Область техники, к которой относится изобретение
Настоящая заявка относится к области биотехнологии, в частности, она касается способа селекции гибридов пшеницы, включающего получение ядерной стерильной линии пшеницы и получение гибридных семян, более конкретно ядерного гена мужской стерильности пшеницы и его мутантов и применения при селекции гибридов.
Уровень техники
Пшеница - самоопыляемая культура, и в основе применения ее гетерозиса лежит создание технической системы для эффективного получения гибридных семян пшеницы. В настоящее время существует три главных способа применения гетерозиса в производстве пшеницы: первый способ - трехлинейная система, то есть для получения гибридных семян используется линия с мужской стерильностью (male sterile line) типа ядерно-цитоплазматического взаимодействия посредством взаимодействия стерильной линии, поддерживающей линии и восстановительной линии; второй способ - использование линии с химически индуцированной мужской стерильностью, то есть для получения гибридных семян применяются средства химической гибридизации, индуцирующие у пшеницы линии с мужской стерильностью; и третий способ - двухлинейная система, то есть для получения гибридных семян используется фототермочувствительная стерильная линия, фертильность которой изменяется в зависимости от фотопериода и температуры. Трехлинейная система исследовалась еще с 1950-х годов, до настоящего времени было выведено более 70 линий пшеницы с мужской стерильностью типа ядерно-цитоплазматического взаимодействия, и наиболее изученными линиями с мужской стерильностью являются линии T-типа, K-типа и Q-типа, цитоплазма которых в основном происходит из различных видов Aegilops, Triticum timopheevii, дикого овса, Haynaldia villosa и др. Хотя были получены комбинации из трех линий для большинства таких стерильных линий, однако из-за присущих им непреодолимых генетических дефектов, таких как меньшее количество восстановительных источников, неблагоприятное влияние чужеродной цитоплазмы, низкий уровень отбора сильно превосходящих комбинаций и низкая чистота семян стерильных линий, эти стерильные линии не получили широкого применения в производстве. В 1980-х годах достигли наивысшей точки исследования по методу химического гибридинга. Поскольку метод химического гибридинга преодолевает различные проблем трехлинейной системы, то он когда-то был известен как перспективная новая технология получения гибридных семян. Однако химические гибридизирующие средства имеют много недостатков, таких как низкая стабильность, отходы химических веществ, токсические и побочные эффекты химических веществ, сильное загрязнение окружающей среды и т.п. Еще предстоит дальнейшая разработка идеальных химических гибридизирующих средств, чтобы их можно было широко использовать в производстве. Поскольку трехлинейная система и метод химического гибридинга сталкиваются со многими трудностями, то быстро развивается двухлинейная система гибридной пшеницы, которая постепенно становится главной тенденцией развития исследований и применения гетерозиса пшеницы в будущем.
В основе двухлинейной системы лежит фототермочувствительная линия мужской стерильности, которая является разновидностью линий с мужской стерильностью, вызванной взаимодействием между генотипом и окружающей средой. Фототермочувствительная линия мужской стерильности обладает двойной функцией (это и линия мужской стерильности, и поддерживающая линия), что упрощает процесс воспроизведения стерильной линии, упрощает получение семян, имеет более широкий диапазон восстановительных линий, облегчает селекцию гибридов с превосходным гетерозисом, поэтому двухлинейная система имеет очень большую степень популяризации. В настоящее время оригинальная двухлинейная система технологии получения гибридной пшеницы в Китае опережает международный уровень, отобранные гибридные сорта пшеницы обладают сильной устойчивостью к стрессам и высокой степенью утилизации воды и удобрений, а повышение урожайности может достигать 15-20%, что оказывает существенное влияние на снижение себестоимости и повышение эффективности производства зерна. Однако мужская стерильность у фототермочувствительной линии стерильности сильно зависит от факторов окружающей среды, а изменение окружающей среды сильно влияет на урожайность, качество и чистоту гибридных семян, поэтому может потребоваться какое-то время, чтобы двухлинейный метод получения гибридной пшеницы пробился в производство, причем ключевым моментом является решение проблемы влияния окружающей среды.
Поскольку вышеуказанные традиционные стандартные способы селекции наталкиваются на узкие места, то ключевым моментом для преодоления узких мест может быть биологическая технология селекции, в которой главное место занимает современная молекулярная биология. В основе применения гетерозиса пшеницы лежит создание эффективной технической системы для получения гибридных семян пшеницы, а ключевым моментом создания эффективной технологии получения гибридных семян пшеницы является получение подходящей линии с мужской стерильностью в качестве женской родительской. Мужская стерильность означает то, что растение не может производить пыльники, пыльцу или мужские гаметы с нормальными функциями в процессе полового размножения, а механизм мужской стерильности является основой для повышения урожайности и качества пшеницы с помощью гетерозиса.
Геном пшеницы огромен и сложен, поэтому до сих пор накоплено мало информации о механизме мужской стерильности у пшеницы. Поэтому применение современной молекулярной биологии и клеточной биологии для исследования механизма мужской стерильности у пшеницы имеет важное теоретическое и практическое значение для расширения исследований и применения гетерозиса пшеницы.
Cущность изобретения
Все ссылки, приведенные в заявке, включены в нее путем ссылки.
Если не указано иначе, все технические и научные термины, используемые в заявке, имеют такие значения, которые понятны рядовым специалистам в данной области. Если не указано иначе, методы, используемые или упомянутые в заявке, представлены стандартными методиками, которые общеизвестны рядовым специалистам в данной области. Материалы, методы и примеры приводятся только для объяснения, а не для ограничения.
Изобретением предусмотрен связанный с фертильностью ген TaMS7, причем нуклеотидная последовательность связанного с фертильностью гена выбрана из одной из следующих групп последовательностей:
(a) нуклеотидных последовательностей, приведенных в SEQ ID NO:1, 2, 3, 4, 5 и 6;
(b) нуклеотидных последовательностей, кодирующих аминокислотные последовательности, приведенные в SEQ ID NO: 7, 8 и 9;
(c) последовательностей ДНК, способных гибридизироваться с приведенными в (a) или (b) последовательностями ДНК в строгих условиях; или
(d) последовательностей ДНК, имеющих сходство на 80% (предпочтительно по меньшей мере на 85%) с любыми из последовательностей (a)-(c) и обладающих связанной с фертильностью функцией; или
(e) последовательностей ДНК, комплементарных любым из последовательностей (a)-(d).
Специалистам в данной области следует отметить, что связанный с фертильностью ген по изобретению также включает последовательность гомологичного гена, который имеет высокую гомологию с нуклеотидной последовательностью или белковой последовательностью гена TaMS7 и обладает такой же функцией регуляции или восстановления фертильности. Гомологичный ген с высокой гомологией и функцией регуляции фертильности включает последовательность ДНК, способную гибридизироваться с последовательностью ДНК, приведенной в SEQ ID NO: 1, 2, 3, 4, 5 или 6, или нуклеотидную последовательность, кодирующую аминокислотную последовательность, который имеет сходство на 85% и более с аминокислотной последовательностью, приведенной в SEQ ID NO: 7, 8 или 9. “Строгие условия”, используемые в изобретении, хорошо известны, к примеру, это гибридизация в течение 12-16 часов при 53-60°C в гибридизационном растворе, содержащем 400 мМ NaCl, 40 мМ PIPES (рН 6,4) и 1 мМ ЭДТА, а затем отмывка промывочным раствором, содержащим 0,5×SSC и 0,1% SDS, в течение 15-60 мин при 62-68°C.
Вышеуказанный гомологичный ген также включает последовательность ДНК, которая имеет сходство по меньшей мере на 80%, 85%, 90%, 95%, 98% или 99% с полноразмерной последовательностью, приведенной в SEQ ID NO: 1, 2, 3, 4, 5 или 6, а также обладает функцией регуляции фертильности и может быть выделена из любого растения. Степень сходства последовательностей можно получить по общедоступному алгоритму биоинформатики, включая алгоритм Myers and Miller, метод глобального совмещения Needleman-Wunsch, метод локального совмещения Smith-Waterman, метод поиска сходства Pearson and Lipman и алгоритм Karlin and Altschul. Они хорошо известны специалистам в данной области.
Изобретением также предусмотрена экспрессионная кассета, которая содержит последовательность ДНК связанного с фертильностью гена, предусмотренного изобретением, причем нуклеотидная последовательность связанного с фертильностью гена выбрана из одной из следующих групп последовательностей:
(a) нуклеотидных последовательностей, приведенных в SEQ ID NO:1, 2, 3, 4, 5 и 6;
(b) нуклеотидных последовательностей, кодирующих аминокислотные последовательности, приведенные в SEQ ID NO: 7, 8 и 9;
(c) последовательностей ДНК, способных гибридизироваться с приведенными в (a) или (b) последовательностями ДНК в строгих условиях; или
(d) последовательностей ДНК, имеющих сходство на 80% (предпочтительно по меньшей мере на 85%) с любыми из последовательностей (a)-(c) и обладающих связанной с фертильностью функцией; или
(e) последовательностей ДНК, комплементарных любым из последовательностей (a)-(d).
В частности, связанный с фертильностью ген в такой экспрессионной кассете также функционально связан с промотором, который может управлять экспрессией связанного с фертильностью гена, причем промоторы включают, без ограничения, конститутивные экспрессирующие промоторы, индуцибельные промоторы, тканеспецифичные промоторы или пространственно-временные промоторы. Более предпочтительно промотор представлен специфичным для пыльников промотором. Предпочтительно нуклеотидная последовательность специфичного для пыльников промотора представлена в SEQ ID NO: 16, 17 или 18.
Экспрессионная кассета по изобретению также включает в себя ген инактивации пыльцы, причем ген инактивации пыльцы может нарушать функцию или образование мужских гамет, содержащих ген инактивации пыльцы, у растений. Гены инактивации пыльцы включают, без ограничения, ген барназы, ген амилазы, ген метилазы DAM и т.п. Более предпочтительно ген инактивации пыльцы представлен геном α-амилазы кукурузы, а нуклеотидная последовательность гена инактивации пыльцы предпочтительно представлена в SEQ ID NO: 25.
Экспрессионная кассета по изобретению также включает в себя отборочный ген, который может использоваться для отбора растений, клеток растительной ткани или вектора, содержащего экспрессионную кассету. Отборочные гены включают, без ограничения, гены устойчивости к антибиотикам или гены устойчивости к гербицидам или гены флуоресцентных белков и т.п. В частности, отборочные гены включают, без ограничения, гены устойчивости к хлорамфениколу, гены устойчивости к гигромицину, гены устойчивости к стрептомицину, гены устойчивости к мирамицину, гены устойчивости к сульфонамидам, гены устойчивости к глифосату, гены устойчивости к фосфинотрицину, ген bar, ген красной флуоресценции DsRED, ген mCherry, ген голубого флуоресцентного белка, ген желтого флуоресцентного белка, гены люциферазы, ген зеленого флуоресцентного белка и т.п.
Изобретением также предусмотрен способ регулирования фертильности растений, причем этот способ означает трансформирование мутанта ms7 с мужской стерильностью связанным с фертильностью геном для восстановления мужской фертильности мутанта ms7 с мужской стерильностью, причем нуклеотидная последовательность связанного с фертильностью гена выбрана из одной из следующих групп последовательностей:
(a) нуклеотидных последовательностей, приведенных в SEQ ID NO:1, 2, 3, 4, 5 и 6;
(b) нуклеотидных последовательностей, кодирующих аминокислотные последовательности, приведенные в SEQ ID NO: 7, 8 и 9;
(c) последовательностей ДНК, способных гибридизироваться с приведенными в (a) или (b) последовательностями ДНК в строгих условиях; или
(d) последовательностей ДНК, имеющих сходство на 80% (предпочтительно по меньшей мере на 85%) с любыми из последовательностей (a)-(c) и обладающих связанной с фертильностью функцией; или
(e) последовательностей ДНК, комплементарных любым из последовательностей (a)-(d).
Изобретением также предусмотрен способ регулирования фертильности растений путем воздействия на экспрессию гена фертильности TaMS7. Мутантный материал с мужской стерильность ms7 получают, без ограничения, методом мутации гена TaMS7; или же ген TaMS7 используется для преобразования фенотипа мужской стерильности, вызванного мутацией TaMS7, посредством комплементации генов с восстановлением фертильности у мутантов с мужской стерильностью ms7. Мутации по изобретению включают замены, делеции или вставки одного или нескольких нуклеотидов в нуклеотидной последовательности гена, регулирующего фертильность. Методы мутации генов включают, без ограничения, физический мутагенез, химический мутагенез, RNAi или редактирование генов типа TALEN, CRISPR-Cas9 и т.п.
Изобретение также включает способ получения мутантов с мужской стерильностью ms7, который представляет собой процесс, который включает мутирование эндогенного гена TaMS7, регулирующего фертильность, или нуклеотидной последовательности сильно гомологичного ему гена, получая растения с мужской стерильностью. Аминокислотная последовательность указанного выше регулирующего фертильность гена TaMS7 представлена в SEQ ID NO: 7, 8 или 9. Нуклеотидная последовательность регулирующего фертильность гена TaMS7 представлена в SEQ ID NO: 1, 2, 3, 4, 5 или 6. “Мутации” включают, без ограничения, мутации генов, вызванные следующими методами типа физических или химических методов, причем химический метод включает мутагенез при помощи таких мутагенов, как EMS и др., а мутации также могут означать точечные мутации либо делеции или вставки ДНК или же глушение генов при помощи RNAi, сайт-направленного мутагенеза и т.д., причем сайт-направленный мутагенез генов включает, без ограничения, методы редактирования генов типа ZFN, TALEN и/или CRISPR/Cas9 и т.п.
Изобретением также предусмотрен способ применения мутантов ms7, причем мутанты вызваны мутацией нуклеотидной последовательности, а растения с мутантной нуклеотидной последовательностью проявляют фенотип мужской стерильности, причем нуклеотидная последовательность представляет собой нуклеотидную последовательность гена TaMS7, предпочтительно приведенную в SEQ ID NO: 1, 2, 3, 4, 5 или 6. Применение мутантов с мужской стерильностью включает, без ограничения, применение в селекции гибридов, более конкретно растения-мутанты ms7 применяются в качестве женских растений с мужской стерильностью и гибридизируются с восстановительной линией для получения гибридных семян.
Изобретением также предусмотрен способ поддержания линий с мужской стерильностью. Способ включает использование мутантов с мужской стерильностью ms7 в качестве акцепторного материала при трансформации и трансформирование акцепторных растений с помощью 3 тесно связанных целевых генов. Эти 3 целевых гена - связанный с фертильностью ген TaMS7, ген инактивации пыльцы и ген селекционного маркера, соответственно. При этом связанный с фертильностью ген TaMS7 может восстанавливать фертильность стерильного акцептора трансформации, ген инактивации пыльцы может инактивировать пыльцу, содержащую трансформированный экзогенный ген, при этом теряется способность к оплодотворению, а селекционный ген может использоваться для отбора трансгенных семян или тканей и нетрансгенных семян или тканей, причем отобранные нетрансгенные семена используются в качестве стерильной линии для получения гибридных семян, а трансгенные семена используются в качестве поддерживающей линии для непрерывного и стабильного получения стерильной линии.
По изобретению, мутанты пшеницы с мужской стерильностью ms7 представляют собой мутанты с мужской стерильностью, вызванной мутацией гена TaMS7. Мутанты с мужской стерильностью по изобретению также могут называться линией стерильности ms7 или линией с мужской стерильностью ms7.
В данном способе поддержания линий с мужской стерильностью гены инактивации пыльцы включают, без ограничения, ген барназы, ген амилазы, ген метилазы DAM и т.п. Более предпочтительно ген инактивации пыльцы представлен геном α-амилазы кукурузы Zm-AA, а его нуклеотидная последовательность предпочтительно представлена в SEQ ID NO: 25. Ген инактивации пыльцы связан со специфичным для мужских гамет промотором. В частности, специфичные для мужских гамет промоторы включают, без ограничения, промотор PG47, промотор Zm13 и т.п. Отборочный ген может применяться для отбора растений или векторов, содержащих экспрессионную кассету. Отборочные гены включают, без ограничения, гены устойчивости к антибиотикам или гены устойчивости к гербицидам или гены флуоресцентных белков и т.п. В частности, отборочные гены включают, без ограничения, гены устойчивости к хлорамфениколу, гены устойчивости к гигромицину, гены устойчивости к стрептомицину, гены устойчивости к мирамицину, гены устойчивости к сульфонамидам, гены устойчивости к глифосату, гены устойчивости к фосфинотрицину, ген bar, ген красной флуоресценции DsRED, ген mCherry, ген голубого флуоресцентного белка, ген желтого флуоресцентного белка, ген люциферазы, ген зеленого флуоресцентного белка и т.п.
Изобретением также предусмотрен способ размножения линий с мужской стерильностью, который включает следующие стадии:
(a) трансформирование линии с мужской стерильностью ms7 следующим вектором для получения поддерживающей линии, содержащей следующий вектор: связанный с фертильностью ген TaMS7, который может восстанавливать мужскую фертильность линии с мужской стерильностью ms7; и ген инактивации пыльцы, экспрессия которого нарушает функцию или образование у растений мужских гамет, содержащих ген инактивации пыльцы, поэтому фертильные мужские гаметы, образующиеся у растений, не содержат вектора; и отборочный ген, который используется для сортировки трансгенных семян или тканей и нетрансгенных семян или тканей; и
(b) самоопыление растений поддерживающей линии, трансформированных данным вектором, для получения семян линии с мужской стерильностью ms7 без вектора и семян поддерживающей линии, содержащих вектор; или же опыление растений линии с мужской стерильностью ms7 пыльцой растений поддерживающей линии для размножения семян линии с мужской стерильностью ms7.
В данном способе размножения линий с мужской стерильностью гены инактивации пыльцы включают, без ограничения, ген барназы, ген амилазы, ген метилазы DAM и т.п. Более предпочтительно ген инактивации пыльцы представлен геном α-амилазы кукурузы Zm-AA, а его нуклеотидная последовательность предпочтительно представлена в SEQ ID NO: 25. Ген инактивации пыльцы связан со специфичным для мужских гамет промотором. В частности, специфичные для мужских гамет промоторы включают, без ограничения, промотор PG47, промотор Zm13 и т.п. Отборочный ген может применяться для отбора растений или векторов, содержащих экспрессионную кассету. Отборочные гены включают, без ограничения, гены устойчивости к антибиотикам или гены устойчивости к гербицидам или гены флуоресцентных белков и т.п. В частности, отборочные гены включают, без ограничения, гены устойчивости к хлорамфениколу, гены устойчивости к гигромицину, гены устойчивости к стрептомицину, гены устойчивости к мирамицину, гены устойчивости к сульфонамидам, гены устойчивости к глифосату, гены устойчивости к фосфинотрицину, ген bar, ген красной флуоресценции DsRED, ген mCherry, ген голубого флуоресцентного белка, ген желтого флуоресцентного белка, ген люциферазы, ген зеленого флуоресцентного белка и т.п.
