Изобретение относится к области промышленной биотехнологии, генетической инженерии и касается получения ауксотрофного штамма-реципиента для генно-инженерных работ, плазмидных векторов для экспрессии генов, штамма-продуцента фосфолипазы А2, который может быть использован пищевой промышленности при получении лизофосфолипидов и поверхностно-активных веществ с эмульгирующими свойствами.
Фосфолипаза А2 - фермент, катализирующий гидролиз сложноэфирной связи во втором положении глицерофосфолипидов [Головач О.А., Таганович А.Д. Роль фосфолипаз А2 в патологии легких // Медицинский журнал. - 2005. - №1. - с. 1-6]. При действии фосфолипазы А2 на фосфолипид образуется 2-лизофосфолипид и жирная кислота. Стерическая и позиционная специфичность фосфолипазы А2 обусловливает ее высокую ценность в биохимии липидов и химии. Фермент используют для установления позиционного распределения жирных кислот при анализе фосфоглицеридов, для разделения рацемических смесей липидов, а также в синтезе липидов для получения фосфоглицеридов со смешанным составом жирных кислот. Таким образом, ферментный препарат фосфолипаза А2 может использоваться в пищевой промышленности в качестве вспомогательного средства для образования лизофосфолипидов и поверхностно-активных веществ с эмульгирующими свойствами [Довжикова И.В. Гистохимическое исследование фосфолипидов в плаценте беременных при герпесе // Бюллетень физиологии и патологии дыхания: Дальневосточный научный центр физиологии и патологии дыхания (Благовещенск). - 2003. - №15. - с. 19-23; Брагина Н.А., Чупин В.В., Булгаков В.Г., Шальнев А.Н. Липидные ингибиторы фосфолипазы А2 // Биоорганическая химия. - 1999. - том 25. - №2. - с. 83-96; Murakami M, Kudo I. Phospholipase A2 // J Biochem. - 2002. - Mar. -131(3):285. - с. 92].
Известен способ получения ферментного препарата фосфолипазы А2 с помощью генномодифицированного плесневого гриба Aspergillus niger PLA-54 (США) с геном, кодирующим фосфолипазу свиной поджелудочной железы из Aspergillus niger GAM-53 и кДНК поджелудочной железы свиньи Aspergillus niger PLA-54 с геном, продуцирующим фосфолипазу А2 из Aspergillus niger GAM-53 (NRRL3122 Aspergillus niger) [GRAS notification of phospholipase A2 from a genetically modified strain of Aspergillus niger // DSM. - 28.10.2005].
Известно также, что ферментный препарат фосфолипазы А2 можно получать с помощью штамма Trichoderma reesei RF8793 (Германия), являющегося носителем гена, кодирующего фосфолипазу А2 плесневого грибка Aspergillus nishimurae [A Lipase Enzyme Preparation Derived from Streptomyces viotaceoruber // Genencor International Inc. - 09.02.2004].
Также в США выпускают ферментный препарат фосфолипазы А2, полученный с помощью Streptomyces violaceoruber JAS-591 [A phospholipase A2 preparation produced by Streptomyces violaceoruber expressing the gene encoding phospholipase A2 from Streptomyces violaceoruber // Nagase ChemteX Corporation. - 17.08.2006].
В Японии выпускают ферментный препарат фосфолипазы А2 Streptomyces violaceoruber, полученный с применением рекомбинантного штамма-продуцента Streptomyces violaceoruber AS-10 [GRAS notification for phospholipase A2 from a genetically modified strain of Trichoderma reesei // AB ENZYMES GmbH. - 12.10.2013].
Основным недостатком данных способов является продолжительное время культивирования штаммов (более 7 суток), высокий риск контаминации, относительно низкая экспрессия целевого продукта и использование трудоемких способов очистки ферментного препарата.
Все вышеизложенное свидетельствует о перспективности создания штаммов-продуцентов фосфолипазы А2 на основе микроорганизмов, лишенных большей части этих недостатков, в частности дрожжей. Среди всех, отличных от Saccharomyces, дрожжей наиболее применяемой гетерологичной системой для производства коммерчески релевантных рекомбинантных белков являются Pichia pastoris (Komagataella phaffii). В сравнении с S. cerevisiae, они достигают более высоких уровней экспрессии, а гипергликозилирование у них не столь высоко, как следствие, получаемый продукт является менее аллергенным и его свойства остаются стабильными. Тем не менее наиважнейшей особенностью Komagataella phaffii (Pichia pastoris) является то, что они могут использовать метанол в качестве единственного источника углерода и энергии. Таким образом, переключение с глюкозной среды на метанольную индуцирует высокий уровень экспрессии рекомбинантного белка в этом хосте. В Komagataella phaffii (Pichia pastoris) рекомбинантные белки могут экспрессироваться внеклеточно, секретируясь в среду или прикрепляясь к клеточной поверхности. Метилотрофные дрожжи слабо секретируют собственные белки в культуральную жидкость, что позволяет получить относительно чистый целевой белок при его секреции.
Изобретение по патенту CN104328095 (МПК A21D 8/04; C11B 3/00; C12N 1/19; C12N 15/55; C12N 15/81; C12N 9/16, опубл. 2015-02-04) относится к созданию ферментного препарата фосфолипазы А2 с наиболее приемлемым рН. Согласно ему оптимизированную последовательность гена фосфолипазы А2, полученнной из S. violaceoruber, клонируют в плазмиду pPIC9K, а затем рекомбинантная плазмида встраивается в хромосому Pichia pastoris (Komagataella phaffii).
