Область техники.
Изобретение относится к биотехнологии, в частности к генетической инженерии, и касается получения термостабильной L-аспарагиназы MrAII и способа ее очистки. Для этого был создан рекомбинантный штамм Escherichia coli - продуцента L- аспарагиназы Melioribacter roseus путем введения в штамм-реципиент гена mrAII в составе рекомбинантной плазмидной ДНК. MrAII представляет собой первый описанный фермент из большой группы близкородственных бактериальных L-АСП II типа, происходящих от представителей группы Bacteroidetes-Chlorobi.
L-аспарагиназа (EC 3.5.1.1; L-аспарагинамидогидролаза) (L-АСП) представляет собой биотехнологически значимый фермент, используемый в фармацевтической, биосенсорной и пищевой промышленности. L-аспарагиназа катализирует превращение аминокислоты L-аспарагина (L-АСП) в L-аспарагиновую кислоту и аммиак. L-АСП фермент известен своими антиканцерогенными свойствами. Фермент оказывает истощающее действие на концентрацию L-АСП во внеклеточной жидкости. Пониженный уровень L-АСП и неспособность чувствительных опухолевых клеток синтезировать собственные L-АСП приводит к ингибированию синтеза белка, остановке клеточного цикла в фазе G1 и, в конечном итоге, апоптозу в лейкемических клетках [1].
L-АСП используется также в пищевой промышленности в качестве технологического агента для снижения уровня акриламида в жареных продуктах. L-АСП катализирует гидролиз L-АСП, не допуская реакции восстанавливающих сахаров с этой аминокислотой с образованием канцерогенного акриламида [2, 3].
Особенностью перечисленных областей биотехнологического использования L-АСП являются повышенные требования к стабильности и активности фермента.
Предшествующий уровень техники.
Усилия по открытию новых перспективных L-аспарагиназ для различных областей биотехнологии превратили эту группу ферментов в растущее семейство с большим разнообразием. L-АСП были выделены из многих архей, бактерий, грибов и дрожжей, тогда как L-АСП растительного и млекопитающего происхождения в настоящее время также известны [3, 4].
Бактериальные L-АСП подразделяются на тип I и тип II [17]. L-АСП типа I участвуют в метаболизме азота и, по-видимому, конститутивно экспрессируются. Эти цитоплазматические ферменты проявляют низкое сродство к L-АСП со значениями KM в миллимолярном диапазоне [5]. С другой стороны, бактериальные L-АСП типа II, по-видимому, участвуют в метаболизме углерода, и их экспрессия жестко регулируется различными факторами. Ферменты II типа расположены в периплазматическом пространстве. Они проявляют высокую специфическую активность в отношении L-АСП со значениями KM на два порядка ниже (микромолярные KM) по сравнению с бактериальными L-АСП I типа [4].
L-АСП растительного типа или L-АСП типа III, по-видимому, играют доминирующую роль в деградации аспарагина путем гидролиза амидной связи боковой цепи L-АСП или ее β-пептидов в значительной части высших растений. Эти ферменты относятся к суперсемейству N-концевых нуклеофильных гидролаз [5]. Высокие значения KM (миллимолярные) этих ферментов ограничивают их применение в биомедицине [5].
Изученные и применяемые на данный момент L-АСП выделены из мезофильных микроорганизмов, имеют ряд недостатков. Проблема стабильности и активности мезофильных L-АСП является одной из ключевых, что определяет направление исследования L-АСП экстремофильного происхождения.
Выявлено, что бактерия Melioribacter roseus, принадлежащая к новому предложенному типу Ignavibacteriae группы Bacteroidetes-Chlorobi [6], обладает двумя L-аспарагиназами: L-АСП класса 2 (растительного типа, MrAIII) и L-АСП класса 1 (бактериального типа II, MrAII), обладающими высоким сходством с соответствующими аналогами, происходящими от других членов группы Bacteroidetes-Chlorobi. Сам M. roseus представляет собой умеренно термофильную, факультативно-анаэробную хемоорганотрофную бактерию [6].
Для гипертермофильных архей Pyrococcus и Thermococcus sp.: Pyrococcus furiosus [7], Pyrococcus yayanosii CH1 [8], Thermococcus kodakaraensis [9], Thermococcus gammatolerans [3] биохимически охарактеризовано несколько L-АСП. Данные нативные L-аспарагиназы гипертермофильных архей превосходят по характеристикам разработанные для пищевых производств и ранее описанные в патентах улучшенные варианты мезофильных L-АСП с повышенной термостабильностью:
- мутантные термостабильные формы L-АСП Aspergillus oryzae и Aspergillus niger при повышенной температуре 70°С характеризуются снижением активности в 2 и более раз в сравнении с активностью при температурном оптимуме 50°С [7];
- мутантные формы L-АСП Aspergillus oryzae, обладающих остаточной активностью, не превышающей 72 % при инкубации в течение 20 минут при 66°С [7];
- штамм Bacillus methylotrophicus MKSY2013 был описан в качестве источника термостабильной L-АСП для биотехнологии в патенте KR101777904B1. Однако выводы о термостабильности данной L-АСП сделаны на основании оценки свойств не очищенного белка, а супернатанта в диапазоне температур 60-95°С в течение 30 минут [10].
При температурном оптимуме 85-95°С нативные L-АСП гипертермофильных архей сохраняют активность в интервале температур 70-90°С до 32 часов. Термостабильность L-АСП гипертермофильных архей также сочетается с повышенной устойчивостью в широком диапазоне рН, в денатурирующих условиях, в присутствии в средах ионов металлов [3].
