Заявление об установлении приоритета
Настоящая заявка испрашивает приоритет на основании заявки PCT № PCT/CN2020/094000 от 02 июня 2020 г., и заявки РСТ № PCT/CN2021/085839 от 7 апреля 2021 г. Полное содержание которой включено в настоящее описание в качестве ссылки.
ОБЛАСТЬ ТЕХНИКИ
Настоящее описание относится к генетически модифицированным животным и клеткам, содержащим локус гуманизированной легкой цепи иммуноглоублина и/или локус гуманизированной тяжелой цепи иммуноглоублина.
УРОВЕНЬ ТЕХНИКИ
Антитела представляют собой иммунологические белки, которые связывают определенный антиген. Как правило, антитела специфичны в отношении мишеней, обладают способностью опосредовать иммунные эффекторные механизмы и имеют длительный период полужизни в сыворотке. Такие свойства делают антитела мощными терапевтическими средствами. Моноклональные антитела используются терапевтически для лечения различных состояний, включая онкологическое заболевание, инфекционное заболевание, аутоиммунное заболевание и воспалительные расстройства. Было предложено много новых форматов антител для различных целей лечения. Например, биспецифические антитела могут связываться с двумя разными мишенями или двумя разными эпитопами на мишени, создавая аддитивный или синергический эффект, превосходящий эффект моноклональных антител. Однако производство биспецифических антител затруднительно из-за проблем с ошибочным спариванием.
Кроме того, эти терапевтические антитела обычно представляют собой человеческие или гуманизированные антитела. Человеческие или гуманизированные антитела могут быть получены путем гуманизации антител грызунов (например, мышиного антитела) или с использованием фаговых библиотек. Антитела, полученные этими методами, часто имеют субоптимальную аффинность связывания и биофизические характеристики, что приводит к трудностям в производстве и плохой фармакокинетике. В частности, процесс гуманизации может неблагоприятно повлиять на аффинность связывания и ввести иммуногенные эпитопы в антитела, а антитела, обнаруженные с использованием фаговых библиотек, демонстрируют ограниченное разнообразие и ненативное спаривание тяжелых и легких цепей иммуноглобулина. Для улучшения свойств часто требуются повторяющиеся и трудоемкие эксперименты. А в некоторых случаях эти антитела также могут быть иммуногенными у пациентов.
Существует потребность в эффективных и рентабельных способах получения гуманизированных антител и, в частности, в платформе для создания гуманизированных биспецифических антител для различных лечебных целей.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к генетически модифицированным животным и клеткам с гуманизированным локусом тяжелой цепи и легкой цепи иммуноглобулина. В некоторых вариантах осуществления генетически модифицированные животные имеют ограниченный набор генов IGKV и IGKJ человека в эндогенном локусе генов легкой цепи иммуноглобулина. В одном аспекте генетически модифицированные животные, как описано в настоящем документе, могут продуцировать вариабельные домены легкой цепи иммуноглобулина, которые могут сочетаться с довольно разнообразным семейством вариабельных доменов тяжелой цепи, включая например, аффинно-зрелые вариабельные домены или соматически мутированные.
В одном аспекте настоящее изобретение относится к генетически модифицированному животному, отличному от человека, содержащему в эндогенном локусе легкой цепи иммуноглобулина экзогенную последовательность генов вариабельной области легкой цепи. В некоторых вариантах осуществления экзогенная последовательность генов вариабельной области легкой цепи содержит не более трех генов IGKV человека и не более двух генов IGKJ человека. В некоторых вариантах осуществления не более трех генов IGKV человека и не более двух генов IGKJ человека функционально связаны с эндогенным геном константного домена легкой цепи.
В некоторых вариантах осуществления не более трех генов IGKV человека выбирают из Таблицы 1, и не более двух генов IGKJ человека выбирают из Таблицы 2.
В некоторых вариантах осущест\вления экзогенная последовательность генов вариабельной области легкой цепи содержит один ген IGKV человека и один ген IGKJ человека.
В некоторых вариантах осуществления экзогенная последовательность генов вариабельной области легкой цепи дополнительно содержит последовательность 3'-UTR IGKJ человека. В некоторых вариантах осуществления экзогенный ген вариабельной области легкой цепи в одной или более клетках животного может подвергаться соматическим гипермутациям.
В некоторых вариантах осуществления соматические гипермутации могут приводить к заменам до одной, двух или трех аминокислот в вариабельных областях легкой цепи в одной или более клетках животного.
В некоторых вариантах осуществления экзогенная последовательность гена вариабельной области легкой цепи содержит один ген IGKV человека и один ген IGKJ человека. В некоторых вариантах осуществления ген IGKV человека выбран из группы, состоящей из IGKV3-20, IGKV3-11 и IGKV1-39. В некоторых вариантах осуществления ген IGKV человека и ген IGKJ человека функционально связаны. В некоторых вариантах осуществления ген IGKV человека представляет собой IGVK3-11. В некоторых вариантах осуществления ген IGKJ человека выбран из группы, состоящей из IGKJ1 и IGKJ4. В некоторых вариантах осуществления ген IGKV человека представляет собой IGKV1-39, и ген IGKJ человека представляет собой IGKJ4. В некоторых вариантах осуществления ген IGKV человека представляет собой IGKV3-11, и ген IGKJ человека представляет собой IGKJ1. В некоторых вариантах осуществления ген IGKV человека представляет собой IGKV3-20, и ген IGKJ человека представляет собой IGKJ1. В некоторых вариантах осуществления животное дополнительно содержит промоторную последовательность, функционально связанную с геном IGKV человека. В некоторых вариантах осуществления последовательность промотора находится в пределах 2500 или 3000 п.н. относительно гена IGKV человека. В некоторых вариантах осуществления промотор представляет собой промотор IGKV3-20, промотор IGKV3-11 или промотор IGKV1-39.
В некоторых вариантах осуществления животное содержит нарушение в эндогенном локусе генов легкой цепи иммуноглобулина животного. В некоторых вариантах осуществления животное представляет собой мышь, и нарушение в эндогенном локусе генов легкой цепи иммуноглобулина животного включает делецию одного или более генов IGKV мыши, представленных в Таблице 3, и одного или более генов IGKJ мыши, представленных в Таблице 4. В некоторых вариантах осуществления животное представляет собой мышь, и нарушение в эндогенном локусе генов легкой цепи иммуноглобулина животного включает делецию последовательности, начиная с IGKV2-137 мыши до IGKJ5 мыши.
В некоторых вариантах осуществления животное содержит эндогенный IGKC. В некоторых вариантах осуществления животное дополнительно содержит интронный энхансер каппа в 5'-области по отношению к эндогенному IGKC и/или 3'-энхансер каппа.
В некоторых вариантах осуществления вариабельная область легкой цепи человека представляет собой реаранжированную последовательность.
В некоторых вариантах осуществления животное является гомозиготным в отношении локуса гена легкой цепи иммуноглобулина. В некоторых вариантах осуществления животное является гетерозиготным в отношении локуса гена легкой цепи иммуноглобулина. В некоторых вариантах осуществления у животного имеется нарушение в эндогенном локусе гена легкой цепи лямбда иммуноглобулина.
В некоторых вариантах осуществления животное представляет собой грызуна (например, мышь).
В некоторых вариантах осуществления животное дополнительно содержит эндогенный локус генов тяжелой цепи иммуноглобулина, один или более генов IGHV человека, один или более генов IGHD человека и один или более генов IGHKJ человека. В некоторых вариантах осуществления гены IGHV человека, гены IGHD человека и гены IGHJ человека функционально связаны и могут подвергаться VDJ-реаранжировке.
В некоторых вариантах осуществления животное содержит по меньшей мере 150 генов IGHV человека, выбранных из Таблицы 5, по меньшей мере 20 генов IGHD человека, выбранных из Таблицы 6, и по меньшей мере 5 генов IGHJ человека, выбранных из Таблицы 7. В некоторых вариантах осуществления животное содержит все гены IGHV человека, все гены IGHD человека и все гены IGHJ человека в эндогенном локусе генов тяжелой цепи иммуноглобулина хромосомы 14 человека. В некоторых вариантах осуществления животное содержит все гены IGHV человека, все гены IGHD человека и все гены IGHJ человека в эндогенном локусе генов тяжелой цепи иммуноглобулина хромосомы 14 человека в клетке человека. В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, полученную из локуса генов тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления немодифицированная последовательность человека составляет по меньшей мере 800 т.п.н.
В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, полученную из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHV(III)-82 человека и до IGHV1-2 человека. В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, полученную из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHV(III)-82 человека и до IGHV6-1 человека. В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, полученную из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHD1-1 человека и до IGHJ6 человека. В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, полученную из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHV(III)-82 человека и до IGHJ6 человека. В некоторых вариантах осуществления животное содержит IGHV(III)-82, IGHV7-81, IGHV4-80, IGHV3-79, IGHV(II)-78-1, IGHV5-78, IGHV7-77, IGHV(III)-76-1, IGHV3-76, IGHV3-75, и IGHV(II)-74-1. В некоторых вариантах осуществления животное содержит IGHV5-10-1 и IGHV3-64D.
В некоторых вариантах осуществления животное дополнительно содержит в эндогенном локусе генов тяжелой цепи иммуноглобулина первую последовательность, содержащую один или более генов IGHV человека;
вторую последовательность, содержащую эндогенную последовательность; и третью последовательность, содержащую один или более генов IGHD человека и один или более генов IGHJ человека. В некоторых вариантах осуществления первая последовательность, вторая последовательность и третья последовательность функционально связаны.
В некоторых вариантах осуществления первая последовательность содержит по меньшей мере 150 генов IGHV человека, выбранных из Таблицы 5. В некоторых вариантах осуществления первая последовательность содержит по меньшей мере 20 генов IGHD человека, выбранных из Таблицы 6. В некоторых вариантах осуществления первая последовательность представляет собой немодифицированную последовательность, полученную из локуса генов тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления первая последовательность составляет по меньшей мере 800 т.п.н. В некоторых вариантах осуществления вторая последовательность содержит эндогенную последовательность, составляющую по меньшей мере 3 т.п.н. В некоторых вариантах осуществления третья последовательность содержит по меньшей мере 20 генов IGHD человека, выбранных из Таблицы 6, и по меньшей мере 5 генов IGHJ человека, выбранных из Таблицы 7.
В некоторых вариантах осуществления третья последовательность содержит все гены IGHD человека, представленные в Таблице 6, и все гены IGHJ человека, представленные в Таблице 7. В некоторых вариантах осуществления третья последовательность представляет собой немодифицированную последовательность, полученную из локуса генов тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления третья последовательность составляет по меньшей мере 50 т.п.н. В некоторых вариантах осуществления животное содержит нарушение в эндогенном локусе генов тяжелой цепи иммуноглобулина животного.
В некоторых вариантах осуществления животное представляет собой мышь, и нарушение в эндогенном локусе генов тяжелой цепи иммуноглобулина животного включает делецию одного или более генов IGHV мыши, представленных в Таблице 8, одного или более генов IGHD мыши, представленных в Таблице 9, и одного или более генов IGHJ мыши, представленных в Таблице 10. В некоторых вариантах осуществления животное представляет собой мышь, и нарушение в эндогенном локусе генов тяжелой цепи иммуноглобулина животного включает делецию последовательности, начиная с IGHV1-85 мыши до IGHJ4 мыши. В некоторых вариантах осуществления животное содержит один или более эндогенных генов IGHM, IGHδ, IGHG3, IGHG1, IGHG2b, IGHG2a, IGHE и IGHA.
В некоторых вариантах осуществления животное является гомозиготным в отношении локуса генов тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления животное является гетерозиготным в отношении локуса генов тяжелой цепи иммуноглобулина.
В одном аспекте настоящее изобретение относится к генетически модифицированному животному, отличному от человека, геном которого содержит эндогенный локус легкой цепи иммуноглобулина, включающий: замену одного или более эндогенных генов IGKV одним или более генами IGKV человека, выбранными из Таблицы 1; и замену одного или более эндогенных генов IGKJ одним или более генами IGKJ человека, выбранными из Таблицы 2. В некоторых вариантах осуществления ген IGKV человека и ген IGKJ человека функционально связаны с эндогенным геном IGKC.
В некоторых вариантах осуществления один или более генов IGKV человека выбраны из группы, состоящей из IGKV3-20, IGKV3-11 и IGKV1-39. В некоторых вариантах осуществления ген IGKV человека представляет собой IGKV3-11. В некоторых вариантах осуществления один или более генов IGKJ человека выбраны из группы, состоящей из IGKJ1 и IGKJ4. В некоторых вариантах осуществления животное дополнительно содержит вставку последовательности 3'-UTR IGKJ человека. В некоторых вариантах осуществления ген IGKV человека представляет собой IGKV1-39, и ген IGKJ человека представляет собой IGKJ4. В некоторых вариантах осуществления ген IGKV человека представляет собой IGKV3-11, и ген IGKJ человека представляет собой IGKJ1. В некоторых вариантах осуществления ген IGKV человека представляет собой IGKV3-20, и ген IGKJ человека представляет собой IGKJ1. В некоторых вариантах осуществления все эндогенные гены IGKV заменены одним или более генами IGKV человека. В некоторых вариантах осуществления все эндогенные гены IGKJ заменены одним или более генами IGKJ человека.
В некоторых вариантах осуществления животное дополнительно содержит последовательность промотора перед геном IGKV человека. В некоторых вариантах осуществления последовательность промотора находится в пределах 3000 п.н. относительно гена IGKV человека. В некоторых вариантах осуществления животное дополнительно содержит интронный энхансер каппа в 5'-области эндогенного IGKC. В некоторых вариантах осуществления животное дополнительно содержит 3'-энхансер каппа.
В некоторых вариантах осуществления геном животного дополнительно содержит эндогенный локус тяжелой цепи иммуноглобулина, включающий: замену одного или более эндогенных генов IGHV, IGHD и IGHJ одним или более генами IGHV, IGHD человека и IGHJ человека. В некоторых вариантах осуществления один или более генов IGHV, IGHD человека и IGHJ человека функционально связаны с одним или более эндогенными генами IGHM, IGHδ, IGHG, IGHE и IGHA. В некоторых вариантах осуществления один или более эндогенных генов IGHV, IGHD и IGHJ заменены по меньшей мере 150 генами IGHV человека, представленными в Таблице 5, по меньшей мере 20 генами IGHD человека, представленными в Таблице 6, и по меньшей мере 5 генами IGHJ человека, представленными в Таблице 7.
В некоторых вариантах осуществления животное представляет собой мышь, и по меньшей мере 180 генов IGHV мыши, представленных в Таблице 8, все гены IGHD мыши, представленные в Таблице 9, и все гены IGHJ мыши, представленные в Таблице 10, заменены.
В некоторых вариантах осуществления у животного отсутствует эндогенный локус вариабельной области тяжелой цепи иммуноглобулина, который способен к реаранжировке и формированию последовательности нуклеиновой кислоты, кодирующей эндогенный вариабельный домен тяжелой цепи (например, вариабельный домен тяжелой цепи мыши). В некоторых вариантах осуществления у животного отсутствует эндогенный локус вариабельной области легкой цепи иммуноглобулина, который способен к реаранжировке и формированию последовательность нуклеиновой кислоты, кодирующую эндогенный вариабельный домен легкой цепи (например, вариабельный домен легкой цепи мыши).
В некоторых вариантах осуществления животное может продуцировать гуманизированное антитело. В некоторых вариантах осуществления антитело содержит вариабельную область легкой цепи, кодируемую геном IGKV и геном IGKJ.
В одном аспекте описание относится к клетке, полученной от животного, как описано в настоящем документе. В некоторых вариантах осуществления клетка представляет собой В-клетку, которая экспрессирует химерную легкую цепь иммуноглобулина, содержащую вариабельный домен легкой цепи иммуноглобулина, который кодируется геном IGKV человека, выбранным из группы, состоящей из IGKV3-20, IGKV3-11 и IGKV1-39, и геном IGKJ человека, выбранным из группы, состоящей из IGKJ1 и IGKJ4. В некоторых вариантах осуществления вариабельный домен легкой цепи иммуноглобулина функционально связан с константной областью легкой цепи, не относящейся к человеку.
В некоторых вариантах осуществления В-клетка, которая экспрессирует химерную тяжелую цепь иммуноглобулина, содержащую вариабельный домен тяжелой цепи иммуноглобулина, полученный в результате реаранжировки одного или более генов IGHV человека, одного или более генов IGHD человека и одного или более генов IGHJ человека. В некоторых вариантах осуществления вариабельный домен тяжелой цепи иммуноглобулина функционально связан с константной областью тяжелой цепи, не относящейся к человеку.
В некоторых вариантах осуществления клетка представляет собой эмбриональную стволовую (ES) клетку.
В одном аспекте описание относится к способу получения химерного антитела, которое специфически связывается с антигеном, причем способ включает экспонирование животного, как описано в настоящем документе, с антигеном; получение гибридомы из клетки, полученной от животного; и сбор химерного антитела, продуцируемого гибридомой.
В некоторых вариантах осуществления способ дополнительно включает секвенирование генома гибридомы.
В одном аспекте описание относится к способу получения антитела, которое специфически связывается с антигеном, причем способ включает экспонирование животного, как описано в настоящем документе, с антигеном; секвенирование нуклеиновых кислот, кодирующих вариабельные области тяжелых и легких цепей иммуноглобулина человека, в клетке, экспрессирующей гибридное антитело, которое специфически связывается с антигеном; и функциональное связывание в клетке нуклеиновой кислоты, кодирующей вариабельную область тяжелой цепи иммуноглобулина человека, с нуклеиновой кислотой, кодирующей константную область тяжелой цепи иммуноглобулина человека, и нуклеиновой кислоты, кодирующей вариабельную область легкой цепи иммуноглобулина человека, с нуклеиновой кислотой, кодирующей константную область легкой цепи иммуноглобулина человека.
В одном аспекте описание относится к способу получения антитела, которое специфически связывается с антигеном, причем указанный способ включает получение последовательности нуклеиновой кислоты, кодирующей вариабельные области тяжелой и легкой цепи иммуноглобулина человека, в клетке, которая экспрессирует гибридное антитело, которое специфически связывается с антигеном, где клетку получают путем экспонирования животного, как описано в настоящем документе, с антигеном; функциональное связывание нуклеиновой кислоты, кодирующей вариабельную область тяжелой цепи иммуноглобулина человека, с нуклеиновой кислотой, кодирующей константную область тяжелой цепи иммуноглобулина человека, и нуклеиновой кислоты, кодирующей вариабельную область легкой цепи иммуноглобулина человека, с нуклеиновой кислотой, кодирующей константную область легкой цепи иммуноглобулина человека; и экспрессию нуклеиновой кислоты в клетке с получением таким образом антитела.
В одном аспекте описание относится к способу получения нуклеиновой кислоты, кодирующей домен связывания антитела, который специфически связывается с антигеном, причем способ включает
экспонирование животного, как описано в настоящем документе, с антигеном; и секвенирование нуклеиновых кислот, кодирующих вариабельные области тяжелой и легкой цепи иммуноглобулина человека, в клетке, которая экспрессирует гибридное антитело, которое специфически связывается с антигеном.
В одном аспекте описание относится к способу получения образца, причем способ включает экспонирование животного, как описано в настоящем документе, с антигеном; и
забор образца у животного. В некоторых вариантах осуществления образец представляет собой ткань селезенки, клетку селезенки или В-клетку.
В одном аспекте описание относится к способу получения биспецифического антитела, причем способ включает экспрессию в клетке последовательности нуклеиновой кислоты, кодирующей полипептид легкой цепи, содержащий VL человека, последовательности нуклеиновой кислоты, кодирующей первый полипептид тяжелой цепи, содержащий первую VH человека, и последовательности нуклеиновой кислоты, кодирующей второй полипептид тяжелой цепи, содержащий вторую VH человека, где последовательность, кодирующая первую VH, получают от животного, как описано в настоящем документе, после его экспонирования с первым антигеном, и последовательность, кодирующая вторую VH, получают от животного или другого животного после его экспонирования со вторым антигеном.
В некоторых вариантах осуществления первые VH и VL образуют первый антигенсвязывающий сайт, который специфически связывается с первым антигеном. В некоторых вариантах осуществления вторые VH и VL образуют второй антигенсвязывающий сайт, который специфически связывается со вторым антигеном.
В одном аспекте описание относится к антителу или его антигенсвязывающему фрагменту, содержащему вариабельную область легкой цепи человека, имеющую последовательность, по меньшей мере на 90%, 95% или 98% идентичную SEQ ID NO: 38, 39 или 40.
В одном аспекте описание относится к множеству антител или их антигенсвязывающих фрагментов, где каждое антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи человека, которая имеет последовательность, по меньшей мере на 90%, 95% или 98% идентичную SEQ ID NO: 38, 39 или 40.
В одном аспекте описание относится к способу получения антитела, которое специфически связывается с интересующим белком, причем способ включает экспонирование животного, описанного в настоящем документе, с интересующим белком, где животное не экспрессирует эндогенный белок, гомологичный интересующему белку; и секвенирование нуклеиновых кислот, кодирующих вариабельные области тяжелых и легких цепей иммуноглобулина человека, в клетке, которая экспрессирует антитело, которое специфически связывается с интересующим белком.
В некоторых вариантах осуществления у животного нарушен ген, кодирующий эндогенный белок. В некоторых вариантах осуществления ген, кодирующий эндогенный белок, нокаутирован.
В некоторых вариантах осуществления эндогенный белок по меньшей мере на 80%, 90%, 95% гомологичен интересующему белку. В некоторых вариантах осуществления эндогенный белок по меньшей мере на 80%, 90% или 95% идентичен интересующему белку.
В некоторых вариантах осуществления интересующий белок представляет собой белок человека. В некоторых вариантах осуществления интересующий белок представляет собой PD-1, CTLA-4, LAG-3, BTLA, PD-L1, CD27, CD28, CD47, CD137, CD154, TIGIT, TIM-3, GITR, SIRPa или OX40.
Если не определено иначе, то все технические и научные термины, использованные в настоящем описании, имеют тот же смысл, который вкладывается в них обычным специалистом в области, к которой принадлежит настоящее изобретение. Материалы и методы описаны в настоящем документе для применения в настоящем изобретении; другие подходящие материалы и методы, известные в данной области, также могут использоваться. Материалы, методы и примеры являются иллюстративными и не предназначены для ограничения. Все публикации, патентные заявки, патенты, последовательности, занесения в базы данных и другие ссылки, упомянутые в настоящем описании, включены ссылкой в полном объеме. В случае противоречий, они будут урегулированы настоящим описанием изобретения, включающим определения.
Другие признаки и преимущества изобретения будут очевидны из следующего подробного описания и чертежей, и из формулы изобретения.
Описание Чертежей
ФИГ. 1А представляет собой блок-схему способа введения генов иммуноглобулина человека в геном мыши.
ФИГ. 1B представляет собой обзор замены вариабельной области тяжелой цепи иммуноглобулина мыши вариабельной областью тяжелой цепи иммуноглобулина человека.
На ФИГ. 2A показаны наиболее часто используемые гены V-области цепи каппа человека.
На ФИГ. 2B показаны наиболее часто используемые гены J-области цепи каппа человека.
ФИГ. 3А представляет собой схематическую диаграмму, показывающую локус легкой цепи иммуноглобулина мыши.
ФИГ. 3B представляет собой обзор замены генов вариабельной области легкой цепи иммуноглобулина мыши реаранжированными генами IGKV и IGKJ человека.
ФИГ. 4 представляет собой схематическую диаграмму, показывающую направленную стратегию гуманизации гена легкой цепи каппа.
ФИГ. 5A представляет собой гистограмму, показывающую массу тела мышей дикого типа (WT) и гетерозиготных мышей hVH/hcVL.
ФИГ. 5B представляет собой гистограмму, показывающую массу печени мышей дикого типа (WT) и гетерозиготных мышей hVH/hcVL.
ФИГ. 5C представляет собой гистограмму, показывающую массу тимуса мышей дикого типа (WT) и гетерозиготных мышей hVH/hcVL.
ФИГ. 5D представляет собой гистограмму, показывающую массу селезенки мышей дикого типа (WT) и гетерозиготных мышей hVH/hcVL.
ФИГ. 5E представляет собой гистограмму, показывающую массу легких мышей дикого типа (WT) и гетерозиготных мышей hVH/hcVL.
ФИГ. 5F представляет собой гистограмму, показывающую массу сердца мышей дикого типа (WT) и гетерозиготных мышей hVH/hcVL.
ФИГ. 5G представляет собой гистограмму, показывающую массу почек мышей дикого типа (WT) и гетерозиготных мышей hVH/hcVL.
ФИГ. 6А представляет собой гистограмму, показывающую процентное содержание иммунных клеток в крови по данным проточной цитометрии. Иммунные клетки крови были получены от мышей дикого типа или мышей hVH/hcVL.
ФИГ. 6B представляет собой гистограмму, показывающую процентное содержание иммунных клеток в селезенке по данным проточной цитометрии. Иммунные клетки были получены от мышей дикого типа или мышей hVH/hcVL.
ФИГ. 7 представляет собой гистограмму, показывающую процентное содержание иммунных клеток в лимфатических узлах по данным проточной цитометрии. Иммунные клетки лимфатических узлов были получены от мышей дикого типа или мышей hVH/hcVL.
На ФИГ. 8А показан процент популяции про-В-клеток (B220lowCD43highIgMlow), пре-В-клеток (B220lowCD43intIgMlow) и незрелых В-клеток (B220highCD43lowIgMhigh) в В-клетках костного мозга. В-клетки костного мозга были получены от мышей дикого типа или мышей hVH/hcVL.
На ФИГ. 8В показан процент популяции плазматических клеток (B220lowIgM-IgD-CD138+) и В-клеток памяти (B220+IgM+IgD-CD38+) в В-клетках костного мозга. В-клетки костного мозга были получены от мышей дикого типа или мышей hVH/hcVL.
На ФИГ. 9 показан процент популяции переходного типа 1 (T1,B220+IgM+IgD-), переходного типа 2 (T2,B220+IgM+IgD+) и зрелых В-клеток (M, B220+IgMlowIgD+) в В-клетках селезенки. В-клетки селезенки были получены от мышей дикого типа или мышей hVH/hcVL.
На ФИГ. 10А показан процент популяции плазматических клеток (B220lowIgM-IgD-CD138+) в В-клетках селезенки. В-клетки селезенки были получены от мышей дикого типа или мышей hVH/hcVL.
На ФИГ. 10В показан процент популяции В-клеток памяти (B220+IgM+IgD-CD38+) в В-клетках селезенки. В-клетки селезенки были получены от мышей дикого типа или мышей hVH/hcVL.
На ФИГ. 10С показан процент популяции В-клеток маргинальной зоны (MZ, B220+CD21+CD23-) и фолликулярных В-клеток (F0, B220+CD21lowCD23+) в В-клетках селезенки. В-клетки селезенки были получены от мышей дикого типа или мышей hVH/hcVL.
На ФИГ. 11А показан результат проточной цитометрии В-клеток селезенки, меченных mB220, у самок мышей дикого типа.
На ФИГ. 11В показан результат проточной цитометрии В-клеток селезенки, меченных mB220, у самок мышей hVH/hcVL.
На ФИГ. 11С показан результат проточной цитометрии В-клеток селезенки, меченных mB220, у самцов мышей дикого типа.
На ФИГ. 11D показан результат проточной цитометрии В-клеток селезенки, меченных с помощью mB220, у самцов мышей hVH/hcVL.
На ФИГ. 11Е показан результат проточной цитометрии В-клеток селезенки, меченных mIgG каппа-FITC и mIgG лямбда-РЕ у самок мышей дикого типа.
НА ФИГ. 11F показан результат проточной цитометрии В-клеток селезенки, меченных mIgG каппа-FITC и mIgG лямбда-PE, у самок мышей hVH/hcVL.
На ФИГ. 11G показан результат проточной цитометрии В-клеток селезенки, меченных mIgG каппа-FITC и mIgG лямбда-РЕ у самцов мышей дикого типа.
На ФИГ. 11H показан результат проточной цитометрии В-клеток селезенки, меченных mIgG каппа-FITC и mIgG лямбда-PE, у самцов мышей hVH/hcVL.
ФИГ. 11I представляет собой гистограмму, показывающую процент В-клеток, меченных mIgG лямбда-PE (mIg лямбда), и В-клеток селезенки, меченных mIgG каппа-FITC (mIg каппа).
На ФИГ. 12A показан процент популяции переходного типа 1 (T1, B220+IgM+IgD-), переходного типа 2 (T2, B220+IgM+IgD+) и зрелых B-клеток (M, B220+IgMlowIgD+) в B-клетках лимфатических узлов. В-клетки лимфатических узлов были получены от мышей дикого типа или мышей hVH/hcVL.
На ФИГ. 12В показан процент популяции плазматических клеток (B220lowIgM-IgD-CD138+) и В-клеток памяти (B220+IgM+IgD-CD38+) в В-клетках лимфатических узлов. В-клетки лимфатических узлов были получены от мышей дикого типа или мышей hVH/hcVL.
На ФИГ. 13А показан результат проточной цитометрии В-клеток лимфатических узлов, меченных mB220, у самок мышей дикого типа.
На ФИГ. 13В показан результат проточной цитометрии В-клеток лимфатических узлов, меченных mB220, у самок мышей hVH/hcVL.
На ФИГ. 13С показан результат проточной цитометрии В-клеток лимфатических узлов, меченных mB220, у самцов мышей дикого типа.
На ФИГ. 13D показан результат проточной цитометрии В-клеток лимфатических узлов, меченных mB220, у самцов мышей hVH/hcVL.
На ФИГ. 13Е показан результат проточной цитометрии В-клеток лимфатических узлов, меченных mIgG каппа-FITC и mIgG лямбда-РЕ у самок мышей дикого типа.
На ФИГ. 13F показан результат проточной цитометрии В-клеток лимфатических узлов, меченных mIgG каппа-FITC и mIgG лямбда-PE, у самок мышей hVH/hcVL.
На ФИГ. 13G показан результат проточной цитометрии В-клеток лимфатических узлов, меченных mIgG каппа-FITC и mIgG лямбда-РЕ, у самцов мышей дикого типа.
На ФИГ. 13H показан результат проточной цитометрии В-клеток лимфатических узлов, меченных mIgG каппа-FITC и mIgG лямбда-PE у самцов мышей hVH/hcVL.
ФИГ. 13I представляет собой гистограмму, показывающую процент В-клеток, меченных mIgG лямбда-PE, (mIg лямбда), и В-клеток лимфатических узлов, меченных mIgG каппа-FITC (mIg каппа).
На ФИГ. 13J показана концентрация подтипов сывороточного иммуноглобулина (Ig) у мышей дикого типа (WT) или мышей hVH/hcVL до иммунизации антигеном. Концентрации подтипа Ig определяли с помощью ИФА.
ФИГ. 14 представляет собой схематическую диаграмму хромосомы 2 человека, на которой выделен локус легкой цепи иммуноглобулина (не в масштабе). VHK представляет собой сегмент для кластера генов IGKV, JHK представляет собой сегмент для кластера генов IGKJ, а CHK представляет собой ген IGKC.
На ФИГ. 15 показан список генов IGKV дистального Vκ-кластера человека и список генов IGKV проксимального Vκ-кластера человека.
ФИГ. 16 представляет собой схематическую диаграмму, показывающую локус тяжелой цепи иммуноглобулина человека (IGH) на хромосоме 14 (14q32. 33).
ФИГ. 17 представляет собой схематическую диаграмму, показывающую локус IGH мыши (Mus musculus) на хромосоме 12 (12F2) (штамм C57BL/6).
ФИГ. 18 представляет собой схематическую диаграмму, показывающую локус цепи каппа иммуноглобулина человека (IGK) на хромосоме 2 (2p11. 2).
ФИГ. 19 представляет собой схематическую диаграмму, показывающую локус IGK мыши (Mus musculus) на хромосоме 6 (6C1).
На ФИГ. 20 представлен репертуар IMGT для локуса тяжелой цепи иммуноглобулина человека (IGH).
На ФИГ. 21 представлен репертуар IMGT для IGH мыши.
На ФИГ. 22 представлен репертуар IMGT для локуса цепи каппа иммуноглобулина человека (IGK).
На ФИГ. 23 представлен репертуар IMGT для IGK мыши.
На ФИГ. 24 показан титр антиген-специфических антител после трех иммунизаций первым тестируемым антигеном у мышей дикого типа и мышей hVHH/H/hcVLK/+.
На ФИГ. 25 показан титр антиген-специфических антител после трех иммунизаций вторым тестируемым антигеном у мышей дикого типа и мышей hVHH/H/hcVLK/+.
На ФИГ. 26A показана частота использования IGHV (частота > 1%) у наивных мышей hVHH/H/hcVLK/+.
На ФИГ. 26B показана частота использования IGHV (частота <1%) у наивных мышей hVHH/H/hcVLK/+.
На ФИГ. 26C показана частота использования IGHD у наивных мышей hVHH/H/hcVLK/+.
На ФИГ. 26D показана частота использования IGHJ у наивных мышей hVHH/H/hcVLK/+.
На ФИГ. 26E показана частота использования IGHV (частота > 1%) у наивных мышей hVHH/H/hcVLK/K.
На ФИГ. 26F показана частота использования IGHV (частота <1%) у наивных мышей hVHH/H/hcVLK/K.
На ФИГ. 26G показана частота использования IGHJ у наивных мышей hVHH/H/hcVLK/K.
ФИГ. 27A представляет собой гистограмму, показывающую распределение длины аминокислотной последовательности CDR3 тяжелой цепи у наивных мышей hVHH/H/hcVLK/+.
ФИГ. 27B представляет собой гистограмму, показывающую распределение длины аминокислотной последовательности CDR3 тяжелой цепи у наивных мышей hVHH/H/hcVLK/K.
На ФИГ. 28А показана частота встречаемости аминокислот в CDR3 тяжелой цепи (длина CDR3 равна 17 аминокислотам) у наивных мышей hVHH/H/hcVLK/+.
На ФИГ. 28B показана частота встречаемости аминокислот в CDR3 тяжелой цепи (длина CDR3 равна 19 аминокислотам) у наивных мышей hVHH/H/hcVLK/+.
На ФИГ. 28C показана частота встречаемости аминокислот в CDR3 тяжелой цепи (длина CDR3 равна 21 аминокислоте) у наивных мышей hVHH/H/hcVLK/+.
На ФИГ. 28D показана частота встречаемости аминокислот в CDR3 тяжелой цепи (длина CDR3 равна 17 аминокислотам) у наивных мышей hVHH/H/hcVLK/K.
На ФИГ. 29 показана частота встречаемости CDR3 тяжелой цепи, которая содержит остатки цистеина, один остаток цистеина и два остатка цистеина в CDR3 с разной длиной у наивных мышей hVHH/H/hcVLK/+.
На ФИГ. 30А показана степень соматических аминокислотных гипермутаций (АА), кодируемых в реаранжированной последовательности IGKV3-11/J1 человека у мышей hVHH/H/hcVLK/+.
На ФИГ. 30В показана степень соматических гипермутаций ДНК в реаранжированной последовательности IGKV3-11/J1 человека у мышей hVHH/H/hcVLK/+.
На ФИГ. 30С показана степень соматических гипермутаций ДНК в реаранжированной последовательности IGKV3-11/J1 человека у мышей hVHH/H/hcVLK/K.
На ФИГ. 31 показаны результаты кластеризации эпитопов для 15 человеческих антител против антигена А.
На ФИГ. 32 представлены последовательности, описанные в настоящем документе.
На ФИГ. 33A показано разнообразие тяжелых цепей с помощью анализа V-D-J-рекомбинации антител, нацеленных на антиген D, у мышей hVHH/H/hcVLK/K.
На ФИГ. 33B показано разнообразие тяжелых цепей с помощью анализа V-D-J-рекомбинации антител, нацеленных на антиген E, у мышей hVHH/H/hcVLK/K.
На ФИГ. 34A-34B показано распределение скорости ассоциации (kon) и скорости диссоциации (koff) антител, нацеленных на антиген D.
На ФИГ. 34C-34D показано распределение скорости ассоциации (kon) и скорости диссоциации (koff) антител, нацеленных на антиген E.
На ФИГ. 35A показано распределение аффинности связывания (KD) антител, нацеленных на антиген D.
На ФИГ. 35B показано распределение аффинности связывания (KD) антител, нацеленных на антиген E.
ПОДРОБНОЕ ОПИСАНИЕ
Моноклональное антитело обычно содержит две тяжелые цепи, где каждый мономер тяжелой цепи связан с идентичной легкой цепью. Из-за целевой специфичности моноклональных антител они обычно могут связываться с одной мишенью. Однако при разработке лекарственных средств необходимы антитела, способные связывать два разных антигена или эпитопа. Например, CD3-специфическое антитело обычно сочетают с различными антиген-специфическими опухолеассоциированными антителами для получения биспецифических антител для лечения онкологических заболеваний. Способность биспецифических антител связывать два разных антигена или эпитопа обеспечивает широкий спектр клинических применений. На сегодняшний день коммерчески доступны по меньшей мере два биспецифических антитела, и большое количество биспецифических антител проходит клинические испытания.
Биспецифическое антитело обычно имеет две разные тяжелые цепи и две разные легкие цепи. Получить их в одной клетке-хозяине сложно, потому что случайное спаривание двух тяжелых и двух легких цепей приводит к экспрессии 10 различных видов IgG, причем только один представляет собой интересующий формат. Было предложено использовать стратегию «выступ-во-впадину» для модификации тяжелых цепей. Некоторые другие модификации также были предложены для легких цепей. Однако эти дополнительные модификации могут неблагоприятно влиять на биохимические и/или биофизические свойства, период полужизни в сыворотке и/или стабильность, приводя к низкой эффективности, нестабильности и высокой иммуногенности. Существует потребность в способах получения биспецифических антител, которые не зависят в значительной степени от конструирования антител.
Настоящее изобретение относится к генетически модифицированным животным и клеткам с гуманизированным локусом легкой цепи иммуноглобулина (например, локусом цепи каппа) и/или гуманизированным локусом тяжелой цепи иммуноглобулина. В одном аспекте гуманизированный локус легкой цепи иммуноглобулина имеет ограниченный набор генов IGKV и генов IGKJ. Генетически сконструированное животное посредством длительного и сложного процесса селекции антител в организме делает биологически приемлемый выбор в сочетании разнообразной коллекции вариабельных доменов тяжелой цепи человека с ограниченным числом вариантов вариабельных доменов легкой цепи человека. Животное сконструировано так, чтобы представлять ограниченное число вариантов вариабельных доменов легкой цепи человека в сочетании с широким разнообразием вариантов вариабельных доменов тяжелой цепи человека. При контрольном заражении иммуногеном у животного вырабатывается антитело к иммуногену, ограниченное в основном или исключительно количеством вариантов легкой цепи в его репертуаре.
В различных вариантах осуществления антитела, продуцируемые у генетически модифицированного животного, имеют тяжелые цепи, которые могут связываться с идентичными или практически идентичными легкими цепями. Это особенно полезно при получении биспецифических антител. Например, такое животное можно иммунизировать первым антигеном для получения В-клетки, которая экспрессирует антитело, которое специфически связывается с первым антигеном. Животное (или животное с такой же модификацией) можно иммунизировать вторым антигеном для получения В-клетки, которая экспрессирует антитело, которое специфически связывается со вторым антигеном. VH можно клонировать из первой В-клетки и второй В-клетки. Два VH могут быть соединены с одним и тем же VL легкой цепи для получения биспецифического антитела. Таким образом, нет необходимости связывать легкую цепь с конкретной тяжелой цепью с помощью конструирования антител (например, внося модификации в последовательности). Это может значительно повысить вероятность успеха разработки биспецифических антител. Фактически, описанные в настоящем документе антитела или последовательности можно дополнительно комбинировать друг с другом для получения полиспецифических антител.
Чтобы получить ограниченный репертуар вариантов вариабельных доменов легкой цепи, животное конструируют так, чтобы ограничить его способность генерировать вариабельные домены легкой цепи с природным разнообразием. Затем эндогенный локус животного может быть модифицирован выбранной подходящей экзогенной последовательностью гена вариабельной области легкой цепи человека, функционально связанной с эндогенным константным доменом легкой цепи животного таким образом, что реаранжированная экзогенная последовательность гена вариабельной области человека может кодировать реаранжированную химерную легкую цепь (человеческая вариабельная, эндогенная константная) или нереаранжированные экзогенные последовательности гена вариабельной области человека с ограниченным разнообразием могут подвергаться реаранжировке и рекомбинации, чтобы кодировать реаранжированную химерную легкую цепь.
Описанные здесь генетически модифицированные животные могут иметь некоторые дополнительные преимущества. Например, в некоторых случаях генетически модифицированные животные, описанные в настоящем документе, имеют полные гены вариабельной области тяжелой цепи человека и реаранжированные гены вариабельной области легкой цепи человека (например, IGKV1-39/IGKJ4 человека). Кроме того, поскольку вся вариабельная область тяжелой цепи в локусе иммуноглобулина человека вводится в геном животного (без модификаций или с ограниченными модификациями), эти гены могут подвергаться VDJ-рекомбинации способом, который очень похож на то, что происходит у человека. Кроме того, продукция антител может быть очень эффективной и иметь скорость, близкую к нормальной, благодаря эффективной VDJ-рекомбинации.
Используемый в настоящем описании термин «антитело» относится к молекуле иммуноглобулина, включающей четыре полипептидные цепи, две тяжелые (Н) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями. Каждая тяжелая цепь содержит вариабельный домен тяжелой цепи (VH) и константную область тяжелой цепи (CH). Каждая легкая цепь содержит вариабельный домен легкой цепи (VL) и константную область легкой цепи (CL). Домены VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые областями, определяющими комплементарность (CDR), перемежающиеся с более консервативными областями, называемыми каркасными областями (FR). Каждый VH и VL содержит три CDR и четыре FR, расположенных от амино-конца к карбоксиконцу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4 (CDR тяжелой цепи могут обозначаться аббревиатурой HCDR1, HCDR2 и HCDR3; CDR легкой цепи могут обозначаться аббревиатурой LCDR1, LCDR2 и LCDR3). Термин «высокоаффинное» антитело относится к антителу, которое имеет KD по отношению к его эпитопу-мишени примерно 10-8 М или ниже (например, примерно или ниже 1×10-8 М, 1×10-9 М, 1×10-10 М или 1×10-11 М). В некоторых вариантах осуществления KD можно измерить с помощью поверхностного плазмонного резонанса, например. BIACORE™ или ИФА.
Используемый в настоящем описании термин «антигенсвязывающий фрагмент» относится к части полноразмерного антитела, где часть антитела способна специфически связываться с антигеном. В некоторых вариантах осуществления антигенсвязывающий фрагмент содержит по меньшей мере один вариабельный домен (например, вариабельный домен тяжелой цепи или вариабельный домен легкой цепи). Неограничивающие примеры фрагментов антител включают, например, фрагменты Fab, Fab', F(ab')2 и Fv.
Используемый в настоящем описании термин «человеческое антитело» относится к антителу, которое кодируется эндогенной нуклеиновой кислотой (например, реаранжированный локус тяжелой или легкой цепи иммуноглобулина человека), присутствующей в организме человека. В некоторых вариантах осуществления человеческое антитело выделяют из тканей человека или продуцируют в культуре человеческих клеток (например, в гибридомных клетках человека). В некоторых вариантах осуществления человеческое антитело продуцируется в клетке, не являющейся человеческой (например, в клеточной линии мыши или хомяка). В некоторых вариантах осуществления человеческое антитело продуцируется в бактериальной или дрожжевой клетке. В некоторых вариантах осуществления человеческое антитело продуцируется у трансгенного животного, не являющегося человеком (например, у мыши), содержащего нереаранжированный или реаранжированный локус иммуноглобулина человека (например, локус тяжелой или легкой цепи иммуноглобулина человека).
Используемый в настоящем описании термин «химерное антитело» относится к антителу, которое содержит последовательность, присутствующую по меньшей мере в двух разных антителах (например, в антителах двух разных видов млекопитающих, таких как антитела человека и мыши). Неограничивающим примером химерного антитела является антитело, содержащее последовательности вариабельного домена (например, всю или часть последовательности вариабельного домена легкой цепи и/или тяжелой цепи) человеческого антитела и константные домены антитела, не являющегося человеческим. Дополнительные примеры химерных антител описаны в настоящем документе и известны в данной области.
Используемый в настоящем описании термин «гуманизированное антитело» относится к антителу, не являющемуся человеческим, которое содержит последовательность, полученную из иммуноглобулина, не являющегося человеческим, (например, из мышиного) и содержит последовательности, полученные из человеческого иммуноглобулина.
Используемые в настоящем описании термины «объект» и «пациент» используются взаимозаменяемо в описании и описывают животное, человека или отличного от человека. Ветеринарные и неветеринарные применения рассматриваются настоящим изобретением. Пациенты могут быть взрослыми людьми или подростками (например, люди моложе 18 лет). Помимо человека, пациенты включают, но не ограничиваются ими, мышей, крыс, хомяков, морских свинок, кроликов, хорьков, кошек, собак и приматов. Включены, например, приматы, отличные от человека (например, обезьяна, шимпанзе, горилла и т. п.), грызуны (например, крысы, мыши, песчанки, хомяки, хорьки, кролики), зайцеобразные, свиньи (например, свинья, миниатюрная свинья), лошадиные, собачьи, кошачьи, крупный рогатый скот и другие домашние, сельскохозяйственные и зоопарковые животные.
Используемые в настоящем описании выражения «специфическое связывание» и «специфически связывается» применительно к антителу означают, что антитело взаимодействует со своей молекулой-мишенью предпочтительно по отношению к другим молекулам, поскольку взаимодействие зависит от присутствия конкретной структуры (т. е. антигенной детерминанты или эпитопа) на молекуле-мишени; другими словами, реагент распознает и связывается с молекулами, которые включают специфическую структуру, а не со всеми молекулами в целом. Антитело, которое специфически связывается с молекулой-мишенью, может называться антителом, специфичным к мишени.
Используемые в настоящем описании термины «полипептид», «пептид» и «белок» используются взаимозаменяемо для обозначения полимеров аминокислот любой длины, по меньшей мере, из двух аминокислот.
Используемые в настоящем описании термины «полинуклеотид», «молекула нуклеиновой кислоты» и «последовательность нуклеиновой кислоты» используются в настоящем описании взаимозаменяемо для обозначения полимеров нуклеотидов любой длины, по меньшей мере, из двух нуклеотидов и включают, без ограничения, ДНК, РНК, ДНК/РНК-гибриды и их модификации.
Используемый в настоящем описании термин «немодифицированная последовательность человека» относится к последовательности, полученной от объекта-человека, клетки человека, культивируемой клетки человека или клеточной линии человека, где последовательность идентична генетической последовательности объекта-человека, клетки человека, культивируемой клетки человека или клеточной линии человека.
Используемый в настоящем описании термин «биспецифическое антитело» включает антитело, способное избирательно связываться с двумя или более эпитопами. Биспецифические антитела обычно содержат две неидентичные тяжелые цепи, причем каждая тяжелая цепь специфически связывается с другим эпитопом либо на двух разных молекулах (например, разные эпитопы на двух разных иммуногенах), либо на одной и той же молекуле (например, разные эпитопы на одном и том же иммуногене). Эпитопы, специфически связанные с биспецифическим антителом, могут быть на одной и той же или на разных мишенях (например, на одном и том же или другом белке). Биспецифические антитела могут быть получены, например, путем объединения тяжелых цепей, которые распознают разные эпитопы одного и того же иммуногена. Например, последовательности нуклеиновых кислот, кодирующие вариабельные последовательности тяжелой цепи, которые распознают разные эпитопы одного и того же иммуногена, могут быть слиты с последовательностями нуклеиновых кислот, кодирующими одинаковые или разные константные области тяжелой цепи, и такие последовательности могут быть экспрессированы в клетке, которая экспрессирует легкую цепь иммуноглобулина. Типичное биспецифическое антитело имеет две тяжелые цепи, каждая из которых имеет три CDR тяжелой цепи, за которыми следуют (от N-конца к С-концу) домен СН1, шарнир, домен СН2 и домен СН3, и легкую цепь иммуноглобулина, которая либо не придает эпитоп-связывающей специфичности, но которая может связываться с каждой тяжелой цепью, или которая может связываться с каждой тяжелой цепью, и которая может связываться с одним или более эпитопами, связанными областями связывания эпитопов тяжелой цепи, или которая может связываться с каждой тяжелой цепью и обеспечивает связывание одной или обеих тяжелых цепей с одним или обоими эпитопами.
Используемый в настоящем описании термин «антитело с общей легкой цепью» относится к антителу, имеющему две или больше двух идентичных легких цепей. В некоторых вариантах осуществления антитело с общей легкой цепью представляет собой биспецифическое антитело с общей легкой цепью.
Локус рестриктированной легкой цепи каппа иммуноглобулина
Локус цепи каппа иммуноглобулина (также известный как IGK или локус каппа иммуноглобулина) представляет собой область на хромосоме (например, хромосома 2 человека), которая содержит гены легких цепей антител человека (или иммуноглобулинов). Аналогично гены легкой цепи иммуноглобулина также могут подвергаться последовательной реаранжировке, которая приводит к продукции зрелой нуклеиновой кислоты легкой цепи иммуноглобулина (например, цепи каппа).
