Изобретение относится к области биотехнологии и ветеринарной вирусологии и представляет собой экспрессирующий плазмидный вектор pET15b-p26-6×His, обеспечивающий высокий уровень экспрессии целевого участка gag-гена и синтез в клетках Е. coli рекомбинантного белка р26 аутентичного капсидному белку р26 вируса инфекционной анемии лощадей (ИНАН). Предлагаемый вектор содержит индуцибельный LacI промотор, ген устойчивости к ампициллину, последовательность размером 717 п.н. капсидного белка р26 вируса ИНАН лощадей, кодирующую белок молекулярной массой 26 кДа с гексагистидиновой меткой (6×His) на его С-концевой части.
Изобретение может быть использовано в научно-исследовательской работе по изучению особенностей репликации вируса ИНАН, патогенеза болезни; прикладной биотехнологии для получения реагентов тест-систем диагностики и контроля ИНАН лошадей: рекомбинантного капсидного белка р26, обладающего антигенными свойствами соответствующего вирусного белка и поли- и моноклональных антител (МКА) к капсидному белку р26 вируса ИНАН. Технология получения рекомбинантного капсидного белка р26 на основе предлагаемого изобретения исключает работы с патогенным вирусом ИНАН, гарантирует биологическую безопасность производства и применения.
Полученные отечественные реагенты обеспечат высокую чувствительность и специфичность разрабатываемых диагностических тест-систем на основе рекомбинантного р26 (rp26) для диагностики ИНАН, которые будут экономичнее импортных коммерческих диагностических наборов для серологической диагностики и мониторинга ИНАН.
Уровень техники
Инфекционная анемия лошадей (ИНАН) - хроническая персистирующая изнуряющая болезнь представителей семейства Equidae. ИНАН имеет большое экономическое значение в конневодстве и попадает под программу нормативного контроля во многих странах мира, в том числе и РФ. Этиологическим агентом ИНАН лошадей является генетически разнообразный, имеющий несколько линий, циркулирующих по всему миру вирус, относящийся к роду Lentivirus семейства Retroviridae [Инфекционная патология животных (2 т.) / Под ред. А.Я. Самуйленко, Б.В. Соловьева, Е.А. Непоклонова, Е.С. Воронина. М., 2006; Сергеев В.А., Непоклонов Е.А., Алипер Т.И. Вирусы и вирусные вакцины. М., 2007; Payne S.L., La Celle K., Pei X.F., Qi X.M., Shao H, Steagall W.K., et all. Long terminal repeat sequences of equine infectious anaemia virus are a major determinant of cell tropism. J. Gen. Virol, 1999, 80: 755-759 (doi: 10.1099/0022-1317-80-3-755], он генетически и антигенно связан с другими лентивирусами, которые характеризуются развитием персистирующей инфекции.
Известными вариантами вируса ИНАН в Российской Федерации являются: штамм «3-К-ВНИИТИБП-ВИЭВ», выделенный в Запорожье в 1967 году К.П. Юровым. Штамм депонирован в коллекции вирусов ВГНКИ, хранится в коллекции ВНИИТИБП [Способ изготовления культуралъного антигена вируса ИНАН лошадей и набор для идентификации АТ/Юров К.П. и др.патент RU2146150 С1, 10.03.2000] и двух современных изолятов, выделенных на территории нашей страны: Нижний Новгород-2011 и Омск-2012. Данные определения нуклеотидных последовательностей и филогенетического анализа позволили отнести штамм «3-К-ВНИИТИБП-ВИЭВ» к группе североамериканского происхождения (гомология 98%), а изоляты Нижний Новгород-2011 и Омск-2012 - к группе европейских (гомология 82-83%) [Юров К.П., Заблоцкий В.Т., Косминков Н.Е. Вирусные болезни лошадей. М., 2010; Филогенетический анализ Российских изолятов вируса ИНАН лошадей с использованием оптимизированной двухраундовой ПЦР/НН Герасимова, О.Л. Колбасова, С.Ж. Цыбанов, Д.В. Колбасов // Селъхозбиология, 2015, том 50, 6, с. 803-809 doi: 10.15389/agrobiology.2015.6.803rus].
Геном вируса ИНАН представлен двумя идентичными молекулами линейной одноцепочечной РНК позитивной полярности [Leroux С., Cadoré J.L., Montelaro R.C. Equine Infectious Anemia Virus (EIAV): what has HIV's country cousin got to tell us? Vet. Res., 2004, 35(4): 485-512 (doi: 10.1051/vetres: 2004020; Cook R.F., Leroux C., Issel C.J. Equine infectious anemia and equine infectious anemia virus in 2013: a review. Vet. Microbiol, 2013, 167(1-2): 181-204. doi: 10.1016/j.vetmic.2013.09.031] и состоит из трех основных структурных генов, расположенных последовательно в направлении 5'→3'. gag-ген, кодирует полипротеин-предшественник Gag (р55), который расщепляется на матриксный (МА) р15, капсидный (СА) р26, нуклеокапсид (NC) p11 и р9, среди которых капсидный белок р26 составляет большую часть вирусной частицы; pol-ген, кодирует полипептиды, обладающие ревертазной, специфической РНК-азной и ДНК-эндонуклеазной активностью и env-ген, кодирует поверхностные вирионные гликопротеины, участвующие в формировании оболочки вириона, специфическом узнавании клетки-хозяина и проникновении вируса в клетку. Гликопротеины оболочки вирионов обладают высокой вариабельностью, что является одним из основных механизмов защиты возбудителя от элиминирующего воздействия факторов иммунитета. Вирусные мутации, приводящие к уклонению от опосредованного клеточного и гуморального иммунного ответа, осложняют диагностику и являются основой неудач при разработке вакцин против ИНАН.
Кодируемый gag-геном белок р26 представляет собой капсидную оболочку вириона, заключенную в вирусную оболочку. Обнаружение антител к антигену р26 является основой для РДП и других коммерческих тестов, используемых в серологических диагностике ИНАН лошадей. Белок р26 обладает высокой антигенностью, стимулирует значительный антительный ответ и обеспечивает определение сероконверсии ИНАН. Тем не менее, антитела к капсидному белку р26 вируса ИНАН не являются протективными.
Для ликвидации и предупреждения распространения болезни применяется тотальный убой лошадей, а также жесткие карантинные мероприятия. С учетом этого лабораторная диагностика имеет решающее значение для принятия решений о ликвидации вспышек и контроля распространения болезни. Контроль ИНАН основан на выявлении носителей вируса, антител к ИНАН с использованием, соответственно, молекулярно-генетических методов ПЦР-диагностики и серологических методов [Герасимова Н.Н., Колбасова О.Л., Янжиева Д.В., Конвисарева Т.С., Еремец В.И. Использование серологических и молекулярно-генетических методов при исследовании на ИНАН проб от однокопытных животных Российской Федерации. В сб.: Научные основы производства и обеспечения качества биологических препаратов для АПК. Щелково, 2014: 96-100].
Современные методы лабораторной диагностики ИНАН, основанные на определении вирусных антигенов или ДНК, не всегда обеспечивают выявление хронически или бессимптомно инфицированных животных. В этих случаях наиболее информативными являются серологические методы, основанные на определении антител к вирусным белкам ИНАН в сыворотках крови животных. Рутинно одобренными и широко используемыми серологическими методами диагностики ИНАН являются иммунодиффузионный тест в агаровом геле (РДП) и методы иммуноферментного анализа (ИФА). Международным тестом для диагностики ИНАН согласно рекомендациям Международного эпизоотического бюро (МЭБ) является РДП, который выявляет преципитирующие антитела против группоспецифического и антигенно стабильного антигена - корового белка р26 вируса. Фактически, капсидный белок р26 ИНАН является надежным реагентом для РДП и ИФА [Shane, В.S., С.J. Issel, and R.С. Montelaro. Enzymelinked immunosorbent assay for the detection of equine infectious anemia virus p26 antigen and antibody. J. Clin. Microbiol. 1984. 19:351-355. 1,2, 11] и в настоящее время включен в коммерческие тесты (EIA AGID test kit [VMRD Inc.] и ViraCHEK / EIA [Synbiotics Corp]. Оба теста - РДП и ИФА являются хорошо зарекомендовавшими себя диагностическими инструментами.
В тест-системах РДП и ИФА в качестве специфического антигена используются препараты вирусных ИНАН антигенов, полученных из селезенки инфицированных лошадей или очищенный культуральный вирусный антиген.