Среди двух аннотированных L-аспарагиназ M. roseus, L-АСП класса 2 (растительного типа) (MrAIII) и L-АСП класса 1 (бактериальный тип II) (MrAII), MrAII была выбрана для дальнейшего детального выяснения, поскольку в настоящее время только бактериальные ферменты типа II одобрены для биомедицинского применения. Кроме этого, L-аспарагиназ M. roseus обладает наибольшей удельной ферментативной активностью, среди изученных экстремофильных бактерий.
Настоящее изобретение касается создания эффективной системы гетерологической экспрессии новой L-АСП из термофильной бактерии Melioribacter roseus, выделенной выделен из микробного мата в потоке горячей воды (46°С) из нефтеразведочной скважины глубиной 2775 м (Россия) [6], разработки способа очистки, а также характеристики фермента для биотехнологического применения.
Раскрытие изобретения.
Задача заявляемой группы изобретений - получение очищенных препаратов новой термостабильной L-АСП MrAII из Melioribacter roseus для биотехнологического применения путем создания эффективной системы гетерологической экспрессии в клетках мезофильной бактерии Escherichia coli.
Задача решена путем:
- конструирования гена mrAII, кодирующего L-АСП из M. roseus, с учетом соответствия требованиям эффективной экспрессии в клетках E. coli;
- клонирования гена mrAII, экспрессии и очистки рекомбинантного фермента;
- определения специфической активности MrAII и кинетических параметров;
- характеристики влияния температуры, рН и ионов металлов на активность ферментов. Определение химической стабильности MrAII.
Для этого:
а) в источнике новой перспективной для биотехнологии L-АСП - термофильной бактерии M. roseus с ранее аннотированной геномной последовательностью GenBank NC_012883 [6] идентифицированы предполагаемые гены MROS_RS06765 и MROS_RS05340 - кодирующие L-АСП M. roseus (MrAIII) и с номером доступа последовательности в GenBank WP_014855981.1 и WP_014855710.1, соответственно. Нуклеотидная последовательность указанных генов представлена в SEQ ID NO: 1 и SEQ ID NO:2.
б) был искусственно синтезирован и клонирован нативный ген L-аспарагиназы II типа из M. roseus (mrAII) в вектор pET-28a(+);
в) сконструированная плазмида была трансформирована в хозяина E. coli BL21 (DE3) для гетерогенной экспрессии MrAII;
г) достигнута ферментативная очистка MrAII с помощью ионообменной хроматографии с конечным выходом 75,5 %;
д) определена удельная активность, субстратная специфичность, Vmax и КМ рекомбинантной L-АСП II типа из M. roseus;
е) определена активность MrAII при различных температурах в диапазоне от 37 до 80°C.
ж) исследовано влияние различных катионов металлов, Ni2+, Cu2+, Mg2+, Zn2+, Ca2+, Fe3+ и ЭДТА на активность MrAII;
з) проверена цитотоксическая активность MrAII на различных типах лейкозных и солидных опухолевых раковых клеток человека вместе с нормальными фибробластами и нормальными CD4+ Т-клетками.
Результаты изучения биохимических, кинетических характеристик показали, что L-АСП M. roseus сочетает высокую специфическую активность, низкую нежелательную глутаминазную активность, высокую термостабильность, устойчивость к химической денатурации, присутствию в среде ионов металлов, что позволяет сделать вывод о перспективности фермента для применения в биомедицине и пищевой промышленности.
Заявляемое изобретение и способ его получения были проиллюстрированы следующими примерами.
Пример 1. Идентификация гена L-аспарагиназы в геноме M. roseus P3M-2
Биоинформационный анализ доступной последовательности всего генома M. roseus показал, что M. roseus P3M-2 обладает двумя ферментами, проявляющими активность L-аспарагиназы - WP_014855981.1 и WP_014855710.1 - кодируется генами MROS_RS06765 и MROS_RS05340, соответственно.
Поиск гомологов показал, что предполагаемая изоаспартилпептидаза/L-аспарагиназа WP_014855981.1 обладает наибольшей идентичностью с ранее охарактеризованной изоаспартилпептидазой/L-аспарагиназой из E. coli - WP_146849425.1 - с 55,2%, классифицированными как L-АСП растительного типа EcAIII. Когда аминокислотная последовательность WP_014855981.1 была сопоставлена с известными L-АСП растительного типа, и были обнаружены все ранее идентифицированные каталитические остатки, включая нуклеофильный остаток треонина, участвующий в авторасщеплении и катализе, за которым следует консервативный пентапептид (VGAVA). Таким образом, фермент WP_014855981.1 M. roseus представляет собой L-аспарагиназу растительного типа (класс 2) - MrAIII. Однако MrAIII примерно на 40% похож с другими охарактеризованными L-АСП растительного типа из Thermococcus kodakarensis (Q5JHT1.1) и Pyrobaculum calidifontis (ABO08395.1) с 43,1% и 39,5%, соответственно.
