Изобретение относится к плазмидной ДНК-матрице pVAX-RBD, молекуле мРНК-RBD, полученной с использованием ДНК-матрицы pVAX-RBD и обеспечивающей синтез и секрецию белка RBD SARS-CoV 2 в организме млекопитающих, а также комплексу, состоящему из наночастиц, содержащих молекулы мРНК-RBD и индуцирующих SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью, и может быть использовано в биотехнологии и генетической инженерии.
Известна фармацевтическая композиция, разработанная американской компанией Moderna ТХ, Inc., состоящая из множества липидных наночастиц со средним размером частиц от 80 до 160 нм и содержащая модифицированную мРНК, кодирующую полипептид. Липидные наночастицы включают катионный липид, нейтральный липид, холестерин и ПЭГ-липид. мРНК содержит 5'-cap, 5'-UTR, N1-метил-псевдоуридин, 3'-UTR и поли-область с не менее чем 100 нуклеотидами (Патент США №10703789, МПК А61К 39/395, опубл. 07.07.2020 г.).
Известен состав полирибонуклеотидной молекулы мРНК, используемый для иммунизации организма млекопитающих и разработанный швейцарской компанией NOVARTIS AG (международная заявка WO, 2018029586, МПК А61К 39/00, опубл. 15.02.2018 г.). Эта молекула мРНК способна непосредственно кодировать и облегчать трансляцию целевого белка(ов), против которого желателен ответ антитела в организме млекопитающего. Молекула содержит несколько компонентов: катионные липиды в виде липосомных наночастиц и открытую рамка считывания, соответствующая аминокислотной последовательности белка(ов), например, целевого белка человека или его фрагмента. Нативная последовательность кодонов оптимизирована для вида-хозяина (например, мыши или кролика) с целью повышения эффективности трансляции и, в конечном счете, уровня экспрессии белка мишени. Дополнительные модификации могут быть внесены в открытую рамку считывания для усиления экспрессии/оборота белка. Для секретируемых или мембранных белков это может включать использование гетерологичных сигнальных пептидов, таких как сигнал секреции интерлейкина 2 (IL-2). В конкретном примере для секретируемых белков молекула мРНК может включать гетерологичный сигнальный пептид, такой как сигнальный пептид человеческого IL-2 или IgG kappa.
Известна мРНК - вакцина, состоящая из последовательности гена эпитопного антигена тримерного спайкового гликопротеина S и/или последовательности генов трансмембранного белка-оболочки Е эпитопного антигена и/или последовательности гена эпитопного антигена мембранного гликопротеина М и/или последовательности гена эпитопа нуклеокапсидного антигена Н, и/или последовательности гена эпитопа антигена рецептор-связывающий домен RBD тримерного спайка S-гликопротеина. Последовательности генов эпитопов антигена разделены соединительными пептидами (патент Китая CN 111821433 А61К 39/215, А61Р 31/14, опубл. 27.10.2020 г.).
Наиболее близким техническим решением (прототипом) является гибридный ген, кодирующий слитый белок RBD нового коронавируса SARS-CoV-2, включающий последовательность кодирующего гена RBD SARS-CoV-2, последовательность кодирующего гена линкера и последовательность кодирующего гена СТВ или CRM197 (заявка Китая CN 111217917, МПК А61К 39/215, C12N 15/62, опубл. 02.06.2020 г.). Гибридный ген входит в состав вектора. Слитый белок RBD содержит: фрагмент RBD нового коронавируса SARS-CoV-2; линкер; фрагмент белка СТВ или CRM197 (А). Слитый белок дополнительно содержит сигнальный пептид и/или молекулярный адъювант. Предпочтительно, сигнальный пептид соединен с N-концом слитого белка, а молекула адъюванта соединена с С-концом слитого белка. Более предпочтительно, сигнальный пептид связан со слитым белком через линкер. Сигнальный пептид представляет собой сигнальный пептид tPA, и/или молекулярный адъювант представляет собой агонист CPG или TLR. Форма вакцины представляет собой вакцину на основе рекомбинантной белковой мРНК-RBD, заключенной в липосомы.
Однако использование липидов для доставки вакцин на основе мРНК в клетки организма имеет свои недостатки. Для решения этих проблем существует множество липосомальных составов, но все они имеют ряд недостатков, связанных, прежде всего, с природой липидов. Положительно заряженные частицы липосом могут связываться с отрицательно заряженными белками и нуклеиновыми кислотами, прилипать к поверхностям клеток, что может дестабилизировать плазматическую мембрану и вызывать побочные реакции у вакцинированных лиц. Использование ионизируемых липидов усложняет состав липидных наночастиц и увеличивает стоимость их производства. Еще один недостаток липидных наночастиц заключается в том, что их необходимо транспортировать и хранить в непрерывном процессе холодовой цепи, что является сложной логистической проблемой. Следует отметить, что LNP представляет собой многокомпонентную структуру для стабилизации которой требуется использование дополнительных компонентов и такие липосомы могут вызывать дополнительные нежелательные реакции на введение разработанной вакцины. Кроме того, приведенные выше патенты-аналоги и прототип обеспечивают недостаточно эффективно секрецию белка RBD из клеток для обеспечения презентации RBD иммунной системе организма.
Техническим результатом заявляемого изобретения является создание менее токсичного и более простого по составу и технологии изготовления средства доставки мРНК-RBD в клетки организма для синтеза в них белка RBD SARS-Cov-2 и повышение уровня секреции указанного белка из клеток для обеспечения презентации белка RBD иммунной системе организма.