Изобретением также предусмотрен способ получения поддерживающих линий, который включает следующие стадии:
(a) трансформирование линии с мужской стерильностью ms7 следующим вектором для получения поддерживающей линии, содержащей следующий вектор: связанный
с фертильностью ген TaMS7, который может восстанавливать мужскую фертильность линии с мужской стерильностью ms7; и ген инактивации пыльцы, экспрессия которого нарушает функцию или образование у растений мужских гамет, содержащих ген инактивации пыльцы, поэтому фертильные мужские гаметы, образующиеся у растений, не содержат вектора; и отборочный ген, который используется для сортировки трансгенных семян или тканей и нетрансгенных семян или тканей; и
(b) самоопыление растений поддерживающей линии, трансформированных данным вектором, для получения семян линии с мужской стерильностью ms7 без вектора и семян поддерживающей линии, содержащих вектор; или же опыление растений линии с мужской стерильностью ms7 пыльцой растений поддерживающей линии для размножения семян линии с мужской стерильностью ms7.
В данном способе получения поддерживающих линий гены инактивации пыльцы включают, без ограничения, ген барназы, ген амилазы, ген метилазы DAM и т.п. Более предпочтительно ген инактивации пыльцы представлен геном α-амилазы кукурузы Zm-AA, а его нуклеотидная последовательность предпочтительно представлена в SEQ ID NO: 25. Ген инактивации пыльцы связан со специфичным для мужских гамет промотором. В частности, специфичные для мужских гамет промоторы включают, без ограничения, промотор PG47, промотор Zm13 и т.п. Отборочный ген может применяться для отбора растений или векторов, содержащих экспрессионную кассету. Отборочные гены включают, без ограничения, гены устойчивости к антибиотикам или гены устойчивости к гербицидам или гены флуоресцентных белков и т.п. В частности, отборочные гены включают, без ограничения, гены устойчивости к хлорамфениколу, гены устойчивости к гигромицину, гены устойчивости к стрептомицину, гены устойчивости к мирамицину, гены устойчивости к сульфонамидам, гены устойчивости к глифосату, гены устойчивости к фосфинотрицину, ген bar, ген красной флуоресценции DsRED, ген mCherry, ген голубого флуоресцентного белка, ген желтого флуоресцентного белка, ген люциферазы, ген зеленого флуоресцентного белка и т.п.
Изобретением также предусмотрен способ размножения поддерживающих линий, который включает следующие стадии:
(a) трансформирование линии с мужской стерильностью ms7 следующим вектором для получения поддерживающей линии, содержащей следующий вектор: связанный
с фертильностью ген TaMS7, который может восстанавливать мужскую фертильность линии с мужской стерильностью ms7; и ген инактивации пыльцы, экспрессия которого нарушает функцию или образование у растений мужских гамет, содержащих ген инактивации пыльцы, поэтому фертильные мужские гаметы, образующиеся у растений, не содержат вектора; и отборочный ген, который используется для сортировки трансгенных семян или тканей и нетрансгенных семян или тканей; и
(b) самоопыление растений поддерживающей линии, трансформированных данным вектором, для получения семян линии с мужской стерильностью ms7 без вектора и семян поддерживающей линии, содержащих вектор, в соотношении 1:1.
Изобретением также предусмотрен способ получения семян, который включает следующие стадии:
(a) трансформирование линии с мужской стерильностью ms7 следующим вектором для получения поддерживающей линии, содержащей следующий вектор: связанный
с фертильностью ген TaMS7, который может восстанавливать мужскую фертильность линии с мужской стерильностью ms7; и ген инактивации пыльцы, экспрессия которого нарушает функцию или образование у растений мужских гамет, содержащих ген инактивации пыльцы, поэтому фертильные мужские гаметы, образующиеся у растений, не содержат вектора;
(b) самоопыление растений поддерживающей линии, полученных при трансформации данным вектором; и
(c) после самоопыления получаются семена линии с мужской стерильностью ms7 без вектора и семена поддерживающей линии, содержащие вектор.
В данном способе размножения или поддержания линий с мужской стерильностью, способе получения или размножения поддерживающих линий и способе получения семян и т.п. по изобретению стадия (a) также может включать: введение вектора, содержащего связанный с фертильностью ген TaMS7, ген инактивации пыльцы и отборочный ген, в общее растение, затем получение трансгенных растений, содержащих вектор, гибридизацию трансгенных растений с линией мужской стерильности ms7, а затем получение, путем направленной селекции, растений поддерживающей линии, содержащих вектор на фоне мужской стерильности ms7.
В данном способе размножения или поддержания линий с мужской стерильностью, способе получения или размножения поддерживающих линий и способе получения семян и т.п. по изобретению нуклеотидную последовательность связанного с фертильностью гена выбирают из одной из следующих групп последовательностей:
(a) нуклеотидных последовательностей, приведенных в SEQ ID NO:1, 2, 3, 4, 5 и 6;
(b) нуклеотидных последовательностей, кодирующих аминокислотные последовательности, приведенные в SEQ ID NO: 7, 8 и 9;
(c) последовательностей ДНК, способных гибридизироваться с приведенными в (a) или (b) последовательностями ДНК в строгих условиях; или
(d) последовательностей ДНК, имеющих сходство на 80% (предпочтительно по меньшей мере на 85%) с любыми из последовательностей (a)-(c) и обладающих связанной с фертильностью функцией; или
(e) последовательностей ДНК, комплементарных любым из последовательностей (a)-(d).
Связанный с фертильностью ген TaMS7 также может быть функционально связан со специфичным для пыльников промотором, который будет управлять экспрессией гена TaMS7 в пыльниках растений. Промотор со специфичностью к экспрессии в пыльниках выбирают из одной из групп, состоящих из промоторов генов, регулирующих фертильность: NP1, MSP1, PAIR1, PAIR2, ZEP1, MELL, PSS1, TDR, UDT1, GAMYB4, PTC1, API5, WDA1, CYP704B2, MS26, MS22, DPW, MADS3, OSC6, RIP1, CSA, AID1, 5126, Ms45 и др. Более предпочтительно нуклеотидная последовательность специфичного для пыльников промотора представлена в SEQ ID NO: 16, 17 или 18. Связанный с фертильностью ген TaMS7 также может быть функционально связан с терминатором, который представлен терминатором любого общеизвестного гена, в частности, нуклеотидная последовательность терминатора представлена в SEQ ID NO: 22, 23 или 24.
В данном способе размножения или поддержания линий с мужской стерильностью, способе получения или размножения поддерживающих линий и способе получения семян и т.п. по изобретению гены инактивации пыльцы включают, без ограничения, ген барназы, ген амилазы, ген метилазы DAM и т.п. Более предпочтительно ген инактивации пыльцы представлен геном α-амилазы кукурузы Zm-AA, а его нуклеотидная последовательность предпочтительно представлена в SEQ ID NO: 25. Ген инактивации пыльцы связан со специфичным для мужских гамет промотором. В частности, специфичные для мужских гамет промоторы включают, без ограничения, промотор PG47, промотор Zm13 и др.
В данном способе размножения или поддержания линий с мужской стерильностью, способе получения или размножения поддерживающих линий и способе получения семян и т.п. по изобретению отборочный ген применяется для отбора растений или векторов, содержащих экспрессионную кассету. Отборочные гены включают, без ограничения, гены устойчивости к антибиотикам или гены устойчивости к гербицидам или гены флуоресцентных белков и т.п. В частности, отборочные гены включают, без ограничения, гены устойчивости к хлорамфениколу, гены устойчивости к гигромицину, гены устойчивости к стрептомицину, гены устойчивости к мирамицину, гены устойчивости к сульфонамидам, гены устойчивости к глифосату, гены устойчивости к фосфинотрицину, ген bar, ген красной флуоресценции DsRED, ген mCherry, ген голубого флуоресцентного белка, ген желтого флуоресцентного белка, ген люциферазы, ген зеленого флуоресцентного белка и т.п.
Изобретением также предусмотрены специфичные для пыльников промоторы, нуклеотидные последовательности которых представлены в SEQ ID NO: 16, 17 или 18. Их вставляют в вектор и соединяют с репортерным геном GUS. С помощью вектора трансформируют рис и пшеницу и в трансгенных растениях выявляют и анализируют активность и профиль экспрессии GUS. При анализе на окрашивание GUS в корнях, стеблях, листьях и цветках трансгенных растений оказывается, что предусмотренный изобретением промотор управляет экспрессией гена GUS в пыльниках растений. Это свидетельствует о том, что SEQ ID NO: 16, 17 или 18 по изобретению является промотором, специфичным для пыльников.
Специфичный для пыльников промотор по изобретению содержит нуклеотидную последовательность по SEQ ID NO: 16, 17 или 18 из списка последовательностей либо содержит нуклеотидную последовательность, которая имеет сходство на 90% с нуклеотидной последовательностью SEQ ID NO: 16, 17 или 18 либо содержит фрагмент из 500 и более чем 500 непрерывных нуклеотидов из последовательности SEQ ID NO: 16, 17 или 18, причем нуклеотидная последовательность, функционально связанная с промотором, управляет экспрессией в пыльниках растений. Экспрессирующие векторы, содержащие данные последовательности, трансгенные линии клеток и бактерии-хозяева и т.п. входят в объем охраны изобретения. Пары праймеров для амплификации любых нуклеотидных фрагментов промоторов по SEQ ID NO: 16, 17 или 18, приведенных в заявке, входят в объем охраны изобретения.
Термин “промотор” в настоящем изобретении означает регуляторный участок ДНК, обычно включающий TATA-бокс, который может направлять РНК-полимеразу II на инициацию синтеза РНК в соответствующем сайте инициации транскрипции определенной кодирующей последовательности. Промотор может включать в себя и другие распознающие последовательности, обычно расположенные по восходящей от блока TATA, которые называют вышележащими элементами промотора с функцией регуляции эффективности транскрипции. Как известно специалистам, хотя в заявке идентифицирована нуклеотидная последовательность области промотора, однако выделение и идентификация других регуляторных элементов, расположенных по восходящей от блока TATA в области данного промотора по изобретению, также входят в объем охраны изобретения. Таким образом, область промотора по изобретению в общем дополнительно определяется как последовательности, включающие вышележащие регуляторные элементы или энхансеры, регулирующие пространственные и временные паттерны экспрессии кодирующей последовательности. Таким же образом могут быть идентифицированы и выделены промоторные элементы, проявляющие тканеспецифическую экспрессию (к примеру, специфичную для мужских тканей), которые можно использовать вместе с другим базовым промотором для изучения преимущественной экспрессии в определенных мужских тканях. Под базовым промотором (core promoter) подразумевается минимальная последовательность, необходимая для инициации транскрипции, к примеру, последовательность, известная как TATA-бокс, которая обычно находится в промоторе кодирующего белок гена. Следовательно, в качестве альтернативы можно использовать вышележащий промотор гена TaMS7 в сочетании с его собственным базовым промотором или базовым промотором из других источников.
Базовым промотором может быть любой известный базовый промотор типа промотора 35S или 19S вируса мозаичности цветной капусты (U.S. Patent No. 5,352,605), промотор убиквитина (U.S. Patent No. 5,510,474), базовый промотор IN2 (U.S. Patent No. 5,364,780) или промотор вируса мозаичности норичника.
Функции промоторов генов можно анализировать следующими методами: нуклеотидную последовательность промотора функционально соединяют с репортерным геном, получая вектор, который можно использовать при трансформации, затем этим вектором трансформируют растения и получают трансгенные растения, а затем изучают экспрессию репортерного гена в каждом органе/ткани растений трансгенного потомства для определения специфичности экспрессии. В качестве альтернативы последовательность промотора, связанную с репортерным геном, субклонируют в экспрессирующий вектор для экспериментов по краткосрочной экспрессии и определяют функцию промотора или других регуляторных участков в экспериментах с краткосрочной экспрессией.
Выбор подходящего экспрессирующего вектора для проверки функции промотора или его регуляторных участков зависит от организма-хозяина и способа введения в него экспрессирующего вектора, причем такие методы хорошо известны рядовым специалистам. В последовательности экспрессирующего вектора для эукариот содержатся участки, контролирующие инициацию транскрипции и контролирующие процессинг. Эти участки функционально связаны с репортерным геном типа YFP, UidA, гена GUS или люциферазы. Экспрессирующий вектор с предполагаемым регуляторным участком из генома может быть введен в целый орган типа пыльцы на определенных стадиях развития или в каллус для функциональной проверки.
Кроме того, промотор по изобретению также может быть связан с другими нуклеотидными последовательностями, помимо гена TaMS7, для управления их экспрессией. Нуклеотидные последовательности, фрагменты и варианты промотора по изобретению могут быть собраны в одну экспрессионную кассету с гетерогенной нуклеотидной последовательностью и использованы для экспрессии в целевых растениях, более предпочтительно для экспрессии в мужских органах растений. Экспрессионная кассета содержит подходящие сайты расщепления для рестрикционных ферментов, которые используются для вставки промотора и гетерогенной нуклеотидной последовательности. Экспрессионные кассеты можно использовать для манипуляций на любых растениях для получения соответствующего ожидаемого фенотипа.
Специфичный для пыльников промотор по изобретению может применяться для управления экспрессией следующей гетерологичной нуклеотидной последовательности для получения трансгенных растений с мужской стерильностью, причем гетерологичная нуклеотидная последовательность может кодировать фермент, способствующий расщеплению углеводов, или модифицирующий фермент, амилазу, деветвящий фермент или пектиназу, более конкретно, к примеру, ген барназы, ген α-амилазы кукурузы, ген ауксина, ген rot B, ген цитотоксина, ген дифтерийного токсина, ген метилазы DAM или доминантный ген мужской стерильности. В некоторых воплощениях нуклеиновая кислота, которая может быть функционально связана с промотором настоящего изобретения по нисходящей, может представлять собой структурный ген, регуляторный ген, антисмысловую последовательность структурного гена, антисмысловую последовательность регуляторного гена или ген малой РНК, способной интерферировать с экспрессией определенного эндогенного гена.
Изобретением также предусмотрены последовательности терминаторов транскрипции, нуклеотидные последовательности которых представлены в SEQ ID NO: 22, 23 или 24, которые имеют функции завершения транскрипции гена.
Изобретением также предусмотрены экспрессионные кассеты, векторы или инженерные штаммы, которые содержат специфичный для пыльников промотор SEQ ID NO: 16, 17 или 18, предусмотренный изобретением. В частности, нуклеотидная последовательность связанного с фертильностью гена TaMS7, предусмотренного изобретением, может быть встроена по нисходящей от промотора SEQ ID NO: 16, 17 или 18, предусмотренного изобретением, для управления экспрессией гена фертильности в акцепторных растениях при трансформации.
Специфичный для пыльников промотор по настоящему изобретению может применяться для специфической экспрессии экзогенного гена в пыльниках, чтобы избежать негативного эффекта, вызванного непрерывной экспрессией экзогенного гена в других тканях растения. Специфичный для пыльников промотор также может применяться для функционального анализа и идентификации генов, связанных с развитием пыльцы растений, а также для получения линий мужской стерильности и поддерживающих линий, а также может применяться для экспериментов по абортированию пыльцы, чтобы избежать проблемы биобезопасности, вызванной переносом трансгенных растений или утечкой пыльцы, а специфичный для пыльников промотор имеет важное значение для создания линий растений с мужской стерильностью и поддерживающих линий.
Нуклеотидная последовательность и последовательность промотора или экспрессионная кассета гена TaMS7, предусмотренного изобретением, может быть вставлена в вектор, плазмиду, искусственную дрожжевую хромосому, искусственную бактериальную хромосому или любой другой вектор, подходящий для трансформации в клетки хозяина. Предпочтительно клетки хозяина представляют собой бактериальные клетки, в особенности клетки для клонирования или содержания полинуклеотида или трансформирования растительных клеток, как-то Escherichia coli, Agrobaterium tumefaciens и Agrobacterium rhizogenes. В случае, когда клетки хозяина представляют собой клетки растений, экспрессионная кассета или вектор могут быть вставлены в геном трансформированных клеток растения, причем вставка может быть сайт-специфичной либо случайной.
Способы введения нуклеотидной последовательности, вектора или экспрессионной кассеты в растения или же трансформации ими растений по изобретению представляют собой стандартные трансгенные методы, с помощью которых нуклеотидная последовательность, вектор или экспрессионная кассета попадает при трансформации в акцепторные клетки или акцепторные растения. Для введения рекомбинантного экспрессирующего вектора в растительные клетки можно использовать любые трансгенные методы, известные специалистам в области биотехнологии растений, при этом получают трансгенные растения по изобретению. Методы трансформации включают прямые и косвенные методы трансформации. Подходящие прямые методы трансформации включают захват ДНК, индуцированный полиэтиленгликолем, трансформацию при помощи липосом, бомбардировки частицами, электропорации и микроинъекции. Методы трансформации также включают метод трансформации растений при помощи Agrobacterium и др. По сравнению с предшествующим уровнем техники изобретение имеет следующие преимущества: оно обеспечивает связанный с фертильностью ген TaMS7 и его промотор, а также способы, в которых ген TaMS7 применяется для размножения и поддержания линий с мужской стерильностью ms7. Связанный с фертильностью ген TaMS7 и способ имеют важное значение для получения, популяризации и применения при селекции гибридов для получения культур. Ген фертильности по изобретению и образующиеся при его мутации стерильные линии обеспечивают ресурсы для селекции гибридов пшеницы, а также обеспечивают необходимые элементы для построения системы селекции гибридов третьего поколения. Линии с мужской стерильностью, образующиеся при мутации гена, применяются для получения гибридных семян, что имеет важное значение для прорыва и улучшения существующих “трехлинейных” и “двухлинейных” технологий селекции гибридов.
Краткое описание фигур
На фиг. 1 представлен анализ экспрессии comp155942_c0_seq4 в пыльниках, когда пыльца находится на стадии мейоза (WT-0), одноядерной стадии (WT-1), двухъядерной стадии (WT-2) и трехъядерной стадии (WT-3). По горизонтальной оси представлены различные стадии развития пыльцы, а по вертикальной - FPKM, который отражает уровень экспрессии гена.
