Эта заявка притязает на приоритет заявки на патент США с серийным № 61/888255, поданной 8 октября 2013 г., и заявки на патент США с серийным № 61/767071, поданной 20 февраля 2013 г., полное содержание каждой из которых включено сюда посредством ссылки.
ЗАЯВЛЕНИЕ О СПОНСИРУЕМОМ ПРАВИТЕЛЬСТВОМ ИССЛЕДОВАНИИ ИЛИ РАЗРАБОТКЕ
Это изобретение было сделано при поддержке правительства по 2R01 NS055140 и 1P01 CA 1322714, присужденных Национальным институтом здравоохранения (NIH). Правительство имеет определенные права на это изобретение.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка содержит список последовательностей, который был представлен в электронном виде в формате ASCII и, таким образом, включен посредством ссылки в его полном объеме. Указанная копия ASCII, созданная 20 февраля 2014 г., названа N2067-7000WO_SL.txt и имеет размер, равный 228605 байтам.
УРОВЕНЬ ТЕХНИКИ
Настоящее изобретение относится в целом к применению T-клеток, созданных так, что они экспрессируют химерный антигенспецифический рецептор (CAR), для лечения заболевания, связанного с экспрессией рецептора III эпидермального фактора роста.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Хотя центральная нервная система (ЦНС) часто считается иммунологически «неприкосновенной» (Okada et al., 2009, Crit Rev Immunol 29: 1-42), недавние исследования вакцины на пациентах со злокачественной глиомой продемонстрировали положительные результаты (Aguilar et al., 2012, Curr Treat Options Oncol 13: 437-450; Ruzevick, et al., 2012, Neurosurg Clin N Am 23: 459-470; 15; и Okada et al., 2011, J Clin Oncol 29: 330-336). Однако эффективность вакцины, которая опирается на активность неповрежденной иммунной системы хозяина, может страдать в результате системного подавления иммунитета вследствие экспрессии в опухоли иммуносупрессивных цитокинов, а также химиотерапии и лучевой терапии. С другой стороны, терапия с использованием адоптивного переноса клеток (АСТ) в виде аутологичных Т-клеток, особенно Т-клеток, трансдуцированных химерными антигенспецифическим рецепторами (CAR), раскрыла перспективы предварительных испытаний на раке системы крови (Kalos et al., 2011, Sci Transl Med 3(95):95ra73; и Porter et al., 2011, New England Journal of Medicine 365: 725-733.
Увеличенная экспрессия рецептора эпидермального фактора роста (EGFR) часто обнаруживается в ряде карцином, включая рак молочной железы, легких, головы и шеи, а также глиобластому. Спонтанные перестройки в гене рецептора EGF были впервые определены в первичных опухолях глиобластомах человека, и почти во всех случаях изменения были зарегистрированы в опухолях с увеличением EGFR. Три различных типа мутантов являются результатом этих перестроек. Самым распространенным из них является характеризующийся делецией мутантный рецептор EGF типа III (EGFRvIII), который характеризуется делецией экзонов 2-7 в мРНК EGFR. Эти делеции соответствуют нуклеотидам 275-1075 кДНК, которые кодируют аминокислоты 6-276, по-видимому, за счет альтернативного сплайсинга или перестроек. Делеция 801 п.о. в пределах экстраклеточного домена гена EGFR вызывает находящееся в рамке усечение нормального белка EGFR, приводящее к рецептору с М.м. 145 кДа, создавая тем самым специфический в отношении опухоли и иммуногенный эпитоп (рассматриваемый в Hatanpaa et al., 2010, Neoplasia 12: 675-684; Mukasa et al., 2010, Proc Natl Acad Sci USA 107: 2616-2621). Экспрессия EGFRvIII отмечалась во многих типах опухолей, в том числе мультиформной глиобластоме (GBM), но редко наблюдается в нормальной ткани. EGFRvIII экспрессируется в 24%-67% случаев GBM, и у пациентов, продолжающих жить ≥1 года, экспрессия EGFRvIII является независимым неблагоприятным прогностическим показателем (Heimberger et al., 2005, Clin. Cancer Res. 11: 1462-1466; Heimberger et al., 2005, J Transl. Med 3: 38).
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящим изобретением обеспечиваются, среди прочего, структуры и способы для управления иммунным ответом у пациентов посредством обеспечения оптимизированных и/или гуманизированных антител или фрагментов антител (например, scFv), которые связывают рецептор III эпидермального фактора роста (EGFRvIII), включенных в конструкцию химерного антигенспецифического рецептора (CAR). В некоторых вариантах осуществления настоящее изобретение относится к применению Т-клеток, которые созданы так, что они экспрессируют антитело или фрагмент антитела, которое(ый) связывает EGFRvIII, например, гуманизированное антитело или фрагмент антитела, которое(ый) связывает EGFRvIII, включенное(ый) в CAR, для лечения рака, связанного с экспрессией EGFRvIII. В некоторых аспектах настоящее изобретение относится к адоптивному переносу клеток, который может быть особенно подходящим для пациентов с глиомой, поскольку гораздо лучше можно манипулировать и контролировать специфичность(ю), количество(м) и функциональный(ым) фенотип(ом) клеток, полученных ex vivo, чем не подвергнутых стимуляции Т-клеток, индуцированных с помощью in vivo иммунизации.
Соответственно, в одном аспекте настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей химерный антигенспецифический рецептор (CAR), причем CAR включает антитело или фрагмент антитела, которое(ый) включает связывающий домен антитела против EGFRvIII (например, гуманизированное антитело или фрагмент антитела, которое(ый) специфически связывается с EGFRvIII), трансмембранный домен и внутриклеточный домен сигнализации (например, внутриклеточный домен сигнализации, включающий костимулирующий домен и/или домен первичной сигнализации). В одном варианте осуществления CAR включает антитело или фрагмент антитела, которое(ый) включает связывающий домен антитела против EGFRvIII, описанный здесь (например, гуманизированное антитело или фрагмент антитела, которое(ый) специфически связывается с EGFRvIII, как описано здесь), трансмембранный домен, описанный здесь, и внутриклеточный домен сигнализации, описанный здесь (например, внутриклеточный домен сигнализации, включающий костимулирующий домен и/или домен первичной сигнализации).
В одном варианте осуществления кодируемый связывающий домен антитела против EGFRvIII включает один или более (например, все три) из определяющего комплементарность участка 1 легкой цепи (CDR1 LC), определяющего комплементарность участка 2 легкой цепи (CDR2 LC) и определяющего комплементарность участка 3 легкой цепи (CDR3 LC) связывающего домена антитела против EGFRvIII, описанного здесь, и один или более (например, все три) из определяющего комплементарность участка 1 тяжелой цепи (CDR1 НС), определяющего комплементарность участка 2 тяжелой цепи (CDR2 НС) и определяющего комплементарность участка 3 тяжелой цепи (CDR3 НС) связывающего домена антитела против EGFRvIII, описанного здесь, например, связывающего домена гуманизированного антитела против EGFRvIII, включающего один или более, например, все три, CDR LC и один или более, например, все три, CDR HC. В одном варианте осуществления кодируемый связывающий домен антитела против EGFRvIII включает вариабельную область легкой цепи, описанную здесь (например, в таблице 2 или SEQ ID NO: 11) и/или вариабельную область тяжелой цепи, описанную здесь (например, в таблице 2 или SEQ ID NO: 11). В одном варианте осуществления кодируемый связывающий домен антитела против EGFRvIII представляет собой scFv, включающий легкую цепь и тяжелую цепь аминокислотной последовательности из таблицы 2 или SEQ ID NO: 11. В одном варианте осуществления связывающий домен антитела против EGFRvIII (например, scFv) включает: вариабельную область легкой цепи, включающую аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 30, 20 или 10 модификаций (например, замен) аминокислотной последовательности вариабельной области легкой цепи, приведенной в таблице 2 или SEQ ID NO: 11, или последовательность, идентичную на 95-99% аминокислотной последовательности, приведенной в таблице 2 или SEQ ID NO: 11; и/или вариабельную область тяжелой цепи, включающую аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 30, 20 или 10 модификаций (например, замен) аминокислотной последовательности вариабельной области тяжелой цепи, приведенной в таблице 2 или SEQ ID NO: 11, или последовательность, идентичную на 95-99% аминокислотной последовательности, приведенной в таблице 2 или SEQ ID NO: 11. В одном варианте осуществления связывающий домен антитела против EGFRvIII включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления последовательность нуклеиновой кислоты, кодирующая связывающий домен антитела против EGFRvIII, включает последовательность SEQ ID NO: 68. В одном варианте осуществления последовательность нуклеиновой кислоты, кодирующая связывающий домен антитела против EGFRvIII, включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 39, SEQ ID NO: 45, SEQ ID NO: 51, SEQ ID NO: 57, SEQ ID NO: 63, SEQ ID NO: 69, SEQ ID NO: 75, SEQ ID NO: 81 и SEQ ID NO: 98, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления кодируемый связывающий домен антитела против EGFRvIII представляет собой scFv, и вариабельная область легкой цепи, включающая аминокислотную последовательность, описанную здесь, например, в таблице 2 или SEQ ID NO: 11, присоединена к вариабельной области тяжелой цепи, включающей аминокислотную последовательность, описанную здесь, например, в таблице 2 или SEQ ID NO: 11, через линкер, например, линкер, описанный здесь. В одном варианте осуществления кодируемый связывающий домен антитела против EGFRvIII включает линкер (Gly4-Ser)n, где n равно 1, 2, 3, 4, 5 или 6, предпочтительно 4 (SEQ ID NO: 110). Вариабельная область легкой цепи и вариабельная область тяжелой цепи scFv могут находиться, например, в любой из следующих ориентаций: вариабельная область легкой цепи-линкер-вариабельная область тяжелой цепи или вариабельная область тяжелой цепи-линкер-вариабельная область легкой цепи.
В одном варианте осуществления кодируемый CAR включает трансмембранный домен, который включает трансмембранный домен белка, выбираемого из группы, состоящей из альфа, бета или дзета-цепи T-клеточного рецептора, CD28, CD3 эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154. В одном варианте осуществления кодируемый трансмембранный домен включает последовательность SEQ ID NO: 15. В одном варианте осуществления кодируемый трансмембранный домен включает аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 20, 10 или 5 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 15, или последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 15. В одном варианте осуществления последовательность нуклеиновой кислоты, кодирующая трансмембранный домен, включает последовательность SEQ ID NO: 8 или последовательность, идентичную ей на 95-99%.
В одном варианте осуществления кодируемый связывающий домен антитела против EGFRvIII соединен с трансмембранным доменом с помощью шарнирной области, например, шарнирной области, описанной здесь. В одном варианте осуществления кодируемая шарнирная область включает SEQ ID NO: 14 или SEQ ID NO: 104 или SEQ ID NO: 106 или SEQ ID NO: 108, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления последовательность нуклеиновой кислоты, кодирующая шарнирную область, включает последовательность SEQ ID NO: 7 или SEQ ID NO: 105, или SEQ ID NO: 107, или SEQ ID NO: 109, или последовательность, идентичную ей на 95-99%.
В одном варианте осуществления выделенная молекула нуклеиновой кислоты, кроме того, включает последовательность, кодирующую костимулирующий домен, например, костимулирующий домен, описанный здесь. В одном варианте осуществления кодируемый костимулирующий домен включает функциональный домен сигнализации белка, выбираемого из группы, состоящей из OX40, CD2, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11а/CD18), ICOS (CD278) и 4-1ВВ (CD137). В одном варианте осуществления кодируемый костимулирующий домен включает последовательность SEQ ID NO: 16. В одном варианте осуществления кодируемый костимулирующий домен включает последовательность SEQ ID NO: 102. В одном варианте осуществления кодируемый костимулирующий домен включает аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 20, 10 или 5 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 16 или SEQ ID NO: 102, или последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 16 или SEQ ID NO: 102. В одном варианте осуществления последовательность нуклеиновой кислоты, кодирующая костимулирующий домен, включает последовательность SEQ ID NO: 9, или последовательность, идентичную ей на 95-99%.
В одном варианте осуществления выделенная молекула нуклеиновой кислоты, кроме того, включает последовательность, кодирующую внутриклеточный домен сигнализации, например, внутриклеточный домен сигнализации, описанный здесь. В одном варианте осуществления кодируемый внутриклеточный домен сигнализации включает функциональный домен сигнализации 4-1BB и/или функциональный домен сигнализации CD3 дзета. В одном варианте осуществления кодируемый внутриклеточный домен сигнализации включает функциональный домен сигнализации CD27 и/или функциональный домен сигнализации CD3 дзета. В одном варианте осуществления кодируемый внутриклеточный домен сигнализации включает последовательность SEQ ID NO: 16 или SEQ ID NO: 102, и/или последовательность SEQ ID NO: 17 или SEQ ID NO: 99. В одном варианте осуществления внутриклеточный домен сигнализации включает аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 20, 10 или 5 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 16 и/или аминокислотной последовательности SEQ ID NO: 17 или SEQ ID NO: 99, или последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 16 или SEQ ID NO: 102 и/или аминокислотной последовательности SEQ ID NO: 17 или SEQ ID NO: 99. В одном варианте осуществления кодируемый внутриклеточный домен сигнализации включает последовательность SEQ ID NO: 16 или SEQ ID NO: 102 и последовательность SEQ ID NO: 17 или SEQ ID NO: 99, причем последовательности, включающие внутриклеточный домен сигнализации, экспрессируются в одной и той же рамке и в виде одной полипептидной цепи. В одном варианте осуществления последовательность нуклеиновой кислоты, кодирующая внутриклеточный домен сигнализации, включает последовательность SEQ ID NO: 9 или SEQ ID NO: 103, или последовательность, идентичную ей на 95-99%, и/или последовательность SEQ ID NO: 10 или SEQ ID NO: 100, или последовательность, идентичную ей на 95-99%.
В другом аспекте настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей конструкцию CAR, включающую лидерную последовательность, например, лидерную последовательность, описанную здесь, например, SEQ ID NO: 13, связывающий домен антитела против EGFRvIII, описанный здесь, например, связывающий домен антитела против EGFRvIII, включающий CDR1 LC, CDR2 LC, CDR3 LC, CDR1 HC, CDR2 HC и CDR3 HC, описанный здесь, например, связывающий домен антитела против EGFRvIII, описанный в таблице 2 или SEQ ID NO: 11, или последовательности, идентичной ей на 95-99%, шарнирную область описанную здесь, например, SEQ ID NO: 14 или SEQ ID NO: 104 или SEQ ID NO: 106 или SEQ ID NO: 108, трансмембранный домен, описанный здесь, например, имеющий последовательность SEQ ID NO: 15, и внутриклеточный домен сигнализации, например, внутриклеточный домен сигнализации, описанный здесь. В одном варианте осуществления кодируемый внутриклеточный домен сигнализации включает костимулирующий домен, например, костимулирующий домен, описанный здесь, например, костимулирующий домен 4-1ВВ, имеющий последовательность SEQ ID NO: 16, и/или домен первичной сигнализации, например, домен первичной сигнализации, описанный здесь, например, стимулирующий домен CD3 дзета, имеющий последовательность SEQ ID NO: 17 или SEQ ID NO: 99. В одном варианте осуществления кодируемый внутриклеточный домен сигнализации включает костимулирующий домен, например, костимулирующий домен, описанный здесь, например, костимулирующий домен CD27, имеющий последовательность SEQ ID NO: 102, и/или домен первичной сигнализации, например, домен первичной сигнализации, описанный здесь, например, стимулирующий домен CD3 дзета, имеющий последовательность SEQ ID NO: 17 или SEQ ID NO: 99. В одном варианте осуществления кодируемый внутриклеточный домен сигнализации включает костимулирующий домен, например, костимулирующий домен, описанный здесь, например, костимулирующий домен 4-1ВВ, имеющий последовательность SEQ ID NO: 16, и домен первичной сигнализации, например, домен первичной сигнализации, описанный здесь, например, стимулирующий домен CD3 дзета, имеющий последовательность SEQ ID NO: 17 или SEQ ID NO: 99. В одном варианте осуществления кодируемый внутриклеточный домен сигнализации включает костимулирующий домен, например, костимулирующий домен, описанный здесь, например, костимулирующий домен CD27, имеющий последовательность SEQ ID NO: 102, и домен первичной сигнализации, например, домен первичной сигнализации, описанный здесь, например, стимулирующий домен CD3 дзета, имеющий последовательность SEQ ID NO: 17 или SEQ ID NO: 99. В одном варианте осуществления выделенная молекула нуклеиновой кислоты, кодирующая конструкцию CAR, включает лидерную последовательность, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO: 6, или последовательностью, идентичной ей на 95-99%. В одном варианте осуществления выделенная молекула нуклеиновой кислоты, кодирующая конструкцию CAR, включает последовательность связывающего домена антитела против EGFR, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO: 39, SEQ ID NO: 45, SEQ ID NO: 51, SEQ ID NO: 57, SEQ ID NO: 63, SEQ ID NO: 69, SEQ ID NO: 75, SEQ ID NO: 81 или SEQ ID NO: 98, или последовательностью, идентичной ей на 95-99%. В одном варианте осуществления выделенная молекула нуклеиновой кислоты, кодирующая конструкцию CAR, включает последовательность связывающего домена антитела против EGFR, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO: 69, или последовательностью, идентичной ей на 95-99%. В одном варианте осуществления выделенная молекула нуклеиновой кислоты, кодирующая конструкцию CAR, включает последовательность связывающего домена антитела против EGFR, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO: 4, или последовательностью, идентичной ей на 95-99%. В одном варианте осуществления выделенная молекула нуклеиновой кислоты, кодирующая конструкцию CAR, включает трансмембранную последовательность, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO: 8, или последовательностью, идентичной ей на 95-99%. В одном варианте осуществления выделенная молекула нуклеиновой кислоты, кодирующая конструкцию CAR, включает последовательность внутриклеточного домена сигнализации, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO: 9 или последовательностью, идентичной ей на 95-99%, и/или последовательностью нуклеиновой кислоты SEQ ID NO: 10, или последовательностью, идентичной ей на 95-99%.
В одном варианте осуществления выделенная молекула нуклеиновой кислоты включает (например, состоит из) нуклеиновую кислоту, кодирующую аминокислотную последовательность CAR - SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 или SEQ ID NO: 90, или аминокислотную последовательность, имеющую по крайней мере одну, две, три, четыре, пять, 10, 15, 20 или 30 модификаций (например, замен), но не более 60, 50 или 40 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 или SEQ ID NO: 90, или аминокислотную последовательность, идентичную на 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 или SEQ ID NO: 90. В одном варианте осуществления выделенная молекула нуклеиновой кислоты включает (например, состоит из) нуклеиновую кислоту, кодирующую аминокислотную последовательность CAR - SEQ ID NO: 1 или SEQ ID NO: 2, или аминокислотную последовательность, имеющую крайней мере одну, две, три, четыре, пять, 10, 15, 20 или 30 модификаций (например, замен), но не более 60, 50 или 40 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 1 или SEQ ID NO: 2, или аминокислотную последовательность, идентичную на 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 1 или SEQ ID NO: 2.
В одном варианте осуществления выделенная молекула нуклеиновой кислоты включает (например, состоит из) последовательность нуклеиновой кислоты SEQ ID NO: 42, SEQ ID NO: 48, SEQ ID NO: 54, SEQ ID NO: 60, SEQ ID NO: 66, SEQ ID NO: 72, SEQ ID NO: 78, SEQ ID NO: 84 или SEQ ID NO: 89, или последовательность нуклеиновой кислоты, идентичную на 85%, 90%, 95%, 96%, 97%, 98% или 99% последовательности нуклеиновой кислоты SEQ ID NO: 42, SEQ ID NO: 48, SEQ ID NO: 54, SEQ ID NO: 60, SEQ ID NO: 66, SEQ ID NO: 72, SEQ ID NO: 78, SEQ ID NO: 84 или SEQ ID NO: 89. В одном варианте осуществления выделенная молекула нуклеиновой кислоты включает (например, состоит из) последовательность нуклеиновой кислоты SEQ ID NO: 18 или SEQ ID NO: 19, или последовательность нуклеиновой кислоты, идентичную на 85%, 90%, 95%, 96%, 97%, 98% или 99% последовательности нуклеиновой кислоты SEQ ID NO: 18 или SEQ ID NO: 19.
В одном аспекте настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей связывающий домен антитела против EGFRvIII, причем связывающий домен антитела против EGFRvIII включает один или более (например, все три) из определяющего комплементарность участка 1 легкой цепи (CDR1 LC), определяющего комплементарность участка 2 легкой цепи (CDR2 LC) и определяющего комплементарность участка 3 легкой цепи (CDR3 LC) связывающего домена антитела против EGFRvIII описанного здесь, и один или более (например, все три) из определяющего комплементарность участка 1 тяжелой цепи (CDR1 НС), определяющего комплементарность участка 2 тяжелой цепи (CDR2 НС) и определяющего комплементарность участка 3 тяжелой цепи (CDR3 НС) связывающего домена антитела против EGFRvIII, описанного здесь, например, связывающего домена гуманизированного антитела против EGFRvIII, включающего один или более, например, все три, CDR LC и один или более, например, все три, CDR НС. В одном варианте осуществления кодируемый связывающий домен антитела против EGFRvIII включает вариабельную область легкой цепи, описанную здесь (например, в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80) и/или вариабельную область тяжелой цепи, описанную здесь (например, в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80). В одном варианте осуществления кодируемый связывающий домен антитела против EGFRvIII представляет собой scFv, включающий легкую цепь и тяжелую цепь аминокислотной последовательности в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80. В варианте осуществления связывающий домен антитела против EGFRvIII (например, scFv) включает: вариабельную область легкой цепи, включающую аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 30, 20 или 10 модификаций (например, замен) аминокислотной последовательности вариабельной области легкой цепи, представленной в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80, или последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80; и/или вариабельную область тяжелой цепи, включающую аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 30, 20 или 10 модификаций (например, замен) аминокислотной последовательности вариабельной области тяжелой цепи, представленной в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80, или последовательность, идентичную на 95-99% аминокислотной последовательности, представленной в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80. В одном варианте осуществления связывающий домен антитела против EGFRvIII включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления последовательность нуклеиновой кислоты, кодирующая связывающий домен антитела против EGFRvIII, включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 39, SEQ ID NO: 45, SEQ ID NO: 51, SEQ ID NO: 57, SEQ ID NO: 63, SEQ ID NO: 69, SEQ ID NO: 75, SEQ ID NO: 81 и SEQ ID NO: 98, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления кодируемым связывающим доменом антитела против EGFRvIII является scFv, и вариабельная область легкой цепи, включающая аминокислотную последовательность, описанную здесь, например, в таблице 2, присоединена к вариабельной области тяжелой цепи, включающей аминокислотную последовательность, описанную здесь, например, в таблице 2, через линкер, например, линкер, описанный здесь. В одном варианте осуществления кодируемый связывающий домен антитела против EGFRvIII включает линкер (Gly4-Ser)n, где n равно 1, 2, 3, 4, 5 или 6, предпочтительно 4 (SEQ ID NO: 110). Вариабельная область легкой цепи и вариабельная область тяжелой цепи scFv могут находиться, например, в любой из следующих ориентаций: вариабельная область легкой цепи-линкер-вариабельная область тяжелой цепи или вариабельная область тяжелой цепи-линкер-вариабельная область легкой цепи.
В другом аспекте настоящее изобретение относится к выделенной молекуле полипептида, кодируемой молекулой нуклеиновой кислоты. В одном варианте осуществления выделенная молекула полипептида включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 и SEQ ID NO: 90, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления выделенный полипептид включает последовательность SEQ ID NO: 73 или последовательность, идентичную ей на 95-99%. В одном варианте осуществления выделенный полипептид включает последовательность SEQ ID NO: 79 или последовательность, идентичную ей на 95-99%.
В другом аспекте настоящее изобретение относится к выделенной молекуле химерного антигенспецифического рецептора (CAR), включающей связывающий домен антитела против EGFRvIII (например, гуманизированное антитело или фрагмент антитела, которое(ый) специфически связывается с EGFRvIII), трансмембранный домен и внутриклеточный домен сигнализации (например, внутриклеточный домен сигнализации, включающий костимулирующий домен и/или домен первичной сигнализации). В одном варианте осуществления CAR включает антитело или фрагмент антитела, которое(ый) включает связывающий домен антитела против EGFRvIII, описанный здесь (например, гуманизированное антитело или фрагмент антитела, которое(ый) специфически связывается с EGFRvIII, как описано здесь), трансмембранный домен, описанный здесь, и внутриклеточный домен сигнализации, описанный здесь (например, внутриклеточный домен сигнализации, включающий костимулирующий домен и/или домен первичной сигнализации, описанный здесь).
В одном варианте осуществления связывающий домен антитела против EGFRvIII включает один или более (например, все три) из определяющего комплементарность участка 1 легкой цепи (CDR1 LC), определяющего комплементарность участка 2 легкой цепи (CDR2 LC) и определяющего комплементарность участка 3 легкой цепи (CDR3 LC) связывающего домена антитела против EGFRvIII, описанного здесь, и один или более (например, все три) из определяющего комплементарность участка 1 тяжелой цепи (CDR1 НС), определяющего комплементарность участка 2 тяжелой цепи (CDR2 НС) и определяющего комплементарность участка 3 тяжелой цепи (CDR3 НС) связывающего домена антитела против EGFRvIII, описанного здесь, например, связывающего домена гуманизированного антитела против EGFRvIII, включающего один или более, например, все три, CDR LC и один или более, например, все три, CDR НС. В одном варианте осуществления связывающий домен антитела против EGFRvIII включает вариабельную область легкой цепи, описанную здесь (например, в таблице 2 или SEQ ID NO: 11), и/или вариабельную область тяжелой цепи, описанную здесь (например, в таблице 2 или SEQ ID NO: 11). В одном варианте осуществления связывающий домен антитела против EGFRvIII представляет собой scFv, включающий легкую цепь и тяжелую цепь аминокислотной последовательности, приведенной в таблице 2 или SEQ ID NO: 11. В одном варианте осуществления связывающий домен антитела против EGFRvIII (например, scFv) включает: вариабельную область легкой цепи, включающую аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 30, 20 или 10 модификаций (например, замен) аминокислотной последовательности вариабельной области легкой цепи, представленной в таблице 2 или SEQ ID NO: 11, или последовательность, идентичную на 95-99% аминокислотной последовательности, представленной в таблице 2 или SEQ ID NO: 11; и/или вариабельную область тяжелой цепи, включающую аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 30, 20 или 10 модификаций (например, замен) аминокислотной последовательности вариабельной области тяжелой цепи, представленной в таблице 2 или SEQ ID NO: 11, или последовательность, идентичную на 95-99% аминокислотной последовательности, представленной в таблице 2 или SEQ ID NO: 11. В одном варианте осуществления связывающий домен антитела против EGFRvIII включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления связывающий домен антитела против EGFRvIII представляет собой scFv, и вариабельная область легкой цепи, включающая аминокислотную последовательность, описанную здесь, например, в таблице 2 или SEQ ID NO: 11, присоединена к вариабельной области тяжелой цепи, включающей аминокислотную последовательность, описанную здесь, например, в таблице 2 или SEQ ID NO: 11, через линкер, например, линкер, описанный здесь. В одном варианте осуществления связывающий домен антитела против EGFRvIII включает линкер (Gly4-Ser)n, где n равно 1, 2, 3, 4, 5 или 6, предпочтительно 4 (SEQ ID NO: 110). Вариабельная область легкой цепи и вариабельная область тяжелой цепи scFv могут находиться, например, в любой из следующих ориентаций: вариабельная область легкой цепи-линкер-вариабельная область тяжелой цепи или вариабельная область тяжелой цепи-линкер-вариабельная область легкой цепи.
В одном варианте осуществления выделенная молекула CAR включает трансмембранный домен белка, выбираемый из группы, состоящей из альфа, бета или дзета-цепи T-клеточного рецептора, CD28, CD3 эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD-134, CD-137 и CD154. В одном варианте осуществления трансмембранный домен включает последовательность SEQ ID NO: 15. В одном варианте осуществления трансмембранный домен включает аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 20, 10 или 5 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 15, или последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 15.
В одном варианте осуществления связывающий домен антитела против EGFRvIII соединен с трансмембранным доменом с помощью шарнирной области, например, шарнирной области, описанной здесь. В одном варианте осуществления кодируемая шарнирная область включает SEQ ID NO: 14 или SEQ ID NO: 104 или SEQ ID NO: 106 или SEQ ID NO: 108, или последовательность, идентичную ей на 95-99%.
В одном варианте осуществления выделенная молекула CAR, кроме того, включает последовательность, кодирующую костимулирующий домен, например, костимулирующий домен, описанный здесь. В одном варианте осуществления костимулирующий домен включает функциональный домен сигнализации белка, выбираемого из группы, состоящей из OX40, CD2, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11а/CD18), ICOS (CD278) и 4-1ВВ (CD137). В одном варианте осуществления костимулирующий домен включает последовательность SEQ ID NO: 16 или SEQ ID NO: 102. В одном варианте осуществления костимулирующий домен включает аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 20, 10 или 5 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 16 или SEQ ID NO: 102, или последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 16 или SEQ ID NO: 102. В одном варианте осуществления выделенная молекула CAR, кроме того, включает последовательность, кодирующую внутриклеточный домен сигнализации, например, внутриклеточный домен сигнализации, описанный здесь. В одном варианте осуществления внутриклеточный домен сигнализации включает функциональный домен сигнализации 4-1BB или CD27 и/или функциональной домен сигнализации CD3 дзета. В одном варианте осуществления внутриклеточный домен сигнализации включает последовательность SEQ ID NO: 16 или SEQ ID NO: 102 и/или последовательность SEQ ID NO: 17. В одном варианте осуществления внутриклеточный домен сигнализации включает последовательность SEQ ID NO: 16 или SEQ ID NO: 102 и/или последовательность SEQ ID NO: 99. В одном варианте осуществления внутриклеточный домен сигнализации включает аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 20, 10 или 5 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 16 или SEQ ID NO: 102 и/или аминокислотной последовательности SEQ ID NO: 17 или SEQ ID NO: 99, или последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 16 или последовательности SEQ ID NO: 102 и/или аминокислотной последовательности SEQ ID NO: 17 или SEQ ID NO: 99. В одном варианте осуществления внутриклеточный домен сигнализации включает последовательность SEQ ID NO: 16 или SEQ ID NO: 102 и последовательность SEQ ID NO: 17 или SEQ ID NO: 99, причем последовательности, включающие внутриклеточный домен сигнализации, экспрессируются в одной и той же рамке и в виде одной полипептидной цепи.
В одном варианте осуществления выделенная молекула CAR, кроме того, включает лидерную последовательность, например, лидерную последовательность, описанную здесь. В одном варианте осуществления лидерная последовательность включает аминокислотную последовательность SEQ ID NO: 13 или последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 13.
В другом аспекте настоящее изобретение относится к выделенной молекуле CAR, включающей лидерную последовательность, например, лидерную последовательность, описанную здесь, например, лидерную последовательность с SEQ ID NO: 13 или с последовательностью, идентичной ей на 95-99%, связывающий домен антитела против EGFRvIII, описанный здесь, например, связывающий домен антитела против EGFRvIII, включающий CDR1 LC, CDR2 LC, CDR3 LC, CDR1 HC, CDR2 HC и CDR3 HC, описанный здесь, например, связывающий домен антитела против EGFRvIII, описанный в таблице 2 или SEQ ID NO: 11, или последовательности, идентичной ей на 95-99%, шарнирную область, например, шарнирную область, описанную здесь, например, шарнирную область с SEQ ID NO: 14 или SEQ ID NO: 104 или SEQ ID NO: 106 или SEQ ID NO: 108, или с 95-99% идентичностью ей, трансмембранный домен, например, трансмембранный домен, описанный здесь, например, трансмембранный домен, имеющий последовательность SEQ ID NO: 15 или последовательность, идентичную ей на 95-99%, внутриклеточный домен сигнализации, например, внутриклеточный домен сигнализации, описанный здесь (например, внутриклеточный домен сигнализации, включающий костимулирующий домен и/или домен первичной сигнализации). В одном варианте осуществления внутриклеточный домен сигнализации включает костимулирующий домен, например, костимулирующий домен, описанный здесь, например, костимулирующий домен 4-1ВВ, имеющий последовательность SEQ ID NO: 16, или костимулирующий домен CD27, имеющий последовательность SEQ ID NO: 102, последовательность, идентичную ей на 95-99%, и/или домен первичной сигнализации, например, домен первичной сигнализации, описанный здесь, например, стимулирующий домен CD3 дзета, имеющий последовательность SEQ ID NO: 17 или SEQ ID NO: 99, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления внутриклеточный домен сигнализации включает костимулирующий домен, например, костимулирующий домен, описанный здесь, например, костимулирующий домен 4-1BB, имеющий последовательность SEQ ID NO: 16, или костимулирующий домен CD27, имеющий последовательность SEQ ID NO: 102, и/или домен первичной сигнализации, например, домен первичной сигнализации, описанный здесь, например, стимулирующий домен CD3 дзета, имеющий последовательность SEQ ID NO: 17 или SEQ ID NO: 99.
В одном варианте осуществления выделенная молекула CAR включает (например, состоит из) аминокислотную последовательность SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 или SEQ ID NO: 90, или аминокислотную последовательность, имеющую по крайней мере одну, две, три, четыре, пять, 10, 15, 20 или 30 модификаций (например, замен), но не более 60, 50 или 40 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 или SEQ ID NO: 90, или аминокислотную последовательность, идентичную на 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 или SEQ ID NO: 90. В одном варианте осуществления выделенная молекула CAR включает (например, состоит из) аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2, или аминокислотную последовательность, имеющую по крайней мере одну, две, три, четыре, пять, 10, 15, 20 или 30 модификаций (например, замен), но не более 60, 50 или 40 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 1 или SEQ ID NO: 2, или аминокислотную последовательность, идентичную на 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 1 или SEQ ID NO: 2. В одном варианте осуществления выделенная молекула CAR включает (например, состоит из) аминокислотную последовательность SEQ ID NO: 73 или аминокислотную последовательность, имеющую по крайней мере одну, две, три, четыре, пять, 10, 15, 20 или 30 модификаций (например, замен), но не более 60, 50 или 40 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 73, или аминокислотную последовательность, идентичную на 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 73. В одном варианте осуществления выделенная молекула CAR включает (например, состоит из) аминокислотную последовательность SEQ ID NO: 79 или аминокислотную последовательность, имеющую по крайней мере одну, две, три, четыре, пять, 10, 15, 20 или 30 модификаций (например, замен), но не более 60, 50 или 40 модификаций (например, замен) аминокислотной последовательности SEQ ID NO: 79, или аминокислотную последовательность, идентичную на 85%, 90%, 95%, 96%, 97%, 98% или 99% аминокислотной последовательности SEQ ID NO: 79.
В одном аспекте настоящее изобретение относится к связывающему домену антитела против EGFRvIII, включающему один или более (например, все три) из определяющего комплементарность участка 1 легкой цепи (CDR1 LC), определяющего комплементарность участка 2 легкой цепи (CDR2 LC) и определяющего комплементарность участка 3 легкой цепи (CDR3 LC) связывающего домена антитела против EGFRvIII, описанного здесь, и один или более (например, все три) из определяющего комплементарность участка 1 тяжелой цепи (CDR1 НС), определяющего комплементарность участка 2 тяжелой цепи (CDR2 НС) и определяющего комплементарность участка 3 тяжелой цепи (CDR3 НС) связывающего домена антитела против EGFRvIII, описанного здесь, например, связывающего домена гуманизированного антитела против EGFRvIII, включающего один или более, например, все три, CDR LC и один или более, например, все три, CDR HC. В одном варианте осуществления связывающий домен антитела против EGFRvIII включает вариабельную область легкой цепи, описанную здесь (например, в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80), и/или вариабельной области тяжелой цепи, описанную здесь (например, в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80). В одном варианте осуществления связывающий домен антитела против EGFRvIII представляет собой scFv, включающий легкую цепь и тяжелую цепь аминокислотной последовательности, представленной в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80. В одном варианте осуществления связывающий домен антитела против EGFRvIII (например, scFv) включает: вариабельную область легкой цепи, включающую аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 30, 20 или 10 модификаций (например, замен) аминокислотной последовательности вариабельной области легкой цепи, представленной в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80, или последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80; и/или вариабельную область тяжелой цепи, включающую аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 30, 20 или 10 модификаций (например, замен) аминокислотной последовательности вариабельной области тяжелой цепи, представленной в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80, или последовательность, идентичную на 95-99% аминокислотной последовательности, представленной в SEQ ID NO: 38, 44, 50, 56, 62, 68, 74 или 80. В одном варианте осуществления связывающий домен антитела против EGFRvIII включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления связывающий домен антитела против EGFRvIII представляет собой scFv, и вариабельная область легкой цепи, включающая аминокислотную последовательность, описанную здесь, например, в таблице 2, присоединена к вариабельной области тяжелой цепи, включающей аминокислотную последовательность, описанную здесь, например, в таблице 2, через линкер, например, линкер, описанный здесь. В одном варианте осуществления связывающий домен антитела против EGFRvIII включает линкер (Gly4-Ser)n, где n равно 1, 2, 3, 4, 5 или 6, предпочтительно 4 (SEQ ID NO: 110). Вариабельная область легкой цепи и вариабельная область тяжелой цепи scFv могут находиться, например, в любой из следующих ориентаций: вариабельная область легкой цепи-линкер-вариабельная область тяжелой цепи или вариабельная область тяжелой цепи-линкер-вариабельная область легкой цепи.
В другом аспекте настоящее изобретение относится к вектору, включающему молекулу нуклеиновой кислоты, описанную здесь, например, молекулу нуклеиновой кислоты, кодирующую CAR, описанный здесь. В одном варианте осуществления вектор выбирают из группы, состоящей из ДНК, РНК, плазмиды, лентивирусного вектора, аденовирусного вектора или ретровирусного вектора.
В одном варианте осуществления вектор представляет собой лентивирусный вектор. В одном варианте осуществления вектор, кроме того, включает промотор. В одном варианте осуществления промотором является промотор EF-1. В одном варианте осуществления промотор EF-1 включает последовательность SEQ ID NO: 97.
В одном варианте осуществления вектором является in vitro транскрибируемый вектор, например, вектор, который транскрибирует РНК с молекулы нуклеиновой кислоты, описанной здесь. В одном варианте осуществления последовательность нуклеиновой кислоты в векторе, кроме того, включает поли(А) хвост, например, поли(А) хвост, описанный здесь, например, включающий приблизительно 150 оснований аденозинов (SEQ ID NO: 111). В одном варианте осуществления последовательность нуклеиновой кислоты в векторе, кроме того, включает 3' UTR, например, 3' UTR, описанный здесь, например, включающий по крайней мере один повтор 3' UTR, происходящего из бета-глобулина человека.
В другом аспекте настоящее изобретение относится к клетке, включающей вектор, описанный здесь. В одном варианте осуществления клетка представляет собой клетку, описанную здесь, например, Т-клетку человека, например, Т-клетку человека, описанную здесь. В одном варианте осуществления Т-клеткой человека является CD8+ Т-клетка.
В другом аспекте настоящее изобретение относится к способу создания клетки, включающему трансдукцию клетки, описанной здесь, например, Т-клетки, описанной здесь, вектором, включающим нуклеиновую кислоту, кодирующую CAR, например, CAR, описанный здесь.
Настоящее изобретение также относится к способу получения популяции созданных с использованием РНК клеток, например, клеток, описанных здесь, например, Т-клеток, транзиторно экспрессирующих экзогенную РНК. Способ включает введение in vitro транскрибированной РНК или синтетической РНК в клетку, причем РНК включает нуклеиновую кислоту, кодирующую молекулу CAR, описанную здесь.
В другом аспекте настоящее изобретение относится к способу обеспечения иммунитета к опухоли у млекопитающего, включающему введение млекопитающему эффективного количества клетки, экспрессирующей молекулу CAR, например, клетки, экспрессирующей молекулу CAR, описанную здесь. В одном варианте осуществления клеткой является аутологичная Т-клетка. В одном варианте осуществления клеткой является аллогенная Т-клетка. В одном варианте осуществления млекопитающим является человек.
В другом аспекте настоящее изобретение относится к способу лечения млекопитающего, имеющего заболевание, связанное с экспрессией EGFRvIII (например, пролиферативного заболевания, предракового состояния и не относящегося к раку симптома, связанного с экспрессией EGFRvIII), включающему введение млекопитающему эффективного количества клеток, экспрессирующих молекулу CAR, например молекулу CAR, описанную здесь.
В одном варианте осуществления заболевание представляет собой заболевание, описанное здесь. В одном варианте осуществления заболеванием, связанным с EGFRvIII, является глиобластома. В одном варианте осуществления заболеванием, связанным с EGFRvIII, является рак, например, рак, выбираемый из группы, состоящей из мультиформной глиобластомы (GBM), анапластической астроцитомы, гигантоклеточной глиобластомы, глиосаркомы, анапластической олигодендроглиомы, анапластической эпендимомы, карциномы сосудистого сплетения, анапластической ганглиоглиомы, пинеобластомы, медуллоэпителиомы, эпендимобластомы, медуллобластомы, супратенториальной примитивной нейроэктодермальной опухоли, атипичной тератоидной/рабдоидной опухоли, рака легкого (например, немелкоклеточного рака легкого), рака молочной железы, предстательной железы, яичников, колоректального рака и рака мочевого пузыря и любой их комбинации, и метастазов любого из раков.
В одном варианте осуществления клетки, экспрессирующие молекулу CAR, например молекулу CAR, описанную здесь, вводят в комбинации с агентом, который увеличивает эффективность клетки, экспрессирующей молекулу CAR, например, с агентом, описанным здесь.
В одном варианте осуществления клетки, экспрессирующие молекулу CAR, например молекулу CAR, описанную здесь, вводят в комбинации с агентом, который уменьшает интенсивность одного или более побочных эффектов, связанных с введением клетки, экспрессирующей молекулу CAR, например, с агентом, описанным здесь.
В одном варианте осуществления клетки, экспрессирующие молекулу CAR, например молекулу CAR, описанную здесь, вводят в комбинации с агентом, который лечит заболевание, связанное с EGFRvIII, например, с агентом, описанным здесь.
В одном варианте осуществления клетки, экспрессирующие молекулу CAR, например молекулу CAR, описанную здесь, вводят в дозе и/или по схеме введения доз, описанной здесь.
В одном варианте осуществления клетки, экспрессирующие молекулу CAR, например молекулу CAR, описанную здесь, вводят в качестве лечения первой линии заболевания, например, рака, например, рака, описанного здесь. В другом варианте осуществления клетки, экспрессирующие молекулу CAR, например молекулу CAR, описанную здесь, вводят в качестве лечения второй, третьей, четвертой линии заболевания, например, рака, например, рака, описанного здесь.
В другом аспекте настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей CAR по настоящему изобретению, выделенной молекуле полипептида CAR по настоящему изобретению, вектору, включающему CAR по настоящему изобретению, и клетке, включающей CAR по настоящему изобретению, для применения в качестве лекарственного средства, например, описанного здесь.
В другом аспекте настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей CAR по настоящему изобретению, выделенной молекуле полипептида CAR по настоящему изобретению, вектору, включающему CAR по настоящему изобретению, и клетке, включающей CAR по настоящему изобретению, для применения в лечении заболевания, экспрессирующего EGFRvIII, например, заболевания, экспрессирующего EGFRvIII, описанного здесь.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1А и 1B представляют собой ряд блок-схем лентивирусных векторов для 3C10 и miR17-92. На фиг. 1А представлен экспрессирующий 3C10-CAR вектор pELNS-3C10-CAR; а на фиг. 1В представлен экспрессирующий miR17-92 лентивирусный вектор.
Фиг. 2A-2C представляют собой ряд изображений, демонстрирующих функциональную экспрессию 3C10-CAR и miR17-92, трансдуцированных с использованием лентивирусного вектора в Т-клетки человека. CD3+ Т-клетки были трансдуцированы только pELNS-3C10-CAR или как pELNS-3C10-CAR, так и FG12-EF1α-miR17-92.
Фиг. 3A-3D представляют собой ряд изображений, демонстрирующих, что коэкспрессия miR17-92 в CAR-Т-клетки придает стойкость к подавляющим эффектам TGF-β и TMZ. CAR-Т-клетки (неокрашенные столбики) и те, которые были котрансдуцированы miR17-92 (окрашенные столбики), сокультивировали с АРС, экспрессирующими EGFRvIII, в присутствии указанных концентраций TGF-β и TMZ.
Фиг. 4А и 4В являются изображениями, демонстрирующими сильные терапевтические эффекты CAR-Т-клеток у мышей, имеющих опухоли U87-EGFRvIII.
Фиг. 5A-5C представляют собой ряд изображений, демонстрирующих, что котрансдукция miR17-92 в CAR-Т-клетки дает улучшение защиты от повторно инокулируемых клеток глиомы.
Фиг. 6 является представлением, демонстрирующим сравнение репрезентативных EGFRvIII CAR (SEQ ID 1, 121 и 2, соответственно, в порядке появления).
Фиг. 7 представляет собой изображение, демонстрирующее, что Т-клетки человека, трансдуцированные EGFRvIII-специфическим CAR, демонстрируют специфический и сильный лизис EGFRvIII-экспрессирующих клеток GBM человека U87 (U87-EGFRvIII).
Фиг. 8 представляет собой график, показывающий, что все экспрессирующие CAR на основе антитела против EGFRvIII Т-клетки устраняют опухолевые клетки, но клеточная структура 3C10.BBz CARТ устраняет опухоли наиболее быстро к дню 7.
Фиг. 9 является таблицей, показывающей последовательности VH и VL гуманизированного антитела против EGFRvIII (SEQ ID NO: 122-127, соответственно, в порядке появления).
Фиг. 10 представляет собой график, демонстрирующий in vitro связывание конструкций растворимого гуманизированного scFv, связывающихся с линией EGFRvIII+ клеток.
Фиг. 11 представляет собой график, демонстрирующий in vitro связывание конструкций растворимого гуманизированного scFv, связывающихся с линией клеток, экспрессирующих EGFR дикого типа, с клоном 73 (также называемым CAR6) и клоном 74 (также называемым CAR7), демонстрируя профиль безопасности.
Фиг. 12 представляет собой график сравнения специфичности мышиного CAR9 и человеческого CAR10 в отношении EGFRvIII и EGFR дикого типа при транзиторной трансфекции клеток Jurkat и обнаружении с использованием слитых с Fc белков.
Фиг. 13 представляет собой график, демонстрирующий трансдукцию первичных Т-клеток донора конструкциями гуманизированного EGFRvIII-специфического CAR mCAR19 (контроль), CAR10, CAR9 и CAR6, окрашенных насыщающим количеством EGFRvIII.
Фиг. 14 представляет собой график, демонстрирующий люциферазную активность конструкций гуманизированного EGFRvIII-специфического CAR, стимулированную ВНК-EGFRvIII клетками, но не клетками дикого типа.
Фиг. 15 представляет собой график, демонстрирующий, что конструкции гуманизированного EGFRvIII-специфического CAR пролиферируют в ответ на стимул U87vIII без фоновой пролиферации на EGFR дикого типа.
Фиг. 16 представляет собой график, демонстрирующий, что конструкции гуманизированного EGFRvIII-специфического CAR пролиферируют in vitro в присутствии стимула U87vIII.
Фиг. 17 представляет собой график, демонстрирующий 4-часовой анализ уничтожения опухоли - высвобождения 51-хрома, в котором конструкция гуманизированного EGFRvIII-специфического CAR, 2173 (CAR6) и CAR9 специфически уничтожают EGFRvIII-экспрессирующие клетки, но не клетки, экспрессирующие EGFR дикого типа.
Фиг. 18 представляет собой график, показывающий изменение размера опухоли (см3, верхняя левая панель) и изменение средней интенсивности эмиссии в опухоли (фотонов/сек/см2/стерадиан, верхняя правая панель), и кривую выживаемости Каплана-Мейера (ниже) in vivo у мышей, получивших CAR+ Т-клетки, трансдуцированные конструкцией гуманизированного EGFRvIII-специфического CAR (CAR6).
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Определения
Кроме особо оговоренных случаев, все технические и научные термины, используемые здесь, имеют такое же значение, в котором их обычно понимает специалист со средним уровнем компетентности в области техники, к которой относится настоящее изобретение.
Артикли «a» и «an» относятся к одному или более чем одному (т.e. к по крайней мере одному) грамматическому подлежащему артикля. В качестве примера, «an element» означает один элемент или более чем один элемент.
Термин «приблизительно», когда относится к измеряемой величине, такой как количество, продолжительность времени и т.п., как подразумевается, охватывает вариации, составляющие ±20% или в некоторых случаях ±10%, или в некоторых случаях ±5%, или в некоторых случаях ±1%, или в некоторых случаях ±0,1% от указанной величины, поскольку такие вариации являются адекватными для выполнения описываемых способов.
Термин «химерный антигенспецифический рецептор» или альтернативно «CAR» относится к рекомбинантной полипептидной конструкции, включающий по крайней мере экстраклеточный антигенсвязывающий домен, трансмембранный домен и цитоплазматический домен сигнализации (также называемый здесь «внутриклеточным доменом сигнализации»), включающий функциональный домен сигнализации, происходящий из стимулирующей молекулы, определенной ниже. В одном аспекте стимулирующей молекулой является дзета цепь, связанная с Т-клеточным рецепторным комплексом. В одном аспекте цитоплазматический домен сигнализации, кроме того, включает один или более функциональных доменов сигнализации, происходящих из по крайней мере одной костимулирующей молекулы, определенной ниже. В одном аспекте костимулирующую молекулу выбирают из 4-1ВВ (т.е. CD137) и/или CD28. В одном аспекте CAR включает химерный слитый белок, включающий экстраклеточный домен распознавания антигена, трансмембранный домен и внутриклеточный домен сигнализации, включающий функциональный домен сигнализации, происходящий из стимулирующей молекулы. В одном аспекте CAR включает химерный слитый белок, включающий экстраклеточный домен распознавания антигена, трансмембранный домен и внутриклеточный домен сигнализации, включающий функциональный домен сигнализации, происходящий из костимулирующей молекулы, и функциональный домен сигнализации, происходящий из стимулирующей молекулы. В одном аспекте CAR включает химерный слитый белок, включающий экстраклеточный домен распознавания антигена, трансмембранный домен и внутриклеточный домен сигнализации, включающий два функциональных домена сигнализации, происходящих из одной или более костимулирующих молекул, и функциональный домен сигнализации, происходящий из стимулирующей молекулы. В одном аспекте CAR включает химерный слитый белок, включающий экстраклеточный домен распознавания антигена, трансмембранный домен и внутриклеточный домен сигнализации, включающий по крайней мере два функциональных домена сигнализации, происходящих из одной или более костимулирующих молекул, и функциональный домен сигнализации, происходящий из стимулирующей молекулы. В одном аспекте CAR включает необязательную лидерную последовательность на амино-конце (N-конце) слитого белка CAR. В одном аспекте CAR, кроме того, включает лидерную последовательность на N-конце экстраклеточного домена распознавания антигена, причем лидерная последовательность необязательно отщепляется от домена распознавания антигена (например, scFv) во время клеточного процессинга и локализации CAR на клеточной мембране.
Термин «домен сигнализации» относится к функциональной части белка, которая действует, передавая информацию в клетке для регулирования клеточной активности через определенные пути передачи сигналов благодаря образованию вторичных мессенджеров или функционированию в качестве эффекторов при ответе на такие мессенджеры.
Термин «EGFR» относится к зрелому полноразмерному рецептору эпидермального фактора роста любого млекопитающего, в том числе человеческим и нечеловеческим формам. Состоящий из 1186 аминокислот EGFR человека описан в Ullrich et al., Nature 309: 418-425 (1984)) и имеет № доступа в GenBank - AF125253 и № доступа в SwissProt - P00533-2.
Термин «EGFRvIII» относится к варианту III рецептора эпидермального фактора роста. EGFRvIII является самым распространенным вариантом EGFR, который наблюдается в опухолях человека, но редко наблюдается в нормальной ткани. Этот белок является результатом делеции в рамке считывания экзонов 2-7 и образования нового остатка глицина на стыке экзонов 1 и 8 в экстраклеточном домене EGFR, в результате чего создается специфический в отношении опухоли эпитоп. EGFRvIII экспрессируется в 24%-67% GBM, но не в нормальных тканях. EGFRvIII также известен как мутант типа III, дельта EGFR, EGFRde2-7 и ΔEGFR и описан в патентах США №№ 6455498, 6127126, 5981725, 5814317, 5710010, 5401828 и 5212290. Экспрессия EGFRvIII может быть результатом хромосомной делеции и может также быть результатом аберрантного альтернативного сплайсинга. Смотрите Sugawa et al., 1990, Proc. Natl. Acad. Sci. 87: 8602-8606.
Используемый здесь термин «антитело» относится к белку или полипептидной последовательности, происходящей из молекулы иммуноглобулина, который специфически связывается с антигеном. Антитела могут быть поликлональными или моноклональными, много- или одноцепочечными, или интактными иммуноглобулинами, и могут быть получены из природных источников или из рекомбинантных источников. Антитела могут представлять собой тетрамеры иммуноглобулиновых молекул.
Термин «фрагмент антитела» относится к по крайней мере одной части интактного антитела, или его рекомбинантных вариантов, и относится к антигенсвязывающему домену, например, определяющей антиген вариабельной области интактного антитела, которая является достаточной для обеспечения распознавания и специфического связывания фрагмента антитела с мишенью, например, антигеном. Примеры фрагментов антитела включают, но без ограничения, Fab-, Fab'-, F(аb')2- и Fv-фрагменты, фрагменты антител scFv, линейные антитела, однодоменные антитела, такие как sdAb (или VL, или VH), VHH-домены семейства верблюдовых и полиспецифические антитела, образованные из фрагментов антител. Термин «scFv» относится к слитому белку, включающему по крайней мере один фрагмент антитела, включающий вариабельную область легкой цепи, и по крайней мере один фрагмент антитела, включающий вариабельную область тяжелой цепи, причем вариабельные области легкой и тяжелой цепей связаны рядом с помощью короткого гибкого полипептидного линкера и могут быть экспрессированы в виде одноцепочечного полипептида, и причем scFv сохраняет специфичность интактного антитела, из которого он происходит. Если не указано иное, как здесь используется, scFv может содержать вариабельные области VL и VH в любом порядке, например, по отношению к N-концу и С-концу полипептида, scFv может включать VL-линкер-VH или может включать VH-линкер-VL.
Часть структуры CAR по настоящему изобретению, включающая антитело или фрагмент антитела, может существовать в различных формах, причем антигенсвязывающий домен экспрессируется в виде части непрерывной полипептидной цепи, включая, например, фрагмент однодоменного антитела (sdAb), одноцепочечное антитело (scFv) и гуманизированное антитело (Harlow et al., 1999, In: Using Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, NY; Harlow et al., 1989, In: Antibodies: A Laboratory Manual, Cold Spring Harbor, New York; Houston et al., 1988, Proc. Natl. Acad. Sci. USA 85: 5879-5883; Bird et al., 1988, Science 242: 423-426). В одном аспекте антигенсвязывающий домен структуры CAR по настоящему изобретению включает фрагмент антитела. В дальнейшем аспекте CAR включает фрагмент антитела, который включает scFv.
Термин «тяжелая цепь антитела» относится к большей из двух типов полипептидных цепей, которая присутствует в молекулах антител в их встречающихся в природе конформациях, и которая обычно определяет класс, к которому относится антитело.
Термин «легкая цепь антитела» относится к меньшей из двух типов полипептидных цепей, присутствующих в молекулах антител в их встречающихся в природе конформациях. Легкие цепи каппа (κ) и лямбда (λ) относятся к двум основным изотипам легких цепей антител.
Термин «рекомбинантное антитело» относится к антителу, которое создают с использованием технологии рекомбинантных ДНК, такому как, например, антитело, экспрессируемое бактериофагом или дрожжевой экспрессионной системой. Термин, как должно также подразумеваться, означает антитело, которое было создано посредством синтеза молекулы ДНК, которая кодирует антитело и которая экспрессирует белок антитело, или аминокислотную последовательность, определяющую антитело, причем ДНК или аминокислотная последовательность были получены, используя технологию рекомбинантных ДНК или аминокислотных последовательностей, которая имеется в распоряжении и хорошо известна в данной области техники.
Используемый здесь термин «антиген» или «Аг(Ag)» определяется как молекула, которая стимулирует иммунный ответ. Этот иммунный ответ может включать или продукцию антител, или активацию специфических иммунологически компетентных клеток, или и то, и другое. Квалифицированному специалисту будет понятно, что любая макромолекула, включая фактически все белки или пептиды, может служить в качестве антигена. Кроме того, антигены могут происходить из рекомбинантной или геномной ДНК. Квалифицированному специалисту будет понятно, что любая ДНК, которая включает нуклеотидную последовательность или частичную нуклеотидную последовательность, кодирующую белок, который индуцирует иммунный ответ, поэтому кодирует «антиген», как этот термин здесь используется. Кроме того, квалифицированному в данной области техники специалисту будет понятно, что не требуется, чтобы антиген кодировался исключительно полноразмерной нуклеотидной последовательностью гена. Со всей очевидностью вытекает, что настоящее изобретение включает, но без ограничения, использование частичных нуклеотидных последовательностей более чем одного гена, и что эти нуклеотидные последовательности организованы в различные комбинации для кодирования полипептидов, которые вызывают желаемый иммунный ответ. Более того, квалифицированному специалисту будет понятно, что совсем не требуется, чтобы антиген кодировался «геном». Со всей очевидностью вытекает, что антиген может быть созданным синтезированным или может происходить из биологического образца, или мог бы быть макромолекулой помимо полипептида. Такой биологический образец может включать, но без ограничения, образец ткани, образец опухоли, клетку или жидкость с другими биологическим компонентами.
Используемый здесь термин «противоопухолевый эффект» относится к биологическому эффекту, который может проявляться по-разному, в том числе, но без ограничения, уменьшением объема опухоли, уменьшением числа опухолевых клеток, уменьшением числа метастазов, увеличением средней ожидаемой продолжительности жизни, уменьшением пролиферации опухолевых клеток, уменьшением выживания опухолевых клеток или уменьшением различных физиологических симптомов, связанных с раковым состоянием. «Противоопухолевый эффект» может также проявляться способностью пептидов, полинуклеотидов, клеток и антител по настоящему изобретению к предотвращению возникновения первичной опухоли.
Термин «аутологичный» относится к любому материалу, происходящему от того же индивидуума, которому он позже должен быть снова введен.
Термин «аллогенный» относится к любому материалу, полученному из другого животного того же вида, что и индивидуум, которому вводят материал. Говорят, что два или более индивидуумов являются аллогенными друг относительно друга, когда гены в одном или более локусов не идентичны. В некоторых аспектах аллогенный материал от индивидуумов одного и того же вида может быть достаточно отличным генетически, чтобы взаимодействовать антигенно.
Термин «ксеногенный» относится к трансплантату, происходящему от животного отличного вида.
Термин «рак» относится к заболеванию, характеризующемуся быстрым и неконтролируемым ростом аберрантных клеток. Раковые клетки могут распространяться локально или через кровяное русло и лимфатическую систему в другие части тела. Примеры различных раков описаны здесь и включают, но без ограничения, глиобластому, рак молочной железы, рак предстательной железы, рак яичника, рак шейки матки, рак кожи, рак поджелудочной железы, колоректальный рак, рак почки, рак печени, рак головного мозга, лимфому, лейкоз, рак легкого и т.п.
Используемый здесь термин «заболевание, связанное с экспрессией EGFRvIII» включает, но без ограничения, заболевание, связанное с экспрессией EGFRvIII, или состояние, связанное с клетками, которые экспрессируют EGFRvIII, включая опухолевые клетки различных раковых заболеваний, таких как, например, глиобластома (в том числе стволовые клетки глиобластомы); рак молочной железы, рак яичников и мелкоклеточный рак легкого; плоскоклеточный рак головы и шеи; медуллобластома, колоректальный рак, рак предстательной железы и рак мочевого пузыря. Без ограничения конкретной теорией или механизмом, полагают, что при вызове антигенспецифического ответа против EGFRvIII, CAR, описанные здесь, обеспечивают одно или более из следующего: нацеленность на EGFRvIlI-экспрессирующие опухолевые клетки и их уничтожение, уменьшение или уничтожение опухолей, облегчение инфильтрации иммунных клеток в опухолевый центр и повышение/расширение противоопухолевых ответов. Поскольку EGFRvIII не экспрессируется на обнаружимых уровнях в нормальной (т.е. не раковой) ткани, предполагается, что преимущественно CAR по настоящему изобретению в основном избегают нацеливания на нормальные ткани и клетки/их уничтожения.
Термин «консервативные модификации последовательности» предназначен для обозначения аминокислотных модификаций, которые существенно не влияют или не изменяют характеристики связывания антитела или фрагмента антитела, содержащего аминокислотную последовательность. Такие консервативные модификации включают замены, добавления и делеции аминокислот. Модификации могут быть введены в антитело или фрагмент антитела по настоящему изобретению с помощью стандартных методов, известных в данной области техники, таких как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Консервативными аминокислотными заменами являются те, в которых аминокислотный остаток заменен аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, были определены в данной области техники. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновую кислоту, глютаминовую кислоту), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глютамин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или более аминокислотных остатков в CAR по настоящему изобретению могут быть заменены другими аминокислотными остатками из того же семейства боковых цепей, и измененный CAR может быть проверен, используя функциональные анализы, описанные здесь.
Термин «стимулирование» относится к первичной ответной реакции, индуцируемой связыванием стимулирующей молекулы (например, TCR/CD3 комплекса) с узнаваемым лигандом, таким образом, опосредуя событие сигнальной трансдукции, такое как, но без ограничения, сигнальная трансдукция с помощью TCR/CD3-комплекса. Стимулирование может опосредовать измененную экспрессию определенных молекул, такую как уменьшение экспрессии TGF-β и/или реорганизация структур цитоскелета и т.п.
Термин «стимулирующая молекула» относится к молекуле, экспрессируемой Т-клеткой, которая обеспечивает цитоплазматическую последовательность(и) первичной сигнализации, регулирующую первичную активацию TCR комплекса стимулирующим образом в по крайней мере некотором аспекте пути передачи сигналов с участием Т-клеток. В одном аспекте первичный сигнал инициируется, например, в результате связывания TCR/CD3 комплекса с молекулой MHC, связанной с пептидом, и которое приводит к опосредованию Т-клеточной реакции, в том числе, но без ограничения, пролиферации, активации, дифференциации и т.п. Цитоплазматическая последовательность первичной сигнализации (также называемая «доменом первичной сигнализации»), которая действует стимулирующим образом, может содержать мотив сигнализации, который известен как иммунорецепторный тирозиновый активирующий мотив или ITAM. Примеры содержащей ITAM цитоплазматической последовательности первичной сигнализации, которая является особенно полезной в настоящем изобретении, включают, но без ограничения, те, которые происходят из TCR дзета, FcR гамма, FcR бета, CD3 гамма, CD3 дельта, CD3 эпсилон, CD5, CD22, CD79a, CD79b, CD278 (также известного как «ICOS») и CD66d. В конкретном CAR по настоящему изобретению внутриклеточный домен сигнализации в любой одном или более CAR по настоящему изобретению включает внутриклеточную последовательность сигнализации, например, последовательность первичной сигнализации CD3-дзета. В конкретном CAR по настоящему изобретению последовательность первичной сигнализации CD3-дзета представляет собой последовательность, представленную как SEQ ID NO: 17, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, обезьяны, примата и т.п. В конкретном CAR по настоящему изобретению последовательность первичной сигнализации CD3-дзета представляет собой последовательность, представленную как SEQ ID NO: 99, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, обезьяны, примата и т.п.
Термин «антигенпрезентирующая клетка» или «АРС» относится к клетке иммунной системы, такой как А-клетка (например, В-клетка, дендритная клетка и т.п.), которая представляет чужеродный антиген в комплексе с главными комплексами гистосовместимости (МНС) на своей поверхности. Т-клетки могут распознавать эти комплексы, используя свои Т-клеточные рецепторы (TCR). АРС процессируют антигены и презентируют их Т-клеткам.
«Внутриклеточный домен сигнализации», как этот термин здесь используется, относится к внутриклеточной части молекулы. Внутриклеточный домен сигнализации порождает сигнал, который стимулирует иммунную эффекторную функцию содержащей CAR клетки, например, CAR-T-клетки. Примеры иммунной эффекторной функции, например, в CAR-T-клетке, включают цитолитическую активность и хэлперную активность, в том числе секрецию цитокинов.
В одном варианте осуществления внутриклеточный домен сигнализации может включать внутриклеточный домен первичной сигнализации. Примеры внутриклеточных доменов первичной сигнализации включают те, которые происходят из молекул, ответственных за первичную стимуляцию, или зависящую от антигена стимуляцию. В одном варианте осуществления внутриклеточный домен сигнализации может включать костимулирующий внутриклеточный домен. Приводимые в качестве примера внутриклеточные домены костимулирующей сигнализации включают те, которые происходят из молекул, ответственных за костимулирующие сигналы, или не зависящую от антигена стимуляцию. Например, в случае CAR-T-клетки внутриклеточный домен первичной сигнализации может включать цитоплазматическую последовательность Т-клеточного рецептора, а внутриклеточный домен костимулирующей сигнализации может включать цитоплазматическую последовательность корецептора или костимулирующей молекулы.
Внутриклеточный домен первичной сигнализации может включать мотив сигнализации, который известен как иммунорецепторный тирозиновый активирующий мотив или ITAM. Примеры содержащих ITAM цитоплазматических последовательностей первичной сигнализации включают, но без ограничения, те, которые происходят из CD3 дзета, FcR гамма, FcR бета, CD3 гамма, CD3 дельта, CD3 эпсилон, CD5, CD22, CD79a, CD79b, и CD66d DAP10 и DAP12.
Термин «дзета» или альтернативно «дзета цепь», «CD3-дзета» или «TCR-дзета» определяется как белок, представленный под входящим № в GenBan - BAG36664.1, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, обезьяны, примата и т.п., а «стимулирующий домен дзета» или альтернативно «стимулирующий домен CD3-дзета» или «стимулирующий домен TCR-дзета» определяется как аминокислотные остатки цитоплазматического домена дзета цепи, которые достаточны для функциональной передачи первичного сигнала, необходимого для активации Т-клеток. В одном аспекте цитоплазматический домен дзета включает остатки с 52 по 164 включительно белка с входящим № в GenBan - BAG36664.1, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, обезьяны, примата и т.п., которые являются их функциональными ортологами. В одном аспекте «стимулирующий домен зета» или «стимулирующий домен CD3-дзета» представляет собой последовательность, представленную как SEQ ID NO: 17. В одном аспекте «стимулирующим доменом зета» или «стимулирующим доменом CD3-дзета» является последовательность, представленная как SEQ ID NO: 99.
Термин «костимулирующая молекула» относится к узнаваемому партнеру по связыванию на T-клетке, который специфически связывается с костимулирующим лигандом, тем самым опосредуя костимуляторную реакцию, проявляемую Т-клеткой, такую как, но без ограничения, пролиферация. Костимулирующими молекулами являются молекулы клеточной поверхности, отличные от антигенспецифических рецепторов или их лигандов, которые необходимы для эффективного иммунного ответа. Костимулирующие молекулы включают, но без ограничения, молекулу МНС класса I, BTLA и Toll-подобный рецептор лиганда, а также OX40, CD2, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11а/CD18) и 4-1ВВ (CD137).
Внутриклеточный домен костимулирующей сигнализации может происходить из внутриклеточной части костимулирующей молекулы. Костимулирующая молекула может быть представлена в следующих семействах белков: белки рецепторы TNF, иммуноглобулин-подобные белки, рецепторы цитокинов, интегрины, сигнальные молекулы активации лимфоцитов (белки SLAM) и NK-клеточные рецепторы активации. Примеры таких молекул включают CD27, CD28, 4-1BB (CD137), OX40, GITR, CD30, CD40, ICOS, BAFFR, HVEM, связанный с функционированием лимфоцитов антиген-1 (LFA-1), CD2, CD7, LIGHT, NKG2C, SLAMF7, NKp80, CD160, B7-Н3 и лиганд, который специфически связывается с CD83, и т.п.
Внутриклеточный домен сигнализации может включать всю внутриклеточную часть, или весь природный внутриклеточный домен сигнализации, молекулы, из которой он происходит, или его функциональный фрагмент.
Термин «4-1ВВ» относится к члену суперсемейства TNFR с аминокислотной последовательностью, представленной как входящий № в GenBank - AAA62478.2, или эквивалентными остатками из не являющегося человеком вида, например, мыши, грызуна, обезьяны, примата и т.п., и «костимулирующий домен 4-1ВВ» определяется как аминокислотные остатки 214-255 белка с входящим № в GenBank - AAA62478.2, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, обезьяны, примата и т.п. В одном аспекте «костимулирующий домен 4-1ВВ» представляет собой последовательность, представленную как SEQ ID NO: 16, или эквивалентные остатки из не являющегося человеком вида, например, мыши, грызуна, обезьяны, примата и т.п.
Термин «кодирование» относится к внутренне присущей способности конкретных последовательностей нуклеотидов в полинуклеотиде, таком как ген, кДНК или мРНК, служить в качестве матриц для синтеза других полимеров и макромолекул в ходе биологических процессов, имеющих или определенную последовательность нуклеотидов (т.е. рРНК, тРНК и мРНК), или определенную последовательность аминокислот и биологические свойства, являющиеся их следствием. Таким образом, ген, кДНК или РНК кодирует белок, если транскрипция и трансляция мРНК, соответствующей этому гену, приводит к образованию белка в клетке или другой биологической системе. И кодирующую цепь, нуклеотидная последовательность которой идентична последовательности мРНК и обычно представлена в списках последовательностей, и некодирующую цепь, используемую в качестве матрицы для транскрипции гена или кДНК, можно назвать кодирующими белок или другой продукт этого гена или кДНК.
Кроме случаев, оговоренных особо, «нуклеотидная последовательность, кодирующая аминокислотную последовательность» включает все нуклеотидные последовательности, которые являются вырожденными вариантами друг друга и которые кодируют одну и ту же аминокислотную последовательность. Выражение «нуклеотидная последовательность, которая кодирует белок или РНК» может также включать интроны в той мере, в которой нуклеотидная последовательность, кодирующая белок, может в каком-либо варианте содержать интрон(ы).
Термины «эффективное количество» и «терапевтически эффективное количество» используются здесь взаимозаменяемо и относятся к количеству соединения, препарата, материала или композиции, как описано здесь, эффективному для достижения конкретного биологического результата.
Термин «эндогенный» относится к любому материалу из или продуцированному внутри организма, клетки, ткани или системы.
Термин «экзогенный» относится к любому материалу, введенному из или продуцированному вне организма, клетки, ткани или системы.
Термин «экспрессия» относится к транскрипции и/или трансляции конкретной нуклеотидной последовательности, управляемой промотором.
Термин «вектор для переноса» относится к смеси химически связанных веществ, которая включает выделенную нуклеиновую кислоту и которая может использоваться для доставки выделенной нуклеиновой кислоты внутрь клетки. Многочисленные векторы известны в данной области, включая, но без ограничения, линейные полинуклеотиды, полинуклеотиды, связанные с ионными или амфифильными соединениями, плазмиды и вирусы. Таким образом, термин «вектор для переноса» включает автономно реплицирующуюся плазмиду или вирус. Этот термин, как будет также подразумеваться, будет, кроме того, включать неплазмидные и невирусные соединения, которые облегчают перенос нуклеиновой кислоты в клетки, такие как, например, соединение полилизин, липосома и т.п. Примеры вирусных векторов для переноса включают, но без ограничения, аденовирусные векторы, аденоассоциированные вирусные векторы, ретровирусные векторы, лентивирусные векторы и т.п.