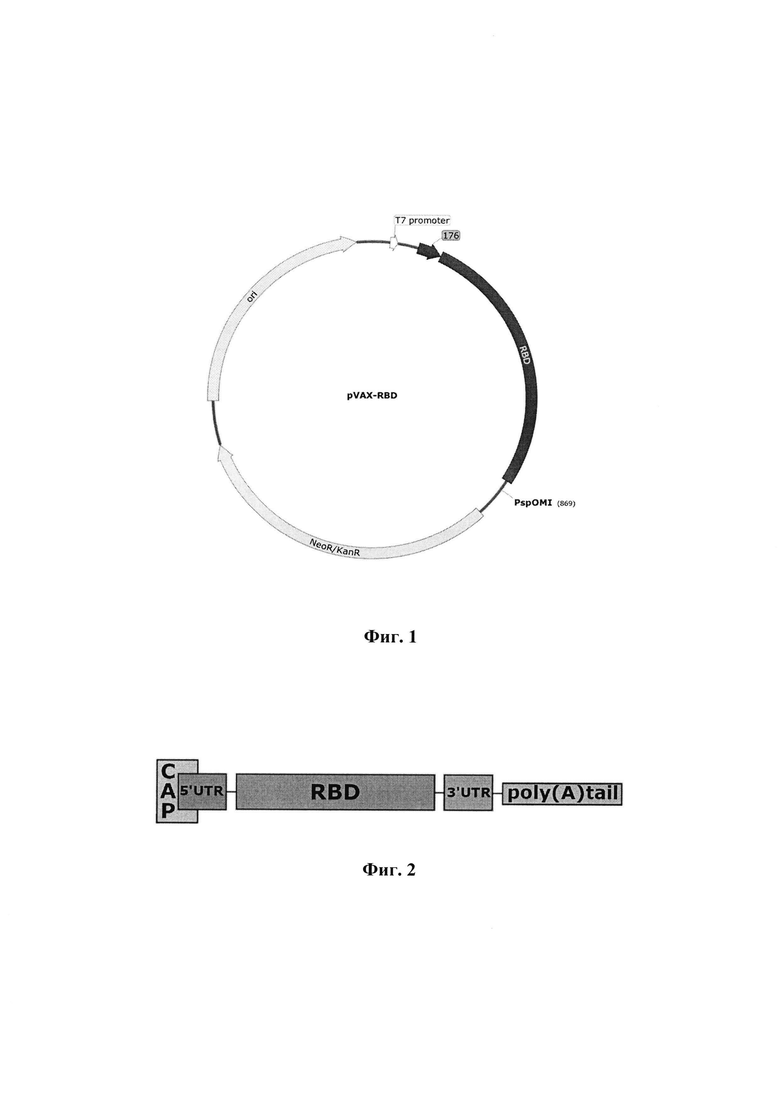
Указанный технический результат достигается созданием плазмидной ДНК-матрицей pVAX-RBD, имеющей нуклеотидную последовательность SEQ ID NO: 1 размером 3710 п.н., молекулярную массу 2.6×103 кДа, содержащая целевой ген, кодирующий химерный белок 176-RBD, перед которым находится промотор РНК-полимеразы фага Т7, обеспечивающий синтез in vitro мРНК-RBD с помощью РНК-полимеразы Т7 и состоящая из следующих фрагментов:
- векторного фрагмента 1484-754 ДНК плазмиды pVAX размером 2981 п.н., содержащего промотор РНК-полимеразы фага Т7 (664-682, размером 19 п.н.);
- ген аминогликозид фосфотрансферазы из Tn5 (1937-2728 размером 702 п.н.),
- ген устойчивости к канамицину (NeoR/KanR) и точку начала репликации ColEl origin (3057-3644 размером 588 п.н.), обеспечивающие селекцию и амплификацию целевой плазмиды в клетках бактерий Escherichia coli;
- фрагмент 755-1483 размером 729 п.н., содержащий искусственный ген 176-RBD с инициирующим кодоном ATG.
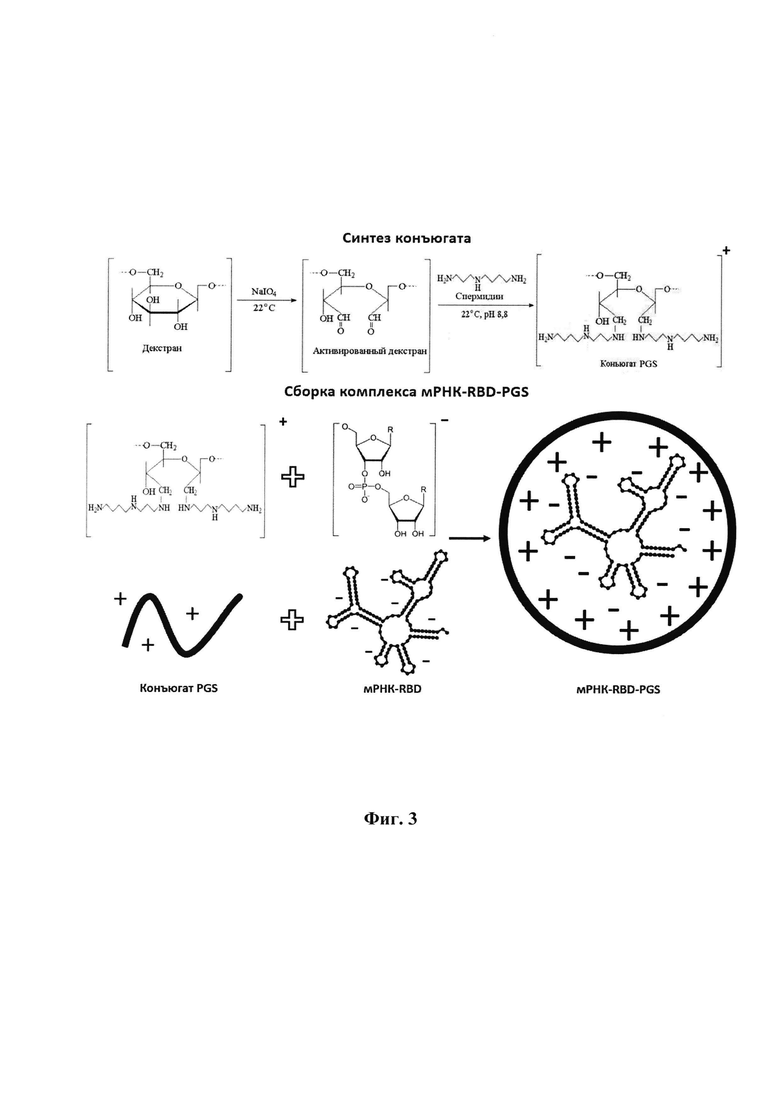
Указанный технический результат достигается также созданием молекулы мРНК-RBD, имеющей нуклеотидную последовательность SEQ ID NO: 2, полученной с использованием ДНК-матрицы pVAX-RBD, содержащей открытую рамку считывания, кодирующую аминокислотную последовательность SEQ ID NO: 3, обеспечивающую синтез и секрецию белка-иммуногена RBD SARS-CoV 2 в организме млекопитающих, и содержащую гибридную сигнальную последовательность 176, консенсусную последовательность Козак, 7-метилгуанозиновый кэп на 5'-конце мРНК, полиаденозиновый (поли А) хвост на 3'-конце мРНК, длиной 150-200 нуклеотидов, и 5'- и 3'-нетранслируемые области (UTR) перед и после открытой рамки считывания.
Указанный технический результат достигается также созданием комплекса в виде наночастиц, индуцирующих SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью, состоящих из молекул конъюгата полиглюкинхпермидин и молекул мРНК-RBD, взятых в разных зарядовых соотношениях от 1:1 до 1:30, соответствующих весовому соотношению от 1:3 до 1:90, имеющих размер в диапазоне 50-900 нанометров, причем каждая наночастица содержит ядро, состоящее из молекул mPHK-RBD и покрытое слоем конъюгата полиглюкинхпермидин, удерживаемого за счет ионного взаимодействия между отрицательно заряженным ядром и положительно заряженным спермидином.