Идентифицированная L-АСП в геноме M. roseus P3M-2 WP_014855710.1 была классифицирована как бактериальная L-АСП II типа (класс 1) (MrAII). Сравнение аминокислотных последовательностей показало, что MrAII демонстрирует высокую гомологию с широкой группой нехарактерных L-АСП из недавно аннотированных геномов бактерий Ignavibacteria (HGR20494.1, HFO52226.1, MBI5730500.1, HET53601.1) - 80.3-79.0 % идентификация, Melioribacter sp. (NJD22110.1, MBS3945884.1) - 80,5-78,3 %, а бактерией Bacteroidetes (MBU0558988.1, MBU1097831.1, MBU0473869.1) - 78,7-76,1 %. Однако уровень гомологии между MrAII и известными L-АСП II типа, включая одобренные FDA антилейкемические средства, довольно низок - ниже, чем между MrAII и MrAIII (44,4 %). Идентичность между MrAII и одобренными FDA L-АСП из E. coli (номер доступа UniProtKB P00805, продается под торговой маркой Elspar, номер доступа GenBank AAA23445.1) и E. chrysanthemi (Dickeya chrysanthemi, Pectobacterium chrysanthemi) (Номер доступа Uni-ProtKB P06608, продаваемый под торговой маркой Erwinaze, номер доступа GenBank AAS67028.1), составляет только 35,2% и 32,4%, соответственно. Интересно, что идентичность между термофильными MrAII и бактериальными L-АСП типа I с известными структурами от других термофилов Thermococcus kodakarensis KOD1 (WP_011250607), Pyrococcus furiosus (WP_011013191.1), Pyrococcus horikoshii (WP_010884185.1) составляет 26,7-26,5. Данные указывают на низкое общее сходство последовательностей MrAII с другими, ранее охарактеризованными L-АСП различного происхождения и типов. MrAII был выбран для дальнейшего детального изучения.
Пример 2. Клонирование гена mrAII: конструирование плазмиды для высокоэффективной гетерологичной экспрессии белка в клетках мезофильной бактерии E. coli.
Ген, который предположительно кодирует L-АСП MROS_RS05340 (mrAII) (последовательность 1199322-1200302 https://www.ncbi.nlm.nih.gov/nuccore/397689003 (по состоянию на 15 октября 2021 г., номер доступа в GenBank белка WP_014855710.1), окруженный сайтами рестрикции NheI/SalI, был искусственно синтезирован компанией TWIST Bioscience (Twist Bioscience, США). Синтезированный нативный ген L-аспарагиназы II типа из M. roseus (mrAII), который состоит из 981 нуклеотида, соответствующего полипептиду из 327 аминокислот, гидролизовали и клонировали в расщепленный NheI/SalI вектор pET-28a(+) под контролем промотора Т7. Сконструированный вектор трансформировали и экспрессировали в E. coli BL21 (DE3).
Пример 3. Экспрессия, выделение и очистка фермента
Сконструированную плазмиду трансформировали в E. coli BL21 (DE3) для гетерологичной экспрессии MrAII. Отобранные рекомбинантные клоны E. coli выращивали, как описано ранее [11]. В среду для культивирования клеток, несущих плазмиды, добавляли 0,05 мг/мл канамицина. Экспрессию целевого белка индуцировали добавлением лактозы к экспрессируемой культуре при плотности А600 1,9 до конечной концентрации 0,2%. Клетки выращивали еще 17-20 ч и осаждали центрифугированием при 4000×g в течение 15 мин.
Все стадии очистки фермента проводили при +4°С. Пять граммов биомассы суспендировали в 50 мл буфера (20 мМ натрий-фосфатного буфера, рН 7,2, 1 мМ глицина, 1 мМ ЭДТА) и разрушали ультразвуковой обработкой [11]. Клеточный дебрис и неразрушенные клетки удаляли центрифугированием (35 000 g, 30 мин). Супернатант, содержащий фермент, наносили на колонку с Q-Sepharose. Белок элюировали линейным градиентом 0-1,0 М NaCl. Собирали фракции колонки, содержащие фермент (0,46-0,7 М NaCl). Ультрафильтрацию, обессоливание и замену буфера проводили с использованием мембран Amicon (Millipore, США). Образцы замораживали и хранили при температуре -20°С.
Концентрацию белка определяли по методу Седмака, используя бычий сывороточный альбумин в качестве стандарта. SDS-PAGE проводили для визуализации и определения чистоты белка, как описано ранее [3].
Ферментативная очистка MrAII была достигнута с помощью ионообменной хроматографии с конечным выходом 75,5 % (таблица 1).
