Изобретение относится к рекомбинантному штамму вируса Сендай, индуцирующему SARS-CoV-2-специфический протективный иммунный ответ при однократной интраназальной вакцинации. Изобретение может быть использовано в биотехнологии, в частности в генетической инженерии для разработки интраназальной векторной вакцины нового поколения против коронавирусной инфекции COVID-19.
Обоснование использования вирусов для доставки «вакцинного гена» заключается в нескольких аспектах. Вакцины с вирусным вектором безопасны и индуцируют как врожденный, так и адаптивный иммунный ответ без вовлечения всего опасного патогена. Более того, вирусным векторам присущи адъювантные свойства благодаря экспрессии различных молекулярных паттернов, ассоциированных с патогеном (PAMPs), и активации врожденного иммунитета [Wang, 2023] [1].
Поскольку входными воротами для SARS-CoV-2 являются слизистые оболочки, для остановки этой инфекции огромное значение приобретает именно мукозальная иммунизация. По сравнению с инъекционными вакцинами, на производстве которых сосредоточены огромные усилия фармацевтических компаний, интраназальные вакцины обеспечивают дополнительные уровни защиты, такие как секреторные димерные IgA (sIgA) и резидентные В- и Т-клетки памяти в слизистой оболочке дыхательных путей [Lund, 2021] [2].
Вирус Сендай (парагрипп мышей I типа, род Respirovirus, семейство Paramyxoviridae) как нельзя лучше подходит для этих целей, поскольку является респираторным вирусом и способен, не вызывая заболевания, ограниченно реплицироваться в клетках бронхиального эпителия человека, а также в некоторых категориях дендритных клеток. В процессе ограниченной репликации рекомбинантные варианты вируса Сендай способны индуцировать в организме человека специфический иммунный ответ на экспрессируемые ими протективно значимые чужеродные вирусные антигены [Зайнутдинов, 2021; Зайчук, 2020] [3; 4]. Использование вируса Сендай в качестве вакцинного вектора также обеспечивает неспецифическую противовирусную защиту организма, поскольку он является одним из лучших природных индукторов интерферонов и применялся для производства человеческого лейкоцитарного интерферона до начала эпохи рекомбинантных белков [Cantell, 1981; Nyman, 1998] [5; 6].
Важным преимуществом вируса Сендай как вакцинного вектора является высокая стабильность его генома. Стабильность связана с необычным свойством генома вируса Сендай, так же, как и других парамиксовирусов, которое называется правилом шести и обозначает строгую полигексамерность длины генома (6n+0, где n - один нуклеотид). Это свойство способствует особенно низкой частоте гомологичной рекомбинации геномных РНК парамиксовирусов [Matsumoto, 2018] [7]. Кроме того, репликация вируса Сендай происходит исключительно в цитоплазме, а не в ядре клетки, и риск генетической интеграции вирусного генома в геном хозяина минимален [Morimoto, 2023] [8].
Вакцинные свойства вируса Сендай активно исследуются в мировой практике. Так, интраназальные вакцины на его основе успешно прошли клинические испытания против парагриппа человека I типа и респираторно-синцитиального вируса [Scaggs Huang, 2021] [9]. Целый ряд рекомбинантных вирусов Сендай проходят доклинические исследования в качестве вакцин против респираторных инфекций человека [Russel, 2021] [10], включая COVID-19 [Morimoto, 2023] [8].
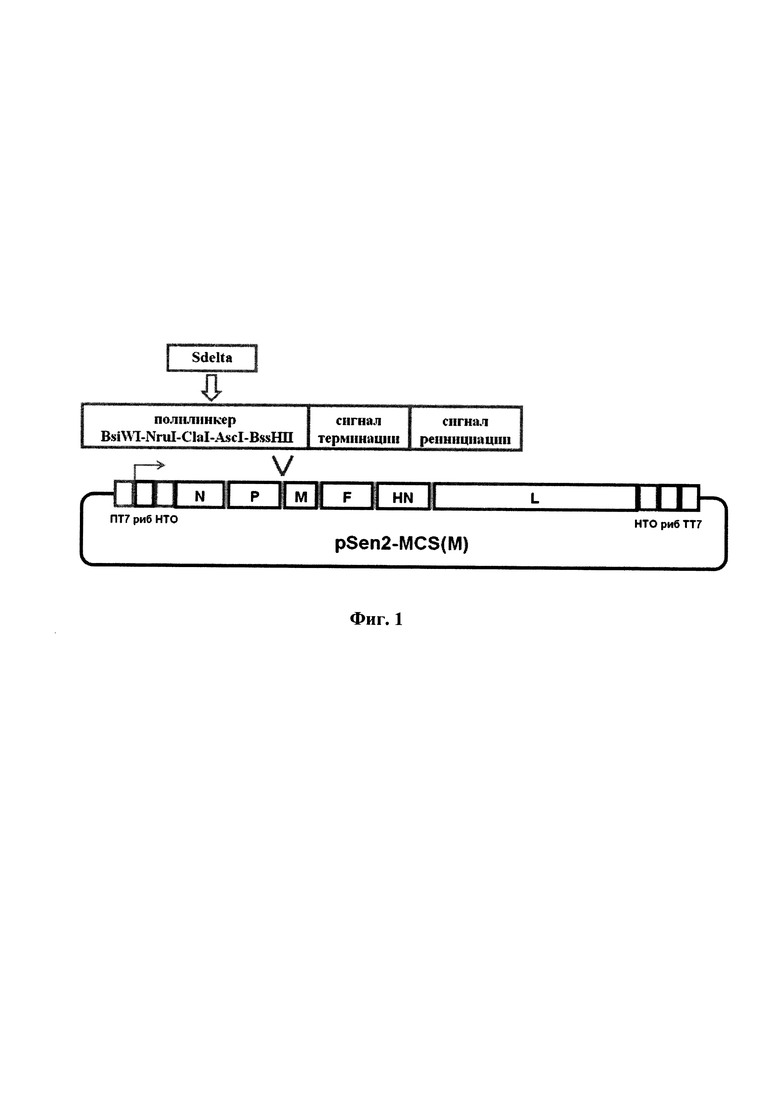
Ранее нами был сконструирован набор рекомбинантных плазмидных ДНК для получения рекомбинантных вирусов Сендай штамма Москва [Патент РФ №2787724] [11]. Для получения рекомбинантного штамма Sen-Sdelta(M) мы использовали четыре плазмидных ДНК, одна из которых содержит полноразмерную ДНК-копию генома вируса с полилинкером для встройки трансгенов между генами Р и M(pSen2-MCS(M)), а три других экспрессируют гены N, Р и L вируса Сендай штамма Москва. Экспрессия генов N, Р и L является необходимым элементом оживления рекомбинантных вариантов вируса, они кодируют белки репликативного комплекса, который синтезирует геномную РНК. Синтез белков N, Р, L и геномной РНК рекомбинантных вариантов вируса Сендай осуществляется с плазмидных ДНК под контролем полимеразы фага Т7. Точность размера генома контролируется рибозимами, встроенными по 3' и 5'концам геномной РНК. Полилинкер для встройки трансгенов в геномных плазмидных ДНК включает пять уникальных сайтов для крупнощепящих эндонуклеаз рестрикции BsiWI, NruI, ClaI, AscI, BssHII, за полилинкером введен сигнал терминации транскрипции трансгена и сигнал реинициации транскрипции последующего гена для РНК-полимеразы вируса Сендай (фиг. 1).
Получение рекомбинантных вариантов вируса Сендай штамма Москва осуществляется в созданной ранее культуре клеток 293-Т7, которая представляет собой дериват культуры клеток почки эмбриона человека 293, конститутивно продуцирующий полимеразу фага Т7 [Кудров, 2023] [12].
По имеющимся к настоящему времени данным, наиболее эффективным иммуногеном в случае инфекции SARS-CoV-2 является полноразмерная копия шиловидного S белка вируса, причем в нативной, не оптимизированной форме [Ilinykh, 2022] [13].