Дрожжи Komagataella phaffii (Pichia pastoris) имеют статус безопасных для человека согласно реестру Федерального управления по контролю за пищевыми продуктами и лекарственными средствами США (GRAS), что позволяет их использовать в качестве продуцентов рекомбинантных ферментов, пригодных для применения в пищевой промышленности.
Известен патент RU 2409671 (МПК C12N 15/00, опубл. 20.01.2011), относящийся к рекомбинантной плазмиде pP10PLC-Вс для экспрессии гена фосфолипазы С РLС-Вс в дрожжах Komagataella phaffii (Pichia pastoris) и штамму дрожжей Komagataella phaffii (Pichia pastoris) PLC-Bc-1 ВКПМ Y-3359 - продуцент фосфолипазы С PLC-Bc.
В патенте RU 2676321 C1 (МПК C12N 1/16, опубл. 27.12.2018), относящемуся к способу получения ферментного препарата фосфолипазы А2 с применением рекомбинантного штамма-продуцента Pichia pastoris X-33/ pPICZαA-PhoA2-StV, описан способ, в котором для селекции дрожжей используются гены антибиотикорезистентности и использован коммерческий плазмидный вектор (Invitrogen), что ограничивает использование полученного штамма.
Технический результат заключается в расширении арсенала средств получения ферментных препаратов фосфолипазы А2.
Технический результат достигается использованием способа получения штамма-продуцента фосфолипазы А2 Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2Sv, включающего в себя:
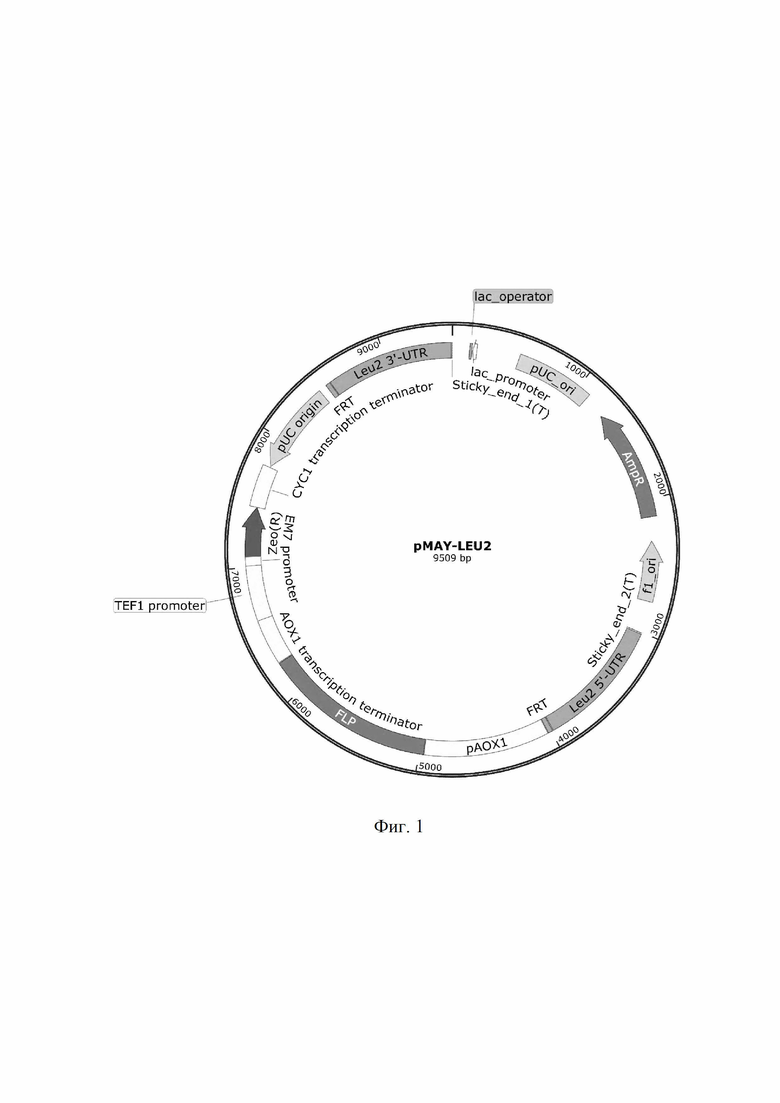
- создание интегративного плазмидного вектора pMAY-LEU2, используемой для получения ауксотрофного по лейцину штамма Komagataella phaffii (Pichia pastoris) YIB Δleu2, который содержит AOX1-промотор индуцибельного типа, ZeoR - ген устойчивости к зеоцину, LEU2 - сайты интеграции в ген, отвечающий за синтез лейцина -, FLP - ген FLP-рекомбиназы, а сконструированная плазмида pMAY-LEU2 имеет размер 9509 п.о.,
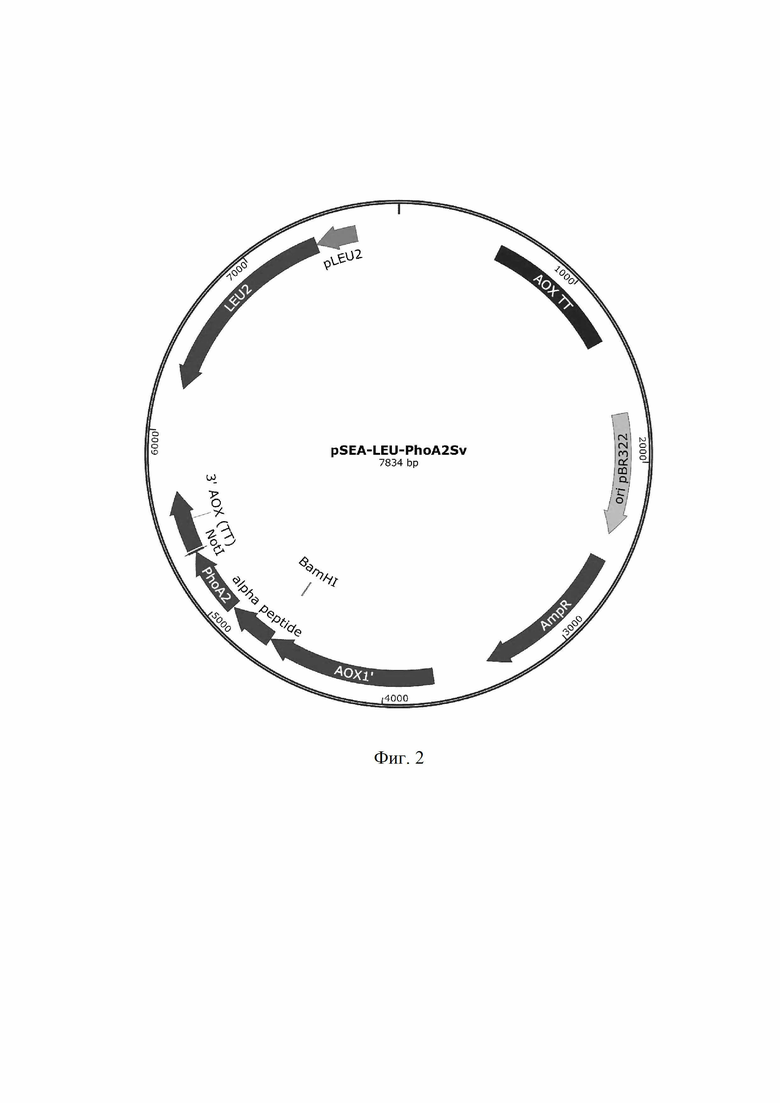
- создание интегративного плазмидного вектора pSEA-LEU-PLA2Sv, для экспрессии генов в дрожжах, содержащего селективный маркер для отбора трансформантов в клетках E.coli, сайт начала репликации, сайт интеграции и экспрессионную кассету, в состав которой входят промотор, терминатор транскрипции и селективный маркер, а в качестве селективного маркера используют последовательность ДНК, кодирующую ген Leu2,
- создание дрожжевого штамма-реципиента Komagataella phaffii (Pichia pastoris) YIB Δleu2, на основе дикого штамма Y-3489, задепонированного в ВКПМ, а данный штамм ауксотрофный по лейцину получен трансформацией вектором pMAY-LEU2 по сайтам интеграции в ген LEU2 с последующим выщеплением последовательности, содержащей гены антибиотикорезистентности,