На фиг. 2 представлен анализ методом ОТ-ПЦР 3 гомологичных генов TaMS7 в различных тканях/органах и пыльниках на разных стадиях развития пшеницы. 1 - корни, 2 - стебли, 3 - листья, 4 - колосья с пыльцой на стадии мейоза, 5 - пыльники с одноядерными микроспорами, 6 - пыльники с двухъядерной пыльцой, 7 - пыльники с трехъядерной пыльцой, 8 - цветочные органы, за исключением пыльников с одноядерными микроспорами, 9 - цветочные органы, за исключением пыльников с двухъядерной пыльцой, и 10 - цветочные органы, за исключением пыльников с трехъядерной пыльцой.
На фиг. 3 представлены генотипы и фенотипы 7 рецессивных ядерных мутантных линий пшеницы с мужской стерильностью, полученных по технологии CRISPR-Cas9: a1 и a2 - два мутантных генотипа TaMS7-A, соответственно, b1 - один мутантный генотип TaMS7-B, d1 и d2 - два мутантных генотипа TaMS7-D, соответственно.
Фиг. 4. Тройной мутант tams7 проявляет признаки полной мужской стерильности. A - колосья дикого типа и тройного мутанта tams7; B - пыльники и пестики растений дикого типа и тройного мутанта tams7; C - зрелые пыльцевые зерна дикого типа и тройного мутанта tams7, окрашенные с помощью I2-KI.
Раскрытие сущности изобретения
Ниже подробно описаны воплощения изобретения, которые реализованы по технической схеме изобретения в качестве предпосылки, а также представлены подробные реализации и конкретный способ работы, но объем защиты изобретения не ограничивается следующими воплощениями.
Воплощение 1. Анализ полногеномных профилей экспрессии в пыльниках пшеницы на разных стадиях развития и получение контигов, экспрессируемых в пыльниках с пыльцой на ранней стадии развития
Собирали пыльники в мейозе и пыльники с одноядерными микроспорами, двухъядерной пыльцой или трехъядерной пыльцой, соответственно. Экстрагировали тотальную РНК с помощью Trizol (Invitrogen) и проводили обработку ДНКазой I (Promega), а затем выделяли мРНК (Ambion). После этого проводили обратную транскрипцию (Invitrogen) очищенной мРНК, обработку ультразвуком (Fisher), приготовление библиотеки (Illumina) и ее амплификацию (Illumina) и секвенирование на установке Illumina.
Прочтения от высокопроизводительного секвенирования транскриптома собирали с помощью программы Trinity, из собранных последовательностей удаляли избыточные последовательности и проводили анализ кластеризации по подобию. Для анализа изменений экспрессии собранных контигов транскриптов сначала совмещали прочтения от высокопроизводительного секвенирования каждого образца с контигами собранных транскриптов с помощью программы TopHat (http://tophat.cbcb.umd.edu/). После этого измеряли нормализованные значения экспрессии совмещаемых контигов транскриптов с помощью программы Cufflink и представляли в виде FPKM (количество фрагментов на 1 т.о. (килобазу) модельного экзона на миллион отображаемых фрагментов).
При анализе полногеномных профилей экспрессии в пыльниках пшеницы на разных стадиях развития оказалось, что 7231 контиг транскриптов сильно экспрессируется в пыльниках с пыльцой на стадии мейоза и одноядерной стадии и не экспрессируется в пыльниках с пыльцой на двухъядерной стадии и трехъядерной стадии. Как видно из фиг. 1, comp155942_c0_seq4 сильно экспрессируется в пыльниках с пыльцой на стадии мейоза и одноядерной стадии и не экспрессируется в пыльниках с пыльцой на двухъядерной стадии и трехъядерной стадии. Ген, соответствующий comp155942_c0_seq4, назвали TaMS7.
Воплощение 2. Проверка специфичности экспрессии гена TaMS7 в различных тканях методом ОТ-ПЦР
Поскольку пшеница является аллогексаплоидом, состоящим из трех комплектов геномов A, B и D, среднее число копий генов составляет 2,8, из которых почти половина (46%) имеют 3-4 копии, 12% имеют 1-2 копии и 42% имеют 5 или более 5 копий. Используя последовательность comp155942_c0_seq4, данные по секвенированию обыкновенной пшеницы, опубликованные CerealsDB и IWGSC (Международный консорциум по секвенированию генома пшеницы), и данные по секвенированию Triticum urartu (донор генома A) и Aegilops tauschii (донор генома D), опубликованные в Nature в 2013 г., мы получили 3 гена TaMS7, которые назвали TaMS7-A, TaMS7-B и TaMS7-D, соответственно.
Последовательности геномной ДНК этих 3 генов TaMS7 представлены в SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3, соответственно, идентичность которых составляет 95-97%; последовательности CDS представлены в SEQ ID NO: 4, SEQ ID NO: 5 и SEQ ID NO: 6, соответственно, идентичность которых составляет 97-98%; а последовательности белков представлены в SEQ ID NO: 7, SEQ ID NO: 8 и SEQ ID NO: 9, соответственно, идентичность которых составляет 98-99%.
Разработали специфичные праймеры для CDS TaMS7-A, TaMS7-B и TaMS7-D, соответственно, и анализировали специфичность экспрессии этих 3 генов во многих тканях пшеницы (корни, стебли и листья, пыльники на разных стадиях развития и другие цветочные органы, помимо пыльников) методом ОТ-ПЦР, а результаты, представленные на фиг. 2, показали, что гены TaMS7 специфически экспрессируются только в колосьях с пыльцой на стадии мейоза и в пыльниках с пыльцой на одноядерной стадии, не экспрессируются в пыльники с пыльцой на двухъядерной стадии и на трехъядерной стадии и не экспрессируются в других цветочных органах, кроме пыльников на разных стадиях развития, и в корнях, стеблях и листьях. Эти результаты показывают, что гены TaMS7 специфически экспрессируются именно в пыльниках, причем специфически экспрессируются только в пыльниках с пыльцой на ранней стадии развития, а промоторы генов TaMS7 являются специфичными для пыльников.
Праймеры для гена TaMS7-A при ОТ-ПЦР:
праймер 1: 5'-ATACTGACACAAGTTTATGGGGCTG-3' (SEQ ID NO: 10),
праймер 2: 5'-AATTACATTCAAATATGGCTCCTTG-3' (SEQ ID NO: 11).
Праймеры для гена TaMS7-B при ОТ-ПЦР:
праймер 3: 5'-GGCCTCGTGAACTCGTCGTATC-3' (SEQ ID NO: 12),
праймер 4: 5'-TGAATTACATGCAAATTTGGCTCCG-3' (SEQ ID NO: 13).
Праймеры для гена TaMS7-D при ОТ-ПЦР:
праймер 5: 5'-ATGTCCAACCAGGAGCACTTCAC-3' (SEQ ID NO: 14),
праймер 6: 5'-GCACAGTTTTTTGAAGCAATGTTG-3' (SEQ ID NO: 15).
Воплощение 3. Получение последовательности промоторов генов TaMS7
Используя последовательности геномной ДНК генов TaMS7-A, TaMS7-B и TaMS7-D, данные по секвенированию обыкновенной пшеницы, опубликованные CerealsDB и IWGSC (Международный консорциум по секвенированию генома пшеницы), и данные по секвенированию Triticum urartu (донор генома A) и Aegilops tauschii (донор генома D), опубликованные в Nature в 2013 г., выделяли промоторы генов TaMS7-A, TaMS7-B и TaMS7-D, которые назвали промотором TaMS7-A, промотором TaMS7-B и промотором TaMS7-D, соответственно, а также называли как pTaMS7-A, pTaMS7-B и pTaMS7-D, соответственно, в настоящей заявке. Длина этих промоторов составляла 2601 п.н., 2635 п.н. и 2821 п.н., соответственно, а их нуклеотидные последовательности представлены в SEQ ID NO: 16, SEQ ID NO: 17 и SEQ ID NO: 18, соответственно.
Воплощение 4. Клонирование промотора TaMS7-D и конструирование экспрессирующего вектора для растений
Для дальнейшей проверки функций указанных выше промоторов проводили эксперимент по проверке функции промотора TaMS7-D по изобретению. Экспрессирующий вектор pBI121 для растений расщепляли двумя рестриктазами HindIII и EcoRI, а полученный фрагмент 35S:GUS лигировали с помощью ДНК-лигазы T4 в вектор pCAMBIA2300 корпорации CAMBIA, который также расщепляли одновременно HindIII и EcoRI, и новый вектор назвали p2300 35S:GUS.
Составляли праймеры для 5'-конца промотора TaMS7-D и по восходящей от ATG:
праймер 7: 5'-aagcttCTGACATAGTACATGTAATCTTTAAATCCATAAC-3' (SEQ ID NO: 19),
праймер 8: 5'-ggatccTTGCGCCGGCGAGCTCGGC-3' (SEQ ID NO: 20).
Последовательность aagctt в праймере 7 является рестрикционным сайтом HindIII, а последовательность ggatcc в праймере 8 является рестрикционным сайтом BamHI.
В качестве матрицы использовали геномную ДНК пшеницы, для амплификации использовали праймер 7 и праймер 8, а условия реакции были следующими: начальная денатурация при 94°C в течение 5 мин; затем 35 циклов, включающих денатурацию при 94°C в течение 30 сек; отжиг при 60°C в течение 30 сек; удлинение при 72°C в течение 2 мин и 30 сек; и окончательное удлинение при 72°C в течение 10 мин. По окончании реакции продукт ПЦР выявляли и извлекали методом электрофореза в 1% агарозном геле. Полученный продукт вставляли в вектор pMD20-T, отбирали и секвенировали положительный клон, последовательность которого представлена в SEQ ID NO: 18. Плазмиду назвали T-pTaMS7-D.
T-pTaMS7-D расщепляли двумя рестриктазами HindIII и BamHI, полученный промотор TaMS7-D лигировали с помощью ДНК-лигазы T4 в вектор p2300 35S:GUS, который также расщепляли одновременно HindIII и BamHI, а затем получали экспрессирующий вектор p2300pTaMS7-D:GUS для растений.
Воплощение 5. Генетическая трансформация риса и гистохимическое выявление экспрессии гена GUS в различных тканях/органах трансгенных растений риса
Экспрессионным вектором p2300TaMS7-D:GUS для растений трансформировали штамм Agrobacterium tumefaciens AGL0 методом теплового шока.
Agrobacterium tumefaciens использовали для инфицирования эмбрионального каллюса риса, а затем Agrobacterium tumefaciens и эмбриональные каллюсы риса совместно культивировали в темноте в течение 2-3 дней. После двух стадий скрининга на устойчивость, предварительной дифференцировки, дифференцировки, укоренения и т.п., наконец, получали трансгенные растения риса p2300TaMS7-D:GUS поколения T0 с устойчивостью к канамицину.
Отбирали трансгенные проростки подходящего размера или определенные ткани, погружали в окрашивающий буфер GUS и инкубировали в течение ночи при 37°C. Затем удаляли реакционный раствор, проводили градиентное обесцвечивание этиловым спиртом и проводили наблюдения под микроскопом. Результаты показали, что экспрессия гена GUS не выявляется в питательных органах трансгенного риса, таких как корни, стебли и листья, и экспрессия гена GUS не выявляется в пыльниках и других цветочных органах с двухъядерной и трехъядерной пыльцой и других цветочных органах, за исключением пыльников с пыльцой на стадии мейоза и на одноядерной стадии, причем промотор TaMS7-D запускал экспрессию гена GUS только в пыльниках с пыльцой на стадии мейоза и на одноядерной стадии, свидетельствуя о том, что промотор TaMS7-D является специфичным для пыльников промотором на ранней стадии развития пыльцы. В соответствии со способом проверки функции промотора TaMS7-D также проверяли функции промотора TaMS7-A и промотора TaMS7-B и оказалось, что промотор TaMS7-A и промотор TaMS7-B тоже являются специфичными для пыльников промоторами, что соответствует результатам эксперимента в воплощении 2.
Воплощение 6. Сайт-направленный нокаут гена фертильности TaMS7 и анализ фенотипа у мутантов
В данном изобретении для сайт-направленного мутагенеза в геноме пшеницы использовали технологию CRISPR-Cas9. В частности, в качестве целевой последовательности в заявке была выбрана последовательность CTGGTGGACCAGCCCATGGT (SEQ ID NO: 21), которая представляет собой консенсусную последовательность генов TaMS7-A, TaMS7-B и TaMS7-D по нуклеотидам 1017-1036 положительной нити TaMS7-A, по нуклеотидам 1035-1054 положительной нити TaMS7-B и по нуклеотидам 1038-1057 положительной нити TaMS7-D. Экспрессионную кассету sgRNA с целевой последовательностью и экспрессионную кассету с Cas9 вставляли в один и тот же вектор pAHC20 и вводили в незрелые зародыши пшеницы с помощью плазмиды, экспрессирующей ген bar, методом бомбардировки частицами, после чего получали трансгенные растения пшеницы.
Проводили молекулярную идентификацию трансгенных растений пшеницы, при этом были идентифицированы мутации в целевых сайтах генов TaMS7-A, TaMS7-B и TaMS7-D: было два типа мутаций в гене TaMS7-A, делеция 5 п.о. и вставка 55 п.о., соответственно; один тип мутации в гене TaMS7-B, делеция 13 п.о. и вставка 54 п.о. одновременно; и два типа мутаций в гене TaMS7-D, один тип - делеция 9 п.о., а другой тип - делеция 7 п.о. и вставка 103 п.о. одновременно (см. фиг. 3).
Осмотр растений, содержащих гомозиготную мутацию или биаллельную мутацию в целевых сайтах генов TaMS7-A, TaMS7-B и TaMS7-D, показал, что растения с таким генотипом проявляли тонкие, мелкие и невыразительные пыльники без пыльцевых зерен с полной мужской стерильностью (см. фиг. 4), их опыляли пыльцой дикого типа для получения семян; тогда как растения, содержащие гомозиготную мутацию или биаллельную мутацию только в одном или двух из генов TaMS7-A, TaMS7-B и TaMS7-D, не проявляли фенотипа мужской стерильности. Это свидетельствует о том, что ген TaMS7 по изобретению является рецессивным геном мужской генной стерильности.
Воплощение 7. Функциональная комплементация трансгенов
В вектор pAHC20 встраивали фрагменты геномной ДНК (SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3) областей, кодирующих TaMS7-A, TaMS7-B и TaMS7-D, соответственно, добавляли спереди 2,5 т.п.н. фрагментов нативного вышележащего промотора (SEQ ID NO: 16, SEQ ID NO: 17 и SEQ ID NO: 18) для управления экспрессией генов и позади 1,4 т.п.н. нижележащих фрагментов (SEQ ID NO: 22, SEQ ID NO: 23 и SEQ ID NO: 24), соответственно, получая векторы, которые были названы pAHC20pTaMS7-A:gTaMS7-A, pAHC20pTaMS7-B:gTaMS7-B и pAHC20pTaMS7-D:gTaMS7-D, соответственно.
Для трансформации или совместной трансформации этими 3 векторами функциональной комплементации мутантов TaMS7-abd пшеницы использовали метод бомбардировки частицами. Пыльцевые зерна положительных трансгенных растений окрашивали с помощью I2-KI и осматривали, показывая, что пыльцевые зерна развивались нормально и были фертильными. Эти анализы также свидетельствуют о том, что гены TaMS7 участвуют в регуляции и контроле развития пыльцы, а мутации генов TaMS7 дают фенотип мужской стерильности.
Воплощение 8. Применение генов TaMS7 в методе селекции гибридов нового поколения
Гены TaMS7 могут применяться в технологии селекции гибридов нового поколения, а основная идея этой технологии заключается в следующем: рецессивные мутанты пшеницы с генной мужской стерильностью используют в качестве материала-акцептора при трансформации, причем стерильных мутантов трансформируют тремя близкородственными целевыми генами. При этом ген-восстановитель фертильности может восстанавливать фертильность акцептора трансформации, ген инактивации пыльцы может инактивировать пыльцевые зерна, содержащие трансгены, то есть у этих зерен пыльцы теряется способность к оплодотворению, ген-маркер семян (отборочный) может использоваться для сортировки трансгенных семян от нетрансгенных семян, а отобранные нетрансгенные семена можно использовать в качестве стерильной линии, тогда как трансгенные семена можно использовать в качестве поддерживающей линии. Поддерживающая линия может опылять стерильную линию для размножения стерильной линии, в то же время поддерживающая линия может самоопыляться, производя потомство в качестве нового поколения поддерживающей линии. В этой технологии применяется биотехнология для получения нетрансгенного продукта, решается проблема искусственного или механического выхолащивания при получении гибридных семян пшеницы, исключаются операции искусственного или механического выхолащивания, получаются семена лучшего качества и чистоты для сеялок и сокращаются трудовые затраты.
Исходя из этих принципов, авторы изобретения использовали ген TaMS7 пшеницы для конструирования экспрессирующего вектора для растений. Перед созданием экспрессирующего вектора для пшеницы авторы изобретения сначала трансформировали пшеницу каждой из трех экспрессионных кассет, ZmBT1-ZmAA, TaMS7 и mCherryW, соответственно, а также проверили функцию каждой экспрессионной кассеты. Результаты показали, что каждая экспрессионная кассета может работать хорошо, как и было задумано, когда ими независимо трансформировали пшеницу.
Далее авторы изобретения сконструировали вектор для трансформации путем сборки следующих ДНК-элементов:
1) вектор pAHC20 использовали в качестве остова, в который операбельно вставляли следующие экспрессионные кассеты;
2) восстанавливающая фертильность экспрессионная кассета гена TaMS7, содержащая связанный с фертильностью ген TaMS7, его промотор и терминатор, которые все были получены из сорта пшеницы CB037, причем три гена TaMS7 из геномов A, B и D пшеницы, а именно TaMS7-A, TaMS7-B и TaMS7-D, могут использоваться для восстановления фертильности стерильной линии ms7 пшеницы одновременно или поодиночке. При этом последовательность геномной ДНК, последовательность промотора и последовательность терминатора гена TaMS7-A представлена в SEQ ID NO: 1, SEQ ID NO: 16 и SEQ ID NO: 22, соответственно; последовательность геномной ДНК, последовательность промотора и последовательность терминатора гена TaMS7-B представлена в SEQ ID NO: 2, SEQ ID NO: 17 и SEQ ID NO: 23, соответственно; а последовательность геномной ДНК, последовательность промотора и последовательность терминатора гена TaMS7-D представлена в SEQ ID NO: 3, SEQ ID NO: 18 и SEQ ID NO: 24, соответственно;
3) экспрессионная кассета гена инактивации пыльцы PG47:ZmBT1-ZmAA-IN2-1: ген инактивации пыльцы ZmAA и транзитный пептид ZmBT1, причем открытую рамку считывания ZmBT1-ZmAA (нуклеотидная последовательность которой представлена в SEQ ID NO: 25) лигировали по нисходящей от промотора PG47 (нуклеотидная последовательность которого представлена в SEQ ID NO: 26) и по восходящей от терминатора IN2-1 (нуклеотидная последовательность которого представлена в SEQ ID NO: 27);
4) экспрессионная кассета гена-маркера семян (отборочного) CaMV35S-LTP2:mCherryW-PINII: открытую рамку считывания гена mCherryW (SEQ ID NO: 28) лигировали между энхансером CaMV35S-промотором LTP2 (SEQ ID NO: 29) и терминатором PINII (SEQ ID NO: 30), получая экспрессионную кассету гена (энхансер CaMV35S-LTP2:mCherryW-PINII).