Термин «экспрессионный вектор» относится к вектору, включающему рекомбинантный полинуклеотид, включающий контролирующие экспрессию последовательности, функционально связанные с экспрессируемой нуклеотидной последовательностью. Экспрессионный вектор включает достаточные для экспрессии действующие в цис-положении элементы; другие элементы для экспрессии могут обеспечиваться клеткой-хозяином или в in vitro экспрессионной системе. Экспрессионные векторы включают все экспрессионные векторы, которые известны в данной области техники, такие как космиды, плазмиды (например, «голые» или содержащиеся в липосомах) и вирусы (например, лентивирусы, ретровирусы, аденовирусы и аденоассоциированные вирусы), которые включают в свой состав рекомбинантный полинуклеотид.
Термин «лентивирус» относится к роду семейства Retroviridae. Лентивирусы являются уникальными среди ретровирусов, являясь способными к инфицированию неделящихся клеток; они могут доставлять значительное количество генетической информации в ДНК клетки-хозяина, а значит они являются одними из самых эффективных средств - векторов для доставки генов. Все из ВИЧ, SIV и FIV являются примерами лентивирусов.
Термин «лентивирусный вектор» относится к вектору, происходящему из по крайней мере части генома лентивируса, в том числе особенно самоинактивирующемуся лентивирусному вектору, представленному в Milone et al. Mol. Ther. 17 (8): 1453-1464 (2009). Другие примеры лентивирусных векторов, которые могут быть использованы в клинике, включают, но без ограничения, например, технологию доставки генов LENTIVECTOR® от Oxford BioMedica, векторную систему LENTIMAX™ от Lentigen и т.п. Неклинические типы лентивирусных векторов также имеются в распоряжении и должны быть известны квалифицированному в данной области техники специалисту.
Используемый здесь термин «гомологичные» или «идентичность» относится к идентичности последовательностей субъединиц между двумя полимерными молекулами, например, между двумя молекулами нуклеиновых кислот, такими как две молекулы ДНК или две молекулы РНК, или между двумя полипептидными молекулами. Когда положение субъединицы в обеих из двух молекул занято одной и той же мономерной субъединицей, например, если положение в каждой из двух молекул ДНК занято аденином, то они являются гомологичными или идентичными в этом положении. Процент гомологии между двумя последовательностями напрямую зависит от числа совпадающих или гомологичных положений, например, если половина (например, пять положений в полимере длиной десять субъединиц) положений в двух последовательностях являются гомологичными, две последовательности гомологичны на 50%; если 90% положений (например, 9 из 10) являются совпадающими или гомологичными, эти две последовательности гомологичны на 90%.
«Гуманизированные» формы нечеловеческих (например, мышиных) антител представляют собой химерные иммуноглобулины, цепи иммуноглобулинов или их фрагменты (такие как Fv, Fab, Fab', F(аb')2 или другие антигенсвязывающие подпоследовательности антител), которые содержат минимальную последовательность, происходящую из нечеловеческого иммуноглобулина. В большинстве случаев гуманизированные антитела и фрагменты антител представляют собой иммуноглобулины человека (антитело-реципиент или фрагмент антитела), в которых остатки из определяющего комплементарность участка (CDR) реципиента заменены остатками из CDR из не являющегося человеком вида (антитела-донора), такого как мышь, крыса или кролик, обладающими желаемой специфичностью, аффинностью и способностью. В некоторых случаях остатки каркасной области (FR) Fv иммуноглобулина человека заменены соответствующими не принадлежащими человеку остатками. Кроме того, гуманизированное антитело/фрагмент антитела может включать остатки, которые не обнаруживаются ни в антителе-реципиенте, ни в импортируемых CDR или каркасных последовательностях. Эти модификации могут дополнительно улучшить и оптимизировать характеристики антитела или фрагмента антитела. В общем, гуманизированное антитело или фрагмент антитела будет включать все или по существу все из по крайней мере одного, а обычно двух вариабельных доменов, в которых по существу все из CDR-участков соответствуют таковым нечеловеческого иммуноглобулина, и все или значительная часть FR-областей принадлежат последовательности иммуноглобулина человека. Гуманизированное антитело или фрагмент антитела могут также включать по крайней мере часть константной области иммуноглобулина (Fc), обычно таковую иммуноглобулина человека. Для получения более подробной информации смотрите Jones et al., Nature, 321: 522-525, 1986; Reichmann et al., Nature, 332: 323-329, 1988; Presta, Curr. Op. Struct. Biol., 2: 593-596, 1992.
Термин антитело «человека» относится к полностью человеческим антителам, а также фактически человеческим антителам. Термин «полностью человеческий» относится к иммуноглобулину, такому как антитело или фрагмента антитела, вся молекула которого имеет человеческое происхождение или состоит из аминокислотной последовательности, идентичной человеческой форме антитела или иммуноглобулина. «Фактически человеческое» антитело представляет собой антитело, которое включает достаточное количество положений аминокислот человека, так что антитело не вызывает иммуногенную реакцию у нормального человека.
Термин «выделенный» означает измененный или удаленный из природного состояния. Например, нуклеиновая кислота или пептид, присутствующий в природе в живом животном, не является «выделенным», но та же нуклеиновая кислота или пептид, которая частично или полностью отделена от сосуществующих материалов ее (его) природного состояния, является выделенной. Выделенная нуклеиновая кислота или белок может существовать в значительной степени очищенной форме или может существовать в неприродном окружении, такой как, например, клетка-хозяин.
В контексте настоящего изобретения используются следующие сокращения для обычно встречающихся нуклеотидных оснований. «A» относится к аденозину, «C» относится к цитозину, «G» относится к гуанозину, «T» относится к тимидину, и «U» относится к уридину.
Термин «функционально связанный» или «транскрипционный контроль» относится к функциональной связи между регуляторной последовательностью и гетерологичной нуклеотидной последовательностью, приводящей к экспрессии последней. Например, первая последовательность нуклеиновой кислоты функционально связана со второй последовательностью нуклеиновой кислоты, когда первая последовательность нуклеиновой кислоты установлена в функциональной связи со второй последовательностью нуклеиновой кислоты. Например, промотор функционально связан с кодирующей последовательностью, если промотор оказывает влияние на транскрипцию или экспрессию кодирующей последовательности. Функционально связанные последовательности ДНК могут быть соседними и, в случае необходимости соединения двух кодирующих белки областей, находятся в одной и той же рамке считывания.
Термин «парентеральное» введение иммуногенной композиции включает, например, подкожную (s.c.), внутривенную (i.v.), внутримышечную (i.m.) или интрастернальную инъекцию, внутрь опухоли, или способы инфузии.
Термин «нуклеиновая кислота» или «полинуклеотид» относится к дезоксирибонуклеиновым кислотам (ДНК) или рибонуклеиновым кислотам (РНК) и их полимерах в или одно-, или двухцепочечной форме. Если специально не ограничивается, термин охватывает нуклеиновые кислоты, содержащие известные аналоги природных нуклеотидов, которые обладают способностями к связыванию, аналогичными таковым ссылочной нуклеиновой кислоты, и которые метаболизируются аналогично со встречающимися в природе нуклеотидами. Если не указано иное, конкретная последовательность нуклеиновой кислоты также подразумеваемым образом охватывает ее консервативно модифицированные варианты (например, замены вырожденными кодонами), аллели, ортологи, SNP и комплементарные последовательности, а также последовательности, указанные в прямой форме. В частности, замены вырожденными кодонами могут быть получены путем создания последовательностей, в которых третье положение одного или более выбранных (или всех) кодонов замещено остатками со смешанными основаниями и/или дезоксиинозиновыми остатками (Batzer et al., Nucleic Acid Res. 19: 5081 (1991); Ohtsuka et al., J. Biol. Chem. 260: 2605-2608 (1985); и Rossolini et al., Mol. Cell. Probes 8: 91-98 (1994)).
Термины «пептид», «полипептид» или «белок» используются взаимозаменяемо и относятся к соединению, состоящему из аминокислотных остатков, ковалентно связанных пептидными связями. Белок или пептид должен содержать по крайней мере две аминокислоты, и не устанавливается ограничение на максимальное число аминокислот, которое может включать последовательность белка или пептида. Полипептиды включают любой пептид или белок, включающий две или более аминокислот, соединенных друг с другом пептидными связями. Используемый здесь термин относится и к коротким цепям, которые обычно называют в данной области техники пептидами, олигопептидами и олигомерами, например, и к более длинным цепям, которые обычно называют в данной области техники белками, множество типов которых существует. «Полипептиды» включают, например, биологически активные фрагменты, по существу гомологичные полипептиды, олигопептиды, гомодимеры, гетеродимеры, варианты полипептидов, модифицированные полипептиды, производные, аналоги, слитые белки, среди прочих. Полипептиды включают природные пептиды, рекомбинантные пептиды или их комбинацию.
Термин «промотор» относится к последовательности ДНК, распознаваемой относящейся к синтезу системой клетки, или введенной, относящейся к синтезу системой, необходимой для инициации специфической транскрипции последовательности полинуклеотида.
Термин «промотор/регуляторная последовательность» относится к последовательности нуклеиновой кислоты, которая необходима для экспрессии продукта гена, функционально связанного с промотором/регуляторной последовательностью. В некоторых случаях эта последовательность может представлять собой базовую промоторную последовательность, а в некоторых случаях эта последовательность может также включать последовательность энхансера и другие регуляторные элементы, которые необходимы для экспрессии продукта гена. Промотором/регуляторной последовательностью может, например, быть промотор/регуляторная последовательность, который экспрессирует продукт гена тканеспецифически.
Термин «конститутивный промотор» относится к нуклеотидной последовательности, которая, после функциональной связи с полинуклеотидом, который кодирует или определяет продукт гена, является причиной продукции продукта гена в клетке в большей части или во всех физиологических условиях клетки.
Термин «индуцируемый промотор» относится к нуклеотидной последовательности, которая, после функциональной связи с полинуклеотидом, который кодирует или определяет продукт гена, является причиной продукции продукта гена в клетке в основном только, когда в клетке присутствует индуктор, который соответствует промотору.
Термин «тканеспецифический промотор» относится к нуклеотидной последовательности, которая, после функциональной связи с полинуклеотидом, который кодирует или определяет продукт гена, является причиной продукции продукта гена в клетке в основном только, если клеткой является клетка типа ткани, соответствующего промотору.
Термин «гибкий полипептидный линкер» или «линкер», используемый в контексте scFv, относится к пептидному линкеру, который состоит из аминокислот, таких как остатки глицина и/или серина, используемому отдельно или в комбинации, чтобы связать вариабельные области тяжелой и легкой цепей вместе. В одном варианте осуществления гибкий полипептидный линкер представляет собой линкер Gly/Ser и включает аминокислотную последовательность (Gly-Gly-Gly-Ser)n (SEQ ID NO: 112), где n - целое положительное число, равное или большее 1. Например, n=1, n=2, n=3, n=4, n=5 и n=6, n=7, n=8, n=9 и n=10. В одном варианте осуществления гибкие полипептидные линкеры включают, но без ограничения, (Gly4 Ser)4 (SEQ ID NO: 113) или (Gly4 Ser)3 (SEQ ID NO: 114). В другом варианте осуществления линкеры включают множественные повторы (Gly2Ser), (GlySer) или (Gly3Ser) (SEQ ID NO: 112). Также в объем настоящего изобретения включены линкеры, описанные в WO2012/138475, который включен сюда в качестве ссылки.
Как здесь используется, 5' кэп-структура (также называемая кэп-структурой РНК, являющейся остатком 7-метилгуанозина кэп-структурой РНК или являющейся м7G кэп-структурой РНК) является модифицированным гуаниновым нуклеотидом, который был добавлен «перед» или на 5'-конец эукариотической мРНК вскоре после начала транскрипции. 5' кэп-структура состоит из концевой группы, которая связана с первым транскрибированным нуклеотидом. Его присутствие важно для распознания рибосомой и защиты от РНКазы. Добавление кэп-структуры связано с транскрипцией и происходит котранскрипционно, так что каждый из этих процессов оказывает влияние на другой процесс. Вскоре после начала транскрипции, 5'-конец синтезируемой мРНК связывается кэп-синтезирующим комплексом, связанным с РНК-полимеразой. Этот ферментативный комплекс катализирует химические реакции, которые необходимы для кэппирования мРНК. Синтез протекает как многоступенчатая биохимическая реакция. Фрагмент кэппирования можно модифицировать для модулирования функциональных возможностей мРНК, таких как ее стабильность или эффективность трансляции.
Как здесь используется, «in vitro транскрибированная РНК» относится к РНК, предпочтительно мРНК, которая была синтезирована in vitro. Как правило, in vitro транскрибированная РНК образуется с вектора для in vitro транскрипции. Вектор для in vitro транскрипции включает матрицу, которая используется для образования in vitro транскрибированной РНК.
Как здесь используется, «поли(А)» представляет собой ряд аденозинов, присоединенных с помощью полиаденилирования к мРНК. В предпочтительном варианте осуществления конструкции для транзиторной экспрессии, поли(А) составляет от 50 до 5000 (SEQ ID NO: 115), предпочтительно больше, чем 64, более предпочтительно больше, чем 100, более предпочтительно больше, чем 300 или 400. Последовательности поли(А) можно модифицировать химически или ферментативно для модулирования функциональных возможностей мРНК, таких как локализация, стабильность или эффективность трансляции.
Как здесь используется, «полиаденилирование» относится к ковалентной связи полиаденилильного фрагмента, или его модифицированного варианта, с молекулой мРНК. В эукариотических организмах большая часть молекул мРНК является полиаденилированной на 3'-конце. 3' поли(А)-хвост является длинной последовательностью из адениновых нуклеотидов (часто несколько сотен), добавленной к пре-мРНК под действием фермента полиаденилатполимеразы. У высших эукариот поли(А)-хвост добавляется на транскрипты, которые содержат специфическую последовательность, сигнал полиаденилирования. Поли(А)-хвост и белок, связанный с ним, помогают в защите мРНК от деградации под действием экзонуклеаз. Полиаденилирование также важно для терминации транскрипции, экспорта мРНК из ядра и трансляции. Полиаденилирование происходит в ядре сразу после транскрипции ДНК в РНК, но, кроме того, может также происходить позднее в цитоплазме. После терминации транскрипции цепь мРНК расщепляется под действием эндонуклеазного комплекса, связанного с РНК-полимеразой. Сайт расщепления, как правило, характеризуются присутствием последовательности оснований AAUAAA вблизи сайта расщепления. После расщепления мРНК остатки аденозина добавляются к свободному 3'-концу в сайте расщепления.
Как здесь используется, «транзиторная» относится к экспрессии не интегрированного трансгена в течение периода, составляющего часы, дни или недели, причем период времени экспрессии короче периода времени экспрессии гена, если он интегрирован в геном или содержится в стабильном плазмидном репликоне в клетке-хозяине.
Термин «путь трансдукции сигнала» относится к биохимической связи между различными молекулами трансдукции сигнала, которые играют роль в передаче сигнала из одной части клетки в другую часть клетки. Выражение «рецептор клеточной поверхности» включает молекулы и комплексы молекул, способные принимать сигнал и передавать сигнал через мембрану клетки.
Термин «субъект», как предполагается, включает живые организмы, в которых может быть вызван иммунный ответ (например, млекопитающих, человека).
Термин «очищенная в значительной степени» клетка относится к клетке, которая по существу не содержит другие типы клеток. Очищенная в значительной степени клетка также относится к клетке, которая была отделена от других типов клеток, с которыми она обычно связана в своем встречающемся в природе состоянии. В некоторых случаях популяция очищенных в значительной степени клеток относится к гомогенной популяции клеток. В других случаях этот термин относится просто к клеткам, которые были отделены от клеток, с которыми они связаны в природе в своем природном состоянии. В некоторых вариантах осуществления клетки подвергают культивированию in vitro. В других вариантах осуществления клетки не подвергают культивированию in vitro.
Термин «терапевтическое», как здесь используется, означает лечение. Терапевтический эффект достигается при ослаблении, подавлении, ремиссии или ликвидации болезненного состояния.
Термин «профилактика», как здесь используется, означает предупреждение или профилактику заболевания или болезненного состояния.
В контексте настоящего изобретения «опухолевый антиген» или «антиген, связанный с гиперпролиферативным нарушением», относится к антигенам, которые характерны для конкретных гиперпролиферативных нарушений. В некоторых аспектах связанные с гиперпролиферативнвми нарушениями антигены настоящего изобретения происходят из раков, включая, но без ограничения, первичную или метастатическую меланому, тимому, лимфому, саркому, рак легкого, рак печени, неходжкинскую лимфому, неходжкинскую лимфому, лейкозы, рак матки, рак шейки рак, рак мочевого пузыря, рак почки и аденокарциномы, такие как рак молочной железы, рак предстательной железы, рак яичников, рак поджелудочной железы и т.п.
Термин «трансфицированная» или «трансформированная, или «трансдуцированная» относится к процессу, с помощью которого экзогенную нуклеиновую кислоту трансфицируют или вводят в клетку-хозяина. «Трансфицированная» или «трансформированная, или «трансдуцированная» клетка является клеткой, которая была подвергнута трансфекции, трансформации или трансдукции экзогенной нуклеиновой кислотой. Клетка включает эмбриональную клетку субъекта и ее потомство.
Термин «специфически связывается» относится к антителу, или лиганду, которое(ый) распознает и связывает узнаваемый белок, являющийся партнером по связыванию (например, стимулирующую и/или костимулирующую молекулу, присутствующую на Т-клетках), присутствующий в образце, но антитело или лиганд практически не распознает или не связывает другие молекулы в образце.
Диапазоны: На протяжении всего этого описания различные аспекты настоящего изобретения могут быть представлены в формате диапазона. Следует понимать, что описание в формате диапазона представлено исключительно для удобства и краткости и не должно рассматриваться как жесткое ограничение объема настоящего изобретения. Соответственно, описание диапазона, как следует рассматривать, содержит конкретно выявляемые все возможные поддиапазоны, а также отдельные численные значения внутри этого диапазона. Например, описание такого диапазона, как от 1 до 6, как следует рассматривать, содержит конкретно выявляемые поддиапазоны, такие как от 1 до 3, от 1 до 4, от 1 до 5, от 2 до 4, от 2 до 6, от 3 до 6 и т.д., а также отдельные численные значения внутри этого диапазона, например, 1, 2, 2,7, 3, 4, 5, 5,3 и 6. В качестве другого примера, такой диапазон, как составляющая 95-99% идентичность, включает что-нибудь с составляющей 95%, 96%, 97%, 98% или 99% идентичностью и включает такие поддиапазоны, как составляющая 96-99%, 96-98%, 96-97%, 97-99%, 97-98% и 98-99% идентичность. Это применимо независимо от ширины диапазона.
Описание
Здесь обеспечиваются смеси химически связанных веществ и способы использования для лечения такого заболевания, как рак, используя химерные антигенспецифические рецепторы (CAR) на основе антитела против EGFRvIII.
В одном аспекте настоящим изобретением обеспечивает ряд химерных антигенспецифических рецепторов, включающих антитело или фрагмент антитела, созданных так, что они специфически связываются с белком EGFRvIII. В одном аспекте настоящим изобретением обеспечивается клетка (например, Т-клетка), созданная для экспрессии CAR, причем CAR-Т-клетка («CART») демонстрирует противоопухолевое свойство. В одном аспекте клетку трансформируют CAR, и CAR экспрессируется на поверхности клетки. В некоторых вариантах осуществления клетку (например, Т-клетку) трансдуцируют с использованием вирусного вектора, кодирующего CAR. В некоторых вариантах осуществления вирусный вектор представляет собой ретровирусный вектор. В некоторых вариантах осуществления вирусный вектор представляет собой лентивирусный вектор. В некоторых таких вариантах осуществления клетка может стабильно экспрессировать CAR. В другом варианте осуществления клетку (например, Т-клетку) трансфицируют нуклеиновой кислотой, например, мРНК, кДНК, ДНК, кодирующей CAR. В некоторых таких вариантах осуществления клетка может транзиторно экспрессировать CAR.
В одном аспекте связывающая белок EGFRvIII часть CAR представляет собой фрагмент антитела scFv. В одном аспекте такие фрагменты антител являются функциональными в том плане, что у них сохраняется эквивалентная аффинность, например, они связывают тот же антиген с эффективностью, сопоставимой с таковой антитела класса IgG, из которого они получены. В одном аспекте такие фрагменты антител являются функциональными в том плане, что они обеспечивают биологическую реакцию, которая может включать, но без ограничения, активацию иммунного ответа, ингибирование вызова трансдукции сигнала от его антигена-мишени, ингибирование активности киназы и т.п., как будет понятно квалифицированному в данной области техники специалисту.
В одном аспекте EGFRvIII-антигенсвязывающий домен CAR представляет собой фрагмент мышиного антитела scFv. В другом аспекте EGFRvIII-антигенсвязывающий домен CAR представляет собой фрагмент антитела scFv, который является гуманизированным по сравнению с мышиной последовательностью scFv, из которого он получен. Образование приводимого в качестве примера родительского мышиного моноклонального антитела против EGFRvIII (3C10) описано в Okamoto et al. (British J. Cancer 1996, 73: 1366-1372). Приводимое в качестве примера полностью человеческое антитело против EGFRvIII (139) описано в Morgan et al. (2012) Human Gene Therapy, 23: 1043-1953, который включен сюда посредством ссылки. В одном аспекте scFv в случае мышиной последовательности включает SEQ ID NO: 11. Гуманизация этого scFv мыши может быть желательной для клинических условий, в которых специфические для мыши остатки могут вызвать ответ против мышиных антигенов у человека (НАМА) у пациентов, получающих лечение против EGFRvIII, например, лечение с использованием Т-клеток, трансдуцированных конструкцией против EGFRvIII.
В одном аспекте связывающий домен антитела против EGFRvIII CAR кодируется трансгеном, последовательность которого была оптимизирована в отношении кодонов для экспрессии в клетке млекопитающего. В одном аспекте вся конструкция CAR по настоящему изобретению кодируется трансгеном, вся последовательность которого была оптимизирована в отношении кодонов для экспрессии в клетке млекопитающего. Оптимизация кодонов относится к открытию того, что частота встречаемости синонимичных кодонов (т.е. кодонов, которые кодируют одну и ту же аминокислоту) в кодирующей ДНК является смещенной у различных видов. Такая вырожденность кодонов делает возможным кодирование идентичных полипептидов рядом нуклеотидных последовательностей. Разнообразные методы оптимизации кодонов известны в данной области техники и включают, например, методы, описанные в по крайней мере патентах США №№ 5786464 и 6114148.
В одном аспекте связывающий домен антитела против EGFRvIII CAR представляет собой связывающий домен гуманизированного антитела против EGFRvIII. Например, в одном варианте осуществления связывающий домен антитела против EGFRvIII включает часть scFv, представленную в SEQ ID NO: 38. В одном аспекте связывающий домен гуманизированного антитела против EGFRvIII включает часть scFv, представленную в SEQ ID NO: 44. В одном аспекте связывающий домен гуманизированного антитела против EGFRvIII включает часть scFv, представленную в SEQ ID NO: 50. В одном аспекте связывающий домен гуманизированного антитела против EGFRvIII включает часть scFv, представленную в SEQ ID NO: 56. В одном аспекте связывающий домен гуманизированного антитела против EGFRvIII включает часть scFv, представленную в SEQ ID NO: 62. В одном аспекте связывающий домен гуманизированного антитела против EGFRvIII включает часть scFv, представленную в SEQ ID NO: 68. В одном аспекте связывающий домен гуманизированного антитела против EGFRvIII включает часть scFv, представленную в SEQ ID NO: 74. В одном аспекте связывающий домен гуманизированного антитела против EGFRvIII включает часть scFv, представленную в SEQ ID NO: 80. В одном аспекте связывающий домен гуманизированного антитела против EGFRvIII включает часть scFv, представленную в SEQ ID NO: 86.
В одном аспекте CAR, описанный здесь, включает антигенсвязывающий домен специфического антитела с внутриклеточным доменом сигнализации. Например, в некоторых аспектах внутриклеточный домен сигнализации включает, но без ограничения, сигнальные модули CD3-дзета цепь, 4-1BB и CD28 и их комбинации.
В одном аспекте антигенсвязывающий домен связывается с EGFRvIII. В одном аспекте CAR включает последовательность, представленную в SEQ ID NO: 43. В одном аспекте CAR включает последовательность, представленную в SEQ ID NO: 49. В одном аспекте CAR включает последовательность, представленную в SEQ ID NO: 55. В одном аспекте CAR включает последовательность, представленную в SEQ ID NO: 61. В одном аспекте CAR включает последовательность, представленную в SEQ ID NO: 67. В одном аспекте CAR включает последовательность, представленную в SEQ ID NO: 73. В одном аспекте CAR включает последовательность, представленную в SEQ ID NO: 79. В одном аспекте CAR включает последовательность, представленную в SEQ ID NO: 85.
В одном аспекте CAR включает по крайней мере один внутриклеточный домен сигнализации, выбираемый из группы, состоящей из домена сигнализации CD137 (4-1BB), домена сигнализации CD28, домена сигнализации CD3 дзета и любой их комбинации. В одном аспекте CAR включает по крайней мере один внутриклеточный домен сигнализации из одной или более костимулирующих молекул, отличных от CD137 (4-1BB) или CD28, домена сигнализации CD3 дзета и любой их комбинации.
Кроме того, настоящим изобретением обеспечиваются структуры CAR и их применение в лекарственных средствах или способах для лечения, среди прочих заболеваний, рака или любой злокачественной опухоли или аутоиммунных заболеваний, в патогенез которых вовлечены клетки или ткани, которые экспрессируют EGFRvIII.
Настоящим изобретением также обеспечиваются структуры и способы для сверхэкспрессии miR-17-92 (микроРНК-17-92), например, в CAR-экспрессирующей клетке, например, Т-клетке. В одном аспекте сверхэкспрессия с трансгена miR-17-92 придает CAR-трансдуцированной Т-клетке повышенную стойкость к индуцированной опухолью иммуносупрессии и химиотерапии, способствуя тем самым продолжительным терапевтическим эффектам.
Химерный антигенспецифический рецептор (CAR)
Настоящее изобретение охватывает рекомбинантную ДНК-конструкцию, включающую последовательности, кодирующие CAR, причем CAR включает фрагмент антитела, который специфически связывается с EGFRvIII, например, фрагмент антитела человека, который специфически связывается с EGFRvIII. В одном аспекте EGFRvIII является EGFRvIII человека, и последовательность нуклеиновой кислоты, кодирующая фрагмент антитела, является смежной с, и находится той же рамке считывания, что и последовательность(ю) нуклеиновой кислоты, кодирующей внутриклеточный домен сигнализации. Внутриклеточный домен сигнализации может включать домен костимулирующей сигнализации и/или домен первичной сигнализации, например, дзета-цепь. Домен костимулирующей сигнализации относится к части CAR, включающей по крайней мере часть внутриклеточного домена костимулирующей молекулы.
В конкретных аспектах конструкция CAR по настоящему изобретению включает scFv-домен, выбираемый из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86, причем scFv может предшествовать необязательная лидерная последовательность, например, представленная в SEQ ID NO: 13, и за ней может следовать необязательная шарнирная последовательность, например, представленная в EQ ID NO: 14 или SEQ ID NO: 104 или SEQ ID NO: 106 или SEQ ID NO: 108, трансмембранная область, например, представленная в SEQ ID NO: 15, внутриклеточной домен сигнализации, который включает SEQ ID NO: 16 или SEQ ID NO: 102, и последовательность CD3 дзета, которая включает SEQ ID NO: 17 или SEQ ID NO: 99, причем домены являются смежными и находятся в одной и той же рамке считывания с образованием одного слитого белка. В настоящее изобретение также включена нуклеотидная последовательность, которая кодирует полипептид каждого из scFv-фрагментов, выбираемых из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86, и каждого из доменов SEQ ID NO: 13-17. В настоящее изобретение также включена нуклеотидная последовательность, которая кодирует полипептид каждого из scFv-фрагментов, выбираемых из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86, и каждого из доменов SEQ ID NO: 13-16 и SEQ ID NO: 99. В одном аспекте конструкция EGFRvIII-специфического CAR включает необязательную лидерную последовательность, экстраклеточный антигенсвязывающий домен, который специфически связывается с EGFRvIII, шарнирную область, трансмембранный домен и внутриклеточный стимулирующий домен. В одном аспекте конструкция EGFRvIII-специфического CAR включает необязательную лидерную последовательность, экстраклеточный антигенсвязывающий домен, который специфически связывается с EGFRvIII, шарнирную область, трансмембранный домен, внутриклеточный домен сигнализации, который включает домен костимулирующей сигнализации и домен первичной стимуляции. Конкретные конструкции EGFRvIII-специфического CAR, содержащие гуманизированный scFv-домен, представлены в SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 и SEQ ID NO: 90. Конкретные конструкции EGFRvIII-специфического CAR, содержащие scFv-домен мыши, представлены в SEQ ID NO: 1 и SEQ ID NO: 2.
Приводимая в качестве примера лидерная последовательность представлена как SEQ ID NO: 13. Приводимая в качестве примера шарнирная/спейсерная последовательность представлена как SEQ ID NO: 14 или SEQ ID NO: 104 или SEQ ID NO: 106 или SEQ ID NO: 108. Приводимая в качестве примера последовательность трансмембранного домена представлена как SEQ ID NO: 15. Приводимая в качестве примера последовательность костимулирующего домена белка 4-1BB предоставляется как SEQ ID NO: 16. Приводимая в качестве примера последовательность костимулирующего домена белка CD27 представлена как SEQ ID NO: 102. Приводимый в качестве примера домен первичной сигнализации из последовательности домена CD3 дзета представлен как SEQ ID NO: 17. Другой приводимый в качестве примера домен первичной сигнализации из последовательности домена CD3 дзета представлен как SEQ ID NO: 99.
В одном аспекте настоящее изобретение охватывает рекомбинантную НК-конструкцию, включающую молекулу нуклеиновой кислоты, кодирующую CAR, причем молекула нуклеиновой кислоты включает последовательность нуклеиновой кислоты, кодирующую связывающий домен антитела против EGFRvIII, например, описанный здесь, который является смежным с, и находится в той же рамке считывания, что и последовательность(ю) нуклеиновой кислоты, кодирующая внутриклеточный домен сигнализации. В одном аспекте связывающий домен антитела против EGFRvIII выбирают из одной или более из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86. В одном аспекте связывающий домен антитела против EGFRvIII кодируется нуклеотидной последовательностью, представленной в последовательности, выбираемой из группы, состоящей из SEQ ID NO: 39, SEQ ID NO: 45, SEQ ID NO: 51, SEQ ID NO: 57, SEQ ID NO: 63, SEQ ID NO: 69, SEQ ID NO: 75, SEQ ID NO: 81 и SEQ ID NO: 98. В одном аспекте связывающий домен антитела против EGFRvIII кодируется SEQ ID NO: 39. В одном аспекте связывающий домен антитела против EGFRvIII кодируется SEQ ID NO: 45. В одном аспекте связывающий домен антитела против EGFRvIII кодируется SEQ ID NO: 51. В одном аспекте связывающий домен антитела против EGFRvIII кодируется SEQ ID NO: 57. В одном аспекте связывающий домен антитела против EGFRvIII кодируется SEQ ID NO: 63. В одном аспекте связывающий домен антитела против EGFRvIII кодируется SEQ ID NO: 69. В одном аспекте связывающий домен антитела против EGFRvIII кодируется SEQ ID NO: 75. В одном аспекте связывающий домен антитела против EGFRvIII кодируется SEQ ID NO: 81.
В одном аспекте настоящее изобретение охватывает рекомбинантную НК-конструкцию, включающую молекулу нуклеиновой кислоты, кодирующую CAR, причем молекула нуклеиновой кислоты включает последовательность нуклеиновой кислоты, кодирующую связывающий домен антитела против EGFRvIII, выбираемый из группы, состоящей из SEQ ID NO: 42, SEQ ID NO: 48, SEQ ID NO: 54, SEQ ID NO: 60, SEQ ID NO: 66, SEQ ID NO: 72, SEQ ID NO: 78, SEQ ID NO: 84 и SEQ ID NO: 90, причем последовательность является смежной с и находится в той же рамке считывания, что и последовательность(ю) нуклеиновой кислоты, кодирующая внутриклеточный домен сигнализации. Приводимый в качестве примера внутриклеточный домен сигнализации, который может использоваться в CAR, включает, но без ограничения, один или более внутриклеточных доменов сигнализации, например, CD3 дзета, CD28, 4-1BB и т.п. В некоторых случаях CAR может включать любую комбинацию внутриклеточных доменов сигнализации из CD3-дзета, CD28, 4-1ВВ и т.п. В одном аспекте НК-конструкция включает SEQ ID NO: 42. В одном аспекте последовательностью нуклеиновой кислоты конструкции CAR является SEQ ID NO: 48. В одном аспекте НК-конструкция включает SEQ ID NO: 54. В одном аспекте НК-конструкция включает SEQ ID NO: 60. В одном аспекте НК-конструкция включает SEQ ID NO: 66. В одном аспекте НК-конструкция включает SEQ ID NO: 72. В одном аспекте НК-конструкция включает SEQ ID NO: 78. В одном аспекте НК-конструкция включает SEQ ID NO: 84.
Последовательности нуклеиновых кислот, кодирующие желаемые молекулы, можно получить, используя рекомбинантные способы, известные в данной области техники, такие как, например, путем скрининга библиотек из клеток, экспрессирующих ген, путем извлечения гена из вектора, который, как известно, включает его, или путем выделения непосредственно из клеток и тканей, содержащих его, используя стандартные методы. Альтернативно, представляющую интерес нуклеиновую кислоту можно синтезировать, а не клонировать.
Настоящее изобретение включает ретровирусные и лентивирусные векторные конструкции, экспрессирующие CAR, которые можно непосредственно трансдуцировать в клетку.
Настоящее изобретение также включает РНК-конструкцию, которую можно непосредственно трансфицировать в клетку. Способ получения мРНК для применения в трансфекции включает in vitro транскрипцию (IVT) матрицы с использованием специально разработанных праймеров, с последующим добавлением поли(А), чтобы получить конструкцию, содержащую 3' и 5' нетранслируемые последовательности («UTR»), 5' кэп и/или участок внутренней посадки рибосомы (IRES), экспрессируемую нуклеиновую кислоту и поли(А)-хвост, обычно длиной 50-2000 оснований. РНК, полученная таким образом, может эффективно трансфицировать различные типы клеток. В одном варианте осуществления матрица включает последовательности для CAR. В одном варианте осуществления РНК-вектор, кодирующий CAR, трансдуцируют в Т-клетку путем электропорации.
Антигенсвязывающий домен
В одном аспекте CAR по настоящему изобретению включает мишень-специфический связывающий элемент, иначе называемый антигенсвязывающим доменом. Выбор этой составляющей зависит от типа и количества лигандов, которые определяют поверхность клетки-мишени. Например, может быть выбран антигенсвязывающий домен, который распознает лиганд, который действует в качестве маркера клеточной поверхности на клетках-мишенях, связанных с конкретным болезненным состоянием. Таким образом, примеры маркеров клеточной поверхности, которые могут действовать в качестве лигандов для антигенсвязывающего домена CAR по настоящему изобретению, включают те, которые связаны с вирусными, бактериальными и паразитарными инфекциями, аутоиммунным заболеванием и раковыми клетками.
В одном аспекте CAR-опосредованная Т-клеточная реакция может быть направлена на представляющий интерес антиген через создание антигенсвязывающего домена, который специфически связывается с желаемым антигеном, в CAR.
В одном из аспектов часть CAR, включающая антигенсвязывающий домен, включает антигенсвязывающий домен, мишенью которого является EGFRvIII. В одном аспекте мишенью антигенсвязывающего домена является EGFRvIII человека. Например, мышиное моноклональное антитело (IgG2b) 3C10 против EGFRvIII было получено путем иммунизации мышей 14-аминокислотным пептидом (LEEKKGNYVVTDHC; SEQ ID NO: 101), включающим EGFRvIII-специфический стык, и продемонстрировало очень специфическое распознавание EGFRvIII без какого-либо обнаруживаемого связывания с EGFR дикого типа (Okamoto et al., British J. Cancer 1996, 73: 1366-1372). Соответственно, в некоторых вариантах осуществления мишенью антигенсвязывающего домена является аминокислотная последовательность, например, аминокислотная последовательность, включающая дополнительный остаток глицина, в области стыка с EGFvIII. В некоторых вариантах осуществления мишенью антигенсвязывающего домена является одна или более аминокислотных последовательностей в аминокислотной последовательности SEQ ID NO: 101.
Антигенсвязывающим доменом может быть любой домен, который связывается с антигеном, включая, но без ограничения, моноклональное антитело, поликлональное антитело, рекомбинантное антитело, антитело человека, гуманизированное антитело и его функциональный фрагмент, в том числе, но без ограничения, однодоменное антитело, такое как вариабельный домен тяжелой цепи (VH), вариабельный домен легкой цепи (VL) и вариабельный домен (VHH) происходящего от семейства верблюдовых нанотела, и альтернативный каркас, который, как известно в данной области техники, функционируют как антигенсвязывающий домен, такой как рекомбинантный домен фибронектина и т.п. В некоторых случаях выгодно, когда антигенсвязывающий домен происходит из того же вида, в котором будет, в конечном счете, использоваться CAR. Например, в случае применения для людей может быть выгодно, когда антигенсвязывающий домен CAR включают человеческие или гуманизированные остатки для антигенсвязывающего домена антитела или фрагмента антитела.
Таким образом, в одном аспекте антигенсвязывающий домен включает антитело человека или фрагмент антитела. В другом аспекте антигенсвязывающий домен включает гуманизированное антитело или фрагмент антитела. В одном варианте осуществления связывающий домен антитела против EGFRvIII включает один или более (например, один, два или все три) из определяющего комплементарность участка 1 легкой цепи (CDR1 LC), определяющего комплементарность участка 2 легкой цепи (CDR2 LC) и определяющего комплементарность участка 3 легкой цепи (CDR3 LC) связывающего домена антитела против EGFRvIII, описанного здесь, и один или более (например, один, два или все три) из определяющего комплементарность участка 1 тяжелой цепи (CDR1 НС), определяющего комплементарность участка 2 тяжелой цепи (CDR2 НС) и определяющего комплементарность участка 3 тяжелой цепи (CDR3 НС) связывающего домена антитела против EGFRvIII, описанного здесь. В одном варианте осуществления связывающий домен антитела против EGFRvIII включает вариабельную область легкой цепи, описанную здесь, и/или вариабельную область тяжелой цепи, описанную здесь. В одном варианте осуществления связывающий домен антитела против EGFRvIII представляет собой scFv, включающий вариабельную область легкой цепи и вариабельную область тяжелой цепи аминокислотной последовательности, например, вариабельную область легкой цепи и вариабельную область тяжелой цепи, описанную здесь. В одном варианте осуществления связывающий домен антитела против EGFRvIII (например, scFv) включает: вариабельную область легкой цепи, включающую аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 30, 20 или 10 модификаций (например, замен) аминокислотной последовательности вариабельной области легкой цепи, представленной здесь, или последовательность, идентичную на 85-99% (например, 90-99% или 95-99%) аминокислотной последовательности, представленной здесь; и/или вариабельную область тяжелой цепи, включающую аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации (например, замены), но не более 30, 20 или 10 модификаций (например, замен) аминокислотной последовательности вариабельной области тяжелой цепи, представленной здесь, или последовательность, идентичную на 85-99% (например, 90-99% или 95-99%) аминокислотной последовательности, представленной здесь. В одном аспекте антигенсвязывающий домен включает одну или более последовательностей, выбираемых из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86. В одном аспекте гуманизированный CAR выбирают из одной или более последовательностей, выбираемых из группы, состоящей из SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 и SEQ ID NO: 90.
В некоторых аспектах нечеловеческое антитело является гуманизированным, причем специфические последовательности или области антитела модифицированы для увеличения сходства с антителом, образуемым в естественным условиях у человека, или его фрагментом. В одном аспекте антигенсвязывающий домен является гуманизированным.
Гуманизированное антитело может быть получено с использованием различных методов, известных в данной области техники, включая, но без ограничения, CDR-прививку (смотрите, например, Европейский патент № ЕР 239400; публикацию международной заявки № WO 91/09967; и патенты США №№ 5225539, 5530101 и 5585089, каждый из которых включен сюда во всей своей полноте посредством ссылки), венерование или изменение поверхности (смотрите, например, Европейский патент № ЕР 592106 и № ЕР 519596; Padlan, 1991, Molecular Immunology, 28 (4/5): 489-498; Studnicka et al., 1994, Protein Engineering, 7(6): 805-814; и Roguska et al., 1994, PNAS, 91: 969-973, каждый из которых включен сюда во всей своей полноте посредством ссылки), перетасовку цепей (смотрите, например, патент США № 5565332, который включен сюда во всей своей полноте посредством ссылки) и методы, описанные, например, в публикации заявки на патент США № US2005/0042664, публикации заявки на патент США № US2005/0048617, патенте США № 6407213, патенте США № 5766886, публикации международной заявки № WO 9317105, Tan et al., 2002, J. Immunol., 169: 1119-25; Caldas et al., 2000, Protein Eng., 13(5): 353-60; Morea et al., 2000, Methods, 20: 267-79; Baca et al., 1997, J. Biol. Chem., 272: 10678-84; Roguska et al., 1996, Protein Eng., 9(10): 895-904; Couto ei al., 1995, Cancer Res., 55: 5973s-5977; Couto et al., 1995, Cancer Res., 55(8): 1717-22; Sandhu 1994 Gene, 150(2): 409-10; и Pedersen et al., 1994, J. Mol. Biol, 235(3): 959-73, каждый из которых включен сюда во всей своей полноте посредством ссылки. Часто каркасные остатки в каркасных областях будут заменены соответствующим остатком из антитела-донора CDR с целью изменения, например, увеличения, связывания с антигеном. Эти замены в каркасных областях идентифицируют способами, хорошо известными в данной области техники, например, путем моделирования взаимодействий остатков CDR и каркасных остатков для идентификации каркасных остатков, важных для связывания с антигеном, и сравнения последовательностей для выявления необычные каркасных остатков в конкретных положениях. (Смотри, например, Queen et al., патент США № 5585089; и Riechmann et al., 1988, Nature, 332: 323, которые включены сюда во всей своей полноте посредством ссылки).
Гуманизированное антитело или фрагмент антитела имеет один или более аминокислотных остатков, остающихся в нем из источника, который является нечеловеческим. Эти нечеловеческие аминокислотные остатки часто называют «импортными» остатками, которые обычно берут из «импортного» вариабельного домена. Как здесь предусмотрено, гуманизированные антитела или фрагменты антител включают один или более CDR из нечеловеческих молекул иммуноглобулинов и каркасные области, причем аминокислотные остатки, составляющие каркасную область, происходят полностью или в основном из зародышевой линии человека. Множество методов гуманизации антител или фрагментов антител хорошо известны в данной области техники и могут быть выполнены по существу в соответствии со способом Winter и сотрудников (Jones et al., Nature, 321: 522-525 (1986); Riechmann et al., Nature, 332: 323-327 (1988); Verhoeyen et al., Science, 239: 1534-1536 (1988)), путем замены CDR грызунов или последовательностями CDR грызунов соответствующих последовательностей антитела человека, т.е. CDR-прививки (EP 239400; публикация РСТ-заявки № WO 91/09967 и патенты США №№ 4816567; 6331415; 5225539; 5530101; 5585089; 6548640, содержание которых включено сюда посредством ссылки во всей их полноте). В таких гуманизированных антителах и фрагментах антител в значительной степени меньше, чем интактный человеческий вариабельный домен был заменен соответствующей последовательностью из не являющегося человеком вида. Гуманизированные антитела часто представляют собой антитела человека, в которых некоторые остатки CDR и, возможно, некоторые остатки каркасных областей (FR) заменены остатками из аналогичных участков в антителах грызунов. Гуманизации антител и фрагментов антител можно также достичь путем венерования или изменения поверхности (EP 592106; EP 519596; Padlan, 1991, Molecular Immunology, 28(4/5): 489-498; Studnicka et al., Protein Engineering, 7(6): 805-814 (1994); и Roguska et al., PNAS, 91:969-973 (1994)) или перетасовки цепей (патент США № 5565332), содержание которых включено сюда посредством ссылки во всей их полноте.
Результатом выбора вариабельных доменов человека, как легких, так и тяжелых цепей, которые будут использоваться при получении гуманизированных антител, является снижение антигенности. В соответствии с так называемым способом «наилучшего соответствия», последовательность вариабельного домена антитела грызуна скринируют по всей библиотеке известных последовательностей вариабельных доменов человека. Последовательность человека, которая наиболее близка к последовательности грызуна, затем принимают в качестве человеческой каркасной области (FR) для гуманизированного антитела (Sims et al., J. Immunol., 151: 2296 (1993); Chothia et al., J. Mol. Biol., 196: 901 (1987), содержание которых включено сюда посредством ссылки во всей их полноте). В другом способе используется конкретная каркасная область, полученная из консенсусной последовательности для всех антител человека конкретной подгруппы легких или тяжелых цепей. Одна и та же каркасная область может использоваться для нескольких различных гуманизированных антител (смотрите, например, Carter et al., Proc. Natl. Acad. Sci. USA, 89: 4285 (1992); Presta et al., J. Immunol., 151: 2623 (1993), содержание которых включено сюда посредством во всей их полноте).
В некоторых аспектах часть структуры CAR по настоящему изобретению, которая включает фрагмент антитела, является гуманизированной с сохранением высокой аффинности к антигену-мишени и других подходящих биологических свойств. В соответствии с одним аспектом настоящего изобретения гуманизированные антитела и фрагменты антител получают с помощью процесса анализа исходных последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются общедоступными и известны квалифицированным в данной области техники специалистам. В наличии имеются компьютерные программы, которые иллюстрируют и демонстрируют возможные трехмерные конформационные структуры выбранных кандидатных последовательностей иммуноглобулина. Осмотр этих демонстраций позволяет проанализировать вероятную роль остатков в функционировании последовательности иммуноглобулина-кандидата, например, проанализировать остатки, которые влияют на способность иммуноглобулина-кандидата к связыванию с антигеном-мишенью. Таким образом, остатки FR могут быть выбраны из реципиентных и импортных последовательностей и объединены, так что достигается требуемая характеристика антитела или фрагмента антитела, такая как увеличенная аффинность к антигену-мишени. В общем, остатки CDR непосредственно и наиболее существенно участвуют во влиянии на связывание антигена.
В одном аспекте связывающим доменом антитела против EGFRvIII является, например, Fv, Fab или (Fab')2, или бифункциональное (например, биспецифическое) гибридное антитело (например, Lanzavecchia et al., Eur. J. Immunol. 17, 105 (1987)). В одном аспекте предусмотренным здесь фрагментом антитела является scFv. В одном аспекте scFv связывает белок EGFRvIII, но не EGFR дикого типа. В некоторых случаях scFv человека может быть также получен из библиотеки дрожжевого дисплея.
В некоторых случаях scFv могут быть приготовлены в соответствии со способом, известным в данной области техники (смотрите, например, Bird et al., (1988) Science 242: 423-426 и Huston et al., (1988) Proc. Natl. Acad. Sci. USA 85: 5879-5883). Молекулы scFv могут быть получены путем связывания VH- и VL-областей вместе с использованием гибких полипептидных линкеров. Молекулы scFv включают линкер (например, линкер Ser-Gly) оптимизированной длины и/или с оптимизированным аминокислотным составом. Длина линкера может сильно влиять на то, каким образом вариабельные области scFv будут укладываться и взаимодействовать. В самом деле, если используется короткий полипептидный линкер (например, 5-10 аминокислот), внутрицепочечная укладка предотвращается. Межцепочечная укладка также требуется для сближения двух вариабельных областей для формирования функционального сайта связывания эпитопа. Для примеров ориентации и размер линкера смотрите, например, Hollinger et al. 1993 Proc Natl Acad. Sci. U.S.A. 90: 6444-6448, публикацию заявки на патент США №№ 2005/0100543, 2005/0175606, 2007/0014794 и публикацию РСТ-заявки №№ WO2006/020258 и WO2007/024715, которые включены сюда посредством ссылки.
ScFv может включать линкер из по крайней мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45, 50 или более аминокислотных остатков между своими VL- и VH-областями. Последовательность линкера может включать любую встречающуюся в природе аминокислоту. В некоторых вариантах осуществления последовательность линкера включает аминокислоты глицин и серин. В другом варианте осуществления последовательность линкера включает ряды повторов глицина и серина, такие как (Gly4Ser)n (SEQ ID NO: 37), где n - положительное целое число, равное или большее 1. В одном варианте осуществления линкером может быть (Gly4Ser)4 (SEQ ID NO: 113) или (Gly4Ser)3 (SEQ ID NO: 114). В результате изменения длины линкера активность может сохраняться или повышаться, что приводит к большей эффективности в исследованиях активности.
Стабильность и мутации
Стабильность связывающего домена антитела против EGFRvIII, например, молекул scFv (например, растворимого scFv), можно оценить относительно биофизических свойств (например, термостабильности) обычной контрольной молекулы scFv или полноразмерного антитела. В одном варианте осуществления гуманизированный scFv имеет термостабильность, превышающую на приблизительно 0,1, приблизительно 0,25, приблизительно 0,5, приблизительно 0,75, приблизительно 1, приблизительно 1,25, приблизительно 1,5, приблизительно 1,75, приблизительно 2, приблизительно 2,5, приблизительно 3, приблизительно 3,5 приблизительно 4, приблизительно 4,5, приблизительно 5, приблизительно 5,5, приблизительно 6, приблизительно 6,5, приблизительно 7, приблизительно 7,5, приблизительно 8, приблизительно 8,5, приблизительно 9, приблизительно 9,5, приблизительно 10 градусов, приблизительно 11 градусов, приблизительно 12 градусов, приблизительно 13 градусов, приблизительно 14 градусов или приблизительно 15 градусов по Цельсию таковую контрольной связывающей молекулы (например, обычной молекулы scFv) в описанных анализах.
Увеличенная термостабильность связывающего домена антитела против EGFRvIII, например, scFv, впоследствии придается всей конструкции EGFRvIII-специфического CAR, что приводит к улучшению терапевтических свойств конструкции EGFRvIII-специфического CAR. Термостабильность связывающего домена антитела против EGFRvIII, например, scFv, может быть увеличена на по крайней мере приблизительно 2°C или 3°C по сравнению с обычным антителом. В одном варианте осуществления связывающий домен антитела против EGFRvIII, например, scFv, имеет увеличенную на 1°C термостабильность по сравнению с обычным антителом. В другом варианте осуществления связывающий домен антитела против EGFRvIII, например, scFv, имеет увеличенную на 2°C термостабильность по сравнению с обычным антителом. В другом варианте осуществления связывающий домен антитела против EGFRvIII, например, scFv, имеет увеличенную на 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15°C термостабильность по сравнению с обычным антителом. Сравнения могут быть сделаны, например, между молекулами scFv, описанными здесь, и молекулами scFv или Fab-фрагментами антитела, из которого происходят VH и VL scFv. Термостабильность может быть измерена с помощью методов, известных в данной области техники. Например, в одном варианте осуществления может быть измерена Tm. Методы измерения Tm и другие методы определения стабильности белков описаны подробнее ниже.
Мутации в scFv (возникающие в результате гуманизации или прямого мутагенеза растворимого scFv) изменяют стабильность scFv и увеличивают общую стабильность scFv и конструкции EGFRvIII-специфического CAR. Стабильность гуманизированного scFv сравнивают с таковой scFv мыши, используя такие измерения, как Tm, температура денатурации и температура агрегации. Способность к связыванию мутантных scFv можно определить, используя анализы, описанных в разделе «Примеры».
В одном варианте осуществления связывающий домен антитела против EGFRvIII, например, scFv, содержит по крайней мере одну мутацию, возникающую в процессе гуманизации, из условия, чтобы мутантный scFv придавал увеличенную стабильность EGFRvIII-специфической конструкции. В другом варианте осуществления связывающий домен антитела против EGFRvIII, например, scFv, содержит по крайней мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 мутаций, возникающих в процессе гуманизации, из условия, чтобы мутантный scFv придавал увеличенную стабильность EGFRvIII-специфической конструкции.
Способы оценки стабильности белков
Стабильность антигенсвязывающего домена можно оценить, используя, например, способы, описанные ниже. Такие способы позволяют определить множество термически инициированных конформационных переходов с разворачиванием, причем наименее стабильный домен или разворачивается первым, или ограничивает порог общей стабильности многодоменного блока, который разворачивается кооперативно (например, многодоменного белка, который демонстрирует один конформационный переход с разворачиванием). Наименее стабильный домен можно идентифицировать рядом дополнительных способов. Мутагенез может быть выполнен, чтобы исследовать, какой домен ограничивает общую стабильность. Кроме того, устойчивость к протеазам многодоменного белка может быть исследована в условиях, когда наименее стабильный домен, как известно благодаря DSC или другим спектроскопическим методам, является по сути развернутым (Fontana, et al., (1997) Fold. Des., 2: R17-26; Dimasi et al. (2009) J. Mol. Biol. 393: 672-692). После идентификации наименее стабильного домена последовательность, кодирующую этот домен (или его часть), можно использовать в качестве тестовой последовательности в способах.
a) Термостабильность
Термостабильность структур можно проанализировать с использованием ряда неограничивающих биофизических или биохимических методов, известных в данной области техники. В некоторых вариантах осуществления термостабильность оценивают с помощью аналитической спектроскопии.
Приводимым в качестве примера методом аналитической спектроскопии является дифференциальная сканирующая калориметрия (DSC). При выполнении DSC используется калориметр, который чувствителен к теплопоглощениям, которые сопровождают развертывание большинства белков или белковых доменов (смотрите, например, Sanchez-Ruiz, et al., Biochemistry, 27: 1648-52, 1988). Для определения термостабильности белка образец белка помещают в калориметр, и температуру повышают до тех пор, пока Fab или scFv не развернется. Температура, при которой белок разворачивается, является показателем общей стабильности белка.
Другим приводимым в качестве примера методом аналитической спектроскопии является спектроскопия кругового дихроизма (CD). В случае CD-спектрометрии измеряется оптическая активность структуры в зависимости от повышения температуры. В случае спектроскопии кругового дихроизма (CD) определяется различия в поглощении света с левовращающей поляризацией и света с правовращающей поляризацией, которые возникают из-за структурной асимметрии. Неупорядоченная или развернутая структура приводит в CD-спектру, очень отличному от такового упорядоченной или свернутой структуры. CD-спектр отражает чувствительность белков к денатурирующим эффектам повышения температуры и, следовательно, является показателем термостабильности белков (смотрите van Mierlo and Steemsma, J. Biotechnol, 79 (3): 281-98, 2000).
Другим приводимым в качестве примера методом аналитической спектроскопии для определения термостабильности является флуоресцентная спектроскопия (смотрите van Mierlo and Steemsma, выше). Еще одним приводимым в качестве примера методом аналитической спектроскопии для определения термостабильности является спектроскопия ядерного магнитного резонанса (ЯМР) (смотрите van Mierlo and Steemsma, выше).
Термостабильность структуры можно измерить биохимически. Приводимым в качестве примера биохимическим методом для оценки термостабильности является анализ теплового испытания. В ходе «анализа теплового испытания» структуру подвергают воздействию диапазона повышенных температур в течение заданного периода времени. Например, в одном варианте осуществления тестовые молекулы scFv или молекулы, включающие молекулы scFv, подвергают воздействию диапазона увеличивающихся температур, например, в течение 1-1,5 часов. Активность белка затем анализируют с помощью соответствующего биохимического анализа. Например, если белком является связывающий белок (например, scFv или scFv-содержащий полипептид), активность связывания связывающего белка можно определить с помощью функционального или количественного ELISA.
Такой анализ может быть осуществлен в формате высокой пропускной способности и форматах, описанных в разделе «Примеры», используя E. coli и скрининг с высокой пропускной способностью. Библиотека вариантов связывающего домена антитела против EGFRvIII, например, scFv, может быть создана, используя способы, известные в данной области техники. Экспрессия связывающего домена антитела против EGFRvIII, например, scFv, может быть индуцирована, и связывающий домен антитела против EGFRvIII, например, scFv, может быть подвергнут тепловому испытанию. Подвергнутые испытанию тестовые образцы можно проанализировать на связывание, и те связывающие домены антитела против EGFRvIII, например, scFvs, которые являются стабильными, могут быть увеличены в масштабе и далее охарактеризованы.
Термостабильность оценивается путем измерения температуры плавления (Tm) структуры с использованием любого из вышеуказанных методов (например, методов аналитической спектроскопии). Температура плавления представляет собой температуру в средней точке кривой теплового перехода, при которой 50% молекул структуры находится в свернутом состоянии (смотри, например, Dimasi et al. (2009) J. Mol Biol. 393: 672-692). В одном варианте осуществления значения Tm для связывающего домена антитела против EGFRvIII, например, scFv, составляют приблизительно 40°C, 41°C, 42°C, 43°C, 44°C, 45°C, 46°C, 47°C, 48°C, 49°C, 50°C, 51°C, 52°C, 53°C, 54°C, 55°C, 56°C, 57°C, 58°C, 59°C, 60°C, 61°C, 62°C, 63°C, 64°C, 65°C, 66°C, 67°C, 68°C, 69°C, 70°C, 71°C, 72°C, 73°C, 74°C, 75°C, 76°C, 77°C, 78°C, 79°C, 80°C, 81°C, 82°C, 83°C, 84°C, 85°C, 86°C, 87°C, 88°C, 89°C, 90°C, 91°C, 92°C, 93°C, 94°C, 95°C, 96°C, 97°C, 98°C, 99°C, 100°C. В одном варианте осуществления значения Tm для IgG составляют приблизительно 40°C, 41°C, 42°C, 43°C, 44°C, 45°C, 46°C, 47°C, 48°C, 49°C, 50°C, 51°C, 52°C, 53°C, 54°C, 55°C, 56°C, 57°C, 58°C, 59°C, 60°C, 61°C, 62°C, 63°C, 64°C, 65°C, 66°C, 67°C, 68°C, 69°C, 70°C, 71°C, 72°C, 73°C, 74°C, 75°C, 76°C, 77°C, 78°C, 79°C, 80°C, 81°C, 82°C, 83°C, 84°C, 85°C, 86°C, 87°C, 88°C, 89°C, 90°C, 91°C, 92°C, 93°C, 94°C, 95°C, 96°C, 97°C, 98°C, 99°C, 100°C. В одном варианте осуществления значения Tm для поливалентного антитела составляют приблизительно 40°C, 41°C, 42°C, 43°C, 44°C, 45°C, 46°C, 47°C, 48°C, 49°C, 50°C, 51°C, 52°C, 53°C, 54°C, 55°C, 56°C, 57°C, 58°C, 59°C, 60°C, 61°C, 62°C, 63°C, 64°C, 65°C, 66°C, 67°C, 68°C, 69°C, 70°C, 71°C, 72°C, 73°C, 74°C, 75°C, 76°C, 77°C, 78°C, 79°C, 80°C, 81°C, 82°C, 83°C, 84°C, 85°C, 86°C, 87°C, 88°C, 89°C, 90°C, 91°C, 92°C, 93°C, 94°C, 95°C, 96°C, 97°C, 98°C, 99°C, 100°C.
Термостабильность также оценивают путем измерения теплоемкости (Cp) структуры, используя аналитический колориметрический метод (например, DSC). Теплоемкость структуры - энергия (например, в ккал/моль), которая требуется для повышения на 1°C температуры 1 моля воды. Так большая Ср является признаком денатурированной или неактивной белковой структуры. Изменение теплоемкости (ΔCp) структуры измеряется путем определения теплоемкости структуры до и после его теплового перехода. Термостабильность может быть также оценена путем измерения или определения других параметров термодинамической стабильности, включая свободную энергию Гиббса разворачивания (ΔG), энтальпию разворачивания (ΔΗ) или энтропию разворачивания (ΔS). Один или более из вышеуказанных биохимических анализов (например, анализ теплового испытания) используются для определения температуры (т.е. значения ТС), при которой 50% структуры сохраняют свою активность (например, активность связывания).
Кроме того, мутации в связывающем домене антитела против EGFRvIII, например, scFv, изменяют термостабильность связывающего домена антитела против EGFRvIII, например, scFv, по сравнению с не мутированным связывающим доменом антитела против EGFRvIII, например, scFv. Когда связывающий домен гуманизированного антитела против EGFRvIII, например, scFv, включен в конструкцию CAR на основе антитела против EGFRvIII, связывающий домен антитела против EGFRvIII, например, гуманизированный scFv, придает термостабильность всей конструкции CAR на основе антитела против EGFRvIII. В одном варианте связывающий домен антитела против EGFRvIII, например, scFv, содержит одну мутацию, которая придает термостабильность связывающему домену антитела против EGFRvIII, например, scFv. В другом варианте связывающий домен антитела против EGFRvIII, например, scFv, содержит множество мутаций, которые придают термостабильность связывающему домену антитела против EGFRvIII, например, scFv. В одном варианте осуществления множество мутации в связывающем домене антитела против EGFRvIII, например, scFv, оказывают аддитивный эффект на термостабильность связывающего домен антитела против EGFRvIII, например, scFv.
b) % Агрегация
Стабильность структуры можно определить путем измерения ее склонности к агрегации. Агрегация может быть определена с помощью ряда неограничивающих биохимических или биофизических методов. Например, агрегация структуры может быть оценена с помощью хроматографии, например, гель-хроматографии (SEC). В ходе SEC молекулы разделяются на основе размера. Колонку заполняют полутвердыми гранулами полимерного геля, которые будут пропускать внутрь себя ионы и небольшие молекулы, но не большие молекулы. Когда белковую композицию вносят в верхнюю часть колонки, компактные свернутые белки (т.е. не агрегированные белки) распределяются по большему объему растворителя, чем тот, который находится в распоряжении крупных белковых агрегатов. Следовательно, крупные агрегаты движутся быстрее через колонку, и таким образом смесь можно разделить или фракционировать на ее компоненты. Каждую фракцию можно отдельно подвергнуть количественному анализу (например, с помощью рассеяния света), по мере того как она элюируется из геля. Соответственно, % агрегации структуры можно определить путем сравнения концентрации фракции с общей концентрацией белка, внесенного в гель. Стабильные структуры элюируют с колонки в виде по существу одной фракции и появляются в виде по существу одного пика в профиле элюции или на хроматограмме.
с) Аффинность
Устойчивость структуры моно оценить путем определения ее аффинности к мишени. Широкий ряд методов определения аффинности известен в данной области техники. В иллюстративном методе определения аффинности используется поверхностный плазмонный резонанс. Поверхностный плазмонный резонанс представляет собой оптическое явление, которое позволяет проводить анализ в режиме реального времени биоспецифических взаимодействий по детекции изменений концентраций белка в матрице биосенсора, например, с использованием системы BIAcore (Pharmacia Biosensor AB, Uppsala, Sweden и Piscataway, N.J.). Для дальнейших описаний смотрите Jonsson, U., et al. (1993) Ann. Biol. Clin. 51: 19-26; Jonsson, U., i (1991) Biotechniques 11: 620-627; Johnsson, B., et al. (1995) J. Mol. Recognit. 8: 125-131; и Johnnson, B., et al. (1991) Anal. Biochem. 198: 268-277.
В одном аспекте антигенсвязывающий домен CAR включает аминокислотную последовательность, которая является гомологичной аминокислотной последовательности антигенсвязывающего домена, описанной здесь, и антигенсвязывающий домен сохраняет желаемые функциональные свойства фрагментов антител против EGFRvIII, описанных здесь. В одном конкретном аспекте структура CAR настоящего изобретения включает фрагмент антитела. В дальнейшем аспекте фрагмент антитела включает scFv.
В различных аспектах антигенсвязывающий домен CAR создают путем модификации одной или более аминокислот в одной или обеих вариабельных областях (например, VH и/или VL), например, в одном или более CDR-участков и/или в одной или более каркасных областей. В одном конкретном аспекте структура CAR настоящего изобретения включает фрагмент антитела. В дальнейшем аспекте фрагмент антитела включает scFv.
Специалисту со средним уровнем компетентности в данной области техники будет понятно, что антитело или фрагмент антитела настоящего изобретения можно подвергнуть дополнительным модификациям из условия, чтобы они отличались по аминокислотной последовательности (например, от дикого типа), но не по желаемой активности. Например, дополнительные нуклеотидные замены, ведущие к заменам аминокислот в положениях «несущественных» аминокислотных остатков, могут быть осуществлены в белке. Например, несущественный аминокислотный остаток в молекуле может быть заменен другим аминокислотным остатком из того же семейства боковых цепей. В другом варианте осуществления ряд аминокислот может быть заменен структурно аналогичным рядом, который отличается порядком и/или составом членов семейства боковых цепей, например, может быть осуществлена консервативная замена, при которой аминокислотный остаток заменяют на аминокислотный остаток, имеющий сходную боковую цепь.
В данной области техники были определены семейства аминокислотных остатков, имеющих сходные боковые цепи, включая основные боковые цепи (например, лизин, аргинин, гистидин), кислые боковые цепи (например, аспарагиновую кислоту, глютаминовую кислоту), незаряженные полярные боковые цепи (например, глицин, аспарагин, глютамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин).
Процент идентичности в контексте двух или более последовательностей нуклеиновых кислот или полипептидных последовательностей относится к двум или более последовательностей, которые являются одинаковыми. Две последовательности являются «по существу идентичными», если две последовательности имеют определенный процент аминокислотных остатков или нуклеотидов, которые являются одинаковыми (например, составляющую 60% идентичность, в некоторых случаях составляющую 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичность по заданной области, или, если не указано, по всей последовательности), при сравнении и совмещении для максимального соответствия в окне сравнения, или на протяжении намеченного района, как определено с помощью одного из следующих алгоритмов сравнения последовательностей или путем совмещения вручную и визуального осмотра. Дополнительно, идентичность существует на протяжении района, длина которого составляет по крайней мере приблизительно 50 нуклеотидов (или 10 аминокислот), или более предпочтительно на протяжении района, длина которого составляет от 100 до 500 или 1000 или более нуклеотидов (или 20, 50, 200 или более аминокислот).
Для сравнения последовательностей, как правило, одна последовательность выполняет функцию контрольной последовательности, с которой сравнивают тестовые последовательности. При использовании алгоритма сравнения последовательностей тестовые и контрольные последовательности вводят в компьютер, задают координаты подпоследовательностей, если необходимо, и задаются параметры программы с использованием алгоритма сравнения последовательностей. Параметры программы по умолчанию могут использоваться, или альтернативные параметры могут быть заданы. Затем алгоритм сравнения последовательностей рассчитывает процент идентичности последовательностей для тестовых последовательностей относительно контрольной последовательности, на основе параметров программы. Методы совмещения последовательностей с целью сравнения хорошо известны в данной области техники. Оптимальное совмещение последовательностей с целью сравнения может быть проведено, например, с помощью алгоритма локальной гомологии Smith и Waterman, (1970) Adv. Appl. Math. 2:482c, с помощью алгоритма совмещения на основе гомологии Needleman и Wunsch, (1970) J. Mol. Biol. 48: 443, с помощью метода поиском подобия Pearson и Lipman, (1988) Proc. Nat'l. Acad. Sci. USA 85: 2444, с использованием компьютерных реализаций этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics, Genetics Computer Group, 575 Science Dr., Madison, WI), или с помощью совмещения вручную и визуального осмотра (смотрите, например, Brent et al., (2003) Current Protocols in Molecular Biology).
Двумя примерами алгоритмов, которые подходят для определения процента идентичности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, описанные в Altschul et al., (1977) Nuc. Acids Res. 25: 3389-3402; и Altschul et al., (1990) J. Mol. Biol. 215: 403-410, соответственно. Программное обеспечение для выполнения анализов BLAST общедоступно через Национальный центр биотехнологической информации.
Процент идентичности между двумя аминокислотными последовательностями может быть также определен с использованием алгоритма Е. Meyers и W. Miller, (1988) Comput. Appl. Biosci. 4: 11-17), который был включен в программу ALIGN (версия 2.0), используя таблицу весов замен остатков РАМ120, штраф на длину гэпа - 12 и штраф за открытие гэпа - 4. Кроме того, процент идентичности между двумя аминокислотными последовательностями можно определить, используя алгоритм Needleman и Wunsch (1970) J. Mol. Biol. 48: 444-453), который был включен в программу GAP в пакете программ GCG (имеющихся на сайте www.gcg.com), используя или матрицу Blossom 62, или матрицу РАМ250 и штраф за открытие гэпа = 16, 14, 12, 10, 8, 6 или 4 и штраф за длину гэпа = 1, 2, 3, 4, 5 или 6.
В одном аспекте настоящим изобретением предусматриваются модификации исходной аминокислотной последовательности антитела или фрагмента (например, scFv), которые создают функционально эквивалентные молекулы. Например, VH или VL связывающего домена антитела против EGFRvIII, например, scFv, в составе CAR, могут быть модифицированы с сохранением составляющей по крайней мере приблизительно 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности каркасной области исходной VH или VL связывающего домена антитела против EGFRvIII, например, scFv. Настоящим изобретением предусматриваются модификации всей конструкции CAR, например, модификации в одной или более аминокислотных последовательностей различных доменов конструкции CAR для создания функционально эквивалентных молекул. Конструкция CAR может быть модифицирована с сохранением составляющей по крайней мере приблизительно 70%, 71%, 72%. 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичности исходной конструкции CAR.
Трансмембранный домен
Что касается трансмембранного домена, в различных вариантах осуществления может быть сконструирован CAR, который включает трансмембранный домен, присоединенный к экстраклеточному домену CAR. Трансмембранный домен может включать одну или более дополнительных аминокислот, примыкающий к трансмембранному району, например, одну или более аминокислот, связанных с экстраклеточным районом белка, из которого был получен трансмембранный белок, (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 до 15 аминокислот экстраклеточного района) и/или одну или более дополнительных аминокислот, связанных с внутриклеточным районом белка, из которого получен трансмембранный белок, (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 до 15 аминокислот внутриклеточного района). В одном аспекте трансмембранным доменом является домен, который связан с одним из других доменов CAR. В некоторых случаях трансмембранный домен может быть выбран или модифицирован путем замены аминокислот во избежание связывания таких доменов с трансмембранными доменами тех же или отличных белков оболочки, например, чтобы свести к минимуму взаимодействия с другими членами рецепторного комплекса. В одном аспекте трансмембранный домен способен к гомодимеризации с другим CAR на поверхности CAR-T-клетки. В отличном аспекте аминокислотная последовательность трансмембранного домена может быть изменена или заменена таким образом, чтобы свести к минимуму взаимодействия со связывающими доменами природного партнера по связыванию, присутствующего в той же CAR-T-клетке.
Трансмембранный домен может быть получен или из природного, или из рекомбинантного источника. Если источник является природным, домен может быть получен из любого мембраносвязанного или трансмембранного белка. В одном аспекте трансмембранный домен способен к передаче сигналов на внутриклеточный домен(ы), всякий раз когда CAR связывается с мишенью. Трансмембранный домен, имеющий конкретное применение в этом изобретении, может включать по крайней мере трансмембранный район(ы), например, альфа, бета или дзета цепи Т-клеточного рецептора, CD28, CD3 эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137, CD154.