Соединение сегмента V (также известного как ген IGKV) и сегмента J (также известного как ген IGKJ) создает непрерывный экзон, который кодирует весь вариабельный домен легкой цепи. В нереаранжированной ДНК сегменты гена V (или кластер гена IGKV) расположены относительно далеко от области С. Сегменты гена J (или кластер генов IGKJ) расположены близко к области C. Соединение сегмента V с сегментом гена J также сближает ген V с последовательностью С-области. Сегмент гена J реаранжированной V-области отделен от последовательности С-области только интроном. Чтобы получить полную информационную РНК легкой цепи иммуноглобулина, экзон V-области соединяется с последовательностью C-области путем сплайсинга РНК после транскрипции.
Локус легкой цепи иммуноглобулина человека расположен на хромосоме 2 человека. В Таблице 1 перечислены гены IGKV и их относительные порядки в этом локусе. Существует несколько различных групп генов IGKV человека, включая гены IGKV1 (включая все гены IGKV, начиная с IGKV1, также известные как VκI), гены IGKV2 (включая все гены IGKV, начиная с IGKV2, также известные как VκII), гены IGKV3 (включая все гены IGKV, начиная с IGKV3, также известные как VκIII), гены IGKV4 (включая все гены IGKV, начиная с IGKV4, также известные как VκIV), гены IGKV5 (включая все гены IGKV, начиная с IGKV5, также известные как VκV), гены IGKV6 (включая все гены IGKV, начиная с IGKV6, также известные как VκVI), и гены IGKV7 (включая все гены IGKV, начиная с IGKV7, также известные как VκVII).
Эти гены IGKV в хромосоме 2 человека также образуют два кластера, проксимальный кластер Vκ и дистальный кластер Vκ (ФИГ. 14). Последовательности в двух кластерах похожи, но не идентичны. Это большое сегментарное дублирование последовательности произошло после дивергенции человеческой линии от самого последнего общего предка с другими человекообразными обезьянами. Соответствующие гены IGVK в каждом кластере суммированы на ФИГ. 15.
В Таблице 2 перечислены все гены IGKJ и их относительные порядки на хромосоме 2 человека. Ген константной области цепи каппа иммуноглобулина (IGKC), который кодирует константные домены легкой цепи иммуноглобулина, расположен после генов IGKV и IGKJ. Эти гены и порядок этих генов также показаны на ФИГ. 18 и ФИГ. 22.
Локус легкой цепи иммуноглобулина мыши расположен на хромосоме 6 мыши. В Таблице 3 перечислены гены IGKV и их относительные порядки в этом локусе.
Gm9728 и Amd-ps2 также расположены в этом локусе. Относительный порядок Gm9728 равен 4, а относительный порядок Amd-ps2 равен 134. В Таблице 4 перечислены все гены IGKJ и их относительные порядки на хромосоме 6 мыши. Ген IGKC, кодирующий константные домены иммуноглобулина легкой цепи, расположен после генов IGKV и IGKJ. Эти гены и порядок этих генов также показаны на ФИГ. 19 и ФИГ. 23.
Настоящее изобретение относится к генетически модифицированному животному, отличному от человека, содержащему один, два, три или не более трех генов IGKV человека и один, два, три или не более трех генов IGKJ человека. В некоторых вариантах осуществления ген IGKV человека и ген IGKJ человека находятся в эндогенном локусе генов легкой цепи иммуноглобулина. В некоторых вариантах осуществления ген IGKV человека и ген IGKJ человека представляют собой реаранжированные последовательности. В некоторых вариантах осуществления они представляют собой нереаранжированные последовательности.
В некоторых вариантах осуществления гены IGKV человека выбраны из любого из генов IGKV, представленных в Таблице 1. В некоторых вариантах осуществления гены IGKJ человека выбраны из любого из генов IGKJ, представленных в Таблице 2.
В некоторых вариантах осуществления животное содержит только один ген IGKV человека и один ген IGKJ человека. В некоторых вариантах осуществления ген IGKV выбран из любого из генов IGKV, представленных в Таблице 1. В некоторых вариантах осуществления ген IGKJ выбран из любого из генов IGKJ, представленных в Таблице 2.
В некоторых вариантах осуществления животное содержит ген IGKV человека, выбранный из IGKV3-20, IGKV1-39, IGKV1D-39, IGKV3-11, IGKV3-15 и IGKV4-1. В некоторых вариантах осуществления животное содержит ген IGKJ человека, выбранный из IGKJ1, IGKJ2 и IGKJ4.
В некоторых вариантах осуществления животное содержит ген IGKV человека, выбранный из IGKV3-20, IGKV3-11, IGKV3-15, IGKV1-39, IGKV1D-39 и IGKV1-12, IGKV1D-12. В некоторых вариантах осуществления животное содержит ген IGKJ человека, выбранный из IGKJ1, IGKJ4 и IGKJ2.
В некоторых вариантах осуществления животное содержит ген IGKV человека, выбранный из IGKV3-20, IGKV3-11 и IGKV1-39. В некоторых вариантах осуществления животное содержит ген IGKJ человека, выбранный из IGKJ1 и IGKJ4.
В некоторых вариантах осуществления животное содержит последовательность промотора перед первым нуклеотидом гена IGKV человека. В некоторых вариантах осуществления последовательность промотора находится в пределах или примерно в пределах 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600, 1700, 1800, 1900, 2000, 2100, 2200, 2300, 2400, 2500, 2600, 2700, 2800, 2900, 3000, 3500, 4000, 4500 или 5000 п.н. перед геном IGKV человека. В некоторых вариантах осуществления промотор представляет собой промотор IGKV3-20 человека, промотор IGKV1-39 человека или промотор IGKV3-11 человека. В некоторых вариантах осуществления последовательность промотора по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентична нуклеотидам 1-2000 SEQ ID NO: 35, нуклеотидам 1-2000 SEQ ID NO: 36 или нуклеотидам 1-2000 SEQ ID NO: 37.
В некоторых вариантах осуществления животное содержит вспомогательную последовательность после последнего нуклеотида гена IGKJ человека. В некоторых вариантах осуществления вспомогательная последовательность содержит последовательность 3'UTR IGKJ мыши или последовательность 3'UTR IGKJ человека. В некоторых вариантах осуществления последовательность по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или на 100% идентична SEQ ID NO: 6 или SEQ ID NO: 7.
В различных вариантах осуществления соответствующий(е) энхансер(ы) может быть сохранен у животного. Например, при модификации локуса каппа для замены эндогенных сегментов гена вариабельной области каппа сегментами гена вариабельной области каппа человека интронный энхансер каппа и 3'-энхансер каппа функционально сохраняются или не разрушаются. В некоторых вариантах осуществления модифицированный локус каппа может подвергаться соматическим гипермутациям. В некоторых вариантах осуществления степень соматических гипермутаций примерно такая же или аналогичная таковой в локусе каппа дикого типа. В некоторых вариантах осуществления по меньшей мере 50%, 60%, 70%, 80%, 90% или 95% вариабельных областей легкой цепи человека в антителах (до или после иммунизации антигеном) имеют по меньшей мере 1 соматическую мутацию. В некоторых вариантах осуществления по меньшей мере 50%, 60%, 70%, 80%, 90% или 95% вариабельных областей легкой цепи человека в антителах (до или после иммунизации антигеном) имеют по меньшей мере 2 соматические мутации. В некоторых вариантах осуществления по меньшей мере 50%, 60%, 70%, 80%, 90% или 95% вариабельных областей легкой цепи человека в антителах (до или после иммунизации антигеном) имеют по меньшей мере 3 соматические мутации.
В некоторых вариантах осуществления животное содержит эндогенный IGKC. В некоторых вариантах осуществления гены IGKV и/или гены IGKJ функционально связаны вместе с геном IGKC (например, с эндогенным геном IGKC).
В некоторых вариантах осуществления ген IGKV и ген IGKJ функционально связаны друг с другом.
В некоторых вариантах осуществления животное содержит эндогенный IGKC. В некоторых вариантах осуществления гены IGKV и/или гены IGKJ функционально связаны друг с другом. Между этими генами может происходить VJ-рекомбинация с образованием функциональных антител. В некоторых вариантах осуществления ген IGKV и ген IGKJ реаранжированы в эндогенном локусе иммуноглобулина с каппа-цепью.
В некоторых вариантах осуществления животное содержит нарушение в эндогенном локусе генов легкой цепи иммуноглобулина животного. В некоторых вариантах осуществления нарушение в эндогенном локусе генов легкой цепи иммуноглобулина животного включает делецию одного или более эндогенных генов IGKV и одного или более эндогенных генов IGKJ.
В некоторых вариантах осуществления животное представляет собой мышь. Нарушение эндогенного локуса генов легкой цепи иммуноглобулина животного включает делецию, составляющую по меньшей мере или примерно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162 или 163 генов IGKV мыши (например, генов, представленных в Таблице 3). В некоторых вариантах осуществления нарушение включает делецию, составляющую примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGKV мыши, выбранных из IGKV2-137, IGKV1-136, IGKV1-135, IGKV14-134-1, IGKV17-134, IGKV1-133, IGKV1-132, IGKV1-131, IGKV14-130, и IGKV9-129. В некоторых вариантах осуществления мышь по-прежнему содержит примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGKV мыши, выбранных из IGKV2-137, IGKV1-136, IGKV1-135, IGKV14-134-1, IGKV17-134, IGKV1-133, IGKV1-132, IGKV1-131, IGKV14-130, и IGKV9-129.
В некоторых вариантах осуществления нарушение включает делецию, составляющую примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGKV мыши, выбранных из IGKV3-10, IGKV3-9, IGKV3-8, IGKV3-7, IGKV3-6, IGKV3-5, IGKV3-4, IGKV3-3, IGKV3-2, и IGKV3-1. В некоторых вариантах осуществления мышь по-прежнему содержит примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGKV мыши, выбранных из IGKV3-10, IGKV3-9, IGKV3-8, IGKV3-7, IGKV3-6, IGKV3-5, IGKV3-4, IGKV3-3, IGKV3-2, и IGKV3-1.
В некоторых вариантах осуществления нарушение включает делецию, составляющую примерно или по меньшей мере 1, 2, 3, 4 или 5 генов IGKJ мыши, выбранных из IGKJ1, IGKJ2, IGKJ3, IGKJ4 и IGKJ5. В некоторых вариантах осуществления мышь все еще содержит примерно или по меньшей мере 1, 2, 3, 4 или 5 генов IGKJ мыши, выбранных из IGKJ1, IGKJ2, IGKJ3, IGKJ4 и IGKJ5 (например, IGKJ5).
В некоторых вариантах осуществления нарушение в эндогенном локусе гена легкой цепи каппа иммуноглобулина животного включает делецию, составляющую примерно или по меньшей мере 500 т.п.н., 600 т.п.н., 700 т.п.н., 800 т.п.н., 900 т.п.н., 1000 т.п.н., 1500 т.п.н., 2000 т.п.н., 2500 т.п.н., 3000 т.п.н. или 3500 т.п.н. эндогенной последовательности.
В некоторых вариантах осуществления удаленная последовательность начинается с IGKV2-137 до IGKJ4, с IGKV1-136 до IGKJ4, с IGKV1-135 до IGKJ4, с IGKV2-137 до IGKJ5, с IGKV1-136 до IGKJ5 или с IGKV1-135 до IGKJ5. (например, с IGKV2-137 до IGKJ5).
В некоторых вариантах осуществления животное содержит примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 последовательностей, которые по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или на 100% идентичны последовательности в локусе генов легкой цепи иммуноглобулина человека. В некоторых вариантах последовательность имеет длину примерно или по меньшей мере 2 или 3 т.п.н. В некоторых вариантах последовательность имеет длину не более 4 т.п.н.
В некоторых вариантах осуществления животное содержит одну или более (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) реаранжированных последовательностей IGKV/IGKJ человека. В некоторых вариантах осуществления реаранжированная последовательность IGKV/IGKJ человека по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или на 100% идентична SEQ ID NO: 35 (например, нуклеотидам 2001-2512 SEQ ID NO: 35). В некоторых вариантах осуществления реаранжированная последовательность IGKV/IGKJ человека по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или на 100% идентична SEQ ID NO: 36 (например, нуклеотидам 2001-2551 SEQ ID NO: 36). В некоторых вариантах осуществления реаранжированная последовательность IGKV/IGKJ человека по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или на 100% идентична SEQ ID NO: 37 (например, нуклеотидам 2001-2572 SEQ ID NO: 37).
В некоторых вариантах осуществления реаранжированная последовательность IGKV/IGKJ человека имеет длину примерно или по меньшей мере 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600, 1700, 1800, 1900, 2000., 2100, 2200, 2300, 2400, 2500, 2600, 2700, 2800, 2900, 3000, 3100, 3200, 3300, 3400, 3500, 3600, 3700, 3800, 3900, 4000, или 50000 п.н. В некоторых вариантах осуществления реаранжированная последовательность IGKV/IGKJ человека имеет длину менее 1500, 1600, 1700, 1800, 1900, 2000, 2100, 2200, 2300, 2400, 2500, 2600, 2700, 2800, 2900, 3000 или 3100 п.н.
В некоторых вариантах осуществления животное может продуцировать иммуноглобулин (например, IgG), содержащий вариабельную область легкой цепи, по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или на 100% идентичную SEQ ID NO: 38. В некоторых вариантах осуществления животное может продуцировать иммуноглобулин (например, IgG), содержащий вариабельную область легкой цепи, по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или на 100% идентичную SEQ ID NO: 39. В некоторых вариантах осуществления животное может продуцировать иммуноглобулин (например, IgG), содержащий вариабельную область легкой цепи, по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или на 100% идентичную SEQ ID NO: 40. В некоторых вариантах осуществления вариабельная область легкой цепи может иметь 1, 2, 3, 4 или 5 мутаций по сравнению с SEQ ID NO: 38, 39 или 40. В некоторых вариантах осуществления константный домен легкой цепи имеет последовательность, по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91% %, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или на 100% идентичную SEQ ID NO: 41.
В некоторых вариантах осуществления животное может иметь одну, две, три, четыре, пять, шесть, семь, восемь, девять или десять немодифицированных последовательностей человека. В некоторых вариантах осуществления немодифицированная последовательность человека имеет длину примерно или по меньшей мере 0,1, 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 т.п.н.
В некоторых вариантах осуществления вариабельная область легкой цепи имеет последовательность SEQ ID NO: 38, содержащую 0, 1 или 2 мутации. В некоторых вариантах осуществления вариабельная область легкой цепи имеет последовательность SEQ ID NO: 39, содержащую 0, 1 или 2 мутации. В некоторых вариантах осуществления вариабельная область легкой цепи имеет последовательность SEQ ID NO: 40, содержащую 0, 1 или 2 мутации.
Более того, в некоторых случаях нокаутируются все мышиные гены IGKV и гены IGKJ (все не псевдогены), и вариабельная область легкой цепи не будет иметь какой-либо последовательности, которая кодируется последовательностью, полученной от мыши, тем самым сводя к минимуму иммуногенность у человека. В различных вариантах осуществления вариабельная область легкой цепи способна подвергаться соматическим мутациям.
Генетически модифицированный локус тяжелой цепи иммуноглобулина
Локус тяжелой цепи иммуноглобулина (также известный как IGH или локус тяжелой цепи иммуноглобулина) представляет собой область на хромосоме (например, на хромосоме 14 человека), которая содержит гены тяжелых цепей антител человека (или иммуноглобулинов).
Эта область представляет зародышевую организацию локуса тяжелой цепи. Локус включает сегменты V (вариабельный), D (разнообразия), J (соединения) и C (константный). Гены в области V образуют кластер генов V (также известный как кластер генов IGHV). Гены в области D образуют кластер генов D (также известный как кластер генов IGHD). Гены в области J образуют кластер генов J (также известный как кластер генов IGHJ).
Во время созревания В-клеток событие рекомбинации на уровне ДНК соединяет один сегмент D (также известный как ген IGHD) с сегментом J (также известный как ген IGHJ); слитый экзон D-J этой частично реаранжированной D-J-области затем соединяется с сегментом V (также известным как ген IGHV). Реаранжированная область V-D-J, содержащая слитый экзон V-D-J, затем транскрибируется и сливается на уровне РНК с константной областью IGHM; этот транскрипт кодирует тяжелую цепь мю. Позже при созревании В-клетки генерируют пре-м-РНК V-D-J-Cmu-Cdelta, которая альтернативно подвергается сплайсингу, чтобы кодировать либо тяжелую цепь мю, либо дельта. Зрелые В-клетки в лимфатических узлах подвергаются рекомбинации с переключением, так что слитый сегмент гена V-D-J приближается к одному из сегментов гена IGHG, IGHA или IGHE, и каждая клетка экспрессирует тяжелую цепь гамма, альфа или эпсилон. Возможная рекомбинация множества различных генов IGHV с несколькими генами IGHJ обеспечивает широкий диапазон распознавания антигена. Дополнительное разнообразие достигается за счет разнообразия соединений, возникающего в результате случайного добавления нуклеотидов терминальной дезоксинуклеотидилтрансферазой, и за счет соматической гипермутации, которая происходит во время созревания В-клеток в селезенке и лимфатических узлах. Известно, что несколько сегментов V, D, J и C не способны кодировать белок и считаются сегментами псевдогенных генов (часто называемыми просто псевдогенами).
Локус тяжелой цепи иммуноглобулина человека расположен на хромосоме 14 человека. В Таблице 5 перечислены гены IGHV и их относительные порядки в этом локусе.
RPS8P1, ADAM6 и KIAA0125 также расположены в этом локусе. Относительный порядок RPS8P1 равен 160, относительный порядок ADAM6 равен 161, а относительный порядок KIA09A0125 равен 164. В Таблице 6 перечислены все гены IGHD и их относительные порядки на хромосоме 14 человека. В Таблице 7 перечислены все гены IGHJ и их относительные порядки на хромосоме 14 человека. Гены константных доменов иммуноглобулинов расположены после генов IGHV, IGHD и IGHJ. Эти гены включают (как показано в следующем порядке): константный домен тяжелой цепи мю иммуноглобулина (IGHM), константный домен тяжелой цепи дельта иммуноглобулина (IGH δ), константный домен тяжелой цепи гамма 3 иммуноглобулина (IGHG3), константный домен тяжелой цепи гамма 1 иммуноглобулина (IGHG1), константный домен тяжелой цепи эпсилон P1 иммуноглобулина (псевдоген) (IGHEP1), константный домен тяжелой цепи альфа 1 иммуноглобулина (IGHA1), константный домен тяжелой цепи гамма P иммуноглобулина (нефункциональный) (IGHGP), константный домен тяжелой цепи гамма 2 иммуноглобулина (IGHG2), константный домен тяжелой цепи гамма 4 иммуноглобулина (IGHG4), константный домен тяжелой цепи эпсилон иммуноглобулина (IGHE) и константный домен тяжелой цепи альфа 2 иммуноглобулина (IGHA2). Эти гены и порядок этих генов также показаны на ФИГ. 16 и ФИГ. 20.
Локус тяжелой цепи иммуноглобулина мыши расположен на хромосоме 12 мыши. В Таблице 8 перечислены гены IGHV и их относительные порядки в этом локусе.
В Таблице 9 перечислены все гены IGHD и их относительные порядки на хромосоме 12 мыши. В Таблице 10 перечислены все гены IGHJ и их относительные порядки на хромосоме 12 мыши. Гены константных доменов иммуноглобулина расположены после генов IGHV, IGHD и IGHJ. Эти гены включают (как показано в следующем порядке): константный домен тяжелой цепи мю иммуноглобулина (IGHM), константный домен тяжелой цепи дельта иммуноглобулина (IGH δ), константный домен тяжелой цепи гамма 3 иммуноглобулина (IGHG3), константный домен тяжелой цепи гамма 1 иммуноглобулина (IGHG1), константный домен тяжелой цепи гамма 2b иммуноглобулина (IGHG2b), константный домен тяжелой цепи гамма 2a иммуноглобулина (IGHG2a), константный домен тяжелой цепи эпсилон иммуноглобулина (IGHE) и константный домен тяжелой цепи альфа иммуноглобулина. Эти гены и порядок этих генов также показаны на ФИГ. 17 и ФИГ. 21.
Настоящее изобретение относится к генетически модифицированному животному, отличному от человека, содержащему один или более генов IGHV человека, один или более генов IGHD человека и/или один или более генов IGHJ человека.
Генетически модифицированные животные могут быть получены путем введения генов иммуноглобулинов человека в геном животных, отличных от человека, для получения животных, которые могут экспрессировать гуманизированные антитела или химерные антитела. На ФИГ. 1А показаны способы получения гуманизированных мышей. В некоторых вариантах осуществления способы сначала включают модификацию области иммуноглобулина человека на хромосоме человека. Затем модифицированные хромосомы человека вводят в реципиентную клетку мыши. Затем вариабельную область иммуноглобулина человека вводят в соответствующую область генома мыши путем прямой замены (например, одностадийной замены). Затем клетки-реципиенты подвергают скринингу, предпочтительно на наличие клеток, не содержащих хромосомы человека. Затем клетки вводят в бластоцисты для получения химерных животных (например, мышей). Последующую селекцию можно проводить для получения животных, содержащих интактный гуманизированный локус иммуноглобулина.
В некоторых вариантах осуществления гены IGHV человека, гены IGHD человека и гены IGHJ человека функционально связаны друг с другом и могут подвергаться VDJ-реаранжировке. В некоторых вариантах осуществления гены IGHV человека, гены IGHD человека и гены IGHJ человека находятся в эндогенном локусе генов тяжелой цепи иммуноглобулина.
В некоторых вариантах осуществления животное содержит примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160 или 161 ген IGHV человека (например, гены, представленные в Таблице 5).
В некоторых вариантах осуществления животное содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов, выбранных из IGHV(III)-82, IGHV7-81, IGHV4-80, IGHV3-79, IGHV(II)-78-1, IGHV5-78, IGHV7-77, IGHV(III)-76-1, IGHV3-76, и IGHV3-75.
В некоторых вариантах осуществления животное содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов, выбранных из IGHV(III)-5-2, IGHV(III)-5-1, IGHV2-5, IGHV7-4-1, IGHV4-4, IGHV1-3, IGHV(III)-2-1, IGHV1-2, IGHV(II)-1-1, и IGHV6-1.
В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, содержащую последовательность, начиная с гена, выбранного из IGHV(III)-82, IGHV7-81, IGHV4-80, IGHV3-79, IGHV(II)-78-1, IGHV5-78, IGHV7-77, IGHV(III)-76-1, IGHV3-76, и IGHV3-75, и заканчивая геном, выбранным из IGHV(III)-5-2, IGHV(III)-5-1, IGHV2-5, IGHV7-4-1, IGHV4-4, IGHV1-3, IGHV(III)-2-1, IGHV1-2, IGHV(II)-1-1, и IGHV6-1. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHV(III)-82 человека и заканчивая IGHV1-2 человека. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHV(III)-82 человека и заканчивая IGHV(II)-1-1 человека. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHV(III)-82 человека и заканчивая IGHV-6-1 человека.
В некоторых вариантах осуществления животное содержит примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26 или 27 генов IGHD человека (например, гены, представленные в Таблице 6). В некоторых вариантах осуществления животное содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов, выбранных из IGHD1-1, IGHD2-2, IGHD3-3, IGHD4-4, IGHD5-5, IGHD4-23, IGHD5-24, IGHD6-25, IGHD1-26 и IGHD7-27.
В некоторых вариантах осуществления животное содержит примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8 или 9 генов IGHJ человека (например, гены, представленные в Таблице 7). В некоторых вариантах осуществления животное содержит 1, 2, 3, 4, 5, 6, 7, 8 или 9 генов IGHJ человека, выбранных из IGHJ1P, IGHJ1, IGHJ2, IGHJ2P, IGHJ3, IGHJ4, IGHJ5, IGHJ3P и IGHJ6.
В некоторых вариантах осуществления животное содержит немодифицированную последовательность человека, содержащую последовательность, начиная с гена, выбранного из IGHD1-1, IGHD2-2, IGHD3-3, IGHD4-4, IGHD5-5, IGHD4-23, IGHD5-24, IGHD6- 25, IGHD1-26 и IGHD7-27, и заканчивая геном, выбранным из IGHJ1P, IGHJ1, IGHJ2, IGHJ2P, IGHJ3, IGHJ4, IGHJ5, IGHJ3P и IGHJ6. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHD1-1 человека и заканчивая IGHJ6 человека.
В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHD1-1 человека и заканчивая IGHD7-27 человека.
В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHJ1P человека и заканчивая IGHJ6 человека. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHJ1 человека и заканчивая IGHJ6 человека.
В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHV(III)-82 человека и заканчивая IGHJ6 человека.
В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHV1-2 человека и заканчивая IGHJ6 человека. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHV(II)-1-1 человека и заканчивая IGHJ6 человека. В некоторых вариантах осуществления немодифицированная последовательность человека получена из локуса генов тяжелой цепи иммуноглобулина человека, начиная с IGHV6-1 человека и заканчивая IGHJ6 человека.
В некоторых вариантах осуществления животное может иметь одну, две, три, четыре, пять, шесть, семь, восемь, девять или десять немодифицированных последовательностей человека. В некоторых вариантах осуществления немодифицированная последовательность человека имеет длину, составляющую примерно или по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 т.п.н.
В некоторых вариантах осуществления животное содержит один или более эндогенных генов, выбранных из группы, состоящей из генов константной области тяжелой цепи мю иммуноглобулина (IGHM), константной области тяжелой цепи дельта иммуноглобулина (IGHδ), константной области тяжелой цепи гамма 3 иммуноглобулина (IGHG3), константной области тяжелой цепи гамма 1 иммуноглобулина (IGHG1), константной области тяжелой цепи гамма 2b иммуноглобулина (IGHG2b), константной области тяжелой цепи гамма 2a иммуноглобулина (IGHG2a), константной области тяжелой цепи эпсилон иммуноглобулина (IGHE) и константной области тяжелой цепи альфа иммуноглобулина (IGHA). В некоторых вариантах осуществления эти эндогенные гены функционально связаны друг с другом. В некоторых вариантах осуществления эти эндогенные гены имеют тот же порядок, что и у животного дикого типа. В некоторых вариантах осуществления у животного может происходить переключение изотипа (переключение класса иммуноглобулина).
В некоторых вариантах осуществления гены IGHV, гены IGHD и/или гены IGHJ функционально связаны друг с другом. VDJ-рекомбинация может происходить среди этих генов и приводить к продуцированию функциональных антител. В некоторых вариантах осуществления эти гены расположены в порядке, сходном с порядком в локусе тяжелой цепи иммуноглобулина человека. Такое расположение предлагает различные преимущества, например, расположение этих генов позволяет получать вариабельные домены тяжелой цепи с разнообразием, которое очень похоже на разнообразие вариабельных доменов тяжелой цепи у человека. Поскольку некоторые случайные последовательности могут быть вставлены в последовательность во время VDJ-рекомбинации, в некоторых вариантах осуществления полный репертуар человеческих антител без модификаций или с минимальными модификациями может снизить вероятность того, что непоследовательность человека будет вставлена во время VDJ-рекомбинации.
В некоторых вариантах осуществления гены IGHV, гены IGHD и/или гены IGHJ функционально связаны вместе с одним или более генами (например, все гены), выбранными из генов IGHM, IGHδ, IGHG3, IGHG1, IGHG2b, IGHG2a, IGHE и IGHA.
В некоторых вариантах осуществления животное содержит нарушение в эндогенном локусе генов тяжелой цепи иммуноглобулина животного. В некоторых вариантах осуществления нарушение в эндогенном локусе генов тяжелой цепи иммуноглобулина животного включает делецию одного или более эндогенных генов IGHV, одного или более эндогенных генов IGHD и одного или более эндогенных генов IGHJ.
В некоторых вариантах осуществления животное представляет собой мышь. Нарушение в эндогенном локусе генов тяжелой цепи иммуноглобулина животного включает делецию, составляющую по меньшей мере или примерно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181 или 182 гена IGHV мыши (например, гены, представленные в Таблице 8). В некоторых вариантах осуществления нарушение включает делецию, составляющую примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 мышиных генов IGHV, выбранных из IGHV1-86, IGHV1-85, IGHV1-84, IGHV1-83, IGHV1-82, IGHV1-81, IGHV1-80, IGHV1-79, IGHV1-78 и IGHV1-77. В некоторых вариантах осуществления мышь по-прежнему содержит примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 мышиных генов IGHV, выбранных из IGHV1-86, IGHV1-85, IGHV1-84, IGHV1-83, IGHV1-82, IGHV1-81, IGHV1-80, IGHV1-79, IGHV1-78 и IGHV1-77 (например, IGHV1-86).
В некоторых вариантах осуществления нарушение включает делецию, составляющую примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGHV мыши, выбранных из IGHV5-6, IGHV5-5, IGHV2-3., IGHV6-1, IGHV5-4, IGHV5-3, IGHV2-2, IGHV5-2, IGHV2-1 и IGHV5-1. В некоторых вариантах осуществления мышь все еще содержит делеции, составляющие примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGHV мыши, выбранных из IGHV5-6, IGHV5-5, IGHV2- 3, IGHV6-1, IGHV5-4, IGHV5-3, IGHV2-2, IGHV5-2, IGHV2-1 и IGHV5-1.
В некоторых вариантах осуществления нарушение в эндогенном локусе генов тяжелой цепи иммуноглобулина животного включает делецию, составляющую по меньшей мере или примерно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 генов IGHD мыши (например, гены, представленные в Таблице 9). В некоторых вариантах осуществления нарушение включает делецию, составляющую примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGHD мыши, выбранных из IGHD5-1, IGHD3-1, IGHD1-1, IGHD6-1, IGHD2-3, IGHD2-7, IGHD2-8, IGHD5-6, IGHD3-2 и IGHD4-1. В некоторых вариантах осуществления мышь по-прежнему содержит примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 генов IGHD мыши, выбранных из IGHD5-1, IGHD3-1, IGHD1-1, IGHD6-1, IGHD2-3, IGHD2-7, IGHD2-8, IGHD5-6, IGHD3-2 и IGHD4-1.
В некоторых вариантах осуществления нарушение включает делецию, составляющую примерно или по меньшей мере 1, 2, 3 или 4 генов IGHJ мыши, выбранных из IGHJ1, IGHJ2, IGHJ3 и IGHJ4. В некоторых вариантах осуществления мышь по-прежнему содержит примерно или по меньшей мере 1, 2, 3 или 4 гена IGHJ мыши, выбранных из IGHJ1, IGHJ2, IGHJ3 и IGHJ4.
В некоторых вариантах осуществления нарушение в эндогенном локусе генов тяжелой цепи иммуноглобулина животного включает делецию, составляющую примерно или по меньшей мере 500 т.п.н., 600 т.п.н., 700 т.п.н., 800 т.п.н., 900 т.п.н., 1000 т.п.н., 1500 т.п.н., 2000 т.п.н., 2500 т.п.н., или 3000 т.п.н. эндогенной последовательности.
В некоторых вариантах осуществления удаленная последовательность начинается с IGHV1-86 до IGHJ4, с IGHV1-85 до IGHJ4, с IGHV1-84 до IGHJ4, с IGHV1-83 до IGHJ4, или с IGHV1-82 до IGHJ4 (например, с IGHV1-85 до IGHJ4).
В некоторых вариантах осуществления животное содержит примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 последовательностей, которые по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или на 100% идентичны последовательности в локусе гена тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления последовательность имеет длину примерно или по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 2500, 3000 или 3500 т.п.н. В некоторых вариантах осуществления последовательность начинается от IGHV(III)-82 человека до IGHV1-2. В некоторых вариантах осуществления последовательность начинается от IGHV7-81 человека до IGHV1-2. В некоторых вариантах осуществления последовательность начинается от IGHV(II)-1-1 человека до IGHVJ6. В некоторых вариантах осуществления последовательность начинается от IGHV6-1 человека до IGHVJ6.
Гены IGHV человека, гены IGHD человека и гены IGHJ человека функционально связаны друг с другом и могут подвергаться VDJ-реаранжировке. В некоторых вариантах осуществления модифицированная мышь имеет полный репертуар генов IGHV, IGHD и IGHJ человека (например, включая все гены человека IGHV, IGHD и IGHJ, не являющиеся псевдогенами). Таким образом, модифицированная мышь может продуцировать полный репертуар человеческих антител.
В некоторых вариантах осуществления можно детектировать использование IGHV3-15, IGHV1-18, IGHV3-21, IGHV5-51, IGHV3-74, IGHV3-30-3, IGHV3-43, IGHV1-24, IGHV3-7, IGHV4-4, IGHV3-53, IGHV4-59, IGHV6-1, IGHV3-23, IGHV3-33, IGHV3-30, IGHV3-48, IGHV4-39, IGHV4-34, или IGHV3-66 (например, с частотой >1% среди реаранжированных последовательностей). В некоторых вариантах осуществления можно детектировать использование IGHV3-38-3, IGHV1-58, IGHV4-38-2, IGHV4-61, IGHV3-NL1, IGHV2-26, IGHV1-2, IGHV7-4-1, IGHV4-28, IGHV3-64, IGHV3-49, IGHV5-10-1, IGHV3-72, IGHV2-5, IGHV2-70, IGHV1-46, IGHV1-3, IGHV3-11, IGHV3-13, IGHV3-20, IGHV3-64D, IGHV1-69, IGHV3-73, IGHV4-30-2, IGHV4-31, или IGHV4-30-4 (например, с частотой <1% среди реаранжированных последовательностей).
В некоторых вариантах осуществления можно детектировать использование IGHD5-24, IGHD2-8, IGHD6-25, IGHD1-14, IGHD4-23, IGHD3-16, IGHD1-20, IGHD2-15, IGHD2-21, IGHD1-1, IGHD5-12, IGHD3-22, IGHD7-27, IGHD4-11, IGHD3-9, IGHD3-3, IGHD2-2, IGHD5-18, IGHD4-17, IGHD3-10, IGHD6-6, IGHD1-26, IGHD1-7, IGHD6-19, или IGHD6-13 (например, с частотой >1% среди реаранжированных последовательностей).
В некоторых вариантах осуществления можно детектировать использование IGHJ1, IGHJ2, IGHJ3, IGHJ4, IGHJ5 или IGHJ6 (например, с частотой >1% среди реаранжированных последовательностей).
В некоторых вариантах осуществления можно детектировать использование IGHV1-24, IGHV4-30-2, IGHV1-18, IGHV3-43, IGHV4-30-4, IGHV5-51, IGHV3-21, IGHV4-31, IGHV3-7, IGHV3-30-3, IGHV3-53, IGHV4-4, IGHV3-74, IGHV3-66, IGHV3-33, IGHV3-23, IGHV6-1, IGHV3-30, IGHV4-34, IGHV3-48, IGHV4-59, или IGHV4-39 (например, с частотой >1% среди реаранжированных последовательностей).
В некоторых вариантах осуществления можно детектировать использование IGHV3-25, IGHV4-38-2, IGHV7-4-1, IGHV3-NL1, IGHV4-61, IGHV1-58, IGHV2-26, IGHV3-72, IGHV5-10-1, IGHV1-46, IGHV3-49, IGHV2-70, IGHV1-2, IGHV3-64, IGHV4-28, IGHV3-20, IGHV1-3, IGHV3-13, IGHV3-73, IGHV3-11, IGHV3-64D, IGHV1-69, IGHV2-5, или IGHV3-15 (например, с частотой <1% среди реаранжированных последовательностей).
Кроме того, поскольку V(D)J-рекомбинация может происходить между эндогенными генами IGHV, IGHD, IGHJ, IGKV и IGKJ и генами человека, если эндогенные гены IGHV, IGHD, IGHJ, IGKV и IGKJ включены в реаранжированный сегмент VDJ тяжелой цепи или реаранжированный сегмент VJ легкой цепи, вполне вероятно, что антитела, генерируемые репертуарами антител, имеют иммуногенные эпитопы у человека. Иммуногенность может привести к выработке антител к лекарственным средствам и может поставить под угрозу эффективность. Здесь, в некоторых вариантах осуществления, эндогенные гены IGHV, IGHD, IGHJ, IGKV и IGKJ были эффективно удалены. В некоторых случаях целые мышиные гены IGHV, IGHD и IGHJ (например, включая все гены, не являющиеся псевдогенами) нокаутированы, и вариабельная область тяжелой цепи не будет иметь какой-либо последовательности, кодируемой последовательностью, полученной от мыши.
Маловероятно, что антитела, генерируемые репертуарами антител, являются иммуногенными для человека. Таким образом, антитела более подходят для использования в качестве терапевтических средств у человека. Следовательно, генетически модифицированные животные обеспечивают выгодную платформу для производства гуманизированных антител.
Генетически модифицированный локус легкой цепи лямбда иммуноглобулина
Локус цепи лямбда иммуноглобулина (также известный как IGL или лямбда-локус иммуноглобулина) представляет собой область на хромосоме (например, хромосоме 22 человека), которая содержит гены легких цепей антител человека (или иммуноглобулинов). Аналогично гены легкой цепи иммуноглобулина также могут подвергаться последовательной реаранжировке, которая приводит к продукции зрелой нуклеиновой кислоты легкой цепи иммуноглобулина (например, лямбда-цепи). У здорового человека общее соотношение каппа и лямбда составляет примерно 2:1 в сыворотке (при измерении интактных цельных антител) или 1:1,5 при измерении свободных легких цепей. У мышей общее соотношение каппа и лямбда составляет примерно 9:1.
В некоторых вариантах осуществления животное содержит локус иммуноглобулина цепи лямбда человека.
В некоторых вариантах осуществления у животного имеется нарушение в эндогенном локусе гена легкой цепи лямбда иммуноглобулина. В некоторых вариантах осуществления нарушение в эндогенном локусе генов легкой цепи иммуноглобулина животного включает делецию одного или более эндогенных генов IGLV, одного или более эндогенных генов IGLJ и/или одного или более генов константной области лямбда-цепи иммуноглобулина (IGLC) (например, IGLC1, IGLC2, IGLC3 и IGLC4).
Локус легкой цепи лямбда иммуноглобулина (локус IGL) мыши расположен на хромосоме 16 мыши. В Таблице 11 перечислены гены IGLV, IGLJ и IGLC и их относительные порядки в этом локусе.
Нарушение эндогенного локуса гена легкой цепи лямбда иммуноглобулина животного включает делецию, составляющую по меньшей мере или примерно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 генов IGLV, IGLJ и IGLC мыши (например, гены, представленные в Таблице 11). В некоторых вариантах осуществления делеция включает примерно или по меньшей мере 1, 2, 3 или 4 гена IGLC мыши, выбранных из IGLC1, IGLC2, IGLC3 и IGLC4. В некоторых вариантах осуществления нарушение включает делецию, составляющую примерно или по меньшей мере 1, 2 или 3 гена IGLV мыши, выбранных из IGLV1, IGLV2 и IGLV3. В некоторых вариантах осуществления нарушение включает делецию, составляющую примерно или по меньшей мере 1, 2, 3, 4 или 5 генов IGLJ мыши, выбранных из IGLJ1, IGLJ2, IGLJ3, IGLJ3P и IGLJ4.
В некоторых вариантах осуществления нарушение в эндогенном локусе генов легкой цепи лямбда иммуноглобулина животного включает делецию, составляющую примерно или по меньшей мере 10 т.п.н., 20 т.п.н., 30 т.п.н., 40 т.п.н., 50 т.п.н., 60 т.п.о., 70 т.п.н., 80 т.п.н., 90 т.п.н., 100 т.п.н., 110 т.п.н., 120 т.п.н., 130 т.п.н., 140 т.п.н., 150 т.п.н., 160 т.п.н., 170 т.п.н., 180 т.п.н., 190 т.п.н., 200 т.п.н., 210 т.п.н., 220 т.п.н., 230 т.п.н., 240 т.п.н., 250 т.п.н., 260 т.п.н., 270 т.п.н., 280 т.п.н., 290 т.п.н., 300 т.п.н., 350 т.п.н., 400 т.п.н., 450 т.п.н., 500 т.п.н. или 1000 т.п.н. В некоторых вариантах осуществления эндогенный ген легкой цепи лямбда иммуноглобулина животного не нарушен.
В некоторых вариантах осуществления удаленная последовательность начинается от IGLV2 до IGLC1, от IGLV3 до IGLC1 или от IGLJ2 до IGLC1.
Подробное описание генетически модифицированных животных и генетически модифицированного локуса иммуноглобулина тяжелой цепи можно найти например, в PCT/CN2020/075698, которая включена в настоящее описание посредством ссылки в полном объеме.
Генетически модифицированные животные
В одном аспекте настоящее изобретение относится к генетически модифицированному животному, отличному от человека, содержащему гуманизированный локус легкой цепи иммуноглобулина и/или гуманизированный локус тяжелой цепи иммуноглобулина. Гуманизированный локус легкой цепи иммуноглобулина содержит ограниченный набор генов IGKV человека и/или один ген IGKJ человека. В некоторых вариантах осуществления эти гены находятся в эндогенном локусе гена иммуноглобулина.
В некоторых вариантах осуществления животное содержит локус иммуноглобулина цепи лямбда человека. В некоторых вариантах осуществления у животного имеется нарушение в эндогенном локусе гена легкой цепи лямбда иммуноглобулина. В некоторых вариантах осуществления у животного нет нарушения эндогенного локуса гена легкой цепи лямбда иммуноглобулина животного.
Генетически модифицированное животное, отличное от человека, может относиться к различным животным, таким как например, мышь, крыса, кролик, свинья, крупный рогатый скот (например, корова, бык, буйвол), олень, овца, коза, курица, кошка, собака, хорек, примат (например, мартышка, макака-резус). Для животных, отличных от человека, для которых подходящие генетически модифицированные эмбриональные стволовые (ES) клетки недоступны, используются другие способы для получения животного, отличного от человека, содержащего генетическую модификацию. Такие методы включают, например, модификацию генома не-ES клеток (например, фибробластов или индуцированных плюрипотентных клеток) и использование ядерного переноса для переноса модифицированного генома в подходящую клетку, например, ооцит и созревание модифицированной клетки (например, модифицированного ооцита) у животного, отличного от человека, в подходящих условиях для формирования эмбриона. Эти способы известны в данной области и описаны, например, в публикации A. Nagy, et al., «Manipulation the Mouse Embryo: A Laboratory Manual (Third Edition)», Cold Spring Harbour Laboratory Press, 2003, которая включена в настоящее описание посредством ссылки в полном объеме. Таким образом, в различных вариантах осуществления сегменты V, D и/или J человека могут быть функционально связаны с последовательностями генов константной области животных, отличных от человека (например, грызуна, мыши, крысы, хомяка). Во время развития В-клеток эти реаранжированные сегменты V, D и/или J человека связаны с константной областью иммуноглобулина животного, отличного от человека.
В одном аспекте животное представляет собой млекопитающее, например, суперсемейства Dipodoidea или Muroidea. В некоторых вариантах осуществления генетически модифицированное животное представляет собой грызуна. Грызун может быть выбран из мыши, крысы и хомяка. В некоторых вариантах осуществления генетически модифицированное животное принадлежит к семейству, выбранному из Calomyscidae (например, мышевидные хомяки), Cricetidae (например, хомяк, крысы и мыши Нового Света, полевки), Muridae (мыши и крысы, песчанки, иглистые мыши, хохлатые крысы), Nesomeyidae (лазающие мыши, скальные крысы, длиннохвостые крысы, малагасийские крысы и мыши), Platacanthomyidae (например, колючие сони) и Spalacidae (например, слепыши, бамбуковые крысы и цокоры). В некоторых вариантах осуществления генетически модифицированный грызун выбран из мыши или крысы (семейство Muridae), песчанки, игольчатой мыши и хохлатой крысы. В некоторых вариантах осуществления отличным от человека животным является мышь.
В некоторых вариантах осуществления животное представляет собой мышь с фоном С57 (например, штамм C57BL, выбранный из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/ 10Cr и C57BL/Ola). В некоторых вариантах осуществления мышь представляет собой штамм 129, выбранный из группы, состоящей из штамма 129P1, 129P2, 129P3, 129X1, 129S1 (например, 129S1/SV, 129S1/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129T1, 129T2. Эти мыши описаны, например, в публикациях Festing et al., Revised nomenclature for strain 129 mice, Mammalian Genome 10: 836 (1999); Auerbach et al., Establishment and Chimera Analysis of 129/SvEv- and C57BL/6-Derived Mouse Embryonic Stem Cell Lines (2000), каждая из которых включена в настоящее описание в качестве ссылки в полном объеме. В некоторых вариантах осуществления генетически модифицированная мышь представляет собой смесь штамма 129 и штамма C57BL/6. В некоторых вариантах осуществления мышь представляет собой смесь штаммов 129 или смесь штаммов BL/6. В некоторых вариантах осуществления мышь представляет собой штамм BALB, например, штамм BALB/c. В некоторых вариантах осуществления мышь представляет собой смесь штамма BALB и другого штамма. В некоторых вариантах осуществления мышь происходит от гибридной линии (например, 50% BALB/c-50% 12954/Sv; или 50% C57BL/6-50% 129).
В некоторых вариантах осуществления животное представляет собой крысу. Крыса может быть выбрана из крыс Wistar, линии LEA, линии Sprague Dawley, линии Fischer, F344, F6 и Dark Agouti. В некоторых вариантах осуществления линия крыс представляет собой смесь двух или более линий, выбранных из группы, состоящей из Wistar, LEA, Sprague Dawley, Fischer, F344, F6 и Dark Agouti.
Животное может иметь одну или более других генетических модификаций и/или других модификаций, подходящих для конкретной цели, для которой создано гуманизированное животное.
Генетически модифицированные животные, отличные от человека, которые содержат модификацию эндогенного локуса гена иммуноглобулина, не относящегося к человеку. В некоторых вариантах осуществления модификация может содержать последовательность нуклеиновой кислоты человека, кодирующую по меньшей мере часть белка человека (например, по меньшей мере, на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или на 99% идентичную последовательности вариабельного домена тяжелой цепи или вариабельного домена легкой цепи человека). Хотя также предложены генетически модифицированные клетки, которые могут содержать модификации, описанные в настоящем документе (например, ES-клетки, соматические клетки), во многих вариантах осуществления генетически модифицированные животные, отличные от человека, включают модификацию эндогенного локуса в зародышевой линии животного.
Генетически модифицированные животные могут экспрессировать гуманизированное антитело и/или химерное антитело из эндогенных мышиных локусов, в которых один или более эндогенных генов иммуноглобулина мыши заменены генами иммуноглобулина человека и/или нуклеотидной последовательностью, по меньшей мере на 10%, 20% 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или на 99% идентичной последовательностям гена иммуноглобулина человека (например, генам IGHV, IGHD, IGHJ, IGKV и/или IGKJ). В различных вариантах осуществления эндогенный локус гена иммуноглобулина, не относящегося к человеку, модифицирован полностью или частично с включением последовательности нуклеиновой кислоты человека.
Может быть выполнен генетический, молекулярный и поведенческий анализ описанных выше млекопитающих, отличных от человека. Настоящее описание также относится к потомству, полученному от млекопитающего, отличного от человека, согласно настоящему описанию, скрещивающегося с теми же или другими генотипами. Млекопитающие, отличные от человека, могут представлять собой любое животное, отличное от человека, известное в данной области и которое можно использовать в способах, описанных в настоящем документе. Предпочтительными млекопитающими, отличными от человека, являются млекопитающие (например, грызуны). В некоторых вариантах осуществления отличным от человека млекопитающим является мышь.
Настоящее описание также относится к клеточной линии или первичной клеточной культуре, полученной из млекопитающего, отличного от человека, или его потомства. Модель на основе клеточной культуры может быть приготовлена, например, следующими способами. Культуры клеток могут быть получены путем выделения из млекопитающего, отличного от человека, альтернативно клетки могут быть получены из культуры клеток, созданной с использованием тех же конструкций и стандартных способов трансфекции клеток. Интеграцию генетических конструкций, содержащих последовательности ДНК, кодирующие человеческие или гуманизированные иммуноглобулины, можно детектировать различными методами.
Существует множество аналитических методов, которые можно использовать для детектирования экзогенной ДНК или модификаций геномной ДНК, включая методы на уровне нуклеиновых кислот (включая подходы количественного определения мРНК с использованием полимеразной цепной реакции с обратной транскриптазой (RT-PCR) или Саузерн-блоттинга, и гибридизация in situ) и методы на уровне белка (включая гистохимию, иммуноблот-анализ и исследования связывания in vitro). Кроме того, уровень экспрессии интересующего гена можно количественно определить с помощью методов ИФА, хорошо известных специалистам в данной области. Для выполнения количественных измерений можно использовать многие стандартные методы анализа. Например, уровни транскрипции можно измерить с помощью ОТ-ПЦР и методов гибридизации, включая защиту от РНКаз, Саузерн-блоттинг, анализ RNA dot (RNAdot). Иммуногистохимическое окрашивание, проточная цитометрия, вестерн-блоттинг также могут быть использованы для оценки присутствия человеческих или гуманизированных белков.
Антитела и антигенсвязывающие фрагменты
В настоящем описании предложены антитела и их антигенсвязывающие фрагменты (например, гуманизированные антитела или химерные антитела), которые получают описанными здесь способами.
