработки реакции.
117. Способ по п. 110 или 111, дополнительно включающий нормализацию:
- первого детектируемого сигнала с применением детектируемого сигнала, генерируемого интактным олигонуклеотидом стебель-петля при первой температуре до указанной амплификации и/или до указанной обработки реакции; и/или
- второго детектируемого сигнала с использованием детектируемого сигнала, генерируемого интактным олигонуклеотидом стебель-петля при дополнительной температуре до указанной амплификации и/или до указанной обработки реакции;
при этом дополнительная температура превышает Tm интактного олигонуклеотида стебель-петля.
118. Способ по любому из пп. 1-117, дополнительно включающий
генерирование сигнала положительного контроля первой мишени с применением известной концентрации первой мишени и/или известной концентрации первого олигонуклеотида после указанной модификации.
119. Способ по любому из пп. 1-118, дополнительно включающий
генерирование положительного контрольного сигнала для первой мишени путем повторения способа на отдельном контрольном образце, содержащем указанную первую мишень.
120. Способ по п. 119, отличающийся тем, что
отдельный контрольный образец, содержащий первую мишень, содержит известную концентрацию первой мишени.
121. Способ по п. 119 или 120, отличающийся тем, что
отдельный контрольный образец, содержащий первую мишень, дополнительно содержит вторую мишень.
122. Способ по любому из пп. 1-121, дополнительно включающий
генерирование сигнала положительного контроля второй мишени с применением известной концентрации второй мишени и/или известной концентрации олигонуклеотида стебель-петля после указанной модификации.
123. Способ по любому из пп. 1-122, дополнительно включающий
генерирование положительного контрольного сигнала для второй мишени путем повторения указанного способа на отдельном контрольном образце, содержащем вторую мишень.
124. Способ по п. 123, отличающийся тем, что
контрольный образец, содержащий вторую мишень, содержит известную концентрацию второй мишени.
125. Способ по п. 123 или 124, отличающийся тем, что
указанный контрольный образец, содержащий вторую мишень, дополнительно содержит указанную первую мишень.
126. Способ по любому из пп. 1-125, дополнительно включающий
генерирование комбинированного положительного контрольного сигнала путем повторения указанного способа на отдельном контрольном образце, содержащем первую и вторую мишени.
127. Способ по п. 126, отличающийся тем, что
комбинированный контрольный образец содержит известную концентрацию первой мишени и/или известную концентрацию второй мишени.
128. Способ по любому из пп. 116-127, дополнительно включающий
нормализацию первого детектируемого сигнала и/или второго детектируемого сигнала с применением любого упомянутого положительного контрольного сигнала.
129. Способ по любому из пп. 116-128, дополнительно включающий
оценку уровней сигнала отрицательного контроля путем повторения способа по любому из пп. 1-115 на отдельном образце отрицательного контроля, который не содержит:
(i) указанной первой мишени; или
(ii) указанной второй мишени; или
(iii) указанной первой мишени или указанной второй мишени.
130. Способ по п. 129, дополнительно включающий
нормализацию первого детектируемого сигнала и/или второго детектируемого сигнала с применением упомянутого отрицательного контрольного сигнала.
131. Способ по любому из пп. 116-130, отличающийся тем, что
любой упомянутый контрольный сигнал является флуоресцентным контрольным сигналом.
132. Способ по любому из пп. 1-131, дополнительно включающий сравнение первого и/или второго детектируемых сигналов с пороговым значением, причем
порог генерируется с применением детектируемых сигналов, полученных из серии образцов, протестированных в соответствии со способом по любому из пп. 1-115 и включающих любой один или более из:
(i) контроль без темплата и первую мишень,
(ii) контроль без темплата и вторую мишень,
(iii) контроль без темплата, первую мишень и вторую мишень,
чтобы таким образом определить упомянутое присутствие или отсутствие первой и второй мишеней в образце.
133. Способ по п. 132, отличающийся тем, что
серию образцов тестируют с применением известной концентрации первого олигонуклеотида и/или известной концентрации интактного олигонуклеотида стебель-петля.