Итак, авторы изобретения создали вектор, содержащий экспрессионную кассету для восстановления фертильности только с одним геном TaMS7, экспрессионную кассету гена инактивации пыльцы и экспрессионную кассету гена-маркера семян. Кроме того, был также сконструирован вектор, содержащий экспрессионную кассету для восстановления фертильности с двумя генами TaMS7, экспрессионную кассету гена инактивации пыльцы и экспрессионную кассету отборочного гена-маркера.
Для трансформации этими 2 векторами мутантов TaMS7-abd пшеницы использовали метод бомбардировки частицами и получили 12 и 9 положительных трансгенных растений, соответственно, в которых трансген представлен единственной копией. Проводили определение активности у пыльцы трансгенных растений и контрольных растений, и результаты показали, что доля стерильных пыльцевых зерен в общей массе пыльцевых зерен у нетрансгенных растений пшеницы дикого типа составляет менее 2%, причем пыльники мутантных растений TaMS7-abd нетрансгенной пшеницы не содержат пыльцевых зерен, тогда как пыльники трансгенных растений содержат пыльцевые зерна, причем доля стерильных пыльцевых зерен в общей массе пыльцевых зерен составляет около 50%. Эти результаты показали, что ген TaMS7 в векторе по изобретению восстанавливает фенотип мужской стерильности у мутантов TaMS7-abd, а ген ZmBT1-ZmAA способен инактивировать пыльцу, как и ожидалось.
Проводили анализ расщепления на флуоресцентные семена и нефлуоресцентные семена у семян поколения T1, вырабатываемых данными трансгенными растениями, который показал разделение в соотношении 1:1, а именно: флуоресцентные семена, несущие экзогенный ген, и нефлуоресцентные семена без экзогенного гена проявляли разделение в соотношении 1:1, указывая на то, что каждый элемент вектора, предусмотренный изобретением, хорошо экспрессируется в целом, а задача создания и селекции стерильной линии может быть реализована; при этом ген TaMS7 может восстанавливать фертильность у мутантного акцептора мужской стерильности, а экспрессия гена ZmBT1-ZmAA и гена mCherryW способна инактивировать пыльцу и маркировать трансгенные семена по флуоресценции, как и ожидалось, соответственно. Таким образом, была установлена технологическая система селекции гибридов нового поколения, а именно: на основе линии пшеницы с генной мужской стерильностью ms7 при трансформации указанным вектором образуется поддерживающая линия, содержащая экзогенный вектор, которая может использоваться для непрерывного получения поддерживающей линии и стерильной линии ms7, причем эта технологическая система решает проблему того, что линия пшеницы с генной мужской стерильностью ms7 не способна размножаться, а полученная линия с мужской стерильностью ms7 может использоваться для получения гибридных семян.
Данная технологическая система также может быть получена следующим способом, а именно: указанным вектором трансформируют обычную пшеницу дикого типа, а трансгенные растения скрещивают с мужской стерильной линией ms7, затем гибридные растения самоопыляются, а семена подвергаются скринингу для получения поддерживающей линии, содержащей экзогенный вектор на фоне гомозиготной мутации ms7, причем поддерживающая линия, содержащая экзогенный вектор, может использоваться для непрерывного получения поддерживающей линии и стерильной линии ms7.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> FRONTIER LABORATORIES OF SYSTEMS CROP DESIGN CO.,LTD.
PEKING UNIVERSITY
<120> Wheat fertility-related gene TaMS7 and application method thereof
<130>
<150> 201710321452.3
<151> 2017-05-09
<160> 30
<170> PatentIn version 3.3
<210> 1
<211> 1923
<212> DNA
<213> Triticum aestivum L.
<400> 1
atggcacttg gccgcgcgag atcgccggcg ctggtgctcg ccgccgccgt ccttggcgcg 60
ctctgcgtcg tcgcgctctc ggaggatggt gcgtgtgttc ggggttttgc atgtcagcta 120
gtacggagta cgtgcctgtg tctgttatca tgatcaatga ccgatggtgg cgtgcgtgtg 180
cagagcaact ggagaacctg cggttcgtgc agcacgcgca ggacgcgccg ctggtgtcgc 240
actacaacta catcgtggtg ggcggcggca cgtccgggtg cccgctggcg gcgacgctgt 300
cggagcactc gcgggtgctg ctgctggagc gcgggggcct cccctaccgc aacatgtcca 360
accaggagca cttcacggac gcgctggccg acacgtcgct ggcgtccccg gcgcagcggt 420
tcgtgtccac ggacggcgtg gtgaacgcgc gggcgcgggt gctgggcggc gggagctgcc 480
tcaacgccgg gttctacacg cgggccagca acgagtacgt gcgcacggcc gggtgggacg 540
ccggcctcgt caactcgtcg taccggtggg tggagcgcgc gctggtgttc cgcccggacg 600
tgccgccgtg gcaggccgcg ctccgggacg cgctgctgga ggccggcgtc acccccgaca 660
acggcttcac cttcgaccac gtcacgggga ccaagatcgg cggcaccatc ttcgacaaca 720
acgggcagcg ccacacggcc gccgacttcc tccggcacgc ccggccgcgg gggctcaccg 780
tggtgctcta cgccacggtg tcgcgggtcc tgttcaggag ccaggagggg gtgccgtacc 840
cggtggcgta cggggtggtg ttcgcggacc cgctgggggt gcagcaccgg gtgtacctcc 900
gggacggggg caagaacgag gtgatcctgt cggcggggac gctggggagc ccgcagctgc 960
tgatgctgag cggcgtgggc ccgcaggtgc acctggaggc gcacagcatc caggtgctgg 1020
tggaccagcc catggtcggg cagggcgtgg ccgacaaccc catgaactcg gtcttcatcc 1080
cgtcgccggt gccggtgggg ctgtccctgg tgcaggtcgt cgggatcacc aagtccggca 1140
gcttcatcga gggcgtgagc ggctccgagt tcggcatccc ggtgtccaac ggcgcccgcc 1200
ggctggccag cttcgggctc ttctccccgc agaccgggca gctcggcacg ctgccgccgg 1260
ggcagaggac gccggaggcg ctgcagcgcg cggcggaggc gatgcggcgg ctggacaggc 1320
gggcgttccg gggcggcttc atcctggaga agatcctggg gccggtgtcg acggggcaca 1380
tcgagctgcg cagcaccgac ccgcgcgcca acccggccgt gaccttcaac tacttccagg 1440
aggcggagga cctggagcgg tgcgtccggg ggatccagac gatcgagcgg gtgatccagt 1500
cgcgcgcctt ctccaacttc acctacgcca acaccaccgt ggagtccatc ttcaccgact 1560
cggccaactt ccccgtcaac ctgctgccgc gccacgtcaa cgactcccgc tcgccggagc 1620
agtactgcag ggagaccgtc atgaccatct ggcactacca cggcgggtgc cacgtcggcg 1680
ccgtcgtcga cgacaactac cgggtgttcg gggtcagggg gctcagggtc atcgacagct 1740
ccaccttcag gtactccccc ggcaccaacc cgcaggccac cgtcatgatg ctcggcaggt 1800
aaacgcaact ctcaaatcac ccccgcaatt atactgatct gaatgaactc aactaacaca 1860
actgttgtaa atctgtacag gtatatgggc ataaagattc aggccgagag atggaggaaa 1920
tga 1923
<210> 2
<211> 1954
<212> DNA
<213> Triticum aestivum L.
<400> 2
atggcacttg gccgcgcgag atcaccggcg ctggtgctcg ccgccgccgt tcttggcgcg 60
ctctgcatcg tcgcactctc ggaggatggt gcgtgtgttc acctgcatgg tttttctggg 120
ggttttgcgt cagctacgtg cgtgtgtctt ttatcatgat caatggacgc tgtgatgatc 180
aatggtgccg tgcgtgtgca gagcaactgg agaacctgcg gttcgtgcag cacgcgcagg 240
acgcgccgct ggtgtcgcac tacaactaca tcgtggtcgg cggcggcacg tccgggtgcc 300
cgctggcggc gacgctgtcg gagcactcgc gggtgctcct gctggagcgc gggggcctcc 360
cctaccgcaa catgtcgaac caggagcact tcacggacgc gctggccgac acgtcgctgg 420
cgtccccggc ccagcggttc atctccacgg acggcgtggt gaacgcgcgg gcgcgggtgc 480
tgggcggcgg gagctgcctc aacgccggat tctacacgcg ggccagcaac gagtacgtgc 540
gcacggccgg gtgggacgcc ggcctcgtga actcgtcgta tcggtgggtg gagcgcgcgc 600
tggtgttccg ccccgacgtg ccgccgtggc aggccgcgct ccgggacgcg ctgctcgagg 660
ccggcgtcac ccccgacaac ggcttcacct tcgatcacgt cacggggacc aagatcggcg 720
gcaccatctt cgacaacaac gggcagcgcc acacggccgc cgacttcctc cggcacgccc 780
ggccccgggg gctcaccgtc gtgctctacg ccacggtgtc ccggatcctg ttcagaagcc 840
aggagggggt gccgtatccg gtggcgtacg gggtggtgtt cgcggacccg ctgggggtgc 900
agcaccgggt gtacctccgg gacgggggga agaacgaggt gatactgtcg gcggggacgc 960
tggggagccc gcagctgctg atgctgagcg gcgtcggccc gcaggcgcac ctcgaggcac 1020
acggcatcca ggtgctggtg gaccagccca tggtcgggca gggcgtggcc gacaacccca 1080
tgaactcggt cttcatcccg tcgccggtgc cggtggggct ctccctggtg caggtcgtcg 1140
ggatcaccaa gtccggcagc ttcatcgagg gcgtgagcgg ctccgagttc ggcatcccgg 1200
tctccgacgg cgcccgccgc ctggccagct tcggcctctt ctccccccag accgggcagc 1260
tcggcacgct gccgccgggg cagaggacgc cggaggcgct gcagcgcgcg gcggaggcga 1320
tgaggcggct ggacaggcgg gcgttccggg gcgggttcat cctggagaag atcctggggc 1380
cggtgtccac gggccacatc gagctgcgca gcaccgaccc gcgcgcgaac ccggcggtga 1440
cgttcaacta cttccaggag gcggaggacc tggagcggtg cgtccggggg atccagacga 1500
tcgagcgggt gatccagtcg cgcgccttct ccaacttcac ctacgccaac accaccgtcg 1560
agtccatctt caccgactcg gccaacttcc ccgtcaacct gctgccgcgg cacgtcaacg 1620
actcccgctc gccggagcag tactgcaggg agaccgtcat gaccatctgg cactaccacg 1680
gcggatgcca cgtcggcgcc gtcgtcgacg acaactaccg ggtgttcggg gtgagggggc 1740
tcagggtgat cgacagctcc accttcaggt actcccccgg caccaacccg caggccaccg 1800
tcatgatgct cggcaggtaa acgcaagtct gaaatgaccc tcgcaattat actgatctga 1860
atgaatgaat caactcaact caactaacac aactatgata aatctgtaca ggtatatggg 1920
cataaagatt caggccgaga gatggaggaa atga 1954
<210> 3
<211> 1952
<212> DNA
<213> Triticum aestivum L.
<400> 3
atggcacttg gccgcgcgag atcgccgacg ctggtgctcg ccgccgcggt ccttggcgcg 60
ctctgcgtcg tcgcactctc ggaggacggt gcgtgtgttc acctgcatgg tttttctggg 120
ggttttgcac gtcagctacg tgcgttgtct cttttatcat gatcactgga cgctgtgatg 180
accaatggtg ccatacgtgt gcagagcaac tggagaacct gcggttcgtg cagcacgcgc 240
aggacgcgcc gctggtgtcg cactacaact acatcgtggt cggcggcggc acgtccgggt 300
gcccgctggc ggcgacgctg tcggagcact cgcgggtgct gctgctggag cgcgggggcc 360
tcccctaccg caacatgtcc aaccaggagc acttcacgga tgcgctggcc gacacgtcgc 420
tggcgtcccc ggcccagcgg ttcgtctcca cggacggcgt ggtgaacgcg cgggcgcggg 480
tgctgggcgg cgggagctgc ctcaacgccg ggttctacac gcgggccagc aacgagtacg 540
tgcgcacggc cgggtgggac gccggcctcg tcaactcgtc gtaccggtgg gtggagcgcg 600
cgctggtgtt ccgccccgac gtgccgccgt ggcaggccgc gctccgggac gcgctgctcg 660
aggccggcgt cacccccgac aacggcttca ccttcgacca cgtcacgggg accaagatcg 720
gcggcaccat cttcgacaac agcgggcagc gccacaccgc ggccgacttc ctccggcacg 780
cccggccccg gcggctcacc gtcgtgctct acgccacggt gtcgcggatc ctgttcagaa 840
gccaggaggg ggtgccgtac ccggtggcgt acggggtggt gttcgcggac ccgctggggg 900
tgcagcaccg ggtgtacctc cgggacggcg ggaagaacga ggtgatactg tcggcgggga 960
cgctggggag cccgcagctg ctgatgctga gcggcgtggg cccgcaggcg cacctggagg 1020
cgcacggcat ccaggtgctg gtggaccagc ccatggtcgg gcagggcgtg gccgacaacc 1080
ccatgaactc ggtcttcatc ccgtcgccgg tgccggtggg gctctccctg gtgcaggtcg 1140
tcgggatcac caagtccggc agcttcatcg agggcgtgag cggctccgag ttcggcatcc 1200
cggtctccga cggcgcccgc cgcctggcca gcttcggcct cttctccccc cagaccgggc 1260
agctcggcac gctgccgccg gggcagagga cgccggaggc gctgcagcgc gcggcggagg 1320
cgatgaggcg gctggacagg cgggcgttcc ggggcggctt catcctggag aagatcctgg 1380
ggccggtgtc gacggggcac atcgagctgc gcagcaccga cccgcgcgcc aacccggccg 1440
tgaccttcaa ctacttccag gaggcggagg acctggagcg gtgcgtccgg gggatccaga 1500
cgatcgagcg ggtgatccag tcgcgcgcct tctccaactt cacctacgcc aacaccaccg 1560
tcgagtccat cttcaccgac tcggccaact tccccgtcaa cctgctgccg cggcacgtca 1620
acgactcccg ctcgccggag cagtactgca gggagaccgt catgaccatc tggcactacc 1680
acggcgggtg ccacgtcggc gccgtcgtcg acgacaacta ccgggtgttc ggggtgaggg 1740
ggctcagggt catcgacagc tccaccttca ggtactcccc cggcaccaac ccacaggcca 1800
ccgtcatgat gctcggcagg taaacgcaag tctgagatga cccccgcaat tatactgatc 1860
tgaatgaatg aatcaactct actaacacaa ctatgataac tctgtacagg tatatgggca 1920
taaagattca ggccgagaga tggaggaaat ga 1952
<210> 4
<211> 1746
<212> DNA
<213> Triticum aestivum L.
<400> 4
atggcacttg gccgcgcgag atcgccggcg ctggtgctcg ccgccgccgt ccttggcgcg 60
ctctgcgtcg tcgcgctctc ggaggatgag caactggaga acctgcggtt cgtgcagcac 120
gcgcaggacg cgccgctggt gtcgcactac aactacatcg tggtgggcgg cggcacgtcc 180
gggtgcccgc tggcggcgac gctgtcggag cactcgcggg tgctgctgct ggagcgcggg 240
ggcctcccct accgcaacat gtccaaccag gagcacttca cggacgcgct ggccgacacg 300
tcgctggcgt ccccggcgca gcggttcgtg tccacggacg gcgtggtgaa cgcgcgggcg 360
cgggtgctgg gcggcgggag ctgcctcaac gccgggttct acacgcgggc cagcaacgag 420
tacgtgcgca cggccgggtg ggacgccggc ctcgtcaact cgtcgtaccg gtgggtggag 480
cgcgcgctgg tgttccgccc ggacgtgccg ccgtggcagg ccgcgctccg ggacgcgctg 540
ctggaggccg gcgtcacccc cgacaacggc ttcaccttcg accacgtcac ggggaccaag 600
atcggcggca ccatcttcga caacaacggg cagcgccaca cggccgccga cttcctccgg 660
cacgcccggc cgcgggggct caccgtggtg ctctacgcca cggtgtcgcg ggtcctgttc 720
aggagccagg agggggtgcc gtacccggtg gcgtacgggg tggtgttcgc ggacccgctg 780
ggggtgcagc accgggtgta cctccgggac gggggcaaga acgaggtgat cctgtcggcg 840
gggacgctgg ggagcccgca gctgctgatg ctgagcggcg tgggcccgca ggtgcacctg 900
gaggcgcaca gcatccaggt gctggtggac cagcccatgg tcgggcaggg cgtggccgac 960
aaccccatga actcggtctt catcccgtcg ccggtgccgg tggggctgtc cctggtgcag 1020
gtcgtcggga tcaccaagtc cggcagcttc atcgagggcg tgagcggctc cgagttcggc 1080
atcccggtgt ccaacggcgc ccgccggctg gccagcttcg ggctcttctc cccgcagacc 1140
gggcagctcg gcacgctgcc gccggggcag aggacgccgg aggcgctgca gcgcgcggcg 1200
gaggcgatgc ggcggctgga caggcgggcg ttccggggcg gcttcatcct ggagaagatc 1260
ctggggccgg tgtcgacggg gcacatcgag ctgcgcagca ccgacccgcg cgccaacccg 1320
gccgtgacct tcaactactt ccaggaggcg gaggacctgg agcggtgcgt ccgggggatc 1380
cagacgatcg agcgggtgat ccagtcgcgc gccttctcca acttcaccta cgccaacacc 1440
accgtggagt ccatcttcac cgactcggcc aacttccccg tcaacctgct gccgcgccac 1500
gtcaacgact cccgctcgcc ggagcagtac tgcagggaga ccgtcatgac catctggcac 1560
taccacggcg ggtgccacgt cggcgccgtc gtcgacgaca actaccgggt gttcggggtc 1620
agggggctca gggtcatcga cagctccacc ttcaggtact cccccggcac caacccgcag 1680
gccaccgtca tgatgctcgg caggtatatg ggcataaaga ttcaggccga gagatggagg 1740
aaatga 1746
<210> 5
<211> 1746
<212> DNA
<213> Triticum aestivum L.