В нашей стране для выявления антител к вирусу ИНАН в реакции диффузии в агаровом геле (РДП) и ИФА в качестве специфического антигена используют адаптированный к росту в культурах клеток кожи и/или мышц, и/или легкого, и/или почек эмбриона лошади штамм «3-К-ВНИИТИБП-ВИЭВ» [Способ изготовления культурального антигена вируса ИНАН лошадей и набор для идентификации АТ / Юров К.П. и др. патент RU 2146150 С1, 10.03.2000]. Методика культивирования тканей или культур клеток инфицированных ИНАН животных предполагает манипуляции с инфекционным агентом, что способствует получению нестабильных противоречивых результатов тестов РДП и ИФА, увеличивает вероятность ложноположительных результатов анализа, поскольку вирус может быть контаминирован другими антигенами или вирусами, отличными от вируса ИНАН, наличием в сыворотке лошадей неспецифических антител к клеточным компонентам или белкам. Кроме того, вирус ИНАН сложно культивировать in vitro, что затрудняет использование этого подхода для крупномасштабного производства антигена.
Применение рекомбинантных белков в качестве специфических антигенов вместо вирусных, гарантирует биологическую безопасность производства и применения диагностических тест-систем и обеспечивает их высокую чувствительность и специфичность.
Darrel & Peisheng, патент США №5427907, раскрывает способ применения синтетического пептида в качестве антигена в иммунологическом анализе для выявления антител против вируса ИНАН в сыворотке крови лошадей. Эта процедура включает в себя только поиск некоторых эпитопов вирусных белков.
Современное производство рекомбинантных белков главным образом базируется на прокариотических системах экспрессии. Преимущество использования Е. coli в качестве продуцента состоит в том, что данный метод является более простым и относительно дешевым, позволяет получать рекомбинантный белок в короткие сроки с использованием непатогенного микроорганизма, особенно когда предполагается крупномасштабное производство.
Известны исследования экспрессии гена корового белка р26 вируса ИНАН в прокариотической экспрессирующей системе, проведенные: Birkett A. J. и др. (1997) [Cloning, expression, purification, and characterization of the major core protein p26 from equine infectious anemia virus ž / Ashley J. Birkett, Belen Yelamos, Ignacio Rodriguez-Crespo, Francisco Gavilanes, Darrell L. Peterson // Biochimica et Biophysica Acta. 1997. - 1339: 62-72]; и его использование для разработки РДП тест-систем серодиагностики ИНАН - ИФА и РДП на основе рекомбинантного р26, синтезированного в компетентных клетках Е. coli Alvarez et al. (2007), Paulo Cesar Peregrino Ferreira et al. (2006), [The amplified DNA was cloned into the pBAD/Thio expression vector, transformed TOP10 E.coli bacterial strain (Invitrogen). Standardization and validation of an agar gel immunodiffusion test for the diagnosis of equine infectious anemia using a recombinant p26 antigen / I. Alvarez, G. Gutierrez, A. Vissani, S. Rodriguez, M. Barrandeguy, K. Trono // Veterinary Microbiology 121 (2007) 344-351; Paulo Cesar Peregrino Ferreira et al. Metod and composition for the diagnosis of EIA virus disease by using the recombinant capsid protein virus (р26) // патент Pub. No.: US 7026133, 04.11.2006]; Piza A. S. Т., (2006) и Singha H. и др., (2013) [Serodiagnosis of equine infectious anemia by agar gel immunodiffusion and ELISA using a recombinant p26 viral protein expressed in Escherichia coli as antigen/A. S.T. Piza, A. Rael Pereira, M.T. Terreran, Otto Mozzerc et all. // Preventive Veterinary Medicine. 2006. V. 78, (3-4), P. 239-245 https://doi.org/10.1016/j.prevetmed. 2006.10.009; Development, evaluation, and laboratory validation of immunoassays for the diagnosis of equine infectious anemia (EIA) using recombinant protein produced from a synthetic p26 gene of EIA virus / Harisankar Singha, Sachin K. Goyal, Praveen Malik, Sandip K. Khurana, Raj K. Singh // Indian J. Virol. 2013. 24(3):349-356. DOI 10.1007/s13337-013-0149-9]. Таким образом, применение rp26 вируса ИНАН в качестве специфического антигена для выявления антител к ИНАН является предпочтительным решением и распространенным подходом, зарекомендовавшим себя во всем мире.
Также для получения специфических к белку р26 вируса ИНАН гипериммунных сывороток и моноклональных антител (МКА) требуется значительное количество соответствующего белка.
Известен штамм клеток E.coli BL21(DE3)pLysS, который используется для высокоэффективной экспрессии генов под контролем Т7 промотора и позволяет достигать высокого уровня экспрессии целевого гена при индукции с IPTG [Алиев А.С., Грудинин М.П., Комиссаров А.Б. и др. Клонирование и экспрессия в E.coli рекомбинантного белка VP1 вируса инфекционной анемии кур // Естественные науки, 2019, №5(83), http://doi.org/10.23670/IRJ.2019.83.5.011]. Клетки этого штамма обладают свойством в присутствии индуктора (IPTG) преимущественно экспрессировать рекомбинантные белки и в минимальном количестве собственные белки.
В качестве прототипа нами взята рекомбинационная кассета на основе аналогичного вектора pET32b(+) системы рЕТ, содержащая ген CP204L вируса АЧС для получения рекомбинантного белка р30 вируса АЧС [Штамм клеток E.coli BL21(DE3)pLysS, клон pTT9/ASFVp30, содержащий рекомбинантную плазмиду со вставкой участка гена CP204L вируса Африканской чумы свиней, кодирующей конформационный эпитоп белка р30, для изготовления диагностических препаратов, Копытов В.О. и др., патент RU 2463343 С1, 2012.10.10].
Технической проблемой является необходимость расширение арсенала источников рекомбинантных антигенов вируса ИНАН, обладающих высокой антигенной и иммуногенной активностью, сохраняющих свои основные иммунобиологические свойства после хроматографической очистки и пригодных для использования в диагностических тест-системах в качестве специфического антигена и получения специфических гипериммунных сывороток и МКА (Моноклинальные антитела) в целях клинической диагностики ИНАН.
Раскрытие сущность изобретения. Техническим результатом является создание нового штамма-продуцента Е. coli BL21(DE3) - клона pET15b-p26AEI/1, содержащего рекомбинантную плазмиду pET15b(+) со вставкой участка gag-гена, кодирующего капсидный белок р26 вируса ИНАН, размером в 717 п.н. (237 а.а) с молекулярной массой 26 кДа, который обеспечивает высокий уровень экспрессии целевого гена и синтез (репродукцию) рекомбинантного белка р26, обладающего антигенными свойствами соответствующего вирусного капсидного белка ИНАН; расширение арсенала источников рекомбинантных антигенов вируса ИНАН, обладающих высокой антигенной и иммуногенной активностью, сохраняющих свои основные иммунобиологические свойства после хроматографической очистки и пригодных для использования в качестве специфического антигена для диагностики ИНАН в клинических образцах животных: выявления антител к вирусу ИНАН в сыворотках крови лошадей и получения контрольных (стандартизованных) гипериммунных моноспецифических сывороток и МКА на лабораторных и целевых животных; для изучения патогенеза болезни, особенностей репродукции вируса ИНАН in vitro и in vivo и иммунного ответа макроорганизма на инвазию вируса ИНАН.
Данный результат достигается получением клона pET15v-p26AEI/1 клеток Е. coli штамма BL21(DE3), который является гиперпродуцентом рекомбинантного белка р26 в растворимой форме. Скринированием полученных рекомбинантных трансформантов на наличие вставки методом ПЦР и изучением культурально-биологических свойств клонов при накоплении в жидкой питательной среде 2YT (рН 6.8-7,2), содержащей 100 мкг/мл ампициллина, оптимальной температуре 37,0±0,5°С и перемешивание при 180 об/мин; определением однородности клонов путем пересева на твердую питательную среду.
Данная разработка связана с использованием рекомбинантной плазмиды с сильным промоторным регионом, обеспечивающим высокий и стабильный уровень транскрипции последовательности целевой кассеты в компетентных клетках Е. coli.
Для получения рекомбинантного белка р26 нами выбран экспрессионный вектор pET15b, содержащий Т7 промотор и энхансер, которые являются наиболее эффективными структурными элементами для экспрессии в прокариотической системе. Кроме того, в его состав входит полилинкер, содержащий сайты эндонуклеаз Hind III и Xho I, что необходимо для клонирования целевой последовательности в вектор.