134. Способ по любому из пп. 1-133, отличающийся тем, что
образец представляет собой биологический образец, полученный от субъекта.
135. Способ по любому из пп. 1-133, отличающийся тем, что способ осуществляют in vitro.
136. Способ по любому из пп. 1-133, отличающийся тем, что способ осуществляют ex vivo.
137. Способ по любому из пп. 1-136, отличающийся тем, что
первая и вторая детектируемые фрагменты излучают в одной и той же цветовой области видимого спектра.
138. Композиция для определения наличия или отсутствия первой и второй мишеней в образце, содержащая:
- первый олигонуклеотид для детектирования первой мишени, причем первая мишень представляет собой нуклеиновую кислоту и комплементарна по меньшей мере части первого олигонуклеотида, и
- первый детектирующий фрагмент, при этом
первый детектирующий фрагмент способен генерировать первый детектируемый сигнал при модификации первого олигонуклеотида и
модификация индуцируется гибридизацией первой мишени с первым олигонуклеотидом путем комплементарного спаривания оснований;
- интактный олигонуклеотид стебель-петля для детектирования второй мишени, содержащий двухцепочечную стеблевую область из гибридизированных нуклеотидов, противоположные цепи которой связаны неразорванной одноцепочечной петлевой областью из негибридизированных нуклеотидов, при этом по меньшей мере одна цепь двухцепочечной стеблевой области содержит второй детектирующий фрагмент; и
первый фермент способен расщеплять один или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля только тогда, когда в образце присутствует вторая мишень, с тем чтобы разрушить одноцепочечную петлевую область и получить разделенный олигонуклеотид стебель-петля;
при этом
второй детектирующий фрагмент способен генерировать второй детектируемый сигнал при диссоциации двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля и
первый и второй детектирующие фрагменты способны генерировать детектируемые сигналы, которые нельзя различить при одной температуре с использованием одного типа детектора.
139. Композиция по п. 138, отличающаяся тем, что
область первого олигонуклеотида, комплементарная первой мишени, имеет другую температуру плавления (Tm) по сравнению с каждой цепью двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля.
140. Композиция по п. 138 или 139, отличающаяся тем, что первый олигонуклеотид отличается по последовательности от:
каждой цепи двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля; и
одноцепочечной петлевой области интактного олигонуклеотида стебель-петля.
141. Композиция по любому из пп. 138-140, отличающаяся тем, что
первый олигонуклеотид представляет собой олигонуклеотид стебель-петля, содержащий двухцепочечную стеблевую область из гибридизированных нуклеотидов на противоположных цепях, которые связаны неразорванной одноцепочечной петлевой областью негибридизированных нуклеотидов, все или часть которых комплементарны первой мишени.
142. Композиция по п. 141, отличающаяся тем, что
первая мишень гибридизуется с первым олигонуклеотидом за счет комплементарного спаривания оснований, вызывающего диссоциацию цепей в двухцепочечной стеблевой области первого олигонуклеотида, что позволяет первой детектирующей молекуле обеспечивать первый детектируемый сигнал.
143. Композиция по любому из пп. 138-140, отличающаяся тем, что
первый олигонуклеотид представляет собой олигонуклеотид стебель-петля, содержащий:
двухцепочечную стеблевую область гибридизированных нуклеотидов, противоположные цепи которой связаны одноцепочечной петлевой областью негибридизированных нуклеотидов, все или часть которых комплементарны первой мишени, и
вторую одноцепочечную область, отходящую от одной из указанных противоположных цепей в 3'-направлении и оканчивающуюся последовательностью, комплементарной области первой мишени, и
блокирующую молекулу, предшествующую указанной последовательности, которая комплементарна области первой мишени.