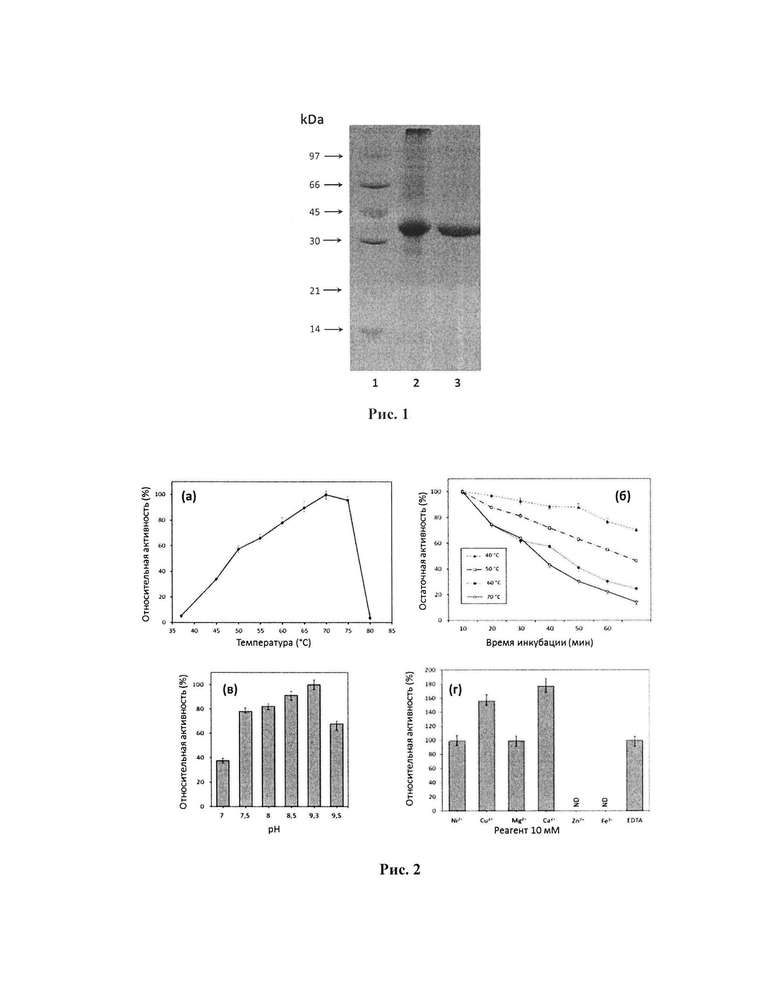
Очищенный фермент показал электрофоретическую однородность с единственной основной полосой на SDS-PAGE (рис. 1). Молекулярная масса очищенного фермента, по оценкам SDS-AGE, составляла приблизительно 34,5 кДа (рис. 1), что соответствовало теоретическому значению, рассчитанному на основе аминокислотной последовательности обработанной формы фермента (35,3 кДа).
Молекулярную массу, теоретический pI и частоты аминокислот анализировали с использованием ProtParam (http://web.expasy.org/protparam/, по состоянию на 11 ноября 2021 г.).
Пример 4. Специфическая активность MrAII и кинетика фермента.
Каталитическую активность L-аспарагиназы измеряли с помощью спектрометра кругового дихроизма Jasco J-815 (Jasco, Япония) с температурно-регулируемой ячейкой. Образцы готовили путем смешивания L-аспарагина 1-40 мМ и раствора MrAII 0,03-0,035 мг/мл. Реакции проводили в кварцевых кюветах (объем 300 мкл, длина оптического пути 1 мм). Зависимую от времени эллиптичность регистрировали при 210 нм. Удельную активность фермента выражали в ЕД/мг белка.
Активности L-глутаминазы и D-аспарагиназы измеряли по той же методике с использованием L-глутамина или D-аспарагина в качестве субстратов соответственно.
Кинетические параметры MrAII определяли в боратном буфере (0,025 М, рН 9,3) при 70°С с L-аспарагином в качестве субстрата. Наблюдаемые данные были подогнаны под уравнение Михаэлиса-Ментен, а кинетические константы KM и Vmax рассчитаны по графикам Лайнуивера-Берка.
Рекомбинантная L-АСП II типа из M. roseus проявляла высокую активность гидролиза в отношении L-АСП с удельной активностью 1530 Ед/мг. Эксперименты по субстратной специфичности показали, что L-АСП II проявляет дополнительную активность L-глутаминазы, но не активность D-аспарагиназы. Фермент мог использовать L-глутамин в качестве субстрата активности фермента с примерно 19 %, когда в качестве субстрата использовался L-АСП.
Были оценены кинетические свойства рекомбинантной L-АСП II типа из M. roseus. Было обнаружено, что Vmax составляет 5573 мкМ/мин. КМ для субстрата L-АСП был оценен в 1,4 мМ для MrAII).
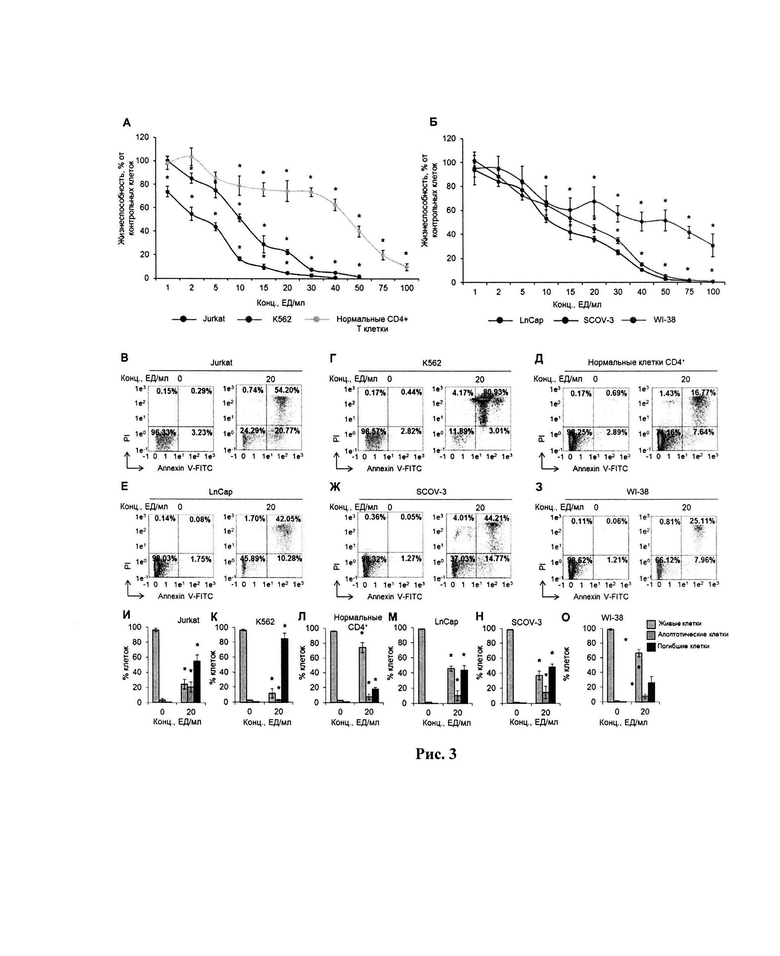
Активность MrAII оценивали при различных температурах в диапазоне от 37 до 80°C. Было показано, что фермент проявляет высокую активность в диапазоне температур от 60 до 75°C с максимумом при 70°C (рис. 2а). Временной ход термической инактивации очищенной L-аспарагиназы показан на рисунке 2б.