<400> 5
atggcacttg gccgcgcgag atcaccggcg ctggtgctcg ccgccgccgt tcttggcgcg 60
ctctgcatcg tcgcactctc ggaggatgag caactggaga acctgcggtt cgtgcagcac 120
gcgcaggacg cgccgctggt gtcgcactac aactacatcg tggtcggcgg cggcacgtcc 180
gggtgcccgc tggcggcgac gctgtcggag cactcgcggg tgctcctgct ggagcgcggg 240
ggcctcccct accgcaacat gtcgaaccag gagcacttca cggacgcgct ggccgacacg 300
tcgctggcgt ccccggccca gcggttcatc tccacggacg gcgtggtgaa cgcgcgggcg 360
cgggtgctgg gcggcgggag ctgcctcaac gccggattct acacgcgggc cagcaacgag 420
tacgtgcgca cggccgggtg ggacgccggc ctcgtgaact cgtcgtatcg gtgggtggag 480
cgcgcgctgg tgttccgccc cgacgtgccg ccgtggcagg ccgcgctccg ggacgcgctg 540
ctcgaggccg gcgtcacccc cgacaacggc ttcaccttcg atcacgtcac ggggaccaag 600
atcggcggca ccatcttcga caacaacggg cagcgccaca cggccgccga cttcctccgg 660
cacgcccggc cccgggggct caccgtcgtg ctctacgcca cggtgtcccg gatcctgttc 720
agaagccagg agggggtgcc gtatccggtg gcgtacgggg tggtgttcgc ggacccgctg 780
ggggtgcagc accgggtgta cctccgggac ggggggaaga acgaggtgat actgtcggcg 840
gggacgctgg ggagcccgca gctgctgatg ctgagcggcg tcggcccgca ggcgcacctc 900
gaggcacacg gcatccaggt gctggtggac cagcccatgg tcgggcaggg cgtggccgac 960
aaccccatga actcggtctt catcccgtcg ccggtgccgg tggggctctc cctggtgcag 1020
gtcgtcggga tcaccaagtc cggcagcttc atcgagggcg tgagcggctc cgagttcggc 1080
atcccggtct ccgacggcgc ccgccgcctg gccagcttcg gcctcttctc cccccagacc 1140
gggcagctcg gcacgctgcc gccggggcag aggacgccgg aggcgctgca gcgcgcggcg 1200
gaggcgatga ggcggctgga caggcgggcg ttccggggcg ggttcatcct ggagaagatc 1260
ctggggccgg tgtccacggg ccacatcgag ctgcgcagca ccgacccgcg cgcgaacccg 1320
gcggtgacgt tcaactactt ccaggaggcg gaggacctgg agcggtgcgt ccgggggatc 1380
cagacgatcg agcgggtgat ccagtcgcgc gccttctcca acttcaccta cgccaacacc 1440
accgtcgagt ccatcttcac cgactcggcc aacttccccg tcaacctgct gccgcggcac 1500
gtcaacgact cccgctcgcc ggagcagtac tgcagggaga ccgtcatgac catctggcac 1560
taccacggcg gatgccacgt cggcgccgtc gtcgacgaca actaccgggt gttcggggtg 1620
agggggctca gggtgatcga cagctccacc ttcaggtact cccccggcac caacccgcag 1680
gccaccgtca tgatgctcgg caggtatatg ggcataaaga ttcaggccga gagatggagg 1740
aaatga 1746
<210> 6
<211> 1746
<212> DNA
<213> Triticum aestivum L.
<400> 6
atggcacttg gccgcgcgag atcgccgacg ctggtgctcg ccgccgcggt ccttggcgcg 60
ctctgcgtcg tcgcactctc ggaggacgag caactggaga acctgcggtt cgtgcagcac 120
gcgcaggacg cgccgctggt gtcgcactac aactacatcg tggtcggcgg cggcacgtcc 180
gggtgcccgc tggcggcgac gctgtcggag cactcgcggg tgctgctgct ggagcgcggg 240
ggcctcccct accgcaacat gtccaaccag gagcacttca cggatgcgct ggccgacacg 300
tcgctggcgt ccccggccca gcggttcgtc tccacggacg gcgtggtgaa cgcgcgggcg 360
cgggtgctgg gcggcgggag ctgcctcaac gccgggttct acacgcgggc cagcaacgag 420
tacgtgcgca cggccgggtg ggacgccggc ctcgtcaact cgtcgtaccg gtgggtggag 480
cgcgcgctgg tgttccgccc cgacgtgccg ccgtggcagg ccgcgctccg ggacgcgctg 540
ctcgaggccg gcgtcacccc cgacaacggc ttcaccttcg accacgtcac ggggaccaag 600
atcggcggca ccatcttcga caacagcggg cagcgccaca ccgcggccga cttcctccgg 660
cacgcccggc cccggcggct caccgtcgtg ctctacgcca cggtgtcgcg gatcctgttc 720
agaagccagg agggggtgcc gtacccggtg gcgtacgggg tggtgttcgc ggacccgctg 780
ggggtgcagc accgggtgta cctccgggac ggcgggaaga acgaggtgat actgtcggcg 840
gggacgctgg ggagcccgca gctgctgatg ctgagcggcg tgggcccgca ggcgcacctg 900
gaggcgcacg gcatccaggt gctggtggac cagcccatgg tcgggcaggg cgtggccgac 960
aaccccatga actcggtctt catcccgtcg ccggtgccgg tggggctctc cctggtgcag 1020
gtcgtcggga tcaccaagtc cggcagcttc atcgagggcg tgagcggctc cgagttcggc 1080
atcccggtct ccgacggcgc ccgccgcctg gccagcttcg gcctcttctc cccccagacc 1140
gggcagctcg gcacgctgcc gccggggcag aggacgccgg aggcgctgca gcgcgcggcg 1200
gaggcgatga ggcggctgga caggcgggcg ttccggggcg gcttcatcct ggagaagatc 1260
ctggggccgg tgtcgacggg gcacatcgag ctgcgcagca ccgacccgcg cgccaacccg 1320
gccgtgacct tcaactactt ccaggaggcg gaggacctgg agcggtgcgt ccgggggatc 1380
cagacgatcg agcgggtgat ccagtcgcgc gccttctcca acttcaccta cgccaacacc 1440
accgtcgagt ccatcttcac cgactcggcc aacttccccg tcaacctgct gccgcggcac 1500
gtcaacgact cccgctcgcc ggagcagtac tgcagggaga ccgtcatgac catctggcac 1560
taccacggcg ggtgccacgt cggcgccgtc gtcgacgaca actaccgggt gttcggggtg 1620
agggggctca gggtcatcga cagctccacc ttcaggtact cccccggcac caacccacag 1680
gccaccgtca tgatgctcgg caggtatatg ggcataaaga ttcaggccga gagatggagg 1740
aaatga 1746
<210> 7
<211> 581
<212> PRT
<213> Triticum aestivum L.
<400> 7
Met Ala Leu Gly Arg Ala Arg Ser Pro Ala Leu Val Leu Ala Ala Ala
1 5 10 15
Val Leu Gly Ala Leu Cys Val Val Ala Leu Ser Glu Asp Glu Gln Leu
20 25 30
Glu Asn Leu Arg Phe Val Gln His Ala Gln Asp Ala Pro Leu Val Ser
35 40 45
His Tyr Asn Tyr Ile Val Val Gly Gly Gly Thr Ser Gly Cys Pro Leu
50 55 60
Ala Ala Thr Leu Ser Glu His Ser Arg Val Leu Leu Leu Glu Arg Gly
65 70 75 80
Gly Leu Pro Tyr Arg Asn Met Ser Asn Gln Glu His Phe Thr Asp Ala
85 90 95
Leu Ala Asp Thr Ser Leu Ala Ser Pro Ala Gln Arg Phe Val Ser Thr
100 105 110
Asp Gly Val Val Asn Ala Arg Ala Arg Val Leu Gly Gly Gly Ser Cys
115 120 125
Leu Asn Ala Gly Phe Tyr Thr Arg Ala Ser Asn Glu Tyr Val Arg Thr
130 135 140
Ala Gly Trp Asp Ala Gly Leu Val Asn Ser Ser Tyr Arg Trp Val Glu
145 150 155 160
Arg Ala Leu Val Phe Arg Pro Asp Val Pro Pro Trp Gln Ala Ala Leu
165 170 175
Arg Asp Ala Leu Leu Glu Ala Gly Val Thr Pro Asp Asn Gly Phe Thr
180 185 190
Phe Asp His Val Thr Gly Thr Lys Ile Gly Gly Thr Ile Phe Asp Asn
195 200 205
Asn Gly Gln Arg His Thr Ala Ala Asp Phe Leu Arg His Ala Arg Pro
210 215 220
Arg Gly Leu Thr Val Val Leu Tyr Ala Thr Val Ser Arg Val Leu Phe
225 230 235 240
Arg Ser Gln Glu Gly Val Pro Tyr Pro Val Ala Tyr Gly Val Val Phe
245 250 255
Ala Asp Pro Leu Gly Val Gln His Arg Val Tyr Leu Arg Asp Gly Gly
260 265 270
Lys Asn Glu Val Ile Leu Ser Ala Gly Thr Leu Gly Ser Pro Gln Leu
275 280 285
Leu Met Leu Ser Gly Val Gly Pro Gln Val His Leu Glu Ala His Ser
290 295 300
Ile Gln Val Leu Val Asp Gln Pro Met Val Gly Gln Gly Val Ala Asp
305 310 315 320
Asn Pro Met Asn Ser Val Phe Ile Pro Ser Pro Val Pro Val Gly Leu
325 330 335
Ser Leu Val Gln Val Val Gly Ile Thr Lys Ser Gly Ser Phe Ile Glu
340 345 350
Gly Val Ser Gly Ser Glu Phe Gly Ile Pro Val Ser Asn Gly Ala Arg
355 360 365
Arg Leu Ala Ser Phe Gly Leu Phe Ser Pro Gln Thr Gly Gln Leu Gly
370 375 380
Thr Leu Pro Pro Gly Gln Arg Thr Pro Glu Ala Leu Gln Arg Ala Ala
385 390 395 400
Glu Ala Met Arg Arg Leu Asp Arg Arg Ala Phe Arg Gly Gly Phe Ile
405 410 415
Leu Glu Lys Ile Leu Gly Pro Val Ser Thr Gly His Ile Glu Leu Arg
420 425 430
Ser Thr Asp Pro Arg Ala Asn Pro Ala Val Thr Phe Asn Tyr Phe Gln
435 440 445
Glu Ala Glu Asp Leu Glu Arg Cys Val Arg Gly Ile Gln Thr Ile Glu
450 455 460
Arg Val Ile Gln Ser Arg Ala Phe Ser Asn Phe Thr Tyr Ala Asn Thr
465 470 475 480
Thr Val Glu Ser Ile Phe Thr Asp Ser Ala Asn Phe Pro Val Asn Leu
485 490 495
Leu Pro Arg His Val Asn Asp Ser Arg Ser Pro Glu Gln Tyr Cys Arg
500 505 510
Glu Thr Val Met Thr Ile Trp His Tyr His Gly Gly Cys His Val Gly
515 520 525
Ala Val Val Asp Asp Asn Tyr Arg Val Phe Gly Val Arg Gly Leu Arg
530 535 540
Val Ile Asp Ser Ser Thr Phe Arg Tyr Ser Pro Gly Thr Asn Pro Gln
545 550 555 560
Ala Thr Val Met Met Leu Gly Arg Tyr Met Gly Ile Lys Ile Gln Ala
565 570 575
Glu Arg Trp Arg Lys
580
<210> 8
<211> 581
<212> PRT
<213> Triticum aestivum L.
<400> 8
Met Ala Leu Gly Arg Ala Arg Ser Pro Ala Leu Val Leu Ala Ala Ala
1 5 10 15
Val Leu Gly Ala Leu Cys Ile Val Ala Leu Ser Glu Asp Glu Gln Leu
20 25 30
Glu Asn Leu Arg Phe Val Gln His Ala Gln Asp Ala Pro Leu Val Ser
35 40 45
His Tyr Asn Tyr Ile Val Val Gly Gly Gly Thr Ser Gly Cys Pro Leu
50 55 60
Ala Ala Thr Leu Ser Glu His Ser Arg Val Leu Leu Leu Glu Arg Gly
65 70 75 80
Gly Leu Pro Tyr Arg Asn Met Ser Asn Gln Glu His Phe Thr Asp Ala
85 90 95
Leu Ala Asp Thr Ser Leu Ala Ser Pro Ala Gln Arg Phe Ile Ser Thr
100 105 110
Asp Gly Val Val Asn Ala Arg Ala Arg Val Leu Gly Gly Gly Ser Cys
115 120 125
Leu Asn Ala Gly Phe Tyr Thr Arg Ala Ser Asn Glu Tyr Val Arg Thr
130 135 140
Ala Gly Trp Asp Ala Gly Leu Val Asn Ser Ser Tyr Arg Trp Val Glu
145 150 155 160
Arg Ala Leu Val Phe Arg Pro Asp Val Pro Pro Trp Gln Ala Ala Leu
165 170 175
Arg Asp Ala Leu Leu Glu Ala Gly Val Thr Pro Asp Asn Gly Phe Thr
180 185 190
Phe Asp His Val Thr Gly Thr Lys Ile Gly Gly Thr Ile Phe Asp Asn
195 200 205
Asn Gly Gln Arg His Thr Ala Ala Asp Phe Leu Arg His Ala Arg Pro
210 215 220
Arg Gly Leu Thr Val Val Leu Tyr Ala Thr Val Ser Arg Ile Leu Phe
225 230 235 240
Arg Ser Gln Glu Gly Val Pro Tyr Pro Val Ala Tyr Gly Val Val Phe
245 250 255
Ala Asp Pro Leu Gly Val Gln His Arg Val Tyr Leu Arg Asp Gly Gly
260 265 270
Lys Asn Glu Val Ile Leu Ser Ala Gly Thr Leu Gly Ser Pro Gln Leu
275 280 285
Leu Met Leu Ser Gly Val Gly Pro Gln Ala His Leu Glu Ala His Gly
290 295 300
Ile Gln Val Leu Val Asp Gln Pro Met Val Gly Gln Gly Val Ala Asp
305 310 315 320
Asn Pro Met Asn Ser Val Phe Ile Pro Ser Pro Val Pro Val Gly Leu
325 330 335
Ser Leu Val Gln Val Val Gly Ile Thr Lys Ser Gly Ser Phe Ile Glu
340 345 350
Gly Val Ser Gly Ser Glu Phe Gly Ile Pro Val Ser Asp Gly Ala Arg
355 360 365
Arg Leu Ala Ser Phe Gly Leu Phe Ser Pro Gln Thr Gly Gln Leu Gly
370 375 380
Thr Leu Pro Pro Gly Gln Arg Thr Pro Glu Ala Leu Gln Arg Ala Ala
385 390 395 400
Glu Ala Met Arg Arg Leu Asp Arg Arg Ala Phe Arg Gly Gly Phe Ile
405 410 415
Leu Glu Lys Ile Leu Gly Pro Val Ser Thr Gly His Ile Glu Leu Arg
420 425 430
Ser Thr Asp Pro Arg Ala Asn Pro Ala Val Thr Phe Asn Tyr Phe Gln
435 440 445
Glu Ala Glu Asp Leu Glu Arg Cys Val Arg Gly Ile Gln Thr Ile Glu
450 455 460
Arg Val Ile Gln Ser Arg Ala Phe Ser Asn Phe Thr Tyr Ala Asn Thr
465 470 475 480
Thr Val Glu Ser Ile Phe Thr Asp Ser Ala Asn Phe Pro Val Asn Leu
485 490 495
Leu Pro Arg His Val Asn Asp Ser Arg Ser Pro Glu Gln Tyr Cys Arg
500 505 510
Glu Thr Val Met Thr Ile Trp His Tyr His Gly Gly Cys His Val Gly
515 520 525
Ala Val Val Asp Asp Asn Tyr Arg Val Phe Gly Val Arg Gly Leu Arg
530 535 540
Val Ile Asp Ser Ser Thr Phe Arg Tyr Ser Pro Gly Thr Asn Pro Gln
545 550 555 560
Ala Thr Val Met Met Leu Gly Arg Tyr Met Gly Ile Lys Ile Gln Ala
565 570 575
Glu Arg Trp Arg Lys
580
<210> 9
<211> 581
<212> PRT
<213> Triticum aestivum L.