Для успешного выделения рекомбинантного белка предложена система очистки с помощью металл-хелатной хроматографии - Ni-NTA. Для этого С-концевую часть белка р26 дополнили меткой - 6×His-tag. Наличие 6×His-tag в белке позволило дополнительно идентифицировать экспрессию и подтвердить подлинность секретируемого рекомбинантного белка в специфичных аналитических методах - ИФА, Western Blot, Dot Blot с применением антител anti-6xHis-tag, HRP (например ab1187, Abcam, фиг. №8, В).
Предлагаемое изобретение предназначено для получение нового штамма-продуцента Е. coli BL21(DE3), клона pET15b-p26AEI/1, содержащего рекомбинантную плазмиду со вставкой участка gag-гена, кодирующего белок р26 вируса ИНАН, размером в 717 пн (237 а.а) с молекулярной массой 26 кДa, который обеспечивает высокий уровень экспрессии целевого гена и синтез (репродукцию) рекомбинантного капсидного белка р26, обладающего антигенными свойствами соответствующего вирусного белка, эффективно очищается методом хроматографии на целлюлозе и пригоден для использования в диагностических тест-системах в качестве специфического антигена для выявления антител к вирусу ИНАН в сыворотках крови лошадей и получения специфических поли- и МКА для разработки и тест-систем выявления антигена вируса ИНАН.
Сущность изобретения состоит в создании эффективного продуцента рекомбинантного белка р26 вируса ИНАН размером 717 п. н с молекулярной массой 26 кДa в растворимой форме на основе штамма клеток Е. coli BL21(DE3).
Получение вектора включает следующие этапы:
1. Определение полной нуклеотидной последовательности основного капсидного белка р26 вируса ИНАН (фиг.1); дизайн конструкции гена p26AEI-6×His, предполагающий расчет и подбор олигонуклеотидных параметров, фланкирующих участка gag-тепа капсидного белка р26 с сайтами рестрикции; амплификацию участка gag-гена, отвечающего за синтез капсидного белка р26 вируса ИНАН (NCBI) с сохранением рамки считывания при клонировании.
2. Выбор штамма Е. coli и реципиентной плазмиды для создания экспрессирующей конструкции.
3. Оптимизация условий экспрессии и хроматографической очистки рекомбинантного белка.
4. Оценка реактивности, специфичности и антигенных свойств рекомбинантного белка методами иммуноблоттинга, ИФА и реакции непрямой гемагглютинации (РИГА).
Плазмидный ДНК вектор pET15b-р26АЕ1 размером 6350 п. о. получают с использованием стандартных методов генной инженерии, коммерчески доступных плазмид и химически синтезированных олигонуклеотидов.
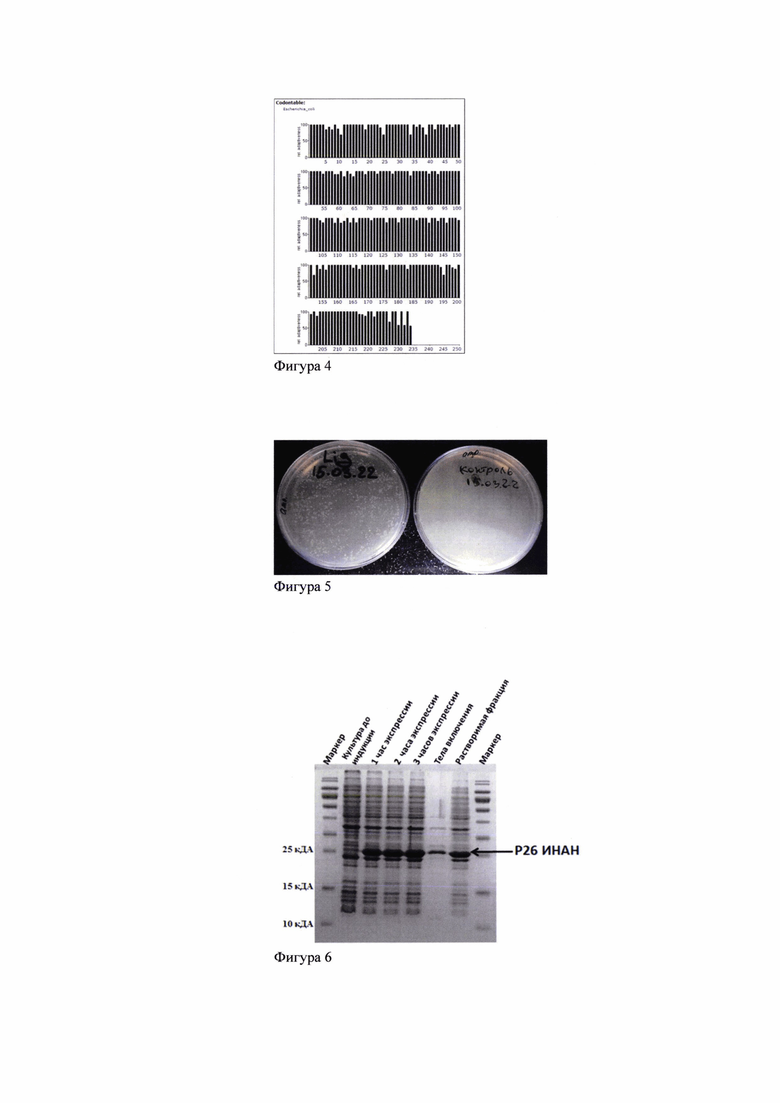
Получение экспрессирующей рекомбинантной плазмиды pET15b(+)-p26AEI (фиг.2) осуществляется путем субклонирования полученной в ПЦР нуклеотидной последовательности гена p26AEI-6×His вируса ИНАН с праймерами F-p26 и R-p26 (табл. 1) в вектор pET15b(+) по сайтам рестрикции NdeI и XhoI в положении 5' и 3'; трансформации клеток Е. coli штамма BL21(DE3) (Promega) плазмидой pET15b(+)-p26AEI; последующий скрининг полученных рекомбинантов на наличие вставки методом ПЦР (фиг.3) и отбор клонов штамма-продуцента Е. coli pET15b-p26AEI/1 (фиг.5) с высоким уровнем экспрессии рекомбинантного белка (rp26) (фиг.6); подбор оптимальных условий: культивирования в жидкой питательной среде 2YT (рН 6.8-7,2), содержащей 100 мкг/мл ампициллина, оптимальной температуре 37,0±0,5°С и перемешивание при 180 об/мин; индукции экспрессии гена (фиг.6) и синтеза целевого белка клоном pET15b-p26AEI/1 клеток Е. coli штамма-продуцента BL21(DE3); очистки (фиг.7); анализа аутентичности (фиг.7), культурально-биологических свойств rp26 вируса ИНАН: специфичности и реактивности в непрямом ТФ ИФА (фиг.8) и РНГА (фиг.10), антигенной активности сыворотки кур (фиг.9). Конечный выход целевого белка составляет не менее 15-20 мг с 1 л бактериальной культуры.
Таким образом, поскольку капсидный белок р26 вируса ИНАН является иммунодоминантным диагностически значимым белком, его рекомбинантный аналог rp26 может быть включен в качестве специфического антигена в состав тест-систем диагностики и контроля ИНАН.
В основе изобретения, лежат разработанные авторами олигонуклеотидные праймеры, представленные в табл. 1.
Примеры иллюстрируют настоящее изобретение и не предназначены для его ограничения по духу или объему. Для того, чтобы это изобретение могло быть лучше понято, описаны следующие примеры.
Осуществление изобретения
ПРИМЕР 1. Иллюстрация возможности получение экспрессионного плазмидного вектора pET15b-p26-6×His, обеспечивающего высокий уровень экспрессии целевого участка gag-гена и синтеза в клетках Е. coli рекомбинантного белка р26 в растворимой форме аутентичного капсидному белку р26 вируса инфекционной анемии лощадей (ИНАН)
Для биоинформатического анализа белка р26, вируса ИНАН лошадей сравнивали сходство его аминокислотной последовательности с помощью Basic Local Alignment Search Tool (BLAST) анализа в Национальном Центр биотехнологической информации (NCBI; http://www.ncbi.nlm.nih.gov), а также в базе данных Uniprot (www.uniprot.org) определили физические данные, такие как теоретический pI, молекулярная масса и коэффициенты экстинкции с использованием Инструмент ProtParam в ExPASy (https://www.expasy.org/).
