Область техники, к которой относится изобретение
Изобретение относится к области молекулярной биологии, медицинской химии, вирусологии и микробиологии, и предназначено для скрининга потенциальных ингибиторов РНК-зависимой РНК-полимеразы (РЗРП) SARS-COV-2 в лабораториях с первым уровнем биобезопасности, не оборудованных для работы с высокопатогенными инфекциями. Более конкретно, изобретение относится к полноразмерным репликонам -искусственно созданным системам, позволяющим проводить тестирование потенциальных ингибиторов РЗРП SARS-COV-2 без использования полноценных вирусных частиц. Предложен новый полноразмерный репликон с двумя блоками репортерных генов, один из которых может транскрибироваться не только вирусной РЗРП, но и клеточными полимеразами, а второй может транскрибироваться только вирусной полимеразой по механизму прерывистой транскрипции, характерной для семейства Coronaviridae.
Уровень техники
Пандемия COVID-19 (COronaVIrus Disease 2019), вызванная бетакоронавирусом SARS-CoV-2, к концу 2023 г. унесла более 7 миллионов жизней [https://covidl9.who.int]. Вирус поражает главным образом легкие и другие сегменты дыхательных путей, вызывая острую пневмонию [Zhu N., Zhang D., Wang W., Li X., Yang В., Song J., Zhao X., Huang В., Shi W., Lu R., Niu P., Zhan F., Ma X., Wang D., Xu W., Wu G., Gao G. F., Tan W., China Novel Coronavirus I., Research T. A Novel Coronavirus from Patients with Pneumonia in China, 2019 // N Engl J Med. - 2020. - T. 382, №8. - C. 727-733. Budinger G. R. S., Misharin A. V., Ridge К. M., Singer B. D., Wunderink R. G. Distinctive features of severe SARS-CoV-2 pneumonia // J Clin Invest. - 2021. - T. 131, №14.]. В отличие от большинства респираторных вирусов, эта коронавирусная инфекция часто вызывает тяжелое воспаление - цитокиновый шторм, что в сочетании с гиперкоагуляцией может привести к развитию острого респираторного дистресс-синдрома [Torres Acosta М. A., Singer В. D. Pathogenesis of COVID-19-induced ARDS: implications for an ageing population // Eur Respir J. - 2020. - T. 56, №3.]. Кроме того, у пациентов с COVID-19 часто наблюдаются признаки внереспираторной патологии, в том числе расстройства желудочно-кишечноготракта, дисфункция печени и поджелудочной железы, неврологические и психиатрические симптомы, признаки патологии сердечно-сосудистой системы [Marjot Т., Webb G. J., Barritt A. S. t, Moon A. M., Stamataki Z., Wong V. W., Barnes E. COVID-19 and liver disease: mechanistic and clinical perspectives // Nat Rev Gastroenterol Hepatol. - 2021. -T. 18, №5. - C. 348-364. Keyhanian K., Umeton R. P., Mohit В., Davoudi V., Hajighasemi F., Ghasemi M. SARS-CoV-2 and nervous system: From pathogenesis to clinical manifestation // J Neuroimmunol. - 2020. - T. 350. - C. 577436. Groff A., Kavanaugh M., Ramgobin D., McClafferty В., Aggarwal C. S., Golamari R., Jain R. Gastrointestinal Manifestations of COVID-19: A Review of What We Know // Ochsner J. - 2021. - T. 21, №2. - C. 177-180. Xie Y., Xu E., Bowe В., Al-Aly Z. Long-term cardiovascular outcomes of COVID-19 // Nat Med. -2022. - T. 28, №3. - C. 583-590. Doroftei В., Ciobica A., Hie O. D., Maftei R., Ilea C. Mini-Review Discussing the Reliability and Efficiency of COVID-19 Vaccines // Diagnostics (Basel). - 2021. - T. 11, №4.]. Интенсивные исследования привели к внедрению в клиническую практику вакцин различных типов [Reynolds С.J., Pade С, Gibbons J. М., Otter A. D., Lin К. М., Munoz Sandoval D., Pieper F. P., Butler D. K., Liu S., Joy G., Forooghi N., Treibel T. A., Manisty C, Moon J. C, sign С.О. I. s., sign С.О. I. C. N. s., Semper A., Brooks Т., McKnight A., Altmann D. M., Boyton R. J., Abbass H., Abiodun A., Alfarih M., Alldis Z., Altmann D. M., Amin О. E., Andiapen M., Artico J., Augusto J. В., Baca G. L., Bailey S. N. L., Bhuva A. N., Boulter A., Bowles R, Boyton R. J., Bracken О. V., O'Brien В., Brooks Т., Bullock N., Butler D. K., Captur G., Carr O., Champion N., Chan C, Chandran A., Coleman Т., Couto de Sousa J., Couto-Parada X., Cross E., Cutino-Moguel Т., D'Arcangelo S., Davies R. H., Douglas В., Di Genova C, Dieobi-Anene K., Diniz M. O., Ellis A., Feehan K., Finlay M., Fontana M., Forooghi N., Francis S., Gibbons J. M., Gillespie D., Gilroy D., Hamblin M., Harker G., Hemingway G., Hewson J., Heywood W., Hickling L. M., Hicks В., Hingorani A. D., Howes L., Itua I., Jardim V., Lee W. J., Jensen M., Jones J., Jones M., Joy G., Kapil V., Kelly C, Kurdi H., Lambourne J., Lin К. M., Liu S., Lloyd A., Louth S., Maini M. K., Mandadapu V., Manisty C, McKnight A., Menacho K., Mfuko C, Mills K., Millward S., Mitchelmore O., Moon C, Moon J., Munoz Sandoval D., Murray S. M., Noursadeghi M., Otter A., Pade C, Palma S., Parker R., Patel K., Pawarova M., Petersen S. E., Piniera В., Pieper F. P., Rannigan L., Rapala A., Reynolds C. J., Richards A., Robathan M., Rosenheim J., Rowe C, Royds M., Sackville West J., Sambile G., Schmidt N. M., Selman H., Semper A., Seraphim A., Simion M., Smit A., Sugimoto M., Swadling L., Taylor S., Temperton N., Thomas S., Thornton G. D., Treibel T. A., Tucker A., Varghese A., Veerapen J., Vijayakumar M., Warner Т., Welch S., White H., Wodehouse Т., Wynne L., Zahedi D., Chain В., Moon J. C. Immune boosting by B. 1.1.529 (Omicron) depends on previous SARS-CoV-2 exposure // Science. - 2022. - T. 377, №6603. - C. eabql841.]. Вцелом они снижают риски заражения и развития тяжелой формы COVID-19, хотя эффективность зависит от их типа. В то же время генетическое разнообразие вируса привело к появлению его новых вариантов, способных обходить ранее существовавший иммунный ответ, сформированный в результате вакцинации или предшествующего инфицирования [Acherjee Т., Behara A., Saad М., Vittorio Т. J. Mechanisms and management of prothrombotic state in COVID-19 disease // Ther Adv Cardiovasc Dis. - 2021. - T. 15. - C. 17539447211053470].
Для терапии COVID-19 к настоящему времени создан только один специфический препарат «Паксловид», являющийся ингибитором основной протеазы вируса. Применяющиеся для лечения коронавирусной инфекции препараты ремдесивир, фавипиравир и некоторые другие аналоги нуклеозидов, направленные на блокирование РНК-полимеразы SARS-CoV-2, не являются специфическими ингибиторами этого фермента и имеют существенное побочное действие. В связи с этим поиск новых специфических ингибиторов РНК-зависимой РНК-полимеразы SARS-CoV-2 (РЗРП) представляется совершенно необходимым.