В некоторых случаях трансмембранный домен может быть присоединен к экстраклеточному району CAR, например, антигенсвязывающему домену CAR, с помощью шарнирной области, например, шарнирной области из белка человека. Например, в одном варианте осуществления шарнирной областью может быть шарнирная область Ig (иммуноглобулина) человека, например, шарнирная область IgG4 или шарнирная область CD8a. В одном варианте осуществления шарнирная область или спейсер включает (например, состоит из) аминокислотную последовательность SEQ ID NO: 14. В одном аспекте трансмембранный домен включает (например, состоит из) трансмембранный домен SEQ ID NO: 15.
В одном аспекте шарнирная область или спейсер включает шарнирную область IgG4. Например, в одном варианте осуществления шарнирная область или спейсер включает шарнирную область с аминокислотной последовательностью ESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGKM (SEQ ID NO: 104). В некоторых вариантах осуществления шарнирная область или спейсер включает шарнирную область, кодируемую нуклеотидной последовательностью GAGAGCAAGTACGGCCCTCCCTGCCCCCCTTGCCCTGCCCCCGAGTTCCTGGGCGGACCCAGCGTGTTCCTGTTCCCCCCCAAGCCCAAGGACACCCTGATGATCAGCCGGACCCCCGAGGTGACCTGTGTGGTGGTGGACGTGTCCCAGGAGGACCCCGAGGTCCAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCACAACGCCAAGACCAAGCCCCGGGAGGAGCAGTTCAATAGCACCTACCGGGTGGTGTCCGTGCTGACCGTGCTGCACCAGGACTGGCTGAACGGCAAGGAATACAAGTGTAAGGTGTCCAACAAGGGCCTGCCCAGCAGCATCGAGAAAACCATCAGCAAGGCCAAGGGCCAGCCTCGGGAGCCCCAGGTGTACACCCTGCCCCCTAGCCAAGAGGAGATGACCAAGAACCAGGTGTCCCTGACCTGCCTGGTGAAGGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAACGGCCAGCCCGAGAACAACTACAAGACCACCCCCCCTGTGCTGGACAGCGACGGCAGCTTCTTCCTGTACAGCCGGCTGACCGTGGACAAGAGCCGGTGGCAGGAGGGCAACGTCTTTAGCTGCTCCGTGATGCACGAGGCCCTGCACAACCACTACACCCAGAAGAGCCTGAGCCTGTCCCTGGGCAAGATG (SEQ ID NO: 105).
В одном аспекте шарнирная область или спейсер включает шарнирную область IgD. Например, в одном варианте осуществления шарнирная область или спейсер включает шарнирную область аминокислотной последовательности RWPESPKAQASSVPTAQPQAEGSLAKATTAPATTRNTGRGGEEKKKEKEKEEQEERETKTPECPSHTQPLGVYLLTPAVQDLWLRDKATFTCFVVGSDLKDAHLTWEVAGKVPTGGVEEGLLERHSNGSQSQHSRLTLPRSLWNAGTSVTCTLNHPSLPPQRLMALREPAAQAPVKLSLNLLASSDPPEAASWLLCEVSGFSPPNILLMWLEDQREVNTSGFAPARPPPQPGSTTFWAWSVLRVPAPPSPQPATYTCVVSHEDSRTLLNASRSLEVSYVTDH (SEQ ID NO: 106). В некоторых вариантах осуществления шарнирная область или спейсер включает шарнирную область, кодируемую нуклеотидной последовательностью AGGTGGCCCGAAAGTCCCAAGGCCCAGGCATCTAGTGTTCCTACTGCACAGCCCCAGGCAGAAGGCAGCCTAGCCAAAGCTACTACTGCACCTGCCACTACGCGCAATACTGGCCGTGGCGGGGAGGAGAAGAAAAAGGAGAAAGAGAAAGAAGAACAGGAAGAGAGGGAGACCAAGACCCCTGAATGTCCATCCCATACCCAGCCGCTGGGCGTCTATCTCTTGACTCCCGCAGTACAGGACTTGTGGCTTAGAGATAAGGCCACCTTTACATGTTTCGTCGTGGGCTCTGACCTGAAGGATGCCCATTTGACTTGGGAGGTTGCCGGAAAGGTACCCACAGGGGGGGTTGAGGAAGGGTTGCTGGAGCGCCATTCCAATGGCTCTCAGAGCCAGCACTCAAGACTCACCCTTCCGAGATCCCTGTGGAACGCCGGGACCTCTGTCACATGTACTCTAAATCATCCTAGCCTGCCCCCACAGCGTCTGATGGCCCTTAGAGAGCCAGCCGCCCAGGCACCAGTTAAGCTTAGCCTGAATCTGCTCGCCAGTAGTGATCCCCCAGAGGCCGCCAGCTGGCTCTTATGCGAAGTGTCCGGCTTTAGCCCGCCCAACATCTTGCTCATGTGGCTGGAGGACCAGCGAGAAGTGAACACCAGCGGCTTCGCTCCAGCCCGGCCCCCACCCCAGCCGGGTTCTACCACATTCTGGGCCTGGAGTGTCTTAAGGGTCCCAGCACCACCTAGCCCCCAGCCAGCCACATACACCTGTGTTGTGTCCCATGAAGATAGCAGGACCCTGCTAAATGCTTCTAGGAGTCTGGAGGTTTCCTACGTGACTGACCATT (SEQ ID NO:107).
В одном аспекте трансмембранный домен может быть рекомбинантный, в этом случае он будет содержать преимущественно гидрофобные остатки, такие как лейцин и валин. В одном аспекте триплет из фенилаланина, триптофана и валина может быть обнаружен на каждом конце рекомбинантного трансмембранного домена.
Необязательно, короткий олиго- или полипептидный линкер, например, длиной от 2 до 10 аминокислот, может формировать связь между трансмембранным доменом и цитоплазматическим районом CAR. Дублет глицин-серин является примером подходящего линкера. Например, в одном аспекте линкер включает аминокислотную последовательность GGGGSGGGGS (SEQ ID NO: 108). В некоторых вариантах осуществления линкер кодируется нуклеотидной последовательностью GGTGGCGGAGGTTCTGGAGGTGGAGGTTCC (SEQ ID NO:109).
Цитоплазматический домен
Цитоплазматический домен или район CAR включает внутриклеточный домен сигнализации. Внутриклеточный домен сигнализации, как правило, ответственен за активацию по крайней мере одной из обычных эффекторных функций иммуноцита, в который был введен CAR. Термин «эффекторная функция» относится к специализированной функции клетки. Эффекторной функцией Т-клетки, например, могут быть цитолитическая активность или хелперная активности, включая секрецию цитокинов. Таким образом, термин «внутриклеточный домен сигнализации» относится к части белка, которая трансдуцирует сигнал для эффекторной функции и предписывает клетке выполнение специализированной функции. Хотя обычно весь внутриклеточный домен сигнализации может использоваться, во многих случаях нет необходимости использовать всю цепь. Если используется усеченная часть внутриклеточного домена сигнализации, такая усеченная часть может использоваться вместо интактной цепи при условии, что она трансдуцирует сигнал для эффекторной функции. Таким образом, термин «внутриклеточный домен сигнализация» как подразумевается, включает любую усеченную часть внутриклеточного домена сигнализации, достаточную для трансдукции сигнала для эффекторной функции.
Примеры внутриклеточных доменов сигнализации для использования в CAR по настоящему изобретению включают цитоплазматические последовательности Т-клеточного рецептора (TCR) и корецепторов, которые действуют совместно, чтобы инициировать сигнальную трансдукцию после задействования антигенспецифического рецептора, а также любое производное или вариант этих последовательностей и любая рекомбинантная последовательность, которая имеет те же функциональные возможности.
Известно, что сигналы, генерируемые благодаря одному только TCR, являются недостаточными для полной активации Т-клетки, и что также требуется вторичный и/или костимулирующий сигнал. Таким образом, можно сказать, что активация Т-клетки осуществляется при посредничестве двух различных классов цитоплазматических сигнальных последовательностей: тех, которые инициируют антигензависимую первичную активацию через TCR, (внутриклеточных доменов первичной сигнализации) и тех, которые действуют независимым от антигена образом, чтобы обеспечить вторичный или костимулирующий сигнал (цитоплазматического домена вторичной сигнализации, например, костимулирующего домена).
Домен первичной сигнализации регулирует первичную активацию TCR комплекса или стимулирующим образом, или ингибирующим образом. Внутриклеточные домены первичной сигнализации, которые действуют стимулирующим образом, могут содержать мотивы сигнализации, которые известны как иммунорецепторные тирозиновые активирующие мотивы или ITAM.
Примеры ITAM-содержащих внутриклеточных доменов первичной сигнализации, которые имеют конкретное применение в настоящем изобретении, включают таковые из TCR дзета, FcR гамма, FcR бета, CD3 гамма, CD3 дельта, CD3 эпсилон, CD5, CD22, CD79a, CD79b, и CD66d. В одном варианте осуществления CAR по настоящему изобретению, например, CAR, выбираемый из группы, состоящей из SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79 и SEQ ID NO: 85, включает внутриклеточный домен сигнализации, например, домен первичной сигнализации CD3-дзета. В одном варианте осуществления домен первичной сигнализации включает модифицированный домен ITAM, например, мутированный домен ITAM, который обладает измененной (например, увеличенной или уменьшенной) активность по сравнению с природным доменом ITAM. В одном варианте осуществления домен первичной сигнализации включает модифицированный, ITAM-содержащий внутриклеточный домен первичной сигнализации, например, оптимизированный и/или усеченный ITAM-содержащий внутриклеточный домен первичной сигнализации. В одном варианте осуществления домен первичной сигнализации включает один, два, три, четыре или более мотивов ITAM.
Внутриклеточный домен сигнализации CAR может включать домен сигнализации CD3-дзета сам по себе, или он может быть объединен с любым другим желаемым внутриклеточным доменом(ами) сигнализации, используемым в связи с CAR по настоящему изобретению. Например, внутриклеточный домен сигнализации CAR может включать часть CD3 дзета-цепи и домен костимулирующей сигнализации. Домен костимулирующей сигнализации относится к части CAR, включающей внутриклеточный домен костимулирующей молекулы. Костимулирующей молекулой является молекула клеточной поверхности, отличная от антигенспецифического рецептора или его лигандов, которая необходима для эффективной ответной реакции лимфоцитов на антиген. Примеры таких молекул включают CD27, CD28, 4-1ВВ (CD137), OX40, CD30, CD40, PD-1, ICOS, связанный с функционированием лимфоцитов антиген-1 (LFA-1), CD2, CD7, LIGHT, NKG2C, В7-H3 и лиганд, который специфически связывается с CD83, и т.п. Например, костимуляция CD27, как было продемонстрировано, приводит к увеличению роста, эффекторной функции и выживания CAR-T-клеток человека in vitro и увеличению выживания Т-клеток человека и противоопухолевой активностью in vivo (Song et al. Blood. 2012; 119 (3): 696-706).
Внутриклеточные сигнальные последовательности в цитоплазматической части CAR по настоящему изобретению могут быть связаны друг с другом в произвольном или определенном порядке. Необязательно, короткий олиго- или полипептидный линкер, например, длиной от 2 до 10 аминокислот (например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот), может формировать связь между внутриклеточной последовательностью сигнализации. В одном варианте осуществления дублет глицин-серин может использоваться в качестве подходящего линкера. В одном варианте осуществления одна аминокислота, например, аланин, глицин, может использоваться в качестве подходящего линкера.
В одном аспекте конструируют внутриклеточный домен сигнализации, который включает два или более, например, 2, 3, 4, 5 или более, доменов костимулирующей сигнализации. В одном варианте осуществления два или более, например, 2, 3, 4, 5 или более, доменов костимулирующей сигнализации разделены линкерной молекулой, например, линкерной молекулой, описанной здесь. В одном варианте осуществления внутриклеточный домен сигнализации включает два домена костимулирующей сигнализации. В некоторых вариантах осуществления линкерной молекулой является остаток глицина. В некоторых вариантах осуществления линкером является остаток аланина.
В одном аспекте конструируют внутриклеточный домен сигнализации, который включает домен сигнализации из CD3-дзета и домен сигнализации из CD28. В одном аспекте конструируют внутриклеточный домен сигнализации, который включает домен сигнализации из CD3-дзета и домен сигнализации из 4-1ВВ. В одном аспекте сигнальный домен 4-1ВВ является сигнальный домен SEQ ID NO: 16. В одном аспекте доменом сигнализации из CD3-дзета является домен сигнализации SEQ ID NO: 17.
В одном аспекте конструируют внутриклеточный домен сигнализации, который включает домен сигнализации из CD3-дзета и домен сигнализации из CD27. В одном аспекте домен сигнализации из CD27 включает аминокислотную последовательность QRRKYRSNKGESPVEPAEPCRYSCPREEEGSTIPIQEDYRKPEPACSP (SEQ ID NO:102). В одном аспекте домен сигнализации из CD27 кодируется последовательностью нуклеиновой кислоты AGGAGTAAGAGGAGCAGGCTCCTGCACAGTGACTACATGAACATGACTCCCCGCCGCCCCGGGCCCACCCGCAAGCATTACCAGCCCTATGCCCCACCACGCGACTTCGCAGCCTATCGCTCC (SEQ ID NO:103).
В одном аспекте CAR-экспрессирующая клетка, описанная здесь, может, кроме того, включать второй CAR, например, второй CAR, который включает отличный антигенсвязывающий домен, например, к той же мишени (EGFRvIII) или отличной мишени.
В другом аспекте настоящим изобретением обеспечивается популяция CAR-экспрессирующих клеток, например, CAR-T-клеток. В некоторых вариантах популяция CAR-экспрессирующих клеток включает смесь клеток, экспрессирующих различных CAR. Например, в одном варианте осуществления популяция CAR-T-клеток может включать первую клетку, экспрессирующую CAR, содержащий описанный здесь связывающий домен антитела против EGFRvIII, и вторую клетку, экспрессирующую CAR, содержащий отличный связывающий домен антитела против EGFRvIII, например, связывающий домен антитела против EGFRvIII, описанный здесь, который отличается от связывающего домена антитела против EGFRvIII в CAR, экспрессируемого первой клеткой. В качестве другого примера, популяция CAR-экспрессирующих клеток может включать первую клетку, экспрессирующую CAR, который включает связывающий домен антитела против EGFRvIII, например, описанный здесь, и вторую клетку, экспрессирующую CAR, который включает антигенсвязывающий домен к мишени, отличной от EGFRvIII. В одном варианте осуществления популяция CAR-экспрессирующих клеток включает, например, первую клетку, экспрессирующую CAR, который включает внутриклеточный домен первичной сигнализации, и вторую клетку, экспрессирующую CAR, который включает домен вторичной сигнализации.
В другом аспекте настоящим изобретением обеспечивается популяция клеток, в которой по крайней мере одна клетка в популяции экспрессирует CAR, содержащий описанный здесь связывающий домен антитела против EGFRvIII, а вторая клетка экспрессирует другой агент, например, агент, который повышает активность CAR-экспрессирующей клетки. Например, в одном варианте осуществления агентом может быть агент, который ингибирует ингибиторную молекулу. Ингибиторные молекулы, например, PD1, могут, в некоторых вариантах осуществления, уменьшать способность CAR-экспрессирующей клетки к индукции эффекторного иммунного ответа. Примеры ингибиторных молекул включают PD1, PD-L1, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2В4 и TGFR бета.
РНК-трансфекция
Здесь описаны способы получения in vitro транскрибированной РНК для CAR. Настоящее изобретение также включает кодирующую CAR РНК-конструкцию, которую можно непосредственно трансфицировать в клетку. Способ получения мРНК для использования в трансфекции может включать in vitro транскрипцию (IVT) матрицы с использованием специально разработанных праймеров, с последующим добавление полиА, чтобы получить конструкцию, содержащую 3' и 5' нетранслируемые последовательности («UTR»), 5' кэп и/или участок внутренней посадки рибосомы (IRES), экспрессируемую нуклеиновую кислоту и поли(А)-хвост, обычно длиной 50-2000 оснований (SEQ ID NO: 116). РНК, полученная таким образом, может эффективно трансфицировать различные типы клеток. В одном аспекте матрица включает последовательности для CAR.
В одном аспекте EGFRvIII-специфический CAR кодируется информационной РНК (мРНК). В одном аспекте мРНК, кодирующую EGFRvIII-специфический CAR, вводят в Т-клетку для получения CAR-T-клетки.
В одном варианте осуществления in vitro транскрибированная РНК для CAR может быть введена в клетку в виде транзиторной трансфекции. РНК получают с помощью in vitro транскрипции, используя созданную с помощью полимеразной цепной реакции (ПЦР) матрицу. Представляющую интерес ДНК из любого источника можно непосредственно превратить с помощью ПЦР в матрицу для in vitro синтеза мРНК, используя соответствующие праймеры и РНК-полимеразу. Источником ДНК может быть, например, геномная ДНК, плазмидная ДНК, фаговая ДНК, кДНК, синтетическая последовательность ДНК или любой другой подходящий источник ДНК. Желаемой матрицей для in vitro транскрипции является CAR настоящего изобретения. Например, матрица для РНК для CAR включает экстраклеточный район, включающий одноцепочечный вариабельный домен противоопухолевого антитела; шарнирную область; трансмембранный домен (например, трансмембранный домен CD8a); и цитоплазматический район, который включает внутриклеточный домен сигнализации, например, включающий домен сигнализации CD3-дзета и домен сигнализации 4-1ВВ.
В одном варианте осуществления ДНК, используемая для ПЦР, содержит открытую рамку считывания. ДНК может происходить из встречающейся в природе последовательности ДНК генома организма. В одном варианте осуществления нуклеиновая кислота может включать некоторые или все 5' и/или 3' нетранслируемые участки (UTR). Нуклеиновая кислота может включать экзоны и интроны. В одном варианте осуществления ДНК, используемая для ПЦР, представляет собой последовательность нуклеиновой кислоты человека. В другом варианте осуществления ДНК, используемая для ПЦР, представляет собой последовательность нуклеиновой кислоты человека, включающую 5' и 3' UTR. Альтернативно, ДНК может быть искусственной последовательностью ДНК, которая в нормальных условиях не экспрессируется во встречающемся в природе организме. Приводимой в качестве примера искусственной последовательностью ДНК является последовательность, которая содержит части генов, которые лигированы вместе с образованием открытой рамки считывания, кодирующей слитый белок. Части ДНК, которые подвергают лигированию, могут происходить из одного организма или из более чем одного организма.
ПЦР используется для создания матрицы для in vitro транскрипции мРНК, которая используется для трансфекции. Способы выполнения ПЦР хорошо известны в данной области техники. Конструируют праймеры для использования в ПЦР, которые содержат участки, по существу комплементарные участкам ДНК, используемой в качестве матрицы для ПЦР. «По существу комплементарные», как здесь используются, относятся к последовательностям нуклеотидов, в которых большая часть или все основания в последовательности праймера являются комплементарными, или одно или более оснований являются некомплементарными, или несовпадающими. По существу комплементарные последовательности способны к отжигу или гибридизации с намеченной ДНК-мишенью в условиях отжига, используемых для ПЦР. Могут быть сконструированы праймеры, которые по существу комплементарны любой части ДНК-матрицы. Например, могут быть сконструированы праймеры для амплификации части нуклеиновой кислоты, которая в нормальных условиях транскрибируется в клетках, (открытой рамки считывания), включающей 5' и 3' UTR. Также могут быть сконструированы праймеры для амплификации части нуклеиновой кислоты, которая кодирует конкретный домен, представляющий интерес. В одном варианте осуществления конструируют праймеры для амплификации кодирующей области кДНК человека, включающей все или части 5' и 3' UTR. Праймеры, применимые для ПЦР, создают с помощью методов синтеза, которые хорошо известны в данной области техники. «Прямыми праймерами» являются праймеры, которые содержат участок нуклеотидов, по существу комплементарных нуклеотидам в ДНК-матрице, которые находятся слева от последовательности ДНК, которая должна быть амплифицирована. «Слева», как здесь используется, относится к положению 5' от последовательности ДНК, которая должна быть амплифицирована, относительно кодирующей цепи. «Обратными праймерами» являются праймеры, которые содержат участок нуклеотидов, по существу комплементарных нуклеотидам в двухцепочечной ДНК-матрице, которые находятся справа от последовательности ДНК, которая должна быть амплифицирована. «Справа», как здесь используется, относится к положению 3' от последовательности ДНК, которая должна быть амплифицирована, относительно кодирующей цепи.
Любая ДНК-полимераза, применимая для ПЦР, может использоваться в описываемых здесь способах. Реагенты и полимераза коммерчески доступны из ряда источников.
Могут также использоваться химические структуры, обладающие способностью поддерживать стабильность и/или эффективность трансляции. Предпочтительно РНК содержит 5' и 3' UTR. В одном варианте осуществления размер 5' UTR находится между 1 и 3000 нуклеотидов. Размер последовательностей 5' и 3' UTR, добавляемых к кодирующей области, может быть изменен с помощью различных способов, в том числе, но без ограничения, конструирования праймеров для ПЦР, которые подвергаются отжигу к различным участкам UTR. Используя этот подход, специалист со средним уровнем компетентности в данной области техники может изменять размеры 5' и 3' UTR, необходимые для достижения оптимальной эффективности трансляции после трансфекции транскрибированной РНК.
5' и 3' UTR могут быть встречающимися в природе, эндогенными 5' и 3' UTR для представляющей интерес нуклеиновой кислоты. Альтернативно, последовательности UTR, которые не являются эндогенными для представляющей интерес нуклеиновой кислоты, могут быть добавлены посредством включения последовательностей UTR в прямой и обратный праймеры или с помощью любых других модификаций матрицы. Последовательности UTR, которые не являются эндогенными для представляющей интерес нуклеиновой кислоты, могут использоваться для изменения стабильности и/или эффективности трансляции РНК. Например, известно, что AU-богатые элементы в последовательностях 3' UTR могут уменьшить стабильность мРНК. Следовательно, можно выбрать или сконструировать 3' UTR, которые увеличивают стабильность транскрибированной РНК, на основе свойств UTR, которые хорошо известны в данной области техники.
В одном варианте осуществления 5' UTR может содержать последовательность Козак эндогенной нуклеиновой кислоты. Альтернативно, когда 5' UTR, который не является эндогенным для представляющей интерес нуклеиновой кислоты, добавляют с помощью ПЦР, как описано выше, консенсусная последовательность Козак может быть переконструирована посредством добавления последовательности 5' UTR. Последовательность Козак может увеличить эффективность трансляции с некоторых РНК-транскриптов, но, по-видимому, не требуется для всех РНК для допуска эффективной трансляции. Необходимость в последовательностях Козак в случае многих мРНК известна в данной области техники. В других вариантах осуществления 5' UTR может быть 5' UTR из РНК-содержащего вируса, геномная РНК которого является стабильной в клетках. В других вариантах осуществления различные аналоги нуклеотидов могут использоваться в 3' или 5' UTR для препятствования деградации мРНК под действием экзонуклеаз.
Чтобы сделать возможным синтез РНК с ДНК-матрицы без необходимости в клонировании гена, промотор транскрипции должен быть присоединен к ДНК-матрице 5' от транскрибируемой последовательности. Когда последовательность, которая функционирует в качестве промотора для РНК-полимеразы, добавлена к 5'-концу прямого праймера, промотор для РНК-полимеразы становится включенным в продукт ПЦР 5' от открытой рамки считывания, которая должна быть транскрибирована. В одном предпочтительном варианте осуществления промотором является промотор для полимеразы T7, описанный здесь где-то в другом месте. Другие применимые промоторы включают, но без ограничения, промоторы для РНК-полимеразы T3 и SP6. Консенсусные нуклеотидные последовательности для промоторов T7, T3 и SP6 известны в данной области техники.
В предпочтительном варианте осуществления мРНК содержит и кэп-структуру на 5'-конце, и 3' поли(A) хвост, которые определяют связывание рибосомы, инициацию трансляции и стабильность мРНК в клетке. На замкнутой в круг ДНК-матрице, например, плазмидной ДНК, РНК-полимераза продуцирует длинный конкатемерный продукт, который не подходит для экспрессии в эукариотических клетках. Транскрипция плазмидой ДНК, линеаризованной на конце 3' UTR, приводит к мРНК с нормальным размером, которая не является эффективной в трансфекции эукариотических клеток, даже если она подвергается полиаденилированию после транскрипции.
На линейной ДНК-матрице РНК-полимераза фага T7 может удлинить 3'-конец транскрипта после последнего основания матрицы (Schenborn and Mierendorf, Nuc Acids Res., 13: 6223-6236 (1985); Nacheva and Berzal-Herranz, Eur. J. Biochem., 270: 1485-1465 (2003).
Традиционным способом интеграции участков полиA/T в ДНК-матрицу является молекулярное клонирование. Однако последовательность полиA/T, интегрированная в плазмидную ДНК, может быть причиной нестабильности плазмиды, и поэтому плазмидные ДНК-матрицы, полученные из бактериальных клеток, часто «испорчены» в высокой степени делециями и другими аберрациями. Это делает процедуры клонирования не только трудными и трудоемкими, но часто ненадежными. Вот почему очень требуется способ, который делает возможным конструирование ДНК-матриц с 3' участком полиA/T без клонирования.
Сегмент полиA/T транскрипционной ДНК-матрицы можно синтезировать во время ПЦР, используя обратный праймер, содержащий полиT-хвост, такой как 100T-хвост (SEQ ID NO: 117) (размер может составлять 50-5000 T (SEQ ID NO: 118)), или создать после ПЦР с помощью любого способа, в том числе, но без ограничения, лигирования ДНК или in vitro рекомбинации. Поли(A)-хвосты также обеспечивают РНК стабильность и уменьшают их деградацию. Как правило, имеется положительная корреляция между размером поли(A)-хвоста и стабильностью транскрибированной РНК. В одном варианте осуществления размер поли(A)-хвоста находится между 100 и 5000 аденозинов (SEQ ID NO: 119).
Поли(A)-хвосты РНК могут быть дополнительно удлинены после in vitro транскрипции с помощью использования поли(A)-полимеразы, такой как поли(А)-полимераза E. coli (E-PAP). В одном варианте осуществления увеличение размера поли(A)-хвоста со 100 нуклеотидов до 300-400 нуклеотидов (SEQ ID NO: 120) приводит к приблизительно двукратному увеличению эффективности трансляции РНК. Кроме того, присоединение различных химических групп к 3'-концу может увеличить стабильность мРНК. Такое присоединение может включать модифицированные/искусственные нуклеотиды, аптамеры и другие соединения. Например, аналоги АТФ могут быть включены в поли(A)-хвост, используя поли(A)-полимеразу. Аналоги АТФ могут дополнительно увеличить стабильность РНК.
5' кэп-структуры также обеспечивают стабильность молекулам РНК. В предпочтительном варианте осуществления РНК, продуцированные с помощью способов, описываемых здесь, включают 5' кэп-структуру. 5' кэп-структуру обеспечивают, используя методы, известные в данной области техники и описанные здесь (Cougot, et al., Trends in Biochem. Sci., 29: 436-444 (2001); Stepinski, et al., RNA, 7: 1468-1495 (2001); Elango, et al., Biochim. Biophys. Res. Commun., 330: 958-966 (2005)).
РНК, продуцированные с помощью способов, описываемых здесь, могут также содержать последовательность участка внутренней посадки рибосомы (IRES). Последовательность IRES может быть любой вирусной, хромосомной или искусственно созданной последовательностью, которая инициирует кэп-независимое связывание рибосомы с мРНК и способствует инициации трансляции. Могут быть включены любые растворенные вещества, подходящие для электропорации в клетки, которые могут содержать факторы, содействующие клеточной проницаемости и жизнеспособности, такие как сахара, пептиды, липиды, белки, антиоксиданты и поверхностно-активные вещества.
РНК можно вести в клетки-мишени, используя любой из множества различных способов, например, коммерчески доступных способов, которые включают, но без ограничения, электропорацию (Amaxa Nucleofector-II (Amaxa Biosystems, Cologne, Германия)), (ECM 830 (BTX) (Harvard Instruments, Boston, Mass.) или Gene Pulser II (BioRad, Denver, Colo.), Multiporator (Eppendort, Hamburg, Германия), трансфекцию с использованием катионных липосом, используя липофекцию, инкапсуляцию полимеров, трансфекцию с использованием пептидов или системы для биолистической доставки частиц, такие как «генные пушки» (смотрите, например, Nishikawa, et al., Hum Gene Ther., 12(8): 861-870 (2001).
НК-конструкции, кодирующие CAR
Настоящим изобретением обеспечиваются молекулы нуклеиновых кислот, кодирующие одну или более конструкций CAR, описанных здесь. В одном аспекте молекула нуклеиновой кислоты обеспечивается в виде мРНК-транскрипта. В одном аспекте молекула нуклеиновой кислоты обеспечивается в виде ДНК-конструкции.
Соответственно, в одном аспекте настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей химерный антигенспецифический рецептор (CAR), причем CAR включает связывающий домен антитела против EGFRvIII (например, связывающий домен гуманизированного антитела против EGFRvIII), трансмембранный домен и внутриклеточный домен сигнализации, включающий стимулирующий домен, например, домен костимулирующей сигнализации и/или домен первичной сигнализации, например, дзета-цепь. В одном варианте осуществления связывающим доменом антитела против EGFRvIII является связывающий домен антитела против EGFRvIII, описанный здесь, например, связывающий домен антитела против EGFRvIII, который включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74 и SEQ ID NO: 80, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления выделенная молекула нуклеиновой кислоты, кроме того, включает последовательность, кодирующую костимулирующий домен. В одном варианте осуществления костимулирующим доменом является функциональный домен сигнализации белка, выбираемого из группы, состоящей из OX40, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11a/CD18), ICOS (CD278) и 4-1BB (CD137). В одном варианте осуществления костимулирующий домен включает последовательность SEQ ID NO: 16, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления трансмембранным доменом является трансмембранный домен белка, выбираемого из группы, состоящей из альфа, бета или дзета цепи T-клеточного рецептора, CD28, CD3 эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154. В одном варианте осуществления трансмембранный домен включает последовательность SEQ ID NO: 15, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления внутриклеточный домен сигнализации включает функциональный домен сигнализации 4-1BB и функциональный домен сигнализации CD3 дзета. В одном варианте осуществления внутриклеточный домен сигнализации включает последовательность SEQ ID NO: 16 или SEQ ID NO: 102, или последовательность, идентичную ей на 95-99%, и последовательность SEQ ID NO: 17 или SEQ ID NO: 99, или последовательность, идентичную ей на 95-99%, причем последовательности, включающие внутриклеточный домен сигнализации, экспрессируются в одной и той же рамке и в виде одной полипептидной цепи. В одном варианте осуществления связывающий домен антитела против EGFRvIII соединен с трансмембранным доменом с помощью шарнирной области, например, шарнирной области, описанной здесь. В одном варианте осуществления шарнирная область включает SEQ ID NO: 14 или SEQ ID NO: 104, или SEQ ID NO: 106, или SEQ ID NO: 108, или последовательность, идентичную ей на 95-99%.
В другом аспекте настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей конструкцию CAR, включающую лидерную последовательность SEQ ID NO: 13, scFv-домен, имеющий последовательность, выбираемую из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86 (или последовательность, идентичную ей на 95-99%), шарнирную область SEQ ID NO: 14 или SEQ ID NO: 104, или SEQ ID NO: 106, или SEQ ID NO: 108 (или последовательности, идентичной ей на 95-99%), трансмембранный домен, имеющий последовательность SEQ ID NO: 15 (или последовательность, идентичную ей на 95-99%), костимулирующий домен 4-1ВВ, имеющий последовательность SEQ ID NO: 16, или костимулирующий домен CD27, имеющий последовательность SEQ ID NO: 102 (или последовательность, идентичную ей на 95-99%), и стимулирующий домен CD3 дзета, имеющий последовательность SEQ ID NO: 17 или SEQ ID NO: 99 (или последовательность, идентичную ей на 95-99%).
В другом аспекте настоящее изобретение относится к выделенной молекуле полипептида, кодируемой молекулой нуклеиновой кислоты. В одном варианте осуществления выделенная молекула полипептида включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 и SEQ ID NO: 90, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления выделенный полипептид включает последовательность SEQ ID NO: 73 или последовательность, идентичную ей на 95-99%. В одном варианте осуществления выделенный полипептид включает последовательность SEQ ID NO: 79 или последовательность, идентичную ей на 95-99%.
В другом аспекте настоящее изобретение относится к молекуле нуклеиновой кислоты, кодирующей молекулу химерного антигенспецифического рецептора (CAR), которая включает связывающий домен антитела против EGFRvIII, трансмембранный домен и внутриклеточный домен сигнализации, включающий стимулирующий домен, и причем указанный связывающий домен антитела против EGFRvIII включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86, или последовательность, идентичную ей на 95-99%.
В одном варианте осуществления кодируемая молекула CAR, кроме того, включает последовательность, кодирующую костимулирующий домен. В одном варианте осуществления костимулирующим доменом является функциональный домен сигнализации белка, выбираемого из группы, состоящей из OX40, CD27, CD28, CDS, ICAM-1, LFA-1 (CD11a/CD18) и 4-1BB (CD137). В одном варианте осуществления костимулирующий домен включает последовательность SEQ ID NO: 16. В одном варианте осуществления трансмембранным доменом является трансмембранный домен белка, выбираемого из группы, состоящей из альфа, бета или дзета цепи T-клеточного рецептора, CD28, CD3 эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154. В одном варианте осуществления трансмембранный домен включает последовательность SEQ ID NO: 15. В одном варианте осуществления внутриклеточный домен сигнализации включает функциональный домен сигнализации 4-1BB и функциональный домен сигнализации дзета. В одном варианте осуществления внутриклеточный домен сигнализации включает последовательность SEQ ID NO: 16 и последовательность SEQ ID NO: 17, причем последовательности, включающие внутриклеточный домен сигнализации, экспрессируются в одной и той же рамке и в виде одной полипептидной цепи. В одном варианте осуществления связывающий домен антитела против EGFRvIII соединен с трансмембранным доменом с помощью шарнирной области. В одном варианте осуществления шарнирная область включает SEQ ID NO: 14. В одном варианте осуществления шарнирная область включает SEQ ID NO: 104 или SEQ ID NO: 106, или SEQ ID NO: 108.
В другом аспекте настоящее изобретение относится к кодируемой молекуле CAR, включающей лидерную последовательность SEQ ID NO: 13, scFv-домен, имеющий последовательность, выбираемую из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 80 и SEQ ID NO: 86, или последовательность, идентичную ей на 95-99%, шарнирную область с SEQ ID NO: 14 или SEQ ID NO: 104, или SEQ ID NO: 106, или SEQ ID NO: 108, трансмембранный домен, имеющий последовательность SEQ ID NO: 15, костимулирующий домен 4-1ВВ, имеющий последовательность SEQ ID NO: 16, или костимулирующий домен CD27, имеющий последовательность SEQ ID NO: 102, и стимулирующий домен CD3 дзета, имеющий последовательность SEQ ID NO: 17 или SEQ ID NO: 99. В одном варианте осуществления кодируемая молекула CAR включает последовательность, выбираемую из группы, состоящей из SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79, SEQ ID NO: 85 и SEQ ID NO: 90, или последовательность, идентичную ей на 95-99%. В одном варианте осуществления кодируемая молекула CAR включает последовательность SEQ ID NO: 73 или последовательность, идентичную ей на 95-99%. В одном варианте осуществления выделенная молекула CAR включает последовательность SEQ ID NO: 79 или последовательность, идентичную ей на 95-99%.
Последовательности нуклеиновых кислот, кодирующие желаемые молекулы, можно получить, используя рекомбинантные методы, известные в данной области техники, такие как, например, посредством скрининга библиотек из клеток, экспрессирующих ген, посредством доставки гена из вектора, который, как известно, включает его, или посредством непосредственного выделения из клеток или тканей, содержащих их, используя стандартные методы. Альтернативно, представляющий интерес ген можно создать синтетически, а не клонировать.
Настоящим изобретением также обеспечиваются векторы, в которые вставлена ДНК настоящего изобретения. Векторы, происходящие из ретровирусов, например, лентивируса, являются подходящими инструментальными средствами для достижения длительного переноса генов, поскольку они делает возможной длительную, стабильную интеграцию трансгена и его воспроизведение в дочерних клетках. Лентивирусные векторы обладают дополнительным преимуществом по сравнению с векторами, происходящими из онко-ретровирусов, таких как вирусы мышиного лейкоза, поскольку они могут трансдуцировать непролиферирующиеся клетки, такие как гепатоциты. Они также обладают дополнительным преимуществом, состоящим в низкой иммуногенности.
Вкратце, экспрессия природных или синтетических нуклеиновых кислот, кодирующих CAR, обычно достигается при функциональной связи нуклеиновой кислоты, кодирующей полипептид CAR или его часть, с промотором и включении конструкции в экспрессионный вектор. Векторы могут быть подходящими для репликации и интеграции в эукариотах. Типичные векторы для клонирования содержат терминаторы транскрипции и трансляции, инициирующие последовательности и промоторы, применимые для регуляции экспрессии желаемой последовательности нуклеиновой кислоты.
Экспрессионные конструкции по настоящему изобретению могут также использоваться для иммунизации нуклеиновой кислотой и генотерапии, используя стандартные протоколы для доставки генов. Методы доставки генов известны в данной области техники. Смотрите, например, патенты США №№ 5399346, 5580859, 5589466, включенные сюда посредством ссылки в их полных объемах. В другом варианте осуществления настоящим изобретением обеспечивается вектор для генотерапии.
Нуклеиновую кислоту можно клонировать во множество типов векторов. Например, нуклеиновую кислоту можно клонировать в вектор, включающий, но без ограничения, плазмиду, фагемиду, производное фага, вирус животного и космиду. Представляющие особый интерес векторы включают экспрессионные векторы, векторы для репликации, векторы для создания зондов и векторы для секвенирования.
Кроме того, экспрессионный вектор может быть привнесен в клетку в форме вирусного вектора. Технология с использованием вирусных векторов хорошо известна в данной области техники и описана, например, в Sambrook и др. (2001, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York) и в других руководствах по вирусологии и молекулярной биологии. Вирусы, которые применимы в качестве векторов, включают, но без ограничения, ретровирусы, аденовирусы, аденоассоциированные вирусы, вирусы герпеса и лентивирусы. Как правило, подходящий вектор содержит начало репликации, функционирующее в по крайней мере одном организме, промоторную последовательность, удобные сайты для эндонуклеаз рестрикции и один или более селектируемых маркеров (например, WO 01/96584; WO 01/29058; и патент США № 6326193).
Было разработано множество систем на основе вирусов для переноса генов в клетки млекопитающих. Например, ретровирусы обеспечивают удобную платформу для систем для доставки генов. Выбранный ген можно вставить в вектор и упаковать в ретровирусные частицы, используя известные в данной области техники методы. Рекомбинантный вирус затем можно выделить и доставить в клетки субъекта или in vivo, или ex vivo. В данной области техники известен ряд ретровирусных систем. В некоторых вариантах осуществления используют аденовирусные векторы. В данной области техники известен ряд аденовирусных векторов. В одном варианте осуществления используют лентивирусные векторы.
Дополнительные элементы промотора, например, энхансеры, регулируют частоту инициации транскрипции. Типично они находятся 5' от сайта инициации в области, находящейся на расстоянии 30-110 п.о. от него, хотя ряд промоторов, как недавно было установлено, также содержит функциональные элементы 3' от сайта инициации. Часто расстояние между элементами промотора является трансформируемым, так что функция промотора сохраняется, когда элементы являются инвертированными или перемещенными относительно друг друга. В промоторе гена тимидинкиназы (tk) расстояние между элементами промотора может быть увеличено до 50 п.о. в отдалении, прежде чем активность начнет уменьшаться. В зависимости от промотора, судя по всему, индивидуальные элементы могут функционировать или кооперативно, или независимо для активации транскрипции.
Одним примером подходящего промотора является промоторная последовательность немедленно-раннего гена цитомегаловируса (CMV). Эта промоторная последовательность является сильной конститутивной промоторной последовательностью, способной к запуску высоких уровней экспрессии любой полинуклеотидной последовательности, функционально связанной с ней. Другим примером подходящего промотора является промотор гена фактора-1α элонгации (EF-1α). Однако могут также использоваться другие конститутивные промоторные последовательности, включая, но без ограничения, ранний промотор вакуолизирующего обезьяньего вируса (SV40), промотор вируса опухоли молочной железы мыши (MMTV), промотор длинного концевого повтора (LTR) вируса иммунодефицита человека (ВИЧ), промотор MoMuLV, промотор вируса птичьего лейкоза, немедленно ранний промотор вируса Эпштейна-Барра, промотор вируса саркомы Рауса, а также промоторы генов человека, такие как, но без ограничения, промотор гена актина, промотор гена миозина, промотор гена гемоглобина и промотор гена креатинкиназы. Кроме того, настоящее изобретение не должно ограничиваться использованием конститутивных промоторов. В качестве части настоящего изобретения также предусматриваются индуцируемые промоторы. Использование индуцируемого промотора обеспечивает молекулярный переключатель, способный включить экспрессию последовательности полинуклеотида, с которой он функционально связан, когда такая экспрессия является желательной, или выключить экспрессию, когда экспрессия нежелательна. Примеры индуцируемых промоторов включают, но без ограничения, промотор гена металлотионина, промотор гена глюкокортикоида, промотор гена прогестерона и тетрациклиновый промотор.
Для оценки экспрессии полипептида CAR или его частей, экспрессионный вектор, который вводят в клетку, может также содержать или ген селектируемого маркера, или ген-репортер, или то и другое для облегчения идентификации и отбора экспрессирующих клеток из популяции клеток, которые пытались трансфицировать или инфицировать посредством вирусных векторов. В других аспектах селектируемый маркер может переноситься на отдельном фрагменте ДНК и использоваться в процедуре котрансфекции. И селектируемые маркеры, и гены-репортеры могут быть фланкированы соответствующими регуляторными последовательностями, чтобы сделать возможной экспрессию в клетках-хозяевах. Применимые селектируемые маркеры включают, например, гены устойчивости к антибиотикам, такие как neo и т.п.
Гены-репортеры используются для идентификации возможно трансфицированных клеток и для оценки функциональности регуляторных последовательностей. Вообще, геном-репортером является ген, который не присутствует в являющемся реципиентом организме или ткани или не экспрессируется в нем и который кодирует полипептид, экспрессия которого обнаруживается по какому-либо легко выявляемому свойству, например, ферментативной активности. Экспрессию гена-репортера анализируют в соответствующий момент времени, после введения ДНК в являющиеся реципиентами клетки. Подходящие гены-репортеры могут включать гены, кодирующие люциферазу, бета-галактозидазу, хлорамфеникол-ацетилтрансферазу, секретируемую щелочную фосфатазу, или ген зеленого флуоресцентного белка (например, Ui-Tei et al., 2000 FEBS Letters 479: 79-82). Подходящие экспрессионные системы хорошо известны и могут быть приготовлены, используя известные методы, или куплены. Вообще, конструкцию с минимальным 5' фланкирующим районом, демонстрирующую наибольший уровень экспрессии гена-репортера, определяют как промотор. Такие промоторные районы могут быть связаны с геном-репортером и использоваться для оценки агентов в отношении способности к модуляции регулируемой промотором транскрипции.
Способы введения и экспрессии генов в клетке известны в данной области техники. В связи с экспрессионным вектором, вектор можно без труда ввести в клетку-хозяина, например, клетку млекопитающего, бактериальную, дрожжевую клетку или клетку насекомого, с помощью любого способа в данной области техники. Например, экспрессионный вектор можно трансфицировать в клетку-хозяина с помощью физического, химического или биологического способа.
Физические способы введения полинуклеотида в клетку-хозяина включают преципитацию с использованием фосфата кальция, липофекцию, бомбардировку частицами, микроинъекцию, электропорацию и т.п. Способы получения клеток, включающих векторы и/или экзогенные нуклеиновые кислоты, хорошо известны в данной области техники. Смотрите, например, Sambrook et al. (2001, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York). Предпочтительным способом введения полинуклеотида в клетку-хозяина является трансфекция с использованием фосфата кальция.
Биологические способы введения представляющего интерес полинуклеотида в клетку-хозяина включают использование ДНК- и РНК-векторов. Вирусные векторы, и в особенности ретровирусные векторы, стали наиболее широко используемым способом введения генов в клетки млекопитающего, например, человека. Другие вирусные векторы могут происходить из лентивируса, поксвирусов, вируса I простого герпеса, аденовирусов и аденоассоциированных вирусов, и т.п. Смотрите, например, патенты США № 5350674 и № 5585362.
Химические способы введения полинуклеотида в клетку-хозяина включают коллоидные дисперсионные системы, такие как макромолекулярные комплексы, нанокапсулы, микросферы, сферы и системы на основе липидов, включая эмульсии типа «масло-в-воде», мицеллы, смешанные мицеллы и липосомы. Приводимой в качестве примера коллоидной системой для применения в качестве носителя для доставки in vitro и in vivo является липосома (например, искусственная мембранная везикула).
В случае использования невирусной системы для доставки, приводимым в качестве примера носителем для доставки является наночастица, например, липосома или другая подходящая система для доставки субмикронного размера. Использование липидных композиций предполагается для введения нуклеиновых кислот в клетку-хозяина (in vitro, ex vivo или in vivo). В другом аспекте нуклеиновая кислота может быть ассоциированной с липидом. Нуклеиновая кислота, ассоциированная с липидом, может быть заключена в водную внутреннюю часть липосомы, распределена в липидном бислое липосомы, присоединена к липосоме через связующую молекулу, которая связывается и с липосомой, и с олигонуклеотидом, захвачена в липосому, образовывать комплекс с липосомой, быть диспергирована в растворе, содержащем липид, смешана с липидом, объединена с липидом, содержаться в виде суспензии в липиде, содержаться или образовывать комплекс с мицеллой, или быть иным образом ассоциирована с липидом. Липидные, липид/ДНК или липид/экспрессионный вектор ассоциированные композиции не ограничиваются какой-либо конкретной структурой в растворе. Например, они могут присутствовать в бислойной структуре, в виде мицелл, или быть с «разрушенной» структурой. Они могут также просто быть распределены в растворе, возможно, образуя агрегаты, которые являются неодинаковыми по размеру или форме. Липиды могут быть жирами, которые могут встречающимися в природе или синтетическими липидами. Например, липиды включают капельки жира, которые встречаются в природе в цитоплазме, а также класс соединений, которые содержат длинноцепочечные алифатические углеводороды и их производные, такие как жирные кислоты, спирты, амины, аминоспирты и альдегиды.
Липиды, подходящие для использования, можно получить из коммерческих источников. Например, димиристилфосфатидилхолин («DMPC») можно получить от Sigma, St. Louis, MO; дицетилфосфат («DCP») можно получить из K & K Laboratories (Plainview, NY); холестерин («Choi») можно получить от Calbiochem-Behring; димиристилфосфатидилглицерин («DMPG») и другие липиды можно получить от Avanti Polar Lipids, Inc. (Birmingham, AL). Маточные растворы липидов в хлороформе или смеси хлороформ/метанол можно хранить при приблизительно -20°C. Хлороформ используется в качестве единственного растворителя, поскольку он легче испаряется, чем метанол. «Липосома» является общим термином, охватывающим множество одно- и многослойных липидных везикул, образованных в результате создания замкнутых липидных бислоев или агрегатов. Липосомы можно охарактеризовать как имеющие везикулярные структуры с мембраной в виде фосфолипидного бислоя и внутренней водной средой. Многослойные липосомы имеют множество липидных слоев, разделенных водной средой. Они образуются самопроизвольно при суспендировании фосфолипидов в большом количестве водного раствора. Липидные компоненты подвергаются самоперестройке до образования замкнутых структур и захватывают воду и растворенные вещества между липидными бислоями (Ghosh et al., 1991 Glycobiology 5: 505-510). Однако также включены композиции, которые имеют структуры в растворе, отличные от нормальной везикулярной структуры. Например, липиды могут принять мицеллярную структуру или просто существовать в виде неоднородных агрегатов липидных молекул. Также предусматриваются комплексы липофектамин-нуклеиновая кислота.
Независимо от способа, используемого для введения экзогенных нуклеиновых кислот в клетку-хозяина или иначе подвергания клетки воздействию ингибитора по настоящему изобретению, для подтверждения присутствия рекомбинантной последовательности ДНК в клетке-хозяине можно выполнить множество анализов. Такие анализы включают, например, «молекулярно-биологические» анализы, хорошо известные квалифицированным в данной области техники специалистам, такие как блоттинг по Саузерну и Нозерн-блоттинг, ОТ-ПЦР и ПЦР; «биохимические» анализы, такие как детектирование присутствия или отсутствия конкретного пептида, например, с помощью иммунологического способа (ELISA и Вестерн-блоттинга) или с помощью описанных здесь анализов для идентификации агентов, которые не оказываются в объеме настоящего изобретения.
Настоящим изобретением, кроме того, обеспечивается вектор, включающий молекулу нуклеиновой кислоты, кодирующую CAR. В одном аспекте кодирующий CAR вектор можно непосредственно трансдуцировать в клетку, например, Т-клетку. В одном аспекте вектором является вектор для клонирования или экспрессионный вектор, например, включающий, но без ограничения, одну или более плазмид (например, экспрессионных плазмид, векторов для клонирования, миниколец, минивекторов, двойных хромосом), ретровирусных и лентивирусных векторных конструкций. В одном аспекте вектор способен к экспрессии конструкции CAR в Т-клетках млекопитающего. В одном аспекте Т-клеткой млекопитающего является Т-клетка человека.
Источники Т-клеток
До размножения и генетической модификации источник T-клеток получают от субъекта. Термин «субъект», как предполагается, включает живые организмы, в которых может быть вызван иммунный ответ (например, млекопитающих). Примеры субъектов включают людей, собак, кошек, мышей, крыс и их трансгенные виды. T-клетки можно получить из ряда источников, включая мононуклеарные клетки периферической крови, костный мозг, ткань лимфатических узлов, пуповинную кровь, ткань вилочковой железы, ткань из места инфекции, асцитические жидкости, плевральный выпот, ткань селезенки и опухоли. В некоторых аспектах настоящего изобретения может использоваться любой ряд линий Т-клеток, имеющихся в данной области техники. В некоторых аспектах настоящего изобретения T-клетки можно получить из дозы крови, полученной от субъекта, используя любой ряд методов, известных квалифицированному специалисту, таких как разделение с использованием Ficoll™. В одном предпочтительном аспекте клетки циркулирующей крови индивидуума получают с помощью афереза. Аферезный продукт типично содержит лимфоциты, включая T-клетки, моноциты, гранулоциты, B-клетки, другие ядросодержащие лейкоциты, эритроциты и тромбоциты. В одном аспекте клетки, полученные с помощью афереза, можно промыть для удаления фракции плазмы и поместить в соответствующий буфер или среду для стадий последующей обработки. В одном аспекте настоящего изобретения клетки промывают забуференным фосфатом солевым раствором (PBS). В альтернативном аспекте в растворе для промывки отсутствует кальций и может отсутствовать магний или могут отсутствовать многие, если не все двухвалентные катионы. Стадии первоначальной активации в отсутствие кальция могут привести к увеличенной активации. Как это будет совершенно понятно специалистам со средним уровнем компетентности в данной области техники, стадия промывки может выполняться с использованием способов, известных квалифицированным в данной области техники специалистам, например, с использованием полуавтоматической «проточной» центрифуги (например, клеточного процессора Cobe 2991, Baxter CytoMate, или Haemonetics Cell Saver 5) в соответствии с инструкциями производителя. После промывки клетки можно ресуспендировать во множестве биосовместимых буферов, таких как, например, не содержащий Ca, не содержащий Mg PBS, PlasmaLyte A, или другой солевой раствор с буфером или без него. Альтернативно, нежелательные компоненты аферезного образца можно удалить, а клетки непосредственно суспендировать в среде для культивирования.
В одном аспекте T-клетки выделяют из лимфоцитов периферической крови посредством лизиса эритроцитов и истощения моноцитов, например, с помощью центрифугирования через градиент PERCOLL™ или с помощью проточного элютрационного центрифугирования. Конкретную субпопуляцию T-клеток, например, CD3+, CD28+, CD4+, CD8+, CD45RA+ и CD45RO+ T-клеток, можно далее выделить с использованием методов позитивного или негативного отбора. Например, в одном аспекте T-клетки выделяют посредством инкубации со сферами, конъюгированными с антителами против CD3/CD28 (например, 3x28), такими как DYNABEADS® M-450 CD3/CD28 T, в течение периода времени, достаточного для позитивного отбора желаемых T-клеток. В одном аспекте период времени составляет приблизительно 30 минут. В дальнейшем аспекте период времени находится в диапазоне от 30 минут до 36 часов или больше и включает все целые числа в нем. В дальнейшем аспекте период времени составляет по крайней мере 1, 2, 3, 4, 5 или 6 часов. Тем не менее, в другом предпочтительном аспекте период времени составляет от 10 до 24 часов. В одном аспекте период времени инкубации составляет 24 часа. Более длинные периоды времени инкубации могут использоваться для выделения T-клеток в любой ситуации, когда существует мало T-клеток по сравнению с другими типами клеток, например, при выделении инфильтрирующих опухоль лимфоцитов (TIL) из опухолевой ткани или от индивидуумов с ослабленным иммунитетом. Кроме того, использование более длинных периодов времени инкубации может увеличить эффективность захвата CD8+ T-клеток. Таким образом, с помощью просто укорочения или удлинения периода времени допускают связывание T-клеток со сферами с антителами против CD3/CD28, и/или посредством увеличения или уменьшения отношения сфер к T-клеткам (описываемого здесь далее), субпопуляции T-клеток можно предпочтительно отбирать в начале культивирования или в другие моменты времени во время процесса. Кроме того, посредством увеличения или уменьшения отношения антител против CD3 и/или CD28 на сферах или других поверхностях, субпопуляции Т-клеток можно предпочтительно отбирать в начале культивирования или в другие моменты времени во время процесса. Квалифицированному специалисту будет понятно, что множество циклов отбора можно также использовать в связи с этим изобретением. В некоторых аспектах может быть желательным выполнение процедуры отбора и использование «не отобранных» клеток в процессе активации и размножения. «Не отобранные» клетки могут быть также подвергнуты дальнейшим циклам отбора.
Обогащение популяции T-клеток с помощью негативного отбора может выполняться с использованием комбинации антител, направленных против маркеров поверхности, присущих только негативно отбираемым клеткам. Одним способом является клеточный сортинг и/или отбор посредством негативной магнитной иммуноадгезии или проточной цитометрии, в случае которой используется смесь моноклональных антител, направленных против маркеров клеточной поверхности, присутствующих на негативно отбираемых клетках. Например, для обогащения в отношении CD4+ клеток с помощью негативного отбора, смесь моноклональных антител типично включает антитела против CD14, CD20, CD1lb, CD16, HLA-DR и CD8. В некоторых аспектах может быть желательным обогащение в отношении или позитивный отбор регуляторных T-клеток, которые типично экспрессируют CD4+, CD25+, CD62Lвысокий уровень, GITR+ и FoxP3+. Альтернативно, в некоторых аспектах регуляторные T-клетки истощают с использованием сфер, конъюгированных с антителами против C25, или другого схожего способа отбора.
В одном варианте осуществления можно отобрать популяцию T-клеток, которая экспрессирует один или более из IFN-γ, TNFα, IL-17A, IL-2, IL-3, IL-4, GM-CSF, IL-10, IL-13, гранзима B и перфорина, или других подходящих молекул, например, других цитокинов. Методы скрининга на предмет клеточной экспрессии могут быть определены, например, с использованием способов, описанных в публикации PCT-заявки № WO 2013/126712.
Для выделения желаемой популяции клеток с помощью позитивного или негативного отбора, концентрация клеток и поверхности (например, частиц, таких как сферы) может варьировать. В некоторых аспектах может быть желательным значительное уменьшение объема, в котором сферы и клетки смешивают вместе, (например, увеличение концентрации клеток) для обеспечения максимального контакта клеток и сфер. Например, в одном аспекте используют концентрацию, составляющую 2 биллиона клеток/мл. В одном аспекте используют концентрацию, составляющую 1 биллион клеток/мл. В дальнейшем аспекте используют концентрацию, превышающую 100 миллионов клеток/мл. В дальнейшем аспекте используют концентрацию клеток, составляющую 10, 15, 20, 25, 30, 35, 40, 45 или 50 миллионов клеток/мл. Тем не менее, в другом аспекте используют концентрацию клеток, выбираемую из 75, 80, 85, 90, 95 или 100 миллионов клеток/мл. В дальнейших аспектах могут использоваться концентрации, составляющие 125 или 150 миллионов клеток/мл. Использование высоких концентраций может привести к увеличению выхода клеток, активации клеток и размножению клеток. Кроме того, использование высоких концентраций клеток создает возможность для более эффективного захвата клеток, которые могут слабо экспрессировать представляющие интерес антигены-мишени, таких как CD28-негативные T-клетки, или из образцов, в которых присутствует множество опухолевых клеток, (т.е. лейкозной крови, ткани опухоли и т.д.). Такие популяции клеток могут иметь терапевтическое значение, и их желательно было бы получить. Например, использование высоких концентраций клеток создает возможность для более эффективного отбора CD8+ T-клеток, которые обычно имеют более слабую экспрессию CD28.
В связанном с этим аспекте может быть желательным использование более низких концентраций клеток. В результате значительного разбавления смеси T-клеток и поверхности (например, частиц, таких как сферы) минимизируются взаимодействия между частицами и клетками. Это позволяет отобрать клетки, которые экспрессируют высокие количества желаемых антигенов, которые связываются с частицами. Например, CD4+ T-клетки экспрессируют CD28 на более высоких уровнях и более эффективно захватываются, чем CD8+ T-клетки в разбавленных концентрациях. В одном аспекте используемая концентрация клеток составляет 5×106/мл. В других аспектах используемая концентрация может составлять от приблизительно 1×105/мл до 1×106/мл и любое целое число в этом диапазоне.
В других аспектах клетки могут подвергаться инкубации на вращающем устройстве в течение переменных промежутков времени при переменных скоростях или при 2-10°C, или при комнатной температуре.
T-клетки для стимуляции могут быть также заморожены после стадии промывки. Без желания ограничиться какой-либо теорией, стадия замораживания и последующего оттаивания обеспечивает более однородный продукт в результате удаления гранулоцитов и в некоторой мере моноцитов в популяции клеток. После стадии промывки, которая удаляет плазму и тромбоциты, клетки можно суспендировать в растворе для замораживания. Хотя множество растворов и параметров для замораживания известно в данной области техники и будет применимо в этой связи, один способ включает использование PBS, содержащего 20% DMSO и 8% сывороточного альбумина человека, или сред для культивирования, содержащих 10% декстрана 40 и 5% декстрозы, 20% сывороточного альбумина человека и 7,5% DMSO, или 31,25% PlasmaLyte-A, 31,25% декстрозы 5%, 0,45% NaCl, 10% декстрана 40 и 5% декстрозы, 20% сывороточного альбумина человека и 7,5% DMSO, или других подходящих сред для замораживания клеток, содержащих, например, Hespan и PlasmaLyte A, клетки затем замораживают до -80°C со скоростью 1° в минуту и хранят в газообразной фазе бака для хранения с жидким азотом. Могут использоваться другие способы контролируемого замораживания, а также неконтролируемое замораживание непосредственно при -20°C или в жидком азоте.
В некоторых аспектах замороженные клетки оттаивают и промывают, как здесь описывается, и допускается их выдерживание в течение одного часа при комнатной температуре до активации, используя способы по настоящему изобретению.
В связи с настоящим изобретением также предусматривается коллекция образцов крови или аферезных продуктов от субъекта в период времени до того момента, когда могли бы понадобится размноженные клетки, описываемые здесь. Как таковой, источник клеток, подвергаемых размножению, может быть получен в любой необходимый момент времени, и быть требуемыми клетками, такими как T-клетки, выделенными и замороженными для применения позже в терапии с использованием T-клеток для любого ряда заболеваний или состояний, в случае которых могла бы полезной терапия с использованием T-клеток, таких как те, которые здесь описаны. В одном аспекте образец крови или аферезный продукт отбирают у здорового в целом субъекта. В некоторых аспектах образец крови или аферезный продукт отбирают у здорового в целом субъекта, который подвержен риску развития заболевания, но у которого еще не развилось заболевание, и представляющие интерес клетки выделяют и замораживают для применения позже. В некоторых аспектах T-клетки могут быть размножены, заморожены и использованы позже. В некоторых аспектах образцы получают от пациента вскоре после диагностирования конкретного заболевания, описанного здесь, но до каких-либо лечений. В дальнейшем аспекте клетки выделяют из образца крови или аферезного продукта от субъекта до какого-либо ряда соответствующих способов лечения, включая, но без ограничения, лечение такими средствами, как натализумаб, эфализумаб, противовирусные средства, химиотерапия, облучение, иммуносупрессанты, такие как циклоспорин, азатиоприн, метотрексат, микофенолят и FK506, антитела или другие иммуноаблативные средства, такие как CAMPATH, антитела против CD3, цитоксан, флударабин, циклоспорин, FK506, рапамицин, микофеноловая кислота, стероиды, FR901228 и облучение.
В дальнейшем аспекте настоящего изобретения T-клетки получают от пациента непосредственно после лечения, которое оставляет субъекта с функциональными Т-клетками. В этой связи было отмечено, что после некоторых лечений рака, в частности, лечений лекарственными средствами, которые повреждают иммунную систему, вскоре после лечения во время периода времени, когда должно бы в норме происходить восстановление пациентов от лечения, качество получаемых T-клеток может быть оптимальным или улучшенным в отношении их способности к размножению ex vivo. Так же, после ex vivo манипуляции, используя описываемые здесь способы, эти клетки могут быть в предпочтительном состоянии для усиленного приживления и in vivo размножения. Таким образом, в связи с настоящим изобретением предусматривается получение клеток крови, включая T-клетки, дендритные клетки или другие клетки гемопоэтической линии, во время этой фазы восстановления. Кроме того, в некоторых аспектах режимы мобилизации (например, мобилизации с использованием GM-CSF) и установления определенного состояния могут использоваться для создания состояния у субъекта, при котором репопуляция, рециркуляция, регенерация и/или размножение конкретных типов клеток является преимущественной, в особенности во время определенного временного окна после терапии. Иллюстративные типы клеток включают T-клетки, B-клетки, дендритные клетки и другие клетки иммунной системы.
Активация и размножение T-клеток
T-клетки можно активировать и размножить, в общем используя способы, описанные, например, в патентах США №№ 6352694, 6534055, 6905680, 6692964, 5858358, 6887466, 6905681, 7144575, 7067318, 7172869, 7232566, 7175843, 5883223, 6905874, 6797514, 6867041; и публикации заявки на патент США № 20060121005.
Как правило, T-клетки по настоящему изобретению размножают посредством контактирования с поверхностью с присоединенным к ней агентом, который стимулирует связанный с комплексом CD3/TCR сигнал, и лигандом, который стимулирует костимулирующую молекулу на поверхности T-клеток. В частности, популяции T-клеток можно стимулировать, как здесь описано, например, посредством контактирования с антителом против CD3, или его антигенсвязывающим фрагментом, или антителом против CD2, иммобилизованным на поверхности, или посредством контактирования с активатором протеинкиназы C (например, бриостатином) вместе с кальциевым ионофором. Для костимуляции вспомогательной молекулы на поверхности T-клетки используют лиганд, который связывается со вспомогательной молекулой. Например, популяцию T-клеток можно привести в контакт с антителом против CD3 и антителом против CD28, в условиях, подходящих для стимуляции пролиферации T-клеток. Для стимуляции пролиферации или CD4+ T-клеток, или CD8+ T-клеток используют антитело против CD3 и антитело против CD28. Примеры антитела против CD28 включают 9.3, B-T3, XR-CD28 (Diaclone, Besancon, Франция), как могут использоваться и другие общеизвестные в данной области техники способы (Berg et al., Transplant Proc. 30(8): 3975-3977, 1998; Haanen et al., J. Exp. Med. 190(9): 1319-1328, 1999; Garland et al., J. Immunol Meth. 227(1-2): 53-63, 1999).
В некоторых аспектах первичный стимулирующий сигнал и костимулирующий сигнал для T-клетки могут быть обеспечены с использованием различных протоколов. Например, агенты, формирующие каждый сигнал, могут находиться в растворе или быть связанными с поверхностью. В случае связывания с поверхностью, агенты могут быть связаны с одной и той же поверхностью (т.е. находиться в «цис»-форме) или с отдельными поверхностями (т.е. находиться в «транс»-форме). Альтернативно, один агент может быть связан с поверхностью, а другой агент находиться в растворе. В одном аспекте агент, формирующий костимулирующий сигнал, связан с поверхностью клетки, а агент, формирующий сигнал первичной активации, находится в растворе или связан с поверхностью. В некоторых аспектах оба агента могут находиться в растворе. В другом аспекте агенты могут находиться в растворимой форме, а затем быть сшиты с поверхностью, например, клеткой, экспрессирующей Fc-рецепторы или антитело или другой связывающий агент, который будет связываться с агентами. В этой связи смотрите, например, публикации заявок на патенты США №№ 20040101519 и 20060034810 для искусственных антигенпрезентирующих клеток (aAPC), которые предусмотрены для применения для активации и размножения T-клеток в настоящем изобретении.
В одном аспекте два агенты иммобилизованы на сферах, или на одной и той же сфере, т.е. «цис», или на отдельных сферах, т.е. «транс». В качестве примера, агентом, формирующим сигнал первичной активации, является антитело против CD3 или его антигенсвязывающий фрагмент, а агентом, формирующим костимулирующий сигнал, является антитело против CD28 или его антигенсвязывающий фрагмент; и оба агента коиммобилизованы на одной и той сфере в эквивалентных молекулярных количествах. В одном аспекте используют составляющее 1:1 отношение каждого антитела, связанного со сферами, для размножения CD4+ T-клеток и роста T-клеток. В некоторых аспектах настоящего изобретения используют отношение антитело против CD3:антитело против CD28, которые связаны со сферами, из условия, чтобы отмечалось увеличение размножения T-клеток по сравнению с размножением, отмечаемым при использовании отношения 1:1. В одном конкретном аспекте отмечается увеличение от приблизительно 1 до приблизительно 3 раз по сравнению с размножением, отмечаемым при использовании отношения 1:1. В одном аспекте отношение антитело против CD3:антитело против CD28, которые связаны со сферами, находится в диапазоне от 100:1 до 1:100 и включает любые целые числа в нем. В одном аспекте настоящего изобретения с частицами связано больше антитела против CD28, чем антитела против CD3, т.е. отношение антитело против CD3:антитело против CD28 меньше 1. В некоторых аспектах настоящего изобретения отношение антитела против CD28 к антителу против CD3, которые связаны со сферами, больше чем 2:1. В одном конкретном аспекте используют составляющее 1:100 отношение CD3:CD28 антител, связанных со сферами. В одном аспекте используют составляющее 1:75 отношение CD3:CD28 антител, связанных со сферами. В дальнейшем аспекте используют составляющее 1:50 отношение CD3:CD28 антител, связанных со сферами. В одном аспекте используют составляющее 1:30 отношение CD3:CD28 антител, связанных со сферами. В одном предпочтительном аспекте используют составляющее 1:10 отношение CD3:CD28 антител, связанных со сферами. В одном аспекте используют составляющее 1:3 отношение CD3:CD28 антител, связанных со сферами. Тем не менее, в другом аспекте используют составляющее 3:1 отношение CD3:CD28 антител, связанных со сферами.
Отношения частиц к клеткам в диапазоне от 1:500 до 500:1 и любые целые числа в нем могут использоваться для стимуляции T-клеток и других клеток-мишеней. Как это совершенно понятно специалистам со средним уровнем компетентности в данной области техники, отношение частиц к клеткам может зависеть от размера частиц по сравнению с клеткой-мишенью. Например, сферы с маленькими размерами могли бы только связать малое число клеток, в то время как более большие сферы могли бы связать большое число. В некоторых аспектах отношение частиц к клеткам находится в диапазоне от 1:100 до 100:1 и включает любые целые числа в нем, и в дальнейших аспектах отношение, которое включает диапазон от 1:9 до 9:1 и любые целые числа в нем, может также использоваться для стимуляции T-клеток. Отношение связанных с антителом против CD3 и антителом против CD28 частиц к T-клеткам, которое приводит к стимуляции T-клеток, может варьировать, как отмечено выше, однако некоторые предпочтительные отношения включают 1:100, 1:50, 1:40, 1:30, 1:20, 1:10, 1:9, 1:8, 1:7, 1:6, 1:5, 1:4, 1:3, 1:2, 1:1, 2:1, 3:1, 4:1, 5:1, 6:1, 7:1, 8:1, 9:1, 10:1 и 15:1, при этом предпочтительное отношение составляет по крайней мере 1:1 частиц на T-клетку. В одном аспекте используют составляющее 1:1 или меньше отношение частиц к клеткам. В одном конкретном аспекте предпочтительное отношение частица:клетка составляет 1:5. В дальнейших аспектах отношение частиц к клеткам может варьировать в зависимости от дня стимуляции. Например, в одном аспекте отношение частиц к клеткам составляет от 1:1 до 10:1 в первый день, и дополнительные частицы добавляют к клеткам каждый день или через день после этого вплоть до 10 дней, в конечных отношениях от 1:1 до 1:10 (на основе подсчета клеток в день добавления). В одном конкретном аспекте отношение частиц к клеткам составляет 1:1 в первый день стимуляции и доводится до 1:5 на третий и пятый день стимуляции. В одном аспекте частицы добавляют ежедневно или через день до конечного отношения 1:1 в первый день и 1:5 на третий и пятый день стимуляции. В одном аспекте отношение частиц к клеткам составляет 2:1 в первый день стимуляции и доводится до 1:10 на третий и пятый день стимуляции. В одном аспекте частицы добавляют ежедневно или через день до конечного отношения 1:1 в первый день и 1:10 на третий и пятый день стимуляции. Квалифицированному в данной области техники специалисту будет понятно, что множество других отношений может быть подходящим для использования в настоящем изобретении. В частности, отношения будут варьировать в зависимости от размера частиц и от размера и типа клеток. В одном аспекте самые типичные отношения для применения находятся вблизи 1:1, 2:1 и 3:1 в первый день.
В дальнейших аспектах настоящего изобретения клетки, такие как T-клетки, объединяют с покрытыми агентами сферами, сферы и клетки впоследствии разделяют, и затем клетки подвергают культивированию. В альтернативном аспекте, до культивирования, покрытые агентами сферы и клетки не разделяют, а культивируют вместе. В дальнейшем аспекте сферы и клетки сначала концентрируют посредством приложения силы, такой как магнитная сила, что приводит к увеличенному лигированию маркеров клеточной поверхности, тем самым индуцируя стимуляцию клеток.
