Область техники
Изобретение относится к области биотехнологии и касается способа получения штаммов дрожжей Komagataella kurtzmanii с улучшенной способностью к усвоению сорбитола. Изобретение может применяться для получения продуцентов рекомбинантных белков на основе K. kurtzmanii, при ферментации которых используют сорбитол.
Уровень техники
В настоящее время биосинтетические платформы (системы экспрессии), разработанные на основе различных видов дрожжей, эффективно используют для производства рекомбинантных белков. В этом качестве дрожжи обеспечивают простоту манипуляций в сочетании с быстрым ростом, способностью осуществлять корректный фолдинг и эффективную секрецию рекомбинантных белков во внеклеточную среду. Более того, дрожжи обладают превосходными ферментационными характеристиками и способны расти до высокой плотности, что увеличивает выход синтезируемого белка и делает производственные процессы более экономичными по сравнению со многими другими альтернативными системами экспрессии [Gellissen, G., Kunze, G., Gaillardin, С., Cregg, J.M., et al. New yeast expression platforms based on methylotrophic Hansenula polymorpha and Pichia pastoris and on dimorphic Arxula adeninivorans and Yarrowia lipolytica - a comparison. FEMS Yeast Res, 2005, 5(11), 1079-96.].
Несмотря на преимущества дрожжевых платформ, они не в состоянии обеспечить одинаково высокий уровень экспрессии любых рекомбинантных белков. Известно, например, что сывороточный альбумин человека секретируется клетками дрожжей до 1,4 г/литр [Kobayashi, K., Kuwae, S., Ohya, Т., Ohda, Т., et al. High-level expression of recombinant human serum albumin from the methylotrophic yeast Pichia pastoris with minimal protease production and activation. J Biosci Bioeng, 2000, 89 (1), 55-61.]. В то же время продукция таких важных белков как, например, интерлейкин человека 2 (hIL2) и фактор роста тромбоцитов (PDGF) чрезвычайно низка [Anzur Lasnik, М., Nemec, М., Romih, R. and Schara, M. An EPR study of the secretion of G-CSF heterologous protein from Pichia pastoris. Biotechnol Bioeng, 2003, 81 (7), 768-74; Guan, В., Chen, F., Lei, J., Li, Y., et al. Constitutive expression of a rhIL-2-HSA fusion protein in Pichia pastoris using glucose as carbon source. Appl Biochem Biotechnol, 2013, 171 (7), 1792-804.]. Как и при использовании других систем, на уровень синтеза целевых рекомбинантных белков в дрожжах коренным образом влияют выбор организма для экспрессии; генетические элементы, управляющие экспрессией целевых генов, такие как сигнальная последовательность, промоторы, терминаторы и различные цис- и транс- регуляторные элементы; условия культивирования рекомбинантного штамма, состав среды [Hohenblum, Н., Gasser, В., Maurer, М., Borth, N., et al. Effects of gene dosage, promoters, and substrates on unfolded protein stress of recombinant Pichia pastoris. Biotechnol Bioeng, 2004, 85 (4), 367-75; Juturu, V. and Wu, J.C. Heterologous Protein Expression in Pichia pastoris: Latest Research Progress and Applications. Chembiochem, 2018, 19 (1), 7-21; Yang, Z. and Zhang, Z. Engineering strategies for enhanced production of protein and bio-products in Pichia pastoris: A review. Biotechnol Adv, 2018, 36 (1), 182-195.].
Для производства белковых молекул одной из первых была разработана и успешно использовалась система экспрессии на основе дрожжей Saccharomyces cerevisiae. Однако ее использование сопровождалось рядом ограничений, таких как нестабильность плазмид, низкий выход белка и неродное гликозилирование, например гипергликозилирование или α-1,3-связанное маннозо-концевое N-связанное гликозилирование белков [Celik, Е. and Calik, P. Production of recombinant proteins by yeast cells. Biotechnol Adv, 2012, 30 (5), 1108-18.]. Решения указанных и ряда других важных проблем удалось частично достичь с помощью альтернативных систем экспрессии, разработанных на основе дрожжей таких видов, как метилотрофные дрожжи Komagataella phaffii (бывш. Pichia pastoris) и Hansenula polymorpha; диморфные дрожжи Arxula adeninivorans и Yarrowia lipolytica; лактозоассимилирующие дрожжи Kluyveromyces lactis, Schizosaccharomyces pombe и некоторые другие почкующиеся дрожжи, такие как Kluyveromyces marxianus, Candida utilis и Zygosaccharomyces bailii. Все они обладают способностью к посттрансляционным модификациям белков, таким как гликозилирование, образование дисульфидных связей, протеолитические модификации и доставке целевых молекул в различные субклеточные компартменты.
