ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение связано со способами получения промышленных продуктов из растительных липидов, особенно из вегетативных частей растений. В частности, в настоящем изобретении предлагаются нефтепродукты, такие как биодизельное топливо и синтетическое дизельное топливо, и способы их получения, а также растения, содержащие повышенный уровень одного или более неполярных липидов, таких как триацилглицериды, и повышенное общее содержание неполярных липидов. В одном конкретном варианте реализации изобретения настоящее изобретение связано с комбинациями модификаций двух или более ферментов, процессирующих липиды, белков масляных включений, снижения уровня ферментов, катализирующих липиды, и/или факторов транскрипции, регулирующих биосинтез липида, с повышением уровня одного или более неполярных липидов и/или общего содержания неполярных липидов и/или содержания мононенасыщенных жирных кислот в растениях или любой их части. В варианте реализации настоящее изобретение связано со способом экстракции липидов. В другом варианте реализации изобретения липид превращается в один или более углеводородных продуктов в собранных вегетативных частях растения, с целью получения алкиловых эфиров жирных кислот, пригодных для применения в качестве возобновляемого биодизельного топлива.
УРОВЕНЬ ТЕХНИКИ
Большая часть энергии в мире, особенно для транспортных целей, поставляется видами топлива, полученными из нефти, запас которой ограничен. Необходимы альтернативные, возобновляемые источники энергии, например, из вырабатываемых биологическим путем масел.
Биосинтез триацилглицеридов
Триацилглицериды (ТАГ) представляют собой основную форму липидов в семенах и состоят из трех ацильных цепей, присоединенных сложноэфирной связью к глицериновому скелету. Жирные кислоты синтезируются в пластиде как промежуточные соединения ацил-ацил-переносящего белка (АПБ), и в их молекуле может образовываться первая кратная связь под действием катализатора. Данная реакция катализируется стеароил-АПБ-десатуразой и дает олеиновую кислоту (C18:1Δ9). Впоследствии ацильные цепи транспортируются к цитозолю и эндоплазматическому ретикулуму (ЭР) как тиоэфиры ацил-Коэнзим (КоА). До вхождения в основной путь биосинтеза ТАГ, также известный как путь Кеннеди или путь глицерил-3-фосфата (G3P), ацильные цепи обычно интегрируются в фосфолипиды мембраны ЭР, где в их молекулах могут образовываться дополнительные кратные связи. Двумя ключевыми ферментами в получении полиненасыщенных жирных кислот являются связанные с мембраной десатуразы FAD2 и FAD3, которые дают линолевую (C18:2Δ9,12) и α-линоленовую кислоту (C18:3Δ9,12,15), соответственно.
Биосинтез ТАГ по пути Кеннеди состоит из серии последовательных реакций ацилирования, в каждой из которых эфиры ацил-КоА используются в качестве доноров ацила. Первая стадия ацилирования обычно происходит в положении sn1 скелета G3P и катализируется глицерин-3-фосфатацилтрансферазой (sn1-GPAT). Продукт, sn1-лизофосфатидиновая кислота (sn1-LPA) служит субстратом для ацилтрансферазы лизофосфатидиновой кислоты, (LPAAT), которая осуществляет присоединение второй цепи ацила в положении sn2, с образованием фосфатидиновой кислоты (ФК). Далее ФК дефосфорилируется до диацилглицерина (ДАГ) фосфатазой фосфатидиновой кислоты (PAP), таким образом, обеспечивая субстрат для заключительной стадии ацилирования. В заключение, третья цепь ацила присоединяется сложноэфирной связью в положении sn3 ДАГ в ходе реакции, катализируемой диацилглицерилацилтрансферазой (DGAT), с образованием ТАГ, который аккумулируется в масляных включениях. Вторая ферментная реакция, осуществляемая фосфатидилглицерилацилтрансферазой (PDAT), также приводит к превращению ДАГ в ТАГ. Данная реакция не связана с DGAT, и в ней используются фосфолипиды в качестве доноров ацила.
Для максимизации выхода с целью коммерческого получения липидов существует потребность в дополнительных средствах повышения уровней липидов, особенно неполярных липидов, например, ДАГ и ТАГ, в трансгенных организмах или их частях, например, растениях, семенах, листьях, водорослях и грибах. Попытки повышения выхода нейтрального липида из растений большей частью были сосредоточены на отдельных критических ферментных стадиях, принимающих участие в биосинтезе жирных кислот или сборке ТАГ. Эти стратегии, однако, привели к скромному повышению содержания масла в семени или листьях. Недавняя работа по конструированию в области метаболизма маслянистых дрожжей Yarrowia lipolytica продемонстрировала, что комбинированный подход к повышению выхода глицерин-3-фосфата и предупреждения разложения ТАГ посредством β-окисления приводил к кумулятивному повышению общего содержания липида (Dulermo c соавт., 2011).
Липиды растений, например, триацилглицериды масла из семян (ТАГ), находят широкое применение, например, в кулинарии (рассыпчатость, текстура, аромат), промышленности (мыло, свечи, духи, косметика, пригодны в качестве агентов для сушки, изолирующих веществ, любрикантов) и обеспечивают питательную ценность. Кроме того, растет интерес к использованию липидов растительного происхождения в качестве биологического топлива.
Для максимизации выхода при коммерческом получении липидов биологическим способом существует потребность в дополнительных средствах для повышения уровней липидов, особенно неполярных липидов, например, ДАГ и ТАГ, в трансгенных организмах или их частях, например, растениях, семенах, листьях, водорослях и грибах.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторами настоящего изобретения идентифицирован способ получения нефтепродукта из вегетативных частей растения.
В первом аспекте настоящего изобретения предлагается способ получения нефтепродукта, включающий стадии:
(i) обработки в реакторе композиции, содержащей:
(a) вегетативные части растения, сухая масса которых составляет по меньшей мере 2 г, и в которых общее содержание неполярных липидов составляет по меньшей мере 5% масс., в пересчете на сухую массу,
(b) растворитель, который содержит воду, спирт или и то, и другое, и
(c) необязательно катализатор,
причем обработка включает нагревание композиции при температуре от около 50°C до около 450°C и давлении от 5 до 350 бар на протяжении периода от 1 до 120 минут, в окислительной, восстановительной или инертной среде,
(ii) извлечение нефтепродукта из реактора с выходом по меньшей мере 35% масс., в пересчете на сухую массу вегетативных частей растения,
с получением таким образом нефтепродукта.
В варианте реализации изобретения сухая масса вегетативных частей растения составляет по меньшей мере 1 кг.
В варианте реализации изобретения общее содержание неполярных липидов в вегетативных частях растения составляет по меньшей мере 10%, по меньшей мере 15%, по меньшей мере 20%, около 25%, около 30%, около 35%, от 10% до 75%, от 20% до 75% или предпочтительно от 30% до 75%, в пересчете на сухую массу.
В варианте реализации изобретения концентрация твердых веществ в композиции составляет от 5% до 90%, предпочтительно от 15% до 50% (сухая масса/масса).
Можно применять любой подходящий катализатор. В варианте реализации изобретения катализатор представляет собой щелочь, кислоту или катализатор на основе драгоценного металла. Например, в варианте реализации изобретения катализатор содержит NaOH или KOH или оба, предпочтительно в концентрации 0,1-2 М.
В варианте реализации изобретения время обработки составляет от 1 до 60 минут, предпочтительно от 10 до 60 минут, более предпочтительно, от 15 до 30 минут. В варианте реализации изобретения, в котором давление составляет менее чем 50 бар, время реакции может составлять до 24 часов или даже до 7 дней. В предпочтительном варианте реализации изобретения температура составляет от 275°C до 360°C, давление составляет от 100 до 200 бар, и длительность реакции составляет 10-60 минут.
В варианте реализации изобретения, в котором растворитель является водой, выход нефтепродукта в результате способа составляет от по меньшей мере 36%, 37%, 38%, 39% или 40% до максимального значения 55% или предпочтительно 60% масс., в пересчете на сухую массу вегетативных частей растения. В данном варианте реализации изобретения нефтепродукт содержит по меньшей мере в 2 раза, предпочтительно по меньшей мере в 3 раза больше углеводородных соединений, чем эфиров жирного ацила. Предпочтительно, нефтепродукт содержит 35%, более предпочтительно, 40% C13-C22 углеводородных соединений.
В другом варианте реализации изобретения, в котором растворитель содержит спирт, предпочтительно метанол, выход нефтепродукта в результате способа составляет от по меньшей мере 36%, 37%, 38%, 39% или 40% до максимального значения 65% или предпочтительно 70% масс., в пересчете на сухую массу вегетативных частей растения. В данном варианте реализации изобретения нефтепродукт содержит по меньшей мере в 1,5 раза, предпочтительно по меньшей мере в 2 раза больше эфиров жирного ацила, чем углеводородных соединений. Предпочтительно, нефтепродукт содержит 40%, более предпочтительно, 50% метиловых эфиров жирных кислот.
В дальнейшем варианте реализации изобретения, в котором растворитель содержит около 80% воды, нефтепродукт содержит около 30% C13-C22 углеводородных соединений, предпочтительно около 35%, более предпочтительно, около 40% C13-C22 углеводородных соединений.
В другом варианте реализации изобретения, в котором растворитель содержит около 50% метанола, нефтепродукт содержит около 50% метиловых эфиров жирных кислот (МЭЖК).
В дальнейшем варианте реализации изобретения содержание воды в извлеченном нефтепродукте составляет менее чем около 15% масс., предпочтительно менее чем 5% масс.
В другом варианте реализации изобретения выход нефтепродукта по меньшей мере на 2% масс. больше, предпочтительно по меньшей мере на 4% масс. больше, чем выход соответствующего способа с применением соответствующих вегетативных частей растения, в которых содержание неполярного липида составляет менее чем 2%, в пересчете на сухую массу.
В варианте реализации изобретения вегетативные части растения на стадии (i)(a) физически обрабатывают одним или больше из сушки, рубки, нарезания, помола, вальцовки, прессования, дробления или перетирания. В альтернативном варианте реализации изобретения вегетативные части растения не подвергают сушке до содержания влаги менее чем 10% до приготовления композиции. Например, вегетативные части растения содержат по меньшей мере 20% или по меньшей мере 30% влаги, или вегетативные части растения сохраняют по меньшей мере 50% от содержания воды, присутствующей в них на момент их сбора.
В варианте реализации изобретения способ дополнительно включает одно или более из:
(i) гидродезоксигенации извлеченного нефтепродукта,
(ii) обработки извлеченного нефтепродукта водородом, с целью снижения уровней кетонов или сахара в нефтепродукте,
(iii) выработки синтетического газа из извлеченного нефтепродукта, и
(iv) фракционирования извлеченного нефтепродукта, с целью получения одного или более из мазута, дизельного топлива, керосина или бензина Например, стадия фракционирования может представлять собой фракционную перегонку.
В варианте реализации изобретения вегетативные части растения включают листья растения, стебли или оба.
В варианте реализации изобретения вегетативные части растения содержат комбинацию экзогенных полинуклеотидов и/или генетических модификаций, как определяется в настоящем документе.
Авторами настоящего изобретения дополнительно продемонстрировано значительное повышение содержания липидов в организмах, особенно в вегетативных частях и семени растений, путем манипулирования биосинтезом жирных кислот, сборкой липидов и путями упаковки липидов, а также снижения катаболизма липидов. Различные комбинации генов и снижение экспрессии генов применяли для достижения значительного повышения содержания масла, что имеет большое значение для выработки биотоплива и других промышленных продуктов, получаемых из масла.
Во втором аспекте настоящего изобретения предлагается рекомбинантная эукариотная клетка, содержащая:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке,
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, и любое одно или два или все три из:
c) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке, по сравнению с соответствующей клеткой, не содержащей генетической модификации,
d) третьего экзогенного полинуклеотида, кодирующего полипептида, который повышает экспорт жирных кислот из пластид клетки, по сравнению с соответствующей клеткой, не содержащей четвертого экзогенного полинуклеотида, и
e) четвертого экзогенного полинуклеотида, кодирующего второй полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клетке.
В варианте реализации изобретения клетка содержит a), b) и c), и необязательно d) или e).
В варианте реализации изобретения клетка содержит a), b) и d), и необязательно c) или e).
В варианте реализации изобретения клетка содержит a), b) и e), и необязательно c) или d).
В варианте реализации изобретения клетка дополнительно содержит один или более или все из:
a) пятого экзогенного полинуклеотида, кодирующего полипептид покрытия масляного включения (ПМВ), предпочтительно связанный с липидной капелькой белок (БСЛК),
b) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды клетки, по сравнению с соответствующей клеткой, не содержащей второй генетической модификации, и
c) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицеридов (ДАГ) в пластиде, по сравнению с соответствующей клеткой, не содержащей третьей генетической модификации.
В варианте реализации изобретения рекомбинантная эукариотная клетка содержит:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке,
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, и
c) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке, по сравнению с соответствующей клеткой, не содержащей генетической модификации,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клетке, и необязательно клетка дополнительно содержит один или более или все из:
d) третьего экзогенного полинуклеотида, кодирующего полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК,
e) четвертого экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид клетки, по сравнению с соответствующей клеткой, не содержащей четвертого экзогенного полинуклеотида,
f) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды клетки, по сравнению с соответствующей клеткой, не содержащей второй генетической модификации, и
g) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицеридов (ДАГ) в пластиде, по сравнению с соответствующей клеткой, не содержащей третьей генетической модификации.
В варианте реализации изобретения клетка является растительной клеткой из или в вегетативной части растения, причем один или более или все промоторы экспрессируются в вегетативной части с более высокими уровнями, чем в семени растения.
В предпочтительном варианте реализации изобретения присутствие c), d) или e), вместе с первым и вторым экзогенными полинуклеотидами повышает общее содержание неполярных липидов в клетке, предпочтительно клетке в вегетативной части растения, такой как лист или стебель, относительно соответствующей клетки, которая содержит первый и второй экзогенные полинуклеотиды, но не содержит каждый из c), d) и e). Более предпочтительно, повышение является синергетическим. Наиболее предпочтительно, по меньшей мере промотор, направляющий экспрессию первого экзогенного полинуклеотида, является промотором, не принадлежащим к конститутивным промоторам.
В варианте реализации изобретения полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT, и полипептид, принимающий участие в катаболизме ТАГ в клетке, является липазой SDP1.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, и полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, и полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, и полипептид, принимающий участие в катаболизме триацилглицеридов (ТАГ) в клетке, является липазой SDP1.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT, и полипептид, принимающий участие в катаболизме триацилглицеридов (ТАГ) в клетке, является липазой SDP1.
В варианте реализации изобретения два фактора транскрипции, если они присутствуют, представляют собой WRI1 и LEC2 или WRI1 и LEC1.
В упомянутых выше вариантах реализации изобретения клетка предпочтительно находится в вегетативной части растения, которое растет на грунте или которое было выращено на грунте, с последующим сбором урожая части растения, причем клетка содержит по меньшей мере 8% масс. ТАГ (% от сухой массы), например, от 8% до 75% или от 8% до 30%. Более предпочтительно, содержание ТАГ составляет по меньшей мере 10%, например, от 10% до 75% или от 10% до 30%. Предпочтительно, указанные уровни ТАГ присутствуют в вегетативных частях до или в ходе стадии цветения или до стадии завязывания семян в ходе развития растения. В указанных вариантах реализации изобретения соотношение содержания ТАГ в листьях к содержанию ТАГ в стеблях растения предпочтительно составляет от 1:1 до 10:1, и/или соотношение повышено по сравнению с соответствующей клеткой, содержащей первый и второй экзогенные полинуклеотиды, но не содержащей первой генетической модификации.
В упомянутых выше вариантах реализации изобретения клетка предпочтительно содержит экзогенный полинуклеотид, кодирующий DGAT, и генетическую модификацию, подавляющую выработку эндогенной липазы SDP1. Более предпочтительно, клетка не содержит экзогенного полинуклеотида, кодирующего PDAT, и/или представляет собой клетка, не являющуюся клеткой Nicotiana benthamiana, и/или WRI1 представляет собой WRI1, не являющийся WRI1 Arabidopsis thaliana (SEQ ID NO: 21 или 22). Наиболее предпочтительно, по меньшей мере один из экзогенных полинуклеотидов в клетке экспрессируется промотором, который не является конститутивным промотором, таким как промотор, который экспрессируется предпочтительно в зеленых тканях или стеблях растения или который активизируется после начала цветения или в ходе старения.
В третьем аспекте настоящего изобретения предлагается рекомбинантная эукариотная клетка, содержащая:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке,
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, и
c) третий экзогенный полинуклеотид, кодирующий полипептид покрытия масляного включения (ПМВ), предпочтительно связанный с липидными капельками полипептид (СЛКП),
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клетке, и, при этом, рекомбинантная эукариотная клетка содержит повышенный уровень одного или более неполярных липидов и/или повышенное количество полипептида ПМВ, относительно соответствующей клетки, которая содержит третий экзогенный полинуклеотид, нуклеотидная последовательность которого комплементарна последовательности, приведенной в SEQ ID NO: 176.
В варианте реализации изобретения клетка согласно упомянутому выше аспекту дополнительно содержит одно или более или все из:
d) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке, по сравнению с соответствующей клеткой, не содержащей первой генетической модификации,
e) четвертого экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид клетки, по сравнению с соответствующей клеткой, не содержащей четвертого экзогенного полинуклеотида,
f) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды клетки, по сравнению с соответствующей клеткой, не содержащей второй генетической модификации, и
g) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицеридов (ДАГ) в пластиде, по сравнению с соответствующей клеткой, не содержащей третьей генетической модификации.
В варианте реализации изобретения полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT и полипептид ПМВ является олеозином. В качестве альтернативы, полипептид ПМВ представляет собой БСЛК.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, и полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, и полипептид ПМВ является олеозином. В качестве альтернативы, полипептид ПМВ представляет собой БСЛК.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT, и полипептид ПМВ является олеозином. В качестве альтернативы, полипептид ПМВ представляет собой БСЛК.
В варианте реализации изобретения клетка содержит два экзогенных полинуклеотида, кодирующие два различных полипептида фактора транскрипции, которые повышают экспрессию в клетке одного или более гликолитических генов и/или генов биосинтеза жирных кислот, такие как WRI1 и LEC2 или WRI1 и LEC1.
В предпочтительном варианте реализации изобретения присутствие третьего экзогенного полинуклеотида, кодирующего полипептид ПМВ, предпочтительно БСЛК, вместе с первым и вторым экзогенными полинуклеотидами, повышает общее содержание неполярных липидов в растительной клетке, предпочтительно клетке в вегетативной части растения, такой как лист или стебель, относительно клетки соответствующего растения, которая содержит первый и второй экзогенные полинуклеотиды, но не содержит третьего экзогенного полинуклеотида. Более предпочтительно, повышение является синергетическим. Наиболее предпочтительно, по меньшей мере промотор, направляющий экспрессию первого экзогенного полинуклеотида, является промотором, не принадлежащим к конститутивным промоторам.
В четвертом аспекте настоящего изобретения предлагается рекомбинантная эукариотная клетка, содержащая пластиды и первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, и один или более или все из:
a) второго экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид клетки, по сравнению с соответствующей клеткой, не содержащей второго экзогенного полинуклеотида,
b) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды клетки, по сравнению с соответствующей клеткой, не содержащей первой генетической модификации, и
c) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде, по сравнению с соответствующей клеткой, не содержащей второй генетической модификации,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клетке.
В варианте реализации изобретения клетка, предпочтительно растительная клетка, содержит a) и необязательно b) или c).
В варианте реализации изобретения клетка согласно упомянутому выше аспекту дополнительно содержит одно или более или все из:
d) третьего экзогенного полинуклеотида, кодирующего полипептид, принимающий участие в биосинтезе одного или более неполярных липидов,
e) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке, по сравнению с соответствующей клеткой, не содержащей третьей генетической модификации, и
f) четвертого экзогенного полинуклеотида, кодирующего полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК.
В предпочтительном варианте реализации изобретения клетка, предпочтительно растительная клетка, содержит первый, второй и третий экзогенные полинуклеотиды и необязательно третью генетическую модификацию или четвертый экзогенный полинуклеотид.
В предпочтительном варианте реализации изобретения присутствие второго экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид клетки, предпочтительно являющегося тиоэстеразой жирного ацила, такой как полипептид FATA, вместе с первым и, если они присутствуют, третьими экзогенными полинуклеотидами повышает общее содержание неполярных липидов в растительной клетке, предпочтительно клетке в вегетативной части растения, такой как лист или стебель, по сравнению с соответствующей растительной клеткой, которая содержит первый и, если они присутствуют, третьи экзогенные полинуклеотиды, но не содержит второго экзогенного полинуклеотида. Более предпочтительно, повышение, обеспечиваемое вторым экзогенным полинуклеотидом, является синергетическим. Наиболее предпочтительно, по меньшей мере промотор, направляющий экспрессию первого экзогенного полинуклеотида, является промотором, не принадлежащим к конститутивным промоторам.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, предпочтительно фактором транскрипции, не являющимся WRI1 Arabidopsis thaliana (SEQ ID NO: 21 или 22), а полипептид, который повышает экспорт жирных кислот из пластид клетки, является тиоэстеразой жирных кислот, предпочтительно полипептидом FATA или FATB, более предпочтительно, полипептидом FATA или тиоэстеразой жирных кислот, не являющейся тиоэстеразой среднецепочечных жирных кислот. На присутствие тиоэстеразы, не являющейся среднецепочечной тиоэстеразой, указывает процент C12:0 и/или C14:0 жирных кислот в общем содержании жирных кислот в клетке, которое является примерно таким же, как и в соответствующей клетке, не содержащей экзогенного полинуклеотида, кодирующего тиоэстеразу. Предпочтительно, клетка дополнительно содержит экзогенный полинуклеотид, кодирующий DGAT, и генетическую модификацию, подавляющую выработку эндогенной липазы SDP1. В варианте реализации изобретения сниженная выработка липазы SDP1 действует синергетически с фактором транскрипции и тиоэстеразой жирных кислот для повышения общего содержания неполярных липидов в клетке. Более предпочтительно, клетка не содержит экзогенного полинуклеотида, кодирующего PDAT, и/или представляет собой клетку, не являющуюся клеткой Nicotiana benthamiana. Наиболее предпочтительно, по меньшей мере один из экзогенных полинуклеотидов в клетке экспрессируется промотором, который не является конститутивным промотором, таким как промотор, который предпочтительно экспрессируется в зеленых тканях или стеблях растения или активизируется в ходе старения.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, и полипептид, принимающий участие в импорте жирных кислот в пластиды клетки, является полипептидом ТГД.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, и полипептид, принимающий участие в выработке диацилглицерида (ДАГ), является пластидной GPAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, который повышает экспорт жирных кислот из пластид клетки, является тиоэстеразой жирных кислот, предпочтительно полипептидом FATA или FATB, и полипептид, принимающий участие в импорте жирных кислот в пластиды клетки, является полипептидом ТГД.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, который повышает экспорт жирных кислот из пластид клетки, является тиоэстеразой жирных кислот, предпочтительно полипептидом FATA или FATB, и полипептид, принимающий участие в выработке диацилглицерида (ДАГ), является пластидной GPAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, принимающий участие в импорте жирных кислот в пластиды клетки, является полипептидом ТГД, и полипептид, принимающий участие в выработке диацилглицерида (ДАГ), является пластидной GPAT.
В варианте реализации изобретения клетка содержит два экзогенных полинуклеотида, кодирующие два различных полипептида фактора транскрипции, которые повышают экспрессию в клетке одного или более гликолитических генов и/или генов биосинтеза жирных кислот, такие как WRI1 и LEC2 или WRI1 и LEC1.
В вариантах реализации второго, третьего и четвертого аспектов изобретения, в которых клетка содержит экзогенный полинуклеотид, кодирующий тиоэстеразу жирных кислот, такую как полипептид FATA или FATB, тиоэстераза предпочтительно является полипептидом FATA или тиоэстеразой жирных кислот, не являющейся среднецепочечной тиоэстеразой жирных кислот.
В пятом аспекте настоящего изобретения предлагается рекомбинантная эукариотная клетка, содержащая:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, предпочтительно фактор транскрипции WRI,
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, которые представляют собой LPAAT с преимущественной активностью в отношении жирных кислот со средней длиной цепи (C8-C14), и
c) третий экзогенный полинуклеотид, кодирующий полипептид, который повышает экспорт C8-C14 жирных кислот из пластид клетки, по сравнению с соответствующей клеткой, не содержащей третьего экзогенного полинуклеотида,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клетке.
В варианте реализации изобретения третий экзогенный полинуклеотид кодирует тиоэстеразу, предпочтительно тиоэстеразу FATB с преимущественной активностью в отношении жирных кислот со средней длиной цепи (C8-C14).
В предпочтительном варианте реализации изобретения присутствие третьего экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт C8-C14 жирных кислот из пластид клетки, вместе с первыми и вторыми экзогенными полинуклеотидами повышает общее содержание СЦЖК в клетке, предпочтительно клетке вегетативной части растения, такой как лист, корень или стебель, по сравнению с соответствующей растительной клеткой, содержащей первый и второй экзогенные полинуклеотиды, но не содержащей третьего экзогенного полинуклеотида. Более предпочтительно, повышение, обеспечиваемое третьим экзогенным полинуклеотидом, является синергетическим. Наиболее предпочтительно, по меньшей мере промотор, направляющий экспрессию первого экзогенного полинуклеотида, является промотором, не принадлежащим к конститутивным промоторам.
В варианте реализации изобретения экзогенный полинуклеотид, кодирующий тиоэстеразу FATB с преимущественной активностью в отношении жирных кислот со средней длиной цепи (C8-C14), содержит аминокислоты, последовательность которых приведена в любой из SEQ ID NO: 193-199, или биологически активный фрагмент любой из них или полипептид, аминокислотная последовательность которого по меньшей мере на 30% идентична любой одной или более из SEQ ID NO: 193-199. Более предпочтительно, экзогенный полинуклеотид, кодирующий тиоэстеразу FATB с преимущественной активностью в отношении жирных кислот со средней длиной цепи (C8-C14), содержит аминокислоты, последовательность которых приведена в SEQ ID NO: 193-199, или биологически активный фрагмент любой из них или полипептид, аминокислотная последовательность которого по меньшей мере на 30% идентична любой одной или обеим из SEQ ID NO: 193-199.
В варианте пятого аспекта фактор транскрипции не является WRI1 Arabidopsis thaliana (SEQ ID NO: 21 или 22).
В варианте пятого аспекта экзогенный полинуклеотид, кодирующий LPAAT, содержит аминокислоты, последовательность которых приведена как SEQ ID NO: 200, или его биологически активный фрагмент или полипептид, аминокислотная последовательность которого по меньшей мере на 30% идентична им.
В варианте пятого аспекта клетка дополнительно содержит одно или более или все из:
d) четвертого экзогенного полинуклеотида, кодирующего дополнительный полипептид, принимающий участие в биосинтезе одного или более неполярных липидов,
e) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке, по сравнению с соответствующей клеткой, не содержащей первой генетической модификации,
f) пятого экзогенного полинуклеотида, кодирующего полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК,
g) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды клетки, по сравнению с соответствующей клеткой, не содержащей второй генетической модификации, и
h) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде, по сравнению с соответствующей клеткой, не содержащей третьей генетической модификации,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клетке.
В варианте пятого аспекта клетка является растительной клеткой из или в вегетативной части растения, и один или более или все из промоторов экспрессируются в вегетативной части с более высокими уровнями, чем в семени растения.
В варианте пятого аспекта жирная кислота со средней длиной цепи является по меньшей мере миристиновой кислотой. В предпочтительном варианте реализации изобретения содержание миристиновой кислоты в клетке составляет по меньшей мере около 8%, по меньшей мере около 10%, по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, от 8% до 25%, от 8% до 20%, от 10% до 25%, от 11% до 25%, от около 15% до 25%, от около 20% до 25% (масс./масс, в пересчете на сухую массу).
В вариантах реализации третьего, четвертого и пятого аспектов клетка предпочтительно находится в вегетативной части растения, которое растет на грунте или которое было выращено на грунте, с последующим сбором урожая частей растения, причем клетка содержит по меньшей мере 8% масс. ТАГ (% от сухой массы), например, от 8% до 75% или от 8% до 30%. Более предпочтительно, содержание ТАГ составляет по меньшей мере 10%, например, от 10% до 75% или от 10% до 30%. Предпочтительно, указанные уровни ТАГ присутствуют в вегетативных частях до или в ходе стадии цветения или до стадии завязывания семян в ходе развития растения. В указанных вариантах реализации изобретения соотношение содержания ТАГ в листьях к содержанию ТАГ в стеблях растения предпочтительно составляет от 1:1 до 10:1, и/или соотношение повышено по сравнению с соответствующей клеткой, содержащей первый и второй экзогенные полинуклеотиды, но не содержащей первой генетической модификации.
В вариантах второго, третьего, четвертого и пятого аспектов клетка предпочтительно содержит экзогенный полинуклеотид, кодирующий DGAT, и генетическую модификацию, подавляющую выработку эндогенной липазы SDP1. В предпочтительном варианте реализации изобретения клетка не содержит экзогенного полинуклеотида, кодирующего PDAT, и/или клетка не является клеткой Nicotiana benthamiana и/или клетка не является клеткой Brassica napus. Наиболее предпочтительно, по меньшей мере один из экзогенных полинуклеотидов в клетке экспрессируется промотором, который не является конститутивным промотором, таким как промотор, который предпочтительно экспрессируется в зеленых тканях или стеблях растения или активизируется в ходе старения.
В варианте реализации изобретения клетка по изобретению (включая второй, третий, четвертый и пятый аспекты) обладает одним или более или всеми из следующих признаков (если это уместно):
i) в клетке повышен синтез общих жирных кислот относительно соответствующей клетки, не содержащей первого экзогенного полинуклеотида, или снижен катаболизм общих жирных кислот относительно соответствующей клетки, не содержащей первого экзогенного полинуклеотида, или присутствуют оба этих признака, таким образом, что клетка содержит повышенный уровень общих жирных кислот относительно соответствующей клетки, не содержащей первого экзогенного полинуклеотида,
ii) в клетке повышена экспрессия и/или активность ацилтрансферазы ацилов жирных кислот, катализирующей синтез ТАГ, ДАГ или МАГ, предпочтительно ТАГ, относительно соответствующей клетки, содержащей первый экзогенный полинуклеотид и не содержащей экзогенного полинуклеотида, который кодирует полипептид, принимающий участие в биосинтезе одного или более неполярных липидов,
iii) в клетке снижена выработка лизофосфатидиновой кислоты (LPA) из ацил-АПБ и G3P в пластидах клетки относительно соответствующей клетки, содержащей первый экзогенный полинуклеотид и не содержащей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде клетки,
iv) в клетке изменено соотношение C16:3 и C18:3 жирных кислот в содержании общих жирных кислот и/или содержании галактолипидов относительно соответствующей клетки, не содержащей экзогенного(ых) полинуклеотида(ов) и/или генетической(их) модификации(й), предпочтительно соотношение снижено,
v) клетка находится в вегетативной части растения, причем общее содержание в ней неполярных липидов составляет по меньшей мере около 8%, по меньшей мере около 10%, по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75% или от около 25% до 50% (масс./масс., в пересчете на сухую массу);
vi) клетка находится в вегетативной части растения, причем содержание ТАГ в ней составляет по меньшей мере около 8%, по меньшей мере около 10%, по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75% или от около 25% до 50% (масс./масс., в пересчете на сухую массу),
vii) полипептид(ы) фактора транскрипции выбран(ы) из группы, состоящей из Wrinkled 1 (WRI1), Leafy Cotyledon 1 (LEC1), LEC1-подобного, Leafy Cotyledon 2 (LEC2), BABY BOOM (BBM), FUS3, ABI3, ABI4, ABI5, Dof4 и Dof11 или из группы, состоящей из MYB73, bZIP53, AGL15, MYB115, MYB118, TANMEI, WUS, GFR2a1, GFR2a2 и PHR1,
viii) олеиновая кислота составляет по меньшей мере 20% (моль %), по меньшей мере 22% (моль %), по меньшей мере 30% (моль %), по меньшей мере 40% (моль %), по меньшей мере 50% (моль %) или по меньшей мере 60% (моль %), предпочтительно около 65% (моль %) или от 20% до около 65% от содержания общих жирных кислот в клетке,
ix) неполярный липид в клетке содержит жирную кислоту, которая содержит гидроксильную группу, эпоксигруппу, циклопропановую группу, двойную углерод-углеродную связь, тройную углерод-углеродную связь, конъюгированные двойные связи, разветвленную цепь, такую как метилированная или гидроксилированная разветвленная цепь, или комбинацию двух или более из них или любую из двух, трех, четырех, пяти или шести вышеупомянутых групп, связей или разветвленных цепей,
x) неполярный липид в клетке содержит одну или более полиненасыщенных жирных кислот, выбранных из эйкозадиеновой кислоты (ЭДК), арахидоновой кислоты (АРК), стеаридоновой кислоты (СДК), эйкозатриеновой кислоты (ЭТрК), эйкозатетраеновой кислоты (ЭТК), эйкозапентаеновой кислоты (ЭПК), докозапентаеновой кислоты (ДПК), докозагексаеновой кислоты (ДГК) или комбинации двух или более из них,
xi) клетка находится в растении или его части, предпочтительно вегетативной части растения, или клетка является клеткой водорослей, например, диатомовых (бацилляриофитов), зеленых водорослей (хлорофитов), зелено-голубых водорослей (цианофитов), золотисто-коричневых водорослей (хризофитов), гаптофитов, коричневых водорослей или водорослей гетероконтов, или клетка получена из или представляет собой микроорганизм, подходящий для ферментации, такой как гриб,
xii) один или более или все из промоторов выбраны промотором, не являющегося конститутивным промотором, предпочтительно тканеспецифического промотора, такого как специфичный для листьев и/или стеблей промотор, регулируемого развитием промотора, такого как специфичный для старения промотор, например, промотор SAG12, индуцибельного промотора или регулируемого циркадным ритмом промотора, причем по меньшей мере один из промоторов, функционально связанных с экзогенным полинуклеотидом, кодирующим полипептид фактора транскрипции, предпочтительно является промотором, не принадлежащим к конститутивным промоторам,
xiii) клетка содержит, в числе общих жирных кислот, среднецепочечные жирные кислоты, предпочтительно C12:0, C14:0 или обе, на уровне по меньшей мере 5% от общего содержания жирных кислот, и необязательно экзогенный полинуклеотид, кодирующий LPAAT, которая проявляет преимущественную активность в отношении жирных кислот со средней длиной цепи (C8-C14), предпочтительно C12:0 или C14:0;
xiv) клетка содержит, в числе общих жирных кислот, уровень олеиновой кислоты и/или уровень пальмитиновой кислоты, по меньшей мере на 2% выше, чем в соответствующей клетке,не содержащей экзогенного(ых) полинуклеотида(ов) и/или генетической(их) модификации(й), и/или уровень α-линоленовой кислоты (АЛК) и/или уровень линолевой кислоты по меньшей мере на 2% ниже, чем в соответствующей клетке, не содержащей экзогенного(ых) полинуклеотида(ов) и/или генетической(их) модификации(й),
xv) неполярный липид в клетке содержит модифицированный уровень общих стеролов, предпочтительно свободных (неэтерифицированных) стеролов, стероильных эфиров, стероилгликозидов, относительно неполярного липида в соответствующей клетке, не содержащей экзогенного(ых) полинуклеотида(ов) и/или генетической(их) модификации(й),
xvi) неполярный липид в клетке содержит воски и/или эфиры восков,
xvii) клетка является одним из членов популяции или коллекции размером по меньшей мере около 1000 таких клеток, предпочтительно в вегетативной части растения или семени,
xviii) клетка содержит экзогенный полинуклеотид, кодирующий супрессор сайленсинга, причем экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клетке,
xix) уровень одного или более неполярных липидов и/или общее содержание неполярных липидов в клетке по меньшей мере на 2% масс. выше, чем в соответствующей клетке, которая содержит экзогенные полинуклеотиды, кодирующие WRI1 Arabidposis thaliana (SEQ ID NO: 21) и DGAT1 Arabidopsis thaliana (SEQ ID NO: 1), и
xx) общее содержание полиненасыщенных жирных кислот (ПНЖК), сниженное относительно общего содержания ПНЖК в соответствующей клетке, не содержащей экзогенного(ых) полинуклеотида(ов) и/или генетической(их) модификации(й).
Следующие варианты реализации изобретения применяются к клетке по изобретению (включая второй, третий, четвертый и пятый аспекты), а также к способам получения клеток и к способам применения клеток. В указанных вариантах реализации изобретения, в которых клетка находится в вегетативной части растения, предпочтительно растение растет на грунте или было выращено на грунте.
В варианте реализации изобретения полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является ацилтрансферазой ацилов жирных кислот, который принимает участие в биосинтезе ТАГ, ДАГ или моноацилглицерида (МАГ) в клетке, предпочтительно ТАГ в клетке, такой как, например, DGAT, PDAT, LPAAT, GPAT или MGAT, предпочтительно DGAT или PDAT.
В варианте реализации изобретения полипептид, принимающий участие в катаболизме триацилглицеридов (ТАГ) в клетке, является липазой SDP1, полипептидом Cgi58, ацил-КоА оксидазой, такой как ACX1 или ACX2 или полипептидом, принимающим участие в β-окислении жирных кислот в клетке, такой как пероксисомальный транспортер АТФ-связывающей кассеты PXA1, предпочтительно липазой SDP1.
В варианте реализации изобретения полипептид покрытия масляного включения (ПМВ) является олеозином, таким как полиолеозин или калеозин, или предпочтительно связанным с липидными капельками белком (БСЛК).
В варианте реализации изобретения полипептид, который повышает экспорт жирных кислот из пластид клетки, является тиоэстеразой C16 или C18 жирных кислот, такой как полипептид FATA или полипептид FATB, переносчиком жирных кислот, таким как полипептид ABCA9, или длинноцепочечной ацил-КоА синтетазой (ДЦАС).
В варианте реализации изобретения полипептид, принимающий участие в импорте жирных кислот в пластиды клетки, является переносчиком жирных кислот или его субъединицей, предпочтительно полипептидом ТГД, таким как, например, полипептид TGD1, полипептид TGD2, полипептид TGD3 или полипептид TGD4.
В варианте реализации изобретения полипептид, принимающий участие в выработке диацилглицерида (ДАГ) в пластиде, является пластидной GPAT, пластидной LPAAT или пластидной PAP.
В одном варианте реализации изобретения клетка получена из или находится в растении 16:3 или в его вегетативной части или семени, причем клетка содержит одно или более или все из следующего:
a) экзогенный полинуклеотид, кодирующий полипептид, который повышает экспорт жирных кислот из пластид клетки, по сравнению с соответствующей клеткой, не содержащей экзогенного полинуклеотида,
b) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды клетки, по сравнению с соответствующей клеткой, не содержащей первой генетической модификации, и
c) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде, по сравнению с соответствующей клеткой, не содержащей второй генетической модификации,
причем экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клетке.
В альтернативном варианте реализации изобретения клетка получена из или находится в растении 18:3 или в его вегетативной части или семени.
В варианте реализации изобретения клетка получена из или находится в листе стебле или корне растения, перед цветением растения, причем общее содержание неполярных липидов в клетке составляет по меньшей мере около 8%, по меньшей мере около 10%, по меньшей мере около 11%, от 8% до 15%, или от 9% до 12% (масс., в пересчете на сухую массу). В варианте реализации изобретения общее содержание неполярных липидов в клетке составляет по меньшей мере на 3%, более предпочтительно по меньшей мере на 5% больше, чем общее содержание неполярных липидов в соответствующей клетке, трансформированной генами, кодирующими WRI1 и DGAT, но не содержащей других экзогенных полинуклеотидов и генетических модификаций, описанных в настоящем документе для второго, третьего, четвертого и пятого аспектов. Более предпочтительно, степень повышения находится в клетке, расположенной в стебле или корне растения.
В варианте реализации изобретения введение одного или более экзогенных полинуклеотидов или генетических модификаций, предпочтительно экзогенного полинуклеотида, кодирующего ПМВ или тиоэстеразу жирного ацила, или генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке, более предпочтительно, экзогенного полинуклеотида, кодирующего тиоэстеразу FATA или БСЛК или снижающего экспрессию эндогенной липазы ТАГ, такой как липаза ТАГ SDP1, в клетке, приводит к синергетическому повышению общего содержания неполярного липида в клетке, при добавлении к паре трансгенов WRI1 и DGAT, особенно перед цветением растения и, даже более конкретно, в стеблях и/или корнях растения. Например, см. Примеры 8, 11 и 15. В предпочтительном варианте реализации изобретения повышение содержания ТАГ в клетке, расположенной в стебле или корне растения, по меньшей мере в 2 раза, более предпочтительно, по меньшей мере в 3 раза больше, чем в соответствующей клетки, трансформированной генами, кодирующими WRI1 и DGAT1, но не содержащей тиоэстеразы FATA, БСЛК и генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке. Наиболее предпочтительно, по меньшей мере промотор, направляющий экспрессию первого экзогенного полинуклеотида, является промотором, не принадлежащим к конститутивным промоторам.
Генетическая модификация может быть любой модификацией природной клетки, которая обеспечивает желательный эффект. Способы генетической модификации клеток хорошо известны из уровня техники. В варианте реализации изобретения каждая из одной или более или всех генетических модификаций представляет собой мутацию эндогенного гена, которая частично или полностью инактивирует ген, предпочтительно введенную мутацию, такую как точечная мутация, инсерция или делеция (или комбинация одного или более из этого). Точечная мутация может представлять собой преждевременный стоп-кодон, мутацию сайта сращивания, мутацию сдвига рамки или мутацию замены аминокислоты, которая снижает активность гена или кодируемого полипептида. Делеция может состоять из одного или более нуклеотидов в пределах транскрибированного экзона или промотора гена, или простираться сквозь или в более чем один экзон, или расширяться до делеции всего гена. Предпочтительно делецию вводят с применением технологий ZF, ПАТЭН или ККППРП. В варианте реализации изобретения одна или более или все генетические модификации представляют собой экзогенный полинуклеотид, кодирующий молекулу РНК, которая подавляет экспрессию эндогенного гена, причем экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клетке. Примеры экзогенного полинуклеотида, снижающего экспрессию эндогенного гена, выбраны из группы, состоящей из антисмыслового полинуклеотида, смыслового полинуклеотида, микроРНК, полинуклеотида, кодирующего полипептид, который связывается с эндогенным ферментом, молекулой двухцепочечной РНК и полученной из нее процессированной молекулой РНК. В варианте реализации изобретения клетка содержит генетические модификации, которые представляют собой введенную мутацию в эндогенном гене и экзогенном полинуклеотиде, кодирующем молекулу РНК, снижающую экспрессию другого эндогенного гена.
В варианте реализации изобретения экзогенный полинуклеотид, кодирующий WRI1, содержит одно или более из следующего:
i) нуклеотиды, кодирующие полипептид, содержащий аминокислоты, последовательность которых приведена в любой из SEQ ID NO: 21-75 или 205-210, или его биологически активный фрагмент или полипептид, аминокислотная последовательность которого по меньшей мере на 30% идентична любой одной или более из SEQ ID NO: 21-75 или 205-210,
ii) нуклеотиды, последовательность которых по меньшей мере на 30% идентична i), и
iii) нуклеотиды, которые гибридизуются с i) и/или ii) в строгих условиях. Предпочтительно, полипептид WRI1 представляет собой полипептид WRI1, не являющийся WRI1 Arabidopsis thaliana (SEQ ID NO: 21 или 22). Более предпочтительно, полипептид WRI1 содержит аминокислоты, последовательность которых приведена в SEQ ID NO: 208, или их биологически активный фрагмент или полипептид, аминокислотная последовательность которого по меньшей мере на 30% идентична им.
В варианте реализации второго, третьего, четвертого или пятого аспектов изобретения рекомбинантная клетка представляет собой клетку клубня картофеля (Solanum tuberosum), клетку клубня или листа сахарной свеклы (Beta vulgaris), клетку сахарного тростника (вид Saccharum) или стебля или листа сорго (Sorghum bicolor), клетку эндосперма однодольного растения, причем клетка содержит повышенное количество общих жирных кислот относительно соответствующей клетки эндосперма дикого типа, такой как, например, клетка зерна пшеницы (Triticum aestivum), зерна риса (вид Oryza) или семени кукурузы (Zea mays), клетка семени вида Brassica, содержащая повышенное количество общих жирных кислот, например, семя рапса, или клетка зерна боба, содержащая повышенное количество общих жирных кислот, например, семени сои (Glycine max).
В шестом аспекте настоящего изобретения предлагается организм, не относящийся к человеческому роду, или его часть, содержащий или состоящий из одной или более клеток по изобретению.
В варианте реализации изобретения часть организма, не относящегося к человеческому роду, представляет собой семя, плод или вегетативную часть растения, такую как воздушная часть растения или зеленая часть, например, лист или стебель.
В другом варианте реализации изобретения организм, не относящийся к человеческому роду, представляет собой фототрофный организм, такой как растение или водоросль или организм, подходящий для ферментации, такой как гриб.
В седьмом аспекте настоящего изобретения предлагается трансгенное растение или его часть, предпочтительно вегетативная часть растения, содержащее:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении,
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, и любое одно или два или все три из:
c) генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в растении, по сравнению с соответствующим растением, не содержащим генетической модификации,
d) третьего экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид клеток растения, по сравнению с соответствующей клеткой, не содержащей четвертого экзогенного полинуклеотида, и
e) четвертого экзогенного полинуклеотида, кодирующего второй полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении.
В варианте реализации изобретения растение или его часть содержит a), b) и c), и необязательно d) или e).
В варианте реализации изобретения растение или его часть содержит a), b) и d), и необязательно c) или e).
В варианте реализации изобретения растение или его часть содержит a), b) и e), и необязательно c) или d).
В предпочтительном варианте реализации изобретения присутствие c), d) или e), вместе с a) и b), повышает общее содержание неполярного липида в растении или его части, предпочтительно вегетативной части растения, такой как лист, корень или стебель, относительно соответствующего растения или его части, которое содержит a) и b), но не содержит каждого из c), d) и e). Более предпочтительно, повышение является синергетическим. Наиболее предпочтительно, по меньшей мере промотор, направляющий экспрессию первого экзогенного полинуклеотида, является промотором, не принадлежащим к конститутивным промоторам.
В варианте реализации изобретения растение или его часть дополнительно содержит одно или более или все из:
a) пятого экзогенного полинуклеотида, кодирующего полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК,
b) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды растения, по сравнению с соответствующим растением, не содержащим второй генетической модификации, и
c) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде, по сравнению с соответствующим растением, не содержащим третьей генетической модификации.
В варианте реализации изобретения трансгенное растение или его часть содержит:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, предпочтительно экспрессирующийся промотором, не являющегося конститутивным промотором,
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, и
c) генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в растении, по сравнению с соответствующим растением, не содержащим генетической модификации,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении, и необязательно растение или его часть дополнительно содержит одно или более или все из:
d) третьего экзогенного полинуклеотида, кодирующего полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК,
e) четвертого экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид растения, по сравнению с соответствующим растением, не содержащим четвертого экзогенного полинуклеотида,
f) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды растения, по сравнению с соответствующим растением, не содержащим второй генетической модификации, и
g) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде, по сравнению с соответствующим растением, не содержащим третьей генетической модификации.
В варианте реализации изобретения часть является вегетативной частью, причем один или более или все из промоторов экспрессируются в вегетативной части с более высокими уровнями, чем в семени растения.
В варианте реализации изобретения полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT, и полипептид, принимающий участие в катаболизме ТАГ в растении, является липазой SDP1.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, и полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, и полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, и полипептид, принимающий участие в катаболизме триацилглицеридов (ТАГ) в растении, является липазой SDP1.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT, и полипептид, принимающий участие в катаболизме триацилглицеридов (ТАГ) в растении, является липазой SDP1.
В варианте реализации изобретения два фактора транскрипции, если они присутствуют, представляют собой WRI1 и LEC2 или WRI1 и LEC1.
В упомянутых выше вариантах реализации изобретения растение предпочтительно растет на грунте или было выращено на грунте, с последующим сбором урожая части растения. Предпочтительно, вегетативная часть растения содержит по меньшей мере 8% масс. ТАГ (% от сухой массы), например, от 8% до 75% или от 8% до 30%. Более предпочтительно, содержание ТАГ составляет по меньшей мере 10%, например, от 10% до 75% или от 10% до 30%. Предпочтительно, указанные уровни ТАГ присутствуют в вегетативной части до или во время стадии цветения или до стадии завязывания семян в ходе развития растения. В указанных вариантах реализации изобретения соотношение содержания ТАГ в листьях к содержанию ТАГ в стеблях растения предпочтительно составляет от 1:1 до 10:1, и/или соотношение повышено по сравнению с соответствующей клеткой, содержащей первый и второй экзогенные полинуклеотиды, но не содержащей первой генетической модификации.
В упомянутых выше вариантах реализации изобретения общее содержание неполярного липида в растении или его части предпочтительно по меньшей мере на 3%, более предпочтительно, по меньшей мере на 5% выше, чем общее содержание неполярного липида в соответствующем растении или его части, трансформированной генами, кодирующими WRI1 и DGAT, но не содержащем других экзогенных полинуклеотидов и генетических модификаций, описанных в настоящем документе. Более предпочтительно, указанная степень повышения присутствует в тканях стебля или корня растения.
В упомянутых выше вариантах реализации изобретения введение одного или более экзогенных полинуклеотидов или генетических модификаций, предпочтительно экзогенного полинуклеотида, кодирующего ПМВ или тиоэстеразу жирных кислот, или генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке, более предпочтительно, экзогенного полинуклеотида, кодирующего тиоэстеразу БСЛК или FATA или снижающего экспрессию эндогенной липазы ТАГ, такой как липаза ТАГ SDP1 в клетке, предпочтительно приводит к синергетическому повышению общего содержания неполярного липида в растении или его части, при добавлении к паре трансгенов WRI1 и DGAT, особенно перед цветением растения, и даже более конкретно, в ткани стебля и/или корня растения. Например, см. Примеры 8, 11 и 15. В предпочтительном варианте реализации изобретения повышение содержания ТАГ в тканях листа, стебля или корня или всех трех частей растения составляет по меньшей мере в 2 раза, более предпочтительно, по меньшей мере в 3 раза, относительно соответствующей части, трансформированной генами, кодирующими WRI1 и DGAT1, но не содержащей экзогенного полинуклеотида, кодирующего ПМВ или тиоэстеразу жирных кислот и генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке.
В упомянутых выше вариантах реализации изобретения растение или его часть предпочтительно содержит второй экзогенный полинуклеотид, кодирующий DGAT, и первую генетическую модификацию, подавляющую выработку эндогенной липазы SDP1 Более предпочтительно, растение или его часть не содержит экзогенного полинуклеотида, кодирующего PDAT, и/или представляет собой растение или его часть, не принадлежащие к Nicotiana benthamiana и/или Brassica napus, и/или WRI1 не является WRI1 Arabidopsis thaliana (SEQ ID NO: 21 или 22). В варианте реализации изобретения растение не является сахарным тростником. Наиболее предпочтительно, по меньшей мере один из экзогенных полинуклеотидов в растении экспрессируется промотором, который не является конститутивным промотором, например, таким как промотор, который предпочтительно экспрессируется в зеленых тканях или стеблях растения или который регулируется после начала цветения или в ходе старения. Предпочтительно, по меньшей мере первый экзогенный полинуклеотид (кодирующий фактор транскрипции) экспрессируется таким промотором.
В восьмом аспекте настоящего изобретения предлагается трансгенное растение или его часть, предпочтительно вегетативная часть растения, содержащее:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении,
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, и
c) третий экзогенный полинуклеотид, кодирующий полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении, и, при этом, растение содержит повышенный уровень одного или более неполярных липидов и/или повышенное количество полипептида ПМВ относительно соответствующего растения, которое содержит третий экзогенный полинуклеотид, нуклеотидная последовательность которого комплементарна последовательности, приведенной в SEQ ID NO: 176.
В предпочтительном варианте реализации изобретения присутствие третьего экзогенного полинуклеотида, кодирующего полипептид ПМВ, вместе с первым и вторым экзогенными полинуклеотидами, повышает общее содержание неполярного липида в растении или его части, предпочтительно вегетативной части растения, такой как лист, корень или стебель, относительно соответствующей части растения, которая содержит первый и второй экзогенные полинуклеотиды, но не содержит третьего экзогенного полинуклеотида. Более предпочтительно, повышение является синергетическим. Наиболее предпочтительно, по меньшей мере промотор, направляющий экспрессию первого экзогенного полинуклеотида, является промотором, не принадлежащим к конститутивным промоторам.
В варианте реализации восьмого аспекта изобретения растение или его часть дополнительно содержит одно или более или все из:
d) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в растении, по сравнению с соответствующим растением, не содержащим первой генетической модификации,
e) четвертого экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид растения, по сравнению с соответствующим растением, не содержащим четвертого экзогенного полинуклеотида,
f) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды растения, по сравнению с соответствующим растением, не содержащим второй генетической модификации, и
g) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде, по сравнению с соответствующим растением, не содержащим третьей генетической модификации.
В варианте реализации изобретения полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT и полипептид ПМВ является олеозином. В качестве альтернативы, полипептид ПМВ представляет собой БСЛК.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, и полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, а полипептид ПМВ является олеозином. В качестве альтернативы, полипептид ПМВ представляет собой БСЛК.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT, и полипептид ПМВ является олеозином. В качестве альтернативы, полипептид ПМВ представляет собой БСЛК.
В варианте реализации изобретения клетка содержит два экзогенных полинуклеотида, кодирующие два различных полипептида фактора транскрипции, которые повышают экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, такие как WRI1 и LEC2 или WRI1 и LEC1.
В девятом аспекте настоящего изобретения предлагается трансгенное растение или его часть, предпочтительно вегетативная часть растения, содержащее первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, и одно или более или все из:
a) второго экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид растения, по сравнению с соответствующим растением, не содержащим второго экзогенного полинуклеотида,
b) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды растения, по сравнению с соответствующим растением, не содержащим первой генетической модификации, и
c) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде, по сравнению с соответствующим растением, не содержащим второй генетической модификации,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении.
В варианте реализации изобретения растение или его часть, предпочтительно вегетативная часть растения, содержит a) и необязательно b) или c)
В варианте реализации девятого аспекта изобретения растение или его часть дополнительно содержит одно или более или все из:
d) третьего экзогенного полинуклеотида, кодирующего полипептид, принимающий участие в биосинтезе одного или более неполярных липидов,
e) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в растении, по сравнению с соответствующим растением, не содержащим третьей генетической модификации, и
f) четвертого экзогенного полинуклеотида, кодирующего полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК.
В предпочтительном варианте реализации изобретения растение или его часть, предпочтительно вегетативная часть растения, содержит первый, второй и третий экзогенные полинуклеотиды и необязательно третью генетическую модификацию или четвертый экзогенный полинуклеотид.
В предпочтительном варианте реализации изобретения присутствие второго экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид растения, предпочтительно являющийся тиоэстеразой жирного ацила, такой как полипептид FATA, вместе с первым и, если они присутствуют, третьими экзогенными полинуклеотидами повышает общее содержание неполярного липида в части растения, предпочтительно вегетативной части растения, такой как лист, корень или стебель, относительно соответствующей части растения, которая содержит первый и, если они присутствуют, третьи экзогенные полинуклеотиды, но не содержит второго экзогенного полинуклеотида. Более предпочтительно, повышение, обеспечиваемое вторым экзогенным полинуклеотидом, является синергетическим. Наиболее предпочтительно, по меньшей мере промотор, направляющий экспрессию первого экзогенного полинуклеотида, является промотором, не принадлежащим к конститутивным промоторам.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, и полипептид, принимающий участие в импорте жирных кислот в пластиды клетки, является полипептидом ТГД.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, а полипептид, принимающий участие в выработке диацилглицерида (ДАГ), является пластидной GPAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, который повышает экспорт жирных кислот из пластид растения, является тиоэстеразой жирных кислот, предпочтительно полипептидом FATA или FATB, и полипептид, принимающий участие в импорте жирных кислот в пластиды растения, является полипептидом ТГД.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, который повышает экспорт жирных кислот из пластид растения, является тиоэстеразой жирных кислот, предпочтительно полипептидом FATA или FATB, и полипептид, принимающий участие в выработке диацилглицерида (ДАГ), является пластидной GPAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, принимающий участие в импорте жирных кислот в пластиды растения, является полипептидом ТГД, и полипептид, принимающий участие в выработке диацилглицерида (ДАГ), является пластидной GPAT.
В варианте реализации изобретения растение содержит два экзогенных полинуклеотида, кодирующие два различных полипептида фактора транскрипции, которые повышают экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, таких как WRI1 и LEC2 или WRI1 и LEC1.
В вариантах реализации седьмого, восьмого и девятого аспектов изобретения, в которых растение содержит экзогенный полинуклеотид, кодирующий тиоэстеразу жирных кислот, такую как, например, полипептид FATA или FATB, тиоэстераза предпочтительно является полипептидом FATA или тиоэстеразой жирных кислот, кроме тиоэстеразы среднецепочечных жирных кислот.
В десятом аспекте настоящего изобретения предлагается трансгенное растение или его часть, предпочтительно вегетативная часть, содержащее:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении, предпочтительно фактора транскрипции WRI,
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, которые представляют собой LPAAT с преимущественной активностью в отношении жирных кислот со средней длиной цепи (C8-C14), и
c) третий экзогенный полинуклеотид, кодирующий полипептид, который повышает экспорт C8-C14 жирных кислот из пластид растения, по сравнению с соответствующим растением, не содержащим третьего экзогенного полинуклеотида,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении.
В предпочтительном варианте реализации изобретения присутствие третьего экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт C8-C14 жирных кислот из пластид растения, вместе с первым и вторым экзогенными полинуклеотидами, повышает общее содержание СЦЖК в части растения, предпочтительно вегетативной части растения, такой как лист, корень или стебель, относительно соответствующей части растения, которая содержит первый и второй экзогенные полинуклеотиды, но не содержит третьего экзогенного полинуклеотида. Более предпочтительно, повышение, обеспечиваемое третьим экзогенным полинуклеотидом, является синергетическим. Наиболее предпочтительно, по меньшей мере промотор, направляющий экспрессию первого экзогенного полинуклеотида, является промотором, не принадлежащим к конститутивным промоторам.
В варианте реализации десятого аспекта изобретения трансгенное растение или его часть дополнительно содержит одно или более или все из:
d) четвертого экзогенного полинуклеотида, кодирующего дополнительный полипептид, принимающий участие в биосинтезе одного или более неполярных липидов,
e) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в растении, по сравнению с соответствующим растением, не содержащим первой генетической модификации,
f) пятого экзогенного полинуклеотида, кодирующего полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК,
g) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды растения, по сравнению с соответствующим растением, не содержащим второй генетической модификации, и
h) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде, по сравнению с соответствующим растением, не содержащим третьей генетической модификации,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении.
В варианте реализации десятого аспекта изобретения фактор транскрипции не является WRI1 Arabidopsis thaliana (SEQ ID NO: 21 или 22), и/или растение не является N. benthamiana.
В варианте реализации десятого аспекта изобретения экзогенный полинуклеотид, кодирующий LPAAT, содержит аминокислоты, последовательность которых приведена в SEQ ID NO: 200, или их биологически активный фрагмент или полипептид LPAAT, аминокислотная последовательность которого по меньшей мере на 30% идентична им.
В варианте реализации десятого аспекта изобретения один или более или все из промоторов экспрессируются в вегетативной части с более высокими уровнями, чем в семени растения, предпочтительно включая по меньшей мере промотор, который экспрессирует первый экзогенный полинуклеотид.
В варианте реализации десятого аспекта изобретения жирная кислота со средней длиной цепи представляет собой по меньшей мере миристиновую кислоту (C14:0). В предпочтительном варианте реализации изобретения содержание миристиновой кислоты в части растения, предпочтительно вегетативной части растения, составляет по меньшей мере около 8%, по меньшей мере около 10%, по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, от 8% до 25%, от 8% до 20%, от 10% до 25%, от 11% до 25%, от около 15% до 25%, от около 20% до 25% (масс./масс., в пересчете на сухую массу).
В вариантах реализации шестого, седьмого, восьмого, девятого и десятого аспектов изобретения растение предпочтительно растет на грунте или было выращено на грунте, с последующим сбором урожая части растения, предпочтительно вегетативной части растения, причем часть растения содержит по меньшей мере 8% масс. ТАГ (% от сухой массы), например, от 8% до 75% или от 8% до 30%. Более предпочтительно, содержание ТАГ составляет по меньшей мере 10%, например, от 10% до 75% или от 10% до 30%. Предпочтительно, указанные уровни ТАГ присутствуют в вегетативной части до или во время стадии цветения или до стадии завязывания семян в ходе развития растения. В указанных вариантах реализации изобретения соотношение содержания ТАГ в листьях к содержанию ТАГ в стеблях растения предпочтительно составляет от 1:1 до 10:1, и/или соотношение повышено по сравнению с соответствующей клеткой, содержащей первый и второй экзогенные полинуклеотиды, но не содержащей первой генетической модификации.
В вариантах реализации шестого, седьмого, восьмого, девятого и десятого аспектов изобретения растение или его часть предпочтительно содержит экзогенный полинуклеотид, кодирующий DGAT, и генетическую модификацию, подавляющую выработку эндогенной липазы SDP1. В предпочтительном варианте реализации изобретения растение или его часть не содержит экзогенного полинуклеотида, кодирующего PDAT, и/или растение не является растением Nicotiana benthamiana. Наиболее предпочтительно, по меньшей мере один из экзогенных полинуклеотидов в растении или его части экспрессируется промотором, который не является конститутивным промотором, например, таким как промотор, который предпочтительно экспрессируется в зеленых тканях или стеблях растения или который регулируется в ходе старения.
В одиннадцатом аспекте настоящего изобретения предлагается растение, содержащее вегетативную часть, или его вегетативная часть, причем общее содержание неполярного липида в вегетативной части составляет по меньшей мере около 18%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 18% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75% или от около 25% до 50% (масс., в пересчете на сухую массу), и, при этом, неполярный липид содержит по меньшей мере 90% триглицеридов (ТАГ).
В предпочтительных вариантах реализации изобретения вегетативная часть растения отличается признаками, описанными в седьмом, восьмом, девятом и десятом аспектах. Растение предпочтительно является растением 18:3.
В варианте реализации упомянутых выше аспектов изобретения растительную клетку или часть растения обрабатывают таким образом, что она становится неспособной размножаться или дать начало живому растению, т.е. ее убивают. Например, растительную клетку или часть растения высушивают и/или перемалывают.
В двенадцатом аспекте настоящего изобретения предлагается растение, содержащее вегетативную часть, или его вегетативная часть, причем содержание ТАГ в вегетативной части составляет по меньшей мере около 18%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 18% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75% или от около 25% до 50% (масс., в пересчете на сухую массу), и, при этом, неполярный липид содержит по меньшей мере 90% триацилглицеридов (ТАГ). Растение предпочтительно является растением 18:3.
В тринадцатом аспекте настоящего изобретения предлагается растение, содержащее вегетативную часть, или его вегетативная часть, причем общее содержание неполярного липида в вегетативной части составляет по меньшей мере 8%, по меньшей мере около 10%, по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс., в пересчете на сухую массу), и, при этом, неполярный липид содержит по меньшей мере 90% триацилглицеридов (ТАГ), при том, что растение является растением 16:3 или его вегетативной частью.
В четырнадцатом аспекте настоящего изобретения предлагается растение, содержащее вегетативную часть, или его вегетативная часть, причем содержание ТАГ в вегетативной части составляет по меньшей мере 8%, по меньшей мере около 10%, по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс., в пересчете на сухую массу), и, при этом, неполярный липид содержит по меньшей мере 90% триацилглицеридов (ТАГ), при том, что растение является растением 16:3 или его вегетативной частью.
В варианте реализации изобретения клетка по изобретению (включая второй, третий, четвертый или пятый аспекты) представляет собой клетку следующих видов или родов, или растение или его часть по изобретению (включая шестой, седьмой, восьмой, девятый, десятый, одиннадцатый, двенадцатый, тринадцатый и четырнадцатый аспекты) представляет собой Acrocomia aculeata (макаубская пальма), Arabidopsis thaliana, Aracinis hypogaea (арахис), Astrocaryum murumuru (мурумуру), Astrocaryum vulgare (тукума), Attalea geraensis (атталия), Attalea humilis (американская масличная пальма), Attalea oleifera (андайя), Attalea phalerata (урикури), Attalea speciosa (бабассу), Avena sativa (овес), Beta vulgaris (сахарная свекла), виды Brassica, например, Brassica carinata, Brassica juncea, Brassica napobrassica, Brassica napus (рапс), Camelina sativa (ложный лен), Cannabis sativa (конопля), Carthamus tinctorius (сафлор красильный), Caryocar brasiliense (пекви), Cocos nucifera (кокосовый орех), Crambe abyssinica (абиссинская капуста), Cucumis melo (дыня), Elaeis guineensis (африканская пальма), Glycine max (соя), Gossypium hirsutum (хлопчатник), виды Helianthu, например, Helianthus annuus (подсолнечник), Hordeum vulgare (ячмень), Jatropha curcas (лечебный орех), Joannesia princeps (лещина арара), виды Lemna (ряска), такие как Lemna aequinoctialis, Lemna disperma, Lemna ecuadoriensis, Lemna gibba (ряска горбатая), Lemna japonica, Lemna minor, Lemna minuta, Lemna obscura, Lemna paucicostata, Lemna perpusilla, Lemna tenera, Lemna trisulca, Lemna turionifera, Lemna valdiviana, Lemna yungensis, Licania rigida (ойтика), Linum usitatissimum (лен), Lupinus angustifolius (люпин), Mauritia flexuosa (пальма бурити), Maximiliana maripa (пальма инайя), виды Miscanthus, такие как Miscanthus x giganteus и Miscanthus sinensis, виды Nicotiana (табак), такие как Nicotiana tabacum или Nicotiana benthamiana, Oenocarpus bacaba (бакаба-до-азейте), Oenocarpus bataua (патайя), Oenocarpus distichus (бакаба-де-лек), виды Oryza (рис), такие как Oryza sativa и Oryza glaberrima, Panicum virgatum (просо прутьевидное), Paraqueiba paraensis (марь), Persea amencana (авокадо), Pongamia pinnata (индийский бук), Populus trichocarpa, Ricinus communis (клещевина обыкновенная), виды Saccharum (сахарный тростник), Sesamum indicum (кунжут), Solanum tuberosum (картофель), виды Sorghum, например, Sorghum bicolor, Sorghum vulgare, Theobroma grandiforum (какао), виды Trifolium, Trithrinax brasiliensis (бразильская игольчатая пальма), виды Triticum (пшеница),Triticum aestivum и Zea mays (кукуруза).
В пятнадцатом аспекте настоящего изобретения предлагается растение картофеля или его часть, предпочтительно клубень, диаметр которого составляет по меньшей мере 2 см, и содержание ТАГ в котором составляет по меньшей мере 0,5%, в пересчете на сухую массу, и/или общее содержание жирных кислот составляет по меньшей мере 1%, предпочтительно по меньшей мере 1,5% или по меньшей мере 2,0%, в пересчете на сухую массу. Картофельный клубень предпочтительно содержит повышенный уровень мононенасыщенных жирных кислот (МНЖК) и/или сниженный уровень полиненасыщенных жирных кислот (ПНЖК), как в общем содержании жирных кислот, так и во фракции ТАГ общего содержания жирных кислот, например, повышенный уровень олеиновой кислоты и сниженный уровень АЛК, по сравнению с соответствующим картофельным клубнем, не содержащим генетических модификаций и/или экзогенного(ых) полинуклеотида(ов). Предпочтительно, уровень АЛК в общем содержании жирных кислот клубня снижен до менее чем 10%, и/или уровень олеиновой кислоты в общем содержании жирных кислот повышен по меньшей мере до 5%, предпочтительно по меньшей мере до 10% или, более предпочтительно, по меньшей мере до 15%, по сравнению с соответствующим картофельным клубнем, не содержащим генетических модификаций и/или экзогенного(ых) полинуклеотида(ов). Кроме того, в варианте реализации изобретения уровень пальмитиновой кислоты в общем содержании жирных кислот клубня повышен, и/или уровни стеариновой кислоты (18:0) снижены в общем содержании жирных кислот клубня, по сравнению с соответствующим картофельным клубнем, не содержащим генетических модификаций и/или экзогенного(ых) полинуклеотида(ов). В варианте реализации изобретения содержание крахмала в клубне составляет от около 90% до 100% масс., по сравнению с клубнем дикого типа, при выращивании в одинаковых условиях.
В варианте реализации изобретения растение картофеля или его часть по изобретению, предпочтительно клубень, содержит:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клубне, и
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клубне в процессе роста растения картофеля.
В предпочтительном варианте реализации изобретения картофельный клубень дополнительно содержит одно или более или все из:
c) третий экзогенный полинуклеотид, кодирующий полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК,
d) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клубне, по сравнению с соответствующим клубнем, не содержащим первой генетической модификации, например, где полипептид является SDP1,
e) четвертого экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид клубня, по сравнению с соответствующим клубнем, не содержащим четвертого экзогенного полинуклеотида,
f) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды клубня, по сравнению с соответствующим клубнем, не содержащим второй генетической модификации, и
g) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиды клубня, по сравнению с соответствующим клубнем, не содержащим третьей генетической модификации.
В дальнейших вариантах реализации изобретения дополнительные генетические модификации в клубне являются такими, как определено в контексте клетки или растения по изобретению.
В шестнадцатом аспекте настоящего изобретения предлагается растение сорго или сахарного тростника или его часть, предпочтительно стебель или лист, общее содержание жирных кислот в котором составляет по меньшей мере 6% или по меньшей мере 8%, в пересчете на сухую массу, и/или содержание ТАГ в стебле составляет по меньшей мере 2% или по меньшей мере 3%, в пересчете на сухую массу, и/или содержание ТАГ повышено по меньшей мере в 50 раз в стебле и/или по меньшей мере в 100 раз в листе, в пересчете на массу. В вариантах реализации изобретения растение сорго или сахарного тростника или его часть, предпочтительно стебель или лист, отличается признаками, определенными в контексте клетки или растения или его части по изобретению.
В семнадцатом аспекте настоящего изобретения предлагается растение сорго или сахарного тростника или его часть, предпочтительно стебель или лист, которое содержит:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении или его части, и
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении или его части в процессе роста растения.
Предпочтительно, промотор, который направляет экспрессию по меньшей мере первого экзогенного полинуклеотида, представляет собой промотор, не являющийся убихитиновым промотором риса (Rubi3). Более предпочтительно, промотор не является убихитиновым промотором или любым другим конститутивным промотором. Предпочтительно, первый и второй экзогенные полинуклеотиды и их соответствующие промоторы объединены в одной генетической конструкции, которая интегрирована в геном растения.
В варианте реализации изобретения содержание сахара в стебле сахарного тростника составляет от около 70% до 100% масс. относительно стебля сахарного тростника дикого типа, при выращивании в одинаковых условиях. В качестве альтернативы, содержание сахара составляет от 50% до 70%.
В варианте реализации изобретения растение сорго или сахарного тростника или его часть, предпочтительно стебель или лист, по изобретению содержит:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в стебле(ях) растения, и
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов,
причем по меньшей мере один из экзогенных полинуклеотидов, предпочтительно по меньшей мере первый экзогенный полинуклеотид функционально связан с промотором, который предпочтительно экспрессируется в стебле(ях) относительно листьев в процессе роста растения.
В варианте реализации изобретения растение сорго или сахарного тростника или его часть по изобретению дополнительно содержит одно или более или все из:
c) третий экзогенный полинуклеотид, кодирующий полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК,
d) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в растении или его части, по сравнению с соответствующим растением или его частью, не содержащими первой генетической модификации,
e) четвертого экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид растения или его части, по сравнению с соответствующим растением или его частью, не содержащими четвертого экзогенного полинуклеотида,
f) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды растения или его части, по сравнению с соответствующим растением или его частью, не содержащими второй генетической модификации, и
g) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластидах растения или его части, по сравнению с соответствующим растением или его частью, не содержащими третьей генетической модификации.
Растение сорго или сахарного тростника или его часть по изобретению предпочтительно содержит повышенный уровень мононенасыщенных жирных кислот (МНЖК) и/или сниженный уровень полиненасыщенных жирных кислот (ПНЖК), как в общем содержании жирных кислот, так и во фракции ТАГ общего содержания жирных кислот, например, повышенный уровень олеиновой кислоты и сниженный уровень АЛК, по сравнению с соответствующим растением или его частью, не содержащими генетических модификаций и/или экзогенного(ых) полинуклеотида(ов).
Предпочтительно, уровень АЛК в общем содержании жирных кислот составляет менее чем 10% и/или уровень олеиновой кислоты в общем содержании жирных кислот составляет по меньшей мере 5%, предпочтительно по меньшей мере 10% или более предпочтительно по меньшей мере 15%, по сравнению с соответствующим растением или его частью, не содержащими генетических модификаций и/или экзогенного(ых) полинуклеотида(ов).
В дальнейших вариантах реализации изобретения дополнительные генетические модификации в растении сорго или сахарного тростника или его части являются такими, как определено в контексте клетки или растения по изобретению.
В восемнадцатом аспекте настоящего изобретения предлагается трансгенное однодольное растение или его часть, предпочтительно лист, семя, стебель, корень или эндосперм, общее содержание жирных кислот или содержание ТАГ в котором повышено по меньшей мере в 5 раз, в пересчете на массу, по сравнению с соответствующим нетрансгенным однодольным растением или его частью. В качестве альтернативы, в изобретении предлагается трансгенное однодольное растение, содержание ТАГ в эндосперме которого составляет по меньшей мере 2,0%, предпочтительно по меньшей мере 3%, более предпочтительно, по меньшей мере 4% или по меньшей мере 5%, масс., или часть растения, предпочтительно лист, стебель, корень, зерно или эндосперм. В варианте реализации изобретения содержание ТАГ в эндосперме составляет по меньшей мере 2%, что по меньшей мере в 5 раз выше, чем в соответствующем нетрансгенном эндосперме. Предпочтительно, растение обладает полной мужской и женской фертильностью, его пыльца по существу является на 100% жизнеспособной, и скорость прорастания его семени составляет от 70% до 100% относительно соответствующего семени дикого типа. В варианте реализации изобретения трансгенное растение представляет собой растение-потомок, по меньшей мере через два поколения от начального трансгенного растения пшеницы, и предпочтительно является гомозиготным по трансгенам. В вариантах реализации изобретения однодольное растение или его часть, предпочтительно лист, стебель, семя или эндосперм, дополнительно отличается одним или более признаками, определенными в контексте клетки или растения по изобретению.
В девятнадцатом аспекте настоящего изобретения предлагается однодольное растение или его часть, предпочтительно лист, зерно, стебель или эндосперм, которое содержит:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении или его части, и
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении или его части в процессе роста растения.
Предпочтительно, промотор, который направляет экспрессию по меньшей мере первого экзогенного полинуклеотида, представляет собой промотор, не являющийся конститутивным промотором.
В варианте реализации изобретения содержание крахмала в семени однодольного растения по изобретению составляет от около 70% до 100% масс. относительно семени дикого типа, если растения, от которых они получены, выращены в одинаковых условиях. Предпочтительными однодольными растениями в упомянутых выше двух аспектах являются пшеница, рис, сорго и кукуруза (маис).
В варианте реализации изобретения однодольное растение или его часть, предпочтительно лист, зерно или эндосперм, по изобретению содержит:
a) первый экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в эндосперме растения, и
b) второй экзогенный полинуклеотид, кодирующий полипептид, принимающий участие в биосинтезе одного или более неполярных липидов,
причем по меньшей мере один из экзогенных полинуклеотидов, предпочтительно по меньшей мере первый экзогенный полинуклеотид, функционально связан с промотором, который в процессе роста растения экспрессируется в эндосперме с более высокими уровнями, чем в листьях.
В предпочтительном варианте реализации изобретения однодольное растение или его часть дополнительно содержит одно или более или все из:
c) третий экзогенный полинуклеотид, кодирующий полипептид покрытия масляного включения (ПМВ), предпочтительно БСЛК,
d) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в растении или его части, по сравнению с соответствующим растением или его частью, не содержащими первой генетической модификации,
e) четвертого экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид растения или его части, по сравнению с соответствующим растением или его частью, не содержащими четвертого экзогенного полинуклеотида,
f) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды растения или его части, по сравнению с соответствующим растением или его частью, не содержащими второй генетической модификации, и
g) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластидах растения или его части, по сравнению с соответствующим растением или его частью, не содержащими третьей генетической модификации.
В варианте реализации изобретения однодольное растение обладает признаками a), b), одним или обоими из d) и e), и необязательно одним из c), f) и g).
Однодольное растение или его часть, предпочтительно лист, семя, стебель или эндосперм по изобретению предпочтительно содержит повышенный уровень мононенасыщенных жирных кислот (МНЖК) и/или сниженный уровень полиненасыщенных жирных кислот (ПНЖК), как в общем содержании жирных кислот, так и во фракции ТАГ общего содержания жирных кислот, например, повышенный уровень олеиновой кислоты и сниженный уровень ЛК (18:2), по сравнению с соответствующим растением или его частью, не содержащими генетических модификаций и/или экзогенного(ых) полинуклеотида(ов).
Предпочтительно, уровень линолевой кислоты (ЛК, 18:2) в общем содержании жирных кислот семени или эндосперма снижен по меньшей мере на 5%, и/или уровень олеиновой кислоты в общем содержании жирных кислот повышен по меньшей мере на 5% относительно соответствующего растения дикого типа или его части, предпочтительно по меньшей мере на 10% или, более предпочтительно, по меньшей мере на 15%, по сравнению с соответствующим растением или его частью, не содержащими генетических модификаций и/или экзогенного(ых) полинуклеотида(ов).
Следующие варианты реализации изобретения применяются к каждому из растений и их частей в соответствии с пятнадцатым, шестнадцатым, семнадцатым, восемнадцатым и девятнадцатым аспектами.
В варианте реализации изобретения полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT и полипептид ПМВ является олеозином. В качестве альтернативы, полипептид ПМВ представляет собой БСЛК.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении или его части, является полипептидом WRI1, и полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении или его части, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, и полипептид ПМВ является олеозином. В качестве альтернативы, полипептид ПМВ представляет собой БСЛК.
В варианте реализации изобретения полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении или его части, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT, и полипептид ПМВ является олеозином. В качестве альтернативы, полипептид ПМВ представляет собой БСЛК.
В варианте реализации изобретения растение или его часть содержит два экзогенных полинуклеотида, кодирующие два различных полипептида фактора транскрипции, которые повышают экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, например, WRI1 и LEC2 или WRI1 и LEC1.
В каждом из вариантов реализации клеток, растений и их частей по изобретению (включая второй, третий, четвертый, пятый, шестой, седьмой, восьмой, девятый, десятый, одиннадцатый, двенадцатый, тринадцатый, четырнадцатый, пятнадцатый, шестнадцатый, семнадцатый, восемнадцатый и девятнадцатый аспекты), предпочтительно полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT, и полипептид, принимающий участие в катаболизме триацилглицеридов (ТАГ) в клетке, является липазой SDP1.
В каждом из вариантов реализации клеток, растений и их частей по изобретению (включая второй, третий, пятый, шестой, седьмой, восьмой, десятый, одиннадцатый, двенадцатый, тринадцатый, четырнадцатый, пятнадцатый, шестнадцатый, семнадцатый, восемнадцатый и девятнадцатый аспекты, но исключая пятый и десятый аспекты) предпочтительно полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является полипептидом WRI1, полипептидом LEC2, полипептидом LEC1 или LEC1-подобным полипептидом, полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT или PDAT, и полипептид, который повышает экспорт жирных кислот из пластид клетки, является тиоэстеразой жирных кислот, предпочтительно полипептидом FATA или FATB, более предпочтительно, полипептидом FATA или тиоэстеразой жирных кислот, кроме тиоэстеразы среднецепочечных жирных кислот.
В каждом из упомянутых выше вариантов реализации клеток, растений и их частей по изобретению (включая второй, третий, четвертый, пятый, шестой, седьмой, восьмой, девятый, десятый, одиннадцатый, двенадцатый, тринадцатый, четырнадцатый, пятнадцатый, шестнадцатый, семнадцатый, восемнадцатый и девятнадцатый аспекты) предпочтительно полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растении или его части, является комбинацией по меньшей мере двух полипептидов, предпочтительно полипептида WRI1 и полипептида LEC2. Более предпочтительно, указанные по меньшей мере два полипептида фактора транскрипции экспрессируются различными промоторами. Наиболее предпочтительно, экзогенные полинуклеотиды, кодирующие указанные по меньшей мере два полипептида, объединены на единой генетической конструкции, интегрированной в геном клетки или растения.
В каждом из упомянутых выше вариантов реализации изобретения, в которых растение является двудольным растением, указанный фактор транскрипции может быть фактором транскрипции однодольного растения. С другой стороны, если растение является однодольным растением, указанный фактор транскрипции может быть фактором транскрипции двудольного растения. Указанный фактор транскрипции предпочтительно представляет собой фактор транскрипции, не являющийся WRI1 A. thaliana (SEQ ID NO: 21 или 22).
В каждом из упомянутых выше вариантов реализации изобретения растение предпочтительно является трансгенным растением-потомком, по меньшей мере через два поколения от начального трансгенного растения, и предпочтительно является гомозиготным по трансгенам.
В дальнейших вариантах реализации изобретения дополнительные генетические модификации в растении или его части являются такими, как определено в контексте клетки по изобретению.
В варианте реализации изобретения растение или его часть по изобретению (включая шестой, седьмой, восьмой, девятый, десятый, одиннадцатый, двенадцатый, тринадцатый, четырнадцатый, пятнадцатый, шестнадцатый, семнадцатый, восемнадцатый и девятнадцатый аспекты) обладает одним или более или всеми из следующих признаков (если это уместно):
i) растение содержит часть, предпочтительно вегетативную часть, в которой повышен синтез общих жирных кислот относительно соответствующей части, не содержащей первого экзогенного полинуклеотида, или снижен катаболизм общих жирных кислот относительно соответствующей части, не содержащей первого экзогенного полинуклеотида, или присутствуют оба этих явления, таким образом, что она содержит повышенный уровень общих жирных кислот относительно соответствующей части, не содержащей первого экзогенного полинуклеотида,
ii) растение содержит часть, предпочтительно вегетативную часть, в которой повышена экспрессия и/или активность ацилтрансферазы ацилов жирных кислот, катализирующей синтез ТАГ, ДАГ или МАГ, предпочтительно ТАГ, относительно соответствующей части, содержащей первый экзогенный полинуклеотид и не содержащей экзогенного полинуклеотида, кодирующего полипептид, принимающий участие в биосинтезе одного или более неполярных липидов,
iii) растение содержит часть, предпочтительно вегетативную часть, в которой снижена выработка лизофосфатидиновой кислоты (LPA) из ацил-АПБ и G3P в пластидах относительно соответствующей части, содержащей первый экзогенный полинуклеотид и не содержащей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластидах части растения,
iv) растение содержит часть, предпочтительно вегетативную часть, которая содержит модифицированное соотношение C16:3 и C18:3 жирных кислот в общем содержании жирных кислот и/или содержании галактолипидов относительно соответствующей части, не содержащей экзогенного(ых) полинуклеотида(ов) и/или генетической(их) модификации(й), предпочтительно соотношение снижено,
v) общее содержание неполярного липида в вегетативной части растения составляет по меньшей мере около 8%, по меньшей мере около 10%, по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу), предпочтительно до цветения,
vi) содержание ТАГ в вегетативной части растения составляет по меньшей мере около 8%, по меньшей мере около 10%, по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу), предпочтительно до цветения,
vii) полипептид(ы) фактора транскрипции выбран(ы) из группы, состоящей из WRI1, LEC1, LEC1-подобного, LEC2, BBM, FUS3, ABI3, ABI4, ABI5, Dof4 и Dof11, предпочтительно WRI1, LEC1 или LEC2, или из группы, состоящей из MYB73, bZIP53, AGL15, MYB115, MYB118, TANMEI, WUS, GFR2a1, GFR2a2 и PHR1,
viii) олеиновая кислота составляет по меньшей мере 20% (моль %), по меньшей мере 22% (моль %), по меньшей мере 30% (моль %), по меньшей мере 40% (моль %), по меньшей мере 50% (моль %), или по меньшей мере 60% (моль %), предпочтительно около 65% (моль %) или от 20% до около 65% от общего содержания жирных кислот в растении или его части,
ix) неполярный липид в растении или его части, предпочтительно вегетативной части, содержит повышенный уровень одной или более жирных кислот, которые содержат гидроксильную группу, эпоксигруппу, циклопропановую группу, двойную углерод-углеродную связь, тройную углерод-углеродную связь, конъюгированные двойные связи, разветвленную цепь, например, метилированную или гидроксилированную разветвленную цепь, или комбинацию двух или более из этого, или любые две, три, четыре, пять или шесть из вышеупомянутых групп, связей или разветвленных цепей,
x) неполярный липид в растении или его части, предпочтительно вегетативной части, содержит одну или более полиненасыщенных жирных кислот, выбранных из эйкозадиеновой кислоты (ЭДК), арахидоновой кислоты (АРК), стеаридоновой кислоты (СДК), эйкозатриеновой кислоты (ЭТрК), эйкозатетраеновой кислоты (ЭТК), эйкозапентаеновой кислоты (ЭПК), докозапентаеновой кислоты (ДПК), докозагексаеновой кислоты (ДГК) или комбинации двух из более из них,
xi) часть является вегетативной частью растения, такой как лист или стебель или его часть,
xii) один или более или все промоторы выбраны промотором, не являющегося конститутивным промотором, предпочтительно тканеспецифического промотора, такого как специфичный для листьев и/или стеблей промотор, регулируемого развитием промотора, такого как специфичный для старения промотор, например, промотор SAG12, индуцибельного промотора или регулируемого циркадным ритмом промотора, причем по меньшей мере один из промоторов, функционально связанных с экзогенным полинуклеотидом, кодирующим полипептид фактора транскрипции, предпочтительно является промотором, не принадлежащим к конститутивным промоторам,
xiii) растение или его часть, предпочтительно вегетативная часть, содержит, в числе общих жирных кислот, среднецепочечные жирные кислоты, предпочтительно C12:0, C14:0 или обе, на уровне по меньшей мере 5% от общего содержания жирных кислот, и необязательно экзогенный полинуклеотид, кодирующий LPAAT, которая проявляет преимущественную активность в отношении жирных кислот со средней длиной цепи (C8-C14), предпочтительно C12:0 или C14:0,
xiv) растение или его часть, предпочтительно вегетативная часть, содержит, в числе общих жирных кислот, уровень олеиновой кислоты и/или уровень пальмитиновой кислоты, по меньшей мере на 2% выше, чем в соответствующем растении или его части, не содержащей экзогенного(ых) полинуклеотида(ов) и/или генетической(их) модификации(й), и/или уровень α-линоленовой кислоты (АЛК) и/или уровень линолевой кислоты, по меньшей мере на 2% ниже, чем в соответствующей клетке, не содержащей экзогенного(ых) полинуклеотида(ов) и/или генетической(их) модификации(й),
xv) неполярный липид в растении или его части, предпочтительно вегетативной части, содержит модифицированный уровень общих стеролов, предпочтительно свободных (неэтерифицированных) стеролов, стероильных эфиров, стероилгликозидов, относительно неполярного липида в соответствующей клетке, не содержащей экзогенного(ых) полинуклеотида(ов) и/или генетической(их) модификации(й),
xvi) неполярный липид в растении или его части содержит воски и/или эфиры восков,
xvii) растение или его часть, предпочтительно вегетативная часть, является членом популяции или коллекции размером по меньшей мере около 1000 таких растений или их частей,
xviii) растение содержит экзогенный полинуклеотид, кодирующий супрессор сайленсинга, причем экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении,
xix) уровень одного или более неполярных липидов и/или общее содержание неполярного липида в растении или его части, предпочтительно вегетативной части растения, по меньшей мере на 2% масс. выше, чем в соответствующей клетке, которая содержит экзогенные полинуклеотиды, кодирующие WRI1 Arabidposis thaliana (SEQ ID NO: 21) и DGAT1 Arabidopsis thaliana (SEQ ID NO: 1), и
xx) общее содержание полиненасыщенных жирных кислот (ПНЖК), сниженное относительно общего содержания ПНЖК в соответствующей клетке, не содержащей экзогенного(ых) полинуклеотида(ов) и/или генетической(их) модификации(й).
xxi) часть растения является клубнем картофеля (Solanum tuberosum), сахарной свеклы (Beta vulgaris), стеблем сахарного тростника (вид Saccharum) или сорго (Sorghum bicolor) семенем однодольного растения с повышенным общим содержанием жирных кислот в эндосперме, например, зерном пшеницы (Triticum aestivum) или зернышком кукурузы (Zea mays), листом вида Nicotiana или семенем боба с повышенным общим содержанием жирных кислот, например, семенем вида Brassica или семенем сои (Glycine max),
xxii) если часть растения является семенем, то семя прорастает в существенной мере с такой же скоростью, как и соответствующее семя дикого типа, или при высевании в грунт дает растение, семена которого прорастают в существенной мере с такой же скоростью относительно соответствующего семени дикого типа, и
xxiii) растение является водорослевым растением, например, из числа диатомовых (бацилляриофитов), зеленых водорослей (хлорофитов), зелено-голубых водорослей (цианофитов), золотисто-коричневых водорослей (хризофитов), гаптофитов, коричневых водорослей или водорослей гетероконтов.
В упомянутых выше вариантах реализации изобретения предпочтительной частью растения является фрагмент листа с площадью поверхности по меньшей мере 1 см2 или фрагмент стебля длиной по меньшей мере 1 см.
В варианте реализации упомянутых выше аспектов изобретения растение или часть растения обрабатывают таким образом, что она становится неспособной размножаться или дать начало живому растению, т.е. ее убивают. Например, растение или часть растения высушивают и/или перемалывают.
В упомянутых выше вариантах реализации изобретения общее содержание неполярного липида в части растения предпочтительно по меньшей мере на 3% выше, более предпочтительно, по меньшей мере на 5% выше, чем общее содержание неполярного липида в соответствующей части растения, трансформированной генами, кодирующими WRI1 и DGAT, но не содержащей других экзогенных полинуклеотидов и генетических модификаций, описанных в настоящем документе для упомянутых выше аспектов. Более предпочтительно, указанная степень повышения относится к стеблю или корню растения.
В варианте реализации изобретения введение одного или более экзогенных полинуклеотидов или генетических модификаций, предпочтительно экзогенного полинуклеотида, кодирующего ПМВ или тиоэстеразу жирного ацила, или генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке, более предпочтительно, экзогенного полинуклеотида, кодирующего тиоэстеразу FATA или БСЛК или снижающего экспрессию эндогенной липазы ТАГ, такой как липаза ТАГ SDP1, в клетке, приводит к синергетическому повышению общего содержания неполярного липида в клетке, при добавлении к паре трансгенов WRI1 и DGAT, особенно перед цветением растения и, даже более конкретно, в стеблях и/или корнях растения. Например, см. Примеры 8, 11 и 15. В предпочтительном варианте реализации изобретения повышение содержания ТАГ в клетке, расположенной в стебле или корне растения, по меньшей мере в 2 раза, более предпочтительно, по меньшей мере в 3 раза выше, чем в соответствующей клетке, трансформированной генами, кодирующими WRI1 и DGAT1, но не содержащей тиоэстеразы FATA, БСЛК и генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в растении. Наиболее предпочтительно, по меньшей мере промотор, направляющий экспрессию первого экзогенного полинуклеотида, является промотором, не принадлежащим к конститутивным промоторам.
В вариантах реализации шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, тринадцатого, четырнадцатого, шестнадцатого, восемнадцатого и девятнадцатого аспектов изобретения растение или его часть предпочтительно является фенотипически нормальным в том смысле, что его способность расти и воспроизводиться в существенной мере не снижена, по сравнению с немодифицированным растением или его частью. Предпочтительно биомасса, скорость роста, скорость прорастания, размер запасающего органа, размер семени и/или количество образованных жизнеспособных семян составляет не менее чем 90% от показателей соответствующего растения дикого типа, при выращивании в идентичных условиях. В варианте реализации изобретения растение обладает мужской и женской фертильностью в той же степени, что и соответствующее растение дикого типа, а его пыльца (если она образуется) так же жизнеспособна, как и пыльца соответствующего растения дикого типа, предпочтительно жизнеспособность составляет около 100%. В варианте реализации изобретения растение образует семя, которому свойственна скорость прорастания по меньшей мере 90% относительно скорости прорастания соответствующего семени растения дикого типа, если вид растений образует семена. В варианте реализации изобретения высота растения по изобретению составляет по меньшей мере 90% относительно высоты соответствующего растения дикого типа, выращенного в таких же условиях. Комбинация каждого из указанных признаков включена. В альтернативном варианте реализации изобретения высота растения по изобретению составляет от 60% до 90% относительно высоты соответствующего растения дикого типа, выращенного в таких же условиях. В варианте реализации изобретения растение или его часть по изобретению, предпочтительно лист растения, не проявляет усиленного некроза, т.е. степень некроза, если он присутствует, такая же, какую демонстрирует соответствующее растение дикого типа или его часть, выращенное в таких же условиях и находящееся на такой же стадии развития растения. Данный признак применяется, в частности, к растению или его части, содержащему экзогенный полинуклеотид, кодирующий тиоэстеразу жирных кислот, такой как тиоэстераза FATB.
Следующие варианты реализации изобретения применяются к растению или его части по изобретению (включая шестой, седьмой, восьмой, девятый, десятый, одиннадцатый, двенадцатый, тринадцатый, четырнадцатый, шестнадцатый, восемнадцатый и девятнадцатый аспекты), а также к способу получения растения или его части или способу их применения. В варианте реализации изобретения полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является ацилтрансферазой ацилов жирных кислот, принимающей участие в биосинтезе ТАГ, ДАГ или моноацилглицерида (МАГ) в растении или его части, предпочтительно ТАГ в растении или его части, такой как DGAT, PDAT, LPAAT, GPAT или MGAT, предпочтительно DGAT или PDAT.
В другом варианте реализации изобретения полипептид, принимающий участие в катаболизме триацилглицеридов (ТАГ) в растении или части растения, является липазой SDP1, полипептидом Cgi58, ацил-КоА оксидазой, такой как ACX1 или ACX2, или полипептидом, принимающим участие в β-окислении жирных кислот в растении, таким как пероксисомальный транспортер АТФ-связывающей кассеты PXA1, предпочтительно липазой SDP1.
В варианте реализации изобретения полипептид покрытия масляного включения (ПМВ) является олеозином, таким как полиолеозин или калеозин, или предпочтительно связанным с липидными капельками белком (БСЛК).
В варианте реализации изобретения полипептид, который повышает экспорт жирных кислот из пластид растения, является тиоэстеразой C16 или C18 жирных кислот, такой как полипептид FATA или полипептид FATB, переносчиком жирных кислот, таким как полипептид ABCA9 или длинноцепочечная ацил-КоА синтетаза (ДЦАС).
В варианте реализации изобретения полипептид, принимающий участие в импорте жирных кислот в пластиды растения, является переносчиком жирных кислот или его субъединицей, предпочтительно полипептидом ТГД, таким как полипептид TGD1, полипептид TGD2, полипептид TGD3 или полипептид TGD4.
В варианте реализации изобретения полипептид, принимающий участие в выработке диацилглицерида (ДАГ) в пластиде, является пластидной GPAT, пластидной LPAAT или пластидной PAP.
В варианте реализации изобретения растение или его часть по изобретению является растением 16:3 или его частью, которая содержит одно или более или все из следующего:
a) экзогенный полинуклеотид, кодирующий полипептид, который повышает экспорт жирных кислот из пластид растения, по сравнению с соответствующим растением, не содержащим экзогенного полинуклеотида,
b) первой генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды растения, по сравнению с соответствующим растением, не содержащим первой генетической модификации, и
c) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде, по сравнению с соответствующим растением, не содержащим второй генетической модификации,
причем экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении, или его части.
В альтернативном варианте реализации изобретения растение или его часть по изобретению является растением 18:3 или его частью.
В варианте реализации изобретения общее содержание неполярного липида в вегетативной части растения перед цветением растения составляет по меньшей мере около 8%, по меньшей мере около 10%, около 11%, от 8% до 15%, или от 9% до 12% (масс./масс., в пересчете на сухую массу)
В варианте реализации изобретения одна или более или все из генетических модификаций представляют собой мутацию эндогенного гена, которая частично или полностью инактивирует ген, такую как точечная мутация, инсерция или делеция (или комбинация одного или более из этого), предпочтительно введенную мутацию. Точечная мутация может представлять собой преждевременный стоп-кодон, мутацию сайта сращивания, мутацию сдвига рамки или мутацию замены аминокислоты, которая снижает активность гена или кодируемого полипептида. Делеция может состоять из одного или более нуклеотидов в пределах транскрибированного экзона или промотора гена, или простираться сквозь или в более чем один экзон, или расширяться до делеции всего гена. Предпочтительно делецию вводят с применением технологий ZF, ПАТЭН или ККППРП. В альтернативном варианте реализации изобретения одна или более или все генетические модификации представляют собой экзогенный полинуклеотид, кодирующий молекулу РНК, которая подавляет экспрессию эндогенного гена, причем экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в растении или его части.
В варианте реализации изобретения экзогенный полинуклеотид, кодирующий WRI1, содержит одно или более из следующего:
i) нуклеотиды, кодирующие полипептид, содержащий аминокислоты, последовательность которых приведена в любой из SEQ ID NO: 21-75 или 205-210, или его биологически активный фрагмент или полипептид, аминокислотная последовательность которого по меньшей мере на 30% идентична любой одной или более из SEQ ID NO: 21-75 или 205-210,
ii) нуклеотиды, последовательность которых по меньшей мере на 30% идентична i), и
iii) нуклеотиды, которые гибридизуются с i) и/или ii) в строгих условиях. Предпочтительно, полипептид WRI1 представляет собой полипептид WRI1, не являющийся WRI1 Arabidopsis thaliana (SEQ ID NO: 21 или 22). Более предпочтительно, полипептид WRI1 содержит аминокислоты, последовательность которых приведена в SEQ ID NO: 208, или их биологически активный фрагмент или полипептид, аминокислотная последовательность которого по меньшей мере на 30% идентична им.
В варианте реализации изобретения общее содержание неполярного липида или одного или более неполярных липидов и/или уровень олеиновой кислоты или ПНЖК в растении или его части определяют методом анализа метиловых эфиров жирных кислот, полученных из растения или его вегетативной части, с применением газовой хроматографии.
В дальнейшем варианте реализации изобретения, в котором часть растения является листом, общее содержание неполярного липида в листе определяют методом анализа с применением ядерного магнитного резонанса (ЯМР).
В варианте реализации изобретения растение или его часть является членом популяции или коллекции размером по меньшей мере около 1000 таких растений или их частей.
В дальнейшем аспекте настоящего изобретения предлагается популяция размером по меньшей мере около 1000 растений, каждое из которых представляет собой растение по изобретению, выращенное в поле.
В другом аспекте настоящего изобретения предлагается коллекция размером по меньшей мере около 1000 вегетативных частей растения, каждая из которых является вегетативной частью растения по изобретению, причем урожай вегетативных частей растения собран с растений, растущих в поле.
В варианте реализации клетки, организма, не относящегося к человеческому роду, растения или его части по изобретению, полипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, является фактором транскрипции WRI1, полипептид, принимающий участие в биосинтезе одного или более неполярных липидов, является DGAT, например, DGAT1 или DGAT2, или PDAT, и полипептид, принимающий участие в катаболизме триацилглицеридов (ТАГ) в клетке, является липазой SDP1. В предпочтительном варианте реализации изобретения полипептид покрытия масляного включения (ПМВ) является олеозином, полипептид, который повышает экспорт жирных кислот из пластид клетки, является тиоэстеразой жирных кислот, например, тиоэстеразой FATA или FATB, полипептид, принимающий участие в импорте жирных кислот в пластиды клетки, является полипептидом ТГД, предпочтительно полипептидом TGD1, и полипептид, принимающий участие в выработке диацилглицерида (ДАГ) в пластиде, является пластидной GPAT. В более предпочтительном варианте реализации изобретения клетка находится в вегетативной части растения, причем содержание ТАГ в вегетативной части растения перед цветением растения составляет по меньшей мере 8% (% от сухой массы).
В варианте реализации изобретения растение, вегетативная часть растения, организм, не относящийся к человеческому роду, или его часть, семя или клубень картофеля содержат первый экзогенный полинуклеотид, кодирующий WRI1, второй экзогенный полинуклеотид, кодирующий DGAT или ПДАТ, предпочтительно DGAT1, третий экзогенный полинуклеотид, кодирующий РНК, которая уменьшает экспрессию гена, кодирующего полипептид SPD1, и четвертый экзогенный полинуклеотид, кодирующий олеозин. В предпочтительных вариантах реализации изобретения, в вегетативной части растения, организме, не относящемся к человеческому роду, или его части, семени или клубне картофеля присутствует один или более, или все из следующих признаков:
i) общее содержание липида по меньшей мере 8%, по меньшей мере 10%, по меньшей мере 12%, по меньшей мере 14% или по меньшей мере 15,5% (%, масс.);
ii) общее содержание липида в вегетативной части растения или организме, не относящемся к человеческому роду, по меньшей мере в 3 раза, по меньшей мере в 5 раз, по меньшей мере в 7 раз, по меньшей мере в 8 раз или по меньшей мере в 10 раз выше относительно соответствующей вегетативной части растения или организма, не относящегося к человеческому роду, в которых отсутствуют экзогенные полинуклеотиды;
iii) общее содержание ТАГ составляет по меньшей мере 5%, по меньшей мере 6%, по меньшей мере 6,5% или по меньшей мере 7% (%, масс. в пересчете на массу сухого вещества или массу семени);
iv) общее содержание ТАГ по меньшей мере в 40 раз, по меньшей мере в 50 раз, по меньшей мере в 60 раз или по меньшей мере в 70 раз, по меньшей мере в 100 раз или по меньшей мере в 120 раз выше, чем в соответствующей вегетативной части растения или организма, не относящегося к человеческому роду, в которых отсутствуют экзогенные полинуклеотиды;
v) олеиновая кислота составляет по меньшей мере 15%, по меньшей мере 19% или по меньшей мере 22% (%, масс., в пересчете на массу сухого семени) всех жирных кислот в ТАГ;
vi) уровень олеиновой кислоты в ТАГ по меньшей мере в 10 раз, по меньшей мере в 15 раз или по меньшей мере в 17 раз превышает уровень в соответствующей вегетативной части растения или организме, не относящемся к человеческому роду, в которых отсутствуют экзогенные полинуклеотиды;
vii) пальмитиновая кислота составляет по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30% или по меньшей мере 33% (%, масс.) жирных кислот в ТАГ;
viii) уровень пальмитиновой кислоты в ТАГ по меньшей мере в 1,5 раза выше относительно соответствующей вегетативной части растения или организма, не относящегося к человеческому роду, в которых отсутствуют экзогенные полинуклеотиды;
ix) линолевая кислота составляет по меньшей мере 22%, по меньшей мере 25%, по меньшей мере 30% или по меньшей мере 34% (%, масс.) жирных кислот в ТАГ;
x) α-линоленовая кислота составляет менее 20%, менее 15%, менее 11% или менее 8% (%, масс.) жирных кислот в ТАГ,
xi) уровень α-линоленовой кислоты в ТАГ по меньшей мере в 5 раз или по меньшей мере в 8 раз ниже, чем в соответствующей вегетативной части растения или организме, не относящемся к человеческому роду, в которых отсутствуют экзогенные полинуклеотиды, и
xii) для картофельного клубня, содержание ТАГ составляет по меньшей мере 0,5%, в пересчете на сухую массу, и/или общее содержание жирных кислот составляет по меньшей мере 1%, предпочтительно по меньшей мере 1,5% или по меньшей мере 2,0%, в пересчете на сухую массу.
Дополнительно предлагается семя растения по изобретению или полученное от такого растения.
В другом аспекте изобретения предлагается трансгенный стебель растения или часть стебля размером по меньшей мере 1 г сухой массы, содержание ТАГ в которой составляет по меньшей мере 5% масс. (сухая масса), предпочтительно по меньшей мере 6%, более предпочтительно по меньшей мере 7%. В варианте реализации изобретения трансгенный стебель растения или часть стебля принадлежит, или предпочтительно ее урожай собран с двудольного растения. В качестве альтернативы, трансгенный стебель растения или часть стебля принадлежит, или предпочтительно ее урожай собран с однодольного растения. В варианте реализации изобретения стебель растения или часть стебля принадлежит или получена от растения, не являющегося сахарным тростником. В вариантах реализации изобретения стебель растения или часть стебля дополнительно отличается одним или более признаками, определенными в контексте клетки или растения по изобретению.
В другом аспекте изобретения предлагается растительная клетка, содержащая:
a) первый экзогенный полинуклеотид, кодирующий PDAT,
b) первую генетическую модификацию, подавляющую эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды клетки, предпочтительно полипептида ТГД, по сравнению с соответствующей клеткой, не содержащей первой генетической модификации, и одно или более из:
c) второй генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке, предпочтительно полипептида SDP1, по сравнению с соответствующей клеткой, не содержащей генетической модификации,
d) второго экзогенного полинуклеотида, кодирующего полипептид, который повышает экспорт жирных кислот из пластид клетки, предпочтительно тиоэстеразу жирного ацила, по сравнению с соответствующей клеткой, не содержащей второго экзогенного полинуклеотида, и
e) третьей генетической модификации, подавляющей эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде, по сравнению с соответствующей клеткой, не содержащей третьей генетической модификации,
причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию полинуклеотида в клетке. В предпочтительном варианте реализации изобретения присутствие в клетке первой, второй или третьей генетической модификации или второго экзогенного полинуклеотида синергетически повышает общее содержание неполярного липида в клетке, по сравнению с соответствующей клеткой, содержащей PDAT, но не содержащей дополнительной генетической модификации или экзогенного полинуклеотида. Более предпочтительно, по меньшей мере один из экзогенных полинуклеотидов экспрессируется промотором, который не принадлежит к конститутивным промоторам.
В другом аспекте настоящего изобретения предлагается способ получения рекомбинантной эукариотной клетки по изобретению, включающий стадии:
i) введения в эукариотную клетку по меньшей мере одного экзогенного полинуклеотида и/или по меньшей мере одной генетической модификации, определенной в настоящем документе, с получением эукариотной клетки, содержащей набор экзогенных полинуклеотидов и/или генетических модификаций, определенных в настоящем документе,
ii) экспрессии экзогенного(ых) полинуклеотида(ов) в клетке или ее клетке-потомке,
iii) анализ содержания липидов в клетке или ее клетке-потомке, и
iv) отбор клетки по изобретению.
В варианте реализации изобретения один или более экзогенные полинуклеотиды стабильно интегрированы в геном клетки или ее клетки-потомка.
В варианте реализации изобретения способ дополнительно включает стадию регенерации трансгенного растения из клетки или клетки-потомка, содержащей один или более экзогенных полинуклеотидов.
В дальнейшем варианте реализации изобретения стадия регенерации трансгенного растения выполняется до стадии экспрессии одного или более экзогенных полинуклеотидов в клетке или ее клетке-потомке и/или до стадии анализа содержимого липида в клетке или ее клетке-потомке и/или до стадии отбора клетки или клетки-потомка, содержащей повышенный уровень одного или более неполярных липидов.
В другом варианте реализации изобретения способ дополнительно содержит стадию получения семени или растения-потомка из трансгенного растения, причем семя или растение-потомок содержит один или более экзогенных полинуклеотидов.
Еще в другом варианте реализации изобретения отобранная клетка или регенерированное из нее растение или вегетативная часть растения или семя регенерированного растения обладает одним или более признаками, определенными в настоящем документе.
В дальнейшем аспекте настоящего изобретения предлагается способ получения растения, в геном которого интегрирован набор экзогенных полинуклеотидов и/или генетических модификаций, определенных в настоящем документе, причем способ включает стадии:
i) скрещивания двух материнских растений, и, при этом, одно растение содержит по меньшей мере один из экзогенных полинуклеотидов и/или по меньшей мере одну из генетических модификаций, определенных в настоящем документе, и другоерастение содержит по меньшей мере один из экзогенных полинуклеотидов и/или по меньшей мере одну из генетических модификаций, определенных в настоящем документе, притом, что два материнских растения содержат набор экзогенных полинуклеотидов и/или генетических модификаций, определенных в настоящем документе,
ii) скрининг одного или более растений-потомков скрещивания на предмет присутствия или отсутствия набора экзогенных полинуклеотидов и/или генетических модификаций, определенных в настоящем документе, и
iii) отбор растения-потомка, содержащего набор экзогенных полинуклеотидов и/или генетических модификаций, определенных в настоящем документе,
с получением таким образом растения.
Дополнительно предложена трансгенная клетка или трансгенное растение, полученные с применением способа по изобретению, или его часть, полученная из него, которая содержит набор экзогенных полинуклеотидов и/или генетических модификаций, определенных в настоящем документе.
Дополнительно предлагается применение набора экзогенных полинуклеотидов и/или генетических модификаций, определенных в настоящем документе, для получения трансгенной клетки, трансгенного организма, не относящегося к человеческому роду, или его части или семени, обладающих повышенной способностью к выработке одного или более неполярных липидов относительно соответствующей клетки, организма, не относящегося к человеческому роду, или его части или семени, не содержащих набора экзогенных полинуклеотидов и/или генетических модификаций, причем каждый экзогенный полинуклеотид функционально связан с промотором, способным направлять экспрессию экзогенного полинуклеотида в трансгенной клетке, трансгенном организме, не относящемся к человеческому роду, или его части или семени.
Предпочтительно, по меньшей мере один из промоторов, функционально связанных с экзогенным полинуклеотидом, который кодирует полипептид фактора транскрипции, представляет собой промотор, не принадлежащий к конститутивным промоторам.
В варианте реализации изобретения трансгенная клетка, организм, не относящийся к человеческому роду, или его часть или семя обладает одним или более признаками, определенными в настоящем документе.
В дальнейшем аспекте настоящего изобретения предлагается способ получения промышленного продукта, включающий стадии:
i) получение рекомбинантной эукариотной клетки по изобретению, трансгенного организма, не относящегося к человеческому роду, или его части по изобретению, трансгенного растения или его части по изобретению, семени по изобретению, или трансгенной клетки или трансгенного растения или его части по изобретению, и
ii) превращение по меньшей мере части липида в клетке, организме, не относящемся к человеческому роду, или его части, растении или его части, или семени в промышленный продукт, с применением нагревания, химических или ферментных средств или любой их комбинации к липиду in situ в организме, не относящемся к человеческому роду, или его части, и
iii) извлечение промышленного продукта,
с получением таким образом промышленного продукта.
В дальнейшем аспекте настоящего изобретения предлагается способ получения промышленного продукта, включающий стадии:
i) получение рекомбинантной эукариотной клетки по изобретению, трансгенного организма, не относящегося к человеческому роду, или его части по изобретению, трансгенного растения или его части по изобретению, семени по изобретению, или трансгенной клетки или трансгенного растения или его части по изобретению, и
ii) физическая обработка клетки, организма, не относящегося к человеческому роду, или его части, растения или его части или семени со стадии i),
iii) одновременное или последующее превращение по меньшей мере части липида в обрабатываемой клетке, организме, не относящемся к человеческому роду, или его части, растении или его части или семени в промышленный продукт, с применением нагревания, химических или ферментных средств или любой их комбинации к липиду в обрабатываемой клетке, организме, не относящемся к человеческому роду, или его части, растении или его части или семени, и
iv) извлечение промышленного продукта,
с получением таким образом промышленного продукта.
В варианте реализации двух упомянутых выше аспектов изобретения часть растения является вегетативной частью растения.
В дальнейшем аспекте настоящего изобретения предлагается способ получения промышленного продукта, включающий стадии:
i) получение вегетативной части растения с общим содержанием неполярного липида по меньшей мере около 18%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 18% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу),
ii) превращение по меньшей мере части липида в вегетативной части растения в промышленный продукт, с применением нагревания, химических или ферментных средств или любой их комбинации к липиду in situ в вегетативной части растения, и
iii) извлечение промышленного продукта,
с получением таким образом промышленного продукта.
В другом аспекте настоящего изобретения предлагается способ получения промышленного продукта, включающий стадии:
i) получение вегетативной части растения с общим содержанием неполярного липида по меньшей мере около 18%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 18% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу),
ii) физическая обработка вегетативной части растения со стадии i),
iii) одновременное или последующее превращение по меньшей мере части липида в обрабатываемой вегетативной части растения в промышленный продукт, с применением нагревания, химических или ферментных средств или любой их комбинации к липиду в обрабатываемой вегетативной части растения, и
iv) извлечение промышленного продукта,
с получением таким образом промышленного продукта.
Еще в другом аспекте настоящего изобретения предлагается способ получения промышленного продукта, включающий стадии:
i) получение вегетативной части растения с общим содержанием неполярного липида по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу), причем растение является растением 16:3 или его вегетативной частью,
ii) превращение по меньшей мере части липида в вегетативной части растения в промышленный продукт, с применением нагревания, химических или ферментных средств или любой их комбинации к липиду in situ в вегетативной части растения, и
iii) извлечение промышленного продукта,
с получением таким образом промышленного продукта.
В другом аспекте настоящего изобретения предлагается способ получения промышленного продукта, включающий стадии:
i) получение вегетативной части растения с общим содержанием неполярного липида по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу), причем растение является растением 16:3 или его вегетативной частью,
ii) физическая обработка вегетативной части растения со стадии i),
iii) одновременное или последующее превращение по меньшей мере части липида в обрабатываемой вегетативной части растения в промышленный продукт, с применением нагревания, химических или ферментных средств или любой их комбинации к липиду в обрабатываемой вегетативной части растения, и
iv) извлечение промышленного продукта,
с получением таким образом промышленного продукта.
В варианте реализации изобретения стадия физической обработки клетки, организма, не относящегося к человеческому роду, или его части, растения или его части, или семени включает одно или более из вальцовки, прессования, дробления или помола клетки, организма, не относящегося к человеческому роду, или его части, растения или его части, или семени.
В варианте реализации изобретения способ включает стадии:
(a) экстракция по меньшей мере части неполярного липида, содержащегося в клетке, организме, не относящемся к человеческому роду, или его части, растении или его части, или семени в форме неполярного липида, и
(b) извлечение экстрагированного неполярного липида,
причем стадии (a) и (b) выполняются до стадии превращения по меньшей мере части липида в клетке, организме, не относящемся к человеческому роду, или его части, растении или его части, или семени в промышленный продукт.
В варианте реализации изобретения экстрагированный неполярный липид содержит триацилглицериды, причем триацилглицериды составляют по меньшей мере 90%, предпочтительно по меньшей мере 95% экстрагированного липида.
В варианте реализации изобретения промышленный продукт является углеводородным продуктом, таким как эфиры жирных кислот, предпочтительно метиловые эфиры жирных кислот и/или этиловые эфиры жирных кислот, алкан, например, метан, этан или длинноцепочечный алкан, смесь алканов с более длинной цепью, алкен, биотопливо, газообразный монооксид углерода и/или водород, биоспирт, например, этанол, пропанол или бутанол, биоуголь или комбинация монооксида углерода, водорода и биоугля. В предпочтительном варианте реализации изобретения общие жирные кислоты в вегетативной части растения содержат по меньшей мере 5% C12:0, C14:0, или сумма C12:0 и C14:0 составляет по меньшей мере 5% от общего содержания жирных кислот, и промышленный продукт, полученный из липида вегетативной части растения, является компонентом авиационного топлива.
В дальнейшем аспекте настоящего изобретения предлагается способ получения экстрагированного липида, включающий стадии:
i) получение рекомбинантной эукариотной клетки по изобретению, трансгенного организма, не относящегося к человеческому роду, или его части по изобретению, трансгенного растения или его части по изобретению, семени по изобретению, или трансгенной клетки или трансгенного растения или его части по изобретению,
ii) экстракция липида из клетки, организма, не относящегося к человеческому роду, или его части, растения или его части, или семени, и
iii) извлечение экстрагированного липида,
с получением таким образом экстрагированного липида.
В дальнейшем аспекте настоящего изобретения предлагается способ получения экстрагированного липида, включающий стадии:
i) получение вегетативной части растения с общим содержанием неполярного липида по меньшей мере около 18%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 18% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу),
ii) экстракцию липида из вегетативной части растения, и
iii) извлечение экстрагированного липида,
с получением таким образом экстрагированного липида.
В дальнейшем аспекте настоящего изобретения предлагается способ получения экстрагированного липида, включающий стадии:
i) получение вегетативной части растения с общим содержанием неполярного липида по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу), причем растение является растением 16:3 или его вегетативной частью,
ii) экстракцию липида из вегетативной части растения, и
iii) извлечение экстрагированного липида,
с получением таким образом экстрагированного липида.
В варианте реализации изобретения способ экстракции включает одно или более из сушки, вальцевания, прессования, дробления или помола клетки, организма, не относящегося к человеческому роду, или его части, растения или его части, или семени, и/или очистки экстрагированного липида или масла из семян.
В варианте реализации изобретения органический растворитель применяется в способе в процессе экстракции, для экстрагирования масла.
В дальнейшем варианте реализации изобретения способ включает извлечение экстрагированного липида или масла путем сбора его в емкость и/или одного или более из дегуммации, дезодорирования, обесцвечивания, сушки, фракционирования экстрагированного липида или масла, удаления по меньшей мере некоторых восков и/или эфиров восков из экстрагированного липида или масла или анализа состава жирных кислот экстрагированного липида или масла.
В варианте реализации изобретения объем экстрагированного липида или масла составляет по меньшей мере 1 литр.
В дальнейшем варианте реализации изобретения применяются один или более или все из следующих признаков:
(i) экстрагированный липид или масло содержит триацилглицериды, причем триацилглицериды составляют по меньшей мере 90%, предпочтительно по меньшей мере 95% или по меньшей мере 96% экстрагированного липида или масла,
(ii) экстрагированный липид или масло содержит свободные стеролы, стероильные эфиры, стероилгликозиды, воски или эфиры восков или любую их комбинацию, и
(iii) общее содержание и/или состав стеролов в экстрагированном липиде или масле значительно отличается от содержания и/или состава стеролов в экстрагированном липиде или масле, полученном из соответствующей клетки, организма, не относящегося к человеческому роду, или его части, растения или его части, или семени.
В варианте реализации изобретения способ дополнительно включает превращение экстрагированного липида или масла в промышленный продукт.
В варианте реализации изобретения промышленный продукт является углеводородным продуктом, таким как эфиры жирных кислот, предпочтительно метиловые эфиры жирных кислот и/или этиловые эфиры жирных кислот, алкан, например, метан, этан или длинноцепочечный алкан, смесь алканов с более длинной цепью, алкен, биотопливо, газообразный монооксид углерода и/или водород, биоспирт, например, этанол, пропанол или бутанол, биоуголь или комбинация монооксида углерода, водорода и биоугля. В предпочтительном варианте реализации изобретения общие жирные кислоты в вегетативной части растения содержат по меньшей мере 5% C12:0, C14:0, или сумма C12:0 и C14:0 составляет по меньшей мере 5% от общего содержания жирных кислот, и промышленный продукт, полученный из липида вегетативной части растения, является компонентом авиационного топлива.
В дальнейшем варианте реализации изобретения часть растения является воздушной частью растения или зеленой частью растения, предпочтительно вегетативной частью растения, такой как лист или стебель растения. В альтернативном варианте реализации изобретения часть растения является клубнем или свеклой, таким как клубень картофеля (Solanum tuberosum) или сахарная свекла.
Еще в другом варианте реализации изобретения способ дополнительно включает стадию сбора урожая клетки, организма, не относящегося к человеческому роду, или его части, растения или его части, такого как клубень или свекла, или семени, предпочтительно с помощью механического уборщика, или посредством процесса, включающего фильтрацию, центрифугирование, осаждение, флотацию или флоккуляцию водорослевых или грибковых организмов.
В другом варианте реализации изобретения уровень липида в клетке, организме, не относящемся к человеческому роду, или его части, растении или его части, или семени и/или в экстрагированном липиде или масле может быть определен методом анализа с применением газовой хроматографии метиловых эфиров жирных кислот, полученных из экстрагированного липида или масла.
Еще в другом варианте реализации изобретения способ дополнительно включает сбор урожая части с растения.
В варианте реализации изобретения часть растения является вегетативной частью растения с общим содержанием неполярного липида по меньшей мере около 18%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 18% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу).
В дальнейшем варианте реализации изобретения часть растения является вегетативной частью растения с общим содержанием ТАГ по меньшей мере около 18%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 18% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу).
В другом варианте реализации изобретения часть растения является вегетативной частью растения с общим содержанием неполярного липида по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу), причем вегетативная часть растения получена от растений 16:3.
Еще в другом варианте реализации изобретения часть растения является вегетативной частью растения с общим содержанием ТАГ по меньшей мере около 11%, по меньшей мере около 12%, по меньшей мере около 15%, по меньшей мере около 20%, по меньшей мере около 25%, по меньшей мере около 30%, по меньшей мере около 35%, по меньшей мере около 40%, по меньшей мере около 45%, по меньшей мере около 50%, по меньшей мере около 55%, по меньшей мере около 60%, по меньшей мере около 65%, по меньшей мере около 70%, от 8% до 75%, от 10% до 75%, от 11% до 75%, от около 15% до 75%, от около 20% до 75%, от около 30% до 75%, от около 40% до 75%, от около 50% до 75%, от около 60% до 75%, или от около 25% до 50% (масс./масс., в пересчете на сухую массу), причем вегетативная часть растения получена от растений 16:3.
Дополнительно предлагается способ получения семени, включающий:
i) выращивание растения по изобретению, и
ii) сбор урожая семени с растения.
В варианте реализации изобретения упомянутый выше способ включает выращивание популяции размером по меньшей мере около 1000 растений, каждое из которых является растением по изобретению, и сбор урожая семени с популяции растений.
Еще в другом аспекте настоящего изобретения предлагается способ ферментации, включающий:
i) обеспечение емкости, содержащей жидкую композицию, которая содержит рекомбинантную эукариотную клетку по изобретению или трансгенный организм, не относящийся к человеческому роду, по изобретению, причем клетка или организм, не относящийся к человеческому роду, пригодны для ферментации, и составляющие, необходимые для ферментации и биосинтеза жирных кислот, и
ii) обеспечение условий, способствующих ферментации жидкой композиции, содержащейся в указанной емкости.
Кроме того, предлагается извлеченный или экстрагированный липид, получаемый из рекомбинантной эукариотной клетки по изобретению, трансгенного организма, не относящегося к человеческому роду, или его части по изобретению, трансгенного растения или его части по изобретению, семени по изобретению или трансгенной клетки или трансгенного растения или его части по изобретению или получаемый с применением способа по изобретению.
В дальнейшем аспекте настоящего изобретения предлагается промышленный продукт, полученный с применением способа по изобретению, который является углеводородным продуктом, таким как эфиры жирных кислот, предпочтительно метиловые эфиры жирных кислот и/или этиловые эфиры жирных кислот, алкан, например, метан, этан или длинноцепочечный алкан, смесь алканов с более длинной цепью, алкен, биотопливо, газообразный монооксид углерода и/или водород, биоспирт, например, этанол, пропанол или бутанол, биоуголь или комбинация монооксида углерода, водорода и биоугля.
Дополнительно предлагается применение рекомбинантной эукариотной клетки по изобретению, трансгенного организма, не относящегося к человеческому роду, или его части по изобретению, трансгенного растения или его части по изобретению, семени по изобретению или трансгенной клетки или трансгенного растения или его части по изобретению или извлеченного или экстрагированного липида по изобретению для производства промышленного продукта.
Примеры промышленных продуктов по изобретению включают, но не ограничиваясь ими, углеводородный продукт, такой как эфиры жирных кислот, предпочтительно метиловые эфиры жирных кислот и/или этиловые эфиры жирных кислот, алкан, например, метан, этан или длинноцепочечный алкан, смесь алканов с более длинной цепью, алкен, биотопливо, газообразный монооксид углерода и/или водород, биоспирт, например, этанол, пропанол или бутанол, биоуголь или комбинация монооксида углерода, водорода и биоугля.
В дальнейшем аспекте настоящего изобретения предлагается способ получения топлива, включающий:
i) приведение в реакцию липида по изобретению со спиртом, необязательно в присутствии катализатора, с получением алкиловых эфиров, и
ii) необязательное смешивание алкиловых эфиров с топливом на основе нефти.
В варианте реализации упомянутого выше способа алкиловые эфиры являются метиловыми эфирами.
Еще в другом аспекте настоящего изобретения предлагается способ получения синтетического дизельного топлива, включающий:
i) превращение липида в рекомбинантной эукариотной клетке по изобретению, трансгенном организме, не относящемся к человеческому роду, или его части по изобретению, трансгенном растении или его части по изобретению, семени по изобретению, или трансгенной клетке или трансгенном растении или его части по изобретению, в бионефть с применением способа, включающего пиролиз или гидротермическую обработку, или в синтетический газ с применением газификации, и
ii) превращение бионефти в синтетическое дизельное топливо с применением способа, включающего фракционирование, предпочтительно с отбором углеводородных соединений, которые конденсируются при температуре от приблизительно 150°C до приблизительно 200°C или от приблизительно 200°C до приблизительно 300°C, или превращение синтетического газа в биотопливо с применением металлического катализатора или микробного катализатора.
В другом аспекте настоящего изобретения предлагается способ получения биотоплива, включающий превращение липида в рекомбинантной эукариотной клетке по изобретению, трансгенном организме, не относящемся к человеческому роду, или его части по изобретению, трансгенном растении или его части по изобретению, семени по изобретению или трансгенной клетке или трансгенном растении или его части по изобретению в бионефть с применением пиролиза, в биоспирт с применением ферментации или в биогаз с применением газификации или анаэробного расщепления.
В варианте реализации упомянутого выше способа часть является вегетативной частью растения.
Кроме того, предлагается способ получения кормов (продуктов питания), включающий смешивание рекомбинантной эукариотной клетки по изобретению, трансгенного организма, не относящегося к человеческому роду, или его части по изобретению, трансгенного растения или его части по изобретению, семени по изобретению или трансгенной клетки или трансгенного растения или его части по изобретению, или получаемых с применением способа по изобретению, или экстракта или его части, по меньшей мере с еще одним питательным ингредиентом.
В дальнейшем аспекте настоящего изобретения предлагаются корма (продукты питания), косметические продукты или химические реактивы, содержащие рекомбинантную эукариотную клетку по изобретению, трансгенный организм, не относящийся к человеческому роду, или его часть по изобретению, трансгенное растение или его часть по изобретению, семя по изобретению или трансгенную клетку или трансгенное растение или его часть по изобретению, или получаемые с применением способа по изобретению, или экстракт или его часть.
В другом аспекте настоящего изобретения предлагается способ кормления животного, включающий предложение животному трансгенного растения или его части по изобретению, семени по изобретению или трансгенного растения или его части по изобретению, или извлеченного или экстрагированного липида по изобретению.
Любой вариант реализации изобретения будет взят для применения соответствующих поправок к любому другому варианту реализации изобретения, если конкретно не указано иное.
Контекст настоящего изобретения не ограничивается конкретными вариантами реализации изобретения, описанными в настоящем документе, которые предназначены только для целей иллюстрации. Функционально равноценные продукты, составы и способы явно находятся в пределах контекста изобретения, как раскрыто в настоящем документе.
В настоящем документе, если конкретно не указано иное или контекст не требует иного, ссылка на одну стадию, состав вещества, группу стадий или группу составов вещества должна интерпретироваться, как включающая единственное и множественное число (т. е., один или более) таких стадий, составов вещества, групп стадий или групп составов вещества.
Изобретение дополнительно раскрыто посредством следующих неограничивающих Примеров и со ссылкой на прилагаемые фигуры.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1. Схема синтеза липида в эукариотных клетках, демонстрирующая экспорт некоторых из жирных кислот, синтезированных в пластидах, в эндоплазматический ретикулум (ЭР) через связанную с пластидой мембрану (СПЛМ), и импорт некоторых из жирных кислот из ЭР в пластиду для эукариотного синтеза галактолипидов. Сокращения:
ацетил-КоА и малонил-КоА: ацетил-кофермент A и малонил-кофермент A;
ACCase: ацетил-КоА карбоксилаза;
FAS: комплекс синтетазы жирных кислот;
16:0-АПБ, 18:0-АПБ и 18:1-АПБ: C16:0-ацилпереносящий белок (АПБ), C18:0-ацилпереносящий белок, C18:1-ацилпереносящий белок;
KAS II: кетоацил-АПБ синтетаза II (EC 2.3.1.41);
PLPAAT: пластидная LPAAT;
PGPAT: пластидная GPAT;
PAP: фосфорилаза ФК (EC 3.1.3.4);
G3P: глицерин-3-фосфат;
LPA: лизофосфатидиновая кислота;
PA: фосфатидиновая кислота;
ДАГ: диацилглицерид;
ТАГ: триацилглицерид;
ацил-КоА и ацил-ФХ: ацил-коэнзим A и ацил-фосфатидилхолин;
ФХ: фосфатидилхолин;
GPAT: глицерил-3-фосфат ацилтрансфераза;
LPAAT: ацилтрансфераза лизофосфатидиновой кислоты (EC 2.3.1.51);
LPCAT: ацил-КоА:лизофосфатидилхолин ацилтрансфераза; или синонимы 1-ацилглицерофосфохолин O-ацилтрансфераза; ацил-КоА:1-ацил-sn-глицеро-3-фосфохолин O-ацилтрансфераза (EC 2.3.1.23),
CPT: холинфосфотрансфераза ХДГ-холин:диацилглицерил холинфосфотрансфераза; или синонимы 1-алкил-2-ацетилглицерил холинфосфотрансфераза; алкилацилглицерил холинфосфотрансфераза; холинфосфотрансфераза; фосфорилхолин-глицерид трансфераза (EC 2.7.8.2),
PDCT: фосфатидилхолин:диацилглицерил холинфосфотрансфераза,
PLC: фосфолипаза C (EC 3.1.4.3),
PLD: фосфолипаза D; холин фосфатаза; лецитиназа D; липофосфодиэстераза II (EC 3.1.4.4),
PDAT: фосфолипид: диацилглисерил ацилтрансфераза; или синоним фосфолипид: 1,2-диацил-sn-глицерил O-ацилтрансфераза (EC 2.3.1.158);
FAD2: Δ 12-десатураза жирных кислот; FAD3, Δ15-десатураза жирных кислот;
UDP-Gal: уридиндифосфатгалактоза;
MGDS: моногалактозилдиацилглицерид синтетаза;
MGDG: моногалактозилдиацилглицерид; ДГДГ: дигалактозилдиацилглицерид
FAD6, 7, 8: пластидная Δ12-десатураза жирных кислот, пластидная ω3-десатураза, пластидная ω3-десатураза, индуцируемые при низкой температуре, соответственно.
Фиг. 2. Схематическая генетическая карта конструкции для повышения содержания масла из семян в двудольных растениях. Сокращения: PRO Pissa-Vicillin, промотор вицилина и 5' НТР Pisum sativum; TMV leader, 5'НТР вируса мозаики табака; Arath-DGAT1, кодирующий белок участок, который кодирует DGAT1 A. thaliana; TER Glyma-Lectin, 3' участок терминатора/полиаденилирования гена лектина G. max; PRO Phavu-Phaseolin, промотор из гена белка фазеолина Phaseolus vulgaris; Arath-WRI1, кодирующий белок участок, который кодирует WRI1 A. thaliana; TER AGRTU-NOS, 3' участок терминатора/полиаденилирования гена НОС Agrobacterium tumefaciens; РRО Phavu-PHA, промотор гена фазеолина Phaseolus vulgaris; Sesin-Oleosin, кодирующий белок участок, который кодирует ген олеозина Sesame indicum; TER Phavu-PHA, 3' участок терминатора/полиаденилирования гена фазеолина Phaseolus vulgaris.
Фиг. 3. Схематическая диаграмма вектора pOIL122. Сокращения: TER Agrtu-NOS, терминатор нопалинсинтетазы Agrobacterium tumefaciens; NPTII, участок кодирования белка неомицинфосфотрансферазы; PRO CaMV35S-Ex2, промотор 35S вируса мозаики цветной капусты с двойным участком энхансера; Arath-DGAT1, участок кодирования белка ацилтрансферазы DGAT1 Arabidopsis thaliana; PRO Arath-Rubisco SSU, промотор маленькой субъединицы Rubisco A. thaliana; Arath-FATA2, участок кодирования белка тиоэстеразы FATA2 A. thaliana; Arath-WRI, участок кодирования белка фактора транскрипции WRI1 A. thaliana; TER Glyma-Lectin, терминатор лектина Glycine max; промотор enTCUP2, криптический конститутивный промотор Nicotiana tabacum; attB1 и attB2, сайты рекомбинации Gateway; фрагмент NB SDP1, участок SDP1 Nicotiana benthamiana, служащий мишенью для сайленсинга шпРНКи; терминатор ОКС, терминатор октопинсинтетазы A. tumefaciens. Признаки скелета за пределами участка T-ДНК получены из pORE04 (Coutu с соавт., 2007).
Фиг. 4. Профили метиловых эфиров общих жирных кислот (% масс.) (МЭЖК), иллюстрирующие влияние опосредованного WRI1+DGAT1 генетического фона с высоким содержанием масла на выработку среднецепочечных жирных кислот (СЦЖК) в листе Nicotiana benthamiana (n=4). Наиболее высокая выработка СЦЖК наблюдалась после добавления Arath-WRI1.
Фиг. 5. Профили общих МЭЖК в листьях (% масс.), проясняющие влияние WRI1 на аккумуляцию СЦЖК (n=4). Добавление Arath-WRI1 значительно повысило выработку соответствующей жирной кислоты (C12:0, C14:0 или C16:0) относительно предыдущего добавления только Cocnu-LPAAT.
Фиг. 6. Уровни общих жирных кислот (ОЖК) (% масс.), уровни ТАГ, уровни СЦЖК (C16:0 и C14:0, % от общих жирных кислот) в ОЖК и СЦЖК в ТАГ (% от общего содержания жирных кислот в ТАГ) в клетках растения после экспрессии комбинаций трех DGAT масличной пальмы с FATB, LPAAT и WRI1. Номера 1-10 такие же, как приведено в тексте (Пример 9).
Фиг. 7. Уровни ТАГ (% сухой массы листа) в ткани листа N. benthamiana, инфильтрованной генами, кодирующими различные полипептиды WRI1, с (прямоугольники справа) или без (прямоугольники слева) кo-экспрессией DGAT1 (n=3). Все образцы дополнительно были инфильтрованы конструкцией P19.
Фиг. 8. Схематическое представление конструкции шпильки SDP1 N. benthamiana. Проиллюстрированные генетические сегменты являются такими, как описано в Примере 11. Сокращения являются такими, как приведено на Фиг. 3. Сайты attB представляют собой сайты рекомбинации из вектора pHELLSGATE12.
Фиг. 9. Содержание ТАГ в образцах зеленых листьев растений табака, трансформированных T-ДНК из pOIL51, линии №61 и №69, собранных перед цветением. Контрольные (родительские) образцы были получены от растений, трансформированных T-ДНК из pJP3502.
Фиг. 10. Уровни ТАГ (% сухой массы) в корне и ткани ствола дикого типа (дт) и трансгенных растений N. tabacum, содержащих только T-ДНК из pJP3502 или дополнительно содержащих T-ДНК из pOIL051.
Фиг. 11. Уровни ТАГ (% сухой массы) в корне и ткани ствола дикого типа (дт) и трансгенных растений N. tabacum, содержащих только T-ДНК из pJP3502 или дополнительно содержащих T-ДНК из pOIL049.
Фиг. 12. Содержание ТАГ в образцах листьев растений табака в стадии завязывания семян, трансформированных T-ДНК из pOIL049, линии №23c и №32b. Контрольные (родительские) образцы были получены от растений, трансформированных T-ДНК из pJP3502. Верхняя линия иллюстрирует процент 18:2 в ТАГ, а нижняя линия иллюстрирует процент 18:3 (АЛК) в содержании жирных кислот.
Фиг. 13. A. Содержание крахмала в листовой ткани растений дикого типа (ДТ) и трансгенных растений, содержащих T-ДНК из pJP3502 (контроль HO) или Т-ДНК как из pJP3502, так и из pOIL051 (pOIL51.61 и pOIL51.69), или как из pJP3502, так и из pOIL049 (pOIL49.32b). Данные представляют объединенные результаты по меньшей мере для трех индивидуальных растений. B. Корреляция между содержанием крахмала и ТАГ в листовой ткани растений дикого типа (ДТ) и трансгенных растений, содержащих T-ДНК из pJP3502 (контроль HO) или Т-ДНК как из pJP3502, так и из pOIL051 (pOIL51.61 и pOIL51.69), или как из pJP3502, так и из pOIL049 (pOIL49.32b). Данные представляют объединенные результаты по меньшей мере для трех индивидуальных растений.
Фиг. 14. Схематическое представление бинарного вектора pTV55. Сокращения: PRO, промотор; TER, 3' участок терминации/полиаденилирования; Arath, A. thaliana; Linus, Linum usitatissimum; Nicta, Nicotiana tabacum; Glyma, G. max; Cnl1, конлинин 1 из льна; Cnl2, конлинин 2 из льна; MAR Nicat-RB7, участок прикрепления к матриксу из RB7 табака, или как на Фиг. 3. Сокращения генов MGAT2, DGAT1, GPAT4, WRI1 такие же, как в тексте.
Фиг. 15. Содержание масла (%) в семенах T2 C. sativa, по данным ЯМР трансформированных pTV55, pTV56 и pTV57. Каждая точка данных представляет среднее содержание масла для трех независимых партий семени по 50 мг от каждой трансгенной линии. Семена отрицательного контроля представляли собой семена C. sativa дикого типа (нетрансформированные), выращенные в таких же условиях в теплице. N указывает на количество независимых трансгенных событий для каждой конструкции.
Фиг. 16. Филогенетическое дерево полипептидов LDAP (Пример 15).
Фиг. 17. Схематическое представление генетической конструкции pJP3506, включая участок T-ДНК между левой и правой границами. Сокращения такие же, как на Фиг. 3, и: Sesin-Oleosin, участок кодирования белка олеозина Sesame indicum.
Фиг. 18. Выход и изменение калорийности для выработки бионефти методом гидротермической обработки (ГТО) из вегетативного растительного материала табака с высоким содержанием масла, дикого типа и трансгенного, в качестве исходного сырья.
КЛЮЧ К ПЕРЕЧНЮ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 полипептид DGAT1 Arabidopsis thaliana (CAB44774.1)
SEQ ID NO: 2 полипептид DGAT2 Arabidopsis thaliana (NP_566952.1)
SEQ ID NO: 3 полипептид DGAT2 Ricinus communis (AAY16324.1)
SEQ ID NO: 4 полипептид DGAT2 Vernicia fordii (ABC94474.1)
SEQ ID NO: 5 полипептид DGAT2 Mortierella ramanniana (AAK84179.1)
SEQ ID NO: 6 полипептид DGAT2 Homo sapiens (Q96PD7.2)
SEQ ID NO: 7 полипептид DGAT2 Homo sapiens (Q58HT5.1)
SEQ ID NO: 8 полипептид DGAT2 Bos taurus (Q70VZ8.1)
SEQ ID NO: 9 полипептид DGAT2 Mus musculus (AAK84175.1)
SEQ ID NO: 10 YFP трипептид-консервативная DGAT2 и/или мотив последовательности MGAT1/2
SEQ ID NO: 11 HPHG тетрапептид-консервативная DGAT2 и/или мотив последовательности MGAT1/2
SEQ ID NO: 12 мотив последовательности EPHS тетрапептид-консервативной растительной DGAT2
SEQ ID NO: 13 RXGFX(K/R)XAXXXGXXX(L/V)VPXXXFG(E/Q) -длинный консервативный мотив последовательности DGAT2, который представляет собой часть предполагаемого глицерил фосфолипидного домена
SEQ ID NO: 14 FLXLXXXN - консервативный мотив последовательности DGAT2 и MGAT1/2 мыши, который представляет собой предполагаемый домен связывания с нейтральным липидом
SEQ ID NO: 15 домен plsC ацилтрансферазы (PF01553) GPAT
SEQ ID NO: 16 домен суперсемейства галогенокислоты дегалогеназы (ГКД)-подобной гидролазы (PF12710) GPAT
SEQ ID NO: 17 домен фосфосерин фосфатазы (PF00702). GPAT4-8 содержит N-концевой участок, гомологичный указанному домену.
SEQ ID NO: 18 Консервативная последовательность аминокислоты GPAT GDLVICPEGTTCREP
SEQ ID NO: 19 Консервативная последовательность аминокислоты GPAT/фосфатазы (Мотив I)
SEQ ID NO: 20 Консервативная последовательность аминокислоты GPAT/фосфатазы (Мотив III)
SEQ ID NO: 21 полипептид WRI1 Arabidopsis thaliana (A8MS57).
SEQ ID NO: 22 полипептид WRI1 Arabidopsis thaliana (Q6X5Y6)
SEQ ID NO: 23 полипептид WRI1 Arabidopsis lyrata подвид lyrata (XP_002876251.1)
SEQ ID NO: 24 полипептид WRI1 Brassica napus (ABD16282.1)
SEQ ID NO: 25полипептид WRI1Brassica napus (ADO16346.1)
SEQ ID NO: 26 полипептид WRI1 Glycine max (XP_003530370.1)
]SEQ ID NO: 27 полипептид WRI1 Jatropha curcas (AEO22131.1)
[SEQ ID NO: 28 полипептид WRI1 Ricinus communis (XP_002525305.1)
SEQ ID NO: 29 полипептид WRI1 Populus trichocarpa (XP_002316459.1)
SEQ ID NO:30 полипептид WRI1 Vitis vinifera (CBI29147.3)
SEQ ID NO: 31 полипептид WRI1 Brachypodium distachyon (XP_003578997.1)
SEQ ID NO: 32 полипептид WRI1 Hordeum vulgare подвид. vulgare (BAJ86627.1)
SEQ ID NO: 33 полипептид WRI1 Oryza sativa (EAY79792.1)
SEQ ID NO: 34 полипептид WRI1 Sorghum bicolor (XP_002450194.1)
SEQ ID NO: 35 полипептид WRI1 Zea mays (ACG32367.1)
SEQ ID NO: 36 полипептид WRI1 Brachypodium distachyon (XP_003561189.1)
SEQ ID NO: 37 полипептид WRI1 Brachypodium sylvaticum (ABL85061.1)
SEQ ID NO: 38 полипептид WRI1 Oryza sativa (BAD68417.1)
SEQ ID NO: 39 полипептид WRI1 Sorghum bicolor (XP_002437819.1)
SEQ ID NO: 40 полипептид WRI1 Sorghum bicolor (XP_002441444.1)
SEQ ID NO: 41 полипептид WRI1 Glycine max (XP_003530686.1)
SEQ ID NO: 42 полипептид WRI1 Glycine max (XP_003553203,1)
SEQ ID NO: 43 полипептид WRI1 Populus trichocarpa (XP_002315794.1)
SEQ ID NO: 44 полипептид WRI1 Vitis vinifera (XP_002270149.1)
SEQ ID NO: 45 полипептид WRI1 Glycine max (XP_003533548.1)
SEQ ID NO: 46 полипептид WRI1 Glycine max (XP_003551723.1)
SEQ ID NO: 47 полипептид WRI1 Medicago truncatula (XP_003621117.1)
SEQ ID NO: 48 полипептид WRI1 Populus trichocarpa (XP_002323836.1)
SEQ ID NO: 49 полипептид WRI1 Ricinus communis (XP_002517474.1)
SEQ ID NO: 50 полипептид WRI1 Vitis vinifera (CAN79925.1)
SEQ ID NO: 51 полипептид WRI1 Brachypodium distachyon (XP_003572236.1)
SEQ ID NO: 52 полипептид WRI1 Oryza sativa (BAD10030.1)
SEQ ID NO: 53 полипептид WRI1 Sorghum bicolor (XP_002444429.1)
SEQ ID NO: 54 полипептид WRI1 Zea mays (NP_001170359.1)
SEQ ID NO: 55 полипептид WRI1 Arabidopsis lyrata подвид lyrata (XP_002889265.1)
SEQ ID NO: 56 полипептид WRI1 Arabidopsis thaliana (AAF68121.1)
SEQ ID NO: 57 полипептид WRI1 Arabidopsis thaliana (NP_178088.2)
SEQ ID NO: 55 полипептид WRI1 Arabidopsis lyrata подвид lyrata (XP_002889265.1)
SEQ ID NO: 59 полипептид WRI1 Thellungiella halophila (BAJ33872.1)
SEQ ID NO: 60 полипептид WRI1 Arabidopsis thaliana (NP_563990.1)
SEQ ID NO: 61 полипептид WRI1Glycine max (XP_003530350.1)
SEQ ID NO: 62 полипептид WRI1 Brachypodium distachyon (XP_003578142.1)
SEQ ID NO: 63 полипептид WRI1 Oryza sativa (EAZ09147.1)
SEQ ID NO: 64 полипептид WRI1 Sorghum bicolor (XP_002460236.1)
SEQ ID NO: 65 полипептид WRI1 Zea mays (NP_001146338.1)
SEQ ID NO: 66 полипептид WRI1 Glycine max (XP_003519167.1)
SEQ ID NO: 67 полипептид WRI1 Glycine max (XP_003550676.1)
SEQ ID NO: 68 полипептид WRI1 Medicago truncatula (XP_003610261.1)
SEQ ID NO: 69 полипептид WRI1 Glycine max (XP_003524030.1)
SEQ ID NO: 70 полипептид WRI1 Glycine max (XP_003525949.1)
SEQ ID NO: 71 полипептид WRI1 Populus trichocarpa (XP_002325111.1)
SEQ ID NO: 72 полипептид WRI1 Vitis vinifera (CBI36586.3)
SEQ ID NO: 73 полипептид WRI1 Vitis vinifera (XP_002273046.2)
SEQ ID NO: 74 полипептид WRI1 Populus trichocarpa (XP_002303866.1)
SEQ ID NO: 75 полипептид WRI1 Vitis vinifera (CBI25261.3)
SEQ ID NO: 76 Sorbi-WRL1
SEQ ID NO: 77 Lupan-WRL1
SEQ ID NO: 78 Ricco-WRL1
SEQ ID NO: 79 полипептид WRI1 Lupin angustifolius
SEQ ID NO: 80 полипептид DGAT1 Aspergillus fumigatus (XP_755172.1)
SEQ ID NO: 81 полипептид DGAT1 Ricinus communis (AAR11479.1)
SEQ ID NO: 82 полипептид DGAT1 Vernicia fordii (ABC94472.1)
SEQ ID NO: 83 полипептид DGAT1 Vernonia galamensis (ABV21945.1)
SEQ ID NO: 84 полипептид DGAT1 Vernonia galamensis (ABV21946.1)
SEQ ID NO: 85 полипептид DGAT1 Euonymus alatus (AAV31083.1)
SEQ ID NO: 86 полипептид DGAT1 Caenorhabditis elegans (AAF82410.1)
SEQ ID NO: 87 полипептид DGAT1 Rattus norvegicus (NP_445889.1)
SEQ ID NO: 88 полипептид DGAT1 Homo sapiens (NP_036211.2)
SEQ ID NO: 89 мотив WRI1 (R G V T/S R H R W T G R)
SEQ ID NO: 90 мотив WRI1 (F/Y E A H L W D K)
SEQ ID NO: 91 мотив WRI1 (D L A A L K Y W G)
SEQ ID NO: 92 мотив WRI1 (S X G F S/A R G X)
SEQ ID NO: 93 мотив WRI1 (H H H/Q N G R/K W E A R I G R/K V)
SEQ ID NO: 94 мотив WRI1 (Q E E A A A X Y D)
SEQ ID NO: 95 полипептид олеозин Brassica napus (CAA57545.1)
SEQ ID NO: 96 полипептид олеозин S1-1Brassica napus (ACG69504.1)
SEQ ID NO: 97 полипептид олеозин S2-1 Brassica napus (ACG69503.1)
SEQ ID NO: 98 полипептид олеозин S3-1 Brassica napus (ACG69513.1)
SEQ ID NO: 99 полипептид олеозин S4-1 Brassica napus (ACG69507.1)
SEQ ID NO: 100 полипептид олеозин S5-1 Brassica napus (ACG69511.1)
SEQ ID NO: 101 полипептид олеозин 1 Arachis hypogaea (AAZ20276.1)
SEQ ID NO: 102 полипептид олеозин 2 Arachis hypogaea (AAU21500.1)
SEQ ID NO: 103 полипептид олеозин 3 Arachis hypogaea (AAU21501.1)
SEQ ID NO: 104 полипептид олеозин 5 Arachis hypogaea (ABC96763.1)
SEQ ID NO: 105 полипептид олеозин 1 Ricinus communis (EEF40948.1)
SEQ ID NO: 106 полипептид олеозин 2 Ricinus communis (EEF51616.1)
SEQ ID NO: 107 полипептид олеозин Glycine max изоформа a (P29530.2)
SEQ ID NO: 108 полипептид олеозин Glycine max изоформа b (P29531.1)
SEQ ID NO: 109 полипептид олеозин Linum usitatissimum низкомолекулярная изоформа (ABB01622.1)
SEQ ID NO: 110 последовательность аминокислоты полипептида олеозин Linum usitatissimum высокомолекулярная изоформа (ABB01624.1)
SEQ ID NO: 111 полипептид олеозин Helianthus annuus (CAA44224.1)
SEQ ID NO: 112 полипептид олеозин Zea mays (NP_001105338.1)
SEQ ID NO: 113 полипептид стеролеозин Brassica napus (ABM30178.1)
SEQ ID NO: 114 полипептид стеролеозин SLO1-1 Brassica napus (ACG69522.1)
SEQ ID NO: 115 полипептид стеролеозин SLO2-1 Brassica napus (ACG69525.1)
SEQ ID NO: 116 полипептид стеролеозин Sesamum indicum (AAL13315.1)
SEQ ID NO: 117 полипептид стеролеозин Zea mays (NP_001152614.1)
SEQ ID NO: 118 полипептид калеозин CLO-1 Brassica napus (ACG69529.1)
SEQ ID NO: 119 полипептид калеозин CLO-3 Brassica napus (ACG69527.1)
SEQ ID NO: 120 полипептид калеозин Sesamum indicum (AAF13743.1)
SEQ ID NO: 121 полипептид калеозин Zea mays (NP_001151906.1)
SEQ ID NO: 122 последовательность Т-ДНК pJP3502 (вставленная в геном)
SEQ ID NO: 123 последовательность вектора pJP3507
SEQ ID NO: 124 последовательность линкера
SEQ ID NO: 125 частичная последовательность CGI-58 Nicotiana benthamiana, выбранная для сайленсинга шпРНКи (pTV46)
SEQ ID NO: 126 частичная последовательность АГФазы N. tabacum, выбранная для сайленсинга шпРНКи (pTV35)
SEQ ID NO: 127 мотив липазы GXSXG
SEQ ID NO: 128 мотив ацилтрансферазы HX(4)D
SEQ ID NO: 129 вероятный мотив связывания с липидом VX(3)HGF
SEQ ID NO: 130 полинуклеотид CGi58 Arabidopsis thaliana (NM_118548.1)
SEQ ID NO: 131 полинуклеотид CGi58 Brachypodium distachyon (XM_003578402.1)
SEQ ID NO: 132 полинуклеотид CGi58 Glycine max (XM_003523590.1)
SEQ ID NO: 133 полинуклеотид CGi58 Zea mays (NM_001155541.1)
SEQ ID NO: 134 полинуклеотид CGi58 Sorghum bicolor (XM_002460493.1)
SEQ ID NO: 135 полинуклеотид CGi58 Ricinus communis (XM_002510439.1)
SEQ ID NO: 136 полинуклеотид CGi58 Medicago truncatula (XM_003603685.1)
SEQ ID NO: 137 полинуклеотид LEC2 Arabidopsis thaliana (NM_102595.2)
SEQ ID NO: 138 полинуклеотид LEC2 Medicago truncatula (X60387.1)
SEQ ID NO: 139 полинуклеотид LEC2 Brassica napus (HM370539.1)
SEQ ID NO: 140 полинуклеотид BBM Arabidopsis thaliana (NM_121749.2)
SEQ ID NO: 141 полинуклеотид BBM Medicago truncatula (AY899909.1)
SEQ ID NO: 142 полипептид LEC2 Arabidopsis thaliana (NP_564304.1)
SEQ ID NO: 143 полипептид LEC2 Medicago truncatula (CAA42938.1)
SEQ ID NO: 144 полипептид LEC2 Brassica napus (ADO16343.1)
SEQ ID NO: 145 полипептид BBM Arabidopsis thaliana (NP_197245.2)
SEQ ID NO: 146 полипептид BBM Medicago truncatula (AAW82334.1)
SEQ ID NO: 147 индуцибельный промотор alcA Aspergilus niger
SEQ ID NO: 148 индуктор AlcR, который активирует промотор AlcA в присутствии этанола
SEQ ID NO: 149 LEC1 Arabidopsis thaliana (AAC39488)
SEQ ID NO: 150 LEC1 Arabidopsis lyrata (XP_002862657)
SEQ ID NO: 151 LEC1 Brassica napus (ADF81045)
SEQ ID NO: 152 LEC1 Ricinus communis (XP_002522740)
SEQ ID NO: 153 LEC1 Glycine max (XP_006582823)
SEQ ID NO: 154 LEC1 Medicago truncatula (AFK49653)
SEQ ID NO: 155 LEC1 Zea mays (AAK95562)
SEQ ID NO: 156 LEC1 Arachis hypogaea (ADC33213)
SEQ ID NO: 157 LEC1-подобный Arabidopsis thaliana (AAN15924)
SEQ ID NO: 158 LEC1-подобный Brassica napus (AHI94922)
SEQ ID NO: 159 LEC1-подобный Phaseolus coccineus (AAN01148)
SEQ ID NO: 160 FUS3 Arabidopsis thaliana (AAC35247)
SEQ ID NO: 161 FUS3 Brassica napus
SEQ ID NO: 162 FUS3Medicago truncatula
SEQ ID NO: 163 последовательность кДНК SDP1 Arabidopsis thaliana, номер доступа NM_120486, 3275 нуклеотидов (нт)
SEQ ID NO: 164 кДНК SDP1 Brassica napus номер доступа GN078290
SEQ ID NO: 165 кДНК SDP1 Brachypodium distachyon, 2670 нуклеотидов (нт)
SEQ ID NO: 166 кДНК SDP1 Populus trichocarpa, 3884 нт
SEQ ID NO: 167 кДНК SDP1 Medicago truncatula; XM_003591377; 2490 нт
SEQ ID NO: 168 кДНК SDP1Glycine max XM_003521103; 2783 нт
SEQ ID NO: 169 кДНК SDP1Sorghum bicolor XM_002458486; 2724 нт
SEQ ID NO: 170 кДНК SDP1Zea mays, NM_001175206; 2985 нт
SEQ ID NO: 171 кДНК SDP1Physcomitrella patens, XM_001758117; 1998 нт
SEQ ID NO: 172 кДНК SDP1 Hordeum vulgare, AK372092; 3439 нт
SEQ ID NO: 173 кДНК SDP1Nicotiana benthamiana, Nbv5tr6404201
SEQ ID NO: 174 участок кДНК SDP1 Nicotiana benthamiana, являющийся мишенью для сайленсинга шпРНКи
SEQ ID NO: 175 промотор гена SDP1 Arabidopsis thaliana, 1,5 тысяч пар оснований
SEQ ID NO: 176 нуклеотидная последовательность, комплементарная гену pSSU-Олеозина в Т-ДНК pJP3502. В порядке (комплементарный последовательности): терминатор лектина Glycine max 348 нт, 3' экзон 255 нт, интрон UBQ10 304 нт, 5' экзон 213 нт, промотор SSU 1751 нт
SEQ ID NO: 177 кДНК пластидной GPAT Arabidopsis thaliana, NM_179407
SEQ ID NO: 178 полипептид пластидной GPAT Arabidopsis thaliana, NM_179407
SEQ ID NO: 179 кДНК пластидной GPAT Populus trichocarpa, XP_006368351
SEQ ID NO: 180 кДНК пластидной GPAT Jatropha curcas, ACR61638
SEQ ID NO: 181 кДНК пластидной GPAT Ricinus communis, XP_002518993
SEQ ID NO: 182 кДНК пластидной GPAT Helianthus annuus, ADV16382
SEQ ID NO: 183 кДНК пластидной GPAT Medicago truncatula, XP_003612801
SEQ ID NO: 184 кДНК пластидной GPAT Glycine max, XP_003516958
SEQ ID NO: 185 кДНК пластидной GPAT Carthamus tinctorius, CAHG3PACTR
SEQ ID NO: 186 кДНК пластидной GPAT Solanum tuberosum, XP_006352898
SEQ ID NO: 187 кДНК пластидной GPAT Oryza sativa, японской, NM_001072027
SEQ ID NO: 188 кДНК пластидной GPAT Sorghum bicolor, XM_002467381
SEQ ID NO: 189 кДНК пластидной GPAT Zea mays, NM_001158637
SEQ ID NO: 190 кДНК пластидной GPAT Hordeum vulgare, AK371419
SEQ ID NO: 191 кДНК пластидной GPAT Physcomitrella patens, XM_001771247
SEQ ID NO: 192 кДНК пластидной GPAT Chlamydomonas reinhardtii, XM_001694925
SEQ ID NO: 193 14:0-АПБ тиоэстераза Cinnamomum camphora (Cinca-TE), хлоропластная, 382 аминокислоты (ак), (номер доступа Q39473.1)
SEQ ID NO: 194 ацил-АПБ тиоэстераза FatB1 Cocos nucifera (Cocnu-TE1; 417 ак, номер доступа AEM72519.1
SEQ ID NO: 195 ацил-АПБ тиоэстераза FatB2 Cocos nucifera (Cocnu-TE2; 423 ак, номер доступа AEM72520.1)
SEQ ID NO: 196 ацил-АПБ тиоэстераза FatB3 Cocos nucifera (Cocnu-TE3; 414 ак, номер доступа AEM72521.1)
SEQ ID NO: 197 ацил-(АПБ) тиоэстераза типа B Cuphea lanceolata (Cupla-TE, 419 ак, номер доступа CAB60830.1)
SEQ ID NO: 198 FatB1 Cuphea viscosissima (Cupvi-TE; 419 ак, номер доступа AEM72522.1)
SEQ ID NO: 199 12:0-АПБ тиоэстераза Umbellularia californica (лауроил-ацилпереносящего белка тиоэстераза) (Umbca-TE, 382 ак; номер доступа Q41635.1)
SEQ ID NO: 200 LPAAT Cocos nucifera (Cocnu-LPAAT, 308 ак, номер доступа Q42670.1)
SEQ ID NO: 201 пластидная LPAAT1 Arabidopsis thaliana (Arath-PLPAAT; 356 ак, номер доступа AEE85783.1)
SEQ ID NO: 202 FATA1Arabidopsis thaliana
SEQ ID NO: 203 FATA2 Arabidopsis thaliana
SEQ ID NO: 204 FATB Arabidopsis thaliana
SEQ ID NO: 205 WRI3 Arabidopsis thaliana
SEQ ID NO: 206 WRI4 Arabidopsis thaliana
SEQ ID NO: 207 WRI1Avena sativa
SEQ ID NO: 208 WRI1 Sorghum bicolor
SEQ ID NO: 209 WRI1 Zea mays
SEQ ID NO: 210 WRI1Triadica sebifera
SEQ ID NO: 211 последовательность промотора пататина B33 S. tuberosum
SEQ ID NO 212-215 и 245-254 олигонуклеотидные праймеры
SEQ ID NO: 216 участок промотора SEE1 Z. mays (1970 нт из номера доступа AJ494982)
SEQ ID NO: 217 последовательность промотора AlSAP A. littoralis, номер доступа DQ885219
SEQ ID NO: 218 последовательность промотора ArRolC A. rhizogenes, номер доступа DQ160187
SEQ ID NO: 219 конструкция шпРНКи, содержащая фрагмент 732 пар оснований (по) пластидной GPAT N. benthamiana
SEQ ID NO: 220 DGAT1 Elaeis guineensis (масличная пальма)
SEQ ID NO: 221 MYB73 G. max, номер доступа ABH02868
SEQ ID NO: 222 bZIP53 A. thaliana, номер доступа AAM14360
SEQ ID NO: 223 AGL15 A. thaliana, номер доступа NP_196883
SEQ ID NO: 224 MYB118 A. thaliana, номер доступа AAS58517
SEQ ID NO: 225 MYB115 A. thaliana, номер доступа AAS10103
SEQ ID NO: 226 TANMEI A. thaliana, номер доступа BAE44475
SEQ ID NO: 227 WUS A. thaliana, номер доступа NP_565429
SEQ ID NO: 228 GFR2a1 B. napus, номер доступа AFB74090
SEQ ID NO: 229 GFR2a2 B. napus, номер доступа AFB74089
SEQ ID NO: 230 PHR1 A. thaliana, номер доступа AAN72198
SEQ ID NO: 231 фрагмент TGD1 N. benthamiana
SEQ ID NO: 232 аминокислоты SDP1 картофеля
SEQ ID NO: 233 нуклеотидная последовательность SDP1 картофеля
SEQ ID NO: 234 малая субъединица АГФ-азы картофеля
SEQ ID NO: 235 нуклеотидная последовательность малой субъединицы АГФ-азы картофеля
SEQ ID NO: 236 нуклеотидная последовательность LDAP-1 Sapium sebiferum
SEQ ID NO: 237 аминокислотная последовательность LDAP-1Sapium sebiferum
SEQ ID NO: 238 нуклеотидная последовательность LDAP-2 Sapium sebiferum
SEQ ID NO: 239 аминокислотная последовательность LDAP-2 Sapium sebiferum
SEQ ID NO: 240 нуклеотидная последовательность LDAP-3 Sapium sebiferum
SEQ ID NO: 241 аминокислотная последовательность LDAP-3 Sapium sebiferum
SEQ ID NO: 242 SDP1 S. bicolor (номер доступа XM_002463620)
SEQ ID NO: 243 нуклеотидная последовательность SDP1 T. aestivum (номер доступа AK334547)
SEQ ID NO: 244 фрагмент SDP1 шпРНКи S. bicolor
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Общие методы
Если конкретно не определено иное, все технические и научные термины, используемые в настоящем документе, следует интерпретировать как имеющие такое же значение, которое им обычно придает средний специалист в данной области (например, культуры клеток, молекулярной генетики, биологии растений, клеточной биологии, химии белка, химии липида и жирной кислоты, получения биотоплива и биохимии).
Если не указано иное, методы химии рекомбинантного белка, культуры клетки и иммунологические методы, применяемые в настоящем изобретении, представляют собой стандартные методики, хорошо известные специалистам из уровня техники. Такие методы описаны и объяснены в литературных источниках, например, J. Perbal, A Practical Guide to Molecular Cloning, John Wiley and Sons (1984), J. Sambrook с соавт., Molecular Cloning: A Laboratory Manual, Cold Spring Harbour Laboratory Press (1989), T.A. Brown (редактор), Essential Molecular Biology: A Practical Approach, Volumes 1 and 2, IRL Press (1991), D.M. Glover and B.D. Glover and B.D. Hames (редакторы), DNA Cloning: A Practical Approach, Volumes 1-4, IRL Press (1995 и 1996), F.M. Ausubel с соавт. (редакторы), Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience (1988, включая все обновления, изданные до настоящего времени), Ed Harlow and David Lane (редакторы) Antibodies: A Laboratory Manual, Cold Spring Harbour Laboratory, (1988), и J.E. Coligan с соавт. (редакторы), Current Protocols in Immunology, John Wiley & Sons (включая все обновления, изданные до настоящего времени).
Некоторые определения
Термин «трансгенный организм, не относящийся к человеческому роду» обозначает, например, цельное растение, водоросль, не относящиеся к человеческому роду животное или организм, пригодный для ферментации, такой как дрожжи или гриб, содержащий один или более экзогенных полинуклеотидов (трансген) или полипептидов. В варианте реализации трансгенный организм, не относящийся к человеческому роду, не является животным или его частью. В одном варианте реализации трансгенный организм, не относящийся к человеческому роду, представляет собой фототрофный организм (например, растение или водоросль), способный к получению из солнечного света энергии для синтеза питательных органических соединений.
Термин «экзогенный» в контексте полинуклеотида или полипептида обозначает полинуклеотид или полипептид, присутствующий в клетке, которая от природы не содержит полинуклеотида или полипептида. Такая клетка в настоящем документе называется «рекомбинантной клеткой» или «трансгенной клеткой». В варианте реализации экзогенный полинуклеотид или полипептид происходит из другого рода относительно клетки, содержащей экзогенный полинуклеотид или полипептид. В другом варианте реализации экзогенный полинуклеотид или полипептид происходит от другого вида. В одном варианте реализации экзогенный полинуклеотид или полипептид экспрессируется в растении-хозяине или растительной клетке, и экзогенный полинуклеотид или полипептид происходит от другого вида или рода. Экзогенный полинуклеотид или полипептид может быть неприродным, например, таким как синтетическая молекула ДНК, которая получена способами рекомбинации ДНК. Молекула ДНК может, часто предпочтительно, содержать кодирующий белок участок, который является кодон-оптимизированным для экспрессии в клетке, с получением таким образом полипептида, который содержит такую же последовательность аминокислот, что и природный полипептид, даже если нуклеотидная последовательность кодирующего белок участка имеет неприродное происхождение. Экзогенный полинуклеотид может кодировать или экзогенный полипептид может представлять собой: диацилглицерил ацилтрансферазу (DGAT), например, DGAT1 или DGAT2, фактор транскрипции Wrinkled 1 (WRI1), на ПМТ, таком как олеозин или, предпочтительно, БСКЛ, тиоэстераза жирной кислоты, такая как полипептид FATA или FATB или полипептид супрессора сайленсинга.
В настоящем документе термин «экстрагированный липид» обозначает состав, экстрагированный из трансгенного организма или его части, который содержит по меньшей мере 60% (масс./масс.) липида.
В настоящем документе термин «неполярный липид» обозначает жирные кислоты и их производные, растворимые в органических растворителях, но не растворимые в воде. Жирные кислоты могут быть жирными кислотами в свободной и/или этерифицированной форме. Примеры этерифицированных форм включают, без ограничений, триацилглицерид (ТАГ), диацилглицерид (ДАГ), моноацилглицерид (МАГ). Неполярные липиды также включают стероиды, эфиры стероидов и эфиры восков. Неполярные липиды также известны, как «нейтральные липиды». При комнатной температуре неполярный липид обычно представляет собой жидкость. Предпочтительно, неполярный липид в основном (> 50%) содержит жирные кислоты, длина которых составляет по меньшей мере 16 атомов углерода. Более предпочтительно, по меньшей мере 50% общих жирных кислот в неполярном липиде представляют собой C18 жирные кислоты, например, олеиновую кислоту. Предпочтительно, по меньшей мере 5% от общих жирных кислот в неполярных липидах представляют собой жирные кислоты C12 или C14 или те и другие. В варианте реализации по меньшей мере 50%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 91%, более предпочтительно по меньшей мере 92%, более предпочтительно по меньшей мере 93%, более предпочтительно по меньшей мере 94%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 96%, более предпочтительно по меньшей мере 97%, более предпочтительно по меньшей мере 98%, более предпочтительно по меньшей мере 99% жирных кислот в неполярном липиде по изобретению могут представлять собой ТАГ. Неполярный липид может быть дополнительно очищен или обработан, например гидролизом с сильным основанием, чтобы высвободить свободную жирную кислоту, или фракционированием, дистилляцией, или подобным методом. Неполярный липид может присутствовать в или быть получен из частей растения, например, семени, листьев, клубней, свеклы или плода, из рекомбинантных клеток или из организмов, не относящихся к человеческому роду, например, дрожжей. Неполярный липид по изобретению может образовывать часть «масла из семян», если он получен из семени.