Пример 5. Влияние температуры, рН и ионов металлов на активность фермента. Химическая стабильность MrAII
Для очищенного фермента исследовали активность при различных температурах и уровнях рН.
Для оптимальной температуры анализировали профиль активности фермента при различных температурах в диапазоне от 45 до 80°С с шагом 5°С. Кроме того, активность MrAII измеряли при 37°C. Смесь анализировали в боратном буфере (0,025 М, рН 9,3).
Термостабильность L-АСП M. roseus определяли путем определения остаточной активности фермента, предварительно инкубированного при различных температурах от 40 до 70°C с шагом 10°C в боратном буфере (0,025 М, рН 9,3).
Оптимальный рН определяли путем оценки ферментативной активности при различных уровнях рН при 70°С в пяти буферных системах. А именно, ацетат натрия (0,05 М, рН 4,0-6,0), фосфат натрия (0,05 М, рН 6,0-7,0), трис-HCl-буфер (0,05 М, рН 7,0-9,0), борат (0,025 М, рН 9,3) и буфер глицин-NaOH (0,05 М, рН 9,5).
MrAII проявлял максимальную активность при рН 9,3 и только 20 % его активности при кислых значениях рН менее 7,0. Профиль активности MrAII, зависящий от рН, в рабочем диапазоне рН 7,0-9,5 показан на рисунке 2в. Согласно экспериментальным данным, рекомбинантный фермент сохранял высокую относительную активность в диапазоне рН 7,5-9,5 в зависимости от буферной системы.
Пример 6. Химические исследования денатурации и влияние ионов различных металлов
Стабильность фермента исследовали через 1 ч инкубации в боратном буфере (0,025 М, рН 9,3) в присутствии 0-8,0 М мочевины. Активность MrAII, исследованного при 70°C в отсутствие мочевины, принимали за 100 %. Измеренную активность сравнивали с активностью фермента без добавления мочевины в тех же условиях.
Влияние ионов металлов на активность MrAII исследовали в присутствии различных катионов (Ni2+, Cu2+, Mg2+, Zn2+, Ca2+, Fe3+) и ЭДТА. Добавляли соли NiCl2, CuSO4, MgCl2, ZnCl2, CaCl2, FeCl3 и ЭДТА в концентрации 10 мМ. Активность фермента анализировали при 70°С и рН 9,3 (в 0,025 М боратном буфере) путем добавления L-аспарагина и соответствующего(их) иона(ов) металла или ЭДТА. Вся процедура повторялась трижды.
Активность без добавления какого-либо иона металла была принята за 100%. Измеренную активность сравнивали с активностью фермента без добавления иона металла или ЭДТА в тех же условиях.
Было исследовано влияние различных катионов металлов, Ni2+, Cu2+, Mg2+, Zn2+, Ca2+, Fe3+ и ЭДТА, добавленных в концентрации 10 мм, на активность MrAII (рисунок 2д). Никаких существенных изменений активности фермента не наблюдалось в присутствии Ni2+, Mg2+ или ЭДТА. Добавление Cu2+ и Ca2+ привело к увеличению активности MrAII на 56 % и 77 %, соответственно. С другой стороны, Zn2+ и Fe3+ полностью ингибировали активность фермента (рис. 3д).
Те же результаты были получены в присутствии 2-8 М растворов мочевины: MrAII полностью терял свою активность при всех испытанных концентрациях.
Пример 7. Определение цитотоксической активности MrAII
Линии клеток хронического миелогенного лейкоза человека K562, острого Т-клеточного лейкоза Jurkat, карциномы предстательной железы LnCap и аденокарциномы яичника SCOV-3 выращивали в среде RPMI-1640. Нормальные человеческие фибробласты WI-38 выращивали в среде DMEM и использовали в качестве нормальных контрольных адгезивных клеток.
Исследование с участием нормальных CD4+ Т-клеток человека было одобрено Этическим комитетом Института биомедицинской химии; письменное информированное согласие было получено от всех участников. Кровь здоровых доноров в возрасте 18-25 лет (n = 4) собирали в пробирки Vacuette K3ЭДТА (Greiner Bio-One, Кремсмюнстер, Австрия). Свежие мононуклеарные клетки периферической крови (РВМС) выделяли с помощью центрифугирования в градиенте плотности Lympholite-H (Cedarlane, Канада). Т-клетки CD4+ очищали от РВМС с использованием набора для выделения CD4+ человека (Miltenyi Biotec, Германия) в соответствии с инструкциями производителя. CD4+ Т-клетки высевали в количестве 5 × 105 клеток/мл и культивировали в колбах объемом 25 см2 в клеточной среде RPMI 1640 с добавлением 10% FBS, 5 мкг/мл анти-CD28, 5 мкг/мл анти-CD3 mAb и 100 ед/мл мл rHu IL-2. Все клеточные линии культивировали в среде 5% CO2/95% воздуха во влажной атмосфере при 37°C и тестировали на заражение микоплазмами перед экспериментом с использованием набора для обнаружения микоплазм PlasmoTest™ (InvivoGen, США).