В настоящее время одним из лидеров среди дрожжевых систем экспрессии признается платформа на основе метилотрофных дрожжей Komagataella phaffii (бывш. Pichia pastoris), которая была сконструирована для производства рекомбинантных белков [Cregg, J.М., Barringer, K.J., Hessler, А.Y. and Madden, K.R. Pichia pastoris as a host system for transformations. Mol Cell Biol, 1985, 5 (12), 3376-85.]. В последнее время платформа K. phaffii используется также для получения небелковых продуктов [Pena, D.A., Gasser, В., Zanghellini, J., Steiger, М.G., et al. Metabolic engineering of Pichia pastoris. Metab Eng, 2018, 50 2-15; Werten, M. W. Т., Eggink, G., Cohen Stuart, M.A. and de Wolf, F.A. Production of protein-based polymers in Pichia pastoris. Biotechnol Adv, 2019, 37 (5), 642-666.]. Высокая эффективность этой биосинтетической платформы K. phaffii обеспечивается способностью этого вида дрожжей к быстрому росту до высокой плотности в полностью синтетической среде [Ergun, В.G. and Calik, P. Lignocellulose degrading extremozymes produced by Pichia pastoris: current status and future prospects. Bioprocess Biosyst Eng, 2016, 39 (1), 1-36.]. Широкому использованию этой платформы способствуют также высокий уровень синтеза рекомбинантных белков, секретируемых во внеклеточную среду, и низкий уровень секреции собственных эндогенных белков.
Экспрессию целевых генов в дрожжах K. phaffii часто осуществляют под контролем промотора PAOX1 гена AOX1. Известно, что PAOX1 является сильным и строго регулируемым промотором, его активность индуцируется метанолом, но подавляется большинством других источников углерода, таких как глюкоза и глицерин, которые являются подходящими субстратами для роста клеток [Cregg, J.М., Madden, K.R., Barringer, K.J., Thill, G.P., et al. Functional characterization of the two alcohol oxidase genes from the yeast Pichia pastoris. Mol Cell Biol, 1989, 9 (3), 1316-23.]. Известно лишь несколько источников углерода, среди которых - сорбитол и маннитол, которые не подавляют активность VAOX1. По этой причине их, в особенности - сорбитол, часто применяют для стимулирования роста дрожжей в процессе индукции метанолом [Inan, М. and Meagher, М.М. Non-repressing carbon sources for alcohol oxidase (AOX1) promoter of Pichia pastoris. J Biosci Bioeng, 2001, 92 (6), 585-9.] [Celik, E., Calik, P. and Oliver, S.G. Fed-batch methanol feeding strategy for recombinant protein production by Pichia pastoris in the presence of co-substrate sorbitol. Yeast, 2009, 26 (9), 473-84.].
Использование сорбитола в качестве дополнительного ростового субстрата для продуцентов на основе K. phaffii приводит к многочисленным позитивным эффектам: к увеличению выхода рекомбинантного белка [Azadi, S., Mahboubi, A., Naghdi, N., Solaimanian, R., et al. Evaluation of Sorbitol-Methanol Co-Feeding Strategy on Production of Recombinant Human Growth Hormone in Pichia Pastoris. Iran J Pharm Res, 2017, 16 (4), 1555-1564; Gao, M.J., Zhan, X.В., Gao, P., Zhang, X., et al. Improving Performance and Operational Stability of Porcine Interferon-alpha Production by Pichia pastoris with Combinational Induction Strategy of Low Temperature and Methanol/Sorbitol Co-feeding. Appl Biochem Biotechnol, 2015, 176 (2), 493-504.], снижению клеточного стресса и смертности [Wang, Z., Wang, Y., Zhang, D., Li, J., et al. Enhancement of cell viability and alkaline polygalacturonate lyase production by sorbitol co-feeding with methanol in Pichia pastoris fermentation. Bioresour Technol, 2010, 101 (4), 1318-23.], снижению потребления кислорода [Niu, H., Jost, L., Pirlot, N., Sassi, H., et al. A quantitative study of methanol/sorbitol co-feeding process of a Pichia pastoris Mut(+)/pAOX1-lacZ strain. Microb Cell Fact, 2013, 12 33.] и к снижению протеолитической деградации [Celik, Е., Calik, P. and Oliver, S.G. Fed-batch methanol feeding strategy for recombinant protein production by Pichia pastoris in the presence of co-substrate sorbitol. Yeast, 2009, 26 (9), 473-84.].
Усвоение сорбитола дрожжами зависит от двух метаболических процессов: от транспорта этого углевода в клетки и от последующего ферментативного превращения во фруктозу под действием фермента сорбитолдегидрогеназы, кодируемого геном SOR1 [Jordan, P., Choe, J.Y., Boles, E. and Oreb, M. Hxt13, Hxt15, Hxt16 and Hxt17 from Saccharomyces cerevisiae represent a novel type of polyol transporters. Sci Rep, 2016, 6 23502; Sarthy, A.V., Schopp, C. and Idler, K.B. Cloning and sequence determination of the gene encoding sorbitol dehydrogenase from Saccharomyces cerevisiae. Gene, 1994, 140 (1), 121-6.]. При этом эффективность усвоения сорбитола непосредственно отражается на эффективности роста дрожжей на средах, содержащих сорбитол в качестве единственного источника углерода. Медленный рост или его отсутствие соответствует низкой эффективности усвоения сорбитола, тогда как увеличение скорости роста свидетельствует об улучшении усвоения сорбитола.