В качестве примера, белки клеточной поверхности можно лигировать посредством допуска контактирования парамагнитных сфер, к которым присоединены антитело против CD3 и антитело против CD28, (3×28 сфер), с T-клетками. В одном аспекте объединяют клетки (например, 104-109 T-клеток) и сферы (например, DYNABEADS® M-450 CD3/CD28 T парамагнитные сферы в соотношении 1:1) в буфере, например, PBS (без двухвалентных катионов, таких как кальция и магния). Снова, специалисты со средним уровнем компетентности в данной области техники могут легко понять, что может использоваться любая концентрация клеток. Например, представляющая интерес клетка-мишень может быть очень редкой в образце и составлять лишь 0,01% от образца, или весь образец (т.е. 100%) может включать представляющую интерес клетку-мишень. Соответственно, любое количество клеток находится в условиях настоящего изобретения. В некоторых аспектах может быть желательным значительное уменьшение объема, в котором смешивают вместе частицы и клетки, (т.е. увеличение концентрации клеток) для обеспечения максимального контакта клеток и частиц. Например, в одном аспекте используют концентрацию, составляющую 2 биллиона клеток/мл. В одном аспекте используют концентрацию, превышающую 100 миллионов клеток/мл. В дальнейшем аспекте используют концентрацию клеток, составляющую 10, 15, 20, 25, 30, 35, 40, 45 или 50 миллионов клеток/мл. Тем не менее, в другом аспекте используют концентрацию клеток, выбираемую из 75, 80, 85, 90, 95 или 100 миллионов клеток/мл. В дальнейших аспектах могут использоваться концентрации, составляющие 125 или 150 миллионов клеток/мл. Использование высоких концентраций может привести к увеличению выхода клеток, активации клеток и размножения клеток. Кроме того, использование высоких концентраций клеток создает возможность для более эффективного захвата клеток, которые могут слабо экспрессировать представляющие интерес антигены-мишени, таких как CD28-негативные T-клетки. Такие популяции клеток могут иметь терапевтическое значение, и их желательно было бы получить в некоторых аспектах. Например, использование высоких концентраций клеток создает возможность для более эффективного отбора CD8+ T-клеток, которые обычно имеют более слабую экспрессию CD28.
В одном аспекте настоящего изобретения смесь можно подвергнуть культивированию в течение интервала от нескольких часов (приблизительно 3 часов) до приблизительно 14 дней или любого часового целого значения в нем. В одном аспекте смесь можно подвергнуть культивированию в течение 21 дня. В одном аспекте настоящего изобретения сферы и T-клетки культивируют вместе в течение приблизительно восьми дней. В одном аспекте сферы и T-клетки культивируют вместе в течение 2-3 дней. Также может быть желательно несколько циклов стимуляции, так что время культивирования T-клеток может составлять 60 дней или более. Подходящие для культивирования T-клеток условия включают соответствующую среду (например, минимальную питательную среду или среду RPMI 1640 или X-vivo 15, (Lonza)), которая может содержать факторы, необходимые для пролиферации и жизнеспособности, включая сыворотку (например, фетальную бычью или человеческую сыворотку), интерлейкин-2 (IL-2), инсулин, IFN-γ, IL-4, IL-7, GM-CSF, IL-10, IL-12, IL-15, TGFβ и TNF-α, или любые другие добавки для роста клеток, известные квалифицированному специалисту. Другие добавки для роста клеток включают, но без ограничения, поверхностно-активное вещество, плазманат и восстанавливающие агенты, такие как N-ацетилцистеин и 2-меркаптоэтанол. Среды могут включать RPMI 1640, AIM-V, DMEM, MEM, α-MEM, F-12, X-Vivo 15 и X-Vivo 20, Optimizer, с добавленными аминокислотами, пируватом натрия и витаминами, или бессывороточные или дополненные соответствующим количеством сыворотки (или плазмы) или определенным рядом гормонов, и/или количеством цитокина(ов), достаточным для роста и размножения T-клеток. Антибиотики, например, пенициллин и стрептомицин, включают только в экспериментальные культуры, но не в культуры клеток, инфузия которых субъекту должна быть проведена. Клетки-мишени поддерживают в условиях, необходимых для поддержания роста, например, при соответствующей температуре (например, 37°C) и атмосферных условиях (например, воздух плюс 5% CO2).
T-клетки, которые были подвергнуты различным периодам времени стимуляции, могут демонстрировать различные характеристики. Например, типичная кровь или аферезные продукты в виде мононуклеарных клеток периферической крови содержат популяцию T-клеток-хелперов (TH, CD4+), которая больше популяции цитотоксических Т-клеток или Т-клеток-супрессоров (TС, CD8+). Ex vivo размножение T-клеток при стимуляции рецепторов CD3 и CD28 порождает популяцию T-клеток, которая до приблизительно дней 8-9 состоит преимущественно из TH-клеток, в то время как через приблизительно 8-9 дней популяция T-клеток включает большую в возрастающей степени популяцию TС-клеток. Соответственно, в зависимости от цели лечения, преимущественной может быть инфузия субъекту популяции T-клеток, состоящей преимущественно из TH-клеток. Так же, если была выделена антигенспецифическая субпопуляция TС-клеток, может быть выгодным размножение этой субпопуляции в большей степени.
Кроме того, помимо маркеров CD4 и CD8, другие фенотипические маркеры могут значительно варьировать, но по большей части, воспроизводимо в ходе процесса размножения клеток. Таким образом, такая воспроизводимость создает возможность для приспособления продукта - активированной T-клетки для конкретных целей.
После конструирования EGFRvIII-специфического CAR, для оценки активности молекулы могут использоваться различные анализы, такие как, но без ограничения, способность к размножению Т-клеток после стимуляции, к поддержанию размножения T-клеток в отсутствие рестимуляции и противораковые активности в соответствующих in vitvo моделях и моделях на животных. Анализы для оценки эффектов EGFRvIII-специфического CAR описаны подробнее ниже.
Анализ с использованием Вестерн-блоттинга экспрессии CAR в первичных Т-клеток может быть использован для обнаружения присутствия мономеров и димеров. Смотрите, например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). Очень кратко, Т-клетки (1:1 смесь CD4+ и CD8+ Т-клеток), экспрессирующие CAR, размножают in vitro в течение более 10 дней с последующим лизисом и электрофорезом в SDS-ПААГ в восстанавливающих условиях. CAR, содержащие полноразмерный цитоплазматический домен TCR-ζ и эндогенную TCR-ζ цепь, детектируют с помощью Вестерн-блоттинга, используя антитело против TCR-ζ цепи. Те же подмножества Т-клеток используются для анализа с использованием электрофореза в SDS-ПААГ в невосстанавливающих условиях, чтобы обеспечить оценку ковалентного образования димера.
In vitro размножение CAR+ Т-клеток после антигенной стимуляции можно измерить с помощью проточной цитометрии. Например, смесь CD4+ и CD8+ Т-клеток стимулируют с использованием αCD3/αCD28 aAPC с последующей трансдукцией лентивирусными векторами, экспрессирующими GFP под контролем анализируемых промоторов. Приводимые в качестве примера промоторы включают промоторы гена IE CMV, EF-1α, убиквитина C или фосфоглицерокиназы (PGK). Флуоресценция GFP оценивается на 6-й день культивирования в подмножествах CD4+ и/или CD8+ Т-клеток с помощью проточной цитометрии. Смотрите, например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). Альтернативно, смесь CD4+ и CD8+ Т-клеток стимулируют покрытыми αCD3/αCD28 магнитными сферами в день 0 и трансдуцируют CAR в день 1, используя бицистронный лентивирусный вектор, экспрессирующий CAR вместе с eGFP с помощью последовательности пропуска рибосомой 2А. Культуры повторно стимулируют с использованием или EGFRvIII+ U-87 клеток (U-87-EGFRvIII), U-87 клеток дикого типа (U-87 дикого типа), или клеток К562, экспрессирующих hCD32 и 4-1BBL, в присутствии антитела против CD3 и СD28 (BBL K562-3/28) после промывки. Экзогенный IL-2 добавляют к культурам через день в количестве 100 МЕ/мл. GFP+ Т-клетки подсчитывают с помощью проточной цитометрии, используя основанный на сферах подсчет. Смотрите, например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009).
Длительное размножение CAR+ Т-клеток в отсутствие повторного стимулирования может также быть измерено. Смотрите, например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). Вкратце, средний объем Т-клеток (фл) измеряется на 8-ой день культивирования, используя счетчик частиц Coulter Multisizer III, после стимуляции покрытыми αCD3/αCD28 магнитными сферами в день 0 и трансдукции указанным CAR в день 1.
Оценка пролиферации клеток и продукции цитокинов была описана ранее, например, в Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). Вкратце, оценка CAR-опосредованной пролиферации осуществляется в титрационных микропланшетах путем смешивания промытых Т-клеток с клетками-мишенями, такими как клетки asU87MG, ВНК или СНО, экспрессирующие EGFRvIII или EGFR дикого типа (wt) или CD32 и CD137 (KT32-BBL), для окончательного соотношения Т-клетки:клетки-мишени=1:1. Моноклональные антитела против CD3 (клон ОКТ3) и против CD28 (клон 9.3) добавляют к культурам с клетками KT32-BBL в качестве положительного контроля для стимулирования пролиферации Т-клеток, поскольку эти сигналы поддерживают длительное размножение CD8+ T-клеток ex vivo. Т-клетки подсчитывают в культурах, используя флуоресцентные сферы CountBright™ (Invitrogen, Carlsbad, CA) и проточную цитометрию, как описано производителем. CAR+ Т-клетки определяют по экспрессии GFP, используя Т-клетки, которые созданы с использованием экспрессирующих связанный с eGFP-2A CAR лентивирусных векторов. В случае CAR+ Т-клеток, не экспрессирующих GFP, CAR+ T-клетки детектируют с помощью биотинилированного рекомбинантного белка EGFRvIII и дополнительного конъюгата авидин-PE. CD4+ и CD8+ экспрессию на Т-клеткам также одновременно детектируют с помощью специфических моноклональных антител (BD Biosciences). Измерения цитокинов выполняются на супернатантах, собранных через 24 часа после рестимулирования, используя набор Cytometric Bead Array для исследования цитокинов Th1/Th2 человека (BD Biosciences, San Diego, CA) в соответствии с инструкциями изготовителя. Флуоресценцию оценивают с помощью проточного цитометра FACScalibur, и данные анализируют в соответствии с инструкциями изготовителя.
Цитотоксичность может быть оценена с помощью стандартного анализа высвобождения 51Cr. Смотрите, например, Milone et al., Molecular Therapy 17(8): 1453-1464 (2009). Вкратце, в клетки-мишени (клетки U87MG, ВНК или СНО, экспрессирующие EGFRvIII или EGFR дикого типа (wt), вводят 51Cr (в виде NaCrО4, New England Nuclear, Boston, MA) при 37°С в течение 2 часов с частым перемешиванием, дважды промывают в полной RPMI и высевают в титрационные микропланшеты. Эффекторные Т-клетки смешивают с клетками-мишенями в лунках, в полной среде RPMI при различных соотношениях эффекторные клетки:клетки-мишени (E:T). Также готовят дополнительные лунки, содержащие только среду (самопроизвольное высвобождение, SR) или 1%-ый раствор детергента Triton-X 100 (полное высвобождение, ТR). Через 4 часа инкубации при 37°С собирают супернатант из каждой лунки. Выводившийся 51Cr затем измеряют с помощью счетчика гамма-частиц (Packard Instrument Co., Waltham, MA). Каждое условие выполняют в по крайней мере трех повторах, и процент лизиса рассчитывают по формуле: % лизиса = (ER-SR)/(ТR-SR), где ER представляет собой среднее значение 51Cr, выводившегося в случае каждого экспериментального условия. Также могут использоваться альтернативные анализы цитотоксичности, такие как анализы цитотоксичности на основе потока, как описано в примере 8.
Люцифераза жука-щелкуна красного и жука-щелкуна зеленого может использоваться, чтобы одновременно следовать за прогрессированием опухоли и направленной миграцией Т-клеток, поскольку в случае каждой из них используется один и тот же субстрат люциферин, но излучается свет на противоположных концах спектра видимого света.
Другие анализы, в том числе те, которые описаны в разделе «Примеры» здесь, а также те, которые известны в данной области техники, также могут использоваться для оценки конструкций EGFRvIII-специфических CAR по настоящему изобретению.
Терапевтическое применение для EGFRvIII-экспрессирующих заболеваний и нарушений
EGFRvIII представляет собой специфический для опухоли, лиганд-независимый, конститутивно активный вариант рецептора эпидермального фактора роста. Настоящим изобретением обеспечиваются композиции и способы для лечения заболеваний и нарушений, связанных с EGFRvIII. Примером заболевания или нарушения, связанного с EGFRvIII, является глиома.
Глиома относится к раку центральной нервной системы, который начинается в глиальных клетках (например, клетках, которые окружают и поддерживают нервные клетки и включают олигодендроциты, астроциты, микроглии и эпендимальные клетки). Глиомы особенно опасны в плане и коэффициента заболеваемости, и злокачественности и подразделяются на семь или более типов, таких как глиобластома и анапластическая астроцитома в соответствии с их детализированным патологическим типом ткани. Стадия заболевания (размер опухоли, наличие дистальных метастазов) и гистологическая злокачественность используются при определении степени злокачественности первичных опухолей головного мозга. Гистологическая злокачественность подразделяется на четыре уровня, т.е. G1-G4 в соответствии с Руководством по лечению опухолей головного мозга ((2002) Kanehara & Co., Ltd.), и они соответствуют WHO1-WHO4, соответственно. Чем больше число, тем выше степень злокачественности. Например, злокачественность глиобластомы равна G4 (WHO4), в то время как злокачественность анапластической астроцитомы равна G3 (WHO3), и как G3, так и G4 определяются как злокачественные. Таким образом, в соответствии с некоторыми вариантами осуществления способы по этому изобретению нацелены на злокачественные глиомы. В других аспектах настоящее изобретение нацелено на мультиформную глиобластому (GBM). В дальнейших вариантах осуществления композиции и способы по настоящему изобретению могут использоваться при лечении других глиом, включая, но без ограничения, анапластическую астроцитому, гигантоклеточную глиобластому, глиосаркому, анапластическую олигодендроглиому, анапластическую эпендимому, карциному сосудистого сплетения, анапластическую ганглиоглиому, пинеобластому, медуллоэпителиому, эпендимобластому, медуллобластому, супратенториальную примитивную нейроэктодермальную опухоль и атипичную тератоидную/рабдоидную опухоль.
Глиобластома является самой распространенной первичной опухолью головного мозга у взрослых. Более половины из 18000 пациентов, у которых диагностируются злокачественные первичные опухоли головного мозга в США каждый год, имеют мультиформную глиобластому. Мультиформная глиобластома представляет собой анапластическую, исключительно клеточную опухоль, с высокими показателями пролиферации, разрастанием мелких сосудов и фокальным некрозом. Признаки и симптомы зависят от нескольких факторов (размера, скорости роста, локализации опухоли в головном мозге) и представлены в основном головной болью, судорогами, неврологические расстройства, изменениями психического состояния. Прогноз мультиформной глиобластомы остается мрачным. Время жизни составляет менее 2 лет для большинства пациентов. Общее состояние по шкале Карнофского (KPS) является одним из самых важных прогностических факторов: пациенты с KPS >70 живы через 18 месяцев в приблизительно 18% случаев, по сравнению с 13% пациентов с более низкими оценками KPS. Первичная мультиформная глиобластома развивается de novo из глиальных клеток, как правило, имеет историю болезни в течение менее шести месяцев, является более распространенной у пожилых пациентов и представляет собой мелкоклеточную опухоль по гистологии. Вторичная мультиформная глиобластома развивается в течение месяцев или лет из уже существующих астроцитом с низкой степенью злокачественности, преимущественно поражает молодых людей и представляет собой гигантоклеточную опухоль по гистологии.
Злокачественные глиомы также известны как глиомы с высокой степенью злокачественности. Они могут поражать головной мозг и спинной мозг. В некоторых аспектах композиции и способы по настоящему изобретению могут использоваться для лечения субъектов, имеющих злокачественную глиому в головном мозге, например, такую, которую выбирают из анапластической астроцитомы (АА), мультиформной глиобластомы (GBM), анапластической олигодендроглиомы (АО) и анапластической олигоастроцитомы (АОА). В некоторых аспектах композиции и способы по настоящему изобретению могут использоваться для лечения субъектов, имеющих мультиформную глиобластому (GBM).
Мультиформная глиобластома является самой злокачественной стадией астроцитомы, со временем дожития менее 2 лет для большинства пациентов. Гистологически эти опухоли характеризуются высокими показателями пролиферации, пролиферацией эндотелиальных клеток и фокальным некрозом. В высокой степени пролиферативной характер этих поражений, по-видимому, является результатом нескольких митогенных эффектов. Один из признаков GBM является пролиферация эндотелиальных клеток. Множество факторов ангиогенеза и их рецепторов обнаруживается в GBM.
Существуют биологические подмножества астроцитом, которые могут отражать клиническую гетерогенность, наблюдаемую в случае этих опухолей. Эти подмножества включают глиомы ствола головного мозга, которые являются формой диффузной, фибриллярной астроцитомы у людей, которые часто развиваются по злокачественному курсу. GBM ствола головного мозга имеют общие генетические признаки с GBM у взрослых, которые поражают молодых пациентов. Плейоморфная ксантоастроцитома (PXA) является поверхностной, астроцитарной опухолью с низкой степенью злокачественности, которая преимущественно поражает молодых взрослых людей. Хотя эти опухоли имеют аномальные гистологические признаки, они, как правило, являются медленно растущими опухолями, которые могут поддаваться хирургическому лечению. Некоторые PXA, однако, могут возвращаться в виде GBM. Волосовидная астроцитома является самой распространенной астроцитарной опухолью у детей и клинически и гистопатологически отличается от диффузной, фибриллярной астроцитомы, которая поражает взрослых. Волосовидные астроцитомы не имеют те же геномные изменения, что и диффузная, фибриллярная астроцитома. Субэпендимальные гигантоклеточные астроцитомы (SEGA) являются перивентрикулярными астроцитарными опухолями с низкой степенью злокачественности, которые, как правило, сопровождаются эпилойэй (TS), и гистологически идентичны так называемым «оплываниями свечи», которые выстилают желудочки пациентов с TS. Подобно другим опухолевым поражениям в ТС, они являются медленно растущими и могут быть больше похожи на гамартомы, чем на истинные новообразования. Десмопластическая астроцитома головного мозга у детей грудного возраста (DCAI) и десмопластическая ганглиоглиома у детей грудного возраста (DIGG) представляют собой большие, поверхностные, обычно кистозные, доброкачественные астроцитомы, которые поражают детей в первый год или два жизни.
Олигодендроглиомы и олигоастроцитомы (смешанные глиомы) представляют собой диффузные, глиальные опухоли в основном ЦНС, которые являются клинически и биологически наиболее близкородственными с диффузными, фибриллярными астроцитомами. Опухоли, однако, гораздо меньше распространены, чем астроцитомы, и имеют, как правило, лучшие прогнозы, чем диффузные астроцитомы. Олигодендроглиомы и олигоастроцитомы могут развиваться или в анапластическую олигодендроглиому, или анапластическую олигоастроцитому WHO степени III, или в GBM WHO степени IV. Таким образом, генетические изменения, которые приводят к олигодендроглиальным опухолям, представляют собой еще один путь к GBM.
Эпендимомы представляют собой клинически разнородную группу глиом, которые варьируют от агрессивных внутрижелудочковых опухолей у детей до доброкачественных опухолей спинного мозга у взрослых. Перерождения эпендимомы в GBM редки. Опухоли сосудистого сплетения также представляют собой разнообразную группу опухолей, которые преимущественно возникают в системе желудочков мозга, начиная от агрессивных супратенториальных внутрижелудочковых опухолей у детей до доброкачественных опухолей мостомозжечкового угла у взрослых. Опухоли сосудистого сплетения иногда регистрировали у пациентов с синдромом Ли-Фраумени и заболеванием Гиппеля-Линдау (VHL).
Медуллобластомы представляют собой злокачественные, примитивные опухоли, которые возникают в задней черепной ямке, в основном у детей. Эти опухоли также встречаются у молодых взрослых людей. Медуллобластомы часто являются хирургически удаляемыми с последующим лечением с использованием химиотерапии и/или облучения. Они могут возвращаться локально или иногда как падающий метастаз из задней черепной ямки в позвоночник. Менингиомы представляют собой распространенные внутричерепные опухоли, которые возникают в мозговых оболочках и сдавливают находящийся ниже мозг. Хотя они, как правило, считаются доброкачественными и лишь редко открыто злокачественными, борьба с этими опухолями часто представляет клинические проблемы. Гистологические степени менингиом варьируют, при этом большинство являются доброкачественными, WHO степень I/IV (82%); реже атипичными, WHO степень II/IV (15%); и редко они встречаются как анапластические или злокачественные, WHO степень III/IV (3%).
Шванномы представляют собой доброкачественные опухоли, которые возникают на периферических нервах. Шванномы могут возникнуть на черепно-мозговых нервах, особенно вестибулярной части восьмого черепного нерва (вестибулярные шванномы, невриномы слухового нерва), где они появляются в виде опухолей мостомозжечкового угла. Гемангиобластомы являются опухолями неясного происхождения, которые состоят из эндотелиальных клеток, перицитов и так называемых стромальных клеток. Эти доброкачественные опухоли чаще всего возникают в мозжечке и спинном мозге молодых взрослых людей. Множественные гемангиобластомы характерны для заболевания Гиппеля-Линдау (VHL). Гемангиоперицитомы (HPC) являются дуральными опухолями, которые могут демонстрировать локально агрессивное поведение и могут метастазировать. Гистогенез дуральной гемангиоперицитомы (HPC) уже давно обсуждается, при этом некоторые авторы определяют ее самостоятельную единицу, а другие определяют ее как подтип менингиомы.
Симптомы как первичных, так и метастатических опухолей головного мозга часто зависят от местоположения в головном мозге и размера опухоли. Поскольку различные отделы мозга отвечают за конкретные функции, клинические симптомы будут сильно варьировать. Опухоли в лобной доле головного мозга могут вызывать слабость и паралич, нарушения настроения, трудности мышления, спутанность сознания и дезориентацию, и широкие эмоциональные перепады настроения. Опухоли в теменной доле могут вызывать судороги, онемение или паралич, трудности в написании от руки, неспособность выполнять простые математические задачи, трудности с определенными движениями и потерю тактильного чувства. Опухоли в затылочной доле могут привести к потере зрения в половине каждого поля зрения, зрительные галлюцинации и судороги. Опухоли в височной доле могут вызывать судороги, нарушения восприятия и нарушения в пространстве и сенсорную афазию. Если опухоль возникает в мозжечке, человек может иметь атаксию, потерю координации, головные боли и рвоту. Опухоли в гипоталамусе могут вызывать эмоциональные изменения и изменения в восприятии горячего и холодного. Кроме того, опухоли в гипоталамусе могут влиять на рост и питание у детей. За исключением мозжечка, опухоль с одной стороны головного мозга вызывает симптомы и повреждения на противоположной стороне тела.
Композиции и способы по настоящему изобретению могут использоваться для лечения субъекта, который был охарактеризован как имеющий клетки или ткани, экспрессирующие EGFRvIII, или у которого подозревается наличие клеток или тканей, экспрессирующих EGFRvIII. Например, субъекты, которым принесет пользу лечение в соответствии с настоящим изобретением, включают субъектов с глиомой или субъектов с подозрением на наличие глиомы, например, о чем свидетельствует наличие одного или более из следующего: головных болей, тошноты и рвоты, судорог, потери зрения, боли, слабости, онемения в конечностях и/или поражения черепно-мозговых нервов в результате повышения внутричерепного давления. В конкретных вариантах осуществления подвергаемой лечению глиомой является мультиформная глиобластома. В соответствии с этим вариантом осуществления мультиформная глиобластома может быть в головном мозге или спинном мозге.
Настоящим изобретением также обеспечиваются способы ингибирования пролиферации или уменьшения популяции EGFRvIII-экспрессирующих клеток, при этом способы включают приведение популяции клеток, включающей EGFRvIII-экспрессирующую клетку, в контакт с экспрессирующей CAR клеткой, описанной здесь, например, Т-клеткой, которая связывается с EGFRvIII-экспрессирующей клеткой. В конкретном варианте осуществления настоящим изобретением обеспечиваются способы ингибирования пролиферации или уменьшения популяции раковых клеток, экспрессирующих EGFRvIII, при этом способы включают приведение популяции EGFRvIII-экспрессирующих раковых клеток в контакт с экспрессирующей CAR клеткой, описанной здесь, например, Т-клеткой, которая связывается с EGFRvIII-экспрессирующей клеткой. В другом варианте осуществления настоящим изобретением обеспечиваются способы ингибирования пролиферации или уменьшения популяции раковых клеток, экспрессирующих EGFRvIII, при этом способы включают приведение популяции EGFRvIII-экспрессирующих раковых клеток в контакт с экспрессирующей EGFRvIII-специфический CAR T-клеткой, описанной здесь, которая связывается с EGFRvIII-экспрессирующей клеткой. В некоторых вариантах осуществления экспрессирующая EGFRvIII-специфический CAR T-клетка по настоящему изобретению уменьшает количество или процент клеток и/или раковых клеток на по крайней мере 25%, по крайней мере 30%, по крайней мере 40%, по крайней мере 50%, по крайней мере 65%, по крайней мере 75%, по крайней мере 85%, по крайней мере 95% или по крайней мере 99% у субъекта с или модели на животном глиомы(ой) или другого(им) рака(ом), связанного(ым) с EGFRvIII-экспрессирующими клетками, по сравнению с отрицательным контролем. В одном аспекте субъектом является человек.
Настоящим изобретением также обеспечиваются способы предотвращения, лечения заболевания, связанного с EGFRvIII-экспрессирующими клетками, (например, глиобластомы), и/или борьбы с ним, при этом способы включают введение субъекту, нуждающемуся в этом, экспрессирующей EGFRvIII-специфический CAR T-клетки, описанной здесь, которая связывается с EGFRvIII-экспрессирующей клеткой. В одном аспекте субъектом является человек.
Настоящим изобретением также обеспечиваются способы предотвращения рецидива рака, связанного с EGFRvIII-экспрессирующими клетками, при этом способы включают введение субъекту, нуждающемуся в этом, экспрессирующей EGFRvIII-специфический CAR T-клетки, описанной здесь, которая связывается с EGFRvIII-экспрессирующей клеткой. В одном варианте осуществления способы включают введение субъекту, нуждающемуся в этом, эффективного количества экспрессирующей EGFRvIII-специфический CAR T-клетки, описанной здесь, которая связывается с EGFRvIII-экспрессирующей клеткой, в комбинации с эффективным количеством другой терапии.
В одном аспекте настоящее изобретение относится к вектору, включающему EGFRvIII-специфический CAR, функционально связанный с промотором для экспрессии в Т-клетках млекопитающих. В одном аспекте настоящим изобретением обеспечивается рекомбинантная T-клетка, экспрессирующая EGFRvIII-специфический CAR, для применения в лечении опухолей, экспрессирующих EGFRvIII. Рекомбинантную Т-клетку, экспрессирующую CAR на основе антитела против EGFRvIII, называют экспрессирующей EGFRvIII-специфический CAR T-клеткой. В одном аспекте экспрессирующая EGFRvIII-специфический CAR T-клетка по настоящему изобретению способна к контактированию опухолевой клетки с по крайней мере одним EGFRvIII-специфическим CAR по настоящему изобретению, представленным на поверхности, таким образом, что экспрессирующая EGFRvIII-специфический CAR T-клетка активируется в ответ на антиген, и CAR-T-клетка воздействует на опухолевую клетку, и рост опухоли ингибируется.
В одном аспекте настоящее изобретение относится к способу ингибирования роста EGFRvIII-экспрессирующей опухолевой клетки, включающему приведение опухолевой клетки в контакт с экспрессирующей EGFRvIII-специфический CAR T-клеткой, описанной здесь, так что CAR-T-клетка активируется в ответ на антиген и воздействует на раковую клетку, при этом рост опухоли ингибируется. В одном аспекте активированная CAR-T-клетка воздействует на раковые клетки и уничтожает их.
В одном аспекте настоящее изобретение относится к способу лечения рака у субъекта. Способ включает введение субъекту экспрессирующей EGFRvIII-специфический CAR Т-клетки, описанной здесь, так что осуществляется лечение рака у субъекта. Примером рака, который поддается лечению с помощью экспрессирующей EGFRvIII-специфический CAR Т-клетки по настоящему изобретению, является рак, связанный с экспрессией EGFRvIII. В одном аспекте раком, связанным с экспрессией EGFRvIII, является глиобластома.
В одном аспекте рак, связанный с EGFRvIII, выбирают из группы, состоящей из мультиформной глиобластомы (GBM), анапластической астроцитомы, гигантоклеточной глиобластомы, глиосаркомы, анапластической олигодендроглиомы, анапластической эпендимомы, карциномы сосудистого сплетения, анапластической ганглиоглиомы, пинеобластомы, медуллоэпителиомы, эпендимобластомы, медуллобластомы, супратенториальной примитивной нейроэктодермальной опухоли и атипичной тератоидной/рабдоидной опухоли, немелкоклеточного рака легкого, рака легкого, молочной железы, предстательной железы, яичников, колоректального рака и рака мочевого пузыря и любой их комбинации.
Настоящее изобретение включает тип клеточной терапии, в случае которой T-клетки генетически модифицируют для экспрессии химерного антигенспецифического рецептора (CAR), и проводят инфузию CAR-Т-клетки реципиенту, нуждающемуся в этом. Клетка, инфузия которой проведена, способна к уничтожению опухолевых клеток у реципиента. В некоторых вариантах осуществления CAR-модифицированные Т-клетки способны к репликации in vivo, что приводит к длительному сохранению, которое может приводить к длительному контролю опухоли. В различных аспектах Т-клетки введенные пациенту, или их потомство сохраняются в организме пациента в течение по крайней мере четырех месяцев, пяти месяцев, шести месяцев, семи месяцев, восьми месяцев, девяти месяцев, десяти месяцев, одиннадцати месяцев, двенадцати месяцев, тринадцати месяцев, четырнадцати месяц, пятнадцати месяцев, шестнадцати месяцев, семнадцати месяцев, восемнадцати месяцев, девятнадцати месяцев, двадцати месяцев, двадцати одного месяца, двадцати двух месяцев, двадцать трех месяцев, двух лет, трех лет, четырех лет или пяти лет после введения Т-клетки пациенту.
В одном аспекте CAR-модифицированные T-клетки, описанные здесь, могут также выступать в качестве типа вакцины для ex vivo иммунизации и/или in vivo терапии для млекопитающего. В одном аспекте млекопитающим является человек.
Что касается ex vivo иммунизации, по крайней мере одно из следующего имеет место in vitro до введения клетки млекопитающему: i) размножение клеток, ii) введение нуклеиновой кислоты, кодирующей CAR, в клетки и/или iii) криоконсервация клеток.
Ex vivo процедуры хорошо известны в данной области техники и подробнее обсуждаются ниже. Вкратце, клетки выделяют от млекопитающего (например, человека) и генетически модифицируют (например, трансдуцируют или трансфицируют in vitro) с использованием вектора, экспрессирующего CAR, описанного здесь. CAR-модифицированную клетку можно ввести млекопитающему-реципиенту для обеспечения терапевтического эффекта. Млекопитающим-реципиентом может быть человек, и CAR- модифицированная клетка может быть аутологичной по отношению к реципиенту. Альтернативно, клетки могут быть аллогенными, сингенными или ксеногенными по отношению к реципиенту.
Процедура для ex vivo размножения гемопоэтических стволовых клеток и клеток-предшественников, описанная в патенте США № 5199942, который включен сюда посредством ссылки, может применяться для клеток по настоящему изобретению. В данной области техники известны другие подходящие способы, по этой причине настоящее изобретение не ограничивается каким-либо конкретным способом ex vivo размножения клеток. Вкратце, ex vivo культивирование и размножение T-клеток включает: (1) получение CD34+ гемопоэтических стволовых клеток и клеток-предшественников от млекопитающего, исходя из сбора периферической крови или эксплантатов костного мозга; и (2) размножение таких клеток ex vivo. Помимо факторов клеточного роста, описанных в патенте США № 5199942, другие факторы, такие как flt3-L, IL-1, IL-3 и c-kit-лиганд, могут использоваться для культивирования и размножения клеток.
Помимо использования вакцины на основе клеток в плане ex vivo иммунизации, настоящим изобретением также обеспечиваются композиции и способы для in vivo иммунизации для вызова иммунного ответа, направленного против антигена у пациента.
В целом, клетки, активированные и размноженные, как здесь описывается, могут использоваться при лечении и предупреждении заболеваний, которые возникают у индивидуумов с ослабленным иммунитетом. В частности, CAR-модифицированные T-клетки по настоящему изобретению используются при лечении заболеваний, нарушений и состояний, связанных с экспрессией EGFRvIII. В некоторых аспектах клетки по настоящему изобретению используются при лечении пациентов, подверженных риску развития заболеваний, нарушений и состояний, связанных с экспрессией EGFRvIII. Таким образом, настоящим изобретением обеспечиваются способы лечения или предупреждения заболеваний, нарушений и состояний, связанных с экспрессией EGFRvIII, включающие введение субъекту, нуждающемуся в этом, терапевтически эффективного количества CAR-модифицированных T-клеток по настоящему изобретению.
CAR-модифицированные T-клетки по настоящему изобретению могут вводиться или отдельно, или в виде фармацевтической композиции в комбинации с разбавителями и/или с другими компонентами, такими как IL-2 или другие цитокины, или популяциями клеток, или другими лекарственными терапиями, например, описанными здесь.
Настоящим изобретением также обеспечиваются способы ингибирования пролиферации или уменьшения популяции EGFRvIII-экспрессирующих клеток, при этом способы включают приведение популяции клеток, включающей EGFRvIII-экспрессирующую клетку, в контакт с экспрессирующей EGFRvIII-специфический CAR T-клеткой, описанной здесь, которая связывается с EGFRvIII-экспрессирующей клеткой. В конкретном аспекте настоящим изобретением обеспечиваются способы ингибирования пролиферации или уменьшения популяции раковых клеток, экспрессирующих EGFRvIII, при этом способы включают приведение популяции EGFRvIII-экспрессирующих раковых клеток в контакт с экспрессирующей EGFRvIII-специфический CAR T-клеткой, описанной здесь, которая связывается с EGFRvIII-экспрессирующей клеткой. В одном аспекте настоящим изобретением обеспечиваются способы ингибирования пролиферации или уменьшения популяции раковых клеток, экспрессирующих EGFRvIII, при этом способы включают приведение популяции EGFRvIII-экспрессирующих раковых клеток в контакт с экспрессирующей EGFRvIII-специфический CAR T-клеткой, описанной здесь, которая связывается с EGFRvIII-экспрессирующей клеткой. В некоторых аспектах экспрессирующая EGFRvIII-специфический CAR T-клетка по настоящему изобретению уменьшает количество или процент клеток и/или раковых клеток на по крайней мере 25%, по крайней мере 30%, по крайней мере 40%, по крайней мере 50%, по крайней мере 65%, по крайней мере 75%, по крайней мере 85%, по крайней мере 95% или по крайней мере 99% у субъекта с или модели на животном глиомы(ой) или другого(им) рака(ом), связанного(ым) с EGFRvIII-экспрессирующими клетками, по сравнению с отрицательным контролем. В одном аспекте субъектом является человек.
Настоящим изобретением также обеспечиваются способы предотвращения, лечения заболевания, связанного с EGFRvIII-экспрессирующими клетками, (например, глиобластомы) и/или борьбы с ним, при этом способы включают введение субъекту, нуждающемуся в этом, экспрессирующей EGFRvIII-специфический CAR T-клетки, описанной здесь, которая связывается с EGFRvIII-экспрессирующей клеткой. В одном аспекте субъектом является человек.
Настоящим изобретением также обеспечиваются способы предотвращения рецидива рака, связанного с EGFRvIII-экспрессирующими клетками, при этом способы включают введение субъекту, нуждающемуся в этом, экспрессирующей EGFRvIII-специфический CAR T-клетки, описанной здесь, которая связывается с EGFRvIII-экспрессирующей клеткой. В одном аспекте способы включают введение субъекту, нуждающемуся в этом, эффективного количества экспрессирующей EGFRvIII-специфический CAR T-клетки, описанной здесь, которая связывается с EGFRvIII-экспрессирующей клеткой, в комбинации с эффективным количеством другой терапии.
Комбинированные терапии
CAR-экспрессирующая клетка, описанная здесь, может использоваться в комбинации с другими известными агентами и терапиями. Вводимые «в комбинации», как здесь используется, означает, что два (или более) различных метода лечения назначаются субъекту во время поражения субъекта заболеванием, например, два или более лечений назначаются после диагностирования у субъекта заболевания и до того, как заболевание было вылечено или устранено, или лечение было прекращено по другим причинам. В некоторых вариантах осуществления назначение одного лечения все еще происходит, когда начинается назначение второго лечения, так что существует перекрывание, что касается введения. Это иногда упоминается здесь как «одновременное» или «параллельное» назначение (доставка). В других вариантах осуществления назначение одного лечения заканчивается до начала назначения другого лечения. В некоторых вариантах осуществления того или другого случая лечение является более эффективным из-за комбинированного введения. Например, второе лечение является более эффективным, например, отмечается эквивалентный эффект при использовании менее интенсивного второго лечения, или второе лечение ослабляет симптомы в большей степени, чем можно было бы увидеть, если бы второе лечение было назначено в отсутствие первого лечения, или аналогичная ситуация наблюдается с первым лечением. В некоторых вариантах осуществления назначение является таким, что ослабление симптома, или уменьшение другого параметра, связанного с заболеванием, больше, чем можно было бы наблюдать в случае одного лечения, назначенного в отсутствие другого. Эффект двух лечений может быть частично аддитивным, полностью аддитивным или более чем аддитивным. Назначение может быть таким, что эффект первого назначенного лечения является все еще выявляемым, когда назначают второе лечение.
CAR-экспрессирующая клетка, описанная здесь, и по крайней мере одно дополнительное терапевтическое средство могут вводиться одновременно, в одной и той же композиции или в отдельных композиций, или последовательно. В случае последовательного введения первой можно вводить CAR-экспрессирующую клетку, описанную здесь, а вторым можно вводить дополнительный агент, или порядок введения может быть обратным.
В других аспектах CAR-экспрессирующую клетку, описанную здесь, можно использовать в схеме лечения в комбинации с хирургическим вмешательством, химиотерапией, облучением, иммунодепрессантами, такими как циклоспорин, азатиоприн, метотрексат, микофенолат и FK506, антителами или другими иммуноаблативными агентами, такими как CAMPATH, антитела против CD3 или другие терапии с использованием антител, цитоксин, флударабин, циклоспорин, FK506, рапамицин, микофенольная кислота, стероиды, FR901228, цитокины, и облучение. Приводимые в качестве примера иммунотерапевтические методы для злокачественной глиомы представлены в Johnson et al. 2010 Curr Neurol Neurosci Rep 10: 259-266. В некоторых вариантах осуществления CAR-экспрессирующую клетку, описанную здесь, можно использовать в схеме лечения в комбинации с агентом, мишенью которого являются экстраклеточные матричные белки, такие как тенасцин, например, антителом против тенасцина, например, 211Аt-меченным антителом против тенасцина. В некоторых вариантах осуществления CAR-экспрессирующую клетку, описанную здесь, можно использовать в схеме лечения в комбинации с иммуномодулирующим агентом, таким как интерферон альфа, интерферон бета, пептидный ингибитор TGF-β2 или поли-ICLC. В некоторых вариантах осуществления CAR-экспрессирующую клетку, описанную здесь, можно использовать в схеме лечения в комбинации с пептидной вакциной против фактора транскрипции WT1, такой как та, которая описана в Izumoto et al. 2008 J Neurosurg 108: 963-971.
В одном варианте осуществления CAR-экспрессирующую клетку, описанную здесь, можно использовать в комбинации с химиотерапевтическим агентом. Приводимые в качестве примера химиотерапевтические агенты включают алкилирующий агент, агент на основе платины, ингибитор ангиогенеза (например, ингибитор пути с участием VEGF, ингибитор тирозинкиназы (например, ингибитор пути с участием EGF), ингибитор mTOR.
Основные химиотерапевтические агенты, рассматриваемые для применения в комбинированной терапии, включают анастрозол (Arimidex®), бикалутамид (Casodex®), блеомицина сульфат (Blenoxane®), бусульфан (Myleran®), бусульфан в инъекционной форме (Busulfex®), капецитабин (Xeloda®), N4-пентоксикарбонил-5-дезокси-5-фторцитидин, карбоплатин (Paraplatin®), кармустин (BiCNU®), хлорамбуцил (Leukeran®), цисплатин (Platinol®), кладрибин (Leustatin®), циклофосфамид (Cytoxan® или Neosar®), цитарабин, цитозинарабинозид (Cytosar-U®), цитарабин в липосомальной форме для инъекции (DepoCyt®), дакарбазин (DTIC-Dome®), дактиномицин (Актиномицин D, Космеган), даунорубицина гидрохлорид (Cerubidine®), даунорубицина цитрат в липосомальной форме для инъекции (DaunoXome®), дексаметазон, доцетаксел (Taxotere®), доксорубицина гидрохлорид (Adriamycin®, Rubex®), этопозид (Vepesid®), флударабина фосфат (Fludara®), 5- фторурацил (Adrucil®, Efudex®), флутамид (Eulexin®), тезацитибин, гемцитабин (дифтордезоксицитидин), гидроксимочевину (Hydrea®), Идарубицин (Idamycin®), ифосфамид (IFEX®), иринотекан (Camptosar®), L-аспарагиназу (ELSPAR®), лейковорина кальция, мелфалан (Alkeran®), 6-меркаптопурин (Purinethol®), метотрексат (Folex®), митоксантрон (Novantrone®), милотарг, паклитаксел (Taxol®), Phoenix (Yttrium90/MX-DTPA), пентостатин, полифепросан 20 с кармустином, имплантат (Gliadel®), тамоксифена цитрат (Nolvadex®), тенипозид (Vumon®), 6-тиогуанин, тиотепу, тирапазамин (Tirazone®), топотекана гидрохлорид для инъекции (Hycamptin®), винбластин (Velban®), винкристин (Oncovin®), и винорелбин (Navelbine®).
Приводимые в качестве примера алкилирующие агенты включают, без ограничения, азотистые иприты, производные этиленимина, алкилсульфонаты, нитрозомочевины и триазены: урациловый иприт (Aminouracil Mustard®, Chlorethaminacil®, Demethyldopan®, Desmethyldopan®, Haemanthamine®, Nordopan®, Uracil nitrogen mustard®, Uracillost®, Uracilmostaza®, Uramustin®, Uramustine®), хлорметин (Mustargen®), циклофосфамид (Cytoxan®, Neosar®, Clafen®, Endoxan®, Procytox®, Revimmune™), ифосфамид (Mitoxana®), мелфалан (Alkeran®), Хлорамбуцил (Leukeran®), пипоброман (Amedel®, Vercyte®), триэтиленмеламин (Hemel®, Hexalen®, Hexastat®), триэтилентиофосфорамин, Темозоломид (Temodar®), тиотепу (Thioplex®), бусульфан (Busilvex®, Myleran®), кармустин (BiCNU®), ломустин (CeeNU®), стрептозоцин (Zanosar®) и Дакарбазин (DTIC-Dome®). Дополнительные приводимые в качестве примера алкилирующие агенты включают, без ограничения, Оксалиплатин (Eloxatin®); Темозоломид (Temodar® и Temodal®); Дактиномицин (также известный как актиномицин-D, Cosmegen®); Мелфалан (также известный как L-PAM, L-сарколизин и фенилаланиновый иприт, Alkeran®); Алтретамин (также известный как гексаметилмеламин (HMM), Hexalen®); Кармустин (BiCNU®); Бендамустин (Treanda®); Бусульфан (Busulfex® и Myleran®); Карбоплатин (Paraplatin®); Ломустин (также известный как CCNU, CeeNU®); Цисплатин (также известный как CDDP, Platinol® и Platinol®-AQ); Хлорамбуцил (Leukeran®); Циклофосфамид (Cytoxan® и Neosar®); Дакарбазин (также известный как DTIC, DIC и имидазолкарбоксамид, DTIC-Dome®); Алтретамин (также известный как гексаметилмеламин (HMM), Hexalen®); Ифосфамид (Ifex®); Преднумустин; Прокарбазин (Matulane®); Мехлоретамин (также известный как азотистый иприт, мустин и мехлоретамина гидрохлорид, Mustargen®); Стрептозоцин (Zanosar®); Тиотепу (также известную как тиофосфороамид, TESPA и TSPA, Thioplex®); Циклофосфамид (Endoxan®, Cytoxan®, Neosar®, Procytox®, Revimmune®) и Бендамустина НС1 (Treanda®).
Приводимые в качестве примера агенты на основе платины включают, без ограничения, карбоплатин, цисплатин и оксалиплатин.
Приводимые в качестве примера ингибиторы ангиогенеза включают, без ограничения, A6 (Angstrom Pharmacueticals), АВТ-510 (Abbott Laboratories), АВТ-627 (Атрасентан) (Abbott Laboratories/Xinlay), АВТ-869 (Abbott Laboratories), Актимид (CC4047, Помалидомид) (Celgene Corporation), AdGVPEDF.11D (GenVec), ADH-1 (Эксгерин) (Adherex Technologies), AEE788 (Novartis), AG-013736 (Акситиниб) (Pfizer), AG3340 (Приномастат) (Agouron Pharmaceuticals), AGX1053 (AngioGenex), AGX51 (AngioGenex), ALN-VSP (ALN-VSP О2) (Alnylam Pharmaceuticals), AMG 386 (Amgen), AMG706 (Amgen), Апатиниб (YN968D1) (Jiangsu Hengrui Medicine), AP23573 (Ридафоролимус/MK8669) (Ariad Pharmaceuticals), AQ4N (Novavea), ARQ 197 (ArQule), ASA404 (Novartis/Antisoma), Атипримод (Callisto Pharmaceuticals), ATN-161 (Attenuon), AV-412 (Aveo Pharmaceuticals), AV-951 (Aveo Pharmaceuticals), Авастин (Бевацизумаб) (Genentech), AZD2171 (Цедираниб/Рецентин) (AstraZeneca), BAY 57-9352 (Телатиниб) (Bayer), BEZ235 (Novartis), BIBF1120 (Boehringer Ingelheim Pharmaceuticals), BIBW 2992 (Boehringer Ingelheim Pharmaceuticals), BMS-275291 (Bristol-Myers Squibb), BMS-582664 (Бриваниб) (Bristol-Myers Squibb), BMS-690514 (Bristol-Myers Squibb), Кальцитриол, CCI-779 (Торисел) (Wyeth), CDP-791 (ImClone Systems), Цефлатонин (Гомогаррингтонин/HHT) (ChemGenex Therapeutics), Целебрекс (Целекоксиб) (Pfizer), CEP-7055 (Цефалон/Sanofi), CHIR-265 (Chiron Corporation), NGR-TNF, COL-3 (Метастат) (Collagenex Pharaceuticals), Комбретастатин (Oxigene), CP-751871 (Фигитумумаб) (Pfizer), CP-547,632 (Pfizer), CS-7017 (Daiichi Sankyo Pharma), CT-322 (Ангиоцепт) (Adnexus), Куркумин, Дальтепарин (Фрагмин) (Pfizer), Дисульфирам (Antabuse), E7820 (Eisai Limited), E7080 (Eisai Limited), EMD 121974 (Циленгитид) (EMD Pharmaceuticals), ENMD-1198 (EntreMed), ENMD-2076 (EntreMed), Эндостар (Simcere), Эрбитукс (ImClone/Bristol-Myers Squibb), EZN-2208 (Enzon Pharmaceuticals), EZN-2968 (Enzon Pharmaceuticals), GC1008 (Genzyme), Генистеин, GSK1363089 (Форетиниб) (GlaxoSmithKline), GW786034 (Пазопаниб) (GlaxoSmithKline), GT-111 (Vascular Biogenics Ltd.), IMC-1121B (Рамуцирумаб) (ImClone Systems), IMC-18F1 (ImClone Systems), IMC-3G3 (ImClone LLC), INCB007839 (Incyte Corporation), INGN 241 (Introgen Therapeutics), Ирессу (ZD1839/Гефитиниб), LBH589 (Фаридак/Панобиностат) (Novartis), Луцентис (Ранибизумаб) (Genentech/Novartis), LY317615 (Энзастаурин) (Eli Lilly and Company), Макуген (Пегаптаниб) (Pfizer), MEDI522 (Абегрин) (MedImmune), MLN518 (Тандутиниб) (Millennium), Неовастат (AE941/Бенефин) (Aeterna Zentaris), Нексавар (Bayer/Onyx), NM-3 (Genzyme Corporation), Носкапин (Cougar Biotechnology), NPI-2358 (Nereus Pharmaceuticals), OSI-930 (OSI), Паломид 529 (Paloma Pharmaceuticals, Inc), Panzem в капсулах (2ME2) (EntreMed), Panzem NCD (2ME2) (EntreMed), PF-02341066 (Pfizer), PF-04554878 (Pfizer), PI-88 (Progen Industries/Medigen Biotechnology), PKC412 (Novartis), Полифенон E (экстракт зеленого чая) (Polypheno E International, Inc), PPI-2458 (Praecis Pharmaceuticals), PTC299 (PTC Therapeutics), PTK787 (Ваталаниб) (Novartis), PXD101 (Белиностат) (CuraGen Corporation), RAD001 (Эверолимус) (Novartis), RAF265 (Novartis), Регорафениб (BAY73-4506) (Bayer), Ревлимид (Celgene), Retaane (Alcon Research), SN38 (липосомальный) (Neopharm), SNS-032 (BMS-387032) (Sunesis), SOM230 (Пасиреотид) (Novartis), Скваламин (Genaera), Сурамин, Сутент (Pfizer), Тарцеву (Genentech), TB-403 (Thrombogenics), Темпостатин (Collard Biopharmaceuticals), Тетратиомолибрат (Sigma-Aldrich), TG100801 (TargeGen), Талидомид (Celgene Corporation), Тинзапарин натрий, TKI258 (Novartis), TRC093 (Tracon Pharmaceuticals Inc.), VEGF Trap (Aflibercept) (Regeneron Pharmaceuticals), VEGF Trap-Eye (Regeneron Pharmaceuticals), Веглин (VasGene Therapeutics), Бортезомиб (Millennium), XL184 (Exelixis), XL647 (Exelixis), XL784 (Exelixis), XL820 (Exelixis), XL999 (Exelixis), ZD6474 (AstraZeneca), Вориностат (Merck) и ZSTK474.
Приводимые в качестве примера ингибиторы рецепторов фактора роста эндотелия сосудов (VEGF) включают, но без ограничения, Бевацизумаб (Avastin®), акситиниб (Inlyta®); Бриваниб аланинат (BMS-582664, (S)-((R)-1-(4-(4-фтор-2-метил-1Н-индол-5-илокси)-5-метилпирроло[2,1-f][1,2,4]триазин-6-илокси)пропан-2-ил)2-аминопропаноат); Сорафениб (Nexavar®); Пазопаниб (Votrient®); Сунитиниб малат (Sutent®); Цедираниб (AZD2171, CAS 288383-20-1); Варгатеф (BIBF1120, CAS 928326-83-4); Форетиниб (GSK1363089); Телатиниб (BAY57-9352, CAS 332012-40-5); Апатиниб (YN968D1, CAS 811803-05-1); Иматиниб (Gleevec®); Понатиниб (AP24534, CAS 943319-70-8); Тивозаниб (AV951, CAS 475108-18-0); Регорафениб (BAY73-4506, CAS 755037-03-7); Ваталаниба дигидрохлорид (РТК787, CAS 212141-51-0); Бриваниб (БМС-540215, CAS 649735-46-6); Вандетаниб (Caprelsa® или AZD6474); Мотесаниба дифосфат (AMG706, CAS 857876-30-3, N-(2,3-дигидро-3,3-диметил-1H-индол-6-ил)-2-[(4-пиридинилметил)амино]-3-пиридинкарбоксамид, описанный в публикации РСТ-заявки № WO 02/066470); Довитиниба ангидрид молочной кислоты (TKI258, CAS 852433-84-2); Линфаниб (ABT869, CAS 796967-16-3); Кабозантиниб (XL184, CAS 849217-68-1); Лестауртиниб (CAS 111358-88-4); N-[5-[[[5-(1,1-диметилэтил)-2-оксазолил]метил]тио]-2-тиазолил]-4-пиперидинкарбоксамид (BMS38703, CAS 345627-80-7); (3R,4R)-4-амино-1-((4-((3-метоксифенил)амино)пирроло[2,1-f][1,2,4]триазин-5-ил)метил)пиперидин-3-ол (BMS690514); N-(3,4-дихлор-2-фторфенил)-6-метокси-7-[[(3aα,5β,6aα)-октагидро-2-метилциклопента[с]пиррол-5-ил]метокси]-4-хиназолинамин (XL647, CAS 781613-23-8); 4-метил-3-[[1-метил-6-(3-пиридинил)-1Н-пиразоло[3,4-d]пиримидин-4-ил]амино]-N-[3-(трифторметил)фенил]бензамид (BHG712, CAS 940310-85-0) и Афлиберцепт (Eylea®).
Приводимые в качестве примера ингибиторы пути с участием EGF включают, без ограничения, тирфостин 46, EKB-569, эрлотиниб (Tarceva®), гефитиниб (IRESSA®), эрбитукс, нимотузумаб, лапатиниб (Tykerb®), цетуксимаб (моноклональное антитело против EGFR), 188Re-меченный нимотузумаб (моноклональное антитело против EGFR) и такие соединения, которые в общем и конкретно раскрыты в WO 97/02266, ЕР 0564409, WO 99/03854, ЕР 0520722, ЕР 0566226, ЕР 0787722, ЕР 0837063, патенте США № 5747498, WO 98/10767, WO 97/30034, WO 97/49688, WO 97/38983 и WO 96/33980. Приводимые в качестве примера антитела против EGFR включают, но без ограничения, Цетуксимаб (Erbitux®); Панитумумаб (Vectibix®); Матузумаб (EMD-72000); Трастузумаб (Herceptin®); Нимотузумаб (hR3); Залутумумаб; TheraCIM h-R3; MDX0447 (CAS 339151-96-1); и ch806 (мАт-806, CAS 946414-09-1). Приводимые в качестве примера ингибиторы рецепторов эпидермального фактора роста (EGFR) включают, но без ограничения, Эрлотиниба гидрохлорид (Tarceva®), Гефитниб (IRESSA®); N-[4-[(3-хлор-4-фторфенил)амино]-7-[[(3"S")-тетрагидро-3-фуранил]окси]-6-хиназолинил]-4(диметиламино)-2-бутенамид, Tovok®); Вандетаниб (Caprelsa®); Лапатиниб (Tykerb®); (3R,4R)-4-амино-1-((4-((3-метоксифенил)амино)пирроло[2,1-f][1,2,4]триазин-5-ил)метил)пиперидин-3-ол (BMS690514); Канертиниба дигидрохлорид (CI-1033); 6-[4-[(4-этил-1-пиперазинил)метил]фенил]-N-[(1R)-1-фенилэтил]-7H-пирроло[2,3-d]пиримидин-4-амин (AEE788, CAS 497839-62-0); Мубритиниб (TAK165); Пелитиниб (EKB569); Афатиниб (BIBW2992); Нератиниб (HKI-272); N-[4-[[1-[(3-фторфенил)метил]-1H-индазол-5-ил]амино]-5-метилпирроло[2,1-f][1,2,4]триазин-6-ил]карбаминовой кислоты, (3S)-3-морфолинилметиловый эфир (BMS599626); N-(3,4-дихлор-2-фторфенил)-6-метокси-7-[[(3аα,5β,6аα)-октагидро-2-метилциклопента[с]пиррол-5-ил]метокси]-4-хиназолинамин (XL647, CAS 781613-23-8); и 4-[4-[[(1R)-1-фенилэтил]амино]-7Н-пирроло[2,3-d]пиримидин-6-ил]фенол (PKI166, CAS 187724-61-4).
Приводимые в качестве примера ингибиторы mTOR включают, без ограничения, рапамицин (Rapamune®) и их аналоги и производные; SDZ-RAD; Темсиролимус (Torisel®; также известен как CCI-779); Ридафоролимус (официально известный как деферолимус, (1R,2R,4S)-4-[(2R)-2[(1R,9S,12S,15R,16E,18R,19R,21R,23S,24E,26E,28Z,30S,32S,35R)-1,18-дигидрокси-19,30-диметокси-15,17,21,23,29,35-гексаметил-2,3,10,14,20-пентаоксо-11,36-диокса-4-азатрицикло[30.3.1.04,9]гексатриаконта-16,24,26,28-тетраен-12-ил]пропил]-2-метоксициклогексилдиметилфосфинат, также известный как АР23573 и MK8669, и описанный в публикации РСТ-заявки № WO 03/064383); Эверолимус (Afinitor® или RAD001); Рапамицин (AY22989, Sirolimus®); Симапимод (CAS 164301-51-3); (5-{2,4-бис[(3S)-3-метилморфолин-4-ил]пиридо[2,3-d]пиримидин-7-ил}-2-метоксифенил)метанол (AZD8055); 2-Амино-8-[транс-4-(2-гидроксиэтокси)циклогексил]-6-(6-метокси-3-пиридинил)-4-метил-пиридо[2,3-d]пиримидин-7(8Н)-он (PF04691502, CAS 1013101-36-4) и N2-[1,4-диоксо-4-[[4-(4-оксо-8-фенил-4H-1-бензопиран-2-ил)морфолини-4-ил]метокси]бутил]-L-аргинилглицил-L-α-аспартил-L-серин, внутренняя соль (SF1126, CAS 936487-67-1).
Приводимые в качестве примера ингибиторы фосфоинозитид-3- киназы (PI3K) включают, но без ограничения, 4-[2-(1Н-индазол-4-ил)-6-[[4-(метилсульфонил)пиперазин-1-ил]метил]тиено[3,2-d]пиримидин-4-ил]морфолин (также известный как GDC 0941 и описанный в публикации РСТ-заявки № WO 09/036082 и WO 09/055730); 2-метил-2-[4-[3-метил-2-оксо-8-(хинолин-3-ил)-2,3-дигидроимидазо[4,5-с]хинолин-1-ил]фенил]пропионитрил (также известный как BEZ 235 или NVP-BEZ 235 и описанный в публикации РСТ-заявки № WO 06/122806); 4-(трифторметил)-5-(2,6-диморфолинопиридин-4-ил)пиридин-2-амин (также известный как BKM120 или NVP-BKM120 и описанный в публикации РСТ-заявки № WO2007/084786); Тозасертиб (VX680 или МК-0457, CAS 639089-54-6); (5Z)-5-[[4-(4-пиридинил)-6-хинолинил]метилен]-2,4-тиазолидиндион (GSK1059615, CAS 958852-01-2); (1Е,4S,4aR,5R,6aS,9aR)-5-(Ацетилокси)-1-[(ди-2-пропениламино)метилен]-4,4a,5,6,6a,8,9,9a-октагидро-11-гидрокси-4-(метоксиметил)-4a,6а-диметил-циклопента[5,6]нафто[1,2-с]пиран-2,7,10(1Н)-трион (PX866, CAS 502632-66-8); и 8-фенил-2-(морфолин-4-ил)хромен-4-он (LY294002, CAS 154447-36-6). Приводимые в качестве примера ингибиторы протеинкиназы В (РКВ) или AKT включают, но без ограничения, 8-[4-(1-аминоциклобутил)фенил]-9-фенил-1,2,4-триазоло[3,4-f][1,6]нафтиридин-3(2Н)-он (МК-2206, CAS 1032349-93-1); Перифосин (KRX0401); 4-додецил-N-1,3,4-тиадиазол-2-ил-бензолсульфонамид (РНТ-427, CAS 1191951-57-1); 4-[2-(4-амино-1,2,5-оксадиазол-3-ил)-1-этил-7-[(3S)-3-пиперидинилметокси]-1Н-имидазо[4,5-с]пиридин-4-ил]-2-метил-3-бутин-2-ол (GSK690693, CAS 937174-76-0); 8-(1-гидроксиэтил)-2-метокси-3-[(4-метоксифенил)метокси]-6H-дибензо[b,d]пиран-6-он (паломид 529, P529 или SG-00529); Трицирбин (6-амино-4-метил-8-(β-D-рибофуранозил)-4H,8H-пирроло[4,3,2-де]пиримидо[4,5-с]пиридазин); (αS)-α-[[[5-(3-метил-1H-индазол-5-ил)-3-пиридинил]окси]метил]-бензолэтанамин (A674563, CAS 552325-73-2); 4-[(4-хлорфенил)метил]-1-(7Н-пирроло[2,3-d]пиримидин-4-ил)-4-пиперидинамин (CCT128930, CAS 885499-61-6); 4-(4-хлорфенил)-4-[4-(1H-пиразол-4-ил)фенил]пиперидин (AT7867, CAS 857531-00-1); и Арчексин (RX-0201, CAS 663232-27-7).
Лекарственные средства, которые или блокируют кальций-зависимую фосфатазу кальциневрин (циклоспорин и FK-506), или ингибируют киназу p70S6, которая важна для индуцируемой фактором роста передачи сигнала, (рапамицин) (Liu et al., Cell 66: 807-815, 1991; Henderson et al., Immun. 73: 316-321, 1991; Bierer et al., Curr. Opin. Immun. 5: 763-773, 1993), могут также использоваться. В дальнейшем аспекте клеточные композиции по настоящему изобретению могут вводиться пациенту в сочетании с (например, до, одновременно или после) трансплантацией костного мозга, Т-клеточной аблативной терапией, используя химиотерапевтические средства, такие как флударабин, наружная дистанционная лучевая терапия (XRT), циклофосфамид и/или антитела, такие как ОКТ3 или САМРАТН. В одном аспекте клеточные композиции по настоящему изобретению вводят после В-клеточной аблативной терапии, такой как агенты, которые вступают в реакцию с CD20, например, Ритуксан. Например, в одном варианте осуществления субъекты могут пройти стандартное лечение высокой дозой химиотерапии с последующей трансплантацией стволовых клеток периферической крови. В некоторых вариантах осуществления, после трансплантации, субъектам осуществляют инфузию размноженных иммунных клеток по настоящему изобретению. В дополнительном варианте осуществления размноженные клетки вводят до или после хирургического вмешательства.
В одном варианте осуществления субъекту может вводиться агент, который уменьшает или облегчает побочный эффект, связанный с введением CAR-экспрессирующей клетки. Побочные эффекты, связанные с введением CAR-экспрессирующей клетки, включают, но без ограничения, CRS и гемофагоцитарный лимфогистиоцитоз (HLH), также называемый синдромом активации макрофагов (MAS). Симптомы CRS включают высокую температуру, тошноту, преходящую гипотензию, гипоксию и т.п. Соответственно, способы, описанные здесь, могут включать введение CAR-экспрессирующей клетки, описанной здесь, субъекту и последующее введение агента для борьбы с повышенными уровнями растворимого фактора в результате лечения CAR-экспрессирующей клеткой. В одном варианте осуществления повышенным у субъекта растворимым фактором является один или более из IFN-γ, TNFα, IL-2 и IL-6. Таким образом, агентом, вводимым с целью лечения этого побочного эффекта, может быть агент, который нейтрализует один или более из этих растворимых факторов. Такие агенты включают, но без ограничения, стероид, ингибитор TNFα и ингибитор IL-6. Примером ингибитора TNFα является энтанерцепт. Примером ингибитора IL-6 является Тоцилизумаб (toc).
В одном варианте осуществления субъекту может вводиться агент, который увеличивает активность CAR-экспрессирующей клетки. Например, в одном варианте осуществления агентом может быть агент, который ингибирует ингибиторную молекулу. Ингибиторные молекулы, например, запрограммированной смерти 1 (PD1), могут, в некоторых вариантах осуществления, уменьшать способность CAR-экспрессирующей клетки к вызову иммунной эффекторной реакции. Примеры ингибиторных молекул включают PD1, PD-L1, CTLA4, TIM3, LAG3, VISTA, BTLA, TIGIT, LAIR1, CD160, 2В4 и TGFR бета. Ингибирование ингибиторной молекулы, например, путем ингибирования на уровне ДНК, РНК или белка, может оптимизировать функционирование CAR-экспрессирующей клетки. В вариантах осуществления ингибиторная нуклеиновая кислота, например, ингибиторная нуклеиновая кислота, например, дсРНК, например, короткая интерферирующая РНК или шпилечная РНК, могут использоваться для ингибирования экспрессии ингибиторной молекулы в CAR-экспрессирующей клетке. В одном варианте осуществления ингибитором является шпилечная РНК. В одном варианте осуществления ингибиторная молекула ингибируется в CAR-экспрессирующей клетке. В этих вариантах осуществления молекула дцРНК, которая ингибирует экспрессию ингибиторной молекулы, связана с нуклеиновой кислотой, кодирующей компонент, например, все компоненты, CAR. В одном варианте осуществления ингибитором ингибиторного сигнала может быть, например, антитело или фрагмент антитела, которое(ый) связывается с ингибиторной молекулой. Например, агент может представлять собой антитело или фрагмент антитела, которое(ый) связывается с PD1, PD-L1, PD-L2 или CTLA4 (например, ипилимумаб (также упоминаемый как MDX-010 и MDX-101 и продаваемый как Yervoy®; Bristol-Myers Squibb; Тремелимумаб (моноклональное антитело изотипа IgG2, доступное от Pfizer, ранее известное как тицилимумаб, СР 675206)).
PD1 является ингибиторным член семейства рецепторов CD28, которое также включает CD28, CTLA-4, ICOS и BTLA. PD-1 представлен на активированных В-клетках, Т-клетках и миелоидных клетках (Agata et al. 1996 Int. Immunol 8: 765-75). Было установлено, что два лиганда для PD1, PD-L1 и PD-L2, подавляют активацию Т-клеток после связывания с PD1 (Freeman et a. 2000 J Exp Med 192: 1027-34; Latchman et al. 2001 Nat Immunol 2: 261-8; Carter et al. 2002 Eur J Immunol 32: 634-43). PD-Ll присутствует в большом количестве в раках человека (Dong et al. 2003 J Mol Med 81: 281-7; Blank et al. 2005 Cancer Immunol. Immunother 54: 307-314; Konishi et al. 2004 Clin Cancer Res 10: 5094). Иммуносупрессия может быть отменена путем ингибирования локального взаимодействия PD1 с PD-L1. Антитела, фрагменты антител и другие ингибиторы PD1, PD-L1 и PD-L2 доступны в данной области техники и могут использоваться в комбинации с CD123-CAR, описанным здесь. Например, ниволумаб (также упоминаемый как BMS-936558 или MDX1106; Bristol-Myers Squibb) представляет собой полностью человеческое моноклональное антитело изотипа IgG4, которое специфически блокирует PD-1. Ниволумаб (клон 5C4) и другие человеческие моноклональные антитела, которые специфически связываются с PD-1, раскрыты в патенте США 8008449 и WO2006/121168. Пидилизумаб (CT-011; Cure Tech) представляет собой гуманизированное моноклональное антитело изотипа IgG1k, которое связывается с PD-1. Пидилизумаб и другие гуманизированные моноклональные антитела против PD1 описаны в WO2009/101611. Ламбролизумаб (также упоминаемый как MK03475; Merck) представляет собой гуманизированное моноклональное антитело изотипа IgG4, которое связывается с PD1. Ламбролизумаб и другие гуманизированные антитела против PD1 описаны в патенте США № 8354509 и WO2009/114335. MDPL3280A (Genentech/Roche) представляет собой моноклональное антитело класса IgG1 с оптимизированным Fc человека, которое связывается с PD-L1. MDPL3280A и другие человеческие моноклональные антитела против PD-L1 описаны в патенте США № 7943743 и публикации заявки на патент США № 20120039906. Другие связывающиеся с PD-L1 агенты включают YW243.55.S70 (вариабельные области тяжелой и легкой цепей приведены в SEQ ID NO: 20 и 21 в WO2010/077634) и MDX-1 105 (также упоминаемый как BMS-936559, и, например, связывающиеся с PD-L1 агенты, раскрытые в WO2007/005874). AMP-224 (В7-DCIg; Amplimmune; например, описанный в WO2010/027827 и WO2011/066342) представляет собой растворимый рецептор в виде слияния с Fc против PD-L1, который блокирует взаимодействие между PD1 и B7-H1. Другие антитела против PD1 включают AMP 514 (Amplimmune), среди прочего, например, антитела против PD1, описанные в патенте США № 8609089, заявках на патенты США 2010028330 и/или 20120114649. Агентом, который увеличивает активность CAR-экспрессирующей клетки, может быть, например, слитый белок, включающий первый домен и второй домен, причем первый домен представляет собой ингибиторную молекулу или ее фрагмент, а второй домен представляет собой полипептид, который ассоциируется с положительным сигналом, например, полипептидом который ассоциируется с положительный сигналом, является CD28, ICOS и их фрагменты, например, внутриклеточный домен сигнализации CD28 и/или ICOS. В одном варианте осуществления слитый белок экспрессируют в той же клетке, которая экспрессировала CAR. В другом варианте осуществления слитый белок экспрессируют в клетке, например, Т-клетке, которая не экспрессирует CAR на основе антитела против EGFRvIII.
В одном варианте осуществления агентом, который увеличивает активность CAR-экспрессирующей клетки, описанной здесь, является miR-17-92.
Фармацевтические композиции и способы лечения
Фармацевтические композиции по настоящему изобретению могут включать CAR-экспрессирующую клетку, например, множество CAR-экспрессирующих клеток, описанных здесь, в сочетании с одним или более фармацевтически или физиологически приемлемыми носителями, разбавителями или наполнителями. Такие композиции могут включать буферы, такие как нейтральный забуференный физиологический раствор, забуференный фосфатом физиологический раствор и т.п.; углеводы, такие как глюкоза, манноза, сахароза или декстраны, маннит; белки; полипептиды или аминокислоты, такие как глицин; антиоксиданты; хелатирующие агенты, такие как EDTA или глутатион; адъюванты (например, гидроксид алюминия); и консерванты. В одном аспекте композиции по настоящему изобретению приготовлены для внутривенного введения.
Фармацевтические композиции по настоящему изобретению могут вводиться способом, подходящим для подвергаемого лечению (или предотвращению) заболевания. Количество и частота введения будет зависеть от таких факторов, как состояние пациента, а также типа и тяжести заболевания пациента, хотя соответствующие дозировки могут быть определены с помощью клинических испытаний.
В одном варианте осуществления фармацевтическая композиция по существу не содержит примесь, например, отсутствуют детектируемые уровни примеси, например, выбираемой из группы, состоящей из эндотоксина, микоплазмы, компетентного по репликации лентивируса (RCL), р24, нуклеиновой кислоты VSV-G, gag ВИЧ, остаточных покрытых антителом против CD3/CD28 сфер, мышиных антител, объединенной сыворотки крови человека, бычьего сывороточного альбумина, бычьей сыворотки, компонентов культуральных сред, клеточных или плазмидных компонентов для упаковки вектора, бактерии и грибка. В одном варианте осуществления бактерией является по крайней мере одна, выбираемая из группы, состоящей из Alcaligenes faecalis, Candida albicans, Escherichia coli, Haemophilus influenza, Neisseria meningitides, Pseudomonas aeruginosa, Staphylococcus aureus, Streptococcus pneumonia и группы А Streptococcus pyogenes.
Когда показано «иммунологически эффективное количество», «противоопухолевое эффективное количество», «опухоль-ингибирующее эффективное количество» или «терапевтическое количество», точное количество композиций по настоящему изобретению для введения может быть определено врачом с учетом индивидуальных различий в возрасте, весе, размере опухоли, степени инфицирования или метастазирования, и состояния пациента (субъекта). Как правило, можно утверждать, что фармацевтическая композиция, включающая Т-клетки, описанные здесь, может вводиться в дозе, составляющей от 104 до 109 клеток/кг веса тела, а в некоторых случаях от 105 до 106 клеток/кг веса тела, в том числе все целые значения в пределах этих диапазонов. Включающие Т-клетки композиции могут также вводиться множество раз в этих дозах. Клетки могут вводиться с помощью методов инфузии, которые общеизвестны в иммунотерапии (смотрите, например, Rosenberg et al., New Eng. J. of Med. 319: 1676, 1988).