Отдельной проблемой во всем мире является нехватка лабораторий для работы со столь высокопатогенным вирусом, как SARS-CoV-2, и, соответственно, весьма актуальна разработка безопасных систем для скрининга соединений без использования вирусных частиц. Такие системы уже созданы для изучения различных стадий жизненного цикла вируса в условиях лаборатории с первым уровнем биобезопасности [Jin Y.-Y., Lin Н., Сао L., Wu W.-C, Ji Y., Du L., Jiang Y., Xie Y., Tong K., Xing F., Zheng F., Shi M., Pan J.-A., Peng X., Guo D. A Convenient and Biosafe Replicon with Accessory Genes of SARS-CoV-2 and Its Potential Application in Antiviral Drug Discovery // Virologica Sinica. - 2021. - T. 36, №5. - C. 913-923. Zhang Q. Y., Deng C. L., Liu J., Li J. Q., Zhang H. Q., Li N., Zhang Y. N., Li X. D., Zhang В., Xu Y., Ye H. Q. SARS-CoV-2 replicon for high-throughput antiviral screening // J Gen Virol. - 2021. - T. 102, №5. Kotaki Т., Xie X., Shi P. Y., Kameoka M. A PCR amplicon-based SARS-CoV-2 replicon for antiviral evaluation // Sci Rep.- 2021. - T. 11, №1. - C. 2229]. В частности, наиболее часто используют модели полноразмерных репликонов: кДНК вируса клонируется в вектор искусственной бактериальной хромосомы [Chiem, К., Ye, С, Martinez-Sobrido, L. Generation of recombinant SARS-CoV-2 using a bacterial artificial chromosome. // Current Protocols in Microbiology. - 2020. - T. 59, №1. -el26]. При трансфекции этого вектора в клетки Vero Е6 клеточной РНК-полимеразой II синтезируется РНК SARS-CoV-2. На данный момент создано несколько полноразмерных репликонов SARS-CoV-2, однако все они кодируют лишь один блок репортерных генов [Chiem К, Morales Vasquez D, Park JG, Piatt RN, Anderson T, Walter MR, Kobie JJ, Ye C, Martinez-Sobrido L. Generation and Characterization of recombinant SARS-CoV-2 expressing reporter genes. // J. Virol. - 2021. - T. 95, №7. - e02209-20; Wang B, Zhang C, Lei X, Ren L, Zhao Z, Wang J, Huang H. Construction of Non-infectious SARS-CoV-2 Replicons and Their Application in Drug Evaluation. // Viro. Sin. - 2021. - T. 36, №5. - C. 890-900; Ye C, Martinez-Sobrido L. Use of a Bacterial Artificial Chromosome to Generate Recombinant SARS-CoV-2 Expressing Robust Levels of Reporter Genes. // Microbiol Spectr. - 2022. - T. 10, №6. -e0273222], что не позволяет всесторонне изучать стадию транскрипции и репликации вируса и однозначно подтвердить РЗРП в качестве мишени действия ингибиторов.
Необходимо подчеркнуть, что для аналогов нуклеозидов необходима именно клеточная система, поскольку механизм их действия включает предварительные модификации в клетке (трифосфорилирование), и для тестирования на индивидуальной РНК-полимеразе аналоги нуклеозидов необходимо предварительно химически превратить в соответствующие 5'-трифосфаты. Это, во-первых, сильно замедляет и удорожает скрининг, а во-вторых, может приводить к искажению результатов.
Таким образом, создание полноразмерного репликона с двумя блоками репортерных генов является весьма актуальной и важной задачей.
Раскрытие сущности изобретения
Техническим результатом изобретения является создание полноразмерного репликона с двумя блоками репортерных генов, для тестирования потенциальных ингибиторов РЗРП SARS-COV-2 без использования полноценных вирусных частиц.
В своем первом воплощении изобретение является генетической конструкцией, представляющей собой кДНК вируса, не содержащую генов структурных белков S, Е и М, с двумя блоками репортерных генов, которая клонирована в вектор искусственной бактериальной хромосомы.
Аспектом воплощения изобретения является то, что первый блок репортерных генов, представленный люциферазой Renilla reniformis и зеленым флуоресцентным белком (Rluc-GFP), расположен под контролем CMV-промотора, находящегося перед 5'UTR SARS-CoV-2.
Другим аспектом воплощения изобретения является то, что второй блок репортерных генов представленный люциферазой светлячка Photinus pyralis и красным флуоресцентным белком (Fluc-RFP) и находится после последовательности, регулирующей прерывистую транскрипцию (TRS-N).
Главным аспектом воплощения изобретения является то, что такое расположение генов и регуляторных последовательностей (CMV - 5'UTR - Rluc-GFP - TRS-N - Flue -RFP - 3'UTR) позволяет оценить уровень ингибирования вирусной РЗРП, поскольку второй блок репортерных генов, находящийся после TRS-N, может транскрибироваться только вирусной полимеразой по механизму прерывистой транскрипции, характерной для семейства Coronaviridae. Первый же блок репортерных генов, находящийся под контролем CMV-промотора, может транскрибироваться не только вирусной РЗРП, но и клеточными полимеразами.
Вторым воплощением изобретения является создание в качестве отрицательного контроля системы полноразмерного репликона, в котором в активном центре полимеразной субъединицы РЗРП (nsp12) произведены замены аминокислот (D760A, D761A) методом сайт-направленного мутагенеза. Данные замены согласно литературным данным приводят к потере каталитической активности РЗРП [Gao Y., Yan L., Huang Y., Liu F., Zhao Y., Cao L., Wang Т., Sun Q., Ming Z., Zhang L., Ge J., Zheng L., Zhang Y., Wang H., Zhu Y., Zhu С., Ни Т., Hua Т., Zhang В., Yang X., Li J., Yang H., Liu Z., Xu W., Guddat L. W., Wang Q., Lou Z., Rao Z. Structure of the RNA-dependent RNA polymerase from COVID-19 virus // Science. - 2020. - T. 368, №6492. - C. 779-782.].
В своем последним воплощении для облегчения поиска нуклеозидных ингибиторов РЗРП без создания их фосфорилированных депо-форм была получена клеточная линия Vero Е6_ТК, экспрессирующая тимидин-киназу вируса простого герпеса, осуществляющую первую стадию фосфорилирования нуклеозидов в клетке.
Таким образом, получена улучшенная безвирусная репортерная система для тестирования ингибиторов РЗРП вируса SARS-CoV-2, проведена ее валидация и показана ее высокая эффективность для тестирования как для ненуклеозидных, так и для нуклеозидных ингибиторов РНК-зависимой РНК-полимеразы SARS-CoV-2. Наличие отрицательного контроля - системы полноразмерного репликона с мутациями в активном центре (D760A, D761А) полимеразной субъединицы РЗРП (nsp12), приводящими к потере каталитической активности, позволяет однозначно идентифицировать РНК-зависимую РНК-полимеразу SARS-COV-2 в качестве мишени антивирусного действия.
Далее изобретение будет раскрыто подробнее со ссылками на фигуры и примеры, которые приводятся исключительно с целью иллюстрации и пояснения сущности заявленного изобретения, но которые не предназначены для ограничения объема притязаний.
Краткое описание фигур и таблиц
Фигура 1. Модель полноразмерного репликона SARS-CoV-2 с двумя блоками репортерных генов. Первый блок репортерных генов Rluc-GFP расположен внутри ORF1a под контролем промотора CMV, второй блок Fluc-RFP расположен между геном белка N и 3'UTR и может транскрибироваться только с помощью механизма прерывистой транскрипции вирусной РЗРП. Ниже приведена схема 6 фрагментов кДНК вируса, из которых производилась сборка полноразмерного репликона (репликон GG1) и клонирование в плазмиду pBAC_lacZ с использованием Golden Gate подхода.
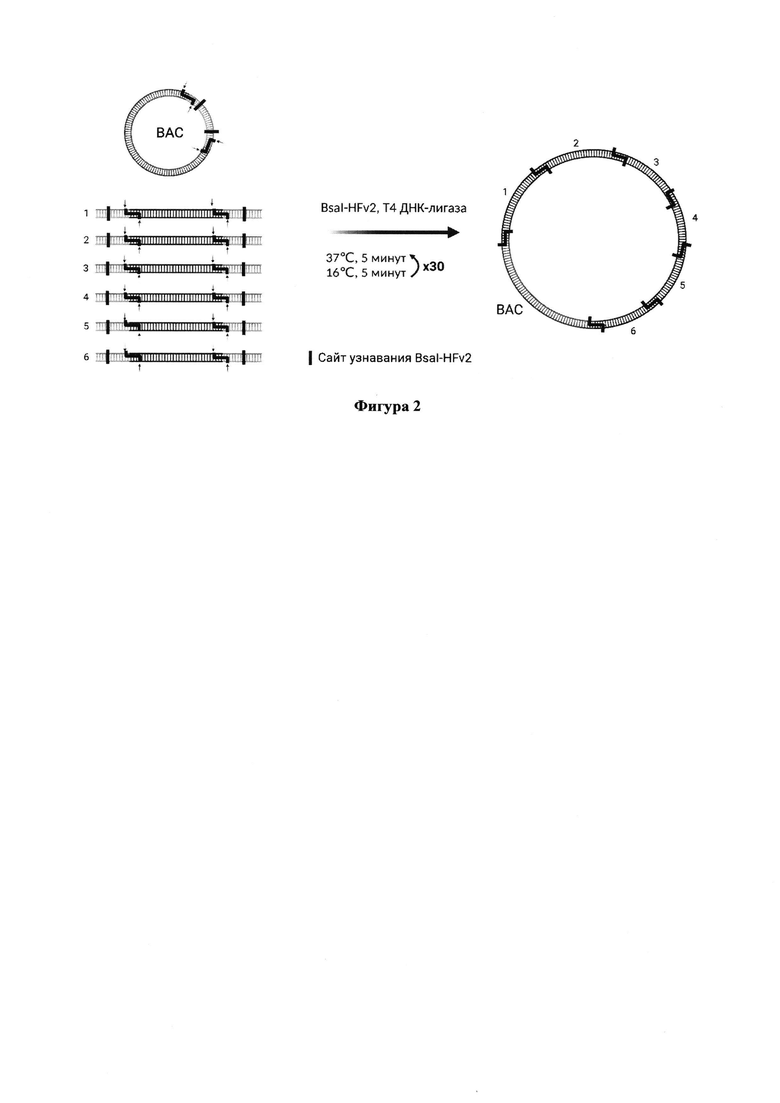
Фигура 2. Схема эксперимента по получению полноразмерного репликона методом Golden Gate клонирования. К смеси pBAC_lacZ и фрагментов кДНК вируса, клонированных в промежуточные вектора pJet 1.2, добавляются эндонуклеаза рестрикции BsaI-HFv2 и Т4 ДНК-лигаза. BsaI-HFv2 вносит два двуцепочечных разрыва в вектор рВАС и вырезает фрагменты 1-6 из промежуточных векторов, не оставляя при этом своих сайтов посадки во фрагментах, необходимых для сборки полноразмерного репликона. Т4 ДНК-лигаза осуществляет лигирование фрагментов с соответствующими липкими концами.