Дрожжи K. kurtzmanii представляют собой отдельный вид, близкородственный K. phaffii [Naumov, G.I., Naumova, E.S., Tyurin, О.V. and Kozlov, D.G. Komagataella kurtzmanii sp.nov., a new sibling species of Komagataella (Pichia) pastoris based on multigene sequence analysis. Antonie Van Leeuwenhoek, 2013, 104 (3), 339-47.]. На основе дрожжей K. kurtzmanii была разработана и уже успешно применяется конкурентоспособная рекомбинантная экспрессионная система. Во многих случаях показатели ее продуктивности превосходят характеристики платформы на основе K. phaffii [Matveeva, A.Yu., Gubaidullin, I.I., Fedorov, A.S. and Kozlov, D.G. Optimization of the Expression of 1,3-1,4-β-glucanase Gene from Rhizomucor miehei in the Komagataella kurtzmanii yeast. Biotechnologiya, 2019, 35 (5), 3-11; Gorbunov, A.A., Akentyev, F.I., Gubaidullin, I.I., Zhiganov, N.I., et al. Biosynthesis and Secretion of Serine Peptidase SerP38 from Tenebrio molitor in the Yeast Komagataella kurtzmanii. Applied Biochemistry and Microbiology, 2021, 57 (9), 917-924.].
Известно, что геномы дрожжей K. kurtzmanii и K. phaffii содержат высокогомологичные аллели гена SOR1, отвечающие за синтез сорбитолдегидрогеназы и усвоение сорбитола [Heistinger, L., Dohm, J.С., Paes, B.G., Koizar, D., et al. Genotypic and phenotypic diversity among Komagataella species reveals a hidden pathway for xylose utilization. Microb Cell Fact, 2022, 21 (1), 70.]. При этом дрожжи K. phaffii способны эффективно усваивать сорбитол, тогда как дрожжи K. kurtzmanii не способны к расту на средах, содержащих сорбитол в качестве единственного источника углерода [Naumov, G.I., Naumova, Е.S., Tyurin, О.V. and Kozlov, D.G. Komagataella kurtzmanii sp.nov., a new sibling species of Komagataella (Pichia) pastoris based on multigene sequence analysis. Antonie Van Leeuwenhoek, 2013, 104 (3), 339-47.]. Это не позволяет использовать сорбитол для подкормки дрожжей K. kurtzmanii в процессе проведения ферментаций и увеличения их эффективности в роли продуцента. В то же время поскольку дрожжи K. kurtzmanii и K. phaffii принадлежат к близкородственным видам, следует ожидать, что усвоение сорбитола дрожжами K. kurtzmanii в процессе ферментации также может сопровождаться положительными эффектами, в частности, повышением продукции целевых гетерологичных продуктов.
Известен пример модификации дрожжей S. cerevisiae для увеличения эффективности усвоения маннитола и сорбитола. Как было показано, решению данной задачи способствует мутация в гене общего транскрипционного корепрессора Tup1-Cyc8 [Tanaka, Н., Murata, K., Hashimoto, W. and Kawai, S. Hsp104-dependent ability to assimilate mannitol and sorbitol conferred by a truncated Cyc8 with a C-terminal polyglutamine in Saccharomyces cerevisiae. PLoS One, 2020, 15 (11), e0242054.].
Таким образом, технической проблемой, на решение которой направлено настоящее изобретение, является совершенствование способов, направленных на получение штаммов дрожжей K. kurtzmanii, способных к усвоению сорбитола.
Раскрытие сущности изобретения
Техническим результатом заявляемого изобретения является получение генетически модифицированных дрожжей K. kurtzmanii, способных к эффективному усвоению сорбитола.
Для достижения технического результата предложен способ получения дрожжей K. kurtzmanii с улучшенной способностью к усвоению сорбитола, заключающийся в том, что клетки дрожжей K. kurtzmanii трансформируют рекомбинантной плазмидной ДНК pPH727-Sdh115, содержащей следующие структурные элементы: фрагмент ДНК, кодирующий ген SOR1 (PAS_chr1-1_0490) дрожжей K. phaffii GS115, включающий последовательность структурного гена сорбитолдегидрогеназы дрожжей K. phaffii размером 1047 н.п. и нативную 3'-UTR (3'-нетранслируемую область) длиной 211 н.п., которая определяется как промотор гена SOR1, терминаторную область гена алкогольоксидазы АОХ1 дрожжей K. kurtzmanii, отвечающую за терминацию транскрипции гена SOR1, ген HIS4 дрожжей K. kurtzmanii, обеспечивающий селекцию трансформантов на среде без добавления гистидина, фрагменты дистальной части промоторной области гена алкогольоксидазы АОХ1 дрожжей K. kurtzmanii для направления интеграции плазмидной ДНК в геном дрожжей, фрагмент ДНК плазмиды pUC18, обеспечивающий амплификацию плазмиды pPH727-Sdh115 в клетках Е. Coli, ген устойчивости ApR, обеспечивающий селекцию плазмидосодержащих клеток Е. coli на среде с антибиотиком ампициллином или клетки дрожжей K. kurtzmanii трансформируют рекомбинантной плазмидной ДНК pPH727-Sdh727, но содержащей вместо гена SOR1 (PAS_chr1-1_0490) дрожжей K. phaffii (SOR1GS115) ген SOR1 дрожжей K. kurtzmanii (SOR1Y-727).
