Группа заявляемых изобретений относится к биотехнологии, в частности к биосинтезу ксилоглюканазы, и представляет собой рекомбинантный штамм дрожжей Komagataella phaffii, несущий на хромосоме ген ксилоглюканазы из Myceliophthora thermophila ВКПМ F-244, содержащей замены (W324A/W325A), направленные на изменение способа действия фермента на субстрат с эндо-процессивного на эндо-диссоциативный, и способный синтезировать эндо-диссоциативную ксилоглюканазу, а также способ микробиологического синтеза эндо-диссоциативной ксилоглюканазы на основе заявленного штамма.
Лигноцеллюлозная биомасса является дешевым возобновляемым источником ферментируемых сахаров являющихся субстратами множества биосинтетических процессов. Ферментативный гидролиз - экологически безопасный способ биоконверсии растительной биомассы в сахара. Сложный полисахаридный состав растительных клеточных стенок, содержащих, помимо целлюлозы, множество разнообразных связующих гликанов, обуславливает необходимость использования широкого спектра ферментов для гидролиза.
Основным структурным полисахаридом первичной клеточной стенки всех двудольных и многих однодольных растений является разновидность бета-глюкана - ксилоглюкан (КГ). Наряду с другими связующими гликанами, он формирует аморфный матрикс, в который погружены микрофибриллы целлюлозы. Поэтому полный гидролиз целлюлозы невозможен без предварительного гидролиза полисахаридов матрикса, в том числе КГ.
Основная цепь молекулы КГ состоит из блоков целлотетраозы, соединенных между собой β-(1,4)-связями. В большинстве случаев каждые три из четырех остатков глюкозы в таком блоке связаны α-(1,6)-связями с остатками D-ксилозы. Некоторые из остатков глюкозы, в свою очередь, могут быть связаны β-(1,2)-связями с остатками D-галактозы или, реже, L-арабинозы. В КГ некоторых растений остатки D-галактозы могут быть связаны α-(1,2)-связями с остатками L-фукозы. Видо- и тканеспецифичность модификаций остатков ксилозы определяет разнообразие типов КГ в растениях.
Ферменты, специфически расщепляющие β-(1,4)-связи основной цепи молекулы КГ, называются ксилоглюканазами.
В соответствии с классификацией Хериссата, карбогидразы разделяются на семейства, объединенные общими структурно-функциональными свойствами, на основании сходства аминокислотной последовательностей (Henrissat and Bairoch, 1996). В настоящее время в базе данных CAZy находится более 140 семейств гликозил-гидролаз (http://www.Cazy.org). Ферменты, гидролизующие ксилоглюкан, обнаружены в семействах гликозил-гидролаз: GH5, GH7, GH12, GH16, GH26, GH44 и GH74. В отличие от других семейств, содержащих помимо ксилоглюканаз, ферменты с иной субстратной специфичностью, семейство GH74 содержит только ферменты, обладающие активностью на ксилоглюкане. В состав семейства GH74 входят:
1) целлюлазы (ЕС 3.2.1.4), расщепляющие β-(1,4)-D-глюкозидные связи в целлюлозе, лихенане и β-D-глюканах злаков;
2) целлобиогидролазы, специфичные к редуцирующим концам олигосахаридов (OXG-RCBH, КФ 3.2.1.150), отщепляющие целлобиозу с восстанавливающего конца молекулы КГ или его олигосахаридов;
3) ксилоглюкан-специфические эндо-β-1,4-глюканазы, ксилоглюканазы (ЕС 3.2.1.151), гидролизующие β-1,4-D-глюкозидные связи у незамещенного остатка глюкозы в основной цепи КГ.
Способ действия ксилоглюканаз на субстрат может быть эндо-процессивным и эндо-диссоциативным. В первом случае ферменты случайным образом прикрепляются к основной цепи молекулы КГ и делают первый разрез, после чего процессивным способом удаляют «строительные блоки» КГ (гептасахариды XXXG, октасахариды XXLG/XLXG и нонасахариды XLLG - классификация Фрая (Fry et al. 1993)) с конца одной из частей молекулы, гидролизуя β-1,4-связи у незамещенного остатка глюкозы. Эндо-диссоциативные ксилоглюканазы открепляются от молекулы КГ после каждого акта гидролиза и затем случайным образом присоединяются к ней снова. В результате действия на КГ эндо-процессивных ксилоглюканаз основными конечными продуктами гидролиза являются «строительные блоки» КГ (XXXG, XXLG, XLXG и XLLG). В процессе гидролиза такими ферментами общая длина полисахаридной цепи, а значит и вязкость растворов КГ, снижаются медленно. Конечными продуктами эндо-диссоциативных ксилоглюканаз являются олигоксилоглюканы различной длины, а вязкость растворов под действием таких ферментов уменьшается очень быстро. Поэтому эндо-диссоциативные ксилоглюканазы могут быть использованы в качестве кормовых добавок для снижения вязкости промышленных субстратов на основе лигноцеллюлозной биомассы, а также кормов для животных.
В источниках информации имеются сведения о микроорганизмах - природных продуцентах ксилоглюканаз: Geotrichum sp. (Yaoi and Mitsuishi, 2004), Aspergillus japonicus, Chrysosporium lucknowense, Trichoderma reesei (Grishutin et al., 2004; Markov et al., 2005; Qi et al., 2013; Lopes et al., 2021), Phanerochaete chrysosporium (Ishida et al., 2007), Cellvibrio japonicus (Attia et al., 2016), Aspergillus clavatus (Damasio et al., 2014), Sporotrichum thermophile, Myceliophthora thermophila и Sporotrichum pruinosum (Krestyanova et al., 2016), Paenibacillus odorifer (Gusakov et al., 2020). Методами селекции и радиационного мутагенеза получены штаммы, синтезирующие повышенные количества ксилоглюканаз: Penicillium verruculosum (RU 2361918), Penicillium funiculosum (RU 2323254), Aspergillus aculeatus (RU 2303057). Подобным образом был получен высокоактивный штамм мицелиального гриба Aspergillus foetidus - продуцента комплекса карбогидраз, содержащего ксилоглюканазу (RU 2323973 C1).
Описаны также полученные методами генной инженерии штаммы Escherichia coli - продуценты рекомбинантных ксилоглюканаз из Geotrichum sp. М128 (US 2004038367; Yaoi and Mitsuishi, 2002), Fusarium graminearum (Habrylo et al., 2012), Rhizomucor miehei (Song et al., 2013), Aspergillus cervinus (RU 2625013 C1), Xanthomonas citri (Feng et al., 2014), Xanthomonas campestris (de Araujo et al., 2013). В качестве реципиентов для гетерологичной экспрессии грибных ксилоглюканаз используют различные штаммы грибов: в Aspergillus oryzae экспрессированы ксилоглюканазы из Malbranchea cinnamomea (US 6500658), в Fusarium venenatum - ксилоглюканаза из Trichoderma reesei (US 2004067569 A1), в Penicillium canescens - ксилоглюканаза из Penicillium canescens (RU 2358756), в Aspergillus niger - ксилоглюканаза из Thielavia australiensis и собственная ксилоглюканаза (WO 2014138983; Hasper et al., 2002), в Aspergillus nidulans - ксилоглюканаза из Aspergillus terreus (Vitcosque et al., 2016).
Также для гетерологичной экспрессии грибных ксилоглюканаз применяют отличные от Е. coli бактериальные реципиенты, например, Acidothermus cellulolyticus использован для гетерологичной экспрессии ксилоглюканаз из Acremonium sp., Cladorrhinum foecundissimum, Humicola insolens, Thielavia terrestris, Trichoderma reesei, Aspergillus aculeatus, Aspergillus aculeatus, Aspergillus fumigatus, Myceliophthora thermophila (Vlasenko et al., 2010).