144. Композиция по п. 143, отличающаяся тем, что:
- первая мишень гибридизуется со второй ее одноцепочечной областью за счет комплементарного спаривания оснований;
композиция дополнительно содержит полимеразу, способную удлинять вторую одноцепочечную область, используя первую мишень в качестве темплатной последовательности, для получения двухцепочечной нуклеиновой кислоты, при этом указанная блокирующая молекула способна предотвращать удлинение полимеразой первой мишени, используя указанную одну противоположную цепь в качестве темплата, и
при денатурации двухцепочечной нуклеиновой кислоты вторая одноцепочечная область, удлиненная полимеразой, способна гибридизоваться с одноцепочечной петлевой областью первого олигонуклеотида за счет комплементарного спаривания оснований с образованием сигнального дуплекса и, таким образом, активируя первый детектирующий фрагмент для обеспечения первого детектируемого сигнала.
145. Композиция по любому из пп. 141-144, отличающаяся тем, что
первый детектирующий фрагмент представляет собой флуорофор.
146. Композиция по п. 145, отличающаяся тем, что
первый олигонуклеотид содержит молекулу гасителя, при этом флуорофор и молекула гасителя расположены на противоположных цепях двухцепочечной стеблевой области первого олигонуклеотида.
147. Композиция по любому из пп. 138-140, отличающаяся тем, что:
- первый олигонуклеотид содержит
первую двухцепочечную область гибридизированных нуклеотидов, первая цепь которой переходит в одноцепочечную область, оканчивающуюся комплементарной последовательностью, способной гибридизоваться с областью первой мишени, при этом первая цепь содержит молекулу-блокатор, предшествующую указанной комплементарной последовательности;
- композиция дополнительно содержит полимеразу.
148. Композиция по п. 147, отличающаяся тем, что:
область первой мишени гибридизуется с указанной комплементарной последовательностью одноцепочечной области путем комплементарного спаривания оснований; и
композиция дополнительно содержит полимеразу, способную удлинять комплементарную последовательность, используя первую мишень в качестве темплатной последовательности, для получения второй двухцепочечной области, при этом указанная блокирующая молекула предотвращает удлинение полимеразой первой мишени с использованием одноцепочечной области в качестве темплата; и,
когда первая и вторая двухцепочечные области денатурированы, комплементарная последовательность, удлиненная полимеразой, способна гибридизоваться с первой цепью первой двухцепочечной области за счет комплементарного спаривания оснований с образованием сигнального дуплекса и, таким образом, активируя первый детектирующий фрагмент для обеспечения первого детектируемого сигнала.
149. Композиция по п. 147 или 148, отличающаяся тем, что
первый детектирующий фрагмент представляет собой флуорофор и модификация увеличивает его расстояние от молекулы гасителя.
150. Композиция по п. 149, отличающаяся тем, что
первый олигонуклеотид содержит молекулу гасителя, при этом флуорофор и молекула гасителя расположены на противоположных цепях первой двухцепочечной области.
151. Композиция по любому из пп. 138-140, отличающаяся тем, что:
- первый олигонуклеотид комплементарен первой области мишени;
- композиция дополнительно содержит дополнительный олигонуклеотид, комплементарный второй области первой мишени, при этом первая и вторая области первой мишени фланкируют друг с другом, но не перекрываются, и каждая способна гибридизоваться с первой мишенью с образованием дуплексной структуры, включающей:
(i) первый двухцепочечный компонент путем гибридизации первого олигонуклеотида с мишенью или путем комплементарного спаривания оснований и
(ii) второй двухцепочечный компонент путем гибридизации дополнительного олигонуклеотида с мишенью путем комплементарного спаривания оснований,
тем самым сближая первый и дополнительные олигонуклеотиды и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
152. Композиция по п. 151, отличающаяся тем, что:
- первый детектирующий фрагмент представляет собой флуорофор и дополнительный олигонуклеотид содержит гаситель;
- указанное формирование дуплексной структуры дополнительно сближает флуорофор и гаситель; и
- указанный детектируемый сигнал представляет собой уменьшение флуоресценции, обеспечиваемое первым детектируемым фрагментом.