Краткое описание чертежей
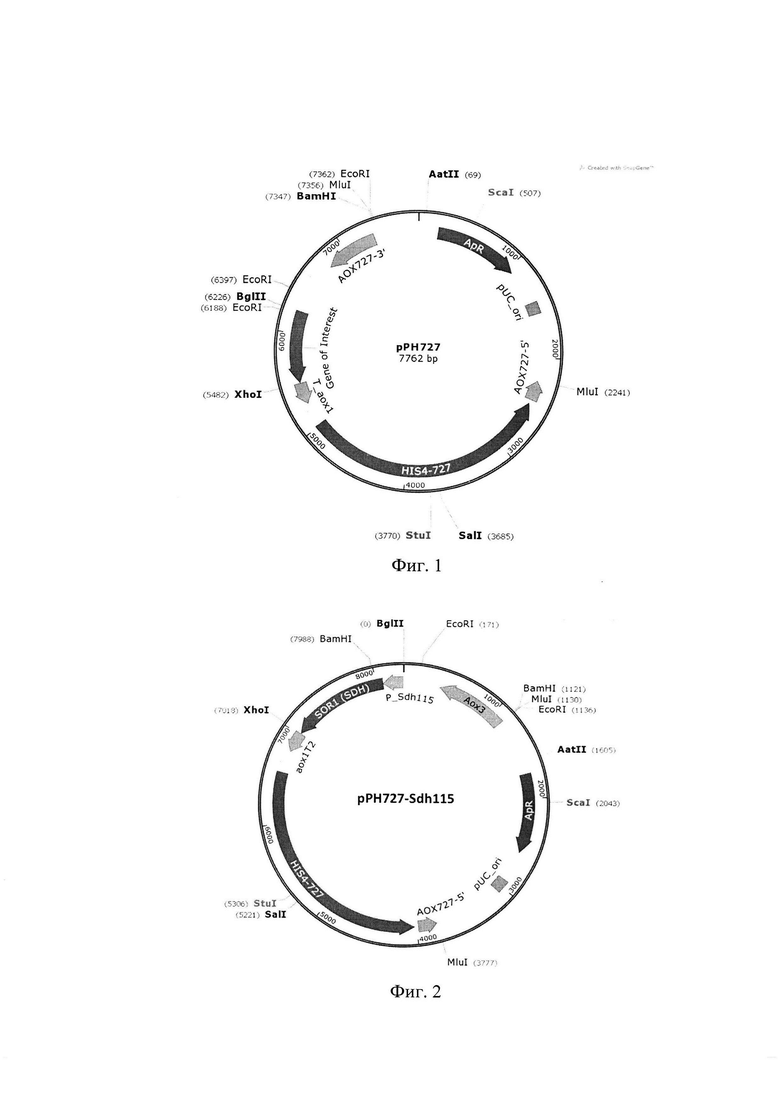
Фиг. 1 - Схема рекомбинантной плазмидной ДНК рРН727
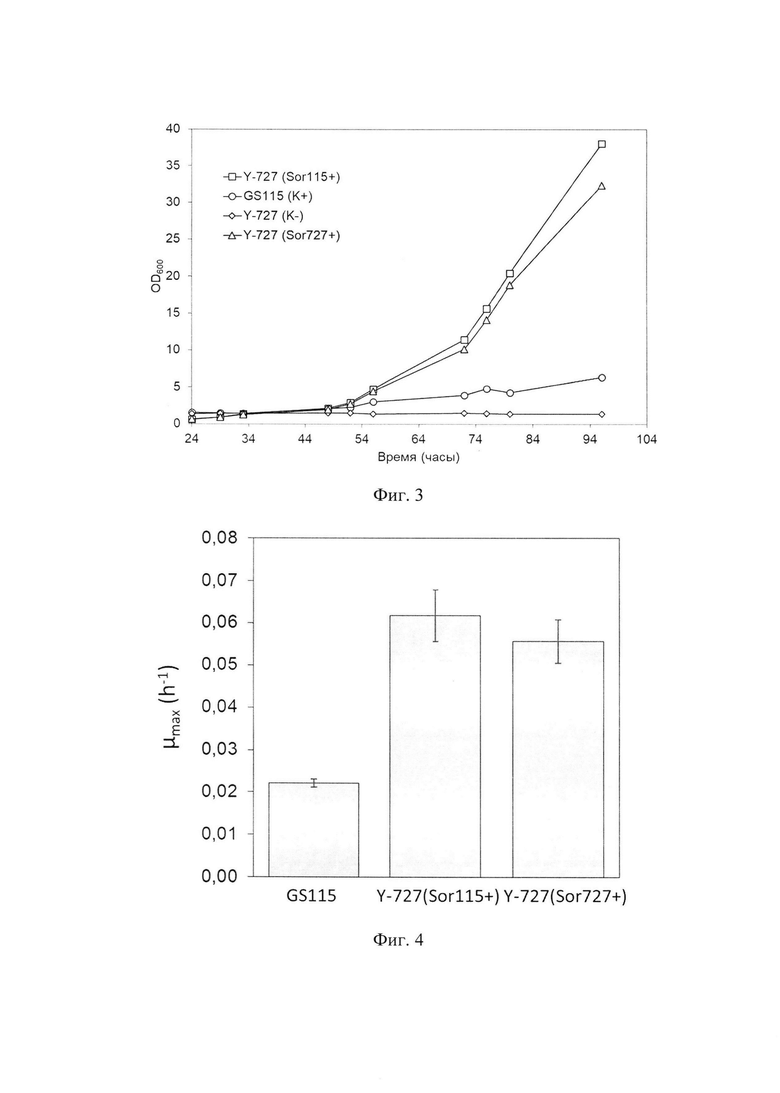
Фиг. 2 - Схема рекомбинантной интегративной плазмидной ДНК pPH727-Sdh115
Фиг. 3 - Динамика роста для четырех отобранных штаммов на минимальной среде с сорбитолом в качестве единственного источника углерода. Y-727 (Sor115+) - штамм K. kurtzmanii, трансформированный вектором pPH727-Sdh115; Y-727 (Sor727+) - штамм K. kurtzmanii, трансформированный вектором pPH727-Sdh727; GS115 (K+) - прототрофный штамм K. phaffii GS115, Y-727 (K-) - штамм K. kurtzmanii Y-727 дикого типа
Фиг. 4 - Анализ удельной скорости роста штаммов K. phaffii GS115, K. kurtzmanii Y-727 (Sor115+) и K. kurtzmanii Y-727 (Sor727+) на минимальной среде с сорбитолом в качестве единственного источника углерода. Доверительный интервал соответствует стандартной ошибке среднего
Осуществление изобретения
Для осуществления заявляемого изобретения решают ключевую задачу по увеличению уровня экспрессии сорбитолдегидрогеназы в дрожжах K. kurtzmanii. Для ее решения используют подходы, известные из уровня техники. К таким подходам относится замена регуляторных элементов в геномном аллеле гена сорбитолдегидрогеназы, внесение дополнительных копий гена сорбитолдегидрогеназы в геном дрожжей K. kurtzmanii, в том числе - гетерологичных (чужеродных), и другие. Данные решения осуществляют с использованием оборудования, реагентов и методик, хорошо известных для специалистов, работающих в данной области техники.