Как правило, антитела (также называемые иммуноглобулинами) состоят из двух классов полипептидных цепей, легких цепей и тяжелых цепей. Неограничивающие примеры антител по настоящему изобретению могут представлять собой интактные антитела с четырьмя цепями иммуноглобулинов, включая две тяжелые цепи и две легкие цепи. Тяжелая цепь антитела может относиться к любому изотипу, включая IgM, IgG, IgE, IgA или IgD, или подклассу, включая IgG1, IgG2, IgG2a, IgG2b, IgG3, IgG4, IgE1, IgE2 и т. д. Легкая цепь может быть легкой цепью каппа или легкой цепью лямбда. Антитело может содержать две идентичные копии легкой цепи и две идентичные копии тяжелой цепи. Тяжелые цепи, каждая из которых содержит один вариабельный домен (или вариабельную область, VH) и множество константных доменов (или константных областей), связываются друг с другом посредством дисульфидной связи в своих константных доменах, образуя «ствол» антитела. Легкие цепи, каждая из которых содержит один вариабельный домен (или вариабельную область, VL) и один константный домен (или константную область), каждая связывается с одной тяжелой цепью посредством дисульфидного связывания. Вариабельная область каждой легкой цепи выровнена с вариабельной областью тяжелой цепи, с которой она связана. Вариабельные области как легких цепей, так и тяжелых цепей содержат три гипервариабельные области, расположенные между более консервативными каркасными областями (FR).
Эти гипервариабельные области, известные как области, определяющие комплементарность, (CDR), образуют петли, которые включают основную антигенсвязывающую поверхность антитела. Четыре каркасных области в основном принимают конформацию бета-листа, а CDR образуют петли, соединяющие, а в некоторых случаях образующие часть структуры бета-листа. CDR в каждой цепи удерживаются в непосредственной близости от каркасных областей и, вместе с CDR из другой цепи, способствуют образованию антигенсвязывающей области.
Способы идентификации областей CDR антитела путем анализа аминокислотной последовательности антитела хорошо известны, и обычно используется ряд определений CDR. Определение Kabat основано на вариабельности последовательности, а определение Chothia основано на расположении структурных петлевых областей. Эти методы и определения описаны, например, в публикациях Martin, «Protein sequence and structure analysis of antibody variable domains,» Antibody engineering, Springer Berlin Heidelberg, 2001. 422-439; Abhinandan, et al. «Analysis and improvements to Kabat and structurally correct numbering of antibody variable domains,» Molecular immunology 45. 14 (2008): 3832-3839; Wu, T.T. and Kabat, E.A. (1970) J. Exp. Med. 132: 211-250; Martin et al., Methods Enzymol. 203: 121-53 (1991); Morea et al., Biophys Chem. 68(1-3):9-16 (Oct. 1997); Morea et al., J. Mol Biol. 275(2):269-94 (Jan. 1998 г.); Chothia et al., Nature 342(6252):877-83 (Dec. 1989); Ponomarenko and Bourne, BMC Structural Biology 7:64 (2007); каждая из которых включена в настоящее описание в качестве ссылки в полном объеме.
CDR важны для распознавания эпитопа антигена. Используемый в настоящем описании термин «эпитоп» представляет собой наименьшую часть молекулы-мишени, способную специфически связываться с антигенсвязывающим доменом антитела. Минимальный размер эпитопа может составлять примерно три, четыре, пять, шесть или семь аминокислот, но эти аминокислоты не обязательно должны быть в последовательной линейной последовательности первичной структуры антигена, поскольку эпитоп может зависеть от трехмерной конфигурации на основе вторичной и третичной структуры антигена.
В некоторых вариантах осуществления антитело представляет собой интактную молекулу иммуноглобулина (например, IgG1, IgG2a, IgG2b, IgG3, IgG4, IgM, IgD, IgE, IgA). Подклассы IgG (IgG1, IgG2, IgG3 и IgG4) высококонсервативны, различаются по своей константной области, особенно по своим шарнирам и верхним доменам CH2. Последовательности и различия подклассов IgG известны в данной области и описаны, например, в публикациях Vidarsson, et al, «IgG subclasses and allotypes: from structure to effector functions.» Frontiers in immunology 5 (2014); Irani, et al. «Molecular properties of human IgG subclasses and their implications for designing therapeutic monoclonal antibodies against infectious diseases.» Molecular immunology 67. 2 (2015): 171-182; Shakib, Farouk, ed. The human IgG subclasses: molecular analysis of structure, function and regulation. Elsevier, 2016; каждая из которых включена в настоящее описание в качестве ссылки в полном объеме.
Антитело также может представлять собой молекулу иммуноглобулина, полученную из любых видов (например, человека, грызуна, мыши, крысы, верблюда). Антитела, раскрытые в настоящем описании, также включают, но не ограничиваются ими, поликлональные, моноклональные, моноспецифичные, полиспецифичные антитела и химерные антитела, которые включают связывающий домен иммуноглобулина, слитый с другим полипептидом. Термин «антигенсвязывающий домен» или «антигенсвязывающий фрагмент» представляет собой часть антитела, которая сохраняет специфическую активность связывания интактного антитела, т. е. любая часть антитела, которая способна специфически связываться с эпитопом на молекуле-мишени интактного антитела. Он включает в себя, например, Fab, Fab', F(ab')2 и варианты этих фрагментов. Таким образом, в некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент могут предсталвять собой, например, scFv, Fv, Fd, dAb, биспецифическое антитело, биспецифическое scFv, диантитело, линейное антитело, молекулу одноцепочечного антитела, полиспецифическое антитело, образованное из фрагментов антител, и любой полипептид который включает связывающий домен, который является доменом связывания антитела или гомологичен ему. Неограничивающие примеры антигенсвязывающих доменов включают например, CDR тяжелой цепи и/или легкой цепи интактного антитела, вариабельные области тяжелой и/или легкой цепи интактного антитела, полноразмерные тяжелые или легкие цепи интактного антитела или отдельные CDR любой из тяжелых цепи или легкой цепи интактного антитела.
В некоторых вариантах осуществления антигенсвязывающий фрагмент может образовывать часть химерного антигенного рецептора (CAR). В некоторых вариантах осуществления химерный антигенный рецептор представляет собой слияния одноцепочечных вариабельных фрагментов (scFv), как описано в настоящем документе, слитых с трансмембранным доменом CD3-дзета и эндодоменом.
В некоторых вариантах осуществления scFV имеет один вариабельный домен тяжелой цепи и один вариабельный домен легкой цепи. В некоторых вариантах осуществления scFV имеет два вариабельных домена тяжелой цепи и два вариабельных домена легкой цепи. В некоторых вариантах осуществления scFV имеет две антигенсвязывающие области, и две антигенсвязывающие области могут связываться с соответствующими антигенами-мишенями.
Антитела и их антигенсвязывающие фрагменты (например, гуманизированные антитела или химерные антитела), которые получают описанными здесь способами, имеют различные преимущества. В некоторых вариантах осуществления не требуется дополнительной оптимизации для получения желаемых свойств (таких как например, аффинность связывания, термостабильность и/или ограниченная агрегация).
В некоторых реализациях антитело (или его антигенсвязывающие фрагменты) специфически связывается с мишенью со скоростью диссоциации (koff) меньше 0,1 с-1, меньше 0,01 с-1, меньше 0,001 с-1, меньше 0,0001 с-1 или меньше 0,00001 с-1. В некоторых вариантах осуществления скорость диссоциации (koff) больше 0,01 с-1, больше 0,001 с-1, больше 0,0001 с-1, больше 0,00001 с-1 или больше 0,000001 с-1. В некоторых вариантах осуществления koff большинства этих антител (например, >50%, >60%, >70% или >80%), составляет от 1×10-2/с до 1×10-3/с.
В некоторых вариантах осуществления кинетическая скорость ассоциации (kon) составляет больше 1×102/мс, больше 1×103/мс, больше 1×104/мс, больше 1×105/мс или больше 1×106/мс. В некоторых вариантах осуществления кинетическая скорость ассоциации (kon) составляет меньше 1×105/мс, меньше 1×106/мс или меньше 1×107/мс. В некоторых вариантах осуществления kon большинства этих антител (например, >50%, >60%, >70% или >80%), составляет от 1×105/мс до 1×106/мс.
Аффинности можно вывести из отношения кинетических констант скорости (KD=koff/kon). В некоторых вариантах осуществления KD составляет меньше 1×10-6 М, меньше 1×10-7 М, меньше 1×10-8 М, меньше 1×10-9 М или меньше 1×10-10 М. В некоторых вариантах осуществления KD составляет меньше 50 нМ, 40 нМ, 30 нМ, 20 нМ, 15 нМ, 10 нМ, 9 нМ, 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ или 1 нМ. В некоторых вариантах осуществления KD больше 1×10-7 М, больше 1×10-8 М, больше 1×10-9 М, больше 1×10-10 М, больше 1×10-11 М или больше 1×10-12 М. В некоторых вариантах осуществления антитело связывается с мишенью с KD, меньшей или равной примерно 0,9 нМ, 0,8 нМ, 0,7 нМ, 0,6 нМ, 0,5 нМ, 0,4 нМ, 0,3 нМ, 0,2 нМ или 0,1 нМ. В некоторых вариантах осуществления KD большинства этих антител (например, >50%, >60%, >70% или >80%) находится в пределах от 0,1 нМ до 100 нМ (например, от 1 нМ до 100 нМ).
В некоторых вариантах осуществления средняя длина CDR3 тяжелой цепи антител, продуцируемых животным, как описано в настоящем документе, составляет 13~15 аминокислот. В некоторых вариантах осуществления длина CDR3 тяжелой цепи большинства этих антител (например, >50%, >60%, >70% или >80%) составляет от 9 до 18 аминокислот или от 10 до 17 аминокислот.
В некоторых вариантах осуществления менее 30%, 20% или 15% CDR3 тяжелой цепи в антителах, продуцируемых животным, как описано в настоящем документе, содержат остаток цистеина (например, один остаток цистеина или два остатка цистеина). В некоторых вариантах осуществления CDR3 тяжелой цепи состоит не более чем из 21 аминокислотного остатка.
В некоторых вариантах осуществления определяют термостабильность. Антитела или антигенсвязывающие фрагменты, как описано в настоящем документе, могут иметь Tm больше 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95 °C.
Поскольку IgG можно описать как многодоменный белок, кривая плавления иногда показывает два перехода или три перехода с первой температурой денатурации Tm D1, второй температурой денатурации Tm D2 и, необязательно, третьей температурой денатурации Tm D3.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, как описано в настоящем документе, имеют Tm D1 больше 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95 °C. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, как описано в настоящем документе, имеют Tm D2 больше 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95 °C. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, как описано в настоящем документе, имеют Tm D3 больше 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75., 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95 °C.
В некоторых вариантах осуществления Tm, Tm D1, Tm D2, Tm D3 составляют меньше 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95 °C.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, как описано в настоящем документе, не образуют агрегации, когда температура составляет меньше 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95 °C.
Методы создания генетически модифицированных животных
Генетически модифицированные животные могут быть получены путем введения генов иммуноглобулинов человека в геном животных, отличных от человека, для получения животных, которые могут экспрессировать гуманизированные антитела или химерные антитела. На ФИГ. 1А показаны способы получения гуманизированных животных. В некоторых вариантах осуществления способы сначала включают модификацию локуса иммуноглобулина человека на хромосоме человека. Затем модифицированные хромосомы человека вводят в реципиентную клетку мыши. Затем вариабельную область иммуноглобулина человека вводят в соответствующую область генома мыши путем прямой замены. Затем клетки-реципиенты скринируют. В некоторых вариантах осуществления клетки не содержат хромосомы человека. Затем клетки вводят в бластоцисты для получения химерных мышей. Последующую селекцию можно проводить для получения мышей, содержащих интактный локус гуманизированного иммуноглобулина.
При создании генетически модифицированных животных можно использовать несколько других методов, в том числе, например, негомологичное соединение концов (NHEJ), гомологичная рекомбинация (HR), нуклеазы с цинковыми пальцами (ZFN), эффекторные нуклеазы, подобные активатору транскрипции (TALEN), и короткие палиндромные повторы, регулярно расположенные группами, (CRISPR)-Cas-система. В некоторых вариантах осуществления используется гомологичная рекомбинация. В некоторых вариантах осуществления редактирование генома CRISPR-Cas9 используется для создания генетически модифицированных животных. Многие из этих методов редактирования генома известны в данной области и описаны, например, в публикации Yin et al., «Delivery technologies for genome editing,» Nature Reviews Drug Discovery 16. 6 (2017): 387-399, которая включена посредством ссылки в полном объеме. Также предусмотрено множество других методов, которые можно использовать при редактировании генома, например, микроинъекцию генетически модифицированного ядра в безъядерный ооцит и слияние безъядерного ооцита с другой генетически модифицированной клеткой.
Процесс генетической модификации может включать замену эндогенной последовательности последовательностью человека путем гомологичной рекомбинации. В некоторых вариантах осуществления расщепление выше и ниже целевого сайта (например, с помощью нуклеаз цинковых пальцев, TALEN или CRISPR) может привести к разрыву двухцепочечной ДНК, и гомологичная рекомбинация используется для замены эндогенной последовательности последовательностью человека.
В некоторых вариантах осуществления способы получения генетически модифицированного гуманизированного животного могут включать стадию замены в эндогенном локусе (или сайте) нуклеиновой кислоты (например, области V, D, J или области V, J) соответствующей областью человеческой последовательности. Последовательность может включать область (например, часть или всю область) генов IGHV, IGHD, IGHJ, IGKV и/или IGKJ. В некоторых вариантах осуществления замена опосредована гомологичной рекомбинацией. В некоторых вариантах осуществления замена опосредована Cre-рекомбиназой.
В некоторых вариантах осуществления модификация локуса легкой цепи иммуноглобулина мыши может быть осуществлена напрямую. В некоторых вариантах осуществления вектор используется непосредственно для замены всей вариабельной области легкой цепи иммуноглобулина мыши. В некоторых вариантах осуществления векторы могут быть вставлены выше V-области и между J-областью и C-областью.
В некоторых вариантах осуществления модификация локуса легкой цепи иммуноглобулина мыши может быть осуществлена более чем за одну стадию. В некоторых вариантах осуществления первый вектор можно использовать для замены всей вариабельной области легкой цепи иммуноглобулина мыши. В некоторых вариантах осуществления вектор включает один или более маркеров селекции. В некоторых вариантах осуществления маркер селекции представляет собой доминантный маркер селекции (например, ген устойчивости к неомицину, или Neo). В некоторых вариантах осуществления маркер селекции представляет собой маркер негативной селекции (например, ген рецептора дифтерийного токсина или DTR). В некоторых вариантах осуществления первый вектор содержит один или более доминантных и/или один или более маркеров негативной селекции. В некоторых вариантах осуществления второй вектор можно использовать для дополнительной замены области, включающей один или более маркеров селекции, как описано в настоящем документе.
На ФИГ. 4 показана направленная стратегия для замены эндогенной вариабельной области легкой цепи иммуноглобулина мыши вариабельной областью легкой цепи человека (например, реаранжированной или нереаранжированной последовательностью вариабельной области легкой цепи человека). Например, вариабельная область легкой цепи человека может включать ген IGKV человека, выбранный из IGKV3-20, IGKV3-11 и IGKV1-39; и ген IGKJ человека, выбранный из IGKJ1 и IGKJ4. В некоторых вариантах осуществления вся вариабельная область легкой цепи мыши заменяется первым вектором, содержащим Neo и ген маркера селекции DTR. В некоторых вариантах осуществления второй вектор содержит от 5' до 3' одно или более из следующего: последовательность гомологического плеча ДНК выше сайта вставки (5'-гомологичное плечо); последовательность промотора перед первым нуклеотидом гена IGKV человека (например, в области по меньшей мере или примерно 2000 п.н. перед геном IGKV человека); ген IGKV человека (например, IGKV3-20, IGKV3-11, или IGKV1-39); ген IGKJ человека (например, IGKJ1 или IGKJ4); вспомогательную последовательность (например, последовательность полиА, последовательность WPRE или последовательность 3'UTR); и последовательность гомологического плеча ДНК ниже сайта вставки (3'-гомологичное плечо).
Эти векторы могут быть интегрированы в геном клеток, и клетки могут быть отобраны по маркерам лекарственной устойчивости или их комбинацией (например, зеоцин, G418 и/или пуромицин). В некоторых вариантах осуществления экспрессируется PB-транспозаза, и генетические элементы между последовательностями-мишенями транспозазы могут быть удалены.
В некоторых вариантах осуществления эти векторы интегрированы в модифицированную хромосому человека. Хромосома человека может быть изменена первой, прежде чем первый и второй векторы будут интегрированы в геном. В некоторых вариантах осуществления один или более дополнительных векторов могут быть добавлены в различные участки хромосомы по мере необходимости. В некоторых вариантах осуществления вектор добавляют между С-областью и центромерой.
Человеческая хромосома может быть получена из клеточных линий человека, опухолевых клеток, первичной клеточной культуры и/или фибробластов человека. В некоторых вариантах осуществления клетку человека вводят с первым вектором, а затем сливают с реципиентной клеткой. Затем модифицированную хромосому отделяют и вводят в другую подходящую клетку-реципиент. Клетки с желаемой устойчивостью отбирают для получения клеток, содержащих только одну хромосому человека. Затем в клетки вводят второй вектор и клетки отбирают по резистентности. Затем, если необходимо, можно ввести третий вектор и/или четвертый вектор. Клетка-реципиент может быть клеткой млекопитающего, клеткой человека или клеткой мыши. В некоторых вариантах осуществления клетка-реципиент представляет собой клетку СНО или, предпочтительно, клетку А9. В некоторых вариантах осуществления модифицированные хромосомы метят флуоресценцией и разделяют. А модифицированные хромосомы вводят в клетки-реципиенты путем микроинъекции хромосом. В некоторых вариантах осуществления донорные клетки индуцируют для образования многоядерных хромосом. Затем эти ядра проталкиваются через клеточную мембрану для создания микроклеток, которые можно слить с клеткой-реципиентом. В некоторых вариантах осуществления также можно использовать опосредованный микроклетками перенос хромосом. Методы манипулирования хромосомами описаны, например, в публикациях CN1200014A; CN109837307A; US20120093785A1; и US2009253902; Kuroiwa et al. «Manipulation of human minichromosomes to carry greater than megabase-sized chromosome inserts.» Nature Biotechnology 18. 10 (2000): 1086-1090; китайском патенте CN1717483A; Paulis, Marianna. «Chromosome Transfer Via Cell Fusion.» Methods in Molecular Biology 738(2011):57; Genes, Chromosomes & Cancer 14: 126127 (1995); Tomizuka et al. «Functional expression and germline atransmission of a human chromosome fragment in chimaeric mice.» Nature Genetics 16. 2 (1997): 133-143; Somatic Cell and Molecular Genetics, Vol. 13, No. 3, 1987, pp. 279-284; каждая из которых включена в настоящее описание посредством ссылки в полном объеме.
Последовательности распознавания LoxP также могут быть добавлены в хромосому человека (например, хромосому человека 2, 14, 22). Клетки также можно обработать ферментом Cre, что приведет к рекомбинации сайтов loxP, что приведет к удалению последовательностей геномной ДНК. В некоторых вариантах осуществления спонтанный разрыв хромосомы также можно использовать для удаления последовательностей геномной ДНК.
Вариабельная область иммуноглобулина мыши может быть заменена вариабельной областью иммуноглобулина человека путем замены (например, гомологичной рекомбинацией или Cre-опосредованной рекомбинацией). В некоторых вариантах осуществления для опосредования замены можно использовать Cre-рекомбинацию. В некоторых вариантах осуществления векторы могут добавлять последовательность распознавания LoxP в хромосому человека. Аналогичные модификации могут быть сделаны в хромосоме мыши, где к хромосоме могут быть добавлены две последовательности распознавания LoxP. Например, Cre-рекомбиназа может затем опосредовать замену областей V, J на хромосоме мыши на области V, J на хромосоме человека или замену областей V, D, J на хромосоме мыши на области V, D, J на хромосоме человека.
Клетки могут быть подвергнуты дополнительному скринингу на наличие клеток, не содержащих хромосомы человека (например, с помощью DT). В некоторых случаях клетки, которые не подвергались скринингу с помощью DT, могут содержать рекомбинантные фрагменты хромосом человека, но эти фрагменты малы и нестабильны в клетках мыши (например, Shinohara et al. (2000) Chromosome Research, 8: 713-725), и естественным образом исчезают во время клеточной пролиферации. В некоторых вариантах осуществления удален большой фрагмент модифицированной хромосомы человека, например, посредством Cre-опосредованной делеции или спонтанного хромосомного разрыва.
5'-концевое плечо гомологии и/или 3'-концевое плечо гомологии может иметь желаемую длину для облегчения гомологичной рекомбинации. В некоторых вариантах осуществления плечо гомологии составляет примерно или по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40 или 50 т.п.н. (например, примерно 3 т.п.н.). В некоторых вариантах осуществления плечо гомологии составляет меньше 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40 или 50 т.п.н.
В некоторых вариантах осуществления вектор может также необязательно включать репортерный белок, например. люциферазу (например, Gluc) или флуоресцентный белок (например, EGFP, BFP и т. д.).
Эти модификации могут быть выполнены в различных клетках. В некоторых вариантах осуществления клетка представляет собой стволовую клетку, эмбриональную стволовую клетку или оплодотворенную яйцеклетку.
Настоящее описание дополнительно относится к способу создания модели гуманизированного животного, включающему следующие стадии:
(а) обеспечение клетки (например, оплодотворенной яйцеклетки) на основе описанных здесь способов;
(b) культивирование клетки в жидкой культуральной среде;
(c) трансплантацию культивируемой клетки в фаллопиеву трубу или матку самки млекопитающего-реципиента, отличного от человека, что позволяет клетке развиваться в матке самки млекопитающего, отличного от человека;
(d) идентификацию передачи по зародышевой линии потомству генетически модифицированного гуманизированного млекопитающего, отличного от человека, от беременной самки на стадии (c).
В некоторых вариантах осуществления отличным от человека млекопитающим в вышеуказанном способе является мышь (например, мышь C57, мышь BALB/c или мышь C57BL/6).
В некоторых вариантах осуществления отличное от человека млекопитающее на стадии (с) представляет собой самку с псевдобеременностью (или ложной беременностью).
В некоторых вариантах осуществления оплодотворенные яйцеклетки для способов, описанных выше, представляют собой оплодотворенные яйцеклетки C57BL/6. Другие оплодотворенные яйцеклетки, которые также можно использовать в описанных здесь способах, включают, но не ограничиваются ими, оплодотворенные яйцеклетки FVB/N, оплодотворенные яйцеклетки BALB/c, оплодотворенные яйцеклетки DBA/1 и оплодотворенные яйцеклетки DBA/2.
Оплодотворенные яйцеклетки могут происходить от любого животного, оличного от человека, например, любого животного, отличного от человека, как описано в настоящем документе. В некоторых вариантах осуществления оплодотворенные яйцеклетки происходят от грызунов. Генетическая конструкция может быть введена в оплодотворенную яйцеклетку путем микроинъекции ДНК. Например, путем культивирования оплодотворенной яйцеклетки после микроинъекции культивируемая оплодотворенная яйцеклетка может быть перенесена ложнобеременному животному, отличному от человека, которое затем рождает млекопитающее, отличное от человека, с получением упомянутого млекопитающего, отличного от человека, в описанных выше методах.
Также предложены клетки, ткани и животные (например, мышь), которые содержат нуклеотидные последовательности, как описано в настоящем документе, а также клетки, ткани и животные (например, мышь), которые экспрессируют гуманизированные или химерные антитела из эндогенного локуса, не относящегося к человеку.
Настоящее описание также относится к различным направленным векторам (например, векторам, которые можно использовать для создания генетически модифицированных животных). В некоторых вариантах осуществления вектор может содержать: а) фрагмент ДНК, гомологичный 5'-концу области, подлежащей изменению (5'-плечо гомологии); b) последовательность, включающая желаемые генетические элементы (например, сайт распознавания LoxP, гены лекарственной устойчивости и/или репортерные гены и т. д.); и c) второй фрагмент ДНК, гомологичный 3'-концу изменяемой области (3'-плечо гомологии). Описание также относится к клетке, содержащей направленные векторы, как описано в настоящем документе.
В некоторых вариантах осуществления гены в клетке являются гетерозиготными. В некоторых вариантах осуществления гены в клетке являются гомозиготными.
В некоторых вариантах осуществления клетка млекопитающего, отличного от человека, представляет собой клетку мыши. В некоторых вариантах осуществления клетка представляет собой оплодотворенную яйцеклетку.
Настоящее описание дополнительно относится к способам создания генетически модифицированной животной модели с двумя или более человеческими или химерными генами. Животное может содержать один или более локусов человеческого или гуманизированного иммуноглобулина и последовательность, кодирующую дополнительный человеческий или химерный белок. В некоторых вариантах осуществления дополнительный человеческий или химерный белок может представлять собой белок 1 запрограммированной клеточной смерти (PD-1), цитотоксический Т-лимфоцит-ассоциированный белок 4 (CTLA-4), белок 3 гена активации лимфоцитов (LAG-3), белок, ассоциированный с В- и Т-лимфоцитами (BTLA), Лиганд 1 белка 1 запрограммированной клеточной смерти (PD-L1), CD27, CD28, CD47, CD137, CD154, Т-клеточный иммунорецептор с доменами Ig и ITIM (TIGIT), Т-клеточный иммуноглобулин и муциновый домен-3 (TIM-3), глюкокортикоид-индуцируемый белок, родственный TNFR (GITR), или член 4 суперсемейства рецепторов TNF (TNFRSF4 или OX40).
Способы создания генетически модифицированных животных моделей с дополнительными человеческими или химерными генами (например, гуманизированными генами) могут включать следующие стадии:
(а) использование описанных здесь способов для получения генетически модифицированного животного, отличного от человека;
(b) скрещивание генетически модифицированного животного, отличного от человека, с другим генетически модифицированным животным, отличным от человека, и затем скрининг потомства для получения генетически модифицированного животного, отличного от человека, с двумя или более человеческими или химерными генами.
В некоторых вариантах осуществления на стадии (b) способа генетически модифицированное животное можно скрещивать с генетически модифицированным животным, отличным от человека, с человеческими или химерными PD-1, CTLA-4, LAG-3, BTLA, PD-L1, CD27, CD28, CD47, CD137, CD154, TIGIT, TIM-3, GITR, SIRPa или OX40. Описаны некоторые из этих генетически модифицированных животных, отличных от человека, например, в документах PCT/CN2017/090320, PCT/CN2017/099577, PCT/CN2017/099575, PCT/CN2017/099576, PCT/CN2017/099574, PCT/CN2017/106024, PCT/CN2017/110494, PCT/CN2017/110435, PCT/CN2017/120388, PCT/CN2018/081628, PCT/CN2018/081629; каждый из которых включен в настоящее описание посредством ссылки в полном объеме.
Настоящее описание дополнительно относится к способам получения нокаутного животного. В некоторых вариантах осуществления, если животное (например, мышь) экспрессирует белок, очень похожий на интересующий антиген, то может быть трудно вызвать иммунный ответ у животного. Это связано с тем, что во время созревания иммунных клеток В-клетки и Т-клетки, которые распознают молекулы MHC, связанные с пептидами собственного происхождения, удаляются из репертуара иммунных клеток. В этих случаях геннетически сконструированное животное может быть дополнительно модифицировано. Соответствующий ген у животного может быть нокаутирован, после чего животное подвергается воздействию интересующего антигена. Поскольку животное не подвергается отрицательной селекции в отношении продукта гена, животное может генерировать антитело, которое может легко специфически связываться с мишенью. Таким образом, в некоторых вариантах осуществления в описании также предложены способы нокаута интересующего гена у генетически модифицированных животных. В некоторых вариантах осуществления ген может быть нокаутирован с помощью различных известных методов редактирования генов, например, системы CRISPR-Cas, TALEN или ZFN. В некоторых вариантах осуществления генетически модифицированное животное можно скрещивать с другим животным, у которого нокаутирован интересующий ген. Как только животное с нокаутным фенотипом создано, оно может подвергаться воздействию интересующего антигена для выработки антитела (например, антитела с общей легкой цепью). В некоторых вариантах осуществления интересующий антиген представляет собой белок человека. В некоторых вариантах осуществления интересующий антиген представляет собой PD-1, CTLA-4, LAG-3, BTLA, PD-L1, CD27, CD28, CD47, CD137, CD154, TIGIT, TIM-3, GITR, SIRPa или OX40.
В некоторых вариантах осуществления генетически модифицированные животные могут иметь человеческий ген ADAM6, эндогенный ген ADAM6 или модифицированный ген ADAM6 (например, в его эндогенном локусе). Белок ADAM6 является членом семейства белков ADAM, где ADAM является аббревиатурой от «A Disintegrin And Metalloprotease». Ген ADAM6 человека, который обычно находится между генами IGHV человека IGHV1-2 и IGHV6-1, представляет собой псевдоген (ФИГ. 16). У мышей есть два гена ADAM6, ADAM6a и ADAM6b. Они расположены в межгенной области между кластерами генов IGHV и IGHD мыши. ADAM6a мыши расположен между IGHV5-1 мыши и IGHD5-1 мыши. ADAM6b мыши расположен между IGHD3-1 мыши и IGHD1-1 мыши. Таким образом, в некоторых вариантах осуществления генетически модифицированные животные могут иметь ген ADAM6 человека. В некоторых вариантах осуществления генетически модифицированные животные не имеют эндогенного гена ADAM6.
В некоторых вариантах осуществления генетически модифицированные животные представляют собой мышей. В некоторых вариантах осуществления мыши модифицированы для включения нуклеотидной последовательности, кодирующей белок ADAM6 (например, ADAM6a или ADAM6b). В некоторых вариантах осуществления последовательность находится в любом подходящем положении. Его можно поместить в межгенную область или в любое подходящее положение в геноме. В некоторых вариантах осуществления нуклеиновая кислота кодирует последовательность, которая по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91% 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или на 100% идентична гену ADAM6a мыши (например, 113539230-113547024 NC_000078.6) или гену ADAM6b мыши (например, 113486188-113492125 NC_000078.6). В некоторых вариантах осуществления нуклеиновая кислота дополнительно включает регуляторные элементы для гена ADAM6a и гена ADAM6b (например, промоторы).
В некоторых вариантах осуществления функциональный локус ADAM6 мыши может быть помещен посреди кластера генов IGHV человека. В некоторых вариантах осуществления локус ADAM6 мыши находится между двумя генами IGVH человека. В некоторых вариантах осуществления псевдоген ADAM6 человека между VH1-2 человека и VH(II)-1-1 человека заменен локусом ADAM6 мыши. В некоторых вариантах осуществления ген ADAM6a и ген ADAM6b расположены между IGHV1-2 человека и VH(II)-1-1 человека в геноме животного. В некоторых вариантах осуществления расположение последовательности ADAM6 мыши в последовательности гена человека может приблизительно соответствовать положению псевдогена ADAM6 человека или может приблизительно соответствовать положению последовательности ADAM6 мыши (например, внутри межгенной области V-D). В некоторых вариантах осуществления генетически модифицированные мыши имеют гуманизированный локус тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления ADAM6a мыши и ADAM6b мыши расположены между генами IGHV1-2 и IGHV6-1 человека. Размещение ADAM6 мыши a и ADAM6 мыши b между генами IGHV1-2 и IGHV6-1 человека может иметь различные преимущества. Например, поскольку эти гены заменяют ген ADAM6 человека в том же локусе, вполне вероятно, что замена гена ADAM6 человека будет иметь ограниченное влияние на VDJ-рекомбинацию, а гены ADAM6a и ADAM6b мыши также могут функционировать должным образом (как в расположении, подобном эндогенному локусу).
Таким образом, в одном аспекте изобретение относится к генетически модифицированному животному, содержащему в эндогенном локусе генов тяжелой цепи иммуноглобулина первую последовательность, содержащую один или более генов IGHV человека; вторую последовательность, содержащую ген ADAM6; и третью последовательность, содержащую один или более генов IGHD человека и один или более генов IGHJ человека. В некоторых вариантах осуществления первая последовательность, вторая последовательность и третья последовательность функционально связаны.
В некоторых вариантах осуществления первая последовательность включает все гены IGHV человека, представленные в Таблице 5, за исключением IGHV2-10, IGHV3-9, IGHV1-8, IGHV(II)-1-1 и IGHV6-1. В некоторых вариантах осуществления первая последовательность включает все гены IGHV человека, представленные в Таблице 5, за исключением IGHV5-10-1 и IGHV3-64D, IGHV(II)-1-1 и IGHV6-1. В некоторых вариантах осуществления первая последовательность содержит IGHV5-10-1 и IGHV3-64D. В некоторых вариантах осуществления первая последовательность представляет собой немодифицированную последовательность, полученную из локуса генов тяжелой цепи иммуноглобулина человека.
В некоторых вариантах осуществления вторая последовательность содержит либо один, либо оба из гена ADAM6a мыши и гена ADAM6b мыши. В некоторых вариантах осуществления животное представляет собой фертильного самца мыши. В некоторых вариантах осуществления вторая последовательность не содержит гена ADAM6a мыши или гена ADAM6b мыши.
В некоторых вариантах осуществления третья последовательность содержит все гены IGHD человека, представленные в Таблице 6, и все гены IGHJ человека, представленные в Таблице 7. В некоторых вариантах осуществления третья последовательность содержит IGHV6-1 человека. В некоторых вариантах осуществления третья последовательность содержит IGHV(II)-1-1 человека. В некоторых вариантах осуществления третья последовательность представляет собой немодифицированную последовательность, полученную из локуса генов тяжелой цепи иммуноглобулина человека.
В некоторых вариантах осуществления AMAM6a и/или ADAM6b представляют собой эндогенные последовательности. В некоторых вариантах осуществления AMAM6a и/или ADAM6b не заменены и/или расположены в своем эндогенном или нативном положении. В некоторых вариантах осуществления гены IGHV мыши перед IGHV1-2 мыши в локусе вариабельной области тяжелой цепи заменены генами IGHV человека. В некоторых вариантах осуществления гены IGHV, IGHD и IGHJ мыши после IGHV6-1 мыши в локусе вариабельной области тяжелой цепи заменены одним или более генами IGHV, IGHD и/или генами IGHJ человека.
Таким образом, в некоторых вариантах осуществления гены IGHV, IGHD и IGHJ мыши могут быть заменены IGHV, IGHD и IGHJ человека путем более чем одной замены. На первой стадии выбранное количество генов IGHV мыши на 5'-стороне ADAM6a (например, все гены IGHV мыши, представленные в Таблице 8) заменены генами IGHV человека. На второй стадии выбранное количество генов IGHD и IGHJ мыши на 3'-стороне ADAM6b (например, все гены IGHD мыши, представленные в Таблице 9, кроме IGHD5-1 и IGHD3-1, и все гены IGHJ, представленные в Таблице 10) заменены генами IGHD человека и генами IGHJ человека. Замена может быть осуществлена путем гомологичной рекомбинации или Cre-опосредованной рекомбинации.
В некоторых вариантах осуществления мыши не имеют генов ADAM6a или ADAM6b мыши. В некоторых вариантах осуществления мыши имеют гены ADAM6 человека.
Для повышения фертильности мышей можно использовать различные методы. В некоторых вариантах осуществления для спаривания можно использовать самок мышей с суперовуляцией. В некоторых вариантах осуществления можно использовать экстракорпоральное оплодотворение. Суперовуляцию можно вызвать введением сывороточного гонадотропина и хорионического гонадотропина (например, CG человека или мыши) половозрелой самке мыши. Взрослых самцов мышей можно умертвить и выделить их cauda epididymides. Проток cauda epididymides разрезают, чтобы высвободить сперму. Затем можно умертвить суперовулирующую зрелую самку мыши и выделить маточные трубы. Комплексы кумулюс-ооцит (COC) могут быть выделены из маточных труб. Затем в COC можно добавить суспензию сперматозоидов и инкубировать для инсеминации. Партеногенетические ооциты, содержащие только один пронуклеус, могут быть удалены. После инкубации эмбрионы на стадии 2 клеток могут быть перенесены самкам-реципиентам. Способы повышения фертильности мышей известны в данной области.
Описание также относится к последовательности нуклеиновой кислоты, которая по меньшей мере на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%., 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентична любой нуклеотидной последовательности, как описано в настоящем документе, и к аминокислотной последовательности, которая по меньшей мере на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентична любой аминокислотной последовательности, как описано в настоящем документе.
В некоторых вариантах осуществления описание относится к нуклеотидным последовательностям, кодирующим любые пептиды, которые описаны в настоящем документе, или любым аминокислотным последовательностям, которые кодируются любыми нуклеотидными последовательностями, описанными в настоящем документе. В некоторых вариантах осуществления последовательность нуклеиновой кислоты составляет менее чем 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 150, 200, 250, 300, 350, 400, 500 или 600 нуклеотидов. В некоторых вариантах осуществления аминокислотная последовательность составляет менее чем 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150., 160, 170, 180, 190, 200, 250, 300, 350 или 400 аминокислотных остатков.
В некоторых вариантах осуществления аминокислотная последовательность (i) содержит аминокислотную последовательность; или (ii) состоит из аминокислотной последовательности, где аминокислотная последовательность представляет собой любую из описанных в настоящем документе последовательностей.
В некоторых вариантах осуществления последовательность нуклеиновой кислоты (i) содержит последовательность нуклеиновой кислоты; или (ii) состоит из последовательности нуклеиновой кислоты, где последовательность нуклеиновой кислоты представляет собой любую из описанных в настоящем документе последовательностей.
Чтобы определить процент идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновых кислот, последовательности выравнивают для оптимальных целей сравнения (например, пробелы могут быть введены в одну или обе из первой и второй последовательностей аминокислот или нуклеиновых кислот для оптимального выравнивания, а негомологичные последовательности могут быть проигнорированы для целей сравнения). Затем сравниваются аминокислотные остатки или нуклеотиды в соответствующих положениях аминокислот или нуклеотидов. Если положение в первой последовательности занято тем же аминокислотным остатком или нуклеотидом что и в соответствующем положении второй последовательности, то молекулы являются идентичными по этому положению. Процент идентичности между двумя последовательностями является функцией числа идентичных положений совместно с последовательностями, принимая во внимание число пробелов и длину каждого пробела, которые должны появляться для оптимального выравнивания двух последовательностей. В иллюстративных целях сравнение последовательностей и определение процента идентичности между двумя последовательностями можно выполнить, например, с помощью матрицы оценки Blossum 62 со штрафом за пробел 12, штрафом за расширение пробела 4 и штрафом за сдвиг пробела 5.
Процент остатков, консервативных по сходным физико-химическими свойствам (процент гомологии), например, лейцина и изолейцина также можно использовать для измерения сходства последовательностей. Семейства аминокислотных остатков, обладающих сходными физико-химическими свойствами, определены в данной области. К этим семействам относятся, например, аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), с кислыми боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), с незаряженными полярные боковыми цепями (например, глицин, аспарагин, глютамин, серин, треонин, тирозин, цистеин), с неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), с бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и с ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Процент гомологии во многих случаях выше, чем процент идентичности. Таким образом, настоящее описание также относится к аминокислотной последовательности, которая имеет по меньшей мере 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% процентов гомологии с любой аминокислотной последовательностью, как описано в настоящем документе, или с нуклеиновой кислотой, кодирующей эти аминокислотные последовательности.
Способы использования генетически модифицированных животных
Генетически модифицированных животных можно использовать для создания гуманизированных или химерных антител, которые могут специфически связываться с мишенью. В некоторых вариантах осуществления мишень (например, белок или фрагмент белка) можно использовать в качестве иммуногена для получения антител у этих животных с использованием стандартных способов получения поликлональных и моноклональных антител. В некоторых вариантах осуществления генетически модифицированное животное подвергается воздействию выбранного антигена в течение времени и в условиях, которые позволяют животному продуцировать антитело, специфичное к антигену.
Поликлональные антитела могут быть получены у животных путем многократных инъекций (например, подкожных или внутрибрюшинных инъекций) антигенного пептида или белка. В некоторых вариантах осуществления антигенный пептид или белок инъецируют по меньшей мере с одним адъювантом. В некоторых вариантах осуществления антигенный пептид или белок можно конъюгировать с агентом, который является иммуногенным для видов, которые должны быть иммунизированы. Животным можно вводить антигенный пептид или белок больше одного раза (например, два, три или четыре раза).
Можно использовать полноразмерный полипептид или белок или, в качестве альтернативы, можно использовать их антигенные пептидные фрагменты в качестве иммуногенов. Антигенный пептид белка содержит по меньшей мере 8 (например, по меньшей мере, 10, 15, 20 или 30) аминокислотных остатков аминокислотной последовательности и включает эпитоп белка, так что антитело, индуцированное против пептида, образует специфический иммунный комплекс с белком.
Иммуноген обычно используется для получения антител путем иммунизации подходящего объекта (например, генетически модифицированного животного, как описано в настоящем документе). Подходящий иммуногенный препарат может содержать, например, рекомбинантно экспрессируемый или химически синтезированный полипептид (например, фрагмент белка). Препарат может дополнительно включать адъювант, такой как полный или неполный адъювант Фрейнда, или подобный иммуностимулирующий агент.
Поликлональные антитела можно получить, как описано выше, путем иммунизации подходящего объекта полипептидом или его антигенным пептидом (таким как например, часть белка) в качестве иммуногена. Титр антител у иммунизированного объекта можно отслеживать с течением времени с помощью стандартных методов, таких как твердофазный иммуноферментный анализ (ИФА) с использованием иммобилизованного полипептида или пептида. При желании молекулы антитела могут быть выделены из млекопитающего (например, из крови) и далее очищены хорошо известными методами, такими как хроматография на протеине A или протеине G, для получения фракции IgG. В надлежащее время после иммунизации, например, когда титры специфических антител самые высокие, клетки, продуцирующие антитела, могут быть получены от объекта и использованы для получения моноклональных антител стандартными методами, такими как метод гибридомы, первоначально описанный Kohler et al. (Nature 256: 495-497, 1975), метод гибридомы B-клеток человека (Kozbor et al., Immunol. Today 4:72, 1983), метод EBV-гибридомы (Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96, 1985), или методы триомы. Технология получения гибридом хорошо известна (см., как правило, Current Protocols in Immunology, 1994, Coligan et al. (Eds.), John Wiley & Sons, Inc., New York, NY). Клетки гибридомы, продуцирующие моноклональное антитело, детектируют путем скрининга супернатантов гибридомной культуры на наличие антител, которые связывают интересующий полипептид или эпитоп, например, с использованием стандартного анализа ИФА.
В одном аспекте настоящее описание относится к мыши, содержащей модификацию эндогенного локуса тяжелой цепи иммуноглобулина, где мышь продуцирует В-клетку, содержащую реаранжированную последовательность иммуноглобулина, функционально связанную с последовательностью гена константной области тяжелой цепи. В некоторых вариантах осуществления реаранжированная последовательность иммуноглобулина, функционально связанная с последовательностью гена константной области тяжелой цепи, содержит последовательность V, D и/или J тяжелой цепи человека. В некоторых вариантах осуществления последовательность гена константной области тяжелой цепи содержит последовательность тяжелой цепи человека или мыши, выбранную из группы, состоящей из СН1, шарнира, СН2, СН3 и их комбинации.
В одном аспекте настоящее изобретение относится к мыши, содержащей модификацию эндогенной легкой цепи иммуноглобулина (например, каппа или лямбда), где мышь продуцирует В-клетку, содержащую реаранжированную последовательность иммуноглобулина, функционально связанную с последовательностью гена константной области легкой цепи. В некоторых вариантах осуществления реаранжированная последовательность иммуноглобулина, функционально связанная с последовательностью гена константной области легкой цепи, содержит последовательность V и/или J легкой цепи человека. В некоторых вариантах осуществления последовательность гена константной области легкой цепи содержит константную область легкой цепи человека или мыши.
В-клетки мыши или клетки селезенки могут содержать реаранжированную последовательность гена вариабельной области иммуноглобулина, не относящегося к мыши, например, функционально связанную с геном константной области иммуноглобулина мыши. Определены последовательности для кодирования вариабельной области тяжелой цепи человека и вариабельной области легкой цепи человека. Последовательности могут быть определены, например, путем секвенирования интересующей гибридомы или В-клеток. В некоторых вариантах осуществления используется скрининг отдельных В-клеток. Он может скринировать природный репертуар антител без необходимости слияния гибридом и комбинаторного дисплея. Например, В-клетки можно смешать с панелью антигенов со штрих-кодом ДНК, так что последовательности штрих-кода (штрих-кодов) антигена и рецептора В-клеток (BCR) отдельных В-клеток восстанавливаются с помощью протоколов секвенирования отдельных клеток.
Антитела можно дополнительно модифицировать для получения гуманизированного антитела или человеческого антитела, например, путем функционального связывания последовательности, кодирующей вариабельную область тяжелой цепи человека, с последовательностью, кодирующей константную область тяжелой цепи человека, и/или функционального связывания последовательности, кодирующей вариабельную область легкой цепи человека, с последовательностью, кодирующей константную область легкой цепи человека.
В описании также предложены способы получения антител, нуклеиновых кислот, клеток, тканей (например, ткани селезенки). В некоторых вариантах осуществления способы включают экспонирование животного, описанного в настоящем документе, с антигеном. Антитела (например, гибридные антитела), нуклеиновые кислоты, кодирующие антитела, клетки и/или ткани (например, ткань селезенки) могут быть получены от животного. В некоторых вариантах осуществления нуклеиновые кислоты, кодирующие вариабельные области тяжелой и легкой цепи иммуноглобулина человека, определяют, например, путем секвенирования. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая вариабельную область тяжелой цепи иммуноглобулина человека, может быть функционально связана с нуклеиновой кислотой, кодирующей константную область тяжелой цепи иммуноглобулина человека. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая вариабельную область легкой цепи иммуноглобулина человека, может быть функционально связана с нуклеиновой кислотой, кодирующей константную область легкой цепи иммуноглобулина человека. В некоторых вариантах осуществления клетки, содержащие нуклеиновые кислоты, как описано в настоящем документе, культивируют и антитела собирают.
В некоторых вариантах осуществления отсутствие генов V, D, J иммуноглобулина мыши (например, генов IGHV, IGHD, IGHJ, IGKV или IGKJ мыши) вносит вклад в последовательность вариабельной области тяжелой цепи и/или легкой цепи. В некоторых вариантах осуществления последовательности вариабельной области тяжелой цепи и/или легкой цепи, продуцируемые животным, являются полностью человеческими и полностью обеспечиваются генами V, D, J иммуноглобулина человека (например, генами IGHV, IGHD, IGHJ, IGKV и IGKJ человека).
Варианты антител или антигенсвязывающих фрагментов, описанных в настоящем документе, могут быть получены путем введения соответствующих нуклеотидных изменений в ДНК, кодирующую человеческое, гуманизированное или химерное антитело, или его антигенсвязывающий фрагмент, описанный в настоящем документе, или путем пептидного синтеза. Такие варианты включают, например, делеции, вставки или замены остатков в аминокислотных последовательностях, которые составляют антигенсвязывающий сайт антитела или антигенсвязывающий домен. В популяции таких вариантов некоторые антитела или антигенсвязывающие фрагменты будут иметь повышенную аффинность к белку-мишени. Любая комбинация делеций, вставок и/или комбинации может быть сделана для получения антитела или его антигенсвязывающего фрагмента, которое обладает повышенной аффинностью связывания с мишенью. Аминокислотные изменения, введенные в антитело или антигенсвязывающий фрагмент, также могут изменять или вводить новые посттрансляционные модификации в антитело или антигенсвязывающий фрагмент, такие как изменение (например, увеличение или уменьшение) количества сайтов гликозилирования, изменение типа сайта гликозилирования (например, изменение аминокислотной последовательности таким образом, что ферменты, присутствующие в клетке, присоединяют другой сахар) или введение новых сайтов гликозилирования.
Антитела, раскрытые в настоящем описании, могут быть получены из любых видов животных, включая млекопитающих. Неограничивающие примеры нативных антител включают антитела, полученные с использованием человека, приматов, например, обезьян и человекообразных обезьян, коров, свиней, лошадей, овец, верблюдовых (например, верблюдов и лам), кур, коз и грызунов (например, крыс, мышей, хомяков и кроликов), включая трансгенных грызунов, генетически сконструированных для выработки человеческих антител.
Человеческие и гуманизированные антитела включают антитела, имеющие вариабельные и константные области, полученные из последовательностей иммуноглобулина зародышевой линии человека (или имеющие ту же аминокислотную последовательность, что и у полученной последовательности). Человеческие антитела могут включать аминокислотные остатки, не кодируемые последовательностями человеческого иммуноглобулина зародышевой линии (например, мутации, введенные случайным или сайт-специфическим мутагенезом in vitro или соматической мутацией in vivo), например, в CDR.
Могут быть сделаны дополнительные модификации антител или антигенсвязывающих фрагментов. Например, остаток(остатки) цистеина может быть введен в Fc-область, тем самым обеспечивая образование межцепочечной дисульфидной связи в этой области. Полученное таким образом гомодимерное антитело может иметь любое увеличенное время полужизни in vitro и/или in vivo. Гомодимерные антитела с увеличенным периодом полужизни in vitro и/или in vivo также могут быть получены с использованием гетеробифункциональных сшивающих агентов, как описано, например, в Wolff et al. (Cancer Res. 53: 2560-2565, 1993). Альтернативно, можно сконструировать антитело, которое имеет двойные Fc-области (см., например, Stevenson et al., Anti-CancerDrug Design 3:219-230, 1989).