В последнее время ведется дискуссия на тему безопасности использования S белка в качестве иммуногена в связи с его возможным участием в развитии коагулопатии, которая является одним из известных осложнений при тяжелой форме COVID-19. В работах, в которых изучалось прямое влияние спайковых белков из вариантов SARS-CoV-2 на активность тромбоцитов и свертываемость крови, были получены противоречивые результаты. Так, в работе Kuhn с соавторами показано, что S белок SARS-CoV-2 через RGD (Arg-Gly-Asp) мотив может слабо взаимодействовать с некоторыми интегринами на поверхности тромбоцитов человека и запускать их стохастическую активацию. Авторы предполагают, что такая активация может быть связана с патогенезом COVID-19 и возникновением коагулопатий [Kuhn, 2023] [14]. Однако в работе Kusudo с соавторами было показано, что спайковые белки из вариантов SARS-CoV-2 (альфа, бета, гамма, дельта) даже в такой высокой дозе как 5 мкг/мл не оказывали влияния на коагуляцию, активность, количество, средний объем тромбоцитов и параметры тромбоэластографии в исследовании ex vivo [Kusudo, 2023][15]. К настоящему времени также накопился большой массив данных по эффективности и безопасности вакцин против COVID-19, в том числе и векторных, включающих спайковый белок SARS-CoV-2 в качестве иммуногена. Крупномасштабный ретроспективный анализ антиковидных вакцин на платформах мРНК, аденовируса, цельновирионной инактивации и субъединичной вакцины на основе полноразмерного S белка не продемонстрировал повышенного риска тромбоэмболии, кровоизлияния и смерти от тромбоэмболии и кровоизлияния после вакцинации [Uaprasert, 2021] [16]. Проведенное в Дании исследование также отмечает, что число случаев тромбоза, зарегистрированных после вакцинации против SARS-CoV-2, остается ниже ожидаемого числа в общей популяции, которое было оценено исходя из уровня заболеваемости "классическим" венозным тромбозом среди всего населения Дании до введения программы вакцинации [∅stergaard, 2021] [17]. Совокупный анализ мировых данных по использованию вакцин против COVID-19 (вакцины BioNTech/Pfizer(mRNA), ModemaTherapeutics(mRNA), Oxford/AstraZenecaChAdOxl-S, Janssen Ad26.COV2-S) показал, что во время заражения SARS-CoV 2 и связанным с ним заболеванием (COVID-19) тромбоз возникает по меньшей мере в 100 раз чаще без вакцинации, чем после нее [Elalamy, 2021] [18]. Опыт вакцинации российской вакциной Спутник V (векторная вакцина на основе аденовируса, включающего S ген SARS-CoV-2) показал, что она предотвращает тяжелое течение COVID-19 с развитием летальных исходов, ТЭЛА (тромбоэмболия легочной артерии), венозных и артериальных тромбозов [Моисеева, 2022] [19].
Принимая во внимание вышеприведенную информацию, в качестве трансгена для получения рекомбинантного вакцинного штамма мы использовали полноразмерную копию ДНК гена S белка природного изолята SARS-CoV-2, относящегося к линии В. 1.617.2. высокопатогенного варианта Дельта (Delta VOC) (фиг.2).
Полученный рекомбинантный штамм Sen-Sdelta(M) депонирован в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора под номером V-2396.
Близких аналогов к заявляемому рекомбинантному штамму Sen-Sdelta(M) не обнаружено.
Известно иммунобиологическое средство для профилактики заболеваний, вызванных вирусом тяжелого респираторного синдрома SARS-CoV-2, на основе рекомбинантного аденовируса человека 5-го серотипа или рекомбинантного аденовируса человека 26-го серотипа, содержащее оптимизированную под экспрессию в клетках млекопитающих последовательность протективного антигена S вируса SARS-CoV-2 с делецией 18 аминокислот на С'-конце гена [Патент РФ №2720614, опубл. 2020 г.] [20].
Однако в качестве вектора используются патогенные для человека аденовирусы, а также двукратное парентеральное введение вакцины. В заявляемом изобретении используется однократное интраназальное введение препарата Sen-Sdelta(M), поскольку такой путь вакцинации является наилучшим в плане простоты использования, наименее травматичным и безопасным в сравнении с парентеральным введением. Кроме того, интраназальное введение наиболее адекватно имитирует природную инфекцию SARS-CoV-2 и обеспечивает формирование первой линии защиты у входных ворот инфекции.
Наиболее близким аналогом (прототипом) является штамм рекомбинантного вируса везикулярного стоматита (VSV), кодирующий белок шипа (S) SARS-CoV-2 или его иммуногенный вариант, который может быть использован для индукции иммунного ответа на SARS-CoV-2, предотвращения заражения вирусом SARS-CoV-2 при одно- или двукратной мукозальной вакцинации (заявка на патент US 2021338804, МПК A61K 39/215; C12N7/01, опубл. 04.11.2021) [21]. Однако в данном прототипе в качестве вектора используется также патогенный для человека вирус везикулярного стоматита, который может вызывать осложнения при вакцинации. В заявляемом изобретении в качестве вектора используется непатогенный для человека вирус Сендай.
Техническим результатом заявляемого изобретения является создание такого иммунобиологического средства для профилактики заболеваний, вызванных вирусом тяжелого респираторного синдрома SARS-CoV-2, в котором в качестве вектора используется не патогенный для человека вирус, который не вызывает осложнения при вакцинации.