Описаны также полученные методами генной инженерии штаммы дрожжей Komagataella phaffii - продуценты рекомбинантных ксилоглюканаз из Phanerochaete chrysosporium (Ishida et al., 2007), Aspergillus cervinus ВКПМ F-612 (RU 2639248 C1) и эндоксилоглюканазы Penicillium oxalicum (CN 104388406 A; Xian et al., 2016). Помимо этого, разработан метод микробиологического синтеза в Komagataella phaffii термостабильной рекомбинантной ксилоглюканазы, кодируемой синтетическим геном на основе нуклеотидной последовательности ксилоглюканазы семейства GH74 из Myceliophthora thermophila ВКПМ F-244 (RU 2605629 C1).
Системы гетерологичной экспрессии на основе бактерий и микроскопических грибов имеют свои преимущества и недостатки. К недостаткам штаммов-продуцентов на основе микроскопических грибов родов Aspergillus и Penicillium относят невысокий уровень синтеза рекомбинантных ферментов по сравнению с E. coli и сложности с проведением генетических манипуляций на штаммах вследствие нестандартизированного и недостаточно разработанного генетического инструментария (Demain and Vaishnav, 2009). Метилотрофные дрожжи (Pichia, Komagataella, Hansenula) являются высокоэффективными продуцентами рекомбинантных белков. Преимуществом метилотрофных дрожжей является то, что их культуру можно выращивать в ферментерах до более высокой плотности, по сравнению с традиционно используемыми дрожжами Saccharomyces cerevisiae, что позволяет получить более высокий уровень продукции целевого белка (Gellissen, 2000). Метилотрофные дрожжи обладают одними из самых мощных в природе промоторов. В отличие от Е. coli, метилотрофные дрожжи обладают способностью ко всем характерным для эукариот посттрансляционным модификациям (гликозилирование, фосфорилирование, образование дисульфидных связей между цистеиновыми остатками, секреция рекомбинантных белков в культивационную среду). Низкий уровень секреции собственных белков существенно облегчает выделение и очистку целевого рекомбинантного продукта (Macauley-Patrick and Fazenda, 2005). Рекомбинантные белки в метилотрофных дрожжах, в отличие от Е. coli, не образуют телец включения.
Ближайшим аналогом заявляемого штамма является рекомбинантный штамм дрожжей Komagataella phaffii ВКПМ Y-4933 - продуцент секретируемой термостабильной ксилоглюканазы семейства GH74 из Myceliophthora thermophila ВКПМ F-244, кодируемой геном mt74sin, интегрированным в хромосому P. pastor is под контролем промотора АОХ1 (RU 2605629, Berezina et al. 2017). Рекомбинантный фермент MtXgh74 является ксилоглюкан-специфической эндо-β-1,4-глюканазой с эндо-процессивным способом действия на субстрат. Деградация полимерной молекулы ксилоглюкана происходит в результате последовательного отщепления ферментом MtXgh74 структурных гепта-, окта- и нонасахаридов ксилоглюкана путем гидролиза β-1,4-гликозидной связи у незамещенных остатков основной цепи полисахарида. В результате фермент MtXgh74 эффективно осахаривает субстрат, продуцируя короткие ксилоглюкан-олигосахариды, но недостаточно эффективно снижает вязкость растворов ксилоглюкана. Снижение динамической вязкости 1% раствора КГ в процессе его гидролиза ферментом MtXgh74 при 50°С составляет 1,79 мПа⋅с в пересчете на 0,01 мкмоль образовавшихся редуцирующих концов.
Ближайшим аналогом заявляемого способа микробиологического синтеза ксилоглюканазы является способ синтеза рекомбинантной секретируемой термостабильной ксилоглюканазы MtXgh74 при ферментации штамма дрожжей Komagataella phaffii ВКПМ Y-4269 (RU 2605629, Berezina et al., 2017). Активность ксилоглюканазы MtXgh74 в конце ферментации в культуральной жидкости ВКПМ Y-4269 составляла не менее 60 ед/мл.
Задача заявляемой группы изобретений состоит в разработке способа микробиологического синтеза ксилоглюканазы на основе рекомбинантного штамма Komagataella phaffii, сочетающего высокий уровень продукции гетерологичного фермента с эндо-диссоциативным способом действия на субстрат, обеспечивающим эффективное снижение вязкости растворов ксилоглюкана и предназначенного для снижения вязкости ксилоглюкан-содержащих субстратов.
Задачу решают путем:
- конструирования рекомбинантного штамма дрожжей Komagataella phaffii ВКПМ Y-4933 - продуцента секретируемой термостабильной ксилоглюканазы GH74, кодируемой мутированный геном mt74sin-mut, соответствующим SEQ ID NO 1,
- разработки способа микробиологического синтеза ксилоглюканазы на основе рекомбинантного штамма дрожжей Komagataella phaffii ВКПМ Y-4933.
Процесс конструирования заявляемого штамма состоит из следующих этапов:
- конструирования мутированного гена mt74sin-mut (SEQ ID NO 1) на основе нуклеотидной последовательности гена mt74sin, кодирующего мутантный вариант ксилоглюканазы семейства GH74 из Myceliophthora thermophila ВКПМ F-244.
- конструирования интегративной плазмидной ДНК (плазмиды) pPIC-mt74sin-mut, содержащей ген mt74sin-mut, являющийся мутантным вариантом гена ксилоглюканазы mt74sin.
- конструирования рекомбинантного штамма дрожжей Komagataella phaffii/mt74sin-mut, способного синтезировать ксилоглюканазу семейства GH74 с эндо-диссоциативным способом действия на субстрат.
- разработки способа микробиологического синтеза ксилоглюканазы на основе рекомбинантного штамма дрожжей Komagataella phaffii/mt74sin-mut, сочетающего высокий уровень синтеза гетерологичной ксилоглюканазы семейства GH74 с эндо-диссоциативным способом действия на субстрат.
Этап 1. Конструирование мутированного гена mt74sin-mut на основе нуклеотидной последовательности гена mt74sin, интегрированного в геном штамма Komagataella phaffii ВКПМ Y-4933 и кодирующего мутантную форму ксилоглюканазы MtXgh74 с эндо-диссоциативным способом действия на субстрат.
Ксилоглюканаза MtXgh74 является эндо-процессивным ферментом. Для изменения способа действия фермента на субстрат с эндо-процессивного на эндо-диссоциативный, конструируют мутантную форму MtXgh74-mut (W324A/W325A). Дизайн нуклеотидной последовательности mt74sin-mut разрабатывают на основании гена mt74sin (GenBank: KY549923.1), интегрированного в геном штамма Komagataella phaffii ВКПМ Y-4933 и кодирующего ксилоглюканазу MtXgh74 из Myceliophthora thermophila ВКПМ F-244 (GenBank: ARR73955.1), (RU 2605629, Berezina et al., 2017). Введение мутаций в ген mt74syn осуществляли методом перекрывающихся ПЦР-продуктов (Overlap PCR) с использованием праймеров и гена mt74syn в качестве матрицы. Для проведения мутагенеза использованы следующие праймеры: F_mtgh74syn_BgIII - AAGAGATCTGGTACCGCTACGGC, R_mtgh74syn_XhoI - GCCCCTCGAGTTATTAATTGGCAGC, SP_II (VATLNSAA) - TAATAGCGCTGCTCCAGATGCCCAGATT, ASP_II (VATLNSAA) - ATCTGGAGCAGCGCTATTAAGGGTAG.
В результате получают фрагмент ДНК, содержащий мутированный ген mt74sin-mut. Корректность введенной мутации подтверждают при помощи секвенирования методом Сэнгера.