В некоторых вариантах осуществления можно осуществить ковалентную модификацию антитела или его антигенсвязывающего фрагмента. Эти ковалентные модификации могут быть сделаны химическим или ферментативным синтезом или ферментативным или химическим расщеплением. Другие типы ковалентных модификаций антитела или фрагмента антитела вводятся в молекулу путем взаимодействия целевых аминокислотных остатков антитела или фрагмента с органическим модифицирующим агентом, который способен реагировать с выбранными боковыми цепями или N- или C-концевыми остатками.
Способы получения биспецифических антител с общими легкими цепями
В настоящем изобретении предложено генетически сконструированное животное, которое экспрессирует ограниченный набор легких цепей, которые могут быть связаны с разнообразными тяжелыми цепями. В различных вариантах осуществления эндогенные гены вариабельной области легкой цепи каппа удалены и заменены одним, двумя, тремя, четырьмя или пятью генами вариабельной области легкой цепи человека, функционально связанными с эндогенным геном константной области цепи каппа. В различных вариантах осуществления животное также содержит нефункциональный локус легкой цепи лямбда или его делецию или делецию, которая делает локус неспособным образовывать легкую цепь лямбда.
В некоторых вариантах осуществления животное содержит локус вариабельной области легкой цепи, в котором отсутствует эндогенный ген вариабельной области легкой цепи и который содержит реаранжированную последовательность V/J человека, функционально связанную с эндогенной константной областью, и где локус экспрессирует легкую цепь, содержащую последовательность V/J человека, связанную с эндогенной константной областью.
В различных вариантах осуществления генетически сконструированное животное при иммунизации интересующим антигеном генерирует В-клетки, которые демонстрируют разнообразие реаранжировок вариабельных областей тяжелой цепи иммуноглобулина человека, которые экспрессируются и функционируют с ограниченным числом (например, 1, 2, 3, 4, 5) реаранжированных легких цепей, включая варианты осуществления, в которых одна или две легкие цепи содержат вариабельные области легкой цепи человека, которые включают, например, от 1 до 5 соматических мутаций. В некоторых вариантах осуществления по меньшей мере 50%, 60%, 70%, 80%, 90% или 95% вариабельных областей легкой цепи человека имеют по меньшей мере 1 соматическую мутацию. В некоторых вариантах осуществления по меньшей мере 50%, 60%, 70%, 80%, 90% или 95% вариабельных областей легкой цепи человека имеют по меньшей мере 2 соматические мутации. В некоторых вариантах осуществления по меньшей мере 50%, 60%, 70%, 80%, 90% или 95% вариабельных областей легкой цепи человека имеют по меньшей мере 3 соматические мутации.
В некоторых вариантах осуществления степень соматических гипермутаций относительно низкая. В некоторых вариантах осуществления не больше 30% или 40% вариабельных областей легкой цепи человека имеют одну или более чем одну соматическую мутацию. В некоторых вариантах осуществления не больше 20% вариабельных областей легкой цепи человека имеют две или более двух соматических мутаций.
В некоторых вариантах осуществления животное, как описано в настоящем документе, может быть иммунизировано первым иммуногеном для получения В-клетки, которая экспрессирует антитело, которое специфически связывается с первым эпитопом. Животное можно иммунизировать вторым иммуногеном для получения В-клетки, экспрессирующей антитело, которое специфически связывается со вторым эпитопом. Вариабельные области тяжелой цепи могут быть клонированы из В-клеток в вектор, который используется для трансфекции клетки для экспрессии реаранжированной вариабельной области тяжелой цепи человека, слитой с константной областью тяжелой цепи человека, и общей вариабельной области легкой цепи, слитой с константной областью легкой цепи человека.
В некоторых вариантах осуществления описанные здесь способы предназначены для получения биспецифического антитела. Биспецифические антитела можно получить, создав интерфейс между парой молекул антител, чтобы максимизировать процент гетеродимеров, выделенных из рекомбинантной клеточной культуры. Например, интерфейс может содержать, по меньшей мере, часть домена СН3 константного домена антитела. В этом способе одну или более небольших боковых цепей аминокислот из интерфейса первой молекулы антитела заменяют более крупными боковыми цепями (как например, у тирозина или триптофана). Компенсаторные «полости» идентичного или сходного размера с крупной боковой цепью (цепями) создаются в интерфейсе второй молекулы антитела путем замены крупных боковых цепей аминокислот более мелкими (как например, у аланина или треонина). Это обеспечивает механизм повышения выхода гетеродимера по отношению к другим нежелательным конечным продуктам, таким как гомодимеры. Этот метод описан, например, в WO 96/27011, которая включена в качестве ссылки в полном объеме.
В некоторых вариантах осуществления можно использовать технологию «выступ-во-впадину» (KIH), которая включает конструирование доменов CH3 для создания либо «выступа», либо «впадины» в каждой тяжелой цепи для стимулирования гетеродимеризации. Технология KIH описана, например, в публикации Xu, Yiren, et al. «Production of bispecific antibodies in 'knobs-into-holes' using a cell-free expression system.» MAbs. Vol. 7. No. 1. Taylor & Francis, 2015, которая включена в качестве ссылки в полном объеме. В некоторых вариантах осуществления одна тяжелая цепь имеет замену T366W и/или S354C (выступ) (нумерация EU), а другая тяжелая цепь имеет замену Y349C, T366S, L368A и/или Y407V (впадина) (нумерация EU). В некоторых вариантах осуществления одна тяжелая цепь имеет одну или более из следующих замен Y349C и T366W (нумерация EU). Другая тяжелая цепь может иметь одну или более следующих замен E356C, T366S, L368A и Y407V (нумерация EU). В некоторых вариантах осуществления одна тяжелая цепь содержит замену T366Y (выступ), а другая тяжелая цепь содержит одну, две или три из этих замен T366S, L368A, Y407V (впадина).
Кроме того, для очистки биспецифических антител можно использовать анионообменную хроматографию. Анионообменная хроматография - это процесс разделения веществ на основе их заряда с использованием ионообменной смолы, содержащей положительно заряженные группы, такие как диэтиламиноэтиловые группы (DEAE). В растворе смола покрыта положительно заряженными противоионами (катионами). Анионообменные смолы связываются с отрицательно заряженными молекулами, вытесняя противоион. Анионообменная хроматография может использоваться для очистки белков на основе их изоэлектрической точки (pI). Изоэлектрическая точка определяется как рН, при котором белок не имеет суммарного заряда. Когда pH > pI, белок имеет суммарный отрицательный заряд, а когда pH < pI, белок имеет суммарный положительный заряд. Таким образом, в некоторых вариантах осуществления в две тяжелые цепи могут быть введены разные аминокислотные замены, так что pI для гомодимера, включающего два плеча A, и pI для гомодимера, включающего два плеча B, различаются. Используемый в настоящем описании термин «плечо» в биспецифическом антителе относится к антигенсвязывающему сайту в биспецифическом антителе, который специфически связывается с конкретным антигеном (или эпитопом). pI для биспецифического антитела, имеющего плечо A и плечо B, будет где-то между двумя pI гомодимеров. Таким образом, два гомодимера и биспецифическое антитело могут высвобождаться при различных условиях рН. Настоящее описание показывает, что несколько замен аминокислотных остатков могут быть введены в тяжелые цепи для корректировки pI. Таким образом, в некоторых вариантах осуществления аминокислотный остаток в VH в положении 83 по Kabat представляет собой лизин, аргинин или гистидин. В некоторых вариантах осуществления аминокислотные остатки в VH в одном или более положениях 1, 6, 43, 81 и 105 (нумерация по Kabat) представляют собой аспарагиновую кислоту или глутаминовую кислоту. В некоторых вариантах осуществления аминокислотные остатки в одном или более положениях 13 и 105 (нумерация по Kabat) представляют собой аспарагиновую кислоту или глутаминовую кислоту. В некоторых вариантах осуществления аминокислотные остатки в одном или более положениях 13 и 42 (нумерация по Kabat) представляют собой лизин, аргинин, гистидин или глицин.
ПРИМЕРЫ
ПРИМЕР 1: Обзор
Были проведены эксперименты по введению генов иммуноглобулина человека в геном мыши для получения мышей, экспрессирующих гуманизированные антитела. На ФИГ. 1А показаны способы получения гуманизированных мышей. Способы включали сначала модификацию области иммуноглобулина человека на хромосоме человека. Затем модифицированные хромосомы человека вводили в реципиентную клетку мыши.
Вариабельную область иммуноглобулина мыши заменяли вариабельной областью иммуноглобулина человека путем прямой замены (например, гомологичной рекомбинацией или Cre-опосредованной рекомбинацией). В некоторых случаях вариабельная область иммуноглобулина человека может быть введена в геном мыши поэтапно. Затем клетки-реципиенты проверяли на корректность замены. Затем клетки вводили в бластоцисты для получения химерных мышей. Последующее разведение проводили для получения мышей, содержащих вариабельные области человеческого или гуманизированного иммуноглобулина.
Поскольку ген тяжелой цепи мыши и два гена легкой цепи расположены на хромосомах 12, 6 и 16, соответственно, мышей, содержащих вариабельную область тяжелой цепи человека или вариабельную область легкой цепи человека, можно получить отдельно. Затем этих мышей можно скрещивать друг с другом для получения мышей, которые могут экспрессировать как вариабельный домен тяжелой цепи человека, так и вариабельный домен общей легкой цепи человека.
ПРИМЕР 2: Модификация локуса тяжелой цепи иммуноглобулина мыши
Локус тяжелой цепи иммуноглобулина расположен на хромосоме 12 мыши. Два сайта рекомбинации вводили по обеим сторонам вариабельной области локуса тяжелой цепи иммуноглобулина.
Проводили также эксперименты по созданию модифицированной хромосомы человека. Два сайта рекомбинации вводили по обеим сторонам вариабельной области локуса тяжелой цепи иммуноглобулина. Затем модифицированную хромосому человека вводили в клетку мыши. Затем клетки подвергали скринингу. Выбирали только клетку, содержащую только одну хромосому человека. Затем Cre-рекомбиназа опосредовала замену областей V, D, J на хромосоме мыши на области V, D, J на хромосоме человека (ФИГ. 1В).
Клетки положительного клона вводили в бластоцисты мышей BALB/c с помощью микроинъекции. Микроинъекцию эмбриона проводили по описанной методике, как например в A. Nagy, et al., «Manipulating the Mouse Embryo: A Laboratory Manual (Third Edition),» Cold Spring Harbor Laboratory Press, 2003. Инъецированные оплодотворенные яйцеклетки затем переносили в питательную среду для кратковременного культивирования, а затем трансплантировали в маточные трубы мыши-реципиента для получения генетически модифицированных гуманизированных мышей (поколение F0). Затем мышей скрещивали с мышами, имеющими фон C57BL/6. ПЦР-анализ проводили на ДНК, полученной из хвоста мышей. Далее мышей несколько раз скрещивали с мышами с фоном BALB/c (например, по меньшей мере 5 раз) для получения гетерозиготных по гуманизированному локусу тяжелых цепей иммуноглобулинов мышей с фоном BALB/c.
Затем гетерозиготных мышей скрещивали друг с другом для получения гомозиготных мышей. Подробное описание того, как получить гуманизированных мышей, гомозиготных по локусу иммуноглобулина тяжелой цепи, представлено в PCT/CN2020/075698, поданной 18, 2020, которая включена в настоящее описание посредством ссылки в полном объеме.
ПРИМЕР 3: Дизайн последовательности VJ-областей легкой цепи
Анализировали использование последовательностей вариабельной области легкой цепи человека в различных антителах.
(1) Данные собирали из базы данных IMGT/GeneFrequency. Статистический анализ данных показал, что пятью самыми часто используемыми генами V-области легкой цепи каппа были следующие: IGKV3-20, IGKV1-39/IGKV1D-39, IGKV3-11, IGKV3-15, IGKV4-1. Тремя наиболее часто используемыми генами J-области легкой цепи каппа были: IGKJ1, IGKJ2, IGKJ4.
2) Анализировали генетическую информацию легкой цепи из 110 молекул антител. Информацию о последовательности получали из базы данных IMGT/mAb-DB. Статистический анализ показал, что 5 топовых генов V-области легкой цепи каппа представляют собой IGKV3-20, IGKV3-11, IGKV3-15, IGKV1-39/IGKV1D-39 и IGKV1-12/IGKV1D-12; и 3 наиболее часто используемых генов J-области легкой цепи каппа представляют собой: IGKJ1, IGKJ4 и IGKJ2 (ФИГ. 2А-2В).
На основании анализа три V-области легкой цепи человека (IGKV3-20, IGKV3-11 и IGKV1-39) и две J-области легкой цепи человека (IGKJ1 и IGKJ4) выбирали для гуманизированного локуса иммуноглобулина легкой цепи.
ПРИМЕР 4: Получение мышей с реаранжированными легкими цепями
Локус легкой цепи иммуноглобулина мыши расположен на хромосоме 6 мыши. ФИГ. 3А представляет собой схематическую диаграмму, показывающую локус легкой цепи иммуноглобулина мыши. Вся VJ-область мыши дикого типа может быть заменена последовательностью реаранжированной VJ-области легкой цепи человека (ФИГ. 3B). В качестве альтернативы процесс может быть осуществлен поэтапно, т.е. сначала может быть нокаутирована VJ-область мыши (например, путем замены VJ-области последовательностью, содержащей Neo и DTR), а затем может быть вставлена последовательность, содержащая реаранжированную VJ-область легкой цепи человека.
Например, хромосому 6 мыши модифицировали путем нокаута всей последовательности вариабельной области локуса легкой цепи каппа иммуноглобулина. Конструировали три различные реаранжированные последовательности VJ легкой цепи человека, которые затем вставляли в VJ-область цепи каппа хромосомы 6 мыши.
Схематическая диаграмма стратегии гуманизации показана на ФИГ. 4. Конструировали три направленных вектора для гомологичной рекомбинации, обладающие общими характеристиками последовательностей, т. е. каждая из которых имеет расположенные выше и ниже последовательности гомологичного плеча и реаранжированные последовательности VJ легкой цепи человека. Для каждого вектора реаранжированную последовательность VJ человека выбирали по-разному. Три вектора содержали реаранжированные последовательности IGKV1-39/J4, IGKV3-11/J1 и IGKV3-20/J1 человека, соответственно, где IGKV1-39 (SEQ ID NO:1) идентична нуклеотидной последовательности 89319625-89320099 номер доступа NCBI. NC_000002.12; IGKV3-11 (SEQ ID NO:2) идентична нуклеотидной последовательности 89027171-89027684 номер доступа NCBI NC_000002.12; IGKV3-20 (SEQ ID NO: 3) идентична нуклеотидной последовательности 89142574-89143108 номер доступа NCBI NC_000002.12; IgKJ1 (SEQ ID NO:4) идентична нуклеотидной последовательности 88861886-88861923 номер доступа NCBI NC_000002.12; IgKJ4 (SEQ ID NO:5) идентична нуклеотидной последовательности 88860886-88860923 номер доступа NCBI NC_000002.12.
Кроме того, каждая реаранжированная последовательность VJ легкой цепи человека также содержала последовательность промотора V человека гена V-области легкой цепи. Последовательность промотора находится примерно в области 2000 п.н. перед геном V-области.
Кроме того, после каждой реаранжированной последовательности VJ человека может быть отдельная вспомогательная последовательность, например, последовательность 3'UTR мыши (SEQ ID NO: 6) или последовательность 3'UTR человека (SEQ ID NO: 7), следующая за VJ-областью легкой цепи человека.
Три вектора вводили в клетки путем редактирования генов, и клетки отбирали с помощью DT-скрининга. Положительные клоны подтверждали ПЦР и Саузерн-блоттингом. ПЦР-анализ проводили с использованием следующих праймеров:
В Таблице 13 L-GT-F расположен на 5'-плече гомологии; R-GT-R расположен на 3'-плече гомологии; L-GT-R и R-GT-F расположены на реаранжированной последовательности VJ легкой цепи человека; Праймеры m-5'loxp-L-GT-F, m-5'loxp-R-GT-R, m-3'lox-L-GT-F и m-3'lox-R-GT-R расположены на хромосоме гуманизированной тяжелой цепи.
Саузерн-блоттинг проводили с использованием следующих зондов:
В Таблице 14 5'-зонд расположен снаружи 5'-плеча гомологии; 3'-зонд расположен снаружи 3'-плеча гомологии; зонд IGKV1-39, зонд IGKV3-11 и зонд IGKV3-20 расположены на гуманизированных фрагментах.
Клетки положительного клона вводили в бластоцисты мышей BALB/c с помощью микроинъекции. Микроинъекцию эмбриона проводили по описанной методике, как например в A. Nagy, et al., «Manipulating the Mouse Embryo: A Laboratory Manual (Third Edition),» Cold Spring Harbor Laboratory Press, 2003. Инъецированные оплодотворенные яйцеклетки затем переносили в питательную среду для кратковременного культивирования, а затем трансплантировали в маточные трубы мыши-реципиента для получения генетически модифицированных гуманизированных мышей (поколение F0). Химерных мышей отбирали для спаривания с гуманизированными гомозиготными мышами с геном тяжелой цепи, получая мышей поколения F1. ПЦР и анализ по Саузерну проводили на ДНК, полученной из хвоста мышей поколения F1, чтобы подтвердить, были ли получены положительные гетерозиготные мыши.
ПРИМЕР 5: Экспрессия общей легкой цепи
Последовательности мРНК вариабельной области легкой цепи у мышей анализировали путем секвенирования. Отбирали неиммунизированных (не экспонированных с определенным антигеном) гуманизированных мышей (гетерозиготных по гуманизированной тяжелой цепи и гетерозиготных с реаранжированной легкой цепью) и экстрагировали РНК из ретроорбитальной крови. Набор 5'-RACE (набор SMARTer RACE 5'/3', Takara Bio USA, Inc., кат. № 634858) использовали для выполнения обратной транскрипции с получением кДНК. Полученную кДНК амплифицировали с использованием праймера IGKC-R и праймера UPM из набора 5'-RACE. Затем секвенировали фрагмент последовательности вариабельной области тяжелой цепи. Последовательность праймера IGKC-R представляла собой 5'-CTAACACTCATTCCTGTTGAAGCTCTTGAC-3' (SEQ ID NO: 34). Результат секвенирования сравнивали с инструментом NCBI Ig Blast для идентификации последовательностей иммуноглобулинов человека, чтобы определить использование генов областей Vκ и Jκ человека. Результаты показали, что 89% из 577 клонов от трех вариантов мышей (выбранных случайным образом) экспрессировали человеческие Vκ и Jκ, а соотношение человеческих Vκ и Jκ у каждой мыши превышало 60%. Это указывает на то, что после того, как эндогенный локус вариабельной области цепи К был полностью заменен реаранжированной последовательностью последовательностей легкой цепи иммуноглобулина человека, гены Vκ и Jκ человека у гуманизированных мышей экспрессировались эффективно и преимущественно. Напротив, цепь κ дикого типа не экспрессировалась или имела относительно низкий уровень экспрессии у гетерозиготных мышей.
ПРИМЕР 6: Созревание В-клеток у мышей hVH/hcVL
Мыши поколения F1 скрещивали друг с другом. Гомозиготный гуманизированный локус тяжелой цепи иммуноглобулина и гетерозиготный гуманизированный локус общей легкой цепи иммуноглобулина (гуманизированные мыши VHH/H/cVLK/+ или обозначенные как мыши hVHH/H/hcVLK/+) использовали для тестирования развития В-клеток. Мышей с гуманизированным локусом VH (гетерозиготным или гомозиготным) и локусом общей легкой цепи иммуноглобулина (гетерозиготным или гомозиготным) также называют мышами hVH/hcVL. Проводили эксперименты для сравнения иммунных систем мышей hVHH/H/hcVLK/+ и мышей дикого типа до иммунизации. Массу тела и массу некоторых органов, например, селезенки, тимуса, печени, сердца, легких и почки измеряли у мышей дикого типа и гетерозиготных мышей hVH/hcVL (ФИГ. 5A-5G). Между мышами дикого типа и гетерозиготными мышами hVH/hcVL не было обнаружено существенных различий в средней массе тела и массе органов (селезенки, тимуса, печени, сердца, легких и почек).
Проточную цитометрию проводили для анализа популяций и распределения лимфоцитов в крови (ФИГ. 6А), селезенке (ФИГ. 6В) и лимфатических узлах (ФИГ. 7) мышей дикого типа и гетерозиготных мышей hVH/hcVL. Результаты показали, что у гетерозиготных мышей hVH/hcVL процент В-клеток, Т-клеток, NK-клеток, CD4+ Т-клеток и CD8+ Т-клеток в крови, селезенке и лимфатических узлах был почти таким же, как у мышей дикого типа. В результатах лейкоциты включали: В-клетки (например, характеризующиеся CD45+, CD19+, TCR-), Т-клетки и клетки натуральные киллеры (NK) (например, характеризующиеся CD45+, TCR-, NK1.1+). Т-клетки дополнительно характеризовались CD45+, CD19-, TCR+. CD4+ Т-клетки (CD4) характеризовались CD45+, CD19-, TCR+, CD4+, CD8-, CD8+ Т-клетки (CD8) характеризовались CD45+, CD19-, TCR+, CD4-, CD8+. В анализ методом проточной цитометрии были включены только интактные, единичные, живые лейкоциты.
На ФИГ. 8А показано процентное содержание В-клеток на разных стадиях созревания в костном мозге. Клетки-предшественники В-клеток в костном мозге анализировали с помощью проточной цитометрии. На основании уровней экспрессии B220 и CD43 клетки-предшественники В-клеток в костном мозге можно разделить на 3 клеточные популяции: про-В-клетки (характеризуемые B220lowCD43highIgMlow), пре-В-клетки (характеризуемые B220lowCD43intIgMlow) и незрелые В-клетки. (характеризуемые B220highCD43lowIgMhigh). Значимых различий между мышами дикого типа и мышами hVH/hcVL не наблюдали. Кроме того, развитие В-клеток также оценивали в костном мозге с помощью проточной цитометрии для селективного окрашивания плазматических клеток (B220lowIgM-IgD-CD138+) и В-клеток памяти (B220+IgM+IgD-CD38+) (ФИГ. 8В). Значимых различий между мышами дикого типа и мышами hVH/hcVL не наблюдали.
На ФИГ. 9 указано процентное содержание В-клеток на разных стадиях созревания. созревания развития В-клеток в селезенке были разделены на Т1 (В-клетки переходного типа 1, характеризующиеся В220+IgM+IgD-), Т2 (В-клетки переходного типа 2, характеризующиеся В220+IgM+IgD+) и зрелые В-клетки (характеризуется B220+IgMlowIgD+). Значимых различий между мышами дикого типа и мышами hVH/hcVL не наблюдали.
Созревание В-клеток также оценивали в селезенке с помощью проточной цитометрии для селективного окрашивания плазматических клеток (B220lowIgM-IgD-CD138+) и В-клеток памяти (B220+IgM+IgD-CD38+) (ФИГ. 10А-10В). Значимых различий между мышами дикого типа и мышами hVH/hcVL не наблюдали. Кроме того, созревание В-клеток также оценивали в маргинальной зоне селезенки (В-клетки маргинальной зоны, MZ-B, характеризующиеся B220+CD21+CD23-) и фолликулярной зоне (фолликулярные В-клетки, обозначаемые как FO-B, характеризующиеся по B220+CD21lowCD23+). На ФИГ. 10C показан процент В-клеток селезенки в маргинальной зоне селезенки (MZ-B) и фолликулярной зоне (FO-B). Значимых различий между мышами дикого типа и мышами hVH/hcVL не наблюдали.
Использование легкой цепи В-клеток оценивали в селезенке с помощью проточной цитометрии. В-клетки сначала селективно метили mB220 (антитело Pacific Blue™ против мышиного/человеческого CD45R/B220, BioLegend, кат. № 103227), а затем mIgG каппа-FITC (антитело FITC против легкой цепи κ Ig мыши, BioLegend, кат. № 409509) и mIgG lambda-PE (PE антитело против легкой цепи λ Ig мыши, BioLegend, кат. № 407307). Значимых различий между мышами дикого типа и мышами hVH/hcVL не наблюдали. Результаты показали, что гуманизация не влияла на соотношение экспрессии цепей κ и λ в селезенке гетерозиготных мышей hVH/hcVL (ФИГ. 11A-11I).
На ФИГ. 12А показано процентное содержание В-клеток на разных стадиях созревания. Стадии созревания В-клеток в лимфатических узлах были разделены на Т1 (В-клетки переходного типа 1, характеризующиеся В220+IgM+IgD-), Т2 (В-клетки переходного типа 2, характеризующиеся В220+IgM+IgD+) и зрелые В-клетки (характеризующиеся B220+IgMlowIgD+). Значимых различий между мышами дикого типа и мышами hVH/hcVL не наблюдали. Созревание В-клеток также оценивали в лимфатических узлах с помощью проточной цитометрии для селективного окрашивания плазматических клеток (B220lowIgM-IgD-CD138+) и В-клеток памяти (B220+IgM+IgD-CD38+) (ФИГ. 12В). Значимых различий между мышами дикого типа и мышами hVH/hcVL не наблюдали.
Использование легкой цепи В-клеток оценивали в лимфатических узлах с помощью проточной цитометрии. В-клетки сначала селективно метили mB220, а затем метили mIgG каппа-FITC и mIgG лямбда-PE. Значимых различий между мышами дикого типа и мышами hVH/hcVL не наблюдали. Результаты показали, что гуманизация не влияла на соотношение экспрессии цепей κ и λ в селезенке гетерозиготных мышей hVH/hcVL (ФИГ. 13A-13I).
Проводили эксперименты для оценки созревания В-клеток у мышей hVHH/H/hcVLK/K. Значимых различий в созревании В-клеток и среднем иммунном органе (например, в селезенке) не детектировали между мышами дикого типа и мышами hVHH/H/hcVLK/K. Как показано на ФИГ. 13J, различные подтипы иммуноглобулина (Ig) в сыворотке мышей hVHH/H/hcVLK/K и мышей дикого типа количественно измеряли с помощью ИФА перед антигенной иммунизацией. Значимых различий не наблюдали.
В другом подобном эксперименте анализировали подтипы Ig мышей hVHH/H/hcVLK/K и мышей дикого типа до и после иммунизации. Значимых различий обнаружено не было (ФИГ. 13H).
ПРИМЕР 7: Продукция антител у мышей hVH/hcVL
Проводили эксперименты для оценки того, нормально ли развивались В-клетки у мышей hVH/hcVL. Измеряли титры антиген-специфических антител в сыворотке иммунизированных мышей hVH/hcVL и мышей дикого типа.
Мышей hVHH/H/hcVLK/+ и мышей дикого типа (C57BL/6) иммунизировали двумя разными антигенами всего три раза. При первой иммунизации вводили полный адъювант Фрейнда (CFA) и 20 мкг антигена. Через две недели вводили неполный адъювант Фрейнда (IFA) и 20 мкг антигена. Через две недели мышам вводили 20 мкг антигена в комбинации с ИФА. Примерно через неделю после третьей иммунизации собирали кровь и анализировали титры антиген-специфических антител мышей дикого типа и мышей hVH/hcVL с помощью ИФА. Мыши hVHH/H/hcVLK/+ доминантно экспрессировали антитела с гуманизированной вариабельной областью общей легкой цепи.
ИФА проводили следующим образом. His-меченый антиген разводили в 1×PBS до 0,5 мкг/мл и добавляли в 96-луночный планшет в количестве 0,1 мл/лунку с последующей инкубацией при 37°С в течение 2 часов. После инкубации каждую лунку трижды промывали 300 мкл 1×PBST, а затем блокировали 250 мкл 1×PBS с добавлением 5% обезжиренного молока при 37°C в течение 1 часа. Затем каждую лунку дважды промывали 300 мкл 1×PBST. Образцы сыворотки мышей hVH/hcVL или мышей дикого типа сначала разбавляли с использованием 1×PBS в соотношении 1:500, 1:2000, 1:8000, 1:32000, 1:128000, 1:512000 или 1:2048000 и затем добавляли в 96-луночный планшет. Образцы сыворотки от неиммунизированных мышей также разбавляли в соотношении 1:500 и добавляли в планшет в качестве пустого контроля. Разведенные образцы сыворотки (120 мкл/лунку) инкубировали при 37°С в 96-луночном планшете в течение 1 часа. После инкубации планшет пять раз промывали 300 мкл/лунку 1×PBST. Затем планшет инкубировали с 0,1 мл/лунку разбавленного 1:20000 козьего анти-мышиного IgG Fc (HRP) при 37°C в течение 1 часа. После инкубации планшет пять раз промывали 300 мкл/лунку 1×PBST. Затем 0,1 мл проявляющего раствора ТМВ добавляли в каждую лунку и планшет выдерживали в темноте при комнатной температуре в течение 10 минут, после чего добавляли 0,1 мл стоп-раствора в каждую лунку. OD450 и OD570 измеряли планшет-ридером, и стандартное значение OD на ФИГ. 24-25 рассчитывали как: стандартное значение OD=OD450 - OD570.
Результаты титров антиген-специфических антител после третьей иммунизации показаны на ФИГ. 24 и ФИГ. 25, соответственно. Результаты показали, что мыши hVH/hcVL могут продуцировать антитела, которые специфически связываются с антигеном, и иммунный ответ у мышей дикого типа и мышей hVH/hcVL был сходным. В частности, титры сывороточных антител были аналогичны антителам дикого типа.
ПРИМЕР 8: Продукция антител у мышей hVH/hcVL
TFR1 и Ova использовали для иммунизации мышей hVHH/H/hcVLK/K и мышей дикого типа (по 3 на каждую группу). Затем с помощью оптофлюидной системы Beacon® выделяли плазматические клетки, которые могут продуцировать антиген-специфические моноклональные антитела. Количество положительных клеток показано ниже.
Антиген А является членом суперсемейства рецепторов фактора некроза опухоли. Антиген А использовали для иммунизации мышей hVHH/H/hcVLK/K. Было получено 42 антитела с общей легкой цепью человека (IGKV3-11/J1). VH и VL этих антител добавляли к константным областям человеческого IgG1 для получения человеческих антител. Затем Biacore использовали для определения аффинности 42 человеческих антител с общей легкой цепью (IGKV3-11/J1), продуцируемых против антигена А.
Очищенные антитела к антигену А разбавляли до 1 мкг/мл и затем вводили в биосенсор Biacore 8K со скоростью 10 мкл/мин в течение примерно 50 секунд для достижения желаемой плотности белка (например, примерно 50 единиц ответа (RU)). Затем меченный His антиген А в концентрации 200 нМ вводили со скоростью 30 мкл/мин в течение 120 секунд. Диссоциацию контролировали в течение 600 секунд. Чип регенерировали после последней инъекции каждого титрования глицином (pH 2, 30 мкл/мин в течение 30 секунд).
Кинетические скорости ассоциации (kon) и скорости диссоциации (koff) были получены одновременно путем аппроксимации данных в глобальном масштабе к модели связывания Ленгмюра 1: 1 (Karlsson, R. Roos, H. Fagerstam, L. Petersson, B., 1994. Methods Enzymology 6. 99-110) с помощью оценочного программного обеспечения Biacore 8K 3. 0, Аффинности выводили из отношения кинетических констант скорости (KD=koff/kon).
Как будет понятно специалисту в данной области техники, тот же способ с соответствующими корректировками параметров (например, концентрация антител) проводили для каждого тестируемого антитела. Результаты показали, что, за исключением A-1C5-IgG1, A-1D10-IgG1 и A-1E1-IgG1, аффинность связывания этих антител достигала или даже превышала 10-8 М, что указывает на то, что методы могут успешно генерировать антитела с высокой аффинностью связывания против антигена.
Анализ корреляции эпитопов
Относительное положение эпитопов анализировали с помощью конкурентного эксперимента поверхностного плазмонного резонанса (SPR). В общей сложности 15 моноклональных антител использовали для изучения эффекта ингибирования связывания (блокировки) друг против друга: A-1B10-IgG1, A-1D7-IgG1, A-1C8-IgG1, A-1F2-IgG1, A-H6L6-IgG1., A-1F9-IgG1, A-1B1-IgG1, A-1D12-IgG1, A-1E7-IgG1, A-1A7-IgG1, A-1B8-IgG1, A-H7L7-IgG1, A-1E12-IgG1, A -H3L3-IgG1 и А-H8L8-IgG1. Другое антитело против антигена А также включали в качестве PC (положительный контроль).
Буфер HBS-EP+ (10 мМ 4-(2-гидроксиэтил)-1-пиперазинэтансульфокислоты (HEPES), 150 мМ NaCl, 3 мМ этилендиаминтетрауксусной кислоты (EDTA) и 0,05% Р20, рН 7,4) разбавляли буфером HBS-EP+ (10×) в качестве рабочего буфера на протяжении всего эксперимента. Анти-His-антитела фиксировали на поверхности сенсорного чипа СМ5 серии S путем связывания аминогрупп с получением анти-His-чипа (т. е., CM5-анти-His-канал 1,8-чип). Затем 1М этаноламин, рН 8,5 вводили для блокирования оставшихся активных карбоксильных групп на поверхности чипа с последующим уравновешиванием с использованием буфера HBS-EP+ в течение 2 часов. Рекомбинантный белок антигена А с His-меткой (1 мкг/мл) вводили в биосенсор Biacore 8K со скоростью 10 мкл/мин в течение 50 секунд и захватывали анти-His-чипом для достижения желаемой плотности белка (т. е., 200 RU.). Пару антител (по 200 нМ каждое) непрерывно вводили на чип со скоростью 30 мкл/мин. Первое вводимое антитело (аналит 1) имело время связывания 250 секунд, а затем вводили второе антитело (аналит 2) со временем связывания 250 секунд. После введения антител в каждом цикле анализа чип дважды регенерировали глициновым буфером (pH 1,7; 30 мкл/мин в течение 30 секунд). Каждая пара моноклональных антител подвергалась одним и тем же экспериментальным стадиям для получения данных об ингибировании связывания, когда каждое моноклональное антитело было соединено с другим антителом.
Величина связывания каждого антитела была получена с использованием программного обеспечения Biacore Insight Evaluation Software. Для количественной оценки интерференции связывания одного антитела с другим рассчитывали коэффициент связывания для сравнения каждой пары антител. Соотношение связывания определяется как значение связывания второго антитела (аналит 2), деленное на значение связывания первого антитела (аналит 1). Для кластерного анализа также использовалось статистическое программное обеспечение. Анализировали корреляцию эпитопов, и 15 антител человека против антигена А были разделены на 4 кластера эпитопов (ФИГ. 31). Таким образом, A-1B10-IgG1, A-1D7-IgG1, A-1C8-IgG1 и A-1F2-IgG1 имеют одинаковые или перекрывающиеся эпитопы; A-1B1-IgG1 и A-1D12-IgG1 имеют одинаковые или перекрывающиеся эпитопы; A-1B8-IgG1, A-H7L7-IgG1 и A-1E12-IgG1 имеют одинаковые или перекрывающиеся эпитопы; A-H3L3-IgG1 и A-H8L8-IgG1 имеют одинаковые или перекрывающиеся эпитопы. Результаты показывают, что мыши hVHH/H/hcVLK/K могут продуцировать антитела, нацеленные на различные эпитопы.
Измерение термостабильности
Термостабильность 17 человеческих антител против антигена А измеряли с помощью набора красителей Protein Thermal Shift™ с использованием систем ПЦР в реальном времени QuantStudio™ 5.
Эксперименты проводили согласно протоколу производителя. Реакции проводили непрерывно в две стадии. В частности, первую стадию выполняли при 1,6°C в секунду при 25°C в течение 2 минут, а вторую стадию проводили при 0,05°C в секунду при 99°C в течение 2 минут. Температуру плавления (Tm) каждого антитела к антигену А определяли, как показано в Таблице ниже.
Результаты показывают, что Tm антител человека против антигена А выше или примерно 70°C, и эти антитела термостабильны.
ПРИМЕР 9: Продукция антител у мышей hVH/hcVL
Антиген В представляет собой трансмембранную сериновую протеазу II типа. Антиген B использовали для иммунизации мышей hVHH/H/hcVLK/K. Затем Biacore использовали для определения аффинности 8 человеческих антител с общей легкой цепью (IGKV3-11/J1), продуцируемых против антигена B.
Пример 10 Блокирование связывания лиганда антигена C с антигеном C
Антиген C представляет собой ингибирующий рецептор на активированных антигеном Т-клетках, который играет критическую роль в индукции и поддержании иммунной толерантности к собственным белкам. Проводили анализы блокирования, чтобы определить, могут ли 129 антител против человеческого Антигена C блокировать связывание между человеческим Антигеном C и его лигандом. Эти антитела были продуцированы мышами hVHH/H/hcVLK/K.
Антитела против Антигена С собирали из супернатанта CHO-S. В каждую лунку планшета добавляли по 30 мкл клеток СНО (примерно 1×105), транзиторно трансфецированных лигандом Антигена С человека. Антитела в супернатанте (примерно 20-200 мкг/мл) собирали и затем разводили в 10 и 100 раз. В каждую лунку добавляли оттитрованные антитела (30 мкл на лунку) и Bitoin-меченный антиген С человека (30 мкл на лунку, с конечной концентрацией 1 мкг/мл в каждой лунке) по 30 мкл на лунку при 4 °С. Клетки с Bitoin-меченным антигеном C человека и антителами инкубировали при 4°С в течение 30 минут.
После двукратной промывки фосфатно-солевым буфером (PBS) в каждую лунку добавляли по 50 мкл Alexa Fluor® 647-меченого стрептавидина (AF647 Streptavidin, Jackson Immuno research, кат. № 016-600-084) в разведении 1:1000 и инкубировали в течение 15 минут при 4°C с последующей промывкой PBS. Сигналы для AF647 определяли с помощью проточной цитометрии (Thermo Attune NX).
Результаты для антител с процентом протестированных клеток, которые имели сигналы стрептавидина в анализе проточной цитометрии, суммированы в Таблице ниже. По мере снижения концентрации антител скорость блокирования снижается, что указывает на то, что эти антитела могут блокировать связывание между Антигеном C и его лигандом.
Отбирали 23 антитела для определения перекрестной реактивности с Антигеном C мыши, Антигеном C обезьяны и Антигеном C собаки с помощью проточной цитометрии. Результаты показаны ниже.
ПРИМЕР 11: Разнообразие антител и распределение аффинности связывания у мышей hVH/hcVL
Антиген D и антиген E представляют собой трансмембранные рецепторные белки типа I. Антиген D и антиген E использовали для иммунизации мышей hVHH/H/hcVLK/K (IGKV3-11/J1) по отдельности. Затем использовали оптофлюидную систему Beacon® для выделения плазматических клеток, которые могут продуцировать антиген-специфические моноклональные антитела.
После выделения РНК из плазматических клеток РНК обратно транскрибировали в кДНК и секвенировали. В группе антигена D было получено в общей сложности 1166 последовательностей вариабельной области тяжелой цепи антитела. В группе антигена Е было получено в общей сложности 1290 последовательностей антител с вариабельной областью тяжелой цепи. Определяли человеческие зародышевые гены этих последовательностей тяжелых цепей антител. Результаты показаны на ФИГ. 33A-33B. Среди них гены V-области всех антител включают в общей сложности 5 подгрупп VH1-VH5 (включая IGHV1, IGHV2, IGHV3, IGHV4 и IGHV5; см. ось X, показанную на ФИГ. 33А-33В). Гены области D включают в общей сложности 6 подгрупп DH1-DH6 (включая IGHD1, IGHD2, IGHD3, IGHD4, IGHD5 и IGHD6; см. левую ось Y, показанную на ФИГ. 33А-33В). Гены области J включают в общей сложности 6 подгрупп JH1-JH6 (включая IGHJ1, IGHJ2, IGHJ3, IGHJ4, IGHJ5 и IGHJ6; см. правую ось Y, показанную на ФИГ. 33А-33В). Результаты показывают, что разнообразие V\D\J тяжелой цепи антитела, продуцируемого против антигена D и антигена E, очень велико. Это демонстрирует, что у гуманизированных мышей с общей легкой цепью разнообразие тяжелых цепей антител не затрагивается.
Затем Biacore использовали для определения аффинности этих антител. Скорости ассоциации (kon) и скорости диссоциации (koff) получали путем аппроксимации данных к модели связывания Ленгмюра 1:1 (Karlsson, R. Roos, H. Fagerstam, L. Petersson, B., 1994. Methods Enzymology 6. 99-110). Аффинности выводили из отношения кинетических констант скорости (KD=koff/kon). Анализировали антитела с KD менее 1×10-8 М. Эти антитела включали 805 антител с общей легкой цепью человека против антигена D и 1026 антител с общей легкой цепью человека против антигена E. Значения аффинности связывания антител (KD), kon и koff были предоставлены и отсортированы на основе KD. Проводили статистический анализ. Распределение KD, kon и koff показано на ФИГ. 34-35.
На ФИГ. 34A-34B представлено распределение скоростей kon и koff для антител против антигена D. На ФИГ. 34C-34D представлено распределение скоростей kon и koff для антител против антигена E. На ФИГ. 35 представлено распределение аффинности связывания антител, которое показывает, что kon-скорость этих антител в основном сконцентрирована между 1×105/мс и 1×106/мс, koff в основном сконцентрирована между 1×10-2/с и 1×10-3/с, а аффинность связывания (KD) в основном сконцентрирована между 0,1 нМ~100 нМ. Эти результаты согласуются с результатами для антител, полученных в результате естественного иммунного ответа. Данные показывают, что мыши hVHH/H/hcVLK/K могут продуцировать разнообразные антитела с высокой аффинностью к различным антигенам.
ПРИМЕР 12: Анализ использования зародышевой линии у мышей hVHH/H/hcVLK/+
Трансгенных мышей с гомозиготным гуманизированным локусом тяжелой цепи иммуноглобулина и гетерозиготным гуманизированным локусом общей легкой цепи иммуноглобулина (гуманизированные мыши VHH/H/cVLK/+ или обозначенные как мыши hVHH/H/hcVLK/+) отбирали для анализа использования зародышевой линии. Мыши hVHH/H/hcVLK/+ имели реаранжированные последовательности IGKV3-11/J1 человека. РНК экстрагировали из спленоцитов мышей перед иммунизацией антигеном. Экстрагированную РНК подвергали обратной транскрипции и вариабельные области тяжелой цепи иммуноглобулина амплифицировали с помощью ПЦР. Более конкретно, верхний праймер, т. е. 5'-праймер быстрой амплификации кДНК-концов (RACE) и нижний праймер, нацеленный на область ниже от генов, кодирующих IgM. Амплифицированный продукт ПЦР очищали, а использование зародышевой линии у наивных мышей hVHH/H/hcVLK/+ определяли с помощью секвенирования следующего поколения (NGS). Всего было получено 131847 достоверных прочтений.
Анализировали использование тяжелых цепей IGHV, IGHD и IGHJ у наивных мышей hVHH/H/hcVLK/+. Результаты показаны на ФИГ. 26A-26D. Были обнаружены транскрипты 46 генов IGHV (включая два псевдогена). Кроме того, были обнаружены транскрипты 25 генов IGHD и 6 генов IGHJ. Например, как показано на ФИГ. 26D, IGHJ3, IGHJ4, IGHJ5 и IGHJ6 часто использовались у наивных мышей hVHH/H/hcVLK/+, в то время как IGHJ1 и IGHJ2 использовались реже. Модель использования зародышевой линии IGHJ в значительной степени согласуется с литературными данными об использовании зародышевой линии IGHJ у человека.
Также определяли использование IGHV и IGHD тяжелой цепи у гомозиготных наивных мышей hVHH/H/hcVLK/K. Всего было получено 123188 достоверных прочтений. Результаты показаны на ФИГ. 26E-26G. Детектировали транскрипты 46 генов IGHV (включая два псевдогена IGHV1-NL1 и IGHV3-38-3). Результаты теста аналогичны результатам гетерозиготных мышей.
Распределение длин CDR3 тяжелой цепи определяли с помощью NGS-секвенирования иммунного репертуара из спленоцитов наивных мышей hVHH/H/hcVLK/+ (n=1). Как показано на ФИГ. 27А, средняя длина CDR3 составляет около 13~15 аминокислот. Результаты для гомозиготных наивных мышей hVHH/H/hcVLK/K показаны на ФИГ. 27B. Результаты также соответствовали средней длине CDR3 тяжелой цепи человека в иммунной системе человека.
Также анализировали типы аминокислот в каждом положении CDR3 тяжелой цепи (HCDR3) (ФИГ. 28А-28С, 28D). Эти паттерны аналогичны аминокислотному составу HCDR3 человека.
Остаток цистеина может образовывать дисульфидную связь. HCDR3 человека может содержать один остаток цистеина или два остатка цистеина, в то время как HCDR3 мыши обычно не содержит цистеина. Результаты на ФИГ. 29 показали, что частота HCDR3 у мышей hVHH/H/hcVLK/+, содержащих остатки цистеина, увеличивается по мере увеличения длины HCDR3. Этот результат согласуется с разнообразием HCDR3 в мононуклеарных клетках периферической крови человека (PBMC). Результаты показывают, что VDJ-рекомбинация у мышей hVH/hcVL работает должным образом.
ПРИМЕР 13: Анализ соматических гипермутаций у мышей hVHH/H/hcVLK/+
Мышей hVHH/H/hcVLK/+ с реаранжированными последовательностями IGKV3-11/J1 человека отбирали для анализа соматических гипермутаций. РНК экстрагировали из спленоцитов мышей перед иммунизацией антигеном. Экстрагированную РНК обратно транскрибировали в кДНК, и реаранжированную область IGKV3-11/J1 человека амплифицировали с помощью ПЦР с использованием праймеров, специфичных для IGKV3-11 и IGJ1 человека. Затем амплифицированный продукт ПЦР обрабатывали для секвенирования следующего поколения (NGS). Всего было получено и проанализировано с помощью инструмента IMGT/HighV-QUEST 48326 достоверных прочтений. Как показано на ФИГ. 30A-30B, результаты показывают, что соматические гипермутации могут возникать в реаранжированной области легкой цепи человека, но большинство соматических гипермутаций ограничены одной или двумя аминокислотными заменами. Результаты для гомозиготных наивных мышей hVHH/H/hcVLK/K представлены на ФИГ. 30С. Всего было получено и проанализировано с помощью инструмента IMGT/HighV-QUEST 48541 достоверное чтение. Как и в случае с гетерозиготными мышами, большинство соматических мутаций ограничивается заменой 1-2 нуклеотидов, то есть заменой 1-2 аминокислот. Это указывает на то, что у мышей hVHH/H/hcVLK/K скорость соматических гипермутаций относительно низка и составляет около 1-2 нуклеотидных (или аминокислотных) изменений.
Другие варианты осуществления
Следует понимать, что хотя изобретение приведено вместе с подробным описанием, представленное выше описание предназначено для иллюстрации и не ограничивает объем изобретения, который определен объемом прилагаемой формулы изобретения. Другие аспекты, преимущества и модификации находятся в пределах представленной ниже формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Biocytogen Pharmaceuticals (Beijing) Co., Ltd.