153. Композиция по любому из пп. 138-140, отличающаяся тем, что:
- первый олигонуклеотид гибридизуется с первой мишенью за счет комплементарного спаривания оснований,
- композиция дополнительно содержит:
праймер, гибридизованный с областью первой мишени путем комплементарного спаривания оснований, и
полимеразу с экзонуклеазной активностью, способную удлинять праймер с использованием первой мишени в качестве темплатной последовательности, тем самым расщепляя первый олигонуклеотид и модифицируя первый олигонуклеотид, позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
154. Композиция по любому из пп. 138-140, отличающаяся тем, что:
- первая мишень гибридизуется с первым олигонуклеотидом путем комплементарного спаривания оснований с образованием таким образом двухцепочечного дуплекса,
- композиция дополнительно содержит рестрикционную эндонуклеазу, способную расщеплять двухцепочечный дуплекс, содержащий первую мишень, тем самым модифицируя первый олигонуклеотид и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
155. Композиция по п. 154, отличающаяся тем, что
рестрикционная эндонуклеаза представляет собой никующую эндонуклеазу, способную связываться с цепью указанного двухцепочечного дуплекса и расщеплять ее, и указанная цепь содержит первый олигонуклеотид.
156. Композиция по любому из пп. 153-155, отличающаяся тем, что
первый детектирующий фрагмент представляет собой флуорофор и указанное изменение первого олигонуклеотида увеличивает расстояние от флуорофора до молекулы гасителя.
157. Композиция по п. 156, отличающаяся тем, что
ервый олигонуклеотид содержит молекулу гасителя.
158. Композиция для определения наличия или отсутствия первой и второй мишеней в образце, содержащая:
- первый олигонуклеотид для детектирования первой мишени, содержащий первый детектирующий фрагмент, при этом
первый детектирующий фрагмент способен генерировать первый детектируемый сигнал при модификации первого олигонуклеотида и
модификация индуцируется первой мишенью;
- интактный олигонуклеотид стебель-петля для детектирования второй мишени, содержащий двухцепочечную стеблевую область из гибридизированных нуклеотидов, противоположные цепи которой связаны неразорванной одноцепочечной петлевой областью из негибридизированных нуклеотидов, при этом по меньшей мере одна цепь двухцепочечной стеблевой области содержит второй детектирующий фрагмент и
первый фермент способен расщеплять один или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля только тогда, когда в образце присутствует вторая мишень, с тем чтобы разрушить одноцепочечную петлевую область и получить разделенный олигонуклеотид стебель-петля;
при этом
второй детектирующий фрагмент способен генерировать второй детектируемый сигнал при диссоциации двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля и
первый и второй детектирующие фрагменты способны генерировать детектируемые сигналы, которые нельзя различить при одной температуре с использованием одного типа детектора.
159. Композиция по п. 158, отличающаяся тем, что первый олигонуклеотид отличается по последовательности от:
каждой цепи двухцепочечной стеблевой области интактного олигонуклеотида стебель-петля; и
одноцепочечной петлевой области интактного олигонуклеотида стебель-петля.
160. Композиция по п. 158 или 159, отличающаяся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты;
- композиция дополнительно содержит:
первый праймер, комплементарный первой последовательности в первой мишени,
второй олигонуклеотид, содержащий компонент, комплементарный второй последовательности в первой мишени, которая отличается от первой последовательности, и область метки, не комплементарную первой мишени,
первую полимеразу, обладающую экзонуклеазной активностью, и
необязательно вторую полимеразу.
161. Композиция по п. 160, отличающаяся тем, что
каждый из первого праймера и второго олигонуклеотида гибридизуется с первой мишенью путем комплементарного спаривания оснований,
первая полимераза способна удлинять первый праймер, используя мишень в качестве темплата, тем самым отщепляя область метки, позволяя отщепленной области метки гибридизоваться с первым олигонуклеотидом путем комплементарного спаривания оснований, и
первая полимераза или необязательная вторая полимераза способны удлинять область метки с использованием первого олигонуклеотида в качестве темплата для создания двухцепочечной последовательности, содержащей первый олигонуклеотид, тем самым модифицируя первый олигонуклеотид и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
162. Композиция по п. 160 или 161, отличающаяся тем, что
первый олигонуклеотид содержит флуорофор и молекулу гасителя.