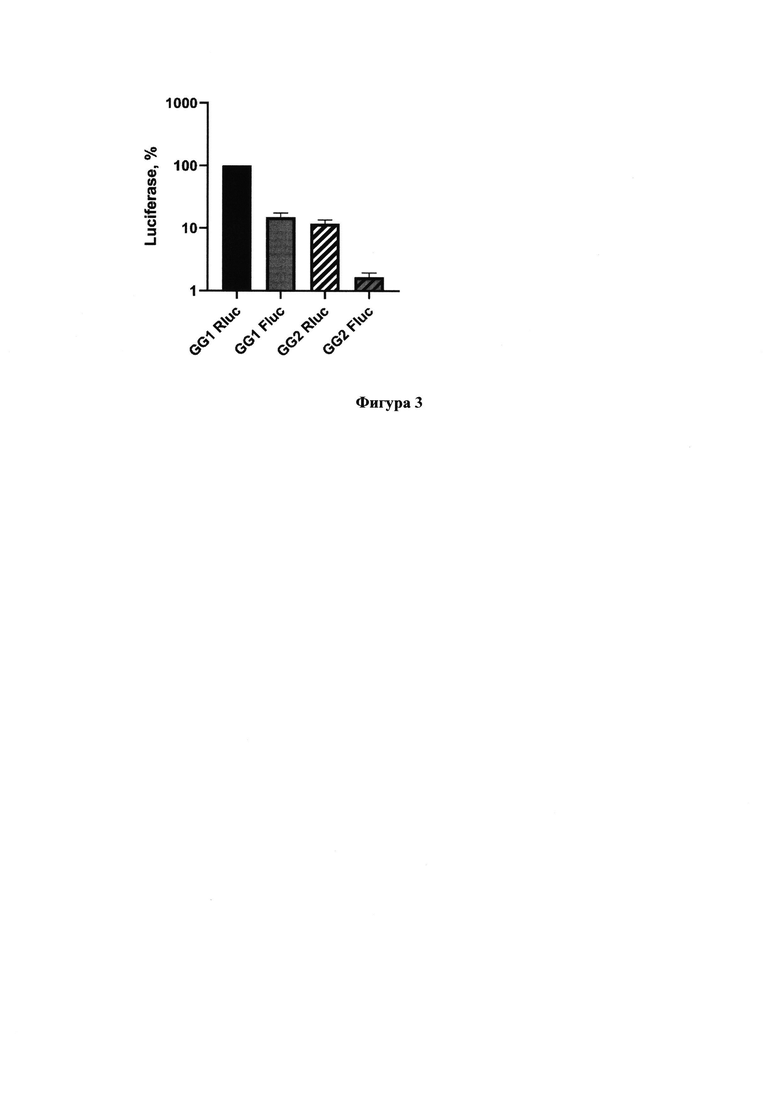
Фигура 3. Сравнение уровней люминесценции люцифераз ренилла (Rluc) и светлячка (Fluc) в клетках Vero Е6, трансфицированных репликонами GG1 или GG2 (репликон, кодирующий каталитически неактивную вирусную полимеразу).
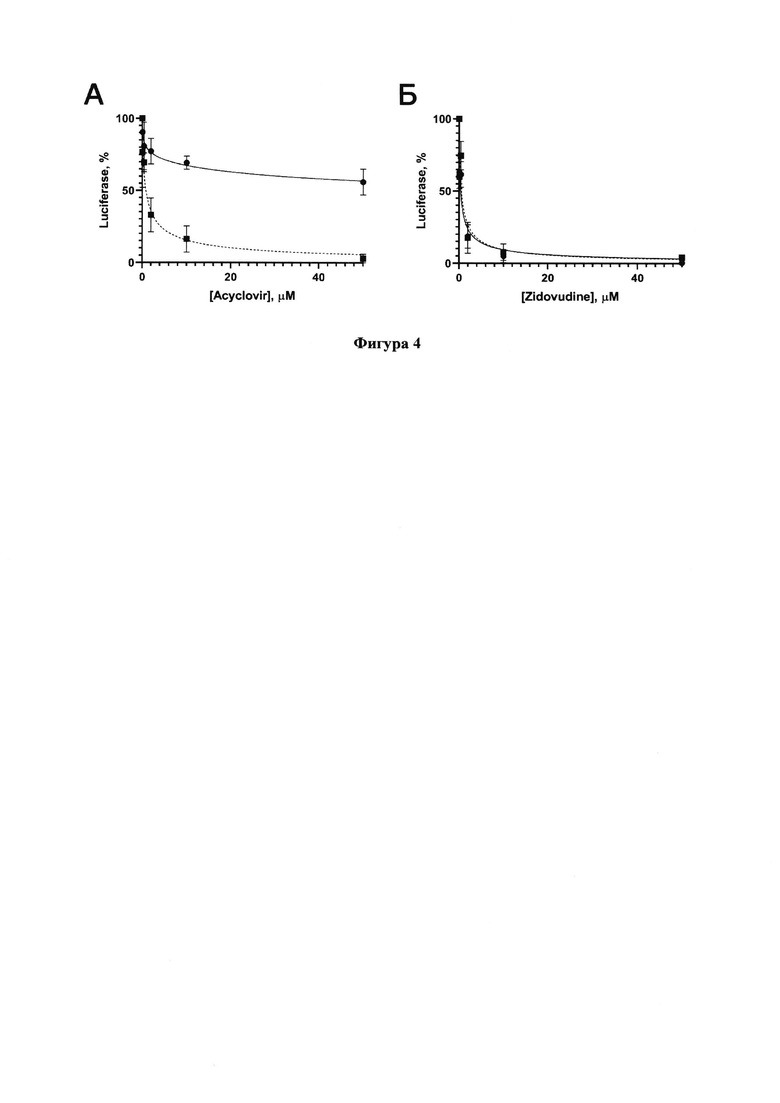
Фигура 4. Проверка ингибирующей активностиацикловира (А) и зидовудина (В) на уровень люминесценции люциферазы в клеточных линиях Vero Е6 (сплошная кривая) и Vero Е6_ТК (пунктирная кривая), транедуцированных реприкативно-некомпетентным вектором на основе ВИЧ-1.
Фигура 5. Структура молнупиравира.
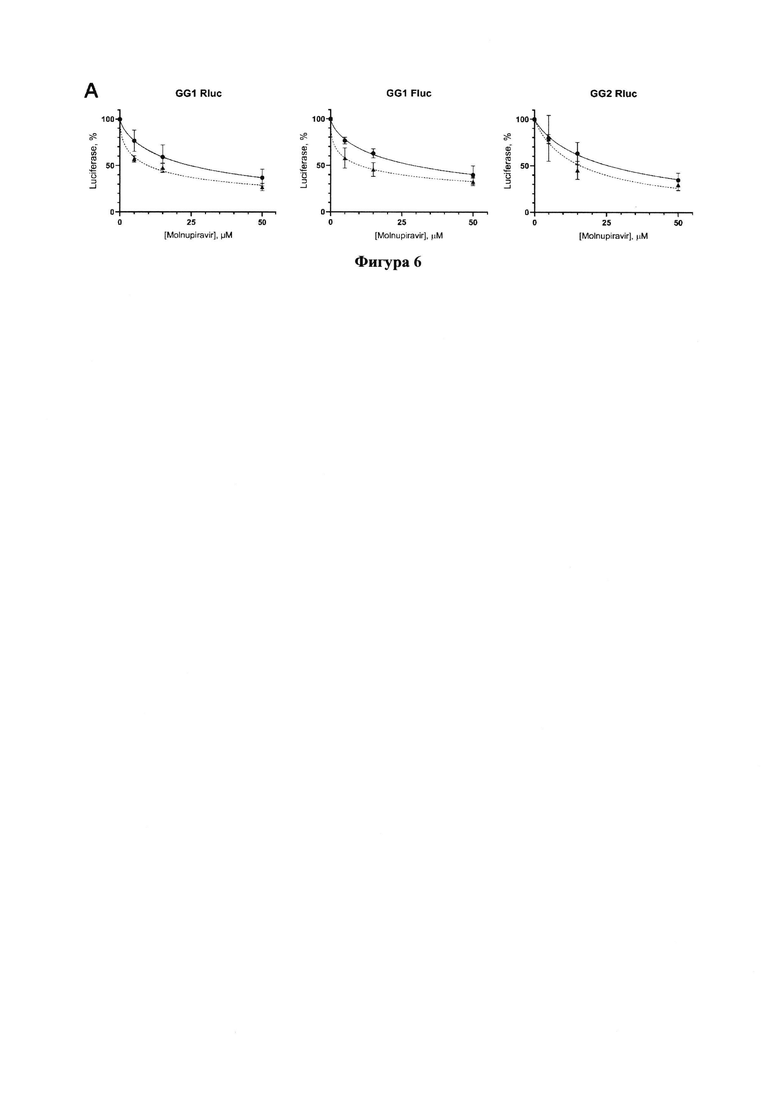
Фигура 6. Результаты тестирования молнупиравира по снижению уровней люцифераз в трансфецированных репортерами GG1 или GG2 клеточных линиях Vero Е6 (сплошная линия) и Vero Е6_ТК (пунктирная).
