ЗАЯВЛЕНИЕ ОБ УСТАНОВЛЕНИИ ПРИОРИТЕТА
По настоящей заявке испрашивается приоритет по PCT/CN 2019/075406, поданной 18 февраля 2019 года, и PCT/CN 2019/106320, поданной 18 сентября 2019 года. Их содержание включено в настоящее описание в качестве ссылки в полном объеме.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к генетически модифицированным животным и клеткам с гуманизированным локусом тяжелой цепи иммуноглобулина и/или гуманизированным локусом легкой цепи иммуноглобулина.
УРОВЕНЬ ТЕХНИКИ
Терапевтические антитела являются одним из самых быстрорастущих классов терапевтических соединений, быстро опережающим рост области низкомолекулярных лекарственных средств. Эти терапевтические антитела, как правило, являются антителами человека или гуманизированными антителами. Антитела человека или гуманизированные антитела можно получать посредством гуманизации антитела грызуна (например, антитела мыши) или с использованием фаговых библиотек. Антитела, получаемые этими способами, зачастую имеют субоптимальные аффинности связывания и биофизические свойства, что приводит к проблемам в производстве и плохой фармакокинетике. В частности, гуманизация может негативно влиять на аффинность связывания и приводить к встраиванию иммуногенных эпитопов в антитела, и антитела, обнаруженные с использованием фаговых библиотек, демонстрируют ограниченное разнообразие и ненативное спаривание тяжелых и легких цепей иммуноглобулин. Для улучшения свойств зачастую необходимы повторяющиеся и времязатратные эксперименты. А в некоторых случаях эти антитела также могут являться иммуногенными для пациентов, что приводит к снижению их эффективности с течением времени.
Одним из возможных подходов для получения полностью человеческих антител является использование трансгенных животных, сконструированных для экспрессии репертуара антител человека. Получение трансгенных животных, таких как мыши, имеющих различные иммуноглобулиновые локусы, делает возможным использование таких трансгенных животных в различных исследованиях и разработках, например, в разработке лекарственных средств и фундаментальных исследованиях в различных биологических системах. Многие из трансгенных животных ранних поколений имели неполные репертуары антител человека, продукцию антител ниже нормальных показателей из-за менее эффективной рекомбинации V(D)J, репертуары эндогенных антител, в которые могли встраиваться иммуногенные эпитопы, и различный другие проблемы. Существует потребность в эффективных и экономичных способах получения антител человека и не являющихся человеком животных, содержащих гуманизированный иммуноглобулиновый локус и обладающих способностью отвечать на антиген с образованием гуманизированных антител.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к генетически модифицированным животным и клеткам с гуманизированным локусом тяжелой цепи и легкой цепи иммуноглобулина.
В некоторых аспектах настоящее изобретение относится к генетически модифицированному, не являющемуся человеком животному, содержащему в эндогенном локусе гена тяжелой цепи иммуноглобулина один или более генов IGHV человека, один или более генов IGHD человека и один или более генов IGHJ человека. В некоторых вариантах осуществления гены IGHV человека, гены IGHD человека и гены IGHJ человека функционально связаны и могут подвергаться реаранжировке VDJ.
В некоторых вариантах осуществления животное содержит приблизительно или по меньшей мере 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160 или 161 ген IGHV человека, выбранный из таблицы 1, приблизительно или по меньшей мере 20, 21, 22, 23, 24, 25, 26 или 27 генов IGHD человека, выбранных из таблицы 2, и приблизительно или по меньшей мере 5, 6, 7, 8 или 9 генов IGHJ человека, выбранных из таблицы 3. В некоторых вариантах осуществления животное содержит все гены IGHV человека из таблицы 1 за исключением IGHV2-10, IGHV3-9 и IGHV1-8, все гены IGHD человека из таблицы 2 и все гены IGHJ человека из таблицы 3. В некоторых вариантах осуществления животное содержит все гены IGHV человека из таблицы 1 за исключением IGHV5-10-1 и IGHV3-64D, все гены IGHD человека из таблицы 2 и все гены IGHJ человека из таблицы 3. В некоторых вариантах осуществления животное содержит все гены IGHV человека, все гены IGHD человека и все гены IGHJ человека в эндогенном локусе гена тяжелой цепи иммуноглобулина на хромосоме 14 человека. В некоторых вариантах осуществления животное содержит все гены IGHV человека, все гены IGHD человека и все гены IGHJ человека в эндогенном локусе гена тяжелой цепи иммуноглобулина на хромосоме 14 человека клетки человека (например, соматической клетки, культивируемой клетки, неимунной клетки, клетки без какой-либо реаранжировки V(D)J).
В некоторых вариантах осуществления животное имеет разрушенный эндогенный локус гена тяжелой цепи иммуноглобулина животного.
В некоторых вариантах осуществления животное является мышью, и разрушение эндогенного локуса гена тяжелой цепи иммуноглобулина животного включает делецию одного или более генов IGHV мыши из таблицы 4, одного или более генов IGHD мыши из таблицы 5 и/или одного или более генов IGHJ мыши из таблицы 6.
В некоторых вариантах осуществления животное являются мышью, и разрушение эндогенного локуса гена тяжелой цепи иммуноглобулина животного включает делецию смежной последовательности, тянущейся от гена IGHV1-85 мыши до гена IGHJ4 мыши.
В некоторых вариантах осуществления животное содержит один или более эндогенных генов IGHM, IGHδ, IGHG3, IGHG1, IGHG2b, IGHG2a, IGHE и IGHA.
В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, полученную из локуса гена тяжелой цепи иммуноглобулина человека.
В некоторых вариантах осуществления немодифицированная последовательность человека составляет приблизительно или по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 т.п.н.
В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, полученную из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHV(III)-82 человека до IGHV1-2 человека. В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, полученную из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHV(III)-82 человека до IGHV6-1 человека. В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, полученную из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHD1-1 человека до человек IGHJ6.
В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, полученную из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHV(III)-82 человека до IGHJ6 человека.
В некоторых вариантах осуществления животное является гомозиготным в отношении локуса гена тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления животное является гетерозиготным в отношении локуса гена тяжелой цепи иммуноглобулина.
В некоторых вариантах осуществления животное дополнительно содержит в эндогенном локусе гена легкой цепи иммуноглобулина один или более человек генов IGKV и один или более генов IGKJ человека.
В некоторых вариантах осуществления животное имеет разрушенный эндогенный локус гена легкой лямбда-цепи иммуноглобулина животного.
В некоторых вариантах осуществления животное является грызуном (например, мышью).
В некоторых аспектах настоящее изобретение относится к генетически модифицированному животному, содержащему в эндогенном локусе гена тяжелой цепи иммуноглобулина, первую последовательность, содержащую один или более генов IGHV человека; вторую последовательность, содержащую эндогенную последовательность; и третью последовательность, содержащую один или более генов IGHD человека, и один или более генов IGHJ человека, где первая последовательность, вторая последовательность и третья последовательность функционально связаны.
В некоторых вариантах осуществления первая последовательность содержит приблизительно или по меньшей мере 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160 или 161 ген IGHV человека, выбранный из таблицы 1. В некоторых вариантах осуществления первая последовательность содержит приблизительно или по меньшей мере 20, 21, 22, 23, 24, 25, 26 или 27 генов IGHD человека, выбранных из таблицы 2.
В некоторых вариантах осуществления первая последовательность является немодифицированной последовательностью, полученной из локуса гена тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления первая последовательность составляет приблизительно или по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 т.п.н.
В некоторых вариантах осуществления вторая последовательность содержит эндогенную последовательность, составляющую приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 т.п.н.
В некоторых вариантах осуществления третья последовательность содержит приблизительно или по меньшей мере 20, 21, 22, 23, 24, 25, 26 или 27 генов IGHD человека, выбранных из таблицы 2. В некоторых вариантах осуществления третья последовательность содержит приблизительно или по меньшей мере 5, 6, 7, 8 или 9 генов IGHJ человека, выбранных из таблицы 3. В некоторых вариантах осуществления третья последовательность содержит все гены IGHD человека из таблицы 2 и все гены IGHJ человека из таблицы 3.
В некоторых вариантах осуществления третья последовательность является немодифицированной последовательностью, полученной из локуса гена тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления третья последовательность составляет приблизительно или по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 т.п.н.
В некоторых вариантах осуществления животное имеет разрушенный эндогенный локус гена тяжелой цепи иммуноглобулина животного.
В некоторых вариантах осуществления животное является мышью, и разрушение эндогенного локуса гена тяжелой цепи иммуноглобулина животного включает делецию одного или более генов IGHV мыши из таблицы 4, одного или более генов IGHD мыши из таблицы 5 и одного или более генов IGHJ мыши из таблицы 6.
В некоторых вариантах осуществления животное является мышью, и разрушение эндогенного локуса гена тяжелой цепи иммуноглобулина животного включает делецию последовательности, тянущейся от IGHV1-85 мыши до IGHJ4 мыши.
В некоторых вариантах осуществления животное содержит один или более эндогенных генов, выбранных из группы, состоящей из генов IGHM, IGHδ, IGHG3, IGHG1, IGHG2b, IGHG2a, IGHE и IGHA.
В некоторых вариантах осуществления животное является гомозиготным в отношении локуса гена тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления животное является гетерозиготным в отношении локуса гена тяжелой цепи иммуноглобулина.
В некоторых вариантах осуществления животное дополнительно содержит в эндогенном локусе гена легкой цепи иммуноглобулина один или более генов IGKV человека и один или более генов IGKJ человека.
В некоторых вариантах осуществления животное имеет разрушенный эндогенный локус гена легкой лямбда-цепи иммуноглобулина животного.
В некоторых вариантах осуществления животное является грызуном (например, мышью).
В некоторых аспектах настоящее изобретение относится к генетически модифицированному, не являющемуся человеком животному, содержащему в эндогенном локусе гена легкой цепи иммуноглобулина один или более генов IGKV человека и один или более генов IGKJ человека.
В некоторых вариантах осуществления животное содержит приблизительно или по меньшей мере 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75 или 76 генов IGKV человека из таблицы 7, и/или содержит приблизительно или по меньшей мере 1, 2, 3, 4 или 5 генов IGKJ человека из таблицы 8.
В некоторых вариантах осуществления животное содержит немодифицированную последовательность, полученную из локуса гена легкой цепи иммуноглобулина человека, тянущегося от IGKV3D-7 человека до IGKJ5 человека.
В некоторых вариантах осуществления животное имеет разрушенный эндогенный локус гена легкой цепи иммуноглобулина животного.
В некоторых вариантах осуществления животное является мышью, и разрушение эндогенного локуса гена легкой цепи иммуноглобулина животного включает делецию одного или более генов IGKV мыши из таблицы 9 и одного или более генов IGKJ мыши из таблицы 10. В некоторых вариантах осуществления животное содержит все гены IGKV человека и все гены IGKJ человека в эндогенном локусе гена каппа-цепи иммуноглобулина на хромосоме 2 человека. В некоторых вариантах осуществления животное содержит все гены IGKV человека и все гены IGKJ человека в эндогенном локусе гена тяжелой цепи иммуноглобулина хромосомы 2 человека клетки человека (например, соматической клетки, культивируемой клетки, неимунной клетки, клетки без какой-либо реаранжировки V(D)J).
В некоторых вариантах осуществления животное является мышью, и разрушение эндогенного локуса гена легкой цепи иммуноглобулина животного включает делецию последовательности, тянущегося от IGKV2-137 мыши до IGKJ5 мыши.
В некоторых вариантах осуществления животное содержит эндогенный IGKC.
В некоторых вариантах осуществления животное является гомозиготным в отношении локуса гена легкой цепи иммуноглобулина. В некоторых вариантах осуществления животное является гетерозиготным в отношении локуса гена легкой цепи иммуноглобулина.
В некоторых вариантах осуществления животное дополнительно содержит в эндогенном локусе гена тяжелой цепи иммуноглобулина один или более генов IGHV человека, один или более генов IGHD человека и один или более генов IGHJ человека.
В некоторых вариантах осуществления животное имеет разрушенный эндогенный локус гена легкой лямбда-цепи иммуноглобулина животного.
В некоторых вариантах осуществления животное является грызуном (например, мышью).
В некоторых аспектах настоящее изобретение относится к генетически модифицированному, не являющемуся человеком животному, геном который содержит эндогенный локус тяжелой цепи иммуноглобулина, содержащий: замену одного или более эндогенных генов IGHV, эндогенных генов IGHD и эндогенных генов IGHJ с одним или более генами IGHV человека, генами IGHD человека и генами IGHJ человека. В некоторых вариантах осуществления гены IGHV человека, гены IGHD человека и гены IGHJ человека функционально связаны с одним или более из эндогенных генов, выбранных из группы, состоящей из генов IGHM, IGHδ, IGHG, IGHE и IGHA.
В некоторых вариантах осуществления один или более эндогенных генов IGHV, эндогенных генов IGHD и эндогенных генов IGHJ заменяют приблизительно или по меньшей мере 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160 или 161 ген IGHV человека из таблицы 1, приблизительно или по меньшей мере 20, 21, 22, 23, 24, 25, 26 или 27 генов IGHD человека из таблицы 2 и приблизительно или по меньшей мере 5, 6, 7, 8 или 9 генов IGHJ человека из таблицы 3.
В некоторых вариантах осуществления животное является мышью, и заменяют приблизительно или по меньшей мере 180 генов IGHV мыши из таблицы 4, все гены IGHD мыши из таблицы 5 и все гены IGHJ мыши из таблицы 6.
В некоторых аспектах настоящее изобретение относится к генетически модифицированному, не являющемуся человеком животному, геном которого содержит эндогенный локус легкой цепи иммуноглобулина содержащий: замену одного или более эндогенных генов IGKV и эндогенных генов IGKJ одним или более генами IGKV человека и генами IGKJ человека. В некоторых вариантах осуществления гены IGKV человека и гены IGKJ человека функционально связаны с эндогенным геном IGKC.
В некоторых вариантах осуществления один или более эндогенных генов IGKV и эндогенных генов IGKJ заменяют приблизительно или по меньшей мере 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75 или 76 генами IGKV человека из таблицы 7 и приблизительно или по меньшей мере 1, 2, 3, 4 или 5 генами IGKJ человека из таблицы 8.
В некоторых вариантах осуществления животное является мышью, и заменяют все гены IGKV мыши из таблицы 9 и все гены IGKJ мыши из таблицы 10.
В некоторых вариантах осуществления у животного отсутствует эндогенный локус вариабельной области тяжелой цепи иммуноглобулина, который может подвергаться реаранжировке и образующий последовательность нуклеиновой кислоты, кодирующую эндогенный вариабельный домен тяжелой цепи (например, вариабельный домен тяжелой цепи мыши).
В некоторых вариантах осуществления у животного отсутствует эндогенный локус вариабельной области легкой цепи иммуноглобулина, который может подвергаться реаранжировке и образующий последовательность нуклеиновой кислоты, кодирующую эндогенный вариабельный домен легкой цепи (например, вариабельный домен легкой цепи мыши).
В некоторых вариантах осуществления животное может продуцировать гуманизированное антитело.
В некоторых аспектах настоящее изобретение относится к клетке, полученной из животного, как представлено в настоящем описании.
В некоторых вариантах осуществления клетка является B-клетка, экспрессирующая химерную тяжелую цепь иммуноглобулина, содержащую вариабельный домен тяжелой цепи иммуноглобулина, полученный посредством реаранжировки одного или более генов IGHV человека, одного или более генов IGHD человека и одного или более генов IGHJ человека. В некоторых вариантах осуществления вариабельный домен тяжелой цепи иммуноглобулина функционально связан с не принадлежащей человеку константной областью тяжелой цепи.
В некоторых вариантах осуществления клетка является B-клеткой, экспрессирующей химерную легкую цепь иммуноглобулина, содержащую вариабельный домен легкой цепи иммуноглобулина, полученный посредством реаранжировки одного или более генов IGKV человека и один или более гены IGKJ человека, и где вариабельный домен легкой цепи иммуноглобулина функционально связан с не принадлежащей человеку константная область легкой цепи.
В некоторых вариантах осуществления клетка является эмбриональной стволовой клеткой (ES).
В некоторых аспектах настоящее изобретение относится к способу получения химерного антитела, специфически связывающегося с антигеном, включающему подвергание животного, представленного в настоящем описании, воздействию антигена; получение гибридомы из клетки, полученной из животного; и сбор химерного антитела, продуцируемого гибридомой. В некоторых вариантах осуществления интересующую клетку выделяют и осуществляют секвенирование для определения последовательности подвергнутой реаранжировке вариабельной области тяжелой цепи и вариабельной области легкой цепи.
В некоторых вариантах осуществления способ дополнительно включает секвенирование генома гибридомы.
В некоторых аспектах настоящее изобретение относится к способу модификации генома клетки, включающему модификацию хромосомы человека, встраивание модифицированной хромосомы человека в клетку животного и индуцирование рекомбинации между модифицированной хромосомой человека и эндогенной хромосомой и, таким образом, замену одного или более эндогенных генов одним или более генами человека.
В некоторых вариантах осуществления модифицированная хромосома человека содержит два или более экзогенных участка рекомбинации.
В некоторых вариантах осуществления эндогенная хромосома содержит два или более экзогенных участка рекомбинации.
В некоторых вариантах осуществления приблизительно или по меньшей мере 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160 или 161 ген IGHV человека, выбранный из таблицы 1, приблизительно или по меньшей мере 20, 21, 22, 23, 24, 25, 26 или 27 генов IGHD человека, выбранных из таблицы 2, и приблизительно или по меньшей мере 5, 6, 7, 8 или 9 генов IGHJ человека, выбранных из таблицы 3, встраивают в эндогенную хромосому посредством рекомбинации.
В некоторых вариантах осуществления приблизительно или по меньшей мере 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75 или 76 генов IGKV человека из таблицы 7 и приблизительно или по меньшей мере 1, 2, 3, 4 или 5 генов IGKJ человека из таблицы 8 встраивают в эндогенную хромосому посредством рекомбинации.
В некоторых вариантах осуществления последовательность человека встраивают в эндогенную хромосому посредством рекомбинации, и последовательность человека составляет приблизительно или по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 т.п.н.
В одном из аспектов настоящее изобретение относится к способу получения антитела, специфически связывающегося с антигеном. Способ включает получение последовательности нуклеиновой кислоты, кодирующей вариабельные области тяжелой и легкой цепи иммуноглобулина человека в клетке, экспрессирующей гибридное антитело, специфически связывающееся с антигеном, где клетку получают посредством подвергания животного, представленного в настоящем описании, воздействию антигена; функциональное связывание нуклеиновой кислоты, кодирующей вариабельную область тяжелой цепи иммуноглобулина человека, с нуклеиновой кислотой, кодирующей константную область тяжелой цепи иммуноглобулина человека, и нуклеиновой кислоты, кодирующей вариабельную область легкой цепи иммуноглобулина человека, с нуклеиновой кислотой, кодирующей константную область легкой цепи иммуноглобулина человека; и экспрессию нуклеиновой кислоты в клетке и, таким образом, получение антитела.
В одном из аспектов настоящее изобретение относится к способу получения нуклеиновой кислоты, кодирующей связывающий домен антитела, специфически связывающийся с антигеном. Способ включает подвергание животного, представленного в настоящем описании, воздействию антигена, и секвенирование нуклеиновых кислот, кодирующих вариабельные области тяжелой и легкой цепи иммуноглобулина человека, в клетке, экспрессирующей гибридное антитело, специфически связывающееся с антигеном.
В одном из аспектов настоящее изобретение относится к способу получения образца, включающему подвергание животного, представленного в настоящем описании, воздействию антигена и получение образца из животного. В некоторых вариантах осуществления образец представляет собой ткань селезенки, клетку селезенки или B-клетку.
В одном из аспектов настоящее изобретение относится к способу получения антитела, специфически связывающегося с антигеном. Способ включает подвергание животного, представленного в настоящем описании, воздействию антигена; получение последовательности (например, посредством секвенирования) нуклеиновых кислот, кодирующих вариабельные области тяжелой и легкой цепи иммуноглобулина человека, в клетке, экспрессирующей гибридное антитело, специфически связывающееся с антигеном; и функциональное связывание в клетке нуклеиновой кислоты, кодирующей вариабельную область тяжелой цепи иммуноглобулина человека, с нуклеиновой кислотой, кодирующей константную область тяжелой цепи иммуноглобулина человека, и нуклеиновой кислоты, кодирующей вариабельную область легкой цепи иммуноглобулина человека, с нуклеиновой кислотой, кодирующей константную область легкой цепи иммуноглобулина человека.
Настоящее изобретение также относится к потомству не являющегося человеком млекопитающего.
В некоторых вариантах осуществления не являющееся человеком млекопитающее является грызуном. В некоторых вариантах осуществления не являющееся человеком млекопитающее является мышью.
Настоящее изобретение также относится к клетке, включающей направленный вектор, как представлено в настоящем описании.
Настоящее изобретение также относится к клетке (например, стволовой клетке, эмбриональной стволовой клетке, иммунной клетке, B-клетке, T-клетке или гибридоме) или линии клеток, или ее культуре первичных клеток, полученных из не являющегося человеком млекопитающего или его потомства. Настоящее изобретение дополнительно относится к ткани, органу или их культуре, полученным из не являющегося человеком млекопитающего или его потомства.
Настоящее изобретение дополнительно относится к применению не являющегося человеком млекопитающего или его потомства, модели на животных, полученной способом, представленным в настоящем описании, в разработке продукта, связанного с иммунизацией, производстве антитела человека или модельной системе для исследований в области фармакологии, иммунологии, микробиологии и медицины.
Если не указано иначе, все технические и научные термины, используемые в настоящем описании, обладают тем же значением, которое общепринято понятным специалисту в области, к которой принадлежит изобретение. Способы и материалы представлены в настоящем описании для использования в настоящем изобретении; также можно использовать другие подходящие способы и материалы, известные в этой области. Материалы, способы и примеры являются исключительно иллюстративными и не предназначены для ограничения. Все публикации, патентные заявки, патенты, последовательности, записи в базах данных и других ссылках, упомянутых в настоящем описании, включены в настоящее описание в качестве ссылки в полном объеме. В случае противоречия настоящее описание, включая определения, будет обладать приоритетом.
Другие признаки и преимущества изобретения будут очевидны из следующего подробного описания, фигур и формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1A является блок-схемой способа встраивания генов иммуноглобулина человека в геном мыши.
Фиг. 1B является общей схемой замены вариабельной области тяжелой цепи иммуноглобулина мыши вариабельной областью тяжелой цепи иммуноглобулина человека.
Фиг. 1C является общей схемой замены вариабельной области легкой цепи иммуноглобулина мыши вариабельной областью легкой цепи иммуноглобулина человека.
Фиг. 2 является схемой, на которой показан локус тяжелой цепи иммуноглобулина мыши.
Фиг. 3A является схемой, на которой показан локус тяжелой цепи иммуноглобулина мыши после встраивания двух участков рекомбинации в геном.
Фиг. 3B является схемой, на которой показан локус тяжелой цепи иммуноглобулина мыши после рекомбинации с направленным вектором.
На фиг. 4 показана стратегия таргетинга для модификации локуса тяжелой цепи иммуноглобулина мыши.
На фиг. 5A показаны результаты анализа ПЦР с использованием пары праймеров mIgHV-5’loxP-L-GT-F/mIGHV-005-L-GT-R2.
На фиг. 5B показаны результаты анализа ПЦР с использованием пары праймеров mIGHV-005-5’loxP-R-GT-F2/mIgHV-5’loxP-R-GT-R.
На фиг. 6A показаны результаты анализа ПЦР с использованием пары праймеров mIGHV-3’lox-L-GT-F2/mIGHV-3’lox-L-GT-R2.
На фиг. 6B показаны результаты анализа ПЦР с использованием пары праймеров mIGHV3’lox-R-GT-F2/mIGHV-3’lox-R-GT-R1.
Фиг. 7 является схемой хромосомы 14 человека, на которой выделен локус тяжелой цепи иммуноглобулина (нарисовано не в масштабе). Локус тяжелой цепи иммуноглобулина содержит вариабельные области (VH, DH, JH) и константную область (CH). VH представляет собой сегмент для кластера генов IGHV, DH представляет собой сегмент для кластера генов IGHD, JH представляет собой сегмент для кластера генов IGHJ, и CH представляет собой кластер генов, экспрессирующий константные домены.
Фиг. 8 является схемой, на которой показана хромосома 14 человека после модификации.
На фиг. 9 показаны модификации хромосомы 14 человека с использованием двух векторов.
На фиг. 10 показана модифицированная хромосома человека 14.
На фиг. 11 показаны результаты анализов ПЦР участка loxP 301 на хромосоме hChr14-mut3 с использованием пары праймеров hIGHV-5’loxP-L-GT-F1 и hIGHV-5’ loxP-R-GT-R. 8-D7 является клоном положительного контроля.
На фиг. 12 показаны результаты анализов ПЦР участка loxP 302 на хромосоме hChr14-mut3. 8-D7 является клоном отрицательного контроля.
Фиг. 13 является изображением флуоресцентной гибридизации in situ (FISH) клеток до модификации хромосомы 14 человека.
Фиг. 14 является изображением FISH клеток после модификации хромосомы человека 14.
Фиг. 15 является схемой, на которой показана модифицированная хромосома 12 мыши.
Фиг. 16 является схемой, на которой показана Cre-опосредованный рекомбинация, посредством которой локус вариабельной области тяжелой цепи мыши заменяют соответствующей последовательностью геномной ДНК человека.
На фиг. 17 показаны результаты анализа ПЦР с использованием пары праймеров M5-L и пары праймеров M5-R.
На фиг. 18 показаны результаты анализа ПЦР с использованием пары праймеров M3.
На фиг. 19 показаны результаты анализа ПЦР с использованием пары праймеров H5-L и пары праймеров H5-R.
На фиг. 20 показаны результаты анализа ПЦР с использованием пары праймеров H3-L и пары праймеров H3-R.
Фиг. 21 является изображением FISH. Белыми стрелками (1) и (2) указана хромосома 12 мыши. Белой стрелкой (3) указан фрагмент хромосомы человека, меченый человек-специфическим зондом IGH Break Apart.
Фиг. 22 является схемой, на которой показана Flp-опосредованная рекомбинация.
Фиг. 23 является схемой, на которой показан локус легкой цепи иммуноглобулина мыши.
Фиг. 24A является схемой, на которой показан локус легкой цепи иммуноглобулина мыши после встраивания двух участков рекомбинации в геном.
Фиг. 24B является схемой, на которой показан локус легкой цепи иммуноглобулина мыши после рекомбинации с направленным вектором.
Фиг. 25 является схемой, на которой показана стратегия генного таргетинга для хромосомы 6 мыши.
На фиг. 26 показаны результаты анализа ПЦР с использованием пары праймеров IGKV-005-C-5G-L-GT-F/IGKV-005-C-5G-L-GT-R2.
На фиг. 27 показаны результаты анализа ПЦР с использованием пары праймеров IGKV-005-C-5G-R-GT-F1/IGKV-005-C-5G-R-GT-R.
Фиг. 28 является схемой хромосомы человека 2, на которой выделен локус легкой цепи иммуноглобулина (нарисован не в масштабе). VHK представляет собой сегмент для кластера генов IGKV, JHK представляет собой сегмент для кластера генов IGKJ, и CHK представляет собой ген IGKC.
Фиг. 29 является схемой, на которой показана модифицированная хромосома 2 человека.
Фиг. 30 является схемой, на которой показана стратегия генного таргетинга для хромосомы 2 человека.
На фиг. 31 показаны результаты анализа ПЦР после первой рекомбинации (встраивание вектора 2702). WT представляет собой клетки H9 дикого типа.
Фиг. 32 является изображением результатов FISH. Белой стрелкой указана модифицированная хромосома 2 человека с правильной рекомбинацией.
На фиг. 33 показаны результаты анализа ПЦР после второй рекомбинации (встраивание вектора 2701).
Фиг. 34 является схемой, на которой показана Cre-опосредованная рекомбинация, при которой последовательность геномной ДНК вариабельной области легкой цепи человека добавляли в соответствующий локус мыши.
Фиг. 35 является изображением флуоресцентной гибридизации in situ (FISH). Стрелками (1) и (3) указана хромосома 6 мыши. Стрелкой (2) указан фрагмент хромосомы человека, меченый человек-специфическим зондом IGK Break Apart.
Фиг. 36 является схемой, на которой показана Flp-опосредованная рекомбинация.
Фиг. 37 является схемой, на которой показан локус тяжелой цепи иммуноглобулина (IGH) человека на хромосоме 14 (14q32.33).
Фиг. 38 является схемой, на которой показан локус IGH мыши (Mus musculus) на хромосоме 12 (12F2) (линия C57BL/6).
Фиг. 39 является схемой, на которой показан локус каппа-цепи иммуноглобулина (IGK) человека на хромосоме 2 (2p11.2).
Фиг. 40 является схемой, на которой показан локус IGK мыши (Mus musculus) на хромосоме 6 (6C1).
Фиг. 41 показан репертуар IMGT для локуса тяжелой цепи иммуноглобулина (IGH) человека.
Фиг. 42 показан репертуар IMGT для IGH мыши.
Фиг. 43 показан репертуар IMGT для локуса каппа-цепи иммуноглобулина (IGK) человека.
Фиг. 44 показан репертуар IMGT для IGK мыши.
На фиг. 45 показаны процентные доли лейкоцитов, определяемых в периферической крои.
На фиг. 46 показаны процентные доли лейкоцитов, определяемых в клетках селезенки.
На фиг. 47 показаны процентные доли лейкоцитов, определяемых в лимфоузлах.
На фиг. 48 показаны процентные доли B-клеток селезенки на разных стадиях развития.
На фиг. 49 показаны процентные доли B-клеток лимфоузлов на разных стадиях развития.
На фиг. 50 показаны процентные доли B-клеток селезенки в краевой зоне (MZ-B) и фолликулярной зоне (FO-B) селезенки.
На фиг. 51A показаны результаты проточной цитометрии B-клеток на разных стадиях развития в костном мозге, полученных из мышей дикого типа. Область 1 соответствует про-B-клеткам, Область 2 соответствует пре-B-клеткам, и Область 3 соответствует незрелым B-клеткам.
На фиг. 51B показаны результаты проточной цитометрии B-клеток на разных стадиях развития в костном мозге, полученных из мышей, гетерозиготных по гуманизированной тяжелой цепи. Область 1 соответствует про-B-клеткам, Область 2 соответствует пре-B-клеткам, и Область 3 соответствует незрелым B-клеткам.
На фиг. 51C показаны результаты проточной цитометрии B-клеток на разных стадиях развития в костном мозге, полученных из мышей, гомозиготных по гуманизированной тяжелой цепи. Область 1 соответствует про-B-клеткам, Область 2 соответствует пре-B-клеткам, и Область 3 соответствует незрелым B-клеткам.
На фиг. 52 показаны уровни изотипов IgA в серийно разведенной сыворотке мыши.
На фиг. 53 показаны уровни изотипов IgG1 в серийно разведенной сыворотке мыши.
На фиг. 54 показаны уровни изотипов IgG2b в серийно разведенной сыворотке мыши.
На фиг. 55 показаны уровни изотипов IgG2c в серийно разведенной сыворотке мыши.
На фиг. 56 показаны уровни изотипов IgG3 в серийно разведенной сыворотке мыши.
На фиг. 57 показаны уровни изотипов IgM в серийно разведенной сыворотке мыши.
На фиг. 58 показано распределение определяемой экспрессии генов IGKV после рекомбинации VJ среди отдельных мышей.
На фиг. 59 показаны результаты проточной цитометрии для мышей дикого типа после иммунизации с помощью BTLA человека.
На фиг. 60 показаны результаты проточной цитометрии для мышей, гомозиготных по гуманизированной тяжелой цепи, после иммунизации с помощью BTLA человека.
На фиг. 61 показаны результаты проточной цитометрии для мышей дикого типа после иммунизации с помощью PD-1 собаки (dPD-1).
На фиг. 62 показаны результаты проточной цитометрии для мышей, гомозиготных по гуманизированной тяжелой цепи, после иммунизации с помощью PD-1 собаки (dPD-1).
На фиг. 63 показаны обобщенные результаты ELISA для мышей дикого типа (черные столбики; мышей помечали как 1-5) и мышей, гомозиготных по гуманизированной тяжелой цепи (серые столбики; мышей помечали как 6-10), после иммунизации овальбумином (OVA).
На фиг. 64 показан список дистального кластера генов Vκ IGKV человека и список проксимального кластера генов Vκ IGKV человека.
На фиг. 65A показана масса тела наивных мышей, мышей дикого типа и мышей hVH/hVL.
На фиг. 65B показана масса селезенки наивных мышей дикого типа и мышей hVH/hVL.
На фиг. 66 показана процентная доля иммунных клеток в селезенке наивных мышей, мышей дикого типа и мышей hVH/hVL.
На фиг. 67A показана процентная доля популяции B-клеток переходного типа 1 (T1, B220+IgM+IgD-), B-клеток переходного типа 2 (T2, B220+IgM+IgD+) и зрелых B-клеток (M, B220+IgMlowIgD+) среди B-клеток селезенки. B-клетки селезенки получают из наивных мышей дикого типа или мышей hVH/hVL.