Концентрация свободного и этерифицированного стероида (например, ситостерола, кампэстрола, стигмастерола, брассикастерола, Δ5-авенастерола, ситостанола, кампестанола и холестерина) в экстрагированном липиде может быть такой, как описано в Phillips с соавт. (2002). Стероиды в растительных маслах присутствуют, как свободные спирты, эфиры с жирными кислотами (этерифицированные стероиды), гликозиды стероидов и ацилированные гликозиды стероидов. Концентрации стероидов в природных растительных маслах (масло из семян) варьируют до максимального содержания приблизительно 1100 мг/100 г. Гидрогенизированное пальмовое масло содержит одну из самых низких концентраций для природных масел, приблизительно 60 мг/100 г. Извлеченные или экстрагированные масла из семян по изобретению предпочтительно содержат от приблизительно 100 до приблизительно 1000 мг общего стероида/100 г масла. В случае применения в качестве продукта питания или корма, предпочтительно, стероиды присутствуют, главным образом, в свободных или этерифицированных формах вместо гликозилированных форм. В маслах из семян по настоящему изобретению, предпочтительно по меньшей мере 50% стероидов в маслах присутствуют, как этерифицированные стероиды, за исключением соевого масла, которое содержит около 25% этерифицированных стероидов. Масло из семени рапса и рапса по изобретению предпочтительно содержат от приблизительно 500 до приблизительно 800 мг общего стероида/100 г, причем основным стероидом является ситостерол, и вторым по распространенности - кампэстрол. Кукурузное масло из семени по изобретению предпочтительно содержит от приблизительно 600 до приблизительно 800 мг общего стероида/100 г, причем основным стероидом является ситостерол. Соевое масло из семени по изобретению предпочтительно содержит от приблизительно 150 до приблизительно 350 мг общего стероида/100 г, причем основным стероидом является ситостерол, и вторым по распространенности - стигмастерол, и, при этом, содержание свободного стероида выше, чем этерицифированного стероида. Хлопковое масло по изобретению предпочтительно содержит от приблизительно 200 до приблизительно 350 мг общего стероида/100 г, причем основным стероидом является ситостерол. Кокосовое масло и пальмовое масло по изобретению предпочтительно содержат от приблизительно 50 до приблизительно 100 мг общего стероида/100 г, причем основным стероидом является ситостерол. Сафлоровое масло из семени по изобретению предпочтительно содержит от приблизительно 150 до приблизительно 250 мг общего стероида/100 г, причем основным стероидом является ситостерол. Арахисовое масло по изобретению предпочтительно содержит от приблизительно 100 до приблизительно 200 мг общего стероида/100 г, причем основным стероидом является ситостерол. Кунжутное масло из семени по изобретению предпочтительно содержит от приблизительно 400 до приблизительно 600 мг общего стероида/100 г, причем основным стероидом является ситостерол. Масло подсолнечника из семени по изобретению предпочтительно содержит от приблизительно 200 до 400 мг общего стероида/100 г, причем основным стероидом является ситостерол. Масла, полученные из вегетативных частей растения по изобретению, предпочтительно содержат менее 200 мг общего стероида/100 г, более предпочтительно менее 100 мг общего стероида/100 г, и наиболее предпочтительно менее 50 мг общего стероида/100 г, причем большая часть стероидов присутствует в форме свободных стероидов.
В настоящем документе термин «масло из семян» обозначает состав, полученный из семени/зерна растения, содержащего по меньшей мере 60% (масс./масс.) липидов, или такой, который можно получить из семени/зерна, если масло из семян все еще присутствует в семени/зерне. Таким образом, масло из семян по изобретению включает масло из семян, которое присутствует в семени/зерне или его части, а также масло из семян, которое экстрагировано из семени/зерна. Масло из семян предпочтительно является экстрагированным маслом из семян. Масло из семян при комнатной температуре обычно представляет собой жидкость. Предпочтительно, общее содержание жирных кислот (ОЖК) в масле из семян в основном (> 50%) включает жирные кислоты, длина которых составляет по меньшей мере 16 атомов углерода. Более предпочтительно, по меньшей мере 50% общих жирных кислот в масле из семян представляют собой C18 жирные кислоты например, олеиновую кислоту. Жирные кислоты обычно присутствуют в этерифицированной форме, например, такой как ТАГ, ДАГ, ацил-КоА или фосфолипид. Жирные кислоты могут быть жирными кислотами в свободной и/или этерифицированной форме. В варианте реализации изобретения по меньшей мере 50%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 91%, более предпочтительно по меньшей мере 92%, более предпочтительно по меньшей мере 93%, более предпочтительно по меньшей мере 94%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 96%, более предпочтительно по меньшей мере 97%, более предпочтительно по меньшей мере 98%, более предпочтительно по меньшей мере 99% жирных кислот в масле из семян по изобретению могут быть найдены в виде ТАГ. В варианте реализации масло из семян по изобретению представляет собой «в существенной мере очищенное» или «очищенное» масло, которое отделено от одного или более других липидов, нуклеиновых кислот, полипептидов или других загрязняющих молекул, с которыми оно связано в семени или в неочищенном экстракте. Предпочтительно, в существенной мере очищенное масло из семян по меньшей мере на 60% свободно, более предпочтительно по меньшей мере на 75% свободно, и более предпочтительно, по меньшей мере на 90% свободно от других компонентов, с которыми оно связано в семени или экстракте. Масло из семян по изобретению может дополнительно содержать молекулы, не относящиеся к классу жирных кислот, например, без ограничений, стероиды. В варианте реализации масло из семян представляет собой масло рапса (вид Brassica, такой как Brassica carinata, Brassica juncea, Brassica napobrassica, Brassica napus), горчичное масло (Brassica juncea), масло других видов Brassica (например, Brassica napobrassica, Brassica camelina), масло подсолнечника (вид Helianthus, такой как Helianthus annuus), льняное масло (Linum usitatissimum), соевое масло (Glycine max), сафлоровое масло (Carthamus tinctorius), кукурузное масло (Zea mays), масло табака (вид Nicotiana, такой как Nicotiana tabacum или Nicotiana benthamiana), арахисовое масло (Arachis hypogaea), пальмовое масло (Elaeis guineensis), хлопковое масло (Gossypium hirsutum), кокосовое масло (Cocos nucifera), масло авокадо (Persea americana), оливковое масло (Olea europaea), масло кешью (Anacardium occidentale), масло макадамии (Macadamia intergrifolia), миндальное масло (Prunus amygdalus), масло из семян овса (Avena sativa), рисовое масло (вид Oryza, такой как Oryza sativa и Oryza glaberrima), масло из семян Arabidopsis (Arabidopsis thaliana) или масло из семян Acrocomia aculeata (макаубская пальма), Aracinis hypogaea (арахис), Astrocaryum murumuru (мурумуру), Astrocaryum vulgare (тукума), Attalea geraensis (атталия), Attalea humilis (американская масличная пальма), Attalea oleifera (андайя), Attalea phalerata (урикури), Attalea speciosa (бабассу), Beta vulgaris (сахарная свекла), Camelina sativa (ложный лен), Caryocar brasiliense (пекви), Crambe abyssinica (абиссинская капуста), Cucumis melo (дыня), Hordeum vulgare (ячмень), Jatropha curcas (лечебный орех), Joannesia princeps (лещина арара), Licania rigida (ойтика), Lupinus angustifolius (люпин), Mauritia flexuosa (пальма бурити), Maximiliana maripa (пальма инайя), вид Miscanthus, такой как Miscanthus x giganteus и Miscanthus sinensis, Oenocarpus bacaba (бакаба-до-азейте), Oenocarpus bataua (патайя), Oenocarpus distichus (бакаба-де-лек), Panicum virgatum (просо прутьевидное), Paraqueiba paraensis (марь), Persea amencana (авокадо), Pongamia pinnata (индийский бук), Populus trichocarpa, Ricinus communis (клещевина), вид Saccharum (сахарный тростник), Sesamum indicum (кунжут), Solanum tuberosum (картофель), вид Sorghum такой как Sorghum bicolor, Sorghum vulgare, Theobroma grandiforum (какао) вид Trifolium, Trithrinax brasiliensis (бразильская игольчатая пальма) и вид Triticum. (пшеница) такой как Triticum aestivum. Масло из семян может быть экстрагировано из семени/зерна любым способом, известным из уровня техники. Такие способы обычно включают экстракцию неполярными растворителями, такими как диэтиловый эфир, петролейный эфир, смеси хлороформ/метанол или бутанол, в общем, ассоциирующимися с первым дроблением семян. Липиды, связанные с крахмалом в зерне, могут быть экстрагированы с помощью насыщенного водой бутанола. Масло из семян может быть «дегуммировано» способами, известными из уровня техники, чтобы удалить полисахариды, или обработано другими способами, чтобы удалить загрязняющие вещества или повысить чистоту, стабильность, или улучшить цвет. ТАГ и другие эфиры в масле из семян могут быть гидролизованы с целью высвобождения свободных жирных кислот, или масло из семян может быть гидрогенизировано, обработано химическим или ферментным способом, как известно из уровня техники.
В настоящем документе термин «жирная кислота» обозначает карбоновую кислоту с алифатическим хвостом размером по меньшей мере 8 атомов углерода, насыщенным или ненасыщенным. Предпочтительные жирные кислоты содержат углеродно-углеродную цепь длиной по меньшей мере 12 атомов углерода. Большинство природных жирных кислот содержат четное количество атомов углерода, поскольку в их биосинтезе участвует ацетат, содержащий два атома углерода. Жирные кислоты могут находиться в свободном состоянии (неэтерифицированные) или в этерифицированной форме, например, как часть ТАГ, ДАГ, МАГ, связи ацил-КоА (тиоэфир), связи ацил-ацилпереносящий белок (АПБ) или другой ковалентно связанной форме. Будучи ковалентно связанной в этерифицированной форме, жирная кислота в настоящем документе обозначается как группа «ацил». Жирная кислота может быть этерифицирована как фосфолипид, например, фосфатидилхолин (ФХ), фосфатидилэтаноламин, фосфатидилсерин, фосфатидилглицерил, фосфатидилинозитол или дифосфатидилглицерил. Насыщенные жирные кислоты не содержат кратных связей или других функциональных групп вдоль цепи. Термин «насыщенный» обозначает водород, в котором все атомы углерода (кроме карбоксильной группы [-COOH]) связаны с максимальным количеством атомов водорода. Иными словами, омега (ω) конец содержит 3 атома водорода (CH3-), и каждый атом углерода в цепи связан с 2 атомами водорода (-CH2-). Ненасыщенные жирные кислоты по форме сходны с насыщенными жирными кислотами, за исключением того, что одна или более алкеновых функциональных групп расположены вдоль цепи, причем каждый алкен заменяет часть одинарной связи «-CH2-CH2-» в цепи частью двойной связи «-CH=CH-» (т. е., двойная связь между двумя атомами углерода). Два последующих атома углерода в цепи, присоединенные к каждому из концов двойной связи, могут находиться в цис- или транс-конфигурации.
В настоящем документе термин "мононенасыщенная жирная кислота" или "МНЖК" обозначает жирную кислоту, содержащую по меньшей мере 12 атомов углерода в углеродной цепи и только одну алкеновую группу (двойная углерод-углеродная связь), которая может находиться в этерифицированной или неэтерифицированной (свободной) форме. В настоящем документе термин "полиненасыщенная жирная кислота" или "ПНЖК" обозначает жирную кислоту, содержащую по меньшей мере 12 атомов углерода в углеродной цепи и по меньшей мере две алкеновые группы (двойные углерод-углеродные связи), которая может находиться в этерифицированной или неэтерифицированной форме.
В настоящем документе жирная кислота со ʺсредней длиной цепиʺ, дополнительно обозначенная как ʺСЦЖКʺ, включает ацильную цепь длиной 8-14 атомов углерода. Ацильная цепь может быть модифицированной (например, она может содержать одну или более двойных связей, гидроксильных групп, эпоксигрупп и т.д) или немодифицированной (насыщенной). Данный термины включают по меньшей мере одну или более или все из каприловой кислоты (C8:0), каприновой кислоты (C10:0), лауриновой кислоты (C12:0) и миристиновой кислоты (C14:0).
«Моноацилглицерид» или «МАГ» представляет собой глицерид, в котором глицерин этерифицирован одной жирной кислотой. В настоящем документе МАГ содержит гидроксильную группу в положении sn-1/3 (также обозначена в настоящем документе как sn-1 МАГ или 1-МАГ или 1/3-МАГ) или sn-2(также обозначена в настоящем документе как 2-МАГ), и, таким образом, МАГ не содержит фосфорилированных молекул, таких как фосфатидиновая кислота (ФК) или фосфатидилхолин (ФХ). МАГ, таким образом, является компонентом нейтральных липидов в клетке.
«Диацилглицерид» или «ДАГ» представляет собой глицерид, в котором глицерин этерифицирован двумя жирными кислотами, которые могут быть одинаковыми или, предпочтительно, разными. В настоящем документе ДАГ содержит гидроксильную группу в положении sn-1,3 или sn-2, и, таким образом, ДАГ не содержит фосфорилированных молекул, таких как ФК или ФХ. ДАГ, таким образом, является компонентом нейтральных липидов в клетке. В пути Кеннеди синтеза ДАГ (Фиг. 1), прекурсор sn-глицерил-3-фосфат (G-3-P) этерифицируется двумя ацильными группами, каждая из которых поступает от сложного эфира жирной кислоты коэнзима A, при этом, первая реакция катализируется глицерин-3-фосфат ацилтрансферазой (GPAT) в положении sn-1 с образованием ЛизоФК, с последующей реакцией присоединения второго ацила в положении sn-2, катализированного ацилтрансферазой лизофосфатидиновой кислоты (LPAAT), с образованием фосфатидиновой кислоты (ФК). Данное промежуточное соединение затем дефосфорилируется PAP с образованием ДАГ. Кроме того, ДАГ может быть образован из ТАГ удалением ацильной группы под действием липазы или из ФХ по существу удалением головной группы холина любым из ферментов карнитинпальмитоилтрансфераза (КПТ), PLC или PLD (Фиг. 1).
«Триацилглицерид» или «ТАГ» представляет собой глицерид, в котором глицерин этерифицирован тремя жирными кислотами, которые могут быть одинаковыми (например, как в триолеине) или, чаще, разными. В пути Кеннеди синтеза ТАГ, ДАГ образуется, как изложено выше, а затем третья ацильная группа присоединяется к основе глицерина под действием DGAT. Альтернативные пути образования ТАГ включают катализированный ферментом PDAT (Фиг. 1) и путь MGAT, описанные в настоящем документе.
В настоящем документе термин "дикий тип" или его вариации обозначает вегетативную часть растения, клетку, семя или не относящийся к человеческому роду организм или его часть, например, клубень или свеклу, которые не были генетически модифицированы, например, введением экзогенного(ых) полинуклеотида(ов) по настоящему изобретению.
Термин "соответствующий" обозначает вегетативную часть растения, клетку, семя или не относящийся к человеческому роду организм или его часть (например, клубень или свекла), которые имеют такой же или подобный генетический фон, как и вегетативная часть растения, клетка, семя или не относящийся к человеческому роду организм или его часть по изобретению, но при этом они не были модифицированы, как описано в настоящем документе (например, вегетативная часть растения, клетка, семя или не относящийся к человеческому роду организм или его часть, в которых отсутствует экзогенный(ые) полинуклеотид(ы) и/или отсутствует генетическая(ие) модификация(и)). В предпочтительном варианте реализации изобретения соответствующая вегетативная часть растения, эукариотная клетка, семя или не относящийся к человеческому роду организм или его часть находятся в той же стадии развития, что и вегетативная часть растения, эукариотная клетка, семя или не относящийся к человеческому роду организм или его часть по изобретению. Например, если не относящийся к человеческому роду организм представляет собой цветущее растение, то соответствующее растение также предпочтительно находится в стадии цветения. Соответствующая вегетативная часть растения, эукариотная клетка, семя или не относящийся к человеческому роду организм или его часть может применяться в качестве контроля для сравнения уровней нуклеиновой кислоты или экспрессии белка или степени и природы модификации признака, например, выработки и/или содержания неполярных липидов, с вегетативной частью растения, эукариотной клеткой, семенем или не относящимся к человеческому роду организмом или его частью по изобретению, которые модифицированы, как описано в настоящем документе. Квалифицированный специалист в данной области с легкостью может определить подходящую "соответствующую" вегетативную часть растения, эукариотную клетку, семя или не относящийся к человеческому роду организм или его часть, ткань, орган или организм для такого сравнения.
В настоящем документе "по сравнению с" или ʺотносительноʺ обозначает сравнение уровней неполярного липида, общего содержания неполярных липидом, содержания жирных кислот или другого параметра вегетативной части растения, эукариотной клетки, семени, не относящегося к человеческому роду организма или его части (такой как клубень или свекла), экспрессирующих один или более экзогенных полинуклеотидов или экзогенных полипептидов, с вегетативной частью растения, эукариотной клеткой, семенем, не относящимся к человеческому роду организма или его частью, в которых отсутствуют один или больше экзогенных полинуклеотидов или полипептидов.
В настоящем документе "повышенная способность к выработке неполярных липидов" является относительным термином, который обозначает повышенное общее количество неполярного липида, выработанного вегетативной частью растения, эукариотной клеткой, семенем или не относящимся к человеческому роду организмом или его частью (например, клубень или свекла) по изобретению, относительно соответствующей вегетативной части растения, эукариотной клетки, семени или не относящегося к человеческому роду организма или его части. В одном варианте реализации изобретения повышено содержание ТАГ и/или полиненасыщенных жирных кислот или содержание олеиновой кислоты в общем содержании жирных кислот неполярного липида, или снижено содержание линоленовой кислоты в общем содержании жирных кислот неполярного липида, например, по меньшей мере на 2% в абсолютных величинах.
В настоящем документе каждый из ʺсинергииʺ, ʺсинергетическогоʺ, ʺдействуя синергетическиʺ и родственных терминов представляет собой сравнительный термин, который означает, что влияние комбинации элементов, присутствующих в клетке, растении или его части по изобретению, например, комбинации элементов A и B, превышает сумму влияния элементов по отдельности в соответствующих клетках, растениях или их частях, например, суммарное влияние A и влияние B. Если более двух элементов присутствуют в клетке, растении или его части, например, элементы A, B и C, это означает, что влияние комбинации всех элементов превышает сумму влияний индивидуальных элементов. В предпочтительном варианте реализации изобретения это означает, что влияние комбинации элементов A, B и C превышает сумму сочетанного влияния элементов A и B и влияния элемента C. В таком случае, может быть указано, что элемент C действует синергетически с элементами A и B. Необходимо понимать, что влияние измеряют в соответствующих клетках, растениях или их частях, например, выращенных в таких же условиях и находящихся в такой же стадии биологического развития.
В настоящем документе "прорастать со скоростью, в существенной мере такой же, как и соответствующее растение дикого типа" обозначает, что семя растения по изобретению относительно способно к прорастанию при сравнении с семенем растения дикого типа, не содержащего определенного(ых) экзогенного(ых) полинуклеотида(ов). Прорастание может быть измерено in vitro на питательной среде для культуры ткани или в грунте, как это происходит в поле. В одном варианте реализации изобретения количество семян, которые прорастают, например, при выращивании в оптимальных условиях теплицы для данного вида растений, составляет по меньшей мере 75%, более предпочтительно, по меньшей мере 90%, по сравнению с соответствующими семенами дикого типа. В другом варианте реализации изобретения семена, которые прорастают, например, при выращивании в оптимальных условиях теплицы для данного вида растений, дают саженцы, растущие со скоростью, которая, в среднем, составляет по меньшей мере 75%, более предпочтительно, по меньшей мере 90%, по сравнению с соответствующими растениями дикого типа. Это обозначается как ʺсила роста саженцевʺ. В варианте реализации изобретения скорость начального роста корней и роста побегов у саженцев по изобретению является по существу такой же, как у соответствующего саженца дикого типа, выращенного в таких же условиях. В варианте реализации изобретения биомасса листьев (сухая масса) растений по изобретению составляет по меньшей мере 80%, предпочтительно по меньшей мере 90% относительно биомассы листьев соответствующих растений дикого типа, выращенных в таких же условиях, предпочтительно в поле. В варианте реализации изобретения высота растений по изобретению составляет по меньшей мере 70%, предпочтительно по меньшей мере 80%, более предпочтительно, по меньшей мере 90% высоты растения относительно соответствующего растения дикого типа, выращенного в таких же условиях, предпочтительно в поле и предпочтительно в состоянии зрелости.
В настоящем документе термин "экзогенный полинуклеотид, который подавляет выработку и/или активность эндогенного полипептида" или его вариации обозначают полинуклеотид, кодирующий молекулу РНК (например, кодирующий искусственную микроРНК (имиРНК) или шпилечную РНК-интерференцию (шпРНКи)), которая подавляет выработку и/или активность или непосредственно подавляет выработку и/или активность (например, представляет собой имиРНК или шпРНК, которая может быть доставлена, например, непосредственно в клетку) эндогенного полипептида, например, SDP1 ТАГ липазы, пластидной GPAT, пластидной LPAAT, полипептида TGD, АГФазы или дельта-12 десатуразы жирных кислот (FAD2), или проявляет комбинацию двух или более из указанных эффектов. Обычно, молекула РНК уменьшает экспрессию эндогенного гена, кодирующего полипептид.
В настоящем документе термин "в пересчете на массу" обозначает массу вещества (например, ТАГ, ДАГ, жирной кислоты) как процент от массы композиции, содержащей вещество (например, семя, лист). Например, если трансгенное семя содержит 25 мкг общих жирных кислот на 120 мкг массы семени, процент от общих жирных кислот в пересчете на массу составляет 20,8%.
В настоящем документе термин "в относительном выражении" обозначает параметр, например, количество вещества в композиции, содержащей вещество, в сравнении с параметром для соответствующей композиции, в процентном выражении. Например, уменьшение с 3 единиц до 2 единиц является уменьшением на 33% в относительном выражении.
В настоящем документе ʺпластидыʺ представляют собой органеллы в растениях, включая морские водоросли, которые являются местом выработки углеродных соединений на основе фотосинтеза, включая сахара, крахмал и жирные кислоты. Пластиды включают хлоропласты, содержащие хлорофилл и осуществляющие фотосинтез, этиопласты, которые являются прекурсорами хлоропластов, а также специализированные пластиды, такие как хромопласты, которые представляют собой окрашенные пластиды для синтеза и хранения пигментов, геронтопласты, которые руководят демонтажем фотосинтетических структур в ходе старения, амилопласты для синтеза и хранения крахмала, элайопласты для хранения липидов и протеинопласты для хранения и модификации белков.
В настоящем документе термин «биологическое топливо» обозначает любой вид топлива, обычно применяемый для выработки энергии машинным оборудованием, например, автомобилями, самолетами, катерами, большими грузовыми машинами или работающими на нефти двигателями, энергия которого происходит от биологической фиксации углерода. Биотопливо включает виды топлива, получаемые в результате превращения биомассы, а также твердой биомассы, жидких видов топлива и биогазов. Примеры биологического топлива включают биоспирты, биодизельное топливо, синтетическое дизельное топливо, вегетативное масло, биоэфиры, биогаз, синтетический газ, твердое биологическое топливо, полученное из водорослей топливо, биоводород, биометанол, 2,5-диметилфуран (ДМФ), биодиметиловый эфир (биоДМЭ), дизельное топливо Фишера-Тропша, биоводородное дизельное топливо, смешанные спирты и древесное дизельное топливо.
В настоящем документе термин «биоспирт» обозначает полученные биологическим способом спирты, например, этанол, пропанол и бутанол. Биоспирты образуются под действием микроорганизмов и/или ферментов посредством ферментации сахаров, гемицеллюлозы или целлюлозы.
В настоящем документе термин «биодизельное топливо» обозначает состав, содержащий метиловые или этиловые эфиры жирной кислоты, полученные из липидов переэтерификацией, причем липиды получены из живых клеток, а не из окаменелых остатков.
В настоящем документе термин «синтетическое дизельное топливо» обозначает форму дизельного топлива, которое получено из возобновляемого сырья вместо окаменелого сырья, используемого в большинстве видов дизельного топлива.
В настоящем документе термин «растительное масло» включает чистое растительное масло (или непосредственно масло) или масло из отходов (побочный продукт других отраслей промышленности), в том числе масло, которое вырабатывается в вегетативной части или семени растения.
В настоящем документе термин «биогаз» обозначает метан или легковоспламеняющуюся смесь метана и других газов, полученные анаэробным расщеплением органического материала анаэробами.
В настоящем документе термин «синтетический газ» обозначает горючую смесь, которая содержит варьирующиеся количества монооксида углерода и водорода, и может содержать другие углеводороды, полученные частичным окислением биомассы. Синтетический газ может быть превращен в метанол в присутствии катализатора (обычно на основе меди), с последующей дегидратацией метанола в присутствии другого катализатора (например, кремния диоксида-алюминия).
В настоящем документе термин «Фишер-Тропш» обозначает набор химических реакций, в ходе которых смесь монооксида углерода и водорода превращается в жидкие углеводороды. Синтетический газ вначале может быть кондиционирован с использованием, например, конверсии водяного газа, чтобы достичь требуемого соотношения H2/CO. Конверсия происходит в присутствии катализатора, обычно железа или кобальта. Температура, давление и катализатор определяют, будет ли образовываться легкая или тяжелая синтетическая нефть. Например, при 330°C образуются в основном бензин и олефины, тогда как при 180-250°C образуются в основном дизельное топливо и воски. Жидкости, полученные из синтетического газа, которые содержат различные углеводородные фракции, представляют собой углеводороды с неразветвленной цепью и высокой степенью чистоты (не содержащие серы).
В настоящем документе термин «биоуголь» обозначает древесный уголь, полученный из биомассы, например, пиролизом биомассы.
В настоящем документе термин «сырье» обозначает материал, например, биомассу или продукт ее превращения (например, синтетический газ), который используют для производства продукта, например, биологического топлива, например, биодизельного топлива или синтетического дизельного топлива.
В настоящем документе термин «промышленный продукт» обозначает углеводородный продукт, который в основном состоит из атомов углерода и водорода, например, метиловые и/или этиловые эфиры жирной кислоты или алканы, такие как метан, смеси алканов с более длинной цепью, которые обычно являются жидкостями при температуре окружающей среды, биологическое топливо, монооксид углерода и/или водорода или биоспирт, например, этанол, пропанол или бутанол или биоуголь. Термин «промышленный продукт» предназначен включать промежуточные продукты, которые могут быть превращены в другие промышленные продукты, например, синтетический газ, который сам по себе рассматривается, как промышленный продукт, может использоваться для синтеза углеводородного продукта, который также рассматривается, как промышленный продукт. Термин «промышленный продукт» в настоящем документе включает обе чистые формы упомянутых выше соединений или, чаще, смесь различных соединений и компонентов, например, углеводородный продукт может содержать соединения с длиной углеродной цепи в определенном интервале, как хорошо известно из уровня техники.
В настоящем документе ʺпотомокʺ подразумевает ближайшее и все последующие поколения потомства, полученные от родительского организма, например, второе, третье или более поздние поколения потомства.
В настоящем документе слово «включают (содержат)» или его вариации, например, «включает (содержит)» или «включающий (содержащий)» следует понимать, как обозначающее включение заявленного элемента, целого числа или стадии, или группы элементов, целых чисел или стадий, но исключение любого другого элемента, целого числа или стадии или группы элементов, целых чисел или стадий.
Термин «и/или», например, «X и/или Y» следует понимать, как обозначающий любое из «X и Y» или «X или Y» и интерпретировать как обеспечивающий явное присутствие обоих значений, или любого из значений.
В настоящем документе термин около (приблизительно), если не указано иное, обозначает +/- 10%, более предпочтительно +/- 5%, более предпочтительно +/- 2%, более предпочтительно +/- 1%, даже более предпочтительно +/- 0,5% от указанного значения.
Выработка неполярных липидов и триацилглицеридов
Настоящее изобретение основано на открытии того факта, что содержание неполярных липидов в рекомбинантных эукариотных клетках может быть повышено с помощью комбинации модификаций, выбранных из обозначенных в настоящем документе как: (A). Push, (B). Pull, (C). Protect, (D). Package, (E). Plastidial export, (F). Plastidial import и (G). Prokariotyc Pathway. Как описано в настоящем документе, клетки без пластид могут содержать различные комбинации A-D, тогда как клетки с пластидами, такие как растительные и водорослевые клетки, могут содержать различные комбинации A-G.
Таким образом, рекомбинантные клетки, трансгенные животные, не относящиеся к человеческому роду, или их часть, а также трансгенные растения или их часть по изобретению содержат целый ряд комбинаций экзогенных полинуклеотидов и/или генетических модификаций, каждая из которых обеспечивает одну из модификаций. Указанные экзогенные полинуклеотиды и/или генетические модификации включают:
(A) экзогенный полинуклеотид, кодирующий полипептид фактора транскрипции, который увеличивает экспрессию одного или боле гликолитических генов и/или генов биосинтеза жирных кислот в клетке, трансгенном животном, не относящемся к человеческому роду, или его части, или трансгенном растении или его части, обеспечивая модификацию ʺPushʺ
(B) экзогенный полинуклеотид, кодирующий полипептид, который принимает участие в биосинтезе одного или более неполярных липидов в клетке, трансгенном животном, не относящемся к человеческому роду, или его части, или трансгенном растении или его части, обеспечивая модификацию ʺPullʺ
(C) генетическая модификация, которая подавляет эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке,трансгенном животном, не относящемся к человеческому роду, или его части, или трансгенном растении или его части, по сравнению с соответствующей клеткой, трансгенным животным, не относящимся к человеческому роду, или его частью, или трансгенным растением или его частью, в которых отсутствует генетическая модификация, обеспечивающая модификацию ʺProtectʺ
(D) экзогенный полинуклеотид, кодирующий полипептид покрытия масляного включения (ПМВ), обеспечивая модификацию ʺPackageʺ,
(E) экзогенный полинуклеотид, кодирующий полипептид, который увеличивает экспорт жирных кислот из пластид клетки, трансгенного животного, не относящегося к человеческому роду, или его части, или трансгенного растения или его части, по сравнению с соответствующей клеткой, трансгенным животным, не относящимся к человеческому роду, или его частью, или трансгенным растением или его частью, в которых отсутствует экзогенный полинуклеотид, обеспечивающий ʺPlastidial exportʺ,
(F) генетическая модификация, подавляющая эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды клетки,трансгенное животное, не относящееся к человеческому роду, или его часть, или трансгенное растение или его часть, по сравнению с соответствующей клеткой,трансгенным животным, не относящимся к человеческому роду, или его частью, или трансгенным растением или его частью, в которых отсутствует генетическая модификация, обеспечивающая ʺPlastidial importʺ, и
(G) генетическая модификация, подавляющая эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицеридов (ДАГ) в пластиде клетки, трансгенном животном, не относящемся к человеческому роду, или его части, или трансгенном растении или его части, по сравнению с соответствующей клеткой,трансгенным животным, не относящимся к человеческому роду, или его частью, или трансгенным растением или его частью, в которых отсутствует генетическая модификация, обеспечивающая модификацию ʺProkaryotic Pathwayʺ.
Предпочтительные комбинации (дополнительно обозначенные в настоящем документе как наборы) экзогенных полинуклеотидов и/или генетические модификации по изобретению представляют собой;
1) A, B и необязательно один из C, D, E, F или G;
2) A, C и необязательно один из D, E, F или G;
3) A, D и необязательно один из E, F или G;
4) A, E и необязательно F или G;
5) A, F и необязательно G;
6) A и G;
7) A, B, C и необязательно один из D, E, F или G;
8) A, B, D и необязательно один из E, F или G;
9) A, B, E и необязательно F или G;
10) A, B, F и необязательно G;
11) A, B, C, D и необязательно один из E, F или G;
12) A, B, C, E и необязательно F или G;
13) A, B, C, F и необязательно G;
14) A, B, D, E и необязательно F или G;
15) A, B, D, F и необязательно G;
16) A, B, E, F и необязательно G;
17) A, C, D и необязательно один из E, F или G;
18) A, C, E и необязательно F или G;
19) A, C, F и необязательно G;
20) A, C, D, E и необязательно F или G;
21) A, C, D, F и необязательно G;
22) A, C, E, F и необязательно пятая модификация G;
23) A, D, E и необязательно F или G;
24) A, D, F и необязательно G;
25) A, D, E, F и необязательно G;
26) A, E, F и необязательно G;
27) шесть из А, B, C, D, E, F и G, за исключением одной из A, B, C, D, E, F или G, и
28) любая из 1-26 выше, где присутствуют два или более экзогенных полинуклеотида, кодирующие два или более различных полипептида фактора транскрипции, которые увеличивают экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке, например, один экзогенный полинуклеотид, кодирующий WRI1, и другой экзогенный полинуклеотид, кодирующий LEC2.
В каждой из приведенных выше предпочтительных комбинаций может присутствовать по меньшей мере два различных экзогенных полинуклеотида, которые кодируют по меньшей мере два различных полипептида фактора транскрипции, увеличивающих экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке,трансгенном животном, не относящемся к человеческому роду, или его части, или трансгенном растении или его части.
Эти модификации описаны, как указано ниже:
A. Модификация ʺPush (Толчок)ʺ отличается повышенным синтезом общих жирных кислот в пластидах эукариотной клетки. В варианте реализации изобретения это происходит благодаря повышенной экспрессии и/или активности фактора транскрипции, который регулирует синтез жирных кислот в пластидах. В одном варианте реализации изобретения это может быть достигнуто путем экспрессии в трансгенной клетке экзогенного полинуклеотида, который кодирует полипептид фактора транскрипции, повышающего экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в клетке. В варианте реализации изобретения повышенный синтез жирных кислот не является вызванным доставкой в клетку модифицированной АККазы, активность которой в меньшей степени ингибируется жирными кислотами, по сравнению с эндогенной АККазой в клетке. В варианте реализации изобретения клетка содержит экзогенный полинуклеотид, кодирующий фактор транскрипции, предпочтительно под контролем промотора, кроме конститутивного промотора. Фактор транскрипции может быть выбран из группы, состоящей из WRI1, LEC1, LEC1-подобного, LEC2, BBM, FUS3, ABI3, ABI4, ABI5, Dof4, Dof11, или из группы, состоящей из MYB73, bZIP53, AGL15, MYB115, MYB118, TANMEI, WUS, GFR2a1, GFR2a2 и PHR1, и предпочтительно представляет собой WRI1, LEC1 или LEC2. В дальнейшем варианте реализации изобретения увеличенный синтез общих жирных кислот является относительным по сравнению с соответствующей клеткой дикого типа. В варианте реализации изобретения присутствует два или более экзогенных полинуклеотида, кодирующие два или более различных полипептида фактора транскрипции.
B. Модификация ʺPull (Притягивание)ʺ отличается повышенной экспрессией и/или активностью в клетке ацилтрансферазы ацилов жирных кислот, которая катализирует синтез ТАГ, ДАГ или МАГ в клетке, такой как DGAT, PDAT, LPAAT, GPAT или MGAT, предпочтительно DGAT или PDAT. В одном варианте реализации изобретения это может быть достигнуто путем экспрессии в трансгенной клетке экзогенного полинуклеотида, который кодирует полипептид, принимающий участие в биосинтезе одного или более неполярных липидов. В варианте реализации изобретения ацилтрансфераза является мембрано-связанной ацилтрансферазой, которая использует субстрат ацил-КоА в качестве донора ацила в случае DGAT, LPAAT, GPAT или MGAT или ацильную группу ФК в качестве донора ацила в случае PDAT. Модификация Pull может быть относительной по сравнению с соответствующей клеткой дикого типа или, предпочтительно, относительной по сравнению с соответствующей клеткой, содержащей модификацию Push. В варианте реализации изобретения клетка содержит экзогенный полинуклеотид, кодирующий ацилтрансферазу ацилов жирных кислот.
C. Модификация ʺProtect (Защита)ʺ отличается снижением катаболизма триацилглицеридов (ТАГ) в клетке. В варианте реализации изобретения это может быть достигнуто путем генетической модификации в клетке, которая подавляет эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов (ТАГ) в клетке, по сравнению с соответствующей клеткой без генетической модификации. В варианте реализации изобретения клетка проявляет сниженную экспрессию и/или активность эндогенной ТАГ липазы в клетке, предпочтительно липазы SDP1, полипептида Cgi58, ацил-КоА оксидазы, такой как ACX1 или ACX2, или полипептида, принимающего участие в β-окислении жирных кислот в клетке, такого как пероксисомальный переносчик ATP-связывающей кассеты PXA1. Это может происходить благодаря экспрессии в клетке экзогенного полинуклеотида, кодирующего молекулу РНК, которая снижает экспрессию, например, эндогенного гена, кодирующего ТАГ липазу, такую как липаза SDP1, ацил-КоА оксидаза или полипептид, принимающий участие в β-окислении жирных кислот в клетке, или путем мутации в эндогенном гене, кодирующем, например, ТАГ липазу, ацил-КоА оксидазу или полипептид, принимающий участие в β-окислении жирных кислот. Это может происходить благодаря экспрессии в клетке экзогенного полинуклеотида, кодирующего молекулу РНК, которая снижает экспрессию, например, эндогенного гена, кодирующего ТАГ липазу, такую как липаза SDP1, ацил-КоА оксидаза или полипептид, принимающий участие в β-окислении жирных кислот в клетке, или путем мутации в эндогенном гене, кодирующем, например, ТАГ липазу, ацил-КоА оксидазу или полипептид, принимающий участие в β-окислении жирных кислот. В варианте реализации изобретения сниженная экспрессия и/или активность является относительной по сравнению с соответствующей клеткой дикого типа или относительной по сравнению с соответствующей клеткой, содержащей модификацию Push.
D. Модификация ʺPackage (Упаковка)ʺ отличается повышенной экспрессией и/или аккумуляцией полипептида покрытия масляного включения (ПМВ). В варианте реализации изобретения это может быть достигнуто путем экспрессии в трансгенной клетке экзогенного полинуклеотида, кодирующего полипептид покрытия масляного включения (ПМВ). Полипептид ПМВ может представлять собой олеозин, такой как полиолеозин, каолеозин или стеролеозин или предпочтительно LDAP. В варианте реализации изобретения уровень олеозина, который аккумулируется в эукариотной клетке, по меньшей мере в 2 раза выше относительно соответствующей клетки, содержащей ген олеозина из T-ДНК pJP3502. В варианте реализации изобретения повышенная экспрессия или аккумуляция полипептида ПМВ не является вызванной исключительно модификацией Push. В варианте реализации изобретения экспрессия и/или аккумуляция является относительной по сравнению с соответствующей клеткой дикого типа или предпочтительно относительно соответствующей клетки, которая содержит модификацию Push.
E. Модификация ʺPlastidial export (Пластидный экспорт)ʺ отличается повышенной скоростью экспорта общих жирных кислот из пластид эукариотной клетки. В одном варианте реализации изобретения это может быть достигнуто путем экспрессии в трансгенной клетке экзогенного полинуклеотида, кодирующего полипептид, который увеличивает экспорт жирных кислот из пластид клетки, по сравнению с соответствующей клеткой, в которой отсутствует экзогенный полинуклеотид. В варианте реализации изобретения это достигается путем повышения экспрессии и/или активности тиоэстеразы (ТЭ) жирных кислот, полипептида-переносчика жирных кислот, такого как полипептид ABCA9, или длинноцепочечной ацил-КоА синтетазы (ДЦАС). В варианте реализации изобретения клетка содержит экзогенный полинуклеотид, кодирующий ТЭ, полипептид-переносчик жирных кислот или ДЦАС. ТЭ может представлять собой полипептид FATB или, предпочтительно, полипептид FATA. В варианте реализации изобретения ТЭ предпочтительно представляет собой ТЭ со специфичностью в отношении СЦЖК. В варианте реализации изобретения модификация Plastidial export является относительной по сравнению с соответствующей клеткой дикого типа или предпочтительно относительной по сравнению с соответствующей клеткой, которая содержит модификацию Push.
F. Модификация ʺPlastidial import (Пластидный импорт)ʺ отличается сниженной скоростью импорта жирных кислот в пластиды клетки из пространства за пределами пластид. В варианте реализации изобретения это может быть достигнуто с помощью генетической модификации в клетке, которая подавляет эндогенную выработку и/или активность полипептида, принимающего участие в импорте жирных кислот в пластиды клетки, по сравнению с соответствующей клеткой, не содержащей генетической модификации. Например, это может происходить благодаря экспрессии в клетке экзогенного полинуклеотида, кодирующего молекулу РНК, которая снижает экспрессию эндогенного гена, кодирующего полипептид-переносчик, такой как полипептид TGD, например полипептид TGD1, TGD2, TGD3 или TGD4, или мутации в эндогенном гене, кодирующем полипептид TGD. В варианте реализации изобретения сниженная скорость импорта является относительной по сравнению с соответствующей клеткой дикого типа или относительной по сравнению с соответствующей клеткой, которая содержит модификацию Push.
G. Модификация ʺProkaryotic Pathway (Прокариотный путь)ʺ отличается сниженным количеством ДАГ или сниженной скоростью выработки ДАГ в пластидах клетки. В варианте реализации изобретения это может быть достигнуто путем генетической модификации в клетке, которая подавляет эндогенную выработку и/или активность полипептида, принимающего участие в выработке диацилглицеридов (ДАГ) в пластиде, по сравнению с соответствующей клеткой, не содержащей генетической модификации. В варианте реализации изобретения сниженное количество или скорость выработки ДАГ является результатом сниженной выработки LPA из ацил-АПБ и G3P в пластидах. Сниженное количество или скорость выработки ДАГ может быть результатом экспрессии в клетке экзогенного полинуклеотида, кодирующего молекулу РНК, которая снижает экспрессию эндогенного гена, кодирующего пластидную GPAT, пластидную LPAAT или пластидную PAP, предпочтительно пластидную GPAT, или мутации в эндогенном гене, кодирующем пластидный полипептид. В варианте реализации изобретения сниженное количество или скорость выработки ДАГ является относительным по сравнению с соответствующей клеткой дикого типа или предпочтительно относительной по сравнению с соответствующей клеткой, которая содержит модификацию Push.
Модификация Push имеет большое значение для изобретения, а модификация Pull является предпочтительной. Модификации Protect и Package могут быть комплементарными, т.е. одной из двух может быть достаточно. Клетка может содержать одну, две или все три из модификаций Plastidial export, Plastidial import и Prokaryotic Pathway. В варианте реализации изобретения по меньшей мере один из экзогенных полинуклеотидов в клетке, предпочтительно по меньшей мере экзогенный полинуклеотид, кодирующий фактор транскрипции, который регулирует синтез жирных кислот в пластидах, экспрессируется под контролем (H) промотора, кроме конститутивного промотора, например, такого как связанный с развитием промотор, промотор, который предпочтительно активен в фотосинтезирующих клетках, тканеспецифичный промотор, промотор, модифицированный путем снижения уровня его экспрессии относительно соответствующего природного промотора, или предпочтительно промотора, представляющего собой специфичный для старения промотор. Более предпочтительно, по меньшей мере экзогенный полинуклеотид, кодирующий фактор транскрипции, который регулирует синтез жирных кислот в пластидах, экспрессируется под контролем промотора, кроме конститутивного промотора, причем экзогенный полинуклеотид, кодирующий молекулу РНК, которая подавляет эндогенную выработку и/или активность полипептида, принимающего участие в катаболизме триацилглицеридов, также экспрессируется под контролем промотора, кроме конститутивного промотора, и, при этом, промоторы могут быть одинаковыми или разными.
Растения вырабатывают некоторые, но не все из их мембранных липидов, такие как MGDG, в пластидах по так называемому прокариотному пути (Фиг. 1). Кроме того, в растениях присутствует эукариотный путь для синтеза галактолипидов и глицеролипидов, по которому ЖК вначале синтезируются в пластиде, а затем в ЭР происходит сборка ЖК в глицеролипиды. MGDG, синтезированный по эукариотному пути, содержит C18:3 жирную кислоту (АЛК), этерифицированную в положении sn-2 MGDG. Сборка скелета ДАГ, содержащего АЛК для синтеза MGDG данным путем, происходит в ЭР, после чего происходит его импорт в пластиду. В противоположность этому, MGDG, синтезированный прокариотным путем, содержит C16:3 жирную кислоту, этерифицированную в положении sn-2 MGDG. Соотношение вклада прокариотного пути относительно эукариотного пути в выработку MGDG (16:3) против MGDG (18:3) представляет собой характерный и отличительный признак различных видов растений (Mongrand с соавт., 1998). Указанный отличительный жирнокислотный состав MGDG позволяет классифицировать все высшие растения (покрытосеменные) на так называемые 16:3 или 18:3 растения. Виды 16:3, примером которых являются Arabidopsis и Brassica napus, в общем содержат активный прокариотный и активный эукариотный пути синтеза MGDG, тогда как виды 18:3, примером которых являются Nicotiana tabacum Pisum sativum и Glycine max, в общем, содержат только (или почти полностью) эукариотный путь синтеза MGDG, с небольшой или отсутствующей аккумуляцией C16:3 жирной кислоты в вегетативных тканях. В настоящем документе ʺрастение 16:3ʺ или ʺвид 16:3ʺ представляет собой такое, которое содержит более чем 2% C16:3 жирной кислоты в общем содержании жирных кислот в его фотосинтезирующих тканях. В настоящем документе ʺрастение 18:3ʺ или ʺвид 18:3ʺ представляет собой такое, которое содержит менее чем 2% C16:3 жирных кислот в общем содержании жирных кислот в его фотосинтетических тканях. Как описано в настоящем документе, растение может быть превращено из растения 16:3 в растение 18:3 путем подходящих генетических модификаций. Соотношение потока между прокариотным и эукариотным путями не является консервативным для разных видов растений или тканей. В видах 16:3 до 40% потока в листьях проходит по прокариотному пути (Browse с соавт., 1986), тогда как в видах 18:3, таких как горох и соя, около 90% ЖК, которые синтезированы в пластиде, экспортируется из пластиды в ЭР, чтобы обеспечить источник ЖК для эукариотного пути (Ohlrogge и Browse, 1995; Somerville с соавт., 2000).
Таким образом, различные количества 18:3 и 16:3 жирных кислот найдены в гликолипидах различных видов растений. Это используется для того, чтобы отличать растения 18:3, в которых жирные кислоты с 3 двойными связями практически полностью являются C18 жирными кислотами, и растения 16:3, которые содержат как C16-, так и C18-жирные кислоты с 3 двойными связями. В хлоропластах растений 18:3 ферментная активность, катализирующая превращение фосфатидата в диацилглицерид и диацилглицерида в моногалактозилдиацилглицерид (МГД), существенно менее выражена, чем в хлоропластах 16:3. В листьях растений 18:3 хлоропласты синтезируют стеароил-ACP2 в строме, вводят первую двойную связь в насыщенную углеводородную цепь, а затем гидролизуют тиоэфир тиоэстеразами (Фиг. 1). Высвобождающийся олеат экспортируется сквозь конверты хлоропластов в мембраны эукариотной части клетки, вероятно, эндоплазматический ретикулум, где он инкорпорируется в ФХ. В этих мембранах в ФХ-связанные олеоильные группы вводятся кратные связи, и далее они перемещаются обратно в хлоропласт. МГД-связанные ацильные группы представляет собой субстраты для введения третьей двойной связи с образованием МГД с двумя линоленоильными остатками. Данный галактолипид является характерным для растений 18:3, например, таких как Asteraceae и Fabaceae. В фотосинтетически активных клетках растений 16:3, которые представлены, например, членами Apiaceae и Brassicaceae, два пути действуют параллельно, чтобы обеспечить тилакоиды МГД.
В одном варианте реализации изобретения вегетативная часть растения, эукариотная клетка, семя или трансгенный организм, не принадлежащий к человеческому роду, или их часть (например, клубень или свекла) по изобретению вырабатывает более высокие уровни неполярных липидов, таких как ТАГ, или имеет более высокое содержание общих жирных кислот (ОЖК), предпочтительно оба, по сравнению с соответствующей вегетативной частью растения, эукариотной клеткой, семенем или организмом, не принадлежащим к человеческому роду, или их частью, в которой отсутствуют генетические модификации или экзогенные полинуклеотиды. В одном примере растения по изобретению дают семена, листья или имеют части листьев с площадью поверхности по меньшей мере 1 см2, стволы и/или клубни с повышенным содержанием неполярных липидов, например, содержанием ТАГ или ОЖК, предпочтительно обоими, по сравнению с соответствующими семенами, листьями, частями листьев с площадью поверхности по меньшей мере 1 см2, стволами или клубнями.
В другом варианте реализации изобретения вегетативная часть растения, трансгенный организм, не относящийся к человеческому роду, или их часть (такая как клубень или свекла), предпочтительно растение, клубень, свекла или семя, вырабатывают ТАГ, обогащенный одной или более конкретных жирных кислот. Широкий спектр жирных кислот может быть инкорпорирован в ТАГ, в том числе насыщенные и ненасыщенные жирные кислоты и короткоцепочечные и длинноцепочечные жирные кислоты. Некоторые неограничивающие примеры жирных кислот, которые могут быть инкорпорированы в ТАГ, и уровень которых может быть повышен, включают: каприновую (10:0), лауриновую (12:0), миристиновую (14:0), пальмитиновую (16:0), пальмитолеиновую (16:1), стеариновую (18:0), олеиновую (18:1), вакценовую (18:1), линолевую (18:2), элеостеариновую (18:3), γ-линоленовую (18:3), α-линоленовую (18:3ω3), стеаридоновую (18:4ω3), арахидоновую (20:0), эйкозадиеновую (20:2), дигомо-γ-линолевую (20:3), эйкозатриеновую (20:3), арахидоновую (20:4), эйкозатетраеновую (20:4), эйкозапентаеновую (20:5ω3), бегеновую (22:0), докозапентаеновую (22:5ω), докозагексаеновую (22:6ω3), лигноцериновую (24:0), ацетэруковую (24:1), церотиновую (26:0) и монтановую (28:0) жирные кислоты. В одном варианте настоящего изобретения вегетативная часть растения, эукариотная клетка, семя или трансгенный организм или их часть (как например, клубень или свекла) обогащены ТАГ, содержащими олеиновую кислоту и/или сниженное содержание линоленовой кислоты (АЛК), предпочтительно по меньшей мере на 2% или по меньшей мере на 5%, в пересчете на абсолютное вещество.
Предпочтительно, вегетативная часть растения, эукариотная клетка, семя или трансгенный организм, не принадлежащий к человеческому роду, или их часть по изобретению трансформированы одной или более химерными ДНК (экзогенные полинуклеотиды). В случае нескольких химерных ДНК, предпочтительно они ковалентно соединены в одну молекулу ДНК, например, такую как единая молекула T-ДНК, и предпочтительно интегрированы в один локус генома клетки-хозяина. В качестве альтернативы, химерные ДНК находятся на двух или более молекулах ДНК, которые могут быть несвязанными с геномом хозяина, или молекула(ы) ДНК не являются интегрированными в геном хозяина, например, как это происходит в экспериментах с временной экспрессией. Растение, вегетативная часть растения, эукариотная клетка, семя или трансгенный организм, не принадлежащий к человеческому роду, или их часть предпочтительно являются гомозиготными по одной молекуле ДНК, инкорпорированной в их геном.
Факторы транскрипции
В эукариотных клетках различные факторы транскрипции принимают участие в синтезе жирных кислот и липидов, содержащих жирные кислоты, такие как ТАГ, и, таким образом, существует возможность манипулировать ими с целью модификации Push. Предпочтительным фактором транскрипции является WRI1. В настоящем документе термин «Wrinkled 1» или «WRI1» или «WRL1» обозначает фактор транскрипции класса AP2/ERWEBP, который регулирует экспрессию нескольких ферментов, принимающих участие в гликолизе и биосинтезе жирных кислот de novo. WRI1 содержит два специфичных для растения (AP2/EREB) домена связывания с ДНК. Дополнительно, WRI1, по меньшей мере в Arabidopsis, также регулирует разложение сахарозы посредством гликолиза, таким образом регулируя поступление прекурсоров для биосинтеза жирных кислот. Иными словами, он управляет потоком углерода от продукта фотосинтеза к запасным липидам. Мутанты wri1, по меньшей мере в Arabidopsis, демонстрируют фенотип сморщенного семени, за счет дефекта во встраивании сахарозы и глюкозы в ТАГ.
Примеры генов, которые транскрибируются WRI1, включают, без ограничений, одну или более, предпочтительно все из генов, кодирующих пируваткиназу (At5g52920, At3g22960), субъединицу E1 альфа (At1g01090) пируватдегидрогеназы (ПДГ), ацетил-КоА карбоксилазу (АЦКазу), субъединицу BCCP2 (At5g15530), еноил-АПБ редуктазу (At2g05990; EAR), фосфоглицерат мутазу (At1g22170), цитозольную фруктокиназу и цитозольную фосфоглицерат мутазу, синтетазу сахарозы (SuSy) (см., например, Liu с соавт., 2010b; Baud с соавт., 2007; Ruuska с соавт., 2002).
WRI1 содержит консервативный домен AP2 (cd00018). AP2 представляет собой домен связывания с ДНК, найденный в регуляторах транскрипции в растениях, например, APETALA2 и EREBP (белок, связывающийся с элементом, который реагирует на этилен). В EREBP домен специфично связывается с боксом 11bp GCC элемента, который реагирует на этилен (ЭРЭ), промоторного элемента, незаменимого для реагирования на этилен. EREBP и фактор связывания с C-повтором CBF1, который принимает участие в ответе на стресс, содержат одну копию домена AP2. APETALA2-подобные белки, которые играют роль в развитии растения, содержат две копии.
Другие мотивы последовательности, которые могут быть найдены в WRI1 и его функциональных гомологах, включают:
1. R G V T/S R H R W T G R (SEQ ID NO:89).
2. F/Y E A H L W D K (SEQ ID NO:90).
3. D L A A L K Y W G (SEQ ID NO:91).
4. S X G F S/A R G X (SEQ ID NO:92).
5. H H H/Q N G R/K W E A R I G R/K V (SEQ ID NO:93).
6. Q E E A A A X Y D (SEQ ID NO:94).
В настоящем документе термин «Wrinkled 1» или «WRI1» также включает «подобные Wrinkled 1» или «WRI1-подобные» белки. Примеры белков WRI1 включают номера доступа: Q6X5Y6, (Arabidopsis thaliana; SEQ ID NO:22), XP_002876251.1 (подвид Arabidopsis lyrata ; Lyrata; SEQ ID NO:23), ABD16282.1 (Brassica napus; SEQ ID NO:24), ADO16346.1 (Brassica napus; SEQ ID NO:25), XP_003530370.1 (Glycine max; SEQ ID NO:26), AEO22131.1 (Jatropha curcas; SEQ ID NO:27), XP_002525305.1 (Ricinus communis; SEQ ID NO:28), XP_002316459.1 (Populus trichocarpa; SEQ ID NO:29), CBI29147.3 (Vitis vinifera; SEQ ID NO:30), XP_003578997.1 (Brachypodium distachyon; SEQ ID NO:31), BAJ86627.1 (Hordeum vulgare подвид. vulgare; SEQ ID NO:32), EAY79792.1 (Oryza sativa; SEQ ID NO:33), XP_002450194.1 (Sorghum bicolor; SEQ ID NO:34), ACG32367.1 (Zea mays; SEQ ID NO:35), XP_003561189.1 (Brachypodium distachyon; SEQ ID NO:36), ABL85061.1 (Brachypodium sylvaticum; SEQ ID NO:37), BAD68417.1 (Oryza sativa; SEQ ID NO:38), XP_002437819.1 (Sorghum bicolor; SEQ ID NO:39), XP_002441444.1 (Sorghum bicolor; SEQ ID NO:40), XP_003530686.1 (Glycine max; SEQ ID NO:41), XP_003553203.1 (Glycine max; SEQ ID NO:42), XP_002315794.1 (Populus trichocarpa; SEQ ID NO:43), XP_002270149.1 (Vitis vinifera; SEQ ID NO:44), XP_003533548.1 (Glycine max; SEQ ID NO:45), XP_003551723.1 (Glycine max; SEQ ID NO:46), XP_003621117.1 (Medicago truncatula; SEQ ID NO:47), XP_002323836.1 (Populus trichocarpa; SEQ ID NO:48), XP_002517474.1 (Ricinus communis; SEQ ID NO:49), CAN79925.1 (Vitis vinifera; SEQ ID NO:50), XP_003572236.1 (Brachypodium distachyon; SEQ ID NO:51), BAD10030.1 (Oryza sativa; SEQ ID NO:52), XP_002444429.1 (Sorghum bicolor; SEQ ID NO:53), NP_001170359.1 (Zea mays; SEQ ID NO:54), XP_002889265.1 (Arabidopsis lyrata подвид lyrata; SEQ ID NO:55), AAF68121.1 (Arabidopsis thaliana; SEQ ID NO:56), NP_178088.2 (Arabidopsis thaliana; SEQ ID NO:57), XP_002890145.1 (Arabidopsis lyrata подвид lyrata; SEQ ID NO:58), BAJ33872.1 (Thellungiella halophila; SEQ ID NO:59), NP_563990.1 (Arabidopsis thaliana; SEQ ID NO:60), XP_003530350.1 (Glycine max; SEQ ID NO:61), XP_003578142.1 (Brachypodium distachyon; SEQ ID NO:62), EAZ09147.1 (Oryza sativa; SEQ ID NO:63), XP_002460236.1 (Sorghum bicolor; SEQ ID NO:64), NP_001146338.1 (Zea mays; SEQ ID NO:65), XP_003519167.1 (Glycine max; SEQ ID NO:66), XP_003550676.1 (Glycine max; SEQ ID NO:67), XP_003610261.1 (Medicago truncatula; SEQ ID NO:68), XP_003524030.1 (Glycine max; SEQ ID NO:69), XP_003525949.1 (Glycine max; SEQ ID NO:70), XP_002325111.1 (Populus trichocarpa; SEQ ID NO:71), CBI36586.3 (Vitis vinifera; SEQ ID NO:72), XP_002273046.2 (Vitis vinifera; SEQ ID NO:73), XP_002303866.1 (Populus trichocarpa; SEQ ID NO:74), и CBI25261.3 (Vitis vinifera; SEQ ID NO:75). Дальнейшие примеры включают Sorbi-WRL1 (SEQ ID NO: 76), Lupan-WRL1 (SEQ ID NO: 77), Ricco-WRL1 (SEQ ID NO: 78), и Lupin angustifolius WRI1 (SEQ ID NO: 79). Предпочтительным WRI1 является WRI1 маиса или WRI1 сорго.
Позже, подмножество WRI1-подобных факторов транскрипции было повторно классифицировано как факторы транскрипции WRI2, WRI3 или WRI4, которые скорее отличаются преимущественной экспрессией в стволах и/или корнях растений, чем в развивающихся семенах (To с соавт., 2012). Несмотря на повторную классификацию, они входят в определение ʺWRI1ʺ в настоящем документе. Предпочтительными WRI1-подобными факторами транскрипции являются те, которые могут дополнить функцию мутации wri1 в растении, особенно функцию в развивающемся семени растения, например, в мутанте wri1 A. thaliana. Кроме того, функция WRI1-подобного полипептида может быть количественно оценена в N. benthamiana при помощи анализов с временной экспрессией, как описано в настоящем документе.
В настоящем документе полипептид ʺЛИСТОВАЯ СЕМЯДОЛЯʺ или ʺLECʺ обозначает фактор транскрипции, который является фактором транскрипции LEC1, LEC1-подобным, LEC2, ABI3 или FUS3 и демонстрирует широкий контроль над созреванием семени и синтезом жирных кислот. LEC2, FUS3 и ABI3 представляют собой родственные полипептиды, каждый из которых содержит ДНК-связывающий домен B3 размером 120 аминокислот (Yamasaki с соавт., 2004), найденный только в растительных белках. Их можно отличить при помощи филогенетического анализа, чтобы определить связь аминокислотной последовательности с членами множества полипептидов A. thaliana, имеющими указанные ниже номера доступа: LEC2, номер доступа AAL12004.1; FUS3 (также известный как FUSCA3), номер доступа AAC35247. LEC1 принадлежит к другому классу полипептидов и гомологичен полипептиду HAP3 класса фактора связывания CBF (Lee с соавт., 2003). Гены LEC1, LEC2 и FUS3 требуются в ходе раннего эмбриогенеза, чтобы предопределить гибель эмбриональных клеток и определить идентичность семядоли, и позже в ходе инициации и поддержания созревания эмбриона (Santos-Mendoza с соавт., 2008). Кроме того, они индуцируют экспрессию генов, кодирующих запасные белки семени, путем связывания с мотивами RY, присутствующими в промоторах, и генов олеозина. Дополнительно, они могут отличаться характером экспрессии в ходе развития семени или своей способностью дополнить соответствующую мутацию в A. thaliana.
В настоящем документе термин ʺЛистовая Семядоля 1ʺ или ʺLEC1ʺ обозначает фактор транскрипции NF-YB-типа, который принимает участие в развитии зиготы и соматическом эмбриогенезе. Эндогенный ген специфично экспрессируется в семени, как в эмбрионе, так и эндосперме. LEC1 активизирует ген, кодирующий WRI1, а также большой класс генов синтеза жирных кислот. Кроме того, эктопическая экспрессия LEC2 вызывает быструю активацию реагирующих на ауксин генов и может вызывать образование соматических эмбрионов. Примеры полипептидов LEC1 включают белки Arabidopsis thaliana (AAC39488, SEQ ID NO:149), Medicago truncatula (AFK49653, SEQ ID NO:154) and Brassica napus (ADF81045, SEQ ID NO:151), A. lyrata (XP_002862657, SEQ ID NO:150), R. communis (XP_002522740, SEQ ID NO:152), G. max (XP_006582823, SEQ ID NO:153), A. hypogaea (ADC33213, SEQ ID NO:156), Z. mays (AAK95562, SEQ ID NO:155).
LEC1-подобный (L1L) является близкородственным LEC1, но отличается другим характером экспрессии гена, который экспрессируется раньше в ходе эмбриогенеза (Kwong с соавт., 2003). Примеры LEC1-подобных полипептидов включают белки Arabidopsis thaliana (AAN15924, SEQ ID NO:157), Brassica napus (AHI94922, SEQ ID NO:158), and Phaseolus coccineus LEC1-like (AAN01148, SEQ ID NO: 159).
В настоящем документе термин ʺЛистовая семядоля 2ʺ или ʺLEC2ʺ обозначает домен B3 фактора транскрипции, который принимает участие в развитии зиготы и соматическом эмбриогенезе и который активизирует экспрессию гена, кодирующего WRI1. Его эктопическая экспрессия облегчает эмбриогенез из вегетативных тканей растения (Alemanno с соавт., 2008). Примеры полипептидов LEC2 включают белки Arabidopsis thaliana (номер доступа NP_564304.1, SEQ ID NO:142), Medicago truncatula (номер доступа CAA42938.1, SEQ ID NO:143) and Brassica napus (номер доступа ADO16343.1, SEQ ID NO:144).
В варианте реализации изобретения экзогенный полинуклеотид по изобретению, который кодирует LEC2, содержит одно или более из следующего:
i) нуклеотиды, кодирующие полипептид, содержащий аминокислоты, последовательность которых приведена в любой из SEQ ID NO: 142-144, или его биологически активный фрагмент или полипептид, аминокислотная последовательность которого по меньшей мере на 30% идентична любой одной или больше из SEQ ID NO: 142-144,
ii) нуклеотиды, последовательность которых по меньшей мере на 30% идентична i), и
iii) полинуклеотид, который гибридизуется с одним или обоими из i) или ii) в строгих условиях.
В настоящем документе термин ʺFUS3ʺ обозначает домен B3 фактора транскрипции, который принимает участие в развитии зиготы и соматическом эмбриогенезе и обнаружен по большей части в протодермальной ткани эмбриона (Gazzarrini с соавт., 2004). Примеры полипептидов FUS3 включают белки Arabidopsis thaliana (AAC35247, SEQ ID NO:160), Brassica napus (XP_006293066.1, SEQ ID NO:161) and Medicago truncatula (XP_003624470, SEQ ID NO:162). Чрезмерной экспрессией любого из LEC1, L1L, LEC2, FUS3 и ABI3 из экзогенного полинуклеотида предпочтительно управляют с помощью регулируемого развитием промотора, такого как специфичный для старения промотор, индуцибельный промотор, или промотор, который сконструирован с целью обеспечения сниженного уровня экспрессии относительно природного промотора, особенно в растениях, кроме Arabidopsis thaliana и B. napus сорт Westar, во избежание аномалий развития, возникающих в ходе развития растения, которые обычно ассоциируются с повышенной экспрессией этих факторов транскрипции (Mu с соавт., 2008).
В настоящем документе термин «BABY BOOM» или «BBM» обозначает фактор транскрипции AP2/ERF, который индуцирует регенерацию в условиях культуры, в норме не поддерживающий регенерацию в растениях дикого типа. Эктопическая экспрессия генов Brassica napus BBM (BnBBM) в B. napus и Arabidopsis индуцирует спонтанный соматический эмбриогенез и органогенез из саженцев, выращиваемых на свободной от гормона основной среде (Boutilier с соавт., 2002). В табаке эктопическая экспрессия BBM является достаточной для индукции дополнительной регенерации побегов и корней на основной среде, но необходим экзогенный цитокинин для образования соматического зародыша (СЭ) (Srinivasan с соавт., 2007). Примеры полипептидов BBM включают белки Arabidopsis thaliana (номер доступа NP_197245.2, SEQ ID NO: 145), маиса (US 7579529), Sorghum bicolor (номер доступа XP_002458927) и Medicago truncatula (номер доступа AAW82334.1, SEQ ID NO:146).
В варианте реализации изобретения экзогенный полинуклеотид по изобретению, который кодирует BBM, содержит, если не указано иное, одно или более из следующего:
i) нуклеотиды, кодирующие полипептид, содержащий аминокислоты, последовательность которых приведена в одной из SEQ ID NO: 145 или 146, или его биологически активный фрагмент или полипептид, аминокислотная последовательность которого по меньшей мере на 30% идентична одной или обеим из SEQ ID NO: 145 или 146,
ii) нуклеотиды, последовательность которых по меньшей мере на 30% идентична i), и
iii) полинуклеотид, который гибридизуется с одним или обоими из i) или ii) в строгих условиях.
Полипептид ABI3 (A. thaliana, номер доступа NP_189108), родственный белку VP1 маиса, экспрессируется с низкими уровнями в вегетативных тканях и влияет на развитие пластид. Полипептид ABI4 A. thaliana, номер доступа NP_181551) принадлежит к семейству факторов транскрипции, которые содержат специфический для растений домен AP2 (Finkelstein с соавт., 1998) и действуют ниже ABI3 в биохимическом пути. ABI5 (A. thaliana номер доступа NP_565840) представляет собой фактор транскрипции семейства bZIP, который влияет на чувствительность ABA и управляет экспрессией некоторых генов LEA в семенах. Он связывает с реагирующим на ABA элементом.
Каждый из следующих факторов транскрипции был выбран на основе того, что они функционируют в ходе эмбриогенеза в растениях. Номера доступа приведены в Таблице 10. Гомологи каждого из них могут быть с легкостью идентифицированы во многих другие видах растений и проанализированы, как описано в Примере 10.
MYB73 представляет собой фактор транскрипции, идентифицированный в сое, который принимает участие в стрессовых реакциях.
bZIP53 представляет собой фактор транскрипции в семействе белка bZIP, идентифицированный в Arabidopsis.
AGL15 (бесполо-подобный 15) представляет собой фактор транскрипции бокса MADS, который нативно экспрессируется в ходе эмбриогенеза. Кроме того, AGL15 нативно экспрессируется в листовом примордии, верхушечном меристеме побегов и молодых цветочных почках, наводя на мысль о том, что AGL15 может дополнительно функционировать в ходе развития после прорастания. AGL15 играет роль в эмбриогенезе и катаболизме гиббереловой кислоты. Он нацелен на домен B3 факторов транскрипции, которые являются ключевыми регуляторами эмбриогенеза.
MYB115 и MYB118 представляют собой факторы транскрипции в семействе MYB из Arabidopsis, принимающем участие в эмбриогенезе.
TANMEI, также известный как EMB2757, кодирует белок повтора WD, необходимый Arabidopsis для эмбрионального развития.
WUS, также известный как Wuschel, представляет собой ген гомеобокса, который управляет пулом стволовых клеток в эмбрионах. Он экспрессируется в стволовой клетке, организующей центр меристема, и необходим для удерживания стволовых клеток в недифференцированном состоянии. Фактор транскрипции связывается с мотивом TAAT компонента ядра.
GFR2a1 и GFR2a2 представляет собой факторы транскрипции по меньшей мере из сои.
Ацилтрансферазы ацилов жирных кислот
В настоящем документе термин "ацилтрансфераза ацилов жирных кислот" обозначает белок, способный к переносу ацильной группы с ацил-КоА, ФХ или ацил-АПБ, предпочтительно ацил-КоА или ФХ, на субстрат, с целью образования ТАГ, ДАГ или МАГ. Указанные ацилтрансферазы включают DGAT, PDAT, MGAT, GPAT и LPAAT.
В настоящем документе термин «диацилглицерил ацилтрансфераза» (DGAT) обозначает белок, который переносит жирную ацильную группу из ацил-КоА к субстрату ДАГ, с образованием ТАГ. Таким образом, термин «активность диацилглицерил ацилтрансферазы» обозначает перенос группы ацила от ацил-КоА к ДАГ, с образованием ТАГ. DGAT может дополнительно выполнять функцию MGAT, но, в основном, функционирует, как DGAT, т. е., обладает более выраженной каталитической активностью в роли DGAT, чем в роли MGAT, если активность фермента выражена в единицах нмоль продукта/мин/мг белка (см. например, Yen с соавт., 2005). Активность DGAT может быть лимитирующей скорость в процессе синтеза ТАГ в семенах (Ichihara с соавт., 1988). DGAT использует субстрат ацил-КоА в качестве донора ацила и переносит его в положение sn-3 ДАГ, с образованием ТАГ. Фермент в своем нативном состоянии функционирует в эндоплазматическом ретикулуме (ЭР) клетки.
Существует три известных типа DGAT, обозначаемых, как DGAT1, DGAT2 и DGAT3, соответственно. Полипептиды DGAT1 являются мембранными белками, которые обычно содержат 10 трансмембранных доменов, полипептиды DGAT2 также являются мембранными белками, но обычно содержат 2 трансмембранных домена, тогда как полипептиды DGAT3 обычно не содержат ни одного и считаются растворимыми в цитоплазме, а не интегрированными в мембраны. Растительные полипептиды DGAT1 обычно содержат около 510-550 остатков аминокислот, в то время как полипептиды DGAT2 обычно содержат около 310-330 остатков. DGAT1 является основным ферментом, ответственным за выработку ТАГ из ДАГ в большинстве развивающихся семян растений, тогда как DGAT2 из таких видов растений, как тунговое дерево (Vernicia fordii) и клещевина (Ricinus communis), которые продуцируют высокие количества необычных жирных кислот, по-видимому, играют важную роль в аккумуляции необычных жирных кислот в ТАГ. Повышенная экспрессия AtDGAT1 в листьях табака приводила к повышению содержания ТАГ в 6-7 раз (Bouvier-Nave с соавт., 2000).
Примеры полипептидов DGAT1 включают белки DGAT1Aspergillus fumigatus (XP_755172.1; SEQ ID NO:80), Arabidopsis thaliana (CAB44774.1; SEQ ID NO:1), Ricinus communis (AAR11479.1; SEQ ID NO:81), Vernicia fordii (ABC94472.1; SEQ ID NO:82), Vernonia galamensis (ABV21945.1 and ABV21946.1; SEQ ID NO:83 and SEQ ID NO:84, соответственно), Euonymus alatus (AAV31083.1; SEQ ID NO:85), Caenorhabditis elegans (AAF82410.1; SEQ ID NO:86), Rattus norvegicus (NP_445889.1; SEQ ID NO:87), Homo sapiens (NP_036211.2; SEQ ID NO:88), а также их варианты и/или мутанты. Примеры полипептидов DGAT2 включают белки, кодируемые генами DGAT2 Arabidopsis thaliana (NP_566952.1; SEQ ID NO: 2), Ricinus communis (AAY16324.1; SEQ ID NO: 3), Vernicia fordii (ABC94474.1; SEQ ID NO: 4), Mortierella ramanniana (AAK84179.1; SEQ ID NO: 5), Homo sapiens (Q96PD7.2; SEQ ID NO: 6) (Q58HT5.1; SEQ ID NO: 7), Bos taurus (Q70VZ8.1; SEQ ID NO: 8), Mus musculus (AAK84175.1; SEQ ID NO: 9), а также их варианты и/или мутанты. Аминокислотные последовательности DGAT1 и DGAT2 демонстрируют незначительную гомологию. Экспрессия в листьях экзогенного DGAT2 была в два раза более эффективной, чем DGAT1 с точки зрения повышения содержания масла (ТАГ). Кроме того, DGAT2 A. thaliana проявляет большее предпочтение в отношении линолеоил-КоА и линоленоил-КоА в качестве доноров ацила, по сравнению с олеоил-КоА, сравнению с DGAT1. Указанное предпочтение в отношении субстрата может применяться, чтобы отличить два класса DGAT, в добавление к их аминокислотным последовательностям.
Примеры полипептидов DGAT3 включают белки, кодируемые генами DGAT3 арахиса (Arachis hypogaea, Saha с соавт., 2006), а также их вариантами и/или мутантами. DGAT обладает незначительной или не обладает обнаружимой активностью MGAT, например, менее 300 пмоль/мин/мг белка, предпочтительно менее 200 пмоль/мин/мг белка, более предпочтительно менее 100 пмоль/мин/мг белка.
В варианте реализации изобретения экзогенный полинуклеотид по изобретению, кодирующий DGAT1, содержит одно или больше из следующего:
i) нуклеотиды, кодирующие полипептид, который содержит аминокислоты, последовательность которых приведена в любой из SEQ ID NO: 1 или 80-88, или его биологически активный фрагмент или полипептид, последовательность аминокислот которого по меньшей мере на 30% идентична любой одной или более из SEQ ID NO: 1 или 80-88,
ii) нуклеотиды, последовательность которых по меньшей мере на 30% идентична i), и
iii) полинуклеотид, который гибридизуется с одним или обоими из i) или ii) в строгих условиях.
В варианте реализации изобретения экзогенный полинуклеотид по изобретению, кодирующий DGAT2, содержит одно или более из следующего
i) нуклеотиды, кодирующие полипептид, который содержит аминокислоты, последовательность которых приведена в любой из SEQ ID NO: 2-9, или его биологически активный фрагмент или полипептид, последовательность аминокислот которого по меньшей мере на 30% идентична любой одной или больше из SEQ ID NO: 2-9,
ii) нуклеотиды, последовательность которых по меньшей мере на 30% идентична i), и
iii) полинуклеотид, который гибридизуется с одним или обоими из i) или ii) в строгих условиях.
В настоящем документе термин ʺфосфолипид:диацилглицерид ацилтрансферазаʺ (PDAT; EC 2.3.1.158) или его синоним ʺфосфолипид:1,2-диацил-sn-глицерид O-ацилтрансферазаʺ обозначает ацилтрансферазу, переносящую ацильную группу с фосфолипида, обычно ФХ, в положение sn-3 ДАГ, с образованием ТАГ. Данная реакция не связана с DGAT, и в ней используются фосфолипиды в качестве доноров ацила. Существует несколько форм PDAT в растительных клетках, включая PDAT1, PDAT2 или PDAT3 (Ghosal с соавт., 2007).
В настоящем документе термин «моноацилглицерил ацилтрансфераза» или «MGAT» обозначает белок, который переносит жирную ацильную группу из ацил-КоА к субстрату МАГ, например, sn-2 МАГ, для образования ДАГ. Таким образом, термин «активность моноацилглицерил ацилтрансферазы» по меньшей мере обозначает перенос ацильной группы от ацил-КоА к МАГ, для образования ДАГ. Термин «MGAT» в настоящем документе включает ферменты, которые действуют на субстраты sn-1/3 МАГ и/или sn-2 МАГ с образованием sn-1,3 ДАГ и/или sn-1,2/2,3-ДАГ, соответственно. В предпочтительном варианте реализации MGAT отдает предпочтение субстрату sn-2 МАГ относительно sn-1 МАГ, или в существенной мере использует в качестве субстрата только sn-2 МАГ. В настоящем документе MGAT не включает ферментов, которые предпочтительно переносят ацильную группу к ЛизоФК относительно МАГ, такие ферменты известны как LPAAT. Таким образом, MGAT предпочтительно использует нефосфорилированные моноацильные субстраты, даже если они также могут обладать низкой каталитической активностью по отношению к ЛизоФК. Предпочтительный MGAT не обладает обнаружимой активностью ацилирования ЛизоФК. Кроме того, MGAT может выполнять функцию DGAT, но, в основном, функционирует как MGAT, т. е., он обладает более выраженной каталитической активностью в роли MGAT, чем в роли DGAT, если активность фермента выражается в единицах нмоль продукта/мин/мг белка (также см. Yen с соавт., 2002). Существует три известных класса MGAT, обозначаемых, как MGAT1, MGAT2 и MGAT3, соответственно. Примеры полипептидов MGAT1, MGAT2 и MGAT3 описаны в WO2013/096993.
В настоящем документе «путь MGAT» обозначает анаболический путь образования ТАГ, отличный от пути Кеннеди, в котором ДАГ образуется ацилированием sn-1 МАГ или, предпочтительно, sn-2 МАГ, катализированным MGAT. В дальнейшем ДАГ может быть использован для образования ТАГ или других липидов. В WO2012/000026 продемонстрировано, во-первых, что листовая ткань растения может синтезировать МАГ из G-3-P таким образом, что МАГ доступен экзогенной MGAT, экспрессирующейся в листовой ткани, во-вторых, что MGAT из различных источников может функционировать в тканях растения, что требует успешного взаимодействия с другими факторами растения, принимающими участие в синтезе липидов, и в-третьих, что ДАГ, образованный под действием экзогенной MGAT, доступен растительной DGAT или экзогенной DGAT для выработки ТАГ. Активность MGAT и DGAT может быть проанализирована путем введения конструкций, кодирующих ферменты (или ферменты-кандидаты) в Saccharomyces cerevisiae, штамм H1246 и демонстрирует аккумуляцию ТАГ.
Некоторые из мотивов, важность которых для каталитической активности DGAT2 продемонстрирована в некоторых DGAT2, также являются консервативными в MGAT ацилтрансферазах. Особый интерес представляет предполагаемый домен связывания с нейтральными липидами с консенсусной последовательностью FLXLXXXN (SEQ ID NO: 14), где каждый X независимо представляет собой любую аминокислоту, кроме пролина, и N представляет собой любую неполярную аминокислоту, расположенную в пределах N-концевого трансмембранного участка, после которого следует предполагаемый домен глицерил/фосфолипид ацилтрансферазы. Мотив FLXLXXXN (SEQ ID NO: 14) найден в DGAT2 мыши (аминокислоты 81-88) и MGAT1/2, но не в DGAT2 дрожжей или растения. Он является важным для активности DGAT2 мыши. Другие мотивы последовательности DGAT2 и/или MGAT1/2 включают:
1. В высокой степени консервативный трипептид YFP (SEQ ID NO: 10) в большинстве полипептидов DGAT2, а также в MGAT1 и MGAT2, например, присутствующий как аминокислоты 139-141 в DGAT2 мыши. Мутация данного мотива в DGAT2 дрожжей посредством неконсервативных замен делает фермент нефункциональным.
2. Тетрапептид HPHG (SEQ ID NO: 11), в высокой степени консервативный в MGAT, а также в последовательностях DGAT2 животных и грибов, например, присутствующий как аминокислоты 161-164 в DGAT2 мыши, и важный для каталитической активности по меньшей мере в DGAT2 дрожжей и мыши. Растительные ацилтрансферазы DGAT2 содержат вместо него консервативную последовательность EPHS (SEQ ID NO: 12), поэтому консервативные модификации в первом и четвертом положениях аминокислот могут быть переносимыми.
3. Более длинный консервативный мотив, который является частью предполагаемого домена фосфолипида глицерина. Примером данного мотива является
RXGFX(K/R)XAXXXGXXX(L/V)VPXXXFG(E/Q) (SEQ ID NO:13), который присутствует как аминокислоты 304-327 в DGAT2 мыши. Последовательность аминокислот в данном мотиве менее консервативна, чем в остальных, как можно было бы ожидать на основании его длины, и гомологи могут быть распознаны путем поиска мотива. Расположение может варьировать для более консервативных аминокислот, т. е., могут присутствовать дополнительные X аминокислоты в пределах мотива или меньшее количество X аминокислот, по сравнению с приведенной выше последовательностью.
Одним важным компонентом в синтезе глицеролипида из жирных кислот, этерифицированных АЦБ или КоА, является фермент, sn-глицерил-3-фосфат ацилтрансфераза (GPAT), которая представляет собой другой полипептид, принимающий участие в биосинтезе неполярных липидов. Указанный фермент принимает участие в различных метаболических путях и осуществлении физиологических функций. Он катализирует следующую реакцию: G3P+жирный ацил-АПБ или -КоА →LPA+свободный-АПБ или -КоА. Катализируемая GPAT реакция происходит в трех отдельных субклеточных структурах растения: пластиде, эндоплазматическом ретикулуме (ЭР) и митохондрии. Указанные реакции катализируются тремя различными видами ферментов GPAT, растворимой формой, которая локализована в строме пластиды, которая использует ацил-АПБ в качестве своего природного ацильного субстрата (ПGPAT на Фиг. 1), и двумя мембраносвязанными формами, локализованными в ЭР и митохондрии, которые используют ацил-КоА и ацил-АПБ в качестве природных доноров ацила, соответственно (Chen с соавт., 2011).
В настоящем документе термин «глицерил-3-фосфат ацилтрансфераза» («GPAT»; EC 2.3.1.15) и его синоним «глицерил-3-фосфат O-ацилтрансфераза» обозначают белок, который ацилирует глицерил-3-фосфат (G-3-P), с образованием ЛизоФК и/или МАГ, причем последний продукт образуется, если GPAT дополнительно обладает активностью фосфатазы по отношению к ЛизоФК. Ацильная группа, которая переносится, происходит от ацил-КоА, если GPAT относится к ЭР-типу GPAT («ацил-КоА:sn-глицерил-3-фосфат 1-O-ацилтрансфераза», дополнительно обозначается как «микросомальная GPAT»), или от ацил-АЦФ, если GPAT является GPAT пластидного типа (GPATПТ). Таким образом, термин «активность глицерил-3-фосфат ацилтрансферазы» обозначает ацилирование G-3-P с образованием ЛизоФК и/или МАГ. Термин «GPAT» включает ферменты, которые ацилируют G-3-P с образованием sn-1 LPA и/или sn-2 LPA, предпочтительно sn-2 LPA. Предпочтительно, GPAT, которая может повышенно экспрессироваться при модификации Pull, представляет собой мембраносвязанную GPAT, которая функционирует в ЭР клетки, более предпочтительно, GPAT9, а пластидная GPAT, которая подавляется при модификации Prokaryotic pathway, представляет собой растворимую GPAT («пластидную GPAT»).В предпочтительном варианте реализации GPAT обладает активностью фосфатазы. В наиболее предпочтительном варианте реализации GPAT представляет собой sn-2 GPAT, обладающий активностью фосфатазы, которая продуцирует sn-2 МАГ.
В настоящем документе термин « sn-1 глицерил-3-фосфат ацилтрансфераза» (sn-1 GPAT) обозначает белок, который ацилирует sn-глицерил-3-фосфат (G-3-P), с предпочтительным образованием 1-ацил -sn-глицерил-3-фосфата (sn-1 LPA). Таким образом, термин «активность sn-1 глицерил-3-фосфат ацилтрансферазы» обозначает ацилирование sn-глицерил-3-фосфата с образованием 1-ацил-sn-глицерил-3-фосфата (sn-1 LPA).
В настоящем документе термин « sn-2 глицерил-3-фосфат ацилтрансфераза» (sn-2 GPAT) обозначает белок, который ацилирует sn-глицерил-3-фосфат (G-3-P) с предпочтительным образованием 2-ацил-sn-глицерил-3-фосфата (sn-2 LPA). Таким образом, термин «активность sn-2 глицерил-3-фосфат ацилтрансферазы» обозначает ацилирование sn-глицерил-3-фосфата с образованием 2-ацил-sn-глицерил-3-фосфата (sn-2 LPA).
Семейство GPAT большое, и все известные члены содержат два консервативных домена, ацилтрансферазный домен plsC (PF01553; SEQ ID NO: 15) и домен суперсемейства галогенкислоты дегалогеназа-подобной гидролазы (PF12710; SEQ ID NO: 16), а также их варианты. В дополнение к этому, по меньшей мере в Arabidopsis thaliana, все GPAT в подклассах GPAT4-GPAT8 содержат N-концевой участок, гомологичный домену фосфосерин фосфатазы (PF00702; SEQ ID NO: 17), и GPAT, которые продуцируют МАГ в качестве продукта, могут быть идентифицированы по наличию такого гомологичного участка. Некоторые GPAT, эндогенно экспрессируемые в ткани листа, содержат консервативную последовательность аминокислот GDLVICPEGTTCREP (SEQ ID NO:18). GPAT4 и GPAT6 содержат консервативные остатки, которые известны как критические для активности фосфатазы, конкретно консервативные аминокислоты в мотиве I (DXDX[T/V][L/V]; SEQ ID NO: 19) и мотиве III (K-[G/S][D/S]XXX[D/N]; SEQ ID NO: 20), расположены на N-конце (Yang с соавт., 2010).
Гомологи Arabidopsis GPAT4 (номер доступа NP_171667.1) и GPAT6 (NP_181346.1) включают AAF02784.1 (Arabidopsis thaliana), AAL32544.1 (Arabidopsis thaliana), AAP03413.1 (Oryza sativa), ABK25381.1 (Picea sitchensis), ACN34546.1 (Zea Mays), BAF00762.1 (Arabidopsis thaliana), BAH00933.1 (Oryza sativa), EAY84189.1 (Oryza sativa), EAY98245.1 (Oryza sativa), EAZ21484.1 (Oryza sativa), EEC71826.1 (Oryza sativa), EEC76137.1 (Oryza sativa), EEE59882.1 (Oryza sativa), EFJ08963.1 (Selaginella moellendorffii), EFJ11200.1 (Selaginella moellendorffii), NP_001044839.1 (Oryza sativa), NP_001045668.1 (Oryza sativa), NP_001147442.1 (Zea mays), NP_001149307.1 (Zea mays), NP_001168351.1 (Zea mays), AFH02724.1 (Brassica napus) NP_191950.2 (Arabidopsis thaliana), XP_001765001.1 (Physcomitrella patens), XP_001769671.1 (Physcomitrella patens), (Vitis vinifera), XP_002275348.1 (Vitis vinifera), XP_002276032.1 (Vitis vinifera), XP_002279091.1 (Vitis vinifera), XP_002309124.1 (Populus trichocarpa), XP_002309276.1 (Populus trichocarpa), XP_002322752.1 (Populus trichocarpa), XP_002323563.1 (Populus trichocarpa), XP_002439887.1 (Sorghum bicolor), XP_002458786.1 (Sorghum bicolor), XP_002463916.1 (Sorghum bicolor), XP_002464630.1 (Sorghum bicolor), XP_002511873.1 (Ricinus communis), XP_002517438.1 (Ricinus communis), XP_002520171.1 (Ricinus communis), ACT32032.1 (Vernicia fordii), NP_001051189.1 (Oryza sativa), AFH02725.1 (Brassica napus), XP_002320138.1 (Populus trichocarpa), XP_002451377.1 (Sorghum bicolor), XP_002531350.1 (Ricinus communis), and XP_002889361.1 (Arabidopsis lyrata).
Были выделены растворимые пластидные GPAT (ПGPAT, также известные как ATS1 в Arabidopsis thaliana), причем кодирующие их гены клонированы из нескольких видов растений, таких как горох (Pisum sativum, номер доступа: P30706.1), шпинат (Spinacia oleraceа, номер доступа: Q43869.1), тыква (Cucurbita moschate, номер доступа: P10349.1), огурец (Cucumis sativus, номер доступа: Q39639.1) и Arabidopsis thaliana (номер доступа: Q43307.2). Растворимая пластидная GPAT представляет собой первую фиксированную стадию того, что известно как прокариотный путь синтеза глицеролипидов и действует только в пластиде (Фиг. 1). Так называемый прокариотный путь расположен исключительно в пластидах растений и обеспечивает сборку ДАГ для синтеза галактолипидов (MGDG и ДГМГ), которые содержат C16:3 жирные кислоты, этерифицированные в положении sn-2 глицеринового скелета.
Консервативные мотивы и/или остатки могут использоваться в качестве основанного на последовательности диагностического признака для идентификации ферментов GPAT. Альтернативно, может применяться более строгий анализ, основанный на функции. Такой анализ включает, например, подкормку меченым глицерил-3-фосфатом клеток или микросом и количественное определение уровней меченых продуктов тонкослойной хроматографией или подобным методом. Активность GPAT приводит к образованию меченой LPA, тогда как активность GPAT/фосфатазы приводит к образованию меченого МАГ.
В настоящем документе термин "ацилтрансфераза лизофосфатидиновой кислоты" (LPAAT; EC 2.3.1.51) и его синонимы ʺ1-ацил-глицерил-3-фосфат ацилтрансферазаʺ, ʺацил-КоА:1-ацил-sn-глицерол-3-фосфат 2-O-ацилтрансферазаʺ и ʺ1-ацилглицерил-3-фосфат O-ацилтрансферазаʺ обозначают белок, который ацилирует лизофосфатидиновую кислоту (LPA) с образованием фосфатидиновой кислоты (ФК, PA). Ацильную группу для переноса получают из ацил-КоА, если LPAAT представляет собой LPAAT ЭР-типа, или из ацил-АПБ, если LPAAT представляет собой LPAAT пластидного типа (ПLPAAT). Таким образом, термин "активность ацилтрансферазы лизофосфатидиновой кислоты" обозначает ацилирование LPA с образованием ФК.
Полипептиды покрытия масляного включения
Семена растения и пыльца аккумулируют ТАГ в субклеточных структурах, называемых масляными включениями, диаметр которых в общем находится в диапазоне 0,5-2,5 мкм. В настоящем документе ʺлипидные капелькиʺ, также называемые ʺмасляными включениямиʺ, представляет собой богатые липидами клеточные органеллы для хранения или обмена нейтральных липидов, в основном содержащие ТАГ. Размер липидных капелек может значительно варьировать, от около 20 нм до 100 мкм. Указанные органеллы содержат ядро ТАГ, окруженное фосфолипидным монослоем, содержащим несколько инкорпорированных белков, принимающих участие в метаболизме и хранении липидов, а также в переносе липидов к другим мембранам, в том числе олеозинов, если масляные включения происходят из растительных семян или тканей цветка (Jolivet с соавт., 2004). В общем, они содержат 0,5-3,5% белка, в то время как остальная часть представлена липидом. Они являются наименее плотными органеллами в большинстве клеток и могут, таким образом, с легкостью быть выделены путем центрифугирования с флотацией. Олеозины представляют самый распространенный (по меньшей мере 80%) белок в мембране масляных включений из семян.
В настоящем документе термин «олеозин» обозначает амфифильный белок, присутствующий в мембране масляных включений в запасных тканях семян (см., например, Huang, 1996; Lin с соавт., 2005; Capuano с соавт., 2007; Lui с соавт., 2009; Shimada и Hara-Nishimura, 2010), и искусственно созданные варианты (см., например WO2011/053169 и WO2011/127118).
Олеозин обладает низкой М.м. (15-26000), что соответствует приблизительно 140-230 аминокислотных остатков, и это позволяет им плотно упаковываться на поверхности масляных включений. В семенах каждого вида обычно представлены два или более олеозинов с различной М.м.. Каждая молекула олеозина содержит относительно гидрофильный, вариабельный N-концевой домен (например, длиной около 48 остатков аминокислоты), центральный полностью гидрофобный домен (например, длиной приблизительно 70-80 остатков аминокислоты), который особенно богат алифатическими аминокислотами, такими как аланин, глицин, лейцин, изолейцин и валин, и амфифильный α-спиральный домен размером около 30-40 остатков аминокислот на C-конце или вблизи него. Обычно центральный гидрофобный домен содержит в центре мотив пролинового узла размером около 12 остатков. В общем, центральный участок гидрофобных остатков вставлен в липидное ядро, и амфифильный N-концевой участок и/или амфифильный C-концевой участок размещен на поверхности масляных включений, причем положительно заряженные остатки инкорпорированы в фосфолипидный монослой, и отрицательно заряженные участки контактируют с внешней средой.
В настоящем документе термин «олеозин» включает полиолеозины, содержащие несколько полипептидов олеозина, соединенных вместе способом «голова к хвосту» в качестве единого полипептида (WO2007/045019), например, 2x, 4x или 6x пептидов олеозина, и калеозины, которые связывают кальций и составляют минорный белковый компонент белков, покрывающих масляные включения в семенах (Froissard с соавт., 2009), а также стеролеозины, которые связываются со стероидами (WO2011/053169). Однако, в общем, значительная часть (по меньшей мере 80%) олеозинов в масляных включениях не будет относиться к калеозинам и/или стеролеозинам. Термин ʺолеозинʺ дополнительно включает полипептиды олеозина, модифицированные искусственным путем, т.е. олеозины, в которых один или более остатков аминокислот природного олеозина искусственно заменены остатками цистеина, как описано в WO2011/053169. Обычно, 4-8 остатков заменены искусственным путем, предпочтительно 6 остатков, но может быть заменено от 2 до 14 остатков. Предпочтительно, амфифильные N-концевой и C-концевой домены содержат замены цистеина. Модификация повышает способность олеозинов к образованию поперечных связей и повышает термостабильность и/или устойчивость белков к разложению протеазами.
Известно значительное количество последовательностей белка олеозина и нуклеотидных последовательностей, кодирующих его, из множества различных видов растений. Примеры включают, без ограничений, олеозины Arabidposis, рапса, кукурузы, риса, арахиса, клещевины, сои, льна, винограда, капусты, хлопчатника, подсолнечника, сорго и ячменя. Примеры олеозинов (с номерами доступа) включают олеозин Brassica napus (CAA57545.1; SEQ ID NO:95), Brassica napus олеозин S1-1 (ACG69504.1; SEQ ID NO:96), Brassica napus олеозин S2-1 (ACG69503.1; SEQ ID NO:97), Brassica napus олеозин S3-1 (ACG69513.1; SEQ ID NO:98), Brassica napus олеозин S4-1 (ACG69507.1; SEQ ID NO:99), Brassica napus олеозин S5-1 (ACG69511.1; SEQ ID NO:100), Arachis hypogaea олеозин 1 (AAZ20276.1; SEQ ID NO:101), Arachis hypogaea олеозин 2 (AAU21500.1; SEQ ID NO:102), Arachis hypogaea олеозин 3 (AAU21501.1; SEQ ID NO:103), Arachis hypogaea олеозин 5 (ABC96763.1; SEQ ID NO:104), Ricinus communis олеозин 1 (EEF40948.1; SEQ ID NO:105), Ricinus communis олеозин 2 (EEF51616.1; SEQ ID NO:106), олеозин Glycine max изоформа a (P29530.2; SEQ ID NO:107), олеозин Glycine max изоформа b (P29531.1; SEQ ID NO:108), олеозин Linum usitatissimum низкомолекулярная изоформа (ABB01622.1; SEQ ID NO:109), олеозин Linum usitatissimum высокомолекулярная изоформа (ABB01624.1; SEQ ID NO:110), олеозин Helianthus annuus (CAA44224.1; SEQ ID NO:111), олеозин Zea mays (NP_001105338.1; SEQ ID NO:112), Brassica napus стеролеозин (ABM30178.1; SEQ ID NO:113), Brassica napus стеролеозин SLO1-1 (ACG69522.1; SEQ ID NO:114), Brassica napus стеролеозин SLO2-1 (ACG69525.1; SEQ ID NO:115), стеролеозин Sesamum indicum (AAL13315.1; SEQ ID NO:116), стеролеозин Zea mays (NP_001152614.1; SEQ ID NO:117), калеозин CLO-1 Brassica napus (ACG69529.1; SEQ ID NO:118), Brassica napus калеозин CLO-3 (ACG69527.1; SEQ ID NO:119), калеозин Sesamum indicum (AAF13743.1; SEQ ID NO:120), калеозин Zea mays (NP_001151906.1; SEQ ID NO:121), калеозин Glycine max (AAB71227). Другими функционально эквивалентными полипептидами липидной инкапсуляции являются пластоглобулины и полипептиды белка жировых капель миокарда (БЖКМ) (WO2011/127118).
В варианте реализации изобретения экзогенный полинуклеотид по изобретению, кодирующий олеозин, содержит, если не указано иное, одно или более из следующего:
i) нуклеотиды, кодирующие полипептид, который содержит аминокислоты, последовательность которых приведена в любой из SEQ ID NO: 95-112, или его биологически активный фрагмент или полипептид, последовательность аминокислот которого по меньшей мере на 30% идентична любой одной или более из SEQ ID NO: 95-112,
ii) нуклеотиды, последовательность которых по меньшей мере на 30% идентична i), и
iii) полинуклеотид, который гибридизуется с одним или обоими из i) или ii) в строгих условиях.
В варианте реализации изобретения экзогенный полинуклеотид по изобретению, кодирующий стеролеозин, содержит, если не указано иное, одно или более из следующего:
i) нуклеотиды, кодирующие полипептид, который содержит аминокислоты, последовательность которых приведена в любой из SEQ ID NO: 113-117, или его биологически активный фрагмент или полипептид, последовательность аминокислот которого по меньшей мере на 30% идентична одной или более из SEQ ID NO: 113-117,
ii) нуклеотиды, последовательность которых по меньшей мере на 30% идентична i), и
iii) полинуклеотид, который гибридизуется с одним или обоими из i) или ii) в строгих условиях.
В настоящем документе ʺбелок, связанный с липидной капелькойʺ или ʺБСЛКʺ обозначает полипептид, который в растениях связан с липидными капельками в тканях или органах, кроме семян, пыльников и пыльцы, например, тканях фруктов, включая перикарпий и мезокарпий. БСЛК могут быть связаны с масляными включениями в семенах, пыльниках или пыльце, а также в тканях или органах, кроме семян, пыльников и пыльцы. Они отличаются от олеозинов, которые являются полипептидами, связанными с поверхностью липидных капелек в тканях семени, пыльниках и пыльце. БСЛК в настоящем документе включают полипептиды БСЛК, образованные природным путем в тканях растения, а также варианты аминокислотной последовательности, полученные искусственным путем. Функция таких вариантов может быть проанализирована, как проиллюстрировано в Примере 15.
Horn с соавт. (2013) идентифицировали два гена БСЛК, которые экспрессируются в перикарпии авокадо. Аминокислотная последовательность кодируемых полипептидов авокадо БСЛК1 и БСЛК2 была на 62% идентичной, а также обладала гомологией к полипептиду, кодируемому At3g05500 Arabidopsis и SRPP-подобному белку резинового дерева. Gidda с соавт. (2013) идентифицировали три гена БСЛК, которые экспрессировались в мезокарпии, но не в ядрах масличной пальмы (Elaeis guineensis) и сделали вывод о том, что гены БСЛК были специфичными для растения и консервативными для всех видов растений. Полипептиды БСЛК могут содержать дополнительные домены (Gidda с соавт., (2013). Гены, кодирующие БСЛК, в общем, активизируются в тканях, не относящихся к семенам, с обильным содержанием липидов, и могут быть идентифицированы таким образом, однако считается, что они экспрессируются во всех клетках, не относящихся к семенам, которые вырабатывают масло, в том числе для временного хранения. Horn с соавт. (2013) иллюстрируют филогенетическое дерево SRPP-подобных белков в растениях. Характерные полипептиды БСЛК описаны в Примере 15 настоящего документа. Гомологи БСЛК в других видах растений могут быть с легкостью идентифицированы специалистами в данной области.
В варианте реализации изобретения экзогенный полинуклеотид по изобретению, кодирующий БСЛК, содержит, если не указано иное, одно или более из следующего:
i) нуклеотиды, кодирующие полипептид, который содержит аминокислоты, последовательность которых приведена в любой из SEQ ID NO: 237, 239 или 241, или его биологически активный фрагмент или полипептид, последовательность аминокислот которого по меньшей мере на 30% идентична любой одной или более из SEQ ID NO: 237, 239 или 241,
ii) нуклеотиды, последовательность которых по меньшей мере на 30% идентична i), и
iii) полинуклеотид, который гибридизуется с одним или обоими из i) или ii) в строгих условиях.
В настоящем документе термин «Полипептид, принимающий участие в биосинтезе крахмала» обозначает любой полипептид, регуляция которого в клетке вниз до уровней ниже нормальных (дикий тип), приводит к снижению уровня синтеза крахмала и снижению уровней крахмала. Примером такого полипептида является АГФаза.
В настоящем документе термин «фосфорилаза АДФ-глюкозы» или «АГФаза» обозначает фермент, который регулирует биосинтез крахмала, катализируя превращение глюкозы-1-фосфата и АТФ в АДФ-глюкозу, которая служит строительным блоком для полимеров крахмала. Активная форма фермента АГФазы состоит из 2 больших и 2 маленьких субъединиц.
Фермент АДФаза в растениях существует, главным образом, в форме тетрамера, который состоит из 2 больших и 2 маленьких субъединиц. Хотя каталитическая и регуляторная роли этих субъединиц отличаются в зависимости от вида (Kuhn с соавт., 2009), в растениях маленькая субъединица проявляет, главным образом, каталитическую активность. Молекулярная масса маленькой субъединицы составляет приблизительно 50-55 кДа. Молекулярная масса большой субъединицы составляет приблизительно 55-60 кДа. Фермент растения интенсивно активизируется 3-фосфоглицератом (ФГА), продуктом фиксации диоксида углерода; в отсутствие ФГА фермент проявляет только около 3% активности. Растительная АГФаза также интенсивно ингибируется неорганическим фосфатом (Фн). Наоборот, бактериальная и водорослевая АГФазы существуют в виде гомотетрамеров размером 50 кДа. Водорослевый фермент, подобно его растительному двойнику, активируется ФГА и ингибируется Фн, тогда как бактериальный фермент активируется фруктозо-1,6-бифосфатом (ФБФ) и ингибируется АМФ и Фн.
ТАГ липазы и бета-окисление
В настоящем документе термин «полипептид, принимающий участие в разложении липида и/или который снижает содержание липида» обозначает любой полипептид, катаболизирующий липид, причем регуляция полипептида в клетке вниз до уровней ниже нормальных (дикий тип) приводит к повышению уровня масла, например, жирных кислот и/или ТАГ, в клетке, предпочтительно клетке вегетативной части, клубне, свекле или семени растения. Примеры таких полипептидов включают, без ограничений, липазы или липазу, такую как полипептид CGi58 (подобный сравнительному идентификатору гена-58), САХАРОЗАВИСИМУЮ триацилглицерил липазу 1 (SDP1) (см., например, Kelly с соавт., 2011) и липазу, описанную в WO 2009/027335.
В настоящем документе термин «ТАГ липаза» (EC.3.1.1.3) обозначает белок, который гидролизует ТАГ до одной или более жирных кислот и любого из ДАГ, МАГ или глицерина. Таким образом, термин «активность ТАГ липазы» обозначает гидролиз ТАГ до глицерина и жирных кислот.
В настоящем документе термин «CGi58» обозначает растворимую ацил-КоА-зависимую ацилтрансферазу лизофосфатидиновой кислоты, кодируемую геном At4g24160 в Arabidopsis thaliana и его гомологами в других растениях, и «Ict1p» в дрожжах и его гомологами. Растительный ген, например, локус гена Arabidopsis At4g24160, экспрессируется как две альтернативные копии: более длинная полноразмерная изоформа (At4g24160.1) и более короткая изоформа (At4g24160.2), в которой отсутствует часть 3' конца (см. James с соавт., 2010; Ghosh с соавт., 2009; US 201000221400). Обе иРНК кодируют белок, гомологичный человеческому белку CGI-58 и другим ортологичным членам данного семейства α/гидролазы (АБГД). В варианте реализации белок CGI58 (At4g24160) содержит три мотива, которые являются консервативными в видах растений: мотив липазы GXSXG (SEQ ID NO: 127), мотив ацилтрансферазы HX(4)D (SEQ ID NO: 128), и VX(3)HGF, вероятный мотив связывания с липидом (SEQ ID NO: 129). Белок CGI-58 человека обладает активностью ацилтрансферазы лизофосфатидиновой кислоты (LPAAT), но не активностью липазы. И наоборот, белки растения и дрожжей содержат канонический мотив последовательности липазы GXSXG (SEQ ID NO: 127), отсутствующий в белках позвоночных (человек, мыши и рыбки данио) и обладающий липазной и фосфолипазной активностью (Ghosh с соавт., 2009). Хотя белки CGI58 растения и дрожжей, похоже, обладают обнаружимыми уровнями активности липазы ТАГ и фосфолипазы А, в дополнение к активности LPAAT, белок человека не проявляет такой активности.
Разрушение гомологичного гена CGI-58 в Arabidopsis thaliana приводит к аккумуляции капелек нейтрального липида в зрелых листьях. Масс-спектроскопический анализ капелек липида, выделенных от мутантов с утратой функции cgi-58,показал, что они содержат триацилглицериды со специфичными жирными кислотами, обычными для листьев. Листья зрелых растений cgi-58 демонстрируют заметное повышение абсолютных уровней триацилглицеридов, более чем в 10 раза превышающих уровни в растениях дикого типа. Уровни липида в запасающих масло семенах cgi-58 оставались неизменными, и, в отличие от мутаций с β-окислением, семена cgi-58 прорастали и росли нормально, не требуя спасения с помощью сахарозы (James с соавт., 2010).
Примеры нуклеотидов, кодирующих полипептиды CGi58, включают нуклеотиды из Arabidopsis thaliana (NM_118548.1 кодирующий NP_194147.2; SEQ ID NO:130), Brachypodium distachyon (XP_003578450.1; SEQ ID NO:131), Glycine max (XM_003523590.1 кодирующий XP_003523638.1; SEQ ID NO:132), Zea mays (NM_001155541.1 кодирующий NP_001149013.1; SEQ ID NO:133), Sorghum bicolor (XM_002460493.1 кодирующий XP_002460538.1; SEQ ID NO:134), Ricinus communis (XM_002510439.1 кодирующий XP_002510485.1; SEQ ID NO:135), Medicago truncatula (XM_003603685.1 кодирующий XP_003603733.1; SEQ ID NO:136), и Oryza sativa (кодирующий EAZ09782.1).
В варианте реализации изобретения генетическая модификация по изобретению подавляет эндогенную выработку CGi58, причем CGi58 кодируется одним или более из следующего:
i) нуклеотиды, содержащие последовательность, приведенную в любой из SEQ ID NO: 130-136,
ii) нуклеотиды, содержащие последовательность, которая по меньшей мере на 30% идентична любой одной или более из SEQ ID NO: 130-136, и
iii) полинуклеотид, который гибридизуется с одним или обоими из i) или ii) в строгих условиях.
Другие липазы, обладающие активностью липазы в отношении ТАГ, включают САХАРО-ЗАВИСИМУЮ триацилглицерид липазу 1 (SDP1, см., например, Eastmond с соавт., 2006; Kelly с соавт., 2011) и SDP1-подобные полипептиды, найденные в растительных видах, а также дрожжах (полипептид TGL4) и животных клетках, которые принимают участие в расщеплении запасных ТАГ. По-видимому, SDP1 и SDP1-подобные полипептиды ответственны за начало расщепления ТАГ в семенах после прорастания (Eastmond с соавт., 2006). Растения с мутантным SDP1, в отсутствие экзогенных WRI1 и DGAT1, демонстрируют повышенные уровни ПНЖК в ТАГ. В настоящем документе ʺполипептиды SDP1ʺ включают SDP1 полипептиды, SDP1-подобные полипептиды и их гомологи в растительных видах. Размер SDP1 и SDP1-подобных полипептидов в растениях составляет 800-910 остатков аминокислот, причем они содержат пататин-подобный домен ацилгидролазы, который может связываться с поверхностями масляных включений и предпочтительно гидролизовать ТАГ относительно ДАГ или МАГ. Считается, что SDP1 проявляет предпочтение к гидролизу ацильной группы в положении sn-2 ТАГ. Arabidopsis содержит по меньшей мере три гена, кодирующих SDP1 липазы, а именно SDP1 (номер доступа NP_196024, нуклеотидная последовательность SEQ ID NO: 163 и гомологи в других видах), SDP1 (номер доступа NM_202720 и гомологи в других видах, Kelly с соавт., 2011) и ATGLL (At1g33270) (Eastmond с соавт., 2006). Особый интерес с точки зрения снижения активности гена представляют гены SDP1, которые экспрессируются в вегетативных тканях растений, например, в листьях, стеблях и корнях. Таким образом, уровни неполярных липидов в вегетативных частях растения могут быть повышены путем снижения активности полипептидов SDP1 в частях растения, например, с помощью мутации эндогенного гена, кодирующего полипептид SDP1, или введением экзогенного гена, который кодирует сайленсинговую РНК молекулу, которая снижает экспрессию эндогенного гена SDP1. Такое снижение приносит конкретную пользу на урожаях клубней, таких как сахарная свекла и картофель, и в ʺвысокосахарозныхʺ растениях, таких как сахарный тростник и сахарная свекла.
Гены, кодирующие гомологи SDP1 (в том числе SDP1-подобные гомологи) в растительных видах выбора, могут быть с легкостью идентифицированы по гомологии к известным последовательностям гена SDP1. Известные нуклеотидные или аминокислотные последовательности SDP1 включают номера доступа: в Brassica napus, GN078290 (SEQ ID NO:164), GN078281, GN078283; Capsella rubella, XP_006287072; Theobroma cacao, XP_007028574.1; Populus trichocarpa, XP_002308909 (SEQ ID NO:166); Prunus persica, XP_007203312; Prunus mume, XP_008240737; Malus domestica, XP_008373034; Ricinus communis, XP_002530081; Medicago truncatula, XP_003591425 (SEQ ID NO:167); Solanum lycopersicum, XP_004249208; Phaseolus vulgaris, XP_007162133; Glycine max, XP_003554141 (SEQ ID NO:168); Solanum tuberosum, XP_006351284; Glycine max, XP_003521151; Cicer arietinum, XP_004493431; Cucumis sativus, XP_004142709; Cucumis melo, XP_008457586; Jatropha curcas, KDP26217; Vitis vinifera, CBI30074; Oryza sativa, Japonica Group BAB61223; Oryza sativa, Indica Group EAY75912; Oryza sativa, Japonica Group NP_001044325; Sorghum bicolor, XP_002458531 (SEQ ID NO:169); Brachypodium distachyon, XP_003567139 (SEQ ID NO:165); Zea mays, AFW85009; Hordeum vulgare, BAK03290 (SEQ ID NO:172); Aegilops tauschii, EMT32802; Sorghum bicolor, XP_002463665; Zea mays, NP_001168677 (SEQ ID NO:170); Hordeum vulgare, BAK01155; Aegilops tauschii, EMT02623; Triticum urartu, EMS67257; Physcomitrella patens, XP_001758169 (SEQ ID NO:171). Предпочтительные последовательности SDP1 для применения в генетических конструкциях с целью подавления экспрессии эндогенных генов, происходят из кДНК, соответствующих генам, которые экспрессируются в наибольшей степени в клетках, вегетативных частях растения или семенах, в зависимости от того, что подлежит модификации. Нуклеотидные последовательности, которые являются в высокой степени консервативными между кДНК, соответствующими всем генам SDP1 в виде растений, являются предпочтительными, если желательно сократить активность всех членов генного семейства в данном виде.
В варианте реализации изобретения генетическая модификация по изобретению подавляет эндогенную выработку SDP1, причем SDP1 кодируется одним или более из следующего:
i) нуклеотиды, последовательность которых приведена в любой из SEQ ID NO: 163-174,
ii) нуклеотиды, последовательность которых по меньшей мере на 30% идентична любой одной или более последовательностям, приведенным как SEQ ID NO: 163-174, и
iii) последовательность нуклеотидов, которая гибридизуется с одним или обоими из i) или ii) в строгих условиях.
Как проиллюстрировано в Примерах, снижение экспрессии и/или активности ТАГ липазы SDP1 в листьях растения значительно повышало содержание ТАГ, как с точки зрения количества ТАГ, которые аккумулируются, так и более раннего наступления периода аккумуляции в ходе развития растения, в контексте co-экспрессии фактора транскрипции WRI1 и ацилтрансферазы ацилов жирных кислот. В частности, повышение наблюдалось в растениях до цветения, и составляло до около 70% масс. (% сухой массы) на момент начала старения. Повышение было относительным по сравнению с уровнями ТАГ, наблюдаемыми в соответствующих листьях растений, трансформированных экзогенными полинуклеотидами, кодирующими WRI1 и ацилтрансферазу ацилов жирных кислот, но без модификации, которая снижает экспрессию и/или активность SDP1.
Кроме того, снижение экспрессии других генов катаболизма ТАГ в частях растения может повысить содержание ТАГ, например, генов ACX, кодирующих ацил-КоА оксидазы, таких как гены Acx1 (At4g16760 и гомологи в других видах растений) или Acx2 (At5g65110 и гомологи в других видах растений). Другим полипептидом, принимающим участие в катаболизме липидов, является PXA1, который представляет собой пероксисомальный переносчик АТФ-связывающей кассеты, необходимый для импорта жирных кислот для β-окисления (Zolman с соавт., 2001).
Экспорт жирных кислот из пластид
В настоящем документе термин "полипептид, который повышает экспорт жирных кислот из пластид клетки" обозначает любой полипептид, который способствует переносу жирных кислот из внутреннего пространства пластид (в клетках, которые содержат пластиды, таких как клетка вегетативной части, клубня, свеклы или семени растения) за пределы пластиды, т.е. в любую другую часть клетки, такую как эндоплазматический ретикулум (ЭР). Примеры таких полипептидов включают, но не ограничиваясь ими, тиоэстеразу C16 или C18 жирных кислот, такую как полипептид FATA или полипептид FATB, тиоэстеразу C8-C14 жирных кислот (которая дополнительно является полипептидом FATB), переносчик жирных кислот, такой как полипептид ABCA9 или длинноцепочечная ацил-КоА синтетаза (ДЦАС).
В настоящем документе термина ʺтиоэстераза жирных кислотʺ или ʺТЖК (FAT)ʺ обозначает фермент, катализирующий гидролиз тиоэфирной связи между ацильным фрагментом и ацилпереносящим белком (АПБ) в ацил-АПБ и высвобождение свободной жирной кислоты. Такие ферменты обычно функционируют в пластидах организма, синтезирующего жирные кислоты de novo. В настоящем документе термин ʺтиоэстераза C16 или C18 жирной кислотыʺ обозначает фермент, катализирующий гидролиз тиоэфирной связи между C16 и/или C18 ацильным фрагментом и АПБ в ацил-АПБ и высвобождение свободной C16 или C18 жирной кислоты в пластиде. Далее свободная жирная кислота переэтерифицируется до КоА в пластидном конверте, и в таком виде она транспортируется из пластиды. Специфичность субстрата фермента тиоэстеразы жирных кислот (ТЖК) в пластиде принимает участие в определении спектра длины цепи и степени насыщенности жирных кислот, экспортируемых из пластиды. Ферменты ТЖК могут быть классифицированы в два класса, на основании их специфичности в отношении субстрата и нуклеотидных последовательностей, FATA и FATB (EC 3.1.2.14) (Jones с соавт., 1995). Полипептиды FATA отдают предпочтение олеоил-АПБ в качестве субстрата, в то время как полипептиды FATB демонстрируют более высокую активность в отношении насыщенных ацил-АПБ с различной длиной цепи, например, воздействуя на пальмитоил-АПБ с образованием свободной пальмитиновой кислоты. Примеры полипептидов FATA, пригодных в соответствии с изобретением, включают, но не ограничиваясь ими, полипептиды из Arabidopsis thaliana (NP_189147), Arachis hypogaea (GU324446), Helianthus annuus (AAL79361), Carthamus tinctorius (AAA33020), Morus notabilis (XP_010104178,1), Brassica napus (CDX77369,1), Ricinus communis (XP_002532744,1) и Camelina sativa (AFQ60946,1). Примеры полипептидов FATB, пригодных в соответствии с изобретением, включают, но не ограничиваясь ими, полипептиды из Zea mays (AIL28766), Brassica napus (ABH11710), Helianthus annuus (AAX19387), Arabidopsis thaliana (AEE28300), Umbellularia californica (AAC49001), Arachis hypogaea (AFR54500), Ricinus communis (EEF47013) и Brachypodium sylvaticum (ABL85052.1).
Подкласс полипептидов FATB составляют тиоэстеразы жирных кислот, которые проявляют гидролитическую активность в отношении насыщенного ацильного фрагмента C8-C14, соединенного тиоэфирной связью с АПБ. Такие ферменты дополнительно обозначают как тиоэстеразы среднецепочечных жирных кислот (СЦЖК) или ферменты СЦ-FAT. Такие ферменты могут дополнительно проявлять активность тиоэстеразы в отношении C16-АПБ, в действительности, они могут проявлять более высокую активность тиоэстеразы в отношении субстрата C16 ацил-АПБ, чем субстрата СЦЖК-АПБ, тем не менее, они рассматриваются в настоящем документе как СЦЖК тиоэстеразы, если при экзогенной экспрессии в растительной клетке они продуцируют по меньшей мере 0,5% СЦЖК в общем содержании жирных кислот. Примеры СЦЖК тиоэстераз предоставлены в Примере 9 настоящего документа.
В настоящем документе термин ʺпереносчик жирной кислотыʺ связан с наличием в мембране пластиды полипептида, который принимает активное участие в переносе жирных кислот из пластиды за ее пределы. Примеры полипептидов ABCA9 (ABC переносчик A член семейства 9), пригодных в соответствии с изобретением, включают, но не ограничиваясь ими, полипептиды из Arabidopsis thaliana (Q9FLT5), Capsella rubella (XP_006279962.1), Arabis alpine (KFK27923.1), Camelina sativa (XP_010457652.1), Brassica napus (CDY23040.1) и Brassica rapa (XP_009136512.1).
В настоящем документе термин ʺацил-КоА синтетазаʺ или ʺАКС (ACS)ʺ (EC 6.2.1.3) обозначает полипептид, который входит в состав семейства лигазы, катализирующего образование жирного ацил-КоА в ходе двухстадийного процесса, проходящего через промежуточное аденилированное соединение, с использованием неэтерифицированной жирной кислоты, КоА и АТФ в качестве субстратов, с образованием эфира ацил-КоА, АМФ и пирофосфата в качестве продуктов. В настоящем документе термин ʺдлинноцепочечная ацил-КоА синтетазаʺ (ДАКС) обозначает АКС, которая проявляет активность по меньшей мере в отношении субстрата C18 свободной жирной кислоты, хотя она может обладать более широкой активностью в отношении любой из свободных C14-C20 жирных кислот. Эндогенные пластидные ферменты ДЦАС локализуются во внешней мембране пластиды и функционируют вместе с тиоэстеразой жирной кислоты с целью экспорта жирных кислот из пластиды (Schnurr с соавт., 2002). В Arabidopsis присутствует по меньшей мере девять идентифицированных генов ДЦАС (Shockey с соавт., 2002). Предпочтительными полипептидами ДЦАС являются принадлежащие к подклассу LACS9, которые в Arabidopsis являются основными пластидными ДЦАС. Примеры полипептидов ДЦАС, пригодных в соответствии с изобретением, включают, но не ограничиваясь ими, полипептиды из Arabidopsis thaliana (Q9CAP8), Camelina sativa (XP_010416710.1), Capsella rubella (XP_006301059.1), Brassica napus (CDX79212.1), Brassica rapa (XP_009104618.1), Gossypium raimondii (XP_012450538.1) и Vitis Vinifera (XP_002285853.1). Гомологи вышеупомянутых полипептидов в других видах могут быть с легкостью идентифицированы специалистами в данной области.
Полипептиды, принимающие участие в выработке диацилглицерида (ДАГ) в пластидах
Уровни неполярных липидов, например, в вегетативных частях растения могут быть дополнительно повышены путем снижения активности полипептидов, принимающих участие в выработке диацилглицерида (ДАГ) в пластиде в частях растения, например, мутацией эндогенного гена, кодирующего такой полипептид, или введением экзогенного гена, который кодирует сайленсинговую молекулу РНК, снижающую экспрессию гена-мишени, принимающего участие в выработке диацилглицерида (ДАГ) в пластиде.
В настоящем документе термин "полипептид, принимающий участие в выработке диацилглицерида (ДАГ) в пластиде" обозначает любой полипептид в пластиде (в клетках, которые содержат пластиды, таких как клетка вегетативной части, клубня, свеклы или семени растения), который непосредственно принимает участие в синтезе диацилглицерида. Примеры таких полипептидов включают, но не ограничиваясь ими, пластидную GPAT, пластидную LPAAT или пластидную PAP.
GPAT описаны в другом месте настоящего документа. Примеры пластидных полипептидов GPAT, которые могут служить мишенью для подавления согласно изобретению, включают, но не ограничиваясь ими, полипептиды из Arabidopsis thaliana (BAA00575), Capsella rubella (XP_006306544.1), Camelina sativa (010499766.1), Brassica napus (CDY43010.1), Brassica rapa (XP_009145198.1), Helianthus annuus (ADV16382.1) и Citrus unshiu (BAB79529.1). Гомологи в других видах могут с легкостью быть идентифицированы специалистами в данной области.
LPAAT описаны в другом месте настоящего документа. Как будет понятно квалифицированному специалисту, пластидные LPAAT, которые служат мишенью для подавления с целью снижения синтеза ДАГ в пластиде, не являются эндогенными LPAAT, которые функционируют за пределами пластиды, например, в ЭР, например, описанных в настоящем документе как пригодные для выработки ТАГ, содержащих среднецепочечные жирные кислоты. Примеры пластидных полипептидов LPAAT, которые могут служить мишенью для подавления согласно изобретению, включают, но не ограничиваясь ими, полипептиды Brassica napus (ABQ42862), Brassica rapa (XP_009137939.1), Arabidopsis thaliana (NP_194787.2), Camelina sativa (XP_010432969.1), Glycine max (XP_006592638.1) и Solanum tuberosum (XP_006343651.1). Гомологи вышеупомянутых полипептидов в других видах могут быть с легкостью идентифицированы специалистами в данной области.
В настоящем документе термин "фосфатаза фосфатидиновой кислоты (PAP) (EC 3.1.3.4)ʺ обозначает белок, осуществляющий гидролиз фосфатной группы на 3-sn-фосфатидате с образованием 1,2-диацил-sn-глицерида (ДАГ) и фосфата. Примеры пластидных полипептидов PAP, которые могут служить мишенью для подавления согласно изобретению, включают, но не ограничиваясь ими, полипептиды из Arabidopsis thaliana (Q6NLA5), Capsella rubella (XP_006288605.1), Camelina sativa (XP_010452170.1), Brassica napus (CDY10405.1), Brassica rapa (XP_009122733.1), Glycine max (XP_003542504.1) и Solanum tuberosum (XP_006361792.1). Гомологи вышеупомянутых полипептидов в других видах могут быть с легкостью идентифицированы специалистами в данной области.
Импорт жирных кислот в пластиды
Уровни неполярных липидов в вегетативных частях растения могут быть дополнительно повышены путем снижения активности полипептидов ТГД в частях растения, например, с помощью мутации эндогенного гена, кодирующего полипептид ТГД, или введения экзогенного гена, кодирующего сайленсинговую молекулу РНК, которая снижает экспрессию эндогенного гена ТГД. В настоящем документе ʺтригалактозилдиацилглицерид полипептидʺ (ТГД, TGD) представляет собой принимающий участие в переносе липидов хлоропластами в ЭР (Xu с соавт., 2010) и принимающий участие в формировании белкового комплекса, осуществляющего функцию пермеазы для липидов. Четыре таких полипептида известны, как формирующие или связанные с ТГД пермеазой, а именно TGD-1 (номер доступа At1g19800 и гомологи в других видах), TGD-2 (номер доступа At2g20320 и гомологи в других видах), TGD-3 (номер доступа NM-105215 и гомологи в других видах) и TGD-4 (At3g06960 и гомологи в других видах) (US 20120237949). Считается, что полипептиды TGD-1, -2 и -3 являются компонентами переносчика АТФ-связывающей кассеты (АСК), связанного с внутренней мембраной конверта хлоропласта. Полипептиды TGD-2 и TGD-4 связываются с фосфатидиновой кислотой, тогда как полипептид TGD-3 действует, как АТФаза в строме хлоропласта. В настоящем документе ʺэндогенный ген ТГДʺ представляет собой ген, кодирующий полипептид ТГД в растении. Мутации в гене TGD-1 A. thaliana вызывали аккумуляцию триацилглицеридов, олигогалактолипидов и фосфатидиновой кислоты (ФК) (Xu с соавт., 2005). Мутации в генах ТГД или генах SDP1, или, в действительности, в любом целевом гене в растении, могут быть введены в сайт-специфической форме с помощью искусственной цинк-пальцевой нуклеазы (ЦПН), эффектора TAL (ПАТЭН) или технологии ККППРП (с применением нуклеазы Cas9 типа), как известно из уровня техники. Предпочтительными экзогенными генами, кодирующими сайленсинговые РНК, являются кодирующие молекулу двухцепочечной РНК, такой как шпилечная РНК или искусственный прекурсор микроРНК.
Ферменты, модифицирующие жирные кислоты
В настоящем документе термин «FAD2» обозначает связанную с мембраной дельта-12 десатуразу жирной кислоты, которая образует кратные связи в олеиновой кислоте (C18:1Δ9)с образованием линолевой кислоты (C18:2Δ9,12).
В настоящем документе термин «эпоксигеназа» или «эпоксигеназа жирной кислоты» обозначает фермент, который вводит эпоксидную группу в жирную кислоту, с получением в результате эпоксижирной кислоты. В предпочтительном варианте реализации эпоксигруппа вводится при 12-м атоме углерода на цепи жирной кислоты, если эпоксигеназа представляет собой Δ12-эпоксигеназу, особенно цепи жирной кислоты C16 или C18. Эпоксигеназа может быть Δ9-эпоксигеназой, Δ15-эпоксигеназой или оказывать свое действие при другом положении в цепи ацила, как известно из уровня техники. Эпоксигеназа может принадлежать к классу P450. Предпочтительные эпоксигеназы принадлежат к классу монооксигеназ, как описано в WO98/46762. Многочисленные эпоксигеназы или предполагаемые эпоксигеназы клонированы и известны в данной области. Другие примеры эпоксигеназ включают белки, содержащие последовательность аминокислоты, представленную в SEQ ID NO: 21 WO 2009/129582, полипептиды, кодируемые генами Crepis paleastina (CAA76156, Lee с соавт., 1998), Stokesia laevis (AAR23815) (монооксигеназный тип), Euphorbia lagascae (AAL62063) (тип P450), CYP2J2 человека (эпоксигеназа арахидоновой кислоты, U37143); CYPIA1 человека (эпоксигеназа арахидоновой кислоты, K03191), а также их варианты и/или мутанты.
В настоящем документе термин «гидроксилаза» или «гидроксилаза жирной кислоты» обозначает фермент, который вводит гидроксильную группу в жирную кислоту, с образованием гидроксилированной жирной кислоты. В предпочтительном варианте реализации гидроксильная группа вводится при 2-м, 12-м и/или 17-м атоме углерода на цепи жирной кислоты C18. Предпочтительно, гидроксильная группа вводится при 12 -м атоме углерода, если гидроксилаза представляет собой Δ12-гидроксилазу. В другом предпочтительном варианте реализации гидроксильная группа вводится при 15-м атоме углерода на цепи жирной кислоты C16. Дополнительно, гидроксилазы могут обладать активностью фермента, такого как десатураза жирной кислоты. Примеры генов, кодирующих Δ12-гидроксилазы включают полученные из Ricinus communis (AAC9010, van de Loo 1995); Physaria lindheimeri, (ABQ01458, Dauk с соавт., 2007); Lesquerella fendleri, (AAC32755, Broun с соавт., 1998); Daucus carota, (AAK30206); гидроксилазы жирных кислот, которые гидроксилируют конец жирных кислот, например: A, thaliana CYP86A1 (P48422, ω-гидроксилаза жирной кислоты); Vicia sativa CYP94A1 (P98188, ω-гидроксилаза жирной кислоты); CYP2E1 мыши (X62595, ω-1 гидроксилаза лауриновой кислоты); CYP4A1 крысы (M57718, ω-гидроксилаза жирной кислоты), а также их варианты и/или мутанты.
В настоящем документе термин «конъюгаза» или «конъюгаза жирной кислоты» обозначает фермент, способный к образованию конъюгационной связи в ацильной цепи жирной кислоты. Примеры конъюгаз включают конъюгазы, кодируемые генами Calendula officinalis (AF343064, Qiu с соавт., 2001); Vernicia fordii (AAN87574, Dyer с соавт., 2002); Punica granatum (AY178446, Iwabuchi с соавт., 2003) и Trichosanthes kirilowii (AY178444, Iwabuchi с соавт., 2003); а также их варианты и/или мутанты.
В настоящем документе термин «ацетиленаза» или «ацетиленаза жирной кислоты» обозначает фермент, который вводит тройную связь в жирную кислоту, что приводит в результате к образованию ацетиленсодержащей жирной кислоты. В предпочтительном варианте реализации тройная связь вводится при 2-м, 6-м, 12-м и/или 17-м атоме углерода на цепи жирной кислоты C18. Примеры ацетиленаз включают полученные из Helianthus annuus (AA038032, ABC59684), а также их варианты и/или мутанты.
Примеры таких модифицирующих жирные кислоты генов включают белки в соответствии со следующими номерами доступа, которые сгруппированы по предполагаемой функции, и гомологи из других видов: Δ12-ацетиленазы ABC00769, CAA76158, AAO38036, AAO38032; Δ12 конъюгазы AAG42259, AAG42260, AAN87574; Δ12-десатуразы P46313, ABS18716, AAS57577, AAL61825, AAF04093, AAF04094; Δ12 эпоксигеназы XP_001840127, CAA76156, AAR23815; Δ12-гидроксилазы ACF37070, AAC32755, ABQ01458, AAC49010; и ферменты Δ12 P450, например, AF406732.
Супрессоры сайленсинга
В варианте реализации изобретения рекомбинантная/трансгенная клетка по изобретению может содержать супрессор сайленсинга.
В настоящем документе ʺсупрессор сайленсингаʺ повышает экспрессию трансгена в клетке по изобретению. Например, присутствие супрессора сайленсинга приводит к более высоким уровням полипептида(ов), продуцируемого экзогенным(и) полинуклеотидом(ами) в клетке по изобретению, по сравнению с соответствующей клеткой, не содержащей супрессора сайленсинга. В варианте реализации изобретения супрессор сайленсинга предпочтительно связывается с молекулой дцРНК, размер которой составляет 21 пару оснований, относительно молекулы дцРНК другой длины. Это является признаком по меньшей мере супрессора сайленсинга типа p19, а именно p19 и его функциональных ортологов. В другом варианте реализации изобретения супрессор сайленсинга предпочтительно связывается с двухцепочечной молекулой РНК с нависающими 5' концами, по сравнению с соответствующей двухцепочечной молекулой РНК с тупыми концами. Это признак супрессора сайленсинга типа V2, а именно V2 и его функциональных ортологов. В варианте реализации изобретения молекула дцРНК или ее процессированный РНК продукт содержит по меньшей мере 19 нуклеотидов подряд, длина которого предпочтительно составляет 19-24 нуклеотида, с 19-24 парами оснований подряд в случае двухцепочечной молекулы шпилечной РНК или процессированного РНК продукта, более предпочтительно, состоит из 20, 21, 22, 23 или 24 нуклеотидов и предпочтительно содержит метилированный нуклеотид, по меньшей мере на 95% идентичный последовательности, комплементарной участку РНК-мишени, причем, участок РНК-мишени находится i) в пределах 5' нетранслируемого участка РНК-мишени, ii) в пределах 5' половины РНК-мишени, iii) в пределах кодирующей белок открытой рамки считывания РНК-мишени, iv) в пределах 3' половины РНК-мишени или v) в пределах 3' нетранслируемого участка РНК-мишени.
Дальнейшие подробности относительно супрессоров сайленсинга хорошо известны из уровня техники и описаны в WO 2013/096992 и WO 2013/096993.
Полинуклеотиды
Термины «полинуклеотид», и «нуклеиновая кислота» используются равнозначно. Они обозначают полимерную форму нуклеотидов любой длины, дезоксирибонуклеотидов или рибонуклеотидов или их аналогов. Полинуклеотид по изобретению может быть геномного, кДНК, полусинтетического или синтетического происхождения, одноцепочечным или двухцепочечным, и вследствие его происхождения или манипуляций с ним: (1) не связан с полноразмерным или частью полинуклеотида, с которым он связан в природе, (2) связан с другим полинуклеотидом, чем тот, с которым он связан в природе, или (3) не встречается в природе. Ниже приведены неограничивающие примеры полинуклеотидов: кодирующие или не кодирующие участки гена или фрагмента гена, экзоны, интроны, информационная РНК (иРНК), транспортная РНК (тРНК), рибосомальная РНК (рРНК), рибозимы, кДНК, рекомбинантные полинуклеотиды, плазмиды, векторы, выделенная ДНК с любой последовательностью, выделенная РНК с любой последовательностью, химерная ДНК с любой последовательностью, зонды нуклеиновой кислоты и праймеры. Для применения in vitro полинуклеотид может включать нуклеотиды, модифицированные, например, посредством конъюгации с меткой.
В настоящем документе «выделенный полинуклеотид» обозначает полинуклеотид, который отделен от полинуклеотидных последовательностей, с которыми он связан или соединен в природном состоянии, или от неприродного полинуклеотида.
В настоящем документе термин «ген» понимается в самом широком контексте и включает дезоксирибонуклеотидные последовательности, содержащие транскрибированный участок и, при условии трансляции, участок кодирования белка структурного гена и, в том числе, последовательности, смежные с кодирующим участком, на 5' и 3' концах, на расстоянии по меньшей мере около 2 тысяч пар оснований на любом конце, которые принимают участие в экспрессии гена. В этом отношении, ген содержит контрольные сигналы, например, промоторы, энхансеры, сигналы терминации и/или полиаденилирования, которые в природе связаны с указанным геном, или гетерологичные контрольные сигналы, и в этом случае ген обозначается, как «химерный ген». Последовательности, которые размещены в направлении 5' по отношению к кодирующему белок участку, и которые присутствуют в иРНК, обозначаются как 5' нетранслируемые последовательности. Последовательности, которые размещены в направлении 3' или по ходу транскрипции относительно кодирующего белок участка, и которые присутствуют в иРНК, обозначаются, как 3' нетранслируемые последовательности. Термин «ген» включает кДНК и геномные формы гена. Геномная форма или клон гена содержит кодирующий участок, который может перемежаться не кодирующими последовательностями под названием «интроны», «вставочные последовательности» или «встроенные последовательности». Интроны представляют собой сегменты гена, которые транскрибированы в ядерную РНК (яРНК). Интроны могут содержать регуляторные элементы, например, энхансеры. Интроны удаляются, или «происходит укорачивание» ядерной или первичной копии; таким образом, интроны отсутствуют в копии иРНК. Ген, содержащий по меньшей мере один интрон, может подвергаться вариабельному сплайсингу с образованием альтернативных иРНК из одного транскрибированного гена и, таким образом, полипептидных вариантов. В природном или химерном гене интроны могут отсутствовать. Функции иРНК в ходе трансляции состоят в определении последовательности или порядка аминокислот в возникающем полипептиде. Термин «ген» включает синтетическую или слитую молекулу, кодирующую полноразмерные белки по изобретению или их части, как раскрыто в настоящем документе, и нуклеотидную последовательность, комплементарную к любому из упомянутого выше.
В настоящем документе «химерная ДНК» обозначает любую молекулу ДНК, которая не найдена в природе; также обозначается в настоящем документе как «конструкция ДНК» или «генетическая конструкция». Обычно, химерная ДНК содержит регуляторную и транскрибированную или кодирующую белок последовательности, которые не найдены вместе в природе. Соответственно, химерная ДНК может содержать регуляторные последовательности и кодирующие последовательности, которые происходят из различных источников или регуляторных последовательностей, и кодирующие последовательности, происходящие из того же источника, но организованные до некоторой степени отличным образом, чем найденные в природе. Открытая рамка считывания может быть или может не быть связанной с ее природными регуляторными элементами, расположенными против хода транскрипции и по ходу транскрипции. Открытая рамка считывания может быть инкорпорирована, например, в геном растения, в неприродном расположении или в репликоне или векторе, где она не найдена в природе, например, бактериальной плазмиде или вирусном векторе. Термин «химерная ДНК» не ограничивается молекулами ДНК, которые являются реплицируемыми в организме-хозяине, но включает ДНК, способную к лигированию в репликон, например, специфическими адаптерными последовательностями.
«Трансген» представляет собой ген, который введен в геном процедурой трансформации. Термин включает ген в клетке-потомке, растении, семени, организме, не относящемся к человеческому роду, или его части, который был введен в геном их родительской клетки. Такие клетки-потомки, и т. п. могут представлять по меньшей мере 3-е или 4-е поколение потомков относительно родительской клетки, которая была первоначально трансформирована, или родительского трансгенного растения (которое в настоящем документе обозначается как растение T0). Потомок может быть получен путем полового воспроизводства или вегетативно, например, из клубней картофеля или отводков сахарного тростника. Термин «генетически модифицированный», «генетическая модификация» и их вариации представляют более широкий термин, который включает введение гена в клетку посредством трансформации или трансдукции, видоизменение гена в клетке и генную модификацию или модуляцию регулирования гена в клетке или потомке любой модифицированной клетки, как изложено выше.
«Геномный участок» в настоящем документе обозначает положение в пределах генома, в котором трансген или группа трансгенов (также обозначается в настоящем документе как кластер) вставлены в клетку или ее предка. Такие участки содержат только нуклеотидны, введенные в результате вмешательства человека, например, способами, раскрытыми в настоящем документе.
«Рекомбинантный полинуклеотид» по изобретению обозначает молекулу нуклеиновой кислоты, которая сконструирована или модифицирована способами искусственной рекомбинации. Рекомбинантный полинуклеотид может присутствовать в клетке в измененном количестве или экспрессироваться с измененной скоростью (например, в случае иРНК), по сравнению с его природным состоянием. В одном варианте реализации полинуклеотид введен в клетку, которая от природы не содержит полинуклеотида. Обычно экзогенная ДНК используется в качестве шаблона для транскрипции иРНК, которая в дальнейшем транслируется в непрерывную последовательность остатков аминокислоты, кодируя полипептид по изобретению в пределах трансформированной клетки. В другом варианте реализации полинуклеотид является для клетки эндогенным, и его экспрессия модифицирована рекомбинантными средствами, например, экзогенная контрольная последовательность введена в направлении относительного целевого эндогенного гена, чтобы предоставить возможность трансформированной клетке экспрессировать полипептид, кодируемый геном, или делеция введена в целевой ген методами ZFN, ПАТЭН или ККППРП.
Рекомбинантный полинуклеотид по изобретению содержит полинуклеотиды, которые не были отделены от других компонентов клеточной или неклеточной системы экспрессии, в которой он присутствует, и полинуклеотиды, образованные в указанных клеточных или неклеточных системах, которые впоследствии очищают по меньшей мере от некоторых других компонентов. Полинуклеотид может быть беспрерывным участком нуклеотидов или может содержать два или более смежных участков нуклеотидов из различных источников (природных и/или синтетических), соединенных таким образом, чтобы сформировать единый полинуклеотид. Обычно, такие химерные полинуклеотиды содержат по меньшей мере открытую рамку считывания, кодирующую полипептид по изобретению, функционально связанную с промотором, пригодным для управления транскрипцией открытой рамки считывания в целевой клетке.
С учетом определенных полинуклеотидов, необходимо понимать, что % идентичности, превышающий приведенные выше, будет охватывать предпочтительные варианты реализации изобретения. Таким образом, если это уместно, в свете минимальных значений % идентичности, предпочтительно, полинуклеотид содержит последовательность полинуклеотида, которая по меньшей мере на 60%, более предпочтительно по меньшей мере на 65%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 91%, более предпочтительно по меньшей мере на 92%, более предпочтительно по меньшей мере на 93%, более предпочтительно по меньшей мере на 94%, более предпочтительно по меньшей мере на 95%, более предпочтительно по меньшей мере на 96%, более предпочтительно по меньшей мере на 97%, более предпочтительно по меньшей мере на 98%, более предпочтительно по меньшей мере на 99%, более предпочтительно по меньшей мере на 99,1%, более предпочтительно по меньшей мере на 99,2%, более предпочтительно по меньшей мере на 99,3%, более предпочтительно по меньшей мере на 99,4%, более предпочтительно по меньшей мере на 99,5%, более предпочтительно по меньшей мере на 99,6%, более предпочтительно по меньшей мере на 99,7%, более предпочтительно по меньшей мере на 99,8%, и даже более предпочтительно по меньшей мере на 99,9% идентична представленной в соответствующей SEQ ID NO.
Полинуклеотид по настоящему изобретению или пригодный для применения в соответствии с настоящим изобретением может селективно гибридизоваться, в строгих условиях, с полинуклеотидом, определенным в настоящем документе. В настоящем документе строгие условия представляют собой: (1) применение в ходе гибридизации денатурирующего агента, такого как формамид, например, 50%, об/об формамида, содержащего 0,1%, масс/об телячьего сывороточного альбумина, 0,1% Фиколла, 0,1% поливинилпирролидона, 50 мМ натрий фосфатного буфера, pH 6,5, содержащего 750 мМ NaCl, 75 мМ натрия цитрата, при 42°C; или (2) применение 50%, формамида, 5 x натрия хлорида+натрия цитрата (0,75 M NaCl, 0,075 M натрия цитрата), 50 мМ натрия фосфата (pH 6,8), 0,1% натрия пирофосфата, 5 x раствор Денхардта, обработанная ультразвуком ДНК спермы лосося (50 г/мл), 0,1% натрия лаурилсульфата и 10% декстрана сульфата при 42°C в 0,2 x натрия хлорида+натрия цитрата и 0,1% натрия лаурилсульфата, и/или (3) применение низкой ионной силы и высокой температуры для промывания, например, 0,015 M NaCl/0,0015 M натрия цитрата/0,1% натрия лаурилсульфата при 50°C.
Полинуклеотиды по изобретению могут содержать, по сравнению с природными молекулами, одну или более мутаций, которые представляют собой делеции, вставки или замены нуклеотидных остатков. Полинуклеотиды, которые содержат мутации относительно референтной последовательности, могут быть природными (т. е., выделенными из природного источника) или синтетическими (например, полученными путем сайт-направленного мутагенеза или тасования ДНК в нуклеиновой кислоте, как изложено выше).
Полинуклеотиды для снижения уровней экспрессии генов
РНК-интерференция
РНК-интерференция (РНКи) особенно пригодна для специфичного снижения экспрессии гена, что ведет к уменьшению выработки конкретного белка, если ген кодирует белок. Не желая ограничиваться теорией, Waterhouse с соавт. (1998) предложили модель механизма, по которому двухцепочечная РНК (дцРНК) может использоваться для снижения выработки белка. Данная технология зависит от присутствия молекул дцРНК, которые содержат последовательность, по существу идентичную иРНК целевого гена или его части. Пригодным образом, дцРНК может быть получена с помощью единственного промотора в рекомбинантном векторе или клетке-хозяине, в которых смысловые и антисмысловые последовательности фланкированы посторонней последовательностью, которая позволяет смысловой и антисмысловой последовательностям гибридизоваться с образованием молекулы дцРНК с посторонней последовательностью, образующей структуру петли. Конструирование и получение подходящих молекул дцРНК находится в пределах компетенции специалиста в данной области, особенно с учетом Waterhouse с соавт. (1998), Smith с соавт. (2000), WO 99/32619, WO 99/53050, WO 99/49029 и WO 01/34815.
В одном из примеров вводят ДНК, которая направляет синтез по меньшей мере частично двухцепочечного продукта(-ов) РНК, гомологичного гену мишени, который подлежит инактивации, например, такого как SDP1, TGD, пластидная GPAT, пластидная LPAAT, пластидная PAP, ген АДФ-глюкозопирофосфорилазы (АГФазы). Таким образом, ДНК содержит смысловые и антисмысловые последовательности, которые при транскрибировании в РНК могут гибридизоваться с образованием двухцепочечного участка РНК. В одном варианте реализации, смысловая и антисмысловая последовательности отделены спейсерным участком, который содержит интрон, отщепляющийся при транскрибировании в РНК. Показано, что такое размещение приводит к более высокой эффективности сайленсинга гена (Smith с соавт., 2000). Двухцепочечный участок может содержать одну или две молекулы РНК, транскрибированные с одного или двух участков ДНК. Считается, что присутствие двухцепочечной молекулы запускает реакцию эндогенной системы, которая разрушает двухцепочечную РНК, а также гомологичную копию РНК, образованную геном-мишенью, эффективно снижая или устраняя активность гена-мишени.
Длина смысловой и антисмысловой последовательностей, которые гибридизуются, должна составлять для каждой по меньшей мере 19 нуклеотидов подряд, предпочтительно по меньшей мере 50 нуклеотидов подряд, более предпочтительно, по меньшей мере 100 или по меньшей мере 200 нуклеотидов подряд. В общем, применяется последовательность размером 100-1000 нуклеотидов, соответствующая участку иРНК гена-мишени. Может использоваться полноразмерная последовательность, соответствующая полному транскрипту гена. Степень идентичности смысловой последовательности целевому транскрипту (и, таким образом, также идентичности антисмысловой последовательности и последовательности, комплементарной транскрипту-мишени) должна составлять по меньшей мере 85%, по меньшей мере 90% или 95-100%. Молекула РНК может, конечно, содержать посторонние последовательности, функция которых может состоять в стабилизации молекулы. Молекула РНК может экспрессироваться под контролем промотора РНК полимеразы II или РНК полимеразы III. Примеры последнего включают промоторы тРНК или мяРНК.
Предпочтительные молекулы малой интерферирующей РНК («миРНК») содержат нуклеотидную последовательность, которая идентична приблизительно 19-25 смежным нуклеотидам иРНК-мишени. Предпочтительно, последовательность миРНК начинается с динуклеотида AA, содержит GC в количестве приблизительно 30-70% (предпочтительно, 30-60%, более предпочтительно 40-60% и более предпочтительно приблизительно 45-55%), и не обладает высокой степенью идентичности ни к одной нуклеотидной последовательности в геноме организма, кроме мишени, в которую она должна быть введена, например, как определяется стандартным поиском BLAST.
МикроРНК
МикроРНК (сокращенно миРНК) в общем представляют собой некодирующие молекулы РНК длиной 19-25 нуклеотидов (в растениях обычно приблизительно 20-24 нуклеотида), происходящие от крупных прекурсоров, которые образуют несовершенные структуры «петля-на-стебле».
миРНК связывают с дополнительными последовательностями на целевых транскриптах информационной РНК (иРНК), что обычно приводит к угнетению трансляции или разложению мишени и сайленсингу гена. Как хорошо известно из уровня техники, на основе природных миРНК могут быть сконструированы искусственные миРНК (имиРНК) для целей уменьшения экспрессии любого целевого гена.
Считается, что в клетках растений процессинг молекул-прекурсоров миРНК в значительной степени происходит в ядре. Процессинг пре-миРНК (содержащих один или более локальных двухцепочечных или «шпилечных» участков, а также обыкновенный 5' «кэп» и полиаденилированный хвост иРНК) происходит до более короткой молекулы-прекурсора миРНК, которая также содержит структуру «петля-на-стебле» или конъюгацию плеч хромосомы и называется «пре-миРНК». В растениях, пре-миРНК расщепляются различными ДАЙСЕР-подобными ферментами (ДСП), образуя дуплексы миРНК:миРНК*. Перед транспортом за пределы ядра, такие дуплексы метилируются.
В цитоплазме, цепь миРНК от спаренной миРНК:миРНК селективно инкорпорируется в активный РНК-индуцированный комплекс сайленсинга (РИКС) для распознавания мишени. Комплексы РИКС содержат конкретное подмножество белков Argonaute, которые осуществляют специфичное в отношении последовательности подавление гена (см., например, Millar и Waterhouse, 2005; Pasquinelli с соавт., 2005; Almeida и Allshire, 2005).
Косупрессия
Гены могут подавлять экспрессию родственных эндогенных генов и/или трансгенов, уже присутствующих в геноме, причем данный признак, назван зависимым от гомологии сайленсингом гена. В большинстве случаев зависимый от гомологии сайленсинг гена относится к одному из двух классов - функционирующий на уровне транскрипции трансгена или действующий посттранскрипционально.
Посттранскрипциональный зависимый от гомологии сайленсинг (т. е., косупрессия) гена описывает прекращение экспрессии трансгена и родственных эндогенных или вирусных генов в трансгенных растениях. Косупрессия часто, но не всегда, возникает, если копий трансгена много, и, в общем, считается запускаемой на уровне процессинга, локализации и/или разложения иРНК. Существуют различные модели для объяснения того, как осуществляется косупрессия (см. Taylor, 1997).
Косупрессия включает введение добавочной копии гена или его фрагмента в растение, в смысловой ориентации относительно промотора, для его экспрессии. Размер смыслового фрагмента, его соответствие участкам гена-мишени, и степень идентичности его последовательности гену-мишени могут быть определены специалистами в данной области. В некоторых случаях дополнительная копия последовательности гена препятствует экспрессии растительного гена-мишени. В WO 97/20936 и EP 0465572 раскрыты способы осуществления подходов косупрессии.
Антисмысловые полинуклеотиды
Термин «антисмысловой полинуклеотид» следует интерпретировать как обозначающий молекулу ДНК или РНК, которая комплементарна по меньшей мере части конкретной молекулы иРНК, кодирующей эндогенный полипептид, и способна препятствовать посттранскрипциональному событию, например, трансляции иРНК. Применение антисмысловых методов хорошо известно из уровня техники (см., например, G. Hartmann and S. Endres, Manual of Antisense Methodology, Kluwer (1999)). Применение антисмысловых методов в растениях рассмотрено Bourque (1995) и Senior (1998). Bourque (1995) перечисляет большое количество примеров того, как антисмысловые последовательности применяются в растительных системах в качестве способа инактивации гена. Bourque также заявляет, что достижение 100% ингибирования любой ферментной активности может не быть необходимым, поскольку частичное ингибирование будет более чем вероятно приводить к измеримому изменению в системе. Senior (1998) указывает, что антисмысловые методы в настоящее время представляют собой хорошо изученную технику для манипуляции экспрессией гена.
В одном варианте реализации антисмысловой полинуклеотид гибридизуется в физиологических условиях, т. е., антисмысловой полинуклеотид (который полностью или частично является одноцепочечным) по меньшей мере способен к образованию двухцепочечного полинуклеотида с иРНК, кодирующей в клетке эндогенный полипептид, например, иРНК SDP1, TGD, пластидной GPAT, пластидной LPAAT, пластидной PAP или АГФазы, в нормальных условиях.
Антисмысловые молекулы могут включать последовательности, которые соответствуют структурным генам или для последовательностей, которые осуществляют контроль над экспрессией гена или событием укорачивания. Например, антисмысловая последовательность может соответствовать целевому кодирующему участку эндогенного гена или 5'-нетранслируемому участку (НТУ) или 3'-НТУ или их комбинации. Она может быть частично комплементарной интронным последовательностям, которые могут быть укорочены в ходе или после транскрипции, предпочтительно только до экзонных последовательностей гена-мишени. Принимая во внимание существенную в общем дивергенцию НТУ, нацеливание данных участков обеспечивает высокую степень специфичности ингибирования гена.
Длина антисмысловой последовательности должна составить по меньшей мере 19 нуклеотидов подряд, предпочтительно по меньшей мере 50 нуклеотидов и более предпочтительно по меньшей мере 100, 200, 500 или 1000 нуклеотидов. Может использоваться полноразмерная последовательность, комплементарная к полноразмерному транскрипту гена. Наиболее предпочтительно, длина составляет 100-2000 нуклеотидов. Степень идентичности антисмысловой последовательности целевому транскрипту должна составлять по меньшей мере 90% и более предпочтительно 95-100%. Антисмысловая молекула РНК, конечно, может содержать посторонние последовательности, функция которых может состоять в стабилизации молекулы.
Рекомбинантные векторы
Один вариант реализации настоящего изобретения включает рекомбинантный вектор, который содержит по меньшей мере один полинуклеотид, раскрытый в настоящем документе, и способен доставлять полинуклеотид в клетку-хозяина. Рекомбинантные векторы включают векторы экспрессии. Рекомбинантные векторы содержат гетерологичные полинуклеотидные последовательности, т. е., полинуклеотидные последовательности, которые в природе не найдены смежно с полинуклеотидом, раскрытым в настоящем документе, и, предпочтительно, получены от другого вида. Вектор может представлять собой РНК или ДНК, и обычно представляет собой вирусный вектор, полученный из вируса, или плазмиду. Плазмидные векторы обычно содержат дополнительные последовательности нуклеиновой кислоты, которые включены для простоты селекции, амплификации и трансформации кассеты экспрессии в прокариотных клетках, например, pUC-полученных векторах, pGEM-полученных векторах или бинарных векторах, содержащих один или более участков T-ДНК. Дополнительные последовательности нуклеиновой кислоты включают источники репликации для автономной репликации вектора, гены селекционного маркера, предпочтительно кодирующие резистентность к антибиотику или гербициду, уникальные множественные сайты клонирования, предусматривающие множественные сайты для вставки последовательностей нуклеиновой кислоты или кодируемых генов в конструкцию нуклеиновой кислоты, и последовательностей, которые увеличивают степень трансформации прокариотных и эукариотных (особенно растительных) клеток.
«Функционально связанный» в настоящем документе обозначает функциональное взаимоотношение двух или более сегментов нуклеиновой кислоты (например, ДНК). Обычно, это выражение обозначает функциональное взаимоотношение элемента регуляции транскрипции (промотор) с транскрибированной последовательностью. Например, промотор функционально связан с кодирующей последовательностью полинуклеотида, раскрытого в настоящем документе, если он стимулирует или модулирует транскрипцию кодирующей последовательности в пригодной клетке. В общем, промоторные элементы регуляции транскрипции, которые функционально связаны с транскрибированной последовательностью, являются физически смежными с транскрибированной последовательностью, т. е., они цис-действующие. Однако, некоторые элементы регуляции транскрипции, например, энхансеры, не должны быть физически смежными или расположенными поблизости кодирующих последовательностей, транскрипцию которых они усиливают.
Если присутствует несколько промоторов, каждый промотор независимо может быть таким же или другим.
Кроме того, рекомбинантные векторы могут содержать одну или более последовательностей сигнального пептида, чтобы позволить удержание экспрессированного полипептида, раскрытого в настоящем документе, в эндоплазматическом ретикулуме (ЭР) клетки или перенос в пластиду, и/или химерные последовательности, которые приводят к экспрессии молекул нуклеиновой кислоты в виде слитых белков. Примеры пригодных сигнальных сегментов включают любой сигнальный сегмент, способный направлять секрецию или локализацию полипептида, раскрытого в настоящем документе.
Чтобы упростить идентификацию трансформантов, рекомбинантный вектор желательно должен включать ген селекционного или пригодного для скрининга маркера. «Ген маркера» обозначает ген, который передает отличный фенотип клеткам, экспрессирующим ген маркера, и, таким образом, позволяет отличать такие трансформированные клетки от клеток, которые не содержат маркера. Ген селекционного маркера обеспечивает признак, по которому может быть осуществлена «селекция» на основе резистентности к агенту селекции (например, гербициду, антибиотику). Ген пригодного для скрининга маркера (или репортерный ген) обеспечивает признак, который можно идентифицировать посредством наблюдения или тестирования, т. е., «скрининга» (например., β-глюкуронидаза, люцифераза, ЗФБ или другая ферментная активность, отсутствующая в нетрансформированных клетках). Примеры селекционных маркеров для селекции трансформантов растения включают, без ограничений, ген hyg, который кодирует резистентность к гигромицину B; ген неомицинфосфотрансферазы (nptII), обеспечивающий резистентность к канамицину, паромомицину; ген глутатион-S-трансферазы из печени крыс, обеспечивающий резистентность к полученным из глутатиона гербицидам, например, описанный в EP 256223; гена глутаминсинтетазы, при повышенной экспрессии обеспечивающий резистентность к ингибиторам глутаминсинтетазы, таким как фосфинотрицин, например, как раскрыто в WO 87/05327; ген ацетилтрансферазы Streptomyces viridochromogenes, обеспечивающий резистентность к агенту селекции фосфинотрицину, например, как раскрыто в EP 275957; ген, кодирующий 5-енолшикимат-3-фосфат синтетазу (ЕШФС), обеспечивающую переносимость N-фосфонометилглицина, например, как раскрыто Hinchee с соавт. (1988); ген bar, обеспечивающий резистентность к биалафосу, например, раскрытый в WO 91/02071; ген нитрилазы, например, bxn Klebsiella ozaenae, который обеспечивает резистентность к бромоксинилу (Stalker с соавт., 1 988); ген дигидрофолатредуктазы (ДГФР), обеспечивающий резистентность к метотрексату (Thillet с соавт., 1988); мутантный ген ацетолактсинтетазы (АЛС), который обеспечивает резистентность к имидазолинону, сульфонилмочевине или другим ингибирующим АЛС химическим веществам (EP 154204); модифицированный ген антранилатсинтетазы, который обеспечивает резистентность к 5-метилтриптофану; или ген далапон дегалогеназы, который обеспечивает резистентность к гербициду.
Предпочтительно, рекомбинантный вектор стабильно инкорпорирован в геном клетки, например, растительные клетки. Соответственно, рекомбинантный вектор может содержать пригодные элементы, которые позволяют вектору инкорпорироваться в геном или хромосому клетки.
Вектор экспрессии
В настоящем документе «вектор экспрессии» представляет собой ДНК вектор, который способен к трансформации клетки-хозяина и эффективной экспрессии одного или более указанных полинуклеотидов. Векторы экспрессии по настоящему изобретению содержат регуляторные последовательности, например, последовательности контроля транскрипции, последовательности контроля трансляции, источники репликации и другие регуляторные последовательности, которые совместимы с клеткой-хозяином и которые управляют экспрессией полинуклеотидов по настоящему изобретению. В частности, векторы экспрессии по настоящему изобретению содержат последовательности контроля транскрипции. Последовательности контроля транскрипции представляют собой последовательности, которые управляют инициацией транскрипции, удлинением транскрипта и окончанием транскрипции. Особенно значимыми последовательностями контроля транскрипции являются такие, которые управляют инициацией транскрипции, например, промоторные, энхансерные, операторные и репрессорные последовательности. Выбор используемых регуляторных последовательностей зависит от организма-мишени, например, растения, и/или органа-мишени или целевой ткани. Такие регуляторные последовательности могут быть получены от любого эукариотного организма, такого как растения или вирусы растения, или могут быть химически синтезированы. Целый ряд векторов, пригодных для стабильной трансфекции растительных клеток или для получения трансгенных растений, раскрыт, например, в Pouwels с соавт., Cloning Vectors: A Laboratory Manual, 1985, supp. 1987, Weissbach and Weissbach, Methods for Plant Molecular Biology, Academic Press, 1989, и Gelvine с соавт., Plant Molecular Biology Manual, Kluwer Academic Publishers, 1990. Обычно, векторы экспрессии растений содержат, например, один или более клонированных генов растения под транскрипциональным контролем 5' и 3' регуляторных последовательностей и доминирующего селекционного маркера. Кроме того, такие векторы экспрессии растений могут содержать регуляторный участок промотора (например, регуляторный участок, управляющий индуцибельной или конститутивной, регулируемой окружающей средой или стадией развития, клеточно- или тканеспецифичной экспрессией), старт-сайт инициации транскрипции, сайт связывания с рибосомой, сайт терминации транскрипции и/или сигнал полиаденилирования.
Раскрыт целый ряд конститутивных промоторов, которые активны в клетках растений. Пригодные промоторы для конститутивной экспрессии в растениях включают, без ограничений, промотор 35S вируса мозаики цветной капусты (CaMV), 35S вируса мозаики норичника (ВМН), индуцируемый светом промотор маленькой субъединицы (МСЕ) рибулозо-1,5-бисфосфат карбоксилазы, рисовый промотор цитозольной триозофосфат изомеразы, промотор аденин фосфорибозилтрансферазы Arabidopsis, промотор гена рисового актина 1, промоторы маннопин синтетазы и октопин синтетазы, промотор Adh, промотор синтетазы сахарозы, промотор комплекса R гена и промотор гена белка, связывающегося с α/βхлорофиллом. Эти промоторы использовались для создания векторов ДНК, которые экспрессируются в растениях, см., например, WO 84/02913. Все указанные промоторы использовались для создания различных видов экспрессируемых в растениях рекомбинантных ДНК векторов.
С целью экспрессии в исходных тканях растения, таких как лист, семя, корень или стебель, промоторы, используемые в настоящем изобретении, предпочтительно демонстрируют относительно высокую экспрессию в указанных конкретных тканях. Для достижения данной цели можно выбирать из целого ряда промоторов для генов с ткане- или клеточноспецифичной или -усиленной экспрессией. Примеры таких промоторов, о которых сообщалось в литературе, включают, промотор глутаминсинтетазы хлоропластов GS2 гороха, промотор фруктозо-1,6-бифосфатазы хлоропласта пшеницы, промотор ядерного фотосинтетического ST-LS1 картофеля, промотор серин/треонинкиназы и промотор глюкоамилазы (CHS) Arabidopsis thaliana. Также сообщалось, что активным в фотосинтетически активных тканях является промотор рибулозо-1,5-бисфосфат карбоксилазы восточной лиственницы (Larix laricinа), промотор гена Cab сосны, Cab6, промотор гена Cab-1 пшеницы, промотора гена Cab-1 шпината, промотор гена Cab-1R риса, промотор пируват, ортофосфат дикиназы (ПФДК) Zea mays, промотор гена Lhcb1*2 табака, промотор симпортера Suc2 Arabidopsis thaliana сахарозы-H30, и промотор для генов тилакоидного белка мембраны шпината (PsaD, PsaF, PsaE, PC, FNR, AtpC, AtpD, Cab, RbcS). Другие промоторы белков, связывающихся с α/βхлорофиллом также могут использоваться в настоящем изобретении, например, промоторы гена LhcB и гена PsbP белой горчицы (Sinapis albа).
Различные промоторы растительных генов, которые регулируются в ответ на экологические, гормональные, химические и/или связанные с развитием сигналы, также могут использоваться для экспрессии генов РНК-связывающего белка в клетках растения, в том числе, промоторы, регулируемые (1) нагреванием, (2) светом (например, промотор RbcS-3A гороха, промотор RbcS маиса), (3) гормонами, например, абсцизиновая кислота, (4) образованием ран (например, WunI) или (5) химическими веществами, такими как метилжасмонат, салициловая кислота, стероидные гормоны, спирт, сафенеры (WO 97/06269), или дополнительно может быть предпочтительным использование (6) специфичных в отношении органа промоторов.
В настоящем документе термин «промотор, специфичный в отношении запасающего органа растения» обозначает промотор, который предпочтительно, по сравнению с другими тканями растения, направляет транскрипцию гена в запасающем органе растения. С целью экспрессии в акцептирующих тканях растения, например, клубня картофельного растения, плода помидора или семени сои, рапса, хлопка, Zea mays, пшеницы, риса и ячменя, промоторам, используемым в настоящем изобретении, предпочтительно свойственна относительно высокая экспрессия в данных конкретных тканях. Может применяться промотор β-конглицинина или другие специфичные для семени промоторы, например, промоторы напина, зеина, линина и фазеолина. Кроме того, могут применяться специфичные в отношении корней промоторы. Примером такого промотора является промотор гена кислой хитиназы. Дополнительно, экспрессия в корневой ткани могла бы быть достигнута с использованием специфичных в отношении корней субдоменов промотора CaMV 35S, которые идентифицированы.
В особенно предпочтительном варианте реализации промотор направляет экспрессию в тканях и органах, в которых имеет место биосинтез липида. Такие промоторы могут действовать на стадии развития семени, в подходящее время для модификации состава липида в семенах. Предпочтительные промоторы для специфичной экспрессии в семени включают: 1) промоторы генов, кодирующих ферменты, принимающие участие в биосинтезе липида и аккумуляции в семенах, например, десатуразы и элонгазы, 2) промоторы генов, кодирующих запасаемые белки семени, и 3) промоторы генов, кодирующих ферменты, принимающие участие в биосинтезе углеводов и аккумуляции в семенах. Пригодные промоторы, специфичные в отношении семени, представляют собой: промотор гена напина масличного рапса (US 5608152), промотор Фарм. USP Vicia faba (Baumlein с соавт., 1991), промотор олеозина Arabidopsis (WO 98/45461), промотор фазеолина Phaseolus vulgaris (US 5504200), промотор Bce4 Brassica (WO 91/13980) или промотор легумина B4 (Baumlein с соавт., 1992), а также промоторы, которые приводят к специфичной экспрессии в семени однодольных, таких как маис, ячмень, пшеница, рожь, рис и т. п. Следует отметить пригодные промоторы гена lpt2 или lpt1 ячменя (WO 95/15389 и WO 95/23230) или промоторы, описанные в WO 99/16890 (промоторы гена гордеина ячменя, гена глютелина риса, гена оризина риса, гена проламина риса, гена глиадина пшеницы, гена глютелина пшеницы, гена зеина маиса, гена глютелина овса, гена казирина сорго, гена секалина ржи). Другие промоторы включают раскрытые Broun с соавт. (1998), Potenza с соавт. (2004), US 20070192902 и US 20030159173. В варианте реализации специфичный для семени промотор предпочтительно экспрессируется в определенных частях семени, например, семядоле(-ях) или эндосперме. Примеры специфичных для семядоли промоторов включают, без ограничений, промотор FP1 (Ellerstrometal.,1996), промотор легумина гороха (Perrin с соавт., 2000) и промотор фитогемагглютинина боба (Perrin с соавт., 2000). Примеры специфичных для эндоспермы промоторов включают, без ограничений, промотор зеина-1 маиса (Chikwamba с соавт., 2003), промотор глютелина-1 риса (Yang с соавт., 2003), промотор D-гордеина ячменя (Horvath с соавт., 2000) и промоторы высокомолекулярного глютенина пшеницы (Alvarez с соавт., 2000). В дальнейшем варианте реализации специфичный для семени промотор не экспрессируется или экспрессируется только на низком уровне в зародыше и/или после того, как семя прорастает.
В другом варианте реализации промотор, специфичный для запасного органа растения, представляет собой специфичный для фрукта промотор. Примеры включают, без ограничений, промотор полигалактуроназы помидора, E8 и Pds, а также промотор аминоциклопропанкарбоксилат оксидазы (АЦК) яблока (обзор см. в Potenza с соавт., 2004). В предпочтительном варианте реализации промотор предпочтительно направляет экспрессию в съедобных частях плода, например, сердцевине плода, относительно кожуры плода или семян в пределах плода.
В варианте реализации индуцибельный промотор представляет собой систему alc Aspergillus nidulans. Примеры индуцибельных систем экспрессии, которые могут использоваться вместо системы Aspergillus nidulans alc, описаны в обзоре Padidam (2003) и Corrado и Karali (2009). В другом варианте реализации индуцибельный промотор представляет собой индуцируемый сафенером промотор, например, такой как промотор ln2-1 или ln2-2 маиса (Hershey и Stoner, 1991), индуцибельный промотор сафенера представляет собой промотор GST-27 маиса (Jepson с соавт., 1994) или соевый промотор GH2/4 (Ulmasov с соавт., 1995).
В другом варианте реализации индуцибельный промотор представляет собой промотор, индуцируемый старением, например, такой как индуцируемый старением промотор SAG (связанный со старением ген) 12 и SAG 13 Arabidopsis (Gan, 1995; Gan и Amasino, 1995) и LSC54 Brassica napus (Buchanan-Wollaston, 1994). Такие промоторы демонстрируют повышенную экспрессию в период около начала старения растительных тканей, особенно листьев.
Для экспрессии в вегетативных тканях могут использоваться специфичные для листьев промоторы, такие как промоторы рибулозобифосфат карбоксилазы (РБФК). Например, гены RBCS1, RBCS2 и RBCS3A помидора экспрессируются в листьях и растущих под действием света саженцах (Meier с соавт., 1997). Могут использоваться промоторы рибулозобифосфат карбоксилазы, экспрессирующиеся почти исключительно в клетках мезофилла листовых пластинок и влагалищ листьев с высокими уровнями, как раскрыто Matsuoka с соавт. (1994). Другой специфичный для листьев промотор представляет собой легко собирающий урожай промотор гена улавливающего свет белка a/b, связывающегося с хлорофиллом (см. Shiina с соавт., 1997). Родственный myb промотор (Atmyb5) гена Arabidopsis thaliana, описанный Li с соавт. (1996), является специфичным для листьев. Промотор Atmyb5 экспрессируется в процессе развития листовых трихом, прилистников и эпидермальных клеток на краях молодой розетки и стеблевых листьях, а также в незрелых семенах. Кроме того, может использоваться листовой промотор, идентифицированный в маисе Busk с соавт. (1997).
В некоторых случаях, например, если LEC2 или BBM рекомбинантно экспрессируется, может быть желательным, чтобы трансген не экспрессировался с высокими уровнями. Примером промотора, который может использоваться в таких случаях, является укороченный промотор напина A, который сохраняет специфичный для семени характер экспрессии, но со сниженным уровнем экспрессии (Tan с соавт., 2011).
5' Нетранслируемые лидерные последовательности могут быть получены промотором, выбранного для экспрессии последовательности гетерологичного гена полинуклеотида по настоящему изобретению, или могут быть гетерологичными относительно участка кодирования фермента, который должен вырабатываться, и могут быть специфично модифицированы, при желании, с целью увеличения трансляции иРНК. Обзор оптимизации экспрессии трансгенов см. в Koziel с соавт. (1996). 5' Нетранслируемые участки также могут быть получены из РНК вирусов растений (среди прочего, вирус мозаики табака, вирус гравировки табака, вирус карликовой мозаики маиса, вирус мозаики люцерны), из пригодных эукариотных генов, растительных генов (лидер гена белка a/b, связывающегося с хлорофиллом, пшеницы и маиса) или из синтетической последовательности гена. Настоящее изобретение не ограничивается конструкциями, в которых нетранслируемый участок получен из 5' нетранслируемой последовательности, которая сопровождает последовательность промотора. Дополнительно, лидерная последовательность может происходить от постороннего промотора или кодирующей последовательности. Лидерные последовательности, пригодные по форме и содержанию для настоящего изобретения, включают лидер Hsp70 маиса (US 5362865 и US 5859347) и элемент омега вируса мозаики табака (ВМТ).