Указанный технический результат достигается созданием рекомбинантного штамма Sen-Sdelta(M) вируса Сендай, используемого для однократной интраназальной вакцинации против COVID-19 и включающего трансген Sdelta, имеющий нуклеотидную последовательность SEQ ID NO: 1, содержащую ген шиловидного S белка дельта-варианта SARS-CoV-2 и депонированный в Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора под номером V-2396.
Штамм Sen-Sdelta(M) характеризуется следующими признаками: Морфологические признаки. Размножение рекомбинантного варианта Sen-Sdelta(M) штамма Москва вируса Сендай происходит в клетках аллантоисной полости 10-11-дневных куриных эмбрионов в течение 72 часов при 37°С. Вирус накапливается в аллантоисной жидкости в титре 4096 ГАЕ, где ГАЕ - гемагглютинирующая единица в РГА с эритроцитами петуха при 4°С. Вирус размножается в культуре клеток почки макаки-резус LLC-MK.2, вызывая формирование бляшкообразных поражений монослоя клеток после инкубации 3 суток при 37°С в атмосфере 5%-ного СО2 под агаровым покрытием при добавлении трипсина в концентрации 10 мкг/мл. Для визуализации бляшек используют 0,1% суспензию эритроцитов морской свинки в растворе PBS с ионами Са2+ и Mg2+.
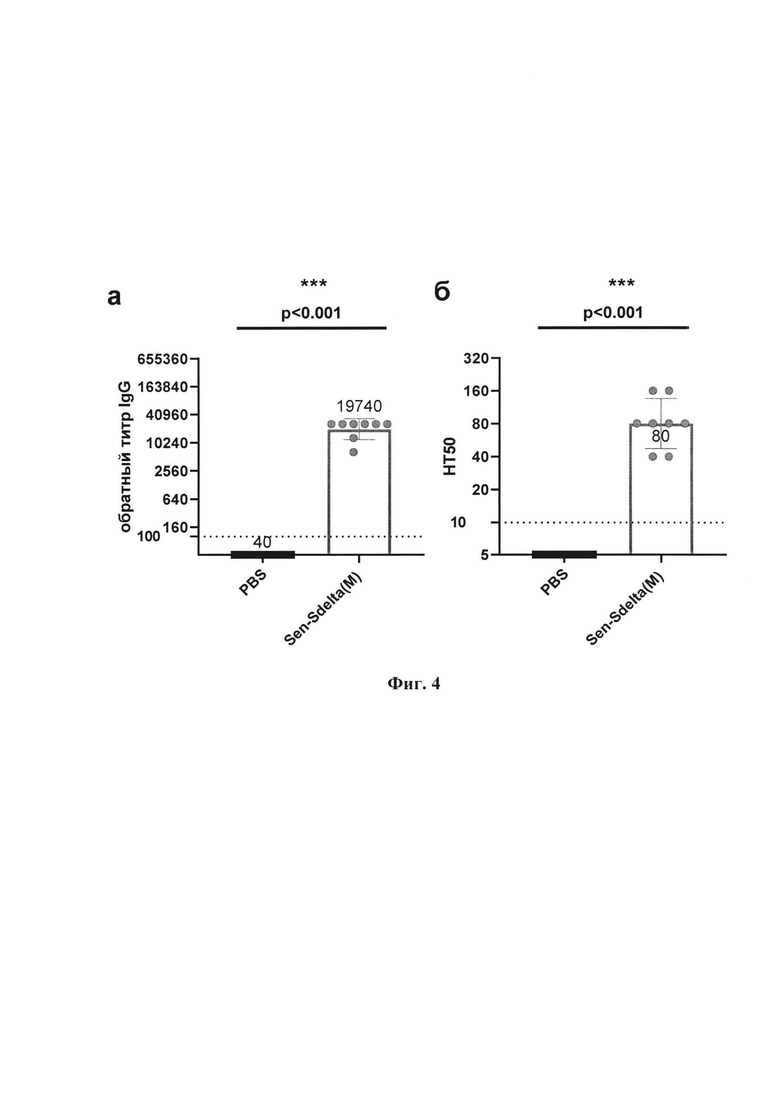
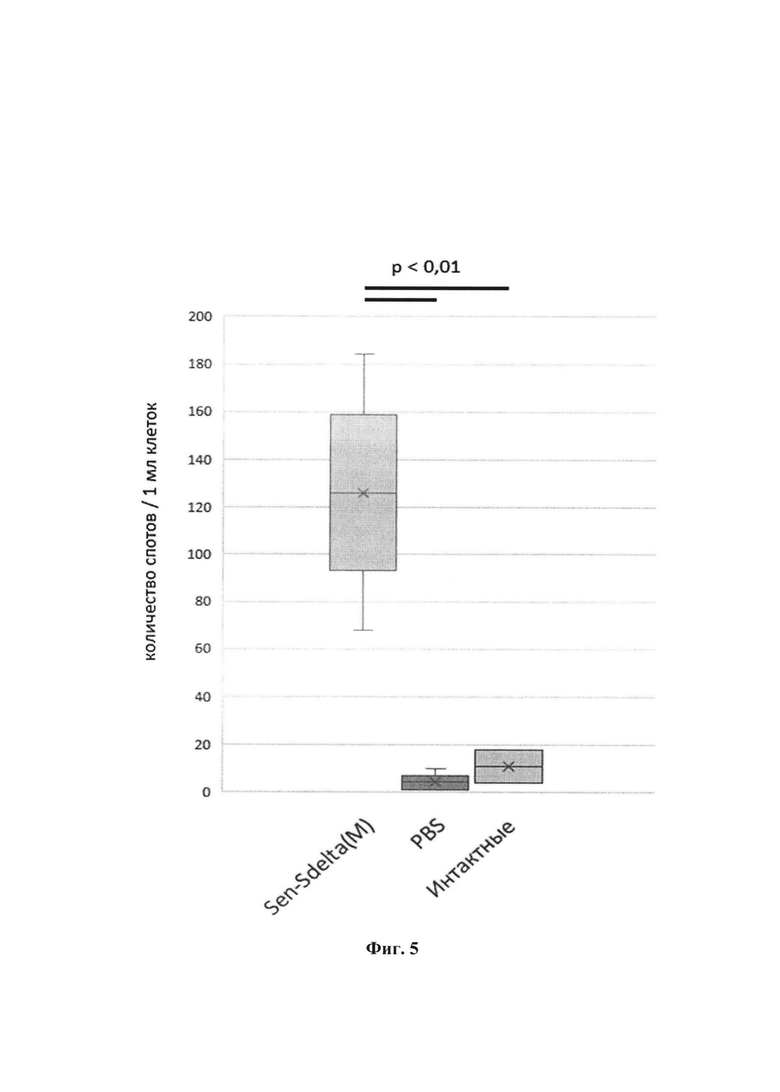
Физиолого-биохимические характеристики и культуральные свойства штамма. Секвенирование генома рекомбинанта выполнено с помощью технологии секвенирования нового поколения (NGS) на платформе MiSEQ. Определена полная нуклеотидная последовательность рекомбинантного штамма Sen-Sdelta(M), длина которой составляет 19254 н. Позиция встройки трансгена S белка: 3669-7485 н. Методом вестерн-блоттинга показана экспрессия трансгена S белка SARS-CoV 2 в лизатах клеток LLC-MK2, инфицированных рекомбинантным штаммом Sen-Sdelta(M) (фиг. 3). При однократной интраназальной вакцинации мышей линии BALB/c и золотистых сирийских хомячков рекомбинантным штаммом Sen-Sdelta(M) происходит индукция иммунного ответа, обеспечивающего значимое снижение репликации SARS-CoV-2 в носовой полости и легких животных после последующего заражения (фиг. 4, 5).
Изобретение иллюстрируется следующими графическими изображениями:
Фиг. 1. Схема конструирования рекомбинантного генома вируса Сендай штамма Москва с введением трансгена Sdelta перед геном М. Синтез геномной РНК рекомбинантных штаммов вируса Сендай осуществляется с плазмидной ДНК под контролем полимеразы фага Т7 - указано стрелкой. ПТ7 - промотор, ТТ7 - терминатор полимеразы Т7, риб - рибозимы.
Фиг. 2. Последовательность трансгена Sdelta с заложенными по краям последовательности сайтами рестрикции BsiWI и BssHII (подчеркнуты). Длина последовательности от стартового до стоп-кодона (помечены цветом) составляет 3816 п.н.
Фиг. 3. Вестерн-блот анализ экспрессии трансгена шиловидного S белка в составе рекомбинантного вируса Сендай Sen-Sdelta(M) в лизатах клеток LLC-MK2. В качестве первичных антител использована сыворотка крови реконвалесцента COVID-19, разведение 1:200. М - контроль молекулярной массы белков (kDa). Дорожки: 1 - лизат неинфицированных клеток LLC-MK2 (отрицательный контроль); 2 - лизат клеток, инфицированных исходным векторным штаммом Москва вируса Сендай; 3 - лизат клеток, инфицированных Sen-Sdelta(M); 4 - S белок, наработанный в трансформированных клетках СНО-K1, 200 нг (положительный контроль).
Фиг. 4. Общие антитела класса IgG и вируснейтрализующие антитела против дельта-штамма SARS-CoV-2 в сыворотках иммунизированных и контрольных мышей линии BALB/c: а - обратные титры IgG, полученные методом ИФА; 6 - титры нейтрализации (НТ50) на культуре клеток Vero Е6. Точки представляют индивидуальные значения, вершины гистограмм показывают средние геометрические значения (СГТ), вертикальные линии отображают границу 95% доверительного интервала (95%ДИ), а горизонтальные пунктирные линии указывают на пороговые значения титра IgG (<1:100) и НТ50 (<1:10) со значениями титров ниже порога для расчета СГТ 1:40 и 1:5 соответственно. Значения р над графиком получены в парном сравнении с применением U-критерия Манна-Уитни (степень достоверности различий *р<0,05; **р<0,01; ***р<0,001).
Фиг. 5. Оценка Т-клеточного ответа методом ELISpot в группах вакцинированных и контрольных мышей линии BALB/c. Число спленоцитов, продуцирующих IFN-γ в ответ на специфическую стимуляцию, на 106 клеток. Средние значения в группах представлены линиями, верхняя и нижняя грани прямоугольников отображают границу 95% доверительного интервала (95%ДИ). Значение р<0,01 над графиком получено в парном сравнении с применением U-критерия Манна-Уитни.
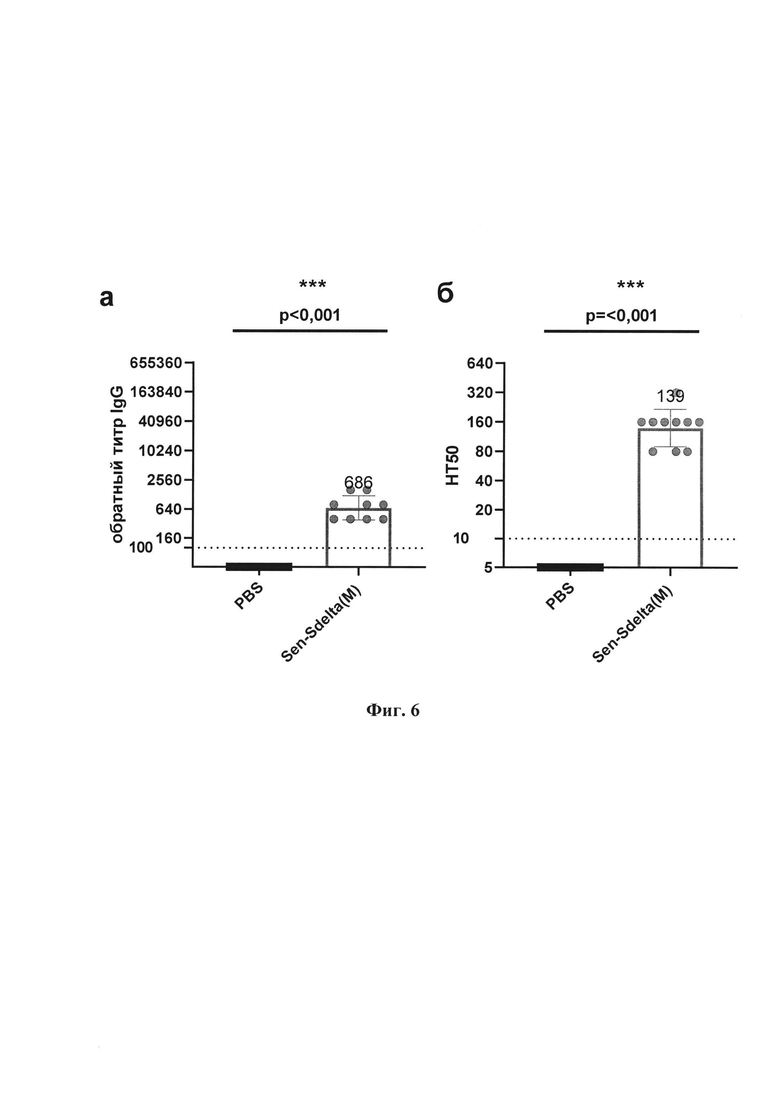
Фиг. 6. Общие антитела IgG и вируснейтрализующие антитела против дельта-штамма SARS-CoV-2 в сыворотках иммунизированных и контрольных хомяков: а - обратные титры IgG, полученные методом ИФА; 6 - титры нейтрализации (НТ50) на культуре клеток Vero Е6. Точки представляют индивидуальные значения, вершины гистограмм показывают средние геометрические значения (СГТ), вертикальные линии отображают границу 95% доверительного интервала (95%ДИ), а горизонтальные пунктирные линии указывают на пороговые значения титра IgG (<1:100) и НТ50 (<1:10) со значениями титров ниже порога для расчета СГТ 1:40 и 1:5 соответственно. Значения р над графиком получены в парном сравнении с применением U-критерия Манна-Уитни (степень достоверности различий *р<0,05; **р<0,01; ***р<0,001).