На фиг. 67B показана процентная доля популяции B-клеток краевой зоны (MZ) и B-клеток фолликулярной зоны (FO) среди B-клеток селезенки. B-клетки селезенки получают из наивных мышей дикого типа или мышей hVH/hVL.
На фиг. 68A показана процентная доля популяция про-B-клеток (B220lowCD43highIgMlow), пре-B-клеток (B220lowCD43intIgMlow) и незрелых B-клеток (B220highCD43lowIgMhigh) среди B-клеток костного мозга. B-клетки костного мозга получают из наивных мышей дикого типа или мышей hVH/hVL.
На фиг. 68B показана процентная доля популяции плазматических клеток (B220lowIgM-IgD-CD138-) и B-клеток памяти (B220+IgM+IgD-CD38+) среди B-клеток костного мозга. B-клетки костного мозга получают из наивных мышей дикого типа или мышей hVH/hVL.
На фиг. 68C показана процентная доля популяции плазматических клеток (B220lowIgM-IgD-CD138-) и B-клеток памяти (B220+IgM+IgD-CD38+) среди B-клеток селезенки. B-клетки селезенки получают из наивных мышей дикого типа или мышей hVH/hVL.
На фиг. 69 показана концентрация подтипов иммуноглобулинов (Ig) в сыворотке у наивных мышей дикого типа или мыши hVH/hVL. Концентрации подтипов Ig количественно измеряли посредством ELISA.
На фиг. 70A показано использование IGHV (частота>1%) у наивных мышей hVH/hVL.
На фиг. 70B показано использование IGHV (частота<1%) у наивных мышей hVH/hVL.
На фиг. 70C показано использование IGHD у наивных мышей hVH/hVL.
На фиг. 70D показано использование IGHJ у наивных мышей hVH/hVL.
На фиг. 71A показано использование IGKV (частота>1%) у наивных мышей hVH/hVL.
На фиг. 71B показано использование IGKV (частота<1%) у наивных мышей hVH/hVL.
На фиг. 71C показано использование IGKJ у наивных мышей hVH/hVL.
Фиг. 72 является гистограммой, на которой показано распределение длин CDR3 тяжелой цепи в аминокислотах у наивных мышей hVH/hVL.
На фиг. 73 показана частота использования аминокислот в CDR3 тяжелой цепи у наивных мышей hVH/hVL.
На фиг. 74 показана частота HCDR3, содержащих остатки цистеина, у мышей hVH/hVL.
Фиг. 75A является изображением гистологического анализа селезенки наивных мышей дикого типа.
Фиг. 75B является изображением гистологического анализа пахового лимфоузла наивных мышей дикого типа.
Фиг. 75C является изображением гистологического анализа пейеровой бляшки наивных мышей дикого типа.
Фиг. 75D является изображением гистологического анализа селезенки наивных мышей hVH/hVL.
Фиг. 75E является изображением гистологического анализа пахового лимфоузла наивных мышей hVH/hVL.
Фиг. 75F является изображением гистологического анализа пейеровой бляшки наивных мышей hVH/hVL.
На фиг. 76A показан титр BCMA-специфических антител после второй и третьей иммунизации с использованием BCMA (антигена созревания B-клеток) человека в качестве антигена для мышей дикого типа и мышей hVH/hVL.
На фиг. 76B показан титр IL4R-специфических антител после второй и третьей иммунизации с использованием IL4R (рецептора интерлейкина-4) человека в качестве антигена для мышей дикого типа и мышей hVH/hVL.
На фиг. 76C показан титр PD-1-специфических антител после второй и третьей иммунизации с использованием PD-1 (белка программируемой гибели клеток 1) человека в качестве антигена для мышей дикого типа и мышей hVH/hVL.
На фиг. 76D показан титр Siglec15-специфических антител после второй и третьей иммунизации с использованием Siglec15 (связывающего сиаловую кислоту, Ig-подобного лектина 15) человека в качестве антигена для мышей дикого типа и мышей hVH/hVL.
На фиг. 76E показан титр SIRPα-специфических антител после второй и третьей иммунизации с использованием SIRPα (сигнального регуляторного белка α) человека в качестве антигена для мышей дикого типа и мышей hVH/hVL.
На фиг. 77A показана масса тела мышей дикого типа и мышей hVH/hVL после иммунизации.
На фиг. 77B показана масса селезенки мышей дикого типа и мышей hVH/hVL после иммунизации.
На фиг. 78 показана процентная доля иммунных клеток в селезенке мышей дикого типа и мышей hVH/hVL после иммунизации.
На фиг. 79A показана процентная доля популяции B-клеток переходного типа 1 (T1, B220+IgM+IgD-), B-клеток переходного типа 2 (T2, B220+IgM+IgD+) и зрелых B-клеток (M, B220+IgMlowIgD+) среди B-клеток селезенки. B-клетки селезенки получают из мышей дикого типа или мышей hVH/hVL после иммунизации.
На фиг. 79B показана процентная доля популяции B-клеток краевой зоны (MZ) и фолликулярной зоны (FO) среди B-клеток селезенки. B-клетки селезенки получают из мышей дикого типа или мышей hVH/hVL после иммунизации.
На фиг. 80A показана процентная доля популяции про-B-клеток (B220lowCD43highIgMlow), про-B-клеток (B220lowCD43intIgMlow) и незрелых B-клеток (B220highCD43lowIgMhigh) среди B-клеток костного мозга. B-клетки костного мозга получают из мышей дикого типа или мышей hVH/hVL после иммунизации.
На фиг. 80B показана процентная доля популяции плазматических клеток (B220lowIgM-IgD-CD138-) и B-клеток памяти (B220+IgM+IgD-CD38+) среди B-клеток костного мозга. B-клетки костного мозга получают из мышей дикого типа или мышей hVH/hVL после иммунизации.
На фиг. 80C показана процентная доля популяции плазматических клеток (B220lowIgM-IgD-CD138-) и B-клеток памяти (B220+IgM+IgD-CD38+) среди B-клеток селезенки. B-клетки селезенки получают из мышей дикого типа или мышей hVH/hVL после иммунизации.
На фиг. 81 показана концентрация подтипов иммуноглобулина (Ig) в сыворотке у наивных мышей дикого типа или мыши hVH/hVL или после третьей иммунизации. Концентрации подтипов Ig определяли посредством ELISA.
На фиг. 82 показана общая концентрация IgG в сыворотке у наивных мышей дикого типа или мыши hVH/hVL или после иммунизации. Концентрации IgG определяли посредством ELISA.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к генетически модифицированным животным и клеткам с гуманизированным локусом тяжелой цепи иммуноглобулина и/или гуманизированным локусом легкой цепи иммуноглобулина (например, локусом каппа-цепи).
Генетически модифицированных животных можно получать посредством встраивания генов иммуноглобулинов человека в геном не являющихся человеком животных для получения животных, которые могут экспрессировать гуманизированные антитела или химерные антитела. На фиг. 1A показаны способы получения гуманизированных мышей. В некоторых вариантах осуществления способы сначала включают модификацию иммуноглобулиновой области человека на хромосоме человека. Затем модифицированные хромосомы человека встраивают в клетку-реципиента мыши. Затем вариабельную область иммуноглобулина человека встраивают в соответствующую область генома мыши посредством прямой замены (например, одностадийной замены). Затем клетки-реципиенты подвергают скринингу, предпочтительно, на клетки, не содержащие хромосомы человека. Затем клетки инъецируют в бластоцисты для получения химерных животных (например, мышей). Для получения животных, содержащих интактный гуманизированный иммуноглобулиновый локус, можно осуществлять последующее скрещивание.
Генетически модифицированные животные, представленные в настоящем описании, могут обладать различные преимущества. Например, в некоторых случаях генетически модифицированные животные, представленные в настоящем описании, обладают полными репертуарами антител человека. Таким образом, вариабельные домены, генерируемые этими животными, могут обладать разнообразием, очень схожим с разнообразием вариабельных доменов у человека. Кроме того, т.к. в геном животного встраивают целую последовательность в иммуноглобулиновом локусе человека (без модификаций или с ограниченными модификациями), эти гены могут подвергаться рекомбинации V(D)J способом, очень схожим с тем, что происходит у человека. Кроме того, продукция антител может быть очень эффективной и иметь скорость, схожую с нормальными скоростями благодаря эффективной рекомбинации V(D)J. Кроме того, т.к. рекомбинация V(D)J может происходить между эндогенными генами IGHV, IGHD, IGHJ, IGKV и IGKJ и генами человека, если эндогенные гены IGHV, IGHD, IGHJ, IGKV и IGKJ встраивают в подвергнутый реаранжировке сегмент тяжелой цепи VDJ или подвергнутый реаранжировке сегмент легкой цепи VJ, вероятно, антитела генерируемые репертуарами антител, содержат иммуногенные эпитопы человека. Иммуногенность может приводить к продукции антител против лекарственных средств и может быть эффективной. В этом случае эндогенные гены IGHV, IGHD, IGHJ, IGKV и IGKJ эффективно подвергаются делеции. Менее вероятно, что антитела, генерируемые репертуарами антител, являются иммуногенными для людей. Таким образом, антитела больше подходят для использования в качестве терапевтических средств для людей. Таким образом, генетически модифицированные животные представляют собой предпочтительную платформу для продукции гуманизированных антител.
В рамках изобретения термин "антитело" относится к молекуле иммуноглобулина, содержащей четыре полипептидные цепи, две тяжелые (H) цепи и две легкие (L) цепи, соединенные друг с другом дисульфидными связями. Каждая тяжелая цепь содержит вариабельный домен тяжелой цепи (VH) и константную область тяжелой цепи (CH). Каждая легкая цепь содержит вариабельный домен легкой цепи (VL) и константную область легкой цепи (CL). Домены VH и VL можно дополнительно разделять на области гипервариабельности, обозначаемые как определяющие комплементарность области (CDR), перемежающиеся с областями, являющимися более консервативными каркасными областями (FR). Каждая VH и VL содержит три CDR и четыре FR, расположены с амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4 (CDR тяжелой цепи можно сокращенно обозначать как HCDR1, HCDR2 и HCDR3; CDR легкой цепи можно сокращенно обозначать как LCDR1, LCDR2 и LCDR3). Термин "высокоаффинное" антитело относится к антителу, имеющему KD в отношении своего целевого эпитопа приблизительно 10-9 M или менее (например, приблизительно 1×10-9 M, 1×10-10 M, 1×10-11 M или 1×10-12 M или менее). В некоторых вариантах осуществления KD можно измерять посредством поверхностного плазмонного резонанса, например, BIACORE™ или ELISA.
В рамках изобретения термин "антигенсвязывающий фрагмент" относится к части полноразмерного антитела, где часть антитела может специфически связываться с антигеном. В некоторых вариантах осуществления антигенсвязывающий фрагмент содержит по меньшей мере один вариабельный домен (например, вариабельный домен тяжелой цепи или вариабельный домен легкой цепи). Неограничивающие примеры фрагментов антител включают, например, Fab-, Fab’-, F(ab’)2- и Fv-фрагменты.
В рамках изобретения термин "антитело человека" относится к антителу, кодируемому нуклеиновой кислотой (например, подвергнутый реаранжировке локус тяжелой или легкой цепи иммуноглобулина человека), присутствующей у человека. В некоторых вариантах осуществления антитело человека получают из человека или продуцируют в культуре клеток человека (например, гибридомных клетках человека). В некоторых вариантах осуществления антитело человека продуцируют в не принадлежащей человеку клетке (например, линии клеток мыши или хомяка). В некоторых вариантах осуществления антитело человека продуцируют в бактериальной или дрожжевой клетке. В некоторых вариантах осуществления антитело человека продуцируют в трансгенном не являющемся человеком животном (например, мыши), содержащем неподвергнутый реаранжировке или подвергнутый реаранжировке иммуноглобулиновый локус человека (например, локус тяжелой или легкой цепи иммуноглобулина человека).
В рамках изобретения термин "химерное антитело" относится к антителу, содержащему последовательность, присутствующую по меньшей мере в двух разных антителах (например, антителах из двух разных видов млекопитающих, таких как антитело человека и мыши). Неограничивающим примером химерного антитела является антитело, содержащее последовательности вариабельных доменов (например, вся или часть последовательности вариабельных доменов легкой цепи и/или тяжелой цепи) антитела человека и константных доменов не принадлежащего человеку антитела. Дополнительные примеры химерных антител представлены в настоящем описании и известны в этой области.
В рамках изобретения термин "гуманизированное антитело" относится к не принадлежащему человеку антителу, содержащему последовательность, полученную из не принадлежащего человеку иммуноглобулина (например, мыши) и содержащую последовательности, полученные из иммуноглобулина человека.
В рамках изобретения термин "одноцепочечные антитело" относится к одному полипептиду, содержащему по меньшей мере два вариабельных домена иммуноглобулина (например, вариабельный домен тяжелой цепи или легкой цепи иммуноглобулина млекопитающего), который может специфически связываться с антигеном.
В рамках изобретения термины "индивидуум" и "пациент" используют взаимозаменяемо на всем протяжении настоящего описания для описания животного, человека или не являющегося человеком животного. В настоящем изобретении предусмотрено ветеринарное и неветеринарное использование. Пациенты-люди могут являться взрослыми людьми или людьми младшего возраста (например, людьми возрастом менее 18 лет). В дополнение к людям, пациенты включают, в качестве неограничивающих примеров, мышей, крыс, хомяков, морских свинок, кроликов, хорьков, кошек, собак и приматов. Включены, например, не являющиеся человеком приматы (например, мартышка, шимпанзе, горилла и т.п.), грызуны (например, крысы, мыши, песчанки, хомяки, хорьки, кролики), зайцеобразные, свиньи (например, свинья, минипиг), лошади, собаки, кошки, крупный рогатый скот и другие домашние, сельскохозяйственные и зоопарковые животные.
В рамках изобретения в отношении антитела фразы "специфическое связывание" и "специфически связывается" означает, что антитело взаимодействует со своей молекулой-мишенью, предпочтительно, с другими молекулами, т.к. взаимодействие зависит от наличия конкретной структуры (т.е. антигенной детерминанты или эпитопа) на молекуле-мишени; другими словами, реагент распознает и связывается с молекулами, включающими специфическую структуру, а не всеми молекулами в целом. Антитело, специфически связывающееся с молекулой-мишенью, можно обозначать как мишене-специфическое антитело.
В рамках изобретения термины "полипептид", "пептид" и "белок" используют взаимозаменяемо в отношении полимеров аминокислот любой длины, по меньшей мере две аминокислоты.
В рамках изобретения термины "полинуклеотид", "молекула нуклеиновой кислоты" и "последовательность нуклеиновой кислоты" используют взаимозаменяемо в настоящем описании в отношении полимеров нуклеотидов любой длины, по меньшей мере два нуклеотида, и они включают, в качестве неограничивающих примеров, ДНК, РНК, гибриды ДНК/РНК и их модификации.
В рамках изобретения термин "немодифицированная последовательность человека" относится к последовательности, полученной из человека, клетки человека, культивируемой клетки человека или линии клеток человека, где последовательность идентична генетической последовательности человека, клетки человека, культивируемой клетки человека или линии клетки человека.
Генетически модифицированный локус тяжелой цепи иммуноглобулина
Локус тяжелой цепи иммуноглобулина (также известный как IGH или локус тяжелой цепи иммуноглобулина) является областью на хромосоме (например, хромосоме 14 человека), содержащей гены тяжелых цепей антител человека (или иммуноглобулинов).
Эта область соответствует организации зародышевой линии локуса тяжелой цепи. Локус включает сегменты V (вариабельный), D (разнообразие), J (соединительный) и C (константный). Гены в области V образуют кластер генов V (также известный как кластер генов IGHV). Гены в области D образуют кластер генов D (также известный как кластер генов IGHD). Гены в области J образуют кластер генов J (также известный как кластер генов IGHJ).
При развитии B-клеток во время рекомбинационного события на уровне ДНК один сегмент D (также известный как ген IGHD) соединяется с сегментом J (также известным как ген IGHJ); затем слитый экзон D-J этой частично подвергнутой реаранжировке области D-J соединяется с сегментом V (также известным как ген IGHV). Затем подвергнутая реаранжировке область V-D-J, содержащая слитый экзон V-D-J, транскрибируется и сливается на уровне РНК с константной областью IGHM; этот транскрипт кодируется тяжелой мю-цепью. Позднее во время развития B-клетки генерируют матричную пре-РНК V-D-J-Cmu-Cdelta, подвергающуюся альтернативному сплайсингу, кодируя тяжелую мю- или дельта-цепь. Зрелые B-клетки в лимфоузлах подвергаются рекомбинации на этапе переключения таким образом, что слитый генный сегмент V-D-J сближается с одним из генных сегментов IGHG, IGHA или IGHE, и каждая клетка экспрессирует тяжелую гамма-, альфа- или эпсилон-цепь. Потенциальная рекомбинация многих разных генов IGHV с несколькими генами IGHJ обеспечивает широкий диапазон распознавания антигенов. Дополнительного разнообразия достигают посредством разнообразия присоединения, что является результатом случайного добавления нуклеотидов с помощью концевой дезоксинуклеотидилтрансферазы и соматической гипермутации, происходящей во время созревания B-клеток в селезенке и лимфоузлах. Известно, что несколько сегментов V, D, J и C не могут кодировать белок, и их считают псевдогенными сегментами (часто просто обозначаемые как псевдогены).
Локус тяжелой цепи иммуноглобулина человека локализован на хромосоме 14 человека. В таблице 1 приведены гены IGHV и их относительный порядок в этом локусе.
Таблица 1. Список генов IGHV на хромосоме 14 человека
RPS8P1, ADAM6, и KIAA0125 также локализованы в этом локусе. Относительный порядок RPS8P1 представляет собой 160, относительный порядок ADAM6 представляет собой 161, и относительный порядок KIAA0125 представляет собой 164. В таблице 2 приведены все гены IGHD и их относительный порядок на хромосоме 14 человека. В таблице 3 приведены все гены IGHJ и их относительный порядок хромосома человека 14. Гены константных доменов иммуноглобулинов расположены после генов IGHV, IGHD и IGHJ. Эти гены включают (показаны в следующем порядке): константную область тяжелой мю-цепи иммуноглобулина (IGHM), константную область тяжелой дельта-цепи иммуноглобулина (IGH δ), константную область тяжелой гамма-цепи иммуноглобулина 3 (IGHG3), константную область тяжелой гамма-цепи иммуноглобулина 1 (IGHG1), константную область тяжелой эпсилон-цепи иммуноглобулина P1 (псевдоген) (IGHEP1), константную область тяжелой альфа-цепи иммуноглобулина 1 (IGHA1), константную область тяжелой гамма-цепи иммуноглобулина P (нефункциональную) (IGHGP), константную область тяжелой гамма-цепи иммуноглобулина 2 (IGHG2), константную область тяжелой гамма-цепи иммуноглобулина 4 (IGHG4), константную область тяжелой эпсилон-цепи иммуноглобулина (IGHE) и константную область тяжелой альфа-цепи иммуноглобулина 2 (IGHA2). Эти гены и порядок этих генов также показаны на фиг. 37 и фиг. 41.
Таблица 2. Список генов IGHD на хромосоме 14 человека
Таблица 3. Список генов IGHJ на хромосоме 14 человека
Локус тяжелой цепи иммуноглобулина мыши локализован на хромосоме 12 мыши. В таблице 4 приведены гены IGHV и их относительный порядок в этом локусе.
Таблица 4. Список генов IGHV на хромосоме 12 мыши
В таблице 5 приведены все гены IGHD и их относительный порядок на хромосоме 12 мыши. В таблице 6 приведены все гены IGHJ и их относительный порядок на хромосоме 12 мыши. Гены константных доменов иммуноглобулинов находятся после генов IGHV, IGHD и IGHJ. Эти гены включают (показаны в следующем порядке): гены константной области тяжелой мю-цепи иммуноглобулина (IGHM), константной области тяжелой дельта-цепи иммуноглобулина (IGH δ), константной области тяжелой гамма-цепи иммуноглобулина 3 (IGHG3), константной области тяжелой гамма-цепи иммуноглобулина 1 (IGHG1), константной области тяжелой гамма-цепи иммуноглобулина 2b (IGHG2b), константной области тяжелой гамма-цепи иммуноглобулина 2a (IGHG2a), константной области тяжелой эпсилон-цепи иммуноглобулина (IGHE) и константной области тяжелой альфа-цепи иммуноглобулина (IGHA). Эти гены и порядок этих генов также показаны на фиг. 38 и фиг. 42.
Таблица 5. Список генов IGHD на хромосоме 12 мыши
Таблица 6. Список генов IGHJ на хромосоме 12 мыши
Настоящее изобретение относится к генетически модифицированному, не являющемуся человеком животному, содержащему один или более генов IGHV человека, один или более генов IGHD человека и/или один или более генов IGHJ человека. В некоторых вариантах осуществления гены IGHV человека, гены IGHD человека и гены IGHJ человека функционально связаны и могут подвергаться реаранжировке VDJ. В некоторых вариантах осуществления гены IGHV человека, гены IGHD человека и гены IGHJ человека находятся в эндогенном локусе гена тяжелой цепи иммуноглобулина.
В некоторых вариантах осуществления животное содержит приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160 или 161 ген IGHV человека (например, гены, приведенные в таблице 1).
В некоторых вариантах осуществления животное содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов, выбранных из IGHV(III)-82, IGHV7-81, IGHV4-80, IGHV3-79, IGHV(II)-78-1, IGHV5-78, IGHV7-77, IGHV(III)-76-1, IGHV3-76 и IGHV3-75.
В некоторых вариантах осуществления животное содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов, выбранных из IGHV(III)-5-2, IGHV(III)-5-1, IGHV2-5, IGHV7-4-1, IGHV4-4, IGHV1-3, IGHV(III)-2-1, IGHV1-2, IGHV(II)-1-1 и IGHV6-1.
В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, содержащую последовательность, начинающуюся с гена, выбранного из IGHV(III)-82, IGHV7-81, IGHV4-80, IGHV3-79, IGHV(II)-78-1, IGHV5-78, IGHV7-77, IGHV(III)-76-1, IGHV3-76 и IGHV3-75, и заканчивающуюся геном, выбранным из IGHV(III)-5-2, IGHV(III)-5-1, IGHV2-5, IGHV7-4-1, IGHV4-4, IGHV1-3, IGHV(III)-2-1, IGHV1-2, IGHV(II)-1-1 и IGHV6-1. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHV(III)-82 человека до IGHV1-2 человека. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHV(III)-82 человека до IGHV(II)-1-1 человека. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHV(III)-82 человека до IGHV-6-1 человека.
В некоторых вариантах осуществления животное содержит приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26 или 27 генов IGHD человека (например, гены, приведенные в таблице 2). В некоторых вариантах осуществления животное содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов, выбранных из IGHD1-1, IGHD2-2, IGHD3-3, IGHD4-4, IGHD5-5, IGHD4-23, IGHD5-24, IGHD6-25, IGHD1-26 и IGHD7-27.
В некоторых вариантах осуществления животное содержит приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8 или 9 генов IGHJ человека (например, гены, приведенные в таблице 3). В некоторых вариантах осуществления животное содержит 1, 2, 3, 4, 5, 6, 7, 8 или 9 генов IGHJ человека, выбранных из IGHJ1P, IGHJ1, IGHJ2, IGHJ2P, IGHJ3, IGHJ4, IGHJ5, IGHJ3P и IGHJ6.
В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, содержащую последовательность, начинающуюся с гена, выбранного из IGHD1-1, IGHD2-2, IGHD3-3, IGHD4-4, IGHD5-5, IGHD4-23, IGHD5-24, IGHD6-25, IGHD1-26 и IGHD7-27, и заканчивающуюся геном, выбранных из IGHJ1P, IGHJ1, IGHJ2, IGHJ2P, IGHJ3, IGHJ4, IGHJ5, IGHJ3P и IGHJ6. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHD1-1 человека до IGHJ6 человека.
В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHD1-1 человека до IGHD7-27 человека.
В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHJ1P человека до IGHJ6 человека. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHJ1 человека до IGHJ6 человека.
В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHV(III)-82 человека до IGHJ6 человека.
В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHV1-2 человека до IGHJ6 человека. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHV(II)-1-1 человека до IGHJ6 человека. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса гена тяжелой цепи иммуноглобулина человека, тянущегося от IGHV6-1 человека до IGHJ6 человека.
В некоторых вариантах осуществления животное может иметь одну, две, три, четыре, пять, шесть, семь, восемь, девять или десять немодифицированных последовательностей человека. В некоторых вариантах осуществления немодифицированная последовательность человека имеет длину приблизительно или по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 т.п.н.
В некоторых вариантах осуществления животное содержит один или более эндогенных генов, выбранных из группы, состоящей из генов константной области тяжелой мю-цепи иммуноглобулина (IGHM), константной области тяжелой дельта-цепи иммуноглобулина (IGHδ), константной области тяжелой гамма-цепи иммуноглобулина 3 (IGHG3), константной области тяжелой гамма-цепи иммуноглобулина 1 (IGHG1), константной области тяжелой гамма-цепи иммуноглобулина 2b (IGHG2b), константной области тяжелой гамма-цепи иммуноглобулина 2a (IGHG2a), константной области тяжелой эпсилон-цепи иммуноглобулина (IGHE) и константной области тяжелой альфа-цепи иммуноглобулина (IGHA). В некоторых вариантах осуществления эти эндогенные гены функционально связаны. В некоторых вариантах осуществления эти эндогенные гены имеют тот же порядок, что и у животного дикого типа. В некоторых вариантах осуществления у животного может происходить переключение изотипа (переключение класса иммуноглобулинов).
В некоторых вариантах осуществления гены IGHV, гены IGHD и/или гены IGHJ функционально связаны. Рекомбинация VDJ может происходить между этими генами и приводить к образованию функциональных антител. В некоторых вариантах осуществления эти гены расположены в порядке, схожем с порядком в локусе тяжелой цепи иммуноглобулина человека. Это расположение обеспечивает различные преимущества, например, расположение этих генов делает возможной продукцию вариабельных доменов тяжелой цепи с разнообразием, очень схожим с разнообразием вариабельных доменов тяжелой цепи у человека. Т.к. некоторые случайные последовательности могут встраиваться в последовательность во время рекомбинации VDJ, в некоторых вариантах осуществления полные репертуары антител человека без модификаций или минимальные модификации могут снижать вероятность того, что не принадлежащая человеку последовательность встраивается во время рекомбинации VDJ.
В некоторых вариантах осуществления гены IGHV, гены IGHD и/или гены IGHJ функционально связаны с одним или более генами (например, всеми генами), выбранными из генов IGHM, IGHδ, IGHG3, IGHG1, IGHG2b, IGHG2a, IGHE и IGHA.
В некоторых вариантах осуществления животное имеет разрушенный эндогенный локус гена тяжелой цепи иммуноглобулина животного. В некоторых вариантах осуществления разрушение эндогенного локуса гена тяжелой цепи иммуноглобулина животного включает делецию одного или более эндогенных генов IGHV, одного или более эндогенных генов IGHD и одного или более эндогенных генов IGHJ.
В некоторых вариантах осуществления животное является мышью. Разрушение эндогенного локуса гена тяжелой цепи иммуноглобулина животного включает делецию по меньшей мере или приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181 или 182 генов мышей IGHV (например, гены, приведенные в таблице 4). В некоторых вариантах осуществления разрушение включает делецию приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGHV мыши, выбранных из IGHV1-86, IGHV1-85, IGHV1-84, IGHV1-83, IGHV1-82, IGHV1-81, IGHV1-80, IGHV1-79, IGHV1-78 и IGHV1-77. В некоторых вариантах осуществления мышь содержит приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGHV мыши, выбранных из IGHV1-86, IGHV1-85, IGHV1-84, IGHV1-83, IGHV1-82, IGHV1-81, IGHV1-80, IGHV1-79, IGHV1-78 и IGHV1-77 (например, IGHV1-86).
В некоторых вариантах осуществления разрушение включает делецию приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGHV мыши, выбранных из IGHV5-6, IGHV5-5, IGHV2-3, IGHV6-1, IGHV5-4, IGHV5-3, IGHV2-2, IGHV5-2, IGHV2-1 и IGHV5-1. В некоторых вариантах осуществления мышь содержит делецию приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGHV мыши, выбранных из IGHV5-6, IGHV5-5, IGHV2-3, IGHV6-1, IGHV5-4, IGHV5-3, IGHV2-2, IGHV5-2, IGHV2-1 и IGHV5-1.
В некоторых вариантах осуществления разрушение эндогенного локуса гена тяжелой цепи иммуноглобулина животного включает делецию по меньшей мере или приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 генов IGHD мыши (например, гены, приведенные в таблице 5). В некоторых вариантах осуществления разрушение включает делецию приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGHD мыши, выбранных из IGHD5-1, IGHD3-1, IGHD1-1, IGHD6-1, IGHD2-3, IGHD2-7, IGHD2-8, IGHD5-6, IGHD3-2 и IGHD4-1. В некоторых вариантах осуществления мышь содержит приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGHD мыши, выбранных из IGHD5-1, IGHD3-1, IGHD1-1, IGHD6-1, IGHD2-3, IGHD2-7, IGHD2-8, IGHD5-6, IGHD3-2 и IGHD4-1.
В некоторых вариантах осуществления разрушение включает делецию приблизительно или по меньшей мере 1, 2, 3 или 4 гена IGHJ мыши, выбранных из IGHJ1, IGHJ2, IGHJ3 и IGHJ4. В некоторых вариантах осуществления мышь содержит приблизительно или по меньшей мере 1, 2, 3 или 4 гена IGHJ мыши, выбранных из IGHJ1, IGHJ2, IGHJ3 и IGHJ4.
В некоторых вариантах осуществления разрушение эндогенного локуса гена тяжелой цепи иммуноглобулина животного включает делецию приблизительно или по меньшей мере 500 т.п.н., 600 т.п.н., 700 т.п.н., 800 т.п.н., 900 т.п.н., 1000 т.п.н., 1500 т.п.н., 2000 т.п.н., 2500 т.п.н. или 3000 т.п.н. эндогенной последовательности.
В некоторых вариантах осуществления делетированная последовательность тянется от IGHV1-86 до IGHJ4, от IGHV1-85 до IGHJ4, от IGHV1-84 до IGHJ4, от IGHV1-83 до IGHJ4 или от IGHV1-82 до IGHJ4 (например, от IGHV1-85 до IGHJ4).
В некоторых вариантах осуществления животное содержит приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 последовательностей, являющихся по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичных последовательности в локусе гена тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления последовательность имеет длину приблизительно или по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 2500, 3000 или 3500 т.п.н. В некоторых вариантах осуществления последовательность тянется от IGHV(III)-82 человека до IGHV1-2. В некоторых вариантах осуществления последовательность начинается от IGHV7-81 человека до IGHV1-2. В некоторых вариантах осуществления последовательность тянется от IGHV(II)-1-1 человека до IGHVJ6. В некоторых вариантах осуществления последовательность тянется от IGHV6-1 человека до IGHVJ6.
Гены IGHV человека, гены IGHD человека и гены IGHJ человека функционально связаны и могут подвергаться реаранжировке VDJ. В некоторых вариантах осуществления модифицированная мышь имеет полные репертуары генов IGHV, IGHD и IGHJ человека (например, включая все непсевдогенные гены IGHV, IGHD и IGHJ человека). Таким образом, модифицированная мышь может продуцировать полный репертуар антител человека. В некоторых вариантах осуществления после рекомбинации VDJ один ген IGHV (например, IGHV3-21 или IGHV3-74) из таблицы 15 вносит вклад в последовательность, кодирующую вариабельную область тяжелой цепи антитела. Один ген IGHD из таблицы 15 вносит вклад в последовательность, кодирующую вариабельную область тяжелой цепи антитела. И один ген IGHJ из таблицы 15 вносит вклад в последовательность, кодирующую вариабельную область тяжелой цепи антитела. В некоторых вариантах осуществления ген IGHV является IGHV3-21 или IGHV3-74.
В некоторых вариантах осуществления один ген IGHV (например, IGHV3-30, IGHV3-33, IGHV4-39 или IGHV4-34) на фиг. 70A и фиг. 70B вносит вклад в последовательность, кодирующую вариабельную область тяжелой цепи антитела. Один ген IGHD (например, IGHD6-19) на фиг. 70C вносит вклад в последовательность, кодирующую вариабельную область тяжелой цепи антитела. И один ген IGHJ (например, IGHJ4 или IGHJ6) на фиг. 70D вносит вклад в последовательность, кодирующую вариабельную область тяжелой цепи антитела. В некоторых вариантах осуществления один ген IGKV (например, IGKV4-1, IGKV1-33, IGKV2-30) на фиг. 71A и фиг. 71B вносит вклад в последовательность, кодирующую вариабельную область легкой цепи антитела. Один ген IGKJ (например, IGKJ1, IGKJ2, или IGKJ4) на фиг. 71C вносит вклад в последовательность, кодирующую вариабельную область легкой цепи антитела.
