Область техники
Настоящее изобретение относится к микроорганизму рода Corynebacterium, продуцирующему L-аминокислоты, и к способу получения L-аминокислот с его применением.
Предшествующий уровень техники
L-аминокислоты использовали в областях кормов для животных, фармацевтических препаратов и косметики и в основном получали посредством ферментации с использованием штаммов, принадлежащих к роду Corynebacterium или Escherichia. Для получения L-аминокислот были проведены различные исследования для разработки штаммов, продуцирующих L-аминокислоты с высокой эффективностью, и технологии процесса ферментации.
Конкретно, в основном использовали подходы, специфичные в отношении целевого вещества, такие как способы увеличения экспрессии гена, кодирующего фермент, вовлеченный в биосинтез L аминокислот, или способы удаления генов, ненужных для биосинтеза (US 8048650 В2 и т.д.).
Описание изобретения
Техническая задача
Авторы настоящего изобретения подтвердили усиление способности микроорганизма рода Corynebacterium, имеющего ослабленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, продуцировать L-аминокислоты, тем самым завершили настоящее изобретение.
Техническое решение
Задача настоящего изобретения заключается в предложении микроорганизма рода Corynebacterium, имеющего ослабленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1.
Другая задача настоящего изобретения заключается в предложении способа получения L-аминокислот, включающего культивирование микроорганизма в культуральной среде.
Другая задача настоящего изобретения заключается в предложении композиции для продуцирования L-аминокислот, содержащей микроорганизм, питательную среду или их комбинацию.
Другая задача настоящего изобретения заключается в предложении способа получения микроорганизма рода Corynebacterium, продуцирующего L-аминокислоты, где способ включает ослабление активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1.
Другая задача настоящего изобретения заключается в предложении применения микроорганизма рода Corynebacterium, имеющего ослабленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, для продуцирования L-аминокислот.
Полезные эффекты
L-аминокислоты могут быть получены с высокой эффективностью с применением микроорганизма по настоящему изобретению.
Лучший вариант
Настоящее изобретение будет подробно описано ниже. Вместе с тем, каждое описание и воплощение, раскрытое в настоящем изобретении, может быть применено к другим описаниям и воплощениям в данном документе. Другими словами, все комбинации различных компонентов, раскрытых в настоящем изобретении, включены в объем настоящего изобретения. Кроме того, объем настоящего изобретения не следует ограничивать конкретными описаниями, представленными ниже.
Специалисты в данной области техники поймут или смогут установить, используя не более чем рутинные эксперименты, множество эквивалентов для конкретных воплощений настоящего изобретения. Предполагается, что такие эквиваленты включены в объем настоящего изобретения.
В одном аспекте настоящего изобретения предложен микроорганизм рода Corynebacterium, имеющий ослабленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1.
Белок по настоящему изобретению может иметь, содержать или по существу состоять из аминокислотной последовательности, представленной в SEQ ID NO: 1.
При использовании здесь термин "белок, содержащий аминокислотную последовательность SEQ ID NO: 1" представляет собой белок, имеющий активность компонента пермеазного ABC-транспортера и может иметь номер NCgl1917 в NCBI (Национальный центр биотехнологической информации).
В настоящем изобретении белок, содержащий аминокислотную последовательность SEQ ID NO: 1, может представлять собой эндогенный белок штамма, но не ограничен этим.
Аминокислотная последовательность белка NCgl1917 может быть получена из GenBank Национального института здравоохранения США (NIH), который представляет собой известную базу данных. Например, информация об аминокислотной последовательности белка NCgl1917 может быть идентифицирована в эталонной последовательности NCBI WP_006284211, но без ограничения этим. Например, белок NCgl1917 может относиться к белку, имеющему пермеазную активность АВС-транспортера, полученному из рода Corynebacterium, но без ограничения этим.
В настоящем изобретении аминокислотная последовательность белка NCgl1917 может содержать аминокислотную последовательность, имеющую по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99.5%, 99,7%, или 99,9% гомологии или идентичности с аминокислотной последовательностью, представленной в SEQ ID NO: 1. Также, специалисту в данной области техники очевидно, что любой белок, имеющий аминокислотную последовательность, содержащую делецию, модификацию, замену, консервативную замену или добавление одной или нескольких аминокислот, попадает в объем настоящего изобретения, если этот белок имеет аминокислотную последовательность, сохраняющую гомологию или идентичность и эффекты, соответствующие белку по настоящему изобретению.
Примеры этого могут включать добавление или делецию последовательности на N-конце, С-конце и/или в середины аминокислотной последовательности и встречающиеся в природе мутации, молчащую мутацию или консервативную замену, не вызывающие изменений в функциях белка.
При использовании здесь термин "консервативная замена" относится к замене одной аминокислоты другой аминокислотой, имеющей аналогичное структурное и/или химическое свойство. Белок может содержать по меньшей мере одну консервативную замену, сохраняя при этом по меньшей мере одну биологическую активность. Такая аминокислотная замена в основном может происходить на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильное™ и/или амфипатической природы остатка. Например, электрически заряженные аминокислоты с боковыми цепями включают положительно заряженные (основные) аминокислоты, такие как аргинин, лизин и гистидин, и отрицательно заряженные (кислые) аминокислоты, такие как глутаминовая кислота и аспарагиновая кислота; а незаряженные аминокислоты с боковыми цепями включают неполярные аминокислоты, такие как глицин, аланин, валин, лейцин, изолейцин, метионин, фенилаланин, триптофан и пролин, и полярные или гидрофильные аминокислоты, такие как серии, треонин, цистеин, тирозин, аспарагин и глутамин. Среди аминокислот ароматические аминокислоты включают фенилаланин, триптофан и тирозин.
При использовании в данном документе термин "гомология" или "идентичность" относится к степени соответствия двух взятых аминокислотных последовательностей или нуклеотидных последовательностей и может быть выражен в процентах. Термины "гомология" и "идентичность" часто могут быть использованы взаимозаменяемо друг с другом.
Гомология последовательностей или идентичность консервативных полинуклеотидов или полипептидов может быть определена с помощью стандартного алгоритма выравнивания и может использоваться со штрафом за пробелы по умолчанию, установленным используемой программой. По существу, гомологичные или идентичные последовательности могут в общем случае гибридизоваться с каждой другой целиком или с ее частью в умеренных или очень жестких условиях. Очевидно, что гибридизация включает гибридизацию полинуклеотида с полинуклеотидом, содержащих обычный кодон или вырожденный кодон.
Гомология, сходство или идентичность двух полинуклеотидных или белковых последовательностей, может быть определена) с использованием любого компьютерного алгоритма, известного в данной области техники, например программы "FASTA" с использованием параметров по умолчанию, как раскрыто в Pearson et al., (1988) Proc. Natl. Acad. Sci. USA 85:2444). Альтернативно, гомология, сходство или идентичность может быть определена(о) с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), который выполняют в программе Needleman из пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277) (версия 5.0.0 или последующие) (включая пакет программ GCG (Devereux, J. et al., Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S.F. et al., J. MOLEC BIOL 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994, и [CARILLO et al.] (1988) ШАМ J Applied Math 48:1073). Например, гомология, сходство или идентичность может быть определена(о) с использованием BLAST из базы данных Национального центра биотехнологической информации (NCBI) или ClustalW.
Гомология, сходство или идентичность полинуклеотидов или белков могут быть определены путем сравнения информации о последовательностях с использованием компьютерной программы GAP (например Needleman et al. (1970), J.Mol Biol. 48: 443), как раскрыто в Smith and Waterman, Adv. Appl. Math (1981) 2:482. В кратком изложении, программа GAP определяет сходство как количество выровненных символов (т.е. нуклеотидов или аминокислот), которые аналогичны, на общее количество символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать (1) двоичную матрицу сравнения (содержащую значение 1 для идентичностей и 0 для неидентичностей) и взвешенную матрицу сравнения Gribskov et al. (1986), Nucl. Acids Res. 14:6745, как описано в Schwartz and Dayhoff, eds., Atlas of Protein Sequence and Structure, Национального фонда биомедицинских исследований, стр. 353-358 (1979) (или матрицу замещения EDNAFULL (EMBOSS version of NCBI NUC4.4)); (2) штраф 3,0 за каждый пробел и дополнительный штраф 0,10 за каждый символ в каждом пробеле (или штраф за открытие пробела в размере 10 и штраф за продление пробела в размере 0,5); и (3) отсутствие штрафа за конечные пробелы.
В настоящем изобретении полинуклеотид, кодирующий белок, содержащий аминокислотную последовательность SEQ ID NO: 1, может быть назван геном Ncgl1917.