Фигура 7. Структура 2'-амино-2'-дезоксиаденозина.
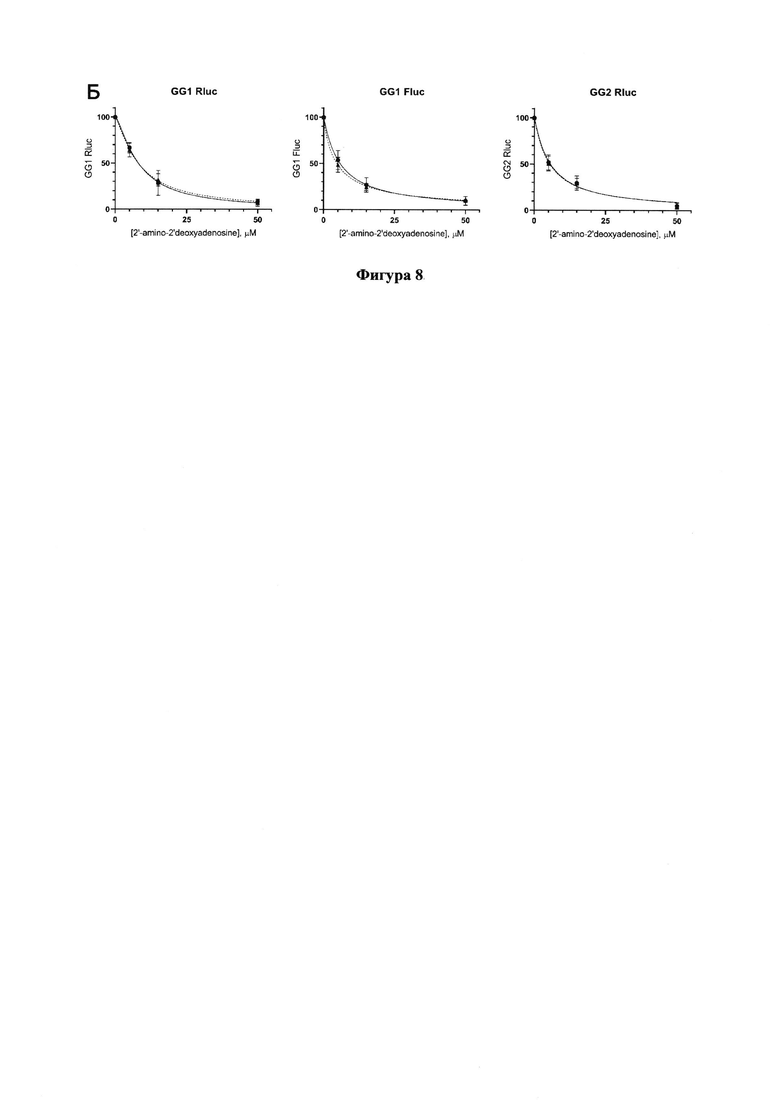
Фигура 8. Результаты тестирования 2'-амино-2'-дезоксиаденозина по снижению уровней люцифераз в трансфецированных репортерами GG1 или GG2 клеточных линиях Vero Е6 (сплошная линия) и Vero Е6_ТК (пунктирная).
Фигура 9. Структура ацикловира.
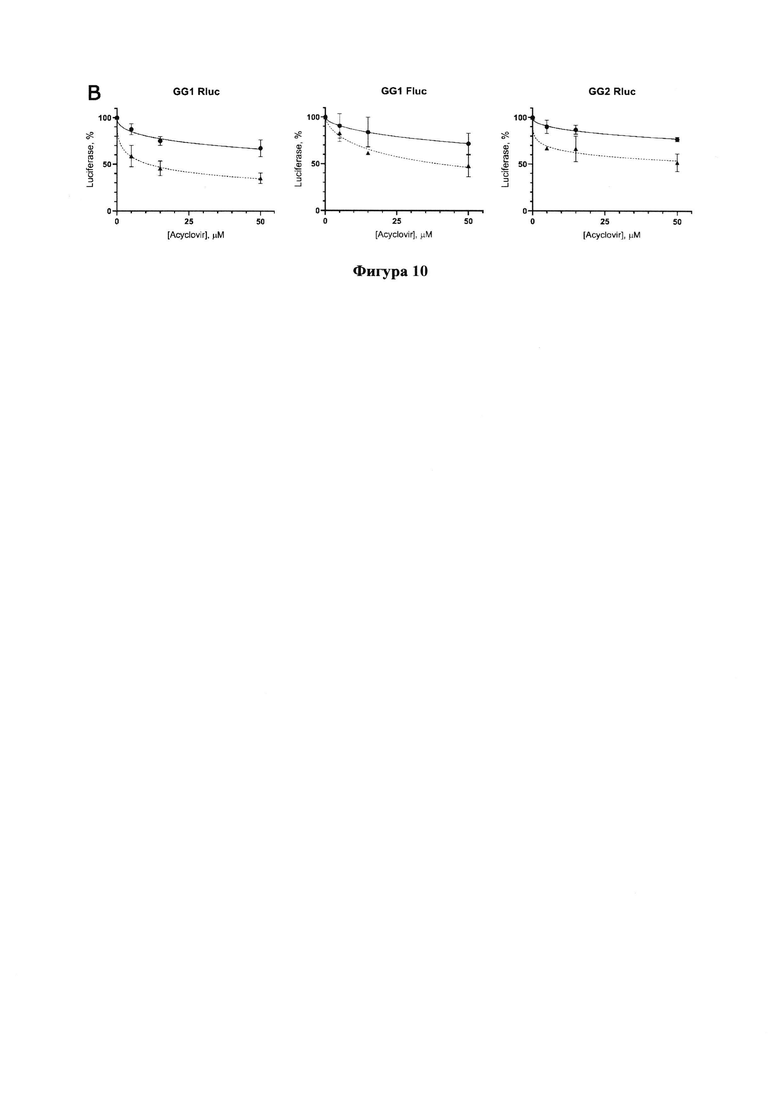
Фигура 10. Результаты тестирования ацикловира по снижению уровней люцифераз в трансфецированных репортерами GG1 или GG2 клеточных линиях Vero Е6 (сплошная линия) и Vero Е6_ТК (пунктирная).
Фигура 11. Структура 3'-О-метиладенозина.
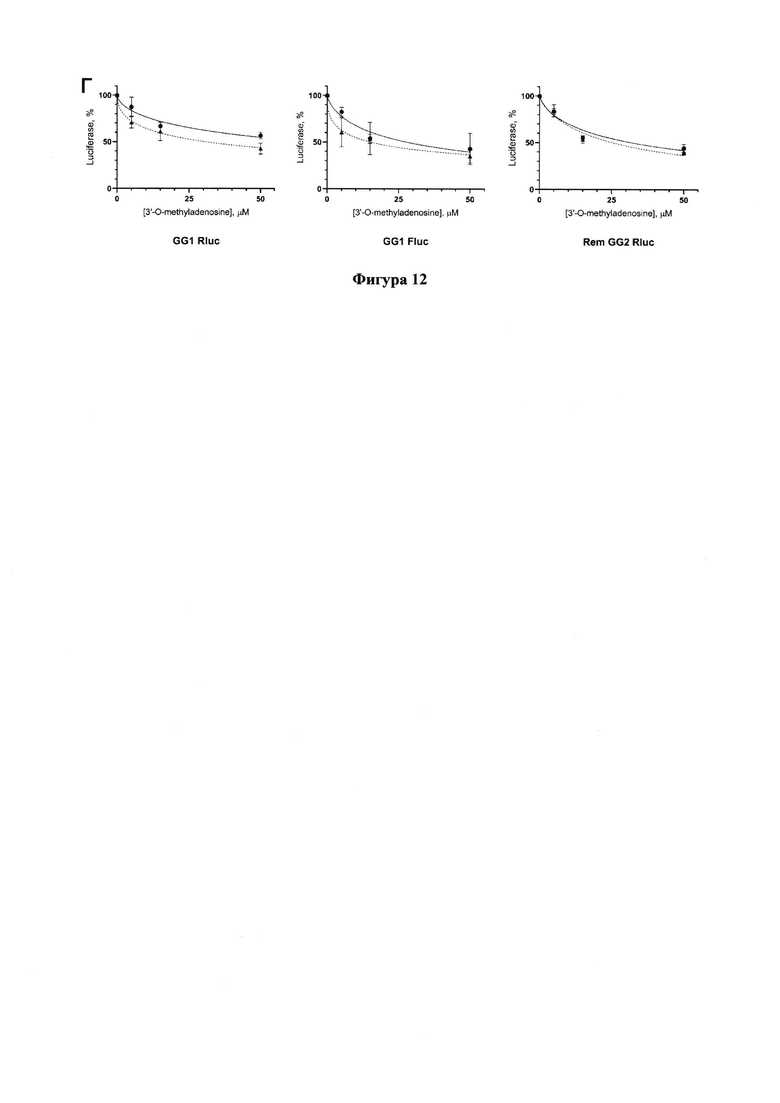
Фигура 12. Результаты тестирования 3'-О-метиладенозина по снижению уровней люцифераз в трансфецированных репортерами GG1 или GG2 клеточных линиях Vero Е6 (сплошная линия) и Vero Е6_ТК (пунктирная).