Для проверки острой токсичности клетки культивировали в течение 72 ч в 96-луночных планшетах в присутствии фермента в диапазоне концентраций 1-100 ЕД/мл и тестировали жизнеспособность клеток путем измерения превращение соли тетразолия, бромида 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолия, в формазан (тест МТТ). Значения IC50 и IC90 (концентрация фермента, при которой ответ снижался на 50% и 90% соответственно) рассчитывали по уравнениям аппроксимации.
Для измерения апоптоза инкубированные клетки диссоциировали с помощью трипсина, ресуспендировали в PBS и инкубировали с аннексином V-FITC и йодидом пропидия (PI) из набора FITC Annexin V/Dead Cell Apoptosis (Life Technologies, США) в соответствии с протоколом производителя. Подсчет 5 × 104 клеток в каждой точке проводили методом проточной цитометрии с помощью MACS Quant Analyzer 10 (Miltenyi Biotec, Германия).
Чтобы проверить цитотоксическую активность MrAII, различные типы лейкозных и солидных опухолевых раковых клеток человека вместе с нормальными фибробластами и нормальными CD4+ Т-клетками культивировали в присутствии различных концентраций фермента. Жизнеспособность клеток и индукцию апоптоза измеряли после 72 ч инкубации (Рисунок 3). В таблице 2 представлены значения IC50 и IC90 для протестированных клеточных линий.
Клеточная линия K562 была наиболее чувствительной среди раковых клеток, и фермент был способен значительно снижать жизнеспособность клеток даже при самой низкой концентрации 1 ЕД/мл (рис. 3а). Нормальные CD4+ Т-клетки были более устойчивы, чем раковые лимфоциты, и при концентрациях выше 10 ЕД/мл наблюдалось значительное снижение жизнеспособности клеток. Обе линии солидных опухолевых клеток LnCap и SCOV-3 продемонстрировали практически одинаковую чувствительность к ферменту (рис. 3б). Нормальные клетки фибробластов человека WI-38 также были чувствительны к низким концентрациям фермента (1 - 15 ЕД/мл); однако они оказались более устойчивыми к более высоким концентрациям (30-100 ЕД/мл).
Результаты показали, что MrAII обладает цитотоксической активностью и может индуцировать апоптоз в раковых клетках, в то время как высокие концентрации необходимы для индукции апоптоза в нормальных клетках.
Стабильная L-аспарагиназа из термофильной бактерии Melioribacter roseus с высоким значением температурного оптимума 70°С может быть использована в биомедицине как противоопухолевый агент, в высокотемпературных процессах в пищевой промышленности для снижения уровня акриламида.
Краткое описание чертежей:
Изобретение проиллюстрировано следующими рисунками.
Рис. 1. Анализ SDS-PAGE:
1 - маркер молекулярной массы белка;
2 - MrAII в бесклеточном гомогенате в концентрации 20 мкг;
3 - очищенный MrAII в концентрации 11 мкг.
Рис. 2. Исследование влияния на активность рекомбинантной L-аспарагиназы TsA температуры, рН, ионов металлов:
(а) влияние температуры на активность и (б) стабильность MrAII. Стабильность выражали в виде остаточной активности (%) после 0-60 мин инкубации;
(в) влияние pH и (г) различных катионов металлов на каталитическую активность MrAII; ND - не определено.
Рис. 3. Цитотоксическая активность MrAII в отношении раковых клеток. Раковые и нормальные клетки культивировали в течение 72 ч в присутствии различных концентраций фермента.
а) жизнеспособность клеток, измеренная с помощью теста МТТ для рака и нормальных Т-лимфоцитов;
б) жизнеспособность клеток, измеренная с помощью МТТ-теста для клеток солидных опухолей и фибробластов; (в-з) репрезентативные диаграммы проточной цитометрии для клеток, инкубированных с 20 ед./мл фермента и меченных аннексином V-FITC и PI. Представлены соотношения живых клеток (нижний левый квадрант), апоптотических клеток (нижний правый квадрант) и мертвых клеток (два верхних квадранта). (и-о) Гистограммы живых, апоптотических и мертвых клеток, инкубированных с 20 ед/мл фермента. Конц. - концентрация. PI, йодид пропидия. n = 4. * p ≤ 0,05 по сравнению с контрольными необработанными клетками.
ЛИТЕРАТУРА
1. Mahajan, R. V.; Kumar, V.; Rajendran, V.; Saran, S.; Ghosh, P.C.; Saxena, R.K. Purification and characterization of a novel and robust L-asparaginase having low-glutaminase activity from bacillus licheniformis: In vitro evaluation of anti-cancerous properties. PLoS One 2014, 9, doi:10.1371/journal.pone.0099037.
2. Nunes, J.C.F.; Cristóvão, R.O.; Freire, M.G.; Santos-Ebinuma, V.C.; Faria, J.L.; Silva, C.G.; Tavares, A.P.M. Recent Strategies and Applications for l-Asparaginase Confinement. Molecules 2020, 25.