Терминация транскрипции достигается с помощью 3' нетранслируемой последовательности ДНК, функционально связанной в векторе экспрессии с целевым полинуклеотидом. 3' Нетранслируемый участок рекомбинантной молекулы ДНК содержит сигнал полиаденилирования, функция которого в растениях состоит в присоединении аденилатных нуклеотидов к 3' концу РНК. 3' Нетранслируемый участок может быть получен из различных генов, которые экспрессируются в клетках растения. В данном случае обычно используются 3' нетранслируемые участки синтетазы нопалина, 3' нетранслируемый участок маленькой субъединицы гена РУБИСКО гороха, 3' нетранслируемый участок соевого гена запасаемого белка 7S семени. Кроме того, пригодными являются 3' транскрибированные, нетранслируемые участки, содержащие полиаденилатный сигнал индуцирующих опухоль (ИО) генов плазмиды Agrobacterium.
Рекомбинантные технологии ДНК могут использоваться для улучшения экспрессии трансформированного полинуклеотида посредством манипуляции, например, эффективностью, с которой транслируются образующиеся в результате транскрипты, путем оптимизации кодонов согласно виду клетки-хозяина или делеции последовательностей, которые дестабилизируют транскрипты, и эффективностью посттрансляционных модификаций.
Транспортные нуклеиновые кислоты
Транспортные нуклеиновые кислоты могут использоваться для доставки экзогенного полинуклеотида в клетку и содержат одну, предпочтительно, две граничные последовательности и один или более целевые полинуклеотиды. Транспортная нуклеиновая кислота может кодировать или не кодировать селекционный маркер. Предпочтительно, транспортная нуклеиновая кислота формирует часть бинарного вектора в клетке бактерии, причем бинарный вектор дополнительно содержит элементы, которые позволяют репликацию вектора у бактерии, селекцию или поддержание бактериальных клеток, содержащих бинарный вектор. При переносе в эукариотную клетку, компонент транспортной нуклеиновой кислоты бинарного вектора способен к интеграции в геном эукариотной клетки или пригоден для экспериментов с временной экспрессией, только с экспрессией в клетке.
В настоящем документе термин «внехромосомная транспортная нуклеиновая кислота» обозначает молекулу нуклеиновой кислоты, которая способна к перемещению из бактерии, например, вида Agrobacterium, в эукариотную клетку, например, клетку листа растения. Внехромосомная транспортная нуклеиновая кислота представляет собой генетический элемент, который известен, как элемент, способный к перемещению, с последующей интеграцией с нуклеотидной последовательностью, содержащейся в пределах его границ, в геном клетки-реципиента. В данном отношении, транспортная нуклеиновая кислота фланкирована обычно двумя «граничными» последовательностями, хотя в некоторых случаях может использоваться одна граница на одном конце, и второй конец транспортной нуклеиновой кислоты генерируется случайным образом в процессе переноса. Целевой полинуклеотид обычно размещается между левой, подобной границе, последовательностью и правой, подобной границе, последовательностью транспортной нуклеиновой кислоты. Полинуклеотид, содержащийся в пределах транспортной нуклеиновой кислоты, может быть функционально связан со множеством других промоторных и терминаторных регуляторных элементов, которые облегчают его экспрессию, т. е., транскрипцию и/или трансляцию полинуклеотида. Транспортные ДНК (Т-ДНК) видов Agrobacterium, например, Agrobacterium tumefaciens или Agrobacterium rhizogenes, и созданные человеком их варианты/мутанты, вероятно, представляют собой лучше всего описанные примеры транспортных нуклеиновых кислот. Другой пример представляет собой Р-ДНК («растительная ДНК»), которая содержит подобные границе последовательности T-ДНК из растений.
В настоящем документе «T-ДНК» обозначает T-ДНК ОИ плазмиды Agrobacterium tumefaciens или из плазмиды Ri Agrobacterium rhizogenes, или их варианты, функция которых состоит в переносе ДНК в растительные клетки. T-ДНК может содержать полноразмерную T-ДНК, включая как правую, так и левую граничные последовательности, но необходимыми являются только минимальные последовательности, необходимые в цис для переноса, т. е., правая T-ДНК граничная последовательность. Т-ДНК по изобретению имеют вставленный в них где-либо между правой и левой граничными последовательностями (если они присутствуют) целевой полинуклеотид. Последовательности, кодирующие факторы, необходимые в транс для переноса T-ДНК в растительную клетку, например, гены vir, могут быть вставлены в T-ДНК или могут присутствовать на том же репликоне, что и T-ДНК, или предпочтительно находятся в транс на совместимом репликоне в хозяине Agrobacterium. Такие «системы бинарного вектора» хорошо известны из уровня техники. В настоящем документе «П-ДНК» обозначает транспортную нуклеиновую кислоту, выделенную из генома растения или ее варианты/мутанты, созданные человеком, и содержит на каждом конце или только на одном конце подобную граничной последовательность T-ДНК.
В настоящем документе «граничная» последовательность транспортной нуклеиновой кислоты может быть выделена из выбранного организма, например, растения или бактерии, или представлять собой ее вариант/мутант, созданный человеком. Граничная последовательность способствует и облегчает перенос полинуклеотида, с которым она связана, и может облегчить его интеграцию в геном клетки-реципиента. В варианте реализации длина граничной последовательности составляет 10-80 пар оснований. Граничные последовательности T-ДНК видов Agrobacterium хорошо известны из уровня техники и включают раскрытые Lacroix с соавт. (2008).
Хотя традиционно использовались только виды Agrobacterium для переноса генов в клетки растений, в настоящее время идентифицировано/разработано большое число систем, которые функционируют подобным видам Agrobacterium способом. В последнее время несколько видов, не относящихся к Agrobacterium, были генетически модифицированы таким образом, чтобы сделать их компетентными для переноса гена (Chung с соавт., 2006; Broothaerts с соавт., 2005). Они включают вид Rhizobium NGR234, Sinorhizobium meliloti и Mezorhizobium loti.
Прямой перенос эукариотных экспрессионных плазмид от бактерий эукариотным хозяевам впервые был осуществлен несколько десятилетий тому назад слиянием клеток млекопитающего и протопластов несущей плазмиду Escherichia coli (Schaffner, 1980). С того времени, количество бактерий, способных доставлять гены в клетки млекопитающего, устойчиво возрастало (Weiss, 2003), будучи обнаружено четырьмя независимыми группами (Sizemore с соавт. 1995; Courvalin с соавт., 1995; Powell с соавт., 1996; Darji с соавт., 1997).
В настоящем документе термины «трансфекция», «трансформация» и их вариации, в общем, используются равнозначно. «Трансфицированными» или «трансформированными» клетками можно манипулировать с целью введения целевого полинуклеотида(ов) или получения из них клеток-потомков.
Рекомбинантные клетки
В изобретении также раскрывается рекомбинантная клетка, например, рекомбинантная растительная клетка или грибковая клетка, которая является клеткой-хозяином, трансформированной одним или более полинуклеотидов или векторов, раскрытых в настоящем документе, или их комбинацией. Пригодные клетки согласно изобретению включают любую клетку, которая может быть трансформирована полинуклеотидом или рекомбинантным вектором по изобретению, кодирующим РНК, полипептид или фермент, раскрытые в настоящем документе. Клетка представляет собой клетку, которая, таким образом, пригодна к использованию для получения липида. Рекомбинантная клетка может быть клеткой в культуре, клеткой in vitro или в организме, например, таком как растение, или в органе, например, таком как семя или лист. Предпочтительно, клетка находится в растении, более предпочтительно в семени растения. В одном варианте реализации рекомбинантная клетка представляет собой клетку, не являющуюся клеткой человека.
Клетки-хозяева, в которые введен(ы) полинуклеотид(ы), могут быть нетрансформированными клетками или клетками, которые уже трансформированы по меньшей мере одной нуклеиновой кислотой. Такие нуклеиновые кислоты могут быть связаны или не связаны с синтезом липида. Клетки-хозяева по настоящему изобретению могут быть эндогенно (т. е., от природы) способны к выработке полипептида(ов), раскрытого в настоящем документе, когда рекомбинантная клетка, полученная из них, обладает повышенной способностью к выработке полипептида(ов) или может быть способна к выработке указанного(ых) полипептида(ов) только после трансформации по меньшей мере одним полинуклеотидом по изобретению. В варианте реализации рекомбинантная клетка по изобретению обладает повышенной способностью к выработке неполярного липида, такого как ТАГ.
Клетки-хозяева по настоящему изобретению могут представлять собой любую клетку, способную к выработке по меньшей мере одного белка, раскрытого в настоящем документе, и включают клетки грибов (в том числе, дрожжей), животных, таких как членистоногие, и растительные клетки, такие как клетки водорослей и растений. В предпочтительном варианте реализации растительная клетка представляет собой клетку семени, в частности, клетку в семядоле или эндосперме семени. Клетки-хозяева могут быть организмом, пригодным для процесса ферментации, например, таким как Yarrowia lipolytica или другие дрожжи. В одном варианте реализации изобретения клетка представляет собой клетку животного. Клетка животного может принадлежать к любому виду животного, например, клетка животного, не относящегося к человеческому роду, клетка позвоночного, не относящегося к человеческому роду, клетка млекопитающего, не относящегося к человеческому роду, или клетки водных животных, например, рыб или ракообразных, беспозвоночных, насекомых и т. п. Примеры клеток водорослей, пригодных в качестве клеток-хозяев по настоящему изобретению, включают, например, виды Chlamydomonas (например, Chlamydomonas reinhardtii), виды Dunaliella, виды Haematococcus, виды Chlorella, виды Thraustochytrium, виды Schizochytrium и виды Volvox.
Трансгенные растения
В изобретении также раскрывается растение, содержащее один или более экзогенных полинуклеотидов или полипептидов и одну или более генетических модификаций по изобретению, клетки по изобретению, вектор по изобретению или их комбинации. Термин «растение» в роли имени существительного обозначает цельные растения, тогда как термин «его часть» обозначает органы растения (например, листья, стебли, корни, цветки, плод), одинарные клетки (например, пыльца), семя, части семени, например, зародыш, эндосперм, щиток зародыша или оболочку семени, ткань растения, например, сосудистую ткань, растительные клетки и их потомки. В настоящем документе части растения включают растительные клетки.
В настоящем документе термин ʺв растенииʺ, в контексте модификации растения означают, что модификация произошла по меньшей мере в одной части растения, включая ситуации, в которых модификация произошла во всем растению, и не исключая ситуаций, в которых модификация происходит только в одной или нескольких, но не во всех частях растения. Например, говорят, что тканеспецифичный промотор экспрессируется ʺв растенииʺ, даже если он может экспрессироваться только в некоторых частях растения. Аналогично, ʺполипептид фактора транскрипции, который повышает экспрессию одного или более гликолитических генов и/или генов биосинтеза жирных кислот в растенииʺ означает, что повышенная экспрессия присутствует по меньшей мере в части растения.
В настоящем документе термин «растение» применяется в самом широком смысле, включая любой организм из царства растений. Кроме того, он включает красные и коричневые водоросли, а также зеленые водоросли. Он включает, без ограничений, любые виды цветковых растений, травы, сельскохозяйственной культуры или злака (например, масличная культура, маис, соя), кормового или силосного, фруктового или овощного растения, травянистого растения, деревянистого растения или дерева. Не подразумевается ограничения растения какой-либо конкретной структурой. Кроме того, он обозначает одноклеточное растение (например, микроводоросль). Термин «его часть» со ссылкой на растение обозначает растительную клетку и ее потомка, множество клеток растения, структуру, которая присутствует на любой стадии развития растения или ткани растения. Такие структуры включают, без ограничений, листья, стебли, цветки, плоды, орехи, корни, семя, оболочку семени, зародыши. Термин «ткань растения» включает дифференцированные и недифференцированные ткани растений, в том числе, присутствующие в листьях, стеблях, цветках, плодах, орехах, корнях, семени, например, зародышевая ткань, эндосперм, ткань кожуры (например, эпидермис, перидерма), сосудистая ткань (например, ксилема, флоэма) или донная ткань (содержащая клетки паренхимы, колленхимы и/или склеренхимы), а также клетки в культуре (например, одинарные клетки, протопласты, каллюс, зародыши, и т. п.). Ткань растения может находиться in planta, в культуре органа, культуре ткани или культуре клеток.
В настоящем документе термин «вегетативная ткань» или «вегетативная часть растения» обозначает любую ткань растения, орган или часть, кроме органов для полового воспроизводства растений. В частности, органы для полового воспроизводства растений представляют собой несущие семена органы, цветки, пыльцу, плоды и семена. Вегетативные ткани и части включают по меньшей мере листья, стебли (в том числе, стрелки и побеги, за исключением головок), клубни и корни растения, за исключением цветков, пыльцы, семени, включая оболочку семени, зародыша и эндосперма, плода, включая ткани мезокарпа, несущие семя стручки и несущие семя головки. В одном варианте реализации изобретения вегетативная часть растения представляет собой воздушную часть растения. В другом или дальнейшем варианте реализации изобретения вегетативная часть растения представляет собой зеленую часть, например, лист или стебель.
«Трансгенное растение» или его вариации обозначают растение, которое содержит трансген, не найденный в растении дикого типа такого же вида, подвида или сорта. Трансгенные растения, раскрытые в контексте настоящего изобретения, включают растения и их потомков, генетически модифицированных с применением рекомбинантных методов, чтобы вызывать выработку по меньшей мере одного полипептида, раскрытого в настоящем документе, в желательном растении или его части. Трансгенные части растения имеют соответствующее значение.
Термины «семя» и «зерно» используются равнозначно в настоящем документе. «Зерно» обозначает зрелое зерно, например, собранное в процессе сбора урожая зерно, или зерно, которое находится на растении, но готово для сбора урожая, а также может обозначать зерно после набухания или прорастания, согласно контексту. Зрелое зерно обычно содержит менее чем приблизительно 18% влаги. В предпочтительном варианте реализации, содержание влаги в зерне находится на уровне, который в общем расценивается как безопасный для хранения, предпочтительно, от 5% до 15%, от 6% до 8%, от 8% до 10% или от 10% до 15%. В настоящем документе «развивающееся семя» обозначает семя до состояния зрелости, обычно найденное в репродуктивных структурах растения после оплодотворения или цветения, но может также обозначать семена до состояния зрелости, которые сняты с растения. Зрелое семя обычно содержит менее чем приблизительно 12% влаги
В настоящем документе термин «запасающий орган растения» обозначает часть растения, специализирующуюся на хранении энергии, например, в форме белков, углеводов, липида. Примерами запасающих органов растения являются семя, плод, клубневидные корни и клубни. Предпочтительным запасающим органом растения по изобретению является семя.
В настоящем документе термин «фенотипически нормальный» обозначает генетически модифицированное растение или его часть, например, трансгенное растение, или запасающий орган, такой как семя, клубень или плод по изобретению, без существенного снижения способности к росту и воспроизводству, по сравнению с немодифицированным растением или его частью. Предпочтительно, биомасса, темпы роста, скорость прорастания, размер запасающего органа, размер семени и/или количество образовавшихся жизнеспособных семян составляет не менее 90% от показателей растения, не содержащего указанных генетических модификаций или экзогенных полинуклеотидов, при выращивании в идентичных условиях. Данный термин не включает признаков растения, которые могут отличаться от растения дикого типа, но которые не влияют на полноценность растения с точки зрения коммерческих целей, например, такие как фенотип балерины листьев саженца. В варианте реализации генетически модифицированное растение или его часть, которые являются фенотипически нормальными, содержат рекомбинантный полинуклеотид, кодирующий подавляющий сайленсинг, функционально связанный с промотором, специфичным для запасающего органа растения, и обладают способностью к росту или воспроизводству, по существу такой же как соответствующее растение или его часть, не содержащие указанного полинуклеотида.
В настоящем документе "начало цветения" или ʺнаступление цветенияʺ со ссылкой на растение обозначает время раскрытия первого цветка на растении или время начала пыления.
В настоящем документе термин "завязывание семян" со ссылкой на несущее семена растение обозначает время, когда первое семя растения достигает зрелости. Обычно это можно определить по цвету семени или содержанию в нем влаги, как хорошо известно из уровня техники.
В настоящем документе термин "старениеʺ со ссылкой на цельное растение обозначает конечную стадию развития растения, которая следует за завершением роста, обычно после того, как воздушная биомасса или высота растения достигает максимума. Старение начинается, когда воздушная биомасса растения достигает максимума, и ее количество начинает уменьшаться, и в общем заканчивается с гибелью большинства тканей растения. В ходе данной стадии растение мобилизует и рециклизует клеточные компоненты из листьев и других частей, которые накапливались в процессе роста, в другие части, чтобы завершить репродуктивное развитие. Старение представляет собой сложный, регулируемый процесс, в который вовлечена экспрессия новых генов или повышенная экспрессия некоторых генов. Часто, некоторые части растения стареют, в то время как другие части того же растения продолжают расти. Таким образом, относительно листа растения или другого зеленого органа, термин ʺстарениеʺ в настоящем документе обозначает время, когда количество хлорофилла в листе или органе начинает снижаться. Старение обычно сопровождается обезвоживанием листа или органа, по большей части в последней стадии старения. Старение обычно можно определить по изменению цвета листа с зеленого по направлению к желтому и, в конечном счете, к коричневому окрашиванию после полного обезвоживания. Считается, что клеточное старение лежит в основе старения растения и органов.
Растения, раскрытые или предусмотренные для применения в практике настоящего изобретения, включают однодольные или двудольные. В предпочтительных вариантах реализации, растения по настоящему изобретению представляют собой сельскохозяйственные растения (например, злаковые и зернобобовые, маис, пшеницу, различные виды картофеля, рис, сорго, пшено, маниок, ячмень), или другие растения из семейства бобовых, такие как соя, бобы или горох. Растения могут выращиваться для производства съедобных корней, клубней, листьев, стеблей, цветков или плодов. Растения могут быть овощными растениями, вегетативные части которых используются в пищу. Растения по изобретению могут представлять собой: Acrocomia aculeata (макаубская пальма), Arabidopsis thaliana, Aracinis hypogaea (арахис), Astrocaryum murumuru (мурумуру), Astrocaryum vulgare (тукума), Attalea geraensis (атталия), Attalea humilis (американская масличная пальма), Attalea oleifera (андайя), Attalea phalerata (урикури), Attalea speciosa (бабассу), Avena sativa (овес), Beta vulgaris (сахарная свекла), виды Brassica, такие как Brassica carinata, Brassica juncea, Brassica napobrassica, Brassica napus (рапс), Camelina sativa (ложный лен), Cannabis sativa (конопля), Carthamus tinctorius (сафлор красильный), Caryocar brasiliense (пекви), Cocos nucifera (кокос), Crambe abyssinica (абиссинская капуста), Cucumis melo (дыня), Elaeis guineensis (африканская пальма), Glycine max (соя), Gossypium hirsutum (хлопчатник), виды Helianthus, такие как Helianthus annuus (подсолнечник), Hordeum vulgare (ячмень), Jatropha curcas (лечебный орех), Joannesia princeps (лещина арара), виды Lemna (ряска), такие как Lemna aequinoctialis, Lemna disperma, Lemna ecuadoriensis, Lemna gibba (ряска горбатая), Lemna japonica, Lemna minor, Lemna minuta, Lemna obscura, Lemna paucicostata, Lemna perpusilla, Lemna tenera, Lemna trisulca, Lemna turionifera, Lemna valdiviana, Lemna yungensis, Licania rigida (ойтика), Linum usitatissimum (лен), Lupinus angustifolius (люпин), Mauritia flexuosa (пальма бурити), Maximiliana maripa (пальма инайя), виды Miscanthus, такие как Miscanthus x giganteus и Miscanthus sinensis, виды Nicotiana (табак), такие как Nicotiana tabacum или Nicotiana benthamiana, Oenocarpus bacaba (бакаба-до-азейте), Oenocarpus bataua (патайя), Oenocarpus distichus (бакаба-де-лек), виды Oryza (рис), такие как Oryza sativa и Oryza glaberrima, Panicum virgatum (просо прутьевидное), Paraqueiba paraensis (марь), Persea amencana (авокадо), Pongamia pinnata (индийский бук), Populus trichocarpa, Ricinus communis (клещевина), виды Saccharum (сахарный тростник), Sesamum indicum (кунжут), Solanum tuberosum (картофель), виды Sorghum, такие как Sorghum bicolor, Sorghum vulgare, Theobroma grandiforum (какао), виды Trifolium, Trithrinax brasiliensis ((бразильская игольчатая пальма), виды Triticum (пшеница), такие как Triticum aestivum, Zea mays (кукуруза), люцерна (Medicago sativa), рожь (Secale cerale), сладкий картофель (Lopmoea batatus), маниок (Manihot esculenta), кофе (виды Cofea), ананас (Anana comosus), цитрусовое дерево (виды Citrus), какао (Theobroma cacao), чай (Camellia senensis), банан (виды Musa), авокадо (Persea americana), инжир (Ficus casica), гуава (Psidium guajava), манго (Mangifer indica), олива (Olea europaea), папайя (Carica papaya), кешью (Anacardium occidentale), макадамия (Macadamia intergrifolia) и миндаль (Prunus amygdalus).
Другие предпочтительные растения включают травы C4, например, в дополнение к упомянутым выше, Andropogon gerardi, Bouteloua curtipendula, B. gracilis, Buchloe dactyloides, Schizachyrium scoparium, Sorghastrum nutans, Sporobolus cryptandrus; травы C3, например, Elymus canadensis, бобы Lespedeza capitata и Petalostemum villosum, полукустарник Aster azureus; и древесные растения, например, Quercus ellipsoidalis и Q. macrocarpa. Другие предпочтительные растения включают травы C3.
В предпочтительном варианте реализации изобретения растение представляет собой покрытосеменное растение.
В варианте реализации изобретения растение представляет собой растение масличной культуры, предпочтительно сельскохозяйственной масличной культуры. В настоящем документе «растение масличной культуры» представляет собой вид растения, используемый для коммерческого получения липида из семян растения. Растение масличной культуры может представлять собой, например, масличный рапс (например, рапс), маис, подсолнечник, сафлор красильный, сою, сорго, лен (семя льна) или сахарную свеклу. К тому же, растение масличной культуры может быть другим представителем Brassica, хлопчатником, арахисом, маком, брюквой, горчицей, клещевиной, кунжутом, сафлором, Jatropha curcas или орехоносным растением. Растение может вырабатывать высокие уровни липида в плоде, например, олива, масличная пальма или кокосовая пальма. Садовые растения, к которым может быть применено настоящее изобретение, представляют собой салат, эндивий или растения вида Brassica, включая капусту, брокколи или цветную капусту. Настоящее изобретение может быть применено к табаку, тыквам, моркови, землянике, помидору или перцу.
В предпочтительном варианте реализации трансгенное растение является гомозиготным по всем и каждому гену, который введен (трансген), таким образом, что его потомок, не отходит от желательного фенотипа. Кроме того, трансгенное растение может быть гетерозиготным по введенному(ым) трансгену(ам), предпочтительно однородно гетерозиготным по трансгену, например, в потомстве F1, выращенном из гибридного семени. Такие растения могут предлагать преимущества, например, силу гибрида, хорошо известную из уровня техники.
Трансформация растений
Трансгенные растения могут быть получены с применением методов, известных из уровня техники, например, раскрытых в общих чертах в Slater et al., Plant Biotechnology - The Genetic Manipulation of Plants, Oxford University Press (2003), и Christou and Klee, Handbook of Plant Biotechnology, John Wiley and Sons (2004).
В настоящем документе термин «стабильно трансформирующий», «стабильно трансформированный» и его вариации обозначают интеграцию полинуклеотида в геном клетки таким образом, что он передается клеткам-потомкам в процессе деления клетки, без необходимости в положительной селекции на предмет их присутствия. Стабильные трансформанты или их потомки могут быть идентифицированы любыми средствами, известными из уровня техники, например, Саузерн-блоттингом на хромосомной ДНК или гибридизацией геномной ДНК in situ, которые позволяют осуществить их отбор.
Опосредованный Agrobacterium перенос представляет собой широко применимую систему для введения генов в растительные клетки, поскольку ДНК может быть введена в клетки цельных тканей растения, органов растения или эксплантов в культуре ткани, для временной экспрессии или стабильной интеграции ДНК в геном растительной клетки. Например, можно применять способы с погружением цветков (в полевых условиях). Применение опосредованных Agrobacterium векторов интеграции в растения с целью введения ДНК в растительные клетки хорошо известно из уровня техники. Участок ДНК для переноса определяется последовательностями границы, и перенесенная ДНК (T-ДНК) обычно вставляется в геном растения. Данный способ является способом выбора вследствие простой и надежной природы переноса гена.
Способы ускорения, которые могут применяться, включают, например, баллистическую трансфекцию и т. п. Одним из примеров способа доставки трансформирующих молекул нуклеиновых кислот в растительные клетки является баллистическая трансфекция. Данный способ рассмотрен Yang et al., Particle Bombardment Technology for Gene Transfer, Oxford Press, Oxford, England (1994). Небиологические частицы (микрочастицы), которые могут быть покрыты нуклеиновыми кислотами и доставлены в клетки, например, незрелых эмбрионов, толкающей силой. Примеры частиц включают состоящие из вольфрама, золота, платины и т.п.
В другом способе пластиды могут быть стабильно трансформированы. Способы, раскрытые для трансформации пластиды в высших растениях, включают доставку с помощью пистолета частицы ДНК, содержащей селекционный маркер, и нацеливание ДНК на геном пластиды посредством гомологичной рекомбинации (US 5451513, US 5545818, US 5877402, US 5932479 и WO 99/05265). Дополнительно, могут применяться другие способы трансформации клетки, которые включают, без ограничений, введение ДНК в растения прямым переносом ДНК в пыльцу, прямой инъекцией ДНК в репродуктивные органы растения или прямой инъекцией ДНК в клетки незрелых зародышей, с последующей регидратацией высушенных зародышей.
Регенерация, развитие и культивирование растений из единичных трансформантов протопласта растения или из различных трансформированных эксплантов хорошо известны из уровня техники (Weissbach с соавт., In: Methods for Plant Molecular Biology, Academic Press, San Diego, Calif., (1988)). Такая регенерация и процесс роста обычно включают стадии селекции трансформированных клеток, культивирования таких индивидуализированных клеток через обыкновенные стадии зародышевого развития, включая стадию укоренившегося сеянца. Трансгенные зародыши и семена регенерируют подобным образом. Полученные трансгенные укоренившиеся побеги в дальнейшем пересаживают в пригодную среду для роста растения, например, грунт.
Развитие или регенерация растений, содержащих чужеродный, экзогенный ген, хорошо известны из уровня техники. Предпочтительно, регенерирующиеся растения самоопыляются, чтобы обеспечить гомозиготные трансгенные растения. В другом случае, пыльцу, полученную от регенерирующих растений, скрещивают с выращенными из семени растениями важных с агрономической точки зрения линий. С другой стороны, пыльца растений таких важных линий используется для опыления регенерированных растений. Трансгенное растение по настоящему изобретению, содержащее желательный полинуклеотид, культивируют с использованием способов, хорошо известных специалисту в данной области.
Чтобы подтвердить присутствие трансгенов в трансгенных клетках и растениях, может быть осуществлена амплификация полимеразной цепной реакцией (ПЦР) или анализ методом Саузерн-блоттинга, с применением методов, известных специалистам из уровня техники. Продукты экспрессии трансгенов могут быть обнаружены любым из множества путей, в зависимости от природы продукта, которые включают Нозерн-блоттинг гибридизацию, Вестерн-блоттинг и ферментный анализ. Как только трансгенные растения получены, они могут выращиваться для получения тканей или частей растения, содержащих желательный фенотип. Урожай ткани растения или частей растения может быть собран, и/или семена могут быть собраны. Семя может служить источником для выращивания дополнительных растений с тканями или частями, обладающими желательными характеристиками. Предпочтительно, урожай вегетативных частей растения собирают в то время, когда выход неполярных липидов находится на самом высоком уровне. В одном варианте реализации урожай вегетативных частей растения собирают во время, близкое к цветению или после начала цветения. Предпочтительно, части растения собирают в период около начала старения, на который обычно указывает пожелтение и высыхание листьев.
Трансгенные растения, полученные с использованием Agrobacterium или других способов трансформации, обычно содержат единственный генетический локус на одной хромосоме. Такие трансгенные растения могут обозначаться как гемизиготные по дополнительному(ым) гену(ам). Более предпочтительным является трансгенное растение, гомозиготное по дополнительному(ым) гену(ам), т. е. трансгенное растение, которое содержит два дополнительных гена, по одному гену в одинаковых локусах на каждой хромосоме из пары хромосом. Гомозиготное трансгенное растение может быть получено путем самооплодотворения гемизиготного трансгенного растения, проращиванием части полученных семян и анализом полученных растений на предмет присутствия целевого гена.
Кроме того, необходимо понимать, что два различных трансгенных растения, которые содержат два независимо разделенных экзогенных гена или локуса, могут быть скрещены (получен гибрид), с целью получения потомства, содержащего оба набора генов или локусов. Самоопыление подходящего потомка F1 может давать растения, гомозиготные по обоим экзогенным генам или локусам. Обратное скрещивание с материнским растением и скрещивание с нетрансгенным растением также предусмотрены, как и вегетативное размножение. Подобным образом, трансгенное растение может быть скрещено со вторым растением, содержащим генетическую модификацию, такую как мутантный ген, с идентификацией потомства, содержащего как трансген, так и генетическую модификацию. Описание других способов разведения, которые обычно применяются для различных признаков и сельскохозяйственных культур, можно найти в Fehr, In: Breeding Methods for Cultivar Development, Wilcox J. ed., American Society of Agronomy, Madison Wis. (1987).
ИНЛПГ
В одном варианте реализации TILLING (Индуцированные Нацеливанием Локальные Повреждения В Геномах, ИНЛПГ) может применяться для получения растений, в которых эндогенные гены содержат мутацию, например, гены, кодирующие полипептид SDP1 или ТГД, пластидную GPAT, пластидную LPAAT, фосфатазу фосфатидиновой кислоты (PAP) или комбинацию двух или более из указанных компонентов. На первой стадии введение мутаций, например новых модификаций одинарных пар оснований, осуществляют в популяции растений посредством обработки семян (или пыльцы) химическим мутагеном, с последующим разведением растений до поколения, в котором мутации будут устойчиво наследоваться. ДНК извлекают и хранят семена всех членов популяции, чтобы создать ресурс, который может быть доступен повторно через некоторое время. Для анализа ИНЛПГ можно применить гетеродуплексные способы с использованием специфичных эндонуклеаз, с целью выявления однонуклеотидного полиморфизма (ОНП). В качестве альтернативы, секвенирование ДНК следующего поколения из пулов мутантных растений можно применять для идентификации мутаций в выбранном гене. Обычно, достигается частота мутаций один мутант на 1000 растений в популяции мутантных растений. С применением данного подхода может быть осуществлен скрининг многих тысяч растений для идентификации любого индивидуума с модификацией единственного основания, а также небольшими инсерциями или делециями (1-30 пар оснований) в любом гене или конкретном участке генома. ИНЛПГ дополнительно описана Slade и Knauf (2005) и Henikoff с соавт. (2004).
В дополнение к возможности эффективного обнаружения мутаций, высокопроизводительная технология ИНЛПГ идеальна для обнаружения природного полиморфизма. Таким образом, детальное исследование неизвестной гомологичной ДНК, образующей гетеродуплекс с известной последовательностью, выявляет количество и расположение полиморфных сайтов. Идентифицированы нуклеотидные модификации, а также инсерции и делеции небольшого размера, в том числе, по меньшей мере некоторые полиморфизмы количества повторов. Указанный признак получил название Ecotilling (Comai с соавт., 2004).
Редактирование генома с применением сайт-специфичных нуклеаз
Редактирование генома с применением сконструированных нуклеаз, таких как направляемые РНК эндонуклеазы или нуклеазы ДНК, состоящие из доменов связывания ДНК, специфичных для последовательности, слитых с неспецифичным модулем расщепления ДНК. Такие сконструированные нуклеазы дают возможность осуществлять эффективные и точные генетические модификации путем индукции направленных разрывов двухцепочечной ДНК, которые побуждают эндогенные клеточные механизмы восстановления ДНК клетки к восстановлению индуцированного разрыва. Такие механизмы включают, например, подверженное ошибкам негомологичное соединение концов (НГСК) и направляемое гомологией восстановление (НГВ).
В присутствии донорской плазмиды с удлиненными гомологичными плечами, НГВ может приводить к введению одного или нескольких трансгенов для коррекции или замены существующих генов. В отсутствии донорской плазмиды, НГСК-опосредованное восстановление дает незначительные инсерционные или делеционные мутации мишени, которые вызывают разрушение гена.
Сконструированные нуклеазы, пригодные для способов по настоящему изобретению, включают цинк-пальцевые нуклеазы (ЦПН), подобные активатору транскрипции (ПАТ) эффекторные нуклеазы (ПАТЭН) и нуклеазы типа CRISPR/Cas9.
Обычно кодирующие нуклеазу гены доставляются в клетки плазмидной ДНК, вирусными векторами или транскрибированной in vitro мРНК.
Цинк-пальцевая нуклеаза (ЦПН) содержит домен связывания с ДНК и домен расщепления ДНК, причем домен связывания с ДНК состоит по меньшей мере из одного «цинкового пальца» и функционально связан с доменом расщепления ДНК. Домен связывания с ДНК цинкового пальца расположен на N-конце белка, а домен расщепления ДНК расположен на C-конце указанного белка.
ЦПН должна содержать по меньшей мере один «цинковый палец». В предпочтительном варианте реализации изобретения ЦПН должна содержать по меньшей мере три «цинковых пальца», чтобы обладать достаточной специфичностью и быть пригодной для направленной генетической рекомбинации в клетке- или организме-хозяине. Обычно, специфичность ЦПН, содержащей более чем три «цинковых пальца», будет постепенно увеличиваться с наличием каждого дополнительного «цинкового пальца».
Домен «цинкового пальца» может происходить из любого класса или вида «цинковых пальцев». В конкретном варианте реализации изобретения домен «цинкового пальца» включает Cis2His2 тип «цинкового пальца», который в самом общем виде представлен, например, факторами транскрипции «цинкового пальца» TFIIIA или Sp1. В предпочтительном варианте реализации изобретения домен «цинкового пальца» включает три «цинковых пальца» типа Cis2His2. Распознавание ДНК и/или специфичность связывания ЦПН могут быть модифицированы с целью достижения целевой генетической рекомбинации на каком-либо выбранном сайте клеточной ДНК. Такая модификация может быть достигнута с применением известных методов молекулярной биологии и/или химического синтеза (см., например, Bibikova с соавт., 2002).
Домен расщепления ДНК ЦПН получают из класса неспецифичных доменов расщепления ДНК, например домен расщепления ДНК рестрикционного фермента типа II, например, FokI (Kim с соавт., 1996). Другие пригодные эндонуклеазы могут содержать, например, HhaI, HindIII, Nod, BbvCI, EcoRI, BglI и AlwI.
Подобная активатору транскрипции (ПАТ) эффекторная нуклеаза (ПАТЭН) содержит домен связывания с ДНК эффектора ПАТ и домен эндонуклеазы.
Эффекторы ПАТ представляют собой белки патогенных бактерий растения, которые впрыскиваются патогеном в растительную клетку, где они перемещаются к ядру и функционируют, как факторы транскрипции для активизации специфических генов растения. Первичная аминокислотная последовательность эффектора ПАТ диктует нуклеотидную последовательность, с которой он связывается. Таким образом, для эффекторов ПАТ могут быть предсказаны сайты-мишени, и эффекторы ПАТ могут быть сконструированы и сгенерированы для целей связывания с конкретными нуклеотидными последовательностями.
Слитые с эффектором ПАТ кодирующие последовательности нуклеиновых кислот представляют собой последовательности, кодирующие нуклеазу или часть нуклеазы, обычно неспецифический домен расщепления рестрикционной эндонуклеазы типа II, например, FokI (Kim с соавт., 1996). Другие пригодные эндонуклеазы могут содержать, например, HhaI, HindIII, Nod, BbvCI, EcoRI, BglI и AlwI. Тот факт, что некоторые эндонуклеазы (например, FokI) функционируют только в виде димеров, может быть с выгодой использован для повышения специфичности эффектора ПАТ в отношении мишени. Например, в некоторых случаях каждый мономер FokI может быть слит с последовательностью эффектора ПАТ, которая распознает другую последовательность мишени ДНК, и только в случае, когда два сайта распознавания находятся в непосредственной близости, неактивные мономеры сближаются настолько, чтобы совместно образовать функционирующий фермент. С включением необходимости в связывании с ДНК для активизации нуклеазы, может быть создан высокоспецифичный в отношении сайта рестрикционный фермент.
Специфичная для последовательности ПАТЭН может распознавать конкретную последовательность в пределах предварительно выбранной нуклеотидной последовательности-мишени, присутствующей в клетке. Таким образом, в некоторых вариантах реализации изобретения нуклеотидная последовательность-мишень может быть просканирована на предмет наличия сайтов для распознавания нуклеазой, и конкретная нуклеаза может быть выбрана на основании последовательности мишени. В других случаях, может быть сконструирована ПАТЭН, нацеленная на конкретную клеточную последовательность.
Редактирование генома с применением программируемых РНК-направляемых ДНК эндонуклеаз
В отличие от сайт-специфичных нуклеаз, описанных выше, система кластеризованных, коротких палиндромных повторов с регулярными промежутками (ККППРП, CRISP)/Cas предлагает альтернативу ЦПН и ПАТЭН для индукции направленных генетических модификаций посредством направляемого РНК расщепления ДНК.
Системы ККППРП зависят от ККППРП РНК (ккРНК) и трансактивирующей химерной РНК (траккРНК) для осуществления специфичного для последовательности расщепления ДНК. Существует три вида систем ККППРП/Cas: в системах II типа Cas9 служит направляемой РНК ДНК эндонуклеазой, которая расщепляет ДНК при распознавании мишени ккРНК-траккРНК. Основание ККППРП РНК спаривается с траккРНК с образованием структуры из двух РНК, которая направляет Cas9 эндонуклеазы к комплементарным сайтам ДНК для расщепления.
Система ККППРП может быть доставлена в растительные клетки путем совместной доставки плазмид, экспрессирующих эндонуклеазу Cas и необходимые компоненты ккРНК. Эндонуклеаза Cas может быть превращена в никазу, чтобы обеспечить дополнительный контроль над механизмом восстановления ДНК (Cong с соавт., 2013).
ККППРП обычно представляют собой короткие, частично палиндромные последовательности размером 24-40 пар оснований, содержащие внутренние и концевые перевернутые повторы размером до 11 пар оснований. Хотя обнаружены изолированные элементы, в общем они организованы в кластеры (в количестве до около 20 или более на геном) повторяющихся фрагментов, разделенных уникальными последовательностями размером 20-58 пар оснований. В общем, ККППРП являются гомогенными в пределах конкретного генома, и большинство из них идентичны. Однако, существуют примеры гетерогенности, например, в Archaea (Mojica с соавт., 2000).
Биомасса растения
Повышение общего содержания липида в биомассе растения эквивалентно более высокому содержанию энергии, что делает ее применение в качестве корма (продукта питания) или фуража или для выработки биологического топлива более экономичным.
Основным компонентом природной биомассы растения являются углеводы (приблизительно 75%, в пересчете на массу сухого вещества) и лигнин (приблизительно 25%), который может варьировать в зависимости от вида растения. Углеводы в основном представляют собой целлюлозные или гемицеллюлозные волокна, которые придают прочность структуре растения, и лигнин, который удерживает волокна вместе. Биомассе растений обычно свойственна низкая плотность энергии в результате двух факторов: физической формы и содержания влаги. Это придает дополнительное неудобство и неэффективность ее хранению и транспортировке без какой-либо предварительной обработки.
Существует целый ряд доступных способов превращения ее в более пригодные формы, в том числе: 1) физическая предварительная обработка (например, размалывание), или 2) трансформация термическими (например, окисление, газификация, пиролиз) или химическими (например, анаэробное расщепление, ферментация, компостирование, переэтерификация) способами. Таким образом, биомассу превращают в то, что может быть описано, как топливная биомасса.
Сжигание
Сжигание представляет собой процесс, в ходе которого легковоспламеняющимся материалам позволяют гореть в присутствии воздуха или кислорода с высвобождением тепла. Основой процесса является окисление. Сжигание представляет собой самый простой способ использования биомассы для получения энергии и применяется для обеспечения теплом. Указанное тепло может непосредственно использоваться множеством способов: 1) нагревание пространства, 2) нагревание воды (или другой жидкости) для центрального или местного нагревания или переработки тепла, 3) получение пара для генерации электричества или движущей силы. Если легковоспламеняющийся горючий материал находится в форме биомассы, окисление касается, в основном, атомов углерода (C) и водорода (H) в целлюлозе, гемицеллюлозе, лигнине и других присутствующих молекулах, с образованием диоксида углерода (CO2) и воды (H2O). Растения по изобретению дают усовершенствованное топливо для сжигания благодаря повышенному содержанию липидов.
Газификация
Газификация представляет собой процесс частичного окисления, посредством которого источник углерода, например, биомасса растения, разлагается на монооксид углерода (CO) и водород (H2), плюс диоксид углерода (CO2) и, возможно, молекулы углеводорода, например, метана (CH4). Если газификация имеет место при относительно низкой температуре, например, от 700°C до 1000°C, газообразный продукт будет содержать относительно высокие уровни углеводородов, по сравнению с высокотемпературной газификацией. В результате, он может использоваться непосредственно для сжигания с целью получения тепла или генерации электричества с помощью паровой турбины или, при условии соответствующего удаления газа, в двигателе внутреннего сгорания для генерации электричества. Камера сжигания для простого парового котла может быть соединена с газификатором, или устройство для получения газа может очищаться от углеводородов с более длинной цепью (смолы), которые хранятся и сжигаются в другом месте. Система газификации может быть тесно связана с газовой турбиной комбинированного цикла для генерации электричества (КЦИГ - комбинированный цикл интегрированной газификации). Высокотемпературная газификация (от 1200°C до 1600°C) приводит к образованию нескольких углеводородов в газообразном продукте и более высокой доле CO и H2. Это известно как синтетический газ (сингаз или биосинтетический газ), который может использоваться для синтеза углеводородов с более длинной цепью, с применением таких методов, как синтез Фишера-Тропша (ФТ). При надлежащем соотношении H2 к CO (2:1), синтез ФТ может применяться для превращения синтетического газа в высококачественное синтетическое биологическое дизельное топливо, совместимое с обычными видами дизельного топлива из окаменелого сырья и дизельными двигателями.
Пиролиз
В настоящем документе термин «пиролиз» подразумевает процесс, в ходе которого используется медленное нагревание в отсутствие кислорода, с целью получения газообразных, нефтяных и угольных продуктов из биомассы. Пиролиз представляет собой термическое или термохимическое превращение материалов на основе липида, особенно на основе триглицерида. Продукты пиролиза включают газ, жидкость и твердый уголь, причем доля каждого зависит от параметров процесса. Более низкие температуры (около 400°C) показывают тенденцию к образованию более твердого угля (медленный пиролиз), тогда как несколько более высокие температуры (около 500°C) дают намного более высокую долю жидкости (бионефть), при условии, что продолжительность обработки паров удерживается на уровне до приблизительно 1 секунды или менее.
Температура от около 275°C до около 375°C может применяться для получения жидкой бионефти с более высоким относительным содержанием углеводородов с большей длиной цепи. Пиролиз включает прямой термической крекинг липидов или комбинацию термического и каталитического крекинга. При температурах приблизительно 400-500°C происходит крекинг, дающий углеводороды с короткой цепью, например, алканы, алкены, алкадиены, ароматические вещества, олефины и карбоновую кислоту, а также монооксид углерода и диоксид углерода.
Могут использоваться четыре основных типа катализатора, в том числе, катализаторы на основе переходного металла, катализаторы типа молекулярного сита, активизированный алюминия оксид и натрия карбонат (Maher с соавт., 2007). Примеры представлены в US 4102938. Алюминия оксид (Al2O3), активированный кислотой, представляет собой эффективный катализатор (US 5233109). Катализаторы типа молекулярного сита представляют собой пористые, высоко кристаллические структуры, которые демонстрируют избирательность в отношении размера, таким образом, что сквозь них могут проникать только молекулы определенных размеров. Они включают катализаторы на основе цеолита, например, ZSM-5 или HZSM-5, которые являются кристаллическими материалами, содержащими AlO4 и SiO4, и другие катализаторы на основе кремния диоксида-алюминия оксида. Активность и селективность указанных катализаторов зависит от кислотности, размера пор и формы пор, и обычно они активны при 300-500°C. Катализаторы на основе переходного металла раскрыты, например, в US 4992605. Катализатор на основе натрия карбоната использовали в пиролизе масел (Dandik и Aksoy, 1998).
В настоящем документе ʺгидротермическая обработкаʺ, ʺГТОʺ, дополнительно называемая ʺтермической деполимеризациейʺ представляет собой вид пиролиза, в ходе которого полученное из растения вещество, конкретно углеродсодержащий материал в полученном растение веществе, вводят в реакцию с водородом для получения продукта в форме бионефти, в основном состоящего из парафиновых углеводородов, наряду с другими газообразными и твердыми веществами. Значительное преимущество ГТО состоит в том, что нет необходимости сушить вегетативный растительный материал перед образованием композиции для введения в реакцию превращения, хотя вегетативный материал растения может быть предварительно высушен, чтобы упростить транспортировку или хранение биомассы. Биомасса может использоваться непосредственно после сбора урожая на поле. Реактор представляет собой любую емкость, способную выдерживать применяемую высокую температуру и давление и устойчивую к коррозии. Растворитель, применяемый для ГТО, содержит воду или полностью является водой или может содержать некоторые углеводородные соединения в форме масла. В общем, растворитель для ГТО не требует добавления спиртов. Реакция превращения может происходить в окислительном, восстановительном или инертном окружении. ʺОкислительныйʺ в настоящем документе означает присутствие воздуха, ʺвосстановительныйʺ означает присутствие восстановительного агента, обычно газообразного водорода или метана, например, 10-15% H2 и остальная часть газообразного N2, и ʺинертныйʺ означает присутствие инертного газа, такого как азот или аргон. Реакцию превращения предпочтительно осуществляют в восстановительных условиях. Углеродсодержащие материалы для превращения включают целлюлозу, гемицеллюлозу, лигнин и белки, а также липиды. Для процесса применяют температуру превращения от 270°C до 400°C и давление от 70 до 350 бар, обычно от 300°C до 350°C и 100-170 бар. В результате процесса образуются пары органических веществ, пиролизные газы и древесный уголь. Пары органических веществ конденсируют для получения бионефти. Извлечение бионефти может быть осуществлено путем охлаждения реактора и снижения давления до атмосферного, что позволяет образование бионефти (органическая) и водной фазы и извлечение бионефти из реактора.
Выход извлеченной бионефти вычисляют как процент от сухой массы исходной биомассы, в пересчете на сухую массу. Его вычисляют по формуле: масса бионефти x 100/сухая масса вегетативных частей растения. Масса бионефти не включает массу воды или твердых веществ, которые могут присутствовать в бионефтяной смеси и с легкостью удаляются фильтрацией или другими известными способами.
Далее бионефть можно разделить на фракции фракционной дистилляцией, с дополнительными процессами очистки или без них. Обычно, фракции, которые конденсируются при этих температурах, носят название: около 370°C, мазут; около 300°C, дизельное масло; около 200°C, керосин; около 150°C, газолин (бензин). Более тяжелые фракции можно расщеплять на более легкие, более желательные фракции, хорошо известные из уровня техники. Дизельное топливо обычно состоит из C13-C22 углеводородных соединений.
В настоящем документе ʺнефтяное дизельное топливоʺ (нефтедиз) обозначает дизельное топливо, полученное из окаменелого топлива, которое регламентируется спецификациями, установленными ASTM D975 в США и EN 590 в Европе. Термин ʺвозобновляемое дизельное топливоʺ в настоящем документе обозначает дизельное топливо, полученное из недавно живой биомассы (не окаменелого топлива), которое удовлетворяет требованиям ASTM D975 и не является моноалкиловыми эфирами. Типичными признаками возобновляемого дизельного топлива являются: цетановое число 75-90, плотность энергии приблизительно 44 МДж/кг, плотность приблизительно 0,78 г/мл, содержание энергии приблизительно 123 кБТЕ/галлон, уровни хлора менее чем 10 пропромилле, точка мутности ниже 0°C.
Переэтерификация
«Переэтерификация» в настоящем документе представляет собой превращение липидов, главным образом, триацилглицеридов, в метиловые эфиры или этиловые эфиры жирных кислот путем приведения в реакцию с короткоцепочечными спиртами, например, метанолом или этанолом, в присутствии катализатора, например, щелочи или кислоты. Метанол применяется чаще за счет низкой стоимости и доступности, однако могут применяться также этанол, пропанол или бутанол или их смеси. Катализаторы могут быть гомогенными катализаторами, гетерогенными катализаторами или ферментными катализаторами. Гомогенные катализаторы включают сульфат железа, и затем KOH. Гетерогенные катализаторы включают CaO, K3PO4 и WO3/ZrO2. Ферментные катализаторы включают Novozyme 435, получаемый из Candida antarctica.
Переэтерификация может осуществляться на экстрагированном масле или предпочтительно непосредственно in situ в вегетативном материале растения. Вегетативные части растения можно высушить и смолоть до использования, чтобы получить композицию для реакции превращения, но это не является необходимым. Преимущество прямого превращения в эфиры жирных кислот, предпочтительно МЭЖК, состоит в том, что для превращения можно применять более низкую температуру и давление, и несмотря на это получать высокий выход продукта, например, содержащего по меньшей мере 50% масс. МЭЖК. Выход извлекаемой посредством переэтерификации бионефти вычисляют так же, как и для процесса ГТО.
Получение неполярных липидов
Методы, которые рутинно применяются в уровне техники, могут применяться для извлечения, обработки, очистки и анализа липидов, таких как ТАГ, выработанных клетками, организмами или их частями по настоящему изобретению. Такие методы раскрыты и к ним дается пояснение в источниках литературы, например, Fereidoon Shahidi, Current Protocols in Food Analytical Chemistry, John Wiley & Sons, Inc. (2001) D1.1.1-D1.1.11, и Perez-Vich с соавт. (1998).
Получение масла из вегетативных частей растения или семян
Обычно семена растения варят, прессуют и/или экстрагируют для получения неочищенного масла из семян, которое в дальнейшем дегуммируют, рафинируют, отбеливают и дезодорируют. В общем, методы измельчения семени известны из уровня техники. Например, семена масличных культур могут быть смягчены обрызгиванием их водой, чтобы увеличить содержание влаги, например, до 8,5%, и вальцуют, используя гладкий валик со щелью 0,23-0,27 мм. В зависимости от вида семени, вода может не добавляться до измельчения. Применение тепла инактивирует ферменты, облегчает дальнейшее разрушение клеток, слияние капелек липида и агломерацию частиц белка, что облегчает процесс экстракции.
В варианте реализации изобретения большая часть масла из семян высвобождается при прохождении через винтовой пресс. Жмых, выходящий из винтового пресса, в дальнейшем экстрагируют растворителем, например гексаном, с применением колонки с подводом тепла. Альтернативно, неочищенное масло из семян, полученное операцией прессования, может быть пропущено сквозь резервуар для отстаивания, оборудованный проволочным дренажным верхом, для удаления твердых веществ, которые сопровождают масло из семян в ходе операции прессования. Осветленное масло из семян может быть пропущено сквозь планшет и рамочный фильтр для удаления любых оставшихся тонких частиц твердого вещества. При желании, масло из семян, полученное в результате процесса экстракции, может быть объединено с осветленным маслом из семян для получения смеси неочищенного масла из семян.
Как только растворитель удаляют из неочищенного масла из семян, прессованные и экстрагированные части объединяют и обрабатывают обычными процедурами обработки липида (т.е., дегуммирование, рафинация каустической содой, обесцвечивание и дезодорирование).
Для экстракции липида из вегетативных частей растений по изобретению применяются способы, аналогичные известным из уровня техники для экстракции масла из семян. Один из путей представляет собой физическую экстракцию, которая часто исключает экстракцию растворителем. Экстракция с помощью шнекового пресса, как и способы экстракции с помощью винтового пресса и гидравлического пресса, представляет собой распространенный вид экстракции. Механическая экстракция обычно менее эффективна, чем экстракция растворителем, при которой органический растворитель (например, гексан) смешивают по меньшей мере с биомассой растения, предпочтительно после высушивания и перемалывания биомассы. Растворитель растворяет липид в биомассе и подобном материале, после чего раствор отделяют от биомассы механической операцией (например, с помощью описанных выше процессов прессования). Данная стадия отделения также может выполняться посредством фильтрации (например, с помощью пресс-фильтра или подобного устройства) или центрифугирования и т. п. В дальнейшем, органический растворитель может быть отделен от неполярного липида (например, дистилляцией). Вторая стадия разделения дает неполярный липид из растения и может давать повторно используемый растворитель, если он пригоден для традиционного сбора в виде паров. В варианте реализации изобретения содержание масла и/или белка в части растения или семени анализируют отражательной спектроскопией в ближней инфракрасной области, как раскрыто в Hom с соавт. (2007), до экстракции.
Если вегетативные части растения не будут использоваться для экстракции липида немедленно, их предпочтительно перерабатывают, чтобы максимально снизить содержание липида (см., например, Christie, 1993), например, сушкой вегетативных частей растения.
Дегуммирование
Дегуммирование является ранней стадией рафинирования масел, и его основная цель состоит в удалении из масла большинства фосфолипидов, которые могут присутствовать в количестве приблизительно 1-2% от общего экстрагированного липида. Добавление к неочищенному маслу ~2% воды, обычно содержащей фосфорную кислоту, при 70-80° C приводит к отделению большинства фосфолипидов, в сопровождении следовых количеств металлов и пигментов. Удаляемый нерастворимый материал, главным образом представляет собой смесь фосфолипидов и триацилглицеридов, и также известен как лецитин. Дегуммирование может осуществляться добавлением концентрированной фосфорной кислоты к неочищенному маслу из семян, чтобы перевести неспособные к гидратации фосфатиды в способную к гидратации форму, и к незначительному количеству присутствующих хелатов металлов. Смолу отделяют от масла из семян центрифугированием. Масло из семян может быть рафинировано добавлением достаточного количества раствора натрия гидроксида для титрования всех жирных кислот, с удалением образованного таким образом мыла.
Щелочная рафинация
Щелочная рафинация представляет собой один из процессов очистки для обработки неочищенного масла, иногда также называемый нейтрализацией. Она обычно следует за дегуммированием и предшествует обесцвечиванию. После дегуммирования масло из семян может быть обработано добавлением достаточного количества раствора щелочи, чтобы оттитровать все жирные кислоты и фосфорные кислоты, с последующим удалением образованного таким способом мыла. Пригодные щелочные материалы включают натрия гидроксид, калия гидроксид, натрия карбонат, лития гидроксид, кальция гидроксид, кальция карбонат и аммония гидроксид. Данный процесс обычно осуществляется при комнатной температуре и позволяет удалить фракцию свободных жирных кислот. Мыло удаляют центрифугированием или экстракцией растворителем мыла, и нейтрализованное масло промывают водой. При необходимости, любая дополнительная щелочь в масле может быть нейтрализована подходящей кислотой, например, хлористоводородной кислотой или серной кислотой.
Отбеливание
Отбеливание представляет собой процесс очистки, в ходе которого масла нагревают до 90-120°C, выдерживая при этой температуре в течение 10-30 минут в присутствии отбеливающей земли (0,2-2,0%) и в отсутствие кислорода, путем проведения операции в атмосфере азота, под струей пара или в вакууме. Данная стадия обработки масла введена с целью удаления нежелательных пигментов (каротиноиды, хлорофилл, госсипол и т.д.), и в ходе данного процесса также удаляются продукты окисления, следовые металлы, соединения серы и следы мыла.
Дезодорирование
Дезодорирование представляет собой обработку масел и жиров при высокой температуре (200-260°C) и низком давлении (0,1-1 мм рт. ст). Это обычно достигается посредством введения пара в масло из семян со скоростью приблизительно 0,1 мл/минуту/100 мл масла из семян. Дезодорирование может выполняться путем нагревания масла из семян до 260°C под вакуумом, с медленным введением пара в масло из семян со скоростью приблизительно 0,1 мл/минуту/100 мл масла из семян. Приблизительно через 30 минут обрызгивания, маслу из семян дают остыть под вакуумом. Масло из семян обычно переносят в стеклянную емкость и продувают аргоном перед хранением при сниженной температуре. Если количество масла из семян ограничено, масло из семян может быть размещено под вакуумом, например, в реакторе Парра, и нагрето до 260°C с выдерживанием в течение такого периода времени, чтобы запах был уничтожен. Такая обработка улучшает цвет масла из семян и удаляет большинство летучих веществ или пахучих соединений, включая любые оставшиеся свободные жирные кислоты, моноацилглицериды и продукты окисления.
Вымораживание
Вымораживание представляет собой процесс, иногда применяемый в коммерческом производстве масел для разделения масел и жиров на твердые (стеарин) и жидкие (олеин) фракции путем кристаллизации при температурах ниже температуры окружающей среды. Впервые он был применен к хлопковому маслу с целью получения свободного от твердых веществ продукта. Его обычно применяют для уменьшения содержания насыщенных жирных кислот в маслах.
Водоросли для получения липидов
За год водоросли могут образовывать массу, в 10-100 раз превышающую массу наземных растений, причем их можно выращивать в открытых водоемах (таких как водоемы по типу водоводов и озер) или в фотобиореакторах. Наиболее распространенные водоросли, вырабатывающие масло, в общем, могут включать диатомовые (бацилляриофиты), зеленые водоросли (хлорофиты), зелено-голубые водоросли (цианофиты) и коричнево-золотистые водоросли (хризофиты). Дополнительно, пятая группа, известная как гаптофитовые водоросли, может быть использована. Группы включают коричневые водоросли и гетероконты. Конкретные неограничивающие примеры водорослей включают Классы: Chlorophyceae, Eustigmatophyceae, Prymnesiophyceae, Bacillariophyceae. Bacillariophytes, способные к выработке масла, включают рода Amphipleura, Amphora, Chaetoceros, Cyclotella, Cymbella, Fragilaria, Hantzschia, Navicula, Nitzschia, Phaeodactylum и Thalassiosira. Конкретные неограничивающие примеры хлорофитов, способных к выработке масла, включают Ankistrodesmus, Botryococcus, Chlorella, Chlorococcum, Dunaliella, Monoraphidium, Oocystis, Scenedesmus и Tetraselmis. В одном из аспектов, хлорофиты могут представлять собой Chlorella или Dunaliella. Конкретные неограничивающие примеры цианофитов, способных к выработке масла, включают Oscillatoria и Synechococcus. Конкретный пример хризофитов, способных к выработке масла, включает Boekelovia. Конкретные неограничивающие примеры гаптофитовых водорослей включают Isochysis и Pleurochysis
Конкретные водоросли, пригодные в соответствии с настоящим изобретением, включают, например, виды Chlamydomonas, например, Chlamydomonas reinhardtii, виды Dunaliella, например, Dunaliella salina, Dunaliella tertiolecta, D. acidophila, D. Lateralis, D. martima, D. parva, D. polmorpha, D. primolecta, D. pseudosalina, D. Quartolecta, D. viridis, виды Haematococcus, виды Chlorella, такие как Chlorella vulgaris, Chlorella sorokiniana или Chlorella protothecoides, виды Thraustochytrium, виды Schizochytrium, виды Volvox, виды Nannochloropsis, Botryococcus braunii, который может содержать более 60%, масс липида, Phaeodactylum tricornutum, Thalassiosira pseudonana, виды Isochrysis, виды Pavlova, виды Chlorococcum, виды Ellipsoidion, виды Neochloris, виды Scenedesmus.
Урожай водорослей по изобретению можно собирать с помощью микросит, центрифугирования, флоккуляции (с применением, например, хитозана, алюминиевых квасцов и хлорида железа) и пенной флотации. Прерывание потока диоксида углерода может вынудить водоросли выпасть хлопьями самим по себе, что называется «аутофлоккуляцией». При пенной флотации, фермер аэрирует воду в пену, а затем снимает водоросли с поверхности. Ультразвуковой и другие методы сбора урожая в настоящий момент разрабатываются.
Липид может быть экстрагирован из водорослей механическим измельчением. При высушивании водорослевая масса сохраняет содержание липида, который в дальнейшем может быть «выжат» с помощью масляного пресса. Кроме того, осмотический шок может быть применен для высвобождения клеточных компонентов, например, липида из водорослей, а извлечение с помощью ультразвука может ускорить процессы экстракции. Химические растворители (например, гексан, бензол, петролейный эфир) часто применяются для экстракции липидов из водорослей. Дополнительно, ферментная экстракция с применением ферментов для разложения клеточных стенок может применяться для извлечения липидов из водорослей. Кроме того, суперкритический CO2 может использоваться в качестве растворителя. В данном способе CO2 превращают в жидкость под давлением и нагревают до точки, в которой он становится суперкритическим (обладает свойствами как жидкости, так и газа), что дает ему возможность выполнять роль растворителя.
В настоящем документе «маслянистый организм» обозначает такой, который аккумулирует по меньшей мере 20% сухой массы в виде триацилглицеридов. В настоящем документе «дрожжи» включают виды Saccharomyces, Saccharomyces cerevisiae, Saccharomyces carlbergensis, виды Candida, виды Kluveromyces, виды Pichia, виды Hansenula, виды Trichoderma, Lipomyces starkey и Yarrowia lipolytica. Предпочтительные дрожжи включают Yarrowia lipolytica или другие маслянистые дрожжи и штаммы видов Saccharomyces.
Применение растительных липидов
Липиды, полученные раскрытыми способами, находят множество применений. В некоторых вариантах реализации липиды используются в качестве пищевых масел. В других вариантах реализации липиды рафинируют и используют в качестве смазки или для других промышленных целей, например, синтеза пластмасс. В некоторых предпочтительных вариантах реализации липиды рафинируют для производства биодизельного топлива. Биодизельное топливо может быть получено из масел, получаемых из растений, водорослей и грибов по изобретению. Применение растительных триацилглицеридов для получения биологического топлива рассмотрено в Durrett с соавт. (2008). Полученное топливо обычно обозначается как биодизельное топливо и обладает динамическим интервалом вязкости 1,9-6,0 мм2с-1 (ASTM D6751). Биоспирт можно получить из сахаров биомассы, не принадлежащих к липидам, которые остаются после экстракции липида. Общие способы получения биологического топлива могут быть найдены, например, в Maher и Bressler, 2007; Greenwell с соавт., 2010; Karmakar с соавт., 2010; Alonso с соавт., 2010; Liu с соавт., 2010a. Gong и Jiang, 2011; Endalew с соавт., 2011; и Semwal с соавт., 2011.
В настоящем изобретении предлагаются способы повышения содержания масла в вегетативных тканях. Растения по настоящему изобретению содержат повышенное количество энергии в листьях и/или стеблях, таким образом, что цельные надземные части растения можно собирать и использовать для получения биотоплива. Кроме того, уровень олеиновой кислоты значительно повышен, притом, что уровень полиненасыщенной жирной альфа-линоленовой кислоты (АЛК) снижен. Таким образом, растения, морские водоросли и грибы по настоящему изобретению снижают затраты на производство биотоплива.
Биодизельное топливо
Производство биодизельного топлива или алкиловых эфиров хорошо известно. Существует три основных способа получения сложного эфира из липидов: 1) катализированная основанием переэтерификация липида с использованием спирта; 2) прямая, катализируемая кислотой этерификация липида с использованием метанола; и 3) превращение липида в жирные кислоты, а затем в алкиловые эфиры с помощью кислотного катализа Может применяться любой способ получения алкиловых эфиров и глицериловых эфиров жирных кислот (в ходе которого этерифицируются одна, две или три гидроксигруппы на глицерине). Жирные кислоты могут быть получены, например, гидролизом или омылением ТАГ с использованием кислотных или основных катализаторов, соответственно, или фермента, например, липазы или эстеразы. Алкиловые эфиры жирных кислот могут быть получены путем введения жирной кислоты в реакцию со спиртом в присутствии кислотного катализатора. Кроме того, алкиловые эфиры жирных кислот могут быть получены путем введения ТАГ в реакцию со спиртом в присутствии кислотного или основного катализатора.
Эфиры глицерина могут быть получены, например, введением глицерина в реакцию с алкилгалогенидом в присутствии основания или с олефином или спиртом в присутствии кислотного катализатора. Алкиловые эфиры могут быть непосредственно смешаны с дизельным топливом или промыты водой или другими водными растворами с целью удаления различных примесей, в том числе катализаторов, перед смешиванием.
Авиационное топливо
Для улучшения характеристик биотоплива разработаны технологии термического и каталитического расщепления (разрыва) химических связей, что позволяет превращать бионефть в биологические альтернативы полученному из нефть дизельному топливу и другим видам топлива, такие как реактивное топливо.
Применение источника среднецепочечных жирных кислот, такого как вырабатывается рекомбинантной эукариотной клеткой по изобретению, трансгенным организмом, не относящимся к человеческому роду, или его частью по изобретению, трансгенным растением или его частью по изобретению, семенем по изобретению или трансгенной клеткой или трансгенным растением или его частью по изобретению, предотвращает потребность в высоких энергиях для расщепления цепи жирной кислоты и получения более коротких молекул, необходимых для реактивного топлива и других видов топлива с требованиями низкой температуры потока. Данный способ включает отщепление одной или более групп среднецепочечных жирных кислотных от глицеридов с получением глицерина и одной или больше свободных жирных кислот. Кроме того, способ включает отщепление одной или более среднецепочечных жирных кислот от глицерина и декарбоксилирование одной или более среднецепочечных жирных кислот с получением одного или более углеводородов для выработки реактивного горючего.
Пищевые (кормовые) продукты
Настоящее изобретение включает составы, которые могут применяться в качестве пищевых (кормовых) продуктов. Для целей настоящего изобретения, «пищевые (кормовые) продукты» включают любой пищевой продукт или препарат для потребления человеком или животным, который служит целям питания или образования тканей или снабжения энергией, и/или сохраняет, восстанавливает или поддерживает соответствующий пищевой статус или метаболическую функцию. Пищевые (кормовые) продукты во изобретению включают питательные составы для младенцев и/или маленьких детей.
Пищевые (кормовые) составы по изобретению включают, например, клетку по изобретению, растение по изобретению, часть растения по изобретению, семя по изобретению, экстракт по изобретению, продукт способа по изобретению или состав вместе с пригодным(и) носителем(ями). Термин «носитель» используется в самом широком смысле и включает любой компонент, который может иметь или не иметь пищевой ценности. Как будет понятно специалисту в данной области, носитель должен быть пригодным для использования (или используемым в достаточно низкой концентрации) в пищевом (кормовом) продукте, таким образом, что он не оказывает вредного влияния на организм, который потребляет пищевой (кормовой) продукт.
Пищевой (кормовой) продукт по настоящему изобретению содержит липид, полученный, прямо или косвенно, с применением способов, клеток или организмов, раскрытых в настоящем документе. Кроме того, состав может находиться в твердой или жидкой форме. Дополнительно, состав может содержать съедобные макронутриенты, витамины и/или минералы в количествах, желательных для конкретного применения. Количества таких ингредиентов будут варьировать, в зависимости от того, предназначен ли состав для применения у нормальных индивидуумов или индивидуумов с особыми потребностями например, индивидуумов, страдающих от метаболических расстройств и т. п.
Примеры пригодных носителей, обладающих пищевой ценностью, включают, без ограничений, макронутриенты, например, съедобные жиры, углеводы и белки. Примеры таких съедобных жиров включают, без ограничений, кокосовое масло, масло бораго, грибное масло, масло черной смородины, соевое масло и моно- и диглицериды. Примеры таких углеводов включают, без ограничений, глюкозу, съедобную лактозу и гидролизованный крахмал. Дополнительно, примеры белков, которые могут применяться в питательном составе по изобретению включают, без ограничений, белки сои, обработанную электродиализом сыворотку, обработанное электродиализом снятое молоко, молочную сыворотку или гидролизаты указанных белков.
Относительно витаминов и минералов, следующее может быть добавлено к составам пищевых (кормовых) продуктов по настоящему изобретению: кальций, фосфор, калий, натрий, хлорид, магний, марганец, железо, медь, цинк, селен, йод и витамины A, E, D, C и группы B. Другие такие витамины и минералы также могут быть добавлены.
Дополнительно, состав пищевого (кормового) продукта по настоящему изобретению может быть добавлен к пище даже в тех случаях, когда нет необходимости в дополнении рациона. Например, состав может быть добавлен к пище любого типа, включая, без ограничений, маргарин, масло, сыры, молоко, йогурт, шоколад, конфеты, легкие закуски, салатные масла, кондитерские масла, кондитерские жиры, мясо, рыбу и напитки.
Дополнительно, липид, полученный в соответствии с настоящим изобретением или клетками-хозяевами, трансформированными таким образом, чтобы содержать и экспрессировать указанные гены, может применяться в качестве пищевых добавок к корму для животного, с целью изменения состава жирных кислот в ткани животного или молоке до более желательного для потребления человеком или животным. Примеры таких животных включают овцу, теленка, лошадей и т. п. Дополнительно, пищевые (кормовые) продукты по изобретению могут использоваться в аквакультуре, с целью повышения уровней жирных кислот в организме рыбы, предназначенной для потребления человеком или животным.
Предпочтительные пищевые (кормовые) продукты по изобретению представляет собой растения, семя и другие части растения, такие как листья, плоды и стебли, которые могут быть использованы непосредственно в качестве пищи или корма человеком или другими животными. Например, животные могут пастись непосредственно на таких растениях, выращенных в поле, или получать более точно отмеренные количества в процессе управляемого кормления. Изобретение включает применение таких растений и частей растения в качестве корма, с целью повышения уровней полиненасыщенных жирных кислот в организме человека и других животных.
Для потребления животными, не принадлежащими к человеческому роду, ком может находиться в любой подходящей форме, например, но не ограничиваясь ими, силос, сено или подножный корм в поле. В варианте реализации изобретения корм для потребления животными представляет собой бобовое растение или его часть, принадлежащее к семейству Fabaceae (или Leguminosae) например люцерна, клевер, горох, люцерна посевная, бобы, чечевица, люпин, мескито, кэроб, соя и арахис.
Составы
Настоящее изобретение также включает составы, особенно фармацевтические составы, содержащие один или более липидов, полученных с применением способов по изобретению.
Фармацевтический состав может содержать один или более липидов, в комбинации со стандартным, известным, нетоксичным фармацевтически приемлемым носителем, адъювантом или растворителем, таким как буферизованный фосфатом раствор соли, вода, этанол, полиолы, растительные масла, увлажнитель или эмульсия, например, эмульсия вода/масло. Состав может находиться в жидкой или твердой форме. Например, состав может находиться в форме таблетки, капсулы, жидкости для приема внутрь, порошка, мази или крема для местного применения. Надлежащая сыпучесть может поддерживаться, например, поддержанием необходимого размера частиц в случае дисперсий и применением поверхностно-активных веществ. Кроме того, может быть желательным введение поддерживающих изотоничность агентов, например, сахаров, натрия хлорида и т. п. Помимо таких инертных разбавителей, состав может включать вспомогательные вещества, например, увлажнители, эмульгаторы и суспендирующие агенты, подсластители, вкусовые добавки и ароматизаторы.
Типичные дозы конкретной жирной кислоты составляют от 0,1 мг до 20 г, для приема от 1 до 5 раз в сутки (до 100 г в сутки) и, предпочтительно, находятся в интервале от приблизительно 10 мг до приблизительно 1, 2, 5 или 10 г в сутки (в виде одной или нескольких доз). Как известно из уровня техники, желательной является доза по меньшей мере около 300 мг/сутки жирной кислоты, особенно полиненасыщенной жирной кислоты. Однако следует понимать, что любое количество жирной кислоты будет полезным для субъекта.
Возможные способы введения фармацевтических составов по настоящему изобретению включают, например, энтеральный и парентеральный. Например, жидкий препарат можно вводить перорально. Дополнительно, гомогенная смесь может быть полностью диспергирована в воде, смешана в стерильных условиях с физиологически приемлемыми разбавителями, консервантами, буферами или пропеллентами, чтобы получить спрей или средство для ингаляций.
Дозы состава при введении субъекту могут быть определены специалистом, обладающим среднем уровнем квалификации в данной области, и зависят от различных факторов, таких как масса тела, возраст, общее состояние здоровья, анамнез, иммунный статус субъекта и т. п.
Дополнительно, составы по настоящему изобретению могут применяться для косметических целей. Составы могут быть добавлены к уже существующим косметическим составам, таким образом, что смесь, или жирная кислота, полученная согласно изобретению, может применяться в качестве единственного «активного» ингредиента в косметическом составе.
Полипептиды
В настоящем документе термины «полипептид» и «белок» в общем используются равнозначно.
Полипептид или класс полипептидов может быть определен по степени идентичности (% идентичности) его последовательности аминокислот референтной последовательности аминокислот или большему % идентичности одной референтной последовательности аминокислот, чем другой. % идентичности полипептида референтной последовательности аминокислот обычно определяется путем анализа GAP (Needleman и Wunsch, 1970; программное обеспечение GCG) с параметрами штрафа на открытие промежутка=5, и штрафа на продление промежутка=0,3. Длина последовательности запроса составляет по меньшей мере 100 аминокислот, и анализ GAP выравнивает две последовательности на протяжении участка длиной по меньшей мере 100 аминокислот. Даже более предпочтительно, длина последовательности запроса составляет по меньшей мере 250 аминокислот, и анализ GAP выравнивает две последовательности на протяжении участка длиной по меньшей мере 250 аминокислот. Даже более предпочтительно, анализ GAP выравнивает две последовательности на протяжении их полной длины. Полипептид или класс полипептидов может обладать такой же ферментной активностью как или отличной активностью от или не обладать активностью референтного полипептида. Предпочтительно, полипептид обладает ферментной активностью, составляющей по меньшей мере 10% от активности референтного полипептида.
В настоящем документе «биологически активный фрагмент» представляет собой часть полипептида по изобретению, которая обладает определенным видом активности полноразмерного референтного полипептида, например, активностью MGAT. Биологически активные фрагменты в настоящем документе исключают полноразмерный полипептид. Биологически активные фрагменты могут быть частью любого размера, до тех пор, пока они сохраняют определенную активность. Предпочтительно, биологически активный фрагмент сохраняет по меньшей мере 10% активности полноразмерного полипептида.
Относительно определенного полипептида или фермента, предусматривается, что более высокий % идентичности, чем раскрытый в настоящем документе, будет включать предпочтительные варианты. Таким образом, если это уместно, в свете минимального % идентичности, предпочтительно, полипептид/фермент содержит последовательность аминокислот, которая по меньшей мере на 60%, более предпочтительно по меньшей мере на 65%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 91%, более предпочтительно по меньшей мере на 92%, более предпочтительно по меньшей мере на 93%, более предпочтительно по меньшей мере на 94%, более предпочтительно по меньшей мере на 95%, более предпочтительно по меньшей мере на 96%, более предпочтительно по меньшей мере на 97%, более предпочтительно по меньшей мере на 98%, более предпочтительно по меньшей мере на 99%, более предпочтительно по меньшей мере на 99,1%, более предпочтительно по меньшей мере на 99,2%, более предпочтительно по меньшей мере на 99,3%, более предпочтительно по меньшей мере на 99,4%, более предпочтительно по меньшей мере на 99,5%, более предпочтительно по меньшей мере на 99,6%, более предпочтительно по меньшей мере на 99,7%, более предпочтительно по меньшей мере на 99,8%, и даже более предпочтительно, по меньшей мере на 99,9% идентична соответствующей указанной SEQ ID NO.
Мутантные последовательности аминокислот полипептидов, раскрытых в настоящем документе, могут быть получены посредством введения подходящих нуклеотидных модификаций в нуклеиновую кислоту, определенную в настоящем документе, или синтезом желательного полипептида in vitro. Такие мутанты включают, например, делеции, вставки или замены остатков в пределах последовательности аминокислот. Комбинация делеций, вставок и замен может быть получена для введения в конечную конструкцию, при условии, что конечный полипептидный продукт обладает желательными характеристиками.
Мутантные (модифицированные) полипептиды могут быть получены с применением любой техники, известной в данной области, например, с применением стратегий направленной эволюции или рационального дизайна (см. ниже). Для продуктов, получаемых из мутантной/модифицированной ДНК, может быть легко проведен скрининг с применением способов, раскрытых в настоящем документе, чтобы определить, обладают ли они активностью фактора транскрипции, ацилтрансферазы жирных кислот или ПМВ.
При конструировании мутантов последовательности аминокислот, расположение сайта мутации и природа мутации будут зависеть от модифицированной характеристики (характеристик). Сайты мутации могут быть модифицированы индивидуально или сериями, например, (1) вначале заменой консервативными альтернативами аминокислот, а затем более радикальными вариантами, в зависимости от полученных результатов, (2) удалением целевого остатка, или (3) вставкой других остатков, смежных с размещенным сайтом.
Длина делеций в последовательности аминокислот, в общем, варьирует от приблизительно 1 до 15 остатков, более предпочтительно, приблизительно 1-10 остатков и обычно приблизительно 1-5 смежных остатков.
В мутантах с заменой по меньшей мере один остаток аминокислоты в полипептиде удален, и другой остаток вставлен на его место. Сайты, представляющие наибольший интерес для мутагенеза с замещением с целью инактивации ферментов, включают сайты, идентифицированные как активный(е) сайт(ы). Другими сайтами, представляющими интерес, являются такие, конкретные остатки которых, полученные от различных штаммов или видов, идентичны. Указанные положения могут быть важны с точки зрения биологической активности. Такие сайты, особенно находящиеся в пределах последовательности по меньшей мере трех других одинаково консервативных сайтов, предпочтительно заменяются относительно консервативным образом. Такие консервативные замены приведены в Таблице 1 под заголовком «примеры замен».
Таблица 1. Примеры замен.
Остаток
Замен
В предпочтительном варианте мутантный/вариантный полипептид содержит только или не более чем один, два, три или четыре консервативных модификации аминокислот, по сравнению с природным полипептидом. Подробности относительно консервативных модификаций аминокислот приведены в Таблице 1. Как будет понятно специалисту, такие незначительные модификации могут быть спрогнозированы на рациональной основе как не влияющие на активность полипептида при экспрессии в рекомбинантной клетке. Мутанты с желательной активностью могут быть сконструированы с применением стандартных методик из уровня техники, например, случайным мутагенезом, направленным мутагенезом или мутагенезом насыщения в известных целевых генах, или путем обработки различных генов «тасованием» ДНК.
ПРИМЕРЫ
Пример 1. Общие материалы и методы
Экспрессия генов в растительных клетках в системе временной экспрессии
Гены экспрессируют в клетках растения с использованием системы временной экспрессии, по существу как раскрыто Voinnet с соавт. (2003) и Wood с соавт. (2009). Бинарные векторы, содержащие кодирующий участок, которые экспрессируются высокоактивным конститутивным промотором e35S, содержащим дуплицированный энхансерный участок, вводят в Agrobacterium tumefaciens штамм AGL1. Химерный бинарный вектор, 35S:p19, для экспрессии вирусного супрессора сайленсинга p19 вводят в AGL1 отдельно, как раскрыто в WO2010/057246. Химерный бинарный вектор, 35S:V2, для экспрессии вирусного супрессора сайленсинга V2 вводят в AGL1 отдельно. Рекомбинантные клетки выращивают до стационарной фазы при температуре 28 °C в среде Лурия-Бертани с добавлением 50 мг/л канамицина и 50 мг/л рифампицина. Затем бактерии гранулируют центрифугированием при 5000 g в течение 5 минут при комнатной температуре, перед ресуспендированием до OD600=1,0 в инфильтрационном буфере, содержащем 10 мМ минимальной эссенциальной среды, pH 5,7, 10 мМ MgCl2 и 100 мкМ ацетосирингона. Далее клетки инкубируют при 28°C со встряхиванием в течение 3 часов, после чего измеряют OD600, и объем каждой культуры, содержащей конструкцию вирусного супрессора 35S:p19 или 35S:V2, необходимый для достижения конечной концентрации OD600=0,125, помещают в свежую пробирку. Доводят до конечного объема упомянутым выше буфером. После этого осуществляют инфильтрацию листьев смесью культуры, и растения обычно выращивают еще в течение 3-5 дней после инфильтрации, перед выделением листовых пластинок для получения очищенного лизата клеток или выделения общего липида.
Трансформация Brassica napus
Семена Brassica napus стерилизуют с применением газообразного хлора, как описано Kereszt с соавт. (2007), и проращивают на питательной среде для культуры тканей. Котиледонные черешки с ножкой 2-4 мм выделяют, как описано Belide с соавт. (2013) и используют в качестве эксплантов. Готовят культуры A. tumefaciens AGL1 (Lazo с соавт., 1991), содержащие бинарный вектор, и котиледонные черешки инокулируют культурами, как описано Belide с соавт. (2013). Инфицированные котиледонные черешки культивируют на среде МС с добавлением 1 мг/л тидиазурона (ТДЗ)+0.1 мг/л 1-нафталинуксусной кислоты (НУК)+3 мг/л AgNO3+250 мг/л цефотаксима, 50 мг/л тиментина и 25 мг/л канамицина и культивируют в течение 4 недель при 24 °C с фотопериодом света-темноты 16 часов/8 часов, с двухнедельными пассажами на такую же среду. Экспланты с зеленым каллюсом переносят в среду для инициации побегов (МС+1 мг/л кинетина+3 мг/л AgNO3+250 мг/л цефотаксима+50 мг/л тиментина+25 мг/л канамицина) и культивируют еще в течение 2-3 недель. Маленькие побеги (~1 см) выделяют из устойчивого каллюса и переносят в среду для удлинения побегов (среда МС с добавлением 0,1 мг/л гибберелловой кислоты+3 мг/л AgNO3+250 мг/л цефотаксима+25 мг/л канамицина) и культивируют еще в течение двух недель. Здоровые побеги с одним или двумя листьями отбирают и переносят в среду для корнеобразования (1/2 МС с добавлением 1 мг/л НУК+20 мг/л аденина сульфата (АДС)+3 мг/л AgNO3+250 мг/л цефотаксима) и культивируют в течение 2-3 недель. ДНК выделяют из маленьких листьев устойчивых побегов с применением набора для выделения ДНК растения (Bioline, Александрия, Новый Южный Уэльс, Австралия), в соответствии с протоколом производителя. Присутствие последовательностей T-ДНК определяют методом ПЦР амплификации на геномной ДНК. Положительные, трансгенные побеги с корнями переносят в горшки, содержащие смесь для выращивания саженцев и выращивают в теплице с дневной температурой 24°C/ночной температурой 16°C (стандартные условия).
Анализ ферментов очищенного лизата листа
Ткани листа Nicotiana benthamiana, предварительно обработанные инфильтрацией, как изложено выше, измельчают в растворе, содержащем 0,1 М калий фосфатного буфера (pH 7,2) и 0,33 М сахарозы, с использованием стеклянного гомогенизатора. Гомогенат листьев центрифугируют при 20 000 g в течение 45 минут при температуре 4° C, после чего собирают каждый супернатант. Содержание белка в каждом супернатанте измеряют согласно Bradford (1976), с помощью мультиметочного счетчика Wallac1420 и реактива красителя для анализа белка Bio-Rad (Bio-Rad Laboratories, Геркулес, США, Калифорния). В анализах ацилтрансферазы используют 100 мкг белка согласно Cao с соавт. (2007), с некоторыми модификациями. Реакционная среда содержит 100 мМ Трис-HCl (pH 7,0), 5 мМ MgCl2, 1 мг/мл альбумина телячьей сыворотки (не содержащего жирных кислот), 200 мМ сахарозы, 40 мМ холодного олеоил-КоА, 16,4 мкМ sn-2 моноолеоилглицерил[14C] (55 мКи/ммоль, American Radiochemicals, Сент-Луис, США, Миссури) или 6,0 мкМ динатриевой соли [14C]глицерил-3-фосфата (G-3-P) (150 мКи/ммоль, American Radiochemicals). Анализ проводят через 7,5, 15 или 30 минут.
Анализ липидов
Анализ содержания масла в семенах Arabidposis
Если содержание масла в семени или состав общих жирных кислот следует определить для семян небольшого размера, например, семян Arabidopsis, то жирные кислоты в семенах метилируют напрямую, без раздавливания семян. Семена сушат в эксикаторе в течение 24 часов, и приблизительно 4 мг семян переносят в стеклянный флакон емкостью 2 мл с навинчивающейся крышкой, покрытой слоем Тефлона. Раствор 0,05 мг тригептадеканоина (ТАГ с тремя C17:0 жирными кислотами) в 0,1 мл толуола помещают во флакон в качестве внутреннего стандарта. Жирные кислоты семени метилируют добавлением 0,7 мл 1 н метанольного HCl (Supelco) во флакон, содержащий материал семян. В дроблении семян для полного метилирования нет необходимости в случае семян небольшого размера, например, семян Arabidopsis. Смесь кратковременно обрабатывают вихревым перемешиванием и инкубируют при 80° C в течение 2 часов. После охлаждения смесей до комнатной температуры, 0,3 мл 0,9% NaCl (масс/об) и 0,1 мл гексана добавляют во флакон и тщательно перемешивают в течение 10 минут в Heidolph Vibramax 110. МЭЖК собирают в стеклянную вставку емкостью 0,3 мл и анализируют методом ГХ с плазменно-ионизационным детектором (ПИД), как описано ниже.
Площадь пика индивидуальных МЭЖК вначале корректируют на основе площади пиков известного количества таких же МЭЖК, присутствующих в коммерческом стандартном GLC-411 (NU-CHEK PREP, INC., США). GLC-411 содержит равные количества 31 жирной кислоты (% масс.), в интервале от C8:0 до C22:6. В случае жирных кислот, которые не присутствовали в стандарте, находят площадь пиков наиболее подобного МЭЖК. Например, площадь пика МЭЖК 16:1d9 используют для 16:1d7, и площадь пика МЭЖК C22:6 используют для C22:5. Скорректированные площади пиков используют для вычисления массы каждого МЭЖК в образце, по сравнению с массой внутреннего стандарта. Масло хранится, в основном, в форме ТАГ, и его массу вычисляют на основании массы МЭЖК. Общее количество моль глицерина определяют путем вычисления количества моль каждого МЭЖК и деления общего количества моль МЭЖК на 3. Содержание ТАГ вычисляют как сумму фрагментов глицерина и жирного ацила, с использованием формулы: %, масс. масла=100 x ((41 x общее количество моль МЭЖК/3)+(общее количество г МЭЖК - (15 x общее количество моль МЭЖК)))/грамм семени, где 41 и 15 представляют собой значения молекулярной массы фрагмента глицерина и метильной группы, соответственно.
Анализ содержания жирной кислоты в семенах Camelina и семенах рапса
Для определения состава жирных кислот в одиночных семенах большего размера, таких как семена рапса и Camelina, выполняют прямое метилирование жирных кислот в семени, как и для семян Arabidopsis, за исключением разрушения оболочек семян. Данный способ позволяет экстрагировать достаточное количество масла из семян, чтобы провести анализ состава жирных кислот. Для определения состава жирных кислот общего липида, экстрагированного из семян, семена раздавливают и липиды экстрагируют с помощью CHCl3/MeOH. Аликвоты экстрагированного липида метилируют и анализируют методом ГХ. Содержание общего липида для объединенного в пул семени рапса (содержание масла в семенах) определяют при помощи двукратной экстракции липида с применением CHCl3/MeOH из известной массы высушенных семян после раздавливания, с последующим метилированием аликвот липида вместе с 17:0 жирными кислотами в качестве внутреннего стандарта. В случае Camelina, липид из известного количества семян метилируют вместе с известным количеством 17:0 жирных кислот, как и в случае анализа масла Arabidopsis, и МЭЖК анализируют методом ГХ. Для количественного определения ТАГ, фракционируют ТАГ из экстрагированного липида с применением ТСХ и непосредственно метилируют на кремния диоксиде с применением 17:0 ТАГ в качестве внутреннего стандарта. Эти способы описаны более подробно ниже.
После сбора урожая в стадии зрелости растения, семена Camelina или рапса обезвоживают путем хранения семян в течение 24 часов при комнатной температуре в эксикаторе, содержащем силикагель в качестве поглотителя влаги. Содержание влаги в семенах обычно составляет 6-8%. Общие липиды экстрагируют из известной массы обезвоженных семян путем измельчения семян с помощью смеси хлороформа и метанола (2/1, об/об) в пробирке Эппендорфа, с использованием гомогенизатора для тканей Reicht (частота 22/секунду в течение 3 минут) и металлического шара. Добавляют один объем 0,1 М раствора KCl, и смесь встряхивают в течение 10 минут. Нижнюю неполярную фракцию собирают после центрифугирования смеси в течение 5 минут при скорости 3000 об/мин. Оставшуюся верхнюю (водную) фракцию промывают 2-мя объемами хлороформа при перемешивании в течение 10 минут. Вторую неполярную фракцию также собирают и объединяют в пул с ранее собранной. Растворитель испаряют из липидов в экстракте в потоке азота и высушенный общий липид растворяют в известном объеме хлороформа.
Для измерения количества липида в экстрагированном материале известное количество 17:0-ТАГ добавляют в качестве внутреннего стандарта и липиды из известного количества семян инкубируют в 1 н метанольной HCl (Supelco) в течение 2 часов при 80°C. Полученный таким образом МЭЖК экстрагируют гексаном и анализируют методом ГХ. Количество отдельного МЭЖК определяют на основании количества МЭЖК 17:0 ТАГ. Массу отдельных МЭЖК, после вычитания массы метильных групп, которыми этерифицированы МЭЖК, переводят в моль путем деления на молекулярную массу отдельных МЭЖК. Общее количество моль всех МЭЖК делят на три, чтобы вычислить количество моль ТАГ, и, таким образом, глицерина. Далее, количество моль ТАГ превращают в массу ТАГ. В конце, процент содержания масла на основе массы семени вычисляют с использованием массы семени, предполагая, что весь экстрагированный липид представляет собой ТАГ или эквивалент ТАГ для целей вычисления содержания масла. Данный способ базируется на Li с соавт., (2006). Другие семена подобного размера, кроме семян Camelina или рапса, также могут быть проанализированы данным способом.
Кроме того, содержание масла в семени рапса и других семенах было измерено методами ядерного магнитного резонанса (Rossell и Pritchard, 1991) пульсирующей волной NMS 100 Minispec (Bruker Pty Ltd Scientific Instruments, Германия), как описано в Примере 14. Метод ЯМР позволяет одновременно измерить содержание влаги. Кроме того, содержание масла в семенах можно измерить отражательной спектроскопией в ближнем ИК-диапазоне, например, с применением монохроматора NIRSystems модель 5000. Кроме того, содержание влаги может быть измерено в образце партии семян, посредством сушки семян в образце в течение 18 часов при температуре приблизительно 100°C, согласно Li с соавт. (2006).
Анализ липидов из образцов лизата листа
Липиды из образцов лизата экстрагируют с использованием смеси хлороформ/метанол/0,1 М раствор KCl (2:1:1) и извлекают. Различные классы липидов в образцах разделяют на пластинках Силикагель 60 для тонкослойной хроматографии (ТСХ) (MERCK, Дармштадт, Германия) (ТСХ), пропитанных 10% раствором борной кислоты. Система растворителей, используемых для фракционирования ТАГ из экстракта липидов, представляет собой хлороформ/ацетон (90:10, об/об). Отдельные классы липидов визуализируют посредством контакта пластинок с парами йода и идентифицируют посредством параллельного анализа аутентичных стандартов на такой же пластинке ТСХ. Пластинки приводят в контакт с фосфорными визуализационными экранами на протяжении ночи и анализируют с помощью визуализатора фосфора Fujifilm FLA-5000 перед жидкостно-сцинтиляционным подсчетом для количественного определения импульсов в минуту.
Выделение общего липида и фракционирование липидов из вегетативных тканей
Состав жирных кислот общего липида в образцах листьев и других вегетативных тканей определяли прямым метилированием жирных кислот в лиофилизированных образцах. Для количественной оценки общего липида, жирные кислоты в известной массе лиофилизированных образцов непосредственно метилируют с 17:0 СЖК. Для определения уровней общих ТАГ в образцах листьев, ТАГ из экстрагированного общего липида фракционируют с помощью ТСХ и метилируют в присутствии внутреннего стандарта 17:0 ТАГ, вследствие присутствия в листьях значительных количеств полярных липидов. Это осуществляют, как указано ниже. Ткани, включая образцы листа, лиофилизируют, взвешивают (сухая масса) и общие липиды, экстрагированные как описано Bligh и Dyer (1959) или с использованием смеси хлороформ/метанол/0,1 М раствор KCl (ХМК; 2:1:1) в качестве растворителя. Общие липиды извлекают из образцов листа N. benthamiana после лиофилизации, добавляя 900 мкл смеси хлороформ/метанол (2:1, об/об) на образец листа диаметром 1 см. Добавляют 0,8 мкг ДАГЭ на 0,5 мг массы сухого листа в качестве внутреннего стандарта, если должен быть проведен анализ ТСХ-ПИД. Образцы гомогенизируют с помощью гомогенизатора для тканей ULTRA-TURRAX производства IKA, после чего добавляют 500 мкл 0,1 М раствора KCl. Образцы обрабатывают вихревым перемешиванием, центрифугируют в течение 5 минут, и нижнюю фракцию собирают. Оставшуюся верхнюю фракцию экстрагируют повторно, добавляя 600 мкл хлороформа, обрабатывают вихревым перемешиванием и центрифугируют в течение 5 минут. Нижнюю фракцию извлекают и объединяют в пул с ранее собранной. Липиды сушат в потоке азота и ресуспендируют в 2 мкл хлороформа на мг сухой массы листа. Общие липиды листьев N. tabacum или образцов листа извлекают, как раскрыто выше, с некоторыми модификациями. Если 4 или 6 листовых дисков (каждый с площадью поверхности приблизительно 1 см2) объединяют, то используют 1,6 мл растворителя ХМК, а если объединяют 3 или менее листовых дисков, то используют 1,2 мл ХМК. Лиофилизированные ткани листа гомогенизируют в пробирке Эппендорфа, содержащей металлический шар, с помощью гомогенизатора для тканей Reicht (Qiagen) в течение 3 минут при частоте 20/секунду.
Отделение нейтральных липидов методами ТСХ и переметилирования
Известные объемы полных экстрактов листа, например, 30 мкл, помещают на пластинку ТСХ силикагель 60 (1×20 см) (Merck KGaA, Германия). Нейтральные липиды фракционируют на различные типы и отделяют от полярных липидов методом ТСХ в уравновешенной емкости для проявления, содержащей систему растворителей гексан/диэтиловый эфир (ДЭЭ)/уксусная кислота (70:30:1, об/об/об). Полосы ТАГ визуализируют посредством обрызгивания примулином, маркируют в УФ-свете, соскабливают с пластины ТСХ, переносят во флаконы для ГХ емкостью 2 мл и сушат с помощью N2. Добавляют 750 мкл 1 н метанольного HCl (Supelco analytical, США) в каждый флакон, вместе с известным количеством C17:0 ТАГ в качестве внутреннего стандарта, в зависимости от количества ТАГ в каждом образце. Обычно, добавляют 30 мкг внутреннего стандарта для образцов с низким содержанием ТАГ, тогда как в случае образцов с высоким содержанием ТАГ применяют до 200 мк г внутреннего стандарта.
Образцы липида для анализа состава жирных кислот методом ГХ переметилируют путем инкубирования смесей при 80°C в течение 2 часов в присутствии метанольного HCl. После охлаждения образцов до комнатной температуры, реакцию останавливают, добавляя 350 мкл H2O. Метиловые эфиры жирных кислот (МЭЖК) извлекают из смеси, добавляя 350 мкл гексана, обрабатывая вихревым перемешиванием и центрифугируя со скоростью 1700 об/мин в течение 5 минут. Верхнюю гексановую фракцию собирают и переносят во флаконы для ГХ с коническими вставками емкостью 300 мкл. После выпаривания, образцы ресуспендируют в 30 мкл гексана. По 1 мкл вводят в прибор для ГХ.
Количество индивидуальных и общих жирных кислот (ОЖК) в липидных фракциях определяют с помощью ГХ, определяя площадь каждого пика, и вычисляя посредством сравнения с площадью пика для известного количества внутреннего стандарта. Содержание ТАГ в листе вычисляют как сумму глицерильных и жирных ацильных фрагментов во фракции ТАГ, с использованием соотношения: %, масс. ТАГ=100 x ((41 x общее количество моль МЭЖК/3)+(общее количество МЭЖК в г - (15 x общее количество моль МЭЖК)))/грамм сухой массы листа, где 41 и 15 представляют собой молекулярную массу глицерильного фрагмента и метильной группы, соответственно.
Капиллярная газожидкостная хроматография (ГХ)
МЭЖК анализируют методом ГХ с использованием ГХ Agilent Technologies 7890A (Пало-Альто, Калифорния, США), оборудованного колонкой SGE BPX70 (70% цианопропил полисилфенилен-силоксан) (30 м x 0,25 мм внутренний диаметр, толщина пленки 0,25 мкм), ПИД, инжектором с расщеплением/без расщепления и аутосемплером с инжектором Agilent Technologies 7693 Series. Гелий используют в качестве газа-носителя. Образцы вводят в режиме расщепления (соотношение 50:1) при температуре печи 150°C. После инжекции температуру печи удерживают на уровне 150°C в течение 1 минуты, затем повышают до 210°C со скоростью 3°C/минуту-1, и в конце до 240°C со скоростью 50°C/минуту-1. Пики количественно измеряют с помощью программного обеспечения Agilent Technologies ChemStation (Rev B.04.03 (16), Пало-Альто, Калифорния, США), на основании ответа известного количества внешнего стандарта GLC-411 (Nucheck) и внутреннего стандарта C17:0-Me.
Количественное определение ТАГ с помощью Iatroscan
1 мкл экстракта липида помещают на один Chromarod-SII для ТСХ-ПИД Iatroscan™ (Mitsubishi Chemical Medience Corporation, Япония). Далее стойку Chromarod переносят в уравновешенную емкость для проявления, содержащую 70 мл системы растворителей гексан/CHCl3 /2-пропанол/муравьиная кислота (85: 10,716:0,567:0,0567, об/об/об/об). После инкубации в течение 30 минут, стойку Chromarod сушат в течение 3 минут при 100° C и немедленно сканируют на анализаторе ТСХ-ПИД Iatroscan MK-6s (Mitsubishi Chemical Medience Corporation. Япония). Площадь пиков внутреннего стандарта ДАГЭ и ТАГ интегрируют с помощью программного обеспечения для интеграции SIC-480II (версия: 7.0-E, SIC System instruments Co., LTD - Япония).
Количественное определение ТАГ осуществляют в две стадии. Вначале во всех образцах сканируют ДАГЭ для коррекции выходов экстракции, после чего концентрированные образцы ТАГ выбирают и разбавляют. Затем, количество ТАГ определяют в разбавленных образцах вторым сканированием в соответствии с внешней калибровкой, используя глицерилтрилинолеат в качестве внешнего стандарта (Sigma-Aldrich).
Количественное определение ТАГ в образцах листа методом ГХ
Площадь пика индивидуального МЭЖК вначале корректируют на основании площади пика известных количеств таких же МЭЖК, присутствующих в коммерческом стандарте GLC-411 (NU-CHEK PREP, Inc., США). Скорректированные значения площади пика используют для вычисления массы каждого МЭЖК в образце, по сравнению с внутренним стандартом. Поскольку масло хранится в основном в форме ТАГ, количество масла вычисляют на основании количества МЭЖК в каждом образце. Общее количество моль глицерина определяют, вычисляя количество моль МЭЖК, с последующим делением общего количества моль МЭЖК на 3 Количество ТАГ вычисляют как сумму глицерильных и жирных ацильных фрагментов, с использованием формулы: %, масс. масла=100 x ((41 x общее количество моль МЭЖК/3)+(общее количество г МЭЖК - (15 x общее количество моль МЭЖК)))/грамм сухой массы листа, где 41 и 15 представляют собой молекулярную массу глицерильного фрагмента и метильной группы, соответственно.
Пример 2. Увеличение содержания липида в вегетативных частях Nicotiana benthamiana
Генетическую конструкцию pJP3502 применяют для получения стабильно трансформированных растений Nicotiana benthamiana с помощью протокола опосредованной Agrobacterium трансформации, как описано в WO2013/096993 для Nicotiana tabacum. Отбирают трансгенные растения с резистентностью к канамицину и выращивают в теплице до состояния зрелости. Образцы листьев собирают на стадии завязывания семян и лиофилизируют. Определяют общее содержание жирных кислот (ОЖК) (% от сухой массы) и состав общего липида в образцах после экстракции в соответствии с Bligh и Dyer (1959), а также содержание и состав фракции триацилглицеридов (ТАГ). Данные приведены в Таблице 2 и Таблице 3. Образец листа с наиболее высоким содержанием масла получен от трансгенного растения №16, содержание ОЖК в котором составило 33% масс. Данный образец содержал 22,5% масс. ТАГ (в пересчете на сухую массу).
Наблюдается выраженная корреляция между изменением состава жирных кислот и содержанием ОЖК или ТАГ. Уровень олеиновой кислоты (C18:1n-9), повышает с увеличением содержания ОЖК и ТАГ, таким образом, что она является преобладающей жирной кислотой в листьях с высоким содержанием ТАГ, например, содержащих 66,8% ОЖК и 66,9% жирных кислот ТАГ в листьях с наиболее высоким содержанием ТАГ. Подобные корреляции наблюдаются для других жирных кислот, например, уровни АЛК снижаются до 4,9% ОЖК и 3,9% ТАГ в листьях с наиболее высоким содержанием ТАГ. Кроме того, наблюдается выраженная корреляция между уровнями C16:3 и содержанием как ОЖК, так и ТАГ, причем содержание C16:3 существенно снижается в образцах с высоким содержанием ОЖК и ТАГ.
Два растения из числа растений с высоким содержанием масла, №14 и №16, были дополнительно проанализированы в ходе фазы старения листьев, когда листья начали желтеть (Таблица 4 и Таблица 5). Несмотря на то, что изменение общего содержания жирных кислот было незначительным в образце с наиболее высокими показателями (32,9% против 33%), количество ТАГ повысилось до 32,6%. В указанных образцах липиды листьев практически полностью состояли из ТАГ.
Таблица 2. Состав и количество общих жирных кислот (ОЖК) (% от сухой массы) в листьях растений Nicotiana benthamiana, стабильно трансформированных T-ДНК из конструкции pJP3502. Образцы дополнительно содержат 0,1-0,3% C14:0 и 0,0-0,7% C20:1
Таблица 3. Состав жирных кислот и количество ТАГ (% от сухой массы) в листьях растений Nicotiana benthamiana, стабильно трансформированных T-ДНК из конструкции pJP3502. Образцы дополнительно содержат 0,1-0,3% C14:0 и 0,0-0,7% C20:1
Таблица 4. Состав и количество (% от сухой массы) общих жирных кислот (ОЖК) в листьях растений Nicotiana benthamiana, стабильно трансформированных T-ДНК из конструкции pJP3502, на стадии желтого листа. Образцы дополнительно содержат 0,1-0,3% C14:0 и 0,0-0,7% C20:1
Таблица 5. Состав жирных кислот и количество (% от сухой массы) ТАГ в листьях растений Nicotiana benthamiana, стабильно трансформированных T-ДНК из конструкции pJP3502. Образцы дополнительно содержат 0,1-0,3% C14:0 и 0,0-0,7% C20:1
Пример 3. Повышение содержания липидов в вегетативных частях растения Nicotiana tabacum
Конструкцию pJP3502 ранее применяли для трансформации Nicotiana tabacum (WO2013/096993). Семя, полученное от гомозиготного растения T1, трансформированного T-ДНК из pJP3502 и содержащее высокий уровень ОЖК и ТАГ, собирают и высевают с целью получения нового поколения растений-потомков T2, однородно гомозиготных по трансгенам. Горшки размещают в теплице таким образом, что зрелые листья растений частично перекрываются в форме типичного купола («купол парашюта»), как это происходило бы при выращивании в поле, или максимально контактируют с прямым солнечным светом (форма без купола). Образцы листьев получают от каждого полностью выросшего растения на стадии завязывания семян и лиофилизируют. Содержание жирных кислот определяют для фракции ТАГ (Таблица 6) после экстракции общих липидов из образцов в соответствии с Bligh и Dyer (1959). Уровни ТАГ в ткани зрелых листьев растений без купола обычно выше, чем в растениях с куполом, с максимально зарегистрированным содержанием ТАГ в листьях 20,6% от сухой массы листа.
Таблица 6. Содержание ТАГ (% от сухой массы) в ткани зрелых листьев трансгенных растений-потомков T2 (Линия 49), трансформированных T-ДНК pJP3502, по сравнению с диким типом (дт).
Пример 4. Повышение содержания масла в вегетативных частях однодольных растений
Химерные конструкции ДНК разработаны таким образом, чтобы повысить содержание масла в однодольных растениях, например, растении C4 S. bicolor (сорго), путем экспрессии комбинации генов, кодирующих WRI1, LEC1 Z. mays (номер доступа AAK95562; SEQ ID NO: 155), DGAT и олеозина в трансгенных растениях. Несколько пар конструкций для биолистической сотрансформации разработаны и получены путем лигирования-клонирования рестрикционного фермента, как указано ниже.
Генетическая конструкция pOIL136 представляет собой бинарный вектор, содержащий три кассеты экспрессии однодольных, а именно ген селекционного маркера, кодирующий фосфинотрицин ацетилтрансферазу (ФАТ) для селекции растения, вторую кассету для экспрессии DGAT и третью для экспрессии олеозина. Вначале получают pJP136 амплификацией промотора гена актина из Oryza sativa (McElroy с соавт., 1990) и инсерции его в качестве фрагмента тупого-ClaI в pORE04 (Coutu с соавт., 2007) с получением pOIL094. Далее получают pOIL095 путем инсерции версии гена олеозина Sesamum indicum, кодон-оптимизированного для экспрессии в однодольных, в pOIL094 на сайте KpnI. pOIL093 получают клонированием версии кодон-оптимизированного для экспрессии в однодольных гена DGAT2a Umbelopsis ramanniana (Lardizabal с соавт., 2008) в качестве фрагмента SmaI-KpnI в вектор, уже содержащий промотор гена убихитина Zea mays. Далее получают pOIL134 клонированием кассеты экспрессии NotI DGAT2a из pOIL093 в pOIL095 на сайтах NotI. pOIL141 получают инсерцией гена селекционного маркера, кодирующего ФАТ в качестве фрагмента BamHI-SacI в вектор, содержащий промотор убихитина Z. mays. В конце получают pOIL136 клонированием кассеты экспрессии убихитина Z. mays::ФАТ в качестве фрагмента тупого-AscI в ZraI-AscI pOIL096. Генетическая конструкция pOIL136, таким образом, содержит следующие кассеты экспрессии: промотор актина O. sativa::олеозин S. indicum, промотор убихитина Z. mays::DGAT2a U. ramanniana и промотор убихитина Z. mays::ФАТ.
Подобный вектор pOIL197, содержащий NPTII вместо ФАТ, конструируют субклонированием кассеты убихитина Z. mays::NPTII из pUKN в качестве фрагмента HindIII-SmaI в сайты AscI (тупые) и HindIII pJP3343. Далее полученный вектор, pOIL196, расщепляют с помощью HindIII (тупые) и AgeI. Полученный фрагмент размером 3358 пар оснований клонируют в сайты ZraI-AgeI pOIL134 с получением pOIL197.
Разработан и получен набор конструкций, содержащих гены, которые кодируют WRI1 Z. mays (ZmWRI) или факторы транскрипции LEC1 (ZmLEC1) под контролем различных промоторов, для биолистической сотрансформации в комбинации с pOIL136, с целью оценки влияния активности промотора и специфичности в отношении клетки на функционирование WRI1 или LEC1, или обоих в случае комбинации, при экспрессии в вегетативных тканях растения C4, такого как сорго. Указанный отдельный набор конструкций не содержит гена селекционного маркера, за исключением pOIL333, который содержит NPTII в качестве селекционного маркера. Различные исследуемые промоторы были такими, как указано ниже. Промотор гена убихитина Z. Mays (pZmUbi) представляет собой активный конститутивный промотор однодольных, в то время как о расширенном промотор CaMV 35S (e35S), содержащем двойной участок энхансера, сообщалось, что он приводит к более низким уровням экспрессии трансгена (обзор в Girijashankar и Swathisree, 2009). Тогда как промотор гена фосфоенолпируваткарбоксилазы (pZmPEPC) Z. mays активен в мезофильных клетках листа (Matsuoka и Minami, 1989), месте фотосинтеза в видах растений C4, промотор гена маленькой субъединицы Z. mays Rubisco (pZmSSU) является специфичным для слоя клеток обкладки сосудистых пучков (Nomura с соавт., 2000; Lebrun с соавт., 1987), т.е. клеток, в которых имеет место фиксация углерода в растениях C4.
Экспрессия гена Z. mays, кодирующего SEE1 цистеинпротеазу (номер доступа AJ494982) идентифицирована, как сходная с экспрессией специфичного для старения промотора SAG12 A. thaliana в ходе развития растения. Таким образом, промотор размером 1970 пар оснований из гена SEE1 (SEQ ID NO: 216) был дополнительно выбран для контроля экспрессии генов, кодирующих факторы транскрипции WRI1 и LEC1 Z. mays. Далее, промоторы из гена, кодирующего белок «цинкового пальца» AlSAP Aeluropus littoralis (Ben Saad с соавт., 2011; номер доступа DQ885219; SEQ ID NO: 217) и промотор из чувствительного к сахарозе гена ArRolC A. rhizogenes (Yokoyama с соавт., 1994; номер доступа DQ160187; SEQ ID NO: 218) дополнительно выбраны для экспрессии ZmWRI1 в ткани стебля. Таким образом, каждый из этих промоторов индивидуально присоединяют в направлении против хода транскрипции к участкам, кодирующим ZmWRI1 или ZmLEC1, как указано ниже.
Промежуточный вектор, pOIL100, вначале получают клонированием последовательности, кодирующей WRI1 Z. mays, и участка терминатора транскрипции/полиаденилирования, фланкированных сайтами AscI-NcоI, в такие же сайты в бинарном векторе pJP3343. Различные версии конструкций для экспрессии WRI1 основаны на данном векторе и получены клонированием различных промоторов в pOIL100. pOIL101 получают клонированием фрагмента XhoI-Sal, содержащего промотор e35S с удвоенным участком энхансера, в сайт XhoI pOIL100. pOIL102 получают клонированием фрагмента HindIII-AvrII, содержащего промотор гена убихитина Z. mays, в сайты HindIII-XbaI pOIL100. pOIL103 получают клонированием фрагмента HindIII-NcoI, содержащего промотор гена PEPC Z. mays, в сайты HindIII-NcoI pOIL100. pOIL104 получают клонированием фрагмента HindIII-AvrII, содержащего промотор гена SSU Z. mays, в сайты HindIII-AvrII pOIL100.
Синтезирован синтетический фрагмент, содержащий участок промотора SEE1 Z. mays, фланкированный уникальными сайтами HindIII-XhoI. Данный фрагмент клонируют в направлении против хода транскрипции по отношению к участку, кодирующему белок WRI1 Z. mays, используя сайты HindIII-XhoI в pOIL100. Результирующий вектор обозначен pOIL329. Синтезирован содержащий синтетический фрагмент участок промотора AlSAP A. littoralis, фланкированный уникальными сайтами XhoI-XbaI. Данный фрагмент клонируют в направлении против хода транскрипции по отношению к участку, кодирующему WRI1 Z. mays, используя сайты XbaI-Xho I в pOIL100. Результирующий вектор обозначен pOIL330. Синтезирован содержащий синтетический фрагмент участок промотора ArRolC A. rhizogenes, фланкированный уникальными сайтами PspOMI-XhoI. Данный фрагмент клонируют в направлении против хода транскрипции по отношению к участку, кодирующему WRI1 Z. mays, используя сайты PspOMI-XhoI в pOIL100. Результирующий вектор обозначен pOIL335. В конце, бинарный вектор (pOIL333), содержащий кассету экспрессии SEE1 Z. mays::ZmLEC1, получают в три стадии. Вначале, вектор экспрессии 35S::GUS конструируют амплификацией участка, кодирующего GUS, с фланкирующими праймерами, содержащими сайты AvrII и KpnI. Далее полученный фрагмент клонируют в сайты SpeI-KpnI pJP3343. Результирующий вектор обозначен pTV111. Затем, участок 35S промотора pTV111 заменяют промотором SEE1Z. mays. На этой стадии последовательность SEE1 Z. mays амплифицируют с применением фланкирующих праймеров, содержащих уникальные сайты HindIII и XhoI. Результирующий фрагмент разрезают соответствующими рестрикционными ферментами и субклонируют в сайты SalI-HindIII pTV111. Результирующий вектор обозначен pOIL332. Далее последовательность, кодирующую ZmLEC1 амплифицируют с применением фланкирующих праймеров, содержащих сайты NotI и EcoRV. Полученный фрагмент субклонируют в соответствующие сайты pOIL332 с получением pOIL333.
Готовят ДНК для биолистической трансформации, расщепляя скелеты векторов pOIL101, pOIL102, pOIL103, pOIL104, pOIL197, pOIL329, pOIL330, pOIL333 и pOIL335 рестрикционными ферментами, с последующим выделением на геле. Затем ДНК pOIL197 смешивают с ДНК pOIL101, pOIL102, pOIL103, pOIL104, pOIL329, pOIL330, pOIL333 или pOIL335 и осуществляют трансформацию эксплантов S. bicolor посредством биолистически опосредованной трансформации. В качестве альтернативы, трансформацию с помощью конструкций для экспрессии таких же комбинаций генов осуществляют по отдельности или в форме сотрансформации путем опосредованной Agrobacterium трансформации (Gurel с соавт., 2009; Wu с соавт., 2014).
Трансгенные растения регенерируют и отбирают резистентные к антибиотикам. В случае, если двумя конструкциями сотрансформируют одно и то же событие, наблюдается повышенное содержание масла в тканях трансгенных растений, не относящихся к семенам.
Химерные конструкции ДНК для опосредованной Agrobacterium трансформации применяются для трансформации Zea mays (кукуруза), как описано Gould с соавт., (1991). Вкратце, экспланты верхушек побегов культивируют с трансгенной Agrobacterium в течение двух дней перед перемещением на соленую среду МС, содержащую канамицин и карбенициллин. После нескольких кругов субкультуривирования, самопроизвольно формируются трансформированные побеги и корни, и экспланты переносят в грунт. Кроме того, конструкции применяются для трансформации Hordeum vulgare (ячмень) и Avena sativa (овес) с применением способов трансформации, известных для указанных видов. Вкратце, в случае ячменя применяют культуры Agrobacterium для трансформации клеток в незрелых эмбрионах ячменя (сорт Golden Promise) в соответствии с опубликованными способами (Tingay с соавт., 1997; Bartlett с соавт., 2008) с некоторыми модификациями в том, что эмбрионы длиной от 1,5 до 2,5 мм выделяют из незрелых зерновок и удаляют эмбриональные оси. Полученные экспланты культивируют в течение 2-3 дней с трансгенной Agrobacterium, а затем культивируют в темноте в течение 4-6 недель на средах, содержащих тиментин и гигромицин для генерации эмбриогенного каллюса, после чего переносят в переходные среды в условиях слабой освещенности на две недели. Затем каллюсы переносят в регенерационные среды, чтобы позволить регенерацию побегов и корней, перед переносом регенерировавших ростков в грунт. Трансформированные растения получают и выращивают до стадии зрелости в теплице.
Пример 5. Повышение содержания масла в двудольных растениях
Содержание масла в двудольных растениях вида Trifolium repens (клевер), бобовых, обычно применяемых в качестве пастбищной культуры, повышается при экспрессии комбинации генов WRI1, DGAT и олеозина в вегетативных частях. Конструкцию pJP3502 применяют для трансформации T. repens путем опосредованной Agrobacterium трансформации (Larkin с соавт., 1996). Вкратце, генетическую конструкцию pJP3502 вводят в A. tumefaciens посредством стандартной процедуры электропорации. Бинарный вектор дополнительно содержит ген селекционного маркера 35S:NptII в пределах T-ДНК. Трансформированные клетки Agrobacterium выращивают на твердых средах ЛБ с добавлением канамицина (50 мг/л) и рифампицина (25 мг/л) и инкубируют при 28°C в течение двух дней. Единичную колонию используют для инициации свежей культуры. После 48 часов энергичного культивирования, клетки Agrobacterium применяют для обработки семядоль T. repens (сорт. Haifa), вырезанных из набухшего семени, как описано Larkin с соавт. (1996). После совместного культивирования в течение трех дней экспланты обрабатывают 25 мг/л канамицина, чтобы отобрать трансформированные побеги, а затем переносят в среду для корнеобразования для формирования корней, перед переносом в грунт.
Шесть трансформированных растений, содержащих T-ДНК из pJP3502, были получены и перенесены в грунт в теплице. Повышенное содержание масла наблюдалось в ткани некоторых из растений, не относящейся к семенам, причем одно растение продемонстрировало повышение уровней ТАГ в листьях более чем в 4 раза. Такие растения пригодны в качестве корма для животных, например, путем выращивания растений на пастбищах, что обеспечивает питание с повышенным содержанием энергии на единицу массы (плотность энергии) и приводит к повышению темпов прироста у животных.
Дополнительно, конструкцию pJP3502 применяют для трансформации других бобовых растений, таких как люцерна посевная (Medicago sativa) и люцерна усеченная (Medicago truncatula), по способу Wright с соавт. (2006), с получением трансгенных растений с повышенным содержанием ТАГ в вегетативных частях. Были получены три предполагаемых трансгенных растения M. truncatula. Трансгенные растения пригодны в качестве пастбищной культуры или в качестве сена или силоса как источника питания для животных, таких как крупный рогатый скот, овцы и лошади, обеспечивая повышенную плотность энергии в корме.
С целью повышения содержания масла в семенах бобовых, синтезирован фрагмент ДНК, содержащий комбинацию двух химерных генов, а именно (a) первого химерного гена, кодирующего WRI1 A. thaliana, экспрессируемый промотором запасаемого белка фазеолина бета-типа Phaseolus vulgaris и 5' НТУ плюс (b) второго химерного гена, кодирующего DGAT1 A. thaliana, экспрессируемый промотором вициллина Pisum sativum и 5' НТУ. Фрагмент ДНК вставляют в бинарный вектор pORE04, содержащий химерный ген, кодирующий олеозин, для получения конструкции T-ДНК, содержащей три химерных гена и ген селекционного маркера (Фиг. 2), которую применяют для трансформации Lupinus angustifolius, другого бобового растения, способом, который описан Pigeaire с соавт. (1997). Вкратце, экспланты верхушек побегов L. angustifolius культивируют с трансгенной Agrobacterium перед тщательным увлажнением раствором канамицина (20 мг/мл) и переносом на регенерирующую среду, не содержащую канамицина. Множественные пазушные побеги, развивающиеся из верхушек побегов, вырезают помещают на среду, содержащую 50 мг/л канамицина, и выжившие побеги переносят на свежую среду, содержащую 50 мг/л канамицина. Затем здоровые побеги переносят в грунт. Гены на T-ДНК экспрессируются в клетках трансформированных растений, повышая содержание масла в вегетативных тканях и семенах. Дополнительно применяют специфичный для семени промотор, управляющий геном WRI1, с целью повышения содержания масла в трансгенных семенах Lupinus.
Кроме того, конструкцию применяют для трансформации Glycine max, как описано Zhang с соавт. (1999), с получением трансгенных растений сои с повышенным содержанием ТАГ в семенах. Были получены трансгенные растения, что продемонстрировано ПЦР на ДНК, полученной из образцов растений. Растения выращивали до стадии зрелости и собирали урожай семян. Ожидается, что, по данным неразрушающего ЯМР, содержание масла в семенах будет повышено.
Вторая генетическая конструкция для повышения содержания масла в семенах люпина и сои сконструирована путем синтеза инсерции ДНК, содержащей три кассеты экспрессии генов, а именно первой, содержащей промотор β-конглицинина Glycine max, экспрессирующий DGAT2A Umbelopsis ramanniana, второй, содержащей промотор KTi3 Glycine max, экспрессирующий WRI1 A. thaliana, и третьей, содержащей промотор β-конглицинина Glycine max, экспрессирующий MGAT2 Mus musculus. Фрагмент SbfI-PstI данной инсерции клонируют в бинарный вектор pORE04 на сайте PstI, с получением pJP3569. Версию без гена MGAT2 получают, клонируя фрагмент SbfI-SwaI меньшего размера в pORE04 на сайтах EcoRV-PstI, с получением pJP3570, Кроме того, получены версии, содержащие только ген WRI1 и только ген DGAT2A. Указанные бинарные векторы применяют для трансформации Glycine max с получением трансгенного семени. Содержание масла в семени первичных трансформантов анализируют неразрушающим ЯМР перед посевом, чтобы получить семя T2.
Версия pJP3569, пригодная для трансформации люпина, получена ПЦР амплификацией кассет экспрессии WRI1 и DGAT2A в единичном ампликоне, адаптированном с помощью рестрикционных сайтов NotI. Фрагмент NotI клонируют в pJP3416 на сайте PspOMI с получением бинарного вектора pJP3678, содержащего ген селекционного маркера ФАТ.
Пример 6. Эксперименты для повышения содержания масла в вегетативных частях рапса
Два бинарных вектора экспрессии применяют для трансформации B. napus (сорт Оскар) с целью изучения влияния на аккумуляцию ТАГ в семени и/или вегетативных тканях. Вначале конструируют плазмиды pJP3414 путем инсерции последовательности, кодирующей кодон-оптимизированный белок WRI1 A. thaliana, в бинарный вектор 35S-pORE04, который содержит пустую кассету экспрессии 35S. Таким образом, T-ДНК pJP3414 содержит кодон-оптимизированную версию фактора транскрипции WRI1 A. thaliana под контролем конститутивного промотора 35S. Ткань листа 11 независимо трансформированных саженцев B. napus T0, трансформированных pJP3414, как описано в Примере 1, каждый образец, которой содержит повышенные уровни ТАГ, сравнивают с растениями, трансформированными пустым вектором (pORE04). Однако, ни в одном из случаев уровень ТАГ не превышал 1%. Максимальные уровни были обнаружены в линии 31, которая содержала до 0,58% ТАГ, в пересчете на сухую массу. Содержание масла в трансгенном семени T1 не было существенно повышенным, по сравнению с семенами дикого типа (Оскар) и трансформированными пустым контрольным вектором. Семена T1 трех линий, демонстрирующих самые высокие уровни ТАГ в ткани листа, регенерированы на среде МС, содержащей 3% сахарозы. Не наблюдалось разницы в прорастании через 5 и 8 дней, по сравнению с нетрансформированным контролем (Оскар).
В попытке дополнительного повышения уровней ТАГ в вегетативных тканях B. napus, применяют второй вектор, pJP3502 (Vanhercke с соавт., 2014), для трансформации B. napus (сорт Оскар). Уровни ТАГ количественно определяют в образцах трансгенных листьев, полученных перед цветением. Однако, содержание ТАГ дополнительно не повышается, и состав жирных кислот не отличается от нетрансформированных контрольных растений в такой же стадии роста.
Результаты экспериментов с B. napus, описанных в данном Примере, которые обеспечивают содержание ТАГ в листьях менее чем 1%, абсолютно противоречат приведенным в Примерах 2 и 3 для видов Nicotiana, которые обеспечивают около 20-30% ТАГ. Авторы изобретения тщательно изучали эти результаты в поиске объяснения разницы между видами. Идентифицировано несколько отличий между видами. Одно из отличий, которое изобретатели приняли как обеспечивающее существенную разницу, состояло в принадлежности Brassica napus к так называемым видам 16:3, тогда как вид Nicotiana относится к так называемым видам 18:3. Это связано с относительным вкладом так называемых прокариотных и эукариотных путей в синтез липида в пластидах (Фиг. 1), и, таким образом, по мнению авторов изобретения, с количеством ДАГ, доступным для синтеза ТАГ. Это навело авторов изобретения на идею о том, что модель с модификацией соотношения синтеза жирных кислот через эукариотный путь относительно прокариотного пути, например, уменьшение аккумуляцию 16:3 относительно 18:3, изменила бы уровень ТАГ, который аккумулируется в растительных клетках или фотосинтезирующих микробных клетках. Они ожидали, что модификация, которая изменит баланс в пользу эукариотного пути, будет обеспечивать преимущество с точки зрения уровней аккумуляции ТАГ, особенно в так называемых растениях 16:3. Вкратце, для превращения клеток ʺ16:3ʺ в более сходные с клетками ʺ18:3ʺ.
Авторами изобретения выдвинута гипотеза о том, что присутствие C18:1-АПБ в пластиде, которое ингибирует АККазу по механизм обратной связи, могло бы быть более выражено в растениях 16:3 за счет синтеза и удерживания жирных кислот в пластиде по прокариотному пути. И наоборот, ими выдвинута гипотеза о том, что растения C18:3 способны к аккумуляции более высоких уровней ТАГ в вегетативных тканях за счет повышенного экспорта C18:1 из пластид для снабжения эукариотного пути. Как показано в Примерах 2-4 настоящего документа, это наблюдалось в таких видах, как N. tabacum и N. benthamiana, в которых присутствует более высокое соотношение пластидных липидов C18:3/C16:3 относительно таких видов, как B. napus, которые содержат низкие уровни C18:3 в пластидных липидах. Данная модель, согласно гипотезе авторов изобретения, объяснила бы, почему стабильная трансформация генами экспрессии WRI+DGAT+олеозина из вектора pJP3502 как в N. tabacum, так и в N. benthamiana приводила к высоким уровням аккумуляции ТАГ и обширным изменениям в составе жирных кислот. И наоборот, трансформация таким же вектором B. napus приводила только к незначительному повышению аккумуляции ТАГ и небольшому изменению состава жирных кислот. Данную модель исследовали, как описано ниже.
Пример 7. Модификация пластидной экспрессии GPAT
Повышенная экспрессия пластидной GPAT в растительных клетках
Целый ряд экспериментов выполнялся для проверки гипотезы о том, что присутствие высокоактивного прокариотного пути 16:3 в растении (т.е. так называемом растении 16:3) обеспечивало бы значительно более низкие уровни ТАГ в вегетативных тканях при введении комбинации генов на pJP3502, относительно растений 18:3. Такие эксперименты описаны в следующих Примерах. Вначале, авторы изобретения оценивали, может ли повышенная экспрессия пластидной GPAT воспрепятствовать высокому уровню аккумуляции ТАГ в трансгенном N. benthamiana, увеличивая поток в прокариотном пути.
Кодирующий участок для экспрессии пластидной GPAT Arabidopsis thaliana, ATS1 (Nishida с соавт., 1993), амплифицируют методом ОТ-ПЦР из общей РНК A. thaliana и клонируют в качестве фрагмента EcoRI-PstI в бинарный вектор экспрессии pJP3343 под контролем промотора 35S, с получением конститутивного вектора экспрессии pOIL098. Влияние повышенной экспрессии пластидной GPAT на генетическом фоне высокого содержания масла в листьях определяли путем инфильтрации ткани листа с высоким содержанием масла химерным вектором pOIL098. Ткань листа с высоким содержанием масла генерируют совместной инфильтрацией бинарными векторами экспрессии WRI1 и DGAT (Пример 1) или инфильтрацией pOIL098 листьев растения Nicotiana, стабильно трансформированного T-ДНК из pJP3502 или другого вектора высокого содержания масла. Ожидается, что содержание масла будет снижено в инфильтрованных пятнах на листьях, соэкспрессирующих ген, кодирующий ATS1. Это определяют методом анализа ОЖК и ТАГ как доли от сухой массы образца. Дополнительно, это определяют путем наблюдения за инкорпорацией меченного ацетата в жирные кислоты, продуцированные микросомами или листовыми лизатами, полученными из инфильтрованных пятен на листьях.
Аккумуляция масла в Arabidopsis thaliana с мутантной пластидной GPAT
Мутант ats1 A. thaliana содержит разрушительную мутацию в гене, кодирующем пластидную GPAT, которая снижает активность пластидной GPAT до уровня только 3,8% от дикого типа (Kunst с соавт., 1988). Уровни аккумуляции ТАГ за пределами семян, по меньшей мере в листьях, стеблях и корнях, как в материнской A. thaliana, так и растении с мутантной ats1 оценивают и сравнивают. T-ДНК конструкции pJP3502 для повышенной экспрессии комбинации генов, кодирующих WRI1, DGAT и олеозин, вводят путем трансформации в растения обоих генотипов. Комбинация генов в T-ДНК pJP3502 повышает синтез жирных кислот в обоих типах генетического фона растений. Однако, ожидается, что в присутствии мутантного ats1 аккумуляция ТАГ в среднем будет существенно выше, чем в трансгенных растениях, полученных от генотипа дикого типа (материнский), за счет снижения активности пластидной GPAT и таким образом уменьшения потока жирных кислот в пластидный прокариотный путь. Соотношение C16:3 и C18:3 жирных кислот существенно снижается в листьях с мутантным ats1, как трансформированных, так и нетрансформированных.
Сайленсинг гена, кодирующего пластидную GPAT в растительных клетках
В дополнение к генетической модификации растения введением мутации в ген, кодирующий пластидную GPAT, поток жирных кислот через прокариотный путь 16:3 может быть уменьшен и, таким образом, повышено содержание масла в вегетативных частях путем сайленсинга пластидной GPAT. Это демонстрируется получением трансгенной кассеты, содержащей конститутивный или специфичный для листьев промотор, экспрессирующий шпилечную РНК, соответствующую участку гена, кодирующего пластидную GPAT выбранного вида. В качестве примера, кассету экспрессии шпилечной РНКи получают с использованием фрагмента SalI-EcoRV последовательности кДНК пластидной GPAT A. thaliana размером 581 пара оснований (NM_179407, SEQ ID NO: 177). Кроме того, участок любого гена, кодирующего пластидную GPAT, последовательность которого в высокой степени идентична нуклеотидной последовательности NM_179407, можно применять для конструирования гена с целью экспрессии шпилечной РНК для сайленсинга эндогенного гена пластидной GPAT. Конструкция шпРНКи, содержащая фрагмент пластидной GPAT N. benthamiana размером 732 пары оснований (SEQ ID NO: 219), фланкированный уникальными сайтами SmaI и KasI, разработана с целью стабильной трансформации N. tabacum. Синтезированный фрагмент пластидной GPAT N. benthamiana субклонируют в сайты SmaI-KasI pJP3303, с получением pOIL113. Ожидается, что уменьшение удерживания жирных кислот в пластидах будет приводить к повышению аккумуляции ТАГ, особенно при объединении с компонентом ʺPushʺ, таким как повышенная экспрессия фактора транскрипции, такого как WRI1, или компонентом ʺPullʺ, таким как DGAT или PDAT и/или сниженная активность SDP1 или ТГД.
Инактивация гена, кодирующего пластидную GPAT или, более того, любого гена может быть достигнута с применением способов ККППРП/Cas9. Например, инактивация гена, кодирующего пластидную GPAT A. thaliana (номер доступа NM_179407) может осуществляться разрушением гена, опосредованным ККППРП/Cas9/короткой направляющей РНК (крРНК), с последующим мутагенезом посредством восстановления ДНК негомологичным соединением концов (НГСК). Перед направленным расщеплением ДНК, Cas9 стимулирует разделение цепей ДНК и позволяет КНРНК гибридизоваться со специфической последовательностью длиной 20 нуклеотидов гена-мишени. Это помещает ДНК-мишень в активный сайт Cas9 в надлежащей ориентации относительно сайта связывания с PAM (тандемные гуанозиновые нуклеотиды). Такое расположение позволяет отдельным нуклеазным доменам Cas9 независимо расщеплять каждую цепь последовательности ДНК-мишени в точке, отстоящей на 3 нуклеотида в направлении против хода транскрипции относительно сайта PAM. Далее происходит восстановление двухцепочечного разрыва ДНК путем подверженного ошибкам НГСК, в ходе которого происходят делеции или инсерции нескольких нуклеотидов, что приводит к инактивации гена пластидной GPAT. Последовательности кнРНК, нацеленные на ген GPAT A. thaliana, идентифицированы и отобраны с применением веб-инструмента CRISPRP (Xie с соавт., 2014). Последовательность-мишень длиной 20 нуклеотидов может быть любой последовательностью длиной 20 нуклеотидов в пределах гена-мишени, в том числе в пределах не кодирующих участков гена, таких как промотор или интрон, при условии, что она является специфической последовательностью в пределах генома. Последовательность может быть вставлена в бинарный вектор, содержащий кассету экспрессии ККППРП/Cas9/кнРНК и селекционный маркер канамицина для растений (Jiang с соавт., 2013) и трансформирована в растительные клетки путем опосредованного Agrobacterium превращения. Можно осуществить скрининг трансгенных растений T1 на предмет мутаций в гене пластидной GPAT методом ПЦР амплификации и секвенирования ДНК.
Пример 8. Повышение экспрессии тиоэстеразы в растительных клетках
Синтез жирных кислот de novo происходит в пластидах эукариотных клеток, где жирные кислоты синтезируются при связывании с ацилпереносящим белком в форме конъюгатов ацил-АПБ. После удлинения цепи до ацильных групп C16:0 и C18:0, а затем введения кратной связи в C18:1 во время связывания с АПБ, жирные кислоты отщепляются от АПБ тиоэстеразами и входят в эукариотный путь посредством экспорта из пластид и транспорта в ЭР, где они принимают участие в биогенезе мембран и запасных липидов. В хлоропластах процесс экспорта включает две стадии: во-первых, ацильные цепи высвобождаются в форме свободных жирных кислот по действием ферментов тиоэстераз (тиоэстераза жирного ацила; ТЖА), во-вторых, введение в реакцию с КоА с образованием эфиров ацил-КоА, что катализируется длинноцепочечными ацил-КоА синтетазами (ДЦАС). A. thaliana содержит 3 тиоэстеразы жирного ацила, которые могут отличаться на основании их специфичности в отношении ацильной цепи. FATA1 и FATA2 преимущественно гидролизуют ненасыщенные ацил-АПБ, в то время как насыщенные цепи ацил-АПБ обычно расщепляются FATB.
С целью изучения влияния соэкспрессии тиоэстеразы и генов, кодирующих полипептиды WRI1 и/или DGAT, на содержание общих жирных кислот, содержание ТАГ и состав жирных кислот получают химерные гены для каждой из трех тиоэстераз A. thaliana путем инсерции кодирующих участков в бинарный вектор экспрессии pJP3343 для временной экспрессии в клетках листьев N. benthamiana промотором 35S. Кодирующие белок участки FATA1 A. thaliana (номер доступа NP_189147.1, SEQ ID NO: 202) и FATA2 (номер доступа NP_193041.1, SEQ ID NO: 203) амплифицируют из кДНК стручков с применением праймеров, содержащих сайты EcoRI и PstI, и далее клонируют в pJP3343 с применением таких же рестрикционных сайтов. Полученные векторы экспрессии обозначены pOIL079 и pOIL080, соответственно. Кодирующий белок участок гена FATB A. thaliana (номер доступа NP_172327.1, SEQ ID NO: 204) амплифицируют с применением праймеров, содержащих фланкированные сайты NotI и SacI, и клонируют в соответствующие рестрикционные сайты pJP3343, с получением pOIL081. Конструкциями pOIL079, pOIL080 и pOIL081 инфильтруют ткань листа N. benthamiana, отдельно или в комбинации с конструкциями, содержащими гены. фактора транскрипции WRI1 A. thaliana (AtWRI1) (pJP3414) и/или DGAT1 ацилтрансферазы (AtDGAT1) (pJP3352). Для сравнения, химерные гены, кодирующие FatB1 Cocos nucifera (CnFATB1) (pJP3630), FatB2 C. nucifera (CnFATB2) (pJP3629) вводят в ткань листа N. benthamiana параллельно с тиоэстеразами Arabidopsis, чтобы сравнить влияние полипептидов FatB, обладающих специфичностью в отношении СЦЖК, с тиоэстеразами Arabidopsis, не обладающими специфичностью в отношении СЦЖК. Все эксперименты с инфильтрацией включают химерный ген для экспрессии супрессора сайленсинга p19, как описано в Примере 1. Отрицательный контроль инфильтруют только T-ДНК p19.
Наблюдалось синергетическое влияние для экспрессии тиоэстеразы и повышенной экспрессии WRI1 и/или DGAT на уровни ТАГ в листьях N. benthamiana. Экспрессия генов тиоэстеразы без генов WRI1 или DGAT существенно повышает уровни ТАГ свыше низкого уровня в отрицательном контроле (только p19). Например, экспрессия тиоэстеразы FATB2 кокоса приводила к повышению уровней ТАГ в листьях в 8,2 раза, по сравнению с отрицательным контролем. Соэкспрессия фактора транскрипции WRI1 A. thaliana с каждой из тиоэстераз дополнительно повышала уровни ТАГ, по сравнению с контролем AtWRI1. Соэкспрессия каждой из тиоэстераз CnFATB1 и CnFATB2 кокоса с WRI1 приводила к более высоким уровням ТАГ, чем каждой из трех тиоэстераз A. thaliana с WRI1. Интересно, что противоположное наблюдалось, если ацилтрансфераза DGAT1 A. thaliana соэкспрессировалась в комбинации с тиоэстеразой и WRI1. Это наводит на мысль о лучшей специфичности в отношении ацильной цепи тиоэстераз A. thaliana и ацилтрансферазы DGAT1 A. thaliana, что приводит к увеличенному потоку ацильных цепей от ацил-АПБ в ТАГ. Кроме того, не-СЦЖК тиоэстеразы были значительно более эффективны с точки зрения увеличения процента олеиновой кислоты в общем содержании жирных кислот в листьях. Соэкспрессия AtWRI1, AtDGAT1 и AtFATA2 приводила к наиболее высокому уровню ТАГ в листьях, обеспечивая уровень, который более чем в 1,6 раза превышал наблюдаемый при соэкспрессии AtWRI1 и AtDGAT1 без тиоэстеразы. Указанные эксперименты подтвердили синергетическое повышение синтезе и аккумуляции масла, при соэкспрессии WRI1 и DGAT, а также продемонстрировали дальнейшее синергетическое увеличение при добавлении тиоэстеразы к комбинации.
Три различных бинарных вектора экспрессии сконструированы для оценки влияния соэкспрессии генов, кодирующих WRI1, DGAT1 и FATA, на уровни ТАГ и состав жирных кислот в листьях стабильно трансформированного N. tabacum. Вектор pOIL121 содержит ген SSU::AtWRI1 для экспрессии AtWRI1 промотором SSU, гена 35S::AtDGAT1 для экспрессии AtDGAT промотором 35S и гена enTCUP2::AtFATA2 для экспрессии AtFATA2 промотором enTCUP2, который является конститутивным промотором. Указанные генетические конструкции получают из pOIL38, вначале расщепляя ДНК с помощью NotI для удаления гена, кодирующего олеозин S. indicum. Кодирующий белок участок гена FATA2 A. thaliana амплифицируют и фланкируют сайтами NotI с применением ДНК pOIL80 в качестве шаблона. Далее этот фрагмент вставляют в сайт NotI pOIL38. Затем pOIL121 служит материнским вектором для pOIL122, который содержит дополнительную кассету шпилечной РНК enTCUP2::SDP1 для опосредованного РНКи сайленсинга эндогенного гена SDP1 в трансгенных растениях. Чтобы этого достичь, полноразмерную кассету шпильки SDP1 N. benthamiana выделяют из pOIL51 (Пример 11) в виде фрагмента SfoI-SmaI и клонируют в сайт SfoI pOIL121, с получением pOIL122 (Фиг. 3). Третий вектор, pOIL123, содержащий гены SSU::WRI1 и 35S::DGAT1 и ген шпилечной РНК enTCUP2::SDP1 получают подобным образом, клонируя кассету шпилечной РНК enTCUP2::SDP1 в виде фрагмента SfoI-SmaI в сайт SfoI pOIL36.
Вкратце, векторы содержали комбинации генов:
pOIL121: SSU::AtWRI1, 35S::AtDGAT1, enTCUP2::AtFATA2.
pOIL122: SSU::AtWRI1, 35S::AtDGAT1, enTCUP2::AtFATA2, enTCUP2::шпилька SDP1.
pOIL123: SSU::AtWRI1, 35S::AtDGAT1, enTCUP2::шпилька SDP1.
Каждую из трех конструкций применяют для получения трансформированных растений N. tabacum (сорт Wi38) путем опосредованной Agrobacterium трансформации. Каждое из соэкспрессии тиоэстеразы FATA2 A. thaliana или сайленсинга эндогенной липазы ТАГ SDP1 в комбинации с экспрессией AtWRI1 и AtDGAT1 приводило к дальнейшему повышению уровней ТАГ, по сравнению с экспрессией AtWRI1 и AtDGAT1 в отсутствие как гена тиоэстеразы, так и гена сайленсинга SDP1. Наиболее высокий выход ТАГ получают с применением pOIL122 посредством сочетанного действия всех четырех химерных генов.
Необходимо отметить, что N. benthamiana является растением 18:3. Такие же конструкции pOIL079, pOIL080 и pOIL081 применяются для трансформации A. thaliana, растения 16:3.
Авторами изобретения была задумана модель, повышающая пластидный экспорт жирных кислот, например, снижения аккумуляции ацил-АПБ в пластидах путем повышения активности тиоэстеразы жирного ацила, таким образом, повышая биосинтеза жирных кислот в результате снижения ингибирования ацетил-КоА карбоксилазы (АККазы) по механизму обратной связи (Andre с соавт., 2012; Moreno-Perez с соавт., 2012). Повышенная экспрессия тиоэстеразы повышает экспорт ацильных цепей из пластид в ЭР, таким образом, предлагая эффективную связь между так называемыми «Push» и «Pull» стратегиями метаболического конструирования.
Пример 9. Выработка среднецепочечных жирных кислот в вегетативных растительных клетках
Eccleston с соавт. (1996) изучали аккумуляцию C12:0 и C14:0 жирных кислот как в семенах, так и в листьях трансгенных растений Brassica napus, трансформированных конститутивно экспрессирующимся геном, кодирующим тиоэстеразу California Bay Laurel 12:0-АПБ (Umbellularia californica). В указанном исследовании сообщалось об аккумуляции значительных уровней C12:0 в зрелых семенах B. napus, но только очень низкие уровни C12:0 наблюдались в ткани листа, несмотря на высокие уровни экспрессии и активности тиоэстеразы 12:0-АПБ. Такие же результаты были получены при трансформации гена в A. thaliana (Voelker с соавт., 1992). Данное исследование было расширено соэкспрессией LPAAT Cocos nucifera и тиоэстеразы Umbellularia californica, что приводило к повышению аккумуляции общей C12:0, в также к увеличению фракции трилаурина в семенах B. napus (Knutzon с соавт., 1999). Таким образом, уровень техники указывает на то, что синтез среднецепочечных жирных кислот (СЦЖК) в вегетативных растительных клетках был проблематичным.
Для оценки влияния введения тиоэстераз, обладающих специфичностью в отношении СЦЖК, в комбинации с другими генами, описанными в настоящем документе, химерные ДНК для экспрессии несколько различных тиоэстераз синтезированы и введены в растительные клетки, отдельно или в комбинациях. Кодирующие белок участки для тиоэстераз из организмов, которые доказанно вырабатывают СЦЖК (Jing с соавт., 2011), синтезируют и вставляют в виде фрагментов EcoRI в бинарный вектор pJP3343, который содержит кассету экспрессии промотора 35S (Vanhercke с соавт., 2013). Тиоэстеразы были следующими: тиоэстераза Cinnamomum camphora14:0-АПБ (обозначена как Cinca-TE) (Yuan с соавт., 1995; номер доступа Q39473.1; SEQ ID NO: 193), ацил-АПБ тиоэстераза FatB1 Cocos nucifera (Cocnu-TE1;номер доступа AEM72519.1 SEQ ID NO: 194), ацил-АПБ тиоэстераза FatB2 Cocos nucifera (Cocnu-TE2; номер доступа AEM72520.1; SEQ ID NO: 195), ацил-АПБ тиоэстераза FatB3 Cocos nucifera (Cocnu-TE3; номер доступа AEM72521.1; SEQ ID NO: 196), ацил-(АПБ) тиоэстераза типа B Cuphea lanceolata (Cupla-TE) (Topfer с соавт., 1995; номер доступа CAB60830.1; SEQ ID NO: 197), Cuphea viscosissima FatB1 (Cupvi-TE; номер доступа AEM72522.1; SEQ ID NO: 198) и 12:0-АПБ тиоэстераза Umbellularia californicа (Umbca-TE) (Voelker с соавт., 1992; номер доступа Q41635.1; SEQ ID NO: 199). Все указанные тиоэстеразы принадлежат к классу FATB и обладают специфичность в отношении СЦЖК. Дополнительно были клонированы кодирующие белок участки для LPAAT C. nucifera (Cocnu-LPAAT, тип СЦЖК) (Knutzon с соавт., 1995; номер доступа Q42670.1; SEQ ID NO: 200) и пластидной LPAAT1 A. thaliana (Arath-PLPAAT; номер доступа AEE85783.1; SEQ ID NO: 201). Ранее было показано, что Cocnu-LPAAT повышает инкорпорацию СЦЖК в положении sn-2 ТАГ в семенах (Knutzon с соавт., 1995), тогда как пластидную LPAAT A. thaliana (Arath-PLPAAT) (Kim с соавт., 2004) применяли в качестве контрольной LPAAT, чтобы определить влияние любой специфичности в отношении СЦЖК, которой Cocnu-LPAAT может обладать. Первая LPAAT использует ацил-КоА в качестве одного субстрата и в своем природном окружении функционирует в ЭР, тогда как последняя ПLPAAT использует ацил-АПБ в качестве субстрата и функционирует в пластиде.
Гены тиоэстеразы вводят в листья Nicotiana benthamiana путем опосредованной Agrobacterium инфильтрации, как описано в Примере 1, наряду с геном для соэкспрессии супрессора сайленсинга p19 и Cocnu-LPAAT или Arath-ПLPAAT, чтобы определить, могут ли СЦЖК вырабатываться в листовой ткани N. benthamiana. Инфильтрованные зоны листьев собирают и лиофилизируют в течение пяти дней после инфильтрации со смесями Agrobacterium, после чего определяют общее содержание и состав жирных кислот методом ГХ, как описано в Примере 1 (Таблица 7). Для данных, приведенных в Таблице 7, ошибка представляет стандартное отклонение для трех инфильтраций. Инфильтрованные зоны контрольных листьев содержали только следовые (< 0,1%) или нулевые уровни C12:0 и C14:0 жирных кислот, тогда как C16:0 присутствовала в количестве 14,9%±0,6 в ОЖК общих липидов листа. Уровни C12:0 значительно повышались только при экспрессии Cocnu-TE3 (1,2%±0,1) и Umbca-TE (1,6%±0,1). Экспрессия каждой из изучаемых тиоэстераз приводила к аккумуляции C14:0 в листьях N. benthamiana, причем Cinca-TE давала наиболее высокий уровень, 11,3%±1,0, Кроме того, экспрессия каждой из тиоэстераз, за исключением Umbca-TE, приводит к повышенным уровням C16:0. Наиболее высокий уровень аккумуляции C16:0 (35,4%±4,7) наблюдается при экспрессии Cocnu-TE1. Выраженный некроз инфильтрованных зон наблюдался в листьях при экспрессии только генов FATB, который, похоже, коррелирует с уровнем выработки СЦЖК. Авторы изобретения считают, что некроз, вероятно, возникает за счет уровней свободных жирных кислот (СЖК) выше оптимальных, а также за счет обширной аккумуляции СЦЖК скорее в фосфолипидных пулах липида, чем в ТАГ.
Таблица 7. Состав общих жирных кислот листа (% от общих жирных кислот листа) для выбранных жирных кислот в листьях Nicotiana benthamiana, инфильтрованных различными тиоэстеразами (ТЭ) и LPAAT. Результаты сгруппированы по соинфильтрованому гену (единичные гены (кроме p19, присутствующего во всех образцах), Arath-LPAAT+различные ТЭ, Cocnu-LPAAT+различные ТЭ). «Контроль» означает неинфильтрованный лист N. benthamiana, тогда как «только p19» содержит только ген супрессора сайленсинга. 16:3 представляет собой 16:3Δ7,10,13; 18:3 представляет собой 18:3Δ9,12,15. Гены идентифицированы, как определено в тексте.
Соинфильтрация химерным геном для экспрессии Arath-PLPAAT с тиоэстеразами демонстрирует тенденцию к снижению аккумуляции как C12:0, так и C14:0, по сравнению с отсутствием LPAAT, при небольшом повышении аккумуляции C16:0. И наоборот, соинфильтрация генами для экспрессии Cocnu-LPAAT или Umbca-TE повышает аккумуляцию C12:0 до 3,3%±0,5, тогда как обнаруженная аккумуляция C14:0 в образце Cinca-TE+Cocnu-LPAAT составляет 14,9%±1,6. Самые высокие уровни C16:0 наблюдались после соэкспрессии Cocnu-TE1 и Cocnu-LPAAT (40,2%±2,8). Добавление LPAAT к каждой зоне инокуляции снижает степень некроза листовой ткани. Неожиданно было обнаружено, что C8:0 и C10:0 жирные кислоты также вырабатывались в растительных клетках в экспериментах с временной экспрессией. Аккумуляция C8:0 и C10:0 не наблюдается при экспрессии только тиоэстеразы. Однако, если экспрессия тиоэстеразы сочетается с соэкспрессией CuphoFatB с CnLPAAT и AtWRI1, обнаруживается присутствие C8:0 в концентрации 0,27 ± 0,09% от общего содержания жирных кислот в растительных клетках. Кроме того, в случае соэкспрессии CuplaFatB с CnLPAAT и AtWRI1, было обнаружено присутствие C10:0 в 0,54 ± 0,16% от общего содержания жирных кислот.
Полученные результаты указывают на то, что специфичность тиоэстераз в отношении ацила, о которой сообщалось ранее, наблюдаемая при экспрессии в семени, по существу сохранялась в листьях N. benthamiana, и указанная система экспрессии была валидной системой для оценки специфичности в отношении ацила. Добавление пластидной ПLPAAT A. thaliana не повышает аккумуляции СЦЖК, хотя иприводит к несколько повышенной аккумуляции C16:0 в клетках A. thaliana. И наоборот, LPAAT C. nucifera повышает аккумуляцию в листьях N. benthamiana, жирных кислот C12:0, C14:0 и C16:0, которые найдены в масле C. nucifera (Laureles с соавт., 2002). Это указывает на то, что природная LPAAT N. benthamiana не экспрессируется с высокими уровнями в листовой ткани или не обладает высокой активностью в отношении субстратов C12:0, C14:0 и C16:0.
Выработка среднецепочечных жирных кислот в вегетативных растительных клетках, аккумулирующих высокие уровни ТАГ
Авторами изобретения ранее была достигнута выработка 15% ТАГ в листьях N. tabacum путем координированной экспрессии химерных генов, кодирующих WRI1 A. thaliana, DGAT1 A. thaliana и олеозин S. indicum (Vanhercke с соавт., 2014). Для оценки возможности достижения или повышения аккумуляция СЦЖК, которая наблюдалась после экспрессии тиоэстераз в комбинации с LPAAT, в растительных клетках, вырабатывающих высокие уровни ТАГ (Vanhercke с соавт., 2013), указанные гены соэкспресируют. Осуществляют инфильтрацию комбинациями C12:0, C14:0 и C16:0 тиоэстеразы/LPAAT с наилучшими результатами (Cocnu-LPAAT плюс тиоэстераза Umbca-TE, Cinca-TE и Cocnu-TE2, соответственно), с предварительно описанными комбинациями Arath-WRI1+DGAT и без них (Vanhercke с соавт., 2013). Данные проиллюстрированы на Фиг. 4.
Аккумуляция соответствующих СЦЖК (C12:0 для Umbca-TE, C14:0 для Cinca-TE и C16:0 для Cocnu-TE2) является систематической и значительно повышается, в основном при добавлении Arath-WRI1 к комбинациям: C12:0 составляет 9,5%±0,9 от общих жирных кислот листа в образцах Umbca-TE+Cocnu-LPAAT+Arath-WRI1, уровень C14:0 составляет 18,5%±2,6 в образцах Cinca-TE+Cocnu-LPAAT+Arath-WRI1, и уровень C16:0 составляет 38,3%±3,0 в образцах Cocnu-TE2+Cocnu-LPAAT+Arath-WRI1. Обнаружено, что инфильтрация тиоэстеразой плюс Arath-WRI1 оказывает существенно более выраженное влияние на C12:0 в присутствии Umbca-TE, на C14:0 в присутствии Cinca-TE и на C16:0 в присутствии Cocnu-TE2, по сравнению с инфильтрацией тиоэстеразой плюс Cocnu-LPAAT в отсутствие WRI1 (Фиг. 5). Добавление Cocnu-LPAAT к смесям тиоэстераза плюс Arath-WRI1 оказывало влияние на состав жирных кислот, с относительно небольшим повышением C12:0 и C14:0, наблюдаемом в наборах Umbca-TE и Cinca-TE, и небольшим снижением C16:0 в наборе Cocnu-TE2. Максимальные зарегистрированные уровни составляли: 8,8%±1,1 C12:0 в общих жирных кислотах листа, наблюдаемые в образцах Umbca-TE+Arath-WRI1+Cocnu-LPAAT, 14,1%±3,5 C14:0 в образцах Cinca-TE+Arath-WRI1+Cocnu-LPAAT и 48,6%±3,7 C16:0 в образцах Cocnu-TE2+Arath-WRI1.
Интересно, что единственной тиоэстеразой, для которой Arath-WRI1 не повышает настолько сильно аккумуляцию СЦЖК, является Cocnu-TE2, хотя аккумуляция по-прежнему значительно возрастает. Добавление только указанного гена приводит к повышению аккумуляции C16:0 с 16,0%±0,4 до 37,3%±0,6, тогда как дальнейшее добавление Arath-WRI1 увеличивает данный показатель только на 48,6%±1,7. Это может происходить благодаря промежуточным соединениям C12:0 и C14:0, относительно недолго существующим в ходе пластидного синтеза жирных кислот, по сравнению с C16:0.
Другие эффекты, которые были отмечены, включают повышение уровней C16:0 и C18:1Δ9 и снижение уровня C18:3Δ9,12,15 в присутствии Arath-WRI1. Дальнейшее добавление Cinca-TE и Cocnu-TE2 все еще дополнительно снижает уровни C18:3Δ9,12,15. В противоположность этому, дополнительное количество C12:0 вырабатывается после добавления Arath-WRI1 к Umbca-TE, по-видимому, скорее за счет C16:0, чем дополнительного количества C18:3Δ9,12,15 (Фиг. 5).
Дополнительно, подмножество образцов было проанализировано методом ЖХ-МС, с целью лучшего понимания аккумуляции СЦЖК. Пластидные галактолипиды моногалактозилдиацилглицерин (MGDG) и дигалактозилдиацилглицерин (ДГДГ) содержат только низкие уровни C12:0 и C14:0 и сниженные уровни C16:0, по сравнению с инфильтрацией контрольным p19. Основным видом C12:0-содержащих MGDG в образцах Umbca-TE является 30:3, указывая на то, что одна C18:3 и одна C12:0 были солокализованы на моногалактозильном скелете. Другим основным видом C12:0-содержащих MGDG является 28:0, указывая на то, что второй жирной кислотой является C16:0. Основными видами C14:0-содержащих MGDG в образцах Cinca-TE являются 28:0 и 30:0, указывая на то, что значительная доля C14:0 в MGDG принадлежит ди-C14:0 или C16:0. C12:0-содержащие и C14:0-содержащие MGDG не были обнаружены в контрольной выборке p19. И наоборот, C16:0-содержащие виды MGDG демонстрируют тенденцию к снижению в образцах Cocnu-TE2. Все основные виды MGDG в образцах дикого типа (C16:3-содержащий 34:6, C18:3-содержащий 34:6 и C18:3-содержащий 36:6) демонстрируют тенденцию к снижению при экспрессии трансгенов. Указанное снижение является наиболее выраженным в присутствии комбинации WRI+DGAT.
Только следовые уровни C12:0-содержащих видов ДГДГ наблюдаются в образцах Umbca-TE. Основными C14:0-содержащими видами, наблюдаемыми в образцах Cinca-TE, являются 28:0 и 30:0, оба из которых отсутствуют в контроле. Кроме того, указанные виды наблюдаются с повышенными уровнями в образцах Cocnu-TE2, но только со следовыми уровнями в образцах Umbca-TE. Все основные виды ДГДГ в образцах дикого типа (C16:0-содержащий 34:3, C18:3-содержащий 34:3 и C18:3-содержащий 36:6) демонстрируют тенденцию к снижению при экспрессии трансгенов. Указанное снижение является наиболее выраженным в присутствии WRI.
Подобным образом, содержание видов ТАГ в общем значительно повышено во всех образцах, содержащих WRI+DGAT, как было описано ранее (Vanherckeetal.,2013) (Vanhercke с соавт., 2013). Обнаружено, что виды C12:0 являются преобладающими в высоком образце Umbca-TE ТАГ, виды C14:0 в образце Cinca-TE с высоким содержанием ТАГ, и виды C16:0 в высоком образце Cocnu-TE2 ТАГ. Анализ фракции ТАГ методом ЖХ-МС демонстрирует, что C12:0-содержащий 36:0 является преобладающим видом ТАГ, в два раза превышая уровень видов ТАГ, содержащих C18:3, во всех образцах Umbca-TE, содержащих фактор транскрипции WRI. Кроме того, C14:0-содержащий 42:0, является преобладающим видом ТАГ в образцах Cinca-TE, сотрансформированных LPAAT, DGAT, WRI или WRI+DGAT, хотя ответ значительно более выражен в случае образцов, содержащих WRI. Содержание нескольких C16:0-содержащих видов ТАГ значительно повышено как в образцах Cinca-TE с высоким содержанием ТАГ (например, 44:0 и 50:3), так и в образцах Cocnu-TE2 (например, 46:0, 48:0, 50:2 и 50:3). Повторимся, наиболее выраженное повышение уровня C16:0 наблюдается в присутствии WRI.
Стабильная трансформация для выработки СЦЖК в вегетативных тканях
Получена серия генетических конструкций в бинарном векторе, с целью стабильной трансформации растений, например, табака комбинациями генов для выработки СЦЖК в вегетативных тканях, чтобы идентифицировать оптимальные комбинации генов. Указанные конструкции содержат ген для экспрессии WRI1 под контролем промотора SSU (см. Пример 8, pOIL121) или специфичного для старения промотора SAG12, ген, кодирующий DGAT масличной пальмы (ниже), ген, кодирующий LPAAT кокоса (CocnuLPAAT, см. выше) под контролем промотора enTCUP, и несколько генов, экспрессирующих различные тиоэстеразы жирного ацила (FATB), экспрессируемые промотором 35S или промотора SAG12. Они описаны ниже.
Клонирование гена, кодирующего DGAT Elaeis guineensis (масличная пальма)
Для начальной оценки различных ферментов DGAT, в том числе характерных ферментов DGAT1, DGAT2 и DGAT3, последовательности-кандидаты DGAT масличной пальмы были идентифицированы на основе опубликованной транскриптомы (Dussert с соавт., 2013) и кодон-оптимизированы для экспрессии в Nicotiana tabacum. Далее кодирующие белок участки по отдельности клонируют в бинарные векторы экспрессии под контролем промотора 35S для оценки в экспериментах с временной экспрессией в листьях N. benthamiana, как описано в Примере 1. Изученные комбинации гена являются такими, как указано ниже:
1 P19 (отрицательный контроль)
2 P19+CnLPAAT+WRI1
3 P19+CnLPAAT+AtWRI1+AtDGAT1
4 P19+CnLPAAT+AtWRI1+EgDGAT1
5 P19+CnLPAAT+AtWRI1+EgDGAT2
6 P19+CnLPAAT+AtWRI1+EgDGAT3
7 P19+CincaFatB
8 P19+CincaFatB+CnLPAAT+WRI1
9 P19+CincaFatB+CnLPAAT+AtWRI1+AtDGAT1
10 P19+CincaFatB+CnLPAAT+AtWRI1+EgDGAT1
11 P19+CincaFatB+CnLPAAT+AtWRI1+EgDGAT2
12 P19+CincaFatB+CnLPAAT+AtWRI1+EgDGAT3
Результаты для уровней ОЖК и ТАГ, а также уровней общих СЦЖК в ОЖК или ТАГ проиллюстрированы на Фиг. 6. По сравнению с AtDGAT1, экспрессия EgDGAT1 приводит к более выраженной аккумуляции общих жирных кислот и повышенных уровней ТАГ. Общее содержание СЦЖК в общем содержании жирных кислот снижается при экспрессии EgDGAT1, по сравнению с AtDGAT1, но уровни СЦЖК, присутствующих в ТАГ, остаются такими же (Фиг. 6).
Подготовка генетических конструкций
Генетические конструкции для стабильной трансформации (Таблица 8) собирают посредством последовательной инсерции кассет генов с применением совместимых сайтов рестрикционных ферментов. Четыре генных конструкции (Таблица 8), каждая из которых содержит ген, кодирующий DGAT1 масличной пальмы (EgDGAT1), который экспрессируется промотором 35S, ген, кодирующий LPAAT C. nucifera (CnLFAAT), который экспрессируется из конститутивного промотора enTCUP2, и ген, кодирующий AtWRI1, который экспрессируется промотором SSU или промотора SAG12, в дополнение к одной из серий генов, кодирующих ферменты FATB.
Кроме того, пять генных конструкций содержат ген для экспрессии шпилечной РНК с целью снижения экспрессии эндогенного гена, кодирующего ацил-активизирующий фермент (ААФ). Шпилька сконструирована на основе сходства последовательности с идентифицированным AAE15 Arabidopsis lyrata (EFH44575.1) и геномом N. benthamiana. Показано, что ААФ принимают участие в повторной активации СЦЖК, и, таким образом, дальнейшем удлинении. Предполагалось, что сайленсинг ААФ может повысить аккумуляцию СЦЖК. Кассету шпильки конструируют в векторе pKANNIBAL, а затем субклонируют в вектор экспрессии pWBVec2 (Wang с соавт., 2004), с экспрессией шпильки под контролем промотора 35S.
Таблица 8. Краткое описание собранных генетических конструкций.
Указанные генетические конструкции применяют для получения трансформированных растений табака сортов Висконсин 38 и линии с высоким содержанием масла, трансформированной T-ДНК из pJP3502. Наблюдается, что растения, трансформированные конструкциями с одним геном FATB, экспрессирующимся промотором 35S, были существенно меньше, чем трансформированные соответствующей конструкцией FATB, экспрессирующейся промотором SAG12 или, конструкциями с четырьмя генами. Считалось, что меньший размер растений вызван синтезом СЦЖК, которые не инкорпорировались эффективно в ТАГ.
Обсуждение
В настоящем исследовании обнаружено, что выработка C12:0 в листовых клетках составляет только около 1,6% от общего содержания жирных кислот после экспрессии только Umbca-TE (Таблица 7). Добавление гена для экспрессии Arath-WRI оказывает более выраженное влияние на аккумуляцию C12:0 и C14:0 в листовой ткани, чем добавление LPAAT кокоса (Фиг. 4 и 5). Это показывает, что WRI1 в комбинации с тиоэстеразой значительно повышает аккумуляцию СЦЖК в листовых клетках, действуя синергетически. Важно отметить, что обнаружена аккумуляция большей части C12:0, C14:0 и C16:0 в листьях в форме ТАГ, липида, который не аккумулируется со значительными уровнями в листьях дикого типа. Данные эксперименты продемонстрировали, что клетки в вегетативных частях растений могут быть модифицированы таким образом, чтобы вырабатывать СЦЖК, особенно C12:0 и C14:0, в ТАГ с высокими уровнями. Дополнительно, уровни C16:0 были значительно повышены.
Пример 10, Влияние различных полипептидов фактора транскрипции на аккумуляцию ТАГ
В экспериментах с WRI1 и DGAT, о которых сообщалось ранее (Vanhercke с соавт., 2013), применяли синтетический ген, кодирующий AtWR I1 A. thaliana (номер доступа AAP80382.1), и синтетический ген, кодирующий AtDGAT1, также из A. thaliana (номер доступа AAF19262; SEQ ID NO: 1). Для сравнения других полипептидов WRI с AtWRI1 на предмет их способности объединяться с DGAT для повышения содержания масла, другие последовательности, кодирующие WRI, были идентифицированы и использованы при генерации конструкций для экспрессии в листьях N. benthamiana. Нуклеотидные последовательности, кодирующие факторы транскрипции A. thaliana WRI3 (номер доступа AAM91814.1, SEQ ID NO: 205) и WRI4 (номер доступа NP_178088.2, SEQ ID NO: 206) (To с соавт., 2012), синтезированы и вставлены в виде фрагментов EcoRI в pJP3343 под контролем промотора 35S. Результирующие бинарные векторы экспрессии обозначены pOIL027 и pOIL028, соответственно. Кодирующую последовательность для WRI1 (AsWRI1, SEQ ID NO: 207) овса (Avena sativa) амплифицируют методом ПЦР из вектора, предложенного проф. Sten Stymne (Шведский университет сельскохозяйственных наук), с применением фланкирующих праймеров, содержащих дополнительные сайты EcoRI. Амплифицированный фрагмент вставляют в pJP3343 с получением pOIL055. Последовательность-кандидат WRI1 из S. bicolor (номер доступа XP_002450194.1, SEQ ID NO: 208) идентифицирована поиском BLASTp на сервере Национального центра биотехнологической информации (НЦБИ) с применением аминокислотной последовательности WRI1 Zea mays (номер доступа NP_001137064.1, SEQ ID NO: 209) в качестве запроса. Кодирующий белок участок гена WRI1S. bicolor (SbWRI1) синтезирован и вставлен в виде фрагмента EcoRI в pJP3343, с получением pOIL056. Ген-кандидат, кодирующий WRI1, идентифицирован из транскриптомы китайского сального дерева (Triadica sebifera; TsWRI1, SEQ ID NO: 210) (Uday с соавт., подана). Кодирующий белок участок синтезирован и вставлен в виде фрагмента EcoRI в pJP3343, с получением pOIL070, Бинарные векторы pJP3414 и pJP3352, содержащие кодирующие последовательности для экспрессии. полипептидов WRI1 и DGAT1 A. thaliana являются такими, как описано Vanhercke с соавт. (2013).
Плазмиды, содержащие последовательности, кодирующие различные WRI, вводят в листовую ткань N. benthamiana для временной экспрессии с применением гена, кодирующего белок вирусного супрессора p19 во всех процедурах инокуляции, как описано в Примере 1. Гены, кодирующие полипептиды WRI, оценивают отдельно или в комбинации с геном ацилтрансферазы DGAT1, причем задача последнего состоит в обеспечении более высокого биосинтеза и аккумуляции ТАГ. Положительным контролем в данном эксперименте служит комбинация генов, кодирующих фактор транскрипции WRI1 A. thaliana и AtDGAT1. Все процедуры инфильтрации осуществляют в трех экземплярах с применением трех различных растений, и уровни ТАГ анализируют, как описано в Примере 1. Экспрессия большинства индивидуальных полипептидов WRI в отсутствие экзогенно введенного DGAT1 приводит к повышенным, но все еще низким уровням ТАГ (< 0,23%, в пересчете на сухую массу) в инфильтрованных зонах листа, по сравнению с контролем, который содержал только конструкцию p19 (Фиг. 7). Исключение составляет TsWRI1, который, сам по себе, по-видимому, не повышает значительно уровней ТАГ. Кроме того, отличия в уровнях ТАГ, полученных при экспрессии различных факторов транскрипции WRI, сами по себе не являются большими. Как AsWRI1, так и SbWRI1 давали уровни ТАГ, подобные собственно AtWRI1. Анализ состава жирных кислот ТАГ выявляет только незначительные изменения, за исключением повышенных уровней C18:1Δ9 при экспрессии AtWRI3 в инфильтрованных листовых тканях (Таблица 9).
Таблица 9. Состав жирных кислот ТАГ в образцах листьев N. benthamiana, инфильтрованных различными химерными генами для экспрессии WRI (n=3).
Все образцы дополнительно инфильтрованы конструкцией P19. Образцы ТАГ дополнительно содержат 0,1-0,4% C14:0; 0,5-1,2% C16:3 и; 0,1-0,7% C18:1Δ11.
В противоположность этому, отличия в выходах ТАГ при экспрессии различных полипептидов WRI являются более выраженными при соэкспрессии с ацилтрансферазой AtDGAT1. Это снова демонстрирует синергетический эффект соэкспрессии WRI1 и DGAT на биосинтез ТАГ в инфильтрованной листовой ткани N. benthamiana, как сообщается Vanhercke с соавт. (2013). Промежуточные уровни ТАГ наблюдаются при соэкспрессии DGAT1 с векторами экспрессии AtWRI3, AtWRI4 и TsWRI1, в то время как уровни, полученные с AsWRI1 и AtWRI1, значительно ниже. В качестве результата, который не мог быть предсказан заранее, наиболее высокие выходы ТАГ получены при соэкспрессии DGAT с SbWRI1, даже если эксперимент проводят на клетках двудольных. Анализ состава жирных кислот обнаруживает повышенные уровни ТАГ C18:1Δ 9 и сниженные уровни C18:3Δ 9,12,15 (АЛК) в случае SbWRI1, AsWRI1 и положительного контроля AtWRI1 (Таблица 9). Однако, в отличие от AtWRI1, экспрессия AsWRI1 и SbWRI1 в обоих случаях продемонстрировала повышенные уровни C16:0, по сравнению с отрицательным контролем p19. Интересно, что инфильтрованные AtWRI3 образцы листьев продемонстрировали отличный профиль ТАГ, обогащенный C18:1Δ 9, в то время как C16:0 и АЛК изменения только слегка коснулись.