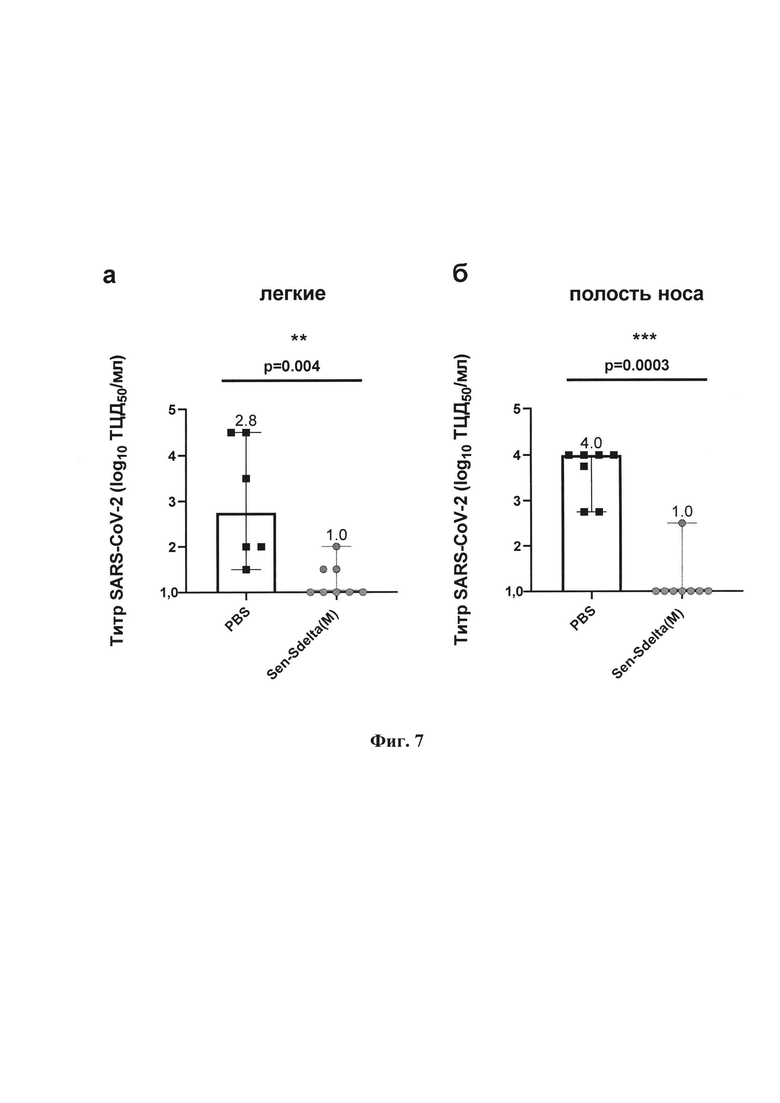
Фиг. 7. Инфекционный титр (lg ТЦД50/мл) на культуре клеток Vero Е6 у мышей BALB/c на 5-е сутки после заражения гамма-штаммом SARS-CoV-2 в гомогенатах тканей: а - легких; 6 - полости носа. Точки представляют индивидуальные значения, вершины гистограмм показывают медианные значения в группе, вертикальные линии отображают границу 95% доверительного интервала (95%ДИ). Значения р над графиком получены в парном сравнении с применением U-критерия Манна-Уитни (степень достоверности различий *р<0,05; **р<0,01; ***р<0,001).
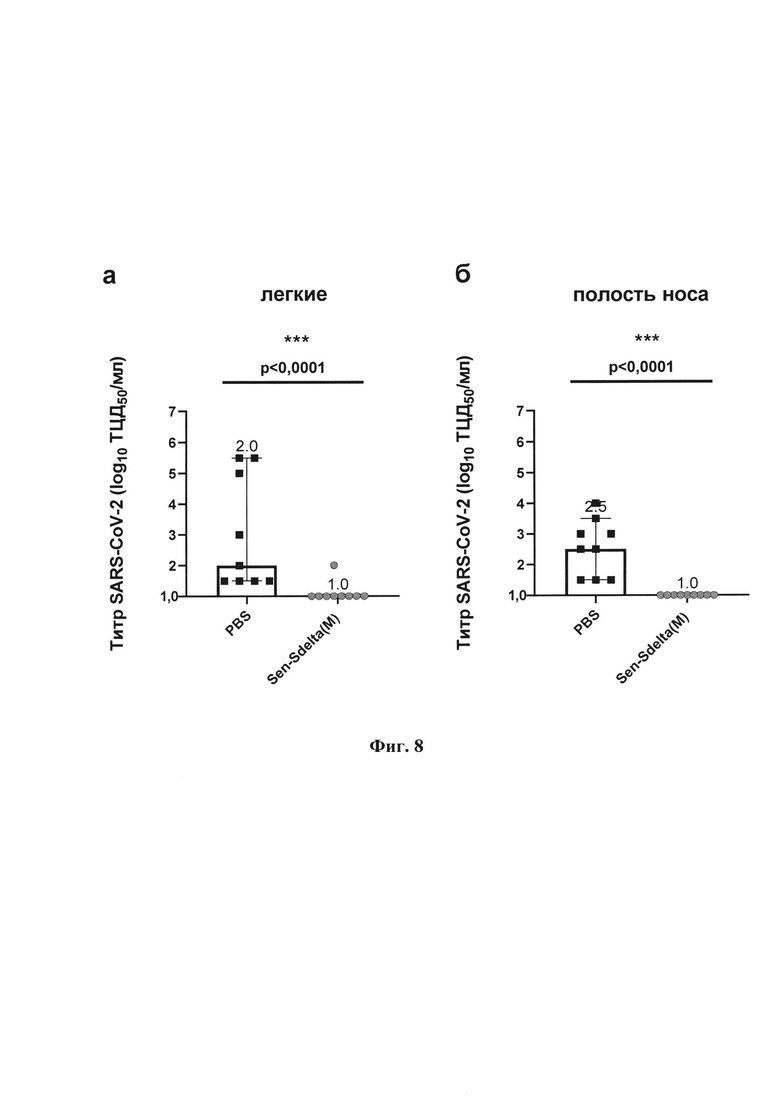
Фиг. 8. Инфекционный титр (lg ТЦД50/мл) на культуре клеток Vero Е6 у хомяков на 6-е сутки после заражения дельта-штаммом SARS-CoV-2 в гомогенатах тканей: а - легких; 6 - полости носа. Точки представляют индивидуальные значения, вершины гистограмм показывают медианные значения в группе, вертикальные линии отображают границу 95% доверительного интервала (95%ДИ). Значения р над графиком получены в парном сравнении с применением U-критерия Манна-Уитни (степень достоверности различий *р<0,05; **р<0,01; ***р<0,001).
Для лучшего понимания сущности изобретения ниже следуют примеры его осуществления.
Пример 1. Конструирование рекомбинантного штамма Sen-Sdelta(M)
Рекомбинантные варианты вируса Сендай получали на основе его российского штамма Москва [Zainutdinov, 2016] [22]. Трансген Sdelta получали методом ОТ-ПЦР с использованием праймеров, представленные в таблице 1, и геномной РНК, полученной из штамма SARS-CoV-2 hCoV-19/Russia/MOS-2406/2021, относящегося к варианту Delta (вариант В. 1.617.2), выделенному от взрослого пациента в июне 2021 г. и депонированному в международной базе GISAID с номером доступа EPI_ISL_7338789 (https://gisaid.org/). Структура S гена представлена на фиг. 2.