Синтез мРНК кодирующей RBD SARS-COV-2 проводили с линеаризованной и очищенной ДНК-матрицы pVAX-RBD (рис. 1), которая содержит промотор РНК-полимеразы бактериофага Т7 и целевой ген RBD. Для конструирования плазмиды использовали фрагмент гена S, соответствующий последовательности RBD (320-542 а.о.) (GenBank MN908947). Оптимизацию кодонного состава последовательности проводили с помощью программы GeneOptimizer (https://www.thermofisher.com/ru/en/home/life-science/cloning/gene-synthesis/geneart-gene-synthesis/geneoptimizer.html). Для обеспечения эффективного транспорта синтезированного белка из клетки к 5'-концу гена был добавлен фрагмент, кодирующий сигнальную последовательность MMRTLILAVLLVYFCATVHC.
Реакцию транскрипции проводили с помощью РНК-полимеразы бактериофага Т7. Реакционная смесь включала в себя 1 мкг линеаризованной матрицы, Т7 полимеразу с буфером, смесь рибонуклеотидов, в которой уридин был заменен на псевдоуридин, ингибитор РНКаз и безнуклеазную воду. Смесь инкубировали в течение 2 часов при температуре 37°C. После инкубации в смесь добавляли 1 мкл ДНКазы, и инкубировали в течение 30 минут при температуре 37°C для разрушения ДНК матрицы. Анализ полученного продукта производили с использованием электрофореза в 2% агарозном геле. Кэпирование и полиаденилирование РНК проводили с использованием соответствующих ферментов. Кэпирование РНК проводили с помощью наборов ScriptCap™ m7G CappingSystem (CellScript, США) по инструкции производителя. Полиаденилирование РНК проводили с использованием набора A-Plus™ Poly(A) PolymeraseTailingKit (CellScript, США), как это рекомендовано производителем.
В результате, была получена мРНК имеющая нуклеотидную последовательность SEQ ID NO: 2, которая была способна обеспечить синтез в эукариотической клетке целевого белка RBD (аминокислотная последовательность SEQ ID NO: 3)
В данном изобретении в качестве препарата для иммунизации используется молекула мРНК. Молекула мРНК должна непосредственно кодировать и обеспечивать трансляцию целевого белка, против которого необходим иммунный ответ. Таким образом, мРНК должна содержать необходимые для ее функционирования компоненты.
Первым компонентом мРНК является открытая рамка считывания, содержащая соответствующую аминокислотную последовательность (SEQ ID NO: 3) белка-антигена RBD SARS-CoV 2. Для повышения эффективности трансляции и уровня синтеза белка проведена оптимизация кодонового состава. Для повышения уровня секреции белка проведена дополнительная модификация. В начале открытой рамки считывания была добавлена гибридная сигнальная последовательность 176, обеспечивающая эффективный транспорт синтезируемого белка из клетки.
Вторым компонентом мРНК является консенсусная последовательность Козак, необходимая для инициации трансляции.
Третьим компонентом мРНК является 7-метилгуанозиновый кэп на 5'-конце мРНК. Этот кэп важен для рекрутирования фактора инициации эукариот elF4E и сборки зрелой рибосомы. Метилгуанозиновый кэп может быть добавлен ферментативно при транскрипции мРНК in vitro.
Четвертым компонентом мРНК является полиаденозиновый (поли А) хвост на 3'-конце мРНК длинной 150-200 нуклеотидов. Известно, что поли А хвост продлевает время полураспада мРНК в клетках, а также способствует эффективной сборке рибосом и трансляции белка. Включение поли А хвоста можно осуществить посредством транскрипции in vitro или ферментативного полиаденилирования с использованием поли (А) полимеразы.
Пятым компонентом молекулы мРНК является включение 5'- и 3'-нетранслируемых областей (UTR) перед и после открытой рамки считывания. Известно, что присутствие UTR может усиливать трансляцию мРНК, а также увеличивать ее период полураспада в клетке.
В предлагаемой заявке на изобретение доставка мРНК-вакцины предлагается осуществлять в клетки организма млекопитающих с помощью конъюгата полиглюкин: спермидин (PGS). В отличие от липосом, PGS содержит только два компонента, полиглюкин и спермидин, что отличает его от липосомных наночастиц. PGS позволяет лиофилизировать нуклеиновую кислоту и хранить ее в течение длительного времени при положительных температурах. Привлекательно то, что компоненты PGS биоразлагаемы и безопасны для человека. Полиглюкин, полимер глюкозы с молекулярной массой 40000, не токсичен для человека и является лицензированным плазмозамещающим препаратом гемодинамического действия, восстанавливающим объем циркулирующей крови. Спермидин - это встречающийся в природе полиамин, содержащийся во всех живых организмах; он важен для поддержания клеточного гомеостаза и участвует во многих биологических процессах, включая рост и пролиферацию клеток, стабилизацию ДНК и РНК, ферментативную модуляцию и регуляцию трансляции. Кроме того, низкая стоимость компонентов вакцины и возможность лиофилизации и хранения конъюгата PGS при +4°C обеспечивают дополнительные технологические преимущества при производстве и транспортировке вакцины.
Изобретение иллюстрируется следующими графическими материалами. На фиг. 1 представлена физическая и генетическая карта ДНК-матрицы pVAX-RBD. На фиг. 2 приведена схема структуры мРНК-RBD. На фиг. 3 представлена химическая формула конъюгата полиглюкин : спермидин (PGS), структура мРНК и комплекса мРНК-PGS. На фиг. 4 представлен электрофорез комплексов мРНК-RBD-PGS в 2% агарозном геле. На фиг. 5 приведена электронная микроскопия наночастиц комплекса мРНК-PGS. На фиг. 6 представлены диаграммы данных, касающихся гуморальных ответов на мышах BALB/C, иммунизированных антигеном SARS-CoV-2: мышей иммунизировали внутримышечно голой мРНК-RBD, комплексами мРНК с конъюгатом полиглюкин : спермидин (мРНК-RBD-PGS или mPHK-GFP-PGS) или физиологический раствор. Фиг. 7 приведены диаграммы оценки жизнеспособности клеток HEK293, обработанных мРНК-RBD-PGS (А) и мРНК-RBD-Lipofectamine 3000 (В), с помощью анализа МТТ.
Ниже приведены примеры конкретного выполнения изобретения.