Данный эксперимент демонстрирует, что фактор транскрипции WRI1 S. bicolor, SbWRI1, превосходит AtWRI1 при соэкспрессии с DGAT с точки зрения повышения уровней ТАГ в вегетативных частях растения. Кроме того, авторами изобретения сделан вывод о том, что фактор транскрипции, например, WRI1, из однодольного растения мог бы надлежащим образом функционировать в клетке двудольного растения, и, в действительности, даже мог бы обладать превосходящей активность, по сравнению с соответствующем фактором транскрипции двудольного растения. Кроме того, фактор транскрипции двудольного растения мог бы надлежащим образом функционировать в клетке однодольного растения.
Применение других факторов транскрипции
Получены генетические конструкции для экспрессии каждого из 14 различных факторов транскрипции в растительных клетках, с целью оценки их способности функционировать в плане повышения уровней ТАГ в комбинации с другими генами, принимающими участие в биосинтезе и аккумуляции ТАГ. Указанные факторы транскрипции были кандидатами в качестве альтернатив WRI1 или для добавления к комбинациям, содержащим один или более из факторов транскрипции WRI1, LEC1 и LEC2, для применения в растительных клетках, особенно в вегетативных частях растения. Их выбор в значительной степени основан на сообщениях об их участии в эмбриогенезе (обзор в Baud и Lepiniec (2010), а также Ikeda с соавт. (2006)), подобно LEC2. Таким образом, проведены эксперименты для оценки их функции с применением системы экспрессии N. benthamiana (Пример 1), как указано ниже.
Нуклеотидные последовательности участков, кодирующих белок для следующих факторов транскрипции кодон-оптимизированы для экспрессии в N. benthamiana и N. tabacum, синтезированы и субклонированы в виде фрагментов NotI-SacI в соответствующие сайты pJP3343: A. thaliana FUS3 (pOIL164) (Luerssen с соавт., 1998; номер доступа AAC35247; SEQ ID NO:160), A. thaliana LEC1L (pOIL165) (Kwong с соавт. 2003; номер доступа AAN15924; SEQ ID NO:157), A. thaliana LEC1 (pOIL166) (Lotan с соавт., 1998; номер доступа AAC39488; SEQ ID NO:149), G. max MYB73 (pOIL167) (Liu с соавт., 2014; номер доступа ABH02868; SEQ ID NO:221), A. thaliana bZIP53 (pOIL168) (Alonso с соавт., 2009; номер доступа AAM14360; SEQ ID NO:222), A. thaliana AGL15 (pOIL169) (Zheng с соавт., 2009; номер доступа NP_196883; SEQ ID NO:223), A. thaliana MYB118 (номер доступа AAS58517; pOIL170; SEQ ID NO:224), MYB115 (Wang с соавт., 2002; номер доступа AAS10103; pOIL171; SEQ ID NO:225), A. thaliana TANMEI (pOIL172) (Yamagishi с соавт., 2005; номер доступа BAE44475; SEQ ID NO:226), A. thaliana WUS (pOIL173) (Laux с соавт., 1996; номер доступа NP_565429; SEQ ID NO:227), A. thaliana BBM (pOIL174) (Boutilier с соавт., 2002; номер доступа AAM33893, SEQ ID NO:145), B. napus GFR2a1 (номер доступа AFB74090; pOIL177; SEQ ID NO:228) и GFR2a2 (номер доступа AFB74089; pOIL178; SEQ ID NO:229) (Liu с соавт. (2012c)). Кроме того, кодон-оптимизированная версия фактора транскрипции PHR1 A. thaliana, принимающего участие в адаптации к условиям фосфатного голодания при высокой освещенности, подобным образом субклонирована в pJP3343 (pOIL189) (Nilsson с соавт. (2012); номер доступа AAN72198; SEQ ID NO: 230). Указанные факторы транскрипции кратко описаны в Таблице 10.
В качестве скринингового теста для определения функции указанных факторов транскрипции, генетические конструкции вводят в клетки листьев N. benthamiana, как описано в Примере 1, с геном, кодирующим DGAT1, или без него, или с другими комбинациями генов, такими как кодирующие WRI1, LEC2, hpSDP1 или тиоэстеразу FATA, и определяют общее содержание липида и состав жирных кислот в клетках листьев. Факторы транскрипции, которые существенно повышают общее содержание липида, идентифицированы и отобраны.
Для стабильной трансформации растений с применением генов, кодирующих альтернативные факторы транскрипции, получены следующие бинарные конструкции. Гены для экспрессии факторов транскрипции используют промотор SSU или промотор SAG12. Показано, что повышенная экспрессия эмбриогенных факторов транскрипции, таких как LEC1 и LEC2, индуцирует различные плеотропные эффекты, нежелательные в настоящем контексте, включая соматический эмбриогенез (Feeney с соавт. (2012); Santos-Mendoza с соавт. (2005); Stone с соавт. (2008); Stone с соавт. (2001); Shen с соавт. (2010)). Для минимизации возможного отрицательного влияния на развитие растений и выход биомассы, специфичные для ткани или стадии развития промоторы являются предпочтительнее конститутивных промоторов с точки зрения управления эктопической экспрессией основных регуляторов эмбриогенеза.
Таблица 10. Дополнительные факторы транскрипции и генетические конструкции для их экспрессии
Пример 11. Сайленсинг ТАГ липазы в растениях, аккумулирующих высокие уровни ТАГ в листовой ткани
Сахарозависимая ТАГ липаза 1 (SDP1) продемонстрировала свою роль в обороте ТАГ в несеменных тканях A. thaliana, а также в ходе прорастания семени (Eastmond с соавт., 2006; Kelly с соавт., 2011; Kelly с соавт., 2013). SDP1 экспрессируется в развивающемся семени, а полипептид SDP1 дополнительно присутствует в зрелом семени в связи с масляными включениями (но не в качестве покрытия). Сайленсинг гена, кодирующего SDP1, приводил к небольшому, но значимому повышению уровней ТАГ в корнях и стеблях A. thaliana (< 0,4%, в пересчете на сухую массу), в то время как еще менее выраженное увеличение наблюдается в листовой ткани (Kelly с соавт., 2013).
Для определения возможности дальнейшего повышения уровней ТАГ в тканях листа и стебля, по сравнению с соэкспрессией AtWRI1 и AtDGAT1, разработан эксперимент с сайленсингом эндогенного гена SDP1 в растениях N. tabacum, которые являются гомозиготными по T-ДНК, содержащей гены для трансгенной экспрессии полипептидов WRI, DGAT1 и олеозина (Vanhercke с соавт., 2014). Поиск BLAST по транскриптоме N. benthamiana (Naim с соавт., 2012) с применением нуклеотидной последовательности AtSDP1 в качестве запроса идентифицировал транскрипт (Nbv5tr6385200, SEQ ID NO: 173) с гомологией в отношении гена SDP1 A. thaliana. Участок (SEQ ID NO: 174) размером 713 пар оснований выбран для опосредованного шпилькой сайленсинга гена. Сконструирован синтетический фрагмент размером 3,903 тыс. пар оснований на основе вектора pHELLSGATE12, который содержит, в приведенном порядке, конститутивный промотор enTCUP2, фрагмент SDP1 N. benthamiana размером 713 пар оснований в смысловой ориентации, фланкированный сайтами attB1 и attB2, интрон Pdk, последовательность интрона cat в обратной ориентации, второй фрагмент SDP1 N. benthamiana размером 713 пар оснований, фланкированный сайтами attB1 и attB2, в обратной (антисмысловой) ориентации, и участок терминатора/сайт полиаденилирования OCS 3' (Фиг. 8). Вставку субклонируют в pJP3303 с применением рестрикционных сайтов SmaI и KasI, причем полученный вектор экспрессии обозначен pOIL051. Данная химерная ДНК содержит ген селекционного маркера, обеспечивающий резистентность к гигромицину.
pOIL051 применяют для получения трансформированных растений N. tabacum путем опосредованной Agrobacterium трансформации. Исходные растительные клетки получают от трансгенных растений, гомозиготных по T-ДНК pJP3502 (Vanhercke с соавт., 2014). Трансгенные растения, содержащие T-ДНК из pOIL051, отбирают по резистентности к гигромицину и переносят в грунт в теплице или в комнату с контролируемой средой для продолжения роста. В ходе развития растения образцы листьев собирают с подтвержденных двойных трансформантов (Растения T0) перед цветением, в период цветения и на стадии завязывания семян, и в каждом определяют уровень ТАГ. Трансгенные растения, содержащие только низкие уровни ТАГ в листьях или содержащие ТАГ на том же уровне, что и контрольные, идентифицируют с помощью экстракции липида из образцов листьев и анализа пятен ТСХ и отбрасывают. Уровни ТАГ в оставшейся популяции трансформантов количественно определяют методом ГХ, как описано в Примере 1. Перед цветением большинство из указанных растений демонстрируют значительно повышенные уровни ТАГ (> 5% от сухой массы листьев) в листовой ткани, в то время как в 4 растениях присутствовали уровни ТАГ выше 10% (Таблица 11). Максимальный уровень ТАГ, наблюдаемый в листьях указанных растений, перед цветением, составил 11,3% в растении 51-13. Для сравнения, трансгенные растения материнской линии N. tabacum, экспрессирующие AtWRI1, AtDGAT1 и олеозин, демонстрирует уровни ТАГ приблизительно 2% перед цветением и около 6% в ходе цветения (Vanhercke с соавт., 2014). Таким образом, добавление SDP1-ингибирующей конструкции к комбинации AtWRI1 плюс AtDGAT1 является синергетическим с точки зрения повышения уровней ТАГ в этих растениях. Неожиданно было обнаружено, что содержание ТАГ в листьях, собранных с двойных трансформированных растений в стадии цветения, было значительно повышено, составляя 30,5%, в пересчете на сухую массу (Таблица 12), что представляет собой увеличение в 5 раз относительно растений без сайленсинга SDP1. К большому удивлению авторов изобретения, уровень ТАГ достиг потрясающего значения 70,7% (% от сухой массы) в образцах стареющих листьев (зеленых и желтых) на стадии завязывания семян (Таблица 13). Применение ЯМР для измерения содержания масла в цельных листьях растений табака на стадии завязывания семян, содержание ТАГ в некоторых зеленых листьях, начавших стареть, составило около 43%, а в некоторых коричневых, дегидратированных листьях составило 42%. Если такие листья зажать между двумя коричневыми бумажными фильтрами, выделяющееся масло впитывается в бумагу и делает ее просвечивающейся, тогда как для листьев контрольного табака этого не наблюдалось, что предлагает простой способ скрининга для обнаружения растений с высоким содержанием масла.
Два первичных трансформанта (№61, №69), каждый из содержал Т-ДНК из pJP3502 и pOIL51 и демонстрировал высокие уровни ТАГ, проанализированы цифровой капельной ПЦР (цкПЦР) с применением специфичной для гена гигромицина пары праймеров, с целью определения количества инсерций T-ДНК pOIL51. Растение, обозначенное №61, содержит одну инсерцию T-ДНК из pOIL51, тогда как растение №69 содержит три инсерции T-ДНК из pOIL51. Проведен повторный скрининг растений-потомков T1 обеих линий методом цкПЦР с целью идентификации гомозиготных, гетерозиготных и нулевых растений. Растения-потомки растения №61, не содержащие инсерций из pOIL51 (нулевые; всего 7) или 2 инсерции T-ДНК (т.е., гомозиготные по указанной T-ДНК; всего 12) отобраны для дальнейшего анализа. Кроме того, растения-потомки линии №69, содержащие инсерции нулевые T-ДНК от pOIL51 (нуль; всего 2) или 2 такие инсерции (всего 15) или 4 или 5 инсерций (всего 5) сохраняли для дальнейшего анализа.
Таблица 11. Уровни ТАГ и состав жирных кислот ТАГ (% от сухой массы листьев) в листовой ткани растений N. tabacum (Поколение T0), экспрессирующих трансгены WRI1, DGAT1 и олеозина и супертрансформированных T-ДНК, кодирующей конструкцию шпильки SDP1 (pOIL051), по сравнению с диким типом (нетрансформированный). Образцы листьев собраны в течение вегетативной стадии (перед цветением). Образцы липида дополнительно содержали 0,0-0,2% C16:3, 0,0-0,4% C20:1; 0,0-0,1% C20:2n-6.
Таблица 12. Уровни ТАГ и состав ТАГ (% от сухой массы листа) в листовой ткани растений N. tabacum (Поколение T0), экспрессирующих трансгены WRI1, DGAT1 и олеозина и супертрансформированных T-ДНК, кодирующей конструкцию шпильки SDP1 (pOIL051). Образцы листьев собраны в период цветения.
Таблица 13. Уровни ТАГ и состав ТАГ (% от сухой массы листа) в листовой ткани растений N. tabacum (Поколение T0), экспрессирующих трансгены WRI1, DGAT1 и олеозина и супертрансформированных T-ДНК, кодирующей конструкцию шпильки SDP1 (pOIL051). Образцы листьев собраны в стадии завязывания семян. Y=желтый лист, G =зеленый лист.
Отобранные растения T1 выращены в теплице в то же время и при тех же условиях, что и контрольные растения. Образцы ткани зеленых листьев растений T1, собранные перед цветением, сушат и определяют содержание общих жирных кислот (ОЖК) и ТАГ методом ГХ. Содержание ОЖК в растениях, содержащих обе Т-ДНК, варьирует от 4,6% до 16,1%, в пересчете на сухую массу, в том числе уровни ТАГ в тех же листьях от 1,2% до 11,8%, в пересчете на сухую массу (Фиг. 9). Это намного больше, чем в растениях, содержащих только T-ДНК из pJP3502, выращенных бок-о-бок в таких же условиях и проанализированных на такой же стадии роста, что снова демонстрирует синергию между снижением активности ТАГ липазы и комбинации WRI1 плюс DGAT. В растениях, содержащих только T-ДНК pJP3502, присутствует от 4,2% до 6,8% ОЖК, в том числе уровни ТАГ от 1,4% до 4,1%, в пересчете на сухую массу (Фиг. 9). Содержание в растениях дикого типа в среднем составляет около 0,8% ОЖК, в том числе менее чем 0,5% ТАГ, в пересчете на сухую массу. Состав жирных кислот в общем содержании жирных кислот и содержание ТАГ в листьях каждой из линий, №61 и №69, является сходным с их составом в листьях, содержащих только T-ДНК из pJP3502 (материнский). По сравнению с контрольными листьями дикого типа, растения, содержащие Т-ДНК из pOIL51 и pJP3502, демонстрируют повышенные уровни C16:0, C18:1 и C18:2 жирных кислот. Такой существенный сдвиг состава жирных кислот в значительной степени происходит за счет C18:3, содержание которой снижается с около 50-55% до около 20-30%, как процент от общего содержания жирных кислот.
Значительное повышение уровней ОЖК, включая уровни ТАГ, между растениями, содержащими только T-ДНК pJP3502, и растениями, содержащими Т-ДНК из pOIL51 и pJP3502, сохраняется на протяжении развития растения. Контрольные растения, содержащие только T-ДНК из pJP3502, содержат от 7,7% до 17,5% ТАГ в ходе цветения, в то время как в период завязывания семян уровни ТАГ варьируют от 14,1% до 20,7%, в пересчете на сухую массу. Содержание ТАГ в листьях растений, содержащих Т-ДНК pJP3502 и pOIL51, варьирует от 6,3% до 23,3% в период цветения и от 12,6% до 33,6% в период завязывания семян. Подобные изменения состава жирных кислот во фракции ТАГ для обеих стадий были обнаружены, как было описано ранее для стадии вегетативного роста.
Кроме того, найдены повышенные уровни ТАГ и в других вегетативных тканях трансгенных растений, таких как корни и стебель. Некоторые ткани корня трансгенных растений N. tabacum, трансформированных T-ДНК pOIL051, содержат 4,4% ТАГ, а некоторые ткани стебля содержат 7,4% ТАГ, в пересчете на сухую массу (Фиг. 10). Растения дикого типа и N. tabacum, содержащие только T-ДНК из pJP3502, демонстрируют намного более низкие уровни ТАГ в обоих видах тканей. Добавление конструкции шпильки SDP1 с целью снижения экспрессии эндогенной липазы ТАГ оказывает очевидное синергетическое влияние с генами, кодирующими фактор транскрипции и биосинтез ТАГ (WRI1 и DGAT), с точки зрения повышения содержания ТАГ в стеблях и корнях. Необходимо отметить, что более низкие уровни ТАГ в корнях сравнимы с тканями стебля в пределах одного и того же растения, в то время как в растениях дикого типа и N. tabacum, содержащие только T-ДНК из pJP3502 наблюдается обратная тенденция. Состав ТАГ в тканях корня и стебля демонстрирует изменения в содержании C18:1 и C18:3 жирных кислот, подобные наблюдаемым ранее в трансгенной листовой ткани. Уровни C18:2 в ТАГ снижены в трансгенной ткани стебля, в то время как содержание C16 жирных кислот обычно снижено в трансгенных тканях корня, по сравнению с контролем дикого типа.
Таким образом, авторами изобретения сделан вывод о том, что добавление экзогенного гена к комбинации WRI1 и DGAT с целью сайленсинга эндогенного гена SDP1 повышает общее содержание жирных кислот, в том числе содержание ТАГ, на всех стадиях роста растения, и оказывает синергетическое влияние с WRI1 и DGAT, особенно в стеблях и корнях. Семена T1 трансгенных растений помещают на питательные среды для тканей in vitro при комнатной температуре, чтобы проверить степень и расчет времени прорастания. Прорастание семени T1 трех независимо трансформированных линий по-прежнему сравнивают с семенем трансгенных растений, трансформированных T-ДНК из pJP3502. Кроме того, ранняя сила саженца, по-видимому, остается интактной. Это было неожиданным, с учетом роли SDP1 в прорастании семени A. thaliana и наблюдаемых дефекты при прорастании мутантов SDP1 (Eastmond с соавт., 2006). Для преодоления любых дефектов прорастания, если они возникнут, разработана вторая конструкция, в которой ингибирующая SDP1 РНК экспрессируется промотором, который в семени по существу не экспрессируется или экспрессируется с низкими уровнями, например, такого как промотор из фотосинтетического гена, например, SSU. Авторы изобретения считают предпочтительным снизить риск вредного влияния на прорастание семени или раннюю силу саженца, чтобы избежать конститутивного промотора для контроля экспрессии РНК, ингибирующей SDP1, или по меньшей мере избежать промотора, экспрессирующегося в семенах.
Необходимо отметить, что растения T0 с наиболее высокими уровнями ТАГ выращены в условиях высокой освещенности в комнате с контролируемой средой (интенсивность света 500 микромоль, световой цикл 16 часов света/26°C-8 часов темноты/18°C), и они выглядели меньше (около 70% высоты относительно растений, трансформированных T-ДНК из pJP3502), чем контрольные растения дикого типа. Авторами изобретения сделан вывод о том, что комбинация трансгенов и/или генетических модификаций для подходов ʺpushʺ, ʺpullʺ, ʺprotectʺ и ʺpackageʺ особенно благоприятна для достижения высоких уровней ТАГ в вегетативных частях растения. В данном примере, WRI1 обеспечивает ʺpushʺ, DGAT обеспечивает ʺpullʺ, сайленсинг SDP1 обеспечивает ʺprotectʺ, и олеозин обеспечивает ʺpackageʺ ТАГ.
Пример 12. Специфическая для старения экспрессия фактора транскрипции
Сообщалось, что эктопическая экспрессия основных регуляторов развития эмбриона и семени, например LEC2, повышает уровни ТАГ в несеменных тканях (Santos-Mendoza с соавт., 2005; Slocombe с соавт., 2009; Andrianov с соавт., 2010). Однако, повышенная экспрессия конститутивного LEC2 в растениях, трансформированных геном 35S-LEC2, приводит к нежелательному плеотропному влиянию на развитие и морфологию растения, включая соматический эмбриогенез и аномальные структура листа (Stone с соавт., 2001; Santos-Mendoza с соавт., 2005). Для оценки того, будет ограничение экспрессии LEC2 стадией старения листьев в ходе развития растения, т.е. после того, как растения полностью выросли и достигли полной биомассы, минимизировать нежелательные фенотипические эффекты, но по-прежнему увеличивать уровни липида в листьях, разработана и получена химерная ДНК для экспрессии LEC2 под контролем специфичного для старения промотора A. thaliana из гена SAG12 (U37336; Gan и Amasino, 1995).
Для получения генетической конструкции, получен синтетический фрагмент ДНК размером 3,635 тыс. пар оснований, содержащий, в указанном порядке, специфичный для старения промотор SAG12 A. thaliana, последовательность, кодирующую белок LEC2 и участок терминатора/полиаденилирования гена лектина Glycine max. Указанный фрагмент вставляют между рестрикционными сайтами SacI и NotI pJP3303. Данная конструкция обозначена как pOIL049 и оценена в листьях растения N. tabacum, стабильно трансформированных генами, кодирующими полипептиды WRI1, DGAT1 и олеозина, содержащими T-ДНК из pJP3502. С применением опосредованных Agrobacterium способов трансформации, конструкцию pOIL049 используют для трансформации клеток растений N. tabacum, гомозиготных по T-ДНК pJP3502 (Пример 3). Трансгенные растения, содержащие гены из pOIL049, отбирают на основе резистентности к гигромицину и выращивают в теплице до стадии зрелости. Образцы ткани трансгенных листьев получены на разных стадиях роста, включая старение листьев, и содержат повышенные уровни ТАГ, по сравнению с материнской линией N. tabacum pJP3502.
Всего получено 149 независимых растений T0 (т.е. первичных трансформантов). Cобраны образцы верхних зеленых листьев всех растений и нижних коричневых, полностью постаревших листьев выбранных событий на стадию завязывания семян в процессе развития растения, и содержание ТАГ количественно определено с помощью ТСХ-ГХ. Количество инсерций T-ДНК pOIL49 в отобранных растениях определено методом цкПЦР, с применением пары праймеров, специфичной в отношении гена гигромицина. Уровень ТАГ 30,2%, в пересчете на сухую массу, наблюдается в ткани зеленых листьев, собранной на стадии завязывания семян. Уровни ТАГ в коричневых листьях ниже в большинстве растений, от которых получены образцы. Однако, три растения (№32b, №8b и №23c) продемонстрировали более высокие уровни ТАГ в ткани постаревших коричневых листьев, чем в зеленых растущих листьях. Указанные растения содержали 1, 2 или 3 инсерции T-ДНК из pOIL49.
Проведен скрининг потомства T1 растений №23c и №32b методом цкПЦР с целью идентификации нулевых, гетерозиготных и гомозиготных по T-ДНК из pOIL049 растений. Растения-потомки растения №23c, содержащего нуль инсерций T-ДНК из pOIL049 (нулевые; всего 7) или две T-ДНК инсерции T-ДНК из pOIL049 (гомозиготные; всего 4) отобран для дальнейшего анализа. Кроме того, растения-потомки растения №32b, содержащие нуль инсерций (нулевые; всего 6) или две инсерции (гомозиготные; всего 9) сохранены для дальнейшего анализа. Образцы ткани зеленых листьев получены перед цветением, и содержание ОЖК и ТАГ определено методом ГХ. Растения дикого типа и растения, трансформированные T-ДНК из pJP3502, были такими же как описано ранее (Пример 11), и были выращен бок-о-бок в одной теплице. Уровни ОЖК перед цветением в листьях трансформантов, содержащих T-ДНК из pOIL049, варьируют от 5,2% до 19,5%, в пересчете на сухую массу (Фиг. 12). Уровни ТАГ в этих же тканях варьируют от 0,8% до 15,4%, в пересчете на сухую массу. Это значительно больше, чем в растениях, содержащих только T-ДНК из pJP3502. Содержание ТАГ в растениях, содержащих Т-ДНК из pJP3502 и pOIL049, дополнительно повышается до уровня от 38,5% до 34,9% в период цветения и завязывания семян, соответственно. Когда состав жирных кислот в общем содержании жирных кислот был проанализирован для листьев, гомозиготных по T-ДНК из pOIL049, наблюдались повышенные уровни C18:2 и сниженные уровни C18:3 (Фиг. 12), в то время как процентное содержание C16:0 и C18:1 остается приблизительно таким же, как и в листьях, трансформированных только T-ДНК из pJP3502. Указанные данные демонстрируют, что добавление второго гена фактора транскрипции под контролем неконститутивного промотора с целью обеспечения регулируемой развитием экспрессии может дополнительно повышать уровни ТАГ в вегетативных тканях растения. Кроме того, данные указывают на некоторую степень экспрессии специфичного для старения промотора SAG12 в зеленой ткани до старения листьев.
Уровни ТАГ значительно повышены в ткани стебля, по сравнению с растениями N. tabacum дикого типа и трансгенными растениями, содержащими только T-ДНК из pJP3502. Некоторые ткани стебля трансгенных растений N. tabacum, трансформированные T-ДНК из pOIL049, содержат 4,9% ТАГ, в пересчете на сухую массу (Фиг. 11). С другой стороны, уровни ТАГ в ткани корня демонстрируют более выраженную вариацию между тремя растениями pOIL049, причем некоторые ткани корня содержат 3,4% ТАГ. Необходимо отметить, что более низкие уровни ТАГ в корнях были сравнимыми с тканью стебля в пределах того же растения, в то время как обратная тенденция наблюдалась в растениях дикого типа и N. tabacum, содержащих только T-ДНК из pJP3502. Состав ТАГ в тканях корня и стебля демонстрирует изменения в содержании C18:1 и C18:3 жирных кислот, подобные наблюдаемым ранее в трансгенной листовой ткани. Уровни C18:2 в ТАГ снижены в трансгенной ткани стебля, в то время как содержание C16 жирных кислот обычно снижено в трансгенных тканях корня, по сравнению с контролем дикого типа.
Получены соответствующие генетические конструкции, кодирующие другие факторы транскрипции под контролем промотора SAG12, а именно LEC1, LEC1like, FUS3, ABI3, ABI4, ABI5 и другие (Пример 10). Например, получены дополнительные конструкции для экспрессии фактора транскрипции однодольных LEC1 Zea mays (Shen с соавт., 2010) или LEC1 Sorghum bicolor (номер доступа Genbank XM_002452582.1) под контролем полученного из однодольных гомолога промотора SAG12 A. thaliana, такого как промотор SEE1 маиса (Robson с соавт., 2004). Дальнейшие конструкции получены для экспрессии факторов транскрипции под контролем регулируемых развитием промоторов, например, таких, которые предпочтительно экспрессируются в период цветения (например, чувствительные к длительности светового дня промоторы), фитохромных промоторов, криптохромных промоторов, или в стеблях растения в ходе вторичного роста, таких как промотор из гена CesA. Указанные конструкции применяются для трансформации растений, причем отбираются растения, которые вырабатывают по меньшей мере 8% ТАГ в вегетативных частях.
Уровни крахмала и сахаров
Уровни крахмала и растворимых сахаров измеряют в листовой ткани, образцы которой получены от растений дикого типа и трансгенных растений, содержащих T-ДНК из pJP3502 или Т-ДНК из pJP3502 и pOIL51 или pJP3502 и pOIL049. В общем, в листьях, содержащих обе Т-ДНК, найдена обратная корреляция между уровнями ТАГ и крахмала в листовой ткани, в пересчете на сухую массу (Фиг. 13). И наоборот, в трансгенных растениях уровни растворимых сахаров в листьях являются таким же, как и в растениях дикого типа, наводя на мысль об отсутствии значимого узкого места в превращении сахаров в ТАГ. Влияние положения листьев в растении наблюдается в растениях дикого типа, в которых уровни крахмала демонстрируют тенденцию к повышению от более низкого к более высокому положению листьев. В трансгенных растениях обнаружено отсутствие такого эффекта.
Пример 13. Специфическая для стебля экспрессия гена, кодирующего фактор транскрипции
Листья растений N. tabacum, экспрессирующих трансгены, которые кодируют WRI1, DGAT и олеозин, на стадии завязывания семян содержат около 16% ТАГ. Однако, уровни ТАГ намного ниже в стеблях (1%) и корнях (1,4%) растений (Vanhercke с соавт., 2014). Авторами изобретения было оценено, являются ли более низкие уровни ТАГ в стеблях и корнях результатом слабой промоторной активности промотора SSU Rubisco, применяемого для экспрессии гена, кодирующего WRI1 в трансгенных растениях. Трансген DGAT в T-ДНК pJP3502 экспрессируется промотором CaMV35S, который более активно экспрессируется в стеблях и корнях, и таким образом он вряд ли является ограничивающим фактором для аккумуляции ТАГ в стеблях и корнях.
В попытке увеличить биосинтез ТАГ в ткани стебля, разработана конструкция, в которой ген, кодирующий WRI1, размещен под контролем. промотора SDP1 A. thaliana. Синтезирован синтетический фрагмент ДНК размером 3,156 тыс. пар оснований, содержащий промотор SDP1 A. thaliana размером 1,5 тыс. пар оснований (SEQ ID NO: 175) (Kelly с соавт., 2013), и далее участок, кодирующий полипептид WRI1 A. thaliana, и участок терминатора/полиаденилирования лектина G. max. Указанный фрагмент вставлен между сайтами SacI и NotI pJP3303. Результирующий вектор обозначен как pOIL050, и в дальнейшем применяется для трансформации клеток растений N. tabacum, гомозиготных по T-ДНК из pJP3502 путем опосредованной Agrobacterium трансформации. Трансгенные растения отобраны на основе резистентности к гигромицину, причем всего 86 независимых трансгенных растений выращены в теплице до стадии зрелости. Получены образцы ткани трансгенного листа и стебля на стадии завязывания семян, которые содержат более высокие уровни ТАГ, чем материнские растения N. tabacum, трансформированные pJP3502.
Пример 14. Повышение содержания масла в семенах
Несколько групп сообщали о повышенных уровнях ТАГ в ткани семени маиса, рапса или Arabidopsis thaliana при повышенной экспрессии индивидуальных генов, кодирующих WRI1 и DGAT1 (Shen с соавт., 2010; Liu с соавт., 2010; Weselake с соавт., 2008; Jako с соавт., 2001; обзор в Liu с соавт., 2012). Недавно, van Erp с соавт. (2014) исследовали влияние соэкспрессии WRI1 и DGAT1 на содержание масла семян в A. thaliana. Абсолютные уровни ТАГ, повышаются с 38% в растениях дикого типа и контроле с пустым вектором до 44% в трансгенных линиях. Сайленсинг липазы ТАГ SDP1 в комбинации с повышенной экспрессией WRI1 и DGAT1 повышал уровни ТАГ до 45%. Необходимо отметить, что, хотя обнаружено увеличение средней массы семян, однако количество семян на растении было ниже, чем в контрольных растениях.
Синтетический фрагмент ДНК размером приблизительно 14,3 тыс. пар оснований, содержащий открытые рамки считывания, кодирующие MGAT2 M. musculus, DGAT1 A. thaliana, WRI1 A. thaliana и полипептиды GPAT4 A. thaliana под контролем специфичных для семени промоторов из генов, кодирующих FAE1, конлинин 1 и конлинин 2, синтезирован и вставлен в виде фрагмента Not I-PstI в pJP3416. Результирующий вектор обозначен как pTV55 (Фиг. 14). Серии производных векторов сконструированы из pTV55 последовательным удалением индивидуальных кассет экспрессии, причем на каждой стадии применялось расщепление рестрикционным ферментом, с последующим самолигированием. Кассету GPAT4 удаляют расщеплением PacI, с получением pTV56. Последующее расщепление SrfI удаляет кассету экспрессии MGAT2, с образованием pTV57. Кассету WRI1 удаляют с применением фланкирующих рестрикционных сайтов SbfI, с получением pTV58. В конце, кассету DGAT1 в pTV58 обменивают на кассету WRI1 путем расщепления Srf I и PacI, с последующей обработкой T4 ДНК полимеразой и лигированием. Кассету WRI1 вырезают из pTV57 с применением SbfI и обрабатывают T4 полимеразой. Лигирование кассеты WRI с тупыми концами в расщепленный SrfI-PacI скелет pTV58 дает pTV59. Каждый вектор содержит e35S (содержащий удвоенный участок энхансера)::ген ФАТ в качестве гена селекционного маркера, обеспечивая резистентность к BASTA.
Вкратце, конструкции содержат следующие комбинации генов:
pTV55: ProCnl1::MGAT2+ProCnl2::DGAT1+ProCnl1::GPAT4+ProFAE1::WRI1;
pTV56: ProCnl1::MGAT2+ProCnl2::DGAT1+ProFAE1::WRI1;
pTV57: ProCnl2::DGAT1+ProFAE1::WRI1;
pTV58; ProCnl2::DGAT1;
pTV59: ProFAE1::WRI1.
Конструкции pTV55-pTV59 по отдельности вводят в штамм AGL1 A. tumefaciens и применяют для трансформации C. sativa (сорт Celine) методом погружения цветков, адаптированным из Liu и другие (2012). Вкратце, недавно открытые цветочные почки погружают в. раствор A. tumefaciens на 15 с, заворачивают в пленку из пластического материала и оставляют на ночь в темноте при 24°C, после чего пластический материал удаляют. Всего выполняют 3-4 погружения цветков, на основании качества доступных цветков. Растения выращивают до уровня зрелости и собирают урожай семян T1. После прорастания семени T1 на грунте, укоренившиеся саженцы T1 (7-10 дней) обрызгивают 0,1% гербицидом BASTA (250 г/л аммония глюфосината; Crop Science Pty Ltd, Виктория, Австралия) для отбора для растений, экспрессирующих ген селекционного маркера ФАТ. Выжившие саженцы отделяют, переносят в свежие горшки с грунтом и выращивают до состояния зрелости в теплице при 22±1°C (день) и 18±1°C (ночь). Урожай семян T2 собирают, и содержание масла (которое по меньшей мере на 95% состоит из ТАГ) для 3 независимых партий семян по 50 мг от каждой линии измеряют методом ЯМР (MQC, Oxford Instruments). Калибровочные образцы готовят с сухой бумагой Kimwipes, содержащей известные количества масла семян C. sativa, в пробирках для ЯМР диаметром 10 мм, чтобы генерировать диапазон содержания масла на основании массы бумаги и масла. Калибровочные образцы опечатывают парафильмом и перед использованием выдерживают при 40°C в нагревательном блоке по меньшей мере в течение 1 часа, чтобы позволить маслу равномерно распределиться в ткани. Калибровочные образцы измеряют трижды с применением зонда с магнитом на 0,55 Тесла диаметром 10 мм, функционирующим на частоте резонанса протона 23,4 МГц в течение 16 секунд. Температуру магнита поддерживают на уровне 40°C. Образцы семян вначале сушат в печи при температуре 105°C на протяжении ночи, чтобы гарантировать содержание влажности менее чем 5%. Затем образцы взвешивают, переносят в стеклянную пробирку диаметром 10 мм и инкубируют при 40°C в течение 1 часа перед анализом методом ЯМР. Содержание масла измеряют методом ЯМР в трех экземплярах с калибровкой на основании массовой компенсации.
Количество вставленных копий T-ДНК в каждой трансформированной линии определяют цифровой ПЦР (цПЦР). Вначале геномную ДНК расщепляют с помощью EcoRI и BamHI, чтобы гарантировать физическое разделение Т-ДНК в случае множественных инсерций. Ген LEAFY C. sativa (lfy) выбран в качестве референтного гена, и селекционный маркер выбран в качестве гена-мишени в реакции мультиплексной цПЦР. Зонды метят с помощью HEX (референтного гена) или FAM (гена-мишени). Условия амплификации были следующими: 95°C в течение 10 минут (повышение со скоростью 2,5°C/с), 39 циклов при 94°C в течение 30 секунд (повышение со скоростью 2,5°C/с) и при 61°C в течение 1 минуты (повышение со скоростью 2,5°C/с), 98°C в течение 10 минут. После ПЦР амплификации, флуоресценцию отдельных капелек измеряют с помощью устройства для считывания капелек QX100 (BioRad), и количество копий вычисляют с применением программного обеспечения QuantaSoft (версия 1.3.2.0, BioRad).
Трансформация C. sativa дает множественные трансгенные события, демонстрирующие повышенные уровни ТАГ в сегрегированных семенах T2, по сравнению с нетрансформированным контролем дикого типа (Фиг. 15). Интересно, что наиболее высокие уровни ТАГ получены с pTV57, который содержит гены, кодирующие WRI1 и DGAT1. Дополнительная инсерция MGAT2 (pTV56) и MGAT2+GPAT4 (pTV55) приводит к несколько более низким уровням ТАГ, по сравнению с pTV55. Определение количества копий выявило 1 или 2 инсерции T-ДНК для линий pTV57, демонстрирующих второе по величине и самое высокое содержание масла, соответственно (Таблица 14).
Гомозиготные растения T2, трансформированные T-ДНК из pTV057, дополнительно были скрещены с растениями C. sativa, трансформированными генами для экспрессии десатураз и элонгаз жирных кислот, необходимых для синтеза и аккумуляции ДГК в семени (WO2013/185184). Семена F1 демонстрируют повышенное содержание масла, по данным ЯМР, по сравнению с семенами C. sativa, трансформированными конструкцией ДГК и без T-ДНК pTV57. Содержание ДГК в семенах определяют путем измерения уровней ДГК во фракции ТАГ, по сравнению с содержанием ДГК C. sativa в материнском растении. Общее содержание ДГК в семенах (мг ДГК/г семян), содержащих обе Т-ДНК, повышено относительно общего содержания ДГК в семенах, содержащих только конструкцию ДГК.
Таблица 14. Содержание масла (%) в семени T2 и количество копий (цифровая ПЦР) в трансгенных событиях pTV57 C. sativa.
Пример 15. Влияние экспрессии белка масляных включений на аккумуляцию и оборот ТАГ
Растения N. tabacum, трансформированные T-ДНК pJP3502 и экспрессирующие трансгены, которые кодируют WRI1, DGAT1 A. thaliana и олеозин S. indicum, содержат повышенные уровни ТАГ в вегетативных тканях. Как проиллюстрировано в Примере 11 выше, при сайленсинге эндогенного гена, кодирующего липазу ТАГ SDP1, в таких растениях, уровни ТАГ в листьях дополнительно повышаются, что указало авторам изобретения на тот факт, что происходит значительный оборот ТАГ в растениях, которые сохранили активность SDP1. Таким образом, был определен уровень экспрессии трансгенов в растениях. Хотя гибридизация методом Нозерн-блоттинга подтвердила выраженную экспрессию WRI1 и DGAT1 и некоторую экспрессию мРНК олеозина, анализ экспрессии методом цифровой ПЦР и кОТ-ПЦР обнаружил только очень низкие уровни транскриптов олеозина. Анализ экспрессии выявил, что ген, кодирующий олеозин, экспрессируется слабо, по сравнению с трансгенами WRI1 и DGAT1. На основании указанных экспериментов, авторами изобретения был сделан вывод о том, что масляные включения в листовой ткани не были полностью защищены от расщепления ТАГ из-за ненадлежащей выработки белка олеозина, кодируемого T-ДНК в pJP3502. Для повышения стабильности аккумуляции ТАГ в процессе развития растения, были разработаны несколько модификаций pJP3502, в которых ген олеозина был заменен. Указанные модифицированные конструкции были такими, как указано ниже.
1. pJP3502 содержит ген (в SEQ ID NO: 176 приведена комплементарная ему последовательность), кодирующий олеозин S. indicum, который слабо экспрессировался. Данный ген содержит внутренний интрон UBQ10, который может снижать уровень экспрессии. Для проверки этого, синтетический фрагмент ДНК размером 502 пары оснований, содержащий ген олеозина S. indicum и не содержащий внутреннего интрона UBQ10, был синтезирован и вставлен в pJP3502 в виде фрагмента NotI, заменяя в pJP3502 ген олеозина, содержащий интрон. Результирующая плазмида обозначена как pOIL040.
2. Промотор маленькой субъединицы (SSU) Rubisco, управляющий экспрессией гена олеозина в pJP3502, был заменен конститутивным промотором enTCUP2. На данный момент, фрагмент размером 2321 пар оснований, содержащий промотор enTCUP2, кодирующий белок олеозин участок, участок терминатора/полиаденилирования лектина G. max и первые 643 пары оснований в направлении по ходу транскрипции промотора SSU, управляющего экспрессией wri1, синтезирован и субклонирован в сайты AscI и SpeI pJP3502, с получением pOIL038.
3. Подобной стратегией следовали для экспрессии сконструированной версии гена олеозина S. indicum, содержащего 6 введенных остатков цистеина (o3-3) под контролем промотора enTCUP2 (Winichayakul с соавт., 2013). Фрагмент размером 2298 пар оснований, содержащий промотор enTCUP2, кодирующий белок олеозин участок o3-3, участок терминатора/полиаденилирования лектина G. max и первые 643 пары оснований в направлении по ходу транскрипции промотора SSU, управляющего экспрессией wri1, синтезирован и субклонирован в сайты AscI и SpeI pJP3502, с получением pOIL037.
4. Сайты NotI, фланкирующие ген олеозина S. indicum в pJP3502, применяются для обмена кодирующего белок участка на участок, кодирующий олеозин 3 арахиса (номер доступа AAU21501.1) (Parthibane с соавт., 2012a; Parthibane с соавт., 2012b). Фрагмент размером 528 пар оснований, содержащий ген олеозина 3, фланкированный сайтами NotI, синтезирован и субклонирован в соответствующий сайт pJP3502. Результирующий вектор обозначен как pOIL041
5. Дополнительно, фланкированный NotI фрагмент размером 1077 пар оснований, который содержит ген, кодирующий стеролеозин A. thaliana (Arab-1) (номер доступа AAM10215.1) (Jolivet с соавт., 2014), синтезирован и субклонирован в сайт NotI pJP3502, с получением pOIL043.
6. Поверхностный белок липидных капелек Nannochloropsis oceanic (ПБЛК) (номер доступа AFB75402.1) (Vieler с соавт., 2012) синтезирован в виде NotI-фланкированного фрагмента размером 504 пары оснований и субклонирован в сайт NotI pJP3502, с получением pOIL044.
7. И наконец, калеозин A. thaliana (CLO3) (номер доступа O22788.1) (Shimada с соавт., 2014) синтезирован в виде фланкированного NotI фрагмента размером 612 пар оснований и субклонирован в pJP3502, с получением pOIL042.
Каждую из этих конструкций вводят в клетки листьев N. benthamiana, как описано в Примере 1. Временная экспрессия как pJP3502, так и pOIL040 в листовой ткани N. benthamiana приводит к повышенным уровням ТАГ и подобным изменениям профиля жирных кислот в ТАГ, однако pOIL040 повышает уровень ТАГ в большей степени (1,3% по сравнению с 0,9%). Каждая из конструкций pOIL037, pOIL038, pOIL041, pOIL042 и pOIL043 применяется для стабильной трансформации растений N. tabacum (сорт W38) с помощью опосредованных Agrobacterium способов. Трансгенные растения отбирают на основании резистентности к канамицину и выращивают в теплице до состояния зрелости. На различных стадиях развития растения отбирают образцы трансгенной листовой ткани, которые содержат повышенные уровни ТАГ, по сравнению с N. tabacum дикого типа и растениями N. tabacum, трансформированными pJP3502.
Клонирование и характеристика полипептидов БСЛК из Sapium sebifera
Олеозины не экспрессируются с высокими уровнями в несеменных тканях растения, аккумулирующих масло, таких как мезокарп оливы, масличной пальмы и авокадо (Murphy, 2012). Вместо этого, в данных тканях идентифицированы белки, связанные с липидными капельками (БСЛК), которые могут играть роль, подобную роли олеозина в тканях семени (Horn с соавт., 2013). Таким образом, авторы изобретения рассмотрели возможность того, что олеозин не является оптимальным упаковочным белком для защиты аккумулированного масла от липазы ТАГ или другого влияния цитозольных ферментов в вегетативных тканях растений. Таким образом, полипептиды БСЛК были идентифицированы и оценены на предмет повышения аккумуляции ТАГ, как указано ниже.
Плод китайского сального дерева Sapium sebifera, члена семейства Euphorbiaceae, представляет особый интерес для авторов изобретения, поскольку он содержит богатую маслом ткань за пределами семени. Недавнее исследование (Divi с соавт., подано для публикации) показало, что указанная маслянистая ткань под названием жировой слой может происходить из мезокарпа плода растения. Таким образом, авторами изобретения подвергнута сомнению транскриптома S. sebifera из-за последовательностей БСЛК. Сравнительный анализ экспрессированных генов в тканях оболочки фрукта и семени выявил группу из трех ранее идентифицированных генов БСЛК, которые экспрессировались с высокими уровнями в жировом слое.
Нуклеотидные последовательности, кодирующие три БСЛК, получены методом ОТ-ПЦР с применением РНК, происходящих из жировой ткани, и трех пар праймеров. Последовательности праймеров основаны на последовательностях ДНК, фланкирующих полный кодирующий участок для каждого из трех генов. Последовательности праймера: для БСЛК1, 5'-TTTTAACGATATCCGCTAAAGG-3' (SEQ ID NO: 245) и 5'-AATGAATGAACAAGAATTAAGTC-3' (SEQ ID NO: 246) AT-3'; БСЛК2, 5'-CTTTTCTCACACCGTATCTCCG-3' (SEQ ID NO: 247) и 5'-AGCATGATATA CTTGTCGAGAAAGC-3' (SEQ ID NO: 248); БСЛК3, 5'-GCGACAGTGTAGCGTTTT-3' (SEQ ID NO: 249) и 5'-ATACATAAAATGAAAACTATTGTGC-3' (SEQ ID NO: 250).
Анализ транскриптомы S. sebifera обнаружил многочисленные ортологи для каждого из генов БСЛК, в том числе восемь БСЛК1, шесть БСЛК2 и шесть БСЛК3 генов, с менее чем 10% дивергенцией последовательностей в пределах каждого семейства гена. Предполагаемые последовательности пептида выровнены, и филогенетическое дерево сконструировано с применением программного обеспечения Genious (Фиг. 16), вместе с гомологами БСЛК из других видов растений, включая два из авокадо (Pam), один из масличной пальмы, один из Parthenium argentatum (Par), два из Arabidopsis (Ath), пять из Taraxacum brevicorniculatum (Tbr), три из Hevea brasiliensis (Hbr), как проиллюстрировано на Фиг. 16. Филогенетическое дерево выявило, что SsLDAP3 разделяет большую степень идентичности аминокислотных последовательностей с полипептидами БСЛК1 и БСЛК2 из авокадо и БСЛК из масличной пальмы, в то время как полипептиды SsLDAP1 и SsLDAP2 были более дивергентными.
Генетические конструкции для повышенной экспрессии БСЛК
Для оценки функции БСЛК из S. sebifera получены векторы экспрессии для экспрессии каждого из указанных полипептидов под контролем промотора 35S в клетках листа. Полноразмерные последовательности кДНК SsLDAP вставляют в вектор назначения pDONR207 посредством реакций рекомбинации, заменяя участки CcdB и Cm(R) вектора назначения фрагментами кДНК SsLDAP. Последующее подтверждение методом анализа рестрикционного расщепления и секвенирования ДНК, конструкции вводят в штамм AGL1 Agrobacterium tumefaciens и применяют для временной экспрессии в клетках листьев N. benthamiana и стабильной трансформации N. tabacum.
Экспрессия каждого из трех генов SsLDAP под транскрипциональным контролем промотора 35S в листьях N. benthamiana в комбинации с экспрессией 35S::AtDGAT1 и 35S::AtWRI1 дает значительно более высокие уровни аккумуляции ТАГ, по сравнению с клетками, инфильтрованными генами 35S::AtDGAT1 и 35S::AtWRI1 без конструкции БСЛК. Уровень ТАГ повышается приблизительно в 2 раза, по сравнению с уровнем ТАГ в контрольных клетках. В клетках, получающих комбинацию генов, наблюдается значительное повышение уровня α-линоленовой кислоты (АЛК) и снижение уровня насыщенных жирных кислот, относительно контрольных клеток.
Солокализация ЖФБ-химерных полипептидов БСЛК с капельками липида в клетках листа
С целью характеристики SsLDAP in vivo и наблюдения за их поведением в динамике, получены экспрессионные конструкции для экспрессии химерных полипептидов, состоящих из полипептидов БСЛК, слитых с желтым флуоресцентным белком (ЖФБ). Для каждого химерного полипептида, ЖФБ был слит в рамке с C-концом SsLDAP. Полная открытая рамка считывания каждого из трех генов БСЛК, без стоп-кодона на 3' конце, слита с последовательностью ЖФБ и химерными генами, вставленными в pDONR207. После подтверждения полученных конструкций методом рестрикционного расщепления и секвенирования ДНК, в конструкции вводят в штамм AGL1 A. tumefaciens и применяют для временной экспрессии в клетках листа N. benthamiana и стабильной трансформации N. tabacum. Через три дня после инфильтрации клеток листа конструкциями БСЛК-ЖФБ, листовые диски из инфильтрованных областей окрашивают Красным Нилом, который положительно окрашивает липидные капельки, и визуализируют под однофокусным микроскопом, чтобы обнаружить красное окрашивание (капельки липида) и флуоресценцию полипептида ЖФБ. Наблюдается солокализация БСЛК-ЖФБ с липидными капельками, указывая на связь БСЛК с липидными капельками в клетках листа.
Пример 16. Модификация жирных кислот в различных пулах липида в листьях, аккумулирующих высокие уровни ТАГ
Авторами изобретения описана выработка повышенных уровней ТАГ в N. tabacum путем соэкспрессии трансгенов, кодирующих WRI1 A. thaliana, DGAT1 A. thaliana и олеозин S. indicum (Vanhercke и другие, 2014). Для изучения возможности совмещения модификаций жирных кислот в различных пулах липида, которые существуют в листьях, с соэкспрессией комбинации гена WRI1 и DGAT1, выполнялись эксперименты с временной экспрессией, чтобы посмотреть, могут ли встречаться жирные кислоты в пулах ацил-КоА и ацил-ФХ. В одном эксперименте экспрессию трансгена, кодирующего элонгазу жирных кислот A. thaliana (AtFae1), которая удлиняет C18:1-КоА до C20:1-КоА, сочетали с генами, кодирующими WRI1 и DGAT, с целью оценки модификаций в пуле ацил-КоА. Во втором эксперименте трансген, кодирующий десатуразу 2 жирных кислот A. thaliana (AtFAD2), которая преобразует C18:1-ФХ в C18:2-ФХ посредством введения кратной связи, сочетали с генами WRI1 и DGAT, с целью оценки модификаций в пуле ФХ. Указанные эксперименты разработаны для оценки доступности ацильных субстратов в ЭР растительных клеток.
Ген, кодирующий AtFAE1, экспрессируется промотором CaMV35S в листьях N. benthamiana, отдельно или в комбинации с WRI1 и DGAT1, как описано в Примере 1, в трех экземплярах. Образцы листьев собирают через 5 дней после инфильтрации клетками Agrobacteriu, содержащими различные комбинации генов. Общий липид экстрагируют из образцов листьев, и в каждом случае фракцию ТАГ отделяют с помощью ТСХ. Определяют состав жирных кислот каждой фракции ТАГ и проводят количественное определение методом ГХ с применением известного количества C17:0-ТАГ в качестве внутреннего стандарта. Как проиллюстрировано в Таблице 15, доля C20:1 в ТАГ значительно повышается при экспрессии AtAFE1. Кроме того, соэкспрессия WRI1 и DGAT1 с AtFAE1 повышает уровень продукта C20:1, по сравнению с контролем, в то время как общее количество ТАГ повышается с 0,8 до 14,9 мкг/мг листа. Продукт C20:1 в ТАГ аккумулируется с таким высоким содержанием, как 0,96 мкг на мг, по сравнению с 0,04 мкг на мг без комбинации WRI1 и DGAT1.
Таблица 15. Модифицированные уровни жирных кислот после трансгенной экспрессии модифицирующих ферментов в листьях N. benthamiana.
Дополнительно, AtFAD2 соэкспрессируют промотором CaMV35S в листьях N. benthamiana, отдельно или в комбинации с WRI1 и DGAT1. Определяют состав жирных кислот фракции ТАГ, и проводят количественное определение, как описано выше. Уровень жирной кислоты C18:2 в ТАГ значительно повышается при соэкспрессии AtFAD2 с WRI1 и DGAT1, с 10,7% до 37,9%, а уровень C18:1 снижается с 19,5% до 7,6%. Кроме того, наблюдаются значительные уровни ТАГ (13,4 мкг/мг листа) при соэкспрессии WRI+DGAT1+AtFAD2 в листьях. Полученные результаты явно демонстрируют, что состав и количество жирных кислот могут быть модифицированы добавлением ферментов модификации жирных кислот в комбинации по меньшей мере с WRI1 и DGAT, и таким образом их можно применять для повышения аккумуляции модифицированных жирнокислотных продуктов, генерируемых в одном или обоих пулах ацил-КоА и ФХ, и в конечном счете, хранения в ТАГ в вегетативных частях растения.
Пример 17. Сайленсинг генов ТГД в растениях
Согласно оценкам Li-Beisson с соавт. (2013), в листьях Arabidopsis (растения 16:3) приблизительно 40% жирных кислот, синтезированных в хлоропластах, поступают в прокариотный путь, тогда как 60% экспортируются для поступления в эукариотный путь. После того десатурации в ЭР, около половины указанных экспортированных жирных кислот возвращается в пластиду, чтобы обеспечить синтез галактолипидов для тилакоидных мембран. В транспорте (импорт) жирных кислот в пластиду, в виде ДАГ или фосфолипидов, принимает участие TGD1, пермеаза-подобный белок внутреннего хлоропластного конверта. Переносчик липидов ABC Arabidopsis, содержащий белки TGD1, 2 и 3, идентифицирован Benning с соавт. (2008 и 2009), и позже Roston с соавт. (2012). Указанный комплекс белка локализован во внутренней мембране хлоропластного конверта, и предполагается, что он опосредует перенос фосфатидата через указанную мембрану. Полипептид TGD2 представляет собой фосфатидил-связывающий белок, и TGD3 представляет собой АТФазу. Новый белок Arabidopsis, TGD4, идентифицирован с помощью генетического подхода (Xu с соавт., 2008), причем инактивация гена TGD4 также блокирует перенос липида из ЭР в пластиды. Последние биохимические данные указывают на то, что TGD4 является фосфатидат-связывающим белком, расположенным во внешней мембране хлоропластного конверта (Wang и Benning, 2012).
Xu с соавт. (2005) описали неплотные аллели tgd1 в A. thaliana, приводящие к снижению роста растения и высокой частоте абортов эмбриона. Листовая ткань мутантов tgd1 A. thaliana содержит повышенные уровни ТАГ, вероятно в форме цитозольных масляных капелек. Кроме того, повышенные уровни ТАГ найдены в корнях мутантов tgd1. Не было обнаружено отличий в содержании масла в семенах. О подобной аккумуляции ТАГ в листовой ткани сообщалось для мутантов A. thaliana tgd2 (Awai с соавт., 2006), tgd3 (Lu с соавт., 2007) и tgd4 (Xu с соавт., 2008). Все мутантные аллели tgd были достаточно неплотными или значительно ослабляли развитие растения.
Сайленсинг TGD1
Конструкция сайленсинга, направленная против пластидного импортера TGD1, сгенерирована на основании полноразмерного транскрипта мРНК, идентифицированного в транскриптоме N. benthamiana. Фрагмент размером 685 пар оснований амплифицируют из кДНК листа N. benthamiana при инкорпорации в сайт PmlI на 5' конце. Вначале фрагмент TGD1 клонируют в pENTR/D-TOPO (Invitrogen), и затем вставляют в вектор назначения pHELLSGATE12 посредством клонирования LR (Gateway). Полученный вектор экспрессии, обозначенный как pOIL025, временно экспрессируют в N. benthamiana, чтобы оценить влияние сайленсинга гена TGD1 на уровни ТАГ в листьях. Конструкцию шпильки TGD1 размещают под контролем индуцибельного промотора alcA A. niger путем субклонирования в виде фрагмента Pml I-EcoRV в сайты NheI (Кленова)-SfoI pOIL020 (ниже). Полученным вектором, обозначенным как pOIL026, супертрансформируют гомозиготную линию N. tabacum pJP3502 с целью дополнительного повышения уровней масла в листьях.
Получены дополнительные конструкции для экспрессии шпилечной РНК с целью снижения экспрессии генов TGD-2, -3 и -4. С применением указанных конструкций получены трансформированные растения, и в трансформантах определено содержание масла. Трансформированные растения скрещены с трансформантами, полученными с применением pJP3502 или других комбинаций генов, как изложено выше.
Пример 18. Экспрессия комбинаций генов в картофельных клубнях
Конструкция pJP3506
Генетическая конструкция, содержащая три гена для экспрессии в картофельных клубнях, получена и применяется для трансформации картофеля. Указанная конструкция обозначена как pJP3506 и базируется на существующем векторе pJP3502 (WO2013/096993) с заменой промоторов, чтобы обеспечить специфическую для клубня экспрессию. pJP3506 содержит (i) ген резистентности к канамицину NPTII под управлением промотора 35S с удвоенным участком энхансера (e35S), в качестве гена селекционного маркера, и три экспрессионные кассеты генов, которые представляют собой (ii) 35S::AtDGAT1, кодирующую DGAT1 Arabidopsis thaliana, (iii) B33::AtWRI1, кодирующую WRI1 Arabidopsis thaliana, и (iv) B33::олеозин кунжута, кодирующую олеозин из Sesame indicum. Нуклеотидные последовательности, кодирующие указанные полипептиды, являются такими же, как в pJP3502. Промотор пататина B33 (B33) представляет собой специфичный для клубня промотор, полученный из Solanum tuberosum, который был предложен д-ром Alisdair Fernie, Институт молекулярной физиологии растений им. Макса Планка, Потсдам, Германия. Круговая карта плазмиды pJP3506 представлена на Фиг. 17.
Последовательность промотора пататина B33 S. tuberosum, применяемая в конструкции pJP3506, представляет собой укороченную версию, в которой 183 нуклеотида удалены на 5' конце и 261 нуклеотид удален на 3' конце, по сравнению с номером доступа GenBank X14483. Нуклеотидная последовательность промотора пататина B33, применяемого в pJP3506, приведена в SEQ ID NO: 211.
Трансформация картофеля
Саженцы картофеля (Solanum tuberosum), сорт Atlantic, выращенные в культуре ткани, в асептических условиях, приобретены у Toolangi Elite, сертифицированного органа семенного картофеля в Виктории (СОСКВА), Австралия. Междоузлия стебля разрезают на куски длиной приблизительно 1 см под суспензией штамма LBA4404 Agrobacterium tumefaciens, содержащего pJP3506. Клетки Agrobacterium выращивают до OD 0,2 и разбавляют равным объемом среды МС. Избыток суспензии Agrobacterium удаляют кратковременным промоканием куском стебля стерильной фильтровальной бумагой, с последующим переносом на среду МС и инкубированием при 24°C в течение двух дней (сокультивирование). Затем междоузлия переносят на свежую среду МС с добавлением 200 мкг/л НУК, 2 мг/л бензиламинопурина (БАП) и 250 мг/л цефотаксима. Селекцию трансгенных каллюсов начинают через 10 дней, после переноса междоузлий на свежую среду МС с добавлением 2 мг/л БАП, 5 мг/л GA3, 50 мг/л канамицина и 250 мг/л цефотаксима. Побеги, регенерирующие из каллюсов, вырезают и помещают на простую среду МС для индукции корней, перед пересадкой в горшок диаметра 15 см, содержащий почвенную смесь и выращиванием в теплице до состояния зрелости растения, включая рост клубня.
Экстракция ДНК и молекулярная идентификация трансгенных растений методом ПЦР
Диски диаметром около 1 см получают из листьев картофеля от растений в теплице. Их помещают в планшет для микротитрования с глубокими лунками и лиофилизируют в течение 48 часов. Далее лиофилизированные образцы листьев измельчают в порошок, добавляя стальной шарик в каждую лунку, и встряхивают планшет в лизаторе для тканей Reicht (Qiagen) с максимальной частотой 28/с в течение 2 минут для каждой стороны планшета для микротитрования. 375 мкл экстракционного буфера, содержащего 0,1 М Трис-HCl, pH 8,0, 0,05 М ЭДТА и 1,25% натрия додецилсульфата (НДС) добавляют в каждую лунку, содержащую порошкообразную листовую ткань. Через 1 час инкубации при 65°C, 187 мкл 6 М ацетата аммония добавляют в каждую лунку, и смеси выдерживают при 4°C в течение 30 минут перед центрифугированием планшетов на протяжении 30 минут при 3000 об/мин. 340 мкл супернатанта из каждой лунки переносят в новый планшет для микротитрования с глубокими лунками, содержащий 220 мкл изопропанола и выдерживают в течение 5 минут при комнатной температуре перед центрифугированием при 3000 об/мин на протяжении 30 минут. Осажденные гранулы ДНК промывают 70% этанолом, сушат на воздухе и ресуспендируют в 225 мкл H2O на образец.
По два мкл каждого препарата ДНК образцов листьев добавляют к 20 мкл реакционной смеси ПЦР с помощью системы ПЦР HotStar (Qiagen). Пару олигонуклеотидных праймеров, основанных на 5' и 3' последовательностях из гена WRI1 Arabidopsis thaliana, кодон-оптимизированного для табака, применяют в реакциях ПЦР. Последовательности праймеров: Nt-Wri-P3: 5'-CACTCGTGCTTTCCATCATC-3' (SEQ ID NO: 212) и Nt-Wri-P1: 5'-GAAGGCTGAGCAACAAGAGG-3'(SEQ ID NO: 213). Кроме того, пару олигонуклеотидных праймеров, основанных на гене DGAT1 Arabidopsis thaliana, кодон-оптимизированном для табака, применяют в отдельной реакции ПЦР для каждого образца ДНК. Последовательности праймеров: Nt-DGAT-P2: 5'-GGCGATTTTGGATTCTGC-3' (SEQ ID NO: 214) и Nt-DGAT-P3: 5'-CCCAACCCTTCCGTATACAT-3' (SEQ ID NO: 215). Амплификацию проводят с начальным циклом при 95°C в течение 15 минут, за которым следуют 40 циклов при 95°C в течение 30 секунд, при 57° C в течение 30 с и при 72°C в течение 60 секунд. Продукты ПЦР обрабатывают электрофорезом на 1% агарозном геле для обнаружения специфичных продуктов амплификации
Анализ липида картофельных клубней
Тонкие ломтики клубней, собранные с регенерированных картофельных растений для подтвержденных трансгенных растений и нетрансформированного контроля, лиофилизируют в течение 72 часов, и анализируют содержание и состав липида. Общие липиды экстрагируют из сухих тканей клубня с применением хлороформа: метанола: 0,1 М KCl (2:1:1 об/об/об), как указано ниже. Лиофилизированные ткани клубня вначале гомогенизируют в хлороформе: метаноле (2:1, об/об) в пробирке Эппендорфа, содержащей металлический шар, с помощью лизатора для тканей Reicht (Qiagen) в течение 3 минут при частоте 29/с. После смешивания каждого гомогената с Vibramax 10 (Heidolph) при 2000 об/мин в течение 15 минут, к каждому образцу добавляют 1/3 объема 0,1 М раствора KCl и дополнительно перемешивают. После центрифугирования при 10000 g в течение 5 минут, из каждого образца отбирают нижнюю фракцию, содержащую липиды, и полностью выпаривают в потоке N2. Каждый препарат липида растворяют в 3 мкл CHCl3 на миллиграмм сухой массы клубня. Аликвоты препаратов липида наносят на пластину для тонкослойной хроматографии (ТСХ) (20 см x 20 см, силикагель 60, Merck) и элюируют гексаном: диэтиловым эфиром: уксусной кислотой (70:30:1, об/об/об). Пластину ТСХ обрызгивают примулином и визуализируют в УФ-свете, чтобы проявить пятна липида. ТАГ и ФХ извлекают, соскабливая кремния диоксид с подходящих полос, и превращают в метиловые эфиры жирных кислот (МЭЖК), инкубируя материал в 1 н метанольной HCl (Supelco, Беллефонт, Пенсильвания) при 80°C в течение 2 часов, вместе с известным количеством тригептадеканоина (Nu-Chek PREP, Inc., США) в качестве внутреннего стандарта для количественного определения липида. МЭЖК анализируют на приборе ГХ-ПИД (7890A ГХ, Agilent Technologies, Пало Альто, Калифорния), оборудованном колонкой BPX70 длиной 30 м (внутренний диаметр 0,25 мм, толщина пленки 0,25 мм, SGE, Остин, США), как было описано ранее (Petrie с соавт., 2012). Пики интегрируют с помощью программного обеспечения Agilent Technologies ChemStation (версия B.04.03).
Среди приблизительно 100 регенерированных индивидуальных трансгенных линий, анализ липидов, полученных из молодых картофельных клубней диаметром приблизительно 2 см обнаружил повышенные уровни общих липидов, ТАГ и фракций фосфолипидов в клубнях многих трансгенных растений, причем наблюдается диапазон от отсутствия повышения до значительного повышения. Первый анализ липидов картофельного клубня показал, что типичный картофельный клубень дикого типа в ранней стадии развития (диаметр около 2 см) содержит около 0,03% ТАГ, в пересчете на сухую массу.
Содержание общих липидов было повышено до 0,5-4,7% масс. (сухая масса) в клубнях 21 индивидуального трансгенного растения, представляющего 16 независимо трансформированных линий (Таблица 16). Клубни линии №69 продемонстрировали наиболее высокую аккумуляцию ТАГ, в среднем 3,3%, в пересчете на сухую массу. Это соответствует повышению приблизительно в 100 раз относительно клубней дикого типа в такой же стадии развития. Дополнительно, клубни этой же трансгенной линии аккумулировали наиболее высокие из наблюдаемые уровней фосфолипидов, 1,0% масс. в молодых клубнях, в пересчете на сухую массу (Таблица 18). Кроме того, повышенная аккумуляция липида сопровождалась модификацией состава жирных кислот в трансгенных клубнях. Трансгенные клубни систематически аккумулировали более высокие проценты насыщенных и мононенасыщенных жирных кислот (МНЖК) и более низкие уровни полиненасыщенных жирных кислот (ПНЖК) в общем содержании жирных кислот и во фракции ТАГ общих жирных кислот (Таблица 17), конкретно, сниженный уровень 18:3 (АЛК), который был уменьшен с около 17% для дикого типа до менее чем 10% в трансгенных клубнях. Уровень олеиновой кислоты (18:1) в общем содержании жирных кислот повышался с около 1% для дикого типа до более чем 5% во многих линиях и более чем 15% в некоторых клубнях. Хотя уровни пальмитиновой кислоты были повышены, уровни стеариновой кислоты (18:0) были сниженные в лучших трансгенных линиях (Таблица 16 и 17).
Трансгенные картофельные растения содержали в теплице, чтобы позволить непрерывный рост клубней. Клубни большего размера линии №69 содержали более высокие уровни ОЖК и ТАГ, чем клубни диаметром приблизительно 2 см.
Дополнительно повышенные уровни ОЖК и ТАГ получены в картофельных клубнях путем добавления химерного гена, кодирующего сайленсинговую РНК для снижения экспрессия эндогенного гена SDP1, в комбинации с генами WRI1 и DGAT.
Дальнейшие комбинации генов для трансформации картофеля
Общую РНК из свежих, развивающихся картофельных (Solanum tuberosum L., сорт Atlantic) клубней экстрагируют способом TRIzol (Invitrogen). Выбранные участки кДНК, кодирующие маленькую субъединицу АГФазы картофеля и SDP1, получены с помощью ОТ-ПЦР с применением следующих праймеров: st-AGPs1: 5'-ACAGACATGTCTAGACCCAGATG-3' (SEQ ID NO: 251), st-AGPa1: 5'-CACTCTCATCCCAAGTGAAGTTGC-3' (SEQ ID NO: 252); st-SDP1-s1: 5'-CTGAGATGGAAGTGAAGCACAGATG-3' (SEQ ID NO: 253), и st-SDP1-a1: 5'-CCATTGTTAGTCCTTTCAGTC-3' (SEQ ID NO: 254). Далее продукты ПЦР очищают и лигируют с pGEMT Easy.
Таблица 16. Общий выход липида (% масс. от сухой массы картофельного клубня) и жирнокислотный состав в характерных молодых клубнях картофеля, трансформированных T-ДНК pJP3506, до цветения растений. Клубни линии 65 были эквивалентны клубням дикого типа (нетрансгенным).
d11
Таблица 17. Выход ТАГ (% масс. от сухой массы картофельного клубня) и его жирнокислотный состав в характерных молодых клубнях картофеля, трансформированных T-ДНК pJP3506, до цветения растений.
d11
Таблица 18. Выход фосфолипидов (% масс. от сухой массы картофельного клубня) и их жирнокислотный состав в характерных молодых клубнях картофеля, трансформированных T-ДНК pJP3506, до цветения.
d11
После верификации секвенированием ДНК, клонированные продукты ПЦР непосредственно применяют в качестве последовательности гена-мишени, с целью получения конструкции шпилечной РНКи, или сливают посредством ПЦР с перекрывающимися праймерами. Три фрагмента ПЦР (SDP1, АГФаза, СЗБ+АГФ) в дальнейшем клонируют в вектор pKannibal, который содержит специфические рестрикционные сайты, для клонирования желательного фрагмента в смысловой и антисмысловой ориентации. Выбранные рестрикционные сайты представляют собой BamHI и HindIII для клонирования фрагмента в смысловой ориентации и KpnI и XhoI для вставки фрагмента в антисмысловой ориентации. Наборы праймеров, применяемые для амплификации трех фрагментов гена-мишени, модифицируют добавлением рестрикционных сайтов, которые направляют фрагмент в клонирующие сайты pKannibal. Экспрессионные кассеты, содержащие фрагмент-мишень ДНК между промотором 35S и терминатором OCS в pKannibal, высвобождают с помощью Not1 и клонируют в бинарный вектор pWBVec2, с гигромицином в качестве селекционного маркера для растений. Такие бинарные векторы вводят в штамм AGL1 A. tumefaciens и применяют для трансформации картофеля, как изложено выше.
Пример 19. Повышение содержания масла в однодольных растениях
Экспрессия в эндосперме
Содержание масла в эндосперме однодольных видов растений Triticum aestivum (пшеница) и, таким образом, в зерне растений, повышают экспрессию комбинации генов, кодирующих WRI1, DGAT и олеозин, в эндосперме в ходе развития семени с применением специфичных для эндоспермы промоторов. Конструкция (обозначенная как pOIL-Endo2) содержит химерные гены: (a) промотор гена Glu1 Brachypodium distachyon::кодирующий белок участок гена Zea mays, кодирующего полипептид ZmWRI1 (SEQ ID NO: 35)::участок терминатора/полиаденилирования из гена лектина Glycine max, (b) промотор гена глютенина Bx17 Triticum aestivum::кодирующий белок участок гена A. thaliana, кодирующего полипептид AtDGAT1 (SEQ ID NO: 1)::участок терминатора/полиаденилирования из гена Nos Agrobacterium tumefaciens, (c) промотор гена GluB4 Oryza sativa::кодирующий белок участок гена Sesame indicum, кодирующего участок полипептида олеозина::участок терминатора/полиаденилирования из гена лектина Glycine max и (d) промотор 35S::участок, кодирующий резистентность к гигромицину, в качестве гена селекционного маркера. Конструкцию применяют для трансформации незрелых эмбрионов T. aestivum (сорт Fielder) путем опосредованной Agrobacterium трансформации. Инокулированные незрелые эмбрионы подвергают воздействию гигромицина, чтобы отобрать трансформированные побеги, а затем переносят в среду корнеобразования для формирования корней, перед переносом в грунт.
Получены тридцать трансформированных растений, на которых завязалось семя T1 и которые содержат T-ДНК из pOIL-Endo2. Урожай зрелых семян собран со всех 30 растений, и 6 семян каждого семейства разрезаны пополам. Половинки, содержащие эмбрион, хранили для последующего проращивания; другую половинку, содержащую по большей части эндосперм, экстрагировали и оценивали содержание масла. T-ДНК, вставленная в геном пшеницы, все еще сегрегируется в семенах T1 указанных растений, поэтому семена T1 представляют собой смесь гомозиготных трансформированных, гетерозиготных трансформированных и нулевых по T-ДНК. Повышенное содержание масла наблюдается в эндосперме некоторых из семян, причем некоторые семена демонстрируют повышение уровней ТАГ более, чем в 5 раз. В половинках с эндоспермом шести семян (сорт Fielder) дикого типа содержание ТАГ составляет около 0,47% масс. (диапазон от 0,37% до 0,60%), по сравнению с содержанием ТАГ 2,5% в некоторых зернах. В некоторых семействах все шесть семян содержат свыше 1,7% ТАГ; другие очевидно сегрегированы между диким типом и повышенным содержанием ТАГ. Кроме того, в эндосперме с повышенным содержанием ТАГ состав жирных кислот модифицирован, демонстрируя повышение процентного содержания олеиновой кислоты и пальмитиновой кислоты и снижение процентного содержания линолевой кислоты (Таблица 19). Семя T1 беспрепятственно прорастало с той же скоростью, что и семя соответствующего дикого типа, причем растения, представляющие индивидуумы как с высоким содержанием масла, так и с низким содержанием масла из 14 семейств T0 выращены до состояния зрелости. Указанные растения обладали полноценной мужской и женской фертильностью.
Зерно пригодно для получения пищевых продуктов для потребления человеком, или в качестве корма для животных, обеспечивая зерно с повышенным содержанием энергии на единицу массы (плотность энергии) и давая повышенные темпы прироста у животных, таких как, например, домашняя птица, свиньи, крупный рогатый скот, овцы и лошади.
Таблица 19. Состав жирных кислот (% от общих жирных кислот) в ТАГ и общее содержание ТАГ (% масс. масла в половинке с эндоспермом) в трансгенном эндосперме пшеницы
Дополнительно, конструкцию pOIL-Endo2 применяют для трансформации кукурузы (Zea mays) и риса (Oryza sativa), с целью получения трансгенных растений с повышенным содержанием ТАГ в эндосперме и, таким образом, в зерне.
Экспрессия в листьях и стеблях
Серия бинарных векторов экспрессии разработана для опосредованной Agrobacterium трансформации сорго (S. bicolor) и пшеницы (Triticum aestivum), с целью повышения содержания масла в вегетативных тканях. Исходными векторами для конструкций являются pOIL093-095, pOIL134 и pOIL100-104 (см. Пример 4). Вначале, фрагмент ДНК, кодирующий опосредованной WRI1 Z. mays, амплифицируют методом ПЦР с применением pOIL104 в качестве шаблона и праймеров, содержащих рестрикционные сайты KpnI. Указанный фрагмент субклонируют в направлении по ходу транскрипции относительно конститутивного промотора Actin1 Oryza sativa в pOIL095, используя сайт KpnI. Результирующий вектор обозначен как pOIL154. Фрагмент ДНК, кодирующий DGAT2a Umbelopsis ramanniana под контролем промотора убихитина Z. mays (pZmUbi), выделяют из pOIL134 в виде фрагмента NotI и вставляют в сайт NotI pOIL154, с получением pOIL155. Экспрессионную кассету, состоящую из кодирующего ФАТ участка под контролем промотора pZmUbi, фланкированного на 3' конце участком терминатора/полиаденилирования NOS A. tumefaciens, конструируют амплификацией кодирующего ФАТ участка с применением pJP3416 в качестве шаблона. Разработаны праймеры для инкорпорации рестрикционных сайтов BamHI и SacI на 5' и 3' конце, соответственно. После двойного расщепления BamHI+SacI, фрагмент ФАТ клонируют в соответствующие сайты pZLUbi1casNK. Полученное промежуточное соединение обозначено как pOIL141. Далее, кассету селекционного маркера ФАТ вводят в скелет pOIL155. В этой точке pOIL141 вначале разрезают с помощью NotI, фрагмента Кленова ДНК полимеразы I с тупыми концами, и далее расщепляют с помощью AscI. Затем полученный фрагмент размером 2622 пар оснований субклонируют в сайты ZraI-AscI pOIL155, с получением pOIL156. В конце, промотор Actin1, управляющий экспрессией WRI1 pOIL156, обменивают на промотор маленькой субъединицы Rubisco Z. mays (pZmSSU), с получением pOIL157. Данный вектор получен ПЦР амплификацией промотора SSU Z. mays, с применением pOIL104 в качестве шаблона и фланкирующих праймеров, содержащих рестрикционные сайты Asi SI и PmlI. Далее полученный ампликон разрезают с помощью SpeI+MluI и субклонируют в соответствующие сайты pOIL156.
Указанные векторы таким образом содержат следующие кассеты экспрессии:
pOIL156: промотор актина 1 O. sativa::WRI1 Z. mays, промотор убихитина Z. mays::DGAT2a U. rammaniana и промотор убихитина Z. mays::ФАТ
pOIL157: промотор SSU Z. mays::WRI1 Z. mays, промотор убихитина Z. mays::DGAT2a U. rammaniana и убихитина Z. mays::ФАТ.
Вторые серии бинарных векторов экспрессии, содержащих промотор старения SEE1 Z. mays (Robson с соавт., 2004, см. Пример 4), фактор транскрипции LEC1 Z. mays (Shen с соавт., 2010) и фрагмент SDP1 шпРНКи S. bicolor, сконструированы, как указано ниже. Вначале участок прикрепления к матриксу (MAR) вводят в pORE04 путем расщепления pDCOT с помощью AatII+ SnaBI, и субклонирования в сайты AatII+ EcoRV pORE04. Полученный промежуточный вектор обозначен как pOIL158. Далее ген селекционного маркера ФАТ под контролем промотора убихитина Z. mays субклонируют в pOIL158. К этому времени, pOIL141 вначале расщеплен NotI, обработан фрагментом Кленова ДНК полимеразы I и в конце расщеплен AscI. Полученный фрагмент вставляют в сайты AscI+ ZraI pOIL158, с получением pOIL159. Оригинальный источник репликации RK2 oriV в pOIL159 заменяют источником RiA4 путем рестрикционного расщепления pJP3416 с помощью SwaI+ SpeI, с последующим субклонированием в сайты SwaI+AvrII pOIL159. Полученный вектор обозначен как pOIL160. Синтезирован фрагмент «part1 старения однодольных» размером 10,019 тыс. пар оснований, содержащий следующие кассеты экспрессии: актин 1 O. sativa::DGAT1 A. thaliana, кодон-оптимизированный для экспрессии в Z. mays, SEE1Z. mays::WRI1Z. mays, SEE1 Z. mays::LEC1Z. mays. Данный фрагмент субклонируют в виде фрагмента SpeI-EcoRV в сайты SpeI-StuI pOIL160, с получением pOIL161. Синтезирован второй фрагмент « part2 старения однодольных» размером 7,967 тыс. пар оснований, который содержит следующие элементы: MAR, убихитин Z. mays::шпРНКи, фрагмент, нацеленный на S. bicolor/SDP1 T. aestivum, пустая кассета под контролем промотора актина 1 O. sativa. Последовательности двух ТАГ липаз SDP1 S. bicolor (номера доступа XM_002463620; SEQ ID NO. 242 и XM_002458486; SEQ ID NO: 169) и одна последовательность SDP1 T. aestivum (номер доступа AK334547) (SEQ ID NO: 243) получены методом поиска BLAST с последовательностью SDP1 A. thaliana (номер доступа NM_120486). Разработана конструкция синтетической шпильки (SEQ ID NO: 244), содержащая четыре фрагмента (67 пар оснований, 90 пар оснований, 50 пар оснований, 59 пар оснований) последовательности XM_002458486 S. bicolor, которая продемонстрировала наиболее высокую идентичность с последовательностью SDP1 T. aestivum. Кроме того, включен фрагмент размером 278 пар оснований, полученный из липазы SDP1 S. bicolor XM_002463620, с целью усиления эффективности сайленсинга в отношении обеих последовательностей SDP1 S. bicolor. Фрагмент «part2 старения однодольных» субклонируют в виде фрагмента BsiWI-EcoRV в сайты BsiWI-FspI pOIL161. Полученный вектор обозначен pOIL162.
Генетические конструкции pOIL156 pOIL157, pOIL162 и pOIL163 применяют для трансформации S. bicolor и T. aestivum с применением опосредованной Agrobacterium трансформации. Трансгенные растения отобраны на основе резистентности к гигромицину и содержат повышенные уровни ТАГ и ОЖК в вегетативных тканях, по сравнению с нетрансформированными контрольными растениями. Такие растения пригодны для питания животных в виде сена или силоса, а также могут применяться для получения зерна или экстракции масла.
Пример 20. Экстракция масла и получение биотоплива
Экстракция липида из листьев
Урожай трансгенных листьев табака, трансформированных T-ДНК из pJP3502, собирают с растений, выращенных в теплице на протяжении летних месяцев. Листья сушат, а затем измельчают до фрагментов размером 1-3 мм перед экстракцией. Измельченный материал обрабатывают экстракцией в аппарате Сокслета (кипячение с обратным холодильником) в течение 24 часов с выбранными растворителями, как описано ниже. 5 г материала сухих листьев табака и 250 мл растворителя применяют в каждом эксперименте с экстракцией.
Экстракция гексаном в качестве растворителя
Гексан обычно находит коммерческое применение в качестве растворителя для экстракции масла из прессованных семян масличных культур, например, рапса, причем гексан экстрагирует нейтральные (неполярные) липиды, и в другой роли он применяется впервые. Масса экстрагированного липида составила 1,47 г из 5 г листового материала, выход липида составил 29% масс. Проведен анализ экстрагированного гексаном липида в ДМСО методом 1H ЯМР. Анализ продемонстрировал сигналы, типичные для длинноцепочечных жирных кислот в триглицериде, без ароматических продуктов. Далее липид обрабатывают ГХМС с целью идентификации основных компонентов. Анализ экстрагированного гексаном липида методом прямой ГХМС провести сложно, поскольку температура кипения слишком высокая, и материал разлагается в ходе ГХМС. В таких ситуациях общая техника анализа включает первоначальное получение метиловых эфиров жирных кислот, которые были получены, как указано ниже: 18 мг липидного экстракта растворяют в 1 мл толуола, добавляют 3 мл сухого 3 н метанольного раствора HCL и перемешивают на протяжении ночи при 60°C. К охлажденному флакону добавляют 5 мл 5% NaCl и 5 мл гексана и встряхивают. Органический слой извлекают, и экстракцию повторяют еще с 5 мл гексана. Объединенные органические фракции нейтрализуют 8 мл 2% KHCO3, отделяют и сушат с помощью Na2SO4. Растворитель выпаривают в потоке N2, после чего концентрацию доводят до 1 мг/мл в гексане для анализа ГХМС. Основными жирными кислотами являются 16:0 (пальмитиновая, 38,9%) и 18:1 (олеиновая, 31,3%).
Экстракция ацетоном в качестве растворителя
Ацетон применяют в качестве растворителя для экстракции, поскольку его свойства растворителя должны обеспечивать экстракцию практически всего липида из листьев, т.е. как неполярных, так и полярных липидов. Экстрагированное ацетоном масло выглядит подобным экстрагированному гексаном липиду. Масса экстрагированного липида составила 1,59 г из 5 г листьев табака, т.е. 31,8% масс. Проведен анализ липида в ДМСО методом 1H ЯМР. Наблюдаются сигналы, типичные для длинноцепочечных жирных кислот в триглицериде, сигналов ароматических продуктов.
Экстракция горячей водой в качестве растворителя
Осуществлена попытка экстракции горячей водой в качестве растворителя для экстракции, с целью определения ее пригодности для получения масла из листьев табака. Экстрагированный водой материал выглядит гелеподобным и желируется при охлаждении. Масса экстракта составила 1,9 г или 38% масс. Полученный материал подобен густому гелю и, вероятно, содержит полярные соединения из листьев, такие как сахара и другие углеводы. Проведен анализ материала в ДМСО методом 1H ЯМР. Анализ продемонстрировал сигналы, типичные для длинноцепочечных жирных кислот в триглицериде, и не показал экстрагированных ароматических продуктов. Лишний твердый материал был экстрагирован гексаном, давая 20% масс. липида, что указывает на неэффективность экстракции неполярных липидов водой.
Экстракция этанолом в качестве растворителя
Этанол применяли в качестве растворителя для экстракции, с целью определения его пригодности для получения масла из листьев табака. Экстрагированный этанолом липид выглядит подобным как липиду, экстрагированному водой, так и липиду, экстрагированному гексаном, он имеет желто-красный цвет, гелеподобный внешний вид и желируется при охлаждении. Масса экстрагированного липида составила 1,88 г из 5 г табака, или 37,6% масс. Кроме того, этанол в качестве растворителя экстрагировал из листьев табака некоторые полярные соединения.
Экстракция эфиром в качестве растворителя
Осуществлена попытка экстракции диэтиловым эфиром в качестве растворителя для экстракции, поскольку предполагалось, что он может экстрагировать меньшее количество примесей, чем другие растворители. В результате экстракции получено 1,4 г или 28% масс. Экстрагированный эфиром липид подобен материалу, экстрагированному гексаном, имеет желтоватый цвет и выглядит немного чище, чем гексановый экстракт. Несмотря на то, что полученный с диэтиловым эфиром экстракт выглядит наиболее чистым маслом, анализ методом ЯМР продемонстрировал смесь большего количества органических соединений.
Получение биодизельного топлива из растений табака
Партия трансгенных растений табака была выращена на протяжении зимы (не являющейся его нормальным сезоном роста), с целью оценки выработки масла в листьях в течение более холодного сезона, с меньшим природным освещением. Листья зрелых растений содержат около 10% масла, в пересчете на сухую массу; это намного ниже, чем в растениях, выращенных в течение летнего периода. Несмотря на это, липид экстрагирован и превращен в биодизельное топливо, как указано ниже. Стадии способа представляли собой: (a) экстракцию неочищенного липида, (b) очистку ТАГ из липида, и (c) превращение очищенных ТАГ в биодизельное топливо.
Гексан (петролейный эфир 40-60C) применяют в качестве растворителя для экстракции, с целью получения масла, в основном содержащего неполярный липид. 500 г листового материала табака сушат, взвешивают, а затем пропитывают гексаном на протяжении ночи при перемешивании. Смесь фильтруют, затем гексановый экстракт сушат с помощью сульфата магния и обрабатывают с активированным углем для дезодорирования масла. Раствор фильтруют, причем полученная жидкость, упаренная на роторном испарителе, дает около 42 граммов неочищенного масла. Оно имеет желтый/зеленый цвет и вязкую консистенцию при охлаждении. Часть указанного масла использовали в попытке получить биодизельное топливо напрямую, однако количество примесей и высокое содержание свободных жирных кислот привело к образованию большого количества мыла, которое препятствовало протеканию реакции метилирования и отделению продукта. Таким образом, в случае получения биодизельного топлива существует необходимость в дальнейшей очистке указанного масла, с целью обогащения фракцией ТАГ до реакции переэтерификации.
Одной из проблем с выращенным зимой образцом является присутствие относительно высоких уровней свободных жирных кислот (СЖК) в экстрагированном материале, что приводит к повышенному образованию мыла, препятствующего отделению метиловых эфиров и глицериновых продуктов. С целью очистки ТАГ в масле исследованы несколько систем растворителя, причем смесь гексана/этилацетата 80:20 выбрана в качестве подходящей для колоночной хроматографии. Выполнено разделение на колонке с кремния диоксидом, с применением гексана:этилацетата (80:20). Более гидрофобные ТАГ элюируются с колонки первыми, в виде оранжевого/желтого масла. Далее элюируется темно-зеленая полоса, содержащая смесь растворимых в гексане компонентов из листьев табака, включая хлорофилл, смешанный с некоторым количеством ТАГ и СЖК. Конечный продукт вымывается из колонки чистым этилацетатом и в основном представляет собой СЖК.
Очищенные ТАГ, обогащенные отдельно от СЖК и фосфатов, а также других примесей, на этой стадии могут быть превращены в биодизельное топливо. Это осуществлено путем введения ТАГ в реакцию с метанолом в присутствии основного катализатора, с получением метилового эфира (биодизельное топливо) и глицерина в качестве побочного продукта. Альтернативный способ, катализируемая кислотой этерификация, может применяться для введения жирных кислот в реакцию со спиртом, с получением биодизельного топлива в присутствии СЖК, с меньшими требованиями к чистоте ТАГ. Другие способы, такие как реакторы с фиксированным слоем, суперкритические реакторы и ультразвуковые реакторы позволяют отказаться или уменьшают применение химических катализаторов, и дополнительно могут применяться для получения биодизельного топлива из липидов. Однако, способы с основным катализом являются наиболее экономичными для превращения очищенных ТАГ, требуя только низких значений температуры и давления, с получением выхода превращения свыше 98% при условии низких влажности и содержания свободных жирных кислот в исходном масле.
Очищенные ТАГ обрабатывают раствором метоксида (NaOH и метанол смешивают до полного растворения) при температуре масла 60°C. Смесь выдерживают при 60°C и перемешивают в течение 2 часов, пока происходит реакция переэтерификации. Реакционную смесь охлаждают до комнатной температуры, при этом образуются две фазы, причем верхний слой представляет собой биодизельное топливо, а нижний слой является глицерином. Далее фазы разделяют с применением делительной воронки, и извлекают биодизельное топливо.
Гидротермическая обработка вегетативных тканей с высоким содержанием масла
Другой, более прямой подход к превращению вегетативных частей растения в промышленные продукты, такие как жидкое горючее, состоит в гидротермической обработке (ГТО). Ее применяют для превращения материала листьев трансгенного табака, содержащего около 30% масс. ОЖК, в возобновляемое биодизельное топливо, которое может быть добавлено в обычную сырую нефть на нефтеперерабатывающем заводе, с целью получения возобновляемого дизельного топлива (парафиновое дизельное топливо). Нефтяное дизельное топливо является смесью большого количества углеводородных соединений, в основном алканов, и определяется как фракция на выходе нефтеперерабатывающего завода в промежутке 200-300°C, обычно содержащая в основном C13-C22 углеводороды. При типичном превращении листа трансгенного табака посредством ГТО, твердый вегетативный растительный материал трансгенного табака смешивают с водой, чтобы получить концентрацию твердых веществ 16-50%. Далее указанную суспензию подвергают воздействию температуры 270-400°C и давления 70-350 бар. Длительность реакции варьирует в промежутке 1-60 минут, причем эксперименты проводились с NaOH и KOH в качестве катализатора и без них.
После окончания ГТО обработки и охлаждения реакционной смеси последнюю разделяют на 3 различных продуктовых потока, а именно газ, твердые вещества и жидкая бионефть. Выход бионефти составляет 25-40%, в пересчете на сухую массу относительно количества исходного материала. На Фиг. 18 проиллюстрировано, что намного больший выход бионефти был получен для материала листьев трансгенного табака, по сравнению с материалом листьев табака соответствующего дикого типа.
Прямое превращение липида в вегетативных частях растения в биодизельное топливо in situ
В другой серии экспериментов водный компонент реакции ГТО был заменен растворителем метанолом. Существует целый ряд причин для попытки применения метанола, причем одна из них состоит в идее непосредственного превращения ТАГ из масла листьев (in situ), в одну стадию, с получением метиловых эфиров жирных кислот (МЭЖК) непосредственно в липиде и прямым получением биодизельного топлива. Применяются такие же условия реакции и оборудование, что и в предыдущих экспериментах с ГТО, с заменой воды метанолом, температура реакции составляет 335°C, и давление 240 бар, с применением NaOH в качестве катализатора. Вегетативные растительные части трансгенного табака дают 47% масс. бионефти, в пересчете на исходную массу, в то время как табак дикого типа дает 35% масс. бионефти. Анализ методом H1 ЯМР двух полученных образцов бионефти продемонстрировал только небольшое количество МЭЖК, причем анализ методом ЯМР бионефти из трансгенного табака продемонстрировал большое количество МЭЖК биодизельного топлива.
Квалифицированным специалистам в данной области будет понятно, что могут быть осуществлены многочисленные вариации и/или модификации изобретения, как проиллюстрировано в конкретных вариантах реализации изобретения, без выхода за пределы контекста изобретения, как было подробно описано. Таким образом, представленные варианты реализации изобретения следует во всех отношениях интерпретировать как иллюстративные, но не ограничивающие.
Все публикации, которые обсуждаются и/или на которые ссылаются в настоящем документе, включены в настоящий документ в полном объеме.
Любое обсуждение документов, действий, материалов, устройств, изделий или подобного, включенное в настоящий документ, служит исключительно цели обеспечения контекста для настоящего изобретения. Это не следует интерпретировать как согласие с тем, что любые или все из этих материалов образуют часть уровня техники или были общедоступным знанием в данной области, относящимся к настоящему изобретению, поскольку они существовали до даты приоритета каждого из пунктов формулы изобретения в данной заявке.
ССЫЛКИ:
Alemanno et al. (2008) Planta 227:853-866.
Almeida and Allshire (2005) TRENDS Cell Biol. 15:251-258.
Alonso et al. (2009) Plant Cell 21: 1747-1761.
Alonso et al. (2010) Green Chem. 12:1493-1513.
Alvarez et al. (2000) Theor. Appl. Genet. 100:319-327.
Andre at al (2012) Proc. Natl. Acad. Sci. U.S.A. 109:10107-10112.
Andrianov et al. (2010) Plant Biotech. J. 8:277-287.
Awai et al (2006) Biochem. Soc. Trans. 34:395-398.
Bartlett et al. (2008) Plant Methods 4:22.
Baud et al. (2007) Plant J. 50:825-838.
Baud and Lepiniec (2010) Progr. Lipid Res. 49: 235-249.
Baumlein et al. (1991) Mol. Gen. Genet. 225:459-467.
Baumlein et al. (1992) Plant J. 2:233-239.
Belide et al. (2013) Plant Cell Tiss. Org. Cult. DOI 10.1007/s11240-013-0295-1.
Ben Saad et al. (2011) Transgenic Res 20: 1003-1018.
Benning et al (2008) Prog. Lipid Res. 47:381-389.
Benning et al (2009) J. Biol. Chem 284:17420-17427.
Bibikova et al. (2002) Genetics 161:1169-1175.
Bligh and Dyer (1959) Canadian Journal of Biochemistry and Physiology 37:911-917.
Bourque (1995) Plant Sci. 105:125-149.
Boutilier et al. (2002) Plant Cell 14:1737-1749.
Bouvier-Nave et al. (2000) European Journal of Biochemistry/FEBS 267:85-96.
Bradford (1976) Anal. Biochem. 72:248-254.
Broothaerts et al. (2005) Nature 433:629-633.
Broun et al. (1998) Plant J. 13:201-210.
Browse et al. (1986) Biochem J 235: 25-31.
Buchanan-Wollaston (1994) Plant Physiol. 105:839-846.
Busk et al. (1997) Plant J. 11:1285-1295.
Cao et al. (2007) J. Lipid Res. 48:583-591.
Capuano et al. (2007) Biotechnol. Adv. 25:203-206.
Chen et al (2011) Plant Physiol. 155:851-865.
Chikwamba et al. (2003) Proc. Natl. Acad. Sci. U.S.A. 100:11127-11132.
Christie (1993) Advances in Lipid Methodology - Two, Oily Press, Dundee, pp195-213.
Chung et al. (2006) BMC Genomics 7:120.
Comai et al. (2004) Plant J 37: 778-786.
Cong et al. (2013) Science 339:819-823.
Corrado and Karali (2009) Biotechnol. Adv. 27:733-743.
Courvalin et al. (1995) Life Sci. 318:1207-1212.
Coutu et al. (2007) Transgenic Res. 16:771-781.
Dandik and Aksoy (1998) Fuel Process Technol. 57: 81-92.
Darji et al. (1997) Cell 91:765-775.
Dauk et al (2007) Plant Sci. 173:43-49.
Dulermo and Nicaud (2011) Metab. Eng. 13:482-491.
Durrett et al. (2008) Plant J. 54:593-607.
Dussert et al. (2013) Plant Physiol.162:1337-1358.
Dyer et al. (2002) Plant Physiol. 130:2027-2038.
Eastmond et al. (2006) Plant Cell 18: 665-675.
Eccleston et al (1996) Planta 198:46-53.
Ellerstrom et al. (1996) Plant Mol. Biol. 32:1019-1027.
Endalew et al. (2011) Biomass and Bioenergy 35:3787-3809.
Feeney et al. (2012) Plant Physiol 162: 1881-1896.
Finkelstein et al. (1998) Plant Cell 10:1043-1054.
Froissard et al. (2009) FEMS Yeast Res 9:428-438.
Gan (1995) Molecular characterization and genetic manipulation of plant senescence. PhD thesis. University of Wisconsin, Madison.
Gan and Amasino (1995) Science 270:1986-1988.
Gazzarrini et al. (2004) Dev. Cell 7:373-385.
Ghosal et al. (2007) Biochimica et Biophysica Acta 1771:1457-1463.
Ghosh et al. (2009) Plant Physiol. 151:869-881.
Gidda et al (2013) Plant Signaling Behav. 8:e27141.
Girijashankar and Swathisree, (2009) Physiol. Mol. Biol. Plants 15: 287-302.
Gong and Jiang (2011) Biotechnol. Lett. 33:1269-1284.
Gould et al. (1991) Plant Physiol. 95:426-434.
Greenwell et al. (2010) J. R. Soc. Interface 7:703-726.
Gurel et al. (2009) Plant Cell Rep. 28:429-444.
Hershey and Stoner (1991) Plant Mol. Biol. 17:679-690.
Hinchee et al. (1988) Biotechnology 6:915-922.
Hom et al. (2007) Euphytica 153:27-34.
Horn et al. (2013). Plant Physiol 162:1926-1936.
Horvath et al. (2000) Proc. Natl. Acad. Sci. U.S.A. 97:1914-1919.
Huang (1996) Plant Physiol. 110:1055-1061.
Ichihara et al (1988) Biochim. Biophys. Acta 958:125-129.
Ikeda et al. (2006) Pl Biotech J. 23: 153-161.
Iwabuchi et al. (2003) J. Biol. Chem. 278:4603-4610.
Jako et al (2001) Plant Physiol. 126:861-874.
James et al. (2010) Proc. Natl. Acad. Sci. USA 107:17833-17838.
Jepson et al. (1994) Plant Mol. Biol. 26:1855-1866.
Jiang, et al. (2013) Nucleic Acids Research 41(20) e188.
Jing et al. (2011) BMC Biochemistry 12:44.
Jolivet et al. (2014) Plant Physiol. Biochem. 42:501-509.
Jones et al. (1995) Plant Cell 7: 359-371.
Karmakar et al. (2010) Bioresource Technology 101:7201-7210.
Kelly et al. (2011) Plant Physiol. 157: 866-875.
Kelly et al (2013a) Plant Biotech. J. 11:355-361.
Kelly et al. (2013b) Plant Physiol. 162:1282-1289.
Kereszt et al. (2007) Nature Protocols 2:948-952.
Kim et al. (1996) Proc. Natl. Acad. Sci. USA 93:1156-1160.
Kim et al. (2004) Plant Physiol. 134:1206-1216.
Knutzon et al. (1995) Plant Physiol. 109:999-1006.
Koziel et al. (1996) Plant Mol. Biol. 32:393-405.
Kuhn et al. (2009) J. Biol. Chem. 284:34092-102.
Kunst et al. (1988) Proc. Natl. Acad. Sci. U.S.A. 85:4143-4147.
Kwong et al. (2003) Plant Cell 15:5-18.
Lacroix et al. (2008) Proc. Natl. Acad. Sci.U.S.A. 105: 15429-15434.
Lardizabal et al. (2008) Plant Physiol. 148: 89-96.
Larkin et al. (1996) Transgenic Res. 5:325-335.
Laureles et al. (2002) J. of Agric. and Food Chem. 50:1581-1586.
Lebrun et al. (1987) Nucl. Acids Res. 15:4360.
Laux et al. (1996) Development 122: 87-96.
Lazo et al. (1991) Bio/Technology 9:963-967.
Lee et al. (1998) Science 280:915-918.
Lee et al., (2003) Proc. Natl. Acad. Sci. U.S.A. 100:2152-2158.
Li-Beisson et al (2013) The Arabidopsis Book, 2013.
Li et al. (1996) FEBS Lett. 379:117-121.
Li et al. (2006) Phytochemistry 67: 904-915.
Lin et al. (2005) Plant Physiol. Biochem. 43:770-776.
Liu et al. (2010a) Fuel 89:2735-2740.
Liu et al. (2010b) Plant Physiol. Biochem. 48: 9-15.
Liu et al (2012a) Prog. Lipid Res. 51:350-377.
Liu et al. (2012b) In Vitro Cellular and Dev. Biol. Plant 48:462-468.
Liu et al. (2012c) J Exp Bot 63: 3727-3740.
Liu et al. (2014) BMC Plant Biol. 14: 73.
Lotan et al. (1998) Cell 93: 1195-1205.
Lu et al (2007) J. Biol. Chem. 282: 35945-35953.
Luerssen et al. (1998) Plant J. 15: 755-764.
Lui et al. (2009) J. Agric. Food Chem. 57: 2308-2313.
Maher and Bressler (2007) Bioresource Technology 98:2351-2368.
Matsuoka et al. (1994) Plant J. 6:311-319.
Matsuoka and Minami (1989) Eur. J. Biochem. 181: 593-598.
McElroy et al. (1990) Plant Cell 2: 163-171.
Meier et al. (1997) FEBS Lett. 415:91-95.
Millar and Waterhouse (2005). Funct Integr Genomics 5:129-135.
Mojica et al. (2000) Mol Microbiol 36:244-246.
Mongrand et al. (1998) Phytochemistry 49:1049-1064.
Moreno-Perez (2012) PNAS 109:10107-10112.
Mu et al. (2008) Plant Physiol. 148:1042-1054.
Murphy et al. (2012). Protoplasma 249:541-585.
Naim et al. (2012) PLoS One 7: e52717.
Needleman and Wunsch (1970) J. Mol Biol. 45: 443-453.
Nilsson et al. (2012) Physiol. Plantarum 144: 35-47.
Nishida et al (1993) Plant Mol. Biol. 21:267-277.
Nomura et al. (2000) Plant Mol. Biol. 44: 99-106.
Ohlrogge and Browse (1995) Plant Cell 7: 957-970.
Padidam (2003) Curr. Opin. Plant Biol. 6:169-77.
Padidam et al. (2003) Transgenic Res. 12:101-9.
Parthibane et al. (2012a) J. Biol. Chem. 287:1946-1965.
Parthibane et al. (2012b) Plant Physiol. 159:95-104.
Pasquinelli et al. (2005). Curr. Opin. Genet. Develop. 15:200-205.
Perez-Vich et al. (1998) J.A.O.C.S. 75:547-555.
Perrin et al. (2000) Mol. Breed. 6:345-352.
Petrie et al. (2012) PLOS One 7: e35214.
Phillips et al. (2002) Journal of Food Composition and Analysis 12:123-142.
Pigeaire et al. (1997) Mol. Breed. 3:341-349.
Potenza et al. (2004) In Vitro Cell Dev. Biol. Plant 40:1-22.
Powell et al. (1996) Vaccines 183, Abstract.
Qiu et al. (2001) J. Biol. Chem. 276:31561-3156.
Robson et al. (2004) Plant Biotechnol J 2:101-112.
Rossell and Pritchard (1991) Analysis of Oilseeds, Fats and Fatty Foods. Elsevier Science Publishers Ltd: London (Chapter 2, pp.48-53).
Roston et al (2012) J. Biol. Chem. 287:21406-21415.
Ruuska et al. (2002) Plant Cell 14:1191-1206.
Saha et al. (2006) Plant Physiol. 141:1533-1543.
Santos-Mendoza et al. (2005) FEBS Lett. 579:4666-4670.
Santos-Mendoza et al. (2008) Plant J. 54:608-620.
Schaffner (1980) Proc. Natl. Acad. Sci. U.S.A. 77:2163-2167.
Schnurr et al. (2002) Plant Physiol 129:1700-1709.
Scott et al. (2010) Plant Biotechnol. J. 8:912-27.
Shen et al. (2010) Plant Phys. 153: 980-987.
Semwal et al. (2011) Bioresource Technology 102:2151-2161.
Senior (1998) Biotech. Genet. Engin. Revs. 15:79-119.
Shen et al. (2010) Plant Physiol. 153:980-987.
Shiina et al. (1997) Plant Physiol. 115:477-483.
Shimada and Hara-Nishimura (2010) Biol. Pharm. Bull. 33:360-363.
Shimada et al. (2014) Plant Physiol. 164:105-118.
Shockey et al. (2002) Plant Physiol 129:1710-1722.
Sizemore et al. (1995) Science 270:299-302.
Slade and Knauf (2005) Transgenic Res. 14: 109-115.
Slocombe et al. (2009) Plant Biotechnol. J. 7: 694-703.
Smith et al. (2000) Nature 407:319-320.
Somerville et al. (2000) Lipids. In BB Buchanan, W Gruissem, RL Jones, eds, Biochemisty and Molecular Biology of Plants. American Society of Plant Physiologists, Rockville, MD, pp 456-527.
Srinivasan et al. (2007) Planta 225:341-51.
Stalker et al. 1988 Science 242: 419-423.
Stone et al. (2001) Proc. Natl. Acad. Sci. U.S.A.98: 11806-11811.
Stone et al. (2008) Proc. Natl. Acad. Sci. U.S.A.105: 3151-3156.
Tan et al. (2011) Plant Physiol. 156:1577-1588.
Taylor (1997) The Plant Cell 9:1245-1249.
Thillet et al. (1988) J. Biol. Chem 263:12500-12508.
Tingay et al. (1997) Plant J. 11:1369-1376.
To et al. (2012) Plant Cell 24:5007-5023.
Topfer et al. (1995) Science 268:681-686.
Ulmasov et al. (1995) Plant Physiol. 108:919-927.
van de Loo et al. (1995) Proc Natl Acad Sci U S A. 92:6743-6747.
Van Erp et al. (2014) Plant Physiol. 165-30-36.
Vanhercke et al. (2013) FEBS Letters 587:364-369.
Vanhercke et al. (2014). Plant Biotech. J. 12:231-239.
Vieler et al. (2012) Plant Physiol. 158:1562-1569.
Voelker et al. (1992) Science 257:72-74.
Voinnet et al. (2003) Plant J. 33:949-956.
Wang and Benning (2012) Plant J 70:614-623.
Wang et al. (2002) Plant J 32:831-843.
Wang et al. (2004) PNAS 101:3275-3280.
Waterhouse et al. (1998). Proc. Natl. Acad. Sci. U.S.A. 95:13959-13964.
Weiss (2003) Int. J. Med. Microbiol. 293:95-106.
Weselake et al. (2008) J. Exp. Botany 59: 3543-3549.
Winichayakul et al. (2013) Plant Physiol. 162:626-639.
Wood et al. (2009). Plant Biotech. J. 7: 914-924.
Wright et al. (2006) Methods Mol Biol. 343:120-135.
Wu et al. (2014) In Vitro Cellular and Dev. Biol.-Plant 50:9-18.
Xie et al. (2014) Mol. Plant 7:923-926.
Xu et al (2010) Plant and Cell Physiol. 51:1019-1028.
Xu et al (2005) Plant Cell 17:3094-3110.
Xu et al (2008) Plant Cell 20:2190-2204.
Yamagishi et al. (2005) Pl Physiol 139: 163-173.
Yamasaki et al. (2004) Plant Cell 16:3448-3459.
Yang et al. (2003) Planta 216:597-603.
Yang et al. (2010)Proc. Natl. Acad. Sci. U.S.A.107:12040-12045.
Yen et al. (2002) Proc. Natl. Acad. Sci. U.S.A. 99:8512-8517.
Yen et al. (2005) J. Lipid Res. 46: 1502-1511.
Yokoyama et al. (1994) Mol Gen Genet 244: 15-22.
Yuan et al. (1995) Proc. Natl. Acad. Sci. U.S.A. 92:10639-10643.
Zhang et al. (1999) Plant Cell, Tissue and Organ Culture 56:37-46.
Zheng et al. (2009) Pl Physiol 21: 2563-2577.
Zolman et al (2001) Plant Physiol. 127:1266-1274.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Commonwealth Scientific and Industrial Research
Organisation
<120> СПОСОБЫ ПОЛУЧЕНИЯ ПРОМЫШЛЕННЫХ ПРОДУКТОВ ИЗ РАСТИТЕЛЬНЫХ ЛИПИДОВ
<130> 517481
<150> AU 2014902617
<151> 2014-07-07
<150> AU 2015900084
<151> 2015-01-13
<150> AU 2015900284
<151> 2015-01-30
<160> 254
<170> PatentIn версия 3.5
<210> 1
<211> 520
<212> Белок
<213> Arabidopsis thaliana
<400> 1
Met Ala Ile Leu Asp Ser Ala Gly Val Thr Thr Val Thr Glu Asn Gly
1 5 10 15
Gly Gly Glu Phe Val Asp Leu Asp Arg Leu Arg Arg Arg Lys Ser Arg
20 25 30
Ser Asp Ser Ser Asn Gly Leu Leu Leu Ser Gly Ser Asp Asn Asn Ser
35 40 45
Pro Ser Asp Asp Val Gly Ala Pro Ala Asp Val Arg Asp Arg Ile Asp
50 55 60
Ser Val Val Asn Asp Asp Ala Gln Gly Thr Ala Asn Leu Ala Gly Asp
65 70 75 80
Asn Asn Gly Gly Gly Asp Asn Asn Gly Gly Gly Arg Gly Gly Gly Glu
85 90 95
Gly Arg Gly Asn Ala Asp Ala Thr Phe Thr Tyr Arg Pro Ser Val Pro
100 105 110
Ala His Arg Arg Ala Arg Glu Ser Pro Leu Ser Ser Asp Ala Ile Phe
115 120 125
Lys Gln Ser His Ala Gly Leu Phe Asn Leu Cys Val Val Val Leu Ile
130 135 140
Ala Val Asn Ser Arg Leu Ile Ile Glu Asn Leu Met Lys Tyr Gly Trp
145 150 155 160
Leu Ile Arg Thr Asp Phe Trp Phe Ser Ser Arg Ser Leu Arg Asp Trp
165 170 175
Pro Leu Phe Met Cys Cys Ile Ser Leu Ser Ile Phe Pro Leu Ala Ala
180 185 190
Phe Thr Val Glu Lys Leu Val Leu Gln Lys Tyr Ile Ser Glu Pro Val
195 200 205
Val Ile Phe Leu His Ile Ile Ile Thr Met Thr Glu Val Leu Tyr Pro
210 215 220
Val Tyr Val Thr Leu Arg Cys Asp Ser Ala Phe Leu Ser Gly Val Thr
225 230 235 240
Leu Met Leu Leu Thr Cys Ile Val Trp Leu Lys Leu Val Ser Tyr Ala
245 250 255
His Thr Ser Tyr Asp Ile Arg Ser Leu Ala Asn Ala Ala Asp Lys Ala
260 265 270
Asn Pro Glu Val Ser Tyr Tyr Val Ser Leu Lys Ser Leu Ala Tyr Phe
275 280 285
Met Val Ala Pro Thr Leu Cys Tyr Gln Pro Ser Tyr Pro Arg Ser Ala
290 295 300
Cys Ile Arg Lys Gly Trp Val Ala Arg Gln Phe Ala Lys Leu Val Ile
305 310 315 320
Phe Thr Gly Phe Met Gly Phe Ile Ile Glu Gln Tyr Ile Asn Pro Ile
325 330 335
Val Arg Asn Ser Lys His Pro Leu Lys Gly Asp Leu Leu Tyr Ala Ile
340 345 350
Glu Arg Val Leu Lys Leu Ser Val Pro Asn Leu Tyr Val Trp Leu Cys
355 360 365
Met Phe Tyr Cys Phe Phe His Leu Trp Leu Asn Ile Leu Ala Glu Leu
370 375 380
Leu Cys Phe Gly Asp Arg Glu Phe Tyr Lys Asp Trp Trp Asn Ala Lys
385 390 395 400
Ser Val Gly Asp Tyr Trp Arg Met Trp Asn Met Pro Val His Lys Trp
405 410 415
Met Val Arg His Ile Tyr Phe Pro Cys Leu Arg Ser Lys Ile Pro Lys
420 425 430
Thr Leu Ala Ile Ile Ile Ala Phe Leu Val Ser Ala Val Phe His Glu
435 440 445
Leu Cys Ile Ala Val Pro Cys Arg Leu Phe Lys Leu Trp Ala Phe Leu
450 455 460
Gly Ile Met Phe Gln Val Pro Leu Val Phe Ile Thr Asn Tyr Leu Gln
465 470 475 480
Glu Arg Phe Gly Ser Thr Val Gly Asn Met Ile Phe Trp Phe Ile Phe
485 490 495
Cys Ile Phe Gly Gln Pro Met Cys Val Leu Leu Tyr Tyr His Asp Leu
500 505 510
Met Asn Arg Lys Gly Ser Met Ser
515 520
<210> 2
<211> 314
<212> Белок
<213> Arabidopsis thaliana
<400> 2
Met Gly Gly Ser Arg Glu Phe Arg Ala Glu Glu His Ser Asn Gln Phe
1 5 10 15
His Ser Ile Ile Ala Met Ala Ile Trp Leu Gly Ala Ile His Phe Asn
20 25 30
Val Ala Leu Val Leu Cys Ser Leu Ile Phe Leu Pro Pro Ser Leu Ser
35 40 45
Leu Met Val Leu Gly Leu Leu Ser Leu Phe Ile Phe Ile Pro Ile Asp
50 55 60
His Arg Ser Lys Tyr Gly Arg Lys Leu Ala Arg Tyr Ile Cys Lys His
65 70 75 80
Ala Cys Asn Tyr Phe Pro Val Ser Leu Tyr Val Glu Asp Tyr Glu Ala
85 90 95
Phe Gln Pro Asn Arg Ala Tyr Val Phe Gly Tyr Glu Pro His Ser Val
100 105 110
Leu Pro Ile Gly Val Val Ala Leu Cys Asp Leu Thr Gly Phe Met Pro
115 120 125
Ile Pro Asn Ile Lys Val Leu Ala Ser Ser Ala Ile Phe Tyr Thr Pro
130 135 140
Phe Leu Arg His Ile Trp Thr Trp Leu Gly Leu Thr Ala Ala Ser Arg
145 150 155 160
Lys Asn Phe Thr Ser Leu Leu Asp Ser Gly Tyr Ser Cys Val Leu Val
165 170 175
Pro Gly Gly Val Gln Glu Thr Phe His Met Gln His Asp Ala Glu Asn
180 185 190
Val Phe Leu Ser Arg Arg Arg Gly Phe Val Arg Ile Ala Met Glu Gln
195 200 205
Gly Ser Pro Leu Val Pro Val Phe Cys Phe Gly Gln Ala Arg Val Tyr
210 215 220
Lys Trp Trp Lys Pro Asp Cys Asp Leu Tyr Leu Lys Leu Ser Arg Ala
225 230 235 240
Ile Arg Phe Thr Pro Ile Cys Phe Trp Gly Val Phe Gly Ser Pro Leu
245 250 255
Pro Cys Arg Gln Pro Met His Val Val Val Gly Lys Pro Ile Glu Val
260 265 270
Thr Lys Thr Leu Lys Pro Thr Asp Glu Glu Ile Ala Lys Phe His Gly
275 280 285
Gln Tyr Val Glu Ala Leu Arg Asp Leu Phe Glu Arg His Lys Ser Arg
290 295 300
Val Gly Tyr Asp Leu Glu Leu Lys Ile Leu
305 310
<210> 3
<211> 340
<212> Белок
<213> Ricinus communis
<400> 3
Met Gly Glu Glu Ala Asn His Asn Asn Asn Asn Asn Asn Ile Asn Ser
1 5 10 15
Asn Asp Glu Lys Asn Glu Glu Lys Ser Asn Tyr Thr Val Val Asn Ser
20 25 30
Arg Glu Leu Tyr Pro Thr Asn Ile Phe His Ala Leu Leu Ala Leu Ser
35 40 45
Ile Trp Ile Gly Ser Ile His Phe Asn Leu Phe Leu Leu Phe Ile Ser
50 55 60
Tyr Leu Phe Leu Ser Phe Pro Thr Phe Leu Leu Ile Val Gly Phe Phe
65 70 75 80
Val Val Leu Met Phe Ile Pro Ile Asp Glu His Ser Lys Leu Gly Arg
85 90 95
Arg Leu Cys Arg Tyr Val Cys Arg His Ala Cys Ser His Phe Pro Val
100 105 110
Thr Leu His Val Glu Asp Met Asn Ala Phe His Ser Asp Arg Ala Tyr
115 120 125
Val Phe Gly Tyr Glu Pro His Ser Val Phe Pro Leu Gly Val Ser Val
130 135 140
Leu Ser Asp His Phe Ala Val Leu Pro Leu Pro Lys Met Lys Val Leu
145 150 155 160
Ala Ser Asn Ala Val Phe Arg Thr Pro Val Leu Arg His Ile Trp Thr
165 170 175
Trp Cys Gly Leu Thr Ser Ala Thr Lys Lys Asn Phe Thr Ala Leu Leu
180 185 190
Ala Ser Gly Tyr Ser Cys Ile Val Ile Pro Gly Gly Val Gln Glu Thr
195 200 205
Phe Tyr Met Lys His Gly Ser Glu Ile Ala Phe Leu Lys Ala Arg Arg
210 215 220
Gly Phe Val Arg Val Ala Met Glu Met Gly Lys Pro Leu Val Pro Val
225 230 235 240
Phe Cys Phe Gly Gln Ser Asn Val Tyr Lys Trp Trp Lys Pro Asp Gly
245 250 255
Glu Leu Phe Met Lys Ile Ala Arg Ala Ile Lys Phe Ser Pro Ile Val
260 265 270
Phe Trp Gly Val Leu Gly Ser His Leu Pro Leu Gln Arg Pro Met His
275 280 285
Val Val Val Gly Lys Pro Ile Glu Val Lys Gln Asn Pro Gln Pro Thr
290 295 300
Val Glu Glu Val Ser Glu Val Gln Gly Gln Phe Val Ala Ala Leu Lys
305 310 315 320
Asp Leu Phe Glu Arg His Lys Ala Arg Val Gly Tyr Ala Asp Leu Thr
325 330 335
Leu Glu Ile Leu
340
<210> 4
<211> 322
<212> Белок
<213> Vernicia fordii
<400> 4
Met Gly Met Val Glu Val Lys Asn Glu Glu Glu Val Thr Ile Phe Lys
1 5 10 15
Ser Gly Glu Ile Tyr Pro Thr Asn Ile Phe Gln Ser Val Leu Ala Leu
20 25 30
Ala Ile Trp Leu Gly Ser Phe His Phe Ile Leu Phe Leu Val Ser Ser
35 40 45
Ser Ile Phe Leu Pro Phe Ser Lys Phe Leu Leu Val Ile Gly Leu Leu
50 55 60
Leu Phe Phe Met Val Ile Pro Ile Asn Asp Arg Ser Lys Leu Gly Gln
65 70 75 80
Cys Leu Phe Ser Tyr Ile Ser Arg His Val Cys Ser Tyr Phe Pro Ile
85 90 95
Thr Leu His Val Glu Asp Ile Asn Ala Phe Arg Ser Asp Arg Ala Tyr
100 105 110
Val Phe Gly Tyr Glu Pro His Ser Val Phe Pro Ile Gly Val Met Ile
115 120 125
Leu Ser Leu Gly Leu Ile Pro Leu Pro Asn Ile Lys Phe Leu Ala Ser
130 135 140
Ser Ala Val Phe Tyr Thr Pro Phe Leu Arg His Ile Trp Ser Trp Cys
145 150 155 160
Gly Leu Thr Pro Ala Thr Arg Lys Asn Phe Val Ser Leu Leu Ser Ser
165 170 175
Gly Tyr Ser Cys Ile Leu Val Pro Gly Gly Val Gln Glu Thr Phe Tyr
180 185 190
Met Lys Gln Asp Ser Glu Ile Ala Phe Leu Lys Ala Arg Arg Gly Phe
195 200 205
Ile Arg Ile Ala Met Gln Thr Gly Thr Pro Leu Val Pro Val Phe Cys
210 215 220
Phe Gly Gln Met His Thr Phe Lys Trp Trp Lys Pro Asp Gly Glu Leu
225 230 235 240
Phe Met Lys Ile Ala Arg Ala Ile Lys Phe Thr Pro Thr Ile Phe Trp
245 250 255
Gly Val Leu Gly Thr Pro Leu Pro Phe Lys Asn Pro Met His Val Val
260 265 270
Val Gly Arg Pro Ile Glu Val Lys Gln Asn Pro Gln Pro Thr Ala Glu
275 280 285
Glu Val Ala Glu Val Gln Arg Glu Phe Ile Ala Ser Leu Lys Asn Leu
290 295 300
Phe Glu Arg His Lys Ala Arg Val Gly Tyr Ser Asp Leu Lys Leu Glu
305 310 315 320
Ile Phe
<210> 5
<211> 355
<212> Белок
<213> Mortierella ramanniana
<400> 5
Met Ala Ser Lys Asp Gln His Leu Gln Gln Lys Val Lys His Thr Leu
1 5 10 15
Glu Ala Ile Pro Ser Pro Arg Tyr Ala Pro Leu Arg Val Pro Leu Arg
20 25 30
Arg Arg Leu Gln Thr Leu Ala Val Leu Leu Trp Cys Ser Met Met Ser
35 40 45
Ile Cys Met Phe Ile Phe Phe Phe Leu Cys Ser Ile Pro Val Leu Leu
50 55 60
Trp Phe Pro Ile Ile Leu Tyr Leu Thr Trp Ile Leu Val Trp Asp Lys
65 70 75 80
Ala Pro Glu Asn Gly Gly Arg Pro Ile Arg Trp Leu Arg Asn Ala Ala
85 90 95
Trp Trp Lys Leu Phe Ala Gly Tyr Phe Pro Ala His Val Ile Lys Glu
100 105 110
Ala Asp Leu Asp Pro Ser Lys Asn Tyr Ile Phe Gly Tyr His Pro His
115 120 125
Gly Ile Ile Ser Met Gly Ser Phe Cys Thr Phe Ser Thr Asn Ala Thr
130 135 140
Gly Phe Asp Asp Leu Phe Pro Gly Ile Arg Pro Ser Leu Leu Thr Leu
145 150 155 160
Thr Ser Asn Phe Asn Ile Pro Leu Tyr Arg Asp Tyr Leu Met Ala Cys
165 170 175
Gly Leu Cys Ser Val Ser Lys Thr Ser Cys Gln Asn Ile Leu Thr Lys
180 185 190
Gly Gly Pro Gly Arg Ser Ile Ala Ile Val Val Gly Gly Ala Ser Glu
195 200 205
Ser Leu Asn Ala Arg Pro Gly Val Met Asp Leu Val Leu Lys Arg Arg
210 215 220
Phe Gly Phe Ile Lys Ile Ala Val Gln Thr Gly Ala Ser Leu Val Pro
225 230 235 240
Thr Ile Ser Phe Gly Glu Asn Glu Leu Tyr Glu Gln Ile Glu Ser Asn
245 250 255
Glu Asn Ser Lys Leu His Arg Trp Gln Lys Lys Ile Gln His Ala Leu
260 265 270
Gly Phe Thr Met Pro Leu Phe His Gly Arg Gly Val Phe Asn Tyr Asp
275 280 285
Phe Gly Leu Leu Pro His Arg His Pro Ile Tyr Thr Ile Val Gly Lys
290 295 300
Pro Ile Pro Val Pro Ser Ile Lys Tyr Gly Gln Thr Lys Asp Glu Ile
305 310 315 320
Ile Arg Glu Leu His Asp Ser Tyr Met His Ala Val Gln Asp Leu Tyr
325 330 335
Asp Arg Tyr Lys Asp Ile Tyr Ala Lys Asp Arg Val Lys Glu Leu Glu
340 345 350
Phe Val Glu
355
<210> 6
<211> 388
<212> Белок
<213> Homo sapiens
<400> 6
Met Lys Thr Leu Ile Ala Ala Tyr Ser Gly Val Leu Arg Gly Glu Arg
1 5 10 15
Gln Ala Glu Ala Asp Arg Ser Gln Arg Ser His Gly Gly Pro Ala Leu
20 25 30
Ser Arg Glu Gly Ser Gly Arg Trp Gly Thr Gly Ser Ser Ile Leu Ser
35 40 45
Ala Leu Gln Asp Leu Phe Ser Val Thr Trp Leu Asn Arg Ser Lys Val
50 55 60
Glu Lys Gln Leu Gln Val Ile Ser Val Leu Gln Trp Val Leu Ser Phe
65 70 75 80
Leu Val Leu Gly Val Ala Cys Ser Ala Ile Leu Met Tyr Ile Phe Cys
85 90 95
Thr Asp Cys Trp Leu Ile Ala Val Leu Tyr Phe Thr Trp Leu Val Phe
100 105 110
Asp Trp Asn Thr Pro Lys Lys Gly Gly Arg Arg Ser Gln Trp Val Arg
115 120 125
Asn Trp Ala Val Trp Arg Tyr Phe Arg Asp Tyr Phe Pro Ile Gln Leu
130 135 140
Val Lys Thr His Asn Leu Leu Thr Thr Arg Asn Tyr Ile Phe Gly Tyr
145 150 155 160
His Pro His Gly Ile Met Gly Leu Gly Ala Phe Cys Asn Phe Ser Thr
165 170 175
Glu Ala Thr Glu Val Ser Lys Lys Phe Pro Gly Ile Arg Pro Tyr Leu
180 185 190
Ala Thr Leu Ala Gly Asn Phe Arg Met Pro Val Leu Arg Glu Tyr Leu
195 200 205
Met Ser Gly Gly Ile Cys Pro Val Ser Arg Asp Thr Ile Asp Tyr Leu
210 215 220
Leu Ser Lys Asn Gly Ser Gly Asn Ala Ile Ile Ile Val Val Gly Gly
225 230 235 240
Ala Ala Glu Ser Leu Ser Ser Met Pro Gly Lys Asn Ala Val Thr Leu
245 250 255
Arg Asn Arg Lys Gly Phe Val Lys Leu Ala Leu Arg His Gly Ala Asp
260 265 270
Leu Val Pro Ile Tyr Ser Phe Gly Glu Asn Glu Val Tyr Lys Gln Val
275 280 285
Ile Phe Glu Glu Gly Ser Trp Gly Arg Trp Val Gln Lys Lys Phe Gln
290 295 300
Lys Tyr Ile Gly Phe Ala Pro Cys Ile Phe His Gly Arg Gly Leu Phe
305 310 315 320
Ser Ser Asp Thr Trp Gly Leu Val Pro Tyr Ser Lys Pro Ile Thr Thr
325 330 335
Val Val Gly Glu Pro Ile Thr Ile Pro Lys Leu Glu His Pro Thr Gln
340 345 350
Gln Asp Ile Asp Leu Tyr His Thr Met Tyr Met Glu Ala Leu Val Lys
355 360 365
Leu Phe Asp Lys His Lys Thr Lys Phe Gly Leu Pro Glu Thr Glu Val
370 375 380
Leu Glu Val Asn
385
<210> 7
<211> 328
<212> Белок
<213> Homo sapiens
<400> 7
Met Ala His Ser Lys Gln Pro Ser His Phe Gln Ser Leu Met Leu Leu
1 5 10 15
Gln Trp Pro Leu Ser Tyr Leu Ala Ile Phe Trp Ile Leu Gln Pro Leu
20 25 30
Phe Val Tyr Leu Leu Phe Thr Ser Leu Trp Pro Leu Pro Val Leu Tyr
35 40 45
Phe Ala Trp Leu Phe Leu Asp Trp Lys Thr Pro Glu Arg Gly Gly Arg
50 55 60
Arg Ser Ala Trp Val Arg Asn Trp Cys Val Trp Thr His Ile Arg Asp
65 70 75 80
Tyr Phe Pro Ile Thr Ile Leu Lys Thr Lys Asp Leu Ser Pro Glu His
85 90 95
Asn Tyr Leu Met Gly Val His Pro His Gly Leu Leu Thr Phe Gly Ala
100 105 110
Phe Cys Asn Phe Cys Thr Glu Ala Thr Gly Phe Ser Lys Thr Phe Pro
115 120 125
Gly Ile Thr Pro His Leu Ala Thr Leu Ser Trp Phe Phe Lys Ile Pro
130 135 140
Phe Val Arg Glu Tyr Leu Met Ala Lys Gly Val Cys Ser Val Ser Gln
145 150 155 160
Pro Ala Ile Asn Tyr Leu Leu Ser His Gly Thr Gly Asn Leu Val Gly
165 170 175
Ile Val Val Gly Gly Val Gly Glu Ala Leu Gln Ser Val Pro Asn Thr
180 185 190
Thr Thr Leu Ile Leu Gln Lys Arg Lys Gly Phe Val Arg Thr Ala Leu
195 200 205
Gln His Gly Ala His Leu Val Pro Thr Phe Thr Phe Gly Glu Thr Glu
210 215 220
Val Tyr Asp Gln Val Leu Phe His Lys Asp Ser Arg Met Tyr Lys Phe
225 230 235 240
Gln Ser Cys Phe Arg Arg Ile Phe Gly Phe Tyr Cys Cys Val Phe Tyr
245 250 255
Gly Gln Ser Phe Cys Gln Gly Ser Thr Gly Leu Leu Pro Tyr Ser Arg
260 265 270
Pro Ile Val Thr Val Val Gly Glu Pro Leu Pro Leu Pro Gln Ile Glu
275 280 285
Lys Pro Ser Gln Glu Met Val Asp Lys Tyr His Ala Leu Tyr Met Asp
290 295 300
Ala Leu His Lys Leu Phe Asp Gln His Lys Thr His Tyr Gly Cys Ser
305 310 315 320
Glu Thr Gln Lys Leu Phe Phe Leu
325
<210> 8
<211> 361
<212> Белок
<213> Bos taurus
<400> 8
Met Lys Thr Leu Ile Ala Ala Tyr Ser Gly Val Leu Arg Gly Thr Gly
1 5 10 15
Ser Ser Ile Leu Ser Ala Leu Gln Asp Leu Phe Ser Val Thr Trp Leu
20 25 30
Asn Arg Ala Lys Val Glu Lys Gln Leu Gln Val Ile Ser Val Leu Gln
35 40 45
Trp Val Leu Ser Phe Leu Val Leu Gly Val Ala Cys Ser Val Ile Leu
50 55 60
Met Tyr Thr Phe Cys Thr Asp Cys Trp Leu Ile Ala Val Leu Tyr Phe
65 70 75 80
Thr Trp Leu Val Phe Asp Trp Asn Thr Pro Lys Lys Gly Gly Arg Arg
85 90 95
Ser Gln Trp Val Arg Asn Trp Ala Val Trp Arg Tyr Phe Arg Asp Tyr
100 105 110
Phe Pro Ile Gln Leu Val Lys Thr His Asn Leu Leu Thr Ser Arg Asn
115 120 125
Tyr Ile Phe Gly Tyr His Pro His Gly Ile Met Gly Leu Gly Ala Phe
130 135 140
Cys Asn Phe Ser Thr Glu Ala Thr Glu Val Ser Lys Lys Phe Pro Gly
145 150 155 160