При использовании здесь термин "полинуклеотид" относится к цепи ДНК или РНК, имеющей определенную минимальную длину, в виде полимера из нуклеотидов, в котором нуклеотидные мономеры связаны друг с другом в виде длинной цепи посредством ковалентных связей. Конкретно, полинуклеотид может представлять собой полинуклеотидный фрагмент, кодирующий белок.
Полинуклеотид, кодирующий белок по настоящему изобретению, может содержать нуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную в SEQ ID NO: 1. В качестве примера настоящего изобретения, полинуклеотид по настоящему изобретению может иметь или содержать последовательность SEQ ID NO: 2. Кроме того, полинуклеотид по настоящему изобретению может содержать последовательность SEQ ID NO: 2 или по существу состоять из нее.
Полинуклеотид по настоящему изобретению может содержать различные модификации, сделанные в кодирующей области в диапазоне, не изменяющем аминокислотную последовательность полипептида, экспрессируемого с кодирующей области, с учетом вырожденности кодона или кодона, предпочтительного для живого организма, в котором должен экспрессироваться полипептид.
Конкретно, полинуклеотид по настоящему изобретению может иметь или содержать нуклеотидную последовательность, имеющую гомологию или идентичность 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более и менее 100%, с последовательностью SEQ ID NO: 2, или может состоять или по существу состоять из нуклеотидной последовательности, имеющей гомологию или идентичность 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более и менее 100%, с последовательностью SEQ ID NO: 2, без ограничения этим.
Кроме того, полинуклеотид может содержать любой зонд, полученный из любых известных последовательностей генов, например нуклеотидной последовательности, гибридизованной в жестких условиях с последовательностью, полностью или частично комплементарной описанной выше полинуклеотидной последовательности, без ограничения. Термин «жесткие условия» относится к условиям, делающим возможной специфическую гибридизацию между полинуклеотидами. Такие условия подробно раскрыты в известных документах (J. Sambrook et al. Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York, 9.50-9.51, 11.7-11.8). Например, жесткие условия могут включать условия, делающие возможной гибридизацию между полинуклеотидами, имеющими высокую гомологию или идентичность, например гомологию или идентичность 70% или более, 75% или более, 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более, или 99% или более, не допуская гибридизацию между полинуклеотидами, имеющими гомологию или идентичность ниже указанных выше гомологий или идентичностей, или промывку один раз, конкретно, два или три раза, в обычных условиях промывки для Саузерн-гибридизации, при концентрации соли и температуре, соответствующих 60°С, 1 × SSC (цитратно-солевой раствор), 0,1% SDS (додецилсульфат натрия), конкретно 60°С, 0,1 × SSC, 0,1% SDS и, более конкретно 68°С, 0,1 × SSC, 0,1% SDS.
Гибридизация требует, чтобы две нуклеиновые кислоты имели комплементарные последовательности, хотя возможны ошибки спаривания между основаниями, в зависимости от жесткости гибридизации. Термин "комплементарный" используют для описания взаимоотношения между нуклеотидными основаниями, которые могут гибридизоваться друг с другом. Например, в контексте ДНК аденин комплементарен тимину и цитозин комплементарен гуанину. Таким образом, полинуклеотид по настоящему изобретению может содержать не только нуклеиновокислотные последовательности, по существу аналогичные последовательностям полинуклеотида, но также выделенные фрагменты нуклеиновой кислоты, комплементарные всей последовательности полинуклеотида.
Конкретно, полинуклеотиды, имеющие гомологию или идентичность с полинуклеотидом по настоящему изобретению, могут быть обнаружены с использованием условий гибридизации, включающих процесс гибридизации при значении Tm 55°С и описанные выше условия. Кроме того, значение Tm может составлять 60°С, 63°С или 65°С, но не ограничено ими, и может быть соответствующим образом скорректировано специалистами в данной области техники в зависимости от предполагаемой цели.
Подходящая жесткость для гибридизации полинуклеотидов может зависеть от длины полинуклеотидов и степени комплементарности, и эти переменные хорошо известны в данной области техники (например Sambrook et al. выше).
При использовании здесь термин "микроорганизм (или штамм)" включает как микроорганизмы дикого типа, так и микроорганизмы, содержащие естественную или искусственную генетическую модификацию, такие как микроорганизмы, имеющие конкретный механизм, ослабленный или усиленный посредством введения экзогенного гена или усиления или инактивации эндогенного гена, и содержащие генетическую модификацию для получения целевого белка, протеина или продукта.
Микроорганизм по настоящему изобретению может представлять собой: микроорганизм, имеющий ослабленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1 по настоящему изобретению; микроорганизм, в котором экспрессия полинуклеотида, кодирующего белок, содержащий аминокислотную последовательность SEQ ID NO: 1 по настоящему изобретению, является ослабленный; или микроорганизм, генетически модифицированный (например рекомбинантный микроорганизм) для ослабления активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1 по настоящему изобретению, с использованием вектора, без ограничения этим.
Например, ослабленная активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, может представлять собой ослабленную экспрессию полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 1 или полинуклеотидa SEQ ID NO: 2. Например, ослабление активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1, может быть достигнуто путем модификации нуклеотидной последовательности полинуклеотида, например вставки, делеции или замены нуклеотидов. Однако данное воплощение не ограничивается этим.
Микроорганизм по настоящему изобретению может представлять собой микроорганизм, имеющий способность продуцировать L-аминокислоты.
Микроорганизм по настоящему изобретению может представлять собой микроорганизм, естественным образом имеющий способность продуцировать L-аминокислоты, или микроорганизм, полученный путем придания родительскому штамму, не способному продуцировать L-аминокислоты, способности продуцирования с ослабленной способностью белка, содержащего аминокислотную последовательность SEQ ID NO: 1 по настоящему изобретению, но без ограничения этим.
Например, микроорганизм по настоящему изобретению может представлять собой штамм или микроорганизм, имеющий ослабленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, посредством трансформации с использованием вектора, сконструированного для ослабления активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1 по настоящему изобретению. С учетом целей настоящего изобретения, микроорганизм может представлять собой микроорганизм, имеющий усиленную способность продуцировать L-аминокислоты, по сравнению с модифицированными микроорганизмами (например встречающийся в природе микроорганизм дикого типа или микроорганизм, в котором активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, не модифицирована), путем ослабления активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1, по настоящему изобретению, во встречающемся в природе микроорганизме дикого типа или микроорганизме, продуцирующем L-аминокислоту, но без ограничения этим.
Например, немодифицированный микроорганизм, в качестве штамма для сравнения с точки зрения усиления способности продуцировать L-аминокислоты, может представлять собой Corynebacterium glutamicum KCCM12502P (KR 102126951 В1), KCCM11222P (US 10590446 B2) или KCCM11248P (KR 101335789 B1), без ограничения этим.
Например, в штамме, имеющем усиленную способность продуцирования L-аминокислоты, способность продуцировать L-аминокислоты может быть усилена примерно на 1% или более, конкретно на 1% или более, примерно на 2,5% или более, примерно на 5% или более, примерно на 6% или более, примерно на 6% или более, примерно на 7% или более, примерно на 8% или более, примерно на 9% или более, примерно на 10% или более, примерно на 10,5% или более, примерно на 11% или более, примерно на 11,5% или более, примерно на 12% или более, примерно на 12,5% или более, примерно на 13% или более, примерно на 13,5% или более, примерно на 14% или более, примерно на 14,5% или более, примерно на 15% или более, примерно на 15,5% или более, примерно на 16% или более, примерно на 16,5% или более, примерно на 17% или более, примерно на 17,5% или более, примерно на 18% или более, примерно на 18,5% или более, примерно на 19% или более, примерно на 19,5% или более примерно 20% или более, примерно на 20,5% или более, примерно на 21% или более, примерно на 21,5% или более, примерно на 22% или более, примерно на 22,5% или более, примерно на 23% или более, примерно на 23,5% или более, примерно на 24% или более, примерно на 24,5% или более, примерно на 25% или более, примерно на 25,5% или более, примерно на 26% или более, примерно на 26,5% или более, примерно на 27% или более, примерно на 27,5% или более, примерно на 28% или более, примерно на 28,5% или более, примерно на 29% или более, примерно на 30% или более, примерно на 30,5% или более, примерно на 31% или более, примерно на 31,5% или более, примерно на 32% или более, примерно на 32,5% или более, примерно на 33% или более, примерно на 33,5% или более, примерно на 34% или более, примерно на 34,5% или более, примерно на 35% или более, по сравнению со способностью продуцирования L-аминокислот у родительского штамма до модификации или немодифицированного микроорганизма, но настоящее изобретение не ограничивается этим, до тех пор, пока получается положительное значение прироста способности продуцировать L-аминокислоты по сравнению со способностью продуцировать L-аминокислоты у родительского штамма до модификации или у немодифицированного микроорганизма. В другом примере в рекомбинантном штамме, имеющем усиленную способность продуцирования L-аминокислоты, способность продуцирования L-аминокислоты может быть усилена примерно в 1,1 раза или более, примерно в 1,12 раза или более, примерно в 1,13 раза или более, примерно в 1,15 раза или более, примерно в 1,16раза или более, примерно в 1,17 раза или более, примерно в 1,18 раза или более примерно в 1,19 раза или менее, примерно в 1,2 раза или более, примерно в 1,25 раза или менее, примерно в 1,3 раза или более, примерно в 1,35 раза или более по сравнению с родительским штаммом до модификации или немодифицированным микроорганизмом, без ограничения этим. Термин "примерно" относится к диапазону, включающему ±0,5, ±0,4, ±0,3, ±0,2, ±0,1 или подобные и включает все числовые значения в диапазоне, равном или близком к числовому значению, следующему за термином "примерно", без ограничения этим.
