Перекрестная ссылка на родственные заявки:
По настоящей заявке испрашивается приоритет патентной заявки Соединенных Штатов No. 61/477414 (поданной 20 апреля 2011 г.; находящейся на рассмотрении), эта заявка включена в настоящее описание в качестве ссылки в полном объеме.
Ссылка на перечень последовательностей:
Заявка включает один или более Перечней последовательностей согласно 37 C.F.R. 1.821 и тому подобное, которые раскрыты как в бумажном, так и в машиночитаемом вариантах, где их описания в бумажном и машиночитаемом вариантах включены в настоящее описание в качестве ссылки в полном объеме.
Известный уровень техники:
Область техники, к которой относится изобретение:
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам и к другим молекулам, которые способны иммуноспецифически связываться с B7-H1 или PD-1. В некоторых вариантах осуществления такие молекулы дополнительно способны модулировать способность B7-H1 или B7-DC связываться с PD-1, или способны влиять на сигнальную активность B7-H1 или PD-1. Изобретение дополнительно относится к применению таких молекул для диагностики и лечения злокачественного новообразования и других заболеваний.
Описание известного уровня техники:
A. Опосредуемые клетками иммунные ответы
Иммунная система человека и других млекопитающих ответственна за обеспечение защиты против инфекций и заболеваний. Такая защита обеспечивается как гуморальным иммунным ответом, так и иммунным ответом, опосредуемым клетками. Гуморальный ответ приводит к продукции антител и других биомолекул, которые способны распознавать и нейтрализовать чужеродные агенты (антигены). В противоположность этому иммунный ответ, опосредуемый клетками, включает активацию Т-клетками макрофагов, клеток-природных киллеров (NK) и антигенспецифических цитотоксических Т-лимфоцитов и секрецию различных цитокинов в ответ на распознавание антигена (Dong, C. et al. (2003) "Immune Regulation by Novel Costimulatory Molecules" Immunolog. Res. 28(1):39-48).
Способность Т-клеток оптимально опосредовать иммунный ответ против антигена требует двух отдельных сигнальных взаимодействий (Viglietta, V. et al. (2007) "Modulating Co-Stimulation" Neurotherapeutics 4:666-675; Korman, A.J. et al. (2007) "Checkpoint Blockade in Cancer Immunotherapy" Adv. Immunol. 90:297-339). Во-первых, антиген, который располагается на поверхности антигенпрезентирующих клеток (APC), должен быть представлен специфичной для антигена наивной CD4+ Т-клетке. При таком представлении сигнал доставляется через рецептор Т-клетки (TCR), что заставляет Т-клетку инициировать иммунный ответ, который должен быть специфичным в отношении представляемого антигена. Во-вторых, серии костимуляторных и ингибиторных сигналов, опосредуемых взаимодействиями между APC и определенными молекулами поверхности Т-клетки, запускают сначала активацию и пролиферацию Т-клеток, и затем их ингибирование. Таким образом, первый сигнал придает специфичность иммунному ответу, тогда как второй сигнал служит для определения природы, степени и продолжительности ответа.
Иммунная система жестко контролируется костимуляторными и коингибиторными лигандами и рецепторами. Эти молекулы являются вторым сигналом для активации Т-клеток и обеспечивают сбалансированной сетью позитивных и негативных сигналов для доведения до максимума иммунных ответов против инфекции, в то же время, ограничивая иммунный ответ на собственные антигены (Wang, L. et al. (March 7, 2011) “VISTA, A Novel Mouse Ig Superfamily Ligand That Negatively Regulates T Cell Responses,” J. Exp. Med. 10.1084/jem.20100619:1-16; Lepenies, B. et al. (2008) “The Role Of Negative Costimulators During Parasitic Infections,” Endocrine, Metabolic & Immune Disorders - Drug Targets 8:279-288). Особую важность приобретает связывание между лигандами B7.1 (CD80) и B7.2 (CD86) антигенпрезентирующей клетки и CD28 и CTLA-4 рецепторами CD4+ Т-лимфоцита (Sharpe, A.H. et al. (2002) “The B7-CD28 Superfamily,” Nature Rev. Immunol. 2:116-126; Dong, C. et al. (2003) “Immune Regulation by Novel Costimulatory Molecules,” Immunolog. Res. 28(1):39-48; Lindley, P.S. et al. (2009) “The Clinical Utility Of Inhibiting CD28-Mediated Costimulation,” Immunol. Rev. 229:307-321). Связывание B7.1 или B7.2 с CD28 стимулирует активацию Т-клеток; связывание B7.1 или B7.2 с CTLA-4 ингибирует такую активацию (Dong, C. et al. (2003) “Immune Regulation by Novel Costimulatory Molecules,” Immunolog. Res. 28(1):39-48; Lindley, P.S. et al. (2009) “The Clinical Utility Of Inhibiting CD28-Mediated Costimulation,” Immunol. Rev. 229:307-321; Greenwald, R.J. et al. (2005) “The B7 Family Revisited,” Ann. Rev. Immunol. 23:515-548). CD28 конститутивно экспрессируется на поверхности Т-клеток (Gross, J., et al. (1992) “Identification And Distribution Of The Costimulatory Receptor CD28 In The Mouse,” J. Immunol. 149:380-388), тогда как экспрессия CTLA4 быстро позитивно регулируется после активации Т-клеток (Linsley, P. et al. (1996) “Intracellular Trafficking Of CTLA4 And Focal Localization Towards Sites Of TCR Engagement,” Immunity 4:535-543). Так как CTLA4 является более высокоаффинным рецептором (Sharpe, A.H. et al. (2002) “The B7-CD28 Superfamily,” Nature Rev. Immunol. 2:116-126), связывание сначала инициирует пролиферацию Т-клеток (через CD28), а затем ингибирует ее (через возникающую экспрессию CTLA4), тем самым ослабляя эффект, когда в пролиферации больше нет необходимости.
Последующие исследования лигандов CD28 рецептора привели к идентификации и характеристики набора молекул, родственных B7 («надсемейства B7») (Coyle, A.J. et al. (2001) “The Expanding B7 Superfamily: Increasing Complexity In Costimulatory Signals Regulating T Cell Function,” Nature Immunol. 2(3):203-209; Sharpe, A.H. et al. (2002) “The B7-CD28 Superfamily,” Nature Rev. Immunol. 2:116-126; Greenwald, R.J. et al. (2005) “The B7 Family Revisited,” Ann. Rev. Immunol. 23:515-548; Collins, M. et al. (2005) “The B7 Family Of Immune-Regulatory Ligands,” Genome Biol. 6:223.1-223.7; Loke, P. et al. (2004) “Emerging Mechanisms Of Immune Regulation: The Extended B7 Family And Regulatory T Cells.” Arthritis Res. Ther. 6:208-214; Korman, A.J. et al. (2007) “Checkpoint Blockade in Cancer Immunotherapy,” Adv. Immunol. 90:297-339; Flies, D.B. et al. (2007) “The New B7s: Playing a Pivotal Role in Tumor Immunity,” J. Immunother. 30(3):251-260; Agarwal, A. et al. (2008) “The Role Of Positive Costimulatory Molecules In Transplantation And Tolerance,” Curr. Opin. Organ Transplant. 13:366-372; Lenschow, D.J. et al. (1996) “CD28/B7 System of T Cell Costimulation,” Ann. Rev. Immunol. 14:233-258; Wang, S. et al. (2004) “Co-Signaling Molecules Of The B7-CD28 Family In Positive And Negative Regulation Of T Lymphocyte Responses,” Microbes Infect. 6:759-766). В настоящее время существует несколько представителей семейства: B7.1 (CD80), B7.2 (CD86), индуцибельный костимуляторный лиганд (ICOS-L), лиганд запрограммированной смерти 1 (PD-L1; B7-H1), лиганд запрограммированной смерти 2 (PD-L2; B7-DC), B7-H3, B7-H4 и B7-H6 (Collins, M. et al. (2005) “The B7 Family Of Immune-Regulatory Ligands,” Genome Biol. 6:223.1-223.7; Flajnik, M.F. et al. (2012) “Evolution Of The B7 Family: Co-Evolution Of B7H6 And Nkp30, Identification Of A New B7 Family Member, B7H7, And Of B7's Historical Relationship With The MHC,” Immunogenetics epub doi.org/10.1007/s00251-012-0616-2).
B. Взаимодействия B7-H1/PD1
1. B7-H1
B7-H1 (PD-L1, CD274) является особенно значимым представителем надсемейства B7, так как он главным образом вовлечен в формирование иммунного ответа на опухоли (Flies, D.B. et al. (2007) “The New B7s: Playing a Pivotal Role in Tumor Immunity,” J. Immunother. 30(3):251-260; патенты Соединенных Штатов No. 6803192, 7794710; публикации патентных заявок Соединенных Штатов No. 2005/0059051, 2009/0055944, 2009/0274666, 2009/0313687; публикации PCT No. WO 01/39722; WO 02/086083). B7-H1 представляет собой трансмембранный белок типа 1 с М.м. приблизительно 33 кДа. Предполагается, что он играет главную роль в подавлении иммунной системы во время определенных состояний, таких как беременность, аллотрансплантаты тканей, аутоиммунное заболевание и другие заболевания, такие как гепатит.
B7-H1 широко экспрессируется в различных тканях человека и мыши, таких как сердце, плацента, мышца, печень плода, селезенка, лимфатические узлы и тимус у обоих видов, а также в печени, легком и почке только у мыши (Martin-Orozco, N. et al. (2007) “Inhibitory Costimulation And Anti-Tumor Immunity,” Semin. Cancer Biol. 17(4):288-298). У человека экспрессия белка B7-H1 найдена в эндотелиальных клетках человека (Chen, Y. et al. (2005) “Expression of B7-H1 in Inflammatory Renal Tubular Epithelial Cells,” Nephron. Exp. Nephrol. 102:e81-e92; de Haij, S. et al. (2005) “Renal Tubular Epithelial Cells Modulate T-Cell Responses Via ICOS-L And B7-H1” Kidney Int. 68:2091-2102; Mazanet, M.M. et al. (2002) “B7-H1 Is Expressed By Human Endothelial Cells And Suppresses T Cell Cytokine Synthesis,” J. Immunol. 169:3581-3588), миокарде (Brown, J.A. et al. (2003) “Blockade Of Programmed Death-1 Ligands On Dendritic Cells Enhances T Cell Activation And Cytokine Production,” J. Immunol. 170:1257-1266), синцитиотрофобластах (Petroff, M.G. et al. (2002) “B7 Family Molecules: Novel Immunomodulators At The Maternal-Fetal Interface,” Placenta 23:S95-S101), в резидентных макрофагах некоторых тканей или в макрофагах, которые активированы интерфероном (IFN)-γ или фактором некроза опухолей (TNF)-α (Latchman, Y. et al. (2001) “PD-L2 Is A Second Ligand For PD-1 And Inhibits T Cell Activation,” Nat. Immunol 2:261-268), and in tumors (Dong, H. (2003) “B7-H1 Pathway And Its Role In The Evasion Of Tumor Immunity,” J. Mol. Med. 81:281-287). У мыши экспрессия белка B7-H1 найдена в эндотелии сердца, в клетках островкового аппарата поджелудочной железы, в тонком кишечнике и плаценте (Martin-Orozco, N. et al. (2007) “Inhibitory Costimulation And Anti-Tumor Immunity,” Semin. Cancer Biol. 17(4):288-298).
2. PD-1
Белок-1 программируемой смерти ("PD-1") представляет собой рецептор B7-H1 и B7-DC. PD-1 представляет собой представитель мембранных белков типа I широкого семейства CD28/CTLA4 регуляторов Т-клеток с М.м. приблизительно 31 кДа (Ishida, Y. et al. (1992) “Induced Expression Of PD-1, A Novel Member Of The Immunoglobulin Gene Superfamily, Upon Programmed Cell Death,” EMBO J. 11:3887-3895; публикации патентных заявок Соединенных Штатов No. 2007/0202100; 2008/0311117; 2009/00110667; патенты Соединенных Штатов No. 6808710; 7101550; 7488802; 7635757; 7722868; публикация PCT No. WO 01/14557). По сравнению с CTLA4, PD-1 более широко негативно регулирует иммунные ответы.
PD-1 экспрессируется на активированных Т-клетках, В-клетках и моноцитах (Agata, Y. et al. (1996) “Expression Of The PD-1 Antigen On The Surface Of Stimulated Mouse T And B Lymphocytes,” Int. Immunol. 8(5):765-772; Yamazaki, T. et al. (2002) “Expression Of Programmed Death 1 Ligands By Murine T Cells And APC,” J. Immunol. 169:5538-5545) и на низких уровнях в Т-клетках природных киллерах (NK) ((Nishimura, H. et al. (2000) “Facilitation Of Beta Selection And Modification Of Positive Selection In The Thymus Of PD-1-Deficient Mice,” J. Exp. Med. 191:891-898; Martin-Orozco, N. et al. (2007) “Inhibitory Costimulation And Anti-Tumor Immunity,” Semin. Cancer Biol. 17(4):288-298).
Внеклеточная область PD-1 состоит из единичного иммуноглобулинового (Ig)V домена с 23% идентичностью эквивалентному домену в CTLA4 (Martin-Orozco, N. et al. (2007) “Inhibitory Costimulation And Anti-Tumor Immunity,” Semin. Cancer Biol. 17(4):288-298). За внеклеточным доменом IgV следует трансмембранная область и внутриклеточный хвост. Внутриклеточный хвост содержит два сайта фосфорилирования, локализованных в иммунорецепторном ингибиторном мотиве на основе тирозина и в иммунорецепторном переключающем мотиве на основе тирозина, что предполагает негативную регуляцию PD-1 сигналов TCR (Ishida, Y. et al. (1992) “Induced Expression Of PD-1, A Novel Member Of The Immunoglobulin Gene Superfamily, Upon Programmed Cell Death,” EMBO J. 11:3887-3895; Blank, C. et al. (Epub 2006 Dec 29) “Contribution Of The PD-L1/PD-1 Pathway To T-Cell Exhaustion: An Update On Implications For Chronic Infections And Tumor Evasion Cancer,” Immunol. Immunother. 56(5):739-745).
Сообщалось об антителах, способных иммуноспецифически связываться с PD-1 (см., например, Agata, T. et al. (1996) “Expression Of The PD-1 Antigen On The Surface Of Stimulated Mouse T And B Lymphocytes,” Int. Immunol. 8(5):765-772).
C. Взаимодействия B7-H1 и PD-1
Взаимодействие B7-H1 и PD-1, как обнаружено, снабжает Т- и В-клетки решающим негативным костимуляторным сигналом (Martin-Orozco, N. et al. (2007) “Inhibitory Costimulation And Anti-Tumor Immunity,” Semin. Cancer Biol. 17(4):288-298) и действует в качестве индуктора клеточной смерти (Ishida, Y. et al. (1992) “Induced Expression Of PD-1, A Novel Member Of The Immunoglobulin Gene Superfamily, Upon Programmed Cell Death,” EMBO J. 11:3887-3895; Subudhi, S.K. et al. (2005) “The Balance Of Immune Responses: Costimulation Verse Coinhibition,” J. Molec. Med. 83:193-202).
Взаимодействие между низкими концентрациями PD-1 рецептора и B7-H1 лиганда приводит к передаче ингибиторного сигнала, который усиленно ингибирует пролиферацию антигенспецифических CD8+ Т-клеток; при более высоких концентрациях взаимодействия с PD-1 не ингибируют пролиферацию Т-клеток, но существенно снижают продукцию множества цитокинов (Sharpe, A.H. et al. (2002) “The B7-CD28 Superfamily,” Nature Rev. Immunol. 2:116-126). Пролиферация Т-клеток и продукция цитокинов как находящимися в покое, так и предварительно активированными CD4 и CD8 Т-клетками, и даже наивными Т-клетками из крови пупочного канатика, как обнаружено, ингибируются растворимыми слитыми белками B7-H1-Fc (Freeman, G.J. et al. (2000) “Engagement Of The PD-1 Immunoinhibitory Receptor By A Novel B7 Family Member Leads To Negative Regulation Of Lymphocyte Activation,” J. Exp. Med. 192:1-9; Latchman, Y. et al. (2001) “PD-L2 Is A Second Ligand For PD-1 And Inhibits T Cell Activation,” Nature Immunol. 2:261-268; Carter, L. et al. (2002) “PD-1:PD-L inhibitory pathway affects both CD4(+) and CD8(+) T cells and is overcome by IL-2,” Eur. J. Immunol. 32(3):634-643; Sharpe, A.H. et al. (2002) “The B7-CD28 Superfamily,” Nature Rev. Immunol. 2:116-126).
Взаимодействия B7-H1 - PD-1 ведут к аресту клеточного цикла на фазе G0-G1, но не увеличивают клеточной смерти (Latchman, Y. et al. (2001) “PD-L2 Is A Second Ligand For PD-1 And Inhibits T Cell Activation,” Nature Immunol. 2:261-268; Carter, L. et al. (2002) “PD-1:PD-L inhibitory pathway affects both CD4(+) and CD8(+) T cells and is overcome by IL-2,” Eur. J. Immunol. 32(3):634-643). Таким образом, комплексирование B7-H1 - PD-1 обладает способностью антагонистически действовать на сигнал B7 - CD28, когда антигенная стимуляция является слабой или лимитирующей, и играет ключевую роль в негативной регуляции ответов Т-клеток.
Передача сигнала, опосредуемого B7-H1 и PD-1, является сложной. Обе молекулы дополнительно связываются с другими белками. B7-H1 способен связываться с B7-1 (CD80) (Butte, M.J. et al. (2008) “Interaction of PD-L1 and B7-1,” Molecular Immunol. 45:3567-3572); PD-1 способен связываться с B7-DC (PD-L2) (Lázár-Molnár, E. et al. (2008) “Crystal Structure Of The Complex Between Programmed Death-1 (PD-1) And Its Ligand PD-L2,” Proc. Natl. Acad. Sci. (USA) 105(30):10483-10488). B7-1 взаимодействует с CD28 для доставки костимуляторного сигнала для активации Т-клеток, что важно на ранних стадиях иммунного ответа (Elloso, M.M. et al. (1999) “Expression and Contribution of B7-1 (CD80) and B7-2 (CD86) in the Early Immune Response to Leishmania major Infection,” J. Immunol. 162:6708-6715). B7-DC является сильным стимулятором Т-клеток, повышая пролиферацию Т-клеток и продукцию IFN-γ. Однако он также проявляет ингибиторное влияние на иммунный ответ через свое взаимодействие с PD-1 (Ishiwata, K. et al. (epub January 10, 2010) “Costimulator Responses Induced by Nippostrongylus brasiliensis,” J. Immunol. 184:2086-2094). Микробы и опухоли, очевидно, используют PD-1 и B7-H1 для ускользания от уничтожения иммунной системой. Различия в связывающей аффинности к различным рецепторам и лигандам, которые взаимодействуют с PD-1 и B7-H1, как предполагается, обусловливают индивидуальные функциональные исходы блокады PD-1 и B7-H1 в моделях заболеваний (Butte, M.J. et al. (2008) “Interaction of PD-L1 and B7-1,” Molecular Immunol. 45:3567-3572). Путь PD-1 вовлечен как играющий ключевую роль также в повреждение иммунной функции в процессе хронической инфекции («Т-клеточное истощение»), и блокада функции PD-1 способна восстановить многие функции Т-клеток (Rodríquez-García, M. et al. (November 19, 2010) “Expression Of PD-L1 And PD-L2 On Human Macrophages Is Up-Regulated By HIV-1 And Differentially Modulated By IL-10,” J. Leukocyte Biol. 89: doi:10.1189/jlb.0610327:1-9).
Роль B7-H1 и PD-1 в ингибировании активации и пролиферации Т-клеток предполагает, что эти молекулы могут служить в качестве терапевтических мишеней для лечения воспаления и злокачественного новообразования. Предлагается применение антител против PD1 для лечения инфекций и опухолей и позитивной модуляции адаптивного иммунного ответа (см. публикации патентных заявок Соединенных Штатов No. 2010/0040614; 2010/0028330; 2004/0241745; 2008/0311117; 2009/0217401; патенты Соединенных Штатов No. 7521051; 7563869; 7595048; публикации PCT No. WO 2004/056875; WO 2008/083174). Наоборот, средства, которые модулируют взаимодействие PD-1 с B7-H1, как предполагается, находят применение в позитивной или негативной модуляции иммунного ответа (см. патенты Соединенных Штатов No. 7029674; 7488802; публикации патентных заявок Соединенных Штатов No. 2007/0122378; 2009/0076250; 2009/0110667; 2009/0263865; 2009/0297518; публикацию PCT No. WO 2006/133396). Подобно этому предлагается использование антител против B7-H1 для лечения инфекций и опухолей и позитивной регуляции адаптивного иммунного ответа (публикации патентных заявок Соединенных Штатов No. 2009/0055944; 2009/0274666; 2009/0317368; патенты Соединенных Штатов No. 6803192; 7794710; публикации PCT No. WO 01/39722; WO 02/086083).
Тем не менее, несмотря на все эти достижения, остается потребность в композициях, способных модулировать взаимодействие между B7-H1 и PD-1. Настоящее изобретение направлено на такие композиции и их применение для лечения злокачественного новообразования и других заболеваний и состояний.
Краткое изложение сущности изобретения:
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам и к другим молекулам, которые способны иммуноспецифически связываться с B7-H1 или PD-1. В некоторых вариантах осуществления такие молекулы дополнительно способны модулировать способность B7-H1 связываться с PD-1 или способны влиять на сигнальную активность B7-H1 или PD-1. Изобретение дополнительно касается применения таких молекул для диагностики и лечения злокачественного новообразования и других заболеваний.
Конкретнее, в изобретении предлагается молекула, содержащая антигенсвязывающий фрагмент антитела, которая иммуноспецифически связывается с B7-H1 или PD-1 и, в особенности, с B7-H1 человека или PD-1 человека, предпочтительно, экспрессирующимися на поверхности живой клетки в эндогенной концентрации или концентрации, полученной в результате трансфекции. Изобретение особенно касается варианта осуществления в виде такой молекулы, в которой антигенсвязывающий фрагмент связывается с B7-H1, и где живая клетка представляет собой опухолевую клетку, клетку, инфицированную патогеном, или антигенпрезентирующую клетку, а также варианта осуществления в виде такой молекулы, в которой антигенсвязывающий фрагмент связывается с PD-1, и где живая клетка представляет собой Т-клетку.
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам и к другим молекулам, которые способны иммуноспецифически связываться с B7-H1 или PD-1. В некоторых вариантах осуществления такие молекулы дополнительно способны модулировать способность B7-H1 связываться с PD-1 или способны влиять на сигнальную активность B7-H1 или PD-1. Изобретение дополнительно касается применения таких молекул для диагностики и лечения злокачественного новообразования и других заболеваний.
Конкретнее, в изобретении предлагается молекула, содержащая антигенсвязывающий фрагмент антитела, которая иммуноспецифически связывается с B7-H1 или PD-1 и, в особенности, с B7-H1 человека или PD-1 человека, предпочтительно, экспрессирующимися на поверхности живой клетки в эндогенной концентрации или концентрации, полученной в результате трансфекции. Изобретение особенно касается варианта осуществления в виде такой молекулы, в которой антигенсвязывающий фрагмент связывается с B7-H1, и где живая клетка представляет собой опухолевую клетку, клетку, инфицированную патогеном, или антигенпрезентирующую клетку, а также варианта осуществления в виде такой молекулы, в которой антигенсвязывающий фрагмент связывается с PD-1, и где живая клетка представляет собой Т-клетку.
Изобретение дополнительно касается варианта осуществления в виде таких молекул, где молекула представляет собой моноклональное антитело, антитело человека, химерное антитело или гуманизированное антитело. Изобретение включает варианты осуществления, где такие антитела являются моноспецифическими, биспецифическими, триспецифическими или мультиспецифическими.
Изобретение дополнительно касается варианта осуществления в виде таких молекул или антител, которые связываются с B7-H1, и где их антигенсвязывающий фрагмент содержит шесть CDR, где CDR содержат по меньшей мере одну консенсусную CDR из CDR антител 1E12, 1F4, 2G11, 3B6 и 3D10 против B7-H1 со всеми оставшимися CDR, выбранными из:
(A) трех CDR легкой цепи и трех CDR тяжелой цепи антитела 1E12 против B7-H1;
(B) трех CDR легкой цепи и трех CDR тяжелой цепи антитела 1F4 против B7-H1;
(C) трех CDR легкой цепи и трех CDR тяжелой цепи антитела 2G11 против B7-H1;
(D) трех CDR легкой цепи и трех CDR тяжелой цепи антитела 3B6 против B7-H1; или
(E) трех CDR легкой цепи и трех CDR тяжелой цепи антитела 3D10 против B7-H1.
Изобретение дополнительно касается варианта осуществления в виде таких молекул или антител, которые связываются с B7-H1, и где их антигенсвязывающий фрагмент содержит шесть CDR, где шесть CDR представляют собой:
(A) три CDR легкой цепи и три CDR тяжелой цепи антитела 1E12 против B7-H1;
(B) три CDR легкой цепи и три CDR тяжелой цепи антитела 1F4 против B7-H1;
(C) три CDR легкой цепи и три CDR тяжелой цепи антитела 2G11 против B7-H1;
(D) три CDR легкой цепи и три CDR тяжелой цепи антитела 3B6 против B7-H1; или
(E) три CDR легкой цепи и три CDR тяжелой цепи антитела 3D10 против B7-H1.
Изобретение дополнительно касается варианта осуществления в виде таких антител, где антитело связывается с B7-H1 и содержит вариабельный домен антитела h3D10 Var 1, h3D10 Var 2, h3D10 Var 3, h3D10 Var 4, h3D10 Var 5, h3D10 Var 6, h3D10 Var 7, h3D10 Var 8, h3D10 Var 9, h3D10 Var 10, h3D10 Var 11, h3D10 Var 12, h3D10 Var 13 или h3D10 Var 14.
Изобретение дополнительно касается варианта осуществления в виде описанных выше молекул или антител, где молекулы или антитела связываются с PD-1, и где антигенсвязывающий фрагмент содержит шесть CDR, где CDR содержат по меньшей мере одну консенсусную CDR из CDR антител 1E3, 1E8 и 1H3 против PD-1 со всеми оставшимися CDR, выбранными из:
(A) трех CDR легкой цепи и трех CDR тяжелой цепи антитела 1E3 против PD-1;
(B) трех CDR легкой цепи и трех CDR тяжелой цепи антитела 1E8 против PD-1; или
(C) трех CDR легкой цепи и трех CDR тяжелой цепи антитела 1H3 против PD-1.
Изобретение дополнительно касается варианта осуществления в виде таких антител, где шесть CDR представляют собой:
(A) три CDR легкой цепи и три CDR тяжелой цепи антитела 1E3 против PD-1;
(B) три CDR легкой цепи и три CDR тяжелой цепи антитела 1E8 против PD-1; или
(C) три CDR легкой цепи и три CDR тяжелой цепи антитела 1H3 против PD-1.
Изобретение дополнительно касается варианта осуществления в виде таких антител, где антитело связывается с PD-1 и содержит вариабельный домен антитела h1H3 Var 1, h1H3 Var 2, h1H3 Var 3, h1H3 Var 4, h1H3 Var 5, h1H3 Var 6, h1H3 Var 7, h1H3 Var 8, h1H3 Var 9, h1H3 Var 10, h1H3 Var 11, h1H3 Var 12, h1H3 Var 13 или h1H3 Var 14.
Изобретение дополнительно касается варианта осуществления в виде описанных выше молекул или антител, где молекула или антитело помечены определяемой меткой или содержит конъюгированный токсин, лекарственное средство, рецептор, фермент, лиганд рецептора.
Изобретение дополнительно касается варианта осуществления в виде описанных выше молекул или антител, где молекула или антитело:
(A) модулирует передачу сигнала, опосредуемую B7-H1 или PD-1;
(B) ослабляет способность B7-H1 связываться с рецептором B7-H1 или ослабляет способность PD-1 связываться с лигандом PD-1;
(C) является агонистом B7-H1 или PD-1 в отношении опосредуемой ими передачи сигнала;
(D) опосредует пролиферацию Т-клеток; или
(E) повышает продукцию IFN-γ.
Изобретение дополнительно касается фармацевтической композиции, содержащей терапевтически эффективное количество любых из описанных выше молекул или антител и физиологически приемлемый носитель или наполнитель. Изобретение дополнительно касается применения такой фармацевтической композиции для лечения злокачественного новообразования, аутоиммунного заболевания, инфекционного заболевания или заболевания, влияющего на количество Т-клеток или на их здоровое состояние. Изобретение дополнительно касается применения такой фармацевтической композиции, где лечение является профилактическим и предлагается перед любым симптомом злокачественного новообразования, аутоиммунного заболевания, инфекционного заболевания или заболевания, влияющего на количество Т-клеток или на их здоровое состояние, или для лечения побочных состояний при трансплантации.
Изобретение дополнительно касается применения любых описанных выше молекул или антител для диагностики злокачественного новообразования, аутоиммунного заболевания (особенно заболевания трансплантат против хозяина), инфекционного заболевания (особенно хронического вирусного заболевания) или заболевания, влияющего на количество Т-клеток или на их здоровое состояние, у индивидуума с помощью анализа клеток индивидуума на их способность связываться с B7-H1 или PD-1.
Изобретение особенно касается варианта осуществления в виде таких молекул, антител и композиций, где B7-H1 представляет собой B7-H1 человека и PD-1 представляет собой PD-1 человека.
Изобретение особенно касается способа диагностики заболевания (особенно злокачественного новообразования) у индивидуума, включающего оценку клеток индивидуума на их способность связываться с любыми из описанных выше молекул, связывающих B7-H1, где в способе предлагается цитологическая оценка для диагностики наличия заболевания у индивидуума.
Изобретение дополнительно касается способа диагностики заболевания (особенно заболевания, влияющего на количество Т-клеток и/или на их здоровое состояние) у индивидуума, включающего оценку клеток индивидуума на их способность связываться с молекулой, связывающей PD-1, где в способе предлагается цитологическая оценка для диагностики наличия и/или прогрессии заболевания у индивидуума, или для оценки ответа индивидуума на лечение.
Краткое описание чертежей
На фигуре 1 представлено тестирование супернатантов гибридом, на связывание антитела с B7-H1. Положительный контроль (PC): 1:1000 разведенные сыворотки от мыши, используемой для создания гибридомы; отрицательный контроль (NC): 5% молоко/PBS. Данные показаны для связывания с B7-H1-Fc и определения с помощью IgG против мыши.
На фигуре 2 представлены результаты экспериментов по определению того, способны ли выделенные антитела против B7-H1 модулировать связывание B7-H1 с PD-1. Положительный контроль: клоны MIH-1 и 29E.2A3 (оба против CD274 человека (B7-H1); два отрицательных контроля: кондиционные среды от неродственной гибридомы (случайные АТ) и векторный контроль (VC).
На фигуре 3 представлена медиана интенсивности флуоресценции (MFI) тестируемых антител против B7-H1 при связывании с CHO-hB7-H1. Как обнаружено, ни один из тестируемых клонов не имел перекрестной реакции с родительской линией CHO, что указывает на то, что экспрессируемое антитело было иммуноспецифическим в отношении B7-H1 человека.
На фигуре 4 представлена медиана интенсивности флуоресценции (MFI), полученная в результате тестирования связывания выбранных антител против B7-H1 человека с клетками CHO, экспрессирующими B7-H1 человека.
На фигуре 5 сравнивается медиана интенсивности флуоресценции (MFI), полученная в результате тестирования связывания антител 5H1 и 1E12 против B7-H1 человека с клетками CHO, экспрессирующими B7-H1 человека.
На фигуре 6 представлено связывание антигена и изотип выделенных антител против PD-1 человека.
На фигурах 7A-7B представлены результаты экспериментов, указывающие на то, что некоторые из выделенных гибридом экспрессировали нейтрализующие антитела против PD-1 человека.
На фигуре 8 представлена медиана интенсивности флуоресценции (MFI) тестируемых антител против PD-1 при связывании с CHO-hPD-1. Как обнаружено, ни один из тестируемых клонов не имел перекрестной реакции с родительской линией CHO, что указывает на то, что экспрессируемое антитело было специфичным в отношении PD-1 человека.
На фигуре 9 представлена медиана интенсивности флуоресценции (MFI), полученная в результате тестирования связывания выбранных антител против PD-1 человека с клетками CHO, экспрессирующими PD-1 человека. Положительный контроль: EH12 (имеющееся в продаже от BioLegend антитело против PD-1 человека); mIgG1: отрицательный контроль - IgG мыши.
На фигуре 10 представлена медиана интенсивности флуоресценции (MFI), полученная в результате основанного на клетках конкурентного анализа при различных концентрациях антител против PD-1 человека.
На фигуре 11 представлена медиана интенсивности флуоресценции (MFI), полученная в результате основанного на клетках конкурентного анализа при концентрации антитела против PD-1 человека 20 мкг/мл.
На фигуре 12 представлены результаты эксперимента, в котором клетки CHO, трансфицированные полноразмерным PD-1 человека, предварительно инкубировали с насыщающей дозой моноклональных антител (mAb) против PD-1 человека или с контрольными Ig перед окрашиванием меченными биотином hB7-H1-FC или hB7-DC mIg.
На фигуре 13, панели A-B, представлено сравнение связывания: (A) имеющегося в продаже антитела против PD-1, EH12 с (B) моноклональным антителом мыши, которое является химерным («ch»), обладая областями Fab антитела мыши против PD-1 человека и областью Fc антитела человека.
На фигуре 14, панели A-B, представлена способность антител против PD-1 по настоящему изобретению проявлять блокирующий эффект в отношении связывания биотинилированного B7-Hl-Fc и биотинилированного B7-DC-Fc с PD-1.
На фигуре 15 показана кривая связывания химерного антитела 1H3 против PD-1 человека с клетками CHO.hPD-1 при определении с помощью антитела против hIg.
На фигурах 16A-16B показаны результаты исследований способности химерного антитела 1H3 против PD-1 человека связываться с первичными Т-клетками человека CD8+ (фигура 16A) и CD4+ (фигура 16B) относительно антител отрицательного контроля (паливизумаб; SYNAGIS®, Medimmune, Inc.).
На фигуре 17 представлена способность антител по настоящему изобретению повышать антигенспецифический ответ Т-клеток при измерении с помощью разведения CFSE при повторном присутствии столбнячного токсина (TT).
На фигурах 18A-18D показана способность гуманизированных вариантов 1H3 (h1H3 Var 1 - h1H3 Var 14) связываться с клетками CHO.hPD1.
На фигурах 19A-19B показана способность гуманизированных антител против PD-1 блокировать взаимодействия между hPD-1-Fc и клетками HEK293, экспрессирующими B7-H1 (фигура 19A) или B7-DC (фигура 19B).
На фигуре 20 представлены кривые связывания h1H3 Var 1 - h1H3 Var 6.
Подробное описание изобретения:
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам и к другим молекулам, которые способны иммуноспецифически связываться с B7-H1 или PD-1. В некоторых вариантах осуществления такие молекулы дополнительно способны модулировать способность B7-H1 или B7-DC в отношении связывания с PD-1, или способны влиять на сигнальную активность B7-H1 или PD-1. Изобретение дополнительно касается применения таких молекул для лечения злокачественного новообразования и других заболеваний.
Молекула, как говорится, способна «иммуноспецифически связываться» со второй молекулой, если такое связывание характеризуется специфичностью и аффинностью антитела к распознаваемому им антигену. Антитела, как говорится, способны «иммуноспецифически связываться» с областью-мишенью или конформационной областью («эпитопом») антигена (и в частности, антигенов: B7-H1 или PD-1), если в такое связывание вовлекается сайт распознавания антигена молекулы иммуноглобулина. Антитело, которое иммуноспецифически связывается с конкретным антигеном, может связываться с другими антигенами с более низкой аффинностью, если другой антиген обладает некоторым сходством последовательности или конформационным сходством, которое распознается сайтом распознавания антигена при определении, например, иммунометодами, методами BIACORE® или другими методами, известными в данной области, но оно не должно связываться с совершенно неродственным антигеном. Однако предпочтительно, чтобы антитела (и их антигенсвязывающие фрагменты) не реагировали перекрестно с другими антигенами. Антитела могут также связываться с другими молекулами путем, который не является иммуноспецифическим, например, с рецепторами FcR с помощью связывающих доменов в других областях/доменах молекулы, которые не входят в сайт распознавания антигена, например, с помощью области Fc.
Как используется в настоящем описании термин «модулировать» относится к способности изменять эффект или результат. В частности, изобретение относится к молекулам (особенно к антителам или их антигенсвязывающим фрагментам, которые иммуноспецифически связываются с B7-H1 человека или PD-1 человека), которые способны модулировать связывание между B7-H1 и PD-1 и/или модулировать передачу сигнала, которая возникает как следствие связывания B7-H1-PD-1. Такая модуляция может приводить к ослаблению или полному блокированию способности B7-H1 связываться с PD-1. В дополнительном варианте осуществления такая модуляция может ослаблять или полностью нейтрализовать способность B7-H1 или PD-1 опосредовать передачу сигнала. В дополнительном варианте осуществления такая модуляция может повышать или другим образом оказывать агонистические эффекты на передачу сигнала через B7-H1 или PD-1 1) либо с помощью усиления взаимодействия между B7-H1 и PD-1 и облегчения связывания B7-H1 - PD-1, ii) либо с помощью прямого связывания с B7-H1 и PD-1 и в результате этого имитирования активности эндогенного лиганда и т.п. В еще одном варианте осуществления такая модуляция может изменять природу взаимодействия между B7-H1 и PD-1 так, что изменяется природа вызываемой передачи сигнала. Например, молекулы по настоящему изобретению могут при связывании с B7-H1 или PD-1 изменять способность таких молекул связываться с другими лигандами и рецептором (например, влияя на способность PD-1 связываться с B7-DC или на способность B7-H1 связываться с B7-1 (CD80)) и тем самым изменяя итоговую активность. Предпочтительно, такая модуляция должна обеспечивать по меньшей мере 10% изменение в измеряемой активности иммунной системы, более предпочтительно, по меньшей мере 50% изменение такой активности, или по меньшей мере 2-кратное, 5-кратное, 10-кратное или, еще более предпочтительно, по меньшей мере 100-кратное изменение такой активности.