Далее приведено описание некоторых вариантов воплощения настоящего изобретения. В то же время настоящее изобретение не следует ограничивать указанными вариантами. Описание деталей, очевидных для специалистов в данной области техники, в данном документе опущены.
В одном из воплощений заявляемого изобретения с целью увеличения уровня экспрессии сорбитолдегидрогеназы клетки дрожжей K. kurtzmanii трансформируют рекомбинантной плазмидной ДНК pPH727-Sdh115 (Пример 1), содержащей следующие структурные элементы:
- фрагмент ДНК, кодирующий ген SOR1 (PAS_chr1-1_0490) дрожжей K. phaffii GS115, включающий последовательность структурного гена сорбитолдегидрогеназы дрожжей K. phaffii размером 1047 н.п. и нативную 3'-UTR (3'-нетранслируемую область) длиной 211 н.п., которая определяется как промотор гена SOR1 [Periyasamy, S., Govindappa, N., Sreenivas, S. and Sastry, K. Isolation, characterization and evaluation of the Pichia pastoris sorbitol dehydrogenase promoter for expression of heterologous proteins. Protein Expr Purif, 2013, 92 (1), 128-33.].
- терминаторную область гена алкогольоксидазы AOX1 дрожжей K. kurtzmanii, отвечающую за терминацию транскрипции гена SOR1;
- ген HIS4 дрожжей K. kurtzmanii, обеспечивающий селекцию трансформантов на среде без добавления гистидина;
- фрагменты дистальной части промоторной области гена алкогольоксидазы АОХ1 дрожжей K. kurtzmanii для направления интеграции плазмидной ДНК в геном дрожжей;
- фрагмент ДНК плазмиды pUC18, обеспечивающий амплификацию плазмиды pPH727-Sdh115 в клетках Е. coli;
- ген устойчивости ApR, обеспечивающий селекцию плазмидосодержащих клеток Е. coli на среде с антибиотиком ампициллином.