Например, микроорганизм, имеющий усиленную способность продуцировать L-аминокислоты, согласно настоящему изобретению, может иметь активность белка, содержащего аминокислотную последовательность SEQ ID NO:l, менее 100%, например, примерно 99,9% или менее, примерно 99% или менее, около 98% или меньше, примерно 97% или менее, примерно 96% или менее, примерно 95% или менее, около 90% или менее, примерно 80% или менее, примерно 70% или менее, примерно 60% или менее, примерно 50% или менее, примерно 40% или менее, примерно 30% или меньше, примерно 20% или менее, примерно 10% или менее, примерно 5% или менее или примерно 0% по сравнению с родительским штаммом до модификации или немодифицированным микроорганизмом, но без ограничения этим.
При использовании здесь термин "немодифицированный микроорганизм" не исключает штаммы, содержащие мутацию, встречающуюся в природе у микроорганизмов, и может относиться к штамму дикого типа или штамму природного типа, или к штамму до трансформации посредством генетической модификации, вызванной естественным или искусственным фактором. Например, немодифицированный микроорганизм может относиться к микроорганизму до ослабления активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1, как описано здесь. "Немодифицированный микроорганизм" может использоваться взаимозаменяемо со "штаммом до модификации", "микроорганизмом до модификации", "немутированным штаммом", "немодифицированным штаммом", "немутированным микроорганизмом" или "эталонным микроорганизмом".
В другом примере настоящего изобретения микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой Corynebacterium glutamicum, Corynebacterium crudilactis, Corynebacterium deserti, Corynebacterium efficiens, Corynebacterium callunae, Corynebacterium stationis, Corynebacterium singulare, Corynebacterium halotolerans, Corynebacterium striatum, Corynebacterium ammoniagenes, Corynebacterium pollutisoli, Corynebacterium imitans, Corynebacterium testudinoris или Corynebacterium flavescens.
При использовании в этом документе термин «ослабление» белка представляет собой расширенное понятие, которое включает как снижение, так и удаление активности, по сравнению с собственной активностью. Термин «ослабление» можно использовать взаимозаменяемо с такими терминами, как инактивация, отсутствие, понижающая регуляция, уменьшение, снижение и аттенуирование.
Ослабление может включать: случай, когда активность белка снижена или удалена по сравнению с собственной активностью белка, проявляемой микроорганизмом, вследствие мутации полинуклеотида, кодирующего белок или подобного; случай, когда активность и/или концентрация (уровень экспрессии) белка в клетках являются более низкими, чем у штамма дикого типа, вследствие подавленной экспрессии полинуклеотида, кодирующего указанное, или подавленной трансляции его в белок; случай, когда полинуклеотид вообще не экспрессируется; и/или случай, когда не получают активность белка, хотя полинуклеотид экспрессируется. Термин "собственная активность" относится к активности белка, которой первоначально обладал родительский штамм до трансформации, дикий тип или немодифицированный микроорганизм, когда микроорганизм трансформируют посредством генетической модификации, вызванной естественным или искусственным фактором. Собственная активность также может использоваться взаимозаменяемо с "активностью до модификации". "Инактивация, дефицит, понижающая регуляция, уменьшение, редуцирование и аттенуирование" активности белка по сравнению с собственной активностью означает снижение активности по сравнению с активностью белка, которой первоначально обладал родительский штамм до трансформации или немодифицированный микроорганизм.
Ослабление активности белка может быть осуществлено посредством, но без ограничения им, любого способа, известного в данной области техники, и может быть достигнуто посредством применения различных способов, хорошо известных в данной области техники (например, Nakashima N et al., Bacterial cellular engineering by genome editing and gene silencing. Int J Mol Sci. 2014;15 (2): 2773-2793, Sambrook et al. Molecular Cloning 2012 и т.д.).
Конкретно, ослабление белка по настоящему изобретению может быть достигнута посредством:
1) делеции гена, кодирующего белок, полностью или частично;
2) модификации области регуляции экспрессии гена (или последовательности регуляции экспрессии) для снижения экспрессии гена, кодирующего белок;
3) модификации аминокислотной последовательности, составляющей белок, для удаления или ослабления активности белка (например делеции/замены/добавления по меньшей мере одной аминокислоты в аминокислотной последовательности);
4) модификации последовательности гена, кодирующего белок, для удаления или ослабления активности белка (например делеции/замены/добавления по меньшей мере одного основания нуклеиновой кислоты в последовательности оснований гена белка для кодирования белка, модифицированного для удаления или ослабления активности этого белка);
5) модификации последовательности оснований, кодирующей инициирующий кодон, или 5'-UTR области транскрипта гена, кодирующего белок;
6) введения антисмыслового олигонуклеотида (например антисмысловой РНК), комплементарно связывающегося с транскриптом гена, кодирующего белок;
7) добавления последовательности, комплементарной последовательности Шайна-Дальгарно гена, кодирующего белок, выше последовательности Шайна-Дальгарно с образованием вторичной структуры, делающей связывание рибосомы невозможным;
8) добавления промотора обратной транскрипции на 3'-конец открытой рамки считывания (ORF) последовательности гена, кодирующего белок (генная инженерия обратной транскрипции, RTE); или
9) любой комбинации из двух или более, выбранных из приведенных выше пп. 1-8, без ограничения этим.
Например, делеция гена, кодирующего белок, полностью или частично, описанная в (1) выше, может быть достигнута путем делеции всего полинуклеотида, кодирующего внутренний целевой белок в хромосоме, или путем замены полинуклеотида на полинуклеотид, в котором один или несколько нуклеотидов удалены, или геном-маркером.
Также модификация области регуляции экспрессии (или последовательности регуляции экспрессии), описанная в (2) выше, может быть достигнута посредством мутации в области регуляции экспрессии (или последовательности регуляции экспрессии) вследствие делеции, вставки, неконсервативной или консервативной замены, или любой их комбинации или замены последовательностью, имеющей более слабую активность. Область регуляции экспрессии содержит промотор, операторную последовательность, последовательность, кодирующую сайта связывания рибосомы, и последовательность регулирования окончания транскрипции и трансляции, без ограничения этим.
Кроме того, модификация последовательности оснований, кодирующей инициирующий кодон, или 5'-UTR-области транскрипта гена, кодирующего белок, описанный в (3) выше, может быть достигнута, например, посредством замены последовательности основание, кодирующей внутренний инициирующий кодон, другой последовательностью оснований, кодирующей инициирующий кодон с более низким уровнем экспрессии белка, без ограничения этим.
Кроме того, модификация аминокислотной последовательности или нуклеотидной последовательности, описанной в (4) и (5) выше, может быть проведена путем индуцирования мутации в аминокислотной последовательности белка или в нуклеотидной последовательности, кодирующей белок, посредством делеции, вставки, неконсервативной или консервативной замены, или любой их комбинации или посредством замены аминокислотной последовательности или нуклеотидной последовательности на аминокислотную последовательность или нуклеотидную последовательность, модифицированную так, чтобы иметь более слабую активность или не иметь активности, для ослабления активности белка, без ограничения этим. Например, экспрессия гена может быть подавлена или ослаблена путем формирования терминирующего кодона посредством введения мутации в нуклеотидную последовательность. В качестве другого примера модификации нуклеотидной последовательности, экспрессия полинуклеотида может быть ослаблена или активность белка, кодируемого полинуклеотидом, может быть ослаблена посредством вставки транспозона в его нуклеотидную последовательность, но воплощение этим не ограничивается.