Кроме того, в некоторых случаях целые гены IGHV мыши, гены IGHD и гены IGHJ (например, включая все непсевдогенные гены) подвергают нокауту, и вариабельная область тяжелой цепи не будет иметь любую последовательность, кодируемую последовательность, полученную из мыши, что, таким образом, минимизирует иммуногенность для человека.
Генетически модифицированный локус легкой каппа-цепи иммуноглобулина
Локус каппа-цепь иммуноглобулина (также известный как IGK или каппа-локус иммуноглобулина) является областью на хромосоме (например, хромосоме 2 человека), содержащей гены легких цепей антител (или иммуноглобулинов) человека. Аналогично, гены легкой цепи иммуноглобулина могут также подвергаться серии реаранжировок, приводящих к продукции зрелой нуклеиновой кислоты легкой цепи (например, каппа-цепи) иммуноглобулина.
Соединение сегмента V (также известного как ген IGKV) и сегмента J (также известного как ген IGKJ) приводит к образованию непрерывного экзона, кодирующего весь вариабельный домен легкой цепи. В неподвергнутой реаранжировке ДНК сегменты гена V (или кластер генов IGKV) локализованы относительно далеко от C-области. Сегменты гена J (или кластер генов IGKJ) локализованы близко к C-области. Соединение сегмента V с сегментом гена J также сближает ген V с последовательностью C-области. Сегмент гена J подвергнутой реаранжировке V-области отделен от последовательности C-области только интроном. Для образования полной матричной РНК легкой цепи иммуноглобулина экзон V-области соединяется с последовательностью C-области посредством сплайсинга РНК после транскрипции.
Локус легкой цепи иммуноглобулина человека локализован на хромосоме 2 человека. В таблице 7 приведены гены IGKV и их относительный порядок в этом локусе. Существует несколько разных групп генов IGKV человека, включая гены IGKV1 (включая все гены IGKV, начинающиеся с IGKV1, также известного как VκI), гены IGKV2 (включая все гены IGKV, начинающиеся с IGKV2, также известного как VκII), гены IGKV3 (включая все гены IGKV, начинающиеся с IGKV3, также известного как VκIII), гены IGKV4 (включая все гены IGKV, начинающиеся с IGKV4, также известного как VκIV), гены IGKV5 (включая все гены IGKV, начинающиеся с IGKV5, также известного как VκV), гены IGKV6 (включая все гены IGKV, начинающиеся с IGKV6, также известного как VκVI) и гены IGKV7 (включая все гены IGKV, начинающиеся с IGKV7, также известного как VκVII).
Эти гены IGKV на хромосоме 2 человека также образуют два кластера, проксимальный кластер Vκ и дистальный кластер Vκ (фиг. 28). Последовательности в двух кластерах схожи, но не идентичны. Эта крупная сегментарная дупликация последовательности произошла из-за дивергенции человеческой линии от наиболее близкого общего предка с другими человекообразными приматами. Соответствующие гены IGVK в каждом кластере показаны на фиг. 64.
Таблица 7. Список генов IGKV на хромосоме 2 человека
В таблице 8 приведены все гены IGKJ и их относительный порядок на хромосоме 2 человека. Ген константной области каппа-цепи иммуноглобулина (IGKC), кодирующий константные домены легкой цепи иммуноглобулинов, локализован после генов IGKV и IGKJ. Эти гены и порядок этих генов также показаны на фиг. 39 и фиг. 43.
Таблица 8. Список генов IGKJ на хромосоме 2 человека
Локус легкой цепи иммуноглобулина мыши локализован на хромосоме 6 мыши. В таблице 9 приведены гены IGKV и их относительный порядок в этом локусе.
Таблица 9. Список генов IGKV на хромосоме 6 мыши
Gm9728 и Amd-ps2 также локализованы в этом локусе. Относительный порядок Gm9728 представляет собой 4, и относительный порядок Amd-ps2 представляет собой 134. В таблице 10 приведены все гены IGKJ и их относительный порядок на хромосоме 6 мыши. Ген IGKC, кодирующий константные домены легкой цепи иммуноглобулинов, находится после генов IGKV и IGKJ. Эти гены и порядок этих генов также показан на фиг. 40 и фиг. 44.
Таблица 10. Список генов IGKJ на хромосоме 6 мыши
Настоящее изобретение относится к генетически модифицированному, не являющемуся человеком животному, содержащему один или более генов IGKV человека и/или один или более генов IGKJ человека. В некоторых вариантах осуществления гены IGKV человека и гены IGKJ человека функционально связаны и могут подвергаться реаранжировке VJ. В некоторых вариантах осуществления гены IGKV человека и гены IGKJ человека находятся в эндогенном локусе гена легкой цепи иммуноглобулина.
В некоторых вариантах осуществления животное содержит приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75 или 76 генов IGKV человека (например, гены, приведенные в таблице 7).
В некоторых вариантах осуществления животное содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов, выбранных из IGKV3D-7, IGKV1D-8, IGKV1D-43, IGKV1D-42, IGKV2D-10, IGKV3D-11, IGKV1D-12, IGKV1D-13, IGKV2D-14 и IGKV3D-15.
В некоторых вариантах осуществления животное содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов, выбранных из IGKV2-10, IGKV1-9, IGKV1-8, IGKV3-7, IGKV1-6, IGKV1-5, IGKV2-4, IGKV7-3, IGKV5-2 и IGKV4-1.
В некоторых вариантах осуществления животное содержит приблизительно или по меньшей мере 1, 2, 3, 4 или 5 генов IGKJ человека (например, гены, приведенные в таблице 3). В некоторых вариантах осуществления животное содержит 1, 2, 3, 4 или 5 генов IGKJ человека, выбранных из IGKJ1, IGKJ2, IGKJ3, IGKJ4 и IGKJ5.
В некоторых вариантах осуществления животное содержит эндогенный IGKC. В некоторых вариантах осуществления гены IGKV и/или гены IGKJ функционально связаны. Рекомбинация VJ может происходить среди этих генов и приводить к образованию функциональных антител. В некоторых вариантах осуществления эти гены расположены в порядке, схожем с порядком в локусе легкой цепи иммуноглобулина человека. Это расположение обеспечивает различные преимущества, например, расположение этих генов делает возможной продукцию вариабельных доменов легких цепей с разнообразием, очень схожим с разнообразием вариабельных доменов легких цепей у человека.
В некоторых вариантах осуществления гены IGKV и/или гены IGKJ функционально связаны с геном IGKC (например, эндогенным геном IGKC).
В некоторых вариантах осуществления животное имеет разрушенный эндогенный локус гена легкой цепи иммуноглобулина животного. В некоторых вариантах осуществления разрушение эндогенного локуса гена легкой цепи иммуноглобулина животного включает делецию одного или более эндогенных генов IGKV и одного или более эндогенных генов IGKJ.
В некоторых вариантах осуществления животное является мышью. Разрушение эндогенного локуса гена тяжелой цепи иммуноглобулина животного включает делецию по меньшей мере или приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162 или 163 гена IGKV мыши (например, гены, приведенные в таблице 9). В некоторых вариантах осуществления разрушение включает делецию приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGKV мыши, выбранных из IGKV2-137, IGKV1-136, IGKV1-135, IGKV14-134-1, IGKV17-134, IGKV1-133, IGKV1-132, IGKV1-131, IGKV14-130 и IGKV9-129. В некоторых вариантах осуществления мышь содержит приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGKV мыши, выбранных из IGKV2-137, IGKV1-136, IGKV1-135, IGKV14-134-1, IGKV17-134, IGKV1-133, IGKV1-132, IGKV1-131, IGKV14-130 и IGKV9-129.
В некоторых вариантах осуществления разрушение включает делецию приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGKV мыши, выбранных из IGKV3-10, IGKV3-9, IGKV3-8, IGKV3-7, IGKV3-6, IGKV3-5, IGKV3-4, IGKV3-3, IGKV3-2 и IGKV3-1. В некоторых вариантах осуществления мышь содержит приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGKV мыши, выбранных из IGKV3-10, IGKV3-9, IGKV3-8, IGKV3-7, IGKV3-6, IGKV3-5, IGKV3-4, IGKV3-3, IGKV3-2 и IGKV3-1.
В некоторых вариантах осуществления разрушение включает делецию приблизительно или по меньшей мере 1, 2, 3, 4 или 5 гены IGKJ мыши, выбранных из IGKJ1, IGKJ2, IGKJ3, IGKJ4 и IGKJ5. В некоторых вариантах осуществления мышь содержит приблизительно или по меньшей мере 1, 2, 3, 4 или 5 генов IGKJ мыши, выбранных из IGKJ1, IGKJ2, IGKJ3, IGKJ4 и IGKJ5 (например, IGKJ5).
В некоторых вариантах осуществления разрушение эндогенного локуса гена легкой каппа-цепи иммуноглобулина животного включает делецию приблизительно или по меньшей мере 500 т.п.н., 600 т.п.н., 700 т.п.н., 800 т.п.н., 900 т.п.н., 1000 т.п.н., 1500 т.п.н., 2000 т.п.н., 2500 т.п.н., 3000 т.п.н. или 3500 т.п.н. эндогенной последовательности.
В некоторых вариантах осуществления делетированная последовательность тянется от IGKV2-137 до IGKJ4, от IGKV1-136 до IGKJ4, от IGKV1-135 до IGKJ4, от IGKV2-137 до IGKJ5, от IGKV1-136 до IGKJ5 или от IGKV1-135 до IGKJ5 (например, от IGKV2-137 до IGKJ5).
В некоторых вариантах осуществления животное содержит приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 последовательностей, являющихся по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичными последовательности локуса гена легкой цепи иммуноглобулина человека. В некоторых вариантах осуществления последовательность имеет длину приблизительно или по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 2500, 3000 или 3500 т.п.н.
В некоторых вариантах осуществления животное может иметь одну, две, три, четыре, пять, шесть, семь, восемь, девять или десять немодифицированных последовательностей человека. В некоторых вариантах осуществления немодифицированная последовательность человека имеет длину приблизительно или по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 2500, 3000 или 3500 т.п.н.
В некоторых вариантах осуществления последовательность тянется от IGKV3D-7 человека до IGKJ5. В некоторых вариантах осуществления последовательность тянется от IGKV3D-7 человека до IGKJ4. В некоторых вариантах осуществления последовательность тянется от IGKV1D-8 человека до IGKJ5. В некоторых вариантах осуществления последовательность тянется от IGKV1D-8 человека до IGKJ4.
Гены IGKV человека и гены IGKJ человека функционально связаны и могут подвергаться реаранжировке VJ. В некоторых вариантах осуществления модифицированная мышь имеет полные репертуары генов IGKV и IGKJ человека (например, включая все непсевдогенные гены IGKV и IGKJ человека). Таким образом, модифицированная мышь может продуцировать полный репертуар антител человека. В некоторых вариантах осуществления после рекомбинации VJ один ген IGKV (например, IGKV1D-43, IGKV1D-13, IGKV1D-16, или IGKV1D-12) из таблицы 16 вносит вклад в последовательность, кодирующую вариабельную область легкой цепи антитела. Один ген IGKJ человека вносит вклад в последовательность, кодирующую вариабельную область легкой цепи антитела. В некоторых вариантах осуществления ген IGKV является IGKV1D-43, IGKV1D-13, IGKV1D-16 или IGKV1D-12. Кроме того, в некоторых случаях целые гены IGKV мыши и гены IGKJ (все непсевдогенные гены) подвергают нокауту, и вариабельная область легкой цепи не будет иметь любую последовательность, кодируемую последовательностью, полученной из мыши, что, таким образом, минимизирует иммуногенность у людей.
В некоторых вариантах осуществления человек гены IGKV проксимального кластера Vκ включены в модифицированную хромосому. В некоторых вариантах осуществления гены IGKV дистального кластера Vκ человека включены в модифицированную хромосому. В некоторых вариантах осуществления и гены IGKV проксимального кластера Vκ человека и гены IGKV дистального кластера Vκ человека включены в модифицированную хромосому.
Генетически модифицированный локус легкой лямбда-цепи иммуноглобулина
Локус лямбда-цепи иммуноглобулина (также известный как IGL или лямбда-локус иммуноглобулина) является областью на хромосоме (например, хромосоме 22 человека), содержащей гены легких цепей антител (или иммуноглобулинов) человека. Аналогично, гены легкой цепи иммуноглобулина также могут подвергаться серии реаранжировок, приводящих к продукции зрелой нуклеиновой кислоты легкой цепи (например, лямбда-цепи) иммуноглобулина. У здоровых людей общее соотношение каппа/лямбда составляет приблизительно 2:1 в сыворотке (при измерении интактных целых антител) или 1:1,5 при измерении свободных легких цепей. У мышей общее соотношение каппа/лямбда составляет приблизительно 9:1.
В некоторых вариантах осуществления животное содержит локус лямбда-цепи иммуноглобулина человека.
В некоторых вариантах осуществления животное имеет разрушенный эндогенный локус гена легкой лямбда-цепи иммуноглобулина животного. В некоторых вариантах осуществления разрушение эндогенного локуса гена легкой цепи иммуноглобулина животного включает делецию одного или более эндогенных генов IGLV, один или более эндогенных генов IGLJ и/или один или более генов константной области иммуноглобулин лямбда-цепи (IGLC) (например, IGLC1, IGLC2, IGLC3 и IGLC4).
Локус легкой лямбда-цепи иммуноглобулина мыши (локус IGL) локализован на хромосоме 16 мыши. В таблице 11 приведены гены IGLV, IGLJ и IGLC и их относительный порядок в этом локусе.
Таблица 11. Список генов в локусе IGL мыши
Разрушение эндогенного локуса гена легкой лямбда-цепи иммуноглобулина животного включает делецию по меньшей мере или приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 генов IGLV, IGLJ и IGLC мыши (например, гены, приведенные в таблице 11). В некоторых вариантах осуществления делеция включает приблизительно или по меньшей мере 1, 2, 3 или 4 гена IGKC мыши, выбранных из IGLC1, IGLC2, IGLC3 и IGLC4. В некоторых вариантах осуществления разрушение включает делецию приблизительно или по меньшей мере 1, 2 или 3 гена IGLV мыши, выбранных из IGLV1, IGLV2 и IGLV3. В некоторых вариантах осуществления разрушение включает делецию приблизительно или по меньшей мере 1, 2, 3, 4 или 5 генов IGLJ мыши, выбранных из IGLJ1, IGLJ2, IGLJ3, IGLJ3P и IGLJ4.
В некоторых вариантах осуществления разрушение эндогенного локуса гена легкой лямбда-цепи иммуноглобулина животного включает делецию приблизительно или по меньшей мере 10 т.п.н., 20 т.п.н., 30 т.п.н., 40 т.п.н., 50 т.п.н., 60 т.п.н., 70 т.п.н., 80 т.п.н., 90 т.п.н., 100 т.п.н., 110 т.п.н., 120 т.п.н., 130 т.п.н., 140 т.п.н., 150 т.п.н., 160 т.п.н., 170 т.п.н., 180 т.п.н., 190 т.п.н., 200 т.п.н., 210 т.п.н., 220 т.п.н., 230 т.п.н., 240 т.п.н., 250 т.п.н., 260 т.п.н., 270 т.п.н., 280 т.п.н., 290 т.п.н., 300 т.п.н., 350 т.п.н., 400 т.п.н., 450 т.п.н., 500 т.п.н. или 1000 т.п.н. В некоторых вариантах осуществления разрушение в эндогенном гене легкой лямбда-цепи иммуноглобулина животного отсутствует.
В некоторых вариантах осуществления делетированная последовательность тянется от IGLV2 до IGLC1, от IGLV3 до IGLC1 или от IGLJ2 до IGLC1.
Генетически модифицированные животные
В одном из аспектов настоящее изобретение относится к генетически модифицированному, не являющемуся человеком животному, содержащему гуманизированный локус тяжелой цепи иммуноглобулина и/или гуманизированный локус легкой цепи иммуноглобулина. В некоторых вариантах осуществления животное содержит один или более генов IGHV человека, один или более генов IGHD человека, один или более генов IGHJ человека, один или более генов IGKV человека и/или один или более генов IGKJ человека. В некоторых вариантах осуществления эти гены находятся в эндогенном локусе генов иммуноглобулинов.
В некоторых вариантах осуществления животное содержит локус лямбда-цепи иммуноглобулина человека. В некоторых вариантах осуществления животное имеет разрушенный эндогенный локус гена легкой лямбда-цепи иммуноглобулина животного. В некоторых вариантах осуществления животное не имеет разрушение эндогенного локуса гена легкой лямбда-цепи иммуноглобулина животного.
Генетически модифицированное, не являющееся человеком животное может являться различными животными, например, мышью, крысой, кроликом, свиньей, крупным рогатым скотом (например, коровой, буйволом, бизоном), оленем, овцой, козой, курицей, кошкой, собакой, хорьком, приматом (например, игрункой, макаком-резусом). Для не являющихся человеком животных, в случае которых недоступны подходящие генетически модифицируемые стволовые клетки (ES), используют другие способы получения не являющегося человеком животного, имеющего генетическую модификацию. Такие способы включают, например, модификацию генома не-ES клетки (например, фибробласта или индуцированной плюрипотентной клетки) и использование ядерного трансфера для переноса модифицированного генома в подходящую клетку, например, ооцит, и вынашивание модифицированной клетки (например, модифицированного ооцита) в не являющемся человеком животном в условиях, подходящих для образования эмбриона. Эти способы известны в этой области и описаны, например, в A. Nagy, et al., "Manipulating Mouse Embryo: A Laboratory Manual (Third Edition)," Cold Spring Harbor Laboratory Press, 2003, включенных в настоящее описание в качестве ссылки в полном объеме. Таким образом, в различных вариантах осуществления сегменты V, D и/или J человека можно функционально связывать с последовательностями генов константной области не являющегося человеком животного (например, грызуна, мыши, крысы, хомяка). Во время развития B-клеток эти подвергнутый реаранжировке сегменты V, D и/или J человека связаны с константной областью иммуноглобулина не являющегося человеком животного.
В одном из аспектов животное является млекопитающим, например, суперсемейства Dipodoidea или Muroidea. В некоторых вариантах осуществления генетически модифицированное животное является грызуном. Грызун может быть выбран из мыши, крысы и хомяка. В некоторых вариантах осуществления генетически модифицированное животное относится к семейству, выбранному из Calomyscidae (например, мышевидные хомячки), Cricetidae (например, хомяк, крысы и мыши Нового света, полевки), Muridae (истинные мыши и крысы, песчанки, иглистые мыши, косматые хомяки), Nesomyidae (рипидомисы, скальные крысы, белохвостые хомяки, малагасийские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). В некоторых вариантах осуществления генетически модифицированный грызун выбран из истинной мыши или крысы (семейство Muridae), песчанки, иглистой мыши и косматого хомяка. В некоторых вариантах осуществления не являющееся человеком животное является мышью.
В некоторых вариантах осуществления животное является мышью с фоном C57 (например, линия C57BL, выбранная из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola). В некоторых вариантах осуществления мышь относится к линии 129, выбранной из группы, состоящей из линии 129P1, 129P2, 129P3, 129X1, 129S1 (например, 129S1/SV, 129S1/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129T1, 129T2. Эти мыши описаны, например, в Festing et al., Revised nomenclature for strain 129 mice, Mammalian Genome 10: 836 (1999); Auerbach et al., Establishment and Chimera Analysis of 129/SvEv- и C57BL/6-Derived Mouse Embryonic Stem Cell Lines (2000), оба из которых включены в настоящее описание в качестве ссылки в полном объеме. В некоторых вариантах осуществления генетически модифицированная мышь представляет собой смешение линии 129 и линии C57BL/6. В некоторых вариантах осуществления мышь представляет собой смешение линий 129 или смешение линий BL/6. В некоторых вариантах осуществления мышь относится к линии BALB, например, линии BALB/c. В некоторых вариантах осуществления мышь представляет собой смешение линии BALB и другой линии. В некоторых вариантах осуществления мышь относится к гибридной линии (например, 50% BALB/c-50% 12954/Sv; или 50% C57BL/6-50% 129).
В некоторых вариантах осуществления животное является крысой. Крыса может быть выбрана из крысы Вистар, линии LEA, линии Спрег-Доули, линии Фишера, F344, F6 и крысы Dark Agouti. В некоторых вариантах осуществления линия крысы представляет собой смешение двух или более линий, выбранных из группы, состоящей из Вистар, LEA, Спрег-Доули, Фишера, F344, F6 и Dark Agouti.
Животное может иметь одну или более других генетических модификаций и/или других модификаций, подходящих для конкретной цели, для которой получают гуманизированное животное.
Генетически модифицированные, не являющиеся человеком животные, содержащие модификацию эндогенного не принадлежащего человеку локуса генов иммуноглобулина. В некоторых вариантах осуществления модификация может включать последовательность нуклеиновой кислоты человека, кодирующую по меньшей мере часть белка человека (например, по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичную последовательности вариабельного домена тяжелой цепи или вариабельного домена легкой цепи человека). Хотя настоящее изображение также относится к генетически модифицированным клеткам, которые могут содержать модификации, представленные в настоящем описании (например, клетки ES, соматическая клетки), во многих вариантах осуществления генетически модифицированные, не являющиеся человеком животные содержат модификацию эндогенного локуса в зародышевой линии животного.
Генетически модифицированные животные могут экспрессировать гуманизированное антитело и/или химерное антитело из эндогенных локусов мыши, где один или более эндогенных генов иммуноглобулинов мыши заменяют генами иммуноглобулинов человека и/или нуклеотидной последовательностью, по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичной последовательностям генов иммуноглобулинов человека (например, генов IGHV, IGHD, IGHJ, IGKV и/или IGKJ). В различных вариантах осуществления эндогенный не принадлежащий человеку локус генов иммуноглобулинов модифицируют полностью или частично так, чтобы они содержали последовательность нуклеиновой кислоты человека.
Можно осуществлять генетические, молекулярные и поведенческие анализы описанных выше не являющихся человеком млекопитающих. Настоящее изобретение также относится к потомству, даваемому не являющимся человеком млекопитающим по изобретению, скрещенным с животным того же или другого генотипа. Не являющиеся человеком млекопитающие могут являться любым не являющимся человеком животным, известным в этой области и которое можно использовать в способах, как представлено в настоящем описании. Предпочтительные не являющиеся человеком млекопитающие являются млекопитающими (например, грызунами). В некоторых вариантах осуществления не являющееся человеком млекопитающее является мышью.
Настоящее изобретение также относится к линии клеток или культуре первичных клеток, полученных из не являющегося человеком млекопитающего или его потомства. Модель на основе культуры клеток можно получать, например, следующими способами. Культуры клеток можно получать посредством выделения из не являющегося человеком млекопитающего, альтернативно, клетку можно получать из культуры клеток, полученной с использованием тех же конструкций и стандартных способов трансфекции клеток. Интеграцию генетических конструкций, содержащих последовательности ДНК, кодирующие иммуноглобулины человека или гуманизированные иммуноглобулины, можно определять различными способами.
Существует множество аналитических способов, которые можно использовать для детекции экзогенной ДНК или модификаций в геномной ДНК, включая способы на уровне нуклеиновой кислоты (включая подходы количественного анализа мРНК с использованием полимеразной цепной реакции с обратной транскрипцией (RT-ПЦР) или Саузерн-блоттинга и гибридизации in situ) и способы на уровне белка (включая гистохимию, иммуноблоттинг и исследования связывания in vitro). Кроме того, уровень экспрессии интересующего гена можно количественно анализировать способами ELISA, хорошо известными специалистам в этой области. Для проведения количественных измерений можно использовать множество стандартных способов анализа. Например, уровни транскрипции можно измерять с использованием RT-ПЦР и способов гибридизации, включая защиту от РНКазы, Саузерн-блоттинг, дот-анализ РНК (RNAdot). Для оценки наличия белков человека или гуманизированных белков также можно использовать иммуногистохимическое окрашивание, проточную цитометрию, анализ вестерн-блоттинга.
Антитела и антигенсвязывающие фрагменты
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам (например, гуманизированным антителам или химерным антителам), получаемым способами, представленными в настоящем описании.
В основном, антитела (также названные иммуноглобулинами) состоят из двух классов полипептидных цепей, легких цепей и тяжелых цепей. Неограничивающее антитело по изобретению может являться интактным антителом из четырех иммуноглобулиновых цепей, содержащим две тяжелые цепи и две легкие цепи. Тяжелая цепь антитела может иметь любой изотип, включая IgM, IgG, IgE, IgA или IgD или подклассы, включая IgG1, IgG2, IgG2a, IgG2b, IgG3, IgG4, IgE1, IgE2 и т.д. Легкая цепь может являться легкой каппа-цепью или легкой лямбда-цепью. Антитело может содержать две идентичные копии легкой цепи и две идентичные копии тяжелой цепи. Тяжелые цепи, каждая из которых содержит один вариабельный домен (или вариабельную область, VH) и множество константных доменов (или константных областей), связываются друг с другом посредством дисульфидной связи в своих константных доменах с образованием "стебля" антитела. Легкие цепи, каждая из которых содержит один вариабельный домен (или вариабельную область, VL) и один константный домен (или константную область), каждый из которых связывается с одной тяжелой цепью с помощью дисульфидной связи. Вариабельная область каждой легкой цепи выравнивается с вариабельной областью тяжелой цепи, с которой она связывается. Вариабельные области обеих легких цепей и тяжелых цепей содержат три гипервариабельные области, расположенные между более консервативными каркасными областями (FR).
Эти гипервариабельные области, известные как определяющие комплементарность области (CDR), образуют петли, содержащие главную антигенсвязывающую поверхность антитела. Четыре каркасные области, главным образом, принимают конформацию бета-листа, и CDR образуют петли, соединяющие и в некоторых случаях образующие часть структуры бета-листа. CDR в каждой цепи удерживаются в непосредственной близости каркасными областями и, вместе с CDR из другой цепи, участвуют в образовании антигенсвязывающей области.
Способы идентификации областей CDR антитела посредством анализа аминокислотной последовательности антитела хорошо известны, и ряд определений CDR является общеупотребительным. Определение по Kabat основано на вариабельности последовательности, и определение по Chothia основано на локализации структурных областей петель. Эти способы и определения описаны, например, в Martin, "Protein sequence and structure analysis of antibody variable domains," Antibody engineering, Springer Berlin Heidelberg, 2001. 422-439; Abhinandan, et al. "Analysis and improvements to Kabat and structurally correct numbering of antibody variable domains," Molecular immunology 45,14 (2008): 3832-3839; Wu, T.T. and Kabat, E.A. (1970) J. Exp. Med. 132: 211-250; Martin et al., Methods Enzymol. 203: 121-53 (1991); Morea et al., Biophys Chem. 68(1-3): 9-16 (Oct. 1997); Morea et al., J Mol Biol. 275(2): 269-94 (Jan.1998); Chothia et al., Nature 342(6252): 877-83 (Dec. 1989); Ponomarenko and Bourne, BMC Structural Biology 7:64 (2007); каждый из которых включен в настоящее описание в качестве ссылки в полном объеме.
CDR важны для распознавания эпитопа антигена. В рамках изобретения "эпитоп" представляет собой наименьшую часть молекулы-мишени, которая может специфически связываться антигенсвязывающим доменом антитела. Минимальный размер эпитопа может составлять приблизительно три, четыре, пять, шесть или семь аминокислот, но эти аминокислоты могут не являться последовательной линейной последовательностью первичной структурой антигена, т.к. эпитоп может зависеть от трехмерной конфигурации антигена на основе вторичной и третичной структуры антигена.
В некоторых вариантах осуществления антитело является интактной молекулой иммуноглобулина (например, IgG1, IgG2a, IgG2b, IgG3, IgG4, IgM, IgD, IgE, IgA). Подклассы IgG (IgG1, IgG2, IgG3 и IgG4) являются высококонсервативными, отличаются своей константной областью, в частности, шарнирной областью и верхними доменами CH2. Последовательности и различия подклассов IgG известны в этой области и описаны, например, в Vidarsson, et al, "IgG subclasses and allotypes: from structure to effector functions." Frontiers in immunology 5 (2014); Irani, et al. "Molecular properties of human IgG subclasses and their implications for designing therapeutic monoclonal antibodies against infectious diseases." Molecular immunology 67,2 (2015): 171-182; Shakib, Farouk, ed. The human IgG subclasses: molecular analysis of structure, function and regulation. Elsevier, 2016; каждый из которых включен в настоящее описание в качестве ссылки в полном объеме.
Антитело также может являться молекулой иммуноглобулина, полученной из любого биологического вида (например, человека, грызуна, мыши, крысы, верблюжьих). Антитела, представленные в настоящем описании, также включают, в качестве неограничивающих примеров, поликлональные, моноклональные, моноспецифические, полиспецифические антитела и химерные антитела, включающие связывающий домен иммуноглобулина, слитый с другим полипептидом. Термин "антигенсвязывающий домен" или "антигенсвязывающий фрагмент" является частью антитела, сохраняющей специфическую связывающую активность интактного антитела, т.е. любой частью антитела, способной специфически связываться с эпитопом на молекуле-мишени интактного антитела. Он включает, например, Fab, Fab', F(ab')2 и варианты этих фрагментов. Таким образом, в некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент может являться, например, scFv, Fv, Fd, dAb, биспецифическим антителом, биспецифическим scFv, диателом, линейным антителом, молекулой одноцепочечного антитела, мультиспецифическим антителом, образованным из фрагментов антител, и любым полипептидом, включающим связывающий домен, являющийся связывающим доменом антитела или гомологичный ему. Неограничивающие примеры антигенсвязывающих доменов включают, например, CDR тяжелой цепи и/или легкой цепи интактного антитела, вариабельные области тяжелой цепи и/или легкой цепи интактного антитела, полноразмерные тяжелые или легкие цепи интактного антитела или отдельную CDR из тяжелой цепи или легкой цепи интактного антитела.
В некоторых вариантах осуществления антигенсвязывающий фрагмент может образовывать часть химерного антигенного рецептора (CAR). В некоторых вариантах осуществления химерный антигенный рецептор представляет собой слитые белки из одноцепочечных вариабельных фрагментов (scFv), как представлено в настоящем описании, слитых с трансмембранным доменом и эндодоменом CD3-дзета.
В некоторых вариантах осуществления scFV содержит один вариабельный домен тяжелой цепи и один вариабельный домен легкой цепи. В некоторых вариантах осуществления scFV содержит два вариабельных домена тяжелой цепи и два вариабельных домена легких цепей. В некоторых вариантах осуществления scFV содержит две антигенсвязывающие области, и две антигенсвязывающие области могут связываться с соответствующими антигенами-мишенями.
Антитела и их антигенсвязывающие фрагменты (например, гуманизированные антитела или химерные антитела), получаемые способами, представленными в настоящем описании, обладают различными преимуществами. В некоторых вариантах осуществления не требуется дополнительная оптимизация для достижения желаемых свойств (например, аффинностей связывания, термической стабильности и/или ограниченной агрегации).
В некоторых вариантах осуществления антитело (или их антигенсвязывающие фрагменты) специфически связывается с мишенью со скоростью диссоциации (koff) менее 0,1 с-1, менее 0,01 с-1, менее 0,001 с-1, менее 0,0001 с-1 или менее 0,00001 с-1. В некоторых вариантах осуществления скорость диссоциации (koff) составляет более 0,01 с-1, более 0,001 с-1, более 0,0001 с-1, более 0,00001 с-1 или более 0,000001 с-1.
В некоторых вариантах осуществления скорость ассоциации(kon) составляет более 1×102/Мс, более 1×103/Мс, более 1×104/Мс, более 1×105/М, или более 1×106/Мс. В некоторых вариантах осуществления скорость ассоциации (kon) составляет менее 1×105/Мс, менее 1×106/Мс или менее 1×107/Мс.
Вывод об аффинностях можно делать из соотношения констант скоростей (KD=koff/kon). В некоторых вариантах осуществления KD составляет менее 1×10-6 M, менее 1×10-7 M, менее 1×10-8 M, менее 1×10-9 M или менее 1×10-10 M. В некоторых вариантах осуществления KD составляет менее 50 нМ, 40 нМ, 30 нМ, 20 нМ, 15 нМ, 10 нМ, 9 нМ, 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ или 1 нМ. В некоторых вариантах осуществления KD составляет более 1×10-7 M, более 1×10-8 M, более 1×10-9 M, более 1×10-10 M, более 1×10-11 M или более 1×10-12 M. В некоторых вариантах осуществления антитело связывается с мишенью с KD приблизительно 0,9 нМ, 0,8 нМ, 0,7 нМ, 0,6 нМ, 0,5 нМ, 0,4 нМ, 0,3 нМ, 0,2 нМ, 0,1 нМ или менее.
В некоторых вариантах осуществления определяют термическую стабильность. Антитела или антигенсвязывающие фрагменты, представленные в настоящем описании, могут иметь Tm более 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95°C.
Т.к. IgG можно описать как мультидоменный белок, кривые плавления иногда демонстрируют два перехода или три перехода с первой температурой денатурации Tm D1 второй температурой денатурации Tm D2 и, необязательно, третьей температурой денатурации Tm D3.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, представленные в настоящем описании, имеют Tm D1 более 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95°C. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, представленные в настоящем описании, имеют Tm D2 более 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95°C. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, представленные в настоящем описании, имеют Tm D3 более 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95°C.