Ампликон трансгена Sdelta был встроен рестриктазно-лигазным методом по сайтам BsiWI - BssHII в полилинкер геномной плазмиды pSen2-MCS(M) (фиг. 1) с получением геномной плазмидной ДНК pSen2-CoVSpike2(M). NGS секвенирование подтвердило ожидаемую первичную структуру полученной плазмидной ДНК.
Рекомбинантный вариант Sen-Sdelta(M) вируса Сендай штамма Москва был получен в результате трансфекции клеток 293-Т7 набором из четырех плазмидных ДНК: геномная pSen2-CoVSpike2(M) и три вспомогательных плазмиды, экспрессирующих гены N, Р и L вируса Сендай [Патент РФ №2787724][11], с последующим переносом трансфекционного материала в аллантоисную полость 11-дневных куриных эмбрионов и выделением рекомбинантного вируса из аллантоисной жидкости после 72 часов инкубации как описано в [Кудров, 2023][12].
Для последующих экспериментов рекомбинантный вирус нарабатывали в аллантоисной жидкости 11-дневных куриных эмбрионов в течение 72 час при 37°С, очищали от клеточных элементов центрифугированием при 1150 × g 10 мин и однократно замораживали при -40°С. После разморозки пробирки с аллантоисной жидкостью центрифугировали при 12000 × g 30 мин при 6°С (осветление). Вирус из надосадочной жидкости концентрировали центрифугированием при 100000 × g 45 мин при 6°С. Полученный осадок суспендировали в фосфатно-солевом буфере (1/10 от исходного объема) с добавлением MgCl2 до конечной концентрации 1 мМ. Полученную суспензию обрабатывали ультразвуком при частоте колебаний наконечника 22 кГц, амплитуде 14-16 мкм и мощности 300 Вт. Обработку ультразвуком проводили три раза по 1 мин с охлаждением после каждой обработки в течение 30 сек во льду. Препарат расфасовывали и хранили при - 40°С.
Титрование вируса Сендай проводили методом бляшек на культуре клеток LLC-MK2 с использованием для визуализации вируса эритроцитов морской свинки как описано в [Tatsumoto, 2018] [23]. Титр вируса выражали в количестве бляшкообразующих единиц (БОЕ) на 1 мл суспензии. Титр очищенного концентрированного препарата вируса Сендай составил 1,0×109 БОЕ/мл.
Пример 2. Оценка экспрессии трансгена S белка SARS-CoV-2 рекомбинантным штаммом Sen-Sdelta(M)
Экспрессию трансгена S белка SARS-CoV-2 оценивали методом вестерн-блоттинга в лизатах клеток почки макаки-резус LLC-MK.2, полученных из Коллекции культур клеток ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора (коллекционный номер 103). Подготовку лизатов клеток LLC-МКг для анализа S белка SARS-CoV-2 проводили по следующей методике. Монослой LLC-MK2 клеток (90% поверхности), выращенный в 6-ти луночных планшетах, инфицировали рекомбинантным штаммом Sen-Sdelta(M) с множественностью 10 БОЕ/кл. После 90 минут сорбции вируса и внесения питательной среды DMEM с добавлением 0,25% раствора трипсина в объеме 1 мкл/мл среды, матрасы инкубировали 72 часа при 37°С. Среду удаляли, лунки дважды промывали холодным PBS с тщательным удалением остатков. В лунки добавляли по 200 мкл лизирующего буфера: RIPA+ коктейль ингибиторов протеаз (Sigma-Aldrich, США), инкубировали на льду, периодически покачивая, до полного лизиса клеток. Лизаты собирали в пробирки и инкубировали на качалке во льду еще 30 мин, далее центрифугировали 14 тыс. об/мин 15 мин при 6°С. Супернатант переносили в новые пробирки и хранили при - 20°С до анализа в вестерн-блоттинге. В качестве отрицательного контроля использовали аналогичным образом подготовленный лизат неинфицированных клеток LLC-MK2 и лизат клеток, инфицированных исходным векторным штаммом Москва вируса Сендай. Пробы для вестерн-блот анализа готовили из расчета 50 мкл лизата клеток + 50 мкл буфера нанесения (0,5М Трис, рН 6,8; 25% глицерин; 10% SDS; 0,5% (w/v) бромфеноловый синий). В качестве первичных антител использовали сыворотку крови человека, ранее вакцинированного Спутник-V и переболевшего в легкой форме SARS-CoV-2. В качестве положительного контрольного белка использовали рекомбинантный белок Sdelta, наработанный в трансформированных клетках СНО-K1 в форме гомотримера [Merkuleva, 2022] [24]. Для проявления использовали соответствующие вторичные антитела, конъюгированные с щелочной фосфатазой (Sigma-Aldrich, США).
Результаты вестерн-блоттинга представлены на фиг.3, из которой следует, что в лизатах клеток, инфицированных рекомбинантным вирусом Sen-Sdelta(M), выявляется негликозилированная (S1, около 100 кДа), мономерная гликозилированная (около 200 кДа), а также тримерная форма белка S с молекулярной массой свыше 260 кДа. Взаимодействие экспрессируемого рекомбинантным вариантом Sen-Sdelta(M) белка S с сывороткой реконвалесцента COVID-19 свидетельствует о его иммуноидентичности вирионному S гликопротеину SARS-CoV-2.
Пример 3. Иммуногенность рекомбинантного штамма Sen-Sdelta(M) на моделях мышей линии BALB/c и золотистых сирийских хомячков
Все работы с животными проводили в помещениях виварного блока лаборатории биологической защиты 2 уровня (BSL-2) и 3 уровня (BSL-3). В работе использовали 18 сирийских хомяков Mesocricetus auratus (самки, возраст 6 недель, вес 80-110 г.) и 36 мышей линии BALB/c Mus musculus strain BALB/c (самки, возраст 8 недель, вес 18-22 г.) свободных от посторонней микрофлоры, которые были получены из питомника лабораторных животных ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора. Хомяков разделяли на две группы (вакцинированная и контрольная) по 9 животных в каждой. Мышей делили на 3 группы: вакцинированная (n=16), контрольная (n=16) и интактные (n=4). Животных содержали на стандартном рационе питания и свободном доступе к воде. Манипуляции с животными, включая вакцинацию, отбор смывов и крови осуществляли после премедикации раствором золетила (Valdepharm, Франция). Вакцинацию штаммом Sen-Sdelta(M) проводили интраназально однократно в дозе 106 БОЕ/хомяк и 105 БОЕ/мышь, объем каждого инокулята составлял 100 мкл для хомяков (по 50 мкл в каждую ноздрю) и 10 мкл для мышей. Контрольным группам вводили аналогичное количество фосфатно-солевого буфера (PBS), на котором проводили разведение вируса. Группе интактных мышей никакие препараты не вводили. Забор крови у животных осуществляли через 28 дней после вакцинации для выявления общих и вируснейтрализующих антител к SARS-CoV-2.
Определение общих антител класса IgG к коронавирусу проводили с использованием тест-системы «Вектор ИФА SARS-CoV-2-АТ» (Вектор-бест, Россия). Для детекции IgG использовали пероксидазный конъюгат «Goat Anti-Mouse IgG» (Biomedicals, США). Нейтрализующую активность сывороток крови определяли на культуре клеток Vero Е6 против вируса hCoV-19/Russia/MOS-2406/2021 (Delta VOC) как описано в [Merkuleva, 2022] [23]. Для оценки клеточного иммунного ответа использовали по 8 мышей из группы вакцинированных и контроля и 4 интактные мыши, у которых на 28 сутки после вакцинации осуществляли забор селезенки. Спленоциты выделяли путем индивидуального последовательного прессования селезенок через клеточный фильтр (Jet BioFil, Китай) с порами 70 и 40 мкм. После удаления эритроцитов с помощью буфера для лизиса эритроцитов (Sigma, США), спленоциты дважды промывали и ресуспендировали в среде RPMI 1640 с добавлением L-глутамина и гентамицина (50 мкг/мл). Интенсивность Т-клеточного иммунного ответа у иммунизированных мышей определяли по числу спленоцитов, продуцирующих IFN-γ с помощью набора ELISpot Plus: Mouse IFN-γ (ALP) (Mabtech, Швеция) в соответствии с инструкцией производителя. Для стимуляции клеток использовали смесь пептидов в концентрации 20 мкг/мл каждый (табл. 2). Пептиды были рассчитаны с помощью инструментов IEDB Analysis Resourse и синтезированы компанией AtaGenix Laboratories (Китай), чистота пептидов составила более 80%.