Введение антисмыслового олигонуклеотида (например антисмысловой РНК), комплементарно связывающегося с транскриптом гена, кодирующего белок, описанный в (6) выше, может быть достигнуто способом, раскрытым в документе (Weintraub, Н. et al., Antisense-RNA as a molecular tool for genetic analysis, Reviews - Trends in Genetics, Vol.1(1) 1986).
Добавление последовательности, комплементарной последовательности Шайна-Дальгарно гена, кодирующего белок выше последовательности Шайна Дальгарно, для формирования вторичной структуры, делающей невозможным связывание рибосомы, описанное в (7) выше, может сделать невозможной трансляцию мРНК или снизить скорость.
Добавление промотора для обратной транскрипции к 3'-концу открытой рамки считывания (ORF) последовательности гена, кодирующего белок (инженерия обратной транскрипции, RTE), описанное в (8) выше, может ослаблять активность посредством образования антисмыслового нуклеотида, комплементарного транскрипту гена, кодирующего белок.
При использовании здесь термин "вектор" может включать конструкцию ДНК, содержащую последовательность оснований полинуклеотида, кодирующего целевой полипептид, и функционально связанную с подходящей областью регуляции экспрессии (последовательностью регуляции экспрессии) так, чтобы иметь возможность экспрессировать целевой полипептид в подходящей клетке-хозяине. Область регуляции экспрессии, может содержать промотор, способный инициировать транскрипцию, любую операторную последовательность для регуляции транскрипции, последовательность, кодирующую подходящий сайт связывания мРНК рибосомы, и последовательность для регуляции окончания транскрипции и трансляции. Когда подходящую клетку-хозяина трансформируют вектором, вектор может реплицироваться или функционировать независимо от генома хозяина или может быть интегрирован в его геном.
Вектор, используемый в настоящем изобретении, конкретно не ограничен, и можно использовать любой вектор, известный в данной области техники. Примеры обычно используемых векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, pWE15, М13, MB L3, MBL4, ГХП, ASHII, API, t10, t11, Charon 4A, Charon 21А и т.д. можно использовать в качестве фагового вектора или космидного вектора. В качестве плазмидного вектора можно использовать векторы на основе pDZ, на основе pBR, на основе pUC, на основе pBluescript II, на основе pGEM, на основе pTZ, на основе pCL, на основе рЕТ и т.д. Конкретно, можно использовать векторы pDZ, pDC, pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и т.д.
Например, полинуклеотид, кодирующий целевой полипептид, может быть встроен в хромосому с использованием вектора для хромосомной вставки в клетки. Встраивание полинуклеотида в хромосому может быть выполнено посредством любого способа, известного в данной области техники, например посредством гомологичной рекомбинации, без ограничения этим. Полинуклеотид может дополнительно содержать селекционный маркер для подтверждения хромосомной вставки. Селекционный маркер используют для отбора клеток, которые трансформированы вектором, то есть для подтверждения встраивания нужных молекул нуклеиновой кислоты, и можно использовать маркеры, обеспечивающие селективные фенотипы, такие как резистентности к лекарственному средству, потребность в питательных веществах, устойчивость к цитотоксическим агентам или поверхностная экспрессия полипептида. Только клетки, экспрессирующие селективный маркер, способны выживать или демонстрировать разные фенотипы в среде, обработанной селективным агентом, и таким образом могут быть отобраны трансформированные клетки.
При использовании здесь термин "трансформация" относится к процессу введения вектора, содержащего полинуклеотид, кодирующий целевой полипептид, в клетку-хозяина или микроорганизм таким образом, что полипептид, кодируемый полинуклеотидом, экспрессируется в клетке-хозяине. Трансформированный полинуклеотид может быть либо в форме, встроенной в хромосому клетки-хозяина, либо в форме, локализованной вне хромосомы, при условии, что полипептид экспрессируется в клетке-хозяине. Кроме того, полинуклеотид содержит ДНК и РНК, кодирующие целевой полипептид. Полинуклеотид может быть введен в клетку-хозяина в любой форме при условии, что полинуклеотид введен в клетку-хозяина и экспрессируется в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, содержащую все основные элементы, необходимые для саморепликации. Экспрессионная кассета в общем случае может содержать промотор, функционально связанный с полинуклеотидом, сигнал окончания транскрипции, сайт связывания рибосомы и сигнал окончания трансляции. Экспрессионная кассета может быть в форме самореплицирующегося вектора экспрессии. Также полинуклеотид может быть введен в клетку-хозяина в его исходной форме и функционально связан с последовательностью, необходимой для экспрессии в клетке-хозяине, без ограничения этим.
Кроме того, при использовании здесь термин "функционально связанный" означает функциональную связь между нуклеотидной последовательностью, кодирующей целевой белок, и промоторной последовательностью, которая инициирует и опосредует транскрипцию полинуклеотида.
В микроорганизме по настоящему изобретению модификация полинуклеотида полностью или частично может быть индуцирована посредством (а) гомологичной рекомбинации с использованием вектора для хромосомной вставки в микроорганизм или редактированием генома с использованием сконструированной нуклеазы (например CRISPR-Cas9) и/или (б) обработки излучением, таким как ультрафиолетовые лучи и радиоактивные лучи, и/или химическим веществом, без ограничения ими. Способ модификации гена полностью или частично может включать технологию рекомбинации ДНК. Например, часть или весь ген может быть делетирован посредством индуцирования гомологичной рекомбинации путем вставки в микроорганизм нуклеотидной последовательности или вектора, содержащего нуклеотидную последовательность, имеющую гомологию с целевым геном. Встроенная нуклеотидная последовательность или вектор могут содержать доминантный селекционный маркер, без ограничения этим.
В другом аспекте настоящего изобретения предлагается способ получения L-аминокислот, включающий культивирование рекомбинантного микроорганизма рода Corynebacterium, имеющего ослабленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, в культуральной среде.
Белок, содержащий аминокислотную последовательность SEQ ID NO: 1, ослабление и микроорганизм являются такими, как описано выше.
Микроорганизм рода Corynebacterium может представлять собой Corynebacterium glutamicum, но не ограничен этим, и является таким, как описано выше.
L-аминокислота может включать по меньшей мере одну аминокислоту, выбранную из L-треонина и L-изолейцина, без ограничения этим.
При использовании здесь термин "культивирование" относится к выращиванию микроорганизма рода Corynebacterium по настоящему изобретению в соответствующим образом подобранном окружении. Процесс культивирования по настоящему изобретению можно выполнять с использованием подходящей культуральной среды и условий культивирования, хорошо известных в данной области техники. Процесс культивирования может быть соответствующим образом подобран специалистом в данной области техники в соответствии с выбранным штаммом. Конкретно, культивирование можно выполнять методом периодического культивирования, методом непрерывного культивирования и методом периодического культивирования с подпиткой, без ограничения ими.
При использовании здесь термин "культуральная среда" относится к веществу, где смешаны, в качестве основных элементов, питательные вещества, необходимые для культивирования микроорганизма рода Corynebacterium, и поставляются питательные вещества и факторы роста, а также вода, которые необходимы для выживания и роста. Конкретно, хотя культуральные среды и другие условия культивирования микроорганизма рода Corynebacterium по настоящему изобретению конкретно не ограничены, если среды обычно используют в культивировании микроорганизмов, микроорганизм рода Corynebacterium по настоящему изобретению можно культивировать в обычной среде, содержащей подходящие источники углерода, источники азота, источники фосфора, неорганические соединения, аминокислоты и/или витамины, в аэробных условиях при регулировании температуры, рН и тому подобного.
Конкретно, культуральная среда для микроорганизма рода Corynebacterium раскрыта в документе ("Manual of Methods for General Bacteriology" by the American Society for Bacteriology (Washington D.C., USA, 1981)).
В настоящем изобретении в качестве источников углерода можно использовать углеводы, такие как глюкоза, сахароза, лактоза, фруктоза и мальтоза; сахарные спирты, такие как маннит и сорбит; органические кислоты, такие как пировиноградная кислота, молочная кислота и лимонная кислота; и аминокислоты, такие как глутаминовая кислота, метионин и лизин. Кроме того, можно использовать натуральные органические питательные вещества, такие как гидролизаты крахмала, меласса, черная меласса, рисовые отруби, маниока, меласса из сахарного тростника и кукурузный сироп, и, в частности, можно использовать углеводы, такие как глюкоза и стерильная предварительно обработанная меласса (т.е. меласса, превращенная в редуцирующие сахара), и подходящие количества из любых других источников углерода также можно использовать без ограничений. Эти источники углерода можно использовать по отдельности или в комбинации из по меньшей мере двух из них, но без ограничения этим.