<400> 9
Met Ala Leu Gly Arg Ala Arg Ser Pro Thr Leu Val Leu Ala Ala Ala
1 5 10 15
Val Leu Gly Ala Leu Cys Val Val Ala Leu Ser Glu Asp Glu Gln Leu
20 25 30
Glu Asn Leu Arg Phe Val Gln His Ala Gln Asp Ala Pro Leu Val Ser
35 40 45
His Tyr Asn Tyr Ile Val Val Gly Gly Gly Thr Ser Gly Cys Pro Leu
50 55 60
Ala Ala Thr Leu Ser Glu His Ser Arg Val Leu Leu Leu Glu Arg Gly
65 70 75 80
Gly Leu Pro Tyr Arg Asn Met Ser Asn Gln Glu His Phe Thr Asp Ala
85 90 95
Leu Ala Asp Thr Ser Leu Ala Ser Pro Ala Gln Arg Phe Val Ser Thr
100 105 110
Asp Gly Val Val Asn Ala Arg Ala Arg Val Leu Gly Gly Gly Ser Cys
115 120 125
Leu Asn Ala Gly Phe Tyr Thr Arg Ala Ser Asn Glu Tyr Val Arg Thr
130 135 140
Ala Gly Trp Asp Ala Gly Leu Val Asn Ser Ser Tyr Arg Trp Val Glu
145 150 155 160
Arg Ala Leu Val Phe Arg Pro Asp Val Pro Pro Trp Gln Ala Ala Leu
165 170 175
Arg Asp Ala Leu Leu Glu Ala Gly Val Thr Pro Asp Asn Gly Phe Thr
180 185 190
Phe Asp His Val Thr Gly Thr Lys Ile Gly Gly Thr Ile Phe Asp Asn
195 200 205
Ser Gly Gln Arg His Thr Ala Ala Asp Phe Leu Arg His Ala Arg Pro
210 215 220
Arg Arg Leu Thr Val Val Leu Tyr Ala Thr Val Ser Arg Ile Leu Phe
225 230 235 240
Arg Ser Gln Glu Gly Val Pro Tyr Pro Val Ala Tyr Gly Val Val Phe
245 250 255
Ala Asp Pro Leu Gly Val Gln His Arg Val Tyr Leu Arg Asp Gly Gly
260 265 270
Lys Asn Glu Val Ile Leu Ser Ala Gly Thr Leu Gly Ser Pro Gln Leu
275 280 285
Leu Met Leu Ser Gly Val Gly Pro Gln Ala His Leu Glu Ala His Gly
290 295 300
Ile Gln Val Leu Val Asp Gln Pro Met Val Gly Gln Gly Val Ala Asp
305 310 315 320
Asn Pro Met Asn Ser Val Phe Ile Pro Ser Pro Val Pro Val Gly Leu
325 330 335
Ser Leu Val Gln Val Val Gly Ile Thr Lys Ser Gly Ser Phe Ile Glu
340 345 350
Gly Val Ser Gly Ser Glu Phe Gly Ile Pro Val Ser Asp Gly Ala Arg
355 360 365
Arg Leu Ala Ser Phe Gly Leu Phe Ser Pro Gln Thr Gly Gln Leu Gly
370 375 380
Thr Leu Pro Pro Gly Gln Arg Thr Pro Glu Ala Leu Gln Arg Ala Ala
385 390 395 400
Glu Ala Met Arg Arg Leu Asp Arg Arg Ala Phe Arg Gly Gly Phe Ile
405 410 415
Leu Glu Lys Ile Leu Gly Pro Val Ser Thr Gly His Ile Glu Leu Arg
420 425 430
Ser Thr Asp Pro Arg Ala Asn Pro Ala Val Thr Phe Asn Tyr Phe Gln
435 440 445
Glu Ala Glu Asp Leu Glu Arg Cys Val Arg Gly Ile Gln Thr Ile Glu
450 455 460
Arg Val Ile Gln Ser Arg Ala Phe Ser Asn Phe Thr Tyr Ala Asn Thr
465 470 475 480
Thr Val Glu Ser Ile Phe Thr Asp Ser Ala Asn Phe Pro Val Asn Leu
485 490 495
Leu Pro Arg His Val Asn Asp Ser Arg Ser Pro Glu Gln Tyr Cys Arg
500 505 510
Glu Thr Val Met Thr Ile Trp His Tyr His Gly Gly Cys His Val Gly
515 520 525
Ala Val Val Asp Asp Asn Tyr Arg Val Phe Gly Val Arg Gly Leu Arg
530 535 540
Val Ile Asp Ser Ser Thr Phe Arg Tyr Ser Pro Gly Thr Asn Pro Gln
545 550 555 560
Ala Thr Val Met Met Leu Gly Arg Tyr Met Gly Ile Lys Ile Gln Ala
565 570 575
Glu Arg Trp Arg Lys
580
<210> 10
<211> 25
<212> DNA
<213> Artificial Synthesis
<400> 10
atactgacac aagtttatgg ggctg 25
<210> 11
<211> 25
<212> DNA
<213> Artificial Synthesis
<400> 11
aattacattc aaatatggct ccttg 25
<210> 12
<211> 22
<212> DNA
<213> Artificial Synthesis
<400> 12
ggcctcgtga actcgtcgta tc 22
<210> 13
<211> 25
<212> DNA
<213> Artificial Synthesis
<400> 13
tgaattacat gcaaatttgg ctccg 25
<210> 14
<211> 23
<212> DNA
<213> Artificial Synthesis
<400> 14
atgtccaacc aggagcactt cac 23
<210> 15
<211> 24
<212> DNA
<213> Artificial Synthesis
<400> 15
gcacagtttt ttgaagcaat gttg 24
<210> 16
<211> 2601
<212> DNA
<213> Triticum aestivum L.
<400> 16
tgactctgtc catcctgaac caaaagaatg ggaaacactt gactaaaact tcaggttcaa 60
ctctccttag caagtaccag tgctactgat atgtgtgatt caagcagcta gtttttgtaa 120
cctcatgatg attacagttg atcttacttt tgtggtgatc acaggtctct tcatatgtga 180
tggcggatgg ggttggccta actttcttgc tctcaagaaa ctcaagaaag gctatgttgt 240
agggtcgagc tgcattgtga aggcagatct cactatcgtt ggtttatcca atgatggcta 300
gatctttacc tcatgccgat ggagataaag ccctttcatc ctcctagcaa gacttatata 360
tgagataatg ttgtggtagc agcactaaaa acttgtgggg tgattagtag taatgagctg 420
gatttgagaa gtgagatgac ttcacgcata tatttctagg tattatatat caagcagaaa 480
tatgttattg gaagtacttg agtttgcatg gatgatgcaa ctatgtcggt cagcttgtct 540
cgctacatgg atgtttgatt cagctgaacc atgttatctc tcgcttggtt tttgtgcacc 600
atggatgtcc ccagttgatg aatcttatgg ttccatgtaa agcactttgc ttctatttat 660
tttgagaagg aaaacggctc ttcccttgtc caccgtccat ttattttgat cccatggtcc 720
aggttttaga actcctagtc ccggaaataa acaactatct tgcaagtgct tcatgtacaa 780
ttgcaacgat gagcaggtga ctaacctaat actccctccg tttctaaata tttgtctttc 840
tagatatttt aacaagtgac tacatacgga gcaaaatgag tggatctaca ctctaaaata 900
tgtctacgta catccgtata tagtagccat ttgaaatgtc tagaaagaca aatatttagg 960
aacagaggga gtatttatta tacttttata aagaatgttg gagtacccat ctctggggag 1020
cctctgcgtg tgaaaatgtc ctccgcgctg tttgttgcac gttggccacc cgcggctggc 1080
tccctggggc gctccgggcc atcgatctag gcggccgcct cgcacccgga ctgcttgcct 1140
ctgggtccat gacgaggggg tgcgggcgcg gttggatcca tcgtgatgtg gtgtttttgg 1200
tcacaccagt ttgtgccctc ttgttggttg tcctgcgcgt gatgggccga ccgaatctgc 1260
ctgtctgtcg tgttgaccac ccgcggctag ctccctgggg cgctccgggc cattgatcta 1320
ggcagccgcc tcgcacctgg actgcttgcc tctgggtcca tgacgagggg tgcgggtgca 1380
gtcggatcca tcgtgatgtg gtgttttcgg tcacaccagt ttgtgccctc ttgttgattg 1440
ccctgcgcgt gatgggctga ccggatctgc ctgttagtgt tgtgttggcc acccgcggct 1500
ggctccttgg ggcgctccag gtcatcgatc taggcggtcg cctcgcaccc ggactgcttg 1560
cctctgggtc catgacgaga ggatgcgggc gcggtcggat ccatcatgat gtggtgtttt 1620
cggtcacacc agtttgtgct ctcttgttgg ttgtcctgcg cgtgatgggc cgaccggatc 1680
tgcctgtcag tgttgtgttg gccacccgcg gctggctcct tggggcgctc cgggctatcg 1740
atctaggcgt ccgcctcgca cccggactgc ttgcctctgg gtccatgacg aggaggtgcg 1800
gacgcggtcg gatccatcat gatgtggtgt tttcggtcac accagtttat gccctcttgt 1860
tggttgatct gcgcgtgatg ggccgactag atctgcctgt cagtgttgtg ttggccaccc 1920
gcggctggct ccatggggcg ctccgcgcca tcgatctagg gggccgcctc gcatccagac 1980
tgcttcagga actgacatcc gttcatttat tttgtcagtt catttatttg aggcaaattt 2040
cttcaggaac tgacatccgt tccatggggc atacgttggg gattggtttc tggcgaacgt 2100
tcgccagatt atctttaccg ctttttttta gtttctaggt ttttcttggt attttattat 2160
tttcttatag tttggaccga tctccttaca tcatagtttt gccttggtac ctgtaatatg 2220
tatttgtgct tgtatataaa aatggaggga gtatagtttt ttcgggaaaa atatagtgca 2280
cgcgcaaaac cgtctcaaga tcggccagta aaccctcccc cttgctgctt catcgcattt 2340
gatatgatat actcaacagc tcctcccgtc taaaaagaaa gttcatcact tctgcatttc 2400
acaaccgatg cattccattt gatataacca ttccctttct atactgacac aagtttatgg 2460
ggctgttctt tgcactgcaa ccgcctccgc attatttaac acatctaccc tcgatctacc 2520
cgcgcgctaa ctcgaggttt ataaaccaag cgaacttttc gcgtccctga agcgcaaagg 2580
atgccgagct cgccggcgca a 2601
<210> 17
<211> 2635
<212> DNA
<213> Triticum aestivum L.
<400> 17
tgctgccaaa gaagaatgga gctttggagg tcaaccatta tcgaccaatc agcttgatcc 60
actctgttgc taagctaatc tccaaagtgc tcgttgcaag gctggcaggg gccctggata 120
aaatcatctc accggcgtag actgcatttc agaaaggcaa atgcagccac gagagctatc 180
agtacgtgca aagctgcgtt cggatgttac ataggacgaa gaaaaaggca cttttcttca 240
aactcgacat agcaaaggct tttgattcgg tgacatggga ggacttgctc gagctcaagc 300
aaaaaaatgg gtttccctcc tagatgggga aactggatag cgctgctact ctcttctacc 360
acctcctcct gcttgctcaa cgaaacgcag agccaatcaa tatctcatgg ttgtggcttt 420
aggcagggtg attcgctatc cctgctgctt ttcatcctgg cgatcgaccc tcttcaccac 480
ctccttcaag ctgctgcaga ggaagagatg atcgctcagc tgacaggccg tgggatcagc 540
atgtggatta gcttatacgc tgatgacgcc gtcattttcg ccaacccggt gaaggaagaa 600
gtggacacgc tgttgttttt gctcagcaga ttcggggagg ccacgggact gaagttaaac 660
caggcaaaat ctgaagtcat cccaattaac tgtgctgatg ttcagttaac tgaggttctg 720
caaaactttg gggggctgca gtccacgttc ccaaccacat acctgggact gccaatctct 780
ccaaggaagc taaggctggt gcatttccaa tttatcattg accggatacg ctcaaggttg 840
gctggttgga aaggcaagct gatgaacatg gcaggtagaa gggtactggt gcgtgctgtg 900
ttgatggcac tgccggtgtt tgcaatgaca gtactcaaag tgccgaagaa gattttaaaa 960
gaggtggaaa aagcaaggag gcagtttcta tgggcacatg atgagaatgt cactggcgcc 1020
aaatgcaagg tggcctgggg caaagtctgt cttccggtgg ccaagggggg gctaggtttg 1080
ttggacctcc atcgtttcag cacagctctc cggctgcgct ggttgtggct ctcctggcaa 1140
cagccacacc ggccctggat gaacttcccg gcccaaggag gacctggagc tgtttgcatc 1200
ggccaccttt gtgcacttgg gcaacggaaa aacagcgcgt ttctggactt gcaaatggat 1260
ctgggcggtg gcgctgcaac acgagttccc atcattgttt cagcatgcca gaagaaaaaa 1320
caggatggtt catgatgcgc tgacaaatga cacatgggta cgtgatctcc ggcatggcaa 1380
catagtagag attgctcacc ggtttttgca aatgtggaga aagattcagg gtgcaggcat 1440
tgttctctcc tccaacgagg accggatcag ctgggtggca accagaggag gaggaagcta 1500
ctcggctaag gctgcttacg atctacaaat cagtgacatt cctccttcag gaggcaaagc 1560
tgcgatttgg agggcttggg cgccaggcac ggtgaagatc ttcgcctggc tgctccacca 1620
ggacaagctt tggtgcaacg atcgcctgca acggcgaggg tggcccaatg ggtatttttg 1680
cgtatgctgc aacagaaatc tggaaagctc aatacacctt ttctgggact gcccttttgc 1740
ccgatctgtc tggccggcag tgtcgcgtag atatggttgc gctgtgctca aagaagcgcg 1800
ggacgcaggt cacagttccc tcagacattg ggaaagactg accgcactaa cgcctcaaag 1860
ttgcagaaaa gggctaaggt cgatgctgtt gctgatcact tgggagtttt ggaaagagag 1920
gaacgcgtgt gtatttagag gaaaaacgcc ccagacggag gatgttcagc gagccatcaa 1980
gaacacgatg gagctctggc gggcagcagg agctcgatgc attgaggccc tgttctgtag 2040
cgagataaaa cgtacggctc atgtagctgg cctagttttg gccattctta gctcttctca 2100
aaccatgatc acttgatcgc ccatgtattt cccactgctt tcttctaaat caatgaagca 2160
ggtgaacccg gatctttcaa aaaaaaacct cctgatgtac tcagtgtcgt ggccggtgat 2220
ttaaccgtgt ccactgatta attggaatgt atggtgttcc aggtcaaaaa agaatgtcaa 2280
atgtaattct ttcggaaaaa atatagtgca cgcgcaaaac cgtctcaaga ttggccagta 2340
aaccctctcg cttgctgctc cgtcgcattt gatatactca acagctcctc ctgtctaaaa 2400
agaaagttca tcacttctgc atttcacgac caatgcattg catttgatat aaccattccc 2460
tttctatagt aacacatgtt tatgggctcc tcgcggctct tctttgcact gcaaccgcct 2520
ccgcattatt taacacatct accctcgatc tacccgcgcg ctaactccag gtttataagc 2580
gaagcgaact tttcgcgtcc ctgaagcgta aaggatgccg agctcgccgg cgcaa 2635
<210> 18
<211> 2821
<212> DNA
<213> Triticum aestivum L.
<400> 18
ctgacatagt acatgtaatc tttaaatcca taaccttctg tttctatcag tgctactagg 60
ttgtaaaatg ttatgtctga caccatcata gatctctcaa tgttgtctct gtcatggaaa 120
tgcatcctca attcccaaac ctcatcatcc agactgcaaa taatataaag cactaatgaa 180
acactaataa aactaataac caaatgtaaa ccctagggca caggacaaac aaagcacaac 240
atcagaacta atctaaaaat gtcaacccta ggcacaggac aaacaaagca cataatctga 300
actaatctaa ataatgtcct agggcacatg acaacaaaat cacaacattt gacacttgta 360
ctgcatttga taggatagaa caatatgaac actaacccta ggggtaccct cgcaacacaa 420
ttgaatggag taccctggca acgcaattga atggagaacg ggaagggaag agcaaagaaa 480
agtggatctc actatacgcc gccgaaggat gacgcgatgt gctggcttcc cgtcgaattg 540
gccttcaccg ccgcggcgaa tgctctggcc gccctgtatt ccatcgaata ttgcgggtcc 600
tcctcctcct cttcctctga cggcttgcta gggttcgaac tcgcgcacag ctcgcctaga 660
tccactgcgg gtcctcctcc actacgtacg atgcgtgcgg cgctgccggg cggagagccg 720
ccaccagcat ctccaccttt cgccggactg cgcatctctg tgtttctatg gaatggcgac 780
ggcaccaatg ggggagggcg acggcagcgg gagcgaggag gagggggaag aagaatgggg 840
gagggcgacg gcggcgggag cgaggaggag ggggaagaag aatgggggag ggcgacggcg 900
gcgggagcga ggaggagggg gaagaagaat gggggagggc gacggcggcg ggagcgagga 960
gggggaaaag agacggatgc gggagtggat gactaaagac gagccctaac ggcgccgtct 1020
aacgccgact gccggctgtc gggacggcct ggagtgccat gtaggcgaaa aacgggccca 1080
acctgtcata aactcactta acacggcccg atccgccgat cagtggtttt tgcaaaacaa 1140
ttacgataga cgtggtgctt tctgcaacaa ttctgtaacc taatgttttc cgcaaaccta 1200
gccctcaaag tggtggtttt atgcaattta ctctgattac agttgatctt acttttgtgg 1260
tgatcacagg tctcttcgta tgtgatggcg gatggggttg gcctaacttt cttggactca 1320
agaaactcaa gaaaggctat gttgtagggt cgagctgcat tgtgaaggca gatctcacta 1380
tcgttggttt atacaatgat ggctagatct ttacctcatg cctatggaga gataaagccc 1440
tttcatcctc ctagcaagac ttatatatga gatgatgttg tggtagcagc actaaaaact 1500
ttttttttag aaaaggagca gcactaaaaa cttggggtga ttagtagtaa tgagctggat 1560
ttgagaagtg agatgacttc acgcatatat ttctaggtat tatatatcaa gcagaaatat 1620
gatattggaa gtacttgtgt ttgcatggat gatgcaacta tgtcggtcag cttgtctcgc 1680
tacatggatg tttgattcag ctaaaccatg ttatctctcg cttggttttt gtgcaccatg 1740
gatgttccag ttgatgaatg gttccatgta aagcgctttg cttctattta ttttgggaag 1800
gaaaacggct ctttccattg tccaccgtcc atttattttg atcccatggg ccagatttta 1860
gaactcccgg tcccggaact aaactaatat ttattacttc tataaagaat gttggagtat 1920
caggtcttcg gggagccttt gcgtgtgaaa atgtcctccg cgttgttcgt tgcatgttgg 1980
ccacccgcgg ttggctccct ggggcgctct gggccgtccg atctaggcag gcgcctcaca 2040
cccggacgac ttgcctatgg gtccatgccg ggggggctgc gggcacggtt gggtccatcg 2100
tgatgtggtg ttttcgatca caccagtttg tgccctcttg ttggttgcct tgcgcgtgac 2160
gggccgaccc tgttggtgta gttcgcttgt gccaggcaag ccgcgcacac ccccggatcg 2220
gggtggcgcg tgttaggaaa ccggagaggg ccatccggcc agccgtcgcg atggcgggcc 2280
agccggccaa ggttcattta tttgaggcaa atttctttag tgacatccgt gctatggggc 2340
gtttgctggg gattggtttc tagcgaacgt tcgccagatt atctttaccg ctttctttta 2400
gtatctaggt ttttcttagt attttattat ttttctcata gtttggaccg atctcctcac 2460
gtcagaatgt caaatgtaat cctttcgaaa aaaatatagt gcacgcgcga aaaccgtctc 2520
aagattggcc agtaaaccct cctgcttact gctccatcgc atttgatata ctccttctgt 2580
gtaaaaagaa tgttcatcac ttttgcattt cacaaccaat gcattgcatt tgatataacc 2640
attccctttc tatagtaaca caagtttatg ggctcctcgc agctgttctt tgcactgcaa 2700
ccgcctccgc attatttaac agatctaccc tcgatctacc cgcgcgctaa ctcgaggttt 2760
ataaaccaag cgaacttttc gcgtccctga agcgcaaagg atgccgagct cgccggcgca 2820
a 2821
<210> 19
<211> 40
<212> DNA
<213> Artificial Synthesis
<400> 19
aagcttctga catagtacat gtaatcttta aatccataac 40
<210> 20
<211> 25
<212> DNA
<213> Artificial Synthesis
<400> 20
ggatccttgc gccggcgagc tcggc 25
<210> 21
<211> 20
<212> DNA
<213> Triticum aestivum L.