3. Dumina, M. V.; Eldarov, M.A.; Zdanov, D.D.; Sokolov, N.N. L-Asparaginases of Extremophilic Microorganisms in Biomedicine. Biochem. Suppl. Ser. B Biomed. Chem. 2020, 14, doi:10.1134/S1990750820040046.
4. Jia, R.; Wan, X.; Geng, X.; Xue, D.; Xie, Z.; Chen, C. Microbial L-asparaginase for application in acrylamide mitigation from food: Current research status and future perspectives. Microorganisms 2021, 9.
5. Michalska, K.; Jaskolski, M. Structural aspects of L-asparaginases, their friends and relations. Acta Biochim. Pol. 2006.
6. Podosokorskaya, O.A.; Kadnikov, V. V.; Gavrilov, S.N.; Mardanov, A. V.; Merkel, A.Y.; Karnachuk, O. V.; Ravin, N. V.; Bonch-Osmolovskaya, E.A.; Kublanov, I. V. Characterization of Melioribacter roseus gen. nov., sp. nov., a novel facultatively anaerobic thermophilic cellulolytic bacterium from the class Ignavibacteria, and a proposal of a novel bacterial phylum Ignavibacteriae. Environ. Microbiol. 2013, 15, doi:10.1111/1462-2920.12067.
7. Bansal, S.; Gnaneswari, D.; Mishra, P.; Kundu, B. Structural stability and functional analysis of L-asparaginase from Pyrococcus furiosus. Biochem. 2010, doi:10.1134/s0006297910030144.
8. Li, X.; Zhang, X.; Xu, S.; Zhang, H.; Xu, M.; Yang, T.; Wang, L.; Qian, H.; Zhang, H.; Fang, H.; et al. Simultaneous cell disruption and semi-quantitative activity assays for high-throughput screening of thermostable L-asparaginases. Sci. Rep. 2018, doi:10.1038/s41598-018-26241-7.
9. Chohan, S.M.; Rashid, N. TK1656, a thermostable L-asparaginase from Thermococcus kodakaraensis, exhibiting highest ever reported enzyme activity. J. Biosci. Bioeng. 2013, doi:10.1016/j.jbiosc.2013.04.005.
10. Novel Bacillus methylotrophicus MKSY2013 producing enhanced thermostable asparaginase and its producing method. Patent KR101777904B1. https://patents.google.com/patent/KR101777904B1/en
11. Dumina, M. V.; Zhgun, A.A.; Pokrovskaya, M. V.; Aleksandrova, S.S.; Zhdanov, D.D.; Sokolov, N.N.; El’darov, M.A. Comparison of Enzymatic Activity of Novel Recombinant L-asparaginases of Extremophiles. Appl. Biochem. Microbiol. 2021, 57, 594-602, doi:10.1134/S0003683821050057
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:1 Аминокислотная последовательность L-аспарагиназы MrAII термофильной бактерии Melioribacter roseus
MKNILIIFTGGTFSMKIDRETNSAVPYYHGEELLKIIPEADKKANISVYEFGNYPGPHMTPELMLELSEIVKKYINQNDTDGIIITHGTDTLEETAYFLDLTVKTEKPIVVIGAMKTSSEPDWDGPKNLLDAISIINNPNSGNMGVLVCLNGEINAASEVTKTHTEDIETFHSLDFGSLGFVENDRVIFNRLPRKLETFETAKINSNVDLIKVYAGMNEKFFKFSADSGADGLVVEAMGVGNVPPKAFEGIKYVIEKGIPVVLVSRCPAGETLDIYGYEGAGKWLKKAGVIFSDYLNGQKARIKLIVCLGITNEIAKLRSMFEDEQ
SEQ ID NO:2 Ген mrAII, кодирующий L-аспарагиназу MrAII термофильной
бактерии Melioribacter roseus
ATGAAAAATATTTTGATAATTTTTACCGGCGGCACCTTTTCGATGAAGATCGACAGGGAAACAAATTCGGCCGTGCCATATTACCATGGCGAAGAGCTCCTCAAAATCATTCCGGAAGCCGATAAGAAAGCCAATATATCCGTCTATGAATTCGGCAACTATCCCGGACCGCACATGACTCCGGAATTAATGCTTGAGTTGTCGGAAATTGTAAAGAAGTATATAAATCAAAACGATACTGATGGAATTATTATTACGCACGGGACAGACACACTCGAAGAGACAGCCTACTTTCTCGATCTCACGGTAAAAACGGAAAAACCGATAGTGGTAATAGGCGCTATGAAAACAAGCTCGGAGCCGGATTGGGACGGGCCGAAGAACCTGCTCGACGCCATATCGATTATTAATAATCCCAACAGCGGAAATATGGGCGTGCTTGTCTGTTTGAACGGCGAAATTAATGCTGCAAGCGAAGTAACGAAAACTCATACCGAAGATATCGAAACATTTCACAGTCTCGATTTCGGTTCGCTCGGATTTGTCGAAAACGACAGGGTGATTTTTAACAGGCTGCCAAGGAAACTTGAAACGTTCGAGACGGCGAAAATCAATTCAAACGTAGATTTGATAAAAGTATATGCCGGAATGAACGAAAAATTTTTCAAATTTTCAGCCGACAGCGGCGCCGACGGGCTGGTAGTCGAAGCTATGGGAGTTGGCAACGTTCCCCCGAAAGCGTTCGAAGGTATAAAGTACGTTATTGAAAAAGGCATCCCGGTGGTATTGGTTTCCAGATGCCCGGCCGGCGAAACTCTCGACATATACGGCTACGAAGGAGCGGGAAAATGGCTAAAAAAGGCGGGAGTTATTTTTTCCGATTACTTGAACGGGCAAAAAGCCAGAATAAAATTGATTGTTTGTCTCGGCATTACTAACGAAATAGCTAAATTGAGATCGATGTTCGAAGATGAACAATAA