Пример 1. Выбор сигнального пептида, обеспечивающего секрецию RBD
Для обеспечения презентации RBD иммунной системе, необходимо чтобы после его синтеза рибосомами клеток он с высокой эффективностью транспортироваля за пределы клетки. В этом случае RBD попадает в межклеточное пространство и доступен для контакта с В-лимфоцитами, обеспечивающих формирование гуморального ответа, выработку антител. Классический путь секреции белков эндоплазматический ретикулум - аппарат Гольджи клеток млекопитающих требует присутствия N-концевой сигнальной последовательности. Так как RBD является фрагментом белка S его секреция из клеток требует введения на N-конец сигнального пептида. В настоящий момент известны аминокислотные последовательности множества пептидов входящих в состав секретируемых белков. Однако анализ этих пептидов при помощи сервера SignalP-5.0 (Armenteros, J. J. A., Tsirigos, K.D.,  , С.K., Petersen, Т.N., Winther, О., Brunak, S., … & Nielsen, H. (2019). SignalP 5.0 improves signal peptide predictions using deep neural networks. Nature biotechnology, 37(4), 420-423.) (таблица 1) показано, что они отличается по вероятной эффективности.
, С.K., Petersen, Т.N., Winther, О., Brunak, S., … & Nielsen, H. (2019). SignalP 5.0 improves signal peptide predictions using deep neural networks. Nature biotechnology, 37(4), 420-423.) (таблица 1) показано, что они отличается по вероятной эффективности.

Для конструирования выбрали наиболее эффективную химерную сигнальную последовательность MMRTLILAVLLVYFCATVHC состоящую из фрагментов сигнальной последовательности люциферазы (Cypridina noctiluca) и фиброина (Dendrolimus spectabilis).
Пример 2. Описание mPHK-RBD
Синтез мРНК кодирующей RBD SARS-COV-2 проводили с линеаризованной и очищенной ДНК-матрицы pVAX-RBD, имеющей нуклеотидную последовательность SEQ ID NO: 1, а также физическую и генетическую карту, представленную на фиг. 1, которая содержит промотор РНК-полимеразы бактериофага Т7 и целевой ген RBD. Для конструирования плазмиды использовали фрагмент гена S, соответствующий RBD (320-542 а.о.) (GenBank MN908947). Оптимизацию кодонного состава последовательности проводили с помощью программы GeneOptimizer (hppts://www.thermofisher.com/ru/en/home/life-science/cloning/gene-synthesis/geneart-gene-synthesis/geneoptimizer.html). Для обеспечения эффективного транспорта синтезированного белка из клетки к 5'-концу гена был добавлен фрагмент, кодирующий сигнальную последовательность MMRTLILAVLLVYFCATVHC.
Реакцию транскрипции проводили с помощью РНК-полимеразы бактериофага Т7. Реакционная смесь включала в себя 1 мкг линеаризованной матрицы, Т7 полимеразу с буфером, смесь рибонуклеотидов, в которой уридин был заменен на псевдоуридин, ингибитор РНКаз и безнуклеазную воду. Смесь инкубировали в течение 2 часов при температуре 37°C. После инкубации в смесь добавляли 1 мкл ДНКазы, и инкубировали в течение 30 минут при температуре 37°C для разрушения ДНК матрицы. Анализ полученного продукта производили с использованием электрофореза в 2% агарозном геле. Кэпирование и полиаденилирование РНК проводили с использованием соответствующих ферментов. Кэпирование РНК проводили с помощью наборов ScriptCap™ m7G CappingSystem (CellScript, США) по инструкции производителя. Полиаденилирование РНК проводили с использованием набора A-Plus™ Poly(A) PolymeraseTailingKit (CellScript, США), как это рекомендовано производителем.
В результате, была получена мРНК имеющая нуклеотидную последовательность SEQ ID NO: 2, которая была способна обеспечить синтез в эукаритической клетке целевого белка RBD (аминокислотная последовательность SEQ ID NO: 3).
Пример 3. Получение и характеризация комплексов мРНК-RBD с конъюгатом полиглюкин-спермидин
3.1. Получение мРНК-RBD. Схематическая структура mPHK-RBD представлена на фиг. 2. мРНК была получены с использованием в качестве матриц линеаризованных плазмид pVAX-RBD и полимеразы Т7. Кэпирование и полиаденилирование РНК проводили с использованием соответствующих ферментов (см. материалы и методы). Об уровне полиаденилирования судили по изменению подвижности РНК с помощью электрофореза в 2% агарозном геле. Полученная РНК сравнивалась с отрицательным контролем без полиаденилирования, наблюдалась разница в полосах от 100 до 200 нуклеотидов.
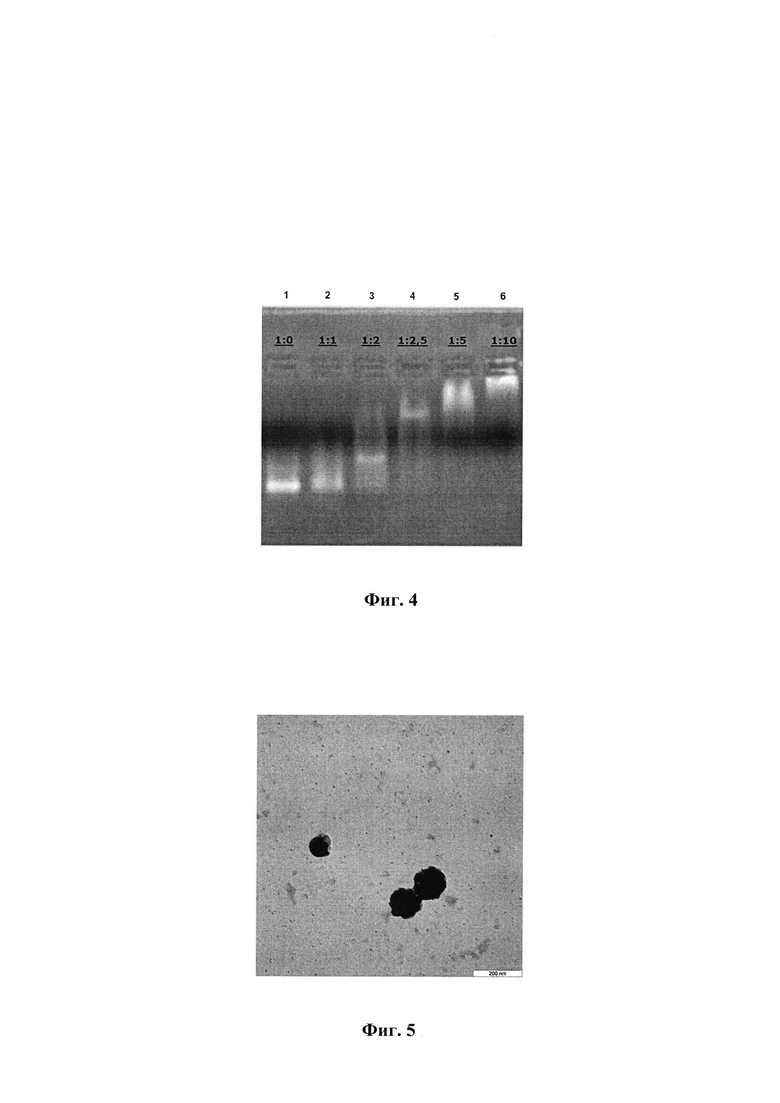
3.2. Получение и характеризация конъюгата полиглюкина со спермидином (PGS). Схема получения конъюгата полиглюкин : спермидин (PGS) представлена на фиг. 3. PGS получали путем конъюгирования полиглюкина (декстран 40000) и спермидина. Декстран 40000 (MP Biomedicals™, США) обрабатывали периодатом натрия (из расчета 50 молекул периодата натрия на 1 молекулу декстрана) в течение 60 мин в Н2О, а затем проводили гель-фильтрацию на колонке Sephadex G-25 в 50 мМ карбонатном буфере (рН 8,8). Процесс активации декстрана контролировали по количеству образовавшихся альдегидных групп, которые определяли фотоколориметрическим методом [Оболенская А.В. Лабораторные работы по химии древесины и целлюлозы. / А.В. Оболенская, З.П. Ельницкая, А.А. Леонович // Учебное пособие для вузов. - М.: «Экология», - 1991. - 320 с.]. Количество альдегидных групп составляло 36-40 на одну молекулу декстрана.