В качестве источников азота можно использовать неорганические источники азота, такие как аммиак, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония и нитрат аммония; и органические источники азота, такие как аминокислоты, например глутаминовая кислота, метионин и глутамин, пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, солодовый экстракт, кукурузный сироп, гидролизат казеина, рыбу или продукты ее разложения и обезжиренный соевый жмых или продукты его разложения. Эти источники азота можно использовать по отдельности или в комбинации из по меньшей мере двух из них, без ограничения этим.
В качестве источников фосфора можно использовать монокалийфосфат, дикалийфосфат или соответствующие им натрийсодержащие соли. В качестве неорганических соединений можно использовать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и тому подобное. Также, можно дополнительно включать аминокислоты, витамины и/или соответствующие предшественники. Эти компоненты и предшественники можно добавлять в питательную среду в периодическом или непрерывном процессе, без ограничения этим.
В настоящем изобретении в процессе культивирования микроорганизма рода Corynebacterium к культурам можно добавлять надлежащим образом такие соединения, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота, для регулирования рН культуральной среды. Также, во время культивирования можно добавлять пеногаситель, такой как полигликолевый эфир жирных кислот, чтобы подавлять образование пены. Кроме того, кислород или кислородсодержащий газ можно вводить в питательную среду для поддержания культуральной среды в аэробном состоянии, или вводить в культуральную среду азот, водород или углекислый газы для поддержания культуры в анаэробных и микроаэробных условиях без введения каких-либо других газов.
В настоящем изобретении температуру культивирования можно поддерживать в диапазоне от 20°С до 45°С, конкретно от 25°С до 40°С, и культивирование можно выполнять в течение примерно от 10 до 160 часов, но без ограничения этим.
L-аминокислоты, полученные посредством культивированием по настоящему изобретению, могут высвобождаться в питательную среду или оставаться в клетках.
Способ получения L-аминокислот по настоящему изобретению может дополнительно включать получение микроорганизма рода Corynebacterium, приготовление культуральной среды для культивирования микроорганизма или любую их комбинацию (независимо от порядка, в любом порядке), например перед стадией культивирования.
Способ получения L-аминокислот по настоящему изобретению может дополнительно включать извлечение L-аминокислот из культуральной среды, полученной после культивирования (культурального бульона, в котором проводят культивирование), или микроорганизма рода Corynebacterium. Стадия извлечения может быть дополнительно включена после стадии культивирования.
Извлечение может представлять собой сбор целевых L-аминокислот с использованием способа культивирования микроорганизма рода Corynebacterium, например подходящего способа, известного в данной области техники, такого как периодический, непрерывный или периодический способ с подпиткой. Например, можно использовать центрифугирование, фильтрацию, обработку осадителем белка (высаливание), экстракцию, ультразвуковую дезинтеграцию, ультрафильтрацию, диализ, различные способы хроматографии, такие как молекулярно-ситовая хроматография (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография и аффинная хроматография, высокоэффективная жидкостная хроматография (ВЭЖХ) и любая их комбинация, и целевые L-аминокислоты могут быть извлечены из культуральной среды или микроорганизма с использованием подходящего способа, хорошо известного в данной области техники.
Кроме того, способ получения L-аминокислот по настоящему изобретению может дополнительно включать стадию очистки. Стадию очистки можно выполнять с использованием подходящего способа, хорошо известного в данной области техники. В одном воплощении, когда способ получения L-аминокислот по настоящему изобретению включает стадии как извлечения, так и очистки, стадии извлечения и очистки можно выполнять непрерывно или с перерывом, независимо от порядка, или можно выполнять одновременно или как одну объединенную стадию, без ограничения этим.
В другом аспекте настоящего изобретения предлагается композиция для получения L-аминокислот, содержащая: микроорганизм рода Corynebacterium, имеющий ослабленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1; культуральную среду, используемую для его культивирования; или их комбинацию.
Белок, содержащий аминокислотную последовательность SEQ ID NO: 1, ослабление, культуральная среда для микроорганизма и L-аминокислоты являются такими, как описано выше.
Композиция по настоящему изобретению может дополнительно содержать любой подходящий эксципиент, обычно используемый в композициях для получения аминокислот. Примеры эксципиента могут включать консервант, увлажнитель, разрыхлитель, суспендирующий агент, буфер, стабилизатор или изотонический агент, без ограничения этим.
В композиции по настоящему изобретению белок, содержащий аминокислотную последовательность SEQ ID NO: 1, ослабление, микроорганизм, питательная среда и L-аминокислоты, являются такими, как описано выше.
В другом аспекте настоящего изобретения предложен способ получения микроорганизма рода Corynebacterium, продуцирующего L-аминокислоты, где способ включает ослабление активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1.
Способ может дополнительно включать модификацию микроорганизма для ослабления активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1.
Ослабление белка, L-аминокислоты и микроорганизм являются такими, как описано выше.
В другом аспекте настоящего изобретения предлагается применение микроорганизма рода Corynebacterium, имеющего ослабленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, для продуцирования L-аминокислот.
Белок, содержащий аминокислотную последовательность SEQ ID NO: 1, ослабление, микроорганизм и L-аминокислоты являются такими, как описано выше.
Вариант осуществления изобретения
Далее настоящее изобретение будет описано более подробно со ссылкой на следующие примеры и экспериментальные примеры. Однако следующие примеры и экспериментальные примеры представлены только для иллюстрации настоящего изобретения, и объем настоящего изобретения не ограничивается ими.
Пример 1. Построение библиотеки случайного мутагенеза с использованием транспозона
Для получения штаммов, имеющих усиленную способность продуцировать L-треонин, библиотеку случайного мутагенеза конструировали следующим образом. Сначала используемый в качестве родительского штамма Corynebacterium glutamicum KCCM12502P (патент Кореи №10-2126951), трансформировали плазмидой, полученной с использованием набора EZ-Tn5™<R6Kγori/KAN-2>TnP Transposome™ (Epicentre) при помощи электроимпульсного способа (Appl. Microbiol. Biotechnol. (1999) 52:541-545) и наносили мазками на комплексную среду для чашек, содержащую канамицин (25 мг/л) и затем получали примерно 20000 колоний.
Комплексная среда для чашек (рН 7,0)
10 г глюкозы, 10 г пептона, 5 г говяжьего экстракта, 5 г дрожжевого экстракта, 18,5 г сердечно-мозговой вытяжки, 2,5 г NaCl, 2 г мочевины, 91 г сорбитола и 20 г агара (из расчета на 1 л дистиллированной воды).
Пример 2. Скрининг библиотеки случайного мутагенеза с использованием транспозона
Примерно 20000 колоний, полученных в Примере 1, инокулировали на селективную среду, содержащую 300 мкл канамицина (25 мг/л), и культивировали в планшете из 96 глубоких лунок при 32°С при 200 об/мин в течение примерно 24 часов.
Селективная среда (рН 8,0)
10 г глюкозы, 5,5 г (NH4)2SO4, 1,2 г MgSO4⋅7H2O, 0,8 г KH2PO4, 16,4 г K2HPO4, 100 мкг биотина, 1000 мкг тиамина HCl, 2000 мкг пантотената кальция и 2000 мкг никотинамида (из расчета на 1 л дистиллированной воды).
Для анализа продуцирования L-треонина, полученного из культурального бульона, использовали нингидриновый метод (Moore, S., Stein, W.Н. Photometric ninghydrin method for use in the chromatography of amino acids. J. Biol. Chem. 1948, 176, 367-388). После завершения культивирования, 10 мкл супернатанта культурального бульона приводили во взаимодействие со 190 мкл раствора нингидрина при 65°С в течение 30 минут, и измеряли поглощение при 570 нм с помощью спектрофотометра, и сравнивали с поглощением контрольного штамма Corynebacterium glutamicum KCCM12502P. Затем отбирали 60 колоний, демонстрирующее более высокое поглощение в качестве мутантных штаммов. Другие колонии демонстрировали поглощение, аналогичное или более низкое, чем у контрольного штамма Corynebacterium glutamicum KCCM12502P.
Отобранные 60 штаммов повторно культивировали таким же образом, как описано выше, и приводили во взаимодействие с раствором нингидрина. В результате отобрали 10 лучших мутантных штаммов, имеющих усиленную способность продуцировать L-треонин по сравнению с родительским штаммом, Corynebacterium glutamicum KCCM12502P, и назвали их 12502P-m1, 12502P-m2, 12502P-m3, 12502P-m4, 12502P-m5, 12502P-m6, 12502P-m7, 12502P-m8, 12502P-m9 и 12502P-m10 соответственно.