Подсчет количества IFN-γ-продуцирующих клеток осуществляли с помощью ELISpot-ридера фирмы Carl Zeiss (Германия).
Результаты оценки иммуногенности штамма Sen-Sdelta(M) представлены на фиг. 4-6.
Как следует из фиг. 4, антитела класса IgG к SARS-CoV-2 обнаружены у всех мышей, вакцинированных Sen-Sdelta(M), со средним геометрическим титром (СГТ) 19740 и 95% доверительным интервалом (95% ДИ) от 12826 до 30381.
Вируснейтрализующие антитела (НТ50) также определены во всех образцах сыворотки крови мышей из группы вакцинированных Sen-Sdelta(M) (СГТ=80; 95% ДИ от 52 до 124). Достоверность сероконверсии (р<0,001) определена в сравнении с контрольной группой, в образцах которой все значения были ниже предела обнаружения метода.
Результаты, представленные на фиг. 5, показывают, что интраназальное введение векторной вакцины Sen-Sdelta(M) приводит к формированию специфического Т-клеточного ответа у мышей. Этот результат значимо отличается от величины иммунного ответа в контрольных группах (р<0,01).
Как следует из фиг. 6, у всех вакцинированных Sen-Sdelta(M) хомяков показана значимая индукция гуморального иммунного ответа в форме образования IgG-антител к антигену дельта-штамма SARS-CoV-2 (фиг.6а), средний геометрическим титр которых составил 686; 95% ДИ [440; 1069]. Также выявлена высокая нейтрализующая активность сывороток крови вакцинированных хомяков к дельта-штамму SARS-CoV-2 (фиг. 6 б), СГТ=152; 95% ДИ [102; 191]. Титры IgG и НТ50 с высокой статистической значимостью (р<0,001, тест Манна-Уитни) отличались от группы контроля, в которой все образцы показали значения титров ниже порога определения.
Таким образом, рекомбинантный штамм вируса Сендай Sen-Sdelta(M) обладает высокой иммуногенностью уже при однократном интраназальном введении и индуцирует формирование как гуморального вируснейтрализующего, так и специфического клеточного иммунного ответа.
Пример 4. Протективность рекомбинантного штамма Sen-Sdelta(M) в отношении инфекции SARS-CoV-2 на моделях мышей линии BALB/c и золотистых сирийских хомячков.
На 30-й день после однократной иммунизации проводили интраназальную инокуляцию опытных и контрольных групп животных, описанных в Примере 3, вирусом SARS-CoV-2 в дозе 50 ИД50 на одно животное, в соответствии с ранее полученными данными о восприимчивости сирийских хомяков [Shipovalov, 2022] [25] и мышей BALB/c [Shipovalov, 2022] [26]. Контроль клинического состояния после заражения проводили ежедневно, у всех животных фиксировали вес и брали смывы из носовой полости. Животных выводили из эксперимента по достижению максимума вирусной нагрузки в тканях легких, на 6-е сутки после заражения хомяков и на 5-е сутки - мышей, извлекали легкие и ткани носовой полости, готовили 10%-ные гомогенаты. Титр SARS-CoV-2 в гомогенатах тканей определяли методом титрования вируса по конечной точке цитопатического действия (ЦПД) на культуре клеток Vero Е6 как описано в [Зайковская, 2022] [27]. Титр вируса выражали в lg ТЦД50/мл. Результаты экспериментов представлены на фиг. 7 и 8.
Как следует из фиг. 7, в результате проведенного исследования выявлено статистически значимое снижение инфекционного титра в тканях легких (р=0,04, фиг. 7а) и полости носа (р=0,003, фиг. 7б) мышей, вакцинированных Sen-Sdelta(M), по сравнению с группой PBS. В частности, относительно контроля, медианный инфекционный титр в тканях легких у вакцинированных животных был снижен на 1,8 lg, а в тканях полости носа - на 3 lg. Кроме того, в 62,5% образцов тканей легких и 87,5% образцов тканей полости носа мышей группы Sen-Sdelta(M) получены значения титра ниже порога определения (титр ≤1 lg ТЦД50/мл). Полученные данные свидетельствуют о высокой эффективности вакцинации в снижении риска заражения и распространения вируса, что подтверждается отсутствием инфекционно- активного вируса в значительном количестве проб из полости носа у мышей, вакцинированных Sen-Sdelta(M).
Аналогичные данные получены при оценке протективности вакцинного штамма Sen-Sdelta(M) на модели золотистого сирийского хомяка (фиг. 8). Обнаружено, что 89% образцов тканей легких и 100% образцов тканей полости носа у вакцинированных хомяков имели титры, находящиеся ниже порога определения (титр ≤1 lg ТЦД50/мл). Достоверное снижение медианного инфекционного титра в группе Sen-Sdelta(M) по сравнению с контролем составило 1 lg в тканях легких (р<0,001, фиг. 8а) и 1,5 lg в тканях полости носа (р <0,001, фиг. 8б). Полученные данные свидетельствуют о высокой эффективности однократной иммунизации Sen-Sdelta(M) в снижении риска заражения и передачи вируса SARS-CoV-2, что подкрепляется отсутствием инфекционно-активного вируса в 100% образцов из полости носа и 89% образцов тканей легких после заражения дельта-штаммом SARS-CoV-2 у вакцинированных Sen-Sdelta(M) сирийских хомячков.
Таким образом, вышеизложенные результаты (примеры 1-4) подтверждают достижение заявляемого технического результата, а именно: на основе российского штамма Москва вируса Сендай создан рекомбинантный штамм Sen-Sdelta(M), экспрессирующий трансген S белка высокопатогенного варианта дельта SARS-CoV-2, который при однократной интраназальной вакцинации индуцирует как гуморальный вируснейтрализующий, так и клеточный иммунный ответ, способный предотвратить развитие инфекции SARS-CoV-2 в организме вакцинированного. Эффективность вакцинации подтверждена для двух видов животных: мыши линии BALB/c и золотистые сирийские хомячки. Штамм Sen-Sdelta(M) может служить прототипом для создания однократных интраназальных вакцин против циркулирующих вариантов SARS-CoV-2, особенно против высокопатогенных, вызывающих озабоченность вариантов (variants of concern, VOC).
Источники информации
1. Wang, S., Liang, В., Wang, W. et al. Viral vectored vaccines: design, development, preventive and therapeutic applications in human diseases. // Sig. Transduct. Target. Ther. - 2023. - V. 8. - 149. https://doi.org/10.1038/s41392-023-01408-5
2. Lund F.E., Randall T.D. Scent of a vaccine. // Science. - 2021. - V. 373(6553). - P. 397-399. https://doi.org/10.1126/science.abg9857
3. Зайнутдинов С.С., Сиволобова Г.Ф., Локтев В.Б., Кочнева Г.В. Мукозальный иммунитет и вакцины против вирусных инфекций. // Вопросы вирусологии. - 2021. - Т. 66 (6). - С. 399-408. https://doi.org/10.36233/0507-4088-82
4. Зайчук Т.А., Нечипуренко Ю.Д., Аджубей А.А., Оникиенко СБ., Черешнев В.А., Зайнутдинов С.С. и др. Проблемы создания вакцин против бетакоронавирусов: антителозависимое усиление инфекции и вирус Сендай как возможный вакцинный вектор // Молекулярная биология. - 2020 - Т. 54 (6) - С. 922-938. https://doi.org/10.31857/S0026898420060154
5. Cantell K., Hirvonen S., Kauppinen H.L., Myllyla G. Production of interferon in human leukocytes from normal donors with the use of Sendai virus // Methods Enzymol. - 1981 - V. 78 (Pt. A). - P. 29-38. https://doi.org/10.1016/0076-6879(81)78094-7
6. Nyman T.A., Tolo H., Parkkinen J., Kalkkinen N. Identification of nine interferon-alpha subtypes produced by Sendai virus-induced human peripheral blood leucocytes // Biochem. J. - 1998 - V. 329 (Pt. 2). - 295-302. https://doi.org/10.1042/bi3290295
7. Matsumoto Y., Ohta K., Kolakofsky D., Nishio M. The control of paramyxovirus genome hexamer length and mRNA editing // RNA. - 2018 - V. 24 (4) - P. 461-467. https://doi.Org/l0.126l/rna.065243.117
8. Morimoto S., Saeki K., Takeshita M., Hirano K., Shirakawa M., Yamada Y. et al. Intranasal Sendai virus-based SARS-CoV-2 vaccine using a mouse model // Genes Cells. - 2023. - V. 28 (1). - P. 29-41. https://doi.org/10.1111/gtc. 12992
9. Scaggs Huang F., Bernstein D.I., Slobod K.S., Portner A., Takimoto Т., Russell C.J. et al. Safety and immunogenicity of an intranasal Sendai virus-based vaccine for human parainfluenza virus type I and respiratory syncytial virus (SeVRSV) in adults // Hum. Vaccin. Immunother. - 2021 - V. 17 (2) - P. 554-559. https://doi.org/10.1080/21645515.2020.1779517
10. Russell C.J., Hurwitz J.L. Sendai virus-vectored vaccines that express envelope glycoproteins of respiratory viruses // Viruses. - 2021 - V. 13 (6). - 1023. https://doi.org/10.3390/v13061023
11. Зайнутдинов С.С., Кочнева Г.В., Сиволобова Г.Ф., Гражданцева А.А. Набор рекомбинантных плазмидных ДНК для получения рекомбинантных вирусов Сендай штамм Москва (варианты). Патент РФ №2787724. Бюл. Изобретений №2 от 12.01.2023 г.