К активированному декстрану добавляли раствор спермидина (Sigma, США) из расчета 15 молекул спермидина на одну молекулу декстрана в 50 мМ карбонатном буфере (рН 8,8), смесь инкубировали 4 ч при температуре (22±2)°C. Затем добавляли боргидрид натрия из расчета 80 молекул боргидрида на 1 молекулу полиглюкина и инкубировали в течение одного часа при температуре (22±2)°C. Непрореагировавшие компоненты удаляли гель-фильтрацией на Sephadex G-25.
3.3. Получение и характеризация комплекса mRNA-PGS.
Схематичное изображение получения комплексов РНК и PGS представлено на фиг. 3. Для формирования комплексов РНК смешивали с PGS в определенных зарядовых соотношениях.
Для формирования комплексов для иммунизации животных мРНК смешивали с PGS в зарядовом соотношении 1:5, что соответствует соотношению по массе 1:15. Смешивали 100 мкл раствора, с концентрацией 0,06 мг/мл мРНК-RBD, с 100 мкл раствора, содержащего 0,93 мг/мл PGS, осторожно перемешивали в течение 2-3 сек., затем смесь инкубировали при комнатной температуре в течение часа. Комплексы mRNA-PGS анализировали с помощью электрофореза в 2% агарозном геле.
Образование комплексов mPHK:PGS было исследовано для определения соотношений устойчивой ассоциации между полимером и мРНК, а также биофизических характеристик комплексов. Комплексообразование оценивали с использованием отношения заряда PGS/мРНК (CR), которое связано с молярным отношением PGS/мРНК (MR). CR представляет собой отношение общего положительного заряда полимера (положительно заряженных периферических аминогрупп в катионном полимере) к общему отрицательному заряду мРНК (фосфатных групп) в образце, которое рассчитывается следующим образом:
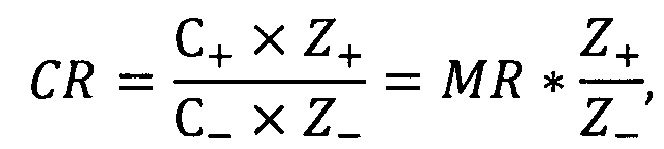
где С+ - итоговая концентрация катионного полимера в образце, С_ - итоговая концентрация нуклеиновой кислоты в образце, Z+ - заряд единичной молекулы катионного полимера, Z- - заряд единичной молекулы нуклеиновой кислоты (Ihnatsyeu-Kachan et al, 2017).
На основе заряда РНК, который соответствует ее длине и нуклеотидному составу, рассчитывали молярную массу полимера, необходимую для нейтрализации отрицательного заряда нуклеиновой кислоты, с учетом того, что на 1 молекулу PGS приходится 45 катионов. Для подбора оптимального соотношения mPHK:PGS для образования комплексов получали серию проб с разными зарядовыми соотношениями (N:P) РНК к полимеру, где (N) отрицательный заряд РНК, а (Р) положительный заряд катионного полимера.
Были взяты следующие N:P PHK:PGS - 1:1, 1:2, 1:2,5, 1:5 и 1:10, что соответствует соотношению по массе 1:3, 1:6, 1:8, 1:15 и 1:30. Так, например, в соотношении по заряду 1:1, брали 1 мкг РНК и смешивали с 3 мкг катионного полимера в равных объемах.
Первоначальной оценкой покрытия мРНК PGS служила подвижность mRNA-PGS в электрическом поле в ходе электрофореза в 2% агарозном геле, степень которой зависит от степени покрытия. На фиг. 4 видно, что при увеличении N:P наблюдается снижение электрофоретической подвижности комплексов мРНК-PGS в геле, что косвенно свидетельствует о формировании комплексов.
Для подтверждения формирования наночастиц была проведена электронная микроскопия (фиг. 5). Размер частиц комплекса mRNA-RBD-PGS при N:P 1:5 составлял 100-200 нм, что соответствует среднему размеру вируса.
Пример 4. Проверка иммуногенных свойств комплексов mPHK:PGS Иммунизация лабораторных животных, оценка вирус-специфических антител.
Работа с животными проводилась в соответствии с «Руководством по уходу и использованию лабораторных животных». Протоколы были одобрены Комитетом по уходу и использованию животных (IACUC) при Государственном научном центре вирусологии и биотехнологии «Вектор» (номер разрешения: НИЦ ВБ «Вектор» / 10.09.2020). Для оценки иммуногенности мРНК-RBD-PGS использовали самок мышей BALB/c массой 16-18 г. Мышей разделили на следующие 4 группы, каждая из которых состояла из 6 животных: Группа 1 (мРНК-RBD-PGS) - мыши, иммунизированные mPHK-RBD-PGS (10 мкг РНК /100 мкл физиологического раствора); Группа 2 (мРНК-RBD) - мыши, иммунизированные голой мРНК-RBD (10 мкг РНК / 100 мкл физиологического раствора); Группа 3 (mPHK-GFP-PGS) - мыши, иммунизированные mPHK-GFP-PGS (10 мкг РНК /100 мкл физиологического раствора); Группа 4 (контроль) - мыши, иммунизированные физиологическим раствором (100 мкл). Мышей иммунизировали трижды внутримышечно (в верхнюю часть бедра задней конечности) в дни 0, 21 и 35. В дни 34 и 48 у мышей брали кровь для анализа сыворотки. Сыворотки отделяли от клеточных элементов центрифугированием (10000 об / мин, 15 минут), нагревали для инактивации системы комплемента (30 минут, 56°C).