В некоторых вариантах осуществления Tm, Tm D1, Tm D2, Tm D3 составляют менее 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95°C.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, представленные в настоящем описании, не образуют агрегаты, когда температура составляет менее 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95°C.
Способы получения генетически модифицированных животных
Генетически модифицированных животных можно получать посредством встраивания генов иммуноглобулинов человека в геном не являющихся человеком животных для получения животных, которые могут экспрессировать гуманизированные антитела или химерные антитела. На фиг. 1A показаны способы получения гуманизированных животных. В некоторых вариантах осуществления способы сначала включают модификацию иммуноглобулинового локуса человека на хромосоме человека. Затем модифицированные хромосомы человека встраивают в клетку-реципиент мыши. Затем вариабельную область иммуноглобулина человека встраивают в соответствующую область генома мыши посредством прямой замены. Затем клетки-реципиенты подвергают скринингу. В некоторых вариантах осуществления клетки не содержат хромосомы человека. Затем клетки инъецируют в бластоцисты для получения химерных мышей. Можно осуществлять последующее скрещивание для получения мышей, содержащих интактный гуманизированный иммуноглобулиновый локус.
Для получения генетически модифицированных животных можно использовать несколько других способов, включая, например, негомологичное соединение концов (NHEJ), гомологичную рекомбинацию (HR), нуклеазы с цинковыми пальцами (ZFN), эффекторные нуклеазы, подобные активаторам транскрипции (TALEN), и систему коротких палиндромных повторов, регулярно расположенных группами (CRISPR)-Cas. В некоторых вариантах осуществления используют гомологичную рекомбинацию. В некоторых вариантах осуществления для получения генетически модифицированных животных используют редактирование генома с помощью CRISPR-Cas9. Многие из этих способов редактирования генома известны в этой области и описаны, например, в Yin et al., "Delivery technologies for genome editing," Nature Reviews Drug Discovery 16,6 (2017): 387-399, включенном в настоящее описание в качестве ссылки в полном объеме. Также известно множество других способов, и их можно использовать в редактировании генома, например, микроинъекцию генетически модифицированного ядра в энуклеированный ооцит и слияние энуклеированного ооцита с другой генетически модифицированной клеткой.
Способ генетической модификации может включать замену эндогенной последовательности последовательностью человека посредством гомологичной рекомбинации. В некоторых вариантах осуществления расщепление выше и ниже целевого участка (например, посредством нуклеаз с цинковыми пальцами, TALEN или CRISPR) может приводить к двухцепочечному разрыву ДНК, и гомологичную рекомбинацию используют для замены эндогенной последовательности последовательностью человека.
В некоторых вариантах осуществления способы получения генетически модифицированного, гуманизированного животного могут включать стадию замены в эндогенном локусе (или участке) нуклеиновой кислоты (например, областей V, D, J или областей V, J) соответствующей областью последовательности человека. Последовательность может включать область (например, часть или всю область) генов IGHV, IGHD, IGHJ, IGKV и/или IGKJ. В некоторых вариантах осуществления замена опосредована гомологичной рекомбинацией. В некоторых вариантах осуществления замена опосредована рекомбиназой Cre.
На фиг. 9 показана стратегия таргетинга для добавления функциональных генетических элементов в хромосому человека. Эти векторы можно встраивать выше V-области, между J-областью и C-областью.
В некоторых вариантах осуществления первый вектор от 5’ до 3’ содержит одно или более из следующего: последовательности ДНК гомологичного плеча выше участка инсерции, промотора PGK, репортерного гена красного флуоресцентного белка (tdTomato), саморасщепляющегося пептида (2A) FMDV (вирусов ящура), гена резистентности к зеоцину (Zeo), последовательности терминации транскрипции/сигнала полиаденилирования (поли(A); "PA"), последовательности распознавания LoxP, гена резистентности к гигромицину (частичной последовательности гигромицинфосфотрансферазы; "3’HygR") и мишени распознавания Flp ("FRT"), нижележащей последовательности ДНК гомологичного плеча и гена DTA.
Второй вектор от 5’ до 3’ содержит одно или более из следующего: последовательности ДНК гомологичного плеча выше участка инсерции, последовательности распознавания LoxP, промотора PGK, частичной последовательности гена резистентности к пуромицину (5’PuroR), экспрессирующего промотора млекопитающего (EF-1a) из фактора элонгации 1 альфа человека, последовательности гена транспозазы piggyBac (PBase), внутренних участков связывания рибосомы (IRES), последовательности гена резистентности к канамицину (Neo), последовательности терминации транскрипции/сигнала полиаденилирования, последовательности ДНК гомологичного плеча ниже участка инсерции и DTA.
Эти векторы можно встраивать в геном клеток и клетки можно подвергать селекции с помощью маркеров резистентности к лекарственному средству или их комбинации (например, зеоцина, G418 и/или пуромицина). В некоторых вариантах осуществления экспрессируется транспозаза PB, и можно осуществлять делецию генетических элементов между последовательностями мишени транспозазы.
В некоторых вариантах осуществления эти векторы встраивают в хромосому человека, подвергнутую модификации. Хромосому человека сначала можно модифицировать перед встраиванием первого и второго векторов в геном. В некоторых вариантах осуществления один или более дополнительных векторов, при необходимости, можно добавлять в различные участки хромосомы. В некоторых вариантах осуществления вектор добавляют между C-областью и центромерой. Третий вектор от 5’ до 3’ содержит одно или более из следующего: последовательности ДНК гомологичного плеча выше участка инсерции, промотора PGK, последовательности гена резистентности к пуромицину (PuroR), последовательности гена тимидинкиназы (TK), последовательности распознавания LoxP, промотора PGK, частичной последовательности гена резистентности к пуромицину (5’PuroR), экспрессирующего промотора млекопитающего (EF-1a), PBase, IRES, Neo, последовательности терминации транскрипции/сигнала полиаденилирования, последовательности ДНК гомологичного плеча ниже участка инсерции и DTA. В некоторых вариантах осуществления эти векторы можно встраивать в ген вариабельной области или константной области. В некоторых вариантах осуществления делеции подвергают часть гена эндогенной вариабельной области или эндогенной константной области. В некоторых вариантах осуществления делеции подвергают крупный фрагмент хромосомы (например, между константной областью и центромерой). Клетки также можно обрабатывать ферментом Cre, что приводит к рекомбинации участков loxP и, таким образом, удалению последовательностей геномной ДНК между J-областью и центромерой на хромосоме 14 человека или между C-областью и центромерой на хромосоме 14 человека. В некоторых вариантах осуществления может происходить спонтанный хромосомный разрыв. Для экспериментов можно выбирать модифицированные хромосомы человека с желаемым хромосомным разрывом.
Хромосому человека можно получать из линий клеток человека, злокачественных клеток, культуры первичных клеток и/или фибробластов человека. В некоторых вариантах осуществления в клетку человека встраивают первый вектор, а затем подвергают ее слиянию с клеткой-реципиентом. Затем модифицированную хромосому отделяют и встраивают в другую подходящую клетку-реципиента. Клетки с желаемой резистентностью подвергают селекции для получения клеток, содержащих только одну хромосому человека. Затем в клетки встраивают второй вектор и клетки подвергают селекции по резистентности. Затем, при необходимости, можно встраивать третий вектор и/или четвертый вектор. Клетка-реципиент может являться клеткой млекопитающего, клеткой человека или клеткой мыши. В некоторых вариантах осуществления клетка-реципиент является клеткой CHO или, предпочтительно, клеткой A9. В некоторых вариантах осуществления модифицированные хромосомы флуоресцентно метят и отделяют. И модифицированные хромосомы инъецируют в клетки-реципиенты посредством микроинъекции хромосом. В некоторых вариантах осуществления донорские клетки индуцируют для мультинуклеации их хромосом. Затем эти ядра проталкивают через клеточную мембрану для получения микроклеток, которые можно подвергать слиянию с клеткой-реципиентом. В некоторых вариантах осуществления также можно использовать опосредованный микроклетками перенос хромосом. Способы манипуляций с хромосомами описаны, например, в CN1200014A, CN109837307A, US20120093785A1 и US2009253902; Kuroiwa et al. "Manipulation of human minichromosomes to carry greater than megabase-sized chromosome inserts." Nature Biotechnology 18,10 (2000)): 1086-1090; патент Китая № CN1717483A; Paulis, Marianna. "Chromosome Transfer Via Cell Fusion." Methods in Molecular Biology 738(2011):57; Genes, Chromosomes & Cancer 14: 126127 (1995); Tomizuka et al. "Functional expression and germline atransmission of a human chromosome fragment in chimaeric mice." Nature Genetics 16,2 (1997): 133-143; Somatic Cell and Molecular Genetics, Vol. 13, No. 3, 1987, pp. 279-284; каждый из которых включен в настоящее описание в качестве ссылки в полном объеме.
В некоторых вариантах осуществления модификацию можно вносить в хромосому мыши. Стратегия таргетинга показана на фиг. 4. Первый вектор может иметь последовательность ДНК гомологичного плеча выше и ниже участка инсерции и последовательность LoxP. В некоторых вариантах осуществления первый вектор от 5’ до 3’ содержит одно или более из следующего: последовательности ДНК гомологичного плеча выше участка инсерции, мишени распознавания Flp (FRT), промотора CAG, гена резистентности к гигромицину (частичной последовательности гигромицинфосфотрансферазы; "5’HygR"), LoxP, FRT, 5-последовательности транспозона PB (PB5’), промотора PGK, репортерного гена синего флуоресцентного белка (BFP), саморасщепляющегося пептида (2A) FMDV, гена резистентности к гигромицину (гигромицинфосфотрансферазы; HygR), 3’-последовательности транспозона PB (PB3’), последовательности ДНК гомологичного плеча ниже участка инсерции и DTA.
Второй вектор может содержать последовательность ДНК гомологичного плеча выше и ниже участка инсерции и последовательность LoxP. В некоторых вариантах осуществления второй вектор от 5’ до 3’ содержит одно или более из следующего: последовательности ДНК гомологичного плеча выше участка инсерции, 5’-последовательности транспозона PB (PB5’), промотора PGK, последовательности репортерного гена зеленого флуоресцентного белка (EGFP), саморасщепляющегося пептида FMDV (2A), последовательности гена резистентности к пуромицину (PuroR), 3’-последовательности транспозона PB (PB3’), мишени распознавания Flp (FRT), частичной последовательности гена резистентности к пуромицину (3’PuroR), саморасщепляющегося пептида (2A) FMDV, рецептора DT (DTR), последовательности распознавания LoxP, последовательности ДНК гомологичного плеча ниже участка инсерции и DTA.
На фиг. 30 показана схожая стратегия таргетинга для локуса легкой каппа-цепи иммуноглобулина. Два вектора можно сначала встраивать в хромосому человека. Первый вектор содержит последовательности ДНК гомологичного плеча выше и ниже участок инсерции, и последовательность распознавания LoxP. В некоторых вариантах осуществления первый вектор от 5’ до 3’ содержит одно или более из следующего: последовательность ДНК гомологичного плеча выше участка инсерции, промотора PGK, tdTomato, саморасщепляющегося пептида (2A) FMDV, Bsr, последовательности терминации транскрипции/сигнала полиаденилирования, последовательности распознавания LoxP, гена резистентности к гигромицину (частичной последовательности гигромицинфосфотрансферазы; "3’HygR"), FRT, последовательности ДНК гомологичного плеча ниже участка инсерции и DTA.
Второй вектор содержит последовательности ДНК гомологичного плеча выше и ниже участка инсерции и последовательность распознавания LoxP. В некоторых вариантах осуществления второй вектор от 5’ до 3’ содержит одно или более из следующего: последовательности ДНК гомологичного плеча выше участка инсерции, последовательности распознавания LoxP, промотора PGK, части последовательности гена резистентности к пуромицину (5’PuroR), EF-1a, PBase, IRES, Neo, последовательности терминации транскрипции/сигнала полиаденилирования, последовательности ДНК гомологичного плеча ниже участка инсерции и DTA.
Последовательности распознавания LoxP также можно добавлять на хромосому человека (например, хромосому 2, 14, 22 человека). Клетки также можно обрабатывать ферментом Cre, что приводит к рекомбинации участков loxP и, таким образом, удалению последовательностей геномной ДНК. В некоторых вариантах осуществления для удаления последовательностей геномной ДНК также можно использовать спонтанный разрыв хромосомы.
Модификацию локуса легкой цепи иммуноглобулина мыши можно осуществлять напрямую. В некоторых вариантах осуществления вектор можно напрямую использовать для замены целой вариабельной области легкой цепи иммуноглобулина мыши. В некоторых вариантах осуществления вектор от 5’ до 3’ содержит: последовательность ДНК гомологичного плеча выше участка инсерции, мишень распознавания Flp (FRT), экспрессирующий промотор млекопитающего (EF-1a) из фактора элонгации 1 альфа человека, ген резистентности к гигромицину (частичную последовательность гигромицинфосфотрансферазы; "5’HygR"), последовательность распознавания LoxP для рекомбиназы Cre, 5’-последовательность транспозона PB (PB5’), репортерный ген синего флуоресцентного белка (BFP), рецептор DT (DTR), саморасщепляющийся пептид (2A) FMDV, последовательность гена резистентности к канамицину (Neo), последовательность терминации транскрипции/сигнала полиаденилирования (поли(A); "PA"), 3’-последовательность транспозона PB (PB3’), частичную последовательность гена резистентности к пуромицину (3’PuroR), саморасщепляющийся пептид (2A) FMDV, рецептор DT (DTR), последовательность распознавания LoxP для рекомбиназы Cre, последовательность ДНК гомологичного плеча ниже участка инсерции и DTA.
Вариабельную область иммуноглобулина мыши можно заменять вариабельной областью иммуноглобулина человека посредством замены (например, гомологичной рекомбинации или Cre-опосредованной рекомбинации). В некоторых вариантах осуществления Cre-рекомбинацию можно использовать для опосредования замены. В некоторых вариантах осуществления с помощью векторов в хромосому человека можно добавлять последовательность распознавания LoxP. Схожие модификации можно вносить в хромосому мыши, где на хромосому можно добавлять две последовательности распознавания LoxP. Затем, например, рекомбиназа Cre может опосредовать замену областей V, J на хромосоме мыши областями V, J на хромосоме человека или замену областей V, D, J на хромосоме мыши областями V, D, J на хромосоме человека.
Клетки дополнительно можно подвергать скринингу на клетки, не имеющие хромосомы человека (например, с помощью DT). В некоторых случаях клетки, не подвергнутые скринингу с помощью DT, могут содержать рекомбинантные фрагменты хромосомы человека, но эти фрагменты являются небольшими и нестабильными в клетках мыши (например, Shinohara et al. (2000) Chromosome Research, 8: 713-725) и естественным образом будут исчезать во время пролиферации клеток. В некоторых вариантах осуществления крупный фрагмент модифицированной хромосомы человека подвергают делеции, например, посредством Cre-опосредованной делеции или спонтанного разрыва хромосомы.
5'-концевое гомологичное плечо и/или 3'-концевое гомологичное плечо может иметь желаемую длину для облегчения гомологичной рекомбинации. В некоторых вариантах осуществления гомологичное плечо составляет приблизительно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40 или 50 т.п.н. (например, приблизительно 3 т.п.н.). В некоторых вариантах осуществления гомологичное плечо составляет менее 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40 или 50 т.п.н.
В некоторых вариантах осуществления вектор также, необязательно, может включать репортерный белок, например, люциферазу (например, Gluc) или флуоресцентный белок (например, EGFP, BFP и т.д.).
Эти модификации можно осуществлять в различных клетках. В некоторых вариантах осуществления клетка является стволовой клеткой, эмбриональной стволовой клеткой или оплодотворенной яйцеклеткой.
Настоящее изобретение также относится к способу получения гуманизированной модели на животных, включающему следующие стадии:
(a) получение клеток (например, оплодотворенной яйцеклетки) на основе способов, представленных в настоящем описании;
(b) культивирование клеток в жидкой среде для культивирования;
(c) трансплантацию культивируемой клетки в фаллопиеву трубу или матку самки-реципиента не являющегося человеком млекопитающего, позволяя клетке развиться в матке самки не являющегося человеком млекопитающего;
(d) идентификацию передачи зародышевой линии потомству генетически модифицированного, гуманизированного, не являющегося человеком млекопитающего от беременной самки на стадии (c).
В некоторых вариантах осуществления не являющееся человеком млекопитающее в указанном выше способе является мышью (например, мышью C57, мышью BALB/c или мышью C57BL/6).
В некоторых вариантах осуществления не являющееся человеком млекопитающее на стадии (c) является самкой с псевдобеременностью (или ложной беременностью).
В некоторых вариантах осуществления оплодотворенные яйцеклетки для описанных выше способов являются оплодотворенными яйцеклетками C57BL/6. Другие оплодотворенные яйцеклетки, которые также можно использовать в способах, представленных в настоящем описании, включают, в качестве неограничивающих примеров, оплодотворенные яйцеклетки FVB/N, оплодотворенные яйцеклетки BALB/c, оплодотворенные яйцеклетки DBA/1 и оплодотворенные яйцеклетки DBA/2.
Оплодотворенные яйцеклетки можно получать из любого не являющегося человеком животного, например, любого не являющегося человеком животного, представленного в настоящем описании. В некоторых вариантах осуществления оплодотворенные яйцеклетки получают из грызунов. Генетическую конструкцию можно встраивать в оплодотворенную яйцеклетку посредством микроинъекции ДНК. Например, с помощью культивирования оплодотворенной яйцеклетки после микроинъекции культивируемую оплодотворенную яйцеклетку можно переносить в ложнобеременное не являющееся человеком животного, которое затем рожает не являющееся человеком млекопитающее, таким образом, получают не являющееся человеком млекопитающее, упомянутое в описанных выше способах.
Настоящее изобретение также относится к клеткам, тканям и животным (например, мыши), содержащим нуклеотидные последовательности, представленные в настоящем описании, а также клеткам, тканям и животным (например, мыши), экспрессирующим гуманизированные или химерные антитела из эндогенного, не принадлежащего человеку локуса.
Настоящее изобретение также относится к различным направленным векторам (например, векторам, которые можно использовать для получения генетически модифицированных животных). В некоторых вариантах осуществления вектор может содержать: a) фрагмент ДНК, гомологичный 5'-концу области, подлежащий изменению (5’-гомологичное плечо); b) последовательность, содержащую желаемые генетические элементы (например, участок распознавания LoxP, гены резистентности к лекарственным средствам и/или репортерным генам и т.д.); и c) второй фрагмент ДНК, гомологичный 3'-концу области, подлежащей изменению (3’-гомологичное плечо). Настоящее изобретение также относится к клетке, содержащей направленные векторы, представленные в настоящем описании.
В некоторых вариантах осуществления гены в клетке являются гетерозиготными. В некоторых вариантах осуществления гены в клетке являются гомозиготными.
В некоторых вариантах осуществления клетка не являющегося человеком млекопитающего является клеткой мыши. В некоторых вариантах осуществления клетка является оплодотворенной яйцеклеткой.
Настоящее изобретение дополнительно относится к способам получения генетически модифицированной модели на животных с двумя или более генами человека или химерными генами. Животное может содержать один или более иммуноглобулиновых локусов человека или гуманизированных иммуноглобулиновых локусов и последовательность, кодирующую дополнительный белок человека или химерный белок. В некоторых вариантах осуществления дополнительный белок человека или химерный белок может являться белком программируемой гибели клеток 1 (PD-1), белком цитотоксических T-лимфоцитов 4 (CTLA-4), белком активации лимфоцитов 3 (LAG-3), B- и T-лимфоцитарным аттенюатором (BTLA), лигандом белка программируемой гибели клеток 1 (PD-L1), CD27, CD28, CD47, CD137, CD154, T-клеточным иммунорецептором с доменами Ig и ITIM (TIGIT), T-клеточным белком 3, содержащим иммуноглобулиновый и муциновый домен (TIM-3), глюкокортикоид-индуцированным TNFR-родственным белком (GITR) или членом 4 суперсемейства рецепторов ФНО (TNFRSF4 или OX40).
Способы получения генетически модифицированной модели на животных с дополнительными генами человека или химерными генами (например, гуманизированными генами) могут включать следующие стадии:
(a) использование способов, представленных в настоящем описании, для получения генетически модифицированного, не являющегося человеком животного;
(b) спаривание генетически модифицированного, не являющегося человеком животного с другим генетически модифицированным, не являющимся человеком животным, а затем скрининг потомства для получения генетически модифицированного, не являющегося человеком животного с двумя или более генами человека или химерными генами.
В некоторых вариантах осуществления на стадии (b) способа генетически модифицированное животное можно спаривать с генетически модифицированным, не являющимся человеком животным с человеческим или химерным PD-1, CTLA-4, LAG-3, BTLA, PD-L1, CD27, CD28, CD47, CD137, CD154, TIGIT, TIM-3, GITR, SIRPa или OX40. Некоторые из эти генетически модифицированных, не являющихся человеком животных описаны, например, в PCT/CN 2017/090320, PCT/CN 2017/099577, PCT/CN 2017/099575, PCT/CN 2017/099576, PCT/CN 2017/099574, PCT/CN 2017/106024, PCT/CN 2017/110494, PCT/CN 2017/110435, PCT/CN 2017/120388, PCT/CN 2018/081628, PCT/CN 2018/081629, каждая из которых включено в настоящее описание в качестве ссылки в полном объеме.
В некоторых вариантах осуществления генетически модифицированные животные могут иметь ген ADAM6 человека, эндогенный ген ADAM6 или модифицированный ген ADAM6. Белок ADAM6 является членом семейства белков ADAM, где ADAM представляет собой аббревиатуру для дизинтегрина и металлопротеазы. Ген ADAM6 человека, как правило, обнаруживаемый между генами IGHV человека IGHV1-2 и IGHV6-1, является псевдогеном (фиг. 37). У мышей есть два гена ADAM6, ADAM6a и ADAM6b. Они находятся в межгенной области между кластерами генов мыши IGHV и IGHD. ADAM6a мыши локализован между IGHV5-1 мыши и IGHD5-1 мыши. ADAM6b мыши локализован между IGHD3-1 мыши и IGHD1-1 мыши. Таким образом, в некоторых вариантах осуществления генетически модифицированные животные могут иметь ген ADAM6 человека. В некоторых вариантах осуществления генетически модифицированные животные не имеют эндогенный ген ADAM6.
В некоторых вариантах осуществления генетически модифицированные животные являются мышами. В некоторых вариантах осуществления мыши модифицированы так, чтобы включать нуклеотидную последовательность, кодирующую белок ADAM6 (например, ADAM6a или ADAM6b). В некоторых вариантах осуществления последовательность помещают в любое подходящее положение. Его можно помещать в межгенную область или в любое подходящее положение в геноме. В некоторых вариантах осуществления нуклеиновая кислота кодирует последовательность, являющуюся по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной гену ADAM6a мыши (например, 113539230-113547024 NC_000078.6; SEQ ID NO: 53) или гену ADAM6b мыши (например, 113486188-113492125 NC_000078.6; SEQ ID NO: 54). В некоторых вариантах осуществления нуклеиновая кислота дополнительно включает регуляторные элементы для гена ADAM6a и гена ADAM6b (например, промоторы).
В некоторых вариантах осуществления функциональный локус ADAM6 мыши можно помещать в середину кластера генов IGHV человека. В некоторых вариантах осуществления локус ADAM6 мыши расположен между двумя генами IGVH человека. В некоторых вариантах осуществления псевдоген ADAM6 человека между VH1-2 человека и VH(II)-1-1 человека заменяют локусом ADAM6 мыши. В некоторых вариантах осуществления ген ADAM6a и ген ADAM6b локализованы между IGHV1-2 человека и VH(II)-1-1 человека в геноме животного. В некоторых вариантах осуществления локализация последовательности ADAM6 мыши в последовательности гена человека может приблизительно соответствовать положению псевдогена ADAM6 человека или положению последовательности ADAM6 мыши (например, в межгенной области V-D). В некоторых вариантах осуществления генетически модифицированные мыши имеют гуманизированный локус тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления ADAM6a мыши и ADAM6b мыши локализованы между генами IGHV1-2 и IGHV6-1 человека. Размещение ADAM6a мыши и ADAM6b мыши между генами IGHV1-2 и IGHV6-1 человека могут иметь различные преимущества. Например, т.к. этими генами заменяют ген ADAM6 человека в том же локусе, вероятно, замена гена ADAM6 человека будет иметь ограниченное влияние на рекомбинацию VDJ, и ген ADAM6a мыши и ген ADAM6b мыши также могут правильно функционировать (т.к. имеет локализацию, схожую с эндогенным локусом).
Таким образом, в одном из аспектов настоящее изобретение относится к генетически модифицированному животному, содержащему в эндогенном локусе гена тяжелой цепи иммуноглобулина первую последовательность, содержащую один или более генов IGHV человека; вторую последовательность, содержащую ген ADAM6; и третью последовательность, содержащую один или более генов IGHD человека и один или более генов IGHJ человека. В некоторых вариантах осуществления первая последовательность, вторая последовательность и третья последовательность функционально связаны.
В некоторых вариантах осуществления первая последовательность содержит все гены IGHV человека из таблицы 1 за исключением IGHV2-10, IGHV3-9, IGHV1-8, IGHV(II)-1-1 и IGHV6-1. В некоторых вариантах осуществления первая последовательность содержит все гены IGHV человека из таблицы 1 за исключением IGHV5-10-1 и IGHV3-64D, IGHV(II)-1-1 и IGHV6-1. В некоторых вариантах осуществления первая последовательность является немодифицированной последовательностью, полученной из локуса гена тяжелой цепи иммуноглобулина человека.
В некоторых вариантах осуществления вторая последовательность содержит один или оба из гена ADAM6a мыши и гена ADAM6b мыши. В некоторых вариантах осуществления животное является фертильным самцом мыши. В некоторых вариантах осуществления вторая последовательность не содержит ген ADAM6a мыши или ген ADAM6b мыши.
В некоторых вариантах осуществления третья последовательность содержит все гены IGHD человека из таблицы 2 и все гены IGHJ человека из таблицы 3. В некоторых вариантах осуществления третья последовательность содержит IGHV6-1 человека. В некоторых вариантах осуществления третья последовательность содержит IGHV(II)-1-1 человека. В некоторых вариантах осуществления третья последовательность является немодифицированной последовательностью, полученной из локуса гена тяжелой цепи иммуноглобулина человека.
В некоторых вариантах осуществления AMAM6a и/или ADAM6b являются эндогенными последовательностями. В некоторых вариантах осуществления AMAM6a и/или ADAM6b не заменяют, и/или они локализованы в своем эндогенном или нативном положении. В некоторых вариантах осуществления гены IGHV мыши перед IGHV1-2 мыши в локусе вариабельной области тяжелой цепи заменяют генами IGHV человека. В некоторых вариантах осуществления гены IGHV, IGHD и IGHJ мыши после IGHV6-1 мыши в локусе вариабельной области тяжелой цепи заменяют одним или более генами IGHV, IGHD и /или IGHJ человека.
Таким образом, в некоторых вариантах осуществления гены IGHV, IGHD и IGHJ мыши можно заменять IGHV, IGHD и IGHJ человека посредством нескольких замен. На первой стадии выбранное количество генов IGHV мыши на 5’-стороне ADAM6a (например, все гены IGHV мыши из таблицы 4) заменяют генами IGHV человека. На второй стадии выбранное количество генов IGHD и IGHJ мыши на 3’-стороне ADAM6b (например, все гены IGHD мыши из таблицы 5, за исключением IGHD5-1 и IGHD3-1, и все гены IGHJ из таблицы 6) заменяют генами IGHD и IGHJ человека. Замену можно осуществлять посредством гомологичной рекомбинации или Cre-опосредованной рекомбинации.
В некоторых вариантах осуществления мыши не имеют гены ADAM6a или ADAM6b мыши. В некоторых вариантах осуществления мыши имеют гены ADAM6 человека.
Для повышения фертильности мышей можно использовать различные способы. В некоторых вариантах осуществления для спаривания можно использовать самок мышей с суперовуляции. В некоторых вариантах осуществления можно использовать оплодотворение in vitro. Суперовуляцию можно индуцировать посредством инъекции сывороточного гонадотропина и хорионического гонадотропина (например, CG человека или мыши) зрелым самкам мышей. Зрелых самцов мышей можно умерщвлять и можно выделять их хвост придатка яичка. Проток хвоста придатка яичка вскрывают для высвобождения спермы. Затем зрелых самок мышей с суперовуляцией можно умерщвлять и можно выделять фаллопиевы трубы. Из фаллопиевой трубы можно высвобождать комплексы кумулюс-ооцит (COC). Затем суспензию спермы можно добавлять к COC и инкубировать для инсеминации. Партеногенетические ооциты, содержащие только один пронуклеус, можно удалять. После инкубации эмбрионы на стадии 2 клеток можно переносить в самок-реципиентов. Способы повышения фертильности мышей известны в этой области.
Настоящее изобретение также относится к последовательности нуклеиновой кислоты, являющейся по меньшей мере на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичной любой нуклеотидной последовательности, представленной в настоящем описании, и аминокислотной последовательности, являющейся по меньшей мере на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичной любой аминокислотной последовательности, представленной в настоящем описании.
В некоторых вариантах осуществления настоящее изобретение относится к нуклеотидным последовательностям, кодирующим любые пептиды, представленные в настоящем описании, или любым аминокислотным последовательностям, кодируемым любыми нуклеотидными последовательностями, представленными в настоящем описании. В некоторых вариантах осуществления последовательность нуклеиновой кислоты содержит менее 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 150, 200, 250, 300, 350, 400, 500 или 600 нуклеотидов. В некоторых вариантах осуществления аминокислотная последовательность содержит менее 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 250, 300, 350 или 400 аминокислотных остатков.
В некоторых вариантах осуществления аминокислотная последовательность (i) содержит аминокислотную последовательность или (ii) состоит из аминокислотной последовательности, где аминокислотная последовательность является любой из последовательностей, представленных в настоящем описании.
В некоторых вариантах осуществления последовательность нуклеиновой кислоты (i) содержит последовательность нуклеиновой кислоты или (ii) состоит из последовательности нуклеиновой кислоты, где последовательность нуклеиновой кислоты является любой из последовательностей, представленных в настоящем описании.
Для определения процента идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновой кислоты последовательности выравнивают в целях оптимального сравнения (например, пропуски можно вносить в одну или обе из первой и второй аминокислотной последовательности или последовательности нуклеиновой кислоты для оптимального выравнивания и при сравнении можно не принимать во внимание негомологичные последовательности). Длина референсной последовательности, выровненной в целях сравнения, составляет по меньшей мере 80% от длины всей референсной последовательности и в некоторых вариантах осуществления составляет по меньшей мере 90%, 95% или 100%. Затем сравнивают аминокислотные остатки или нуклеотиды в соответствующих положениях аминокислот или положениях нуклеотидов. Если положение в первой последовательности занято тем же аминокислотным остатком или нуклеотидом, что и соответствующее положение во второй последовательности, то молекулы идентичны в этом положении (в рамках изобретения "идентичность" аминокислоты или нуклеиновой кислоты эквивалентна "гомологии" аминокислоты или нуклеиновой кислоты). Процент идентичности между двумя последовательностями является функцией количества идентичных положений, общих для последовательностей, с учетом количества пропусков и длины каждого пропуска, который необходимо внести для оптимального выравнивания двух последовательностей. В целях по настоящему изобретению сравнение последовательностей и определение процента идентичности между двумя последовательностями можно осуществлять с использованием матрицы весов Blossum 62 со штрафом за пропуск 12, штрафом за увеличение пропуска 4 и штрафом за пропуск со сдвигом рамки считывания 5.
Для измерения сходства последовательностей также можно использовать процентную долю остатков, консервативных в отношении схожих физико-химических свойств (процент гомологии), например, лейцина и изолейцина. В этой области определены семейства аминокислотных остатков, имеющих схожие физико-химические свойства. Эти семейства включают, например, аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновую кислоту, глутаминовую кислоту), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Во многих случаях процент гомологии выше процента идентичности. Таким образом, настоящее изобретение также относится к аминокислотной последовательности, имеющей по меньшей мере 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% процент гомологии в отношении любой аминокислотной последовательности, представленной в настоящем описании, или нуклеиновой кислотой, кодирующей эти аминокислотные последовательности.
Способы использования генетически модифицированных животных
Генетически модифицированных животных можно использовать для получения гуманизированных или химерных антител, которые могут специфически связываться с мишенью. В некоторых вариантах осуществления мишень (например, белок или фрагмент белка) можно использовать в качестве иммуногена для получения антител в этих животных стандартными способами получения поликлональных и моноклональных антител. В некоторых вариантах осуществления генетически модифицированное животное подвергают воздействию выбранного антигена в течение периода времени и в условиях, позволяющих животному продуцировать антитело, специфическое в отношении антигена.
Поликлональные антитела можно индуцировать у животных посредством множества инъекций (например, подкожных или интраперитонеальных инъекций) антигенного пептида или белка. В некоторых вариантах осуществления антигенный пептид или белок инъецируют по меньшей мере с одним адъювантом. В некоторых вариантах осуществления антигенный пептид или белок можно конъюгировать со средством, иммуногенным для биологического вида, подлежащего иммунизации. Животным можно инъецировать антигенный пептид или белок несколько раз (например, два раза, три раза или четыре раза).