163. Композиция по п. 162, отличающаяся тем, что:
- первый олигонуклеотид содержит флуорофор и молекулу гасителя и
- указанное удлинение области метки увеличивает расстояние между флуорофором и молекулой гасителя.
164. Композиция по п. 158 или 159, отличающаяся тем, что:
- первая мишень является кофактором каталитической активности ферментов;
- композиция дополнительно содержит ДНКзим или рибозим, требующий кофактор для каталитической активности, и
- ДНКазим или рибозим способен связываться с первой мишенью и гибридизоваться с первым олигонуклеотидом путем комплементарного спаривания оснований, тем самым расщепляя и модифицируя первый олигонуклеотид, позволяя первому детектирующему фрагменту генерировать первый детектируемый сигнал.
165. Композиция по п. 164, отличающаяся тем, что кофактор представляет собой ион металла или ион металла, выбранный из Mg2+, Mn2+, Ca2+, Pb2+.
166. Композиция по п. 158 или 159, отличающаяся тем, что:
- первый олигонуклеотид является субстратом для многокомпонентного фермента нуклеиновой кислоты (МНКзима);
- композиция дополнительно содержит МНКзим, способный расщеплять первый олигонуклеотид, когда первая мишень присутствует в образце; и
при этом МНКзим способен связываться с первой мишенью и гибридизоваться с первым олигонуклеотидом путем комплементарного спаривания оснований через его субстратные плечи и указанная гибридизация облегчает расщепление первого олигонуклеотида, тем самым модифицируя его и позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
167. Композиция по п. 166, отличающаяся тем, что:
- первая мишень представляет собой последовательность нуклеиновой кислоты; и
- первая мишень гибридизуется с сенсорными плечами МНКзима путем комплементарного спаривания оснований, чтобы тем самым облегчить сборку МНКзима.
168. Композиция по п. 158 или 159, отличающаяся тем, что:
- первая мишень является аналитом, белком, соединением или молекулой;
- первый олигонуклеотид является субстратом для аптазима; и
- композиция дополнительно содержит аптазим, содержащий аптамерную область, способную связываться с первой мишенью, и область нуклеиновой кислоты с ферментативной активностью, способную расщеплять первый олигонуклеотид и тем самым модифицировать его, позволяя первому детектирующему фрагменту обеспечивать первый детектируемый сигнал.
169. Композиция по п. 168, отличающаяся тем, что
первая мишень связывается с аптамерной областью аптазима и первый олигонуклеотид гибридизуется с активной областью, имеющей ферментную активность по отношению к нуклеиновой кислоте за счет комплементарного спаривания оснований, облегчающего расщепление первого олигонуклеотида и, таким образом, модифицируя его, что позволяет первому детектирующему фрагменту обеспечить первый детектируемый сигнал.
170. Композиция по любому из пп. 166-169, отличающаяся тем, что
первый детектирующий фрагмент представляет собой флуорофор и указанное изменение первого олигонуклеотида увеличивает расстояние от флуорофора до молекулы гасителя.
171. Композиция по п. 170, отличающаяся тем, что
первый олигонуклеотид содержит молекулу гасителя.
172. Композиция по любому из пп. 138-144, 147, 148, 151, 153-155, 158-161 и 164-169, отличающаяся тем, что:
- первый обнаруживающий фрагмент представляет собой наночастицу, наночастицу металла, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которым связан первый олигонуклеотид; и
- первый детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета и/или
(iii) изменение спектра поглощения,
возникающий из первого детектирующего фрагмента после указанной модификации первого олигонуклеотида.
173. Композиция по п. 172, отличающаяся тем, что:
- первый детектирующий фрагмент представляет собой электрохимический агент, с которым связан первый олигонуклеотид;
- первый детектируемый сигнал представляет собой изменение электрохимического сигнала.