ИФА сыворотки. В качестве антигенов для ELISA использовали эукариотические белки RBD и S-тример, любезно предоставленные лабораторией иммунохимии ФБУН ГНЦ ВБ Вектор Роспотребнадзора (Россия). Белок RBD (1 мкг / мл) и S-тример (1 мкг / мл) адсорбировали в лунках 96-луночного планшета в PBS (Greiner_bio one, Германия) при 4°C в течение 12 часов, затем промывали PBST и блокировали с 1% раствором казеина в промывочном буфере в течение 60 минут при комнатной температуре. После этого образцы сыворотки добавляли в трехкратном серийном разведении, начиная с 1:50, и инкубировали в течение 60 минут при 37°C. После промывания добавляли кроличьи антитела против мышиного IgG, конъюгированные с пероксидазой хрена (Sigma, США), и инкубировали в течение 60 минут при 37°C. Планшет промывали и добавляли к нему раствор субстрата ТМВ (Amresco, США). После остановки реакции стоп-раствором оптическую плотность измеряли при длине волны 450 нм с помощью считывающего устройства для ELISA (ChroMate Awareness Technology inc., США). Графики построены с использованием GraphPad Prism 6.0 и Excel 2016. Нейтрализующая активность иммунных сывороток. В работе использован штамм коронавируса 2019-nCoV nCoV / Victoria / 1/2020 (Государственная коллекция возбудителей вирусных инфекций и риккетсиозов ФБУН, ГНЦ БВ «Вектор» Роспотребнадзора, РФ). Рабочая доза вируса составила 100 CPD50. Для оценки эффективности нейтрализующей активности сывороток готовили их серийные двукратные разведения, начиная с разведения от 1:10 до 1:1000. Для разведения сывороток использовали среду MEM с глутамином и добавлением антибиотиков (пенициллин 100 ЕД / мл, стрептомицин-100 мкг / мл). Смесь разведений сыворотки и рабочего разведения вируса готовили в равных объемах. Смесь инкубировали 1 ч при комнатной температуре, затем добавляли в лунки 96-луночного планшета с монослоем культуры клеток Vero Е6. Чашки инкубировали в течение 4 дней при 37°C, 5% CO2. Для окрашивания клеток в каждую лунку планшета добавляли 150 мкл 0,2% раствора генцианвиолета (добавляли 1 г генцианвиолета в 20 мл 96% этанола, 120 мл 40% формалина и 350 мл раствора Хенкса). Через 30 минут жидкость из лунок удалили и промыли водой. Результаты регистрировались визуально. Любое специфическое повреждение клеточной культуры в лунке считалось цитопатическим эффектом вируса (CPV). Нейтрализующую активность сывороток иммунизированных животных оценивали по титру (разведению) сывороток, при котором регистрировали защиту в 50% лунок с культурой клеток от цитопатического действия вируса. Титр нейтрализующих антител рассчитывали по формуле Рида-Мюнха [Reed, L.J.; Muench, Н.A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493-197, doi:10.1093/oxfordjournals.aje.a118408.].
Для оценки иммуногенности вакцинных конструкций мРНК-RBD-PGS и мРНК-RBD мышей иммунизировали трижды в дни 0, 21 и 35. Через пять и семь недель после начала иммунизации у мышей брали сыворотку, которую были протестированы с помощью ELISA на наличие антител, специфически распознающих RBD и S-тример, а также была исследована способность сывороток нейтрализовать вирус. В качестве контролей использовали сыворотку от мышей, иммунизированных мРНК-GFP-PGS, и физиологический раствор.
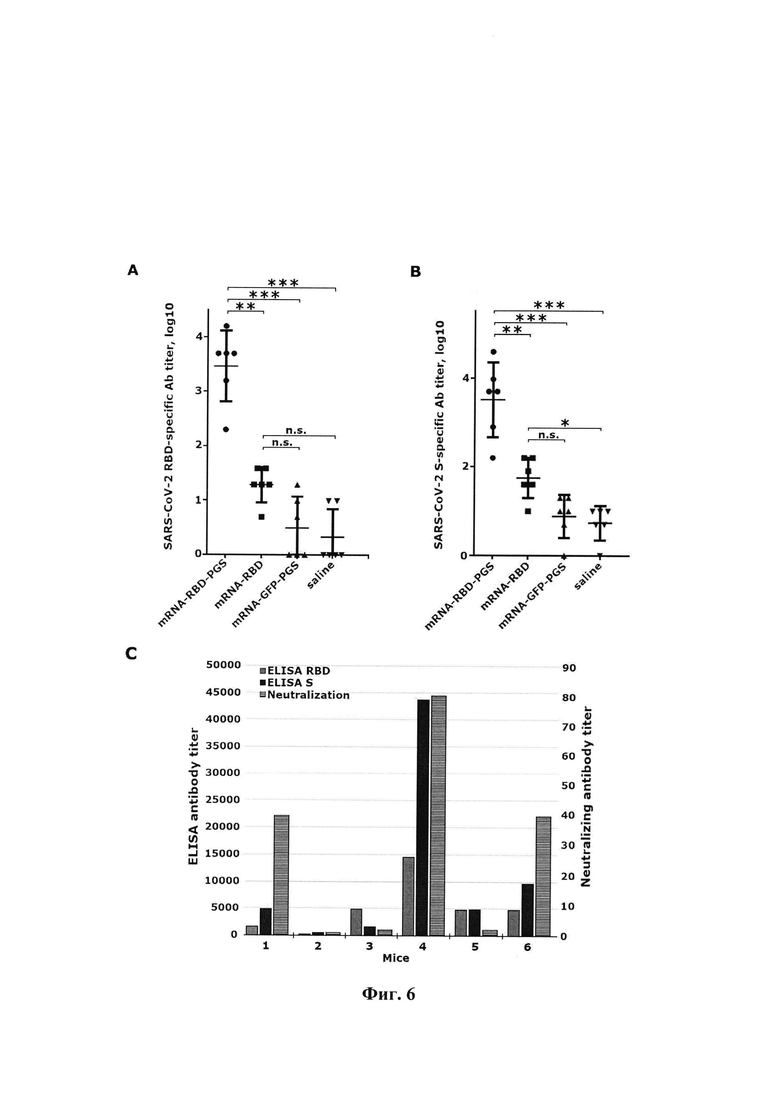
Данные ELISA представлены на фиг. 6 (гуморальные ответы на мышах BALB/C, иммунизированных антигеном SARS-CoV-2). Мышей иммунизировали внутримышечно голой мРНК-RBD, комплексами мРНК с конъюгатом полиглюкин : спермидин (мРНК-RBD-PGS или mPHK-GFP-PGS) или физиологический раствор трижды через 0, 21 и 35 дней (10 мкг мРНК на мышь). Образцы сыворотки крови мышей собирали через 48 дней после первой иммунизации. (А) Титры специфических IgG антител к SARS-CoV-2 RBD и (В) титры специфических антител IgG к SARS-CoV-2 S определяли с помощью ELISA. Статистический анализ проводился с использованием программного обеспечения GraphPad Prism 6.0. Данные представлены как среднее значение ± стандартная ошибка среднего. Достоверность рассчитывалась с использованием двухфакторного дисперсионного анализа с множественными сравнительными тестами (n.s., статистически недостоверно; * р<0,05, ** р<0,01, *** р<0,001). (С) Данные ELISA и вируснейтрализующей активности сывороток мышей, иммунизированных мРНК-RBD-PGS. Ось абсцисс: 1-6-число мыши. Левая ось ординат представляет собой обратный титр антител, полученный с помощью ELISA, правая ось ординат - обратное значение титра нейтрализующей сыворотки: серая полоса - специфические антитела к RBD, белая полоса - специфические антитела против RBD, черная полоса - вирус нейтрализующие антитела.