Как используется в настоящем описании термин «антитело» предназначен для обозначения молекулы иммуноглобулина, которая обладает антигенраспознающим сайтом «вариабельной области». Термин «вариабельная область» предназначен для отличия такого домена иммуноглобулина от доменов, которые широко используются антителами (таких как домен Fc). Вариабельная область содержит «гипервариабельную область», остатки которой отвечают за связывание антигена. Гипервариабельная область содержит аминокислотные остатки из «области, определяющей комплементарность» или «CDR» (т.е. обычно приблизительно остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и приблизительно остатки 27-35 (H1), 50-65 (H2) и 95-102 (H3) в вариабельном домене тяжелой цепи; Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)) и/или такие остатки из «гипервариабельной петли» (т.е. остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (H1), 53-55 (H2) и 96-101 (H3) в вариабельном домене тяжелой цепи; Chothia, C. et al. (1987) “Canonical Structures For The Hypervariable Regions Of Immunoglobulins,” J. Mol. Biol. 196:901-917). Остатки «области каркаса» или «FR» представляют собой те остатки вариабельного домена, которые отличны от остатков гипервариабельной области, как определено в настоящем описании. Термин антитело включает моноклональные антитела, мультиспецифические антитела, антитела человека, гуманизированные антитела, синтетические антитела, химерные антитела, приближенные к верблюдным антитела (см., например, Muyldermans et al., 2001, Trends Biochem. Sci. 26:230; Nuttall et al., 2000, Cur. Pharm. Biotech. 1:253; Reichmann and Muyldermans, 1999, J. Immunol. Meth. 231:25; международные публикации No. WO 94/04678 и WO 94/25591; патент США No. 6005079), одноцепочечные Fv (scFv) (см., например, Pluckthun in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds. Springer-Verlag, New York, pp. 269-315 (1994)), одноцепочечные антитела, связанные дисульфидными связями Fv (sdFv), интратела и антиидиотипические (анти-Id) антитела (включая, например, анти-Id и анти-анти-Id антитела к антителам по изобретению). В частности, такие антитела включают молекулы иммуноглобулинов любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса.
Как используется в настоящем описании термин «антигенсвязывающий фрагмент» антитела относится к одной или более частям антитела, которые содержат области, определяющие комплементарность («CDR»), и необязательно остатки каркаса, которые содержат антигенраспознающий сайт «вариабельной области» антитела, и проявляют способность к иммуноспецифическому связыванию антигена. Такие фрагменты включают Fab', F(ab')2, Fv, одноцепочечные антитела (ScFv) и их мутанты, природные варианты и слитые белки, содержащие антигенраспознающий сайт «вариабельной области» антитела, и гетерологичный белок (например, токсин, антигенраспознающий сайт для отличного антигена, фермент, рецептор, лиганд рецептора и т.п.). Как используется в настоящем описании термин «фрагмент» относится к пептиду или полипептиду, содержащему аминокислотную последовательность по меньшей мере из 5 последовательных аминокислотных остатков, по меньшей мере из 10 последовательных аминокислотных остатков, по меньшей мере из 15 последовательных аминокислотных остатков, по меньшей мере из 20 последовательных аминокислотных остатков, по меньшей мере из 25 последовательных аминокислотных остатков, по меньшей мере из 40 последовательных аминокислотных остатков, по меньшей мере из 50 последовательных аминокислотных остатков, по меньшей мере из 60 последовательных аминокислотных остатков, по меньшей мере из 70 последовательных аминокислотных остатков, по меньшей мере из 80 последовательных аминокислотных остатков, по меньшей мере из 90 последовательных аминокислотных остатков, по меньшей мере из 100 последовательных аминокислотных остатков, по меньшей мере из 125 последовательных аминокислотных остатков, по меньшей мере из 150 последовательных аминокислотных остатков, по меньшей мере из 175 последовательных аминокислотных остатков, по меньшей мере из 200 последовательных аминокислотных остатков или по меньшей мере из 250 последовательных аминокислотных остатков.
Антитела человека, химерные или гуманизированные антитела особенно предпочтительны для использования in vivo у человека, однако антитела мыши или антитела других видов могут быть успешно применены для многих вариантов использования (например, для методов определения in vitro или in situ, экстренного использования in vivo и т.п.). Полностью человеческие антитела особенно желательны для терапевтического лечения индивидуумов, являющихся человеком.
Антитела человека могут быть получены с помощью разнообразных методов, известных в данной области техники, включая методы фагового дисплея, описанные выше, с использованием библиотек антител, происходящих от последовательностей иммуноглобулинов человека (см. патенты США No. 4444887 и 4716111; и международные публикации No. WO 98/46645, WO 98/50433, WO 98/24893, WO 98/16654, WO 96/34096, WO 96/33735 и WO 91/10741). Антитела человека могут быть получены с использованием трансгенных мышей, которые не способны экспрессировать функционирующие эндогенные иммуноглобулины, но которые могут экспрессировать гены иммуноглобулинов человека. Например, комплексы генов тяжелых и легких цепей иммуноглобулинов могут быть введены случайным образом или с помощью гомологичной рекомбинации в эмбриональные стволовые клетки мыши. Альтернативно, вариабельная область, константная область и отличительная область человека могут быть введены в эмбриональные стволовые клетки мыши в дополнение к генам тяжелых и легких цепей человека. Генам тяжелых и легких цепей иммуноглобулинов мыши может быть придано отсутствие функционирования отдельно или одновременно с введением локусов иммуноглобулинов человека с помощью гомологичной рекомбинации. В частности, гомозиготная делеция области JH препятствует продукции эндогенных антител. Модифицированные эмбриональные стволовые клетки развиваются и вводятся в бластоцисты с помощью микроинъекций с получением химерных мышей. Химерные мыши затем скрещиваются с получением гомозиготного потомства, которое экспрессирует антитела человека. Трансгенных мышей иммунизируют с использованием общепринятых методов выбранным антигеном, например, целым полипептидом по изобретению или его частью. Моноклональные антитела, направленные против антигена, могут быть получены от иммунизированных трансгенных мышей с использованием традиционных гибридомных методов (см., например, патент США No. 5916771). Трансгены иммуноглобулинов человека, которые несут трансгенные мыши, реорганизуются в процессе дифференцировки В-клеток и в последующем происходит переключение их классов и соматическая мутация. Таким образом, при использовании такого метода можно получить терапевтически подходящие антитела IgG, IgA, IgM и IgE. Для общего представления об этом методе получения антител человека см. Lonberg and Huszar (1995, Int. Rev. Immunol. 13:65-93, включенную в настоящее описание в качестве ссылки в полном объеме). Для подробного обсуждения этого метода получения антител человека и моноклональных антител человека и протоколов получения таких антител см., например, международные публикации No. WO 98/24893, WO 96/34096 и WO 96/33735; и патенты США No. 5413923, 5625126, 5633425, 5569825, 5661016, 5545806, 5814318 и 5939598, которые включены в настоящее описание в качестве ссылки в полном объеме. Кроме того, такие компании, как Abgenix, Inc. (Freemont, CA) и Medarex (Princeton, NJ), могут быть привлечены для снабжения антителами человека, направленными против выбранного антигена, с использованием метода, сходного с описанным выше.
«Химерное антитело» представляет собой молекулу, в которой различные части антитела происходят от различных молекул иммуноглобулинов, такую как антитела, имеющие вариабельную область, происходящую от антитела, не относящегося к человеку, и константную область иммуноглобулина человека. Методы получения химерных антител известны в данной области. См., например, Morrison, 1985, Science 229:1202; Oi et al., 1986, BioTechniques 4:214; Gillies et al., 1989, J. Immunol. Methods 125:191-202; и патенты США No. 6311415, 5807715, 4816567 и 4816397. Химерные антитела, содержащие одну или более CDR от видов, не относящихся к человеку, и области каркаса от молекулы иммуноглобулина человека, могут быть получены с использованием разнообразных методов, известных в данной области, включая, например, пересадку CDR (EP 239400; международная заявка No. WO 91/09967; и патенты США No. 5225539, 5530101 и 5585089), винирование или перекладку (патенты EP 592106; EP 519596; Padlan, 1991, Molecular Immunology 28(4/5):489-498; Studnicka et al., 1994, Protein Engineering 7:805; и Roguska et al., 1994, Proc. Natl. Acad. Sci. USA 91:969), и перетасовку цепей (патент США No. 5565332).
Изобретение особенно касается «гуманизированных антител» (см., например, европейские патенты No. EP 239400, EP 592106 и EP 519596; международные публикации No. WO 91/09967 и WO 93/17105; патенты США No. 5225539, 5530101, 5565332, 5585089, 5766886 и 6407213; и Padlan, 1991, Molecular Immunology 28(4/5):489-498; Studnicka et al., 1994, Protein Engineering 7(6):805-814; Roguska et al., 1994, PNAS 91:969-973; Tan et al., 2002, J. Immunol. 169:1119-25; Caldas et al., 2000, Protein Eng. 13:353-60; Morea et al., 2000, Methods 20:267-79; Baca et al., 1997, J. Biol. Chem. 272:10678-84; Roguska et al., 1996, Protein Eng. 9:895-904; Couto et al., 1995, Cancer Res. 55 (23 Supp):5973s-5977s; Couto et al., 1995, Cancer Res. 55:1717-22; Sandhu, 1994, Gene 150:409-10; Pedersen et al., 1994, J. Mol. Biol. 235:959-73; Jones et al., 1986, Nature 321:522-525; Reichmann et al., 1988, Nature 332:323-329; и Presta, 1992, Curr. Op. Struct. Biol. 2:593-596). Как используется в настоящем описании термин «гуманизированное антитело» относится к иммуноглобулину, содержащему область каркаса от человека и одну или более CDR от иммуноглобулина, происходящего не от человека (обычно от мыши или крысы). Иммуноглобулин, происходящий не от человека, предоставляющий CDR, называют «донором», а иммуноглобулин человека, предоставляющий каркас, называют «акцептором». Нет необходимости в присутствии константных областей, но если они присутствуют, они должны быть по существу идентичными константным областям иммуноглобулина человека, т.е. иметь идентичность по меньшей мере приблизительно 85-90%, предпочтительно, приблизительно 95% или более. Следовательно, все части гуманизированного иммуноглобулина за исключением по возможности CDR являются по существу идентичными соответствующим частям природных последовательностей иммуноглобулинов человека. Гуманизированное антитело представляет собой антитело, содержащее гуманизированную легкую цепь и гуманизированную тяжелую цепь иммуноглобулина. Например, гуманизированное антитело не должно охватывать обычное химерное антитело, потому что, например, вся вариабельная область химерного антитела не происходит от человека. Говорится, что антитело-донор «гуманизировано» с помощью процесса «гуманизации», потому что полученное гуманизированное антитело, как ожидается, связывается с тем же самым антигеном, что и антитело-донор, которое предоставляет CDR. Большей частью гуманизированные антитела представляют собой иммуноглобулины человека (антитело-реципиент), в которых остатки гипервариабельных областей реципиента заменены остатками гипервариабельных областей (антитела-донора) от видов, не относящихся к человеку, таких как мышь, крыса, кролик или нечеловекообразный примат, имеющими желаемые специфичность, аффинность и емкость. В некоторых случаях остатки области каркаса (FR) иммуноглобулина человека заменяют соответствующими остатками не от человека. Более того, гуманизированные антитела могут содержать остатки, которых не находят в антителе-реципиенте или в антителе-доноре. Эти модификации делают для дополнительного улучшения работы антитела. В целом, гуманизированное антитело должно содержать по существу все, по меньшей мере, из одного и обычно из двух вариабельных доменов, в которых все или по существу все гипервариабельные области соответствуют областям иммуноглобулина, происходящего не от человека, и все или по существу все FR представляют собой FR последовательности иммуноглобулина человека. Гуманизированное антитело необязательно может также содержать по меньшей мере часть константной области (Fc) иммуноглобулина, обычно иммуноглобулина человека, которая иммуноспецифически связывается с полипептидом FcγRIIB, который изменен введением замен, делеций или добавок аминокислотных остатков (т.е. введением мутаций).
Антитела, применяемые в способах по настоящему изобретению, могут быть моноспецифическими. Представляют интерес также биспецифические антитела, триспецифические антитела или антитела более высокой мультиспецифичности, которые характеризуются специфичностью к различным мишеням в дополнение к B7-H1 или PD-1. В предпочтительном варианте осуществления такие мультиспецифические антитела должны характеризоваться специфичностью к различным мишеням иммунных клеток. Например, такие антитела могут связываться как с B7-H1, так и с B7-DC, и таким образом модулировать оба зависимых от PD-1 ответа. Наоборот, такие антитела могут связываться с PD-1 и B7-1 и препятствовать обоим зависимым от B7-H1 ответам. В другом варианте осуществления мультиспецифическое антитело связывается с молекулами (рецепторами или лигандами), вовлеченными в альтернативные иммуномодуляторные пути, с такими как CTLA4, TIM3, TIM4, OX40, CD40, GITR, 4-1-BB, B7-H4, LIGHT или LAG3, для усиления иммуномодуляторных эффектов. Более того, мультиспецифическое антитело может связываться с эффекторными молекулами, такими как цитокины (например, IL-7, IL-15, IL-12, IL-4 TGF-бета, IL-10, IL-17, IFNg, Flt3, BLys) и хемокины (например, CCL21), которые особенно могут подходить для модуляции как острого, так и хронического иммунных ответов.
Кроме того, мультиспецифические антитела могут связываться с антигеном, который важен для направленной доставки антитела к конкретному клеточному типу или ткани. Например, те антитела, которые связывают как PD-1, так и CD27 (или B7-H1 и CD27) могут способствовать колокализации активированных В-клеток памяти (mBAct) и антигенпрезентирующих клеток (APC), так что PD-1, представленный такими APC, может взаимодействовать с лигандами на поверхности В-клеток mBAct, стимулируя выживание mBAct В-клеток. Так как потеря mBAct В-клеток является ускоряющим фактором в прогрессии инфекции ВИЧ до СПИДа (Titanji, K. et al. (2010) “Acute Depletion Of Activated Memory B Cells Involves The PD-1 Pathway In Rapidly Progressing SIV-Infected Macaques,” J. Clin. Invest. 120(11):3878-3890), антитела, которые связываются как с PD-1, так и с CD27, находят применение в лечении инфекции ВИЧ и в профилактики или задержке развития СПИДа. Как обсуждалось выше, путь PD-1 вовлечен как играющий ключевую роль в нарушение иммунной функции при хронической инфекции ВИЧ («истощение Т-клеток») (Khaitan, A. et al. (2011) “Revisiting Immune Exhaustion During HIV Infection,” Curr. HIV/AIDS Rep. 8:4-11; Rodríquez-García, M. et al. (November 19, 2010) “Expression Of PD-L1 And PD-L2 On Human Macrophages Is Up-Regulated By HIV-1 And Differentially Modulated By IL-10,” J. Leukocyte Biol. 89: doi:10.1189/jlb.0610327:1-9; Grabmeier-Pfistershammer, K. et al. (2011) “Identification of PD-1 as a Unique Marker for Failing Immune Reconstitution in HIV-1-Infected Patients on Treatment,” J Acquir. Immune Defic. Syndr. 56(2):118-124). Макрофаги, как показано, вносят существенный вклад на ранних стадиях инфекции ВИЧ (Carter, C. A. et al. (2008) “Cell Biology Of HIV-1 Infection Of Macrophages,” Ann. Rev. Microbiol. 62:425-443; Noursadeghi, M. et al. (2006) “HIV-1 Infection Of Mononuclear Phagocytic Cells: The Case For Bacterial Innate Immune Deficiency In AIDS,” Lancet Infect. Dis. 6:794-804). Соответственно, антитела (особенно, если они конъюгированы с токсином), которые связываются с PD-1 и со специфическим маркером макрофагов (таким, как CD14, CD68, CD163, TLR2 и т.п.) находят применение в профилактики инфекции ВИЧ. Кроме того, антитела, которые связываются со множеством маркеров истощения Т-клеток (например, с PD-1 и с любым или со всеми из: CTLA4, TIM3, TIM4 или LAG-3), находят применение для лечения или диагностики иммунологической реактивности. Другие представляющие интерес антигены-мишени включают маркеры раковых клеток.
Кроме того, обнаружено, что PD-1+ CD8+ клетки обладают активностью против ВИЧ (Killian, M.S. et al. (2011) “Natural Suppression of Human Immunodeficiency Virus Type 1 Replication Is Mediated by Memory CD8+ T Cells,” J. Virol. 85(4):1696-1705). Таким образом, антитела, которые связываются как с PD-1, так и с CD8, находят применение, например, в способах ex vivo для выделения и получения обогащенной популяции таких клеток для конечного использования при лечении инфекции ВИЧ и СПИДа у больных.
Другие маркеры, которые могут быть использованы в таких биспецифических, триспецифических или мультиспецифических антителах против PD-1 или против B7-H1, включают CD4, CD8, CD25 и CTLA-4 (см. статьи De Keersmaecker, B. et al. (2011) (“Fighting with the Enemy’s Weapons? The Role of Costimulatory Molecules in HIV,” Curr. Molec. Med. 566-5240/11: 1-25; and Sarikonda, G. (2011) “Immunosuppressive Mechanisms During Viral Infectious Diseases;” Methods in Molec. Biol. 677:431-447, обе включены в настоящее описание в качестве ссылки).
Аналогично, хотя CD4 Т-клетки требуются, чтобы замедлить рост и распространение M. tuberculosis, опосредуемое PD-1 ингибирование также требуется для предотвращения стимуляции CD4+ Т-клетками тяжелого заболевания (Barber, D.L. et al. (2011) “CD4 T Cells Promote Rather than Control Tuberculosis in the Absence of PD-1-Mediated Inhibition,” J. Immunol. 186:1598-1607; Sakai, S. et al. (2010) “PD-1 - PD-L1 pathway impairs Th1 immune response in the late stage of infection with Mycobacterium bovis bacillus Calmette-Guérin,” Intl. Immunol. 22(12):915-925; Lázár-Molnár, E. et al. (2010) “Programmed Death-1 (PD-1)-Deficient Mice Are Extraordinarily Sensitive To Tuberculosis,” Proc. Natl. Acad. Sci. (USA) 107(30):13402-13407). Таким образом, антитела, которые связываются с CD4 и PD-1, находят применение в лечении туберкулеза и в профилактики или задержке развития туберкулеза.
Последовательности ДНК, кодирующие предпочтительные последовательности каркаса акцептора от человека, включают, но не ограничиваются этим, сегменты FR от VH сегмента VH1-18 и JH6 зародышевой линии человека и VL сегмент VK-A26 и JK4 зародышевой линии человека. В конкретном варианте осуществления одну или более CDR вставляют в области каркаса с использованием общепринятых методов рекомбинантной ДНК. Области каркаса могут представлять собой природные области или консенсусные области каркаса и, предпочтительно, области каркаса от человека (см., например, Chothia et al., 1998, J. Mol. Biol. 278:457-479 для перечня областей каркаса от человека).
Гуманизированное или химерное антитело по изобретению может включать по существу все, по меньшей мере, из одного и обычно двух вариабельных доменов, в которых все или по существу все области CDR соответствуют областям иммуноглобулина, происходящего не от человека (т.е. от антитела-донора), и все или по существу все области каркаса представляют собой области консенсусной последовательности иммуноглобулина человека. Предпочтительно, антитело по изобретению содержит также по меньшей мере часть константной области (Fc) иммуноглобулина, обычно иммуноглобулина человека. Константные домены антител по изобретению могут быть выбраны в соответствии с предполагаемой функцией антитела, в частности, с эффекторной функцией, которая может потребоваться. В некоторых вариантах осуществления константные домены антител по изобретению представляют собой (или включают) домены IgA, IgD, IgE, IgG или IgM человека. В конкретном варианте осуществления используются константные домены IgG человека, особенно изотипов IgG1 и IgG3, в том случае, когда гуманизированные антитела по изобретению предназначены для терапевтического использования и требуются эффекторные функции антител, такие как зависимая от антител клеточная цитотоксичность (ADCC) и зависимая от комплемента цитотоксичность (CDC). Например, PD-1 интенсивно экспрессируется на Т-клетках также как на редких периферических Т-клеточных лимфомах, таких как ангиоиммунобластная Т-клеточная лимфома (AITL). Антитела против PD-1 с ADCC или CDC активностью особенно подходят в качестве терапевтических средств для лечения таких типов злокачественных новообразований. В альтернативных вариантах осуществления изотипы IgG2 и IgG4 используются, когда антитело по изобретению предназначено для терапевтических целей и эффекторная функция антитела не требуется. Например, если желательно повысить активность Т-клеток с помощью направленного действия на PD-1 на поверхности Т-клеток, то эффекторные функции, которые будут уничтожить Т-клетку, могут быть нежелательными. Изобретение охватывает константные домены Fc, содержащие одну или более аминокислотных модификаций, которые изменяют эффекторные функции антитела как это раскрыто в публикациях патентных заявок США No. 2005/0037000 и 2005/0064514.
В некоторых вариантах осуществления антитело по изобретению содержит как легкую цепь, так и, по меньшей мере вариабельный домен тяжелой цепи. В других вариантах осуществления антитело по изобретению может дополнительно содержать одну или более из областей CH1, шарнира, CH2, CH3 и CH4 тяжелой цепи. Антитело может быть выбрано из любого класса иммуноглобулинов, включая IgM, IgG, IgD, IgA и IgE, и из любого изотипа, включая IgG1, IgG2, IgG3 и IgG4. В некоторых вариантах осуществления константный домен представляет собой фиксирующий комплемент константный домен, когда желательно, чтобы антитело проявляло цитотоксическую активность, и классом антитела обычно является IgG1. В других вариантах осуществления, когда цитотоксическая активность нежелательна, константный домен может относиться к классу IgG2. Антитело по изобретению может содержать последовательности от более одного класса или изотипа, и выбор конкретных константных доменов для оптимизации желаемых эффекторных функций находится в рамках знаний специалистов в данной области.
Не требуется, чтобы каркас и области CDR гуманизированного антитела точно соответствовали родительским последовательностям, например, CDR донора или консенсусный каркас могут подвергаться мутации с помощью замены, вставки или делеции, по меньшей мере, одного остатка, так что остаток CDR или каркаса в этом месте не будет соответствовать либо консенсусной, либо донорской последовательности антитела. Предпочтительно, однако, чтобы такие мутации не были распространенными. Обычно по меньшей мере 75% остатков гуманизированного антитела должно соответствовать остаткам родительской области каркаса (FR) и последовательностям CDR, чаще 90% и, наиболее предпочтительно, более 95%. Гуманизированные антитела могут быть получены с использованием разнообразных методов, известных в данной области, включая, но ими не ограничиваясь, пересадку CDR (европейский патент No. EP 239400; международная публикация No. WO 91/09967; и патенты США No. 5225539, 5530101 и 5585089), винирование или перекладку (европейские патенты No. EP 592106 и EP 519596; Padlan, 1991, Molecular Immunology 28(4/5):489-498; Studnicka et al., 1994, Protein Engineering 7(6):805-814; and Roguska et al., 1994, Proc. Natl. Acad. Sci. 91:969-973), перетасовку цепей (патент США No. 5565332), и методы, раскрытые, например, в патентах США No. 6407213, 5766886, 5585089, международной публикации No. WO 9317105, Tan et al., 2002, J. Immunol. 169:1119-25, Caldas et al., 2000, Protein Eng. 13:353-60, Morea et al., 2000, Methods 20:267-79, Baca et al., 1997, J. Biol. Chem. 272:10678-84, Roguska et al., 1996, Protein Eng. 9:895-904, Couto et al., 1995, Cancer Res. 55 (23 Supp):5973s-5977s, Couto et al., 1995, Cancer Res. 55:1717-22, Sandhu, 1994, Gene 150:409-10, Pedersen et al., 1994, J. Mol. Biol. 235:959-73, Jones et al., 1986, Nature 321:522-525, Riechmann et al., 1988, Nature 332:323, and Presta, 1992, Curr. Op. Struct. Biol. 2:593-596. Часто остатки каркаса в областях каркаса должны быть заменены соответствующим остатком из CDR антитела-донора для изменения, предпочтительно, улучшения, связывания антигена. Эти замены в каркасе идентифицируются с помощью методов, хорошо известных в данной области, например, с помощью моделирования взаимодействий CDR и остатков каркаса для идентификации остатков каркаса, важных для связывания антигена, и сравнения последовательностей для идентификации необычных остатков каркаса в конкретных положениях (см., например, Queen et al., патент США No. 5585089; публикации США No. 2004/0049014 и 2003/0229208; патенты США No. 6350861; 6180370; 5693762; 5693761; 5585089; и 5530101 и Riechmann et al., 1988, Nature 332:323).
Антитела по настоящему изобретению могут быть получены с помощью любого метода, известного в данной области, подходящего для получения полипептидов, например, с помощью синтеза in vitro, получения с помощью рекомбинантной ДНК и тому подобного. Предпочтительно, гуманизированные антитела получают с помощью метода рекомбинантной ДНК. Антитела по изобретению могут быть получены с использованием метода рекомбинантной экспрессии иммуноглобулинов. Рекомбинантное получение молекул иммуноглобулинов, включая гуманизированные антитела, описано в патенте США No. 4816397 (Boss et al.), патентах США No. 6331415 и 4816567 (оба от Cabilly et al.), патенте U.K. GB 2188638 (Winter et al.) и патенте U.K. GB 2209757. Методы рекомбинантной экспрессии иммуноглобулинов, включая гуманизированные иммуноглобулины, могут быть также найдены в Goeddel et al., Gene Expression Technology Methods in Enzymology Vol. 185 Academic Press (1991), and Borreback, Antibody Engineering, W. H. Freeman (1992). Дополнительная информация, касающаяся создания, дизайна и экспрессии рекомбинантных антител может быть найдена в Mayforth, Designing Antibodies, Academic Press, San Diego (1993).
Иллюстративный метод получения рекомбинантных химерных антител по изобретению может включать следующее: a) конструирование с помощью общепринятых методов молекулярной биологии вектора экспрессии, который кодирует и экспрессирует тяжелую цепь антитела, в которой CDR и вариабельная область моноклонального антитела мыши против B7-H1 (или против PD-1) соединены с областью Fc, происходящей от иммуноглобулина человека, в результате чего получают вектор для экспрессии тяжелой цепи химерного антитела; b) конструирование с помощью общепринятых методов молекулярной биологии вектора экспрессии, который кодирует и экспрессирует легкую цепь моноклонального антитела мыши против B7-H1 (или против PD-1), в результате чего получают вектор для экспрессии легкой цепи химерного антитела; c) перенос векторов экспрессии в клетку-хозяина с помощью общепринятых методов молекулярной биологии с получением трансфицированной клетки-хозяина для экспрессии химерных антител; и d) культивирование трансфицированной клетки с помощью общепринятых методов культивирования клеток, так что получают химерные антитела.
Иллюстративный метод получения рекомбинантных гуманизированных антител по изобретению может включать следующее: a) конструирование с помощью общепринятых методов молекулярной биологии вектора экспрессии, который кодирует и экспрессирует тяжелую цепь антитела, в которой CDR и минимальная часть каркаса вариабельной области, которые требуются для сохранения специфичности связывания антитела-донора, происходят от иммуноглобулина, происходящего не от человека, например, от моноклонального антитела мыши против B7-H1 (или против PD-1), и оставшаяся часть антитела происходит от иммуноглобулина человека, в результате чего получают вектор для экспрессии тяжелой цепи гуманизированного антитела; b) конструирование с помощью общепринятых методов молекулярной биологии вектора экспрессии, который кодирует и экспрессирует легкую цепь антитела, в которой CDR и минимальная часть каркаса вариабельной области, которые требуются для сохранения специфичности связывания антитела-донора, происходят от иммуноглобулина, происходящего не от человека, например, от моноклонального антитела мыши против B7-H1 (или против PD-1), и оставшаяся часть антитела происходит от иммуноглобулина человека, в результате чего получают вектор для экспрессии легкой цепи гуманизированного антитела; c) перенос векторов экспрессии в клетку-хозяина с помощью общепринятых методов молекулярной биологии с получением трансфицированной клетки-хозяина для экспрессии гуманизированных антител; и d) культивирование трансфицированной клетки с помощью общепринятых методов культивирования клеток, так что получают гуманизированные антитела.
В случае любого иллюстративного метода клетки-хозяева могут быть котрансфицированы такими векторами экспрессии, которые могут содержать различные маркеры селекции, но за исключением того случая, когда последовательности, кодирующие тяжелую и легкую цепи, являются, предпочтительно, идентичными. Этот метод обеспечивает эквивалентную экспрессию полипептидов тяжелой и легкой цепей. Альтернативно, может быть использован один вектор, который кодирует полипептиды как тяжелой, так и легкой цепей. Кодирующие последовательности тяжелой и легкой цепей могут содержать кДНК или геномную ДНК, или обе. Клетка-хозяин для экспрессии рекомбинантного антитела по изобретению может представлять собой либо бактериальную клетку, такую как Escherichia coli, или, более предпочтительно, эукариотную клетку (например, клетку яичника китайского хомячка (CHO) или клетку HEK-293). Выбор вектора экспрессии зависит от выбора клетки-хозяина, и он может быть выбран так, чтобы иметь желаемую экспрессию и регуляторные характеристики в выбранной клетке-хозяине. Другие клеточные линии, которые могут быть использованы, включают, но не ограничиваются этим, CHO-K1, NSO и PER.C6 (Crucell, Leiden, Netherlands). Более того, использование кодонов может быть оптимизировано, когда клетку-хозяина выбирают с учетом случайности видового использования кодонов и повышения экспрессии белка. Например, для экспрессии в клетках CHO ДНК, кодирующая антитела, может включать кодоны, используемые, предпочтительно, Cricetulus griseus (от которого происходят клетки яичников китайского хомячка). Методы оптимизации кодонов могут быть использованы для содействия улучшенной экспрессии желаемой клеткой-хозяином (см., например, Wohlgemuth, I. et al. (2011) “Evolutionary Optimization Of Speed And Accuracy Of Decoding On The Ribosome,” Philos. Trans. R. Soc. Lond. B Biol. Sci. 366(1580):2979-2986; Jestin, J.L. et al. (2009) “Optimization Models And The Structure Of The Genetic Code,” J. Mol. Evol. 69(5):452-457; Bollenbach, T. et al. (2007) “Evolution And Multilevel Optimization Of The Genetic Code,” Genome Res. 17(4):401-404; Kurland, C.G. et al. (1984) “Optimization Of Translation Accuracy,” Prog. Nucleic Acid Res. Mol. Biol. 31:191-219; Grosjean, H. et al. (1982) “Preferential Codon Usage In Prokaryotic Genes: The Optimal Codon-Anticodon Interaction Energy And The Selective Codon Usage In Efficiently Expressed Genes,” Gene 18(3):199-209).
Любое из описанных выше антител может быть использовано для создания антиидиотипических антител с использованием методов, хорошо известных специалистам в данной области (см., например, Greenspan, N.S. et al. (1989) “Idiotypes: Structure And Immunogenicity,” FASEB J. 7:437-444; and Nisinoff, A. (1991) “Idiotypes: Concepts And Applications,” J. Immunol. 147(8):2429-2438).
Связывающие свойства любого из описанных выше антител, если это желательно, могут быть дополнительно улучшены с помощью скрининга вариантов, которые проявляют такие желаемые характеристики. Например, такие антитела могут быть созданы с использованием различных методов фагового дисплея, известных в данной области. В методах фагового дисплея функциональные домены антител помещают на поверхность фаговых частиц, которая несет кодирующие их полинуклеотидные последовательности. В конкретном варианте осуществления такой фаг может быть использован для представления антигенсвязывающих доменов, таких как Fab и Fv или стабилизированный дисульфидной связью Fv, экспрессируемых из репертуарной или комбинаторной библиотеки антител (например, человека или мыши). Фаг, экспрессирующий антигенсвязывающий домен, который связывается с представляющим интерес антигеном, может быть выбран или идентифицирован с помощью антигена, например, с использованием меченого антигена или антигена, связанного с твердой подложкой или шариком или захваченного ими. Фаг, используемый в этих методах, обычно представляет собой нитевидный фаг, включая fd и M13. Антигенсвязывающие домены экспрессируют в виде рекомбинантного слитого белка с белком гена III или гена VIII фага. Примеры методов фагового дисплея, которые могут быть использованы для создания иммуноглобулинов или их фрагментов по настоящему изобретению, включают описанные в Brinkman, U. et al. (1995) “Phage Display Of Disulfide-Stabilized Fv Fragments,” J. Immunol. Methods, 182:41-50, 1995; Ames, R.S. et al. (1995) “Conversion Of Murine Fabs Isolated From A Combinatorial Phage Display Library To Full Length Immunoglobulins,” J. Immunol. Methods, 184:177-186; Kettleborough, C.A. et al. (1994) “Isolation Of Tumor Cell-Specific Single-Chain Fv From Immunized Mice Using Phage-Antibody Libraries And The Re-Construction Of Whole Antibodies From These Antibody Fragments,” Eur. J. Immunol., 24:952-958, 1994; Persic, L. et al. (1997) “An Integrated Vector System For The Eukaryotic Expression Of Antibodies Or Their Fragments After Selection From Phage Display Libraries,” Gene, 187:9-18; Burton, D.R. et al. (1994) “Human Antibodies From Combinatorial Libraries,” Adv. Immunol. 57:191-280; публикациях PCT WO 92/001047; WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; WO 95/15982; WO 95/20401; и патентах США No. 5698426; 5223409; 5403484; 5580717; 5427908; 5750753; 5821047; 5571698; 5427908; 5516637; 5780225; 5658727; 5733743 и 5969108.
Как описано в представленных выше ссылках, после выбора фага области, кодирующие антитело, могут быть выделены из фага и использованы для создания целых антител, включая гуманизированные антитела или любые другие желаемые фрагменты, и экспрессированы в любом желаемом хозяине, включая клетки млекопитающих, клетки насекомых, клетки растений, дрожжи и бактерии, например, как подробно описано ниже. Например, методы рекомбинантного получения фрагментов Fab, Fab' и F(ab')2 также могут применяться с использованием методов, известных в данной области, таких как описанные в публикации PCT WO 92/22324; Mullinax, R.L. et al. (1992) “Expression Of A Heterodimeric Fab Antibody Protein In One Cloning Step,” BioTechniques, 12(6):864-869; and Sawai et al. (1995) “Direct Production Of The Fab Fragment Derived From The Sperm Immobilizing Antibody Using Polymerase Chain Reaction And cDNA Expression Vectors,” Am. J. Reprod. Immunol. 34:26-34; and Better, M. et al. (1988) “Escherichia coli Secretion Of An Active Chimeric Antibody Fragment,” Science 240:1041-1043). Примеры методов, которые могут быть использованы для получения одноцепочечных Fv и антител, включают описанные в патентах США No. 4946778 и 5258498; Huston, J.S. et al. (1991) “Protein Engineering Of Single-Chain Fv Analogs And Fusion Proteins,” Methods in Enzymology 203:46-88; Shu, L. et al., “Secretion Of A Single-Gene-Encoded Immunoglobulin From Myeloma Cells,” Proc. Natl. Acad. Sci. (USA) 90:7995-7999; and Skerra. A. et al. (1988) “Assembly Of A Functional Immunoglobulin Fv Fragment In Escherichia coli,” Science 240:1038-1040.
Метод фагового дисплея может быть использован для повышения аффинности антитела по изобретению к B7-H1 и/или PD-1. Этот метод должен быть пригоден для получения высокоаффинных антител, которые могут быть использованы в комбинаторных методах по изобретению. Этот метод, обозначаемый как созревание аффинности, использует мутагенез или прогулку CDR и повторный отбор с использованием таких рецепторов или лигандов (или их внеклеточных доменов) или их антигенного фрагмента для идентификации антител, которые связываются с антигеном с более высокой аффинностью по сравнению с исходным или родительским антителом (см., например, Glaser, S.M. et al. (1992) “Antibody Engineering By Codon-Based Mutagenesis In A Filamentous Phage Vector System,” J. Immunol. 149:3903-3913). Подвергшиеся мутагенезу полные кодоны в отличие от одиночных нуклеотидов приводят к полурандомизированному репертуару аминокислотных мутаций. Библиотеки могут быть сконструированы как состоящие из пула вариантов клонов, каждый из которых отличается изменением одной аминокислоты в одной CDR и который содержит варианты, представляющие каждую возможную аминокислотную замену для каждого остатка CDR. Может быть проведен скрининг мутантов на повышенную связывающую аффинность к антигену путем приведения в контакт иммобилизованных мутантов с меченым антигеном. Любой метод скрининга, известный в данной области техники, может быть использован для идентификации мутантных антител с повышенной авидностью к антигену (например, ELISA) (см., например, Wu, H. et al. (1998) “Stepwise In Vitro Affinity Maturation Of Vitaxin, An Alphav Beta3-Specific Humanized Mab,” Proc. Natl. Acad. Sci. (USA) 95(11):6037-6042; Yelton, D.E. et al. (1995) “Affinity Maturation Of The BR96 Anti-Carcinoma Antibody By Codon-Based Mutagenesis,” J. Immunol. 155:1994-2004). Можно при возможности использовать прогулку CDR, которая рандомизирует легкую цепь (см. Schier et al. (1996) “Isolation Of Picomolar Affinity Anti-C-Erbb-2 Single-Chain Fv By Molecular Evolution Of The Complementarity Determining Regions In The Center Of The Antibody Binding Site,” J. Mol. Biol. 263:551-567).
Таким образом, в изобретении рассматривается применение случайного мутагенеза совместно с методами фагового дисплея для идентификации улучшенных CDR и/или вариабельных областей. Метод фагового дисплея альтернативно может быть использован для повышения (или снижения) аффинности CDR с помощью направленного мутагенеза (например, созревания аффинности или «прогулки CDR»). Этот метод использует антиген-мишень или его антигенный фрагмент для идентификации антител, имеющих CDR, которые связываются с более высокой (или более низкой) аффинностью с антигеном по сравнению с исходным или родительским антителом (см., например, Glaser, S.M. et al. (1992) “Antibody Engineering By Codon-Based Mutagenesis In A Filamentous Phage Vector System,” J. Immunol. 149:3903-3913). Подвергшиеся мутагенезу полные кодоны в отличие от одиночных нуклеотидов приводят к полурандомизированному репертуару аминокислотных мутаций. Библиотеки могут быть сконструированы как состоящие из пула вариантов клонов, каждый из которых отличается изменением одной аминокислоты в одной CDR и который содержит варианты, представляющие каждую возможную аминокислотную замену для каждого остатка CDR. Может быть проведен скрининг мутантов с повышенной (или пониженной) связывающей аффинностью к антигену путем приведения в контакт иммобилизованных мутантов с меченым антигеном. Любой метод скрининга, известный в данной области техники, может быть использован для идентификации мутантных антител с повышенной (или пониженной) авидностью к антигену (например, ELISA) (см., например, Wu, H. et al. (1998) “Stepwise In Vitro Affinity Maturation Of Vitaxin, An Alphav Beta3-Specific Humanized Mab,” Proc. Natl. Acad. Sci. (USA) 95(11):6037-6042; Yelton, D.E. et al. (1995) “Affinity Maturation Of The BR96 Anti-Carcinoma Antibody By Codon-Based Mutagenesis,” J. Immunol. 155:1994-2004). Можно при возможности использовать прогулку CDR, которая рандомизирует легкую цепь (см. Schier et al. (1996) “Isolation Of Picomolar Affinity Anti-C-Erbb-2 Single-Chain Fv By Molecular Evolution Of The Complementarity Determining Regions In The Center Of The Antibody Binding Site,” J. Mol. Biol. 263:551-567).