В некоторых аспектах может быть желательным введение субъекту активированных Т-клеток и затем впоследствии повторный забор крови (или выполнение афереза), активирование Т-клеток из него в соответствии с настоящим изобретением и подвергание пациента реинфузии этих активированных и размноженных Т-клеток. Этот процесс может осуществляться несколько раз каждые несколько недель. В некоторых аспектах Т-клетки могут быть активированы из заборов крови от 10 см3 до 400 см3. В некоторых аспектах Т-клетки активируются от заборов крови, составляющих 20 см3, 30 см3, 40 см3, 50 см3, 60 см3, 70 см3, 80 см3, 90 см3 или 100 см3.
Введение рассматриваемых композиций может осуществляться любым удобным способом, в том числе путем аэрозольной ингаляции, инъекции, приема внутрь, переливания, имплантации или трансплантации. Композиции, описанные здесь, могут вводиться пациенту трансартериально подкожно, внутрикожно, внутрь опухоли, внутрь узлов, интрамедуллярно, внутримышечно, с помощью внутривенной (IV) инъекции или внутрибрюшинно. В одном аспекте включающую Т-клетки композицию по настоящему изобретению вводят пациенту с помощью внутрикожной или подкожной инъекции. В одном аспекте включающую Т-клетки композицию по настоящему изобретению вводят с помощью внутривенной инъекции. Композиции Т-клеток могут вводиться непосредственно в опухоль, лимфатический узел или места инфекции.
В конкретном приводимом в качестве примера аспекте субъекты могут подвергаться лейкаферезу, в случае которого лейкоциты собирают, обогащают или истощают ex vivo с целью отбора и/или изоляции представляющей интерес клетки, например, Т-клетки. Эти изоляты Т-клеток можно размножить с помощью методов, известных в данной области техники, и обработать из условия, чтобы могли быть введены одна или более конструкций CAR по настоящему изобретению, тем самым создавая CAR-Т-клетку по настоящему изобретению. Субъекты, нуждающиеся в этом, могут затем пройти стандартное лечение высокой дозой химиотерапии с последующей трансплантацией стволовых клеток периферической крови. В некоторых аспектах после или одновременно с трансплантацией, субъектов подвергают инфузии размноженных CAR-Т-клеток по настоящему изобретению. В дополнительном аспекте размноженные клетки вводят до или после хирургического вмешательства.
Доза указанных выше лечений, назначаемых пациенту, будет варьировать в зависимости от точной природы состояния, подлежащего лечению, и реципиента лечения. Увеличение или уменьшение дозы для введения человеку может выполняться в соответствии с принятой в данной области техники практикой.
ПРИМЕРЫ
Настоящее изобретение далее подробно описано со ссылкой на следующие экспериментальные примеры. Эти примеры представлены лишь с целью иллюстрации и, как предполагается, не являются ограничением, если не предусмотрено иное. Таким образом, настоящее изобретение не следует никоим образом рассматривать как ограниченное следующими примерами, а точнее оно, как должно подразумеваться, охватывает всякие и все вариации, которые станут очевидны в результате представленных здесь идей.
Без дальнейшего описания, полагают, что специалист со средним уровнем компетентности в данной области техники может, используя предшествующее описание и следующие иллюстративные примеры, приготовить и использовать соединения по настоящему изобретению и осуществить на практике заявленные способы. Следующие рабочие примеры конкретно показывают различные аспекты настоящего изобретения и не должны рассматриваться как ограничение никоим образом остальной части описания.
Пример 1: Перенаправленные аутологичные Т-клетки, созданные для экспрессии направленного на EGFRvIII химерного антигенспецифического рецептора у пациентов с диагнозом EGFRvIII+ глиобластома
Следующие эксперименты были разработаны для разрешения вопроса, будут ли Т-клетки человека, перенаправленные на поверхностный белок EGFRvIII с помощью основанного на антителе химерного антигенспецифического рецептора (CAR), эффективными в уничтожении EGFRvIII+ модели глиобластомы у мышей NSG. Кроме того, были разработаны эксперименты для оценки приживления и сохранения этих клеток. Были проверены три различных формы CAR, включающие два различных одноцепочечных вариабельных фрагмента (часть CAR, связывающуюся с антигеном EGFRvIII) и внутриклеточные домены сигнализации (4-1BB и CD3 дзета с и без CD28).
Мышь NOD/scid/γcnulll (NSG) с иммунодефицитом является отличной моделью для ксенотрансплантации - приживления линий опухолевых клеток человека (линия опухоли головного мозга U87, которая является EGFR+ и имеет варианты, созданные, чтобы быть EGFRvIII+) и Т-клеток человека. После приживления Т-клетки человека могут сохраняться у мышей NSG в течение приблизительно 2 месяцев, или до развития смертельной ксеногенной GVHD (гомологичной болезни) (xGVHD), что зависит от дозы и донора вводимых с помощью инфузии Т-клеток человека.
Вкратце, был создан новый CAR (CAR на основе 3C10) на основе лентивируса, включающий scFv, происходящий из моноклонального антитела против EGFRvIII - 3C10. Этот CAR был протестирован in vitro и в ксеногенных моделях на мышах. Модели на мышах NOD/scid/γc(-/-) (NSG) широко использовались для доклинических оценок терапии с использованием CAR, в том числе оценки длительного сохранения введенных с помощью инфузии Т-клеток человека. Мыши NSG, несущие опухоли U87-EGFRvIII дня 7 в головном мозге, получали внутрибрюшинные инъекции темозоломида (1 мг/дозу) ежедневно в дни 7-11 и внутривенные инфузии: 2×106 Т-клеток человека, трансдуцированных ex vivo вектором, кодирующим CAR на основе NSG, или контрольным вектором, кодирующим улучшенный зеленый флуоресцентный белок (EGFP), в дни 7 и 17. В день 21 BLI-сигналы были не детектируемыми у всех мышей, получивших CAR-трансдуцированные Т-клетки, в то время как мыши, подвергнутые лечению трансдуцированными контролем Т-клетками, демонстрируют возобновление роста опухоли у 4 из 5 мышей после транзиторного противоопухолевого эффекта, вызванного темозоломидом. В отдельном эксперименте мышей, подвергнутых лечению CAR-Т-клетками, умерщвляли в день 21, и инфильтрацию CAR-трансдуцированными Т-клетками оценивали с помощью иммуногистохимии, используя конъюгированное с биотином мАт против F(аb')2 (специфичное в отношении 3C10CAR) и стрептавидин-фикоэритрин (РЕ). Введенные с помощью внутривенной инфузии CAR-Т-клетки, как представлялось на основе интенсивных PE-сигналов, сильно инфильтрировали опухоль, в то время как контрольная ткань, окрашенная стрептавидином-PE, но без мАт против F(аb')2, продемонстрировала лишь фоновые сигналы.
Теперь представляются материалы и методы, используемые в этих экспериментах.
Материалы и методы
Модель на мышах NSG
Недавно была создана колония мышей NOD/scid/γcnulll (NSG) с иммунодефицитом. У мышей NSG отсутствуют Т- и В-клетки, клетки-природные киллеры, а также ослаблена функция дендритных клеток. Было подтверждено, что приживление активированных Т-клеток было лучше у мышей NSG, чем в предыдущей модели на мышах NOD/scid/β2Mnull. По этой причине модель NSG использовали в экспериментах по ксенотрансплантации материала человека.
Структура и характеристики биологической системы
Хотя многие из моноклональных и поликлональных Ат, направленных против EGFRvIII, обладают перекрестной реактивностью с EGFR дикого типа или другими неспецифическими белками, моноклональное антитело (мАт) 3C10, которое было первоначально создано путем иммунизации мышей 14-аминокислотным пептидом, включающим EGFRvIII-специфический стык, продемонстрировало очень специфическое распознавание EGFRvIII с незначительным выявляемым связыванием EGFR дикого типа (Okamoto et al., 1996 Br J Cancer 73: 1366-1372). Лентивирусный вектор высокой степени чистоты использовали для трансдукции Т-клеток.
Приготовление клеток для инфузии мышам
Клетками для инфузии мышам являются Т-клетки человека. Обогащенные мононуклеарными клетками человека аферезные продукты получают лейкаферезом, проводимым здоровым донорам-добровольцам в University of Pennsylvania Human Immunology Core. Все образцы получают в соответствии с протоколом, утвержденным Экспертным советом университета, и информированное письменное согласие получают от каждого донора. Т-клетки подвергают негативному отбору, используя смесь с обогащением Т-клетками человека RosetteSep (Stemcell Technologies, Vancouver, Канада). Т-клетки передаются в лабораторию TRP, где их активируют с использованием CD3/28 сфер высокой степени чистоты и размножают в RPMI с глутамином, 10% FBS, 20 мМ Hepes, 100 Е/мл пенициллина и 100 мкг/мл стрептомицина. Трансдукция вектора происходит через 24 часа, при этом упакованные лентивирусные векторы добавляют непосредственно к активированным культурам. От клеток отделяют сферы в день 5, и рост контролируется с помощью Coulter Multisizer 3 (Beckman Coulter, Fullerton, CA) в отношении изменений размера (фл) и общего числа клеток, сохраняя концентрацию от 0,7E6 (0,7×106) до 2E6 клеток/мл. Эффективность трансдукции для CAR-трансдуцированных Т-клеток проверяют с помощью проточной цитометрии путем окрашивания или козьим антимышиным антителом (GAM, для CAR на основе 3C10), или козьим античеловеческим антителом (GAH, для CAR на основе 139). Мышам осуществляют инфузию 1 млн CAR+ Т-клеток на мышь с помощью введения в хвостовую вену в день 0 исследования.
Лечение темозоломидом (TMZ)
Мыши, несущие внутрикожно опухоль U87-EGFRvIII и получающие CAR+ Т-клетки в день 0, впоследствии получают внутрибрюшинные инъекции (i.p.) TMZ на дни 0-4 (ежедневно в течение 5 дней): TMZ растворяют в DMEM в концентрации = 6,67 мг/мл. Каждая мышь получает 50 мкл раствора TMZ (333 мкг/дозу; приблизительно 17 мг/кг/доза) с помощью внутрибрюшинных инъекций.
CAR-T-клетки для клинического применения
EGFRvIII-CAR-Т-клетки получают в лаборатории по продукции клеток и вакцин для клинический исследований (CVPF), и клеточный продукт представляет собой аутологичные Т-лимфоциты. Обогащение CD3+ Т-клетками продукта лейкафереза осуществляют с помощью истощения моноцитов путем проточного элютриационного центрифугирования. В день 0 процесс продукции начинают с активации обогащенных Т-клеток, используя покрытые мАт против CD3/CD28 магнитные сферы. Культуру Т-клеток подвергают воздействию кодирующего EGFRvIII-специфический CAR лентивирусного вектора и размножению. Процесс продукции Т-клеток инициируется в статической тканевой культуре (в день 0-5) с последующим переносом в биореактор Wave, если это необходимо для дополнительного размножения в соответствии с условиями перфузии. В конце культивирования от клеток отделяют магнитные сферы, клетки промывают, концентрируют и подвергают криоконсервации. Модифицированный Т-клеточный продукт подвергают криоконсервации в криомешках в объеме, зависящем от числа клеток (в конечной концентрации, составляющей максимум 108/мл), используя морозильник с контролируемой скоростью. Замороженные, экспрессирующие EGFRvIII-специфический CAR Т-клеточные продукты хранят в морозильной камере, поставленной на контроль, при ≤-130°С. Результаты экспериментов описаны ниже.
Искоренение внутричерепных EGFRvIII-экспрессирующих глиобластом с помощью CAR-Т-клеток
Глиобластома (GBM) является самой распространенной и самой злокачественной первичной опухолью головного мозга и обуславливает приблизительно 12000 связанных с раком смертельных случаев в США каждый год. Медиана выживаемости пациентов с GBM короче 15 месяцев после лечения комбинацией химиотерапии (темозоломидом) с лучевой терапией (RT). Терапия с использованием адоптивного переноса клеток (АСТ) в виде аутологичных Т-клеток, особенно Т-клеток, трансдуцированных химерными антигенспецифическими рецепторами (CAR), раскрыла перспективы предварительных испытаний на раке системы крови. АСТ с использованием CAR-T-клеток может быть особенно подходящей для пациентов с GBM, поскольку гораздо лучше можно манипулировать и контролировать специфичность(ю), количество(м) и функциональный(ым) фенотип(ом) клеток, полученных ex vivo, чем природных Т-клеток, индуцированных с помощью in vivo иммунизации.
Вариант III рецептора эпидермального фактора роста (EGFRvIII) является самым распространенным вариантом EGFR, который наблюдается в опухолях человека, но редко наблюдается в нормальной ткани. Этот белок является результатом делеции в рамке считывания экзонов 2-7 и образования нового остатка глицина на стыке экзонов 1 и 8 в экстраклеточном домене EGFR, в результате чего создается специфический в отношении опухоли эпитоп. EGFRvIII экспрессируется в 24%-67% GBM, но не в нормальных тканях.
Для разработки эффективной CAR-терапии для GBM были созданы три новые лентивирусные конструкции CAR, направленные на EGFRvIII. Каждый из этих векторов кодирует одноцепочечный вариабельный фрагмент (scFv), происходящий из EGFRvIII-специфических мышиных моноклональных антител (мАт) 3C10 или EGFRvIII-специфических гуманизированных моноклональных антител (мАт), названных «139» (фиг. 6). ScFv 3C10 был связан с шарнирной областью CD8a, 4-1BB- и CD3ζ-доменами с или без трансмембранных и внутриклеточных доменов CD28 (3C10BBz28-CAR и 3C10BBz-CAR, соответственно). ScFv 139 был связан с шарнирной областью CD8a, 4-1BB- и CD3ζ-доменами (139BBz-CAR). Т-клетки человека, трансдуцированные каждым из этих CAR, продемонстрировали специфический и сильный лизис EGFRvIII-экспрессирующих клеток GBM - U87 (U87-EGFRvIII); смотрите фиг. 7. Мыши с ослабленным иммунитетом NOD/scid/γc(-/-) (NSG), имеющие опухоли U87-EGFRvIII дня 7 в головном мозге, получили внутривенные инфузии из 1×106 Т-клеток человека, трансдуцированных ex vivo: 1) 139BBz-CAR; 2) 3C10BBz-CAR; 3) 3C10BBz28-CAR; 4) контрольным CD19BBz-CAR, мишенью которого является CD19 человека. Эти мыши также получали внутрибрюшинные инъекции темозоломида (330 мкг/доза) ежедневно в дни 7-11. Рост опухоли контролировали с помощью биолюминесцентной визуализации (BLI), поскольку клетки U87-EGFRvIII также экспрессируют люциферазу. Все мыши, подвергнутые лечению только физиологическим раствором, умерли в день 21 из-за быстрого роста опухоли, и лечение темозоломидом без ACT приводило к подавлению, но не искоренению опухолей U87-EGFRvIII. Мыши, получающие CD19BBz-CAR-T-клетки и темозоламид, продемонстрировали некоторые аллогенные реакции против U87-EGFRvIII, но опухоли продолжали расти у этих мышей. С другой стороны, у всех мышей, получающих 139BBz-CAR-, 3C10BBZ-CAR- или 3C10BBz28-CAR-трансдуцированные Т-клетки, BLI-сигналы уменьшилась до уровней, меньше исходных, к дню 21, что говорит о полном искоренении опухолей (фиг. 8). Важно отметить, что мыши, получающие 3C10BBz-CAR-T-клетки, устраняли опухоль быстрее, чем или 3C10BBz28- или 139BBz-CAR-T-клетки, что говорит о том, что комбинация 3C10 с BBz может обеспечить лучшую ответную реакцию у пациентов. Рост опухоли и периферические иммунные реакции контролировали для определения, является ли какой-либо из трех векторов, кодирующих EGFRvIII-CAR, лучше других по длительным противоопухолевым эффектам.
Представленные здесь результаты всецело поддерживают разработку клинического испытания фазы I ACT с использованием Т-клетки, экспрессирующей EGFRvIII-специфический CAR, у пациентов с GBM, которые одновременно получают стандарт лечения - химиотерапию с использованием темозоломида.
План клинического испытания
Открытое предварительное исследование с использованием одной группы было разработано для определения безопасности, переносимость и возможности приживления EGFRvIII-CAR-T-клеток у пациентов с вновь диагностированными EGFRvIII+ GBM. Как правило, всем субъектам вводят дозы аутологичных EGFRvIII-CAR-Т-клеток. Приемлемых субъектов подвергают лейкаферезу для получения большого количества мононуклеарных клеток периферической крови (РВМС) для получения EGFRvIII-CART-клеток. Т-клетки очищают от РВМС, трансдуцируют кодирующим CAR на основе гуманизированного 3C10 лентивирусным вектором, размножают in vitro и подвергают криоконсервации в соответствующих аликвотах-дозах. Клетки, подвергаемые инфузии, оттаивают у постели непосредственно перед инфузией в день 0.
Субъектов подвергают анализам крови для оценки безопасности и приживления и сохранения EGFRvIII-CAR-T-клеток регулярно по неделю 4 включительно (день 28). Субпопуляции циркулирующих Т-клеток, которые содержат вектор, кодирующий 3C10-CAR, оценивают в различные моменты времени после инфузии и сравнивают с образцом на исходном уровне. После 28 дня субъекты оцениваются ежемесячно до 6 месяцев с использованием истории болезни, физикального обследования, ЯМР-томографии головного мозга и анализов крови или в соответствии со стандартом лечения.
Анализы крови с целью научных исследований проводятся одновременно с этим посещениями. Спустя шесть месяцев пациентов контролируют каждые 2 месяца в течение двух лет. После момента времени, равного двум годам, субъекты входят в пролонгированное исследование для ежегодного контролирования по телефону и с помощью вопросника в течение еще тринадцати лет для оценки с целью диагностирования длительных проблем со здоровьем, таких, как развитие новых злокачественностей, в соответствии с правилами FDA, относящимися к исследованиям по переносам генов.
Без желания ограничиться какой-либо конкретной теорией, полагают, что из-за очень ограниченной экспрессии белка EGFRvIII не предвидят какой-либо вид эффективной активации Т-клеток вне опухоли. Предпочтительно, когда осуществляют лишь одну инфузию EGFRvIII-CART-клеток, и поэтому также не предвидят развитие реакций аллергического типа. Однако одной токсичностью, с которой можно столкнуться, является воспаление в результате неспецифической активации Т-клеток в месте опухоли. Симптомы и признаки отека головного мозга будут тщательно контролироваться, и с ним будет осуществляться борьба. В некоторых вариантах осуществления воспаление в результате неспецифической активации Т-клеток можно лечить путем введения противовоспалительного агента, такого как стероидный агент.
Пример 2: Котрансдукция miR-17-92 увеличивает противоопухолевую активность Т-клеток, трансдуцированных химерным антигенспецифическим рецептором на основе антитела против EGFRvIII, у мышей с ксенотрансплантатами глиобластомы человека
Экспрессия miR-17-92 придает фенотип 1-го типа и увеличивает выживаемость Т-клеток. Сообщалось, что экспрессия miR-17-92 подавлена в Т-клетках, полученных из пациентов с глиобластомой (GBM). Для увеличения эффективности терапии с использованием адоптивного переноса клеток против GBM, используя Т-клетки, трансдуцированные химерными антигенспецифическими рецепторами, (CAR-T-клетки) был сконструирован новый лентивирусный вектор для miR-17-92 и CAR, состоящего из специфического в отношении варианта III рецептора эпидермального фактора роста (EGFRvIII) одноцепочечного вариабельного фрагмента (scFv), связанного последовательно с модулем сигнализации - CD3ζ цепью Т-клеточного рецептора и костимулирующими мотивами CD137 (4-1BB) и CD28 (pELNS-3C10-CAR). Помимо антигенспецифических и сильных цитотоксических активностей против клеток GBM U87, стабильно экспрессирующих EGFRvIII (U87-EGFRvIII), CAR-T-клетки, котрансдуцированные miR-17-92, продемонстрировали увеличенную стойкость к подавляющим Т-клетки эффектам трансформирующего фактора роста (TGF)-β и темозоломида по сравнению с CAR-Т-клетками без котрансдукции miR-17-92. У мышей, несущих внутричерепные ксенотрансплантаты U87-EGFRvIII, CAR-Т-клетки с или без экспрессией(и) miR-17-92 с трансгена продемонстрировали схожие уровни сильных терапевтических эффектов, не демонстрируя какой-либо неконтролируемый рост CAR-T-клеток. Однако, когда эти мыши были повторно подвергнуты инъекции клеток U87-EGFRvIII в головном мозге, мыши, получающие котрансдуцированные CAR-T-клетки, продемонстрировали улучшенную защиту по сравнению с мышами, подвергнутыми лечению CAR-Т-клетками без котрансдукции miR-17-92. Эти данные подтверждают, что miR-17-92 может быть включена в CAR для увеличения эффективности у пациентов с GBM.
Результаты экспериментов описаны ниже.
Конструирование лентивирусных векторов для EGFRvIII-специфических CAR и miR-17-92
Был создан лентивирусный вектор для CAR, который распознает EGFRvIII через одноцепочечный вариабельный фрагмент (scFv), происходящий из специфического в отношении EGFRvIII моноклонального антитела (мАт) человека 3C10 (pELNS-3C10-CAR) (смотрите фиг. 1А). В этой конструкции промотор EF-1α управляет экспрессией слитого белка CAR, включающего происходящий из 3C10 scFv, трансмембранный домен (ТМ) CD28, а также 4-1ВВ и внутриклеточные домены (ICD) и CD3ζ домены. Также была создана лентивирусная конструкция для miR-17-92, используя самоинактивирующийся вектор (SIN) на основе FG12, (FG12-EF1α-miR-17-92) (смотрите фиг. 1B). В этом векторе промотор EF1α управляет экспрессией miR-17-92, и промотор UbiC человека управляет экспрессией гена улучшенного зеленого флуоресцентного белка (EGFP), гена-маркера для отслеживания трансдуцированных клеток. Сокращения, используемые на схеме: RSV/5'LTR ВИЧ-1: гибрид промотор-R RSV/длинный концевой повтор U5, EF-1α: промотор 1α-субъединицы фактора элонгации человека, VH: вариабельная область тяжелой цепи иммуноглобулина 3С10, VL: вариабельная область легкой цепи иммуноглобулина 3C10, Δ-3'- LTR ВИЧ-1: 3' длинный концевой повтор с самоинактивацией с делецией в районе U3, CMV/5' LTR ВИЧ-1: гибрид промотор-R LTR/длинный концевой повтор U5, UbiC: промотор гена убиквитина С.
In vitro характеристики Т-клеток человека, трансдуцированных CAR и miR-17-92
Полученные от здоровых доноров CD3+ Т-клетки трансдуцировали pELNS-3C10-CAR, и клетки оценивали в отношении уровней экспрессии трансгена с помощью проточной цитометрии для экспрессии 3C10-CAR и miR-17-92 с помощью антитела против (Fab')2 мыши и EGFP, соответственно (фиг. 2А, слева). Используя Ат против F(аb')2 мыши, которое является специфическим для происходящего из 3C10 scFv на Т-клетках человека, почти половина (48,9%) Т-клеток, экспрессирующих происходящий из 3C10 scFv на своей поверхности, была обнаружена.
Для получения Т-клеток человека, экспрессирующих как CAR, так и экспрессируемый с трансгена miR-17-92, CD3+ Т-клетки котрансдуцировали pELNS-3C10-CAR и FG12-EF1α-miR-17-92 путем последовательного инфицирования двумя лентивирусными векторами. Через 24 часа после первоначальной трансдукции pELNS-3C10-CAR, Т-клетки трансдуцировали FG12-EF1α-miR-17-92. Было отмечено, что приблизительно четверть (23,6%) от общего количества Т-клеток экспрессировала как CAR, так и EGFP (фиг. 2А, справа). Для последующих in vitro исследований, CAR-трансдуцированные Т-клетки (CAR-Т-клетки) были обогащены, используя биотинилированное Ат против F(аb')2 и анти-биотин MACS. Основываясь на эффективности котрансфекции (фиг. 2А, справа), по крайней мере 50% CAR-Т-клеток также экспрессировали EGFR (следовательно, экспрессируемый с трансгена miR-17-92). В соответствии с ПЦР в режиме реального времени, в подвергнутых обогащению с использованием Ат против F(аb')2, miR-17-92-котрансдуцированых CAR-Т-клетках отмечалась экспрессия miR-17-92, превышающая в 3-4 раза таковую в Т-клетках, трансдуцированных только CAR (фиг. 2b). На фиг. 2В представлены уровни экспрессии членов кластера miR-17-92, miR-17-3р, miR-17-5p и miR-92a-1, в трансдуцированных Т-клетках, измеренные с помощью количественной ПЦР в режиме реального времени. Представлены средние значения + среднеквадратическое отклонение 3 повторных измерений одного из трех экспериментов со схожими результатами. * означает р<0,05 между двумя группами, используя коэффициент Стьюдента. На фиг. 2C представлены EGFRvIII-специфические цитотоксические активности трансдуцированных Т-клеток, оцененные с помощью 12-часового анализа высвобождения 51Cr при различных соотношениях E:T, в отношении 51Cr-меченных U87-EGFRvIII или контрольных U87 клеток. Контрольные клетки были контроль(EGFP)-трансдуцированными Т-клетками. Значения означают среднее значение + среднеквадратическое отклонение в лунках в виде трех повторов.
В то время как контроль-трансдуцированные Т-клетки продемонстрировали лишь фоновые уровни лизиса обеих линий родительских клеток U87 (EGFRvIII-негативных) и U87-EGFRvIII, Т-клетки, трансдуцированные CAR, продемонстрировали сильный и специфический лизис EGFRvIII-экспрессирующих клеток GBM человека - U87 (U87-EGFRvIII) с лишь фоновыми уровнями цитотоксических эффектов против родительских клеток U87 (фиг. 2С). В этих 12-часовых анализах высвобождения 51Cr, котрансдукция CAR-Т-клеток, используя miR-17-92, не повышала значительно их специфическую цитотоксическую активность в отношении клеток-мишеней U87-EGFRvIII.
Котрансдукция miR-17-92 дает увеличенное высвобождение IFN-γ и придает стойкость к подавляющим эффектам, вызываемым TGF-β и темозоломидом (TMZ - стандартом лечения)
В предыдущем исследовании (Sasaki et al., 2010, J. Transl Med 8: 17) CD4+ Т-клетки, полученные от трансгенных по miR-17-92 мышей, продемонстрировали увеличенную продукцию IFN-γ по сравнению с аналогами, полученными от мышей дикого типа; и трансфекция Т-клеток человека Jurkat, используя miR-17-92, приводит к повышенной стойкости к индуцируемой активацией гибели клеток (AICD).
Были проведены эксперименты для оценки, обеспечивает ли котрансдукция CAR-Т-клеток, используя miR-17-92, увеличенную продукцию IFN-γ, пролиферацию клеток и меньшую степень апоптоза, когда они подвергаются воздействию химиотерапевтического агента TMZ или иммуносупрессивного цитокина TGF-β.
Когда CAR-T-клетки стимулировали, используя EGFRvIII-трансдуцированные искусственные антигенпрезентирующие клетки (aAPC) без TGF-β или TMZ, клетки экспрессировали схожие уровни IFN-γ с или без котрансдукции(ей). Однако, когда клетки подвергали воздействию увеличивающихся доз TGF-β или TMZ, CAR-Т-клетки без котрансдукции miR-17-92 продуцировали IFN-γ на значительно пониженных уровнях, в то время как в котрансдуцированных CAR-Т-клетках сохранялась продукция IFN-γ на высоком уровне (фиг. 3а). Неокрашенные столбики и окрашенные столбики представляют результаты от CAR-Т-клеток (без miR-17-72) и котрансдуцированных miR-17-92 CAR-Т-клеток, соответственно. На фиг. 3А показан IFN-γ, продуцированный трансдуцированными Т-клетками в течение 25 часа в конце 24 часа 96-часового сокультивирования. На фиг. 3В показано, что относительные уровни пролиферации между группами оценивали с помощью анализа WST1 после 3-дневного курса сокультивирования. На фиг. 3C и 3D показан апоптоз CAR-Т-клеток, оцененный с использованием Аннексина-V и PI. На фиг. 3С представлена средняя интенсивность флуоресценции для Аннексина-V на CAR-Т-клетках, подвергнутых воздействию TMZ. Значения означают среднее значение + среднеквадратическое отклонение в лунках в виде трех повторов. (* означает р<0,05). На фиг. 3D представлены гистограммы для Аннексина-V+ и/или PI+, построенные на основе данных проточной цитометрии в одном из трех экспериментов со схожими результатами.
Были проведены эксперименты для оценки эффектов котрансдукции miR-17-92 на пролиферацию CAR-Т-клеток в присутствии TMZ в культуре. Эксперименты были разработаны с индукцией пролиферации CAR-Т-клеток с помощью EGFRvIII-экспрессирующей аAPC, и пролиферацию оценивали с помощью анализа WST-1 (фиг. 3B). Без TMZ, miR-17-92-котрансдуцированные CAR-T-клетки продемонстрировали тенденцию к более быстрой скорости пролиферации по сравнению с контрольными CAR-Т-клетками, но различие не было значимым. Чтобы конкретно оценить эффект TMZ на пролиферацию CAR-Т-клеток, на фиг. 3В, скорость пролиферации клеток в каждой группе представлена относительно пролиферации тех же клеток без TMZ. При добавлении в культуру увеличивающихся концентраций TMZ, степень подавления роста была значительно меньше в случае miR-17-92-котрансдуцированных CAR-Т-клеток, чем в контрольных CAR-Т-клетках.
Были проведены эксперименты для оценки, будет ли котрансдукция miR-17-92 делать CAR-T-клетки более устойчивыми к TMZ-индуцированному апоптозу. С этой целью проводились оценки с помощью проточной цитометрии Аннексин V+ и йодид пропидия (PI)+ CAR-T-клеток в увеличивающихся концентрациях TMZ (фиг. 3С и 3D). Было отмечено дозозависимое увеличение как характеризующихся апоптозом на ранних стадиях (Аннексин V+ PI-) клеток, апоптозных/некротических клеток (Аннексин V+ PI+), так и некротических (Аннексин V- PI+) клеток, и miR-17-92-котрансдуцированные CAR-T-клетки продемонстрировали меньшую степень апоптозных изменения по сравнению с контрольными CAR-Т-клетками.
Внутривенная инъекция CAR-Т-клеток в комбинации с TMZ приводит к полной ремиссии укоренившихся опухолей U87-EGFRvIII у мышей NSG
Были проведены эксперименты для оценки эффективности CAR-Т-клеток у мышей с ослабленным иммунитетом NOD/scid/γc(-/-) (NSG), имеющих укоренившиеся (день 7) внутричерепные опухоли U87-EGFRvIII. Мыши получали однократные внутривенные инфузии miR-17-92-котрансдуцированных CAR-T-клеток, CAR-T-клеток без котрансдукции miR-17-9, или контроль-трансдуцированных Т-клеток (2×106/мышь) через хвостовую вену. Поскольку пациенты с недавно диагностированной GBM обычно получают терапию с использованием TMZ, эксперименты были разработаны с назначением внутрибрюшинных (i.p.) ежедневных инъекций TMZ в течение 5 дней, начиная со дня инфузии Т-клеток (фиг. 4А). На фиг. 4В представлен анализ Каплана-Мейера. Медиана выживаемости мышей, получивших CAR-Т-клетки (с или без котрансдукции miR-17-92), была значительно больше таковой для мышей с контроль-трансдуцированными Т-клетками (р<0,05). Само по себе лечение TMZ было неэффективным, поскольку все контрольные мыши, получающие TMZ и контроль-трансдуцированные Т-клетки, умерли в пределах 3-х недель (21 день) после инфузии Т-клеток (фиг. 4В). Хотя одна из пяти мышей с CAR-Т-клетками и две из пяти мышей с miR-17-92-котрансдуцированными CAR-Т-клетками умерли из-за прогрессирования опухоли к дню 22, все остальные мыши в этих группах жили больше 40 дней. Представлены результаты одного из двух независимых экспериментов со схожими результатами. Не было статистически значимого различия в выживаемости мышей, получающих miR-17-92-котрансдуцированные CAR-T-клетки в сравнение с CAR-Т-клетками без котрансдукции miR-17-92 (критерий Кокса: р=0,5485).
miR-17-92-котрансдуцированные CAR-T-клетки обеспечивают постоянную защиту от опухолей U87-EGFRvIII у мышей
Для определения того, могут ли CAR-T-клетки, инфузия которых была осуществлена мышам в эксперименте, представленном на фиг. 4, обеспечить длительную защиту хозяев от опухолей U87- EGFRvIII, оставшимся в живых повторно инокулировали клетки U87-EGFRvIII в контралатеральное полушарие головного мозга в день 49 (фиг. 5). Хотя повторно инокулированные опухолевые клетки росли у всех трех мышей, получавших CAR-Т-клетки, ни одна из мышей, получавших miR-17-92-котрансдуцированные CAR-Т-клетки, не продемонстрировала BLI-сигналы, выходящие за рамки фонового уровня. Эти результаты дают веские основания предполагать, что котрансдукции кластера miR-17-92 обеспечивает длительную сохраняемость CAR-Т-клеток, тем самым обеспечивая длительную защиту хозяина от роста опухоли. Продольные измерения среднего потока фотонов опухолевого происхождения + среднеквадратическое отклонение от 2-х групп мышей. Фоновый уровень люминесценции (до 103 фотонов в секунду) был определен на основе уровней, отмечаемых у мышей без опухолей, изображения которых получали параллельно с мышами с опухолями в группах лечения.
miR-17-92 можно включить в CAR для увеличения эффективности
Представленные здесь результаты демонстрируют эффекты коэкспрессии miR-17-92 в Т-клетках, трансдуцированных новым CAR на основе антитела против EGFRvIII (3C10-CAR), включающим scFv 3C10 с CD3ζ цепью, CD137 (4-1BB) и CD28. Представленные результаты показывают, что коэкспрессия miR-17-92 придает увеличенную стойкость к подавляющим рост Т-клеток эффектам TGF-β и темозоломида. In vivo Т-клетки, котрансдуцированные как 3C10-CAR, так и miR-17-92, продемонстрировали более длительные терапевтические эффекты, чем Т-клетки, трансдуцированные только 3C10-CAR.
Лентивирусная трансдукция miR-17-92 в настоящем исследовании обеспечивает эктопическую сверхэкспрессию miR-кластера в трансдуцированных Т-клетках. Однако в физиологических условиях уровни экспрессии эндогенной miR-17-92 в Т-клетках, по-видимому, жестко регулируются. В CD8+ Т-клетках человека экспрессия miR-17-92 выявляется на высоких уровнях в «необученных» клетках, но уменьшается по мере дифференцировки клеток (Salaun et al., 2011, J Transl Med 9: 44). В модели вирусной инфекции лимфоцитарного хориоменингита на мышах, экспрессия miR-17-92 сильно увеличивается после активации Т-клеток, однако уменьшается после клонального размножения и далее заглушается во время развития памяти (Wu et al., 2012, Proc Natl Acad Sci USA 109: 9965-9970). В этом исследовании, на которое ссылаются, miR-17-92 является необходимой для быстрого размножения Т-клеток и экспрессии IFN-γ в них. Однако сверхэкспрессия miR-17-92 смещает дифференциацию в сторону короткоживущих терминальных эффекторных клеток. Неспособность уменьшить экспрессию miR-17-92 приводит к постепенной потере клеток памяти и развитию дефектных центральных клеток памяти (Wu et al., 2012, Proc Natl Acad Sci USA 109: 9965-9970). Эти наблюдения необязательно согласуются с результатами, представленными здесь, поскольку отмечалась сохраняемость miR-17-92-котрансдуцированных CAR-Т-клеток и их практическая способность защитить хозяев от повторного инокулированных клеток U87-EGFRvIII. Без желания быть ограниченными какой-либо конкретной теорией, полагают, что это наблюдение связано с комбинаторными эффектами костимулирующих молекул, предусмотренных в CAR, и miR-17-92.
Хотя miR-17-92 была описана как онкогенная микроРНК (van Haaften and Agami, 2010, Genes & Development 24: 1-4), сама по себе сверхэкспрессия miR-17-92, как известно, не является онкогенной в лимфоцитах (Xiao et al., 2008, Nat Immunol 9: 405-414). Действительно, неконтролируемая пролиферация miR-17-92-трансдуцированных Т-клеток в настоящем исследовании не отмечалась. Тем не менее, в качестве альтернативного подхода для лучшего обеспечения безопасности, транзиторная трансдукция Т-клеток самой miR-17-92, вместо лентивирусного стабильного переноса, и многократная инъекция этих Т-клеток может представлять собой рациональный подход без связанных опасений по поводу безопасности интеграции вирусных векторов (Zhao et al., 2010, Cancer Research 70: 9053-9061).
Что касается CAR, мишенью которого является EGFRvIII, для терапии GBM, недавно Morgan и др. оценили последовательности scFv, происходящие из семи различных мАт против EGFRvIII, в том числе 3C10 и 139 человека, в γ-ретровирусных CAR (Morgan et al., 2012, Hum Gene Ther 23: 1043-1053). Определение in vitro характеристик этих CAR выявило 3C10 и 139 в качестве двух из трех клонов, которые приводили к специфической продукции IFN-γ в ответ на EGFRvIII-экспрессирующие клетки-мишени, но не на клетки, экспрессирующие ген EGFR дикого типа.
Важно также понимать, что EGFRvIII экспрессируется только в выборке пациентов с GBM и части клеток GBM даже в случаях «EGFRvIII-позитивных» клеток (Heimberger et al., 2005, Clin. Cancer Res. 11: 1462-1466). Иммунотерапия, мишенью которой является EGFRvIII в качестве одной мишени, скорее всего, приведет к разрастанию клеток GBM, в которых была уменьшена экспрессия антигена, на который направлена иммунотерапия (Sampson et al., 2010, J Clin Oncol 28: 4722-4729). В ряде предыдущих исследований были разработаны CAR к GBM-связанным антигенам, таким как IL-13Rα2 (Kong et al., 2012, Clin Cancer Res 18: 5949-5960; Kahlon et al., 2004, Cancer Res. 64: 9160- 9166), HER-2 (Ahmed et al., 2010, Clinical Cancer Research 16: 474-485) и EphA2 (Chow, K.K. et al. T Cells Redirected to EphA2 for the Immunotherapy of Glioblastoma. Mol Ther (2012). Без желания ограничиться какой-либо конкретной теорией, полагают, что в эффективной терапии с использованием CAR должны, в конечном счете, использоваться Т-клетки, которые способны противостоять GBM-индуцированным механизмам подавления и быть направленными на несколько антигенов, так что подвергнутые инфузии Т-клетки будут проявлять эффективные и устойчивые терапевтические эффекты против GBM с гетерогенными профилями экспрессии антигена.
Представленные здесь результаты демонстрируют преимущества использования T-клеток, котрансдуцированных pELNS-3C10-CAR и FG12-EF1а-miR-17-92. В качестве альтернативного подхода для достижения коэкспрессии CAR и трансгена miR-17-92, был сконструирован лентивирусный вектор на основе pELNS, который экспрессирует как ген 3C10-CAR, так и ген miR-17-92 в виде одного транскрипта. Использование этого одного «сдвоенного» вектора может иметь преимущество в плане относительно простых процедур трансдукции и прямых регуляторных процессов по сравнению с подходом на основе двух векторов. Кроме того, все Т-клетки, которые экспрессируют CAR, должны также экспрессировать miR-17-92. Однако было установлено, что эффективность трансдукции «сдвоенного» вектора ниже, чем при использовании подхода на основе двух векторов, по-видимому, поскольку титр лентивируса уменьшается по мере увеличения размера вставки. Как обсуждалось здесь в другом месте, лентивирусная трансдукция гена 3C10-CAR и электропорации miR-17-92 в комбинации могут представлять собой подходящую стратегию.
В настоящем исследовании было также установлено, что от 40% до 60% CD3+ CAR-Т-клеток были CD4+, и что CD4+ CAR-Т-клетки эффективно лизировали клетки U87-EGFRvIII EGFRvIII-специфическим образом. Сообщалось, что Перфорин+ CD4+ Т-клетки опосредуют цитотоксические активности через путь с участием перфорина/гранзима В, но не через путь с участием Fas/FasL (Porakishvili et al., 2004, Haematologica 89: 435-443). Поэтому полагают, что CD4+ CAR-T-клетки в данном исследовании экспрессировали перфорин и гранзим для опосредования отмечаемых литических активностей в отношении клеток U87-EGFRvIII.
В заключение, данное исследование дает прочную основу для оценки включающей miR-17-92 терапии с использованием CAR.
Пример 3: Последовательности CAR
Мышиное моноклональное антитело (мАт) 3C10 было первоначально получено путем иммунизации мышей 14-аминокислотным пептидом (PEP3), включающим EGFRvIII-специфический стык, и продемонстрировало очень специфическое распознавание EGFRvIII без какого-либо обнаруживаемого связывания с EGFR дикого типа (Okamoto et al., British J. Cancer 1996, 73: 1366-1372). Впоследствии был получен одноцепочечный вариабельный фрагмент (scFv) мАт 3C10, и была получен кДНК для scFv 3C10. Хотя авидность и/или антиген-специфичность исходных мАт может часто теряется в формах scFv, scFv 3C10 сохранил свою избирательную реактивность с EGFRvIII-специфическим эпитопом (Nakayashiki et al., Jpn. J.Cancer Res. 2000, 91: 1035-1043).
EGFRvIII-специфический CAR был создан путем клонирования 3C10scFv (мыши) с CD28, 4-1ВВ и CD3 дзета в лентивирусную плазмиду pELNS в качестве остова (с промотором EF1). Другой EGFRvIII-специфический CAR был создан путем клонирования 3C10scFv в лентивирусный остов pELNS с шарнирной областью CD8/CD8TM/4-1ВВ/CD3 дзета, экспрессией которого управляет промотор EFLA.
3C10scFv-CD28BBzeta CAR (аминокислотная последовательность) (SEQ ID NO: 1)
MALPVTALLLPLALLLHAARPGSEIQLQQSGAELVKPGASVKLSCTGSGFNIEDYYIHWVKQRTEQGLEWIGRIDPENDETKYGPIFQGRATITADTSSNTVYLQLSSLTSEDTAVYYCAFRGGVYWGPGTTLTVSSGGGGSGGGGSGGGGSHMDVVMTQSPLTLSVAIGQSASISCKSSQSLLDSDGKTYLNWLLQRPGQSPKRLISLVSKLDSGVPDRFTGSGSGTDFTLRISRVEAEDLGIYYCWQGTHFPGTFGGGTKLEIKASTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDFWVLVVVGGVLACYSLLVTVAFIIFWVRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
3C10scFv-BBz CAR (аминокислотная последовательность) (SEQ ID NO: 2)
MALPVTALLLPLALLLHAARPGSEIQLQQSGAELVKPGASVKLSCTGSGFNIEDYYIHWVKQRTEQGLEWIGRIDPENDETKYGPIFQGRATITADTSSNTVYLQLSSLTSEDTAVYYCAFRGGVYWGPGTTLTVSSGGGGSGGGGSGGGGSHMDVVMTQSPLTLSVAIGQSASISCKSSQSLLDSDGKTYLNWLLQRPGQSPKRLISLVSKLDSGVPDRFTGSGSGTDFTLRISRVEAEDLGIYYCWQGTHFPGTFGGGTKLEIKASTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
3C10scFv-CD28BBzeta CAR (последовательность нуклеиновой кислоты) (SEQ ID NO: 18)
atggccttaccagtgaccgccttgctcctgccgctggccttgctgctccacgccgccaggccgggatccgagattcagctgcagcaatctggggcagaacttgtgaagccaggggcctcagtcaagctgtcctgcacaggttctggcttcaacattgaagactactatattcactgggtgaagcagaggactgaacagggcctggaatggattggaaggattgatcctgagaatgatgaaactaaatatggcccaatattccagggcagggccactataacagcagacacatcctccaacacagtctacctgcaactcagcagcctgacatctgaggacactgccgtctattactgtgcctttcgcggtggagtctactgggggccaggaaccactctcacagtctcctcaggaggtggtggttccggtggtggtggttccggaggtggtggttcacatatggatgttgtgatgacccagtctccactcactctatcggttgccattggacaatcagcctccatctcttgcaagtcaagtcagagcctcttagatagtgatggaaagacatatttgaattggttgttacagaggccaggccagtctccaaagcgcctaatctctctggtgtctaaactggactctggagtccctgacaggttcactggcagtggatcagggacagatttcacactgagaatcagcagagtggaggctgaggatttgggaatttattattgctggcaaggtacacattttcctgggacgttcggtggagggaccaagctggagataaaagctagcaccacgacgccagcgccgcgaccaccaacaccggcgcccaccatcgcgtcgcagcccctgtccctgcgcccagaggcgtgccggccagcggcggggggcgcagtgcacacgagggggctggacttcgcctgtgatttttgggtgctggtggtggttggtggagtcctggcttgctatagcttgctagtaacagtggcctttattattttctgggtgaggagtaagaggagcaggctcctgcacagtgactacatgaacatgactccccgccgccccgggcccacccgcaagcattaccagccctatgccccaccacgcgacttcgcagcctatcgctccaaacggggcagaaagaaactcctgtatatattcaaacaaccatttatgagaccagtacaaactactcaagaggaagatggctgtagctgccgatttccagaagaagaagaaggaggatgtgaactgagagtgaagttcagcaggagcgcagacgcccccgcgtacaagcagggccagaaccagctctataacgagctcaatctaggacgaagagaggagtacgatgttttggacaagagacgtggccgggaccctgagatggggggaaagccgagaaggaagaaccctcaggaaggcctgtacaatgaactgcagaaagataagatggcggaggcctacagtgagattgggatgaaaggcgagcgccggaggggcaaggggcacgatggcctttaccagggtctcagtacagccaccaaggacacctacgacgcccttcacatgcaggccctgccccctcgc
3C10scFv-BBz CAR (последовательность нуклеиновой кислоты) (SEQ ID NO: 19)
Atggccttaccagtgaccgccttgctcctgccgctggccttgctgctccacgccgccaggccgggatccgagattcagctgcagcaatctggggcagaacttgtgaagccaggggcctcagtcaagctgtcctgcacaggttctggcttcaacattgaagactactatattcactgggtgaagcagaggactgaacagggcctggaatggattggaaggattgatcctgagaatgatgaaactaaatatggcccaatattccagggcagggccactataacagcagacacatcctccaacacagtctacctgcaactcagcagcctgacatctgaggacactgccgtctattactgtgcctttcgcggtggagtctactgggggccaggaaccactctcacagtctcctcaggaggtggtggttccggtggtggtggttccggaggtggtggttcacatatggatgttgtgatgacccagtctccactcactctatcggttgccattggacaatcagcctccatctcttgcaagtcaagtcagagcctcttagatagtgatggaaagacatatttgaattggttgttacagaggccaggccagtctccaaagcgcctaatctctctggtgtctaaactggactctggagtccctgacaggttcactggcagtggatcagggacagatttcacactgagaatcagcagagtggaggctgaggatttgggaatttattattgctggcaaggtacacattttcctgggacgttcggtggagggaccaagctggagataaaagctagcaccacgacgccagcgccgcgaccaccaacaccggcgcccaccatcgcgtcgcagcccctgtccctgcgcccagaggcgtgccggccagcggcggggggcgcagtgcacacgagggggctggacttcgcctgtgatatctacatctgggcgcccttggccgggacttgtggggtccttctcctgtcactggttatcaccctttactgcaaacggggcagaaagaaactcctgtatatattcaaacaaccatttatgagaccagtacaaactactcaagaggaagatggctgtagctgccgatttccagaagaagaagaaggaggatgtgaactgagagtgaagttcagcaggagcgcagacgcccccgcgtacaagcagggccagaaccagctctataacgagctcaatctaggacgaagagaggagtacgatgttttggacaagagacgtggccgggaccctgagatggggggaaagccgagaaggaagaaccctcaggaaggcctgtacaatgaactgcagaaagataagatggcggaggcctacagtgagattgggatgaaaggcgagcgccggaggggcaaggggcacgatggcctttaccagggtctcagtacagccaccaaggacacctacgacgcccttcacatgcaggccctgccccctcgc
ScFv-фрагмент, названный «139», является антителом человека против EGFRvIII (Morgan et al. 2012 Hum Gene Ther 23 (10): 1043-53). EGFRvIII-специфический CAR, включающий scFv 139, был создан с помощью первоначального синтеза scFv 139. Последовательность scFv 139 клонировали с лидерной последовательностью, шарнирной областью CD8, трансмембранным доменом (ТМ) и желаемыми доменами сигнализации. Например, последовательность scFv 139 клонировали с доменами сигнализации для 4-1ВВ и CD3 дзета. Конструкцию CAR (139scFv-BBZ) экспрессировали с вектора pELNS в случае продукции лентивируса.
139scFv-BBz CAR (аминокислотная последовательность) (SEQ ID NO: 3)
MALPVTALLLPLALLLHAARPGSDIQMTQSPSSLSASVGDRVTITCRASQGIRNNLAWYQQKPGKAPKRLIYAASNLQSGVPSRFTGSGSGTEFTLIVSSLQPEDFATYYCLQHHSYPLTSGGGTKVEIKRTGSTSGSGKPGSGEGSEVQVLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTNYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAGSSGWSEYWGQGTLVTVSSASTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIYIWAPLAGTCGVLLLSLVITLYCKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
139scFv-BBz CAR (последовательность нуклеиновой кислоты) (SEQ ID NO: 20)
Atggccttaccagtgaccgccttgctcctgccgctggccttgctgctccacgccgccaggccgggatccgacatccagatgacccagagccctagcagcctgagcgccagcgtgggcgacagagtgaccatcacctgtcgggccagccagggcatcagaaacaacctggcctggtatcagcagaagcccggcaaggcccccaagagactgatctacgctgccagcaatctgcagagcggcgtgcccagcagattcaccggaagcggctccggcaccgagttcaccctgatcgtgtccagcctgcagcccgaggacttcgccacctactactgcctgcagcaccacagctaccctctgaccagcggcggaggcaccaaggtggagatcaagcggaccggcagcaccagcggcagcggcaagcctggcagcggcgagggaagcgaggtccaggtgctggaatctggcggcggactggtgcagcctggcggcagcctgagactgagctgtgccgccagcggcttcaccttcagcagctacgccatgtcttgggtccggcaggctcctggaaagggcctggaatgggtgtccgccatcagcggctctggcggctccaccaactacgccgacagcgtgaagggccggttcaccatcagccgggacaacagcaagaacaccctgtatctgcagatgaacagcctgagagccgaggacaccgccgtgtactactgtgccggcagcagcgggtggagcgagtactggggccagggcacactggtcacagtgtctagcgctagcaccacgacgccagcgccgcgaccaccaacaccggcgcccaccatcgcgtcgcagcccctgtccctgcgcccagaggcgtgccggccagcggcggggggcgcagtgcacacgagggggctggacttcgcctgtgatatctacatctgggcgcccttggccgggacttgtggggtccttctcctgtcactggttatcaccctttactgcaaacggggcagaaagaaactcctgtatatattcaaacaaccatttatgagaccagtacaaactactcaagaggaagatggctgtagctgccgatttccagaagaagaagaaggaggatgtgaactgagagtgaagttcagcaggagcgcagacgcccccgcgtacaagcagggccagaaccagctctataacgagctcaatctaggacgaagagaggagtacgatgttttggacaagagacgtggccgggaccctgagatggggggaaagccgagaaggaagaaccctcaggaaggcctgtacaatgaactgcagaaagataagatggcggaggcctacagtgagattgggatgaaaggcgagcgccggaggggcaaggggcacgatggcctttaccagggtctcagtacagccaccaaggacacctacgacgcccttcacatgcaggccctgccccctcgct
КОМПОНЕНТЫ CAR
Последовательности нуклеиновых кислот:
• Нуклеотидная последовательность (мыши) scFv 3C10; (SEQ ID NO: 4)
GAGATTCAGCTGCAGCAATCTGGGGCAGAACTTGTGAAGCCAGGGGCCTCAGTCAAGCTGTCCTGCACAGGTTCTGGCTTCAACATTGAAGACTACTATATTCACTGGGTGAAGCAGAGGACTGAACAGGGCCTGGAATGGATTGGAAGGATTGATCCTGAGAATGATGAAACTAAATATGGCCCAATATTCCAGGGCAGGGCCACTATAACAGCAGACACATCCTCCAACACAGTCTACCTGCAACTCAGCAGCCTGACATCTGAGGACACTGCCGTCTATTACTGTGCCTTTCGCGGTGGAGTCTACTGGGGGCCAGGAACCACTCTCACAGTCTCCTCAGGAGGTGGTGGTTCCGGTGGTGGTGGTTCCGGAGGTGGTGGTTCACATATGGATGTTGTGATGACCCAGTCTCCACTCACTCTATCGGTTGCCATTGGACAATCAGCCTCCATCTCTTGCAAGTCAAGTCAGAGCCTCTTAGATAGTGATGGAAAGACATATTTGAATTGGTTGTTACAGAGGCCAGGCCAGTCTCCAAAGCGCCTAATCTCTCTGGTGTCTAAACTGGACTCTGGAGTCCCTGACAGGTTCACTGGCAGTGGATCAGGGACAGATTTCACACTGAGAATCAGCAGAGTGGAGGCTGAGGATTTGGGAATTTATTATTGCTGGCAAGGTACACATTTTCCTGGGACGTTCGGTGGAGGGACCAAGCTGGAGATAAAA
• Нуклеотидная последовательность (гуманизированная) scFv 139; (SEQ ID NO: 5)
GACATCCAGATGACCCAGAGCCCTAGCAGCCTGAGCGCCAGCGTGGGCGACAGAGTGACCATCACCTGTCGGGCCAGCCAGGGCATCAGAAACAACCTGGCCTGGTATCAGCAGAAGCCCGGCAAGGCCCCCAAGAGACTGATCTACGCTGCCAGCAATCTGCAGAGCGGCGTGCCCAGCAGATTCACCGGAAGCGGCTCCGGCACCGAGTTCACCCTGATCGTGTCCAGCCTGCAGCCCGAGGACTTCGCCACCTACTACTGCCTGCAGCACCACAGCTACCCTCTGACCAGCGGCGGAGGCACCAAGGTGGAGATCAAGCGGACCGGCAGCACCAGCGGCAGCGGCAAGCCTGGCAGCGGCGAGGGAAGCGAGGTCCAGGTGCTGGAATCTGGCGGCGGACTGGTGCAGCCTGGCGGCAGCCTGAGACTGAGCTGTGCCGCCAGCGGCTTCACCTTCAGCAGCTACGCCATGTCTTGGGTCCGGCAGGCTCCTGGAAAGGGCCTGGAATGGGTGTCCGCCATCAGCGGCTCTGGCGGCTCCACCAACTACGCCGACAGCGTGAAGGGCCGGTTCACCATCAGCCGGGACAACAGCAAGAACACCCTGTATCTGCAGATGAACAGCCTGAGAGCCGAGGACACCGCCGTGTACTACTGTGCCGGCAGCAGCGGGTGGAGCGAGTACTGGGGCCAGGGCACACTGGTCACAGTGTCTAGC
• Лидерная последовательность (последовательность нуклеиновой кислоты); (SEQ ID NO: 6)
ATGGCCTTACCAGTGACCGCCTTGCTCCTGCCGCTGGCCTTGCTGCTCCACGCCGCCAGGCCG
• Шарнирная область (последовательность нуклеиновой кислоты); (SEQ ID NO: 7)
ACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGAT
• Трансмембранный домен (последовательность нуклеиновой кислоты); (SEQ ID NO: 8)
ATCTACATCTGGGCGCCCTTGGCCGGGACTTGTGGGGTCCTTCTCCTGTCACTGGTTATCACCCTTTACTGC
• Внутриклеточный домен 4-1ВВ (последовательность нуклеиновой кислоты); (SEQ ID NO: 9)
AAACGGGGCAGAAAGAAACTCCTGTATATATTCAAACAACCATTTATGAGACCAGTACAAACTACTCAAGAGGAAGATGGCTGTAGCTGCCGATTTCCAGAAGAAGAAGAAGGAGGATGTGAACTG
• CD3 дзета (последовательность нуклеиновой кислоты); (SEQ ID NO: 10)
AGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACAAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCCCCTCGC
• CD3 дзета (последовательность нуклеиновой кислоты; эталонная последовательность в NCBI - NM_000734.3); (SEQ ID NO: 100)
agagtgaagttcagcaggagcgcagacgcccccgcgtaccagcagggccagaaccagctctataacgagctcaatctaggacgaagagaggagtacgatgttttggacaagagacgtggccgggaccctgagatggggggaaagccgagaaggaagaaccctcaggaaggcctgtacaatgaactgcagaaagataagatggcggaggcctacagtgagattgggatgaaaggcgagcgccggaggggcaaggggcacgatggcctttaccagggtctcagtacagccaccaaggacacctacgacgcccttcacatgcaggccctgccccctcgc
Аминокислотные последовательности:
• Аминокислотная последовательность (мыши) scFv 3C10; (SEQ ID NO: 11)
EIQLQQSGAELVKPGASVKLSCTGSGFNIEDYYIHWVKQRTEQGLEWIGRIDPENDETKYGPIFQGRATITADTSSNTVYLQLSSLTSEDTAVYYCAFRGGVYWGPGTTLTVSSGGGGSGGGGSGGGGSHMDVVMTQSPLTLSVAIGQSASISCKSSQSLLDSDGKTYLNWLLQRPGQSPKRLISLVSKLDSGVPDRFTGSGSGTDFTLRISRVEAEDLGIYYCWQGTHFPGTFGGGTKLEIK
• Аминокислотная последовательность (человека) scFv 139; (SEQ ID NO: 12)
DIQMTQSPSSLSASVGDRVTITCRASQGIRNNLAWYQQKPGKAPKRLIYAASNLQSGVPSRFTGSGSGTEFTLIVSSLQPEDFATYYCLQHHSYPLTSGGGTKVEIKRTGSTSGSGKPGSGEGSEVQVLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTNYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAGSSGWSEYWGQGTLVTVSS
• Лидерная последовательность (аминокислотная последовательность) (SEQ ID NO: 13)
MALPVTALLLPLALLLHAARP
• Шарнирная область (аминокислотная последовательность) (SEQ ID NO: 14)
TTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACD
• Трансмембранный домен (аминокислотная последовательность) (SEQ ID NO: 15)
IYIWAPLAGTCGVLLLSLVITLYC
• Внутриклеточный домен 4-1ВВ (аминокислотная последовательность) (SEQ ID NO: 16)
KRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCEL
• CD3 дзета-домен (аминокислотная последовательность) (SEQ ID NO: 17)
RVKFSRSADAPAYKQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
• CD3 дзета-домен (аминокислотная последовательность; эталонная последовательность в NCBI - NM_000734.3) (SEQ ID NO: 99)
RVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 11, предоставлена как SEQ ID NO: 4. Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 12, предоставлена как SEQ ID NO: 5. Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 13, предоставлена как SEQ ID NO: 6. Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 14, предоставлена как SEQ ID NO: 7. Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 15, предоставлена как SEQ ID NO: 8. Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 16, предоставлена как SEQ ID NO: 9. Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 17, предоставлена как SEQ ID NO: 10. Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 1, предоставлена как SEQ ID NO: 18. Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 2, предоставлена как SEQ ID NO: 19. Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 3, предоставлена как SEQ ID NO: 20. Нуклеотидная последовательность, кодирующая полипептид SEQ ID NO: 99, предоставлена как SEQ ID NO: 100.
Пример 4: Прогнозные указания CDR для EGFRvIIICAR
Прогнозные указания CDR для EGFRvIII-специфического CAR в соответствии с Kabat являются следующими:
• VH:
EIQLQQSGAELVKPGASVKLSCTGSGFNIEDYYIHWVKQRTEQGLEWIGRIDPENDETKYGPIFQGRATITADTSSNTVYLQLSSLTSEDTAVYYCAFRGGVYWGPGTTLTVSS; (SEQ ID NO: 21);
где CDR1 представляет собой DYYIH (SEQ ID NO: 22), CDR2 представляет собой RIDPENDETKYGPIFQG (SEQ ID NO: 23), и CDR3 представляет собой RGGVY (SEQ ID NO: 24).
• VL:
DVVMTQSPLTLSVAIGQSASISCKSSQSLLDSDGKTYLNWLLQRPGQSPKRLISLVSKLDSGVPDRFTGSGSGTDFTLRISRVEAEDLGIYYCWQGTHFPGTFGGGTKLEIK; (SEQ ID NO: 25);
где CDR1 представляет собой KSSQSLLDSDGKTYLN (SEQ ID NO: 26), CDR2 представляет собой LVSKLDS (SEQ ID NO: 27), и CDR3 представляет собой WQGTHFPGT (SEQ ID NO: 28).
Прогнозные указания CDR для EGFRvIII-специфического CAR в соответствии с Chothia являются следующими:
• VH:
EIQLQQSGAELVKPGASVKLSCTGSGFNIEDYYIHWVKQRTEQGLEWIGRIDPENDETKYGPIFQGRATITADTSSNTVYLQLSSLTSEDTAVYYCAFRGGVYWGPGTTLTVSS; (SEQ ID NO: 29);
где CDR1 представляет собой GFNIEDY (SEQ ID NO: 30), CDR2 представляет собой DPENDE (SEQ ID NO: 31), и CDR3 представляет собой RGGVY (SEQ ID NO: 32).
• VL:
DVVMTQSPLTLSVAIGQSASISCKSSQSLLDSDGKTYLNWLLQRPGQSPKRLISLVSKLDSGVPDRFTGSGSGTDFTLRISRVEAEDLGIYYCWQGTHFPGTFGGGTKLEIK; (SEQ ID NO: 33);
где CDR1 представляет собой SQSLLDSDGKTY (SEQ ID NO: 34), CDR2 представляет собой LVS (SEQ ID NO: 35), и CDR3 представляет собой GTHFPG (SEQ ID NO: 36).
Пример 5: Гуманизация мышиного антитела против EGFRvIII
Гуманизация мышиного антитела против EGFRvIII желательна для клинических условий, в которых специфические для мыши остатки могут индуцировать реакцию против мышиного антигена у людей (НАМА) - у пациентов, получающих лечение Т-клетками, трансдуцированными мышиной CAR-конструкцией. Последовательности VH и VL происходящего из гибридомы антитела против EGFRvIII мыши были извлечены из опубликованной литературы (Morgan et al. (2012) Human Gene Therapy, 23: 1043-1953, выше). Гуманизация была достигнута путем пересадки CDR-участков из антитела против EGFRvIII мыши в акцепторные каркасные области зародышевой линии человека VH1_1-f или VH5_5a а также VK2_A17 или VK4_B3 (баз данных vBASE). Помимо CDR-участков, несколько остатков каркасных областей, т.е. VK2 #36, #49, VK4 #2, #36, #46, #49, VH1 #2, #24, #76, #94 и VH5 #2, #24, #73, #76, #94, которые, как полагают, поддерживают структурную целостность CDR-участков, были сохранены из мышиной последовательности. Кроме того, человеческие J-элементы JH6 и JK4 были использованы для тяжелой и легкой цепи, соответственно. Полученные в результате аминокислотные последовательности гуманизированного антитела были обозначены VK2_A17/Hz1 и VK4_B3/Hz1 для легких цепей и VH1_1-f/Hz1, VH5_5-a/Hz1 для тяжелых цепей, показанных на фиг. 9. Нумерация остатков подчиняется Kabat (Kabat E.A. et al., 1991, выше). Для определения CDR были использованы как Kabat, так и Chothia и др., 1987 (выше). Остатки каркасных областей, сохраненные из мышиного антитела против EGFRvIII, показаны жирным шрифтом/курсивом в рамочке, остатки CDR подчеркнуты.
На основе последовательностей гуманизированных легкой и тяжелой цепей, продемонстрированных на фиг. 9, в общей сложности 8 комбинаций каркасных областей были использованы для создания растворимых scFv для дальнейшей валидации. Порядок, в котором VH- и VL-домены появляются в scFv, изменялся (т.е. ориентация VL-VH или VH-VL), и четыре копии субъединицы «G4S» (SEQ ID NO: 37), которая включает последовательность GGGGS (SEQ ID NO: 37), использовали для соединения каркасных областей. На фиг. 9 представлены CDR в последовательностях VH и VL, рассчитанные в соответствии с Kabat и др. и Chothia и др. (выше).
Клонирование:
Были получены последовательности ДНК, кодирующие мышиные и гуманизированные VL- и VH-домены, и кодоны конструкций были оптимизированы для экспрессии в клетках Homo sapiens.
Последовательности, кодирующие VL- и VH-домены, субклонировали в экспрессионные векторы, подходящие для секреции в клетках млекопитающих. Элементы экспрессионного вектора включают промотор (промотор-энхансер цитомегаловируса (CMV)), сигнальную последовательность для облегчения секреции, сигнал полиаденилирования и терминатор транскрипции (гена бычьего гормона роста (BGH)), элемент, делающий возможной репликацию эписомы и репликацию в прокариотах (например, начало репликации SV40 и ColE1 или другие известные в данной области техники) и элементы, которые делают возможным отбор (ген устойчивости к ампициллину и маркер зеоцин).
Пример 6: Характеристики гуманизированных растворимых scFv-фрагментов антитела против EGFRvIII
Были получены растворимые scFv-фрагменты, описанные выше, используя стандартные методы молекулярной биологии. Эти растворимые scFv были использованы в исследованиях описания свойств для изучения стабильности, экспрессии на клеточной поверхности и связывающих свойств scFv.
Экспрессия и очистка scFv
Для трансфекции каждой конструкции scFv приблизительно 3е8 клеток 293F трансфицировали 100 мкг плазмиды, используя PEI в качестве реагента для трансфекции в соотношении 3:1 (PEI:ДНК). Клетки выращивали в 100 мл среды ΕΧΡi293 Expression Media (Invitrogen) в шейкерной колбе при 37°С, 125 оборотов в минуту, 8% CО2. Культуру собирали через шесть дней и использовали для очистки белка.
Клетки 293F собирали путем осаждения при 3500×g в течение 20 минут. Супернатант собирали и фильтровали через фильтрующее устройство VacuCap90 PF Filter Unit (w/0,8/0,2 мкм Super Membrane, PALL). Приблизительно 400 мкл агарозных сфер Ni-NTA (Qiagen) добавляли к супернатанту. Смесь вращали и инкубировали в течение 4 ч при 4°С. Ее загружали в колонку для очистки и промывали промывочным буфером с 20 мМ гистидина. Белок элюировали, используя 500 мкл буфера для элюции с 300 мМ гистидина. Образцы подвергали диализу против буфера PBS в течение ночи при 4°С. Образцы белка подвергали количественному анализу, используя спектрофотометр NanoDrop 2000, микроячейка.
ЕС50 при определяемом с помощью FACS связывании очищенных scFv с клетками, экспрессирующими или EGFR человека дикого типа, или EGFRvIII
Были проведены следующие эксперименты, чтобы показать, что все гуманизированные варианты scFv против EGFRvIII характеризуются сравнимым связыванием с EGFRvIII, но не связыванием с EGFR дикого типа.
Суспензию клеток HEK293F транзиторно трансфицировали или hEGFR дикого типа, или hEGFRvIII и собирали через 2 дня после трансфекции. Приблизительно 5e5 клеток/на лунку переносили в 96-луночный планшет BD Falcon. Клетки центрифугировали при 900 оборотов в минуту (в центрифуге Sorval Legend XT) в течение 3 минут. Супернатант удаляли. Образцы белка scFv против EGFRvIII разводили в DPBS с 5% FBS. Образцы добавляли в лунки, перемешивали и инкубировали в течение 1 часа. Клетки промывали дважды в DPBS с 5% FBS. Клетки инкубировали с анти-поли(His) PE (R&D) в течение 1 часа, дважды промывали перед анализом FACS (LSRII от BD Biosciences).
Было определено, что ЕС50 scFv мыши (m3C10) для hEGFRvIII составляет ~5 нМ, как показано на фиг. 10. Все гуманизированные варианты scFv против EGFRvIII показали значения ЕС50 в диапазоне ЕС50 от однозначного числа до небольшого двухзначного числа нМ (5-50 нМ). Кроме того, не отмечалось заметное связывание конструкций 2173 и 2174 с экспрессирующими EGFR дикого типа клеточными линиями, что означает улучшенный профиль безопасности по сравнению с мышиным 3C10, как показано на фиг. 11. На основании этих исследований, клон 2173 был отобран для дальнейшей клинической характеристики, как показано в примере 8.
Пример 7: Конструкции гуманизированных EGFRvIII-специфических CAR
ScFv, используемые в конечных конструкциях CAR, были получены из гуманизированных каркасных последовательностей, описанных в примере 1. Порядок, в котором VH- и VL-домены появляются в scFv, изменялся (т.е. ориентация VL-VH или VH-VL). Линкер (G4S)4 (SEQ ID NO: 37) использовали для соединения вариабельных доменов для создания scFv, представленных в таблице 1.
Конструкции гуманизированных scFv против EGFRvIII с показом ориентации VH и VL и длины линкера (Таблица раскрывает «G4S» как SEQ ID NO: 37)
mEGFRvIII 3C10
Последовательности гуманизированных scFv-фрагментов, представлены ниже в таблице 2 (SEQ ID NO: 38, SEQ ID NO: 44, SEQ ID NO: 50, SEQ ID NO: 56, SEQ ID NO: 62, SEQ ID NO: 68, SEQ ID NO: 74 и SEQ ID NO: 80). Эти scFv-фрагменты были использованы с дополнительными последовательностями, SEQ ID NO: 13-17, для образования полных конструкций CAR с SEQ ID NO: SEQ ID NO: 43, SEQ ID NO: 49, SEQ ID NO: 55, SEQ ID NO: 61, SEQ ID NO: 67, SEQ ID NO: 73, SEQ ID NO: 79 и SEQ ID NO: 85.
Все эти клоны содержали изменение остатка Q/K в домене сигнализации костимулирующего домена, происходящего из CD3 дзета цепи.
Конструкции гуманизированных EGFRvIII-специфических CAR
нуклеотиды
ScFv-фрагменты CAR затем клонировали в лентивирусные векторы для создания конструкции полноразмерной CAR в одной рамке считывания, и используя промотор EF1 альфа для экспрессии (SEQ ID NO: 97).