Ile Arg Pro Tyr Leu Ala Thr Leu Ala Gly Asn Phe Arg Met Pro Val
165 170 175
Leu Arg Glu Tyr Leu Met Ser Gly Gly Ile Cys Pro Val Asn Arg Asp
180 185 190
Thr Ile Asp Tyr Leu Leu Ser Lys Asn Gly Ser Gly Asn Ala Ile Ile
195 200 205
Ile Val Val Gly Gly Ala Ala Glu Ser Leu Ser Ser Met Pro Gly Lys
210 215 220
Asn Ala Val Thr Leu Arg Asn Arg Lys Gly Phe Val Lys Leu Ala Leu
225 230 235 240
Arg His Gly Ala Asp Leu Val Pro Thr Tyr Ser Phe Gly Glu Asn Glu
245 250 255
Val Tyr Lys Gln Val Ile Phe Glu Glu Gly Ser Trp Gly Arg Trp Val
260 265 270
Gln Lys Lys Phe Gln Lys Tyr Ile Gly Phe Ala Pro Cys Ile Phe His
275 280 285
Gly Arg Gly Leu Phe Ser Ser Asp Thr Trp Gly Leu Val Pro Tyr Ser
290 295 300
Lys Pro Ile Thr Thr Val Val Gly Glu Pro Ile Thr Ile Pro Arg Leu
305 310 315 320
Glu Arg Pro Thr Gln Gln Asp Ile Asp Leu Tyr His Ala Met Tyr Val
325 330 335
Gln Ala Leu Val Lys Leu Phe Asp Gln His Lys Thr Lys Phe Gly Leu
340 345 350
Pro Glu Thr Glu Val Leu Glu Val Asn
355 360
<210> 9
<211> 388
<212> Белок
<213> Mus musculus
<400> 9
Met Lys Thr Leu Ile Ala Ala Tyr Ser Gly Val Leu Arg Gly Glu Arg
1 5 10 15
Arg Ala Glu Ala Ala Arg Ser Glu Asn Lys Asn Lys Gly Ser Ala Leu
20 25 30
Ser Arg Glu Gly Ser Gly Arg Trp Gly Thr Gly Ser Ser Ile Leu Ser
35 40 45
Ala Leu Gln Asp Ile Phe Ser Val Thr Trp Leu Asn Arg Ser Lys Val
50 55 60
Glu Lys Gln Leu Gln Val Ile Ser Val Leu Gln Trp Val Leu Ser Phe
65 70 75 80
Leu Val Leu Gly Val Ala Cys Ser Val Ile Leu Met Tyr Thr Phe Cys
85 90 95
Thr Asp Cys Trp Leu Ile Ala Val Leu Tyr Phe Thr Trp Leu Ala Phe
100 105 110
Asp Trp Asn Thr Pro Lys Lys Gly Gly Arg Arg Ser Gln Trp Val Arg
115 120 125
Asn Trp Ala Val Trp Arg Tyr Phe Arg Asp Tyr Phe Pro Ile Gln Leu
130 135 140
Val Lys Thr His Asn Leu Leu Thr Thr Arg Asn Tyr Ile Phe Gly Tyr
145 150 155 160
His Pro His Gly Ile Met Gly Leu Gly Ala Phe Cys Asn Phe Ser Thr
165 170 175
Glu Ala Thr Glu Val Ser Lys Lys Phe Pro Gly Ile Arg Pro Tyr Leu
180 185 190
Ala Thr Leu Ala Gly Asn Phe Arg Met Pro Val Leu Arg Glu Tyr Leu
195 200 205
Met Ser Gly Gly Ile Cys Pro Val Asn Arg Asp Thr Ile Asp Tyr Leu
210 215 220
Leu Ser Lys Asn Gly Ser Gly Asn Ala Ile Ile Ile Val Val Gly Gly
225 230 235 240
Ala Ala Glu Ser Leu Ser Ser Met Pro Gly Lys Asn Ala Val Thr Leu
245 250 255
Lys Asn Arg Lys Gly Phe Val Lys Leu Ala Leu Arg His Gly Ala Asp
260 265 270
Leu Val Pro Thr Tyr Ser Phe Gly Glu Asn Glu Val Tyr Lys Gln Val
275 280 285
Ile Phe Glu Glu Gly Ser Trp Gly Arg Trp Val Gln Lys Lys Phe Gln
290 295 300
Lys Tyr Ile Gly Phe Ala Pro Cys Ile Phe His Gly Arg Gly Leu Phe
305 310 315 320
Ser Ser Asp Thr Trp Gly Leu Val Pro Tyr Ser Lys Pro Ile Thr Thr
325 330 335
Val Val Gly Glu Pro Ile Thr Val Pro Lys Leu Glu His Pro Thr Gln
340 345 350
Lys Asp Ile Asp Leu Tyr His Ala Met Tyr Met Glu Ala Leu Val Lys
355 360 365
Leu Phe Asp Asn His Lys Thr Lys Phe Gly Leu Pro Glu Thr Glu Val
370 375 380
Leu Glu Val Asn
385
<210> 10
<211> 3
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<400> 10
Tyr Phe Pro
1
<210> 11
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<400> 11
His Pro His Gly
1
<210> 12
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<400> 12
Glu Pro His Ser
1
<210> 13
<211> 24
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<220>
<221> X
<222> (2)..(2)
<223> любая аминокислота
<220>
<221> X
<222> (5)..(5)
<223> любая аминокислота
<220>
<221> X
<222> (6)..(6)
<223> Лизин (K) или Аргинин (R)
<220>
<221> X
<222> (7)..(7)
<223> любая аминокислота
<220>
<221> X
<222> (9)..(11)
<223> любая аминокислота
<220>
<221> X
<222> (13)..(15)
<223> любая аминокислота
<220>
<221> X
<222> (16)..(16)
<223> Лейцин (L) или Валин (V)
<220>
<221> X
<222> (19)..(21)
<223> любая аминокислота
<220>
<221> X
<222> (24)..(24)
<223> Глютаминовая кислота (E) или Глютамин (Q)
<400> 13
Arg Xaa Gly Phe Xaa Xaa Xaa Ala Xaa Xaa Xaa Gly Xaa Xaa Xaa Xaa
1 5 10 15
Val Pro Xaa Xaa Xaa Phe Gly Xaa
20
<210> 14
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<220>
<221> X
<222> (3)..(3)
<223> любая аминокислота
<220>
<221> X
<222> (5)..(7)
<223> любая аминокислота
<400> 14
Phe Leu Xaa Leu Xaa Xaa Xaa Asn
1 5
<210> 15
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<400> 15
Ala Leu Val Val Ala Asn His Gln Ser Phe Leu Asp Pro Leu Val Leu
1 5 10 15
Ser Ala Leu Leu Pro Arg Lys Gly Gly Arg Val Arg Phe Val Ala Lys
20 25 30
Lys Glu Leu Phe Tyr Val Pro Leu Leu Gly Trp Leu Leu Arg Leu Leu
35 40 45
Gly Ala Ile Phe Ile Asp Arg Glu Asn Gly Arg Leu Ala Arg Ala Ala
50 55 60
Leu Arg Glu Ala Val Arg Leu Leu Arg Asp Gly Gly Trp Leu Leu Ile
65 70 75 80
Phe Pro Glu Gly Thr Arg Ser Arg Pro Gly Lys Leu Leu Pro Phe Lys
85 90 95
Lys Gly Ala Ala Arg Leu Ala Leu Glu Ala Gly Val Pro Ile Val Pro
100 105 110
Val Ala Ile Arg Gly Thr
115
<210> 16
<211> 187
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<220>
<221> X
<222> (15)..(15)
<223> любая аминокислота
<220>
<221> X
<222> (18)..(18)
<223> любая аминокислота
<220>
<221> X
<222> (23)..(23)
<223> любая аминокислота
<220>
<221> X
<222> (25)..(26)
<223> любая аминокислота
<220>
<221> X
<222> (28)..(30)
<223> любая аминокислота
<220>
<221> X
<222> (32)..(33)
<223> любая аминокислота
<220>
<221> X
<222> (35)..(38)
<223> любая аминокислота
<220>
<221> X
<222> (41)..(41)
<223> любая аминокислота
<220>
<221> X
<222> (46)..(48)
<223> любая аминокислота
<220>
<221> X
<222> (53)..(53)
<223> любая аминокислота
<220>
<221> X
<222> (55)..(57)
<223> любая аминокислота
<220>
<221> X
<222> (61)..(61)
<223> любая аминокислота
<220>
<221> X
<222> (67)..(67)
<223> любая аминокислота
<220>
<221> X
<222> (72)..(72)
<223> любая аминокислота
<220>
<221> X
<222> (74)..(77)
<223> любая аминокислота
<220>
<221> X
<222> (79)..(79)
<223> любая аминокислота
<220>
<221> X
<222> (114)..(114)
<223> любая аминокислота
<220>
<221> X
<222> (127)..(128)
<223> любая аминокислота
<220>
<221> X
<222> (136)..(136)
<223> любая аминокислота
<220>
<221> X
<222> (139)..(142)
<223> любая аминокислота
<220>
<221> X
<222> (144)..(144)
<223> любая аминокислота
<220>
<221> X
<222> (150)..(150)
<223> любая аминокислота
<220>
<221> X
<222> (164)..(165)
<223> любая аминокислота
<220>
<221> X
<222> (167)..(172)
<223> любая аминокислота
<400> 16
Ala Val Phe Asp Lys Asp Gly Thr Leu Thr Glu Asp Asp Thr Xaa Phe
1 5 10 15
Leu Xaa Tyr Leu Leu Lys Xaa Leu Xaa Xaa Leu Xaa Xaa Xaa Leu Xaa
20 25 30
Xaa Asp Xaa Xaa Xaa Xaa Gly Ser Xaa Leu Thr Leu Ser Xaa Xaa Xaa
35 40 45
Asp Leu Leu Glu Xaa Leu Xaa Xaa Xaa Gly Gly Ile Xaa Val Ile Gly
50 55 60
Leu Ala Xaa Arg Tyr Leu Glu Xaa Leu Xaa Xaa Xaa Xaa Glu Xaa Ala
65 70 75 80
Lys Leu Phe Glu Gly Phe Ile Lys Pro Asp Ala Ala Glu Leu Leu Lys
85 90 95
Glu Leu His Glu Ala Gly Leu Arg Val Val Val Leu Thr Gly Asp Pro
100 105 110
Arg Xaa Ile Ala Lys Pro Val Ala Lys Glu Leu Gly Ile Asp Xaa Xaa
115 120 125
Asn Val Leu Ala Thr Glu Leu Xaa Asp Glu Xaa Xaa Xaa Xaa Val Xaa
130 135 140
Gly Arg Ile Thr Gly Xaa Leu Asp Lys Ala Arg Ala Val Glu Arg Leu
145 150 155 160
Val Val Leu Xaa Xaa Lys Xaa Xaa Xaa Xaa Xaa Xaa Val Val Ala Ile
165 170 175
Gly Asp Ser Ala Asn Asp Leu Pro Ala Leu Lys
180 185
<210> 17
<211> 190
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<400> 17
Ile Lys Ala Val Val Phe Asp Lys Asp Gly Thr Leu Thr Asp Gly Glu
1 5 10 15
Pro Pro Ile Ala Glu Ala Ile Val Glu Ala Ala Ala Glu Leu Gly Leu
20 25 30
Pro Leu Leu Leu Pro Leu Glu Glu Val Glu Lys Leu Leu Gly Arg Gly
35 40 45
Val Glu Gly Ile Glu Arg Ile Leu Leu Glu Gly Gly Leu Thr Ala Glu
50 55 60
Leu Leu Leu Glu Leu Glu Gly Glu Leu Ala Ala Gly Lys Thr Ala Val
65 70 75 80
Leu Val Ala Leu Asp Gly Glu Val Leu Gly Leu Ile Ala Leu Ala Asp
85 90 95
Lys Leu Tyr Pro Gly Ala Arg Glu Ala Leu Lys Ala Leu Lys Glu Arg
100 105 110
Gly Ile Lys Val Ala Ile Leu Thr Asn Gly Asp Arg Ala Asn Ala Glu
115 120 125
Ala Val Leu Glu Ala Leu Gly Leu Ala Asp Leu Phe Asp Val Ile Val
130 135 140
Asp Ser Asp Asp Val Gly Pro Val Lys Pro Lys Pro Glu Ile Phe Leu
145 150 155 160
Lys Ala Leu Glu Arg Leu Gly Val Lys Pro Glu Glu Val Leu Met Val
165 170 175
Gly Asp Gly Val Asn Asp Ala Pro Ala Leu Ala Ala Ala Gly
180 185 190
<210> 18
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<400> 18
Gly Asp Leu Val Ile Cys Pro Glu Gly Thr Thr Cys Arg Glu Pro
1 5 10 15
<210> 19
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<220>
<221> X
<222> (2)..(2)
<223> любая аминокислота
<220>
<221> X
<222> (4)..(4)
<223> любая аминокислота
<220>
<221> X
<222> (5)..(5)
<223> Треонин (T) или Валин (V)
<220>
<221> X
<222> (6)..(6)
<223> Лейцин (L) или Валин (V)
<400> 19
Asp Xaa Asp Xaa Xaa Xaa
1 5
<210> 20
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<220>
<221> X
<222> (2)..(2)
<223> 17-20 аминокислот; где аминокислоты могут быть любыми аминокислотами
<220>
<221> X
<222> (3)..(3)
<223> Глицин (G) или Серин (S)
<220>
<221> X
<222> (4)..(4)
<223> Аспарагиновая кислота (D) или Серин (S)
<220>
<221> X
<222> (5)..(7)
<223> любая аминокислота
<220>
<221> X
<222> (8)..(8)
<223> Аспарагиновая кислота (D) или Аспарагин (N)
<400> 20
Lys Xaa Xaa Xaa Xaa Xaa Xaa Xaa
1 5
<210> 21
<211> 356
<212> Белок
<213> Arabidopsis thaliana
<400> 21
Met Asp Trp Glu Ile Arg Gly Ser Ser Leu Gly Gln Lys Leu Leu Glu
1 5 10 15
Phe Asp Ser Glu Gln Glu Arg Gln Thr Arg Phe Arg Ala Tyr Asp Ser
20 25 30
Glu Glu Ala Ala Ala His Thr Tyr Asp Leu Ala Ala Leu Lys Tyr Trp
35 40 45
Gly Pro Asp Thr Ile Leu Asn Phe Pro Ala Glu Thr Tyr Thr Lys Glu
50 55 60
Leu Glu Glu Met Gln Arg Val Thr Lys Glu Glu Tyr Leu Ala Ser Leu
65 70 75 80
Arg Arg Gln Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly
85 90 95
Val Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg
100 105 110
Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Asn Thr Gln Glu
115 120 125
Glu Ala Ala Ala Ala Tyr Asp Met Ala Ala Ile Glu Tyr Arg Gly Ala
130 135 140
Asn Ala Val Thr Asn Phe Asp Ile Ser Asn Tyr Ile Asp Arg Leu Lys
145 150 155 160
Lys Lys Gly Val Phe Pro Phe Pro Val Asn Gln Ala Asn His Gln Glu
165 170 175
Gly Ile Leu Val Glu Ala Lys Gln Glu Val Glu Thr Arg Glu Ala Lys
180 185 190
Glu Glu Pro Arg Glu Glu Val Lys Gln Gln Tyr Val Glu Glu Pro Pro
195 200 205
Gln Glu Glu Glu Glu Lys Glu Glu Glu Lys Ala Glu Gln Gln Glu Ala
210 215 220
Glu Ile Val Gly Tyr Ser Glu Glu Ala Ala Val Val Asn Cys Cys Ile
225 230 235 240
Asp Ser Ser Thr Ile Met Glu Met Asp Arg Cys Gly Asp Asn Asn Glu
245 250 255
Leu Ala Trp Asn Phe Cys Met Met Asp Thr Gly Phe Ser Pro Phe Leu
260 265 270
Thr Asp Gln Asn Leu Ala Asn Glu Asn Pro Ile Glu Tyr Pro Glu Leu
275 280 285
Phe Asn Glu Leu Ala Phe Glu Asp Asn Ile Asp Phe Met Phe Asp Asp
290 295 300
Gly Lys His Glu Cys Leu Asn Leu Glu Asn Leu Asp Cys Cys Val Val
305 310 315 320
Gly Arg Glu Ser Pro Pro Ser Ser Ser Ser Pro Leu Ser Cys Leu Ser
325 330 335
Thr Asp Ser Ala Ser Ser Thr Thr Thr Thr Thr Thr Ser Val Ser Cys
340 345 350
Asn Tyr Leu Val
355
<210> 22
<211> 430
<212> Белок
<213> Arabidopsis thaliana
<400> 22
Met Lys Lys Arg Leu Thr Thr Ser Thr Cys Ser Ser Ser Pro Ser Ser
1 5 10 15
Ser Val Ser Ser Ser Thr Thr Thr Ser Ser Pro Ile Gln Ser Glu Ala
20 25 30
Pro Arg Pro Lys Arg Ala Lys Arg Ala Lys Lys Ser Ser Pro Ser Gly
35 40 45
Asp Lys Ser His Asn Pro Thr Ser Pro Ala Ser Thr Arg Arg Ser Ser
50 55 60
Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe Glu Ala
65 70 75 80
His Leu Trp Asp Lys Ser Ser Trp Asn Ser Ile Gln Asn Lys Lys Gly
85 90 95
Lys Gln Val Tyr Leu Gly Ala Tyr Asp Ser Glu Glu Ala Ala Ala His
100 105 110
Thr Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Asp Thr Ile Leu
115 120 125
Asn Phe Pro Ala Glu Thr Tyr Thr Lys Glu Leu Glu Glu Met Gln Arg
130 135 140
Val Thr Lys Glu Glu Tyr Leu Ala Ser Leu Arg Arg Gln Ser Ser Gly
145 150 155 160
Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His
165 170 175
Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr
180 185 190
Leu Tyr Leu Gly Thr Tyr Asn Thr Gln Glu Glu Ala Ala Ala Ala Tyr
195 200 205
Asp Met Ala Ala Ile Glu Tyr Arg Gly Ala Asn Ala Val Thr Asn Phe
210 215 220
Asp Ile Ser Asn Tyr Ile Asp Arg Leu Lys Lys Lys Gly Val Phe Pro
225 230 235 240
Phe Pro Val Asn Gln Ala Asn His Gln Glu Gly Ile Leu Val Glu Ala
245 250 255
Lys Gln Glu Val Glu Thr Arg Glu Ala Lys Glu Glu Pro Arg Glu Glu
260 265 270
Val Lys Gln Gln Tyr Val Glu Glu Pro Pro Gln Glu Glu Glu Glu Lys
275 280 285
Glu Glu Glu Lys Ala Glu Gln Gln Glu Ala Glu Ile Val Gly Tyr Ser
290 295 300
Glu Glu Ala Ala Val Val Asn Cys Cys Ile Asp Ser Ser Thr Ile Met
305 310 315 320
Glu Met Asp Arg Cys Gly Asp Asn Asn Glu Leu Ala Trp Asn Phe Cys
325 330 335
Met Met Asp Thr Gly Phe Ser Pro Phe Leu Thr Asp Gln Asn Leu Ala
340 345 350
Asn Glu Asn Pro Ile Glu Tyr Pro Glu Leu Phe Asn Glu Leu Ala Phe
355 360 365
Glu Asp Asn Ile Asp Phe Met Phe Asp Asp Gly Lys His Glu Cys Leu
370 375 380
Asn Leu Glu Asn Leu Asp Cys Cys Val Val Gly Arg Glu Ser Pro Pro
385 390 395 400
Ser Ser Ser Ser Pro Leu Ser Cys Leu Ser Thr Asp Ser Ala Ser Ser
405 410 415
Thr Thr Thr Thr Thr Thr Ser Val Ser Cys Asn Tyr Leu Val
420 425 430
<210> 23
<211> 430
<212> Белок
<213> Arabidopsis lyrata
<400> 23
Met Lys Arg Arg Leu Thr Thr Ser Thr Ser Ser Ser Ser Pro Ser Ser
1 5 10 15
Ser Val Ser Ser Ser Thr Thr Thr Ser Ser Pro Ile Gln Ser Glu Ala
20 25 30
Pro Arg Pro Lys Arg Ala Lys Arg Ala Lys Lys Ser Ser Pro Ser Gly
35 40 45
Asp Lys Ser His Asn Pro Thr Ser Pro Ala Ser Thr Arg Arg Ser Ser
50 55 60
Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe Glu Ala
65 70 75 80
His Leu Trp Asp Lys Ser Ser Trp Asn Ser Ile Gln Asn Lys Lys Gly
85 90 95
Lys Gln Gly Ala Tyr Asp Ser Glu Glu Ala Ala Ala His Thr Tyr Asp
100 105 110
Leu Ala Ala Leu Lys Tyr Trp Gly Pro Asp Thr Ile Leu Asn Phe Pro
115 120 125
Ala Glu Thr Tyr Thr Lys Glu Leu Glu Glu Met Gln Arg Val Thr Lys
130 135 140
Glu Glu Tyr Leu Ala Ser Leu Arg Arg Gln Ser Ser Gly Phe Ser Arg
145 150 155 160
Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg
165 170 175
Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu
180 185 190
Gly Thr Tyr Asn Thr Gln Glu Glu Ala Ala Ala Ala Tyr Asp Met Ala
195 200 205
Ala Ile Glu Tyr Arg Gly Ala Asn Ala Val Thr Asn Phe Asp Ile Ser
210 215 220
Asn Tyr Ile Asp Arg Leu Lys Lys Lys Gly Val Phe Pro Phe Pro Val
225 230 235 240
Asn Gln Pro Asn His Gln Glu Ala Ile Leu Val Glu Ala Lys Gln Glu
245 250 255
Ile Glu Thr Arg Glu Ala Lys Glu Glu Pro Arg Glu Glu Val Lys Gln
260 265 270
Gln Tyr Val Glu Glu Pro Pro Gln Glu Glu Lys Glu Glu Glu Lys Ala
275 280 285
Glu Gln Gln Glu Ala Glu Phe Val Gly Tyr Lys Asp Glu Gly Ala Val
290 295 300
Val Asn Cys Cys Ile Asp Ser Ser Ala Ile Met Glu Met Asn Arg Cys
305 310 315 320
Gly Asp Asn Asn Glu Leu Ala Trp Asn Phe Cys Met Met Asp Ser Gly
325 330 335
Phe Ala Pro Phe Leu Thr Asp Gln Asn Leu Ser Asn Glu Asn Pro Ile
340 345 350
Glu Tyr Pro Glu Leu Phe Asn Glu Leu Ala Phe Glu Asp Asn Ile Asp
355 360 365
Phe Met Phe Asp Glu Ala Lys Asn Asp Cys Leu Ser Leu Glu Asn Leu
370 375 380
Asp Cys Cys Val Val Gly Arg Glu Ser Pro Thr Ser Ser Ser Ser Pro
385 390 395 400
Leu Ser Cys Phe Ser Thr Asp Ser Ala Ser Ser Thr Thr Thr Thr Thr
405 410 415
Ser Val Ser Cys Asn Tyr Leu Gly Leu Phe Val Gly Ser Glu
420 425 430
<210> 24
<211> 413
<212> Белок
<213> Brassica napus
<400> 24
Met Lys Arg Pro Leu Thr Thr Ser Pro Ser Ser Ser Ser Ser Thr Ser
1 5 10 15
Ser Ser Ala Cys Ile Leu Pro Thr Gln Ser Glu Thr Pro Arg Pro Lys
20 25 30
Arg Ala Lys Arg Ala Lys Lys Ser Ser Leu Arg Ser Asp Val Lys Pro
35 40 45
Gln Asn Pro Thr Ser Pro Ala Ser Thr Arg Arg Ser Ser Ile Tyr Arg
50 55 60
Gly Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp
65 70 75 80
Asp Lys Ser Ser Trp Asn Ser Ile Gln Asn Lys Lys Gly Lys Gln Val
85 90 95
Tyr Leu Gly Ala Tyr Asp Ser Glu Glu Ala Ala Ala His Thr Tyr Asp
100 105 110
Leu Ala Ala Leu Lys Tyr Trp Gly Pro Asn Thr Ile Leu Asn Phe Pro
115 120 125
Val Glu Thr Tyr Thr Lys Glu Leu Glu Glu Met Gln Arg Cys Thr Lys
130 135 140
Glu Glu Tyr Leu Ala Ser Leu Arg Arg Gln Ser Ser Gly Phe Ser Arg
145 150 155 160
Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg
165 170 175
Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu
180 185 190
Gly Thr Tyr Asn Thr Gln Glu Glu Ala Ala Ala Ala Tyr Asp Met Ala
195 200 205
Ala Ile Glu Tyr Arg Gly Ala Asn Ala Val Thr Asn Phe Asp Ile Gly
210 215 220
Asn Tyr Ile Asp Arg Leu Lys Lys Lys Gly Val Phe Pro Phe Pro Val
225 230 235 240
Ser Gln Ala Asn His Gln Glu Ala Val Leu Ala Glu Thr Lys Gln Glu
245 250 255
Val Glu Ala Lys Glu Glu Pro Thr Glu Glu Val Lys Gln Cys Val Glu
260 265 270
Lys Glu Glu Ala Lys Glu Glu Lys Thr Glu Lys Lys Gln Gln Gln Glu
275 280 285
Val Glu Glu Ala Val Ile Thr Cys Cys Ile Asp Ser Ser Glu Ser Asn
290 295 300
Glu Leu Ala Trp Asp Phe Cys Met Met Asp Ser Gly Phe Ala Pro Phe
305 310 315 320
Leu Thr Asp Ser Asn Leu Ser Ser Glu Asn Pro Ile Glu Tyr Pro Glu
325 330 335
Leu Phe Asn Glu Met Gly Phe Glu Asp Asn Ile Asp Phe Met Phe Glu
340 345 350
Glu Gly Lys Gln Asp Cys Leu Ser Leu Glu Asn Leu Asp Cys Cys Asp
355 360 365
Gly Val Val Val Val Gly Arg Glu Ser Pro Thr Ser Leu Ser Ser Ser
370 375 380
Pro Leu Ser Cys Leu Ser Thr Asp Ser Ala Ser Ser Thr Thr Thr Thr
385 390 395 400
Ala Thr Thr Val Thr Ser Val Ser Trp Asn Tyr Ser Val
405 410
<210> 25
<211> 415
<212> Белок
<213> Brassica napus
<400> 25
Met Lys Arg Pro Leu Thr Thr Ser Pro Ser Thr Ser Ser Ser Thr Ser
1 5 10 15
Ser Ser Ala Cys Ile Leu Pro Thr Gln Pro Glu Thr Pro Arg Pro Lys
20 25 30
Arg Ala Lys Arg Ala Lys Lys Ser Ser Ile Pro Thr Asp Val Lys Pro
35 40 45
Gln Asn Pro Thr Ser Pro Ala Ser Thr Arg Arg Ser Ser Ile Tyr Arg
50 55 60
Gly Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp
65 70 75 80
Asp Lys Ser Ser Trp Asn Ser Ile Gln Asn Lys Lys Gly Lys Gln Val
85 90 95
Tyr Leu Gly Ala Tyr Asp Ser Glu Glu Ala Ala Ala His Thr Tyr Asp
100 105 110
Leu Ala Ala Leu Lys Tyr Trp Gly Pro Asp Thr Ile Leu Asn Phe Pro
115 120 125
Ala Glu Thr Tyr Thr Lys Glu Leu Glu Glu Met Gln Arg Cys Thr Lys
130 135 140
Glu Glu Tyr Leu Ala Ser Leu Arg Arg Gln Ser Ser Gly Phe Ser Arg
145 150 155 160
Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg
165 170 175
Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu
180 185 190
Gly Thr Tyr Asn Thr Gln Glu Glu Ala Ala Ala Ala Tyr Asp Met Ala
195 200 205
Ala Ile Glu Tyr Arg Gly Ala Asn Ala Val Thr Asn Phe Asp Ile Ser
210 215 220
Asn Tyr Ile Asp Arg Leu Lys Lys Lys Gly Val Phe Pro Phe Pro Val
225 230 235 240
Ser Gln Ala Asn His Gln Glu Ala Val Leu Ala Glu Ala Lys Gln Glu
245 250 255
Val Glu Ala Lys Glu Glu Pro Thr Glu Glu Val Lys Gln Cys Val Glu
260 265 270
Lys Glu Glu Pro Gln Glu Ala Lys Glu Glu Lys Thr Glu Lys Lys Gln
275 280 285
Gln Gln Gln Glu Val Glu Glu Ala Val Val Thr Cys Cys Ile Asp Ser
290 295 300
Ser Glu Ser Asn Glu Leu Ala Trp Asp Phe Cys Met Met Asp Ser Gly
305 310 315 320
Phe Ala Pro Phe Leu Thr Asp Ser Asn Leu Ser Ser Glu Asn Pro Ile
325 330 335
Glu Tyr Pro Glu Leu Phe Asn Glu Met Gly Phe Glu Asp Asn Ile Asp
340 345 350
Phe Met Phe Glu Glu Gly Lys Gln Asp Cys Leu Ser Leu Glu Asn Leu
355 360 365
Asp Cys Cys Asp Gly Val Val Val Val Gly Arg Glu Ser Pro Thr Ser
370 375 380
Leu Ser Ser Ser Pro Leu Ser Cys Leu Ser Thr Asp Ser Ala Ser Ser
385 390 395 400
Thr Thr Thr Thr Thr Ile Thr Ser Val Ser Cys Asn Tyr Ser Val
405 410 415
<210> 26
<211> 285
<212> Белок
<213> Glycine max
<400> 26
Met Lys Arg Ser Pro Ala Ser Ser Cys Ser Ser Ser Thr Ser Ser Val
1 5 10 15
Gly Phe Glu Val His His Pro Ile Glu Lys Arg Arg Pro Lys His Pro
20 25 30
Arg Arg Asn Asn Leu Lys Ser Gln Lys Cys Lys Gln Asn Gln Thr Thr
35 40 45
Thr Gly Gly Arg Arg Ser Ser Ile Tyr Arg Gly Val Thr Arg His Arg
50 55 60
Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Ser Ser Trp Asn
65 70 75 80
Asn Ile Gln Ser Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala Tyr Asp
85 90 95
Thr Glu Glu Ser Ala Ala Arg Thr Tyr Asp Leu Ala Ala Leu Lys Tyr
100 105 110
Trp Gly Lys Asp Ala Thr Leu Asn Phe Pro Ile Glu Thr Tyr Thr Lys
115 120 125
Asp Leu Glu Glu Met Asp Lys Val Ser Arg Glu Glu Tyr Leu Ala Ser
130 135 140
Leu Arg Arg Gln Ser Ser Gly Phe Ser Arg Gly Ile Ser Lys Tyr Arg
145 150 155 160
Gly Val Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly
165 170 175
Arg Val Cys Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Lys Thr Gln
180 185 190
Glu Glu Ala Ala Val Ala Tyr Asp Met Ala Ala Ile Glu Tyr Arg Gly
195 200 205
Val Asn Ala Val Thr Asn Phe Asp Ile Ser Asn Tyr Met Asp Lys Ile
210 215 220
Lys Lys Lys Asn Asp Gln Thr Leu Gln Gln Gln Gln Thr Glu Val Gln
225 230 235 240
Thr Glu Thr Val Pro Asn Ser Ser Asp Ser Glu Glu Ala Glu Val Glu
245 250 255
Gln Gln His Thr Thr Thr Ile Thr Thr Pro Pro Pro Ser Glu Asn Leu
260 265 270
His Met Leu Pro Gln Glu His Gln Val Gly Gly Trp Val
275 280 285
<210> 27
<211> 417
<212> Белок
<213> Jatropha curcas
<400> 27
Met Lys Arg Ser Ser Ala Ser Ser Cys Ser Ser Ser Ser Ser Ser Ser
1 5 10 15
Ser Ser Pro Ser Ser Ser Ser Ser Ser Ala Cys Ser Ala Ser Ser Ser
20 25 30
Cys Leu Asp Ser Val Ser Pro Pro Asn His His Gln Leu Arg Ser Glu
35 40 45
Lys Ser Lys Ser Lys Arg Ile Arg Lys Ile Gln Thr Lys Gln Asp Lys
50 55 60
Cys Gln Thr Thr Ala Thr Thr Thr Ser Pro Ser Gly Gly Gly Arg Arg
65 70 75 80
Ser Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe
85 90 95
Glu Ala His Leu Trp Asp Lys Ser Ser Trp Asn Asn Ile Gln Asn Lys
100 105 110
Lys Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Asn Glu Glu Ala Ala
115 120 125
Ala His Thr Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Gln Asp Thr
130 135 140
Thr Leu Asn Phe Pro Ile Glu Thr Tyr Ser Lys Glu Leu Glu Glu Met
145 150 155 160
Gln Lys Met Ser Lys Glu Glu Tyr Leu Ala Ser Leu Arg Arg Arg Ser
165 170 175
Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His
180 185 190
His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn
195 200 205
Lys Tyr Leu Tyr Leu Gly Thr Tyr Asn Thr Gln Glu Glu Ala Ala Ala
210 215 220
Ala Tyr Asp Met Ala Ala Ile Glu Tyr Arg Gly Ala Asn Ala Val Thr
225 230 235 240
Asn Phe Asp Val Ser His Tyr Ile Asp Arg Leu Lys Lys Lys Gly Ile
245 250 255
Pro Leu Asp Lys Ile Leu Pro Glu Thr Leu Ser Lys Gly Ser Lys Glu
260 265 270
Ser Glu Glu Ile Glu Arg Thr Ser Pro Leu Pro Leu Pro Ser Pro Pro
275 280 285
Ser Pro Ser Ile Thr Pro Leu His Glu Glu Ile Val Ser Pro Gln Leu
290 295 300
Leu Glu Thr Glu Cys Pro Gln His Pro Pro Cys Met Asp Thr Cys Thr
305 310 315 320
Met Ile Val Met Asp Pro Ile Glu Glu His Glu Leu Thr Trp Ser Phe
325 330 335
Cys Leu Asp Ser Gly Leu Val Pro Leu Pro Val Pro Asp Leu Pro Leu
340 345 350
Ala Asn Gly Cys Glu Leu Pro Asp Leu Leu Asp Asp Thr Gly Phe Glu
355 360 365
Asp Asn Ile Asp Leu Ile Phe Asp Ala Cys Cys Phe Gly Asn Asp Ala
370 375 380
Asn Pro Ala Asp Glu Asn Gly Lys Glu Arg Leu Ser Ser Ala Ser Thr
385 390 395 400
Ser Pro Ser Cys Ser Thr Thr Leu Thr Ser Val Ser Cys Asn Tyr Ser
405 410 415
Val
<210> 28
<211> 443
<212> Белок
<213> Ricinus communis
<400> 28
Met Lys Arg Ser Pro Thr Ser Pro Cys Ser Ser Ser Ser Ser Ser Ser
1 5 10 15
Tyr Ser Ser Ser Ser Ala Ser Ser Ser Cys Val Gly Pro Asp Asp Thr
20 25 30
Pro Val Ala Pro Gly Ser His His His His Asp His His Gln Leu Arg
35 40 45
Ser Gln Lys Ser Ser Lys Arg Ile Arg Lys Val Lys Lys Lys Gln Gln
50 55 60
Asn His Asn Ile Asp Gln Asn Asn Thr Asn Thr Thr Ile Thr Ala Pro
65 70 75 80
Thr Ser Ala Arg Arg Ser Ser Ile Tyr Arg Gly Val Thr Arg His Arg
85 90 95
Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Ser Ser Trp Asn
100 105 110
Asn Ile Gln Asn Lys Lys Gly Arg Gln Gly Ala Tyr Asp Asn Glu Glu
115 120 125
Ala Ala Ala His Thr Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro
130 135 140
Glu Thr Thr Leu Asn Phe Pro Ile Glu Thr Tyr Pro Lys Glu Leu Glu
145 150 155 160
Glu Met Gln Lys Met Ser Lys Glu Glu Tyr Leu Ala Ser Leu Arg Arg
165 170 175
Gln Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala
180 185 190
Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe
195 200 205
Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Asn Thr Gln Glu Glu Ala
210 215 220
Ala Ala Ala Tyr Asp Met Ala Ala Ile Glu Tyr Arg Gly Ala Asn Ala
225 230 235 240
Val Thr Asn Phe Asp Ile Ser Asn Tyr Ile Asp Arg Leu Lys Lys Lys
245 250 255
Gly Ile Leu Leu Asp Gln Ile Leu Pro Asp Gln Pro Leu Arg Lys Cys
260 265 270
Ser Ser Glu Ser Glu Glu Ala Glu Ala Glu Ala Glu Val Glu Arg Leu
275 280 285
Pro Ser Leu Pro Ser Ser Ile Leu Pro Gln Glu Gln Asp Thr Ile Ser
290 295 300
Pro Gln Leu Gln Cys Thr Gln Leu Leu Pro Ser Met Asp Ser Cys Thr
305 310 315 320
Met Ile Asn Met Asp Pro Ile Glu Asp Asn Glu Leu Thr Trp Ser Phe
325 330 335
Cys Leu Asp Ser Gly Leu Thr Leu Phe Ser Val Pro Glu Leu Pro Leu
340 345 350
Glu Asn Ala Cys Glu Leu Pro Asp Leu Phe Asp Asp Thr Gly Phe Glu
355 360 365
Asp Asn Ile Asp Leu Ile Phe Asp Gly Cys Cys Phe Gly Asn Asp Asp
370 375 380
Asp Gly Gly Gly Gly Ala Asn His Gln Glu Phe Met Val Glu Ser Arg
385 390 395 400
Gly Cys Arg Val Gly Glu Val Gly Ile Ser Gly Ser Met Glu Glu Glu
405 410 415
Asn Gly Lys Glu Met Cys Cys Ser Ser Ser Ser Pro Ser Cys Ser Thr
420 425 430
Thr Thr Ser Val Ser Cys Cys Asn Tyr Ser Val
435 440
<210> 29
<211> 402
<212> Белок
<213> Populus trichocarpa
<400> 29
Met Lys Arg Ser Ser Ser Cys Ser Ser Ser Ser Ser Ser Ser Ser Ser
1 5 10 15
Ser Cys Val Ala Ser Glu Ser Ile His Lys Pro Lys Ala Lys Arg Ile
20 25 30
Arg Lys Asn Gln Lys Ser Asn Gln Gly Lys Ser Gln Asn Ala Ala Ala
35 40 45
Ala Ala Ala Asn Asn Ser His Asn Ser Gly Lys Arg Ser Ser Ile Tyr
50 55 60
Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe Glu Ala His Leu
65 70 75 80
Trp Asp Lys Ser Ser Trp Asn Ser Ile Gln Asn Lys Lys Gly Lys Gln
85 90 95
Gly Ala Tyr Asp Asn Glu Glu Ala Ala Ala His Thr Tyr Asp Leu Ala
100 105 110
Ala Leu Lys Tyr Trp Gly Ser Glu Thr Thr Leu Asn Phe Pro Ile Glu
115 120 125
Thr Tyr Thr Lys Glu Ile Glu Glu Met Gln Lys Val Thr Lys Glu Glu
130 135 140
Tyr Leu Ala Ser Leu Arg Arg Gln Ser Ser Gly Phe Ser Arg Gly Val
145 150 155 160
Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu
165 170 175
Ala Arg Ile Gly Arg Val Tyr Gly Asn Lys Tyr Leu Tyr Leu Gly Thr
180 185 190
Tyr Asn Thr Gln Glu Glu Ala Ala Ala Ala Tyr Asp Met Ala Ala Ile
195 200 205
Gln Tyr Arg Gly Ala Asn Ala Val Thr Asn Phe Asp Val Ser Asn Tyr
210 215 220
Ile Glu Arg Leu Arg Lys Lys Gly Ile Pro Ile Asp Arg Ile Leu Gln
225 230 235 240
Glu Gln Gln Leu Leu Asn Asn Ser Val Asp Ser Ser Val Glu Val Glu
245 250 255
Val Glu Gln Pro Thr Pro Pro Pro Gln Gln Gln Gln Glu Glu Gln Glu
260 265 270
Gln Lys Ile Val Ser Ser Ser Ser Gln Leu Gln Cys Ser Gln Leu Asn
275 280 285
Ser Ser Leu Asp Gly Thr Pro Pro Met Val Ile Met Asp Thr Ile Glu
290 295 300
Glu His Glu Leu Ala Trp Ser Phe Cys Met Asp Ser Gly Leu Ser Leu
305 310 315 320
Thr Met Pro Asp Leu Pro Leu Glu Asn Ser Cys Glu Leu Pro Asp Leu
325 330 335
Phe Asp His Thr Gly Phe Glu Asp Asn Ile Asp Leu Ile Phe Asp Ala
340 345 350
Cys Cys Tyr Gly Lys Glu Ala Asn Pro Ala Gly Tyr Thr Leu Glu Asp
355 360 365
Asn Ser Thr Gly Gly Val Glu Glu Asp Arg Leu Ser Ser Asp Ser Val
370 375 380
Ser Asn Ser Pro Thr Ser Ser Thr Thr Thr Ser Val Ser Cys Asn Tyr
385 390 395 400
Ser Val
<210> 30
<211> 409
<212> Белок
<213> Vitis vinifera
<400> 30
Met Val Lys Arg Ser Ser Pro Gly Ser Ser Ser Ser Pro Ser Ser Ser
1 5 10 15
Ser Thr Ser Ser Asp Ala Ala Ser Arg Pro Ala Pro Pro Ser Gly Gly
20 25 30
Lys Pro Lys Ser Arg Lys Lys Glu Ala Lys Lys Asn Ser Asn Gly Asn
35 40 45
Gly Ser Asn Ser Lys Asn Lys Arg Thr Ser Ile Tyr Arg Gly Val Thr
50 55 60
Lys His Arg Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Ser
65 70 75 80
Ser Trp Asn Asp Ile Ser Asn Lys Arg Gly Arg Gln Gly Ala Tyr Tyr
85 90 95
Asn Glu Glu Ala Ala Ala Arg Thr Tyr Asp Leu Ala Ala Leu Lys Tyr
100 105 110
Trp Gly Pro Thr Thr Pro Leu Asn Phe Pro Leu Glu Thr Tyr Gln Lys
115 120 125
Asp Ala Glu Glu Met Glu Lys Met Ser Lys Glu Glu Tyr Leu Ala Leu
130 135 140
Leu Arg Arg Gln Ser Asn Gly Phe Ser Arg Gly Val Ser Lys His His
145 150 155 160
His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Leu Gly Asn Lys
165 170 175
Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr Gln Glu Glu Ala Ala Ala Ala
180 185 190
Tyr Asp Met Ala Ala Ile Glu Tyr Arg Gly Leu Asn Ala Val Thr Asn
195 200 205
Phe Asp Ile Ser Asn Tyr Val Lys Leu Gly Arg Val Glu Ala Gln Val
210 215 220
Gln Glu Leu Ala Gln Gln Leu Gln Pro Asn Thr Pro Ile Gly Pro Gln
225 230 235 240
Asn Glu Leu Gln Lys Glu Glu Glu Glu Gln Leu Gln Glu Pro Val Leu
245 250 255
Ser Ser Ser Gln His Leu Pro Ser Met Asp Ser Ser Ala Met Glu Ile
260 265 270
Met Asp Pro Ala Asp Asp Pro Asp Leu Pro Trp Asn Phe Cys Ala Tyr
275 280 285
Ser Thr Leu Leu Val Pro Asp Val Pro Leu Gly Lys Gly Gly Glu Leu
290 295 300
Ser Asp Leu Phe Tyr Glu Lys Gly Phe Glu Asp Asn Ile Asp Tyr Met
305 310 315 320
Phe Glu Gly Ala Ala Gly Asn Glu Glu Glu Ser Asn Ser Ala Glu Asn
325 330 335
Gly Val Lys Glu Asn Gly Phe Met His Glu Leu Glu Val Asp Gly Lys
340 345 350
Leu Gln Asn Val Val Gly Phe Phe Phe Leu Ser Phe Phe Phe Leu Pro
355 360 365
Lys Arg Ala Gly Ile Arg Lys Arg Gly Val Asp Ser Cys Met Gln Leu
370 375 380
Phe Leu Tyr Phe Val Phe Leu Phe Tyr Pro Phe Leu Pro Glu Val Ser
385 390 395 400
Lys Phe Leu Phe His Leu Ser Leu Asp
405
<210> 31
<211> 420
<212> Белок
<213> Brachypodium distachyon
<400> 31
Met Lys Arg Ser Pro Pro Gln Pro Ser Pro Ser Pro Ser Ser Ser Pro
1 5 10 15
Ala Ser Ser Ser Ser Ser Pro Ser Ser Ser Asp Ser Ser Ser Ser Ile
20 25 30
Ala Ile Pro Arg Lys Arg Ala Arg Thr Ala Ala Ala Ala Ala Gly Gly
35 40 45
Gly Lys Ala Arg Ala Ala Ala Ala Lys Arg Pro Lys Lys Asp Gly Lys
50 55 60
Asp Ser Gly Ser Ser Ser Asn Gly Gly Gly Gly Gly Gly Gly Lys Arg
65 70 75 80
Ser Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe
85 90 95
Glu Ala His Leu Trp Asp Lys Asn Cys Phe Thr Ser Leu Gln Asn Lys
100 105 110
Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Thr Glu Glu Ala
115 120 125
Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Glu
130 135 140
Thr Thr Leu Asn Phe Ser Ala Asp Asp Tyr Gly Lys Glu Arg Ser Glu
145 150 155 160
Met Glu Ala Val Ser Arg Glu Glu Tyr Leu Ala Ala Leu Arg Arg Arg
165 170 175
Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg
180 185 190
His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Leu Gly
195 200 205
Asn Lys Tyr Leu Tyr Leu Gly Thr Phe Asp Thr Gln Glu Glu Ala Ala
210 215 220
Arg Ala Tyr Asp Leu Ala Ala Ile Gln Tyr Arg Gly Ala Asn Ala Val
225 230 235 240
Thr Asn Phe Asp Ile Ser Arg Tyr Leu Asp Gln Pro Gln Leu Leu Glu
245 250 255
Gln Leu Gln Gln Gln Gln Gly Pro Gln Val Val Ala Ala Leu Gln Glu
260 265 270
Glu Ala Gln Arg Asp His Gln Ser Asp Asn Ala Val Gln Glu Leu Asn
275 280 285
Ser Gly Glu Ala Gln Thr Pro Gly Gly Ile Asp Glu Pro Ile Ala Ile
290 295 300
Gly Asp Ser Thr Glu Asp Ile Asn Thr Ser Leu Thr Val Asp Asp Ile
305 310 315 320
Ile Glu Glu Ser Leu Trp Ser Pro Tyr Glu Phe Asp Ile Met Ala Gly
325 330 335
Val Asn Val Ser Asn Ser Met Asn Leu Ser Glu Leu Phe Ser Asp Val
340 345 350
Ala Phe Glu Gly Asn Ile Gly Cys Leu Phe Glu Glu Cys Ser Gly Ile
355 360 365
Asp Asp Cys Ser Ser Arg His Gly Ala Gly Leu Ala Ala Phe Gly Leu
370 375 380
Phe Thr Glu Gly Asp Asp Lys Leu Lys Asp Val Ser Glu Met Glu Met
385 390 395 400
Glu Val Asn Pro Gln Ala Asn Asp Val Ser Cys Pro Pro Lys Met Ile
405 410 415
Thr Val Cys Asn
420
<210> 32
<211> 423
<212> Белок
<213> Hordeum vulgare
<400> 32
Met Lys Arg Ser Pro Pro Pro Gln Pro Ser Pro Ser Ser Ser Pro Ala
1 5 10 15
Cys Ser Pro Ser Pro Ser Ser Pro Ser Ser Ser Asp Ser Ser Ser Ile
20 25 30
Ala Ile Pro Arg Lys Arg Ala Arg Thr Gln Lys Ala Gly Ser Ala Lys
35 40 45
Ala Lys Ala Ala Pro Lys Arg Ala Lys Lys Asp Ser Gly Arg Ser Thr
50 55 60
Lys Asp Ser Asp Ala Ser Ala Asn Gly Ala Ala Ala Ser Gly Lys Arg
65 70 75 80
Ser Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe
85 90 95
Glu Ala His Leu Trp Asp Lys Asn Cys Phe Thr Ser Ile Gln Asn Lys
100 105 110
Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Thr Glu Glu Ala
115 120 125
Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Glu
130 135 140
Thr Thr Leu Asn Phe Thr Val Asp Glu Tyr Ala Lys Glu Arg Ser Glu
145 150 155 160
Met Glu Ala Val Ser Arg Glu Glu Tyr Leu Ala Ala Leu Arg Arg Arg
165 170 175
Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg
180 185 190
His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Leu Gly
195 200 205
Asn Lys Tyr Leu Tyr Leu Gly Thr Phe Asp Thr Gln Glu Glu Ala Ala
210 215 220
Arg Ala Tyr Asp Leu Ala Ala Ile Glu Tyr Arg Gly Ala Asn Ala Val
225 230 235 240
Thr Asn Phe Asp Ile Ser Arg Tyr Leu Asp Gln Pro Gln Leu Leu Ala
245 250 255
Gln Leu Glu Gln Gly Pro Gln Val Val Pro Ala Leu Gln Glu Glu Leu
260 265 270
Gln His Asp His Gln Ser Asp Asn Ala Val Gln Glu Leu Asn Ser Gly
275 280 285
Glu Ala Gln Lys Pro Gly Ser Val Ser Glu Pro Ile Ala Val Asp Asp
290 295 300
Thr Asp Asn Thr Gly Asp Ile Gly Ala Pro Leu Val Phe Asp Ser Gly
305 310 315 320
Val Glu Glu Asn Leu Trp Ser Pro Cys Met Asp Tyr Asp Val Asp Pro
325 330 335
Ile Phe Gly Pro Asn Ile Ser Ser Ser Met Asn Leu Ser Glu Trp Phe
340 345 350
Asn Asp Pro Ala Phe Glu Ser Asn Ile Gly Tyr Met Phe Glu Gly Cys
355 360 365
Ser Asp Val Asp Asp Cys Ser Thr Arg His Gly Ala Gly Leu Ser Ala
370 375 380
Leu Gly Phe Leu Lys Glu Gly Asp Asp Lys Leu Lys Asp Gly Ser Asp
385 390 395 400
Met Glu Ala Glu Ile Thr Pro Gln Ala Asn Asp Val Ser Cys Pro Pro
405 410 415
Lys Met Ile Thr Val Cys Asn
420
<210> 33
<211> 443
<212> Белок
<213> Oryza sativa
<400> 33
Met Ala Lys Arg Ser Ser Pro Asp Pro Ala Ser Ser Ser Pro Ser Ala
1 5 10 15
Ser Ser Ser Pro Ser Ser Pro Ser Ser Ser Ser Ser Glu Asp Ser Ser
20 25 30
Ser Pro Met Ser Met Pro Cys Lys Arg Arg Ala Arg Pro Arg Thr Asp
35 40 45
Lys Ser Thr Gly Lys Ala Lys Arg Pro Lys Lys Glu Ser Lys Glu Val
50 55 60
Val Asp Pro Ser Ser Asn Gly Gly Gly Gly Gly Lys Arg Ser Ser Ile
65 70 75 80
Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe Glu Ala His
85 90 95
Leu Trp Asp Lys Asn Cys Ser Thr Ser Leu Gln Asn Lys Lys Lys Gly
100 105 110
Arg Gln Gly Ala Tyr Asp Ser Glu Glu Ala Ala Ala Arg Ala Tyr Asp
115 120 125
Leu Ala Ala Leu Lys Tyr Trp Gly Pro Glu Thr Val Leu Asn Phe Pro
130 135 140
Leu Glu Glu Tyr Glu Lys Glu Arg Ser Glu Met Glu Gly Val Ser Arg
145 150 155 160
Glu Glu Tyr Leu Ala Ser Leu Arg Arg Arg Ser Ser Gly Phe Ser Arg
165 170 175
Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg
180 185 190
Trp Glu Ala Arg Ile Gly Arg Val Leu Gly Asn Lys Tyr Leu Tyr Leu
195 200 205
Gly Thr Phe Asp Thr Gln Glu Glu Ala Ala Lys Ala Tyr Asp Leu Ala
210 215 220
Ala Ile Glu Tyr Arg Gly Ala Asn Ala Val Thr Asn Phe Asp Ile Ser
225 230 235 240
Cys Tyr Leu Asp Gln Pro Gln Leu Leu Ala Gln Leu Gln Gln Glu Pro
245 250 255
Gln Leu Leu Ala Gln Leu Gln Gln Glu Pro Gln Val Val Pro Ala Leu
260 265 270
His Glu Glu Pro Gln Asp Asp Asp Arg Ser Glu Asn Ala Val Gln Glu
275 280 285
Leu Ser Ser Ser Glu Ala Asn Thr Ser Ser Asp Asn Asn Glu Pro Leu
290 295 300
Ala Ala Asp Asp Ser Ala Glu Cys Met Asn Glu Pro Leu Pro Ile Val
305 310 315 320
Asp Gly Ile Glu Glu Ser Leu Trp Ser Pro Cys Leu Asp Tyr Glu Leu
325 330 335
Asp Thr Met Pro Gly Ala Tyr Phe Ser Asn Ser Met Asn Phe Ser Glu
340 345 350
Trp Phe Asn Asp Glu Ala Phe Glu Gly Gly Met Glu Tyr Leu Phe Glu
355 360 365
Gly Cys Ser Ser Ile Thr Glu Gly Gly Asn Ser Met Asp Asn Ser Gly
370 375 380
Val Thr Glu Tyr Asn Leu Phe Glu Glu Cys Asn Met Leu Glu Lys Asp
385 390 395 400
Ile Ser Asp Phe Leu Asp Lys Asp Ile Ser Asp Phe Leu Asp Lys Asp
405 410 415
Ile Ser Ile Ser Asp Gly Glu Arg Ile Ser Pro Gln Ala Asn Asn Ile
420 425 430
Ser Cys Pro Gln Lys Met Ile Ser Val Cys Asn
435 440
<210> 34
<211> 420
<212> Белок
<213> Sorghum bicolor
<400> 34
Met Asp Met Glu Arg Ser Gln Gln Gln Lys Ser Pro Thr Glu Ser Pro
1 5 10 15
Pro Pro Pro Ser Pro Ser Ser Ser Ser Ser Ser Val Ser Ala Asp Thr
20 25 30
Val Leu Pro Pro Pro Gly Lys Arg Arg Arg Ala Ala Thr Thr Ala Lys
35 40 45
Ala Lys Ala Gly Ala Lys Pro Lys Arg Ala Arg Lys Asp Ala Ala Ala
50 55 60
Ala Ala Asp Pro Pro Pro Pro Pro Ala Ala Ala Ala Ala Gly Lys Arg
65 70 75 80
Ser Ser Val Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe
85 90 95
Glu Ala His Leu Trp Asp Lys His Cys Leu Ala Ala Leu His Asn Lys
100 105 110
Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Ser Glu Glu Ala
115 120 125
Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Glu
130 135 140
Thr Leu Leu Asn Phe Pro Val Glu Asp Tyr Ser Ser Glu Met Pro Glu
145 150 155 160
Met Glu Gly Val Ser Arg Glu Glu Tyr Leu Ala Ser Leu Arg Arg Arg
165 170 175
Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg
180 185 190
His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly
195 200 205
Asn Lys Tyr Leu Tyr Leu Gly Thr Phe Asp Thr Gln Glu Glu Ala Ala
210 215 220
Lys Ala Tyr Asp Leu Ala Ala Ile Glu Tyr Arg Gly Val Asn Ala Val
225 230 235 240
Thr Asn Phe Asp Ile Ser Cys Tyr Leu Asp His Pro Leu Phe Leu Ala
245 250 255
Gln Leu Gln Gln Glu Pro Gln Val Val Pro Ala Leu Asn Gln Glu Ala
260 265 270
Gln Pro Asp Gln Ser Glu Thr Glu Thr Ile Ala Gln Glu Ser Val Ser
275 280 285
Ser Glu Ala Lys Thr Pro Asp Asp Asn Ala Glu Pro Asp Asp Asn Ala
290 295 300
Glu Pro Asp Asp Ile Ala Glu Pro Leu Ile Thr Val Asp Asp Ser Ile
305 310 315 320
Glu Glu Ser Leu Trp Ser Pro Cys Met Asp Tyr Glu Leu Asp Thr Met
325 330 335
Ser Arg Ser Asn Phe Gly Ser Ser Ile Asn Leu Ser Glu Trp Phe Asn
340 345 350
Asp Ala Asp Phe Asp Ser Asn Ile Gly Cys Leu Phe Asp Gly Cys Ser
355 360 365
Ala Val Asp Glu Gly Gly Lys Asp Gly Val Gly Leu Ala Asp Phe Ser
370 375 380
Leu Leu Glu Asp Phe Ser Leu Phe Glu Ala Gly Asp Gly Gln Leu Lys
385 390 395 400
Asp Val Leu Ser Asp Met Glu Glu Gly Ile Gln Pro Pro Thr Met Ile
405 410 415
Ser Val Cys Asn
420
<210> 35
<211> 395
<212> Белок
<213> Zea mays
<400> 35
Met Glu Arg Ser Gln Arg Gln Ser Pro Pro Pro Pro Ser Pro Ser Ser
1 5 10 15
Ser Ser Ser Ser Val Ser Ala Asp Thr Val Leu Val Pro Pro Gly Lys
20 25 30
Arg Arg Arg Ala Ala Thr Ala Lys Ala Gly Ala Glu Pro Asn Lys Arg
35 40 45
Ile Arg Lys Asp Pro Ala Ala Ala Ala Ala Gly Lys Arg Ser Ser Val
50 55 60
Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe Glu Ala His
65 70 75 80
Leu Trp Asp Lys His Cys Leu Ala Ala Leu His Asn Lys Lys Lys Gly
85 90 95
Arg Gln Val Tyr Leu Gly Ala Tyr Asp Ser Glu Glu Ala Ala Ala Arg
100 105 110
Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Glu Thr Leu Leu
115 120 125
Asn Phe Pro Val Glu Asp Tyr Ser Ser Glu Met Pro Glu Met Glu Ala
130 135 140
Val Ser Arg Glu Glu Tyr Leu Ala Ser Leu Arg Arg Arg Ser Ser Gly
145 150 155 160
Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His
165 170 175
Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr
180 185 190
Leu Tyr Leu Gly Thr Phe Asp Thr Gln Glu Glu Ala Ala Lys Ala Tyr
195 200 205
Asp Leu Ala Ala Ile Glu Tyr Arg Gly Val Asn Ala Val Thr Asn Phe
210 215 220
Asp Ile Ser Cys Tyr Leu Asp His Pro Leu Phe Leu Ala Gln Leu Gln
225 230 235 240
Gln Glu Pro Gln Val Val Pro Ala Leu Asn Gln Glu Pro Gln Pro Asp
245 250 255
Gln Ser Glu Thr Gly Thr Thr Glu Gln Glu Pro Glu Ser Ser Glu Ala
260 265 270
Lys Thr Pro Asp Gly Ser Ala Glu Pro Asp Glu Asn Ala Val Pro Asp
275 280 285
Asp Thr Ala Glu Pro Leu Ser Thr Val Asp Asp Ser Ile Glu Glu Gly
290 295 300
Leu Trp Ser Pro Cys Met Asp Tyr Glu Leu Asp Thr Met Ser Arg Pro
305 310 315 320
Asn Phe Gly Ser Ser Ile Asn Leu Ser Glu Trp Phe Ala Asp Ala Asp
325 330 335
Phe Asp Cys Asn Ile Gly Cys Leu Phe Asp Gly Cys Ser Ala Ala Asp
340 345 350
Glu Gly Ser Lys Asp Gly Val Gly Leu Ala Asp Phe Ser Leu Phe Glu
355 360 365
Ala Gly Asp Val Gln Leu Lys Asp Val Leu Ser Asp Met Glu Glu Gly
370 375 380
Ile Gln Pro Pro Ala Met Ile Ser Val Cys Asn
385 390 395
<210> 36
<211> 413
<212> Белок
<213> Brachypodium distachyon
<400> 36
Met Glu Ala Tyr Cys Ser Thr Leu Val Lys Asp Glu Leu Ile Asn Gly
1 5 10 15
Gly Gly Gly Gly Ser Ala Gly Gly Met Arg Tyr Cys Glu Ala Ala Pro
20 25 30
Arg Val Ser Pro Pro Val Ala Ile Lys Ser Val Lys Arg Arg Lys Arg
35 40 45
Glu Pro Pro Ala Val Ser Gly Met Thr Thr Val Ser Gly Gly Gly Lys
50 55 60
Asp Gly Asp Lys Ser Ala Gly Asn Ala Ala Ala Lys Arg Ser Ser Arg
65 70 75 80
Phe Arg Gly Val Ser Arg His Arg Trp Thr Gly Arg Phe Glu Ala His
85 90 95
Leu Trp Asp Lys Gly Thr Trp Asn Pro Thr Gln Lys Lys Lys Gly Lys
100 105 110
Gln Val Tyr Leu Gly Ala Tyr Asn Glu Glu Glu Ala Ala Ala Arg Ala
115 120 125
Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Thr Thr Tyr Thr Asn
130 135 140
Phe Pro Val Val Asp Tyr Glu Lys Glu Leu Lys Val Met Gln Gly Val
145 150 155 160
Ser Arg Glu Glu Tyr Leu Ala Ser Ile Arg Arg Lys Ser Asn Gly Phe
165 170 175
Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn
180 185 190
Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu
195 200 205
Tyr Leu Gly Thr Tyr Ser Thr Gln Glu Glu Ala Ala Arg Ala Tyr Asp
210 215 220
Ile Ala Ala Ile Glu Tyr Arg Gly Ile Asn Ala Val Thr Asn Phe Asp
225 230 235 240
Leu Ser Ser Tyr Ile Arg Trp Leu Lys Pro Asn Ser Thr Ile Asn Thr
245 250 255
Asn Thr Pro Ala Ala Glu Leu Ala Ile Leu Gly Gly Gly Gly Thr Pro
260 265 270
Ala Ala Leu Ile Thr Pro Pro Pro Thr Met His Val Pro Arg Leu Leu
275 280 285
Pro Pro Leu Val Lys Gly Arg Gly Ser Ser Ile Ala Asp Asp Val Ser
290 295 300
Ala Gly Ser Cys Val Phe Gly Gly Pro Ser Pro Ser Pro Ser Pro Thr
305 310 315 320
Thr Thr Ala Leu Ser Leu Leu Leu Arg Ser Ser Val Phe Gln Glu Leu
325 330 335
Val Ala Gln Gln Gln Pro Pro Ser Thr Val Asp Asp Asp Asp Asp Ile
340 345 350
Gly Gly His Ala Ala Val Ser Asp Ala Ala Gln Arg Ala Ala Glu Glu
355 360 365
Asn Glu Glu Ser Phe Gly Glu Val Leu Tyr Gly Ala Gly Glu Gly Glu
370 375 380
Ala Ala Thr Ala Phe Ser Cys Ser Met Tyr Glu Leu Gly Leu Asp Asp
385 390 395 400
Asn Phe Ala Arg Ile Glu Glu Ser Leu Trp Gly Cys Leu
405 410
<210> 37
<211> 423
<212> Белок
<213> Brachypodium sylvaticum
<400> 37
Met Glu Ala Tyr Cys Ser Ser Leu Val Lys Asp Glu Leu Ile Asn Gly
1 5 10 15
Gly Gly Gly Gly Ala Gly Gly Met Arg Tyr Cys Glu Ala Ala Pro Arg
20 25 30
Val Ser Pro Pro Val Ala Ile Lys Ser Val Lys Arg Arg Lys Arg Glu
35 40 45
Pro Pro Ala Val Ser Gly Met Thr Thr Val Ser Gly Gly Gly Gly Gly
50 55 60
Asn Gly Lys Asp Gly Asp Lys Ser Ala Gly Asn Ala Ala Ala Ala Lys
65 70 75 80
Arg Ser Ser Arg Phe Arg Gly Val Ser Arg His Arg Trp Thr Gly Arg
85 90 95
Phe Glu Ala His Leu Trp Asp Lys Gly Thr Trp Asn Pro Thr Gln Lys
100 105 110
Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala Tyr Asn Glu Glu Glu Ala
115 120 125
Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Thr
130 135 140
Thr Tyr Thr Asn Phe Pro Val Val Asp Tyr Glu Lys Glu Leu Lys Val
145 150 155 160
Met Gln Gly Val Ser Arg Glu Glu Tyr Leu Ala Ser Ile Arg Arg Lys
165 170 175
Ser Asn Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg
180 185 190
His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly
195 200 205
Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr Gln Glu Glu Ala Ala
210 215 220
Arg Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg Gly Ile Asn Ala Val
225 230 235 240
Thr Asn Phe Asp Leu Ser Ser Tyr Ile Arg Trp Leu Lys Pro Asn Ser
245 250 255
Ala Ala Asn Thr Asn Thr Pro Pro Ala Ala Ala Ala Glu Leu Ala Ile
260 265 270
Leu Gly Gly Ala Pro Ala Ala Leu Ile Ser Pro Ala Pro Ala Pro Thr
275 280 285
Thr Met Arg Val Pro Arg Leu Leu Pro Pro Leu Val Arg Gly Arg Gly
290 295 300
Gly Ser Ile Pro Asp Asp Val Ser Ala Gly Gly Ser Cys Val Phe Gly
305 310 315 320
Ser Pro Ser Pro Ser Pro Ser Pro Thr Thr Thr Ser Ala Leu Ser Leu
325 330 335
Leu Leu Arg Ser Ser Val Phe Gln Glu Leu Val Ala Gln Gln Gln Pro
340 345 350
Pro Ser Ile Val Asp Asp Asp Asp Gly Val Gly Gly Gln Glu Ala Val
355 360 365
Ser Asp Ala Ala Glu Arg Ala Ala Glu Glu Asn Glu Glu Ser Phe Gly
370 375 380
Glu Val Leu Tyr Gly Ala Gly Glu Gly Glu Ala Ala Ala Ala Phe Ser
385 390 395 400
Cys Ser Met Tyr Glu Leu Gly Leu Asp Asp Ser Phe Ala Arg Ile Glu
405 410 415
Glu Ser Leu Trp Gly Cys Leu
420
<210> 38
<211> 399
<212> Белок
<213> Oryza sativa
<400> 38
Met Glu Thr Tyr Gly Leu Val Lys Asp Glu Leu Leu His Gly Ile Gly
1 5 10 15
Gly Gly Gln Gly Arg Leu Tyr Cys Glu Val Lys Pro Thr Ala Ala Pro
20 25 30
Ala Val Ile Thr Ala Ala Gly Gly Gly Ala Lys Ser Val Lys Arg Arg
35 40 45
Lys Arg Glu Pro Ser Ala Ala Ala Met Ser Ala Val Thr Val Ala Gly
50 55 60
Asn Gly Lys Glu Ala Gly Gly Ser Asn Ala Ala Asn Lys Arg Ser Ser
65 70 75 80
Arg Phe Arg Gly Val Ser Arg His Arg Trp Thr Gly Arg Phe Glu Ala
85 90 95
His Leu Trp Asp Lys Gly Thr Trp Asn Pro Thr Gln Lys Lys Lys Gly
100 105 110
Lys Gln Val Tyr Leu Gly Ala Tyr Asn Glu Glu Asp Ala Ala Ala Arg
115 120 125
Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Thr Thr Tyr Thr
130 135 140
Asn Phe Pro Val Ala Asp Tyr Glu Lys Glu Leu Lys Leu Met Gln Gly
145 150 155 160
Val Ser Lys Glu Glu Tyr Leu Ala Ser Ile Arg Arg Lys Ser Asn Gly
165 170 175
Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His
180 185 190
Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr
195 200 205
Leu Tyr Leu Gly Thr Tyr Ser Thr Gln Glu Glu Ala Ala Arg Ala Tyr
210 215 220
Asp Ile Ala Ala Ile Glu Tyr Arg Gly Ile Asn Ala Val Thr Asn Phe
225 230 235 240
Asp Leu Ser Thr Tyr Ile Arg Trp Leu Lys Pro Pro Ser Ser Ser Ser
245 250 255
Ala Ala Gly Thr Pro His His His Gly Gly Gly Met Val Val Gly Ala
260 265 270
Asp Arg Val Leu Ala Pro Ala Gln Ser Tyr Pro Ile Ser Ala Ala Ala
275 280 285
Asp Asp Asp Val Ala Gly Cys Trp Arg Pro Leu Pro Ser Pro Ser Ser
290 295 300
Ser Thr Thr Thr Ala Leu Ser Leu Leu Leu Arg Ser Ser Met Phe Gln
305 310 315 320
Glu Leu Val Ala Arg Gln Pro Val Val Glu Gly Asp Asp Gly Gln Leu
325 330 335
Ala Val Val Ser Gly Asp Asp Ala Asp Ala Asp Ala Asp Ser Asp Val
340 345 350
Lys Glu Pro Pro Pro Glu Ser Glu Tyr Gly Glu Val Phe Ala Ser Asp
355 360 365
Glu Ala Ala Ala Ala Ala Ala Tyr Gly Cys Ser Met Tyr Glu Leu Asp
370 375 380
Asp Ser Phe Ala Leu Ile Asp Asp Ser Val Trp Asn Cys Leu Ile
385 390 395
<210> 39
<211> 488
<212> Белок
<213> Sorghum bicolor
<400> 39
Met Glu Thr Tyr Ser Leu Gln Val Lys Asp Glu Leu His Gly Gly Gly
1 5 10 15
Ile Gly Ile Gly Gly Gly Gly Gln Gly Leu Tyr Cys Gly Ala Thr Pro
20 25 30
Arg Pro Ala Ala Pro Ala Ala Thr Gly Gly Gly Gly Gly Gly Gly Asp
35 40 45
Gly Ala Val Lys Ser Asn Lys Arg Ser Arg Lys Arg Glu Pro Pro Pro
50 55 60
Pro Pro Pro Ser Ser Leu Val Thr Met Ser Asn Gly Gly Lys Asp Glu
65 70 75 80
Ala Val Ala Gly Ser Gly Asp Lys Ser Ala Ser Ser Asn Ser Asn Ala
85 90 95
Ser Lys Arg Ser Ser Arg Phe Arg Gly Val Ser Arg His Arg Trp Thr
100 105 110
Gly Arg Phe Glu Ala His Leu Trp Asp Lys Gly Thr Trp Asn Pro Thr
115 120 125
Gln Lys Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala Tyr Asn Glu Glu
130 135 140
Asp Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly
145 150 155 160
Pro Thr Thr Tyr Thr Asn Phe Pro Val Val Asp Tyr Glu Arg Glu Leu
165 170 175
Lys Val Met Gln Asn Val Ser Lys Glu Glu Tyr Leu Ala Ser Ile Arg
180 185 190
Arg Lys Ser Asn Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val
195 200 205
Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val
210 215 220
Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr Gln Glu Glu
225 230 235 240
Ala Ala Arg Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg Gly Ile Asn
245 250 255
Ala Val Thr Asn Phe Asp Leu Ser Thr Tyr Ile Arg Trp Leu Lys Pro
260 265 270
Gly Gly Gly Val Glu Asp Ser Ala Ala Gly Thr Pro Thr Ser Gly Val
275 280 285
Arg Ala Pro Gly Ile Pro Pro Ala Ser Leu Ser Leu Gln Ala Gly Gly
290 295 300
Leu Leu Gln His Pro His Gly Ala Ala Ala Gly Met Leu Gln Val Asp
305 310 315 320
Val Asp Asp Leu Tyr Arg Gly Gln Leu Ala Ala Ala Arg Gly Ala Ala
325 330 335
Leu Phe Ser Gly Gly Ile Asp Asp Val Gly Ser Val Tyr Ala Ala Gly
340 345 350
Ser Ala Gly Pro Ser Pro Thr Ala Leu Cys Ala Gly Arg Pro Ser Pro
355 360 365
Ser Pro Ser Pro Ser Ser Ser Thr Thr Ala Leu Ser Leu Leu Leu Arg
370 375 380
Ser Ser Val Phe Gln Glu Leu Val Ala Arg Asn Ala Gly Gly Gly Ala
385 390 395 400
Ala Gln Gln Gln Gln Leu Val Val Ala Asp Asp Asp Gly Ala Val Ser
405 410 415
Pro Ala Asp Val Val Asp Ala Lys Val Glu Gln Pro Glu Ala Glu Gly
420 425 430
Glu Leu Gly Arg His Gly Asp Gln Leu Tyr Gly Ala Ala Arg Ala Asp
435 440 445
Glu Asp Glu Asp Ala Phe Ala Cys Ser Met Tyr Glu Leu Asp Asp Ser
450 455 460
Phe Ala Arg Met Glu Gln Ser Leu Trp Gly Cys Leu Arg Ser Ser Asp
465 470 475 480
Ala Pro Asp Asn Met Asn Asn Leu
485
<210> 40
<211> 443
<212> Белок
<213> Sorghum bicolor
<400> 40
Met Glu Ser Ser Gly Met Met Met Val Lys Ser Glu Ile Glu Ser Cys
1 5 10 15
Gly Tyr Pro Gly Pro Ser Ser Ser Thr Ala Pro Ala Ala Gly Val Val
20 25 30
Ile Gly Gly Ser Ala Thr Thr Glu Arg Gly Glu Gly Gly His His His
35 40 45
His His His Gln Val Val Val Arg Arg Arg Arg Arg Glu Pro Pro Leu
50 55 60
Leu Ala Pro Ile Ala Gly Gly Gly Ile Gly Lys Pro Leu Pro Ser Ile
65 70 75 80
Thr Val Lys Arg Ser Ser Arg Phe Arg Gly Val Ser Arg His Arg Trp
85 90 95
Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Asn Ser Trp Asn Pro
100 105 110
Thr Gln Arg Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala Tyr Asp Glu
115 120 125
Glu Glu Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp
130 135 140
Gly Pro Thr Thr Tyr Thr Asn Phe Pro Val Met Asp Tyr Glu Lys Glu
145 150 155 160
Leu Lys Ile Met Glu Asn Leu Thr Lys Glu Glu Tyr Leu Ala Ser Leu
165 170 175
Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly
180 185 190
Val Ala Arg His His Gln Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg
195 200 205
Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr Gln Glu
210 215 220
Glu Ala Ala Arg Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Lys Gly Val
225 230 235 240
Asn Ala Val Thr Asn Phe Asp Leu Arg Ser Tyr Ile Thr Trp Leu Lys
245 250 255
Pro Ser Gly Ala Pro Ala Ala Phe Asn Pro Glu Ala Ala Leu Leu Met
260 265 270
Gln Ala Ala Pro Ala Glu Gln Leu Leu His Pro Ala Glu Thr Ala Gln
275 280 285
Met Leu Pro Arg Val Gly Asn Pro Phe Leu Leu Asp His Gly Ala Ala
290 295 300
Pro Pro Gly Ser Ser Gly Gly Gly Gly Gln Asp Ala Ser Met Ser Ser
305 310 315 320
Met Val Ser Pro Gly Ala Gly Gly Gly Met Arg Arg Arg Gly Ser Ser
325 330 335
Thr Ala Leu Ser Leu Leu Leu Lys Ser Ser Met Phe Arg Gln Leu Val
340 345 350
Glu Lys Asn Ser Asp Ala Glu Glu Gly Val Arg Asp Arg Glu Asp Ala
355 360 365
Ala Ala Ala Ala Ala Ala Ala His Pro Ala Gly Pro Gly Asp Ala Tyr
370 375 380
Glu Tyr His Asn Phe Phe Gln Gly Glu Ala Pro Pro Asp Met Cys Asp
385 390 395 400
Leu Phe Ser Ser Gly Gly Gly Gly Asp His Ala Arg Asn Ala Gly Phe
405 410 415
His Gly Glu Ile Ala Ala Cys Tyr Asp Asp Gly Glu Gly Leu Asp Gly
420 425 430
Trp Asn Gly Phe Gly Asn Met Ser Ser Leu Gln
435 440
<210> 41
<211> 407
<212> Белок
<213> Glycine max
<400> 41
Met Glu Leu Ala Pro Val Lys Ser Glu Leu Ser Pro Arg Ser His Arg
1 5 10 15
Leu Leu Met Ile Asp Gly Ser Glu Val Ile Gly Thr Lys Cys Val Lys
20 25 30
Arg Arg Arg Arg Asp Ser Ser Thr Ala Val Leu Gly Gly Asn Gly Gln
35 40 45
Gln Gly Glu Gln Leu Glu Glu Gln Lys Gln Leu Gly Gly Gln Ser Thr
50 55 60
Ala Thr Thr Val Lys Arg Ser Ser Arg Phe Arg Gly Val Ser Arg His
65 70 75 80
Arg Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Gly Thr Trp
85 90 95
Asn Pro Thr Gln Lys Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala Tyr
100 105 110
Asn Asp Glu Glu Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys
115 120 125
Tyr Trp Gly Ile Ser Thr Phe Thr Asn Phe Pro Val Ser Asp Tyr Glu
130 135 140
Lys Glu Ile Glu Ile Met Lys Thr Val Thr Lys Glu Glu Tyr Leu Ala
145 150 155 160
Ser Leu Arg Arg Arg Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr
165 170 175
Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile
180 185 190
Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr
195 200 205
Gln Glu Glu Ala Ala Arg Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg
210 215 220
Gly Ile Asn Ala Val Thr Asn Phe Asp Leu Ser Thr Tyr Ile Arg Trp
225 230 235 240
Leu Arg Pro Gly Thr His Pro Thr Ala Ser His Asp Gln Lys Pro Ser
245 250 255
Thr Asp Ala Gln Pro Phe Ala Thr Ser Asn Ser Met Gln Ala Arg Gly
260 265 270
Asn Ile Glu Val Ser Asn Ser Asn Lys Asn Ser Phe Pro Ser Gly Lys
275 280 285
Leu Asp Ser Thr Lys Lys Arg Asp Phe Ser Lys Tyr Met Asn Pro Leu
290 295 300
Ser Pro Cys Asn Lys Pro Ser Ser Pro Thr Ala Leu Gly Leu Leu Leu
305 310 315 320
Lys Ser Ser Val Phe Arg Glu Leu Met Gln Arg Asn Leu Asn Ser Ser
325 330 335
Ser Glu Glu Ala Glu Glu Val Glu Leu Lys Tyr Pro His Glu Gly Asn
340 345 350
Asp Gly Val Gly Gly Ile Tyr Asp Asn Glu Asn Thr Asn Asn Ser Tyr
355 360 365
Phe Cys Ser Ser Asn Ile Ser Arg Leu Pro Asn Leu Glu Ser Ser Glu
370 375 380
Glu Ser Pro Leu Pro Met Tyr His Gly Thr Val Gln Ser Leu Trp Asn
385 390 395 400
Ser Ala Phe Asn Met Ser Asn
405
<210> 42
<211> 406
<212> Белок
<213> Glycine max
<400> 42
Met Glu Leu Ala Pro Val Lys Ser Glu Leu Ser Pro Arg Ser His Arg
1 5 10 15
Leu Val Ile Ile Asp Gly Ser Asp Val Ile Ser Thr Lys Cys Ala Lys
20 25 30
Arg Arg Arg Arg Asp Ser Ser Met Ala Val Leu Gly Gly Asn Gly Gln
35 40 45
Gln Gly Glu Gln Leu Glu Glu Gln Lys Gln Leu Gly Gly Gln Ser Thr
50 55 60
Ala Thr Thr Val Lys Arg Ser Ser Arg Phe Arg Gly Val Ser Arg His
65 70 75 80
Arg Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Gly Thr Trp
85 90 95
Asn Pro Thr Gln Lys Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala Tyr
100 105 110
Asn Asp Glu Glu Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys
115 120 125
Tyr Trp Gly Thr Ser Thr Phe Thr Asn Phe Pro Val Ser Asp Tyr Glu
130 135 140
Lys Glu Ile Glu Ile Met Lys Thr Val Thr Lys Glu Glu Tyr Leu Ala
145 150 155 160
Ser Leu Arg Arg Arg Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr
165 170 175
Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile
180 185 190
Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr
195 200 205
Gln Glu Glu Ala Ala Arg Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg
210 215 220
Gly Ile Asn Ala Val Thr Asn Phe Asp Leu Ser Thr Tyr Ile Arg Trp
225 230 235 240
Leu Arg Pro Gly Thr His Pro Thr Ala Ser His Asp Gln Lys Pro Ser
245 250 255
Thr Asp Ala Gln Leu Phe Ala Thr Ser Asn Ser Met Gln Thr Arg Gly
260 265 270
Asn Ile Glu Val Ser Asn Ser Asn Met His Ser Phe Pro Ser Gly Glu
275 280 285
Leu Asp Ser Thr Lys Lys Arg Asp Phe Ser Lys Tyr Met Asn Pro Leu
290 295 300
Ser Pro Cys Asn Lys Pro Ser Ser Pro Thr Ala Leu Gly Leu Leu Leu
305 310 315 320
Lys Ser Ser Val Phe Arg Glu Leu Met Gln Arg Asn Leu Asn Ser Ser
325 330 335
Ser Glu Glu Ala Asp Val Glu Leu Lys Tyr Pro Gln Glu Gly Asn Asp
340 345 350
Gly Val Gly Gly Ile Tyr Asp Asn Asp Asn Thr Ser Asn Ser Tyr Phe
355 360 365
Cys Ser Ser Asn Ile Ser Arg Leu Pro Asn Leu Glu Ser Ser Glu Glu
370 375 380
Cys Pro Leu Pro Met Tyr His Gly Thr Met Gln Ser Leu Trp Asn Ser
385 390 395 400
Ala Phe Asn Met Ser Asn
405
<210> 43
<211> 418
<212> Белок
<213> Populus trichocarpa
<400> 43
Met Glu Met Thr Arg Asn Thr Gly Asp Gln Ile Ser Leu Gly Arg Arg
1 5 10 15
Arg Leu Cys Met Ile Glu Glu Glu Arg Arg Ala Gly Glu Ala Gly Lys
20 25 30
Cys Ile Lys Arg Arg Arg Arg Asp Pro Ser Thr Phe Ala Leu Ser Cys
35 40 45
Asn Ile Asn Asp Gln Gln Ser Asp Gln Gln Gln Gln Gln Gln Ser Leu
50 55 60
Gly Asp Arg Thr Ala Ala Val Ala Thr Thr Val Lys Arg Ser Ser Arg
65 70 75 80
Phe Arg Gly Val Ser Arg His Arg Trp Thr Gly Arg Phe Glu Ala His
85 90 95
Leu Trp Asp Lys Gly Thr Trp Asn Pro Thr Gln Arg Lys Lys Gly Lys
100 105 110
Gln Gly Ala Tyr Asp Glu Glu Glu Ser Ala Ala Arg Ala Tyr Asp Leu
115 120 125
Ala Ala Leu Lys Tyr Trp Gly Thr Ser Thr Phe Thr Asn Phe Pro Ala
130 135 140
Ser Asp Tyr Glu Lys Glu Ile Glu Ile Met Lys Thr Val Thr Lys Glu
145 150 155 160
Glu Tyr Leu Ala Ser Leu Arg Arg Arg Ser Ser Gly Phe Ser Arg Gly
165 170 175
Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg Trp
180 185 190
Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly
195 200 205
Thr Tyr Ser Thr Gln Glu Glu Ala Ala His Ala Tyr Asp Ile Ala Ala
210 215 220
Ile Glu Tyr Arg Gly Ile Asn Ala Val Thr Asn Phe Asp Leu Ser Thr
225 230 235 240
Tyr Ile Arg Trp Leu Lys Pro Glu Ala Ser Leu Pro Ala Pro Gln Thr
245 250 255
Gln Glu Ser Lys Pro Ala Ser Asp Pro Leu Pro Met Ala Thr Phe Ser
260 265 270
Asn His Leu Pro Ser Glu Lys Pro Thr Gln Leu Ser Val Leu Gln Met
275 280 285
Asp Pro Ser Leu Met Asp Asn Leu Asn Thr Pro Lys Asn Glu Asp Ile
290 295 300
Phe His Arg Lys Thr Leu Pro Val Ser Pro Leu Thr Arg Ser Ser Ser
305 310 315 320
Ser Thr Ala Leu Ser Leu Leu Phe Lys Ser Ser Ile Phe Lys Glu Leu
325 330 335
Val Glu Lys Asn Leu Asn Thr Thr Ser Glu Glu Ile Glu Glu Asn Asp
340 345 350
Ser Lys Asn Pro His Asn Gly Asn Asn Asn Ala Gly Glu Ala Phe Tyr
355 360 365
Asp Gly Leu Ser Pro Ile Pro His Thr Gly Thr Ser Thr Glu Asp Pro
370 375 380
Phe Leu Cys Ser Glu Gln Gly Glu Thr Asn Thr Leu Pro Pro Tyr Ser
385 390 395 400
Gly Met Glu Gln Ser Leu Trp Asn Gly Ala Leu Ser Met Pro Ser Arg
405 410 415
Phe His
<210> 44
<211> 336
<212> Белок
<213> Vitis vinifera
<400> 44
Met Glu Met Thr Thr Val Lys Ser Glu Leu Gly Leu Glu Arg Gly Arg
1 5 10 15
Leu Cys Thr Ala Glu Thr Asp Ala Leu Glu Val Thr Lys Cys Val Lys
20 25 30
Arg Arg Arg Arg Asp Pro Ser Ala Val Thr Pro Gly Cys Ser Lys Gln
35 40 45
Gly Glu Gln Gln Lys Gln Val Leu Leu Gln Ala Gly Gln Ser Ile Thr
50 55 60
Ala Ile Ala Thr Thr Met Lys Arg Ser Ser Arg Phe Arg Gly Val Ser
65 70 75 80
Arg His Arg Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Gly
85 90 95
Ser Trp Asn Val Thr Gln Arg Lys Lys Gly Lys Gln Val Tyr Leu Gly
100 105 110
Ala Tyr Asp Glu Glu Glu Ser Ala Ala Arg Ala Tyr Asp Leu Ala Ala
115 120 125
Leu Lys Tyr Trp Gly Pro Ser Thr Phe Thr Asn Phe Pro Val Ser Asp
130 135 140
Tyr Glu Lys Glu Ile Glu Ile Met Gln Gly Leu Thr Lys Glu Glu Tyr
145 150 155 160
Leu Ala Ser Leu Arg Arg Arg Ser Ser Gly Phe Ser Arg Gly Val Ser
165 170 175
Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu Ala
180 185 190
Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr
195 200 205
Ser Thr Gln Glu Glu Ala Ala His Ala Tyr Asp Ile Ala Ala Ile Glu
210 215 220
Tyr Arg Gly Ile Asn Ala Val Thr Asn Phe Glu Leu Ser Thr Tyr Val
225 230 235 240
Arg Trp Leu Arg Pro Arg Ala Thr Ala Leu Thr Pro Gln Glu Pro Arg
245 250 255
Ser Asn Ser Ile Met Gln Ala Ser Ser Asn Cys Leu Pro Asn Glu Glu
260 265 270
Val Glu Leu Ser Phe Leu Ser Pro Asn Pro Phe Thr Val Asp Asp Leu
275 280 285
Ala Thr Pro Leu Lys Gln Glu Lys Phe Gln Arg Glu Val Ser Ile Ser
290 295 300
Pro Cys Thr Lys Ser Ser Ser Pro Thr Ala Leu Ser Leu Leu His Arg
305 310 315 320
Ser Ser Val Phe Arg Gln Leu Val Glu Lys Asn Ser Asn Ser Ile Glu
325 330 335
<210> 45
<211> 389
<212> Белок
<213> Glycine max
<400> 45
Met Ala Met Met Lys Glu Asn Ile Ile Glu Val Ser Leu Gly Arg Arg
1 5 10 15
Gln Met Ser Met Thr Glu Gly Glu Phe Gln Gly Thr Arg Ser Val Lys
20 25 30
Arg Arg Arg Arg Glu Val Ala Ala Ala Ala Gly Ser Gly Asp Asp Asn
35 40 45
His Gln Gln Gln Leu Pro Gln Gln Glu Val Gly Glu Asn Thr Thr Val
50 55 60
Asn Thr Thr Lys Arg Ser Ser Arg Phe Arg Gly Val Ser Arg His Arg
65 70 75 80
Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp Lys Leu Ser Trp Asn
85 90 95
Ile Thr Gln Lys Lys Lys Gly Lys Gln Gly Ala Tyr Asp Glu Glu Glu
100 105 110
Ser Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Thr
115 120 125
Ser Thr Phe Thr Asn Phe Pro Ile Ser Asp Tyr Glu Lys Glu Ile Gln
130 135 140
Ile Met Gln Thr Met Thr Lys Glu Glu Tyr Leu Ala Thr Leu Arg Arg
145 150 155 160
Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala
165 170 175
Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe
180 185 190
Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr Gln Glu Glu Ala
195 200 205
Ala Arg Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg Gly Ile His Ala
210 215 220
Val Thr Asn Phe Asp Leu Ser Thr Tyr Ile Lys Trp Leu Lys Pro Ser
225 230 235 240
Gly Gly Gly Thr Leu Glu Ala Asn Leu Glu Ser His Ala Ala Leu Glu
245 250 255
His Gln Lys Val Ala Ser Pro Ser Asn Tyr Ala Leu Thr Glu Glu Ser
260 265 270
Lys Ser Leu Ala Leu His Asn Ser Phe Phe Ser Pro Tyr Ser Leu Asp
275 280 285
Ser Pro Val Lys His Glu Arg Phe Gly Asn Lys Thr Tyr Gln Phe Ser
290 295 300
Ser Asn Lys Ser Ser Ser Pro Thr Ala Leu Gly Leu Leu Leu Arg Ser
305 310 315 320
Ser Leu Phe Arg Glu Leu Val Glu Lys Asn Ser Asn Val Ser Gly Glu
325 330 335
Glu Asp Asp Gly Glu Ala Thr Lys Asp Gln Gln Thr Gln Ile Ala Thr
340 345 350
Asp Asp Asp Leu Gly Gly Ile Phe Phe Asp Ser Phe Ser Asp Ile Pro
355 360 365
Phe Val Cys Asp Pro Asn Arg Tyr Asp Leu Glu Leu Gln Glu Arg Asp
370 375 380
Leu His Ser Ile Phe
385
<210> 46
<211> 389
<212> Белок
<213> Glycine max
<400> 46
Met Val Met Met Lys Glu Asn Ile Ile Glu Glu Lys Leu Gly Arg Ser
1 5 10 15
Gln Met Ser Met Val Glu Gly Glu Phe Gln Gly Thr Trp Gly Val Lys
20 25 30
Arg Arg Arg Arg Glu Val Ala Ala Ala Ala Ser Ser Gly Asp Asp Asn
35 40 45
His His Gln Gln Leu Pro Gln Gln Glu Val Gly Glu Asn Ser Ser Ile
50 55 60
Ser Thr Thr Lys Arg Ser Ser Arg Phe Arg Gly Val Ser Arg His Arg
65 70 75 80
Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp Lys Leu Ser Trp Asn
85 90 95
Ile Thr Gln Lys Lys Lys Gly Lys Gln Gly Ala Tyr Asp Glu Glu Glu
100 105 110
Ser Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Asn
115 120 125
Ser Thr Phe Thr Asn Phe Pro Ile Ser Asp Tyr Glu Lys Glu Ile Glu
130 135 140
Ile Met Gln Thr Met Thr Lys Glu Glu Tyr Leu Ala Thr Leu Arg Arg
145 150 155 160
Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala
165 170 175
Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe
180 185 190
Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr Gln Glu Glu Ala
195 200 205
Ala Arg Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg Gly Ile His Ala
210 215 220
Val Thr Asn Phe Asp Leu Ser Thr Tyr Ile Lys Trp Leu Lys Pro Ser
225 230 235 240
Gly Gly Gly Thr Pro Glu Glu Asn Leu Glu Ser His Ala Val Leu Glu
245 250 255
His Gln Lys Leu Ala Ser Pro Ser Asn Tyr Ala Leu Thr Glu Glu Ser
260 265 270
Lys Ser Leu Val Leu Pro Asn Ser Phe Ile Ser Pro Asp Ser Leu Asp
275 280 285
Ser Pro Val Lys His Glu Ser Phe Gly Asn Lys Thr Tyr Gln Phe Ser
290 295 300
Arg Asn Lys Ser Ser Ser Pro Thr Ala Leu Gly Leu Leu Leu Arg Ser
305 310 315 320
Ser Leu Phe Arg Glu Leu Val Glu Lys Asn Ser Asn Val Ser Gly Glu
325 330 335
Glu Ala Asp Gly Glu Val Thr Lys Asp Gln Gln Pro Gln Leu Ala Ser
340 345 350
Asp Asp Asp Leu Asp Gly Ile Phe Phe Asp Ser Phe Gly Asp Ile Pro
355 360 365
Phe Val Cys Asp Pro Thr Arg Tyr Asn Leu Glu Leu Gln Glu Arg Asp
370 375 380
Leu His Ser Ile Phe
385
<210> 47
<211> 392
<212> Белок
<213> Medicago truncatula
<400> 47
Met Ala Met Leu Ile Glu Asn Glu Val Met Cys Leu Gly Lys Ser Gln
1 5 10 15
Arg Ser Met Asp Gly Lys Glu Val Lys Gly Ala Arg Arg Val Lys Arg
20 25 30
Gln Arg Arg Asp Ala Ile Val Pro Lys Ile Gly Asp Asp Ala Asn Lys
35 40 45
Met Ala Gln Lys Gln Val Gly Glu Asn Ser Thr Thr Asn Thr Ser Lys
50 55 60
Arg Ser Ser Arg Phe Arg Gly Val Ser Arg His Arg Trp Thr Gly Arg
65 70 75 80
Phe Glu Ala His Leu Trp Asp Lys Leu Ser Trp Asn Thr Thr Gln Lys
85 90 95
Lys Lys Gly Lys Gln Gly Ala Tyr Asp Glu Glu Glu Ser Ala Ala Arg
100 105 110
Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Thr Ser Thr Phe Thr
115 120 125
Asn Phe Pro Ile Ser Asp Tyr Asp Lys Glu Ile Glu Ile Met Asn Thr
130 135 140
Met Thr Lys Glu Glu Tyr Leu Ala Thr Leu Arg Arg Lys Ser Ser Gly
145 150 155 160
Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His
165 170 175
Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr
180 185 190
Leu Tyr Leu Gly Thr Tyr Ser Thr Gln Glu Glu Ala Ala Arg Ala Tyr
195 200 205
Asp Ile Ala Ala Ile Glu Tyr Arg Gly Ile His Ala Val Thr Asn Phe
210 215 220
Glu Leu Ser Ser Tyr Ile Lys Trp Leu Lys Pro Glu Thr Thr Thr Glu
225 230 235 240
Glu Asn His Glu Ser Gln Ile Leu Gln Lys Glu Ser Arg Thr Leu Ala
245 250 255
Pro Pro Asn Asn Ser Thr Leu Leu Gln Glu Ser Lys Leu Leu Ala Leu
260 265 270
Gln Lys Ser Phe Phe Ile Pro Asn Asp Leu Asn Ser Thr Glu Lys Gln
275 280 285
Glu Ser Ser Phe Glu Asn Lys Asn Tyr His Phe Leu Ser Asn Lys Ser
290 295 300
Thr Ser Pro Thr Ala Leu Ser Leu Leu Leu Arg Ser Ser Leu Phe Arg
305 310 315 320
Glu Leu Leu Glu Lys Asn Ser Asn Val Ser Glu Asp Glu Val Thr Lys
325 330 335
Glu Gln Gln Gln Gln Gln Ile Thr Ser Asp Asp Glu Leu Gly Gly Ile
340 345 350
Phe Tyr Asp Gly Ile Asp Asn Ile Ser Phe Asp Phe Asp Pro Asn Ser
355 360 365
Cys Asn Ile Glu Leu Gln Glu Arg Asp Leu His Ser Ile Ser Cys Leu
370 375 380
Tyr Gln Tyr Leu Asn Phe Gly Gln
385 390
<210> 48
<211> 386
<212> Белок
<213> Populus trichocarpa
<400> 48
Met Met Met Ile Lys Asn Glu Glu Asn Pro Gly Arg Arg Arg Gly Cys
1 5 10 15
Ile Ala Asp Ser Glu Ala Gln Val Ala Arg Cys Val Lys Arg Arg Arg
20 25 30
Arg Asp Pro Ala Ile Val Ala Leu Gly Ser Asp Asp Asn Gln Ser Gln
35 40 45
Gln Gln Met Pro Gln Lys Gln Thr Asp Gln Thr Ser Ala Ala Thr Thr
50 55 60
Val Lys Arg Ser Ser Arg Phe Arg Gly Val Ser Arg His Arg Trp Thr
65 70 75 80
Gly Arg Phe Glu Ala His Leu Trp Asp Lys Leu Ser Trp Asn Val Thr
85 90 95
Gln Lys Lys Lys Gly Lys Gln Gly Ala Tyr Asp Glu Glu Glu Ser Ala
100 105 110
Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Thr Ser Thr
115 120 125
Phe Thr Asn Phe Pro Ile Ser Asp Tyr Glu Lys Glu Ile Glu Ile Met
130 135 140
Gln Thr Val Thr Lys Glu Glu Tyr Leu Ala Ser Leu Arg Arg Lys Ser
145 150 155 160
Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His
165 170 175
His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn
180 185 190
Lys Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr Gln Glu Glu Ala Ala Arg
195 200 205
Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg Gly Ile Asn Ala Val Thr
210 215 220
Asn Phe Asp Leu Ser Thr Tyr Ile Arg Trp Ile Lys Pro Gly Val Ala
225 230 235 240
Ala Gln Ala Ala Ala Asn Glu Leu Gln Thr Val Thr Asp Pro Gln Thr
245 250 255
Ala Ala Thr Leu Thr Asp Thr Tyr Thr Pro Arg Glu Glu Thr Lys Pro
260 265 270
Ser Leu Phe Leu Pro Asn Gln Phe Thr Ala Asp Tyr Leu Asn Ser Pro
275 280 285
Pro Lys Leu Asp Ala Phe Gln Asn Asn Ile Phe Val Asp Ser Ser Asn
290 295 300
Lys Thr Ser Ser Pro Thr Ala Leu Ser Leu Leu Leu Arg Ser Ser Val
305 310 315 320
Phe Arg Glu Leu Val Glu Lys Asn Ser Asn Val Cys Glu Glu Glu Thr
325 330 335
Asp Gly Asn Glu Ile Lys Asn Gln Pro Met Ala Gly Ser Asp Asp Glu
340 345 350
Tyr Gly Gly Ile Phe Tyr Asp Gly Ile Gly Asp Ile Pro Phe Val Tyr
355 360 365
Ser Ser Asn Lys Tyr Ser Leu Gly Leu Glu Glu Arg Glu Leu Gln Phe
370 375 380
Val Leu
385
<210> 49
<211> 372
<212> Белок
<213> Ricinus communis
<400> 49
Met Glu Met Met Met Val Lys Asn Glu Glu Ile Ser Gly Arg Arg Arg
1 5 10 15
Ala Ser Val Thr Glu Ser Glu Ala Tyr Val Ala Arg Cys Val Lys Arg
20 25 30
Arg Arg Arg Asp Ala Ala Val Val Thr Val Gly Gly Asp Asp Ser Gln
35 40 45
Ser His Gln Gln Gln Gln Gln Gln Gln Pro Glu Gln Gln Ala His Gln
50 55 60
Ile Ser Ala Ala Thr Thr Val Lys Arg Ser Ser Arg Tyr Arg Gly Val
65 70 75 80
Ser Arg His Arg Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys
85 90 95
Leu Ser Trp Asn Val Thr Gln Lys Lys Lys Gly Lys Gln Gly Ala Tyr
100 105 110
Asp Glu Glu Glu Ser Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys
115 120 125
Tyr Trp Gly Thr Ser Thr Phe Thr Asn Phe Pro Ile Ser Asp Tyr Glu
130 135 140
Lys Glu Ile Glu Ile Met Gln Thr Val Thr Lys Glu Glu Tyr Leu Ala
145 150 155 160
Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr
165 170 175
Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile
180 185 190
Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr
195 200 205
Gln Glu Glu Ala Ala Arg Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg
210 215 220
Gly Ile Asn Ala Val Thr Asn Phe Asp Leu Ser Thr Tyr Ile Arg Trp
225 230 235 240
Leu Lys Pro Glu Val Ala Ala Gln Val Ala Ala Asn Glu Pro Gln Thr
245 250 255
Val Ala Glu Ser Arg Met Leu Pro Ser Ile Asn Asn Arg Ile Ala Arg
260 265 270
Glu Glu Ser Lys Pro Ser Phe Phe Ser Ala Thr Pro Phe Ser Leu Asp
275 280 285
Cys Trp Ser Tyr Pro Arg Lys Gln Glu Glu Phe Gln Asn Arg Thr Pro
290 295 300
Ile Thr Pro Cys Ser Lys Thr Ser Ser Pro Thr Ala Leu Ser Leu Leu
305 310 315 320
Leu Arg Ser Ser Ile Phe Arg Glu Leu Val Glu Lys Asn Ser Asn Val
325 330 335
Ser Glu Asp Glu Asn Glu Gly Glu Glu Thr Lys Asn Gln Ser Gln Ile
340 345 350
Gly Ser Asp Asp Glu Phe Gly Gly Leu Phe Tyr Glu Arg Ile Gly Asp
355 360 365
Ile Pro Phe Ile
370
<210> 50
<211> 404
<212> Белок
<213> Vitis vinifera
<400> 50
Met Glu Met Met Arg Val Lys Ser Glu Glu Asn Leu Gly Arg Arg Arg
1 5 10 15
Met Cys Val Ala Asp Ala Glu Ala Gln Gly Thr Arg Cys Val Lys Arg
20 25 30
Arg Arg Arg Asp Pro Ala Ile Val Thr Leu Gly Cys Asp Asp Gln Ser
35 40 45
Gln Gln Gln Gln Leu Pro Asn Gln Gln Pro Asp Gln Ala Ser Ala Ala
50 55 60
Thr Thr Val Lys Arg Ser Ser Arg Phe Arg Gly Val Ser Arg His Arg
65 70 75 80
Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Phe Ser Trp Asn
85 90 95
Val Thr Gln Lys Lys Lys Gly Lys Gln Gly Ala Tyr Asp Glu Glu Glu
100 105 110
Ser Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Ala
115 120 125
Ser Thr Phe Thr Asn Phe Pro Val Ser Asp Tyr Glu Lys Glu Ile Glu
130 135 140
Ile Met Gln Ser Val Thr Lys Glu Glu Tyr Leu Ala Cys Leu Arg Arg
145 150 155 160
Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala
165 170 175
Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe
180 185 190
Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ser Thr Gln Glu Glu Ala
195 200 205
Ala Arg Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg Gly Ile Asn Ala
210 215 220
Val Thr Asn Phe Asp Leu Ser Thr Tyr Ile Arg Trp Leu Asn Pro Ala
225 230 235 240
Ala Asn Asn Pro Val Val Pro His Glu Ser Arg Ala Asn Thr Glu Pro
245 250 255
Gln Ala Leu Ala Ser Ser Asn Phe Val Leu Ser Glu Glu Ser Glu Pro
260 265 270
Leu Phe Phe His Ser Asn Ser Phe Thr Met Asp Asp Leu Asn Pro Pro
275 280 285
His Lys Gln Glu Val Phe Gln Thr Lys Ile Pro Ile Glu Pro Cys Ser
290 295 300
Lys Ser Ser Ser Pro Thr Ala Leu Gly Leu Leu Leu Arg Ser Ser Ile
305 310 315 320
Phe Arg Glu Leu Val Glu Lys Asn Ser Asn Ala Pro Glu Asp Glu Thr
325 330 335
Asp Ala Glu Asp Thr Lys Asn Gln Gln Gln Val Gly Ser Asp Asp Glu
340 345 350
Tyr Gly Ile Phe Tyr Asp Gly Ile Gly Asp Ile Pro Phe Val Cys Pro
355 360 365
Ser Asn Gly Asp Arg Asn Glu Leu Gln Glu Arg Leu Pro Leu Pro Phe
370 375 380
Thr Ile Ser Gln Gly Asn Pro Tyr Gly Thr Ala Val Leu Thr Ser Met
385 390 395 400
Gln Ser Ile Asn
<210> 51
<211> 378
<212> Белок
<213> Brachypodium distachyon
<400> 51
Met Ala Lys Gln Arg Thr Asp Ser Ala Gly Thr Asp Ala Ala Ala Val
1 5 10 15
Gln Leu Thr Lys Pro Lys Arg Thr Arg Lys Ser Val Pro Arg Arg Glu
20 25 30
Ser Pro Ser Arg Arg Thr Ser Ala Tyr Arg Gly Val Thr Arg His Arg
35 40 45
Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Asn Thr Trp Thr
50 55 60
Gln Ser Gln Arg Lys Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala Tyr
65 70 75 80
Gly Gly Glu Glu Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys
85 90 95
Tyr Trp Gly Arg Asp Thr Val Leu Asn Phe Pro Leu Ser Asn Tyr Asp
100 105 110
Glu Glu Trp Lys Glu Met Glu Gly Gln Ser Arg Glu Glu Tyr Ile Gly
115 120 125
Ser Leu Arg Arg Lys Ser Thr Gly Phe Ser Arg Gly Val Ser Lys Tyr
130 135 140
Arg Gly Val Ala Arg His His His Asn Gly Lys Trp Glu Ala Arg Ile
145 150 155 160
Gly Arg Val Tyr Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Gly Thr
165 170 175
Gln Glu Glu Ala Ala Met Ala Tyr Asp Ile Ala Ala Ile Glu His Arg
180 185 190
Gly Leu Asn Ala Val Thr Asn Phe Asp Val Ser Arg Tyr Ile Asp Trp
195 200 205
His Arg Arg Leu Cys Arg Asp Leu Gly Asp Asn Ile Ile Thr Pro Leu
210 215 220
Thr Asn Pro Thr Val Asp Leu Glu Glu Ala Met Ala Gly Asp Asp Asp
225 230 235 240
Asp Gly Gln Phe Leu Leu Pro Ser Gln Ala Thr Thr Pro Pro Ser Thr
245 250 255
Ser Ser Ala Leu Gly Leu Leu Leu Leu Ser Pro Arg Leu Lys Glu Val
260 265 270
Ile Glu Gly Ser Gly Ala Ala Ser Ala Met Ala Ala Ser Thr Ser Glu
275 280 285
Ser Ser Ala Ala Gly Ser Pro Pro Pro Ser Trp Ser Ser Ser Ser Cys
290 295 300
Ser Pro Ser Pro Pro Ser Pro Ser His Ser Pro Pro Glu Thr Gln Gln
305 310 315 320
Lys Gln Gln Gln Gln Glu Tyr Gly Ala Ser Ala Ala Ala Ala Arg Cys
325 330 335
Ser Phe Pro Asp Asp Val Gln Thr Tyr Phe Gly Cys Glu Asp Gly Cys
340 345 350
Ala Glu Val Asp Thr Phe Leu Phe Gly Asp Leu Ser Ala Tyr Ala Ala
355 360 365
Pro Met Phe Gln Phe Glu Leu Leu Asp Val
370 375
<210> 52
<211> 416
<212> Белок
<213> Oryza sativa
<400> 52
Met Ala Lys Arg Arg Ser Asn Gly Glu Thr Ala Ala Ala Ser Ser Asp
1 5 10 15
Asp Ser Ser Ser Gly Val Cys Gly Gly Gly Gly Gly Gly Glu Val Glu
20 25 30
Pro Arg Arg Arg Gln Lys Arg Pro Arg Arg Ser Ala Pro Arg Asp Cys
35 40 45
Pro Ser Gln Arg Ser Ser Ala Phe Arg Gly Val Thr Arg His Arg Trp
50 55 60
Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Asn Thr Trp Asn Glu
65 70 75 80
Ser Gln Ser Lys Lys Gly Arg Gln Gly Ala Tyr Asp Gly Glu Glu Ala
85 90 95
Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly His Asp
100 105 110
Thr Val Leu Asn Phe Pro Leu Ser Thr Tyr Asp Glu Glu Leu Lys Glu
115 120 125
Met Glu Gly Gln Ser Arg Glu Glu Tyr Ile Gly Ser Leu Arg Arg Lys
130 135 140
Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg
145 150 155 160
His His His Asn Gly Lys Trp Glu Ala Arg Ile Gly Arg Val Phe Gly
165 170 175
Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala Thr Gln Glu Glu Ala Ala
180 185 190
Val Ala Tyr Asp Ile Ala Ala Ile Glu His Arg Gly Leu Asn Ala Val
195 200 205
Thr Asn Phe Asp Ile Asn Leu Tyr Ile Arg Trp Tyr His Gly Ser Cys
210 215 220
Arg Ser Ser Ser Ala Ala Ala Ala Thr Thr Ile Glu Asp Asp Asp Phe
225 230 235 240
Ala Glu Ala Ile Ala Ala Ala Leu Gln Gly Val Asp Glu Gln Pro Ser
245 250 255
Ser Ser Pro Ala Thr Thr Arg Gln Leu Gln Thr Ala Asp Asp Asp Asp
260 265 270
Asp Asp Leu Val Ala Gln Leu Pro Pro Gln Leu Arg Pro Leu Ala Arg
275 280 285
Ala Ala Ser Thr Ser Pro Ile Gly Leu Leu Leu Arg Ser Pro Lys Phe
290 295 300
Lys Glu Ile Ile Glu Gln Ala Ala Ala Ala Ala Ala Ser Ser Ser Gly
305 310 315 320
Ser Ser Ser Ser Ser Ser Thr Asp Ser Pro Ser Ser Ser Ser Ser Ser
325 330 335
Ser Leu Ser Pro Ser Pro Leu Pro Ser Pro Pro Pro Gln Gln Gln Pro
340 345 350
Thr Val Pro Lys Asp Asp Gln Tyr Asn Val Asp Met Ser Ser Val Ala
355 360 365
Ala Ala Arg Cys Ser Phe Pro Asp Asp Val Gln Thr Tyr Phe Gly Leu
370 375 380
Asp Asp Asp Gly Phe Gly Tyr Pro Glu Val Asp Thr Phe Leu Phe Gly
385 390 395 400
Asp Leu Gly Ala Tyr Ala Ala Pro Met Phe Gln Phe Glu Leu Asp Val
405 410 415
<210> 53
<211> 440
<212> Белок
<213> Sorghum bicolor
<400> 53
Met Ala Arg Pro Arg Lys Asn Ala Gly Thr Asp Glu Asp Asn Pro Asn
1 5 10 15
Ala Ala Thr Gly Val Ser Val Thr Gly Lys Pro Pro Lys Leu Lys Arg
20 25 30
Val Arg Arg Lys Gly Glu Pro Arg Glu Ser Ser Thr Pro Ser Gln Arg
35 40 45
Ser Ser Ala Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe
50 55 60
Glu Ala His Leu Trp Asp Lys Asp Ala Arg Asn Gly Ser Arg Asn Lys
65 70 75 80
Lys Gly Lys Gln Gly Ala Tyr Asp Asp Glu Glu Ala Ala Ala Arg Ala
85 90 95
His Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Ala Thr Val Leu Asn
100 105 110
Phe Pro Leu Cys Gly Tyr Asp Glu Glu Leu Arg Glu Met Glu Ala Gln
115 120 125
Pro Arg Glu Glu Tyr Ile Gly Ser Leu Arg Arg Arg Ser Ser Gly Phe
130 135 140
Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn
145 150 155 160
Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Leu Gly Asn Lys Tyr Leu
165 170 175
Tyr Leu Gly Thr Phe Ala Thr Gln Glu Glu Ala Ala Val Ala Tyr Asp
180 185 190
Ile Ala Ala Ile Glu His Arg Gly Leu Asn Ala Val Thr Asn Phe Asp
195 200 205
Ile Ser His Tyr Val Asn His Trp His Arg His Cys His Gly Pro Ser
210 215 220
Asp Asp Ser Leu Gly Val Val Val Asp Asp Val Ala Ala Phe Gln Leu
225 230 235 240
Pro Asp Asp Leu Pro Glu Cys Pro Ala Ala Ala Ile Gly Val Glu Glu
245 250 255
Thr Thr Gly Gly Asp Ala Glu Phe His Asn Gly Glu Glu Gly Tyr Leu
260 265 270
Gln His His Thr Ser Gly Pro Phe Gly Ala Gln Gln Gln Leu Pro Asp
275 280 285
Glu Thr Gly Ala Leu Ala Ala His Gln Met Ala Pro Asn Ser Ser Ala
290 295 300
Leu Asp Met Val Leu Gln Ser Pro Lys Phe Lys Glu Leu Met Glu Gln
305 310 315 320
Val Ser Ala Ala Ala Ala Ala Val Ala Ser Glu Ser Ser Ile Gly Gly
325 330 335
Ser Met Ser Ser Ser Ser Pro Ser Pro Ser Leu Ser Ser Phe Ser Pro
340 345 350
Ser Pro Leu Gln Leu Pro Ser Pro Ser Ser Leu Ser Ser Phe Ser Pro
355 360 365
Ser Ser Pro Leu Gln Gln Pro Ser Pro Pro Leu Gln Gln Pro Glu Phe
370 375 380
Val Glu Gly Ala Pro Ala Ala Arg Cys Ser Phe Pro Asp Asp Val Gln
385 390 395 400
Thr Phe Phe Asp Phe Glu Asn Glu Ser Asp Met Ser Phe Met Tyr Ala
405 410 415
Glu Val Asp Thr Phe Leu Phe Gly Asp Leu Gly Ala Tyr Ala Ala Pro
420 425 430
Ile Phe His Phe Asp Leu Asp Val
435 440
<210> 54
<211> 408
<212> Белок
<213> Zea mays
<400> 54
Met Ala Arg Pro Arg Lys Asn Gly Gly Thr Asp Glu Asp Asp Ala Asn
1 5 10 15
Ala Ala Thr Gly Ala Thr Gly Lys Pro Lys Lys Leu Met Lys Arg Ala
20 25 30
Arg Arg Lys Ser Glu Ser Pro Ser Pro Arg Ser Ser Ala Tyr Arg Gly
35 40 45
Val Thr Arg His Arg Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp
50 55 60
Lys Asp Ala Arg Asn Gly Ser Arg Ser Lys Lys Gly Lys Gln Val Tyr
65 70 75 80
Leu Gly Ala Tyr Asp Asp Glu Asp Ala Ala Ala Arg Ala His Asp Leu
85 90 95
Ala Ala Leu Lys Tyr Trp Gly Pro Ala Gly Thr Val Leu Asn Phe Pro
100 105 110
Leu Ser Gly Tyr Asp Glu Glu Arg Arg Glu Met Glu Gly Gln Pro Arg
115 120 125
Glu Glu Tyr Val Ala Ser Leu Arg Arg Arg Ser Ser Gly Phe Ala Arg
130 135 140
Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg
145 150 155 160
Trp Glu Ala Arg Ile Gly Arg Val Leu Gly Asn Lys Tyr Leu Tyr Leu
165 170 175
Gly Thr Tyr Ala Thr Gln Glu Glu Ala Ala Val Ala Tyr Asp Met Ala
180 185 190
Ala Ile Glu His Arg Gly Phe Asn Ala Val Thr Asn Phe Asp Ile Ser
195 200 205
His Tyr Ile Asn His Trp His Arg His Cys His Gly Pro Cys Asp Gly
210 215 220
Ser Leu Gly Ala Met Asp Val Ala Pro Asn Val Ser Leu Glu Leu Asp
225 230 235 240
Leu Leu Glu Cys Pro Ala Thr Val Gly Leu Gly Leu Glu Glu Thr Thr
245 250 255
Gly Asp Asp Glu Phe His Asn Arg Glu Asp Tyr Leu Gly His Leu Phe
260 265 270
Gly Val Gln Gln Leu Pro Asp Glu Met Gly Pro Pro Ala His Gln Met
275 280 285
Ala Pro Ala Ser Ser Ala Leu Asp Leu Val Leu Gln Ser Pro Arg Phe
290 295 300
Lys Glu Leu Met Gln Gln Val Ser Ala Ala Gly Ala Ser Glu Thr Asn
305 310 315 320
Gly Gly Ser Met Arg Ser Ser Pro Ser Thr Ser Leu Cys Ser Phe Ser
325 330 335
Pro Ser Pro Leu Glu Leu Pro Ser Pro Pro Leu Gln Gln Pro Thr Glu
340 345 350
Phe Ile Asp Gly Ala Pro Pro Arg Cys Ser Phe Pro Asp Asp Val Gln
355 360 365
Ser Phe Phe Asp Phe Lys Asn Asp Asn Asp Met Ser Phe Val Tyr Ala
370 375 380
Glu Val Asp Thr Phe Leu Phe Gly Asp Leu Gly Ala Tyr Ala Pro Pro
385 390 395 400
Met Phe Asp Phe Asp Leu Tyr Glu
405
<210> 55
<211> 304
<212> Белок
<213> Arabidopsis lyrata
<400> 55
Met Ala Lys Val Ser Arg Arg Ser Lys Lys Thr Ile Val Glu Asp Glu
1 5 10 15
Ile Ser Asp Lys Thr Ala Ser Ala Ser Glu Ala Ala Ser Ile Val Phe
20 25 30
Lys Ser Lys Arg Lys Arg Lys Ser Pro Pro Arg Asn Ala Pro Pro Gln
35 40 45
Arg Ser Ser Pro Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg
50 55 60
Tyr Glu Ala His Leu Trp Asp Lys Asn Ser Trp Asn Glu Thr Gln Thr
65 70 75 80
Lys Lys Gly Arg Gln Val Tyr Ile Gly Ala Tyr Asp Glu Glu Glu Ala
85 90 95
Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Arg Asp
100 105 110
Thr Leu Leu Asn Phe Pro Leu Leu Ile Tyr Asp Glu Asp Val Lys Glu
115 120 125
Met Glu Gly Gln Ser Lys Glu Glu Tyr Ile Gly Ser Leu Arg Arg Lys
130 135 140
Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg
145 150 155 160
His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly
165 170 175
Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala Thr Gln Glu Glu Ala Ala
180 185 190
Ile Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg Gly Leu Asn Ala Val
195 200 205
Thr Asn Phe Asp Val Ser Arg Tyr Leu Asn Pro Asp Ala Ala Asp Ser
210 215 220
Lys Pro Ile Arg Asn Asp Pro Glu Ser Ser Asp Asp Asn Lys Cys Pro
225 230 235 240
Lys Ser Glu Glu Ile Ile Glu Pro Ser Thr Ser Pro Glu Ala Ile Thr
245 250 255
Thr Arg Arg Ser Phe Pro Asp Asp Ile Gln Thr Tyr Phe Gly Cys Gln
260 265 270
Asp Ser Gly Lys Leu Ala Thr Glu Glu Asp Val Ile Phe Gly Gly Leu
275 280 285
Asn Ser Phe Ile Asn Pro Gly Phe Tyr Asn Glu Phe Asp Tyr Gly Pro
290 295 300
<210> 56
<211> 308
<212> Белок
<213> Arabidopsis thaliana
<400> 56
Met Ala Lys Val Ser Gly Arg Ser Lys Lys Thr Ile Val Asp Asp Glu
1 5 10 15
Ile Ser Asp Lys Thr Ala Ser Ala Ser Glu Ser Ala Ser Ile Ala Leu
20 25 30
Thr Ser Lys Arg Lys Arg Lys Ser Pro Pro Arg Asn Ala Pro Leu Gln
35 40 45
Arg Ser Ser Pro Tyr Arg Gly Val Thr Arg Trp Thr Gly Arg Tyr Glu
50 55 60
Ala His Leu Trp Asp Lys Asn Ser Trp Asn Asp Thr Gln Thr Lys Lys
65 70 75 80
Gly Arg Gln Gly Ala Tyr Asp Glu Glu Glu Ala Ala Ala Arg Ala Tyr
85 90 95
Asp Leu Ala Ala Leu Lys Tyr Trp Gly Arg Asp Thr Leu Leu Asn Phe
100 105 110
Pro Leu Pro Ser Tyr Asp Glu Asp Val Lys Glu Met Glu Gly Gln Ser
115 120 125
Lys Glu Glu Tyr Ile Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser
130 135 140
Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly
145 150 155 160
Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr
165 170 175
Leu Gly Thr Tyr Ala Thr Gln Glu Glu Ala Ala Ile Ala Tyr Asp Ile
180 185 190
Ala Ala Ile Glu Tyr Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Val
195 200 205
Ser Arg Tyr Leu Asn Pro Asn Ala Ala Ala Asp Lys Ala Asp Ser Asp
210 215 220
Ser Lys Pro Ile Arg Ser Pro Ser Arg Glu Pro Glu Ser Ser Asp Asp
225 230 235 240
Asn Lys Ser Pro Lys Ser Glu Glu Val Ile Glu Pro Ser Thr Ser Pro
245 250 255
Glu Val Ile Pro Thr Arg Arg Ser Phe Pro Asp Asp Ile Gln Thr Tyr
260 265 270
Phe Gly Cys Gln Asp Ser Gly Lys Leu Ala Thr Glu Glu Asp Val Ile
275 280 285
Phe Asp Cys Phe Asn Ser Tyr Ile Asn Pro Gly Phe Tyr Asn Glu Phe
290 295 300
Asp Tyr Gly Pro
305
<210> 57
<211> 303
<212> Белок
<213> Arabidopsis thaliana
<400> 57
Met Ala Lys Val Ser Gly Arg Ser Lys Lys Thr Ile Val Asp Asp Glu
1 5 10 15
Ile Ser Asp Lys Thr Ala Ser Ala Ser Glu Ser Ala Ser Ile Ala Leu
20 25 30
Thr Ser Lys Arg Lys Arg Lys Ser Pro Pro Arg Asn Ala Pro Leu Gln
35 40 45
Arg Ser Ser Pro Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg
50 55 60
Tyr Glu Ala His Leu Trp Asp Lys Asn Ser Trp Asn Asp Thr Gln Thr
65 70 75 80
Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Glu Glu Glu Ala
85 90 95
Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Arg Asp
100 105 110
Thr Leu Leu Asn Phe Pro Leu Pro Ser Tyr Asp Glu Asp Val Lys Glu
115 120 125
Met Glu Gly Gln Ser Lys Glu Glu Tyr Ile Gly Ser Leu Arg Arg Lys
130 135 140
Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg
145 150 155 160
His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Ala
165 170 175
Thr Gln Glu Glu Ala Ala Ile Ala Tyr Asp Ile Ala Ala Ile Glu Tyr
180 185 190
Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Val Ser Arg Tyr Leu Asn
195 200 205
Pro Asn Ala Ala Ala Asp Lys Ala Asp Ser Asp Ser Lys Pro Ile Arg
210 215 220
Ser Pro Ser Arg Glu Pro Glu Ser Ser Asp Asp Asn Lys Ser Pro Lys
225 230 235 240
Ser Glu Glu Val Ile Glu Pro Ser Thr Ser Pro Glu Val Ile Pro Thr
245 250 255
Arg Arg Ser Phe Pro Asp Asp Ile Gln Thr Tyr Phe Gly Cys Gln Asp
260 265 270
Ser Gly Lys Leu Ala Thr Glu Glu Asp Val Ile Phe Asp Cys Phe Asn
275 280 285
Ser Tyr Ile Asn Pro Gly Phe Tyr Asn Glu Phe Asp Tyr Gly Pro
290 295 300
<210> 58
<211> 332
<212> Белок
<213> Arabidopsis lyrata
<400> 58
Met Glu Glu Ile Thr Arg Lys Ser Lys Lys Thr Ser Val Glu Asn Glu
1 5 10 15
Thr Gly Asp Asp Gln Ser Ala Thr Ser Val Val Val Lys Ala Lys Arg
20 25 30
Lys Arg Arg Ser Gln Pro Arg Asp Ala Pro Pro Gln Arg Ser Ser Val
35 40 45
His Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala His
50 55 60
Leu Trp Asp Lys Asn Ser Trp Asn Glu Thr Gln Ser Lys Lys Gly Arg
65 70 75 80
Gln Gly Ala Tyr Asp Glu Glu Asp Ala Ala Ala Arg Ala Tyr Asp Leu
85 90 95
Ala Ala Leu Lys Tyr Trp Gly Arg Asp Thr Ile Leu Asn Phe Pro Leu
100 105 110
Cys Asn Tyr Glu Glu Asp Ile Lys Glu Met Glu Ser Gln Ser Lys Glu
115 120 125
Glu Tyr Ile Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly
130 135 140
Val Ser Lys Tyr Arg Gly Val Ala Lys His His His Asn Gly Arg Trp
145 150 155 160
Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly
165 170 175
Thr Tyr Ala Thr Gln Glu Glu Ala Ala Ile Ala Tyr Asp Ile Ala Ala
180 185 190
Ile Glu Tyr Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Ile Ser Arg
195 200 205
Tyr Met Lys Leu Pro Val Pro Glu Asn Pro Ile Asp Ala Ala Asn Asn
210 215 220
Leu Leu Glu Ser Pro His Ser Asp Ser Ser Pro Phe Ile Asn Pro Thr
225 230 235 240
His Glu Ser Asp Leu Ser Gln Ser Gln Ser Ser Ser Asp Asp Asn Asp
245 250 255
Asp Arg Lys Thr Lys Leu Leu Lys Ser Ser Pro Leu Asn Ala Glu Glu
260 265 270
Val Ile Gly Pro Ser Thr Pro Pro Glu Ile Ala Pro Pro Arg Arg Ser
275 280 285
Phe Pro Glu Asp Ile Gln Thr Tyr Phe Gly Cys Gln Asn Ser Gly Lys
290 295 300
Leu Thr Thr Glu Glu Asp Asp Val Ile Phe Gly Asp Leu Asp Ser Phe
305 310 315 320
Leu Thr Pro Asp Phe Tyr Ser Glu Leu Asn Asp Cys
325 330
<210> 59
<211> 328
<212> Белок
<213> Thellungiella halophila
<400> 59
Met Ala Lys Val Ser Gln Arg Ser Lys Lys Thr Ile Val Asn Asp Glu
1 5 10 15
Ile Ser Asp Lys Lys Ala Val Ala Val Ala Ser Val Ser Ser Ser Ala
20 25 30
Phe Leu Lys Ser Lys Arg Lys Arg Lys Leu Pro Pro Gln Asn Ala Pro
35 40 45
Pro Gln Arg Ser Ser Ser Tyr Arg Gly Val Thr Arg His Arg Trp Thr
50 55 60
Gly Arg Tyr Glu Ala His Leu Trp Asp Lys Asn Cys Trp Asn Glu Thr
65 70 75 80
Gln Thr Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Glu Glu
85 90 95
Glu Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly
100 105 110
Arg Asp Thr Leu Leu Asn Phe Pro Leu Pro Thr Tyr Glu Glu Asp Val
115 120 125
Lys Glu Met Glu Gly His Ser Arg Glu Glu Tyr Ile Gly Ser Leu Arg
130 135 140
Arg Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val
145 150 155 160
Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val
165 170 175
Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala Thr Gln Glu Glu
180 185 190
Ala Ala Arg Ala Tyr Asp Ile Ala Ala Ile Glu Tyr Arg Gly Leu Asn
195 200 205
Ala Val Thr Asn Phe Asp Val Ser Arg Tyr Leu Asn Leu Pro Glu Ser
210 215 220
Lys Asn Pro Ser Ala Ala Ala Asn His Leu Pro Asp Glu Ser Asp Tyr
225 230 235 240
Tyr Asp Ser Met Pro Val Arg Asn Pro Asn His Glu Pro Arg Ser Pro
245 250 255
Asp Gly Gln Thr Ser Ser Glu Asp Asn Asp Tyr Thr Lys Thr Glu Glu
260 265 270
Thr Leu Asp Pro Glu Ala Ile Pro Ser Arg Arg Ser Phe Pro Asp Asp
275 280 285
Ile Gln Thr Tyr Phe Gly Cys Gln Asp Ser Gly Lys Leu Ala Thr Glu
290 295 300
Glu Asp Val Ile Phe Gly Gly Phe Asn Ser Phe Ile Asn Pro Gly Phe
305 310 315 320
Tyr Asn Asp Phe Asp Tyr Ala Pro
325
<210> 60
<211> 345
<212> Белок
<213> Arabidopsis thaliana
<400> 60
Met Phe Ile Ala Val Glu Val Ser Pro Val Met Glu Asp Ile Thr Arg
1 5 10 15
Gln Ser Lys Lys Thr Ser Val Glu Asn Glu Thr Gly Asp Asp Gln Ser
20 25 30
Ala Thr Ser Val Val Leu Lys Ala Lys Arg Lys Arg Arg Ser Gln Pro
35 40 45
Arg Asp Ala Pro Pro Gln Arg Ser Ser Val His Arg Gly Val Thr Arg
50 55 60
His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp Lys Asn Ser
65 70 75 80
Trp Asn Glu Thr Gln Thr Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala
85 90 95
Tyr Asp Glu Glu Asp Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu
100 105 110
Lys Tyr Trp Gly Arg Asp Thr Ile Leu Asn Phe Pro Leu Cys Asn Tyr
115 120 125
Glu Glu Asp Ile Lys Glu Met Glu Ser Gln Ser Lys Glu Glu Tyr Ile
130 135 140
Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys
145 150 155 160
Tyr Arg Gly Val Ala Lys His His His Asn Gly Arg Trp Glu Ala Arg
165 170 175
Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala
180 185 190
Thr Gln Glu Glu Ala Ala Ile Ala Tyr Asp Ile Ala Ala Ile Glu Tyr
195 200 205
Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Ile Ser Arg Tyr Leu Lys
210 215 220
Leu Pro Val Pro Glu Asn Pro Ile Asp Thr Ala Asn Asn Leu Leu Glu
225 230 235 240
Ser Pro His Ser Asp Leu Ser Pro Phe Ile Lys Pro Asn His Glu Ser
245 250 255
Asp Leu Ser Gln Ser Gln Ser Ser Ser Glu Asp Asn Asp Asp Arg Lys
260 265 270
Thr Lys Leu Leu Lys Ser Ser Pro Leu Val Ala Glu Glu Val Ile Gly
275 280 285
Pro Ser Thr Pro Pro Glu Ile Ala Pro Pro Arg Arg Ser Phe Pro Glu
290 295 300
Asp Ile Gln Thr Tyr Phe Gly Cys Gln Asn Ser Gly Lys Leu Thr Ala
305 310 315 320
Glu Glu Asp Asp Val Ile Phe Gly Asp Leu Asp Ser Phe Leu Thr Pro
325 330 335
Asp Phe Tyr Ser Glu Leu Asn Asp Cys
340 345
<210> 61
<211> 299
<212> Белок
<213> Glycine max
<400> 61
Met Ala Lys Lys Ser Gln Lys Ser Leu Lys Asn Asn Asn Asn Asn Asn
1 5 10 15
Thr Thr Arg Lys Arg Thr Arg Lys Ser Val Pro Arg Asp Ser Pro Pro
20 25 30
Gln Arg Ser Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly
35 40 45
Arg Tyr Glu Ala His Leu Trp Asp Lys Asn Cys Trp Asn Glu Ser Gln
50 55 60
Ser Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Asp Glu Glu
65 70 75 80
Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Gln
85 90 95
Asp Thr Ile Leu Asn Phe Pro Leu Ser Asn Tyr Glu Glu Lys Leu Lys
100 105 110
Glu Met Glu Gly Gln Ser Lys Glu Glu Tyr Ile Gly Ser Leu Arg Arg
115 120 125
Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala
130 135 140
Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe
145 150 155 160
Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala Thr Gln Glu Glu Ala
165 170 175
Ala Ala Ala Tyr Asp Met Ala Ala Ile Glu Tyr Arg Gly Leu Asn Ala
180 185 190
Val Thr Asn Phe Asp Leu Ser Arg Tyr Ile Asn Trp Pro Arg Pro Lys
195 200 205
Thr Glu Glu Asn His Gln Asn Thr Pro Ser Asn Gln Asn Val Asn Ser
210 215 220
Asn Ala Glu Leu Glu Leu Gly Ser Ala Ser Asp Glu Ile Thr Glu Glu
225 230 235 240
Gly Val Ala Arg Ser Ser Glu Ser Glu Ser Asn Pro Ser Arg Arg Thr
245 250 255
Phe Pro Glu Asp Ile Gln Thr Ile Phe Glu Asn Asn Gln Asp Ser Gly
260 265 270
Ile Tyr Ile Glu Asn Asp Asp Ile Ile Phe Gly Asp Leu Gly Ser Phe
275 280 285
Gly Ala Pro Ile Phe His Phe Glu Leu Asp Val
290 295
<210> 62
<211> 393
<212> Белок
<213> Brachypodium distachyon
<400> 62
Met Ala Lys Pro Arg Lys Asn Ser Ala Ala Ala Asn Asn Asn Asn Asn
1 5 10 15
Asp Asn Ser Thr Asn Ala Asn Asn Ala Val Ala Glu Ala Ala Ala Ala
20 25 30
Asp Val Arg Ala Lys Pro Lys Lys Arg Thr Arg Lys Ser Val Pro Arg
35 40 45
Glu Ser Pro Ser Gln Arg Ser Ser Ile Tyr Arg Gly Val Thr Arg His
50 55 60
Arg Trp Thr Gly Arg Phe Glu Ala His Leu Trp Asp Lys Asn Ser Trp
65 70 75 80
Asn Glu Ser Gln Asn Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala Tyr
85 90 95
Asp Glu Glu Glu Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys
100 105 110
Tyr Trp Gly Pro Asp Thr Ile Leu Asn Phe Pro Leu Ser Val Tyr Asp
115 120 125
Asp Glu Leu Lys Glu Met Glu Gly Gln Ser Arg Glu Glu Tyr Ile Gly
130 135 140
Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr
145 150 155 160
Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile
165 170 175
Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala Thr
180 185 190
Gln Glu Glu Ala Ala Met Ala Tyr Asp Met Ala Ala Ile Glu Tyr Arg
195 200 205
Gly Leu Asn Ala Val Thr Asn Phe Asp Leu Ser Arg Tyr Ile Lys Trp
210 215 220
Leu Arg Pro Gly Gly Gly Val Asp Ser Ala Ala Ala Ala Ala Ala Arg
225 230 235 240
Asn Pro His Pro Met Leu Ala Gly Leu Ala Thr Gln Glu Glu Leu Pro
245 250 255
Ala Ile Asp His Leu Leu Asp Gly Met Ala Phe Gln Gln His Gly Leu
260 265 270
His Ser Ser Ser Ala Ala Ala Ala Ala Ala Gln Glu Phe Pro Leu Pro
275 280 285
Pro Ala Leu Gly His Ala Pro Thr Thr Ser Ala Leu Ser Leu Leu Leu
290 295 300
Gln Ser Pro Lys Phe Lys Glu Met Ile Glu Arg Thr Ser Ala Ala Glu
305 310 315 320
Thr Thr Thr Thr Ala Thr Thr Thr Ser Ser Ser Ser Ser Pro Arg Pro
325 330 335
Ala Ala Ser Pro Gln Cys Ser Phe Pro Glu Asp Ile Gln Thr Phe Phe
340 345 350
Gly Cys Asp Asp Gly Val Gly Val Gly Val Gly Ala Val Gly Tyr Thr
355 360 365
Asp Val Asp Gly Leu Phe Phe Gly Asp Leu Ser Ala Tyr Ala Ser Ser
370 375 380
Thr Ala Phe His Phe Glu Leu Asp Leu
385 390
<210> 63
<211> 379
<212> Белок
<213> Oryza sativa
<400> 63
Met Ala Lys Pro Arg Lys Asn Ser Thr Thr Thr Asn Thr Ser Ser Ser
1 5 10 15
Gly Val Ala Ala Ala Ala Ala Ala Ala Ala Val Lys Pro Lys Arg Thr
20 25 30
Arg Lys Ser Val Pro Arg Glu Ser Pro Ser Gln Arg Ser Ser Val Tyr
35 40 45
Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe Glu Ala His Leu
50 55 60
Trp Asp Lys Asn Ser Trp Asn Glu Ser Gln Asn Lys Lys Gly Lys Gln
65 70 75 80
Val Tyr Leu Gly Ala Tyr Asp Asp Glu Glu Ala Ala Ala Arg Ala Tyr
85 90 95
Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Asp Thr Ile Leu Asn Phe
100 105 110
Pro Leu Ser Ala Tyr Glu Gly Glu Leu Lys Glu Met Glu Gly Gln Ser
115 120 125
Arg Glu Glu Tyr Ile Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser
130 135 140
Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly
145 150 155 160
Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr
165 170 175
Leu Gly Thr Tyr Ala Thr Gln Glu Glu Ala Ala Met Ala Tyr Asp Met
180 185 190
Ala Ala Ile Glu Tyr Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Leu
195 200 205
Ser Arg Tyr Ile Lys Trp Leu Arg Pro Gly Ala Asp Gly Ala Gly Ala
210 215 220
Ala Gln Asn Pro His Pro Met Leu Gly Ala Leu Ser Ala Gln Asp Leu
225 230 235 240
Pro Ala Ile Asp Leu Asp Ala Met Ala Ser Ser Phe Gln His Asp Gly
245 250 255
His Gly Ala Ala Ala Ala Ala Ala Gln Leu Ile Pro Ala Arg His Ser
260 265 270
Leu Gly His Thr Pro Thr Thr Ser Ala Leu Ser Leu Leu Leu Gln Ser
275 280 285
Pro Lys Phe Lys Glu Met Ile Glu Arg Thr Ser Ala Ala Glu Thr Thr
290 295 300
Thr Thr Ser Ser Thr Thr Thr Ser Ser Ser Ser Pro Ser Pro Pro Gln
305 310 315 320
Ala Thr Lys Asp Asp Gly Ala Ser Pro Gln Cys Ser Phe Pro Lys Asp
325 330 335
Ile Gln Thr Tyr Phe Gly Cys Ala Ala Glu Asp Gly Ala Ala Gly Ala
340 345 350
Gly Tyr Ala Asp Val Asp Gly Leu Phe Phe Gly Asp Leu Thr Ala Tyr
355 360 365
Ala Ser Pro Ala Phe His Phe Glu Leu Asp Leu
370 375
<210> 64
<211> 398
<212> Белок
<213> Sorghum bicolor
<400> 64
Met Ala Lys Pro Arg Lys Asn Ser Ala Ala Ala Asn Asn Asn Asn Ser
1 5 10 15
Ser Ser Asn Gly Ala Gly Asp Leu Thr Pro Arg Ala Lys Pro Lys Arg
20 25 30
Thr Arg Lys Ser Val Pro Arg Glu Ser Pro Thr Gln Arg Ser Ser Val
35 40 45
Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe Glu Ala His
50 55 60
Leu Trp Asp Lys Asn Ser Trp Asn Glu Ser Gln Asn Lys Lys Gly Lys
65 70 75 80
Gln Val Tyr Leu Gly Ala Tyr Asp Asp Glu Glu Ala Ala Ala Arg Ala
85 90 95
Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Asp Thr Ile Leu Asn
100 105 110
Phe Pro Ala Ser Ala Tyr Glu Gly Glu Met Lys Gly Met Glu Gly Gln
115 120 125
Ser Arg Glu Glu Tyr Ile Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe
130 135 140
Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn
145 150 155 160
Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu
165 170 175
Tyr Leu Gly Thr Tyr Ala Thr Gln Glu Glu Ala Ala Met Ala Tyr Asp
180 185 190
Met Ala Ala Ile Glu Tyr Arg Gly Leu Asn Ala Val Thr Asn Phe Asp
195 200 205
Leu Ser Arg Tyr Ile Lys Trp Leu Arg Pro Gly Ala Gly Gly Met Ala
210 215 220
Ala Ala Ala Ala Ala Ala Gln Asn Pro His Pro Met Leu Gly Gly Leu
225 230 235 240
Ala Gln Gln Leu Leu Leu Pro Pro Pro Ala Asp Thr Thr Thr Thr Asp
245 250 255
Gly Ala Gly Ala Ala Ala Phe Gln His Asp His His Gly Ala Glu Ala
260 265 270
Phe Pro Leu Pro Pro Arg Thr Ser Leu Gly His Thr Pro Thr Thr Ser
275 280 285
Ala Leu Ser Leu Leu Leu Gln Ser Pro Lys Phe Lys Glu Met Ile Gln
290 295 300
Arg Thr Glu Ser Gly Thr Thr Thr Thr Thr Thr Thr Thr Ser Ser Leu
305 310 315 320
Ser Ser Ser Pro Pro Pro Thr Pro Ser Pro Ser Pro Pro Arg Arg Ser
325 330 335
Pro Ala Pro Thr Gln Pro Pro Val Gln Ala Ala Ala Arg Asp Ala Ser
340 345 350
Pro His Gln Arg Gly Phe Pro Glu Asp Val Gln Thr Phe Phe Gly Cys
355 360 365
Glu Asp Thr Ala Gly Ile Asp Val Glu Ala Leu Phe Phe Gly Asp Leu
370 375 380
Ala Ala Tyr Ala Thr Pro Ala Phe His Phe Glu Met Asp Leu
385 390 395
<210> 65
<211> 396
<212> Белок
<213> Zea mays
<400> 65
Met Ala Arg Pro Arg Lys Asn Ser Ala Ala Ala Ala Asn Asn Asn Asn
1 5 10 15
Ser Asn Thr Thr Asn Ala Gly Asn Ala Ala Val Asp Leu Ala Ala Arg
20 25 30
Val Lys Pro Lys Arg Thr Arg Lys Ser Val Pro Arg Glu Ser Pro Ser
35 40 45
Gln Arg Ser Ser Val Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly
50 55 60
Arg Phe Glu Ala His Leu Trp Asp Lys Asn Ser Trp Asn Glu Ser Gln
65 70 75 80
Asn Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala Tyr Asp Asp Glu Asp
85 90 95
Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro
100 105 110
Asp Thr Ile Leu Asn Phe Pro Ala Ser Ala Tyr Glu Ala Glu Leu Lys
115 120 125
Glu Met Glu Gly Gln Ser Arg Glu Glu Tyr Ile Gly Ser Leu Arg Arg
130 135 140
Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala
145 150 155 160
Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe
165 170 175
Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Gly Thr Gln Glu Glu Ala
180 185 190
Ala Met Ala Tyr Asp Met Ala Ala Ile Glu Tyr Arg Gly Leu Asn Ala
195 200 205
Val Thr Asn Phe Asp Leu Ser Arg Tyr Ile Lys Trp Leu Arg Pro Gly
210 215 220
Ala Gly Ala Ala Gln Asn Pro His Pro Met Leu Asp Gly Leu Ala Gln
225 230 235 240
Gln Leu Leu Leu Ser Pro Glu Gly Thr Ile Asp Gly Ala Ala Phe His
245 250 255
Gln Gln Gln His Asp His Arg Gln Gln Gly Ala Ala Glu Leu Pro Leu
260 265 270
Pro Pro Arg Ala Ser Leu Gly His Thr Pro Thr Thr Ser Ala Leu Gly
275 280 285
Leu Leu Leu Gln Ser Ser Lys Phe Lys Glu Met Ile Gln Arg Ala Ser
290 295 300
Ala Ala Glu Ser Gly Thr Thr Thr Val Thr Thr Thr Ser Ser Ser Ser
305 310 315 320
Ser Gln Pro Pro Thr Pro Thr Pro Thr Pro Ser Pro Ser Pro Pro Pro
325 330 335
Thr Pro Pro Val Gln Pro Ala Arg Asp Ala Ser Pro Gln Cys Ser Phe
340 345 350
Pro Glu Asp Ile Gln Thr Phe Phe Gly Cys Glu Asp Val Ala Gly Val
355 360 365
Gly Ala Gly Val Asp Val Asp Ala Leu Phe Phe Gly Asp Leu Ala Ala
370 375 380
Tyr Ala Ser Pro Ala Phe His Phe Glu Met Asp Leu
385 390 395
<210> 66
<211> 362
<212> Белок
<213> Glycine max
<400> 66
Met Ala Lys Lys Ser Gln Leu Arg Thr Gln Lys Asn Asn Ala Thr Asn
1 5 10 15
Asp Asp Ile Asn Leu Asn Ala Thr Asn Thr Val Ile Thr Lys Val Lys
20 25 30
Arg Thr Arg Arg Ser Val Pro Arg Asp Ser Pro Pro Gln Arg Ser Ser
35 40 45
Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala
50 55 60
His Leu Trp Asp Lys His Cys Trp Asn Glu Ser Gln Asn Lys Lys Gly
65 70 75 80
Arg Gln Gly Ala Tyr Asp Asn Glu Glu Ala Ala Ala His Ala Tyr Asp
85 90 95
Leu Ala Ala Leu Lys Tyr Trp Gly Gln Asp Thr Ile Leu Asn Phe Pro
100 105 110
Leu Ser Asn Tyr Leu Asn Glu Leu Lys Glu Met Glu Gly Gln Ser Arg
115 120 125
Glu Glu Tyr Ile Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg
130 135 140
Gly Ile Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg
145 150 155 160
Trp Glu Ala Arg Ile Gly Lys Val Phe Gly Asn Lys Tyr Leu Tyr Leu
165 170 175
Gly Thr Tyr Ala Thr Gln Glu Glu Ala Ala Thr Ala Tyr Asp Leu Ala
180 185 190
Ala Ile Glu Tyr Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Leu Ser
195 200 205
Arg Tyr Ile Lys Trp Leu Lys Pro Asn Asn Thr Asn Ser Asn Asn Asp
210 215 220
Gln Ile Ser Ile Asn Leu Thr Asn Ile Asn Asn Asn Cys Thr Asn Asn
225 230 235 240
Phe Ile Pro Asn Pro Asp Gln Glu Gln Glu Val Ser Phe Phe His Asn
245 250 255
Gln Asp Ser Leu Asn Asn Thr Ile Val Glu Glu Ala Thr Leu Val Pro
260 265 270
His Gln Pro Arg Pro Ala Ser Ala Thr Leu Ala Leu Glu Leu Leu Leu
275 280 285
Gln Ser Ser Lys Phe Lys Glu Met Val Glu Met Thr Ser Val Ala Asn
290 295 300
Leu Ser Thr Gln Met Glu Ser Asp Gln Leu Pro Gln Cys Thr Phe Pro
305 310 315 320
Asp His Ile Gln Thr Tyr Phe Glu Tyr Glu Asp Ser Asn Lys Tyr Glu
325 330 335
Glu Gly Asp Asp Leu Leu Phe Lys Phe Ser Glu Phe Ser Ser Ile Val
340 345 350
Pro Phe Tyr His Cys Asp Glu Phe Glu Ser
355 360
<210> 67
<211> 370
<212> Белок
<213> Glycine max
<400> 67
Met Ala Lys Lys Ser Gln Leu Arg Thr Gln Lys Asn Asn Val Thr Thr
1 5 10 15
Asn Asp Asp Asn Asn Leu Asn Val Thr Asn Thr Val Thr Thr Lys Val
20 25 30
Lys Arg Thr Arg Arg Ser Val Pro Arg Asp Ser Pro Pro Gln Arg Ser
35 40 45
Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu
50 55 60
Ala His Leu Trp Asp Lys His Cys Trp Asn Glu Ser Gln Asn Lys Lys
65 70 75 80
Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Asn Glu Glu Ala Ala Ala
85 90 95
His Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Gln Asp Thr Ile
100 105 110
Leu Asn Phe Pro Leu Ser Asn Tyr Leu Asn Glu Leu Lys Glu Met Glu
115 120 125
Gly Gln Ser Arg Glu Glu Tyr Ile Gly Ser Leu Arg Arg Lys Ser Ser
130 135 140
Gly Phe Ser Arg Gly Ile Ser Lys Tyr Arg Gly Val Ala Arg His His
145 150 155 160
His Asn Gly Arg Trp Glu Ala Arg Ile Gly Lys Val Phe Gly Asn Lys
165 170 175
Tyr Leu Tyr Leu Gly Thr Tyr Ala Thr Gln Glu Glu Ala Ala Thr Ala
180 185 190
Tyr Asp Leu Ala Ala Ile Glu Tyr Arg Gly Leu Asn Ala Val Thr Asn
195 200 205
Phe Asp Leu Ser Arg Tyr Ile Lys Trp Leu Lys Pro Asn Asn Asn Thr
210 215 220
Asn Asn Val Ile Asp Asp Gln Ile Ser Ile Asn Leu Thr Asn Ile Asn
225 230 235 240
Asn Asn Asn Asn Cys Thr Asn Ser Phe Thr Pro Ser Pro Asp Gln Glu
245 250 255
Gln Glu Ala Ser Phe Phe His Asn Lys Asp Ser Leu Asn Asn Thr Ile
260 265 270
Val Glu Glu Val Thr Leu Val Pro His Gln Pro Arg Pro Ala Ser Ala
275 280 285
Thr Ser Ala Leu Glu Leu Leu Leu Gln Ser Ser Lys Phe Lys Glu Met
290 295 300
Met Glu Met Thr Ser Val Ala Asn Leu Ser Ser Thr Gln Met Glu Ser
305 310 315 320
Glu Leu Pro Gln Cys Thr Phe Pro Asp His Ile Gln Thr Tyr Phe Glu
325 330 335
Tyr Glu Asp Ser Asn Arg Tyr Glu Glu Gly Asp Asp Leu Met Phe Lys
340 345 350
Phe Asn Glu Phe Ser Ser Ile Val Pro Phe Tyr Gln Cys Asp Glu Phe
355 360 365
Glu Ser
370
<210> 68
<211> 356
<212> Белок
<213> Medicago truncatula
<400> 68
Met Ala Lys Lys Ser Gln Lys Gln Ile Glu Lys Asp Asp Asn Ala Ser
1 5 10 15
Asn Asp Asn Asp Asn Leu Asn Pro Ser Asn Thr Val Thr Thr Lys Ala
20 25 30
Lys Arg Thr Arg Lys Ser Val Pro Arg Thr Ser Pro Pro Gln Arg Ser
35 40 45
Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu
50 55 60
Ala His Leu Trp Asp Lys Asn Cys Trp Asn Glu Ser Gln Asn Lys Lys
65 70 75 80
Gly Arg Gln Gly Ala Tyr Asp Asn Glu Glu Thr Ala Ala His Ala Tyr
85 90 95
Asp Leu Ala Ala Leu Lys Tyr Trp Gly Gln Asp Thr Ile Ile Asn Phe
100 105 110
Pro Leu Ser Asn Tyr Gln Lys Glu Leu Ile Glu Met Glu Ser Gln Ser
115 120 125
Arg Glu Glu Tyr Ile Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser
130 135 140
Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly
145 150 155 160
Arg Trp Glu Ala Arg Ile Gly Lys Val Phe Gly Asn Lys Tyr Leu Tyr
165 170 175
Leu Gly Thr Tyr Ala Thr Gln Glu Glu Ala Ala Thr Ala Tyr Asp Met
180 185 190
Ala Ala Ile Glu Tyr Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Leu
195 200 205
Ser Arg Tyr Ile Lys Trp Leu Lys Pro Asn Asn Asn Asn Asn Asp Asp
210 215 220
Asn Asn Lys Ser Asn Ile Asn Leu Cys Asp Ile Asn Ser Asn Ser Ser
225 230 235 240
Ala Asn Asp Ser Asn Ser Asn Glu Glu Leu Glu Phe Ser Leu Val Asp
245 250 255
Asn Glu Ile Ser Leu Asn Asn Ser Ile Asp Glu Ala Thr Leu Val Gln
260 265 270
Pro Arg Pro Thr Ser Ala Thr Ser Ala Leu Glu Leu Leu Leu Gln Ser
275 280 285
Ser Lys Phe Lys Glu Met Val Glu Met Ala Ser Met Thr Ser Asn Val
290 295 300
Ser Thr Thr Leu Glu Ser Asp Gln Leu Ser Gln Cys Ala Phe Pro Asp
305 310 315 320
Asp Ile Gln Thr Tyr Phe Glu Tyr Glu Asn Phe Asn Asp Thr Met Leu
325 330 335
Glu Asp Leu Asn Ser Ile Met Pro Thr Phe His Tyr Asp Phe Glu Gly
340 345 350
Ala Glu Val Leu
355
<210> 69
<211> 347
<212> Белок
<213> Glycine max
<400> 69
Met Ala Lys Gln Gln Thr His Lys Ile Asn Ala Ser Thr Asn Asn Asn
1 5 10 15
Ile Ser Thr Thr Asn Thr Val Thr Ala Lys Val Lys Arg Thr Arg Arg
20 25 30
Ser Val Pro Arg Asp Ser Pro Pro Gln Arg Ser Ser Ile Tyr Arg Gly
35 40 45
Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp
50 55 60
Lys Asn Cys Trp Asn Glu Ser Gln Asn Lys Lys Gly Arg Gln Gly Ala
65 70 75 80
Tyr Asp Asp Glu Glu Ala Ala Ala His Ala Tyr Asp Leu Ala Ala Leu
85 90 95
Lys Tyr Trp Gly Gln Asp Thr Ile Leu Asn Phe Pro Leu Ser Thr Tyr
100 105 110
Gln Asn Glu Leu Lys Glu Met Glu Gly Gln Ser Arg Glu Glu Tyr Ile
115 120 125
Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys
130 135 140
Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg
145 150 155 160
Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala
165 170 175
Thr Gln Glu Glu Ala Ala Thr Ala Tyr Asp Met Ala Ala Ile Glu Tyr
180 185 190
Arg Gly Val Asn Ala Val Thr Asn Phe Asp Leu Ser Arg Tyr Ile Lys
195 200 205
Trp Leu Lys Pro Asn Asn Asn Asn Thr Thr Val Asn Ser Asn Leu Ile
210 215 220
Asp Ser Asn Pro Asn Cys Glu Thr Asn Phe Thr Ser Asn Ser Asn Gln
225 230 235 240
Gln Gln Gly Phe Asn Phe Phe Asn Arg Gln Glu Ser Phe Asn Asn Glu
245 250 255
Glu Ala Ala Met Thr Gln Pro Arg Pro Ala Val Ala Thr Ser Ala Leu
260 265 270
Gly Leu Leu Leu Gln Ser Ser Lys Phe Lys Glu Met Met Glu Met Thr
275 280 285
Ser Ala Thr Asp Leu Ser Thr Pro Pro Ser Glu Ser Glu Leu Pro Ser
290 295 300
Cys Thr Phe Pro Asp Asp Ile Gln Thr Tyr Phe Glu Cys Glu Asp Ser
305 310 315 320
His Arg Tyr Gly Glu Gly Asp Asp Ile Met Phe Ser Val Leu Asn Gly
325 330 335
Phe Val Pro Pro Met Phe His Cys Asp Asp Phe
340 345
<210> 70
<211> 351
<212> Белок
<213> Glycine max
<400> 70
Met Ala Lys Gln Gln Thr His Glu Ile Asn Ala Ser Thr Asn Asn Asn
1 5 10 15
Ile Asn Thr Thr Lys Thr Val Thr Thr Lys Val Lys Arg Thr Arg Arg
20 25 30
Ser Val Pro Arg Asn Ser Pro Pro Gln Arg Ser Ser Ile Tyr Arg Gly
35 40 45
Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp
50 55 60
Lys Asn Cys Trp Asn Glu Ser Gln Asn Lys Lys Gly Arg Gln Gly Ala
65 70 75 80
Tyr Asp Asp Glu Glu Ala Ala Ala His Ala Tyr Asp Leu Ala Ala Leu
85 90 95
Lys Tyr Trp Gly Gln Asp Thr Ile Leu Asn Phe Pro Leu Ser Thr Tyr
100 105 110
Gln Asn Glu Leu Lys Glu Met Glu Gly Gln Ser Arg Glu Glu Tyr Ile
115 120 125
Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys
130 135 140
Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg
145 150 155 160
Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala
165 170 175
Thr Gln Glu Glu Ala Ala Thr Ala Tyr Asp Met Ala Ala Ile Glu Tyr
180 185 190
Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Leu Ser Arg Tyr Ile Lys
195 200 205
Trp Leu Lys Pro Asn Asn Asn Asn Asn Lys Val Asn Ser Asn Asn Leu
210 215 220
Ile Val Ser Ile Pro Asn Cys Ala Thr Asn Phe Thr Pro Asn Ser Asn
225 230 235 240
Gln Gln Gln Gly Phe Asn Phe Phe Asn Ser Gln Glu Ser Phe Asn Asn
245 250 255
Asn Glu Glu Ala Ala Met Thr Gln Pro Arg Pro Ala Ala Ala Thr Ser
260 265 270
Ala Leu Gly Leu Leu Leu Gln Ser Ser Lys Phe Lys Glu Met Met Glu
275 280 285
Met Thr Ser Ala Ile Asp Leu Ser Thr Pro Pro Ser Glu Ser Glu Leu
290 295 300
Pro Pro Cys Thr Phe Pro Asp Asp Ile Gln Thr Tyr Phe Glu Cys Glu
305 310 315 320
Asp Ser His Arg Tyr Gly Glu Gly Asp Asp Ile Met Phe Ser Glu Leu
325 330 335
Asn Gly Phe Val Pro Pro Met Phe His Cys Asp Asp Phe Glu Ala
340 345 350
<210> 71
<211> 353
<212> Белок
<213> Populus trichocarpa
<400> 71
Met Ala Lys Leu Ser Gln Lys Asn Thr Lys Asn Thr Ala Ser Asn Asn
1 5 10 15
Asn Asn Thr Thr Asn Gly Val Thr Lys Val Lys Arg Thr Arg Arg Ser
20 25 30
Val Pro Arg Asp Ser Pro Pro Gln Arg Ser Ser Ile Tyr Arg Gly Val
35 40 45
Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp Lys
50 55 60
Asn Cys Trp Asn Glu Ser Gln Asn Lys Lys Gly Arg Gln Gly Ala Tyr
65 70 75 80
Asp Asp Glu Glu Ala Ala Ala His Ala Tyr Asp Leu Ala Ala Leu Lys
85 90 95
Tyr Trp Gly Pro Glu Thr Ile Leu Asn Phe Pro Leu Ser Thr Tyr Gln
100 105 110
Asn Glu Leu Lys Glu Met Glu Gly Gln Ser Arg Glu Glu Cys Ile Gly
115 120 125
Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr
130 135 140
Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile
145 150 155 160
Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala Thr
165 170 175
Gln Glu Glu Ala Ala Thr Ala Tyr Asp Met Ala Ala Ile Glu Tyr Arg
180 185 190
Gly Leu Asn Ala Val Thr Asn Phe Asp Leu Ser Arg Tyr Ile Lys Trp
195 200 205
Leu Lys Pro Asn Gln Asn Asn Thr Asp Asn Asn Asn Gly Leu Asp Leu
210 215 220
Pro Asn Pro Ile Ile Gly Thr Asp Asn Ser Thr His Pro Asn Pro Asn
225 230 235 240
Gln Glu Leu Gly Thr Thr Phe Leu Gln Ile Asn Gln Gln Thr Tyr Gln
245 250 255
Pro Ser Glu Thr Thr Leu Thr Gln Pro Arg Pro Ala Thr Asn Pro Ser
260 265 270
Ser Ala Leu Gly Leu Leu Leu Gln Ser Ser Lys Phe Lys Glu Met Met
275 280 285
Glu Met Thr Ala Val Thr Asp Cys Pro Pro Thr Pro Pro Ser Gly Leu
290 295 300
Asp Pro Thr Pro Cys Ser Phe Leu Glu Asp Val Gln Thr Tyr Phe Asp
305 310 315 320
Cys Leu Asp Ser Ser Asn Tyr Gly Asp Gln Gly Asp Asp Met Ile Phe
325 330 335
Gly Asp Leu Asn Ser Phe Val Pro Pro Met Phe Gln Cys Asp Phe Glu
340 345 350
Thr
<210> 72
<211> 323
<212> Белок
<213> Vitis vinifera
<400> 72
Met Ala Lys Leu Ser Gln Gln Asn His Lys Asn Ser Ala Asn Ser Asn
1 5 10 15
Ala Thr Asn Thr Thr Leu Ser Val Thr Lys Val Lys Arg Thr Arg Lys
20 25 30
Thr Val Pro Arg Asp Ser Pro Pro Gln Arg Ser Ser Ile Tyr Arg Gly
35 40 45
Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp
50 55 60
Lys Asn Cys Trp Asn Glu Ser Gln Asn Lys Lys Gly Arg Gln Val Tyr
65 70 75 80
Leu Gly Ala Tyr His Asp Glu Glu Ala Ala Ala His Ala Tyr Asp Leu
85 90 95
Ala Ala Leu Lys Tyr Trp Gly Pro Glu Thr Ile Leu Asn Phe Pro Leu
100 105 110
Ser Thr Tyr Glu Lys Glu Leu Lys Glu Met Glu Gly Leu Ser Arg Glu
115 120 125
Glu Tyr Ile Gly Ser Leu Arg Arg Arg Ser Ser Gly Phe Ser Arg Gly
130 135 140
Val Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg Trp
145 150 155 160
Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly
165 170 175
Thr Tyr Ala Thr Gln Glu Glu Ala Ala Thr Ala Tyr Asp Met Ala Ala
180 185 190
Ile Glu Tyr Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Leu Ser Arg
195 200 205
Tyr Ile Lys Trp Leu Lys Pro Asn Gln Asn Asn Pro Cys Glu Gln Pro
210 215 220
Asn Asn Pro Asn Leu Asp Ser Asn Leu Thr Pro Asn Pro Asn His Asp
225 230 235 240
Phe Gly Ile Ser Phe Leu Asn His Pro Gln Thr Ser Gly Thr Ala Ala
245 250 255
Cys Lys Met Met Glu Met Thr Thr Ala Ala Asp His Leu Ser Thr Pro
260 265 270
Pro Glu Ser Glu Leu Pro Arg Cys Ser Phe Pro Asp Asp Ile Gln Thr
275 280 285
Tyr Phe Glu Cys Gln Asp Ser Gly Ser Tyr Glu Glu Gly Asp Asp Val
290 295 300
Ile Phe Ser Glu Leu Asn Ser Phe Ile Pro Pro Met Phe Gln Cys Asp
305 310 315 320
Phe Ser Ala
<210> 73
<211> 347
<212> Белок
<213> Vitis vinifera
<400> 73
Met Ala Lys Leu Ser Gln Gln Asn His Lys Asn Ser Ala Asn Ser Asn
1 5 10 15
Ala Thr Asn Thr Thr Leu Ser Val Thr Lys Val Lys Arg Thr Arg Lys
20 25 30
Thr Val Pro Arg Asp Ser Pro Pro Gln Arg Ser Ser Ile Tyr Arg Gly
35 40 45
Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp
50 55 60
Lys Asn Cys Trp Asn Glu Ser Gln Asn Lys Lys Gly Arg Gln Gly Ala
65 70 75 80
Tyr His Asp Glu Glu Ala Ala Ala His Ala Tyr Asp Leu Ala Ala Leu
85 90 95
Lys Tyr Trp Gly Pro Glu Thr Ile Leu Asn Phe Pro Leu Ser Thr Tyr
100 105 110
Glu Lys Glu Leu Lys Glu Met Glu Gly Leu Ser Arg Glu Glu Tyr Ile
115 120 125
Gly Ser Leu Arg Arg Arg Ser Ser Gly Phe Ser Arg Gly Val Ser Lys
130 135 140
Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg
145 150 155 160
Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala
165 170 175
Thr Gln Glu Glu Ala Ala Thr Ala Tyr Asp Met Ala Ala Ile Glu Tyr
180 185 190
Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Leu Ser Arg Tyr Ile Lys
195 200 205
Trp Leu Lys Pro Asn Gln Asn Asn Pro Cys Glu Gln Pro Asn Asn Pro
210 215 220
Asn Leu Asp Ser Asn Leu Thr Pro Asn Pro Asn His Asp Phe Gly Ile
225 230 235 240
Ser Phe Leu Asn His Pro Gln Thr Ser Gly Thr Ala Ala Cys Ser Glu
245 250 255
Pro Pro Leu Thr Gln Thr Arg Pro Pro Ile Ala Ser Ser Ala Leu Gly
260 265 270
Leu Leu Leu Gln Ser Ser Lys Phe Lys Glu Met Met Glu Met Thr Thr
275 280 285
Ala Ala Asp His Leu Ser Thr Pro Pro Glu Ser Glu Leu Pro Arg Cys
290 295 300
Ser Phe Pro Asp Asp Ile Gln Thr Tyr Phe Glu Cys Gln Asp Ser Gly
305 310 315 320
Ser Tyr Glu Glu Gly Asp Asp Val Ile Phe Ser Glu Leu Asn Ser Phe
325 330 335
Ile Pro Pro Met Phe Gln Cys Asp Phe Ser Ala
340 345
<210> 74
<211> 275
<212> Белок
<213> Populus trichocarpa
<400> 74
Met Gly Lys Thr Ser Lys Gln Ser Leu Lys Asn Ser Ala Asn Thr Ser
1 5 10 15
Ile Asn Pro Ala Thr Lys Val Lys Arg Thr Arg Lys Ser Val Pro Arg
20 25 30
Asp Ser Pro Pro Gln Arg Ser Ser Ile Tyr Arg Gly Val Thr Arg His
35 40 45
Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp Lys Asn Cys Trp
50 55 60
Asn Glu Ser Gln Asn Lys Lys Gly Arg Gln Gly Ala Tyr Asp Asp Glu
65 70 75 80
Glu Ala Ala Gly His Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly
85 90 95
Gln Asp Thr Ile Leu Asn Phe Pro Leu Ser Thr Tyr Glu Glu Glu Phe
100 105 110
Lys Glu Met Glu Gly His Ser Lys Glu Glu Tyr Ile Gly Ser Leu Arg
115 120 125
Arg Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val
130 135 140
Ala Arg His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val
145 150 155 160
Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala Thr Gln Glu Glu
165 170 175
Ala Ala Thr Ala Tyr Asp Met Ala Ala Ile Glu Tyr Arg Gly Leu Asn
180 185 190
Ala Val Thr Asn Phe Asp Leu Ser Arg Tyr Ser Ser Lys Phe Lys Glu
195 200 205
Met Leu Glu Arg Thr Ser Ala Ser Asp Cys Pro Leu Thr Pro Pro Glu
210 215 220
Ser Asp Arg Asp Pro Pro Arg Arg Ser Phe Pro Asp Asp Ile Gln Thr
225 230 235 240
Tyr Phe Asp Cys Gln Asp Ser Ser Ser Tyr Thr Asp Gly Asp Asp Ile
245 250 255
Ile Phe Gly Asp Leu His Ser Phe Ala Ser Pro Ile Phe His Cys Glu
260 265 270
Leu Asp Gly
275
<210> 75
<211> 304
<212> Белок
<213> Vitis vinifera
<400> 75
Met Ala Lys Thr Ser Gln Lys Ser Gln Lys Thr Thr Gly Asn Ser Thr
1 5 10 15
Asn Asn Asn Gly Gly Ser Val Ala Lys Val Lys Arg Thr Arg Lys Ser
20 25 30
Val Pro Arg Asp Ser Pro Pro Gln Arg Ser Ser Ile Phe Arg Gly Val
35 40 45
Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp Lys
50 55 60
Asn Cys Trp Asn Glu Ser Gln Asn Lys Lys Gly Arg Gln Val Tyr Leu