Фигура 13. Структура соединения LN-140.
Фигура 14. Результаты тестирования ингибитора LN-140 по снижению уровней люцифераз в трансфецированных репортерами GG1 или GG2 клеточных линиях Vero Е6 (сплошная линия) и Vero Е6_ТК (пунктирная).
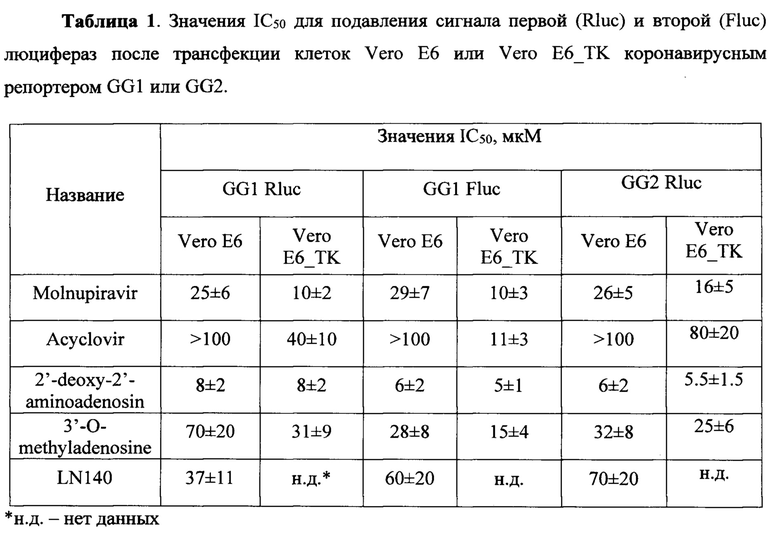
Таблица 1. Значения IC50 для подавления сигнала первой (Rluc) и второй (Fluc) люцифераз после трансфекции клеток Vero Е6 или Vero Е6_ТК коронавирусным репортером GG1 или GG2.
Осуществление изобретения
Предложена модель полноразмерного репликона (Фигура 1). Под контролем промотора CMV находится последовательность модифицированного генома SARS-CoV-2, несодержащего генов структурных белков S, Е и М, начинающаяся с 5'UTR. Затем следует последовательность полипротеина ORFlab, включающая первый блок репортерных генов, размещенный между NSP1 и NSP2. Данный блок включает в себя ген устойчивости к бластицидину и слитый ген Rluc-GFP, а также последовательности Р2А, Т2А и Е2А, вызывающие пропуск образования пептидной связи в процессе трансляции. Последние необходимы для правильного синтеза как NSP1 и NSP2, так и белков первого репортерного блока. Сразу после последовательности полипротеина следует ген белка N под контролем TRS-N. Несмотря на то, что в геноме вируса после ORFlab находится ген белка S под контролем TRS orflab/S, в литературе показано, что подобное размещение на этом месте гена белка N не нарушает процесс эффективного образования субгеномных РНК [Almazan F., Dediego М. L., Galan С, Escors D., Alvarez E., Ortego J., Sola I., Zuniga S., Alonso S., Moreno J. L., Nogales A., Capiscol C, Enjuanes L. Construction of a severe acute respiratory syndrome coronavirus infectious cDNA clone and a replicon to study coronavirus RNA synthesis // J Virol. - 2006. - T. 80, №21. - C. 10900-6]. За геном белка N размещается последовательность второго репортерного блока, представленная слитым геном Fluc-RFP. Второй блок также отделен от гена белка N последовательностью Р2А. После гена Fluc-RFP находятся последовательности 3'UTR, рибозим вируса гепатита D (Rz) и сигнал полиаденилирования SV40 poly (А). Наличие двух блоков репортерных генов позволяет однозначно определять уровень ингибирования РНК-зависимой РНК-полимеразы (РЗРП), а кроме того, оценивать эффективность протекания прерывистой транскрипции и искать клеточные факторы, участвующие в процессе репликации и транскрипции SARS-CoV-2.
Для получения предлагаемого репликона (далее GG1) использован метод Golden Gate клонирования (Фигура 2). Метод основан на использовании эндонуклеаз рестрикции IIS типа, которые узнают и связывают ассиметричные последовательности ДНК и осуществляют расщепление обеих цепей ДНК на определенном расстоянии от сайта посадки. Благодаря этим особенностям эндонуклеаз рестрикции типа IIS становится возможным одновременное клонирование нескольких фрагментов. В результате предварительных ПЦР с коронавирусной кДНК, а также генов, необходимых репортерных белков, были получены 6 фрагментов для сборки полноразмерного репликона (Фигура 1), которые затем каждый по отдельности был клонирован в промежуточный вектор pJet 1.2. Сборка производилась с использованием 7 фрагментов, т.к. помимо описанных 6 фрагментов кДНК вируса в качестве вектора использовалась плазмида pBAC_lacZ (Фигура 2).
В качестве вектора была выбрана искусственная бактериальная хромосома (ВАС), поскольку такой тип вектора позволяет нарабатывать большие генетические конструкции, содержащие токсичные для клеток E.coli последовательности. Для получения полноразмерного репликона в бактериальных клетках был использован специальный штамм E.coli - TransforMax EPI300, в геноме которого содержится индуцибельный мутантный ген trfA, необходимый для инициации увеличения числа копий вектора ДНК с ориджина репликации oriV. Соответственно в этом штамме можно не только нарабатывать большие генетические конструкции, но и индуцированно увеличивать число копий вектора на клетку.
Для получения однозначных данных по ингибированию именно РЗРП SARS-CoV-2 была получена контрольная конструкция с каталитически неактивной РЗРП, содержащей мутации в активном центре (далее GG2). Для этого в последовательности nsp12 полимеразной субъединицы РЗРП SARS-CoV-2 в составе фрагмента 4 два аминокислотных остатка каталитической триады активного центра nspl2 были заменены остатками аланина: D760A и D761A. Потеря каталитической активности РЗРП SARS-CoV-2 в результате этих замен была показана ранее [Gao Y., Yan L., Huang Y., Liu F., Zhao Y., Cao L., Wang Т., Sun Q., Ming Z., Zhang L., Ge J., Zheng L., Zhang Y., Wang H., Zhu Y., Zhu С, Ни Т., Hua Т., Zhang В., Yang X., Li J., Yang H., Liu Z., Xu W., Guddat L. W., Wang Q., Lou Z., Rao Z. Structure of the RNA-dependent RNA polymerase from COVID-19 virus // Science. - 2020. - T. 368, №6492. - C. 779-782.]. Наличие мутаций подтверждали секвенированием ПЦР-продукта соответствующего участка гена nsp12.
Для корректного сопоставления результатов тестирования потенциальных ингибиторов в предлагаемой системе с данными, полученными при исследованиях наклетках, инфицированных SARS-CoV-2, была выбрана клеточная линия Vero Е6. Эта линия является наиболее широко используемой в работах по изучению различных вирусов из-за отсутствия интерферонового ответа в клетках при заражении вирусом. Клетки трансфицировали полученными репортерами (GG1 или GG2), а спустя 48 часов измеряли уровень люминесценции первой люциферазы (Rluc) и второй люциферазы (Fluc) (Фигура 3). Уровень люминесценции первой люциферазы значительно превосходил уровень второй люциферазы, которая не могла синтезироваться клеточными РНК-полимеразами, а только РЗРП SARS-CoV-2. Помимо этого уровень люминесценции обеих люцифераз при трансфекции мутантным репортером GG2 был значительно ниже, чем в случае репортера GG1 с полностью функциональной РЗРП. Уровень второй люциферазы в случае мутантного репортера был незначительно выше фона, поэтому при дальнейших исследованиях ингибиторов его не измеряли.