Пример 3. Анализ способности штаммов со случайными мутациями продуцировать L-треонин
Для окончательного отбора штаммов с воспроизводимо усиленной способностью продуцировать L-треонин, из 10 лучших мутантов, отобранных в Примере 2, выполняли культивирование в колбе с использованием следующей культуральной среды. Продуцирование L-треонина измеряли согласно следующему способу. Сначала каждый из штаммов инокулировали в колбу 250 мл с угловой перегородкой, содержащую 25 мл посевной среды с добавлением канамицина (25 мг/л), и культивировали при встряхивании при 30°С в течение 20 часов при 200 об/мин. Затем 1 мл посевной культуры инокулировали в колбу 250 мл с угловой перегородкой, содержащую 24 мл культуральной среды с добавлением канамицина (25 мг/л), и культивировали при встряхивании при 32°С в течение 72 часов при 200 об/мин. Составы культуральной среды для посева и среды для продуцирования являются следующими и результаты культивированных штаммов приведены в Таблице 1.
Среда для посева (рН 7,0)
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 0,1 мг биотина, 1 мг тиамина HCl, 22 мг пантотената кальция и 2 мг никотинамида (из расчета на 1 л дистиллированной воды).
Среда для продуцирования (рН 7,0)
63 г глюкозы, 28 г (NH4)2SO4, 20 г соевого белка, 14 г мелассы, 1,1 г KH2PO4, 1,2 г MgSO4⋅7H2O, 1,8 мг биотина, 9 мг тиамина HCl, 9 мг пантотената кальция, 180 мг MnSO4, 200 мг FeSO4, 1 мг ZnSO4, 1 мг CuSO4 и 30 г СаСО3 (из расчета на 1 л дистиллированной воды).
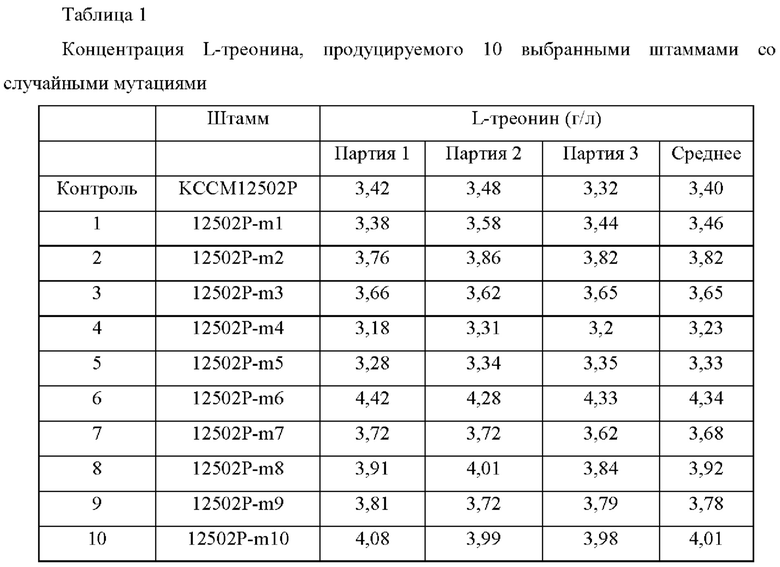
Из отобранных 10 мутантных штаммов в конечном счете был выбран 12502Р-m6 в качестве штамма, у которого способность продуцировать L-треонин была значительно усилена.
Пример 4. Выявление причины усиления способности продуцировать L-треонин у отобранных в конечном счете штаммов
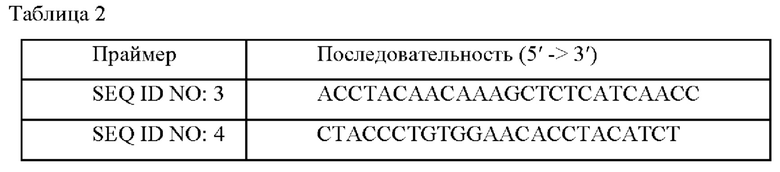
В данном примере пытались идентифицировать ген, удаленный посредством случайной вставки транспозона, у мутантных штаммов, отобранных в конечном счете в Примере 3. Геномную ДНК 12502Р-т6 экстрагировали, расщепляли и лигировали для трансформации Е. coli DH5α, и этот штамм наносили мазками на твердую среду LB, содержащую канамицин (25 мг/л). После отбора 20 трансформированных колоний получали плазмиду, содержащую часть неизвестного гена. В результате анализа его нуклеотидной последовательности с использованием праймеров, содержащих SEQ ID NOS: 3 и 4 из набора EZ-Tn5™<R6Kγori/KAN-2>TnP Transposome™, было подтверждено, что активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, была ослаблена, так как ген, содержащий нуклеотидную последовательность SEQ ID NO:2 (Ncgl1917), был инактивирован на основании нуклеотидных последовательностей, представленных в GenBank Национального института здравоохранения США (NIH).
Пример 5. Получение рекомбинантного вектора и штамма с ослабленной активностью белка, содержащего аминокислотную последовательность SEP ID NO: 1
Рекомбинантный вектор, способный делетировать ген, имеющий нуклеотидную последовательность SEQ ID NO:2, которая была подтверждена в Примере 4, из хромосомы штаммов, принадлежащих к роду Corynebacterium, конструировали в соответствии со следующим способом. Сначала геномную ДНК, выделенную из штамма WT, использовали в качестве матрицы, и сайт распознавания фермента рестрикции Smal встраивали в 5'-фрагмент, расположенный на 249 п. н. выше TQU&Ncgl1917, для синтеза праймеров SEQ ID NOS: 5 и 6. Кроме того, сайт распознавания фермента рестрикции Smal вставляли в 3'-фрагмент, расположенный на 410 п. н. ниже гена Ncgl1917, для синтеза праймеров SEQ ID NOS:7 и 8 (Таблица 3).

Конкретно, выполняли ПЦР с использованием хромосомной ДНК Corynebacterium glutamicum дикого типа АТСС 13032 в качестве матрицы (Sambrook et al., Molecular Cloning Laboratory Manual (1989), Cold Spring Harbor Laboratories). Из указанного выше получали фрагменты ДНК длиной 249 п.н. выше и длиной 410 п.н. ниже участка, который кодирует белок, кодируемый нуклеотидной последовательностью SEQ ID NO: 2. В этом случае ПЦР выполняли в следующих условиях: денатурация при 94°С в течение 2 минут, 30 циклов денатурации при 94°С в течение 1 минуты, отжиг при 56°С в течение 1 минуты и полимеризация при 72°С в течение 40 секунд, и затем полимеризация при 72°С в течение 10 минут. При этом вектор pDCM2 (KR 10-2020-0136813А), который обрабатывали ферментом рестрикции Smal и обрабатывали нагревом при 65°С в течение 20 минут, лигировали для встраивания фрагмента ДНК, амплифицированного посредством ПЦР, используя набор Infusion Cloning, и затем им трансформировали Е. coli DH5α. Штамм наносили мазками на твердую среду LB, содержащую канамицин (25 мг/л). Отбирали колонии, трансформированные вектором, в который посредством ПЦР был встроен ген-мишень, используя праймеры SEQ ID NO: 5 и 8, и получали плазмиду, используя известный способ выделения плазмид. Плазмида была названа pDCM2-ANcgl1917. Штамм Corynebacterium glutamicum KCCM12502P, продуцирующий треонин, трансформировали полученным вектором посредством электроимпульсного метода. Штамм KCCM12502P, в котором был инактивирован ген Ncgl1917, был назван KCCM12502P::А1917.
Пример 6. Оценка способности продуцировать L-треонин штамма, имеющего ослабленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, полученного из Corynebacterium glutamicum KCCM125Q2P
Для анализа способности продуцировать L-треонин полученного штамма Corynebacterium glutamicum KCCM12502P::А1917 выполняли культивирование в колбе с использованием следующей культуральной среды. Исходный штамм, Corynebacterium glutamicum KCCM12502P, использовали в качестве контроля, и продуцирование L-треонина измеряли согласно следующему способу. Каждый из штаммов инокулировали в колбу 250 мл с угловой перегородкой, содержащую 25 мл среды для посева, и культивировали при встряхивании при 30°С в течение 20 часов при 200 об/мин. Затем 1 мл посевной культуры инокулировали в колбу 250 мл с угловой перегородкой, содержащую 24 мл культуральной среды, и культивировали при встряхивании при 32°С в течение 72 часов при 200 об/мин. Составы среды для посева и среды для продуцирования являются следующими, и результаты культивирования штаммов приведены в Таблице 4 Среда для посева (рН 7.0)
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г K2HPO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 0,1 мг биотина, 1 мг тиамина HCl, 22 мг пантотената кальция и 2 мг никотинамида (из расчета на 1 л дистиллированной воды).