65 70 75 80
Gly Ala Tyr Asp Asp Glu Glu Ala Ala Ala His Ala Tyr Asp Leu Ala
85 90 95
Ala Leu Lys Tyr Trp Gly Gln Glu Thr Ile Leu Asn Phe Pro Leu Ser
100 105 110
Ala Tyr Gln Glu Glu Leu Lys Glu Met Glu Gly Gln Ser Lys Glu Glu
115 120 125
Tyr Ile Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly Val
130 135 140
Ser Lys Tyr Arg Gly Val Ala Arg His His His Asn Gly Arg Trp Glu
145 150 155 160
Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr
165 170 175
Tyr Ala Thr Gln Glu Glu Ala Ala Thr Ala Tyr Asp Met Ala Ala Ile
180 185 190
Glu Tyr Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Leu Ser Arg Tyr
195 200 205
Ile Asn Ser Pro Ala Pro Asn Pro Asn Pro Ser Asp His Glu Leu Gly
210 215 220
Leu Ser Phe Leu Gln Gln Gln His Gly Ser Asp Ala Thr Glu Leu Pro
225 230 235 240
Leu Ser His Ala Arg Ser Asp Cys Pro Leu Thr Pro Pro Asp Gln Ile
245 250 255
Glu Met Pro Arg Ser Ser Phe Pro Asp Asp Ile Gln Thr Tyr Phe Asp
260 265 270
Cys Gln Glu Thr Asn Ser Tyr Gly Glu Ser Asp Asp Ile Ile Phe Gly
275 280 285
Asp Leu Lys Tyr Phe Ser Ser Pro Met Phe Gln Cys Glu Leu Asp Thr
290 295 300
<210> 76
<211> 393
<212> Белок
<213> Sorghum bicolor
<400> 76
Met Ala Ser Pro Asn Pro Glu Ala Ala Ala Gly Leu Gln Thr Val Ala
1 5 10 15
Val Ala Ala Gly Gly Gly Glu Gly Gly Ser Ser Ser Ser Leu Gly Ala
20 25 30
Val Ala Gly Ala Ala Ala Val Ser Ser Ser Gly Glu Leu Val Pro Arg
35 40 45
Arg Ser Leu Ala Val Arg Lys Glu Arg Val Cys Thr Ala Lys Glu Arg
50 55 60
Ile Ser Arg Met Pro Pro Cys Ala Ala Gly Lys Arg Ser Ser Ile Tyr
65 70 75 80
Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu
85 90 95
Trp Asp Lys Ser Thr Trp Asn Gln Asn Gln Asn Lys Lys Gly Lys Gln
100 105 110
Gly Ala Tyr Asp Asp Glu Glu Ala Ala Ala Arg Ala Tyr Asp Leu Ala
115 120 125
Ala Leu Lys Tyr Trp Gly Ala Gly Thr Gln Ile Asn Phe Pro Val Ser
130 135 140
Asp Tyr Ala Arg Asp Leu Glu Glu Met Gln Met Ile Ser Lys Glu Asp
145 150 155 160
Tyr Leu Val Ser Leu Arg Arg Gln Leu His Asn Ser Arg Trp Asp Thr
165 170 175
Ser Leu Gly Leu Gly Asn Asp Tyr Met Ser Leu Ser Cys Gly Lys Asp
180 185 190
Ile Met Leu Asp Gly Lys Phe Ala Gly Ser Phe Gly Leu Glu Arg Lys
195 200 205
Ile Asp Leu Thr Asn Tyr Ile Arg Trp Trp Leu Pro Lys Lys Thr Arg
210 215 220
Gln Ser Asp Thr Ser Lys Thr Glu Glu Ile Ala Asp Glu Ile Arg Ala
225 230 235 240
Ile Glu Ser Ser Met Gln Gln Thr Glu Pro Tyr Lys Leu Pro Ser Leu
245 250 255
Gly Leu Gly Ser Pro Ser Lys Pro Ser Ser Val Gly Leu Ser Ala Cys
260 265 270
Ser Ile Leu Ser Gln Ser Asp Ala Phe Lys Ser Phe Leu Glu Lys Ser
275 280 285
Thr Lys Leu Ser Glu Glu Cys Thr Leu Ser Lys Glu Ile Val Glu Gly
290 295 300
Lys Thr Val Ala Ser Val Pro Ala Thr Gly Tyr Asp Thr Gly Ala Ile
305 310 315 320
Asn Ile Asn Met Asn Glu Leu Leu Val Gln Arg Ser Thr Tyr Ser Met
325 330 335
Ala Pro Val Met Pro Thr Pro Met Lys Thr Thr Trp Ser Pro Ala Asp
340 345 350
Pro Ser Val Asp Pro Leu Phe Trp Ser Asn Phe Val Leu Pro Ser Ser
355 360 365
Gln Pro Val Thr Met Ala Thr Ile Thr Thr Thr Thr Asn Glu Val Ser
370 375 380
Ser Ser Asp Pro Phe Gln Ser Gln Glu
385 390
<210> 77
<211> 428
<212> Белок
<213> Lupinus angustifolius
<400> 77
Met Ala Ser Ser Ser Ser Asp Pro Gly Lys Ser Glu Ile Gly Gly Gly
1 5 10 15
Ala Ala Glu Thr Ser Glu Ala Ala Ala Val Ala Val Ala Val Thr Asn
20 25 30
Asp Gln Ser Leu Leu Tyr Arg Gly Leu Lys Lys Ala Lys Lys Glu Arg
35 40 45
Gly Cys Thr Ala Lys Glu Arg Ile Ser Lys Met Pro Pro Cys Ala Ala
50 55 60
Gly Lys Arg Ser Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr
65 70 75 80
Gly Arg Tyr Glu Ala His Leu Arg Asp Lys Ser Thr Trp Asn Gln Asn
85 90 95
Gln Asn Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala Tyr Asp Asp Glu
100 105 110
Glu Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly
115 120 125
Pro Gly Thr Leu Ile Asn Phe Pro Val Thr Asp Tyr Thr Arg Asp Leu
130 135 140
Glu Glu Met Gln Asn Val Ser Arg Glu Glu Tyr Leu Ala Ser Leu Arg
145 150 155 160
Arg Lys Ser Ser Gly Phe Ser Arg Gly Ile Ser Lys Tyr Arg Ala Leu
165 170 175
Ser Ser Arg Trp Glu Pro Ser Tyr Ser Arg Phe Ala Gly Ser Asp Tyr
180 185 190
Phe Asn Ser Met His Tyr Gly Ala Gly Asp Asp Ser Ala Ala Glu Ser
195 200 205
Glu Tyr Ala Ser Gly Phe Cys Ile Glu Arg Lys Ile Asp Leu Thr Gly
210 215 220
His Ile Lys Trp Trp Gly Ser Asn Lys Ser Arg Gln Pro Asp Ala Gly
225 230 235 240
Thr Arg Leu Ser Glu Glu Lys Arg His Gly Phe Ala Gly Asp Ile Cys
245 250 255
Ser Glu Pro Lys Thr Leu Glu Gln Lys Val Gln Pro Thr Glu Pro Tyr
260 265 270
Gln Met Pro Glu Leu Gly Arg Ser His Asn Glu Lys Lys His Arg Ser
275 280 285
Ser Ala Val Ser Ala Leu Ser Ile Leu Ser Gln Ser Ala Ala Tyr Lys
290 295 300
Ser Leu Gln Glu Lys Ala Ser Lys Lys Gln Glu Asn Ser Thr Asp Asn
305 310 315 320
Asp Glu Asn Glu Asn Lys Asn Thr Val Asn Lys Leu Asp His Gly Lys
325 330 335
Ala Val Glu Lys Ser Ser Asn His Asp Gly Gly Ser Asp Arg Val Asp
340 345 350
Ile Glu Ile Gly Thr Thr Gly Ala Leu Ser Leu Gln Arg Asn Ile Tyr
355 360 365
Pro Leu Thr Pro Phe Leu Ser Ala Pro Leu Leu Thr Ala Tyr Asn Thr
370 375 380
Val Asp Pro Ser Leu Val Asp Pro Val Leu Trp Thr Ser Leu Val Pro
385 390 395 400
Met Leu Ser Ala Gly Leu Ser Cys Pro Thr Gln Val Thr Lys Thr Glu
405 410 415
Thr Ser Ser Ser Tyr Thr Ile Phe Gln Pro Glu Gly
420 425
<210> 78
<211> 440
<212> Белок
<213> Ricinus communis
<400> 78
Met Ala Ser Ser Ser Ser Asp Pro Gly Leu Lys Pro Glu Leu Gly Gly
1 5 10 15
Gly Ser Gly Gly Glu Ser Ser Glu Ala Val Ile Ala Asn Asp Gln Leu
20 25 30
Leu Leu Tyr Arg Gln Leu Lys Lys Pro Lys Lys Glu Arg Gly Cys Thr
35 40 45
Ala Lys Glu Arg Ile Ser Lys Met Pro Pro Cys Thr Ala Gly Lys Arg
50 55 60
Ser Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Tyr
65 70 75 80
Glu Ala His Leu Trp Asp Lys Ser Thr Trp Asn Gln Asn Gln Asn Lys
85 90 95
Lys Gly Lys Gln Gly Ala Tyr Asp Asp Glu Glu Ala Ala Ala Arg Ala
100 105 110
Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Gly Thr Leu Ile Asn
115 120 125
Phe Pro Val Thr Asp Tyr Ser Arg Asp Leu Glu Glu Met Gln Asn Val
130 135 140
Ser Arg Glu Glu Tyr Leu Ala Ser Leu Arg Arg Lys Ser Ser Gly Phe
145 150 155 160
Ser Arg Gly Ile Ser Lys Tyr Arg Gly Leu Ser Ser Gln Trp Asp Ser
165 170 175
Ser Phe Gly Arg Met Pro Gly Ser Glu Tyr Phe Ser Ser Ile Asn Tyr
180 185 190
Gly Ala Ala Asp Asp Pro Ala Ala Glu Ser Glu Tyr Val Gly Ser Leu
195 200 205
Cys Phe Glu Arg Lys Ile Asp Leu Thr Ser Tyr Ile Arg Trp Trp Gly
210 215 220
Phe Asn Lys Thr Arg Glu Ser Val Ser Lys Ser Ser Asp Glu Arg Lys
225 230 235 240
His Gly Tyr Gly Glu Asp Ile Ser Glu Leu Lys Ser Ser Glu Trp Ala
245 250 255
Val Gln Ser Thr Glu Pro Tyr Gln Met Pro Arg Leu Gly Met Pro Asp
260 265 270
Asn Gly Lys Lys His Lys Cys Ser Lys Ile Ser Ala Leu Ser Ile Leu
275 280 285
Ser His Ser Ala Ala Tyr Lys Asn Leu Gln Glu Lys Ala Ser Lys Lys
290 295 300
Gln Glu Asn Cys Thr Asp Asn Asp Glu Lys Glu Asn Lys Lys Thr Asn
305 310 315 320
Lys Met Asp Tyr Gly Lys Ala Val Glu Lys Ser Thr Ser His Asp Gly
325 330 335
Ser Asn Glu Arg Leu Gly Ala Ala Leu Gly Met Ser Gly Gly Leu Ser
340 345 350
Leu Gln Arg Asn Ala Tyr Gln Leu Ala Pro Phe Leu Ser Ala Pro Leu
355 360 365
Leu Thr Asn Tyr Asn Ala Ile Asp Pro Leu Val Asp Pro Ile Leu Trp
370 375 380
Thr Ser Leu Val Pro Val Leu Pro Ala Gly Phe Ser Arg Asn Ser Glu
385 390 395 400
Val Gly Met Gly Leu Gln Ile Val Ser Cys His Lys Asp Arg Asp Lys
405 410 415
Phe Asn Leu Tyr Leu Leu Ser Ala Gly Gly Val Ser Thr Phe Leu Leu
420 425 430
Leu Val Val His Trp Arg Phe Cys
435 440
<210> 79
<211> 428
<212> Белок
<213> Lupinus angustifolius
<400> 79
Met Ala Ser Ser Ser Ser Asp Pro Gly Lys Ser Glu Ile Gly Gly Gly
1 5 10 15
Ala Ala Glu Thr Ser Glu Ala Ala Ala Val Ala Val Ala Val Thr Asn
20 25 30
Asp Gln Ser Leu Leu Tyr Arg Gly Leu Lys Lys Ala Lys Lys Glu Arg
35 40 45
Gly Cys Thr Ala Lys Glu Arg Ile Ser Lys Met Pro Pro Cys Ala Ala
50 55 60
Gly Lys Arg Ser Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr
65 70 75 80
Gly Arg Tyr Glu Ala His Leu Arg Asp Lys Ser Thr Trp Asn Gln Asn
85 90 95
Gln Asn Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala Tyr Asp Asp Glu
100 105 110
Glu Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly
115 120 125
Pro Gly Thr Leu Ile Asn Phe Pro Val Thr Asp Tyr Thr Arg Asp Leu
130 135 140
Glu Glu Met Gln Asn Val Ser Arg Glu Glu Tyr Leu Ala Ser Leu Arg
145 150 155 160
Arg Lys Ser Ser Gly Phe Ser Arg Gly Ile Ser Lys Tyr Arg Ala Leu
165 170 175
Ser Ser Arg Trp Glu Pro Ser Tyr Ser Arg Phe Ala Gly Ser Asp Tyr
180 185 190
Phe Asn Ser Met His Tyr Gly Ala Gly Asp Asp Ser Ala Ala Glu Ser
195 200 205
Glu Tyr Ala Ser Gly Phe Cys Ile Glu Arg Lys Ile Asp Leu Thr Gly
210 215 220
His Ile Lys Trp Trp Gly Ser Asn Lys Ser Arg Gln Pro Asp Ala Gly
225 230 235 240
Thr Arg Leu Ser Glu Glu Lys Arg His Gly Phe Ala Gly Asp Ile Cys
245 250 255
Ser Glu Pro Lys Thr Leu Glu Gln Lys Val Gln Pro Thr Glu Pro Tyr
260 265 270
Gln Met Pro Glu Leu Gly Arg Ser His Asn Glu Lys Lys His Arg Ser
275 280 285
Ser Ala Val Ser Ala Leu Ser Ile Leu Ser Gln Ser Ala Ala Tyr Lys
290 295 300
Ser Leu Gln Glu Lys Ala Ser Lys Lys Gln Glu Asn Ser Thr Asp Asn
305 310 315 320
Asp Glu Asn Glu Asn Lys Asn Thr Val Asn Lys Leu Asp His Gly Lys
325 330 335
Ala Val Glu Lys Ser Ser Asn His Asp Gly Gly Ser Asp Arg Val Asp
340 345 350
Ile Glu Ile Gly Thr Thr Gly Ala Leu Ser Leu Gln Arg Asn Ile Tyr
355 360 365
Pro Leu Thr Pro Phe Leu Ser Ala Pro Leu Leu Thr Ala Tyr Asn Thr
370 375 380
Val Asp Pro Ser Leu Val Asp Pro Val Leu Trp Thr Ser Leu Val Pro
385 390 395 400
Met Leu Ser Ala Gly Leu Ser Cys Pro Thr Gln Val Thr Lys Thr Glu
405 410 415
Thr Ser Ser Ser Tyr Thr Ile Phe Gln Pro Glu Gly
420 425
<210> 80
<211> 508
<212> Белок
<213> Aspergillus fumigatus
<400> 80
Met Lys Met Ser Ala Ser Lys Thr Val Thr Ser Ser Ala Ser Ala Val
1 5 10 15
Ser Thr Ser Ser Gly Arg Ser Thr Pro Ser Lys Leu Val Asn Gly Ala
20 25 30
Thr Arg Asn Gly Ser Ala Ala Ala Gly Asn Gly Ser Thr Gly Thr Ala
35 40 45
Lys Gly Lys Arg Arg Ser Lys Tyr Arg His Val Ala Ala Tyr His Ser
50 55 60
Glu Leu Arg His Ser Ser Leu Ser Arg Glu Thr Ser Val Val Pro Ser
65 70 75 80
Phe Leu Gly Phe Arg Asn Leu Met Val Ile Val Leu Val Ala Met Asn
85 90 95
Leu Arg Leu Ile Ile Glu Asn Phe Met Lys Tyr Gly Val Leu Ile Cys
100 105 110
Ile Lys Cys His Asp Tyr Arg Lys Gln Asp Val Val Leu Gly Ser Ile
115 120 125
Leu Phe Ala Leu Val Pro Cys His Leu Phe Leu Ala Tyr Ile Ile Glu
130 135 140
Leu Val Ala Ala Gln Gln Ser Lys Lys Thr Val Gly Arg Gln Lys Lys
145 150 155 160
Asp Leu Ser Thr Glu Glu Arg Glu Arg Glu Gln Gln Ala Phe Arg Ser
165 170 175
Thr Trp Arg Tyr Thr Ala Phe Phe His Thr Val Asn Ala Thr Leu Cys
180 185 190
Leu Ala Val Thr Ser Phe Val Val Tyr Phe Tyr Ile Asn His Pro Gly
195 200 205
Ile Gly Thr Ile Cys Glu Leu His Ala Ile Ile Val Trp Leu Lys Asn
210 215 220
Cys Ser Tyr Ala Phe Thr Asn Arg Asp Leu Arg Gln Ala Met Val Asp
225 230 235 240
Pro Ser Ala Glu Ser Ala Leu Pro Glu Ile Tyr Ser Thr Cys Pro Tyr
245 250 255
Pro Arg Asn Ile Thr Leu Gly Asn Leu Thr Tyr Phe Trp Leu Ala Pro
260 265 270
Thr Leu Val Tyr Gln Pro Val Tyr Pro Arg Ser Ser His Ile Arg Trp
275 280 285
Ser Phe Val Ala Lys Arg Leu Ala Glu Phe Phe Gly Leu Ala Val Phe
290 295 300
Ile Trp Leu Leu Ser Ala Gln Tyr Ala Ala Pro Val Leu Arg Asn Ser
305 310 315 320
Ile Asp Lys Ile Ala Val Met Asp Ile Ala Ser Ile Leu Glu Arg Val
325 330 335
Met Lys Leu Ser Thr Ile Ser Leu Val Ile Trp Leu Ala Gly Phe Phe
340 345 350
Ala Leu Phe Gln Ser Leu Leu Asn Ala Leu Ala Glu Val Met Arg Phe
355 360 365
Gly Asp Arg Glu Phe Tyr Thr Asp Trp Trp Asn Ser Pro Ser Leu Gly
370 375 380
Ala Tyr Trp Arg Ser Trp Asn Arg Pro Val Tyr Leu Phe Met Lys Arg
385 390 395 400
His Val Phe Ser Pro Leu Val Gly Arg Gly Trp Ser Pro Phe Ala Ala
405 410 415
Ser Phe Met Val Phe Ser Leu Ser Ala Val Leu His Glu Met Leu Val
420 425 430
Gly Ile Pro Thr His Asn Leu Ile Gly Val Ala Phe Ala Gly Met Met
435 440 445
Phe Gln Leu Pro Leu Ile Ala Val Thr Ala Pro Phe Glu Lys Val Asn
450 455 460
Asp Ala Leu Gly Lys Ile Val Gly Asn Ser Ile Phe Trp Val Ser Phe
465 470 475 480
Cys Leu Val Gly Gln Pro Leu Gly Ala Leu Leu Tyr Phe Phe Ala Trp
485 490 495
Gln Ala Lys Tyr Gly Ser Val Ser Lys Ile His Val
500 505
<210> 81
<211> 521
<212> Белок
<213> Ricinus communis
<400> 81
Met Thr Ile Leu Glu Thr Pro Glu Thr Leu Gly Val Ile Ser Ser Ser
1 5 10 15
Ala Thr Ser Asp Leu Asn Leu Ser Leu Arg Arg Arg Arg Thr Ser Asn
20 25 30
Asp Ser Asp Gly Ala Leu Ala Asp Leu Ala Ser Lys Phe Asp Asp Asp
35 40 45
Asp Asp Val Arg Ser Glu Asp Ser Ala Glu Asn Ile Ile Glu Asp Pro
50 55 60
Val Ala Ala Val Thr Glu Leu Ala Thr Ala Lys Ser Asn Gly Lys Asp
65 70 75 80
Cys Val Ala Asn Ser Asn Lys Asp Lys Ile Asp Ser His Gly Gly Ser
85 90 95
Ser Asp Phe Lys Leu Ala Tyr Arg Pro Ser Val Pro Ala His Arg Ser
100 105 110
Leu Lys Glu Ser Pro Leu Ser Ser Asp Leu Ile Phe Lys Gln Ser His
115 120 125
Ala Gly Leu Phe Asn Leu Cys Ile Val Val Leu Val Ala Val Asn Ser
130 135 140
Arg Leu Ile Ile Glu Asn Leu Met Lys Tyr Gly Trp Leu Ile Lys Thr
145 150 155 160
Gly Phe Trp Phe Ser Ser Arg Ser Leu Arg Asp Trp Pro Leu Phe Met
165 170 175
Cys Cys Leu Ser Leu Pro Val Phe Pro Leu Ala Ala Tyr Leu Val Glu
180 185 190
Lys Ala Ala Tyr Arg Lys Tyr Ile Ser Pro Pro Ile Val Ile Phe Leu
195 200 205
His Val Ile Ile Thr Ser Ala Ala Val Leu Tyr Pro Ala Ser Val Ile
210 215 220
Leu Ser Cys Glu Ser Ala Phe Leu Ser Gly Val Thr Leu Met Glu Leu
225 230 235 240
Ala Cys Met Val Trp Leu Lys Leu Val Ser Tyr Ala His Thr Asn Tyr
245 250 255
Asp Met Arg Ala Ile Ala Asp Thr Ile His Lys Glu Asp Ala Ser Asn
260 265 270
Ser Ser Ser Thr Glu Tyr Cys His Asp Val Ser Phe Lys Thr Leu Ala
275 280 285
Tyr Phe Met Val Ala Pro Thr Leu Cys Tyr Gln Pro Ser Tyr Pro Arg
290 295 300
Thr Ala Phe Ile Arg Lys Gly Trp Val Phe Arg Gln Phe Val Lys Leu
305 310 315 320
Ile Ile Phe Thr Gly Phe Met Gly Phe Ile Ile Glu Gln Tyr Ile Asn
325 330 335
Pro Ile Val Gln Asn Ser Gln His Pro Leu Lys Gly Asp Leu Leu Tyr
340 345 350
Ala Ile Glu Arg Val Leu Lys Leu Ser Val Pro Asn Leu Tyr Val Trp
355 360 365
Leu Cys Leu Phe Tyr Cys Phe Phe His Leu Trp Leu Asn Ile Val Ala
370 375 380
Glu Leu Leu Arg Phe Gly Asp Arg Glu Phe Tyr Lys Asp Trp Trp Asn
385 390 395 400
Ala Lys Thr Val Glu Glu Tyr Trp Arg Met Trp Asn Met Pro Val His
405 410 415
Lys Trp Met Val Arg His Ile Tyr Phe Pro Cys Leu Arg Arg Lys Ile
420 425 430
Pro Arg Gly Val Ala Ile Val Ile Ala Phe Phe Val Ser Ala Val Phe
435 440 445
His Glu Leu Cys Ile Ala Val Pro Cys His Met Phe Lys Leu Trp Ala
450 455 460
Phe Phe Gly Ile Met Phe Gln Ile Pro Leu Val Val Ile Thr Asn Tyr
465 470 475 480
Phe Gln Arg Lys Phe Arg Ser Ser Met Val Gly Asn Met Ile Phe Trp
485 490 495
Phe Phe Phe Cys Ile Leu Gly Gln Pro Met Cys Val Leu Leu Tyr Tyr
500 505 510
His Asp Leu Met Asn Arg Asp Gly Asn
515 520
<210> 82
<211> 526
<212> Белок
<213> Vernicia fordii
<400> 82
Met Thr Ile Pro Glu Thr Pro Asp Asn Ser Thr Asp Ala Thr Thr Ser
1 5 10 15
Gly Gly Ala Glu Ser Ser Ser Asp Leu Asn Leu Ser Leu Arg Arg Arg
20 25 30
Arg Thr Ala Ser Asn Ser Asp Gly Ala Val Ala Glu Leu Ala Ser Lys
35 40 45
Ile Asp Glu Leu Glu Ser Asp Ala Gly Gly Gly Gln Val Ile Lys Asp
50 55 60
Pro Gly Ala Glu Met Asp Ser Gly Thr Leu Lys Ser Asn Gly Lys Asp
65 70 75 80
Cys Gly Thr Val Lys Asp Arg Ile Glu Asn Arg Glu Asn Arg Gly Gly
85 90 95
Ser Asp Val Lys Phe Thr Tyr Arg Pro Ser Val Pro Ala His Arg Ala
100 105 110
Leu Lys Glu Ser Pro Leu Ser Ser Asp Asn Ile Phe Lys Gln Ser His
115 120 125
Ala Gly Leu Phe Asn Leu Cys Ile Val Val Leu Val Ala Val Asn Ser
130 135 140
Arg Leu Ile Ile Glu Asn Ile Met Lys Tyr Gly Trp Leu Ile Lys Thr
145 150 155 160
Gly Phe Trp Phe Ser Ser Arg Ser Leu Arg Asp Trp Pro Leu Leu Met
165 170 175
Cys Cys Leu Thr Leu Pro Ile Phe Ser Leu Ala Ala Tyr Leu Val Glu
180 185 190
Lys Leu Ala Cys Arg Lys Tyr Ile Ser Ala Pro Thr Val Val Phe Leu
195 200 205
His Ile Leu Phe Ser Ser Thr Ala Val Leu Tyr Pro Val Ser Val Ile
210 215 220
Leu Ser Cys Glu Ser Ala Val Leu Ser Gly Val Ala Leu Met Leu Phe
225 230 235 240
Ala Cys Ile Val Trp Leu Lys Leu Val Ser Tyr Ala His Thr Asn Phe
245 250 255
Asp Met Arg Ala Ile Ala Asn Ser Val Asp Lys Gly Asp Ala Leu Ser
260 265 270
Asn Ala Ser Ser Ala Glu Ser Ser His Asp Val Ser Phe Lys Ser Leu
275 280 285
Val Tyr Phe Met Val Ala Pro Thr Leu Cys Tyr Gln Pro Ser Tyr Pro
290 295 300
Arg Thr Ala Ser Ile Arg Lys Gly Trp Val Val Arg Gln Phe Val Lys
305 310 315 320
Leu Ile Ile Phe Thr Gly Phe Met Gly Phe Ile Ile Glu Gln Tyr Ile
325 330 335
Asn Pro Ile Val Gln Asn Ser Gln His Pro Leu Lys Gly Asp Leu Leu
340 345 350
Tyr Ala Ile Glu Arg Val Leu Lys Leu Ser Val Pro Asn Leu Tyr Val
355 360 365
Trp Leu Cys Met Phe Tyr Cys Phe Phe His Leu Trp Leu Asn Ile Leu
370 375 380
Ala Glu Leu Leu Arg Phe Gly Asp Arg Glu Phe Tyr Lys Asp Trp Trp
385 390 395 400
Asn Ala Arg Thr Val Glu Glu Tyr Trp Arg Met Trp Asn Met Pro Val
405 410 415
His Lys Trp Met Val Arg His Ile Tyr Phe Pro Cys Leu Arg His Lys
420 425 430
Ile Pro Arg Gly Val Ala Leu Leu Ile Thr Phe Phe Val Ser Ala Val
435 440 445
Phe His Glu Leu Cys Ile Ala Val Pro Cys His Ile Phe Lys Leu Trp
450 455 460
Ala Phe Ile Gly Ile Met Phe Gln Ile Pro Leu Val Gly Ile Thr Asn
465 470 475 480
Tyr Leu Gln Asn Lys Phe Arg Ser Ser Met Val Gly Asn Met Ile Phe
485 490 495
Trp Phe Ile Phe Cys Ile Leu Gly Gln Pro Met Cys Leu Leu Leu Tyr
500 505 510
Tyr His Asp Leu Met Asn Arg Lys Gly Thr Thr Glu Ser Arg
515 520 525
<210> 83
<211> 523
<212> Белок
<213> Vernonia galamensis
<400> 83
Met Ala Leu Leu Asp Thr Pro Gln Ile Gly Glu Ile Thr Thr Thr Ala
1 5 10 15
Thr Thr Thr Ile Arg Arg Arg Thr Thr Val Lys Pro Asp Ala Gly Ile
20 25 30
Gly Asp Gly Leu Phe Asp Ser Ser Ser Ser Ser Lys Thr Asn Ser Ser
35 40 45
Phe Glu Asp Gly Asp Ser Leu Asn Gly Asp Phe Asn Asp Lys Phe Lys
50 55 60
Glu Gln Ile Gly Ala Gly Asp Glu Ser Lys Asp Asp Ser Lys Gly Asn
65 70 75 80
Gly Gln Lys Ile Asp His Gly Gly Val Lys Lys Gly Arg Glu Thr Thr
85 90 95
Val Val His Tyr Ala Tyr Arg Pro Ser Ser Pro Ala His Arg Arg Ile
100 105 110
Lys Glu Ser Pro Leu Ser Ser Asp Ala Ile Phe Lys Gln Ser His Ala
115 120 125
Gly Leu Phe Asn Leu Cys Ile Val Val Leu Val Ala Val Asn Gly Arg
130 135 140
Leu Ile Ile Glu Asn Leu Met Lys Tyr Gly Leu Leu Ile Asn Ser Asn
145 150 155 160
Phe Trp Phe Ser Ser Arg Ser Leu Arg Asp Trp Pro Leu Leu Met Cys
165 170 175
Cys Leu Thr Pro Ser Asp Phe Pro Leu Ala Ala Tyr Ile Val Glu Lys
180 185 190
Leu Ala Trp Lys Lys Arg Ile Ser Asp Pro Val Val Ile Thr Leu His
195 200 205
Val Ile Ile Thr Thr Thr Ala Ile Leu Tyr Pro Val Phe Met Ile Leu
210 215 220
Arg Phe Asp Ser Val Val Leu Ser Gly Val Ser Leu Met Leu Cys Ala
225 230 235 240
Cys Ile Asn Trp Leu Lys Leu Val Ser Phe Val His Thr Asn Tyr Asp
245 250 255
Met Arg Ser Leu Leu Asn Ser Thr Asp Lys Gly Glu Val Glu Pro Met
260 265 270
Ser Ser Asn Met Asp Tyr Phe Tyr Asp Val Asn Phe Lys Ser Leu Val
275 280 285
Tyr Phe Met Val Ala Pro Thr Leu Cys Tyr Gln Ile Ser Tyr Pro Arg
290 295 300
Thr Ala Phe Ile Arg Lys Gly Trp Val Leu Arg Gln Leu Ile Lys Leu
305 310 315 320
Val Ile Phe Thr Gly Phe Met Gly Phe Ile Ile Glu Gln Tyr Ile Asn
325 330 335
Pro Ile Val Lys Asn Ser Arg His Pro Leu Lys Gly Asp Phe Leu Tyr
340 345 350
Ala Ile Glu Arg Val Leu Lys Leu Ser Val Pro Asn Leu Tyr Val Trp
355 360 365
Leu Cys Met Phe Tyr Cys Phe Phe His Leu Trp Leu Asn Ile Leu Ala
370 375 380
Glu Leu Leu Cys Phe Gly Asp Arg Glu Phe Tyr Lys Asp Trp Trp Asn
385 390 395 400
Ala Gln Thr Ile Glu Glu Tyr Trp Arg Leu Trp Asn Met Pro Val His
405 410 415
Lys Trp Ile Val Arg His Leu Tyr Phe Pro Cys Leu Arg Asn Gly Ile
420 425 430
Pro Lys Gly Ala Ala Ile Leu Val Ala Phe Phe Met Ser Ala Val Phe
435 440 445
His Glu Leu Cys Ile Ala Val Pro Cys His Ile Phe Lys Phe Trp Ala
450 455 460
Phe Ile Gly Ile Met Phe Gln Val Pro Leu Val Leu Leu Thr Asn Tyr
465 470 475 480
Leu Gln His Lys Phe Gln Asn Ser Met Val Gly Asn Met Ile Phe Trp
485 490 495
Cys Phe Phe Ser Ile Phe Gly Gln Pro Met Cys Val Leu Leu Tyr Tyr
500 505 510
His Asp Val Met Asn Gln Lys Gly Lys Ser Lys
515 520
<210> 84
<211> 517
<212> Белок
<213> Vernonia galamensis
<400> 84
Met Ala Leu Leu Asp Thr Pro Gln Ile Gly Glu Ile Thr Thr Thr Ala
1 5 10 15
Thr Thr Thr Ile Arg Gln His Pro Leu Gly Lys Pro Asp Ala Gly Ile
20 25 30
Gly Asp Gly Leu Phe Ser Ser Ser Ser Ser Lys Thr Asn Ser Ser Phe
35 40 45
Glu Asp Gly Asp Ser Leu Asn Gly Asp Phe Asn Asp Lys Phe Lys Glu
50 55 60
Gln Ile Gly Ala Gly Asp Glu Ser Lys Lys Gly Asn Gly Lys Ile Asp
65 70 75 80
His Gly Gly Val Lys Lys Gly Arg Glu Thr Thr Val Val His Tyr Ala
85 90 95
Tyr Arg Pro Ser Ser Pro Ala His Arg Arg Ile Lys Glu Ser Pro Leu
100 105 110
Ser Ser Asp Ala Ile Phe Lys Gln Ser His Ala Gly Leu Phe Asn Leu
115 120 125
Cys Ile Val Val Leu Val Ala Val Asn Gly Arg Leu Ile Ile Glu Asn
130 135 140
Leu Met Lys Tyr Gly Leu Leu Ile Asn Ser Lys Phe Trp Phe Ser Ser
145 150 155 160
Arg Ser Leu Arg Asp Trp Pro Leu Leu Met Cys Trp Leu Thr Pro Ser
165 170 175
Asp Phe Pro Leu Ala Ala Tyr Ile Val Glu Lys Leu Ala Trp Lys Lys
180 185 190
Arg Ile Ser Asp Pro Val Val Ile Thr Leu His Val Val Ile Thr Thr
195 200 205
Thr Ala Ile Leu Tyr Pro Ile Phe Met Ile Leu Arg Phe Asp Ser Val
210 215 220
Val Leu Leu Gly Val Ser Leu Met Leu Cys Ala Cys Ile Asn Trp Leu
225 230 235 240
Lys Leu Val Ser Phe Val His Thr Asn Tyr Asp Met Arg Ser Leu Leu
245 250 255
Asn Ser Thr Gly Lys Gly Glu Val Glu Pro Met Ser Ser Asn Met Asp
260 265 270
Tyr Phe Tyr Asp Ile Asn Phe Lys Ser Leu Val Tyr Phe Met Val Ala
275 280 285
Pro Thr Leu Cys Tyr Gln Ile Ser Tyr Pro Arg Thr Ala Phe Ile Arg
290 295 300
Lys Gly Trp Val Phe Arg Gln Leu Ile Lys Leu Val Ile Phe Thr Gly
305 310 315 320
Phe Met Gly Phe Ile Ile Glu Gln Tyr Ile Asn Pro Ile Val Lys Asn
325 330 335
Ser Arg His Pro Leu Asn Gly Asp Phe Leu Tyr Ala Ile Glu Arg Val
340 345 350
Leu Lys Val Ser Val Pro Asn Leu Tyr Val Trp Leu Cys Met Phe Tyr
355 360 365
Cys Phe Phe His Leu Trp Leu Asn Ile Leu Ala Glu Leu Leu Trp Phe
370 375 380
Gly Asp Arg Glu Phe Tyr Lys Asp Trp Trp Asn Thr Gln Thr Ile Glu
385 390 395 400
Glu Tyr Trp Arg Leu Trp Asn Met Pro Val His Lys Trp Ile Val Arg
405 410 415
His Leu Tyr Phe Pro Cys Leu Arg Asn Gly Ile Ser Lys Gly Ala Ala
420 425 430
Ile Leu Val Ala Phe Phe Met Ser Ala Val Phe His Glu Leu Cys Ile
435 440 445
Ala Val Pro Cys His Ile Leu Lys Phe Trp Ala Phe Ile Gly Ile Met
450 455 460
Phe Gln Val Pro Leu Val Leu Leu Thr Asn Tyr Leu Gln His Lys Phe
465 470 475 480
Gln Asn Ser Met Val Gly Asn Met Ile Phe Trp Cys Phe Phe Ser Ile
485 490 495
Phe Gly Gln Pro Met Cys Val Phe Leu Tyr Tyr His Glu Val Asn Gln
500 505 510
Lys Gly Lys Ser Lys
515
<210> 85
<211> 507
<212> Белок
<213> Euonymus alatus
<400> 85
Met Ala Ala Asn Leu Asn Glu Ala Ser Asp Leu Asn Phe Ser Leu Arg
1 5 10 15
Arg Arg Thr Gly Gly Ile Ser Ser Thr Thr Val Pro Asp Ser Ser Ser
20 25 30
Glu Thr Ser Ser Ser Glu Ala Asp Tyr Leu Asp Gly Gly Lys Gly Ala
35 40 45
Ala Asp Val Lys Asp Arg Gly Asp Gly Ala Val Glu Phe Gln Asn Ser
50 55 60
Met Lys Asn Val Glu Arg Ile Glu Lys His Glu Ser Arg Val Gly Leu
65 70 75 80
Asp Ser Arg Phe Thr Tyr Arg Pro Ser Val Pro Ala His Arg Thr Ile
85 90 95
Lys Glu Ser Pro Leu Ser Ser Asp Ala Ile Phe Lys Gln Ser His Ala
100 105 110
Gly Leu Phe Asn Leu Cys Ile Val Val Leu Val Ala Val Asn Ser Arg
115 120 125
Leu Ile Ile Glu Asn Leu Met Lys Tyr Gly Trp Leu Ile Arg Ser Gly
130 135 140
Phe Trp Phe Ser Ser Arg Ser Leu Arg Asp Trp Pro Leu Phe Met Cys
145 150 155 160
Cys Leu Thr Leu Pro Val Phe Pro Leu Ala Ala Phe Leu Phe Glu Lys
165 170 175
Leu Ala Gln Lys Asn Leu Ile Ser Glu Pro Val Val Val Leu Leu His
180 185 190
Ile Val Asn Thr Thr Ala Ala Val Leu Tyr Pro Val Leu Val Ile Leu
195 200 205
Arg Cys Asp Ser Ala Phe Met Ser Gly Val Thr Leu Met Leu Phe Ala
210 215 220
Cys Ile Val Trp Leu Lys Leu Val Ser Tyr Ala His Thr Asn Tyr Asp
225 230 235 240
Met Arg Ala Leu Thr Lys Ser Val Glu Lys Gly Asp Thr Pro Leu Ser
245 250 255
Ser Gln Asn Met Asp Tyr Ser Phe Asp Val Asn Ile Lys Ser Leu Ala
260 265 270
Tyr Phe Met Val Ala Pro Thr Leu Cys Tyr Gln Ile Ser Tyr Pro Arg
275 280 285
Thr Pro Tyr Val Arg Lys Gly Trp Val Val Arg Gln Phe Val Lys Leu
290 295 300
Ile Ile Phe Thr Gly Leu Met Gly Phe Ile Ile Glu Gln Tyr Ile Asn
305 310 315 320
Pro Ile Val Gln Asn Ser Gln His Pro Leu Lys Gly Asn Phe Leu Tyr
325 330 335
Ala Ile Glu Arg Val Leu Lys Leu Ser Val Pro Asn Leu Tyr Val Trp
340 345 350
Leu Cys Met Phe Tyr Cys Leu Phe His Leu Trp Leu Asn Ile Leu Ala
355 360 365
Glu Leu Leu Cys Phe Gly Asp Arg Glu Phe Tyr Lys Asp Trp Trp Asn
370 375 380
Ala Lys Thr Val Glu Glu Tyr Trp Arg Met Trp Asn Met Pro Val His
385 390 395 400
Lys Trp Met Val Arg His Ile Tyr Phe Pro Cys Leu Arg Asn Gly Ile
405 410 415
Pro Lys Gly Val Ala Phe Val Ile Ser Phe Leu Val Ser Ala Val Phe
420 425 430
His Glu Leu Cys Ile Ala Val Pro Cys His Ile Phe Lys Leu Trp Ala
435 440 445
Phe Phe Gly Ile Met Leu Gln Val Pro Leu Val Leu Ile Thr Ser Tyr
450 455 460
Leu Gln Asn Lys Phe Arg Ser Ser Met Val Gly Asn Met Met Phe Trp
465 470 475 480
Phe Ser Phe Cys Ile Phe Gly Gln Pro Met Cys Leu Leu Leu Tyr Tyr
485 490 495
His Asp Leu Met Asn Arg Asn Gly Lys Met Glu
500 505
<210> 86
<211> 498
<212> Белок
<213> Caenorhabditis elegans
<400> 86
Met Gln Met Arg Gln Gln Thr Gly Arg Arg Arg Arg Gln Pro Ser Glu
1 5 10 15
Thr Ser Asn Gly Ser Leu Ala Ser Ser Arg Arg Ser Ser Phe Ala Gln
20 25 30
Asn Gly Asn Ser Ser Arg Lys Ser Ser Glu Met Arg Gly Pro Cys Glu
35 40 45
Lys Val Val His Thr Ala Gln Asp Ser Leu Phe Ser Thr Ser Ser Gly
50 55 60
Trp Thr Asn Phe Arg Gly Phe Phe Asn Leu Ser Ile Leu Leu Leu Val
65 70 75 80
Leu Ser Asn Gly Arg Val Ala Leu Glu Asn Val Ile Lys Tyr Gly Ile
85 90 95
Leu Ile Thr Pro Leu Gln Trp Ile Ser Thr Phe Val Glu His His Tyr
100 105 110
Ser Ile Trp Ser Trp Pro Asn Leu Ala Leu Ile Leu Cys Ser Asn Ile
115 120 125
Gln Ile Leu Ser Val Phe Gly Met Glu Lys Ile Leu Glu Arg Gly Trp
130 135 140
Leu Gly Asn Gly Phe Ala Ala Val Phe Tyr Thr Ser Leu Val Ile Ala
145 150 155 160
His Leu Thr Ile Pro Val Val Val Thr Leu Thr His Lys Trp Lys Asn
165 170 175
Pro Leu Trp Ser Val Val Met Met Gly Val Tyr Val Ile Glu Ala Leu
180 185 190
Lys Phe Ile Ser Tyr Gly His Val Asn Tyr Trp Ala Arg Asp Ala Arg
195 200 205
Arg Lys Ile Thr Glu Leu Lys Thr Gln Val Thr Asp Leu Ala Lys Lys
210 215 220
Thr Cys Asp Pro Lys Gln Phe Trp Asp Leu Lys Asp Glu Leu Ser Met
225 230 235 240
His Gln Met Ala Ala Gln Tyr Pro Ala Asn Leu Thr Leu Ser Asn Ile
245 250 255
Tyr Tyr Phe Met Ala Ala Pro Thr Leu Cys Tyr Glu Phe Lys Phe Pro
260 265 270
Arg Leu Leu Arg Ile Arg Lys His Phe Leu Ile Lys Arg Thr Val Glu
275 280 285
Leu Ile Phe Leu Ser Phe Leu Ile Ala Ala Leu Val Gln Gln Trp Val
290 295 300
Val Pro Thr Val Arg Asn Ser Met Lys Pro Leu Ser Glu Met Glu Tyr
305 310 315 320
Ser Arg Cys Leu Glu Arg Leu Leu Lys Leu Ala Ile Pro Asn His Leu
325 330 335
Ile Trp Leu Leu Phe Phe Tyr Thr Phe Phe His Ser Phe Leu Asn Leu
340 345 350
Ile Ala Glu Leu Leu Arg Phe Ala Asp Arg Glu Phe Tyr Arg Asp Phe
355 360 365
Trp Asn Ala Glu Thr Ile Gly Tyr Phe Trp Lys Ser Trp Asn Ile Pro
370 375 380
Val His Arg Phe Ala Val Arg His Ile Tyr Ser Pro Met Met Arg Asn
385 390 395 400
Asn Phe Ser Lys Met Ser Ala Phe Phe Val Val Phe Phe Val Ser Ala
405 410 415
Phe Phe His Glu Tyr Leu Val Ser Val Pro Leu Lys Ile Phe Arg Leu
420 425 430
Trp Ser Tyr Tyr Gly Met Met Gly Gln Ile Pro Leu Ser Ile Ile Thr
435 440 445
Asp Lys Val Val Arg Gly Gly Arg Thr Gly Asn Ile Ile Val Trp Leu
450 455 460
Ser Leu Ile Val Gly Gln Pro Leu Ala Ile Leu Met Tyr Gly His Asp
465 470 475 480
Trp Tyr Ile Leu Asn Phe Gly Val Ser Ala Val Gln Asn Gln Thr Val
485 490 495
Gly Ile
<210> 87
<211> 498
<212> Белок
<213> Rattus norvegicus
<400> 87
Met Gly Asp Arg Gly Gly Ala Gly Ser Ser Arg Arg Arg Arg Thr Gly
1 5 10 15
Ser Arg Val Ser Val Gln Gly Gly Ser Gly Pro Lys Val Glu Glu Asp
20 25 30
Glu Val Arg Glu Ala Ala Val Ser Pro Asp Leu Gly Ala Gly Gly Asp
35 40 45
Ala Pro Ala Pro Ala Pro Ala Pro Ala His Thr Arg Asp Lys Asp Arg
50 55 60
Gln Thr Ser Val Gly Asp Gly His Trp Glu Leu Arg Cys His Arg Leu
65 70 75 80
Gln Asp Ser Leu Phe Ser Ser Asp Ser Gly Phe Ser Asn Tyr Arg Gly
85 90 95
Ile Leu Asn Trp Cys Val Val Met Leu Ile Leu Ser Asn Ala Arg Leu
100 105 110
Ser Leu Glu Asn Leu Ile Lys Tyr Gly Ile Leu Val Asp Pro Ile Gln
115 120 125
Val Val Ser Leu Phe Leu Lys Asp Pro Tyr Ser Trp Pro Ala Pro Cys
130 135 140
Leu Ile Ile Ala Ser Asn Ile Phe Ile Val Ala Thr Phe Gln Ile Glu
145 150 155 160
Lys Arg Leu Ser Val Gly Ala Leu Thr Glu Gln Met Gly Leu Leu Leu
165 170 175
His Val Val Asn Leu Ala Thr Ile Ile Cys Phe Pro Ala Ala Val Ala
180 185 190
Leu Leu Val Glu Ser Ile Thr Pro Val Gly Ser Leu Phe Ala Leu Ala
195 200 205
Ser Tyr Ser Ile Ile Phe Leu Lys Leu Ser Ser Tyr Arg Asp Val Asn
210 215 220
Leu Trp Cys Arg Gln Arg Arg Val Lys Ala Lys Ala Val Ser Ala Gly
225 230 235 240
Lys Lys Val Ser Gly Ala Ala Ala Gln Asn Thr Val Ser Tyr Pro Asp
245 250 255
Asn Leu Thr Tyr Arg Asp Leu Tyr Tyr Phe Ile Phe Ala Pro Thr Leu
260 265 270
Cys Tyr Glu Leu Asn Phe Pro Arg Ser Pro Arg Ile Arg Lys Arg Phe
275 280 285
Leu Leu Arg Arg Val Leu Glu Met Leu Phe Phe Thr Gln Leu Gln Val
290 295 300
Gly Leu Ile Gln Gln Trp Met Val Pro Thr Ile Gln Asn Ser Met Lys
305 310 315 320
Pro Phe Lys Asp Met Asp Tyr Ser Arg Ile Ile Glu Arg Leu Leu Lys
325 330 335
Leu Ala Val Pro Asn His Leu Ile Trp Leu Ile Phe Phe Tyr Trp Leu
340 345 350
Phe His Ser Cys Leu Asn Ala Val Ala Glu Leu Leu Gln Phe Gly Asp
355 360 365
Arg Glu Phe Tyr Arg Asp Trp Trp Asn Ala Glu Ser Val Thr Tyr Phe
370 375 380
Trp Gln Asn Trp Asn Ile Pro Val His Lys Trp Cys Ile Arg His Phe
385 390 395 400
Tyr Lys Pro Met Leu Arg Leu Gly Ser Asn Lys Trp Met Ala Arg Thr
405 410 415
Gly Val Phe Trp Ala Ser Ala Phe Phe His Glu Tyr Leu Val Ser Ile
420 425 430
Pro Leu Arg Met Phe Arg Leu Trp Ala Phe Thr Ala Met Met Ala Gln
435 440 445
Val Pro Leu Ala Trp Ile Val Asn Arg Phe Phe Gln Gly Asn Tyr Gly
450 455 460
Asn Ala Ala Val Trp Val Thr Leu Ile Ile Gly Gln Pro Val Ala Val
465 470 475 480
Leu Met Tyr Val His Asp Tyr Tyr Val Leu Asn Tyr Asp Ala Pro Val
485 490 495
Gly Ala
<210> 88
<211> 488
<212> Белок
<213> Homo sapiens
<400> 88
Met Gly Asp Arg Gly Ser Ser Arg Arg Arg Arg Thr Gly Ser Arg Pro
1 5 10 15
Ser Ser His Gly Gly Gly Gly Pro Ala Ala Ala Glu Glu Glu Val Arg
20 25 30
Asp Ala Ala Ala Gly Pro Asp Val Gly Ala Ala Gly Asp Ala Pro Ala
35 40 45
Pro Ala Pro Asn Lys Asp Gly Asp Ala Gly Val Gly Ser Gly His Trp
50 55 60
Glu Leu Arg Cys His Arg Leu Gln Asp Ser Leu Phe Ser Ser Asp Ser
65 70 75 80
Gly Phe Ser Asn Tyr Arg Gly Ile Leu Asn Trp Cys Val Val Met Leu
85 90 95
Ile Leu Ser Asn Ala Arg Leu Phe Leu Glu Asn Leu Ile Lys Tyr Gly
100 105 110
Ile Leu Val Asp Pro Ile Gln Val Val Ser Leu Phe Leu Lys Asp Pro
115 120 125
Tyr Ser Trp Pro Ala Pro Cys Leu Val Ile Ala Ala Asn Val Phe Ala
130 135 140
Val Ala Ala Phe Gln Val Glu Lys Arg Leu Ala Val Gly Ala Leu Thr
145 150 155 160
Glu Gln Ala Gly Leu Leu Leu His Val Ala Asn Leu Ala Thr Ile Leu
165 170 175
Cys Phe Pro Ala Ala Val Val Leu Leu Val Glu Ser Ile Thr Pro Val
180 185 190
Gly Ser Leu Leu Ala Leu Met Ala His Thr Ile Leu Phe Leu Lys Leu
195 200 205
Phe Ser Tyr Arg Asp Val Asn Ser Trp Cys Arg Arg Ala Arg Ala Lys
210 215 220
Ala Ala Ser Ala Gly Lys Lys Ala Ser Ser Ala Ala Ala Pro His Thr
225 230 235 240
Val Ser Tyr Pro Asp Asn Leu Thr Tyr Arg Asp Leu Tyr Tyr Phe Leu
245 250 255
Phe Ala Pro Thr Leu Cys Tyr Glu Leu Asn Phe Pro Arg Ser Pro Arg
260 265 270
Ile Arg Lys Arg Phe Leu Leu Arg Arg Ile Leu Glu Met Leu Phe Phe
275 280 285
Thr Gln Leu Gln Val Gly Leu Ile Gln Gln Trp Met Val Pro Thr Ile
290 295 300
Gln Asn Ser Met Lys Pro Phe Lys Asp Met Asp Tyr Ser Arg Ile Ile
305 310 315 320
Glu Arg Leu Leu Lys Leu Ala Val Pro Asn His Leu Ile Trp Leu Ile
325 330 335
Phe Phe Tyr Trp Leu Phe His Ser Cys Leu Asn Ala Val Ala Glu Leu
340 345 350
Met Gln Phe Gly Asp Arg Glu Phe Tyr Arg Asp Trp Trp Asn Ser Glu
355 360 365
Ser Val Thr Tyr Phe Trp Gln Asn Trp Asn Ile Pro Val His Lys Trp
370 375 380
Cys Ile Arg His Phe Tyr Lys Pro Met Leu Arg Arg Gly Ser Ser Lys
385 390 395 400
Trp Met Ala Arg Thr Gly Val Phe Leu Ala Ser Ala Phe Phe His Glu
405 410 415
Tyr Leu Val Ser Val Pro Leu Arg Met Phe Arg Leu Trp Ala Phe Thr
420 425 430
Gly Met Met Ala Gln Ile Pro Leu Ala Trp Phe Val Gly Arg Phe Phe
435 440 445
Gln Gly Asn Tyr Gly Asn Ala Ala Val Trp Leu Ser Leu Ile Ile Gly
450 455 460
Gln Pro Ile Ala Val Leu Met Tyr Val His Asp Tyr Tyr Val Leu Asn
465 470 475 480
Tyr Glu Ala Pro Ala Ala Glu Ala
485
<210> 89
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<220>
<221> X
<222> (4)..(4)
<223> Треонин (T) или Серин (S)
<400> 89
Arg Gly Val Xaa Arg His Arg Trp Thr Gly Arg
1 5 10
<210> 90
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<220>
<221> X
<222> (1)..(1)
<223> Фенилаланин (F) или Тирозин (Y)
<400> 90
Xaa Glu Ala His Leu Trp Asp Lys
1 5
<210> 91
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<400> 91
Asp Leu Ala Ala Leu Lys Tyr Trp Gly
1 5
<210> 92
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<220>
<221> misc_feature
<222> (2)..(2)
<223> Xaa может быть любой природной аминокислотой
<220>
<221> X
<222> (5)..(5)
<223> Серин (S) или Аланин (A)
<220>
<221> X
<222> (8)..(8)
<223> любая аминокислота
<400> 92
Ser Xaa Gly Phe Xaa Arg Gly Xaa
1 5
<210> 93
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<220>
<221> X
<222> (3)..(3)
<223> Гистидин (H) или Гутамин (Q)
<220>
<221> X
<222> (6)..(6)
<223> Аргинин (R) или Лизин (K)
<220>
<221> X
<222> (12)..(12)
<223> Аргинин (R) или Лизин (K)
<220>
<221> misc_feature
<222> (13)..(13)
<223> Xaa может быть любой природной аминокислотой
<400> 93
His His Xaa Asn Gly Xaa Trp Glu Ala Arg Ile Gly Xaa Val
1 5 10
<210> 94
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> консервативная последовательность
<220>
<221> X
<222> (7)..(7)
<223> любая аминокислота
<400> 94
Gln Glu Glu Ala Ala Ala Xaa Tyr Asp
1 5
<210> 95
<211> 165
<212> Белок
<213> Brassica napus
<400> 95
Met Gly Ile Leu Arg Lys Lys Lys His Glu Arg Lys Pro Ser Phe Lys
1 5 10 15
Ser Val Leu Thr Ala Ile Leu Ala Thr His Ala Ala Thr Phe Leu Leu
20 25 30
Leu Ile Ala Gly Val Ser Leu Ala Gly Thr Ala Ala Ala Phe Ile Ala
35 40 45
Thr Met Pro Leu Phe Val Val Phe Ser Pro Ile Leu Val Pro Ala Gly
50 55 60
Ile Thr Thr Gly Leu Leu Thr Thr Gly Leu Ala Ala Ala Gly Gly Ala
65 70 75 80
Gly Ala Thr Ala Val Thr Ile Ile Leu Trp Leu Tyr Lys Arg Ala Thr
85 90 95
Gly Lys Ala Pro Pro Lys Val Leu Glu Lys Val Leu Lys Lys Ile Ile
100 105 110
Pro Gly Ala Ala Ala Ala Pro Ala Ala Ala Pro Gly Ala Ala Pro Ala
115 120 125
Ala Ala Pro Ala Ala Ala Pro Ala Val Ala Pro Ala Ala Ala Pro Ala
130 135 140
Ala Ala Pro Ala Pro Lys Pro Ala Ala Pro Pro Ala Pro Lys Pro Ala
145 150 155 160
Ala Ala Pro Ser Ile
165
<210> 96
<211> 193
<212> Белок
<213> Brassica napus
<400> 96
Met Ala Asp Val Arg Thr His Ala His Gln Val Gln Val His Pro Leu
1 5 10 15
Arg Gln Gln Glu Gly Gly Ile Lys Val Val Tyr Pro Gln Ser Gly Pro
20 25 30
Ser Ser Thr Gln Val Leu Ala Val Ile Ala Gly Val Pro Val Gly Gly
35 40 45
Thr Leu Leu Thr Leu Ala Gly Leu Thr Leu Ala Gly Ser Val Ile Gly
50 55 60
Leu Met Leu Ala Phe Pro Leu Phe Leu Ile Phe Ser Pro Val Ile Val
65 70 75 80
Pro Ala Ala Phe Val Ile Gly Leu Ala Met Thr Gly Phe Met Ala Ser
85 90 95
Gly Ala Ile Gly Leu Thr Gly Leu Ser Ser Met Ser Trp Val Leu Asn
100 105 110
His Ile Arg Arg Val Arg Glu Arg Met Pro Asp Glu Leu Glu Glu Ala
115 120 125
Lys Gln Arg Leu Ala Asp Met Ala Glu Tyr Val Gly Gln Arg Thr Lys
130 135 140
Asp Ala Gly Gln Thr Ile Glu Glu Lys Ala His Asp Val Arg Glu Ser
145 150 155 160
Lys Thr Tyr Asp Val Arg Asp Arg Asp Thr Lys Gly His Thr Ala Thr
165 170 175
Gly Gly Asp Arg Asp Thr Lys Thr Thr Arg Glu Val Arg Val Ala Thr
180 185 190
Thr
<210> 97
<211> 188
<212> Белок
<213> Brassica napus
<400> 97
Met Ala Asn Val Asp Arg Arg Val Asn Val Asp Arg Thr Asp Lys Gly
1 5 10 15
Leu Gln Leu Gln Pro Gln Tyr Glu Asp Arg Val Gly Tyr Gly Tyr Gly
20 25 30
Tyr Gly Gly Asn Thr Asp Tyr Lys Ser Cys Gly Pro Ser Thr Asn Gln
35 40 45
Ile Val Ala Leu Ile Ala Gly Val Pro Ile Gly Gly Ser Leu Leu Ala
50 55 60
Leu Ala Gly Leu Thr Leu Ala Gly Ser Val Ile Gly Phe Met Leu Ser
65 70 75 80
Ile Pro Leu Phe Leu Leu Phe Ser Pro Val Ile Val Pro Ala Ala Leu
85 90 95
Thr Ile Gly Leu Ala Val Thr Gly Ile Leu Ala Ser Gly Leu Phe Gly
100 105 110
Leu Thr Gly Leu Ser Ser Val Ser Trp Val Leu Asn Tyr Ile Arg Gly
115 120 125
Arg Ser Asp Thr Val Pro Glu Gln Leu Asp Tyr Ala Lys Arg Arg Met
130 135 140
Ala Asp Ala Val Gly Tyr Ala Gly Gln Lys Gly Lys Glu Met Gly Gln
145 150 155 160
Tyr Val Gln Asp Lys Ala His Glu Ala His Asp Thr Ser Leu Thr Thr
165 170 175
Glu Thr Asn Gly Lys Thr Arg Arg Ala His Ile Ala
180 185
<210> 98
<211> 180
<212> Белок
<213> Brassica napus
<400> 98
Met Ala Asp Thr Ala Arg Thr His His Asp Ile Thr Ser Arg Asp Gln
1 5 10 15
Tyr Pro Ile Leu Gly Arg Asp Arg Asp Gln Tyr Pro Tyr Gly Arg Ser
20 25 30
Asp Tyr Gln Thr Ser Gly Gln Asp Tyr Ser Lys Thr Arg Gln Ile Ala
35 40 45
Lys Ala Ala Thr Ala Val Thr Ala Gly Gly Ser Leu Leu Val Leu Ser
50 55 60
Ser Leu Thr Leu Val Gly Thr Val Ile Ala Leu Thr Val Ala Thr Thr
65 70 75 80
Leu Leu Val Ile Phe Ser Pro Ile Leu Val Pro Ala Leu Ile Thr Val
85 90 95
Ala Leu Leu Ile Thr Gly Phe Leu Ser Ser Gly Gly Phe Gly Ile Ala
100 105 110
Asp Ile Thr Val Phe Ser Trp Ile Tyr Lys Tyr Ala Thr Gly Glu His
115 120 125
Pro Gln Gly Ser Asp Lys Leu Asp Ser Ala Arg Met Lys Leu Gly Thr
130 135 140
Lys Ala Gln Asp Ile Lys Asp Arg Ala Gln Tyr Tyr Gly Gln Gln His
145 150 155 160
Thr Gly Gly Glu His Asp Arg Asp Arg Thr Arg Gly Thr His His Thr
165 170 175
Thr Thr Thr Thr
180
<210> 99
<211> 210
<212> Белок
<213> Brassica napus
<400> 99
Met Ala Asp Thr His Arg Val Asp Arg Thr Asp Arg His Leu Gln Phe
1 5 10 15
Gln Ser Pro Tyr Glu Gly Gly Arg Val Ser Ile Gln Tyr Glu Gly Gly
20 25 30
Gly Gly Ala Gly Gly Tyr Gly Gly Arg Gly Gly Gly Tyr Gly Ala Glu
35 40 45
Gly Tyr Lys Ser Met Met Pro Glu Arg Gly Pro Ser Ser Thr Gln Val
50 55 60
Leu Ser Phe Leu Val Gly Val Pro Ile Val Gly Ser Leu Leu Ala Ile
65 70 75 80
Ala Gly Leu Leu Leu Ala Gly Ser Val Ile Gly Leu Leu Ile Ser Ile
85 90 95
Pro Leu Phe Leu Leu Phe Ser Pro Val Ile Val Pro Ala Ala Leu Thr
100 105 110
Ile Gly Leu Ala Ala Thr Gly Phe Leu Ala Ser Gly Met Phe Gly Leu
115 120 125
Thr Gly Leu Ser Ser Val Ser Trp Val Leu Asn Tyr Leu Arg Gly Thr
130 135 140
Arg Lys Ser Ser Val Pro Glu Gln Leu Glu Tyr Ala Lys Lys Arg Met
145 150 155 160
Ala Asp Ala Val Gly Tyr Ala Gly Gln Lys Gly Lys Gly Met Gly Gln
165 170 175
His Val Gln Asn Lys Ala Gln Glu Ala Lys Gln Tyr Asp Ile Ser Lys
180 185 190
Thr His Asp Thr Thr Thr Lys Gly His Glu Thr Thr Gln Arg Thr Ala
195 200 205
Ala Ala
210
<210> 100
<211> 149
<212> Белок
<213> Brassica napus
<400> 100
Met Ala Asn Gln Thr Arg Thr His Gln Asp Ile Ile Val Arg Asp Ser
1 5 10 15
Arg Ile Thr Leu Asp Arg Asp His Pro Lys Thr Gly Ala Gln Met Val
20 25 30
Lys Val Ala Thr Gly Val Ala Ala Gly Gly Ser Leu Leu Val Leu Ser
35 40 45
Gly Leu Thr Leu Ala Gly Thr Val Ile Ala Phe Ala Val Ala Thr Pro
50 55 60
Leu Leu Ile Ile Phe Ser Pro Val Leu Val Pro Ala Val Ile Thr Val
65 70 75 80
Val Leu Ile Ile Thr Gly Phe Leu Ala Ser Gly Gly Phe Gly Ile Ala
85 90 95
Ala Ile Thr Ala Phe Ser Trp Leu Tyr Arg His Met Thr Gly Ser Gly
100 105 110
Ser Asp Gln Lys Ile Glu Ser Ala Arg Met Lys Val Gly Ser Arg Gly
115 120 125
Tyr Asp Thr Lys Tyr Gly Gln His Asn Ile Gly Val His Gln Gln His
130 135 140
Gln Gln Ala Ala Ser
145
<210> 101
<211> 137
<212> Белок
<213> Arachis hypogaea
<400> 101
Met Ala Glu Ala Leu Tyr Tyr Gly Gly Arg Gln Arg Gln Glu Gln Pro
1 5 10 15
Arg Ser Thr Gln Leu Val Lys Ala Thr Thr Ala Val Val Ala Gly Gly
20 25 30
Ser Leu Leu Ile Leu Ala Gly Leu Val Leu Ala Gly Thr Val Ile Gly
35 40 45
Leu Thr Thr Ile Thr Pro Leu Phe Val Ile Phe Ser Pro Val Leu Val
50 55 60
Pro Ala Val Ile Thr Val Ala Leu Leu Gly Leu Gly Phe Leu Ala Ser
65 70 75 80
Gly Gly Phe Gly Val Ala Ala Ile Thr Val Leu Thr Trp Ile Tyr Arg
85 90 95
Tyr Val Thr Gly Lys His Pro Pro Gly Ala Asn Gln Leu Asp Thr Ala
100 105 110
Arg His Lys Leu Met Gly Lys Ala Arg Glu Ile Lys Asp Phe Gly Gln
115 120 125
Gln Gln Thr Ser Gly Ala Gln Ala Ser
130 135
<210> 102
<211> 150
<212> Белок
<213> Arachis hypogaea
<400> 102
Met Thr Asp Arg Thr Gln Pro His Ala Val Gln Val His Thr Thr Ala
1 5 10 15
Gly Arg Phe Gly Asp Thr Ala Ala Gly Thr Asn Arg Tyr Ala Asp Arg
20 25 30
Gly Pro Ser Thr Ser Lys Val Ile Ala Val Ile Thr Gly Leu Pro Ile
35 40 45
Gly Gly Thr Leu Leu Leu Phe Ala Gly Leu Ala Leu Ala Gly Thr Leu
50 55 60
Leu Gly Leu Ala Val Thr Thr Pro Leu Phe Ile Leu Phe Ser Pro Val
65 70 75 80
Ile Val Pro Ala Thr Ile Val Val Gly Leu Ser Val Ala Gly Phe Leu
85 90 95
Thr Ser Gly Ala Cys Gly Leu Thr Gly Leu Ser Ser Phe Ser Trp Val
100 105 110
Met Asn Tyr Ile Arg Gln Thr His Gly Ser Val Pro Glu Gln Leu Glu
115 120 125
Met Ala Lys His Arg Met Ala Asp Val Ala Gly Tyr Val Gly Gln Lys
130 135 140
Thr Lys Asp Val Gly Gln
145 150
<210> 103
<211> 166
<212> Белок
<213> Arachis hypogaea
<400> 103
Met Ser Asp Gln Thr Arg Thr Gly Tyr Gly Gly Gly Gly Ser Tyr Gly
1 5 10 15
Ser Ser Tyr Gly Gly Gly Gly Thr Tyr Gly Ser Ser Tyr Gly Thr Ser
20 25 30
Tyr Asp Pro Ser Thr Asn Gln Pro Ile Arg Gln Ala Ile Lys Phe Met
35 40 45
Thr Ala Ser Thr Ile Gly Val Ser Phe Leu Ile Leu Ser Gly Leu Ile
50 55 60
Leu Thr Gly Thr Val Ile Gly Leu Ile Ile Ala Thr Pro Leu Leu Val
65 70 75 80
Ile Phe Ser Pro Ile Leu Val Pro Ala Ala Ile Thr Leu Ala Leu Ala
85 90 95
Ala Gly Gly Phe Leu Phe Ser Gly Gly Cys Gly Val Ala Ala Ile Ala
100 105 110
Ala Leu Ser Trp Leu Tyr Ser Tyr Val Thr Gly Lys His Pro Ala Gly
115 120 125
Ser Asp Arg Leu Asp Tyr Ala Lys Gly Val Ile Ala Asp Lys Ala Arg
130 135 140
Asp Val Lys Asp Arg Ala Lys Asp Tyr Ala Gly Ala Gly Arg Ala Gln
145 150 155 160
Glu Gly Thr Pro Gly Tyr
165
<210> 104
<211> 176
<212> Белок
<213> Arachis hypogaea
<400> 104
Met Ala Thr Ala Thr Asp Arg Ala Pro His Gln Val Gln Val His Thr
1 5 10 15
Pro Thr Thr Gln Arg Val Asp Val Pro Arg Arg Gly Tyr Asp Val Ser
20 25 30
Gly Gly Gly Ile Lys Thr Leu Leu Pro Glu Arg Gly Pro Ser Thr Ser
35 40 45
Gln Ile Ile Ala Val Leu Val Gly Val Pro Thr Gly Gly Thr Leu Leu
50 55 60
Leu Leu Ser Gly Leu Ser Leu Leu Gly Thr Ile Ile Gly Leu Ala Ile
65 70 75 80
Ala Thr Pro Val Phe Ile Phe Phe Ser Pro Val Ile Val Pro Ala Val
85 90 95
Val Thr Ile Gly Leu Ala Val Thr Gly Ile Leu Thr Ala Gly Ala Cys
100 105 110
Gly Leu Thr Gly Leu Met Ser Leu Ser Trp Met Ile Asn Phe Ile Arg
115 120 125
Gln Val His Gly Thr Thr Val Pro Asp Gln Leu Asp Ser Val Lys Arg
130 135 140
Arg Met Ala Asp Met Ala Asp Tyr Val Gly Gln Lys Thr Lys Asp Ala
145 150 155 160
Gly Gln Glu Ile Gln Thr Lys Ala Gln Asp Val Lys Arg Ser Ser Ser
165 170 175
<210> 105
<211> 153
<212> Белок
<213> Ricinus communis
<400> 105
Met Ala Asp Arg Pro Gln Pro His Gln Val Gln Val His Arg Tyr Asp
1 5 10 15
Pro Thr Thr Gly Tyr Lys Gly Gln Gln Lys Gly Pro Ser Ala Ser Lys
20 25 30
Val Leu Ala Val Leu Thr Phe Leu Pro Val Gly Gly Gly Leu Leu Ser
35 40 45
Leu Ser Gly Ile Thr Leu Thr Asn Thr Leu Ile Gly Met Ala Ile Ala
50 55 60
Thr Pro Leu Phe Ile Leu Phe Gly Pro Ile Ile Leu Pro Ala Ala Val
65 70 75 80
Val Ile Gly Leu Ala Met Met Ala Phe Met Val Ala Gly Ala Leu Gly
85 90 95
Leu Ser Gly Leu Thr Ser Gln Ser Trp Ala Leu Lys Tyr Phe Arg Glu
100 105 110
Gly Thr Ala Met Pro Glu Ser Leu Asp Gln Ala Lys Lys Arg Met Gln
115 120 125
Asp Met Ala Gly Tyr Val Gly Met Lys Thr Lys Glu Val Gly Gln Asp
130 135 140
Ile Gln Arg Lys Ala Gln Glu Gly Lys
145 150
<210> 106
<211> 138
<212> Белок
<213> Ricinus communis
<400> 106
Met Ala Glu His Gln Gln Ser Pro Val Val Ser His Arg Pro Arg Val
1 5 10 15
Asn Gln Leu Val Lys Ala Gly Thr Ala Ala Thr Ala Gly Ser Ser Leu
20 25 30
Leu Phe Leu Ser Gly Leu Thr Leu Thr Gly Thr Val Ile Ala Leu Ala
35 40 45
Leu Ala Thr Pro Leu Met Val Leu Phe Ser Pro Val Leu Leu Pro Ala
50 55 60
Val Ile Ile Ile Ser Leu Ile Gly Ala Gly Phe Leu Thr Ser Gly Gly
65 70 75 80
Phe Gly Phe Gly Ala Ile Leu Val Leu Ser Trp Ile Tyr Arg Tyr Val
85 90 95
Thr Gly Lys Gln Pro Pro Gly Ala Glu Ser Leu Asp Gln Ala Arg Leu
100 105 110
Lys Leu Ala Gly Lys Ala Arg Glu Met Lys Asp Arg Ala Glu Gln Phe
115 120 125
Gly Gln His Val Thr Gly Gln Gln Thr Ser
130 135
<210> 107
<211> 226
<212> Белок
<213> Glycine max
<400> 107
Met Thr Thr Gln Val Pro Pro His Ser Val Gln Val His Thr Thr Thr
1 5 10 15
Thr His Arg Tyr Glu Ala Gly Val Val Pro Pro Gly Ala Arg Phe Glu
20 25 30
Thr Ser Tyr Glu Ala Gly Val Lys Ala Ala Ser Ile Tyr His Ser Glu
35 40 45
Arg Gly Pro Thr Thr Ser Gln Val Leu Ala Val Leu Ala Gly Leu Pro
50 55 60
Val Gly Gly Ile Leu Leu Leu Leu Ala Gly Leu Thr Leu Ala Gly Thr
65 70 75 80
Leu Thr Gly Leu Ala Val Ala Thr Pro Leu Phe Val Leu Phe Ser Pro
85 90 95
Val Leu Val Pro Ala Thr Val Ala Ile Gly Leu Ala Val Ala Gly Phe
100 105 110
Leu Thr Ser Gly Ala Phe Gly Leu Thr Ala Leu Ser Ser Phe Ser Trp
115 120 125
Ile Leu Asn Tyr Ile Arg Glu Thr Gln Pro Ala Ser Glu Asn Leu Ala
130 135 140
Ala Ala Ala Lys His His Leu Ala Glu Ala Ala Glu Tyr Val Gly Gln
145 150 155 160
Lys Thr Lys Glu Val Gly Gln Lys Thr Lys Glu Val Gly Gln Asp Ile
165 170 175
Gln Ser Lys Ala Gln Asp Thr Arg Glu Ala Ala Ala Arg Asp Ala Arg
180 185 190
Glu Ala Ala Ala Arg Asp Ala Arg Glu Ala Ala Ala Arg Asp Ala Lys
195 200 205
Val Glu Ala Arg Asp Val Lys Arg Thr Thr Val Thr Ala Thr Thr Ala
210 215 220
Thr Ala
225
<210> 108
<211> 223
<212> Белок
<213> Glycine max
<400> 108
Met Thr Thr Val Pro Pro His Ser Val Gln Val His Thr Thr Thr His
1 5 10 15
Arg Tyr Glu Ala Gly Val Val Pro Pro Ala Arg Phe Glu Ala Pro Arg
20 25 30
Tyr Glu Ala Gly Ile Lys Ala Pro Ser Ser Ile Tyr His Ser Glu Arg
35 40 45
Gly Pro Thr Thr Ser Gln Val Leu Ala Val Val Ala Gly Leu Pro Val
50 55 60
Gly Gly Ile Leu Leu Leu Leu Ala Gly Leu Thr Leu Ala Gly Thr Leu
65 70 75 80
Thr Gly Leu Val Val Ala Thr Pro Leu Phe Ile Ile Phe Ser Pro Val
85 90 95
Leu Ile Pro Ala Thr Val Ala Ile Gly Leu Ala Val Ala Gly Phe Leu
100 105 110
Thr Ser Gly Val Phe Gly Leu Thr Ala Leu Ser Ser Phe Ser Trp Ile
115 120 125
Leu Asn Tyr Ile Arg Glu Thr Gln Pro Ala Ser Glu Asn Leu Ala Ala
130 135 140
Ala Ala Lys His His Leu Ala Glu Ala Ala Glu Tyr Val Gly Gln Lys
145 150 155 160
Thr Lys Glu Val Gly Gln Lys Thr Lys Glu Val Gly Gln Asp Ile Gln
165 170 175
Ser Lys Ala Gln Asp Thr Arg Glu Ala Ala Ala Arg Asp Ala Arg Asp
180 185 190
Ala Arg Glu Ala Ala Ala Arg Asp Ala Arg Asp Ala Lys Val Glu Ala
195 200 205
Arg Asp Val Lys Arg Thr Thr Val Thr Ala Thr Thr Ala Thr Ala
210 215 220
<210> 109
<211> 155
<212> Белок
<213> Linum usitatissimum
<400> 109
Met Asp Gln Thr His Gln Thr Tyr Ala Gly Thr Thr Gln Asn Pro Ser
1 5 10 15
Tyr Gly Gly Gly Gly Thr Met Tyr Gln Gln Gln Gln Pro Arg Ser Tyr
20 25 30
Gln Ala Val Lys Ala Ala Thr Ala Ala Thr Ala Gly Gly Ser Leu Ile
35 40 45
Val Leu Ser Gly Leu Ile Leu Thr Ala Thr Val Ile Ser Leu Ile Ile
50 55 60
Ala Thr Pro Leu Leu Val Ile Phe Ser Pro Val Leu Val Pro Ala Leu
65 70 75 80
Ile Thr Val Gly Leu Leu Ile Thr Gly Phe Leu Ala Ser Gly Gly Phe
85 90 95
Gly Val Ala Ala Val Thr Val Leu Ser Trp Ile Tyr Arg Tyr Val Thr
100 105 110
Gly Gly His Pro Ala Gly Gly Asp Ser Leu Asp Gln Ala Arg Ser Lys
115 120 125
Leu Ala Gly Lys Ala Arg Glu Val Lys Asp Arg Ala Ser Glu Phe Ala
130 135 140
Gln Gln His Val Thr Gly Gly Gln Gln Thr Ser
145 150 155
<210> 110
<211> 180
<212> Белок
<213> Linum usitatissimum
<400> 110
Met Ala Asp Arg Thr Thr Gln Pro His Gln Val Gln Val His Thr Gln
1 5 10 15
His His Tyr Pro Thr Gly Gly Ala Phe Gly Arg Tyr Glu Gly Val Leu
20 25 30
Lys Gly Gly Pro Tyr His Gln Gln Gly Thr Gly Ser Gly Pro Ser Ala
35 40 45
Ser Lys Val Leu Ala Val Met Thr Ala Leu Pro Ile Gly Gly Thr Leu
50 55 60
Leu Ala Leu Ala Gly Ile Thr Leu Ala Gly Thr Met Ile Gly Leu Ala
65 70 75 80
Ile Thr Thr Pro Ile Phe Val Ile Cys Ser Pro Val Leu Val Pro Ala
85 90 95
Ala Leu Leu Ile Gly Phe Ala Val Ser Ala Phe Leu Ala Ser Gly Met
100 105 110
Ala Gly Leu Thr Gly Leu Thr Ser Leu Ser Trp Phe Ala Arg Tyr Leu
115 120 125
Gln Gln Ala Gly Gln Gly Val Gly Val Gly Val Pro Asp Ser Phe Asp
130 135 140
Gln Ala Lys Arg Arg Met Gln Asp Ala Ala Gly Tyr Met Gly Gln Lys
145 150 155 160
Thr Lys Glu Val Gly Gln Glu Ile Gln Arg Lys Ser Gln Asp Val Lys
165 170 175
Ala Ser Asp Lys
180
<210> 111
<211> 181
<212> Белок
<213> Helianthus annuus
<400> 111
Thr Thr Thr Thr Tyr Asp Arg His Phe Thr Thr Thr Gln Pro His Tyr
1 5 10 15
Arg Gln Asp Asp Arg Ser Arg Tyr Asp Gln Gln Thr His Ser Gln Ser
20 25 30
Thr Ser Arg Thr Leu Ala Ile Ile Ala Leu Leu Pro Val Gly Gly Ile
35 40 45
Leu Leu Gly Leu Ala Ala Leu Thr Phe Ile Gly Thr Leu Ile Gly Leu
50 55 60
Ala Leu Ala Thr Pro Leu Phe Val Ile Phe Ser Pro Ile Ile Val Pro
65 70 75 80
Ala Val Leu Thr Ile Gly Leu Ala Val Thr Gly Phe Leu Ala Ser Gly
85 90 95
Thr Phe Gly Leu Thr Gly Leu Ser Ser Leu Ser Tyr Leu Phe Asn Met
100 105 110
Val Arg Gln Thr Ala Gly Ser Val Pro Glu Ser Leu Asp Tyr Val Lys
115 120 125
Gly Thr Leu Gln Asp Ala Gly Glu Tyr Ala Gly Gln Lys Thr Lys Asp
130 135 140
Phe Gly Gln Lys Ile Gln Ser Thr Ala His Glu Met Gly Asp Gln Gly
145 150 155 160
Gln Val Gly Val His Ala Gln Val Gly Gly Gly Lys Glu Gly Arg Lys
165 170 175
Ser Gly Asp Arg Thr
180
<210> 112
<211> 156
<212> Белок
<213> Zea mays
<400> 112
Met Ala Asp His His Arg Gly Ala Thr Gly Gly Gly Gly Gly Tyr Gly
1 5 10 15
Asp Leu Gln Arg Gly Gly Gly Met His Gly Glu Ala Gln Gln Gln Gln
20 25 30
Lys Gln Gly Ala Met Met Thr Ala Leu Lys Ala Ala Thr Ala Ala Thr
35 40 45
Phe Gly Gly Ser Met Leu Val Leu Ser Gly Leu Ile Leu Ala Gly Thr
50 55 60
Val Ile Ala Leu Thr Val Ala Thr Pro Val Leu Val Ile Phe Ser Pro
65 70 75 80
Val Leu Val Pro Ala Ala Ile Ala Leu Ala Leu Met Ala Ala Gly Phe
85 90 95
Val Thr Ser Gly Gly Leu Gly Val Ala Ala Leu Ser Val Phe Ser Trp
100 105 110
Met Tyr Lys Tyr Leu Thr Gly Lys His Pro Pro Ala Ala Asp Gln Leu
115 120 125
Asp His Ala Lys Ala Arg Leu Ala Ser Lys Ala Arg Asp Val Lys Asp
130 135 140
Ala Ala Gln His Arg Ile Asp Gln Ala Gln Gly Ser
145 150 155
<210> 113
<211> 244
<212> Белок
<213> Brassica napus
<400> 113
Val Ser Lys Pro Asp Asp Cys Arg Arg Ile Val Asp Glu Thr Ile Ser
1 5 10 15
His Phe Gly Arg Leu Asp His Leu Val Asn Asn Ala Gly Ile Met Gln
20 25 30
Ile Ser Met Phe Glu Asn Ile Glu Glu Ile Thr Arg Thr Arg Ala Val
35 40 45
Met Asp Thr Asn Phe Trp Gly Ser Val Tyr Thr Thr Arg Ala Ala Leu
50 55 60
Pro Tyr Leu Arg Gln Ser Asn Gly Lys Ile Val Ala Met Ser Ser Ser
65 70 75 80
Ala Ala Trp Leu Thr Ala Pro Arg Met Ser Phe Tyr Asn Ala Ser Lys
85 90 95
Ala Ala Leu Leu Asn Phe Phe Glu Thr Leu Arg Ile Glu Leu Gly Ser
100 105 110
Asp Val His Ile Thr Ile Val Thr Pro Gly Tyr Ile Glu Ser Glu Leu
115 120 125
Thr Gln Gly Lys Tyr Phe Ser Gly Glu Gly Glu Leu Val Val Asn Gln
130 135 140
Asp Ile Arg Asp Val Gln Ile Gly Ala Phe Pro Val Thr Ser Val Ser
145 150 155 160
Gly Cys Ala Lys Gly Ile Val Lys Gly Val Cys Arg Lys Gln Arg Tyr
165 170 175
Val Thr Glu Pro Ser Trp Phe Lys Val Thr Tyr Leu Trp Lys Val Phe
180 185 190
Cys Pro Glu Leu Ile Glu Trp Gly Cys Arg Leu Leu Phe Leu Ser Gly
195 200 205
His Gly Thr Ser Glu Lys Asn Ala Leu Asn Lys Lys Ile Leu Asp Ile
210 215 220
Pro Gly Val Arg Ser Ala Leu Tyr Pro Glu Ser Ile Arg Thr Pro Glu
225 230 235 240
Ile Lys Ser Glu
<210> 114
<211> 349
<212> Белок
<213> Brassica napus
<400> 114
Met Glu Leu Ile Asn Asp Phe Leu Asn Leu Thr Ala Pro Phe Phe Thr
1 5 10 15
Phe Phe Gly Leu Cys Phe Phe Leu Pro Pro Phe Tyr Phe Phe Lys Phe
20 25 30
Val Gln Ser Ile Phe Ser Thr Ile Phe Ser Glu Asn Val Tyr Gly Lys
35 40 45
Val Val Leu Ile Thr Gly Ala Ser Ser Gly Ile Gly Glu Gln Leu Ala
50 55 60
Tyr Glu Tyr Ala Ser Lys Gly Ala Cys Leu Ala Leu Thr Ala Arg Arg
65 70 75 80
Lys Asn Arg Leu Glu Glu Val Ala Glu Ile Ala Arg Glu Val Gly Ser
85 90 95
Pro Asn Val Val Thr Val His Ala Asp Val Ser Lys Pro Asp Asp Cys
100 105 110
Arg Arg Ile Val Asp Glu Thr Ile Ser His Phe Gly Arg Leu Asp His
115 120 125
Leu Val Asn Asn Ala Gly Ile Met Gln Ile Ser Met Phe Glu Asn Ile
130 135 140
Glu Glu Ile Thr Arg Thr Arg Ala Val Met Asp Thr Asn Phe Trp Gly
145 150 155 160
Ala Val Tyr Thr Thr Arg Ala Ala Leu Pro Tyr Leu Arg Gln Ser Asn
165 170 175
Gly Lys Ile Val Ala Met Ser Ser Ser Ala Ala Trp Leu Thr Ala Pro
180 185 190
Arg Met Ser Phe Tyr Asn Ala Ser Lys Ala Ala Leu Leu Asn Phe Phe
195 200 205
Glu Thr Leu Arg Ile Glu Leu Gly Ser Asp Val His Ile Thr Ile Val
210 215 220
Thr Pro Gly Tyr Ile Glu Ser Glu Leu Thr Gln Gly Lys Tyr Val Ser
225 230 235 240
Gly Glu Gly Glu Leu Val Val Asn Gln Asp Ile Arg Asp Val Gln Ile
245 250 255
Gly Ala Phe Pro Val Thr Ser Val Ser Gly Arg Ala Lys Gly Ile Val
260 265 270
Lys Gly Val Cys Arg Lys Glu Arg Tyr Val Thr Glu Pro Ser Trp Phe
275 280 285
Lys Val Thr Tyr Leu Trp Lys Val Phe Cys Pro Glu Leu Ile Glu Trp
290 295 300
Gly Cys Arg Leu Met Phe Leu Ser Gly His Gly Thr Pro Glu Glu Asn
305 310 315 320
Ala Leu Asn Lys Lys Ile Leu Asp Ile Pro Gly Val Arg Ser Ala Leu
325 330 335
Tyr Pro Glu Pro Ile Arg Thr Pro Glu Ile Lys Ser Glu
340 345
<210> 115
<211> 456
<212> Белок
<213> Brassica napus
<400> 115
Met Val Asp Leu Leu Asn Ser Val Met Asn Leu Val Ala Pro Pro Ala
1 5 10 15
Thr Met Val Val Met Ala Phe Ser Trp Pro Leu Leu Cys Phe Ile Thr
20 25 30
Phe Ser Glu Arg Leu Tyr Asn Ser Tyr Phe Val Thr Glu Asp Met Glu
35 40 45
Asp Lys Val Val Val Ile Thr Gly Ala Ser Pro Ala Ile Gly Glu Gln
50 55 60
Ile Ala Tyr Glu Tyr Ala Lys Arg Gly Ala Asn Leu Val Leu Val Ala
65 70 75 80
Arg Arg Glu Gln Arg Leu Arg Val Val Ser Asn Asn Ala Arg Gln Ile
85 90 95
Gly Ala Asn His Val Ile Ile Ile Ala Ala Asp Val Val Lys Glu Asp
100 105 110
Asp Cys Arg Arg Phe Ile Thr Gln Ala Val Asn Tyr Tyr Gly Arg Val
115 120 125
Asp His Leu Val Asn Ser Ala Ser Leu Gly His Thr Phe Tyr Phe Asp
130 135 140
Glu Val Ser Asp Thr Thr Val Phe Pro His Leu Leu Asp Ile Asn Phe
145 150 155 160
Trp Gly Asn Val Tyr Pro Thr Tyr Val Ala Leu Pro His Leu Gln Lys
165 170 175
Thr Asn Gly Arg Ile Val Val Asn Ala Ser Val Glu Asn Trp Leu Pro
180 185 190
Leu Pro Arg Met Ser Leu Tyr Ser Ala Ala Lys Ala Ala Leu Val Asn
195 200 205
Phe Tyr Glu Thr Leu Arg Phe Glu Leu Asn Gly Asp Val Gly Ile Thr
210 215 220
Ile Ala Thr His Gly Trp Ile Gly Ser Glu Met Ser Arg Gly Lys Phe
225 230 235 240
Met Leu Glu Glu Gly Ala Glu Met Gln Trp Lys Glu Glu Arg Glu Val
245 250 255
Pro Ala Asn Gly Gly Pro Leu Glu Glu Phe Ala Lys Met Ile Val Ala
260 265 270
Gly Ala Cys Arg Gly Asp Ala Tyr Val Lys Phe Pro Asn Trp Tyr Asp
275 280 285
Val Phe Leu Leu Tyr Arg Val Phe Thr Pro Asn Val Leu Arg Trp Thr
290 295 300
Phe Lys Leu Leu Leu Ser Ser Glu Gly Ser Arg Gln Ser Ser Leu Val
305 310 315 320
Gly Val Gly Gln Gly Leu Pro Pro Glu Glu Ser Ser Ser Gln Met Lys
325 330 335
Leu Met Leu Glu Gly Gly Ser Pro Arg Val Thr Ala Ser Pro Pro Arg
340 345 350
Tyr Thr Pro Ser Pro Ser Pro Pro His His Thr Ala Ser Pro Pro Arg
355 360 365
Tyr Thr Pro Ser Pro Ser Pro Pro His His Thr Ser Ser Pro Gln Arg
370 375 380
Tyr Thr Pro Ser Pro Ser Pro Pro His Tyr Thr Ser Ser Arg His Arg
385 390 395 400
Tyr Thr Pro Ser Pro Ser Pro Pro His Tyr Thr Glu Ser Pro Pro Leu
405 410 415
Tyr Thr Glu Ser Pro Pro His Tyr Thr Thr Ser Pro Asn Trp Tyr Thr
420 425 430
Glu Ser Pro Pro Arg Tyr Thr Pro Ser Pro Ser Pro Pro Arg Phe Ser
435 440 445
Arg Phe Asn Ile Gln Glu Leu Pro
450 455
<210> 116
<211> 348
<212> Белок
<213> Sesamum indicum
<400> 116
Met Asp Leu Ile His Thr Phe Leu Asn Leu Ile Ala Pro Pro Phe Thr
1 5 10 15
Phe Phe Phe Leu Leu Phe Phe Leu Pro Pro Phe Gln Ile Phe Lys Phe
20 25 30
Phe Leu Ser Ile Leu Gly Thr Leu Phe Ser Glu Asp Val Ala Gly Lys
35 40 45
Val Val Val Ile Thr Gly Ala Ser Ser Gly Ile Gly Glu Ser Leu Ala
50 55 60
Tyr Glu Tyr Ala Lys Arg Gly Ala Cys Leu Val Leu Ala Ala Arg Arg
65 70 75 80
Glu Arg Ser Leu Gln Glu Val Ala Glu Arg Ala Arg Asp Leu Gly Ser
85 90 95
Pro Asp Val Val Val Val Arg Ala Asp Val Ser Lys Ala Glu Asp Cys
100 105 110
Arg Lys Val Val Asp Gln Thr Met Asn Arg Phe Gly Arg Leu Asp His
115 120 125
Leu Val Asn Asn Ala Gly Ile Met Ser Val Ser Met Leu Glu Glu Val
130 135 140
Glu Asp Ile Thr Gly Tyr Arg Glu Thr Met Asp Ile Asn Phe Trp Gly
145 150 155 160
Tyr Val Tyr Met Thr Arg Phe Ala Ala Pro Tyr Leu Arg Asn Ser Arg
165 170 175
Gly Arg Ile Val Val Leu Ser Ser Ser Ser Ser Trp Met Pro Thr Pro
180 185 190
Arg Met Ser Phe Tyr Asn Ala Ser Lys Ala Ala Ile Ser Gln Phe Phe
195 200 205
Glu Thr Leu Arg Val Glu Phe Gly Pro Asp Ile Gly Ile Thr Leu Val
210 215 220
Thr Pro Gly Phe Ile Glu Ser Glu Leu Thr Gln Gly Lys Phe Tyr Asn
225 230 235 240
Ala Gly Glu Arg Val Ile Asp Gln Asp Met Arg Asp Val Gln Val Ser
245 250 255
Thr Thr Pro Ile Leu Arg Val Glu Ser Ala Ala Arg Ser Ile Val Arg
260 265 270
Ser Ala Ile Arg Gly Glu Arg Tyr Val Thr Glu Pro Ala Trp Phe Arg
275 280 285
Val Thr Tyr Trp Trp Lys Leu Phe Cys Pro Glu Val Met Glu Trp Val
290 295 300
Phe Arg Leu Met Tyr Leu Ala Ser Pro Gly Glu Pro Glu Lys Glu Thr
305 310 315 320
Phe Gly Lys Lys Val Leu Asp Tyr Thr Gly Val Lys Ser Leu Leu Tyr
325 330 335
Pro Glu Thr Val Gln Val Pro Glu Pro Lys Asn Asp
340 345
<210> 117
<211> 350
<212> Белок
<213> Zea mays
<400> 117
Met Leu Gly Met Ser Arg Thr Gly Leu Ala Gly Ala Ala Leu Arg Val
1 5 10 15
Ala Leu Thr Ala Leu Leu Pro Leu Val Leu Pro Ala Tyr Tyr Val Tyr
20 25 30
Lys Leu Thr Thr Tyr Leu Leu Gly Ala Val Phe Pro Glu Asp Val Ala
35 40 45
Gly Lys Val Val Leu Ile Thr Gly Ala Ser Ser Gly Ile Gly Glu His
50 55 60
Leu Ala Tyr Glu Tyr Ala Lys Arg Gly Ala Tyr Leu Ala Leu Val Ala
65 70 75 80
Arg Arg Glu Ala Ser Leu Arg Glu Val Gly Asp Val Ala Leu Gly Leu
85 90 95
Gly Ser Pro Gly Val Leu Val Leu Pro Ala Asp Val Ser Lys Pro Arg
100 105 110
Asp Cys Glu Gly Phe Ile Asp Asp Thr Ile Ser Tyr Phe Gly Arg Leu
115 120 125
Asp His Leu Val Asn Asn Ala Ser Ile Trp Gln Val Cys Lys Phe Glu
130 135 140
Glu Ile Gln Asp Val Arg His Leu Arg Ala Leu Met Asp Ile Asn Phe
145 150 155 160
Trp Gly His Val Tyr Pro Thr Arg Leu Ala Ile Pro His Leu Arg Arg
165 170 175
Ser Arg Gly Arg Ile Val Gly Val Thr Ser Asn Ser Ser Tyr Ile Phe
180 185 190
Ile Gly Arg Asn Thr Phe Tyr Asn Ala Ser Lys Ala Ala Ala Leu Ser
195 200 205
Phe Tyr Asp Thr Leu Arg Met Glu Leu Gly Ser Asp Ile Arg Ile Thr
210 215 220
Glu Val Val Pro Gly Val Val Glu Ser Glu Ile Thr Lys Gly Lys Met
225 230 235 240
Leu Thr Lys Gly Gly Glu Met Lys Val Asp Gln Asp Glu Arg Asp Ala
245 250 255
Ile Leu Gly Pro Thr Pro Ala Glu Pro Val Gly Asp Phe Ala Arg Thr
260 265 270
Val Val Arg Asp Val Cys Arg Gly Ala Arg Tyr Val Phe Glu Pro Arg
275 280 285
Trp Tyr Met Gly Val Tyr Leu Leu Arg Ala Cys Leu Pro Glu Val Leu
290 295 300
Ala Trp Asn Ser Arg Leu Leu Thr Val Asp Thr Val Gly Ala Ser Thr
305 310 315 320
Thr Asp Thr Leu Gly Lys Trp Leu Val Glu Leu Pro Gly Val Arg Arg
325 330 335
Val Val Gln Pro Pro Ser Leu Arg Ser Pro Glu Ile Lys Asp
340 345 350
<210> 118
<211> 245
<212> Белок
<213> Brassica napus
<400> 118
Met Gly Thr Ala Thr Glu Ile Met Glu Arg Asp Ala Met Ala Thr Val
1 5 10 15
Ala Pro Tyr Ala Pro Val Thr Phe His Arg Arg Ala Arg Val Asp Leu
20 25 30
Asp Asp Arg Leu Pro Lys Pro Tyr Met Pro Arg Ala Leu Gln Ala Pro
35 40 45
Asp Arg Glu His Pro Tyr Gly Thr Pro Gly His Lys Asn Tyr Gly Leu
50 55 60
Ser Val Leu Gln Gln His Val Ala Phe Phe Asp Ile Asp Asp Asn Gly
65 70 75 80
Ile Ile Tyr Pro Trp Glu Thr Tyr Ser Gly Leu Arg Met Ile Gly Phe
85 90 95
Asn Ile Ile Gly Ser Leu Ile Ile Ala Ala Val Ile Asn Leu Ala Leu
100 105 110
Ser Tyr Ala Thr Leu Pro Gly Trp Leu Pro Ser Pro Phe Phe Pro Ile
115 120 125
Tyr Ile His Asn Ile His Lys Ser Lys His Gly Ser Asp Ser Arg Thr
130 135 140
Tyr Asp Asn Glu Gly Arg Phe Met Pro Val Asn Leu Glu Leu Ile Phe
145 150 155 160
Ser Lys Tyr Ala Lys Thr Leu Pro Asp Lys Leu Ser Leu Gly Glu Leu
165 170 175
Trp Asp Met Thr Glu Gly Gln Arg Asp Ala Trp Asp Ile Phe Gly Trp
180 185 190
Phe Ala Ser Lys Ile Glu Trp Gly Leu Leu Tyr Leu Leu Ala Arg Asp
195 200 205
Glu Glu Gly Phe Leu Ser Lys Glu Ala Ile Arg Arg Cys Phe Asp Gly
210 215 220
Ser Leu Phe Glu Tyr Cys Ala Lys Ile Tyr Val Gly Ile Asn Glu Asp
225 230 235 240
Lys Thr Ala Tyr Tyr
245
<210> 119
<211> 244
<212> Белок
<213> Brassica napus
<400> 119
Met Val Arg Glu Ser Met Gly Glu Glu Ser Glu Ala Phe Ala Thr Thr
1 5 10 15
Ala Pro Leu Ala Pro Val Thr Gly Glu Arg Lys Val Arg Asn Asp Leu
20 25 30
Glu Glu Thr Leu Pro Lys Pro Tyr Leu Ala Arg Ala Leu Val Ala Pro
35 40 45
Asp Thr Glu His Pro Asn Gly Ser Glu Gly His Asp Ser Lys Gly Met
50 55 60
Ser Val Thr Gln Gln His Val Ala Phe Phe Asp Gln Asn Gly Asp Gly
65 70 75 80
Ile Val Tyr Pro Trp Glu Thr Tyr Ala Gly Phe Arg Asp Leu Gly Phe
85 90 95
Asn Pro Ile Ser Ser Val Phe Trp Ala Ile Phe Ile Asn Phe Ala Phe
100 105 110
Ser Tyr Val Thr Leu Pro Ser Trp Leu Pro Ser Pro Leu Leu Pro Val
115 120 125
Tyr Ile Asp Asn Ile His Lys Ala Lys His Gly Ser Asp Ser Ser Thr
130 135 140
Tyr Asp Thr Glu Gly Arg Tyr Val Pro Val Asn Leu Glu Asn Ile Phe
145 150 155 160
Ser Lys Tyr Ala Leu Thr Ala Pro Asn Lys Ile Thr Leu Lys Glu Leu
165 170 175
Trp Asn Leu Thr Glu Gly Asn Arg Met Ala Ile Asp Pro Phe Gly Trp
180 185 190
Leu Ala Asn Lys Val Glu Trp Leu Leu Val Tyr Leu Leu Ala Lys Asp
195 200 205
Glu Glu Gly Phe Val Ser Lys Glu Ala Val Arg Gly Val Phe Asp Ala
210 215 220
Ser Phe Phe Glu Tyr Cys Ala Lys Lys Asn Lys Glu Lys Ala Asp Ser
225 230 235 240
Arg Lys Gln Asp
<210> 120
<211> 245
<212> Белок
<213> Sesamum indicum
<400> 120
Met Ala Thr His Val Leu Ala Ala Ala Ala Glu Arg Asn Ala Ala Leu
1 5 10 15
Ala Pro Asp Ala Pro Leu Ala Pro Val Thr Met Glu Arg Pro Val Arg
20 25 30
Thr Asp Leu Glu Thr Ser Ile Pro Lys Pro Tyr Met Ala Arg Gly Leu
35 40 45
Val Ala Pro Asp Met Asp His Pro Asn Gly Thr Pro Gly His Val His
50 55 60
Asp Asn Leu Ser Val Leu Gln Gln His Cys Ala Phe Phe Asp Gln Asp
65 70 75 80
Asp Asn Gly Ile Ile Tyr Pro Trp Glu Thr Tyr Ser Gly Leu Arg Gln
85 90 95
Ile Gly Phe Asn Val Ile Ala Ser Leu Ile Met Ala Ile Val Ile Asn
100 105 110
Val Ala Leu Ser Tyr Pro Thr Leu Pro Gly Trp Ile Pro Ser Pro Phe
115 120 125
Phe Pro Ile Tyr Leu Tyr Asn Ile His Lys Ala Lys His Gly Ser Asp
130 135 140
Ser Gly Thr Tyr Asp Thr Glu Gly Arg Tyr Leu Pro Met Asn Phe Glu
145 150 155 160
Asn Leu Phe Ser Lys His Ala Arg Thr Met Pro Asp Arg Leu Thr Leu
165 170 175
Gly Glu Leu Trp Ser Met Thr Glu Ala Asn Arg Glu Ala Phe Asp Ile
180 185 190
Phe Gly Trp Ile Ala Ser Lys Met Glu Trp Thr Leu Leu Tyr Ile Leu
195 200 205
Ala Arg Asp Gln Asp Gly Phe Leu Ser Lys Glu Ala Ile Arg Arg Cys
210 215 220
Tyr Asp Gly Ser Leu Phe Glu Tyr Cys Ala Lys Met Gln Arg Gly Ala
225 230 235 240
Glu Asp Lys Met Lys
245
<210> 121
<211> 243
<212> Белок
<213> Zea mays
<400> 121
Met Ser Ser Tyr Ser Pro Pro Pro Pro Pro Pro Arg Asp Gln Ser Met
1 5 10 15
Asp Thr Glu Ala Pro Asn Ala Pro Ile Thr Arg Glu Arg Arg Leu Asn
20 25 30
Pro Asp Leu Gln Glu Gln Leu Pro Lys Pro Tyr Leu Ala Arg Ala Leu
35 40 45
Glu Ala Val Asp Pro Ser His Pro Gln Gly Thr Lys Gly Arg Asp Pro
50 55 60
Arg Gly Met Ser Val Leu Gln Gln His Ala Ala Phe Phe Asp Arg Asn
65 70 75 80
Gly Asp Gly Val Ile Tyr Pro Trp Glu Thr Phe Gln Gly Leu Arg Ala
85 90 95
Ile Gly Cys Gly Leu Thr Val Ser Phe Ala Phe Ser Ile Leu Ile Asn
100 105 110
Leu Phe Leu Ser Tyr Pro Thr Gln Pro Gly Trp Leu Pro Ser Pro Leu
115 120 125
Leu Ser Ile Arg Ile Asp Asn Ile His Lys Gly Lys His Gly Ser Asp
130 135 140
Ser Glu Thr Tyr Asp Thr Glu Gly Arg Phe Asp Pro Ser Lys Phe Asp
145 150 155 160
Ala Ile Phe Ser Lys Tyr Gly Arg Thr His Pro Asn Ala Ile Thr Arg
165 170 175
Asp Glu Leu Ser Ser Met Leu Gln Gly Asn Arg Asn Thr Tyr Asp Phe
180 185 190
Leu Gly Trp Leu Ala Ala Ala Gly Glu Trp Leu Leu Leu Tyr Ser Leu
195 200 205
Ala Lys Asp Lys Asp Gly Leu Leu Gln Arg Glu Thr Val Arg Gly Leu
210 215 220
Phe Asp Gly Ser Leu Phe Glu Arg Leu Glu Asp Asp Asn Asn Lys Lys
225 230 235 240
Lys Ser Ser
<210> 122
<211> 11142
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ТДНК
<400> 122
tcctgtggtt ggcatgcaca tacaaatgga cgaacggata aaccttttca cgccctttta 60
aatatccgat tattctaata aacgctcttt tctcttaggt ttacccgcca atatatcctg 120
tcaaacactg atagtttaaa ctgaaggcgg gaaacgacaa tctgctagtg gatctcccag 180
tcacgacgtt gtaaaacggg cgccctagaa tctaattatt ctattcagac taaattagta 240
taagtatttt tttaatcaat aaataataat taataattta ttagtaggag tgattgaatt 300
tataatatat tttttttaat catttaaaga atcttatatc tttaaattga caagagtttt 360
aaatggggag agtgttatca tatcacaagt aggattaatg tgttatagtt tcacatgcat 420
tacgataagt tgtgaaagat aacattatta tatataacaa tgacaatcac tagcgatcga 480
gtagtgagag tcgtcttatt acactttctt ccttcgatct gtcacatggc ggcggcccga 540
attctcacac aaggtagttg caagacactg aagtggtggt agtggtagta gaagaagcag 600
aatcggtaga aaggcaagac aatggagaag atgaagatgg tggagattct cttcccacaa 660
cgcagcaatc aaggttttca aggttaaggc actcgtgctt tccatcatcg aacatgaagt 720
cgatgttatc ctcgaaagca agctcgttga agagttctgg gtactcaatt gggttctcgt 780
tagcaaggtt ttgatcggta aggaatgggg agaatccagt atccatcatg cagaagttcc 840
aagcaagttc gttgttatct ccgcacctat ccatttccat gatggtggaa gaatcaatgc 900
agcagttaac aacggcagct tcctcagaat atcccacaat ttcagcctct tgttgctcag 960
ccttctcttc ctctttttct tcttcctctt gaggtggttc ctcaacgtat tgttgcttaa 1020
cctcttccct aggttcctct ttagcttctc tagtctcaac ctcttgctta gcctcaacaa 1080
gaataccctc ttgatggtta gcctggttaa ctgggaatgg gaaaacgccc ttcttcttaa 1140
gcctgtcgat gtagttggag atatcgaagt tggtaacagc gttagcacct ctgtactcaa 1200
tagcagccat atcataagca gctgcagcct cttcttgagt gttgtaagtt ccgaggtaga 1260
ggtacttgtt tccgaaaact cttccaatcc tagcttccca tcttccgtta tgatgatgcc 1320
tagcaactcc cctatactta gaaactcccc tagagaatcc agatgactgc cttctaaggg 1380
aagcaagata ctcttctttg gtcaccctct gcatctcttc aagttctttg gtgtaagtct 1440
cagctgggaa gttaagaatg gtatctgggc cccaatactt aagagcagca agatcatagg 1500
tatgagcagc agcctcttca gaatcataag ctccaaggta aacctgcttg cccttcttgt 1560
tttggatgga gttccaagag gacttatccc aaaggtgagc ttcgaatctt ccagtccatc 1620
tatgcctagt aacacctctg tagatagatg accttctggt agaagctgga gaagttgggt 1680
tatgagactt atcgccagat ggagatgact tcttagccct cttagctctc tttggtcttg 1740
gagcttcaga ttgaattggg ctagaggtag tagtagaaga ggacactgaa gaagatggag 1800
aactagagca ggtagaggta gtgagcctct tcttcatgaa ttctgttctt ctttactctt 1860
tgtgtgactg aggtttggtc tagtgctttg gtcatctata tataatgata acaacaatga 1920
gaacaagctt tggagtgatc ggagggtcta ggatacatga gattcaagtg gactaggatc 1980
tacaccgttg gattttgagt gtggatatgt gtgaggttaa ttttacttgg taacggccac 2040
aaaggcctaa ggagaggtgt tgagaccctt atcggcttga accgctggaa taatgccacg 2100
tggaagataa ttccatgaat cttatcgtta tctatgagtg aaattgtgtg atggtggagt 2160
ggtgcttgct cattttactt gcctggtgga cttggccctt tccttatggg gaatttatat 2220
tttacttact atagagcttt catacctttt ttttaccttg gatttagtta atatataatg 2280
gtatgattca tgaataaaaa tgggaaattt ttgaatttgt actgctaaat gcataagatt 2340
aggtgaaact gtggaatata tatttttttc atttaaaagc aaaatttgcc ttttactaga 2400
attataaata tagaaaaata tataacattc aaataaaaat gaaaataaga actttcaaaa 2460
aacagaacta tgtttaatgt gtaaagatta gtcgcacatc aagtcatctg ttacaatatg 2520
ttacaacaag tcataagccc aacaaagtta gcacgtctaa ataaactaaa gagtccacga 2580
aaatattaca aatcataagc ccaacaaagt tattgatcaa aaaaaaaaaa cgcccaacaa 2640
agctaaacaa agtccaaaaa aaacttctca agtctccatc ttcctttatg aacattgaaa 2700
actatacaca aaacaagtca gataaatctc tttctgggcc tgtcttccca acctcctaca 2760
tcacttccct atcggattga atgttttact tgtacctttt ccgttgcaat gatattgata 2820
gtatgtttgt gaaaactaat agggttaaca atcgaagtca tggaatatgg atttggtcca 2880
agattttccg agagctttct agtagaaagc ccatcaccag aaatttacta gtaaaataaa 2940
tcaccaatta ggtttcttat tatgtgccaa attcaatata attatagagg atatttcaaa 3000
tgaaaacgta tgaatgttat tagtaaatgg tcaggtaaga cattaaaaaa atcctacgtc 3060
agatattcaa ctttaaaaat tcgatcagtg tggaattgta caaaaatttg ggatctacta 3120
tatatatata atgctttaca acacttggat ttttttttgg aggctggaat ttttaatcta 3180
catatttgtt ttggccatgc accaactcat tgtttagtgt aatactttga ttttgtcaaa 3240
tatatgtgtt cgtgtatatt tgtataagaa tttctttgac catatacaca cacacatata 3300
tatatatata tatatattat atatcatgca cttttaattg aaaaaataat atatatatat 3360
atagtgcatt ttttctaaca accatatatg ttgcgattga tctgcaaaaa tactgctaga 3420
gtaatgaaaa atataatcta ttgctgaaat tatctcagat gttaagattt tcttaaagta 3480
aattctttca aattttagct aaaagtcttg taataactaa agaataatac acaatctcga 3540
ccacggaaaa aaaacacata ataaatttgg ggcccctaga atctaattat tctattcaga 3600
ctaaattagt ataagtattt ttttaatcaa taaataataa ttaataattt attagtagga 3660
gtgattgaat ttataatata ttttttttaa tcatttaaag aatcttatat ctttaaattg 3720
acaagagttt taaatgggga gagtgttatc atatcacaag taggattaat gtgttatagt 3780
ttcacatgca ttacgataag ttgtgaaaga taacattatt atatataaca atgacaatca 3840
ctagcgatcg agtagtgaga gtcgtcttat tacactttct tccttcgatc tgtcacatgg 3900
cggcggcccg cggccgcttc attactcgag ccaggaggat ggatcgatgc tggtctgaga 3960
ccctgctacc ggttgctgac tgaactgctc ggcacggtcc ttcatttcac gggccttgct 4020
cgccaacttt gtcttggccg actccaactg atccgctccg ggtggatgtt tccccgtcag 4080
gtaacggtag atccaggaca gcacagacag agcggcaaca ccaaatcccc cgcttgccag 4140
aaaacccgct cccaacagga agatggtgat gactgcagat cagaaaaact cagattaatc 4200
gacaaattcg atcgcacaaa ctagaaacta acaccagatc tagatagaaa tcacaaatcg 4260
aagagtaatt attcgacaaa actcaaatta tttgaacaaa tcggatgata tctatgaaac 4320
cctaatcgag aattaagatg atatctaacg atcaaaccca gaaaatcgtc ttcgatctaa 4380
gattaacaga atctaaacca aagaacatat acgaaattgg gatcgaacga aaacaaaatc 4440
gaagattttg agagaataag gaacacagaa atttacctgc agggaccagt acaggcgaga 4500
agatcaccag gagaggtgtg gcgattgtca gcgcaatgac cgttccagcc agggtcaacc 4560
cggataacac caacaggcta cctccggcag taaccgcggt cgctgccttt acaacacgct 4620
gagcacgcgg ttgcagttgc aagtgggggg cacgtgtttg ttgctgctgc ccgtagtgct 4680
ctgccatggt tttttttaac ggagcaagcg gccgctgttc ttctttactc tttgtgtgac 4740
tgaggtttgg tctagtgctt tggtcatcta tatataatga taacaacaat gagaacaagc 4800
tttggagtga tcggagggtc taggatacat gagattcaag tggactagga tctacaccgt 4860
tggattttga gtgtggatat gtgtgaggtt aattttactt ggtaacggcc acaaaggcct 4920
aaggagaggt gttgagaccc ttatcggctt gaaccgctgg aataatgcca cgtggaagat 4980
aattccatga atcttatcgt tatctatgag tgaaattgtg tgatggtgga gtggtgcttg 5040
ctcattttac ttgcctggtg gacttggccc tttccttatg gggaatttat attttactta 5100
ctatagagct ttcatacctt ttttttacct tggatttagt taatatataa tggtatgatt 5160
catgaataaa aatgggaaat ttttgaattt gtactgctaa atgcataaga ttaggtgaaa 5220
ctgtggaata tatatttttt tcatttaaaa gcaaaatttg ccttttacta gaattataaa 5280
tatagaaaaa tatataacat tcaaataaaa atgaaaataa gaactttcaa aaaacagaac 5340
tatgtttaat gtgtaaagat tagtcgcaca tcaagtcatc tgttacaata tgttacaaca 5400
agtcataagc ccaacaaagt tagcacgtct aaataaacta aagagtccac gaaaatatta 5460
caaatcataa gcccaacaaa gttattgatc aaaaaaaaaa aacgcccaac aaagctaaac 5520
aaagtccaaa aaaaacttct caagtctcca tcttccttta tgaacattga aaactataca 5580
caaaacaagt cagataaatc tctttctggg cctgtcttcc caacctccta catcacttcc 5640
ctatcggatt gaatgtttta cttgtacctt ttccgttgca atgatattga tagtatgttt 5700
gtgaaaacta atagggttaa caatcgaagt catggaatat ggatttggtc caagattttc 5760
cgagagcttt ctagtagaaa gcccatcacc agaaatttac tagtaaaata aatcaccaat 5820
taggtttctt attatgtgcc aaattcaata taattataga ggatatttca aatgaaaacg 5880
tatgaatgtt attagtaaat ggtcaggtaa gacattaaaa aaatcctacg tcagatattc 5940
aactttaaaa attcgatcag tgtggaattg tacaaaaatt tgggatctac tatatatata 6000
taatgcttta caacacttgg attttttttt ggaggctgga atttttaatc tacatatttg 6060
ttttggccat gcaccaactc attgtttagt gtaatacttt gattttgtca aatatatgtg 6120
ttcgtgtata tttgtataag aatttctttg accatataca cacacacata tatatatata 6180
tatatatatt atatatcatg cacttttaat tgaaaaaata atatatatat atatagtgca 6240
ttttttctaa caaccatata tgttgcgatt gatctgcaaa aatactgcta gagtaatgaa 6300
aaatataatc tattgctgaa attatctcag atgttaagat tttcttaaag taaattcttt 6360
caaattttag ctaaaagtct tgtaataact aaagaataat acacaatctc gaccacggaa 6420
aaaaaacaca taataaattt gggcgcgccg cgtattggct agagcagctt gccaacatgg 6480
tggagcacga cactctcgtc tactccaaga atatcaaaga tacagtctca gaagaccaaa 6540
gggctattga gacttttcaa caaagggtaa tatcgggaaa cctcctcgga ttccattgcc 6600
cagctatctg tcacttcatc aaaaggacag tagaaaagga aggtggcacc tacaaatgcc 6660
atcattgcga taaaggaaag gctatcgttc aagatgcctc tgccgacagt ggtcccaaag 6720
atggaccccc acccacgagg agcatcgtgg aaaaagaaga cgttccaacc acgtcttcaa 6780
agcaagtgga ttgatgtgat aacatggtgg agcacgacac tctcgtctac tccaagaata 6840
tcaaagatac agtctcagaa gaccaaaggg ctattgagac ttttcaacaa agggtaatat 6900
cgggaaacct cctcggattc cattgcccag ctatctgtca cttcatcaaa aggacagtag 6960
aaaaggaagg tggcacctac aaatgccatc attgcgataa aggaaaggct atcgttcaag 7020
atgcctctgc cgacagtggt cccaaagatg gacccccacc cacgaggagc atcgtggaaa 7080
aagaagacgt tccaaccacg tcttcaaagc aagtggattg atgtgatatc tccactgacg 7140
taagggatga cgcacaatcc cactatcctt cgcaagacct tcctctatat aaggaagttc 7200
atttcatttg gagaggacac gctgaaatca ccagtctctc tctacaaatc tatctctgcg 7260
atcgcatggc gattttggat tctgctggcg ttactacggt gacggagaac ggtggcggag 7320
agttcgtcga tcttgatagg cttcgtcgac ggaaatcgag atcggattct tctaacggac 7380
ttcttctctc tggttccgat aataattctc cttcggatga tgttggagct cccgccgacg 7440
ttagggatcg gattgattcc gttgttaacg atgacgctca gggaacagcc aatttggccg 7500
gagataataa cggtggtggc gataataacg gtggtggaag aggcggcgga gaaggaagag 7560
gaaacgccga tgctacgttt acgtatcgac cgtcggttcc agctcatcgg agggcgagag 7620
agagtccact tagctccgac gcaatcttca aacagagcca tgccggatta ttcaacctct 7680
gtgtagtagt tcttattgct gtaaacagta gactcatcat cgaaaatctt atgaagtatg 7740
gttggttgat cagaacggat ttctggttta gttcaagatc gctgcgagat tggccgcttt 7800
tcatgtgttg tatatccctt tcgatctttc ctttggctgc ctttacggtt gagaaattgg 7860
tacttcagaa atacatatca gaacctgttg tcatctttct tcatattatt atcaccatga 7920
cagaggtttt gtatccagtt tacgtcaccc taaggtgtga ttctgctttt ttatcaggtg 7980
tcactttgat gctcctcact tgcattgtgt ggctaaagtt ggtttcttat gctcatacta 8040
gctatgacat aagatcccta gccaatgcag ctgataaggc caatcctgaa gtctcctact 8100
acgttagctt gaagagcttg gcatatttca tggtcgctcc cacattgtgt tatcagccaa 8160
gttatccacg ttctgcatgt atacggaagg gttgggtggc tcgtcaattt gcaaaactgg 8220
tcatattcac cggattcatg ggatttataa tagaacaata tataaatcct attgtcagga 8280
actcaaagca tcctttgaaa ggcgatcttc tatatgctat tgaaagagtg ttgaagcttt 8340
cagttccaaa tttatatgtg tggctctgca tgttctactg cttcttccac ctttggttaa 8400
acatattggc agagcttctc tgcttcgggg atcgtgaatt ctacaaagat tggtggaatg 8460
caaaaagtgt gggagattac tggagaatgt ggaatatgcc tgttcataaa tggatggttc 8520
gacatatata cttcccgtgc ttgcgcagca agataccaaa gacactcgcc attatcattg 8580
ctttcctagt ctctgcagtc tttcatgagc tatgcatcgc agttccttgt cgtctcttca 8640
agctatgggc ttttcttggg attatgtttc aggtgccttt ggtcttcatc acaaactatc 8700
tacaggaaag gtttggctca acggtgggga acatgatctt ctggttcatc ttctgcattt 8760
tcggacaacc gatgtgtgtg cttctttatt accacgacct gatgaaccga aaaggatcga 8820
tgtcatgagc gatcgcgatc gttcaaacat ttggcaataa agtttcttaa gattgaatcc 8880
tgttgccggt cttgcgatga ttatcatata atttctgttg aattacgtta agcatgtaat 8940
aattaacatg taatgcatga cgttatttat gagatgggtt tttatgatta gagtcccgca 9000
attatacatt taatacgcga tagaaaacaa aatatagcgc gcaaactagg ataaattatc 9060
gcgcgcggtg tcatctatgt tactagatcc ctgcagggcg tattggctag agcagcttgc 9120
caacatggtg gagcacgaca ctctcgtcta ctccaagaat atcaaagata cagtctcaga 9180
agaccaaagg gctattgaga cttttcaaca aagggtaata tcgggaaacc tcctcggatt 9240
ccattgccca gctatctgtc acttcatcaa aaggacagta gaaaaggaag gtggcaccta 9300
caaatgccat cattgcgata aaggaaaggc tatcgttcaa gatgcctctg ccgacagtgg 9360
tcccaaagat ggacccccac ccacgaggag catcgtggaa aaagaagacg ttccaaccac 9420
gtcttcaaag caagtggatt gatgtgataa catggtggag cacgacactc tcgtctactc 9480
caagaatatc aaagatacag tctcagaaga ccaaagggct attgagactt ttcaacaaag 9540
ggtaatatcg ggaaacctcc tcggattcca ttgcccagct atctgtcact tcatcaaaag 9600
gacagtagaa aaggaaggtg gcacctacaa atgccatcat tgcgataaag gaaaggctat 9660
cgttcaagat gcctctgccg acagtggtcc caaagatgga cccccaccca cgaggagcat 9720
cgtggaaaaa gaagacgttc caaccacgtc ttcaaagcaa gtggattgat gtgatatctc 9780
cactgacgta agggatgacg cacaatccca ctatccttcg caagaccttc ctctatataa 9840
ggaagttcat ttcatttgga gaggacacgc tgaaatcacc agtctctctc tacaaatcta 9900
tctctctcga gatgattgaa caagatggat tgcacgcagg ttctccggcc gcttgggtgg 9960
agaggctatt cggctatgac tgggcacaac agacaatcgg ctgctctgat gccgccgtgt 10020
tccggctgtc agcgcagggg aggccggttc tttttgtcaa gaccgacctg tccggtgccc 10080
tgaatgaact tcaagacgag gcagcgcggc tatcgtggct ggccacgacg ggcgttcctt 10140
gcgcagctgt gctcgacgtt gtcactgaag cgggaaggga ctggctgcta ttgggcgaag 10200
tgccggggca ggatctcctg tcatctcacc ttgctcctgc cgagaaagta tccatcatgg 10260
ctgatgcaat gcggcggctg catacgcttg atccggctac ctgcccattc gaccaccaag 10320
cgaaacatcg catcgagcga gcacgtactc ggatggaagc cggtcttgtc gatcaggatg 10380
atctggacga agagcatcag gggctcgcgc cagccgaact gttcgccagg ctcaaggcgc 10440
gcatgcccga cggcgaggat ctcgtcgtga ctcatggcga tgcctgcttg ccgaatatca 10500
tggtggaaaa tggccgcttt tctggattca tcgactgtgg ccggctgggt gtggcggacc 10560
gctatcagga catagcgttg gctacccgtg atattgctga agagcttggc ggcgaatggg 10620
ctgaccgctt cctcgtgctt tacggtatcg ccgctcccga ttcgcagcgc atcgccttct 10680
atcgccttct tgacgagttc ttctgaaacg cgtgatcgtt caaacatttg gcaataaagt 10740
ttcttaagat tgaatcctgt tgccggtctt gcgatgatta tcatataatt tctgttgaat 10800
tacgttaagc atgtaataat taacatgtaa tgcatgacgt tatttatgag atgggttttt 10860
atgattagag tcccgcaatt atacatttaa tacgcgatag aaaacaaaat atagcgcgca 10920
aactaggata aattatcgcg cgcggtgtca tctatgttac tagatcgacg tccgtacggt 10980
taaaaccacc ccagtacatt aaaaacgtcc gcaatgtgtt attaagttgt ctaagcgtca 11040
atttgtttac accacaatat atcctgccac cagccagcca acagctcccc gaccggcagc 11100
tcggcacaaa atcaccactc gatacaggca gcccatcagt cc 11142
<210> 123
<211> 16749
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность вектора
<400> 123
gcttcctcgt gctttacggt atcgccgctc ccgattcgca gcgcatcgcc ttctatcgcc 60
ttcttgacga gttcttctga aacgcgtgat cgttcaaaca tttggcaata aagtttctta 120
agattgaatc ctgttgccgg tcttgcgatg attatcatat aatttctgtt gaattacgtt 180
aagcatgtaa taattaacat gtaatgcatg acgttattta tgagatgggt ttttatgatt 240
agagtcccgc aattatacat ttaatacgcg atagaaaaca aaatatagcg cgcaaactag 300
gataaattat cgcgcgcggt gtcatctatg ttactagatc gacgtccgta cggttaaaac 360
caccccagta cattaaaaac gtccgcaatg tgttattaag ttgtctaagc gtcaatttgt 420
ttacaccaca atatatcctg ccaccagcca gccaacagct ccccgaccgg cagctcggca 480
caaaatcacc actcgataca ggcagcccat cagtccacta gacgctcacc gggctggttg 540
ccctcgccgc tgggctggcg gccgtctatg gccctgcaaa cgcgccagaa acgccgtcga 600
agccgtgtgc gagacaccgc agccgccggc gttgtggata cctcgcggaa aacttggccc 660
tcactgacag atgaggggcg gacgttgaca cttgaggggc cgactcaccc ggcgcggcgt 720
tgacagatga ggggcaggct cgatttcggc cggcgacgtg gagctggcca gcctcgcaaa 780
tcggcgaaaa cgcctgattt tacgcgagtt tcccacagat gatgtggaca agcctgggga 840
taagtgccct gcggtattga cacttgaggg gcgcgactac tgacagatga ggggcgcgat 900
ccttgacact tgaggggcag agtgctgaca gatgaggggc gcacctattg acatttgagg 960
ggctgtccac aggcagaaaa tccagcattt gcaagggttt ccgcccgttt ttcggccacc 1020
gctaacctgt cttttaacct gcttttaaac caatatttat aaaccttgtt tttaaccagg 1080
gctgcgccct gtgcgcgtga ccgcgcacgc cgaagggggg tgccccccct tctcgaaccc 1140
tcccggcccg ctctcgcgtt ggcagcatca cccataattg tggtttcaaa atcggctccg 1200
tcgatactat gttatacgcc aactttgaaa acaactttga aaaagctgtt ttctggtatt 1260
taaggtttta gaatgcaagg aacagtgaat tggagttcgt cttgttataa ttagcttctt 1320
ggggtattta aatactgtag aaaagaggaa ggaaataata aatggctaaa atgagaatat 1380
caccggaatt gaaaaaactg atcgaaaaat accgctgcgt aaaagatacg gaaggaatgt 1440
ctcctgctaa ggtatataag ctggtgggag aaaatgaaaa cctatattta aaaatgacgg 1500
acagccggta taaagggacc acctatgatg tggaacggga aaaggacatg atgctatggc 1560
tggaaggaaa gctgcctgtt ccaaaggtcc tgcaccttga acggcatgat ggctggagca 1620
atctgctcat gagtgaggcc gatggcgtcc tttgctcgga agagtatgaa gatgaacaaa 1680
gccctgaaaa gattatcgag ctgtatgcgg agtgcatcag gctctttcac tccatcgaca 1740
tatcggattg tccctatacg aatagcttag acagccgctt agccgaattg gattacttac 1800
tgaataacga tctggccgat gtggattgcg aaaactggga agaagacacc ccatttaaag 1860
atccgcgcga gctgtatgat tttttaaaga cggaaaagcc cgaagaggaa cttgtctttt 1920
cccacggcga cctgggagac agcaacatct ttgtgaaaga tggcaaagta agtggcttta 1980
ttgatcttgg gagaagcggc agggcggaca agtggtatga cattgccttc tgcgtccggt 2040
cgatcaggga ggatattggg gaagaacagt atgtcgagct attttttgac ttactgggga 2100
tcaagcctga ttgggagaaa ataaaatatt atattttact ggatgaattg ttttagtacc 2160
tagatgtggc gcaacgatgc tggcgacaag caggagcgca ccgacttctt ccgcatcaag 2220
tgttttggct ctcaggccga ggcccacggc aagtatttgg gcaaggggtc gctggtattc 2280
gtgcagggca agattcggaa taccaagtac gagaaggacg gccagacggt ctacgggacc 2340
gacttcattg ccgataaggt ggattatctg gacaccaagg caccaggcgg atcaaatcag 2400
gaataagggc acattgcccc ggcgtgagtc ggggcaatcc cgcaaggagg gtgaatgaat 2460
cggacgtttg accggaaggc atacaggcaa gaactgatcg acgcggggtt ttccgccgag 2520
gatgccgaaa ccatcgcaag ccgcaccgtc atgcgtgcgc cccgcgaaac cttccagtcc 2580
gtcggctcga tggcccagca agctacggcc aagatcgagc gcgacagcgt gcaactggct 2640
ccccctgccc tgcccgcgcc atcggccgcc gtggagcgtt cgcgtcgtct cgaacaggag 2700
gcggcaggtt tggcgaagtc gatgaccatc gacacgcgag gaactatgac gaccaagaag 2760
cgaaaaaccg ccggcgagga cctggcaaaa caggtcagcg aggccaagca agccgcgttg 2820
ctgaaacaca cgaagcagca gatcaaggaa atgcagcttt ccttgttcga tattgcgccg 2880
tggccggaca cgatgcgagc gatgccaaac gacacggccc gctctgccct gttcaccacg 2940
cgcaacaaga aaatcccgcg cgaggcgctg caaaacaagg tcattttcca cgtcaacaag 3000
gacgtgaaga tcacctacac cggcgtcgag ctgcgggccg acgatgacga actggtgtgg 3060
cagcaggtgt tggagtacgc gaagcgcacc cctatcggcg agccgatcac cttcacgttc 3120
tacgagcttt gccaggacct gggctggtcg atcaatggcc ggtattacac gaaggccgag 3180
gaatgcctgt cgcgcctaca ggcgacggcg atgggcttca cgtccgaccg cgttgggcac 3240
ctggaatcgg tgtcgctgct gcaccgcttc cgcgtcctgg accgtggcaa gaaaacgtcc 3300
cgttgccagg tcctgatcga cgaggaaatc gtcgtgctgt ttgctggcga ccactacacg 3360
aaattcatat gggagaagta ccgcaagctg tcgccgacgg cccgacggat gttcgactat 3420
ttcagctcgc accgggagcc gtacccgctc aagctggaaa ccttccgcct catgtgcgga 3480
tcggattcca cccgcgtgaa gaagtggcgc gagcaggtcg gcgaagcctg cgaagagttg 3540
cgaggcagcg gcctggtgga acacgcctgg gtcaatgatg acctggtgca ttgcaaacgc 3600
tagggccttg tggggtcagt tccggctggg ggttcagcag ccagcgcttt actgagatcc 3660
tcttccgctt cctcgctcac tgactcgctg cgctcggtcg ttcggctgcg gcgagcggta 3720
tcagctcact caaaggcggt aatacggtta tccacagaat caggggataa cgcaggaaag 3780
aacatgtgag caaaaggcca gcaaaaggcc aggaaccgta aaaaggccgc gttgctggcg 3840
tttttccata ggctccgccc ccctgacgag catcacaaaa atcgacgctc aagtcagagg 3900
tggcgaaacc cgacaggact ataaagatac caggcgtttc cccctggaag ctccctcgtg 3960
cgctctcctg ttccgaccct gccgcttacc ggatacctgt ccgcctttct cccttcggga 4020
agcgtggcgc tttctcatag ctcacgctgt aggtatctca gttcggtgta ggtcgttcgc 4080
tccaagctgg gctgtgtgca cgaacccccc gttcagcccg accgctgcgc cttatccggt 4140
aactatcgtc ttgagtccaa cccggtaaga cacgacttat cgccactggc agcagccact 4200
ggtaacagga ttagcagagc gaggtatgta ggcggtgcta cagagttctt gaagtggtgg 4260
cctaactacg gctacactag aagaacagta tttggtatct gcgctctgct gaagccagtt 4320
accttcggaa aaagagttgg tagctcttga tccggcaaac aaaccaccgc tggtagcggt 4380
ggtttttttg tttgcaagca gcagattacg cgcagaaaaa aaggatctca agaagatcct 4440
ttgatctttt ctacggggtc tgacgctcag tggaacgaaa actcacgtta agggattttg 4500
gtcatgagat tatcaaaaag gatcttcacc tagatccttt tggatctcct gtggttggca 4560
tgcacataca aatggacgaa cggataaacc ttttcacgcc cttttaaata tccgattatt 4620
ctaataaacg ctcttttctc ttaggtttac ccgccaatat atcctgtcaa acactgatag 4680
tttaaactga aggcgggaaa cgacaatctg ctagtggatc tcccagtcac gacgttgtaa 4740
aacgggcgtc tgcgatcgct gaagttccta tacttttcag agaataggaa cttcggaata 4800
ggaacttccc atgggatcta gtaacataga tgacaccgcg cgcgataatt tatcctagtt 4860
tgcgcgctat attttgtttt ctatcgcgta ttaaatgtat aattgcggga ctctaatcat 4920
aaaaacccat ctcataaata acgtcatgca ttacatgtta attattacat gcttaacgta 4980
attcaacaga aattatatga taatcatcgc aagaccggca acaggattca atcttaagaa 5040
actttattgc caaatgtttg aacgatcacg ctagcggata acaatttcac acagggatat 5100
cactagtaaa aggtaccgag ctcctgcagt atcgatgcgg ccgcaaagtc gacgaattct 5160
cattagcaga actcaagatg ctgatcctct ggaacgttga acttgagctt gtgttcctcg 5220
aaaagcttgc acaactcttt gatgtaacgc tggtgaagtc tatcaacttc ctctctagaa 5280
ggctgaggag tcatttgaac ctcgataggc tttccaacga tagtagtgat aggctgtctg 5340
aaaggcatga gtccgaaaga gtattggaaa actccccttc catggaaaag tggaaggctg 5400
attcccataa tcttttggag tctgttctgg atccatctaa gccaagttcc aggagtgttc 5460
tcaacctggt tgaagaggtt gttctctccg aatgagaaga taggaacaag agcagcacca 5520
tgcataagag caagtctgat gaatccctta cggttcttca agagaagtct gtaagcacca 5580
ggtctagcat caagagcctc ttgagcacct ccaacgatga tagcaagaag gtttccacca 5640
ccctttctgc taaggatgtg atcagcagaa actttctcgc tagacacgag tccaccagac 5700
atgatgtaat ctctgaagaa tggagccctg aaccaaacgg taagcatcat aaggtaggat 5760
ctgattccag ggaacaaaga ggtgaatcca gtagactcag tacagaggtt aaggaaagca 5820
ccagcagcaa gaacaccatg aggatggaat ccagcaatgt agttacggct aggatcaagc 5880
tcagcagtct taacgagaga cacagggaag taatccttca tgtacttcca gatggccaat 5940
cttctgaaga attggatagg tctaccacct tgtctaggct tatcccaatc caagtaccac 6000
caggtagcgt aaagaacaga gaaaagccag aacctggtga acaagagtcc aacgaagata 6060
acgatgcaga gttgagcaag agcaaggaat gagaaaaccc actgaagaac agcgaaagtc 6120
tgcaatcttc tctcccaagg aacaagaagt ggagcgaact cgaccatgaa ttcagtcccc 6180
cgtgttctct ccaaatgaaa tgaacttcct tatatagagg aagggtcttg cgaaggatag 6240
tgggattgtg cgtcatccct tacgtcagtg gagatatcac atcaatccac ttgctttgaa 6300
gacgtggttg gaacgtcttc tttttccacg atgctcctcg tgggtggggg tccatctttg 6360
ggaccactgt cggcagaggc atcttcaacg atggcctttc ctttatcgca atgatggcat 6420
ttgtaggagc caccttcctt ttccactatc ttcacaataa agtgacagat agctgggcaa 6480
tggaatccga ggaggtttcc ggatattacc ctttgttgaa aagtctcaat tgccctttgg 6540
tcttctgaga ctgtatcttt gatatttttg gagtagacaa gtgtgtcgtg ctccaccatg 6600
ttgacgaaga ttttcttctt gtcattgagt cgtaagagac tctgtatgaa ctgttcgcca 6660
gtctttacgg cgagttctgt taggtcctct atttgaatct ttgactccat gggatccaag 6720
ggccctagaa tctaattatt ctattcagac taaattagta taagtatttt tttaatcaat 6780
aaataataat taataattta ttagtaggag tgattgaatt tataatatat tttttttaat 6840
catttaaaga atcttatatc tttaaattga caagagtttt aaatggggag agtgttatca 6900
tatcacaagt aggattaatg tgttatagtt tcacatgcat tacgataagt tgtgaaagat 6960
aacattatta tatataacaa tgacaatcac tagcgatcga gtagtgagag tcgtcttatt 7020
acactttctt ccttcgatct gtcacatggc ggcggcccga attctcacac aaggtagttg 7080
caagacactg aagtggtggt agtggtagta gaagaagcag aatcggtaga aaggcaagac 7140
aatggagaag atgaagatgg tggagattct cttcccacaa cgcagcaatc aaggttttca 7200
aggttaaggc actcgtgctt tccatcatcg aacatgaagt cgatgttatc ctcgaaagca 7260
agctcgttga agagttctgg gtactcaatt gggttctcgt tagcaaggtt ttgatcggta 7320
aggaatgggg agaatccagt atccatcatg cagaagttcc aagcaagttc gttgttatct 7380
ccgcacctat ccatttccat gatggtggaa gaatcaatgc agcagttaac aacggcagct 7440
tcctcagaat atcccacaat ttcagcctct tgttgctcag ccttctcttc ctctttttct 7500
tcttcctctt gaggtggttc ctcaacgtat tgttgcttaa cctcttccct aggttcctct 7560
ttagcttctc tagtctcaac ctcttgctta gcctcaacaa gaataccctc ttgatggtta 7620
gcctggttaa ctgggaatgg gaaaacgccc ttcttcttaa gcctgtcgat gtagttggag 7680
atatcgaagt tggtaacagc gttagcacct ctgtactcaa tagcagccat atcataagca 7740
gctgcagcct cttcttgagt gttgtaagtt ccgaggtaga ggtacttgtt tccgaaaact 7800
cttccaatcc tagcttccca tcttccgtta tgatgatgcc tagcaactcc cctatactta 7860
gaaactcccc tagagaatcc agatgactgc cttctaaggg aagcaagata ctcttctttg 7920
gtcaccctct gcatctcttc aagttctttg gtgtaagtct cagctgggaa gttaagaatg 7980
gtatctgggc cccaatactt aagagcagca agatcatagg tatgagcagc agcctcttca 8040
gaatcataag ctccaaggta aacctgcttg cccttcttgt tttggatgga gttccaagag 8100
gacttatccc aaaggtgagc ttcgaatctt ccagtccatc tatgcctagt aacacctctg 8160
tagatagatg accttctggt agaagctgga gaagttgggt tatgagactt atcgccagat 8220
ggagatgact tcttagccct cttagctctc tttggtcttg gagcttcaga ttgaattggg 8280
ctagaggtag tagtagaaga ggacactgaa gaagatggag aactagagca ggtagaggta 8340
gtgagcctct tcttcatgaa ttcactagtg attaaatttt gttggtgctt tgagcatata 8400
acaagcatgg tatatatagg cacgtaaaca agttgagaaa ttttactttg agtttgacat 8460
aaccaataaa agttagtgct gtttattacc tcactcagtt tgcaccgcaa ctgtcgttag 8520
tgatgtttac ctttcctttt tctattattt attagtatta tataatatat atatatgtgt 8580
gatgagactt gaaattgttt agcaccgcaa atgtccttct tgaggggagg ttttcttttg 8640
ctgaggttgg ggtgtcacat acacccccct ctatggactc aacgtccttg ctgaggttta 8700
ccccacacta catgagattt ttctagactc aatactatga tatttctcgc cttatcggaa 8760
ttggttaaac tcagttgaag ttagggtcat atcgataaaa ttgacacatg atcgactctg 8820
atattaaaca gattctctcc ctcgaacctc actcactttc ctttttctat tctttattag 8880
tattatataa tagatccgtt ccaaccattc acgtacataa gaagagagat attttttttt 8940
aatggactaa catgacaaat aaaacaaaca aaggagtaat gatcactaca acaaattaga 9000
ttatgaggga caaataattt catcatctat aaatcatgtt tcgtcactaa aaattttgtg 9060
tgacgaaaaa gatttcgtca atcagttgtc actaaaaata tacaaagacg atttaatgat 9120
gtttaccttt ccttttctat tctttattag tattatataa taaatatatg tgtgatgaga 9180
cttgaaattg tttagcaccg caaatgtcct tgttgaaggg aggttttctt ttgctgaggt 9240
tggggtgtca catacacccc ctctatggac tgaacgtcct ttttgaggtt tattttacac 9300
tgcatgagat ttttctagat tcaacattat gatttctaga ctcaacacta cgatcgtcac 9360
taaagactat tttttatata taaaaaaaat actttgtcct taaatgtata aattagggat 9420
aaatttatta ttataaaaaa ggttaataat tttgtgatta aatctattat tttgtcactg 9480
aaagtgtttg cttttaccga cgacatatat gtcactaaat attatcataa gtagtgacaa 9540
ttacaattgt cacaaaataa aaaaaattat tcatattcaa caaaaaaggg tactacgaca 9600
atacattttt tgtcactgaa agtaatcaag ttgtgataaa ttaatttatt taatgacaaa 9660
aatatttgta tcaaaattca cccatgatca tataataaaa ataactaaaa ttatactaaa 9720
gcataaatga caagaaaatc taactaaaac atatcaaata ttactcctaa acaaagacat 9780
ataagtaaaa atttcttcca aagtatcaat aacgtggtga cacatagctt gcaatcaatc 9840
ttgcttcaat tttcaccttt tatacctgta aaaagaaaga gaaaataaaa caatgattta 9900
aaaatcgaat tcccgcggcc cctagaatct aattattcta ttcagactaa attagtataa 9960
gtattttttt aatcaataaa taataattaa taatttatta gtaggagtga ttgaatttat 10020
aatatatttt ttttaatcat ttaaagaatc ttatatcttt aaattgacaa gagttttaaa 10080
tggggagagt gttatcatat cacaagtagg attaatgtgt tatagtttca catgcattac 10140
gataagttgt gaaagataac attattatat ataacaatga caatcactag cgatcgagta 10200
gtgagagtcg tcttattaca ctttcttcct tcgatctgtc acatggcggc ggcccgcggc 10260
cgcttcatta ctcgagccag gaggatggat cgatgctggt ctgagaccct gctaccggtt 10320
gctgactgaa ctgctcggca cggtccttca tttcacgggc cttgctcgcc aactttgtct 10380
tggccgactc caactgatcc gctccgggtg gatgtttccc cgtcaggtaa cggtagatcc 10440
aggacagcac agacagagcg gcaacaccaa atcccccgct tgccagaaaa cccgctccca 10500
acaggaagat ggtgatgact gcagatcaga aaaactcaga ttaatcgaca aattcgatcg 10560
cacaaactag aaactaacac cagatctaga tagaaatcac aaatcgaaga gtaattattc 10620
gacaaaactc aaattatttg aacaaatcgg atgatatcta tgaaacccta atcgagaatt 10680
aagatgatat ctaacgatca aacccagaaa atcgtcttcg atctaagatt aacagaatct 10740
aaaccaaaga acatatacga aattgggatc gaacgaaaac aaaatcgaag attttgagag 10800
aataaggaac acagaaattt acctgcaggg accagtacag gcgagaagat caccaggaga 10860
ggtgtggcga ttgtcagcgc aatgaccgtt ccagccaggg tcaacccgga taacaccaac 10920
aggctacctc cggcagtaac cgcggtcgct gcctttacaa cacgctgagc acgcggttgc 10980
agttgcaagt ggggggcacg tgtttgttgc tgctgcccgt agtgctctgc catggaaatt 11040
ttgttggtgc tttgagcata taacaagcat ggtatatata ggcacgtaaa caagttgaga 11100
aattttactt tgagtttgac ataaccaata aaagttagtg ctgtttatta cctcactcag 11160
tttgcaccgc aactgtcgtt agtgatgttt acctttcctt tttctattat ttattagtat 11220
tatataatat atatatatgt gtgatgagac ttgaaattgt ttagcaccgc aaatgtcctt 11280
cttgagggga ggttttcttt tgctgaggtt ggggtgtcac atacaccccc ctctatggac 11340
tcaacgtcct tgctgaggtt taccccacac tacatgagat ttttctagac tcaatactat 11400
gatatttctc gccttatcgg aattggttaa actcagttga agttagggtc atatcgataa 11460
aattgacaca tgatcgactc tgatattaaa cagattctct ccctcgaacc tcactcactt 11520
tcctttttct attctttatt agtattatat aatagatccg ttccaaccat tcacgtacat 11580
aagaagagag atattttttt ttaatggact aacatgacaa ataaaacaaa caaaggagta 11640
atgatcacta caacaaatta gattatgagg gacaaataat ttcatcatct ataaatcatg 11700
tttcgtcact aaaaattttg tgtgacgaaa aagatttcgt caatcagttg tcactaaaaa 11760
tatacaaaga cgatttaatg atgtttacct ttccttttct attctttatt agtattatat 11820
aataaatata tgtgtgatga gacttgaaat tgtttagcac cgcaaatgtc cttgttgaag 11880
ggaggttttc ttttgctgag gttggggtgt cacatacacc ccctctatgg actgaacgtc 11940
ctttttgagg tttattttac actgcatgag atttttctag attcaacatt atgatttcta 12000
gactcaacac tacgatcgtc actaaagact attttttata tataaaaaaa atactttgtc 12060
cttaaatgta taaattaggg ataaatttat tattataaaa aaggttaata attttgtgat 12120
taaatctatt attttgtcac tgaaagtgtt tgcttttacc gacgacatat atgtcactaa 12180
atattatcat aagtagtgac aattacaatt gtcacaaaat aaaaaaaatt attcatattc 12240
aacaaaaaag ggtactacga caatacattt tttgtcactg aaagtaatca agttgtgata 12300
aattaattta tttaatgaca aaaatatttg tatcaaaatt cacccatgat catataataa 12360
aaataactaa aattatacta aagcataaat gacaagaaaa tctaactaaa acatatcaaa 12420
tattactcct aaacaaagac atataagtaa aaatttcttc caaagtatca ataacgtggt 12480
gacacatagc ttgcaatcaa tcttgcttca attttcacct tttatacctg taaaaagaaa 12540
gagaaaataa aacaatgatt taaaggcgcg ccgcgtattg gctagagcag cttgccaaca 12600
tggtggagca cgacactctc gtctactcca agaatatcaa agatacagtc tcagaagacc 12660
aaagggctat tgagactttt caacaaaggg taatatcggg aaacctcctc ggattccatt 12720
gcccagctat ctgtcacttc atcaaaagga cagtagaaaa ggaaggtggc acctacaaat 12780
gccatcattg cgataaagga aaggctatcg ttcaagatgc ctctgccgac agtggtccca 12840
aagatggacc cccacccacg aggagcatcg tggaaaaaga agacgttcca accacgtctt 12900
caaagcaagt ggattgatgt gataacatgg tggagcacga cactctcgtc tactccaaga 12960
atatcaaaga tacagtctca gaagaccaaa gggctattga gacttttcaa caaagggtaa 13020
tatcgggaaa cctcctcgga ttccattgcc cagctatctg tcacttcatc aaaaggacag 13080
tagaaaagga aggtggcacc tacaaatgcc atcattgcga taaaggaaag gctatcgttc 13140
aagatgcctc tgccgacagt ggtcccaaag atggaccccc acccacgagg agcatcgtgg 13200
aaaaagaaga cgttccaacc acgtcttcaa agcaagtgga ttgatgtgat atctccactg 13260
acgtaaggga tgacgcacaa tcccactatc cttcgcaaga ccttcctcta tataaggaag 13320
ttcatttcat ttggagagga cacgctgaaa tcaccagtct ctctctacaa atctatctct 13380
gcgatcgcat ggcgattttg gattctgctg gcgttactac ggtgacggag aacggtggcg 13440
gagagttcgt cgatcttgat aggcttcgtc gacggaaatc gagatcggat tcttctaacg 13500
gacttcttct ctctggttcc gataataatt ctccttcgga tgatgttgga gctcccgccg 13560
acgttaggga tcggattgat tccgttgtta acgatgacgc tcagggaaca gccaatttgg 13620
ccggagataa taacggtggt ggcgataata acggtggtgg aagaggcggc ggagaaggaa 13680
gaggaaacgc cgatgctacg tttacgtatc gaccgtcggt tccagctcat cggagggcga 13740
gagagagtcc acttagctcc gacgcaatct tcaaacagag ccatgccgga ttattcaacc 13800
tctgtgtagt agttcttatt gctgtaaaca gtagactcat catcgaaaat cttatgaagt 13860
atggttggtt gatcagaacg gatttctggt ttagttcaag atcgctgcga gattggccgc 13920
ttttcatgtg ttgtatatcc ctttcgatct ttcctttggc tgcctttacg gttgagaaat 13980
tggtacttca gaaatacata tcagaacctg ttgtcatctt tcttcatatt attatcacca 14040
tgacagaggt tttgtatcca gtttacgtca ccctaaggtg tgattctgct tttttatcag 14100
gtgtcacttt gatgctcctc acttgcattg tgtggctaaa gttggtttct tatgctcata 14160
ctagctatga cataagatcc ctagccaatg cagctgataa ggccaatcct gaagtctcct 14220
actacgttag cttgaagagc ttggcatatt tcatggtcgc tcccacattg tgttatcagc 14280
caagttatcc acgttctgca tgtatacgga agggttgggt ggctcgtcaa tttgcaaaac 14340
tggtcatatt caccggattc atgggattta taatagaaca atatataaat cctattgtca 14400
ggaactcaaa gcatcctttg aaaggcgatc ttctatatgc tattgaaaga gtgttgaagc 14460
tttcagttcc aaatttatat gtgtggctct gcatgttcta ctgcttcttc cacctttggt 14520
taaacatatt ggcagagctt ctctgcttcg gggatcgtga attctacaaa gattggtgga 14580
atgcaaaaag tgtgggagat tactggagaa tgtggaatat gcctgttcat aaatggatgg 14640
ttcgacatat atacttcccg tgcttgcgca gcaagatacc aaagacactc gccattatca 14700
ttgctttcct agtctctgca gtctttcatg agctatgcat cgcagttcct tgtcgtctct 14760
tcaagctatg ggcttttctt gggattatgt ttcaggtgcc tttggtcttc atcacaaact 14820
atctacagga aaggtttggc tcaacggtgg ggaacatgat cttctggttc atcttctgca 14880
ttttcggaca accgatgtgt gtgcttcttt attaccacga cctgatgaac cgaaaaggat 14940
cgatgtcatg agcgatcgcg atcgttcaaa catttggcaa taaagtttct taagattgaa 15000
tcctgttgcc ggtcttgcga tgattatcat ataatttctg ttgaattacg ttaagcatgt 15060
aataattaac atgtaatgca tgacgttatt tatgagatgg gtttttatga ttagagtccc 15120
gcaattatac atttaatacg cgatagaaaa caaaatatag cgcgcaaact aggataaatt 15180
atcgcgcgcg gtgtcatcta tgttactaga tccctgcagg gcgtattggc tagagcagct 15240
tgccaacatg gtggagcacg acactctcgt ctactccaag aatatcaaag atacagtctc 15300
agaagaccaa agggctattg agacttttca acaaagggta atatcgggaa acctcctcgg 15360
attccattgc ccagctatct gtcacttcat caaaaggaca gtagaaaagg aaggtggcac 15420
ctacaaatgc catcattgcg ataaaggaaa ggctatcgtt caagatgcct ctgccgacag 15480
tggtcccaaa gatggacccc cacccacgag gagcatcgtg gaaaaagaag acgttccaac 15540
cacgtcttca aagcaagtgg attgatgtga taacatggtg gagcacgaca ctctcgtcta 15600
ctccaagaat atcaaagata cagtctcaga agaccaaagg gctattgaga cttttcaaca 15660
aagggtaata tcgggaaacc tcctcggatt ccattgccca gctatctgtc acttcatcaa 15720
aaggacagta gaaaaggaag gtggcaccta caaatgccat cattgcgata aaggaaaggc 15780
tatcgttcaa gatgcctctg ccgacagtgg tcccaaagat ggacccccac ccacgaggag 15840
catcgtggaa aaagaagacg ttccaaccac gtcttcaaag caagtggatt gatgtgatat 15900
ctccactgac gtaagggatg acgcacaatc ccactatcct tcgcaagacc ttcctctata 15960
taaggaagtt catttcattt ggagaggaca cgctgaaatc accagtctct ctctacaaat 16020
ctatctctct cgagatgatt gaacaagatg gattgcacgc aggttctccg gccgcttggg 16080
tggagaggct attcggctat gactgggcac aacagacaat cggctgctct gatgccgccg 16140
tgttccggct gtcagcgcag gggaggccgg ttctttttgt caagaccgac ctgtccggtg 16200
ccctgaatga acttcaagac gaggcagcgc ggctatcgtg gctggccacg acgggcgttc 16260
cttgcgcagc tgtgctcgac gttgtcactg aagcgggaag ggactggctg ctattgggcg 16320
aagtgccggg gcaggatctc ctgtcatctc accttgctcc tgccgagaaa gtatccatca 16380
tggctgatgc aatgcggcgg ctgcatacgc ttgatccggc tacctgccca ttcgaccacc 16440
aagcgaaaca tcgcatcgag cgagcacgta ctcggatgga agccggtctt gtcgatcagg 16500
atgatctgga cgaagagcat caggggctcg cgccagccga actgttcgcc aggctcaagg 16560
cgcgcatgcc cgacggcgag gatctcgtcg tgactcatgg cgatgcctgc ttgccgaata 16620
tcatggtgga aaatggccgc ttttctggat tcatcgactg tggccggctg ggtgtggcgg 16680
accgctatca ggacatagcg ttggctaccc gtgatattgc tgaagagctt ggcggcgaat 16740
gggctgacc 16749
<210> 124
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность линкера
<400> 124
atttaaatgc ggccgcgaat tcgtcgattg aggacgtccc tactagacct gctggacctc 60
ctcctgctac ttactacgat tctctcgctg tgcatatggt cagtcatgcc cgggcctgca 120
ggcggccgca tttaaat 137
<210> 125
<211> 434
<212> ДНК
<213> Искусственная последовательность
<220>
<223> шпРНКи
<400> 125
gtgagcaatg aaccaagatt tatcaatacc gttacttttg atagcaaaga gggttctcct 60
actcttgtta tggtccacgg atatggtgcc tctcagggtt tcttctttcg gaatttttat 120
gcccttgcga ggcatttcaa agttattgct attgatcagc ttggctgggg tggttcaagc 180
aggcctgact tcacatgcag aagtacagaa gagactgaag attggtttat tgattccttt 240
gaggagtggc gcaaagccaa aaaccttagc aactttattt tgcttgggca ctcctttgga 300
gggtatgtcg ctgcaaaata tgctctcaag catccagagc atgttcagca gttgattctg 360
gtaggaccag ctggatttac atcagagact gaacatatgt ccgagcggct tacccagttt 420
agagcaacat ggaa 434
<210> 126
<211> 593
<212> ДНК
<213> Искусственная последовательность
<220>
<223> шпРНКи
<400> 126
actgctgatg ctgtcaggca gtatctatgg ttgtttgagg agcataatgt tcttgaattc 60
ctcgtacttg ctggagatca tctatatcga atggattatg aaaagttcat tcaagcccac 120
agagaaacag atgctgatat tactgttgcc gcactgccaa tggatgaaaa gcgagccact 180
gcatttggtc tcatgaagat tgacgaagaa ggacgcatta ttgaatttgc agagaaaccg 240
aaaggagagc aattgaaagc aatgaaagtg gatactacca ttttaggtct tgatgatgag 300
agagctaaag agatgccttt tatcgcaagt atgggtatat atgtcattag caaagatgtg 360
atgttaaact tacttcgtga taagttccct ggtgccaatg attttggcag tgaagttatt 420
cctggtgcaa cttcgcttgg gatgagagtg caagcttatt tatatgatgg atactgggaa 480
gatattggta ccatcgaagc tttctacaat gccaatttgg gcattaccaa aaagccagtc 540
ccagatttta gcttctatga ccgatcagct ccaatctaca cccaacctcg ata 593
<210> 127
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> мотив липазы
<220>
<221> misc_feature
<222> (2)..(2)
<223> Xaa может быть любой природной аминокислотой
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa может быть любой природной аминокислотой
<400> 127
Gly Xaa Ser Xaa Gly
1 5
<210> 128
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> мотив ацилтрансферазы
<220>
<221> X
<222> (2)..(5)
<223> любая аминокислота
<400> 128
His Xaa Xaa Xaa Xaa Asp
1 5
<210> 129
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> вероятный мотив связывания с липидом
<220>
<221> X
<222> (2)..(4)
<223> любая аминокислота
<400> 129
Val Xaa Xaa Xaa His Gly Phe
1 5
<210> 130
<211> 1224
<212> ДНК
<213> Arabidopsis thaliana
<400> 130
atggcggaag aaatctcaaa gacgaaggtg ggatcttctt ctactgcttc ggtggctgat 60
tcatctgctg ctgcgtcggc tgcaacgaat gcggccaaat caagatggaa aattttgtgg 120
cctaattcgc tccggtggat tcctacgtcc accgattaca tcatcgccgc cgagaaacgt 180
cttctctcca tcctcaagac gccttatgta caagagcaag tcagtattgg ttcaggacca 240
ccaggttcta aaatcaggtg gtttaggtct acgagcaatg agtcacgtta catcaacact 300
gttacatttg atgccaagga gggagctcct acactcgtca tggttcatgg ttatggtgct 360
tctcaagggt ttttcttccg taattttgat gctcttgcca gtcgatttag agtgatcgct 420
attgatcaac ttgggtgggg tggttcaagt aggcctgatt ttacatgtag aagcacagaa 480
gaaactgagg catggtttat cgactccttt gaggaatggc gtaaagccca gaatctcagt 540
aactttattc tattaggaca ttcttttgga ggctatgttg ctgctaaata cgcgcttaag 600
catcctgaac atgttcaaca cttaattctg gtgggatctg ctgggttctc agcagaagca 660
gatgccaaat cagaatggct cactaaattt agagcaacat ggaaaggtgc agtcctaaat 720
catttatggg agtcaaattt cactcctcag aagctggtta gaggattagg tccttggggt 780
ccaggtcttg taaatcggta tacaactgca agatttggtg cacattcgga gggaactggg 840
ctaacagaag aggaagccaa attgctaacc gattatgtgt accatacttt ggctgcaaag 900
gctagtggag agttatgctt gaaatacatc ttctcatttg gagcatttgc taggaagccc 960
ctcttacaaa gtgcatcaga gtggaaagtg ccaacaacgt ttatctatgg aatgaatgat 1020
tggatgaact atcaaggtgc ggtggaagcg aggaaatcca tgaaggtccc ttgcgaaatc 1080
attcgggttc cacagggtgg tcattttgtg ttcatagaca acccaattgg ttttcattct 1140
gcagtgcttt atgcttgccg caagtttata tctcaagact cctctcatga tcaacaactc 1200
ctagatggtc tacgattggt ttag 1224
<210> 131
<211> 1700
<212> ДНК
<213> Brachypodium distachyon
<400> 131
tccgcgcccg aaacgatccc aacagaagct ctaatctcca aagccgccgc gcgtgttgag 60
ggtggtgcgg ggaaaagctt ggtgtcgtga gcccccgtgt cgcatgcgcc gcgctgccgc 120
cgtgacgagg atggcagcga ccgaggagat gaggcaggcg tccgccgccg ccgccgccac 180
ggtgaccgag gcctcggcgt cggcggcccc gcccgcgggg tccaggtggg cgcgggtgtg 240
gccggccgcg ctgcgctgga tccccacctc caccgagcgc atcatcgccg ccgagaagcg 300
cctcctctcc gtactcaaaa ctgggtatgt ccaagaacaa gttaacattg gctcggctcc 360
acccgggtca aaagtaagat ggtttagatc atcaagtgat gagccaaggt tcatcaatac 420
agtgacattt gatagcaagg agaatgctcc cactcttgtc atggtccatg gttatggtgc 480
ttcacagggt ttcttcttta gaaattttga tgcccttgca agccgtttcc gagtgattgc 540
cattgatcag cttggttggg gtggatcaag tagacctgac ttcacctgta aaagtaccga 600
agaaactgag gcttggttca tagattcttt agaggaatgg cgtaaagcaa agaacctcag 660
taattttata ttgctcggtc attctttcgg aggatatgtt gcagcaaaat atgccttgca 720
gcatcctgaa cacgtgcagc acttaatttt ggtcggttct gctgggtttt catcagaaac 780
agatcatagc tctgagtggt taaccaagtt tcgagcaaca tggaaaggca tgctagtgaa 840
ccaactatgg gagtccaatt ttactcctca aagaattgta agaggattgg gtccttgggg 900
cccagatttg gttcgcagat ataccactgc taggtttggc tcatattcaa caggtgaatt 960
actaacagaa catgagtccg gcttgctgac agattacatt taccatacat tagctgccaa 1020
agctagtgga gagctgtgct tgaaatatat tttttccttg ggggcatttg caaggaaacc 1080
tcttctgcag agtgcatctg actggaaagt gccgaccact ttcatatatg gccatgacga 1140
ttggatgaaa taccaggggg cacagcaagc acgcaaggat atgaaagttc cttgcgaaat 1200
catcagagtc ccacagggag gacattttgt gttcatagat aacccttccg ggttccattc 1260
ggcagtcttc tatgcgtgcc ggaaattttt atctggagat gcagaggagg gtctctctct 1320
tcctgatggc ctgatatctg catgacagca tgaggcgcga tgtcatacca attagcggta 1380
tgaacacaaa gcaaagctat acggagctag gaaatgttac aaatgtcacg actcaccaga 1440
aatgttacaa atgtcaccac tcaccagttt cctttttgta tgtatgaatt gtgtgaatat 1500
acacgtcatt catatttgcc ggcgtatcag tacttcaata gtgataaaac atgatcatat 1560
atatatgtat gatttctcta gtcggttctc atcaagtcaa gttattgtga ttggtgaatg 1620
atatactttc caggtcaact ttgtgtttgc atgtacaaac tatcatggaa catatcagta 1680
tagtttatga tttgtcttcc 1700
<210> 132
<211> 1484
<212> ДНК
<213> Glycine max
<400> 132
gtcatggatg cgcgtcactg ctcgcgttca ttataatggc ggaagagata accaagaacg 60
acgtcggagt aacctccaaa accaccagaa gcagctccag gttctggcct cgttggattc 120
ccacttccac cgatcacatc atcgctgccg agaagcgcct tctttccgtc gtcaagactg 180
gttatgttca agagcatgtt aacattggct ctggtcctcc tggctccaaa gtgaggtggt 240
tccgttcatc cagcaacgag ccgcggttta ttaacaccgt tacatttgac agtaaacccc 300
attctccaac gcttgtcatg attcatggtt atgctgcttc acaggggttc ttttttcgca 360
attttgacgc gcttgcgtct cgatttagag tcattgctgt tgatcaactt ggatggggtg 420
gatcgagcag acctgatttc acatgcaaaa gcactgaaga aactgaggca tggtttattg 480
attcttttga ggaatggaga aaagccaaaa acttgagcaa ttttatactg ctcggacatt 540
cttttggtgg ttatgttgct gccaaatatg cgctcaagca ccctgagcat gtacaacact 600
tgattctggt tggatctgct ggattttcat ctgaatcaga tgcaaagtct gagtggataa 660
caaggtttcg agcaacatgg aagggggcag ttttgaacca tctttgggaa tcaaatttca 720
cacctcagaa acttgtcagg ggtttaggtc cttggggtcc caacatagtc cgcaagtata 780
caagtgctag gtttggtaca cattcaactg gggaaatact gactgaagag gaatcaacat 840
tgctgacaga ctatgtttac cacacattgg cggccaaagc tagtggagag ctgtgcttaa 900
aatatatttt ttcatttgga gcatttgcta ggatgcccct tcttctcagt gcctcagagt 960
ggaaggtgcc caccactttc atgtatggtt tccaagactg gatgaattat caaggtgccc 1020
aagaagctcg caagcatatg aaggttccat gcgaaatcat caggattcct cagggtgggc 1080
actttgcgtt cattgacaac ccaactgcct tccattcagc tgttttttat gcttgtcgaa 1140
ggtttcttac acctgatcca gacaatgaat ctcttcctaa agggctaacc tctgcatagg 1200
ttaggtctta attttgtgct attcctgtct atatgtattt taatattttt ttttactaat 1260
taaatttcat aattgaatga aatcatatgt atattgtttc agtaaagtgg aatttactga 1320
aaatatttgt aatagcaact tcaacaaaaa tcgatttgta ggagaaattt cttccctgga 1380
aattgttcta ttttaaatct tgttgctcat aagatattat gacttcattc aactaataat 1440
tcatgtcgtt taggaaaagt agttagttat attaaatttg tcaa 1484
<210> 133
<211> 1662
<212> ДНК
<213> Zea mays
<400> 133
accatacggg cggggccgca ccgaccgaac ctaaccgaga gcacgagcat acccgtcccg 60
actccgactg cagagcatca gccgaggaga aaagtcggga gaaacgcgcg tgacgtctgc 120
ccgcgtcgta tgcgccgcgc tgccgtcgcc gcgacgacga cgacgaccag gatggcagcc 180
gaggagatga gacgggcctc cgcctcaacg gccacggcgg agatgccggc gtcgccggcg 240
ccggcgcaag cggggtcgag gtgggcgcgg gtgtggccgc gcgcgctccg gtggatcccc 300
acctccacgg accgcatcat cgccgccgag aagcgactcc tcacgatagt caaaactgga 360
tatgtccagg aacgagtcaa cattggctct gctccacctg ggtcaaaagt aagatggttt 420
aggtcagcaa gtgatgaacc aaggttcatt aatactgtaa catttgatag caaggagaat 480
gcccccaccc tggttatggt ccatggctat ggagcttcac aggggttctt ctttcgaaac 540
tttgatgccc ttgcaagccg ttttagggtg attgccattg atcagcttgg ctggggtggt 600
tcaagcagac ctgacttcac atgtaaaagt accgaagaaa ctgaggcatg gttcatagat 660
tctttcgagg agtggcgcaa ggccaagaac ctcagtaatt ttatattgct tggtcactct 720
tttggaggat atgttgctgc aaaatatgcg ctaaagcacc ctgaacacgt tcaacagttg 780
attttggttg gtcctgctgg cttctcatca gaaacagagc atagctctga gtggttaacc 840
aagtttcgag caacatggaa aggcatgcta atgaatcgtc tttgggagtc caattttact 900
ccccaaaggg ttattagggg attgggtcct tggggtccag gtctagtaca gagatatacc 960
agtgccaggt ttggtacaag ttctactggt gaattactaa cagatgaaga atcggcattg 1020
atgacagatt atatgtacca tacgttagct gccaaagcta gtggagagct gtgcttgaaa 1080
tatatatttt ccttcggggc atttgcaagg aaacctcttc tgcagtgcgc gtccgattgg 1140
aaagtgccga ctactttcat atatgggcag caagattgga tgaactacca aggcgctcag 1200
caagcacgga aggacatgaa agttccttgt gaaataatca gggtgccgca gggtggacat 1260
tttgtgttca tagacaaccc ttcagggttc cactcggctg tcttctatgc gtgccgtaat 1320
cttctatcag taaatggaga ggagggattc acatttcctg atggcctaat atctgcgtga 1380
agtggcatgt tcaacaagct tgctcaacaa cagtttacat aaagcaaaga tatacgattg 1440
tggaaatcat tgcccatttc caccaatttg cttgtatacg gattatgctg tgtatatatt 1500
acataacaaa tgtattagta tcatttaatg cacgatttgt gaaagggcct gagtttgtat 1560
ttagcgaatt ttaggttggt ttttttccct ttttcttctt tcagtgcgct tgctagtcaa 1620
tcccatacta taagccgtga tcatttaaaa aaaaaaaaaa aa 1662
<210> 134
<211> 1763
<212> ДНК
<213> Sorghum bicolor
<400> 134
actgcagatg cgcggtcgtc ggctccggct cgcggaggcg agaacggcga accagcccgt 60
gtctctgttc cctttcttcc ctttaaaaac acggcaaaaa aaaagctagc cggttacgct 120
accgaaccga acggctcggc acgcgggcac gggcgcgggg tcgcaccgga aaagcacgag 180
cagagcagac ctgacgtctc cagactgcag gagcatcatc agtcgaggag gaggaagtgt 240
ggggggggga aagggaaacg tgcgcgtcgt atgcgcctcg ctgccgtcgc caggacgacc 300
aggatggcag ccgaggagat gaggcgggcc tcggcctccg cggcggtcgc ggccacgacg 360
gaggcggcgc cggcgccggc gcaagcgggg tccaggtggg cgcgggtgtg gccgagcgcg 420
ctccggtgga tccccacctc cacggatcgc atcatcgccg cggagaagcg gctcctctcg 480
atagtcaaaa ctgggtatgt ccaggaacaa gtcaacattg gctcagctcc acctgggtca 540
aaagtaagat ggtttaggtc agcgagtgat gaaccaaggt tcattaatac tgtaacattt 600
gatggcaagg agaacgcccc caccctggtt atggtccatg gctatggagc ttcacagggg 660
ttcttctttc gaaactttga cgcccttgca agtcgtttta gggtgattgc cattgatcag 720
cttggctggg gtggttcaag cagacctgac ttcacatgta aaagtaccga agaaactgag 780
gcatggttca tagattcttt tgaggagtgg cgcaaggcca agaacctcag taattttata 840
ttgcttggtc actcctttgg aggatatgtt gcggcaaagt atgccctaaa gcaccctgaa 900
cacattcagc acttagtttt ggttggtcct gctggcttct cgtcagaaac agaccatagc 960
tctgagtggt taaccaagtt tcgagcaaca tggaaaggca tgctagtgaa tcatctttgg 1020
gagtccaatt ttactcccca aagagttatt agaggattgg gcccttgggg tccaggtcta 1080
gtacaaagat ataccagtgc caggtttggt acacgttcaa ctggtgatat actaacagat 1140
caagaatcaa cattgttgac agattatatt taccatacct tagctgccaa agctagtgga 1200
gagctgtgct tgaaatatat attttccttc ggggcatttg caaggaaacc tcttctgcag 1260
tgcgcatccg attggaaagt gccgactact ttcatatatg gtcaggaaga ttggatgaac 1320
taccaagggg ctcagcaagc acggaaggac atgaaagttc cttgtgaaat aatcagggtg 1380
ccacagagtg gacattttgt gtttatagac aacccttcag ggttccactc ggctgtcttc 1440
tacgcgtgcc gtaatctttt atcccaaaat ggggaggagg gcttcacatt tcctggtggc 1500
ctaatatctg catgaagtgg catgttcaac aatcttatcg tgcccaacaa tagtttatat 1560
gaagcaaaga tatacgatgg tggaaatctt tgctcatttc caccaatctg gaaatatttg 1620
tgccctcttc caccaatttg tttgtatacg gattatgccg tgtatatatt ctgtgttgac 1680
tgtaagaaac ataatgtatt aacattatgt aatgtatgta cgattcttta tttgattttc 1740
aacttgcaat acgcaagaac cac 1763
<210> 135
<211> 1399
<212> ДНК
<213> Ricinus communis
<400> 135
cgccttttta ccagtcaatt tccattttta tatataagtg cttttgctta atttaagact 60
aactacagcg acgaattcgc gtttatgaaa ttgcttcgcc tacgactgct acgagtatct 120
agctcctcaa tatcatcaat aatggcggaa ggggctgctg ccacatcagc atcagcatca 180
gcgtcagcgt cagcgtcatg ggcaaaaaca agatctctac ggccatctgc tctccgttgg 240
atcccaactt caaccgatca catcatcgcc gccgaaaagc gtcttctctc cctcgtcaag 300
actccctatg ttgtggaaca agtgaatata gggtctggcc caccggggtc gaaggtgagg 360
tggtttcgtt ctaaaagcga cgaggcacgg tttattaaca cggttacttt tgatagcaaa 420
gaggaggatt ctcctacact ggttatggtt catggatatg ctgcttctca aggcttcttc 480
tttcgcaatt ttgatgctct tgcttctcgt ttcaggctca ttgctattga tcagctcggt 540
tggggtggat caagtagacc tgattttacg tgtaagagca ctgaagaaac tgaggcatgg 600
ttcattgact cctttgaggc ttggcgtaaa gagaaaaacc tcagtaactt catcttactt 660
ggacattctt tcggagggta tattgcagct aaatatgcac tcaagcatcc tgagcatgtt 720
caacatctga ttttagtggg atctgctgga ttttcatcag aatctgaaga caaatctgag 780
cagcttactc ggttcagagc aacatggaag ggagcagttt tgaatcattt atgggagtct 840
aattttactc ctcagaaggt tattagaggt ttaggtcctt ggggtccaga tctcgtacgc 900
aagtacacaa ctgctagatt tggttcatat tcaactggtg agatattaaa ggaggaggag 960
tccaaattgc ttacagacta tgtgtaccat accttagccg ccaaagctag tggagagcta 1020
tgcttgaaat atatattttc ttttggagca tttgctcgga tgccccttct acaaagcgcg 1080
tcacaatgga aagtgccaac tactttcata tatgggatgc aagattggat gaattatcaa 1140
ggggcccaaa gagctcgcaa agatatgaat gtcccatgtg aaatcattag ggttcctcag 1200
ggcgggcact tcgttttcat agacaaccca actgggtttc attcagctgt gttatatgcc 1260
tgccggagat ttctctcacc cgatcctgat aatgaatctc ttcctgaagg tctgatatct 1320
gcgtaggaag tgtggtttgt aattatttct tttttatttg ttgtgtataa tttatctgag 1380
aatttccaat tctttcaat 1399
<210> 136
<211> 1480
<212> ДНК
<213> Medicago truncatula
<400> 136
ggttggctca tagttccttt tacctgttga aaacaaaaca tatggagtaa cattttagtc 60
agaaattcaa agctacgcac ttgattaaac taattatcga aaaatggcgg aagaaattag 120
acaaaaggac gacgtcgatt catcttcgaa atctaaaagc ttctggtctt cactccgttg 180
gattcccact tctaccgatc atatcatcgc cgctgagaaa cgccttcttt ccattatcaa 240
gactgggtat gctcaagagc atgttaatat aggttctggt cctcctggct ctaaagttag 300
atggttccgt tcaaccagta acgagccacg ctttctcaac actgttacat ttgatagtaa 360
acccgattct cctacacttg ttatggttca tggatacgct gcttctcagg gtttcttctt 420
tcgcaatttt gatgctctcg cctctcgttt cagaatcatt gctgttgatc aacttggttg 480
gggaggatca agcagacctg atttcacatg caaaagtacc gaagaaactg aggcatggtt 540
cattgattct ttcgaggaat ggagaaaagc caaaaatctt accaatttca tactgcttgg 600
acattctttt ggtggttatg ttgcttccaa atacgcgctc aagcaccctc agcacgtaca 660
acacttaatt ttggtgggac ctgccgggtt tacagaagaa acagatccaa agactgagtt 720
tgttactaag tttcgagcaa catggaaggg agcagttctg aaccatctat gggaatctaa 780
ttttacacct cagaaaattg tcagaggttt aggtccttgg ggtcctaaca tggtccgcaa 840
atatacaagt gctaggtttg gtacacattc aaccgggcaa aaactgattg acgaggaatc 900
aagtctgctg actgattatg tttatcatac attggcggcc aaagctagtg gggagctgtg 960
tttaaaatat atttttgcat ttggagcatt tgctaggatg ccccttcttc aaagtgctca 1020
agagtggaag gtgcccacca cattcatata tggttacgaa gattggatga attatgaagg 1080
tgcccaagaa gctcgcaagc atatgaaggt tccatgtgaa attatcaggg tccctaaggc 1140
cggccatttt gtgttcattg acaacccaag tggcttccat tcagctgtgt tttatgcttg 1200
tcgaaggttt cttaccccaa attcggacaa tgaatctctt cccgaagggc tatcgtctgc 1260
ttaggattta attttgcatc aatccagtgt atattaatat ggttattaat ttttttttac 1320
ttcataactg aatgaagccg tgtcttgttt ctcagtgaag tggaatataa tggaaatata 1380
tgtaattgta ataacaataa tattgatttg ttggggaact ttgaggacaa aaacatattc 1440
tggtaaaatt ttgttgcaca tgcgacaaac atatgctgtg 1480