Методы достижения такого созревания аффинности описаны, например, в: Krause, J.C. et al. (2011) “An Insertion Mutation That Distorts Antibody Binding Site Architecture Enhances Function Of A Human Antibody,” MBio. 2(1) pii: e00345-10. doi: 10.1128/mBio.00345-10; Kuan, C.T. et al. (2010) “Affinity-Matured Anti-Glycoprotein NMB Recombinant Immunotoxins Targeting Malignant Gliomas And Melanomas,” Int. J. Cancer 10.1002/ijc.25645; Hackel, B.J. et al. (2010) “Stability And CDR Composition Biases Enrich Binder Functionality Landscapes,” J. Mol. Biol. 401(1):84-96; Montgomery, D.L. et al. (2009) “Affinity Maturation And Characterization Of A Human Monoclonal Antibody Against HIV-1 gp41,” MAbs 1(5):462-474; Gustchina, E. et al. (2009) “Affinity Maturation By Targeted Diversification Of The CDR-H2 Loop Of A Monoclonal Fab Derived From A Synthetic Naïve Human Antibody Library And Directed Against The Internal Trimeric Coiled-Coil Of Gp41 Yields A Set Of Fabs With Improved HIV-1 Neutralization Potency And Breadth,” Virology 393(1):112-119; Finlay, W.J. et al. (2009) “Affinity Maturation Of A Humanized Rat Antibody For Anti-RAGE Therapy: Comprehensive Mutagenesis Reveals A High Level Of Mutational Plasticity Both Inside And Outside The Complementarity-Determining Regions,” J. Mol. Biol. 388(3):541-558; Bostrom, J. et al. (2009) “Improving Antibody Binding Affinity And Specificity For Therapeutic Development,” Methods Mol. Biol. 525:353-376; Steidl, S. et al. (2008) “In Vitro Affinity Maturation Of Human GM-CSF Antibodies By Targeted CDR-Diversification,” Mol. Immunol. 46(1):135-144; and Barderas, R. et al. (2008) “Affinity maturation of antibodies assisted by in silico modeling,” Proc. Natl. Acad. Sci. (USA) 105(26):9029-9034.
В изобретении особенно рассматриваются получение и использование «производных» любых из описанных выше антител и их антигенсвязывающих фрагментов. Термин «производное» относится к антителу или его антигенсвязывающему фрагменту, которые иммуноспецифически связываются с антигеном, но которые содержат одну, две, три, четыре, пять или более аминокислотных замен, добавок, делеций или модификаций относительно «родительской» молекулы (или молекулы дикого типа). Такие аминокислотные замены или добавки могут быть введены в виде природных аминокислотных остатков (т.е. кодируемых ДНК) или неприродных аминокислотных остатков. Такие аминокислоты могут быть гликозилированными (например, содержать измененную маннозу, 2-N-ацетилглюкозамин, галактозу, фукозу, глюкозу, сиаловую кислоту, 5-N-ацетилнейраминовую кислоту, 5-гликольнейраминовую кислоту и т.п.), ацетилированными, пэгилированными, фосфорилированными, амидированными, дериватизированными с помощью известных защитных/блокирующих групп, протеолитически расщепляемыми, связанными с клеточным лигандом или другим белком, и т.д. В некоторых вариантах осуществления измененные углеводные модификации модулируют одно или более из следующего: солюбилизацию антитела, облегчение субклеточного транспорта и секреции антитела, стимуляцию сборки антител, конформационную целостность и опосредуемую антителом эффекторную функцию. В конкретном варианте осуществления измененные углеводные модификации повышают опосредуемую антителом эффекторную функцию по сравнению с антителом, не имеющим углеводной модификации. Углеводные модификации, которые ведут к измененной опосредуемой антителом эффекторной функции, хорошо известны в данной области (например, см. Shields, R.L. et al. (2002) “Lack Of Fucose On Human IgG N-Linked Oligosaccharide Improves Binding To Human Fcgamma RIII And Antibody-Dependent Cellular Toxicity.,” J. Biol. Chem. 277(30): 26733-26740; Davies J. et al. (2001) “Expression Of GnTIII In A Recombinant Anti-CD20 CHO Production Cell Line: Expression Of Antibodies With Altered Glycoforms Leads To An Increase In ADCC Through Higher Affinity For FC Gamma RIII,” Biotechnology & Bioengineering 74(4): 288-294). Методы изменения содержания углеводов известны специалистам в данной области, см., например, Wallick, S.C. et al. (1988) “Glycosylation Of A VH Residue Of A Monoclonal Antibody Against Alpha (1----6) Dextran Increases Its Affinity For Antigen,” J. Exp. Med. 168(3): 1099-1109; Tao, M.H. et al. (1989) “Studies Of Aglycosylated Chimeric Mouse-Human IgG. Role Of Carbohydrate In The Structure And Effector Functions Mediated By The Human IgG Constant Region,” J. Immunol. 143(8): 2595-2601; Routledge, E.G. et al. (1995) “The Effect Of Aglycosylation On The Immunogenicity Of A Humanized Therapeutic CD3 Monoclonal Antibody,” Transplantation 60(8):847-53; Elliott, S. et al. (2003) “Enhancement Of Therapeutic Protein In Vivo Activities Through Glycoengineering,” Nature Biotechnol. 21:414-21; Shields, R.L. et al. (2002) “Lack Of Fucose On Human IgG N-Linked Oligosaccharide Improves Binding To Human Fcgamma RIII And Antibody-Dependent Cellular Toxicity.,” J. Biol. Chem. 277(30): 26733-26740).
В некоторых вариантах осуществления гуманизированное антитело представляет собой производное. Такое гуманизированное антитело содержит замены, делеции или добавки аминокислотных остатков в одной или более CDR, происходящих не от человека. Производное гуманизированного антитела может иметь по существу такое же связывание, лучшее связывание или худшее связывание по сравнению с недериватизированным гуманизированным антителом. В конкретных вариантах осуществления один, два, три, четыре или пять аминокислотных остатков CDR подвергаются заменам, делециям или добавкам (т.е. мутируют).
Производное антитела или фрагмента антитела может быть модифицировано с помощью химических модификаций с использованием методов, известных специалистам в данной области техники, включая, но ими не ограничиваясь, специфическое химическое расщепление, ацетилирование, составление, метаболический синтез туникамицина и т.п. В одном из вариантов осуществления производное антитела должно обладать функцией, сходной или идентичной родительскому антителу. В другом варианте осуществления производное антитело будет демонстрировать измененную активность по сравнению с родительским антителом. Например, производное антитела (или его фрагмента) может связываться со своим эпитопом более прочно или быть более устойчивым к протеолизу, чем родительское антитело.
Замены, добавки или делеции в дериватизированных антителах могут находиться в области Fc антитела и могут тем самым служить для модификации связывающей аффинности антитела к одному или более FcγR. Методы модификации антител с модифицированным связыванием с одним или более FcγR известны в данной области, см., например, публикации PCT No. WO 04/029207, WO 04/029092, WO 04/028564, WO 99/58572, WO 99/51642, WO 98/23289, WO 89/07142, WO 88/07089 и патенты США No. 5843597 и 5642821. В некоторых вариантах осуществления изобретение охватывает антитела, которые имеют измененную аффинность к активирующему FcγR, например, FcγRIIIA. Предпочтительно, такие модификации также имеют измененную опосредуемую Fc эффекторную функцию. Модификации, которые влияют на опосредуемую Fc эффекторную функцию, хорошо известны в данной области (см. патент США No. 6194551 и WO 00/42072). В одном конкретном варианте осуществления модификация области Fc приводит к антителу с измененной опосредуемой антителом эффекторной функцией, с измененным связыванием с другими рецепторами Fc (например, активирующими рецепторами Fc), с измененной зависимой от антител клеточной цитотоксической активностью (ADCC), с измененной связывающей C1q активностью, с измененной зависимой от комплемента цитотоксической активностью (CDC), с фагоцитарной активностью или с любыми их сочетаниями.
Дериватизированные антитела могут быть использованы для изменения периода полужизни (например, периода полужизни в сыворотке) родительских антител у млекопитающего, предпочтительно, у человека. Предпочтительно, такое изменение должно приводить к периоду полужизни выше 15 дней, предпочтительно, выше 20 дней, выше 25 дней, выше 30 дней, выше 35 дней, выше 40 дней, выше 45 дней, выше 2 месяцев, выше 3 месяцев, выше 4 месяцев или выше 5 месяцев. Повышенные периоды полужизни гуманизированных антител или их фрагментов по настоящему изобретению у млекопитающего, предпочтительно, у человека, приводят к более высокому титру указанных антител или фрагментов антител в сыворотке млекопитающего и, таким образом, снижают частоту введения указанных антител или фрагментов антител и/или снижают концентрацию указанных антител или фрагментов антител при введении. Антитела или их фрагменты, имеющие повышенные периоды полужизни in vivo, могут быть созданы с помощью методов, известных специалистам в данной области техники. Например, антитела или их фрагменты с повышенными периодами полужизни in vivo могут быть созданы путем модификации (например, замены, делеции или добавления) аминокислотных остатков, идентифицированных как вовлеченные во взаимодействие между доменом Fc и рецептором FcRn. Гуманизированные антитела по изобретению могут быть сконструированы для повышения биологической полужизни (см., например, патент США No. 6277375). Например, гуманизированные антитела по изобретению могут быть сконструированы с изменением в Fc-шарнирном домене для повышения периода полужизни in vivo или в сыворотке.
Антитела или их фрагменты с повышенными периодами полужизни in vivo могут быть созданы путем присоединения к указанным антителам или фрагментам антител полимерных молекул, таких как полиэтиленгликоль (ПЭГ) высокой молекулярной массы. ПЭГ может быть присоединен к указанным антителам или фрагментам антител с мультифункциональным линкером или без него, либо с помощью сайт-специфической конъюгации ПЭГ с N- или C-концом указанных антител или фрагментов антител, либо через эпсилон-аминогруппы, присутствующие на остатках лизина. Могут быть использованы линейные или разветвленные дериватизированные полимеры, что ведет к минимальной потере биологической активности. Степень конъюгации должна быть тщательно прослежена с помощью SDS-PAGE и масс-спектрометрии для того, чтобы убедиться в надлежащей конъюгации молекул ПЭГ с антителами. Непрореагировавший ПЭГ может быть отделен от конъюгатов антитело-ПЭГ, например, гель- или ионно-обменной хроматографией.
Антитела по изобретению можно также модифицировать с помощью методов и агентов присоединения, описанных Davis et al. (см. патент США No. 4179337) для того, чтобы снабжать композициями, которые можно вводить в циркуляторное русло млекопитающих по существу с отсутствием иммунного ответа.
Изобретение охватывает модификацию остатков каркаса гуманизированных антител по изобретению. Остатки каркаса в областях каркаса могут быть заменены соответствующим остатком из CDR антитела-донора для изменения, предпочтительно, улучшения, связывания антигена. Эти замены в каркасе идентифицируются с помощью методов, хорошо известных в данной области, например, с помощью моделирования взаимодействий CDR и остатков каркаса с идентифицируемыми остатками каркаса, важными для связывания антигена, и сравнения последовательностей с идентифицируемыми необычными остатками каркаса в конкретных положениях. (См., например, патент США No. 5585089; и Riechmann, L. et al. (1988) "Reshaping Human Antibodies For Therapy," Nature 332:323-327).
Настоящим изобретением также охватываются антитела против B7-H1 человека и против PD-1 человека (и более предпочтительно, гуманизированные антитела) и их антигенсвязывающие фрагменты, которые рекомбинантно слиты или химически конъюгированы (включая как ковалентную, так и нековалентную конъюгацию) с гетерологичной молекулой (т.е. с неродственной молекулой). Соединение необязательно должно быть прямым, но может осуществляться через линкерные последовательности.
Часть Fc слитого белка может варьироваться по изотипу или подклассам, может быть химерной или гибридной, и/или может быть модифицированной, например, для улучшения эффекторных функций, контроля периода полужизни, тканевой доступности, усиления биофизических характеристик, таких как стабильность, и улучшения эффективности продукции (и меньшей стоимости). Многие модификации, пригодные для конструирования раскрытых слитых белков, и методы их получения известны в данной области, см., например, Mueller, J.P. et al. (1997) “Humanized Porcine VCAM-Specific Monoclonal Antibodies With Chimeric Igg2/G4 Constant Regions Block Human Leukocyte Binding To Porcine Endothelial Cells,” Mol. Immun. 34(6):441-452, Swann, P.G. (2008) “Considerations For The Development Of Therapeutic Monoclonal Antibodies,” Curr. Opin. Immun. 20:493-499 (2008), и Presta, L.G. (2008) “Molecular Engineering And Design Of Therapeutic Antibodies,” Curr. Opin. Immun. 20:460-470. В некоторых вариантах осуществления область Fc представляет собой нативную область Fc IgG1, IgG2 или IgG4. В некоторых вариантах осуществления область Fc представляет собой гибрид, например, гибрид, состоящий из константных областей Fc IgG2/IgG4. Модификации области Fc включают, но не ограничиваются этим, IgG4, модифицированный для предотвращения связывания с рецепторами Fc гамма и комплементом, IgG1, модифицированный для улучшения связывания с одним или более рецепторами Fc гамма, IgG1, модифицированный для сведения к минимуму эффекторной функции (изменения аминокислот), IgG1 с измененным гликаном/его отсутствием (обычно путем изменения экспрессии хозяином), и IgG1 с измененным рН-зависимым связыванием с FcRn. Область Fc может включать полную область шарнира или менее полную области шарнира.
Терапевтический исход у больных, которые получали лечение ритуксимабом (химерным моноклональным антителом IgG1 мыши/человека против CD20) при неходжкинской лимфоме или макроглобулинемии Вальденстрема, коррелировал с индивидуальной экспрессией аллельных вариантов рецепторов Fcγ с индивидуально присущей им аффинностью к домену Fc IgG1 человека. В частности, больные с высокоаффинными аллелями низкоаффинного активирующего рецептора Fc CD16A (FcγRIIIA) демонстрировали более высокие уровни ответа и в случаях неходжкинской лимфомы улучшенную выживаемость без прогрессирования заболевания. В других вариантах осуществления домен Fc может содержать одну или более аминокислотных вставок, делеций или замен, которые снижают связывание с низкоаффинным ингибиторным рецептором Fc CD32B (FcγRIIB) и удерживают связывание на уровне дикого типа или повышают связывание с низкоаффинным активирующим рецептором Fc CD16A (FcγRIIIA).
Другие варианты осуществления включают гибриды IgG2-4 и мутанты IgG4, которые характеризуются пониженным связыванием с FcR, что повышает их период полужизни. Примеры гибридов IG2-4 и мутантов IgG4 описаны у Angal, S. et al. (1993) “A Single Amino Acid Substitution Abolishes The Heterogeneity Of Chimeric Mouse/Human (IgG4) Antibody,” Molec. Immunol. 30(1):105-108; Mueller, J.P. et al. (1997) “Humanized Porcine VCAM-Specific Monoclonal Antibodies With Chimeric IgG2/G4 Constant Regions Block Human Leukocyte Binding To Porcine Endothelial Cells,” Mol. Immun. 34(6):441-452; и в патенте США No. 6982323. В некоторых вариантах осуществления в домене IgG1 и/или IgG2 осуществляют делеции, например, Angal et al. описывают IgG1 и IgG2, имеющие замену серина 241 на пролин.
В предпочтительном варианте осуществления домен Fc содержит аминокислотные вставки, делеции или замены, которые усиливают связывание с CD16A. Большое количество замен в домене Fc IgG1 человека, которые усиливают связывание с CD16A и снижают связывание с CD32B, известно в данной области и описано у Stavenhagen, J.B. et al. (2007) “Fc Optimization Of Therapeutic Antibodies Enhances Their Ability To Kill Tumor Cells In Vitro And Controls Tumor Expansion In Vivo Via Low-Affinity Activating Fcgamma Receptors,” Cancer Res. 57(18):8882-8890. Иллюстративные варианты доменов Fc IgG1 человека с пониженным связыванием с CD32B и/или повышенным связыванием с CD16A содержат замены F243L, R929P, Y300L, V305I или P296L. Эти аминокислотные замены могут присутствовать в домене Fc IgG1 человека в любом сочетании. В одном из вариантов осуществления вариант домена Fc IgG1 человека содержит замены F243L, R929P и Y300L. В другом варианте осуществления вариант домена Fc IgG1 человека содержит замены F243L, R929P, Y300L, V305I и P296L. В другом варианте осуществления вариант домена Fc IgG1 человека содержит замену N297Q, так как эта мутация аннулирует связывание с FcR.
В одном из вариантов осуществления такие гетерологичные молекулы представляют собой полипептиды, имеющие по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90 или по меньшей мере 100 аминокислот. Такие гетерологичные молекулы альтернативно могут представлять собой ферменты, гормоны, рецепторы клеточной поверхности, части лекарственного средства, такие как: токсины (такие как абрин, рицин А, экзотоксин синегнойной палочки (т.е. PE-40), дифтерийный токсин, рицин, гелонин или противовирусный белок фитолакки американской), белки (такие как фактор некроза опухолей, интерферон (например, α-интерферон, β-интерферон), фактор роста нервов, фактор роста из тромбоцитов, активатор тканевого плазминогена или апоптотический фактор (например, фактор-α некроза опухолей, фактор-β некроза опухолей), средства, модифицирующие биологический ответ (такие как, например, лимфокины (например, интерлейкин-1 ("IL-1"), интерлейкин-2 ("IL-2"), интерлейкин-6 ("IL-6")), гранулоцит-макрофаг-колониестимулирующий фактор ("GM-CSF"), гранулоцит-колониестимулирующий фактор ("G-CSF") или макрофаг-колониестимулирующий фактор ("M-CSF")), или факторы роста (например, гормон роста ("GH"))), цитотоксины (например, цитостатическое или цитолитическое средство, такое как паклитаксол, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксатрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, монометилауристатин F (MMAF), монометилауристатин E (MMAE; например, ведотин) и пуромицин и его аналоги или гомологи), антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил, декарбазин), алкилирующие средства (например, мехлорэтамин, тиоепа, хлорамбуцил, мелфалан, BiCNU® (кармустин; BSNU) и ломустин (CCNU), циклотосфамид, бусульфан, дибромоманнит, стрептозотоцин, митомицин C и цисдихлородиамин платины (II) (DDP) цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (AMC)), или антимитогенные средства (например, винкристин и винбластин).
Методы конъюгации таких терапевтических частей с антителами хорошо известны; см., например, Arnon et al., “Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy”, in Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), 1985, pp. 243-56, Alan R. Liss, Inc.); Hellstrom et al., “Antibodies For Drug Delivery”, in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), 1987, pp. 623-53, Marcel Dekker, Inc. ); Thorpe, “Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review”, in Monoclonal Antibodies ‘84: Biological And Clinical Applications, Pinchera et al. (eds.), 1985, pp. 475-506); “Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy”, in Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), 1985, pp. 303-16, Academic Press; Thorpe et al. (1982) “The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates,” Immunol. Rev. 62:119-158; Carter, P.J. et al. (2008) “Antibody-Drug Conjugates for Cancer Therapy,” Cancer J. 14(3):154-169; Alley, S.C. et al. (2010) “Antibody-Drug Conjugates: Targeted Drug Delivery For Cancer,” Curr. Opin. Chem. Biol. 14(4):529-537; Carter, P. et al. (2005) “Designer Antibody-Based Therapeutics For Oncology,” Amer. Assoc. Cancer Res. Educ. Book. 2005(1):147-154; Carter, P.J. et al. (2008) “Antibody-Drug Conjugates For Cancer Therapy,” Cancer J. 14(3):154-169; Chari, R.V.J. (2008) “Targeted Cancer Therapy: Conferring Specificity To Cytotoxic Drugs,” Acc. Chem Res. 41(1):98-107; Doronina, S.O. et al. (2003) “Development Of Potent Monoclonal Antibody Auristatin Conjugates For Cancer Therapy,” Nat. Biotechnol. 21(7):778-784; Ducry, L. et al. (2010) “Antibody-Drug Conjugates: Linking Cytotoxic Payloads To Monoclonal Antibodies,” Bioconjug Chem. 21(1):5-13; Senter, P.D. (2009) “Potent Antibody Drug Conjugates For Cancer Therapy,” Curr. Opin. Chem. Biol. 13(3):235-244; and Teicher, B.A. (2009) “Antibody-Drug Conjugate Targets,” Curr Cancer Drug Targets. 9(8):982-1004.
Любая из молекул по настоящему изобретению может быть слита с маркерными последовательностями, такими как пептидные, для облегчения очистки. В предпочтительных вариантах осуществления маркерная аминокислотная последовательность представляет собой пептид гексагистидин, метку гемагглютинина «HA», которая соответствует эпитопу, происходящему от белка гемагглютинина гриппа (Wilson, I.A. et al. (1984) “The Structure Of An Antigenic Determinant In A Protein,” Cell, 37:767-778) и «хвостовую» метку (Knappik, A. et al. (1994) “An Improved Affinity Tag Based On The FLAG Peptide For The Detection And Purification Of Recombinant Antibody Fragments,” Biotechniques 17(4):754-761).
В настоящем изобретении охватываются также антитела или их антигенсвязывающие фрагменты, которые конъюгированы с диагностическим или терапевтическим средством или любой другой молекулой, для которой желательно увеличить период полужизни в сыворотке. Антитела могут использоваться в диагностических целях (in vivo, in situ или in vitro), например, для мониторинга развития или прогрессирования заболевания, нарушения или инфекции, в виде части клинической процедуры тестирования, например, для определения эффективности данного режима лечения. Определение может быть облегчено путем присоединения антитела к детектируемому веществу. Примеры детектируемых веществ включают различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества, биолюминесцентные вещества, радиоактивные вещества, позитронно-эмиссионные металлы и нерадиоактивные парамагнитные ионы металлов. Детектируемое вещество может быть соединено или конъюгировано либо прямо с антителом, либо непрямо через промежуточное соединение (такое как, например, линкер, известный в данной области техники), с использованием методов, известных в данной области техники. См., например, патент США No. 4741900 в отношении ионов металлов, которые могут быть конъюгированы с антителами для применения в диагностических целях в соответствии с настоящим изобретением. Такие диагностика и определение могут осуществляться путем присоединения антитела к детектируемым веществам, включая, но ими не ограничиваясь, различные ферменты, ферменты, включая, но ими не ограничиваясь, пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу; комплексы простетических групп, такие как, но ими не ограничиваясь, стрептавидин/биотин и авидин/биотин; флуоресцентные вещества, такие как, но ими не ограничиваясь, умбеллиферон, флуоресцин, флуоресцина изотиоцианат, родамин, дихлортриазиниламина флуоресцин, данзилхлорид или фикоэритрин; люминесцентное вещество, такое как, но ими не ограничиваясь, люминол; биолюминесцентные вещества, такие как, но ими не ограничиваясь, люцифераза, люциферин и экворин; радиоактивные вещества, такие как, но ими не ограничиваясь, висмут (213Bi), углерод (14C), хром (51Cr), кобальт (57Co), фтор (18F), гадолиний (153Gd, 159Gd), галлий (68Ga, 67Ga), германий (68Ge), гольмий (166Ho), индий (115In, 113In, 112In, 111In), йод (131I, 125I, 123I, 121I), лантаний (140La), лютеций (177Lu), марганец (54Mn), молибден (99Mo), палладий (103Pd), фосфор (32P), празеодимий (142Pr), прометий (149Pm), рений (186Re, 188Re), родий (105Rh), рутемий (97Ru), самарий (153Sm), скандий (47Sc), селен (75Se), стронций (85Sr), сера (35S), технеций (99Tc), таллий (201Ti), олово (113Sn, 117Sn), тритий (3H), ксенон (133Xe), иттербий (169Yb, 175Yb), иттрий (90Y), цинк (65Zn); позитронно-эмиссионные металлы, используемые при различных видах позитронно-эмиссионной томографии и нерадиоактивные парамагнитные ионы металлов.
Молекулы по настоящему изобретению могут быть конъюгированы со вторым антителом с образованием гетероконъюгата антител, как описано Segal в патенте США No. 4676980. Такие гетероконъюгаты антител могут дополнительно связываться с гаптенами (таким как флуоресцеин и т.п.) или с маркерами клеток (например, 4-1-BB, B7-H4, CD4, CD8, CD14, CD25, CD27, CD40, CD68, CD163, CTLA4, GITR, LAG-3, OX40, TIM3, TIM4, TLR2, LIGHT, ICOS, B7-H3, B7-H7, B7-H7CR, CD70, CD47 и т.п.) или с цитокинами (например, IL-7, IL-15, IL-12, IL-4 TGF-бета, IL-10, IL-17, IFNγ, Flt3, BLys) или с хемокинами (например, CCL21) и т.п.
Молекулы по настоящему изобретению могут быть присоединены к твердым подложкам, которые особенно подходят для иммуноанализов или очистки антигена-мишени или других молекул, способных связываться с антигеном-мишенью, который иммобилизован на подложке путем связывания с антителом или антигенсвязывающим фрагментом по настоящему изобретению. Такие твердые подложки включают, но ими не ограничиваются, стекло, целлюлозу, полиакриламид, нейлон, полистирол, поливинилхлорид или полипропилен.
Настоящее изобретение дополнительно включает молекулы нуклеиновой кислоты (ДНК или РНК), которые кодируют любое из таких антител, слитых белков или фрагментов, а также векторные молекулы (такие как плазмиды), которые способны переносить или реплицировать такие молекулы нуклеиновой кислоты. Нуклеиновые кислоты могут быть одноцепочечными, двухцепочечными, могут содержать как одноцепочечные, так и двухцепочечные части.
A. Предпочтительные модуляторные композиции по настоящему изобретению
Изобретение особенно касается антител, которые иммуноспецифически связываются с B7-H1 или с PD-1 и/или модулируют способность B7-H1 связываться с PD-1 у индивидуума. Как используется в настоящем описании «индивидуум», предпочтительно, представляет собой млекопитающее, такое как не относящееся к приматам (например, коровы, свиньи, лошади, кошки, собаки, крысы и т.п.), и приматы (например, обезьяны и человек), наиболее предпочтительно, человек. Изобретение, таким образом, особенно относится к гуманизированным антителам и их антигенсвязывающим фрагментам, которые иммуноспецифически связываются с B7-H1 человека или с PD-1 человека и модулируют способность B7-H1 связываться с PD-1 у человека или в ткани человека (in situ или ex vivo).
Наиболее предпочтительно, чтобы такие антитела и антигенсвязывающие фрагменты могли обладать достаточной авидностью для модуляции способности B7-H1 (особенно при его экспрессии в эндогенной концентрации) и его локализации на поверхности APC индивидуума для связывания PD-1 (особенно при его экспрессии в эндогенной концентрации) и его локализации на поверхности Т-клетки такого индивидуума и наоборот. Термин «эндогенная концентрация» относится к уровню, при котором эндогенная молекула экспрессируется в природе (т.е. в отсутствие векторов экспрессии или рекомбинантных промоторов) в нормальной, злокачественной или инфицированной клетке.
В одном из вариантов осуществления такая модуляция должна включать ингибирование или другое вмешательство в связывание такого (предпочтительно, экспрессируемого эндогенно) и представленного B7-H1 и такого (предпочтительно, экспрессируемого эндогенно) и представленного PD-1. В альтернативном варианте осуществления такая модуляция должна включать повышение или другое облегчение связывания эндогенно экспрессируемого и представленного B7-H1, и эндогенно экспрессируемого и представленного PD-1. В еще одном варианте осуществления такая модуляция включает прямую агонистическую активность, в результате чего связывание антитела против B7-H1 или против PD-1 запускает передачу сигнала через соответствующий рецептор.
(1) Предпочтительные антитела против B7-H1 человека и их CDR
В соответствии с настоящим изобретением такие молекулы могут быть получены с помощью скрининга линий гибридом на те, которые продуцируют антитело, являющееся иммуноспецифическим для B7-H1 человека, и последующего необязательного скрининга среди таких линий тех, которые проявляют модулирующую активность (например, нейтрализующую активность, агонистическую активность, активность в отношении изменения передачи сигнала и т.п.). В изобретении особенно предлагаются клоны против B7-H1 человека: 1E12, 1F4, 2G11, 3B6 и 3D10.
Антитела, экспрессируемые клонами против B7-H1 человека, были секвенированы для выявления их вариабельных доменов. Последовательности CDR представлены жирным шрифтом и подчеркнуты:
Клон 1E12 против B7-H1 человека
Вариабельная область легкой цепи:
DIVMTQSHKL MSTSVGDRVS ITCKASQDVG TAVAWYQQKP GQSPKLLIYW
ASTRHTGVPD RFTGSGSGTD FTLTISNVQS EDLADYFCQQ DSSYPLTFGA
GTKVELK (SEQ ID NO:1)
Вариабельная область тяжелой цепи:
EVKLQESGPS LVKPSQTLSL TCSVTGYSIT SDYWNWIRKF PGNKLEYVGY
ISYTGSTYYN PSLKSRISIT RDTSKNQYYL QLNSVTSEDT ATYYCARYGG
WLSPFDYWGQ GTTLTVSS (SEQ ID NO:2)
Клон 1F4 против B7-H1 человека
Вариабельная область легкой цепи:
DIVTTQSHKL MSTSVGDRVS ITCKASQDVG TAVAWYQQKP GQSPKLLIYW
ASTRHTGVPD RFTGSGSGTD FTLTISNVQS EDLADYFCQQ DSSYPLTFGA
GTKVELK (SEQ ID NO:3)
Вариабельная область тяжелой цепи:
EVQLQESGPG LVAPSQSLSI TCTVSGFSLT TYSINWIRQP PGKGLEWLGV
MWAGGGTNSN SVLKSRLIIS KDNSKSQVFL KMNSLQTDDT ARYYCARYYG
NSPYYAIDYW GQGTSVTVSS (SEQ ID NO:4)
Клон 2G11 против B7-H1 человека
Вариабельная область легкой цепи:
DIVMTQSPSS LAVSVGEKVS MGCKSSQSLL YSSNQKNSLA WYQQKPGQSP
KLLIDWASTR ESGVPDRFTG SGSGTDFTLT ISSVKAEDLA VYYCQQYYGY
PLTFGAGTKL ELK (SEQ ID NO:5)
Вариабельная область тяжелой цепи:
EVKLQESGPS LVKPSQTLSL TCSVTGYSII SDYWNWIRKF PGNKLEYLGY
ISYTGSTYYN PSLKSRISIT RDTSKNQYYL QLNSVTTEDT ATYYCARRGG
WLLPFDYWGQ GTTLTVSS (SEQ ID NO:6)
Клон 3B6 против B7-H1 человека
Вариабельная область легкой цепи:
DIVMTQSPAI MSASPGEKVT MTCSASSSIR YMHWYQQKPG TSPKRWISDT
SKLTSGVPAR FSGSGSGTSY ALTISSMEAE DAATYYCHQR SSYPWTFGGG
TKLEIK (SEQ ID NO:7)
Вариабельная область тяжелой цепи:
EVKLQESGPS LVKPGASVKL SCKASGYTFT SYDINWVKQR PGQGLEWIGW
IFPRDNNTKY NENFKGKATL TVDTSSTTAY MELHSLTSED SAVYFCTKEN
WVGDFDYWGQ GTTLTLSS (SEQ ID NO:8)
Клон 3D10 против B7-H1 человека
Вариабельная область легкой цепи:
QIVLSQSPAI LSASPGEKVT MTCRASSSVS YIYWFQQKPG SSPKPWIYAT
FNLASGVPAR FSGSGSGTSY SLTISRVETE DAATYYCQQW SNNPLTFGAG
TKLELK (SEQ ID NO:9)
Вариабельная область тяжелой цепи:
EVQLQQSGPD LVTPGASVRI SCQASGYTFP DYYMNWVKQS HGKSLEWIGD
IDPNYGGTTY NQKFKGKAIL TVDRSSSTAY MELRSLTSED SAVYYCARGA
LTDWGQGTSL TVSS (SEQ ID NO:10)
(2) Предпочтительные антитела против PD-1 человека и их CDR
Альтернативно такие антитела могут быть получены с помощью скрининга линий гибридом на те, которые продуцируют антитело, являющееся иммуноспецифическим для PD-1 человека, и последующего скрининга среди таких линий тех, которые проявляют модулирующую активность (например, нейтрализующую активность, агонистическую активность, активность в отношении изменения передачи сигнала и т.д.). В изобретении особенно предлагаются клоны против PD-1 человека: 1E3, 1E8 и 1H3.
Антитела, экспрессируемые клонами против PD-1 человека, были секвенированы для выявления их вариабельных доменов. Последовательности CDR представлены жирным шрифтом и подчеркнуты:
Клон 1E3 против PD-1 человека
Вариабельная область легкой цепи:
DIQMTQFPSS LCASQGGKVT VTCKASQDIN NYMAWYQHKP GKGPRLLIHY
TSTLLSGIPS RFSGSGSGRD YSFSISNLEP EDIATYYCLQ YDNLWTFGGG
TKLEIK (SEQ ID NO:11)
Вариабельная область тяжелой цепи:
EVQLQQSGPV LVKPGASVKM SCKASGYTFT DYYMNWVKQS HGKSLEWIGN
INPYNGGTTY NQKFKGKATL TVDKSSRTAY MEINSLTSED SAVYYCARGR
IYDGSLDYWG QGTALTVSS (SEQ ID NO:12)
Клон 1E8 против PD-1 человека
Вариабельная область легкой цепи:
DIVMTQSQKF MSTSVGDRVS VTCKASQSVD TNVAWYQQKP GQSPKALIFS
ASYRYSGVPD RFTGSGSGTD FTLTINSVQS EDLAEYFCQQ YNSYPYTFGS
GTKLEIK (SEQ ID NO:13)
Вариабельная область тяжелой цепи:
QVQLQQSGAE LAKPGASVRL SCKASGYTFT NYWMHWVKQR PGQGLEWIGH
INPSSGFTTY NQNFKDKATL TADKSSNTAY MQLSSLTYED SAVYFCARED
YDVDYWGQGT TLTVSS (SEQ ID NO:14)
Клон 1H3 против PD-1 человека
Вариабельная область легкой цепи:
QIVLTQSPAL MSASPGEKVT MTCSASSSVS YMYWYQQKPR SSPKPWIYLT
SNLASGVPAR FSGSGSGTSY SLTISSMEAE DAATYYCQQW SSNPFTFGSG
TKLEIK (SEQ ID NO:15)
Вариабельная область тяжелой цепи:
EVQLVESGGG LVKPGGSLKL SCAASGFTFS DYGMHWVRQA PEKGLEWVAY
ISSGSYTIYY TDTVKGRFTI SRDNAKNTLF LQMTSLRSED TAMYYCARRG
YGSFYEYYFD YWGQGTTLTV SS (SEQ ID NO: 16)
(3) Консенсусные CDR антител против B7-H1 человека и против PD-1 человека по настоящему изобретению
Проводили анализ CDR идентифицированных антител для того, чтобы идентифицировать последовательности консенсусных CDR и вероятных вариантов последовательностей CDR, которые должны обеспечивать сходными связывающими свойствами. Такие варианты CDR подвергали компьютерной обработке с использованием анализа Blosum62.iij в соответствии с таблицей 1. В таблице 1 представлены баллы за замены. Чем больше баллов, тем более консервативна замена и, следовательно, более вероятно, что замена не будет влиять на функцию.
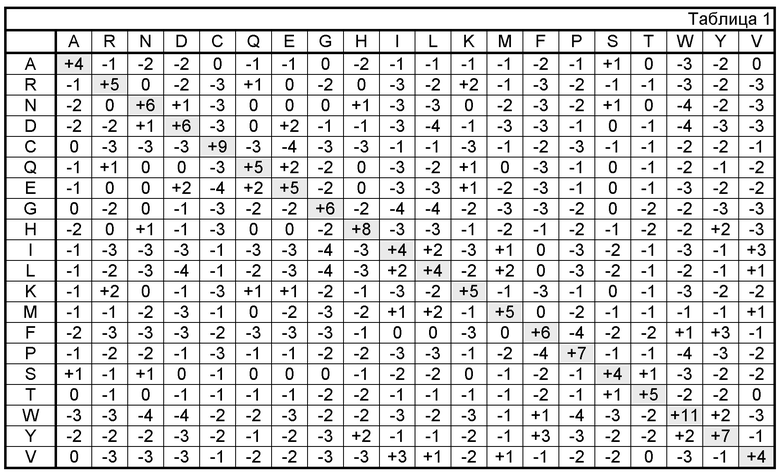
Настоящее изобретение позволяет составлять новые антитела и антигенсвязывающие фрагменты, имеющие 1, 2, 3, 4, 5 или 6 вариантов CDR. Так как методами по настоящему изобретению идентифицировано существенное количество различных CDR, изобретение дает возможность распознавания остатков CDR, которые, вероятно, будут требоваться в любом варианте конкретной идентифицированной CDR. Такие остатки представлены жирным шрифтом в таблице 2, таблице 3, таблице 4 и таблице 5. Эти остатки, как обнаружено, варьируются среди сравниваемых CDR, баллы за замены из таблицы 1 предоставляют средство для определения идентичностей позволяемых замен. Например, если конкретный остаток конкретной CDR, как обнаруживается, варьируется как R или S, то, так как R и S имеют балл за замену -1, любая замена R или S, имеющая балл за замену -1 или выше, является также подходящей как и наблюдаемые варианты (R или S) (или более подходящей, чем R или S) для создания варианта CDR, имеющего связывающие свойства, которые достаточно сходны со свойствами конкретной CDR, что позволяет варианту CDR применяться вместо нее, так чтобы образовывать функционирующее антитело против B7-H1 или против PD-1 или его антигенсвязывающий фрагмент. Для каждого положения выбор остатка, имеющего более высокий балл за замену, является предпочтительным по сравнению с остатком, имеющим более низкий балл за замену.
В таблице 2 представлен анализ CDR легких цепей антител против B7-H1 и предлагается консенсусная последовательность наблюдаемых и предпочтительных вариантов CDR легких цепей антител против B7-H1 по настоящему изобретению.