Среда для продуцирования (рН 7.0)
63 г глюкозы, 28 г (NH4)2SO4, 20 г соевого белка, 14 г мелассы, 1,1 г KH2PO4, 1,2 г MgSO4⋅7H2O, 1,8 мг биотина, 9 мг тиамина HCl, 9 мг пантотената кальция, 180 мг MnSO4, 200 мг из FeSO4, 1 мг ZnSO4, 1 мг CuSO4 и 30 г СаСО3 (из расчета на 1 л дистиллированной воды).
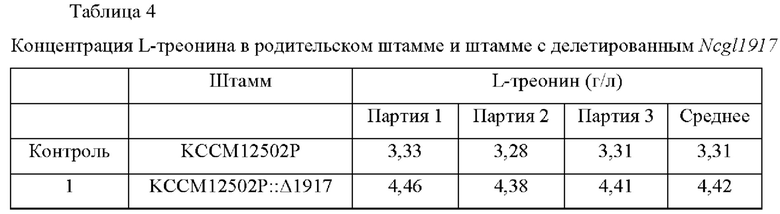
Как описано выше, в случае, когда ген, содержащий нуклеотидную последовательность SEQ ID NO: 2, был делетирован у штамма, продуцирующего L-треонин, Corynebacterium glutamicum KCCM12502P, было подтверждено, что способность продуцировать L-треонин улучшалась в среднем на 35% по сравнению с родительским штаммом.
Таким образом, было подтверждено, что способность продуцировать L-треонин может быть улучшена посредством делетирования гена, содержащего нуклеотидную последовательность SEQ ID NO: 2, у микроорганизма рода Corynebacterium.
На основании вышеописанных результатов было подтверждено, что инактивация белка, содержащего аминокислотную последовательность SEQ ID NO: 1 в штамме, продуцирующем L-треонин, является эффективной для усиления способности продуцировать L-треонин, и штамм KCCM12502P::Δ1917 был назван СА09-0907 и депонирован в Корейском культурном центре микроорганизмов (KCCM) и получил номер регистрации KCCM12946P 1 февраля 2021 года.
Пример 7. Получение штамма, обладающего ослабленной активностью белка, содержащего аминокислотную последовательность SEP ID NO: 1, полученного из Corynebacterium glutamicum KCCM11222P, и оценка его способности продуцировать L-треонин
Для того чтобы определить, достигается ли такой же эффект у другого штамма, принадлежащего к Corynebacterium glutamicum, продуцирующему L-треонин, был получен штамм путем удаления гена, содержащего нуклеотидную последовательность SEQ ID NO: 2, из штамма, продуцирующего L-треонин, Corynebacterium glutamicum KCCM11222P (US 10590446 В2), таким же образом, как в Примере 5, и он был назван KCCM11222P::Δ1917.
Для анализа способности продуцировать L-треонина полученного штамма Corynebacterium glutamicum KCCM11222P::Δ 1917, этот штамм культивировали таким же образом, как в Примере 6. В качестве контроля использовали исходный штамм Corynebacterium glutamicum KCCM11222P. Результаты культивирования штамма приведены в Таблице 5.
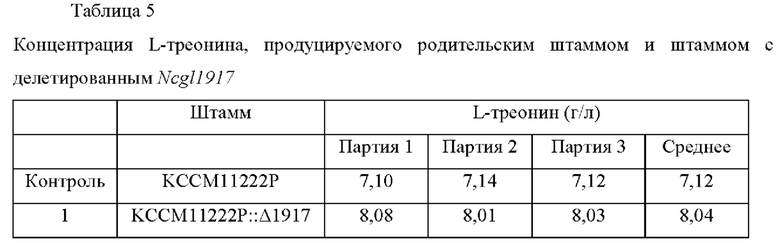
Как описано выше, в случае, когда ген, содержащий нуклеотидную последовательность SEQ ID NO: 2, был делетирован у продуцирующего L-треонин штамма Corynebacterium glutamicum KCCM11222P, было подтверждено, что способность продуцировать L-треонин улучшалась в среднем на 12%.
При объединении результатов Примеров 5-7 было подтверждено, что способность продуцировать L-треонин может быть усилена посредством ослабления активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1, у микроорганизма рода Corynebacterium.
Пример 8. Получение штамма, имеющего ослабленную активность белка, содержащего аминокислотную последовательность SEP ID NO: 1, произведенного из Corynebacterium glutamicum KCCM11248P, и оценка его способности продуцировать L-изолейцин
Для определения, получается ли тот же эффект усиления способности продуцировать L-аминокислоты таким же способом в штамме, принадлежащем к Corynebacterium glutamicum, продуцирующему L-изолейцин, получали штамм посредством делеции гена, содержащего нуклеотидную последовательность SEQ ID NO:2, из штамма, продуцирующего L-изолейцин, Corynebacterium glutamicum KCCM11248P (патент Кореи №10-1335789), таким же способом, как в Примере 5, и он был назван KCCM11248P::Δ1917. Для анализа способности продуцировать L-изолейцин у полученного штамма Corynebacterium glutamicum KCCM11248P::Δ1917 выполняли культивирование в колбе с использованием следующей культуральной среды. В качестве контроля использовали исходный штамм Corynebacterium glutamicum KCCM11248P. Продуцирование L-изолейцина измеряли посредством следующего способа. Сначала каждый из штаммов инокулировали в колбу 250 мл с угловой перегородкой, содержащую 25 мл среды для посева, и культивировали при встряхивании при 30°С в течение 20 часов при 200 об/мин. Затем 1 мл среды для посева инокулировали в колбу 250 мл с угловой перегородкой, содержащую 24 мл культуральной среды, и культивировали при встряхивании при 32°С в течение 72 часов при 200 об/мин. Составы культуральной среды для посева и среды для продуцирования являются следующими, и результаты культивирования штаммов приведены в Таблице 6.
Среда для посева (рН 7.0)
20 г глюкозы, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2HPO4, 0,5 г MgSO4⋅7H2O, 0,1 мг биотина, 1 мг тиамина HCl, 22 мг пантотената кальция и 2 мг никотинамида (из расчета на 1 л дистиллированной воды).
Среда для продуцирования (рН 7.2)
100 г глюкозы, 5 г дрожжевого экстракта, 16 г (NH4)2SO4, 1 г KH2PO4, 1 г MgSO4⋅7H2O, 10 мг FeSO4⋅7H2O, 10 мг MnSO4⋅H2O и 200 мкг биотина (из расчета на 1 л дистиллированной воды).
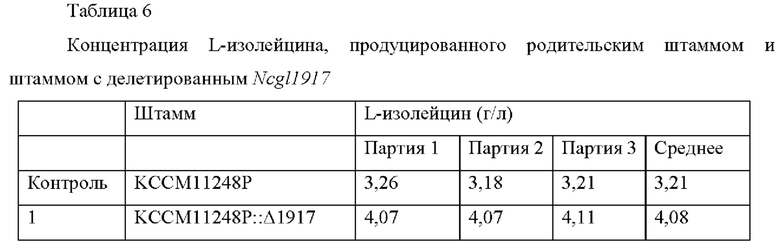
Как описано выше, было подтверждено, что способность продуцировать L-изолейцин улучшалась в среднем на 27% по сравнению со штаммом, продуцирующим L-изолейцин, Corynebacterium glutamicum KCCM11248P, когда был делетирован ген, содержащий нуклеотидную последовательность SEQ ID NO: 2.
Таким образом, было подтверждено, что способность продуцировать L-изолейцин может быть усилена путем ослабления активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1 в микроорганизме рода Corynebacterium.