Нуклеозидные ингибиторы, в отличие от ненуклеозидных, для подтверждения РНК-зависимой РНК-полимеразы SARS-CoV-2 в качестве мишени в экспериментах на выделенном ферменте необходимо предварительно превратить в соответствующие 5'-трифосфаты. Это довольно трудоемкий синтез, который сильно ограничивает возможности скрининга. Вместе с тем, при тестировании самих модифицированных нуклеозидов в качестве ингибиторов есть риск ложноотрицательных результатов, что сужает спектр потенциальных ингибиторов. Для решения этой проблемы была создана клеточная линия, в геноме которой содержится ген тимидин-киназы вируса простого герпеса человека первого типа (ТК). Наличие ТК в клетках существенно увеличивает возможность протекания первого акта фосфорилирования тестируемых нуклеозидных соединений, поскольку данный фермент обладает относительно низкой субстратной специфичностью [Guettari N, Loubiere L, Brisson E, Klatzmann D. Use of herpes simplex virus thymidine kinase to improve the antiviral activity of zidovudine. Virology. 1997 Sep l;235(2):398-405].
Получены клетки Vero E6_ТК, стабильно экспрессирующие ТК. В качестве способа доставки гена ТК в целевые клетки была выбрана транедукция клеток лентивирусным вектором. В геноме вектора лентивирусные гены заменены на ген ТК, слитый с геном устойчивости к пуромицину и геном зеленого флуоресцентного белка (GFP). После транедукции клеток лентивирусным вектором осуществляли селекцию клеток на среде с пуромицином (6 мкг/мл) для отбора ТК-экспрессирующих клеток (Vero Е6_ТК). Для дополнительной очистки ТК-позитивных клонов клетки подвергали сортировке по наличию флуоресценции GFP, слитого с ТК.
Способность аналогов нуклеозидов фосфорилироваться в полученных клетках проверяли с помощью функционального теста. Для этого клетки линии Vero Е6 и Vero Е6_ТК трансдуцировали VSV-G-псевдотипированным репликативно-некомпетентным лентивирусным вектором на основе ВИЧ-1, кодирующим люциферазу светлячка под контролем промотора цитомегаловируса (CMV). Затем к клеткам добавляли ацикловир или зидовудин в различных концентрациях и через 24 часа анализировали уровень люминесценции (Фигура 4). Зидовудин был практически одинаково активен на обеих клеточных линиях (IC50 составили порядка 0,4 мкМ в обоих случаях), в то время как ацикловир проявлял значительно более высокую ингибирующую активность в присутствии тимидинкиназы в клетках Vero Е6_ТК, чем в Vero (IC50 составили 0,8 мкМ и более 100 мкМ, соответственно). Способность ацикловира ингибировать ВИЧ-1 в тканях, инфицированных вирусом простого герпеса, была продемонстрирована ранее [Lisco А, Vanpouille С, Tchesnokov ЕР, Grivel JC, Biancotto A, Brichacek В, Elliott J, Fromentin E, Shattock R, Anton P, Gorelick R, Balzarini J, McGuigan C, Derudas M, Gone M, Schinazi RF, Margolis L. Acyclovir is activated into a HIV-1 reverse transcriptase inhibitor in herpesviras-infected human tissues. // Cell Host Microbe. 2008. - T. 4, №3. - C. 260-270]. Полученные с ацикловиром данные подтвердили то, что нам удалось получить линию, стабильно экспрессирующую ТК. Отсутствие различий в значениях IC50 для зидовудина подтверждает его эффективное фосфорилирование с помощью клеточных киназ в клетках Vero Е6. Тестирование потенциальных ингибиторов РЗРП SARS-CoV-2 проводилось нами не только на клеточной линии Vero Е6, но и на специально полученной линии Vero Е6_ТК.
Тестирование соединений в качестве потенциальных ингибиторов РЗРП SARS-CoV-2 проводилось на клеточных линиях Vero Е6 и Vero Е6_ТК. В качестве потенциальных ингибиторов РЗРП SARS-CoV-2 были выбраны соединения с разными механизмами действия. Помимо исследования влияния данных веществ на уровень люцифераз в трансфицированных клетках был проведен МТТ-тест, который показал, что в исследованных концентрациях соединения не оказывают значительного цитопатического действия на клетки Vero Е6 и Vero Е6_ТК.
В качестве отрицательного контроля использовалась система полноразмерного репликона, содержащего мутации (D760A, D761A) в активном центре полимеразной субъединицы РЗРП (nsp12), приводящие к потере каталитической активности РЗРП.
Таким образом, нами была получена улучшенная безвирусная репортерная система для поиска нуклеозидных и ненуклеозидных ингибиторов РЗРП вируса SARS-CoV-2. Дляоблегчения поиска нуклеозидных ингибиторов РЗРП без создания фосфорилированных депо-форм была создана клеточная линия Vero Е6_ТК, экспрессирующая тимидин-киназу вируса простого герпеса, осуществляющую первую стадию фосфорилирования нуклеозидов в клетке.
Далее приведены примеры, иллюстрирующие данное изобретение. Варианты и модификации осуществления изобретения, которые могут быть воспроизведены, не отходя от общей концепции настоящего изобретения и без привлечения собственной изобретательской деятельности, также будут входить в объем притязаний настоящего изобретения.
Пример 1. Получение полноразмерного репликона GG1, клонированного в искусственную бактериальную хромосому.
Реакцию клонирования методом Golden Gate проводят с помощью набора NEBridge Golden Gate Assembly Kit. В реакционную смесь добавляют 6 фрагментов генома SARS-CoV-2, находящихся в промежуточных векторах pJET1.2 (Thermo Fisher Scientific, США), и вектор pBAC-lacZ (Addgene, #13422), смесь эндонуклеазы BsaI-HFv2 и Т4 ДНК лигазы, после чего смесь помещают в термоциклер. В термоциклере устанавливают следующую программу: {37°С, 5 минут→16°С, 5 минут}×30→55°С, 5 минут. После термоциклирования смесь используют для трансформации специального штамма TransforMax EPI300 E.coli, в котором возможно индуцированное увеличение копийности вектора. После трансформации клетки рассеивают на твердую среду (2.5% LB, 1.5% агар) с хлорамфениколом (12.5 мкг/мл) и оставляют на 36-48 часов при 30°С.Далее материал некоторых колоний переносят в жидкую среду (2.5% LB, 12.5 мкг/мл хлорамфеникол) и растят 12 часов при 30°С, затем разбавляют до оптической плотности 0.3-0.4, после чего добавляют индуктор (L-арабинозу) до итоговой концентрации 2 мМ. Культуры растят 6 часов при 30°С. Конструкции выделяют в соответствии с протоколами наборов для выделения плазмидной ДНК. Выделенную ДНК далее используют для трансфекции клеток Vero Е6 или Vero Е6_ТК.
Пример 2. Получение полноразмерного репликона GG2, содержащего мутации в активном центре РЗРП, клонированного в искусственную бактериальную хромосому.
Перед клонированием методом Golden Gate проводят сайт-направленный мутагенез в гене, кодирующем полимеразную субъединицу РЗРП (nsp12), во фрагменте 4. К фрагменту 4 добавляют полимеразу Q5 High-Fidelity DNA Polymerase (NEB, США) и 5'-фосфорилированные праймеры SEQ ID NO: 1 и SEQ ID NO: 2 (Перечень последовательностей). Дизайн праймеров проводят с использованием онлайн ПО NEBaseChanger.neb.com. Для того, чтобы избавиться от исходного метилированного вектора используют эндонуклеазу рестрикции DpnI. Для проверки правильности протекания мутагенеза проводят секвенирование ДНК в компании ЦКП «Геном». Далее содержащий необходимые мутации фрагмент используют для получения репликона GG2, как описано в Примере 1.
Пример 3. Получение клеточной линии, стабильно экспрессирующей тимидин-киназу вируса простого герпеса человека первого типа.
Клетки линии Vero Е6 инфицируют псевдовирусными частицами, кодирующими в своем геноме зеленый флуоресцентный белок (GFP), ген устойчивости к пуромицину и тимидин-киназу вируса простого герпеса человека первого типа (HSV-TK). Через 24 часа после инфекции, культуральную среду заменяют на среду, содержащую пуромицин в концентрации 8.5 мкг/мл. Эффективность селекции определяют по доле клеток, экспрессирующих GFP на приборе MACSQuant Analyzer 10. Ввиду низкой эффективности селекции прибегают к помощи сортировки GFP+клеток на проточном цитофлуориметре BD F ACS Aria III. Полученную клеточную линию Vero Е6_ТК используют для дальнейших экспериментов.
Пример 4. Тестирование контрольного соединения молнупиравира с использованием клеточной системы для изучения репликации SARS-CoV-2.