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЙ ГЕН tsA, КОДИРУЮЩИЙ ТЕРМОСТАБИЛЬНУЮ L-АСПАРАГИНАЗУ ИЗ АРХЕИ THERMOCOCCUS SIBIRICUS, РЕКОМБИНАНТНЫЙ ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ ТЕРМОСТАБИЛЬНОЙ L-АСПАРАГИНАЗЫ И СПОСОБ ОЧИСТКИ ЭТОГО ФЕРМЕНТА | 2022 |
|
RU2794225C1 |
| СПОСОБ УЛУЧШЕНИЯ КАТАЛИТИЧЕСКИХ СВОЙСТВ ГИПЕРТЕРМОФИЛЬНЫХ L-АСПАРАГИНАЗ I ТИПА, L-АСПАРАГИНАЗА I ТИПА ИЗ ГИПЕРТЕРМОФИЛЬНОЙ АРХЕИ THERMOCOCCUS SIBIRICUS, ПОЛУЧЕННАЯ ТАКИМ СПОСОБОМ | 2023 |
|
RU2836049C1 |
| МУТАНТНАЯ РЕКОМБИНАНТНАЯ L-АСПАРАГИНАЗА Wolinella succinogenes (ВАРИАНТЫ) | 2014 |
|
RU2545722C1 |
| ПРОДУЦЕНТ L-АСПАРАГИНАЗЫ E. COLI И ЭКСПРЕССИОННАЯ ПЛАЗМИДА PET28A-ASNSYN, КОДИРУЮЩАЯ L-АСПАРАГИНАЗУ | 2023 |
|
RU2817891C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pHisTevTSIB0821, ТРАНСФОРМИРОВАННЫЙ ЕЮ ШТАММ Escherichia coli Rosetta(DE3)/pHisTevTSIB0821 И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ПРОЛИДАЗЫ TSIB_0821 | 2012 |
|
RU2509154C1 |
| Рекомбинантный штамм бактерий Escherichia coli Rosetta 2(DE3)/pET28c-TstP36H-Sso7d - продуцент химерной термостабильной ДНК-полимеразы TstP36H-Sso7d из Thermococcus stetteri | 2024 |
|
RU2840236C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pACYC-LANS(KM), ШТАММ Escherichia coli BL21(DE3), ТРАНСФОРМИРОВАННЫЙ РЕКОМБИНАНТНОЙ ДНК pACYC-LANS(KM), И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ L-АСПАРАГИНАЗЫ Erwinia carotovora | 2010 |
|
RU2441916C1 |
| ГИБРИДНЫЙ БЕЛОК НА ОСНОВЕ L-АСПАРАГИНАЗЫ Wolinella succinogenes, ШТАММ Escherichia coli - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ГИБРИДНОГО БЕЛКА, ОБЛАДАЮЩЕГО ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТЬЮ | 2014 |
|
RU2562166C1 |
| СПОСОБ ВЫЯВЛЕНИЯ ШТАММОВ БАКТЕРИЙ-ПРОДУЦЕНТОВ L-АСПАРАГИНАЗЫ | 2008 |
|
RU2398876C2 |
| Модифицированный ген рас бактерий Escherichia coli, кодирующий предшественник фермента с активностью пенициллин G ацилазы, рекомбинантный штамм Escherichia coli - продуцент пенициллин G ацилазы и способ микробиологического синтеза этого фермента | 2015 |
|
RU2624022C1 |
Изобретение относится к биотехнологии. Предложен рекомбинантный штамм Escherichia coli BL21(DE3) - продуцент термостабильной L-аспарагиназы II типа из Meliobacter roseus (MrAII), полученный путем введения синтезированного гена mrAII, соответствующего последовательности SEQ ID NO: 2, в составе вектора рЕТ-28а(+) под контролем промотора Т7 в штамм-реципиент Escherichia coli BL21(DE3). Изобретение обеспечивает получение термостабильной L-аспарагиназы II типа из Meliobacter roseus (MrAII), проявляющей активность гидролиза в отношении L-аспарагина с удельной активностью 1530 Ед/мг. 3 ил., 2 табл., 7 пр.
Рекомбинантный штамм Escherichia coli BL21(DE3) - продуцент термостабильной L-аспарагиназы II типа из Meliobacter roseus (MrAII), полученный путем введения синтезированного гена mrAII, соответствующего последовательности SEQ ID NO: 2, в составе вектора рЕТ-28а(+) под контролем промотора Т7 в штамм-реципиент Escherichia coli BL21(DE3).
| ДУМИНА М.В | |||
| и др | |||
| Сравнительная активность новых рекомбинантных L-аспарагиназ экстремофилов | |||
| Прикладная биохимия и микробиология, 2021, том 57, N 5, с | |||
| Детекторный радиоприемник гетеродин | 1923 |
|
SU467A1 |
| база данных NCBI Reference Sequence: WP_014855710.1, 19.05.2013 | |||
| Аsparaginase/glutaminase [Melioribacter roseus] | |||
| Найдено онлайн: | |||