Участок gag-гена p26AEI-6×His, кодирующего капсидный белок р26 вируса ИНАН лошадей (фиг.1) был сконструирован таким образом, чтобы нести сайты рестрикции NdeI и XhoI с целью направленного клонирования в прокариотическом векторе экспрессии pET15b, с включением гексагистидиновой метки на С - конце белка для проведения металл-хелатной хроматографии на этапе выделения белка rp26.
Для амплификации синтезированной последовательности гена p26AEI-6×His использовали олигонуклкеотидные праймеры, представленные в табл. 1. Синтезированный продукт выделяли из геля, обрабатывали смесью рестриктаз NdeI и XhoI в положении 5' и 3', соответственно и полученную вставку с геном p26AEI-6×His лигировали в вектор pET15b+, содержащий Т7-промотор В результате была сконструирована плазмида pET15b/p26×6His, кодирующая рекомбинантный белок р26 (фиг 2). Полученный в результате плазмидный конструкт и являлся конечным продуктом для клонирования.
Клонированный ген р26 выделяли из плазмиды pET15b с помощью расщепления ферментами NdeI и XhoI и высвобожденную вставку элюировали из агарозного геля с использованием набора MinElute Gel Extraction Kit (Qiagen, Hilden, Германия) в соответствии с инструкциями производителя. После этого элюированную вставку р26 субклонировали в рестрицированный ВатHI и HindIII прокариотический вектор экспрессии pET15b (Qiagen, Hilden, Германия). Далее образец ДНК плазмиды pET15b-р26АЕ1 концентрировали для получения высоко эффективного препарата для экспрессии в клетках Е. coli.
Полученная рекомбинантная плазмида pET15b/p26×6His использована для трансформации Е.coli BL21 (DE3) компетентных клеток. Экспрессия р26 проверена с помощью электрфореза в ПААГ и ELISA.
Трансформацию компетентных клеток плазмидой pET15b/p26×6His осуществляли методом электропорации.
Трансформация клеток штамма-реципиента Escherichia coli BL21 (DE3). Подготовленные компетентные клетки Е. coli BL21 Rosetta (DE3) помещали в ледяную баню, после чего к 100 мкл клеток добавляли раствор рекомбинантной плазмидной ДНК (5 нг) аккуратно перемешали содержимое легким встряхиванием и инкубировали на льду в течение 1 ч. Затем клетки опускали в водяную баню при 42°С на 30 с и повторно инкубировали на льду в течение 2 мин. Далее к клеткам добавляли 400 мкл среды SOB (2% триптон, 0,5% дрожжевой экстракт, 5М NaCl, 1М KCl), инкубировали при температуре 37°С на орбитальном шейкере при 180 об/мин. в течение 40-60 мин и высевали на на чашки Петри, содержащие LB-агаризованную среду с селективный антибиотиком (ампициллин 100 мкг/мл), инкубировали в суховоздушном термостате при 37°С в течение 14-16 ч.
Для подтверждения наличия вставки целевого гена в плазмидах проводили выравнивание полученной нуклеотидной последовательности стандартными методами (Blast, GenBank), которое показало, что рассчитанная аминокислотная последовательность целевого гена p26AEI-6×His в клоне №1 конструкции pET15b-p26AEI полностью совпадает с аминокислотным участком gag-гена капсидного белка р26 вируса ИНАН (NCBI) с 1 по 226 аминокислоту. Учитывая все критерии стратегии получения рекомбинантного белка р26 с 1 по 226 аминокислоту, финальная аминокислотная последовательность выглядит следующим образом: к указанной последовательности был добавлен сигнальный пептид и на С- концевом участке молекулы - 6×His-tag (фиг.3).
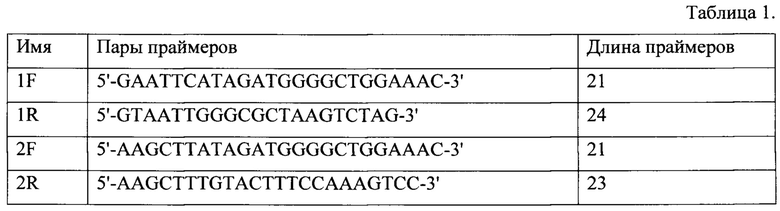
Для увеличения эффективности трансляции каждого кодона белка р26 вируса ИНАН была проведена кодоновая оптимизация с подбором кодона при транскрипции в клетках E.coli. при экспрессии в прокариотической системе (фиг.4).
Положительные трансформанты подвергали скринингу путем отбора одной колонии (фиг.5), экспрессирующей рекомбинантный белок р26 (rp26), с помощью электрофореза в 15% полиакриламидном геле (SDS-PAGE). Идентичность клонированной ДНК была подтверждена методом автоматического секвенирования ДНК, выполненного в компании АО Евроген, Москва.
Получен клон pET15b-p26AEI/1, содержащий рекомбинантную плазмиду со вставкой участка gag-гена, кодирующего коровый белок р26 вируса ИНАН.
Затем проводили индукцию синтеза белка путем внесения изопропил-β-D-тиогалактопиранозида (IPTG).
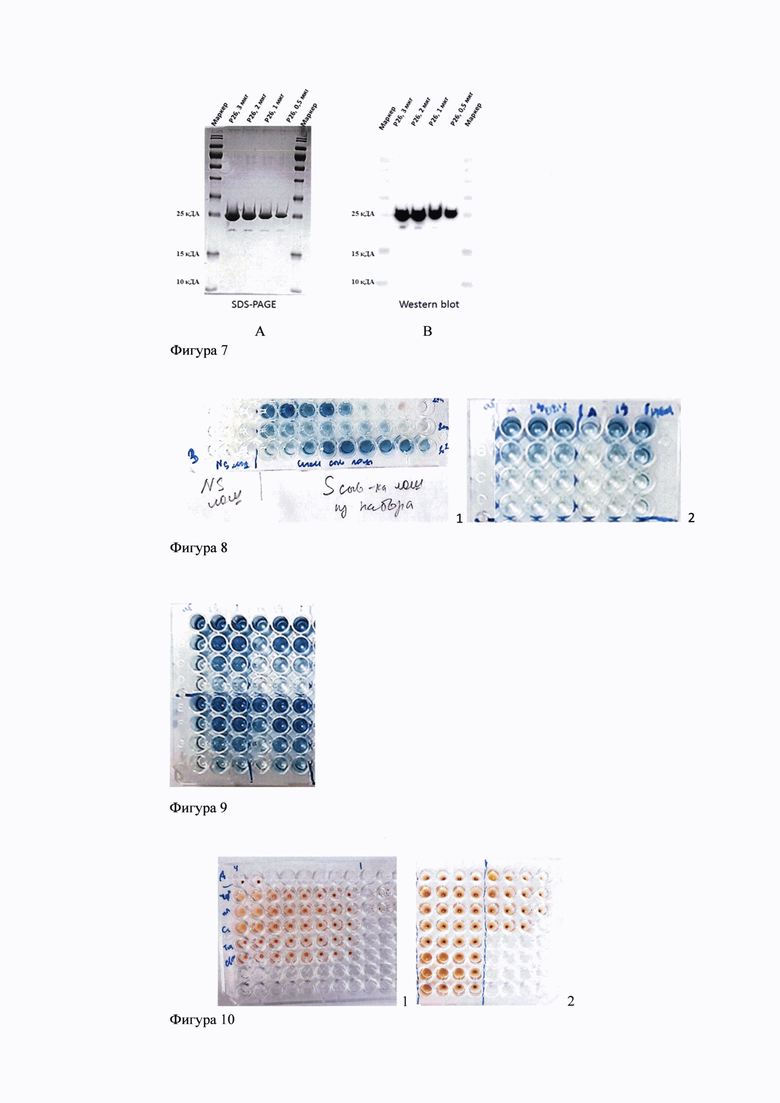
Индукция экспрессии, гена p26AEI-6×His. Использовали штамм-продуцент рекомбинантного белка р26 ИНАН (rp26 ИНАН) на основе клеток Escherichia coli BL21(DE3). Для определения оптимальной концентрации индуктора, обеспечивающей получение высокого уровня экспрессии гена p26AEI-6×His, кодирующего rp26 ИНАН использовали IPTG в трех концентрациях (0,1; 0,5 или 1 мМ). Компетентные клетки BL21(DE3) инкубировали при +37°С в термостатированном шейкере роторного типа при 180 об/мин в течение ночи в LB среде (1% триптон, 1% дрожжевой экстракт и 1% натрий хлористый), содержащей ампициллин в концентрации 100 мкг/мл. Разводили культуру свежей LB средой в 50 раз и выращивали в термостатированном шейкере роторного типа при 180 об/мин +37°С до достижения культурой клеток оптической плотности 0,6-0,8 при длине волны 600 нм. Индукцию экспрессии рекомбинантного гена проводили в течение 18 ч для после добавления IPTG к культуре до конечной концентрации 1 мМ. Наличие и уровень синтеза rp26 ИНАН оценивали методом электрофоретического разделения белков в 12%-ном полиакриламидном геле. Биосинтез rp26 ИНАН с гистидиновой меткой наблюдали на высоком уровне (20±5 мг на 1 л культуры клеток штамма-продуцента), при этом белок накапливался в растворимой форме.