<210> 137
<211> 1317
<212> ДНК
<213> Arabidopsis thaliana
<400> 137
gatctctctc cctctctctc tctctctctc cgggaaaaat ggataacttc ttaccctttc 60
cctcttctaa cgcaaactct gtccaagaac tctctatgga tcctaacaac aatcgctcgc 120
acttcacaac agtccctact tatgatcatc atcaggctca gcctcatcac ttcttgcctc 180
cgttttcata cccggtggag cagatggcgg cggtgatgaa tcctcagccg gtttacttat 240
cggagtgtta tcctcagatc ccggttacgc aaaccggaag tgaattcggt tctctggttg 300
gtaatccttg tttgtggcaa gagagaggtg gttttcttga tccgcgtatg acgaagatgg 360
caaggatcaa caggaaaaac gccatgatga gatcaagaaa caactctagc cctaattcta 420
gtccaagtga gttggttgat tcaaagagac agctgatgat gcttaacttg aaaaataacg 480
tgcagatctc cgacaagaaa gatagctacc aacagtccac atttgataac aagaagctta 540
gggttttgtg tgagaaggaa ttgaagaaca gcgatgttgg gtcactcggg aggatagttc 600
taccaaagag agatgcagaa gcaaatcttc cgaagctatc tgataaagaa ggaatcgttg 660
tacagatgag agatgttttc tctatgcagt cttggtcttt caaatacaag ttttggtcca 720
ataacaagag cagaatgtat gtcctcgaga acacaggaga atttgtgaag caaaatggag 780
ctgagatagg agacttttta acaatatacg aggacgaaag caagaatctc tacttcgcca 840
tgaatggaaa ttcgggaaaa caaaatgaag gaagagaaaa tgagtcgagg gaaaggaacc 900
actacgaaga ggcaatgctt gattacatac caagagacga agaggaagct tccattgcaa 960
tgctcatcgg aaatctaaac gatcactatc ccatccctaa cgatctcatg gacctcacca 1020
ctgaccttca gcaccatcaa gccacgtcct catcaatgcc acctgaggat cacgcgtacg 1080
tgggttcatc cgatgatcag gtgagcttta acgactttga gtggtggtga tatggtggtg 1140
gaagttctca agttcataac cccctttatg aaaatagacc ttaagatata caaaagagat 1200
taaaagaaaa aaaagttagt atatttcatc atatctctca ttgaagatga gatttatatc 1260
tataattgtt taatagtgtt tttattactt ttctatcaat atattaaagt tttaatt 1317
<210> 138
<211> 1439
<212> ДНК
<213> Medicago truncatula
<400> 138
tttcatcctt acatattttg catattgaaa cacgtaggat ggaataagat tgataacaaa 60
aattgcattg tttgcatatt gaaaacatgg gacaattgca tgggttcatg tgcttcatta 120
taagccacac attaggaaac acaggttgat attcaccact atttaacata agaatatctc 180
atgtgtaagc attcatacaa atatcacaat tgaattaaaa accaaagaaa tgtcttcctc 240
taacttctct tgtatcctat ccatctcctt aacattcttc atcttgctac tgaacaaggt 300
gaattcagca gaaacaactt ccttttccat cacaaaattt gtcccagatc aaaagaatct 360
catcttccaa ggcgatgcga aaactgcctc aacagggaag ttagaactct ccaaggcagt 420
caagaactct attggtagag ctctttattc cgcccctatt cacatttggg atagcaaaac 480
cggtagtgtg gctaactttc aaactacctt cacctttaca ataacggcgc ctaatactta 540
taatgttgca gacggtcttg cattcttcat tgcaccaatt gatactaagc cgaaatcaat 600
tcatcatgga ggataccttg gagttttcga tagcaaaact tacaaaaaat caattcaaac 660
tgttgcagtt gaaattgaca ctttctataa tgctcaatgg gatccaaatc ccggaaatat 720
aagtagcact ggtcgacata ttggaatcga tgtaaactct atcaaatcaa taagcaccgt 780
gccgtggagt ttggaaaaca ataaaaaggc taatgttgcg atagggttta atggtgcaac 840
aaatgtgttg agtgttgatg tggaatatcc tttgattcgt cattataccc taagtcatgt 900
tgtgcctttg aaggatgttg ttcctgagtg ggtaaggatt ggtttctctt cttctactgg 960
agccgaatat tcagcacatg atattttatc gtggtctttt gattcaaagt tgaacctagg 1020
ttttgagaac aatatcaatg ccaatgtttc aagctctact caagctgcat agttgaaaac 1080
ttatccatta tgtatgtgtg agtgtaacca accagtctaa gaaaactata ataagatacc 1140
tgaaataatg gttcattatc gtgtagtaga aatatggtca caccatatct tctttttttt 1200
ttaataaatt atggaataat gctatttctc gcgagagtta tgtttcggaa agattcatga 1260
atagatgtta atcaattaga tctatatata tatatatata tatatatata tatatatata 1320
tagcattttc ttaaattatg catatgtaat atcgtgtaat gctattgttt atatcaatga 1380
atggtgtttt gtagtcacat aattcgtaat ttctctccat gagaacagcg aaccaatta 1439
<210> 139
<211> 1393
<212> ДНК
<213> Brassica napus
<400> 139
gagatgggta tccctataag gtgcagcatc gaaccatctg caacattttg actcgttttc 60
ttttgtgttt ttataacatc tgtctcttct tcactcgatc tccctctctt ttctttttca 120
atctccccaa cgaacctccc ttcataactc tctttctctc cccgggaaat atggataact 180
tcttgccctt ttcctcttct aacgcaaact ctgtccaaga actctccatg gatcttaaca 240
agaatcgctc gcacttctcc atggcgcagc ctcagcactt gttgccgcct tactcgtacg 300
ttgcatgtcc ggcacttgat cagacgggga ccatgaatca tcagcctctt cactcatcgg 360
atgcttttcc tcagatcccg gttgtacaaa ccggaggtga attcggctat ttggtttgta 420
agcccggtgt gaggcaggaa cgaggtggat ttcttgatcc acactccact aagatggcta 480
ggatcaacag gaagaaggcg atgctaagat caagaaacaa ctctaaccct aattctagtt 540
cgaatgagtt ggttgattca aggagacaag tggctcttac catgaaaaat aatgccgaga 600
ttgctgctag aaaagatttt tatcgattct cctcattcga taacaagaaa cttagggttt 660
tgttggtgaa gcacttgaag aacagcgatg ttgggtcact tggaaggatt gttctaccaa 720
agagagaagc agaaggaaat cttccggagc tatctgataa agaaggaatg gtattagaga 780
tgagagatgt tgactctgtg cagtcttggt ctttcaaata caagtactgg tccaataaca 840
agagcagaat gtatgtcctc gaaaacacag gagaatttgt gaagaaaaat ggagtattga 900
tgggagacta tctaacaatc tacgaggacg aaagcaagaa tctctacttc tccatcagaa 960
agcacccaca caaacaaaat gatggaagag aggatgagtc gatggaagtt atcgagatga 1020
acttctatga agatataatg tttgattaca taccaaatga tgaagacgat tccattgcaa 1080
tgctcctcgg aaatctaaac gagcactatc cctacccaaa tgatcttatg gatctcactg 1140
tcaatcttga tcagcatcag caagccacct cctcgtcgcc acctgctgat cacatgagct 1200
cgaacgattt cttatggtga tgtgatggac gttgatatgg attccctttg agatgatata 1260
caagggatga aaagaaaaga gtatcatatt catatccata tttgtttgat aaaatgtgtt 1320
tgttcccaat ctattattta tgaaaaactt atttgtgttt aactccagat taattaaata 1380
tttttcattt gac 1393
<210> 140
<211> 1755
<212> ДНК
<213> Arabidopsis thaliana
<400> 140
atgaactcga tgaataactg gttaggcttc tctctctctc ctcatgatca aaatcatcac 60
cgtacggatg ttgactcctc caccaccaga accgccgtag atgttgccgg agggtactgt 120
tttgatctgg ccgctccctc cgatgaatct tctgccgttc aaacatcttt tctttctcct 180
ttcggtgtca ccctcgaagc tttcaccaga gacaataata gtcactcccg agattgggac 240
atcaatggtg gtgcatgcaa taacattaac aataacgaac aaaatggacc aaagcttgag 300
aatttcctcg gccgcaccac cacgatttac aataccaacg agaccgttgt agatggaaat 360
ggcgattgtg gaggaggaga cggtggtggt ggcggctcac taggcctttc gatgataaaa 420
acatggctga gtaatcattc ggttgctaat gctaatcatc aagacaatgg taacggtgca 480
cgaggcttgt ccctctctat gaattcatct actagtgata gcaacaacta caacaacaat 540
gatgatgtcg tccaagagaa gactattgtt gatgtcgtag aaactacacc gaagaaaact 600
attgagagtt ttggacaaag gacgtctata taccgcggtg ttacaaggca tcggtggaca 660
ggtagatacg aggcacattt atgggacaat agttgcaaaa gagaaggcca gactcgcaaa 720
ggaagacaag tttatctggg aggttatgac aaagaagaaa aagcagctag ggcttacgat 780
ttagccgcac taaagtattg gggaaccacc actactacta acttcccctt gagtgaatat 840
gagaaagagg tagaagagat gaagcacatg acgaggcaag agtatgttgc ctctctgcgc 900
aggaaaagta gtggtttctc tcgtggtgca tcgatttatc gaggagtaac aaggcatcac 960
caacatggaa ggtggcaagc taggatcgga agagtcgccg gtaacaaaga cctctacttg 1020
ggaactttcg gcacacagga agaggctgct gaggcttatg acattgcagc cattaaattc 1080
agaggattaa gcgcagtgac taacttcgac atgaacagat acaatgttaa agcaatcctc 1140
gagagcccga gtctacctat tggtagttct gcgaaacgtc tcaaggacgt taataatccg 1200
gttccagcta tgatgattag taataacgtt tcagagagtg caaataatgt tagcggttgg 1260
caaaacactg cgtttcagca tcatcaggga atggatttga gcttattgca gcaacagcag 1320
gagaggtacg ttggttatta caatggagga aacttgtcta ccgagagtac tagggtttgt 1380
ttcaaacaag aggaggaaca acaacacttc ttgagaaact cgccgagtca catgactaat 1440
gttgatcatc atagctcgac ctctgatgat tctgttaccg tttgtggaaa tgttgttagt 1500
tatggtggtt atcaaggatt cgcaatccct gttggaacat cggttaatta cgatcccttt 1560
actgctgctg agattgctta caacgcaaga aatcattatt actatgctca gcatcagcaa 1620
caacagcaga ttcagcagtc gccgggagga gattttccgg tggcgatttc gaataaccat 1680
agctctaaca tgtactttca cggggaaggt ggtggagaag gggctccaac gttttcagtt 1740
tggaacgaca cttag 1755
<210> 141
<211> 2061
<212> ДНК
<213> Medicago truncatula
<400> 141
atgaacttgt taggtttctc tctatctcca caagaacaac atccatcaac acaagatcaa 60
acggtggctt cccgttttgg gttcaaccct aatgaaatct caggctctga tgttcaagga 120
gatcactgct atgatctctc ttctcacaca actcctcatc attcactcaa cctttctcat 180
cctttttcca tttatgaagc tttccacaca aataacaaca ttcacaccac tcaagattgg 240
aaggagaact acaacaacca aaacctacta ttgggaacat catgcatgaa ccaaaatgtg 300
aacaacaaca accaacaagc acaaccaaag ctagaaaact tcctcggtgg acactctttc 360
accgaccatc aagaatacgg tggtagcaac tcatactctt cattacacct cccacctcat 420
cagccggaag catcctgtgg cggtggtgat ggtagtacaa gtaacaataa ctcaataggt 480
ttatctatga taaaaacatg gctcagaaac caaccaccac caccagaaaa caacaacaat 540
aacaacaatg aaagtggtgc acgtgtgcag acactatcac tttctatgag tactggctca 600
cagtcaagtt catctgtgcc tcttctcaat gcaaatgtga tgagtggtga gatttcctca 660
tcggaaaaca aacaaccacc cacaactgca gttgtacttg atagcaacca aacaagtgtc 720
gttgaaagtg ctgtgcctag aaaatccgtt gatacatttg gacaaagaac ttccatttac 780
cgtggtgtaa caaggcatag atggacaggg agatatgaag ctcacctttg ggataatagt 840
tgtagaagag aggggcagac tcgcaaagga aggcaagttt acttgggagg ttatgacaaa 900
gaagaaaaag cagctagagc ctatgatttg gcagcactaa aatattgggg aacaactact 960
acaacaaatt ttccaattag ccattatgaa aaagaagtgg aagaaatgaa gcatatgaca 1020
aggcaagagt acgttgcgtc attgagaagg aaaagtagtg gtttttcacg aggtgcatcc 1080
atttaccgag gagtaacaag acatcatcaa catggtagat ggcaagctag gattggaaga 1140
gttgcaggca acaaagatct ctacctagga actttcagca ctcaagaaga ggcagcagag 1200
gcatatgatg tggcagcaat aaaattcaga ggactgagtg cagttacaaa ctttgacatg 1260
agcagatatg atgtcaaaac catacttgag agcagcacat taccaattgg tggtgctgca 1320
aagcgtttaa aagacatgga gcaagttgaa ttgaatcatg tgaatgttga tattagccat 1380
agaactgaac aagatcatag catcatcaac aacacttccc atttaacaga acaagccatc 1440
tatgcagcaa caaatgcatc taattggcat gcactttcat tccaacatca acaaccacat 1500
catcattaca atgccaacaa catgcagtta cagaattatc cttatggaac tcaaactcaa 1560
aagctttggt gcaaacaaga acaagattct gatgatcata gtacttatac tactgctact 1620
gatattcatc aactacagtt agggaataat aataacaata ctcacaattt ctttggttta 1680
caaaatatca tgagtatgga ttctgcttcc atggataata gttctggatc taattctgtt 1740
gtttatggtg gtggagatca tggtggttat ggaggaaatg gtggatatat gattccaatg 1800
gctattgcaa atgatggtaa ccaaaatcca agaagcaaca acaattttgg tgagagtgag 1860
attaaaggat ttggttatga aaatgttttt gggactacta ctgatcctta tcatgcacag 1920
gcagcaagga acttgtacta tcagccacaa caattatctg ttgatcaagg atcaaattgg 1980
gttccaactg ctattccaac acttgctcca aggactacca atgtctctct atgtcctcct 2040
ttcactttgt tgcatgaata g 2061
<210> 142
<211> 363
<212> Белок
<213> Arabidopsis thaliana
<400> 142
Met Asp Asn Phe Leu Pro Phe Pro Ser Ser Asn Ala Asn Ser Val Gln
1 5 10 15
Glu Leu Ser Met Asp Pro Asn Asn Asn Arg Ser His Phe Thr Thr Val
20 25 30
Pro Thr Tyr Asp His His Gln Ala Gln Pro His His Phe Leu Pro Pro
35 40 45
Phe Ser Tyr Pro Val Glu Gln Met Ala Ala Val Met Asn Pro Gln Pro
50 55 60
Val Tyr Leu Ser Glu Cys Tyr Pro Gln Ile Pro Val Thr Gln Thr Gly
65 70 75 80
Ser Glu Phe Gly Ser Leu Val Gly Asn Pro Cys Leu Trp Gln Glu Arg
85 90 95
Gly Gly Phe Leu Asp Pro Arg Met Thr Lys Met Ala Arg Ile Asn Arg
100 105 110
Lys Asn Ala Met Met Arg Ser Arg Asn Asn Ser Ser Pro Asn Ser Ser
115 120 125
Pro Ser Glu Leu Val Asp Ser Lys Arg Gln Leu Met Met Leu Asn Leu
130 135 140
Lys Asn Asn Val Gln Ile Ser Asp Lys Lys Asp Ser Tyr Gln Gln Ser
145 150 155 160
Thr Phe Asp Asn Lys Lys Leu Arg Val Leu Cys Glu Lys Glu Leu Lys
165 170 175
Asn Ser Asp Val Gly Ser Leu Gly Arg Ile Val Leu Pro Lys Arg Asp
180 185 190
Ala Glu Ala Asn Leu Pro Lys Leu Ser Asp Lys Glu Gly Ile Val Val
195 200 205
Gln Met Arg Asp Val Phe Ser Met Gln Ser Trp Ser Phe Lys Tyr Lys
210 215 220
Phe Trp Ser Asn Asn Lys Ser Arg Met Tyr Val Leu Glu Asn Thr Gly
225 230 235 240
Glu Phe Val Lys Gln Asn Gly Ala Glu Ile Gly Asp Phe Leu Thr Ile
245 250 255
Tyr Glu Asp Glu Ser Lys Asn Leu Tyr Phe Ala Met Asn Gly Asn Ser
260 265 270
Gly Lys Gln Asn Glu Gly Arg Glu Asn Glu Ser Arg Glu Arg Asn His
275 280 285
Tyr Glu Glu Ala Met Leu Asp Tyr Ile Pro Arg Asp Glu Glu Glu Ala
290 295 300
Ser Ile Ala Met Leu Ile Gly Asn Leu Asn Asp His Tyr Pro Ile Pro
305 310 315 320
Asn Asp Leu Met Asp Leu Thr Thr Asp Leu Gln His His Gln Ala Thr
325 330 335
Ser Ser Ser Met Pro Pro Glu Asp His Ala Tyr Val Gly Ser Ser Asp
340 345 350
Asp Gln Val Ser Phe Asn Asp Phe Glu Trp Trp
355 360
<210> 143
<211> 280
<212> Белок
<213> Medicago truncatula
<400> 143
Met Ser Ser Ser Asn Phe Ser Cys Ile Leu Ser Ile Ser Leu Thr Phe
1 5 10 15
Phe Ile Leu Leu Leu Asn Lys Val Asn Ser Ala Glu Thr Thr Ser Phe
20 25 30
Ser Ile Thr Lys Phe Val Pro Asp Gln Lys Asn Leu Ile Phe Gln Gly
35 40 45
Asp Ala Lys Thr Ala Ser Thr Gly Lys Leu Glu Leu Ser Lys Ala Val
50 55 60
Lys Asn Ser Ile Gly Arg Ala Leu Tyr Ser Ala Pro Ile His Ile Trp
65 70 75 80
Asp Ser Lys Thr Gly Ser Val Ala Asn Phe Gln Thr Thr Phe Thr Phe
85 90 95
Thr Ile Thr Ala Pro Asn Thr Tyr Asn Val Ala Asp Gly Leu Ala Phe
100 105 110
Phe Ile Ala Pro Ile Asp Thr Lys Pro Lys Ser Ile His His Gly Gly
115 120 125
Tyr Leu Gly Val Phe Asp Ser Lys Thr Tyr Lys Lys Ser Ile Gln Thr
130 135 140
Val Ala Val Glu Ile Asp Thr Phe Tyr Asn Ala Gln Trp Asp Pro Asn
145 150 155 160
Pro Gly Asn Ile Ser Ser Thr Gly Arg His Ile Gly Ile Asp Val Asn
165 170 175
Ser Ile Lys Ser Ile Ser Thr Val Pro Trp Ser Leu Glu Asn Asn Lys
180 185 190
Lys Ala Asn Val Ala Ile Gly Phe Asn Gly Ala Thr Asn Val Leu Ser
195 200 205
Val Asp Val Glu Tyr Pro Leu Ile Arg His Tyr Thr Leu Ser His Val
210 215 220
Val Pro Leu Lys Asp Val Val Pro Glu Trp Val Arg Ile Gly Phe Ser
225 230 235 240
Ser Ser Thr Gly Ala Glu Tyr Ser Ala His Asp Ile Leu Ser Trp Ser
245 250 255
Phe Asp Ser Lys Leu Asn Leu Gly Phe Glu Asn Asn Ile Asn Ala Asn
260 265 270
Val Ser Ser Ser Thr Gln Ala Ala
275 280
<210> 144
<211> 349
<212> Белок
<213> Brassica napus
<400> 144
Met Asp Asn Phe Leu Pro Phe Ser Ser Ser Asn Ala Asn Ser Val Gln
1 5 10 15
Glu Leu Ser Met Asp Leu Asn Lys Asn Arg Ser His Phe Ser Met Ala
20 25 30
Gln Pro Gln His Leu Leu Pro Pro Tyr Ser Tyr Val Ala Cys Pro Ala
35 40 45
Leu Asp Gln Thr Gly Thr Met Asn His Gln Pro Leu His Ser Ser Asp
50 55 60
Ala Phe Pro Gln Ile Pro Val Val Gln Thr Gly Gly Glu Phe Gly Tyr
65 70 75 80
Leu Val Cys Lys Pro Gly Val Arg Gln Glu Arg Gly Gly Phe Leu Asp
85 90 95
Pro His Ser Thr Lys Met Ala Arg Ile Asn Arg Lys Lys Ala Met Leu
100 105 110
Arg Ser Arg Asn Asn Ser Asn Pro Asn Ser Ser Ser Asn Glu Leu Val
115 120 125
Asp Ser Arg Arg Gln Val Ala Leu Thr Met Lys Asn Asn Ala Glu Ile
130 135 140
Ala Ala Arg Lys Asp Phe Tyr Arg Phe Ser Ser Phe Asp Asn Lys Lys
145 150 155 160
Leu Arg Val Leu Leu Val Lys His Leu Lys Asn Ser Asp Val Gly Ser
165 170 175
Leu Gly Arg Ile Val Leu Pro Lys Arg Glu Ala Glu Gly Asn Leu Pro
180 185 190
Glu Leu Ser Asp Lys Glu Gly Met Val Leu Glu Met Arg Asp Val Asp
195 200 205
Ser Val Gln Ser Trp Ser Phe Lys Tyr Lys Tyr Trp Ser Asn Asn Lys
210 215 220
Ser Arg Met Tyr Val Leu Glu Asn Thr Gly Glu Phe Val Lys Lys Asn
225 230 235 240
Gly Val Leu Met Gly Asp Tyr Leu Thr Ile Tyr Glu Asp Glu Ser Lys
245 250 255
Asn Leu Tyr Phe Ser Ile Arg Lys His Pro His Lys Gln Asn Asp Gly
260 265 270
Arg Glu Asp Glu Ser Met Glu Val Ile Glu Met Asn Phe Tyr Glu Asp
275 280 285
Ile Met Phe Asp Tyr Ile Pro Asn Asp Glu Asp Asp Ser Ile Ala Met
290 295 300
Leu Leu Gly Asn Leu Asn Glu His Tyr Pro Tyr Pro Asn Asp Leu Met
305 310 315 320
Asp Leu Thr Val Asn Leu Asp Gln His Gln Gln Ala Thr Ser Ser Ser
325 330 335
Pro Pro Ala Asp His Met Ser Ser Asn Asp Phe Leu Trp
340 345
<210> 145
<211> 584
<212> Белок
<213> Arabidopsis thaliana
<400> 145
Met Asn Ser Met Asn Asn Trp Leu Gly Phe Ser Leu Ser Pro His Asp
1 5 10 15
Gln Asn His His Arg Thr Asp Val Asp Ser Ser Thr Thr Arg Thr Ala
20 25 30
Val Asp Val Ala Gly Gly Tyr Cys Phe Asp Leu Ala Ala Pro Ser Asp
35 40 45
Glu Ser Ser Ala Val Gln Thr Ser Phe Leu Ser Pro Phe Gly Val Thr
50 55 60
Leu Glu Ala Phe Thr Arg Asp Asn Asn Ser His Ser Arg Asp Trp Asp
65 70 75 80
Ile Asn Gly Gly Ala Cys Asn Thr Leu Thr Asn Asn Glu Gln Asn Gly
85 90 95
Pro Lys Leu Glu Asn Phe Leu Gly Arg Thr Thr Thr Ile Tyr Asn Thr
100 105 110
Asn Glu Thr Val Val Asp Gly Asn Gly Asp Cys Gly Gly Gly Asp Gly
115 120 125
Gly Gly Gly Gly Ser Leu Gly Leu Ser Met Ile Lys Thr Trp Leu Ser
130 135 140
Asn His Ser Val Ala Asn Ala Asn His Gln Asp Asn Gly Asn Gly Ala
145 150 155 160
Arg Gly Leu Ser Leu Ser Met Asn Ser Ser Thr Ser Asp Ser Asn Asn
165 170 175
Tyr Asn Asn Asn Asp Asp Val Val Gln Glu Lys Thr Ile Val Asp Val
180 185 190
Val Glu Thr Thr Pro Lys Lys Thr Ile Glu Ser Phe Gly Gln Arg Thr
195 200 205
Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Tyr Glu
210 215 220
Ala His Leu Trp Asp Asn Ser Cys Lys Arg Glu Gly Gln Thr Arg Lys
225 230 235 240
Gly Arg Gln Val Tyr Leu Gly Gly Tyr Asp Lys Glu Glu Lys Ala Ala
245 250 255
Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Thr Thr Thr
260 265 270
Thr Asn Phe Pro Leu Ser Glu Tyr Glu Lys Glu Val Glu Glu Met Lys
275 280 285
His Met Thr Arg Gln Glu Tyr Val Ala Ser Leu Arg Arg Lys Ser Ser
290 295 300
Gly Phe Ser Arg Gly Ala Ser Ile Tyr Arg Gly Val Thr Arg His His
305 310 315 320
Gln His Gly Arg Trp Gln Ala Arg Ile Gly Arg Val Ala Gly Asn Lys
325 330 335
Asp Leu Tyr Leu Gly Thr Phe Gly Thr Gln Glu Glu Ala Ala Glu Ala
340 345 350
Tyr Asp Ile Ala Ala Ile Lys Phe Arg Gly Leu Ser Ala Val Thr Asn
355 360 365
Phe Asp Met Asn Arg Tyr Asn Val Lys Ala Ile Leu Glu Ser Pro Ser
370 375 380
Leu Pro Ile Gly Ser Ser Ala Lys Arg Leu Lys Asp Val Asn Asn Pro
385 390 395 400
Val Pro Ala Met Met Ile Ser Asn Asn Val Ser Glu Ser Ala Asn Asn
405 410 415
Val Ser Gly Trp Gln Asn Thr Ala Phe Gln His His Gln Gly Met Asp
420 425 430
Leu Ser Leu Leu Gln Gln Gln Gln Glu Arg Tyr Val Gly Tyr Tyr Asn
435 440 445
Gly Gly Asn Leu Ser Thr Glu Ser Thr Arg Val Cys Phe Lys Gln Glu
450 455 460
Glu Glu Gln Gln His Phe Leu Arg Asn Ser Pro Ser His Met Thr Asn
465 470 475 480
Val Asp His His Ser Ser Thr Ser Asp Asp Ser Val Thr Val Cys Gly
485 490 495
Asn Val Val Ser Tyr Gly Gly Tyr Gln Gly Phe Ala Ile Pro Val Gly
500 505 510
Thr Ser Val Asn Tyr Asp Pro Phe Thr Ala Ala Glu Ile Ala Tyr Asn
515 520 525
Ala Arg Asn His Tyr Tyr Tyr Ala Gln His Gln Gln Gln Gln Gln Ile
530 535 540
Gln Gln Ser Pro Gly Gly Asp Phe Pro Val Ala Ile Ser Asn Asn His
545 550 555 560
Ser Ser Asn Met Tyr Phe His Gly Glu Gly Gly Gly Glu Gly Ala Pro
565 570 575
Thr Phe Ser Val Trp Asn Asp Thr
580
<210> 146
<211> 686
<212> Белок
<213> Medicago truncatula
<400> 146
Met Asn Leu Leu Gly Phe Ser Leu Ser Pro Gln Glu Gln His Pro Ser
1 5 10 15
Thr Gln Asp Gln Thr Val Ala Ser Arg Phe Gly Phe Asn Pro Asn Glu
20 25 30
Ile Ser Gly Ser Asp Val Gln Gly Asp His Cys Tyr Asp Leu Ser Ser
35 40 45
His Thr Thr Pro His His Ser Leu Asn Leu Ser His Pro Phe Ser Ile
50 55 60
Tyr Glu Ala Phe His Thr Asn Asn Asn Ile His Thr Thr Gln Asp Trp
65 70 75 80
Lys Glu Asn Tyr Asn Asn Gln Asn Leu Leu Leu Gly Thr Ser Cys Met
85 90 95
Asn Gln Asn Val Asn Asn Asn Asn Gln Gln Ala Gln Pro Lys Leu Glu
100 105 110
Asn Phe Leu Gly Gly His Ser Phe Thr Asp His Gln Glu Tyr Gly Gly
115 120 125
Ser Asn Ser Tyr Ser Ser Leu His Leu Pro Pro His Gln Pro Glu Ala
130 135 140
Ser Cys Gly Gly Gly Asp Gly Ser Thr Ser Asn Asn Asn Ser Ile Gly
145 150 155 160
Leu Ser Met Ile Lys Thr Trp Leu Arg Asn Gln Pro Pro Pro Pro Glu
165 170 175
Asn Asn Asn Asn Asn Asn Asn Glu Ser Gly Ala Arg Val Gln Thr Leu
180 185 190
Ser Leu Ser Met Ser Thr Gly Ser Gln Ser Ser Ser Ser Val Pro Leu
195 200 205
Leu Asn Ala Asn Val Met Ser Gly Glu Ile Ser Ser Ser Glu Asn Lys
210 215 220
Gln Pro Pro Thr Thr Ala Val Val Leu Asp Ser Asn Gln Thr Ser Val
225 230 235 240
Val Glu Ser Ala Val Pro Arg Lys Ser Val Asp Thr Phe Gly Gln Arg
245 250 255
Thr Ser Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Tyr
260 265 270
Glu Ala His Leu Trp Asp Asn Ser Cys Arg Arg Glu Gly Gln Thr Arg
275 280 285
Lys Gly Arg Gln Val Tyr Leu Gly Gly Tyr Asp Lys Glu Glu Lys Ala
290 295 300
Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Thr Thr Thr
305 310 315 320
Thr Thr Asn Phe Pro Ile Ser His Tyr Glu Lys Glu Val Glu Glu Met
325 330 335
Lys His Met Thr Arg Gln Glu Tyr Val Ala Ser Leu Arg Arg Lys Ser
340 345 350
Ser Gly Phe Ser Arg Gly Ala Ser Ile Tyr Arg Gly Val Thr Arg His
355 360 365
His Gln His Gly Arg Trp Gln Ala Arg Ile Gly Arg Val Ala Gly Asn
370 375 380
Lys Asp Leu Tyr Leu Gly Thr Phe Ser Thr Gln Glu Glu Ala Ala Glu
385 390 395 400
Ala Tyr Asp Val Ala Ala Ile Lys Phe Arg Gly Leu Ser Ala Val Thr
405 410 415
Asn Phe Asp Met Ser Arg Tyr Asp Val Lys Thr Ile Leu Glu Ser Ser
420 425 430
Thr Leu Pro Ile Gly Gly Ala Ala Lys Arg Leu Lys Asp Met Glu Gln
435 440 445
Val Glu Leu Asn His Val Asn Val Asp Ile Ser His Arg Thr Glu Gln
450 455 460
Asp His Ser Ile Ile Asn Asn Thr Ser His Leu Thr Glu Gln Ala Ile
465 470 475 480
Tyr Ala Ala Thr Asn Ala Ser Asn Trp His Ala Leu Ser Phe Gln His
485 490 495
Gln Gln Pro His His His Tyr Asn Ala Asn Asn Met Gln Leu Gln Asn
500 505 510
Tyr Pro Tyr Gly Thr Gln Thr Gln Lys Leu Trp Cys Lys Gln Glu Gln
515 520 525
Asp Ser Asp Asp His Ser Thr Tyr Thr Thr Ala Thr Asp Ile His Gln
530 535 540
Leu Gln Leu Gly Asn Asn Asn Asn Asn Thr His Asn Phe Phe Gly Leu
545 550 555 560
Gln Asn Ile Met Ser Met Asp Ser Ala Ser Met Asp Asn Ser Ser Gly
565 570 575
Ser Asn Ser Val Val Tyr Gly Gly Gly Asp His Gly Gly Tyr Gly Gly
580 585 590
Asn Gly Gly Tyr Met Ile Pro Met Ala Ile Ala Asn Asp Gly Asn Gln
595 600 605
Asn Pro Arg Ser Asn Asn Asn Phe Gly Glu Ser Glu Ile Lys Gly Phe
610 615 620
Gly Tyr Glu Asn Val Phe Gly Thr Thr Thr Asp Pro Tyr His Ala Gln
625 630 635 640
Ala Ala Arg Asn Leu Tyr Tyr Gln Pro Gln Gln Leu Ser Val Asp Gln
645 650 655
Gly Ser Asn Trp Val Pro Thr Ala Ile Pro Thr Leu Ala Pro Arg Thr
660 665 670
Thr Asn Val Ser Leu Cys Pro Pro Phe Thr Leu Leu His Glu
675 680 685
<210> 147
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> индуцибельный промотор
<400> 147
tcgatagttg tgatagttcc cacttgtccg tccgcatcgg catccgcagc tcgggatagt 60
tccgacctag gattggatgc atgcggaacc gcacgagggc ggggcggaaa ttgacacacc 120
actcctctcc acgcaccgtt caagaggtac gcgtatagag ccgtatagag cagagacgga 180
gcactttctg gtactgtccg cacgggatgt ccgcacggag agccacaaac gagcggggcc 240
ccgtacgtgc tctcctaccc caggatcgca tccccgcata gctgaacatc tatataaaga 300
cccccaaggt tctcagtctc accaacatca tcaacc 336
<210> 148
<211> 2466
<212> ДНК
<213> Искусственная последовательность
<220>
<223> индуктор
<400> 148
atggccgaca ctagaagaag gcagaaccac tcttgtgacc catgccgtaa gggcaagaga 60
agatgtgatg ctccagagaa ccgtaacgag gctaatgaga acggatgggt gtcatgctct 120
aactgcaaga ggtggaacaa ggactgcacc ttcaactggc ttagctccca aaggtctaag 180
gctaagggtg ctgctccaag agctaggact aagaaggcta ggactgctac tactacctcc 240
gagccttcta cttccgctgc tactattcca actcccgagt ccgataatca cgatgctcca 300
ccagtgatca actcccacga tgctttgcca tcttggactc agggacttct ttctcaccct 360
ggcgatctct tcgacttctc ccattctgct attccagcta acgctgagga tgctgctaac 420
gtgcaatctg atgctccatt cccatgggat cttgctatcc caggcgattt ctctatggga 480
cagcaacttg agaagcccct ctccccattg tctttccagg ctgttcttct tccaccacac 540
tccccaaaca ctgatgatct cattcgtgag cttgaggaac agactaccga tccagattcc 600
gtgactgaca ctaactccgt tcagcaagtt gctcaggatg gctctctttg gtctgatagg 660
cagtctccac tcctcccaga aaacagtttg tgcatggctt ccgactctac cgctagaagg 720
tatgctaggt ccaccatgac caagaacctc atgaggatct accacgactc catggaaaac 780
gccctttctt gctggcttac tgagcacaac tgcccatact ccgaccagat ttcttacctc 840
ccaccaaagc aaagggctga gtggggacca aattggtcta acaggatgtg cattagggtg 900
tgcaggctcg atagggtgtc aacttctctt agaggaaggg ctctctccgc tgaagaagat 960
aaggctgctg ctagggcact tcaccttgct attgtggctt tcgcttctca gtggactcaa 1020
catgctcaaa ggggagctgg acttaacgtc ccagctgata ttgctgctga cgagcgttct 1080
attaggcgta acgcttggaa tgaggctagg catgcacttc agcacactac tggaatccca 1140
tccttcaggg tgatcttcgc caacatcatc ttcagcctca ctcagtccgt gctcgatgat 1200
gatgagcaac atggaatggg agctaggctc gataagcttc tcgagaatga tggtgctcca 1260
gtgttcctcg agactgctaa taggcagctc tacaccttca ggcacaagtt cgctaggatg 1320
cagagaaggg gtaaggcttt caataggctt cctggtggat ccgtggcttc tactttcgct 1380
ggaattttcg agactcccac cccctcatct gagtctccac aacttgatcc agtggtggct 1440
tctgaggaac acaggtctac tctgtctctc atgttctggc tcgggatcat gttcgacact 1500
ctgtctgctg ctatgtacca gaggccactt gttgtgtccg atgaggactc ccagatctct 1560
tctgcttctc caccaagaag aggtgccgag actcctatta accttgattg ctgggagcca 1620
ccaaggcagg tcccatctaa tcaagagaag tctgatgtgt ggggcgacct gttccttagg 1680
acttctgatt ctttgcccga ccacgagtcc cacactcaaa tttctcaacc agctgctagg 1740
tggccatgca cttatgaaca agctgctgct gctctctcct ctgctactcc tgttaaggtg 1800
ttgctttaca ggcgtgtgac tcagctccag actttgttgt ataggggagc ttctccagct 1860
aggcttgagg ctgctattca gaggactctc tacgtgtaca accactggac tgctaagtac 1920
cagccattca tgcaggattg cgttgccaac catgagcttc tcccatccag gatccagtct 1980
tggtacgtga tccttgatgg acactggcac cttgctgcta tgcttttggc tgatgtgctc 2040
gagtccatcg acagggattc ctactccgat atcaaccaca tcgacctcgt gactaagctc 2100
aggcttgata acgctcttgc tgtgtctgct ctcgctaggt catctcttag aggccaagaa 2160
ctcgatccag gcaaggcttc tccaatgtac aggcacttcc acgactccct tactgaggtt 2220
gcattccttg ttgagccatg gactgtggtg ctcatccact catttgctaa ggctgcttac 2280
atcctcctcg attgccttga tcttgatggt cagggaaacg ctctcgctgg ataccttcaa 2340
cttaggcaga actgcaacta ctgcatcagg gctctccagt tccttggccg taagtctgat 2400
atggctgctc tcgtggctaa ggatcttgag aggggactca acggaaaggt cgacagcttc 2460
ctctaa 2466
<210> 149
<211> 208
<212> Белок
<213> Arabidopsis thaliana
<400> 149
Met Thr Ser Ser Val Ile Val Ala Gly Ala Gly Asp Lys Asn Asn Gly
1 5 10 15
Ile Val Val Gln Gln Gln Pro Pro Cys Val Ala Arg Glu Gln Asp Gln
20 25 30
Tyr Met Pro Ile Ala Asn Val Ile Arg Ile Met Arg Lys Thr Leu Pro
35 40 45
Ser His Ala Lys Ile Ser Asp Asp Ala Lys Glu Thr Ile Gln Glu Cys
50 55 60
Val Ser Glu Tyr Ile Ser Phe Val Thr Gly Glu Ala Asn Glu Arg Cys
65 70 75 80
Gln Arg Glu Gln Arg Lys Thr Ile Thr Ala Glu Asp Ile Leu Trp Ala
85 90 95
Met Ser Lys Leu Gly Phe Asp Asn Tyr Val Asp Pro Leu Thr Val Phe
100 105 110
Ile Asn Arg Tyr Arg Glu Ile Glu Thr Asp Arg Gly Ser Ala Leu Arg
115 120 125
Gly Glu Pro Pro Ser Leu Arg Gln Thr Tyr Gly Gly Asn Gly Ile Gly
130 135 140
Phe His Gly Pro Ser His Gly Leu Pro Pro Pro Gly Pro Tyr Gly Tyr
145 150 155 160
Gly Met Leu Asp Gln Ser Met Val Met Gly Gly Gly Arg Tyr Tyr Gln
165 170 175
Asn Gly Ser Ser Gly Gln Asp Glu Ser Ser Val Gly Gly Gly Ser Ser
180 185 190
Ser Ser Ile Asn Gly Met Pro Ala Phe Asp His Tyr Gly Gln Tyr Lys
195 200 205
<210> 150
<211> 236
<212> Белок
<213> Arabidopsis lyrata
<400> 150
Met Glu Arg Gly Ala Pro Phe Ser His Tyr Gln Leu Pro Lys Ser Ile
1 5 10 15
Ser Glu Leu Asn Leu Asp Gln His Ser Asn Pro Asn Pro Met Thr Ser
20 25 30
Ser Val Val Val Ala Asp Ala Ser Asp Asn Asn Lys Gly Ile Val Ala
35 40 45
Gln Gln Gln Pro Pro Cys Met Ala Arg Glu Gln Asp Gln Tyr Met Pro
50 55 60
Ile Ala Asn Val Ile Arg Ile Met Arg Lys Ile Leu Pro Ser His Ala
65 70 75 80
Lys Ile Ser Asp Asp Ala Lys Glu Thr Ile Gln Glu Cys Val Ser Glu
85 90 95
Tyr Ile Ser Phe Val Thr Gly Glu Ala Asn Glu Arg Cys Gln Arg Glu
100 105 110
Gln Arg Lys Thr Ile Thr Ala Glu Asp Ile Leu Trp Ala Met Ser Lys
115 120 125
Leu Gly Phe Asp Asn Tyr Val Asp Pro Leu Thr Val Phe Ile Asn Arg
130 135 140
Tyr Arg Glu Ile Glu Thr Asp Arg Gly Ser Ala Leu Arg Glu Pro Pro
145 150 155 160
Ser Leu Arg Gln Ala Tyr Gly Gly Asn Gly Ile Gly Phe His Gly Pro
165 170 175
Ser His Gly Leu Pro Pro Pro Gly Pro Tyr Gly Tyr Gly Met Leu Asp
180 185 190
Gln Ser Met Val Met Gly Gly Gly Arg Tyr Tyr Gln Asn Gly Ser Ser
195 200 205
Gly Gln Asp Glu Ser Ser Ala Gly Gly Gly Ser Ser Ser Ser Ile Asn
210 215 220
Gly Met Pro Ala Phe Asp Ser Tyr Gly Gln Tyr Lys
225 230 235
<210> 151
<211> 230
<212> Белок
<213> Brassica napus
<400> 151
Met Glu Arg Gly Ala Pro Leu Ser His Tyr Gln Leu Pro Lys Ser Asn
1 5 10 15
Ser Gly Leu Asn Leu Asp Gln His Asn Asn Ser Ile Pro Thr Met Thr
20 25 30
Gly Ser Ile Ser Ala Cys Asp Asp Lys Asn Lys Thr Ile Leu Pro Gln
35 40 45
Gln Gln Pro Ser Met Pro Arg Glu Gln Asp Gln Tyr Met Pro Ile Ala
50 55 60
Asn Val Ile Arg Ile Met Arg Lys Ile Leu Pro Pro His Ala Lys Ile
65 70 75 80
Ser Asp Asp Ala Lys Glu Thr Ile Gln Glu Cys Val Ser Glu Tyr Ile
85 90 95
Ser Phe Val Thr Gly Glu Ala Asn Glu Arg Cys Gln Arg Glu Gln Arg
100 105 110
Lys Thr Ile Thr Ala Glu Asp Ile Leu Trp Ala Met Ser Lys Leu Gly
115 120 125
Phe Asp Asp Tyr Val Gly Pro Leu Asn Val Phe Ile Asn Arg Tyr Arg
130 135 140
Glu Phe Glu Thr Asp Arg Gly Cys Ser Leu Arg Gly Glu Ser Ser Phe
145 150 155 160
Lys Pro Val Tyr Gly Gly Ser Gly Met Gly Phe His Gly Pro Pro Pro
165 170 175
Pro Gly Ser Tyr Gly Tyr Gly Met Leu Asp Gln Ser Met Val Met Gly
180 185 190
Gly Gly Arg Tyr Tyr His Asn Gly Ser Gly Gln Asp Gly Ser Val Ser
195 200 205
Gly Gly Gly Gly Ser Ser Ser Ser Met Asn Gly Met Pro Val Tyr Asp
210 215 220
Gln Tyr Gly Gln Tyr Lys
225 230
<210> 152
<211> 252
<212> Белок
<213> Ricinus communis
<400> 152
Met Glu Arg Gly Gly Arg Val His Arg Tyr Arg Arg His Ala Lys Gln
1 5 10 15
Pro Thr Pro Thr Thr Ser Ala Thr Ala Ser Thr Ser Pro Gly Met Ser
20 25 30
Ser Val Gln Thr Thr Ile Cys Ser Asn Ile Asn Leu Pro Ser Thr Leu
35 40 45
Ser Leu Ser Asn Ser Thr Ala Ala Pro Gln Ala Pro Gln Gln Gln Gln
50 55 60
Leu Gln Pro Ser Gln Cys Leu Val Arg Glu Gln Asp Gln Tyr Met Pro
65 70 75 80
Ile Ala Asn Val Ile Arg Ile Met Arg Arg Ile Leu Pro Pro His Ala
85 90 95
Lys Ile Ser Asp Asp Ala Lys Glu Thr Ile Gln Glu Cys Val Ser Glu
100 105 110
Tyr Ile Ser Phe Ile Thr Gly Glu Ala Asn Asp Arg Cys Gln Arg Glu
115 120 125
Gln Arg Lys Thr Ile Thr Ala Glu Asp Val Leu Trp Ala Met Gly Lys
130 135 140
Leu Gly Phe Asp Asp Tyr Val Glu Pro Leu Thr Leu Phe Leu Asn Arg
145 150 155 160
Tyr Arg Glu Met Glu Asn Glu Arg Ser Thr Ile Arg Asp Pro Ile Leu
165 170 175
Lys Arg Ser Ser Val Gly Val Val Asp Tyr Gly Asn Leu Gly Met Asn
180 185 190
Pro Phe Met Pro Thr Phe Pro Met Ile Pro Pro Pro Gln Gly Tyr Phe
195 200 205
Asp Ser Asn Met Leu Gly Gly Tyr Tyr Arg Asp Ala Pro Asp Gly Ala
210 215 220
Ser Gly Ala Ala Ser Gly Ser Asn Leu Ala Ala Ser Ser Ala Pro Asn
225 230 235 240
Ser Leu Leu His Phe Asp Pro Phe Ala Gln Phe Lys
245 250
<210> 153
<211> 198
<212> Белок
<213> Glycine max
<400> 153
Met Asn Met Asn Met Arg Gln Gln Gln Val Ala Ser Ser Asp Gln Asn
1 5 10 15
Cys Ser Asn His Ser Ala Ala Gly Glu Glu Asn Glu Cys Thr Val Arg
20 25 30
Glu Gln Asp Arg Phe Met Pro Ile Ala Asn Val Ile Arg Ile Met Arg
35 40 45
Lys Ile Leu Pro Pro His Ala Lys Ile Ser Asp Asp Ala Lys Glu Thr
50 55 60
Ile Gln Glu Cys Val Ser Glu Tyr Ile Ser Phe Ile Thr Gly Glu Ala
65 70 75 80
Asn Glu Arg Cys Gln Arg Glu Gln Arg Lys Thr Ile Thr Ala Glu Asp
85 90 95
Val Leu Trp Ala Met Ser Lys Leu Gly Phe Asp Asp Tyr Ile Glu Pro
100 105 110
Leu Thr Met Tyr Leu His Arg Tyr Arg Glu Leu Glu Gly Asp Arg Thr
115 120 125
Ser Met Arg Gly Glu Pro Leu Gly Lys Arg Thr Val Glu Tyr Ala Thr
130 135 140
Leu Ala Thr Ala Phe Val Pro Pro Pro Phe His His His Asn Gly Tyr
145 150 155 160
Phe Gly Ala Ala Met Pro Met Gly Thr Tyr Val Arg Glu Thr Pro Pro
165 170 175
Asn Ala Ala Ser Ser His His His His Gly Ile Ser Asn Ala His Glu
180 185 190
Pro Asn Ala Arg Ser Ile
195
<210> 154
<211> 240
<212> Белок
<213> Medicago truncatula
<400> 154
Met Glu Thr Gly Gly Gly Phe His Gly Tyr Arg Lys Leu Pro Thr Asn
1 5 10 15
Thr Asn Ser Ser Ala Val Ala Gly Thr Leu Lys Leu Ser Ser Val Ser
20 25 30
Glu Met Asn Thr Arg Gln Gln Val Gly Glu Gln Asn Asn Asn Gly Thr
35 40 45
Glu Gln Asp Asn Glu Cys Ile Val Arg Glu Gln Asp Arg Phe Met Pro
50 55 60
Ile Ala Asn Val Ile Arg Ile Met Arg Lys Ile Leu Pro Pro His Ala
65 70 75 80
Lys Ile Ser Asp Asp Ala Lys Glu Thr Ile Gln Glu Cys Val Ser Glu
85 90 95
Tyr Ile Ser Phe Ile Thr Gly Glu Ala Asn Glu Arg Cys Gln Arg Glu
100 105 110
Gln Arg Lys Thr Ile Thr Ala Glu Asp Val Leu Trp Ala Met Ser Lys
115 120 125
Leu Gly Phe Asp Asp Tyr Ile Glu Pro Leu Thr Met Tyr Leu His Arg
130 135 140
Tyr Arg Glu Leu Glu Gly Asp Arg Thr Ser Met Arg Val Glu Pro Leu
145 150 155 160
Gly Lys Arg Gly Met Glu Tyr Gly Asn Leu Gly Gly Phe Val Pro Gln
165 170 175
Phe His Ile Gly His Pro Asn Gly Gly Tyr Tyr Gly Asn Ala Ala Pro
180 185 190
Thr Tyr Met Met Arg Asp Gly Asn Asn Asn Asn Asn Asn Asn Asn Asn
195 200 205
Ala Pro Asn Ala Ala Asn Ala Ala Gly Gly Ser Ser His Ser Gln Ala
210 215 220
Leu Ala Asn Ala Glu Ala Asn Gly His His His His His Gln Tyr Lys
225 230 235 240
<210> 155
<211> 278
<212> Белок
<213> Zea mays
<400> 155
Met Asp Ser Ser Ser Phe Leu Pro Ala Ala Gly Ala Glu Asn Gly Ser
1 5 10 15
Ala Ala Gly Gly Ala Asn Asn Gly Gly Ala Ala Gln Gln His Ala Ala
20 25 30
Pro Ala Ile Arg Glu Gln Asp Arg Leu Met Pro Ile Ala Asn Val Ile
35 40 45
Arg Ile Met Arg Arg Val Leu Pro Ala His Ala Lys Ile Ser Asp Asp
50 55 60
Ala Lys Glu Thr Ile Gln Glu Cys Val Ser Glu Tyr Ile Ser Phe Ile
65 70 75 80
Thr Gly Glu Ala Asn Glu Arg Cys Gln Arg Glu Gln Arg Lys Thr Ile
85 90 95
Thr Ala Glu Asp Val Leu Trp Ala Met Ser Arg Leu Gly Phe Asp Asp
100 105 110
Tyr Val Glu Pro Leu Gly Ala Tyr Leu His Arg Tyr Arg Glu Phe Glu
115 120 125
Gly Asp Ala Arg Gly Val Gly Leu Val Pro Gly Ala Ala Pro Ser Arg
130 135 140
Gly Gly Asp His His Pro His Ser Met Ser Pro Ala Ala Met Leu Lys
145 150 155 160
Ser Arg Gly Pro Val Ser Gly Ala Ala Met Leu Pro His His His His
165 170 175
His His Asp Met Gln Met His Ala Ala Met Tyr Gly Gly Thr Ala Val
180 185 190
Pro Pro Pro Ala Gly Pro Pro His His Gly Gly Phe Leu Met Pro His
195 200 205
Pro Gln Gly Ser Ser His Tyr Leu Pro Tyr Ala Tyr Glu Pro Thr Tyr
210 215 220
Gly Gly Glu His Ala Met Ala Ala Tyr Tyr Gly Gly Ala Ala Tyr Ala
225 230 235 240
Pro Gly Asn Gly Gly Ser Gly Asp Gly Ser Gly Ser Gly Gly Gly Gly
245 250 255
Gly Ser Ala Ser His Thr Pro Gln Gly Ser Gly Gly Leu Glu His Pro
260 265 270
His Pro Phe Ala Tyr Lys
275
<210> 156
<211> 225
<212> Белок
<213> Arachis hypogaea
<400> 156
Met Glu Thr Gly Gly Gly Phe His Gly Tyr Arg Asn Leu Pro Thr Thr
1 5 10 15
Thr Ser Gly Leu Lys Leu Ser Val Ser Glu Met Asn Met Arg Ala Val
20 25 30
Glu Asn Asn Thr Gly Ser Ser Asn Asn Asn His Thr Asp Asp Asn Glu
35 40 45
Cys Thr Val Arg Glu Gln Asp Arg Phe Met Pro Ile Ala Asn Val Ile
50 55 60
Arg Ile Met Arg Lys Ile Leu Pro Pro His Ala Lys Ile Ser Asp Asp
65 70 75 80
Ala Lys Glu Thr Ile Gln Glu Cys Val Ser Glu Tyr Ile Ser Phe Ile
85 90 95
Thr Gly Glu Ala Asn Glu Arg Cys Gln Arg Glu Gln Arg Lys Thr Ile
100 105 110
Thr Ala Glu Asp Val Leu Trp Ala Met Ser Lys Leu Gly Phe Asp Asp
115 120 125
Tyr Ile Glu Pro Leu Thr Met Tyr Leu His Arg Tyr Arg Glu Leu Glu
130 135 140
Gly Asp Arg Thr Ser Met Arg Gly Glu Pro Leu Gly Lys Arg Thr Val
145 150 155 160
Asp Tyr Gly Thr Leu Gly Val Ala Ala Ala Ser Thr Phe Val Pro Pro
165 170 175
Phe His Ile Gly His His His His His Pro His Pro Ser Ser Tyr Tyr
180 185 190
Gly Thr Pro Met Gly Asn Tyr Ile Arg Asp Ala Pro Asn Ala Gly Ser
195 200 205
Ser Leu Gln Pro Pro Ser Leu Ala His Ala Glu Pro Asn Thr Gln Tyr
210 215 220
Lys
225
<210> 157
<211> 234
<212> Белок
<213> Arabidopsis thaliana
<400> 157
Met Glu Arg Gly Gly Phe His Gly Tyr Arg Lys Leu Ser Val Asn Asn
1 5 10 15
Thr Thr Pro Ser Pro Pro Gly Leu Ala Ala Asn Phe Leu Met Ala Glu
20 25 30
Gly Ser Met Arg Pro Pro Glu Phe Asn Gln Pro Asn Lys Thr Ser Asn
35 40 45
Gly Gly Glu Glu Glu Cys Thr Val Arg Glu Gln Asp Arg Phe Met Pro
50 55 60
Ile Ala Asn Val Ile Arg Ile Met Arg Arg Ile Leu Pro Ala His Ala
65 70 75 80
Lys Ile Ser Asp Asp Ser Lys Glu Thr Ile Gln Glu Cys Val Ser Glu
85 90 95
Tyr Ile Ser Phe Ile Thr Gly Glu Ala Asn Glu Arg Cys Gln Arg Glu
100 105 110
Gln Arg Lys Thr Ile Thr Ala Glu Asp Val Leu Trp Ala Met Ser Lys
115 120 125
Leu Gly Phe Asp Asp Tyr Ile Glu Pro Leu Thr Leu Tyr Leu His Arg
130 135 140
Tyr Arg Glu Leu Glu Gly Glu Arg Gly Val Ser Cys Ser Ala Gly Ser
145 150 155 160
Val Ser Met Thr Asn Gly Leu Val Val Lys Arg Pro Asn Gly Thr Met
165 170 175
Thr Glu Tyr Gly Ala Tyr Gly Pro Val Pro Gly Ile His Met Ala Gln
180 185 190
Tyr His Tyr Arg His Gln Asn Gly Phe Val Phe Ser Gly Asn Glu Pro
195 200 205
Asn Ser Lys Met Ser Gly Ser Ser Ser Gly Ala Ser Gly Ala Arg Val
210 215 220
Glu Val Phe Pro Thr Gln Gln His Lys Tyr
225 230
<210> 158
<211> 231
<212> Белок
<213> Brassica napus
<400> 158
Met Glu Arg Gly Gly Phe His Gly Tyr Arg Lys Phe Ser Leu Asn Thr
1 5 10 15
Thr Asn Pro Ser Glu Pro Ala Arg Phe Leu Met Ala Glu Gly Ser Met
20 25 30
Gln Leu Ala Glu Pro Asn Gln Thr Asn Lys Thr Ala Asn Gly Gly Glu
35 40 45
Glu Glu Cys Val Val Arg Glu Gln Asp Arg Phe Met Pro Ile Ala Asn
50 55 60
Val Ile Arg Ile Met Arg Arg Ile Leu Pro Ala His Ala Lys Ile Ser
65 70 75 80
Asp Asp Ser Lys Glu Thr Ile Gln Glu Cys Val Ser Glu Tyr Ile Ser
85 90 95
Phe Ile Thr Gly Glu Ala Asn Glu Arg Cys Gln Arg Glu Gln Arg Lys
100 105 110
Thr Ile Thr Ala Glu Asp Val Leu Trp Ala Met Ser Lys Leu Gly Phe
115 120 125
Asp Asp Tyr Ile Glu Pro Leu Thr Leu Tyr Leu His Arg Tyr Arg Glu
130 135 140
Leu Glu Gly Asp Arg Gly Val Gly Tyr Asn Ala Gly Ser Val Gly Met
145 150 155 160
Thr Ser Gly Met Val Val Lys Arg Pro Asn Gly Thr Met Gly Glu Tyr
165 170 175
Gly Ala Tyr Gly Val Val Pro Gly Met His Met Ala Pro Tyr His Tyr
180 185 190
Arg His Gln Asn Gly Tyr Ala Tyr Ser Gly Asn Glu Pro Asp Ser Lys
195 200 205
Met Gly Gly Pro Ser Ser Ala Ala Asn Gly Ser Arg Val Glu Leu Phe
210 215 220
Pro Thr Gln Gln His Lys Tyr
225 230
<210> 159
<211> 216
<212> Белок
<213> Phaseolus coccineus
<400> 159
Met Glu Ser Gly Gly Phe His Gly Tyr Arg Lys Leu Pro Asn Thr Thr
1 5 10 15
Ser Pro Gly Leu Lys Leu Ser Val Ser Asp Met Asn Asn Val Asn Thr
20 25 30
Ser Arg Gln Val Ala Gly Asp Asn Asn His Thr Ala Asp Glu Ser Asn
35 40 45
Glu Cys Thr Val Arg Glu Gln Asp Arg Phe Met Pro Ile Ala Asn Val
50 55 60
Ile Arg Ile Met Arg Lys Ile Leu Pro Pro His Ala Lys Ile Ser Gly
65 70 75 80
Asp Ala Lys Glu Thr Ile Gln Glu Cys Val Ser Glu Tyr Ile Ser Phe
85 90 95
Ile Thr Gly Glu Ala Asn Glu Arg Cys Gln Arg Glu Gln Arg Lys Thr
100 105 110
Ile Thr Ala Glu Asp Val Leu Trp Ala Met Ser Lys Leu Gly Phe Asp
115 120 125
Asp Tyr Met Glu Pro Leu Thr Met Tyr Leu His Arg Tyr Arg Glu Leu
130 135 140
Glu Gly Asp Arg Thr Ser Met Arg Gly Glu Ser Leu Gly Lys Arg Thr
145 150 155 160
Ile Glu Tyr Ala Pro Met Gly Val Gly Val Ala Thr Ala Phe Val Pro
165 170 175
Pro Gln Phe His Pro Asn Gly Tyr Tyr Gly Pro Ala Met Gly Ala Tyr
180 185 190
Val Ala Pro Pro Asn Ala Ala Ser Ser His His His Gly Met Pro Asn
195 200 205
Thr Glu Pro Asn Ala Arg Ser Met
210 215
<210> 160
<211> 312
<212> Белок
<213> Arabidopsis thaliana
<400> 160
Met Val Asp Glu Asn Val Glu Thr Lys Ala Ser Thr Leu Val Ala Ser
1 5 10 15
Val Asp His Gly Phe Gly Ser Gly Ser Gly His Asp His His Gly Leu
20 25 30
Ser Ala Ser Val Pro Leu Leu Gly Val Asn Trp Lys Lys Arg Arg Met
35 40 45
Pro Arg Gln Arg Arg Ser Ser Ser Ser Phe Asn Leu Leu Ser Phe Pro
50 55 60
Pro Pro Met Pro Pro Ile Ser His Val Pro Thr Pro Leu Pro Ala Arg
65 70 75 80
Lys Ile Asp Pro Arg Lys Leu Arg Phe Leu Phe Gln Lys Glu Leu Lys
85 90 95
Asn Ser Asp Val Ser Ser Leu Arg Arg Met Ile Leu Pro Lys Lys Ala
100 105 110
Ala Glu Ala His Leu Pro Ala Leu Glu Cys Lys Glu Gly Ile Pro Ile
115 120 125
Arg Met Glu Asp Leu Asp Gly Phe His Val Trp Thr Phe Lys Tyr Arg
130 135 140
Tyr Trp Pro Asn Asn Asn Ser Arg Met Tyr Val Leu Glu Asn Thr Gly
145 150 155 160
Asp Phe Val Asn Ala His Gly Leu Gln Leu Gly Asp Phe Ile Met Val
165 170 175
Tyr Gln Asp Leu Tyr Ser Asn Asn Tyr Val Ile Gln Ala Arg Lys Ala
180 185 190
Ser Glu Glu Glu Glu Val Asp Val Ile Asn Leu Glu Glu Asp Asp Val
195 200 205
Tyr Thr Asn Leu Thr Arg Ile Glu Asn Thr Val Val Asn Asp Leu Leu
210 215 220
Leu Gln Asp Phe Asn His His Asn Asn Asn Asn Asn Asn Asn Ser Asn
225 230 235 240
Ser Asn Ser Asn Lys Cys Ser Tyr Tyr Tyr Pro Val Ile Asp Asp Val
245 250 255
Thr Thr Asn Thr Glu Ser Phe Val Tyr Asp Thr Thr Ala Leu Thr Ser
260 265 270
Asn Asp Thr Pro Leu Asp Phe Leu Gly Gly His Thr Thr Thr Thr Asn
275 280 285
Asn Tyr Tyr Ser Lys Phe Gly Thr Phe Asp Gly Leu Gly Ser Val Glu
290 295 300
Asn Ile Ser Leu Asp Asp Phe Tyr
305 310
<210> 161
<211> 321
<212> Белок
<213> Brassica napus
<400> 161
Met Met Ala Asp Glu Asn Val Glu Thr Lys Ala Ser Thr Leu Ile Ala
1 5 10 15
Ser Val Gly His Gln Gly His Gly Phe Gly Ser Gly Ser Gly Gly His
20 25 30
His Gly Leu Ser Ala Ser Val Pro Leu Leu Gly Val Asn Ser Lys Lys
35 40 45
Arg Arg Met Pro Arg Gln Arg Arg Ser Ser Ser Ser Phe Asn Leu Leu
50 55 60
Ser Leu Pro Pro Pro Met Pro Leu Ser Pro His Val Pro Thr Pro Leu
65 70 75 80
Ser Ala Arg Lys Ile Asp Pro Arg Lys Leu Arg Phe Leu Phe Gln Lys
85 90 95
Glu Leu Lys Asn Ser Asp Val Ser Ser Leu Arg Arg Met Ile Leu Pro
100 105 110
Lys Lys Ala Ala Glu Ala His Leu Pro Ala Leu Glu Cys Lys Glu Gly
115 120 125
Ile Pro Ile Arg Met Glu Asp Leu Asp Gly Leu His Val Trp Thr Phe
130 135 140
Lys Tyr Arg Tyr Trp Pro Asn Asn Asn Ser Arg Met Tyr Val Leu Glu
145 150 155 160
Asn Thr Gly Asp Phe Val Asn Ala His Gly Leu Gln Leu Gly Asp Phe
165 170 175
Ile Met Val Tyr Leu Asp Leu Asp Ser Asn Asn Tyr Val Ile Gln Ala
180 185 190
Arg Lys Ala Ser Glu Glu Glu Glu Glu Glu Glu Asp Val Thr Ile Ile
195 200 205
Glu Glu Asp Asp Val Tyr Thr Asn Leu Thr Lys Ile Glu Asn Thr Val
210 215 220
Val Asn Asp Leu Leu Ile Gln Asp Phe Asn His His Asn Asp Asn Ser
225 230 235 240
Ser Asn Asn Asn Ser Asn Asn Asn Ile Asn Asn Asn Lys Cys Ser Tyr
245 250 255
Tyr Tyr Pro Val Ile Asp Asp Ile Thr Thr Asn Thr Ala Ser Phe Val
260 265 270
Tyr Asp Thr Thr Thr Leu Thr Ser Asn Asp Ser Pro Leu Asp Phe Leu
275 280 285
Gly Gly His Thr Thr Thr Thr Thr Asn Thr Tyr Tyr Ser Lys Phe Gly
290 295 300
Ser Phe Glu Gly Leu Gly Ser Val Glu Asn Ile Ser Leu Asp Asp Phe
305 310 315 320
Tyr
<210> 162
<211> 314
<212> Белок
<213> Medicago truncatula
<400> 162
Met Met Met Asp Glu Gly Glu Gly Lys Lys Lys Val Val Val Gln Lys
1 5 10 15
Thr Glu Ala Cys Gly Phe Met Ala Gly Val Glu Asp Glu Leu Gly Phe
20 25 30
Val Asn Val Lys Gly Asp Asn Asn Asn Gly Ser Gly Gln Arg Ile His
35 40 45
His Asp His Gly Phe Val Ala Ala Ala Phe Gly Thr Val His Arg Lys
50 55 60
Lys Arg Met Ala Arg Gln Arg Arg Ser Ser Ser Ser Thr Ile Thr Ile
65 70 75 80
His Leu Lys Asn Leu Pro Ser Ser Thr Thr Thr Thr Thr Thr Thr Thr
85 90 95
Thr Ser His Val Pro Ile Ser Pro Ile Pro Pro Leu Phe His Ser Leu
100 105 110
Pro Pro Ala Arg Glu Ile Asp His Arg Arg Leu Arg Phe Leu Phe Gln
115 120 125
Lys Glu Leu Lys Asn Ser Asp Val Ser Ser Leu Arg Arg Met Val Leu
130 135 140
Pro Lys Lys Ala Ala Glu Ala Phe Leu Pro Val Leu Glu Ser Lys Glu
145 150 155 160
Gly Ile Leu Leu Ser Met Asp Asp Leu Asp Gly Leu His Val Trp Ser
165 170 175
Phe Lys Tyr Arg Phe Trp Pro Asn Asn Asn Ser Arg Met Tyr Val Leu
180 185 190
Glu Asn Thr Gly Asp Phe Val Ser Thr His Gly Leu Arg Phe Gly Asp
195 200 205
Ser Ile Met Val Tyr Gln Asp Asn Gln Asn His Asn Tyr Val Ile Gln
210 215 220
Ala Lys Lys Ala Cys Asp Gln Asp Glu Tyr Met Glu Glu Ala Asn Asp
225 230 235 240
Thr Ile Asn His Ile Phe Val Asp Asp Tyr Glu Val Asn Lys Ser Cys
245 250 255
Phe Asp Val Ala Tyr Pro Ala Met Asn Asp Thr Ser Met Ser Phe Ile
260 265 270
Tyr Asp Thr Thr Ile Ser Asn Asp Ser Pro Leu Asp Phe Leu Gly Gly
275 280 285
Ser Met Thr Asn Tyr Ser Arg Ile Gly Ser Val Glu Thr Phe Gly Ser
290 295 300
Val Glu Asn Leu Ser Leu Asp Asp Phe Tyr
305 310
<210> 163
<211> 3275
<212> ДНК
<213> Arabidopsis thaliana
<400> 163
ggttggctat atggtccaaa ttttgatttg caatatgaga ttgcacagag agaacaatct 60
ttcattatga ttaattattg tacaagtaac aaacaccaat ctccgatata ctttggctct 120
ttagcacatt gttatgctag aagttagcgg aaatctatat gttgttaaac gcagcgttta 180
aattgaacag tgtaatttac cttgaaattt taagactaca tgctgtttag aatttcagat 240
gaaaacatct tgatgtttta gaaatccacg tgggaatagc gtaaaatctt atccaacgaa 300
cttattttgg ttttgttgta tttgtgcaag tcgtcacgct aatcgaaaaa agaaaagaaa 360
aaaagaagcc gtcatgatcg gccatttctc ggccgagtct gagtctgact ctgcgtccgt 420
gtcaccatta tcagatcgag cctgtcttat ctcgttgcga ttccctatgc aaaaatcttc 480
ttcttttttt tattccccca tttatctctg atctcttctc tcttctcaag taaacctctc 540
tgcttcacgt ctcttctttt cttgtcgatt ttccccagat aatcagttga aaacacaccc 600
aaattcatct tcgaatcaat aatggatata agtaatgagg ctagtgtcga tcccttttcg 660
attggaccat catctatcat gggtcgaacc attgctttca gagtcttgtt ctgtagatca 720
atgtcacagc ttaggcgtga tctctttcgg ttcttgttgc attggtttct tagatttaag 780
ctgaccgttt caccgtttgt gtcgtggttt catcctcgga accctcaagg gattttagcg 840
gtggttacaa tcattgcctt tgtgttgaaa cgatacacga atgtgaaaat aaaggcggaa 900
atggcttacc ggaggaagtt ttggaggaat atgatgcgga cggctttgac ttatgaggaa 960
tgggctcatg ctgctaagat gttagagaag gaaacaccaa agatgaatga atctgatctt 1020
tatgatgaag agttggttaa gaacaagctt caggagcttc gtcatcgtcg ccaagaaggc 1080
tcacttagag acattatgtt ttgtatgaga gctgatttgg tgaggaatct cggtaatatg 1140
tgtaattcgg agcttcataa aggtagactt caggttccta gacatatcaa agagtacatt 1200
gatgaggtgt ctactcagtt gagaatggtt tgtaactctg attcagagga gctttcttta 1260
gaagagaagc tttcttttat gcatgaaaca cggcatgcct ttggtagaac ggctttgctt 1320
ttgagtggtg gggcttctct tggtgcgttt catgttggtg tggttaggac tttggttgag 1380
cataagcttt tacctcgaat aattgctggt tctagtgttg gatccatcat ttgtgctgtt 1440
gtggcctcaa ggtcttggcc agaactacag agtttctttg agaattcttt gcattcttta 1500
cagttctttg atcagctcgg aggcgtgttc tcaatagtga aacgggtaat gacacaaggg 1560
gctctacacg atatcagaca gttgcaatgt atgcttagaa acctcacaag caatctcaca 1620
ttccaagaag cttatgacat gacaggaagg attctcggga tcaccgtttg ctccccaaga 1680
aagcatgaac ctcctcggtg tcttaactat ttgacttcgc ctcatgtggt tatatggagc 1740
gcagtgactg cttcttgtgc ttttcctggt ctctttgaag ctcaagagct aatggctaaa 1800
gatcgaagtg gagagatcgt accgtatcat ccacctttca atttggatcc agaagtaggc 1860
actaaatcat catctggacg ccggtggaga gatggtagtt tggaggttga tttaccaatg 1920
atgcagctta aagaactgtt caatgtcaat cattttattg tgagccaagc caatcctcac 1980
attgctccat tactgcgtct aaaggattta gttcgagctt atggtggtag attcgcagct 2040
aagctcgcgc atctagtgga gatggaggtc aaacatagat gcaaccaggt attagagctc 2100
ggttttcctc tcggtggact cgcaaagctt tttgctcagg agtgggaagg tgatgttaca 2160
gttgtaatgc ctgctactct tgctcagtac tcgaagatta tacaaaatcc gactcatgtc 2220
gagcttcaga aagcggctaa ccaaggaaga agatgcactt gggagaagct ctcagccata 2280
aaatcaaact gcgggatcga gcttgcgctt gatgattctg tagctattct taaccatatg 2340
cggaggctca agaaaagtgc ggagagagcc gccactgcca cgtcttcgtc tcatcacgga 2400
ttggcttcaa ccaccagatt caatgcttca agaagaatcc catcttggaa cgtccttgcc 2460
agagagaact caacaggctc actggatgat ctagtcactg acaataacct ccacgcttct 2520
tcgggcagga atttaagcga cagtgaaaca gagagcgtgg agttgagttc ttggacaaga 2580
actggtggac ctttaatgag aacagcttct gctaataagt tcattgattt tgttcagagt 2640
cttgatatcg acattgcatt ggtcagagga tttagtagca gtcccaattc tccagcagtt 2700
cctcctggtg gctcgtttac tccaagcccg agatccatag cggctcattc ggatatcgaa 2760
tcaaacagca atagcaacaa tcttggaaca agcacttcaa gcataacagt tactgaaggt 2820
gatcttctac agcctgagag aacgagtaac ggatttgtgt taaacgtcgt taaaagagag 2880
aacttgggaa tgccatcgat tgggaaccaa aatacagagt taccagagag tgtacagctc 2940
gatataccgg agaaggagat ggattgtagc tctgtatcag aacacgaaga agatgataac 3000
gacaatgaag aagaacataa cggctcgagt ctggttactg tttcttcaga agattccggt 3060
ttacaagaac cggtgtctgg tagtgttata gatgcttaga gtgtgattga ttcaagtgag 3120
tatagattct taattaaatt tgcagagttt ccaaagggtt tagtgcacca cttgtgtatg 3180
tttgtattgc ttattgtttg aaattcattt gtgaaatcga aatatatctg taaattcaga 3240
aaatattctc tcatccatta caaaatattt gagtc 3275
<210> 164
<211> 2795
<212> ДНК
<213> Brassica napus
<400> 164
tcccacgctc aggttctaat tgcaaaaaag gatcatactt tccttattaa aatcaatttc 60
ctgtgcttga tttctatctt aggaagctcg tagtagtttc tctgatagtg aatttgatga 120
aacaaagaaa aaaatgctga cttggtctca gattctaatt aaacacacac acacacataa 180
cctccaatgg atataagcaa cgaggccaat gtcgatccct tctcaatcgg accaacctcc 240
atcctcggcc gaaccatcgc cttcagagtc ctcttctgca aatcaatgct ccagctccgc 300
cgcgacctct tccgcttcct cctccactgg ttcctcacac tcaagctcgc cgtctccccc 360
tttgtctcct ggttccaccc ccggaacccc caggggatcc tcgccgtcgt cacgatcatc 420
gccttcgtcc tgaaacgcta caccaacgtg aaggccaagg ccgagatggc ctaccgtaga 480
aagttctgga ggaacatgat gcgcgcggcg ttgacttacg aggaatgggc tcacgccgct 540
aagatgttgg ataaagagac tccgaagatg aacgagtccg atctttacga tgaagagttg 600
gttaagaaca agctaatgga gcttcgtcat cgacgtcatg agggctctct tagagacatt 660
attttctgta tgagagctga tcttgtgaga aatctcggta atatgtgtaa ccctgagctt 720
cacaagggaa ggcttcacgt gccgagactc atcaaagagt atatcgatga ggtctctaca 780
cagcttagga tggtttgcga catggacact gaagagcttt ctctggagga gaaactttct 840
tttatgcatg agaccagaca cgcgtatgga agaacagctc tacttctcag tggaggagct 900
tctcttgggg ctttccatct tggtgtggtc aagacgcttg tggaacataa gctattgcca 960
agaattatag ctggttcaag cgtggggtct gtaatgtgtg cggttgtggg gacaaggtca 1020
tggcccgagt tgcagagctt ctttgaaggg tcctggcatg ctctgcagtt ctttgatcag 1080
atgggaggaa ttttcactac tgtgaagcgg gttatgactc aaggcgcagt ccatgagatc 1140
cggcatctgc aatggaagtt gaggaatctc accaacaatc tcacagtccg gaatttccgg 1200
gtcgacgact tcgaggatac tcgggataac ggtttgctca ccgacgaagc actagccgcc 1260
tcggtgctta actatctcac ttctcctcac gtggtgatat ggagcgcggt gactgcttct 1320
tgcgctttcc ctggtttgtt tgaagctcag gagctgatgg ctaaagatag gagtggggag 1380
atagtgccgt atcatccgcc ttttaatttg gaaccggagg aaggtgggga taagtcgtct 1440
acgaggaggt ggagagatgg gagtttggag gttgatttgc cgatgatgca gcttaaggag 1500
ctgtttaatg ttaatcattt tattgtgagc caggctaatc ctcacattgc tccgttgctg 1560
cgtttgaagg atatagttag agcttatgga ggtcgatttg cagcaaagct cgcgcaactc 1620
gcggagatgg aagtgaagca tagatgtaat caagtactag aactcgggct tcctctaaga 1680
gaagtagctt cactatttgc tcaagaatgg gaaggcgatg tcacaattgt catgccagct 1740
actttttctc agtacttgaa gatcatacaa gtcgacgatt tcgtcgagct tcaaaaagcc 1800
gctaaccaag gaaggagatg cacttgggag aagctatcag ccataaaagc aaactgtggg 1860
atcgagcttg cgcttgatga gtgtgtaact aatcttaacc atatgcgtag gctcaacaga 1920
agcgctgaga gagccgctgc tgctgctggc acgtcctcct cgtctcatca cggattagct 1980
tcaacgacaa gattcaatgc ttctagaaga atcccgtctt ggaacgtcat cgctagagag 2040
aactcaactg gctcactgga cgacctcgtc actgacagta acaataataa tctccacgcg 2100
gggaggaacc taagcgacag cgaaacggag agcgtggaga tgagttcttg gacgaggact 2160
ggtggaccgt tgatgagaac agcttctgct aataggttca ctgactttgt ccatggtctt 2220
gacgtggaca ttgcgttgac aagagggttt actagcagcc ctaactctcc agcggttcct 2280
ggcccggtta gtccgagttt tagtccaaga tcgagatcct tggcggctca atccgagagc 2340
gaatctgaca agagggaaag tagcaacagt tctagtatat cagctactga aggtgatctt 2400
ctgcagcctg agagaacgag taacggtttt gttttgaacg ttgttagaag agagaacttg 2460
gggatgcctg tggagaacca gagcggtgag ctgccggaga gtgtacagat agatatacct 2520
gagagggaga tggataatag ctctgtctca ggacatgaag atgataatga tgataatgat 2580
gatgaagaag aagaacataa gggctcggtt ccggttaaag attccggttt acaagattct 2640
tgtagtgtaa tagatgctta gactgatttg atccgagtga agagattctt gttcagcaaa 2700
gatcttggag tgttttagtg ctttgtaaat agtacaacta taggccgcaa gtaaggtgca 2760
tgttgtgtat gtttgcagtg attatgttga aaatt 2795
<210> 165
<211> 2670
<212> ДНК
<213> Brachypodium distachyon
<400> 165
atggaagaat ccggagaagc gagtattggg gccttcagga tcgggccgtc gacgcttctc 60
ggccgcggcg tcgcgcttcg cgtgctcctg ttcagctcgc tctggcgtct ccgggcgcgg 120
gcgcgcgccg ctgtgtcgcg cgtgcgcagg gccacgctgc caatggccgc gtcctggctt 180
cacctcagga acacccatgg cgtcctcctg attctcgtgc tcttcgggtt gctcctcagg 240
aagctctccg gtgcgcggtc gcggctggcg ctggcgcgcc ggcgtaggct gtgcaagagc 300
gcgatgcgct acgcggcgac gtatgagcag tgggtgcgtg ccgccaaggt gctcgacaga 360
atgtctgagc aggtgaacga gtctgatttt tacgacgagg agctgatcaa gagtaggctt 420
gaggagctcc ggaggcggag ggaggaaggg tcgctccggg atgtggtgtt ctgtatgcgc 480
ggcgatctcg tgaggaactt ggggaacatg tgtaatcctg agcttcataa gggcaggctc 540
gaggtgccca ggctgataaa agatttcatt gatgaggttt caactcagct gaaaatggtg 600
tgtgaatctg acaccgatgc gttatttttg gaagagaagc ttgcctttgt tcaggaaacc 660
aggcatgcct atgggaggac agcactactc ttaagtgggg gcgcttcact gggctctttc 720
catgtaggtg tagtgaaaac attagttgag cataagcttc tgcctcggat aatagcaggg 780
tctagcgttg gttccattat atgttcaatt gttgctactc gaacatggcc tgagattgag 840
agcttcttca tagactcatt acaaatctta cagttcttcg gtaggatagg tggaattttt 900
gctgtgacca aacgggttat gacttatggt gcacttcatg acattagcca gatgcaaagg 960
cttttgaggg atctcacaag taacttaaca tttcaagagg cttacgatat aactggccgt 1020
gttcttgggg tcactgtttg ctctcccaga aaaaacgagc cacctcgctg cctcaactac 1080
ctgacatcac cacatgttgt tatctggagt gctgtaactg cttcgtgtgc attccctggc 1140
ctctttgaag ctcaggaatt gatggcaaag gataggtttg gccacatagt tcccttccat 1200
gcgccctttt ccacagatcc agaacaaggt cctggagcat caaagcggcg atggagggac 1260
ggaagtttgg agatggattt accgatgatg caactaaagg agttattcaa tgtgaatcat 1320
ttcatcgtca gccaagctaa tcctcacatc tctccactcc tccgaatgaa ggagattgtc 1380
agatcctatg gaggtcgctt tgcgggaaag ctcgctcgtc ttgctgagat ggaggtgaag 1440
tatcgatgta accaagttct agaagttggc ctcccactgg gaggacttgc aaagttgttt 1500
gctcaggact gggagggtga tgtcactatg gttatgccag caacagtagc tcagtacttg 1560
aagattatac aagatccaac atatgcagaa ctccaaatgg ctgccaatca gggtcaaaga 1620
tgcacgtggg agaagctctc agcgatcaga gcaaactgtg caattgaact tgcattggat 1680
gaatccattg cggttctcaa ccacaaacga aggttaagaa gaagcacaag ggcagcagct 1740
tcttcccagg aatataccag caatgttcga ctcagaacac caaggagggt accctcatgg 1800
agctgcatca gtcgagagaa ttcgtcagga tctctctcag aagatcactt tgcggtcgct 1860
atttcatcca gtcaccaagg tactatacga gttgatggcg caccaaacat gcctcatcat 1920
gttcgtcaca gttcacacga tggaagtgag agcgaatcag aaaccattga cttaaattca 1980
tggaccagga gtggtgggcc tctaatgagg acttcatcag ctgatcagtt catcagtttt 2040
atccagaatc tcgagattga atctgagttc gatagggttc gtactacaga ggatgacaat 2100
acaggtattt tatcaggatc tacattttca aaagatccat acccaaacat tagttctaga 2160
gtcactacac cagatagatg cacagaagtt tctgaaacag agtcgtgcaa cgccggcaac 2220
acaagcatca ctgtttctga aggagatttg ctacaacctg agaggactac caccggaatt 2280
ctactcaatt ttgtcagaag agaagatctg cttggtcagc ataacagtga tgctgacatg 2340
accgaaagct ccttagccga agcatatgtg gacacatcac atttggaatc ttgtgatgcc 2400
atctcagcct ctgacagttc tgaaggtaac aaagacgcag ctgactcaga gaatctcttg 2460
gtttctcatg cagatttagt aacttcgcat caatcttcag ttgatgataa caaaggtggc 2520
tagattttga aagaattctt ttagtggctt gctaagtcga tgctgtacag gaaaaactgt 2580
agtgtctccg tttcgtgagc actactgctg gtagcatagt gaatattgta ctttgtacca 2640
gatactaaat aaatttgatt gcttgccatt 2670
<210> 166
<211> 3884
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Populus trichocarpa
<400> 166
gttttctttc ccttatccgc ctttgattgc aaaagtcaat gtcagagcca tcaccccctc 60
cttgctcaat ctttacgtaa ccgtatgtat atccttatct tcttctaaca tttcccaaga 120
ctccgatcct gtatttattg tattcacttc acccttcttc tctctttcct tcccgaacga 180
aaacaaagtc tcaatctttc attctctgtt tgtctaaagt ctgtacattc ttcactttct 240
cgagttgggt ttctttcttg aatttggttt cttgggtttg attttgtttt tcaagtggat 300
attgctattt attgggtggt gatattgaga cccttttgtt agttttgtat attggttttt 360
gaggtggatg tagttttttt aggggtttta gggtttggtt attgaaaact catatggcaa 420
ggttggcttc tggcaatctg gatttataag attctgtttt tcttgttgac acagtacagg 480
atcaaaaggg ttggattttt gttacttgtc aatatcttct tattttgtga tagctagtcc 540
ttttgcatta ggattgcata tctttattct atctacttca ttgtctctct atatattgcc 600
atcctatccg gggagagaca gattcaattg ttttattgtc cttctcattc tcattagaat 660
caaagtcttg acatacaatc ctttcacaat tgtgaaattt gattccttag tgaccatcta 720
ttgtagctgt ttcatatttg tttcgttcaa gctaattctg ttgttagatt tgagacaaaa 780
gaaggccccg cttccaatta cagaccactt tcttgttttg gttttagcta agatatggat 840
ataagcaatg aggccagtgt tgaccctttc aaaatcggac cttcatcgat cattggtagg 900
acaattgctt tcagagttct gttctgtaaa tcaatctcac atttgaggca aaaaatcttt 960
catgtgttgt tgaattacat ttatagagtt ggtgaatttg tggcgcctat gttatcatgg 1020
tttcatccaa ggaatccaca agggatattg gccatgatga cgataattgc atttttattg 1080
aaaagatatg cgaatgttaa attgagggcc gaaacagcgt ataggaggaa attttggagg 1140
aatacgatga gaactgcgtt gacatacgag gagtggtttc atgctgctaa aatgcttgat 1200
aaagagaccc caaagatgca tgaatgtgat ctctatgatg aagaactagt caggaacaag 1260
cttcaagagc tccaccaccg tcgccaagag ggatgtctta gagatataat cttttttatg 1320
agagccgatc ttgtaagaaa tctcggtaat atgtgtaacc ctgagcttca caagggtagg 1380
cttcaagtgc ccaagctcat aaaggaatat attgacgagg tctcaactca gttaagaatg 1440
gtttgtgact ccgattcaga ggagctttcg ttggaagaaa agcttgcttt catgcatgaa 1500
acgagacatg cttttgggag aacagctttg cttctgagtg gaggtgcttc acttggagcg 1560
tttcatgtgg gtgtggttaa aacactggtg gagcacaagc ttatgccccg aataattgct 1620
ggttctagtg tggggtcaat tatgtgttca gttgttgcca ccagatcgtg gccagagctg 1680
caaagttttt ttgaggattc ctggcactcg tttcagtttt ttgaccaatt gggtggaatt 1740
ttcacagttg tgaagagggt catgagacaa ggagctgttc atgaaatccg gcagttgcaa 1800
tggatgttaa ggcatcttac aagtaatctt acatttcaag aagcttatga catgactggt 1860
cgaattcttg ggatcacagt ttgctcacct aggaagcatg agccccctag atgccttaat 1920
taccttactt cccctcatgt tgttatatgg agtgcagtca ctgcttcttg tgcttttcct 1980
ggcctttttg aagcccagga actaatggca aaggacagaa gtggggaact tgtgccttat 2040
cacccaccct ttaatctgga tcctgaagaa ggatctgatg cacctatgcg taggtggagg 2100
gatggtagcc tggagattga tttaccaatg atacaattga aggaactatt caatgtcaat 2160
cattttattg taagtcaagc gaatcctcac attgctccat tgttgagact gaaggatata 2220
gtcagggcat atgggggtag ctttgctgcc aagcttgctc atctcgctga gatggaggta 2280
aaacatagat gcaatcaggt attggaactt ggttttccat taggtggact tgccaagctt 2340
tttgctcaag aatgggaagg tgatgttact gttgttatgc ctgccacact cgctcagtac 2400
tcaaaaatta ttcaaaaccc aaatcacttg gagcttcaaa aggcatcaaa ccaaggcaga 2460
aggtgcacat gggagaagct ttctgccata aaagctaatt gtggtattga gcttgctctt 2520
gatgagtgtg tttctgttct gaaccacatg cgtagactca aaaggagtgc tgagagagct 2580
gctgctgctt ctcatggcca agcaagctct gcgagcacat tgagatttag tgcttcaaaa 2640
agaattcctt cttggaattg catcgcaaga gaaaactcaa caggctcact tgaagaagac 2700
ttccttgctg atgttgcttc aacattccat caaggagttg gtgtggctgg aggaacttct 2760
actggtagga atttgagaac acaacgcaac ctacatcatg atggaagtga tagtgaatct 2820
gaaagtgtag atttgaattc ttggacaaga tctggcgggc ctttgatgag gactgcttct 2880
gcaaataagt tcattgactt tgtccaaagt ctggatgttg attctgagct aaggaaaggc 2940
ttcatggctc atcctaactc gcctggggct cagatgggag gcagggatcc atataatcag 3000
atctcaagag tgacaacccc agatagaaat tcagaaagtg agtttgatca gagagatttt 3060
agcaatagaa attctactgg tggttctagc attacagtca ccgaaggaga ttttttgcag 3120
cctgaaagaa tccataacgg gtttgtgctg aatattgtaa agaaagaaga tttggcacat 3180
cccaatagga tccatgattt ggagaattac aatagtgaag ttcctgaatg tgttcagctt 3240
gattgtcctg aaaaggacat ggatgctagc tcagaatcgg actatgctgc agaggaagac 3300
gactcccctg caacagattc cttgcataaa tcagcttcca ctcttgatca cacagatgat 3360
tctgtcgttc atgacattca ggagaagcat gtcgtggatg gttaactttg agtttcttct 3420
gcattactgt accaaaatat tgggtggagt tgattcccgg gttactgtca atcaaaggtt 3480
tccgactttc cgtcacaact ggagtatcat agacgagatt tagaatctgt ttatttttta 3540
ttttaaaaat atttttgaaa aaaattttga ttttattttg attttatttt tgttttaaat 3600
taatattttt ttggtgtttt tcatattatt ttgatatgtt gatattaaaa ataaattttt 3660
aatatcaatt attcaatcag atatattttt aagtaaaaca agacagtttg aaaagtaatc 3720
ggaactttta aaaggttgct cttagtagtg aattataaaa aacaattgaa agcaatctgg 3780
cagcgtcagg ctattgctgt tgtaaactaa ttttgtgcgc atactatgca acaattgtaa 3840
tccacatgct tagatttcag ccaacgagat ggaatttgac cctc 3884
<210> 167
<211> 2490
<212> ДНК
<213> Medicago truncatula
<400> 167
atggatcgta taagtaatga agccactgtt gatctttttc caatcggtcc ttcaggaatt 60
cttgcccgaa caattgcatt cagagtcctt ttctgcaaat ccatttcaca tttaaggtat 120
caattattct taactttatt cgattcgttt catagattta gaaaattctg gggacccatt 180
atatcatcct tgcatccaaa aaaccctcaa gggatattag ccatcatcac cattctcgct 240
ttcttgttaa aacgttacag taatgttaaa gtaagagctg aattagcata caggagaaaa 300
ttttggagaa atatgatgag atcagctttg acttatgagg agtgggctca tgcagctaag 360
atgcttgata aagagacgac attgaagacg atgaatgaat ccgattttta cgatgtagaa 420
ttggttagga ataaggttca agagttacga catcgtagac aagaggggtc tcttagagat 480
attatctttt gtatgagagc tgatcttgtt agaaatttag gtaatatgtg taaccctcag 540
cttcataaag gtaggcttca tgtgccgaga cagattaagg agtatattga tgaggtggcg 600
atgcagttga gaatggtttg tcattctgat tccgaggagc tttctttgga agaaaagctt 660
gctttcatgc atgaaactag acacgcgttt gggaggacgg ctttgttgtt gagtggtggt 720
gcttctcttg gagcttttca tgtcggtgta gttaaaacct tggtggaaca taaacttatg 780
ccgaggataa tttctggttc gagtgtagga tccattatgt gctctattgt tgctactagg 840
tcttggcctg agcttcaaag cttttttgaa gattcgttgc actcgttaca gttttttgat 900
caaatgggtg ggatttttac gattgtcaag agggttacaa catttggtgc agttcatgag 960
atcagacagt tgcagattat gttgaggcat ctaacgagca atcttacatt tcaagaagct 1020
tacgacatga caggtcgagt tcttgggatt acagtttgct ccccaaggaa gcatgaaccg 1080
cctagatgtc ttaactactt gacttcaccc catgttgtta tatggagtgc agtcacagct 1140
tcttgtgcct ttcctggtct ttttgaggct caggaattga tggcaaagga tagaagtgga 1200
gagattgttc cttaccatcc tccatttaat ttgggtcctg aagagggttc ctcacaagtg 1260
cggcgttgga gggatggtag cttggagatc gatctaccta tgatgcagtt gaaagaattg 1320
ttcaatgtca atcattttat tgttagtcag gccaatcctc atattgcgcc attattgaga 1380
ttaaaagaat ttgtacgagc ttatggaggt aattttgctg ccaagctggc tcatctggta 1440
gagatggagg ttaaacatcg atgtaatcaa atactggaac ttggttttcc attaggtgga 1500
cttgccaagc tgtttgctca ggactgggaa ggtgatgtga cagttgttat gcctgctact 1560
cttgctcagt actcaaaaat tatccagaac ccttcttatg tggagcttca gaaggcagct 1620
aaccaaggga gaagatgcac ttgggagaag ctttcagcca ttaaagcaaa ttgtggaatt 1680
gagcttgctc ttgatgagtg tgttgcaatt ctcaatcata tgagaagact caaaagaagt 1740
gccgagagag ctgcttctgc ttctcatggt ctttctagta ctgtcaaatt tagtgcttca 1800
aaaagaattc catcatggaa tgtcattgcg cgagagaatt ctacaggatc tcttgaagac 1860
tttcttgcag acactgctgc ttcatttcat cacggggtta gtagttccag tggagccacg 1920
ggtaaaaatt ccaagcacca ccgcagcatg catgatgtaa gtgacagtga atccgaaagt 1980
gctgaattga atacctggac cagatctggt ggtcctttga tgagaactgc ttcggcagat 2040
atgttcaccg actttgtcca aaacttagaa gttgatactg aactaaacag aggaatggga 2100
actaatttta gccctcgtga ttcccagtat cacagtccca gattaacaac accggataga 2160
tgctccgaga actcagaacc cgatcagaga gaaaatggca acaaggttgt catgaatgga 2220
tctagcataa tggtaactga aggtgatctt ttgcagcctg agagaatcca taatggaatt 2280
gtgtttaatg ttgtcaagaa agaagactta acaccttcaa gtaggagtca tgattatgat 2340
agtgaaattg ctgagtgtct ccaaattgaa tgtccaggga aggagatgga tgatgctgct 2400
agctcagctt cagaaaacgg agatgacgat tctgcaacag ctaggcccct aactgaaaca 2460
ccagactcta atcctacaga taattcctga 2490
<210> 168
<211> 2783
<212> ДНК
<213> Glycine max
<400> 168
atggatcata ttagtaatga ggccagtgtt gaccgttttc caattggtcc ttctggcatt 60
cttggtagga caattgcttt cagggttctt ttttgcaagt ctatctcaca ttttaggcac 120
cacatattta ttgtgttgtt agatctcttc tataggttta gggggggttt ggcatccttt 180
atatcatggt tgcatcccag gaaccctcaa gggatattgg caatgatgac aattgttgct 240
ttcttgttga aacgatacac aaatgtgaaa tcaagggctg aaatggcata taggaggaag 300
ttttggagaa acatgatgag aagtgctttg acctatgagg agtgggctca tgcagctaag 360
atgcttgata aagagacaac aaagatgaat gaatcagacc tttatgatgt ggaattggtg 420
aggaacaagc ttcaagagct ccgccaccgt cgacaagagg gatctctcgg agatataatg 480
ttttttatgc gtgccgatct tattagaaat ttaggtaata tgtgtaaccc tgaactacac 540
aagggtaggc ttcaggtgcc taaattaatc aaggagtaca ttgatgaagt aacgactcaa 600
ttgagaatgg tctgtgattc tgattcagag gagctatcat tggaagaaaa gcttgctttc 660
atgcatgaaa ctaggcatgc atttgggagg actgctttgc tgttaagtgg gggtgcctct 720
cttggagctt ctcatgtggg tgtagttaaa acactggtag aacataaact catgcctagg 780
ataattgctg gttcaagtgt gggatccatt atgtgtgctg ttgttgccac taggacttgg 840
cctgagctcc agagcttttt tgaggattca tggcactcat tgcaattttt tgatcaaatg 900
ggtgggattt ttgcagttgt taagagagtc acaacattgg gtgctgttca tgagatcaga 960
cagttgcaaa tgatgttgag gcatctaaca agcaacctta catttcaaga agcttatgac 1020
atgacaggca gaattcttgg gattactgtt tgttccccaa ggaagcatga accgcctaga 1080
tgtcttaact acttgacttc accccatgtg gttatatgga gtgcagtaac cgcttcttgt 1140
gcctttcctg gcctttttga ggctcaagaa ttgatggcaa aggatagaag tggagagatt 1200
gttccttacc accctccttt taacttaggc cctgaagagg gctccacacc agtgcgccgt 1260
tggagggatg gtagcttgga gattgattta cctatgatgc agttgaaaga actattcaat 1320
gtcaatcatt ttatagttag tcaggccaac cctcatattg caccactatt gagattgaaa 1380
gaatttgtgc ggacttatgg gggcaacttt gctgccaagc ttgctcatct tgtggagatg 1440
gaggtgaaac ataggtgtca tcaaatactg gaacttggtt ttccattagg tggacttgct 1500
aaattgtttg ctcaagactg ggaaggtgat gtgactgttg ttattcccgc aactcttgct 1560
cagtacacca aaattataca gaacccttct tatggagagc ttcaaaaggc agccaaccaa 1620
gggagaagat gtacctggga gaaactttca gccataaaag caaattgtgg cattgagctt 1680
gctcttgatg agtgtgttgt gattctcaat catatgagaa gactaaagag aattgctgag 1740
agagctgctt ctgcctctca tggtttgtcc agcactgtca ggttcagtgc ttcaaaaaga 1800
attccttcgt ggaattgcat tgcacgagag aattcgaccg gctcccttga ggaccttact 1860
gatgttgcct cctcattgca tcaaggcatc ggcagttcca gcagagccaa tggcaaaact 1920
tggaagaccc accgtggcat acatgatgga agtgacagtg actctgaaag tgttgatttg 1980
cattcttgga caagaactgg cgggcctttg atgagaacta cttcagcaaa tatgttcgtt 2040
gattttctcc aaaacttaga ggttgatacg gatcctaata aaggcttagt gagtcacact 2100
atccataatg attttcagta tcatagcccc aggctcacaa cactagatag gaactctgat 2160
agcacagaat ctgagccaag ggaaactggc aacagggttg tcaatgtgtc cagcatactt 2220
gtgaccgaag gtgatcttct gcagcctgaa aggatccata atgggattgt gtttaatgtt 2280
gtcaagaaag aagacttgtc acccttaagt agtagcagtc atggttttga aaattacaac 2340
attgaagttg ctgaatgtgt ccaagatgag tgtccaggga aggagataga tgctgctagc 2400
tctgcatctg aacacggaga tgatgaagaa tccatgccag ccaggtcctt aactgacatg 2460
ccagattaca attccattga tcatcattcg ggcacagatt cgggtatgga tcaaagcatt 2520
gttgacagtt agtgtcaagt atcagttctt ttccagtgac attttaatat tttgttccta 2580
ttgccctcca tattgtaaat agtactcatt ctagacttgg agaggtcttt attcatgatt 2640
ttgatgggaa tagcccacca atttggtttg ctcataaatg taacaaagat aaagagtttg 2700
tatacataaa ttccacgaca acattgatat ttcttggtta ccacttctca gatgaatgaa 2760
atggagacat ggttttcata att 2783
<210> 169
<211> 2724
<212> ДНК
<213> Sorghum bicolor
<400> 169
atggatgaca tcgccagcga ggcgccggtg ggggcgttcg ccatcggccc gtccacggcg 60
ctgggccgcg ccgtcgcgct ccgggtgctg ctctgcggct ccgcggcgcg cctgcggcac 120
cgcctggccg cggcgctccg cgccgcgctg cccgtcgcgg cggcgtggct gcacccgcgc 180
gacaacacgc gcgggatcct gctcgccgtc tgcgccgtcg cgctcctgct gcggggccga 240
cgcggcaggg ccgggctgcg ggcgagggtg cagtccgcct accgccgcaa gttctggcgg 300
aacatgatgc gcgccgcgct cacctacgag gagtgggcgc acgcggcgcg gatgctggag 360
cgcgaggccg ccccgcgccg cgccagcgac gccgacctct acgacgagga gctcgtccgc 420
aataagctcc gcgagctcag gcaccggcgc cacgagggat cgctcaggga catcgtcttc 480
tgcatgcgcg cggacctgct caggaacctc ggcaatatgt gcaaccccga actgcacaaa 540
gggaggctgc aggtgcctag actcataaag gaatacattg aggaagtatc tactcaactg 600
aaaatggtct gtgattctga ttcagatgag ttgcctcttg aagagaaact cgcatttatg 660
catgagacaa ggcatgcctt tggtagaaca gccttgctgc taagtggagg tgcttcattg 720
ggatcctttc atgtgggtgt tgttaaaacc ttggtagagc ataaactttt gccaaggata 780
atttcaggat caagtgttgg ctcgataatg tgttctatag tagcaacaag atcatggcct 840
gagctggaga gcttttttga agagtggcat tccctgaaat tttttgatca gatgggtgga 900
atctttcctg tggttaaaag aattttgacg caaggcgctg ttcatgatat aaggcacttg 960
caggtgcttt tgagaaacct taccagcaat ttgacatttc aagaagctta tgacatgact 1020
ggtcggattc ttgttgtcac cgtgtgttct ccaaggaagc atgagccgcc tcgatgccta 1080
aactatttaa catcacctca tgttcttatc tggagtgcag taacagcttc ctgtgctttt 1140
cctggacttt ttgaggccca agaattgatg gcaaaagata gatttggtca aaccattcct 1200
ttccatgctc cattcttatt aggcatagaa gaacgaactg ttgctccaac ccgccgctgg 1260
agagatggga gcttagaaag cgatttaccc atgaagcaat tgaaggaact attcaatgtg 1320
aatcatttca tagtaagcca agccaatcct cacatagctc cgctgttgag actaaaggaa 1380
atcgtcaggg cttatggagg cagcttcgct gccaagcttg ctgaacttgc tgagatggaa 1440
gtcaaacata ggtgtaatca agttttggaa cttggatttc ctctaggagg attagctaaa 1500
ttatttgctc aagattggga aggcgatgtt acagttgtta tgccagccac tcttgcgcag 1560
tattccaaga tgatacagaa cccatcttat gctgagcttc agaaggctgc gaatcaaggt 1620
aggagatgca cttgggaaaa gctatcagcc atcagggcaa attgtgctat tgagcttgca 1680
ctggatgaat gtgttgccct cctgaaccac ttgcgtaggc taaagaggag tgcagaaaga 1740
gcatccgcat cgcaaggata tggtccagca atcaggttct gcccatctag gaggattcca 1800
tcctggaatc tcatagcaag agaaaattca actggttctc ttgaagaaga aatgcttaca 1860
tctcctcaag gacctggagg agttgctgga acatctacca gaaaccagta tcctcagaga 1920
agtgcacatg agagcagcga cagtgaatct gagagtattg atttacactc ttggacaaga 1980
agtggtggcc ctcttatgag gacaacctca gccaataaat tcatcagctt tgttcagaat 2040
cttgagatcg acacagaatc cagaacaatt ccatcgaggg aagacataac tgatcttgtg 2100
acaccaaatg ctggtacctt ggcagctcat gcagtgagta gagaagcaat cgataggagc 2160
ttggacaatt cagctttaga tatccatgat accagtaccc ctagatcgac atttggccct 2220
tcaacaagta ttgtggtttc tgaaggtgac ttgttgcagc ctgaaaagat tgaaaatggt 2280
attttgttta atgttgtaag gagggatact ctgctcgggt ctagtagtgg agttgagtct 2340
caaggatctc ctcgggaacc agatgttgaa acagtacaga cggagtgcct tgatggcgtg 2400
tctacttctg atgatgatga tgacaaggaa ctaaatgcca ttgatgatgg aggaactagt 2460
cccatgagca gaaataatct acaacatcag gggtcctcac tggaagaaaa attataccat 2520
ccctcttcct taaattctga agacgagaca aacacaaaca aaccagaagc tgcatcgatt 2580
tttgatatat gtacagatat gcatccggca tctattagcc tacctgaagg gtcttcagaa 2640
aagacagaac tggaaacaac aaagattcct gatgacaatt cagctgttat gaatgatgaa 2700
gttgcctcag gtgctggtaa ctaa 2724
<210> 170
<211> 2985
<212> ДНК
<213> Zea mays
<400> 170
cattctctct ctctctctcc cttttcaatt tcgggggctt tatttctctc ctcccacgcc 60
ttgcatcttc ttgatgcgtt gcgtcccgac tcgaggcaga catcggaggc gcgccactga 120
cttagctcgc gatttctaga tccgcaaccc tgcgctgctc aactccgatt cttttagttc 180
ggattcgggt agcagcaggg cttcggtggc gatgacttga ttgatacgaa ttggggattt 240
cttgctgttt gcgcctctct tcgtatcggc gctgaagggc tcagctcgag attagaacaa 300
tttgggtttt ggggtctttt cttcttctac tactgctggt caagttcaca gaaatcgctg 360
gctccgtggt ccaatggacg agtccgggga agcgagcgtc ggctccttca ggatcgggcc 420
gtcgacgctg ctgggccgcg gggtggcgtt ccgcgtgctc ctcttcagct cgctgtggcg 480
cctgcgggcg cgcgcgtacg cggccatctc gcgcgtgcgc agcgcggcgc tgccggtggc 540
ggcgtcctgg ctgcacctca ggaacagcca cggcgtcctc ctcatggccg tgctcctcgc 600
cctcttcctg aggaaactct cggccgcgcg gtcgcgggcg gcgctcgcgc gccggcgcag 660
gcagcacgag aaggccatgc tgcatgccgg gacgtacgag gtctgggcgc gcgccgccaa 720
ggtgctcgac aagatgtctg agcaggtcca cgaggcggat ttctacgacg aggagctcat 780
caggaatagg ctcgaggaac tccggagacg gagggaggac gggtcgctcc gggacgtggt 840
gttctgtatg cgcggcgatc ttgttaggaa cttggggaac atgtgcaacc ctgaacttca 900
caagggcagg ctagaggttc ctaagcttat aaaggagtac attgaagagg tttctactca 960
actaagaatg gtgtgcgaat ctgacactga cgagttgctg ttggaagaga aacttgcctt 1020
tgttcaggag accaggcatg cctttgggag gacagcgcta ctcttaagtg ggggtgcttc 1080
actcgggtct ttccatgtag gtgtagtgaa aacattggtt gagcataagc ttctgcctcg 1140
gattatagca ggatcaagcg ttggttccat catatgttcg atcgttgcta cccgtacatg 1200
gcctgagatt gagagcttct tcacagactc attacagacc ttgcagtttt tcgacaggat 1260
gggcggaatt tttgcagtga tgaggcgtgt caccacttat ggtgcactgc atgacattag 1320
ccagatgcaa aggcttttga gggatctcac aagtaactta acatttcaag aggcttatga 1380
catgaccggc cgtgttcttg ggatcaccgt ttgctctcct agaaaaaatg agccaccccg 1440
ctgcctcaac tacctgacgg caccacatgt tgttatttgg agtgcagtaa ccgcctcttg 1500
tgcatttcct gggctctttg aagctcagga actgatggca aaggataggt tcggcaacat 1560
cgttcccttc catgcaccct ttgccacaga tcctgaacaa ggtcctggag catcgaagcg 1620
caggtggaga gacgggagct tggagatgga tttacccatg atgagactta aggagttgtt 1680
taatgtaaac catttcattg tgagccaaac taaccctcac atttctccac tcctccggat 1740
gaaagagctt gttagagcct atggagggcg ctttgctgga aagcttgctc gtcttgctga 1800
aatggaggtt aagtatcgat gcaaccaaat cctagagatc ggtcttccaa tgggaggact 1860
tgcaaaattg tttgcccagg attgggaggg tgatgtgacc atggttatgc cagcaacact 1920
tgctcagtac ttgaagatca ttcagaatcc aacatacgcg gagctccaaa tggctgccaa 1980
ccaaggccgc aggtgcacat gggagaagct ctccgcaatc agagcgaact gcgccattga 2040
acttgcactt gacgaatcca tagcggttct aaaccacaaa cggaggctaa aacgaagcat 2100
ggagaggacg gcggcggctt cgcagggtca ctctaactat gtccgaccca agactccgag 2160
gaggataccg tcatggagcc gcatcagtcg agagaactct ttggagtctc tctcggaaga 2220
gatctctgcg gttgctgctt cgtccatgca gcaaggcgct gctcttgttg tcggcgcacc 2280
accaacgact ctttctcagc atgttcggcg cagttctcat gacggaagtg agagtgagtc 2340
agaaaccatt gaccttaatt cctggaccag gagtggaggg cctctaatga ggacagcgtc 2400
cgccgacagg ttcatcagtt tcatccataa cattgagatt gacacagaat taagtaggcc 2460
ctgtgctgtg gaaggtgatg ctgcaggtat tttgtcagaa tctaccttcc caaacggtcc 2520
acgaccgaac aatagctcaa gtgttagtat gccaggtaga tgcacagaaa attctgggac 2580
cgagtcgtgc aacactgtca acaccagagc ttctactccc acaagcatgg ctgttcgtga 2640
aggagatttg ctgccgcctg aaagcactac tgataatgtc ctacttaaca ttgtgaaaag 2700
agacgccctg caggatggtg taactgaatt ggcggaaagc tcctgcgctg aaggatatgc 2760
ggcaaactgt gacaccgtct cagggctaga ctgctgaagg taacaagacg ctcgctgctg 2820
acttgagcaa tcaacaatta gctgatgatt agattcttct tgattttgat gatgaaaggt 2880
catttatatg tagctcacta cagcaacgca gtgtaggaaa attgtacctg ctcgatttaa 2940
actttaaaga gcatgccatg agtagctttg ttaatgttaa tattc 2985
<210> 171
<211> 1998
<212> ДНК
<213> Physcomitrella patens
<400> 171
atgaattact tagacactga cgccgacgct gcgctagagc atttcggcat tggacctctg 60
actttggcgc aaagagttgt ggcctttcgc gtcctatttt gtcgttgggt gaaagagctt 120
cgtgttgccc tcgcaaagag gctgcagcgg acacggaggg tatggagaca ggtgttctat 180
atgtggtttg ggtggttgaa ccctcgaaat cccagcgtcc ttctgttagc tgccgttgta 240
gcaaccatgc tcatgagaag agcgaaggca gggtctcaga aagcagagat tgcgtacaga 300
cggaagttct ggtccaattt aatgagggca gctttgacgt atgaggaatg ggctcatgcg 360
gcgcggatgc tagagaagga gcagaatcgg aggaaagatt cagacttgta cgatgaggat 420
ttggtgcgtt cgaagctcaa cgatcttcga ttgcgtcgtt tggagggtgg tgtggaggac 480
attcttttct gcattagggc cgatttagtg cgtaatttgg gtaacatgtg caatcccgaa 540
ctgcacaaag gccggctaca aactcccccc ctcatccagg aatacatcaa cgaagtgaga 600
taccatcttc gagctgtgtg tgggagcgac tcggacagct tcacacttga cgaaaaaatt 660
gcttttattc atgaaacccg ccatggtttt ggtcgcactg cacttcttct gagtggtgga 720
gcagctcttg gagcgtttca tcttggggtt gttcgaaccc ttgtcgagca tcgtttactt 780
ccccgagtga ttgccggtgc cagtgtggga tctgtcatat gctcatttgc tgcaactcga 840
acttggacag agctccagag ctttttcgaa gacaccatgc cccccatgca ctttttcgaa 900
aacatgggga gcatttttgc tattgcgcac aggcttctga ctcgaggtgc tgtgcatgaa 960
attggtatgc tgcaaaggaa aatgagacag ctcattgggg atttgacctt tcaggaagct 1020
tacgatctat ctggccgcgt gcttggaatc tctgtatgct cacctcggag actcgagcct 1080
ccgagatgtt taaattattt aacttctccc catgtagtca tttggagcgc agtcactgca 1140
tcctgcgcat tcccaggcct ttttgaagca caggagctga tggcgaagga tcgaactggt 1200
caacttgtac cctatcattc gccacctcag gttggccccg aggacaagga catggaaaag 1260
gggattggga agcggcgatg gcgagacggc agtctggaaa gcgatttgcc aatgatgcag 1320
ttgaaggaac tgtttaatgt gaatcatttc attgtcagcc aggcgaatcc gcatattaca 1380
ccatttttga ggttcaagga ttttgttcgt gcatatggag gagatttcgc tggaaaattg 1440
gcacacttag cggagatgga ggttaagcac cggtgcaagc agatgatgga gatgggcttt 1500
gaggtgtttg gattggctaa gctcttcgca caagattggg aaggagatgt cacgattgtg 1560
atgccggcca cttttgccca gtttgccaag atcatcacga acctgacagc cacagatctt 1620
cgcaaggcag tgatgcaagg ccgacgctgc acctgggcga agctatcagc cattcaggcc 1680
aactgtggca tagaattgat gctagacgaa tgtgtctctg aattaaaccg tcgtaggaaa 1740
gccctgcgtg aaatagagcg cagcgcaatg cagagcagcc atggtgggat gcgcgggtta 1800
tcaggaacaa agcgtatccc atcctggaac atcatcgccc gagagaattc ctgcggttcg 1860
ctagatgaag agagtcttca cgaggtgcgg atcccacatg atggtagcga cagcgacgat 1920
aatctggacc aaaatcagct ttcgtggacg agagcaggtg gcccgctcat gcggaccgca 1980
tcagcagcca aattcgtg 1998
<210> 172
<211> 3439
<212> ДНК
<213> Hordeum vulgare
<400> 172
gatcgcagtt agtttggctt gtacgtcgcg ttccccttcc acccttatct ccttctccgg 60
ctgaccggga cgccgcattt gtcccatcca cggcacggca cggcacgggc acgggaggga 120
gaagaagaag cccagctcga ctcctcctcc gcctcctcct ttcctctgat cccctccgtt 180
tgcccattcc ccagatccca gcacgccatg cccgggcgcg caggcgccaa gccgcaccgc 240
gcgcatttct cttccgccct gctccgatcc aaggccgcgg aggtgaccca gtgagctctc 300
ccgccacgcc cgtccgtccg ccggttcatc ggtcgcccat ggacgtcatc accaacgagg 360
cgcgcgtggg ggcgttcgcg atcggcccgt ccacggcggc gggccgggcg ctcgcgctgc 420
gcgtgctcct ctgcggctcc ctggcgcggc tgcggcaccg cctcgccgcc gcgctgcgcg 480
ccgcggcgcc cctggcggcg gcctggctgc acccgcgcca caacacgcgg gggatcctgc 540
tggccgtctg cgccgtcgcg ctcctgctgc gcggccgcgg gggccgcgcc ggggtgcgcg 600
cgcgcgtgca gtccgcctac cgccgcaagt tctggcgcaa catgatgcgc gccgcgctca 660
cctacgagga gtgggcgcac gccgcgcgga tgctcgagcg agagacgccg cgccgcgcca 720
ccgacgccga cctctacgac gaggagctcg tgcgcaacaa gctccgcgag ctcaggcacc 780
gtcgccagga gggctcgctc agggacatcg tcttctgcat gcgcgccgac ctgctcagga 840
accttggtaa catgtgcaac cccgagctcc acaagttgag gctgcaggtg cctaaactca 900
tcaaggaata cattgaggag gtatctactc aactgaaaat ggtttgcaat tctgattcag 960
acgagttacc tctcgaggag aaactggcat ttatgcatga gacaaggcat gcctttggta 1020
gatctgcctt actgctaagt ggaggagctt catttgggtc tttccatgta ggtgttgtga 1080
aaaccttggt agagcataag cttctaccta ggattatttc aggatcaagc gttggcgcaa 1140
taatgtgtgc tattgtcgcc acaaggtcat ggccagaact ggagagtttt tttgaggagt 1200
ggcattcctt gaaattcttt gaccaaatgg gtgggatctt tcctgtattt aaaagaattt 1260
tgacgcatgg ggctgttcat gacattaggc acttgcagac gcaattgaga aatcttacaa 1320
gcaacttaac atttcaagag gcatatgaca tgactggccg ggttctcgtt gttaccgtgt 1380
gttctccaag aaaacatgag ccacctcgat gcctgaacta tttgacgtca cctcacgttc 1440
tcatctggag tgcggtaact gcttcctgtg ctttccctgg actttttgag gcccaggagt 1500
tgatggccaa agatagattc ggagaaacag ttccttttca tgctccattc ttgttgggcg 1560
tggaggaacg agctgatgct gctacacggc gatggagaga tgggagctta gaaagtgatt 1620
tgcccatgaa gcagttgaag gaattattca acgtaaatca cttcatagta agccaagcca 1680
atcctcacat tgctccatta ctgagactaa aggagatcat cagggcttat ggaggcagct 1740
ttgctgcaaa gcttgctgaa cttgctgaga tggaagttaa gcataggttc aatcaagttc 1800
tggaacttgg atttccatta ggaggaatag ctaagttatt tgctcaacat tgggaaggtg 1860
atgtgacaat tgttatgcca gccactcttg ctcagtattc gaagatcata cagaatcctt 1920
cgtattctga gcttcagaaa gcagcaagtc agggtaggcg atgcacttgg gaaaagctct 1980
ctgccatcag ggcaaactgc gctattgagc ttgcattaga tgaatgtgtt gccctcctga 2040
accacatgcg taggctgaag agaagtgcag aaagagcagc cgcttcacaa ggatatggtg 2100
ctacaattag actctgtcca tctagaagga ttccgtcatg gaatctcata gcaagagaaa 2160
attcaactgg ttctctcgat gaggagatgc tcacatctcc cactgttaca agccatcaag 2220
cagttggagg gactgctggg ccatctaaca gaaatcacca tctccaacat agtatacatg 2280
atagcagtga cagtgaatct gagagtatag acttgaactc atggacgaga agtggtggcc 2340
ctctcatgag gacagcctcg gctaataaat tcatcagctt tgttcagaac cttgagattg 2400
acccagagtt cagaacaatt tcaccaaagg ggagtgaagg tgatattttg acaccgaata 2460
gtaacttgtt tgctggtcac ccaattggta gagagccagt tgataatcat ccaaggcctg 2520
ttactcctgg taggacctca ggcaatacag gttccgatcc tcatgatact cctgttccta 2580
ggtctccatt tggtctttcc gcgagtatca tggtccctga aggtgacttg ctgcagcctg 2640
aaaagattga gaatggtatt ttattcaatg ttgtccgaag ggatactctc ctagcgtcta 2700
ctagcggagt tgaacctcat ggatcttcac atgaggcaga tgtggaaact gtaccgaccg 2760
agtgccttta tggtgcttcg gatgacgacg acaacgtgga actgaatgcc aatgatgaag 2820
cgctatctga tcgtggagat cagagatctt cagttgcagg aaatctagat tcgtccgctt 2880
ccatggactg tcaagctgaa gcaagtacta ctcgatcaga agctccatct ctctttgata 2940
tctgtgtgga gattcctcca gcaaccatga ccacagaaaa tagtcggcct gacgagcctt 3000
cttcagacat aagactggag actgtaaaga cagaatgccc tgatgagaat tctgctgctg 3060
ggaatgctga agttgactca gttcctgcca gtaaagaatc ttcctattgg tctcagacat 3120
cagaaattgg acagcagcat caagtggata tgggatctgt gaactcctgt actgtttcat 3180
tttcagaaga tgatagacat gtgagcctta tttcgaacga gaaaccggtc actacttcca 3240
gtggcggagc tgagagtatg acatctggaa gaagtgaagc tgactagcat agaacttgcc 3300
tgttgaccga cctaatgttt ttctgtgttg ggacttggta gtttgaacaa ttcagcttga 3360
tctgatccat gctatgtgtg caatttaaac tcgtgtcacg atcaaactga attgtgtcta 3420
tatgtaggtg ttgtaatcc 3439
<210> 173
<211> 3470
<212> ДНК
<213> Nicotiana benthamiana
<400> 173
gttatctgat ccaaacttct gactttttct attttccgaa tccctatgtt ttttaataaa 60
tccatctctg ccattgcagt gatatattca tttattgtta tcaccttctt catttattgg 120
tccctctgtg ttttccatat attgaaggag aaaacattaa ctttatgcga ttttgtagtt 180
tttctggttg attcctacaa ccccttttga cattgatctt gtgggttaca aaaaacattg 240
aatctttatg tcaaaatttg atctttgtat ttcattttaa attgaaattt gatttttggg 300
ggtattaagg attcttttgt cggttgattt tgtgcctttt ttgccaagtt cttgtcggtc 360
tctgagctga atttccataa tttgacaaaa agaaaaggct aaagcagaaa ggttgggagt 420
ttctttcttt gactttcaga aactaaggta ttttctttga tctaattctt gttaatatct 480
ggttcaatct gattccgttg aatcttgtga atagcctttg tttccctatt gtcagaaaat 540
tatttccttt tcactttcct cgactctcag aagttagtac aatctttgtt ctgctaaatc 600
ttgtgaataa cctttagctt agagttttag gtatctgtat attgggttct cttaacattt 660
agcctagaag ccttctctag gattagtccc ccttttcatt gagatggata taagtaatga 720
ggctacaatt gacttctttt ccattggacc tactacgata ttgggtcgaa caatcgcctt 780
tagagtgttg ttctgtaaat caatttcaca attgaagcat cacctatttc atttcttgat 840
atattacttg tacaaattca agaatggttt gtcatactac ttgacaccct tgatctcgtg 900
gttgcaccct cgtaatccac aaggaatatt ggcattggta acgcttctcg ccttcttgtt 960
gaggcgatac acgaatgtaa aaatcaaggc tgagatggcc tataggagga agttttggag 1020
gaatatgatg agatctgcat tgacttatga ggagtgggct catgctgcca agatgctaga 1080
taaagagacc cctaaaatga atgaggcaga tctttatgat gtagaattag ttcgaaataa 1140
actccaagag cttcgacatc gtaggcaaga gggttctatg agggatatca tattctgtat 1200
gagagctgac cttgttagga atcttggtaa tatgtgtaat ccagaacttc acaagggaag 1260
gcttcatgtg cctagactga ttaaggatta tattgatgag gtttcaactc agttgagaat 1320
ggtatgcgac tctgattcgg aggagcttct cttggaagag aagcttgctt tcatgcatga 1380
aacaagacat gcctttggta ggacagcttt gcttttaagt ggaggtgctt ctttaggagc 1440
tttccatgtg ggcgtggtga aaacacttgt agaacacaaa ctgatgccac ggataattgc 1500
tggttcaagt gtcggctcga ttatgtgctc catagttgca actcgatctt ggcctgagct 1560
ccagagtttt ttcgaggact cctggcactc tttgcaattt ttcgatcagt tgggtgggat 1620
ttttactatt ttcaggaggg tcatgaccca gggtgctgta catgagatca gacagctgca 1680
ggtgctgtta cgtaatctca cgaataatct tactttccaa gaagcctatg acatgactgg 1740
tagagttctg gggattactg tttgctcgcc taggaaacat gaacctccta gatgcttgaa 1800
ctacttgact tcacctcatg ttgttatatg gagtgccgtt accgcttctt gtgcctttcc 1860
tggtctcttc gaagctcaag aacttatggc aaaggataga agtggagatc ttgttccata 1920
tcacccacca tttcatttgg gtcctgatgc cacttctagt gcatctgctc gtcgttggag 1980
ggatggtagc ttggaggttg atttgccaat gatgcagcta aaggagctct tcaatgtcaa 2040
tcactttatt gtgagccagg cgaatccgca tattgctcca ctgctgagga tcaaagagtt 2100
tgtaagagct tatggaggca actttgctgc caagcttgct caacttacgg aaatggaggt 2160
gaagcacaga tgcaatcagg tattagaact tggttttccc ttgggaggat tagcaaagct 2220
ttttgctcaa gaatgggagg gtgatgtaac tgttgtaatg cctgccactc tagctcagta 2280
ctcaaaaatc atacagaatc cctcgactct ggagctgcaa aaagcagcaa atcaaggaag 2340
aaggtgcact tgggaaaaac tctcagccat gaaagcaaac tgtggaattg agcttgcact 2400
tgatgaatgc gttgctatac tgaatcacat gcgtagactg aaaaggagtg ctgagagggc 2460
ggctgctgct tcacatggct tggcaagcac tgtcagattt aacacttcca gaagaattcc 2520
ttcttggaac tgcattgcac gagagaactc aacaggctcc cttgaagatt ttcttgcgga 2580
tgttgctgct tcacatcatc aaggaggcag tggttcgggg gcgcatgtta accgtagttg 2640
gcgaacgcac cggaatgcac atgatggtag tgacagtgag ccggaaaatg tggaccttaa 2700
ttcttggaca agatcgggtg gtcctttgat gaggacaaca tcagctgata agtttattga 2760
ctttgtccag aacttggaaa ttggttcgcg attgaacaaa ggattgacta ttgacctcaa 2820
caatattatt cctcagatgg caagcaggga ccatttctcc ccgagcccaa gggtaacaac 2880
acctgataga agttcagata cagaatttga tcaaagagat tttagttaca gggtccctgc 2940
gagtagttca agcattatgg taggcgaagg tgaccttctg cagcctgaaa ggactaacag 3000
cggtattgtc ttcaatgtgg taaggaaagg agacttgacc ccatcgaaca gaagccttga 3060
ttcagaaaat aatagttccg tgcaggatgc agttgctgag tgcgtgcaac ttgaaagtcc 3120
agaaaaggag atggatatta gctcagtatc ggaggatggt gagaatgatg ttgggcaagg 3180
aagtagggta aatgaagttg attgtagtaa aaatcgttca tcaatcggtg atggcaacga 3240
taagcaagtt attgatactt gagagtttag ctttgattat tctacacagg ccattcgaat 3300
tattttttat actcaaatgg agcttctttc agagctaaca cactcagaat tggggttgta 3360
aatagtgcaa gtagcaaatc tgtaataaat gtttagtgta gtcatcaccc ttctactagt 3420
tcaaagtggc tcagttcaat tcaaattcag aacttcgata attcatgttt 3470
<210> 174
<211> 713
<212> ДНК
<213> Nicotiana benthamiana
<400> 174
tgtatgagag ctgaccttgt taggaatctt ggtaatatgt gtaatccaga acttcacaag 60
ggaaggcttc atgtgcctag actgattaag gattatattg atgaggtttc aactcagttg 120
agaatggtat gcgactctga ttcggaggag cttctcttgg aagagaagct tgctttcatg 180
catgaaacaa gacatgcctt tggtaggaca gctttgcttt taagtggagg tgcttcttta 240
ggagctttcc atgtgggcgt ggtgaaaaca cttgtagaac acaaactgat gccacggata 300
attgctggtt caagtgtcgg ctcgattatg tgctccatag ttgcaactcg atcttggcct 360
gagctccaga gttttttcga ggactcctgg cactctttgc aatttttcga tcagttgggt 420
gggattttta ctattttcag gagggtcatg acccagggtg ctgtacatga gatcagacag 480
ctgcaggtgc tgttacgtaa tctcacgaat aatcttactt tccaagaagc ctatgacatg 540
actggtagag ttctggggat tactgtttgc tcgcctagga aacatgaacc tcctagatgc 600
ttgaactact tgacttcacc tcatgttgtt atatggagtg ccgttaccgc ttcttgtgcc 660
tttcctggtc tcttcgaagc tcaagaactt atggcaaagg atagaagtgg aga 713
<210> 175
<211> 1500
<212> ДНК
<213> Arabidopsis thaliana
<400> 175
cgaaaaaaga agtagaatat atatatatat atatatatat atatatatat atatatattc 60
gtgtggacat cataaatgcc taaatgataa tagttgattt cgagttttat tttcgttact 120
tccaatcaaa ttctccttgc accatattta tttttttact gtgagaacat atataagtat 180
atattggaat tacgtatccg agaggttttt gcatatttcg tttatttatt ttcgatatcc 240
acactactgt attattaaaa atttgaaaaa ttcaactagg gcttttcatc ttctctagaa 300
ttattcgttt atttatgtcg atgtccacac tattattaaa ataaaacgag aggatatggt 360
tggatcatcc aagtttcgtt tatgactctt tgttcattta caaacgttta gttttccact 420
taagttttga aaagagttaa tttccaatat attcggcaca gtttttcaag tgtattcatc 480
tgtttttttt ttttttggtt ggctatatgg tccaaatttt gatttgcaat atgagattgc 540
acagagagaa caatctttca ttatgattaa ttattgtaca agtaacaaac accaatctcc 600
gatatacttt ggctctttag cacattgtta tgctagaagt tagcggaaat ctatatgttg 660
ttaaacgcag cgtttaaatt gaacagtgta atttaccttg aaattttaag actacatgct 720
gtttagaatt tcagatgaaa acatcttgat gttttagaaa tccacgtggg aatagcgtaa 780
aatcttatcc aacgaactta ttttggtttt gttgtatttg tgcaagtcgt cacgctaatc 840
gaaaaaagaa aagaaaaaaa gaagccgtca tgatcggcca tttctcggcc gagtctgagt 900
ctgactctgc gtccgtgtca ccattatcag atcgagcctg tcttatctcg ttgcgattcc 960
ctatgcaaaa atcttcttct tttttttatt cccccattta tctctgatct cttctctctt 1020
ctcaagtaaa cctctctgct tcacgtctct tcttttcttg tcgattttcc ccagataatc 1080
aggtaaataa ggctactttc ttatttgatc tggtggtctt tgtgttgaaa tctctgggtt 1140
ttctctgttg atttcaaagt tctctctttt tttttttgtt tactgggtgc tgtgaaaaat 1200
gatcttgtca aagtctcctc ttttcatcga attgaaactc taattagaaa aaagatcata 1260
acttttatta aaaaaatgag tttgctttgc ttaattttgc gaattgcttc atagattcat 1320
tgattagcct atttggggta acaaaaaaaa gctgacacgg tttcagattc caaaaataga 1380
tcatgactct gtttcttctc tgcagaggtt ttaataaata tatgcttctt ctcatgagtt 1440
ctcgtttttt ttgtcacctt cgcagttgaa aacacaccca aattcatctt cgaatcaata 1500
<210> 176
<211> 2871
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, комплементарная гену pSSU-олеозина
В Т-ДНК pJP3502. В порядке (комплементарная последовательность):
Терминатор лектина Glycine max 348 нт, 3' экзон 255 нт, Интрон UBQ10
304 нт, 5' экзон 213 нт, промотор SSU
<400> 176
ggcccctaga atctaattat tctattcaga ctaaattagt ataagtattt ttttaatcaa 60
taaataataa ttaataattt attagtagga gtgattgaat ttataatata ttttttttaa 120
tcatttaaag aatcttatat ctttaaattg acaagagttt taaatgggga gagtgttatc 180
atatcacaag taggattaat gtgttatagt ttcacatgca ttacgataag ttgtgaaaga 240
taacattatt atatataaca atgacaatca ctagcgatcg agtagtgaga gtcgtcttat 300
tacactttct tccttcgatc tgtcacatgg cggcggcccg cggccgcttc attactcgag 360
ccaggaggat ggatcgatgc tggtctgaga ccctgctacc ggttgctgac tgaactgctc 420
ggcacggtcc ttcatttcac gggccttgct cgccaacttt gtcttggccg actccaactg 480
atccgctccg ggtggatgtt tccccgtcag gtaacggtag atccaggaca gcacagacag 540
agcggcaaca ccaaatcccc cgcttgccag aaaacccgct cccaacagga agatggtgat 600
gactgcagat cagaaaaact cagattaatc gacaaattcg atcgcacaaa ctagaaacta 660
acaccagatc tagatagaaa tcacaaatcg aagagtaatt attcgacaaa actcaaatta 720
tttgaacaaa tcggatgata tctatgaaac cctaatcgag aattaagatg atatctaacg 780
atcaaaccca gaaaatcgtc ttcgatctaa gattaacaga atctaaacca aagaacatat 840
acgaaattgg gatcgaacga aaacaaaatc gaagattttg agagaataag gaacacagaa 900
atttacctgc agggaccagt acaggcgaga agatcaccag gagaggtgtg gcgattgtca 960
gcgcaatgac cgttccagcc agggtcaacc cggataacac caacaggcta cctccggcag 1020
taaccgcggt cgctgccttt acaacacgct gagcacgcgg ttgcagttgc aagtgggggg 1080
cacgtgtttg ttgctgctgc ccgtagtgct ctgccatggt tttttttaac ggagcaagcg 1140
gccgctgttc ttctttactc tttgtgtgac tgaggtttgg tctagtgctt tggtcatcta 1200
tatataatga taacaacaat gagaacaagc tttggagtga tcggagggtc taggatacat 1260
gagattcaag tggactagga tctacaccgt tggattttga gtgtggatat gtgtgaggtt 1320
aattttactt ggtaacggcc acaaaggcct aaggagaggt gttgagaccc ttatcggctt 1380
gaaccgctgg aataatgcca cgtggaagat aattccatga atcttatcgt tatctatgag 1440
tgaaattgtg tgatggtgga gtggtgcttg ctcattttac ttgcctggtg gacttggccc 1500
tttccttatg gggaatttat attttactta ctatagagct ttcatacctt ttttttacct 1560
tggatttagt taatatataa tggtatgatt catgaataaa aatgggaaat ttttgaattt 1620
gtactgctaa atgcataaga ttaggtgaaa ctgtggaata tatatttttt tcatttaaaa 1680
gcaaaatttg ccttttacta gaattataaa tatagaaaaa tatataacat tcaaataaaa 1740
atgaaaataa gaactttcaa aaaacagaac tatgtttaat gtgtaaagat tagtcgcaca 1800
tcaagtcatc tgttacaata tgttacaaca agtcataagc ccaacaaagt tagcacgtct 1860
aaataaacta aagagtccac gaaaatatta caaatcataa gcccaacaaa gttattgatc 1920
aaaaaaaaaa aacgcccaac aaagctaaac aaagtccaaa aaaaacttct caagtctcca 1980
tcttccttta tgaacattga aaactataca caaaacaagt cagataaatc tctttctggg 2040
cctgtcttcc caacctccta catcacttcc ctatcggatt gaatgtttta cttgtacctt 2100
ttccgttgca atgatattga tagtatgttt gtgaaaacta atagggttaa caatcgaagt 2160
catggaatat ggatttggtc caagattttc cgagagcttt ctagtagaaa gcccatcacc 2220
agaaatttac tagtaaaata aatcaccaat taggtttctt attatgtgcc aaattcaata 2280
taattataga ggatatttca aatgaaaacg tatgaatgtt attagtaaat ggtcaggtaa 2340
gacattaaaa aaatcctacg tcagatattc aactttaaaa attcgatcag tgtggaattg 2400
tacaaaaatt tgggatctac tatatatata taatgcttta caacacttgg attttttttt 2460
ggaggctgga atttttaatc tacatatttg ttttggccat gcaccaactc attgtttagt 2520
gtaatacttt gattttgtca aatatatgtg ttcgtgtata tttgtataag aatttctttg 2580
accatataca cacacacata tatatatata tatatatatt atatatcatg cacttttaat 2640
tgaaaaaata atatatatat atatagtgca ttttttctaa caaccatata tgttgcgatt 2700
gatctgcaaa aatactgcta gagtaatgaa aaatataatc tattgctgaa attatctcag 2760
atgttaagat tttcttaaag taaattcttt caaattttag ctaaaagtct tgtaataact 2820
aaagaataat acacaatctc gaccacggaa aaaaaacaca taataaattt g 2871
<210> 177
<211> 1578
<212> ДНК
<213> Arabidopsis thaliana
<400> 177
gtcacacaca cataaacact ccacacgctc tgcttcgtcc aatcaccaaa cacgctttaa 60
tgactctcac gttttcctcc tccgccgcaa ccgttgccgt tgctgctgca accgtaacct 120
cctccgctag ggttccggtt tatccactcg cttcgtcgac tcttcgtgga ttagtatctt 180
tcagattaac cgcgaagaag ctgtttctgc cgcctcttcg ttctcgcggc ggcgttagtg 240
tgagagccat gtctgagctt gttcaggata aagaatcgtc cgtcgcggcg agcattgctt 300
tcaatgaagc cgccggtgag acgccgagtg agcttagtca ttcccgtact ttcttggatg 360
cgcgaagtga acaagatctt ttatctggta tcaagaagga agctgaagct ggaaggttgc 420
cagcaaatgt tgcagcagga atggaagaat tgtattggaa ctacaaaaat gcagttttaa 480
gtagtggagc ttccagggca gatgaaactg ttgtatcaaa catgtctgtt gcttttgatc 540
gcatgcttct tggtgtggag gatccttata cttttaatcc atatcataaa gcagtcagag 600
aaccatttga ctactacatg tttgtccata catacatccg tcctcttatt gatttcaaaa 660
attcgtacgt tggaaatgct tctatattct ctgagctgga agacaagatt cgacagggac 720
acaatatcgt gttgatatca aaccatcaaa gtgaagctga tccggctgtc atttctctat 780
tgcttgaagc acaatctcct ttcataggag agaacattaa atgtgtggct ggtgatcgag 840
tcatcactga tcctctttgt aagccgttca gtatgggaag gaacctcata tgtgtttact 900
cgaaaaagca catgaatgat gatcctgagc ttgttgacat gaaaagaaaa gcaaacacac 960
gaagcttaaa ggagatggct acaatgctaa ggtctggcgg tcaacttata tggattgcac 1020
caagcggtgg aagggaccgc ccgaatcctt ctactgggga atggtttcct gcaccctttg 1080
atgcttcttc ggtagacaac atgagaagac tggttgaaca ttctggcgct cctggacata 1140
tatatccaat gtctttgctt tgctatgaca tcatgccccc tccaccccag gttgagaaag 1200
aaatcggaga gaaaagatta gttgggtttc acggtactgg actatcaatt gctcctgaaa 1260
tcaacttctc agacgtcaca gcagactgcg agagccctaa tgaggcgaaa gaagcataca 1320
gccaagcttt gtacaagtcg gtgaatgaac aatacgagat cttaaactct gcgattaaac 1380
acagaagagg agtagaagca tcaacttcaa gggtctcttt gtcacaacct tggaattagt 1440
ctctcgtttt agggtaacac tttcaaaact cataaatctt ctgtctcaga agttttgttg 1500
caactgtata tatattgaga gagagagcat tgttctttca tttgcaggat acacaaacac 1560
aatcaatgga aaatactc 1578
<210> 178
<211> 459
<212> Белок
<213> Arabidopsis thaliana
<400> 178
Met Thr Leu Thr Phe Ser Ser Ser Ala Ala Thr Val Ala Val Ala Ala
1 5 10 15
Ala Thr Val Thr Ser Ser Ala Arg Val Pro Val Tyr Pro Leu Ala Ser
20 25 30
Ser Thr Leu Arg Gly Leu Val Ser Phe Arg Leu Thr Ala Lys Lys Leu
35 40 45
Phe Leu Pro Pro Leu Arg Ser Arg Gly Gly Val Ser Val Arg Ala Met
50 55 60
Ser Glu Leu Val Gln Asp Lys Glu Ser Ser Val Ala Ala Ser Ile Ala
65 70 75 80
Phe Asn Glu Ala Ala Gly Glu Thr Pro Ser Glu Leu Ser His Ser Arg
85 90 95
Thr Phe Leu Asp Ala Arg Ser Glu Gln Asp Leu Leu Ser Gly Ile Lys
100 105 110
Lys Glu Ala Glu Ala Gly Arg Leu Pro Ala Asn Val Ala Ala Gly Met
115 120 125
Glu Glu Leu Tyr Trp Asn Tyr Lys Asn Ala Val Leu Ser Ser Gly Ala
130 135 140
Ser Arg Ala Asp Glu Thr Val Val Ser Asn Met Ser Val Ala Phe Asp
145 150 155 160
Arg Met Leu Leu Gly Val Glu Asp Pro Tyr Thr Phe Asn Pro Tyr His
165 170 175
Lys Ala Val Arg Glu Pro Phe Asp Tyr Tyr Met Phe Val His Thr Tyr
180 185 190
Ile Arg Pro Leu Ile Asp Phe Lys Asn Ser Tyr Val Gly Asn Ala Ser
195 200 205
Ile Phe Ser Glu Leu Glu Asp Lys Ile Arg Gln Gly His Asn Ile Val
210 215 220
Leu Ile Ser Asn His Gln Ser Glu Ala Asp Pro Ala Val Ile Ser Leu
225 230 235 240
Leu Leu Glu Ala Gln Ser Pro Phe Ile Gly Glu Asn Ile Lys Cys Val
245 250 255
Ala Gly Asp Arg Val Ile Thr Asp Pro Leu Cys Lys Pro Phe Ser Met
260 265 270
Gly Arg Asn Leu Ile Cys Val Tyr Ser Lys Lys His Met Asn Asp Asp
275 280 285
Pro Glu Leu Val Asp Met Lys Arg Lys Ala Asn Thr Arg Ser Leu Lys
290 295 300
Glu Met Ala Thr Met Leu Arg Ser Gly Gly Gln Leu Ile Trp Ile Ala
305 310 315 320
Pro Ser Gly Gly Arg Asp Arg Pro Asn Pro Ser Thr Gly Glu Trp Phe
325 330 335
Pro Ala Pro Phe Asp Ala Ser Ser Val Asp Asn Met Arg Arg Leu Val
340 345 350
Glu His Ser Gly Ala Pro Gly His Ile Tyr Pro Met Ser Leu Leu Cys
355 360 365
Tyr Asp Ile Met Pro Pro Pro Pro Gln Val Glu Lys Glu Ile Gly Glu
370 375 380
Lys Arg Leu Val Gly Phe His Gly Thr Gly Leu Ser Ile Ala Pro Glu
385 390 395 400
Ile Asn Phe Ser Asp Val Thr Ala Asp Cys Glu Ser Pro Asn Glu Ala
405 410 415
Lys Glu Ala Tyr Ser Gln Ala Leu Tyr Lys Ser Val Asn Glu Gln Tyr
420 425 430
Glu Ile Leu Asn Ser Ala Ile Lys His Arg Arg Gly Val Glu Ala Ser
435 440 445
Thr Ser Arg Val Ser Leu Ser Gln Pro Trp Asn
450 455
<210> 179
<211> 2455
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Populus trichocarpa
<400> 179
agtgcgggtg attgggtgag gagtgaagac gctgatttta gaggttttga gagagtggca 60
gtctgcagag aataggaatc cgaccatatc ctccaaaacc cgcgctggac tcagtcaccg 120
ccaatcatca atcagccacc catcaacacc aaaaatcccc gtccttttga tttccaccac 180
ataaaaatag cacactgctc ctccttcact ccattcctat cttaataata ataataataa 240
taaagctcaa ctcttctctt ctaagtcaag acatgatcct ttccattcct gctccttcgt 300
cggcattctt cacaactact aaaccgtctc caccttttcc tagggtttct aaactctgct 360
tcttaacccc ctcatattct ctttcccttc gttttagatc cactgctcga cgctccactt 420
cttttccttg tgtcctctct tctctcaacc ttcacgcaat ggctgaactc gttcaggata 480
aagaagtctt cgcttctgct gaagttgatt acagcaagaa gaaaaacagg actcgttctc 540
gctcgtttct tgatgcaaca actgaacaag agttactgtc gggaatcagg aaggaatcag 600
aagcaggaaa acttccttca aatgttgctg caggaatgaa agatctgtat cagaactaca 660
aaaccgcagt tttgcaaagt ggaattccca acgcacatga gattgtattg gaaaatatgg 720
ctgctgcatt ggatcttata ttctttgatg ttgaggaccc gtttatcttc tcaccttatc 780
acaaagcttt gagaaagcca tatgactact ttgaatttgg tcaaaagtat atccgtccat 840
tgattgattt tagaaattca tatgtaggca atgtttccat tttcaatgaa attcaagaga 900
agcttcggca gggtcacaat attgtcttga tatcaaacca ccaaactgaa gcagatccag 960
ctgtcattgc actgttgctt gaaacatcaa gccctcacat tgctgaaaac ttgatctatg 1020
ttgctgggga tagagttgtc acagatcctc tttgcaagcc attcagcatg ggaaggaatc 1080
ttatatgtgt atactcaaaa aagcacatga atgatgaccc tgaacattca gaggagaaga 1140
gaaaagcaaa tatccgaagt ttgaaagaga tggctttgct tttaaggggt ggctcacaaa 1200
tagtctggat tgcaccaagt ggtggcaggg accgtccaga tcccttgtca ggagagtggt 1260
atccggcaca ctttgatgct tcttcagtag acaacatgag aaggcttgct gaacattctg 1320
gagctccagg acatgtttat cctctggcac tattatgcca tgacatcatg ccccctccgc 1380
ctcaggtgga aaaggaaatt ggagagagaa gagttatttc atttcatgga gttggattat 1440
cagttgcacc agaaatcagc ttctctgaag ttacagcggc atatgaaaat cctgaagagg 1500
ctaaggaggt atatacagag gctctgtata agtctgtgac tgagcaatac aatgtgctta 1560
aatctgctgt acatggaaaa caagggctag gggcgtccat tccaactgtt tctttgtcac 1620
agccatggaa ttagtcaacc ttttctatac ttgattaggc caatagtttt gttatatagt 1680
tctgcaactc ctggaccaca attctagcgg tccttctagt caagtatgtg ccaggagaag 1740
cttctctctc catgatgata tggatggctt tttctggaga tgcaatctaa gctacaagtt 1800
tttgctgtgc ttacattcta tcaaagccaa atctcacaca atatcttgaa gccaaattca 1860
tctgaaacgc gagctgttcc agaggttcaa tttcaggtgt gcagataaca gttcctagta 1920
aacacaagag ctagtcgtct gaggcgatat acatgtatat tttctcaatt ttttggtggc 1980
cgatcatatt ctttttacac caattgctca attgctactc atttttctcc ctcgttcacc 2040
ttcaataact agaagttttc atgctataac acttgcacac agaagtacta tgaacagagt 2100
tggagcacat tttgcctctt gactaaacaa gacttgtttt tagctgccac accaaacttt 2160
ttatatgatg caattatggt agtcgttttc tcttgttttg gtcaaaaccc aaaccagcta 2220
tagttgctac agccaatcga gagtggtgca tgtttgtttg tttttttttt tttttttgtc 2280
ctcagttata gtaaccatgt tcaactgaac tatgcatctc ttaggacacc acctcttaag 2340
ccccgtgatc taaccgtgtt ttcgaatttt tttttttttt ttgggctttt ggtttattta 2400
aacgcagcag ctttgaccca agttaaaaca aaaaaatcta ttaaaaaaat tgtag 2455
<210> 180
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Jatropha curcas
<400> 180
atgacacttt ctgcttttcc ttccacattc ctctttagaa tacaatcgcc atcaacgcct 60
agggtttcca tttccctccc ttccttatct tcaaagctct gtttggttcc tccctctttt 120
tctcctcctt cgcttgctct taaatcgagt gcgcgaagga ccatttgtcc ttgcttgctc 180
tcttctctca acgccaacgt ggctcacctt ctcaaggagg aaaaagaagt tgtggcttcg 240
gcttccggct gcgagaagga ggaggaaaag aagatggaac agcctagtca ctcccgcact 300
ttcctgcatg ccagaacgga acaagatttg ctgtctggaa ttagaaaaga agcagaagca 360
gggaggttgc cttcaaatgt tgcagcaggg atggaagaat tgtatcagaa ttatagaaat 420
gcagtgatac aaagtggaac ccccaatgca gaagagatca tactgtcaaa tatggccgtt 480
gctttggatc gtataagctt ggatgttgag gacccttttg tcttctcaca ttatcacaga 540
gcattgagag agccgtttga ctactataac ttcggtcaaa attatattcg tcctttggtt 600
gattttagaa attcttatgt tggcaatatt tcccttttcc atgaagtgga agagaagctt 660
cagcagggtc ataatattgt cttgatgtca aatcaccaaa ctgaagcaga cccggctata 720
attgcattgc tgcttgagaa aacaaagccc tatattgctg agaatttgat ctatatagca 780
ggtggtagag tcataacaga tcctctttgc aagccattca gcatgggaag gaatcttata 840
tgcgtgtact caaaaaaaca catgaatgat gttcctgagc ttactgagat gaagaaaaga 900
gcaaacatac ggagtttgaa ggagatggcc attccattaa ggggtgggtc acgaatagtg 960
tggattgccc caagtggtgg tagggaccgc ccagatcatc tgactggaga atggtatcca 1020
gcaccatttg atgcttcttc agtggataac atgagaaggc ttgctgaaca ttctggtgct 1080
cctgggcata tttatccatt ggcattatta tgccatgaca taatgccccc tccccttcag 1140
gtgcaaaagg aaattggaga gaaacgagtg atctcctttc atggggttgg attatcaatt 1200
gcaccgggaa tcagcttctc tgaaattgcg ggtagttgtg aaaatcctga agaggcaaag 1260
aacatttatt cacaacttct gtatgattca gtgactgcgc aatacaacgt gcttaaatct 1320
gccataaatg gcaaacgagg gctagaggct tcaattccaa ctgtctcttt gtcacaacca 1380
tggaattaa 1389
<210> 181
<211> 1368
<212> ДНК
<213> Ricinus communis
<400> 181
atgattcttt ccattctttc ccctacacta ccatcgccta gggtttgtat ttccatttct 60
tctgtatctt caaagctctc tctagtccct gtctcttctt tttctcttcc tcctcctttg 120
gccatagtaa gatggtcatc aaggtcctcc atttgtcctt gtttcttctc ttcttctctc 180
aacgccaatc cagtccccga actcctcaac gatgataaga agaagaacaa caacaacaac 240
aagagcaaga agggaaagtg tactcctcac tcccgcactt ttcttgatgc aagaactgaa 300
caagagttgc tgtatggaat taggaaggaa gcagatgcag ggaggttgcc tttaaacatt 360
gcagcaggga tggaagaagt ttatcggaat tatagaaatg cagttttgca aagtggaatt 420
ccaaatgcaa aagaaatcat actgtcaaat atggctgttg cgttagatcg tatgtgcttg 480
gatgttgagg acccttttgt cttctcacct tatcataaag cactaagaga accattcgat 540
tactataatt ttggtcaaaa ttatatccgt cctctgattg attttaggaa ttcatatgtt 600
ggcaacattt cgcttttcca tgaagttgag cagaagcttc agcagggtca caatattatt 660
ttgatgtcaa accaccagac tgaagcagat ccagctgtca ttgcattgtt gcttgaaaaa 720
acaaatccct acattgctga gaatttgatc tacgttgcag gtgatagagt tgtaacagat 780
actctatgca agccattcag catgggaagg aatcttatat gtgtgtactc gaaaaaacac 840
atggctgatg ttcctgagct tactgagatg aagaaaaaag caaacattcg cagtttaaag 900
gagatggtca tgattttaag ggatgggtct caaattgttt ggattgctcc aagtggtggc 960
agggaccgcc cagattcttt gactggagaa tggtgtccag caccctttga tgcttcttca 1020
gtggataaca tgagaaggat tactgaacat tctggcgctc caggacatat ttttccatta 1080
gcgttgttat gccacgatat catgccccct ccacctgagg tacaaaagga aattggagaa 1140
agaagaatga tctcctttca tggagctgga ttatctattg cacccgaaat cagcttctct 1200
gaaattgctg ttgcttgcga agatcatgaa gaggctaaga acgcatatgc acaggtttta 1260
tatgattctg tgactgagca atacaatgtg cttaaatctg ccatacatgg aaaacaagga 1320
ctagaggcat caacttctac cgtctcattg tcgcaaccat gggattag 1368
<210> 182
<211> 1344
<212> ДНК
<213> Helianthus annuus
<400> 182
atgtcgattc tcccgtcttc ttctcctact ctcttcttct ccaccgcaaa ccctagggtt 60
tctgtttctc tttcacttac ttctacagtt tctacatctt catccgtgcg cagtcgctcg 120
attttccggc attttccgta cctagcgttt tctagggcag cgaatgccgc cgcggagacg 180
tttgaaggca agaagtggtc gtcgtcctcc gctacacaac cgatctccgg atccgagctc 240
ggttactcgc atacattcat cgatgctctg tctgaacaag atcttctttc tgtaattcaa 300
agagaggtag aagctggagc actgccaaaa catatcgctc actcaatgga ggaactctat 360
cagaactaca aaaatgcggt tttccaaagt ggtaatccct gtgcagaaga tactgtattg 420
tcaaacatgc gtgtagcatt tgatcgaatg ttcttggatg tgaaggagcc tttcgaattt 480
tcaccgtatc atgaagctat tcgagagcct tttaattact atatgtttgg tcaaaattat 540
attcgtcctc tgatcaattt cagggaatca tatgttggca acgtctctct tttcagtgaa 600
atggaagaac aactgaagca gggtgaaaat gtaattttga tctcaaacca ccaatccgaa 660
gcagatccag ctgtcattgc cttgttgctt gaaacaacaa atccttatat ttccgagaac 720
ataatctatg tggcagggga cagagttata acggatcctc tttgtaagcc tttcagcatg 780
ggaaggaact tgctgtgcgt atattcaaaa aaacatatga acgatgttcc tgagcttgct 840
gatatgaaaa ggagagcaaa tacaagaagt ttaaaagaga tggctttgct tttgaggggt 900
ggatcaaaaa taatatggat tgcaccaagt ggtggaaggg acaggcctga tcccgtcaca 960
aatcaatggt ttccagcacc attcgatgcc agttctctgg acaacatgag aaggcttgtg 1020
gaccatgctg gtgtggtggg tcatatatat cctttagcca tactatgcca tgacatcatg 1080
ccccctcctc ctcaggttga gaaagaaatt ggagagaaaa ggttgatatc ttttcatggc 1140
actggaatat cagttgcacc tgaagttgat ttccaaaacg ccactgcttc ttgtggatcc 1200
cccgaggagg ccaaggcagt ttattcacag gcactttatg attcagtgtg cgagcaatac 1260
aacgtgctac aatccgccat aaatggagca aaaggcttag aagcatcaac atcaagtgtc 1320
tcattgtcgc aacctgttga ctag 1344
<210> 183
<211> 1374
<212> ДНК
<213> Medicago truncatula
<400> 183
atgtttacaa caccattttc ttctccttca accgcatttt tctctccacc taaagcctca 60
tattcttctt cttcttcttc ttcttcttct tcttcttcgt tacctcttcg tagttctttc 120
actttttatc atcttcgatt taatgcaaca acttcttctt cttctgtaac aacttctgga 180
acttcttctt cttcatattg ttctcctctt gctttcaatt ctaataataa aaaacctaaa 240
gaaatttctg ctaatatggc ggcttcttct gtttcttctc gcactttcct caatgccaga 300
aatgaacaag atgttctttc tggaattaag aaggaagtag aagccggaac tttgcccccc 360
actattgctg aagggatgga agaattgtac cttaactata aaagtgcagt tgttaaaagt 420
ggagatccca aagcagatga gattgtattg tcaaatatga ctgctttatt agatcgcata 480
tttttggatg tgaaggagcc ttttgtcttt gaagcacacc ataaagcaaa gagagagcct 540
tttgactact acatgtttgg ccaaaattat attcgtccct tagttgattt caacacttct 600
tacgttggca acatgcccct tttcatacaa atggaagagc aacttaagca gggacacaat 660
attatcttga tgtcaaacca ccaaagtgaa gctgatccag ctattattgc attgctgctt 720
gaaatgcgac ttccacatat tgctgaaaac ttgatttatg tggcaggaga tagagttata 780
accgatcctc tatgcaagcc cttcagtatt ggcaggaatc tgatctgtgt ttattcaaaa 840
aagcacatgc ttgatgatcc agcacttgta gagacgaaaa gaaaagcaaa tacacgaagt 900
ctgaaggaaa tggccacgct tttaaggagt ggatcacaaa taatttggat tgccccaagc 960
ggtggtaggg atcgaccagt tgccaactct ggggaatggg caccggcacc ctttgattct 1020
tcttcagtgg acaatatgcg aaggcttgtc gatcattcag gtccaccagg tcatatctat 1080
cctatggcaa tactgtgcca tgacataatg ccccctccac taaaggttga aaaagaaatt 1140
ggggagaaaa gaattatatc atatcatggg actggcatat cacttgctcc agaaataagc 1200
ttttccgaca tcactgcttc ttgtgaaaat cctgaaaagg ctaaagaagc atactcgaaa 1260
gccttgtatg attctgtgac tagtcaatat gatgtgctgg agtctgccat acacggcaaa 1320
aaaggattag aagcatcaac tcccgcagtt tccttgtcgc agccatggaa gtag 1374
<210> 184
<211> 1967
<212> ДНК
<213> Glycine max
<400> 184
ggctgagact gaggagcgga tcctatctct ctttcacaca ctctccttct ctttcgtatg 60
aagaatgagc acgaccggtt cttcggctta ccactgtgtg gcacacctcc caaataataa 120
gactatgttt atgctctcta cgccgccaac aaccacattc ttcgctacgc ctagggttct 180
tccgtttctc tcttcaaaac tttcttcttc ttcttcttct tctactgcgt cgtcctcgcc 240
ttgttgctcc tccatcactc ccaaggttaa atccaaagat aacaacaatt gctacctcgt 300
ctccgctaaa cattctcccg ctaacatgtc cgcttcggtt tcgtcacgca ccttcctcaa 360
cgctcggaac gaacaagagc ttctagctgg aatcaggaaa gaagtagaag ctggatctct 420
gcctgctaat gttgctgcag gaatggaaga agtgtacaat aactataaaa gtgcagttat 480
ccaaagtgga gatcccaagt caaaggagat tgtattgtcg aatatgattg ctttattgga 540
tcgcatattc ttggatgtga cggatccttt tgtctttcaa ccacaccaca aagcaaagag 600
agagcctttt gactactacg tgtttggtca gaattatatc cgtcctttag ttgatttcaa 660
aaattcttat gttggcaaca tgcccctttt cattgaaatg gaagagaaac ttaagcaggg 720
acacaacatc atcttgatgt caaatcacca aactgaagct gatccagcca tcattgcttt 780
gctgctcgaa acacgactcc catatattgc tgaaaacatg acctatgtag caggagatag 840
agttataact gatcctctgt ccaaaccatt cagtattggc aggaatctca tttgtgttta 900
ctctaaaaag cacatgcttg atgatccagc tcttgtagag atgaaaagaa atgcaaatat 960
acgagctctg aaggaaatgg ctatgctttt aaggagtgga tcacaaatag tctggattgc 1020
cccaagtggt ggaagggatc gcccagatcc ccacaccgga gaatgggcac cggcaccctt 1080
tgatacttct tcggtagata atatgagaag acttgttgaa cattctggtc caccgggcca 1140
tgtatatcct ttggcgatat tgtgccatga tataatgccc cctccactaa aggttgagaa 1200
agaaattggg gagaaaagaa ttatatcctt tcatgggact ggcatatcag tggctccagc 1260
attaagcttt tctgaaacta ctgctactag tgaaaatcct gaaaaggcta aggaggtatt 1320
cacaaaagcc ctgtatgatt ctgtgacgga gcaatataat gtgctgaaat ctgcaataca 1380
tggcaaaaaa ggatttgaag catcaactcc agtagtttct ttgtcacagt catggaagta 1440
gatgaaatct gcatttcttc attgcaattt gctctgatgc agaagcaagt tacaagactt 1500
cagtcaaaca atttcaactg attcacttct gagggactgc ctattactac accggtcacc 1560
gaatgattta gcttgttgga agtttgcagt caaatacata tttttcattt catttttcct 1620
tttgctcttg gttgccgtta tcagcattca attcatctgg aatctgtttc agttcagaag 1680
gttcaaattc tgctgcttac tgtacaggtc tctcttagtt cggtgtcaga tttggttcgt 1740
tgactgataa aatactaaat tttttaccta caattttgtg atcaggctta gctagctgaa 1800
tagataaaat ataattggtt ccatttgtat tttaagtcaa ctttgttcca ttatagatga 1860
atagatgtta gtattacatg ttcagacggg gtcagtgaat aaactggtcc aaatgctaat 1920
gcaaaattat tcatattggt aaaataaaag ctctacagtt accgtta 1967
<210> 185
<211> 1674
<212> ДНК
<213> Carthamus tinctorius
<400> 185
tctctctctc tcacacacaa cacacaaaac acacactact gctactttct ctctctacta 60
cactctcctc tcgctatgtc gatcttcttc tctccttcct cccctactct cttcttctcc 120
accacaaacg caaatcctag ggtttctcct tcatcttcac cttcttctgc cttcactcct 180
cctctgtctt cttctcgcct ccgcccgatt ctccgggggt ttccgtgcct cgcgttctct 240
gcgccggcga atgccgccca tggcacggcg gagaccgtcc acggcaataa gtggccgtca 300
ccgtcgtcct cctcctctgc tgctacgcaa ccgtccgctg gatccgacca cggtcactct 360
cgtacattca tcgatgctcg ttccgaacaa gatcttcttt ctggaattca aagagagttg 420
gaagctggaa cactgccaaa acatattgct caagcaatgg aggagctata tcagaactac 480
aaaaatgcag ttctccaaag tgcggctcct catgcagaag atattgtgtt gtcaaacatg 540
cgtgtagcgt ttgatcgtat gttcttggat gtgaaggagc cgtttgaatt ttcaccatat 600
catgaagcta ttttggaacc ttttaactac tatatgtttg gtcaaaatta tattcggcct 660
ttggtcaatt tcagggaatc atacgttggc aatgtctccg ttttcggtgt aatggaagag 720
cagcttaagc agggtgacaa ggtggttttg atctcaaacc atcaaacaga agcagatcca 780
gctgttattg ccttgatgct tgaaacaaca aacccccata tttctgagaa cataatctac 840
gtggcagggg atagagtaat aacagatcct ctttgcaagc ctttcagcat gggaaggaat 900
ctgttgtgcg tgtattcaaa aaagcatatg aatgatgttc ctgagcttgc tgagatgaaa 960
aaaagatcaa atacaagaag tttaaaaggg aggatggctt tgcttttgag gggcggatct 1020
aaaataatat ggattgcgcc aagtggtggc agggacaggc cagatcctat cacaaatcag 1080
tggtttccgg caccgtttga tgccacttcg cttgacaaca tgagaaggct cgtggaccat 1140
gctggtttgg tgggtcacat atatccttta gccatattgt gccatgacat catgccccct 1200
cctcttcagg ttgagaaaga aattggagag aagagttgga tctcttttca tggcaccgga 1260
atatcagtgg caccggaaat taatttccaa gaagttactg cctcttgtgg gtcccccgag 1320
gaggcgaagg cagcttattc acaggcactc tatgattccg tgtgtgaaca atacaaggtg 1380
ctacattctg cggtacatgg aggaaaaggg ttagaagcat caacaccaag tgtctcgttg 1440
tcacaaccct tgcagtttct cgattagtct cttggtttag aggaggtgaa agcatattct 1500
tttgtttaga tgacataggt gtatagatga taccgaagaa tagatgtaca aacaagtgat 1560
agaaagatgt atgtctaatc aaaaaatgtt ttctgcatct tgtaaaggga tcttcaaaac 1620
agacctttta ttttagctgc agcaaccaat atatcaaaac aggtttttct tttt 1674
<210> 186
<211> 1893
<212> ДНК
<213> Solanum tuberosum
<400> 186
cgcacatatt catttcactc actttctttc ccgacccttt ctctctctaa agctctccag 60
tctgtggtga tgttgatcct ctcagcggct tcgtcttctt cttcctcctt catgctttct 120
tccgcttcgt cttcttctgc acgcattccg aggcagttat cttcattttc aacttgtgtt 180
ccagtagtag taacaactgt ttcttctgca gcaacttcga ctctatttcc gatttcctgc 240
ttcggtgtga aatcgaggac tgttgggatt cggaagctgc ggtgtgccgt tttttgtgct 300
tcgaaggtac gtggaatggc agaaatgatt gaagatgcca tgacggtttc tgcttctgag 360
agccatgagc ttccgcagtc ccgagacttc cttgacgcac gcactggaga agacttgcta 420
tctgctgttc aaaaagctgt ggaagatgaa aaactgccgc ttaatgttgc tgaaggaatg 480
gaggatttgt atcagaacta tcggaatgca gttttacaaa gtggagtccc caaagcagat 540
gaggccactt tgtataacat ggctcttgta tttgatcgtg tttttgtgga tgtgaaggat 600
ccttttgaat tctcgccata tcataaggcc attcgtgaac cttttgacta ttacaagttt 660
ggtcaaaatt atatccgcca gctagttgat ttcaggagtt cttatgttgg gaatatctca 720
gttttcggtg aaatggcaga gaagcttaaa cagggtgata atgttgtctt gatgtcaaac 780
catcaaagtg aagccgatcc tgcgattatt gcactcttga ttgaatcaaa gctcccagat 840
attgctgaga acattattta tgttgctgga gatagagtta ttactgatcc tctttgcaag 900
ccattcagca tgggaaggaa tctcctgtgt gtttattcga aaaaacatat gaatgatgac 960
cccgaacttg ctgagatgaa aaagagagca aacacaagaa gcttgaagga gatggctttg 1020
ctattgaggg gtggatcaaa aataatatgg attgctccta gtggtggaag agataggcca 1080
gaccctgtta caaacgaatg gtatccagca ccatttgatg cttccgcgac agacaacatg 1140
aggaggcttg tacaacatgc tggtgtccct ggtcacattt atcctctagc aattttgtgc 1200
catgatatta tgccccctcc cgcccaggtt gagaaaaata tcggggagaa aagagttgta 1260
tcttttcatg gagctggcat atctgtggca cccaaaattg attttcatga ggttgctggt 1320
gctttggagg accctgaggc taagatggta tatacaaagg cactttatga ctctgtaagc 1380
cagcagtaca atgtgctaaa ttctgctata catggcaaac aaggactgaa ggcatcaata 1440
cctagtgttt cattatcaca accatggcag tagcttctct tccaacttta tttttcatat 1500
cttgttgctg tagtcagttt tgcagatgtt tgtttggcag ttacaatcaa atcacaagga 1560
ttacactcac aatctttcca cataccacgc ttgcatgtgg ttagtctatg cagaaagttg 1620
atacaaacaa agtaattctc gaagttacag caaacataac ctgaaggaat ttttttggca 1680
gggttagata attcttttga cacgaatgta cagttgcttt acattgtatt tataccaaat 1740
gttagatcca aatttgttag taatgatagc tttcaagtac tcaattctga ctttttaagg 1800
tcaagtgtta gtagctatcc tagattgctg ctcatcttgc ctttgaagtg gtaatccaat 1860
ttgttgagaa atataataaa tgatgctctg cta 1893
<210> 187
<211> 2016
<212> ДНК
<213> Oryza sativa
<400> 187
gggctggaga tggagatgga gatggagatg gagggtgggt ttggcacaaa tcccgaagcg 60
ctccggcgac cactcccaac ccagtcccca ctagggtaac aacccccttt cggattaggt 120
ttctagaagc ttcttctatg caggcgccgc cgctcgcctc ctcgccgtcg ccggcgtgga 180
ccgccatcct gcccgcgccg gcgaggctct gctgctcccg ccgcggcgcc ctccgcctcg 240
aagccaaggc cgcctggagg ccggcggccc gagggccgcg ggtgccggcc aagggcgccg 300
tcctcgcctc cgaggtggtg ggcccctctc ccctcctcga cgcgcgcaac gagcaagagc 360
tcattttgca tatcagaaag gaagtggaga aagggaagct gcctgcagat gtcgctgcca 420
atctagaaga gctatactac aactacaagg acgcggttat gcagagcagg gatccaaatg 480
cacacgacat cgtgctttca aacatggtgg ccctgttcga ttgtgttctg ctcgatgtag 540
agaatccgtt tacctttccg ccttatcaca aagctgtcag ggaaccattc gactattaca 600
tgtttggtca gaactacatt aggccccttg tagactatag aaattcatat gttggtaata 660
tatccatttt ccaagacatg gaacagaagc tccaacaggg ccataatgtt gttctgatgt 720
ctaaccatca gacagaagca gatccagcaa tcattgcttt gctgcttgaa agaagcaacc 780
catggatcag cgaaaacata gtttatgttg ctggtgatag ggttgttaca gatcctctct 840
gcaagccatt tagtatggga agaaacctca tttgtgtgta ctcaaaaaag catatgaatg 900
attttcctga gctagttgat atgaagagga gggcaaatac tcgtagtctg aaggaaatgg 960
ctttactttt acgtggcggt tcacagataa tttggatagc accaagtggt ggtagagatc 1020
gtccggatcc tttgacagga gaatggcatc cggcaccatt tgatgcatct gcagtggaca 1080
acatgaggag gcttctggag cattctggtg ttcctgggca catatatcca ctctcactgc 1140
tctgctatga ggttatgcct ccaccacaga aggttgagaa agagattggt gagcaaaggg 1200
ttatatcctt ccatggtgta ggcttgtcag taactgaaga gataaagtac agcgacatta 1260
cggttcatac ccaaaatgtc gacgagtgca gagagaaatt ctcagagtca ttgtacaact 1320
cagtcgttga tcagtataat gcgctcaaat ctgctatctt tagaggtcga ggagcagatt 1380
catcggacag tgccatctca ctctcacaac catggcgatg aaactccgct ttctcagttt 1440
tgttctgtct ggatttctca atgaagttac cttcatttct tttcgacaca gcagatgaac 1500
tgctgccgac attgcaattt ttcctggcag aaccttttaa acttcggtat cctaacccat 1560
actaatcatg aaggggaggc tgttactgtc atgcaaatct tgcctagtat gatgatttta 1620
cccagctgaa tcccagccac acatgatgcg ttcgttcatt gtttgcacac aaatattatt 1680
gcgtcatatg agtattcttt gggtcagaac tgcacagcaa cgcggcctgg gcactcaatc 1740
tggcatgttg tctatggggt gcatgcttgt taacagaaga agcccaacat gtgggatttt 1800
gttttttgcg gttaattttt ttcctgtttt ccttttgttc catgtatata tattcgattt 1860
tgatctccag gtttggagat acaatggtca aagtgttatg atagtctctt agtttgttgc 1920
ctcgaagtta tactcgggcg caacatgtct gactgatatt ctgatgatgt tactcgtttc 1980
tgaacttcct gacgccaata tggtgcttgg atgttg 2016
<210> 188
<211> 1888
<212> ДНК
<213> Sorghum bicolor
<400> 188
atgcacgcgc cgccgctggt cgcgttcgca gggggcgcct gccccgccac cgccgcctcg 60
tcctcgccgt cgccctggct ggcctcgccg cgggccgcca tcctcgccgc gccggcgagg 120
ctcctacggt cccgccgcgg ggcacttcgg ctggaagcca aggccgcgtg gagggctgcc 180
ggagggggac ggggcccgag ggtcccggcc aagggcgctg tgctcgcctc ctatatgggc 240
gccgaggagg tggtgggacc ttcgtcgctg ctcgacgagg aagagctcat ttcacatatc 300
agaaaggaac tggataatgg aaaactccct gcagatgttg ccagtaatct ggaggagttg 360
tattataatt acaggaatgc tgttctgcaa aatggagatc ctaatgcata tgagatcatg 420
ctttcaaata tgacggcttt gtttgatcgt gttctactgg atgtacagaa tccatttacc 480
tttccacctt atcacaaagc tgtgcgagaa ccgttcgact attacatgtt tggtcagaac 540
tacattaggc ctctggtaga tttcaggaac tcctatgttg gcaacatttc actttttcat 600
gacatggaag agaagctgca ccagggccac aatgttgttt tgatgtctaa ccatcagaca 660
gaagcagatc cagcaattat ctccttgctt cttgaaaaaa ccaatccatg gattagtgaa 720
aacatagttt atgttgctgg agatagggtt gttatggatc cactttgcaa gccatttagc 780