В качестве иммуногенов можно использовать полноразмерный полипептид или белок или, альтернативно, его антигенные пептидные фрагменты. Антигенный пептид из белка содержит по меньшей мере 8 (например, по меньшей мере 10, 15, 20 или 30) аминокислотных остатков из аминокислотной последовательности и включает такой эпитоп белка, что антитело, индуцированное против пептида, образует специфический иммунный комплекс с белком.
Иммуноген, как правило, используют для получения антител посредством иммунизации подходящего индивидуума (например, генетически модифицированного животного, представленного в настоящем описании). Подходящий иммуногенный препарат может содержать, например, рекомбинантно экспрессируемый или химически синтезированный полипептид (например, фрагмент белка). Получение может дополнительно включать адъювант, такой как полный или неполный адъювант Фрейнда, или схожее иммуностимулирующее средство.
Поликлональные антитела можно получать, как описано выше, посредством иммунизации подходящего индивидуума с использованием полипептида или его антигенного пептида (например, части белка) в качестве иммуногена. Титр антител у иммунизированного индивидуума можно подвергать мониторингу с течением времени стандартными способами, такими как твердофазный иммуноферментный анализ (ELISA) с использованием иммобилизованного полипептида или пептида. При желании, молекулы антител можно выделять из млекопитающего (например, из крови) и дополнительно очищать хорошо известными способами, такими как хроматография с протеином A или протеином G, для получения фракции IgG. В подходящее время после иммунизации, например, когда титр специфических антител является наиболее высоким, из индивидуума можно получать антителопродуцирующие клетки и использовать их для получения моноклональных антител стандартными способами, такими как гибридомный способ, исходно описанный Kohler et al. (Nature 256:495-497, 1975), способ с использованием гибридомы из B-клеток человека (Kozbor et al., Immunol. Today 4:72, 1983), способ с использованием EBV-гибридомы (Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96, 1985) или триомный способ. Технология получения гибридом хорошо известна (в целом, см. Current Protocols in Immunology, 1994, Coligan et al. (Eds.), John Wiley & Sons, Inc., New York, NY). Гибридомные клетки, продуцирующие моноклональное антитело, определяют посредством скрининга супернатантов культур гибридом на антитела, связывающиеся с интересующим полипептидом или эпитопом, например, с использованием стандартного анализа ELISA.
В одном из аспектов настоящее изобретение относится к мыши, содержащей модификацию локуса эндогенной тяжелой цепи иммуноглобулина, где мышь продуцирует B-клетку, содержащую подвергнутую реаранжировке последовательность иммуноглобулина, функционально связанную с последовательностью гена константной области тяжелой цепи. В некоторых вариантах осуществления подвергнутая реаранжировке последовательность иммуноглобулина, функционально связанная с последовательностью гена константной области тяжелой цепи, содержит последовательность V, D и/или J тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена константной области тяжелой цепи содержит последовательность тяжелой цепи человека или мыши, выбранную из группы, состоящей из CH1, шарнирной области, CH2, CH3 и их комбинации.
В одном из аспектов настоящее изобретение относится к мыши, содержащей модификацию локуса эндогенной легкой цепи иммуноглобулина (например, каппа или лямбда), где мышь продуцирует B-клетку, содержащую подвергнутую реаранжировке последовательность иммуноглобулина, функционально связанную с последовательностью гена константной области легкой цепи. В некоторых вариантах осуществления подвергнутая реаранжировке последовательность иммуноглобулина, функционально связанная с последовательностью гена константной области легкой цепи, содержит последовательность V и/или J легкой цепи человека. В некоторых вариантах осуществления последовательность гена константной области легкой цепи содержит константную область легкой цепи человека или мыши.
B-клетки или клетки селезенки мыши могут содержать подвергнутую реаранжировке немышиную последовательность гена вариабельной области иммуноглобулина, например, функционально связанную с геном константной области иммуноглобулина мыши. Определяют последовательности, кодирующие вариабельную область тяжелой цепи человека и вариабельную область легкой цепи человека. Последовательности можно определять, например, посредством секвенирования интересующей гибридомы или B-клеток. В некоторых вариантах осуществления используют скрининг отдельных B-клеток. Можно осуществлять скрининг природного репертуара антитела без необходимости слияния гибридомы и комбинаторного дисплея. Например, B-клетки можно смешивать с панелью антигенов с ДНК-штрихкодами таким образом, что и последовательности штрихкодов антигена, и последовательности штрихкодов B-клеточного рецептора (BCR) отдельных B-клеток выделяют способами секвенирования отдельных клеток.
Антитела можно дополнительно модифицировать для получения гуманизированного антитела или антитела человека, например, посредством функционального связывания последовательности, кодирующей вариабельную область тяжелой цепи человека, с последовательностью, кодирующей константную область тяжелой цепи человека, и/или функционального связывания последовательности, кодирующей вариабельную область легкой цепи человека, с последовательностью, кодирующей константную область легкой цепи человека.
В некоторых вариантах осуществления, если мышь экспрессирует белок, очень схожий с интересующим антигеном, может быть трудным вызвать иммунный ответ у мыши. Это является результатом того, что во время развития иммунных клеток B-клетки и T-клетки, распознающие молекулы MHC, связанные с пептидами собственного происхождения, удаляются из репертуара иммунных клеток. В этих случаях гуманизированную мышь можно дополнительно модифицировать. Соответствующий ген у мыши можно подвергать нокауту, а затем мышь подвергают воздействию интересующего антигена. Т.к. мышь не подвергается отрицательной селекции на продукт гена, мышь может генерировать антитело, которое легко может специфически связываться с мишенью.
Настоящее изобретение также относится к способам получения антител, нуклеиновых кислот, клеток, тканей (например, ткани селезенки). В некоторых вариантах осуществления способы включают подвергание животного, представленного в настоящем описании, воздействию антигена. Антитела (например, гибридные антитела), нуклеиновые кислоты, кодирующие антитела, клетки и/или ткани (например, ткань селезенки) можно получать из животного. В некоторых вариантах осуществления нуклеиновые кислоты, кодирующие вариабельные области тяжелой и легкой цепи иммуноглобулина человека, определяют, например, посредством секвенирования. В некоторых вариантах осуществления нуклеиновую кислоту, кодирующую вариабельную область тяжелой цепи иммуноглобулина человека, можно функционально связывать с нуклеиновой кислотой, кодирующей константную область тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления нуклеиновую кислоту, кодирующую вариабельную область легкой цепи иммуноглобулина человека, можно функционально связывать с нуклеиновой кислотой, кодирующей константную область легкой цепи иммуноглобулина человека. В некоторых вариантах осуществления клетки, содержащие нуклеиновые кислоты, представленные в настоящем описании, культивируют и собирают антитела.
В некоторых вариантах осуществления ни один из генов иммуноглобулинов V, D, J мыши (например, ни один из генов IGHV, IGHD, IGHJ, IGKV или IGKJ мыши) вносит вклад в последовательность вариабельной области тяжелой цепи и/или легкой цепи. В некоторых вариантах осуществления последовательность вариабельной области тяжелой цепи и/или легкой цепи, продуцируемая животным, является полностью человеческой и полностью образована генами V, D, J иммуноглобулинов человека (например, генами IGHV, IGHD, IGHJ, IGKV и IGKJ человека).
Варианты антител или антигенсвязывающих фрагментов, представленные в настоящем описании, можно получать посредством встраивание подходящих изменений нуклеотидов в ДНК, кодирующую человеческое, гуманизированное или химерное антитело или его антигенсвязывающий фрагмент, представленный в настоящем описании, или посредством пептидного синтеза. Такие варианты включают, например, делеции, инсерции или замены остатков в аминокислотных последовательностях, составляющих антигенсвязывающий участок антитела или антигенсвязывающего домена. В популяции таких вариантов некоторые антитела или антигенсвязывающие фрагменты будут иметь повышенную аффинность для белка-мишени. Можно получать любую комбинацию делеций, инсерций и/или комбинаций для получения антитела или его антигенсвязывающего фрагмента, имеющего повышенную аффинность связывания с мишенью. Изменения аминокислот, встроенные в антитело или антигенсвязывающий фрагмент, также могут изменять или вносить новые посттрансляционные модификации в антитело или антигенсвязывающий фрагмент, такие как изменение (например, повышение или снижение) количество участков гликозилирования, изменение типа участка гликозилирования (например, изменение аминокислотной последовательности таким образом, что разные сахара присоединяются ферментами, присутствующими в клетке) или встраивание новых участков гликозилирования.
Антитела, представленные в настоящем описании, можно получать из любого вида животного, включая млекопитающих. Неограничивающие примеры нативных антител включают антитела, полученный из людей, приматов, например, мартышек и человекообразных приматов, коров, свиней, лошадей, овец, верблюжьих (например, верблюдов и лам), кур, коз и грызунов (например, крыс, мышей, хомяков и кроликов), включая трансгенных грызунов, генетически сконструированные для получения антител человека.
Антитела человека и гуманизированные антитела включают антитела, содержащие вариабельные и константные области, полученные из (или имеющие ту же аминокислотную последовательность, что и антитела, полученные из) последовательностей иммуноглобулинов зародышевой линии человека. Антитела человека могут включать аминокислотные остатки, некодируемые последовательностями иммуноглобулинов зародышевой линии человека (например, мутации, встраиваемые посредством случайного или сайт-специфического мутагенеза in vitro или в результате соматической мутации in vivo), например, в CDR.
Можно вносить дополнительные модификации в антитела или антигенсвязывающие фрагменты. Например, можно встраивать остатки цистеина в Fc-область, таким образом, делая возможным образованием межцепочечной дисульфидной связи в этой области. Полученное таким образом гомодимерное антитело может иметь повышенное время полужизни in vitro и/или in vivo. Гомодимерные антитела с повышенным временем полужизни in vitro и/или in vivo также можно получать с использованием гетеробифункциональных кросс-линкеров, как описано, например, в Wolff et al. (Cancer Res. 53:2560-2565, 1993). Альтернативно, можно конструировать антитело, имеющее двойные Fc-области (см., например, Stevenson et al., Anti-Cancer Drug Design 3:219-230, 1989).
В некоторых вариантах осуществления можно вносить ковалентную модификацию в антитело или его антигенсвязывающий фрагмент. Эти ковалентные модификации можно осуществлять посредством химического или ферментативного синтеза или ферментативного или химического расщепления. Другие типы ковалентных модификаций антитела или фрагмента антитела вносят в молекулу посредством реакции целевых аминокислотных остатков антитела или фрагмента с органическим дериватизирующим средством, которое может реагировать с выбранными боковыми цепями или N- или C-концевыми остатками.
ПРИМЕРЫ
Далее настоящее изобретение описано в следующих примерах, не ограничивающих объем изобретения, описанный в формуле изобретения.
ПРИМЕР 1: Обзор
Эксперименты осуществляли для встраивания генов иммуноглобулина человека в геном мыши для получения мышей, экспрессирующих гуманизированные антитела. На фиг. 1A показаны способы получения гуманизированных мышей. Способы сначала включают модификацию иммуноглобулиновой области человека на хромосоме человека. Затем модифицированные хромосомы человека встраивали в клетку-реципиент мыши.
Вариабельную область иммуноглобулина мыши заменяли вариабельной областью иммуноглобулина человека посредством прямой замены (например, гомологичной рекомбинации или Cre-опосредованной рекомбинации). В некоторых случаях вариабельную область иммуноглобулина человека можно встраивать в геном мыши с помощью постадийного подхода. Затем клетки-реципиенты подвергали скринингу на правильную замену. Затем клетки инъецировали в бластоцисты для получения химерных мышей. Затем осуществляли скрещивание для получения мышей, содержащих интактные вариабельные области иммуноглобулинов человека.
Т.к. ген тяжелой цепи и два гена легкой цепи мыши локализованы на хромосомах 12, 6 и 16, соответственно, мышей, содержащих вариабельную область тяжелой цепи человека или вариабельную область легкой цепи человека, можно получать раздельно (фиг. 1B и 1C). Затем этих мышей можно спаривать друг с другом для получения мышей, которые могут экспрессировать и вариабельный домен тяжелой цепи человека, и вариабельный домен легкой цепи человека.
ПРИМЕР 2: Модификация локуса тяжелой цепи иммуноглобулина мыши
Локус тяжелой цепи иммуноглобулина локализован на хромосоме мыши 12. Фиг. 2 является схемой, на которой показан локус тяжелой цепи иммуноглобулина мыши. Два участка рекомбинации (1301, 1302) встраивали на обеих сторонах вариабельной области из локуса тяжелой цепи иммуноглобулина, и полученная модифицированная хромосома показана на фиг. 3A-3B. Один из них является участком loxP дикого типа, другой является гетероспецифическим мутантным участком lox (lox2272). Рекомбинация не может происходить между участком loxP дикого типа и гетероспецифическим мутантным участком lox. Модификацию осуществляли на эмбриональных стволовых клетках мыши. Общая схема стратегии таргетинга показана на фиг. 4. Вектор (V1401) от 5’ до 3’ содержит: последовательность ДНК гомологичного плеча выше участка инсерции, мишень распознавания Flp (FRT), промотор CAG, ген резистентности к гигромицину (частичную последовательность гигромицинфосфотрансферазы; "5’HygR"), LoxP (1301), FRT, 5’-последовательность транспозона PB (PB5’), промотор PGK, репортерный ген синего флуоресцентного белка (BFP), саморасщепляющийся пептид (2A) FMDV, ген резистентности к гигромицину (гигромицинфосфотрансферазу; HygR), 3’-последовательность транспозона PB (PB3’), последовательность ДНК гомологичного плеча ниже участка инсерции и DTA.
Вектор (V1402) от 5’ до 3’ содержит: последовательность ДНК гомологичного плеча выше участка инсерции, 5’-последовательность транспозона PB (PB5’), промотор PGK, последовательность репортерного гена зеленого флуоресцентного белка (EGFP), саморасщепляющийся пептид (2A) FMDV, последовательность гена резистентности к пуромицину (PuroR), 3’-последовательность транспозона PB (PB3’), мишень распознавания Flp (FRT), частичную последовательность гена резистентности к пуромицину (3’PuroR), саморасщепляющийся пептид (2A) FMDV, рецептор DT (DTR), последовательность распознавания LoxP (1302), последовательность ДНК гомологичного плеча ниже участка инсерции и DTA.
Векторы (V1401 и V1402) встраивали в эмбриональные стволовые клетки мыши. Затем клетки подвергали скринингу с помощью гигромицина B и пуромицина. Интеграцию экзогенных генов в геномы мышей подтверждали посредством ПЦР. Результаты показаны на фиг. 5A-5B и фиг. 6A-6B. Подтверждали, что клоны под номерами 030, 035, 036 и 037 являлись положительными.
Анализ ПЦР осуществляли с использованием следующих праймеров:
mIgHV-5’loxP-L-GT-F: 5’-gccaaggaatttaaaaggggattgaaagcaa-3’ (SEQ ID NO: 1),
mIGHV-005-L-GT-R2: 5’-gccctccatgtacagcttcatgtgc-3’ (SEQ ID NO: 2);
mIGHV-005-5’loxP-R-GT-F2: 5’-actgggcttgtcgagacagagaaag-3’ (SEQ ID NO: 3),
mIgHV-5’loxP-R-GT-R: 5’-ccacagcccgatctacttggctttt-3’ (SEQ ID NO: 4);
mIGHV-3’lox-L-GT-F2: 5’-gcaaggttttgactaagcggagcac-3’ (SEQ ID NO: 5);
mIGHV-3’lox-L-GT-R2: 5’-tgacgcatgtgttttatcggtctgt-3’ (SEQ ID NO: 6);
mIGHV3’lox-R-GT-F2: 5’-gtgcctgacacgtgctacgagattt-3’ (SEQ ID NO: 7);
mIGHV-3’lox-R-GT-R1: 5’-ttcaacaataagcagggccagaggg-3’ (SEQ ID NO: 8).
Из этих праймеров mIgHV-5’loxP-L-GT-F и mIgHV-5’loxP-R-GT-R локализованы на хромосоме мыши, mIGHV-005-L-GT-R2 и mIGHV-005-5’loxP-R-GT-F2 локализованы на векторе 1401, mIGHV-3’lox-L-GT-F2 и mIGHV-3’lox-R-GT-R1 локализованы на хромосоме мыши, mIGHV-3’lox-L-GT-R2 и mIGHV3’lox-R-GT-F2 локализованы на векторе 1402.
ПРИМЕР 3: Модификация хромосомы человека 14
Целью эксперимента является получение модифицированной хромосомы человека по меньшей мере с двумя участками рекомбинации. Два участка рекомбинации встраивали на обеих сторонах вариабельной области из локуса тяжелой цепи иммуноглобулина.
Локус тяжелой цепи иммуноглобулина локализован на хромосоме человека 14. Фиг. 7 является схемой хромосомы человека 14, на которой выделен локус тяжелой цепи иммуноглобулина.
Модифицированная вариабельная область человека показана на фиг. 8. Общая схема стратегии таргетинга показана на фиг. 9. Как показано на фиг. 9, участки 301 и 302 являются участками рекомбинации. Участок рекомбинации 1301 и участок рекомбинации 301 идентичны. Участок рекомбинации 1302 и участок рекомбинации 302 идентичны.
Эксперименты осуществляли для встраивания векторов в хромосому человека 14 выше V-области и между J-областью и C-областью. Первый направленный вектор (V401) от 5’ до 3’ содержит последовательность ДНК гомологичного плеча выше участка инсерции, промотор PGK, репортерный ген красного флуоресцентного белка (tdTomato), саморасщепляющийся пептид (2A) из FMDV (вирусов ящура), последовательность гена резистентности к зеоцину (Zeo), последовательность терминации транскрипции/сигнала полиаденилирования (поли(A); "PA"), последовательность распознавания LoxP (301) для рекомбиназы Cre, ген резистентности к гигромицину (частичную последовательность гигромицинфосфотрансферазы; "3’HygR") и мишень распознавания Flp ("FRT"), нижележащую последовательность ДНК гомологичного плеча и ген субъединицы A дифтерийного токсина (DTA).
Второй вектор (V402) от 5’ до 3’ содержит следующее: последовательность ДНК гомологичного плеча выше участка инсерции, последовательность распознавания LoxP (302) для рекомбиназы Cre, промотор PGK, частичную последовательность гена резистентности к пуромицину (5’PuroR), экспрессирующий промотор млекопитающего (EF-1a) из фактора элонгации 1 альфа человека, последовательность гена транспозазы piggyBac (PBase), внутренние участки связывания рибосомы (IRES), последовательность гена резистентности к канамицину (Neo), последовательность терминации транскрипции/сигнала полиаденилирования (поли(A); "PA"), последовательность ДНК гомологичного плеча ниже участка инсерции и DTA.
В некоторых экспериментах векторы (V401, V402) встраивали в клетки и клетки подвергали селекции с помощью соответствующих маркеров резистентности к лекарственным средствам или их комбинации (зеоцин, G418).
Существует множество способов встраивания интересующих векторов в хромосому человека. Хромосому человека можно получать из линий клеток человека, злокачественных клеток, первичной культуры клеток и/или фибробластов человека. В одном из экспериментов первый вектор встраивают в хромосому. Модифицированную хромосому можно добавлять в клетку-реципиента, а затем второй вектор можно встраивать в модифицированную хромосому. В некоторых экспериментах V401 сначала встраивали в клетки человека, а затем хромосому флуоресцентно метили, а затем разделяли, а затем модифицированную хромосому инъецировали в клетки-реципиенты посредством микроинъекции. Затем V402 встраивали в клетки. В другом эксперименте фибробласты человека подвергали селекции и встраивали в них вектор 402. Затем фибробласты человека подвергали слиянию с клетками-реципиентами (клетками A9 или клетками CHO).
В некоторых экспериментах один или более векторов можно встраивать в хромосому 14 человека в желаемые места посредством гомологичной рекомбинации, например, одновременно. Векторы могут содержать маркеры резистентности к лекарственным средствам (зеоцину, G418), а затем клетки подвергают скринингу. Хромосому метят, а затем отделяют, а затем модифицированную хромосому инъецируют в клетки-реципиенты посредством микроинъекций хромосом.
В некоторых экспериментах встраивают один или более дополнительных векторов. Эти дополнительные векторы можно встраивать в разные места хромосом 14 человека, при необходимости. В одном из экспериментов третий вектор от 5’ до 3’ может содержать следующее: последовательность ДНК гомологичного плеча выше участка инсерции, промотор PGK, последовательность гена резистентности к пуромицину (PuroR), последовательность гена тимидинкиназы (TK), последовательность распознавания LoxP (302), промотор PGK, частичную последовательность гена резистентности к пуромицину (5’PuroR), экспрессирующий промотор млекопитающего (EF-1a) из фактора элонгации 1 альфа человека, последовательность гена транспозазы piggyBac (PBase), внутренние участки связывания рибосомы (IRES), последовательность гена резистентности к канамицину (Neo), последовательность терминации транскрипции/сигнала полиаденилирования (поли(A); "PA"), последовательность ДНК гомологичного плеча ниже участка инсерции и DTA. Вектор встраивают в C-область.
В одном из экспериментов третий вектор (V403) встраивали между C-областью и кинетохором. Вектор от 5’ до 3’ содержит следующее: последовательность ДНК гомологичного плеча выше участка инсерции, промотор PGK, последовательность гена резистентности к пуромицину (PuroR), последовательность гена тимидинкиназы (TK), последовательность распознавания LoxP (302), промотор PGK, частичную последовательность гена резистентности к пуромицину (5’PuroR), экспрессирующий промотор млекопитающего (EF-1a) из фактора элонгации 1 альфа человека, последовательность гена транспозазы piggyBac (PBase), внутренние участки связывания рибосомы (IRES), последовательность гена резистентности к канамицину (Neo), последовательность терминации транскрипции/сигнала полиаденилирования (поли(A); "PA"), последовательность ДНК гомологичного плеча ниже участка инсерции и DTA.
В одном из экспериментов фибробласты человека подвергали селекции и встраивали в них вектор 402. Затем фибробласты человека подвергали слиянию с клетками-реципиентами (клетками A9 или клетками CHO). Модифицированную хромосому отделяли и встраивали в другую подходящую клетку-реципиента. Затем клетки подвергали селекции по резистентности к G418 для получения клеток, содержащих только одну хромосому человека. Затем вектор 401 встраивали в клетки и клетки подвергали селекции по резистентности к зеоцину. После этого вектор 403 встраивали в клетки и клетки подвергали селекции по резистентности к пуромицину. Полученные клоны, выбранные после скрининга, обрабатывали ферментом Cre. Хромосомные способы описаны например, в Kuroiwa et al. "Manipulation of human minichromosomes to carry greater than megabase-sized chromosome inserts." Nature Biotechnology 18,10 (2000)): 1086-1090; CN1200014A; CN109837307A; US20120093785A1; US2009253902; CN1717483A; Paulis, Marianna. "Chromosome Transfer Via Cell Fusion." Methods in Molecular Biology 738(2011):57; Genes, Chromosomes & Cancer 14: 126127 (1995); Tomizuka et al. "Functional expression and germline atransmission of a human chromosome fragment in chimaeric mice." Nature Genetics 16,2 (1997): 133-143; и Somatic Cell and Molecular Genetics, Vol. 13, No. 3, 1987, pp. 279-284; каждый из которых включен в настоящее описание в качестве ссылки в полном объеме.
Осуществляли ПЦР для подтверждения наличия 5’-концевого участка рекомбинации 301 и 3’-концевого участка рекомбинации 302 на хромосоме. Клетки без случайной инсерции подтверждали посредством Саузерн-блоттинга и анализировали посредством флуоресцентной гибридизации in situ (FISH).
На фиг.10 показана модифицированная хромосома 14 человека. На фиг. 11 показаны результаты ПЦР-идентификации участка loxP 301 на хромосоме hChr14-mut3. На фиг. 12 показаны результаты ПЦР-идентификации участка loxP 302 на хромосоме hChr14-mut3. Как показано на фигурах, 12 клонов (под номерами 1-B2, 1-B8, 1-D6, 1-D10, 1-F11, 1-G11, 2-A2, 3-E5, 3-G5, 3-H4, 5-C3 и 6-F11) являлись положительными клонами.
В экспериментах использовали следующие праймеры для ПЦР:
hIGHV-5’loxP-L-GT-F1: 5’-TCAAAGTCAATTTCCTCAGCGAGGCT-3’ (SEQ ID NO: 9),
hIGHV-5’loxP-R-GT-R: 5’-AGGGAGGGAATGGAATGAGGGTGAT-3’ (SEQ ID NO: 10);
hIGHV-3’loxP-L-GT-F1: 5’-CCATGTGACCCATTCGAGTGTCCTG-3’ (SEQ ID NO: 11),
hIGHV-3’loxP-R-GT-R: 5’-TTGTGAGGGCTCAAGTTCAGTGCAT-3’ (SEQ ID NO: 12).
Анализ FISH осуществляли с использованием положительных клонов с использованием зонда для FISH CCP14 (CytoTest Inc., Rockville, MD, кат. № CT-CCP014). Типичные изображения FISH для клона 1-D10 показаны на фиг. 13 и 14. На фиг. 13 белой стрелкой указаны полноразмерные хромосомы 14 человека (до модификации). На фиг. 14 белой стрелкой указан фрагмент модифицированной хромосомы 14 человека.
ПРИМЕР 4: Встраивание хромосом человека или фрагмента в клетки ES мыши
Модифицированную хромосому, полученную в примере 3, встраивали в клетку, полученную в примере 2, описанными выше способами. Затем клетки подвергали скринингу с помощью G418. Выбирали только клетку, содержащую только одну хромосому человека. На фиг.15 показана модифицированная хромосома 12 мыши.
Затем рекомбиназа Cre опосредовала замену областей V, D, J на хромосоме mChr12-mut2 мыши областями V, D, J на хромосоме hChr14-mut3 человека (фиг. 16). Последовательность ДНК хромосомы человека заменяли последовательностью между участками рекомбинации 1301 и 1302. Гигромицин и пуромицин использовали для скрининга положительных клеток. Далее клетки подвергали скринингу с помощью DT для получения клеток мыши, не содержащих хромосомы человека, перед инъекцией в бластоцисты мыши. В некоторых случаях клетки напрямую инъецировали в бластоцисты без скрининга с помощью DT.
После Cre-рекомбинации клетки тестировали для подтверждения встраивания последовательностей гена человека в геном мыши. Результаты ПЦР показаны на фиг. 17, 18, 19 и 20. Все результаты ПЦР показали, что клетки под номерами 1-B4, 1-B10 и 2-A7 имели правильную рекомбинацию, и хромосомы человека в клетках 1-B10 исчезали. Зонды Murine Whole Chromosome Painting Probes (Cytocell Ltd, Cambridge, UK; кат. № AMP12R) и человек-специфические зонды IGH Break Apart (Cytocell Ltd, Cambridge, UK; кат. № LPH 014) использовали для тестирования клеток 1-B10 посредством FISH. Результат показан на фиг. 21, где подтверждено наличие фрагментов хромосомы человека в хромосоме мыши. Эти праймеры показаны из таблицы ниже.
Таблица 12
ПРИМЕР 5: Получение мышей, содержащих гуманизированный локус тяжелой цепи иммуноглобулина
Клетки положительных клонов инъецировали в бластоцисты мышей BALB/c посредством микроинъекции. Микроинъекцию эмбриона осуществляли способом, описанным, например, в A. Nagy, et al., "Manipulating Mouse Embryo: A Laboratory Manual (Third Edition)," Cold Spring Harbor Laboratory Press, 2003. Затем подвергнутые инъекции оплодотворенные яйцеклетки переносили в среду для культивирования для кратковременного культивирования, а затем трансплантировали в фаллопиеву трубу мыши-реципиента для получения генетически модифицированных гуманизированных мышей (поколение F0). Затем мышей спаривали с мышами, имеющими фон C57BL/6. Для спаривания с рабочими мышами Flp выбирали черное потомство (фиг. 22). Анализ ПЦР осуществляли с использованием ДНК, полученной из хвоста мыши. Далее мышей скрещивали с мышами с фоном BALB/c несколько раз (например, по меньшей мер, 5 раз) для получения мышей, гетерозиготных по гуманизированному локусу тяжелой цепи иммуноглобулина, с фоном BALB/c.
Для подтверждения экспрессии мышью тяжелой цепи антитела человека получали кровь химерной мыши (поколение F0) и черной мыши (поколение F1). РНК выделяли и подвергали обратной транскрипции для получения кДНК. Следующие праймеры для ПЦР использовали для амплификации последовательности, а затем последовательности секвенировали.
Таблица 13. Праймеры для ПЦР
ПРИМЕР 6: Модификация локуса легкой цепи иммуноглобулина мыши
Локус легкой цепи иммуноглобулина локализован на хромосоме мыши 6. Фиг. 23 является схемой, на которой показан локус легкой цепи иммуноглобулина мыши. Два участка рекомбинации встраивали на обеих сторонах вариабельной области из локуса легкой цепи иммуноглобулина, и полученная модифицированная хромосома показана на фиг. 24A-24B. Подробности стратегии таргетинга показаны на фиг. 25.
Модификацию осуществляли на эмбриональных стволовых клетках мыши. Вектор (V3901) от 5’ до 3’ содержал: последовательность ДНК гомологичного плеча выше участка инсерции, мишень распознавания Flp (FRT), экспрессирующий промотор млекопитающего (EF-1a) из фактора элонгации 1 альфа человека, ген резистентности к гигромицину (частичную последовательность гигромицинфосфотрансферазы; "5’HygR"), последовательность распознавания LoxP (1101) для рекомбиназы Cre, 5’-последовательность транспозона PB (PB5’), репортерный ген синего флуоресцентного белка (BFP), рецептор DT (DTR), саморасщепляющийся пептид (2A) FMDV, последовательность гена резистентности к канамицину (Neo), последовательность терминации транскрипции/сигнала полиаденилирования (поли(A); "PA"), 3’-последовательность транспозона PB (PB3’), частичную последовательность гена резистентности к пуромицину (3’PuroR), саморасщепляющийся пептид (2A) FMDV, рецептор DT (DTR), последовательность распознавания LoxP (1102) для рекомбиназы Cre, последовательность ДНК гомологичного плеча ниже участка инсерции и DTA.
Векторы (V3901) встраивали в эмбриональные стволовые клетки мыши. Клетки подвергали скринингу с помощью соответствующих генетических маркеров резистентности к антибиотикам или их комбинаций. Интеграцию вектора V3901 в правильный локус в геномы мышей подтверждали посредством ПЦР. Результаты показаны на фиг. 26-27. Два объединенных результата ПЦР подтверждали, что клетки под номерами 208, 209, 215, 217 и 269 являлись положительными клонами.
Анализ ПЦР осуществляли с использованием следующих праймеров:
IGKV-005-C-5G-L-GT-F: 5'-TCACACACTACAGCTTCCACCACAA-3' (SEQ ID NO: 34);
IGKV-005-C-5G-L-GT-R2: 5'-CGGGGAAAAGTCGACTCTAGAACGG-3' (SEQ ID NO: 35);
IGKV-005-C-5G-R-GT-F1: 5'-ACTGCATTCTAGTTGTGGTTTGTCCA-3' (SEQ ID NO: 36);
IGKV-005-C-5G-R-GT-R: 5'-GGCCTGGAAAACTCAGCTATCCTTT-3' (SEQ ID NO: 37).
Из этих праймеров IGKV-005-C-5G-L-GT-F и IGKV-005-C-5G-R-GT-R локализованы на хромосоме мыши, IGKV-005-C-5G-L-GT-R2 и IGKV-005-C-5G-R-GT-F1 локализованы на векторе V3901.
Таким образом, два участка рекомбинации встраивали в хромосому 6 мыши в эмбриональных стволовых клетках мыши.
ПРИМЕР 7: Модификация хромосомы 2 человека
Локус легкой цепи иммуноглобулина человека локализован на хромосоме 2 человека. Фиг. 28 является схемой хромосомы человека 2, на которой выделен локус легкой цепи иммуноглобулина.
Два участка рекомбинации встраивали на обеих сторонах вариабельной области из локуса легкой цепи иммуноглобулина. Область между VHK и центромерой (кинетохором) подвергали делеции для получения более короткой искусственной хромосомы для последующих экспериментов. Схожие участки рекомбинации встраивали в вариабельную область иммуноглобулинового локуса мыши на хромосоме 6. Затем хромосому человека встраивали в клетку-реципиента мыши для получения гуманизированного локуса легкой цепи иммуноглобулина.
Модифицированная хромосома человека 2 показана на фиг. 29. Стратегия таргетинга показана на фиг. 30. Вектор (V2701) от 5’ до 3’ содержит: последовательность ДНК гомологичного плеча выше участка инсерции, промотор PGK, последовательность репортерного гена красного флуоресцентного белка (tdTomato), саморасщепляющийся пептид (2A) FMDV, бластицидин-S-дезаминазу (Bsr) из Aspergillus terreus, последовательность терминации транскрипции/сигнал полиаденилирования (поли(A); "PA"), последовательность распознавания LoxP 2601, ген резистентности к гигромицину (частичную последовательность гигромицинфосфотрансферазы; "3’HygR"), мишень распознавания Flp (FRT), последовательность ДНК гомологичного плеча ниже участка инсерции и субъединицу A дифтерийного токсина (DTA).