174. Композиция по п. 173, отличающаяся тем, что
электрохимический агент выбирают из любой одной или более наночастиц, метиленового синего, толуолового синего, орасетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
175. Композиция по любому из пп. 172-174, отличающаяся тем, что:
- второй детектирующий фрагмент представляет собой наночастицу, металлическую наночастицу, наночастицу благородного металла, наночастицу щелочного металла, наночастицу золота или наночастицу серебра; с которой связана по меньшей мере одна цепь двухцепочечной области второго олигонуклеотида, и
- второй детектируемый сигнал представляет собой:
(i) изменение показателя преломления,
(ii) изменение цвета и/или
(iii) изменение спектра поглощения,
возникающие в результате указанной диссоциации двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
176. Композиция по любому из пп. 172-174, отличающаяся тем, что:
- второй детектирующий фрагмент представляет собой электрохимический агент, с которым связан второй олигонуклеотид; и
- второй детектируемый сигнал представляет собой изменение электрохимического сигнала, возникающее при указанной диссоциации двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля.
177. Композиция по п. 176, отличающаяся тем, что
электрохимический агент выбирают из любой одной или более наночастиц, метиленового синего, толуолового синего, орасетового синего, Hoechst 33258, [Ru(phen)3]2+, ферроцена и/или дауномицина.
178. Композиция по любому из пп. 145, 146, 149, 150, 152, 156, 157, 162, 163, 170 и 171, отличающаяся тем, что:
- второй детектирующий фрагмент представляет собой флуорофор и
- второй детектируемый сигнал, обеспечиваемый указанным вторым детектирующим фрагментом при диссоциации двухцепочечной стеблевой области разделенного олигонуклеотида стебель-петля, увеличивает расстояние от флуорофора до молекулы гасителя.
179. Композиция по п. 178, отличающаяся тем, что
флуорофор и молекула гасителя расположены на противоположных цепях двухцепочечной стеблевой области олигонуклеотида стебель-петля.
180. Композиция по любому из пп. 138-179, отличающаяся тем, что:
- первый фермент представляет собой первый МНКзим,
- первый МНКзим связан со второй мишенью,
- плечи субстрата указанного первого МНКзима гибридизуются путем комплементарного спаривания оснований с областью одной петли интактного олигонуклеотида стебель-петля, тем самым облегчая расщепление одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля и обеспечивая разделенный олигонуклеотид стебель-петля.
181. Композиция по п. 180, отличающаяся тем, что:
- вторая мишень или представляет собой последовательность нуклеиновой кислоты; и
- вторая мишень гибридизуется с сенсорными плечами первого МНКзима путем комплементарного спаривания оснований, чтобы тем самым облегчить сборку первого МНКзима.
182. Композиция по любому из пп. 138-179, отличающаяся тем, что:
- вторая мишень является аналитом, белком, соединением или молекулой;
- первый фермент представляет собой аптазим, содержащий аптамер, способный связываться со второй мишенью; и
- аптамер связывается со второй мишенью, тем самым делая первый фермент каталитически активным.
183. Композиция по п. 182, отличающаяся тем, что
первый фермент является любым из следующих: апта-ДНКзим, апта-рибозим, апта-МНКзим.
184. Композиция по любому из пп. 138-179, отличающаяся тем, что:
- вторая мишень является аналитом, белком, соединением или молекулой;
- одноцепочечная петлевая область интактного олигонуклеотида стебель-петля является субстратом для аптазима; и
- композиция дополнительно содержит аптазим, содержащий область аптамера, способную связываться со второй мишенью, и область фермента нуклеиновой кислоты, способную расщеплять один или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля с образованием, таким образом, разделенного олигонуклеотида стебель-петля.
185. Композиция по п. 184, отличающаяся тем, что
вторая мишень связана с аптамерной областью аптазима и одноцепочечная петлевая область интактного олигонуклеотида стебель-петля гибридизуется с активной областью, имеющей ферментную активность по отношению к нуклеиновой кислоте, за счет комплементарного спаривания оснований, облегчая расщепление одного или более негибридизованных нуклеотидов интактного олигонуклеотида стебель-петля с образованием разделенного олигонуклеотида стебель-петля.