- создание на его основе штамма дрожжей Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2Sv, полученного трансформацией штамма Komagataella phaffii (Pichia pastoris) YIB Δleu2 плазмидным вектором pSEA-LEU-PLA2Sv, с последующим отбором на среде YNB; при этом штамм, обеспечивающий синтез рекомбинантной фосфолипазы А2, не содержащий генов антибиотикорезистентности, задепонирован в ВКПМ под номером Y-4513.
Способ получения штамма-продуцента ферментного препарата фосфолипазы А2 обеспечивает высокую экспрессию целевого продукта, не содержащего гены атибиотикорезистентности.
Технический результат достигается созданием генетической конструкции в виде рекомбинантного вектора pSEA-LEU-PLA2Sv и штамма Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2Sv, обеспечивающих индуцируемый синтез с высоким и стабильным выходом рекомбинантной фосфолипазы А2 в культуральную среду.
Экспрессия гена фосфолипазы А2 в составе плазмиды pSEA-LEU-PLA2Sv находится под контролем промотора гена АОХ1, содержащего области, обеспечивающие активацию транскрипции в присутствии метанола в культуральной среде, а также область инициации транскрипции. Промотор гена АОХ1 относится к числу наиболее сильных дрожжевых промоторов. Уровень экспрессии генов, находящихся под контролем АОХ1 промотора, эффективно регулируется источниками углерода. Транскрипция гена АОХ1 полностью блокирована при выращивании дрожжей на среде с глюкозой, на среде с глицерином наблюдается только базальный уровень экспрессии гена. Использование метанола в качестве единственного источника углерода значительно усиливает экспрессию гена АОХ1 и, следовательно, генов, находящихся под контролем АОХ1 промотора. Это позволяет регулировать синтез фосфолипазы А2 в клетках дрожжей. Регулируемая экспрессия клонированного гена позволяет существенно снизить метаболическую нагрузку на клетку дрожжей.
Для получения на основе Komagataella phaffii (Pichia pastoris) фермента фосфолипаза А2 создан ауксотрофный штамм YIB Δleu2 и сконструированы векторы pMAY-LEU2 для создания ауксотрофного штамма-реципиента, который содержит AOX1-промотор индуцибельного типа,ZeoR - ген устойчивости к зеоцину, LEU2 - сайты интеграции в ген отвечающий за синтез лейцина -, FLP - ген FLP-рекомбиназы. Сконструированная плазмида pMAY-LEU2 имеет размер 9509 п.о. (Фиг. 1) и состоит из следующих элементов:
1) фрагмент 5' AOX1 промотора: 4028-4962 п.о.
2) ген FLP-рекомбиназы: 4968-6251 п.о.
3) FRT - сайты рекомбинации: 3989-4022 п.о и 8517-8550 п.о.
4) терминатор транскрипции 3-AOX (TT): 6258-6599 п.о.
5) сайт интеграции LEU2: 3016-3982 п.о. и 8557-9501 п.о.
6) ori pUC: 511-1099 п.о. и 7783-8456 п.о.
7) ген резистентности к ампициллину AmpR: 1273-2130 п.о.