Этап 2. Конструирование интегративной плазмидной конструкции pPIC-mt74sin-mut
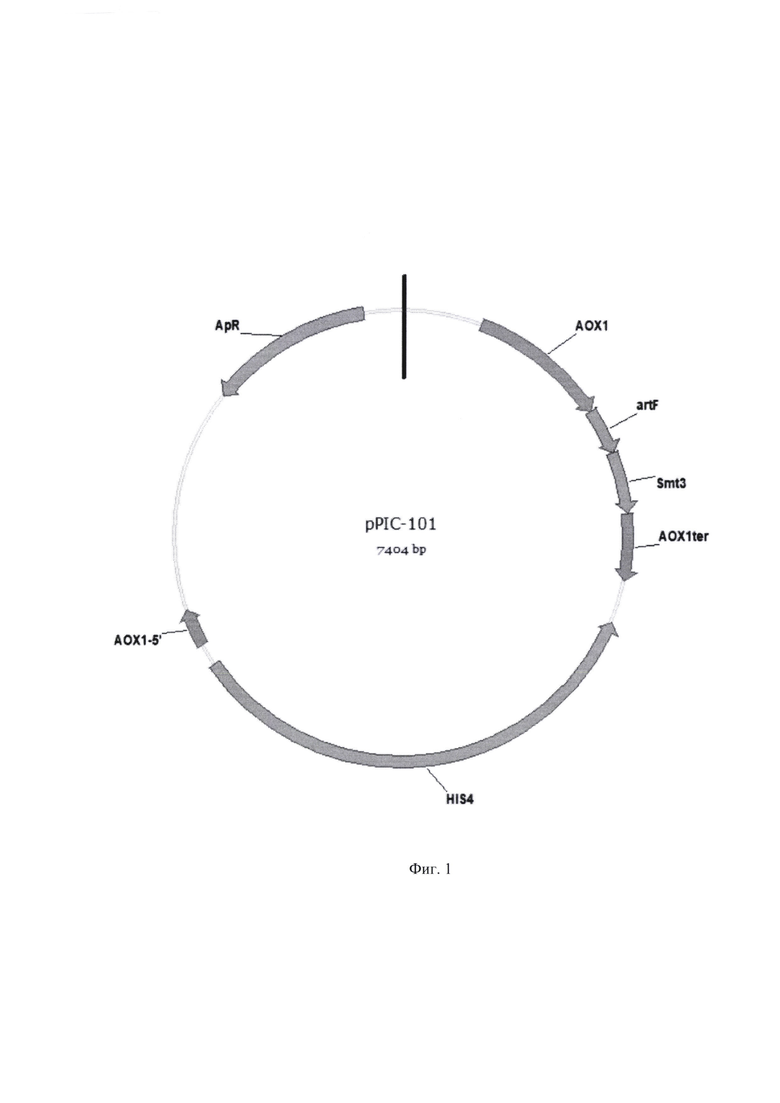
Плазмиду pPIC-mt74sin-mut конструируют путем клонирования полученного на этапе 1 фрагмента ДНК, содержащего мутированнвый ген mt74sin-mut, в вектор pPIC-101 разработанный для интеграции в штаммы Komagataella phaffii (his 4, mut+). Вектор pPIC-101 размером 7404 пары оснований содержит промотор АОХ1; терминатор АОХ1; кодирующую область гена HIS4; кодирующую область гена bla, обеспечивающего устойчивость штаммов Е. coli к ампициллину; репликон pBR-322; последовательность, кодирующую сигнальный пептид α-фактор дрожжей.
Плазмида pPIC-mt74sin-mut размером 9285 пар оснований, наряду с генами вектора pPIC-101, содержит кодирующую область мутированного гена mt74sin-mut ксилоглюканазы под контролем промотора АОХ1. (Фиг. 2).
Этап 3. Получение рекомбинантного штамма Komagataella phajfii/mt74sin-mut
В качестве штамма-реципиента используют штамм Komagataella phaffii (his 4, mut+), не синтезирующий ксилоглюканазу. Компетентные клетки данного штамма трансформируют плазмидой pPIC-mt74sin-mut. В результате получают рекомбинантный штамм Komagataella phaffii/mt74sin-mut, способный синтезировать ксилоглюканазу семейства GH74 с эндо-диссоциативным способом действия на субстрат.
Штамм Komagataella phaffii/mt74sin-mut депонирован во Всероссийской коллекции промышленных микроорганизмов как Komagataella phaffii ВКПМ Y-4933;
Заявляемый штамм Komagataella phaffii ВКПМ Y-4933 имеет следующие морфологические и физиолого-биохимические характеристики:
Морфологические признаки
При культивировании при температуре 28°С в течение 48 часов на агаризованной среде YPD следующего состава (мас.%):
пептон - 2
дрожжевой экстракт - 1
глюкоза - 2
агар - 2
вода - остальное.
Клетки имеют овальную форму, 3-4 мкм в диаметре; почкуются; почкование истинное, многостороннее; истинного мицелия не образуют. На агаризованной среде YPD формируются колонии светло-бежевого цвета с ровным краем, матовой поверхностью, линзовидным профилем и пастообразной консистенцией. При росте в жидкой среде YPD при 28°С в течение 24 ч культивирования - жидкость мутная, осадок белый, коагуляции не наблюдается, пристеночных пленок культура не образует.
Физиолого-биохимические признаки
Штамм является факультативным анаэробом. Температура роста - 20-33°С (оптимум - 28°С). рН среды культивирования - 4,8-7,4 (оптимум - 6,0). В качестве источников углерода штамм может использовать глюкозу, глицерин, метанол, олеат, сорбитол, рамнозу. Не утилизирует галактозу, ксилозу, арабинозу. В качестве источников азота штамм может использовать аминокислоты, сернокислый аммоний, азотнокислый аммоний. Существенными признаками штамма является отсутствие потребности в гистидине. Штамм Komagataella phaffii ВКПМ Y-4933 синтезирует рекомбинантную секретируемую ксилоглюканазу с эндо-диссоциативным способом действия на субстрат.
Способ микробиологического синтеза ксилоглюканазы в общем виде
Посевной материал, представляющий собой клетки рекомбинантного штамма-продуцента, подготавливают путем инкубации в течение 15-24 часов при температуре 29°С на среде YPD при постоянной аэрации на термостатируемой качалке (250 об/мин). Затем выросшую культуру переносят в соотношении 1:200 (по объему) в среду YPgM следующего состава (мас.%):
пептон - 2
дрожжевой экстракт - 1
глицерин 0,5
метанол - 0,5
вода - остальное.
Процесс биосинтеза ведут в колбах Эрленмейера, содержащих 100 мл среды YPgM, в течение 62 часов в ротационном шейкере-термостате (250 об/мин), при температуре 28°С. Каждые 24 часа проводят индукцию метанолом, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%. По истечении 62 часов биомассу отделяют центрифугированием. Наличие рекомбинантной ксилоглюканазы в культуральной среде определяют при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия с последующей зимограммой (Маниатис и др., 1984). Количественную оценку специфической активности рекомбинантной ксилоглюканазы осуществляют при помощи ДНС-метода (Miller, 1959).
Уровень синтеза ксилоглюканазы заявляемым способом составляет не менее 60 ед/мл культуральной жидкости, что сопоставимо с ближайшим аналогом.
Изобретение проиллюстрировано следующими фигурами графических изображений:
Фиг. 1 - Схема плазмидной ДНК pPIC-101. Плазмидный вектор pPIC-101: промотор АОХ1; терминатор АОХ1; ген HIS4; ген bla, обеспечивающий устойчивость к ампициллину (ApR).
Фиг. 2 - Схема рекомбинантной плазмидной ДНК pPIC-mt74sin-mut. Плазмидный вектор pPIC-mt74sin-mut: промотор АОХ1; терминатор АОХ1; ген HIS4; ген bla, обеспечивающий устойчивость к ампициллину (ApR); mt74syn-mut.