186. Композиция по любому из пп. 138-179, отличающаяся тем, что:
- вторая мишень представляет собой последовательность нуклеиновой кислоты; и
- первый фермент представляет собой первую рестрикционную эндонуклеазу, и
- вторая мишень гибридизуется с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований с образованием двухцепочечной последовательности для связывания первой рестрикционной эндонуклеазы с одним или более негибридизированными нуклеотидами интактного олигонуклеотида стебель-петля с образованием разделенного олигонуклеотида стебель-петля.
187. Композиция по п. 186, отличающаяся тем, что
первая рестрикционная эндонуклеаза представляет собой эндонуклеазу первого разрыва, способную связываться и расщеплять цепь указанной двухцепочечной последовательности для первой рестрикционной эндонуклеазы, и указанная цепь включает интактный олигонуклеотид стебель-петля.
188. Композиция по любому из пп. 138-179, отличающаяся тем, что:
- первый фермент включает полимеразу с экзонуклеазной активностью,
- вторую мишень гибридизуют с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля путем комплементарного спаривания оснований с образованием первой двухцепочечной последовательности, содержащей область второй мишени,
- композиция дополнительно содержит первый олигонуклеотид-праймер, гибридизованный за счет комплементарного спаривания оснований со второй мишенью с образованием второй двухцепочечной последовательности, расположенной выше относительно первой двухцепочечной последовательности, содержащей область второй мишени, и
- праймер можно удлинить, используя полимеразу с экзонуклеазной активностью и вторую мишень в качестве темплатной последовательности, расщепляя одноцепочечную петлевую область первой двухцепочечной последовательности и тем самым образуя разделенный олигонуклеотид стебель-петля.
189. Композиция по любому из пп. 138-179, отличающаяся тем, что:
- первый фермент представляет собой экзонуклеазу и
- вторая мишень гибридизуется путем комплементарного спаривания оснований с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля, образуя первую двухцепочечную последовательность, содержащую область второй мишени, с которой может ассоциироваться первый фермент, обладающий экзонуклеазной активностью, и, таким образом, расщеплять одноцепочечную петлевую область первой двухцепочечной последовательности, содержащей вторую мишень, с образованием разделенного олигонуклеотида стебель-петля.
190. Композиция по любому из пп. 138-179, отличающаяся тем, что:
- первый фермент представляет собой ДНКзим или рибозим, требующий кофактора для каталитической активности, и
- вторая мишень является кофактором и связана с ДНКзимом или рибозимом,
- ДНКзим или рибозим гибридизуется с одноцепочечной петлевой областью интактного олигонуклеотида стебель-петля за счет комплементарного спаривания оснований, что позволяет ему расщеплять один или более негибридизированных нуклеотидов одноцепочечной петли интактного олигонуклеотида стебель-петля и тем самым образовать разделенный олигонуклеотид стебель-петля.
191. Композиция по п. 190, отличающаяся тем, что кофактор представляет собой ион металла или ион металла, выбранный из Mg2+, Mn2+, Ca2+, Pb2+.
192. Композиция по любому из пп. 138-150, 153, 156-158, 166 или 167, отличающаяся тем, что:
- первый олигонуклеотид выбран из любого одного или более из: Молекулярный Маяк®, праймера Scorpion®, праймера TaqMan® или субстрата МНКзима.
193. Композиция по любому из пп. 138-192, отличающаяся тем, что
первая мишень и/или вторая мишень представляют собой ампликон нуклеиновой кислоты.
| WO 2016025452 A1, 18.02.2016 | |||
| БЕСКОНТАКТНЫЕ СПОСОБЫ ОБНАРУЖЕНИЯ МОЛЕКУЛЯРНЫХ КОЛОНИЙ, НАБОРЫ РЕАГЕНТОВ И УСТРОЙСТВО ДЛЯ ИХ ОСУЩЕСТВЛЕНИЯ | 2006 |
|
RU2394915C2 |
| KUANG et al., Molecular beacon nano-sensors for probing living cancer cells | |||
| Trends in Biotechnology, 2017, vol | |||
| Скоропечатный станок для печатания со стеклянных пластинок | 1922 |
|
SU35A1 |