Данные на фиг. 6 показали, что через две недели после второй иммунизации наблюдалось небольшое увеличение титра специфических антител, но не было обнаружено значительных различий с группами отрицательного контроля (данные не показаны). Через две недели после третьей иммунизации в группе мРНК-RBD-PGS наблюдалось значительное увеличение (в 100-1000 раз) титра специфических антител как к RBD, так и к S-тримеру по сравнению с сыворотками мышей, иммунизированных мРНК.-RBD, мРНК-GFP-PGS и физиологический раствор (фиг. 6А, В).
Анализ нейтрализующей активности сывороток проводили с использованием живого вируса SARS-CoV-2 штамма nCoV / Victoria / 1/2020 и культуры клеток Vero Е6. Антитела, нейтрализующие вирус, были обнаружены только в группе животных, иммунизированных мРНК-RBD-PGS, а сыворотка животных с высокими титрами антител против RBD обладала вируснейтрализующей активностью (фиг. 6, С).
Пример 5. Исследование цитотоксичности комплексов mPHK-RBD-PGS. Анализ жизнеспособности клеток (МТТ)
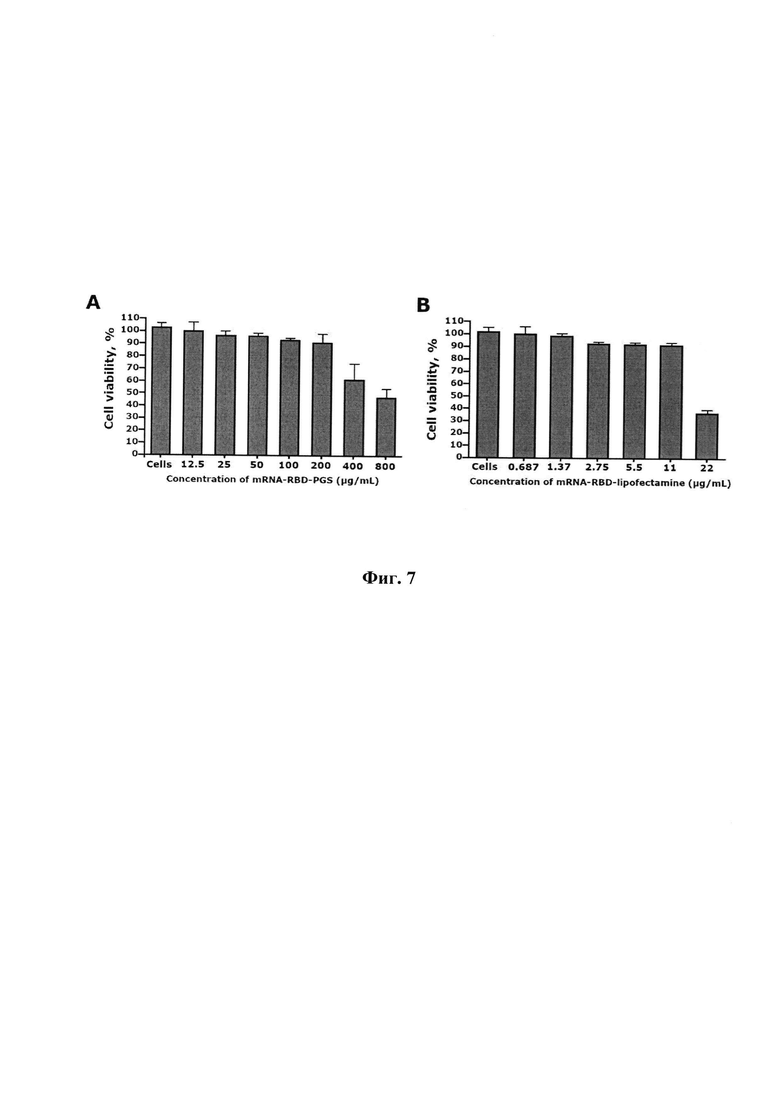
Цитотоксичность комплексов мРНК-RBD-PGS и mPHK-RBD-липофектамин 3000 оценивали с помощью МТТ-теста. Клетки НЕК293 (1×105 клеток / мл) в 100 мкл культуральной среды высевали в 96-луночные планшеты и инкубировали в течение ночи. На следующий день к клеткам добавляли комплексы мРНК-RBD-PGS и MPHK-RBD-липофектамин 3000 и инкубировали в течение 72 часов. После 72 часов инкубации клеток с тестируемыми комплексами в каждую лунку добавляли 20 мкл раствора МТТ (5 мг / мл) и инкубировали в течение 2 часов в CO2-инкубаторе. После инкубации среду отсасывали и добавляли диметилсульфоксид (50 мкл / лунку), чтобы остановить реакцию. Оптическую плотность определяли на микропланшетном ридере Varioskan LUX (Thermo Fisher Scientific, США) при длине волны 570 нм. Жизнеспособность клеток в присутствии комплексов рассчитывали по формуле: (оптическая плотность тестовых лунок / оптическая плотность контрольных лунок) × 100%.