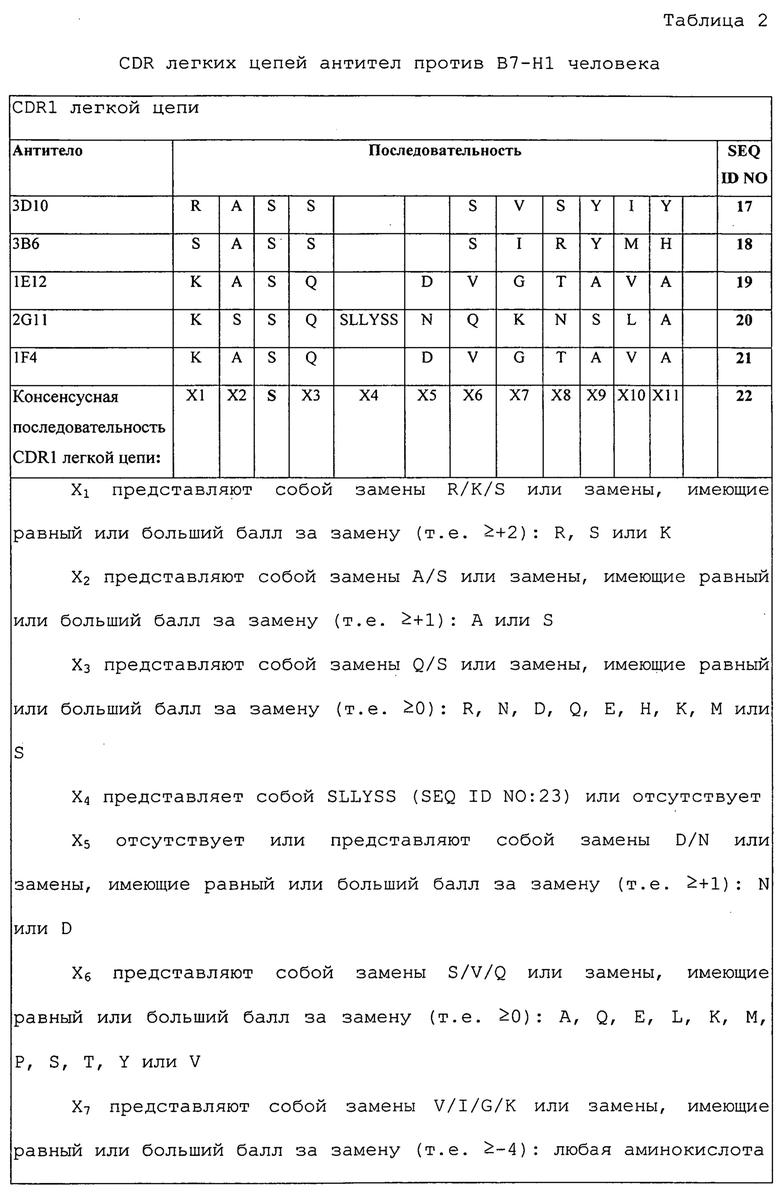

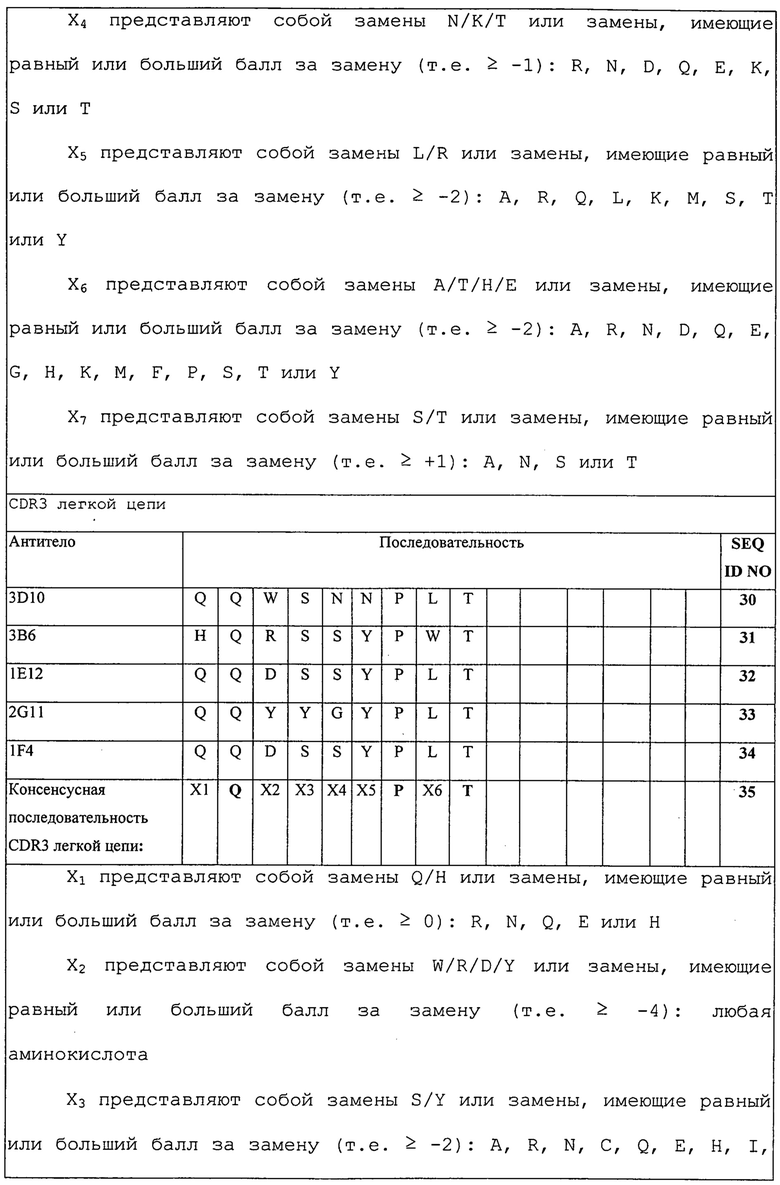

В таблице 3 представлен анализ CDR тяжелых цепей антител против B7-H1 и предлагается консенсусная последовательность наблюдаемых и предпочтительных вариантов CDR тяжелых цепей антител против B7-H1 по настоящему изобретению.
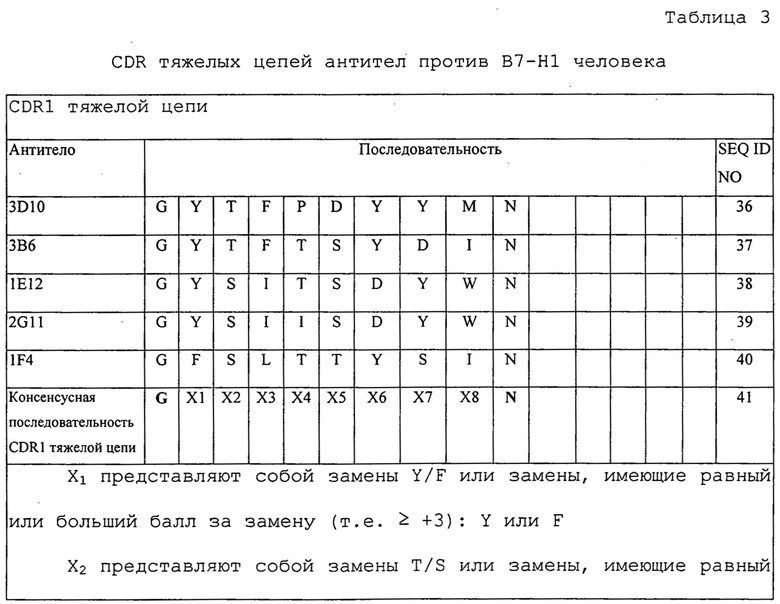




Таким образом, в дополнение к антителам и их антигенсвязывающим фрагментам, которые обладают CDR антител против B7-H1: 1E12, 1F4, 2G11, 3B6 и 3D10, в изобретении дополнительно предлагаются антитела и их антигенсвязывающие фрагменты, которые обладают CDR, имеющими описанные выше консенсусные последовательности легких и/или тяжелых цепей.
В таблице 4 представлен анализ CDR легких цепей антител против PD-1 и предлагается консенсусная последовательность наблюдаемых и предпочтительных вариантов CDR легких цепей антител против PD-1 по настоящему изобретению.
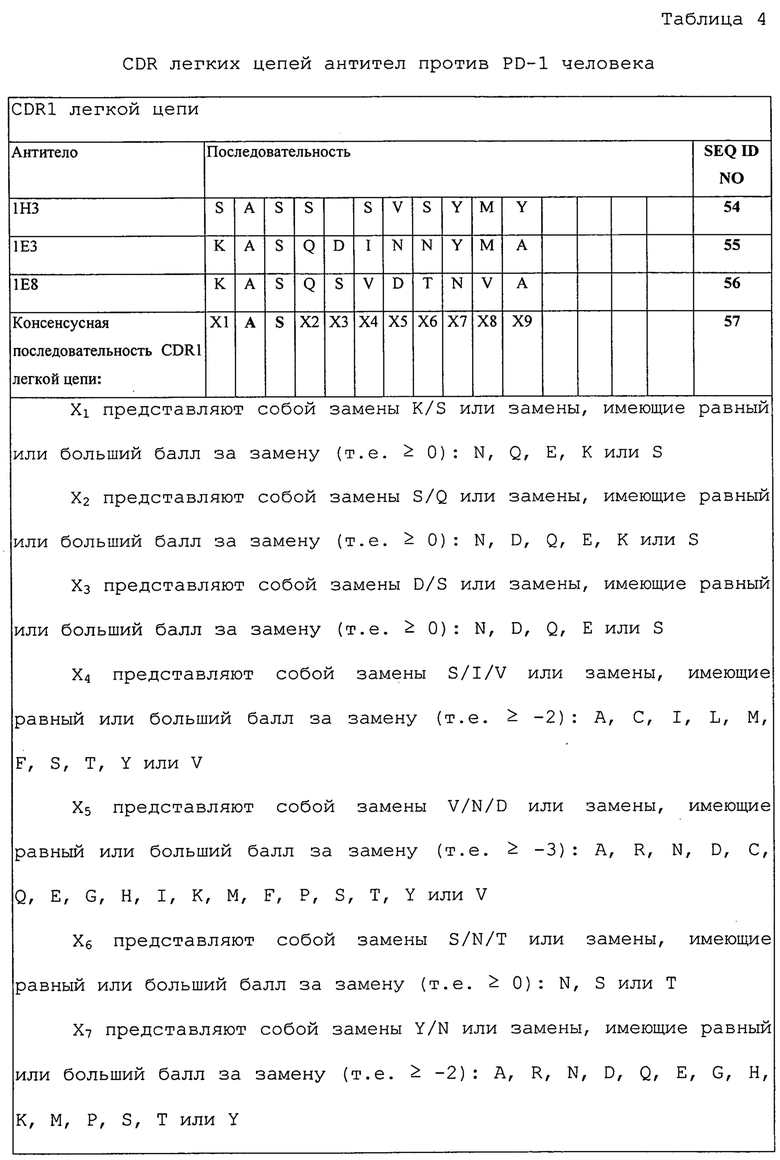
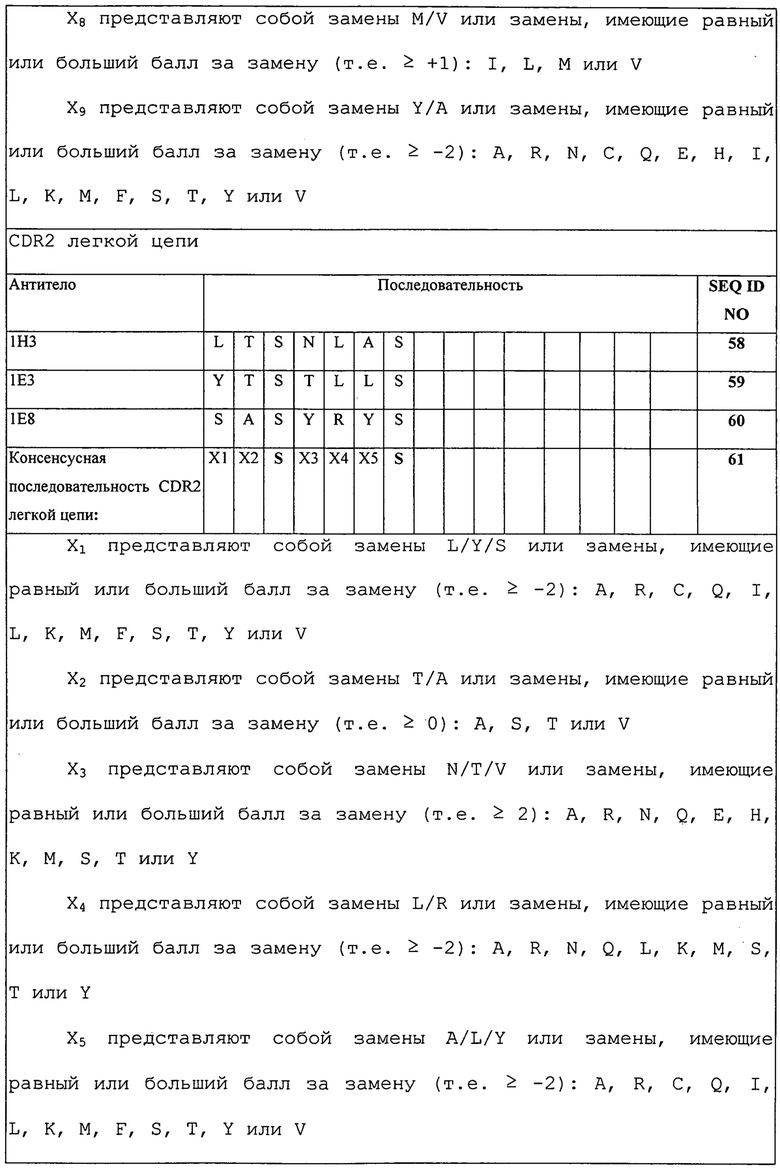
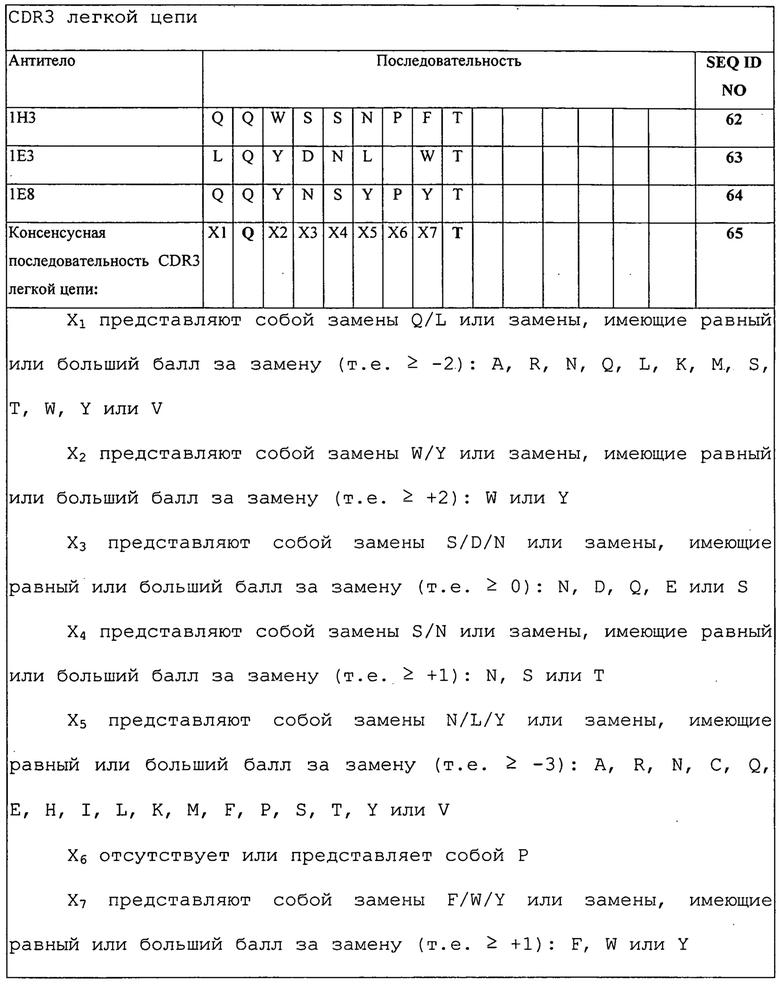
В таблице 5 представлен анализ CDR тяжелых цепей антител против PD-1 и предлагается консенсусная последовательность наблюдаемых и предпочтительных вариантов CDR тяжелых цепей антител против PD-1 по настоящему изобретению.


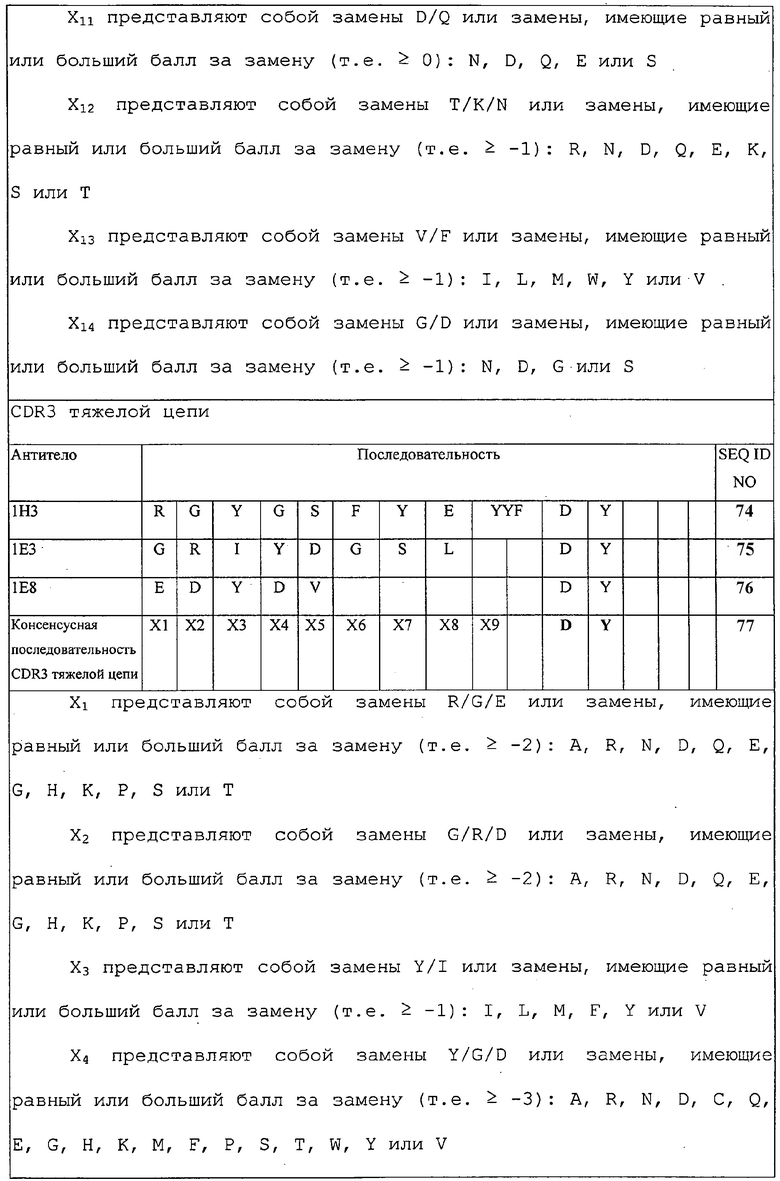

Таким образом, в дополнение к антителам и их антигенсвязывающим фрагментам, которые обладают CDR антител против PD-1: 1E3, 1E8 и 1H3, в изобретении дополнительно предлагаются антитела и их антигенсвязывающие фрагменты, которые обладают CDR, имеющими описанные выше консенсусные последовательности легких и/или тяжелых цепей.
В настоящем изобретении охватываются антитела или их фрагменты, содержащие аминокислотную последовательность вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, которая по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99% идентична аминокислотной последовательности вариабельной области тяжелой цепи и/или легкой цепи мышиного моноклонального антитела, продуцируемого указанными выше клонами, и которая характеризуется иммуноспецифическим связыванием с B7-H1 или PD-1. В настоящем изобретении дополнительно охватываются антитела или их фрагменты, которые содержат CDR, которая по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99% идентична аминокислотной последовательности CDR перечисленных выше клонов и которая характеризуется иммуноспецифическим связыванием с B7-H1 или PD-1. Определение процента идентичности двух аминокислотных последовательностей может быть осуществлено при сравнении белков с помощью программы BLAST.
В конкретном варианте осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению должны содержать одну, две, три, четыре, пять или, более предпочтительно, все шесть CDR описанных выше предпочтительных антител и проявлять способность к связыванию B7-H1 или PD-1 человека.
B. Терапевтическое и профилактическое применение предпочтительных композиций по настоящему изобретению
Изобретение особенно относится к терапевтическому и/или профилактическому применению молекул (особенно антител или их антигенсвязывающих фрагментов), которые иммуноспецифически связываются с B7-H1 человека или PD-1 человека и/или которые способны модулировать связывание между B7-H1 и PD-1, в качестве таковых эти молекулы (B7-H1 или PD-1) экспрессируются эндогенно и локализуются у индивидуума (например, у больного человека), и/или которые способны модулировать передачу сигнала через B7-H1 или PD-1.
Как используется в настоящем описании термины «лечить», «лечение», «обработка» и «терапевтическое использование» относятся к устранению, снижению или облегчению одного или более симптомов заболевания или нарушения, усиливающихся в результате взаимодействий B7-H1 и PD-1. Как используется в настоящем описании термин «терапевтически эффективное количество» относится к количеству терапевтического средства, достаточному для опосредующей роли в клинически выраженных устранении, снижении или облегчении таких симптомов. Эффект является клинически выраженным, если его величина достаточна для вклада в здоровье или в прогноз у индивидуума-реципиента. Терапевтически эффективное количество может относиться к количеству терапевтического средства, достаточному для задержки или сведению к минимуму наступления болезни, например, для задержки или сведению к минимуму распространения злокачественного новообразования. Терапевтически эффективное количество может также относиться к количеству терапевтического средства, которое обеспечивает терапевтическое преимущество в лечении или управлении заболеванием. Кроме того, терапевтически эффективное количество в отношении терапевтического средства по изобретению обозначает то количество терапевтического средства, одного или в сочетании с другими видами терапии, которое обеспечивает терапевтическое преимущество в лечении или управлении заболеванием, например, достаточно для повышения терапевтической эффективности терапевтического антитела, достаточного для лечения или управления заболеванием.
Как используется в настоящем описании термин «профилактическое средство» относится к средству, которое может быть использовано для профилактики нарушения или заболевания перед выявлением каких-либо симптомов такого нарушения или заболевания. «Профилактически эффективное» количество представляет собой количество профилактического средства, достаточное для осуществления такой профилактики. Профилактически эффективное количество может также относиться к количеству профилактического средства, которое обеспечивает профилактическое преимущество в предотвращении заболевания. Кроме того, профилактически эффективное количество в отношении профилактического средства по изобретению обозначает то количество профилактического средства, одного или в сочетании с другими средствами, которое обеспечивает профилактическое преимущество в профилактике заболевания.
Количество доз и частота введения, предлагаемые в настоящем описании, охватываются терминами терапевтически эффективные и профилактически эффективные. Дозировка и частота введения дополнительно обычно должны варьироваться в соответствии с факторами, специфичными для каждого больного в зависимости от вводимых конкретных терапевтических или профилактических средств, тяжести и типа злокачественного новообразования, пути введения, а также от возраста, массы тела, восприимчивости и прошлой истории болезни больного. Подходящие схемы введения могут быть выбраны специалистом в данной области техники путем рассмотрения таких факторов и следуя, например, дозировкам, описанным в литературе и рекомендуемым в Physician 's Desk Reference (56th ed., 2002).
1. Применение положительных модуляторов иммунной системы
В предпочтительном варианте осуществления такие антитела и фрагменты связываются с этими антигенами в одном или более сайтах B7-H1 или PD-1, проксимальных по отношению к сайту связывания B7-H1 - PD-1 и нарушая это связывание. Как обсуждалось выше, взаимодействия между PD-1 и B7-H1 ингибирует пролиферацию Т-клеток и снижает продукцию многих цитокинов (см., например, Sharpe, A.H. et al. (2002) "The B7-CD28 Superfamily," Nature Rev. Immunol. 2:116-126). Таким образом, в предпочтительном варианте осуществления введение молекул по настоящему изобретению индивидууму оказывает положительный модулирующий эффект на иммунную систему индивидуума в результате антагонистического действия на связывание B7-H1 - PD-1. В другом варианте осуществления авидность и/или аффинность антитела против PD-1 могут быть такими, что оно только связывается (и блокирует связывание B7-H1) с Т-клетками, которые экспрессируют очень высокие уровни PD-1, в результате чего Т-клетки истощаются или нарушается их функционирование, и благодаря этому можно специфически направленно действовать на данную клеточную популяцию. Таким образом, изобретение относится к применению антител по настоящему изобретению как опосредующих повышение продукции IFN-γ. Таким образом, оно относится к применению таких антител для лечения заболеваний и состояний, которые отвечают на лечение IFN-γ, таких как рак яичников и другие формы рака, хроническое грануломатозное заболевание, остеопетроз, атаксия Фридриха и т.д. Дополнительно изобретение особенно относится к применению антител по настоящему изобретению как опосредующих повышение пролиферации Т-клеток. Таким образом, оно относится к применению таких антител для лечения заболеваний и состояний, которые можно лечить с помощью повышения пролиферации Т-клеток, таких как: СПИД; тяжелый сочетанный иммунодефицит (SCID); синдром Оменна; гипоплазия хрящей-волос; потеря Т-клеток или случаи их удаления при трансплантации органа или ткани, или при химиотерапии; гипогаммаглобулинемия; X-сцепленная агаммаглобулинемия; временная гипогаммаглобулинемия; дисгаммаглобулинемия; дефицит IgA; дефицит IgG; дефицит IgM; синдром гипер IgM; синдром Вискотта-Олдрича; синдром гипер IgE; общий вариабельный иммунодефицит; синдром ICF; гипоплазия тимуса (например, синдром Ди Георга; синдром Незелофа; синдром атаксии-телангиэктазии); дефицит нуклеозидфосфорилазы пуринов; дефицит аденозиндеаминазы; дефицит ZAP70; синдром голых лимфоцитов; лейкопения; лимфоцитопения (например, идиопатическая CD4+ лимфоцитопения); или дефицит комплемента. Изобретение особенно относится к применению антител по настоящему изобретению как опосредующих и повышенную продукцию IFN-γ, и повышенную пролиферацию Т-клеток.
Положительная модуляция иммунной системы особенно желательна при лечении разных типов злокачественных новообразований и хронических инфекций, и, таким образом, настоящее изобретение находит применение для лечения таких нарушений. Как PD-1, так и B7-H1, сверхэкспрессируются при инфицировании ВИЧ (Xu, Huanbin et al. (2010) “Increased B7-H1 Expression on Dendritic Cells Correlates with Programmed Death 1 Expression on T Cells in Simian Immunodeficiency Virus-Infected Macaques and May Contribute to T Cell Dysfunction and Disease Progression,” J. Immunol. 185:7340-7348; Grabmeier-Pfistershammer, K. et al. (2011) “Identification of PD-1 as a Unique Marker for Failing Immune Reconstitution in HIV-1-Infected Patients on Treatment,” J Acquir. Immune Defic. Syndr. 56(2):118-124). Таким образом, экспрессия B7-H1 на таких клетках может быть использована для диагностики ВИЧ у людей. Следовательно, антитела против PD-1 и против B7-H1 по настоящему изобретению находят особое применение в качестве терапевтических средств для лечения инфицирования ВИЧ и СПИДа. Как используется в настоящем описании термин «злокачественное новообразование» относится к неоплазме или опухоли, возникающих в результате аномального неконтролируемого роста клеток. Как используется в настоящем описании злокачественное новообразование однозначно включает лейкозы и лимфомы. Термин «злокачественное новообразование» относится к заболеванию, касающемуся клеток, которые обладают способностью к метастазированию в отдаленных местах и проявляют фенотипические черты, которые отличают их от черт нераковых клеток, например, образование колоний в трехмерном субстрате, таком как мягкий агар, или образование тубулярных сетей или паутиноподобных матриксов в трехмерной базальной мембране или в препарате внеклеточного матрикса. Нераковые клетки не образуют колоний в мягком агаре и образуют отдельные сфероподобные структуры в трехмерной базальной мембране или в препаратах внеклеточного матрикса.
Типы злокачественного новообразования и относящиеся к нему заболевания, которые можно лечить или предотвращать с помощью способов и композиций по настоящему изобретению, включают, но ими не ограничиваются, следующие: лейкозы, включая, но, не ограничиваясь этим, острый лейкоз, острый лимфоцитный лейкоз, острые миелоцитные лейкозы, такие как миелобластный, промиелоцитный, миеломоноцитный, моноцитный эритролейкемический лейкоз, и миелодиспластический синдром, хронические лейкозы, такие как, но ими не ограничиваясь, хронический миелоцитный (гранулоцитный) лейкоз, хронический лимфоцитный лейкоз, волосатоклеточный лейкоз, истинная полицитемия; лимфомы, такие как, но ими не ограничиваясь, лимфомы типа болезни Ходжкина или неходжкинской лимфомы (например, диффузная анапластическая лимфома, негативная по киназе (ALK), В-крупноклеточная лимфома (DLBCL); диффузная анапластическая лимфома, позитивная по киназе (ALK), В-крупноклеточная лимфома (DLBCL); диффузная анапластическая лимфома, позитивная по киназе (ALK), ALK+ анапластическая крупноклеточная лимфома (ALCL), острая миелоидная лимфома (AML)); множественные миеломы, такие как, но ими не ограничиваясь, вялотекущая множественная миелома, несекреторная миелома, остеосклеротическая миелома, плазмаклеточный лейкоз, солитарная плазмоцитома и экстрамедуллярная плазмацитома; макроглобулинемию Вальденстрема; моноклональную гаммапатию неопределенного значения; доброкачественную моноклональную гаммапатию; заболевание тяжелых цепей; саркомы кости и соединительной ткани, такие как, но ими не ограничиваясь, саркома кости, остеосаркома, хондросаркома, саркома Эвинга, злокачественная гигантоклеточная опухоль, фибросаркома кости, хордома, периостальная саркома, саркомы мягких тканей, ангиосаркома (гемангиосаркома), фибросаркома, саркома Капоши, леймиосаркома, липосаркома, лимфангиосаркома, неврилеммома, рабдомиосаркома, синовиальная саркома; опухоли мозга, включая, но, не ограничиваясь этим, глиому, астроцитому, глиому ствола мозга, эпендимому, олигодендроглиому, неглиальную опухоль, неврилеммому слухового нерва, краниофарингиому, медуллобластому, менингиому, пинеоцитому, пинеобластому, первичную лимфому мозга; рак молочной железы, включая, но, не ограничиваясь этим, аденокарциному, лобулярную (мелкоклеточную) карциному, внутрипротоковую карциному, медуллярный рак молочной железы, слизистый рак молочной железы, протоковый рак молочной железы, папиллярный рак молочной железы, болезнь Педжета и воспалительный рак молочной железы; рак надпочечников, включая, но, не ограничиваясь этим, феохромоцитому и адренокортикальную карциному; рак щитовидной железы, включая, но, не ограничиваясь этим, папиллярный или фолликулярный рак щитовидной железы, медуллярный рак щитовидной железы и анапластический рак щитовидной железы; рак поджелудочной железы, включая, но, не ограничиваясь этим, инсулиному, гастриному, глюкагоному, випому, секретирующую соматостатин опухоль, и карциноидную опухоль или опухоль островковых клеток; типы рака гипофиза, включая, но, не ограничиваясь этим, болезнь Кушинга, секретирующую пролактин опухоль, акромегалию и несахарный диабет; типы рака глаза, включая, но, не ограничиваясь этим, меланому глаза, такую как меланома радужки, хориоидальная меланома и меланома цилиарного тела и ретинобластома; типы рака влагалища, включая, но, не ограничиваясь этим, сквамозноклеточную карциному, аденокарциному и меланому; рак вульвы, включая, но, не ограничиваясь этим, сквамозноклеточную карциному, меланому, аденокарциному, базальноклеточную карциному, саркому и болезнь Педжета; типы рака шейки матки, включая, но, не ограничиваясь этим, сквамозноклеточную карциному и аденокарциному; типы рака матки, включая, но, не ограничиваясь этим, карциному эндометрия и саркому матки; типы рака яичника, включая, но ими не ограничиваясь, карциному эпителия яичника, пограничную опухоль, опухоль зародышевых клеток, и стромальную опухоль; типы рака пищевода, включая, но, не ограничиваясь этим, сквамозный рак, аденокарциному, аденоидную кистозную карциному, мукоэпидермоидную карциному, аденосквамозную карциному, саркому, меланому, плазмацитому, бородавчатый рак и овсяноклеточную (мелкоклеточную) карциному; типы рака желудка, включая, но, не ограничиваясь этим, аденокарциному, грибовидную (полипоидную), изъязвляющую, поверхностно распространяющуюся, малигнизированную лимфому, липосаркому, фибросаркому и саркокарциному; типы рака ободочной кишки; типы рака прямой кишки; типы рака печени, включая, но, не ограничиваясь этим, гепатоклеточную карциному и гепатобластому, типы рака желчного пузыря, включая, но, не ограничиваясь этим, аденокарциному; холангиокарциномы, включая, но, не ограничиваясь этим, папиллярную, нодулярную и диффузную; типы рака легкого, включая, но, не ограничиваясь этим, немелкоклеточный рак легкого, сквамозноклеточную карциному (эпидермоидную карциному), аденокарциному, крупноклеточную карциному и мелкоклеточный рак легкого; типы рака семенников, включая, но, не ограничиваясь этим, герминативную опухоль, семиному, анапластическую, классическую (типичную), сперматоцитную несеминому, эмбриональную карциному, тератомную карциному, хориокарциному (опухоль желточного мешка), типы рака простаты, включая, но, не ограничиваясь этим, аденокарциному, лейомиосаркому и рабдомиосаркому; типы рака полового члена; типы рака ротовой полости, включая, но, не ограничиваясь этим, сквамозноклеточную карциному; типы базального рака; типы рака слюнной железы, включая, но ими не ограничиваясь, аденокарциному, мукоэпидермоидную карциному и аденоидно-кистозную карциному; типы рака гортани, включая, но ими не ограничиваясь, сквамозноклеточный рак и бородавчатый рак; типы рака кожи, включая, но ими не ограничиваясь, базальноклеточную карциному, сквамозноклеточную карциному и меланому, поверхностно распространяющуюся меланому, нодулярную меланому, меланому типа злокачественного лентиго, акральную лентигинозную меланому; типы рака почки, включая, но ими не ограничиваясь, почечноклеточный рак, аденокарциному, гипернефрому, фибросаркому, переходно-клеточный рак (почечной лоханки и/или мочеточника); опухоль Вильмса; типы рака мочевого пузыря, включая, но ими не ограничиваясь, переходно-клеточную карциному, сквамозноклеточный рак, аденокарциному, саркокарциному. Кроме того типы злокачественных новообразований включают миксосаркому, остеогенную саркому, эндотелиосаркому, лимфангиоэндотелиосаркому, мезотелиому, синовиому, гемангиобластому, эпителиальную карциному, цисаденокарциному, бронхогенную карциному, карциному потовых желез, карциному сальных желез, папиллярную карциному и папиллярные аденокарциномы (в качестве обзора таких нарушений см. Fishman et al., 1985, Medicine, 2d Ed., J.B. Lippincott Co., Philadelphia and Murphy et al., 1997, Informed Decisions: The Complete Book of Cancer Diagnosis, Treatment, and Recovery, Viking Penguin, Penguin Books U.S.A., Inc., United States of America)).
Соответственно, способы и композиции по изобретению также подходят для лечения или профилактики разнообразных типов злокачественных новообразований или других заболеваний с аномальной пролиферацией, включая (но, не ограничиваясь этим) следующее: карциному, включая карциному мочевого пузыря, молочной железы, ободочной кишки, почки, печени, легкого, яичника, поджелудочной железы, желудка, шейки матки, щитовидной железы и кожи; включая сквамозноклеточную карциному; гематопоэтические опухоли лимфоидного ряда, включая лейкоз, острый лимфоцитный лейкоз, острый лимфобластный лейкоз, В-клеточную лимфому, Т-клеточную лимфому, лимфому Беркетта; гематопоэтические опухоли миелоидного ряда, включая острый и хронический миелогенный лейкозы и промиелоцитный лейкоз; опухоли мезенхимального происхождения, включая фибросаркому и рабдомиосаркому; другие опухоли, включая меланому, семиному, тератокарциному, нейробластому и глиому; опухоли центральной и периферической нервной системы, включая астроцитому, нейробластому, глиому и шванномы; опухоли мезенхимального происхождения, включая фибросаркому, рабдомиосаркому и остеосаркому; и другие опухоли, включая меланому, пигментную ксеродерму, кератоакантому, семиному, фолликулярный рак щитовидной железы и тератокарциному. Рассматривается также, что типы злокачественных новообразований, которые обусловлены аберрациями апоптоза, также должны подвергаться лечению с помощью методов и композиций по изобретению. Такие типы злокачественного новообразования могут включать, но ими не ограничиваться, фолликулярные лимфомы, карциномы с мутациями p53, гормонзависимые опухоли молочной железы, простаты и яичника, и предраковые нарушения, такие как семейный аденоматозный полипоз и миелодиспластические синдромы. В конкретных вариантах осуществления злокачественные изменения или изменения с нарушением пролиферации (такие как метаплазии и дисплазии) или гиперпролиферативные нарушения лечат или предотвращают с помощью способов и композиций по изобретению в яичнике, мочевом пузыре, молочной железе, ободочной кишке, легком, коже, поджелудочной железе или матке. В других конкретных вариантах осуществления с помощью способов и композиций по изобретению лечат или предотвращают саркому, меланому или лейкоз.
Раковые клетки приобретают характерный набор функциональных способностей в процессе их развития, хотя и с помощью различных механизмов. Такие способности включают ускользание от апоптоза, самодостаточность в отношении ростовых сигналов, отсутствие чувствительности к ингибирующим рост сигналам, тканевую инвазию/метастазирование, неограниченный потенциал пополнения и поддерживаемый ангиогенез. Термин «раковая клетка» предназначен для охвата как предраковых клеток, так и малигнизированных раковых клеток. В некоторых вариантах осуществления к раку относится доброкачественная опухоль, которая сохраняет свою локализацию. В других вариантах осуществления к раку относится злокачественная опухоль, которая способна к инвазии и разрушению соседних структур организма и к распространению в отдаленные места. В еще одних вариантах осуществления рак связан со специфическим раковым антигеном (например, антигеном панкарциномы (KS 1/4), антигеном карциномы яичника (CA125), простатспецифическим антигеном (PSA), канцероэмбриональным антигеном (CEA), CD19, CD20, HER2/neu и т.п.).
Антитела и фрагменты антител по настоящему изобретению особенно подходят для лечения типов злокачественных новообразований, которые связаны с клетками (например, истощенными Т-клетками, В-клетками, моноцитами и т.д.), которые экспрессируют аномально высокие уровни PD-1 (Youngblood, B. (2011) “Chronic Virus Infection Enforces Demethylation Of The Locus That Encodes PD-1 In Antigen-Specific CD8(+) T Cells,” Immunity 35(3):400-412; Spahn, J. et al. (2011) “Ineffective CD8(+) T-Cell Immunity To Adeno-Associated Virus Can Result In Prolonged Liver Injury And Fibrogenesis,” Amer. J. Pathol. 179(5):2370-2381; Wang, C. et al. (2011) “Phenotype, Effector Function, And Tissue Localization Of PD-1-Expressing Human Follicular Helper T Cell Subsets,” BMC Immunol. 12:53, 1-15; Eichbaum, Q. (2011) “PD-1 Signaling In HIV And Chronic Viral Infection Potential For Therapeutic Intervention?” Curr. Med. Chem. 18(26):3971-3980; Hallett, W.H. et al. (2011) “Immunosuppressive Effects Of Multiple Myeloma Are Overcome By PD-L1 Blockade,” Biol Blood Marrow Transplant. 17(8):1133-1145; Ni, L. et al. (2010) “PD-1 Modulates Regulatory T Cells And Suppresses T-Cell Responses In HCV-Associated Lymphoma,” Immunol. Cell. Biol. 89(4):535-539; Inozume, T. et al. (2010) “Selection Of CD8+PD-1+ Lymphocytes In Fresh Human Melanomas Enriches For Tumor-Reactive T Cells,” J. Immunother. 33(9):956-964; and Jin, H.T. et al. (2010) “Cooperation Of Tim-3 And PD-1 In CD8 T-Cell Exhaustion During Chronic Viral Infection,” Proc. Natl. Acad. Sci. (USA) 107(33):14733-14738).