<120> ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА,
СОДЕРЖАЩИЕ ЛОКУС ОБЩЕЙ ЛЕГКОЙ ЦЕПИ ИММУНОГЛОБУЛИНА
<130> 44835-0059WO3
<150> PCT/CN2021/097652
<151> 2021-06-01
<150> PCT/CN2020/094000
<151> 2020-06-02
<150> PCT/CN2021/085839
<151> 2021-04-07
<160> 41
<170> PatentIn version 3.5
<210> 1
<211> 477
<212> ДНК
<213> Искусственная последовательность
<220>
<223> IGKV1-39
<400> 1
atggacatga gggtccccgc tcagctcctg gggctcctgc tactctggct ccgaggtaag 60
gatggagaac actaggaatt tactcagcca gtgtgctcag tactgactgg aacttcaggg 120
aagttctctg ataacatgat taatagtaag aatatttgtt tttatgtttc caatctcagg 180
tgccagatgt gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga 240
cagagtcacc atcacttgcc gggcaagtca gagcattagc agctatttaa attggtatca 300
gcagaaacca gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg 360
ggtcccatca aggttcagtg gcagtggatc tgggacagat ttcactctca ccatcagcag 420
tctgcaacct gaagattttg caacttacta ctgtcaacag agttacagta cccctcc 477
<210> 2
<211> 516
<212> ДНК
<213> Искусственная последовательность
<220>
<223> IGKV3-11
<400> 2
atggaagccc cagctcagct tctcttcctc ctgctactct ggctcccagg tgaggggaac 60
atgaggtggt tttgcacatt agtgaaaact cttgccacct ctgctcagca agaaatataa 120
ttaaaattca aagtatatca acaattttgg ctctactcaa agacagttgg tttgatcttg 180
attacatgag tgcatttctg ttttatttcc aatttcagat accaccggag aaattgtgtt 240
gacacagtct ccagccaccc tgtctttgtc tccaggggaa agagccaccc tctcctgcag 300
ggccagtcag agtgttagca gctacttagc ctggtaccaa cagaaacctg gccaggctcc 360
caggctcctc atctatgatg catccaacag ggccactggc atcccagcca ggttcagtgg 420
cagtgggtct gggacagact tcactctcac catcagcagc ctagagcctg aagattttgc 480
agtttattac tgtcagcagc gtagcaactg gcctcc 516
<210> 3
<211> 537
<212> ДНК
<213> Искусственная последовательность
<220>
<223> IGKV3-20
<400> 3
atggaaaccc cagcgcagct tctcttcctc ctgctactct ggctcccagg tgaggggaac 60
atgggatggt tttgcatgtc agtgaaaacc ctctcaagtc ctgttacctg gcaactctgc 120
tcagtcaata caataattaa agctcaatat aaagcaataa ttctggctct tctgggaaga 180
caatgggttt gatttagatt acatgggtga cttttctgtt ttatttccaa tctcagatac 240
caccggagaa attgtgttga cgcagtctcc aggcaccctg tctttgtctc caggggaaag 300
agccaccctc tcctgcaggg ccagtcagag tgttagcagc agctacttag cctggtacca 360
gcagaaacct ggccaggctc ccaggctcct catctatggt gcatccagca gggccactgg 420
catcccagac aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcagcag 480
actggagcct gaagattttg cagtgtatta ctgtcagcag tatggtagct cacctcc 537
<210> 4
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> IGKJ1
<400> 4
gtggacgttc ggccaaggga ccaaggtgga aatcaaac 38
<210> 5
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> IGKJ4
<400> 5
gctcactttc ggcggaggga ccaaggtgga gatcaaac 38
<210> 6
<211> 1189
<212> ДНК
<213> Mus musculus
<400> 6
gtaagtacac ttttctcatc tttttttatg tgtaagacac aggttttcat gttaggagtt 60
aaagtcagtt cagaaaatct tgagaaaatg gagagggctc attatcagtt gacgtggcat 120
acagtgtcag attttctgtt tatcaagcta gtgagattag gggcaaaaag aggctttagt 180
tgagaggaaa gtaattaata ctatggtcac catccaagag attggatcgg agaataagca 240
tgagtagtta ttgagatctg ggtctgactg caggtagcgt ggtcttctag acgtttaagt 300
gggagatttg gaggggatga ggaatgaagg aacttcagga tagaaaaggg ctgaagtcaa 360
gttcagctcc taaaatggat gtgggagcaa actttgaaga taaactgaat gacccagagg 420
atgaaacagc gcagatcaaa gaggggcctg gagctctgag aagagaagga gactcatccg 480
tgttgagttt ccacaagtac tgtcttgagt tttgcaataa aagtgggata gcagagttga 540
gtgagccgta ggctgagttc tctcttttgt ctcctaagtt tttatgacta caaaaatcag 600
tagtatgtcc tgaaataatc attaagctgt ttgaaagtat gactgcttgc catgtagata 660
ccatggcttg ctgaataatc agaagaggtg tgactcttat tctaaaattt gtcacaaaat 720
gtcaaaatga gagactctgt aggaacgagt ccttgacaga cagctcaagg ggtttttttc 780
ctttgtctca tttctacatg aaagtaaatt tgaaatgatc ttttttatta taagagtaga 840
aatacagttg ggtttgaact atatgtttta atggccacgg ttttgtaaga catttggtcc 900
tttgttttcc cagttattac tcgattgtaa ttttatatcg ccagcaatgg actgaaacgg 960
tccgcaacct cttctttaca actgggtgac ctcgcggctg tgccagccat ttggcgttca 1020
ccctgccgct aagggccatg tgaacccccg cggtagcatc ccttgctccg cgtggaccac 1080
tttcctgagg cacagtgata ggaacagagc cactaatctg aagagaacag agatgtgaca 1140
gactacacta atgtgagaaa aacaaggaaa gggtgactta ttggagatt 1189
<210> 7
<211> 500
<212> ДНК
<213> Homo sapiens
<400> 7
gtaagtaatt tttcactatt gtcttctgaa atttgggtct gatggccagt attgactttt 60
agaggcttaa ataggagttt ggtaaagatt ggtaaatgag ggcatttaag atttgccatg 120
ggttgcaaaa gttaaactca gcttcaaaaa tggatttgga gaaaaaaaga ttaaattgct 180
ctaaactgaa tgacacaaag taaaaaaaaa aagtgtaact aaaaaggaac ccttgtattt 240
ctaaggagca aaagtaaatt tatttttgtt cactcttgcc aaatattgta ttggttgttg 300
ctgattatgc atgatacaga aaagtggaaa aatacatttt ttagtctttc tcccttttgt 360
ttgataaatt attttgtcag acaacaataa aaatcaatag cacgccctaa gaaaaatcag 420
ggaaaagtga agtgtaccta tttgctatgt agaagaggca gcttacttga aaatcagcag 480
caatgttgtt tttagagtct 500
<210> 8
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 8
tcacacacta cagcttccac cacaa 25
<210> 9
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 9
tgggcgcgcc agactctaaa 20
<210> 10
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 10
tgggtctgat ggccagtatt gact 24
<210> 11
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 11
ggcctggaaa actcagctat ccttt 25
<210> 12
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 12
gccaaggaat ttaaaagggg attgaaagca a 31
<210> 13
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 13
agggagggaa tggaatgagg gtgat 25
<210> 14
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 14
ccatgtgacc cattcgagtg tcctg 25
<210> 15
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 15
cttaccattt gcggtgcctg gtttc 25
<210> 16
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 16
tcacacacta cagcttccac cacaa 25
<210> 17
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 17
tgggcgcgcc agactctaaa 20
<210> 18
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 18
tgggtctgat ggccagtatt gact 24
<210> 19
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 19
ggcctggaaa actcagctat ccttt 25
<210> 20
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 20
tcacacacta cagcttccac cacaa 25
<210> 21
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 21
tgggcgcgcc agactctaaa 20
<210> 22
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 22
tgggtctgat ggccagtatt gact 24
<210> 23
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 23
ggcctggaaa actcagctat ccttt 25
<210> 24
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 24
agacttgatg gtgtggagtg gggta 25
<210> 25
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 25
tgggccctgt actttgcttg aacat 25
<210> 26
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 26
tgatgggtca accatgttcc tgtgg 25
<210> 27
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 27
tcagccattg cttctgcttt ctcct 25
<210> 28
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 28
gaaaagtggc tttgatggtg caggg 25
<210> 29
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 29
tgcccatttt tctgcccttg ggtat 25
<210> 30
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 30
aggcattcct tatgccagtc agcat 25
<210> 31
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 31
tttcccatgt cctgctgctt tcctt 25
<210> 32
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 32
catttaggga gctgactggg cacaa 25
<210> 33
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 33
tctgggtcct aactgagcag ctctt 25
<210> 34
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 34