Исследовалось влияние комплексов мРНК-RBD-PGS и липосом мРИК-RBD-липофектамин 3000 на жизнеспособность клеток FIEK293T с помощью анализа МТТ. Тест МТТ показал, что при инкубации клеток HEK293 с комплексами mPFIK-RBD-PGS в диапазоне концентраций от 800 до 12,5 мкг / мл доля жизнеспособных клеток относительно контроля составляла от 51,3±12,8% до 98,5±3,3% соответственно (фиг. 7, А). Контроль необработанных клеток HEK293 составил 99,1±2,4%. Когда мРНК-RBD-Lipofectamine 3000 добавляли к культуре клеток в диапазоне концентраций от 22 до 0,687 мкг / мл, процент жизнеспособности клеток составлял от 36,8±2,6 до 99,9±4,8% соответственно (фиг. 7, В). Значения СС50 мРНК-RBD-PGS и mPHK-RBD-Lipofectamine 3000 против клеток HEK293 составляли 625 мкг / мл и 20,4 мкг / мл соответственно.
Таким образом, заявляемый комплекс в виде наночастиц, содержащих молекулы мРНК-RBD, покрытых конъюгатом полиглюкин-спермидин, индуцируют SARS-CoV-специфические антитела, обладают вируснейтрализующей активностью и являются менее токсичными, чем липосомальные формы мРНК-RBD в прототипе.
Приложение



Изобретение относится к биотехнологии и генетической инженерии. Создана плазмидная ДНК-матрица pVAX-RBD, имеющая нуклеотидную последовательность SEQ ID NO: 1 размером 3710 п.н., молекулярную массу 2.6×103 кДа, содержащая целевой ген, кодирующий химерный белок 176-RBD, перед которым находится промотор РНК-полимеразы фага Т7, обеспечивающий синтез in vitro мРНК-RBD с помощью РНК-полимеразы Т7. Молекула мРНК-RBD, имеющая нуклеотидную последовательность SEQ ID NO: 2, получена с использованием ДНК-матрицы pVAX-RBD, содержит открытую рамку считывания, кодирующую аминокислотную последовательность SEQ ID NO: 3, обеспечивающую синтез и секрецию белка-иммуногена RBD SARS-CoV 2 в организме млекопитающих, и содержащую гибридную сигнальную последовательность 176, консенсусную последовательность Козак, 7-метилгуанозиновый кэп на 5'-конце мРНК, полиаденозиновый (поли А) хвост на 3'-конце мРНК, длиной 150-200 нуклеотидов, и 5'- и 3'-нетранслируемые области (UTR) перед и после открытой рамки считывания. Комплекс в виде наночастиц, индуцирующих SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью, состоящих из молекул конъюгата полиглюкинхпермидин и молекул мРНК-RBD, взятых в разных зарядовых соотношениях от 1:1 до 1:30, соответствующих весовому соотношению от 1:3 до 1:90, имеющих размер в диапазоне 50-900 нанометров, причем каждая наночастица содержит ядро, состоящее из молекул мРНК-RBD и покрытое слоем конъюгата полиглюкинхпермидин, удерживаемого за счет ионного взаимодействия между отрицательно заряженным ядром и положительно заряженным спермидином. Техническим результатом является создание более эффективного средства доставки мРНК-RBD в клетки организма для синтеза в них белка RBD SARS-Cov-2 и повышение уровня секреции указанного белка из клеток для обеспечения презентации белка RBD иммунной системе организма. 3 н.п. ф-лы, 7 ил., 1 табл., 5 пр.
1. Плазмидная ДНК-матрица pVAX-RBD для получения молекулы мРНК-RBD, имеющая нуклеотидную последовательность SEQ ID NO: 1 размером 3710 п.н., молекулярную массу 2.6×103 кДа, содержащая целевой ген, кодирующий химерный белок 176-RBD, перед которым находится промотор РНК-полимеразы фага Т7, обеспечивающий синтез in vitro мРНК-RBD с помощью РНК-полимеразы Т7 и состоящая из следующих фрагментов:
- векторного фрагмента 1484-754 ДНК плазмиды pVAX размером 2981 п.н., содержащего промотор РНК-полимеразы фага Т7 (664-682, размером 19 п.н.);
- ген аминогликозид фосфотрансферазы из Tn5 (1937-2728 размером 702 п.н.),
- ген устойчивости к канамицину (NeoR/KanR) и точку начала репликации ColE1 origin (3057-3644 размером 588 п.н.), обеспечивающие селекцию и амплификацию целевой плазмиды в клетках бактерий Escherichia coli;
- фрагмент 755-1483 размером 729 п.н., содержащий искусственный ген 176-RBD с инициирующим кодоном ATG.
2. Молекула мРНК-RBD для получения комплекса в виде наночастиц, индуцирующих SARS-CoV 2-специфичные антитела, имеющая нуклеотидную последовательность SEQ ID NO: 2, полученная с использованием ДНК-матрицы pVAX-RBD по п.1, содержащая открытую рамку считывания, кодирующую аминокислотную последовательность SEQ ID NO: 3, обеспечивающую синтез и секрецию белка-иммуногена RBD SARS-CoV 2 в организме млекопитающих, и содержащую гибридную сигнальную последовательность 176, консенсусную последовательность Козак, 7-метилгуанозиновый кэп на 5'-конце мРНК, полиаденозиновый (поли А) хвост на 3'-конце мРНК, длиной 150-200 нуклеотидов, и 5'- и 3'-нетранслируемые области (UTR) перед и после открытой рамки считывания.
3. Комплекс в виде наночастиц, индуцирующих SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью, состоящих из молекул конъюгата полиглюкинхпермидин и молекул мРНК-RBD по п.2, взятых в разных зарядовых соотношениях от 1:1 до 1:30, соответствующих весовому соотношению от 1:3 до 1:90, имеющих размер в диапазоне 50-900 нанометров, причем каждая наночастица содержит ядро, состоящее из молекул мРНК-RBD и покрытое слоем конъюгата полиглюкин : спермидин, удерживаемого за счет ионного взаимодействия между отрицательно заряженным ядром и положительно заряженным спермидином.
| Искусственный ген Stbl_RBD_TrM_SC2, кодирующий бицистронную структуру, образованную последовательностями рецепторсвязывающего домена гликопротеина S коронавируса SARS-CoV-2, трансмембранного региона, P2A-пептида и гликопротеина G VSV, рекомбинантная плазмида pStem-rVSV-Stbl_RBD_TrM_SC2, обеспечивающая экспрессию искусственного гена, и рекомбинантный штамм вируса везикулярного стоматита rVSV-Stbl_RBD_TrM_SC2, используемый для создания вакцины против коронавируса SARS-CoV-2 | 2020 |
|
RU2733832C1 |
| Рекомбинантные ДНК pG4223 и плазмидная ДНК pQE 30-pG4223, штамм Escherichia coli M 15-G4223, обеспечивающие получение полипептида G4223, селективно связывающего IgG, и его применение в аффинной хроматографии для выделения IgG | 2017 |
|
RU2715672C2 |
| CN 111676248 A, 18.09.2020. | |||