Сходно с их применением в отношении опухолей, как обсуждалось выше, антитела и антигенсвязывающие фрагменты по настоящему изобретению могут быть использованы сами по себе или в виде адъюванта в сочетании с вакцинами или с антибактериальными средствами для стимуляции иммунного ответа против токсинов или собственных антигенов, или против патогенов (например, вирусов, таких как ВИЧ, HTLV, вирус гепатита, вирус гриппа, респираторный синцитиальный вирус, вирус коровьей оспы, вирус бешенства; бактерий, таких как Mycobacteria, Staphylococci, Streptococci, Pneumonococci, Meningococci, Conococci, Klebsiella, Proteus, Serratia, Pseudomonas, Legionella, Corynebacteria, Salmonella, Vibrio, Clostridia, Bacilli, Pasteurella, Leptospirosis, Bordatella и особенно против таких патогенов, которые связаны с холерой, столбняком, ботулизмом, сибирской язвой, чумой и болезнью Лайма; или против грибковых или паразитарных патогенов, таких как Candida (albicans, krusei, glabrata, tropicalis и т.п.), Cryptococcus, Aspergillus (jumigatus, niger, и т.п.), Genus Mucorales (mucor, absidia, rhizophus), Sporothrix (schenkii), Blastomyces (dermatitidis), Paracoccidioides (brasiliensis), Coccidioides (immitis) и Histoplasma (capsulatum), Entamoeba, histolytica, Balantidium coli, Naegleria fowleri, Acanthamoeba sp., Giardia lambia, Cryptosporidium sp., Pneumocystis carinii, Plasmodium vivax, Babesia microti, Trypanosoma brucei, Trypanosoma cruzi, Toxoplasma gondi и т.п.), Sporothrix, Blastomyces, Paracoccidioides, Coccidioides, Histoplasma, Entamoeba, Histolytica, Balantidium, Naegleria, Acanthamoeba, Giardia, Cryptosporidium, Pneumocystis, Plasmodium, Babesia или Trypanosoma и т.п. Таким образом, антитела и антигенсвязывающие фрагменты по настоящему изобретению находят применение при лечении инфекционного заболевания.
2. Применение негативных модуляторов иммунной системы
В альтернативном варианте осуществления антитела по настоящему изобретению против B7-H1 или против PD-1 применяются для получения антиидиотипических пептидов или антител (Wallmann, J. et al. (2010) “Anti-Ids in Allergy: Timeliness of a Classic Concept,” World Allergy Organiz. J. 3(6):195-201; Nardi, M. et al. (2000) “Antiidiotype Antibody Against Platelet Anti-Gpiiia Contributes To The Regulation Of Thrombocytopenia In HIV-1-ITP Patients,” J. Exp. Med. 191(12):2093-2100) или миметиков (Zang, Y.C. et al. (2003) “Human Anti-Idiotypic T Cells Induced By TCR Peptides Corresponding To A Common CDR3 Sequence Motif In Myelin Basic Protein-Reactive T Cells,” Int. Immunol. 15(9):1073-1080; Loiarro, M. et al. (Epub 2010 Apr 8) “Targeting TLR/IL-1R Signalling In Human Diseases,” Mediators Inflamm. 2010:674363). Такие молекулы служат в качестве заменителей B7-H1 или PD-1 и, следовательно, их введение индивидууму негативно регулирует иммунную систему такого индивидуума путем имитации или облегчения связывания B7-H1 - PD-1. Такие молекулы находят применение при лечении заболевания трансплантат против хозяина. Аналогично, антитела-агонисты, которые i) повышают связывание между такими антителами и таким рецептором/лигандом или ii) запускают передачу сигнала, когда прямо связываются с B7-H1 или PD-1, находят применение в качестве агонистов сигнального пути B7-H1-PD-1 и в результате этого находят применение для лечения воспаления и аутоиммунного заболевания с помощью прямого или непрямого действия в качестве агонистов рецепторной активности.
Биспецифические антитела, характеризующиеся иммуноспецифическим связыванием как с PD-1, так и с B7-H1, способны связываться как с APC, так и с Т-клетками и, таким образом, способствуют колокализации APC и Т-клеток. Такая колокализация содействует способности таких клеток связываться друг с другом через молекулы B7-H1 и PD-1, которые не связаны с антителом или с коингибиторными молекулами. Такое связывание вызывает негативную модуляцию иммунной системы реципиента.
Негативная модуляция иммунной системы желательна при лечении воспалительных и аутоиммунных заболеваний и болезни трансплантат против хозяина (GvHD). Примеры аутоиммунных заболеваний, которые можно лечить путем введения антител по настоящему изобретению, включают, но не ограничиваются этим, очаговую алопецию, анкилозирующий спондилоартрит, антифосфолипидный синдром, аутоиммунную болезнь Аддисона, аутоиммунные заболевания надпочечника, аутоиммунную гемолитическую анемию, аутоиммунный гепатит, аутоиммунный оофорит и орхит, аутоиммунную тромбоцитопению, болезнь Бехчета, буллезный пемфигоид, кардиомиопатию, дерматит целиакия спру, синдром хронической усталости и иммунной дисфункции (CFIDS), хроническую воспалительную демиелинизирующую полинейропатию, синдром Черджа-Стросса, рубцовый пемфигоид, синдром CREST, болезнь холодовых агглютининов, болезнь Крона, дискоидную красную волчанку, эссенциальную смешанную криоглобулинемию, фибромиалгию-фибромиозит, гломерулонефрит, болезнь Грейвса, синдром Гийена-Барре, тиреоидит Хашимото, идиопатический легочный фиброз, идиопатическую тромбоцитопеническую пурпуру (ITP), нейропатию IgA, ювенильный артрит, красный плоский лишай, красную волчанку, болезнь Меньера, смешанное заболевание соединительной ткани, множественный склероз, нейромиелит зрительного нерва (NMO), сахарный диабет типа 1 или опосредуемый иммунной системой сахарный диабет, миастению гравис, обыкновенную пузырчатку, пернициозную анемию, нодозный полиартериит, полихондрию, полигландулярные синдромы, ревматическую полимиалгию, полимиозит и дерматомиозит, первичную агаммаглобулинемию, первичный билиарный цирроз, псориаз, псориазный артрит, феномен Рейно, синдром Рейтера, ревматоидный артрит, саркоидоз, склеродерму, синдром Шегрена, синдром мышечной скованности, системную красную волчанку, красную волчанку, артериит Такаясу, временный артериит/гигантоклеточный артериит, поперечный миелит, язвенный колит, увеит, васкулиты, такие как дерматит - герпетиформный васкулит, витилиго и грануломатоз Вегенера.
Примеры воспалительных нарушений, которые можно предотвращать, лечить или которыми можно управлять, в соответствии с методами по изобретению, включают, но не ограничиваются этим, астму, энцефалит, воспалительное заболевание кишечника, хроническую обструктивную болезнь легких (COPD), аллергические нарушения, септический шок, легочный фиброз, недифференцированную спондилоартропатию, недифференцированную артропатию, артрит, воспалительный остеолизис и хроническое воспаление из-за хронической вирусной или бактериальной инфекции.
Таким образом, антитела и антигенсвязывающие фрагменты по настоящему изобретению находят применение при лечении воспалительных и аутоиммунных заболеваний.
C. Способы введения
Известны различные системы доставки, например, инкапсулирование в липосомы, микрочастицы, микрокапсулы, рекомбинантные клетки, способные экспрессировать антитело или слитый белок, опосредуемый рецепторами эндоцитоз (см., например, Wu and Wu, 1987, J. Biol. Chem. 262:4429-4432), конструкция нуклеиновой кислоты в виде части ретровирусного или другого вектора и т.п., и они могут быть использованы для введения терапевтических или профилактических композиций по настоящему изобретению.
Способы введения гуманизированного антитела по изобретению включают, но не ограничиваются этим, инъекцию в виде парентерального введения (например, внутрикожного, внутримышечного, внутрибрюшинного, внутривенного и подкожного), эпидурального введения и введения в слизистую оболочку (например, в виде интраназального и перорального пути). В конкретном варианте осуществления антитела по изобретению вводят внутримышечно, внутривенно или подкожно. Композиции можно вводить с помощью любого подходящего пути, например, с помощью инфузии или болюсной инъекции, с помощью всасывания через эпителиальную или кожно-слизистую выстилку (например, слизистую оболочку ротовой полости, слизистую оболочку прямой кишки и кишечника и т.п.) и их можно вводить совместно с другими биологически активными средствами. Введение может быть системным или местным. Кроме того, может также применяться легочное введение, например, путем использования ингалятора или небулайзера и состава с распыляющим средством. См., например, патенты США No. 6019968; 598520; 5985309; 5934272; 5874064; 5855913; 5290540 и 4880078; и публикации PCT No. WO 92/19244; WO 97/32572; WO 97/44013; WO 98/31346 и WO 99/66903. В конкретном варианте осуществления может быть желательным введение фармацевтических композиций по изобретению местно в область, которая нуждается в лечении; это может быть достигнуто, например, но без ограничения, с помощью местной инфузии, инъекции или посредством имплантата, причем указанный имплантат бывает пористым, непористым или желатиновым материалом, включая мембраны, такие как силиконовые мембраны или волокна. Предпочтительно, при введении антитела по изобретению с осторожностью должны использоваться материалы, на которых не абсорбируется антитело или слитый белок.
В некоторых вариантах осуществления гуманизированные или химерные антитела по изобретению составляются в липосомы для направленной доставки антител по изобретению. Липосомы представляют собой везикулы, содержащие концентрически организованные фосфолипидные бислои, которые инкапсулируют водную фазу. Липосомы обычно содержат различные типы липидов, фосфолипидов и/или поверхностно-активных веществ. Компоненты липосом организованы в виде бислойной конфигурации, сходной с липидной организацией биологических мембран. Липосомы являются особенно предпочтительными носителями для доставки, частично, из-за их биосовместимости, низкой иммуногенности и низкой токсичности. Способы получения липосом известны в данной области техники и охватываются изобретением, см., например, Epstein et al., 1985, Proc. Natl. Acad. Sci. USA, 82: 3688; Hwang et al., 1980 Proc. Natl. Acad. Sci. USA, 77: 4030-4; патенты США No. 4485045 и 4544545.
Изобретением также охватываются способы получения липосом с продолжительным временем полужизни в сыворотке, т.е. с повышенным временем существования в циркуляторном русле, такие как описанные в патенте США No. 5013556. Предпочтительные липосомы, используемые в способах по изобретению, не исчезают быстро из циркуляторного русла, т.е. не захватываются системой мононуклеарных фагоцитов (MPS). Изобретением охватываются стерически стабилизированные липосомы, которые получают с использованием обычных методов, известных специалисту в данной области. Хотя не рассматривается связь с конкретным механизмом действия, стерически стабилизированные липосомы содержат липидные компоненты с объемными и крайне гибкими гидрофильными частями, что снижает нежелательную реакцию липосом с сывороточными белками, снижает опсонизацию с компонентами сыворотки и снижает распознавание MPS. Стерически стабилизированные липосомы, предпочтительно, получают с использованием полиэтиленгликоля. Для получения липосом и стерически стабилизированной липосомы, см., например, Bendas et al., 2001 BioDrugs, 15(4): 215-224; Allen et al., 1987 FEBS Lett. 223: 42-6; Klibanov et al., 1990 FEBS Lett., 268: 235-7; Blum et al., 1990, Biochim. Biophys. Acta., 1029: 91-7; Torchilin et al., 1996, J. Liposome Res. 6: 99-116; Litzinger et al., 1994, Biochim. Biophys. Acta, 1190: 99-107; Maruyama et al., 1991, Chem. Pharm. Bull., 39: 1620-2; Klibanov et al., 1991, Biochim Biophys Acta, 1062; 142-8; Allen et al., 1994, Adv. Drug Deliv. Rev, 13: 285-309. Изобретением также охватываются липосомы, которые адаптированы для направленной доставки к конкретному органу, см., например, патент США No. 4544545, или для направленной доставки к конкретной клетке, см., например, публикацию патентной заявки США No. 2005/0074403. Особенно подходящие липосомы для использования в композициях и методах по изобретению могут быть получены методом выпаривания с обратной фазой липидной композиции, содержащий фосфатидилхолин, холестерин и дериватизированный ПЭГ фосфатидилэтаноламин (PEG-PE). Липосомы продавливаются через фильтры с определенным размером пор с получением липосом желаемого диаметра. В некоторых вариантах осуществления фрагмент антитела по изобретению, например, F(ab'), может быть конъюгирован с липосомами с использованием ранее описанных методов, см., например, Martin et al., 1982, J. Biol. Chem. 257: 286-288.
Гуманизированные или химерные антитела по изобретению могут быть составлены в виде иммунолипосом. Иммунолипосомы относятся к липосомальным композициям, в которых антитело по изобретению или его фрагмент соединен ковалентно или нековалентно с поверхностью липосомы. Химия присоединения антитела к поверхности липосомы известна в данной области техники и охватывается изобретением, см., например, патент США No. 6787153; Allen et al., 1995, Stealth Liposomes, Boca Rotan: CRC Press, 233-44; Hansen et al., 1995, Biochim. Biophys. Acta, 1239: 133-144. В наиболее предпочтительных вариантах осуществления иммунолипосомы для применения в методах и композициях по изобретению дополнительно стерически стабилизированы. Предпочтительно, чтобы гуманизированные антитела по изобретению были связаны ковалентно или нековалентно с гидрофобным якорем, который стабильно укореняется в липидном бислое липосомы. Примеры гидрофобных якорей включают, но ими не ограничиваются, фосфолипиды, например, фосфатидилэтаноламин (PE), фосфатидилинозитол (PI). Для достижения ковалентного связывания между антителом и гидрофобным якорем могут быть использованы любые из биохимических стратегий, известных в данной области техники, см., например, J. Thomas August, ed., 1997, Gene Therapy: Advances in Pharmacology, Volume 40, Academic Press, San Diego, CA, p. 399-435. Например, функциональная группа молекулы антитела может взаимодействовать с активной группой липосомы, связанной с гидрофобным якорем, например, аминогруппа боковой цепи лизина антитела может быть соединена со связанным с липосомой N-глутарилфосфатидилэтаноламином, активированным растворимым в воде карбодиимидом; или тиоловая группа восстановленного антитела может быть соединена с липосомами через реактивные в отношении тиолов якоря, такие как пиридилтиопропионилфосфатидилэтаноламин. См., например, Dietrich et al., 1996, Biochemistry, 35: 1100-1105; Loughrey et al., 1987, Biochim. Biophys. Acta, 901: 157-160; Martin et al., 1982, J. Biol. Chem. 257: 286-288; Martin et al., 1981, Biochemistry, 20: 4429-38. Хотя не рассматривается связь с конкретным механизмом действия, иммунолипосомные составы, включающие антитело по изобретению, особенно эффективны в качестве терапевтических средств, так как они доставляют антитело в цитоплазму клетки-мишени, т.е. клетки, содержащей рецептор, с которым связывается антитело. Иммунолипосомы, предпочтительно, имеют повышенный период полужизни в крови, специфически направлены на клетки и могут интернализоваться в цитоплазму клеток-мишеней, тем самым избегая потери терапевтического средства или его деградации по эндолизосомальному пути.
Иммунолипосомные композиции по изобретению содержат один или более липидов, образующих везикулу, антитело по изобретению или его фрагмент или производное, и необязательно гидрофильный полимер. Липид, образующий везикулу, предпочтительно, представляет собой липид с двумя углеродными цепями, такими как ацильные цепи, и группу с полярной головкой. Примеры липидов, образующих везикулу, включают фосфолипиды, например, фосфатидилхолин, фосфатидилэтаноламин, фосфатидную кислоту, фосфатидилинозитол, сфингомиелин, и гликолипиды, например, цереброзиды, ганглиозиды. Дополнительные липиды, подходящие для составов по изобретению, известны специалисту в данной области и охватываются изобретением. В некоторых вариантах осуществления иммунолипосомные композиции дополнительно содержат гидрофильный полимер, например, полиэтиленгликоль и ганглиозид GMI, который повышает период полужизни липосомы в сыворотке. Методы конъюгации гидрофильных полимеров с липосомами хорошо известны в данной области и охватываются изобретением. Для обзора иммунолипосом и методов их получения см., например, публикацию патентной заявки США No. 2003/0044407; международную публикацию PCT No. WO 97/38731, Vingerhoeads et al., 1994, Immunomethods, 4: 259-72; Maruyama, 2000, Biol. Pharm. Bull. 23(7): 791-799; Abra et al., 2002, Journal of Liposome Research, 12(1&2): 1-3; Park, 2002, Bioscience Reports, 22(2): 267-281; Bendas et al., 2001 BioDrugs, 14(4): 215-224, J. Thomas August, ed., 1997, Gene Therapy: Advances in Pharmacology, Volume 40, Academic Press, San Diego, CA, p. 399-435.
В изобретении также предлагаются гуманизированные или химерные антитела по изобретению, которые упакованы в герметически запаянный контейнер, такой как ампула или пакет-саше, с указанием количества антитела. В одном из вариантов осуществления антитела по изобретению предоставляются в виде сухого, стерильного, лиофилизованного порошка или свободного от воды концентрата в герметически запаянном контейнере, и они могут быть восстановлены, например, водой или физиологическим раствором, до концентрации, подходящей для введения индивидууму. Предпочтительно, антитела по изобретению предоставляются в виде сухого, стерильного, лиофилизованного порошка в герметически запаянном контейнере в единице лекарственной формы по меньшей мере 5 мг, более предпочтительно, по меньшей мере 10 мг, по меньшей мере 15 мг, по меньшей мере 25 мг, по меньшей мере 35 мг, по меньшей мере 45 мг, по меньшей мере 50 мг или по меньшей мере 75 мг. Лиофилизованные антитела по изобретению должны храниться при температуре от 2 до 8°С в их исходном контейнере, и антитела должны вводиться в пределах 12 часов, предпочтительно, в пределах 6 часов, в пределах 5 часов, в пределах 3 часов или в пределах 1 часа после реконституции. В альтернативном варианте осуществления антитела по изобретению предоставляются в жидкой форме в герметически запаянном контейнере с указанием количества и концентрации антитела, слитого белка или конъюгированной молекулы. Предпочтительно, жидкая форма антител предоставляется в герметически запаянном контейнере с концентрацией антител по меньшей мере 1 мг/мл, более предпочтительно, по меньшей мере 2,5 мг/мл, по меньшей мере 5 мг/мл, по меньшей мере 8 мг/мл, по меньшей мере 10 мг/мл, по меньшей мере 15 мг/мл, по меньшей мере 25 мг/мл, по меньшей мере 50 мг/мл, по меньшей мере 100 мг/мл, по меньшей мере 150 мг/мл, по меньшей мере 200 мг/мл.
Точная доза, которая будет применяться в составе, должна также зависеть от пути введения и тяжести состояния и должна быть выбрана в соответствии с решением практикующего врача и состоянием каждого больного. Эффективные дозы могут быть экстраполированы на основе кривых дозозависимости, полученных в тест-системах на животных моделях in vitro. Для антител, охватываемых изобретением, доза, вводимая больному, обычно составляет от 0,01 мг/кг до 100 мг/кг массы тела больного. Предпочтительно, доза, вводимая больному, обычно составляет от 0,01 мг/кг до 20 мг/кг, от 0,01 мг/кг до 10 мг/кг, от 0,01 мг/кг до 5 мг/кг, от 0,01 до 2 мг/кг, от 0,01 до 1 мг/кг, от 0,01 мг/кг до 0,75 мг/кг, от 0,01 мг/кг до 0,5 мг/кг, от 0,01 мг/кг до 0,25 мг/кг, от 0,01 до 0,15 мг/кг, от 0,01 до 0,10 мг/кг, от 0,01 до 0,05 мг/кг или от 0,01 мг/кг до 0,025 мг/кг массы тела больного. В частности, изобретением охватывается доза, вводимая больному, составляющая 0,2 мг/кг, 0,3 мг/кг, 1 мг/кг, 3 мг/кг, 6 мг/кг или 10 мг/кг. Такая низкая доза как 0,01 мг/кг, как прогнозируется, способна проявлять заметные фармакодинамические эффекты. Уровень доз 0,10-1 мг/кг, как прогнозируется, является наиболее подходящим. Более высокие дозы (например, 1-30 мг/кг) также, как ожидается, должны быть действенными. Обычно антитела человека имеют более длительный период полужизни в организме человека, чем антитела от других видов, из-за иммунного ответа на чужеродные полипептиды. Таким образом, часто можно использовать более низкие дозы антител человека и более редкое введение. Кроме того, дозировка и частота введения антител по изобретению или их фрагментов может быть снижена путем повышения захвата антител и проникновения их в ткани с помощью таких модификаций, как липидизация.
В еще одном варианте осуществления композиции могут быть доставлены с помощью системы контролируемого высвобождения или поддерживаемого высвобождения. Любой способ, известный специалисту в данной области, может быть использован для получения составов с поддерживаемым высвобождением, содержащих антитела по изобретению. См., например, патент США No. 4526938; публикацию PCT WO 91/05548; публикацию PCT WO 96/20698; Ning et al., 1996, “Intratumoral Radioimmunotheraphy of a Human Colon Cancer Xenograft Using a Sustained-Release Gel,” Radiotherapy & Oncology 39:179-189, Song et al., 1995, “Antibody Mediated Lung Targeting of Long-Circulating Emulsions,” PDA Journal of Pharmaceutical Science & Technology 50:372-397; Cleek et al., 1997, “Biodegradable Polymeric Carriers for a bFGF Antibody for Cardiovascular Application,” Pro. Int’l. Symp. Control. Rel. Bioact. Mater. 24:853-854; and Lam et al., 1997, “Microencapsulation of Recombinant Humanized Monoclonal Antibody for Local Delivery,” Proc. Int’l. Symp. Control Rel. Bioact. Mater. 24:759-760. В одном из вариантов осуществления в системе для контролируемого высвобождения может быть использована помпа (см. Langer, выше; Sefton, 1987, CRC Crit. Ref. Biomed. Eng. 14:20; Buchwald et al., 1980, Surgery 88:507; and Saudek et al., 1989, N. Engl. J. Med. 321:574). В другом варианте осуществления для достижения контролируемого высвобождения антител могут быть использованы полимерные материалы (см., например, Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Florida (1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York (1984); Ranger and Peppas, 1983, J., Macromol. Sci. Rev. Macromol. Chem. 23:61; см. также Levy et al., 1985, Science 228:190; During et al., 1989, Ann. Neurol. 25:351; Howard et al., 1989, J. Neurosurg. 7 1:105); патент США No. 5679377; патент США No. 5916597; патент США No. 5912015; патент США No. 5989463; патент США No. 5128326; публикацию PCT No. WO 99/15154; и публикацию PCT No. WO 99/20253). Примеры полимеров, используемых в составах для поддерживаемого высвобождения, включают, но не ограничиваются этим, поли(2-гидроксиэтилметакрилат), поли(метилметакрилат), поли(акриловую кислоту), поли(этилен-совинилацетат), поли(метакриловую кислоту), полигликолиды (PLG), полиангидриды, поли(N-винилпирролидон), поли(виниловый спирт), полиакриламид, поли(этиленгликоль), полилактиды (PLA), поли(лактид-согликолиды) (PLGA) и полиортоэфиры. В еще одном варианте осуществления система с контролируемым высвобождением может быть помещена вблизи терапевтической мишени (например, легких), в результате чего требуется только часть системной дозы (см., например, Goodson, in Medical Applications of Controlled Release, выше, vol. 2, pp. 115-138 (1984)). В другом варианте осуществления полимерные композиции, пригодные в качестве имплантатов с контролируемым высвобождением, используются в соответствии с Dunn et al. (см. патент США 5945155). Этот конкретный метод основан на терапевтическом эффекте контролируемого высвобождения биологически активного вещества из полимерной системы in situ. Имплантация обычно может быть осуществлена в любом участке организма больного, нуждающегося в терапевтическом лечении. В другом варианте осуществления используются неполимерные системы доставки с поддерживаемым высвобождением, в соответствии с чем в организме индивидуума в качестве системы доставки лекарства используется неполимерный имплантат. При имплантации в организм органический растворитель имплантата должен рассеиваться, диспергироваться или вымываться из композиции в окружающую тканевую жидкость, и неполимерный материал должен постепенно коагулировать или преципитировать с образованием твердого, микропористого матрикса (см. патент США 5888533). Системы с контролируемым высвобождением обсуждаются в обзоре Langer (1990, Science 249: 1527-1533). Любой метод, известный специалисту в данной области техники, может быть использован для получения составов с поддерживаемым высвобождением, содержащих один или более терапевтических средств по изобретению. См., например, патент США No. 4526938; международные публикации No. WO 91/05548 и WO 96/20698; Ning et al., 1996, Radiotherapy & Oncology 39:179-189; Song et al., 1995, PDA Journal of Pharmaceutical Science & Technology 50:372-397; Cleek et al., 1997, Pro. Int’l. Symp. Control. Rel. Bioact. Mater. 24:853-854; and Lam et al., 1997, Proc. Int’l. Symp. Control Rel. Bioact. Mater. 24:759-760.
В конкретном варианте осуществления, когда терапевтическая или профилактическая композиция по изобретению представляет собой нуклеиновую кислоту, кодирующую антитело по изобретению или его антигенсвязывающий фрагмент, нуклеиновую кислоту можно вводить in vivo для стимуляции экспрессии кодируемого ею антитела с помощью конструирования ее в виде части подходящего вектора экспрессии, содержащего нуклеиновую кислоту, и введения его так, чтобы он становился внутриклеточным, например, при использовании ретровирусного вектора (см. патент США No. 4980286), или путем прямого введения с использованием бомбардировки микрочастицами (например, с помощью генной пушки; Biolistic, Dupont), или с помощью покрытия липидами или рецепторами клеточной поверхности, или трансфицирующими средствами, или путем введения ее в связи с гомеобокс-подобным пептидом, который известен как проникающий в ядро (см., например, Joliot et al., 1991, Proc. Natl. Acad. Sci. USA 88:1864-1868) и т.д. Альтернативно, нуклеиновая кислота может быть введена внутрь клетки и включена в ДНК клетки-хозяина для экспрессии с помощью гомологичной рекомбинации.
Лечение индивидуума терапевтически или профилактически эффективным количеством антител по изобретению может включать однократное лечение или, предпочтительно, может включать серии лечебного воздействия.
D. Фармацевтические композиции
Композиции по изобретению включают основную массу композиций лекарственного средства, подходящего для получения фармацевтических композиций (т.е. композиций, которые подходят для введения индивидууму или больному), которые могут быть использованы для получения единиц лекарственных форм. Такие композиции содержат профилактически или терапевтически эффективное количество профилактического и/или терапевтического средства, раскрытого в настоящем описании, или сочетание этих средств и фармацевтически приемлемый носитель. Предпочтительно, композиции по изобретению содержат профилактически или терапевтически эффективное количество гуманизированных антител по изобретению и фармацевтически приемлемый носитель.
В конкретном варианте осуществления термин «фармацевтически приемлемый» обозначает одобренный регулирующим ведомством федерального или государственного правительства или включенный в список фармакопеи США или другой общепризнанной фармакопеи как разрешенный для использования у животных и более конкретно у человека. Термин «носитель» относится к разбавителю, адъюванту (например, (полному или неполному) адъюванту Фрейнда), наполнителю, поверхностно-активному веществу, криозащитному средству или носителю, с которыми вводят терапевтическое средство. Такие фармацевтические носители могут представлять собой стерильные жидкости, такие как вода и масла, включая вазелин, масла животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и тому подобное. Вода является предпочтительным носителем, когда фармацевтическую композицию вводят внутривенно. Физиологические растворы и растворы водной декстрозы и глицерина могут применяться в качестве жидких носителей, особенно для вводимых растворов. Подходящие фармацевтические наполнители включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое снятое молоко, глицерин, пропиленгликоль, воду, этанол, полисорбат-80 и тому подобное. Композиция, если это желательно, может также содержать минорные количества увлажняющих или эмульгирующих средств или средств, забуферивающих рН. Эти композиции могут иметь форму растворов, суспензий, эмульсий, таблеток, пилюль, капсул, порошков, составов с поддерживаемым высвобождением и тому подобного.
Обычно ингредиенты композиций по изобретению предлагаются либо по отдельности, либо смешанными друг с другом в единицу лекарственной формы, например, в виде сухого лиофилизованного порошка или свободного от воды концентрата в герметически запаянном контейнере, таком как ампула или саше, с указанием количества активного средства. Когда композиция предназначена для введения с помощью инфузии, она может быть дозирована в инфузионной бутыли, содержащей стерильную воду или физиологический раствор фармацевтической степени чистоты. Когда композицию вводят с помощью инъекции, ампула стерильной воды или физиологического раствора для инъекций может быть предоставлена так, чтобы ингредиенты могли смешиваться перед введением.
Композиции по изобретению могут быть составлены в виде нейтральных или солевых форм. Фармацевтически приемлемые соли включают, но не ограничиваются этим, соли, образованные с анионами, такие как происходящие от хлористоводородной, фосфорной, уксусной, щавелевой, винной кислот и т.п., и соли, образованные с катионами, такими как происходящие от натрия, калий, аммония, кальция, гидроксидов железа, изопропиламина, триэтиламина, 2-этиламиноэтанола, гистидина, прокаина и т.д.
E. Наборы
В изобретении предлагается фармацевтическая упаковка или набор, содержащий один или более контейнеров, наполненных гуманизированными антителами по изобретению. Дополнительно в фармацевтическую упаковку или набор могут быть также включены один или более других профилактических или терапевтических средств, подходящих для лечения заболевания. В изобретении также предлагается фармацевтическая упаковка или набор, содержащий один или более контейнеров, наполненных одним или более ингредиентами фармацевтических композиций по изобретению. Необязательно в сочетании с таким(ми) контейнером(рами) может находиться аннотация в форме предписания государственного агентства, регулирующего производство, применение или продажу фармацевтических или биологических продуктов, причем аннотация отражает одобрение агентством производства, применения или продажи для введения человеку.
В настоящем изобретении предлагается набор, который может быть использован в указанных выше методах. В одном варианте осуществления набор включает одно или более гуманизированных антител по изобретению. В другом варианте осуществления набор дополнительно включает один или более других профилактических или терапевтических средств, подходящих для лечения злокачественных новообразований в одном или более контейнерах. В другом варианте осуществления набор дополнительно включает одно или более цитотоксических антител, которые связываются с одним или более раковыми антигенами, связанными с раком. В определенных вариантах осуществления другое профилактическое или терапевтическое средство представляет собой химиотерапевтическое средство. В других вариантах осуществления профилактическое или терапевтическое средство представляет собой биологическое или гормональное терапевтическое средство.
F. Методы диагностики
Антитела по изобретению и их антигенсвязывающие фрагменты могут быть использованы для диагностических целей, например, для определения, диагностики или мониторинга заболеваний, нарушений или инфекций, связанных с экспрессией B7-H1 или PD-1. Для определения или диагностики заболевания, нарушения или инфекции, особенно аутоиммунного заболевания, в изобретении предлагается включение: (a) определения экспрессии B7-H1 или PD-1 в клетках или в образце ткани индивидуума с использованием одного или более антител (или их фрагментов), которые иммуноспецифически связываются с такими антигенами; и (b) сравнения уровня антигена с контрольным уровнем, например, с уровнем в образце нормальной ткани или перед лечением, где повышение или понижение определяемого уровня антигена по сравнению с контрольным уровнем антигена указывает на ответ индивидуума на лечение заболевания, нарушения или инфекции. Таким образом, в изобретении предлагается также мониторинг прогрессии заболевания, нарушения или инфекции, включающий: (a) определение экспрессии B7-H1 или PD-1 в клетках или в образце ткани индивидуума в одну временную точку с использованием одного или более антител (или их фрагментов), которые иммуноспецифически связываются с такими антигенами; и (b) сравнение уровня экспрессии B7-H1 или PD-1 в клетках или в образце ткани индивидуума в другую временную точку или с течением времени, где повышение или понижение определяемого уровня антигена указывает на прогрессию заболевания, нарушения или инфекции. В изобретении дополнительно предлагается мониторинг ответа на лечение, включающий: (a) определение экспрессии B7-H1 или PD-1 в клетках или в образце ткани индивидуума перед лечением одним или более антителами (или их фрагментами), которые иммуноспецифически связываются с такими антигенами; и (b) определение экспрессии B7-H1 или PD-1 в клетках или в образце ткани индивидуума перед лечением и в одну или более временные точки после лечения и сравнение уровня антигена в течение этого периода времени, где повышение или понижение определяемого уровня антигена по сравнению с уровнем антигена до лечения указывает на ответ на лечение. Такие антитела и фрагменты, предпочтительно, применяются в иммунотестах, таких как твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ (RIA) и сортинг активированных флуоресценцией клеток (FACS).
В одном из аспектов изобретение относится к применению таких антител и фрагментов, и особенно таких антител и фрагментов, которые связываются с B7-H1 человека, в качестве реагентов для IHC анализа клеток in vitro или образцов ткани in situ или in vivo. Например, так как B7-H1 экспрессируется раковыми клетками, но не нормальной тканью (Dong, H. (2003) “B7-H1 Pathway And Its Role In The Evasion Of Tumor Immunity,” J. Mol. Med. 81:281-287), определение его присутствия на клетке с помощью такого связывания клетки с такими антителами или фрагментами указывает и диагностирует раковую клетку. Таким образом, в изобретении предлагается цитологический тест, диагностирующий наличие рака у индивидуума.
Присутствие B7-H1 на опухолевых клетках, как обнаружено, усиливает апоптоз Т-клеток, которые отвечают на опухоль (патент Соединенных Штатов No. 7794710). Таким образом, с помощью определения распространения или степени, с которой опухолевые клетки больного раком экспонируют B7-H1 на своей поверхности, в настоящем изобретении предлагается средство для определения клинической значимости рака и степени, до которой он будет рефрактерен к иммунному ответу.
Сходно, B7-H1 экспрессируется на лимфоидных клетках и дендритных клетках слизистой оболочки (как на миелоидных, так и на плазмацитоидных дендритных клетках), и его экспрессия существенно повышается после инфицирования SIV (Xu, Huanbin et al. (2010) “Increased B7-H1 Expression on Dendritic Cells Correlates with Programmed Death 1 Expression on T Cells in Simian Immunodeficiency Virus-Infected Macaques and May Contribute to T Cell Dysfunction and Disease Progression,” J. Immunol. 185:7340-7348). Таким образом, экспрессия B7-H1 на таких клетках может быть использована для диагностики ВИЧ у человека. Соответственно, обнаружено, что экспрессия PD-1 на CD8+ клетках повышается в условиях инфекции ВИЧ (Killian, M.S. et al. (2011) “Natural Suppression of Human Immunodeficiency Virus Type 1 Replication Is Mediated by Memory CD8+ T Cells,” J. Virol. 85(4):1696-1705). Таким образом, антитела, которые связываются как с PD-1, так и с CD8, находят особое применение для диагностики инфицирования ВИЧ и прогрессии СПИДа.
В дополнительном аспекте изобретение относится к применению таких антител и фрагментов, и особенно таких антител и фрагментов, которые связываются с PD-1 человека. PD-1 находит особое применение в качестве маркера хронической активации иммунной системы и истощения Т-клеток. Его экспрессия повышается на Т-клетках виремического больного, инфицированного ВИЧ, и коррелирует с вирусной нагрузкой у этих больных (Khaitan, A. et al. (2011) “Revisiting Immune Exhaustion During HIV Infection,” Curr. HIV/AIDS Rep. 8:4-11; Grabmeier-Pfistershammer, K. et al. (2011) “Identification of PD-1 as a Unique Marker for Failing Immune Reconstitution in HIV-1-Infected Patients on Treatment,” J Acquir. Immune Defic. Syndr. 56(2):118-124). Таким образом, PD-1 находит особое применение в качестве маркера прогрессии ВИЧ. Наиболее предпочтительно, чтобы экспрессия PD-1 оценивалась с использованием проточной цитометрии. С помощью использования таких антител и фрагментов Т-клетки (которые экспрессируют PD-1) могут быть проанализированы по множеству параметров (например, количеству клеток, клеточному размеру, фенотипу и здоровому состоянию клеток и т.д.) даже при присутствии в гетерогенных препаратах. Таким образом, совместно с антителами к PD-1 и их антигенсвязывающими фрагментами такие методы могут быть использованы для диагностики степени или тяжести СПИДа, лейкозов и другого заболевания, влияющего на количество Т-клеток и их здоровое состояние. Методы проточной цитометрии, которые можно адаптировать для целей настоящего изобретения, раскрыты в Peters, J.M. et al. (2011) “Multiparameter Flow Cytometry In The Diagnosis And Management Of Acute Leukemia,” Arch. Pathol. Lab. Med. 135(1):44-54; Meyerson, H.J. (2010) “A Practical Approach To The Flow Cytometric Detection And Diagnosis Of T-Cell Lymphoproliferative Disorders,” Lab. Hematol. 16(3):32-52; Vandewoestyne, M. et al. (Epub 2010 Aug 3) Laser Capture Microdissection In Forensic Research: A Review,” Int. J. Legal. Med. 124(6):513-521; Ornatsky, O. et al. (Epub 2010 Jul 21) “Highly Multiparametric Analysis By Mass Cytometry,” J. Immunol. Meth. 361(1-2):1-20; Mach, W.J. et al. (Epub 2010 Jul 13) “Flow Cytometry And Laser Scanning Cytometry, A Comparison Of Techniques,” J. Clin. Monit. Comput. 24(4):251-259; and Chattopadhyay, P.K. et al. (2010) “Good Cell, Bad Cell: Flow Cytometry Reveals T-Cell Subsets Important In HIV Disease,” Cytometry A. 77(7):614-622; и в патентах Соединенных Штатов No. 7876436; 7847923; 7842244; 7746466; 7590500; 7527978; 7507548; 7491502; 7486387; 7479630; 7465543; 7354773; 6794152 и 6784981.