ctaacactca ttcctgttga agctcttgac 30
<210> 35
<211> 3012
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Реарранжированная последовательность нуклеиновой кислоты IGKV1-39+J4 человека
<400> 35
gcaggcagca tatttataat gcacagtaaa cactaggagg aaacaaggca gtgagagagg 60
aaagagagca gcgataccga aaatgtcctc agcgagaagc taccacagag gatgaatgga 120
gatcaagccc acgtggaaac atgggaaaat gtctcagtat ttttccacct aagaagggag 180
ggagatgggg tatgtataca cctccctgtc ctcactgatt gagggctttc cgagaggatg 240
ctcattccag gtgctgtgat aggccatgtg tacaggcagg gctgccttct ccagcttcag 300
atagagcagt gaggaaaaga tatggccatg gggggtcagc agaagtacag caaagggaaa 360
agggaaaggg tagcaagagt gacaactata ttcacccccc ccacacacac acacacacac 420
acgaaattgt gtattgcaat ccagaactgc ttctctctga acctaaatct tagcaagcag 480
tttaccagta actgcccttg aaattcaggc ccctggaaag gagcaggggg ttgtgtacag 540
gctataccac agcagtctgc ccacccttag tgatgcatga gtaatgctcc ctggactccc 600
caggttctag tcttctcatg tcgatgtagt tgattccact tcccttgctg cacaaccagg 660
ctgggatgcc tgggcagagg cagacatgtg aggtataggg gttcaaatct gtttccaagt 720
tttatccagc ttcaaagcat ttctccgtgt acatgagcgg tggcttgaca ggagatggag 780
actctctttc ctggatgtga ggcaaggagg caggcgtctg agtcaggatg atgtccctac 840
tcactgctaa agagaaaagt ggctttgatg gtgcagggca gggaaatgca ctgagtggtc 900
gccaccctca cagaagagaa agtgttcact gacctggcct ttccccaggg cctctccctc 960
ccattgcttt ccagaaagcc atgatttttg agagccacac ctgaacactc acaaacatta 1020
tggtgggaaa agcagatcag agcattaggc aagttgcatt accttggcct tcttcctttg 1080
gagacaattg atgtggggtt ctagattgac ccagagtttc aagtttatcc tgattcaggc 1140
ttcaacagct ggaggaagaa acagagatgt tttttgaagt aaacagatct agcattacta 1200
atcaaccctt catactgatg acctatggga aataataccc aagggcagaa aaatgggcag 1260
aataagggga gccccaaacc aagacgaagc tgctgcccat tgagaccctg ggtattacag 1320
agacctatag ctctggataa tggaagatct atgagtggca caggcgctga ggaatcacag 1380
catcattatc gtgcatctgc agggaattgc ttgtaaatat actggtaatt acaaatgttt 1440
aaggtcacta caaatacttt ggagtgtatt aaatatgctt ctgataaaga ctgtttttct 1500
cacatgaaac aatgggaacc atgtgacaat cacagaggtg ttgttactat agcaaaaggg 1560
attgttactc tccacatccc tttaagtaac ttgaaggcct gatagaccca ccctctaaga 1620
cttcattaga cattccctac gaatggttat actctcctgt atactcccaa tacaactcta 1680
aaatatatta ttccatatag tccttaggtt tgtattaaag tttgactttt ttccttcaaa 1740
atatctcttg tcacaacagc ggctctagag agaaatacat tccctccagg caaatctatg 1800
ctgcgctggt ctgacctggg accctgggga cattgcccct gtgctgagtt actaagatga 1860
gccagccctg cagctgtgct cagcctgccc catgccctgc tgattgattt gcatgttcca 1920
gagcacagcc ccctgccctg aagacttttt tatgggctgg tcgcaccctg tgcaggagtc 1980
agtctcagtc aggacacagc atggacatga gggtccccgc tcagctcctg gggctcctgc 2040
tactctggct ccgaggtaag gatggagaac actaggaatt tactcagcca gtgtgctcag 2100
tactgactgg aacttcaggg aagttctctg ataacatgat taatagtaag aatatttgtt 2160
tttatgtttc caatctcagg tgccagatgt gacatccaga tgacccagtc tccatcctcc 2220
ctgtctgcat ctgtaggaga cagagtcacc atcacttgcc gggcaagtca gagcattagc 2280
agctatttaa attggtatca gcagaaacca gggaaagccc ctaagctcct gatctatgct 2340
gcatccagtt tgcaaagtgg ggtcccatca aggttcagtg gcagtggatc tgggacagat 2400
ttcactctca ccatcagcag tctgcaacct gaagattttg caacttacta ctgtcaacag 2460
agttacagta cccctctcac tttcggcgga gggaccaagg tggagatcaa acgtaagtaa 2520
tttttcacta ttgtcttctg aaatttgggt ctgatggcca gtattgactt ttagaggctt 2580
aaataggagt ttggtaaaga ttggtaaatg agggcattta agatttgcca tgggttgcaa 2640
aagttaaact cagcttcaaa aatggatttg gagaaaaaaa gattaaattg ctctaaactg 2700
aatgacacaa agtaaaaaaa aaaagtgtaa ctaaaaagga acccttgtat ttctaaggag 2760
caaaagtaaa tttatttttg ttcactcttg ccaaatattg tattggttgt tgctgattat 2820
gcatgataca gaaaagtgga aaaatacatt ttttagtctt tctccctttt gtttgataaa 2880
ttattttgtc agacaacaat aaaaatcaat agcacgccct aagaaaaatc agggaaaagt 2940
gaagtgtacc tatttgctat gtagaagagg cagcttactt gaaaatcagc agcaatgttg 3000
tttttagagt ct 3012
<210> 36
<211> 3051
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Реарранжированная последовательность нуклеиновой кислоты IGKV3-11+J1 человека
<400> 36
caggagtgag cctggtttga tgcctctctc cccaacacga tagaagtgtg gcacaaatct 60
ttgaaaaatt cagtttcccc gtggcaacaa caactacctg ggactgaaaa cttcttccct 120
cgctctagtc ctttcttcta cacccacttc cacctcatct gtgactcata caatacttgt 180
caggaaagat tctggaaaaa gcaaagagac ttccttagag gtgtcagaga ttcctgtacc 240
agcatctgtc catctctaga gggggttgtg agtatgagga agagcagagc ttgtcaatct 300
tctacttgct ttcactccca ctgtatttcc taacaacagc aaccagagca acagccatat 360
catcacagga cgaacctctc ctacttccaa ggcttttatt tcagtaaatc tgctctacct 420
ctatctcagg cagctagaag ttttgatact catacaaata ctactgcagc tttctgttca 480
taattggaaa agtagacaag actcagtgta atgcaggcat tccttatgcc agtcagcatt 540
cagtttttgg atcatcattg cacacatata cccaccatgt gtctaatata tatgtaaaaa 600
tccatgaagc aagagtcata atagctagca tttgatattg tattgtattt tcctcttata 660
tcatcttctc cttttcgtcc ttaaaaaaaa tctgttcaag tcagtctaaa ttaattattg 720
gatgataagt agataaaatc ttttatttga taacacattg acccaatgaa tatgtttctt 780
tgcaagacat agtcctcact tccaagataa caagcctgac aaaattatac tggagcaagt 840
ccacaagtaa tgatggtagc ttttccttat tgtcagtcct ggggcaaaaa taagacaaaa 900
gataacaagg tagactaaag attatgtaag aaagaaggaa agcagcagga catgggaaac 960
tttcatagga taacattttg ataatggatg atgagaatta atgcgttaga cagagatggg 1020
cgggaatgat tgaaggtctg agcattttag cacagattaa gaccaaatca ttaggatttt 1080
aagagttgtg tacagttagt gaagaaaaag ccctagaatt taatttgact gttgataaaa 1140
cattcttgga ttagattgaa gactcttttc tgtgccaagt aagtatattt atgataatga 1200
tgatgactgt agtgctaaat atttaatcaa taaaaacaaa aataattgcc gcatacataa 1260
tgtcctgagt actactgtaa atgttttatc ttattttctt taaactgtct acagcactgt 1320
aaggtaggca ccagtattgt cacagttaca cagatatgga aactgagaca cagggaagtt 1380
aagttagttg gtcaatttca agcaatcggc aagccatgga gcatctatgt cagggctgcc 1440
aggacatgtg actgtaaaca gaagttttaa ctttttaact caaagagggt atgtgtctgg 1500
gttaatggaa agtttcagga ccctcagaaa acattactaa caagcaaatg aaaggtgtat 1560
ctggaagatt aagttctaac agactcctca tttccatcga tccaataatg cacttaggga 1620
gatgactggg catattgagg ataggaagag agaaatgaaa acacagcctt ttatattgtt 1680
cttaacaggc ttgtgccaaa catcatctgg gtgaatttag gtgattgagg agaagaaaga 1740
cataggaatg aaattctctg agcacaaggg agaagttcta cactcagact gagccaacag 1800
acttttctgg cctgacaacc agggtggcgc aggatgctca gtgcagagag gaagaagcag 1860
gtggtctctg cagctggaag ctcagctccc acccagctgc tttgcatgtc cctcccagct 1920
gccctacctt ccagagccca tatcaatgcc tgtgtcagag ccctggggag gaactgctca 1980
gttaggaccc agagggaacc atggaagccc cagctcagct tctcttcctc ctgctactct 2040
ggctcccagg tgaggggaac atgaggtggt tttgcacatt agtgaaaact cttgccacct 2100
ctgctcagca agaaatataa ttaaaattca aagtatatca acaattttgg ctctactcaa 2160
agacagttgg tttgatcttg attacatgag tgcatttctg ttttatttcc aatttcagat 2220
accaccggag aaattgtgtt gacacagtct ccagccaccc tgtctttgtc tccaggggaa 2280
agagccaccc tctcctgcag ggccagtcag agtgttagca gctacttagc ctggtaccaa 2340
cagaaacctg gccaggctcc caggctcctc atctatgatg catccaacag ggccactggc 2400
atcccagcca ggttcagtgg cagtgggtct gggacagact tcactctcac catcagcagc 2460
ctagagcctg aagattttgc agtttattac tgtcagcagc gtagcaactg gcctccgacg 2520
ttcggccaag ggaccaaggt ggaaatcaaa cgtaagtaat ttttcactat tgtcttctga 2580
aatttgggtc tgatggccag tattgacttt tagaggctta aataggagtt tggtaaagat 2640
tggtaaatga gggcatttaa gatttgccat gggttgcaaa agttaaactc agcttcaaaa 2700
atggatttgg agaaaaaaag attaaattgc tctaaactga atgacacaaa gtaaaaaaaa 2760
aaagtgtaac taaaaaggaa cccttgtatt tctaaggagc aaaagtaaat ttatttttgt 2820
tcactcttgc caaatattgt attggttgtt gctgattatg catgatacag aaaagtggaa 2880