Из фиг.6 видно, что максимальный синтез rp26 ИНАН наблюдался при индукции в течение 3-х часов с использованием IPTG в концентрации 1 мМ. При дальнейшем культивировании синтез rp26 ИНАН снижается. Таким образом, оптимальными условиями культивирования для получения rp26 ИНАН в штамме Е. coli BL21(DE3) являются: культивирование бактерий при 37°С в течение 3-х часов после индукции 1 мМ IPTG (фиг.6).
Далее, проводили препаративное наращивание клеток клон pET15b-p26AEI/1, штамма Е. coli BL21(DE3).
Накопление биомассы клеток, содержащих рекомбинантный белок.
Клетки штамма-продуцента rp26 ИНАН pET15b-p26AEI/1 Е. coli BL21(DE3) культивировали в 500 мл среды YT*2 (1,6% триптон, 1% дрожжевой экстракт, 0,5% NaCl), содержащей ампициллин (100 мкг/мл) при 37°С на орбитальном шейкере при 180 об/мин в течение 4 часов до оптической плотности, достигающей 0,6-0,8 оптических единиц при длине волны - 600 нм.
По окончании культивирования для отделения дебриса, нерастворимых белков биомассу центрифугировали 10 мин. при 5000 g; для концентрирования осадок ресуспендировали в фосфатно-солевом буфере (ФС-буфер:10 мМ Na2HPO4, 1,8 мМ KH2PO4, 15 мМ NaCl, 2,7 мМ KCl) и центрифугировали в течение 1 ч с ускорением 7000 g при 4°С.
Полученную биомассу клеток хранили при -20°С или осадок клеток ресуспендировали в лизирующем буфере, содержащем 50 мМ NaH2PO4, 300 мМ NaCl и 20 мМ имидазол (рН 8,0). Ультразвуковую дезинтеграцию клеток осуществляли, используя режимы: мощность 0,5 кВт, температура 4°С, продолжительность 5 мин. Наличие целевых белков подтверждали методом электрофоретического разделения белков в 12%-м полиакриламидном геле.
После центрифугирования образца при 20000 g в течение 30 мин проводили очистку рекомбинантного белка р26 ИНАН (rp26 ИНАН).
Очистку рекомбинантного белка р26 (rp26) в растворимой форме, полученного в прокариотической (Е. coli) экспрессирующей системе до электрофоретической гомогенности проводили обычными методами разделения белков. Первичную очистку белка rp26, несущего His-tag осуществляли методом колоночной Ni-NTA хроматографии в соответствии с инструкциями производителя (Qiagen, Hilden, Германия) в денатурирующих условиях. Полученный препарат дополнительно подвергали Superdex-200 гельфильтрации. Далее удаляли гистидиновую метку путем инкубации с энтерокиназой в соответствующем буфере.
Хроматографическую очистку проводили методом металл-аффинной хроматографии. Супернатант наносили на хроматографическую колонку со смолой Ni-NTA ("Qiagen", США). Колонку промывали буфером, содержащим 20 мМ Трис-HCl, 0,5 М NaCl и 20 мМ имидазол (рН 7,9). Белок элюировали буфером «В», содержащим 20 мМ Трис-HCl, 0,5 М NaCl и 1 М имидазол (рН 7,9). Раствор белка подвергали диализу против 1000-кратного объема 10 мМ К-фосфатного буфера (рН 6,4). Собирали фракции по 1 мл, концентрацию белка в них определяли по методу Лоури.
Визуально чистоту полученного ферментного препарата контролировали с помощью полиакриламидного SDS-PAGE электрофореза с окрашиванием бриллиантовым синим Кумасси G-250 (фиг.7 позиция А). При анализе в SDS-PAGE в восстанавливающих и денатурирующих условиях очищенный rp26 ИНАН белок мигрировал в виде мономера 26 кДа (фиг. 6 и 7 позиция А). Результаты гель-фильтрационной хроматографии показали, что белок р26 находится в растворе в виде мономера.
Наличие 6×His-tag в белке р26 позволило дополнительно идентифицировать экспрессию и подтвердить подлинность секретируемого белка rp26 ИНАН в Е. coli с помощью специфичных аналитических методов - Western Blot с применением антител anti-6×His-tag, HRP (ab1187, Abcam), (фиг.7) и ТФ ИФА (фиг.8).
Аликвоты очищенного белка объединяли, подвергали диализу против фосфатно-солевого буфера (PBS, рН 7,2) и концентрацию белка измеряли по методу Лоури с использованием коммерческого набора для оценки белка (Merck Bioscience, Бангалор, Индия). Очищенный белок rp26 хранили при -70°С в виде исходных растворов с концентрацией 0,5 мг/мл в аликвотах по 0,5 мл.
Анализ активности и специфичности рекомбинантного белка р26 (rp26) проводили методами иммуноблоттинга (фиг. №7, позиция В), ИФА и реакции непрямой гемагглютинации (РНГА) с использованием сывороток лошадей - нормальной (отрицательной) и специфичных к вирусу ИНАН и кур, интактных и иммунизированных rh26 вируса ИНАН.
ПРИМЕР 2. Определение специфичности рекомбинантного белка р26 (антигена)
Специфическую реактивность белка rp26 определяли вестерн-блоттингом с использованием специфической положительной и отрицательной сывороток лошадей. Мембраны с перенесенным белком блокировали 5% обезжиренным молоком в PBS-T (PBS, содержащий 0,05% Tween-20) в течение ночи при 4°С, дважды промывали PBS-T, нарезали полосками и инкубировали с разведением 1:200 эталонной-положительной к ИНАН сывороткой, полевой сывороткой лошади, инфицированной ИНАН и эталонной отрицательной сывороткой лошади в течение 2 часов при 37°С. Трижды промывали PBS-T и инкубировали с пероксидазным конъюгатом анти-лошадиных IgG в разведении 1:10000 (Sigma-Aldrich, Сент-Луис, США) в течение 1 ч на качалке при 37°С. Затем полоски промывали PBS-T, инкубировали с субстратным Трис-буфером (рН 7,6) с добавлением диаминобензидина (6 мг/10 мл) в присутствии 30 мкл 30% Н2О2. Учет результатов проводили в сравнении с отрицательным (К-) и положительным (К+) контрольными образцами. Реакцию считали положительной при окрашивании исследуемого иммунострипа (полоски) различной интенсивности и ширины на уровне положительного контроля (соответствующей мол. массе белка р26) и отсутствии специфического окрашивания отрицательного контроля (фиг.7).
Белковые полосы образца сравнивали с контролем или маркерами размера и определяли молекулярную массу в дальтонах для идентификации целевого белка.