Таким образом, антитела и фрагменты по настоящему изобретению находят применение при определении и диагностике заболевания, нарушения или инфекции у человека. В одном варианте осуществления такая диагностика включает: a) введение индивидууму (например, парентерально, подкожно или внутрибрюшинно) эффективного количества меченого антитела или антигенсвязывающего фрагмента, которые иммуноспецифически связываются с B7-H1 или PD-1; b) ожидание в течение временного интервала, следующего за введением, возможности концентрирования меченых молекул в тех местах индивидуума, где экспрессируется B7-H1 или PD-1 (и удаления несвязанных меченых молекул до фонового уровня); c) определение фонового уровня; и d) определение меченого антитела у индивидуума, так что определение меченого антитела выше фонового уровня указывает на то, что у индивидуума есть заболевание, нарушение или инфекция. В соответствии с этим вариантом осуществления антитело метят визуализирующейся частью, которая поддается определению с использованием визуализирующей системы, известной специалисту в данной области. Фоновый уровень может быть определен с помощью различных методов, включая сравнение определяемого количества меченых молекул со стандартной величиной, ранее определенной для конкретной системы.
В данной области должно быть понятно, что размер индивидуума и используемая система визуализации должны определять количество визуализирующейся части, необходимое для получения диагностических изображений. Визуализация опухолей in vivo описана у S.W. Burchiel et al., “Immunopharmacokinetics of Radiolabeled Antibodies and Their Fragments.” (Chapter 13 in Tumor Imaging: The Radiochemical Detection of Cancer, S.W. Burchiel and B. A. Rhodes, eds., Masson Publishing Inc. (1982).
В зависимости от нескольких вариабельных, включая тип используемой метки и способ введения, временной интервал после введения дающий возможность меченым молекулам концентрироваться в местах индивидуума и несвязанным меченым молекулам исчезать до фонового уровня составляет от 6 до 48 часов или от 6 до 24 часов, или от 6 до 12 часов. В другом варианте осуществления временной интервал после введения составляет от 5 до 20 дней или от 5 до 10 дней.
В одном из вариантов осуществления мониторинг заболевания, нарушения или инфекции осуществляется с помощью повторения метода диагностики заболевания, нарушения или инфекции, например, через один месяц после первоначального диагноза, через шесть месяцев после первоначального диагноза, через один год после первоначального диагноза и т.п.
Присутствие меченой молекулы может быть определено у индивидуума с использованием методов, известных в данной области техники для сканирования in vivo. Эти методы зависят от типа используемой метки. Специалисты в данной области техники способны выбрать подходящий метод определения конкретной метки. Методы и устройства, которые могут быть использованы для диагностических методов по изобретению включают, но не ограничиваются этим, компьютерную томографию (CT), сканирование всего организма с помощью, например, позитронно-эмиссионной томографии (PET), магнитно-резонансной визуализации (MRI) и сонографии.
В конкретном варианте осуществления молекулу метят радиоизотопом и определяют у больного с использованием чувствительного к излучению хирургического инструмента (Thurston et al., патент США No. 5441050). В другом варианте осуществления молекулу метят флуоресцентным соединением и определяют у больного с использованием чувствительного к флуоресценции сканирующего прибора. В другом варианте осуществления молекулу метят испускающим позитроны металлом и определяют у больного с использованием позитронно-эмиссионной томографии. В еще одном варианте осуществления молекулу метят парамагнитной меткой и определяют у больного с использованием магнитно-резонансной визуализации.
Имея в данный момент описание изобретения в целом, то же самое должно быть с большей легкостью понято при ссылке на последующие примеры, которые предлагаются с целью иллюстрации и не предназначены для ограничения настоящего изобретения, пока это не описано.
Пример 1
Выделение и характеристика антител против B7-H1 человека
Для того, чтобы выделить высокоаффинные нейтрализующие антитела против B7-H1 человека, мышей сначала иммунизировали и затем делали бустерное введение B7-H1-Fc человека. Спленоциты от животных, позитивных в отношении антител против B7-H1, сливали с клетками миеломы, следуя стандартному протоколу. Проводили скрининг полученных гибридом мыши на те гибридомы, которые экспрессируют моноклональные антитела, иммунореактивные в отношении B7-H1. Антитела дополнительно оценивали для определения того, являются ли они антителами IgG или IgM. Соответственно, B7-H1-Fc или негативный контроль иммобилизовали на твердой подложке. Супернатанты гибридом затем вводили в контакт с подложкой, и определяли присутствие антитела против B7-H1 с использованием меченых антимышиных IgG или антимышиных IgM. На фигуре 1 представлены результаты по тестируемым супернатантам гидридом и показано выделение множественных линий гибридом, которые экспрессируют антитело, иммунореактивное в отношении B7-H1 человека.
Проводили скрининг идентифицированных гибридом для определения того, являются ли экспрессируемые ими антитела нейтрализующими антителами и по существу способны блокировать связывание между B7-H1 и PD-1. PD-1-Fc иммобилизовали на твердой подложке, которую затем инкубировали в присутствии разведенных кондиционных сред, содержащих биотинилированный B7-H1-Fc. Способность B7-H1 и PD-1 связываться друг с другом определяли путем оценки связывания стрептавидина-пероксидазы хрена (SA-HRP) с твердой подложкой. Антитело против B7-H1, способное модулировать связывание B7-H1 с PD-1, таким образом, опосредует снижение связывания SA-HAS в этом тесте. Результаты эксперимента представлены на фигуре 2 и указывают на то, что некоторые из выделенных гибридом экспрессируют нейтрализующие антитела против B7-H1 человека. Антитело MIH-1 (против CD274 человека (B7-H1) (Chen, Y. et al. (Epub 2005 Nov 11) “Expression Of B7-H1 In Inflammatory Renal Tubular Epithelial Cells,” Nephron. Exp. Nephrol. 102(3-4):e81-e92) использовали в качестве положительного контроля. 29E.2AE представляет собой антитело против PD-1, но в этом тесте было показано, что оно не является нейтрализующим. Кондиционные среды от неродственных гибридом (случайное Ab) и векторный контроль (VC) использовали в качестве негативных контролей.
Для того, чтобы определить, способны ли экспрессируемые нейтрализующие антитела связываться с B7-H1, который локализуется на поверхности клетки, проводили анализ связывания клеток. Супернатант от каждого клона гибридомы разводили 1:4 и инкубировали в присутствии родительских клеток CHO и клона линии CHO, который гиперэкспрессирует полноразмерный B7-H1 человека. После осуществления возможности связывания клетки отмывали, и присутствие остаточного связывания антитела против B7-H1 с клетками определяли с использованием флуоресцентно-меченных антимышиных антител IgG. Медиана интенсивности флуоресценции (MFI) для связывания CHO-hB7-H1 показана на фигуре 3. Ни один из тестируемых клонов, как обнаружено, не давал перекрестной реакции с родительской линией CHO, указывая на то, что экспрессируемое антитело было специфичным для B7-H1 человека.
В качестве дополнительной оценки различные концентрации трех клонов сравнивали с антителом MIH-1 против B7-H1. Антитело очищали с использованием протеина G и оценивали на способность связываться с B7-H1 при присутствии на эндогенных уровнях и локализации на поверхности APC. Клетки CHO, которые экспрессировали B7-H1, инкубировали с различными концентрациями (10, 1 или 0,1 мкг/мл) антитела против B7-H1 с последующей инкубацией с APC-конъгированными антителами осла против антител мыши. Связывание определяли путем измерения медианы интенсивности флуоресценции. Результаты показывают, что тестируемые антитела проявляли более высокую авидность в отношении B7-H1, чем контрольное антитело (MIH1) (фигура 4) и что антитело 1E12 проявляло более высокую авидность по отношению к B7-H1, чем контрольное антитело (5H1) против В7-Н1 (Dong, H. et al. (2002) “Tumor-Associated B7-H1 Promotes T-Cell Apoptosis: A Potential Mechanism Of Immune Evasion,” Nature Med. 8(8):793-800) (фигура 5).
В заключение, данные показывают, что получено много гибридом, экспрессирующих антитела против B7-H1 человека. Все клоны продуцируют антитела IgG, которые распознают B7-H1-Fc. Клоны 1B3, 1D11, 1E2, 1E4, 1E10, 2A6, 2E12, 2F2, 2F5, 2F11, 3A4 и 3B1 являются слабыми при скрининге по анализу связывания. Низкий сигнал может быть обусловлен низкими уровнями экспрессии и/или слабой аффинностью.
Существенно, что несколько клонов (например, клоны: 1D5, 1E12, 1F4, 2A7, 2G11, 3B6, 3D10) проявляли сильную нейтрализующую активность. Все они являются нейтрализующими при всех тестируемых концентрациях и, очевидно, хорошо связываются с антигеном. MFI для связывания с клетками CHO-B7-H1 составляет следующее: 1D5 = 50,821; 1E12 = 56,152; 1F4 = 62,015; 2A7 = 49,008; 2G11 = 55,947; 3B6 = 59,638; 3D10 = 53,114.
Пример 2
Выделение и характеристика антител против PD-1 человека
Для того, чтобы выделить высокоаффинные нейтрализующие антитела против PD-1 человека, мышей сначала иммунизировали и затем делали бустерное введение PD-1-Fc человека. Спленоциты от животных, позитивных в отношении антител против PD-1, сливали с клетками миеломы, следуя стандартному протоколу. Проводили скрининг полученных гибридом мыши на те гибридомы, которые экспрессируют высокоаффинные моноклональные антитела, иммунореактивные в отношении PD-1 человека. Антитела дополнительно оценивали для определения того, являются ли они антителами IgG или IgM. Соответственно, PD-1-Fc или негативный контроль (B7-H4-Fc) иммобилизовали на твердой подложке. Супернатанты гибридом затем вводили в контакт с подложкой, и определяли присутствие антитела против PD-1 с использованием меченых антимышиных IgG или антимышиных IgM. На фигуре 6 показано связывание антигена и изотип выделенных антител против PD-1 человека и показано выделение множественных линий гибридом, которые экспрессируют антитело, иммунореактивное в отношении PD-1 человека.
Проводили скрининг идентифицированных гибридом для определения того, являются ли экспрессируемые ими антитела нейтрализующими антителами и по существу способны блокировать связывание между B7-DC и PD-1. B7-DC-Fc, слитый белок, который связывает PD-1, иммобилизовали на твердой подложке, которую затем инкубировали в присутствии разведенных кондиционных сред, содержащих биотинилированный PD-1-Fc. Способность B7-DC и PD-1 связываться друг с другом определяли путем оценки связывания стрептавидина-пероксидазы хрена (SA-HRP) с твердой подложкой. Антитело против PD-1, способное блокировать связывание B7-DC с PD-1, таким образом, опосредует снижение связывания SA-HRP в этом тесте. Результаты эксперимента представлены на фигурах 7A и 7B и указывают на то, что некоторые из выделенных гибридом экспрессируют нейтрализующие антитела против PD-1 человека (на фигуре 7B графически показано то же самое, что и на фигуре 7A, но в другом масштабе).
Для того, чтобы определить способны ли экспрессируемые нейтрализующие антитела связываться с PD-1, который локализуется на клеточной поверхности, проводили анализ связывания клеток. Супернатант от каждого клона гибридомы разводили 1:4 и инкубировали в присутствии родительских клеток CHO и клона линии CHO, который гиперэкспрессирует полноразмерный PD-1 человека. После осуществления возможности связывания клетки отмывали, и присутствие остаточного связывания антитела против PD-1 с клетками определяли с использованием флуоресцентно-меченных антимышиных антител IgG. Медиана интенсивности флуоресценции (MFI) для связывания CHO-hPD-1 показана на фигуре 8. Результаты показывают, что антитела 1E3, 1E8 и 1H3 особенно способны связываться с PD-1, экспрессируемым на клеточной поверхности. Ни один из тестируемых клонов, как обнаружено, не давал перекрестной реакции с родительской линией CHO, указывая на то, что экспрессируемое антитело было специфичным для PD-1 человека. J116 представляет собой имеющееся в продаже контрольное антитело против PD-1 человека (eBioscience, Inc.). Изотипирование выявило, что 1E3, 1E8, 1H3 представляют собой IgG1/каппа.
Два клона (1E3 и 1H3) были выбраны для дополнительного исследования. В качестве дополнительной оценки различные концентрации двух клонов сравнивали с антителами M3 и EH-12 против PD-1. Антитело очищали с использованием протеина G и оценивали на способность связываться с PD-1, экспрессируемым на поверхности клеток CHO. Клетки CHO-hPD1 окрашивали, инкубируя с немеченым антителом против PD-1 (10, 1 и 0,1 мкг/мл) и затем с APC-конъюгированными антителами осла против антител мыши, и представляли медианы интенсивности флуоресценции. Результаты теста представлены на фигуре 9. Негативный контроль представляет собой IgG мыши (mIgG1); позитивными контролями являются M3 (нейтрализующее моноклональное антитело против PD-1 человека (Wu, K. et al. (2009) “Kupffer Cell Suppression of CD8+ T Cells in Human Hepatocellular Carcinoma Is Mediated by B7-H1/Programmed Death-1 Interactions,” Cancer Res 69(20):8067-8075) and anti-PD-1 antibody EH12 (Dorfman, D.M. et al. (2006) “Programmed Death-1 (PD-1) Is A Marker Of Germinal Center-Associated T Cells And Angioimmunoblastic T-Cell Lymphoma,” Am. J. Surg. Pathol. 30(7):802-810).
Конкурентный анализ на основе клеток осуществляли путем инкубации 50000 клеток CHO-hPD1 в течение 30 минут с антителом против PD-1 (присутствующим в диапазоне концентраций 0-0,1 мг/мл). Затем добавляли 10 мкг/мл APC-меченного B7-DC-Fc, и инкубацию продолжали в течение дополнительных 30 минут, после этого измеряли флуоресценцию связанного B7-DC-Fc. Медиана интенсивности флуоресценции представлена на фигуре 10. Конкурентный анализ повторяли при фиксированной концентрации антитела 20 мкг/мл (фигура 11).
В заключение, результаты показывают, что клоны 1E3, 1E8 и 1H3 проявляют нейтрализующую активность и хорошо распознают антиген. Клон 1E6 способен перекрестно связываться с PD-1 без его нейтрализации и тем самым демонстрируя повышенное связывание. Однако этот клон, очевидно, не связывается с PD-1 на клеточной поверхности.
Пример 3
Получение гуманизированных антител: общая методология
Как указано выше, для определенных целей, включая, например, применение in vivo для лечения заболевания человека, предпочтительно, использовать гуманизированное производное описанных выше антител против B7-H1 человека и/или против PD-1 человека.
Для образования таких производных последовательности каркаса антител 3D10 или 1H3 («родительские» последовательности) сначала выравнивали с последовательностями каркаса набора «акцепторных» антител человека для того, чтобы идентифицировать различия в последовательностях каркаса. Гуманизация осуществлялась путем замены остатков каркаса, несовпадающих в родительской и акцепторной последовательностях. Замены в потенциально важных положениях, таких как положения в зоне Верньера, положения в области VH/VL межцепочечного взаимодействия или в определяющих положениях канонического класса CDR, анализировали в плане ожидания обратных мутаций (см. Foote, J. et al. (1992) “Antibody Framework Residues Affecting The Conformation Of The Hypervariable Loops,” J. Molec. Biol. 224:487-499). Идентифицировано всего 14 вариантов гуманизированных последовательностей.
Базу данных по консервативному домену (COD) (Marchler-Bauer, et al. (2011) “COD: A Conserved Domain Database For The Functional Annotation Of Proteins,” Nucleic Acids Res. 39:D225-D229) использовали для определения содержания доменов каждой аминокислотной цепи и аппроксимации границ каждого домена. Были точно определены границы вариабельных доменов наряду с границами областей, определяющих комплементарность (CDR), в соответствии с некоторыми обычно используемыми определениями (Kabat, E.A. et al. (1991) “Sequences of Proteins of Immunological Interest,” Fifth Edition. NIH Publication No. 91-3242; Chothia, C. et al. (1987) “Canonical Structures For The Hypervariable Regions Of Immunoglobulins,” J. Mol. Biol. 196:901-917); Honegger, A. et al. (2001) “Yet Another Numbering Scheme For Immunoglobulin Variable Domains: An Automatic Modeling And Analysis Tool,” J. Molec. Biol. 309(3):657-670; определения CDR по Chothia, C. et al. (1987) “Canonical Structures For The Hypervariable Regions Of Immunoglobulins,” J. Mol. Biol. 196:901-917) будут использоваться ниже в отношении таких гуманизированных последовательностей.
Множественные выравнивания родительской последовательности с последовательностями мыши и зародышевой линии человека создавали с использованием MAFFT (Katoh, K. et al. (2002) “MAFFT: A Novel Method For Rapid Multiple Sequence Alignment Based On Fast Fourier Transform,” Nucleic Acids Res. 30: 3059-3066), и вводы в каждое выравнивание упорядочивали в соответствии с идентичностью последовательностей родительской последовательности. Референсные наборы были редуцированы до уникального набора последовательностей путем кластеризации последовательностей со 100% идентичностью и исключения резервных вводов.
Выбор оптимальных акцепторных каркасов был основан на идентичности последовательностей всех родительских антител акцептору вдоль каркаса обеих цепей; однако положения, которые включают VH/VL межцепочечное взаимодействие, представляют особый интерес. Кроме того, длины петель CDR и положения CDR, ответственные за дискретный набор канонических структур, которые были определены для 5 CDR (Chothia, C. et al. (1987) “Canonical Structures For The Hypervariable Regions Of Immunoglobulins,” J. Mol. Biol. 196:901-917; Martin, A.C. et al. (1996) “Structural Families In Loops Of Homologous Proteins: Automatic Classification, Modelling And Application To Antibodies,” J. Molec. Biol 263:800-815; Al-Laziniki, B. et al. (1997) “Standard Conformations For The Canonical Structures Of Immunoglobulins,” J. Molec. Biol. 273:927-948), сравнивали с зародышевыми линиями для определения того, какие каркасы зародышевых линий имеют такие же остатки в пограничной области и, как известно, поддерживают сходные конформации петель CDR. В таблице 6 и в таблице 7 представлены консервативные положения в пределах взаимодействия VH/VL и положения, которые определяют канонический класс CDR (соответственно), с нумерацией в соответствии с определением Chrothia.
Консервативные положения в пределах взаимодействия VH/VL антитела 1H3
Положения, определяющие канонический класс CDR антитела 1H3
На основе выравнивания последовательностей родительских антител с последовательностями зародышевых линий человека были идентифицированы точные соответствия вводов. Выбор предпочтительной зародышевой линии человека основывался на требуемых критериях: (1) идентичности последовательностей вдоль каркаса; (2) идентичности или совместимости остатков межцепочечного взаимодействия; (3) поддержке петель в канонических конформациях родительских CDR; (4) сочетании зародышевых линий тяжелых и легких цепей, найденных в экспрессируемых антителах; и (5) присутствии сайтов N-гликозилирования, которые должны быть удалены.
Была создана структурная модель области Fv антитела 1H3. Кандидатные структурные фрагменты матрицы каркаса (FR) и областей, определяющих комплементарность (CDR), а также полных Fv были сопоставлены, ранжированы и выбраны из базы данных антител, основанной на идентичности их последовательностей последовательности-мишени, а также на качественных кристаллографических измерениях структуры матрицы, таких как разрешение в ангстремах (Å).
Для того, чтобы провести структурное выравнивание CDR с матрицами FR 5 остатков на каждой стороне CDR включали в матрицу CDR. Выравнивание фрагментов осуществляли на основе перекрывающихся сегментов и выравнивания созданных структурных последовательностей. Фрагменты матриц наряду с выравниванием обрабатывали с помощью MODELLER (Sali, A. et al. (1993) “Comparative Protein Modelling By Satisfaction Of Spatial Restraints,” J. Molec. Biol. 234:779-815). Этот протокол создает конформационные ограничения, возникающие из набора выровненных структурных матриц. Группа структур, которая удовлетворяла ограничениям, была создана с помощью процедур оптимизации с использованием сопряженных градиентов и симулированных закалок. Модельные структуры были выбраны из этой группы на основе показателя энергии, выведенного из показателя структуры белка и удовлетворения конформационным константам. Модели были проверены, и боковые цепи положений, которые отличаются между мишенью и матрицей, были оптимизированы с использованием алгоритма оптимизации боковых цепей и сведения к минимуму энергии. Подходящие средства визуализации и компьютеризации использовали для оценки конформационной вариабельности, локальной упаковки и анализа поверхности CDR для выбора одной или более предпочтительных моделей.
Структурная модель родительского антитела была сконструирована и проверена на недостатки структуры, такие как плохая упаковка атомов, напряженность в длине связей, углы связей или двугранные углы. Эти недостатки структуры могут указывать на потенциальные проблемы в отношении структурной стабильности антитела. По протоколу моделирования проводится поиск для сведения к минимуму таких недостатков. Первоначальная структурная модель гуманизированного Fv содержит все безопасные замены (т.е. замены, которые не должны влиять на связывающую аффинность или стабильность) и осмотрительные замены (т.е. замена положения осуществляется, но положение может быть важным для связывающей аффинностью). Замены в положениях, которые рассматриваются как связанные с риском снижения связывающей аффинности или снижения стабильности, не вносятся. Поиск и выбор матрицы осуществляли отдельно от поиска родительской матрицы для того, чтобы создать хорошую автономную модель, а не вариант, близко подогнанный к родительской модели. После осуществления оценки потенциальных замен модель обновляли для отражения предпочтительных замен и эффекта обратных мутаций.
Пример 4
Получение гуманизированных антител против B7-H1 человека
Для иллюстрации продукции таких гуманизированных производных, гуманизированные производные антитела 3D10 против B7-H1 человека получали в соответствии с описанным выше методом.
Выравнивания последовательностей при сравнении вариабельных доменов легких цепей 3D10 с зародышевыми линиями человека создавали, используя: легкие цепи зародышевых линий IGKV3 (IGKV3-11*01, IGKV3-11*02, IGKV3-NL5*01, IGKV3D-11*01, IGKV3-NL4*01, IGKV3D-7*01, IGKV3D-20*01, IGKV3-20*01, IGKV3-20*02 и IGKV3-15*01), легкие цепи зародышевых линий IGKV1 (IGKV1-9*01, IGKV1-39*01, IGKV1D-13*01, IGKV1-16*01, IGKV1-8*01, IGKV1-13*02, IGKV1-NL1*01, IGKV1D-43*01, IGKV1-27*01 и IGKV1-12*01) и легкие цепи зародышевых линий IGKJ (IGKJ4*01, IGKJ2*02, IGKJ2*01, IGKJ2*04, IGKJ2*03, IGKJ5*01, IGKJ1*01 и IGKJ3*01).
Выравнивания последовательностей при сравнении вариабельных доменов тяжелых цепей 3D10 с зародышевыми линиями человека создавали, используя: тяжелые цепи зародышевых линий IGHV1 (IGHV1-2*02, IGHV1-2*04, IGHV1-f*01, IGHV1-48*01, IGHV1-2*03, IGHV1-2*01, IGHV1-46*02, IGHV1-2*05, IGHV1-3*01 и IGHV1-8*01), тяжелые цепи зародышевых линий IGHV3 (IGHV3-49*04, IGHV3-49*01, IGHV3-49*02, IGHV3-49*03, IGHV3-64*01, IGHV3-64*02, IGHV3-72*01, IGHV3-66*01 и IGHV3-23*01), и тяжелые цепи зародышевых линий IGHJ (IGHJ3*02, IGHJ6*01, IGHJ3*01, IGHJ6*03, IGHJ5*02, IGHJ5*01, IGHJ4*01, IGHJ1*01, IGHJ6*04 и IGHJ2*01).
(A) Гуманизация легкой цепи
На основе указанных выше критериев легкая цепь антитела 3D10, как обнаружено, является наиболее сходной с легкой цепью зародышевой линии IGKV3-11*01 (SEQ ID NO:78):
EIVLTQSPAT LSLSPGERAT LSCRASQSVS SYLAWYQQKP GQAPRLLIYD ASNRATGIPA RFSGSGSGTD FTLTISSLEP EDFAVYYCQQ RSNWP
и IGKV1-9*01 (SEQ ID NO:79):
DIQLTQSPSF LSASVGDRVT ITCRASQGIS SYLAWYQQKP GKAPKLLIYA ASTLQSGVPS RFSGSGSGTE FTLTISSLQP EDFATYYCQQ LNSYP
причем IGKV3-11*01 является предпочтительной, как представлено в таблице 8, (остатки CDR антитела 3D10 показаны курсивом, идентичные выровненные остатки представлены подчеркнутыми):
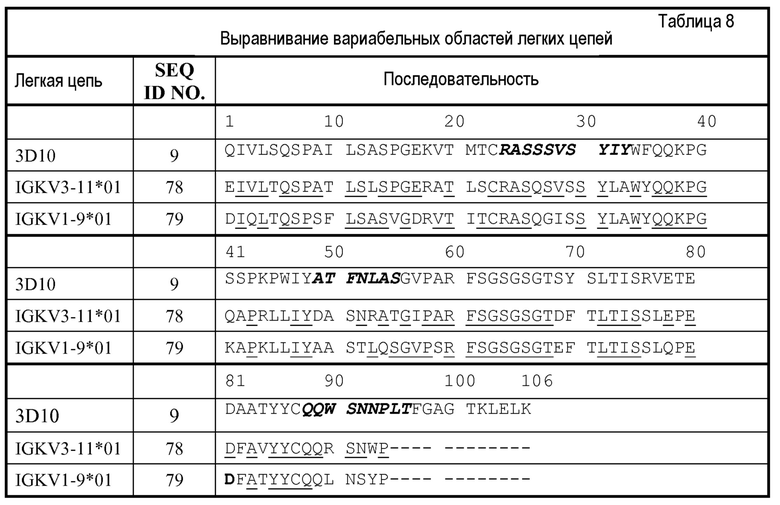
J-сегмент генов сравнивали с родительской последовательностью выше FR4, и J-сегмент IGKJ4*01 (SEQ ID NO:80: LTFGGGTKVEIK) был выбран для легкой цепи.
Как показано выше, легкая цепь антитела 3D10 имеет короткую L1 CDR из десяти остатков, сходную с каноническим классом I петель. Зародышевые линии человека не имеют такой короткой L1 CDR. Каждая из выбранных зародышевых линий, IGKV3-11*01 и IGKV1-9*01, имеет более короткие петли L1, содержащие остатки каркаса, корректные для поддержания этого типа петли. Хорошее сходство последовательностей в целом наблюдается между обоими каркасами акцептора и родительской последовательностью, однако наблюдаются важные различия в пограничных положениях Y33 и P45. Остаток Y33 находится в L1 CDR, и в то время как два выбранных семейства акцепторов не содержат тирозина в этом положении, последовательности других зародышевых линий содержат его. Различие между каркасами акцепторов и родительской последовательностью в положении P45 является следствием того, что родительская зародышевая линия обладает FR2 не сходным с любой зародышевой линией человека. В результате изменения в этой области предполагались независимо от выбранного каркаса акцептора, так что IGKV3-11*01 и IGKV1-9*01 были лучшими в качестве каркасов акцепторов.
Были созданы три гуманизированных цепи для каждого из двух предпочтительных каркасов акцептора, IGKV3-11*01 и IGKV1-9*01. Три LC1 цепи происходят от IGKV3-11*01; три LC2 цепи происходят от IGKV1-9*01. Первая гуманизированная цепь для каждого каркаса акцептора содержала все гуманизирующие замены, считающиеся возможными, и была наиболее гуманизированной из трех цепей. Вторая гуманизированная цепь для каждого каркаса акцептора содержала несколько обратных мутаций в положениях, которые изменяют заряд, потенциально вмешиваются в упаковку остова или могут влиять на конформацию CDR. Третья цепь для каждого из каркасов акцептора содержала большинство обратных мутаций, включая замены, которые изменяют заряд и могут потенциально изменять связывающую аффинность. Последовательности этих шести гуманизированных цепей представлены ниже.
Последовательности этих шести гуманизированных цепей представлены в таблице 9 с различиями относительно легкой цепи родительского антитела 3D10, показанными жирным шрифтом и подчеркнутыми.
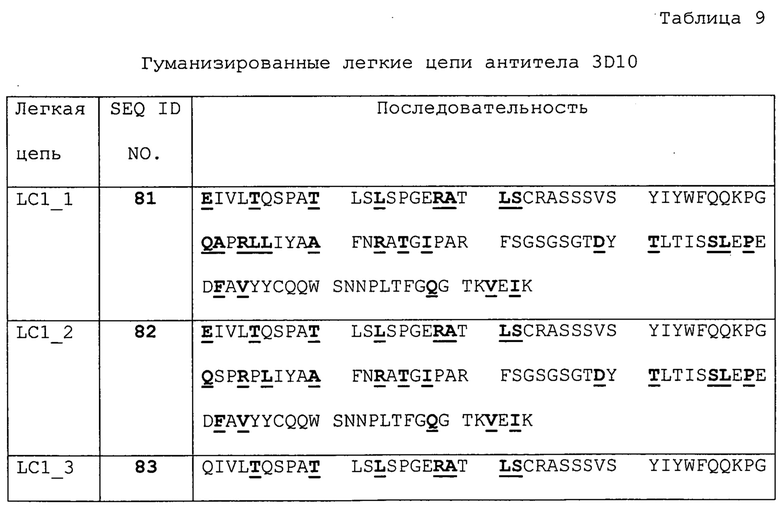
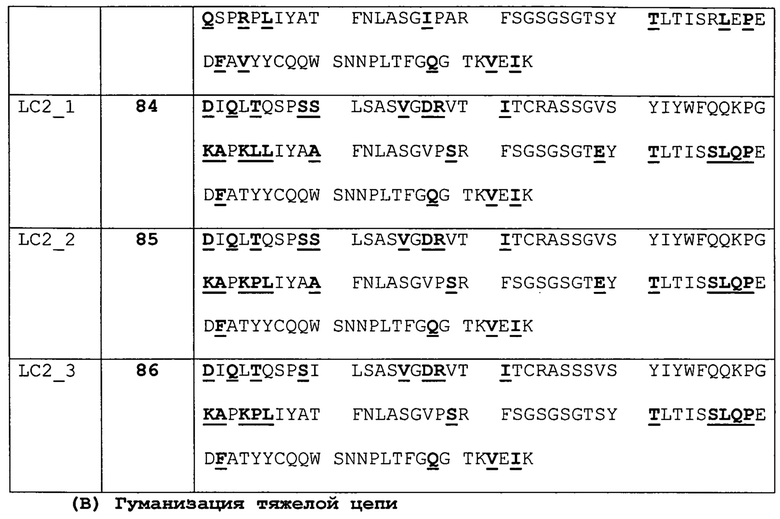
В свете обсуждаемых выше критериев два кандидатных каркаса акцептора зародышевых линий: IGHV1-2*02 (зародышевая линия IGHV1) и IGHV3-49*04 (зародышевая линия IGHV3) были выбраны для гуманизации тяжелой цепи антитела 3D10.
Каркас акцептора IGHV 1-2*02 был выбран из-за сходства его последовательности с родительской последовательностью и того факта, что он содержал очень сходные остатки, вовлеченные в упаковку остова домена. Каркас акцептора IGHV3-49*04 был выбран после рассмотрения и отказа от других последовательностей зародышевых линий, сходных с IGHV 1-2*02. В качестве каркаса акцептора IGHV3-49*04 несколько больше отличается от родительской последовательности, чем IGHV 1-2*02, так как требуется большее количество замен, однако каркас акцептора этой зародышевой линии может поддерживать CDR родительского антитела. Последовательности каркаса акцептора IGHV 1-2*02 и каркаса акцептора IGHV3-49*04 представлены ниже:
Каркас акцептора IGHV 1-2*02 (SEQ ID NO:87):
QVQLVQSGAE VKKPGASVKV SCKASGYTFT GYYMHWVRQA PGQGLEWMGW INPNSGGTNY AQKFQGRVTM TRDTSISTAY MELSRLRSDD TAVYYCAR
Каркас акцептора IGHV3-49*04 (SEQ ID NO:88):
EVQLVESGGG LWQPGRSLRL SCTASGFTFG DYAMSWVRQA PGKGLEWVGF IRSKAYGGTT EYAASVKGRF TISRDDSKSI AYLQMNSLKT EDTAVYYCTR
В таблице 10 представлено выравнивание этих последовательностей с тяжелой цепью антитела 3D10 (остатки CDR антитела 3D10 показаны курсивом, идентичные выровненные остатки представлены подчеркнутыми):
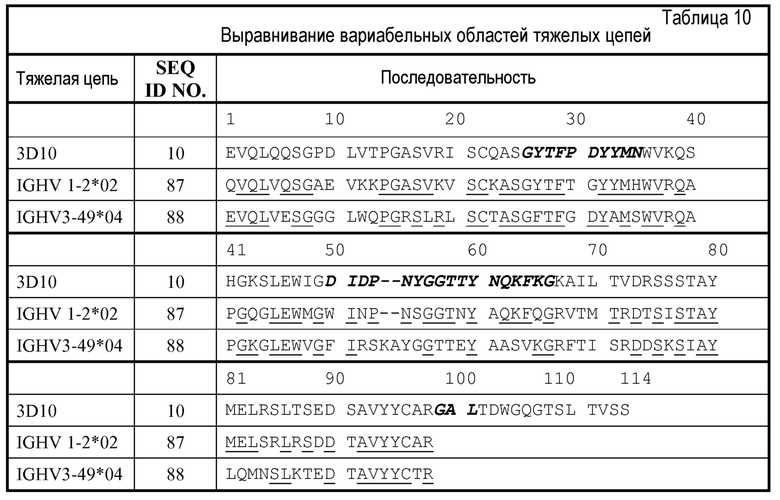
J-сегмент генов сравнивали с родительской последовательностью выше FR4, и J-сегмент IGHJ3*02 (SEQ ID NO:89: DAFDIWGQGTMVTVSS) был выбран для тяжелой цепи.
Были созданы три гуманизированных цепи для каждого из двух предпочтительных каркасов акцептора, IGHV1-2*02 и IGHV3-49*04. Три HC1 цепи происходят от IGHV1-2*02; три HC2 цепи происходят от IGHV3-49*04). Первая гуманизированная цепь для каждого каркаса акцептора содержала все гуманизирующие замены, считающиеся возможными, и была наиболее гуманизированной из трех цепей. Вторая гуманизированная цепь для каждого каркаса акцептора содержала несколько обратных мутаций в положениях, которые изменяют заряд, потенциально вмешиваются в упаковку остова или могут влиять на конформацию CDR. Третья цепь для каждого из каркасов акцептора содержала большинство обратных мутаций, включая замены, которые изменяют заряд и могут потенциально изменять связывающую аффинность. Последовательности этих шести гуманизированных цепей представлены ниже:
DYYMNWVRQA PGKGLEWVGD IDPNYGGTTY AASVKGRFTI SVDRSKSIAY LQMSSLKTED TAVYYCTRGA LTDWGQGTMV TVSS
Последовательности этих шести гуманизированных цепей представлены в таблице 11 с различиями относительно тяжелой цепи родительского антитела 3D10, показанными жирным шрифтом и подчеркнутыми.
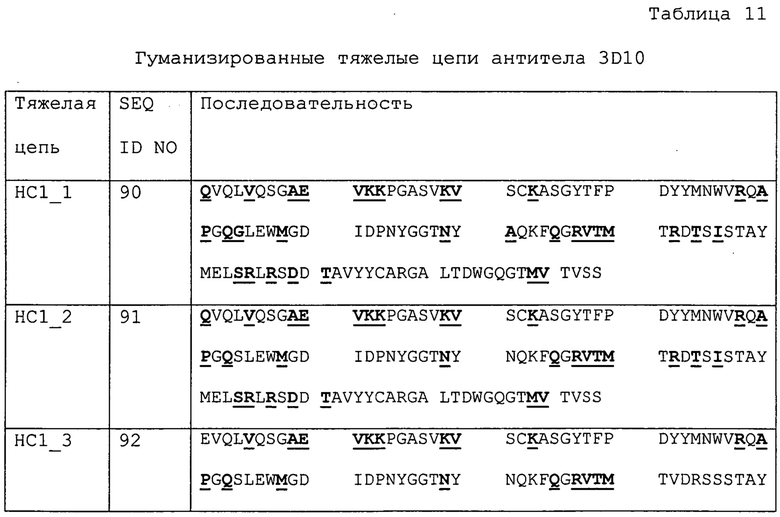

Поиск подтвердил, что существовали антитела с сочетанием зародышевых линий, сходных со спариванием IGKV3-11*01 с IGHV1-2*02. Образованная пара отмечена как акцептор 1. Дополнительно найдены антитела с сочетанием зародышевых линий, сходных со спаренными IGKV 1-9*01 и IGHV3-49*04. Образованная пара отмечена как акцептор 2.
Сочетания описанных выше гуманизированных легких и тяжелых цепей привело к созданию 14 вариантов гуманизированных антител, чьи последовательности описаны в таблице 12.
Гуманизированные антитела 3D10
Пример 5
Получение гуманизированных антител против PD-1 человека
Для иллюстрации продукции таких гуманизированных производных, гуманизированные производные антитела 1H3 против PD-1 человека получали в соответствии с описанным выше методом (химерное антитело, содержащее вариабельную область 1H3 и область Fc IgG1 человека, использовали в качестве родительского антитела).
Выравнивания последовательностей при сравнении вариабельных доменов легких цепей 1H3 с зародышевыми линиями человека создавали, используя: легкие цепи зародышевых линий IGKV3 (IGKV3-11*01, IGKV3-11*02, IGKV3D-11*01, IGKV3D-20*01, IGKV3-NL4*01, IGKV3D-7*01, IGKV3-20*01, IGKV3-NL5*01, IGKV3-15*01, IGKV3-NL1*01, IGKV3-20*01, IGKV3-NL2*01, IGKV3-NL3*01), легкие цепи зародышевых линий IGKV1 (IGKV1-9*01, IGKV1D-43*01, IGKV1-39*01, IGKV1D-13*02, IGKV1-8*01, IGKV1D-13*01, IGKV1-12*01, IGKV1D-16*01, IGKV1-5*01 и IGKV1-NL1*01) и легкие цепи зародышевых линий IGKJ (IGKJ2*02, IGKJ2*01, IGKJ2*04, IGKJ2*03, IGKJ5*01, IGKJ4*01, IGKJ3*01 и IGKJ1*01).
Выравнивания последовательностей при сравнении вариабельных доменов тяжелых цепей 1H3 с зародышевыми линиями человека создавали, используя: тяжелые цепи зародышевых линий IGHV3 (IGHV3-48*01, IGHV3-48*02, IGHV3-48*03, IGHV3-11*01, IGHV3-21*01, IGHV3-11*03, IGHV3-30*03, IGHV3-9*01, IGHV3-7*01 и IGHV3-30*10), тяжелые цепи зародышевых линий IGHV1 (IGHV1-3*01, IGHV1-69*08, IGHV1-69*11, IGHV1-46*01, IGHV1-69*05, IGHV1-69*06, IGHV1-69*01, IGHV1-46*02, IGHV1-69*02 и IGHV1-69*10), и тяжелые цепи зародышевых линий IGHJ (IGHJ6*01, IGHJ6*03, IGHJ4*01, IGHJ6*04, IGHJ5*02, IGHJ3*02, IGHJ5*01, IGHJ3*01, IGHJ2*01 и IGHJ1*01).