8) ген резистентности к зеоцину ZeoR: 7080-7454 п.о.
9) Tef-промотор: 6600-7011 п.о.
10) EM7-промотор: 7012-7079 п.о.
11) терминатор транскрипции СYC1: 7455-7772 п.о.
Вектор pSEA-LEU2-PLA2Sv для экспрессии рекомбинантной фосфолипазы А2, который содержит AOX1-промотор индуцибельного типа, сигнал секреции α-фактора Saccharomyces cerevisiae, а также ген синтеза лейцина LEU2. В вектор pSEA-LEU2 по сайтам EcoRI и NotI под сигнальным пептидом α-фактора был заклонирован оптимизированный ген фосфолипазы А2 Streptomyces violaceoruber. Сконструированная плазмида pSEA-LEU-PLA2Sv имеет размер 7834 п.о. (Фиг. 2) и состоит из следующих элементов:
1) фрагмент 5' AOX1 промотора: 3726-4673 п.о.
2) сигнальный пептид α-фактора Saccharomyces cerevisiae: 4674-4940 п.о.
3) ген фосфолипазы PLA2 Sv: 4947-5315 п.о.
4) терминатор транскрипции 3-AOX (TT): 5335-5667 п.о. и 572-1328 п.о.
5) ген синтеза лейцина LEU2: 6242-7369 п.о.
6) ori pBR322: 1176-2410 п.о.
7) ген резистентности к ампициллину AmpR: 2555-3415 п.о.
Полученный производственный штамм Pichia pastoris YIB Δleu2-PLA2Sv производит фосфолипазу А2 Streptomyces violaceoruber с помощью глубинной ферментации. Фермент секретируется с молекулярной массой 14 кДа.
Рекомбинантный штамм Komagataella phaffii (Pichia pastoris) YIB-leu2/ pSEA-LEU-PLA2Sv характеризуется следующими признаками.
Морфологические признаки. Клетки округлой, слегка овальной формы, размером 3-7 мкм, часть клеток имеет на поверхности почки или соединена с дочерними клетками.
Культуральные признаки. Клетки хорошо растут на полной органической среде YPD - дрожжевой экстракт-1%, пептон - 1 %, глюкоза - 2%. Кроме того, клетки хорошо растут на минеральной среде SC: 1,34% Yeast Nitrogen Base («Difco», США), а также на других синтетических средах для дрожжей, содержащих в качестве источника углерода глюкозу (2%) или глицерин (1%). При росте на твердых средах клетки образуют гладкие, круглые колонии с матовой поверхностью, светло-кремового цвета, край неровный. При росте в жидких средах образуют интенсивную однородную суспензию. Культура имеет характерный запах метилотрофных дрожжей.
Физиолого-биохимические признаки. Клетки растут в пределах от 4 до 37°С. Оптимальной температурой выращивания является 30°С. При росте в аэробных условиях клетки незначительно закисляют среду. Оптимум pH для роста составляет 4,5-6,5. В качестве источника углерода клетки могут использовать многие простые соединения, такие как глюкоза, глицерин, метанол. В качестве источника азота клетки могут использовать минеральные соли в аммонийной форме, аминокислоты, мочевину.
Штамм хранится обычным способом в суспензии с глицерином (15%) при -80°С.
В результате трансформации линеаризованной по сайтам BglII плазмидой и последующей гомологичной рекомбинации происходит встраивание экспрессионной кассеты в хромосому дрожжей Komagataella phaffii (Pichia pastoris) YIB-Δleu2, что обеспечивает стабильное поддержание клонированного гена фосфолипазы А2.
На основе штамма Pichia pastoris Y-3489 методами генной инженерии получили ауксотрофный по лейцину штамм Pichia pastoris YIB-Δleu2. Для этого собрали генетическую конструкцию pMAY-LEU2 (Фиг.1) с генами устойчивости к зеоцину, FLP-рекомбиназы, фланкированными FRT-сайтами.
Исходный штамм P.pastoris Y-3489 дикого типа трансформируют фрагментом 5'leu2-FRT-FLP-ZeoR-FRT-3'leu2, полученным при обработке плазмиды pMay-LEU2 рестриктазой AvrII. Трансформация проводится методом электропорации (Shixuan Wu and Geoffrey J. Letchworth, 2004). Полученные трансформанты YIB Δleu2::[+FLP +ZeoR] отбирают на полной среде YPD с добавлением 250 мкг/мл зеоцина.
Далее устойчивые к зеоцину трансформанты анализируют на минимальной среде YNB (Yeast Nitrogen Base), отбирая неспособные к росту клоны.
Далее отобранные клоны YIB Δleu2::[+FLP +ZeoR] культивируют в жидкой полной среде и затем переносят в среду с метанолом для индукции синтеза FLP-рекомбиназы. В результате рекомбиназа выщепляет кассету [+FLP +ZeoR] между FRT-сайтами и клоны теряют устойчивость к зеоцину. Полученные клоны YIB Δleu2 высевают на среду с зеоцином и отбирают потерявшие устойчивость.
Отсутствие генов устойчивости к антибиотику подтверждают методом ПЦР. Для этого клоны, высевают на жидкую питательную среду YPD и культивируют в течение суток, после чего из полученной суспензии клеток с использованием набора Yeast Preparation Kit (Jena Bioscience) выделяют геномную ДНК, которая используется в качестве матрицы для проведения реакции. На основе сиквенса гена устойчивости к зеоцину из использованной для получения штамма конструкции сконструировали праймеры для наработки фрагмента гена:
прямой - 5'-GTTGACCAGTGCCGTTCCGG-3';
обратный - 5'- TGCTCCTCGGCCACGAAGTG -3'.