Вектор (V2702) от 5’ до 3’ содержит: последовательность ДНК гомологичного плеча выше участка инсерции, последовательность распознавания LoxP 2602, промотор PGK, часть последовательности гена резистентности к пуромицину (5’PuroR), EF-1a, PBase, IRES, последовательность гена резистентности к канамицину (Neo), последовательность терминации транскрипции/сигнала полиаденилирования (поли(A); "PA"), последовательность ДНК гомологичного плеча ниже участка инсерции и DTA.
Последовательность вектора (V2702) подтверждали посредством секвенирования. Вектор встраивали в клетки H9 человека посредством трансфекции. Затем клетки подвергали скринингу по резистентности к G418 и уабаину. Интеграцию генов в геномы человека подтверждали посредством ПЦР. Результаты показаны на фиг. 31. Подтверждали, что клоны под номерами 01, 02, 03 и 04 являются положительными клонами.
Анализ ПЦР осуществляли с использованием следующих праймеров:
3’ L-L-GT-F: 5’-AAGGTGACTCTGCAATCAGCCTCTG-3’ (SEQ ID NO: 38),
3’ L-L-GT-R1: 5’-TCATCTACAGCCACAACGTGAGCAG-3’ (SEQ ID NO: 39);
3’ L-R-GT-F1: 5’-CCCATGTACAGGTTCCGCATGAACT-3’ (SEQ ID NO: 40),
3’ L-R-GT-R: 5’-CTCCGTCCGCTTTTATTTCCCCTGT-3’ (SEQ ID NO: 41).
Выбирали клетки с модифицированными хромосомами, подходящие для дальнейших экспериментов. Модифицированную хромосому человека встраивали в клетки-реципиенты хромосомным способом. Клетки-реципиенты A9 подвергали скринингу на резистентность к G418. Для дальнейшего редактирования генома выбирали клетки, содержащие только одну хромосому человека.
Во время скрининга клон под номером 03, показанный на фиг. 31, имел правильную рекомбинацию, и его мерили с помощью зонда для подсчета хромосомы 2 человека (зонда для FISH CCP2) (CytoTest Inc., Rockville, MD, кат. № CT-CCP002). Результат подтверждал, что в клетке находилась модифицированная хромосома 2 человека (фиг. 32).
Затем вектор (V2701) встраивали в клетки. Клетки подвергали скринингу по резистентности к G418 и бластицидину S. Рекомбинацию подтверждали посредством ПЦР. Праймеры приведены в таблице ниже. Результаты свидетельствовали о том, что 5-C3, 1-H2, 1-H9 и 1-G5 являлись положительными клонами (фиг. 33).
Таблица 14
ПРИМЕР 8: Получение мышей, которые могут продуцировать гуманизированные антитела
Эмбриональные стволовые клетки мыши и клетки, полученные в примере 7, подвергали слиянию и в эмбриональную стволовую клетку мыши, полученную в примере 6, встраивали модифицированную хромосому 2 человека. Выбирали клетки ES мыши, содержащие только одну модифицированную хромосому 2 человека, и после Cre-опосредованной рекомбинации клетки подвергали скринингу, как показано на фиг. 34. После Cre-рекомбинации клетки тестировали для подтверждения встраивания последовательностей генов человека в геном мыши. Зонды Mouse Whole Chromosome Painting Probes (Cytocell Ltd, Cambridge, UK; Кат. № AMP06G) и человек-специфические зонды IGK Break Apart (Cytocell Ltd, Cambridge, UK; Кат. № LPH 034) использовали для проверки ПЦР-подтвержденных положительных клонов посредством флуоресцентной гибридизации in situ (FISH). Результат показан на фиг. 35 и подтверждает наличие фрагментов хромосомы человека в хромосоме мыши. Клетки инъецировали в бластоцисты. Можно получать мышей, содержащих гуманизированный локус легкой цепи иммуноглобулина.
Выбирали химерных мышей для спаривания с мышами C57BL/6, получая серое и черное потомство (поколение F1). Анализ ПЦР осуществляли на ДНК, полученной из хвоста черных мышей, и выбирали положительных мышей поколения F1 для спаривания с рабочими мышами Flp. На фиг. 36 показана схема Flp-опосредованной рекомбинации. Мыши, полученные способами, имели фон C57BL/6. Мыши с разным фоном могут обладать разными преимуществами, и гетерозиготных или гомозиготных мышей, полученных способами по изобретению, можно использовать для получения мышей с некоторым другим фоном посредством возвратного скрещивания (например, мыши BALB/c имеют преимущество гуморального иммунитета) в течение нескольких поколений для получения мышей с желаемым фоном.
Несколько мышей выбирали и скрещивали с мышами BALB/c несколько раз для получения гетерозиготных мышей с фоном BALB/c. Затем гетерозиготных мышей скрещивали друг с другом для получения гомозиготных мышей.
Мышей с гуманизированным локусом легкой цепи иммуноглобулина и мышей с гуманизированным локусом тяжелой цепи иммуноглобулина скрещивали друг с другом для получения мышей и с гуманизированным локусом тяжелой цепи иммуноглобулина, и с гуманизированным локусом легкой цепи иммуноглобулина.
ПРИМЕР 9: Развитие B-клеток у трансгенных мышей
Эксперименты осуществляли для сравнения иммунных систем гуманизированных мышей и мышей дикого типа. Выбирали трех мышей дикого типа (WT) возрастом 9-10 недель, трех мышей, гетерозиготных по гуманизированному локусу тяжелой цепи иммуноглобулина, и трех мышей, гомозиготных по гуманизированному локусу тяжелой цепи иммуноглобулина. Среди них гетерозиготные мыши и гомозиготные мыши имели схожую массу тела, внешний вид и жизнеспособность по сравнению с мышами дикого типа. Получали периферическую кровь, селезенку, лимфоузлы и костный мозг ткани этих мышей, и не наблюдали очевидных анатомических изменений (например, не наблюдали различий размера, морфологии и массы селезенки среди трех групп мышей). Осуществляли проточную цитометрию для анализа популяций лимфоцитов и их распределения в периферической крови, селезенки и лимфоузлах (фиг. 45-47) и популяции B-клеток в селезенке, лимфоузлах и костном мозге (фиг. 48-50) мышей. В результате лейкоциты включали: B-клетки (например, охарактеризованные как CD45+, CD19+, TCR-), T-клетки и естественные киллеры (NK) (например, охарактеризованные как CD45+, TCR-, NK1.1+). T-клетки охарактеризованы как CD45+, CD19-, TCR+. CD4+ T-клетки (CD4) охарактеризованы как CD45+, CD19-, TCR+, CD4+, CD8-. И CD8+ T-клетки (CD8) охарактеризованы как CD45+, CD19-, TCR+, CD4-, CD8+. Только интактные, отдельные, живые лейкоциты включали в анализ проточной цитометрии.
Стадии развития B-клеток в лимфоузлах и селезенке разделяют на T1 (B-клетки переходного типа 1, охарактеризованные как B220+IgM+IgD-), T2 (B-клетки переходного типа 2, охарактеризованные как B220+IgM+IgD+) и зрелые B-клетки (охарактеризованные как B220+IgMlowIgD+). На фиг. 45-47 показаны процентные доли лейкоцитов в разных образцах ткани. На фиг. 48-49 показаны процентные доли B-клеток на разных стадиях развития.
Кроме того, развитие B-клеток также оценивали в краевой зоне (B-клетки краевой зоны, MZ-B, охарактеризованные как B220+CD21+CD23-) и фолликулярной зоне селезенки (фолликулярные B-клетки, обозначаемые как FO-B, охарактеризованные как B220+CD21lowCD23+). На фиг. 50 показаны процентные доли B-клеток селезенки в краевой зоне (MZ-B) и фолликулярной зоне (FO-B) селезенки.
С учетом разных стадий развития B-клетки в костном мозге классифицировали как про-B-клетки (охарактеризованные как B220lowCD43highIgMlow), пре-B-клетки (охарактеризованные как B220lowCD43intIgMlow) и незрелые B-клетки (охарактеризованные как B220highCD43lowIgMhigh). На фиг. 51A-51C показаны процентные доли B-клеток на разных стадиях развития в костном мозге.
Если сравнивать с мышами дикого типа, процентные доли иммунных клеток и B-клеток у гуманизированных мышей были схожими, и не наблюдали статистически значимых различий среди разных групп. Не наблюдали значимых дефектов дифференцировки B-клеток у гетерозиготных по гуманизированной тяжелой цепи мышей (гетерозигот) или гомозиготных по гуманизированной тяжелой цепи мышей (гомозигот).
ПРИМЕР 10: Анализ изотипов иммуноглобулинов в сыворотке
Кроме того, анализировали уровни различных иммуноглобулинов в сыворотке неиммунизированных мышей в примере выше. Мыши включали мышей WT, мышей, гетерозиготных по гуманизированному локусу тяжелой цепи иммуноглобулина, и мышей, гомозиготных по гуманизированному локусу тяжелой цепи иммуноглобулина.
Эксперименты осуществляли с использованием набора Clonotyping System-B6/C57J-HRP (Southern Biotech, кат. №5300-05B). Сначала антитело захвата козы против Ig мыши, ads-UNLB человека разводили PBS (Solarbio, кат. № P1020) до 10 мкг/мл. Затем 0,1 мл разведенного антитела добавляли в каждую лунку планшета для твердофазного иммуноферментного анализа (ELISA) и инкубировали при 37°C в течение 2 часов. Затем планшет промывали и блокировали при 4°C в течение 12 часов. Образцы сыворотки серийно разводили 1% BSA (Cell Signaling, кат. №9998). В каждую лунку добавляли 0,1 мл разведенных образцов с последующей инкубацией при 37°C в течение 1 часа. В лунку добавляли 1% BSA в качестве пустой контрольной точки.
Затем планшет промывали PBS, содержащим 0,05% Tween-20 (Amresco, кат. № M147). Добавляли HRP-конъюгированное вторичное антитело (антитело козы против IgA, IgG (1, 2b, 2c, 3), IgM мыши) (разведенное в 300 раз 1% BSA, 0,1 мл на лунку) и инкубировали с образцом в течение 1 часа при 37°C. Затем планшет промывали и проявляли посредством добавления 0,1 мл раствора хромогена TMB (Beyotime Biotechnology, кат. № P0209) в каждую лунку. После инкубации в темноте при комнатной температуре в течение 8 минут в каждую лунку добавляли 0,1 мл реакционного раствора (Beijing Dingguo Changsheng Biotechnology Co. LTD., кат. № EIA-0032). Оптическую плотность при 450 нм и 570 нм измеряли с использованием спектрофотометра для чтения микропланшетов (Thermo MULTISKAN GO, Thermo Fisher Scientific), и вычисляли стандартное значение OD.
Результаты свидетельствовали о том, что мыши с гуманизированным локусом тяжелой цепи иммуноглобулина могли продуцировать изотипы антител IgA, IgG1, IgG2b, IgG2c, IgG3 и IgM, и мыши имели схожие уровни экспрессии для каждого изотипа по сравнению с мышами дикого типа (фиг. 52-57). Это свидетельствует о том, что гуманизация сегментов генов вариабельной области тяжелой цепи не имела значимого неблагоприятного эффекта в отношении переключения класса, экспрессии или секреции антитела различных изотипов.
ПРИМЕР 11: Рекомбинация V(D)J сегментов генов вариабельной области человека у мышей
Последовательности мРНК вариабельной области тяжелой цепи и вариабельной области легкой цепи у мышей анализировали посредством секвенирования нового поколения.
Выбирали одну неиммунизированную (не подвергнутую воздействую конкретного антигена) мышь, гомозиготную по гуманизированной тяжелой цепи. Клетки селезенки собирали из мыши для выделения РНК. Для осуществления обратной транскрипции для получения кДНК использовали набор 5' RACE (SMARTer RACE 5'/3' Kit, Takara Bio USA, Inc., кат. №634858). Полученную кДНК подвергали ПЦР-амплификации с использованием праймера, специфического для константной области IgM, и праймера UPM из набора 5' RACE для получения последовательности фрагмента вариабельной области тяжелой цепи с последующим секвенированием. Последовательность праймера, специфического для константной области IgM, представляла собой 5'-ccaagcttacgagggggaagacatttgggaa-3' (SEQ ID NO: 50).
В другом эксперименте выбирали одиннадцать мышей, гетерозиготных по гуманизированной легкой цепи, и из ретроорбитальной крови выделяли РНК. После обратной транскрипции способом, описанным выше, праймеры VKF1 и IgKC-tag использовали для амплификации генов легкой цепи семейства VκI с последующим секвенированием. Использовали следующие последовательности праймеров:
Последовательность VKF1: 5'-cataagatctcgmcatccrgwtgacccagt-3’ (SEQ ID NO: 51);
Последовательность праймера IgKC-tag: 5'-ctaacactcattcctgttgaagctcttgac-3' (SEQ ID NO: 52).
Результаты секвенирования сравнивали с поиском Ig в NCBI Blast на последовательности иммуноглобулинов человека для определения экспрессии генов VH, DH, JH и Vκ, Jκ человека после рекомбинации V(D)J. Для 135 анализированных клонов предварительные результаты свидетельствовали об экспрессии некоторых сегментов генов VH, DH и всех сегментов генов JH (таблица 15). Некоторые из этих сегментов генов локализованы близко к участку модификации на гуманизированном фрагменте и на некотором расстоянии от участка модификации. Это свидетельствует о том, что гены VH, DH и JH человека на фрагменте хромосомы человека, встроенном в мышей, можно рекомбинировать для экспрессии тяжелой цепи человека после замены фрагмента эндогенной хромосомы последовательностью тяжелой цепи иммуноглобулина человека.
Большинство генов легкой цепи семейства VκI определяли в 441 клоне, полученном из мышей, гетерозиготных по гуманизированной легкой цепи (таблица 16). Аналогично результатам детекции тяжелой цепи, некоторые из этих генов в гуманизированном фрагменте локализованы очень близко к участку модификации и на некотором расстоянии от участка модификации. Это свидетельствует о том, что после замены локуса эндогенной вариабельной области легкой цепи иммуноглобулина локусом вариабельной области легкой цепи иммуноглобулина человека гены Vκ и Jκ человека, встроенные в геном мыши, можно рекомбинировать для экспрессии легкой цепи с вариабельной областью легкой цепи человека. Дополнительный анализ результатов для одиннадцати мышей показал, что определенное распределение генов IGKV не отличалось значимо среди тестируемых мышей (фиг. 58).
Таблица 15. Список определенных генов IGHV, генов IGHD и IGHJ, экспрессирующихся после рекомбинации VDJ
Таблица 16. Список определенных генов IGKV, экспрессирующихся после рекомбинации VJ
ПРИМЕР 12: Иммунизация и продукция антител в гуманизированных мышах
Пять мышей дикого типа (WT) и пять мышей, гомозиготных по гуманизированной тяжелой цепи (возрастом 9-10 недель), выбирали случайным образом и иммунизировали экзогенными антигенами. Мышей повторно иммунизировали раз в две недели все за три иммунизации. После второй иммунизации и третьей иммунизации получали ретроорбитальную кровь. Получали сыворотку, а затем измеряли титр в сыворотке посредством ELISA или FACS для определения и анализа антиген-специфического гуморального ответа. В исследовании использовали три антигена, представлявшие собой hBTLA, dPD1 и OVA (фиг. 59-63). Результаты свидетельствовали о том, что после второй иммунизации большинство мышей дикого типа (WT) и мышей, гомозиготных по гуманизированной тяжелой цепи, продуцировали антиген-специфические антитела. После третьей иммунизации титр антител повышался до 1×104-1×105. Результаты теста иммуногенной потенции являлись, по существу, такими же у гуманизированных мышей, что и у мышей дикого типа, что свидетельствует о том, что локус гуманизированной вариабельной области иммуноглобулина у мышей является функциональным и может приводить к продукции антиген-специфических антител.
ПРИМЕР 13: Развитие B-клеток у мышей hVH/hVL
Мышей, гомозиготных по гуманизированному локусу тяжелой цепи иммуноглобулина (мышей с гуманизированным VH или мышей hVH), и мышей с гуманизированным локусом легкой цепи иммуноглобулина (мышей с гуманизированным VL или мышей hVL) скрещивали друг с другом для получения мышей, гомозиготных по гуманизированному локусу тяжелой цепи иммуноглобулина и гомозиготных по гуманизированному локусу легкой цепи иммуноглобулина (мышей с гуманизированными VH/VL или мышей hVH/hVL). Мышей hVH/hVL можно использовать для получения гуманизированного моноклонального антитела in vivo.
Эксперименты осуществляли для сравнения иммунных систем наивных мышей с гуманизированными VH/VL и наивных мышей дикого типа. Измеряли массу тела и массу селезенки у мышей дикого типа и мышей hVH/hVL (фиг. 65A-65B). Не наблюдали значимых различий средней массы тела и массы селезенки между мышами дикого типа и мышами hVH/hVL.
Осуществляли проточную цитометрию для анализа популяций и распределения лимфоцитов в селезенке (фиг. 66) и популяций B-клеток в селезенке и костном мозге (фиг. 67A-67B, 68A-68C) мышей. Результаты свидетельствовали о том, что у мышей hVH/hVL процентные доли B-клеток, T-клеток, NK-клеток, CD4+ T-клеток и CD8+ T-клеток в селезенке являлись почти идентичными процентам у мышей дикого типа. В результате лейкоциты включали: B-клетки (например, охарактеризованные как CD45+, CD19+, TCR-), T-клетки, и естественные киллеры (NK) (например, охарактеризованные как CD45+, TCR-, NK1.1+). T-клетки охарактеризованы как CD45+, CD19-, TCR+. CD4+ T-клетки (CD4) охарактеризованы как CD45+, CD19-, TCR+, CD4+, CD8-. И CD8+ T-клетки (CD8) охарактеризованы как CD45+, CD19-, TCR+, CD4-, CD8+. В анализ проточной цитометрии включали только интактные, отдельные, живые лейкоциты.
На фиг. 67A показаны процентные доли B-клеток на разных стадиях развития. Стадии развития B-клеток в селезенке классифицировали как T1 (B-клетки переходного типа 1, охарактеризованные как B220+IgM+IgD-), T2 (B-клетки переходного типа 2, охарактеризованные как B220+IgM+IgD+) и зрелые B-клетки (охарактеризованные как B220+IgMlowIgD+). Кроме того, развитие B-клеток также оценивали в краевой зоне (B-клетки краевой зоны, MZ-B, охарактеризованные как B220+CD21+CD23-) и фолликулярной зоне селезенки (фолликулярные B-клетки, обозначаемый как FO-B, охарактеризованные как B220+CD21lowCD23+). На фиг. 67B показаны процентные доли B-клеток селезенки в краевой зоне (MZ-B) и фолликулярной зоне селезенки (FO-B). Не наблюдали значимых различий между мышами дикого типа и мышами hVH/hVL.
На фиг. 68A показаны процентные доли B-клеток на разных стадиях развития в костном мозге. Клетки-предшественники B-клеток в костном мозге анализировали посредством проточной цитометрии. Учитывая уровни экспрессии B220 и CD43, клетки-предшественники B-клеток в костном мозге можно разделять на 3 популяции клеток: про-B-клетки (охарактеризованные как B220lowCD43highIgMlow), пре-B-клетки (охарактеризованные как B220lowCD43intIgMlow) и незрелые-B-клетки (охарактеризованные как B220highCD43lowIgMhigh). Не наблюдали значимых различий между мышей дикого типа и мышей hVH/hVL.
Кроме того, развитие B-клеток также оценивали в костном мозге или селезенке посредством проточной цитометрии для селективного окрашивания плазматических клеток (B220lowIgM-IgD-CD138-) и B-клеток памяти (B220+IgM+IgD-CD38+) (фиг. 68B-68C). Не наблюдали значимых различий между мышами дикого типа и мышами hVH/hVL.
Разные подтипы иммуноглобулинов (Ig) в сыворотке мышей hVH/hVL и мышей дикого типа количественно измеряли посредством ELISA. Всего шесть мышей выбирали для каждой группы. Не наблюдали значимых различий уровней IgA, IgG1, IgG2b, IgG2c, IgG3 и IgM (фиг.69).
Эти эксперименты показали, что иммунная система мышей hVH/hVL является функциональной, и гуманизированный иммуноглобулиновый локус у мышей hVH/hVL может правильно взаимодействовать с константными областями иммуноглобулинов мыши.
ПРИМЕР 14: Анализ использования зародышевой линии у мышей hVH/hVL
Анализировали использование IGHV, IGHD и IGHJ тяжелой цепи у наивных мышей hVH/hVL (без стимуляции антигеном). Результаты показаны на фиг. 70A-70D. Кроме того, также анализировали использование IGKV и IGKJ каппа-цепи. Результаты показаны на фиг. 71A-71C.
Использование зародышевой линии у наивных мышей hVH/hVL определяли посредством секвенирования нового поколения (NGS). Например, как показано на фиг. 71C, IGKJ1, IGKJ2 и IGKJ4 часто использовались у наивных мышей hVH/hVL, в то время как IGKJ3 и IGKJ5 наблюдали реже. Такой профиль использования IGKJ зародышевой линии соответствует описанному в литературе для использования IGKJ зародышевой линии человека.
Распределение длины CDR3 тяжелой цепи определяли посредством секвенирования NGS иммунного репертуара из спленоцитов наивных мышей hVH/hVL (n=2). Как показано на фиг. 72, медиана длины CDR3 составляла 14 аминокислот. Результаты соответствовали медиане длины CDR3 тяжелой цепи человека в иммунной системе человека.
Анализировали тип аминокислот в каждом положении CDR3 тяжелой цепи (HCDR3) (фиг. 73). Наблюдали множество профилей, включая повышение частоты использования тирозина и повышение использование DH2 (IGHD2) семейства зародышевой линии. Эти профили схожи с композицией аминокислот в HCDR3 человека.
Остаток цистеина может образовывать дисульфидную связь. HCDR3 человека может содержать один остаток цистеина или два остатка цистеина, в то время как HCDR3 мыши, как правило, не содержит цистеин. В результатах на фиг. 74 показана частота HCDR3 мышей hVH/hVL, содержащих остатки цистеина, и частота повышается с увеличением длины HCDR3. Эти результаты соответствуют разнообразию HCDR3 в мононуклеарных клетках периферической крови человека (PBMC).
ПРИМЕР 15: Гистологический анализ лимфоидных органов
Селезенку, паховый лимфоузел и пейерову бляшку из наивных мышей дикого типа или наивных мышей hVH/hVL окрашивали гематоксилином-эозином. Типичные срезы показаны на фиг. 75A-75F. Мыши дикого типа (C57BL/6) и мыши hVH/hVL демонстрировали нормальную структуру с хорошо определенными фолликулами, и не наблюдали значимых различий гистологической морфологии.
ПРИМЕР 16: Образование антител у мышей hVH/hVL
После второй и третьей иммунизации антигенами BCMA, IL4R, PD-1, Siglec15 и SIRPα получали кровь и анализировали титры антиген-специфических антител у мышей дикого типа (C57BL/6) и мышей hVH/hVL посредством ELISA (фиг. 76A-76E). Результаты свидетельствовали о том, что мыши hVH/hVL могут продуцировать антитела, специфически связывающиеся с антигеном, и иммунный ответ у мышей дикого типа и мышей hVH/hVL был схожим.
ПРИМЕР 17: Развитие B-клеток у мышей hVH/hVL
Эксперименты осуществляли для сравнения иммунных систем мышей с гуманизированными VH/VL и мышей дикого типа после иммунизации. Измеряли массу тела и массу селезенки у мышей дикого типа и мышей hVH/hVL (фиг. 77A-77B). Не наблюдали значимых различий средней массы тела и массы селезенки между мышами дикого типа и мышами hVH/hVL.
Проточную цитометрию осуществляли для анализа популяций и распределение лимфоцитов в селезенке (фиг. 78) и популяции B-клеток в селезенке и костном мозге (фиг. 79A-79B, 80A-80C) мышей.
На фиг. 79A показаны процентные доли B-клеток на разных стадиях развития в селезенке. Кроме того, развитие B-клеток также оценивали в краевой зоне и фолликулярной зоне селезенки. На фиг. 79B показаны процентные доли B-клеток селезенки в краевой зоне (MZ-B) и фолликулярной зоне (FO-B) селезенки. Не наблюдали значимых различий между мышами дикого типа и мышами hVH/hVL.
На фиг. 80A показаны процентные доли B-клеток на разных стадиях развития в костном мозге. Клетки-предшественники B-клеток в костном мозге анализировали посредством проточной цитометрии. Не наблюдали значимых различий между мышами дикого типа и мышами hVH/hVL.
Кроме того, развитие B-клеток также оценивали в костном мозге или селезенке посредством проточной цитометрии для селективного окрашивания плазматических клеток (B220lowIgM-IgD-CD138-) и B-клеток памяти (B220+IgM+IgD-CD38+) (фиг. 80B-80C). Не наблюдали значимых различий между мышами дикого типа и мышами hVH/hVL.
Различные подтипы иммуноглобулинов (Ig) в сыворотке мышей hVH/hVL и мышей дикого типа количественно измеряли посредством ELISA. Всего шесть мышей выбирали для каждой группы. Не наблюдали значимых различий уровней IgA, IgG1, IgG2b, IgG2c, IgG3 и IgM (фиг. 81). Кроме того, общее количество IgG в сыворотке мышей hVH/hVL и мышей дикого типа количественно измеряли посредством ELISA. Не наблюдали значимых различий (фиг. 82).
ДРУГИЕ ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ
Следует понимать, что, хотя настоящее изобретение представлено в комбинации с его подробным описанием, изложенное выше описание приведено в иллюстративных целях и не ограничивает объем изобретения, определяемый объемом формулы изобретения. Другие аспекты, преимущества и модификации входят в объем формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Beijing Biocytogen Co., Ltd.