На основе общей идентичности последовательностей, спаривания пограничных положений и сходства канонических положений классических CDR было идентифицировано два семейства зародышевых линий для каждой легкой и тяжелой цепи в качестве возможных каркасов акцептора (IGKV3 и IGKV1 для легкой цепи и 1GHV3 и IGHV1 для тяжелой цепи). Антитело 1H3, как обнаружено, является наиболее сходным с легкой цепью зародышевой линии 1GKV3-11*01 и тяжелой цепью IGHV3-48*01. На основе общей идентичности последовательностей, спаривания пограничных положений и сходства канонических положений классических CDR было идентифицировано два семейства зародышевых линий для каждой легкой и тяжелой цепи в качестве возможных каркасов акцептора (IGKV3 и IGKV1 для легкой цепи и 1GHV3 и IGHV1 для тяжелой цепи).
(A) Гуманизация легкой цепи
Антитело 1H3 имеет короткую L1 CDR из 10 остатков и входит в канонический класс I. Ни одна зародышевая линия человека не имеет такой короткой L1 CDR, но наиболее близкие зародышевые линии в каждом выбранном семействе (IGKV3-II*01 и IGKV1-9*01 имеют короткие L1 петли и содержат корректные остатки каркаса для поддержания L1 петель класса I. Общее сходство последовательностей было хорошим для обоих каркасов акцептора; однако наблюдались различия в двух пограничных положениях (Y33 и P45). Y33 находится в L1 CDR, и в то время как два семейства акцепторов не содержат тирозина в этом положении, другие семейства потенциальных акцепторов содержат его. Различие в P45 было связано с количеством других различий в FR2 родительской легкой цепи. Вкратце, родительская зародышевая линия принадлежит к семейству зародышевых линий мыши с FR2, не сходным с любой зародышевой линией человека.
Зародышевые линии 1GKV3-11 *01 и IGKV 1-9*01 были выбраны в качестве каркасов легких цепей акцептора. Последовательности легких цепей 1GKV3-11 *01 и IGKV 1-9*01 представлены как SEQ ID NO:78 и SEQ ID NO:79, соответственно. Выравнивания этих последовательностей с легкой цепью антитела 1H3 представлено в таблице 13, (остатки CDR антитела 1H3 показаны курсивом, идентичные выровненные остатки представлены подчеркнутыми):
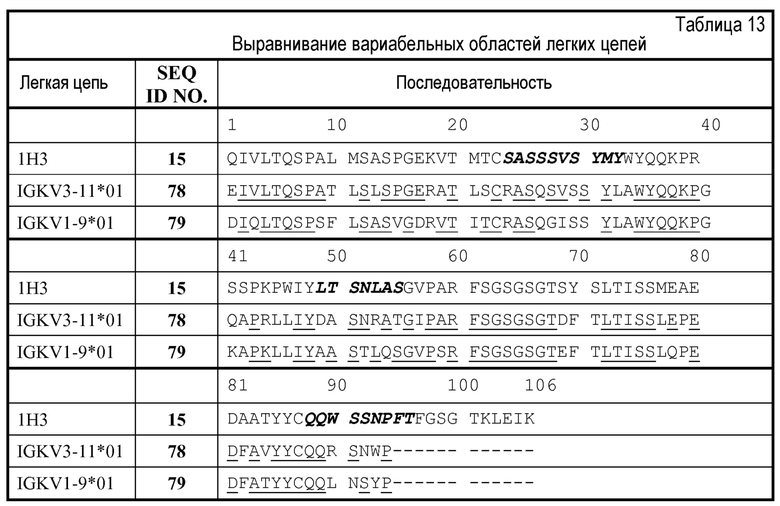
J-сегмент генов сравнивали с родительской последовательностью выше FR4, и J-сегмент IGKJ2*02 (SEQ ID NO:96: CTFGQGTKLEIK) был выбран для легкой цепи.
Было создано две гуманизированных цепи для каждого из двух предпочтительных каркасов акцептора, IGKV3-11*01 и IGKV1-9*01. Две LC1 цепи происходят от IGKV3-11*01; две LC2 цепи происходят от IGKV1-9*01. Первая гуманизированная цепь для каждого каркаса акцептора содержала все гуманизирующие замены, считающиеся возможными, и была наиболее гуманизированной из трех цепей. Вторая гуманизированная цепь для каждого каркаса акцептора содержала несколько обратных мутаций в положениях, которые изменяют заряд, потенциально вмешиваются в упаковку остова или могут влиять на конформацию CDR. Последовательности этих четырех гуманизированных цепей представлены ниже:
Последовательности этих четырех гуманизированных цепей представлены в таблице 14 с различиями относительно легкой цепи родительского антитела 1H3 Var1, показанными жирным шрифтом и подчеркнутыми.


В свете обсуждаемых выше критериев два кандидатных каркаса акцептора зародышевых линий: IGHV3-48*01 (зародышевая линия IGHV3) и IGHV1-3*01 (зародышевая линия IGHV1) были выбраны для гуманизации тяжелой цепи антитела 1H3 Var1.
Каркас акцептора IGHV3-48*01 был выбран в качестве главного каркаса тяжелой цепи акцептора из-за общего сходства последовательностей. Пограничные остатки спарены за исключением H35, положения, в котором допускается некоторая вариабельность, и он содержит корректные остатки для определения канонического класса H1 CDR. H2 CDR не попадает в какой-либо канонический класс последовательностей из-за тирозина в положении 56. Выбор каркаса второй тяжелой цепи акцептора был сделан после удаления любой зародышевой линии, близкородственной IGHV3-48*01. В результате наиболее близкой зародышевой линией стала IGHV1-3*01. Количество различий, принимаемых во внимание, становится более высоким, так как упаковка нижнего остова отличается, однако зародышевая линия должна поддерживать CDR и функционировать в качестве подходящего каркаса акцептора. Было отмечено, что родительская последовательность содержит цистеин в консервативном положении 60, где все зародышевые линии человека содержат тирозин. Последовательности каркаса акцептора 1GHV3-48*01 и каркаса акцептора IGHV1-3*01 представлены ниже:
Каркас акцептора 1GHV3-48*01 (SEQ ID NO:101):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS SYSMNWVRQA PGKGLEWVSY ISSSSSTIYY ADSVKGRFTI SRDNAKNSLY LQMNSLRAED TAVYYCAR
Каркас акцептора IGHV1-3*01 (SEQ ID NO:102):
QVQLVQSGAE VKKPGASVKV SCKASGYTFT SYAMHWVRQA PGQRLEWMGW INAGNGNTKY SQKFQGRVTI TRDTSASTAY MELSSLRSED TAVYYCAR
В таблице 15 представлено выравнивание этих последовательностей с тяжелой цепью антитела 1H3 (остатки CDR антитела 1H3 показаны курсивом, идентичные выровненные остатки представлены подчеркнутыми):


J-сегмент генов сравнивали с родительской последовательностью выше FR4, и J-сегмент IGHJ6*01 (SEQ ID NO:103: WGQGTTVTV) был выбран для тяжелой цепи.
Было создано три гуманизированных цепи для каждого из двух предпочтительных каркасов акцептора, IGHV3-48*01 и IGHV1-3*01. Три HC1 цепи происходят от IGHV3-48*01; три HC2 цепи происходят от IGHV1-3*01). Первая гуманизированная цепь для каждого каркаса акцептора содержала все гуманизирующие замены, считающиеся возможными, и была наиболее гуманизированной из трех цепей. Вторая гуманизированная цепь для каждого каркаса акцептора содержала несколько обратных мутаций в положениях, которые изменяют заряд, потенциально вмешиваются в упаковку остова или могут влиять на конформацию CDR. Третья цепь для каждого из каркасов акцептора содержала большинство обратных мутаций, включая замены, которые изменяют заряд и могут потенциально изменять связывающую аффинность. Последовательности этих шести гуманизированных цепей представлены ниже:
DYGMHWVRQA PGKGLEWVAY ISSGSYTIYS ADSVKGRFTI SRDNAKNTLY LQMSSLRAED TAVYYCARRG YGSFYEYYFD YWGQGTTVTV SS
Последовательности этих шести гуманизированных цепей представлены в таблице 16 с различиями относительно тяжелой цепи родительского антитела 1H3, показанными жирным шрифтом и подчеркнутыми.
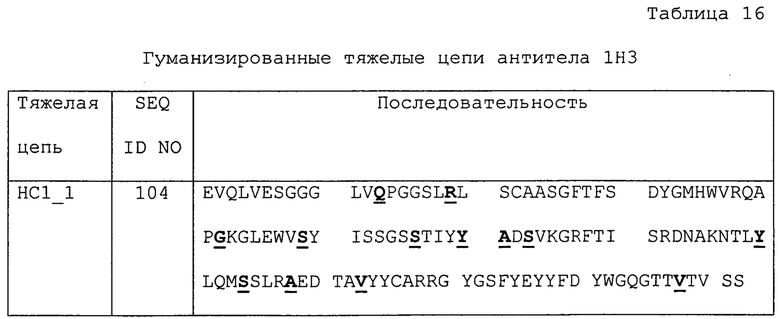
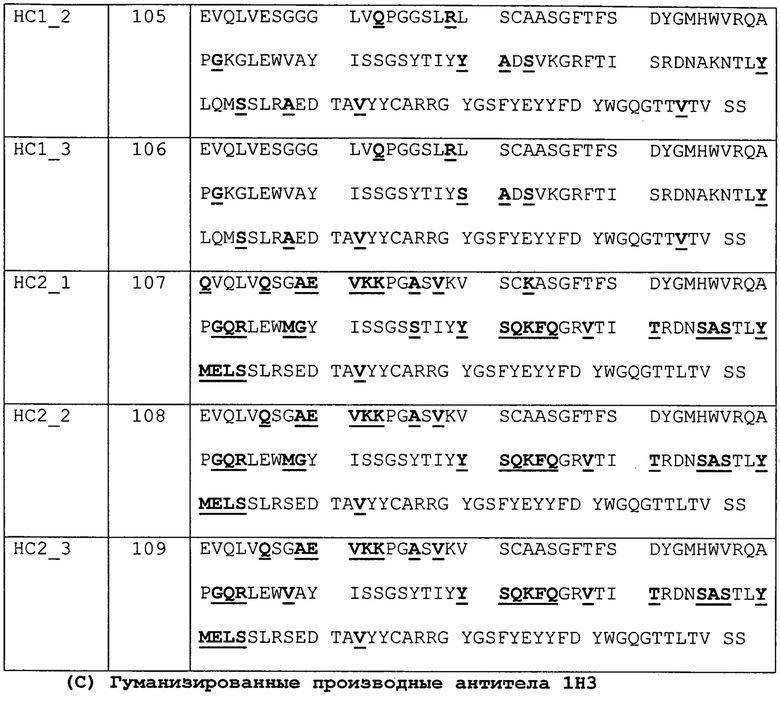
Поиск подтвердил, что существуют антитела с сочетанием зародышевых линий, сходных со спариванием IGKV3-11*01 с IGHV3-48*01. Образованная пара отмечена как акцептор 1. Впоследствии найдены антитела со сходным спариванием IGKVI-9*01 с IGHVI-3*01. Образованная пара отмечена как акцептор 2.
Сочетания описанных выше гуманизированных легких и тяжелых цепей привело к созданию 14 вариантов гуманизированных антител, чьи последовательности описаны в таблице 17.
Гуманизированные антитела 1H3
Пример 6
Характеристика антитела 1H3 против PD-1 человека
Для того? чтобы оценить свойства антител против PD-1 по настоящему изобретению получали конструкт, который обладал химерными («ch») мышиными областями Fab антитела 1H3 против PD-1 человека и областью Fc IgG1 человека (конструкт 1H3). Конструкт тестировали на его способность связываться с PD-1 человека.
На фигуре 12 представлены результаты эксперимента, в котором клетки CHO, трансфицированные полноразмерным PD-1 человека, предварительно инкубировали с насыщающей дозой mAb против PD-1 перед окраской меченными биотином антителами против hB7-H1-Fc или против mFc hB7-DC. На фигуре 12 показано, что конструкт мышиного моноклонального антитела, который обладает химерными («ch») областями Fab мыши антитела 1H3 против PD-1 человека и областью Fc IgG1 человека, блокирует связывание B7-H1-Fc и B7-DC-Fc с клетками, экспрессирующими PD-1 человека, в результате связывания антитела с клетками. M1, m3 и 1E8 все представляют собой антитела против PD-1 позитивного контроля. На невосстанавливающем геле конструкт мигрирует в виде одной полосы с М.м. приблизительно 200; эта полоса заменяется полосой с М.м. приблизительно 52 в восстанавливающих условиях.
Конструкт 1H3 имел аффинность с KD 2,19 нМ, Ka «скорости ассоциации» 0,734×10-5/Мсек, и Kd «скорости диссоциации» 1,61×10-4/сек. EC50 конструкта, как обнаружено, составляла 75 нг. На фигуре 13 сравнивается связывание, полученное с этим конструктом, со связыванием имеющегося в продаже антитела EH12 против PD-1. Конструкт, как показано, способен полностью блокировать способность hPD-1 (экспрессируемого клетками CHO) связываться с B7-H1-Fc или B7-DC-Fc, как показано на фигуре 14. На фигуре 15 показано, что химерный конструкт 1H3 способен связываться с hPD-1-Fc и блокировать связывание такого hPD-1-Fc с hB7-H1 экспрессируемым клетками CHO.
Способность конструкта 1H3 связываться с основными Т-клетками человека оценивали относительно контрольного антитела (паливизумаба; SYNAGIS®, Medimmune, Inc.); 1H3 продемонстрировал повышенное связывание с клетками как CD8, так и CD4 (фигуры 16A-16B).
Для иллюстрации функциональных характеристик антител по настоящему изобретению оценивали способность конструкта 1H3 и конструктов химерных антител, имеющих области FAB антитела 1H3 («конструкт 1H3»), 1E3 («конструкт 1E3»), и антитела 3D10 («конструкт 3D10») повышать активность Т-клеток. Незрелые дендритные клетки (DC) экспонировали с TNFα и PGE2 в течение двух дней (клетки инкубировали в присутствии 50 мкг/мл токсоида столбняка (TT) в течение ночи на второй день созревания). Полученные в результате клетки, как обнаружено, стали зрелыми дендритными клетками как определено по их приобретенной способности экспрессировать B7-H1 и B7-DC. Зрелые DC клетки затем инкубировали в течение двух недель в присутствии меченных карбоксифлуоресцеинсукцинимидиловым эфиром (CFSE) аутологичных Т-клеток и 100 нг/мл TT, и описанных выше конструктов антител. Как показано на фигуре 17, антитела по настоящему изобретению были способны блокировать взаимодействия B7-H1-PD-1 при измерении по экспансии антигенспецифических Т-клеток памяти. На 7 день проводили анализ цитокинов, присутствующих в супернатантах клеток (таблица 18).
Как показано в таблице 18, оба конструкта антител против hPD-1 стимулировали ответы как Th1, так и Th2. Очень низкое количество IL-1β, IL-2, IL-4, IL-7, IL-10 и G-CSF было выявлено в супернатантах. Все mAb имели очень низкие уровни эндотоксина, менее 0,01 EU/мг; кроме того, DC и Т-клетки содержались в бессывороточных средах. Внутриклеточное окрашивание клеток на IFNγ на 7 день выявило, что тогда как только 0,15% контрольных клеток было IFN-γ+, 1,9% клеток, которые были проинкубированы с 1H3 конструктом, 0,91% клеток, которые были проинкубированы с 1E3 конструктом, и 3,2% клеток, которые были проинкубированы с 3D10 конструктом, было IFN-γ+.
Таким образом, суммируя, 1H3 конструкт, как обнаружено, опосредует приблизительно 7-кратное повышение пролиферации Т-клеток и приблизительно 12-кратное увеличение продукции IFN-γ на клетку. Кумулятивный эффект такого действия заключается приблизительно в 100-кратном увеличении секреции IFNγ.
В качестве дополнительной характеристики DC, происходящим из моноцитов, давали созревать путем инкубации с TNFα и PGE2. Клетки затем пульсировали в течение 2 часов в присутствии пула пептидов CEF, ограниченного смешанными классом I и классом II (т.е. пептидов цитомегаловируса, вируса Эпштейна-Барра и вируса гриппа), и инкубировали с меченными CFSE аутологичными Т-клетками (колонка LD, 95% чистота) в течение двух недель. Обработанные клетки затем инкубировали в течение двух недель в присутствии меченных CFSE аутологичных Т-клеток (колонка LD, 95% чистота) и описанных выше конструктов антител. На 7 день процент разведенных CFSE Т-клеток, как было найдено, составлял 40% для клеток, проинкубированных с контрольным антителом, 37% для клеток, проинкубированных с конструктом 1H3, 50% для клеток, проинкубированных с конструктом 3D10, и 57% для клеток, проинкубированных с антителом CA-18C3 (моноклональное антитело, специфичное против IL-1α). На 11 день опять оценивали процент разведенных CFSE Т-клеток. Процент разведенных CFSE Т-клеток, как затем было найдено, составлял 17% для клеток, проинкубированных с контрольным антителом, и 38% для клеток, проинкубированных с конструктом 1H3. Процент разведенных CFSE Т-клеток, как затем было найдено, составлял 27% для клеток, проинкубированных с антителом CA-18C3.
Супернатанты обработанных клеток также анализировали на цитокины IL-2 и IFN-γ на 7 день (таблица 19).
Супернатанты обработанных клеток также анализировали на цитокины IFN-γ, TNFα и GM-CSF на 11 день. Конструкт 1H3, как обнаружено, опосредует увеличение всех трех цитокинов относительно контрольного антитела (таблица 20).
В качестве дополнительной функциональной характеристики DC, происходящим из моноцитов (полученным от позитивного по HLA-A2 донора PBMC), давали созревать путем экспозиции с TNFα и PGE2, и зрелые DC затем пульсировали с ограниченными HLA-A2 пептидами MART-1 и Flu M1 в течение 2 часов. После чего клетки инкубировали в течение двух недель в присутствии меченных CFSE аутологичных Т-клеток (колонка LD, 95% чистота) и описанного выше конструкта 1H3 или конструкта 3D10. Влияние пептидов MART-1 и M1 на продукцию цитокинов показано в таблице 21.
Пример 7
Характеристика гуманизированных антител против PD-1
Описанные выше антитела 1H3_var 1-1H3_var 14 (см. таблицу 17) оценивали для подтверждения их способности связываться с PD-1 человека и их терапевтических возможностей. Полипептиды, кодирующие антитела, экспрессировали в клетках CHO и титры функциональных антител определяли с помощью ELISA (таблица 22).
Для уверенности в том, что связывание было специфичным для PD-1 человека, связывание оценивали с использованием клеток CHO, которые были трансфицированы для экспрессии PD-1 человека. Результаты таких экспериментов по связыванию представлены на фигурах 18A-18D. С помощью повторения таких экспериментов по связыванию в присутствии 1, 3, 10, 30, 100, 300, 1000 или 3000 нг варианта h1H3 было обнаружено, что способность таких гуманизированных антител связываться с PD-1 во всех случаях зависела от концентрации антитела.
Для того, чтобы продемонстрировать способность гуманизированных антител против PD-1 по настоящему изобретению блокировать взаимодействия между PD-1 и его природными лигандами, экспрессирующие PD-1 клетки HEK293 инкубировали в присутствии B7-H1 (или B7-DC) и выбранных вариантов h1H3. Варианты антител h1H3, как обнаружено, были способны блокировать связывание B7-H1 с клетками HEK293 (фигура 19A (B7-H1); фигура 19B (B7-DC) (Ctl = синагис, WT = химерное 1H3).