Для ПЦР используют Taq-полимеразу в составе ScreenMix от компании Евроген. Амплификацию всех фрагментов проводят на приборе Т100 (BioRad) при следующих параметрах: денатурация при 95°С 30 сек, отжиг праймеров при 56°С 30 сек, элонгация при 72°С 30 сек. Расчетный размер ампликона - 359 пар оснований, в качестве положительного контроля используют исходный вектор.
Далее в целях проверки отобранные клоны YIB Δleu2 трансформируют методом электропорации полученной далее плазмидой pSEA2 -LEU2 (линеризованной по сайту PmeI в промоторе AOX1) и высевают на минимальную среду YNB.
В качестве проверки у полученных клонов YIB Δleu2 выделяют хромосомную ДНК в качестве матрицы ПЦР с праймерами, фланкирующими ген LEU2:
прямой 5'-CTGACAAGACACAGCAAGA-3';
обратный 5'-CACCAAGATCGTCAAGAA-3'.
В качестве контроля проводят ПЦР с теми же праймерами по хромосомной ДНК дрожжей дикого типа. Полученные ПЦР продукты проверяют методом электрофореза в агарозном геле. Длина ПЦР продукта штамма дикого типа - 2705 п.о., при делеции LEU2 теряется участок его рамки считывания размером 793 п.о. и итоговый фрагмент составляет 1912 п.о.
Таким образом, получили штамм-реципиент Pichia pastoris YIB Δleu2 ауксотрофный по лейцину и не содержащий генов атибиотикорезистентности.
Данный штамм-реципиент используют для вставки гена фосфолипазы А2.
Экспрессионный вектор pSEA-LEU получают на основе вектора pPic9 (invitrogen). Для этого штамм дрожжей Pichia pastoris GS115 культивируют на жидкой питательной среде YPD в течение суток, после чего с использованием набора Yeast Preparation Kit (Jena Bioscience) из полученной суспензии клеток выделяют геномную ДНК и используют в качестве матрицы для ПЦР-амплификации участка гена синтеза лейцина. Для амплификации используют праймеры:
прямой 5'-agcatctagaAGGAGGTATCTTACTGGTGGTATTTA-3'
обратный 5'-agttgtacaTGGTATTGTTTGTATGGAGGATATTAT-3'
На 5'-конце каждого праймера находится сайт рестрикции для эндонуклеазы, образующей липкие концы: XbaI для прямого праймера и BsrGI - для обратного (в приведенных последовательностях выделены жирным шрифтом).
Для ПЦР используют Phusion полимеразу от компании New England Biolabs. Амплификацию всех фрагментов проводят на приборе Т100 (BioRad) при следующих параметрах: денатурация при 98°С 10 сек, отжиг праймеров при 61°С 30 сек, элонгация при 72°С 60 сек, 30 циклов. Расчетный размер ампликона - 1733 пары оснований. Полученный фрагмент очищают с использованием набора CleanUp Standart (Евроген).
Полученный таким образом ампликон гидролизуют эндонуклеазами рестрикции XbaI и BsrGI очищают в агарозном геле и лигируют с вектором рPic9 (invitrogen), расщепленным ферментами рестрикции по сайтам XbaI и BsrGI и обработанным щелочной фосфатазой, используя ДНК-лигазу фага Т4.
Лигазной смесью трансформируют компетентные клетки E.coli Top10, приготовленные накануне обработкой хлористым кальцием. После стандартной процедуры трансформации (0°С - 30 мин, 42°С - 45 секунд, 0°С - 2 мин) клетки разводят в 3-10 раз средой SOB (Hanahan, D. (1983). "Studies on transformation of Escherichia coli with plasmids". Journal of Molecular Biology. 166 (4): 557-580), подращивают в течение одного часа и высевают на агар LB, содержащий ампициллин в концентрации 100 мкг/мл. Посевы инкубируют при 37°С.
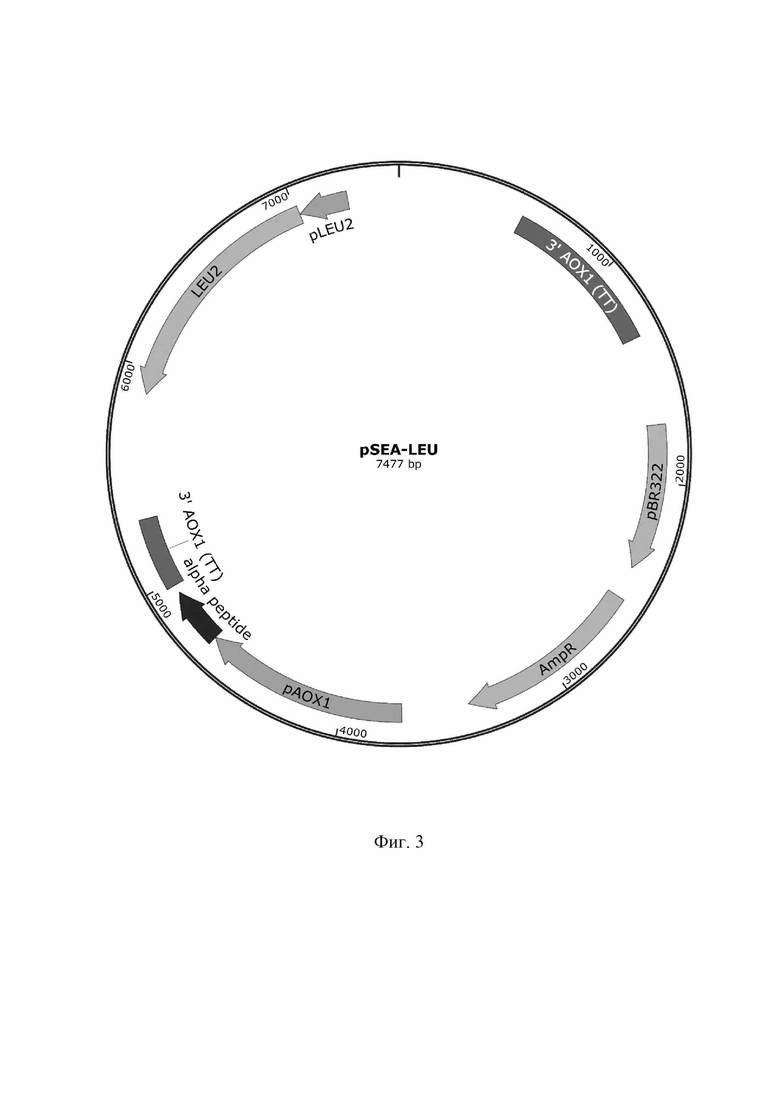
На следующие сутки выросшие ампициллинрезистентные колонии тестируют с помощью праймеров использованных ранее для амплификации гена LEU2 и отбирают позитивные клоны, из которых с использованием набора Plasmid Miniprep (Евроген) выделяют плазмидную ДНК. Выделенные плазмидные ДНК гидролизуют эндонуклеазами рестрикции XbaI и BsrGI и проверяют на наличие вставок, соответствующих по размеру последовательности, кодирующей ген LEU2. Из клонов, плазмидная ДНК которых содержит данную последовательность, выделяют вектор pSEA-LEU (Фиг. 3).