<400> 21
ctggtggacc agcccatggt 20
<210> 22
<211> 1453
<212> DNA
<213> Triticum aestivum L.
<400> 22
tatttcaagg agccatattt gaatgtaatt catggcataa tattgtattt ggggaacctg 60
gtttagacag attaatttgt ttttttccac ttattctcta ggcatagtct tttagtgcct 120
gtttgttgat tattatgaaa cccagtcatc cctttaacat gcatttctct gcattgcaaa 180
ataagaagga aaaacttatg ttgcaaaggc atgacggtgg ttcaagcaac aatacttcga 240
tcaaaaaaat tggaaaccgg ttgccacatc cacctaccaa tttccattga taacaagagt 300
agcaaaccag aatactgtgc aggatgcatt tacagttttg tactagaaac ttaatttcat 360
gaaattacat accactattt tacttggtgg ttgttacatg ctacaaacgc caggttccct 420
tcagagggat ccagcaagcc atcatcttat ctatacaggg accgagtaca tgtcaaagga 480
acatgatgag ttaagaatgg aacagccaaa ggcagaaatg acttgaacat tttttttcct 540
tgtttccaac ttgccctact tgcctgctac ctatatattg gcttgcgaaa ttctctccct 600
acctatgtta tcaggaacca gaaaatgcta cttcggcatc tctgttgtcc agaataccca 660
agaagtcacc aacaattgca tgcttgcata agaataattc atatagtgag gttttcttgg 720
atggccgtga aggagccaca aatgtttccg cccatgtctg ggaatgtcgc acatcctggc 780
aagggtttca ccttgccttt acggtcaaca cttacagggt acttgaggag gtggcctctc 840
atctcagtca cctcatttgc aaagaactga tcccagttat gttgcccgag ttgacgaacc 900
cgcctcatgc actctaggct ctctggatgg ttaaagtctt cctcgataat tccaatatgc 960
tcagcccaga gggacattct gtagccataa atctacataa aaaatcacca aacttgattt 1020
aagagtataa tccagaaata ccagctgcgt cataactaga ttattaccct aagagatgag 1080
atgatatacc ccgacaacct ttttcacttg ctgttttaga atgcacatat tagtatttat 1140
attcacataa ctgtacttgt agaattgcca ccaacagtat ttatgtgtca ttacatgtct 1200
atagtttagt gtatatataa ttgaaaaaaa ggtaaatgtt aaatgggtgt agttggaggc 1260
agtctctcta aataaggtga aacaattgta atgaagcagc agttctaagc aacgcataac 1320
atcaggaaga caagtacaaa atctgcaggc acaatctcaa aatgcaagct catgaaaaca 1380
cctgtcttct gcaggtacca aaagatgtga ttcagcagct tgaacttgaa acatgaggca 1440
ttacctgtcc acg 1453
<210> 23
<211> 1446
<212> DNA
<213> Triticum aestivum L.
<400> 23
tatttcacgg agccaaattt gcatgtaatt catggcataa tattgtattt gggggacctg 60
gtttagaaca gattaatttg gtttttccac ttattctcta ggcatagtct tttagtgcct 120
gtttgttgat tattatgaaa cccatacatc cctttaacat gcatttctct gcactgtaaa 180
ataagaagga aaacttatgt cccaaaggca tgatggtggt tcaagcaaca atacttcggt 240
aaaaaaatgg aaccggttgc cacatccacc caccaatttc cattgatgac aagaggagca 300
aaccaaggcc caacattgct tcagaatagt gtgaaggatg catttacagt tttgcactag 360
aaactttgtt tcatcaaatt acatataaca tactactatt tctcttggtg gttgtcactt 420
gctacaaacg ccaggttccc ttcagaggga tccagcatgt aatcatctta catatacagg 480
gaccgaatac atgtaaaagg aacatgatta gttaagatgg gaacaaccaa aaggcagaaa 540
tgacttgatt tttttttcct tgtttccaac ttgccctact tgcctgctac ctacatattg 600
gcttgcgaaa ttctctccct acctatgtta tcaggaacca gaaagtgcta cttcggcatc 660
tctgttgtcc ataataccca agaagtcacc aacaattgca tgcttgcata agcagaattc 720
atattgtgag gttttcctgg atggccgtga aggagccaca aatgtttccg cccatgtctg 780
ggaatgtcgc acatcctggc aagggtttca ccttgccttt acggtcaaca cttacagggt 840
acttgaggag gtggcctctc atctcagtca cctcatttgc aaagaactga tcccagttat 900
gttgcccgag ttgacgaacc cgcctcatgc actctaggct ctctggatgg ttaaagtctt 960
cctcgataat tccaatatgc tcagcccaga gggacattct gtagccataa atctacataa 1020
aaaatcacca aacttgattt agagtataat ccagaaatac cagctgcgtc ataagtcata 1080
actagattat taccctaaga gatgagataa tatacccaga caaccttttt cactgctgtt 1140
ttagaatgca catattagta tttaattcac ataagaactg tacttgtaga attgccacca 1200
acagtatgta tgtgtcatta cgtgtctata gtttagtgta tatataatta aaaaaaggta 1260
aatgttaaat gggcgtagtt ggaggcagtc tctctgaaaa cgtaaaaaat aagtaaggtg 1320
aaacaattgt aatgaagcag tggttctaag caacgcataa catcaggaag acaagtacaa 1380
aatctgcagg cacaatctca aaatgcaagc tcatgaaaac acctgtcttc tgcaggtacc 1440
aaaaaa 1446
<210> 24
<211> 1472
<212> DNA
<213> Triticum aestivum L.
<400> 24
tatttcacgg agccaaattt gcatgtaatt cgtggcataa tattgtattt gggggacctg 60
gtttagaaca gattaatttg gtttttccac ttattctcta ggcatagtct tttagtgcct 120
gtttgttgat tactatgaaa cccagacatc cctttaacat gcatttctct gcattgcaaa 180
ataagaaaga aaacttatgt tgcaaaggca tgatggtggt tcaagcaaca atacttcggt 240
aaaaaaatgg aaaccggttg caacatccac ccaccaattt ccattgatga caagagtagc 300
aaaccaaggc ccaacattgc ttcaaaaaac tgtgcaggat gcatttacag ttttgcacta 360
gaaacttagt ttcatcaaat tacatataac atactactat ttctcttggt ggttgtcact 420
tgctacaaac gccaggttcc cttcagaggg atccagcatg taatcatctt acctatacag 480
ggaccgaata catgtaaaag gaacatgatt agttaagacg ggaacaacca aaaggcagaa 540
atgacttgaa ttttttttcc ttgtttccaa cttgccctac ttgcctgcta cctacatatt 600
ggcttgcgaa attctctccc tacctatgtt atcaggaacc agaaagtgct acttcggcat 660
ctctgttgtc cagaataccc aagaagccaa caattgcatt cttgcataag cagaattcat 720
attgtgaggt tttcctggat ggccgtgaag gagccacaaa tgtttccgcc catgtctggg 780
aatgtcgcac atcctggcaa gggtttcacc ttgcctttac ggtcaacact tacagggtac 840
ttgaggaggt ggcctctcat ctcagtcacc tcattggcaa agaactgatc ccagttatgt 900
tgcccgagtt gacgaacccg cctcatgcac tctaggctct ctggatggtt aaagtcttcc 960
tcgataattc caatatgctc agcccagagg gacattctgt agccataaat ctacataaaa 1020
gatcaccaaa cttgatttag agtataatcc agaaatacca gctgcgtcat aactagatta 1080
ttaccctata agagatgaga tgatataccc cgacaacctt tttcacttgc tgttttagaa 1140
tgcacatatt agtatttata ttcacataag aactgtactt gtataattgc caccaacagt 1200
atttatgtgc cattacgtgt ctatagttta gtgtatatat aataaaaaag ggtaaatgtt 1260
aaataggtgt agttggaggc agtctctctg aaaacgtaaa aaataagtta aaacaattgt 1320
aatgaagcag cggttctaag caacgcataa catcaggaag acaagtacaa aatctgcagg 1380
cacaatctca aaatgcaagc tcatgagaac acctgtcttc tgcaggtacc aaaaaatgtg 1440
attcagcagc ttgaacttga aacatgaggc at 1472
<210> 25
<211> 1487
<212> DNA
<213> Zea mays
<400> 25
atggcggcga caatggcagt gacgacgatg gtgacgagga gcaaggagag ctggtcgtca 60
ttgcaggtcc cggcggtggc attcccttgg aagccacgag gtggcaagac cggcggcctc 120
gagttccctc gccgggcgat gttcgccagc gtcggcctca acgtgtgccc gggcgtcccg 180
gcggggcgcg acccgcggga gcccgatccc aaggtcgtcc gggcggcctg cggcctggtc 240
caggcacaag tcctcttcca ggggtttaac tgggagtcgt gcaagcagca gggaggctgg 300
tacaacaggc tcaaggccca ggtcgacgac atcgccaagg ccggcgtcac gcacgtctgg 360
ctgcctccac cctcgcactc cgtctcgcca caaggctaca tgccaggccg cctatacgac 420
ctggacgcgt ccaagtacgg cacggcggcg gagctcaagt ccctgatagc ggcgttccac 480
ggcaggggcg tgcagtgcgt ggcggacatc gtcatcaacc accggtgcgc ggaaaagaag 540
gacgcgcgcg gcgtgtactg catcttcgag ggcgggactc ccgacgaccg cctggactgg 600
ggccccggga tgatctgcag cgacgacacg cagtactcgg acgggacggg gcaccgcgac 660
cgggcgaggg gttcgcggcg gcgcccgaca tcgaccacct caacccgcgc gtgcagcggg 720
agctctccgc ctggctcaac tggctcaggt ccgacgccgt ggggttcgac ggctggcgcc 780
tcgacttcgc caagggctac tcgccggccg tcgccagaat gtacgtggag agcacggggc 840
cgccgagctt cgtcgtcgcg gagatatgga actcgctgag ctacagcggg gacggcaagc 900
cggcgcccaa ccaggaccag tgccggcagg agctgctgga ctggacgcgg gccgtcggcg 960
ggcccgccat ggcgttcgac ttccccacca agggcctgct gcaggcgggc gtgcaggggg 1020
agctgtggcg gctgcgcgac agctccggca acgcggccgg cctgatcggg tgggcgcccg 1080
agaaggccgt caccttcgtc gacaaccatg acaccgggtc gacgcagaag ctctggccgt 1140
tcccatccga caaggtcatg cagggctacg cctacatcct cacccatcca ggagtcccct 1200
gcattttcta cgaccacatg ttcgactgga acctgaagca ggagatatcc acgctgtctg 1260
ccatcagggc gcggaacggc atccgcgccg ggagcaagct gcggatcctc gtggcggacg 1320
cggacgcgta cgtggccgtc gtcgacgaga aggtcatggt gaagatcggg acaaggtacg 1380
gcgtgagcag cgtggtcccg tcggatttcc acccggcggc gcacggcaag gactactgcg 1440
tctgggagaa agcgagcctc cgcgtcccgg cggggcgcca cctctag 1487
<210> 26
<211> 2736
<212> DNA
<213> Zea mays
<400> 26
tgcaccggac actgtctggt ggcataccag acagtccggt gtgccagatc agggcaccct 60
tcggttcctt tgctcctttg cttttgaacc ctaactttga tcgtttattg gtttgtgttg 120
aacctttatg cacctgtgga atatataatc tagaacaaac tagttagtcc aatcatttgt 180
gttgggcatt caaccaccaa aattatttat aggaaaaggt taaaccttat ttccctttca 240
atctccccct ttttggtgat tgatgccaac acaaaccaaa gaaaatatat aagtgcagaa 300
ttgaactagt ttgcataagg taagtgcata ggttacttag aattaaatca atttatactt 360
ttacttgata tgcatggttg ctttctttta ttttaacatt ttggaccaca tttgcaccac 420
ttgttttgtt ttttgcaaat ctttttggaa attctttttc aaagtctttt gcaaatagtc 480
aaaggtatat gaataagatt gtaagaagca ttttcaagat ttgaaatttc tccccctgtt 540
tcaaatgctt ttcctttgac taaacaaaac tccccctgaa taaaattctc ctcttagctt 600
tcaagagggt tttaaataga tatcaattgg aaatatattt agatgctaat tttgaaaata 660
taccaattga aaatcaacat accaatttga aattaaacat accaatttaa aaaatttcaa 720
aaagtggtgg tgcggtcctt ttgctttggg cttaatattt ctcccccttt ggcattaatc 780
gccaaaaacg gagactttgt gagccattta tactttctcc ccattggtaa atgaaatatg 840
agtgaaagat tataccaaat ttggacagtg atgcggagtg acggcgaagg ataaacgata 900
ccgttagagt ggagtggaag ccttgtcttc gccgaagact ccatttccct ttcaatctac 960
gacttagcat agaaatacac ttgaaaacac attagtcgta gccacgaaag agatatgatc 1020
aaaggtatac aaatgagcta tgtgtgtaat gtttcaatca aagtttcgag aatcaagaat 1080
atttagctca ttcctaagtt tgctaaaggt tttatcatct aatggtttgg taaagatatc 1140
gactaattgt tctttggtgc taacataagc aatctcgata tcaccccttt gttggtgatc 1200
cctcaaaaag tgataccgaa tgtctatgtg cttagtgcgg ctgtgttcaa cgggattatc 1260
cgccatgcag atagcactct cattgtcaca taggagaggg actttgctca atttgtagcc 1320
atagtcccta aggttttgcc tcatccaaag taattgcaca caacaatgtc ctgcggcaat 1380
atacttggct tcggcggtag aaagagctat tgagttttgt ttctttgaag tccaagacac 1440
cagggatctc cctagaaact gacaagtccc tgatgtgctc ttcctatcaa ttttacaccc 1500
tgcccaatcg gcatctgaat atcctattaa atcaaaggtg gatcccttgg ggtaccaaag 1560
accaaattta ggagtgtaaa ctaaatatct catgattctt ttcacggccc taaggtgaac 1620
ttccttagga tcggcttgga atcttgcaca catgcatata gaaagcatac tatctggtcg 1680
agatgcacat aaatagagta aagatcctat catcgaccgg tatacctttt ggtctacgga 1740
tttacctccc gtgtcgaggt cgagatgccc attagttccc atgggtgtcc tgatgggctt 1800
ggcatccttc attccaaact tgttgagtat gtcttgaatg tactttgttt ggctgatgaa 1860
ggtgccatct tggagttgct tgacttgaaa tcctagaaaa tatttcaact tccccatcat 1920
agacatctcg aatttcggaa tcatgatcct actaaactct tcacaagtag atttgttagt 1980
agacccaaat ataatatcat caacataaat ttggcataca aacaaaactt ttgaaatggt 2040
tttagtaaag agagtaggat cggctttact gactctgaag ccattagtga taagaaaatc 2100
tcttaggcat tcataccatg ctgttggggc ttgcttgagc ccataaagcg cctttgagag 2160
tttataaaca tggttagggt actcactatc ttcaaagccg agaggttgct caacatagac 2220
ctattcaccc catttgatca cttttttggt ccttcaggat ctaatagtta tgtataattt 2280
agagtctctt gtttaatggc cagatatttc taattaatct aagaatttat gatatttttt 2340
aattttttat catgtctgat gagaattaac ataaaggctc aattgggtcc tgaattaata 2400
atagagtgaa aattaatcca gaggctctat tagaaccttc aattagtaat accaagatat 2460
atataagata gtagagtata gtttaaatgt tggcattgtt cattctttct tttgttattt 2520
aatttatgct ttccacggtg gttagtggtt acttctgaag ggtccaaata atgcatgaag 2580
agtttgagga caagaagtct gccctaaaaa tagcgatgca aaggcatggt gtccaagcca 2640
tacatatagc gcactaattt tatcagcaga acaatggtat ttataggtcc tagtgcccag 2700
gcaacaagag acacgaataa agcatcgatc acgaca 2736
<210> 27
<211> 344
<212> DNA
<213> Zea mays
<400> 27
tgacaaagca gcattagtcc gttgatcggt ggaagaccac tcgtcagtgt tgagttgaat 60
gtttgatcaa taaaatacgg caatgctgta agggttgttt tttatgccat tgataataca 120
ctgtactgtt cagttgttga actctatttc ttagccatgc caagtgcttt tcttattttg 180
aataacatta cagcaaaaag ttgaaagaca aaaaaaaaaa cccccgaaca gagtgctttg 240
ggtcccaagc tactttagac tgtgttcggc gttcccccta aatttctccc cctatatctc 300
actcacttgt cacatcagcg ttctctttcc cctatatctc cacg 344
<210> 28
<211> 711
<212> DNA
<213> Artificial Synthesis
<400> 28
atggtgtcca agggcgagga ggacaacatg gccatcatca aggagttcat gcgcttcaag 60
gtgcacatgg agggctccgt gaacggccac gagttcgaga tcgagggcga gggcgagggc 120
cgcccctacg agggcaccca aaccgccaag ctcaaggtga ccaagggtgg ccccctcccc 180
ttcgcctggg acatcctctc cccacaattc atgtacggct ccaaggccta cgtgaagcac 240
cccgccgaca tccccgacta cctcaagctc tccttccccg agggcttcaa gtgggagcgc 300
gtgatgaact tcgaggacgg cggcgtggtg accgtgaccc aagactcctc cctccaagac 360
ggcgagttca tctacaaggt gaagctccgc ggcaccaact tcccctccga cggccccgta 420
atgcaaaaga agaccatggg ctgggaggcc tcctccgagc ggatgtaccc cgaggacggc 480
gccctcaagg gcgagatcaa gcaaaggctc aagctcaagg acggcggcca ctacgacgcc 540
gaggtgaaga ccacctacaa ggccaagaag cccgtgcaac tccccggcgc ctacaacgtg 600
aacatcaagc tcgacatcac ctcccacaac gaggactaca ccatcgtgga gcaatacgag 660