В таблице 23 представлены результаты, полученные при процедуре очистки и временной экспрессии в масштабе 500 мл. Невосстанавливающие гели показали, что экспрессируемые антитела преимущественно мигрируют в виде одной полосы приблизительно 160 кДа; при анализе в восстанавливающем геле эта полоса заменялась полосами приблизительно в 60 кДа и 30 кДа. Результаты показывают, что h1H3 варианты акцептора 1: h1H3 Var1, h1H3 Var3, h1H3 Var4 и h1H3 Var6 характеризовались хорошим связыванием с PD-1 человека, тогда как h1H3 варианты акцептора 2: h1H3 Var7 - h1H3 Var14 характеризовались плохим связыванием с PD-1 человека. Соответственно, тяжелую и легкую цепи из h1H3 вариантов акцептора 1: h1H3 Var1 - h1H3 Var6, клонировали в вектор двойных генов (DGV; Lonza Biologics, Berkshire, UK; Bebbington, C.R. et al. (1992) “High-Level Expression Of A Recombinant Antibody From Myeloma Cells Using A Glutamine Synthetase Gene As An Amplifiable Selectable Marker,” Biotechnology (NY) 10(2):169-175) и трансфицировали в клетки CHO для возможности получения стабильных клеточных линий, продуцирующих антитела.
Анализ связывания при концентрации антитела 0,5, 1,5, 5, 15, 50, 150 и 500 нг/мл, а также 1,5, 5, 15 и 50 мкг/мл выявил, что связывание зависело от концентрации. Определяли специфическую связывающую способность вариантов в отношении PD-1 при концентрациях антитела в диапазоне от 0 до приблизительно 350 нМ. На фигуре 20 представлены полученные кривые для h1H3 Var1 - h1H3 Var6, которые указывают, что эти антитела связываются с PD-1. Антитела h1H3 Var1 и h1H3 Var4 проявляют пониженное связывание относительно родительского антитела. Напротив, h1H3 Var 2, h1H3 Var 3, h1H3 Var 5 и h1H3 Var 6 характеризуются связыванием, которое сравнимо со связыванием родительского антитела. Данные по EC50 этих антител представлены в таблице 23.
Все публикации и патенты, указанные в настоящем описании, включены в настоящий документ в качестве ссылки в той же степени как если бы каждая индивидуальная публикация или патентная заявка была бы специально и индивидуально указана как включенная в качестве ссылки в полном объеме. Несмотря на то, что изобретение описано в связи с конкретными вариантами его осуществления, должно быть понятно, что возможны дополнительные его модификации, и эта заявка рассматривается как покрывающая любые варианты, способы применения или адаптаций изобретения, следуя в целом принципам изобретения и включая такие отступления от настоящего раскрытия, как подпадающие под известную или обычную практику в той области техники, к которой принадлежит изобретение, и как возможные для применения к существенным признакам, изложенным в настоящем описании ранее.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА | 2016 |
|
RU2731202C2 |
| АНТИТЕЛА К H7CR | 2013 |
|
RU2691428C2 |
| АНТИТЕЛА К H7CR | 2013 |
|
RU2650756C2 |
| АНТИТЕЛА ПРОТИВ SIGLEC-15 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2759334C2 |
| СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ ПРОТИВ В7-Н1 | 2010 |
|
RU2571204C2 |
| ЛЕЧЕНИЕ РАКА КОМБИНАЦИЕЙ АНТАГОНИСТА PD-1 И ДИНАЦИКЛИБА | 2014 |
|
RU2705795C2 |
| Антитело против PD-1 и его применение | 2017 |
|
RU2739610C1 |
| СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ ПРОТИВ В7-Н1 | 2010 |
|
RU2706200C2 |
| АНТИ-PD-L1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2783685C2 |
| АНТИТЕЛА К БЕЛКУ ПРОГРАММИРУЕМОЙ КЛЕТОЧНОЙ СМЕРТИ 1 | 2018 |
|
RU2812915C2 |


















































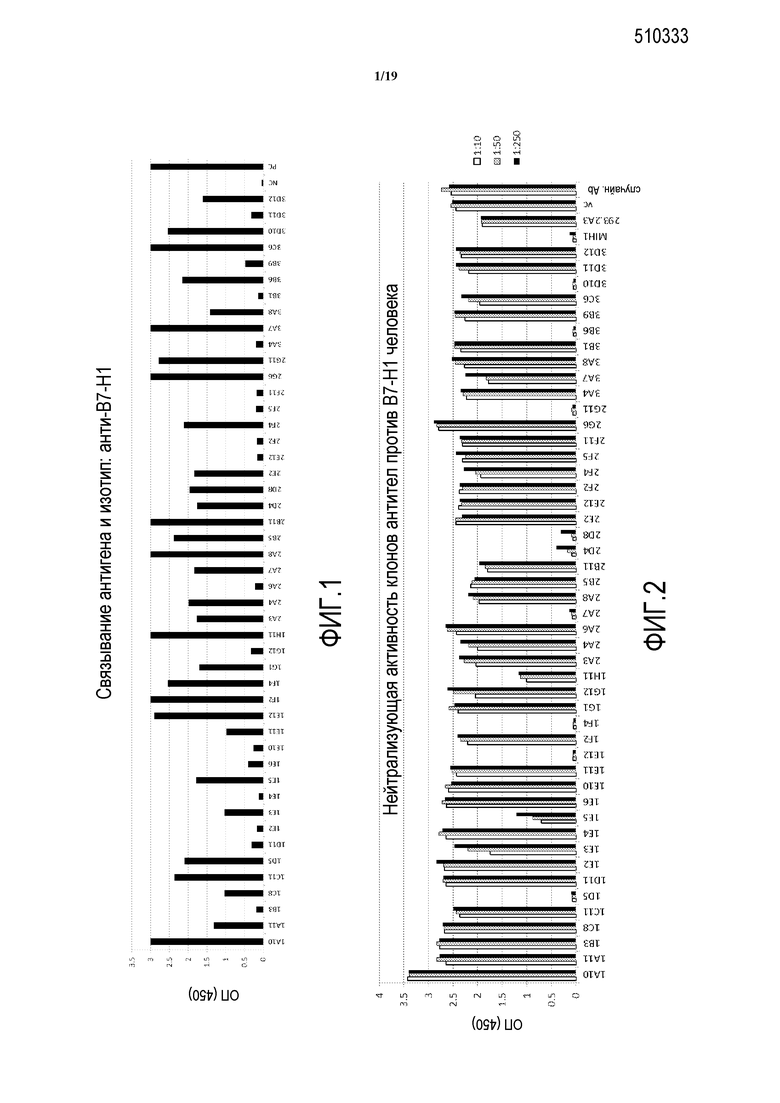
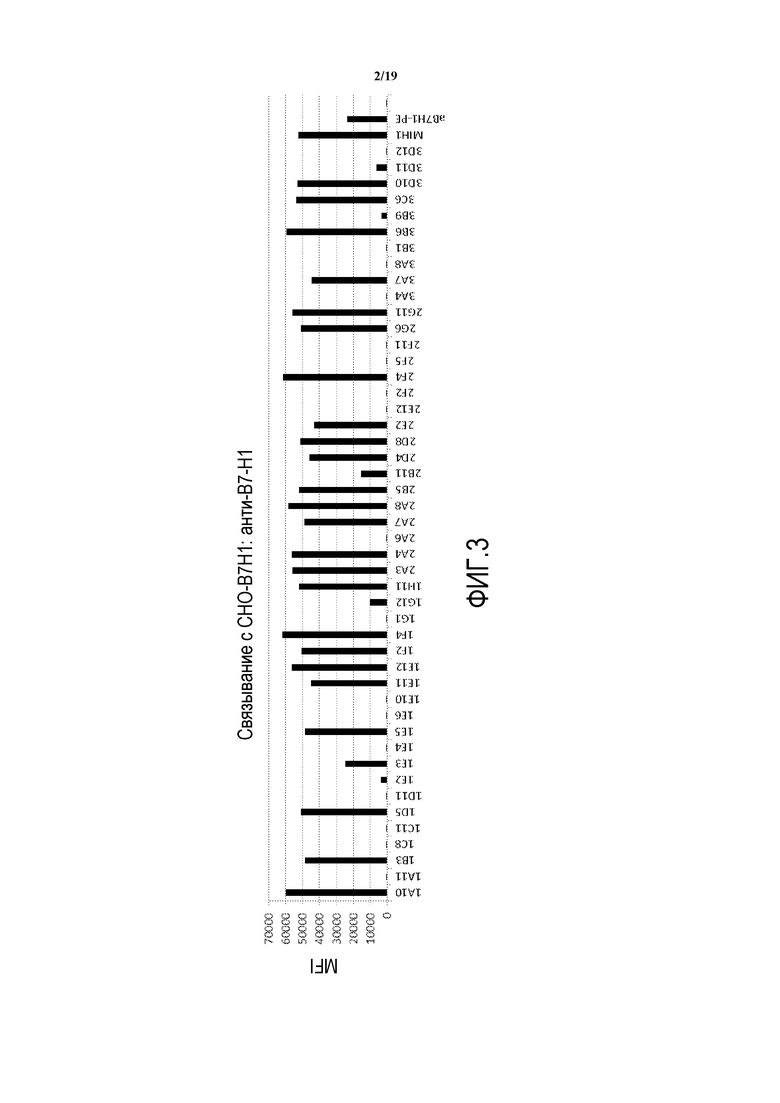
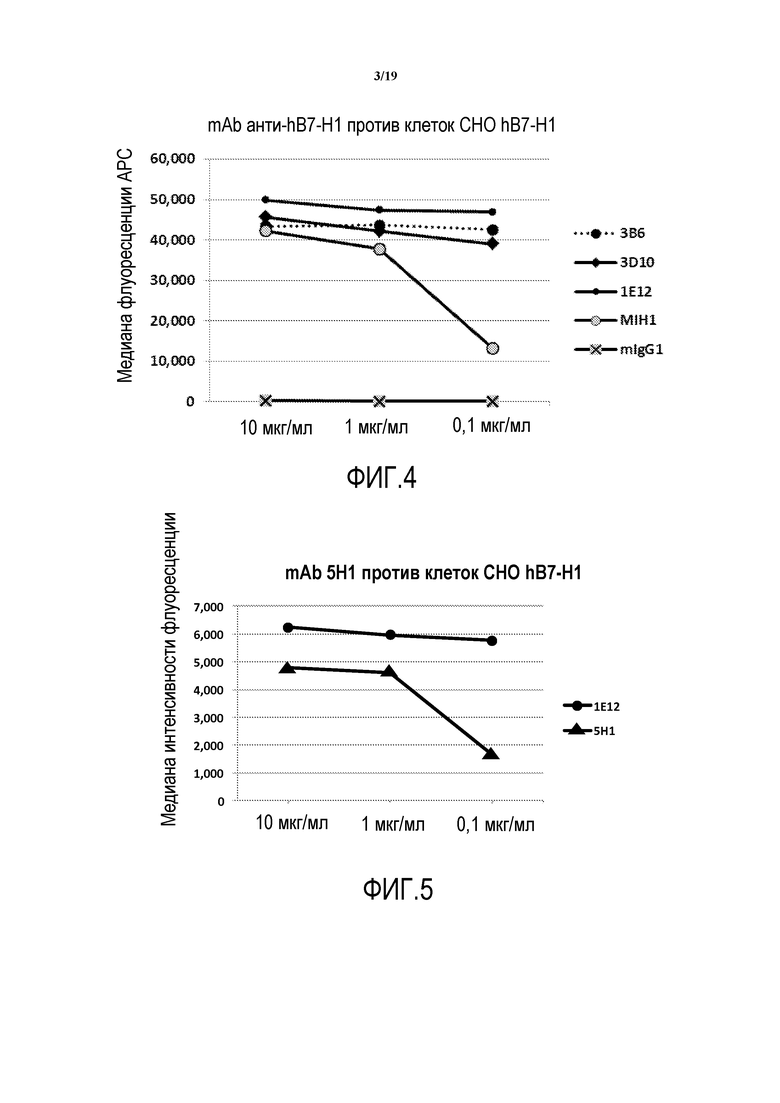
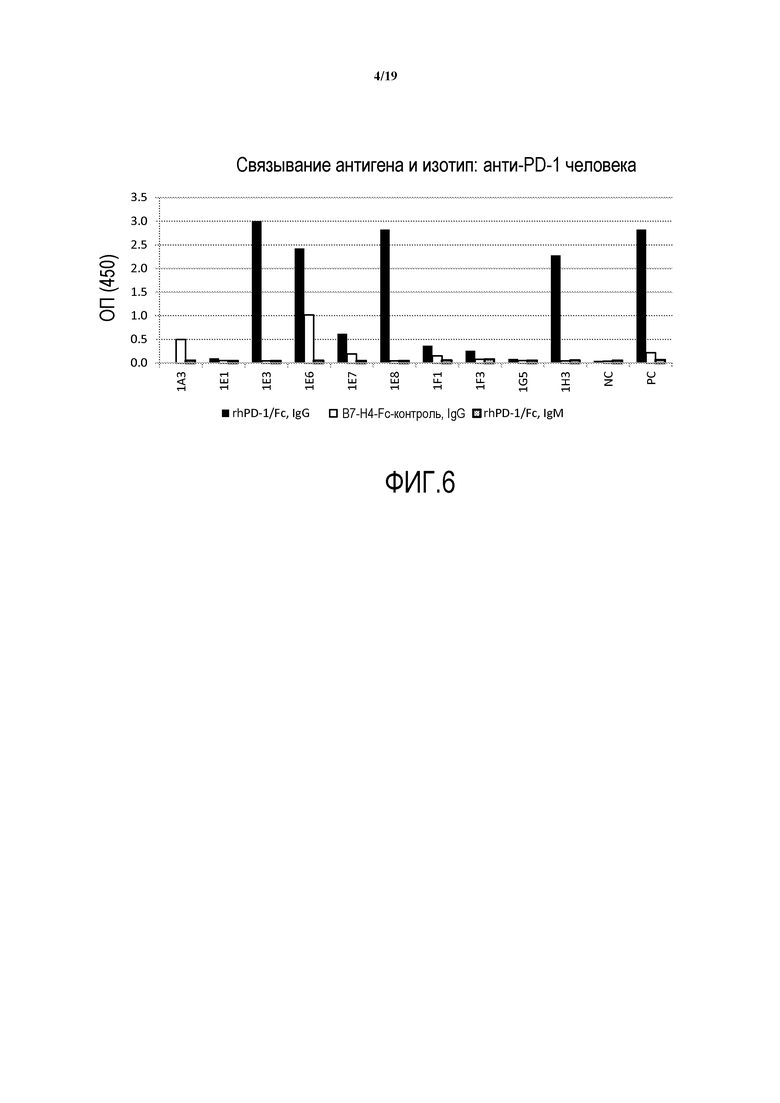
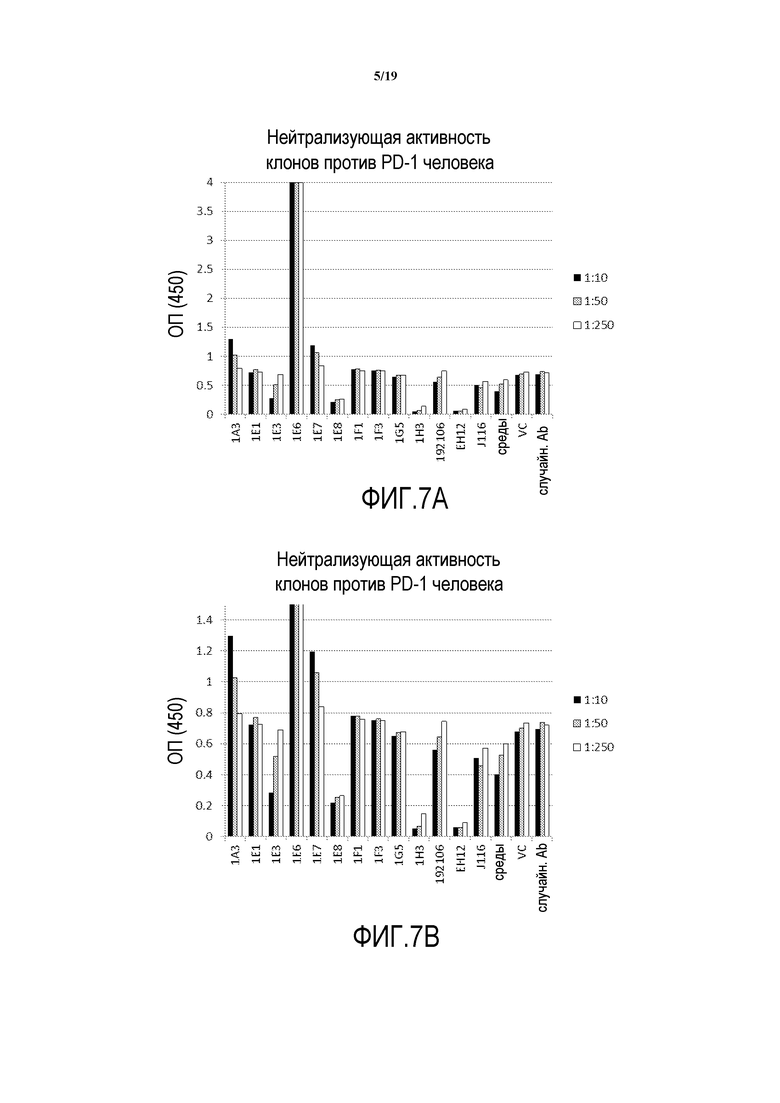


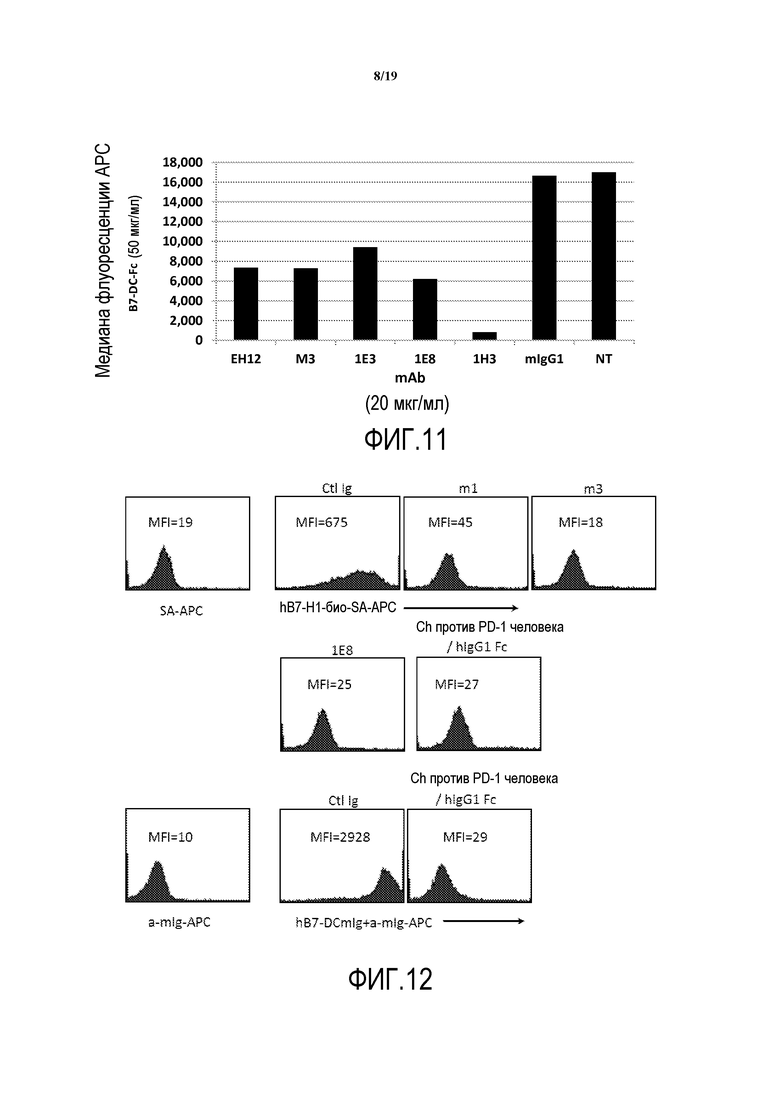
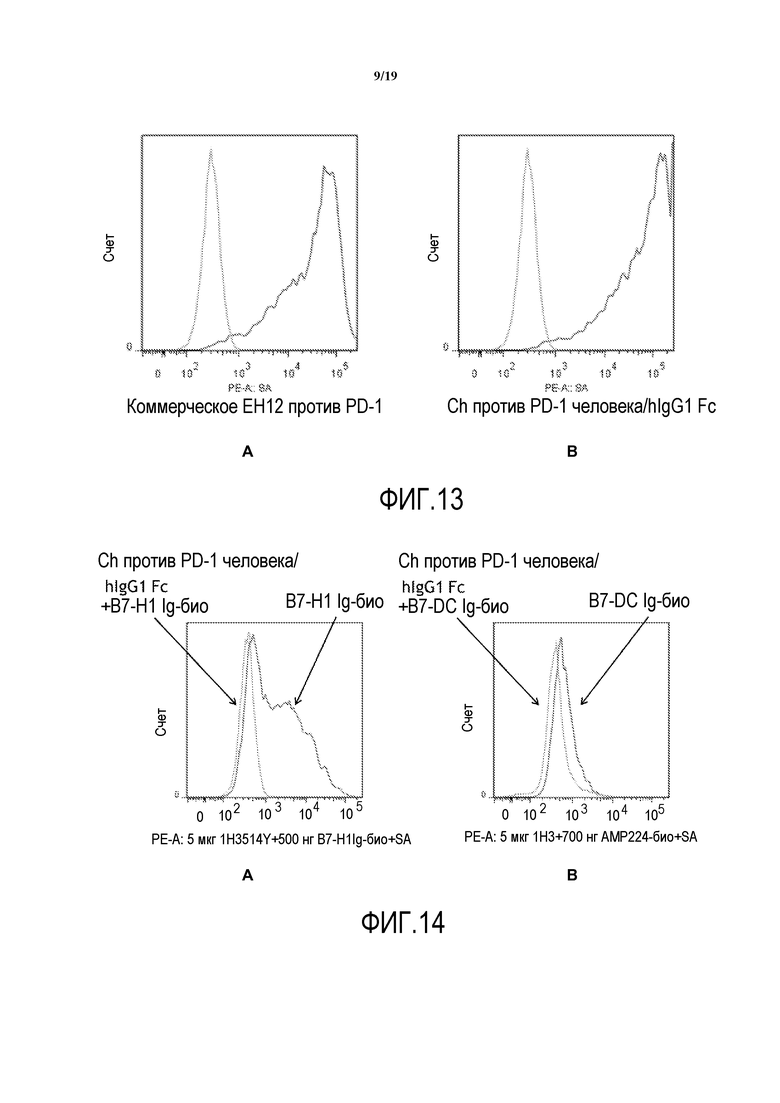
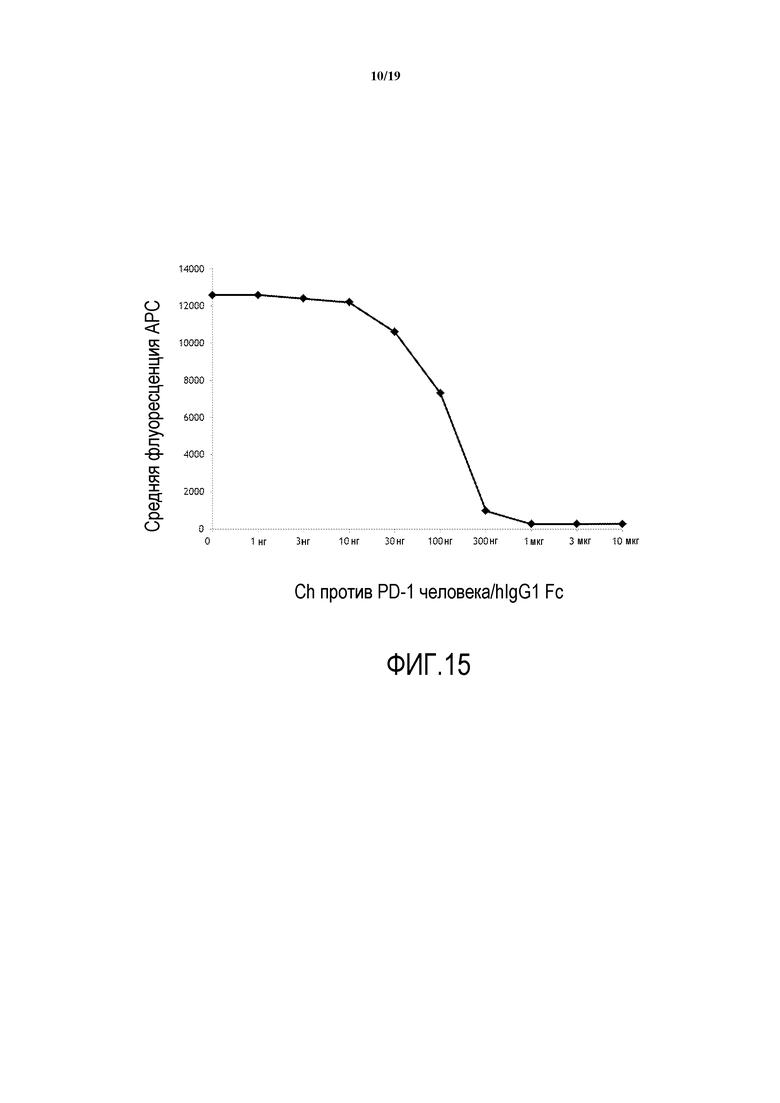
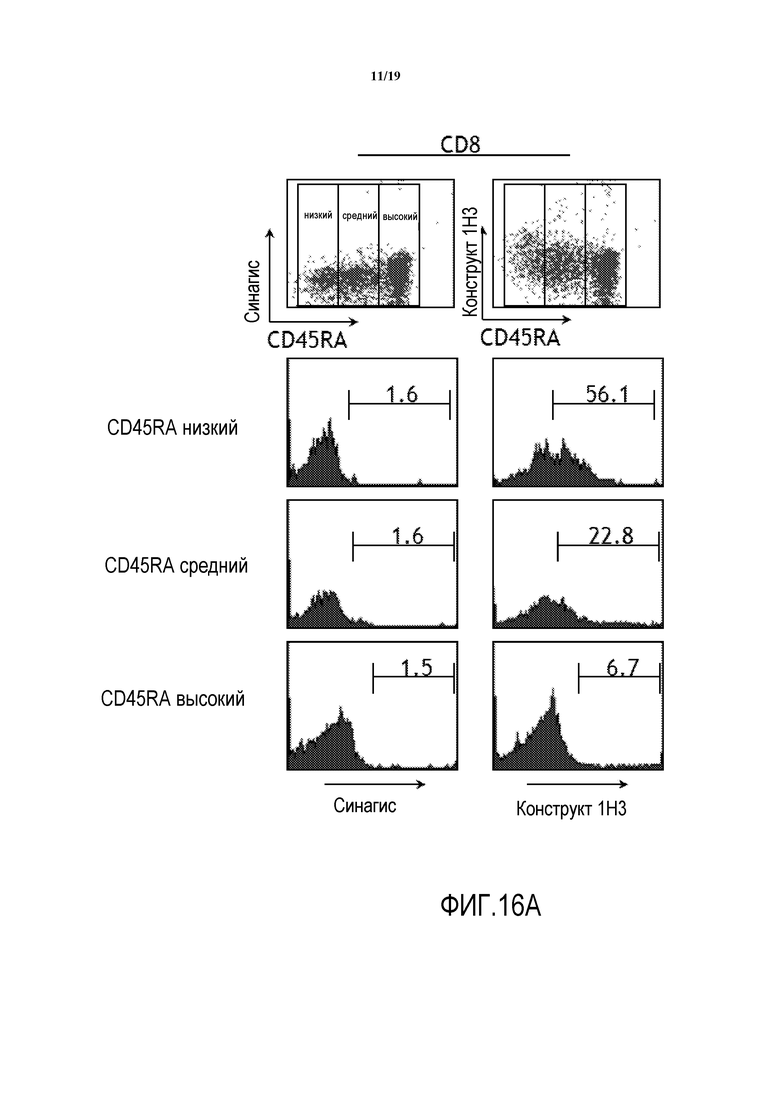
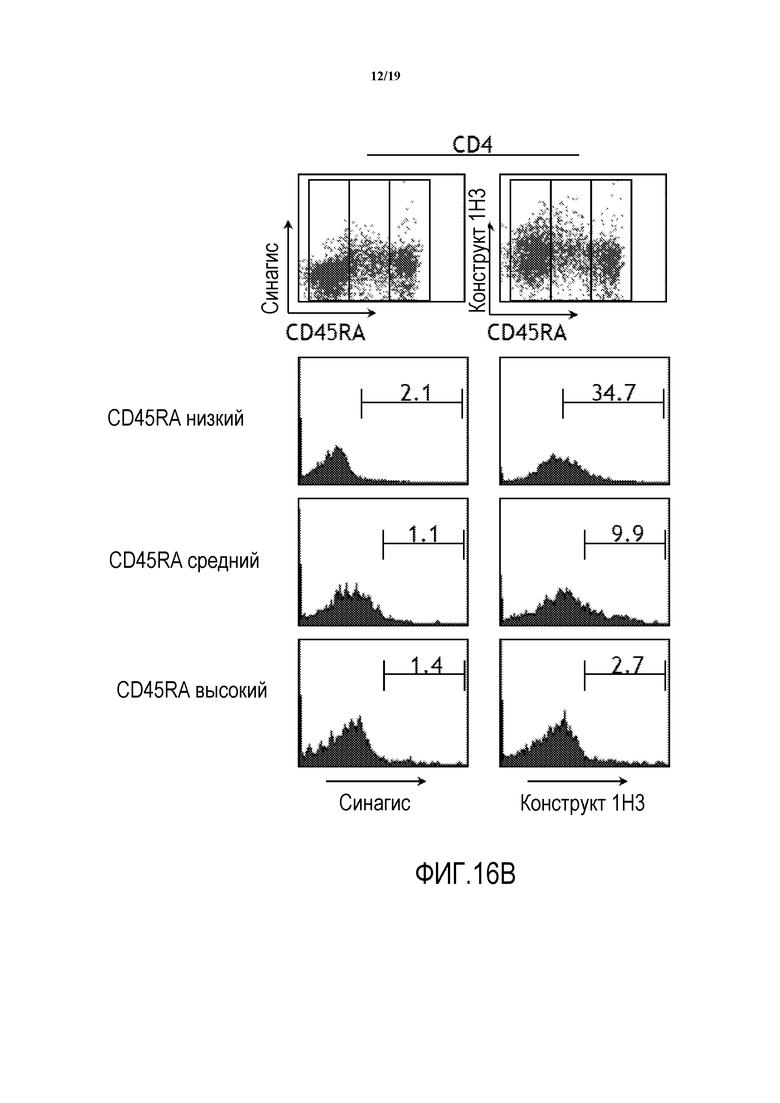
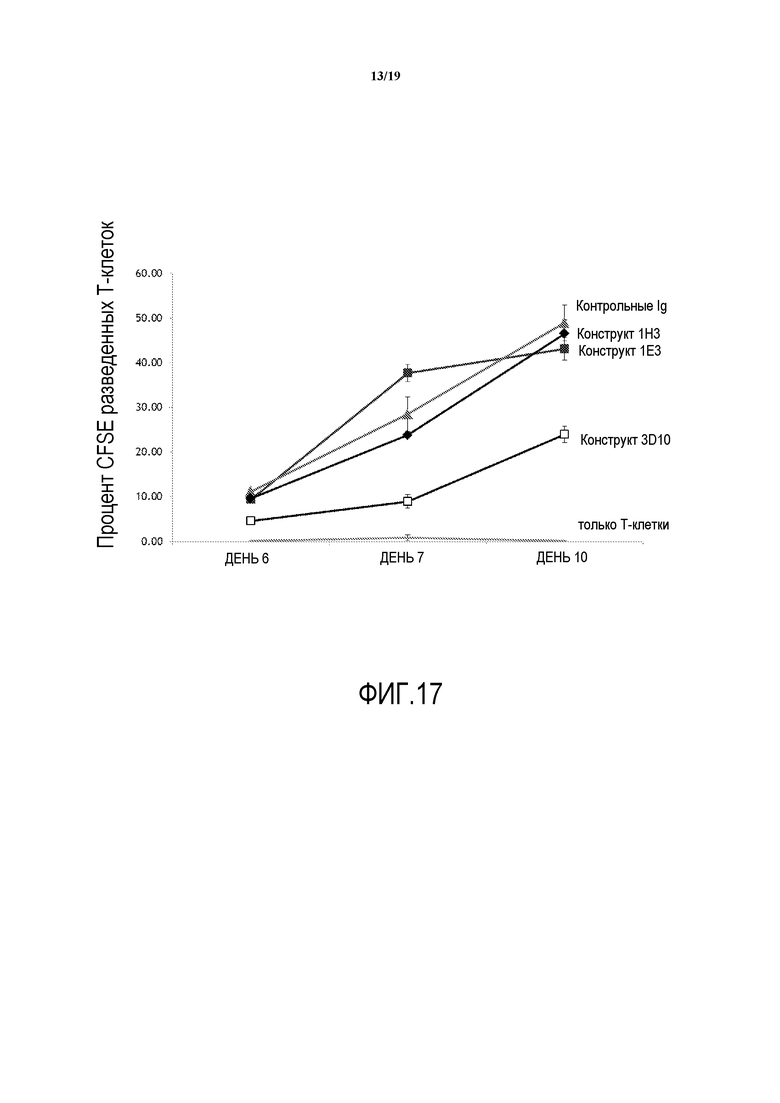
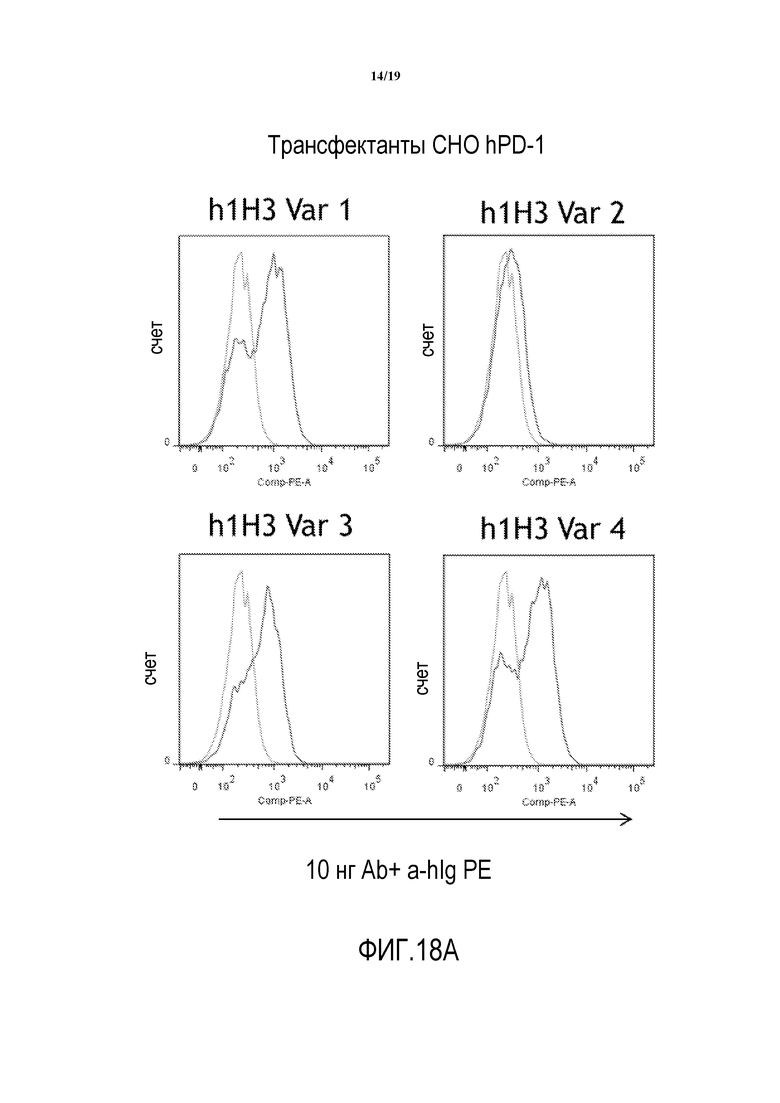

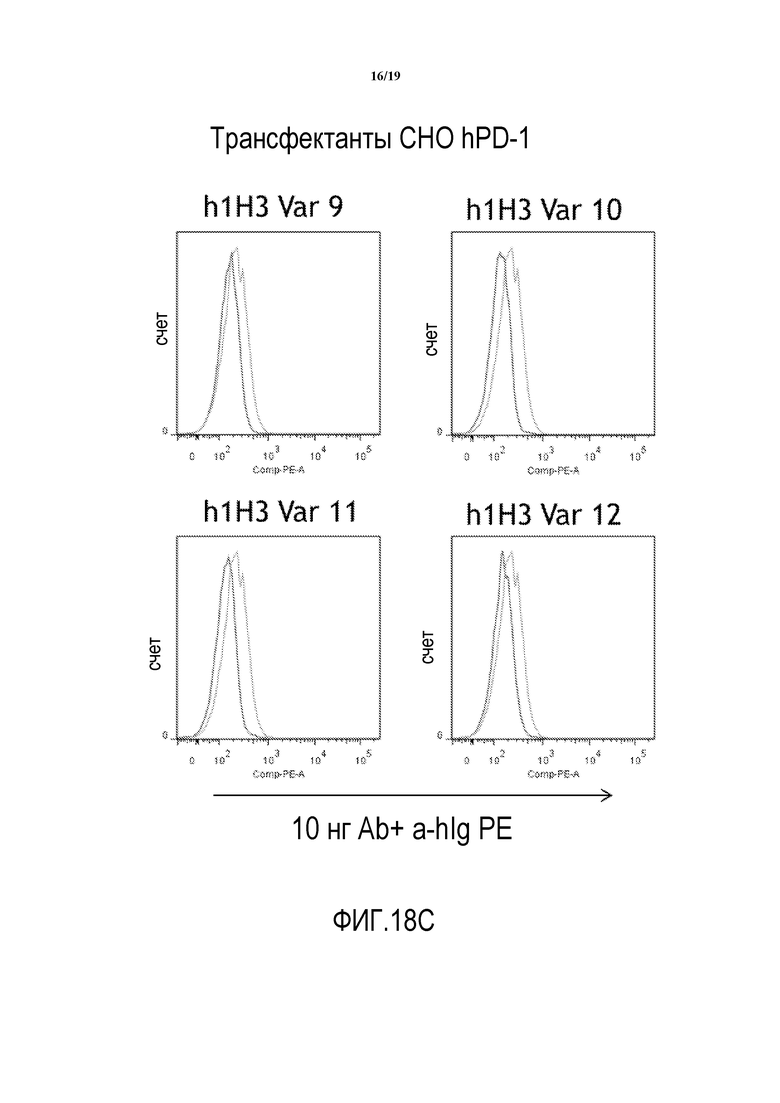
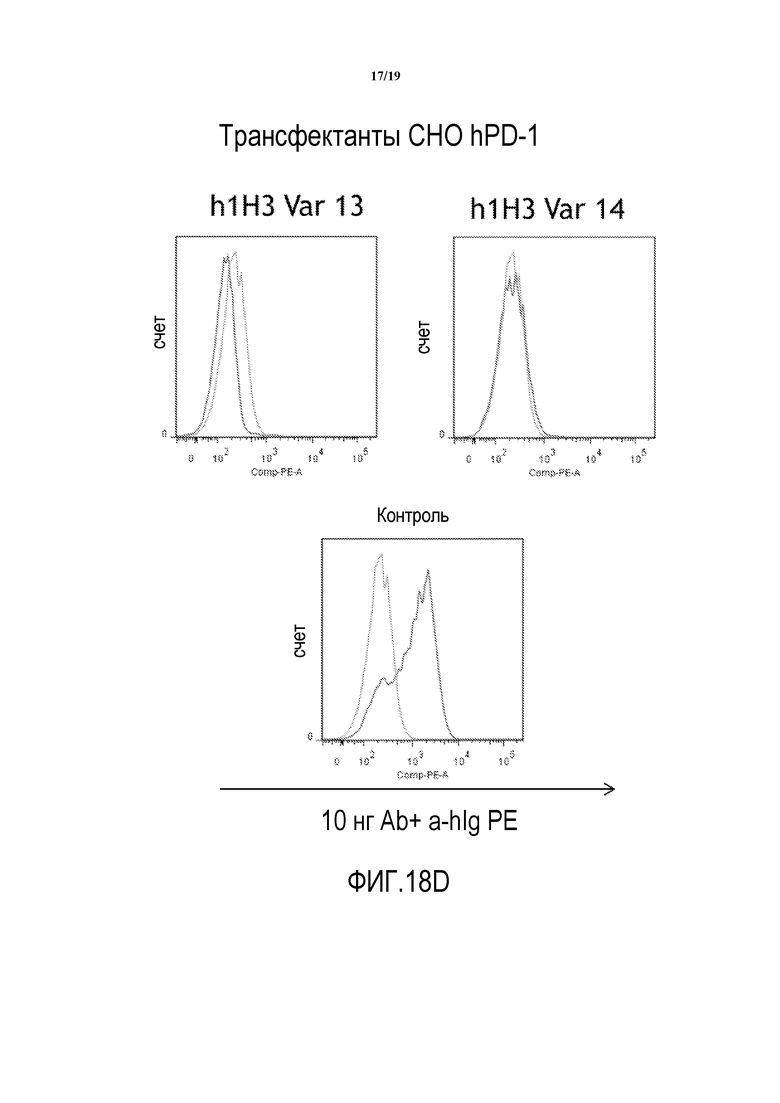
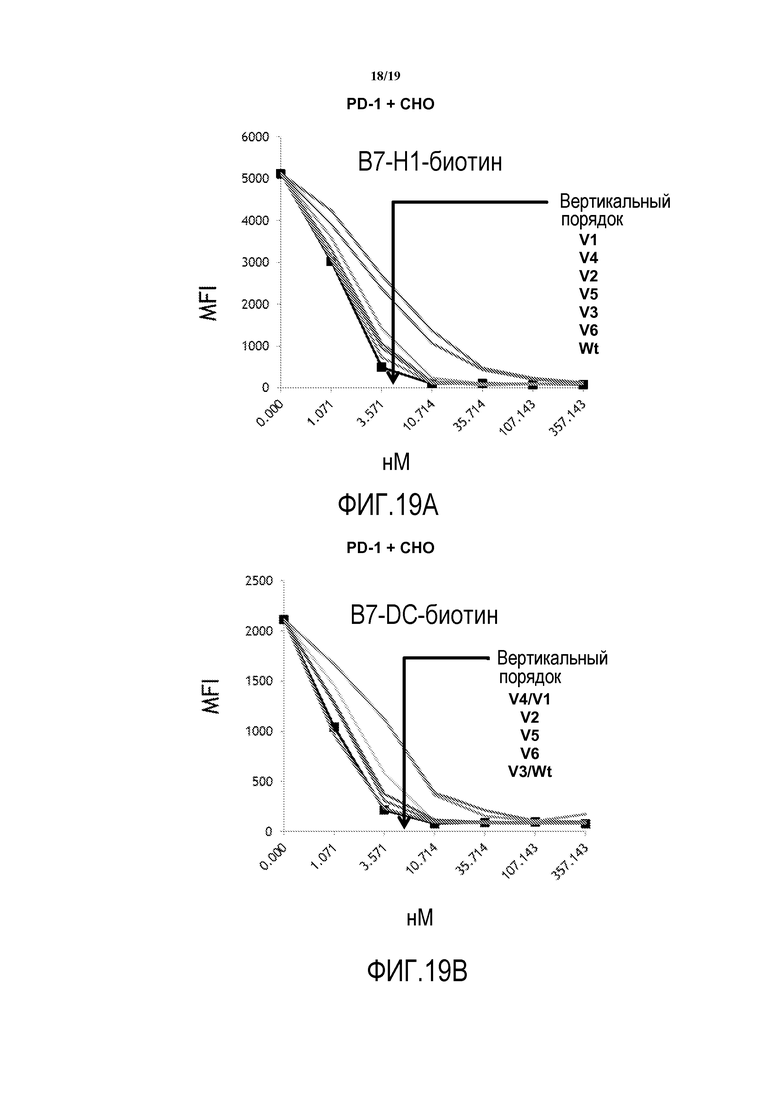
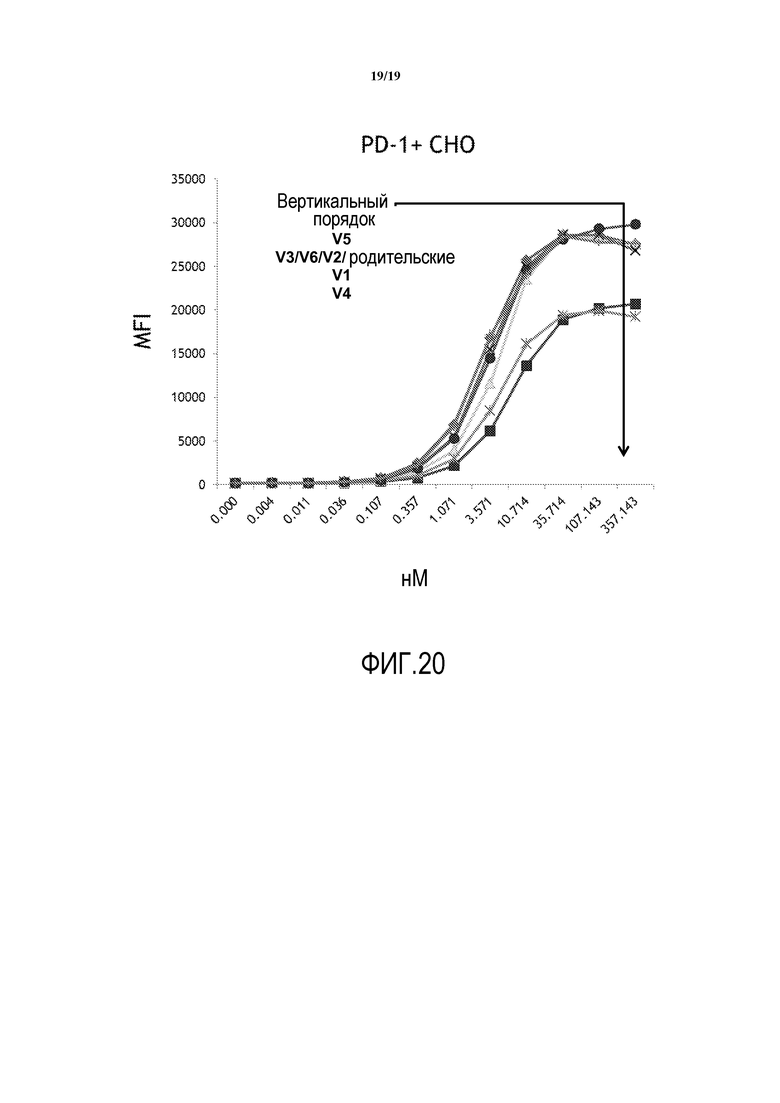
Изобретение относится к области биохимии. Описана группа изобретений, включающая антитело или его антигенсвязывающий фрагмент, которые связываются с PD-1; гуманизированное антитело или его антиген-связывающий фрагмент; фармацевтическая композиция для лечения заболевания, связанного со взаимодействием В7-Н1 с PD-1; способ лечения злокачественной опухоли у индивидуума, а также способ лечения инфекционного заболевания, связанного со взаимодействием В7-Н1 с PD-1, где вышеуказанные способы включают введение фармацевтической композиции; применение вышеуказанного антитела или антиген-связывающего фрагмента для получения лекарственного средства для лечения злокачественной опухоли, а также для получения лекарственного средства для лечения инфекционного заболевания, связанного со взаимодействием В7-Н1 с PD-1; способ обнаружения или диагностики заболевания, нарушения или инфекции, связанного (ой) со взаимодействием В7-Н1 с PD-1, способ контроля за прогрессированием заболевания, нарушения или инфекции, способ контроля ответа на лечение заболевания, нарушения или инфекции, способ лечения заболевания, отличающегося повышенной экспрессией PD-1, способ лечения заболевания, отличающегося повышенной экспрессией В7-Н1, где способы включают в себя использование вышеуказанного антитела или его антиген-связывающего фрагмента. Изобретение расширяет арсенал антител, связывающихся с PD-1. 12 н. и 25 з.п. ф-лы, 20 ил., 23 табл., 7 пр.
1. Антитело или его антиген-связывающий фрагмент, содержащее шесть гипервариабельных областей (CDR),
где CDR включают три CDR легкой цепи последовательности SEQ ID NO: 98 и три CDR тяжелой цепи последовательности SEQ ID NO: 106, и
где антитело или антиген-связывающий фрагмент связывается с PD-1.
2. Антитело или антиген-связывающий фрагмент по п. 1, где три CDR легкой цепи содержат первую CDR легкой цепи, содержащую аминокислоты 24-33 последовательности SEQ ID NO: 98, вторую CDR легкой цепи, содержащую аминокислоты 49-55 последовательности SEQ ID NO: 98, и третью CDR легкой цепи, содержащую аминокислоты 88-96 последовательности SEQ ID NO: 98.
3. Антитело или антиген-связывающий фрагмент по п. 1, где три CDR тяжелой цепи содержат первую CDR тяжелой цепи, содержащую аминокислоты 26-35 последовательности SEQ ID NO: 106, вторую CDR тяжелой цепи, содержащую аминокислоты 50-66 последовательности SEQ ID NO: 106, и третью CDR тяжелой цепи, содержащую аминокислоты 99-111 последовательности SEQ ID NO: 106.
4. Антитело или антиген-связывающий фрагмент по п. 1, содержащее вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 98.
5. Антитело или антиген-связывающий фрагмент по п. 1, содержащее вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 106.
6. Антитело или антиген-связывающий фрагмент по п. 1, содержащее вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 98, и вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 106.
7. Антитело или антиген-связывающий фрагмент по п. 1, где связанный PD-1 экспрессируется на поверхности живой клетки при эндогенной или трансфецирующей концентрации.
8. Антитело или антиген-связывающий фрагмент по п. 7, где живой клеткой является Т-клетка.
9. Антитело или антиген-связывающий фрагмент по п. 1, где PD-1 представляет собой PD-1 человека.
10. Антитело или антиген-связывающий фрагмент по п. 1, где антитело или антиген-связывающий фрагмент
(A) ослабляет способность лиганда PD-1 связываться с PD-1;
(B) оказывают антагонистическое действие на PD-1-опосредованную передачу сигнала;
(C) повышает Т-клеточную пролиферацию;
(D) повышает продукцию IFN-γ; или
(E) оказывает комбинацию этих действий.
11. Антитело или антиген-связывающий фрагмент по п. 1, содержащее константный домен из константной области иммуноглобулина (Fc).
12. Антитело или антиген-связывающий фрагмент по п. 11, где константный домен представляет собой константный домен человека.
13. Антитело или антиген-связывающий фрагмент по п. 12, где константный домен человека представляет собой домен IgA, IgD, IgE, IgG или IgM.
14. Антитело или антиген-связывающий фрагмент по п. 13, где константный домен IgG человека представляет собой домен IgGl, IgG2, IgG3 или IgG4.
15. Антитело или антиген-связывающий фрагмент по п. 1, где антитело или антиген-связывающий фрагмент является меченым или конъюгированным с токсином, лекарственным средством, рецептором, ферментом, лигандом рецептора.
16. Антитело или антиген-связывающий фрагмент по п. 1, где антитело представляет собой моноклональное антитело, антитело человека, химерное антитело или гуманизированное антитело.
17. Гуманизированное антитело или его антиген-связывающий фрагмент, содержащее вариабельную область легкой цепи, содержащей аминокислотную последовательность SEQ ID NO: 98, вариабельную область тяжелой цепи, содержащей аминокислотную последовательность SEQ ID NO: 106, и константный домен IgG4 человека, причем антитело или его антиген-связывающий фрагмент связывают PD-1.
18. Фармацевтическая композиция для лечения заболевания, связанного со взаимодействием В7-Н1 с PD-1, содержащая терапевтически эффективное количество антитела или его антиген-связывающего фрагмента по любому из пп. 1-17, и физиологически приемлемый носитель или эксципиент.
19. Способ лечения злокачественной опухоли у индивидуума, где способ включает введение индивидууму терапевтически эффективного количества фармацевтической композиции по п. 18.
20. Способ по п. 19, где фармацевтическую композицию вводят до появления каких-либо симптомов злокачественной опухоли.
21. Способ лечения инфекционного заболевания, связанного со взаимодействием В7-Н1 с PD-1, у индивидуума, где способ включает введение индивидууму терапевтически эффективного количества фармацевтической композиции по п. 18.
22. Способ по п. 21, где фармацевтическую композицию вводят до появления каких-либо симптомов заболевания.
23. Способ по п. 22, где инфекционное заболевание представляет собой хроническое вирусное заболевание.
24. Применение антитела или его антиген-связывающего фрагмента по любому из пп. 1-17 для получения лекарственного средства для лечения злокачественной опухоли.
25. Применение антитела или антиген-связывающего фрагмента по любому из пп. 1-17 для получения лекарственного средства для лечения инфекционного заболевания, связанного со взаимодействием В7-Н1 с PD-1.
26. Способ обнаружения или диагностики заболевания, нарушения или инфекции, связанного(ой) со взаимодействием В7-Н1 с PD-1, включающий:
(a) оценку экспрессии PD-1 в клетках или в образце ткани индивидуума, используя антитело или его антиген-связывающий фрагмент по любому из пп. 1-17; и
(b) сравнение уровня PD-1 с контрольным уровнем, где повышение в исследуемом уровне PD-1 по сравнению с контрольным уровнем является показателем заболевания, нарушения или инфекции.
27. Способ по п. 26, где экспрессию PD-1 оценивают твердофазным иммуноферментным анализом (ELISA), радиоиммунологическим анализом (RIA) или сортировкой клеток с активированной флуоресценцией (FACS).
28. Способ контроля за прогрессированием заболевания, нарушения или инфекции, связанного(ой) со взаимодействием В7-Н1 с PD-1, включающий:
(a) оценку экспрессии PD-1 в клетках или в образце ткани индивидуума в первый момент времени, используя антитело или его антиген-связывающий фрагмент по любому из пп. 1-17; и
(b) сравнение уровня PD-1 в клетках или в образце ткани индивидуума во второй момент времени или в течение времени, где повышение в исследуемом уровне PD-1 является показателем прогрессирования заболевания, нарушения или инфекции.
29. Способ по п. 28, где экспрессию PD-1 оценивают твердофазным иммуноферментным анализом (ELISA), радиоиммунологическим анализом (RIA) или сортировкой клеток с активированной флуоресценцией (FACS).
30. Способ контроля ответа на лечение заболевания, нарушения или инфекции, связанного(ой) со взаимодействием В7-Н1 с PD-1, включающий:
(a) оценку экспрессии PD-1 в клетках или в образце ткани индивидуума перед лечением с использованием антитела или его антиген-связывающего фрагмента по любому из пп. 1-17; и
(b) оценку экспрессии PD-1 в клетках или в образце ткани индивидуума в одну или более временные точки после лечения, и сравнение уровня PD-1 в течение времени, где повышение исследуемого уровня PD-1 по сравнению с уровнем PD-1 до лечения указывает на ответ на лечение.
31. Способ по п. 30, где экспрессию PD-1 оценивают твердофазным иммуноферментным анализом (ELISA), радиоиммунологическим анализом (RIA) или сортировкой клеток с активированной флуоресценцией (FACS).
32. Способ лечения заболевания, отличающегося повышенной экспрессией PD-1, у индивидуума, где способ включает стадии:
(i) определения, страдает ли индивидуум заболеванием, отличающимся повышенной экспрессией PD-1 путем
(a) оценки экспрессии PD-1 в клетках или в образце ткани индивидуума, используя антитело или его антиген-связывающий фрагмент по любому из пп. 1-17;
(b) сравнения уровня PD-1 с контрольным уровнем, где повышение исследуемого уровня PD-1 по сравнению с контрольным уровнем указывает на то, что индивидуум страдает заболеванием, отличающимся повышенной экспрессией PD-1; и
(ii) введения индивидууму терапевтически эффективного количества фармацевтической композиции по п. 18, если индивидуум страдает заболеванием, отличающимся повышенной экспрессией PD-1.
33. Способ по п. 32, где заболевание, отличающееся повышенной экспрессией PD-1, представляет собой злокачественную опухоль.
34. Способ по п. 32, где заболевание, отличающееся повышенной экспрессией PD-1, представляет собой инфекцию.
35. Способ лечения заболевания, отличающегося повышенной экспрессией В7-Н1, у индивидуума, где способ включает стадии
(i) определения, страдает ли индивидуум заболеванием, отличающимся повышенной экспрессией В7-Н1 путем
(a) оценки экспрессии В7-Н1 в клетках или в образце ткани индивидуума, используя антитело или антиген-связывающий фрагмент;
(b) сравнения уровня В7-Н1 с контрольным уровнем, где повышение исследуемого уровня В7-Н1 по сравнению с контрольным уровнем указывает на то, что индивидуум страдает заболеванием, отличающимся повышенной экспрессией В7-Н1; и
(ii) введения индивидууму терапевтически эффективного количества фармацевтической композиции по п. 18, если индивидуум страдает заболеванием, отличающимся повышенной экспрессией В7-Н1.
36. Способ по п. 35, где заболевание, отличающееся повышенной экспрессией В7-Н1, представляет собой злокачественную опухоль.
37. Способ по п. 35, где заболевание, отличающееся повышенной экспрессией В7-Н1, представляет собой инфекцию.
| WO2004056875 A1 08.07.2004 | |||
| US 7488802 B2 10.02.2009 | |||
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1 (PD-1) И СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТИ-PD-1-АНТИТЕЛ САМОСТОЯТЕЛЬНО ИЛИ В КОМБИНАЦИИ С ДРУГИМИ ИММУНОТЕРАПЕВТИЧЕСКИМИ СРЕДСТВАМИ | 2006 |
|
RU2406760C2 |
| WO 2006121168 A1 16.11.2006. | |||