За сутки до тестирования, клетки линий Vero Е6 или Vero Е6_ТК рассевают на 48-луночный планшет в количестве 6*104 клеток на лунку. На следующий день проводят трансфекцию клеток полноразмерным репликоном GG1 или GG2 с использованием реагента Lipofectamine 3000 (Thermo Fisher Scientific, США) согласно протоколу производителя. Через 6 часов после трансфекции исходную среду заменяют на среду, содержащую молнупиравир (Фигура 5) в концентрации 0, 5, 15, 50 мкМ при этом концентрация диметилсульфоксида (DMSO) в культуральной среде составляла 1%. Через 48 часов после трансфекции эффективность ингибирования оценивают по изменению люминесцентного сигнала от люцифераз рениллы (Rluc) и светлячка (Fluс). Активность люциферазы рениллы (Renilla reniformis) и люциферазы светлячка (Photinus pyralis) измеряют с помощью набора Dual-Luciferase Reporter Assay System (Promega, США) на приборе BioTek Synergy HI Multimode Reader (Agilent, США).
Молнупиравир проявляет ингибирующую активность относительно РЗРП, причем на линии клеток Vero Е6_ТК (пунктирная кривая) ингибирование происходит более эффективно (Фигура 6). Значения IC50 приведены в Таблице 1.
Пример 5. Тестирование 2'-амино-2'-дезоксиаденозина с использованием клеточной системы для изучения репликации SARS-CoV-2.
За сутки до тестирования, клетки линий Vero Е6 или Vero Е6_ТК рассевают на 48-луночный планшет в количестве 6*104 клеток на лунку. На следующий день проводят трансфекцию клеток полноразмерным репликоном GG1 или GG2 с использованием реагента Lipofectamine 3000 (Thermo Fisher Scientific, США) согласно протоколу производителя. Через 6 часов после трансфекции исходную среду заменяют на среду, содержащую 2'-амино-2'-дезоксиаденозин (Фигура 7) в концентрации 0, 5, 15, 50 мкМ при этом концентрация диметилсульфоксида (DMSO) в культуральной среде составляла 1%. Через 48 часов после трансфекции эффективность ингибирования оценивают по изменению люминесцентного сигнала от люцифераз рениллы (Rluc) и светлячка (Fluс). Активность люциферазы рениллы (Renilla reniformis) и люциферазы светлячка (Photinus pyralis) измеряют с помощью набора Dual-Luciferase Reporter Assay System (Promega, США) на приборе BioTek Synergy HI Multimode Reader (Agilent, США).
2'-Амино-2'-дезоксиаденозин проявляет ингибирующую активность относительно РЗРП, причем в обеих линиях клеток, Vero Е6 и Vero Е6_ТК, ингибирование происходит одинаково (Фигура 8), что говорит о том, что этот нуклеозид эффективно фосфорилируется клеточными ферментами в клетках Vero Е6. Значения IC50 приведены в Таблице 1.
Пример 6. Тестирование ацикловира с использованием клеточной системы для изучения репликации SARS-CoV-2.
За сутки до тестирования, клетки линий Vero Е6 или Vero Е6_ТК рассевают на 48-луночный планшет в концентрации 6*104 клеток на лунку. На следующий день ставят трансфекцию клеток полноразмерным репликоном GG1 или GG2 с использованием реагента Lipofectamine 3000 (Thermo Fisher Scientific, США) согласно протоколу производителя. Через 6 часов после трансфекции исходную среду заменяют на среду, содержащую ацикловир (фигура 9) в концентрации 0, 5, 15, 50 мкМ при этом концентрация диметилсульфоксида (DMSO) в культуральной среде составляла 1%. Через 48 часов после трансфекции эффективность ингибирования оценивают по изменениюлюминесцентного сигнала от люцифераз рениллы (Rluc) и светлячка (Fluс). Активность люциферазы рениллы (Renilla reniformis) и люциферазы светлячка (Photinus pyralis) измеряют с помощью набора Dual-Luciferase Reporter Assay System (Promega, США) на приборе BioTek Synergy HI Multimode Reader (Agilent, США).
Ацикловир проявляет ингибирующую активность относительно РЗРП, причем на линии клеток Vero Е6_ТК (пунктирная кривая) ингибирование происходит более эффективно, что подтверждает необходимость предварительного фосфорилирования ацикловира ТК вируса простого герпеса для проявления его ингибирующего действия (Фигура 10). Значения IC50 приведены в Таблице 1.
Пример 7. Тестирование 3'-О-метиладенозина с использованием клеточной системы для изучения репликации SARS-CoV-2.
За сутки до тестирования, клетки линий Vero Е6 или Vero Е6_ТК рассевают на 48-луночный планшет в концентрации 6*104 клеток на лунку. На следующий день ставят трансфекцию клеток полноразмерным репликоном GG1 или GG2 с использованием реагента Lipofectamine 3000 (Thermo Fisher Scientific, США) согласно протоколу производителя. Через 6 часов после трансфекции исходную среду заменяют на среду, содержащую 3'-О-метиладенозина (Фигура 11) в концентрации 0, 5, 15, 50 мкМ при этом концентрация диметилсульфоксида (DMSO) в культуральной среде составляла 1%. Через 48 часов после трансфекции эффективность ингибирования оценивают по изменению люминесцентного сигнала от люцифераз рениллы (Rluc) и светлячка (Fluc). Активность люциферазы рениллы (Renilla reniformis) и люциферазы светлячка (Photinus pyralis) измеряют с помощью набора Dual-Luciferase Reporter Assay System (Promega, США) на приборе BioTek Synergy HI Multimode Reader (Agilent, США).
3'-О-Метиладенозин проявляет ингибирующую активность относительно РЗРП, причем на линии клеток Vero Е6_ТК (пунктирная кривая) ингибирование происходит чуть более эффективно (Фигура 12). Значения IC50 приведены в Таблице 1.
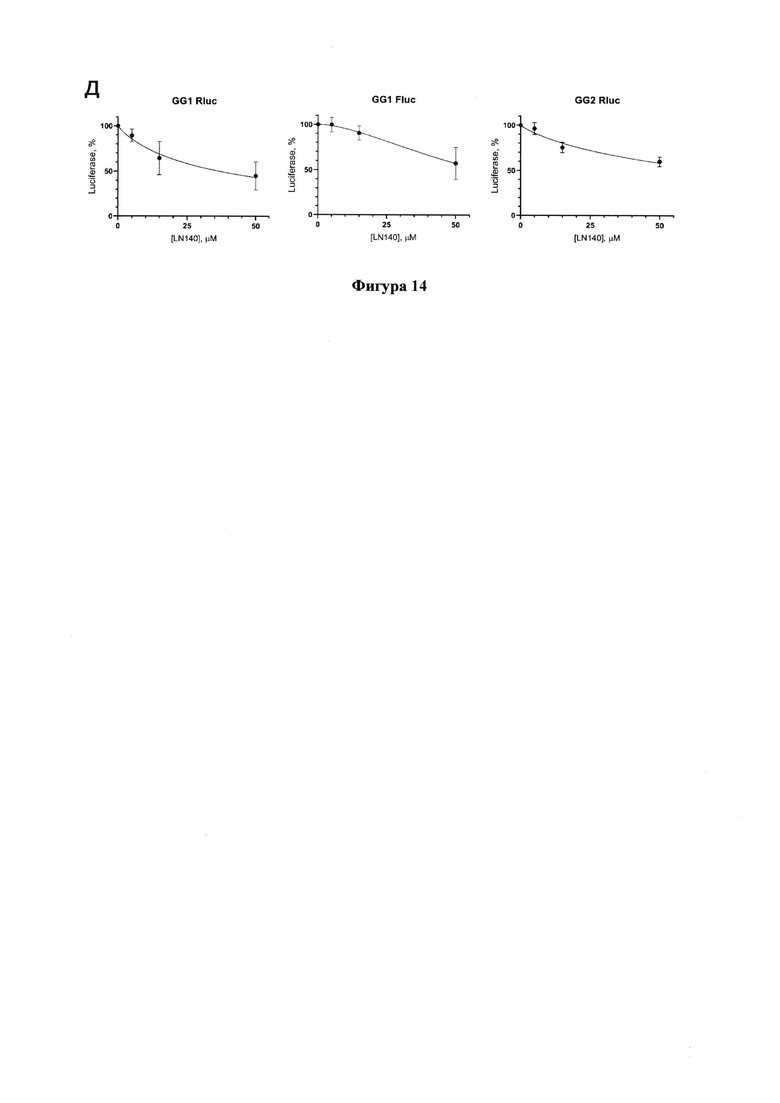
Пример 8. Тестирование соединения LN140 с использованием клеточной системы для изучения репликации SARS-CoV-2.