<120> ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ ЖИВОТНЫЕ
С ГУМАНИЗИРОВАННЫМ ИММУНОГЛОБУЛИНОВЫМ ЛОКУСОМ
<130> 1
<160> 54
<170> PatentIn version 3.5
<210> 1
<211> 31
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 1
gccaaggaat ttaaaagggg attgaaagca a
31
<210> 2
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 2
gccctccatg tacagcttca tgtgc
25
<210> 3
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 3
actgggcttg tcgagacaga gaaag
25
<210> 4
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 4
ccacagcccg atctacttgg ctttt
25
<210> 5
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 5
gcaaggtttt gactaagcgg agcac
25
<210> 6
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 6
tgacgcatgt gttttatcgg tctgt
25
<210> 7
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 7
gtgcctgaca cgtgctacga gattt
25
<210> 8
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 8
ttcaacaata agcagggcca gaggg
25
<210> 9
<211> 26
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 9
tcaaagtcaa tttcctcagc gaggct
26
<210> 10
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 10
agggagggaa tggaatgagg gtgat
25
<210> 11
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 11
ccatgtgacc cattcgagtg tcctg
25
<210> 12
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 12
ttgtgagggc tcaagttcag tgcat
25
<210> 13
<211> 31
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 13
gccaaggaat ttaaaagggg attgaaagca a
31
<210> 14
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 14
cgagagctgt ggagagaaag gcaaa
25
<210> 15
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 15
tatgtcctgc gggtaaatag ctgcg
25
<210> 16
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 16
agggagggaa tggaatgagg gtgat
25
<210> 17
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 17
ccatgtgacc cattcgagtg tcctg
25
<210> 18
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 18
cttaccattt gcggtgcctg gtttc
25
<210> 19
<211> 26
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 19
tcaaagtcaa tttcctcagc gaggct
26
<210> 20
<211> 28
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 20
aggaattgta tcccataggc tagcacgt
28
<210> 21
<211> 24
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 21
gacgacgtga ccctgttcat cagc
24
<210> 22
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 22
ccacagcccg atctacttgg ctttt
25
<210> 23
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 23
gcaaggtttt gactaagcgg agcac
25
<210> 24
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 24
cactagtctc gtgcagatgg acagc
25
<210> 25
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 25
gttagaagac ttcctctgcc ctcgg
25
<210> 26
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 26
ttgtgagggc tcaagttcag tgcat
25
<210> 27
<211> 20
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 27
caggtscagc tggtrcagtc
20
<210> 28
<211> 20
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 28
agggatccag agttccaggt
20
<210> 29
<211> 20
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 29
cagrtcacct tgaaggagtc
20
<210> 30
<211> 20
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 30
saggtgcagc tggtggagtc
20
<210> 31
<211> 20
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 31
caggtgcagc tgcaggagtc
20
<210> 32
<211> 20
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 32
gargtgcagc tggtgcagtc
20
<210> 33
<211> 20
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 33
caggtacagc tgcagcagtc
20
<210> 34
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 34
tcacacacta cagcttccac cacaa
25
<210> 35
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 35
cggggaaaag tcgactctag aacgg
25
<210> 36
<211> 26
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 36
actgcattct agttgtggtt tgtcca
26
<210> 37
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 37
ggcctggaaa actcagctat ccttt
25
<210> 38
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 38
aaggtgactc tgcaatcagc ctctg
25
<210> 39
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 39
tcatctacag ccacaacgtg agcag
25
<210> 40
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 40
cccatgtaca ggttccgcat gaact
25
<210> 41
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 41
ctccgtccgc ttttatttcc cctgt
25
<210> 42
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 42
gttataacac ggggagtgcg tgtgc
25
<210> 43
<211> 30
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 43
gtttggacaa accacaacta gaatgcagtg
30
<210> 44
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 44
gcaacggcta caatcaacag catcc
25
<210> 45
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 45
tgggtctggg acagacttca ctctc
25
<210> 46
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 46
aaggtgactc tgcaatcagc ctctg
25
<210> 47
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 47
tcatctacag ccacaacgtg agcag
25
<210> 48
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 48
cccatgtaca ggttccgcat gaact
25
<210> 49
<211> 25
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 49
ctccgtccgc ttttatttcc cctgt
25
<210> 50
<211> 31
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 50
ccaagcttac gagggggaag acatttggga a
31
<210> 51
<211> 30
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 51
cataagatct cgmcatccrg wtgacccagt
30
<210> 52
<211> 30
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 52
ctaacactca ttcctgttga agctcttgac
30
<210> 53
<211> 7795
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 53
gggatgacag attctctgtt cagtgcactc agggtctgcc tccacgagaa tcaccatgtc
60
ctttctcaag actgtgttct gtgcagtgcc ctgtcagtgg aaatctggag agcatgcttc
120
catgagcttg tgagtagtat atctagtaag ccatggcttt gtgttaatgg tgatgttcta
180
catatcagtt ctctggctta ataatgaggt gatgattcta tgttcctgta acgcttcctc
240
aactgggtcc taagtctttc ttcactccat ctattcctct aaggaatgat cctgaaaatc
300
ccatcacaaa ctataggaga tgggaaccat caaaaaacac agtgacaaag aggtgggaac
360
gcatcagggt tcaggaacca tattttaaaa agatatcgta aataacttct taaaagagat
420
atagacaaat ctccattaat acggagacca gaggcctaag gctaagaacc aatggtggct
480
caaggtctcc tgctacccga ggagcaaacg tagagcagtt tctaatgatt tatttaaaat
540
atagaatcaa aagtaccagt ttgcaatttt gaaagattta tttcagcaat gcaacaacat
600
caggtggtgc cgagtccaac acgtcttatg tcccatgata taaacaaagg ccatccagaa
660
ctgtggactg gagttctacc ttgtccccta atgacattca gatttttttt ccattctctt
720
tatcttagag gagacagggg gctaactcat tttacttgtc ctttgcttgt tcttgccaag
780
aacgtaaagc agcttgcaag tcttcaaacc taaatatctt agtaactcct acacgagtgg
840
caatgccaaa gagcagtgca acaaagagga agtaaatacg accaaagagt attcttaaat
900
acactactgg ctctaggttc tgttttatta tgcgcctttg aaccggaggg gacccactgt
960
ctatgctccc actgtgtccc tcttctttgc actttggagg gctccaacca aaatggcaat
1020
ggcaattccg acgattgtta cacactcctc tgaaattgca tttttctggg gtgcagtcat
1080
aacccaaacg agataaactt ccattgcaag ctcctcgatc acagaactta ccccttgaac
1140
acggggtacc atgtctcacc aatccagcat ctgctgtttc tgtcccacga tgttcatcaa
1200
gcccaaagca ggtaacccca gagataaccg attgatggaa tgaaacatgt tcttgcaaaa
1260
atggaagatt ggtgacattg gtacactgca accttccaca cagcttgtcc tgatcagcac
1320
aagcattgaa tgtgaggctt tcttctgctc tagtacaatg cccaaatcga aaccgttgtt
1380
tgttgatgtc atagcactta atattagcat tcttagcact tacaccaaag atttccatgc
1440
attgtatgtt gcgatcagtg cagttacctt tatagcagta accctcttct gagcatggtg
1500
tcccatcttg cagataagtg tcatctgggc aaatgaactt agagccacta cagtactctg
1560
gaagatcaca tatgttctgg ataggtctgc agagtgtccc agaaggactg taagtgcaat
1620
ttgcacagca taattcttta tcacaaatgc taccaggtgt taacctgcaa tcatttccac
1680
agcagggatc tgaataacat gccttttggg agccacagtc acactgctca ttgttatcta
1740
ctttgaagtt tccacaaaac ttataagtca atgatgtatt ataataaaca tgacggtcat
1800
agaaaagaca tggcatcaga tcaggagtat taagtatgtt gcttatctct gcaagggaac
1860
aattgctgaa agcatctgtt aattgaggat ttttgaacat gatgcaggtg ttccttctct
1920
ggcagataca gtacccctca tcatgtttta ggcctaaact ccttccaaca cgattggtta
1980
ttataataga taaaaataaa ggatttcgac catgttgacc aagacaaatt agggctgagg
2040
gagaacatat actcctctca gctggattaa cagcatcatc tcctggcgaa ttcttgttaa
2100
ttatagctcc tgcatcaggc ctaaaatgag cataaaatac tctctcatag aaagtatgag
2160
cctgccctcc tggaactcga aaatcttgtg aaaatggatc agcctcggta tacacagtca
2220
tgagaaagac atagtaccgc atatgaagat tggtcagata ggtgtccatt aaactaatga
2280
ctttaaacaa atactcaaca gtagatgaaa gtttgtcacc tccagaagca ctatatacag
2340
aatgggttgc ttgaaagtgg ccttttatag cagctggatg tgtagcgtaa ttcttactag
2400
atagtctggg agctccatct gcatattcca atctggagga gggagaacct gtattatggc
2460
tccagtgctt ccatgcattc ataggccctg tgtcatcaga ctcagatact atctgagaaa
2520
caaggtgttc aaagctctgt gaatcattga ggggtttgat ttcataggta aggttatcca
2580
actttatgac ccctgacagg cccccataac aagtatccac agtgaccatg gattgcagga
2640
tcccctccag gtagccaata tagtaacaat ctacaggaaa aaaggggtac tccatctgta
2700
aggctccttg gtcatcttga gttgtcagca acaagtgtct gggccaaatg agtgtctttc
2760
tccgcaggtg gatgatatgt ctctggcccc gaaaacgcaa gctatacgag agcagtcttt
2820
gtgcttgaag tcctttggta tggtagatct ccttccgagg aataaccacc tccgatgaga
2880
tgtaacgcca agtgggatgg ccttgagaac accagactgg aaccaggagg agcagccaga
2940
gtgcaaatag caagaggagg accctgggga ccacaggtct ttccactagc ctcatgcccc
3000
aggtcagaga taacatcctg ggtggagcta actccctctg ctgtggccac tgcctggtct
3060
agaaaatact gacagaggac taaaaacctc ctcaggctcc caacctaagt ggttacccag
3120
acaactggag ttaggtaaca gtcactgggt gtggcaggaa ttgagtctga atgtgttagc
3180
tgaggttgag gttaaatatt gtcaaaaggg atgtctataa atgtgcctgg acaagaaaag
3240
tcagaagcag caaggagtgt ctctgacagg ctcaatcctt tcttttcttt ttttgaagtt
3300
caaaatatca tttccacgtg aatgtatttg gttcccagtg tgactctggg tctctttcta
3360
ggagtcaata tttctttata tcttggctca tgtttttcac agttgttcta acttcttgtt
3420
ttgttttgtt tgtttgtttg tttgaaagtt agaagtaaat actgtctata ttagcctttt
3480
agctataaat gattgttttt atttcttcta atcatgtttt gtttgagttt tggttaaact
3540
atttacaaat gagttttttt tttccttttg ggtgttgctc gaaagtttgg agctttctgt
3600
taatattgtg ttgttgtttc tccaatatta ttagacctga gaattctacc tgggtacctg
3660
tgaactccag aatttttaaa aattccatct cttgggaaca ttatctctga ccccgtctga
3720
ggccgaagtg gctgtccccc tccaaccttt agtatctttc tttcctgact attgggattt
3780
cttcaagcaa tcaggctgat gggttctcag cagtgagacc agtagactgt cggtatgaac
3840
gtcgaagagt ctgccacaca ctccgggttc atcaacagtg ctttcgcgtc tcttactttt
3900
gtagaaggaa atgcagcctc tgagttttct ccaagaaatc attgatgaaa gggtgaaaag
3960
atgggtatca cccggagttc atgacaagcc ctggctcaga cacgtgagca aggtctacag
4020
ccccaaagat aggctgccct gcaacatgta tttataagat aggagaaaaa aatgggtagt
4080
tggagggttg atcaacttac ttcctctcaa acatatatat ctcatctaag tgtgcagggg
4140
aaaactctgt agaactactg ggatacctgc tcacccccag gagcctcatg aataagtctc
4200
tgcttctgcc ttgtagccat gagcattact gcacctgata cccctgcagc ttcctaggga
4260
agagggagga agtgacttgg cccctgtctg gttaaggtaa gaggagataa atcccttctc
4320
attgattagg gtgagagggg tcatgtgctc tatcattggt gacccagttg ggacatgggt
4380
ttataccaaa gtcatcactc tgaggttctg tgtaccacca ggctgaactc ccatatccta
4440
catggacata ggacaacacc aagcagaagg aggttttagg actaaactga aggacagaga
4500
tgcggtttct aaacaactag ggagtgccag ggccagcctc tctaaccact ataggacact
4560
gtggagtctg gttacaaaga gagattactc aaggtcctta gcactgatta cagagcatat
4620
ctcagatgcc ttctgctgac cagatgtatc tttgcataat ctgcctatcc agattcagaa
4680
aattgatgcc acatagccaa gtggactttc aggaacagac gatttaaaaa caggcagaga
4740
gatgtgagag aaaggagaag gagagagaga agggagaggg agagaagaga gagggagacg
4800
gagaaggaaa gagggagaag gagaaggaga gaaggggcat ggacagaggg agggacagaa
4860
ggagagagga gatagagagg gggataagga agaagggagg gagggagaga gagagaaggc
4920
taagtctttc catacctggg tcccaatacc tcttataacc caagcacatg gtttcacata
4980
tcacaatgcg gttgggatat agataactgt aaatacttgt gaaaataatg gggctgagat
5040
ctggggtttt catgatagtt tcaaagtcac cgtactgact aaaaccttcc actggcccat
5100
ctccagcttc ctaatctgag ggtatcaaat ttcccactaa gtgtgtttag aaagatctcc
5160
acctttttgc ccttgtcttc cagtgcccca cctacgttct ggtctcccac atctgatgtc
5220
ttctcagtga ttctggccct gcctgctcca cagctacaaa ccccttccta taatgagctc
5280
tgtgctgagc catcatcctg aatcaatcca ccttaagcag atgttttgct tatttttcct
5340
gtgtccatac tacagaggaa aggtaggcat gtagaagctg aagcatctca cctcattcca
5400
agcaccctca gtctctaaat gtgccccctt gtttccagaa gtgcaacctc aagcatcttt
5460
tattcattca tcttagaggg ccacatgtgc tgtagtgtta taagatgaaa tttaaagcat
5520
taattattcc taacaagcca attaaacaag ccaaaaacat tcatcagtca ttcccatgga
5580
acctctgaag catcttcctg ctctaacctt gggttttcca gggctgctct gggatcacag
5640
gagctgtcct gtctaccagc catataaagg cagacctatc agaattacac cagacttctc
5700
accatagact ataaaagcca gaatatcctg gacagatgtt atacagaaac taagagaaca
5760
caaatgccag cccaggctac tatacccagc aaaactctca attaccatcg atgaagaaac
5820
caagatattc cattacaagt ccaaatttac acaatatctt tccataaatc cagccctaca
5880
aaggatagca gatggaaaac tccaacacag gtaggaaaac tacaccctag aaagagcact
5940
aaagtaatca tctttcaaca cactcaaaag aagataacca cacaaacata attccacctc
6000
taacaacaaa aataaagtag gcaacaatca ctattcctta atatctcttt taacatcaat
6060
ggactcaatt ctccaataaa aagacataga ctaacagact gaatacataa acaggacaca
6120
gcattttgct gcataaagca aacacagcgt tacttttttt tttctaaatg acatttttta
6180
ttagatattg tctttattga catttcaaat gttatcccct ttcctggttt accctctgaa
6240
atcccctatc tcctccccct ccccctgctc accaatccac ccactcccac ttccaggccc
6300
tggcaatccc ctatatttgg gcatagagcc ttcacaggac caaggtactc tccttgcatt
6360
gatgaccaac tagtccattc tctgctacaa atgcagctag atctatgagt cccaccatgt
6420
tttcttttgt tggtggtttc atgccaggga gctcttggag tactgattgg ttcatattgt
6480
tgttctccct atggggttac aaaacccttc aacttcttgg gtcctttctc tggctgcctc
6540
attggggacc ttgtgcgaag tccaatggat gactgtgagc atccacttct gtatttgcca
6600
ggcactggca gagcctctca gaagacagct atatcaagat cctggcagca agctcttgtt
6660
ggtatccaca aaagtgtctg gtggttgtct atgggatgga tccccaaagg ggcagtctct
6720
ggatggtcat tccttcagtc tctgttccac actttgtctc tttaactcct tccatgacta
6780
ttttattcct ccctctaaga aggaccgaag tattcatact ttggtcttcc ttcttgaaat
6840
tcatgtgttt tgtgaattgt atctttgata ttccgaactt ctgggctaat atccacttat
6900
cagtgagtga atatcatgtg tgttcttatg tgattgagtt acctcactca ggatgatatc
6960
ctccagaacc atccatttgt ctaagaattt aatgaattca ttgtttttaa tagctgagga
7020
gtactccatt gtgtaaatgt accacatttt ctgtacccat tgttctcttg agggacatct
7080
gggttcttta aagcttctgg acattaaata taaggctgct atggaaatag tggagaatgt
7140
gtccttatta catgttggag catcttctgg gtatatgccc aggagtgcta ttgctggatc
7200
ctctgatagt actatgtcca attttctgag gaactgccaa actgatttac agagtggttg
7260
taccagcttg caattccacc agcaatggag aaatgttccc cttcctccac atcctcacca
7320
acatctgctg tcacctcaat ttgttcttag tgattcagac aggtgtgagg tggaatatca
7380
gggttgtttg gcatttccct gatgactagt gatattgaaa aaaattttaa gtgtttctca
7440
gccattcagt attcttcagt tgagaattca ctgtttagct ctgtactcag gtttttttaa
7500
tagggttatt tggttttctg gagtctaacg tcttgaattc tttctatata ttggatatta
7560
gccctctgtc atatttagga ttggtaaaga tctttcccaa tatgttggct gcctttttgt
7620
gtcctttgcc ttacagaacc tttttaattt tatgaggtcc catttgctaa ttcttcattt
7680