Промотор EF1 альфа
GTGAGGCTCCGGTGCCCGTCAGTGGGCAGAGCGCACATCGCCCACAGTCCCCGAGAAGTTGGGGGGAGGGGTCGGCAATTGAACCGGTGCCTAGAGAAGGTGGCGCGGGGTAAACTGGGAAAGTGATGTCGTGTACTGGCTCCGCCTTTTTCCCGAGGGTGGGGGAGAACCGTATATAAGTGCAGTAGTCGCCGTGAACGTTCTTTTTCGCAACGGGTTTGCCGCCAGAACACAGGTAAGTGCCGTGTGTGGTTCCCGCGGGCCTGGCCTCTTTACGGGTTATGGCCCTTGCGTGCCTTGAATTACTTCCACCTGGCTGCAGTACGTGATTCTTGATCCCGAGCTTCGGGTTGGAAGTGGGTGGGAGAGTTCGAGGCCTTGCGCTTAAGGAGCCCCTTCGCCTCGTGCTTGAGTTGAGGCCTGGCCTGGGCGCTGGGGCCGCCGCGTGCGAATCTGGTGGCACCTTCGCGCCTGTCTCGCTGCTTTCGATAAGTCTCTAGCCATTTAAAATTTTTGATGACCTGCTGCGACGCTTTTTTTCTGGCAAGATAGTCTTGTAAATGCGGGCCAAGATCTGCACACTGGTATTTCGGTTTTTGGGGCCGCGGGCGGCGACGGGGCCCGTGCGTCCCAGCGCACATGTTCGGCGAGGCGGGGCCTGCGAGCGCGGCCACCGAGAATCGGACGGGGGTAGTCTCAAGCTGGCCGGCCTGCTCTGGTGCCTGGCCTCGCGCCGCCGTGTATCGCCCCGCCCTGGGCGGCAAGGCTGGCCCGGTCGGCACCAGTTGCGTGAGCGGAAAGATGGCCGCTTCCCGGCCCTGCTGCAGGGAGCTCAAAATGGAGGACGCGGCGCTCGGGAGAGCGGGCGGGTGAGTCACCCACACAAAGGAAAAGGGCCTTTCCGTCCTCAGCCGTCGCTTCATGTGACTCCACGGAGTACCGGGCGCCGTCCAGGCACCTCGATTAGTTCTCGAGCTTTTGGAGTACGTCGTCTTTAGGTTGGGGGGAGGGGTTTTATGCGATGGAGTTTCCCCACACTGAGTGGGTGGAGACTGAAGTTAGGCCAGCTTGGCACTTGATGTAATTCTCCTTGGAATTTGCCCTTTTTGAGTTTGGATCTTGGTTCATTCTCAAGCCTCAGACAGTGGTTCAAAGTTTTTTTCTTCCATTTCAGGTGTCGTGA (SEQ ID NO: 97)
Экспрессия на поверхности CAR9, CAR10 и выбранные конструкции гуманизированных EGFRvIII-специфических CAR и окрашивание, определяемое с помощью FACS
Следующие эксперименты показали, что, по-видимому, существует различие в аффинности к EGFRvIII, на основе in vitro исследований связывания в случае как клеток Jurkat, так и первичных Т-клеток.
В клетки Jurkat E6 электропорировали или вектор для CAR9, или вектор для CAR10, используя набор Amaxa Cell Line Nucleofector Kit V (Lonza, Colgne AG, Германия) и программу X-001. Через день после трансфекции 0,5×106 клеток в 0,2 мл буфера FACS (DPBS буфера, содержащего 5% FBS) помещали в каждую лунку 96-луночного планшета V-образной формы (Greiner Bio-One, Германия) и инкубировали в течение 10 минут при комнатной температуре. Затем клетки центрифугировали и ресуспендировали в 0,2 мл буфера FACS с различными концентрациями EGFRvIII-Fc или EGFRwt-Fc и инкубировали при 4°С в течение 30 минут. Затем клетки промывали буфером FACS три раза и инкубировали с 0,2 мл буфера FACS с 2 мкл конъюгированного с PE Fc против IgG человека (Jackson ImmunoResearch Laboratories, West Grove, PA) в течение 30 минут при 4°С в темноте. После промывания 0,2 мл буфера FACS три раза клетки анализировали в устройстве LSRII (BD Biosciences, San Jose, CA), используя программное обеспечение FACSDiva (BD Biosciences, San Jose, CA). Иммунофлуоресцентное окрашивание было проанализировано как относительная логарифмическая флуоресценции живых клеток, и определяли процент PE-позитивных клеток.
Как показано на фиг. 12, связывание CAR9, экспрессируемого в клетках Jurkat, со слитым белком EGFRvIII-Fc приблизительно в 1000 раз сильнее, чем с EGFR-Fc дикого типа. Кроме того, CART-клеточная структура, экспрессирующая CAR10, проявляет значительно меньшее (в ~40 раз) связывание с EGFRvIII, чем с CAR9. Это говорит о том, что, хотя мышиный CAR9 связывается с EGFRvIII, он по-прежнему сохраняет некоторое связывания с EGFR дикого типа. Кроме того, это убедительно свидетельствует о том, что CAR9 имеет более высокую аффинность к EGFRvIII, чем конструкция CAR10.
Дальнейшие эксперименты на первичных Т-клетках дали схожие результаты. Вкратце, первичные CD3+ Т-клетки человека стимулировали, используя покрытые антителом против CD3/CD28 сферы, в течение 24 часов, а затем трансдуцировали лентивирусными векторами, кодирующими или CAR9, CAR10, CAR6, или контрольный CAR, с множественностью заражения (MOI) 3:1. В эксперимент была также включена популяция контроль-трансдуцированных Т-клеток. Эти клетки размножали в течение приблизительно 8-9 дней в культуре, пока они не начали переходить в состояние покоя. В этот момент времени 0,5×106 клеток помещали в каждую 96-луночного планшета V-образной формы. Клетки промывали один раз PBS и окрашивали реагентом, позволяющим отличить живые клетки от мертвых, (1:1000 в PBS), в течение 30 мин на льду. Затем клетки дважды промывали буфером FACS буфера и инкубировали с 1 мкг/мл биотинилированного белка EGFRvIII или EGFR дикого типа в течение 30 мин на льду. Затем клетки промывали два раза и инкубировали с 0,2 мл буфера FACS с использованием 1:1000 разведения стрептавидина-РЕ в течение 15 мин на льду. После двукратной промывки буфером FACS клетки анализировали в LSRII. Иммунофлуоресцентное окрашивание было проанализировано как относительная логарифмическая флуоресценция живых клеток, и процент PE-позитивных клеток определяли в сочетании с геометрическим средним позитивной популяции.
Как показано на фиг. 13, CAR9 и CAR6 демонстрируют среднее геометрическое (21K для CAR9, 27К для CAR6) для связывания с EGFRvIII, превышающее в 10 раз таковое для CAR10 (лишь 2K), когда насыщающие количества белка EGFRvIII используются для детектирования, даже если все конструкции трансдуцированы эквивалентно (~50% эффективность трансдукции для всех). Аналогичным образом, специфичность в отношении белка EGFR дикого типа приблизительно в 10 раз меньше, что отображает логарифмический сдвиг в меньшую сторону для окрашивания белком EGFR дикого типа. Это обеспечивает дополнительную поддержку данных, полученных с использованием клеток Jurkat, представленных выше, которые указывают на то, что CAR9 и CAR6 обладают большей аффинностью к белку EGFRvIII, чем CAR10, при экспрессии в первичных Т-клетках, и предполагает, что они будут более эффективными в клинике.
Функциональный анализ панели конструкций гуманизированных CAR был проведен, как описано в примере 8.
Пример 8: Анализ конструкций гуманизированных EGFRvIII-специфических CAR в Т-клетках
Для оценки возможности нацеливания на EGFRvIII с помощью CAR- технологии, гуманизированные scFv-фрагменты против EGFRvIII были клонированы в лентивирусный вектор для экспрессии CAR с использованием CD3 дзета-цепи и костимулирующей молекулы 4-1ВВ в двух различных конфигурациях. Оптимальная конструкция выбирается на основе количества и качества эффекторной Т-клеточной реакции EGFRvIII-CAR-трансдуцированных Т-клеток в ответ на мишени EGFRvIII+ и EGFR дикого типа. Эффекторные Т-клеточные реакции включают, но без ограничения, размножение, пролиферацию, удвоение клеток, продукцию цитокинов и уничтожение клеток-мишеней или цитолитическую активность (дегрануляцию).
Материалы и методы
Создание линии клеток-репортеров Jurkat для первоначальной характеристики функции CAR
В качестве альтернативы трансдукции и активации первичных T-клеток, линия клеток-репортеров Jurkat-NFAT может использоваться для оценки функциональной активности конструкций CAR. Линию Т-клеток Jurkat (E6-1) трансфицировали репортерной конструкцией NFAT-люцифераза, и стабильная, клональная клеточная линия (JNL) была отобрана для дальнейшей характеристики на основе сильной индукции репортера NFAT после стимуляции РМА и иономицином. Клетки JNL трансдуцировали лентивирусными векторами с MOI 5:1 и затем размножали в течение 5-7 дней. Перед использованием в анализе определяли процент трансдуцированных клеток (экспрессирующих EGFRvIII-специфический CAR с представлением на клеточной поверхности) и относительную интенсивность флуоресценции этой экспрессии в них с помощью анализа с использованием проточной цитометрии в LSRII. Исходя из гистограмм, относительные уровни экспрессии CAR могут быть проверены путем сравнения процента трансдуцированных клеток с их относительной интенсивностью флуоресценции.
Оценка Т-клеточной активации экспрессирующих EGFRvIII-специфический гуманизированный CAR клеток JNL
Для оценки активации Т-клеток в линии клеток-репортеров JNL, клетки JNL или CAR-трансдуцированные клетки JNL высевают в количестве 50000 клеток на лунку в 96-луночный черный планшет с прозрачным дном. Клетки-мишени (родительские клетки ВНК или клетки ВНК, созданные для экспрессии или EGFRvIIII, или EGFR дикого типа) добавляют в лунки с получением соотношений эффектор:мишень (Е:Т)=1:2, 1:1, 1:0,3, 1:0,1, 1:0,03, 1:01 и 1:0,003. РМА и иономицин используются в качестве положительного контроля для активации. Клетки подвергают инкубации при 37°С в течение 16-24 часов. В конце инкубации равный объем реагента для анализы люциферазы Bright-Glo добавляют в каждую лунку. Планшет инкубируют при комнатной температуре в течение 10 минут, а затем люминесценцию измеряют с помощью люминометра.
Создание перенаправленных, экспрессирующих EGFRvIII-специфический гуманизированный CAR Т-клеток
Лентивирусные векторы для переноса гуманизированного EGFRvIII-специфического CAR используются для получения геномного материала, который упаковывается в псевдотипированные лентивирусные частицы VSVg. ДНК лентивирусного вектора для переноса смешивают с тремя компонентами для упаковки VSVg, gag/pol и rev в сочетании с реагентом липофектамином для трансфекции их вместе в Т-клетки 293. Через 24 и 48 ч среду собирают, фильтруют и концентрируют с помощью ультрацентрифугирования или хроматографии. Полученный в результате вирусный препарат хранят при -80°С. Количество трансдуцирующих единиц определяют титрованием на SupT1-клетках.
Перенаправленные, экспрессирующие EGFRvIII-специфический CAR Т-клетки получают путем активации свежих Т-клеток при введении в контакт с CD3×28 сферами в течение 24 часов, с последующим добавлением соответствующего количества трансдуцирующих единиц для получения желаемого процента трансдуцированных Т-клеток. Этим модифицированным Т-клеткам позволяют размножаться, пока они не станут покоящимися и не уменьшаться в размере (~300 фл), после чего их подвергают криоконсервации для последующего анализа. Количества и размеры клеток определяют, используя Coulter multisizer III. Перед криоконсервацией определяют процент трансдуцированных клеток (экспрессирующих EGFRvIII-специфический CAR с представлением на клеточной поверхности) и относительную интенсивность флуоресценции этой экспрессии в них с помощью анализа с использованием проточной цитометрии в LSRII. Исходя из гистограмм, относительные уровни экспрессии CAR могут быть исследованы путем сравнения процента трансдуцированных клеток с их относительной интенсивностью флуоресценции.
Оценка цитолитической активности, способностей к пролиферации и секреции цитокинов перенаправленных на EGFRvIII, экспрессирующих гуманизированный CAR Т-клеток.
Для оценки функциональных способностей экспрессирующих гуманизированный EGFRvIII-специфический CAR Т-клеток к уничтожению, пролиферации и секреции цитокинов, клетки оттаивали и оставляли для восстановления в течение ночи. Помимо гуманизированных конструкций, мышиный CAR9 использовали с целью сравнения, в то время как SS1-BBz использовали в качестве экспрессируемого CAR без нацеливания для определения фонового CAR/T-клеточного эффекта. Для этого анализа цитотоксичности на основе проточной цитометрии, клетки-мишени окрашивали CSFE для количественного анализа их присутствия. Клетки-мишени также окрашивали на предмет выявления экспрессии EGFRvIII для подтверждения схожих уровней антигенов-мишеней. Цитолитические активности EGFRvIII-CAR-Т-клеток определяли при титровании соотношений эффектор:клетка-мишень = 10:1, 3:1, 1:1, 0,3:1 и 0: 1, где эффекторы были определены как Т-клетки, экспрессирующие CAR на основе антитела против EGFRvIII. Анализы начинали путем смешивания соответствующего количества Т-клеток с постоянным количеством клеток-мишеней. Через 4 или 16 ч общий объем каждой смеси удаляли, и каждую лунку промывали. Т-клетки окрашивали на CD3, и все клетки окрашивали маркером, позволяющим отличить живые клетки от мертвых, 7AAD. После последней промывки осажденные клетки ресуспендировали в определенном объеме с заданным числом сфер для подсчета. Данные по окрашиванию клеток собирали с помощью проточной цитометрии в LSRII и анализировали с использованием программного обеспечения FlowJo, используя сферы для количественного анализа результатов.
Для измерения клеточной пролиферации и продукции цитокинов Т-клеток(ами) с гуманизированным CAR, специфическим в отношении EGFRvIII, клетки размораживали и предоставляли возможность восстановиться в течение ночи. Помимо гуманизированного CAR, специфического в отношении EGFRvIII, мышиный CAR9 использовали с целью сравнения, в то время как SS1-BBz использовали в качестве экспрессируемого CAR без нацеливания для определения фонового CAR/T-клеточного эффекта. Т-клетки были направлены на U87, линию клеток глиобластомы, порожденной астроцитомой, экспрессирующей или не экспрессирующей EGFRvIII. Кроме того, CD3×28 сферы использовали для оценки возможности Т-клеток отвечать на второй ряд иммунологических эндогенных сигналов. Для анализа пролиферации Т-клетки окрашивали CSFE. Пролиферация представляла собой разведение красящего вещества CSFE, отражающее разделение родительских меток теперь на две дочерние клетки. В ходе анализа были проверены лишь соотношения эффектор:мишень = 1:1 и 1:0, где эффекторы были определены как все Т-клеток (CD4 и 8), приведенные к представлению химерного рецептора к EGFRvIII в одинаковых процентах. Анализ проводили в двух повторах и через 24 часа после смешивания клеток. Супернатант удаляли для определения продукции цитокинов. Через 5 дней Т-клетки окрашивали на предмет выявления живых/мертвых клеток, используя Live/Dead Violet (Invitrogen), затем окрашивали на экспрессию CAR и фенотипировали как или CD4, или CD8 клетки. После последней промывки осажденные клетки ресуспендировали в определенном объеме с заданным числом сфер для подсчета BD. Данные по окрашиванию клеток собирали с помощью проточной цитометрии в LSRII и анализировали с использованием программного обеспечения FlowJo, используя сферы для количественного анализа результатов. Общее количество клеток определяли по количеству подсчитанных клеток относительно определенного количества сфер, умноженному на долю сфер, еще не подсчитанных.
Результаты
Анализ с использованием клеток-репортеров Jurkat для проверки способности экспрессирующих гуманизированный EGFRvIII-специфический CAR Т-клеток распознавать экспрессирующие EGFRvIII клетки-мишени.
Способность CART-клеточных структур к индукции активации после связи с мишенью определяли с использованием линии клеток-репортеров JNL. Линия клеток JNL была создана с использованием репортерной конструкции NFAT-люцифераза, которая индуцируется после связи с CAR. Клетки JNL трансдуцировали различными CAR-EGFRvIII-конструкциями (CAR9, CAR3, CAR6, CAR8 и CAR10). Эффективность трансдукции оценивали с помощью проточной цитометрии, и она, как было установлено, составляет приблизительно 45-52% для всех конструкций. Клетки JNL-CAR-EGFRvIII затем стимулировали с использованием семи различных соотношений Е:Т, используя три различных линии клеток-мишеней (родительские клетки ВНК, ВНК-EGFRvIII или ВНК-EGFR WT). Родительские клетки JNL и клетки JNL, экспрессирующие контрольный CAR, были включены в качестве дополнительных контролей. Результаты, представленные на фиг. 14, показывают, что значительная, мишень-индуцированная активации может произойти уже при соотношениях 1:0,01 в случае всех проверенных конструкций, и CAR6 и CAR10 вызывают наибольшую активацию при более высоких соотношениях E:T. Никакой значительной активации не отмечалось в случае экспрессирующих EGFR дикого типа клеток или с помощью экспрессирующих контрольный CAR клеток JNL. Эти данные демонстрируют специфичность CAR-конструкций в отношении мишени EGFRvIII и отсутствие перекрестной реактивности с мишенью в виде EGFR дикого типа.
Трансдукция первичных T-клеток человека конструкциями гуманизированных EGFRvIII-специфических CAR и их размножение
CD3+ Т-клетки были получены из продуктов афереза или цельной крови от здоровых доноров. Как описано выше, Т-клетки стимулировали покрытыми антителом против CD3×CD28 сферами в течение 24 часов, а затем трансдуцировали, используя сконцентрированные супернатанты, содержащие лентивирусы, с множественность заражения = 3. Клетки размножали в культуре в течение 8-10 дней.
Представленности на клеточной поверхности гуманизированных EGFRvIII-специфических CAR являются сопоставимыми, и их уровни экспрессии очень схожи с таковыми мышиного CAR9. Наложение гистограмм, на которых представлен характер окрашивания на представленность на поверхности клеток - трандуцированных каждым из гуманизированных EGFRvIII-специфических CAR Т-клеток и средняя интенсивность флуоресценции (MFI), рассчитанная, исходя из этих профилей, хорошо коррелирует с процентом трансдуцированных клеток.
Анализ пролиферации для проверки способности экспрессирующих EGFRvIII клеток-мишеней к стимуляции экспрессирующих гуманизированный, EGFRvIII-специфический CAR Т-клеток.
Способность экспрессирующих EGFRvIII-специфический CAR Т-клеток к пролиферации в ответ на связь с мишенью оценивали в анализе на пролиферацию. Субпопуляции были подсчитаны с помощью проточной цитометрии. Донорские Т-клетки трансдуцировали или гуманизированными CAR, мышиным CAR9, или SS1 (мишенью которого является мезотелин). CAR смешивали в соотношении 1:1 или 1:0 с клетками-мишенями и сокультивировали в течение 5 дней. На фиг. 15 продемонстрирована способность ND407 EGFRvIII-CAR-Т-клеток к пролиферации антигенспецифическим образом. Столбик с пунктирной линией указывает на количество посеянных Т-клеток, и сравнительно, увеличение количества Т-клеток не было обнаружено при связи с U87, в то время как связь с U87-EGFRvIII индуцировала пролиферацию, которая была специфической в отношении популяции экспрессирующих EGFRvIII-специфический CAR Т-клеток. Относительный ответ на ND407 указывал на то, что CAR6 и CAR8 являются более функциональными, чем CAR9 или CAR3. Результат, полученный с использованием покрытых Ат против CD3×28 сфер, указывает на то, что их стимуляция не была достаточной, чтобы стимулировать пролиферацию во второй цикле активации, аналогично отсутствию стимуляции вообще.
ND407 Т-клетки были использованы для скрининга различных huEGFRvIII-специфических CAR на их способность к размножению преимущественно CAR+ Т-клеток. На фиг. 16 показано, что CAR5 и CAR6 последовательно обладают самой большой способностью к размножению CAR+ Т-клеток у каждого донора. Увеличение CAR+ Т-клеток является результатом успешной связи с мишенью, пролиферации и пережития индуцированной активацией гибели клеток из-за распознавания антигена.
Анализ уничтожения для проверки способности экспрессирующих гуманизированный EGFRvIII-специфический CAR Т-клеток к уничтожению экспрессирующих EGFRvIII клеток-мишеней
Способность экспрессирующих EGFRvIII-специфический CAR Т-клеток к уничтожению мишеней проверяли в анализе высвобождения хрома. Была создана линия клеток глиобластомы человек, U-87MG, которая экспрессирует или рецептор EGFR дикого типа, или мутант EGFRvIII. Эти созданные линии клеток служили в качестве мишеней в случае анализа уничтожения. Три эффекторные CAR-Т-клетки использовали для определения специфичности уничтожения клеток-мишеней: 1) Т-клетки человека, трансдуцированные для экспрессии 3C10 мыши (CAR9), 2) Т-клетки человека, трансдуцированные для экспрессии гуманизированного варианта 3C10 мыши, названного CAR6, и 3) Т-клетки человека, трансдуцированные CAR, специфическим для мезотелина, SS1. Все эффекторные клетки были приведены к представлению 30% CAR+ трансдукции. Клетки-мишени метили хромом-51 и промывали непосредственно перед сокультивированием. Эффекторы и мишени смешивали вместе в указанных соотношениях (Е:Т) и допускали инкубацию в течение 4 часов.
Результаты, представленные на фиг. 17(А), показывает, что CAR-Т-клетки, смешанные с клетками U-87, экспрессирующими рецептор EGFR дикого типа, продемонстрировали отсутствие уничтожения клеток выше фона до Е:Т=50: 1. Однако результаты, представленные на фиг. 17(В), показывают, что в отличие от этого экспрессирующие EGFRvIII-специфический CAR Т-клетки, CAR9 или CAR6, смешанные с клетками U-87, экспрессирующими EGFRvIII, продемонстрировали специфическое уничтожение при соотношениях Е:Т от 6,25:1 до 50: 1. Не наблюдалось значительное уничтожение, когда экспрессирующие специфический для мезотелина CAR Т-клетки использовали в качестве эффекторов. Эти данные показывают специфическое в отношении мишени уничтожение клеток-мишеней, экспрессирующих EGFRvIII, Т-клетками, экспрессирующими CAR9 и CAR6, но отсутствие уничтожения клеток, экспрессирующих EGFR дикого типа, или экспрессирующей неспецифический CAR T-клеткой, SSI.
Анализ цитокинов для проверки способности экспрессирующих гуманизированный EGFRvIII-специфический CAR T-клеток к содействию противоопухолевому ответу и демонстрации специфичности
Способность экспрессирующих EGFRvIII-специфический CAR T-клеток к индукции цитокина в ответ на связь с мишенью оценивали в анализе сокультивирования. CAR-Т-клетки сокультивировали с экспрессирующими мишень клетками в течение 18-24 ч при различных соотношениях мишень:эффектор (0,3:1, 1:1, 3:1 и 10:1). Клетки-мишени включали клетки U87, экспрессирующие эндогенный белок EGFR дикого типа (U87), клетки U87, сверхэкспрессирующие EGFRvIII (U87-VIII), родительские клетки ВНК (клетки почки детеныша хомяка), клетки ВНК, сверхэкспрессирующие белок человека EGFR дикого типа (ВНК wt), или клетки ВНК, сверхэкспрессирующие белок человека EGFRvIII (ВНК-VIII). Спустя 18-24 часов супернатанты удаляли из культур, и цитокины анализировали, используя Cytometric Bead Assay (CBA). Результаты ясно показали, что 1) CAR6 и CAR9 Т-клетки индуцировали схожие уровни IFNγ в ответ на EGFRvIII-экспрессирующие клетки, и 2) что ни та, ни другая популяция CAR-Т-клеток не индуцировала IFNγ в ответ на экспрессирующие EGFR дикого типа клетки. Важно отметить, что эти данные вместе с данными, касающимися уничтожения и пролиферации, указывают на то, что CAR6 и CAR9 демонстрируют функциональную специфичность в отношении EGFRvIII и обладают способностью к содействию противоопухолевому иммунному ответу.
Пример 9: Экспрессирующие CAR на основе гуманизированного антитела против EGFRvIII Т-клетки уменьшают массу опухоли у мышей
Было установлено, что экспрессирующие CAR на основе гуманизированного антитела против EGFRvIII Т-клетки уменьшают массу опухоли in vivo у мышей. Например, трансдуцированные лентивирусным вектором, кодирующим #2173 (CAR6) - химерный антигенспецифический рецептор (CAR) на основе гуманизированного антитела против EGFRvIII, Т-лимфоциты человека доставляли внутривенно ксеногенным, с ослабленным иммунитетом NOD/SCID/common-gamma chain-/- мышам, получившим установившиеся опухоли глиомы U87vIII in vivo. В случае контрольных мышей с установившимися к дню 5 подкожными боковыми опухолями U87vIII, получающих подобранные в соответствии с донором Т-клетки, трансдуцированные не CAR, опухоли росли быстро, и в соответствии с прямым измерением подкожной опухоли, используя циркуль (максимальная ширина × максимальную длина), и в соответствии с эмиссией фотонов, измеряемой с помощью Spectrum в in vivo системе визуализации (IVIS). В случае подвергнутых лечению мышей, получающих даже небольшое количество (0,5-1×106) CAR6-трансдуцированных клеток, рост опухоли был заметно сниженным у мышей в зависимости от дозы.
В этом примере 1×106 клеток глиомы U87vIII человека, экспрессирующих EGFRvIII, GFP+Luc+ промывали и вводили подкожно в 100 мкл физиологического раствора в бока 30 мышей с ослабленным иммунитетом NSG (N=10/группа). Т-клетки человека стимулировали, используя покрытые антителом против CD3/28 сферы, и трансдуцировали с использованием лентивирусного вектора гуманизированным scFv #2173 против EGFRvIII CAR (CAR6). После трансдукции, ex vivo размножения и удаление сфер, CAR-трансдуцированные Т-клетки (~50% CAR+ в соответствии с проточной цитометрией) промывали и вводили в 100 мкл физиологического раствора в хвостовую вену через 5 дней после имплантации опухоли. Рост опухоли оценивали путем измерения с помощью циркуля (вверху слева) и люциферин-индуцированной эмиссии фотонов (справа вверху). Измерения начинали через 7 дней после переноса Т-клеток и через 12 дней после инъекции опухоли. SEM (сканирующая электронная микроскопия) представлена на фиг. 18 (N=10 мышей/группу). Выживание каждой группы наносили на график - кривые Каплана Мейера на фиг. 18 (ниже). Все мыши, получающие контрольные Т-клетки, умерли к дню 26, при этом выживаемость для группы, получающей 0,5×106 и 1,0×106 CAR6 Т-клеток, составляла 30% и 90%, соответственно, на дату - экспериментальный день 30.
ЭКВИВАЛЕНТЫ
Описания каждого и любого патента, заявки на патент и публикации, приведенного(ой) здесь, таким образом включены сюда посредством ссылки в их полном объеме. Хотя это изобретение было описано со ссылкой на конкретные аспекты, очевидно, что другие аспекты и вариации этого изобретения могут быть разработаны квалифицированными в данной области техники специалистами без отступа от фактического существа и объема настоящего изобретения. Прилагаемая формула изобретения, как предполагается, включает все такие варианты осуществления и эквивалентные вариации.
--->
Список последовательностей
<110> NOVARTIS AG
TRUSTEES OF THE UNIVERSITY OF PENNSYLVANIA
UNIVERSITY OF PITTSBURGH - OF THE COMMONWEALTH SYSTEM
OF HIGHER EDUCATION
<120> Лечение рака с использованием химерного антигенспецифического
рецептора на основе гуманизированного антитела против EGFRvIII
<130> N2067-7000WO
<140>
<141>
<150> 61/888,255
<151> 2013-10-08
<150> 61/767,071
<151> 2013-02-20
<160> 127
<170> Патент в версии 3.5
<210> 1
<211> 535
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 1
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Gly Ser Glu Ile Gln Leu Gln Gln Ser Gly Ala
20 25 30
Glu Leu Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Thr Gly Ser
35 40 45
Gly Phe Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Lys Gln Arg Thr
50 55 60
Glu Gln Gly Leu Glu Trp Ile Gly Arg Ile Asp Pro Glu Asn Asp Glu
65 70 75 80
Thr Lys Tyr Gly Pro Ile Phe Gln Gly Arg Ala Thr Ile Thr Ala Asp
85 90 95
Thr Ser Ser Asn Thr Val Tyr Leu Gln Leu Ser Ser Leu Thr Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly
115 120 125
Pro Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser His Met Asp Val Val Met Thr Gln
145 150 155 160
Ser Pro Leu Thr Leu Ser Val Ala Ile Gly Gln Ser Ala Ser Ile Ser
165 170 175
Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu
180 185 190
Asn Trp Leu Leu Gln Arg Pro Gly Gln Ser Pro Lys Arg Leu Ile Ser
195 200 205
Leu Val Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Thr Gly Ser
210 215 220
Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser Arg Val Glu Ala Glu
225 230 235 240
Asp Leu Gly Ile Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr
245 250 255
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Ala Ser Thr Thr Thr Pro
260 265 270
Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu
275 280 285
Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His
290 295 300
Thr Arg Gly Leu Asp Phe Ala Cys Asp Phe Trp Val Leu Val Val Val
305 310 315 320
Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile
325 330 335
Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr
340 345 350
Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln
355 360 365
Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Lys Arg Gly
370 375 380
Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val
385 390 395 400
Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu
405 410 415
Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg Ser Ala Asp
420 425 430
Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
435 440 445
Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg
450 455 460
Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly
465 470 475 480
Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu
485 490 495
Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu
500 505 510
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His
515 520 525
Met Gln Ala Leu Pro Pro Arg
530 535
<210> 2
<211> 491
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 2
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Gly Ser Glu Ile Gln Leu Gln Gln Ser Gly Ala
20 25 30
Glu Leu Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Thr Gly Ser
35 40 45
Gly Phe Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Lys Gln Arg Thr
50 55 60
Glu Gln Gly Leu Glu Trp Ile Gly Arg Ile Asp Pro Glu Asn Asp Glu
65 70 75 80
Thr Lys Tyr Gly Pro Ile Phe Gln Gly Arg Ala Thr Ile Thr Ala Asp
85 90 95
Thr Ser Ser Asn Thr Val Tyr Leu Gln Leu Ser Ser Leu Thr Ser Glu
100 105 110
Asp Thr Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly
115 120 125
Pro Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser His Met Asp Val Val Met Thr Gln
145 150 155 160
Ser Pro Leu Thr Leu Ser Val Ala Ile Gly Gln Ser Ala Ser Ile Ser
165 170 175
Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu
180 185 190
Asn Trp Leu Leu Gln Arg Pro Gly Gln Ser Pro Lys Arg Leu Ile Ser
195 200 205
Leu Val Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Thr Gly Ser
210 215 220
Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser Arg Val Glu Ala Glu
225 230 235 240
Asp Leu Gly Ile Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr
245 250 255
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Ala Ser Thr Thr Thr Pro
260 265 270
Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu
275 280 285
Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His
290 295 300
Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu
305 310 315 320
Ala Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr
325 330 335
Cys Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe
340 345 350
Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg
355 360 365
Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser
370 375 380
Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr
385 390 395 400
Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys
405 410 415
Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn
420 425 430
Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu
435 440 445
Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly
450 455 460
His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr
465 470 475 480
Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
485 490
<210> 3
<211> 488
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 3
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser
20 25 30
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala
35 40 45
Ser Gln Gly Ile Arg Asn Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly
50 55 60
Lys Ala Pro Lys Arg Leu Ile Tyr Ala Ala Ser Asn Leu Gln Ser Gly
65 70 75 80
Val Pro Ser Arg Phe Thr Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu
85 90 95
Ile Val Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu
100 105 110
Gln His His Ser Tyr Pro Leu Thr Ser Gly Gly Gly Thr Lys Val Glu
115 120 125
Ile Lys Arg Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly
130 135 140
Glu Gly Ser Glu Val Gln Val Leu Glu Ser Gly Gly Gly Leu Val Gln
145 150 155 160
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
165 170 175
Ser Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
180 185 190
Glu Trp Val Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Asn Tyr Ala
195 200 205
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
210 215 220
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
225 230 235 240
Tyr Tyr Cys Ala Gly Ser Ser Gly Trp Ser Glu Tyr Trp Gly Gln Gly
245 250 255
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Thr Thr Pro Ala Pro Arg
260 265 270
Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg
275 280 285
Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly
290 295 300
Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr
305 310 315 320
Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Lys Arg
325 330 335
Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro
340 345 350
Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu
355 360 365
Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg Ser Ala
370 375 380
Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu
385 390 395 400
Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly
405 410 415
Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu
420 425 430
Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser
435 440 445
Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly
450 455 460
Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu
465 470 475 480
His Met Gln Ala Leu Pro Pro Arg
485
<210> 4
<211> 729
<212> ДНК
<213> Mus sp.
<400> 4
gagattcagc tgcagcaatc tggggcagaa cttgtgaagc caggggcctc agtcaagctg 60
tcctgcacag gttctggctt caacattgaa gactactata ttcactgggt gaagcagagg 120
actgaacagg gcctggaatg gattggaagg attgatcctg agaatgatga aactaaatat 180
ggcccaatat tccagggcag ggccactata acagcagaca catcctccaa cacagtctac 240
ctgcaactca gcagcctgac atctgaggac actgccgtct attactgtgc ctttcgcggt 300
ggagtctact gggggccagg aaccactctc acagtctcct caggaggtgg tggttccggt 360
ggtggtggtt ccggaggtgg tggttcacat atggatgttg tgatgaccca gtctccactc 420
actctatcgg ttgccattgg acaatcagcc tccatctctt gcaagtcaag tcagagcctc 480
ttagatagtg atggaaagac atatttgaat tggttgttac agaggccagg ccagtctcca 540
aagcgcctaa tctctctggt gtctaaactg gactctggag tccctgacag gttcactggc 600
agtggatcag ggacagattt cacactgaga atcagcagag tggaggctga ggatttggga 660
atttattatt gctggcaagg tacacatttt cctgggacgt tcggtggagg gaccaagctg 720
gagataaaa 729
<210> 5
<211> 720
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 5
gacatccaga tgacccagag ccctagcagc ctgagcgcca gcgtgggcga cagagtgacc 60
atcacctgtc gggccagcca gggcatcaga aacaacctgg cctggtatca gcagaagccc 120
ggcaaggccc ccaagagact gatctacgct gccagcaatc tgcagagcgg cgtgcccagc 180
agattcaccg gaagcggctc cggcaccgag ttcaccctga tcgtgtccag cctgcagccc 240
gaggacttcg ccacctacta ctgcctgcag caccacagct accctctgac cagcggcgga 300
ggcaccaagg tggagatcaa gcggaccggc agcaccagcg gcagcggcaa gcctggcagc 360
ggcgagggaa gcgaggtcca ggtgctggaa tctggcggcg gactggtgca gcctggcggc 420
agcctgagac tgagctgtgc cgccagcggc ttcaccttca gcagctacgc catgtcttgg 480
gtccggcagg ctcctggaaa gggcctggaa tgggtgtccg ccatcagcgg ctctggcggc 540
tccaccaact acgccgacag cgtgaagggc cggttcacca tcagccggga caacagcaag 600
aacaccctgt atctgcagat gaacagcctg agagccgagg acaccgccgt gtactactgt 660
gccggcagca gcgggtggag cgagtactgg ggccagggca cactggtcac agtgtctagc 720
<210> 6
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
олигонуклеотид»
<400> 6
atggccttac cagtgaccgc cttgctcctg ccgctggcct tgctgctcca cgccgccagg 60
ccg 63
<210> 7
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 7
accacgacgc cagcgccgcg accaccaaca ccggcgccca ccatcgcgtc gcagcccctg 60
tccctgcgcc cagaggcgtg ccggccagcg gcggggggcg cagtgcacac gagggggctg 120
gacttcgcct gtgat 135
<210> 8
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
олигонуклеотид»
<400> 8
atctacatct gggcgccctt ggccgggact tgtggggtcc ttctcctgtc actggttatc 60
accctttact gc 72
<210> 9
<211> 126
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 9
aaacggggca gaaagaaact cctgtatata ttcaaacaac catttatgag accagtacaa 60
actactcaag aggaagatgg ctgtagctgc cgatttccag aagaagaaga aggaggatgt 120
gaactg 126
<210> 10
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 10
agagtgaagt tcagcaggag cgcagacgcc cccgcgtaca agcagggcca gaaccagctc 60
tataacgagc tcaatctagg acgaagagag gagtacgatg ttttggacaa gagacgtggc 120
cgggaccctg agatgggggg aaagccgaga aggaagaacc ctcaggaagg cctgtacaat 180
gaactgcaga aagataagat ggcggaggcc tacagtgaga ttgggatgaa aggcgagcgc 240
cggaggggca aggggcacga tggcctttac cagggtctca gtacagccac caaggacacc 300
tacgacgccc ttcacatgca ggccctgccc cctcgc 336
<210> 11
<211> 243
<212> БЕЛОК
<213> Mus sp.
<400> 11
Glu Ile Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Thr Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Lys Gln Arg Thr Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr Val Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Pro Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser His Met Asp Val Val Met Thr Gln Ser Pro Leu Thr Leu Ser Val
130 135 140
Ala Ile Gly Gln Ser Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu
145 150 155 160
Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro
165 170 175
Gly Gln Ser Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser
180 185 190
Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr
195 200 205
Leu Arg Ile Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys
210 215 220
Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Leu
225 230 235 240
Glu Ile Lys
<210> 12
<211> 240
<212> БЕЛОК
<213> Homo sapiens
<400> 12
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Ile Val Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His His Ser Tyr Pro Leu
85 90 95
Thr Ser Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Gly Ser Thr
100 105 110
Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser Glu Val Gln Val
115 120 125
Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile Ser
165 170 175
Gly Ser Gly Gly Ser Thr Asn Tyr Ala Asp Ser Val Lys Gly Arg Phe
180 185 190
Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Gly Ser Ser
210 215 220
Gly Trp Ser Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
225 230 235 240
<210> 13
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 13
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro
20
<210> 14
<211> 45
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 14
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp
35 40 45
<210> 15
<211> 24
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 15
Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu
1 5 10 15
Ser Leu Val Ile Thr Leu Tyr Cys
20
<210> 16
<211> 42
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 16
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
35 40
<210> 17
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 17
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 18
<211> 1605
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 18
atggccttac cagtgaccgc cttgctcctg ccgctggcct tgctgctcca cgccgccagg 60
ccgggatccg agattcagct gcagcaatct ggggcagaac ttgtgaagcc aggggcctca 120
gtcaagctgt cctgcacagg ttctggcttc aacattgaag actactatat tcactgggtg 180
aagcagagga ctgaacaggg cctggaatgg attggaagga ttgatcctga gaatgatgaa 240
actaaatatg gcccaatatt ccagggcagg gccactataa cagcagacac atcctccaac 300
acagtctacc tgcaactcag cagcctgaca tctgaggaca ctgccgtcta ttactgtgcc 360
tttcgcggtg gagtctactg ggggccagga accactctca cagtctcctc aggaggtggt 420
ggttccggtg gtggtggttc cggaggtggt ggttcacata tggatgttgt gatgacccag 480
tctccactca ctctatcggt tgccattgga caatcagcct ccatctcttg caagtcaagt 540
cagagcctct tagatagtga tggaaagaca tatttgaatt ggttgttaca gaggccaggc 600
cagtctccaa agcgcctaat ctctctggtg tctaaactgg actctggagt ccctgacagg 660
ttcactggca gtggatcagg gacagatttc acactgagaa tcagcagagt ggaggctgag 720
gatttgggaa tttattattg ctggcaaggt acacattttc ctgggacgtt cggtggaggg 780
accaagctgg agataaaagc tagcaccacg acgccagcgc cgcgaccacc aacaccggcg 840
cccaccatcg cgtcgcagcc cctgtccctg cgcccagagg cgtgccggcc agcggcgggg 900
ggcgcagtgc acacgagggg gctggacttc gcctgtgatt tttgggtgct ggtggtggtt 960
ggtggagtcc tggcttgcta tagcttgcta gtaacagtgg cctttattat tttctgggtg 1020
aggagtaaga ggagcaggct cctgcacagt gactacatga acatgactcc ccgccgcccc 1080
gggcccaccc gcaagcatta ccagccctat gccccaccac gcgacttcgc agcctatcgc 1140
tccaaacggg gcagaaagaa actcctgtat atattcaaac aaccatttat gagaccagta 1200
caaactactc aagaggaaga tggctgtagc tgccgatttc cagaagaaga agaaggagga 1260
tgtgaactga gagtgaagtt cagcaggagc gcagacgccc ccgcgtacaa gcagggccag 1320
aaccagctct ataacgagct caatctagga cgaagagagg agtacgatgt tttggacaag 1380
agacgtggcc gggaccctga gatgggggga aagccgagaa ggaagaaccc tcaggaaggc 1440
ctgtacaatg aactgcagaa agataagatg gcggaggcct acagtgagat tgggatgaaa 1500
ggcgagcgcc ggaggggcaa ggggcacgat ggcctttacc agggtctcag tacagccacc 1560
aaggacacct acgacgccct tcacatgcag gccctgcccc ctcgc 1605
<210> 19
<211> 1473
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 19
atggccttac cagtgaccgc cttgctcctg ccgctggcct tgctgctcca cgccgccagg 60
ccgggatccg agattcagct gcagcaatct ggggcagaac ttgtgaagcc aggggcctca 120
gtcaagctgt cctgcacagg ttctggcttc aacattgaag actactatat tcactgggtg 180
aagcagagga ctgaacaggg cctggaatgg attggaagga ttgatcctga gaatgatgaa 240
actaaatatg gcccaatatt ccagggcagg gccactataa cagcagacac atcctccaac 300
acagtctacc tgcaactcag cagcctgaca tctgaggaca ctgccgtcta ttactgtgcc 360
tttcgcggtg gagtctactg ggggccagga accactctca cagtctcctc aggaggtggt 420
ggttccggtg gtggtggttc cggaggtggt ggttcacata tggatgttgt gatgacccag 480
tctccactca ctctatcggt tgccattgga caatcagcct ccatctcttg caagtcaagt 540
cagagcctct tagatagtga tggaaagaca tatttgaatt ggttgttaca gaggccaggc 600
cagtctccaa agcgcctaat ctctctggtg tctaaactgg actctggagt ccctgacagg 660
ttcactggca gtggatcagg gacagatttc acactgagaa tcagcagagt ggaggctgag 720
gatttgggaa tttattattg ctggcaaggt acacattttc ctgggacgtt cggtggaggg 780
accaagctgg agataaaagc tagcaccacg acgccagcgc cgcgaccacc aacaccggcg 840
cccaccatcg cgtcgcagcc cctgtccctg cgcccagagg cgtgccggcc agcggcgggg 900
ggcgcagtgc acacgagggg gctggacttc gcctgtgata tctacatctg ggcgcccttg 960
gccgggactt gtggggtcct tctcctgtca ctggttatca ccctttactg caaacggggc 1020
agaaagaaac tcctgtatat attcaaacaa ccatttatga gaccagtaca aactactcaa 1080
gaggaagatg gctgtagctg ccgatttcca gaagaagaag aaggaggatg tgaactgaga 1140
gtgaagttca gcaggagcgc agacgccccc gcgtacaagc agggccagaa ccagctctat 1200
aacgagctca atctaggacg aagagaggag tacgatgttt tggacaagag acgtggccgg 1260
gaccctgaga tggggggaaa gccgagaagg aagaaccctc aggaaggcct gtacaatgaa 1320
ctgcagaaag ataagatggc ggaggcctac agtgagattg ggatgaaagg cgagcgccgg 1380
aggggcaagg ggcacgatgg cctttaccag ggtctcagta cagccaccaa ggacacctac 1440
gacgcccttc acatgcaggc cctgccccct cgc 1473
<210> 20
<211> 1465
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 20
atggccttac cagtgaccgc cttgctcctg ccgctggcct tgctgctcca cgccgccagg 60
ccgggatccg acatccagat gacccagagc cctagcagcc tgagcgccag cgtgggcgac 120
agagtgacca tcacctgtcg ggccagccag ggcatcagaa acaacctggc ctggtatcag 180
cagaagcccg gcaaggcccc caagagactg atctacgctg ccagcaatct gcagagcggc 240
gtgcccagca gattcaccgg aagcggctcc ggcaccgagt tcaccctgat cgtgtccagc 300
ctgcagcccg aggacttcgc cacctactac tgcctgcagc accacagcta ccctctgacc 360
agcggcggag gcaccaaggt ggagatcaag cggaccggca gcaccagcgg cagcggcaag 420
cctggcagcg gcgagggaag cgaggtccag gtgctggaat ctggcggcgg actggtgcag 480
cctggcggca gcctgagact gagctgtgcc gccagcggct tcaccttcag cagctacgcc 540
atgtcttggg tccggcaggc tcctggaaag ggcctggaat gggtgtccgc catcagcggc 600
tctggcggct ccaccaacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 660
aacagcaaga acaccctgta tctgcagatg aacagcctga gagccgagga caccgccgtg 720
tactactgtg ccggcagcag cgggtggagc gagtactggg gccagggcac actggtcaca 780
gtgtctagcg ctagcaccac gacgccagcg ccgcgaccac caacaccggc gcccaccatc 840
gcgtcgcagc ccctgtccct gcgcccagag gcgtgccggc cagcggcggg gggcgcagtg 900
cacacgaggg ggctggactt cgcctgtgat atctacatct gggcgccctt ggccgggact 960
tgtggggtcc ttctcctgtc actggttatc accctttact gcaaacgggg cagaaagaaa 1020
ctcctgtata tattcaaaca accatttatg agaccagtac aaactactca agaggaagat 1080
ggctgtagct gccgatttcc agaagaagaa gaaggaggat gtgaactgag agtgaagttc 1140
agcaggagcg cagacgcccc cgcgtacaag cagggccaga accagctcta taacgagctc 1200
aatctaggac gaagagagga gtacgatgtt ttggacaaga gacgtggccg ggaccctgag 1260
atggggggaa agccgagaag gaagaaccct caggaaggcc tgtacaatga actgcagaaa 1320
gataagatgg cggaggccta cagtgagatt gggatgaaag gcgagcgccg gaggggcaag 1380
gggcacgatg gcctttacca gggtctcagt acagccacca aggacaccta cgacgccctt 1440
cacatgcagg ccctgccccc tcgct 1465
<210> 21
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 21
Glu Ile Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Thr Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Lys Gln Arg Thr Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr Val Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Pro Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser
<210> 22
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 22
Asp Tyr Tyr Ile His
1 5
<210> 23
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 23
Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe Gln
1 5 10 15
Gly
<210> 24
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 24
Arg Gly Gly Val Tyr
1 5
<210> 25
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 25
Asp Val Val Met Thr Gln Ser Pro Leu Thr Leu Ser Val Ala Ile Gly
1 5 10 15
Gln Ser Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 26
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 26
Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn
1 5 10 15
<210> 27
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 27
Leu Val Ser Lys Leu Asp Ser
1 5
<210> 28
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 28
Trp Gln Gly Thr His Phe Pro Gly Thr
1 5
<210> 29
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 29
Glu Ile Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Thr Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Lys Gln Arg Thr Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr Val Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Pro Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser
<210> 30
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 30
Gly Phe Asn Ile Glu Asp Tyr
1 5
<210> 31
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 31
Asp Pro Glu Asn Asp Glu
1 5
<210> 32
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 32
Arg Gly Gly Val Tyr
1 5
<210> 33
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 33
Asp Val Val Met Thr Gln Ser Pro Leu Thr Leu Ser Val Ala Ile Gly
1 5 10 15
Gln Ser Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 34
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 34
Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
1 5 10
<210> 35
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 35
Leu Val Ser
1
<210> 36
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 36
Gly Thr His Phe Pro Gly
1 5
<210> 37
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 37
Gly Gly Gly Gly Ser
1 5
<210> 38
<211> 246
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 38
Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asn Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr Gln Ser Pro Asp Ser
130 135 140
Leu Ala Val Ser Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser
145 150 155 160
Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln
165 170 175
Gln Lys Pro Gly Gln Pro Pro Lys Arg Leu Ile Ser Leu Val Ser Lys
180 185 190
Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr
195 200 205
Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val
210 215 220
Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly
225 230 235 240
Thr Lys Val Glu Ile Lys
245
<210> 39
<211> 738
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 39
gaaatccagc tggtccaatc gggagctgag gtcaagaagc cgggagccac cgtcaagatc 60
tcatgcaagg ggtcgggatt caacatcgag gactactaca ttcactgggt gcagcaagct 120
ccgggaaaag gcctggaatg gatgggcaga atcgacccag aaaacgacga aactaagtac 180
ggaccgattt tccaaggaag agtgactatc accgccgata cttcaaccaa taccgtctac 240
atggaactga gctcgctccg gtccgaagat actgcagtgt attactgtgc ctttcgcgga 300
ggggtgtact ggggccaagg aactactgtc actgtctcgt caggaggcgg agggtcggga 360
ggaggcggga gcggaggcgg tggctcgggt ggcggaggaa gcgacgtggt gatgacccag 420
tccccggact ccctcgccgt gagcctcgga gagagggcga ctatcaattg caagtcgtcc 480
cagtcacttc tggattccga tggtaaaacg tacctcaact ggctgcagca aaagccaggg 540
cagccaccca aacggttgat ctcccttgtg tccaaactgg atagcggagt gcctgaccgc 600
ttctcgggtt ccggtagcgg gaccgacttc accctgacga tcagctcact gcaggcggag 660
gacgtggcag tgtactactg ctggcaggga acccacttcc ctggcacctt tggaggtggc 720
accaaggtgg agatcaag 738
<210> 40
<211> 831
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 40
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgaaatcc agctggtcca atcgggagct gaggtcaaga agccgggagc caccgtcaag 120
atctcatgca aggggtcggg attcaacatc gaggactact acattcactg ggtgcagcaa 180
gctccgggaa aaggcctgga atggatgggc agaatcgacc cagaaaacga cgaaactaag 240
tacggaccga ttttccaagg aagagtgact atcaccgccg atacttcaac caataccgtc 300
tacatggaac tgagctcgct ccggtccgaa gatactgcag tgtattactg tgcctttcgc 360
ggaggggtgt actggggcca aggaactact gtcactgtct cgtcaggagg cggagggtcg 420
ggaggaggcg ggagcggagg cggtggctcg ggtggcggag gaagcgacgt ggtgatgacc 480
cagtccccgg actccctcgc cgtgagcctc ggagagaggg cgactatcaa ttgcaagtcg 540
tcccagtcac ttctggattc cgatggtaaa acgtacctca actggctgca gcaaaagcca 600
gggcagccac ccaaacggtt gatctccctt gtgtccaaac tggatagcgg agtgcctgac 660
cgcttctcgg gttccggtag cgggaccgac ttcaccctga cgatcagctc actgcaggcg 720
gaggacgtgg cagtgtacta ctgctggcag ggaacccact tccctggcac ctttggaggt 780
ggcaccaagg tggagatcaa gggatcgcac caccatcacc atcatcatca c 831
<210> 41
<211> 277
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 41
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val
20 25 30
Lys Lys Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Gly Ser Gly Phe
35 40 45
Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Gln Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys
65 70 75 80
Tyr Gly Pro Ile Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser
85 90 95
Thr Asn Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr
100 105 110
Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly
115 120 125
Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr
145 150 155 160
Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly Glu Arg Ala Thr Ile
165 170 175
Asn Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
180 185 190
Leu Asn Trp Leu Gln Gln Lys Pro Gly Gln Pro Pro Lys Arg Leu Ile
195 200 205
Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly
210 215 220
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
225 230 235 240
Glu Asp Val Ala Val Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly
245 250 255
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Ser His His His
260 265 270
His His His His His
275
<210> 42
<211> 1470
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 42
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgagatcc agctggtgca gtcgggagct gaagtcaaaa agcctggcgc aaccgtcaag 120
atctcgtgca aaggatcagg gttcaacatc gaggactact acatccattg ggtgcaacag 180
gcacccggaa aaggcctgga gtggatgggg aggattgacc cagaaaatga cgaaaccaag 240
tacggaccga tcttccaagg acgggtgacc atcacggctg acacttccac taacaccgtc 300
tacatggaac tctcgagcct tcgctcggaa gataccgcgg tgtactactg cgcctttaga 360
ggtggagtct actggggaca agggactacc gtcaccgtgt cgtcaggtgg cggaggatca 420
ggcggaggcg gctccggtgg aggaggaagc ggaggaggtg gctccgacgt ggtgatgacg 480
cagtcaccgg actccttggc ggtgagcctg ggtgaacgcg ccactatcaa ctgcaagagc 540
tcccagagct tgctggactc cgatggaaag acttatctca attggctgca acagaagcct 600
ggccagccgc caaagagact catctcactg gtgagcaagc tggatagcgg agtgccagat 660
cggttttcgg gatcgggctc aggcaccgac ttcaccctga ctatttcctc cctccaagcc 720
gaggatgtgg ccgtctacta ctgttggcag gggactcact tcccggggac cttcggtgga 780
ggcactaagg tggagatcaa aaccactacc ccagcaccga ggccacccac cccggctcct 840
accatcgcct cccagcctct gtccctgcgt ccggaggcat gtagacccgc agctggtggg 900
gccgtgcata cccggggtct tgacttcgcc tgcgatatct acatttgggc ccctctggct 960
ggtacttgcg gggtcctgct gctttcactc gtgatcactc tttactgtaa gcgcggtcgg 1020
aagaagctgc tgtacatctt taagcaaccc ttcatgaggc ctgtgcagac tactcaagag 1080
gaggacggct gttcatgccg gttcccagag gaggaggaag gcggctgcga actgcgcgtg 1140
aaattcagcc gcagcgcaga tgctccagcc tacaagcagg ggcagaacca gctctacaac 1200
gaactcaatc ttggtcggag agaggagtac gacgtgctgg acaagcggag aggacgggac 1260
ccagaaatgg gcgggaagcc gcgcagaaag aatccccaag agggcctgta caacgagctc 1320
caaaaggata agatggcaga agcctatagc gagattggta tgaaagggga acgcagaaga 1380
ggcaaaggcc acgacggact gtaccaggga ctcagcaccg ccaccaagga cacctatgac 1440
gctcttcaca tgcaggccct gccgcctcgg 1470
<210> 43
<211> 490
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 43
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val
20 25 30
Lys Lys Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Gly Ser Gly Phe
35 40 45
Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Gln Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys
65 70 75 80
Tyr Gly Pro Ile Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser
85 90 95
Thr Asn Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr
100 105 110
Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly
115 120 125
Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr
145 150 155 160
Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly Glu Arg Ala Thr Ile
165 170 175
Asn Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
180 185 190
Leu Asn Trp Leu Gln Gln Lys Pro Gly Gln Pro Pro Lys Arg Leu Ile
195 200 205
Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly
210 215 220
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
225 230 235 240
Glu Asp Val Ala Val Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly
245 250 255
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
420 425 430
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
435 440 445
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
450 455 460
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
465 470 475 480
Ala Leu His Met Gln Ala Leu Pro Pro Arg
485 490
<210> 44
<211> 246
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 44
Asp Val Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln Lys Pro Gly Gln Pro
35 40 45
Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys
130 135 140
Lys Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Gly Ser Gly Phe Asn
145 150 155 160
Ile Glu Asp Tyr Tyr Ile His Trp Val Gln Gln Ala Pro Gly Lys Gly
165 170 175
Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr
180 185 190
Gly Pro Ile Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr
195 200 205
Asn Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala
210 215 220
Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly Thr
225 230 235 240
Thr Val Thr Val Ser Ser
245
<210> 45
<211> 738
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 45
gatgtcgtga tgacccagtc cccagactcc ctcgcagtgt ccttgggaga acgggccacc 60
atcaactgca aatcgagcca gtcactgctg gactcagacg gaaagaccta cctcaactgg 120
ctgcagcaga agcctggcca gccaccgaag cgcctgatct ccctggtgtc caagctggac 180
tcgggcgtcc cggacaggtt tagcggtagc ggctcgggaa ccgacttcac tctgaccatt 240
agctcgctcc aagctgaaga tgtggcggtc tactactgct ggcaggggac ccacttcccc 300
gggacctttg gcggaggaac taaagtcgaa atcaaaggag gaggcggatc aggtggagga 360
ggcagcggag gaggagggag cggcggtggc ggctccgaaa ttcaacttgt gcaatccggt 420
gccgaggtga agaaacctgg tgccactgtc aagatctcgt gtaagggatc gggattcaat 480
atcgaggact actacatcca ctgggtgcaa caggcgccag gaaagggatt ggagtggatg 540
ggtcgcatcg acccggaaaa cgatgagact aagtacggac cgatcttcca aggccgggtc 600
acgatcactg cggatacctc cactaatacc gtgtatatgg agctctcgtc actgagaagc 660
gaagatacgg ccgtgtacta ctgcgcattc agaggaggtg tgtactgggg ccagggaact 720
actgtgaccg tgtcgtcg 738
<210> 46
<211> 831
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 46
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgatgtcg tgatgaccca gtccccagac tccctcgcag tgtccttggg agaacgggcc 120
accatcaact gcaaatcgag ccagtcactg ctggactcag acggaaagac ctacctcaac 180
tggctgcagc agaagcctgg ccagccaccg aagcgcctga tctccctggt gtccaagctg 240
gactcgggcg tcccggacag gtttagcggt agcggctcgg gaaccgactt cactctgacc 300
attagctcgc tccaagctga agatgtggcg gtctactact gctggcaggg gacccacttc 360
cccgggacct ttggcggagg aactaaagtc gaaatcaaag gaggaggcgg atcaggtgga 420
ggaggcagcg gaggaggagg gagcggcggt ggcggctccg aaattcaact tgtgcaatcc 480
ggtgccgagg tgaagaaacc tggtgccact gtcaagatct cgtgtaaggg atcgggattc 540
aatatcgagg actactacat ccactgggtg caacaggcgc caggaaaggg attggagtgg 600
atgggtcgca tcgacccgga aaacgatgag actaagtacg gaccgatctt ccaaggccgg 660
gtcacgatca ctgcggatac ctccactaat accgtgtata tggagctctc gtcactgaga 720