aaatacattt tttagtcttt ctcccttttg tttgataaat tattttgtca gacaacaata 2940
aaaatcaata gcacgcccta agaaaaatca gggaaaagtg aagtgtacct atttgctatg 3000
tagaagaggc agcttacttg aaaatcagca gcaatgttgt ttttagagtc t 3051
<210> 37
<211> 3072
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Реарранжированная последовательность нуклеиновой кислоты IGK V3-20+J1
человека
<400> 37
agccagtgga tgactatttt ctttcctgag actgataatg gtcattccac ccaatggtcc 60
aaactacatg cagtggtaat ggtcatgcag gcctttctgc cacctcttgc tatattttta 120
ccaactcatg ggccactgcc aacagtctag ctatctgatc aggaaaatag caaccgagtg 180
actggactat taaagcatcc cctctgtgga gaaaaggact atggcaacag cttgccacct 240
ataagagaca aatatgtcac tcagctggat gctgggtcta ccatgaccac ccttgagatg 300
aacttatgcc atgtttttgg atgcccactg agactttgct ttgaccaagg aaagttcttt 360
actgcccaaa tgacgtcaat ggacacactc tcatgggaca tgatggattt tccatacacc 420
ttgttatcca aaggccaatg gatctattta atgctagaac agctgactca cacggcaact 480
gaagagtgta catcagggca acctgctagt agagtggtac cccatctgac tacagcaatt 540
ggacattaaa cactgcactg cagagcaagg gaaagacggc acggaggtgc aggttgaaaa 600
acactgagtt gagtgaggaa gaagttgggc aaggccagtt tcctgataga gctgccactg 660
tgaaatccca aactcagtgt ttccaatcat ttttttcctt ttatagtgat gttcttgggg 720
gtgttgcagt ttaggccacc atgataaccc agacaggcct cctaactcta attggatgga 780
atgattcccc tgggtgcctt ctatgggttc cacaggatca agaaatatca gggtgttaat 840
ttccttctgc tggtaggagg gtcatctgat tcctccatta gggtggatga cgttatcaga 900
catgtcaatt ttctacagta cttgacctgc cttctagaac tggacaaccg agggtgaaag 960
gactggatca agcaacaatg gcaatgggtg cccaaagagg tagtagtctc agaacaggga 1020
gagacagact gagtccctcc actaactcag cccaacacct gtggatgagt aggggatgcc 1080
tgagatcctg gggtggatgg gaggtggggc actgatctgt caatctgctt tttcttcaag 1140
gatcaggcag cagagaccca gaagcttcat gtctttgtaa ggctcttcca agccaacaca 1200
gataatttga caaaacactg tatctgcatc ccagacctca ccctgtcaca gactgaagtg 1260
gttgtttcat cctactaagg gtaaactata ccagctatac agaataaaaa gactggatag 1320
tttagaggat cacccaagaa atagttcttt gcctgcatgg acaaaaccat cttctgtctt 1380
tagggaaatg gtatcactac cctgaggatt tggagcccag gtctcactta tctgtgcagt 1440
tgtgaaagtc ctcacaccca cagtgctgag gttaattgaa tgttactctt ttaatttctg 1500
caaagaatga gacagcttct ggaccctcag gaaagatcac taacaagtaa atacaagtat 1560
atccggaaga taaagttgta atagactctt cctttcaacc tgatccatca tgcatttagg 1620
gagctgactg ggcacaagtt ggagcagaaa gagaaaaatg aaaccacagc cttctatttt 1680
gtttctaaca gacttgtacc aaacattctg tggctcaatc taggtgatgg tgagacaaga 1740
ggacacaggg gttaaattct gtggccgcag gggagaagtt ctaccctcag actgagccaa 1800
cggccttttc tggcctgatc acctgggcat gggctgctga gagcagaaag gggaggcaga 1860
ttgtctctgc agctgcaagc ccagcacccg ccccagctgc tttgcatgtc cctcccagcc 1920
gccctgcagt ccagagccca tatcaatgcc tgggtcagag ctctggagaa gagctgctca 1980
gttaggaccc agagggaacc atggaaaccc cagcgcagct tctcttcctc ctgctactct 2040
ggctcccagg tgaggggaac atgggatggt tttgcatgtc agtgaaaacc ctctcaagtc 2100
ctgttacctg gcaactctgc tcagtcaata caataattaa agctcaatat aaagcaataa 2160
ttctggctct tctgggaaga caatgggttt gatttagatt acatgggtga cttttctgtt 2220
ttatttccaa tctcagatac caccggagaa attgtgttga cgcagtctcc aggcaccctg 2280
tctttgtctc caggggaaag agccaccctc tcctgcaggg ccagtcagag tgttagcagc 2340
agctacttag cctggtacca gcagaaacct ggccaggctc ccaggctcct catctatggt 2400
gcatccagca gggccactgg catcccagac aggttcagtg gcagtgggtc tgggacagac 2460
ttcactctca ccatcagcag actggagcct gaagattttg cagtgtatta ctgtcagcag 2520
tatggtagct caccttggac gttcggccaa gggaccaagg tggaaatcaa acgtaagtaa 2580
tttttcacta ttgtcttctg aaatttgggt ctgatggcca gtattgactt ttagaggctt 2640
aaataggagt ttggtaaaga ttggtaaatg agggcattta agatttgcca tgggttgcaa 2700
aagttaaact cagcttcaaa aatggatttg gagaaaaaaa gattaaattg ctctaaactg 2760
aatgacacaa agtaaaaaaa aaaagtgtaa ctaaaaagga acccttgtat ttctaaggag 2820
caaaagtaaa tttatttttg ttcactcttg ccaaatattg tattggttgt tgctgattat 2880
gcatgataca gaaaagtgga aaaatacatt ttttagtctt tctccctttt gtttgataaa 2940
ttattttgtc agacaacaat aaaaatcaat agcacgccct aagaaaaatc agggaaaagt 3000
gaagtgtacc tatttgctat gtagaagagg cagcttactt gaaaatcagc agcaatgttg 3060
tttttagagt ct 3072
<210> 38
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> общая легкая цепь VL, кодируемая IGKV1-39/IGKJ4 человека
<400> 38
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 39
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> общая легкая цепь VL, кодируемая IGKV3-11/IGKJ1 человека
<400> 39
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 40
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> общая легкая цепь VL, кодируемая IGKV3-20/IGKJ1 человека
<400> 40
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 41
<211> 107
<212> Белок
<213> Mus musculus
<400> 41
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
1 5 10 15
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
35 40 45
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
65 70 75 80
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
85 90 95
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМ ИММУНОГЛОБУЛИНОВЫМ ЛОКУСОМ | 2020 |
|
RU2822367C2 |
| КОЛЛЕКЦИЯ И СПОСОБЫ ЕЕ ПРИМЕНЕНИЯ | 2010 |
|
RU2569187C2 |
| АНТИТЕЛА И ДРУГИЕ МОЛЕКУЛЫ, КОТОРЫЕ СВЯЗЫВАЮТ В7-Н1 И PD-1 | 2012 |
|
RU2625034C2 |
| КОЛЛЕКЦИЯ И СПОСОБЫ ЕЕ ПРИМЕНЕНИЯ | 2011 |
|
RU2603080C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ ДЛЯ ТЕСТИРОВАНИЯ ТЕРАПЕВТИЧЕСКИХ АГЕНТОВ | 2019 |
|
RU2805212C2 |
| ПРОДУЦИРУЮЩИЕ АНТИТЕЛА МЛЕКОПИТАЮЩИЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ | 2020 |
|
RU2820926C2 |
| ПРОДУЦИРУЮЩИЕ АНТИТЕЛА МЛЕКОПИТАЮЩИЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ | 2009 |
|
RU2559524C2 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА К ОНКОСТАТИНУ М И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2600444C2 |
| ПРОДУЦИРУЮЩИЕ АНТИТЕЛА МЛЕКОПИТАЮЩИЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ | 2009 |
|
RU2731084C2 |
| ИДЕНТИФИКАЦИЯ АССОЦИИРОВАННЫХ С ОБРАЗЦОМ ПОЛИНУКЛЕОТИДОВ | 2012 |
|
RU2644681C2 |
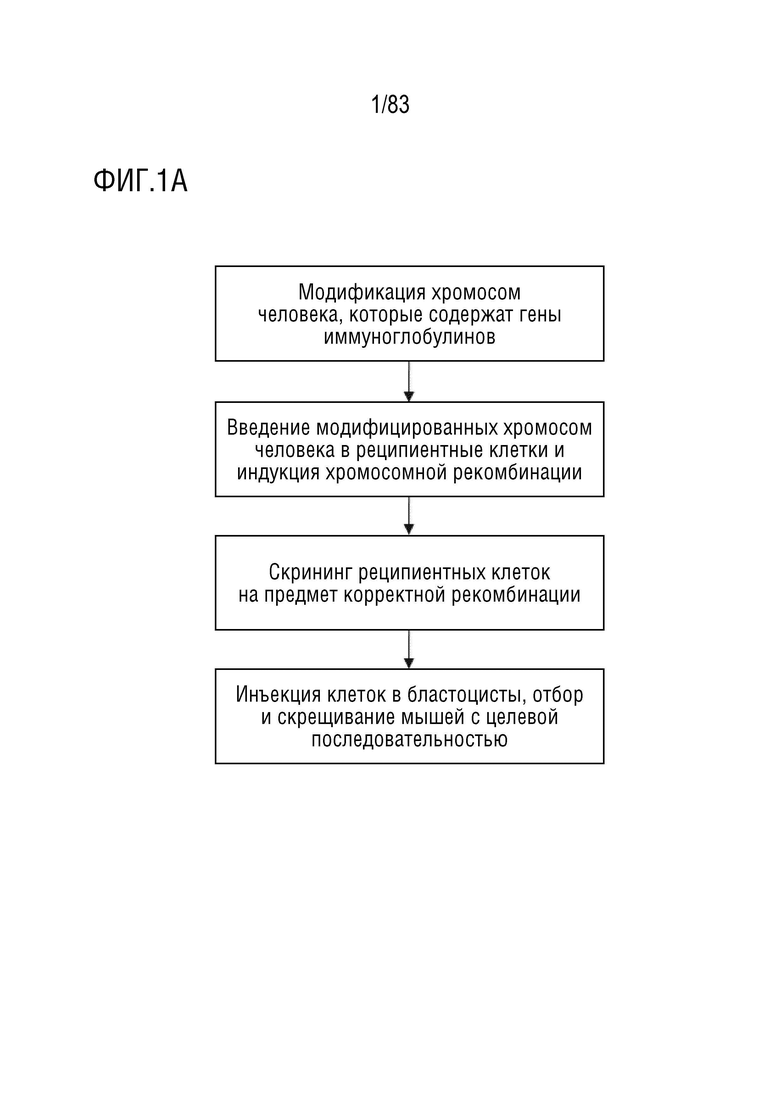
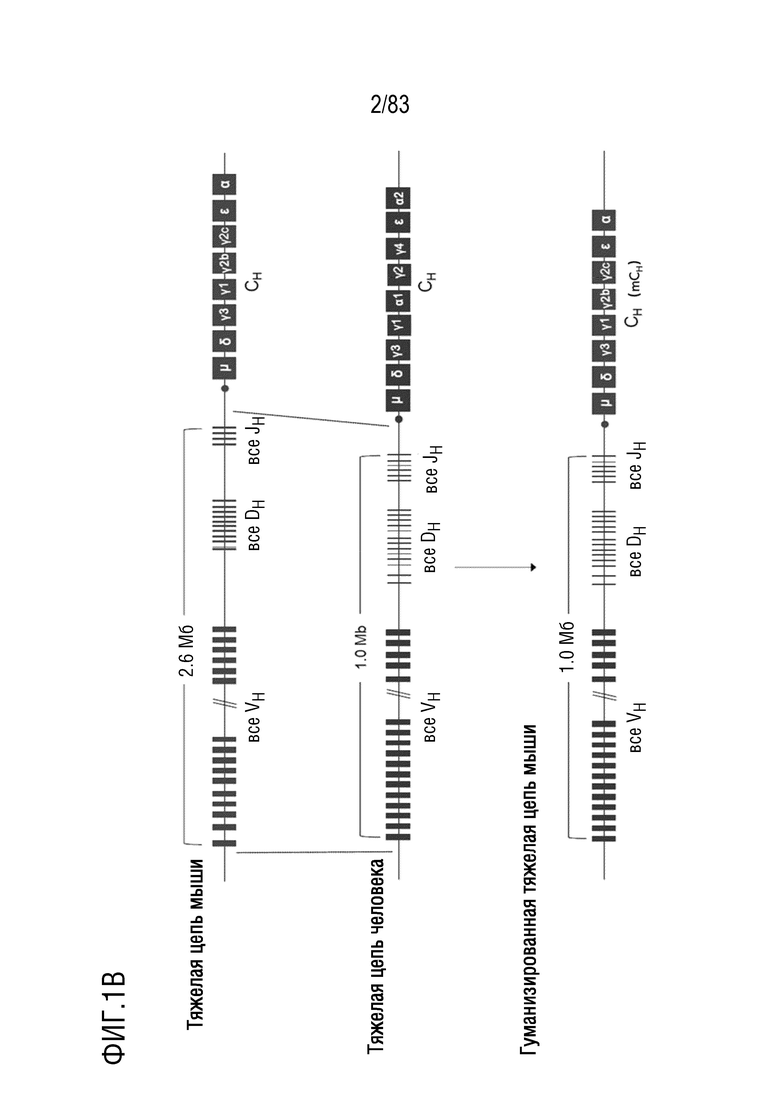
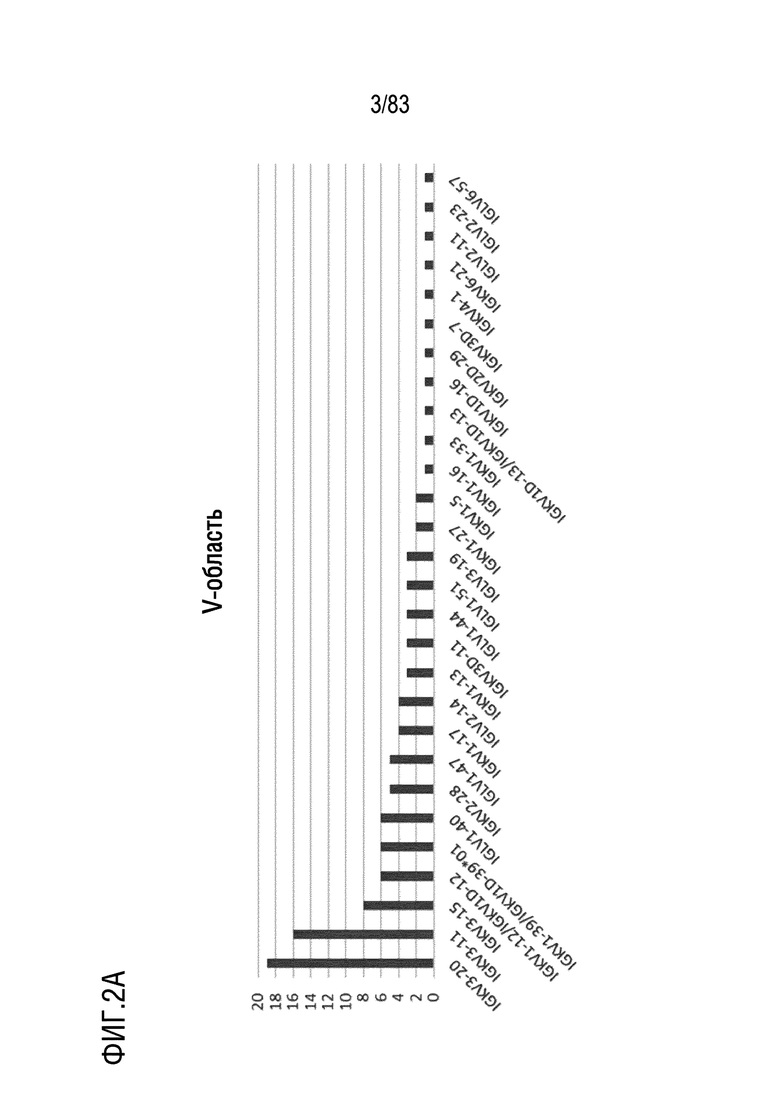
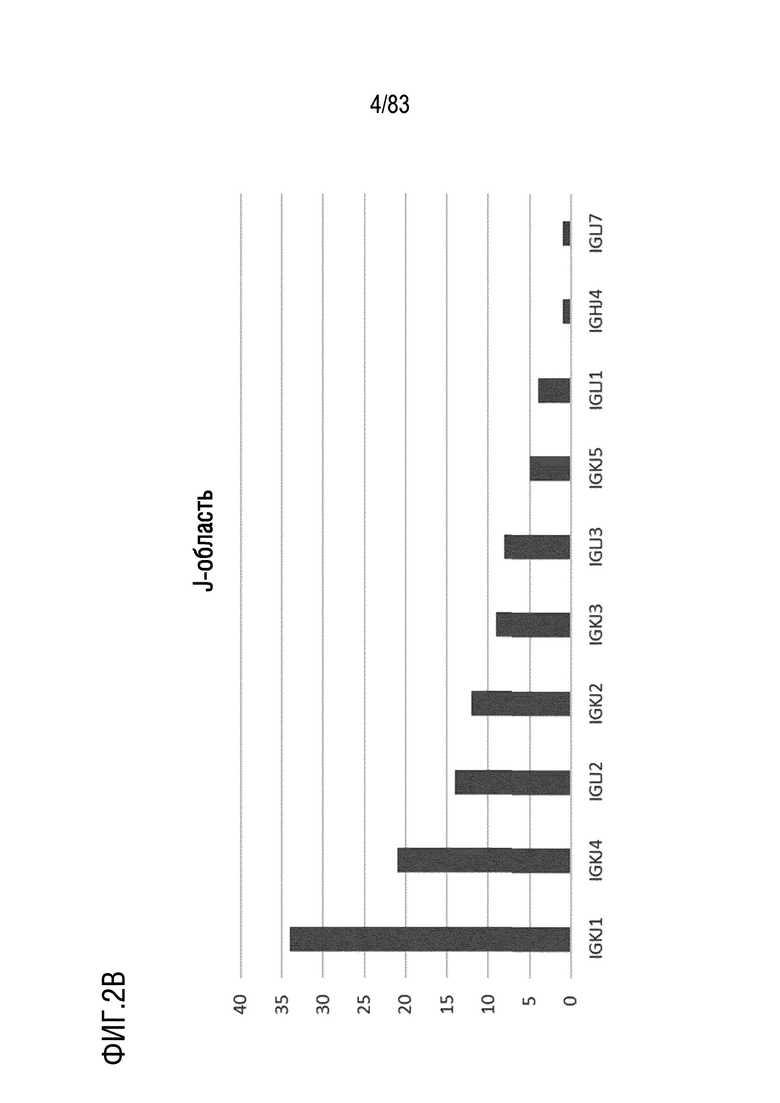
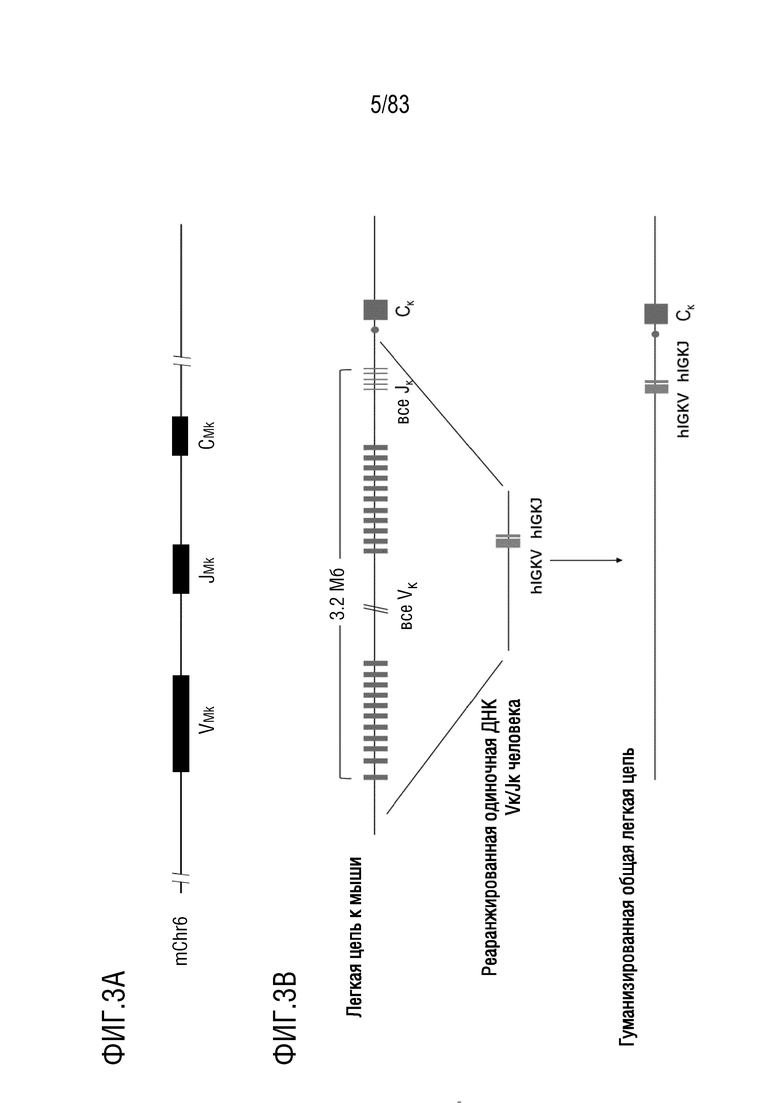
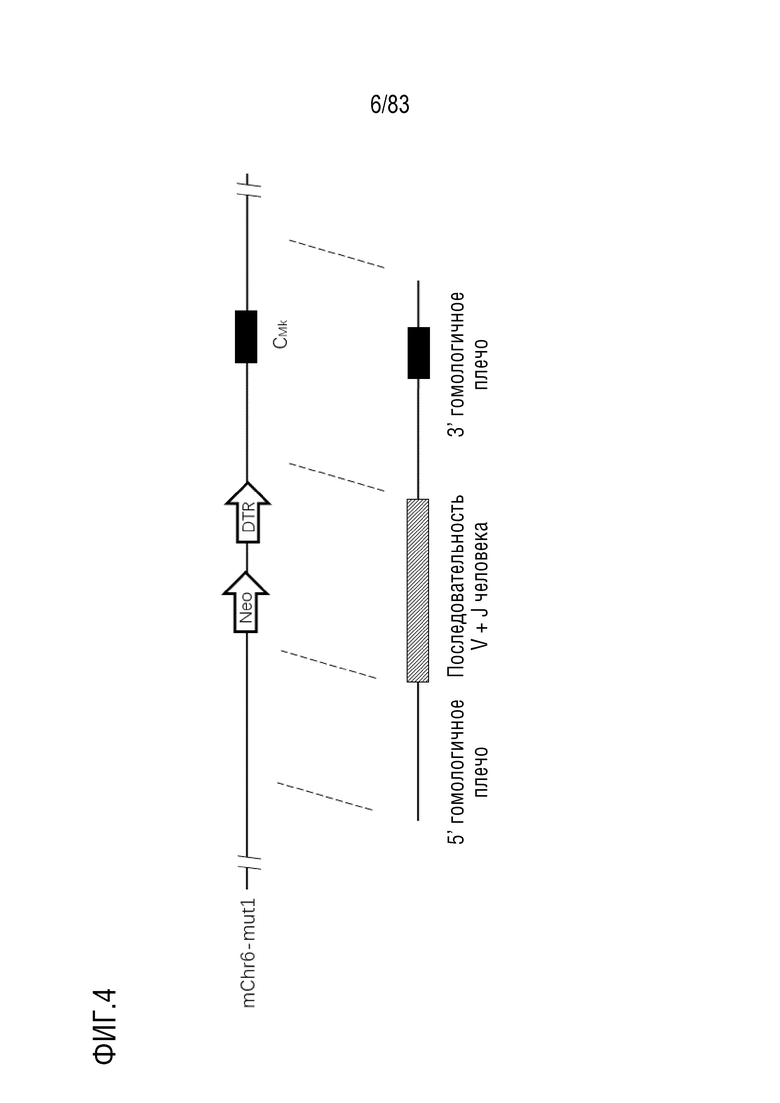
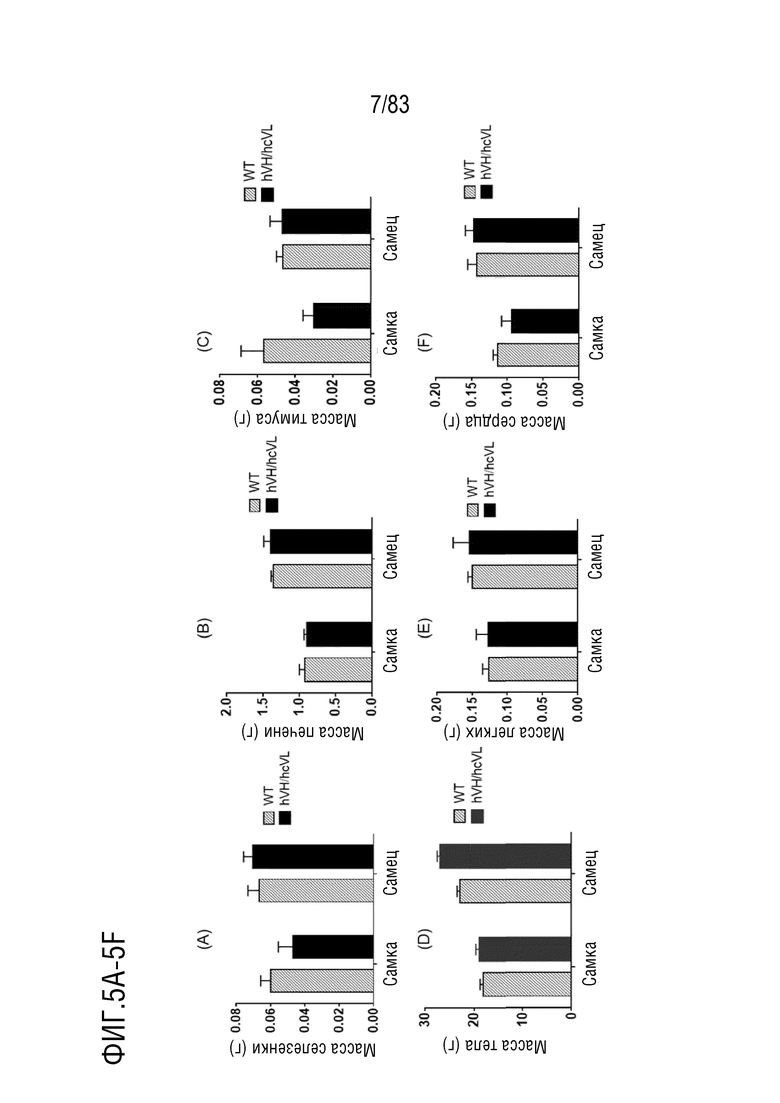
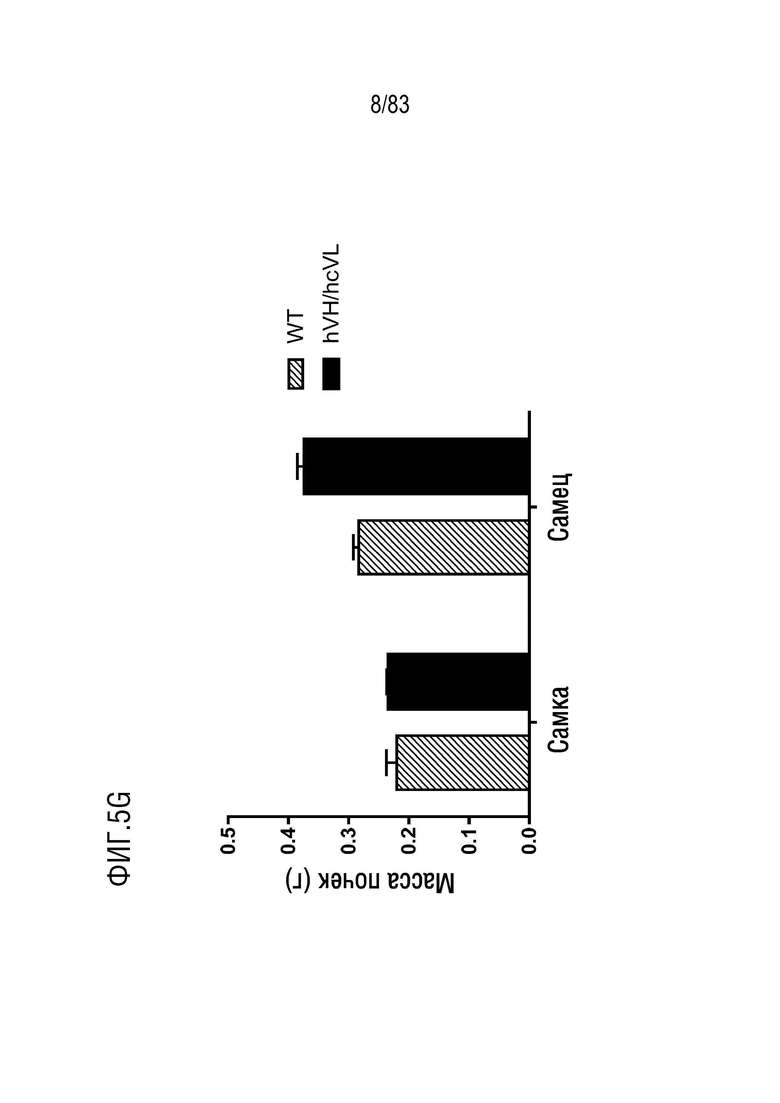
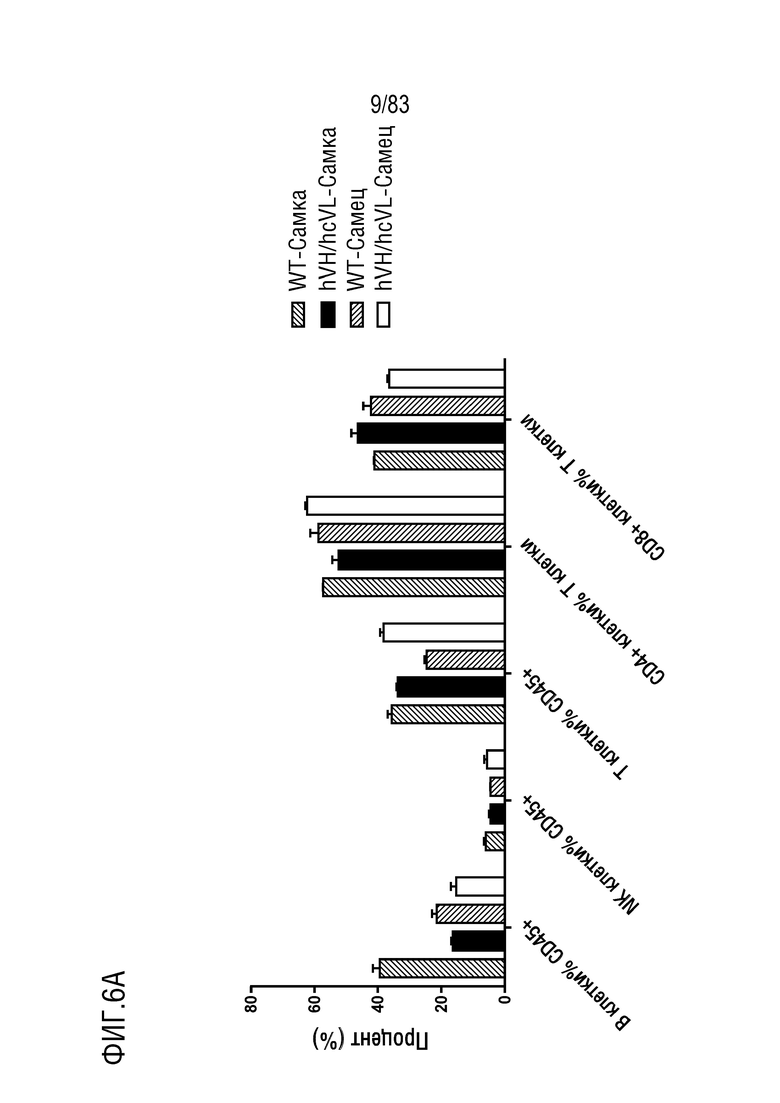
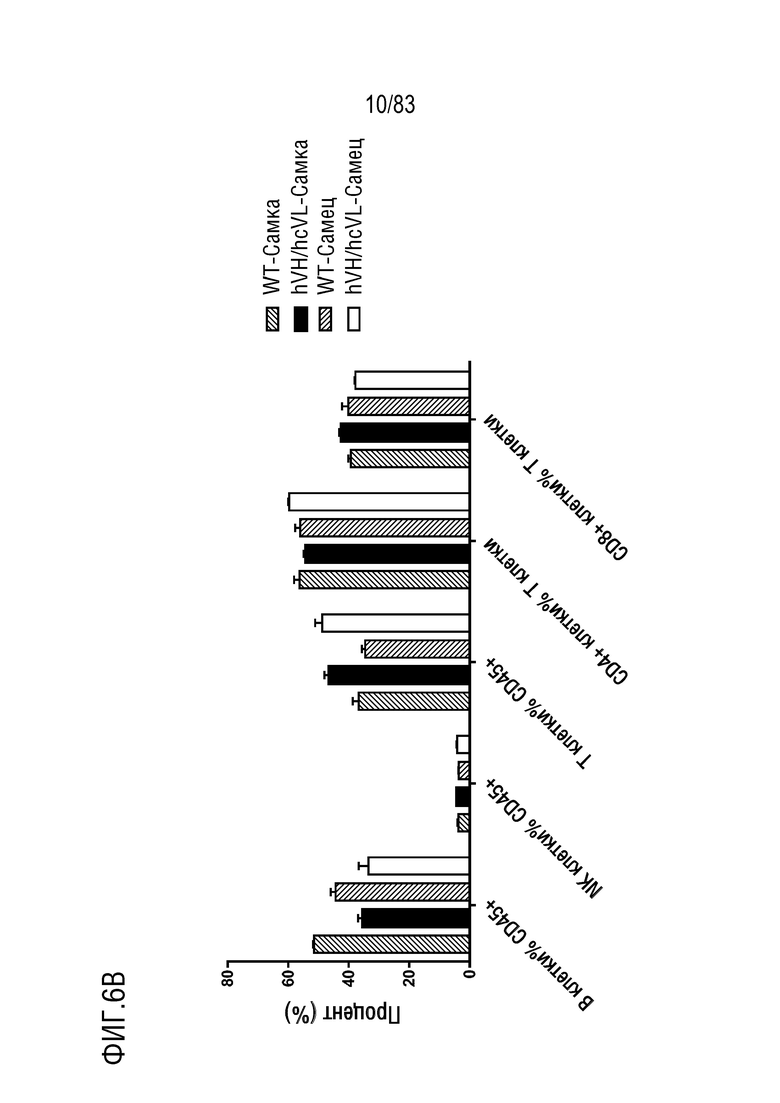
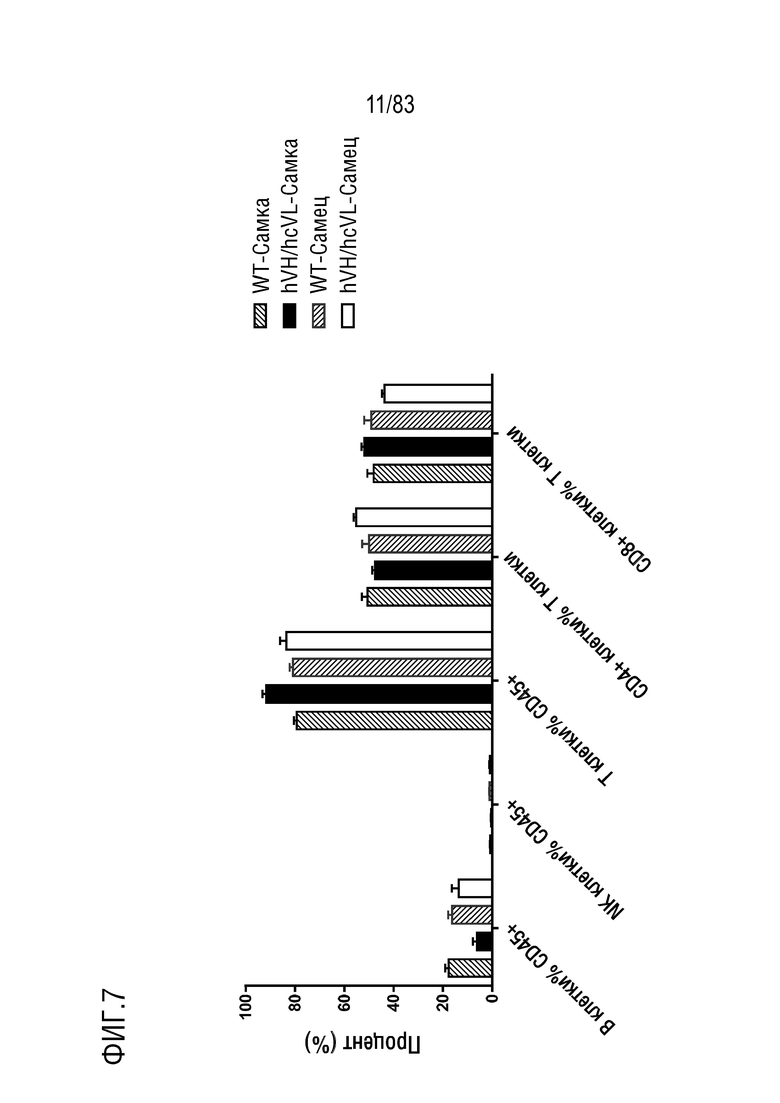
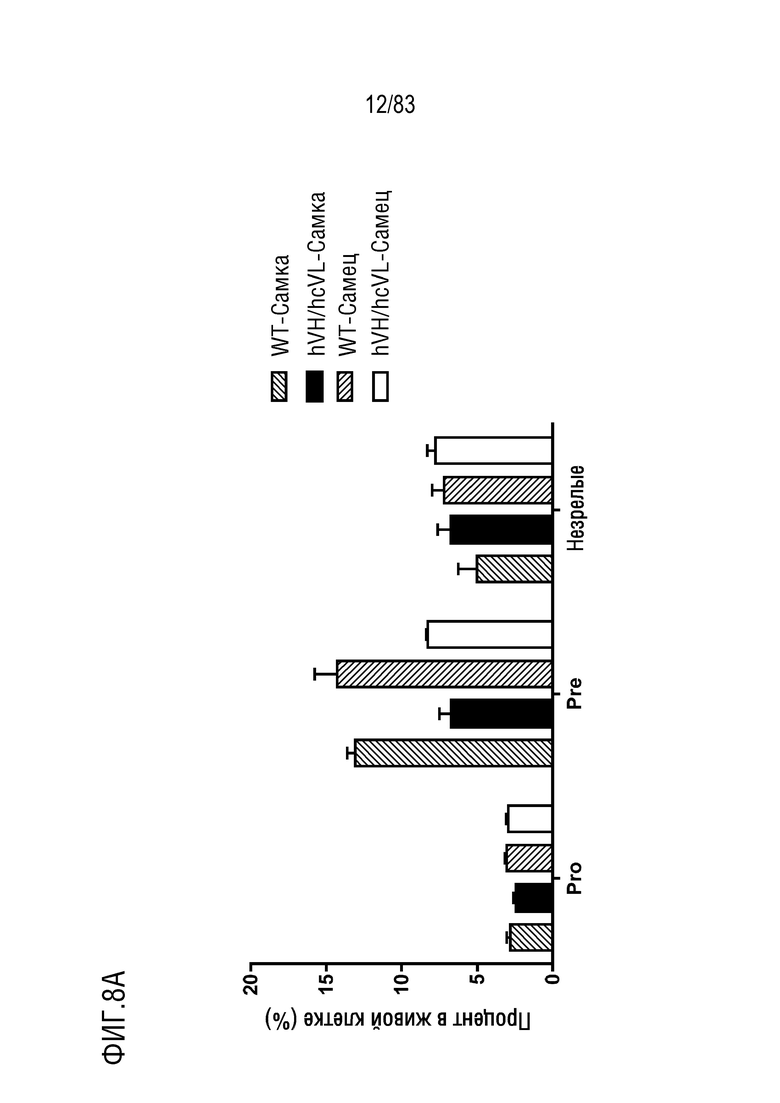
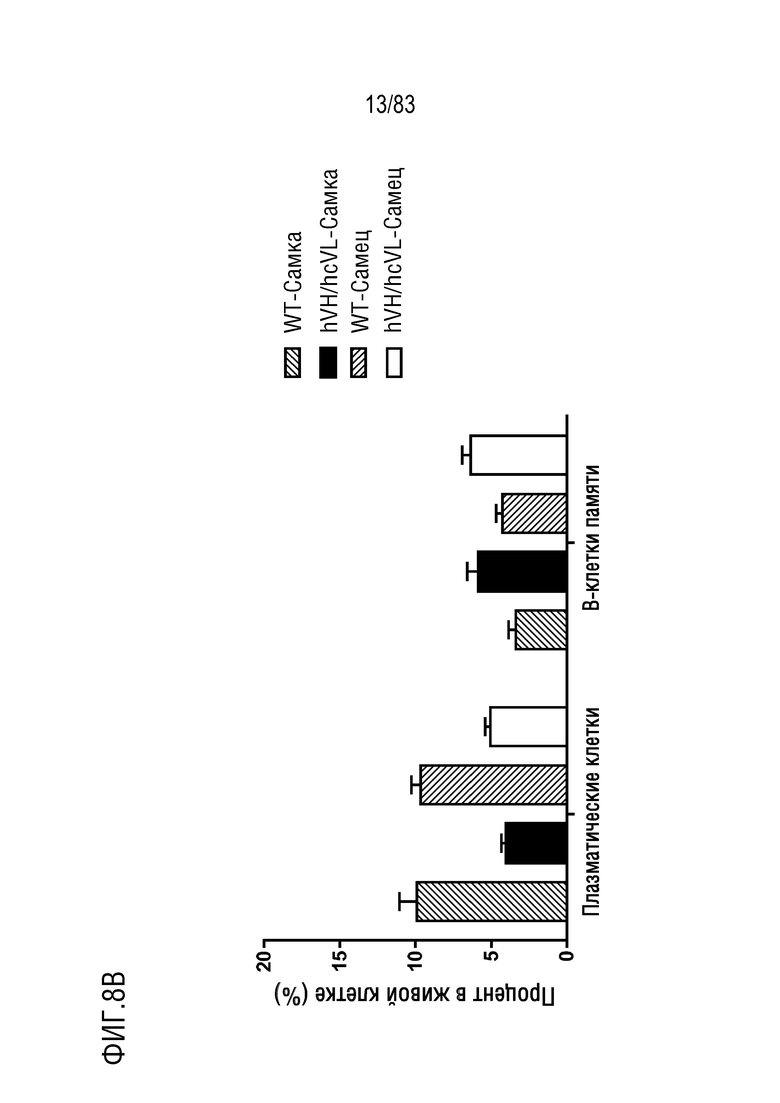
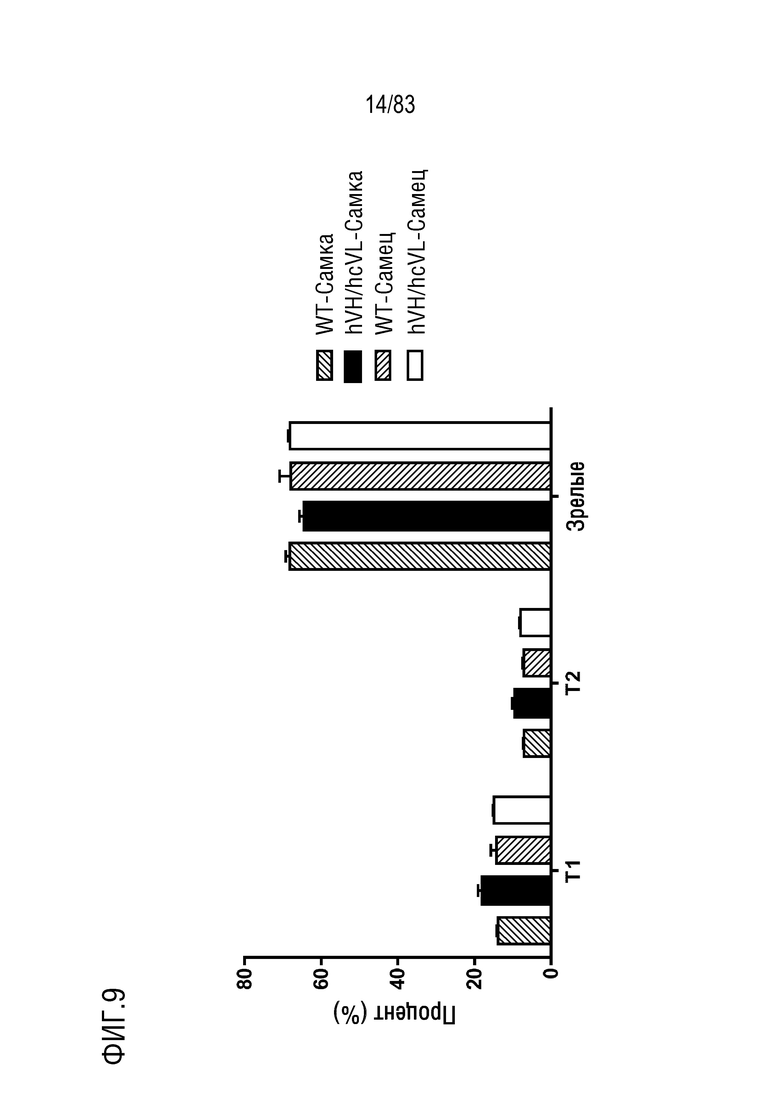
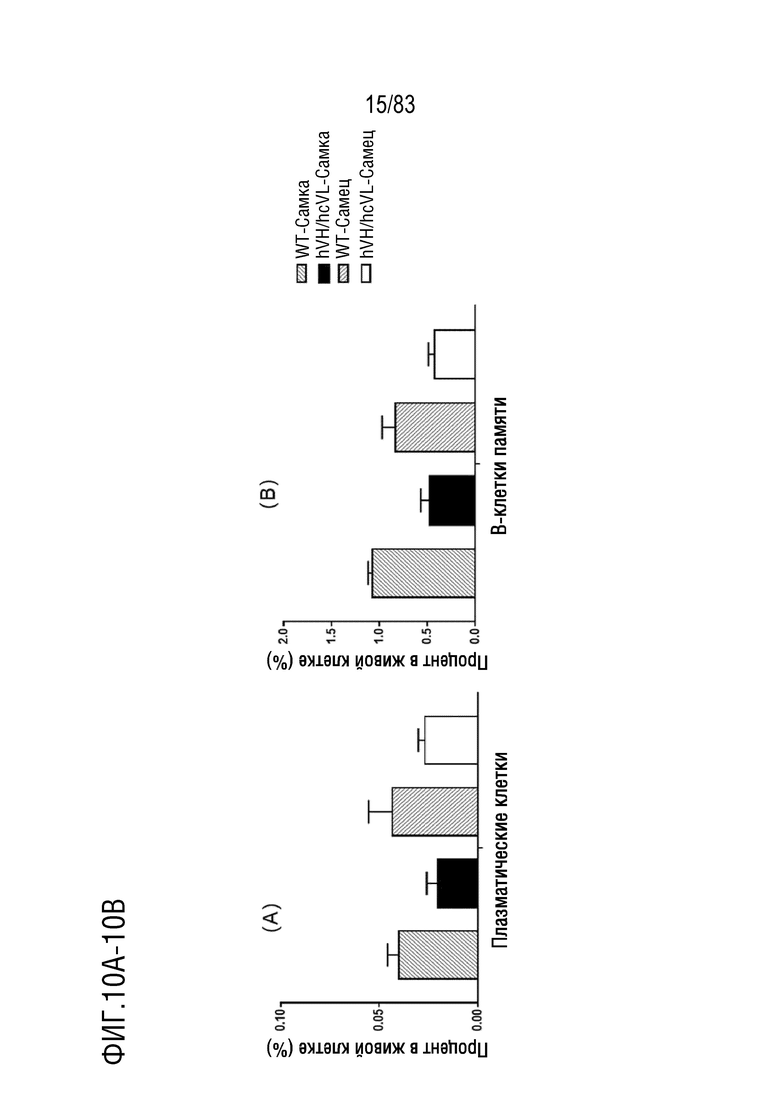
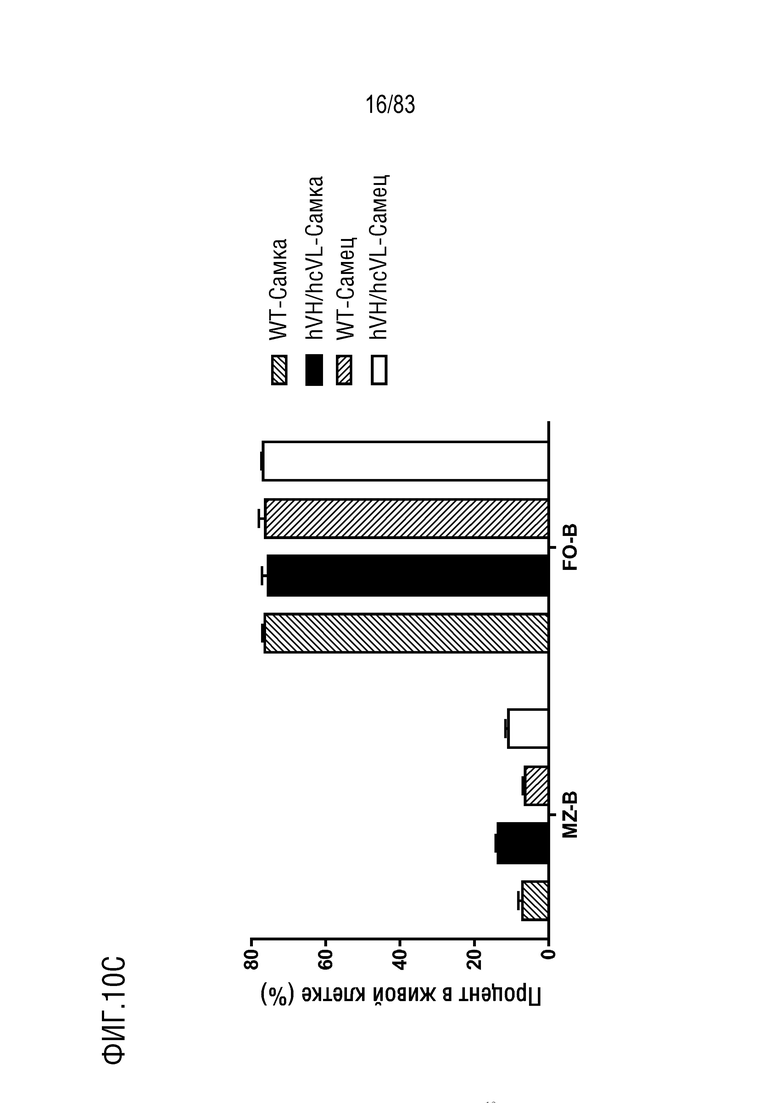
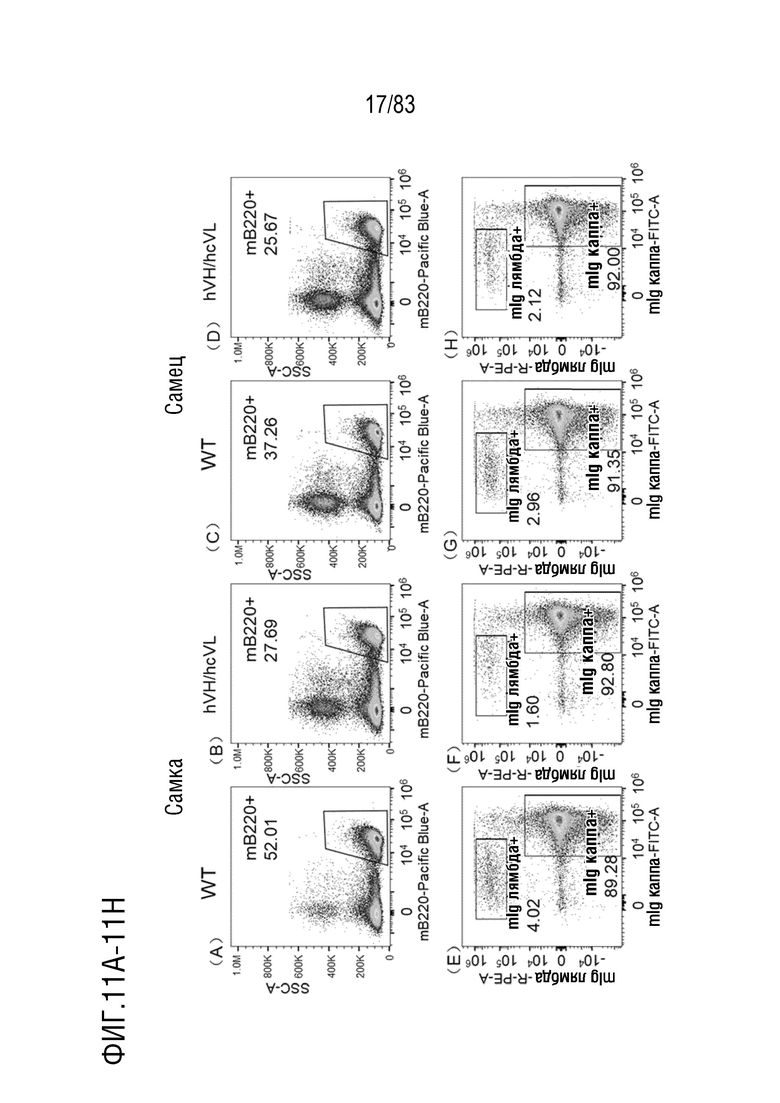
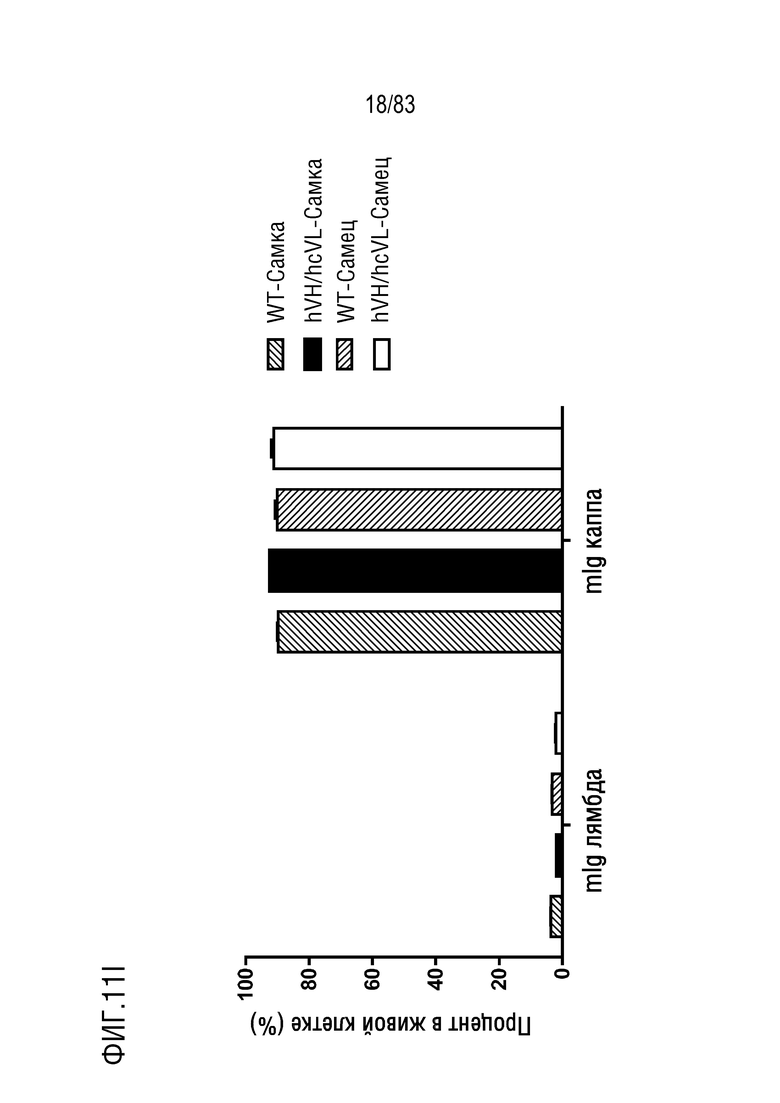
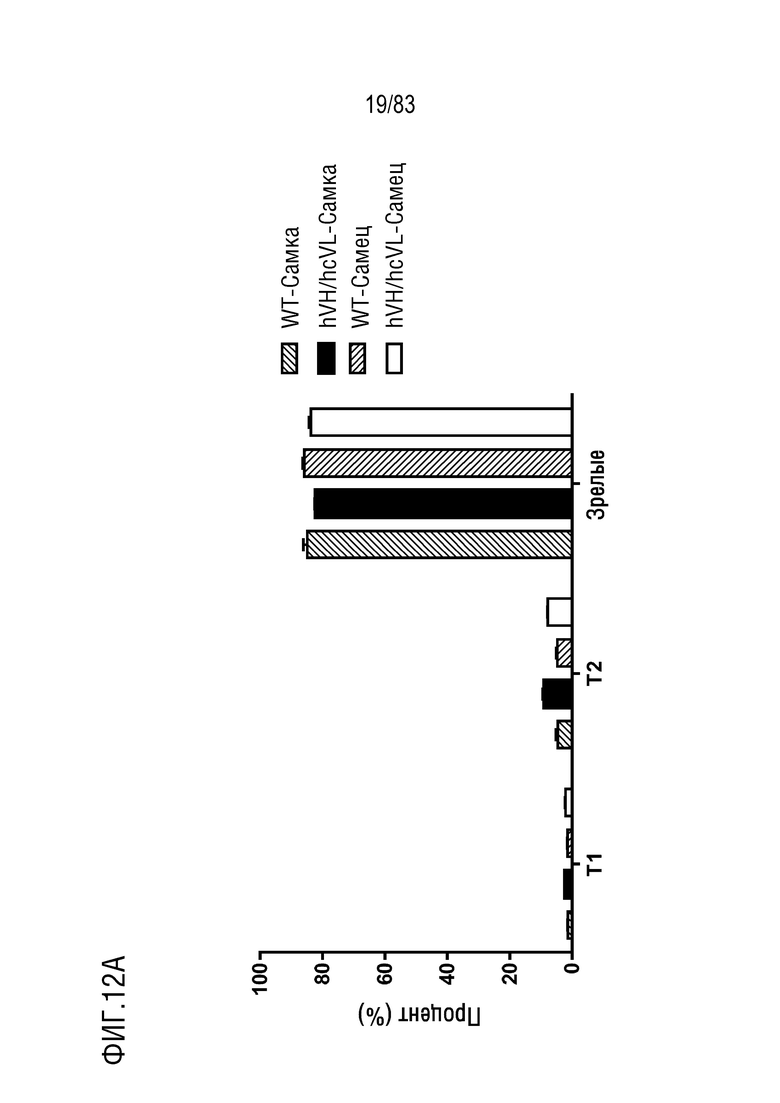
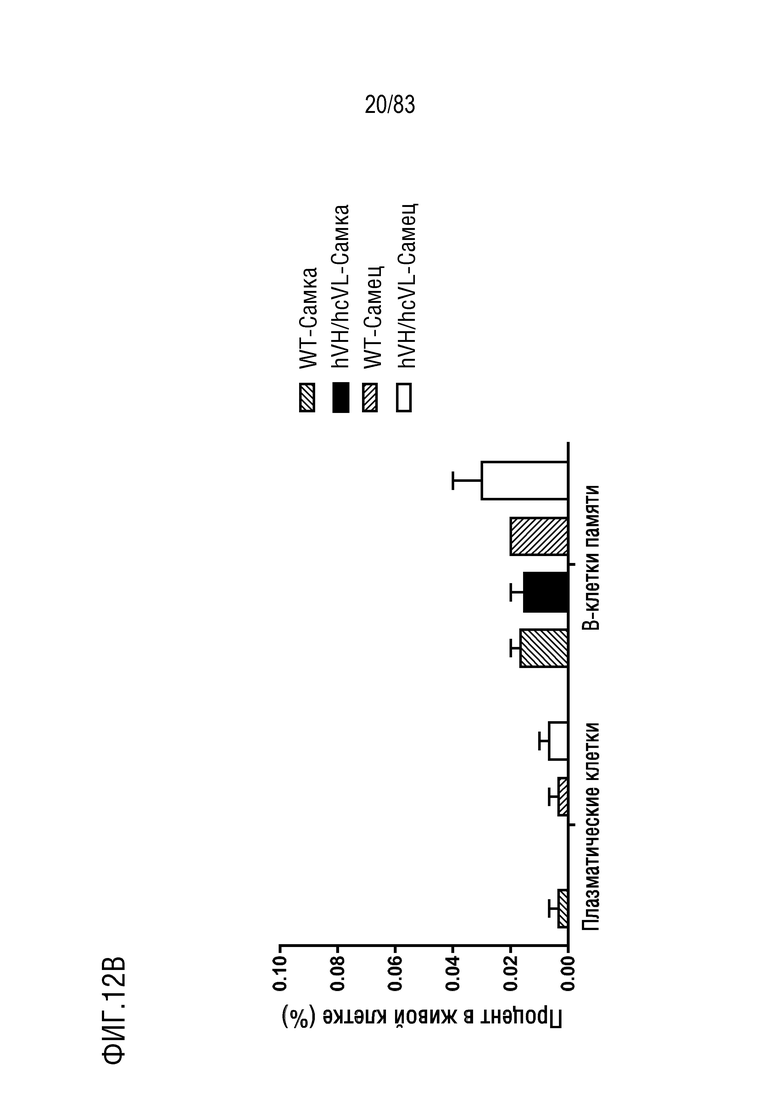
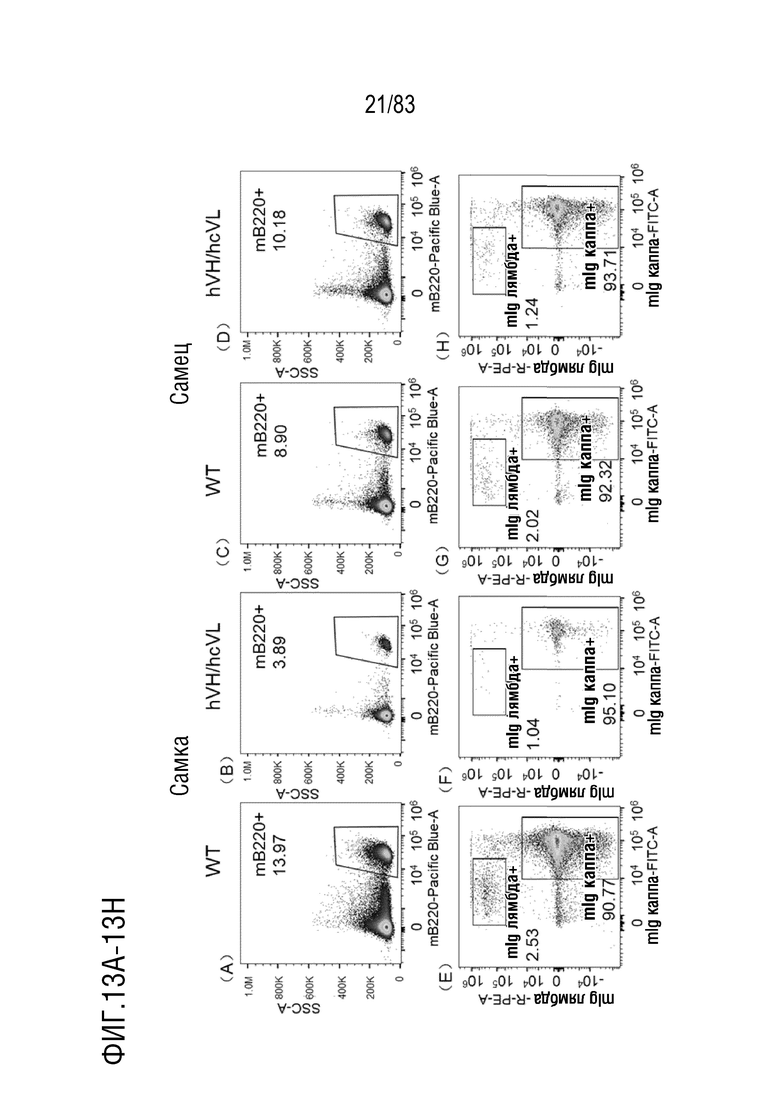
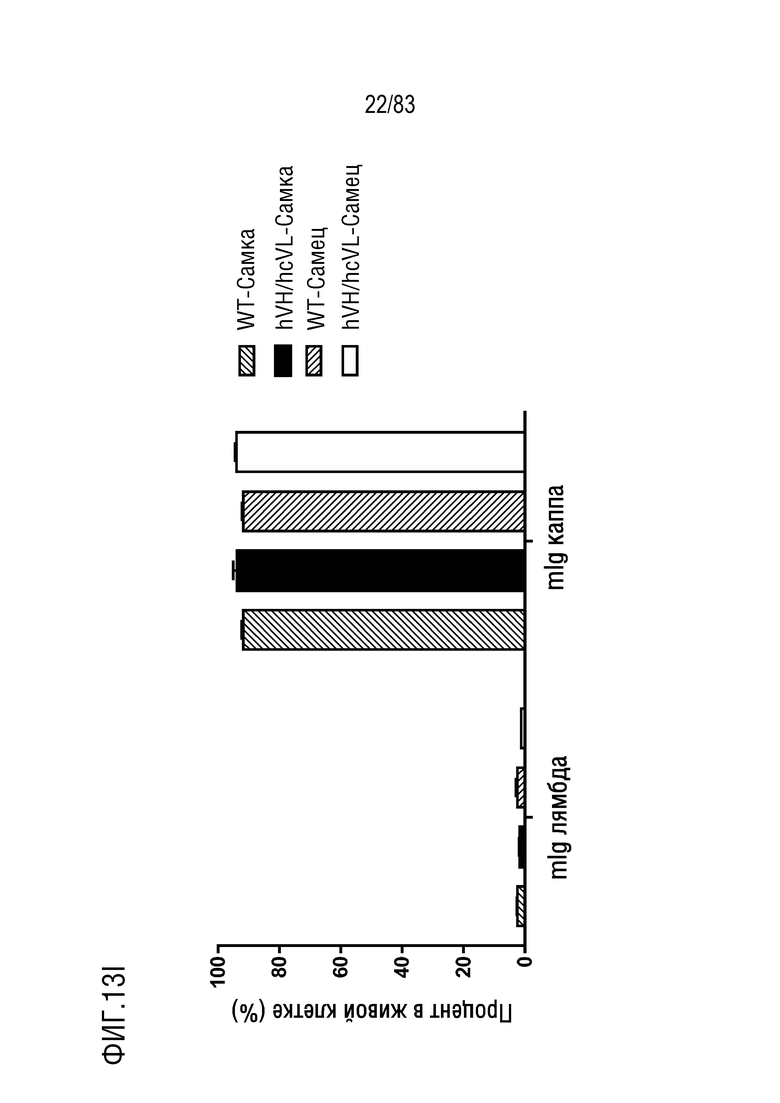
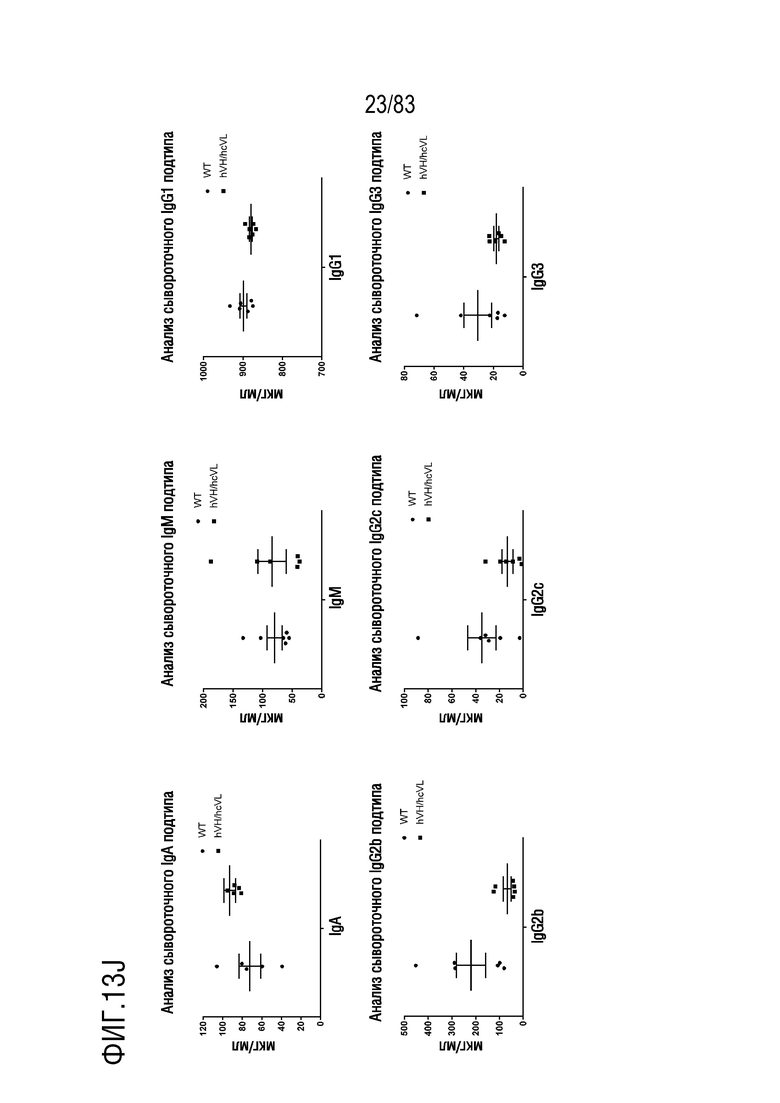

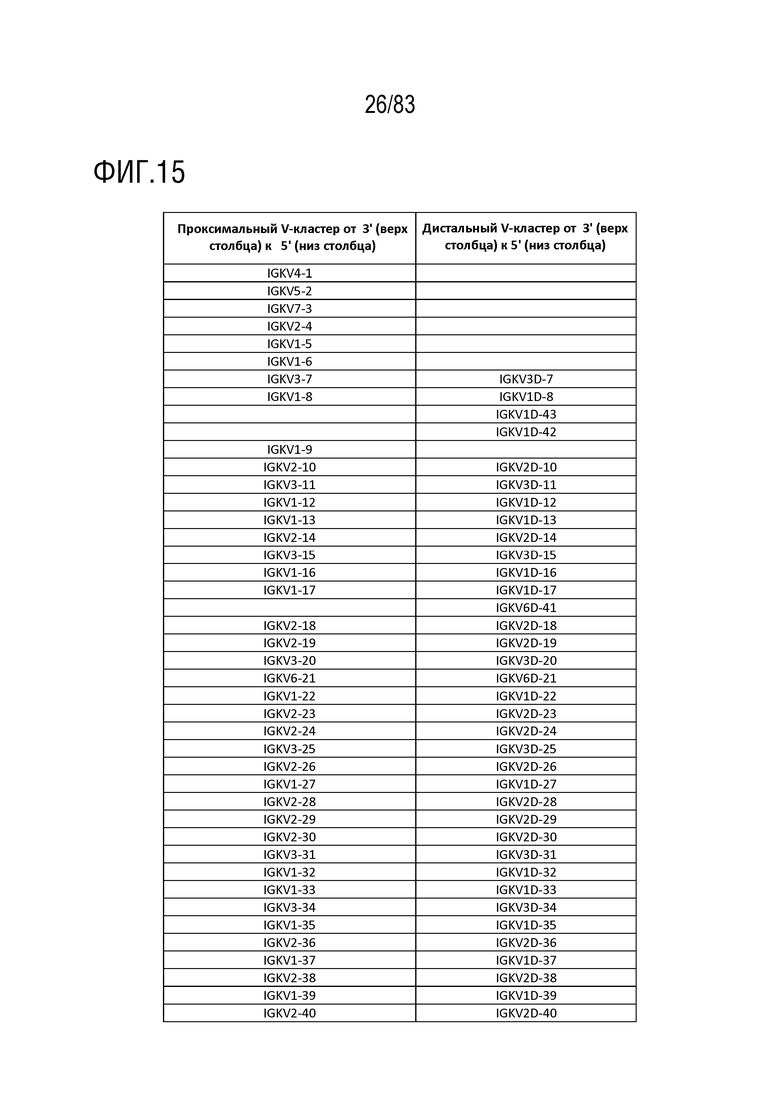
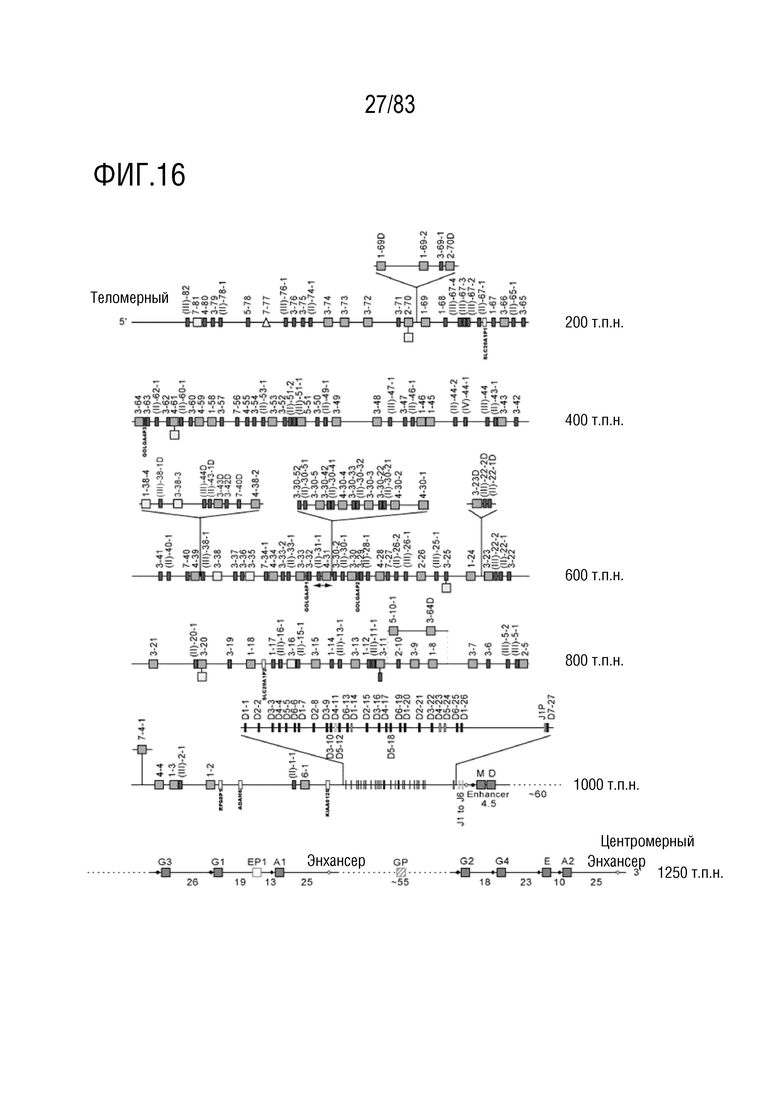

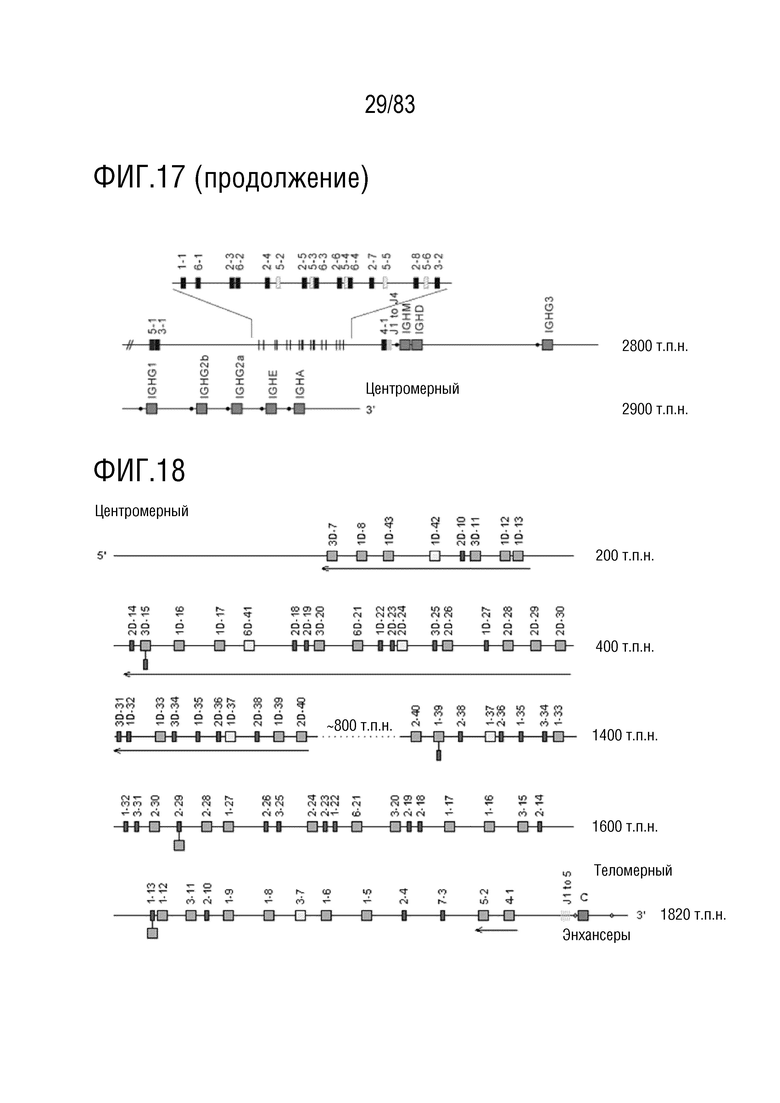
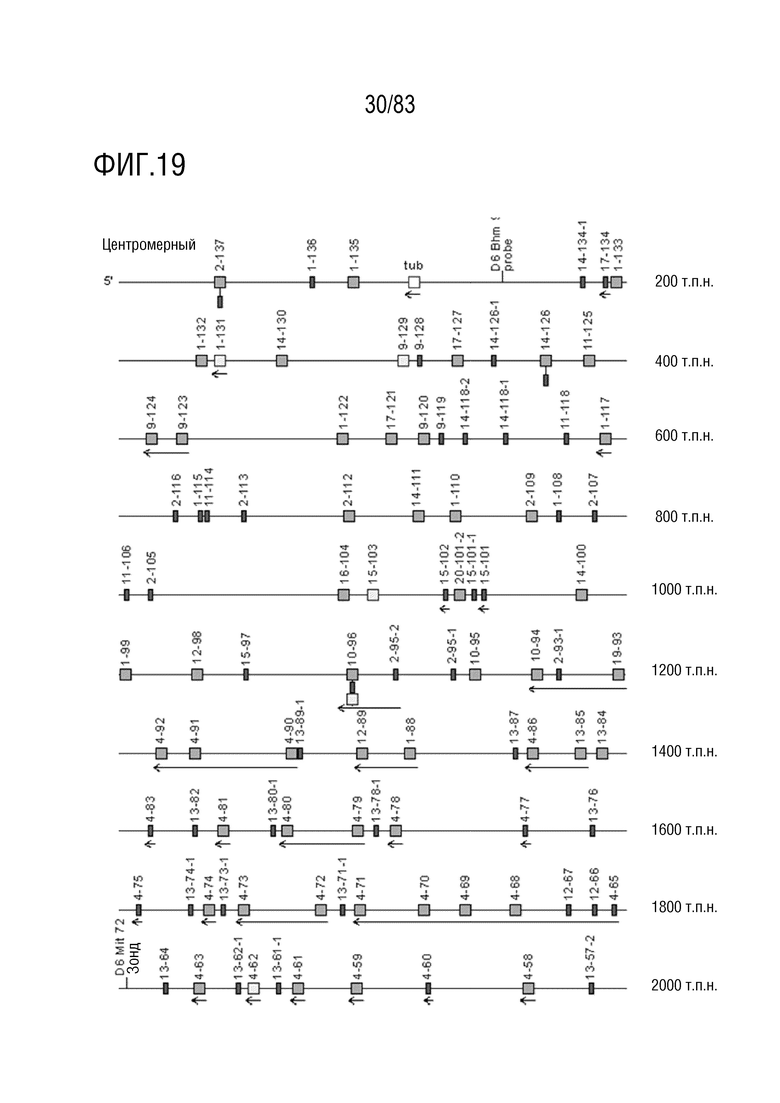
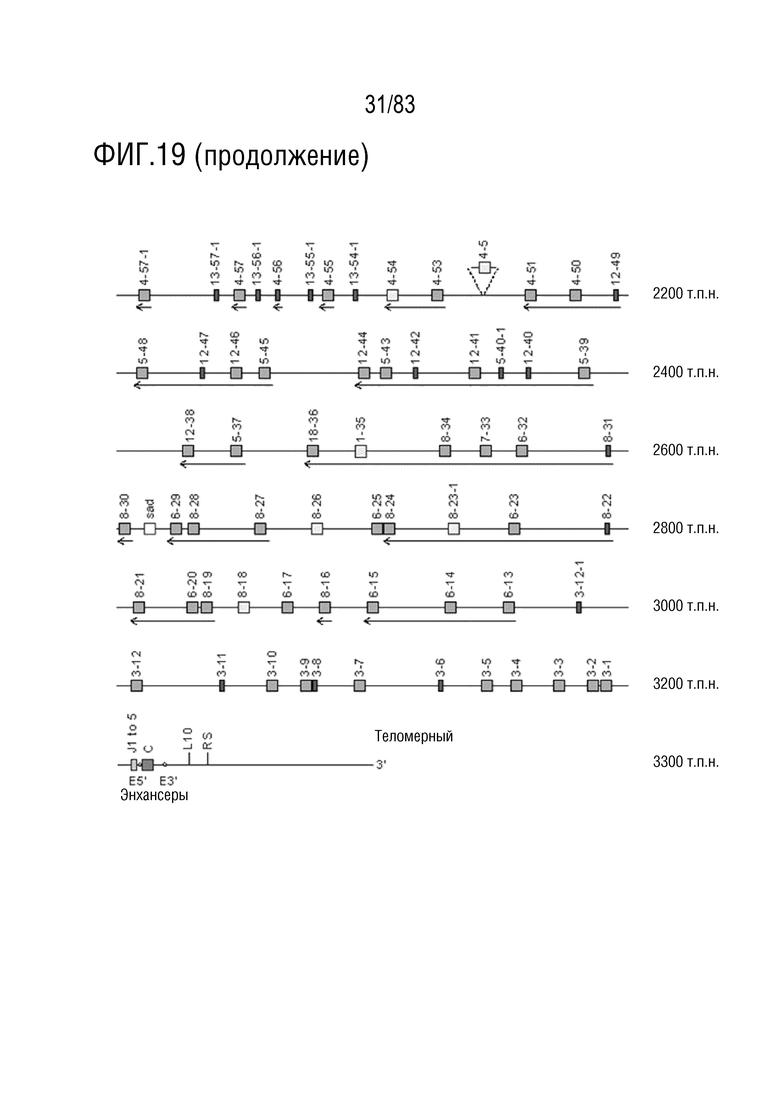
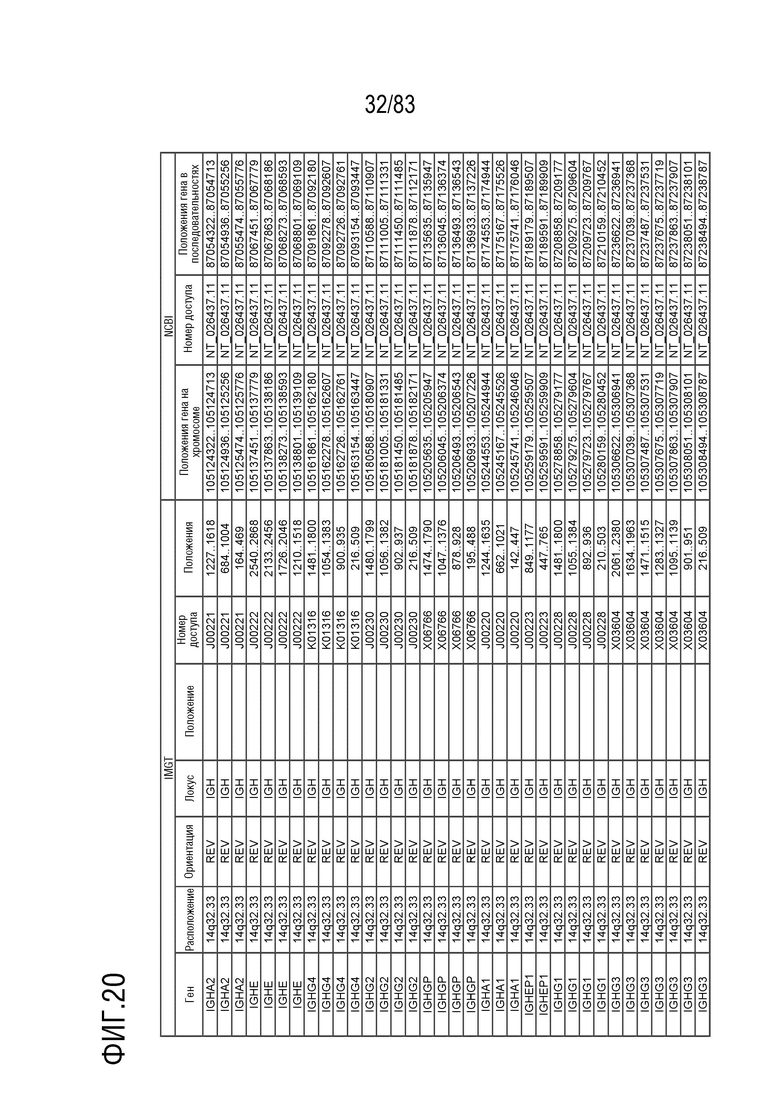





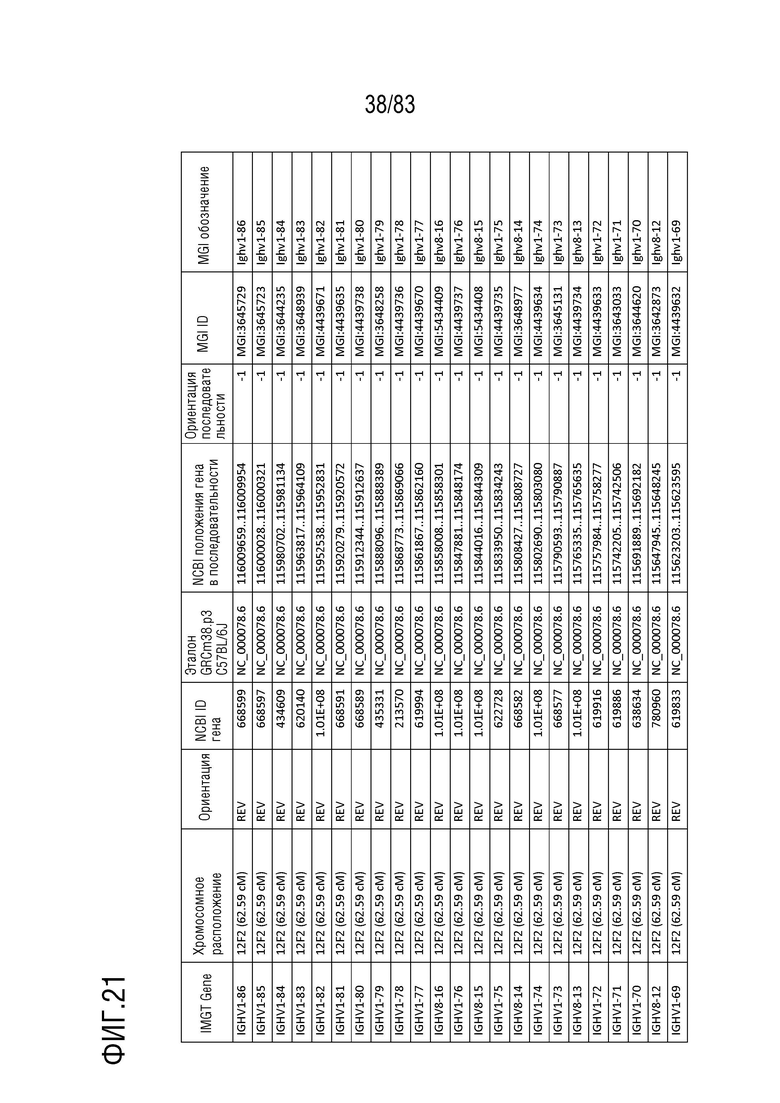
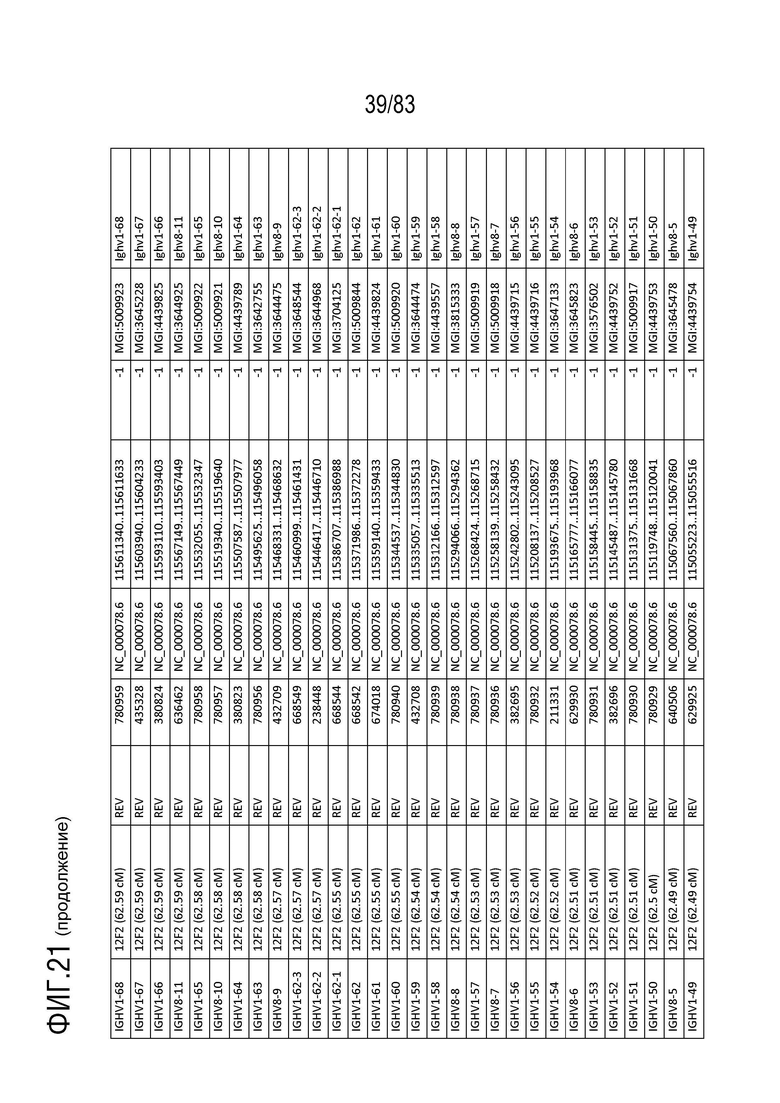
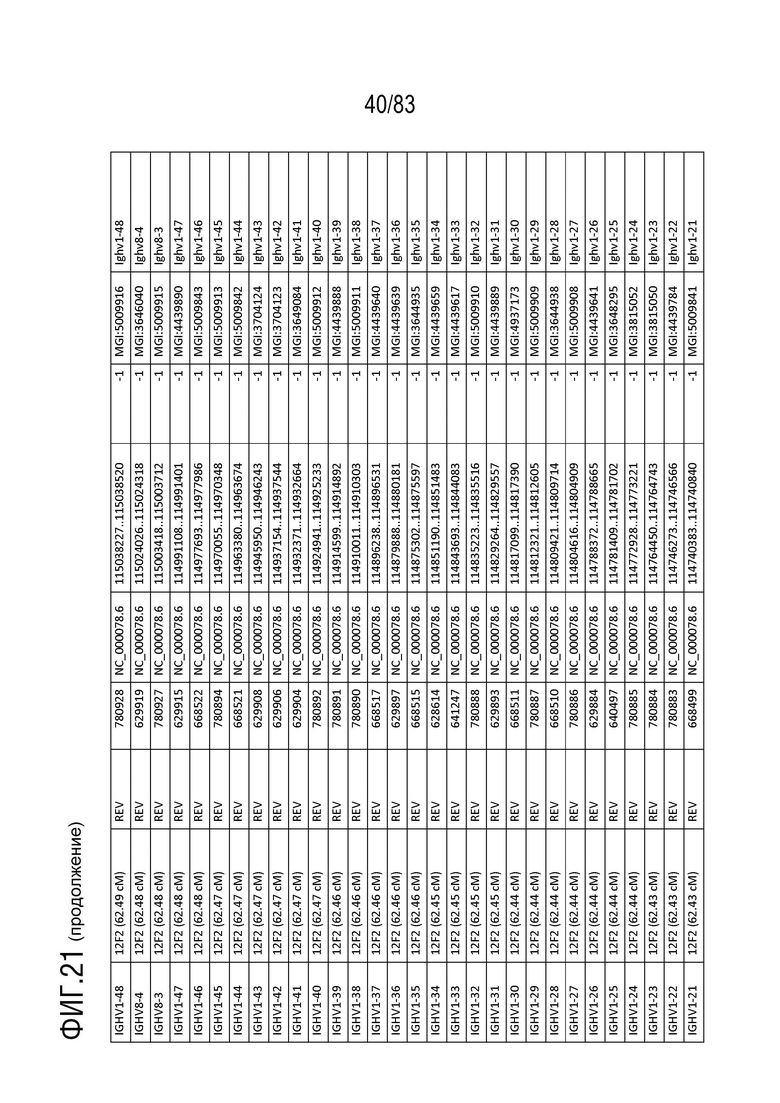
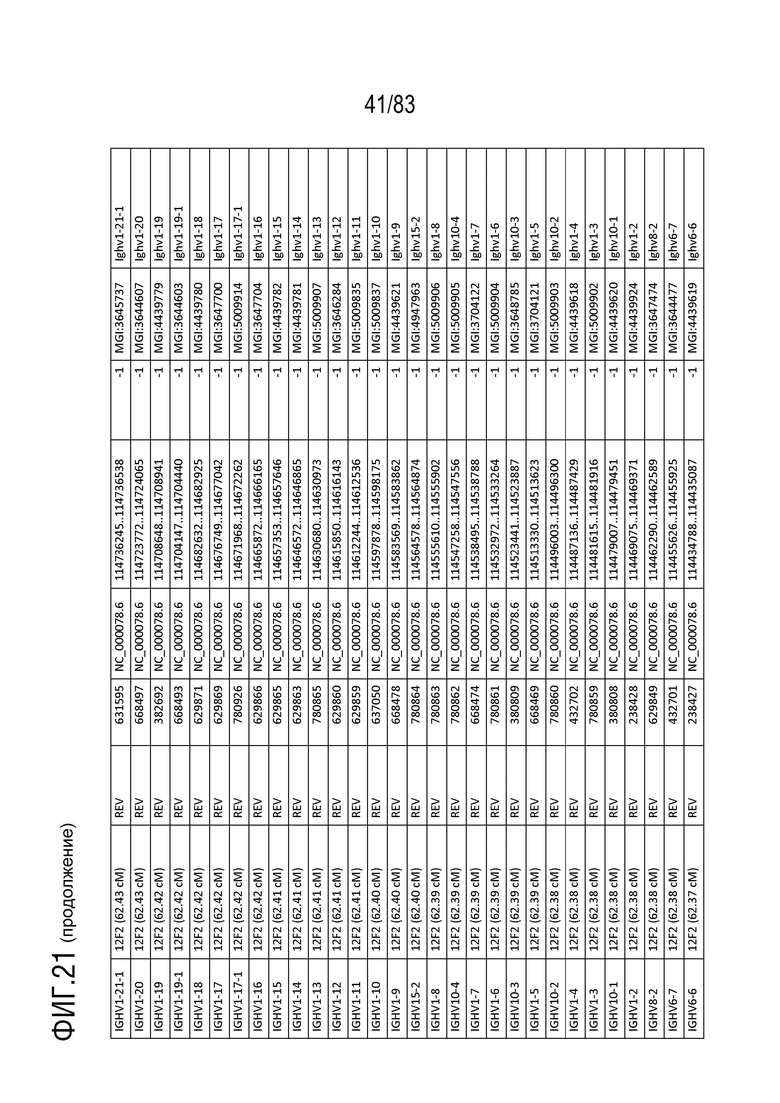
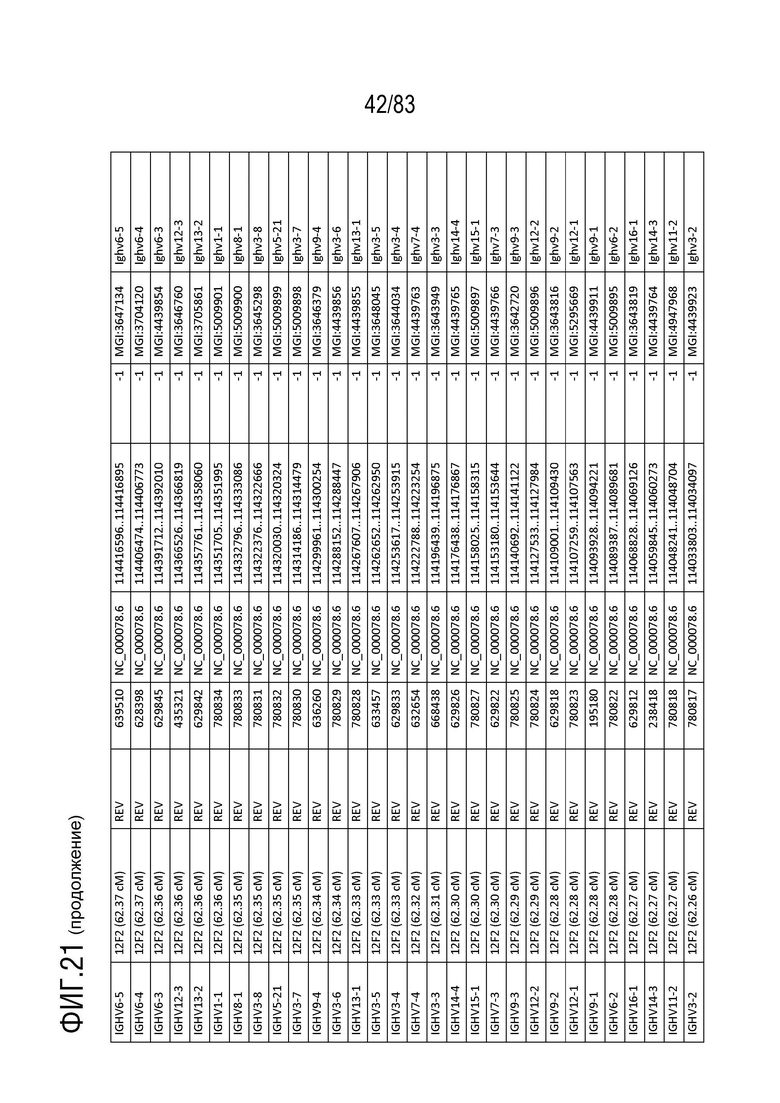
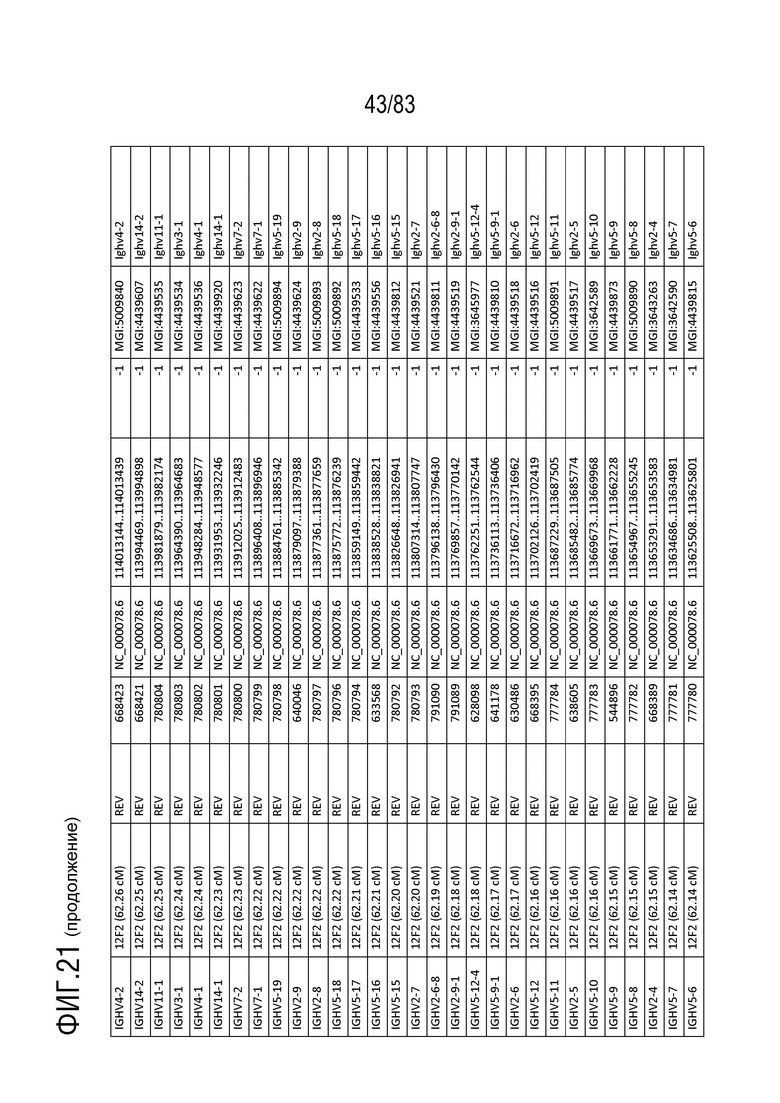