Ген фосфолипазы А2 был получен путем химико-ферментативного синтеза нуклеотидной последовательности, кодирующей фосфолипазу А2 в Streptomyces violaceoruber, полученной из открытых баз данных.
Наработку фрагмента ДНК, содержащего полноразмерный ген PhoA2-StV, получают при помощи полимеразной цепной реакции с использованием в качестве матрицы плазмиды, содержащей синтезированный ген PhoA2-StV SEC ID No1 и праймеров:
прямой 5'-atatgaattcGCTCCAGCAGATAAACC-3'
обратный 5'- tcgcggccgcTCAACCAAAAATCTTTA-3'
На 5'-конце каждого праймера находится сайт рестрикции для эндонуклеазы, образующей липкие концы: EcoRI для прямого праймера и NotI - для обратного (в приведенных последовательностях выделены жирным шрифтом).
Реакцию амплификации гена PhoA2 проводят используя Phusion полимеразу от компании New England Biolabs. Амплификацию всех фрагментов проводят на приборе Т100 (BioRad) при следующих параметрах: денатурация при 98°С 10 сек, отжиг праймеров при 61°С 30 сек, элонгация при 72°С 30 сек, 30 циклов. Расчетный размер ампликона - 389 пар оснований. Полученный фрагмент очищают с использованием набора CleanUp Standart (Евроген), используя в качестве матрицы плазмиду с геном PhoA2 (50 нг) с помощью праймеров PhoA2 EcoRI for и PhoA2 SalI rev mut. Процесс амплификации состоит из 15 циклов со следующими параметрами: денатурация 95°С - 20 сек, отжиг 60°С - 20 сек, элонгация 68°С - 30 сек.
После амплификации очищеный фрагмент ДНК обрабатывают рестриктазами EcoRI и NotI, чистят с использованием набора CleanUp Standart (Евроген) и клонируют в вектор pSEA-LEU, обработанный теми же рестриктазами.
Полученной лигазной смесью трансформируют компетентные клетки E. coli Top10. Выделенные из клонов плазмиды анализируются на присутствие вставки картированием; отсутствие мутаций подтверждается секвенированием. Из клонов, плазмидная ДНК которых содержит последовательность гена PhoA2, выделяют вектор pSEA-LEU-PLA2Sv (Фиг. 2).
Конструкцию pSEA-LEU-PLA2Sv линеаризовали по сайту BglII. Полученный фрагмент 5'pAOX1-a-factor signal-PLA2Sv-3'AOX1(TT)-LEU2-3'AOX1(TT) длиной 5436 п.о., не содержащий ori pBR322 и гена резистентности к ампициллину AmpR, очищен в агарозном геле и набором Cleanup standart и подготовлен к трансформации методом электропорации на приборе MicroPulser (BioRad).
Суспензию компетентных клеток Komagataella phaffii (Pichia pastoris)YIB Δleu2 в количестве 40 мкл помещают в охлажденные пробирки. Добавляют к компетентным клеткам 5-20 мкг линеаризованной плазмиды pSEA-LEU-PLA2Sv и осторожно перемешивают. Помещают пробирки в ледяную баню на 5 мин. Переносят смесь в охлажденную стерильную кювету для электропоратора с шириной зазора 2 мм. Трансформацию методом электропорации клеток проводят в режиме Fungi -> Pic. После проведения трансформации в кюветы вносят 1 мл 1 М ледяного сорбита. Переносят смесь в стерильную пробирку с 1 мл среды YPD и инкубируют при 30 0С в течение 1-3 часов.
Аликвоты трансформированных клеток Komagataella phaffii (Pichia pastoris) YIB Δleu2 100 и/или 200 мкл пересевают на твердую питательную среду YNB и инкубируют в термостате при +30°С в течение 72 ч.
Проросшие колонии трансформированных клеток Komagataella phaffii (Pichia pastoris) YIB Δleu2 пересевают в культуральные планшеты, содержащие по 2 мл среды YPG: дрожжевой экстракт-1%, пептон - 1 %, глицерин - 0,5 %. В засеянные культуральные планшеты в стерильных условиях необходимо вносить по 1% метанола от объема питательной среды в каждую засеянную лунку каждые 24 часа с момента засева. Планшеты размещают в шейкере-инкубаторе и устанавливают следующие условия: скорость перемешивания - 250 об/мин, температурный режим - 30°С, продолжительность культивирования - 72 ч.