agcgaagata cggccgtgta ctactgcgca ttcagaggag gtgtgtactg gggccaggga 780
actactgtga ccgtgtcgtc ggggtcacat caccaccatc atcatcacca c 831
<210> 47
<211> 277
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 47
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Val Val Met Thr Gln Ser Pro Asp Ser Leu
20 25 30
Ala Val Ser Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln
35 40 45
Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln
50 55 60
Lys Pro Gly Gln Pro Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu
65 70 75 80
Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr
100 105 110
Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr
115 120 125
Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser
145 150 155 160
Gly Ala Glu Val Lys Lys Pro Gly Ala Thr Val Lys Ile Ser Cys Lys
165 170 175
Gly Ser Gly Phe Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Gln Gln
180 185 190
Ala Pro Gly Lys Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn
195 200 205
Asp Glu Thr Lys Tyr Gly Pro Ile Phe Gln Gly Arg Val Thr Ile Thr
210 215 220
Ala Asp Thr Ser Thr Asn Thr Val Tyr Met Glu Leu Ser Ser Leu Arg
225 230 235 240
Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr
245 250 255
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Ser His His His
260 265 270
His His His His His
275
<210> 48
<211> 1470
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 48
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgacgtgg tcatgactca aagcccagat tccttggctg tctcccttgg agaaagagca 120
acgatcaatt gcaaaagctc gcagtccctg ttggactccg atggaaaaac ctacctcaac 180
tggctgcagc agaagccggg acaaccacca aagcggctga tttccctcgt gtccaagctg 240
gacagcggcg tgccggatcg cttctcgggc agcggctcgg gaaccgattt tactctcact 300
atttcgtcac tgcaagcgga ggacgtggcg gtgtattact gctggcaggg cactcacttc 360
ccgggtactt ttggtggagg taccaaagtc gaaatcaagg gtggaggcgg gagcggagga 420
ggcgggtcgg gaggaggagg atcgggtggc ggaggctcag aaatccagct ggtgcagtca 480
ggtgccgaag tgaagaagcc tggggccacg gtgaagatct cgtgcaaggg gagcggattc 540
aacatcgagg attactacat ccattgggtg caacaggccc ctggcaaagg gctggaatgg 600
atgggaagga tcgaccccga gaatgacgag actaagtacg gcccgatctt ccaaggacgg 660
gtgaccatca ctgcagacac ttcaaccaac accgtctaca tggaactctc ctcgctgcgc 720
tccgaggaca ccgccgtgta ctactgtgct ttcagaggag gagtctactg gggacaggga 780
acgaccgtga ccgtcagctc aaccactacc ccagcaccga ggccacccac cccggctcct 840
accatcgcct cccagcctct gtccctgcgt ccggaggcat gtagacccgc agctggtggg 900
gccgtgcata cccggggtct tgacttcgcc tgcgatatct acatttgggc ccctctggct 960
ggtacttgcg gggtcctgct gctttcactc gtgatcactc tttactgtaa gcgcggtcgg 1020
aagaagctgc tgtacatctt taagcaaccc ttcatgaggc ctgtgcagac tactcaagag 1080
gaggacggct gttcatgccg gttcccagag gaggaggaag gcggctgcga actgcgcgtg 1140
aaattcagcc gcagcgcaga tgctccagcc tacaagcagg ggcagaacca gctctacaac 1200
gaactcaatc ttggtcggag agaggagtac gacgtgctgg acaagcggag aggacgggac 1260
ccagaaatgg gcgggaagcc gcgcagaaag aatccccaag agggcctgta caacgagctc 1320
caaaaggata agatggcaga agcctatagc gagattggta tgaaagggga acgcagaaga 1380
ggcaaaggcc acgacggact gtaccaggga ctcagcaccg ccaccaagga cacctatgac 1440
gctcttcaca tgcaggccct gccgcctcgg 1470
<210> 49
<211> 490
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 49
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Val Val Met Thr Gln Ser Pro Asp Ser Leu
20 25 30
Ala Val Ser Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln
35 40 45
Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln
50 55 60
Lys Pro Gly Gln Pro Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu
65 70 75 80
Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr
100 105 110
Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr
115 120 125
Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser
145 150 155 160
Gly Ala Glu Val Lys Lys Pro Gly Ala Thr Val Lys Ile Ser Cys Lys
165 170 175
Gly Ser Gly Phe Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Gln Gln
180 185 190
Ala Pro Gly Lys Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn
195 200 205
Asp Glu Thr Lys Tyr Gly Pro Ile Phe Gln Gly Arg Val Thr Ile Thr
210 215 220
Ala Asp Thr Ser Thr Asn Thr Val Tyr Met Glu Leu Ser Ser Leu Arg
225 230 235 240
Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr
245 250 255
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
420 425 430
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
435 440 445
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
450 455 460
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
465 470 475 480
Ala Leu His Met Gln Ala Leu Pro Pro Arg
485 490
<210> 50
<211> 246
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 50
Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly His Val Thr Ile Ser Ala Asp Thr Ser Ile Asn Thr Val Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr Gln Ser Pro Leu Ser
130 135 140
Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser
145 150 155 160
Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln
165 170 175
Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile Ser Leu Val Ser Lys
180 185 190
Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr
195 200 205
Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val
210 215 220
Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly
225 230 235 240
Thr Lys Val Glu Ile Lys
245
<210> 51
<211> 738
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 51
gagattcagc tggtccaaag cggcgcagaa gtgaaaaagc caggggaatc gttgcgcatc 60
agctgtaaag gttccggctt caacatcgag gactattaca tccattgggt gcggcagatg 120
ccaggaaagg ggctggaatg gatgggacgg attgacccgg agaacgacga aaccaagtac 180
ggaccgatct ttcaaggaca cgtgactatc tccgccgaca ccagcatcaa tacggtgtac 240
ctccaatggt cctcactcaa ggcctcggat accgcgatgt actactgcgc gttcagagga 300
ggcgtctact ggggacaagg gactactgtg actgtctcat caggaggtgg aggaagcgga 360
ggaggtggct cgggcggagg tggatcggga ggaggagggt ccgatgtggt gatgacccag 420
tccccactgt cgctcccggt gaccctcgga cagcctgcta gcatctcgtg caaatcctcg 480
caatccctgc tggactcgga cggaaaaacg tacctcaatt ggctgcagca gcgccctggc 540
cagagcccga gaaggcttat ctcgctggtg tcaaagctgg atagcggtgt gcccgaccgg 600
ttcagcggct cagggtcagg aaccgatttc accttgaaga tctcccgcgt ggaagccgaa 660
gatgtcggag tctactactg ctggcagggt actcacttcc cggggacctt tggtggcggc 720
actaaggtcg agattaag 738
<210> 52
<211> 831
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 52
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgagattc agctggtcca aagcggcgca gaagtgaaaa agccagggga atcgttgcgc 120
atcagctgta aaggttccgg cttcaacatc gaggactatt acatccattg ggtgcggcag 180
atgccaggaa aggggctgga atggatggga cggattgacc cggagaacga cgaaaccaag 240
tacggaccga tctttcaagg acacgtgact atctccgccg acaccagcat caatacggtg 300
tacctccaat ggtcctcact caaggcctcg gataccgcga tgtactactg cgcgttcaga 360
ggaggcgtct actggggaca agggactact gtgactgtct catcaggagg tggaggaagc 420
ggaggaggtg gctcgggcgg aggtggatcg ggaggaggag ggtccgatgt ggtgatgacc 480
cagtccccac tgtcgctccc ggtgaccctc ggacagcctg ctagcatctc gtgcaaatcc 540
tcgcaatccc tgctggactc ggacggaaaa acgtacctca attggctgca gcagcgccct 600
ggccagagcc cgagaaggct tatctcgctg gtgtcaaagc tggatagcgg tgtgcccgac 660
cggttcagcg gctcagggtc aggaaccgat ttcaccttga agatctcccg cgtggaagcc 720
gaagatgtcg gagtctacta ctgctggcag ggtactcact tcccggggac ctttggtggc 780
ggcactaagg tcgagattaa gggctcacac catcatcacc atcaccacca c 831
<210> 53
<211> 277
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 53
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val
20 25 30
Lys Lys Pro Gly Glu Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Phe
35 40 45
Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Arg Gln Met Pro Gly Lys
50 55 60
Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys
65 70 75 80
Tyr Gly Pro Ile Phe Gln Gly His Val Thr Ile Ser Ala Asp Thr Ser
85 90 95
Ile Asn Thr Val Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr
100 105 110
Ala Met Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly
115 120 125
Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr
145 150 155 160
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
165 170 175
Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
180 185 190
Leu Asn Trp Leu Gln Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile
195 200 205
Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly
210 215 220
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
225 230 235 240
Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly
245 250 255
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Ser His His His
260 265 270
His His His His His
275
<210> 54
<211> 1470
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 54
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgaaatcc agctggtgca aagcggagcc gaggtgaaga agcccggaga atccctgcgc 120
atctcgtgta agggttccgg ctttaacatc gaggattact acatccactg ggtgagacag 180
atgccgggca aaggtctgga atggatgggc cgcatcgacc cggagaacga cgaaaccaaa 240
tacggaccaa tcttccaagg acatgtgact atttccgcgg atacctccat caacactgtc 300
tacttgcagt ggagctcgct caaggcgtcg gataccgcca tgtactactg cgcattcaga 360
ggaggtgtgt actggggcca gggcactacg gtcaccgtgt cctcgggagg tggagggtca 420
ggaggcggag gctcgggcgg tggaggatca ggcggaggag gaagcgatgt ggtcatgact 480
caatccccac tgtcactgcc tgtcactctg gggcaaccgg cttccatctc atgcaagtca 540
agccaatcgc tgctcgactc cgacggaaaa acctacctca attggcttca gcagcgccca 600
ggccagtcgc ctcggaggct gatctcactc gtgtcgaagc ttgactccgg ggtgccggat 660
cggtttagcg gaagcggatc ggggaccgac ttcacgttga agattagccg ggtggaagcc 720
gaggacgtgg gagtctatta ctgctggcag gggacccact tcccggggac tttcggagga 780
ggcaccaaag tcgagattaa gaccactacc ccagcaccga ggccacccac cccggctcct 840
accatcgcct cccagcctct gtccctgcgt ccggaggcat gtagacccgc agctggtggg 900
gccgtgcata cccggggtct tgacttcgcc tgcgatatct acatttgggc ccctctggct 960
ggtacttgcg gggtcctgct gctttcactc gtgatcactc tttactgtaa gcgcggtcgg 1020
aagaagctgc tgtacatctt taagcaaccc ttcatgaggc ctgtgcagac tactcaagag 1080
gaggacggct gttcatgccg gttcccagag gaggaggaag gcggctgcga actgcgcgtg 1140
aaattcagcc gcagcgcaga tgctccagcc tacaagcagg ggcagaacca gctctacaac 1200
gaactcaatc ttggtcggag agaggagtac gacgtgctgg acaagcggag aggacgggac 1260
ccagaaatgg gcgggaagcc gcgcagaaag aatccccaag agggcctgta caacgagctc 1320
caaaaggata agatggcaga agcctatagc gagattggta tgaaagggga acgcagaaga 1380
ggcaaaggcc acgacggact gtaccaggga ctcagcaccg ccaccaagga cacctatgac 1440
gctcttcaca tgcaggccct gccgcctcgg 1470
<210> 55
<211> 490
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 55
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val
20 25 30
Lys Lys Pro Gly Glu Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Phe
35 40 45
Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Arg Gln Met Pro Gly Lys
50 55 60
Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys
65 70 75 80
Tyr Gly Pro Ile Phe Gln Gly His Val Thr Ile Ser Ala Asp Thr Ser
85 90 95
Ile Asn Thr Val Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr
100 105 110
Ala Met Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly
115 120 125
Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr
145 150 155 160
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
165 170 175
Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
180 185 190
Leu Asn Trp Leu Gln Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile
195 200 205
Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly
210 215 220
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
225 230 235 240
Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly
245 250 255
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
420 425 430
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
435 440 445
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
450 455 460
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
465 470 475 480
Ala Leu His Met Gln Ala Leu Pro Pro Arg
485 490
<210> 56
<211> 246
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 56
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys
130 135 140
Lys Pro Gly Glu Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Phe Asn
145 150 155 160
Ile Glu Asp Tyr Tyr Ile His Trp Val Arg Gln Met Pro Gly Lys Gly
165 170 175
Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr
180 185 190
Gly Pro Ile Phe Gln Gly His Val Thr Ile Ser Ala Asp Thr Ser Ile
195 200 205
Asn Thr Val Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala
210 215 220
Met Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly Thr
225 230 235 240
Thr Val Thr Val Ser Ser
245
<210> 57
<211> 738
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 57
gacgtcgtca tgacccagag cccgctgtca ctgcctgtga ccctgggcca gccggcgtcc 60
attagctgca aatcctcgca atccctgctc gactcagacg gaaaaacgta cttgaactgg 120
ctccaacagc gccctgggca atccccaagg cggcttatct cactcgtcag caagctcgat 180
agcggtgtcc cagacagatt ttcgggctcg ggatcgggca ctgatttcac tctgaagatc 240
tcgcgggtgg aagccgagga tgtgggagtg tactattgct ggcagggcac tcacttcccc 300
gggacgtttg gcggaggaac taaggtcgag atcaaaggag gaggtggatc aggcggaggt 360
gggagcggag gaggaggaag cggtggtgga ggttccgaaa tccagctggt gcaatcagga 420
gccgaggtga agaagccggg agaatccctg cgcatctcgt gcaagggctc gggcttcaac 480
atcgaggatt actacatcca ctgggtgcgg cagatgccgg gaaaggggtt ggaatggatg 540
ggacgcattg acccggaaaa tgatgaaacc aaatacgggc caatcttcca aggccacgtg 600
accattagcg ctgacacttc catcaacacc gtgtaccttc agtggtcctc actgaaggcg 660
tcggacactg ccatgtacta ctgtgcattc agaggagggg tctactgggg acagggcacc 720
accgtgaccg tgagctcc 738
<210> 58
<211> 831
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 58
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgacgtcg tcatgaccca gagcccgctg tcactgcctg tgaccctggg ccagccggcg 120
tccattagct gcaaatcctc gcaatccctg ctcgactcag acggaaaaac gtacttgaac 180
tggctccaac agcgccctgg gcaatcccca aggcggctta tctcactcgt cagcaagctc 240
gatagcggtg tcccagacag attttcgggc tcgggatcgg gcactgattt cactctgaag 300
atctcgcggg tggaagccga ggatgtggga gtgtactatt gctggcaggg cactcacttc 360
cccgggacgt ttggcggagg aactaaggtc gagatcaaag gaggaggtgg atcaggcgga 420
ggtgggagcg gaggaggagg aagcggtggt ggaggttccg aaatccagct ggtgcaatca 480
ggagccgagg tgaagaagcc gggagaatcc ctgcgcatct cgtgcaaggg ctcgggcttc 540
aacatcgagg attactacat ccactgggtg cggcagatgc cgggaaaggg gttggaatgg 600
atgggacgca ttgacccgga aaatgatgaa accaaatacg ggccaatctt ccaaggccac 660
gtgaccatta gcgctgacac ttccatcaac accgtgtacc ttcagtggtc ctcactgaag 720
gcgtcggaca ctgccatgta ctactgtgca ttcagaggag gggtctactg gggacagggc 780
accaccgtga ccgtgagctc cggctcgcat caccatcatc accaccatca c 831
<210> 59
<211> 277
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 59
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu
20 25 30
Pro Val Thr Leu Gly Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln
35 40 45
Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln
50 55 60
Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile Ser Leu Val Ser Lys Leu
65 70 75 80
Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr
100 105 110
Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr
115 120 125
Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser
145 150 155 160
Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Arg Ile Ser Cys Lys
165 170 175
Gly Ser Gly Phe Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Arg Gln
180 185 190
Met Pro Gly Lys Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn
195 200 205
Asp Glu Thr Lys Tyr Gly Pro Ile Phe Gln Gly His Val Thr Ile Ser
210 215 220
Ala Asp Thr Ser Ile Asn Thr Val Tyr Leu Gln Trp Ser Ser Leu Lys
225 230 235 240
Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr
245 250 255
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Ser His His His
260 265 270
His His His His His
275
<210> 60
<211> 1470
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 60
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgacgtcg tcatgaccca atcccctctc tccctgccgg tcaccctggg tcagccggcg 120
tcgatctcat gcaaaagctc acagtccctg ctggattcgg acggaaaaac ctacttgaac 180
tggctccaac agaggccggg tcagtcccct cgcagactga tctcgctggt gagcaagctc 240
gactcgggtg tgccggatcg gttctccggg tcaggatcgg gcaccgactt tacgctcaag 300
atttcgagag tggaggccga ggatgtggga gtgtactatt gctggcaggg cacgcatttc 360
cccgggacct ttggaggcgg gactaaggtg gaaatcaagg gaggtggcgg atcaggcgga 420
ggaggcagcg gcggaggtgg atcaggaggc ggagggtcag agatccagct ggtccaaagc 480
ggagcagagg tgaagaagcc aggcgagtcc cttcgcattt cgtgcaaagg gagcggcttc 540
aacattgaag attactacat ccactgggtg cggcaaatgc caggaaaggg tctggaatgg 600
atgggacgga tcgacccaga aaatgatgaa actaagtacg gaccgatctt ccaaggacac 660
gtcactatct ccgcggacac ttcgatcaac accgtgtacc tccagtggag cagcttgaaa 720
gcctccgaca ccgctatgta ctactgtgcc ttccgcggag gagtctactg gggacagggg 780
actactgtga ccgtgtcgtc caccactacc ccagcaccga ggccacccac cccggctcct 840
accatcgcct cccagcctct gtccctgcgt ccggaggcat gtagacccgc agctggtggg 900
gccgtgcata cccggggtct tgacttcgcc tgcgatatct acatttgggc ccctctggct 960
ggtacttgcg gggtcctgct gctttcactc gtgatcactc tttactgtaa gcgcggtcgg 1020
aagaagctgc tgtacatctt taagcaaccc ttcatgaggc ctgtgcagac tactcaagag 1080
gaggacggct gttcatgccg gttcccagag gaggaggaag gcggctgcga actgcgcgtg 1140
aaattcagcc gcagcgcaga tgctccagcc tacaagcagg ggcagaacca gctctacaac 1200
gaactcaatc ttggtcggag agaggagtac gacgtgctgg acaagcggag aggacgggac 1260
ccagaaatgg gcgggaagcc gcgcagaaag aatccccaag agggcctgta caacgagctc 1320
caaaaggata agatggcaga agcctatagc gagattggta tgaaagggga acgcagaaga 1380
ggcaaaggcc acgacggact gtaccaggga ctcagcaccg ccaccaagga cacctatgac 1440
gctcttcaca tgcaggccct gccgcctcgg 1470
<210> 61
<211> 490
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 61
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu
20 25 30
Pro Val Thr Leu Gly Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln
35 40 45
Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln
50 55 60
Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile Ser Leu Val Ser Lys Leu
65 70 75 80
Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr
100 105 110
Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr
115 120 125
Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser
145 150 155 160
Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Arg Ile Ser Cys Lys
165 170 175
Gly Ser Gly Phe Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Arg Gln
180 185 190
Met Pro Gly Lys Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn
195 200 205
Asp Glu Thr Lys Tyr Gly Pro Ile Phe Gln Gly His Val Thr Ile Ser
210 215 220
Ala Asp Thr Ser Ile Asn Thr Val Tyr Leu Gln Trp Ser Ser Leu Lys
225 230 235 240
Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr
245 250 255
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
420 425 430
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
435 440 445
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
450 455 460
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
465 470 475 480
Ala Leu His Met Gln Ala Leu Pro Pro Arg
485 490
<210> 62
<211> 246
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 62
Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asn Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr Gln Ser Pro Leu Ser
130 135 140
Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser
145 150 155 160
Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln
165 170 175
Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile Ser Leu Val Ser Lys
180 185 190
Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr
195 200 205
Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val
210 215 220
Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly
225 230 235 240
Thr Lys Val Glu Ile Lys
245
<210> 63
<211> 738
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 63
gaaatccagc tcgtgcagag cggagccgag gtcaagaaac cgggtgctac cgtgaagatt 60
tcatgcaagg gatcgggctt caacatcgag gattactaca tccactgggt gcagcaggca 120
ccaggaaaag gacttgaatg gatgggccgg atcgacccgg aaaatgacga gactaagtac 180
ggccctatct tccaaggacg ggtgacgatc accgcagaca ctagcaccaa caccgtctat 240
atggaactct cgtccctgag gtccgaagat actgccgtgt actactgtgc gtttcgcgga 300
ggtgtgtact ggggacaggg taccaccgtc accgtgtcat cgggcggtgg aggctccggt 360
ggaggagggt caggaggcgg tggaagcgga ggaggcggca gcgacgtggt catgactcaa 420
tcgccgctgt cgctgcccgt cactctggga caacccgcgt ccatcagctg caaatcctcg 480
cagtcactgc ttgactccga tggaaagacc tacctcaact ggctgcagca acgcccaggc 540
caatccccaa gacgcctgat ctcgttggtg tcaaagctgg actcaggggt gccggaccgg 600
ttctccggga gcgggtcggg cacggatttc actctcaaga tctccagagt ggaagccgag 660
gatgtgggag tctactactg ctggcaggga acccatttcc ctggaacttt tggcggagga 720
actaaggtcg agattaaa 738
<210> 64
<211> 831
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 64
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgaaatcc agctcgtgca gagcggagcc gaggtcaaga aaccgggtgc taccgtgaag 120
atttcatgca agggatcggg cttcaacatc gaggattact acatccactg ggtgcagcag 180
gcaccaggaa aaggacttga atggatgggc cggatcgacc cggaaaatga cgagactaag 240
tacggcccta tcttccaagg acgggtgacg atcaccgcag acactagcac caacaccgtc 300
tatatggaac tctcgtccct gaggtccgaa gatactgccg tgtactactg tgcgtttcgc 360
ggaggtgtgt actggggaca gggtaccacc gtcaccgtgt catcgggcgg tggaggctcc 420
ggtggaggag ggtcaggagg cggtggaagc ggaggaggcg gcagcgacgt ggtcatgact 480
caatcgccgc tgtcgctgcc cgtcactctg ggacaacccg cgtccatcag ctgcaaatcc 540
tcgcagtcac tgcttgactc cgatggaaag acctacctca actggctgca gcaacgccca 600
ggccaatccc caagacgcct gatctcgttg gtgtcaaagc tggactcagg ggtgccggac 660
cggttctccg ggagcgggtc gggcacggat ttcactctca agatctccag agtggaagcc 720
gaggatgtgg gagtctacta ctgctggcag ggaacccatt tccctggaac ttttggcgga 780
ggaactaagg tcgagattaa agggagccac catcatcatc accaccacca c 831
<210> 65
<211> 277
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 65
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val
20 25 30
Lys Lys Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Gly Ser Gly Phe
35 40 45
Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Gln Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys
65 70 75 80
Tyr Gly Pro Ile Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser
85 90 95
Thr Asn Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr
100 105 110
Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly
115 120 125
Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr
145 150 155 160
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
165 170 175
Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
180 185 190
Leu Asn Trp Leu Gln Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile
195 200 205
Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly
210 215 220
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
225 230 235 240
Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly
245 250 255
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Ser His His His
260 265 270
His His His His His
275
<210> 66
<211> 1470
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 66
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgaaatcc agctcgtgca gagcggagcc gaggtcaaga aaccgggtgc taccgtgaag 120
atttcatgca agggatcggg cttcaacatc gaggattact acatccactg ggtgcagcag 180
gcaccaggaa aaggacttga atggatgggc cggatcgacc cggaaaatga cgagactaag 240
tacggcccta tcttccaagg acgggtgacg atcaccgcag acactagcac caacaccgtc 300
tatatggaac tctcgtccct gaggtccgaa gatactgccg tgtactactg tgcgtttcgc 360
ggaggtgtgt actggggaca gggtaccacc gtcaccgtgt catcgggcgg tggaggctcc 420
ggtggaggag ggtcaggagg cggtggaagc ggaggaggcg gcagcgacgt ggtcatgact 480
caatcgccgc tgtcgctgcc cgtcactctg ggacaacccg cgtccatcag ctgcaaatcc 540
tcgcagtcac tgcttgactc cgatggaaag acctacctca actggctgca gcaacgccca 600
ggccaatccc caagacgcct gatctcgttg gtgtcaaagc tggactcagg ggtgccggac 660
cggttctccg ggagcgggtc gggcacggat ttcactctca agatctccag agtggaagcc 720
gaggatgtgg gagtctacta ctgctggcag ggaacccatt tccctggaac ttttggcgga 780
ggaactaagg tcgagattaa aaccactacc ccagcaccga ggccacccac cccggctcct 840
accatcgcct cccagcctct gtccctgcgt ccggaggcat gtagacccgc agctggtggg 900
gccgtgcata cccggggtct tgacttcgcc tgcgatatct acatttgggc ccctctggct 960
ggtacttgcg gggtcctgct gctttcactc gtgatcactc tttactgtaa gcgcggtcgg 1020
aagaagctgc tgtacatctt taagcaaccc ttcatgaggc ctgtgcagac tactcaagag 1080
gaggacggct gttcatgccg gttcccagag gaggaggaag gcggctgcga actgcgcgtg 1140
aaattcagcc gcagcgcaga tgctccagcc tacaagcagg ggcagaacca gctctacaac 1200
gaactcaatc ttggtcggag agaggagtac gacgtgctgg acaagcggag aggacgggac 1260
ccagaaatgg gcgggaagcc gcgcagaaag aatccccaag agggcctgta caacgagctc 1320
caaaaggata agatggcaga agcctatagc gagattggta tgaaagggga acgcagaaga 1380
ggcaaaggcc acgacggact gtaccaggga ctcagcaccg ccaccaagga cacctatgac 1440
gctcttcaca tgcaggccct gccgcctcgg 1470
<210> 67
<211> 490
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 67
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val
20 25 30
Lys Lys Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Gly Ser Gly Phe
35 40 45
Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Gln Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys
65 70 75 80
Tyr Gly Pro Ile Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser
85 90 95
Thr Asn Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr
100 105 110
Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly
115 120 125
Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr
145 150 155 160
Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly Gln Pro Ala Ser Ile
165 170 175
Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
180 185 190
Leu Asn Trp Leu Gln Gln Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile
195 200 205
Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly
210 215 220
Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser Arg Val Glu Ala
225 230 235 240
Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly
245 250 255
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
420 425 430
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
435 440 445
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
450 455 460
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
465 470 475 480
Ala Leu His Met Gln Ala Leu Pro Pro Arg
485 490
<210> 68
<211> 246
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 68
Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly His Val Thr Ile Ser Ala Asp Thr Ser Ile Asn Thr Val Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr Gln Ser Pro Asp Ser
130 135 140
Leu Ala Val Ser Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser
145 150 155 160
Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln
165 170 175
Gln Lys Pro Gly Gln Pro Pro Lys Arg Leu Ile Ser Leu Val Ser Lys
180 185 190
Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr
195 200 205
Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val
210 215 220
Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly
225 230 235 240
Thr Lys Val Glu Ile Lys
245
<210> 69
<211> 738
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 69
gaaatccagc tggtgcagtc aggcgccgag gtcaagaagc cgggagagtc gctgagaatc 60
tcgtgcaagg gctcggggtt caacatcgag gactactaca ttcactgggt caggcagatg 120
ccgggaaagg gactggaatg gatgggccgg atcgacccag aaaatgacga aaccaaatac 180
gggccgattt ttcaaggcca cgtgactatc agcgcagaca cgagcatcaa cactgtctac 240
ctccagtggt cctcgcttaa ggccagcgat accgctatgt actactgcgc attcagaggc 300
ggggtgtact ggggacaagg aaccactgtg accgtgagca gcggaggtgg cggctcggga 360
ggaggtggga gcggaggagg aggttccggc ggtggaggat cagatgtcgt gatgacccag 420
tccccggact ccctcgctgt ctcactgggc gagcgcgcga ccatcaactg caaatcgagc 480
cagtcgctgt tggactccga tggaaagact tatctgaatt ggctgcaaca gaaaccagga 540
caacctccca agcggctcat ctcgcttgtg tcaaaactcg attcgggagt gccagaccgc 600
ttctcggggt ccgggagcgg aactgacttt actttgacca tttcctcact gcaagcggag 660
gatgtggccg tgtattactg ttggcagggc acgcatttcc ctggaacctt cggtggcgga 720
actaaggtgg aaatcaag 738
<210> 70
<211> 834
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 70
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgaaatcc agctggtgca gtcaggcgcc gaggtcaaga agccgggaga gtcgctgaga 120
atctcgtgca agggctcggg gttcaacatc gaggactact acattcactg ggtcaggcag 180
atgccgggaa agggactgga atggatgggc cggatcgacc cagaaaatga cgaaaccaaa 240
tacgggccga tttttcaagg ccacgtgact atcagcgcag acacgagcat caacactgtc 300
tacctccagt ggtcctcgct taaggccagc gataccgcta tgtactactg cgcattcaga 360
ggcggggtgt actggggaca aggaaccact gtgaccgtga gcagcggagg tggcggctcg 420
ggaggaggtg ggagcggagg aggaggttcc ggcggtggag gatcagatgt cgtgatgacc 480
cagtccccgg actccctcgc tgtctcactg ggcgagcgcg cgaccatcaa ctgcaaatcg 540
agccagtcgc tgttggactc cgatggaaag acttatctga attggctgca acagaaacca 600
ggacaacctc ccaagcggct catctcgctt gtgtcaaaac tcgattcggg agtgccagac 660
cgcttctcgg ggtccgggag cggaactgac tttactttga ccatttcctc actgcaagcg 720
gaggatgtgg ccgtgtatta ctgttggcag ggcacgcatt tccctggaac cttcggtggc 780
ggaactaagg tggaaatcaa gggatcacac caccatcatc accatcacca ccat 834
<210> 71
<211> 278
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 71
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val
20 25 30
Lys Lys Pro Gly Glu Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Phe
35 40 45
Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Arg Gln Met Pro Gly Lys
50 55 60
Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys
65 70 75 80
Tyr Gly Pro Ile Phe Gln Gly His Val Thr Ile Ser Ala Asp Thr Ser
85 90 95
Ile Asn Thr Val Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr
100 105 110
Ala Met Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly
115 120 125
Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr
145 150 155 160
Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly Glu Arg Ala Thr Ile
165 170 175
Asn Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
180 185 190
Leu Asn Trp Leu Gln Gln Lys Pro Gly Gln Pro Pro Lys Arg Leu Ile
195 200 205
Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly
210 215 220
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
225 230 235 240
Glu Asp Val Ala Val Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly
245 250 255
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Ser His His His
260 265 270
His His His His His His
275
<210> 72
<211> 1470
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 72
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgagattc agctcgtgca atcgggagcg gaagtcaaga agccaggaga gtccttgcgg 120
atctcatgca agggtagcgg ctttaacatc gaggattact acatccactg ggtgaggcag 180
atgccgggga agggactcga atggatggga cggatcgacc cagaaaacga cgaaactaag 240
tacggtccga tcttccaagg ccatgtgact attagcgccg atacttcaat caataccgtg 300
tatctgcaat ggtcctcatt gaaagcctca gataccgcga tgtactactg tgctttcaga 360
ggaggggtct actggggaca gggaactacc gtgactgtct cgtccggcgg aggcgggtca 420
ggaggtggcg gcagcggagg aggagggtcc ggcggaggtg ggtccgacgt cgtgatgacc 480
cagagccctg acagcctggc agtgagcctg ggcgaaagag ctaccattaa ctgcaaatcg 540
tcgcagagcc tgctggactc ggacggaaaa acgtacctca attggctgca gcaaaagcct 600
ggccagccac cgaagcgcct tatctcactg gtgtcgaagc tggattcggg agtgcccgat 660
cgcttctccg gctcgggatc gggtactgac ttcaccctca ctatctcctc gcttcaagca 720
gaggacgtgg ccgtctacta ctgctggcag ggaacccact ttccgggaac cttcggcgga 780
gggacgaaag tggagatcaa gaccactacc ccagcaccga ggccacccac cccggctcct 840
accatcgcct cccagcctct gtccctgcgt ccggaggcat gtagacccgc agctggtggg 900
gccgtgcata cccggggtct tgacttcgcc tgcgatatct acatttgggc ccctctggct 960
ggtacttgcg gggtcctgct gctttcactc gtgatcactc tttactgtaa gcgcggtcgg 1020
aagaagctgc tgtacatctt taagcaaccc ttcatgaggc ctgtgcagac tactcaagag 1080
gaggacggct gttcatgccg gttcccagag gaggaggaag gcggctgcga actgcgcgtg 1140
aaattcagcc gcagcgcaga tgctccagcc tacaagcagg ggcagaacca gctctacaac 1200
gaactcaatc ttggtcggag agaggagtac gacgtgctgg acaagcggag aggacgggac 1260
ccagaaatgg gcgggaagcc gcgcagaaag aatccccaag agggcctgta caacgagctc 1320
caaaaggata agatggcaga agcctatagc gagattggta tgaaagggga acgcagaaga 1380
ggcaaaggcc acgacggact gtaccaggga ctcagcaccg ccaccaagga cacctatgac 1440
gctcttcaca tgcaggccct gccgcctcgg 1470
<210> 73
<211> 490
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 73
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val
20 25 30
Lys Lys Pro Gly Glu Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Phe
35 40 45
Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Arg Gln Met Pro Gly Lys
50 55 60
Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys
65 70 75 80
Tyr Gly Pro Ile Phe Gln Gly His Val Thr Ile Ser Ala Asp Thr Ser
85 90 95
Ile Asn Thr Val Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr
100 105 110
Ala Met Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly
115 120 125
Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Val Val Met Thr
145 150 155 160
Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly Glu Arg Ala Thr Ile
165 170 175
Asn Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr
180 185 190
Leu Asn Trp Leu Gln Gln Lys Pro Gly Gln Pro Pro Lys Arg Leu Ile
195 200 205
Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Ser Gly
210 215 220
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
225 230 235 240
Glu Asp Val Ala Val Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly
245 250 255
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
420 425 430
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
435 440 445
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
450 455 460
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
465 470 475 480
Ala Leu His Met Gln Ala Leu Pro Pro Arg
485 490
<210> 74
<211> 246
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 74
Asp Val Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln Lys Pro Gly Gln Pro
35 40 45
Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys
130 135 140
Lys Pro Gly Glu Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Phe Asn
145 150 155 160
Ile Glu Asp Tyr Tyr Ile His Trp Val Arg Gln Met Pro Gly Lys Gly
165 170 175
Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr
180 185 190
Gly Pro Ile Phe Gln Gly His Val Thr Ile Ser Ala Asp Thr Ser Ile
195 200 205
Asn Thr Val Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala
210 215 220
Met Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly Thr
225 230 235 240
Thr Val Thr Val Ser Ser
245
<210> 75
<211> 738
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 75
gacgtggtga tgacccaatc gccagattcc ctggcagtgt ccctgggcga acgcgccact 60
attaactgca aatcgtcaca gtccttgctt gattccgacg gaaagaccta cctcaattgg 120
ctccagcaga agccaggaca accgccaaag agactgatct ccctggtgtc aaagctggac 180
tcgggagtgc ctgatcggtt ctcgggtagc gggagcggca ccgacttcac tctgaccatc 240
tcgtcactcc aggctgagga cgtggccgtg tattactgtt ggcagggtac tcactttccg 300
ggcactttcg gaggcggcac caaggtggag attaaaggag gaggcggaag cggaggtgga 360
ggatcgggag gtggtgggag cggcggagga gggagcgaga tccagctcgt ccaatcggga 420
gcggaagtga agaagcccgg agagtcactt agaatctcat gcaaggggtc gggcttcaac 480
atcgaggatt actacatcca ttgggtccgc cagatgcctg gtaaaggact ggaatggatg 540
gggaggattg acccggaaaa cgacgaaact aagtacggac cgatctttca agggcacgtg 600
actatctccg ctgatacctc aatcaatact gtctacctcc agtggtcctc gctgaaagca 660
agcgacaccg cgatgtacta ctgcgccttc cggggaggag tgtactgggg ccaaggcacc 720
acggtcacgg tcagctcc 738
<210> 76
<211> 834
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 76
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgacgtgg tgatgaccca atcgccagat tccctggcag tgtccctggg cgaacgcgcc 120
actattaact gcaaatcgtc acagtccttg cttgattccg acggaaagac ctacctcaat 180
tggctccagc agaagccagg acaaccgcca aagagactga tctccctggt gtcaaagctg 240
gactcgggag tgcctgatcg gttctcgggt agcgggagcg gcaccgactt cactctgacc 300
atctcgtcac tccaggctga ggacgtggcc gtgtattact gttggcaggg tactcacttt 360
ccgggcactt tcggaggcgg caccaaggtg gagattaaag gaggaggcgg aagcggaggt 420
ggaggatcgg gaggtggtgg gagcggcgga ggagggagcg agatccagct cgtccaatcg 480
ggagcggaag tgaagaagcc cggagagtca cttagaatct catgcaaggg gtcgggcttc 540
aacatcgagg attactacat ccattgggtc cgccagatgc ctggtaaagg actggaatgg 600
atggggagga ttgacccgga aaacgacgaa actaagtacg gaccgatctt tcaagggcac 660
gtgactatct ccgctgatac ctcaatcaat actgtctacc tccagtggtc ctcgctgaaa 720
gcaagcgaca ccgcgatgta ctactgcgcc ttccggggag gagtgtactg gggccaaggc 780
accacggtca cggtcagctc cggctcccat caccaccacc atcaccatca tcac 834
<210> 77
<211> 278
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 77
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Val Val Met Thr Gln Ser Pro Asp Ser Leu
20 25 30
Ala Val Ser Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln
35 40 45
Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln
50 55 60
Lys Pro Gly Gln Pro Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu
65 70 75 80
Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr
100 105 110
Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr
115 120 125
Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser
145 150 155 160
Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Arg Ile Ser Cys Lys
165 170 175
Gly Ser Gly Phe Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Arg Gln
180 185 190
Met Pro Gly Lys Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn
195 200 205
Asp Glu Thr Lys Tyr Gly Pro Ile Phe Gln Gly His Val Thr Ile Ser
210 215 220
Ala Asp Thr Ser Ile Asn Thr Val Tyr Leu Gln Trp Ser Ser Leu Lys
225 230 235 240
Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr
245 250 255
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Ser His His His
260 265 270
His His His His His His
275
<210> 78
<211> 1470
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 78
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgacgtgg tgatgactca gtcgcctgac tcgctggctg tgtcccttgg agagcgggcc 120
actatcaatt gcaagtcatc ccagtcgctg ctggattccg acgggaaaac ctacctcaat 180
tggctgcagc aaaaaccggg acagcctcca aagcggctca tcagcctggt gtccaagttg 240
gacagcggcg tgccagaccg cttctccggt tcgggaagcg gtactgattt cacgctgacc 300
atctcatccc tccaagcgga ggatgtggca gtctactact gttggcaggg cacgcatttt 360
ccgggcactt ttggaggagg gaccaaggtc gaaatcaagg gaggaggtgg ctcgggcgga 420
ggaggctcgg gaggaggagg atcaggaggc ggtggaagcg agattcaact ggtccagagc 480
ggcgcagaag tcaagaagcc gggtgaatcg ctcagaatct cgtgcaaagg atcgggattc 540
aacatcgagg actactacat tcactgggtc agacaaatgc cgggcaaagg gctggaatgg 600
atggggagga tcgaccccga aaacgatgaa accaagtacg gaccaatctt ccaagggcac 660
gtgaccattt cggcggacac ctcaatcaac actgtgtacc tccagtggag ctcacttaag 720
gccagcgata ccgccatgta ctattgcgct ttccgcggag gggtgtactg gggacagggc 780
actactgtga ccgtgtcatc caccactacc ccagcaccga ggccacccac cccggctcct 840
accatcgcct cccagcctct gtccctgcgt ccggaggcat gtagacccgc agctggtggg 900
gccgtgcata cccggggtct tgacttcgcc tgcgatatct acatttgggc ccctctggct 960
ggtacttgcg gggtcctgct gctttcactc gtgatcactc tttactgtaa gcgcggtcgg 1020
aagaagctgc tgtacatctt taagcaaccc ttcatgaggc ctgtgcagac tactcaagag 1080
gaggacggct gttcatgccg gttcccagag gaggaggaag gcggctgcga actgcgcgtg 1140
aaattcagcc gcagcgcaga tgctccagcc tacaagcagg ggcagaacca gctctacaac 1200
gaactcaatc ttggtcggag agaggagtac gacgtgctgg acaagcggag aggacgggac 1260
ccagaaatgg gcgggaagcc gcgcagaaag aatccccaag agggcctgta caacgagctc 1320
caaaaggata agatggcaga agcctatagc gagattggta tgaaagggga acgcagaaga 1380
ggcaaaggcc acgacggact gtaccaggga ctcagcaccg ccaccaagga cacctatgac 1440
gctcttcaca tgcaggccct gccgcctcgg 1470
<210> 79
<211> 490
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 79
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Val Val Met Thr Gln Ser Pro Asp Ser Leu
20 25 30
Ala Val Ser Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln
35 40 45
Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln
50 55 60
Lys Pro Gly Gln Pro Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu
65 70 75 80
Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr
100 105 110
Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr
115 120 125
Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser
145 150 155 160
Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Arg Ile Ser Cys Lys
165 170 175
Gly Ser Gly Phe Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Arg Gln
180 185 190
Met Pro Gly Lys Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn
195 200 205
Asp Glu Thr Lys Tyr Gly Pro Ile Phe Gln Gly His Val Thr Ile Ser
210 215 220
Ala Asp Thr Ser Ile Asn Thr Val Tyr Leu Gln Trp Ser Ser Leu Lys
225 230 235 240
Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr
245 250 255
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
420 425 430
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
435 440 445
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
450 455 460
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
465 470 475 480
Ala Leu His Met Gln Ala Leu Pro Pro Arg
485 490
<210> 80
<211> 246
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 80
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys
130 135 140
Lys Pro Gly Ala Thr Val Lys Ile Ser Cys Lys Gly Ser Gly Phe Asn
145 150 155 160
Ile Glu Asp Tyr Tyr Ile His Trp Val Gln Gln Ala Pro Gly Lys Gly
165 170 175
Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr
180 185 190
Gly Pro Ile Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr
195 200 205
Asn Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala
210 215 220
Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly Thr
225 230 235 240
Thr Val Thr Val Ser Ser
245
<210> 81
<211> 738
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 81
gatgtggtca tgacgcagtc accactgtcc ctccccgtga cccttggaca gccagcgtcg 60
attagctgca agtcatccca atccctgctc gattcggatg gaaagaccta tctcaactgg 120
ctgcagcaaa gacccggtca gagccctagg agactcatct cgttggtgtc aaagctggac 180
agcggagtgc cggaccggtt ttccggttcg ggatcgggga cggacttcac tctgaagatt 240
tcacgggtgg aagctgagga tgtgggagtg tactactgct ggcagggaac ccatttccct 300
ggcacttttg gcggaggaac taaggtcgaa atcaagggag gaggtggctc gggaggaggc 360
ggatcgggcg gaggcgggag cggcggagga gggtccgaaa tccaacttgt ccagtcagga 420
gccgaagtga agaaaccggg agccaccgtc aaaatcagct gtaagggatc gggattcaat 480
atcgaggact actacatcca ctgggtgcag caagctccgg gcaaaggact ggagtggatg 540
gggcgcatcg acccagagaa cgacgaaacc aaatacggcc cgatcttcca agggcgggtg 600
accatcaccg cggacacctc aactaacact gtgtacatgg agctgagctc cctgcgctcc 660
gaagatactg cagtctacta ctgcgccttc cgcggtggtg tgtactgggg acagggcacc 720
actgtgactg tcagctcg 738
<210> 82
<211> 831
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 82
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgatgtgg tcatgacgca gtcaccactg tccctccccg tgacccttgg acagccagcg 120
tcgattagct gcaagtcatc ccaatccctg ctcgattcgg atggaaagac ctatctcaac 180
tggctgcagc aaagacccgg tcagagccct aggagactca tctcgttggt gtcaaagctg 240
gacagcggag tgccggaccg gttttccggt tcgggatcgg ggacggactt cactctgaag 300
atttcacggg tggaagctga ggatgtggga gtgtactact gctggcaggg aacccatttc 360
cctggcactt ttggcggagg aactaaggtc gaaatcaagg gaggaggtgg ctcgggagga 420
ggcggatcgg gcggaggcgg gagcggcgga ggagggtccg aaatccaact tgtccagtca 480
ggagccgaag tgaagaaacc gggagccacc gtcaaaatca gctgtaaggg atcgggattc 540
aatatcgagg actactacat ccactgggtg cagcaagctc cgggcaaagg actggagtgg 600
atggggcgca tcgacccaga gaacgacgaa accaaatacg gcccgatctt ccaagggcgg 660
gtgaccatca ccgcggacac ctcaactaac actgtgtaca tggagctgag ctccctgcgc 720
tccgaagata ctgcagtcta ctactgcgcc ttccgcggtg gtgtgtactg gggacagggc 780
accactgtga ctgtcagctc ggggtcccac catcatcacc accaccatca c 831
<210> 83
<211> 277
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 83
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu
20 25 30
Pro Val Thr Leu Gly Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln
35 40 45
Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln
50 55 60
Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile Ser Leu Val Ser Lys Leu
65 70 75 80
Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr
100 105 110
Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr
115 120 125
Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser
145 150 155 160
Gly Ala Glu Val Lys Lys Pro Gly Ala Thr Val Lys Ile Ser Cys Lys
165 170 175
Gly Ser Gly Phe Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Gln Gln
180 185 190
Ala Pro Gly Lys Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn
195 200 205
Asp Glu Thr Lys Tyr Gly Pro Ile Phe Gln Gly Arg Val Thr Ile Thr
210 215 220
Ala Asp Thr Ser Thr Asn Thr Val Tyr Met Glu Leu Ser Ser Leu Arg
225 230 235 240
Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr
245 250 255
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Ser His His His
260 265 270
His His His His His
275
<210> 84
<211> 1470
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 84
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgatgtgg tcatgacgca gtcaccactg tccctccccg tgacccttgg acagccagcg 120
tcgattagct gcaagtcatc ccaatccctg ctcgattcgg atggaaagac ctatctcaac 180
tggctgcagc aaagacccgg tcagagccct aggagactca tctcgttggt gtcaaagctg 240
gacagcggag tgccggaccg gttttccggt tcgggatcgg ggacggactt cactctgaag 300
atttcacggg tggaagctga ggatgtggga gtgtactact gctggcaggg aacccatttc 360
cctggcactt ttggcggagg aactaaggtc gaaatcaagg gaggaggtgg ctcgggagga 420
ggcggatcgg gcggaggcgg gagcggcgga ggagggtccg aaatccaact tgtccagtca 480
ggagccgaag tgaagaaacc gggagccacc gtcaaaatca gctgtaaggg atcgggattc 540
aatatcgagg actactacat ccactgggtg cagcaagctc cgggcaaagg actggagtgg 600
atggggcgca tcgacccaga gaacgacgaa accaaatacg gcccgatctt ccaagggcgg 660
gtgaccatca ccgcggacac ctcaactaac actgtgtaca tggagctgag ctccctgcgc 720
tccgaagata ctgcagtcta ctactgcgcc ttccgcggtg gtgtgtactg gggacagggc 780
accactgtga ctgtcagctc gaccactacc ccagcaccga ggccacccac cccggctcct 840
accatcgcct cccagcctct gtccctgcgt ccggaggcat gtagacccgc agctggtggg 900
gccgtgcata cccggggtct tgacttcgcc tgcgatatct acatttgggc ccctctggct 960
ggtacttgcg gggtcctgct gctttcactc gtgatcactc tttactgtaa gcgcggtcgg 1020
aagaagctgc tgtacatctt taagcaaccc ttcatgaggc ctgtgcagac tactcaagag 1080
gaggacggct gttcatgccg gttcccagag gaggaggaag gcggctgcga actgcgcgtg 1140
aaattcagcc gcagcgcaga tgctccagcc tacaagcagg ggcagaacca gctctacaac 1200
gaactcaatc ttggtcggag agaggagtac gacgtgctgg acaagcggag aggacgggac 1260
ccagaaatgg gcgggaagcc gcgcagaaag aatccccaag agggcctgta caacgagctc 1320
caaaaggata agatggcaga agcctatagc gagattggta tgaaagggga acgcagaaga 1380
ggcaaaggcc acgacggact gtaccaggga ctcagcaccg ccaccaagga cacctatgac 1440
gctcttcaca tgcaggccct gccgcctcgg 1470
<210> 85
<211> 490
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 85
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu
20 25 30
Pro Val Thr Leu Gly Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln
35 40 45
Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln
50 55 60
Arg Pro Gly Gln Ser Pro Arg Arg Leu Ile Ser Leu Val Ser Lys Leu
65 70 75 80
Asp Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Lys Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr
100 105 110
Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr
115 120 125
Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Gln Leu Val Gln Ser
145 150 155 160
Gly Ala Glu Val Lys Lys Pro Gly Ala Thr Val Lys Ile Ser Cys Lys
165 170 175
Gly Ser Gly Phe Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Gln Gln
180 185 190
Ala Pro Gly Lys Gly Leu Glu Trp Met Gly Arg Ile Asp Pro Glu Asn
195 200 205
Asp Glu Thr Lys Tyr Gly Pro Ile Phe Gln Gly Arg Val Thr Ile Thr
210 215 220
Ala Asp Thr Ser Thr Asn Thr Val Tyr Met Glu Leu Ser Ser Leu Arg
225 230 235 240
Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr
245 250 255
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Thr Thr Thr Pro Ala
260 265 270
Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser
275 280 285
Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr
290 295 300
Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala
305 310 315 320
Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys
325 330 335
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
340 345 350
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
355 360 365
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg
370 375 380
Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn
385 390 395 400
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
405 410 415
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
420 425 430
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
435 440 445
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
450 455 460
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
465 470 475 480
Ala Leu His Met Gln Ala Leu Pro Pro Arg
485 490
<210> 86
<211> 243
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 86
Glu Ile Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Thr Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Lys Gln Arg Thr Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr Val Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Pro Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser His Met Asp Val Val Met Thr Gln Ser Pro Leu Thr Leu Ser Val
130 135 140
Ala Ile Gly Gln Ser Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu
145 150 155 160
Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro
165 170 175
Gly Gln Ser Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser
180 185 190
Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr
195 200 205
Leu Arg Ile Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys
210 215 220
Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Leu
225 230 235 240
Glu Ile Lys
<210> 87
<211> 822
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 87
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgagatcc agctccaaca gagcggagcc gaactggtca aaccgggagc gtcggtgaag 120
ttgtcatgca ctggatcggg cttcaacatc gaggattact acatccactg ggtcaagcaa 180
cgcaccgagc aggggctgga atggatcgga cggatcgacc ccgaaaacga tgaaaccaag 240
tacgggccta tcttccaagg acgggccacc attacggctg acacgtcaag caataccgtc 300
tacctccagc tttccagcct gacctccgag gacactgccg tgtactactg cgccttcaga 360
ggaggcgtgt actggggacc aggaaccact ttgaccgtgt ccagcggagg cggtggatca 420
ggaggaggag gctcaggcgg tggcggctcg cacatggacg tggtcatgac tcagtccccg 480
ctgaccctgt cggtggcaat tggacagagc gcatccatct cgtgcaagag ctcacagtcg 540
ctgctggatt ccgacggaaa gacttatctg aactggctgc tccaaagacc agggcaatca 600
ccgaaacgcc ttatctccct ggtgtcgaaa ctcgactcgg gtgtgccgga tcggtttacc 660
ggtagcgggt ccggcacgga cttcactctc cgcatttcga gggtggaagc ggaggatctc 720
gggatctact actgttggca gggaacccac ttccctggga cttttggagg cggaactaag 780
ctggaaatca agggtagcca tcaccatcac caccaccatc at 822
<210> 88
<211> 274
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 88
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Glu Ile Gln Leu Gln Gln Ser Gly Ala Glu Leu
20 25 30
Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Thr Gly Ser Gly Phe
35 40 45
Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Lys Gln Arg Thr Glu Gln
50 55 60
Gly Leu Glu Trp Ile Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys
65 70 75 80
Tyr Gly Pro Ile Phe Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser
85 90 95
Ser Asn Thr Val Tyr Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr
100 105 110
Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Pro Gly
115 120 125
Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser His Met Asp Val Val Met Thr Gln Ser Pro
145 150 155 160
Leu Thr Leu Ser Val Ala Ile Gly Gln Ser Ala Ser Ile Ser Cys Lys
165 170 175
Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp
180 185 190
Leu Leu Gln Arg Pro Gly Gln Ser Pro Lys Arg Leu Ile Ser Leu Val
195 200 205
Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
210 215 220
Gly Thr Asp Phe Thr Leu Arg Ile Ser Arg Val Glu Ala Glu Asp Leu
225 230 235 240
Gly Ile Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly
245 250 255
Gly Gly Thr Lys Leu Glu Ile Lys Gly Ser His His His His His His
260 265 270
His His
<210> 89
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 89
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgagatcc agctccaaca gagcggagcc gaactggtca aaccgggagc gtcggtgaag 120
ttgtcatgca ctggatcggg cttcaacatc gaggattact acatccactg ggtcaagcaa 180
cgcaccgagc aggggctgga atggatcgga cggatcgacc ccgaaaacga tgaaaccaag 240
tacgggccta tcttccaagg acgggccacc attacggctg acacgtcaag caataccgtc 300
tacctccagc tttccagcct gacctccgag gacactgccg tgtactactg cgccttcaga 360
ggaggcgtgt actggggacc aggaaccact ttgaccgtgt ccagcggagg cggtggatca 420
ggaggaggag gctcaggcgg tggcggctcg cacatggacg tggtcatgac tcagtccccg 480
ctgaccctgt cggtggcaat tggacagagc gcatccatct cgtgcaagag ctcacagtcg 540
ctgctggatt ccgacggaaa gacttatctg aactggctgc tccaaagacc agggcaatca 600
ccgaaacgcc ttatctccct ggtgtcgaaa ctcgactcgg gtgtgccgga tcggtttacc 660
ggtagcgggt ccggcacgga cttcactctc cgcatttcga gggtggaagc ggaggatctc 720
gggatctact actgttggca gggaacccac ttccctggga cttttggagg cggaactaag 780
ctggaaatca agaccactac cccagcaccg aggccaccca ccccggctcc taccatcgcc 840
tcccagcctc tgtccctgcg tccggaggca tgtagacccg cagctggtgg ggccgtgcat 900
acccggggtc ttgacttcgc ctgcgatatc tacatttggg cccctctggc tggtacttgc 960
ggggtcctgc tgctttcact cgtgatcact ctttactgta agcgcggtcg gaagaagctg 1020
ctgtacatct ttaagcaacc cttcatgagg cctgtgcaga ctactcaaga ggaggacggc 1080
tgttcatgcc ggttcccaga ggaggaggaa ggcggctgcg aactgcgcgt gaaattcagc 1140
cgcagcgcag atgctccagc ctacaagcag gggcagaacc agctctacaa cgaactcaat 1200
cttggtcgga gagaggagta cgacgtgctg gacaagcgga gaggacggga cccagaaatg 1260
ggcgggaagc cgcgcagaaa gaatccccaa gagggcctgt acaacgagct ccaaaaggat 1320
aagatggcag aagcctatag cgagattggt atgaaagggg aacgcagaag aggcaaaggc 1380
cacgacggac tgtaccaggg actcagcacc gccaccaagg acacctatga cgctcttcac 1440
atgcaggccc tgccgcctcg g 1461
<210> 90
<211> 487
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 90
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Glu Ile Gln Leu Gln Gln Ser Gly Ala Glu Leu
20 25 30
Val Lys Pro Gly Ala Ser Val Lys Leu Ser Cys Thr Gly Ser Gly Phe
35 40 45
Asn Ile Glu Asp Tyr Tyr Ile His Trp Val Lys Gln Arg Thr Glu Gln
50 55 60
Gly Leu Glu Trp Ile Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys
65 70 75 80
Tyr Gly Pro Ile Phe Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser
85 90 95
Ser Asn Thr Val Tyr Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr
100 105 110
Ala Val Tyr Tyr Cys Ala Phe Arg Gly Gly Val Tyr Trp Gly Pro Gly
115 120 125
Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser His Met Asp Val Val Met Thr Gln Ser Pro
145 150 155 160
Leu Thr Leu Ser Val Ala Ile Gly Gln Ser Ala Ser Ile Ser Cys Lys
165 170 175
Ser Ser Gln Ser Leu Leu Asp Ser Asp Gly Lys Thr Tyr Leu Asn Trp
180 185 190
Leu Leu Gln Arg Pro Gly Gln Ser Pro Lys Arg Leu Ile Ser Leu Val
195 200 205
Ser Lys Leu Asp Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
210 215 220
Gly Thr Asp Phe Thr Leu Arg Ile Ser Arg Val Glu Ala Glu Asp Leu
225 230 235 240
Gly Ile Tyr Tyr Cys Trp Gln Gly Thr His Phe Pro Gly Thr Phe Gly
245 250 255
Gly Gly Thr Lys Leu Glu Ile Lys Thr Thr Thr Pro Ala Pro Arg Pro
260 265 270
Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro
275 280 285
Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu
290 295 300
Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys
305 310 315 320
Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Lys Arg Gly
325 330 335
Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val
340 345 350
Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu
355 360 365
Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg Ser Ala Asp
370 375 380
Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
385 390 395 400
Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg
405 410 415
Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly
420 425 430
Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu
435 440 445
Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu
450 455 460
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His
465 470 475 480
Met Gln Ala Leu Pro Pro Arg
485
<210> 91
<211> 240
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 91
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Ile Val Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His His Ser Tyr Pro Leu
85 90 95
Thr Ser Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Gly Ser Thr