Тестирование соединения LN140, которое не может выступать в качестве нуклеозидного ингибитора РНК-полимеразы и не нуждается поэтому в фосфорилировании, проводят только на линии Vero Е6. За сутки до тестирования, клетки линий Vero Е6 или Vero Е6_ТК рассевают на 48-луночный планшет в концентрации 6*104клеток на лунку. На следующий день ставят трансфекцию клеток полноразмерным репликоном GG1 или GG2 с использованием реагента Lipofectamine 3000 (Thermo Fisher Scientific, США) согласно протоколу производителя. Через 6 часов после трансфекции исходную среду заменяют на среду, содержащую LN140 (Фигура 13) в концентрации 0, 5, 15, 50 мкМ при этом концентрация диметилсульфоксида (DMSO) в культуральной среде составляла 1%. Через 48 часов после трансфекции эффективность ингибирования оценивают по изменению люминесцентного сигнала от люцифераз рениллы (Rluc) и светлячка (Flue). Активность люциферазы рениллы (Renilla reniformis) и люциферазы светлячка (Photinus pyralis) измеряют с помощью набора Dual-Luciferase Reporter Assay System (Promega, США) на приборе BioTek Synergy HI Multimode Reader (Agilent, США).
Соединение LN140 проявляет некоторую ингибирующую активность по отношению к обеим люциферазам, однако его действие на люциферазу ренилла более выражено, чем на люциферазу светлячка (Фигура 14). Значения IC50 приведены в Таблице 1.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="БЕЗОПАСНАЯ
КЛЕТОЧНАЯ РЕПОРТЕРНАЯ СИСТЕМА ДЛЯ ПОИСКА НУКЛЕОЗИДНЫХ И
НЕНУКЛЕОЗИДНЫХ ИНГИБИТОРОВ РНК-ЗАВИСИМОЙ РНК-ПОЛИМЕРАЗЫ
SARS-COV-2.xml" softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2024-01-24">
<ApplicantFileReference>24.12.2024</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное учреждение науки Институт молекулярной биологии им. В.А.
Энгельгардта Российской академии наук (ИМБ РАН)</ApplicantName>
<ApplicantNameLatin>Engelhardt Institute of Molecular Biology,
Russian Academy of Sciences</ApplicantNameLatin>
<InventionTitle languageCode="ru">БЕЗОПАСНАЯ КЛЕТОЧНАЯ РЕПОРТЕРНАЯ
СИСТЕМА ДЛЯ ПОИСКА НУКЛЕОЗИДНЫХ И НЕНУКЛЕОЗИДНЫХ ИНГИБИТОРОВ
РНК-ЗАВИСИМОЙ РНК-ПОЛИМЕРАЗЫ SARS-COV-2</InventionTitle>
<SequenceTotalQuantity>2</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>24</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..24</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>misc_difference</INSDFeature_key>
<INSDFeature_location>1..24</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q3">
<INSDQualifier_name>standard_name</INSDQualifier_name>
<INSDQualifier_value>nsp12_D760A_D761A_fw</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atactctctgccgctgctgttgtg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q5">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>misc_difference</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q6">
<INSDQualifier_name>standard_name</INSDQualifier_name>
<INSDQualifier_value>nsp12_D760A_D761A_rev</INSDQualifier_value
>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>catcattgagaaatgtttacg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Средство для ингибирования репликации вируса SARS-CoV-2, опосредованного РНК-интерференцией | 2020 |
|
RU2733361C1 |
| Комбинированное лекарственное средство, обладающее противовирусным эффектом в отношении нового коронавируса SARS-CoV-2 | 2021 |
|
RU2746362C1 |
| ПРОТИВОВИРУСНАЯ КОМПОЗИЦИЯ | 2020 |
|
RU2740660C1 |
| ТРАНС-РЕПЛИЦИРУЮЩАЯ РНК | 2017 |
|
RU2752580C2 |
| СОСТАВ ДЛЯ ВВЕДЕНИЯ РНК | 2017 |
|
RU2746118C2 |
| РЕЦЕПТУРА ДЛЯ ВВЕДЕНИЯ РНК | 2019 |
|
RU2797147C2 |
| ПРОТИВОВИРУСНАЯ КОМПОЗИЦИЯ | 2020 |
|
RU2740657C1 |
| Рекомбинантный штамм Sen-Sdelta(M) вируса Сендай, продуцирующий S белок SARS-CoV-2 и используемый для получения интраназальной вакцины против COVID-19 | 2024 |
|
RU2837536C1 |
| Модифицированный антисмысловой олигонуклеотид против вируса SARS-CoV-2 | 2020 |
|
RU2750584C1 |
| Плазмидная ДНК-матрица pVAX-RBD, молекул мРНК-RBD, полученная с использованием ДНК-матрицы pVAX-RBD, обеспечивающая синтез и секрецию белка RBD SARS-CoV 2 в клетках организма млекопитающих и комплекс в виде наночастиц, содержащих молекулы мРНК-RBD, индуцирующих SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью | 2021 |
|
RU2754230C1 |





Изобретение относится к области молекулярной биологии, медицинской химии, вирусологии и микробиологии и предназначено для скрининга потенциальных ингибиторов РНК-зависимой РНК-полимеразы (РЗРП) SARS-CоV-2 в лабораториях с первым уровнем биобезопасности, не оборудованных для работы с высокопатогенными инфекциями. Более конкретно, изобретение относится к полноразмерным репликонам - искусственно созданным системам, позволяющим проводить тестирование потенциальных ингибиторов РЗРП SARS-CоV-2 без использования полноценных вирусных частиц. В качестве безопасной клеточной системы для тестирования ингибиторов РНК-зависимой РНК-полимеразы SARS-CoV-2 получена генетическая конструкция, представляющая собой кДНК вируса, не содержащую генов структурных белков S, Е и М, с двумя блоками репортерных генов, которая клонирована в вектор искусственной бактериальной хромосомы. Первый блок репортерных генов, представленный люциферазой Renilla reniformis и зеленым флуоресцентным белком (Rluc-GFP), находится под контролем CMV-промотора, находящегося перед 5'UTR SARS-CoV-2. Второй блок репортерных генов представлен люциферазой светлячка Photinus pyralis и красным флуоресцентным белком (Fluc-RFP) и находится после последовательности, регулирующей транскрипцию (TRS-N). Такое расположение генов и регуляторных последовательностей (CMV-5'UTR-Rluc-GFP-TRS-N-Fluc-RFP-3'UTR) позволяет оценить уровень ингибирования вирусной РЗРП, поскольку второй блок репортерных генов, находящийся после TRS-N, может транскрибироваться только вирусной полимеразой по механизму прерывистой транскрипции, характерной для семейства Coronaviridae. Первый же блок репортерных генов, находящийся под контролем CMV-промотора, может транскрибироваться не только вирусной РЗРП, но и клеточными полимеразами. Для использования в качестве отрицательного контроля создана система полноразмерного репликона, в котором в активном центре полимеразной субъединицы РЗРП (nsp12) произведены замены аминокислот (D760A, D761A), приводящие к потере каталитической активности РЗРП. Для облегчения поиска нуклеозидных ингибиторов РЗРП без создания фосфорилированных депо-форм создана клеточная линия Vero Е6_ТК, экспрессирующая тимидин-киназу вируса простого герпеса, осуществляющую первую стадию фосфорилирования нуклеозидов в клетке. 3 н.п. ф-лы, 14 ил., 1 табл., 8 пр.
1. Полноразмерный репликон, позволяющий проводить тестирование потенциальных ингибиторов РНК-зависимой РНК-полимеразы SARS-CоV-2 без использования полноценных вирусных частиц, представляющий собой кДНК вируса, не содержащую генов структурных белков S, Е и М, с двумя блоками репортерных генов, клонированную в вектор искусственной бактериальной хромосомы, при этом один из блоков репортерных генов состоит из последовательности Е2А, гена устойчивости к бластицидину, гена последовательности Т2А, слитого гена люциферазы Renilla reniformis и зеленого флуоресцентного белка Rluc-GFP, а также из последовательности Р2А и находится под контролем CMV-промотора, находящегося перед 5'UTR SARS-CoV-2, второй блок репортерных генов состоит из последовательности Р2А, слитого гена люциферазы светлячка Photinus pyralis и красного флуоресцентного белка Fluc-RFP и находится после последовательности, регулирующей транскрипцию TRS-N.
2. Блок репортерных генов для получения полноразмерного репликона по п. 1, состоящий из последовательности Е2А, гена устойчивости к бластицидину, гена последовательности Т2А, слитого гена люциферазы Renilla reniformis и зеленого флуоресцентного белка Rluc-GFP, а также из последовательности Р2А.
3. Блок репортерных генов для получения полноразмерного репликона по п. 1, состоящий из последовательности Р2А, слитого гена люциферазы светлячка Photinus pyralis и красного флуоресцентного белка Fluc-RFP.
| CHENGJIN YE, KEVIN CHIEM et al., Analysis of SARS-CoV-2 infection dynamic in vivo using reporter-expressing viruses, Proc Natl Acad Sci U S A, 2021, vol.118, n.41, реферат | |||
| ANDREAS MARKUS LOENING, TIMOTHY DAVID FENN et al., Crystal Structures of the Luciferase and Green Fluorescent Protein from Renilla reniformis, J Mol Biol | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |