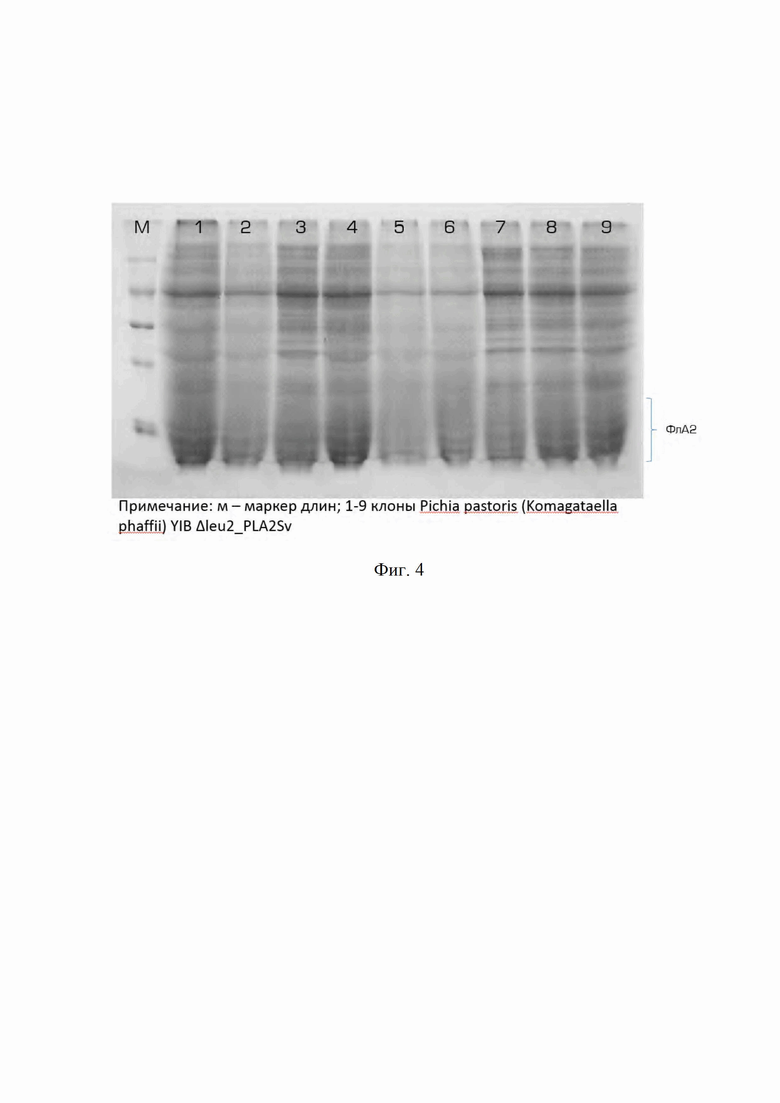
Для определения продуктивности штамма культуральную жидкость из планшетов, отделенную от биомассы, анализируют электрофорезом в 15% полиакриламидном геле. Гель окрашивают Кумасси R-250 по стандартной методике и определяют относительное количество белка в полосе целевого продукта (результаты ПААГ-электрофореза приведены на Фиг. 4).
Штамм Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2Sv, отличающийся наибольшей продукцией целевого белка, задепонирован во Всероссийской коллекции промышленных микроорганизмов под номером Y-4513.
Как следует из приведенных примеров, заявляемое изобретение позволяет получить штамм-продуцент Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2Sv, производящий активную рекомбинантную фосфолипазу А2, секретируемую в питательную среду.
Перечень последовательностей
Оптимизированная нуклеотидная последовательность гена фосфолипазы А2 из Streptomyces violaceoruber
SEQ ID No:1
ATGGCTCCAGCAGATAAACCTCAAGTTTTGGCATCATTCACCCAG
ACATCCGCATCATCCCAGAACGCATGGTTGGCCGCTAACAGAAAC
CAATCTGCTTGGGCTGCTTACGAATTTGATTGGTCTACTGATTT
GTGTACTCAAGCTCCAGATAACCCTTTTGGTTTCCCATTCAACACT
GCTTGTGCTAGACATGATTTCGGTTACAGAAAC
TATAAGGCTGCTGGTTCTTTTGATGCTAACAAGTCCAGAATTGAT
TCTGCTTTCTATGAGGATATGAAGAGAGTCTGCACTGGTTATAC
CGGAGAGAAGAACACTGCCTGTAATTCCACTGCCTGGAC
CTACTATCAAGCCGTAAAGATTTTTGGTT
| название | год | авторы | номер документа |
|---|---|---|---|
| Получение гена фосфолипазы А2 с измененным оптимумом рН путем удаления сайтов гликозилирования | 2019 |
|
RU2766448C2 |
| Способ получения ферментного препарата фосфолипазы А2 с применением рекомбинантного штамма-продуцента Pichia pastoris X-33/ pPICZαA-PhoA2-StV | 2018 |
|
RU2676321C1 |
| Способ получения секреторного рекомбинантного белка ORF2 цирковируса свиней второго типа в дрожжах Komagataella phaffii BG11/ pPic-CV2-7 в форме вирусоподобных частиц | 2023 |
|
RU2832292C1 |
| Способ получения дрожжей K. kurtzmanii с улучшенной способностью к усвоению сорбитола | 2023 |
|
RU2814987C1 |
| Фосфолипаза А2 для экспрессии в дрожжах (варианты) | 2018 |
|
RU2716087C1 |
| Рекомбинантная плазмидная ДНК, способ ее конструирования и штамм дрожжей Komagataella pastoris - продуцент фрагмента белка VP2 вируса инфекционной бурсальной болезни птиц | 2021 |
|
RU2800472C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В ДРОЖЖАХ PICHIA PASTORIS ГЕНА ФОСФОЛИПАЗЫ, ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS - ПРОДУЦЕНТ ФОСФОЛИПАЗЫ | 2009 |
|
RU2409671C1 |
| Рекомбинантная плазмида, способ её конструирования и штамм дрожжей Komagataella pastoris - продуцент иммунного интерферона-гамма собаки | 2020 |
|
RU2756852C2 |
| ИНТЕГРАТИВНЫЙ ПЛАЗМИДНЫЙ ВЕКТОР ДЛЯ ЭКСПРЕССИИ ГЕНОВ В ДРОЖЖАХ | 2008 |
|
RU2388823C1 |
| Штамм Komagataella phaffii T07/pPZL-4x-OA-xyl-AsOr, обладающий способностью продуцировать ксиланазу из грибов вида Aspergillus oryzae | 2022 |
|
RU2796447C1 |
Изобретение относится к области промышленной биотехнологии, генетической инженерии и касается получения ауксотрофного штамма-реципиента для генно-инженерных работ, плазмидных векторов для экспрессии генов, штамма-продуцента фосфолипазы А2, который может быть использован пищевой промышленности при получении лизофосфолипидов и поверхностно-активных веществ с эмульгирующими свойствами. Заявлен способ получения штамма-продуцента фосфолипазы А2 Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2Sv, включающий в себя: создание интегративного плазмидного вектора pMAY-LEU2, используемой для получения ауксотрофного по лейцину штамма Komagataella phaffii (Pichia pastoris) YIB Δleu2, который содержит AOX1-промотор индуцибельного типа, ZeoR - ген устойчивости к зеоцину, LEU2 - сайты интеграции в ген отвечающий за синтез лейцина, FLP - ген FLP-рекомбиназы, а сконструированная плазмида pMAY-LEU2 имеет размер 9509 п.