cgcgccgagg gccgccactc caccggcggc atggacgagc tgtacaagtg a 711
<210> 29
<211> 1149
<212> DNA
<213> Artificial Synthesis
<400> 29
cgtcaacatg gtggagcacg acacgcttgt ctactccaaa aatatcaaag atacagtctc 60
agaagaccaa agggcaattg agacttttca acaaagggta atatccggaa acctcctcgg 120
attccattgc ccagctatct gtcactttat tgtgaagata gtggaaaagg aaggtggctc 180
ctacaaatgc catcattgcg ataaaggaaa ggccatcgtt gaagatgcct ctgccgacag 240
tggtcccaaa gatggacccc cacccacgag gagcatcgtg gaaaaagaag acgttccaac 300
cacgtcttca aagcaagtgg attgatgtga tgaattcaac cgtctcttcg tgagaataac 360
cgtggcctaa aaataagccg atgaggataa ataaaatgtg gtggtacagt acttcaagag 420
gtttactcat caagaggatg cttttccgat gagctctagt agtacatcgg acctcacata 480
cctccattgt ggtgaaatat tttgtgctca tttagtgatg ggtaaatttt gtttatgtca 540
ctctaggttt tgacatttca gttttgccac tcttaggttt tgacaaataa tttccattcc 600
gcggcaaaag caaaacaatt ttattttact tttaccactc ttagctttca caatgtatca 660
caaatgccac tctagaaatt ctgtttatgc cacagaatgt gaaaaaaaac actcacttat 720
ttgaagccaa ggtgttcatg gcatggaaat gtgacataaa gtaacgttcg tgtataagaa 780
aaaattgtac tcctcgtaac aagagacgga aacatcatga gacaatcgcg tttggaaggc 840
tttgcatcac ctttggatga tgcgcatgaa tggagtcgtc tgcttgctag ccttcgccta 900
ccgcccactg agtccgggcg gcaactacca tcggcgaacg acccagctga cctctaccga 960
ccggacttga atgcgctacc ttcgtcagcg acgatggccg cgtacgctgg cgacgtgccc 1020
ccgcatgcat ggcggcacat ggcgagctca gaccgtgcgt ggctggctac aaatacgtac 1080
cccgtgagtg ccctagctag aaacttacac ctgcaactgc gagagcgagc gtgtgagtgt 1140
agccgagta 1149
<210> 30
<211> 310
<212> DNA
<213> Solanum tuberosum
<400> 30
ctagacttgt ccatcttctg gattggccaa cttaattaat gtatgaaata aaaggatgca 60
cacatagtga catgctaatc actataatgt gggcatcaaa gttgtgtgtt atgtgtaatt 120
actagttatc tgaataaaag agaaagagat catccatatt tcttatccta aatgaatgtc 180
acgtgtcttt ataattcttt gatgaaccag atgcatttca ttaaccaaat ccatatacat 240
ataaatatta atcatatata attaatatca attgggttag caaaacaaat ctagtctagg 300
tgtgttttgc 310
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВОССТАНОВЛЕНИЕ ФЕРТИЛЬНОСТИ У РАСТЕНИЙ | 2019 |
|
RU2831789C2 |
| ГЕН-ВОССТАНОВИТЕЛЬ RF4 ДЛЯ ЦИТОПЛАЗМАТИЧЕСКОЙ МУЖСКОЙ СТЕРИЛЬНОСТИ (CMS) C-ТИПА КУКУРУЗЫ, МОЛЕКУЛЯРНЫЕ МАРКЕРЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2747979C2 |
| ГЕН-ВОССТАНОВИТЕЛЬ RF4 ДЛЯ ЦИТОПЛАЗМАТИЧЕСКОЙ МУЖСКОЙ СТЕРИЛЬНОСТИ (CMS) C-ТИПА КУКУРУЗЫ, МОЛЕКУЛЯРНЫЕ МАРКЕРЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2729107C2 |
| СНИЖЕНИЕ ПРЕВРАЩЕНИЯ НИКОТИНА В НОРНИКОТИН В РАСТЕНИЯХ | 2015 |
|
RU2733837C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ РЕДУЦИРУЮЩИХ САХАРОВ В РАСТЕНИИ | 2019 |
|
RU2801948C2 |
| КОМПОЗИЦИИ ДЛЯ ИНДУКЦИИ ГАПЛОИДИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2816649C1 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ САХАРОВ И АМИНОКИСЛОТ В РАСТЕНИИ (SULTR3) | 2020 |
|
RU2826107C1 |
| ТРАНСГЕННОЕ РАСТЕНИЕ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2018 |
|
RU2758722C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ АМИНОКИСЛОТ В РАСТЕНИИ | 2019 |
|
RU2799785C2 |
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ РЕКОМБИНАНТНЫЕ КЛЕТКИ BACILLUS И ДРУГОЙ АГЕНТ БИОЛОГИЧЕСКОЙ БОРЬБЫ | 2015 |
|
RU2736827C2 |
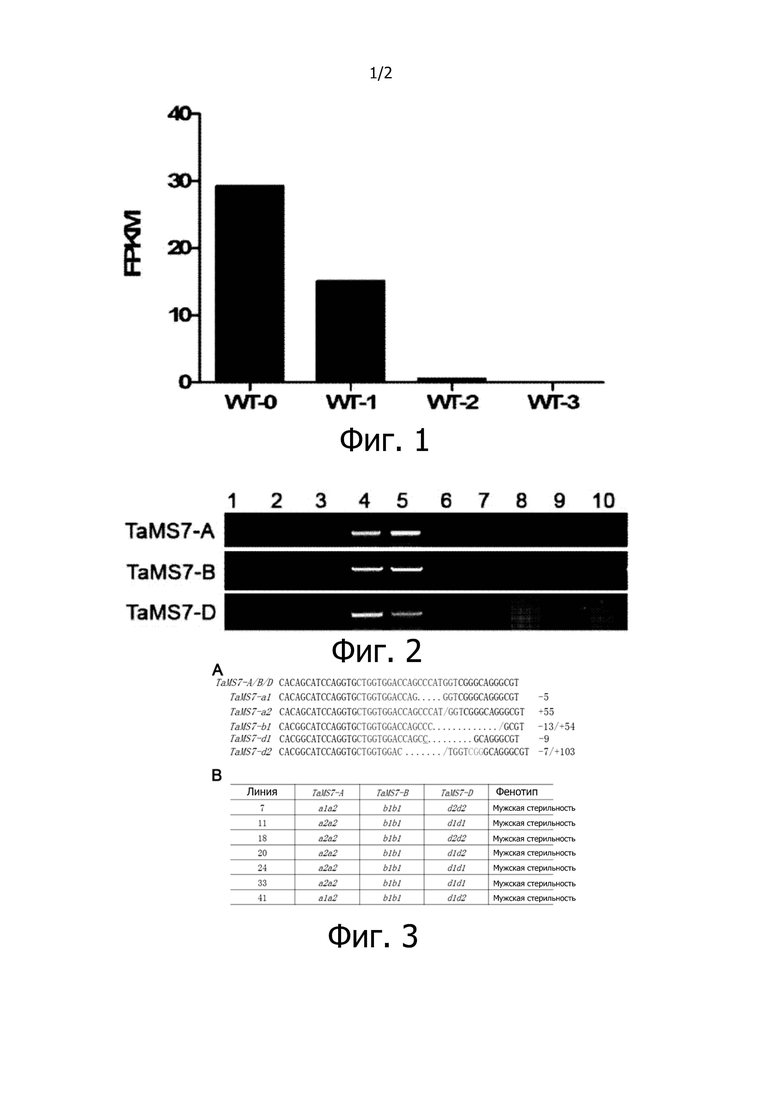
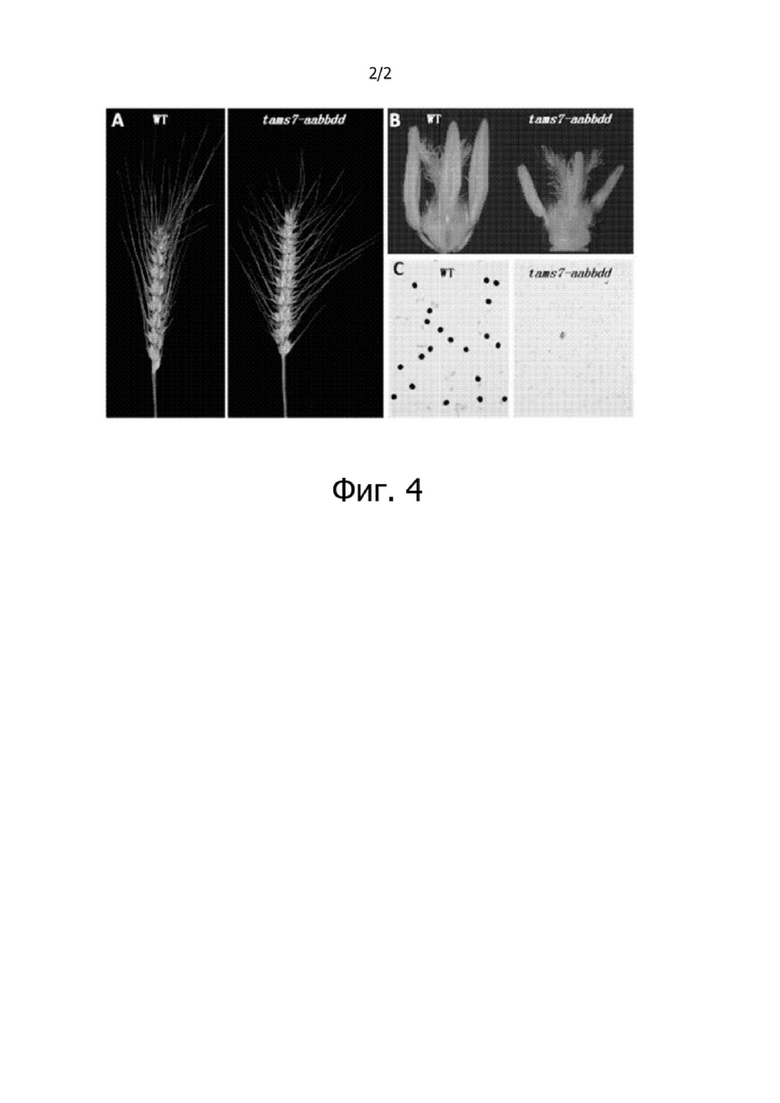
Изобретение относится к области биохимии, в частности к восстанавливающему фертильность гену TaMS7. Также раскрыты экспрессионная кассета, экспрессирующий вектор, генетически модифицированная бактерия, содержащая указанный восстанавливающий фертильность ген TaMS7. Раскрыты применение указанного гена, способ регуляции фертильности растений с помощью указанного гена. Способ размножения линии с мужской стерильностью с помощью указанного гена. Изобретение позволяет эффективно получать растение пшеницы с восстанавливающим фертильность геном TaMS7. 9 н. и 13 з.п. ф-лы, 8 пр., 4 ил.
1. Восстанавливающий фертильность ген TaMS7, нуклеотидная последовательность которого выбрана из нуклеотидных последовательностей, приведенных в SEQ ID NO: 1, 2 или 3.
2. Экспрессионная кассета, содержащая восстанавливающий фертильность ген TaMS7 по п. 1.
3. Экспрессирующий вектор, содержащий ген восстановления фертильности TaMS7 по п. 1.
4. Генетически модифицированная бактерия, содержащая ген восстановления фертильности TaMS7 по п. 1, для клонирования или хранения полинуклеотида или для трансформации растительной клетки.
5. Применение восстанавливающего фертильность гена TaMS7, экспрессионной кассеты, экспрессирующего вектора или генетически-модифицированной бактерии для увеличения уровня экспрессии гена восстановления фертильности TaMS7, в котором связанный с фертильностью ген TaMS7, экспрессионная кассета, экспрессирующий вектор или генетически-модифицированная бактерия включают одну из нуклеотидных последовательностей, приведенных в SEQ ID NO: 1, 2 или 3.
6. Способ регуляции фертильности растений, который увеличивает уровень экспрессии гена восстановления фертильности TaMS7, тем самым регулируя фертильность растений, в котором нуклеотидная последовательность восстанавливающего фертильность гена TaMS7 выбрана из нуклеотидных последовательностей, приведенных в SEQ ID NO: 1, 2 или 3.
7. Способ по п. 6, в котором нуклеотидная последовательность восстанавливающего фертильность гена включает замену, делецию или вставку одного или нескольких нуклеотидов.
8. Способ по п. 6 или 7, в котором мутирование включает мутации генов при помощи следующих методов: мутагенеза при помощи мутагенов, сайленсинга генов или редактирования генов, причем редактирование генов предпочтительно включает сайт-направленные мутации генов, а сайт-направленные мутации генов включают методы редактирования генов типа ZFN, TALEN и/или CRISPR/Cas9.
9. Способ по п. 6, который включает комплементацию фенотипа мужской стерильности, вызванного мутацией гена TaMS7, с помощью нуклеотидной последовательности гена TaMS7, с восстановлением фертильности у линии с мужской стерильностью ms7.
10. Применение способа, приведенного в любом из пп. 6-9, для регуляции фертильности растений путем увеличения уровня экспрессии гена восстановления фертильности TaMS7.
11. Способ размножения линии с мужской стерильностью, который включает следующие стадии:
(a) трансформирование линии с мужской стерильностью ms7 следующим вектором для получения поддерживающей линии, содержащей следующий вектор, причем вектор включает: восстанавливающий фертильность ген TaMS7 по п. 1, который может восстанавливать мужскую фертильность линии с мужской стерильностью ms7, и ген инактивации пыльцы, экспрессия которого нарушает функцию или образование у растений мужских гамет, содержащих ген инактивации пыльцы, поэтому фертильные мужские гаметы, образующиеся у растений, не содержат вектора; и отборочный ген, который используется для сортировки трансгенных семян и нетрансгенных семян; и
(b) самоопыление растений поддерживающей линии, трансформированных данным вектором, для получения линии с мужской стерильностью ms7 без вектора и семян поддерживающей линии, содержащих вектор; либо опыление растений линии с мужской стерильностью ms7 зернами пыльцы растений поддерживающей линии для размножения семян линии с мужской стерильностью ms7;
в котором мужская стерильная линия представляет собой мужскую стерильную линию пшеницы.
12. Способ по п. 11, в котором нуклеотидная последовательность восстанавливающего фертильность гена TaMS7 выбрана из нуклеотидных последовательностей, приведенных в SEQ ID NO: 1, 2 или 3 .
13. Способ по п. 12, в котором экспрессия восстанавливающего фертильность гена TaMS7 управляется специфичным для пыльцы промотором экспрессии, причем нуклеотидная последовательность специфичного для пыльцы промотора экспрессии предпочтительно представлена в SEQ ID NO: 16, 17 или 18.
14. Способ по любому из пп. 11-13, в котором ген инактивации пыльцы включает ген барназы, ген амилазы или метилазы DAM, а предпочтительно ген инактивации пыльцы представлен геном α-амилазы кукурузы, более предпочтительно нуклеотидная последовательность которого представлена в SEQ ID NO: 25.
15. Способ по п. 14, в котором ген инактивации пыльцы связан с промотором, который специфически экспрессируется в мужских гаметах, причем промотор предпочтительно представлен промотором PG47 или промотором Zm13.
16. Способ по любому из пп. 11-15, в котором отборочный ген включает ген устойчивости к антибиотикам или ген устойчивости к гербицидам или ген флуоресцентного белка; предпочтительно отборочный ген включает ген устойчивости к хлорамфениколу, ген устойчивости к гигромицину, ген устойчивости к стрептомицину, ген устойчивости к мирамицину, ген устойчивости к сульфонамидам, ген устойчивости к глифосату, ген устойчивости к фосфинотрицину, ген bar, ген красной флуоресценции DsRED, ген mCherry, ген голубого флуоресцентного белка, ген желтого флуоресцентного белка, ген люциферазы или ген зеленого флуоресцентного белка.
17. Способ получения поддерживающей линии, который включает следующие стадии:
(a) трансформирование линии с мужской стерильностью ms7 следующим вектором для получения поддерживающей линии, содержащей следующий вектор: восстанавливающий фертильность ген TaMS7 по п. 1, который может восстанавливать мужскую фертильность линии с мужской стерильностью ms7, и ген инактивации пыльцы, экспрессия которого нарушает функцию или образование у растений мужских гамет, содержащих ген инактивации пыльцы, поэтому мужские гаметы, образующиеся у растений, не содержат вектора; и отборочный ген, который используется для сортировки трансгенных семян и нетрансгенных семян; и
(b) самоопыление растений поддерживающей линии, трансформированных данным вектором, для получения линии с мужской стерильностью ms7 без вектора и семян поддерживающей линии, содержащих вектор;
в котором мужская стерильная линия представляет собой мужскую стерильную линию пшеницы.
18. Способ по п. 17, в котором нуклеотидная последовательность восстанавливающего фертильность гена TaMS7 выбрана из нуклеотидных последовательностей, приведенных в SEQ ID NO: 1, 2 или 3.
19. Способ по п. 18, в котором восстанавливающий фертильность ген TaMS7 управляется специфичным для пыльцы промотором, причем нуклеотидная последовательность специфичного для пыльцы промотора предпочтительно представлена в SEQ ID NO: 16, 17 или 18.
20. Способ по любому из пп. 17-19, в котором ген инактивации пыльцы включает ген барназы, ген амилазы или метилазы DAM, а предпочтительно ген инактивации пыльцы представлен геном α-амилазы кукурузы, более предпочтительно нуклеотидная последовательность которого представлена в SEQ ID NO: 25.
21. Способ по п. 20, в котором ген инактивации пыльцы связан с промотором, который специфически экспрессируется в мужских гаметах, причем промотор предпочтительно представлен промотором PG47 или промотором Zm13.
22. Способ по любому из пп. 17-21, в котором отборочный ген включает ген устойчивости к антибиотикам или ген устойчивости к гербицидам или ген флуоресцентного белка; предпочтительно отборочный ген включает ген устойчивости к хлорамфениколу, ген устойчивости к гигромицину, ген устойчивости к стрептомицину, ген устойчивости к мирамицину, ген устойчивости к сульфонамидам, ген устойчивости к глифосату, ген устойчивости к фосфинотрицину, ген bar, ген красной флуоресценции DsRED, ген mCherry, ген голубого флуоресцентного белка, ген желтого флуоресцентного белка, ген люциферазы или ген зеленого флуоресцентного белка.
| Matsumoto T | |||
| and et.al | |||
| Пишущая машина для тюркско-арабского шрифта | 1922 |
|
SU24A1 |
| DUAN and et.al | |||
| Optimizing de novo common wheat transcriptome assembly using short-read RNA-Seq data, BMC Genomics, 2012, V.13, N.392, pp.1-12 | |||
| ПРИМЕНЕНИЕ ДЕНДРИМЕРНОЙ НАНОТЕХНОЛОГИИ ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ | 2010 |
|
RU2571928C2 |