ПРИМЕР 3. Активность и специфичность полученного rp26 белка ИНАН определяли в непрямом ТФ ИФА (фиг.8) и РИГА (фиг.10)
Метод непрямого ИФА. Постановку непрямого ИФА проводили по следующей схеме. В лунках 96-луночных планшетов для ИФА (NUNC MaxiSorps, Дания) сорбировали на в 0,05 М карбонатно-бикарбонатном буфере, рН 9,5: специфический антиген ИНАН лошадей из коммерческого «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината из расчета 2 мкг/лунку; полученный очищенный рекомбинантный белок р26 ИНАН лошадей в концентрации 2,5-5 мкг/мл, инкубировали в течение 18 ч при 4°С, промывали ФСБ-Т (ФСБ, содержащий 0,05% Твин-20) и высушивали. В качестве блокирующего раствора использовали ФСБ-Т буфер с 5% обезжиренного молока. Далее в лунки последовательно вносили по 100 мкл исследуемых сывороток (использовали серию двукратных разведений 1:100, 1:200 и т.д.) в ФСБТ: отрицательная (К-) и специфическая к ИНАН (К+) сыворотки лошади из коммерческого «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината; нормальные полевые сыворотки лошадей; полевые специфические к ИНАН сыворотки лошади с активностью в РДП 1:4-1:8. Затем вносили вторичные антитела - IgG кролика к Ig лошади (1:500), пероксидазный конюгат протеина А (1:800). На каждом этапе планшеты инкубировали 1 ч при комнатной температуре с последующей трехкратной отмывкой ФСБ-Т. Далее в лунки вносили по 100 мкл субстратного раствора на основе АБТС («ABTS», «Sigma») на 0,05М цитратном буфере, инкубировали в течение 20-30 мин при комнатной температуре. Интенсивность окрашивания в лунках определяли на планшетном фотометре Multiskan EX (Thermo Scientific, США) при длине волны 405 нм (А405). Положительными считали сыворотки со значением А405 в 2,1 и более раза превышающие значения А405 отрицательной сыворотки.
Результаты. Нормальные сыворотки интактных лошадей не взаимодействовали со специфическим коммерческим антигеном вируса ИНАН и его рекомбинантным аналогом rp26. Титр специфических к ИНАН сывороток лошадей со специфическим коммерческим антигеном при дозе сенсибилизации 2 мкг/лунку составил 1:100-1:300; с рекомбинантным белком р26 при дозе сенсибилизации 250 нг/лунку - 1:300-1:12800.
Определение активности и специфичности рекомбинантного белка р26 вируса ИНАН (rp26) и сывороток животных в реакции непрямой гемагглютинации (РНГА).
Реакция непрямой гемагглютинации. Эритроциты барана, сенсибилизированные рекомбинантным белком р26 вируса ИНАН из расчета 1,0 мкг/мл. Сыворотки кур: сыворотка SPF цыплят, кур кросса Доминант до иммунизации и гипериммунные к rp26 вируса ИНАН; сыворотки лошадей: отрицательная (К-) и специфическая к ИНАН (К+) сыворотки лошади из коммерческого «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината, нормальные полевые сыворотки лошадей, полевые специфические к ИНАН сыворотки лошади с активностью в РДП 1:4-1:8. Сыворотки использовали в разведении с 1:4 двукратным шагом.
Постановку и учет результатов реакции проводили согласно общепринятой методике. В 96-луночных планшетах с U-образным дном готовили последовательные 2-кратные разведения исследуемых сывороток на физиологичсеком растворе (рН7,2-7,4) в объеме 50 мкл/лунку. Затем вносили по 50 мкл/лунку 0,5% взвесь эритроцитов барана, сенсибилизированных рекомбинантным белком р26 вируса ИНАН. Встряхивали иинкубировали при комнатной температуре до момента осаждения эритроцитов в виде пуговки в контрольных лунках (контроль спонтанной агглютинации). Результаты реакции учитывали в крестах.
Результаты: титр специфических антигемагглютининов к ИНАН в сыворотках кур составил 1:8-1:16, при отсутствии признаков спонтанной агглютинации эритроцитов в лунках с сыворотками SPF цыплят и кур до иммунизации.
Титр антигемагглютининов в сыворотках лошадей, специфичных к ИНАН составил 1:4-1:8 (что соответствовало их активности в реакции дифузионной преципитации), при отсутствии признаков спонтанной агглютинации эритроцитов в лунках с отрицательными сыворотками лошадей.
ПРИМЕР 4. Антигенность рекомбинантного белка, полученного на основе экспрессионный вектор pET15b (+), продуцируемого клоном pET15b-p26AEI/1 штамма Escherichia coli BL21(DE3). (фиг.9). Антигенная активность рекомбинантного белка р26 вируса ИНАН (rp26)
Для определения антигенности рекомбинантного белка р26 вируса ИНАН (rp26 ИНАН) использовали молодок кур-несушек кросса Доминант массой 1,5-2,0 кг в возрасте не более 6 мес., которых иммунизировали подкожно в области 2/3 шеи препаратом очищенного rp26 ИНАН из расчета 100 мкг/голову. Первое введение препарата проводили с полным адъювантом Фрейнда. В последующем животным инъецировали 3-кратно с интервалом 14 суток препарат rp26 ИНАН с неполным адъювантом Фрейнда. Для контроля продукции специфических антител у животных отбирали пробы крови на 10 сутки после третьей инъекции препарата. Через 10 сут после 4-ой иммунизации обескровливали животных согласно «Методическим рекомендациям по обескровливанию лабораторных животных».
Специфичность и активность полученных сывороток оценивали в непрямом варианте ТФ ИФА (по общепринятой методике, принцип описан в примере 3) на 96 луночных планшетах, сенсибилизированных, соответственно, специфическим и отрицательным контрольными антигенами вируса ИНАН (из расчета 20-40 мкг/мл), препаратом очищенного rp26 ИНАН (из расчета 2,5-5 мкг/мл); в качестве контролей использовали: лизат интактной культуры клеток кожи лошади и лизат клеток Е. coli штамма BL21(DE3), эталонные положительную к ИНАН (К+) и отрицательную (К-) сыворотки лошади из «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината; вторичные антитела: IgG кролика к Ig лошади, IgG мыши к IgM кур, соответствующие пероксидазные конъюгаты протеина А и моноклональных антител к IgG(H+L) кур и IgG мыши.
Полученная гипериммунная моноспецифическая сыворотка крови кур к rp26 вируса ИНАН в непрямом варианте ТФ ИФА специфически реагировала как с вирусным антигеном ИНАН, так и с хроматографически очищенным rp26 ИНАН и не взаимодействовала с отрицательными антигенами - с лизатом интактной культуры клеток кожи лошади и лизатом клеток Е. coli штамма BL21(DE3). Титр полученной гипериммунной моноспецифической сыворотки крови птиц к хроматографически очищенному rp26 ИНАН составил: со специфическим коммерческим антигеном - 1:100-1:300 IgG(Y); 1:300 IgM кур; с rp26 ИНАН - 1:900-1:2700 IgG(Y); 1:2700-5400 IgM кур.
Таким образом, полученные результаты свидетельствуют о высокой антигенной активности rp26 ИНАН, полученного с применением заявляемого экспрессионного плазмидного вектора pET15b-p26-6×His, обеспечивающего синтез в клетках Е. coli рекомбинантного белка р26 аутентичного капсидному белку р26 вируса инфекционной анемии лощадей (ИНАН).
В результате использованного нами подхода получен препарат rp26 ИНАН, который обладает высокой антигенностью, активностью, специфичностью и может быть использован в качестве специфического антигена в диагностических тест-системах серодиагностики ИНАН лошадей и пригоден в качестве антигена для получения поли- и MICA, специфичных к rp26 вируса ИНАН.
ПРИМЕР 5 Характеристика штамма продуцента
1. Вид микроорганизма: штамм Е. coli BL21(DE3)/pET15b-p26AEI/1 продуцент рекомбинантного белка р26 вируса инфекционной анемии лошадей (ИНАН).
2. Номер в других коллекциях: нет.
3. Основание для депонирования: рекомбинантная ДНК плазмида полученная с помощью стандартных методов генной инженерии, коммерчески доступных плазмид и химически синтезированных олигонуклеотидов.
4. Родословная: штамм-реципиент Е. coli BL21(DE3). Генотип штамма - F-ompT gal dcm lon hsdSB(rB-mB-) λ(DE3[lacI lacUV5-T7 gene 1 ind1 sam7 nin5]).
5. Патогенность: относится к III группе ПБА в соответствии с классификацией микроорганизмов - возбудителей инфекционных заболеваний человека, простейших, гельминтов и ядов биологического происхождения по группам патогенности санитарных правил 1.3.2322-08 «Безопасность работы с микроорганизмами III-IV групп патогенности (опасности) и возбудителями паразитарных болезней».
6. Заключение о группе патогенности: не определялось.
Дата, источник и место выделения: Штамм-продуцент получен в 2022 году в лаборатории иммунологии Федерального научного центра - Всероссийский научно-исследовательский институт экспериментальной ветеринарии имени К.И. Скрябина и Я.Р. Ковленко Российской академии наук (ФГБНУ ФНЦ ВИЭВ РАН).
7. Где идентифицирована культура: ФГБНУ ФНЦ ВИЭВ РАН.
8. Методы идентификации: SDS-PAAG, Western-Blott, секвенирование.