В другом воплощении заявляемого изобретения дрожжи K. kurtzmanii трансформируют рекомбинантной плазмидной ДНК pPH727-Sdh727 (Пример 2), аналогичной плазмиде pPH727-Sdh115, но содержащей вместо гена SOR1 (PAS_chr1-1_0490) дрожжей K. phaffii (SOR1GS115) ген SOR1 дрожжей K. kurtzmanii {SOR1Y-727).
В одном из вариантов осуществления заявляемого изобретения трансформации указанными плазмидами подвергают реципиентный штамм дрожжей K. kurtzmanii Y-727 Δhis4 [Matveeva, A.Yu., Gubaidullin, I.I., Fedorov, A.S. and Kozlov, D.G. Optimization of the Expression of 1,3-1,4-β-glucanase Gene from Rhizomucor miehei in the Komagataella kurtzmanii yeast. Biotechnologiya, 2019, 35 (5), 3-11.], а процедуру трансформации осуществляют в 3 этапа (Пример 3):
1. Перед началом трансформации плазмидную ДНК pPH727-Sdh115 или pPH727-Sdh727 обрабатывают рестриктазой MluI, открывая гомологичные фланги, направляющие интеграцию в локус гена AOX1 генома дрожжей;
2. Приготавливают компетентные клетки дрожжей и осуществляют их трансформацию подготовленной плазмидной ДНК, для чего используют любые подходящие протоколы, известные из уровня техники, применяемые для приготовления компетентных клеток и трансформации дрожжей K. phaffii;
3. По завершении процедуры трансформации трансформанты выращивают на селективной среде без добавления гистидина.
В результате осуществления процедуры трансформации получают модифицированные штаммы дрожжей K. kurtzmanii с увеличенным уровнем экспрессии сорбитолдегидрогеназы.
Для подтверждения способности модифицированных штаммов к эффективному усвоению сорбитола полученные трансформанты выращивают на синтетической среде, содержащей необходимые для роста дрожжей соли, витамины, нуклеотиды и аминокислоты, а также сорбитол в качестве единственного источника углерода. В качестве отрицательного контроля выращивают исходный нетрансформированный реципиентный штамм. Для всех штаммов определяют параметры скорости роста на минимальной среде с сорбитолом любым способом, известным из уровня техники (Пример 4).
В процессе поэтапного выполнения представленного варианта технического решения в рамках патентных исследований осуществляют:
- разработку рекомбинантных плазмид pPH727-Sdh115 и рРН727-Sdh727, содержащих нуклеотидные последовательности генов SOR1GS115 (Пример 1) и SOR1Y-727 (Пример 2), соответственно.
- трансформацию дрожжей K. kurtzmanii сконструированными плазмидами с последующим получением трансформированных штаммов Y-727 (Sor115+) и Y-727 (Sor727+) соответственно (Пример 3).
- Измерение удельной скорости роста для полученных трансформантов на минимальной среде с сорбитолом (Пример 4).
Пример 1. Конструирование вектора pPH727-Sdh115, содержащего ген SOR1 дрожжей K. phaffii
Базовым вектором для конструирования служит плазмида рРН727 (фиг. 1), которая содержит следующие элементы:
- нативный терминатор гена алкогольоксидазы АОХ1 дрожжей Komagataella kurtzmanii Y-727
- последовательность гена HIS4 штамма Komagataella kurtzmanii Y-727
- фрагменты дистальной части промотора гена алкогольоксидазы АОХ1 штамма Komgataella kurtzmanii Y-727
- точку начала репликации плазмид pUCori
- ген устойчивости к ампициллину (ApR), продукт которого обеспечивает отбор трансформантов на среде с антибиотиком
Фрагмент гена SOR1GS115 получают путем амплификации на матрице хромосомной ДНК штамма K. phaffii GS115 с использованием праймеров 5'-agatagatcttgttaaaaagttgtatattattaatgg и 5'-atgtctcgagttactctgggccgtcaatgatag. Полученный фрагмент обрабатывают рестриктазами BglII и XhoI, и затем лигируют в вектор рРН727, предварительно гидролизованный по тем же сайтам. В результате клонирования получают вектор pPH727-Sdh115 (фиг. 2), в котором кодирующая последовательность гена SOR1GS115 окружена промотором гена SOR1GS115 и терминатором гена АОХ1 штамма K. kurtzmanii Y-727.
Вектор pPH727-Sdh115 в дальнейшем используют для интеграции гена SOR1GS115 из K. phaffii в геном дрожжей K. kurtzmanii, в результате чего получают генетически модифицированные дрожжи K. kurtzmanii Y-727 (Sor115+)
Пример 2. Конструирование вектора pPH727-Sdh727, содержащего ген SOR1 дрожжей K. kurtzmanii
Вектор pPH727-Sdh727 получают также, как и вектор pPH727-Sdh115 из Примера 1, за исключением того, что в качестве дополнительного гена сорбитолдегидрогеназы используют SOR1 дрожжей K. kurtzmanii Y-727. SOR1Y-727 получают также путем амплификации на матрице хромосомной ДНК штаммов K. kurtzmanii по праймерам, указанным в Примере 1.