12. Кудров Г.А., Зайнутдинов С.С., Гражданцева А.А., Шиповалов А.В., Сиволобова Г.Ф., Семенова А.В. и др. Интраназальная вакцина против COVID-19 на основе рекомбинантного варианта вируса Сендай (Paramyxoviridae: Respirovirus) штамма Москва // Вопросы вирусологии. - 2023. - Т. 68(3). - С. 42-54. https://doi.org/10.36233/0507-4088-172
13. Ilinykh P.A., Periasamy S., Huang, K. et al. A single intranasal dose of human parainfluenza virus type 3-vectored vaccine induces effective antibody and memory T cell response in the lungs and protects hamsters against SARS-CoV-2 // npj Vaccines. - 2022. - V. 7: 47. https://doi.org/10.1038/s41541 -022-00471-3
14. Kuhn C.C., Basnet N., Bodakuntla S., Alvarez-Brecht P., Nichols S., Martinez-Sanchez A., et al. Direct Cryo-ET observation of platelet deformation induced by SARS-CoV-2 spike protein // Nat. Commun. - 2023. - V. 14. - 620. https://doi.org/10.1038/s41467-023-36279-5
15. Kusudo E., Murata Y., Kawamoto S., Egi M. Variant-derived SARS-CoV-2 spike protein does not directly cause platelet activation or hypercoagulability // Clinical and Experimental Medicine - 2023. - V. 23. - P. 3701-3708. https://doi.org/10.1007/s10238-023-01091-4
16. Uaprasert N. Panrong K., Rojnuckarin P., Chiasakul T. Thromboembolic and hemorrhagic risks after vaccination against SARS-CoV-2: a systematic review and meta-analysis of randomized controlled trials // Thrombosis Journal. - 2021. - V. 19:86 https://doi.org/10.1186/s 12959-021-00340-4
17. ∅stergaard S.D., Schmidt M., Horváth-Puhó E., Thomsen R.W., Sørensen H.T. Thromboembolism and the Oxford-AstraZeneca COVID-19 vaccine: side-effect or coincidence? // Lancet. - 2021. - V. 397 (10283). - P. 1441-1443. https://doi.org/10.1016/S0140-673 6(21)00762-5
18. Elalamy I., Gerotziafas G., Alamowitch S., Laroche J.P., Van Dreden P., Ageno W. et al. Scientific Reviewer Committee. SARS-CoV-2 Vaccine and Thrombosis: An Expert Consensus on Vaccine-Induced Immune Thrombotic Thrombocytopenia // Thromb. Haemost. - 2021. - V. 121 (8). - P. 982-991. https://doi.org/10.1055/a-1499-0119
19. Моисеева А.Г., Серова H.Ю., Шайхутдинова Р.А., Жарова А.С., Коротких А.В., Лидер Р.Ю., Казанцев А.Н. Тромбозы у невакцинированных и вакцинированных пациентов вакциной "Спутник V" (Гам-КОВИДВак), инфицированных COVID-19 // Российский кардиологический журнал. - 2022. - Т. 27 (7): 5091. https://doi.org/10.15829/1560-4071-2022-5091
20. Зубкова О.В., Ожаровская Т.А. Должикова И.В., Попова О., Щебляков Д.В., Гроусова Д.М., Джаруллаева А.Ш., и др. Иммунобиологическое средство и способ его использования для индукции специфического иммунитета против вируса тяжелого острого респираторного синдрома SARS-CoV-2 (варианты). Патент РФ №2720614. Опубл. 12.05.2020. Бюл. №14.
21. Заявка на патент US 2021338804, МПК A61K 39/215; C12N7/01, опубл. 04.11.2021 г. (прототип).
22. Zainutdinov S.S., Tikunov A.Y., Matveeva O.V., Netesov S.V., Kochneva G.V. Complete genome sequence of Sendai virus oncolytic strain Moscow // Genome Announcements. - 2016. - V. 4 (4): e00818-16. https://doi.org/10.1128/genomeA.00818-16
23. Tatsumoto N., Miyauchi Т., Arditi M., Yamashita M. Quantification of infectious Sendai virus using plaque assay. // Bio. Protoc - 2018. - V. 8 (21). https://doi.org/10.21769/BioProtoc.3068
24. Merkuleva I.A., Shcherbakov D.N., Borgoyakova M.B., Shanshin D.V., Rudometov A.P., Karpenko L.I. et al. Comparative immunogenicity of the recombinant receptor-binding domain of protein S SARS-CoV-2 obtained in prokaryotic and mammalian expression systems // Vaccines. - 2022. - V. 10: 96. https://doi.org/10.3390/vaccines 10010096
25. Shipovalov A.V., Kudrov G.A., Tomilov A.A., Bodnev S.A., Boldyrev N.D., Ovchinnikova A.S. et al. Pathogenicity of the SARS-CoV-2 virus variants of concern for the Syrian golden hamster // Problems of Particularly Dangerous Infections. - 2022. - V. 3. - P. 164-169. (In Russ.). https://doi.org/10.21055/0370-1069-2022-3-164-169
26. Shipovalov A.V., Kudrov G.A., Tomilov A.A., Bodnev S.A., Boldyrev N.D., Ovchinnikova A.S. et al. Susceptibility to SARS-CoV-2 virus variants of concern in mouse models // Problems of Particularly Dangerous Infections. - 2022. - V. 1. - P. 148-155. (In Russ.). https://doi.org/10.21055/0370-1069-2022-1-148-155
27. Зайковская A.B., Гладышева A.B., Карташов М.Ю., Таранов О.С., Овчинникова А.С., Шиповалов А.В., Пьянков О.В. Изучение в условиях in vitro биологических свойств штаммов коронавируса SARS-CoV-2, относящихся к различным генетическим вариантам // Проблемы особо опасных инфекций. - 2022. - Т. 1. - С.94-100. https://doi.org/10.21055/0370-1069-2022-1-94-100
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3"
fileName="Приложение_Рекомбинантный штамм Sen-Sdelta(M) для
вакцинации против COVID-19.xml" softwareName="WIPO Sequence"
softwareVersion="2.3.0" productionDate="2024-03-20">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>1234567</ApplicationNumberText>
<FilingDate>2024-03-09</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>1234</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>12345678</ApplicationNumberText>
<FilingDate>2024-03-01</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное бюджетное учреждение
науки «Государственный научный центр вирусологии и биотехнологии
«Вектор» Федеральной службы по надзору в сфере защиты прав
потребителей и благополучия человека (ФБУН ГНЦ ВБ «Вектор»
Роспотребнадзора)</ApplicantName>
<ApplicantNameLatin>Federalnoe byudzhetnoe uchrezhdenie nauki
"Gosudarstvennyj nauchnyj tsentr virusologii i biotekhnologii
"Vektor" Federalnoj sluzhby po nadzoru v sfere zashchity
prav potrebitelej i blagopoluchiya cheloveka (FBUN GNTS VB
"Vektor" Rospotrebnadzora) (RU)</ApplicantNameLatin>
<InventionTitle languageCode="ru">Рекомбинантный штамм Sen-Sdelta(M)
вируса Сендай, продуцирующий S белок SARS-CoV-2 и используемый для
получения интраназальной вакцины против COVID-19</InventionTitle>
<SequenceTotalQuantity>1</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>3832</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..3832</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cgtacgccaccatgtttgtttttcttgttttattgccactagtctctag
tcagtgtgttaatcttagaaccagaactcaattaccccctgcatacactaattctttcacacgtggtgtt
tattaccctgacaaagttttcagatcctcagttttacattcaactcaggacttgttcttacctttctttt
ccaatgttacttggttccatgctatacatgtctctgggaccaatggtactaagaggtttgataaccctgt