9. Морфологические признаки: Клетки палочковидной формы, грамотрицательные, неспороносные.
10. Культуральные особенности: на агаре "Дифко" - колонии круглые, гладкие, мутные, блестящие серые, край ровный. При росте на жидких средах (на минимальной среде с глюкозой или среде Лурье-Бертрана) образуют интенсивную ровную муть. Клетки растут при температуре 37°С при оптимуме рН от 6,8 до 7,5.
Индукцию белкового биосинтеза вызывают путем добавления в культуральную среду индуктора изопропилтио-β-D-галактозида (ИПТГ). Выращивание проводят до достижения наибольшего содержания рекомбинантного белка относительного суммарного белка клетки (6 часов при 37°С после внесения ИПТГ).
Способность трансформированных штаммов гидролизовать хромогенный субстрат (сине-белый тест). В плазмидах с таким маркером бета-галактозидаза расщепляет субстрат X-gal (5-бромо-4-хлоро3-индоил-бета-D-галактопиранозид), превращая его в нерастворимый окрашенный продукт. При вставке в плазмиду целевого гена последовательность гена lacZ нарушается и клетка, трансформированная данной плазмидой, теряет способность к модификации субстрата и остается неокрашенной.
11. Физико-биологические признаки.
12. Биохимические свойства: не определялось.
13. Серологические свойства: не определялось.
14. Продукция антигенов, ферментов, токсинов: продуцирует рекомбинантный белок р26 - аналог основного капсидного белка р26 вируса инфекционной анемии лошадей.
15. Вирулентность для лабораторных животных: не определялось.
16. Устойчивость к антибактериальным препаратам: устойчивость к ампициллину.
17. Отношение к гомо- и гетерологичным фагам (профаги): не определялось.
18. Генетические характеристики: штамм-продуцент Е. coli BL21(DE3)/pET15b-p26AEI/1 отличается от штамма-реципиента Е. coli BL21(DE3) наличием рекомбинантной плазмиды pET15b(+) со вставкой участка gag-гена, кодирующего капсидный белок р26 вируса ИНАН, размером в 717 п.н. (237 а.а), которая и придает ему устойчивость к ампицилину.
19. Дополнительные сведения:
штамм-продуцент Е. coli BL21(DE3)/pET15b-p26AEI/1, содержащий рекомбинантную плазмиду рЕТ15и(+) со вставкой участка gag-гена p26AEI-6×His, кодирующего основной капсидный белок р26 вируса ИНАН, размером в 717 пн (237 а.а) с молекулярной массой 26 кДa, который обеспечивает высокий уровень экспрессии целевого гена и синтез (репродукцию) рекомбинантного белка р26 в растворимой форме, обладающего антигенными свойствами соответствующего вирусного капсидного белка ИНАН. Конечный выход целевого белка составляет не менее 15-20 мг с 1 л бактериальной культуры.
Прочие:
20. Условия культивирования: клетки штамма-продуцента Е. coli BL21(DE3)pET15b-p26AEI/1 культивировали в 500 мл среды YTx2 (1,6% триптон, 1% дрожжевой экстракт, 0,5% NaCl), содержащей ампициллин (100 мкг/мл) при 37°С на орбитальном шейкере при 180 об/мин в течение 4 часов после индукции белкового биосинтеза изопропилтио-β-D-галактозида (ИПТГ), до оптической плотности, достигающей 0,6-0,8 оптических единиц при длине волны - 600 нм.
21. Условия хранения: штамм хранится в пробирках типа эппендорф в нативном виде при минус 80°С среде LB, содержащей 15% глицерина. Объем составляет 0.3 мл, который содержит 106 КОЕ/мл.
22. Литературные ссылки: нет.
23. Авторы: Каторкин С.А., Капустина О.В., Алексеенкова С.В., Гулюкин А.М. Гулюкин Е.А.
24. Форма депонирования: ответственное хранение.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ ИЗОБРАЖЕНИЙ
Эти и другие объекты, признаки и многие сопутствующие преимущества изобретения будут лучше поняты при чтении следующего подробного описания при рассмотрении в связи с прилагаемыми чертежами, в которых:
Фиг. 1. Последовательность фрагмента gag-гена лентивируса ИНАН размером 717 п.н., кодирующего полную нуклеотидную последовательность основного капсидного белка р26, нуклеотидная последовательность которого доступа под номером GenBank: DQ452090.1 - Equine infectious anemia virus p26 recombinant protein gene, partial cds.
Фиг. 2. Схематическое изображение плазмидной ДНК. Экспрессионный вектор pET15b-p26AEI; индуцибельный LacI промотор, ген устойчивости к ампициллину. Наличие 6×His-tag на С - концевом участке молекулы. Получена и использована в дальнейшей работе экспрессирующая генно-инженерная конструкция pET15b-p26AEI.
Фиг. 3. Результаты выравнивания аминокислотной последовательности целевого гена p26AEI-6×His в клоне №1 конструкции pET15b-p26AEI. Учитывая все критерии стратегии получения рекомбинантного белка р26 с 1 по 226 аминокислоту, финальная аминокислотная последовательность выглядит следующим образом: к указанной последовательности был добавлен сигнальный пептид и на С-концевом участке молекулы - 6×His-tag. Рассчитанная аминокислотная последовательность целевого гена в клоне №1 конструкции pET15b-p26AEI полностью совпадает с аминокислотным участком gag-гена капсидного белка р26 вируса ИНАН (NCBI) с 1 по 226 аминокислоту.
Фиг. 4. Анализ кодоновой оптимизации белка р26 вируса ИНАН при экспрессии в прокариотической системе экспрессии. Для увеличения эффективности трансляции каждого кодона белка р26 вируса ИНАН была проведена кодоновая оптимизация с подбором кодона при транскрипции в клетках E.coli.
Фиг. 5. Колонии положительных клонов (Lig) после этапа трансформации E.coli штамм BL-21 DE3 (культивирование термостате в течение 16 часов при 37°С).
Фиг. 6. Экспрессия рекомбинантного белка р26 вируса ИНАН лошадей в клетках Е. coli штамм BL-21 DE3. Индукция 1 мМ IPTG. Экспрессия три часа после индукции. Уровень экспрессии составил около 30% от тотального белка. Локализация в цитоплазме в растворимой форме.
Фиг. 7. Результаты очистки, анализа чистоты и подлинности синтезируемого рекомбинантного белка р26 вируса ИНАН лошадей в клетках Е. coli штамм BL-21 DE3
Позиция А. Электрофоретическая чистота рекомбинантного белка р26 составила более 95% (по данным электрофореза SDS-PAGE); при анализе в SDS-PAGE как в восстанавливающих, так и в денатурирующих условиях очищенный белок мигрировал в виде мономера 26 кДа. Гель-фильтрационная хроматография также указывает на то, что белок находится в растворе в виде мономера.
Позиция В. Специфическое связывание с антителами против 6×His-tag в формате Western Blot.
Фиг. 8. Реактивность и специфичность рекомбинантного белка р26 вируса ИНАН (rp26) и сывороток лошадей в непрямом ТФ ИФА.
Плашка 1. На подложке рекомбинантный белок р26 ИНАН (полноразмерный) 500 нг/лунку. 2-кратные разведения сывороток лошадей с 1:100: F1-3 - отрицательная (К-) сыворотка лошади из коммерческого «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината; G,H1-3 - нормальные полевые сыворотки лошадей; F4-12 - специфическая к ИНАН (К+) из коммерческого «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината с активностью в РДП 1:8; G4-12 - специфическая к ИНАН сыворотка лошади 1 с активностью в РДП 1:4; Н1-4 -специфическая к ИНАН сыворотка лошади 2, полевая.
Результаты. Нормальные сыворотки интактных лошадей не взаимодействуют с rp26 вируса ИНАН. Титр специфических к ИНАН сывороток лошадей составил IgG - 1:1600-1:12800.
Плашка 2. На подложке рекомбинантный белок р26 (из расчета 250 нг/лунку) и специфический антиген ИНАН (из расчета 2 мкг/лунку), ряды: 1,4 - специфический антиген ИНАН из коммерческого «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината; 2,5 - rp26 (фрагмент); 3,6 - rp26 (полноразмерный). Специфические к ИНАН сыворотки лошадей в разведении с 1:100 3-х кратным шагом: AD1-3 - сыворотка лошади из коммерческого «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината с активностью в РДП 1:8; AD4-6 - слабо положительная сыворотка лошади полевая 1 с активностью в РДП 1:4.