100 105 110
Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser Glu Val Gln Val
115 120 125
Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ala Ile Ser
165 170 175
Gly Ser Gly Gly Ser Thr Asn Tyr Ala Asp Ser Val Lys Gly Arg Phe
180 185 190
Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Gly Ser Ser
210 215 220
Gly Trp Ser Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
225 230 235 240
<210> 92
<211> 720
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 92
gatatccaaa tgactcagag cccttcatcc ctgagcgcca gcgtcggaga cagggtgacc 60
atcacgtgcc gggcatccca aggcattaga aataacttgg cgtggtatca gcaaaaacca 120
ggaaaggccc cgaagcgcct gatctacgcg gcctccaacc ttcagtcagg agtgccctcg 180
cgcttcaccg ggagcggtag cggaactgag tttaccctta tcgtgtcgtc cctgcagcca 240
gaggacttcg cgacctacta ctgcctccag catcactcgt acccgttgac ttcgggaggc 300
ggaaccaagg tcgaaatcaa acgcactggc tcgacgtcag ggtccggtaa accgggatcg 360
ggagaaggat cggaagtcca agtgctggag agcggaggcg gactcgtgca acctggcggg 420
tcgctgcggc tcagctgtgc cgcgtcgggt tttactttca gctcgtacgc tatgtcatgg 480
gtgcggcagg ctccgggaaa ggggctggaa tgggtgtccg ctatttccgg ctcgggtgga 540
agcaccaatt acgccgactc cgtgaaggga cgcttcacca tctcacggga taactccaag 600
aatactctgt acctccagat gaactcgctg agagccgagg acaccgcagt gtactactgc 660
gcagggtcaa gcggctggtc cgaatactgg ggacagggca ccctcgtcac tgtcagctcc 720
<210> 93
<211> 807
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 93
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgatatcc aaatgactca gagcccttca tccctgagcg ccagcgtcgg agacagggtg 120
accatcacgt gccgggcatc ccaaggcatt agaaataact tggcgtggta tcagcaaaaa 180
ccaggaaagg ccccgaagcg cctgatctac gcggcctcca accttcagtc aggagtgccc 240
tcgcgcttca ccgggagcgg tagcggaact gagtttaccc ttatcgtgtc gtccctgcag 300
ccagaggact tcgcgaccta ctactgcctc cagcatcact cgtacccgtt gacttcggga 360
ggcggaacca aggtcgaaat caaacgcact ggctcgacgt cagggtccgg taaaccggga 420
tcgggagaag gatcggaagt ccaagtgctg gagagcggag gcggactcgt gcaacctggc 480
gggtcgctgc ggctcagctg tgccgcgtcg ggttttactt tcagctcgta cgctatgtca 540
tgggtgcggc aggctccggg aaaggggctg gaatgggtgt ccgctatttc cggctcgggt 600
ggaagcacca attacgccga ctccgtgaag ggacgcttca ccatctcacg ggataactcc 660
aagaatactc tgtacctcca gatgaactcg ctgagagccg aggacaccgc agtgtactac 720
tgcgcagggt caagcggctg gtccgaatac tggggacagg gcaccctcgt cactgtcagc 780
tcccatcacc atcaccacca ccatcac 807
<210> 94
<211> 269
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 94
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
20 25 30
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln
35 40 45
Gly Ile Arg Asn Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala
50 55 60
Pro Lys Arg Leu Ile Tyr Ala Ala Ser Asn Leu Gln Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Thr Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Ile Val
85 90 95
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His
100 105 110
His Ser Tyr Pro Leu Thr Ser Gly Gly Gly Thr Lys Val Glu Ile Lys
115 120 125
Arg Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly
130 135 140
Ser Glu Val Gln Val Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
145 150 155 160
Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser
165 170 175
Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
180 185 190
Val Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Asn Tyr Ala Asp Ser
195 200 205
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu
210 215 220
Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr
225 230 235 240
Cys Ala Gly Ser Ser Gly Trp Ser Glu Tyr Trp Gly Gln Gly Thr Leu
245 250 255
Val Thr Val Ser Ser His His His His His His His His
260 265
<210> 95
<211> 1452
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 95
atggccctcc ctgtcaccgc cctgctgctt ccgctggctc ttctgctcca cgccgctcgg 60
cccgatatcc aaatgactca gagcccttca tccctgagcg ccagcgtcgg agacagggtg 120
accatcacgt gccgggcatc ccaaggcatt agaaataact tggcgtggta tcagcaaaaa 180
ccaggaaagg ccccgaagcg cctgatctac gcggcctcca accttcagtc aggagtgccc 240
tcgcgcttca ccgggagcgg tagcggaact gagtttaccc ttatcgtgtc gtccctgcag 300
ccagaggact tcgcgaccta ctactgcctc cagcatcact cgtacccgtt gacttcggga 360
ggcggaacca aggtcgaaat caaacgcact ggctcgacgt cagggtccgg taaaccggga 420
tcgggagaag gatcggaagt ccaagtgctg gagagcggag gcggactcgt gcaacctggc 480
gggtcgctgc ggctcagctg tgccgcgtcg ggttttactt tcagctcgta cgctatgtca 540
tgggtgcggc aggctccggg aaaggggctg gaatgggtgt ccgctatttc cggctcgggt 600
ggaagcacca attacgccga ctccgtgaag ggacgcttca ccatctcacg ggataactcc 660
aagaatactc tgtacctcca gatgaactcg ctgagagccg aggacaccgc agtgtactac 720
tgcgcagggt caagcggctg gtccgaatac tggggacagg gcaccctcgt cactgtcagc 780
tccaccacta ccccagcacc gaggccaccc accccggctc ctaccatcgc ctcccagcct 840
ctgtccctgc gtccggaggc atgtagaccc gcagctggtg gggccgtgca tacccggggt 900
cttgacttcg cctgcgatat ctacatttgg gcccctctgg ctggtacttg cggggtcctg 960
ctgctttcac tcgtgatcac tctttactgt aagcgcggtc ggaagaagct gctgtacatc 1020
tttaagcaac ccttcatgag gcctgtgcag actactcaag aggaggacgg ctgttcatgc 1080
cggttcccag aggaggagga aggcggctgc gaactgcgcg tgaaattcag ccgcagcgca 1140
gatgctccag cctacaagca ggggcagaac cagctctaca acgaactcaa tcttggtcgg 1200
agagaggagt acgacgtgct ggacaagcgg agaggacggg acccagaaat gggcgggaag 1260
ccgcgcagaa agaatcccca agagggcctg tacaacgagc tccaaaagga taagatggca 1320
gaagcctata gcgagattgg tatgaaaggg gaacgcagaa gaggcaaagg ccacgacgga 1380
ctgtaccagg gactcagcac cgccaccaag gacacctatg acgctcttca catgcaggcc 1440
ctgccgcctc gg 1452
<210> 96
<211> 484
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 96
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
20 25 30
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln
35 40 45
Gly Ile Arg Asn Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala
50 55 60
Pro Lys Arg Leu Ile Tyr Ala Ala Ser Asn Leu Gln Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Thr Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Ile Val
85 90 95
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His
100 105 110
His Ser Tyr Pro Leu Thr Ser Gly Gly Gly Thr Lys Val Glu Ile Lys
115 120 125
Arg Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly
130 135 140
Ser Glu Val Gln Val Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
145 150 155 160
Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser
165 170 175
Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
180 185 190
Val Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Asn Tyr Ala Asp Ser
195 200 205
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu
210 215 220
Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr
225 230 235 240
Cys Ala Gly Ser Ser Gly Trp Ser Glu Tyr Trp Gly Gln Gly Thr Leu
245 250 255
Val Thr Val Ser Ser Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro
260 265 270
Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys
275 280 285
Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala
290 295 300
Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu
305 310 315 320
Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Lys Arg Gly Arg Lys Lys
325 330 335
Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr
340 345 350
Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly
355 360 365
Gly Cys Glu Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala
370 375 380
Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg
385 390 395 400
Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu
405 410 415
Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
420 425 430
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
435 440 445
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
450 455 460
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
465 470 475 480
Leu Pro Pro Arg
<210> 97
<211> 1183
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 97
gtgaggctcc ggtgcccgtc agtgggcaga gcgcacatcg cccacagtcc ccgagaagtt 60
ggggggaggg gtcggcaatt gaaccggtgc ctagagaagg tggcgcgggg taaactggga 120
aagtgatgtc gtgtactggc tccgcctttt tcccgagggt gggggagaac cgtatataag 180
tgcagtagtc gccgtgaacg ttctttttcg caacgggttt gccgccagaa cacaggtaag 240
tgccgtgtgt ggttcccgcg ggcctggcct ctttacgggt tatggccctt gcgtgccttg 300
aattacttcc acctggctgc agtacgtgat tcttgatccc gagcttcggg ttggaagtgg 360
gtgggagagt tcgaggcctt gcgcttaagg agccccttcg cctcgtgctt gagttgaggc 420
ctggcctggg cgctggggcc gccgcgtgcg aatctggtgg caccttcgcg cctgtctcgc 480
tgctttcgat aagtctctag ccatttaaaa tttttgatga cctgctgcga cgcttttttt 540
ctggcaagat agtcttgtaa atgcgggcca agatctgcac actggtattt cggtttttgg 600
ggccgcgggc ggcgacgggg cccgtgcgtc ccagcgcaca tgttcggcga ggcggggcct 660
gcgagcgcgg ccaccgagaa tcggacgggg gtagtctcaa gctggccggc ctgctctggt 720
gcctggcctc gcgccgccgt gtatcgcccc gccctgggcg gcaaggctgg cccggtcggc 780
accagttgcg tgagcggaaa gatggccgct tcccggccct gctgcaggga gctcaaaatg 840
gaggacgcgg cgctcgggag agcgggcggg tgagtcaccc acacaaagga aaagggcctt 900
tccgtcctca gccgtcgctt catgtgactc cacggagtac cgggcgccgt ccaggcacct 960
cgattagttc tcgagctttt ggagtacgtc gtctttaggt tggggggagg ggttttatgc 1020
gatggagttt ccccacactg agtgggtgga gactgaagtt aggccagctt ggcacttgat 1080
gtaattctcc ttggaatttg ccctttttga gtttggatct tggttcattc tcaagcctca 1140
gacagtggtt caaagttttt ttcttccatt tcaggtgtcg tga 1183
<210> 98
<211> 729
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 98
gagatccagc tccaacagag cggagccgaa ctggtcaaac cgggagcgtc ggtgaagttg 60
tcatgcactg gatcgggctt caacatcgag gattactaca tccactgggt caagcaacgc 120
accgagcagg ggctggaatg gatcggacgg atcgaccccg aaaacgatga aaccaagtac 180
gggcctatct tccaaggacg ggccaccatt acggctgaca cgtcaagcaa taccgtctac 240
ctccagcttt ccagcctgac ctccgaggac actgccgtgt actactgcgc cttcagagga 300
ggcgtgtact ggggaccagg aaccactttg accgtgtcca gcggaggcgg tggatcagga 360
ggaggaggct caggcggtgg cggctcgcac atggacgtgg tcatgactca gtccccgctg 420
accctgtcgg tggcaattgg acagagcgca tccatctcgt gcaagagctc acagtcgctg 480
ctggattccg acggaaagac ttatctgaac tggctgctcc aaagaccagg gcaatcaccg 540
aaacgcctta tctccctggt gtcgaaactc gactcgggtg tgccggatcg gtttaccggt 600
agcgggtccg gcacggactt cactctccgc atttcgaggg tggaagcgga ggatctcggg 660
atctactact gttggcaggg aacccacttc cctgggactt ttggaggcgg aactaagctg 720
gaaatcaag 729
<210> 99
<211> 112
<212> БЕЛОК
<213> Homo sapiens
<400> 99
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 100
<211> 336
<212> ДНК
<213> Homo sapiens
<400> 100
agagtgaagt tcagcaggag cgcagacgcc cccgcgtacc agcagggcca gaaccagctc 60
tataacgagc tcaatctagg acgaagagag gagtacgatg ttttggacaa gagacgtggc 120
cgggaccctg agatgggggg aaagccgaga aggaagaacc ctcaggaagg cctgtacaat 180
gaactgcaga aagataagat ggcggaggcc tacagtgaga ttgggatgaa aggcgagcgc 240
cggaggggca aggggcacga tggcctttac cagggtctca gtacagccac caaggacacc 300
tacgacgccc ttcacatgca ggccctgccc cctcgc 336
<210> 101
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 101
Leu Glu Glu Lys Lys Gly Asn Tyr Val Val Thr Asp His Cys
1 5 10
<210> 102
<211> 48
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 102
Gln Arg Arg Lys Tyr Arg Ser Asn Lys Gly Glu Ser Pro Val Glu Pro
1 5 10 15
Ala Glu Pro Cys Arg Tyr Ser Cys Pro Arg Glu Glu Glu Gly Ser Thr
20 25 30
Ile Pro Ile Gln Glu Asp Tyr Arg Lys Pro Glu Pro Ala Cys Ser Pro
35 40 45
<210> 103
<211> 123
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 103
aggagtaaga ggagcaggct cctgcacagt gactacatga acatgactcc ccgccgcccc 60
gggcccaccc gcaagcatta ccagccctat gccccaccac gcgacttcgc agcctatcgc 120
tcc 123
<210> 104
<211> 230
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 104
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys Met
225 230
<210> 105
<211> 690
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 105
gagagcaagt acggccctcc ctgcccccct tgccctgccc ccgagttcct gggcggaccc 60
agcgtgttcc tgttcccccc caagcccaag gacaccctga tgatcagccg gacccccgag 120
gtgacctgtg tggtggtgga cgtgtcccag gaggaccccg aggtccagtt caactggtac 180
gtggacggcg tggaggtgca caacgccaag accaagcccc gggaggagca gttcaatagc 240
acctaccggg tggtgtccgt gctgaccgtg ctgcaccagg actggctgaa cggcaaggaa 300
tacaagtgta aggtgtccaa caagggcctg cccagcagca tcgagaaaac catcagcaag 360
gccaagggcc agcctcggga gccccaggtg tacaccctgc cccctagcca agaggagatg 420
accaagaacc aggtgtccct gacctgcctg gtgaagggct tctaccccag cgacatcgcc 480
gtggagtggg agagcaacgg ccagcccgag aacaactaca agaccacccc ccctgtgctg 540
gacagcgacg gcagcttctt cctgtacagc cggctgaccg tggacaagag ccggtggcag 600
gagggcaacg tctttagctg ctccgtgatg cacgaggccc tgcacaacca ctacacccag 660
aagagcctga gcctgtccct gggcaagatg 690
<210> 106
<211> 282
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 106
Arg Trp Pro Glu Ser Pro Lys Ala Gln Ala Ser Ser Val Pro Thr Ala
1 5 10 15
Gln Pro Gln Ala Glu Gly Ser Leu Ala Lys Ala Thr Thr Ala Pro Ala
20 25 30
Thr Thr Arg Asn Thr Gly Arg Gly Gly Glu Glu Lys Lys Lys Glu Lys
35 40 45
Glu Lys Glu Glu Gln Glu Glu Arg Glu Thr Lys Thr Pro Glu Cys Pro
50 55 60
Ser His Thr Gln Pro Leu Gly Val Tyr Leu Leu Thr Pro Ala Val Gln
65 70 75 80
Asp Leu Trp Leu Arg Asp Lys Ala Thr Phe Thr Cys Phe Val Val Gly
85 90 95
Ser Asp Leu Lys Asp Ala His Leu Thr Trp Glu Val Ala Gly Lys Val
100 105 110
Pro Thr Gly Gly Val Glu Glu Gly Leu Leu Glu Arg His Ser Asn Gly
115 120 125
Ser Gln Ser Gln His Ser Arg Leu Thr Leu Pro Arg Ser Leu Trp Asn
130 135 140
Ala Gly Thr Ser Val Thr Cys Thr Leu Asn His Pro Ser Leu Pro Pro
145 150 155 160
Gln Arg Leu Met Ala Leu Arg Glu Pro Ala Ala Gln Ala Pro Val Lys
165 170 175
Leu Ser Leu Asn Leu Leu Ala Ser Ser Asp Pro Pro Glu Ala Ala Ser
180 185 190
Trp Leu Leu Cys Glu Val Ser Gly Phe Ser Pro Pro Asn Ile Leu Leu
195 200 205
Met Trp Leu Glu Asp Gln Arg Glu Val Asn Thr Ser Gly Phe Ala Pro
210 215 220
Ala Arg Pro Pro Pro Gln Pro Gly Ser Thr Thr Phe Trp Ala Trp Ser
225 230 235 240
Val Leu Arg Val Pro Ala Pro Pro Ser Pro Gln Pro Ala Thr Tyr Thr
245 250 255
Cys Val Val Ser His Glu Asp Ser Arg Thr Leu Leu Asn Ala Ser Arg
260 265 270
Ser Leu Glu Val Ser Tyr Val Thr Asp His
275 280
<210> 107
<211> 847
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 107
aggtggcccg aaagtcccaa ggcccaggca tctagtgttc ctactgcaca gccccaggca 60
gaaggcagcc tagccaaagc tactactgca cctgccacta cgcgcaatac tggccgtggc 120
ggggaggaga agaaaaagga gaaagagaaa gaagaacagg aagagaggga gaccaagacc 180
cctgaatgtc catcccatac ccagccgctg ggcgtctatc tcttgactcc cgcagtacag 240
gacttgtggc ttagagataa ggccaccttt acatgtttcg tcgtgggctc tgacctgaag 300
gatgcccatt tgacttggga ggttgccgga aaggtaccca cagggggggt tgaggaaggg 360
ttgctggagc gccattccaa tggctctcag agccagcact caagactcac ccttccgaga 420
tccctgtgga acgccgggac ctctgtcaca tgtactctaa atcatcctag cctgccccca 480
cagcgtctga tggcccttag agagccagcc gcccaggcac cagttaagct tagcctgaat 540
ctgctcgcca gtagtgatcc cccagaggcc gccagctggc tcttatgcga agtgtccggc 600
tttagcccgc ccaacatctt gctcatgtgg ctggaggacc agcgagaagt gaacaccagc 660
ggcttcgctc cagcccggcc cccaccccag ccgggttcta ccacattctg ggcctggagt 720
gtcttaaggg tcccagcacc acctagcccc cagccagcca catacacctg tgttgtgtcc 780
catgaagata gcaggaccct gctaaatgct tctaggagtc tggaggtttc ctacgtgact 840
gaccatt 847
<210> 108
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 108
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 109
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
олигонуклеотид»
<400> 109
ggtggcggag gttctggagg tggaggttcc 30
<210> 110
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<220>
<221> различный признак
<222> (1)..(30)
<223> /примечание=«Эта последовательность может включать 1, 2, 3, 4, 5 или 6
повторяющихся единиц «Gly Gly Gly Gly Ser»
<220>
<221> ВАРИАНТ
<222> (6)..(30)
<223> /замена = « »
<220>
<221> различный признак
<222> (1)..(30)
<223> /примечание = «Вариантные остатки, указанные в последовательности,
не являются предпочтительными относительно остатков в
комментариях для различных положений»
<220>
<221> различный признак
<222> (1)..(30)
<223> /примечание = «Смотрите поданное описание изобретения для подробного
описания замен и предпочтительных вариантов осуществления»
<400> 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25 30
<210> 111
<211> 150
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 111
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 150
<210> 112
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 112
Gly Gly Gly Ser
1
<210> 113
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 113
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 114
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
пептид»
<400> 114
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 115
<211> 5000
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<220>
<221> различный признак
<222> (1)..(5000)
<223> /примечание = «Эта последовательность может включать от 50 до 5000
нуклеотидов»
<220>
<221> вариация
<222> (51)..(5000)
<223> /замена = « »
<220>
<221> различный признак
<222> (1)..(5000)
<223> /примечание = «Вариантные основания, указанные в последовательности,
не являются предпочтительными относительно оснований в комментариях
для различных положений»
<220>
<221> различный признак
<222> (1)..(5000)
<223> /примечание = «Смотрите поданное описание изобретения для подробного
описания замен и предпочтительных вариантов осуществления»
<400> 115
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 420
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 480
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 600
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 720
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 780
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 840
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 960
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1020
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1080
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1140
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1200
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1260
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1320
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1380
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1440
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1500
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1560
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1620
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1680
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1860
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1980
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2040
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2100
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2160
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2220
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2280
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2340
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2400
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2460
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2520
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2580
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2640
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2700
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2760
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2820
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2880
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2940
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3000
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3060
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3420
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3480
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3600
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3720
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3780
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3840
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3960
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4020
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4080
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4140
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4200
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4260
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4320
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4380
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4440
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4500
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4560
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4620
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4680
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4860
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4980
aaaaaaaaaa aaaaaaaaaa 5000
<210> 116
<211> 2000
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<220>
<221> различный признак
<222> (1)..(2000)
<223> /примечание = «Эта последовательность может включать от 50 до 2000
нуклеотидов»
<220>
<221> вариация
<222> (51)..(2000)
<223> /замена = « »
<220>
<221> различный признак
<222> (1)..(2000)
<223> /примечание = «Вариантные основания, указанные в последовательности,
не являются предпочтительными относительно оснований в комментариях
для различных положений»
<400> 116
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 420
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 480
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 600
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 720
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 780
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 840
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 960
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1020
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1080
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1140
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1200
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1260
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1320
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1380
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1440
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1500
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1560
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1620
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1680
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1860
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1980
aaaaaaaaaa aaaaaaaaaa 2000
<210> 117
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<400> 117
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 60
tttttttttt tttttttttt tttttttttt tttttttttt 100
<210> 118
<211> 5000
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<220>
<221> различный признак
<222> (1)..(5000)
<223> /примечание = «Эта последовательность может включать от 50 до 5000
нуклеотидов»
<220>
<221> вариация
<222> (51)..(5000)
<223> /замена = « »
<220>
<221> различный признак
<222> (1)..(5000)
<223> /примечание = «Вариантные основания, указанные в последовательности,
не являются предпочтительными относительно оснований в комментариях
для различных положений»
<400> 118
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 60
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 120
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 180
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 240
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 300
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 360
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 420
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 480
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 540
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 600
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 660
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 720
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 780
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 840
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 900
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 960
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1020
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1080
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1140
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1200
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1260
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1320
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1380
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1440
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1500
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1560
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1620
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1680
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1740
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1800
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1860
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1920
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 1980
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2040
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2100
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2160
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2220
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2280
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2340
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2400
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2460
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2520
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2580
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2640
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2700
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2760
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2820
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2880
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 2940
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3000
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3060
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3120
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3180
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3240
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3300
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3360
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3420
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3480
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3540
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3600
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3660
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3720
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3780
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3840
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3900
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 3960
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4020
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4080
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4140
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4200
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4260
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4320
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4380
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4440
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4500
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4560
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4620
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4680
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4740
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4800
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4860
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4920
tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt tttttttttt 4980
tttttttttt tttttttttt 5000
<210> 119
<211> 5000
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<220>
<221> различный признак
<222> (1)..(5000)
<223> /примечание = «Эта последовательность может включать от 100 до 5000
нуклеотидов»
<220>
<221> вариация
<222> (101)..(5000)
<223> /замена = « »
<220>
<221> различный признак
<222> (1)..(5000)
<223> /примечание = «Вариантные основания, указанные в последовательности,
не являются предпочтительными относительно оснований в комментариях
для различных положений»
<400> 119
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 420
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 480
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 600
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 720
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 780
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 840
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 960
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1020
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1080
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1140
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1200
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1260
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1320
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1380
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1440
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1500
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1560
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1620
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1680
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1860
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 1980
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2040
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2100
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2160
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2220
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2280
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2340
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2400
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2460
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2520
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2580
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2640
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2700
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2760
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2820
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2880
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 2940
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3000
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3060
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3420
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3480
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3540
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3600
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3660
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3720
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3780
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3840
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3900
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 3960
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4020
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4080
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4140
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4200
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4260
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4320
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4380
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4440
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4500
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4560
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4620
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4680
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4740
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4800
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4860
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4920
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 4980
aaaaaaaaaa aaaaaaaaaa 5000
<210> 120
<211> 400
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полинуклеотид»
<220>
<221> различный признак
<222> (1)..(400)
<223> /примечание = «Эта последовательность может включать от 100 до 400
нуклеотидов»
<220>
<221> вариация
<222> (101)..(400)
<223> /замена = « »
<220>
<221> различный признак
<222> (1)..(400)
<223> /примечание = «Вариантные основания, указанные в последовательности,
не являются предпочтительными относительно оснований в комментариях
для различных положений»
<220>
<221> различный признак
<222> (1)..(400)
<223> /примечание = «Смотрите поданное описание изобретения для подробного
описания замен и предпочтительных вариантов осуществления»
<400> 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 120
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 180
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 240
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 300
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 360
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 400
<210> 121
<211> 487
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 121
Met Val Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Ile Ile Pro Asp Ile Gln Met Thr Gln Ser Pro Ser Ser
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Ile Ile Ile Cys Arg Ala Ser
35 40 45
Gln Gly Ile Arg Asn Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Arg Leu Ile Tyr Ala Ala Ser Asn Leu Gln Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Thr Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Ile
85 90 95
Val Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln
100 105 110
His His Ser Tyr Pro Leu Thr Ser Gly Gly Gly Thr Lys Val Glu Ile
115 120 125
Lys Phe Thr Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu
130 135 140
Gly Ser Glu Val Gln Val Leu Glu Ser Gly Gly Gly Leu Val Gln Pro
145 150 155 160
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
165 170 175
Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
180 185 190
Trp Val Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Asn Tyr Ala Asp
195 200 205
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr
210 215 220
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
225 230 235 240
Tyr Cys Ala Gly Ser Ser Gly Trp Ser Glu Tyr Trp Gly Gln Gly Thr
245 250 255
Leu Val Thr Val Ser Ser Ala Ser Thr Thr Thr Pro Ala Pro Arg Pro
260 265 270
Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro
275 280 285
Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu
290 295 300
Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys
305 310 315 320
Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Lys Arg Gly
325 330 335
Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val
340 345 350
Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu
355 360 365
Glu Glu Gly Gly Cys Glu Leu Arg Val Lys Phe Ser Arg Ser Ala Asp
370 375 380
Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
385 390 395 400
Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg
405 410 415
Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly
420 425 430
Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu
435 440 445
Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu
450 455 460
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His
465 470 475 480
Met Gln Ala Leu Pro Pro Arg
485
<210> 122
<211> 114
<212> БЕЛОК
<213> Mus musculus
<400> 122
Glu Ile Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Thr Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Lys Gln Arg Thr Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly Arg Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr Val Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Pro Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser
<210> 123
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 123
Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asn Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 124
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 124
Glu Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Phe Asn Ile Glu Asp Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asn Asp Glu Thr Lys Tyr Gly Pro Ile Phe
50 55 60
Gln Gly His Val Thr Ile Ser Ala Asp Thr Ser Ile Asn Thr Val Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Phe Arg Gly Gly Val Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 125
<211> 112
<212> БЕЛОК
<213> Mus musculus
<400> 125
Asp Val Val Met Thr Gln Ser Pro Leu Thr Leu Ser Val Ala Ile Gly
1 5 10 15
Gln Ser Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 126
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 126
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 127
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание = «Описание искусственной последовательности: синтетический
полипептид»
<400> 127
Asp Val Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln Lys Pro Gly Gln Pro
35 40 45
Pro Lys Arg Leu Ile Ser Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Gly Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИЗБИРАТЕЛЬНОЙ ЭКСПРЕССИИ БЕЛКА | 2017 |
|
RU2795467C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР КЛЕТОК | 2015 |
|
RU2751362C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) ПРОТИВ CD123 ДЛЯ ИСПОЛЬЗОВАНИЯ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2015 |
|
RU2724999C2 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ С ИСПОЛЬЗОВАНИЕМ ГУМАНИЗИРОВАННОГО ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА ПРОТИВ ВСМА | 2015 |
|
RU2751660C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА CLL-1 | 2015 |
|
RU2741120C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА ПРОТИВ МЕЗОТЕЛИНА ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2714902C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ЛЕЧЕНИЯ РАКА | 2017 |
|
RU2826270C2 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО РЕЦЕПТОРА АНТИГЕНА ПРОТИВ CD19 | 2015 |
|
RU2815417C2 |
| CD20 ТЕРАПИЯ, CD22 ТЕРАПИЯ И КОМБИНИРОВАННАЯ ТЕРАПИЯ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) K CD19 | 2016 |
|
RU2752918C2 |
| ВИДЫ КОМБИНИРОВАННОЙ ТЕРАПИИ С ИСПОЛЬЗОВАНИЕМ ХИМЕРНЫХ АНТИГЕННЫХ РЕЦЕПТОРОВ И ИНГИБИТОРОВ PD-1 | 2017 |
|
RU2809160C2 |
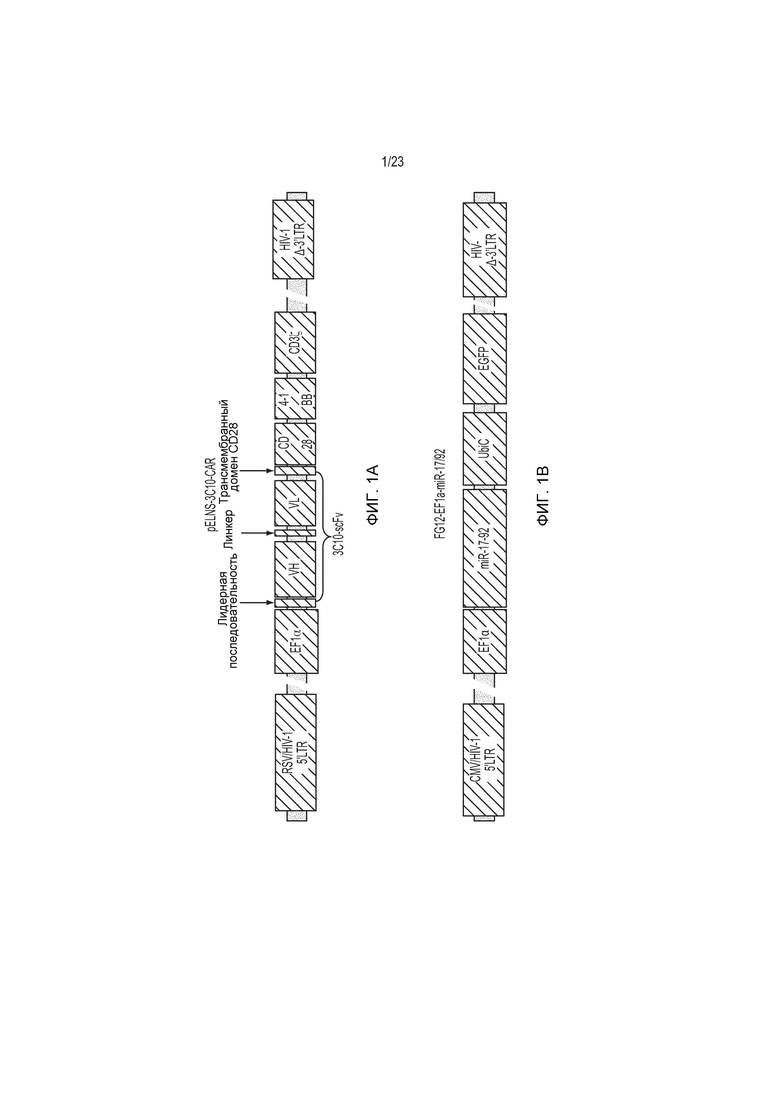
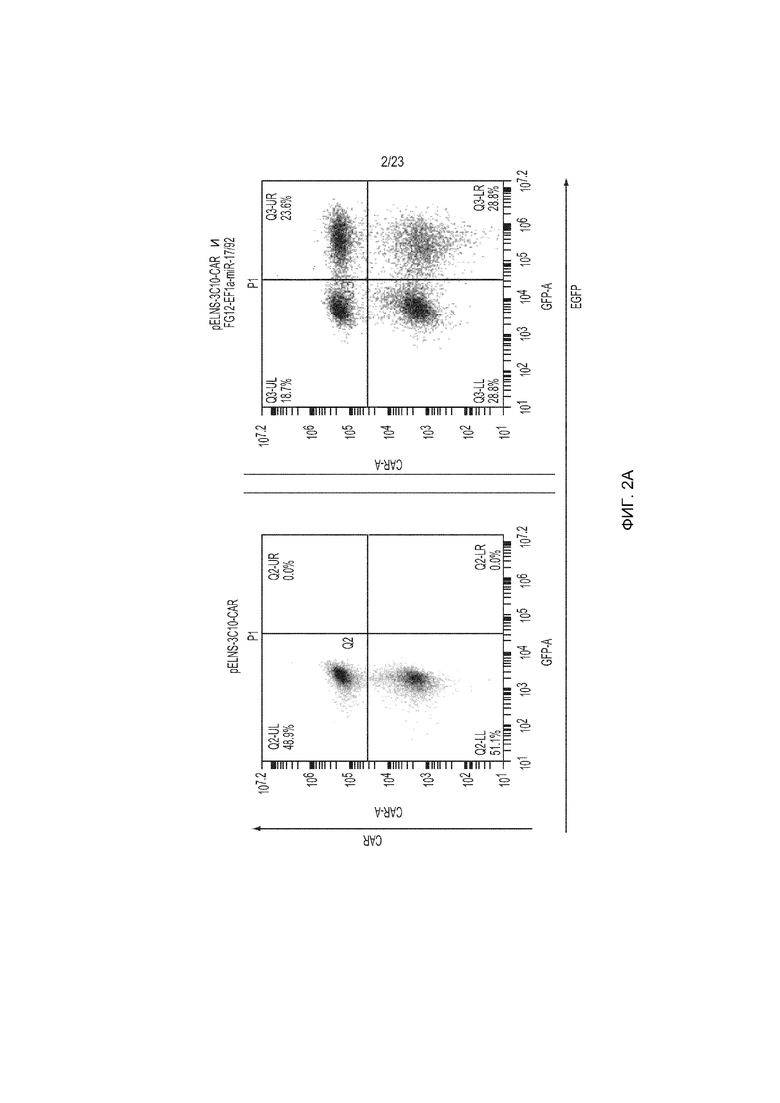

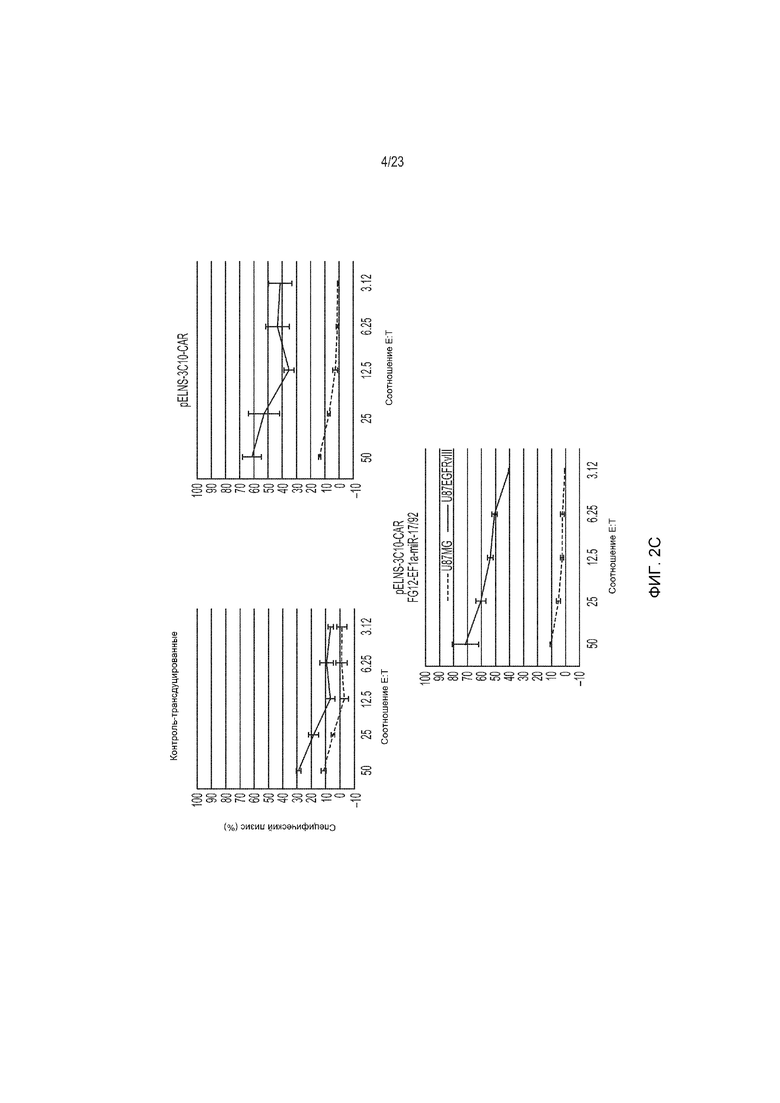
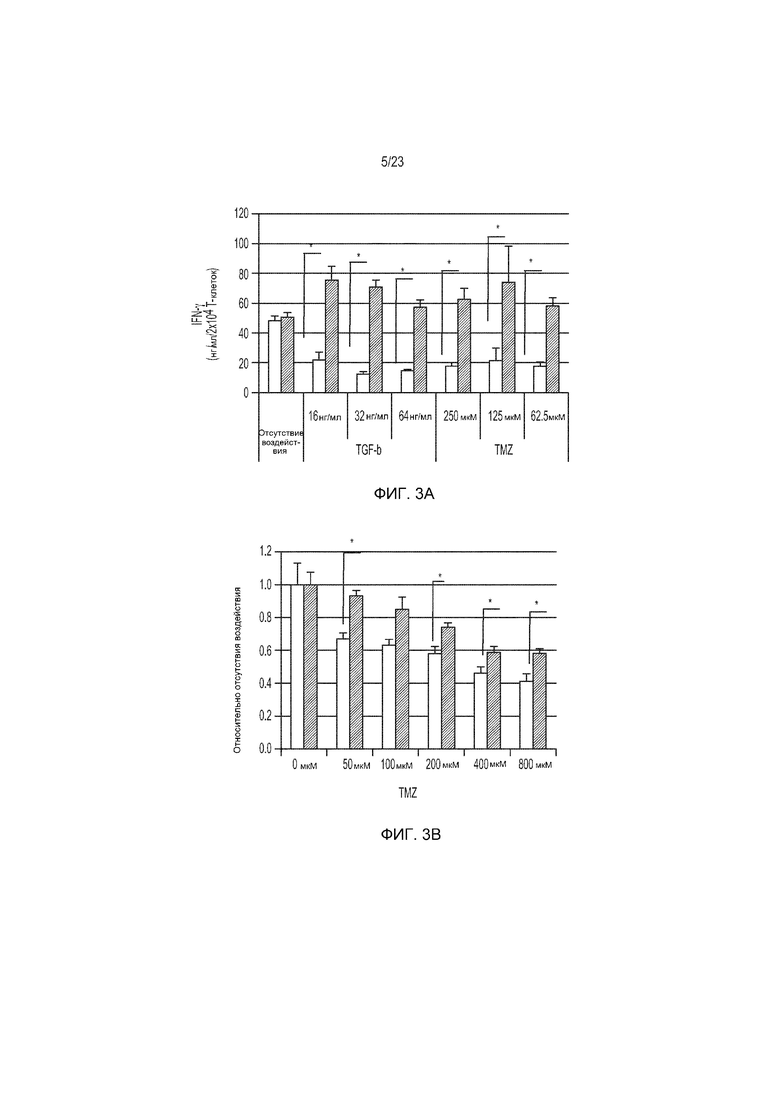
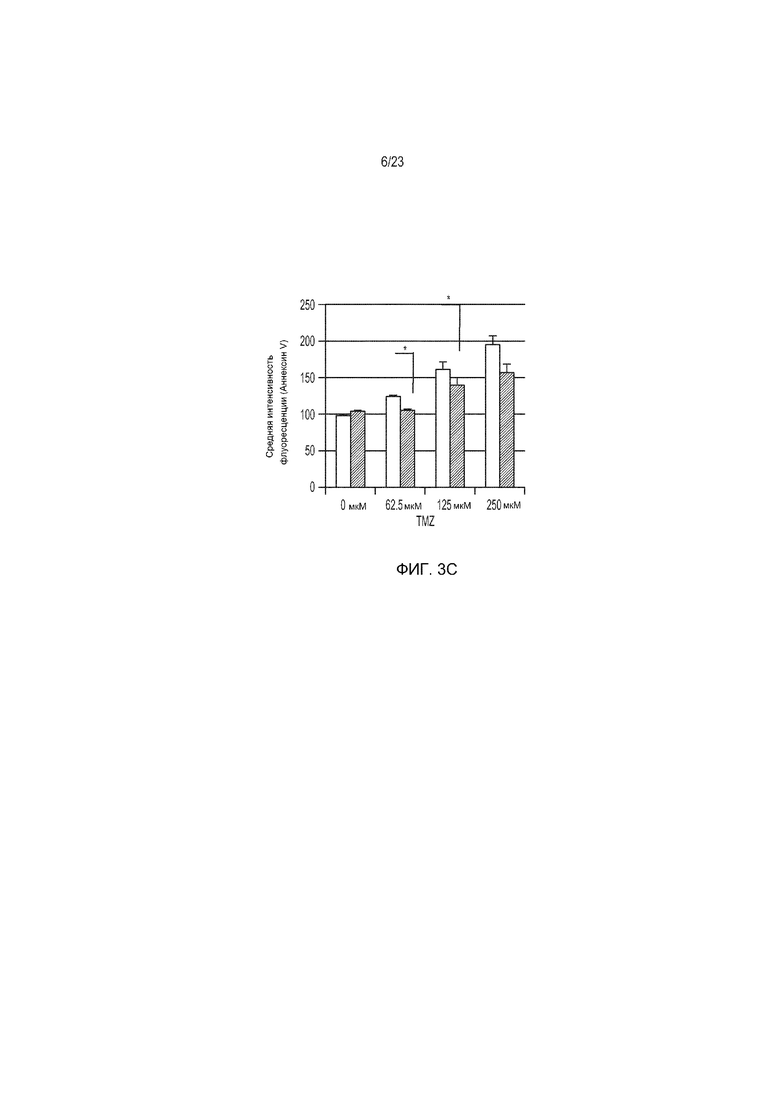

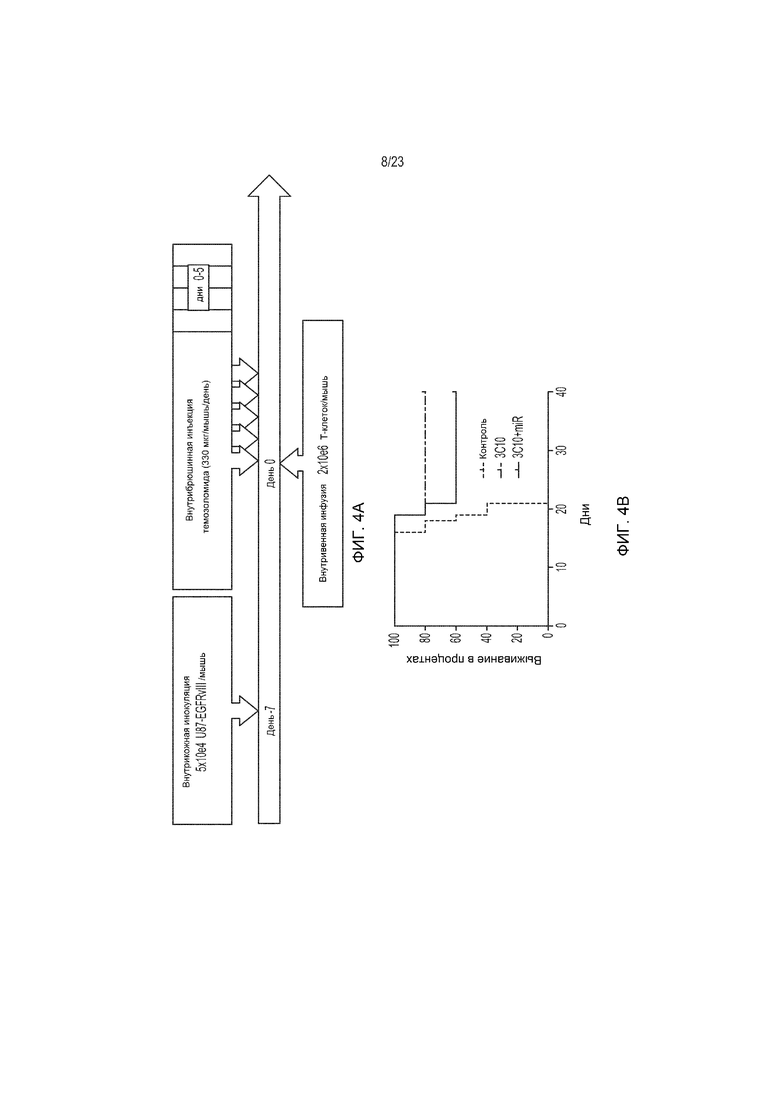
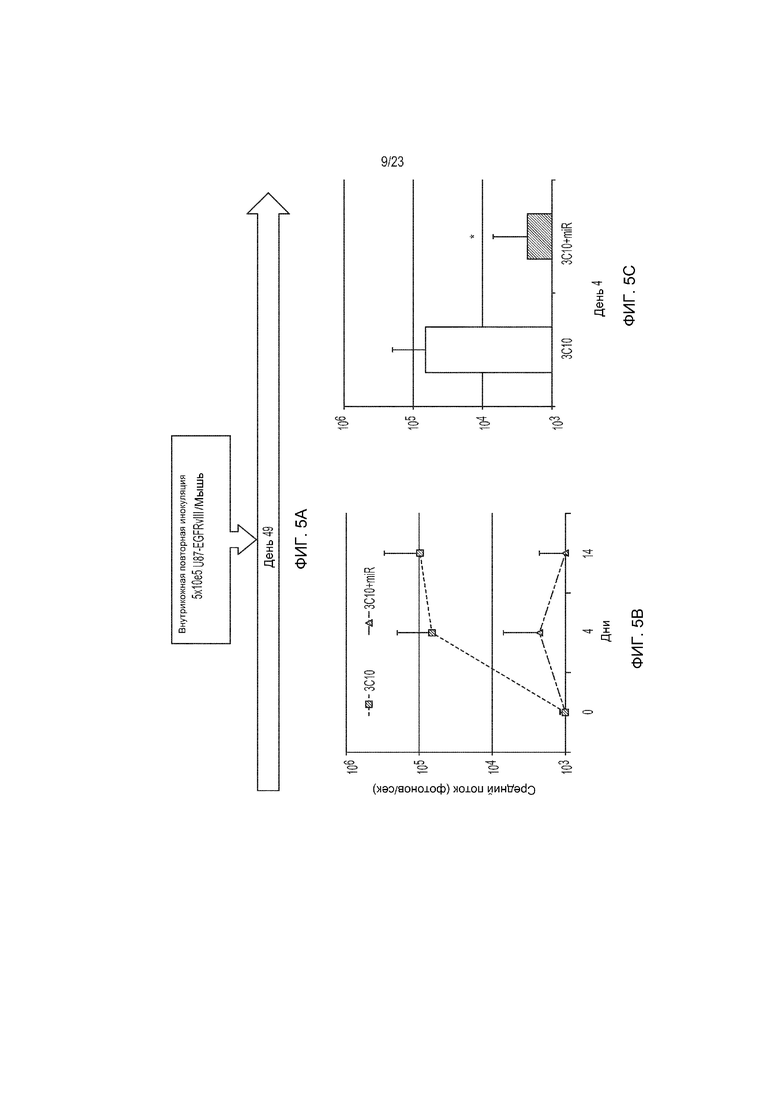
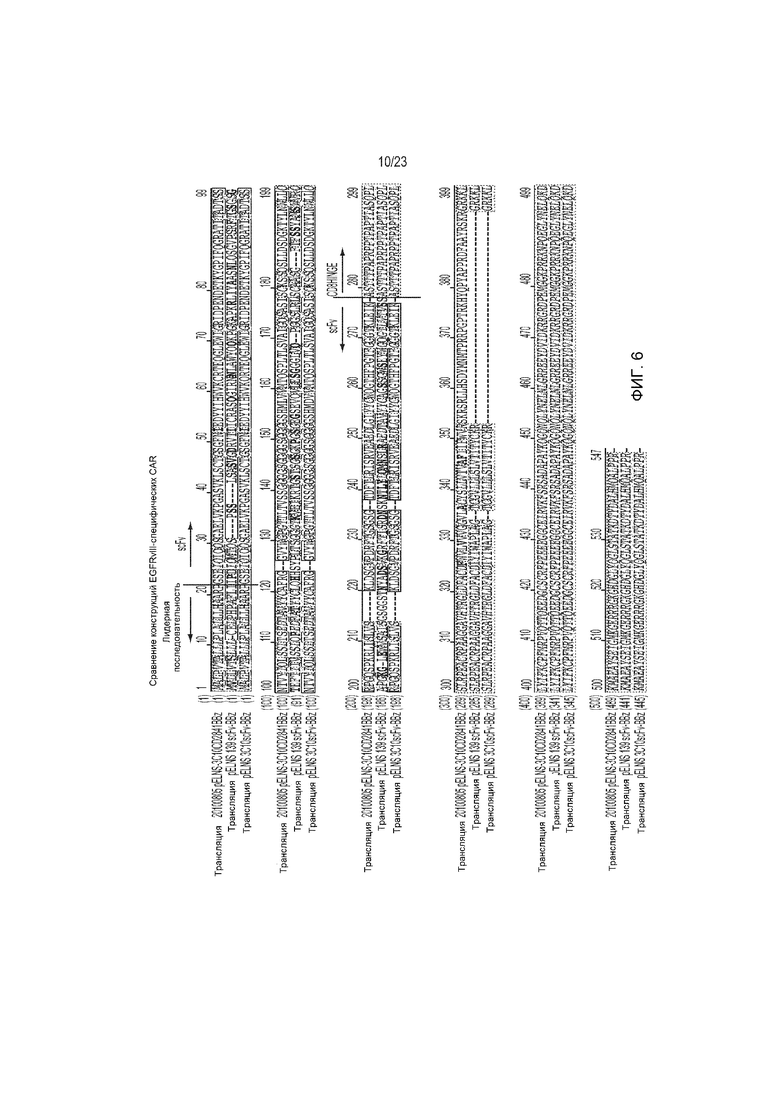
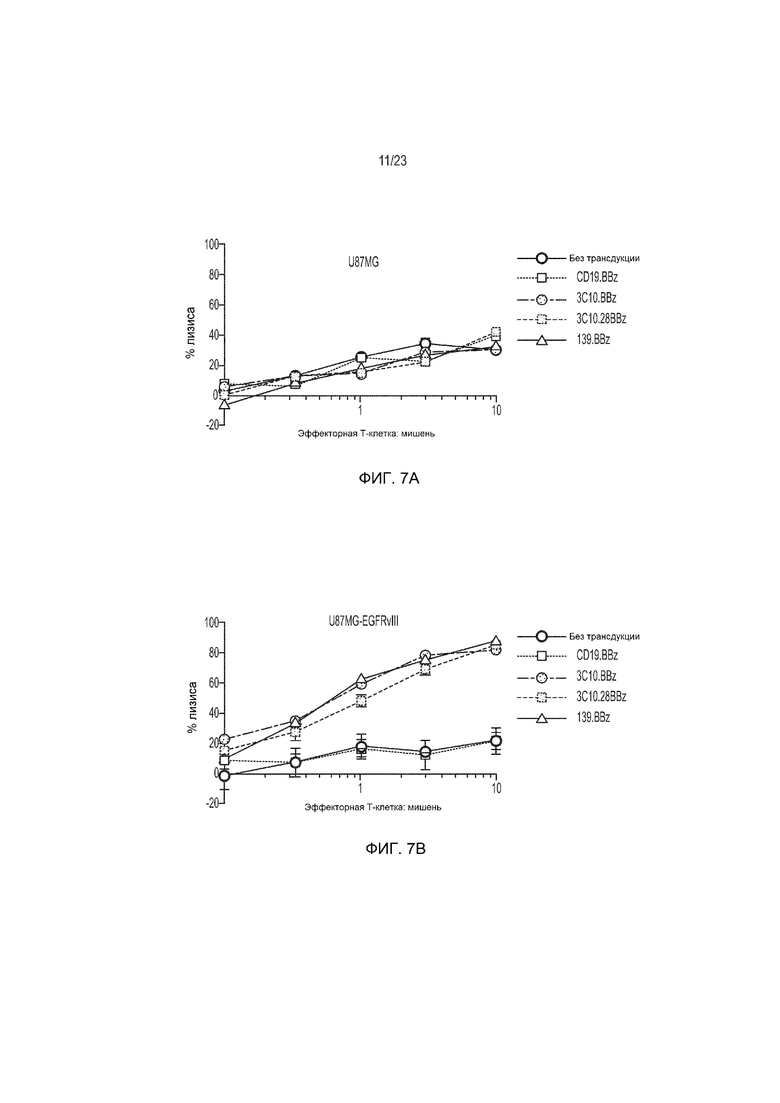
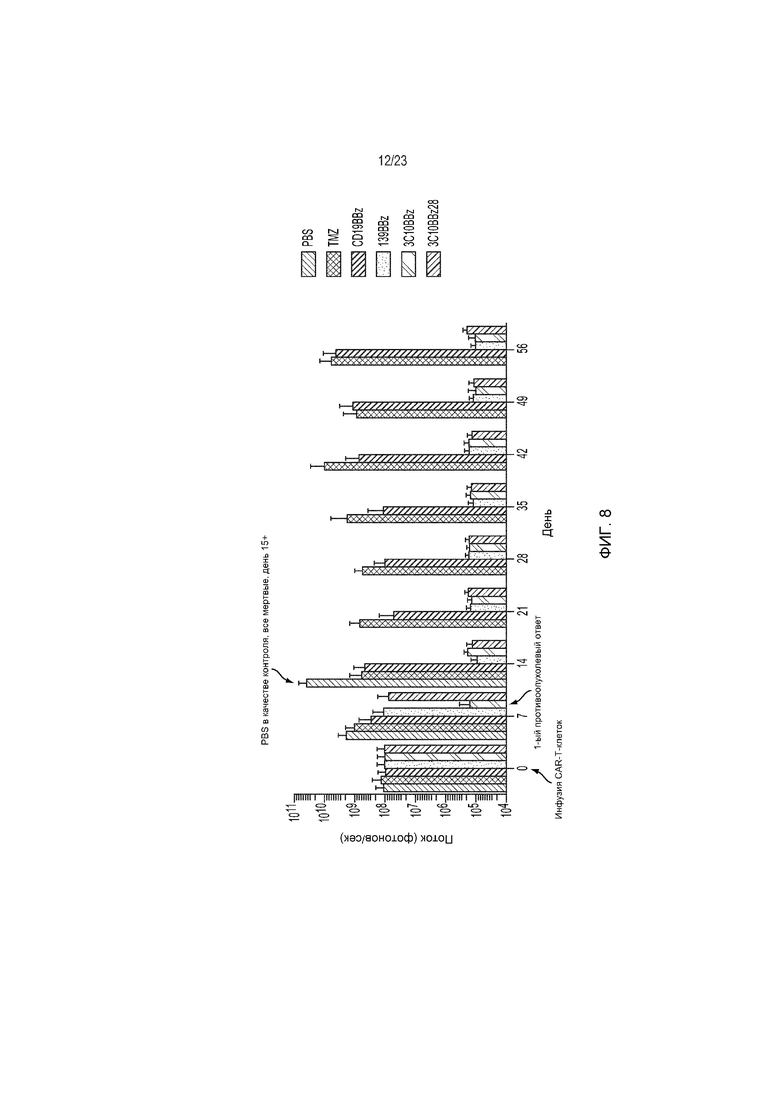
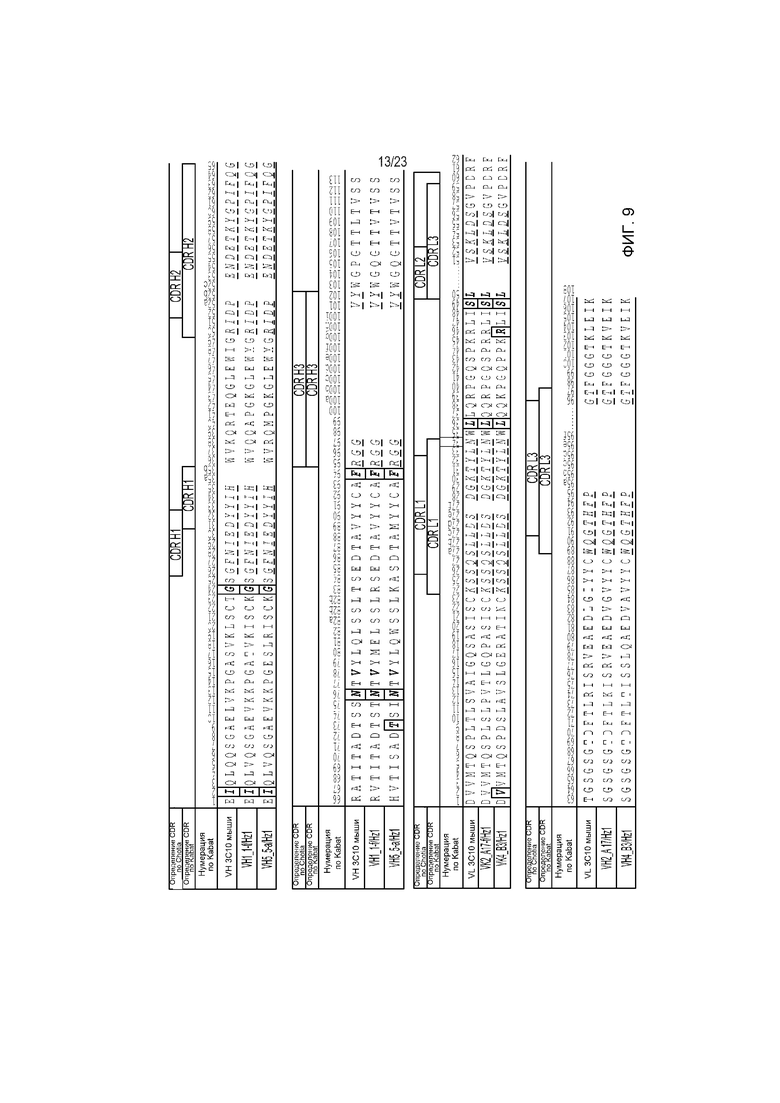
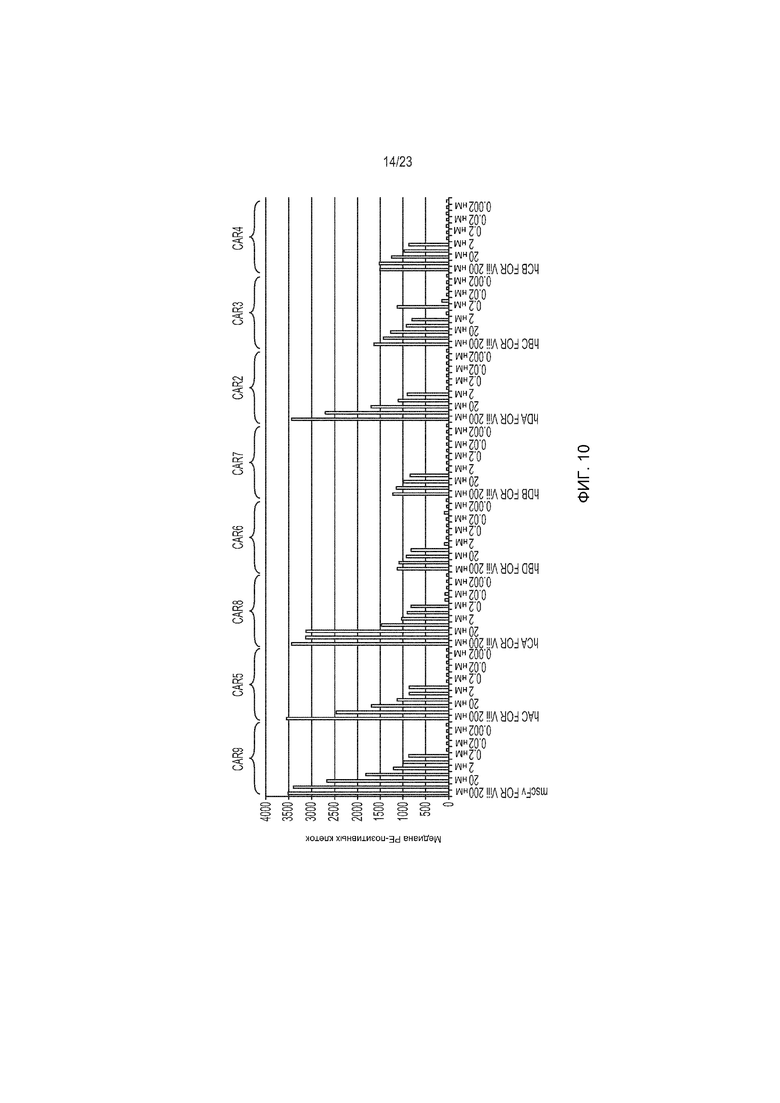
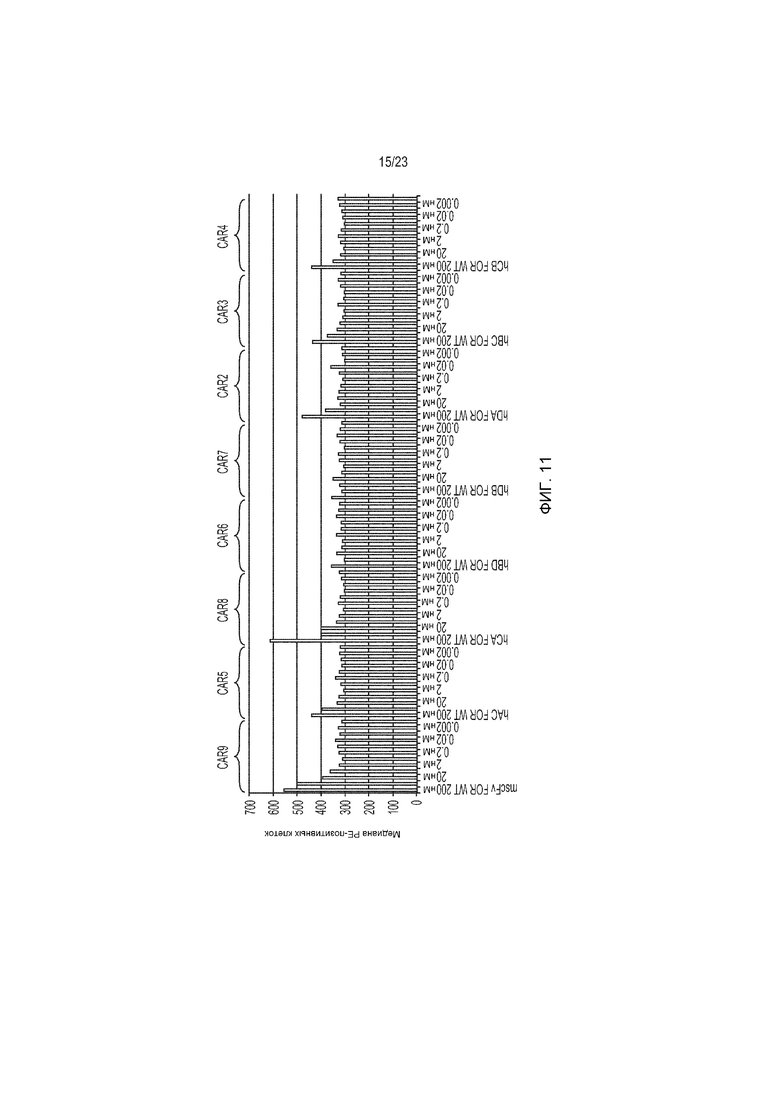
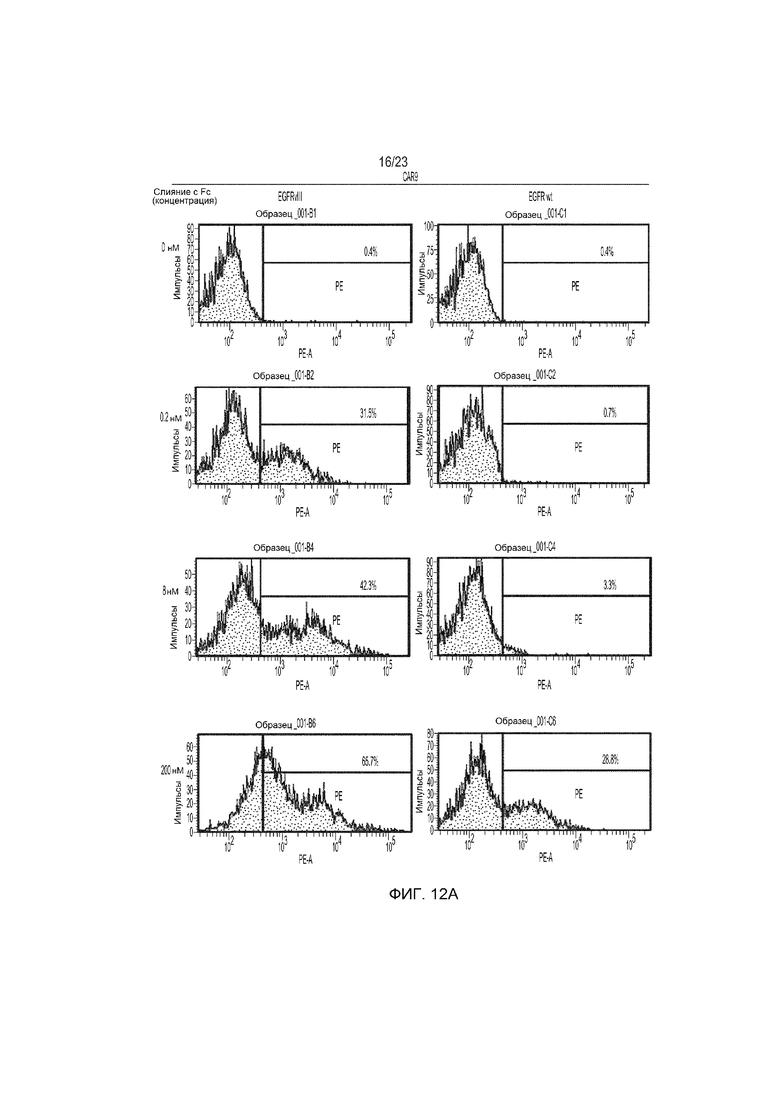
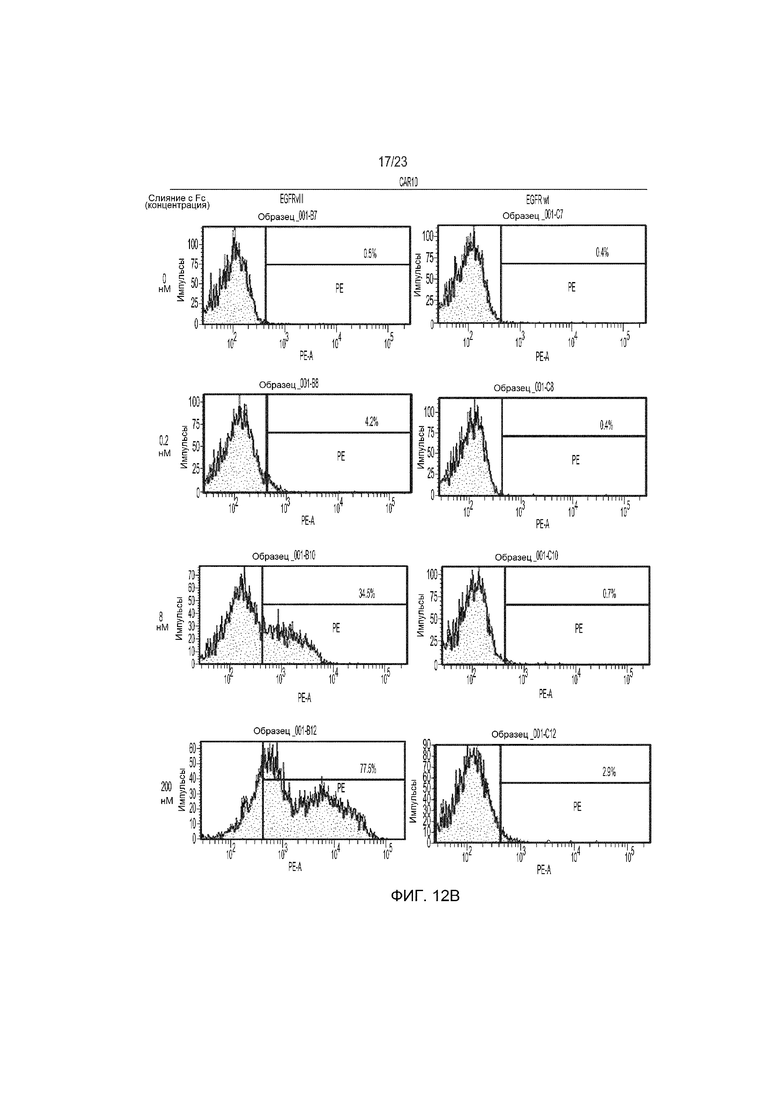

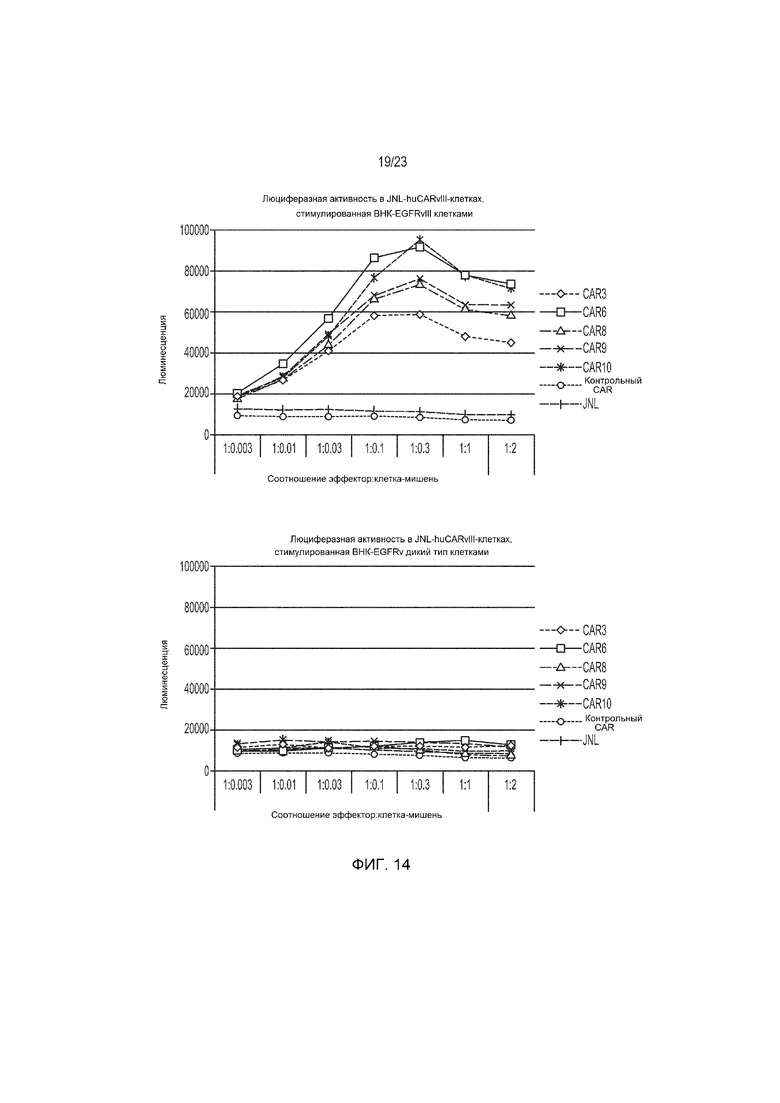
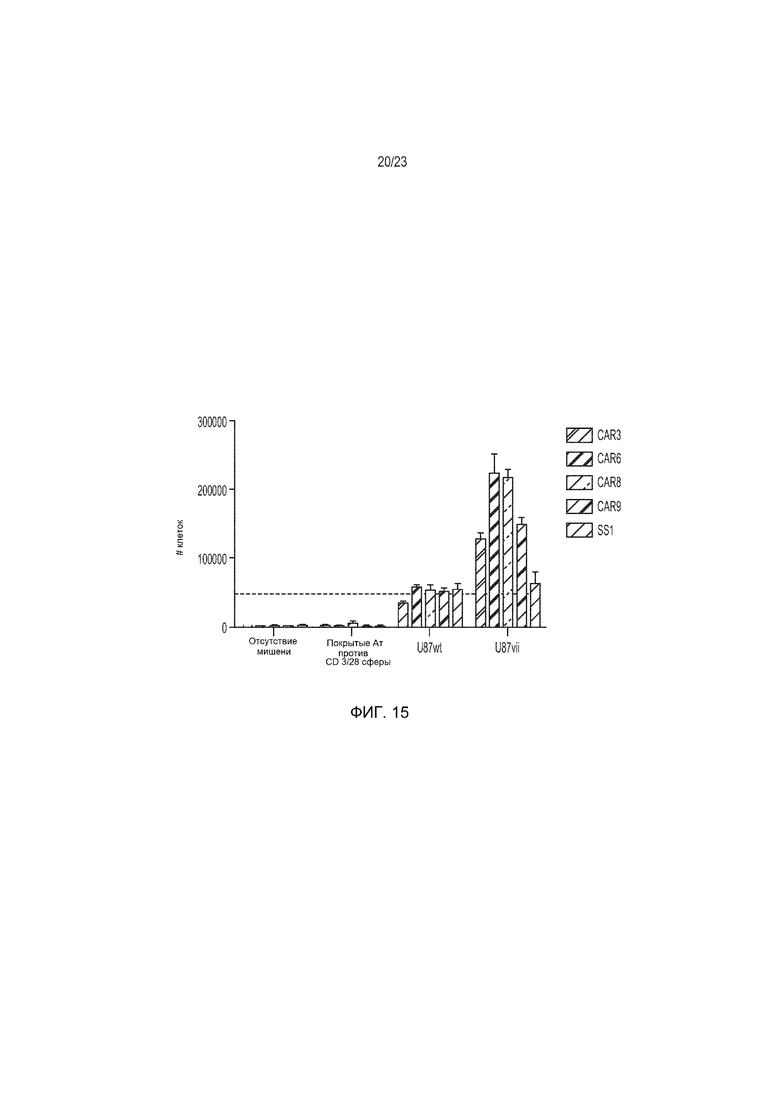
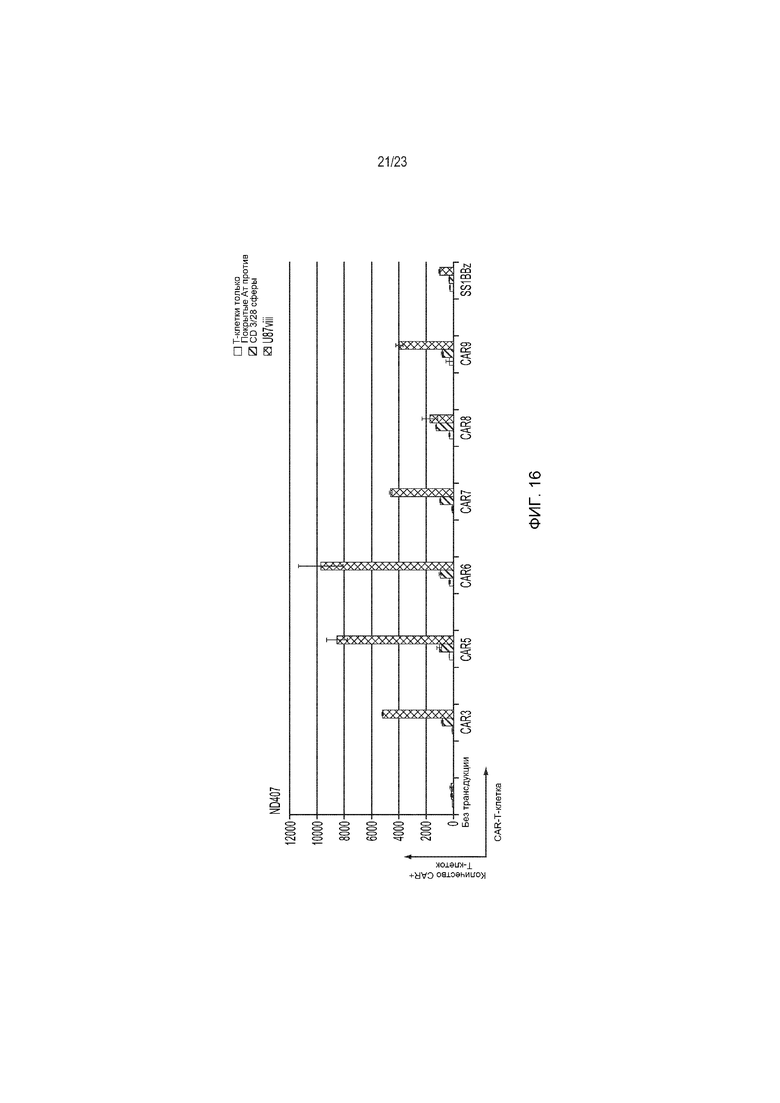
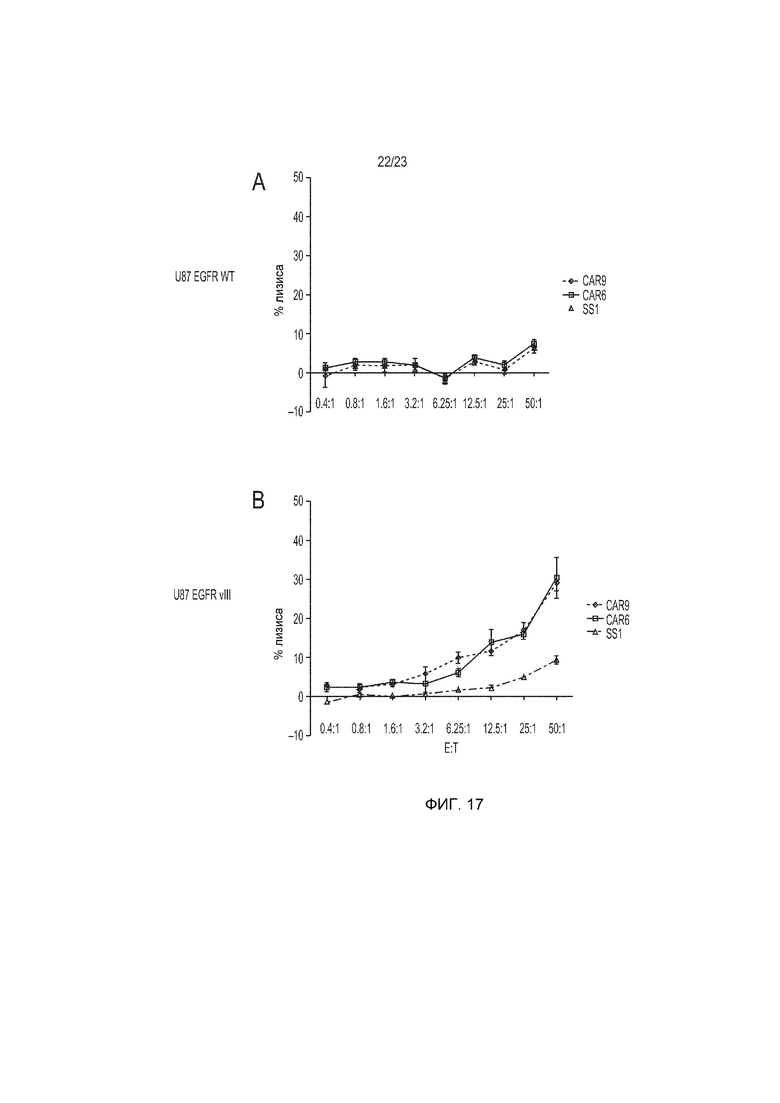
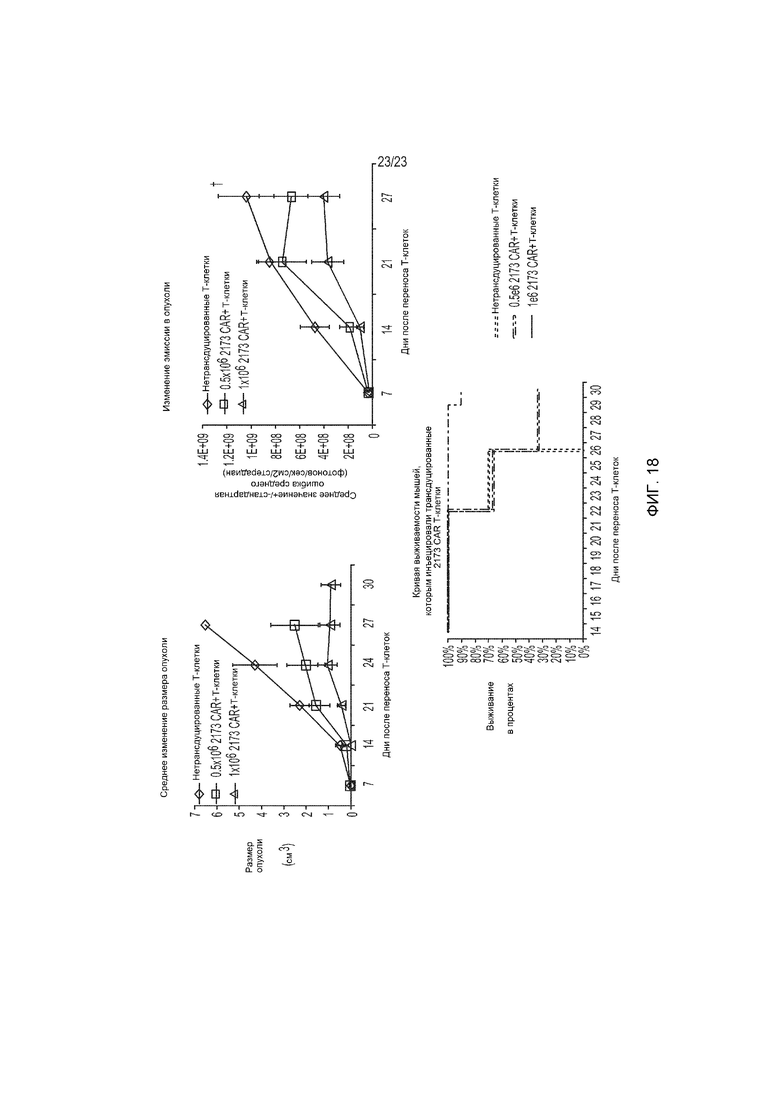
Группа изобретений относится к иммунологии. Предложен антигенсвязывающий домен, который связывает EGFRvIII и содержит CDR1 LC, CDR2 LC, CDR3 LC, CDR1 HC, CDR2 HC и CDR3 HC в соответствии с SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 22, SEQ ID NO: 23 и SEQ ID NO: 24. Также предложен содержащий указанный домен химерный антигенный рецептор, кодирующие нуклеиновые кислоты, вектор, клетка, а также применения указанных изобретений в производстве лекарственного средства, способы создания клетки и получения популяции клеток, способ обеспечения иммунитета, способ лечения заболевания, ассоциированного с экспрессией EGFRvIII. Изобретения могут быть использованы в терапии заболевания, ассоциированного с экспрессией EGFRvIII, с использованием адоптивного переноса клеток. 17 н. и 50 з.п. ф-лы, 18 ил., 2 табл., 9 пр.
1. Антигенсвязывающий домен, который связывает EGFRvIII, где указанный антигенсвязывающий домен содержит вариабельную область легкой цепи и вариабельную область тяжелой цепи, и где:
(i) вариабельная область легкой цепи содержит:
(a) определяющий комплементарность участок 1 легкой цепи (CDR1 LC), содержащий аминокислотную последовательность SEQ ID NO: 26;
(b) определяющий комплементарность участок 2 легкой цепи (CDR2 LC), содержащий аминокислотную последовательность SEQ ID NO: 27; и
(c) определяющий комплементарность участок 3 легкой цепи (CDR3 LC), содержащий аминокислотную последовательность SEQ ID NO: 28;
(ii) вариабельная область тяжелой цепи содержит:
(a) определяющий комплементарность участок 1 тяжелой цепи (CDR1 НС), содержащий аминокислотную последовательность SEQ ID NO: 22;
(b) определяющий комплементарность участок 2 тяжелой цепи (CDR2 НС), содержащий аминокислотную последовательность SEQ ID NO: 23; и
(c) определяющий комплементарность участок 3 тяжелой цепи (CDR3 НС), содержащий аминокислотную последовательность SEQ ID NO: 24; и
(iii) антигенсвязывающий домен содержит аминокислотную последовательность, имеющую не более 30 аминокислотных модификаций по сравнению с аминокислотной последовательностью вариабельной области легкой цепи и вариабельной области тяжелой цепи SEQ ID NO: 80, где указанные аминокислотные модификации не содержатся в CDR1 LC, CDR2 LC, CDR3 LC, CDR1 HC, CDR2 HC и CDR3 HC.
2. Антигенсвязывающий домен по п.1, где указанные аминокислотные модификации: (i) находятся в каркасной области указанного антигенсвязывающего домена против EGFRvIII, и (ii) представляют собой точечные мутации.
3. Антигенсвязывающий домен по п.1 или 2, где:
(a) вариабельная область легкой цепи содержит:
(i) аминокислотную последовательность с не более чем 20 или 10 аминокислотными модификациями аминокислотной последовательности вариабельной области легкой цепи SEQ ID NO: 80, где указанные аминокислотные модификации не содержатся в CDR1 LC, CDR2 LC или CDR3 LC;
(ii) аминокислотную последовательность с 95-99% идентичностью аминокислотной последовательности вариабельной области легкой цепи SEQ ID NO: 80, где указанные аминокислотные модификации не содержатся в CDR1 LC, CDR2 LC или CDR3 LC; или
(iii) аминокислотную аминокислотную последовательность вариабельной области легкой цепи SEQ ID NO: 80; и
(b) вариабельная область тяжелой цепи содержит:
(i) аминокислотную последовательность с не более чем 20 или 10 аминокислотными модификациями аминокислотной последовательности вариабельной области тяжелой цепи SEQ ID NO: 80, где указанные аминокислотные модификации не содержатся в CDR1 HC, CDR2 HC или CDR3 HC;
(ii) аминокислотную последовательность с 95-99% идентичностью аминокислотной последовательности вариабельной области тяжелой цепи SEQ ID NO: 80, где указанные аминокислотные модификации не содержатся в CDR1 HC, CDR2 HC или CDR3 HC; или
(iii) аминокислотную последовательность вариабельной области тяжелой цепи SEQ ID NO: 80.
4. Антигенсвязывающий домен против EGFRvIII по любому из пп.1-3, содержащий аминокислотную последовательность, имеющую по меньшей мере одну, две или три аминокислотные модификации, но не более чем 30, 20 или 10 аминокислотных модификаций аминокислотной последовательности SEQ ID NO: 80, где указанные аминокислотные модификации не содержатся в CDR1 LC, CDR2 LC, CDR3 LC, CDR1 HC, CDR2 HC и CDR3 HC.
5. Антигенсвязывающий домен против EGFRvIII по любому из пп.1-3, содержащий аминокислотную последовательность с 95-99% идентичностью аминокислотной последовательности SEQ ID NO: 80, где указанные аминокислотные модификации не содержатся в CDR1 LC, CDR2 LC, CDR3 LC, CDR1 HC, CDR2 HC и CDR3 HC.
6. Антигенсвязывающий домен против EGFRvIII по любому из пп.1-3, содержащий аминокислотную последовательность SEQ ID NO: 80.
7. Выделенная молекула нуклеиновой кислоты, кодирующая антигенсвязывающий домен по любому из пп.1-6.
8. Молекула химерного антигенспецифического рецептора (CAR), включающая антигенсвязывающий домен по любому из пп.1-6, трансмембранный домен и внутриклеточный домен сигнализации.
9. Молекула CAR по п.8, где внутриклеточный домен сигнализации включает костимулирующий домен и/или домен первичной сигнализации.
10. Молекула CAR по п.8 или 9, где антигенсвязывающий домен против EGFRvIII включает антитело или фрагмент антитела.
11. Молекула CAR по любому из пп.8-10, где антигенсвязывающий домен против EGFRvIII представляет собой scFv.
12. Молекула CAR по любому из пп.8-11, где указанный трансмембранный домен содержит трансмембранный домен белка, выбираемого из группы, состоящей из альфа, бета или дзета-цепи T-клеточного рецептора, CD28, CD3 эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD-134, CD-137 и CD154.
13. Молекула CAR по п.12, где трансмембранный домен включает аминокислотную последовательность SEQ ID NO: 15.
14. Молекула CAR по п.12, где трансмембранный домен включает аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации, но не более 20, 10 или 5 модификаций аминокислотной последовательности SEQ ID NO: 15, или аминокислотную последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 15.
15. Молекула CAR по любому из пп.8-14, где антигенсвязывающий домен антитела против EGFRvIII соединен с трансмембранным доменом с помощью шарнирной области.
16. Молекула CAR по п.15, где шарнирная область включает аминокислотную последовательность SEQ ID NO: 14 или аминокислотную последовательность, идентичную ей на 95-99%.
17. Молекула CAR по любому из пп.8-16, где указанный внутриклеточный домен сигнализации включает костимулирующий домен.
18. Молекула CAR по п.17, где костимулирующий домен включает функциональный домен сигнализации белка, выбираемого из группы, состоящей из OX40, CD27, CD28, ICAM-1, LFA-1 (CD11а/CD18), ICOS (CD278) и 4-1ВВ (CD137).
19. Молекула CAR по п.17, где костимулирующий домен включает аминокислотную последовательность SEQ ID NO: 16.
20. Молекула CAR по п.17, где костимулирующий домен включает аминокислотную последовательность, содержащую по крайней мере одну, две или три модификации, но не более 20, 10 или 5 модификаций аминокислотной последовательности SEQ ID NO: 16, или аминокислотную последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 16.
21. Молекула CAR по любому из пп.8-20, где указанный внутриклеточный домен сигнализации включает функциональный домен сигнализации.
22. Молекула CAR по п.21, где внутриклеточный домен сигнализации включает функциональный домен сигнализации 4-1BB и/или функциональный домен сигнализации CD3 дзета.
23. Молекула CAR по п.21, где внутриклеточный домен сигнализации включает аминокислотную последовательность SEQ ID NO: 16 и/или аминокислотную последовательность SEQ ID NO: 17.
24. Молекула CAR по п.21, где внутриклеточный домен сигнализации включает аминокислотную последовательность SEQ ID NO: 16 и/или аминокислотную последовательность SEQ ID NO: 99.
25. Молекула CAR по п.21, где внутриклеточный домен сигнализации включает аминокислотную последовательность, имеющую по крайней мере одну, две или три модификации, но не более 20, 10 или 5 модификаций аминокислотной последовательности SEQ ID NO: 16 и/или аминокислотной последовательности SEQ ID NO: 17 или SEQ ID NO: 99, или последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 16 и/или аминокислотной последовательности SEQ ID NO: 17 или SEQ ID NO: 99.
26. Молекула CAR по п.21, где внутриклеточный домен сигнализации включает аминокислотную последовательность SEQ ID NO: 16 и аминокислотную последовательность SEQ ID NO: 17 или SEQ ID NO: 99, где аминокислотные последовательности, включающие внутриклеточный домен сигнализации, экспрессируются в одной и той же рамке и в виде одной полипептидной цепи.
27. Молекула CAR по любому из пп.8-26, дополнительно включающая лидерную последовательность.
28. Молекула CAR по п.27, где лидерная последовательность включает аминокислотную последовательность SEQ ID NO: 13 или аминокислотную последовательность, идентичную на 95-99% аминокислотной последовательности SEQ ID NO: 13.
29. Молекула CAR по любому из пп.8-28, содержащая аминокислотную последовательность SEQ ID NO: 85 или аминокислотную последовательность, идентичную ей на 95-99%.
30. Выделенная молекула нуклеиновой кислоты, кодирующая молекулу CAR по любому из пп.8-29.
31. Выделенная молекула нуклеиновой кислоты по п.30, где последовательность нуклеиновой кислоты, кодирующая антигенсвязывающий домен, включает последовательность нуклеиновой кислоты SEQ ID NO: 80 или последовательность нуклеиновой кислоты, идентичную ей на 95-99%, где указанные модификации не содержатся в последовательностях нуклеиновой кислоты, кодирующих CDR1 LC, CDR2 LC, CDR3 LC, CDR1 HC, CDR2 HC и CDR3 HC.
32. Выделенная молекула нуклеиновой кислоты по п.30 или 31, где последовательность нуклеиновой кислоты, кодирующая трансмембранный домен, включает последовательность нуклеиновой кислоты SEQ ID NO: 8 или последовательность, идентичную ей на 95-99%.
33. Выделенная молекула нуклеиновой кислоты по любому из пп.30-32, где кодируемый антигенсвязывающий домен против EGFRvIII соединен с трансмембранным доменом шарнирной областью, и где последовательность нуклеиновой кислоты, кодирующая шарнирную область, включает последовательность нуклеиновой кислоты SEQ ID NO: 7 или последовательность нуклеиновой кислоты, идентичную ей на 95-99%.
34. Выделенная молекула нуклеиновой кислоты по любому из пп.30-33, где кодируемый внутриклеточный домен сигнализации содержит костимулирующий домен, и где последовательность нуклеиновой кислоты, кодирующая костимулирующий домен, включает последовательность нуклеиновой кислоты SEQ ID NO: 9 или последовательность нуклеиновой кислоты, идентичную ей на 95-99%.
35. Выделенная молекула нуклеиновой кислоты по любому из пп.30-34, где последовательность нуклеиновой кислоты, кодирующая внутриклеточный домен сигнализации, включает последовательность нуклеиновой кислоты SEQ ID NO: 9 или последовательность нуклеиновой кислоты, идентичную ей на 95-99%, и/или последовательность нуклеиновой кислоты SEQ ID NO: 10 или SEQ ID NO: 100, или последовательность нуклеиновой кислоты, идентичную ей на 95-99%.
36. Экспрессионный вектор, включающий молекулу нуклеиновой кислоты, кодирующую молекулу CAR по любому из пп.8-29.
37. Вектор по п.36, где вектор выбирают из группы, состоящей из ДНК, РНК, плазмиды, лентивирусного вектора, аденовирусного вектора или ретровирусного вектора.
38. Вектор по п.36 или 37, дополнительно включающий промотор.
39. Вектор по п.38, где промотор представляет собой промотор EF-1.
40. Вектор по п.39, где промотор EF-1 включает последовательность нуклеиновой кислоты SEQ ID NO: 97.
41. Вектор по любому из пп.36-40, где вектор представляет собой in vitro транскрибируемый вектор.
42. Вектор по любому из пп.36-41, где последовательность нуклеиновой кислоты в векторе дополнительно включает поли(А) хвост.
43. Вектор по любому из пп.36-42, где последовательность нуклеиновой кислоты в векторе дополнительно включает 3' UTR.
44. Клетка для экспрессии молекулы CAR по любому из пп.8-29, включающая вектор по любому из пп.36-43.
45. Клетка по п.44, где клетка представляет собой Т-клетку.
46. Клетка по п.45, где Т-клетка представляет собой CD8+ Т-клетку.
47. Клетка по любому из пп.44-46, где клетка представляет собой клетку человека.
48. Способ создания T-клетки, включающий трансдукцию Т-клетки вектором по любому из пп.36-43.
49. Способ получения популяции созданных с использованием РНК клеток, включающий введение in vitro транскрибированной РНК или синтетической РНК в клетку, где РНК включает нуклеиновую кислоту, кодирующую молекулу CAR по любому из пп.8-29.
50. Способ обеспечения иммунитета к опухоли у млекопитающего, имеющего заболевание, ассоциированное с экспрессией EGFRvIII, включающий введение млекопитающему эффективного количества популяции Т-клеток, экспрессирующих молекулу CAR по любому из пп.8-29.
51. Способ лечения млекопитающего, имеющего заболевание, связанное с экспрессией EGFRvIII, включающий введение млекопитающему эффективного количества популяции Т-клеток, экспрессирующих молекулу CAR по любому из пп.8-29.
52. Способ по п.50 или 51, где Т-клетки представляют собой аутологичные Т-клетки или аллогенные Т-клетки.
53. Способ по любому из пп.50-52, где Т-клетки представляют собой CD8+ Т-клетки.
54. Способ по любому из пп.50-53, где указанное млекопитающее представляет собой человека.
55. Способ по любому из пп.50-54, где заболевание, связанное с EGFRvIII, представляет собой глиобластому.
56. Способ по любому из пп.50-54, где заболевание, связанное с EGFRvIII, представляет собой рак, выбираемый из группы, состоящей из мультиформной глиобластомы (GBM), анапластической астроцитомы, гигантоклеточной глиобластомы, глиосаркомы, анапластической олигодендроглиомы, анапластической эпендимомы, карциномы сосудистого сплетения, анапластической ганглиоглиомы, пинеобластомы, медуллоэпителиомы, эпендимобластомы, медуллобластомы, супратенториальной примитивной нейроэктодермальной опухоли, атипичной тератоидной/рабдоидной опухоли, рака легкого, карциномы молочной железы, карциномы предстательной железы, карциномы яичников, колоректальной карциномы и карциномы мочевого пузыря и любой их комбинации, и метастазов любого из раков.
57. Способ по п.56, где рак легкого представляет собой немелкоклеточную карциному легкого.
58. Способ по любому из пп.50-57, где клетки, экспрессирующие молекулу CAR, вводят в комбинации с агентом, который увеличивает эффективность клетки, экспрессирующей молекулу CAR.
59. Способ по любому из пп.50-58, где клетки, экспрессирующие молекулу CAR, вводят в комбинации с агентом, который уменьшает интенсивность одного или более побочных эффектов, связанных с введением клетки, экспрессирующей молекулу CAR.
60. Способ по любому из пп.50-59, где клетки, экспрессирующие молекулу CAR, вводят в комбинации с агентом, который лечит заболевание, связанное с EGFRvIII.
61. Применение молекулы нуклеиновой кислоты по любому из пп.7 или 30-35 в производстве лекарственного средства для лечения заболевания, ассоциированного с экспрессией EGFRvIII.
62. Применение молекулы CAR по любому из пп.8-29 в производстве лекарственного средства для лечения заболевания, ассоциированного с экспрессией EGFRvIII.
63. Применение антигенсвязывающего домена по любому из пп.1-6 в производстве лекарственного средства для лечения заболевания, ассоциированного с экспрессией EGFRvIII.
64. Применение Т-клетки, содержащей CAR, содержащий антигенсвязывающий домен по любому из пп.1-6, в качестве лекарственного средства для лечения заболевания, ассоциированного с экспрессией EGFRvIII.
65. Применение Т-клетки, содержащей CAR, содержащий антигенсвязывающий домен по любому из пп.1-6, в производстве лекарственного средства для лечения заболевания, ассоциированного с экспрессией EGFRvIII.
66. Применение вектора по любому из пп.36-43 в производстве лекарственного средства для лечения заболевания, ассоциированного с экспрессией EGFRvIII.
67. Применение Т-клетки по п.45 или 46 в производстве лекарственного средства для лечения заболевания, ассоциированного с экспрессией EGFRvIII.
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| OHNO M | |||
| et al | |||
| Retrovirally engineered T-cell-based immunotherapy targeting type III variant epidermal growth factor receptor, a glioma-associated antigen | |||
| Cancer science, 2010, v.101, no.12, p.2518-2524 | |||
| NAKAYASHIKI N | |||
| et al | |||
| Production of a Single-chain Variable Fragment Antibody Recognizing Type III Mutant | |||