cctaccatttaatgatggtgtttattttgcttccattgagaagtctaacataataagaggctggattttt
ggtactactttagattcgaagacccagtccctacttattgttaataacgctactaatgttgttattaaag
tctgtcaatttcaattttgtaatgatccatttttggatgtttattaccacaaaaacaacaaaagttggat
ggaaagtggagtttattctagtgcgaataattgcacttttgaatatgtctctcagcctttccttatggac
cttgaaggaaaacagggtaatttcaaaaatcttagggaatttgtgtttaagaatattgatggttatttta
aaatatattctaagcacacgcctattaatttagtgcgtgatctccctcagggtttttcggctttagaacc
attggtagatttgccaataggtattaacatcactaggtttcaaactttacttgctttacatagaagttat
ttgactcctggtgattcttcttcaggttggacagctggtgctgcagcttattatgtgggttatcttcaac
ctaggacttttctattaaaatataatgaaaatggaaccattacagatgctgtagactgtgcacttgaccc
tctctcagaaacaaagtgtacgttgaaatccttcactgtagaaaaaggaatctatcaaacttctaacttt
agagtccaaccaacagaatctattgttagatttcctaatattacaaacttgtgcccttttggtgaagttt
ttaacgccaccagatttgcatctgtttatgcttggaacaggaagagaatcagcaactgtgttgctgatta
ttctgtcctatataattccgcatcattttccacttttaagtgttatggagtgtctcctactaaattaaat
gatctctgctttactaatgtctatgcagattcatttgtaattagaggtgatgaagtcagacaaatcgctc
cagggcaaactggaaagattgctgattataattataaattaccagatgattttacaggctgcgttatagc
ttggaattctaacaatcttgattctaaggttggtggtaattataattaccggtatagattgtttaggaag
tctaatctcaaaccttttgagagagatatttcaactgaaatctatcaggccggtagcaaaccttgtaatg
gtgttgaaggttttaattgttactttcctttacaatcatatggtttccaacccactaatggtgttggtta
ccaaccatacagagtagtagtactttcttttgaacttctacatgcaccagcaactgtttgtggacctaaa
aagtctactaatttggttaaaaacaaatgtgtcaatttcaacttcaatggtttaacaggcacaggtgttc
ttactgagtctaacaaaaagtttctgcctttccaacaatttggcagagacattgctgacactactgatgc
tgtccgtgatccacagacacttgagattcttgacattacaccatgttcttttggtggtgtcagtgttata
acaccaggaacaaatacttctaaccaggttgctgttctttatcagggtgttaactgcacagaagtccctg
ttgctattcatgcagatcaacttactcctacttggcgtgtttattctacaggttctaatgtttttcaaac
acgtgcaggctgtttaataggggctgaacatgtcaacaactcatatgagtgtgacatacccattggtgca
ggtatatgcgctagttatcagactcagactaattctcgtcggcgggcacgtagtgtagctagtcaatcca
tcattgcctacactatgtcacttggtgcagaaaattcagttgcttactctaataactctattgccatacc
cacaaattttactattagtgttaccacagaaattctaccagtgtctatgaccaagacatcagtagattgt
acaatgtacatttgtggtgattcaactgaatgcagcaatcttttgttgcaatatggcagtttttgtacac
aattaaaccgtgctttaactggaatagctgttgaacaagacaaaaacacccaagaagtttttgcacaagt
caaacaaatttacaaaacaccaccaattaaagattttggtggttttaatttttcacaaatattaccagat
ccatcaaaaccaagcaagaggtcatttattgaagatctacttttcaacaaagtgacacttgcagatgctg
gcttcatcaaacaatatggtgattgccttggtgatattgctgctagagacctcatttgtgcacaaaagtt
taacggccttactgttttgccacctttgctcacagatgaaatgattgctcaatacacttctgcactgtta
gcgggtacaatcacttctggttggacctttggtgcaggtgctgcattacaaataccatttgctatgcaaa
tggcttataggtttaatggtattggagttacacagaatgttctctatgagaaccaaaaattgattgccaa
ccaatttaatagtgctattggcaaaattcaagactcactttcttccacagcaagtgcacttggaaaactt
caaaatgtggtcaaccaaaatgcacaagctttaaacacgcttgttaaacaacttagctccaattttggtg
caatttcaagtgttttaaatgatatcctttcacgtcttgacaaagttgaggctgaagtgcaaattgatag
gttgatcacaggcagacttcaaagtttgcagacatatgtgactcaacaattaattagagctgcagaaatc
agagcttctgctaatcttgctgctactaaaatgtcagagtgtgtacttggacaatcaaaaagagttgatt
tttgtggaaagggctatcatcttatgtccttccctcagtcagcacctcatggtgtagtcttcttgcatgt
gacttatgtccctgcacaagaaaagaacttcacaactgctcctgccatttgtcatgatggaaaagcacac
tttcctcgtgaaggtgtctttgtttcaaatggcacacactggtttgtaacacaaaggaatttttatgaac
cacaaatcattactacagacaacacatttgtgtctggtaactgtgatgttgtaataggaattgtcaacaa
cacagtttatgatcctttgcaacctgaattagactcattcaaggaggagttagataaatattttaagaat
catacatcaccagatgttgatttaggtgacatctctggcattaatgcttcagttgtaaacattcaaaaag
aaattgaccgcctcaatgaggttgccaagaatttaaatgaatctctcatcgatctccaagaacttggaaa
gtatgagcagtatataaaatggccatggtacatttggctaggttttatagctggcttgattgccatagta
atggtgacaattatgctttgctgtatgaccagttgctgtagttgtctcaagggctgttgttcttgtggat
cctgctgcaaatttgatgaagacgactctgagccagtgctcaaaggagtcaaattacattacacatagcg
cgc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЙ ВИРУС ГРИППА, ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ПРОФИЛАКТИКИ COVID-19 И ГРИППА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2022 |
|
RU2802058C1 |
| Вакцина на основе вирусоподобных частиц (VLP) для профилактики COVID-19 для парентерального применения | 2024 |
|
RU2834060C1 |
| ПОЛИПЕПТИД С АНТИ-SARS-CoV-2 АКТИВНОСТЬЮ И ПРОЛОНГИРОВАННЫМ ВРЕМЕНЕМ ЦИРКУЛЯЦИИ В КРОВОТОКЕ | 2023 |
|
RU2822355C1 |
| Пептидные иммуногены и вакцинная композиция против коронавирусной инфекции COVID-19 с использованием пептидных иммуногенов | 2020 |
|
RU2743593C1 |
| Способ создания живой вакцины против коронавирусной инфекции COVID-19 на основе пробиотического штамма Enterococcus faecium L3 и живая вакцина Enterococcus faecium L3-pentF-covid-19 | 2020 |
|
RU2745626C1 |
| Вакцинная композиция против коронавирусной инфекции COVID-19 | 2020 |
|
RU2743595C1 |
| Пептидные иммуногены, используемые в качестве компонентов вакцинной композиции против коронавирусной инфекции COVID-19 | 2020 |
|
RU2743594C1 |
| Пептидные иммуногены и вакцинная композиция против коронавирусной инфекции COVID-19 с использованием пептидных иммуногенов | 2020 |
|
RU2738081C1 |
| Вакцина против гриппа типа А, гриппа типа B и COVID-19 | 2021 |
|
RU2751485C1 |
| Способ создания рекомбинантного штамма энтерококка L3-SARSN1 на основе биологически активного штамма Enterococcus faecium L3 | 2022 |
|
RU2820058C1 |


Изобретение относится к биотехнологии. Предложен рекомбинантный штамм Sen-Sdelta(M) вируса Сендай, используемый для получения одноразовой интраназальной векторной вакцины против COVID-19, продуцирующий S белок SARS-CoV-2, включающий трансген Sdelta, имеющий нуклеотидную последовательность SEQ ID NO: 1, содержащую ген шиповидного S белка дельта-варианта SARS-CoV-2. Изобретение может эффективно использоваться для получения одноразовой интраназальной векторной вакцины против COVID-1. 8 ил., 2 табл., 4 пр.
Рекомбинантный штамм Sen-Sdelta(M) вируса Сендай, используемый для получения одноразовой интраназальной векторной вакцины против COVID-19, продуцирующий S белок SARS-CoV-2, включающий трансген Sdelta, имеющий нуклеотидную последовательность SEQ ID NO: 1, содержащую ген шиповидного S белка дельта-варианта SARS-CoV-2.
| КУДРОВ Г.А | |||
| и др | |||
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| Вопросы вирусологии, 2023, т | |||
| Способ получения смеси хлоргидратов опийных алкалоидов (пантопона) из опийных вытяжек с любым содержанием морфия | 1921 |
|
SU68A1 |
| Кузнечный горн | 1921 |
|
SU215A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Опубл | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| ВЕТРЯНОЙ ДВИГАТЕЛЬ | 1927 |
|
SU7167A1 |