Пример 1. Конструирование рекомбинантной плазмидной ДНК pPIC-mt74sin-mut, содержащей синтетический ген mt74sin-mut, кодирующий ксилоглюканазу семейства GH74 с эндо-диссоциативным способом действия на субстрат
Все стандартные генно-инженерные и микробиологические манипуляции проводят по известным методикам (Маниатис с соавт, 1984).
Синтетический ген mt74sin-mut (SEQ ID NO 1) получают путем введения мутаций в ген mt74syn методом перекрывающихся ПЦР-продуктов.
Плазмиду pPIC-mt74sin-mut конструируют путем клонирования фрагмента, содержащего ген mt74sin-mut в интегративный вектор pPIC-101 (Фиг. 1).
Гидролиз плазмиды pPIC-101 и фрагмента ДНК, содержащего ген mt74sin-mut, проводят рестриктазами BglII и XhoI (Fermentas, Латвия). Реакционную смесь очищают набором для выделения ДНК из реакционных смесей (Биосилика, Россия) согласно инструкции производителя. Лигирование проводят в реакционной смеси объемом 20 мкл, содержащей 20 нг ДНК вектора, 1,5 нг ДНК фрагмента и 5 ед. Т4 ДНК лигазы (Fermentas, Латвия), согласно методике производителя. Полученной лигазной смесью в количестве 5 мкл трансформируют компетентные клетки штамма Е. coli XL1 blue MRF' (Stratagene, США) (генотип - Δ(mcrΔ)183 Δ(mcrCB-hsdSMR-mrr)173 end A1 supE44 thi-1 recA1 gyrA96 relA1 lac [F' proAB lacIqZΔM15 Tn10 (Tetr)]). Плазмидную ДНК полученных трансформантов анализируют путем гидролиза рестриктазами BglII и XhoI. В результате отбирают клоны, содержащие BglII/XhoI фрагменты размером, равным исходному фрагменту вставки. Правильность клонирования подтверждают секвенированием каждой из рекомбинантных вставок методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology, США). В результате получают рекомбинантную плазмиду pPIC-mt74sin-mut размером 9285 пар оснований, содержащую наряду с генами вектора pPIC-101, также мутированный ген mt74sin-mut, кодирующий секретируемую ксилоглюканазу семейства GH74 с эндо-диссоциативным способом действия на субстрат (SEQ ID NO 2) (Фиг. 2).
Пример 2. Получение заявляемого штамма Komagataella phaffii ВКПМ Y-4933 - продуцента эндо-диссоциативной ксилоглюканазы семейства GH74
С целью получения рекомбинантного штамма Komagataella phaffii ВКПМ Y-4933 - продуцента ксилоглюканазы, клетки штамма Komagataella phaffii GS115 (his 4, mut+) (Invitrogen) трансформируют плазмидой pPIC-mt74sin-mut. Трансформацию проводят 5 мг линеаризованной плазмидной ДНК, для получения которой исходную плазмиду обрабатывают эндонуклеазой рестрикции MluI.
Культуру дрожжей штамма Komagataella phaffii GS115 (his 4, mut+) выращивают на среде YPD, в аэробных условиях (250 об/мин) до оптической плотности 1-2 единицы. Клетки биомассы, отделенные центрифугированием при 3000g в течение 5 минут, ресуспендируют в буфере, содержащем 1 мМ дитиотрейтол и 100 мМ HEPES, затем инкубируют на шейкере в течение 30 минут и после двукратного промывания холодной деионизированной водой ресуспендируют в 1 мл 1 М сорбитола. Суспензию разделяют на аликвоты по 40 мкл. Электропорацию проводят на приборе GenePulser Xcell (Biorad) в кюветах с зазором 2 мм при 1500 В, 25 мкФ, 600 Ом. После электропорации быстро добавляют 1 мл холодного раствора 1 М сорбитола, инкубируют 1 час при температуре 30°С и высевают на селективную среду для отбора трансформантов. Отбор трансформантов проводят на минимальной среде М9, содержащей (мас.%):
КН2РО4 - 0,1
MgSO4 - 0,05
NaCl - 0,01
CaCl2 - 0,01
(NH4)2SO4 - 0,35
глюкоза - 2
тиамин - 0,02
рибофлавин - 0,02
никотиновая кислота - 0,02
п-аминобензойная кислота - 0,02
пантотенат кальция - 0,02
биотин - 0,0002
пиридоксин - 0,02
инозит - 1
фолиевая кислота - 0,02
вода - остальное.
Клетки инкубируют при 29°С в течение 3 суток. Трансформанты выявляют методом ПЦР-скрининга. В результате получают заявляемый штамм Komagataella phaffii ВКПМ Y-4933, продуцирующий секретируемую ксилоглюканазу с эндо-диссоциативным способом действия на субстрат.
Пример 3. Микробиологический синтез термостабильной секретируемой ксилоглюканазы заявляемым штаммом Komagataella phaffii ВКПМ Y-4933
Посевной материал выращивают путем инкубации клеток штамма Komagataella phaffii ВКПМ Y-4933 в течение 15-24 часов при температуре 29°С на среде YPD при постоянной аэрации (250 об/мин). Среду YPgM засевают подготовленным посевным материалом и ведут процесс биосинтеза в аэробных условиях при температуре 28°С, проводя индукцию метанолом.
Уровень синтеза ксилоглюканазы заявляемым способом составляет не менее 60 Ед/мл, что сопоставимо с таковым у ближайшего аналога - штамма Komagataella phaffii ВКПМ Y-4269. При инкубации препарата фермента в течение часа при 50°С, рН 5.0 сохраняется 100% ферментативной активности, что не уступает ближайшему аналогу.
Конечными продуктами гидролиза КГ ферментом MtXgh74-mut являются олигоксилоглюканы различной длины, в то время как конечными продуктами гидролиза КГ ближайшим аналогом MtXgh74 являются «строительные блоки» КГ (XXXG, XXLG, XLXG и XLLG).
Кинетические константы Vmax и kcat для MtXgh74-mut составляют 76 Ед/мг белка и 97 с-1, что превышает данные показатели для ближайшего аналога, ксилоглюканазы MtXgh74 (44 Ед/мг белка и 56 с-1, соответственно). Кинетическая константа Km для MtXgh74-mut и ближайшего аналога MtXgh74 составляет 24 и 0,45 мг КГ в мл, соответственно, что объясняет различие в способе действия ферментов на субстрат: эндо-диссоциатитвный для MtXgh74-mut и эндо-процессивный для MtXgh74.
Снижение динамической вязкости 1% раствора КГ в процессе его гидролиза эндо-диссоциативной ксилоглюканазой MtXgh74-mut при 50°С составляет 3,55 мПа⋅с в пересчете на 0,01 мкмоль образовавшихся редуцирующих концов, что в два раза превышает это значение для ближайшего аналога - эндо-процессивной ксилоглюканазы MtXgh74 (1,79 мПа⋅с в пересчете на 0,01 мкмоль образовавшихся редуцирующих концов).
Таким образом, получен рекомбинантный штамм дрожжей Komagataella phaffii ВКПМ Y-4933, способный к биосинтезу секретируемой эндо-диссоциативной ксилоглюканазы семейства GH74, кодируемой мутированным геном. При культивировании заявляемым способом уровень синтеза целевого продукта у заявляемого штамма сопоставим с таковым у ближайшего аналога, но в отличие от ближайшего аналога, продуцируемый фермент является является эндо-диссоциативной ксилоглюканазой и эффективно снижает вязкость растворов ксилоглюкана.
Источники информации
1. Berezina OV, Herlet J, Rykov SV, Kornberger P, Zavyalov A, Kozlov D, Sakhibgaraeva L, Krestyanova I, Schwarz WH, Zverlov VV, Liebl W, Yarotsky SV. Thermostable multifunctional GH74 xyloglucanase from Myceliophthora thermophila: high-level expression in Komagataella phaffii and characterization of the recombinant protein. Appl Microbiol Biotechnol. 2017 Jul; 101 (14): 5653-5666. doi: 10.1007/s00253-017-8297-2.
2. RU 2605629 // Рекомбинантный штамм дрожжей Komagataella phaffii - продуцент секретируемой термостабильной ксилоглюканазы, кодируемой синтетическим геном, и способ микробиологического синтеза секретируемой термостабильной ксилоглюканазы на основе этого штамма // Дата приоритета: 16.12.2015.
3. Маниатис Т., Фрич Э., Сэмбрук Дж., Молекулярное клонирование, М.: Мир, 1984; Клонирование ДНК. Методы. Под ред. Д. Гловера, Пер. с англ., Москва, Мир, 1988.
4. Fry SC, York WS, Albersheim P, Darvill A, Hayashi T, Joseleau JP, Kato Y, Lorences EP, Maclachlan GA, McNeil M, Mort AJ, Reid JSG, Seitz HU, Selvendran RR, Voragen AGJ, White AR (1993) An unambiguous nomenclature for xyloglucan-derived oligosaccharides. Physiol Plant, 89 (1): 1-3. doi: 10.1111/j.1399-3054.1993.tb01778.x.
5. Demain AL, Vaishnav P. // Production of recombinant proteins by microbes and higher organisms // Biotechnol Adv., 2009.
6. Grishutin S.G., Gusakov A.V., Markov A.V., Ustinov B.B., Semenova M.V., Sinitsyn A.P. // Specific xyloglucanases as a new class of polysaccharide-degrading enzymes.// Biochim Biophys Acta, 2004.
7. Habrylo O., Song X., Forster A., Jeltsch J.M., Phalip V. // Characterization of the four GH12 Endoxylanases from the plant pathogen Fusarium graminearum. // J. Microbiol Biotechnol., 2012.
8. Hasper A.A., Dekkers E., van Mil M., van de Vondervoort P.J., de Graaff L.H. // EglC, a new endoglucanase from Aspergillus niger with major activity towards xyloglucan. // Appl Environ Microbiol., 2002.
9. Ishida Т., Yaoi K., Hiyoshi A., Igarashi K., Samejima M. // Substrate recognition by glycoside hydrolase family 74 xyloglucanase from the basidiomycete Phanerochaete chrysosporium. // FEBS J., 2007.
10. Markov A.V., Gusakov A.V., Kondratyeva E.G., Okunev O.N., Bekkarevich A.O., Sinitsyn A.P. // New effective method for analysis of the component composition of enzyme complexes from Trichoderma reesei. // Biochemistry (Mosc), 2005.
11. Miller, G.L. // Use of dinitrosalicylic acid reagent for determination of reducing sugar // Analytical Chemistry. 1959.
12. Qi H., Bai F., Liu A.// Purification and characteristics of xyloglucanase and five other cellulolytic enzymes from Trichoderma reesei QM9414. // Biochemistry (Mosc), 2013.
13. Yaoi K., Mitsuishi Y.// Purification, characterization, cloning, and expression of a novel xyloglucan-specific glycosidase, oligoxyloglucan reducing end-specific cellobiohydrolase.// J Biol Chem., 2002.
14. Yaoi K., Mitsuishi Y. // Purification, characterization, cDNA cloning, and expression of a xyloglucan endoglucanase from Geotrichum sp. M128. // FEBS Lett., 2004.
15. Song S., Tang Y., Yang S., Yan Q., Zhou P., Jiang Z. // Characterization of two novel family 12 xyloglucanases from the thermophilic Rhizomucor miehei. // Appl Microbiol Biotechnol., 2013.
16. Vlasenko E.,  M., Cherry J., Xu F. // Substrate specificity of family 5, 6, 7, 9, 12, and 45 endoglucanases. // Bioresour Technol., 2010.
M., Cherry J., Xu F. // Substrate specificity of family 5, 6, 7, 9, 12, and 45 endoglucanases. // Bioresour Technol., 2010.
17. Macauley-Patrick S, Fazenda ML (2005) Heterologous protein production using the Komagataella phaffii expression system. Yeast 22: 249-270.
18. Gellissen G (2000) Heterologous protein production in methylotrophic yeasts. Appl. Microbiol. Biotechnol. 54(6): 741-750.
19. RU 2358756 // Способ получения ферментного препарата для расщепления гемицеллюлозных гетерополисахаридов клеточной стенки растений и ферментный препарат (варианты) // Дата приоритета: 26.11.2007.
20. RU 2303057 // Штамм мицелиального гриба Aspergillus aculeatus - продуцент комплекса карбогидраз, содержащего ксиланазы, бета-глюканазы, пектиназы и ксилоглюканазы // Дата приоритета: 14.11.2005.
21. RU 2323254 // Штамм мицелиального гриба Penicillium funiculosum - продуцент комплекса карбогидраз, содержащего целлюлазы, глюканазы, глюкозидазы, ксиланазы и ксилоглюканазы, и способ получения ферментного препарата комплекса карбогидраз для осахаривания лигноцеллюлозных материалов // Дата приоритета: 04.04.2006.
22. RU 2361918 // Штамм мицелиального гриба Penicillium verruculosum - продуцент комплекса целлюлаз, ксиланазы и ксилоглюканазы и способ получения ферментного препарата комплекса целлюлаз, ксиланазы и ксилоглюканазы для гидролиза целлюлозы и гемицеллюлозы // Дата приоритета: 26.02.2008.
23. US 6500658 // Xyloglucanase from Malbranchea // Priority date: US 20000653778 20000901; WO 2000DK00450 20000811; US 19990149397 P 19990817.
24. US 2004038367 // Novel xyloglucan oligosaccharide-degrading enzyme, polynucleotide encoding the enzyme, and method of preparing the enzyme // Priority date: JP 20020083433 20020325.
25. US 2004067569 A1 // Polypeptides having xyloglucanase activity and nucleic acids encoding same // Priority date: US 20030420191 20030418; US 20020373987 P 20020419.
26. WO 2014138983 A1 // Novel cell wall deconstruction enzymes of Malbranchea cinnamomea, Thielavia australiensis, and Paecilomyces byssochlamydoides, and uses thereof // Priority date: US 201361783222 P 20130314; US 201361783313 P 20130314; US 201361783485 P 20130314.
--->
Перечень последовательностей
<110> Федеральное государственное бюджетное учреждение "Государственный
научно-исследовательский институт генетики и селекции промышленных
микроорганизмов Национального Исследовательского Центра
«Курчатовский Институт»
<120> Рекомбинантный штамм дрожжей Komagataella phaffii – продуцент
секретируемой эндо-диссоциативной ксилоглюканазы семейства GH74,
кодируемой мутированным геном, и способ микробиологического синтеза
секретируемой эндо-диссоциативной ксилоглюканазы на основе этого штамма
<160> 2
<170> PatentIn version 3.1
<210> 1
<211> 2336
<212> DNA
<213> Artificial sequence: mt74syn-mut
<220>
<221> gene
<222> (37)..(2242)
<223> mt74syn-mut
<400> 1
agtgcaccaa ttggtcgacc tcgagttaat taacgtaaga tctggtaccg ctacggcttg 60
gaagaatgtc aacattggag gcggtggtgg ttttgtccca ggtataatct ttcaccctaa 120
agagaagggt gttgcctatg ctagaactga cataggaggc ttgtacagac tgaacccaga 180
tgatgactct tggacaccat tgacggataa tttgggcact aatgagaagt ggggaagatg 240
gggtattgac gcagttgcag ttgatccgca agatgctgac agagtctacg ctgctgtagg 300
catgtataca aacgattggg accccaatcc aggctctatt atccgtagtt ctgatcgagg 360
agctacatgg gaggttaccg aattaccctt caaagtggga ggtaatatgc ctggaagagg 420
tatgggtgag agattggctg tagatcctgc caacagtgac atcttgttct ttggtgctag 480
gtctggtaat ggtctgtgga agtctgcaga cggaggtgtt acttgggcaa gagtagaatc 540
cttcaccaat gtcggaactt atgctcctga cccatctgat gctaccggtt tgaactctga 600
cttgataggt ctgacttttg ttaccttcga ttcgacttca aacgttgttg gtggagcaac 660
ttcccgtata tttgtcggta ctgcagacaa caagactgca tcggtttacg tttccgagga 720
tgctggagcc acatggaaac cagtagaagg acaaccaggt gcattctttc ctcacaaatg 780
cgtgttgcaa ccagaggaaa aggccctata tctatcctat tctaatggag ctggccctta 840
tgacggaacg ttgggagctg tctaccgata cgatttggct gcacgaacct ggacggacat 900
cacaccagct tcaggagggg acctttactt tgggttcggt ggactatcag ttgacttgag 960
gaaacccggt actcttatgg tagctaccct taatagctgg tggccagatg cccagattta 1020
cagaagtacg gattcgggcg ccacatggag taagatttgg gagtgggctg cttacccgga 1080
tatgaactgg tactacggtt tgtacacaga caaagcccct tggataaacg ccggtttcat 1140
ctctcaggac accaaaagac tgggttggat gattgaggca ttagagattg acccacatga 1200
ttccgatcac tggctatatg gcaccggtct aactttgtat ggtggtcatg accttaccaa 1260
atgggataca gttacaagga acgttactat ttccagcctt gctgctggaa ttgaagaaat 1320
ggctgtctta ggcttagcat ctccacctaa tggaagtgaa ctgctggctg ctgttggtga 1380
tgactgtgga ttcacttttc gtgaatctca ggatttaggt acgtcaccgc aaaccccttg 1440
gatgaatcct atttggactt caactactga tgtggattat gccggaaacg aacccgacca 1500
tgttgtgaga gtcggtaatt ctcaaggtgc acctcaagta gctgtttcgg aagatggtgg 1560
agtaacttgg tcagcacatc caggggcaga cgggaccacc aacagtggaa ctttagcata 1620
ttcagccgat gccgatacta tcgtctggag ttcaggttcc gcaggggtgc tgaggtccca 1680
aaaccaggga gcttttgctg ctgtcgggtc tttgccaagc ggcgccacag tggctgccga 1740
tagaagaaat aacacagtct tttacgctgc aagtggagct tccttttaca gatccactga 1800
cactggcgcc acatttgcca aagttgccag cgcctttgga tcgaaggttg ccgctgtcaa 1860
ggctattgct gcacatcccg tggttgctgg agaagtatgg gttgccactg acgctggatt 1920
gttccgttcc atcgattatg gggccacttt ctctgctgct tctggttcca ttaccgatgc 1980
aattcaggtg tctttaggaa aaggagatgg ttcagcttgg aacgtgtacg tatttgggac 2040
cggtcctgaa gggaccaagt tgtatgcaag cgctgatgaa ggtgccacat gggtggatat 2100
ccaaggagag cagggcttcg gttctttgtc agctaacaga cttgttggtt caggaaatgt 2160
ggctggtcaa gtctacgttg gcacaaatgg tagaggtgtt ttctatgcaa aggttagtct 2220
ttctgctgct gccaattaat aactcgaggt gggcgatcgc tctagagcta gcgaattcct 2280
gtgtgaaatt gttatccgct cacaattcca cacaacatac gagccggaag cataaa 2336
//
<210> 2
<211> 9285
<212> DNA
<213> Artificial sequence: pPIC-mt74sin-mut
<220>
<221> promoter
<222> (6100)..( 6834)
<223> AOX1
<224> gene
<225> (585)..(3119)
<226> HIS4, complement
<227> terminator
<228> (6)..( 356)
<229> AOX1ter
<230> gene
<231> (4627)..( 5487)
<232> ApR, complement
<233> gene
<234> (7071)..( 9284)
<235> mt74sin-mut, complement
<400> 1
tcgaggggcc gcgaattaat tcgccttaga catgactgtt cctcagttca agttgggcac 60
ttacgagaag accggtcttg ctagattcta atcaagagga tgtcagaatg ccatttgcct 120
gagagatgca ggcttcattt ttgatacttt tttatttgta acctatatag tataggattt 180
tttttgtcat tttgtttctt ctcgtacgag cttgctcctg atcagcctat ctcgcagctg 240
atgaatatct tgtggtaggg gtttgggaaa atcattcgag tttgatgttt ttcttggtat 300
ttcccactcc tcttcagagt acagaagatt aagtgagaag ttcgtttgtg caagctatcg 360
tccattccga cagcatcgcc agtcactatg gcgtgctgct agcgctatat gcgttgatgc 420
aatttctatg cgcacccgtt ctcggagcac tgtccgaccg ctttggccgc cgcccagtcc 480
tgctcgcttc gctacttgga gccactatcg actacgcgat catggcgacc acacccgtcc 540
tgtggatcta tcgaatctaa atgtaagtta aaatctctaa ataattaaat aagtcccagt 600
ttctccatac gaaccttaac agcattgcgg tgagcatcta gaccttcaac agcagccaga 660
tccatcactg cttggccaat atgtttcagt ccctcaggag ttacgtcttg tgaagtgatg 720
aacttctgga aggttgcagt gttaactccg ctgtattgac gggcatatcc gtacgttggc 780
aaagtgtggt tggtaccgga ggagtaatct ccacaactct ctggagagta ggcaccaaca 840
aacacagatc cagcgtgttg tacttgatca acataagaag aagcattctc gatttgcagg 900
atcaagtgtt caggagcgta ctgattggac atttccaaag cctgctcgta ggttgcaacc 960
gatagggttg tagagtgtgc aatacacttg cgtacaattt caacccttgg caactgcaca 1020
gcttggttgt gaacagcatc ttcaattctg gcaagctcct tgtctgtcat atcgacagcc 1080
aacagaatca cctgggaatc aataccatgt tcagcttgag acagaaggtc tgaggcaacg 1140
aaatctggat cagcgtattt atcagcaata actagaactt cagaaggccc agcaggcatg 1200
tcaatactac acagggctga tgtgtcattt tgaaccatca tcttggcagc agtaacgaac 1260
tggtttcctg gaccaaatat tttgtcacac ttaggaacag tttctgttcc gtaagccata 1320
gcagctactg cctgggcgcc tcctgctagc acgatacact tagcaccaac cttgtgggca 1380
acgtagatga cttctggggt aagggtacca tccttcttag gtggagatgc aaaaacaatt 1440
tctttgcaac cagcaacttt ggcaggaaca cccagcatca gggaagtgga aggcagaatt 1500
gcggttccac caggaatata gaggccaact ttctcaatag gtcttgcaaa acgagagcag 1560
actacaccag ggcaagtctc aacttgcaac gtctccgtta gttgagcttc atggaatttc 1620
ctgacgttat ctatagagag atcaatggct ctcttaacgt tatctggcaa ttgcataagt 1680
tcctctggga aaggagcttc taacacaggt gtcttcaaag cgactccatc aaacttggca 1740
gttagttcta aaagggcttt gtcaccattt tgacgaacat tgtcgacaat tggtttgact 1800
aattccataa tctgttccgt tttctggata ggacgacgaa gggcatcttc aatttcttgt 1860
gaggaggcct tagaaacgtc aattttgcac aattcaatac gaccttcaga agggacttct 1920
ttaggtttgg attcttcttt aggttgttcc ttggtgtatc ctggcttggc atctcctttc 1980
cttctagtga cctttaggga cttcatatcc aggtttctct ccacctcgtc caacgtcaca 2040
ccgtacttgg cacatctaac taatgcaaaa taaaataagt cagcacattc ccaggctata 2100
tcttccttgg atttagcttc tgcaagttca tcagcttcct ccctaatttt agcgttcaac 2160
aaaacttcgt cgtcaaataa ccgtttggta taagaacctt ctggagcatt gctcttacga 2220
tcccacaagg tggcttccat ggctctaaga ccctttgatt ggccaaaaca ggaagtgcgt 2280
tccaagtgac agaaaccaac acctgtttgt tcaaccacaa atttcaagca gtctccatca 2340
caatccaatt cgatacccag caacttttga gttcgtccag atgtagcacc tttataccac 2400
aaaccgtgac gacgagattg gtagactcca gtttgtgtcc ttatagcctc cggaatagac 2460
tttttggacg agtacaccag gcccaacgag taattagaag agtcagccac caaagtagtg 2520
aatagaccat cggggcggtc agtagtcaaa gacgccaaca aaatttcact gacagggaac 2580
tttttgacat cttcagaaag ttcgtattca gtagtcaatt gccgagcatc aataatgggg 2640
attataccag aagcaacagt ggaagtcaca tctaccaact ttgcggtctc agaaaaagca 2700
taaacagttc tactaccgcc attagtgaaa cttttcaaat cgcccagtgg agaagaaaaa 2760
ggcacagcga tactagcatt agcgggcaag gatgcaactt tatcaaccag ggtcctatag 2820
ataaccctag cgcctgggat catcctttgg acaactcttt ctgccaaatc taggtccaaa 2880
atcacttcat tgataccatt attgtacaac ttgagcaagt tgtcgatcag ctcctcaaat 2940
tggtcctctg taacggatga ctcaacttgc acattaactt gaagctcagt cgattgagtg 3000
aacttgatca ggttgtgcag ctggtcagca gcatagggaa acacggcttt tcctaccaaa 3060
ctcaaggaat tatcaaactc tgcaacactt gcgtatgcag gtagcaaggg aaatgtcata 3120
cttgaagtcg gacagtgagt gtagtcttga gaaattctga agccgtattt ttattatcag 3180
tgagtcagtc atcaggagat cctctacgcc ggacgcatcg tggccggcat caccggcgat 3240
ctaacatcca aagacgaaag gttgaatgaa acctttttgc catccgacat ccacaggtcc 3300
attctcacac ataagtgcca aacgcaacag gaggggatac actagcagca gaccgttgca 3360
aacgcaggac ctccactcct cttctcctca acacccactt ttgccatcga aaaaccagcc 3420
cagttattgg gcttgattgg cacgcgtgag cttggcgtaa tcatggtcat agctgtttcc 3480
tgtgtgaaat tgttatccgc tcacaattcc acacaacata cgagccggaa gcataaagtg 3540
taaagcctgg ggtgcctaat gagtgagcta actcacatta attgcgttgc gctcactgcc 3600
cgctttccag tcgggaaacc tgtcgtgcca gctgcattaa tgaatcggcc aacgcgcggg 3660
gagaggcggt ttgcgtattg ggcgctcttc cgcttcctcg ctcactgact cgctgcgctc 3720
ggtcgttcgg ctgcggcgag cggtatcagc tcactcaaag gcggtaatac ggttatccac 3780
agaatcaggg gataacgcag gaaagaacat gtgagcaaaa ggccagcaaa aggccaggaa 3840
ccgtaaaaag gccgcgttgc tggcgttttt ccataggctc cgcccccctg acgagcatca 3900
caaaaatcga cgctcaagtc agaggtggcg aaacccgaca ggactataaa gataccaggc 3960
gtttccccct ggaagctccc tcgtgcgctc tcctgttccg accctgccgc ttaccggata 4020
cctgtccgcc tttctccctt cgggaagcgt ggcgctttct catagctcac gctgtaggta 4080
tctcagttcg gtgtaggtcg ttcgctccaa gctgggctgt gtgcacgaac cccccgttca 4140
gcccgaccgc tgcgccttat ccggtaacta tcgtcttgag tccaacccgg taagacacga 4200
cttatcgcca ctggcagcag ccactggtaa caggattagc agagcgaggt atgtaggcgg 4260
tgctacagag ttcttgaagt ggtggcctaa ctacggctac actagaagga cagtatttgg 4320
tatctgcgct ctgctgaagc cagttacctt cggaaaaaga gttggtagct cttgatccgg 4380
caaacaaacc accgctggta gcggtggttt ttttgtttgc aagcagcaga ttacgcgcag 4440
aaaaaaagga tctcaagaag atcctttgat cttttctacg gggtctgacg ctcagtggaa 4500
cgaaaactca cgttaaggga ttttggtcat gagattatca aaaaggatct tcacctagat 4560
ccttttaaat taaaaatgaa gttttaaatc aatctaaagt atatatgagt aaacttggtc 4620
tgacagttac caatgcttaa tcagtgaggc acctatctca gcgatctgtc tatttcgttc 4680
atccatagtt gcctgactcc ccgtcgtgta gataactacg atacgggagg gcttaccatc 4740
tggccccagt gctgcaatga taccgcgaga cccacgctca ccggctccag atttatcagc 4800
aataaaccag ccagccggaa gggccgagcg cagaagtggt cctgcaactt tatccgcctc 4860
catccagtct attaattgtt gccgggaagc tagagtaagt agttcgccag ttaatagttt 4920
gcgcaacgtt gttgccattg ctacaggcat cgtggtgtca cgctcgtcgt ttggtatggc 4980
ttcattcagc tccggttccc aacgatcaag gcgagttaca tgatccccca tgttgtgcaa 5040
aaaagcggtt agctccttcg gtcctccgat cgttgtcaga agtaagttgg ccgcagtgtt 5100
atcactcatg gttatggcag cactgcataa ttctcttact gtcatgccat ccgtaagatg 5160
cttttctgtg actggtgagt actcaaccaa gtcattctga gaatagtgta tgcggcgacc 5220
gagttgctct tgcccggcgt caatacggga taataccgcg ccacatagca gaactttaaa 5280
agtgctcatc attggaaaac gttcttcggg gcgaaaactc tcaaggatct taccgctgtt 5340
gagatccagt tcgatgtaac ccactcgtgc acccaactga tcttcagcat cttttacttt 5400
caccagcgtt tctgggtgag caaaaacagg aaggcaaaat gccgcaaaaa agggaataag 5460
ggcgacacgg aaatgttgaa tactcatact cttccttttt caatattatt gaagcattta 5520
tcagggttat tgtctcatga gcggatacat atttgaatgt atttagaaaa ataaacaaat 5580
aggggttccg cgcacatttc cccgaaaagt gccacctgac gtctaagaaa ccattattat 5640
catgacatta acctataaaa ataggcgtat cacgaggccc tttcgtctcg cgcgtttcgg 5700
tgatgacggt gaaaacctct gacacatgca gctcccggag acggtcacag cttgtctgta 5760
agcggatgcc gggagcagac aagcccgtca gggcgcgtca gcgggtgttg gcgggtgtcg 5820
gggctggctt aactatgcgg catcagagca gattgtactg agagtgcacc atatgcggtg 5880
tgaaataccg cacagatgcg taaggagaaa ataccgcatc aggcgccatt cgccattcag 5940
gctgcgcaac tgttgggaag ggcgatcggt gcgggcctct tcgctattac gccagctggc 6000
gaaaggggga tgtgctgcaa ggcgattaag ttgggtaacg ccagggtttt cccagtcacg 6060
acgttgtaaa acgacggcca gtgaattaat tcacgcgtgc gctcattcca attccttcta 6120
ttaggctact aacaccatga ctttattagc ctgtctatcc tggcccccct ggcgaggttc 6180
atgtttgttt atttccgaat gcaacaagct ccgcattaca cccgaacatc actccagatg 6240
agggctttct gagtgtgggg tcaaatagtt tcatgttccc caaatggccc aaaactgaca 6300
gtttaaacgc tgtcttggaa cctaatatga caaaagcgtg atctcatcca agatgaacta 6360
agtttggttc gttgaaatgc taacggccag ttggtcaaaa agaaacttcc aaaagtcgcc 6420
ataccgtttg tcttgtttgg tattgattga cgaatgctca aaaataatct cattaatgct 6480
tagcgcagtc tctctatcgc ttctgaaccc cggtgcacct gtgccgaaac gcaaatgggg 6540
aaacacccgc tttttggatg attatgcatt gtctccacat tgtatgcttc caagattctg 6600
gtgggaatac tgctgatagc ctaacgttca tgatcaaaat ttaactgttc taacccctac 6660
ttgacagcaa tatataaaca gaaggaagct gccctgtctt aaaccttttt ttttatcatc 6720
attattagct tactttcata attgcgactg gttccaattg acaagctttt gattttaacg 6780
acttttaacg acaacttgag aagatcaaaa aacaactaat tattcgaagg atcaaaatga 6840
gattgttact gttgctgtta ttgctgttgc ctgcagcatt aggtgctcca gtcaacacta 6900
caacagaaga tgaaacggca caaattccgg ctgaagctgt catcggttac tcagatttag 6960
aaggggattt cgatgttgct gttttgccat tttccaacag cacaaataac gggttattgt 7020
ttataaatac tactattgcc agcattgctg ctaaagaaga aggggtatcc atggaaaaga 7080
gatctggtac cgctacggct tggaagaatg tcaacattgg aggcggtggt ggttttgtcc 7140
caggtataat ctttcaccct aaagagaagg gtgttgccta tgctagaact gacataggag 7200
gcttgtacag actgaaccca gatgatgact cttggacacc attgacggat aatttgggca 7260
ctaatgagaa gtggggaaga tggggtattg acgcagttgc agttgatccg caagatgctg 7320
acagagtcta cgctgctgta ggcatgtata caaacgattg ggaccccaat ccaggctcta 7380
ttatccgtag ttctgatcga ggagctacat gggaggttac cgaattaccc ttcaaagtgg 7440
gaggtaatat gcctggaaga ggtatgggtg agagattggc tgtagatcct gccaacagtg 7500
acatcttgtt ctttggtgct aggtctggta atggtctgtg gaagtctgca gacggaggtg 7560
ttacttgggc aagagtagaa tccttcacca atgtcggaac ttatgctcct gacccatctg 7620
atgctaccgg tttgaactct gacttgatag gtctgacttt tgttaccttc gattcgactt 7680
caaacgttgt tggtggagca acttcccgta tatttgtcgg tactgcagac aacaagactg 7740
catcggttta cgtttccgag gatgctggag ccacatggaa accagtagaa ggacaaccag 7800
gtgcattctt tcctcacaaa tgcgtgttgc aaccagagga aaaggcccta tatctatcct 7860
attctaatgg agctggccct tatgacggaa cgttgggagc tgtctaccga tacgatttgg 7920
ctgcacgaac ctggacggac atcacaccag cttcaggagg ggacctttac tttgggttcg 7980
gtggactatc agttgacttg aggaaacccg gtactcttat ggtagctacc cttaatagct 8040
ggtggccaga tgcccagatt tacagaagta cggattcggg cgccacatgg agtaagattt 8100
gggagtgggc tgcttacccg gatatgaact ggtactacgg tttgtacaca gacaaagccc 8160
cttggataaa cgccggtttc atctctcagg acaccaaaag actgggttgg atgattgagg 8220
cattagagat tgacccacat gattccgatc actggctata tggcaccggt ctaactttgt 8280
atggtggtca tgaccttacc aaatgggata cagttacaag gaacgttact atttccagcc 8340
ttgctgctgg aattgaagaa atggctgtct taggcttagc atctccacct aatggaagtg 8400
aactgctggc tgctgttggt gatgactgtg gattcacttt tcgtgaatct caggatttag 8460
gtacgtcacc gcaaacccct tggatgaatc ctatttggac ttcaactact gatgtggatt 8520
atgccggaaa cgaacccgac catgttgtga gagtcggtaa ttctcaaggt gcacctcaag 8580
tagctgtttc ggaagatggt ggagtaactt ggtcagcaca tccaggggca gacgggacca 8640
ccaacagtgg aactttagca tattcagccg atgccgatac tatcgtctgg agttcaggtt 8700
ccgcaggggt gctgaggtcc caaaaccagg gagcttttgc tgctgtcggg tctttgccaa 8760
gcggcgccac agtggctgcc gatagaagaa ataacacagt cttttacgct gcaagtggag 8820
cttcctttta cagatccact gacactggcg ccacatttgc caaagttgcc agcgcctttg 8880
gatcgaaggt tgccgctgtc aaggctattg ctgcacatcc cgtggttgct ggagaagtat 8940
gggttgccac tgacgctgga ttgttccgtt ccatcgatta tggggccact ttctctgctg 9000
cttctggttc cattaccgat gcaattcagg tgtctttagg aaaaggagat ggttcagctt 9060
ggaacgtgta cgtatttggg accggtcctg aagggaccaa gttgtatgca agcgctgatg 9120
aaggtgccac atgggtggat atccaaggag agcagggctt cggttctttg tcagctaaca 9180
gacttgttgg ttcaggaaat gtggctggtc aagtctacgt tggcacaaat ggtagaggtg 9240
ttttctatgc aaaggttagt ctttctgctg ctgccaatta ataac 9285
//
<---

Группа изобретений относится к биотехнологии. Предложен рекомбинантный штамм дрожжей Komagataella phaffii ВКПМ Y-4933, продуцирующий секретируемую эндо-диссоциативную ксилоглюканазу семейства GH74, кодируемую мутантным геном, соответствующим SEQ ID NO 1. Также предложен способ микробиологического синтеза ксилоглюканазы с использованием указанного штамма. Группа изобретений обеспечивает высокий уровень продукции ксилоглюканазы семейства GH74 с эндо-диссоциативным способом действия на субстрат, обеспечивающим эффективное снижение вязкости растворов ксилоглюкана. 2 н.п. ф-лы, 3 пр., 2 ил.
1. Рекомбинантный штамм дрожжей Komagataella phaffii ВКПМ Y-4933 - продуцент секретируемой эндо-диссоциативной ксилоглюканазы семейства GH74, кодируемой мутантным геном, соответствующим SEQ ID NO 1.
2. Способ микробиологического синтеза ксилоглюканазы, предусматривающий культивирование рекомбинантных микроорганизмов, содержащих ген ксилоглюканазы, в аэробных условиях в питательной среде, включающей источники углерода, азота и минеральные добавки до максимального накопления целевого продукта, отличающийся тем, что в качестве продуцента используют рекомбинантный штамм по п. 1.
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Pichia pastoris - ПРОДУЦЕНТ СЕКРЕТИРУЕМОЙ ТЕРМОСТАБИЛЬНОЙ КСИЛОГЛЮКАНАЗЫ, КОДИРУЕМОЙ СИНТЕТИЧЕСКИМ ГЕНОМ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОЙ ТЕРМОСТАБИЛЬНОЙ КСИЛОГЛЮКАНАЗЫ НА ОСНОВЕ ЭТОГО ШТАММА | 2015 |
|
RU2605629C1 |
| База данных GenBank: KY549923.1, 21.05.2017 | |||
| Приспособление в центрифугах для регулирования количества жидкости или газа, оставляемых в обрабатываемом в формах материале, в особенности при пробеливании рафинада | 0 |
|
SU74A1 |
| Найдено онлайн: https://www.ncbi.nlm.nih.gov/nucleotide/KY549923.1?report=genbank&log$=nuclalign&blast_rank=1&RID=09TEX1F4013 Дата обращения 10.02.2022 | |||
| XIAN L | |||
| ET AL | |||
| Identification and characterization of an acidic | |||