Вектор pPH727-Sdh727 используют для интеграции гена SOR1Y-727 из K. kurtzmanii в геном дрожжей K. kurtzmanii, в результате чего получают генетически модифицированные дрожжи K. kurtzmanii Y-727 (Sor727+)
Пример 3. Получение генетически модифицированных дрожжей K. kurtzmanii Y-727 (Sor115+) и Y-727 (Sor727+)
Перед трансформацией векторы pPH727-Sdh115 и pPH727-Sdh727 линеаризуют рестриктазой MluI. 1 мкг полученной ДНК каждого вектора используют для трансформации электрокомпетентных клеток K. kurtzmanii Y-727 Δhis4 методом электропорации. Селекцию трансформантов проводят на агаризованной среде YNBD (1,7 г/л дрожжевая азотная добавка, 5 г/л сульфат аммония, 20 г/л глюкоза, 20 г/л агар). Свежие колонии полученных трансформантов однократно рассеивают до единичных колоний на свежей чашке с YNBD. Отдельную колонию трансформанта после рассева переносят на агаризованную среду YPD (10 г/л дрожжевой экстракт, 20 г/л пептон, 20 г/л глюкоза, 20 г/л агар) и хранят при +4°С.
Отобранные отдельные трансформанты называют штаммами K. kurtzmanii Y-727 (Sor115+) и K. kurtzmanii Y-727 (Sor727+) в соответствии с типом плазмиды, которой последние были трансформированы. Отобранные штаммы являются генетически модифицированными, их геномы содержат содержат дополнительные гены SOR1GS115 и SOR1Y-727, кодирующие ферменты Sdh115 и Sdh727, соответственно.
Пример 4. Анализ роста штаммов K. kurtzmanii Y-727 (Sor115+) и K. kurtzmanii Y-727 (Sor727+) на минимальной среде с сорбитолом
Свежие культуры полученных штаммов K. kurtzmanii Y-727 (Sor115+) и Y-727 (Sor727+), а также штаммов K. phaffii GS115 и K. kurtzmanii Y-727 дикого типа засевают в стеклянные пробирки с 5 мл YNBS (1,7 г/л дрожжевая азотная добавка, 5 г/л сульфат аммония, 20 г/л сорбитол, 100 mM калий-фосфатный буфер рН 6.0) до конечной оптической плотности (OD600) от 0,1 до 0,2. Пробирки инкубируют в шейкере (30°С, 250 об/мин) в течение 96 часов. Регулярно стерильно отбирают аликвоту из каждой пробирки и измеряют оптическую плотность при λ=600 нм. Для полученных значений строят зависимость значения оптической плотности от времени культивирования (фиг. 3). На основе полученных данных строят графики роста клеток анализируемых штаммов. С использованием построенных графиков в программе Excel аналитически определяют уравнение линии тренда, описывающей экспоненциальную зависимость роста для штаммов в заданных условиях, из которого получают значение удельных скоростей роста (фиг. 4).
Полученные данные демонстрируют, что родительский штамм K. kurtzmanii Y-727 дикого типа практически не способен к росту на среде с сорбитолом. В отличие от него генетически модифицированные штаммы K. kurtzmanii Y-727 (Sor115+) и Y-727 (Sor727+) растут на среде с сорбитолом, причем скорость их роста в 3 раза превышает показатель контрольного штамма K. phaffii GS115.
Таким образом, представленные данные доказывают эффективность заявляемого способа получения дрожжей K. kurtzmanii с улучшенной способностью к усвоению сорбитола.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения штамма-продуцента фосфолипазы А2 Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2S | 2020 |
|
RU2746817C1 |
| ПРИМЕНЕНИЕ ШТАММА ДРОЖЖЕЙ Komagataella pastoris В КАЧЕСТВЕ РЕЦИПИЕНТА ДЛЯ КОНСТРУИРОВАНИЯ ПРОДУЦЕНТОВ ЦЕЛЕВОГО БЕЛКА | 2013 |
|
RU2522479C1 |
| Трансформант дрожжей Komagataella kurtzmanii, продуцирующий бета-глюканазу | 2019 |
|
RU2722563C1 |
| Трансформант Komagataella phaffii - продуцент рекомбинантного химозина в активной форме | 2022 |
|
RU2805486C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS - ПРОДУЦЕНТ СЕКРЕТИРУЕМОЙ КСИЛОГЛЮКАНАЗЫ СЕМЕЙСТВА GH12, КОДИРУЕМОЙ МУТИРОВАННЫМ ГЕНОМ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОЙ КСИЛОГЛЮКАНАЗЫ НА ОСНОВЕ ЭТОГО ШТАММА | 2023 |
|
RU2833908C1 |
| Трансформант Komagataella phaffii, содержащий ген HAC1, продуцент рекомбинантного химозина Vicugna pacos в активной форме | 2023 |
|