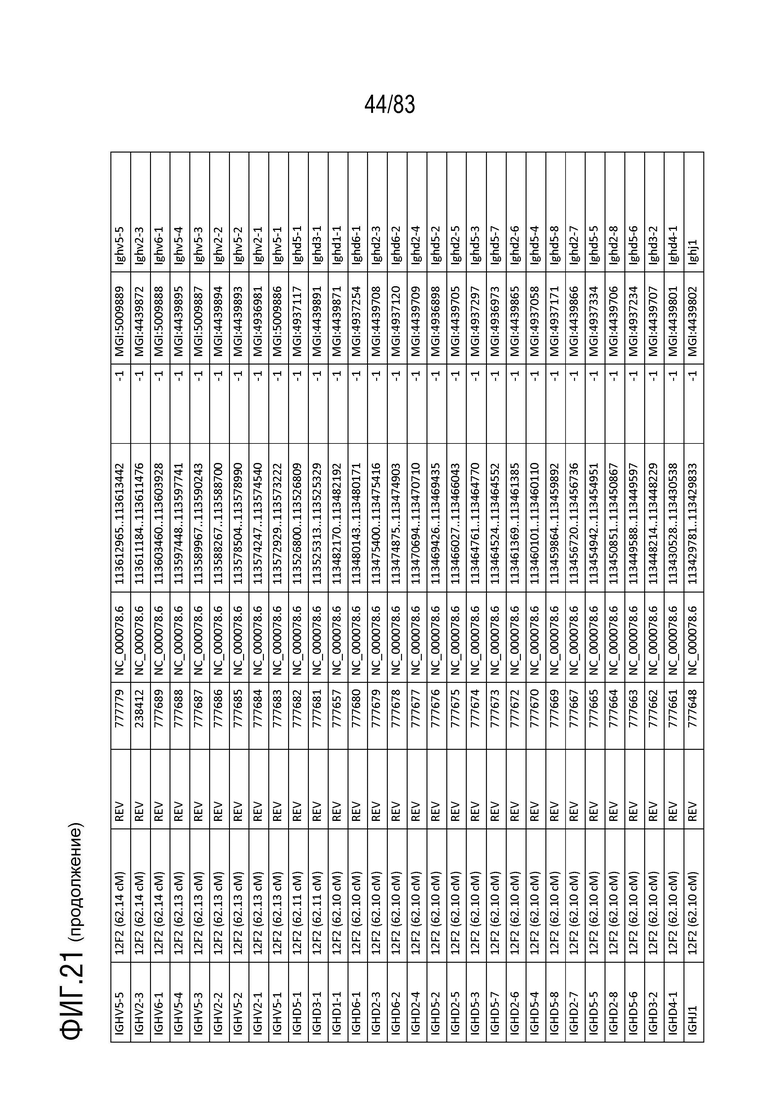




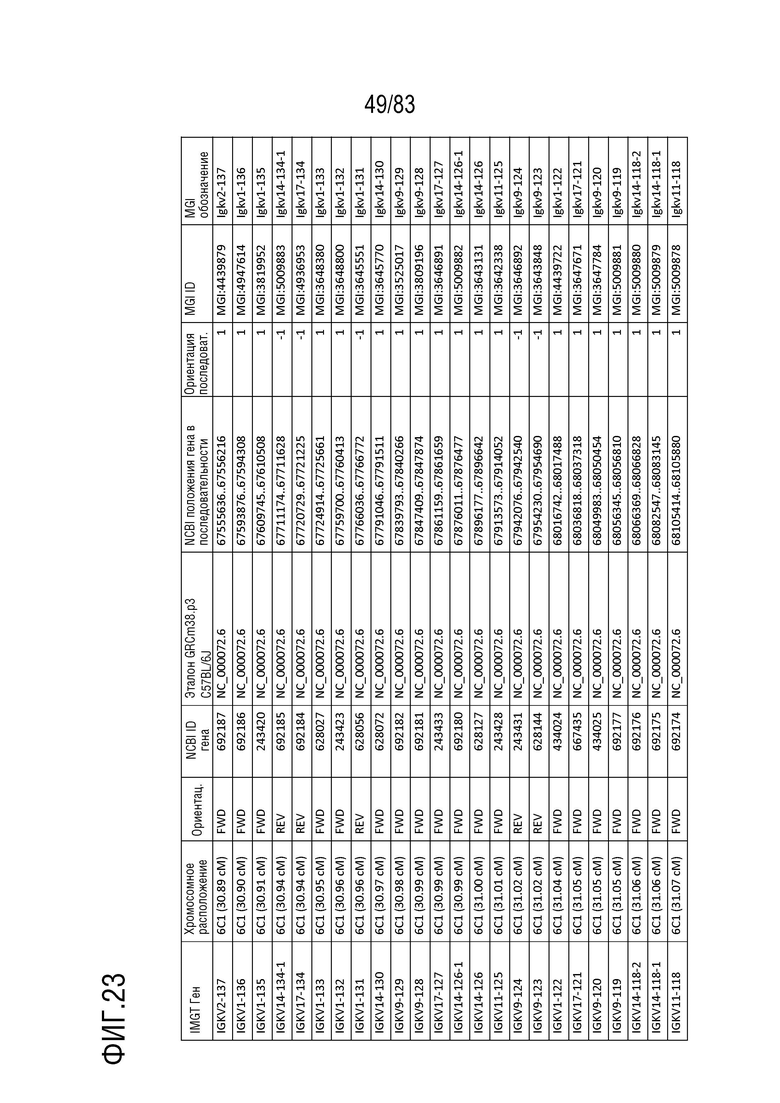
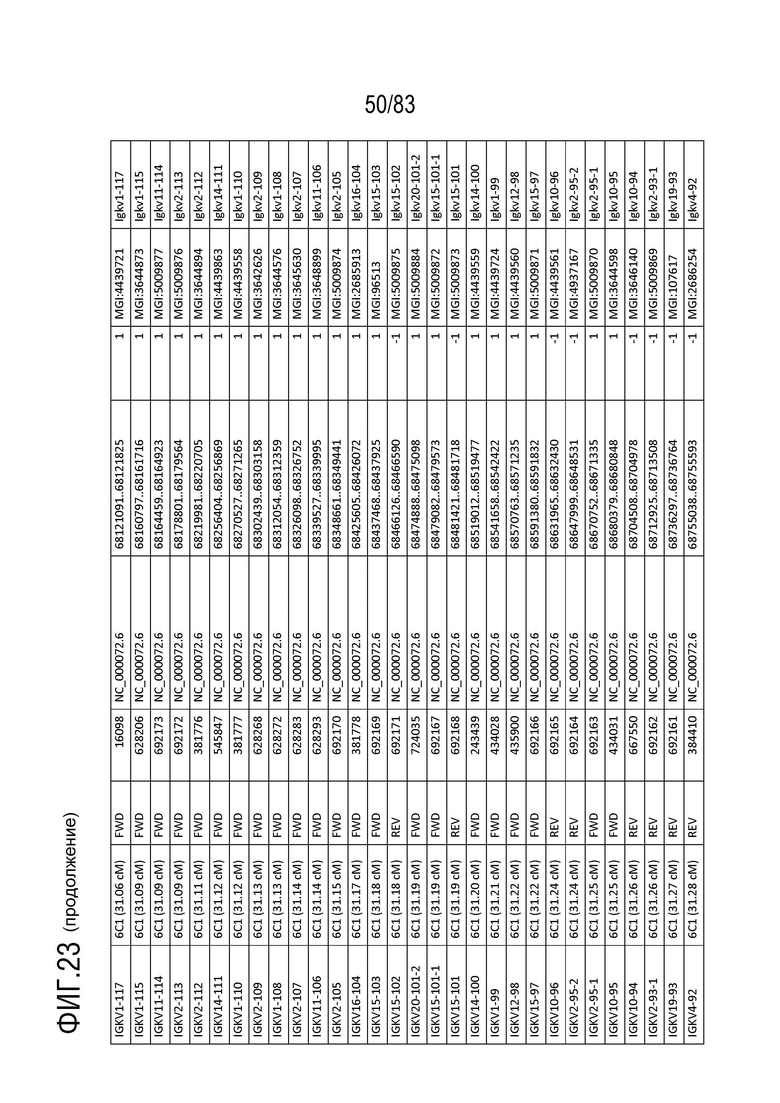
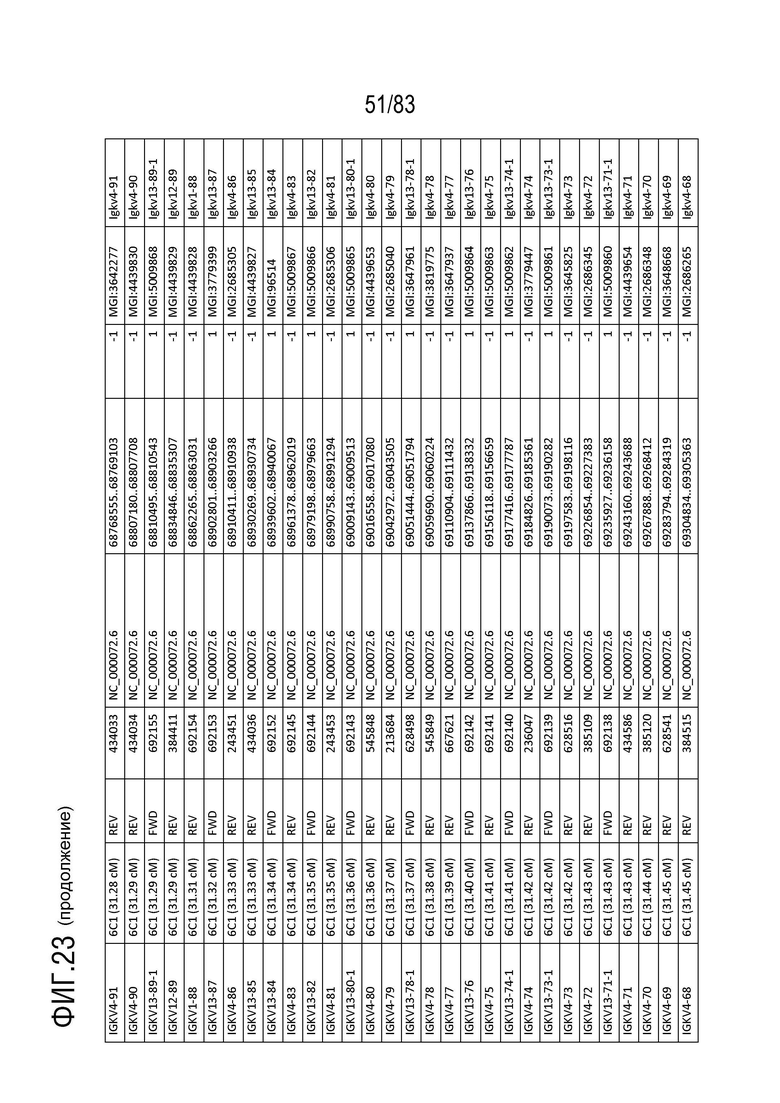
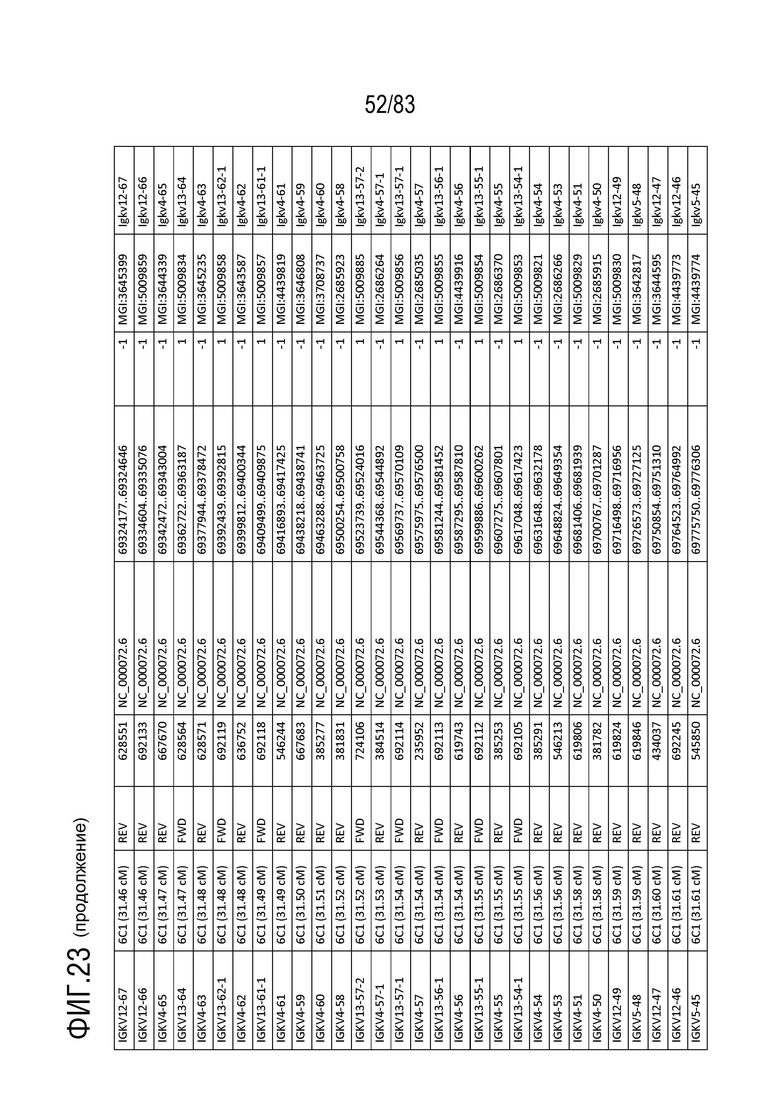
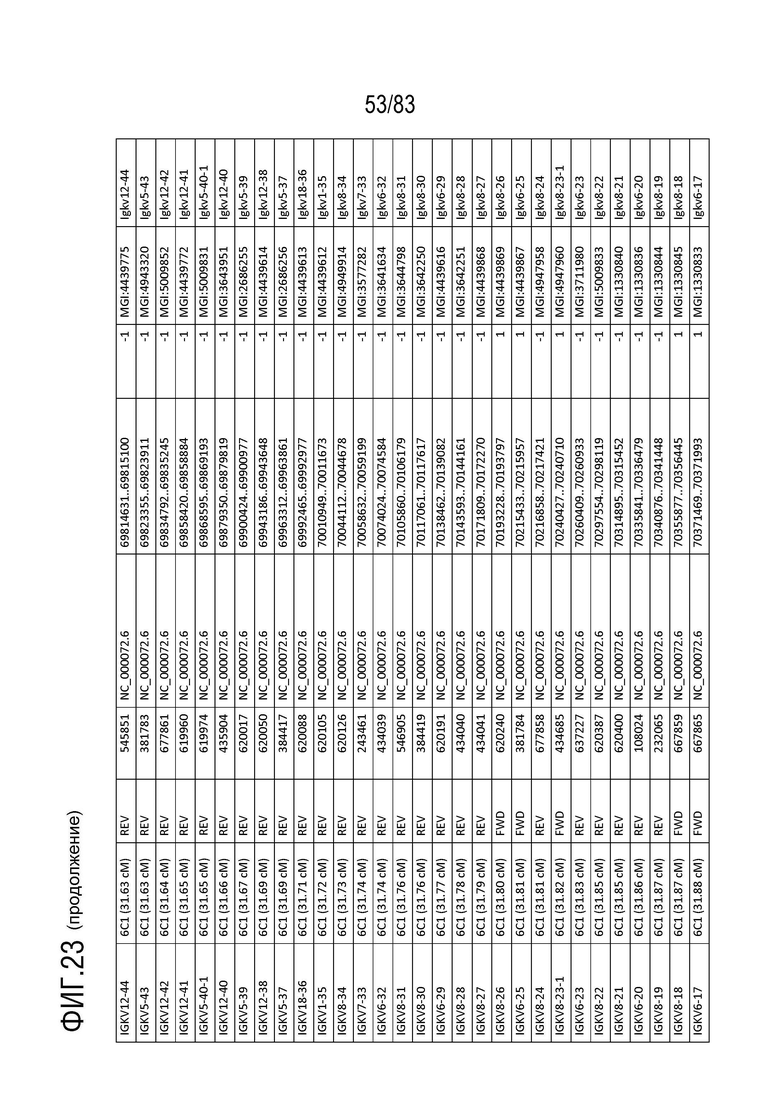
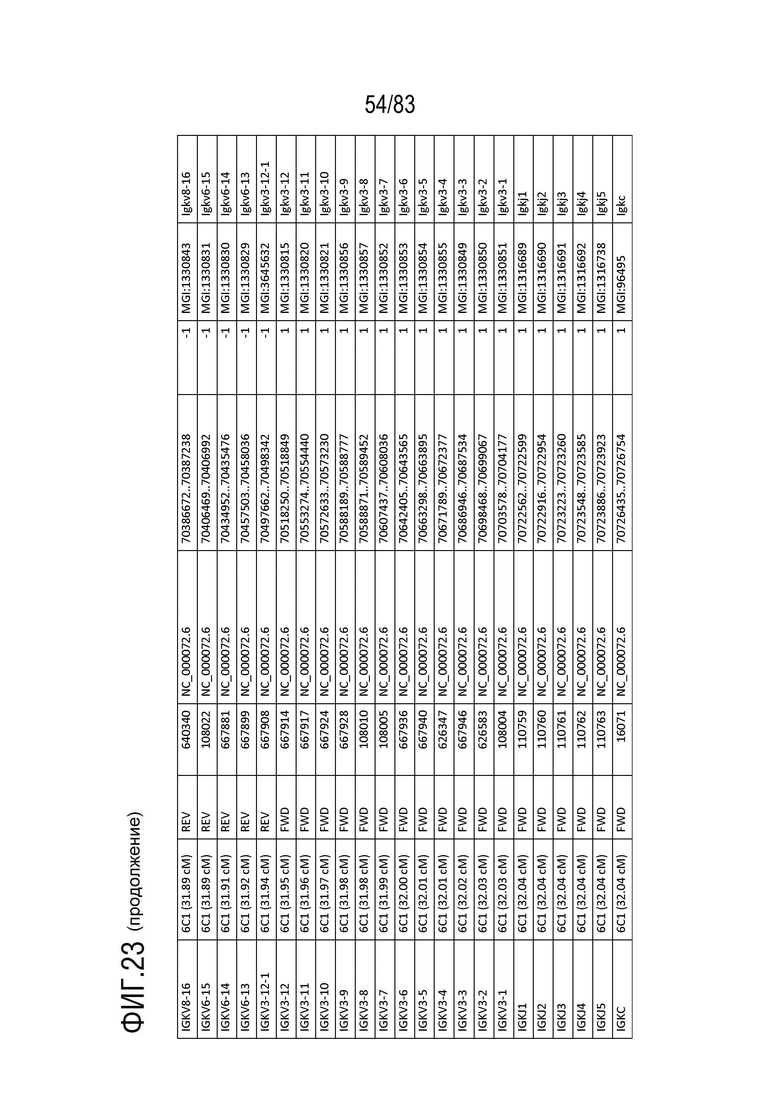
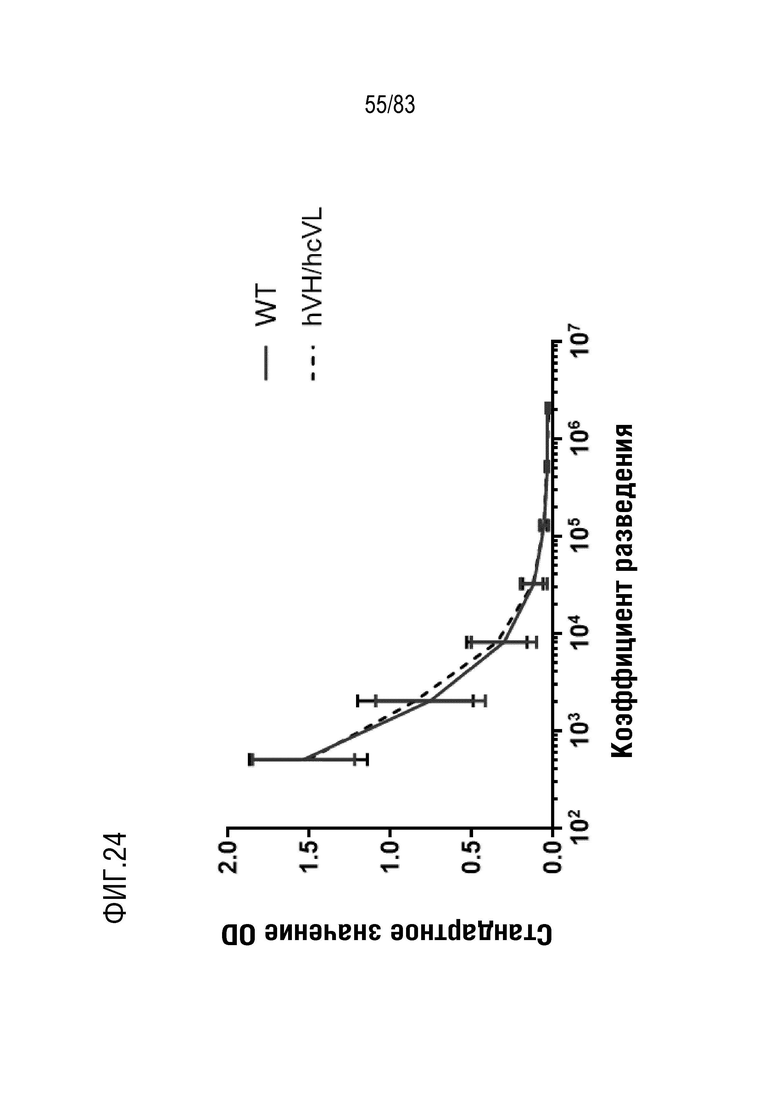
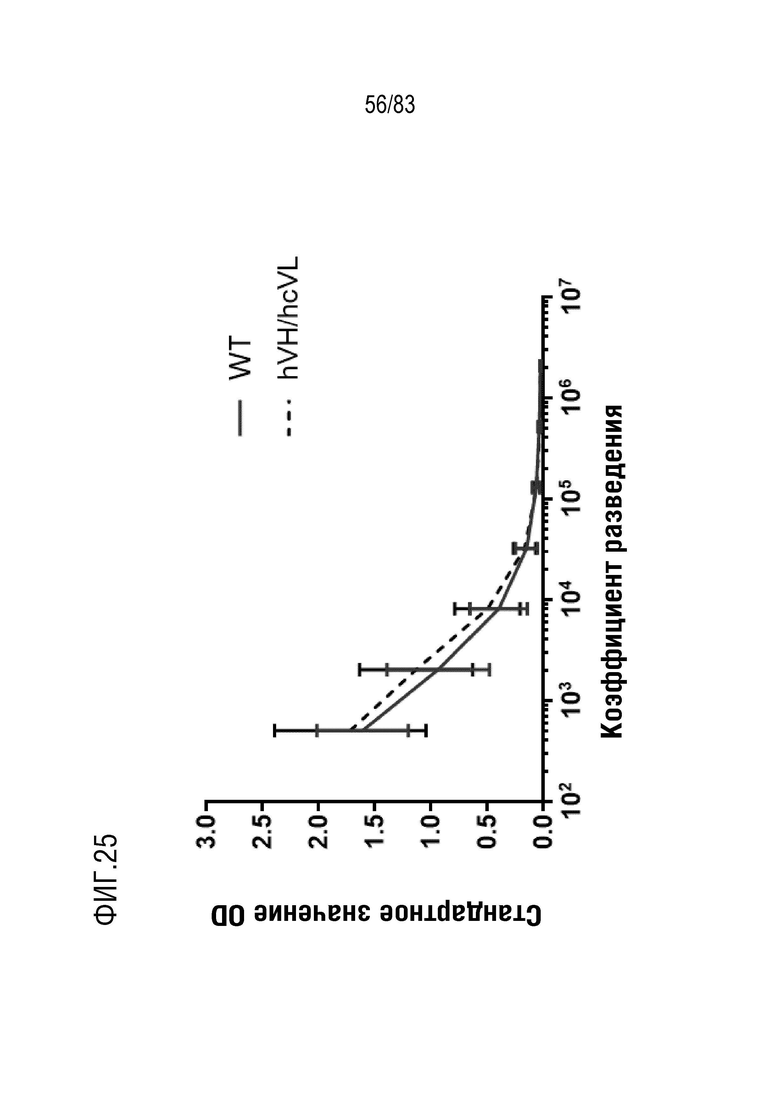
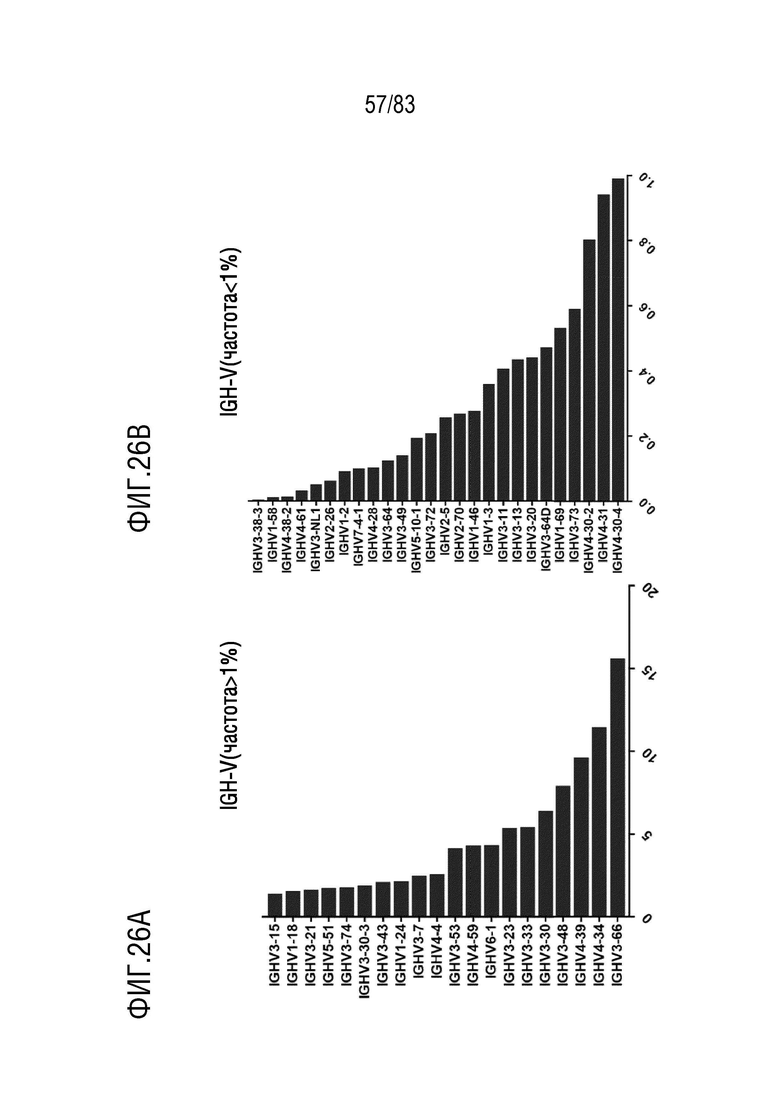
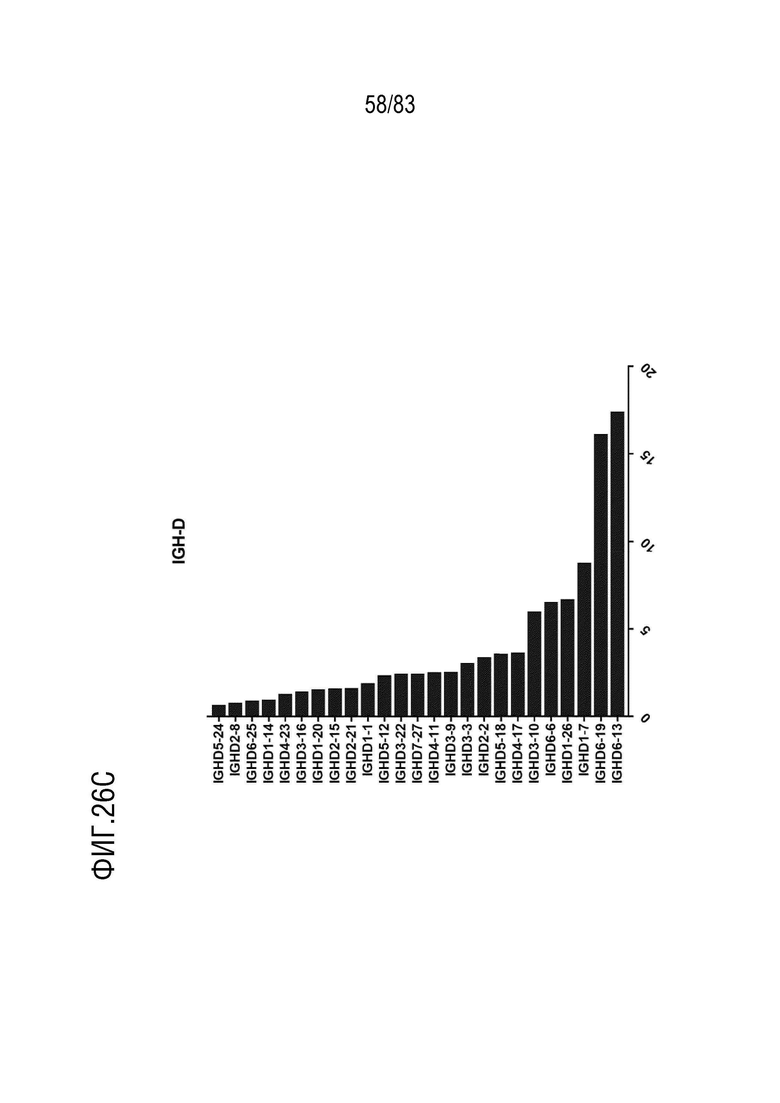
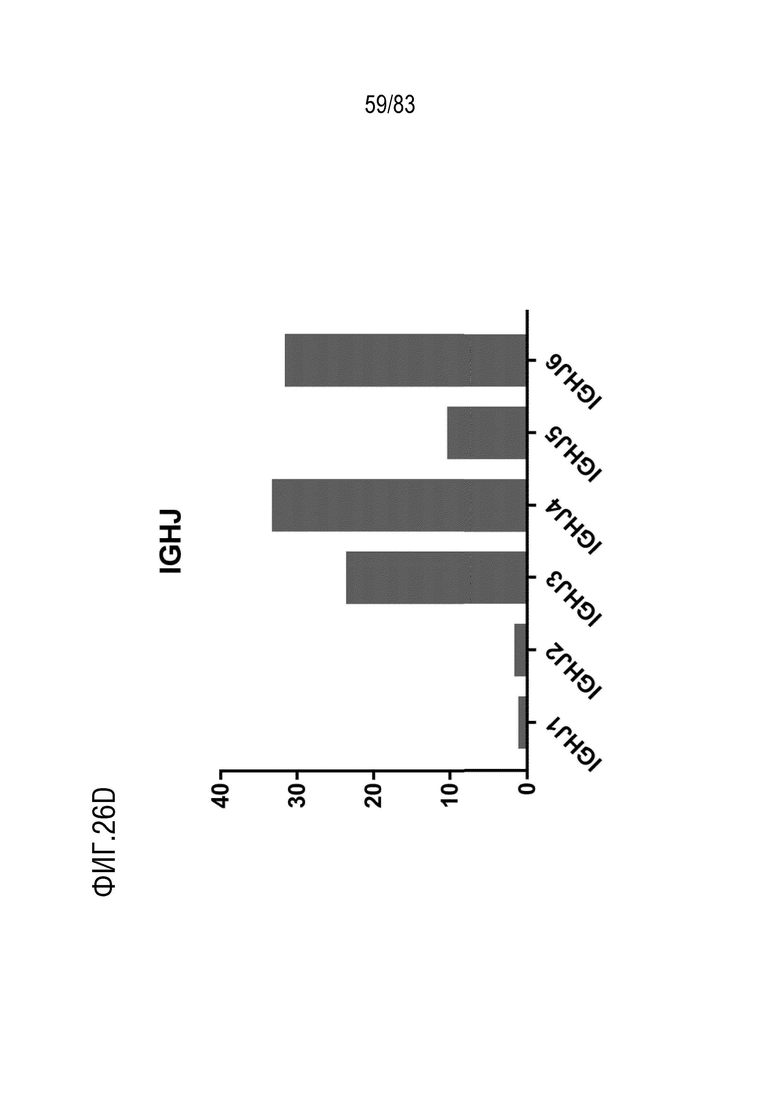
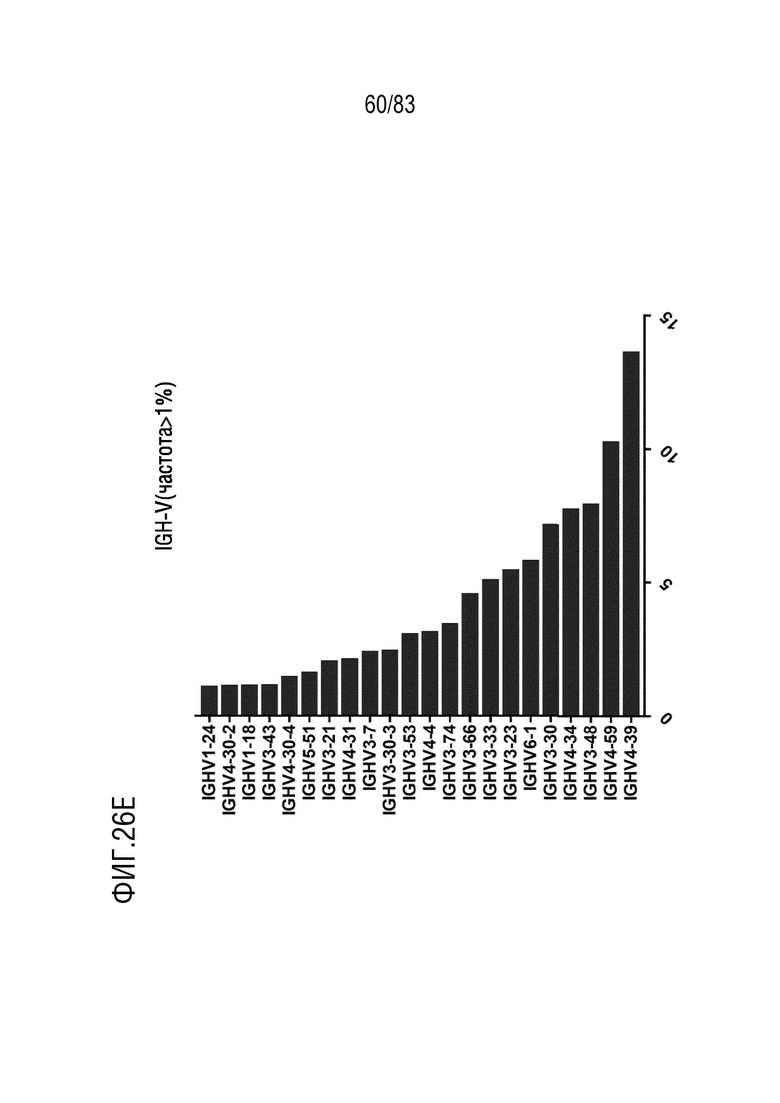
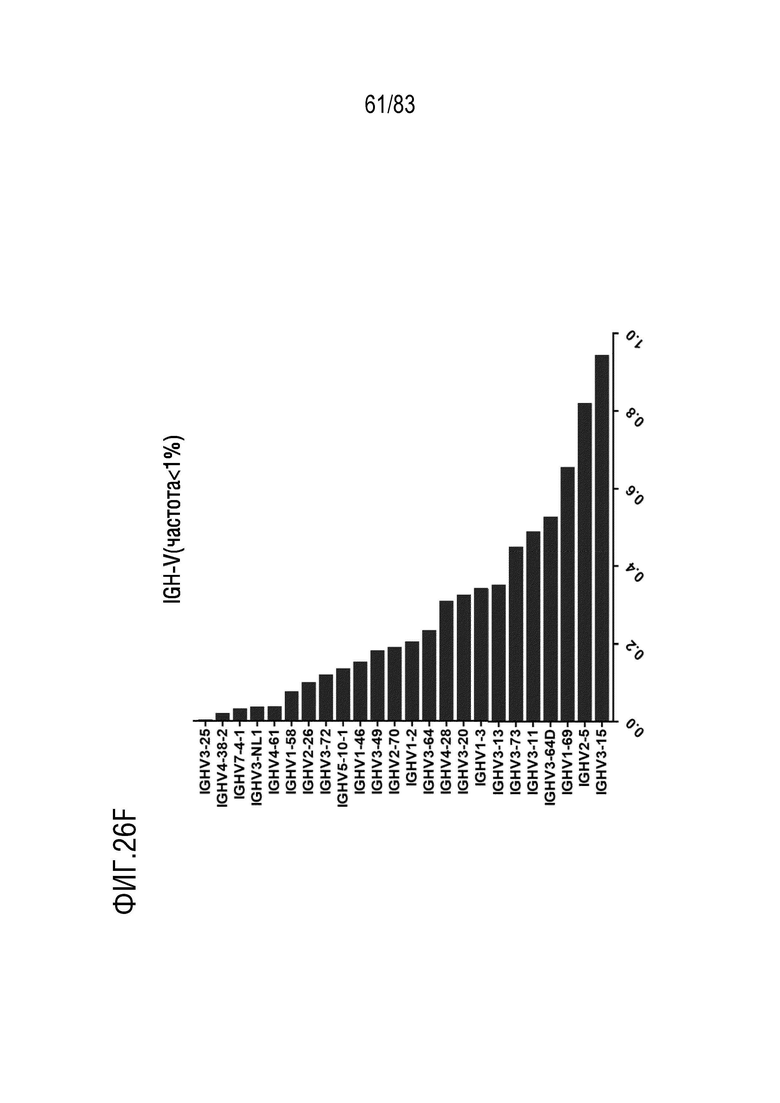
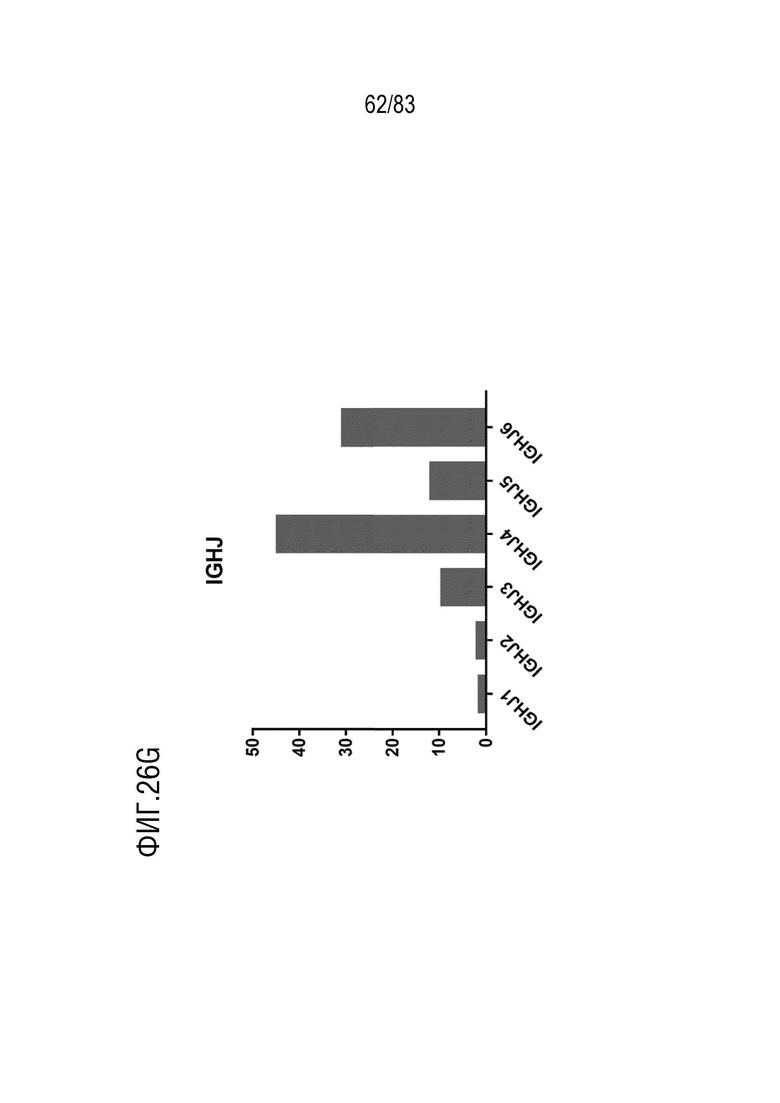
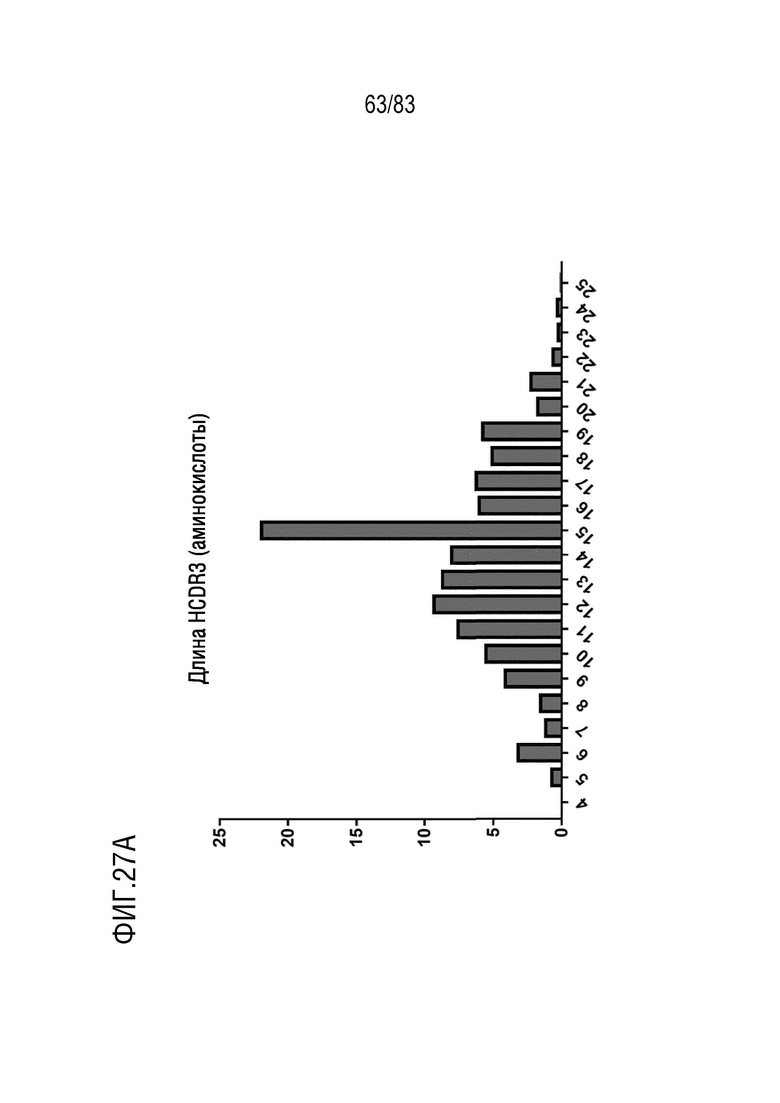
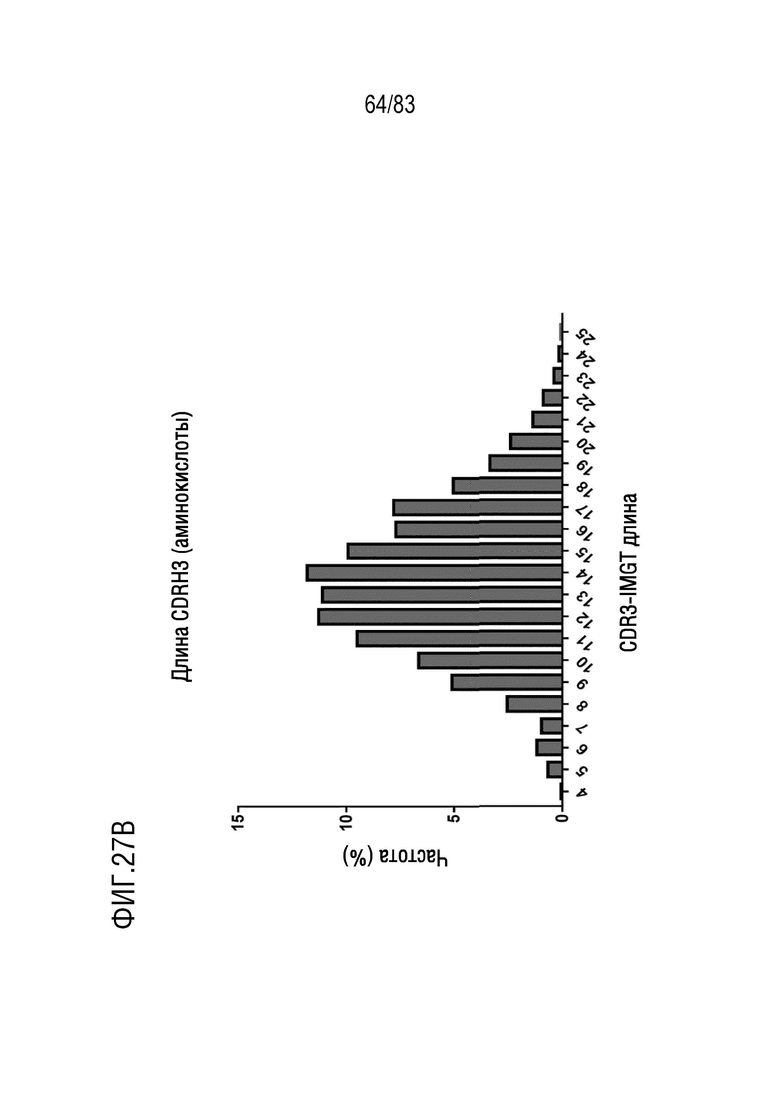




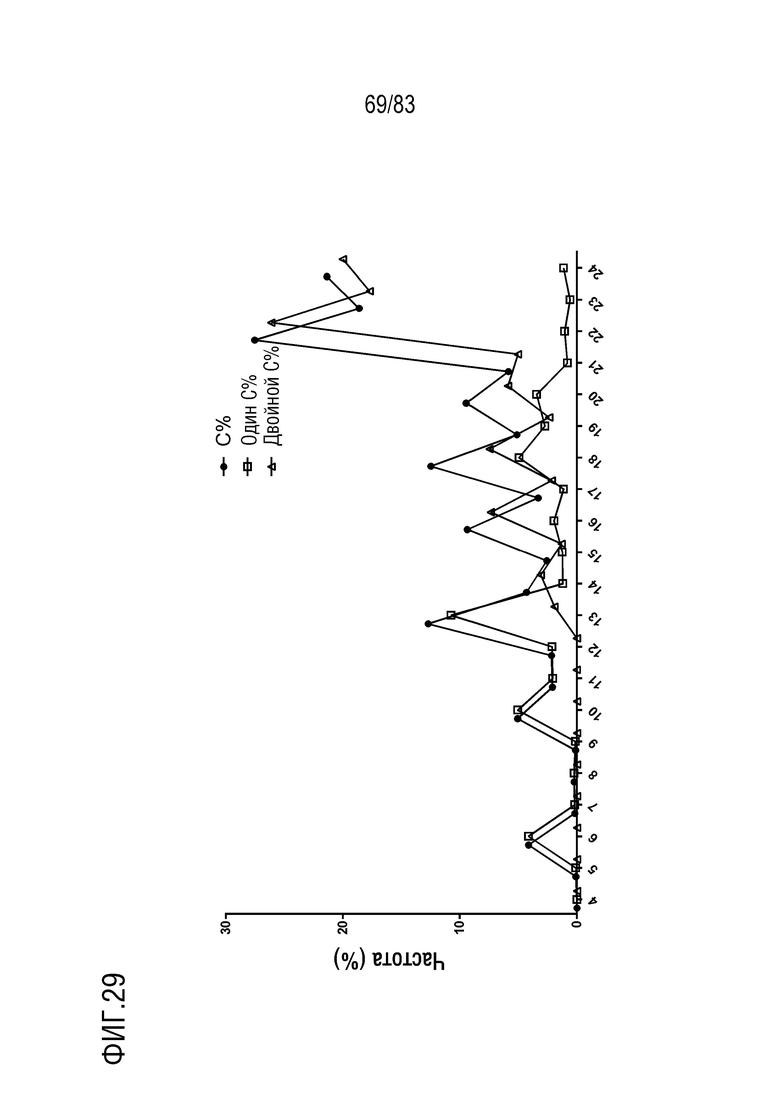
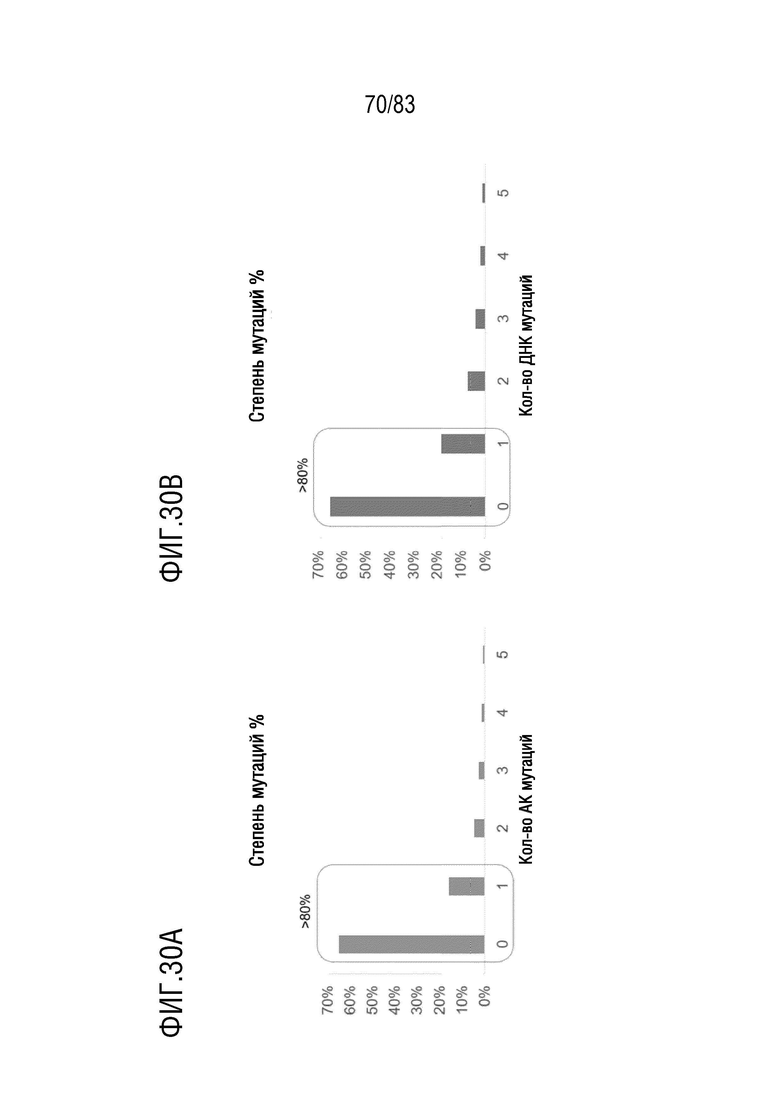
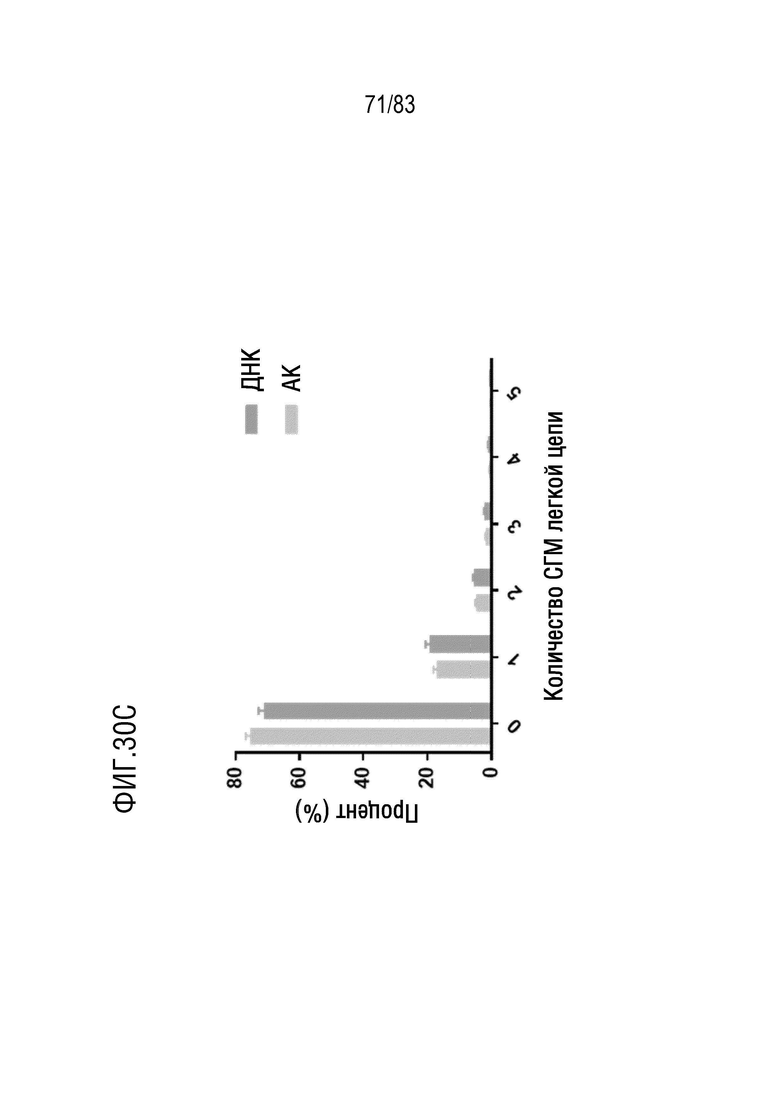
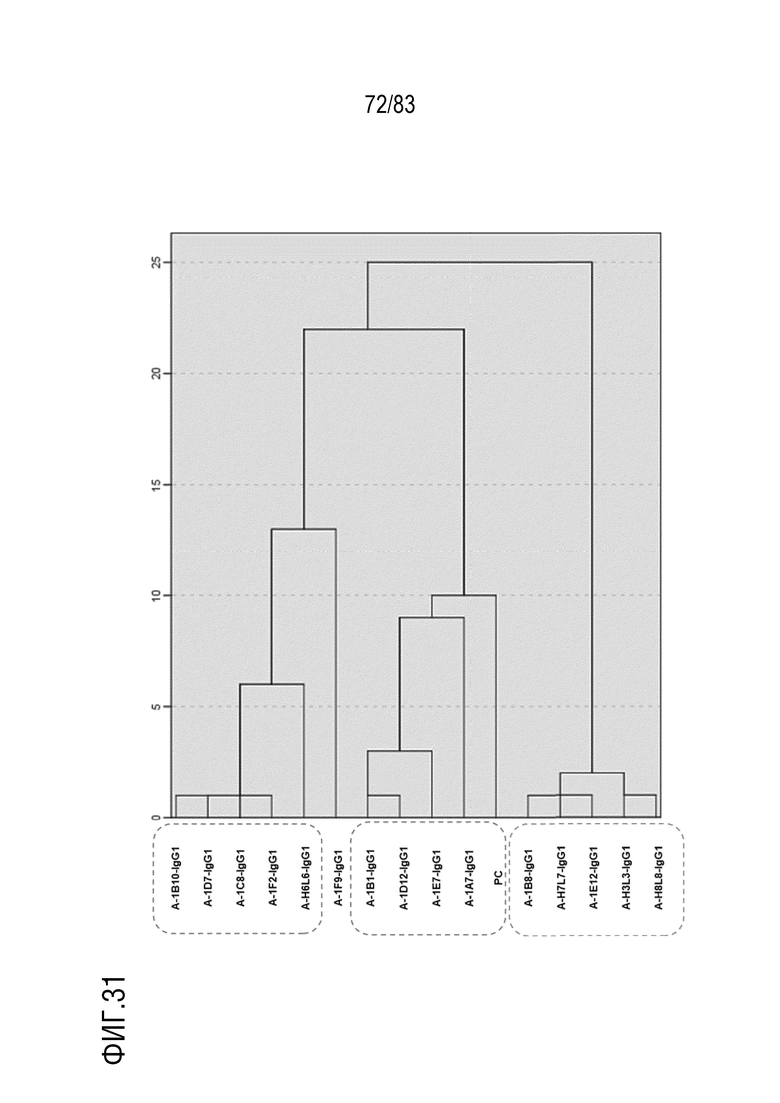






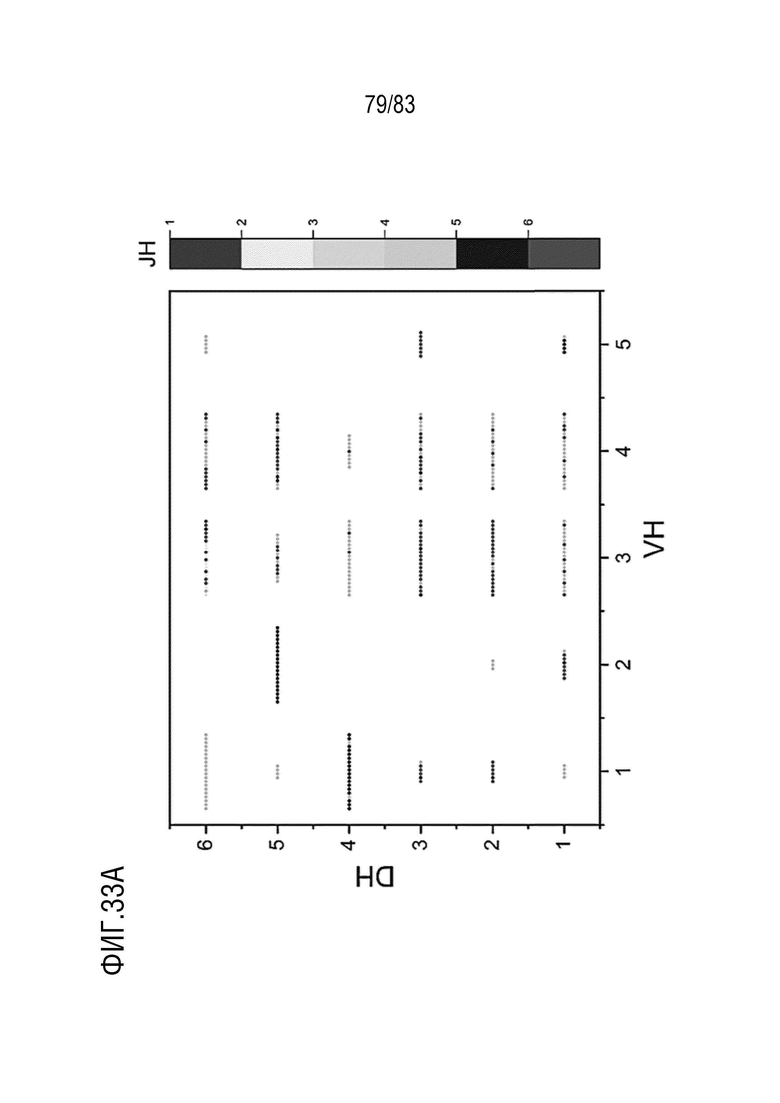
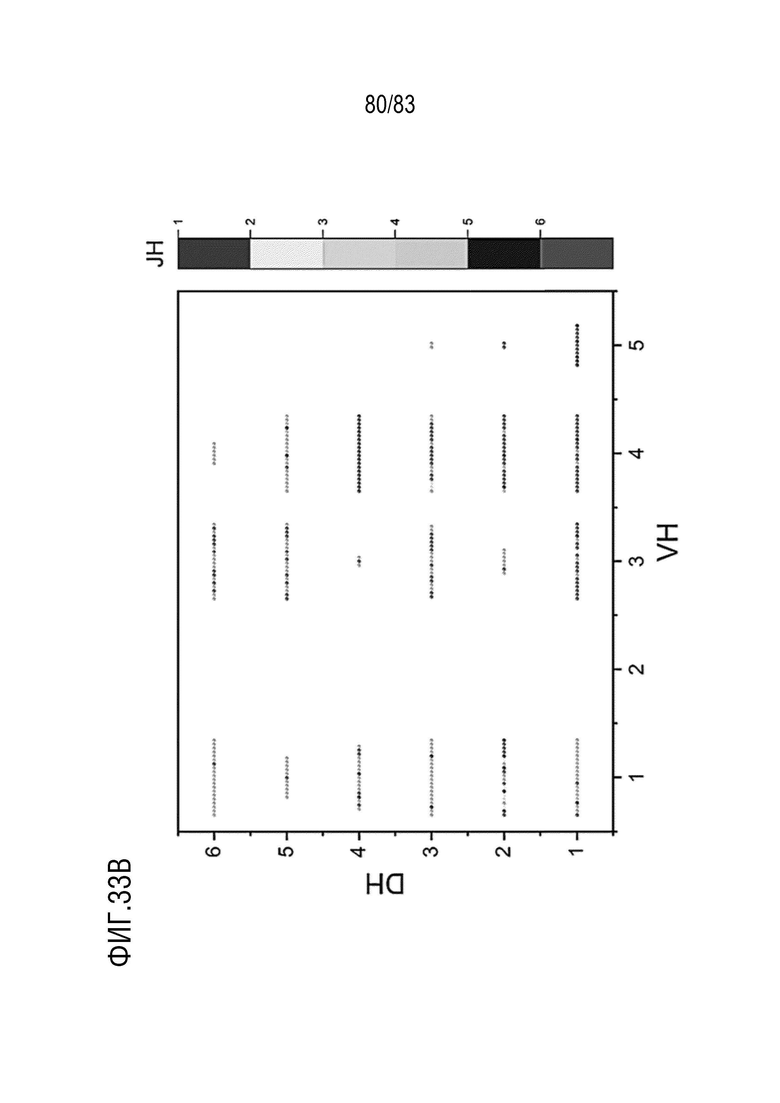
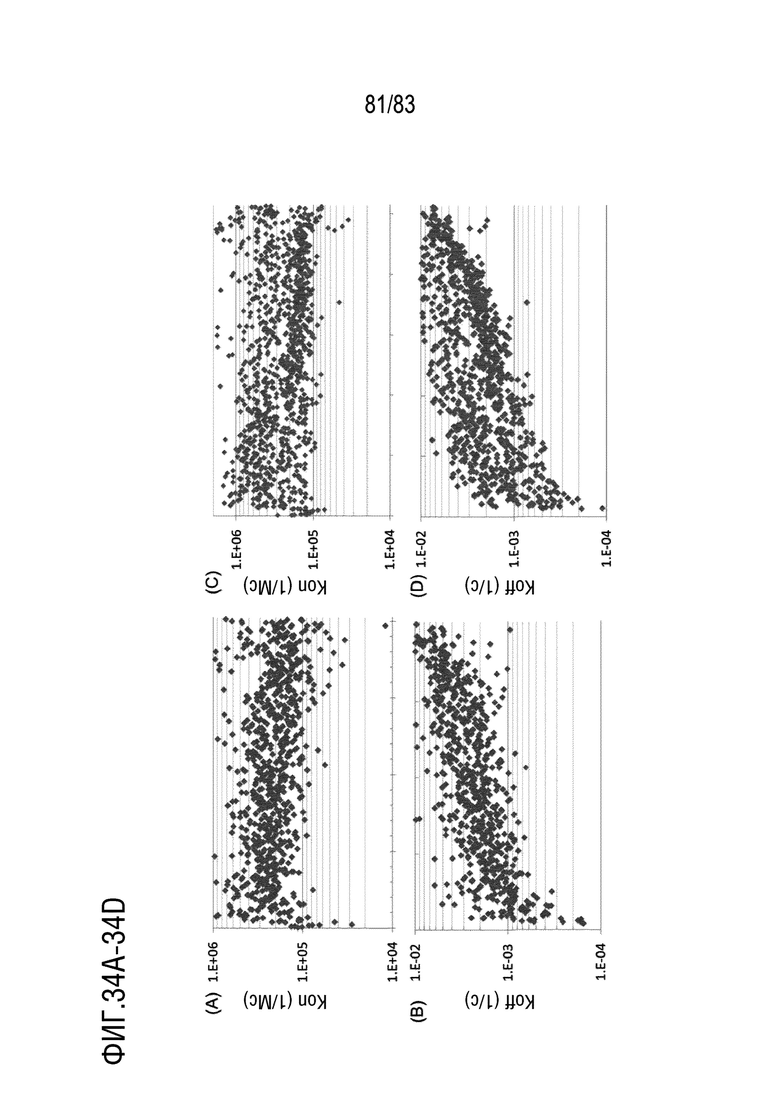
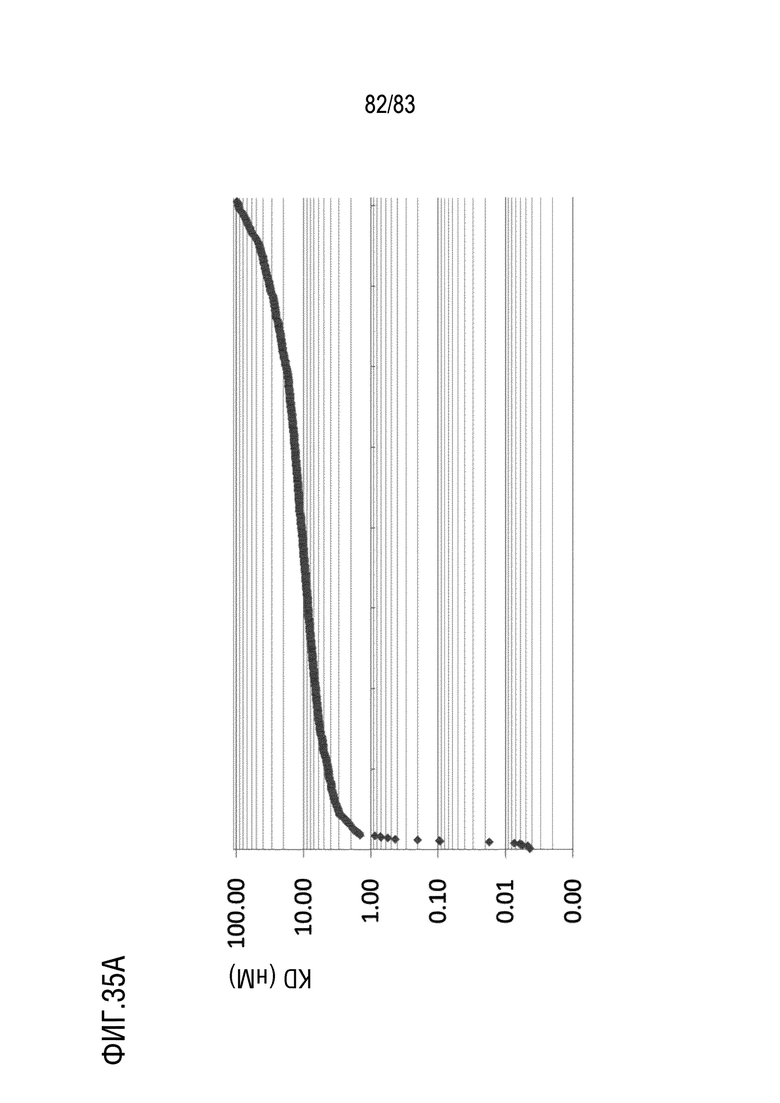
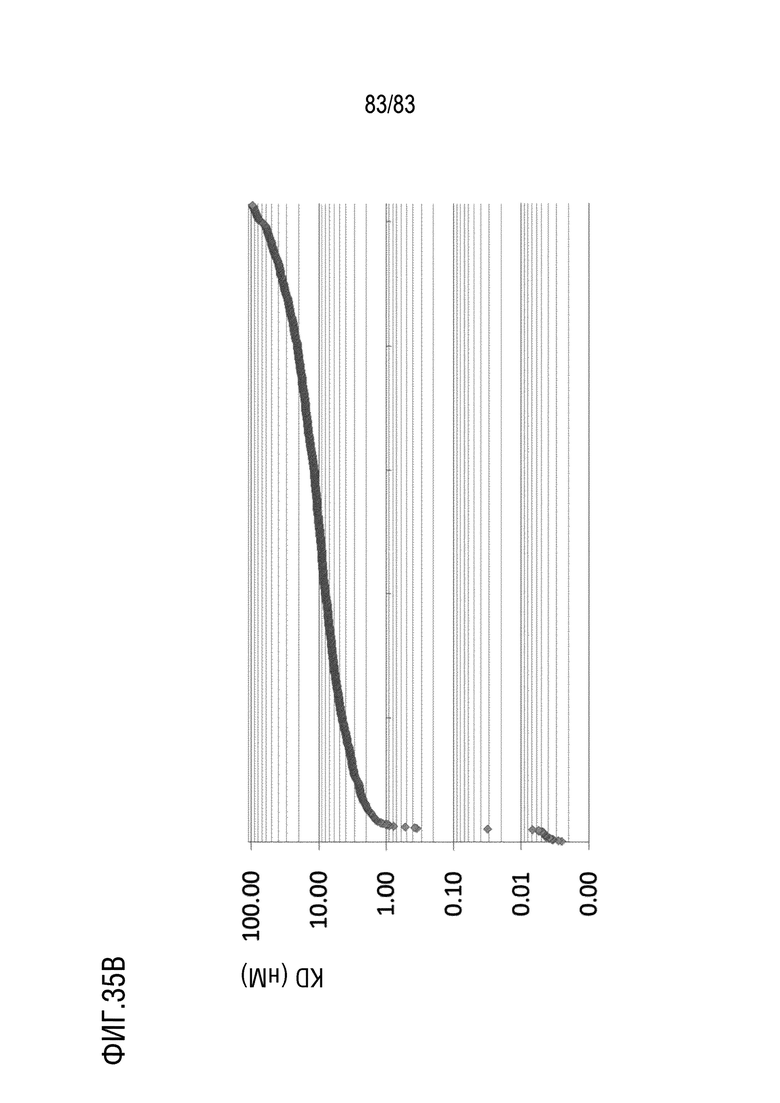
Изобретение относится к области биотехнологии, в частности к генетически модифицированной мыши, геном которой содержит в эндогенном локусе легкой цепи каппа иммуноглобулина экзогенную генную последовательность вариабельной области легкой цепи каппа, причем экзогенная генная последовательность вариабельной области легкой цепи каппа содержит только один ген IGKV человека и только один ген IGKJ человека, а также к полученной из него клетке. Изобретение эффективно для продукции антитела. 2 н. и 17 з.п. ф-лы, 35 ил., 22 табл., 13 пр.
1. Генетически модифицированная мышь, геном которой содержит в эндогенном локусе легкой цепи каппа иммуноглобулина экзогенную генную последовательность вариабельной области легкой цепи каппа, причем экзогенная генная последовательность вариабельной области легкой цепи каппа содержит только один ген IGKV человека и только один ген IGKJ человека,
причем ген IGKV человека и ген IGKJ человека функционально связаны с эндогенным геном константного домена легкой цепи, причем
(1) ген IGKV человека представляет собой IGKV1-39, а ген IGKJ человека представляет собой IGKJ4; или
(2) ген IGKV человека представляет собой IGKV3-11, а ген IGKJ человека представляет собой IGKJ1,
причем мышь продуцирует антитело, содержащее вариабельную область легкой цепи (VL) человека, в ответ на антиген.
2. Генетически модифицированная мышь по п. 1, где экзогенная генная последовательность вариабельной области легкой цепи дополнительно содержит последовательность 3'-UTR IGKJ человека.
3. Генетически модифицированная мышь по п. 1 или 2, где экзогенный ген вариабельной области легкой цепи в одной или более клетках мыши подвержен соматическим гипермутациям.
4. Генетически модифицированная мышь по любому из пп. 1-3, где ген IGKV человека представляет собой IGKV1-39, а ген IGKJ человека представляет собой IGKJ4.
5. Генетически модифицированная мышь по любому из пп. 1-3, где ген IGKV человека представляет собой IGKV3-11, а ген IGKJ человека представляет собой IGKJ1.
6. Генетически модифицированная мышь по любому из пп. 1-5, где мышь дополнительно содержит последовательность промотора, которая функционально связана с геном IGKV человека, причем последовательность промотора находится в пределах 3000 п.н. гена IGKV человека.
7. Генетически модифицированная мышь по любому из пп. 1-6, где мышь содержит эндогенный IGKC.
8. Генетически модифицированная мышь по любому из пп. 1-7, где мышь является гомозиготной или гетерозиготной по локусу гена легкой цепи иммуноглобулина.
9. Генетически модифицированная мышь по любому из пп. 1-8, где мышь дополнительно содержит в эндогенном локусе гена тяжелой цепи иммуноглобулина один или более генов IGHV человека, один или более генов IGHD человека и один или более генов IGHKJ человека, где гены IGHV человека, гены IGHD человека и гены IGHJ человека функционально связаны и могут подвергаться реаранжировке VDJ.
10. Генетически модифицированная мышь по любому из пп. 1-9, где мышь продуцирует антитело или его антигенсвязывающий фрагмент, содержащий вариабельную область легкой цепи человека, имеющую аминокислотную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO: 38 или 39.
11. Генетически модифицированная мышь по любому из пп. 1-10, где мышь продуцирует антитело или его антигенсвязывающий фрагмент, содержащий вариабельную область легкой цепи человека, которая содержит аминокислотную последовательность, представленную в SEQ ID NO: 38.
12. Генетически модифицированная мышь по любому из пп. 1-10, где мышь продуцирует антитело или его антигенсвязывающий фрагмент, содержащий вариабельную область легкой цепи человека, которая содержит аминокислотную последовательность, представленную в SEQ ID NO: 39.
13. Генетически модифицированная мышь по любому из пп. 1-12, где экзогенная генная последовательность вариабельной области легкой цепи может быть обнаружена с помощью ПЦР с использованием одной или более пар праймеров, выбранных из группы, состоящей из:
(a) SEQ ID NO: 8 и SEQ ID NO: 9;
(b) SEQ ID NO: 10 и SEQ ID NO: 11;
(c) SEQ ID NO: 16 и SEQ ID NO: 17; и
(d) SEQ ID NO: 18 и SEQ ID NO: 19.
14. Генетически модифицированная мышь по любому из пп. 1-13, где мышь содержит в эндогенном локусе легкой цепи иммуноглобулина последовательность нуклеиновой кислоты, которая на 95% идентична SEQ ID NO: 35.
15. Генетически модифицированная мышь по любому из пп. 1-14, где мышь содержит в эндогенном локусе легкой цепи иммуноглобулина последовательность нуклеиновой кислоты, идентичную SEQ ID NO: 35.
16. Генетически модифицированная мышь по любому из пп. 1-13, где мышь содержит в эндогенном локусе легкой цепи иммуноглобулина последовательность нуклеиновой кислоты, которая на 95% идентична SEQ ID NO: 36.
17. Генетически модифицированная мышь по любому из пп. 1-13, где мышь содержит в эндогенном локусе легкой цепи иммуноглобулина последовательность нуклеиновой кислоты, идентичную SEQ ID NO: 36.
18. Генетически модифицированная мышь по любому из пп. 1-17, где эндогенная вариабельная генная последовательность легкой цепи каппа нефункциональна.
19. Выделенная иммунная клетка, полученная из генетически модифицированной мыши по любому из пп. 1-18, для получения последовательностей нуклеиновых кислот, кодирующих антитело, где клетка содержит в эндогенном локусе легкой цепи каппа иммуноглобулина экзогенную генную последовательность вариабельной области легкой цепи каппа, причем экзогенная генная последовательность вариабельной области легкой цепи каппа содержит только один ген IGKV человека и только один ген IGKJ человека, причем ген IGKV человека и ген IGKJ человека функционально связаны с эндогенным геном константного домена легкой цепи, причем (1) ген IGKV человека представляет собой IGKV1-39, а ген IGKJ человека представляет собой IGKJ4; или (2) ген IGKV человека представляет собой IGKV3-11, а ген IGKJ человека представляет собой IGKJ1.
| WO 2011097603 A1, 11.08.2011 | |||
| WO 2009157771 A2, 30.12.2009 | |||
| LISA D.TAYLOR et al | |||
| A transgenic mouse that expresses a diversity of human sequence heavy and light chain immunoglobulins, Nucleic Acids Research, 1992, Vol | |||
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| ТРАНСГЕННЫЕ ЖИВОТНЫЕ И СПОСОБЫ ПРИМЕНЕНИЯ | 2011 |
|
RU2580017C2 |