о., создание интегративного плазмидного вектора pSEA-LEU-PLA2Sv, для экспрессии генов в дрожжах, содержащего селективный маркер для отбора трансформантов в клетках E.coli, сайт начала репликации, сайт интеграции и экспрессионную кассету, в состав которой входят промотор, терминатор транскрипции и селективный маркер, а в качестве селективного маркера используют последовательность ДНК, кодирующую ген Leu2, создание дрожжевого штамма-реципиента Komagataella phaffii (Pichia pastoris) YIB Δleu2, на основе дикого штамма Y-3489, задепонированного в ВКПМ, а данный штамм ауксотрофный по лейцину получен трансформацией вектором pMAY-LEU2 по сайтам интеграции в ген LEU2 с последующим выщеплением последовательности, содержащей гены антибиотикорезистентности, создание на его основе штамма дрожжей Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2Sv, полученного трансформацией штамма Komagataella phaffii (Pichia pastoris) YIB Δleu2 плазмидным вектором pSEA-LEU-PLA2Sv, с последующим отбором на среде YNB; при этом штамм, обеспечивающий синтез рекомбинантной фосфолипазы А2, не содержащий генов антибиотикорезистентности, задепонирован в ВКПМ под номером Y-4513. 4 ил.
Способ получения штамма-продуцента фосфолипазы А2 Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2Sv, включающий в себя:
- создание интегративного плазмидного вектора pMAY-LEU2, используемой для получения ауксотрофного по лейцину штамма Komagataella phaffii (Pichia pastoris) YIB Δleu2, который содержит AOX1-промотор индуцибельного типа, ZeoR - ген устойчивости к зеоцину, LEU2 - сайты интеграции в ген, отвечающий за синтез лейцина, FLP - ген FLP-рекомбиназы, а сконструированная плазмида pMAY-LEU2 имеет размер 9509 п.о.,
- создание интегративного плазмидного вектора pSEA-LEU-PLA2Sv, для экспрессии генов в дрожжах, содержащего селективный маркер для отбора трансформантов в клетках E.coli, сайт начала репликации, сайт интеграции и экспрессионную кассету, в состав которой входят промотор, терминатор транскрипции и селективный маркер, а в качестве селективного маркера используют последовательность ДНК, кодирующую ген Leu2,
- создание дрожжевого штамма-реципиента Komagataella phaffii (Pichia pastoris) YIB Δleu2, на основе дикого штамма Y-3489, задепонированного в ВКПМ, а данный штамм ауксотрофный по лейцину получен трансформацией вектором pMAY-LEU2 по сайтам интеграции в ген LEU2 с последующим выщеплением последовательности, содержащей гены антибиотикорезистентности,
- создание на его основе штамма дрожжей Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2Sv, полученного трансформацией штамма Komagataella phaffii (Pichia pastoris) YIB Δleu2 плазмидным вектором pSEA-LEU-PLA2Sv, с последующим отбором на среде YNB; при этом штамм, обеспечивающий синтез рекомбинантной фосфолипазы А2, не содержащий генов антибиотикорезистентности, задепонирован в ВКПМ под номером Y-4513.
| Фосфолипаза А2 для экспрессии в дрожжах (варианты) | 2018 |
|
RU2716087C1 |
| RU 2009111842 A, 10.10.2010 | |||
| CN 104328095 A, 04.02.2015 | |||
| Способ получения ферментного препарата фосфолипазы А2 с применением рекомбинантного штамма-продуцента Pichia pastoris X-33/ pPICZαA-PhoA2-StV | 2018 |
|
RU2676321C1 |