RU2828277C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Pichia pastoris - ПРОДУЦЕНТ СЕКРЕТИРУЕМОЙ ТЕРМОСТАБИЛЬНОЙ КСИЛОГЛЮКАНАЗЫ, КОДИРУЕМОЙ СИНТЕТИЧЕСКИМ ГЕНОМ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОЙ ТЕРМОСТАБИЛЬНОЙ КСИЛОГЛЮКАНАЗЫ НА ОСНОВЕ ЭТОГО ШТАММА | 2015 |
|
RU2605629C1 |
| Интегративная кассета для экспрессии генов и ее использование при конструировании трансформанта дрожжей Komagataella mondaviorum | 2023 |
|
RU2836330C1 |
| Способ получения секреторного рекомбинантного белка ORF2 цирковируса свиней второго типа в дрожжах Komagataella phaffii BG11/ pPic-CV2-7 в форме вирусоподобных частиц | 2023 |
|
RU2832292C1 |
| Рекомбинантный штамм дрожжей Komagataella phaffii - продуцент секретируемой эндо-диссоциативной ксилоглюканазы семейства GH74, кодируемой мутированным геном, и способ микробиологического синтеза секретируемой эндо-диссоциативной ксилоглюканазы на основе этого штамма | 2021 |
|
RU2771581C1 |
Изобретение относится к биотехнологии. Предложен способ получения дрожжей Komagataella kurtzmanii с улучшенной способностью к усвоению сорбитола, включающий трансформацию клеток дрожжей K. kurtzmanii рекомбинантной плазмидной ДНК pPH727-Sdh115, содержащей: фрагмент ДНК, кодирующий ген SOR1 (PAS_chr1-1_0490) дрожжей K. phaffii GS115, включающий последовательность структурного гена сорбитолдегидрогеназы дрожжей K. phaffii размером 1047 н.п. и нативную 3'-UTR (3'-нетранслируемую область) длиной 211 н.п., определяемую как промотор гена SOR1, терминаторную область гена алкогольоксидазы АОХ1 дрожжей K. kurtzmanii, ген HIS4 дрожжей K. kurtzmanii, фрагменты дистальной части промоторной области гена алкогольоксидазы АОХ1 дрожжей K. kurtzmanii, фрагмент ДНК плазмиды pUC18, ген устойчивости ApR, обеспечивающий селекцию плазмидосодержащих клеток Е. coli на среде с ампициллином; либо трансформацию клеток дрожжей K. kurtzmanii рекомбинантной плазмидной ДНК pPH727-Sdh727, содержащей вместо гена SOR1 (PAS_chr1-1_0490) дрожжей K. phaffii (SOR1GS115) ген SOR1 дрожжей K. kurtzmanii (SOR1Y-727). Изобретение обеспечивает расширение арсенала продуцентов рекомбинантных белков на основе K. kurtzmanii, при ферментации которых используют сорбитол. 4 ил., 4 пр.
Способ получения дрожжей K. kurtzmanii с улучшенной способностью к усвоению сорбитола, заключающийся в том, что клетки дрожжей K. kurtzmanii трансформируют рекомбинантной плазмидной ДНК pPH727-Sdh115, содержащей следующие структурные элементы: фрагмент ДНК, кодирующий ген SOR1 (PAS_chr1-1_0490) дрожжей K. phaffii GS115, включающий последовательность структурного гена сорбитолдегидрогеназы дрожжей K. phaffii размером 1047 н.п. и нативную 3'-UTR (3'-нетранслируемую область) длиной 211 н.п., которая определяется как промотор гена SOR1, терминаторную область гена алкогольоксидазы АОХ1 дрожжей K. kurtzmanii, отвечающую за терминацию транскрипции гена SOR1, ген HIS4 дрожжей K. kurtzmanii, обеспечивающий селекцию трансформантов на среде без добавления гистидина, фрагменты дистальной части промоторной области гена алкогольоксидазы АОХ1 дрожжей K. kurtzmanii для направления интеграции плазмидной ДНК в геном дрожжей, фрагмент ДНК плазмиды pUC18, обеспечивающий амплификацию плазмиды pPH727-Sdh115 в клетках Е. Coli, ген устойчивости ApR, обеспечивающий селекцию плазмидосодержащих клеток Е. coli на среде с антибиотиком ампициллином или клетки дрожжей K. kurtzmanii трансформируют рекомбинантной плазмидной ДНК pPH727-Sdh727, но содержащей вместо гена SOR1 (PAS_chr1-1_0490) дрожжей K. phaffii (SOR1GS115) ген SOR1 дрожжей K. kurtzmanii (SOR1Y-727).
| AZADI SAEED et al | |||
| "Evaluation of sorbitol-methanol co-feeding strategy on production of recombinant human growth hormone in Pichia pastoris"; Iranian journal of pharmaceutical research, 2017, N 16(4), p.1555-1564 | |||
| ТЮРИН О.В | |||
| "Разработка системы экспрессии генов на основе метилотрофных дрожжей Komagataella kurtzmanii"; Диссертация на соискание |