Приведенное выше описание настоящего изобретения представлено для иллюстрации, и специалисту в данной области техники должно быть понятно, что могут быть внесены различные изменения и модификации без изменения технической концепции и существенных признаков настоящего изобретения. Таким образом, очевидно, что описанные выше воплощения являются иллюстративными во всех аспектах и не ограничивают настоящее изобретение. Таким образом, объем изобретения определяется не подробным описанием, а формулой изобретения и ее эквивалентами, и все изменения в пределах объема формулы изобретения и ее эквивалентов следует считать включенными в настоящее изобретение.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> MICROORGANISM OF THE GENUS CORYNEBACTERIUM PRODUCING L-AMINO ACID
AND METHOD FOR PRODUCING L-AMINO ACID USING SAME
<130> OPA21296
<150> KR 10-2021-0051233
<151> 2021-04-20
<160> 8
<170> KoPatentIn 3.0
<210> 1
<211> 344
<212> PRT
<213> Artificial Sequence
<220>
<223> NCgl1917
<400> 1
Met Pro Asn Asn Glu Phe His Thr Asn His Ser Leu Gly Gln Asp Asp
1 5 10 15
Gln Thr Pro Asp Gln Ala His Phe Phe Pro Gln Gly Arg Gly Glu Ala
20 25 30
Leu Val Arg Pro Gly Gln Glu His Phe Ile Ala Ala Thr Asp Glu Thr
35 40 45
Gly Leu Gly Ala Val Asp Ala Val Ala Asp Asp Ser Ala Pro Thr Ser
50 55 60
Met Trp Gly Glu Ala Trp Arg Asp Leu Arg Arg Arg Pro Leu Phe Trp
65 70 75 80
Val Ser Ala Val Leu Ile Ile Leu Ala Leu Leu Leu Ala Ala Val Pro
85 90 95
Gln Leu Phe Thr Ser Thr Asp Pro Gln Phe Cys Val Leu Ala Asn Ser
100 105 110
Leu Asp Gly Pro Gln Ser Gly His Pro Phe Gly Phe Asp Arg Gln Gly
115 120 125
Cys Asp Ile Phe Ala Arg Thr Val Tyr Gly Ala Arg Ala Ser Val Ala
130 135 140
Val Gly Val Leu Thr Thr Leu Leu Val Ala Leu Ile Gly Thr Val Phe
145 150 155 160
Gly Ala Leu Ala Gly Phe Phe Gly Gly Ile Met Asp Thr Ile Leu Ser
165 170 175
Arg Ile Thr Asp Met Phe Phe Ala Ile Pro Leu Val Leu Ala Ala Ile
180 185 190
Val Val Met Gln Met Phe Lys Glu His Arg Thr Ile Val Thr Val Val
195 200 205
Leu Val Leu Gly Leu Phe Gly Trp Thr Asn Ile Ala Arg Ile Thr Arg
210 215 220
Gly Ala Val Met Thr Ala Lys Asn Glu Glu Tyr Val Thr Ser Ala Arg
225 230 235 240
Ala Leu Gly Ala Ser Lys Ala Lys Ile Leu Leu Ser His Ile Met Pro
245 250 255
Asn Ala Ala Ala Pro Ile Ile Val Tyr Ala Thr Val Ala Leu Gly Thr
260 265 270
Phe Ile Val Ala Glu Ala Thr Leu Ser Phe Leu Gly Ile Gly Leu Pro
275 280 285
Pro Ser Ile Val Ser Trp Gly Ala Asp Ile Ala Lys Ala Gln Thr Ser
290 295 300
Leu Arg Thr Gln Pro Met Val Leu Phe Tyr Pro Ala Met Ala Leu Ala
305 310 315 320
Leu Thr Val Leu Ser Phe Ile Met Met Gly Asp Val Val Arg Asp Ala
325 330 335
Leu Asp Pro Lys Ser Arg Lys Arg
340
<210> 2
<211> 1035
<212> DNA
<213> Artificial Sequence
<220>
<223> Ncgl1917
<400> 2
atgcctaata atgaattcca cacaaaccac tcgttgggcc aagatgatca aaccccagat 60
caggctcatt tcttcccaca aggacgaggc gaggctctag ttcgaccagg tcaagagcac 120
ttcatcgcag ccactgatga aaccggactt ggtgccgtcg atgctgttgc tgatgactct 180
gcaccaacct ccatgtgggg cgaagcgtgg cgagaccttc gtcgtcgacc actgttctgg 240
gtctctgcgg tgttgattat tttggcgctt ctcctggccg cagttccgca gctgtttacc 300
tcaacggatc cccagttctg tgtgctggca aactctcttg atggtccaca gtctggacat 360
cccttcggat tcgaccgtca aggttgcgat atttttgctc gtaccgtcta cggtgctcgt 420
gcctcggtcg ccgtcggtgt gttgaccacg ttactggtcg ccctcatcgg tactgtattt 480
ggtgctttgg ctggcttctt tggtggcatc atggatacca tcctctcccg catcaccgac 540
atgttcttcg ccattccact ggttctggca gccatcgttg tgatgcagat gttcaaggaa 600
caccgcacca tcgtcaccgt ggttttggtg cttgggcttt tcggctggac caacattgcg 660
cgtattaccc gtggagcggt gatgaccgca aagaatgaag agtatgtcac ctccgcacgt 720
gcgcttggtg catcaaaagc caagatactg ctgtctcaca tcatgccaaa cgccgcagca 780
cccatcattg tgtatgcaac tgtggcactg ggaacattca tcgtggcaga ggcgacgctc 840
tccttcctgg gcattggcct tccaccatca attgtctcct ggggtgctga tatcgcgaag 900
gcacaaacct cccttcgtac ccaacccatg gtgctgttct accccgcaat ggcacttgca 960
ctaaccgttt tgagcttcat catgatgggc gatgtcgtcc gcgacgctct ggatcctaag 1020
tcgaggaagc gatga 1035
<210> 3
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 3
acctacaaca aagctctcat caacc 25
<210> 4
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 4
ctaccctgtg gaacacctac atct 24
<210> 5
<211> 48
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 5
acggccagtg aattcgagct cggtacccat gcctaataat gaattcca 48
<210> 6
<211> 47
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 6
tggtccagcc gaaaagccca agcaccaccg cagagaccca gaacagt 47
<210> 7
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 7
gtggtgcttg ggcttttcgg 20
<210> 8
<211> 45
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер
<400> 8
tgcaggtcga ctctagagga tcccctcatc gcttcctcga cttag 45
<---
Изобретение относится к биотехнологии. Предложен микроорганизм рода Corynebacterium, продуцирующий L-аминокислоту и имеющий удаленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, при этом L-аминокислота включает по меньшей мере одну, выбранную из L-треонина и L-изолейцина. Также предложены способ получения L-аминокислоты с использованием указанного микроорганизма и композиция для получения L-аминокислоты, содержащая указанный микроорганизм. Изобретение обеспечивает продуцирование увеличенного выхода L-аминокислота, где L-аминокислота включает по меньшей мере одну, выбранную из L-треонина и L-изолейцина. 3 н. и 4 з.п. ф-лы, 6 табл., 8 пр.
1. Микроорганизм рода Corynebacterium, продуцирующий L-аминокислоту и имеющий удаленную активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1,
где L-аминокислота включает по меньшей мере одну, выбранную из L-треонина и L-изолейцина.
2. Микроорганизм по п. 1, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
3. Микроорганизм по п. 1, который имеет усиленную способность продуцировать L-аминокислоты по сравнению с микроорганизмом, у которого активность белка, содержащего аминокислотную последовательность SEQ ID NO: 1, не удалена.
4. Способ получения L-аминокислоты, включающий культивирование в среде микроорганизма рода Corynebacterium по любому из пп. 1-3, где L-аминокислота включает по меньшей мере одну, выбранную из L-треонина и L-изолейцина.
5. Способ по п. 4, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
6. Способ по п. 4, дополнительно включающий извлечение L-аминокислоты из микроорганизма или среды.
7. Композиция для получения L-аминокислоты, содержащая: микроорганизм по любому из пп. 1-3 и эксципиент, где L-аминокислота включает по меньшей мере одну, выбранную из L-треонина и L-изолейцина.
| KR 100816472 B1, 26.03.2008 | |||
| База данных NCBI Reference Sequence: WP_006284211.1, 17.04.2020 | |||
| ABC transporter permease [Corynebacterium glutamicum] | |||
| Найдено онлайн: https://www.ncbi.nlm.nih.gov/protein/WP_006284211.1/ Дата обращения 18.04.2024 | |||
| НЕЛИНЕЙНО-ОПТИЧЕСКАЯ СРЕДА | 2003 |
|
RU2267145C2 |
| WO 2016172341 A2, 27.10.2016 | |||
| МИКРООРГАНИЗМ, ПРОДУЦИРУЮЩИЙ О-ФОСФОСЕРИН, И СПОСОБ ПОЛУЧЕНИЯ L-ЦИСТЕИНА ИЛИ ЕГО ПРОИЗВОДНЫХ ИЗ О-ФОСФОСЕРИНА С ЕГО ИСПОЛЬЗОВАНИЕМ | 2011 |
|
RU2536250C1 |