atgggaagaa atctcatttg cgtgtactcg aaaaagcata tgaatgattt tcctgagcta 840
gttgagatga agaggagatc aaatacccga agtctcaagg aaatggcctt gcttttacgt 900
ggtggctcac agttaatttg gattgcacca agtggtggta gagaccgccc aaatccttca 960
acaggagaat ggtacccggc gccattcgat tcatctgcag tggacaatat gaggaggctt 1020
ctggagcatg ctggcgttcc tgggcacata tatccactat cattgctgtg ctatgaggtt 1080
atgcctccac cacaacaggt tgagaaagag attggtgagc agagggtgat atccttccat 1140
ggagtaggct tgtcagtaac tgaagaaata aaatatgggg atattactgc tcataccaag 1200
aatgctgatg agggaaggga gctattcaca aatactttgt acaactcagt tgttaatcag 1260
tacgatgtgc tcaaatctgc tatctttaga gatcgtggag cagctgtatc aaacaatgtc 1320
atctcattgt cacaaccatg gagatgaatg ttagctttct cagtttgggt ccagatttat 1380
tactgaagtt accttttcag aagagcaggt gaactgccat tgtgcaattt cactggagaa 1440
actcttgaac tttaatcttt ttgataccac tcgactttat cagtcatggt ggagcctgtc 1500
attgtcatgc agatccttgc taagaagtct gtggacaact gttggttggt caagggtgac 1560
tggtgattct gcacataggg atcctcgtaa ctgttgcatg cggtcgtccg caaattactg 1620
gttgctcagc aacgtgctgg ttgggcactg aggaatccgt caggttgcat cctttttgcc 1680
ttgacgtcaa tttgtgtagt tgaaggttga agtgataaat tgttttatct tgtcttgtca 1740
tcatgtatat aggctcgagt cttttttggc tccacatttt tttggagata taaaagcagc 1800
aggagttatg acatgccctc agtcggccct ccttgttgaa accctttgga tgtaacctgt 1860
ctatttctta tatatactca ctgaaagt 1888
<210> 189
<211> 2046
<212> ДНК
<213> Zea mays
<400> 189
gaggattcat ctcgtgtcga cgacgccttc gccctctccg agtctccgtc cgtcttccgc 60
gtcctccgca gctggactcg tgcgctattc cccaggacgc tactcccact agggttttcg 120
gattaggttt ctagaacctt ccaccgccgc ctctccatgc acgcgccgcc gctggtcacg 180
ttcgcagggg gcgcctgccc caccaccgcc tccgcttcgc cgtcgccctg gttggcctcg 240
ccgcgggacg ccatctttgc cgcgccggcg aggcccctac ggtcccgccg cgggacactc 300
cggctggaag ctaaggccgc gtggagggct gccggagggg gacggggccc gcgggtcccg 360
gccaagggcg ctgtgctcgc ctcctatatg ggcgccgagg aggtggtggg accatcgtcg 420
ctgcttgacg aggaagagtt catttctcac atcagaaagg aactggataa tggaaaactt 480
cctgcagatg ttgccagtaa cctggaggag ctgtattata attacaggaa tgcggttctg 540
caaaatggag atccaaatgc atatgaggtc atgctttcaa atatgatgac cttgtttgat 600
cgtgttctac tggatgtaca gaatccattt aactttccac cttatcataa agctttgcga 660
gaaccgttcg actattacat gtttggtcag aactacatta ggcctctggt agatttcagg 720
aactcctatg ttggcaacat ttcccttttc catgatatcg aagagaatct ccaccagggc 780
cacaatgttg ttttgatgtc taaccatcag tcagaagcag atccagcaat tattgccttg 840
cttcttgaaa aaaccaatcc ttggattagt gaaaacatag tttatgttgc tggcgatagg 900
gttgttaccg atccgctttg caagccattt agcatgggaa gaaatctcat ttgcgtgtac 960
tcgaaaaagc atatgaatga tttccctgag ctaattgaga tgaagaggag atcaaatact 1020
cgaagtctca aggaaatggc attgctttta cgtggtggtt cacagttaat ttggattgca 1080
ccgagtggtg gtagagaccg cccaaatccc tcatcaggag aatggtaccc ggcaccattc 1140
gattcatctg cagtggacaa tatgaggagg cttctggagc atgctggtgt tcctgggcac 1200
atatatccac tatcattgct gtgctatgag gttatgcctc caccacaaca ggttgagaag 1260
gagattggtg agcagagggt gatatccttc catggagcag gcttgtcagt aactgaagaa 1320
ataaactatg gagacattac tgctcatacc aagaatgctg atgagggaag ggagctattc 1380
acaaatacct tgtacaactc agttgttaac cagtacaatg tgctcaaatc tgctatcttt 1440
agagatcgtg gagcagctgt atcaaacaat gtcatctcac tgtcacaacc atggcgatga 1500
atgtccagtt tcgttactga agttaccttt tcaaaagagc aggtgaacta tcattgtgca 1560
attttgctgg gagaaactct tgaacttaaa tctttttgat atcactagac ttcatcaatc 1620
atggtggagc ctgttattgt catgcagatg ctcgctaaga agtctgcaac tgttggttgg 1680
tcaagggtga ttggcgattt tgcacatacg gatccgcagt tactgctgcc tcaggttgct 1740
cagcaatgtg ctgctggttg ggcaccgagg aatccatcag gttgcatcct ttttgcccgg 1800
acgtcgattt gtgcagttga agtggtaaac gtttttttat cttgtttcgt catcatgtat 1860
atacgtaggc ttgagtcatt gttggctcca cattttttgg agatataaat gcggtaggag 1920
ttatgagatg tccacagtcg gccctccctt gttgaattcc tttggatgta tcctctctct 1980
ttcgtacatg cactcactga aagtcaatgc aaatatctcg tgtttctagt aaaaaaaaaa 2040
aaaaaa 2046
<210> 190
<211> 1994
<212> ДНК
<213> Hordeum vulgare
<400> 190
gagaaaccaa gaagcgagaa tcggcaccgt tcccgtcgcc gcctcgtcgc ctccgtcgcc 60
cgtcttcctt ttccgccgat tcgtgcccac caccaccact ctccttccca ctggctaggg 120
ttttcggttt ctagaacctc acgcccgccc gctccatgca agccccgccg ctcgccgcgc 180
tcgccggagg cgcctgggcc tctcaccgac ctgccatact agcggcgccg gcgggcctcc 240
gccgtcccag gcgctgcgcc ctccggctgc ccgcgtggag ggcggccgga ggcggccggg 300
ccccgcggct accggtcaag ggcgccgtgc tcgcctccga cacgggggcg gacgaggagg 360
tcgcggggcc atcgcccctg ctcgacgtgc gcagcgagca agagttcgtt ctacgcgtca 420
ggaaggaagt ggagagaggg aagttgcgtc cagatgttgc tgacaacttt gaaaacctgt 480
actgcaatta caagaatgcg gtgctacaaa atggggatcc aaatgcatat cagatcatgc 540
tttccaacat gatggattta tttgaccgcg ttctgctaga tgcagagaat ccatttacgt 600
ttcagcctta tcacaaggcc atcagagaac cgtttgacta ttacactttc ggtcagaact 660
acattaggcc actggtagat tttaggaact cttatgtcgg taacatttct gtattcagtg 720
atatggagaa gcagctccgg cagggtcata atgttgttct gatgtctaat catcagacag 780
aagcggatcc agcagttatt gccttgtcac ttgaaagaag caatccgtgg attagcgaga 840
acatagttta tgttgctggg gatagggttc ttacagatcc tctttgcaag ccatttagca 900
tgggaagaaa cctcctttgt gtgtactcaa aaaagcatat gaatgatttt cctgagctaa 960
ttgagatgaa gaggagggca aacactcgaa gtctcaagga aatggctttg cttttacgtg 1020
ggggttcaca tataatttgg atagctccga gtggcggtag agaccgtcct gaccccttga 1080
ctggagaatg gcacccggcg ccatttgatg catctgcagt ggataatatg aggaggcttc 1140
tggagcattc tggcgttcct gggcacatat atccattatc attgctatgc tatgagatta 1200
tgcctccacc acaacagatt gagaaagaga ttggtgagca aagggtgata tccttccatg 1260
gtgtaggctt gtcagtagct gaagaaataa agtatgggga tgttactgct caatctcaga 1320
atgctgatga ggcaaggggg aacttctcag aggctctgta cagttcagtt gttgatcaat 1380
ataatgtcct caagtctgct atctttagag accgtggagc agtttcgtcg aaccctgcca 1440
tctcactctc gcaaccatgg cggtgaaact aagctttctc aggcctggat ttctcatttc 1500
ttttcgacag agcagatgaa ctgctatagt gcaacgttgt ggttttttgc tgggatggcc 1560
ttaaactttg atgtcgtcac agttaggatg aggccctgca gatcctgtaa gttgttgaag 1620
tcgcgggaag gaaaaaccgt gtgatatgct gctacaccgt gttcatgtag tgacaggaag 1680
tctgcggctg ttgtcaggtc taaatcctaa atagcacggc ggaacccagc agcagatgat 1740
gcatgtgttc atcttttgtg aacagctact gctgtatcag atggctatca tctgggccag 1800
attggtccag caaatacaga ttggcccctg gatcctggca gtcgtctgga tcaaaatgct 1860
gatatttctt tttgtgctcg tcttattttt ttgattagtt ttgtgtacat attaattctt 1920
ttgctccaaa atttggagac acatgacatg atatatacag agcagagccc aatatgtgtc 1980
gccttcctgt taac 1994
<210> 191
<211> 1936
<212> ДНК
<213> Physcomitrella patens
<400> 191
ggacgagcgg agtggagagc tatggcggca gcagctggtt ccgctggcgt ggtatgttgg 60
tctagggcag agaagcagca tgccccggtc agggggggtg gaactagtgt taccagtagt 120
accagtggca gcggccatgc gtcgttgaaa gggagcttcg atcggctcca aggtaaccgc 180
cttctgccgc aagccttgac tatgccgtcg ctgtttcggg cgaaacgcaa tggcagaagg 240
acgccgggga atgccgtgac caatttcggg aaatctgaat tccatcgtga aattagtggg 300
agtacgcggg cgaccacgca ggtggctgaa gccaccacag ctggtcttag ggagaccatt 360
gaggaccgcg ctattatcga cggtcattct cacagttttg aaggaattca atcggaagaa 420
gagttgatgc aggtaattga aaaggaggtg gaatccggtc ggctgccgaa gcgtgctggc 480
gcgggaatgg tagagttgta tcgcaattat cgagatgctg tagtgagcag tggcgtagaa 540
aatgcgatgg atattgttgt gaaagtcatg tcaactgtgt tggaccggat tcttctgcag 600
ttcgaggagc cattcacatt tggatcgcac cacaagagaa tggtggagcc gtatgattac 660
tacacatttg gtcagaacta tgtgcgtcct ctcctagatt tcaggaactc ttaccttggg 720
aacttaaaga tctttgacca gatagagaag aacctgaaag aggggcacaa cgtcattttt 780
ctatccaatc accagactga ggcagatcct gctgttatgg cgctgttgct tgagcactct 840
cacccctatt tggcagagaa cttgacctat gtggctggag acagggttgt gctggatcca 900
ttctgcaaac cttttagtat gggcaggaat ctcttgtgcg tgtattcaaa aaagcacatt 960
cacgatgtac cggaccttgc tgaaatgaaa atcaaagcta atgcgaagac tttgagacag 1020
atgacgatcc tgctgaggca gggaggtcaa ttattatggg tagcacccag tggtggacgc 1080
gatcgccctg atcctgagac caacgaatgg gttcctgcac attttgactc gtctgctgtg 1140
gagaatatga agcgactatc tgacattgtc cgagtacctg ctcatttaca tgccctatca 1200
ttactatgtt ttgagattat gccacctcct gtccaggtac aaaaggagct aggagagcga 1260
agagcagtag gatttagcgg agttggtcta gccgtttccg agcaactaga ttatgattcc 1320
attgcgaagt tagtcgacga ttccaaaaat gcgaaggatg ccttttcgga tgcggcatgg 1380
agcgaagtca atgatatgta taacgtgtta aaagaagcaa tttatggtga ccaaggttgt 1440
gctgttagca cagattcctt gagactggaa cagccctggt ttgatggaag caggcgaact 1500
gattgaaaat aggtcatttg aagttttatg taaaagtatg aagcatcctt attgcttttt 1560
acgctgtcta agctccaagg atgtaagaat tcagcagcgt gtataatggc tacattgtca 1620
tgtgatattc tttctgattc gtgcgacacg atggccatgc ctgctcaatc cttgtcacca 1680
ggcgtctcag taggaaacgg tggtactgat tgctgtctgt ccgacttgat ttagtagctc 1740
ggattctgcg tactggataa cttggtctgg taatagggac cgatcctatc ggtgaggagt 1800
ttgtgatata gatcaatact gcactttgtt acaatcggaa tagatgcatt cattattcat 1860
ccaagccaac acatcctgag ttggagcata agttgaagca ctcctcaact tcattgaaag 1920
gagatttctc actacg 1936
<210> 192
<211> 2260
<212> ДНК
<213> Chlamydomonas reinhardtii
<400> 192
gaagaatgct gcacgcgact cagcagcgcg cggtcgctgg ccgtcgcccg ttctcgggtg 60
cgcgcgcgtc gaaccgcgtt gttgctcacg cggctgcgac cgtcgccacc agtctgccga 120
ccgttgacgt ccagttccac cagcctaagc tggcgggcgt gaccaacgag cagcagttca 180
aggcggtaat caaggggctg gtcgctcagg gcaagttccc tccgcagctg gagcccgctt 240
gggattactt ctatgacaac tacaagaagg ctgtcaccag cagtggcgtc gctggggccg 300
atgagaagct tgtcacccag gtgcaagcca gcattctgga caatgtcctg aaccaggcgg 360
tgaaccccta caccttcccc tctttccaca cccgcctaat tgagccctac aactactatg 420
acttcggtca gcgctacgtc gcgaccctca tcgacttcca gaactccgtg ctgggtttcc 480
gcgagcgttt cgaccgcgtt caggagctgc tggaccagaa gcacaacgtt gttatcctcg 540
cgaaccacca gacggaggcc gaccccggtg tgtttgccca tatgctggcg aagacgcacc 600
ctaagctggc gacggatgtg atctacgtcg ctggcgaccg cgttgtcacc gatccgatgt 660
gcaagccctt ctccatgggc cgcaacctct tctgcgtgca ctccaagaag cacatggacg 720
acgctccgga gctgaaggcc gcaaagatgg agaccaaccg caagacgctg gtcgccatgc 780
aacgcaagct gaacgagggc ggcacgctca tgtggatcgc ccccagcggc ggccgcgacc 840
gccccaacgc caacgacgag tgggtgcccg ataactttga tcccgccgcc gtggagctga 900
tgcgcaacct ggtgcagcgc gccaagcagc cgggccacct gatgcccatg tccatgttca 960
gctaccccat gatgccgccg cccaagaccg tggacaagtc cattggcgag cgccgcctca 1020
cggccttcac gggcgtgggc atctccctgt gcgaggagct ggacgtggcg gccatcatcg 1080
cggccagcgg ctcggaggag aaggagcaga aggctctggc caaggccgcg cacgacgcgg 1140
tgaaggagtc gtacgcggtg ctgtccaagg ccatccagga tcccgccttc cgcgccaccc 1200
gcaaggagtt cacacagccc tggatggcgt aaggaggcgg gagcagcagc ggcagtggcg 1260
gcagcgacag cagtggcgga tcttgggggc acgaggcagc agctagcaca cgcggggccg 1320
gtggcggcag cggacgggag aggtgctggt gcggatgctg gtgccaggag cagtgcgtct 1380
atgcctggcg gcggcgccga ccggtgctga agctgttgtc ggcagcagca gcgggaactt 1440
gcgctggcga tggctgaagg tgatgtggct ggccgtacag caaatgctgc tgtcacgcat 1500
tgtcagcggc ggccgctggg tccgctggga tgtgaggagt gcgagatcag cataggccag 1560
gcgggtgggt tgcggtgccg ctgccgacac gtcgcacagg aaggagcagg aagggtgcgg 1620
ctgagcacca gggtcacttg gggcctgtgt tagcgtgacc gggcctgaaa ggggtatttg 1680
tctggggagc agctcgacgc acattcgtgg gattgcttag gaaggagcgt tgggatggct 1740
gtgccgcgcg gtgtgcccag cacgttgact ggctctgacc cgggtcaacc aatgcgttgc 1800
ggccgttgca acgatgcttc taatcagcgt gagatggagt gtacgaatgc aggtatgacg 1860
atgaccatag acgagtgtga gcgtgtgtat ctgtggattg gacacggttc attcaatcca 1920
ttcgtaccgg acatatgact atacaagacc gtgtggagtg tgtttgtgcg ctcaagactg 1980
gacgaagtgg gcgctcccga tgtggagcgc ggtgctcgcg ggttgtgtgt cttgccgcaa 2040
cgcaagcagc gtggcgtggt gtggatgatt cttgcattat gactggggtt ttaccggcgg 2100
cccagagcgt gtgtggagca aaagcaaaaa caggaacaag gagccgtgtg cacaattcgc 2160
gcggatggtg gtggctgggg atctgacagg agagtgccaa cggcggccgg gtgctagtgc 2220
ttgaacatca tgtgatattg tgattgtaca aatggactgc 2260
<210> 193
<211> 382
<212> Белок
<213> Cinnamomum camphora
<400> 193
Met Ala Thr Thr Ser Leu Ala Ser Ala Phe Cys Ser Met Lys Ala Val
1 5 10 15
Met Leu Ala Arg Asp Gly Arg Gly Met Lys Pro Arg Ser Ser Asp Leu
20 25 30
Gln Leu Arg Ala Gly Asn Ala Gln Thr Ser Leu Lys Met Ile Asn Gly
35 40 45
Thr Lys Phe Ser Tyr Thr Glu Ser Leu Lys Lys Leu Pro Asp Trp Ser
50 55 60
Met Leu Phe Ala Val Ile Thr Thr Ile Phe Ser Ala Ala Glu Lys Gln
65 70 75 80
Trp Thr Asn Leu Glu Trp Lys Pro Lys Pro Asn Pro Pro Gln Leu Leu
85 90 95
Asp Asp His Phe Gly Pro His Gly Leu Val Phe Arg Arg Thr Phe Ala
100 105 110
Ile Arg Ser Tyr Glu Val Gly Pro Asp Arg Ser Thr Ser Ile Val Ala
115 120 125
Val Met Asn His Leu Gln Glu Ala Ala Leu Asn His Ala Lys Ser Val
130 135 140
Gly Ile Leu Gly Asp Gly Phe Gly Thr Thr Leu Glu Met Ser Lys Arg
145 150 155 160
Asp Leu Ile Trp Val Val Lys Arg Thr His Val Ala Val Glu Arg Tyr
165 170 175
Pro Ala Trp Gly Asp Thr Val Glu Val Glu Cys Trp Val Gly Ala Ser
180 185 190
Gly Asn Asn Gly Arg Arg His Asp Phe Leu Val Arg Asp Cys Lys Thr
195 200 205
Gly Glu Ile Leu Thr Arg Cys Thr Ser Leu Ser Val Met Met Asn Thr
210 215 220
Arg Thr Arg Arg Leu Ser Lys Ile Pro Glu Glu Val Arg Gly Glu Ile
225 230 235 240
Gly Pro Ala Phe Ile Asp Asn Val Ala Val Lys Asp Glu Glu Ile Lys
245 250 255
Lys Pro Gln Lys Leu Asn Asp Ser Thr Ala Asp Tyr Ile Gln Gly Gly
260 265 270
Leu Thr Pro Arg Trp Asn Asp Leu Asp Ile Asn Gln His Val Asn Asn
275 280 285
Ile Lys Tyr Val Asp Trp Ile Leu Glu Thr Val Pro Asp Ser Ile Phe
290 295 300
Glu Ser His His Ile Ser Ser Phe Thr Ile Glu Tyr Arg Arg Glu Cys
305 310 315 320
Thr Met Asp Ser Val Leu Gln Ser Leu Thr Thr Val Ser Gly Gly Ser
325 330 335
Ser Glu Ala Gly Leu Val Cys Glu His Leu Leu Gln Leu Glu Gly Gly
340 345 350
Ser Glu Val Leu Arg Ala Lys Thr Glu Trp Arg Pro Lys Leu Thr Asp
355 360 365
Ser Phe Arg Gly Ile Ser Val Ile Pro Ala Glu Ser Ser Val
370 375 380
<210> 194
<211> 417
<212> Белок
<213> Cocos nucifera
<400> 194
Met Val Ala Ser Val Ala Ala Ser Ala Phe Phe Pro Thr Pro Ser Phe
1 5 10 15
Ser Ser Thr Ala Ser Ala Lys Ala Ser Lys Thr Ile Gly Glu Gly Ser
20 25 30
Glu Ser Leu Asp Val Arg Gly Ile Val Ala Lys Pro Thr Ser Ser Ser
35 40 45
Ala Ala Met Gln Gly Lys Val Lys Ala Gln Ala Val Pro Lys Ile Asn
50 55 60
Gly Thr Lys Val Gly Leu Lys Thr Glu Ser Gln Lys Ala Glu Glu Asp
65 70 75 80
Ala Ala Pro Ser Ser Ala Pro Arg Thr Phe Tyr Asn Gln Leu Pro Asp
85 90 95
Trp Ser Val Leu Leu Ala Ala Val Thr Thr Ile Phe Leu Ala Ala Glu
100 105 110
Lys Gln Trp Thr Leu Leu Asp Trp Lys Pro Arg Arg Pro Asp Met Leu
115 120 125
Thr Asp Ala Phe Ser Leu Gly Lys Ile Val Gln Asp Gly Leu Ile Phe
130 135 140
Arg Gln Asn Phe Ser Ile Arg Ser Tyr Glu Ile Gly Ala Asp Arg Thr
145 150 155 160
Ala Ser Ile Glu Thr Leu Met Asn His Leu Gln Glu Thr Ala Leu Asn
165 170 175
His Val Arg Asn Ala Gly Leu Leu Gly Asp Gly Phe Gly Ala Thr Pro
180 185 190
Glu Met Ser Lys Arg Asn Leu Ile Trp Val Val Thr Lys Met Gln Val
195 200 205
Leu Val Glu His Tyr Pro Ser Trp Gly Asp Val Val Glu Val Asp Thr
210 215 220
Trp Val Gly Ala Ser Gly Lys Asn Gly Met Arg Arg Asp Trp His Val
225 230 235 240
Arg Asp Tyr Arg Thr Gly Gln Thr Ile Leu Arg Ala Thr Ser Val Trp
245 250 255
Val Met Met Asn Lys His Thr Arg Lys Leu Ser Lys Met Pro Glu Glu
260 265 270
Val Arg Ala Glu Ile Gly Pro Tyr Phe Val Glu His Ala Ala Ile Val
275 280 285
Asp Glu Asp Ser Arg Lys Leu Pro Lys Leu Asp Asp Asp Thr Ala Asp
290 295 300
Tyr Ile Lys Trp Gly Leu Thr Pro Arg Trp Ser Asp Leu Asp Val Asn
305 310 315 320
Gln His Val Asn Asn Val Lys Tyr Ile Gly Trp Ile Leu Glu Ser Ala
325 330 335
Pro Ile Ser Ile Leu Glu Asn His Glu Leu Ala Ser Met Thr Leu Glu
340 345 350
Tyr Arg Arg Glu Cys Gly Arg Asp Ser Val Leu Gln Ser Leu Thr Ala
355 360 365
Ile Ser Asn Asp Cys Thr Gly Gly Leu Pro Glu Ala Ser Ile Glu Cys
370 375 380
Gln His Leu Leu Gln Leu Glu Cys Gly Ala Glu Ile Val Arg Gly Arg
385 390 395 400
Thr Gln Trp Arg Pro Arg Arg Ala Ser Gly Pro Thr Ser Ala Gly Ser
405 410 415
Ala
<210> 195
<211> 423
<212> Белок
<213> Cocos nucifera
<400> 195
Met Val Ala Ser Ile Ala Ala Ser Ala Phe Phe Pro Thr Pro Ser Ser
1 5 10 15
Ser Ser Ser Ala Ala Ser Ala Lys Ala Ser Lys Thr Ile Gly Glu Gly
20 25 30
Pro Gly Ser Leu Asp Val Arg Gly Ile Val Ala Lys Pro Thr Ser Ser
35 40 45
Ser Ala Ala Met Gln Glu Lys Val Lys Val Gln Pro Val Pro Lys Ile
50 55 60
Asn Gly Ala Lys Val Gly Leu Lys Val Glu Thr Gln Lys Ala Asp Glu
65 70 75 80
Glu Ser Ser Pro Ser Ser Ala Pro Arg Thr Phe Tyr Asn Gln Leu Pro
85 90 95
Asp Trp Ser Val Leu Leu Ala Ala Val Thr Thr Ile Phe Leu Ala Ala
100 105 110
Glu Lys Gln Trp Thr Leu Leu Asp Trp Lys Pro Arg Arg Pro Asp Met
115 120 125
Leu Ala Asp Ala Phe Gly Leu Gly Lys Ile Val Gln Asp Gly Leu Val
130 135 140
Phe Lys Gln Asn Phe Ser Ile Arg Ser Tyr Glu Ile Gly Ala Asp Arg
145 150 155 160
Thr Ala Ser Ile Glu Thr Leu Met Asn His Leu Gln Glu Thr Ala Leu
165 170 175
Asn His Val Lys Ser Ala Gly Leu Met Gly Asp Gly Phe Gly Ala Thr
180 185 190
Pro Glu Met Ser Lys Arg Asn Leu Ile Trp Val Val Thr Lys Met Arg
195 200 205
Val Leu Ile Glu Arg Tyr Pro Ser Trp Gly Asp Val Val Glu Val Asp
210 215 220
Thr Trp Val Gly Pro Thr Gly Lys Asn Gly Met Arg Arg Asp Trp His
225 230 235 240
Val Arg Asp His Arg Ser Gly Gln Thr Ile Leu Arg Ala Thr Ser Val
245 250 255
Trp Val Met Met Asn Lys Asn Thr Arg Lys Leu Ser Lys Val Pro Glu
260 265 270
Glu Val Arg Ala Glu Ile Gly Pro Tyr Phe Val Glu Arg Ala Ala Ile
275 280 285
Val Asp Glu Asp Ser Arg Lys Leu Pro Lys Leu Asp Glu Asp Thr Thr
290 295 300
Asp Tyr Ile Lys Lys Gly Leu Thr Pro Arg Trp Gly Asp Leu Asp Val
305 310 315 320
Asn Gln His Val Asn Asn Val Lys Tyr Ile Gly Trp Ile Leu Glu Ser
325 330 335
Ala Pro Ile Ser Ile Leu Glu Asn His Glu Leu Ala Ser Met Ser Leu
340 345 350
Glu Tyr Arg Arg Glu Cys Gly Arg Asp Ser Val Leu Gln Ser Leu Thr
355 360 365
Ala Val Ser Asn Asp Leu Thr Asp Gly Leu Val Glu Ser Gly Ile Glu
370 375 380
Cys Gln His Leu Leu Gln Leu Glu Cys Gly Thr Glu Leu Val Lys Gly
385 390 395 400
Arg Thr Glu Trp Arg Pro Lys His Ser Pro Ala Leu Gly Asn Met Gly
405 410 415
Pro Thr Pro Gly Gly Ser Ala
420
<210> 196
<211> 414
<212> Белок
<213> Cocos nucifera
<400> 196
Met Val Ala Ser Val Ala Ala Ser Ser Ser Phe Phe Pro Val Pro Ser
1 5 10 15
Ser Ser Ser Ser Ala Ser Ala Lys Ala Ser Arg Gly Ile Pro Asp Gly
20 25 30
Leu Asp Val Arg Gly Ile Val Ala Lys Pro Ala Ser Ser Ser Gly Trp
35 40 45
Met Gln Ala Lys Ala Ser Ala Arg Ala Ile Pro Lys Ile Asp Asp Thr
50 55 60
Lys Val Gly Leu Arg Thr Asp Val Glu Glu Asp Ala Ala Ser Thr Ala
65 70 75 80
Arg Arg Thr Ser Tyr Asn Gln Leu Pro Asp Trp Ser Met Leu Leu Ala
85 90 95
Ala Ile Arg Thr Ile Phe Ser Ala Ala Glu Lys Gln Trp Thr Leu Leu
100 105 110
Asp Ser Lys Lys Arg Gly Ala Asp Ala Val Ala Asp Ala Ser Gly Val
115 120 125
Gly Lys Met Val Lys Asn Gly Leu Val Tyr Arg Gln Asn Phe Ser Ile
130 135 140
Arg Ser Tyr Glu Ile Gly Val Asp Lys Arg Ala Ser Val Glu Ala Leu
145 150 155 160
Met Asn His Phe Gln Glu Thr Ser Leu Asn His Cys Lys Cys Ile Gly
165 170 175
Leu Met His Gly Gly Phe Gly Cys Thr Pro Glu Met Thr Arg Arg Asn
180 185 190
Leu Ile Trp Val Val Ala Lys Met Leu Val His Val Glu Arg Tyr Pro
195 200 205
Trp Trp Gly Asp Val Val Gln Ile Asn Thr Trp Ile Ser Ser Ser Gly
210 215 220
Lys Asn Gly Met Gly Arg Asp Trp His Val His Asp Cys Gln Thr Gly
225 230 235 240
Leu Pro Ile Met Arg Gly Thr Ser Val Trp Val Met Met Asp Lys His
245 250 255
Thr Arg Arg Leu Ser Lys Leu Pro Glu Glu Val Arg Ala Glu Ile Thr
260 265 270
Pro Phe Phe Ser Glu Arg Asp Ala Val Leu Asp Asp Asn Gly Arg Lys
275 280 285
Leu Pro Lys Phe Asp Asp Asp Ser Ala Ala His Val Arg Arg Gly Leu
290 295 300
Thr Pro Arg Trp His Asp Phe Asp Val Asn Gln His Val Asn Asn Val
305 310 315 320
Lys Tyr Val Gly Trp Ile Leu Glu Ser Val Pro Val Trp Met Leu Asp
325 330 335
Gly Tyr Glu Val Ala Thr Met Ser Leu Glu Tyr Arg Arg Glu Cys Arg
340 345 350
Met Asp Ser Val Val Gln Ser Leu Thr Ala Val Ser Ser Asp His Ala
355 360 365
Asp Gly Ser Pro Ile Val Cys Gln His Leu Leu Arg Leu Glu Asp Gly
370 375 380
Thr Glu Ile Val Arg Gly Gln Thr Glu Trp Arg Pro Lys Gln Gln Ala
385 390 395 400
Arg Asp Leu Gly Asn Met Gly Leu His Pro Thr Glu Ser Lys
405 410
<210> 197
<211> 419
<212> Белок
<213> Cuphea lanceolata
<400> 197
Met Val Ala Ala Ala Ala Thr Ser Ala Phe Phe Pro Val Pro Ala Pro
1 5 10 15
Gly Thr Ser Pro Lys Pro Gly Lys Ser Gly Asn Trp Pro Ser Ser Leu
20 25 30
Ser Pro Thr Phe Lys Pro Lys Ser Ile Pro Asn Ala Gly Phe Gln Val
35 40 45
Lys Ala Asn Ala Ser Ala His Pro Lys Ala Asn Gly Ser Ala Val Asn
50 55 60
Leu Lys Ser Gly Ser Leu Asn Thr Gln Glu Asp Thr Ser Ser Ser Pro
65 70 75 80
Pro Pro Arg Ala Phe Leu Asn Gln Leu Pro Asp Trp Ser Met Leu Leu
85 90 95
Thr Ala Ile Thr Thr Val Phe Val Ala Ala Glu Lys Gln Trp Thr Met
100 105 110
Leu Asp Arg Lys Ser Lys Arg Pro Asp Met Leu Val Asp Ser Val Gly
115 120 125
Leu Lys Ser Ile Val Arg Asp Gly Leu Val Ser Arg Gln Ser Phe Leu
130 135 140
Ile Arg Ser Tyr Glu Ile Gly Ala Asp Arg Thr Ala Ser Ile Glu Thr
145 150 155 160
Leu Met Asn His Leu Gln Glu Thr Ser Ile Asn His Cys Lys Ser Leu
165 170 175
Gly Leu Leu Asn Asp Gly Phe Gly Arg Thr Pro Gly Met Cys Lys Asn
180 185 190
Asp Leu Ile Trp Val Leu Thr Lys Met Gln Ile Met Val Asn Arg Tyr
195 200 205
Pro Thr Trp Gly Asp Thr Val Glu Ile Asn Thr Trp Phe Ser Gln Ser
210 215 220
Gly Lys Ile Gly Met Ala Ser Asp Trp Leu Ile Ser Asp Cys Asn Thr
225 230 235 240
Gly Glu Ile Leu Ile Arg Ala Thr Ser Val Trp Ala Met Met Asn Gln
245 250 255
Lys Thr Arg Arg Phe Ser Arg Leu Pro Tyr Glu Val Arg Gln Glu Leu
260 265 270
Thr Pro His Phe Val Asp Ser Pro His Val Ile Glu Asp Asn Asp Gln
275 280 285
Lys Leu His Lys Phe Asp Val Lys Thr Gly Asp Ser Ile Arg Lys Gly
290 295 300
Leu Thr Pro Arg Trp Asn Asp Leu Asp Val Asn Gln His Val Ser Asn
305 310 315 320
Val Lys Tyr Ile Gly Trp Ile Leu Glu Ser Met Pro Ile Glu Val Leu
325 330 335
Glu Thr Gln Glu Leu Cys Ser Leu Thr Val Glu Tyr Arg Arg Glu Cys
340 345 350
Gly Met Asp Ser Val Leu Glu Ser Val Thr Ala Val Asp Pro Ser Glu
355 360 365
Asn Gly Gly Arg Ser Gln Tyr Lys His Leu Leu Arg Leu Glu Asp Gly
370 375 380
Thr Asp Ile Val Lys Ser Arg Thr Glu Trp Arg Pro Lys Asn Ala Gly
385 390 395 400
Thr Asn Gly Ala Ile Ser Thr Ser Thr Ala Lys Thr Ser Asn Gly Asn
405 410 415
Ser Ala Ser
<210> 198
<211> 419
<212> Белок
<213> Искусственная последовательность
<220>
<223> Cuphea viscosissima
<400> 198
Met Val Ala Ala Ala Ala Thr Ser Ala Phe Phe Pro Val Pro Ala Pro
1 5 10 15
Gly Thr Ser Pro Lys Pro Gly Lys Ser Gly Asn Trp Pro Ser Ser Leu
20 25 30
Ser Pro Thr Phe Lys Pro Lys Ser Ile Pro Asn Gly Gly Phe Gln Val
35 40 45
Lys Ala Asn Ala Ser Ala His Pro Lys Ala Asn Gly Ser Ala Val Asn
50 55 60
Leu Lys Ser Gly Ser Leu Asn Thr Gln Glu Asp Thr Ser Ser Ser Pro
65 70 75 80
Pro Pro Arg Ala Phe Leu Asn Gln Leu Pro Asp Trp Ser Met Leu Leu
85 90 95
Thr Ala Ile Thr Thr Val Phe Val Ala Ala Glu Lys Gln Trp Thr Met
100 105 110
Leu Asp Arg Lys Ser Lys Arg Pro Asp Met Leu Val Asp Ser Val Gly
115 120 125
Leu Lys Ser Ile Val Arg Asp Gly Leu Val Ser Arg His Ser Phe Ser
130 135 140
Ile Arg Ser Tyr Glu Ile Gly Ala Asp Arg Thr Ala Ser Ile Glu Thr
145 150 155 160
Leu Met Asn His Leu Gln Glu Thr Thr Ile Asn His Cys Lys Ser Leu
165 170 175
Gly Leu His Asn Asp Gly Phe Gly Arg Thr Pro Gly Met Cys Lys Asn
180 185 190
Asp Leu Ile Trp Val Leu Thr Lys Met Gln Ile Met Val Asn Arg Tyr
195 200 205
Pro Thr Trp Gly Asp Thr Val Glu Ile Asn Thr Trp Phe Ser Gln Ser
210 215 220
Gly Lys Ile Gly Met Ala Ser Asp Trp Leu Ile Ser Asp Cys Asn Thr
225 230 235 240
Gly Glu Ile Leu Ile Arg Ala Thr Ser Val Trp Ala Met Met Asn Gln
245 250 255
Lys Thr Arg Arg Phe Ser Arg Leu Pro Tyr Glu Val Arg Gln Glu Leu
260 265 270
Thr Pro His Phe Val Asp Ser Pro His Val Ile Glu Asp Asn Asp Gln
275 280 285
Lys Leu Arg Lys Phe Asp Val Lys Thr Gly Asp Ser Ile Arg Lys Gly
290 295 300
Leu Thr Pro Arg Trp Asn Asp Leu Asp Val Asn Gln His Val Ser Asn
305 310 315 320
Val Lys Tyr Ile Gly Trp Ile Leu Glu Ser Met Pro Ile Glu Val Leu
325 330 335
Glu Thr Gln Glu Leu Cys Ser Leu Thr Val Glu Tyr Arg Arg Glu Cys
340 345 350
Gly Met Asp Ser Val Leu Glu Ser Val Thr Ala Val Asp Pro Ser Glu
355 360 365
Asn Gly Gly Arg Ser Gln Tyr Lys His Leu Leu Arg Leu Glu Asp Gly
370 375 380
Thr Asp Ile Val Lys Ser Arg Thr Glu Trp Arg Pro Lys Asn Ala Gly
385 390 395 400
Thr Asn Gly Ala Ile Ser Thr Ser Thr Ala Lys Thr Ser Asn Gly Asn
405 410 415
Ser Val Ser
<210> 199
<211> 382
<212> Белок
<213> Umbellularia californica
<400> 199
Met Ala Thr Thr Ser Leu Ala Ser Ala Phe Cys Ser Met Lys Ala Val
1 5 10 15
Met Leu Ala Arg Asp Gly Arg Gly Met Lys Pro Arg Ser Ser Asp Leu
20 25 30
Gln Leu Arg Ala Gly Asn Ala Pro Thr Ser Leu Lys Met Ile Asn Gly
35 40 45
Thr Lys Phe Ser Tyr Thr Glu Ser Leu Lys Arg Leu Pro Asp Trp Ser
50 55 60
Met Leu Phe Ala Val Ile Thr Thr Ile Phe Ser Ala Ala Glu Lys Gln
65 70 75 80
Trp Thr Asn Leu Glu Trp Lys Pro Lys Pro Lys Leu Pro Gln Leu Leu
85 90 95
Asp Asp His Phe Gly Leu His Gly Leu Val Phe Arg Arg Thr Phe Ala
100 105 110
Ile Arg Ser Tyr Glu Val Gly Pro Asp Arg Ser Thr Ser Ile Leu Ala
115 120 125
Val Met Asn His Met Gln Glu Ala Thr Leu Asn His Ala Lys Ser Val
130 135 140
Gly Ile Leu Gly Asp Gly Phe Gly Thr Thr Leu Glu Met Ser Lys Arg
145 150 155 160
Asp Leu Met Trp Val Val Arg Arg Thr His Val Ala Val Glu Arg Tyr
165 170 175
Pro Thr Trp Gly Asp Thr Val Glu Val Glu Cys Trp Ile Gly Ala Ser
180 185 190
Gly Asn Asn Gly Met Arg Arg Asp Phe Leu Val Arg Asp Cys Lys Thr
195 200 205
Gly Glu Ile Leu Thr Arg Cys Thr Ser Leu Ser Val Leu Met Asn Thr
210 215 220
Arg Thr Arg Arg Leu Ser Thr Ile Pro Asp Glu Val Arg Gly Glu Ile
225 230 235 240
Gly Pro Ala Phe Ile Asp Asn Val Ala Val Lys Asp Asp Glu Ile Lys
245 250 255
Lys Leu Gln Lys Leu Asn Asp Ser Thr Ala Asp Tyr Ile Gln Gly Gly
260 265 270
Leu Thr Pro Arg Trp Asn Asp Leu Asp Val Asn Gln His Val Asn Asn
275 280 285
Leu Lys Tyr Val Ala Trp Val Phe Glu Thr Val Pro Asp Ser Ile Phe
290 295 300
Glu Ser His His Ile Ser Ser Phe Thr Leu Glu Tyr Arg Arg Glu Cys
305 310 315 320
Thr Arg Asp Ser Val Leu Arg Ser Leu Thr Thr Val Ser Gly Gly Ser
325 330 335
Ser Glu Ala Gly Leu Val Cys Asp His Leu Leu Gln Leu Glu Gly Gly
340 345 350
Ser Glu Val Leu Arg Ala Arg Thr Glu Trp Arg Pro Lys Leu Thr Asp
355 360 365
Ser Phe Arg Gly Ile Ser Val Ile Pro Ala Glu Pro Arg Val
370 375 380
<210> 200
<211> 308
<212> Белок
<213> Cocos nucifera
<400> 200
Met Asp Ala Ser Gly Ala Ser Ser Phe Leu Arg Gly Arg Cys Leu Glu
1 5 10 15
Ser Cys Phe Lys Ala Ser Phe Gly Met Ser Gln Pro Lys Asp Ala Ala
20 25 30
Gly Gln Pro Ser Arg Arg Pro Ala Asp Ala Asp Asp Phe Val Asp Asp
35 40 45
Asp Arg Trp Ile Thr Val Ile Leu Ser Val Val Arg Ile Ala Ala Cys
50 55 60
Phe Leu Ser Met Met Val Thr Thr Ile Val Trp Asn Met Ile Met Leu
65 70 75 80
Ile Leu Leu Pro Trp Pro Tyr Ala Arg Ile Arg Gln Gly Asn Leu Tyr
85 90 95
Gly His Val Thr Gly Arg Met Leu Met Trp Ile Leu Gly Asn Pro Ile
100 105 110
Thr Ile Glu Gly Ser Glu Phe Ser Asn Thr Arg Ala Ile Tyr Ile Cys
115 120 125
Asn His Ala Ser Leu Val Asp Ile Phe Leu Ile Met Trp Leu Ile Pro
130 135 140
Lys Gly Thr Val Thr Ile Ala Lys Lys Glu Ile Ile Trp Tyr Pro Leu
145 150 155 160
Phe Gly Gln Leu Tyr Val Leu Ala Asn His Gln Arg Ile Asp Arg Ser
165 170 175
Asn Pro Ser Ala Ala Ile Glu Ser Ile Lys Glu Val Ala Arg Ala Val
180 185 190
Val Lys Lys Asn Leu Ser Leu Ile Ile Phe Pro Glu Gly Thr Arg Ser
195 200 205
Lys Thr Gly Arg Leu Leu Pro Phe Lys Lys Gly Phe Ile His Ile Ala
210 215 220
Leu Gln Thr Arg Leu Pro Ile Val Pro Met Val Leu Thr Gly Thr His
225 230 235 240
Leu Ala Trp Arg Lys Asn Ser Leu Arg Val Arg Pro Ala Pro Ile Thr
245 250 255
Val Lys Tyr Phe Ser Pro Ile Lys Thr Asp Asp Trp Glu Glu Glu Lys
260 265 270
Ile Asn His Tyr Val Glu Met Ile His Ala Leu Tyr Val Asp His Leu
275 280 285
Pro Glu Ser Gln Lys Pro Leu Val Ser Lys Gly Arg Asp Ala Ser Gly
290 295 300
Arg Ser Asn Ser
305
<210> 201
<211> 356
<212> Белок
<213> Arabidopsis thaliana
<400> 201
Met Asp Val Ala Ser Ala Arg Ser Ile Ser Ser His Pro Ser Tyr Tyr
1 5 10 15
Gly Lys Pro Ile Cys Ser Ser Gln Ser Ser Leu Ile Arg Ile Ser Arg
20 25 30
Asp Lys Val Cys Cys Phe Gly Arg Ile Ser Asn Gly Met Thr Ser Phe
35 40 45
Thr Thr Ser Leu His Ala Val Pro Ser Glu Lys Phe Met Gly Glu Thr
50 55 60
Arg Arg Thr Gly Ile Gln Trp Ser Asn Arg Ser Leu Arg His Asp Pro
65 70 75 80
Tyr Arg Phe Leu Asp Lys Lys Ser Pro Arg Ser Ser Gln Leu Ala Arg
85 90 95
Asp Ile Thr Val Arg Ala Asp Leu Ser Gly Ala Ala Thr Pro Asp Ser
100 105 110
Ser Phe Pro Glu Pro Glu Ile Lys Leu Ser Ser Arg Leu Arg Gly Ile
115 120 125
Phe Phe Cys Val Val Ala Gly Ile Ser Ala Thr Phe Leu Ile Val Leu
130 135 140
Met Ile Ile Gly His Pro Phe Val Leu Leu Phe Asp Pro Tyr Arg Arg
145 150 155 160
Lys Phe His His Phe Ile Ala Lys Leu Trp Ala Ser Ile Ser Ile Tyr
165 170 175
Pro Phe Tyr Lys Ile Asn Ile Glu Gly Leu Glu Asn Leu Pro Ser Ser
180 185 190
Asp Thr Pro Ala Val Tyr Val Ser Asn His Gln Ser Phe Leu Asp Ile
195 200 205
Tyr Thr Leu Leu Ser Leu Gly Lys Ser Phe Lys Phe Ile Ser Lys Thr
210 215 220
Gly Ile Phe Val Ile Pro Ile Ile Gly Trp Ala Met Ser Met Met Gly
225 230 235 240
Val Val Pro Leu Lys Arg Met Asp Pro Arg Ser Gln Val Asp Cys Leu
245 250 255
Lys Arg Cys Met Glu Leu Leu Lys Lys Gly Ala Ser Val Phe Phe Phe
260 265 270
Pro Glu Gly Thr Arg Ser Lys Asp Gly Arg Leu Gly Ser Phe Lys Lys
275 280 285
Gly Ala Phe Thr Val Ala Ala Lys Thr Gly Val Ala Val Val Pro Ile
290 295 300
Thr Leu Met Gly Thr Gly Lys Ile Met Pro Thr Gly Ser Glu Gly Ile
305 310 315 320
Leu Asn His Gly Asn Val Arg Val Ile Ile His Lys Pro Ile His Gly
325 330 335
Ser Lys Ala Asp Val Leu Cys Asn Glu Ala Arg Ser Lys Ile Ala Glu
340 345 350
Ser Met Asp Leu
355
<210> 202
<211> 362
<212> Белок
<213> Arabidopsis thaliana
<400> 202
Met Leu Lys Leu Ser Cys Asn Val Thr Asp Ser Lys Leu Gln Arg Ser
1 5 10 15
Leu Leu Phe Phe Ser His Ser Tyr Arg Ser Asp Pro Val Asn Phe Ile
20 25 30
Arg Arg Arg Ile Val Ser Cys Ser Gln Thr Lys Lys Thr Gly Leu Val
35 40 45
Pro Leu Arg Ala Val Val Ser Ala Asp Gln Gly Ser Val Val Gln Gly
50 55 60
Leu Ala Thr Leu Ala Asp Gln Leu Arg Leu Gly Ser Leu Thr Glu Asp
65 70 75 80
Gly Leu Ser Tyr Lys Glu Lys Phe Val Val Arg Ser Tyr Glu Val Gly
85 90 95
Ser Asn Lys Thr Ala Thr Val Glu Thr Ile Ala Asn Leu Leu Gln Glu
100 105 110
Val Gly Cys Asn His Ala Gln Ser Val Gly Phe Ser Thr Asp Gly Phe
115 120 125
Ala Thr Thr Thr Thr Met Arg Lys Leu His Leu Ile Trp Val Thr Ala
130 135 140
Arg Met His Ile Glu Ile Tyr Lys Tyr Pro Ala Trp Gly Asp Val Val
145 150 155 160
Glu Ile Glu Thr Trp Cys Gln Ser Glu Gly Arg Ile Gly Thr Arg Arg
165 170 175
Asp Trp Ile Leu Lys Asp Ser Val Thr Gly Glu Val Thr Gly Arg Ala
180 185 190
Thr Ser Lys Trp Val Met Met Asn Gln Asp Thr Arg Arg Leu Gln Lys
195 200 205
Val Ser Asp Asp Val Arg Asp Glu Tyr Leu Val Phe Cys Pro Gln Glu
210 215 220
Pro Arg Leu Ala Phe Pro Glu Glu Asn Asn Arg Ser Leu Lys Lys Ile
225 230 235 240
Pro Lys Leu Glu Asp Pro Ala Gln Tyr Ser Met Ile Gly Leu Lys Pro
245 250 255
Arg Arg Ala Asp Leu Asp Met Asn Gln His Val Asn Asn Val Thr Tyr
260 265 270
Ile Gly Trp Val Leu Glu Ser Ile Pro Gln Glu Ile Val Asp Thr His
275 280 285
Glu Leu Gln Val Ile Thr Leu Asp Tyr Arg Arg Glu Cys Gln Gln Asp
290 295 300
Asp Val Val Asp Ser Leu Thr Thr Thr Thr Ser Glu Ile Gly Gly Thr
305 310 315 320
Asn Gly Ser Ala Thr Ser Gly Thr Gln Gly His Asn Asp Ser Gln Phe
325 330 335
Leu His Leu Leu Arg Leu Ser Gly Asp Gly Gln Glu Ile Asn Arg Gly
340 345 350
Thr Thr Leu Trp Arg Lys Lys Pro Ser Ser
355 360
<210> 203
<211> 367
<212> Белок
<213> Arabidopsis thaliana
<400> 203
Met Leu Lys Leu Ser Cys Asn Val Thr Asp His Ile His Asn Leu Phe
1 5 10 15
Ser Asn Ser Arg Arg Ile Phe Val Pro Val His Arg Gln Thr Arg Pro
20 25 30
Ile Ser Cys Phe Gln Leu Lys Lys Glu Pro Leu Arg Ala Ile Leu Ser
35 40 45
Ala Asp His Gly Asn Ser Ser Val Arg Val Ala Asp Thr Val Ser Gly
50 55 60
Thr Ser Pro Ala Asp Arg Leu Arg Phe Gly Arg Leu Met Glu Asp Gly
65 70 75 80
Phe Ser Tyr Lys Glu Lys Phe Ile Val Arg Ser Tyr Glu Val Gly Ile
85 90 95
Asn Lys Thr Ala Thr Ile Glu Thr Ile Ala Asn Leu Leu Gln Glu Val
100 105 110
Ala Cys Asn His Val Gln Asn Val Gly Phe Ser Thr Asp Gly Phe Ala
115 120 125
Thr Thr Leu Thr Met Arg Lys Leu His Leu Ile Trp Val Thr Ala Arg
130 135 140
Met His Ile Glu Ile Tyr Lys Tyr Pro Ala Trp Ser Asp Val Val Glu
145 150 155 160
Ile Glu Thr Trp Cys Gln Ser Glu Gly Arg Ile Gly Thr Arg Arg Asp
165 170 175
Trp Ile Leu Lys Asp Cys Ala Thr Gly Glu Val Ile Gly Arg Ala Thr
180 185 190
Ser Lys Trp Val Met Met Asn Gln Asp Thr Arg Arg Leu Gln Arg Val
195 200 205
Thr Asp Glu Val Arg Asp Glu Tyr Leu Val Phe Cys Pro Pro Glu Pro
210 215 220
Arg Leu Ala Phe Pro Glu Glu Asn Asn Ser Ser Leu Lys Lys Ile Pro
225 230 235 240
Lys Leu Glu Asp Pro Ala Gln Tyr Ser Met Leu Gly Leu Lys Pro Arg
245 250 255
Arg Ala Asp Leu Asp Met Asn Gln His Val Asn Asn Val Thr Tyr Ile
260 265 270
Gly Trp Val Leu Glu Ser Ile Pro Gln Glu Ile Ile Asp Thr His Glu
275 280 285
Leu Lys Val Ile Thr Leu Asp Tyr Arg Arg Glu Cys Gln Gln Asp Asp
290 295 300
Ile Val Asp Ser Leu Thr Thr Ser Glu Thr Pro Asn Glu Val Val Ser
305 310 315 320
Lys Leu Thr Gly Thr Asn Gly Ser Thr Thr Ser Ser Lys Arg Glu His
325 330 335
Asn Glu Ser His Phe Leu His Ile Leu Arg Leu Ser Glu Asn Gly Gln
340 345 350
Glu Ile Asn Arg Gly Arg Thr Gln Trp Arg Lys Lys Ser Ser Arg
355 360 365
<210> 204
<211> 412
<212> Белок
<213> Arabidopsis thaliana
<400> 204
Met Val Ala Thr Ser Ala Thr Ser Ser Phe Phe Pro Val Pro Ser Ser
1 5 10 15
Ser Leu Asp Pro Asn Gly Lys Gly Asn Lys Ile Gly Ser Thr Asn Leu
20 25 30
Ala Gly Leu Asn Ser Ala Pro Asn Ser Gly Arg Met Lys Val Lys Pro
35 40 45
Asn Ala Gln Ala Pro Pro Lys Ile Asn Gly Lys Lys Val Gly Leu Pro
50 55 60
Gly Ser Val Asp Ile Val Arg Thr Asp Thr Glu Thr Ser Ser His Pro
65 70 75 80
Ala Pro Arg Thr Phe Ile Asn Gln Leu Pro Asp Trp Ser Met Leu Leu
85 90 95
Ala Ala Ile Thr Thr Ile Phe Leu Ala Ala Glu Lys Gln Trp Met Met
100 105 110
Leu Asp Trp Lys Pro Arg Arg Ser Asp Met Leu Val Asp Pro Phe Gly
115 120 125
Ile Gly Arg Ile Val Gln Asp Gly Leu Val Phe Arg Gln Asn Phe Ser
130 135 140
Ile Arg Ser Tyr Glu Ile Gly Ala Asp Arg Ser Ala Ser Ile Glu Thr
145 150 155 160
Val Met Asn His Leu Gln Glu Thr Ala Leu Asn His Val Lys Thr Ala
165 170 175
Gly Leu Leu Gly Asp Gly Phe Gly Ser Thr Pro Glu Met Phe Lys Lys
180 185 190
Asn Leu Ile Trp Val Val Thr Arg Met Gln Val Val Val Asp Lys Tyr
195 200 205
Pro Thr Trp Gly Asp Val Val Glu Val Asp Thr Trp Val Ser Gln Ser
210 215 220
Gly Lys Asn Gly Met Arg Arg Asp Trp Leu Val Arg Asp Cys Asn Thr
225 230 235 240
Gly Glu Thr Leu Thr Arg Ala Ser Ser Val Trp Val Met Met Asn Lys
245 250 255
Leu Thr Arg Arg Leu Ser Lys Ile Pro Glu Glu Val Arg Gly Glu Ile
260 265 270
Glu Pro Tyr Phe Val Asn Ser Asp Pro Val Leu Ala Glu Asp Ser Arg
275 280 285
Lys Leu Thr Lys Ile Asp Asp Lys Thr Ala Asp Tyr Val Arg Ser Gly
290 295 300
Leu Thr Pro Arg Trp Ser Asp Leu Asp Val Asn Gln His Val Asn Asn
305 310 315 320
Val Lys Tyr Ile Gly Trp Ile Leu Glu Ser Ala Pro Val Gly Ile Met
325 330 335
Glu Arg Gln Lys Leu Lys Ser Met Thr Leu Glu Tyr Arg Arg Glu Cys
340 345 350
Gly Arg Asp Ser Val Leu Gln Ser Leu Thr Ala Val Thr Gly Cys Asp
355 360 365
Ile Gly Asn Leu Ala Thr Ala Gly Asp Val Glu Cys Gln His Leu Leu
370 375 380
Arg Leu Gln Asp Gly Ala Glu Val Val Arg Gly Arg Thr Glu Trp Ser
385 390 395 400
Ser Lys Thr Pro Thr Thr Thr Trp Gly Thr Ala Pro
405 410
<210> 205
<211> 345
<212> Белок
<213> Arabidopsis thaliana
<400> 205
Met Phe Ile Ala Val Glu Val Ser Pro Val Met Glu Asp Ile Thr Arg
1 5 10 15
Gln Ser Lys Lys Thr Ser Val Glu Asn Glu Thr Gly Asp Asp Gln Ser
20 25 30
Ala Thr Ser Val Val Leu Lys Ala Lys Arg Lys Arg Arg Ser Gln Pro
35 40 45
Arg Asp Ala Pro Pro Gln Arg Ser Ser Val His Arg Gly Val Thr Arg
50 55 60
His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp Lys Asn Ser
65 70 75 80
Trp Asn Glu Thr Gln Thr Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala
85 90 95
Tyr Asp Glu Glu Asp Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu
100 105 110
Lys Tyr Trp Gly Arg Asp Thr Ile Leu Asn Phe Pro Leu Cys Asn Tyr
115 120 125
Glu Glu Asp Ile Lys Glu Met Glu Ser Gln Ser Lys Glu Glu Tyr Ile
130 135 140
Gly Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly Val Ser Lys
145 150 155 160
Tyr Arg Gly Val Ala Lys His His His Asn Gly Arg Trp Glu Ala Arg
165 170 175
Ile Gly Arg Val Phe Gly Asn Lys Tyr Leu Tyr Leu Gly Thr Tyr Ala
180 185 190
Thr Gln Glu Glu Ala Ala Ile Ala Tyr Asp Ile Ala Ala Ile Glu Tyr
195 200 205
Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Ile Ser Arg Tyr Leu Lys
210 215 220
Leu Pro Val Pro Glu Asn Pro Ile Asp Thr Ala Asn Asn Leu Leu Glu
225 230 235 240
Ser Pro His Ser Asp Leu Ser Pro Phe Ile Lys Pro Asn His Glu Ser
245 250 255
Asp Leu Ser Gln Ser Gln Ser Ser Ser Glu Asp Asn Asp Asp Arg Lys
260 265 270
Thr Lys Leu Leu Lys Ser Ser Pro Leu Val Ala Glu Glu Val Ile Gly
275 280 285
Pro Ser Thr Pro Pro Glu Ile Ala Pro Pro Arg Arg Ser Phe Pro Glu
290 295 300
Asp Ile Gln Thr Tyr Phe Gly Cys Gln Asn Ser Gly Lys Leu Thr Ala
305 310 315 320
Glu Glu Asp Asp Val Ile Phe Gly Asp Leu Asp Ser Phe Leu Thr Pro
325 330 335
Asp Phe Tyr Ser Glu Leu Asn Asp Cys
340 345
<210> 206
<211> 303
<212> Белок
<213> Arabidopsis thaliana
<400> 206
Met Ala Lys Val Ser Gly Arg Ser Lys Lys Thr Ile Val Asp Asp Glu
1 5 10 15
Ile Ser Asp Lys Thr Ala Ser Ala Ser Glu Ser Ala Ser Ile Ala Leu
20 25 30
Thr Ser Lys Arg Lys Arg Lys Ser Pro Pro Arg Asn Ala Pro Leu Gln
35 40 45
Arg Ser Ser Pro Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg
50 55 60
Tyr Glu Ala His Leu Trp Asp Lys Asn Ser Trp Asn Asp Thr Gln Thr
65 70 75 80
Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Glu Glu Glu Ala
85 90 95
Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Arg Asp
100 105 110
Thr Leu Leu Asn Phe Pro Leu Pro Ser Tyr Asp Glu Asp Val Lys Glu
115 120 125
Met Glu Gly Gln Ser Lys Glu Glu Tyr Ile Gly Ser Leu Arg Arg Lys
130 135 140
Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg
145 150 155 160
His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Ala
165 170 175
Thr Gln Glu Glu Ala Ala Ile Ala Tyr Asp Ile Ala Ala Ile Glu Tyr
180 185 190
Arg Gly Leu Asn Ala Val Thr Asn Phe Asp Val Ser Arg Tyr Leu Asn
195 200 205
Pro Asn Ala Ala Ala Asp Lys Ala Asp Ser Asp Ser Lys Pro Ile Arg
210 215 220
Ser Pro Ser Arg Glu Pro Glu Ser Ser Asp Asp Asn Lys Ser Pro Lys
225 230 235 240
Ser Glu Glu Val Ile Glu Pro Ser Thr Ser Pro Glu Val Ile Pro Thr
245 250 255
Arg Arg Ser Phe Pro Asp Asp Ile Gln Thr Tyr Phe Gly Cys Gln Asp
260 265 270
Ser Gly Lys Leu Ala Thr Glu Glu Asp Val Ile Phe Asp Cys Phe Asn
275 280 285
Ser Tyr Ile Asn Pro Gly Phe Tyr Asn Glu Phe Asp Tyr Gly Pro
290 295 300
<210> 207
<211> 445
<212> Белок
<213> Avena sativa
<400> 207
Met Lys Arg Ser Pro Pro Pro Ala Pro Pro Ala Ala Pro Pro Pro Pro
1 5 10 15
Gln Pro Ser Pro Ser Ser Ser Ser Pro Ala Cys Ser Pro Ser Pro Ser
20 25 30
Ser Ser Ser Cys Pro Ser Ser Ser Asp Ser Ser Ser Ile Val Ile Pro
35 40 45
Arg Lys Arg Ala Arg Thr Gln Lys Ala Ala Ser Gly Lys Pro Lys Ala
50 55 60
Lys Ala Ser Ala Lys Arg Pro Lys Lys Asp Ala Ser Arg Ser Ser Lys
65 70 75 80
Glu Thr Asp Ala Asn Gly Ala Ala Ala Ala Ala Gly Lys Arg Ser Ser
85 90 95
Ile Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe Glu Ala
100 105 110
His Leu Trp Asp Lys Asn Cys Phe Thr Ser Val Gln Asn Lys Lys Lys
115 120 125
Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Thr Glu Asp Ala Ala Ala
130 135 140
Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Ser Glu Thr Ile
145 150 155 160
Leu Asn Phe Ser Val Glu Asp Tyr Ala Lys Glu Met Pro Glu Met Glu
165 170 175
Ala Val Ser Arg Glu Glu Tyr Leu Ala Ala Leu Arg Arg Arg Ser Ser
180 185 190
Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His
195 200 205
His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Leu Gly Asn Lys
210 215 220
Tyr Leu Tyr Leu Gly Thr Phe Asp Thr Gln Glu Glu Ala Ala Lys Ala
225 230 235 240
Tyr Asp Leu Ala Ala Ile Glu Tyr Arg Gly Ala Asn Ala Val Thr Asn
245 250 255
Phe Asp Ile Ser Cys Tyr Leu Asp Gln Pro Gln Leu Leu Ala Gln Leu
260 265 270
Gln Gln Gly Pro Gln Val Val Pro Ala Leu Gln Glu Glu Leu Gln His
275 280 285
Asp Val Gln His Asp Leu Gln Asn Asp Asn Ala Val Gln Glu Leu Asn
290 295 300
Ser Gly Glu Val Gln Met Pro Gly Ala Met Asp Glu Pro Ile Ala Leu
305 310 315 320
Asp Asp Ser Thr Glu Cys Ile Asn Thr Pro Phe Glu Phe Asp Phe Ser
325 330 335
Val Glu Glu Asn Leu Trp Ser Pro Cys Met Asp Tyr Glu Leu Asp Ala
340 345 350
Ile Leu Gly Asn Asn Thr Ser Asn Ser Ala Asn Met Asn Glu Trp Phe
355 360 365
Asn Asp Ser Thr Phe Glu Ser Asn Ile Gly Cys Leu Phe Glu Gly Cys
370 375 380
Ser Asn Ile Asp Asp Cys Ser Ser Ser Lys His Cys Ala Asp Leu Ala
385 390 395 400
Ala Phe Asp Phe Phe Lys Glu Gly Asp Asp Asn Asp Phe Ser Asn Met
405 410 415
Glu Met Glu Ile Thr Pro Gln Ala Asn Asp Val Ser Cys Pro Pro Asn
420 425 430
Asp Val Ser Cys Pro Pro Lys Met Ile Thr Val Cys Asn
435 440 445
<210> 208
<211> 420
<212> Белок
<213> Sorghum bicolor
<400> 208
Met Asp Met Glu Arg Ser Gln Gln Gln Lys Ser Pro Thr Glu Ser Pro
1 5 10 15
Pro Pro Pro Ser Pro Ser Ser Ser Ser Ser Ser Val Ser Ala Asp Thr
20 25 30
Val Leu Pro Pro Pro Gly Lys Arg Arg Arg Ala Ala Thr Thr Ala Lys
35 40 45
Ala Lys Ala Gly Ala Lys Pro Lys Arg Ala Arg Lys Asp Ala Ala Ala
50 55 60
Ala Ala Asp Pro Pro Pro Pro Pro Ala Ala Ala Ala Ala Gly Lys Arg
65 70 75 80
Ser Ser Val Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe
85 90 95
Glu Ala His Leu Trp Asp Lys His Cys Leu Ala Ala Leu His Asn Lys
100 105 110
Lys Lys Gly Arg Gln Val Tyr Leu Gly Ala Tyr Asp Ser Glu Glu Ala
115 120 125
Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Glu
130 135 140
Thr Leu Leu Asn Phe Pro Val Glu Asp Tyr Ser Ser Glu Met Pro Glu
145 150 155 160
Met Glu Gly Val Ser Arg Glu Glu Tyr Leu Ala Ser Leu Arg Arg Arg
165 170 175
Ser Ser Gly Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg
180 185 190
His His His Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly
195 200 205
Asn Lys Tyr Leu Tyr Leu Gly Thr Phe Asp Thr Gln Glu Glu Ala Ala
210 215 220
Lys Ala Tyr Asp Leu Ala Ala Ile Glu Tyr Arg Gly Val Asn Ala Val
225 230 235 240
Thr Asn Phe Asp Ile Ser Cys Tyr Leu Asp His Pro Leu Phe Leu Ala
245 250 255
Gln Leu Gln Gln Glu Pro Gln Val Val Pro Ala Leu Asn Gln Glu Ala
260 265 270
Gln Pro Asp Gln Ser Glu Thr Glu Thr Ile Ala Gln Glu Ser Val Ser
275 280 285
Ser Glu Ala Lys Thr Pro Asp Asp Asn Ala Glu Pro Asp Asp Asn Ala
290 295 300
Glu Pro Asp Asp Ile Ala Glu Pro Leu Ile Thr Val Asp Asp Ser Ile
305 310 315 320
Glu Glu Ser Leu Trp Ser Pro Cys Met Asp Tyr Glu Leu Asp Thr Met
325 330 335
Ser Arg Ser Asn Phe Gly Ser Ser Ile Asn Leu Ser Glu Trp Phe Asn
340 345 350
Asp Ala Asp Phe Asp Ser Asn Ile Gly Cys Leu Phe Asp Gly Cys Ser
355 360 365
Ala Val Asp Glu Gly Gly Lys Asp Gly Val Gly Leu Ala Asp Phe Ser
370 375 380
Leu Leu Glu Asp Phe Ser Leu Phe Glu Ala Gly Asp Gly Gln Leu Lys
385 390 395 400
Asp Val Leu Ser Asp Met Glu Glu Gly Ile Gln Pro Pro Thr Met Ile
405 410 415
Ser Val Cys Asn
420
<210> 209
<211> 395
<212> Белок
<213> Zea mays
<400> 209
Met Glu Arg Ser Gln Arg Gln Ser Pro Pro Pro Pro Ser Pro Ser Ser
1 5 10 15
Ser Ser Ser Ser Val Ser Ala Asp Thr Val Leu Val Pro Pro Gly Lys
20 25 30
Arg Arg Arg Ala Ala Thr Ala Lys Ala Gly Ala Glu Pro Asn Lys Arg
35 40 45
Ile Arg Lys Asp Pro Ala Ala Ala Ala Ala Gly Lys Arg Ser Ser Val
50 55 60
Tyr Arg Gly Val Thr Arg His Arg Trp Thr Gly Arg Phe Glu Ala His
65 70 75 80
Leu Trp Asp Lys His Cys Leu Ala Ala Leu His Asn Lys Lys Lys Gly
85 90 95
Arg Gln Val Tyr Leu Gly Ala Tyr Asp Ser Glu Glu Ala Ala Ala Arg
100 105 110
Ala Tyr Asp Leu Ala Ala Leu Lys Tyr Trp Gly Pro Glu Thr Leu Leu
115 120 125
Asn Phe Pro Val Glu Asp Tyr Ser Ser Glu Met Pro Glu Met Glu Ala
130 135 140
Val Ser Arg Glu Glu Tyr Leu Ala Ser Leu Arg Arg Arg Ser Ser Gly
145 150 155 160
Phe Ser Arg Gly Val Ser Lys Tyr Arg Gly Val Ala Arg His His His
165 170 175
Asn Gly Arg Trp Glu Ala Arg Ile Gly Arg Val Phe Gly Asn Lys Tyr
180 185 190
Leu Tyr Leu Gly Thr Phe Asp Thr Gln Glu Glu Ala Ala Lys Ala Tyr
195 200 205
Asp Leu Ala Ala Ile Glu Tyr Arg Gly Val Asn Ala Val Thr Asn Phe
210 215 220
Asp Ile Ser Cys Tyr Leu Asp His Pro Leu Phe Leu Ala Gln Leu Gln
225 230 235 240
Gln Glu Pro Gln Val Val Pro Ala Leu Asn Gln Glu Pro Gln Pro Asp
245 250 255
Gln Ser Glu Thr Gly Thr Thr Glu Gln Glu Pro Glu Ser Ser Glu Ala
260 265 270
Lys Thr Pro Asp Gly Ser Ala Glu Pro Asp Glu Asn Ala Val Pro Asp
275 280 285
Asp Thr Ala Glu Pro Leu Thr Thr Val Asp Asp Ser Ile Glu Glu Gly
290 295 300
Leu Trp Ser Pro Cys Met Asp Tyr Glu Leu Asp Thr Met Ser Arg Pro
305 310 315 320
Asn Phe Gly Ser Ser Ile Asn Leu Ser Glu Trp Phe Ala Asp Ala Asp
325 330 335
Phe Asp Cys Asn Ile Gly Cys Leu Phe Asp Gly Cys Ser Ala Ala Asp
340 345 350
Glu Gly Ser Lys Asp Gly Val Gly Leu Ala Asp Phe Ser Leu Phe Glu
355 360 365
Ala Gly Asp Val Gln Leu Lys Asp Val Leu Ser Asp Met Glu Glu Gly
370 375 380
Ile Gln Pro Pro Ala Met Ile Ser Val Cys Asn
385 390 395
<210> 210
<211> 430
<212> Белок
<213> Triadica sebifera
<400> 210
Met Ala Ser Ser Ser Ser Asp Pro Val Leu Lys Ala Glu Leu Gly Ser
1 5 10 15
Ser Gly Gly Gly Cys Ser Ser Gly Gly Gly Gly Glu Ser Ser Glu Ala
20 25 30
Val Ile Ala Asn Asp Gln Leu Leu Leu Tyr Arg Gly Leu Lys Lys Pro
35 40 45
Lys Lys Glu Arg Gly Cys Thr Ala Lys Glu Arg Ile Ser Lys Met Pro
50 55 60
Pro Cys Thr Ala Gly Lys Arg Ser Ser Ile Tyr Arg Gly Val Thr Arg
65 70 75 80
His Arg Trp Thr Gly Arg Tyr Glu Ala His Leu Trp Asp Lys Ser Thr
85 90 95
Trp Asn Gln Asn Gln Asn Lys Lys Gly Lys Gln Val Tyr Leu Gly Ala
100 105 110
Tyr Asp Asp Glu Glu Ala Ala Ala Arg Ala Tyr Asp Leu Ala Ala Leu
115 120 125
Lys Tyr Trp Gly Pro Gly Thr Leu Ile Asn Phe Pro Val Thr Asp Tyr
130 135 140
Thr Arg Asp Leu Glu Glu Met Gln Asn Met Ser Arg Glu Glu Tyr Leu
145 150 155 160
Ala Ser Leu Arg Arg Lys Ser Ser Gly Phe Ser Arg Gly Ile Ser Lys
165 170 175
Tyr Arg Gly Leu Ser Ser Arg Trp Glu Ser Ser Val Gly Arg Met Pro
180 185 190
Gly Ser Glu Tyr Phe Ser Ser Ile Asn Tyr Val Asp Asp Pro Ala Ala
195 200 205
Glu Ser Glu Tyr Val Gly Ser Leu Cys Phe Glu Arg Lys Ile Asp Leu
210 215 220
Thr Ser Tyr Ile Lys Trp Trp Gly Leu Asn Lys Thr Arg Gln Ala Glu
225 230 235 240
Ser Ile Ser Lys Ser Ala Glu Glu Thr Lys Pro Gly Cys Ala Glu Asp
245 250 255
Ile Gly Gly Glu Leu Lys Thr Thr Glu Trp Ala Ile Gln Pro Thr Glu
260 265 270
Pro Tyr Gln Met Pro Arg Leu Gly Met Pro Val His Val Lys Lys His
275 280 285
Lys Gly Ser Lys Ile Ser Ala Leu Ser Val Leu Ser Gln Ser Ala Ala
290 295 300
Phe Lys Ser Leu Gln Glu Lys Ala Ser Lys Lys Gln Glu Asn Ser Thr
305 310 315 320
Asp Asn Asp Glu Asn Glu Asn Lys Asn Thr Asn Thr Asn Lys Ile Asp
325 330 335
Tyr Gly Lys Ala Val Glu Thr Ser Ala Ser His Asp Ser Ser Asn Glu
340 345 350
Arg Pro Val Thr Ala Leu Gly Met Ser Gly Gly Leu Ser Leu Lys Arg
355 360 365
Asn Val Tyr Gln Leu Thr Pro Phe Leu Ser Ala Pro Leu Leu Thr Asn
370 375 380
Tyr Gly Thr Ile Asp Gln Leu Val Asp Pro Ile Leu Trp Ala Ser Leu
385 390 395 400
Val Pro Val Leu Pro Thr Gly Leu Ser Arg Asn Pro Glu Val Thr Lys
405 410 415
Thr Glu Thr Ser Ser Thr Tyr Thr Phe Phe Arg Pro Glu Glu
420 425 430
<210> 211
<211> 1531
<212> ДНК
<213> Solanum tuberosum
<400> 211
ttttaaatca ttgttttatt ttctctttct ttttacaggt ataaaaggtg aaaattgaag 60
caagattgat tgcaagctat gtgtcaccac gttattgata ctttggaaga aatttttact 120
tatatgtctt tgtttaggag taatatttga tatgttttag ttagattttc ttgtcattta 180
tgctttagta taattttagt tatttttatt atatgatcat gggtgaattt tgatacaaat 240
atttttgtca ttaaataaat taatttatca caacttgatt actttcagtg acaaaaaatg 300
tattgtcgta gtaccctttt ttgttgaata tgaataattt tttttatttt gtgacaattg 360
taattgtcac tacttatgat aatatttagt gacatatatg tcgtcggtaa aagcaaacac 420
tttcagtgac aaaataatag atttaatcac aaaattatta acctttttta taataataaa 480
tttatcccta atttatacat ttaaggacaa agtatttttt ttatatataa aaaatagtct 540
ttagtgacga tcgtagtgtt gagtctagaa atcataatgt tgaatctaga aaaatctcat 600
gcagtgtaaa ataaacctca aaaaggacgt tcagtccata gagggggtgt atgtgacacc 660
ccaacctcag caaaagaaaa cctcccttca acaaggacat ttgcggtgct aaacaatttc 720
aagtctcatc acacatatat ttattatata atactaataa agaatagaaa aggaaaggta 780
aacatcatta aatcgtcttt gtatattttt agtgacaact gattgacgaa atctttttcg 840
tcacacaaaa tttttagtga cgaaacatga tttatagatg atgaaattat ttgtccctca 900
taatctaatt tgttgtagtg atcattactc ctttgtttgt tttatttgtc atgttagtcc 960
attaaaaaaa aatatctctc ttcttatgta cgtgaatggt tggaacggat ctattatata 1020
atactaataa agaatagaaa aaggaaagtg agtgaggttc gagggagaga atctgtttaa 1080
tatcagagtc gatcatgtgt caattttatc gatatgaccc taacttcaac tgagtttaac 1140
caattccgat aaggcgagaa atatcatagt attgagtcta gaaaaatctc atgtagtgtg 1200
gggtaaacct cagcaaggac gttgagtcca tagagggggg tgtatgtgac accccaacct 1260
cagcaaaaga aaacctcccc tcaagaagga catttgcggt gctaaacaat ttcaagtctc 1320
atcacacata tatatatatt atataatact aataaataat agaaaaagga aaggtaaaca 1380
tcactaacga cagttgcggt gcaaactgag tgaggtaata aacagcacta acttttattg 1440
gttatgtcaa actcaaagta aaatttctca acttgtttac gtgcctatat ataccatgct 1500
tgttatatgc tcaaagcacc aacaaaattt a 1531
<210> 212
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный праймер
<400> 212
cactcgtgct ttccatcatc 20
<210> 213
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный праймер
<400> 213
gaaggctgag caacaagagg 20
<210> 214
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный праймер
<400> 214
ggcgattttg gattctgc 18
<210> 215
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Олигонуклеотидный праймер
<400> 215
cccaaccctt ccgtatacat 20
<210> 216
<211> 1970
<212> ДНК
<213> Zea mays
<400> 216
ggtacctttt ttcccagaga taaatgtgga atagctctac aaacaaacgg catgatgctg 60
acacttggat ggcgaccttg caatcccaag aactattgca tacggttgcc agtcgacaaa 120
tatctacgcc atgcatggct acggtcggaa tacaccgtag cggcgggtaa ctcgccgata 180
ccgtccacgt gtccttggat gcccggtcgc tgatacttct ggtcttctgg acatgcacca 240
agacaatcaa gtgattcaac cttaatttaa cataatataa ataatacgta acatccaact 300
gacgtgttca cctatagaga atattccttc tgattctact ttcagaatga tgccgttgcc 360
gtgtatcgag caagtactct cactcgaagt atcttatctc ccacatccag cacaaaaatc 420
ttctgttcgt ggcaaatctt gtggcggttg aacgaaagaa tgctatataa gtagctatag 480
agaacgtatt atgtgtaaac caaccgttca gtgtaaatcg tgtgtaaata gtcatgttaa 540
ttttttggcg gcagatcaag tacaaactgt atgcctcgga taaacatgta caaaccacaa 600
cactggccac tagatctata tccaacgttc ataaccatcc atccctctct gctgcactct 660
gcaaacaagc acccccatct cgtagcaaca tcttgtctcc gacaagctct cgatgtagtg 720
gaggccctcc accgcaatat cctagtgtat gatgttggag aagcgactcc taaataatgg 780
tggcaagatg ttgctaggtt tgtagccata gcctcaatct aagatcatcc caagccatgg 840
gacctgattc tacgaggcct acaaccaggc atgacacgtc gtctacccac tcttgtgcat 900
catcggtcac ttgatctgac ttggttccta accacttacc ctaggttcca aagccctaag 960
tttctcgtat attgttagtc attcttagtg ggagttttat gtgtatttca ttcctgttaa 1020
atagcatgcc aactaagcaa acatgatgat ataatatgca atctaataaa aagatatatg 1080
agtgggtttc ataaaaaagg gagagagttt catgaggagt gaaactctga atacagatac 1140
tgatatgaca gctttaaaag tagtgttatg aaatcatcat tgagaaatgg tattagcact 1200
caatcgattt ctacgctgtc aattgtcatg agcacaattt tcacccaaag aggcacacca 1260
gcaatgtccg cttgtagtgt ccgagacgtt gctccatcgc cgtcgtcttg tttctgtgcg 1320
ctccattcaa tgcggcaagt ggctcaatcc caagcggtcg tcgcctccca gccccagcag 1380
caaaatatct tcccatgcgg ccatgccttg aaaattggaa tagattctct agattcaccg 1440
ccgcgtcatc ttcactactt tctcactggc ccaatcagca tctccttctc cgagctcaat 1500
catgctcagt caagcgtcac caatggcgtc acggttggtt ttgtcactgt ctgcatgcaa 1560
gggtattttg cttcgcaagt gtaaatggaa aatggatcta aacaactgca ctgcaccaat 1620
tttggaacgc ggaaccgaga gtctgtttgg gttcgtttga aacgcgctga tgtttctcat 1680
tttttaatag atgtagttac ctgatactat ttaagttgga cgatcaaacg actgtgtcaa 1740
gtgtgattaa gaaaagcatc gaaaataaaa tttatcgcca taaaaagtta aaaacagtgg 1800
ataatagtag gacctcataa tagaaaaaat tatcaaacgg aatggagggg cccaacgcag 1860
tatatagcag ccgggtggtg ccggacatcc gacgctcgtg ccagcaggcc attcttctcg 1920
ccttactccc tcacagaacc cagtaaaata tcgccagtcc cgccgtcgag 1970
<210> 217
<211> 584
<212> ДНК
<213> Aeluropus littoralis
<400> 217
cccaagcttg accgatgcac acgctacctg ccaaggctcc ctccatccgc actctgcatc 60
gtcgcttcgg cgtaaacttc cacgtagtac ttgtacgatt ctagctagac ccagtgcgcc 120
caccctaccg ccggcgagcg ggcccccatc tcgcgccagg cttccatgcg ggtccaccgt 180
ggaccagccc tacgccgaac cgagcccatc cctccaccct ttcaccgcca agcgggaccc 240
gcgttggacc tttccgcttg gctggccccc accagcgtcc acgcgggcca acggcctcgc 300
gaaatggatc tccacacgac aaaccaaaac gagaagaaaa taaatggaaa ggaaagaaac 360
ggatcgccac gcgttccaga ggcgtccgct aaccacccga ttatgcttgc gcagcgtgcg 420
taacctcgtc gtggggttaa tccgggtggc cggatcggga aagccacggc ctttataacc 480
catccctgcc ggatcgaacc ggtaccggaa acaaaaacag ggggagaaaa aaagttcttc 540
gcgaggaagg aaaaggaaaa gtcgcgtgcc gtcctcgccc acag 584
<210> 218
<211> 928
<212> ДНК
<213> Agrobacterium rhizogenes
<400> 218
ttagcgaaag gatgtcaaaa aaggatgccc ataattggga ggagtggggt aaagcttaaa 60
gttggcccgc tattggattt cgcgaaagcg gcattggcaa acgtggagat tgctgcattc 120
aagatacttt ttctattttc tggttaagat gtaaagtatt gccacaatca tattaattac 180
taacattgta tatgtaatat agtgcggaaa ttatctatgc caaaatgatg tattaataat 240
agcaataata atatgtgtta atctttttca atcgggaata cgtttaagcg attatcgtgt 300
tgaataaatt attccaaaag gaaatacatg gttttggaga acctgctata gatatatgcc 360
aaatttacac tagtttagtg ggtgcaaaac tattatctct gtttctgagt ttaataaaaa 420
ataaataagc agggcgaata gcagttagcc taagaaggaa tggtggccat gtacgtgctt 480
ttaagagacc ctataataaa ttgccagctg tgttgctttg gtgccgacag gcctaacgtg 540
gggtttagct tgacaaagta gcgcctttcc gcagcataaa taaaggtagg cgggtgcgtc 600
ccattattaa aggaaaaagc aaaagctgag attccataga ccacaaacca ccattattgg 660
aggacagaac ctattccctc acgtgggtcg ctagctttaa acctaataag taaaaacaat 720
taaaagcagg caggtgtccc ttctatattc gcacaacgag gcgacgtgga gcatcgacag 780
ccgcatccat taattaataa atttgtggac ctatacctaa ctcaaatatt tttattattt 840
gctccaatac gctaagagct ctggattata aatagtttgg atgcttcgag ttatgggtac 900
aagcaacctg tttcctactt tgttacca 928
<210> 219
<211> 732
<212> ДНК
<213> Искусственная
<220>
<223> конструкция шпРНКи, содержащая фрагмент размером 732 пары оснований
<400> 219
aatgcagttt tacaaagtgg agtacccaaa gcagatgaga tcattttgta taacatggct 60
cttgtgttgg atcgtatttt tgtggatgtg aaggatgctt ttgagttctc accacatcat 120
aaggccattc gtgaaccttt tgactattac aagtttggcc aaaattatat ccgcccttta 180
cttgatttca ggagttctta tgttggcaat atatcagttt ttggtgaaat agaagagaag 240
ctcaagcagg gcgttaatgt tgttttgatg tcaaaccacc aaagtgaagc agatccagcg 300
gttattgctc tgttgcttga atcgaggcac ccatacattg ctgagaacat aatttatgtt 360
gcaggagata gagttattac tgatcctctt tgcaagccat tcagcatggg aaggaatctc 420
ctgtgtgttt attcgaaaaa acatatgggt gatgacccca aacttgtcga gaagaaaagg 480
agagcaaaca caagaagctt gaaggagatg gctgtgctat tgaggggtgg atcaaaacta 540
atatggattg ctcctagtgg tggaagagat aggccaaacc ctgttacaaa agaatggtat 600
ccagcgccat ttgatgcttc ttcaacagac aacatgagaa ggcttgtaga acatgctggt 660
gtccctggtc acatttatcc tctagcaata ttatgctatg atattatgcc ccctccgccc 720
caggttgaga aa 732
<210> 220
<211> 512
<212> Белок
<213> Elaeis guineensis
<400> 220
Met Ala Val Ser Lys Asn Pro Glu Thr Leu Ala Pro Asp Gln Glu Pro
1 5 10 15
Ser Lys Glu Ser Asp Leu Arg Arg Arg Pro Ala Ser Ser Pro Ser Ser
20 25 30
Thr Ala Ala Ser Pro Ala Val Pro Asp Ser Ser Ser Arg Thr Ser Ser
35 40 45
Ser Ile Thr Gly Ser Trp Thr Thr Ala Leu Asp Gly Asp Ser Gly Ala
50 55 60
Gly Ala Val Arg Ile Gly Asp Pro Lys Asp Arg Ile Gly Glu Ala Asn
65 70 75 80
Asp Ile Gly Glu Lys Lys Lys Ala Cys Ser Gly Glu Val Pro Val Gly
85 90 95
Phe Val Asp Arg Pro Ser Ala Pro Val His Val Arg Val Val Glu Ser
100 105 110
Pro Leu Ser Ser Asp Thr Ile Phe Gln Gln Ser His Ala Gly Leu Leu
115 120 125
Asn Leu Cys Val Val Val Leu Ile Ala Val Asn Ser Arg Leu Ile Ile
130 135 140
Glu Asn Leu Met Lys Tyr Gly Leu Leu Ile Gly Ser Gly Phe Phe Phe
145 150 155 160
Ser Ser Arg Leu Leu Arg Asp Trp Pro Leu Leu Ile Cys Ser Leu Thr
165 170 175
Leu Pro Val Phe Pro Leu Gly Ser Tyr Met Val Glu Lys Leu Ala Tyr
180 185 190
Lys Lys Phe Ile Ser Glu Pro Val Val Val Ser Leu His Val Ile Leu
195 200 205
Ile Ile Ala Thr Ile Met Tyr Pro Val Phe Val Ile Leu Arg Cys Asp
210 215 220
Ser Pro Ile Leu Ser Gly Ile Asn Leu Met Leu Phe Val Ser Ser Ile
225 230 235 240
Cys Leu Lys Leu Val Ser Tyr Ala His Ala Asn Tyr Asp Leu Arg Ser
245 250 255
Ser Ser Asn Ser Ile Asp Lys Gly Ile His Lys Ser Gln Gly Val Ser
260 265 270
Phe Lys Ser Leu Val Tyr Phe Ile Met Ala Pro Thr Leu Cys Tyr Gln
275 280 285
Pro Ser Tyr Pro Arg Thr Thr Cys Ile Arg Lys Gly Trp Val Ile Cys
290 295 300
Gln Leu Val Lys Leu Val Ile Phe Thr Gly Val Met Gly Phe Ile Ile
305 310 315 320
Glu Gln Tyr Ile Asp Pro Ile Ile Lys Asn Ser Gln His Pro Leu Lys
325 330 335
Gly Asn Val Leu Asn Ala Met Glu Arg Val Leu Lys Leu Ser Ile Pro
340 345 350
Thr Leu Tyr Val Trp Leu Cys Val Phe Tyr Cys Thr Phe His Leu Trp
355 360 365
Leu Asn Ile Leu Ala Glu Leu Leu Cys Phe Gly Asp Arg Glu Phe Tyr
370 375 380
Lys Asp Trp Trp Asn Ala Lys Thr Ile Glu Glu Tyr Trp Arg Met Trp
385 390 395 400
Asn Met Pro Val His Lys Trp Met Leu Arg His Val Tyr Leu Pro Cys
405 410 415
Ile Arg Asn Gly Ile Pro Lys Gly Val Ala Met Val Ile Ser Phe Phe
420 425 430
Ile Ser Ala Ile Phe His Glu Leu Cys Ile Gly Ile Pro Cys His Ile
435 440 445
Phe Lys Phe Trp Ala Phe Ile Gly Ile Met Phe Gln Val Pro Leu Val
450 455 460
Ile Leu Thr Lys Tyr Leu Gln Asn Lys Phe Lys Ser Ala Met Val Gly
465 470 475 480
Asn Met Ile Phe Trp Phe Phe Phe Ser Ile Tyr Gly Gln Pro Met Cys
485 490 495
Val Leu Leu Tyr Tyr His Asp Val Met Asn Arg Lys Val Gly Thr Glu
500 505 510
<210> 221
<211> 74
<212> Белок
<213> Glycine max
<400> 221
Met Ala Asp Ile Asp Arg Ser Phe Asp Asn Asn Val Ser Ala Val Ser
1 5 10 15
Thr Glu Lys Ser Ser Gln Val Ser Asp Val Glu Phe Ser Glu Ala Glu
20 25 30
Glu Ile Leu Ile Ala Met Val Tyr Asn Leu Val Gly Glu Arg Trp Ser
35 40 45
Leu Ile Ala Gly Arg Ile Pro Gly Arg Thr Ala Glu Glu Ile Glu Lys
50 55 60
Tyr Trp Thr Ser Arg Phe Ser Thr Ser Gln
65 70
<210> 222
<211> 146
<212> Белок
<213> Arabidopsis thaliana
<400> 222
Met Gly Ser Leu Gln Met Gln Thr Ser Pro Glu Ser Asp Asn Asp Pro
1 5 10 15
Arg Tyr Ala Thr Val Thr Asp Glu Arg Lys Arg Lys Arg Met Ile Ser
20 25 30
Asn Arg Glu Ser Ala Arg Arg Ser Arg Met Arg Lys Gln Lys Gln Leu
35 40 45
Gly Asp Leu Ile Asn Glu Val Thr Leu Leu Lys Asn Asp Asn Ala Lys
50 55 60
Ile Thr Glu Gln Val Asp Glu Ala Ser Lys Lys Tyr Ile Glu Met Glu
65 70 75 80
Ser Lys Asn Asn Val Leu Arg Ala Gln Ala Ser Glu Leu Thr Asp Arg
85 90 95
Leu Arg Ser Leu Asn Ser Val Leu Glu Met Val Glu Glu Ile Ser Gly
100 105 110
Gln Ala Leu Asp Ile Pro Glu Ile Pro Glu Ser Met Gln Asn Pro Trp
115 120 125
Gln Met Pro Cys Pro Met Gln Pro Ile Arg Ala Ser Ala Asp Met Phe
130 135 140
Asp Cys
145
<210> 223
<211> 268
<212> Белок
<213> Arabidopsis thaliana
<400> 223
Met Gly Arg Gly Lys Ile Glu Ile Lys Arg Ile Glu Asn Ala Asn Ser
1 5 10 15
Arg Gln Val Thr Phe Ser Lys Arg Arg Ser Gly Leu Leu Lys Lys Ala
20 25 30
Arg Glu Leu Ser Val Leu Cys Asp Ala Glu Val Ala Val Ile Val Phe
35 40 45
Ser Lys Ser Gly Lys Leu Phe Glu Tyr Ser Ser Thr Gly Met Lys Gln
50 55 60
Thr Leu Ser Arg Tyr Gly Asn His Gln Ser Ser Ser Ala Ser Lys Ala
65 70 75 80
Glu Glu Asp Cys Ala Glu Val Asp Ile Leu Lys Asp Gln Leu Ser Lys
85 90 95
Leu Gln Glu Lys His Leu Gln Leu Gln Gly Lys Gly Leu Asn Pro Leu
100 105 110
Thr Phe Lys Glu Leu Gln Ser Leu Glu Gln Gln Leu Tyr His Ala Leu
115 120 125
Ile Thr Val Arg Glu Arg Lys Glu Arg Leu Leu Thr Asn Gln Leu Glu
130 135 140
Glu Ser Arg Leu Lys Glu Gln Arg Ala Glu Leu Glu Asn Glu Thr Leu
145 150 155 160
Arg Arg Gln Val Gln Glu Leu Arg Ser Phe Leu Pro Ser Phe Thr His
165 170 175
Tyr Val Pro Ser Tyr Ile Lys Cys Phe Ala Ile Asp Pro Lys Asn Ala
180 185 190
Leu Ile Asn His Asp Ser Lys Cys Ser Leu Gln Asn Thr Asp Ser Asp
195 200 205
Thr Thr Leu Gln Leu Gly Leu Pro Gly Glu Ala His Asp Arg Arg Thr
210 215 220
Asn Glu Gly Glu Arg Glu Ser Pro Ser Ser Asp Ser Val Thr Thr Asn
225 230 235 240
Thr Ser Ser Glu Thr Ala Glu Arg Gly Asp Gln Ser Ser Leu Ala Asn
245 250 255
Ser Pro Pro Glu Ala Lys Arg Gln Arg Phe Ser Val
260 265
<210> 224
<211> 437
<212> Белок
<213> Arabidopsis thaliana
<400> 224
Met Glu Phe Glu Ser Val Phe Lys Met His Tyr Pro Tyr Leu Ala Ala
1 5 10 15
Val Ile Tyr Asp Asp Ser Ser Thr Leu Lys Asp Phe His Pro Ser Leu
20 25 30
Thr Asp Asp Phe Ser Cys Val His Asn Val His His Lys Pro Ser Met
35 40 45
Pro His Thr Tyr Glu Ile Pro Ser Lys Glu Thr Ile Arg Gly Ile Thr
50 55 60
Pro Ser Pro Cys Thr Glu Ala Phe Gly Ala Cys Phe His Gly Thr Ser
65 70 75 80
Asn Asp His Val Phe Phe Gly Met Ala Tyr Thr Thr Pro Pro Thr Ile
85 90 95
Glu Pro Asn Val Ser His Val Ser His Asp Asn Thr Met Trp Glu Asn
100 105 110
Asp Gln Asn Gln Gly Phe Ile Phe Gly Thr Glu Ser Thr Leu Asn Gln
115 120 125
Ala Met Ala Asp Ser Asn Gln Phe Asn Met Pro Lys Pro Leu Leu Ser
130 135 140
Ala Asn Glu Asp Thr Ile Met Asn Arg Arg Gln Asn Asn Gln Val Met
145 150 155 160
Ile Lys Thr Glu Gln Ile Lys Lys Lys Asn Lys Arg Phe Gln Met Arg
165 170 175
Arg Ile Cys Lys Pro Thr Lys Lys Ala Ser Ile Ile Lys Gly Gln Trp
180 185 190
Thr Pro Glu Glu Asp Lys Leu Leu Val Gln Leu Val Asp Leu His Gly
195 200 205
Thr Lys Lys Trp Ser Gln Ile Ala Lys Met Leu Gln Gly Arg Val Gly
210 215 220
Lys Gln Cys Arg Glu Arg Trp His Asn His Leu Arg Pro Asp Ile Lys
225 230 235 240
Lys Asp Gly Trp Thr Glu Glu Glu Asp Ile Ile Leu Ile Lys Ala His
245 250 255
Lys Glu Ile Gly Asn Arg Trp Ala Glu Ile Ala Arg Lys Leu Pro Gly
260 265 270
Arg Thr Glu Asn Thr Ile Lys Asn His Trp Asn Ala Thr Lys Arg Arg
275 280 285
Gln His Ser Arg Arg Thr Lys Gly Lys Asp Glu Ile Ser Leu Ser Leu
290 295 300
Gly Ser Asn Thr Leu Gln Asn Tyr Ile Arg Ser Val Thr Tyr Asn Asp
305 310 315 320
Asp Pro Phe Met Thr Ala Asn Ala Asn Ala Asn Ile Gly Pro Arg Asn
325 330 335
Met Arg Gly Lys Gly Lys Asn Val Met Val Ala Val Ser Glu Tyr Asp
340 345 350
Glu Gly Glu Cys Lys Tyr Ile Val Asp Gly Val Asn Asn Leu Gly Leu
355 360 365
Glu Asp Gly Arg Ile Lys Met Pro Ser Leu Ala Ala Met Ser Ala Ser
370 375 380
Gly Ser Ala Ser Thr Ser Gly Ser Ala Ser Gly Ser Gly Ser Gly Val
385 390 395 400
Thr Met Glu Ile Asp Glu Pro Met Thr Asp Ser Trp Met Val Met His
405 410 415
Gly Cys Asp Glu Val Met Met Asn Glu Ile Ala Leu Leu Glu Met Ile
420 425 430
Ala His Gly Arg Leu
435
<210> 225
<211> 359
<212> Белок
<213> Arabidopsis thaliana
<400> 225
Met Tyr His Gln Asn Leu Ile Ser Ser Thr Pro Asn Gln Asn Ser Asn
1 5 10 15
Pro His Asp Trp Asp Ile Gln Asn Pro Leu Phe Ser Ile His Pro Ser
20 25 30
Ala Glu Ile Pro Ser Lys Tyr Pro Phe Met Gly Ile Thr Ser Cys Pro
35 40 45
Asn Thr Asn Val Phe Glu Glu Phe Gln Tyr Lys Ile Thr Asn Asp Gln
50 55 60
Asn Phe Pro Thr Thr Tyr Asn Thr Pro Phe Pro Val Ile Ser Glu Gly
65 70 75 80
Ile Ser Tyr Asn Met His Asp Val Gln Glu Asn Thr Met Cys Gly Tyr
85 90 95
Thr Ala His Asn Gln Gly Leu Ile Ile Gly Cys His Glu Pro Val Leu
100 105 110
Val His Ala Val Val Glu Ser Gln Gln Phe Asn Val Pro Gln Ser Glu
115 120 125
Asp Ile Asn Leu Val Ser Gln Ser Glu Arg Val Thr Glu Asp Lys Val
130 135 140
Met Phe Lys Thr Asp His Lys Lys Lys Asp Ile Ile Gly Lys Gly Gln
145 150 155 160
Trp Thr Pro Thr Glu Asp Glu Leu Leu Val Arg Met Val Lys Ser Lys
165 170 175
Gly Thr Lys Asn Trp Thr Ser Ile Ala Lys Met Phe Gln Gly Arg Val
180 185 190
Gly Lys Gln Cys Arg Glu Arg Trp Arg Asn His Leu Arg Pro Asn Ile
195 200 205
Lys Lys Asn Asp Trp Ser Glu Glu Glu Asp Gln Ile Leu Ile Glu Val
210 215 220
His Lys Ile Val Gly Asn Lys Trp Thr Glu Ile Ala Lys Arg Leu Pro
225 230 235 240
Gly Arg Ser Glu Asn Ile Val Lys Asn His Trp Asn Ala Thr Lys Arg
245 250 255
Arg Leu His Ser Val Arg Thr Lys Arg Ser Asp Ala Phe Ser Pro Arg
260 265 270
Asn Asn Ala Leu Glu Asn Tyr Ile Arg Ser Ile Thr Ile Asn Asn Asn
275 280 285
Ala Leu Met Asn Arg Glu Val Asp Ser Ile Thr Ala Asn Ser Glu Ile
290 295 300
Asp Ser Thr Arg Cys Glu Asn Ile Val Asp Glu Val Met Asn Leu Asn
305 310 315 320
Leu His Ala Thr Thr Ser Val Tyr Val Pro Glu Gln Ala Val Leu Thr
325 330 335
Trp Gly Tyr Asp Phe Thr Lys Cys Tyr Glu Pro Met Asp Asp Thr Trp
340 345 350
Met Leu Met Asn Gly Trp Asn
355
<210> 226
<211> 386
<212> Белок
<213> Arabidopsis thaliana
<400> 226
Met Ser Lys Arg Pro Pro Pro Asp Pro Val Ala Val Leu Arg Gly His
1 5 10 15
Arg His Ser Val Met Asp Val Ser Phe His Pro Ser Lys Ser Leu Leu
20 25 30
Phe Thr Gly Ser Ala Asp Gly Glu Leu Arg Ile Trp Asp Thr Ile Gln
35 40 45
His Arg Ala Val Ser Ser Ala Trp Ala His Ser Arg Ala Asn Gly Val
50 55 60
Leu Ala Val Ala Ala Ser Pro Trp Leu Gly Glu Asp Lys Ile Ile Ser
65 70 75 80
Gln Gly Arg Asp Gly Thr Val Lys Cys Trp Asp Ile Glu Asp Gly Gly
85 90 95
Leu Ser Arg Asp Pro Leu Leu Ile Leu Glu Thr Cys Ala Tyr His Phe
100 105 110
Cys Lys Phe Ser Leu Val Lys Lys Pro Lys Asn Ser Leu Gln Glu Ala
115 120 125
Glu Ser His Ser Arg Gly Cys Asp Glu Gln Asp Gly Gly Asp Thr Cys
130 135 140
Asn Val Gln Ile Ala Asp Asp Ser Glu Arg Ser Glu Glu Asp Ser Gly
145 150 155 160
Leu Leu Gln Asp Lys Asp His Ala Glu Gly Thr Thr Phe Val Ala Val
165 170 175
Val Gly Glu Gln Pro Thr Glu Val Glu Ile Trp Asp Leu Asn Thr Gly
180 185 190
Asp Lys Ile Ile Gln Leu Pro Gln Ser Ser Pro Asp Glu Ser Pro Asn
195 200 205
Ala Ser Thr Lys Gly Arg Gly Met Cys Met Ala Val Gln Leu Phe Cys
210 215 220
Pro Pro Glu Ser Gln Gly Phe Leu His Val Leu Ala Gly Tyr Glu Asp
225 230 235 240
Gly Ser Ile Leu Leu Trp Asp Ile Arg Asn Ala Lys Ile Pro Leu Thr
245 250 255
Ser Val Lys Phe His Ser Glu Pro Val Leu Ser Leu Ser Val Ala Ser
260 265 270
Ser Cys Asp Gly Gly Ile Ser Gly Gly Ala Asp Asp Lys Ile Val Met
275 280 285
Tyr Asn Leu Asn His Ser Thr Gly Ser Cys Thr Ile Arg Lys Glu Ile
290 295 300
Thr Leu Glu Arg Pro Gly Val Ser Gly Thr Ser Ile Arg Val Asp Gly
305 310 315 320
Lys Ile Ala Ala Thr Ala Gly Trp Asp His Arg Ile Arg Val Tyr Asn
325 330 335
Tyr Arg Lys Gly Asn Ala Leu Ala Ile Leu Lys Tyr His Arg Ala Thr
340 345 350
Cys Asn Ala Val Ser Tyr Ser Pro Asp Cys Glu Leu Met Ala Ser Ala
355 360 365
Ser Glu Asp Ala Thr Val Ala Leu Trp Lys Leu Tyr Pro Pro His Lys
370 375 380
Ser Leu
385
<210> 227
<211> 292
<212> Белок
<213> Arabidopsis thaliana
<400> 227
Met Glu Pro Pro Gln His Gln His His His His Gln Ala Asp Gln Glu
1 5 10 15
Ser Gly Asn Asn Asn Asn Asn Lys Ser Gly Ser Gly Gly Tyr Thr Cys
20 25 30
Arg Gln Thr Ser Thr Arg Trp Thr Pro Thr Thr Glu Gln Ile Lys Ile
35 40 45
Leu Lys Glu Leu Tyr Tyr Asn Asn Ala Ile Arg Ser Pro Thr Ala Asp
50 55 60
Gln Ile Gln Lys Ile Thr Ala Arg Leu Arg Gln Phe Gly Lys Ile Glu
65 70 75 80
Gly Lys Asn Val Phe Tyr Trp Phe Gln Asn His Lys Ala Arg Glu Arg
85 90 95
Gln Lys Lys Arg Phe Asn Gly Thr Asn Met Thr Thr Pro Ser Ser Ser
100 105 110
Pro Asn Ser Val Met Met Ala Ala Asn Asp His Tyr His Pro Leu Leu
115 120 125
His His His His Gly Val Pro Met Gln Arg Pro Ala Asn Ser Val Asn
130 135 140
Val Lys Leu Asn Gln Asp His His Leu Tyr His His Asn Lys Pro Tyr
145 150 155 160
Pro Ser Phe Asn Asn Gly Asn Leu Asn His Ala Ser Ser Gly Thr Glu
165 170 175
Cys Gly Val Val Asn Ala Ser Asn Gly Tyr Met Ser Ser His Val Tyr
180 185 190
Gly Ser Met Glu Gln Asp Cys Ser Met Asn Tyr Asn Asn Val Gly Gly
195 200 205
Gly Trp Ala Asn Met Asp His His Tyr Ser Ser Ala Pro Tyr Asn Phe
210 215 220
Phe Asp Arg Ala Lys Pro Leu Phe Gly Leu Glu Gly His Gln Glu Glu
225 230 235 240
Glu Glu Cys Gly Gly Asp Ala Tyr Leu Glu His Arg Arg Thr Leu Pro
245 250 255
Leu Phe Pro Met His Gly Glu Asp His Ile Asn Gly Gly Ser Gly Ala
260 265 270
Ile Trp Lys Tyr Gly Gln Ser Glu Val Arg Pro Cys Ala Ser Leu Glu
275 280 285
Leu Arg Leu Asn
290
<210> 228
<211> 453
<212> Белок
<213> Brassica napus
<400> 228
Met Asp Leu Gly Ser Val Thr Gly Asn Val Asn Gly Ser Pro Ser Leu
1 5 10 15
Lys Glu Leu Arg Glu Ser Lys Gln Asp Arg Ser Glu Phe Asp Gly Glu
20 25 30
Asp Cys Leu Gln Gln Ser Ser Lys Leu Ala Arg Thr Ile Ala Glu Asp
35 40 45
Lys His Leu Pro Ser Ser Tyr Ala Ala Ala Tyr Ser Arg Pro Met Ser
50 55 60
Phe His Gln Gly Ile Pro Leu Ala Arg Ser Ala Ser Leu Leu Ser Ser
65 70 75 80
Asp Ser Arg Arg Gln Glu His Met Leu Ser Phe Ser Asp Lys Pro Glu
85 90 95
Ala Phe Asp Phe Ser Lys Tyr Val Gly Leu Asp Asn Asn Lys Asn Ser
100 105 110
Leu Ser Pro Phe Leu His Gln Leu Pro Pro Pro Tyr Cys Arg Thr Pro
115 120 125
Gly Gly Gly Tyr Gly Ser Gly Gly Met Met Met Ser Met Gln Gly Lys
130 135 140
Gly Pro Phe Thr Leu Thr Gln Trp Ala Glu Leu Glu Gln Gln Ala Leu
145 150 155 160
Ile Tyr Lys Tyr Ile Thr Ala Asn Val Pro Val Pro Ser Ser Leu Leu
165 170 175
Ile Ser Ile Gln Lys Ser Phe Tyr Pro Tyr Arg Ser Phe Pro Pro Ser
180 185 190
Ser Phe Gly Trp Gly Thr Phe His Leu Gly Phe Ala Gly Gly Lys Met
195 200 205
Asp Pro Glu Pro Gly Arg Cys Arg Arg Thr Asp Gly Lys Lys Trp Arg
210 215 220
Cys Ser Lys Asp Ala Val Pro Asp Gln Lys Tyr Cys Glu Arg His Ile
225 230 235 240
Asn Arg Gly Arg His Arg Ser Arg Lys Pro Val Glu Val Gln Pro Gly
245 250 255
Gln Thr Ala Ala Ser Lys Ala Ala Ala Val Ala Ser Arg Asn Thr Ala
260 265 270
Ser Gln Ile Pro Asn Asn Arg Val Gln Asn Val Ile Tyr Pro Ser Thr
275 280 285
Val Asn Leu Pro Pro Lys Glu Gln Arg Asn Asn Asn Asn Ser Ser Phe
290 295 300
Gly Phe Gly His Val Thr Ser Pro Ser Leu Leu Thr Ser Ser Tyr Leu
305 310 315 320
Asp Phe Ser Ser Asn Gln Asn Lys Pro Glu Glu Leu Lys Ser Asp Trp
325 330 335
Thr Gln Leu Ser Met Ser Ile Pro Val Ala Ser Ser Ser Pro Ser Ser
340 345 350
Thr Ala Gln Asp Lys Thr Thr Leu Ser Pro Leu Arg Leu Asp Leu Pro
355 360 365
Ile Gln Ser Gln Gln Glu Thr Leu Glu Ala Val Arg Lys Val Asn Thr
370 375 380
Trp Ile Pro Ile Ser Trp Gly Asn Ser Leu Gly Gly Pro Leu Gly Glu
385 390 395 400
Val Leu Asn Ser Thr Thr Ser Ser Pro Thr Leu Gly Ser Ser Pro Thr
405 410 415
Gly Val Leu Gln Lys Ser Thr Phe Cys Ser Leu Ser Asn Ser Ser Ser
420 425 430
Val Thr Ser Pro Val Ala Asp Asn Asn Arg Asn Asn Asn Val Asp Tyr
435 440 445
Phe His Tyr Thr Thr
450
<210> 229
<211> 461
<212> Белок
<213> Brassica napus
<400> 229
Met Asp Leu Gly Ser Val Thr Gly Asn Val Asn Gly Ser Pro Gly Leu
1 5 10 15
Lys Glu Leu Arg Gly Ser Lys Gln Asp Arg Ser Gly Phe Asp Gly Glu
20 25 30
Asp Cys Leu Gln Gln Ser Ser Lys Leu Ala Arg Thr Ile Ala Glu Asp
35 40 45
Lys His Leu Pro Ser Ser Tyr Ala Ala Tyr Ser Arg Pro Met Ser Phe
50 55 60
His Gln Gly Ile Pro Leu Thr Arg Ser Ala Ser Leu Leu Ser Ser Asp
65 70 75 80
Ser Arg Arg Gln Glu His Met Leu Ser Phe Ser Asp Lys Pro Glu Ala
85 90 95
Phe Asp Phe Ser Lys Tyr Val Gly Leu Asp Asn Asn Lys Asn Ser Leu
100 105 110
Ser Pro Phe Leu His Gln Leu Pro Pro Pro Tyr Cys Arg Ser Ser Gly
115 120 125
Gly Gly Tyr Gly Ser Gly Gly Met Met Met Ser Met Gln Gly Lys Gly
130 135 140
Pro Phe Thr Leu Thr Gln Trp Ala Glu Leu Glu Gln Gln Ala Leu Ile
145 150 155 160
Tyr Lys Tyr Ile Thr Ala Asn Val Pro Val Pro Ser Ser Leu Leu Ile
165 170 175
Ser Ile Gln Lys Ser Phe Tyr Pro Tyr Arg Ser Phe Pro Pro Ser Ser
180 185 190
Phe Gly Trp Gly Thr Phe His Leu Gly Phe Ala Gly Gly Lys Met Asp
195 200 205
Pro Glu Pro Gly Arg Cys Arg Arg Thr Asp Gly Lys Lys Trp Arg Cys
210 215 220
Ser Lys Asp Ala Val Pro Glu Gln Lys Tyr Cys Glu Arg His Ile Asn
225 230 235 240
Arg Gly Arg His Arg Ser Arg Lys Pro Val Glu Val Gln Pro Gly Gln
245 250 255
Thr Ala Ala Ser Lys Ala Val Ala Ser Arg Asp Thr Ala Ser Gln Ile
260 265 270
Pro Ser Asn Arg Val Gln Asn Val Ile Tyr Pro Ser Asn Val Asn Leu
275 280 285
Gln Pro Lys Glu Gln Arg Asn Asn Asp Asn Ser Pro Phe Gly Phe Gly
290 295 300
His Val Thr Ser Ser Ser Leu Leu Thr Ser Ser Tyr Leu Asp Phe Ser
305 310 315 320
Ser Asn Gln Glu Lys Pro Ser Gly Asn His His Asn Gln Ser Ser Trp
325 330 335
Pro Glu Glu Leu Lys Ser Asp Trp Thr Gln Leu Ser Met Ser Ile Pro
340 345 350
Val Ala Ser Ser Ser Pro Ser Ser Thr Ala Gln Asp Lys Thr Ala Leu
355 360 365
Ser Pro Leu Arg Leu Asp Leu Pro Ile Gln Ser Gln Gln Glu Thr Leu
370 375 380
Glu Ser Ala Arg Lys Val Asn Thr Trp Ile Pro Ile Ser Trp Gly Asn
385 390 395 400
Ser Leu Gly Gly Pro Leu Gly Glu Val Leu Asn Ser Thr Thr Ser Ser
405 410 415
Pro Thr Leu Gly Ser Ser Pro Thr Gly Val Leu Gln Lys Ser Thr Phe
420 425 430
Cys Ser Leu Ser Asn Ser Ser Ser Val Thr Ser Pro Ile Ala Asp Asn
435 440 445
Asn Arg Asn Asn Asn Val Asp Tyr Phe His Tyr Thr Thr
450 455 460
<210> 230
<211> 409
<212> Белок
<213> Arabidopsis thaliana
<400> 230
Met Glu Ala Arg Pro Val His Arg Ser Gly Ser Arg Asp Leu Thr Arg
1 5 10 15
Thr Ser Ser Ile Pro Ser Thr Gln Lys Pro Ser Pro Val Glu Asp Ser
20 25 30
Phe Met Arg Ser Asp Asn Asn Ser Gln Leu Met Ser Arg Pro Leu Gly
35 40 45
Gln Thr Tyr His Leu Leu Ser Ser Ser Asn Gly Gly Ala Val Gly His
50 55 60
Ile Cys Ser Ser Ser Ser Ser Gly Phe Ala Thr Asn Leu His Tyr Ser
65 70 75 80
Thr Met Val Ser His Glu Lys Gln Gln His Tyr Thr Gly Ser Ser Ser
85 90 95
Asn Asn Ala Val Gln Thr Pro Ser Asn Asn Asp Ser Ala Trp Cys His
100 105 110
Asp Ser Leu Pro Gly Gly Phe Leu Asp Phe His Glu Thr Asn Pro Ala
115 120 125
Ile Gln Asn Asn Cys Gln Ile Glu Asp Gly Gly Ile Ala Ala Ala Phe
130 135 140
Asp Asp Ile Gln Lys Arg Ser Asp Trp His Glu Trp Ala Asp His Leu
145 150 155 160
Ile Thr Asp Asp Asp Pro Leu Met Ser Thr Asn Trp Asn Asp Leu Leu
165 170 175
Leu Glu Thr Asn Ser Asn Ser Asp Ser Lys Asp Gln Lys Thr Leu Gln
180 185 190
Ile Pro Gln Pro Gln Ile Val Gln Gln Gln Pro Ser Pro Ser Val Glu
195 200 205
Leu Arg Pro Val Ser Thr Thr Ser Ser Asn Ser Asn Asn Gly Thr Gly
210 215 220
Lys Ala Arg Met Arg Trp Thr Pro Glu Leu His Glu Ala Phe Val Glu
225 230 235 240
Ala Val Asn Ser Leu Gly Gly Ser Glu Arg Ala Thr Pro Lys Gly Val
245 250 255
Leu Lys Ile Met Lys Val Glu Gly Leu Thr Ile Tyr His Val Lys Ser
260 265 270
His Leu Gln Lys Tyr Arg Thr Ala Arg Tyr Arg Pro Glu Pro Ser Glu
275 280 285
Thr Gly Ser Pro Glu Arg Lys Leu Thr Pro Leu Glu His Ile Thr Ser
290 295 300
Leu Asp Leu Lys Gly Gly Ile Gly Ile Thr Glu Ala Leu Arg Leu Gln
305 310 315 320
Met Glu Val Gln Lys Gln Leu His Glu Gln Leu Glu Ile Gln Arg Asn
325 330 335
Leu Gln Leu Arg Ile Glu Glu Gln Gly Lys Tyr Leu Gln Met Met Phe
340 345 350
Glu Lys Gln Asn Ser Gly Leu Thr Lys Gly Thr Ala Ser Thr Ser Asp
355 360 365
Ser Ala Ala Lys Ser Glu Gln Glu Asp Lys Lys Thr Ala Asp Ser Lys
370 375 380
Glu Val Pro Glu Glu Glu Thr Arg Lys Cys Glu Glu Leu Glu Ser Pro
385 390 395 400
Gln Pro Lys Arg Pro Lys Ile Asp Asn
405
<210> 231
<211> 685
<212> ДНК
<213> Nicotiana benthamiana
<400> 231
aaagtccact ggaagaatac tcttcaacag ttggaaagag ttggacctaa gtcggttggt 60
gtctgtctgt taacagcagc ttttgttggc atggccttca ctatccaatt tgttagagaa 120
ttcactagat tagggttaaa tagatctgtt ggtggggtgt tggcccttgc cttttcaaga 180
gagctaagtc cagttgtcac atcaattgta gttgctgggc gtatcggtag tgcatttgct 240
gcggaactgg gcactatgca ggtatctgag cagactgaca cgttgagagt tcttggtgca 300
aatcctgttg attatttggt gacaccaaga gtgattgctt cttgcgttgc attaccattt 360
ttaaccctaa tgtgctttac agttggaatg gcatccagcg cccttttggc agatggtgtt 420
tatggaatta gcataaacat aatcttagat tctgctctga gagctcttag atcatgggac 480
cttattagtg caatgattaa atcaggggtg tttggtgcta ttatatccat cataagctgt 540
gcttgggggg tcaccacgct gggaggtgcc aaaggggttg gagagtcgac tacttcagca 600
gtagttttat ctcttgttgg catattcata gctgactttg ctctctcttg ctgtttcttc 660
cagggtgctg gcgattccct gaaga 685
<210> 232
<211> 824
<212> Белок
<213> Solanum tuberosum
<400> 232
Met Asp Ile Ser Asn Glu Ala Lys Val Glu Phe Ile Ser Ile Gly Pro
1 5 10 15
Ser Ser Ile Val Gly Arg Thr Ile Ala Phe Arg Val Leu Phe Cys Lys
20 25 30
Ser Ile Ser Arg Leu Arg His Asn Ile Phe His Phe Leu Ile Tyr Tyr
35 40 45
Leu Tyr Lys Ile Lys Asn Cys Leu Ser Tyr Tyr Leu Thr Pro Leu Ile
50 55 60
Lys Trp Phe His Pro Arg Asn Pro Gln Gly Ile Leu Ala Leu Val Thr
65 70 75 80
Leu Leu Ala Phe Leu Leu Arg Arg Tyr Thr Asn Val Lys Ile Arg Ala
85 90 95
Asp Met Val Tyr Lys Arg Lys Phe Trp Arg Asn Met Met Lys Ser Ala
100 105 110
Leu Thr Tyr Glu Glu Trp Ala His Ala Ala Lys Met Leu Glu Lys Glu
115 120 125
Thr Pro Lys Met Asn Glu Ala Glu Phe Tyr Asp Glu Glu Leu Val Val
130 135 140
Asn Lys Leu Gln Glu Leu Gln His Arg Arg Asn Glu Gly Ser Leu Arg
145 150 155 160
Asp Ile Met Phe Phe Met Arg Ala Asp Leu Val Arg Asn Leu Gly Asn
165 170 175
Met Cys Asn Pro Gln Leu His Lys Gly Arg Leu His Val Pro Lys Leu
180 185 190
Ile Lys Glu Tyr Ile Asp Glu Val Ser Thr Gln Leu Lys Met Val Cys
195 200 205
Asp Tyr Asp Ser Asp Glu Ile Leu Leu Glu Glu Lys Leu Ala Phe Met
210 215 220
His Glu Thr Arg His Ala Phe Gly Arg Thr Ala Leu Leu Leu Ser Gly
225 230 235 240
Gly Ala Ser Leu Gly Ala Phe His Val Gly Val Val Lys Thr Leu Val
245 250 255
Glu His Lys Leu Met Pro Arg Ile Ile Ala Gly Ser Ser Val Gly Ser
260 265 270
Ile Met Cys Ser Val Val Ala Thr Arg Ser Trp Pro Glu Leu Gln Ser
275 280 285
Phe Phe Glu Asn Phe Trp His Val Leu Gln Pro Phe Glu Gln Met Gly
290 295 300
Gly Ile Leu Thr Val Phe Arg Arg Ile Met Arg Gln Gly Ala Val His
305 310 315 320
Glu Ile Arg Gln Leu Gln Val Met Leu Arg His Leu Thr Asn Asn Leu
325 330 335
Thr Phe Gln Glu Ala Tyr Asp Met Thr Gly Arg Val Leu Gly Ile Thr
340 345 350
Val Cys Ser Pro Arg Lys His Glu Pro Pro Arg Cys Leu Asn Tyr Leu
355 360 365
Thr Ser Pro His Val Val Ile Trp Ser Ala Val Thr Ala Ser Cys Ala
370 375 380
Phe Pro Gly Leu Phe Glu Ala Gln Glu Leu Met Ala Lys Asp Arg Ser
385 390 395 400
Gly Asn Leu Val Pro Tyr His Pro Pro Phe His Leu Glu Pro Asp Gln
405 410 415
Ala Ala Ala Ser Gly Ser Ser Ala Arg Arg Trp Arg Asp Gly Ser Leu
420 425 430
Glu Ile Asp Leu Pro Met Met Gln Leu Lys Glu Leu Phe Asn Val Asn
435 440 445
His Phe Ile Val Ser Gln Ala Asn Pro His Ile Ala Pro Leu Leu Arg
450 455 460
Ile Lys Glu Phe Val Arg Ala Tyr Gly Gly Asn Phe Ala Ala Lys Leu
465 470 475 480
Ala His Leu Thr Glu Met Glu Val Lys His Arg Cys Asn Gln Val Leu
485 490 495
Glu Leu Gly Phe Pro Leu Arg Gly Leu Ala Lys Leu Phe Ala Gln Asp
500 505 510
Trp Glu Gly Asp Val Thr Val Val Met Pro Ala Thr Leu Ala Gln Tyr
515 520 525
Leu Lys Ile Ile Gln Asn Pro Ser Thr Leu Glu Val Gln Lys Ala Ala
530 535 540
Asn Gln Gly Arg Arg Cys Thr Trp Glu Lys Leu Ser Ala Ile Lys Ala
545 550 555 560
Asn Cys Gly Ile Glu Leu Ala Leu Asp Glu Cys Val Ala Ile Leu Asn
565 570 575
His Met Arg Arg Leu Lys Arg Ser Ala Glu Arg Ala Ala Ala Ala Ser
580 585 590
Gln Gly Met Ser Ser Ser Thr Val Lys Leu Asn Ala Ser Arg Arg Ile
595 600 605
Pro Ser Trp Asn Cys Ile Ala Arg Glu Asn Ser Thr Gly Ser Leu Glu
610 615 620
Glu Asp Phe His Ala Asp Ala Ser Ser Ser Leu His His His Asn Ala
625 630 635 640
Gly Arg Asn Trp Arg Cys Asn Asn Lys Asn Ala Ala His Asp His His
645 650 655
Gly Ser Asp Ser Glu Ser Glu Asn Ala Asp Asn Asn Ser Trp Thr Arg
660 665 670
Ser Gly Gly Pro Leu Met Arg Thr Thr Ser Ala Asp Lys Phe Ile Asp
675 680 685
Tyr Val Gln Asn Leu Glu Met His Pro Ser Gln Arg Ser Ser Arg Gly
690 695 700
Leu Ser Ile Asp Leu Asn Asn Val Val Val Arg Glu Pro Leu Ser Pro
705 710 715 720
Ser Pro Arg Val Thr Thr Pro Ala Arg Arg Ser Asp Thr Glu Phe Asp
725 730 735
Gln Arg Asp Ile Arg Ile Ile Val Ala Glu Gly Asp Leu Leu Gln Thr
740 745 750
Glu Arg Thr Asn Asn Gly Ile Val Phe Asn Val Val Arg Arg Gly Asp
755 760 765
Leu Thr Pro Ser Asn Arg Ser Leu Asp Ser Glu Asn Asn Ser Cys Phe
770 775 780
His Asp Pro Val Ala Glu Cys Val Gln Leu Glu Asn Pro Glu Lys Asp
785 790 795 800
Met Asp Ile Ser Ser Ala Ser Glu Asp Gly Glu Asn Ala Val Leu Asp
805 810 815
Glu Val Thr Lys Asn Gln Ile Ile
820
<210> 233
<211> 2475
<212> ДНК
<213> Solanum tuberosum
<400> 233
atggatataa gtaatgaggc taaagtagag ttcatttcca taggaccttc ttcaattgta 60
ggtcgaacaa tagcctttcg agttttgttt tgcaaatcaa tatcgcggtt gaggcacaac 120
atttttcatt tcttgatata ttacttgtac aagatcaaga attgtctgtc atactacttg 180
acacctttga tcaaatggtt tcacccgcgt aatccacagg ggatattagc attagtaaca 240
cttctagcct tcttgttgag gcgatatacg aatgtaaaaa tcagggctga tatggtttat 300
aagaggaaat tttggaggaa tatgatgaaa tctgcattaa cttatgagga atgggctcat 360
gctgcgaaaa tgttggagaa agagacacct aaaatgaatg aagcagagtt ttatgatgaa 420
gagttagttg taaataaact tcaagaactt caacatcgtc gtaatgaagg atctttaaga 480
gatattatgt tctttatgag agctgatctt gtgagaaatc tgggtaatat gtgtaatcca 540
cagcttcata agggtaggct tcatgtgcct aaacttatta aggagtatat tgatgaggtt 600
tcaactcagt tgaaaatggt atgtgattat gattcagatg agattttgtt ggaggagaag 660
cttgctttta tgcatgaaac aagacatgct tttggtagga cagcattgct tttaagcggg 720
ggcgcgtctt tgggagcttt tcatgttggt gtggttaaga cattggttga gcacaagctt 780
atgccaagga taattgctgg ttcgagtgtt ggatcgatta tgtgttctgt agttgcaact 840
cggtcttggc ctgagctgca gagttttttt gagaattttt ggcatgtgtt gcagccgttt 900
gaacagatgg gtggaattct aactgttttc aggaggatca tgagacaagg ggctgtacat 960
gagattaggc agttgcaggt gatgttacgc catctcacga ataatcttac tttccaagaa 1020
gcttacgata tgactggtcg agttctaggg attactgttt gctcccctag aaaacatgaa 1080
cctcctagat gtttgaacta cttgacttca cctcatgttg ttatatggag tgctgtgact 1140
gcttcttgcg cgtttcctgg tctgtttgaa gctcaagaac tgatggcaaa ggatagaagt 1200
ggtaatcttg ttccttatca tccaccattt catttggaac ctgatcaggc tgcagcttct 1260
ggttcatctg ctcgtcgatg gagggatggt agcttggaga tcgatctacc tatgatgcag 1320
ctaaaagagc tattcaacgt aaaccacttt atcgtgagcc aggcgaatcc acatattgct 1380
cctttactca ggatcaaaga gtttgtaaga gcttatggag gcaactttgc tgccaagctt 1440
gctcatctta ctgagatgga agtgaagcac agatgcaatc aggtactgga acttggtttt 1500
cccttgaggg gattagccaa gctatttgct caagattggg aaggcgatgt caccgttgta 1560
atgccagcca ctcttgctca gtacttgaag atcatacaga atccctctac tttggaggtt 1620
caaaaagcag caaatcaagg gaggagatgc acttgggaga aactatcagc cattaaggca 1680
aattgtggaa ttgagcttgc tcttgatgag tgtgtagcaa tactcaacca tatgcgtaga 1740
ctaaaaagga gcgcggagag agcagctgct gcttcacaag gcatgtcaag cagcacagtc 1800
aaactcaatg cttctagacg tattccttct tggaattgca ttgcaagaga gaactcaaca 1860
ggctcccttg aagaagactt tcacgcggat gcttcttcct ctcttcatca tcacaatgct 1920
ggtcgaaact ggcgttgtaa taacaagaat gctgcacatg atcatcatgg tagtgacagt 1980
gagtctgaaa acgcggataa taattcttgg acaagatcag gtggtccatt gatgaggaca 2040
acatcagctg ataagtttat tgactatgta caaaacttgg aaatgcatcc ttcacaacga 2100
tcgagcagag gactgagtat tgacctcaac aatgttgtag tcagggagcc tctttctccg 2160
agtccacgag tgacaacacc tgctaggaga tcagatacag aatttgatca aagagacatc 2220
agaattatcg tcgctgaagg tgatttacta cagactgaaa ggactaacaa tgggattgta 2280
ttcaatgtgg taaggagagg agacttaact ccatcaaaca ggagtcttga ttcagaaaac 2340
aacagttgct ttcatgatcc agtggccgaa tgcgtgcaac tcgaaaatcc tgagaaggat 2400
atggatataa gttcagcatc agaagatgga gaaaatgcag tactagatga agtaacaaaa 2460
aatcagatca tataa 2475
<210> 234
<211> 521
<212> Белок
<213> Solanum tuberosum
<400> 234
Met Ala Ala Ser Ile Gly Ala Leu Lys Ser Ser Pro Ser Ser Asn Asn
1 5 10 15
Cys Ile Asn Glu Arg Arg Asn Asp Ser Thr Arg Ala Val Ser Ser Arg
20 25 30
Asn Leu Ser Phe Ser Ser Ser His Leu Ala Gly Asp Lys Leu Met Pro
35 40 45
Ile Ser Ser Leu Arg Ser Gln Gly Val Arg Phe Asn Val Arg Arg Ser
50 55 60
Ser Leu Ile Val Pro Pro Lys Ala Val Ser Asp Ser Gln Asn Ser Gln
65 70 75 80
Thr Cys Leu Asp Pro Asp Ala Ser Arg Ser Val Leu Gly Ile Ile Leu
85 90 95
Gly Gly Gly Ala Gly Thr Arg Leu Tyr Pro Leu Thr Lys Lys Arg Ala
100 105 110
Lys Pro Ala Val Pro Leu Gly Ala Asn Tyr Arg Leu Ile Asp Ile Pro
115 120 125
Val Ser Asn Cys Leu Asn Ser Asn Ile Ser Lys Ile Tyr Val Leu Thr
130 135 140
Gln Phe Asn Ser Ala Ser Leu Asn Arg His Leu Ser Arg Ala Tyr Ala
145 150 155 160
Ser Asn Met Gly Gly Tyr Lys Asn Glu Gly Phe Val Glu Val Leu Ala
165 170 175
Ala Gln Gln Ser Pro Glu Asn Pro Asp Trp Phe Gln Gly Thr Ala Asp
180 185 190
Ala Val Arg Gln Tyr Leu Trp Leu Phe Glu Glu His Thr Val Leu Glu
195 200 205
Tyr Leu Ile Leu Ala Gly Asp His Leu Tyr Arg Met Asp Tyr Glu Lys
210 215 220
Phe Ile Gln Ala His Arg Glu Thr Asp Ala Asp Ile Thr Val Ala Ala
225 230 235 240
Leu Pro Met Asp Glu Lys Arg Ala Thr Ala Phe Gly Leu Met Lys Ile
245 250 255
Asp Glu Glu Gly Arg Ile Ile Glu Phe Ala Glu Lys Pro Gln Gly Glu
260 265 270
Gln Leu Gln Ala Met Lys Val Asp Thr Thr Ile Leu Gly Leu Asp Asp
275 280 285
Lys Arg Ala Lys Glu Met Pro Phe Ile Ala Ser Met Gly Ile Tyr Val
290 295 300
Ile Ser Lys Asp Val Met Leu Ser Leu Leu Arg Asp Lys Phe Pro Gly
305 310 315 320
Ala Asn Asp Phe Gly Ser Glu Val Ile Pro Gly Ala Thr Ser Leu Gly
325 330 335
Met Arg Val Gln Ala Tyr Leu Tyr Asp Gly Tyr Trp Glu Asp Ile Gly
340 345 350
Thr Ile Glu Ala Phe Tyr Asn Ala Asn Leu Gly Ile Thr Lys Lys Pro
355 360 365
Val Pro Asp Phe Ser Phe Tyr Asp Arg Ser Ala Pro Ile Tyr Thr Gln
370 375 380
Pro Arg Tyr Leu Pro Pro Ser Lys Met Leu Asp Ala Asp Val Thr Asp
385 390 395 400
Ser Val Ile Gly Glu Gly Cys Val Ile Lys Ser Cys Lys Ile His His
405 410 415
Ser Val Val Gly Leu Arg Ser Cys Ile Ser Glu Gly Ala Ile Ile Glu
420 425 430
Asp Ser Leu Leu Met Gly Ala Asp Tyr Tyr Glu Thr Asp Ala Asp Arg
435 440 445
Lys Leu Leu Ala Ala Lys Gly Ser Val Pro Ile Gly Ile Gly Lys Asn
450 455 460
Cys His Ile Lys Arg Ala Ile Ile Asp Lys Asn Ala Arg Ile Gly Asp
465 470 475 480
Asn Val Lys Ile Ile Asn Lys Asp Asn Val Gln Glu Ala Ala Arg Glu
485 490 495
Thr Asp Gly Tyr Phe Ile Lys Ser Gly Ile Val Thr Val Ile Lys Asp
500 505 510
Ala Leu Ile Pro Ser Gly Ile Ile Ile
515 520
<210> 235
<211> 1819
<212> ДНК
<213> Solanum tuberosum
<400> 235
ctagtgattg caatcacact ctaccacaca ctctctagta gagagatcag ttgataacaa 60
gctttgttaa caatggcggc ttccattgga gccttaaaat cttcaccttc ttctaacaat 120
tgcatcaatg agagaagaaa tgattctaca cgtgcagtat ccagcagaaa tctctcattt 180
tcgtcttctc atctcgccgg agacaagttg atgcctatat cgtccttacg ttcccaagga 240
gtccgattca atgtgagaag aagttcattg attgtgccgc ctaaggctgt ttctgattcg 300
cagaattcac agacatgtct agacccagat gctagccgga gtgttttggg aattattctt 360
ggaggtggag ctgggacccg actttatcct ctaactaaaa aaagagcaaa gccagctgtt 420
ccacttggag caaattatcg tctgattgac attcctgtaa gcaactgctt gaacagtaac 480
atatccaaga tctatgttct cacacaattc aactctgcct ctctgaatcg ccacctttca 540
cgagcatatg ctagcaacat gggaggatac aaaaacgagg gctttgtgga agttcttgct 600
gctcaacaaa gtccagagaa ccccgattgg ttccagggca cggctgatgc tgtcagacaa 660
tatctgtggt tgtttgagga gcatactgtt cttgaatacc ttatacttgc tggagatcat 720
ctgtatcgaa tggattatga aaagtttatt caagcccaca gagaaacaga tgctgatatt 780
accgttgccg cactgccaat ggacgagaag cgtgccactg cattcggtct catgaagatt 840
gacgaagaag gacgcattat tgaatttgca gagaaaccgc aaggagagca attgcaagca 900
atgaaagtgg atactaccat tttaggtctt gatgacaaga gagctaaaga aatgcctttc 960
attgccagta tgggtatata tgtcattagc aaagacgtga tgttaagcct acttcgtgac 1020
aagttccctg gggccaatga ttttggtagt gaagttattc ctggtgcaac ttcacttggg 1080
atgagagtgc aagcttattt atatgatggg tactgggaag atattggtac cattgaagct 1140
ttctacaatg ccaatttggg cattacaaaa aagccggtgc cagattttag cttttacgac 1200
cgatcagccc caatctacac ccaacctcga tatctaccac catcaaaaat gcttgatgct 1260
gatgtcacag atagtgtcat tggtgaaggt tgtgtgatca agagctgtaa gattcatcat 1320
tccgtggttg gactcagatc atgcatatca gagggagcaa ttatagaaga ctcacttttg 1380
atgggggcag attactatga gactgatgct gacaggaagt tgctggctgc aaagggcagt 1440
gtcccaattg gcatcggcaa gaattgtcac attaaaagag ccattatcga caagaatgcc 1500
cgtatagggg acaatgtgaa gatcattaac aaagacaacg ttcaagaagc ggctagggaa 1560
acagatggat acttcatcaa gagtgggatt gtcaccgtca tcaaggatgc tttgattcca 1620
agtggaatca tcatctgaag gaatgcgttt taacttggtt gtcctccaag attttggcta 1680
aacagccatg aggtagaaac gtgctgaact tttattttcc tgagctgtag aaatctagtg 1740
tacatctttc tgttatgata cttctcatta cccctacaag agaagactgg atgctgtaaa 1800
aattattcgt ctagaataa 1819
<210> 236
<211> 1173
<212> ДНК
<213> Sapium sebiferum L.
<400> 236
tgccaatagc cagccaataa aacatctaca cgttttcaca cggcttttca tcacagccgt 60
tgtttttctc atctcactcc gtgccttcat cttcatcctc ttctcctctc tctctgtctc 120
tatatgtata gaagcgttag atgtcttgcg ttgttaacca attcattttt cgctttctgc 180
ttcttctaat attataagaa agtttgattc ttcttcttgt caatctttgt tcgcggcttt 240
taacgatatc cgctaaagga aatttgaaat ttcaattatg gccgatggaa acgtcaattc 300
gcaagaacag atggctaagc aggaggaaca gaggctgaag tatttggagt ttgtacaagt 360
ggctgcaata catgctgtgg tgaccttcac aaacctctat gtttatgcca aaaacaagtc 420
gggtccattg aagcccggtg ttgagactgt tgaaggtacg gtcaagagtg tggttggacc 480
tgtttatggc aagttccatg atgttcccat tgaggttctc aagtttgtcg atcgcaagat 540
tgatcaatct gtaagcagcc tagacagccg tgtgcctcca gttgtgaagc agttatcggc 600
ccaagcattt tcagtggctc gcgaagcccc agtggctgct cgtgctgtgg cttctgaagt 660
gcagactgct ggagtgaagg aaactgcatc tgggttggca agaactctgt acttcaaata 720
tgaacccaag gccaaggagc tatacaccaa gtatgaacca aaagcggaac agtgtgctgc 780
ctctgcctgg cgtaagctca atcaactccc agtcttccct catgtagctc aggttgttat 840
gccaacagca gcttattgtt ctgaaaagta caaccaggca gtacttacca ccgctgagaa 900
aggatacaga gtgtcctctt atttgccttt tgtgcccact gagagaattg ctaagttgtt 960
taggaatgag gcacctgaat ctaccccttt cctttccaat tgagcaagat gctgataaat 1020
gattcacaat ggacatgtgg acagaataaa aatctttgga tattatatgg tactgtgtat 1080
ttcaaggttc aagattactc tctacaatgt gtgaattttt gtttcagatg acttaattct 1140
tgttcattca ttatatatat atatatatat ata 1173
<210> 237
<211> 241
<212> Белок
<213> Sapium sebiferum L.
<400> 237
Met Ala Asp Gly Asn Val Asn Ser Gln Glu Gln Met Ala Lys Gln Glu
1 5 10 15
Glu Gln Arg Leu Lys Tyr Leu Glu Phe Val Gln Val Ala Ala Ile His
20 25 30
Ala Val Val Thr Phe Thr Asn Leu Tyr Val Tyr Ala Lys Asn Lys Ser
35 40 45
Gly Pro Leu Lys Pro Gly Val Glu Thr Val Glu Gly Thr Val Lys Ser
50 55 60
Val Val Gly Pro Val Tyr Gly Lys Phe His Asp Val Pro Ile Glu Val
65 70 75 80
Leu Lys Phe Val Asp Arg Lys Ile Asp Gln Ser Val Ser Ser Leu Asp
85 90 95
Ser Arg Val Pro Pro Val Val Lys Gln Leu Ser Ala Gln Ala Phe Ser
100 105 110
Val Ala Arg Glu Ala Pro Val Ala Ala Arg Ala Val Ala Ser Glu Val
115 120 125
Gln Thr Ala Gly Val Lys Glu Thr Ala Ser Gly Leu Ala Arg Thr Leu
130 135 140
Tyr Phe Lys Tyr Glu Pro Lys Ala Lys Glu Leu Tyr Thr Lys Tyr Glu
145 150 155 160
Pro Lys Ala Glu Gln Cys Ala Ala Ser Ala Trp Arg Lys Leu Asn Gln
165 170 175
Leu Pro Val Phe Pro His Val Ala Gln Val Val Met Pro Thr Ala Ala
180 185 190
Tyr Cys Ser Glu Lys Tyr Asn Gln Ala Val Leu Thr Thr Ala Glu Lys
195 200 205
Gly Tyr Arg Val Ser Ser Tyr Leu Pro Phe Val Pro Thr Glu Arg Ile
210 215 220
Ala Lys Leu Phe Arg Asn Glu Ala Pro Glu Ser Thr Pro Phe Leu Ser
225 230 235 240
Asn
<210> 238
<211> 1252
<212> ДНК
<213> Sapium sebiferum L.
<400> 238
ctacttttcc ctagcattag tattctaggc cccactctgt agattcctcc agctgcctga 60
tctaattttt tatcaactct tgaccgttcg atcatcccaa cggctcagat tcactagtac 120
ttttctcaca ccgtatctcc gattctccat gactccatcg atataaatcg cagtgctcat 180
caactgaatt ctcgaaattg cggttacaag ctgctataag aagcgaaaag aaacgctgag 240
aaacaggatc cgttcctcct ccctcgtttt ttactcctta caagatggag accgagaaga 300
agattcctga attgaagcac ttagggttcg tgaggatggc tgctattcag tcactgattt 360
gcgtctcgaa tctctacgat tacgcgaagc ataactcagg acctttgaga tccactgttg 420
gaaccgtgga gggtgccgta accaccgtag taggtccagt ttaccagaaa ttcaaagacc 480
ttcctgatga tcttcttgta tatgttgata agaaggtgga tgaaggaaca cacaagtttg 540
ataagcatgc tccacctatt gctaagaagg ctgcgagcca agcccatagt ttgtttcata 600
tagccttgga gaaggtcgaa aaactcgtgc aggaggctcg tgcaggagga cctcgtgctg 660
ctctgcattt tgtggctaca gagtcgaagc acttggcgtt gacccaatct gtgaagctgt 720
atagtaaact taatcagttc cctgtcattc acactgttac agatgtaacc cttcccacag 780
ctactcactg gtcagataag tataaccata cccttatgga cctgacccgg aagggttata 840
cgatctttgg ttatttgcct ttggttccta ttgatgacat atctaagaca tttaaacaaa 900
gtaaagcaga ggagaaagaa aatgcaacta cgcataaatc tgattcatcg gattccgact 960
aaacggttgc catcatgtct aatgggtgtg gtttgttaag tatagtggtt tgcgaaaatg 1020
ttctagggtt tatgagcctg ctcgaaagat gctgagaaat ggaaatctgt actatttagg 1080
agtttttccg tactataata atgagtatga atggtttgta aattctgcct tgtgctttct 1140
cgacaagtat atcatgcttc tattttttac tactacttac tggactactg aattgtctca 1200
taattgtccc tagtgtctaa ttaaatatca cctccaaaat attattgaaa aa 1252
<210> 239
<211> 225
<212> Белок
<213> Sapium sebiferum L.
<400> 239
Met Glu Thr Glu Lys Lys Ile Pro Glu Leu Lys His Leu Gly Phe Val
1 5 10 15
Arg Met Ala Ala Ile Gln Ser Leu Ile Cys Val Ser Asn Leu Tyr Asp
20 25 30
Tyr Ala Lys His Asn Ser Gly Pro Leu Arg Ser Thr Val Gly Thr Val
35 40 45
Glu Gly Ala Val Thr Thr Val Val Gly Pro Val Tyr Gln Lys Phe Lys
50 55 60
Asp Leu Pro Asp Asp Leu Leu Val Tyr Val Asp Lys Lys Val Asp Glu
65 70 75 80
Gly Thr His Lys Phe Asp Lys His Ala Pro Pro Ile Ala Lys Lys Ala
85 90 95
Ala Ser Gln Ala His Ser Leu Phe His Ile Ala Leu Glu Lys Val Glu
100 105 110
Lys Leu Val Gln Glu Ala Arg Ala Gly Gly Pro Arg Ala Ala Leu His
115 120 125
Phe Val Ala Thr Glu Ser Lys His Leu Ala Leu Thr Gln Ser Val Lys
130 135 140
Leu Tyr Ser Lys Leu Asn Gln Phe Pro Val Ile His Thr Val Thr Asp
145 150 155 160
Val Thr Leu Pro Thr Ala Thr His Trp Ser Asp Lys Tyr Asn His Thr
165 170 175
Leu Met Asp Leu Thr Arg Lys Gly Tyr Thr Ile Phe Gly Tyr Leu Pro
180 185 190
Leu Val Pro Ile Asp Asp Ile Ser Lys Thr Phe Lys Gln Ser Lys Ala
195 200 205
Glu Glu Lys Glu Asn Ala Thr Thr His Lys Ser Asp Ser Ser Asp Ser
210 215 220
Asp
225
<210> 240
<211> 938
<212> ДНК
<213> Sapium sebiferum L.
<400> 240
gagtattcac actctggcct gattgggttt gctataaagg gcgatcgttg caacgctcca 60
tattgtctac ttggttttgt ttcaaatctc atcattttgt aaatttgcga cagtgtagcg 120
ttttctagga aaaaggttgc taaaggaaag tagttatcaa accgcagaaa tggcggaatc 180
cgaacttaat caacacacag atatggttca agatgatgat aaaaaactca agtatctaga 240
ttttgtacaa gtggccgcga tctatgttgt ggtttgtttc tctagtatct atgaatatgc 300
taaggaaaac tccggtccac taaaaccagg ggtccaagcc gttgagtgta ccgtcaaaac 360
tgtaataagt ccggtttacg agaagtttcg cgacgtacct tttgaactcc ttaaattcgt 420
cgatcgtaaa gttgacaact ctctaggcga gttggacagg cacgtgccgt cgctggtgaa 480
gcaggcatca agccaagctc gagctgtggc tagtgaaatt caacatgctg gattggtaga 540
cgcaactaag aacattgcga agacgatgta tacaaagtat gaactgacgg cttggcagct 600
ctactgcaaa tacaagccgg tggctaagcg ttacgcggtg tcgacctggc gctcattgaa 660
ccagcttcct ctgtttcctc aagcggctca gattgcaatc ccaactgctg cttcgtggtc 720
tgagaaatac aataagatgg ttcgttacac gaaagataga ggatatccag cggcggtgta 780
tctgccattg atctcggttg agaggattgc caaggtgttc aatgaagact taaacgggcc 840
caccgtccct accaatggat catccgccgc agcacaatag ttttcatttt atgtatttat 900
gtcagattga agacgctccg gagattttga aaacctga 938
<210> 241
<211> 194
<212> Белок
<213> Sapium sebiferum L.
<400> 241
Met Ala Glu Ser Glu Leu Asn Gln His Thr Asp Met Val Gln Asp Asp
1 5 10 15
Asp Lys Lys Leu Lys Tyr Leu Asp Phe Val Gln Val Ala Ala Ile Tyr
20 25 30
Val Val Val Cys Phe Ser Ser Ile Tyr Glu Tyr Ala Lys Glu Asn Ser
35 40 45
Gly Pro Leu Lys Pro Gly Val Gln Ala Val Glu Cys Thr Val Lys Thr
50 55 60
Val Ile Ser Pro Val Tyr Glu Lys Phe Arg Asp Val Pro Phe Glu Leu
65 70 75 80
Leu Lys Phe Val Asp Arg Lys Val Asp Asn Ser Leu Gly Glu Leu Asp
85 90 95
Arg His Val Pro Ser Leu Val Lys Gln Ala Ser Ser Gln Ala Arg Ala
100 105 110
Val Ala Ser Glu Ile Gln His Ala Gly Leu Val Asp Ala Thr Lys Asn
115 120 125
Ile Ala Lys Thr Met Tyr Thr Lys Tyr Glu Leu Thr Ala Trp Gln Leu
130 135 140
Tyr Cys Lys Tyr Lys Pro Val Ala Lys Arg Tyr Ala Val Ser Thr Trp
145 150 155 160
Arg Ser Leu Asn Gln Leu Pro Leu Phe Pro Gln Ala Ala Gln Ile Ala
165 170 175
Ile Pro Thr Ala Ala Ser Trp Ser Glu Lys Tyr Asn Lys Met Val Arg
180 185 190
Tyr Thr
<210> 242
<211> 2526
<212> ДНК
<213> Sorghum bicolor
<400> 242
atggacgagt ccggggaagc gagcgtcggc tccttcagga tcggcccgtc gacgctgctg 60
ggccgcgggg tggcgctccg cgtgcttctc ttcagctcgc tgtggcgcct gcgggcgcgc 120
gcgtacgccg ccatctcgcg cgtgcgcagc gcggtgctgc cggtggcggc gtcctggctt 180
cacctcagga acacccacgg cgtcctcctc atggtcgtcc tcttcgccct ctccctgagg 240
aagctctccg gcgcgcggtc gcgggcggcg ctcgcgcgcc ggcgcaggca gtacgagaag 300
gccatgctgc atgccgggac gtacgaggtc tgggcccgcg ccgccaatgt gctcgacaag 360
atgtctgatc aggtccatga ggcggatttc tatgacgagg agctgatcag gaacaggctt 420
gaggacctcc ggaggcggag ggaggacggg tcgctgcggg acgtggtgtt ctgtatgcgc 480
ggcgatcttg ttaggaactt ggggaacatg tgcaatcctg aacttcacaa gggcaggcta 540
gaggttccta agcttataaa ggaatacatt gaagaggttt ctattcaact aagaatggtg 600
tgcgaatctg acactgatga gttgctattg ggagagaagc ttgcctttgt tcaggagacc 660
aggcatgcct ttgggaggac agccctactc ttaagtgggg gtgcttcact ggggtctttc 720
catgtaggtg tagtgaaaac attggttgag cataagcttc tgcctcggat tatagcagga 780
tcaagcgttg gttccattat atgttcgatt gttgctaccc ggacatggcc tgagattgag 840
agcttcttca cagactcatt acagaccttg cagttctttg ataggatggg tggaattttt 900
gcagtgatga ggcgagtcac cactcatggt gcactgcatg acattagcca gatgcaaagg 960
cttctgaggg atctcacaag taacttaaca tttcaagagg cttatgacat gactggccgt 1020
gtccttggga tcaccgtttg ctctcctaga aaaaatgagc caccccgctg cctcaactat 1080
ctgacgtcgc cgcacgttgt tatttggagt gctgtaactg cctcttgtgc atttcctggg 1140
ctctttgaag ctcaggaact gatggcgaag gatagattcg gcaacatagt tcccttccat 1200
gcaccctttg ccacagatcc tgaacaaggt cctggagcat caaagcgccg gtggagagat 1260
gggagcctgg aaatggattt gcccatgatg agactcaagg agttgtttaa tgtaaaccat 1320
ttcattgtga gccaaactaa tcctcacatt tctcccctcc tccgaatgaa agagcttgtt 1380
agagtctatg gagggcgctt tgctggaaag cttgctcgtc ttgctgagat ggaggttaag 1440
tatcgatgta accaaatcct agagattggt cttccaatgg gaggacttgc aaaattgttt 1500
gctcaggact gggagggtga tgtcaccatg gttatgccgg caacagtagc tcagtatttg 1560
aagattattc agaatccaac atatgcggag ctccaaatgg ctgccaacca aggccgcagg 1620
tgtacatggg agaagctctc tgcaatcaga gcaaactgtg ccatcgaact tgcattggat 1680
gaatctatag cagttttaaa ccacaaacgg aggctaaaaa gaagcatgga gaggacagag 1740
gctgctttgc agggtcattc taactatgtt cgactcaaaa ctccaaggag ggtaccatca 1800
tggagctgca tcagtcgaga gaattcttca gaatctctct cggaagagat ttcagcagtt 1860
gctacttcaa ccgcgcagca aggtgctgct cttgttgtcg gcacagccac tctttctcac 1920
catgttcgac gcaattctca tgacggaagt gagagtgaat cagaaaccat tgaccttaat 1980
tcctggacca ggagtggtgg gcctctaatg aggacagcat ctgctgacat gttcatcagt 2040
ttcatccata accttgagat tgacacggaa ttaagtaggc cctgtactgt ggagggtggt 2100
actgcaggta tttcgtcaga atctaccttc ccaaatgatc cacaaccgaa caatggctca 2160
agtgttacta ctccaggtag atgcacagaa aattctgaga ccgaggcata cgacactgtc 2220
aacaccagag ccagtcaggc ttctactccc acaagcatcg ctgtttctga aggagatttg 2280
ctgcagcctg aaagcattgc tgacggtatc ctgcttaaca ttgtgaaaag agatgccttg 2340
caggctcaaa atgacagcgt aactgaattg gccgaaagct cctgcactga aacatatgcg 2400
gaaacttgtg acaccatctc agggtctggc actgctgaag ataacaagga tactgctgac 2460
tcaagcaatc actcacttga tattgatgct tttgtagttt cgcatcaacc ttcagctgat 2520
gattag 2526
<210> 243
<211> 3099
<212> ДНК
<213> Triticum aestivum
<400> 243
atgcccgcgc ctgcaggtgc gtgcagccaa gccccaccgc tcgccttcta ttccgcgtcc 60
cctagcttgg cccggccctg ctccgatcca aggccgcggc ggtggcccag tgccctctcc 120
ctcctgccac gccgtccgcc gcccatggac gtcatcacca acgaggcgcg cgtgggggcg 180
ttcgcgatcg gcccgtccac ggcggcgggg cgcgcgctcg cgctgcgcgt gctcctctgc 240
ggctcgctgg cgcggctgcg gcaccgcctc gccgccgcgc tgcgcgccgc ggcgccgctg 300
gcggcggcct ggctgcaccc gcgccacaac acgcggggga tcctgctcgc cgtctgcgcc 360
gtcgcgctgc tgctgcgcgg ccgcgggggc cgcgccgggg tgcgcgcgcg ggtgcagtcc 420
gcctaccgcc gcaagttctg gcggaacatg atgcgcgccg cgctcaccta cgaggagtgg 480
gcgcacgccg cgcggatgct cgagcgggag acgccgcgcc gcgtcaccga cgccgacctc 540
tacgacgagg agctcgtgtg caacaagctc cgtgagctca ggcaccgccg tcaggagggc 600
tcgctcaggg acatcgtctt ctgcatgcgc gccgatctgc tcaggaacct tggtaacatg 660
tgcaaccccg agctccacaa gttgaggctg caggtgccta aaaccatcaa ggagtacatt 720
gaggaggtat ctactcaact gaaaatggtt tgcaattctg attcggacga gttacccctt 780
gaagagaaac tggcatttat gcatgagaca agacatgcct ttggtagatc ggccctactg 840
ctaagtggag gtgcttcatt tggctctttc catgtgggtg ttgtgaaaac cttggtagag 900
cataagcttc tacctaggat tatttcagga tcaagcgttg gcgcaataat gtgtgctatt 960
gtagccacac ggtcatggcc agaactagag agtttttttg aggagtggca ttccttgaaa 1020
ttctttgacc agatgggtgg gatctttcct gtatttaaaa gaattttgac gcatggagcg 1080
gttcatgaca ttaggcactt gcagacgcag ttgagaaatc ttacaagcaa tttgacattt 1140
caagaggcat atgacatgac tggccgggtt ctcgttgtta ctgtgtgttc tccaagaaaa 1200
catgagccac cacgatgcct gaactatttg acatcacctc atgttctcat ttggagtgca 1260
gtaactgctt cctgtgcttt tcctggactt tttgaggccc aggagttgat ggccaaagat 1320
agattcggag aaacagttcc ttttcatgct ccattcttgt tgggtgtgga ggaacgagct 1380
gacgctgcta cacggcgctg gagagatggc agcttagaaa gtgatttacc catgaagcaa 1440
ttgaaggaat tattcaacgt aaatcacttc atagtaagcc aagccaatcc tcacattgct 1500
ccattactga gactaaagga gatcatcagg gcttacggag gcagctttgc tgcaaagctt 1560
gctgaacttg ctgagatgga agttaagcat aggttcaatc aagttctgga acttggattt 1620
ccattaggag gaatagctaa gttgtttgct caacattggg aaggtgatgt gacaatcgtt 1680
atgccagcca cacttgctca gtattcgaag atcatacaga atccttcgta ttctgagctt 1740
cagaaagccg caagtcaggg taggcgatgc acttgggaaa agctctctgc tatcagggca 1800
aactgcgcta ttgagcttgc attagatgaa tgtgttgccc tcctgaacca catgcgtagg 1860
ctgaagagaa gtgcagaaag agcagctgct tcacaaggat atggtgctac aattagactc 1920
tgtccatcta gaaggattcc atcatggaat ctcatagcaa gagaaaattc aactggttct 1980
ctcgatgagg aaatgctcac atgtcccact gttacgagcc atcaagcagt tggagggact 2040
gctgggccat ctaacagaaa tcaccatctc caacatagta tgcatgatag cagtgacagt 2100
gaatctgaga gtatagactt gaactcatgg acgagaagtg gtggccctct catgagaaca 2160
gcctcagcta ataaattcat cagctttgtt cagaaccttg agattgacac agaattcaga 2220
acaatttcac caagggggag cgaaggtgat attgttacac cgaatagtaa cttatttgct 2280
ggtcacccaa ttggtagaga gccagttgat aaccatccag ggcctgctac tcctggtagg 2340
acctcaggca attcaggttg cgatcctcat gatactcctg ttcctaggtc tccatttggt 2400
ctttccacaa gtatcatggt ccctgaaggt gacttgctgc agccggaaaa gattgagaat 2460
ggtattttat tcaatgttgt gagaagggat gctcttgtag cgactactag cggagttgaa 2520
cctcatggat cttcacagga agcagatgtg gaaactgtac cgaccgagtg cctttatggt 2580
gcttcggatg acgacgacga caacgtggaa ctgaatgctg atcatgaagc attatctgac 2640
cctggagatc agagatcctc agttgcagga aacctagatc cgtccacttc catggattgt 2700
caagctgatg aaacaagtac tactcgatca gaagctccat ctctctttaa tatctgtgtg 2760
gagattcctc cagcaaccat gatcagagaa aatagtcggc ccgacgagcc ttcttcagac 2820
ataagactgg agattgtaaa gacagaatgc cctgatgaga attcagctgc tgggaacgat 2880
gaagttggct cagttcctgc caataaagaa tcttcctatt gttctcagac agctgaaaat 2940
agacagcagc atcaagttga tatgggatct gtgaactcct gtagtgtttc agtttcagaa 3000
gatgataggc atgtcagcct catttcgaac gagaaaccag ttactacttc cagtggcgga 3060
gcggagagta tgacatctgg aagaaatgaa gctgactag 3099
<210> 244
<211> 2198
<212> ДНК
<213> Искусственная последовательность
<220>
<223> фрагмент шпРНКи SDP1 S. bicolor
<400> 244
gcggcggcgt ggctgcaccc gcgcgacaac acgcgcggga tcctgctcgc cgtctgcgcc 60
gtcgcgctgg gtgcagtccg cctaccgccg caagttctgg cggaacatga tgcgcgccgc 120
gctcacctac gaggagtggg cgcacgcggc gcggatgctt ggagtgcagt aacagcttcc 180
tgtgcttttc ctggactttt tgaggcccac catctaggag gattccatcc tggaatctca 240
tagcaagaga aaattcaact ggttctctat gtgcaatcct gaacttcaca agggcaggct 300
agaggttcct aagcttataa aggaatacat tgaagaggtt tctattcaac taagaatggt 360
gtgcgaatct gacactgatg agttgctatt gggagagaag cttgcctttg ttcaggagac 420
caggcatgcc tttgggagga cagccctact cttaagtggg ggtgcttcac tggggtcttt 480
ccatgtaggt gtagtgaaaa cattggttga gcataagctt ctgcctcgga ttatagcagg 540
atcaagaagg gtggacccag ctttcttgta caaagtggtc tcgaggaatt cggtacccca 600
gcttggtaag gaaataatta ttttcttttt tccttttagt ataaaatagt taagtgatgt 660
taattagtat gattataata atatagttgt tataattgtg aaaaaataat ttataaatat 720
attgtttaca taaacaacat agtaatgtaa aaaaatatga caagtgatgt gtaagacgaa 780
gaagataaaa gttgagagta agtatattat ttttaatgaa tttgatcgaa catgtaagat 840
gatatactag cattaatatt tgttttaatc ataatagtaa ttctagctgg tttgatgaat 900
taaatatcaa tgataaaata ctatagtaaa aataagaata aataaattaa aataatattt 960
ttttatgatt aatagtttat tatataatta aatatctata ccattactaa atattttagt 1020
ttaaaagtta ataaatattt tgttagaaat tccaatctgc ttgtaattta tcaataaaca 1080
aaatattaaa taacaagcta aagtaacaaa taatatcaaa ctaatagaaa cagtaatcta 1140
atgtaacaaa acataatcta atgctaatat aacaaagcgc aagatctatc attttatata 1200
gtattatttt caatcaacat tcttattaat ttctaaataa tacttgtagt tttattaact 1260
tctaaatgga ttgactatta attaaatgaa ttagtcgaac atgaataaac aaggtaacat 1320
gatagatcat gtcattgtgt tatcattgat cttacatttg gattgattac agttgggaag 1380
ctgggttcga aatcgataag cttgcgctgc agttatcatc atcatcatag acacacgaaa 1440
taaagtaatc agattatcag ttaaagctat gtaatatttg cgccataacc aatcaattaa 1500
aaaatagatc agtttaaaga aagatcaaag ctcaaaaaaa taaaaagaga aaagggtcct 1560
aaccaagaaa atgaaggaga aaaactagaa atttacctgc acaagcttgg atcctctaga 1620
ccactttgta caagaaagct gggtccaccc ttcttgatcc tgctataatc cgaggcagaa 1680
gcttatgctc aaccaatgtt ttcactacac ctacatggaa agaccccagt gaagcacccc 1740
cacttaagag tagggctgtc ctcccaaagg catgcctggt ctcctgaaca aaggcaagct 1800
tctctcccaa tagcaactca tcagtgtcag attcgcacac cattcttagt tgaatagaaa 1860
cctcttcaat gtattccttt ataagcttag gaacctctag cctgcccttg tgaagttcag 1920
gattgcacat agagaaccag ttgaattttc tcttgctatg agattccagg atggaatcct 1980
cctagatggt gggcctcaaa aagtccagga aaagcacagg aagctgttac tgcactccaa 2040
gcatccgcgc cgcgtgcgcc cactcctcgt aggtgagcgc ggcgcgcatc atgttccgcc 2100
agaacttgcg gcggtaggcg gactgcaccc agcgcgacgg cgcagacggc gagcaggatc 2160
ccgcgcgtgt tgtcgcgcgg gtgcagccac gccgccgc 2198
<210> 245
<211> 22
<212> ДНК
<213> Искусственная
<220>
<223> Олигонуклеотидный праймер
<400> 245
ttttaacgat atccgctaaa gg 22
<210> 246
<211> 23
<212> ДНК
<213> Искусственная
<220>
<223> Олигонуклеотидный праймер
<400> 246
aatgaatgaa caagaattaa gtc 23
<210> 247
<211> 22
<212> ДНК
<213> Искусственная
<220>
<223> Олигонуклеотидный праймер
<400> 247
cttttctcac accgtatctc cg 22
<210> 248
<211> 25
<212> ДНК
<213> Искусственная
<220>
<223> Олигонуклеотидный праймер
<400> 248
agcatgatat acttgtcgag aaagc 25
<210> 249
<211> 18
<212> ДНК
<213> Искусственная
<220>
<223> Олигонуклеотидный праймер
<400> 249
gcgacagtgt agcgtttt 18
<210> 250
<211> 25
<212> ДНК
<213> Искусственная
<220>
<223> Олигонуклеотидный праймер
<400> 250
atacataaaa tgaaaactat tgtgc 25
<210> 251
<211> 23
<212> ДНК
<213> Искусственная
<220>
<223> Олигонуклеотидный праймер
<400> 251
acagacatgt ctagacccag atg 23
<210> 252
<211> 24
<212> ДНК
<213> Искусственная
<220>
<223> Олигонуклеотидный праймер
<400> 252
cactctcatc ccaagtgaag ttgc 24
<210> 253
<211> 25
<212> ДНК
<213> Искусственная
<220>
<223> Олигонуклеотидный праймер
<400> 253
ctgagatgga agtgaagcac agatg 25
<210> 254
<211> 21
<212> ДНК
<213> Искусственная
<220>
<223> Олигонуклеотидный праймер
<400> 254
ccattgttag tcctttcagt c 21
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СНИЖЕНИЕ ПРЕВРАЩЕНИЯ НИКОТИНА В НОРНИКОТИН В РАСТЕНИЯХ | 2015 |
|
RU2733837C2 |
| РАСТЕНИЯ С МОДИФИЦИРОВАННЫМИ ПРИЗНАКАМИ | 2017 |
|
RU2809117C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2020 |
|
RU2833880C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2018 |
|
RU2809244C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2740312C2 |
| СПОСОБЫ УСОВЕРШЕНСТВОВАНИЯ ОДНОДОЛЬНЫХ РАСТЕНИЙ | 2015 |
|
RU2727428C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ САХАРОВ И АМИНОКИСЛОТ В РАСТЕНИИ (SULTR3) | 2020 |
|
RU2826107C1 |
| СПОСОБЫ УСОВЕРШЕНСТВОВАНИЯ РАСТЕНИЙ | 2015 |
|
RU2727424C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2817591C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2816526C2 |
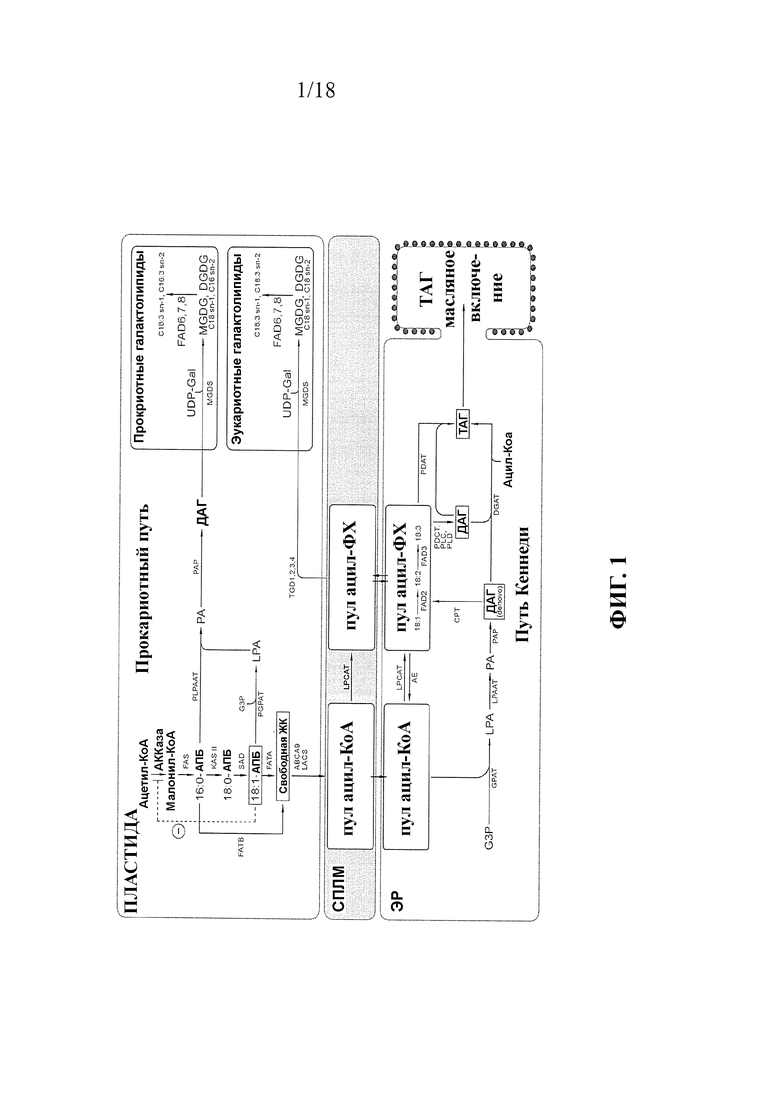

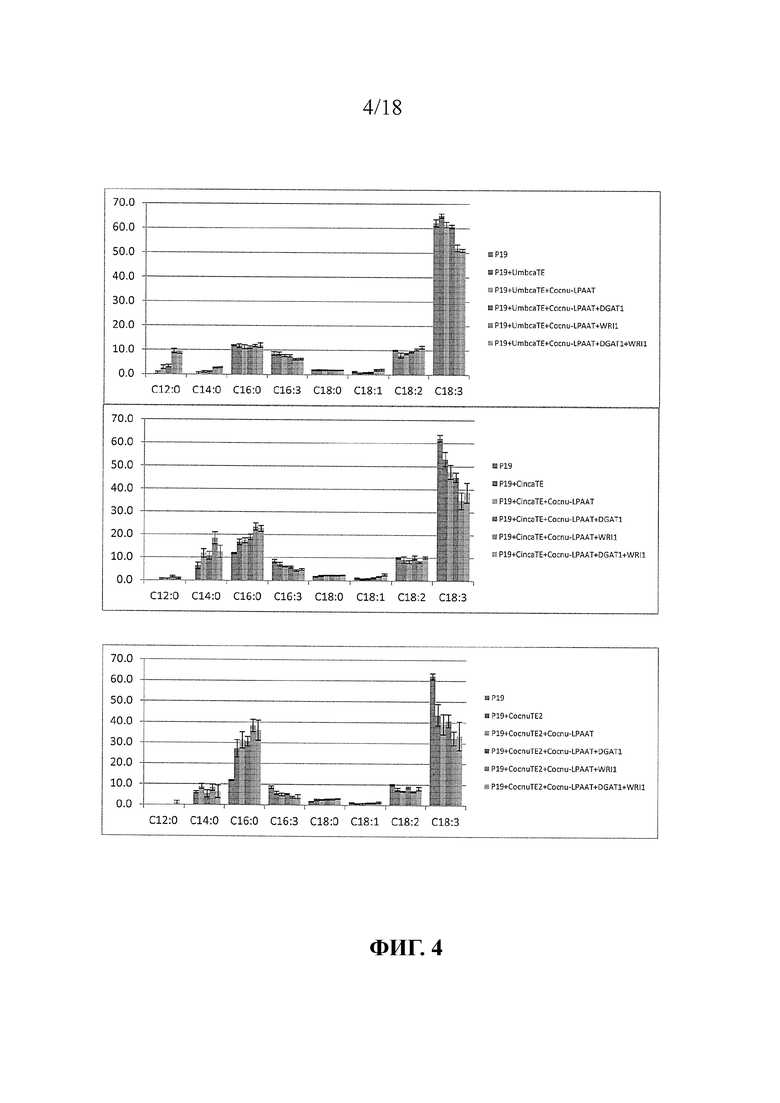
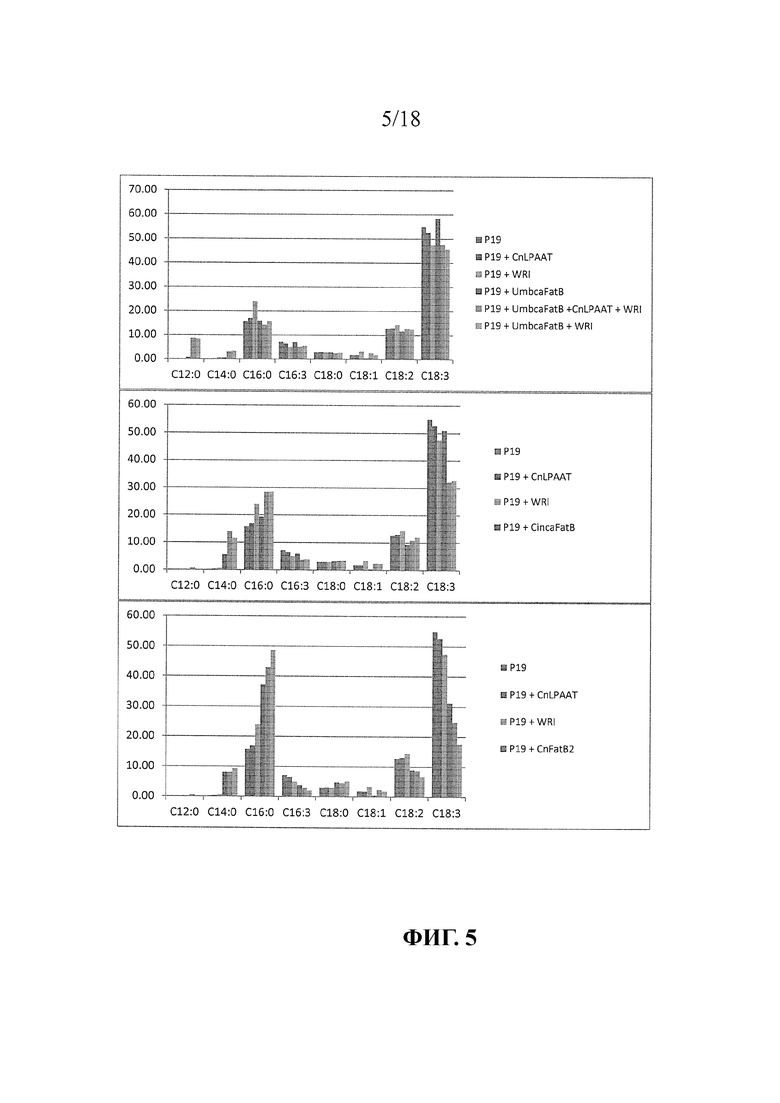

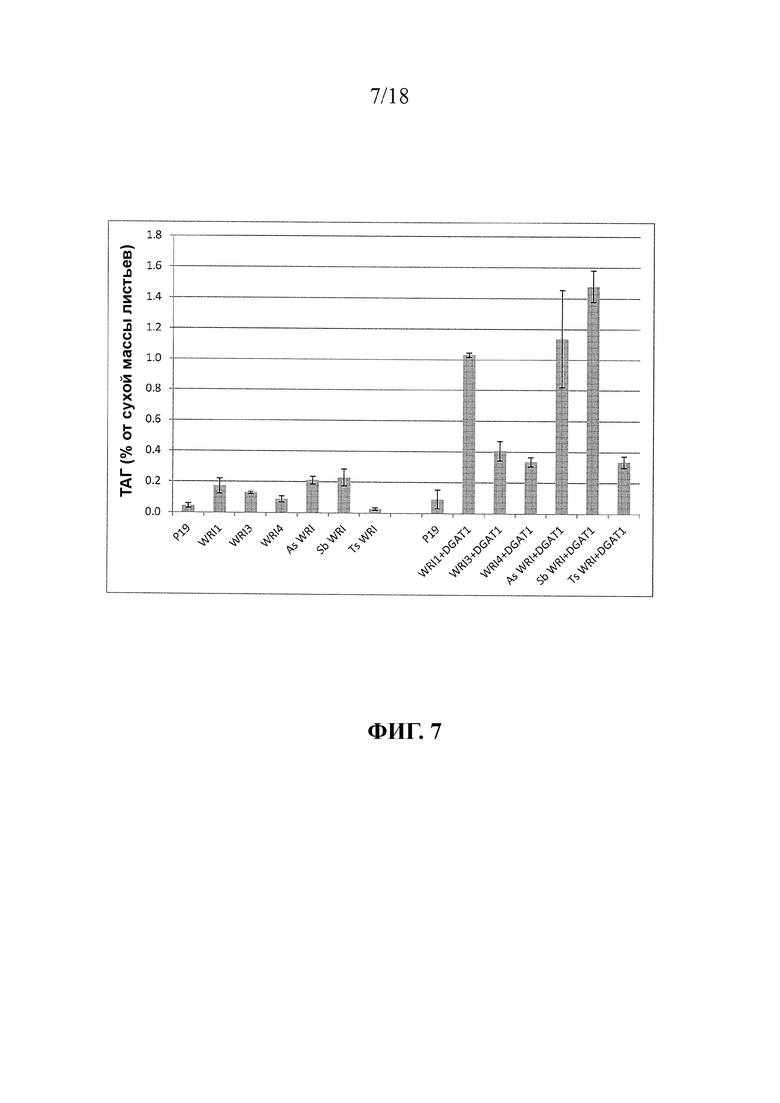

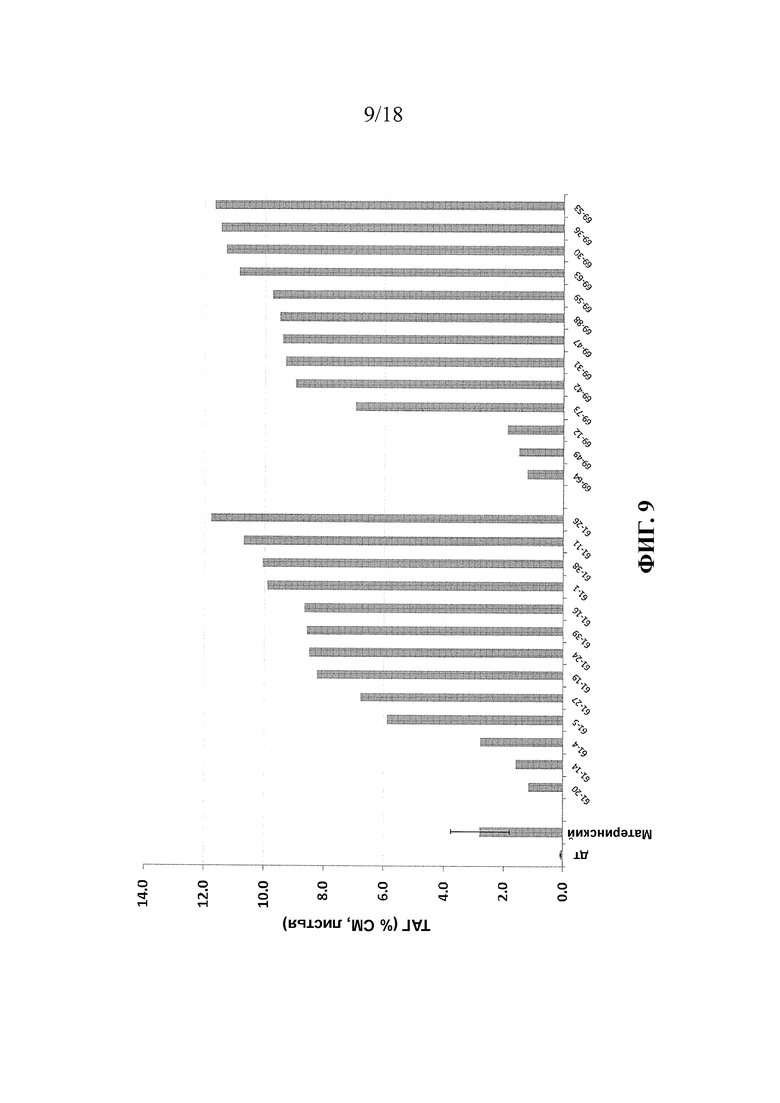
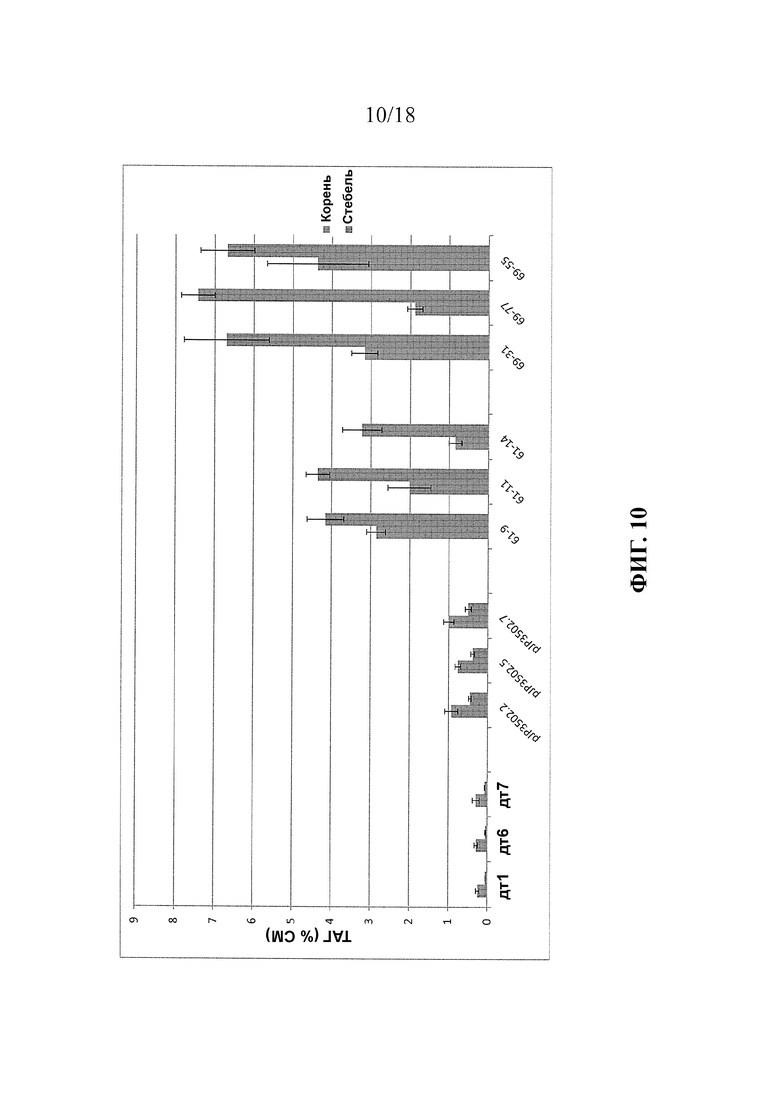
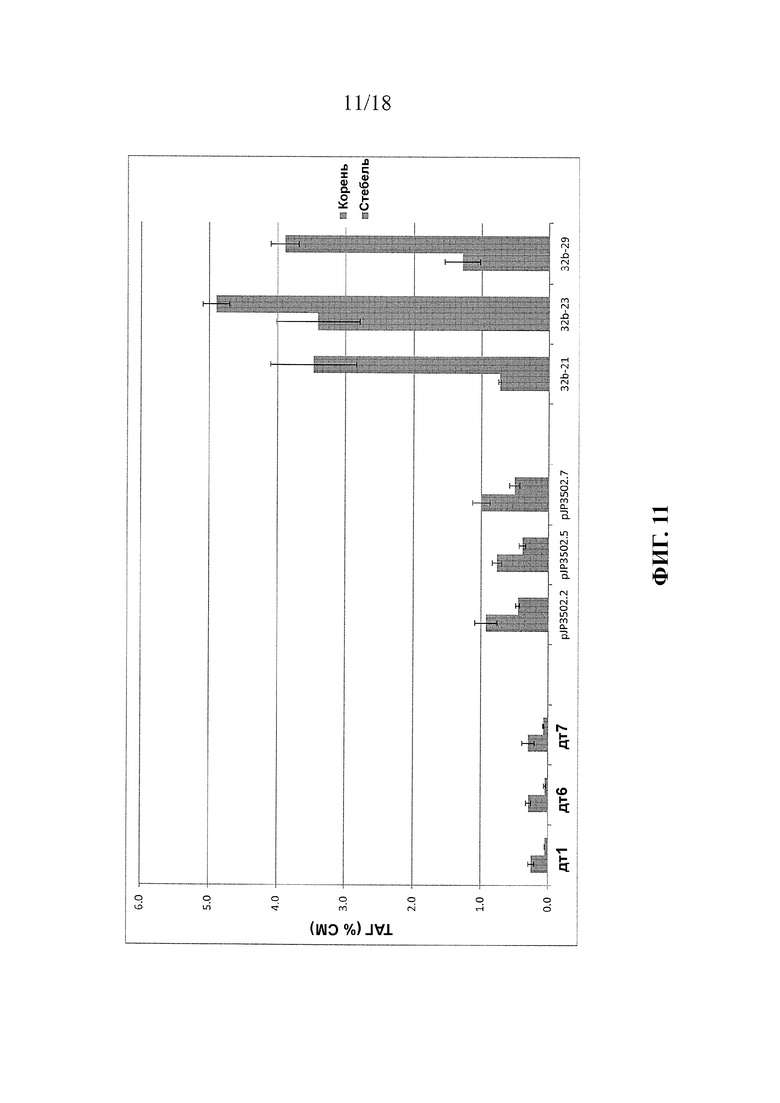
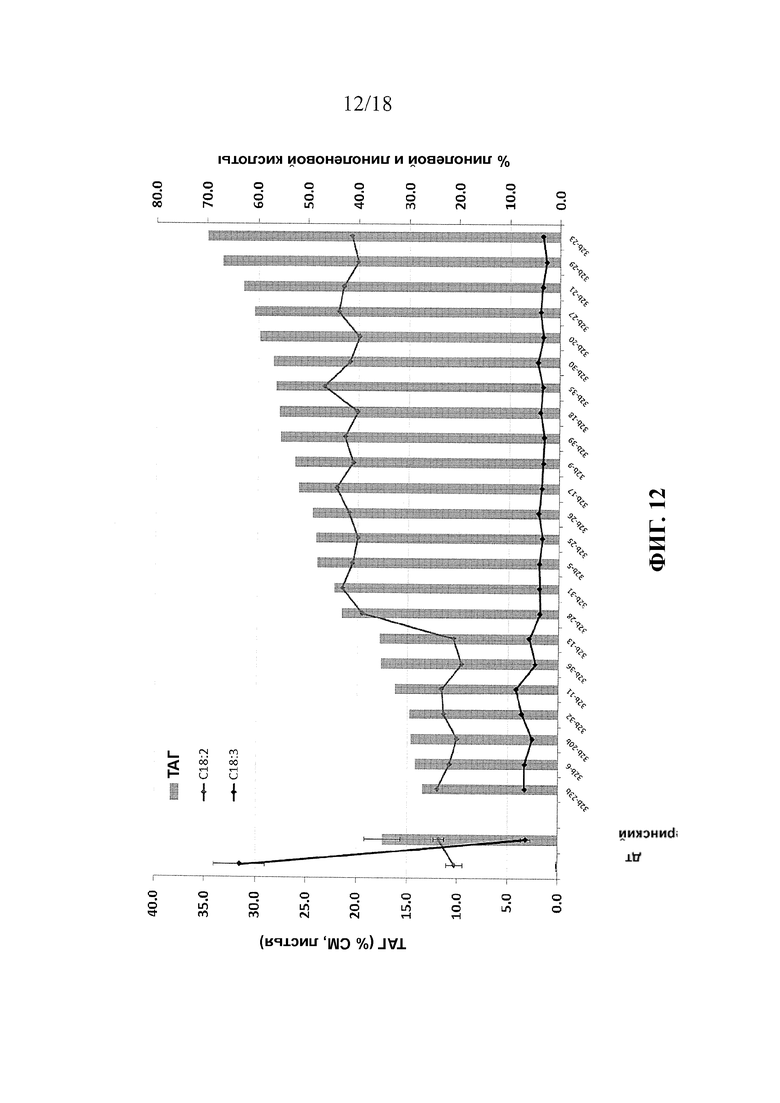
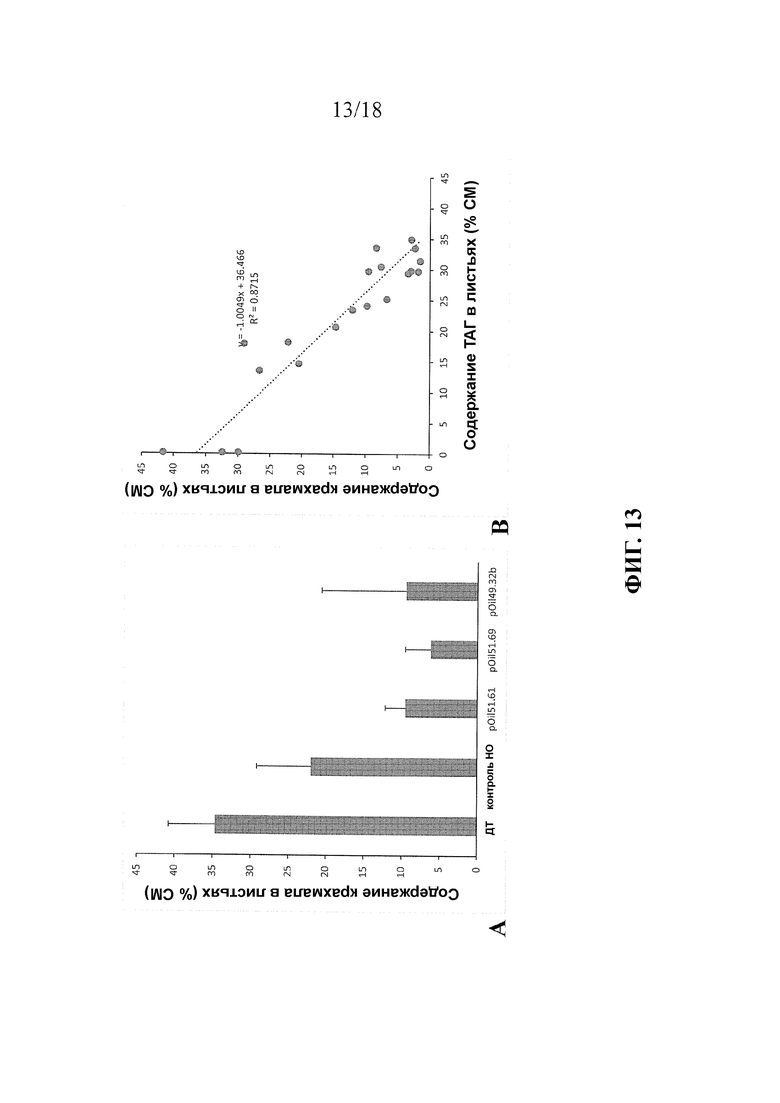


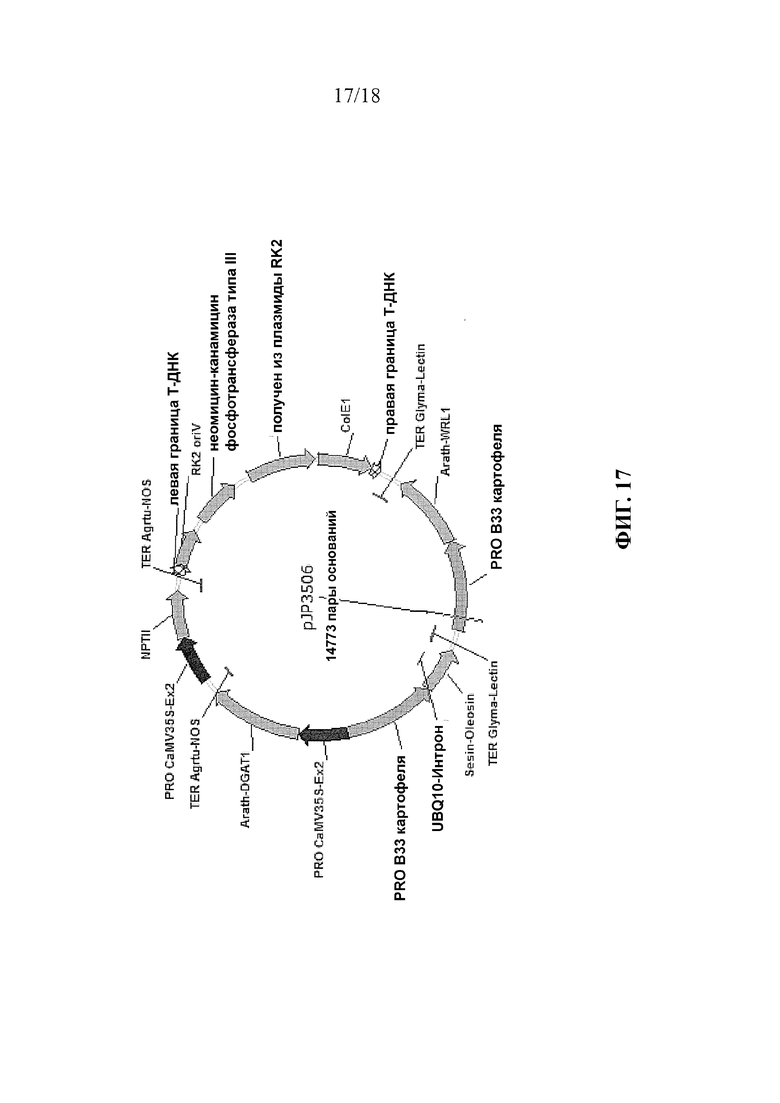
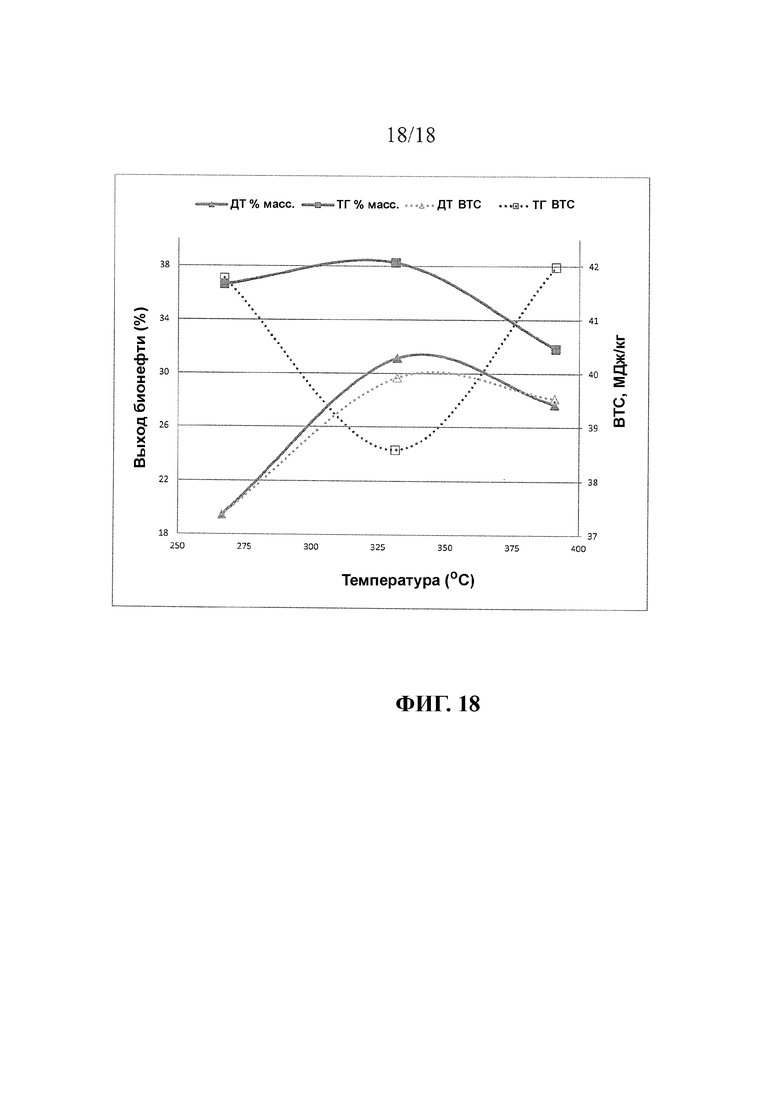
Изобретение относится к биотехнологии и представляет собой способ получения нефтепродукта, включающий стадии:(i) обработки в реакторе композиции, содержащей (a) вегетативные части растения, сухая масса которых составляет по меньшей мере 2 г и в которых общее содержание неполярных липидов составляет по меньшей мере 5% масс., в пересчете на сухую массу,(b) растворитель, который содержит воду, спирт или и то, и другое, и (c) необязательно катализатор, причем обработка включает нагревание композиции при температуре от около 270°C до около 400°C и давлении от 70 до 350 бар на протяжении периода от 1 до 120 минут в окислительной, восстановительной или инертной среде,(ii) извлечение нефтепродукта из реактора с выходом по меньшей мере 35% масс., в пересчете на сухую массу вегетативных частей растения, с получением таким образом нефтепродукта, где нефтепродукт является углеводородным продуктом, включающим эфиры жирных кислот, один или несколько алканов, один или несколько алкенов или комбинацию любых двух или нескольких из них. Изобретение позволяет получать нефтепродукты с высокой степенью эффективности. 9 н. и 15 з.п. ф-лы, 18 ил., 19 табл., 18 пр.
1. Способ получения нефтепродукта, включающий стадии:
(i) обработки в реакторе композиции, содержащей
(a) вегетативные части растения, сухая масса которых составляет по меньшей мере 2 г и в которых общее содержание неполярных липидов составляет по меньшей мере 5% масс., в пересчете на сухую массу,
(b) растворитель, который содержит воду, спирт или и то, и другое, и
(c) необязательно катализатор,
причем обработка включает нагревание композиции при температуре от около 270°C до около 400°C и давлении от 70 до 350 бар на протяжении периода от 1 до 120 минут в окислительной, восстановительной или инертной среде,
(ii) извлечение нефтепродукта из реактора с выходом по меньшей мере 35% масс., в пересчете на сухую массу вегетативных частей растения,
с получением таким образом нефтепродукта, где нефтепродукт является углеводородным продуктом, включающим эфиры жирных кислот, один или несколько алканов, один или несколько алкенов или комбинацию любых двух или нескольких из них.
2. Способ по п. 1, в котором сухая масса вегетативных частей растения составляет по меньшей мере 1 кг.
3. Способ по п. 1 или 2, в котором общее содержание неполярных липидов в вегетативных частях растения составляет по меньшей мере 10%, по меньшей мере 15%, по меньшей мере 20%, около 25%, около 30%, около 35% или от 30% до 75%, в пересчете на сухую массу.
4. Способ по любому из пп. 1-3, в котором части вегетативного растения получены из или находятся в листе или стебле растения до цветения растения и части вегетативного растения включают общее содержание неполярных липидов по меньшей мере приблизительно 8%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 11 %, от 8% до 15% или от 9% до 12% масс., в пересчете на сухую массу.
5. Способ по любому из пп. 1-4, в котором концентрация твердых веществ в композиции составляет от 5% до 90%.
6. Способ по любому из пп. 1-5, в котором катализаторы содержат NaOH, или KOH, или оба предпочтительно в концентрации 0,1-2 М.
7. Способ по любому из пп. 1-6, в котором время обработки составляет от 1 до 60 минут, предпочтительно от 10 до 60 минут, более предпочтительно от 15 до 30 минут.
8. Способ по любому из пп. 1-7, в котором, если растворитель является водой, выход нефтепродукта в результате способа составляет от по меньшей мере 36%, 37%, 38%, 39% или 40% до максимального значения 55% или 60% масс., в пересчете на сухую массу вегетативных частей растения.
9. Способ по любому из пп. 1-8, в котором, если растворитель содержит около 80% воды, нефтепродукт содержит около 30% углеводородных соединений С13-С22.
10. Способ по любому из пп. 1-9, в котором если растворитель содержит спирт, то выход нефтепродукта в результате способа составляет от по меньшей мере 36%, 37%, 38%, 39% или 40% до максимального значения 65% или 70% масс., в пересчете на сухую массу вегетативных частей растения.
11. Способ по любому из пп. 1-10, в котором если растворитель содержит около 50% метанола, то нефтепродукт содержит около 50% метиловых эфиров жирных кислот (МЭЖК).
12. Способ по любому из пп. 1-11, в котором содержание воды в извлеченном нефтепродукте составляет менее чем около 15% масс.
13. Способ по любому из пп. 1-12, в котором выход нефтепродукта по меньшей мере на 2% масс. больше, чем выход соответствующего способа с применением соответствующих вегетативных частей растения, в которых содержание неполярного липида составляет менее чем 2%, в пересчете на сухую массу.
14. Способ по любому из пп. 1-13, в котором вегетативные части растения на стадии (i)(a) физически обрабатывают одним или более из сушки, рубки, нарезки, помола, вальцовки, прессования, дробления или перетирания.
15. Способ по любому из пп. 1-14, в котором вегетативные части растения включают листья растения, стебли или и то, и другое.
16. Способ получения гидрогенизированного нефтепродукта, включающий стадии
(i) получения нефтепродукта с использованием способа по любому из пп. 1-15, и
(ii) гидрогенизацию извлеченного нефтепродукта.
17. Способ снижения уровней кетонов или сахаров в нефтепродукте, включающий стадии
(i) получения нефтепродукта с использованием способа по любому из пп. 1-15, и
(ii) обработку извлеченного нефтепродукта водородом для снижения уровней кетонов или сахаров в нефтепродукте.
18. Способ получения синтетического газа, включающий стадии
(i) получения нефтепродукта с использованием способа по любому из пп. 1-15, и
(ii) превращение извлеченного нефтепродукта в синтетический газ.
19. Способ получения мазута, включающий стадии
(i) получения нефтепродукта с использованием способа по любому из пп. 1-15, и
(ii) фракционирование извлеченного нефтепродукта для получения мазута.
20. Способ получения дизельного топлива, включающий стадии
(i) получения нефтепродукта с использованием способа по любому из пп. 1-15, и
(ii) фракционирование извлеченного нефтепродукта для получения дизельного топлива.
21. Способ получения керосина, включающий стадии
(i) получения нефтепродукта с использованием способа по любому из пп. 1-15, и
(ii) фракционирование извлеченного нефтепродукта для получения керосина.
22. Способ получения бензина, включающий стадии
(i) получения нефтепродукта с использованием способа по любому из пп. 1-15, и
(ii) фракционирование извлеченного нефтепродукта для получения бензина.
23. Способ получения промышленного продукта, включающий стадии
(i) получения нефтепродукта с использованием способа по любому из пп. 1-15, и
(ii) превращение извлеченного нефтепродукта в промышленный продукт с применением нагревания, химических или ферментных средств или любой их комбинации, где промышленный продукт представляет собой биоспирт.
24. Способ по п. 23, в котором биоспирт представляет собой этанол, пропанол, бутанол или комбинацию одного или более или всех из них.
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| СПОСОБ ПОЛУЧЕНИЯ НЕФТЕПРОДУКТОВ | 2004 |
|
RU2261264C1 |