Вторичные антитела IgG кролика к Ig лошади (1:500), пероксидазный конюгат протеина А (1:800).
Результаты. Титр специфических к ИНАН IgG сывороток лошадей составил: со специфическим коммерческим антигеном - 1:100-1:300; с рекомбинантным белком р26 - 1:300-1:900.
Фиг. 9. Антигенная активность рекомбинантного белка р26 вируса ИНАН (rp26).
На подложке рекомбинантный белок р26 (rp26) и специфический антиген ИНАН из расчета 2 мкг/лунку: 1,4 - специфический антиген ИНАН из коммерческого «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината; 2,5 - rp26 (фрагмент); 3,6 - rp26 (полноразмерный). Сыворотки кур, иммунизированные rp26 в разведении с 1:100 3-х кратным шагом: AD1-6 - IgM и Е-Н1-6 - IgG(Y).
Конъюгат вторичных антител: AD1-6 - IgG мыши к IgG(Y) (H+L) кур; Е-Н1-6 - IgG мыши к IgM кур.
Результаты. Титр специфических антител кур к rp26 ИНАН: со специфическим коммерческим антигеном - 1:100-1:300 IgG(Y); 1:300 IgM кур; с рекомбинантным белком р26 - 1:900-1:2700 IgG(Y); 1:2700-5400 IgM кур.
Фиг. 10. Активность и специфичность, рекомбинантного белка р26 вируса ИНАН (гр26) и сывороток животных в реакции непрямой гемагглютинации (РНГА).
Реакция непрямой гемагглютинации. Эритроциты барана, сенсибилизированные рекомбинантным белком р26 вируса ИНАН из расчета 1,0 мкг/мл. Сыворотки кур кросса Доминант, иммунизированные rp26 (100 мкг/гол.), сыворотки лошадей разведение с 1:4 двукратным шагом.
Плашка 1. Сыворотки кур: А1-2 - сыворотка SPF цыплят; В - F1-8 - сыворотки кур кросса Доминант, иммунизированные rp26 вируса ИНАН.
Результаты: титр специфических антигемагглютининов к ИНАН в сыворотках кур составил 1:8-1:16, при отсутствии признаков спонтанной агглютинации эритроцитов в лунках с сыворотками SPF цыплят (А1-2) и кур до иммунизации (Е, F1-8);
Плашка 2. Сыворотки кур и лошадей. A - G5-8 сыворотки кур: А5-8 - сыворотка SPF-цыплят, В - G5-8 - сыворотки кур кросса Доминант, иммунизированные rp26 вируса ИНАН.
Сыворотки лошадей Н5-8 и A - D9-12: Н5-8 - специфическая к ИНАН сыворотка лошади 1 с активностью в РДП 1:4; А9-12 - специфическая к ИНАН (К+) из коммерческого «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината с активностью в РДП 1:8; В9-12 - специфическая к ИНАН сыворотка лошади 2, полевая, С9-12 - нормальная сыворотка интактной лошади; D9-12 - (К-) отрицательная сыворотка лошади из коммерческого «Набора для диагностики инфекционной анемии лошадей в реакции диффузионной преципитации (РДП)» Щелковского биокомбината.
Результаты: титр антигемагглютининов в сыворотках кур, иммунизированных rp26 составил 1:4 (В, С5-8), 1:8-1:16 (F, G5-8), при отсутствии признаков агглютинации эритроцитов в лунках с сыворотками SPF-цыплят и кур до иммунизации (D, E5-8); титр антигемагглютининов в сыворотках лошадей, специфичных к ИНАН составил 1:4-1:8 (Н5-8, А39-12,), при отсутствии признаков спонтанной агглютинации эритроцитов в лунках с отрицательными сыворотками лошадей (С9-12 и D9-12).
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="ЭКСПРЕССИОННЫЙ
ПЛАЗМИДНЫЙ ВЕКТОР ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА Р26
АУТЕНТИЧНОГО ОСНОВНОМУ КАПСИДНОМУ БЕЛКУ Р26 ВИРУСА ИНФЕКЦИОННОЙ
АНЕМИИ ЛОШАДЕЙ В ПРОКАРИОТИЧЕСКОЙ СИСТЕМЕ..xml" softwareName="WIPO
Sequence" softwareVersion="2.3.0" productionDate="2024-09-04">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2024108390</ApplicationNumberText>
<FilingDate>2024-03-29</FilingDate>
</ApplicationIdentification>
<ApplicantName languageCode="ru">ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ
БЮДЖЕТНОЕ НАУЧНОЕ УЧРЕЖДЕНИЕ "ФЕДЕРАЛЬНЫЙ НАУЧНЫЙ ЦЕНТР -
ВСЕРОССИЙСКИЙ НАУЧНО-ИССЛЕДОВАТЕЛЬСКИЙ ИНСТИТУТ ЭКСПЕРИМЕНТАЛЬНОЙ
ВЕТЕРИНАРИИ ИМЕНИ К.И. СКРЯБИНА И Я.Р. КОВАЛЕНКО РОССИЙСКОЙ АКАДЕМИИ
НАУК" (ФГБНУ ФНЦ ВИЭВ РАН)</ApplicantName>
<ApplicantNameLatin>Federal Scientific Centre VIEV (FSC
VIEV)</ApplicantNameLatin>
<InventorName languageCode="ru">Каторкин Сергей
Александрович</InventorName>
<InventorNameLatin>Katorkin Sergey Alexandrovich
</InventorNameLatin>
<InventionTitle languageCode="ru">ЭКСПРЕССИОННЫЙ ПЛАЗМИДНЫЙ ДНК
ВЕКТОР pET15b-p26AEI РАЗМЕРОМ 6350 П.О ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО
БЕЛКА Р26 АУТЕНТИЧНОГО ОСНОВНОМУ КАПСИДНОМУ БЕЛКУ Р26 ВИРУСА
ИНФЕКЦИОННОЙ АНЕМИИ ЛОШАДЕЙ В ПРОКАРИОТИЧЕСКОЙ
СИСТЕМЕ.</InventionTitle>
<SequenceTotalQuantity>4</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>24</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..24</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>infectious anemia of
horses</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gaattcatagatggggctggaaac</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>infectious anemia of
horses</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gtaattgggcgctaagtctag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>24</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..24</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>infectious anemia of
horses</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aagcttatagatggggctggaaac</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>23</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..23</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q8">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>infectious anemia of
horses</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aagctttgtactttccaaagtcc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---

Изобретение относится к области биотехнологии, в частности к экспрессионному плазмидному ДНК вектору pET15b-p26AEI размером 6350 п.о. Указанный вектор предназначен для получения рекомбиантного белка р26 вируса инфекционной анемии лошадей (ИНАН) в прокариотической системе и включает: индуцибельный Lac промотор; последовательность устойчивости к ампициллину; последовательность фрагмента gag-гена, кодирующего основной капсидный белок р26, размером 717 п.н.; шесть полигистидиновых последовательностей (6×His-tag) на С-конце белка; pUC ori - сайт начала репликации плазмиды; bla- ген β-лактамазы для селекции целевых клонов в прокариотических клетках. Настоящее изобретение обеспечивает высокий уровень экспрессии целевого гена и синтез (репродукцию) рекомбинантного белка р26. 10 ил., 1 табл., 5 пр.
Экспрессионный плазмидный ДНК вектор pET15b-p26AEI размером 6350 п.о. для получения рекомбиантного белка р26 вируса инфекционной анемии лошадей в прокариотической системе, включающий в себя следующие элементы: индуцибельный Lac промотор; последовательность устойчивости к ампициллину; последовательность фрагмента gag-гена, кодирующего основной капсидный белок р26 вируса инфекционной анемии лощадей (ИНАН), размером 717 п.н.; шесть полигистидиновых последовательностей (6×His-tag) на С-конце белка; pUC ori - сайт начала репликации плазмиды; bla- ген β-лактамазы для селекции целевых клонов в прокариотических клетках.
| US 2003143205 A1, 31.07.2003 | |||
| WO 2019246056 A1, 26.12.2019 | |||
| DE ARRUDA COUTINHO, L.C | |||
| et al | |||
| Production of Equine Infectious Anemia Virus (EIAV) antigen in Pichia pastoris | |||
| Journal of virological methods, 2013, 191(2), 95-100 | |||
| ZHANG, W | |||
| et al | |||
| Gag protein epitopes recognized by ELA-A-restricted cytotoxic T lymphocytes from horses with long-term |