tacagcacaa gccattggtg ttctgttcaa aaatctttcc ccctgaaccc tatcttcgag
7740
gatcttcccc actttctcct ctataagttt cagtgtctct attattgtgc tgagg
7795
<210> 54
<211> 5938
<212> ДНК/РНК
<213> Искусственная последовательность
<400> 54
tgatgtggga acgcttcagt gttcaggaac catatgattt atttaaaata tagaatcaaa
60
agtaccaatt tgcagttttg aaagatttat tccagtgtaa gcattagcaa tgcaccaaca
120
tcaggtgatt tctgaatcca acacgtctta tgtcctcatg atattaaaaa aaaaaaaagg
180
ccatccagaa ctgtgaactt gagttctacc ttgttcccta ctgacattca gattttcttt
240
tttgcattct ctttatctta caggagacag gaggggaggg ctaactcatt ttactttggc
300
ttgtcccttg ctggtccttg cccagaacgt aaagtagctt gcaagtcttc aaatctaaaa
360
atcttagtaa ctcctacacg agtggcaatg ccaaagagca gtgcaacaaa gaggaagtaa
420
atacgaccaa agagtattct taaatacacc actggctctt gtttttgttt tattgtgtgc
480
ctttgaactg gaggggaccc actgtctatg ctcccactta gtccctcttc tttgcactct
540
ggaggcttcc aaccaaaatg acaatggcaa ttccgatgat tgttacacac tcctctaaaa
600
ctgcattttt ctggggtgca gtcataaccc aaatgagata aacttccact gcaagctcct
660
tgatcacaga acttactttt ggagcagggg gtaccatgtc tcaccattcc agcatctgtt
720
gtttctgtcc cacgatgttc atcaagccca aagcaggtaa acccagagat aatcgattga
780
tggaatgaaa catgttcttg caaatatgga agattggtga cattggtaca ctgcaacctt
840
ccacacagct tgtcctgatc agcacaagca ttgaatgtga ggctttcttc tgctctagta
900
caatgcccaa atcgaaaccg ttgtttgttg atgtcatagc acttaatatt agcattctta
960
gcacttacac caaagatttc catgcattgt atgttgcgat cagtgcagtt acctttatag
1020
cagtaaccat cttctgagca tggtgtccca tcttgcagat aagtgtcatc tgggcaaatg
1080
tatttagtcc cattacagta ctctggaaga tcacatatgt tctggatagg tctgcagagt
1140
gtcccagaag gactgtaagt gcaatttgca cagcataatt ctttatcaca aatgctacca
1200
ggtgttaacc tgcaatcatt tccacagcag ggatctgaat aacatgcctt ttgggagcca
1260
cagtcacact gctcatcgtt atctactttg aagtttccac aaaacttata agtcaatgat
1320
gtattataat aaacatgacg gtcatagaaa agacatggca tcagaccagg agtattaagt
1380
atgttgctta tctctgcaag ggaacaattg ctgaaagcat ctgttaattg aggatgtctg
1440
aacataatgc aggtgttcct tctctggcag acacagtacc cctcatcata ttttaagcct
1500
aaactccttc caacacgatt ggttattata ggagataaaa ataaaggatt tcgatcatat
1560
ttaccaatac aaattagggc taaggaagaa catatactcc tctcagctgg attaacctgg
1620
ttatcttgtg gcccatactt attaagtaaa actcctgcat caggcttaaa tttattataa
1680
aagactgaca catagtaatt ataagccgac cctcctggaa ctgcaaactc aagtcgaaat
1740
ggatcagaat tggtgtacac agtcatgaga aagacatagt accgcatatg aagattggtc
1800
agataggtgt ccattaaact aatgacttga aacaaatacc caacagtaga tgaaagtttg
1860
tcacctgcag cagaattata tacagaattg gttgcttgaa agtggccttt tatagcagct
1920
ggatgtgtag cgtagttctt actagatatt ctgggagctc catctgcata ttccaatctg
1980
gaggagggag aacctgtatt atggctccag tgcttccatg cattcatagg ccctgtgtca
2040
tcagactcag atactatctg agaaacaagg tgttcaaagc tctgtgaatc attgaggggt
2100
ttgatttcat aggtaaggtc atctaacttc atgacccctg acaggccccc ataacaagta
2160
tccacagtga ccatggattg tgggatcccc tccaggtagc caatatagta acaatctaca
2220
ggaaaaaagg ggtaatccat ctgtaaggct ccttggtcat cttgagttgt cagcaacaag
2280
tgtctgggcc aaatgagtgt ctttctccgc aggtggatga tatgtctctg gccccgaaaa
2340
tgcaagctat atgagagcag tctttgtgct tgaagtcctt tggtatggta gatctccttc
2400
cgaggaataa ccacctccga tgagatgtaa cgccaagtag gatggccttg agaacaccag
2460
actggaacca ggaggagcag ccagagtgca aatagcaaga ggaggaccct ggggaccaca
2520
ggtctttcca ctagcctcat gccccaggtc agagataaca tcctgggtgg agctaaatcc
2580
ctctgctgtg gccactgcct ggtctagaaa atactgacag aggactaaaa acctcctcag
2640
gctcccaacc taagtggtta cccagacaac tggagttagg taacagtcac tgggtgtggc
2700
aggaattgag tctgaatgtg ttagctgagg ttgaggttaa atattgtcaa aagggatgtc
2760
tataaatgtg cctggacaag aaaagtcaga agcagcaagg agtgtctctg acaggctcaa
2820
tcctttcttt tctttttttg aagttcaaaa tatcatttcc acgtgaatgt atttggttcc
2880
cagtgtgact ctgggtctct ttctaggagt caatatttct ttatatcttg gctcatgttt
2940
ctcacagttg ttctaatttc ttgttttgtt ttgtttgttt gtttgaacgt tagtagtaaa
3000
tactgtctat attagccttt tagctataaa tgattgtttt tatttcttct aatcatattt
3060
tgtttgagtt ttggttaaac tatttacaaa tgagtttttt ttttttcctt ttgggtgttg
3120
ctcgaaagtt tggagctttc tgttaatatt gtgttgttat ttttccaata ttattagacc
3180
tgagaattct atctgggtac ctgtgaactc tagaattttt aaaaattcca tctcttggga
3240
acattacctc tgaccccgtc tgaggccgaa gtggctgtcc ccctccaacc tttagtatct
3300
ttctttcctg actattggga tttcttcaag caatcaggct gatgggttct cagcagtgag
3360
accagtagac tgccggtatg aacgtcgaag agactgccac acactccagg ttcatcaaca
3420
gtgctttcgc gtctcttact tttgtagaag gaaaagcagc ctctgagtta tctccaagaa
3480
atcattaatg aaagagttaa aagatgggta tcacccggag ttcatgacaa gccctggctc
3540
agacacgtga gcaaggtcta cagccccaaa gataggctgc cctgcaacat gtatttataa
3600
gatagaagaa aaaaatgggt ggttggaggg ttgatcaact tacttcctct caaacatata
3660
tatctcatct aagtgtgcag gggaaaactc tgtaggacta ctgggattgt tattatcatt
3720
attattatta ttattattat tattattatt attattatta ttaacttaag gcattttatt
3780
agatattttc ttcatttagt tttcaaatgt tatccccgga acctcctata ctctctccct
3840
gccctgctcc ccaacccacc cactcctaca tcctggccct ggcattcccc tatactgtgg
3900
cagatgatct tcgtaagacc aagagccttt cctcccattg atggcctact aggctatcct
3960
cttttacata tgcaactaga gtcacagctc tggggaggta ttgcttagtt catattgttt
4020
ttcctcctat agggttgcag atccctttag ctccttgggt actttctcta gctcctccat
4080
tgggggccct gtgttccatc caatagatga ctgtgagcat ccacttctgt atttgccagg
4140
tattggcatg gatcttactg caccttctga actctctaag cagctttcct ggtcacctcc
4200
aggagcctca tgaataagtc tctgcttccc ccttgtggct atgagcatta ctgcacctga
4260
tacaccctgc agcttcctag ggaagaggga ggaagtggct tggcccctgt ctggttaagg
4320
taagaggaga taaatccctt ctcatgaatt agggtgagaa gggtcatgtg ctctatcatt
4380
ggtgaccaac ttggggacat gggcttatac agtcatcact ctgaggctct gtgtaccacc
4440
agactgaact cccatatcct acatgcacat aggacaacac caagtagaag gaggttttag
4500
gactaaactg aaggacagag atggggtttc taaacaacta gggagtgcca gggccagcct
4560
ctctaaccac tataggacac tatggagtct ggttacaaag agagattact caaggtcctt
4620
agcactgatt acagagcata tctcagatgc cttctgctga ccagatgtat ctttgcataa
4680
tctgcctatc cagattcaga aaattgatgc cacatagcca agtggacttt caggaacaga
4740
cgatttaaaa acaggcagag agatgtgaga gaaaggagaa ggagagagag aagggagagg
4800
gagagaagag agagggagac ggagaaggaa agagggagaa ggagaaggag agaaggggca
4860
tggacagagg gagggacaga aggagagagg agatagagag ggggataagg aagaaaggag
4920
ggagggagag agagagaagg ctaagtcttt ccatacctgg gtcccaatac ctcttataac
4980
ccaagcacat ggtttcagat atcacaatgc ggttgggata tagataactg taaatacttg
5040
tgaaaataat ggggctgaga tctggggttt tcatgatagt ttcaaagtca ctgtactgac
5100
taaaaccttc cactggccca tctccagctt gttaatctga gggtatcaaa tttcccacta
5160
agtgtgttta gaaagatctc cacctttttg ccctagtctt ccagtgcccc acctacgttc
5220
tggtctccca catctgatgt cttctcagtg attctggccc tgcctgctcc acagctacaa
5280
accccttcct ataatgagct ctgtgctgag ccatcatcct gaatcaatcc accttaagca
5340
gatgttttgc ttatttttcc tgtgtccata ctacagagga agggtaggca tgtagaagct
5400
gaggcatctc atctcactct aagcaccctc agtctctaaa tgtgcccctt tgtttccagc
5460
agttcagcct caagcatctt ttattcactc gtcttagagg gacacatgtg ctgtagtgtt
5520
ataagatgaa atttaaagca ttagttattc ccaacaagcc aattaaacaa gccaaaaaca
5580
ttcatcagtc attcccatgg aacctctgaa gcatcttcct gctctaacct tgagtttcct
5640
agggctgctg tgggatcaca ggagctgtcc tgtttaccag cctatcctgt cccacgggat
5700
tcagttatta gtgggtgcga gggggaccgc aaacctggaa gaaaatggga ttggaagaga
5760
aaagagaaac gaagaccaag tagatctttt cctatcaagg tcttcgttta ttaggctgag
5820
gtgcctggtg taaagcatgc atcgcgggga ataggaaggg gtcgaggggg aattttacaa
5880
agaacaaaga agcgggcatc tgctgacatg agggccgaag tcaggctcca ggcagcgg
5938
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, СОДЕРЖАЩИЕ ЛОКУС ОБЩЕЙ ЛЕГКОЙ ЦЕПИ ИММУНОГЛОБУЛИНА | 2021 |
|
RU2841160C1 |
| ПРОДУЦИРУЮЩИЕ АНТИТЕЛА МЛЕКОПИТАЮЩИЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ | 2009 |
|
RU2559524C2 |
| ПРОДУЦИРУЮЩИЕ АНТИТЕЛА МЛЕКОПИТАЮЩИЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ | 2020 |
|
RU2820926C2 |
| ПРОДУЦИРУЮЩИЕ АНТИТЕЛА МЛЕКОПИТАЮЩИЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ | 2009 |
|
RU2731084C2 |
| КОЛЛЕКЦИЯ И СПОСОБЫ ЕЕ ПРИМЕНЕНИЯ | 2010 |
|
RU2569187C2 |
| АНТИТЕЛА И ДРУГИЕ МОЛЕКУЛЫ, КОТОРЫЕ СВЯЗЫВАЮТ В7-Н1 И PD-1 | 2012 |
|
RU2625034C2 |
| ГУМАНИЗИРОВАННЫЕ НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕКУ ЖИВОТНЫЕ С ОГРАНИЧЕННЫМИ ЛОКУСАМИ ТЯЖЕЛОЙ ЦЕПИ ИММУНОГЛОБУЛИНА | 2013 |
|
RU2656155C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2761639C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ЛЕГКУЮ ЦЕПЬ С ВАРИАБЕЛЬНОЙ ОБЛАСТЬЮ АЛЬФА ЧЕЛОВЕКА И КОНСТАНТНОЙ ОБЛАСТЬЮ МЫШИ | 2011 |
|
RU2590594C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ ДЛЯ ТЕСТИРОВАНИЯ ТЕРАПЕВТИЧЕСКИХ АГЕНТОВ | 2019 |
|
RU2805212C2 |




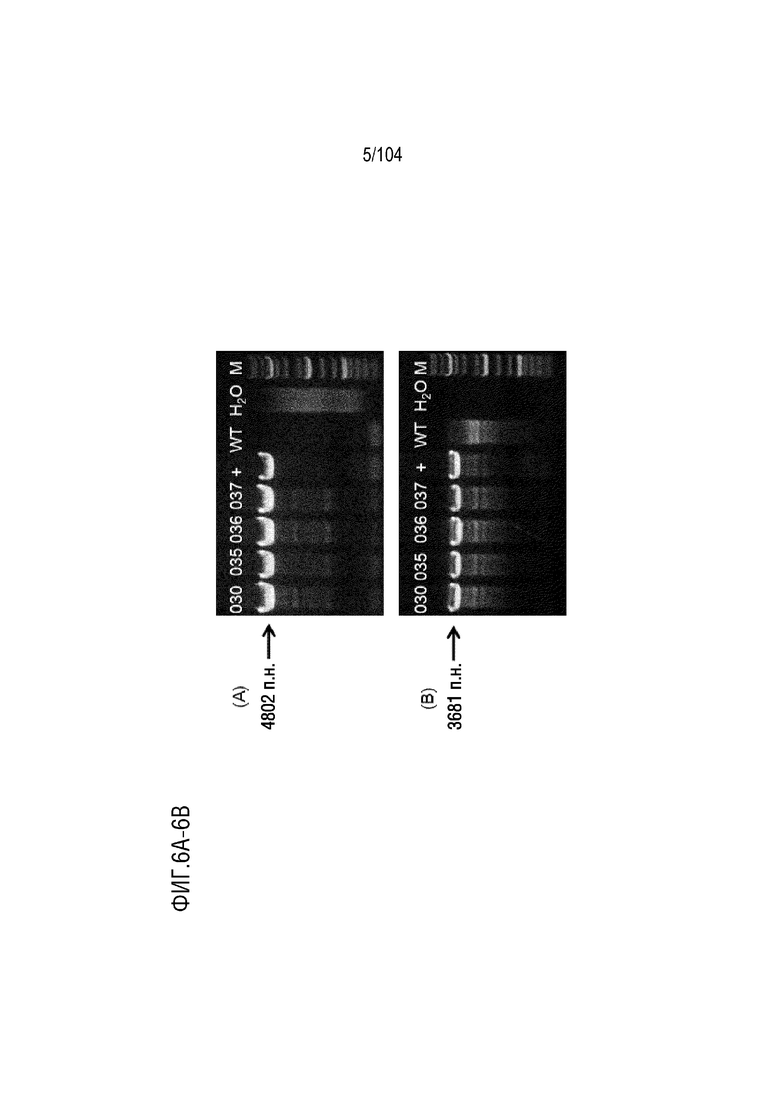

















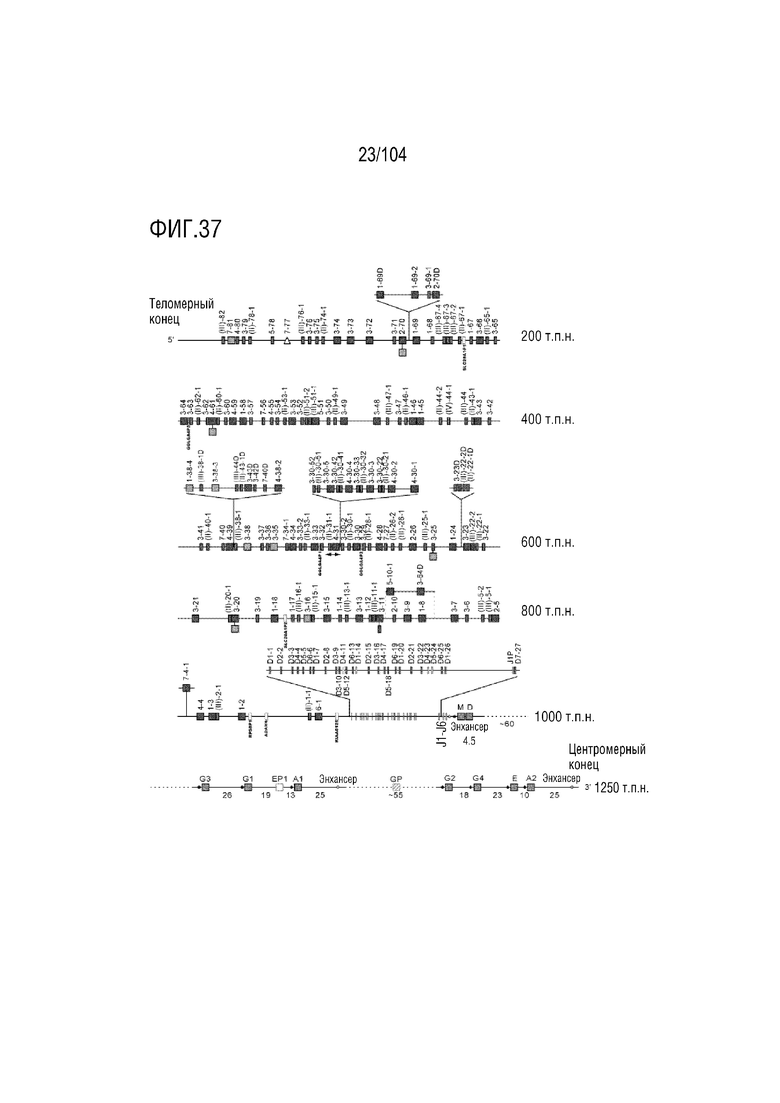






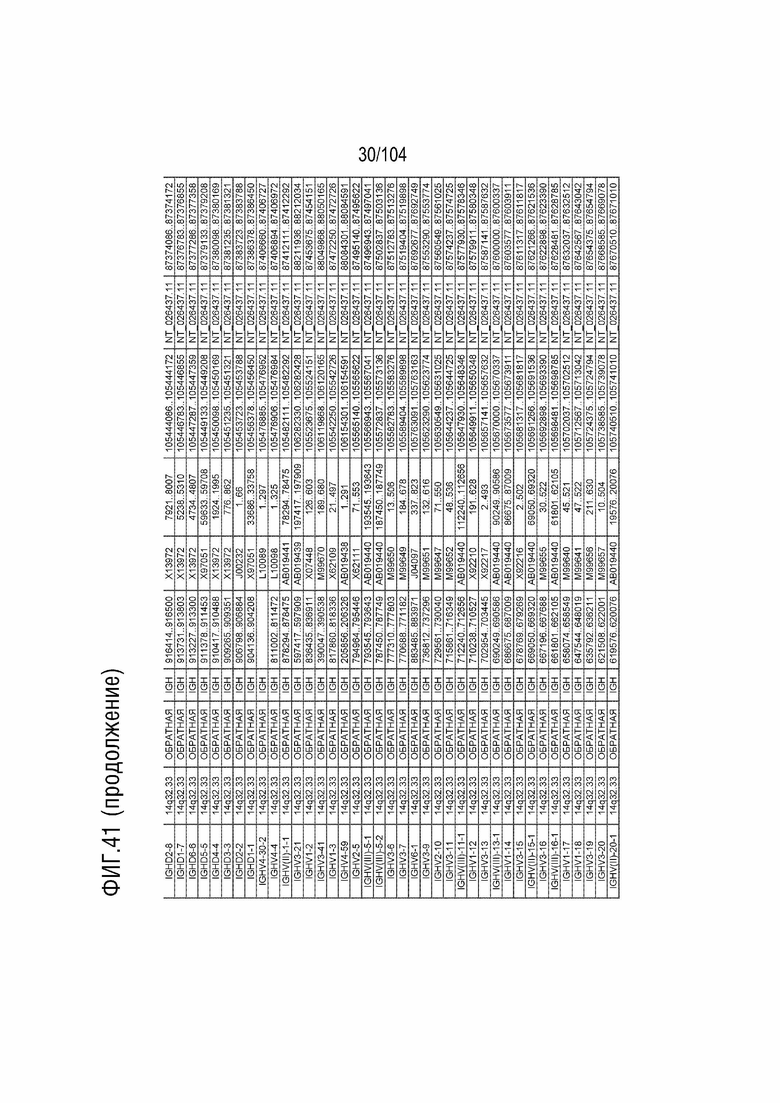


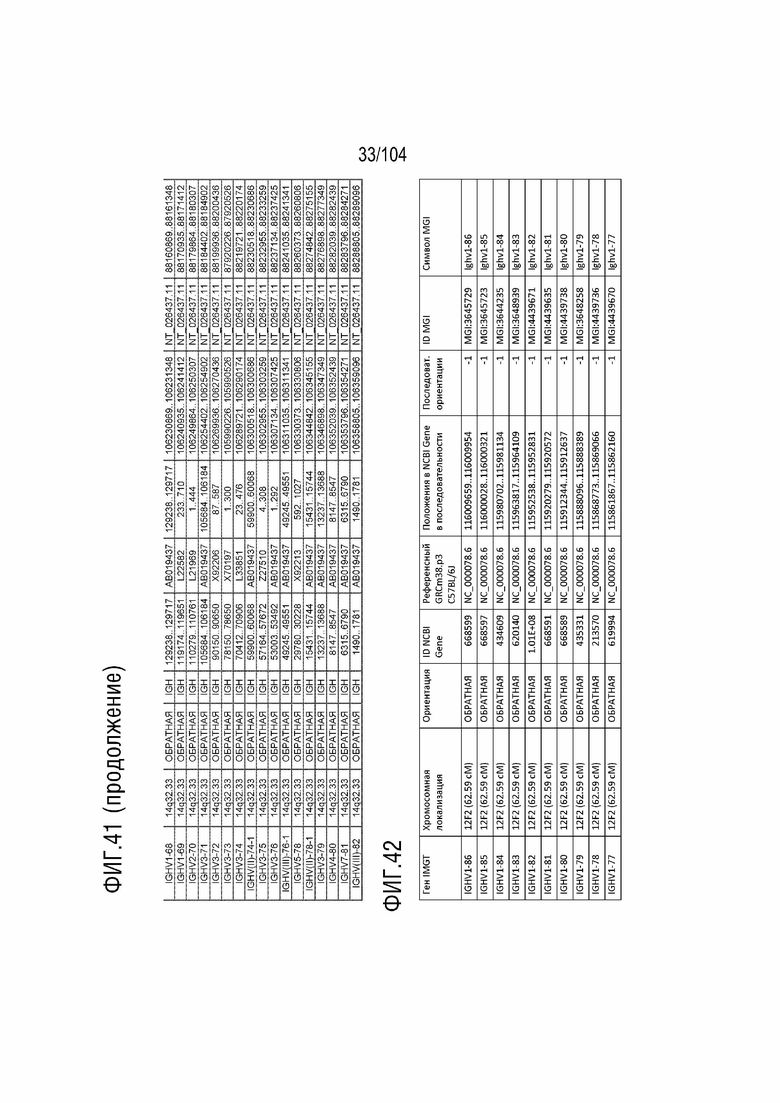
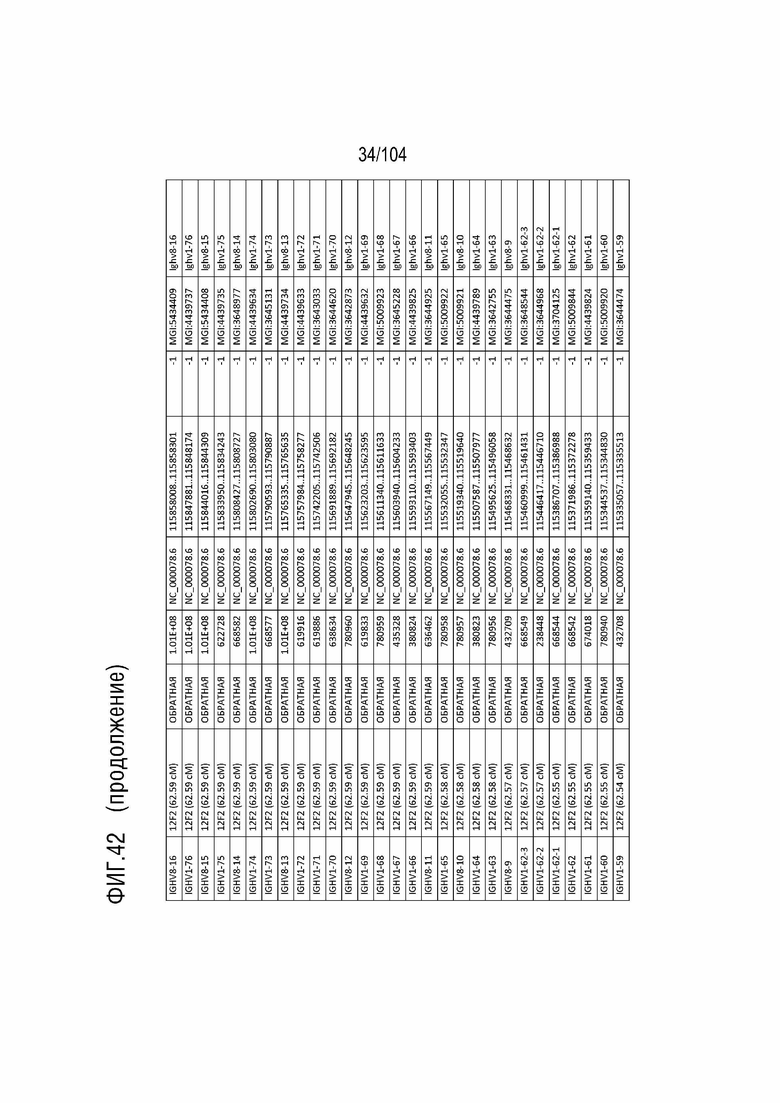












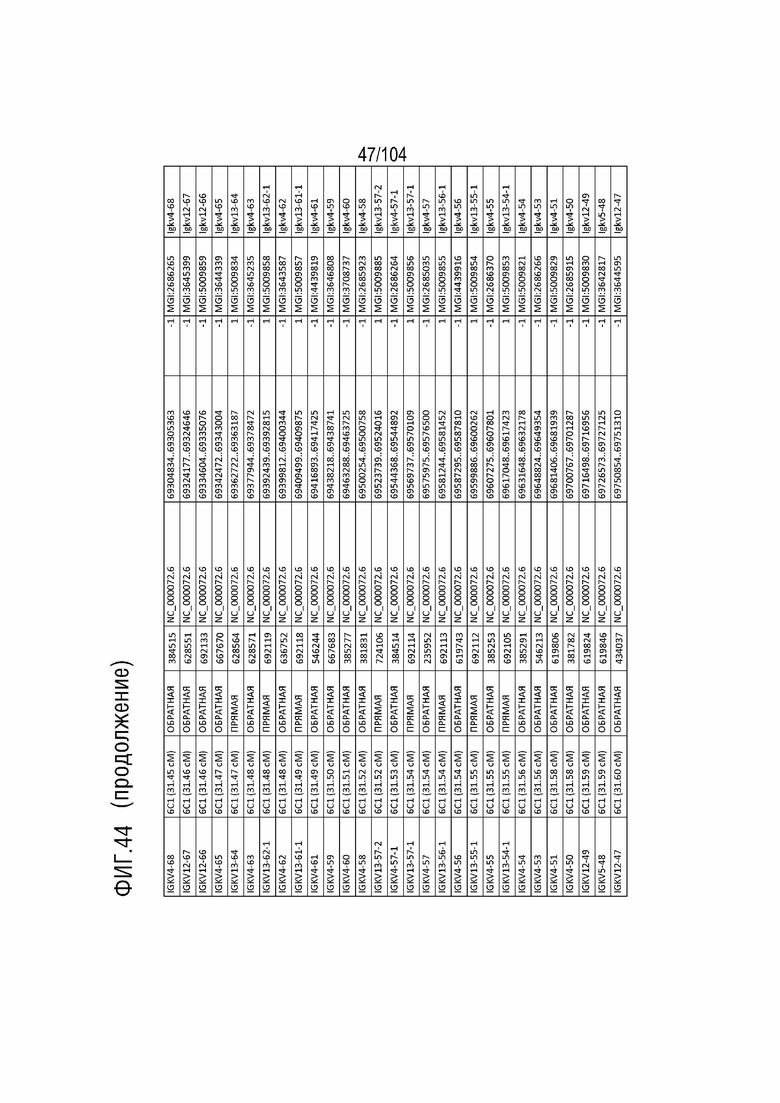




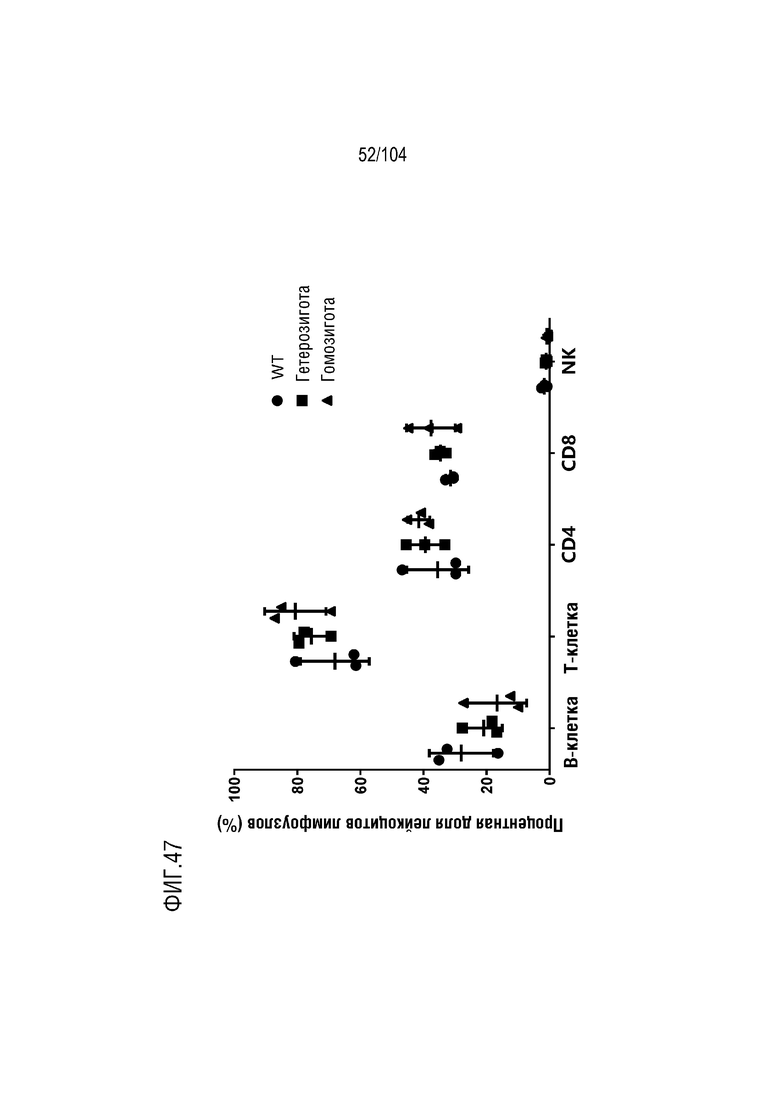









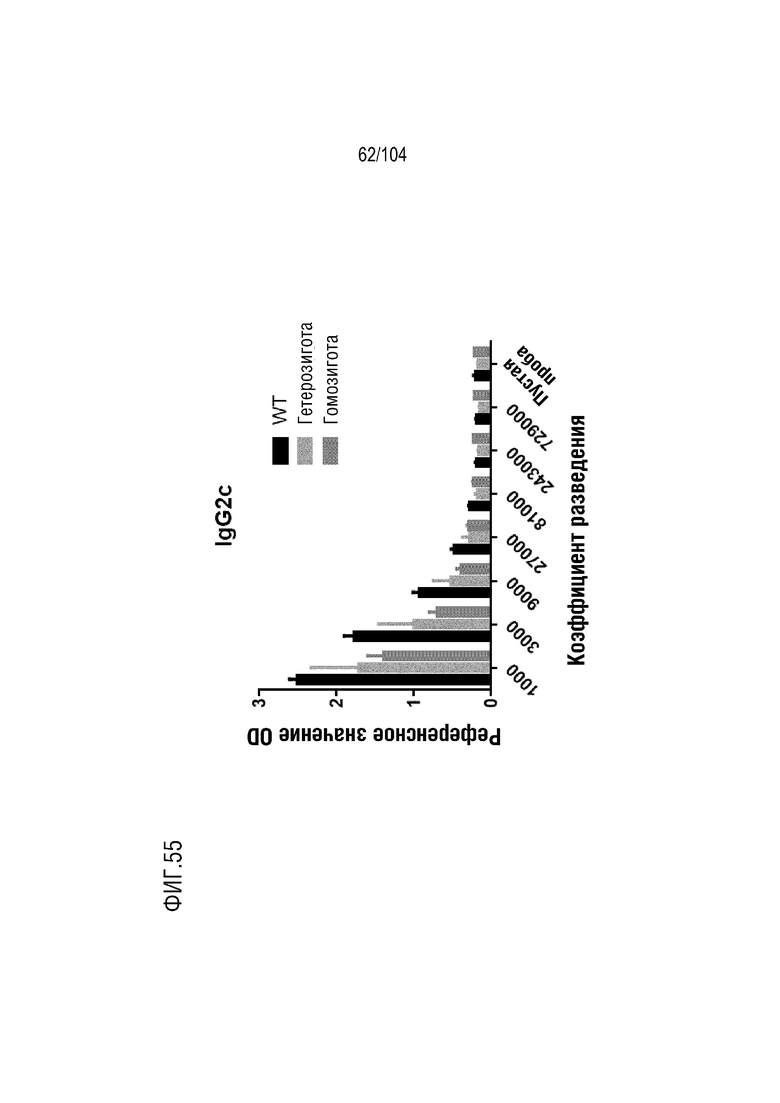







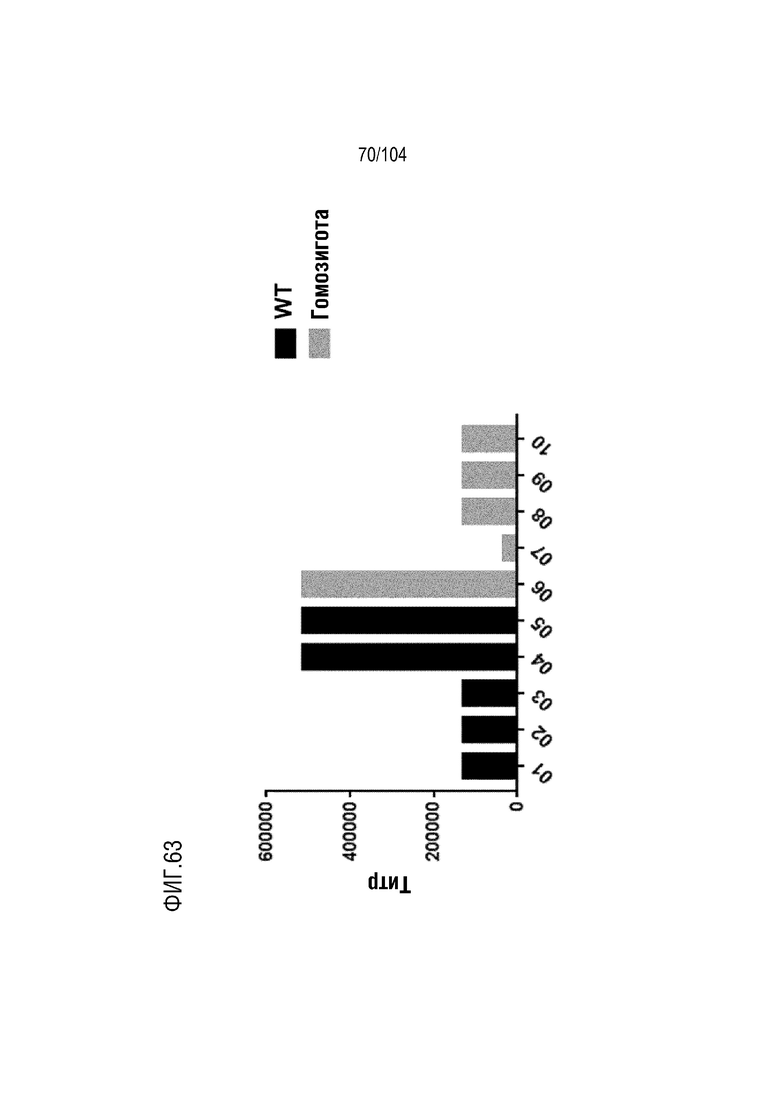
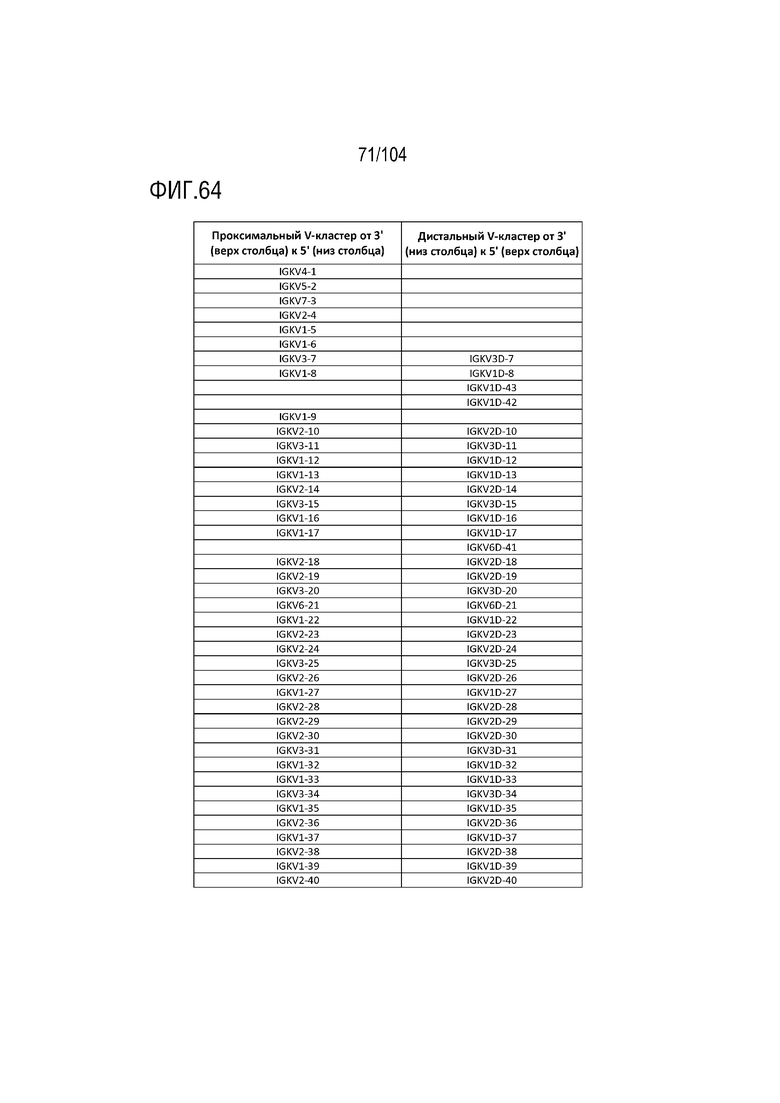














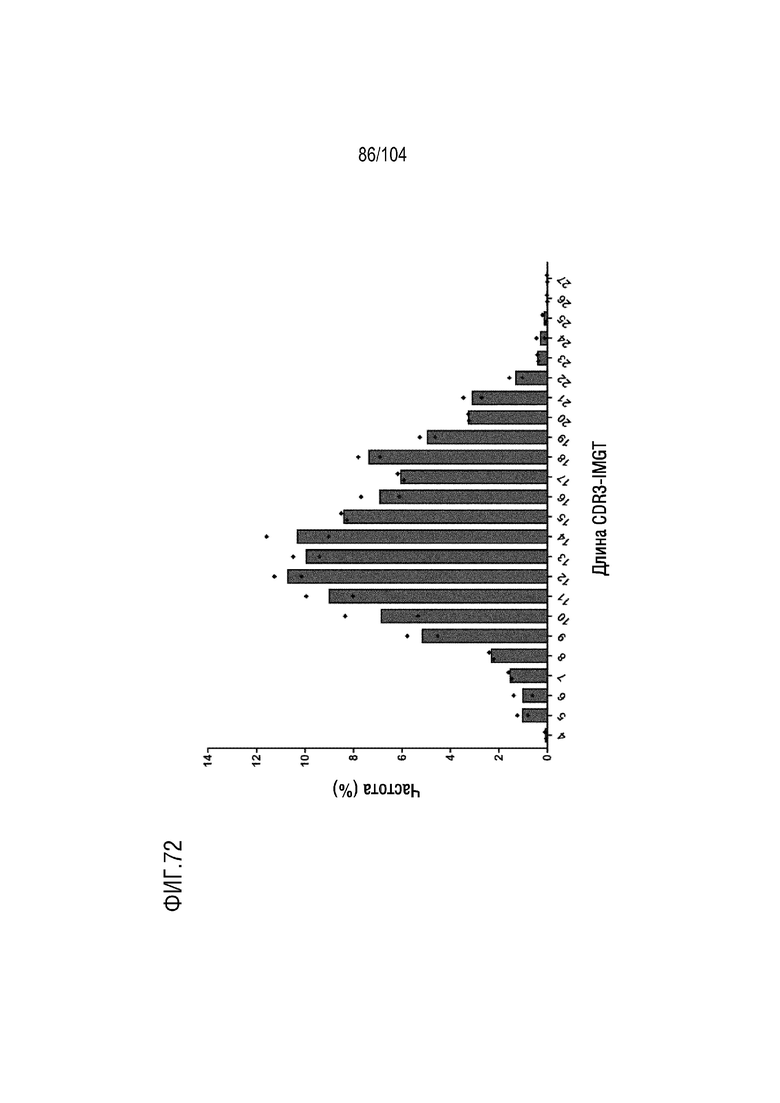




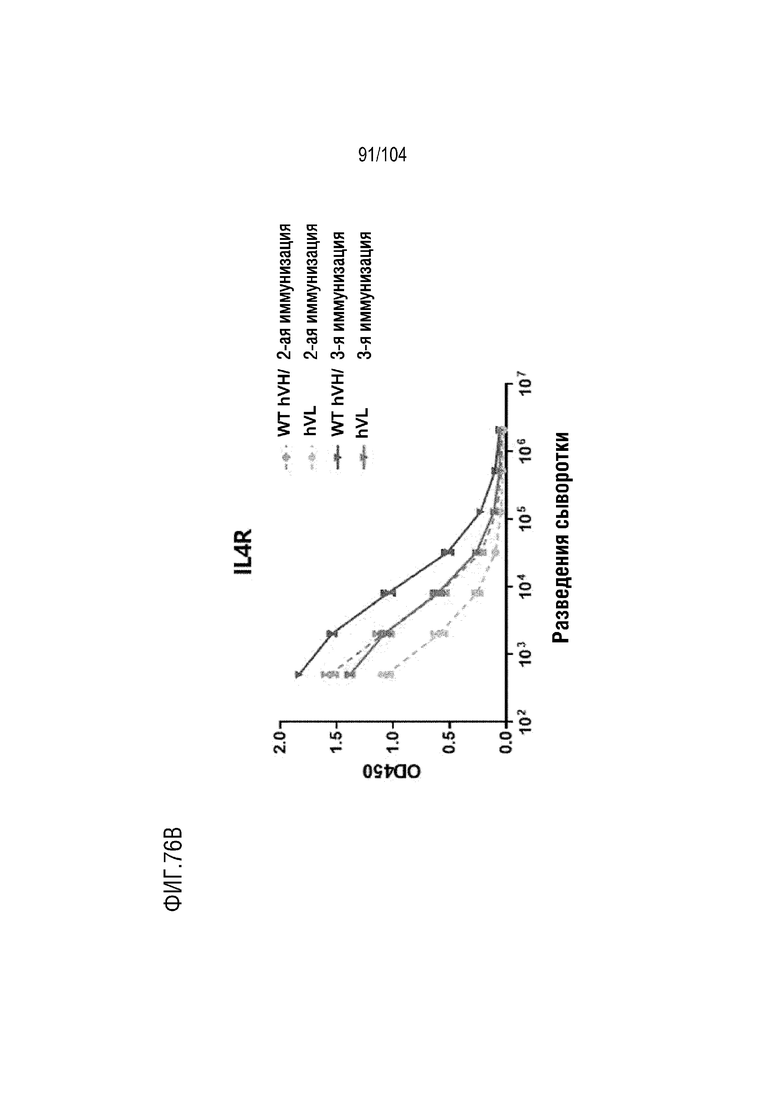


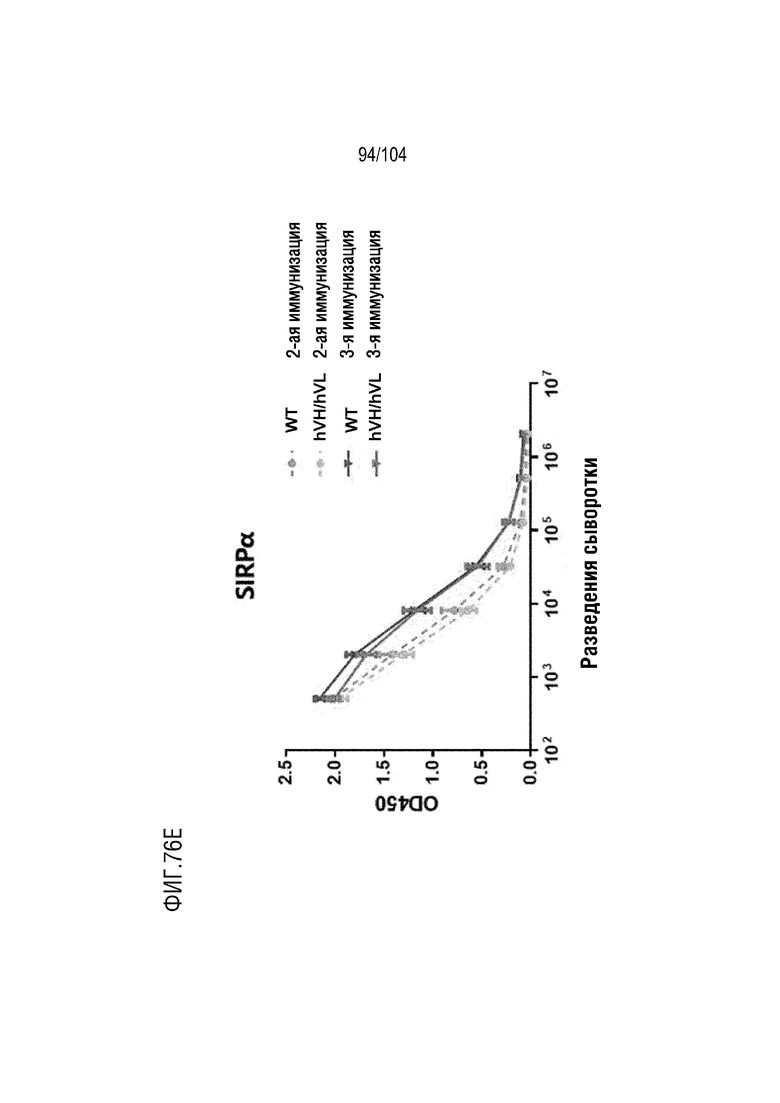










Изобретение относится к области биотехнологии, в частности к генетически модифицированной мыши, содержащей в эндогенном локусе гена тяжелой цепи иммуноглобулина все гены IGHV человека, все гены IGHD человека и все гены IGHJ человека в локусе гена тяжелой цепи иммуноглобулина хромосомы 14 человека из клетки человека, а также к его клетке. Изобретение эффективно для получения гуманизированных антител. 2 н. и 12 з.п. ф-лы, 82 ил., 16 табл., 17 пр.
1. Генетически модифицированная мышь для получения гуманизированных антител, где мышь содержит в эндогенном локусе гена тяжелой цепи иммуноглобулина все гены вариабельных участков тяжелой цепи иммуноглобулинов (IGHV) человека, все гены разнообразия тяжелой цепи иммуноглобулинов (IGHD) человека и все гены стыка тяжелой цепи иммуноглобулинов (IGHJ) человека в локусе гена тяжелой цепи иммуноглобулина хромосомы 14 человека из клетки человека, где гены IGHV человека, гены IGHD человека и гены IGHJ человека функционально связаны и могут подвергаться реаранжировке VDJ, где мышь содержит IGHV(III)-82, IGHV7-81, IGHV4-80, IGHV3-79, IGHV(II)-78-1, IGHV5-78, IGHV7-77, IGHV(III)-76-1, IGHV3-76, IGHV3-75 и IGHV(II)-74-1 человека в эндогенном локусе гена тяжелой цепи иммуноглобулина, где мышь содержит делецию в эндогенном локусе гена тяжелой цепи иммуноглобулина мыши.
2. Мышь по п. 1, где делеция содержит делецию одного или более эндогенных генов IGHV.
3. Мышь по п. 2, где разрыв в эндогенном локусе гена тяжелой цепи иммуноглобулина содержит делецию смежной последовательности, начиная от гена IGHV1-85 до гена IGHJ4.
4. Мышь по п. 1, где мышь содержит немодифицированную последовательность человека, выделенную из локуса гена тяжелой цепи иммуноглобулина человека, начиная от IGHV(III)-82 человека до IGHV1-2 человека.
5. Мышь по п. 1, где мышь содержит немодифицированную последовательность человека, выделенную из локуса гена тяжелой цепи иммуноглобулина человека, начиная от IGHD1-1 человека до IGHJ6 человека.
6. Мышь по п. 1, где мышь является гомозиготным по отношению к локусу гена тяжелой цепи иммуноглобулина.
7. Мышь по п. 1, где мышь содержит в эндогенном локусе гена легкой цепи иммуноглобулина один или более генов вариабельных участков каппа-цепи иммуноглобулинов (IGKV) человека и один или более генов стыка каппа-цепи иммуноглобулинов(IGKJ) человека.
8. Мышь по п. 1, где мышь может продуцировать гуманизированное антитело.
9. Мышь по п.1, где модификация может быть выявлена с помощью ПЦР с использованием одной или более пар праймеров, выбранных из группы, состоящей из:
(a) SEQ ID NO: 13 и SEQ ID NO: 14;
(b) SEQ ID NO: 15 и SEQ ID NO: 16;
(c) SEQ ID NO: 17 и SEQ ID NO: 18;
(d) SEQ ID NO: 19 и SEQ ID NO: 20;
(e) SEQ ID NO: 21 и SEQ ID NO: 22;
(f) SEQ ID NO: 23 и SEQ ID NO: 24; и
(g) SEQ ID NO: 25 и SEQ ID NO: 26.
10. Мышь по любому из пп. 1-8, где мышь содержит все гены IGKV человека и все гены IGKJ человека в эндогенном локусе гена легкой цепи иммуноглобулина.
11. Мышь по любому из пп. 1-8, где мышь обладает полным репертуаром антител человека.
12. Мышь по любому из пп. 1-8, где мышь может в выявляемом количестве экспрессировать IGHV3-15, IGHV3-53, IGHV3-66, IGHV5-51, IGHV1-24, IGHV1-18, IGHV1-69, IGHV3-7, IGHV3-74, IGHV3-23, IGHV3-43, IGHV3-21, IGHV3-30-3, IGHV4-59, IGHV6-1, IGHV3-48, IGHV4-34, IGHV4-39, IGHV3-33, IGHV3-30, IGHV4-61, IGHV3-47, IGHV3-NL1, IGHV3-69-1, IGHV7-4-1, IGHV1-58, IGHV4-28, IGHV2-26, IGHV2-5, IGHV3-64, IGHV3-20, IGHV2-70, IGHV3-11, IGHV4-30-2, IGHV3-13, IGHV3-49, IGHV5-10-1, IGHV3-72, IGHV1-2, IGHV4-30-4, IGHV1-46, IGHV3-64D, IGHV1-3, IGHV3-73, IGHV4-4 и IGHV4-31.
13. Мышь по любому из пп. 1-8, где мышь может в выявляемом количестве экспрессировать IGKV1D-12, IGKV2D-30, IGKV1-27, IGKV6-21, IGKV1-6, IGKV1-39, IGKV1-16, IGKV1-17, IGKV2D-29, IGKV3-11, IGKV1-5, IGKV1-12, IGKV1-9, IGKV3-20, IGKV3-15, IGKV2-28, IGKV2-30, IGKV1-33, IGKV4-1, IGKV6D-41, IGKV3D-11, IGKV2D-24, IGKV1D-43, IGKV1D-17, IGKV7-3, IGKV2D-26, IGKV1-37, IGKV2-29, IGKV1-13, IGKV5-2, IGKV2-40, IGKV3-7, IGKV3D-20, IGKV1D-8, IGKV6D-21, IGKV1-8, IGKV2-24 и IGKV3D-15.
14. Клетка, полученная от мыши по любому из пп. 1-8, для получения гуманизированных антител, где клетка содержит в эндогенном локусе гена тяжелой цепи иммуноглобулина все гены вариабельных участков тяжелой цепи иммуноглобулинов (IGHV) человека, все гены разнообразия тяжелой цепи иммуноглобулинов (IGHD) человека и все гены стыка тяжелой цепи иммуноглобулинов (IGHJ) человека в локусе гена тяжелой цепи иммуноглобулина хромосомы 14 человека из клетки человека, где гены IGHV человека, гены IGHD человека и гены IGHJ человека функционально связаны и могут подвергаться реаранжировке VDJ, где клетка содержит IGHV(III)-82, IGHV7-81, IGHV4-80, IGHV3-79, IGHV(II)-78-1, IGHV5-78, IGHV7-77, IGHV(III)-76-1, IGHV3-76, IGHV3-75 и IGHV(II)-74-1 человека в эндогенном локусе гена тяжелой цепи иммуноглобулина, где клетка содержит делецию в эндогенном локусе гена тяжелой цепи иммуноглобулина мыши.
| US 20140331343 A1, 06.04.2014 | |||
| E-CHIANG LEE et al., Complete humanization of the mouse immunoglobulin loci enables efficient therapeutic antibody discovery, Nature Biotechnology, 2014, vol | |||
| Способ образования коричневых окрасок на волокне из кашу кубической и подобных производных кашевого ряда | 1922 |
|
SU32A1 |
| Приспособление для постепенного включения и выключения фрикционных муфт в самодвижущихся экипажах и т.п. | 1919 |
|
SU356A1 |
| WO 2013041844 A1, 31.07.2014 | |||
| LYNN E | |||
| MACDONALD et al., Precise and in situ genetic humanization of 6 Mb of mouse immunoglobulin genes, PNAS, | |||

